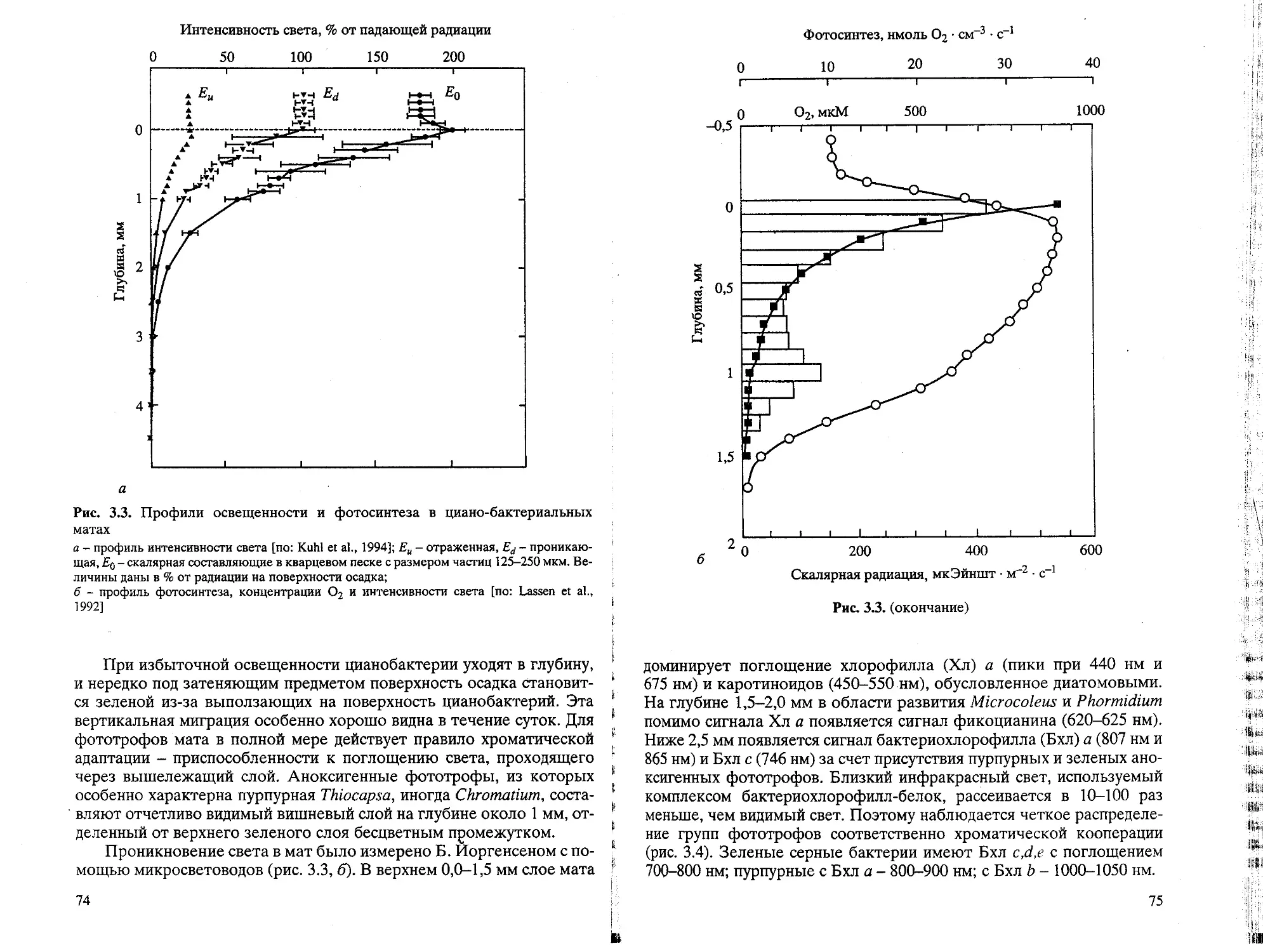
/































































































































































Author: Заварзин Г.А.
Tags: микробиология биология природоведческая микробиология
ISBN: 5-02-006454-8
Year: 2003




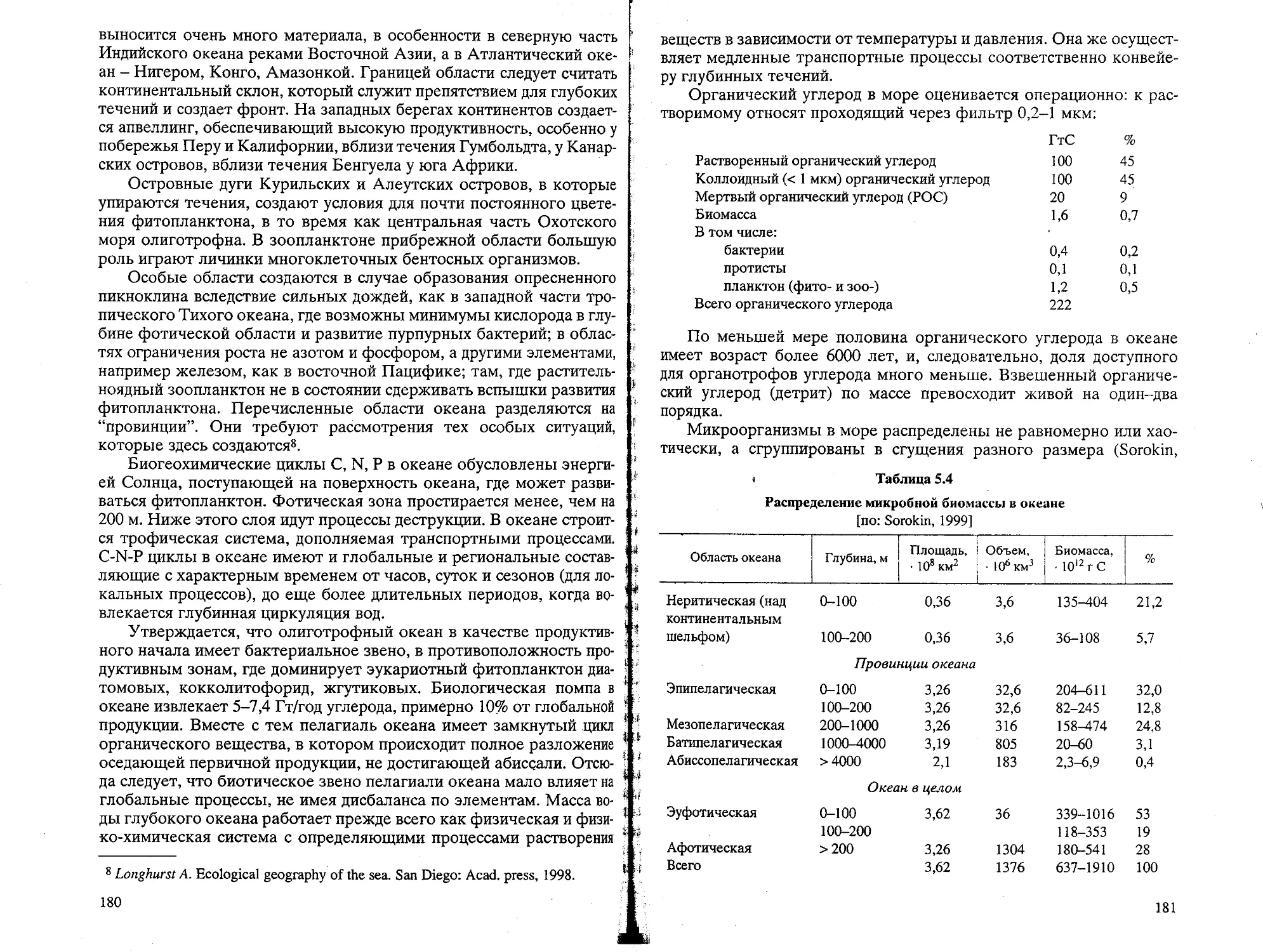
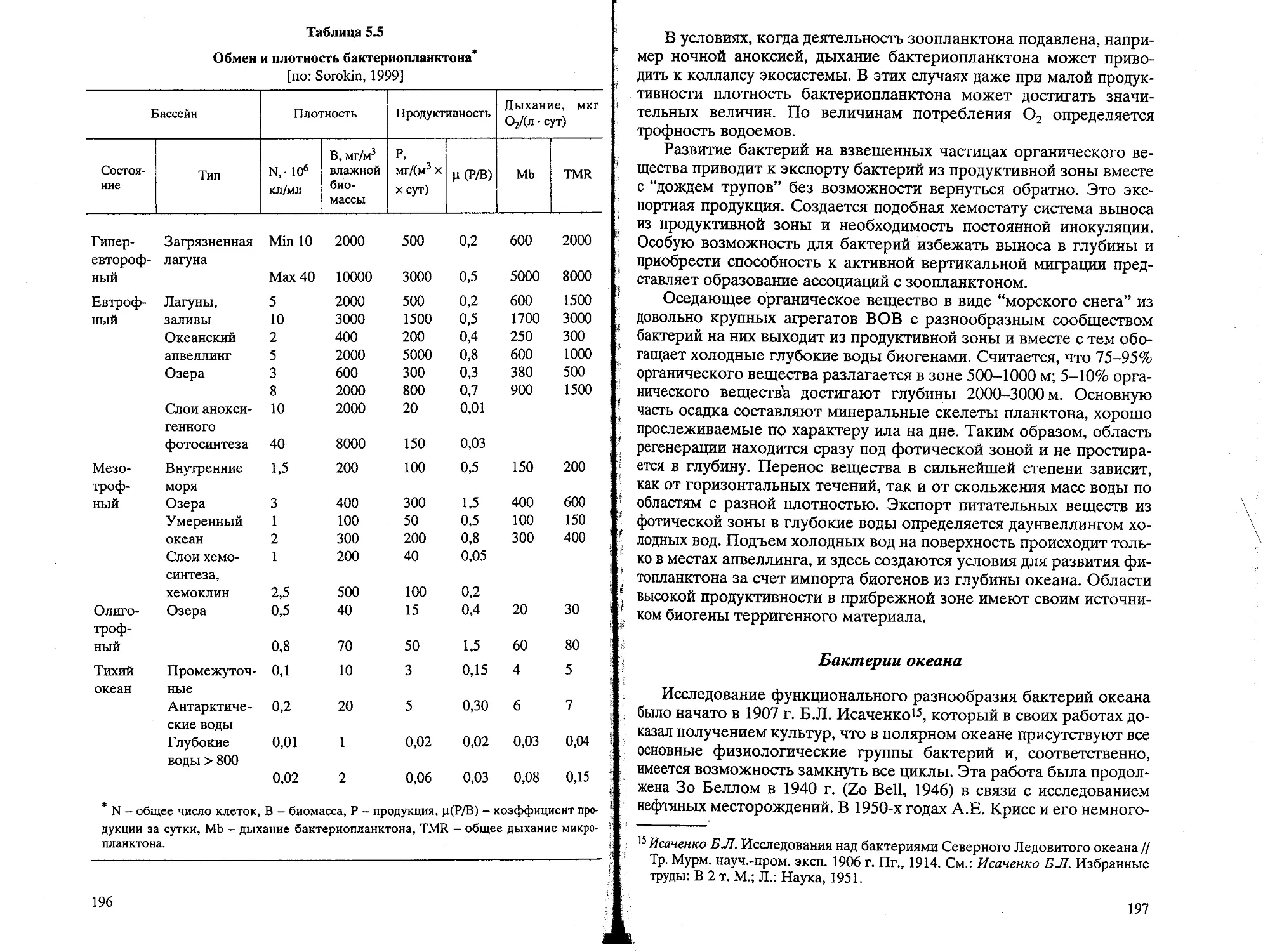
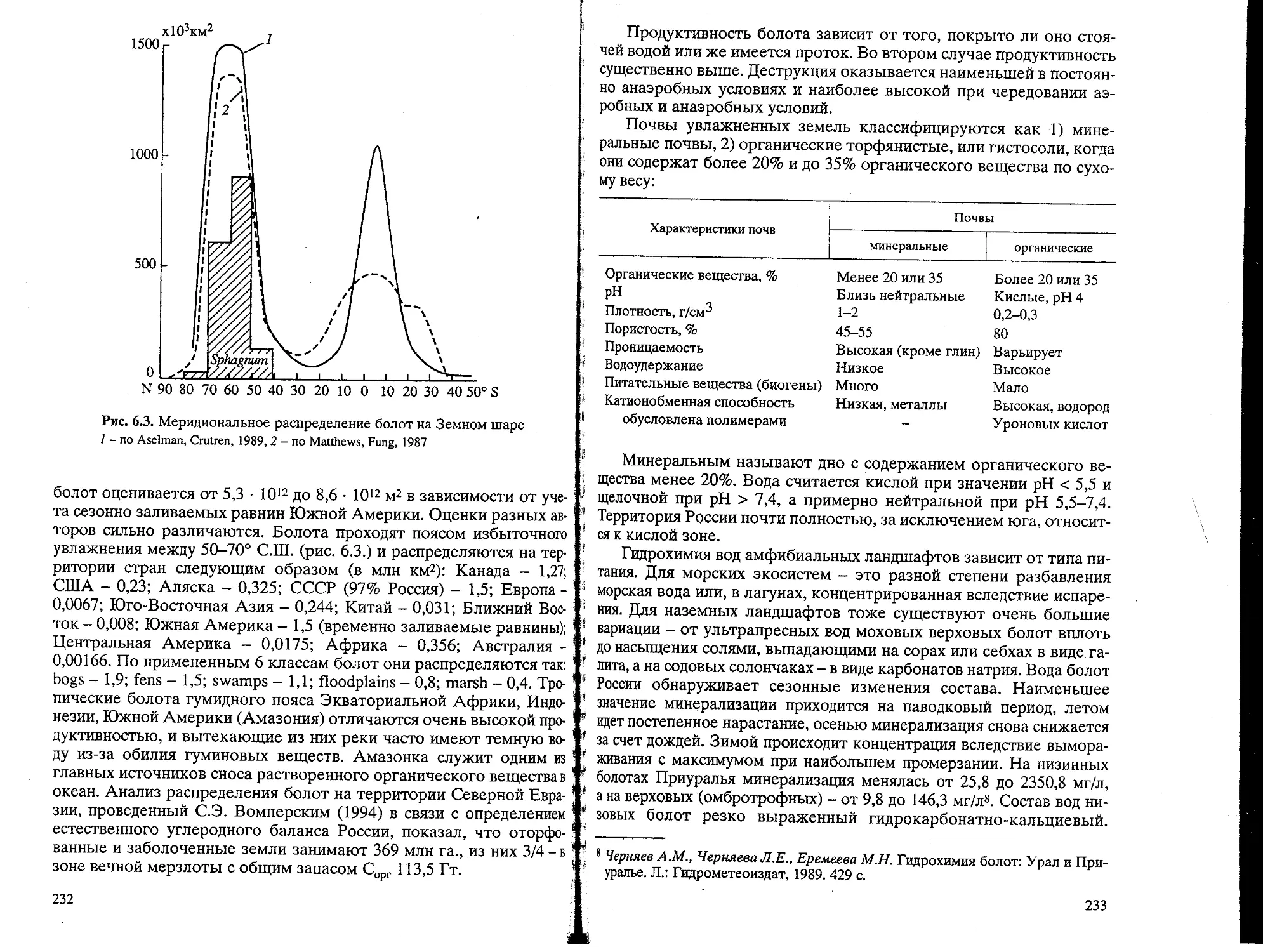
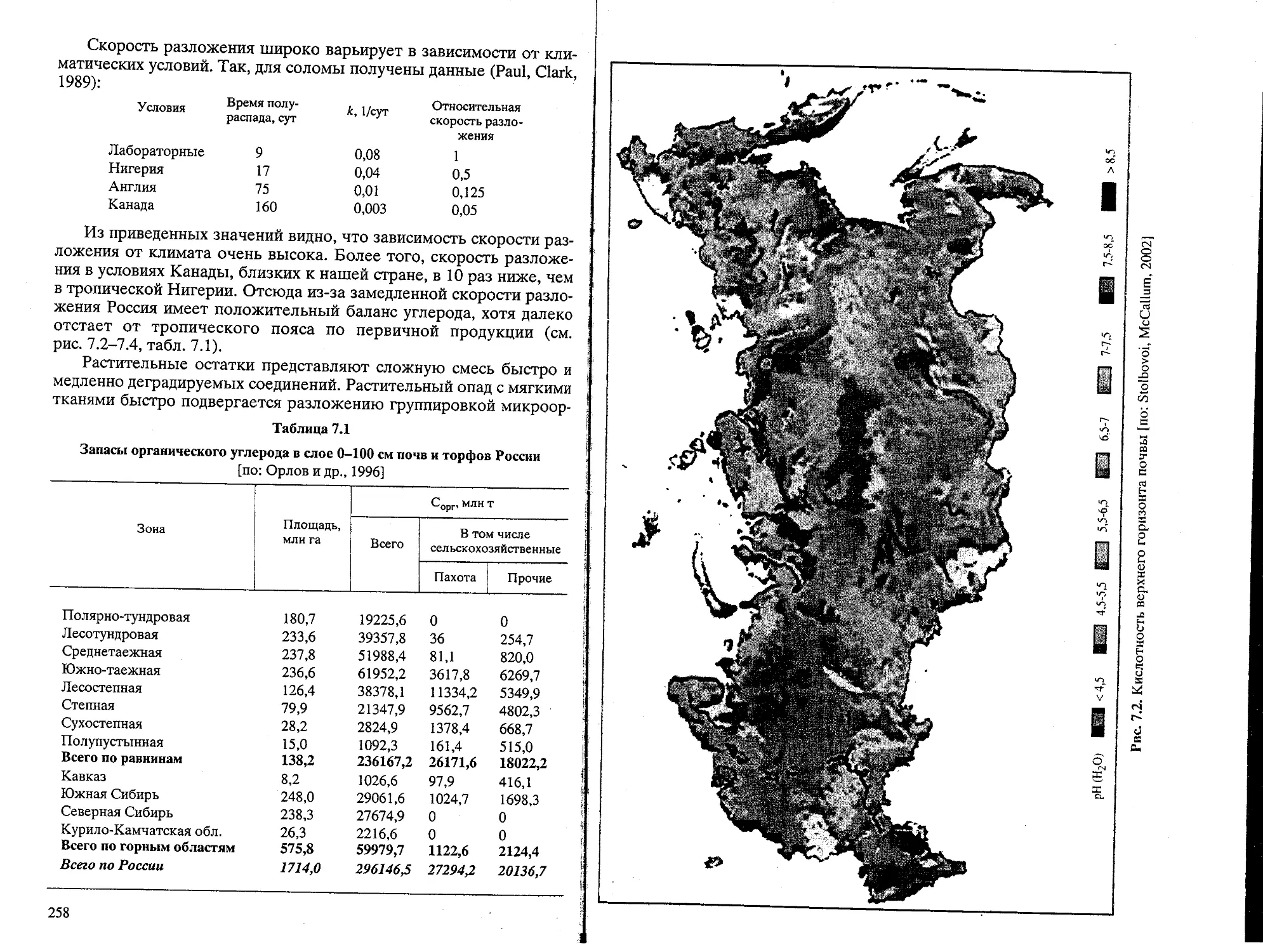
Text
ГА.ЗАВАРЗИН
УДК 579
ББК 28.4
313
Ответственный редактор
кандидат биологических наук Н.Н. Колотилова
Рецензенты:
член-корреспондент РАН А.Ю. Розанов,
доктор биологических наук В.М. Горленко
Заварзин Г.А.
Лекции по природоведческой микробиологии / Г.А. Заварзин; Отв.
ред. Н.Н. Колотилова; Ин-т микробиологии. - М.: Наука, 2003. - 348 с.
ISBN 5-02-006454-8 (в пер.)
Книга основана на курсе лекций, прочитанных в МГУ им. М.В. Ломоносова в
1995-2002 гг. Основное внимание уделено анализу природной среды обитания в рамках
больших биосферных систем. Бактерии — катализаторы биогеохимических циклов, сфор-
сформировали биосферу в течение первых двух третей ее существования и остаются основой
биогеохимической машины планеты.
Для широкого круга естествоиспытателей.
ТП-2003-1-88
ISBN 5-02-006454-8 (в пер.)
© Российская академия наук, 2003
© Издательство "Наука"
(художественное оформление), 2003
ПРЕДИСЛОВИЕ
Предлагаемая вниманию читателя книга основана на материале
лекций, которые автор в течение нескольких лет читал в МГУ
студентам-микробиологам 4-го курса. При этом выяснилось, что уг-
углубленное изучение физиологии и биохимии микроорганизмов на
основе чистых культур, дополненное геномикой и протеомикой, за-
заставляет студентов забывать о больших системах, в которых дейст-
действуют микроорганизмы. Для того чтобы вписать знания микробиоло-
микробиологов в понимание среды обитания микроорганизмов и вызываемых
их деятельностью изменений, приходилось открывать окно в боль-
большой мир геосферных и биосферных систем. Приходится говорить
о месте микробов в естествознании, а для этого хотя бы бегло рас-
рассказывать о природных системах. Главными оказались понятия о
географической оболочке Земли с ландшафтом как средой обита-
обитания микробных сообществ. Аутоэкология микроорганизмов оказа-
оказалась подчиненным разделом.
В центре изложения лежит идея о кооперативном микробном
сообществе, взаимодействующем со своей средой обитания на осно-
основе трофических связей, включающих химические трансформации
веществ, служащих прямым источником энергии для деятельности
микроорганизмов. Главное внимание уделено циклу органического
углерода с фотоавтотрофными продуцентами и органотрофными
деструкторами. Однако этот цикл вызывает множество сопряжен-
сопряженных реакций в геосфере, обозначаемых как биологически опосредо-
опосредованные. Эти реакции не необходимы для жизнедеятельности мик-
микробного сообщества и иногда даже вредны для него, вызывая
сукцессию со сменой сообществ, но без воздействия биоты они не
происходят. Задачей изложения было сформировать приоритеты
микробиологических исследований на основе иерархии природных
систем. В результате складывается мировоззрение, отличное от
того, которое мы находим в учебниках общей микробиологии.
Материал, касающийся организации сообщества, и необходи-
необходимые предварительные сведения о бактериальной клетке и разнооб-
разии микроорганизмов, изложены в кратком "Введении в природо-
природоведческую микробиологию" (Г.А. Заварзин, Н.Н. Колотилова; М.:
Книжный дом "Университет", 2001) и здесь не повторяются, хотя не-
необходимы для понимания.
Студенты очень многому научили меня, затрудняясь в ответах на
вопросы о выводах из прямых наблюдений в природе. Для выводов
необходимы хотя бы поверхностные, но широкие знания, позволяю-
позволяющие избежать ошибок, которые, к сожалению, часто возникают из-
за движения от частного к общему, от элемента к системе. Пожалуй,
в наибольшей степени на мое изложение здесь повлияли взгляды
совсем забытого нами А. Гумбольдта - в их современном преломле-
преломлении. Лекции более или менее самостоятельны, и поэтому были неиз-
неизбежные повторы. Мир микробов, представленный здесь, рассмат-
рассматривается в проекции на то время, когда на Земле не было высших
организмов. При этом выясняется, что этот мир был самодостато-
самодостаточен. Природоведческая микробиология оказалась необъятным
предметом, и многое осталось за рамками изложения. Упорядоче-
Упорядочение изложения заставило вынести за рамки книги почти весь мате-
материал по биогеохимическим циклам, катализируемым микробами.
Книга не могла бы быть издана, если бы не усилия ответствен-
ответственного редактора Н.Н. Колотиловой привести в порядок тексты моих
лекций, написанных в разное время и менявшихся, по мере того как
я расширял свои знания о предмете. Вклад редактора неоценим, и ее
сотрудничество было необходимым условием подготовки книги.
Лекция 1
БИОСФЕРА И МИКРОБЫ
1.1. БИОСФЕРНЫЙ ПОДХОД
Естествознание во второй половине XX в. развивалось в двух на-
направлениях. С одной стороны, все более углублялось понимание
элементарных процессов путем редукционизма, проявившееся в об-
области биологии в формировании молекулярной биологии и молеку-
молекулярной генетики, но, с другой стороны, росло сознание необходимо-
необходимости интегративного подхода, поскольку игнорирование системных за-
закономерностей прямо угрожает существованию как всего человече-
человечества в целом, так и отдельных государств и наций. Эмоциональной
причиной этого широко обсуждаемого изменения в приоритетах,
обозначаемого как смена парадигмы, стало всеобщее осознание ог-
ограниченности земного пространства, обусловленное развитием воз-
воздушного транспорта и возможностью наблюдения Земли из космоса.
Рост в ограниченном пространстве ведет к исчерпанию ресурсов и
гибели популяции подобно тому, как это происходит в культуре ми-
микроорганизмов. Особенно быстро исчерпание ресурсов происходит
в связи с экспоненциальным ростом, который создает впечатление
порогового перехода. На основе этих представлений были разрабо-
разработаны математические модели, претендовавшие на прогностическое
значение и, во всяком случае, сумевшие определить порядок при-
приоритетов.
В ответ на беспокойство, обусловленное представлением об ог-
ограниченности экологических ресурсов, в которые попадало общече-
общечеловеческое достояние, была разработана отражавшая идеологию
системного анализа международная геосферно-биосферная про-
программа, целью которой стало, по сути, возрождение естествознания
как науки, изучающей именно природу, а не понятие "окружающей
среды" или "среды обитания". Системный подход потребовал иных
подходов и приоритетов, чем молекулярная биология, генетика,
идеология выживания наиболее приспособленных. Возникло
множество морально-философских рассуждений на тему "эко-
"экология", включая "экологию сознания", полностью вытеснивших
первоначальный биологический смысл понятия и заменивших его
Редукционизм
1896 \ С. Н. Виноградский
Системный
подход
1859 / Ч. Дарвин
1799
1738
К. Линней
Рис 1.1. Преобладание редукционистского и системного подхода в истории
естествознания
Имена выдающихся выразителей тенденций выбраны условно для обозначения периодов
перехода
экодемагогией. Именно поэтому в лекциях по природоведческой ми-
микробиологии термин "экология" употребляется в значении "аутэко-
логия".
Основным орудием новой эпохи стала возможность компъютер-
ной обработки очень больших баз данных и анализа очень больших
систем уравнений. При этом оказалось, что иногда изменение в от-
относительно малом или, казалось бы, второстепенном параметре
сильно влияет на систему в целом. Отсюда возникла необходимость
в целостном подходе к природе, а не в мозаичном.
Не следует думать, что переход от редукционизма к интегратиз-
му (или холизму) как способу познания окружающего мира представ-
представляет особенность конца XX в. На самом деле, между крайними
взглядами всегда были колебания (рис. 1.1). Замечательным иссле-
исследователем, хорошо владевшим суммой знаний своего времени, был
в конце XVIII - начале XIX в. Александр Гумбольдт, написавший
эмоциональные "Ansichten der Natur" ("Картинки с природы") по
впечатлениям от своих путешествий, которые затем, в конце его
жизни, вылились в книгу "Космос" - величественную картину приро-
природы как единой системы. Начальные посылки А. Гумбольдта осно-
основывались на сознательном противопоставлении изучения системы
одновременно взаимодействующих явлений природы описанию мно-
множества видов, систематизированному за полвека до него К. Линнеем.
Именно системному мировоззрению, составляющему суть естество-
естествознания, следует современный исследователь глобальных изменений
природной среды. Но современный профессионал знает много того,
что неизвестно простому наблюдателю, и это знание позволяет ему
связать проявления природы в систему и видеть "Солнце в малой ка-
капле вод", как писал Державин в оде "Бог", отражавшей целостное
религиозное мировоззрение XVIII в. Взгляды Гумбольдта на единую
систему Земли с сильным воздействием климата на живой мир, от-
отразившиеся прежде всего в биогеографии, суть которой составляет
зональное распределение растительного покрова в зависимости от
климатических зон, во второй половине XIX в. уступили место исто-
истории происхождения (филогении) как единственно заслуживающему
внимания научному объяснению явлений природы. Система уступи-
уступила место хаосу случайных изменений, самопроизвольно приводяще-
приводящему к порядку, в котором выживают наиболее приспособленные.
Замечательная по своей логичности и отсутствию необходимости
каких бы то ни было внешних сил для достижения устойчивого со-
состояния теория Ч. Дарвина стала не только биологической теорией,
но и концепцией в мировоззрении, давшей универсальное объясне-
объяснение существующему введением понятия естественного отбора.
Но она только косвенно отвечала на вопрос, поставленный А. Гум-
Гумбольдтом, заменив взаимодействие происхождением.
Геосферно-биосферные взаимодействия с акцентом на баланс
элементов широко изучались В.И. Вернадским и позволили ему
сформулировать понятие "Биосферы" как взаимодействующей сис-
системы, понимание которой может быть достигнуто через понимание
биогеохимических циклов. Основной мерой стало количество ато-
атомов вещества, находящихся в составе определенных соединений и
пород. В продолжение этой линии рассуждения было введено мета-
метафорическое понятие "живого вещества". В биосферной концепции
большое внимание уделялось влиянию биоты на поверхностные
оболочки Земли, главным образом за счет концентрационной функ-
функции организмов. Изменение масштаба исследований вернуло тео-
теорию от индивидуальных изменений к рассмотрению целостной сис-
системы в рамках космизма.
В последней трети XX в. интерес вызвала концепция "Геи",
предложенная англичанином Дж. Ловелоком почти в виде научного
памфлета, в котором планета Земля рассматривалась как единая си-
стема, как "живой организм", способный приспосабливать окружа-
окружающую среду к своему существованию в ней, превращая ее в свою
среду обитания. Суть этой концепции составила гипотеза регуляции
климата живыми организмами за счет обратных связей в атмосфер-
но-биосферной системе. Гипотеза Геи привлекла особое внимание в
связи с угрозой глобального потепления из-за вызванного челове-
человеком изменения состава атмосферы.
Во всех этих научных течениях на втором плане оставался "мир
невидимых". Целостная биосферная концепция В.И. Вернадского,
основанная на биогеохимических циклах, странным образом обош-
обошла катализаторов этих циклов, хотя важная роль микроорганизмов
и декларировалась им. Простой и ясный взгляд С.Н. Виноградского
на природу и роль в ней микробного мира как "ферментов" (в пер-
первоначальном французском значении слова) остался невостребо-
невостребованным.
Анализ упомянутых выше общих проблем естествознания при-
привел нас к выводу, что деятельность микроорганизмов не служит про-
простым дополнением к выработанной геохимиками системе взглядов.
Напротив, в концептуальном отношении бактерии оказываются ос-
основным двигателем биосферной системы биогеохимических циклов,
катализируя их ключевые реакции. В отношении историческо-
исторического аспекта, объясняющего состояние природы через историю воз-
возникновения ее комподентов, микробиология имеет особое значение
для естествознания, потому что микробы были первыми обитателя-
обитателями Земли и они сформировали ту биогеохимическую систему, кото-
которая осталась основой процессов, происходящих на поверхности Зем-
Земли. Исходным послужил цикл органического углерода и сопряженных
с ним циклов других элементов. Бактерии сформировали и продук-
продукционную фотосинтетическую ветвь цикла, и деструкционную,
сопряженную с циклами других элементов. Эта система была перво-
первоначальной и обусловила устойчивое развитие биосферы, не исклю-
исключающее катастрофические сукцессионные перестройки. Последую-
Последующие формы эволюционно вписывались в уже существующую систему
и лишь затем трансформировали ее.
Чтобы обозначить область знания, которая рассматривается ни-
ниже, удобно ввести термин "природоведческая микробиология". Она
входит в естествознание, возврат к которому как синтезу знаний о
природе необходим для человечества. Термин представляет кальку с
немецкого Naturwissenschaftlichen Mikrobiologie, понимая естество-
естествознание в тех рамках, как оно употреблялось в прошлом в смысле
изучения скорее "природы", чем "естества", и противопоставляя его
"natural history", как это декларировал в 1799 г. А. Гумбольдт. У нас
слово "естествознание" заменили в школьных учебниках "природо-
"природоведением", понимая под природой нечто противоположное лабора-
лаборатории. В литературе, посвященной юридическим аспектам эколо-
экологии, употребляются латинские термины "in situ" и "ex situ", обозна-
обозначающие соответственно "на месте" (в смысле "в местообитании") и
"вне места" (в смысле "в лаборатории"). В более широком смысле тер-
термины обозначают изучение объекта в составе большей системы или
вне ее. Деятельность микроорганизмов, преимущественно прокари-
отных, изучает общая микробиология, противопоставляемая
медицинской и другим специальным областям. В общую микробио-
микробиологию традиционно входит изучение разнообразия бактерий, преж-
прежде всего физиологического. К ней пытаются также относить те
общебиологические процессы, которые изучаются на примере бак-
бактерий как моделей живого существа и на самом деле относятся к об-
области биохимии, энзимологии, молекулярной биологии - короче,
дисциплин, изучающих внутриклеточные процессы, и которые, не-
несомненно, нужно знать микробиологу. Критическим для микробио-
микробиологии стало развитие геномики, позволившей изучать геном как
"паспорт" организма и отдельные гены вне их реализации в актив-
активности in situ.
В природоведческой микробиологии изучаются функциональ-
функциональное разнообразие микроорганизмов с аутэкологией видов, их взаи-
взаимодействие в составе микробных сообществ, деятельность микроб-
микробных сообществ в составе биосферы. Для последнего аспекта нами
был ранее предложен условный термин "глобальная микробиоло-
микробиология", который можно было бы заменить термином "биосферная
микробиология", не меняя сути рассматриваемых предметов.
Угол зрения в природоведческой микробиологии оказывается
существенно отличным от того исследования бинарных отноше-
отношений паразит-хозяин, которое доминирует в медицинской микро-
микробиологии, или концентрации внимания на одном объекте, которо-
которого требует биотехнология. Природоведческая микробиология
поневоле оказывается под сильным влиянием междисциплинарных
аспектов, прежде всего проистекающих из комплекса наук о
Земле.
Перед обсуждением деятельности микроорганизмов в природе
нужно рассмотреть те аспекты геосферно-биосферной системы, ко-
которые важны для понимания взаимодействий. К сожалению, одним
из явных пробелов в знаниях микробиологов (да и других биологов-
"экспериментаторов") оказывается география с ее центральным по-
понятием - ландшафтом (акваторией). Поэтому приходится давать
краткие пояснения, которые покажутся излишними естествоиспы-
естествоиспытателям-натуралистам. В географии большое значение имеет на-
наблюдение и сравнительный анализ явлений в противоположность
эксперименту. К сожалению, экспериментаторы, относя наблюде-
наблюдение к описательным дисциплинам, забывают, что именно наблюде-
наблюдение нового явления, вне зависимости от сложности применяемой для
наблюдения аппаратуры, служит первоначальным импульсом иссле-
исследования, а не запланированный эксперимент, представляющий
разработку на основе комбинации известных положений. Другим
аспектом, проистекающим из наблюдения, служит история событий,
необходимая для их реконструкции, поскольку время нельзя модели-
моделировать в лаборатории.
1.2. СИСТЕМА БИОГЕОХИМИЧЕСКИХ ЦИКЛОВ
1.2.1. Цикл органического углерода
Биогеохимическая машина планеты представляется системой
взаимосвязанных циклов элементов. Эти циклы действуют как в
планетарном масштабе, так и в конкретных ландшафтах-экосисте-
ландшафтах-экосистемах. Общим правилом служит тезис "циклы в циклах". Он действу-
действует на всех иерархических уровнях. Определяющим для биосферы
служит цикл органического углерода, полностью преобразовавший
состояние поверхностных оболочек Земли, движущей силой кото-
которого служит энергия Солнца, использованная фотоавтотрофами.
Биогеохимические циклы сопряжены друг с другом. Система био-
биогеохимических циклов определяется ведущим циклом органическо-
органического углерода Сорг и сопряженными с ним в эквимолекулярном отно-
отношении 1:1:1 циклами углекислоты СО2 и кислорода О2 (рис. 1.2).
Цикл органического углерода обусловлен, во-первых, первичной
продукцией за счет использования внешней солнечной энергии фо-
тоавтотрофными оксигенными организмами: цианобактериями, во-
водорослями, растениями и в небольшой степени хемоавтотрофам» за
счет поступления эндогенного водорода в газогидротермах и, во-
вторых, деструкцией органотрофными организмами, аэробными и
анаэробными. Деструкционная ветвь цикла органического углерода
наиболее сложна, и поэтому ее следует рассмотреть подробнее, оз-
ознакомившись с общими принципами организации участия сообществ
микробов в циклах. Конечным продуктом деструкции служит угле-
углекислота, замыкающая цикл органического углерода и сопрягающая
его с циклом неорганического углерода и циклом кислорода. Цикл
углерода суммируется двумя уравнениями: продукции СО2 + Н2О =
= [СН2О] + О2 и деструкции [СН2О] + О2 = СО2 + Н2О, где символ
[СН2О] означает органический углерод, или Сорг. В реакциях суще-
существуют эквимолярные отношения между углекислотой, кислородом
и [СН2О]. Следовательно, молярное нетто-количество О2 равно ко-
количеству Сорг, выведенному из цикла и обусловленному дибалансом
между продукцией и деструкцией. Резервуары Сорг определяются ди-
динамическим резервуаром биомассы, суммирующим биомассу проду-
продуцентов и деструкторов; резервуаром мортмассы, которая не успела
разложиться под действием деструкторов; резервуаром остаточного
органического вещества, трудно поддающегося разложению. Дина-
Динамический резервуар биомассы различен для организмов с коротким
жизненным циклом и длительным циклом, как у деревьев.
10
CaSiO3 + СО2 = СаСО3 + SiO2
MeSiAl
= [СН2О]+О2
[СН2О] + О2 = СО2 + Н2О
SiAl
0,7
Атмосфера
СО2
38
Океан
1280
Океан
S
7440
Породы
Рис. 1.2. Сопряжение биогеохимических циклов. (Резервуары в 1018 г.)
Ведущим является цикл органического углерода с первичной продукцией, обусловленной
фотоавтотрофной ассимиляцией СО2 и органотрофной деструкцией. С ним сопряжены ана-
анаболические циклы азота и фосфора. Катаболический цикл серы и в прошлом, по-видимому,
цикл железа обусловливают деструкцию Сорг Химическое выветривание изверженных по-
пород ускоряется биотически опосредованными реакциями с ведущей ролью углекислоты
Предположение, что суммарный запас органического вещества
в биомассе должен составлять биомассу первичных продуцентов
плюс биомассу деструкторов, равную примерно половине биомассы
продуцентов, оказывается неверным, потому что продуценты с ко-
коротким жизненным циклом, как фитопланктон океана, могут пере-
перекачивать углерод в организмы с длительным жизненным циклом,
как зоопланктон, и всю последующую трофическую пирамиду в
океане. Таким образом, необходимо оценивать не резервуары, а по-
потоки в них. Пребывание углерода в мортмассе определяется кинети-
кинетикой ее деструкции. Наконец, пребывание углерода в остаточных ус-
устойчивых соединениях, начальной формой которых можно считать
гумус, определяется в значительной степени геологическими факто-
11
рами, прежде всего седиментогенезом, так как устойчивый углерод
поступает в осадочные породы.
Отсюда видно, что в состоянии системы Земли первоочередную
роль играет дисбаланс между продукцией и деструкцией. Кислород
атмосферы представляет недоиспользованный в деструкции остаток
кислорода фотосинтеза. Важны не столько абсолютные величины
продукции и деструкции, сколько разность между ними. Экосистемы
с низкой продуктивностью могут оказаться в сумме накопителями
органического углерода, если деструкция в них еще более замедле-
замедлена. Такая картина действительно наблюдается в холодной тундре и
в анаэробных болотах. Деструкция не может в целом превышать
продукцию, и поэтому система должна неизбежно двигаться в на-
направлении накопления кислорода и органического вещества. При
этом деструкция зависит от содержания кислорода и существенно
различна в аноксических и аэробных условиях. Разность между про-
продукцией и деструкцией определяется наличием ловушек, в приведен-
приведенном примере холодной и анаэробной, к которым надо добавить
обычную геологическую седиментационную ловушку.
Первичную продукцию можно считать пропорциональной содер-
содержанию хлорофилла. В свою очередь, содержание хлорофилла прямо
зависит от освещаемой увлажненной поверхности, которую можно
считать приблизительно постоянной в истории Земли. Этому прямо-
прямолинейному подходу приходится противопоставить ряд ограничений.
Солнечная постоянная в геологическом масштабе времени не была
постоянной, и светимость Солнца изменялась, а для меньших перио-
периодов времени имело значение закономерное изменение положения
Земли относительно Солнца. Соотношение увлажненной поверхно-
поверхности менялось в эпохи высокого и низкого стояния материков, в та-
лассократические и теократические эпохи. Как уже упоминалось,
могла существенно изменяться скорость деструкции. Однако важ-
важнейшим фактором, обусловливающим продукцию, является ее огра-
ограничение биогенными элементами. Вместо упрощенной формулы
продукта первичной реакции фотоавтотрофной ассимиляции угле-
углекислоты следует написать обобщенную формулу биомассы с соот-
соотношением элементов C:N:P = 106:16:1. В этом случае синтез биомас-
биомассы может быть ограничен, как действительно наблюдается, доступ-
доступностью биогенов.
1.2.2. Сопряжение циклов элементов
с циклом Сер,.
Синтез биомассы требует участия кроме углерода и воды других
биогенных элементов, обусловливая их количественное сопряжение
в анаболических реакциях. Цикл азота сопряжен с циклом органи-
органического углерода соотношением Сорг: Nopr = 6:1 для синтеза биомас-
биомассы; в нем происходят также превращения неорганических форм азота.
12
Цикл азота с его этапами: азотфиксацией, аммонификацией, нитри-
нитрификацией, денитрификацией - целиком определяется деятельностью
бактерий. Продукция определяется наличием резервуара доступно-
доступного азота, который пополняется только за счет азотфиксации и
уменьшается за счет денитрификации и экспортной продукции морт-
массы, выводимой из цикла. Способность к азотфиксации широко
распространена среди бактерий, особенно анаэробных, но связана с
большой затратой энергии. Аэробные организмы, в том числе циа-
нобактерии, ассимилируют N2, создавая локально или временно ано-
ксические условия. Ассимиляция азота идет из его связанных мине-
минеральных форм, аммиака или нитратов. На примере цикла азота видно,
что эукариотные организмы могут существовать только в условиях
действующей бактериальной системы, обеспечивающей поступле-
поступление связанного азота.
Цикл фосфора стехиометрически связан с циклом органическо-
органического углерода в отношении С : Р > 100 : 1 в реакциях анаболизма. В цик-
цикле фосфора, в отличие от других биогенных элементов, отсутствует
стадия воздушной миграции, обеспечивающая равномерное распре-
распределение по всему земному шару, за исключением второстепенной по
значимости миграции с аэрозолем. Первичным источником попол-
пополнения доступного фосфора служит его выщелачивание из извержен-
изверженных пород и поступление в состав Рорг биомассы. Из мортмассы в ре-
результате деструкции фосфор поступает в регенерационный цикл,
частично захоранивается с мортмассой, частично переходит в неус-
неусвояемые соединения, прежде всего фосфаты кальция. Содержащие
фосфаты осадки переходят в осадочные породы, из которых фос-
фосфор может быть мобилизован микроорганизмами либо поступает в
геологический рецикл. В результате фосфор в большей степени,
чем азот, служит лимитирующим биогеном, ограничивая развитие
не только эукариот, но и прокариот.
Цикл кальция тесно связан с циклом фосфора образованием фо-
фосфатов кальция, но в гораздо большем масштабе он имеет значение
для цикла неорганического углерода, который приходится рассмат-
рассматривать совместно с циклами щелочноземельных элементов Са и Mg.
Поступление кальция обусловлено углекислотным выщелачивани-
выщелачиванием изверженных пород, приводящим в конечном итоге к образова-
образованию глин как остаточных алюмосиликатных компонентов и кар-
карбонатов кальция и магния. Этот процесс в общем представляет
однонаправленную цепь реакций образования осадочных пород и
обращается лишь с выносом карбонатных пород на поверхность
геологическими процессами и началом рецикла. В геологическом
масштабе реакция выщелачивания очень значительна, но она мед-
медленная и ускоряется в 10-1000 раз под действием биоты, обуслов-
обусловленным локальной генерацией СО2 из аккумулированного углерода
мортмассы при деструкции, но в еще большей степени образовани-
образованием органических кислот. Основная реакция представлена обрати-
13
мым растворением углекислоты и ее диссоциацией:
СО2+Н2О = [Н2СО3 ] <± Н++НСОз = 2 Н+ f
Реакция установления равновесия ускоряется в тысячи раз под дей-
действием универсального по распространенности цинк-содержащего
фермента карбоангидразы:
Н2О
Эта реакция определяет кислотно-щелочную реакцию среды че-
через бикарбонатное равновесие, которое прямо связано с рН. Цикл
неорганического углерода смыкается с циклом органического угле-
углерода через углекислоту воздуха и ее растворенные формы в гидро-
гидросфере, концентрация которых сильно зависит от температуры. В ли-
литосфере неорганический углерод представлен в основном карбонат-
карбонатными породами, прежде всего карбонатом кальция. Обратимая
реакция образования карбоната кальция может быть сокращенно
записана как
Са
2+
= СаСО3 + СО2+Н2О.
Отсюда ясно, что в сочетании с реакцией бикарбонатного равно-
равновесия эта реакция определяет щелочность среды. Осаждение карбо-
карбоната кальция организмами может быть вызвано нарушением равно-
равновесия за счет удаления СО2 с образованием органического углерода
или же дегазации при повышении температуры, и тогда приведенная
выше реакция сдвигается вправо. Термодинамические условия рав-
равновесия бикарбоната и кальция очень подробно описаны, но в при-
природных условиях большую роль играет кинетика реакций, в резуль-
результате которой океан оказывается пересыщенным по кальцию. В при-
присутствии Са2+ щелочность не может возрастать, поскольку карбо-
карбонат- и бикарбонат-ионы удаляются с образованием кальцита. Под-
кисление среды не может происходить, пока присутствует карбонат
кальция. В результате область развития нейтрофильных организ-
организмов обусловлена кальцием, и их можно назвать "кальциефилами".
В этом отношении осаждение карбонатов кальция в геологической
истории привело к формированию "нормальных" условий.
Современный цикл кальция определяется прежде всего биоло-
биологически опосредованными реакциями растворения (выщелачива-
(выщелачивания) и осаждения карбонатов, а также биоминералогией образова-
образования скелета протестами и макроорганизмами. Образование в начале
протерозоя мощных толщ биогенных карбонатных пород обуслов-
обусловлено опосредованным действием циано-бактериальных сообществ,
а с кембрия - прямым действием известковых планктонных организ-
14
мов, как кокколитофориды и формаминиферы, и бентосных рифо-
строителей.
Первичное поступление кальция и магния в биологические ци-
циклы связано с выщелачиванием силикатных изверженных пород и
циклом кремния в биологически опосредованных процессах. Вы-
Выщелачивание обусловливает попутное поступление в водную фазу
микроэлементов. Цикл кремния в фанерозое обусловлен его
использованием как скелетного материала диатомовыми, радиоля-
радиоляриями, губками. В результате кремний ведет себя в океане как био-
биогенный элемент, и это связано с появлением скелетной функции у
эукариот.
Цикл серы сопрягается с циклом органического углерода в реак-
реакциях сульфидогенеза, катализируемых только прокариотами, суль-
сульфат- и сероредуцирующими организмами. Окисление соединений
серы осуществляется в двух направлениях: анаэробного окисления
аноксигенными фототрофными и аэробного - хемолитотрофными
организмами. В биогеохимическом цикле серы участвуют следую-
следующие формы соединений серы, создающие значительные резервуа-
резервуары: 1) сульфаты, преимущественно сульфаты моря; 2) сульфиды в
виде растворенного H2S и нерастворимых сульфидов металлов, час-
частично эндогенного (вулканического) и в основном экзогенного (био-
(биогенного) происхождения; 3) сера, в значительной части эндогенного
происхождения. Разнообразные промежуточные соединения непол-
неполного окисления серы, как тиосульфат или SO2, появляются в тран-
транзитных формах и незначительной концентрации, не образуя резер-
резервуары. Подобно тому как конечным результатом сопряженных цик-
циклов углекислоты, органического углерода, кислорода оказывается
накопление кислорода атмосферы, конечным продуктом серного
цикла оказываются сульфаты океана.
Цикл железа сопрягается с циклом кислорода деятельностью аэ-
аэробных железобактерий, окисляющих закисное железо в гидрат
окиси железа, и с циклом углерода деятельностью железоредуциру-
ющих бактерий, образующих восстановленное железо Fe(II) и маг-
магнетит. Цикл железа связан с циклом серы через образование суль-
сульфидов железа и их окисление бактериями. Хотя железобактерии от-
относятся к наиболее давно известным микробам, роль которых в
области геологической микробиологии была признана, цикл железа
и катализирующие его организмы исследованы только в последнее
время. В геологическом прошлом цикл железа играл гораздо ббль-
шую роль, приведя в раннем протерозое к формированию значимых
для баланса кислорода мощных отложений окислов железа.
Кратко рассмотрев связь между биогеохимическими циклами в
биосфере, следует признать, что они действуют как сопряженная си-
система реакций, и изменения в условиях протекания одного цикла ска-
скажутся на условиях протекания другого (рис. 1.3). На основе термоди-
термодинамики можно рассчитать соотношения этих циклов, но большая не-
15
I Атмосфера:
{Выветривание]
Новообразованные
- (карбонаты) •.'
Остаточные
(глины)
Рис. 1.3. Связь геосферных процессов с биотическими
-»- потоки, —> - каталитическое воздействие, -»- опосредованные реакции
Геологические процессы, обусловленные эндогенной энергией и в зоне гипергенеза - сол-
солнечной энергией, создают среду для биоты и выступают как независимые переменные
определенность обусловливается ведущим циклом органического
углерода, баланс в котором определяется соотношением продукции
и деструкции. Оба компонента могут варьировать, причем вариации
в них зависят от разных факторов. При этом большую роль играет
кинетика процессов. Следовательно, стехиометрические соотноше-
соотношения могут служить скорее для полуколичественной оценки приори-
приоритетов. Вместе с тем простейшее рассмотрение циклов убеждает в су-
существовании определенных тенденций в развитии системы, обуслов-
обусловленных неполной замкнутостью задающего цикла органического
углерода и соответственно движения в область более окисленного со-
состояния биосферы. Далее очевидно, что вся система сопряженных ци-
циклов катализируется бактериями, которые являются необходимым и
достаточным условием для ее действия. Сопряжение циклов требует
согласованной работы групп бактерий, катализирующих определен-
определенные этапы превращений. Эти группы организованы в сообщества.
Поэтому природоведческая микробиология действительно оказыва-
оказывается базовой дисциплиной для естествознания, и центральной пробле-
проблемой в ней оказывается деятельность сообществ микроорганизмов.
1.3. ГЕОБИОФИЗИКА
Биогеохимические циклы описывают планетарную машину Зем-
Земли в абстрактных химических терминах. Реализация циклов осуще-
осуществляется в конкретной географической среде. Масштабы рассмат-
рассматриваемой среды могут быть различными: от планеты в целом до
ландшафта и малой экосистемы. Ключевым понятием в географии
служит ландшафт. Ландшафты объединяются в ландшафтные зо-
зоны, определяющим фактором которых служит гидротермический
режим, характеризуемый климатом. В свою очередь, климат опреде-
определяется прежде всего количеством энергии, получаемой участком
земной поверхности, а затем - состоянием атмосферы над этим уча-
участком, обусловливаемым циркуляцией воздушных масс как в мас-
масштабе всей планеты, так и регионально. Гидротермический режим
описывается влажностью (гумидностью) или сухостью (аридностью)
и температурой, холодной или теплой. Для обозначения климата
есть ряд терминов (полярный, бореальный, тропический и т.д.). Кли-
Климат определяет растительные зоны.
Ландшафтно-климатические зоны явились основой биогеогра-
биогеографии. Для микробиологии, ориентированной на эксперименты с чис-
чистыми лабораторными культурами, вопросы биогеографии не имели
значения. Если для зоологов и ботаников большое значение имеет
проблема географической изоляции, то для микробиологов этот во-
вопрос не возникал - "микробы есть всюду, где есть условия для их
развития". Однако в природоведческой микробиологии, ключевым
объектом которой является микробное сообщество, проблемой слу-
служит взаимодействие микроорганизмов с растительным покровом,
которое не может рассматриваться вне связи с климатом.
Что такое погода и климат? Погода - атмосферные условия,
господствующие в регионе в определенное время, зависящие от теп-
тепла или холода, облачности, влажности, ветра. Климат - "синтез по-
погодных условий в данной области, определяемый долгосрочной ста-
статистикой вариаций состояния атмосферы". Поэтому, рассматривая
биосферные проблемы, следует в первую очередь обратить внима-
внимание на атмосферу.
Климат обусловлен энергетическими механизмами. Разное ко-
количество энергии, получаемой разными регионами от Солнца, при-
приводит к перемещению масс вещества, обусловливаемому быстрой
циркуляцией атмосферы и медленной циркуляцией в океане. Цирку-
Циркуляция относится к области физики и создает сложную климатиче-
климатическую систему, где взаимодействуют воздушные массы атмосферы и
водные - океана. Отсюда происходит перераспределение вещества,
и продукты одной зоны переносятся в другую, где они могут под-
подвергнуться превращениям. Вследствие переноса вещества создается
взаимодействие между различными регионами, ландшафтами, внут-
внутри ландшафтов на разных пространственно-временных уровнях,
16
17
вплоть до формирования микрозон микроорганизмами. Геохи-
Геохимическая трактовка оказывается недостаточной, и реализация био-
биогеохимических циклов в конкретных природных условиях требует
понимания физических процессов. Гетерогенность системы оказы-
оказывается важнейшим условием ее существования, и связь между раз-
разными частями ее осуществляется материальными потоками - транс-
транспортом.
Транспорт вещества представляет первостепенную проблему
для экологии микроорганизмов, которая рассматривается в рамках
геобиофизики. Эта проблема оказалась ключевой при построении
реальных математических моделей для природных процессов, так
как часто именно перенос вещества составляет лимитирующее зве-
звено в реакции. Геобиофизика описывает физические условия сущест-
существования сообществ прежде всего в динамических характеристиках
потоков вещества. Подробно некоторые ее разделы разрабатыва-
разрабатывались как агрофизика культивируемых ландшафтов. Обычно она
связана с процессами макропереноса вещества в масштабах ланд-
ландшафта и геосферы. Однако для микробных сообществ имеет значе-
значение перенос вещества не только в пределах элементарного ланд-
ландшафта с его микроклиматом, но и внутри сообщества на расстояни-
расстояниях в доли миллиметра, когда транспорт обеспечивается только
молекулярной диффузией. Для экологии микроорганизмов геобио-
геобиофизика имеет принципиальное значение. В течение почти полусто-
полустолетия внимание биологов было сконцентрировано на биологии
клетки и процессах, происходящих в ней. Но экология микроорга-
микроорганизмов имеет дело прежде всего с тем, что находится вне клетки.
В результате микробиологи вступили в область систем высшего
ранга по сравнению с клеткой и вышли из приложения разработок
биохимии к прокариотным организмам.
Для географической среды обитания микробных сообществ
принципиальное значение имеет ее неоднородность с разнонаправ-
разнонаправленными процессами в разных областях. Неоднородность среды
обитания создает возможность обмена веществом как по горизонта-
горизонтали, так и по вертикальному профилю. Именно за счет неоднородно-
неоднородности сред обитания оказывается возможным развитие системы в це-
целом. Поэтому наивны попытки представить Землю на любом этапе
ее развития как однородную среду с определенной температурой,
химическим составом. В особенности это относится к микробным
сообществам, местообитания которых могут быть пространственно
ограниченны.
Особенности условий существования организмов в географиче-
географических средах заставляют последовательно рассмотреть атмосферу,
водные системы с Мировым океаном, почву, горные породы. Но все
эти части взаимодействуют друг с другом через подвижные формы
вещества. Например, для газового состава атмосферы первостепенное
значение имеет ее взаимодействие с растворенными газами океана,
18
для состава вод определяющее значение имеет выщелачивание по-
пород, обусловливаемое атмосферным гидрологическим циклом. По-
Поэтому отдельное рассмотрение каждой из сред до известной степени
условно, как условно и рассмотрение циклов элементов по отдель-
отдельности.
Природоведческая микробиология призвана объяснить, каким
образом сообщество микроорганизмов способствует формирова-
формированию ландшафтов прежде всего через их геохимию и как ландшафты
определяют существование сообществ. Если в протерозое микро-
микроорганизмы полностью определяли биотическую составляющую
ландшафтов, то начиная с силура они действуют опосредованно че-
через взаимодействие с растительным покровом на суше, а с неопро-
неопротерозойской революции около миллиарда лет назад, - с фитопланк-
фитопланктоном океана и водорослями мелководий.
1.4. ЛАНДШАФТ
Природоведческая микробиология требует понимания условий,
в которых микроорганизмы взаимодействуют с природой, а это по-
понимание, в свою очередь, должно опираться на науки о Земле, пре-
прежде всего на знание географической оболочки. Центральное гео-
географическое понятие здесь - ландшафт. Абстрактная характеристи-
характеристика среды обитания организмов носит название экосистемы и чаще
всего употребляется экологами. В экологии микроорганизмов упот-
употребляются два понятия: аутэкология, описывающая поведение
отдельного вида в среде обитания, и синэкология, относящаяся к со-
сообществу или экосистеме. Местообитание организмов называется
биотопом, а сумма организмов, обитающих в нем, биоценозом. Для
макроорганизмов эти понятия хорошо разработаны на основе опре-
определения биоразнообразия как флористического или фаунистическо-
го описания и на основе определения численности видов. В основу
современного понимания биоразнообразия положено понятие "эко-
"экосистемы". В микробиологии термины "биотоп" и "биоценоз" упот-
употребляются крайне редко. У микробиологов флористические описания
сейчас заменены перечислением генетических клонов, получаемых
методами молекулярной биологии. Однако главная особенность ми-
микробиологии, в отличие, например, от зоологии, состоит в том, что
бактерии являются мощными геохимическими агентами и взаимо-
взаимодействуют с геосферой. Поэтому для микробиологов определяю-
определяющим условия обитания микробиоты является геохимия ландшафта.
Термин "ландшафт" происходит от немецкого das Land - страна,
земля - и der Schaft - общность. Под ландшафтом понимается про-
пространство земной поверхности, занятое однородной растительно-
растительностью и обладающее однородным почвенным покровом. Ландшафты,
в свою очередь, объединяются в ландшафтные зоны, обусловленные
19
климатом и рельефом. Климат определяет температуру и влаж-
влажность: тропический, умеренный, бореальный, арктический, арид-
аридный, гумидный, семиаридный. Рельеф дает равнинные и горные
ландшафты с их вертикальной зональностью. Для микробиолога,
однако, важен и микроклимат в ландшафте, относящийся иногда к
отдельному валуну. Разница в микроклимате наглядно иллюстриру-
иллюстрируется здесь пестрым ковром лишайников. Микроорганизмы развива-
развиваются в мозаичных местах обитания, иногда неточно называемых
экосистемами, но сумма этих мест определяет процессы, происходя-
происходящие в ландшафте. К водным местам обитания, особенно морским,
термин "ландшафт" обычно не применяется, и здесь говорят либо
об акваториях, либо о конкретных местообитаниях. Нельзя гово-
говорить и о ландшафте применительно к глубинной микрофлоре гор-
горных пород. В этих случаях термин "экосистема" оказывается более
удобным.
Внутри каждого ландшафта действует устоявшийся круговорот
веществ, в той или иной мере замкнутый. Для микробиолога состо-
состояние ландшафта определяется деятельностью в нем микробного со-
сообщества как важнейшего компонента единого продукционно-дест-
рукционного цикла. Местообитанием для микроорганизмов служит
зона в элементе ландшафта - например, донные отложения в водо-
водоеме, ризосфера. В состав ландшафта в качестве подсистем входят
растительный покров и связанные с ним живые организмы (биота),
почва, кора выветривания, поверхностные и грунтовые воды, состав-
составляющие водоемы и водотоки, приземная атмосфера (аэротоп).
Ландшафт представляет гетерогенную систему, включающую эле-
элементарные ландшафты или фации и биогеоценозы. Элементарный
ландшафт представляет такой компонент системы, который мо-
может быть увеличен до больших размеров без существенного изме-
изменения характера происходящих в нем процессов. На этом основано
применение метода мезокосма, т.е. изолированного крупного об-
образца экосистемы для изучения происходящих в ландшафте про-
процессов в полностью контролируемых условиях. Элементарные
ландшафты сами по себе гетерогенны и включают элементы ланд-
ландшафта; например, для верхового болота это кочки и мочажины.
Для элементарного ландшафта характерна вертикальная диффе-
дифференциация, которая включает ярусность наземной части, почвен-
почвенные горизонты в подземной. Вертикальная дифференциация
позволяет упрощать рассмотрение до одномерной модели - разре-
разреза профиля.
В ландшафте происходит объединение его частей в систему
вследствие переноса вещества. Круговорот вещества в ландшафте
определяется превращениями вещества, вызванными поступлением
веществ извне, трансформацией в ландшафте под действием физи-
физических, химических, биологических факторов, выносом вещества из
ландшафта. Циклический характер процессов в ландшафте опреде-
20
ляется в первую очередь биотическим циклом углерода с продук-
продукционной и деструкционной ветвями. Циклические процессы в ланд-
ландшафте определяют его устойчивость как системы. Устойчивость си-
систем трактуется как расширенный принцип Ле Шателье: система
способна уменьшать внешние воздействия за счет внутренней пере-
перестройки до пределов, за которыми наступает разрушение. Внутрен-
Внутренние ресурсы ландшафта определяют его буферную емкость. Это
свойство ландшафтов и аналогичных водных систем обусловлено
саморегуляцией и называется самоочищением и самовозобновлени-
самовозобновлением. Процесс самоочищения связан в первую очередь с деятельно-
деятельностью микроорганизмов-деструкторов.
Из однородности растительного покрова и почвы следует, что
в пределах ландшафта и микробное сообщество должно быть бо-
более или менее однородным. Однако из-за широкого различия в раз-
размерах общую характеристику микробного сообщества трудно дать
даже для элементарного ландшафта, в элементах которого для ми-
микробного сообщества условия оказываются различными. В зависи-
зависимости от дели исследования необходимо подразумевать границы
для рассматриваемого сообщества, которые могут быть на уровне
взаимодействующих колоний или элементов ландшафта в шкале
дециметров-метров. С точки зрения микробиолога ландшафт
представляет собой мозаику условий для развития микроорганиз-
микроорганизмов. Вместе с тем каждый ландшафт имеет характерный для
него геохимический круговорот, и можно говорить о характерных
круговоротах для ландшафтов тундровой или тропической зоны.
В интерпретации результатов вопрос стоит о степени обобщения и
возможной детализации в каждом отдельном случае. Например,
можно обобщать результаты для донных отложений глубоковод-
глубоководного морского бассейна, хотя и в этих отложениях действуют
сообщества с определенной микрозональностью, для толщи
воды олиготрофного океана или же для агроэкосистем. Мозаич-
ность экосистем обусловливает необходимость иерархического
подхода с четким пониманием допустимой степени усреднения и
способа усреднения с тем, чтобы от кочки не перейти ко всему
болоту.
Ландшафты подразделяются на автономные, как, например,
верховое болото, и подчиненные, как предгорные равнины (педи-
плен). Они различаются по степени притока аллохтонного вещества
и автохтонного внутреннего цикла. Тогда первая задача для микро-
микробиолога при решении задач экологии микроорганизмов в конкрет-
конкретной ландшафтной обстановке состоит в том, чтобы определить
направления движения вещества. Обычно для этого используются
стоковые серии ландшафтов, или катены, определяющие горизон-
горизонтальный перенос веществ.
Ландшафты объединяются в ландшафтные зоны на основе био-
биогеографии, где руководящим признаком служит климатическая
21
U
н
|
Средняя годовая температура, °С
-30 -20 -10 0
10
20
30
Разность осадков и испарения, мм/год
-400 -300 -200 -100 0
200
300 400
Площадь, океан/суша
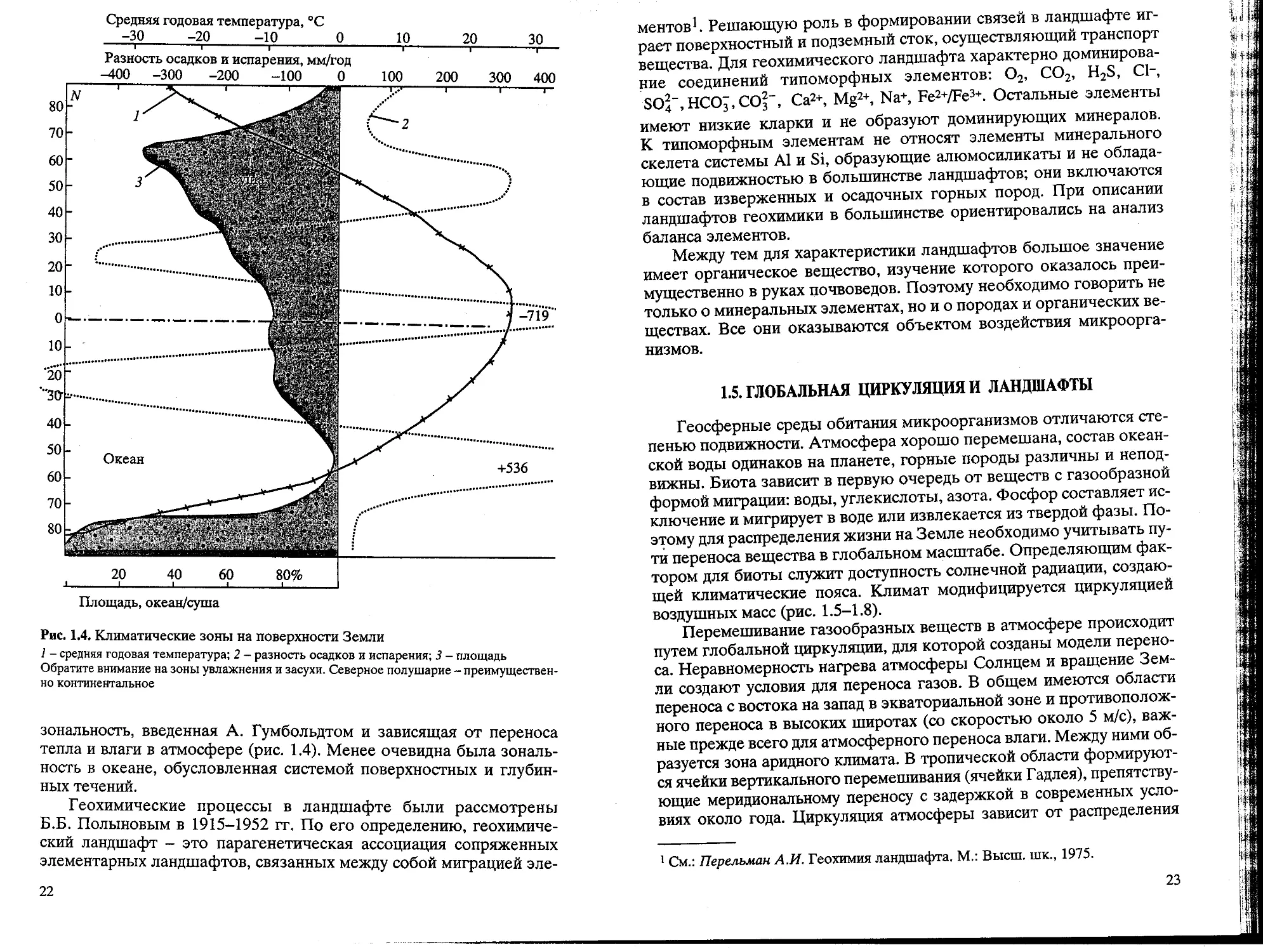
Рис. 1.4. Климатические зоны на поверхности Земли
1 - средняя годовая температура; 2 - разность осадков и испарения; 3 - площадь
Обратите внимание на зоны увлажнения и засухи. Северное полушарие - преимуществен-
преимущественно континентальное
зональность, введенная А. Гумбольдтом и зависящая от переноса
тепла и влаги в атмосфере (рис. 1.4). Менее очевидна была зональ-
зональность в океане, обусловленная системой поверхностных и глубин-
глубинных течений.
Геохимические процессы в ландшафте были рассмотрены
Б.Б. Полыновым в 1915-1952 гг. По его определению, геохимиче-
геохимический ландшафт - это парагенетическая ассоциация сопряженных
элементарных ландшафтов, связанных между собой миграцией эле-
22
ментов1. Решающую роль в формировании связей в ландшафте иг-
играет поверхностный и подземный сток, осуществляющий транспорт
вещества. Для геохимического ландшафта характерно доминирова-
доминирование соединений типоморфных элементов: О2, СО2, H2S, C1-,
8С>4~,НСОз,СОз"", Са2+, Mg2+, Na+, Fe2+/Fe3+. Остальные элементы
имеют низкие кларки и не образуют доминирующих минералов.
К типоморфным элементам не относят элементы минерального
скелета системы А1 и Si, образующие алюмосиликаты и не облада-
обладающие подвижностью в большинстве ландшафтов; они включаются
в состав изверженных и осадочных горных пород. При описании
ландшафтов геохимики в большинстве ориентировались на анализ
баланса элементов.
Между тем для характеристики ландшафтов большое значение
имеет органическое вещество, изучение которого оказалось преи-
преимущественно в руках почвоведов. Поэтому необходимо говорить не
только о минеральных элементах, но и о породах и органических ве-
веществах. Все они оказываются объектом воздействия микроорга-
микроорганизмов.
1.5. ГЛОБАЛЬНАЯ ЦИРКУЛЯЦИЯ И ЛАНДШАФТЫ
Геосферные среды обитания микроорганизмов отличаются сте-
степенью подвижности. Атмосфера хорошо перемешана, состав океан-
океанской воды одинаков на планете, горные породы различны и непод-
неподвижны. Биота зависит в первую очередь от веществ с газообразной
формой миграции: воды, углекислоты, азота. Фосфор составляет ис-
исключение и мигрирует в воде или извлекается из твердой фазы. По-
Поэтому для распределения жизни на Земле необходимо учитывать пу-
пути переноса вещества в глобальном масштабе. Определяющим фак-
фактором для биоты служит доступность солнечной радиации, создаю-
создающей климатические пояса. Климат модифицируется циркуляцией
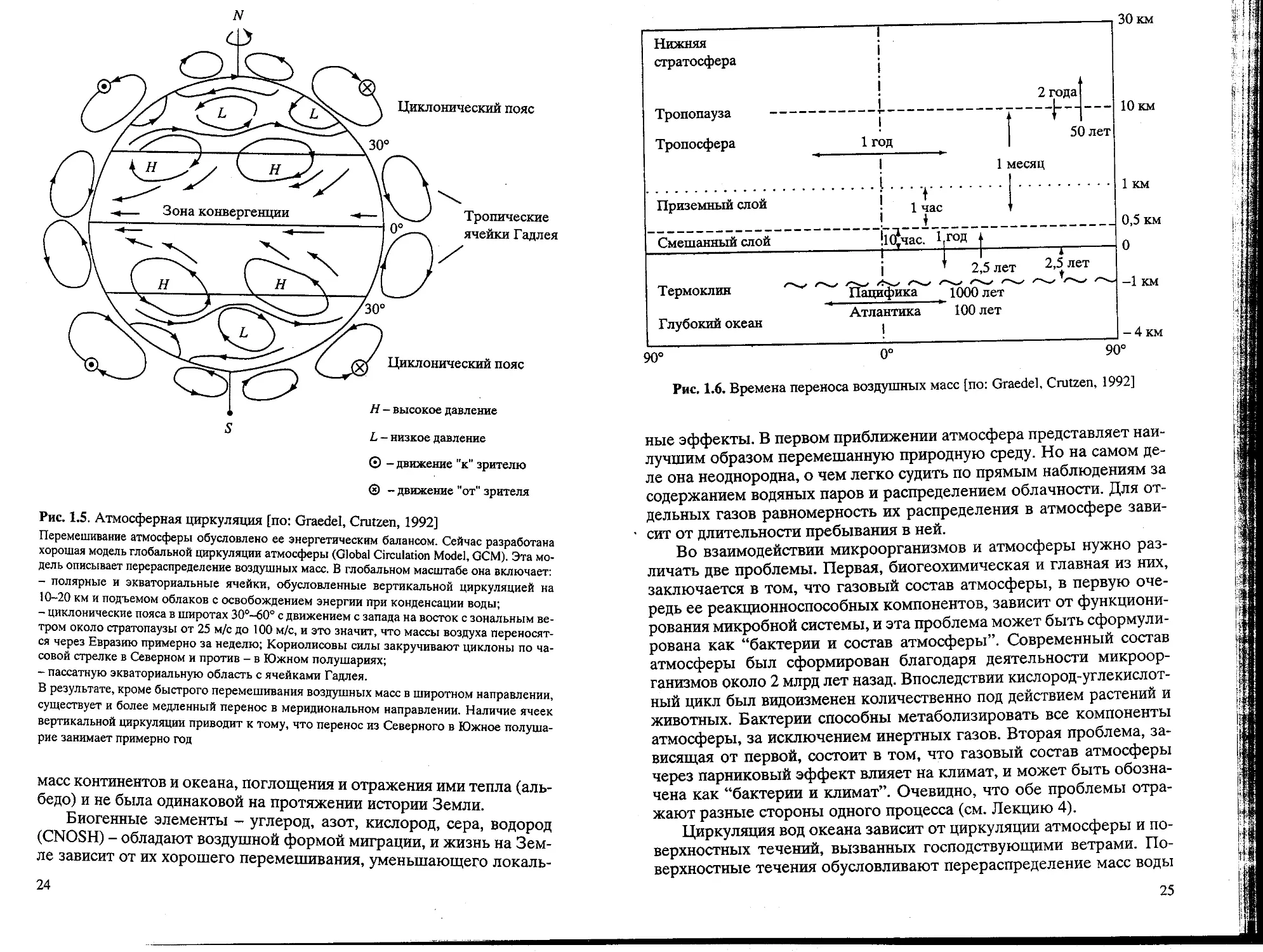
воздушных масс (рис. 1.5-1.8).
Перемешивание газообразных веществ в атмосфере происходит
путем глобальной циркуляции, для которой созданы модели перено-
переноса. Неравномерность нагрева атмосферы Солнцем и вращение Зем-
Земли создают условия для переноса газов. В общем имеются области
переноса с востока на запад в экваториальной зоне и противополож-
противоположного переноса в высоких широтах (со скоростью около 5 м/с), важ-
важные прежде всего для атмосферного переноса влаги. Между ними об-
образуется зона аридного климата. В тропической области формируют-
формируются ячейки вертикального перемешивания (ячейки Гадлея), препятству-
препятствующие меридиональному переносу с задержкой в современных усло-
условиях около года. Циркуляция атмосферы зависит от распределения
1 См.: Перельман А.И. Геохимия ландшафта. М.: Высш. шк., 1975.
23
Циклонический пояс
_ Тропические
°°-—- ячейки Гадлея
30°
( (Ы Циклонический пояс
Я - высокое давление
L - низкое давление
© - движение "к" зрителю
© - движение "от" зрителя
Рис. 1.5. Атмосферная циркуляция [по: Graedel, Crutzen, 1992]
Перемешивание атмосферы обусловлено ее энергетическим балансом. Сейчас разработана
хорошая модель глобальной циркуляции атмосферы (Global Circulation Model, GCM). Эта мо-
модель описывает перераспределение воздушных масс. В глобальном масштабе она включает:
- полярные и экваториальные ячейки, обусловленные вертикальной циркуляцией на
10-20 км и подъемом облаков с освобождением энергии при конденсации воды;
- циклонические пояса в широтах 30°-60° с движением с запада на восток с зональным ве-
ветром около стратопаузы от 25 м/с до 100 м/с, и это значит, что массы воздуха переносят-
переносятся через Евразию примерно за неделю; Кориолисовы силы закручивают циклоны по ча-
часовой стрелке в Северном и против - в Южном полушариях;
- пассатную экваториальную область с ячейками Гадлея.
В результате, кроме быстрого перемешивания воздушных масс в широтном направлении,
существует и более медленный перенос в меридиональном направлении. Наличие ячеек
вертикальной циркуляции приводит к тому, что перенос из Северного в Южное полуша-
полушарие занимает примерно год
масс континентов и океана, поглощения и отражения ими тепла (аль-
(альбедо) и не была одинаковой на протяжении истории Земли.
Биогенные элементы - углерод, азот, кислород, сера, водород
(CNOSH) - обладают воздушной формой миграции, и жизнь на Зем-
Земле зависит от их хорошего перемешивания, уменьшающего локаль-
24
Нижняя
стратосфера
Тропопауза
Тропосфера
1год
Приземный слой
Смешанный слой
2 года!
50 лет
1 месяц
1 час
Термоклин
Глубокий океан
Паци<
ацифика
Атлантика
2,5 лет
"Тбо^лет
" 100 лет
2,5 лет
30 км
10 км
1 км
0,5 км
0
-1 км
-4 км
90° 0° 90°
Рис. 1.6. Времена переноса воздушных масс [по: Graedel, Crutzen, 1992]
ные эффекты. В первом приближении атмосфера представляет наи-
наилучшим образом перемешанную природную среду. Но на самом де-
деле она неоднородна, о чем легко судить по прямым наблюдениям за
содержанием водяных паров и распределением облачности. Для от-
отдельных газов равномерность их распределения в атмосфере зави-
зависит от длительности пребывания в ней.
Во взаимодействии микроорганизмов и атмосферы нужно раз-
различать две проблемы. Первая, биогеохимическая и главная из них,
заключается в том, что газовый состав атмосферы, в первую оче-
очередь ее реакционноспособных компонентов, зависит от функциони-
функционирования микробной системы, и эта проблема может быть сформули-
сформулирована как "бактерии и состав атмосферы". Современный состав
атмосферы был сформирован благодаря деятельности микроор-
микроорганизмов около 2 млрд лет назад. Впоследствии кислород-углекислот-
ный цикл был видоизменен количественно под действием растений и
животных. Бактерии способны метаболизировать все компоненты
атмосферы, за исключением инертных газов. Вторая проблема, за-
зависящая от первой, состоит в том, что газовый состав атмосферы
через парниковый эффект влияет на климат, и может быть обозна-
обозначена как "бактерии и климат". Очевидно, что обе проблемы отра-
отражают разные стороны одного процесса (см. Лекцию 4).
Циркуляция вод океана зависит от циркуляции атмосферы и по-
поверхностных течений, вызванных господствующими ветрами. По-
Поверхностные течения обусловливают перераспределение масс воды
25
глубинными течениями и предполагают области опускания поверх-
поверхностных вод (даунвеллинг) и подъема глубинных (апвеллинг). Эти
области зависят от распределения континентов. Для биоты течения
в океане являются определяющими вследствие поступления пита-
питательных веществ с глубинными водами, создающими области повы-
повышенной продуктивности. Большее значение для биоты имеет верти-
вертикальное распределение водных масс, стратификация вод океана,
обусловленная разным удельным весом теплой и холодной воды.
Поверхностные воды океана находятся в равновесии с газами атмо-
атмосферы, и благодаря вертикальной циркуляции газы переносятся
вниз. Если бы не было обусловленной поверхностными течениями
циркуляции, то нагрев поверхностных вод Солнцем вызвал бы со-
состояние стратификации с крайне медленным вертикальным перено-
переносом. Такие состояния океана наблюдались в геологическом прошлом,
а сейчас моделью их считается Черное море.
Циркуляция вод океана определяет поступление питательных
веществ и соответственно развитие фитопланктона. Центральные
области океанов в середине циклонических течений обеднены био-
той и представляют олиготрофные зоны. Помимо горизонтальной
циркуляции с поверхностными течениями для океана первостепен-
первостепенное значение имеет вертикальный перенос с глубинными течения-
течениями. Подъем глубинных вод - апвеллинг - обогащает продукцион-
продукционную фотическую зону биогенными элементами, в первую очередь
нитратом. Поэтому биологическая продуктивность океана весьма
неравномерна. В ней существуют зоны повышенной продуктивно-
продуктивности и бедные зоны. Они подробнее рассмотрены в Лекции 5.
Химический состав вод океана нормируется по солености, под
которой подразумевается содержание С1-иона. Биота контролирует
Рис. 1.7. Распределение ландшафтов
Ландшафт определяется, во-первых, климатом, во-вторых, тектоникой. Быстрые текто-
тектонические процессы идут на границах плит. Внутри плит тектоника определяется рифто-
генезом и горячими точками подъема (up-lift) с вулканической активностью. Примерами
рифтогенеза служат Восточно-Африканский Рифт или область Байкала. Кратоны пред-
представляют тектонически спокойные и стабильные континентальные области. К ним отно-
относятся щиты, где эрозия обнажила старую кристаллическую основу; платформы, где пло-
плоские щиты перекрыты осадочной оболочкой или же интракратонными бассейнами с тол-
толстыми отложениями осадков; пассивные окраины континентов. Между кратонами и гор-
горными поясами располагаются предгорные бассейны седиментации. Согласно Ронову
A980) интракратонные бассейны, пассивные окраины, предгорные области представляют
места долговременного накопления осадков. Базу для седиментации дает уровень океана.
Если бы время было достаточным, то континенты уплощились бы до уровня океана. Од-
Однако уровень океана претерпевает эвстатические колебания. Уровень океана отмечается
террасами. Короткопериодичные колебания связаны с оледенением и климатом и имеют
амплитуду около 100 м с перераспределением воды между океаном и гляциосферой. Ин-
Интракратонные бассейны Северной Америки, Русской платформы, Бразилии указывают
на отложение с амплитудой в 100-200 м и периодичностью 10-80 млн лет. Длительные
низкие стояния океана в 300-400 м были в кембрии и триасе-юре. В геологии аналогом
ландшафта служат фации, и фациальный анализ дает исходную базу для реконструкции
ландшафта
27
ОКЕАН
Склон
3,6 км 0,00002
Склон
0,0002
Рис. 1.8. Схема различия между эпиконтинентальными и окраинными морями
как мелководными местообитаниями
Эпиконтинентальные моря образуются на внутриконтинентальных равнинах в периоды
высокого стояния океана (талассократические эпохи). Они сохраняют связь с океаном и в
аридном поясе могут служить областью отложения эвапоритов аналогично Кара-Богаз-Го-
Кара-Богаз-Голу на Каспии. Эпиконтинентальные моря отличаются от внутриконтинентальных бессточ-
бессточных водоемов (как Каспий) тем, что сохраняют связь с морем. Окраинные моря имеют
связь с морем, океаном, которая может быть ограничена островными дугами, но не более
в основном содержание минорных компонентов океанской воды,
прежде всего идущих на образование скелета - Са, Si, Sr, и в свою
очередь зависит от биогенных элементов, лимитирующими из кото-
которых являются прежде всего фосфор, азот и затем железо.
Главным следствием глобальной циркуляции для биоты служит
формирование климатических зон как на суше, так и в океане. Зо-
Зональный климат меняется в зависимости от рельефа и перераспре-
перераспределения воздушных потоков, создавая региональные климатические
условия. Зависимое гь биоты от климата была установлена и описа-
описана А. Гумбольдтом. Она находит отражение в ландшафтных зонах.
В последние десятилетия большое внимание уделяется модифика-
модификации климата биотой через состав атмосферы и зависящий от него
парниковый эффект. Этот вторичный по отношению к первичному
радиационному балансу эффект стал центральным в гипотезе "Геи",
выдвинутой Дж. Ловелоком. По его представлениям, биота модифи-
модифицирует окружающую среду в благоприятном для себя направлении в
глобальном масштабе.
В действии биоты следует различать две группы явлений: пря-
прямые и опосредованные. Например, состав атмосферы прямо зависит
от деятельности биоты, продуцирующей и использующей углекис-
углекислоту, кислород, метан. Вместе с тем изменения в содержании СО2
28
вызывают изменения и в карбонатно-силикатном цикле выветрива-
выветривания, и в климате, и в циклах других элементов в качестве вторичных,
опосредованных следствий. Вторичные опосредованные результаты
действия биоты могут быть не менее значительными, чем прямые,
но до широкого применения анализа систем они ускользали от вни-
внимания исследователей.
Рекомендуемая литература
1. АндрузДж., Бримблекумб П., Джикелз Т., Лисе П. Введение в химию окру-
окружающей среды. М: Мир, 1999. 271 с.
2. Вернадский В.И. Биосфера. Л., 1926.
3. Виноградский С.Н. О роли микробов в общем круговороте жизни. Речь,
произнесенная С.Н. Виноградским на общем собрании членов император-
императорского Института экспериментальной медицины 8 декабря 1896 г. СПб., 1896.
4. Глобальный биогеохимический цикл серы и влияние на него деятельности
человека / Ред. М.В. Иванов, Дж. Фриней. М.: Наука, 1979.
5. Добровольский В.В. Основы биогеохимии. М.: Высш. шк., 1998.
6. Заварзин Г.А. Смена парадигмы в биологии // Вест. РАН. 1995. Т. 65. № 1.
С. 8-23.
7. Кузнецов СИ., Иванов М.В.,Ляликова Н.Н. Введение в геологическую ми-
микробиологию. М., 1962.
8. ПерельманА.И., Касимов Н.С. Геохимия ландшафта. М.: Астрея-2000,1999.
768 с.
9. Ронов А.Б. Осадочная оболочка Земли: количественные закономерности
строения, состава и эволюции. М: Наука, 1980. 79 с. (чтения им. В.И. Вер-
Вернадского).
10. Федоров В Д., Гильманов Т.Г. Экология. М.: Изд-во МГУ, 1980. 464 с.
11. Эволюция глобального биогеохимического цикла серы / Ред. М.В. Иванов.
М.: Наука, 1989.
12. Martinus Willem Bejerinck: His life and his work / Ed. C.van Iterson, L.E. den
Dooren de Jons, A.J. Kluyver. Madison, (WI.): Publ. of Sci. Techn., 1983.
13. Fenchel Т., King G-M., Blackburn Т.Н. Bacterial biogeochemistry: The ecophysiol-
ogy of mineral cycling. San Diego etc.: Acad. press, 1998.
14. Graedel ТА., Crutzen PJ. Atmospheric change: An Earth system perspective. N.Y.:
Freeman, 1992. 448 p.
15. Lovelock J. The ages of Gaia: A biography of our living Earth. Oxford: Oxford Univ.
press, 1995. 256 p.
16. Microbial biogeochemistry / Ed. W.E. Krumbein. Publications, Oxford etc.:
Blackwell, 1983. 330 p.
17. Schlesinger W.H. Biochemistry. An analysis of global change. San Diego etc.: Acad.
press, 1997.
Лекция 2
МИКРОБНОЕ СООБЩЕСТВО
2.1. ТРОФИЧЕСКАЯ СТРУКТУРА СООБЩЕСТВА
Микробное сообщество представляет совокупность взаимодей-
взаимодействующих между собой функционально различных микроорганиз-
микроорганизмов. Они связаны единством времени и пространства. Микробное
сообщество подчиняется системным закономерностям. Организация
сообщества следует задаче обеспечить наибольшую устойчивость в
рамках естественного отбора. Устойчивость обеспечивается адапта-
адаптационной динамикой с заменой одних видов другими в зависимости от
внутренних причин при развитии сообщества в экосистеме (сукцес-
(сукцессия). Помимо топических ограничений - соответствия внешним ус-
условиям температуры, солености, подвижности среды - взаимодейст-
взаимодействие в сообществе ограничено внутренним транспортом и скоростью
передачи сигнала от одного компонента к другому. Для микробного
сообщества главную роль при этом играет молекулярная диффузия.
Виды в бактериальном сообществе существуют за счет катализиру-
катализируемых ими химических энергодающих реакций.
Из имеющегося набора функционально сходных организмов до-
доминируют те из них, чьи кинетические характеристики более всего со-
соответствуют условиям, складывающимся в сообществе. Сообщество с
химической точки зрения определяется термодинамикой и кинетикой
осуществляемых окислительно-восстановительных реакций. Но физи-
физико-химический подход дает только первую приблизительную картину
возможностей, реализация которых зависит от биологических особен-
особенностей организмов. К таким особенностям относятся, например, спо-
способность к выживанию, сопротивление выносу из системы, выеданию
и другие свойства, которые могут обеспечить процветание в экологи-
экологических нишах, входящих в абстрактные "фундаментальные ниши".
2.1.1. Кооперация в сообществе
Основу связей в сообществе, объединяющих его в единое целое,
представляют трофические связи, обусловленные образованием и
использованием веществ микроорганизмами. Если оставить в стороне
30
первичную продукцию фотоавтотрофных организмов, сообщество
которых организовано таким образом, чтобы обеспечить полное ис-
использование фотосинтетически активной радиации (ФАР), то тро-
трофическая структура деструктивной ветви подчиняется задаче полно-
полного использования энергии поступающих органических веществ. Для
бактерий использование термодинамически выгодных химических
реакций представляет главную возможность существования и обес-
обеспечивает соответствующую нишу. Естественный отбор заполняет
все вакансии в такой термодинамически детерминированной систе-
системе. Полнота использования энергии химических реакций и есть
главнейшее правило в организации трофической системы микроб-
микробного сообщества. Трофическая система микробного сообщества
организуется таким образом, чтобы любая возможность получения
доступной энергии была использована. По закону Гесса разница ме-
между начальным и конечным уровнем не зависит от пути достижения
этого состояния. Поэтому в сообществе может быть множество раз-
различных маршрутов, реализуемых в зависимости от внешних усло-
условий, однако все они должны дать возможность существовать орга-
организмам, катализирующим отдельные реакции. Эта возможность
определяется термодинамической достаточностью реакции для
обеспечения энергетики вида бактерий (см. ниже).
Трофическая организация бактериального сообщества основы-
основывается на специализации организмов по используемым субстратам и
образуемым продуктам. Тогда достаточно составить список орга-
организмов, образующих продукты, и сопоставить его со списком орга-
организмов, использующих эти вещества, чтобы получить картину тро-
трофических взаимодействий в сообществе (рис. 2.1). Для ее построения
используются три множества: веществ, организмов, взаимодейст-
взаимодействий. Она представлена ориентированным графом, в котором стрел-
стрелки изображают потоки вещества, узлы, изображенные кружками, -
пулы веществ, другие узлы в виде блоков — функциональные груп-
группировки организмов. Поскольку организмы образуют различные
продукты и могут использовать различные субстраты, то в один и
тот же узел могут входить и выходить из него несколько стрелок.
Граф трофических отношений в сообществе представляет транс-
транспортную сеть, аналогичную графу производства какого-либо изде-
изделия. Такая схема дает представление о качественном взаимодейст-
взаимодействии в системе и наиболее продуктивна для ее концептуального ана-
анализа. Переход к количественному описанию может быть получен,
если приписать группам организмов внутри блоков закономерности
роста в зависимости от поступления субстрата (например, по уравне-
уравнению Моно), для субстратов использовать текущие значения концен-
концентрации, стрелкам приписать характеристики транспорта, зависящие
от диффузии. В результате получается детерминированая система.
Однако вследствие разнообразия организмов она оказывается очень
изменчивой и плохо укладывающейся в жесткие рамки кинетических
31
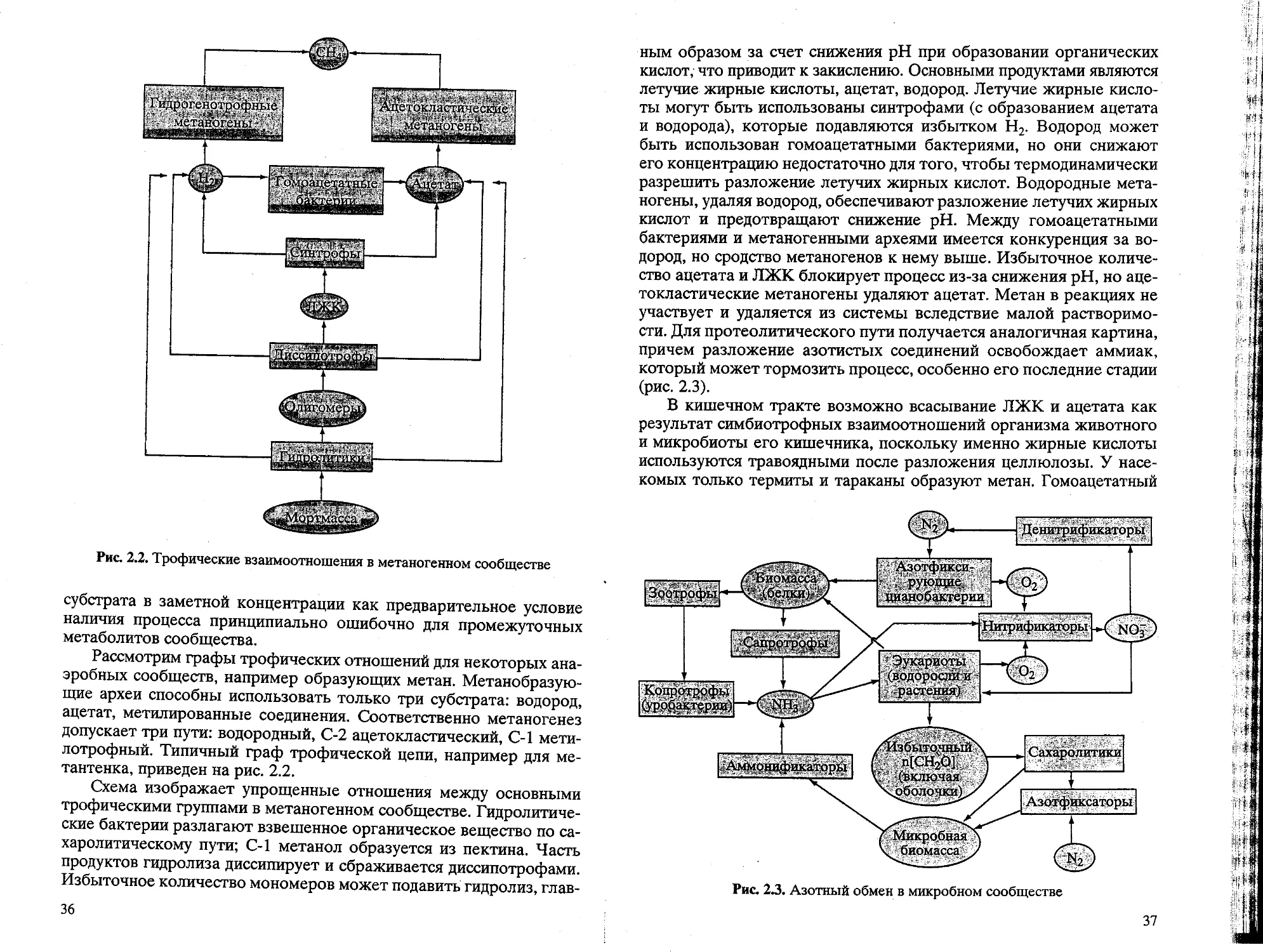
Зеленые, растения
1- ¦ и водоросли" ¦
Цианобактерии
Гидролйтики
>^ Олигомеры J)
Синтрофы| -{ Ацетат ) + ( Н2
Рис. 2.1. Трофические взаимоотношения в микробном сообществе
ограничений. Характеристики компонентов системы меняются в за-
зависимости от двух процессов: адаптации вида организма или же от
адаптивной динамики со сменой видов в функциональной группи-
группировке. Важно, что сообщество микроорганизмов функционирует
как единое целое с кооперативными трофическими связями, опреде-
определяющими план химических взаимодействий.
Ведущим в микробной системе служит цикл органического уг-
углерода. Для него важно построить карту метаболических связей с
указанием ключевых функциональных группировок. При этом в
сообществе бактерий одни организмы используют продукты дру-
других, и такая ступенчатая организация наиболее наглядна для разло-
разложения вещества. Продукт-субстратные взаимодействия между
компонентами сообщества лучше рассмотреть на примере анаэ-
анаэробного сообщества. Здесь последовательно действуют группы ор-
организмов, взаимодействие которых обеспечивает полное разложе-
разложение поступающего вещества в несколько этапов с максимальным
использованием энергии. Каждая группа отличается характерны-
характерными субстратами и продуктами обмена, что позволяет следить за их
деятельностью по результатам химического анализа. Функцио-
32
нальная группа микроорганизмов представляет компонент систе-
системы сообщества.
Поступающее органическое вещество физически разделяется на
две категории: 1) растворимое органическое вещество - РОВ (по-ан-
(по-английски DOC - dissolved organic compounds) и 2) взвешенное органи-
органическое вещество частиц - ВОВ (по-английски РОС - particulated
organic compounds). Растворенное органическое вещество наиболее
доступно и быстро используется бродилыциками, например молоч-
молочнокислыми или маслянокислыми бактериями при компостировании
растительных остатков. Основную часть мортмассы продуцентов
составляет нерастворимое органическое вещество, в частности обо-
оболочки клеток продуцентов. Примером их может служить целлюло-
целлюлоза. Вначале нерастворимое органическое вещество частиц должно
быть превращено в растворимое, так как прокариотная клетка в
принципе не обладает способностью к захвату частичек вещества
через клеточную стенку. Именно в этом состоит основное экофи-
зиологическое отличие прокариотных микробов от эукариотных
фаготрофных протестов.
Первый этап представляет гидролиз, осуществляемый гидроли-
тиками. Они проводят процесс, как правило, находясь в контакте с
твердой фазой гидролизуемых веществ, хотя действие обусловлено
внеклеточными гидролазами. Организмы должны обеспечить мини-
минимальное рассеяние продуктов гидролиза. Это может быть обеспече-
обеспечено физическим расположением клеток. Наиболее наглядно процесс
идет у целлюлозоразлагающих клостридий, которые имеют "цел-
люлосому" - экзоферментный комплекс, находящийся на слизевом
канате и способный тесно прилегать к поверхности целлюлозного
волокна, осуществляя его гидролиз. Образующийся в результате
гидролиза дисахарид целлобиоза уже может быть усвоен клеткой.
Условием работы экзофермента оказывается невозможность под-
подвода к нему метаболической энергии. Экзоферменты нескольких
клеток имеют кооперативное действие, и продукты гидролиза обоб-
обобществляются и могут быть использованы любой клеткой. Поэтому
гидролитические организмы образуют микроколонию на поверхно-
поверхности гидролизуемого субстрата, что снижает потери гидролизата от
рассеяния в среде. По другому варианту клетки целлюлозолитиче-
ских бактерий, например цитофаг, плотно прилегают к волокнам
целлюлозы, опять-таки с целью минимизировать потери на синтез
экзофермента и рассеяние гидролизата. Стратегией гидролитиков
является заселение поверхности нерастворимого субстрата, и их об-
обмен пропорционален доступной поверхности. Гидролйтики сущест-
существуют в условиях избытка субстрата, недоступного другим организ-
организмам, пока он не подвергнут гидролизу. Гидролйтики в особенности
специализированы по используемым веществам. В них следует раз-
различать группировки сахаролитических, протеолитических, липо-
литических организмов, использующих соответственно полимеры
2. Г. А. Заварзин
33
углеводов, азотистых соединений, липидов и продукты их гидроли-
гидролиза. Специализация выступает очень четко, например, для разложе-
разложения гетероциклических соединений. Она создает основание для
функционального биоразнообразия соответственно разнообразию
субстратов. Специализация по субстратам дает возможность обна-
обнаружить организм, приспособленный к его использованию в качест-
качестве предпочтительного источника, путем применения элективной ме-
методики С.Н. Виноградского.
Рассеиваемый при гидролизе полимера олигомерный продукт мо-
может быть использован "микрофлорой рассеяния", теперь называемой
диссипотрофамих. Они представлены в основном одиночными клет-
клетками, часто подвижными. В анаэробном сообществе их наглядными
представителями служат спирохеты с очень выгодным отношением
поверхности к объему. Зависимость диссипотрофов от гидролитиков
очевидна. Менее ясна зависимость гидролитиков от диссипотрофов.
Связана она в числе прочего с тем, что диссипотрофы снижают кон-
концентрацию продуктов гидролиза ниже порогового уровня, регулиру-
регулирующего синтез гидролаз, находящийся под метаболическим контро-
контролем. Диссипотрофам противоположны копиотрофы, использующие
высокие концентрации доступных веществ (при автолизе).
И гидролитики, и диссипотрофы в анаэробном сообществе не
нуждаются во внешнем стоке электронов и осуществляют броже-
брожение. Большинство из них способно к выбросу молекулярного водо-
водорода за счет выделяющей гидрогеназы. Особенности брожений свя-
связаны, как известно, с вариациями в использовании разных внутри-
внутриклеточных акцепторов водорода. Почти все продукты брожений не
могут быть сброжены далее и представлены, помимо водорода, низ-
низшими жирными кислотами, спиртами и аналогичными продуктами.
Анаэробные гидролитики и диссипотрофы объединяются в группи-
группировку первичных анаэробов, четко просматриваемую при анаэроб-
анаэробных процессах как бродилыцики, отвечающие за кислотогенную,
или иначе водородную фазу. Важнейшим их продуктом помимо Н2
является ацетат, а также летучие жирные кислоты (ЛЖК).
Группировка вторичных анаэробов включает организмы с ана-
анаэробным окислительным обменом, обусловленным использованием
несбраживаемых продуктов первичных анаэробов как доноров
электронов и внешних неорганических акцепторов электронов в
окислительно-восстановительных реакциях, приводящих к образо-
образованию полностью окисленного продукта разложения органического
вещества - СО2 - и восстановленного неорганического соединения.
Для неорганических соединений четко соблюдается термодинамиче-
термодинамическая последовательность окислительно-восстановительных потен-
потенциалов как предпочитаемых акцепторов; наименее выгодные тер-
модинамически акцепторы используются в последнюю очередь. По-
Поэтому развитие вторичных анаэробов возможно лишь в поле термо-
термодинамической устойчивости восстановленного продукта реакции в
координатах Eh-pU.
Важнейшими донорами электрона для вторичных анаэробов
служат: водород и/или НСООН; ацетат и/или этанол; летучие жир-
жирные кислоты (ЛЖК). Эти соединения лежат на главных трофиче-
трофических маршрутах. Последовательность использования акцепторов
электронов включает: A) восстановление Fe(III), B) восстановление
нитратов, денитрификацию, анаэробное образование аммония, C)
восстановление соединений серы в сероводород, или сульфидогенез,
D) восстановление углекислоты в метан, или метаногенез. Послед-
Последний процесс не нуждается во внешних источниках окислителя, по-
поскольку углекислота образуется при брожении. Каждый из этих
процессов наблюдается в природе в определенных условиях в зави-
зависимости от поступления акцептора. Их последовательность четко
просматривается во времени, например при затоплении почвы, и в
пространстве, например по профилю иловых осадков океана.
Вторичные анаэробы используют все продукты первичных ана-
анаэробов. Поэтому получается трофическая структура сообщества с
продукт-субстратным взаимодействием как основным принципом
организации. Продукты обобществляются, образуя пулы. Посколь-
Поскольку разные организмы образуют сходные продукты, то ребра от не-
нескольких узлов сходятся вместе и вместо цепей получаются сети.
Важнейшим узлом служит образование ацетата, или ацетогенез,
представляющий промежуточный этап между первичными и вто-
вторичными анаэробами. Точно так же в зависимости от доступного ак-
акцептора электрона сходятся вместе и ребра на единый продукт вос-
восстановления неорганического акцептора, например H2S при сульфи-
догенезе. Для того чтобы сообщество существовало сколько-нибудь
длительное время и было автономным, необходимо, чтобы дест-
деструкция была полной и не накапливалось бы никаких продуктов. На
самом деле это требование выполняется лишь отчасти, и происходит
накопление не полностью разложенных или устойчивых (как гуму-
гумусовые вещества) продуктов обмена сообщества, что ведет к сукцес-
сукцессии - последовательной смене сообществ.
Для автономного сообщества можно написать цикл, где заглав-
заглавными буквами обозначены организмы, а строчными — их продукты.
Начинается цикл с первичных продуцентов автотрофов и заканчи-
заканчивается полным разложением ("минерализацией", если использовать
старый, очень неудобный термин):
СО2
Z -> CO2.
1 Васильева Л.В., Заварзин Г.А. Диссипотрофы в микробном сообществе //
Микробиология. 1995. Т. 64, № 2. С. 239-244.
34
Заметим, кстати, что высокая концентрация какого-либо проме-
промежуточного вещества в сообществе означает, что эффективность
его потребления недостаточна. Требование определять наличие
г* 35
Рис 2.2. Трофические взаимоотношения в метаногенном сообществе
субстрата в заметной концентрации как предварительное условие
наличия процесса принципиально ошибочно для промежуточных
метаболитов сообщества.
Рассмотрим графы трофических отношений для некоторых ана-
анаэробных сообществ, например образующих метан. Метанобразую-
щие археи способны использовать только три субстрата: водород,
ацетат, метилированные соединения. Соответственно метаногенез
допускает три пути: водородный, С-2 ацетокластический, С-1 мети-
лотрофный. Типичный граф трофической цепи, например для ме-
тантенка, приведен на рис. 2.2.
Схема изображает упрощенные отношения между основными
трофическими группами в метаногенном сообществе. Гидролитиче-
Гидролитические бактерии разлагают взвешенное органическое вещество по са-
харолитическому пути; С-1 метанол образуется из пектина. Часть
продуктов гидролиза диссипирует и сбраживается диссипотрофами.
Избыточное количество мономеров может подавить гидролиз, глав-
36
ным образом за счет снижения рН при образовании органических
кислот, что приводит к закислению. Основными продуктами являются
летучие жирные кислоты, ацетат, водород. Летучие жирные кисло-
кислоты могут быть использованы синтрофами (с образованием ацетата
и водорода), которые подавляются избытком Н2. Водород может
быть использован гомоацетатными бактериями, но они снижают
его концентрацию недостаточно для того, чтобы термодинамически
разрешить разложение летучих жирных кислот. Водородные мета-
ногены, удаляя водород, обеспечивают разложение летучих жирных
кислот и предотвращают снижение рН. Между гомоацетатными
бактериями и метаногенными археями имеется конкуренция за во-
водород, но сродство метаногенов к нему выше. Избыточное количе-
количество ацетата и ЛЖК блокирует процесс из-за снижения рН, но аце-
токластические метаногены удаляют ацетат. Метан в реакциях не
участвует и удаляется из системы вследствие малой растворимо-
растворимости. Для протеолитического пути получается аналогичная картина,
причем разложение азотистых соединений освобождает аммиак,
который может тормозить процесс, особенно его последние стадии
(рис. 2.3).
В кишечном тракте возможно всасывание ЛЖК и ацетата как
результат симбиотрофных взаимоотношений организма животного
и микробиоты его кишечника, поскольку именно жирные кислоты
используются травоядными после разложения целлюлозы. У насе-
насекомых только термиты и тараканы образуют метан. Гомоацетатный
Денитрификаторы
Зоотрофы
збытонный
„. п[СН2О].
\ (включая
оболочки
.Л
i.
I.J.
Рис. 2.3. Азотный обмен в микробном сообществе
37
путь, хотя и дает ацетат, вместе с тем приводит к потере сообщест-
сообществом части водорода.
Трофическая система органотрофов определяется в первую
очередь правилом Виноградского A896): для каждого природного
вещества есть микроорганизмы, способные его разложить. Отсю-
Отсюда можно представить себе совокупность органотрофов, располо-
расположенных на маршрутах, ведущих от компонентов мортмассы к СО2
как конечному продукту. Соответственно можно обозначить груп-
группы маршрутов как протеолитический, сахаролитический, липолити-
ческий пути. Помимо бактерий к аэробным органотрофам относят-
относятся грибы. Микромицеты составляют трофические группировки,
аналогичные группировкам бактерий, превосходя их в отношении
ксерофитизма и связанной с ним возможности обитать в среде влаж-
влажного воздуха.
В отношении набора используемых веществ органотрофы раз-
разделяются на политрофов (полифагов) и монотрофов (монофагов).
Первые способны использовать широкий набор веществ, вторые ог-
ограничены по пищевым возможностям. Такое деление не абсолютно
жестко, но для организмов есть предпочтительные субстраты.
Например, организмы мочки льна ("Bacillus felsineus") предпочита-
предпочитают пектин, хотя растут и на растворимых углеводах. Маннит пред-
почителен для Azotobacter, хотя он растет и на других веществах.
Практика выделения микроорганизмов оставила рецептуру для пре-
преимущественного роста тех или иных видов на определенных
субстратах. Для выделения политрофов подходят "универсальные"
среды, обычно отвары вроде мясо-пептонного, соевого или дрожже-
дрожжевого агара с добавлением глюкозы. Легкодоступные вещества выхо-
выходят из клеток при превращении биомассы в мортмассу в результате
автолиза. Они составляют примерно 20% органического вещества
биомассы. Их используют зямотетл-копиотрофы (от лат. copia -
обилие, множество). Политрофы находятся в состоянии общей кон-
конкуренции, в то время как специализация монотрофов выводит их из
конкуренции в самостоятельные экологические ниши.
Используемые вещества делятся на легко- и труднодоступные. К
первым относятся мономеры и растворимые полимеры. К труднодос-
труднодоступным относятся нерастворимые полимеры. Доступность органиче-
органических веществ зависит от их агрегатного состояния. Нерастворимые в
воде липиды относятся к труднодоступным веществам, хотя они
могут содержать и мономеры. В большей степени это относится к ве-
веществам структурных компонентов клеток, по существу предназна-
предназначенных противостоять разрушению. Они составляют основную часть
мортмассы и в природе описываются как взвешенные органические
вещества (ВОВ), часто состоящие из остатков организмов.
Для использования нерастворимых полимеров необходим их
гидролиз, который, очевидно, зависит от поверхности, доступной
для заселения бактериями. Гидролитические организмы достаточно
38
Трофические
группы
Продуценты
Гидролитики
Диссшютрофы
Синтрофы
Вторичные
анаэробы
Газотрофы
Автохтоны
Копиотрофы Социалисты Автохтоны
Зимогены Кооператоры Олиготрофы
Время сукцессии
t°
Топические
характеристики
местообитания
(комбинаторная
матрица)
Рис. 2.4. Функциональное разнообразие микробных сообществ
специфичны по используемым субстратам: одни гидролизуют цел-
целлюлозу, другие - хитин, третьи способны разлагать (хотя и с трудом)
кератин. Гидролитическим организмам сопутствуют диссипотрофы,
использующие низкие концентрации рассеиваемого вещества.
Наконец, остаточное органическое вещество, наиболее труднодос-
труднодоступное, как лигнин, и возникший в результате поликонденсации гумус,
разлагают автохтонные организмы, часто в процессе соокисления.
Общую картину разложения мортмассы продуцентов аэробны-
аэробными деструкторами можно представить в виде части схемы экологи-
экологических вакансий в цикле углерода. Функциональное разнообразие
микроорганизмов при этом дополняется топическим разнообразием
мест обитания: каждому местообитанию соответствует свой набор
организмов ацидофильных или алкалофильных, галофильных или
обитателей ультрапресных вод. Основная масса принадлежит орга-
организмам "нормальных" мест обитания, в которых растительность да-
дает больше органического вещества. Именно для них культуральные
методы дали больше всего представителей (рис. 2.4).
Таким образом, в сообществе принципиально важны обратные
связи, и оно действует как метаболическое целое.
39
2.1.2. Энергетика сообщества
Микроорганизмы получают энергию для своей жизнедеятель-
жизнедеятельности в результате окислительно-восстановительных реакций, пред-
представляющих перенос электрона от донора к акцептору. Каждый
отдельный организм представляет электрохимический элемент,
осуществляя две полуреакции, разделенные мембраной. Отсюда,
кстати, следует вывод, что жизнь может существовать только в дис-
дискретной форме и никакого "живого вещества" быть не может в
принципе.
По сути, в энергетике микроорганизмов мы имеем дело с элект-
электрохимической реакцией. Микробиологам термодинамические рас-
расчеты в координатах Eh-pH особенно удобны для выяснения обмена
литотрофных организмов. Общее правило такое: хемолитотроф-
ный организм может развиваться в области термодинамической
устойчивости продукта энергодающей реакции.
Возможность развития бактерий обусловливается термодинами-
термодинамикой реакции катаболизма. Урожай определяют как УАТР - число
граммов биомассы, синтезированной за счет 1 моля АТФ, образо-
образовавшегося в метаболизме. У большого числа анаэробов эта величи-
величина оказалась близкой к 10 г абсолютно сухой биомассы (а.с.б.) на
1 моль АТФ. ГАТР - это не константа и зависит от условий роста, ха-
характера синтезируемых запасных продуктов, скорости роста. При
росте на глюкозе теоретический урожай на 1 АТФ может составлять
27 г/моль; на ацетате с углекислотой - 15 г/моль; на углекислоте -
5 г/моль. Экспериментально полученный урожай составляет около
50% от теоретического. Остальное расходуется на "энергию поддер-
поддержания".
Анализ термодинамики микроорганизмов был проведен Р. Тау-
Тауэром2. При этом энзиматический механизм получения энергии —
биоэнергетика клетки — остается за рамками общей картины, одна-
однако важен конечный результат — образование АТФ. Формально
можно представить энергетический обмен организмов как образо-
образование и потребление водорода в окислительно-восстановительной
реакции, где Н+ - акцептор, а Н2 - донор электрона. Синтез АТФ из
АДФ требует 32 кДж на моль при стандартных условиях. В клетке
содержание АТФ и фосфата около 10 мМ, АДФ - около 1 мМ, и фи-
физиологический синтез требует 49 кДж. Необратимые затраты соста-
составляют 20 кДж. Следовательно, суммарные затраты составляют при-
примерно 70 кДж/моль и даже при понижении энергетического заряда
клетки не становятся ниже 60 кДж. На синтез 1 АТФ требуется пе-
перенос трех зарядов через клеточную мембрану. Отсюда предполо-
предположение, что наименьшим используемым квантом энергии в катабо-
катаболизме является эквивалент АТФ, оказывается неверным. Мини-
Минимальное количество энергии соответствует 20 кДж/моль субстрата в
энергетической реакции, связанной с переносом протона. Для того
чтобы реакция могла быть энергодающей для популяции вида орга-
организмов, плотность потока вещества должна быть достаточной, что-
чтобы обеспечить поддержание популяции организмов вопреки их есте-
естественному отмиранию и гибели. Если же поток энергии от реакции
определенного вещества через локальное место обитания организ-
организмов недостаточен для поддержания популяции, то маловероятно и
существование специализированной группы организмов. Вопрос со-
состоит в том, насколько общие закономерности биоэнергетики, уста-
установленные для отдельного организма, могут быть применены для
сообщества в целом. В начале 1960-х годов Баас-Бекинг, Каплан и
Мур3 составили диаграммы распространения основных физиологи-
физиологических групп в координатах ?h-pH, аналогичные диаграммам полей
устойчивости минералов. На этих диаграммах приведены поля
встречаемости биологических реакций, свойственных характерным
группам процессов: сульфидогенезу, метаногенезу, окислению же-
железа. Различные реакции наблюдаются практически по всему полю
наблюдаемых в природе значений Eh-pH, совместимых с существо-
существованием воды.
Подобно тому как внутри клетки микроорганизма сопрягаются
энергодающие и энергопотребляющие реакции, в химической сис-
системе сопряженных окислительно-восстановительных реакций, осу-
осуществляемых микробным сообществом, возможно такое их сочета-
сочетание, которое в сумме дает энергетический выигрыш, достаточный
для существования сообщества. Но в отличие от клетки, где есть пе-
переносчик энергии, в сообществе условия должны быть таковы, чтобы
обеспечить выигрыш, достаточный для существования каждой от-
отдельной группы или вида организмов.
2.1.3. Межвидовой перенос водорода.
Синтрофия
Принципиальным открытием в области энергетики взаимо-
взаимодействия между организмами было открытие катаболической
синтрофии М. Брианом и Е. Волиным в 1970-х годах на основе
межвидового переноса водорода. Катаболическая синтрофия не
позволяет ставить знак равенства между метаболизмом чистой
культуры и возможностями питания организмов в сообществе.
Явление катаболической синтрофии означает, что сообщество
микроорганизмов можно и следует рассматривать как биологиче-
2 Thauer R., Jungerman К., Decker К. Energy conservation in anaerobic chemotrophic
bacteria//Bacteriol. Rev. 1977. Vol. 41. P. 100-180.
40
3 Басс-Бекинг Л.Г.М., Каплан И.Р., Мур Д. Пределы колебаний рН и окисли-
окислительно-восстановительных потенциалов природных сред // Геохимия литоге-
литогенеза. М., 1963. С. 11-84.
41
ское единство, обусловленное термодинамическими возможностя-
возможностями. Термин "синтрофия" применялся для обозначения взаимодей-
взаимодействия организмов, окисляющих жирные кислоты, с водородными
метаногенами и взаимодействия фототрофных зеленых серных
бактерий с сульфатредукторами. В первом случае работал водо-
водородный цикл, во втором - серный. Ключевую проблему представ-
представляет концентрация переносчика, которая должна быть ниже пода-
подавляющей для первой бактерии и выше нижнего предела для
потребляющей.
Вопрос состоит в том, может ли концентрационный член в
уравнении Гиббса служить достаточным источником энергии для
системы из двух организмов? Концентрационный член приобрета-
приобретает определяющее значение, если продукт эффективно удаляется
из сферы реакции и его концентрация поддерживается малой по
отношению к субстрату. Выход энергии может быть достаточно
малым, если используется не брожение, а хемиосмотический
механизм.
В 1936 г. Г. Баркер, работая в Дельфте, описал под названием
"Methanobacillus omeljanskii" споровый организм, который образо-
образовывал метан из этанола. Потом в Калифорнии ученица Баркера
Т. Штадтман описала еще ряд культур метаногенов, которые пре-
превращали в метан органические кислоты. Казалось бы, была решена
задача описания биологического процесса метанового сбраживания
органических кислот, которое с очевидностью наблюдалось на пра-
практике в метаногенном сообществе. В 1970-е годы с развитием стро-
строго анаэробной техники было показано, что большинство метаноге-
метаногенов использует водород и есть только два организма, способные ис-
использовать ацетат: Methanosarcina и Methanothrix (=Methanosaeta).
Бриан и Волин доказали, что на самом деле культура
"Methanobacillus omeljanskiF содержит два организма: один - споро-
споровый, близкий к Clostridium kluyveri, который разлагает этанол с об-
образованием ацетата и выделением Н2, и второй - M.o.h. - водород-
водородную метанобразующую бактерию, которая превращает Н2 в
инертный метан. Реакция выделения Н2 из этанола термодинами-
термодинамически невыгодна и становится возможной лишь при глубоком уда-
удалении Н2 из сферы реакции. В результате суммарная реакция тер-
термодинамически возможна и способна обеспечить оба организма
необходимой энергией. Реакция была названа межвидовым пере-
переносом водорода.
За этой работой последовала целая серия других, и был разо-
разобран ряд реакций с бинарными культурами. Организмы, способные
к развитию при условии, что Н2 удаляется из сферы реакции, полу-
получили название синтрофов и составили особую функциональную
группу в сообществе. Межвидовой перенос водорода составляет
промежуточное звено между первичными и вторичными анаэроба-
анаэробами. Синтрофы развиваются совместно с Н2-использующими анаэ-
42
ровными организмами гидрогенотрофами. Среди них важны две
группы: литотрофные метаногены и литотрофные сульфидогены.
Был описан целый ряд культур, в которых анаэробное окисление
субстрата обусловлено присутствием Н2-использующего компонен-
компонента. Для термодинамической разрешенности за счет удаления Н2 из
сферы реакции важен тот нижний предел концентрации, до которо-
которого тот или иной организм способен удалять Н2. Довольно быстро
было установлено, что сульфатвосстанавливающие бактерии в этом
отношении эффективнее метаногенов. Но для независящего от
внешних акцепторов сообщества важны метаногены, поскольку
СО2, служащий акцептором для них, образуется первичными анаэ-
анаэробами. Такая работа была выполнена с чистыми культурами мета-
метаногенов на основе использования высокочувствительного твердо-
электролитного детектора Н2 у нас в лаборатории Института мик-
микробиологии РАН. Концентрацию Н2 измеряли в газовой фазе, кото-
которая предположительно находится в равновесии с жидкой по закону
Генри (концентрация растворенного газа находится в равновесии с
газовой фазой пропорционально парциальному давлению). Содер-
Содержание газа измеряется в ppm (partes pro million), т.е. 1О6 по объему.
Оказалось, что водородные метаногены имеют определенные поро-
пороги, до которых они могут снижать содержание Н2. Для мезофильных
Methanobacterium эта величина была около 5 ppm, а для
Methanosarcina - около 50 ppm. Для термофильных Methanobacterium
пороговая величина была существенно выше, чем для мезофильных,
и составляла 20-30 ppm. Для метаногенного сообщества навоза (а
оно представляет богатейший набор разных видов) пороговые кон-
концентрации точно соответствовали пороговым значениям для чистых
культур водородных метаногенов. Повышение концентрации Н2
служит сигналом разбалансированности в работе сообщества вслед-
вследствие какого-либо стресса. Определение нижнего порога концент-
концентрации Н2 стало стандартным приемом и осуществляется очень про-
просто - определяют концентрацию Н2 в культуре в конце стационар-
стационарной фазы роста при лимите по водороду.
Выделение и потребление Н2 осуществляется гидрогеназами,
катализирующими обратимую реакцию Н2 = 2Н+ + 2е-. Следова-
Следовательно, можно ожидать, что устанавливаются некоторые равно-
равновесные концентрации Н2. Впервые такая равновесная концентра-
концентрация была установлена при разложении метанола Methanosarcina.
При выделении чистой культуры метаносарцины Т.Н. Жилина
обнаружила в качестве упорного спутника Desulfovibrio vulgaris,
который метанол не использует, а на Н2 растет. Оказалось, что
метилотрофные метаногены при росте на метиламинах поддержи-
поддерживают стационарную концентрацию Н2 за счет его выделения.
Таким образом, в бинарной культуре Methanosarcina +
Desulfovibrio метаноген выполнял роль синтрофа по отношению к
сульфидогену.
43
Сейчас известен ряд пороговых значений для разных групп вто-
вторичных Н2-использующих литотрофных анаэробов. Они располага-
располагаются в следующем порядке:
Гомоацетатные Около 1000 ррт
Метанобразующие 100-10
Сульфатвосстанавливающие 10-1
Железовосстанавливающие 1-0,1
Аэробные водородные Менее 0,1
Межвидовой перенос водорода подвел черту под эпохой исклю-
исключительного господства чистых культур в бактериологии, юридиче-
юридически закрепленного Иерусалимским кодексом номенклатуры бакте-
бактерий, по которому узаконивались только "чистые культуры в между-
международно признанных коллекциях". Этот перелом проходил в 1980-х
годах и до сих пор еще не вполне осознан представителями физиоло-
го-биохимического направления и клеточной биологии. Для приро-
природоведческого направления общей микробиологии наряду с чистой
культурой объектом исследования стало микробное сообщество.
Термодинамический выигрыш при удалении продукта реакции
может быть получен не только для Н2, но и для других соединений.
Из них особенное внимание привлек формиат, который обладает
высокой растворимостью и не переходит в газовую фазу. Фор-
Формиат образуется в ряде смешанных брожений, особенно у энтеробак-
терий и клостридий. Он легко разлагается формиатлиазой по реак-
реакции НСООН = СО2 + Н2. Чувствительность и легкость методик оп-
определения формиата не так удобны, как для Н2, и отчасти поэтому
ясности здесь нет. Мнение, что формиат может быть объектом меж-
межвидового переноса, сомнению не подвергается, но значение этого
процесса в природе оспаривается.
2.1.4. С-2 путь и ацетогенез
Главным продуктом первичных анаэробов и таких вторичных
анаэробов, как гомоацетогены и сульфатредукторы с неполным
окислением, является ацетат, и проблема его метаболизма оказы-
оказывается центральной для сообщества. Концентрация ацетата в анаэ-
анаэробных осадках довольно высока и достигает миллимолярных зна-
значений. Ацетат очень хорошо используется аэробными бактериями,
если он выходит в область поступления кислорода. Формально
можно указать на целый ряд анаэробных организмов, способных
использовать ацетат. К ним относятся факультативно анаэробные
бактерии, восстанавливающие железо или нитрат, некоторые
сульфатредукторы, как Desulfobacter, железоредукторы. В фоти-
ческой зоне ацетат и другие жирные кислоты хорошо используют
различные аноксигенные фототрофы. В метантенках 70% метана
образуются через ацетат, но пока, как уже отмечалось, известно
44
только два ацетокластических метаногена - Methanosarcina и
Methanothrix, причем второй обладает на порядок большим сродст-
сродством к ацетату. Далее, ацетат могут использовать сульфатредукто-
сульфатредукторы с полным окислением жирных кислот. В накопительных куль-
культурах сульфатредукторов давно показано, что ацетат служит
лучшим субстратом для сульфидогенеза и, вероятно, хорошо ис-
используется в природе.
Наконец, серовосстанавливающие сульфидогены ориентирова-
ориентированы на ацетат, и известны два характерных представителя - мезо-
фильный Desulfobulbus и умеренно термофильная Desulfurella. Не-
Недавно открытые железовосстанавливающие бактерии, например
Geobacter, используют ацетат как предпочтительный субстрат, но
опять-таки пороговая концентрация для них не определена. Ацетат
может использоваться синтрофно с выделением Н2 по образцу дру-
других жирных кислот, как это показал Вигель для термофильного ме-
таногенного сообщества. Такая же ситуация имеет место для алка-
лофилов.
Межвидовой перенос веществ и термодинамические ограниче-
ограничения, обусловленные концентрацией метаболитов в анаэробном со-
сообществе, демонстрируют значение кооперации в трофической
системе, собранной из организмов с разной специализацией. Эти ор-
организмы не только не конкурируют друг с другом, но жестко взаи-
взаимозависимы. Рассматривая парные взаимодействия в трофической
системе, удалось установить, что, как правило, наиболее тесные вза-
взаимодействия наблюдаются у организмов крайне удаленных друг от
друга филогенетически групп. Например, археи-метаногены зависят
от грамположительных первичных анаэробов. Диссипотрофы-спи-
рохеты зависят от грамположительных гидролитиков. Анаэробное
окисление метана, установленное геохимиками и долгое время не
поддававшееся микробиологическому решению с поиском чистых
культур, оказалось зависящим от консорциума микроорганизмов, в
котором археи-метаногены из Methanosarcinales занимают централь-
центральное положение в микроколонии, а протеобактерии сульфатредукто-
сульфатредукторы Desulfosarcina — периферическое. Отсюда следует, что сообще-
сообщество возникает комбинаторным путем по требованиям кооперации,
а не обусловленной конкуренцией дивергенцией (Заварзин, 2000).
Представленная картина возникновения сообществ на основе
кооперативного взаимодействия между специализированными орга-
организмами, в предельном случае монотрофами, основана на традиции,
восходящей к С.Н. Виноградскому и М.В. Бейеринку. Существует и
противоположный взгляд, восходящий к Л. Пастеру и Р. Коху, осно-
основанный на представлении о широкой приспособляемости организ-
мов-политрофов. Он был сформулирован Г.А. Надсоном4: "...мик-
4 Надсон Г.А. Микроорганизмы как геологические деятели. СПб. Типография
Т.П. Сойкин, 1903. 98 с. См.: Г.А. Надсон. Избр. тр. М.: Наука, 1967. Т. 1.
45
робы одинаковы, а условия различны; поэтому различны и резуль-
результаты". Для аэробных органотрофов почвы такой взгляд развил
Д.Г. Звягинцев5. Он исходит из того, что в почве содержится боль-
большой пул организмов, которые могут приспособиться к использова-
использованию разных веществ, когда они поступают в почву. Например, ба-
бациллы или псевдомонады способны расти на десятках или сотнях
разных веществ, и, прежде чем специалист успеет размножиться,
многочисленные универсалы используют поступившие субстраты.
Кроме того, в почве микроорганизмы разделены пространственно и
растут в виде микроколоний как чистые культуры. Гидролитиче-
Гидролитические ферменты обильны и устойчивы в почве, и продукты гидроли-
гидролиза общедоступны. Поэтому нельзя выделять организмы сахаролити-
ческого, протеолитического, липолитического и т.д. путей. Почвен-
Почвенные организмы представляют не кооперативное сообщество, а ком-
комплекс (ассоциацию) независимых видов. Такие комплексы приуро-
приурочены к определенным условиям и географическим зонам. Основной
материал для исследования комплексов был получен в традициях са-
санитарно-гигиенической школы высевом на плотные среды для ин-
индикаторных организмов. Другим методом служит мультисубстрат-
ный анализ почвенной болтушки по системе "Эколог", разработан-
разработанной П.А. Кожевиным. Метод заключается в исследовании немед-
немедленного использования почвенной микробиотой набора из 30-70
субстратов по образованию окрашенного формазана из тетразолия.
При этом получаются статистически достоверные различия между
разными образцами, которые, однако, трудно интерпретировать
концептуально. Эти крайние взгляды противопоставлены подходу
С.Н. Виноградского и традиции общей микробиологии. Они основа-
основаны на материале доминирующей в почве группировки аэробных ор-
органотрофов. Для такой группировки возможен выбор между адапта-
адаптацией уже накопившихся организмов и адаптивной динамикой раз-
размножающихся.
2.1.5. Экологические ниши
Функциональная группа в сообществе соответствует фундамен-
фундаментальной экологической нише. Внутри каждой ниши (прямоугольника
на наших схемах) действует правило Гаузе: только один вид может
доминировать в данных условиях. Это правило совершенно анало-
аналогично законам рыночной экономики, где устойчивым конечным со-
состоянием является монополия на товар. Конкуренция продолжается
до вытеснения конкурентов, как это происходит в накопительной
культуре. Уцелевают спутники, которые составляют интересней-
интереснейшую часть группировки функциональной ниши, осуществляя либо
5 Д.Г. Звягинцев. Почва и микроорганизмы. М.: Изд-во МГУ, 1987. 256 с.
46
удаление токсического побочного продукта, например альдегидов
в метанотрофном сообществе, либо снабжая их факторами роста.
Здесь рассмотрена функциональная ниша в сообществе, которая
определяется взаимодействием с другими компонентами сообщест-
сообщества. Для взаимодействия существенны лишь некоторые внешние
характеристики организмов, в то время как другие, обращенные во
внутреннюю сферу клетки, второстепенны для деятельности сооб-
сообщества.
Другую категорию составляют внешние, топические, факторы,
важные для всего сообщества и каждого вида в нем. Они определя-
определяются экологией сообщества и вида и характеризуются как экологи-
экологические ниши. Для бактерий аутэкологические характеристики
обычно описываются как их экофизиология в зависимости от внеш-
внешних факторов. Для описания свойств организма недостаточно
физиологии, потому что выигрыш для вида во многом зависит от
морфологии, например подвижности, способности к прикреплению,
образованию гликокаликса, мицелиального роста. Многие из этих при-
признаков контролируются внешними условиями, достаточно вспом-
вспомнить разнообразие морфологии цианобактерий, относящихся к
одной трофической группе, и разнообразие форм роста одной и той
же цианобактерий в зависимости от условий. Ряд морфологических
признаков определяет нахождение организма внутри сообщества с
его определенной архитектурой, как в биопленках.
Экологическая ниша находится под воздействием осциллирую-
осциллирующих внешних факторов, к которым относится, например, смена дня
и ночи с разным поступлением кислорода и экзометаболитов фото-
фотосинтетиков. На примере циано-бактериального мата можно проил-
проиллюстрировать, к каким разнообразным воздействиям приводит это
неизбежное изменение внешних условий, о котором склонны забы-
забывать исследователи лабораторных культур. Организмы циано-бак-
циано-бактериального сообщества должны переходить от крайних условий ги-
пероксии к восстановительным условиям сульфидогенеза.
Экологическая ниша находится под флуктуирующим воздейст-
воздействием среднесрочных внешних факторов - смены сезонов и погоды
с колебаниями осадков и температуры, стрессовых штормовых
явлений.
Экологическая ниша находится под воздействием долгосрочных
закономерных сукцессионных изменений в сообществе, наиболее
наглядным проявлением которых является изменение концентрации
субстрата, поступающего к функциональной группировке, а также
накопление продуктов обмена сообщества, прямых или обусловлен-
обусловленных биологически опосредованными изменениями в местообитании,
например цементацией порового пространства.
Экологическая ниша приурочена к определенным ландшафт-
ландшафтным условиям, например гидротермам, гиперсоленым лагунам, ще-
щелочным озерам, не говоря об очень специализированных местах
47
обитания вроде областей окисления сульфидных руд. При этом раз-
разнообразие экстремофилов ограничено доминированием немногих
видов ("правило Тинемана"). Вместе с тем "нормальные" места оби-
обитания дают возможность существования большому числу особей,
конкуренция между которыми становится очень жесткой.
Выход из проблемы находится в биоразнообразии представите-
представителей одной функциональной группы. В главе 4 "Происхождения ви-
видов" Ч.Дарвин писал6, что биоразнообразие есть путь gjn конкурен-
конкуренции, т.е. фактически уход из-под действия естественного отбора. Ес-
Естественный отбор на самом деле представляет путь к унификации,
принадлежности всех к одной партии победителей. Для "нормаль-
"нормальных" условий, например в почве, возможности конкуренции шире,
так как приток субстратов велик, и соответственно возможно более
широкое разнообразие организмов. Это разнообразие связано с на-
набором используемых субстратов. Многие органотрофные организ-
организмы, как псевдомонады, например, способны использовать в качест-
качестве единственного источника углерода и энергии сотню органических
веществ, лишь бы продукты разложения были совместимы с цент-
центральным метаболизмом. Они универсальны по своим пищевым по-
потребностям и оказываются первыми, которые используют необыч-
необычный субстрат, например ксенобиотик. Такого рода потенции дают
уникальную пищевую нишу. Организмы этого типа легче всего вы-
выделяются в лабораторную культуру. Другие организмы, особенно
среди жестко лимитированных по энергии анаэробов, оказываются
способными использовать лишь ограниченный набор субстратов.
Итак, имеются две стратегии: либо использовать возможности орга-
организма для потенциально широкого круга субстратов -
политрофная стратегия, либо сконцентрировать их на развитии
транспортных систем с высоким сродством к узкому кругу субстра-
субстратов - стратегия монотрофов.
Исследование поведения вида в разных экологических условиях
называется аутэкологией и часто обозначается просто как эколо-
экология вида. В значительной степени подход связан с обнаружением ви-
вида в определенных условиях, обычно регистрируемым в флористи-
флористических списках, например для цианобактерий, грибов. Для бактерий
в связи с их повсеместным распространением и микроочаговостью
мест обитания этот подход обычно рассматривается лишь как тен-
тенденция. Описание вида обычно включает важные для приуроченно-
приуроченности к месту обитания топические характеристики. В противополож-
противоположность аутэкологии поведение сообщества организмов обозначается
как синэкология.
Для классификации поведения разных видов в местах их обита-
обитания применяются разные подходы. Они имеют значение главным
образом для наиболее многочисленных функциональных групп в со-
сообществе: во-первых, первичных продуцентов, представленных ок-
сигенными фотоавтотрофами, и, во-вторых, аэробных деструкто-
деструкторов с органогетеротрофным типом обмена. Между ними велика
конкуренция. Для относительно немногочисленных представителей
высокоспециализированных групп большее значение имеет их поло-
положение в трофической системе, определяемое спецификой катабо-
катаболизма и морфо-физиологическими критериями, а также кинетикой
роста.
По стратегиям роста в соответствии с логистическим уравне-
уравнением Ферхюльста A838):
t = rN(l-NIK),
6 Дарвин Ч. Происхождение видов. Сочинения Чарльза Дарвина. СПб., 1896.
Т. 1, ч. 1.
48
где N - число особей, г - коэффициент прироста, К - емкость среды,
a t - время, выделяют два типа:
r-стратеги, которые обладают высокой удельной скоростью ро-
роста, приближающейся к максимальной при освоении субстрата или
места обитания; это колонизаторы, пионерные формы, зимогены,
по терминологии Виноградского;
Х-стратеги, приспособленые к существованию при насыщении
емкости среды обитания; это представители климаксного, или зре-
зрелого, состояния сообщества.
Эти две стратегии сопоставляются с обилием субстрата. Орга-
Организмы, развивающиеся при обилии субстрата, получили название
копиотрофов, а при недостатке - олиготрофов. Количественная
оценка притока субстрата для водных копиотрофов составляет бо-
более 50 мг С/(л • сут), а для олиготрофов - менее 1 мг С/(л • сут). (Коли-
(Количественные оценки получены из хемостатных опытов.) Оба понятия
при всем их удобстве в практическом употреблении не определены
четко. Казалось бы, среди Х-стратегов должны были бы доминиро-
доминировать олиготрофы, но это не совсем так. В зрелом сообществе боль-
большее значение приобретают межвидовые связи, в то время как
r-стратеги менее зависят друг от друга. Лабораторные чистые куль-
культуры по определению должны быть представлены r-стратегами, не
зависящими от других членов сообщества и способными развивать-
развиваться в свежей среде, колонизовать ее. Поэтому в коллекциях живых
культур организмов преобладают именно копиотрофы. Приведен-
Приведенные жизненные стратегии обусловлены временем развития сообще-
сообщества в данном месте обитания, или сукцессией, последовательностью
смены видов в нем.
Если в целом сообщество организмов представляет кооператив-
кооперативную систему из взаимодействующих функциональных (физиологи-
(физиологических) групп, то внутри каждой группы предполагаются конку-
конкурентные отношения в соответствии с правилом Гаузе о стремлении
к доминированию и монополизации ниши. Конкуренция бактерий со
сходными пищевыми потребностями основана на кинетике роста.
49
2.1.6. Терминология
Взаимодействия в микробных сообществах описываются с по-
помощью терминов, которые имеют неодинаковое понимание у раз-
разных авторов. М. Александер (Alexander, 1997) попытался предста-
представить в общей форме современное понимание ключевых терминов
(environment, niche, ecosystem, community, population, consortium,
parameter, succession), которые хаотически употребляются, особенно
в англоязычной литературе. Популяция означает собрание индиви-
индивидуальных организмов с общими характеристиками и имеющих
близкое общее происхождение, т.е. принадлежащих к одному виду.
Термин "популяция" имеет генетическое значение. Совершенно
неверно употребление слова "популяция" для обозначения, напри-
например, микробного населения почвы. Сообщество (community, а не
society!) в английской литературе означает всю совокупность орга-
организмов, находящихся вместе. В русской литературе для этого поня-
понятия употребляется слово "биоценоз", и Александер настойчиво
рекомендует пользоваться им. Здесь я употребляю слово "сообще-
"сообщество" для совокупности взаимодействующих организмов, образую-
образующих систему связей между собой, а не для всего множества орга-
организмов биценоза. Для взаимодействующих организмов Александер
рекомендует термин "консорциум" как собрания организмов, кото-
которые имеют функциональную связь друг с другом, что совпадает с
"сообществом" в употребленном здесь смысле. Однако в практиче-
практическом словоупотреблении бактериологов консорциум обозначает
весьма тесно взаимодействующие друг с другом организмы не-
немногих видов, образующие физически связанные агрегаты. Для
множества объектов, объединенных общей морфологической
структурой, представленной гликокаликсом, как в биопленке,
очень подошло бы слово "симпозиум", если бы оно не было приня-
принято в употребление в ином смысле - как общее словоговорение.
Далее отмечается смешение понятий местообитания (habitat) и
экологической ниши. Если первое относится к географически или
микроструктурно (microhabitat) определяемому положению биоце-
биоценоза в пространстве (биотоп), то второе относится к абстрактной
характеристике свойств организмов в местообитании. В этом
смысле термин "экологическая ниша" совпадает с понятием
функциональной, или физиологической, группы организмов со
сходными свойствами. Этому пониманию близко соответствовал
"морфофизиологический род" золотого века микробиологии.
В условиях замены функциональной систематики филогенетиче-
филогенетической с соответствующими способами идентификации объектов прихо-
приходится использовать формулировку "морфофизиологическая группа"
или употреблять обозначение старых родов в тривиальном
смысле, например для спорообразующих анаэробов говорить
"клостридии".
50
Сообщество микроорганизмов, несмотря на повсеместность их
зародышей, представляет определенные ограничения для видов, за-
зависящие не только от абиотических факторов местообитания, слу-
служащих селективным фактором. Чтобы вступить в сообщество,
привнесенный извне (allien) зародыш должен успешно включиться в
существующую в сообществе систему взаимодействий. При этом
рассматриваются два этапа: 1) колонизация с малыми взаимодейст-
взаимодействиями организмов между собой при освоении нового пространства
или питательного субстрата и 2) климактерическое сообщество, на-
находящееся в состоянии гомеостаза, т.е. способности поддерживать
свою устойчивость и интегрированность в местообитании, несмотря
на внешние воздействия и потенциальные внутренние изменения со-
состава с течением времени (сукцессию). Для характеристики свойств
организмов сообщества в состоянии гомеостаза можно применить
название "социалистов" в смысле, использованном еще А. Гум-
Гумбольдтом для социальных растений (тростниковые заросли, сфагно-
сфагновое болото и т.п.). Возможность проникновения извне в такое сооб-
сообщество крайне ограниченна: оно обладает свойством гетерофобии,
т.е. отторжения чужеродного. Проще говоря, оно не дает ему места.
С таким пониманием особенностей сообщества согласно большин-
большинство микробиологов, имеющих опыт экологической работы. Про-
Простое введение зародышей несвойственных местообитанию организ-
организмов редко бывает достаточным, чтобы изменить сообщество. В спе-
специально практическом смысле это общее положение, которое
может быть нарушено в определенных условиях, ограничивает
применение "бактериальных удобрений" и "бактериальных биоре-
медиаторов" и вызывает живую дискуссию при обсуждении воз-
возможной роли генетически измененных организмов. Таким обра-
образом, большинство микробиологов понимают, что микробное сооб-
сообщество представляет нечто внутренне согласованное, а не простое
собрание зародышей в одном месте, из которого существующие
культуральные методики могут выделить не только составляю-
составляющие, незаменимые для сообщества компоненты, но и нечто прив-
привнесенное.
Сообщество с сильными кооперативными связями может быть
устойчивым лишь в относительно стабильной обстановке и оказы-
оказывается подверженным разрушению под действием внешних стрес-
стрессов. Стрессы вызывают развитие быстро адаптирующихся и коло-
колонизующих организмов. Иную группировку составляют сообщества,
находящиеся в условиях постоянных и резких стрессов, исключаю-
исключающих все нерезистентные организмы, кроме стресс-толерантных.
Для таких организмов А.А. Горбушиной предложено название
"пойкилофильных", т.е. предпочитающих разные (от греч. poiki-
los - различный), резко изменяющиеся условия, устраняющие их
конкурентов. Примером таких сообществ могут служить сообще-
сообщества литофильных лишайников.
51
2.2. ФИЗИЧЕСКАЯ ОРГАНИЗАЦИЯ СООБЩЕСТВА
2.2.1. Парагистология сообществ
Микробиологи обычно игнорируют организацию микробных
сообществ и их расположение в пространстве, ограничиваясь мор-
морфологией клеток в гомогенной культуре. Структура колоний чис-
чистых культур на плотных средах, обусловленная возрастом клеток и
разными условиями обмена с окружающим пространством, рассма-
рассматривается как чисто лабораторное явление. Однако в природе при-
приходится наблюдать массовое развитие микроорганизмов в виде раз-
разного рода скоплений и биопленок. Крайним примером колониально-
колониального роста служит развитие лихенизированных грибов на поверхности
камней, где литофильные лишайники создают пестрый покров. Для
природоведческой микробиологии физическая организация микроб-
микробных сообществ разнородных организмов определяет взаимодейст-
взаимодействие микроорганизмов в пространстве.
Микроорганизмы могут развиваться только в водной фазе, за
исключением воздушного мицелия актиномицетов, грибов. По от-
отношению к водной фазе они могут находиться в объеме жидкости,
как организмы планктона, на поверхности раздела вода-газ, как
неполон, на поверхности твердых частиц в виде пленок обрастания
или в виде бентоса. Наконец, следует отметить развитие микроор-
микроорганизмов на поверхности макроорганизмов, как у эпифитов, или на
слизистых поверхностях животных.
В графе трофических взаимодействий в сообществе (рис. 2.1)
стрелки-ребра соответствуют не только использованию вещества,
но и его транспорту между компонентами сообщества. Перенос ве-
вещества будет тем быстрее, чем меньше расстояние. Чтобы удержи-
удерживаться в фиксированном положении друг относительно друга, ор-
организмы должны иметь физические приспособления. В результате
возникают агрегаты микроорганизмов вплоть до консорциумов с
очень тесным, почти мембрана к мембране, расположением кле-
клеток синтрофных организмов, как у "Chlorochromatium aggregatum".
Многокомпонентное сообщество анаэробных организмов в метан-
тенках может образовывать гранулы размером в гречневую крупу,
в которых метаноген образует либо пористую псевдопаренхиму,
как метаносарцина, или же клубок нитей, как метанотрикс, а в сво-
свободном пространстве располагаются другие организмы сообщест-
сообщества (рис. 2.5). В результате метаногенное сообщество в грануле спо-
способно осуществлять всю трофическую цепь преобразований от рас-
растворимого органического вещества (РОВ) до СН4 и СО2. Еще более
сложную организацию имеют биопленки и циано-бактериальные
маты, рассматриваемые в отдельной лекции (см. Лекцию 3).
Физическая организация сообщества в агрегатах подчиняется
правилу минимальньа диффузионных расстояний, обеспечивающему
52
Рис. 2.5. Микоплазма в культурах метаносарцины (фото Т.Н. Жилиной, Г. А. За-
варзина)
Ультратонкий срез через макроцисту метаносарцины. Слева — неразрушенный агрегат, в
котором клетки метаносарцины образуют псевдопаренхиму /, справа - часть разрушен-
разрушенного агрегата, замещенная микоплазмами 2. Общая оболочка макроцисты - 3
наиболее быстрый обмен веществами между взаимодействующими
организмами.
Противоположная ситуация возникает при конкуренции за суб-
субстрат, например О2, когда организмы должны иметь свободное диф-
диффузионное пространство между клетками, представляющее каналы
проводимости. В этом случае слизь между клетками отстраняет их
друг от друга и создает возможность максимального использования
пространства.
В почвенных комочках-агрегатах взаимное расположение груп-
группировок организмов позволяет сосуществовать аэробам и анаэро-
анаэробам. Наружный слой слагается аэробами, обращенными в сторону
почвенного раствора и пор аэрации. Интенсивное поглощение ими
О2 позволяет развиваться внутри агрегата анаэробам, восстановлен-
восстановленные продукты обмена которых используются аэробами. Такой агре-
агрегат заведомо образуется при диаметре комочка около 10 мм, но в за-
зависимости от доступности органического вещества может достигать
и 1 мм.
В подвижной среде сообщества должны осуществлять "антихе-
мостатную" стратегию накопления биомассы в требуемой зоне пу-
путем обрастания твердых субстратов. В биотехнологии была развита
53
целая область работы с иммобилизованными клетками. Аналогич-
Аналогичный процесс происходит в природе. Механизм формирования обрас-
обрастаний и закрепления на субстрате был исследован Д.Г. Звягинцевым
для почвы, а К. Маршаллом - для водной среды. Начальный этап
формирования биопленки основывается на гидрофобном взаимо-
взаимодействии с твердым субстратом. Для этого некоторые организмы
имеют "прикрепительные диски" - выделение адгезивного материа-
материала, видимого при микроскопии как пятно у полюса клетки. Отчетли-
Отчетливо виден прикрепительный диск у каулобактеров на конце стебелька,
а у гифомикробов - у основания клетки. Целая группа "стебелько-
"стебельковых бактерий" характеризуется наличием "неклеточных стебель-
стебельков" - тяжей экскретируемого материала, позволяющего им
прикрепляться к поверхности и вместе с тем выносить клетку за
пределы пограничного слоя.
Следующая группа приспособлений обусловлена образованием
ворса из прикрепленных жестких нитей. Начальный этап представ-
представляет Leptothrix sideropus - псевдомонада, образующая цепочку
клеток, причем базальная клетка выделяет нечто вроде зонтика,
распространяющегося по границе раздела вода-воздух в нейстоне.
Более развитый тип представляет образование трубчатых чехлов,
как у Sphaerotilus, переходящих в ожелезненные трубки, как у
Leptothrix ochracea. Рост такого типа с высокой механической устой-
устойчивостью нитей на разрыв особенно свойственен кренофилам, раз-
развивающимся как неподвижная фаза в подвижной среде, например в
виде обрастаний в быстротекущих ручьях, откуда они и получили
свое название, или в прибойной зоне. Поток вещества даже при низ-
низкой его концентрации оказывается достаточным для развития об-
обрастаний. Типичным примером может служить термофильная серо-
серобактерия Thermothrix, образующая ослепительно белые от серы
космы длиной в метры в быстротекущих термальных ручьях. Клет-
Клетки Thermothrix слегка изогнуты и, переплетаясь друг с другом, увели-
увеличивают прочность косм на разрыв.
Есть две экологические стратегии для механической устойчиво-
устойчивости бентосных сообществ, давно получившие название "войлок"
(или "ворс" для начальной стадии) и "кожа". Войлок приобретает
механическую устойчивость за счет переплетения противодейству-
противодействующих разрыву нитей в "ткань" (в текстильном смысле). При этом
промежутки между нитями свободны и не создают диффузионных
ограничений, за исключением искривленного пути. Такую структу-
структуру ткани создает большинство нитчатых водорослей и некоторые
цианобактерии, как Oscillatoria и Phormidium (см. подробнее Лек-
Лекцию 3).
"Кожа" представляет механизм формирования биопленки за
счет гликокаликса - выделяемой микроорганизмами слизи. Биоп-
Биопленка состоит на более 95% из воды и может рассматриваться как
иммобилизованная вода в матрице полимера. Ограничения для диф-
54
фузии в таком полимере более значительны. Типичным представи-
представителем таких плотных структур оказывается циано-бактериальный
мат Microcoleus. Механическая плотность "кожи" очень высока и со-
соответствует примерно 1-2% агару.
В почве в субаэральных условиях возникает другая структура в
виде "паутины", или "сети", образуемой мицелиальными организ-
организмами, как стрептомицеты и грибы. Сеть гиф может охватывать
значительное пространство и в крайнем случае представляет ре-
результат развития одной колонии, как у лесных базидиомицетов, за-
занимающих сотни метров с протянувшимися в подстилке ризомор-
фами.
Физические приспособления связаны с локализацией клеток
друг относительно друга и обусловлены формированием слизей за
счет секретируемых клеткой иногда очень обильных экзополиме-
ров. Экзополимеры 1) удерживают организмы внутри локального
пространства и обеспечивают макростабильность по отношению к
физическим факторам, прежде всего вымыванию; 2) обеспечивают
макроструктуру сообщества с оптимальными диффузионными рас-
расстояниями; 3) ограничивают диффузию веществ молекулярной
диффузией и создают транспортные колодцы для проникновения
питательных веществ; 4) связывают питательные вещества; 5) огра-
ограничивают проникновение вредных факторов как химической приро-
природы, так и питающихся бактериями мелких хищников-протист. Тер-
Термин "гликокаликс" применяется для полисахаридных структур, ле-
лежащих кнаружи от наружной мембраны грамотрицательной клетки
или пептидогликана грамположительной. Гликокаликс может быть
в разных состояниях - от крайне рыхлой слизи до ограниченной
сверху капсулы - и переходить в жесткий чехол или обвертки, как у
некоторых цианобактерии. Экзополимеры в сообществе представ-
представляют смесь продуцируемых разными организмами и отчасти (!)
обобществленных полимеров. Таким образом, микробное сообще-
сообщество за счет образования экзополимеров создает нечто вроде ткани,
которая может изучаться методами парагистологии. Она особенно
важна для бентосных организмов, но и взвешенные организмы соз-
создают агрегаты-хлопья, как, например, Zoogloea ramigera - в аэротен-
ках или "морской снег" - в океане.
2.2.2. Межвидовой транспорт в сообществе
Поскольку в сообществе происходит обмен продуктами и суб-
субстратами между организмами, то значение приобретает перенос ве-
вещества от одного организма к другому. Этот перенос осуществляет-
осуществляется путем молекулярной диффузии. Перенос от точечного источника
пропорционален квадрату расстояния. Поэтому для взаимодейству-
взаимодействующих организмов важно сближение на минимальное диффузионное
расстояние. На самом деле источник является не единичным точеч-
55
ным, а некоторое число их распределено в трехмерном пространст-
пространстве. Значит, оптимальным будет некое подобие кристаллической ре-
решетки.
Понять значение правила минимального диффузионного рас-
расстояния удалось на основе метаногенного сообщества в очистных
сооружениях. Для очистки жидких стоков сахароваренных заводов
в Голландии в начале 1970-х годов Г. Леттинга спроектировал ме-
тантенки с восходящим потоком. Установка сначала работала не-
немногим лучше обычных ферментеров с полным смешением. Но
постепенно работа установки улучшалась, и, наконец, ее эффек-
эффективность многократно превысила проектную. Оказалось, что в
ферментере накопились гранулы миллиметрового размера. Эти
гранулы были сложены метаногенным сообществом. Именно так
Н. Зёнген в 1906 г. накопил метаногенов на ацетате, используя
слив культуры с накапливавшегося на дне осадка. В то время как
Г. Леттинга конструировал свои ферментеры, Т.Н. Жилина вос-
воспроизводила методику Зёнгена, выделяя метаносарцину в проточ-
проточной установке с восходящим потоком. На дне накапливался желто-
желтоватый осадок, напоминавший по виду пшенную или гречневую кру-
крупу. Осадок был тяжелым с удельным весом около двух, а жидкость
над осадком оставалась прозрачной. Когда гранулы этого осадка
разрезали и посмотрели под микроскопом, оказалось, что они
представляют что-то вроде гаструлы и сложены рыхлой псевдопа-
псевдопаренхимой метаносарцины. В межклеточном пространстве распола-
располагались разнообразные палочковидные бактерии, среди которых
можно было опознать клостридии. Гранулы стали объектом тща-
тщательных исследований и морфологических, и физиологических.
Оказалось, что они содержат полный набор анаэробных организ-
организмов, необходимых для разложения различных органических ве-
веществ. В зрелом иле матрицу гранулы представляли сплетенные в
клубок нити метанотрикса, в петлях которого располагались дру-
другие организмы.
Другим примером подобных отношений могут служить хлопья
в аэробной зоне. Постоянной загадкой для исследователей являет-
является развитие анаэробных организмов в зоне фотосинтеза. Так про-
происходит образование метана в слое развития цианобактерий, на-
например в культуре из термофильного мата. Подобный результат
был получен с выделением метана из клубков Trichodesmium в
океане. Обычным является нахождение анаэробов в аэрируемом
слое почвы. Наружный слой сферической гранулы сложен аэроб-
аэробными организмами. При достаточном поступлении растворимого
субстрата из внутренней зоны, где разлагается нерастворимое ор-
органическое вещество, например растительные остатки, и вследст-
вследствие медленной диффузии кислорода - в 1000 раз более медленной,
чем в газовой фазе, - внутри образуется анаэробная зона, где
может развиваться анаэробное сообщество. Размеры такой грану-
56
лы укладываются в миллиметровую шкалу, составляя менее 1см
для комка почвы.
Еще более наглядный пример представляют бентосные сообще-
сообщества. В них последовательные этапы превращения веществ и после-
последовательно действующих функциональных групп организмов распо-
располагаются послойно. В результате фронтального одностороннего
поступления субстрата временная последовательность реакций пе-
переходит в пространственную. Трехмерная организация планктонного
сообщества трансформируется в одномерную бентосного. Особенно
тщательно организация сообщества была исследована на примере
циано-бактериальных матов, которые дальше будут разобраны бо-
более подробно.
Модель молекулярной диффузии является не единственной, хо-
хотя и наиболее важной. Кроме молекулярной диффузии значение
имеет конвекционный перенос вещества вследствие, например, тем-
температурного перепада днем и ночью. Конвекционный перенос заме-
заметен уже в небольших водоемах. Особые условия возникают, когда в
среде имеется турбуленция и возможно перемешивание. При рас-
рассмотрении транспортных процессов особенно важно понимание
масштаба. Так, для одиночных микробных клеток турбуленция не
имеет значения, поскольку вихри существенно больше их размеров
и определяют перемешивание вне диффузионного слоя, но для мик-
микробных хлопьев в несколько миллиметров-сантиметр поток у по-
поверхности уже значителен.
При современной тенденции исследовать природные процессы с
помощью математического моделирования оказалось, что физиче-
физический перенос вещества в пределах исследуемой реальной системы
представляет лимитирующее звено в работе сообщества и далее
вплоть до биосферы. Для обозначения этого подхода было предло-
предложено название "геобиофизика", и слово это постепенно привилось,
чтобы отличать от биофизики с интересом к квантовому выходу
фотореакции или проводимости нервного импульса.
Особенно привлекательны для анализа так называемые "мик-
"микробные консорциумы", т.е. сообщества тесно связанных, вплоть до
соприкосновения мембран, разнородных синтрофирующих микро-
микроорганизмов. В таких сообществах действуют "мини-циклы", не до-
допускающие выхода промежуточных продуктов вовне сообщества.
Они явились подтверждением ранее установленной закономерности
трофического взаимодействия филогенетически удаленных групп
организмов7. Например, анаэробное окисление метана осуществля-
осуществляется при взаимодействии архей-метаногенов и протеобактерий-
сульфатредукторов с формированием консорциума из расположен-
расположенных в центре метаногенов и окружающих их сульфатредукторов,
7 Заварзин Г.А. Микробное сообщество в прошлом и настоящем // Микробиол.
журн. 1989. Т. 51, № 6. С. 3-14.
57
причем за счет межвидового переноса становится возможным обра-
обращение реакции метаногенеза8. Нитрификаторы-протеобактерии
Nitrosomonas и Nitrobacter принадлежат к разным филогенетическим
ветвям, но тесно связаны трофически продукцией и окислением ни-
нитрита. В биопленках нитрозомонас образует ядро шаровидного аг-
агрегата в 0,1 мм, окруженного слоем грушевидных нитробактеров
или тонких нитроспир. Сложная структура уже упоминавшегося ме-
таногенного агрегата метаносарцин с внутренней полостью была
сначала установлена Т.Н. Жилиной на ультратонких срезах с харак-
характерной размерностью 100 мкм. Применение флуоресцентной гибри-
гибридизации к аналогичным гранулам дало структуру концентрического
строения агрегата: наружный слой состоит из органотрофных бро-
дилыциков, под ним располагаются ацетотрофные метаногены и
тесные ассоциации синтрофных организмов с гидрогенотрофными
метаногенами. Если же присутствует сульфат, то на поверхности
пропионат-использующего многовидового агрегата располагается
сульфидоген Desulfobulbus.
Установлено, что состояние организма в суспензионной культу-
культуре резко отличается от его состояния в биопленке: по-разному экс-
прессируются гены ответа на высокое осмотическое давление, из-
избыточную плотность клеток, ограничение кислородом. Структура
сообщества создается за счет усиленного синтеза внеклеточных по-
полимеров. Биопленки организованы таким способом, что имеются
микроколонии организмов с большими каналами между микроколо-
микроколониями. Считается, что эти каналы обеспечивают поступление суб-
субстратов и продуктов к микроколониям на поверхности и в глубине
биопленки за счет как диффузии, так и тока жидкости по каналам.
О скорости роста в разных частях биопленки можно судить по со-
содержанию рибосом в клетках, что видно даже в фазовом контрасте.
Обычно во внутренних частях колонии или агрегата клетки более
прозрачны, чем снаружи. Тот же вывод был получен с флуоресцент-
флуоресцентными пробами на состояние клетки. В биопленках с многими вида-
видами удается получить ответ о состоянии клетки, находящейся в близ-
близком соседстве с продуцентом. Физическое взаимодействие может
быть не слишком тесным, причем кооперация может сменяться кон-
конкуренцией. Пространственная организация многовидового сообще-
сообщества отражает синтрофные взаимодействия и состояние микроколо-
микроколоний в обитаемых микронишах на уровне 10-100 мкм. Гетерогенные
микробные сообщества самоорганизуются при условиях ограниче-
Boetius A., Raveschlag К., Schubert CJ. et al. A marine microbial consortium appar-
apparently mediating anaerobic oxidation of methane // Nature. 2000. Vol. 407, № 5.
P. 623-626; Леин А.Ю., Иванов М.В., Пименов М.В., Гулин М.Б. Геохимиче-
Геохимические особенности карбонатных построек, образующихся при микробном
окислении метана в анаэробных условиях // Микробиология. 2002. Т. 71, № 1.
С. 89-102.
ния транспорта субстрата и возникновения градиентов за счет го-
меоморфного морфогенеза в духе Д'Арси Томпсона за счет эгоисти-
эгоистического поведения клеток. По другой гипотезе, сообщество про-
пространственно организуется под генетическим контролем с воздейст-
воздействием ацилированного гомосерин лактона как межвидового сигнала
кворум-фактора, включая альтруистический синтез эдификатором
внеклеточных структурных компонентов для всего сообщества.
Исследование морфологии и пространственной организации сооб-
сообществ получило мощное обоснование с применением флуоресцент-
флуоресцентной окраски с гибридизацией, специфической для видов организмов.
2.2.3. Градиенты и транспорт в сообществе
Разобраться с процессами в пространственно структурирован-
структурированном сообществе удалось с помощью микроэлектродной техники,
развитой датчанином Н. Ревсбеком9 и широко примененной Б. Йор-
генсеном10 для природных сообществ на примере микробных матов
или биопленок.
Время диффузии для молекулы составляет t = z2/2D, где z - рас-
расстояние, a D - молекулярная диффузия в воде (D = 2,0 • 10~5 см2 • сг1).
Для молекулярной диффузии О2 получены следующие значе-
значения:
Расстояние, z l мкм 10 мкм 100 мкм 1 мм 1 см 10 см 1м
Время, t О, мс 30 мс 3 с 4 мин 7 час 25 сут 8 лет
Если исходить из представления о клетке как "мешке с фермен-
ферментами", то из диффузионных ограничений можно понять разницу ме-
между скоростями процессов в прокариотной клетке с характерными
размерами 1 мкм и эукариотной с размерами 10-100 мкм. Эукарио-
ты явно нуждаются в компартментализации процессов, чтобы лока-
локализовать процессы в пространстве, где транспортные ограничения
не действуют. Дальнейшее увеличение размеров организмов связа-
связано с решением транспортных проблем и локализацией специфиче-
специфических процессов в тканях и органах. Как решают транспортную про-
проблему мицелиальные организмы, например грибы? Здесь следует
различать транспорт по внешней поверхности - апопласту - и по ци-
цитоплазме - перипласту.
Микробные сообщества решают транспортную задачу для раз-
разнородных организмов, создавая "псевдоткань". Поверхность микроб-
микробного мата слизистая. В результате турбуленция Е вблизи поверхности
58
9 Reevsbech N.P. Analysis of microbial mats by use of electrochemical microsensors:
recent advances // Microbial mats / Ed. L.D. Stal, P. Caumette. B. ets.: Springer, 1994.
P.135-148.
10 J0rgensen B.B. Diffusion processes and boundary layers in microbial mats //
Microbial mats / Ed. L.D. Stal, P. Caumette. B. ets.: Springer, 1994. P. 243-254.
59
Турбулентная
диффузия
Турбулентный
поток -Е dC/dz
Поток через
пограничный
диффузионный слой-?)(См - C0)/Zg
Поток в осадке -фД dc/dz
с.
Вода
о2—
сс
f Осадок
-.0
Рис. 2.6. Диффузионный пограничный слой [по: J0rgensen, Reevsbech, 1985]
мата затухает и на расстоянии долей миллиметра становится мень-
меньше молекулярной диффузии D. Над поверхностью образуется диф-
диффузионный пограничный слой. Состояние пограничного слоя в мас-
масштабе 100 мкм удалось экспериментально измерить с помощью ми-
микроэлектродов. Увеличение скорости потока над матом приводит к
уменьшению пограничного слоя от > 1 мм до 0,1 мм при 8 см/с, бо-
более резкому градиенту, более глубокому проникновению О2 в мат.
Это особенно важно для микроаэробов, как Beggiatoa, и других гра-
градиентных организмов. Над неровной поверхностью мата погранич-
пограничный слой следует рельефу мата, но возникают микрозавихрения.
Со стороны потока вещество глубже проникает в гребень, в то вре-
время как со стороны понижения оно высасывается из структуры. В ре-
результате, казалось бы, однородное дно океанических осадков ока-
оказывается мозаичным из-за придонных течений.
Внутри мата значения D, оказывается, составляют только
50-60% от молекулярной диффузии. Для кислорода (Со - концентра-
концентрация в среде) получается картина, представленная на рис. 2.6.
Отсюда по наклону кривой концентрации вычисляется дыхание.
Градиенты могут быть очень крутыми и зависят от концентрации
используемых веществ, в первую очередь от доступного органиче-
органического вещества, используемого для дыхания. Например, для залива
у берегов Дании до оседания планктона кислород проникал на глу-
глубину 4 мм в осадок, а после цветения - лишь на 1 мм. В циано-бакте-
риальных матах градиенты еще более крутые и составляют доли
1 мм. Таким образом, переход между аэробной и анаэробной зрнами
при достаточном количестве органического вещества составляет до-
доли миллиметра. В этих условиях сосуществование аэробных организ-
организмов, поглощающих кислород, и находящихся под их защитой анаэро-
анаэробов вполне возможно в сообществах, имеющих размеры > 1 мм.
60
Микроэлектроды были изготовлены для разных неорганиче-
неорганических веществ, и это позволило экспериментально проследить за про-
процессами внутри слизистой матрицы сообщества. Аналогично О2 из-
измеряются градиенты H2S. В результате оказывается, что в морских
осадках восстановление сульфатов с образованием H2S происходит
только ниже зоны с кислородом и нитратом на глубине 1,1 мм и пре-
прекращается на глубине 2,1 мм вследствие исчерпания сульфата. Пол-
Полное окисление H2S происходило в слое 0,3 мм с временем оборота в
несколько секунд. Окисление H2S создавало дополнительное к ды-
дыханию поглощение кислорода на органических веществах.
Из этого примера можно вывести по меньшей мере два следст-
следствия: 1) серный цикл сульфатредукции - окисления H2S - может слу-
служить дополнением к окислительной системе донных осадков с вре-
временем оборота в секунды-десяток секунд; 2) градиент в осадке дос-
достаточно резкий, чтобы мог существовать микробный тиобиос, т.е.
организмы, пересекающие градиент от H2S-3OHbi до кислородной
зоны, т.к. для этого достаточно иметь длину в сотню мкм. Такой дли-
длины достигают нитчатые серобактерии, как Thioploca, образующие
маты у берегов Чили. Они окисляют H2S за счет накопленного в ва-
вакуоли нитрата.
Денитрификация была измерена в биопленках на поверхности
орошаемого биофильтра в очистных сооружениях с помощью ком-
комбинированного микросенсора на О2 и N2O, причем редукция подав-
подавлялась ацетиленом. Денитрификация отсутствовала при концентра-
концентрации О2> 10-20 мкМ. При концентрации нитрата в воде 125 мкМ зо-
зона денитрификации, располагавшаяся на глубине 0,2 мм, где О2 был
полностью исчерпан, составляла 0,3 мм, а при концентрации нитра-
нитрата 1250 мкМ уходила в глубину более 1 мм. В присутствии аммония
в слое с О2 происходит нитрификация с активным поглощением ки-
кислорода, и в результате нитрат диффундирует вниз в бескислород-
бескислородный слой, где он используется.
Из приведенных примеров видно, что существование очень рез-
резких градиентов в структурированном сообществе позволяет взаимо-
взаимодействовать противоположно направленным процессам и замыкать
циклы внутри одного сообщества.
2.2.4. Коллоидная среда обитания микроорганизмов
Организмы, находящиеся в органическом геле, можно рассмат-
рассматривать как коллоидные системы с важными для них электростати-
электростатическими закономерностями. При обсуждении условий существова-
существования микроорганизмов в природе обычно ограничиваются вопросами
химических реакций, определяющих в первую очередь катаболизм
бактерий. Между тем на самом деле микроорганизмы представляют
собой дисперсные системы, находящиеся в воде как дисперсионной
среде. Распространение микроорганизмов в виде аэрозолей в возду-
61
хе с пылевыми частицами не связано с их жизнедеятельностью.
Клетки бактерий, имеющие размеры 0,3-10 мкм, представляют осо-
особый размерный класс твердых частиц на границе между высокодис-
высокодисперсными системами с размерами 1-100 нм и грубодисперсными
> 10 мкм. Среднедисперсные системы изучены хуже других, так как
они гетерогенны и обладают свойствами некоторых высокодисперс-
высокодисперсных систем, которые входят в их состав в виде частей клеток и в то
же время слишком велики, чтобы подчиняться свойствам собствен-
собственно коллоидных растворов. Они представляют собой суспензии.
Высокодисперсные системы седиментационно-устойчивы и об-
образуют коллоидные растворы - золи, часто представленные органи-
органическими высокомолекулярными соединениями, в том числе и слизя-
ми, образуемыми микроорганизмами. В них протекают молекуляр-
но-кинетические явления в виде диффузии, осмоса, высока скорость
химических реакций на поверхности частиц. Фактически средой
обитания таких организмов является гель, образованный каркасом
из слизи, внутри которого находятся клетки разных взаимосвязан-
взаимосвязанных между собой видов организмов, образующих сообщество. Обра-
Образующиеся агрегаты могут быть сопоставлены с коагулировавшими
коллоидами и достигают размерности грубодисперсных частиц.
Не следует ограничивать внимание органическими дисперсными си-
системами - в среде обитания микроорганизмы сталкиваются с высо-
высокодисперсными минеральными системами глинистых минералов,
гидроокислов, силикатов, способных образовывать устойчивые кол-
коллоидные растворы.
Грубодисперсные системы, как правило, седиментационно неус-
неустойчивы и оседают, создавая осадки с поровым пространством и ин-
терстициальным пространством между поровыми частицами, кото-
которое достаточно по размерам для развития бактерий. В таких поли-
полидисперсных гетерогенных системах протекает деятельность орга-
организмов в иловых осадках, содержащих воду и разные твердые фазы,
и в почве, где имеется и газовая фаза.
Дисперсная фаза оценивается либо численно, по числу частиц в
единице объема системы пч (число клеток в единице объема, кл/мл),
либо по массовой концентрации пм (абсолютно сухая биомасса,
а.с.б., г/мл), либо по объему поб. Последняя величина дает ограниче-
ограничение для численности микроорганизмов размером около 1 мкм по-
порядка 1012 кл/мл. В химии обычно границей для коллоидных частиц
принимается размер < 0,1 мкм, но почвоведы в связи с поведением
глинистой фракции принимают границу < 2 мкм в связи с возраста-
возрастанием роли поверхностных явлений у плоских частиц глинистых
минералов. В этой размерной категории находятся и бактерии, для
которых можно выделить три состояния: планктонной суспензии,
взвешенных хлопьев, осадка. Для палочковидных бактерий, как и
для частиц глинистых минералов, можно наблюдать четыре разные
состояния: независимое (пептизированное) расположение, агрегиро-
62
ванное бок о бок (палисадное), агрегированное полюс к боку ("лам-
("ламповый ерш" на поверхности трихома), сетчатое (например,
Thiodyction), определяемое распределением зарядов на поверхности.
Увеличение поверхности раздела дисперсной фазы, обусловлен-
обусловленное поверхностной энергией, приводит к интенсификации поверхно-
поверхностных явлений. В результате химические и физические процессы в
дисперсных системах отличаются от обычной химии растворов, с
помощью которой излагается микробиология в общих курсах. Час-
Частицы удерживаются в коллоидном растворе благодаря поверхност-
поверхностному двойному слою, который препятствует их сближению и слипа-
слипанию. Двойной слой уменьшается, когда в растворе возрастает содер-
содержание электролитов. Слипание частиц происходит, когда они сбли-
сближаются на расстояние менее 15 А. В результате происходит коагуля-
коагуляция коллоидного раствора, и укрупненные хлопья оседают. Напро-
Напротив, при низком содержании электролитов частицы переходят во
взвешенное состояние, происходит пептизация. Даже такие тяже-
тяжелые частицы, как глинистые минералы, в дистиллированной воде
могут образовывать очень устойчивую взвесь.
Поверхностные явления самопроизвольно ведут к уменьшению
поверхностной энергии. Они обусловливаются снижением поверх-
поверхности раздела фаз и/или поверхностного натяжения. Снижение по-
поверхности раздела фаз ведет к укрупнению частиц в результате ко-
коагуляции и коалесценции (слиянию капель эмульсии). Поверхност-
Поверхностное натяжение определяется s - удельной свободной поверхностной
энергией. Изменение поверхностного натяжения ведет к адсорбции,
адгезии и электрическим процессам с образованием двойного слоя и
электрокинетическим явлениям. Применительно к прикреплению
бактерий, как уже отмечалось, процесс был подробно изучен
Д.Г. Звягинцевым. Во многом поведение суспензии бактерий проте-
протекает аналогично адсорбции на поверхности. Образование биопле-
биопленок начинается с контакта клеток с поверхностью.
Адгезия (прилипание, смачивание) обусловлена связью между
разнородными телами при их контакте, допускающем молекуляр-
молекулярные взаимодействия. У микроорганизмов прикрепление к поверхно-
поверхности обеспечивают гидрофобные взаимодействия с твердой фазой.
Синтез соответствующих соединений находится под генетическим
контролем. Гидрофобные взаимодействия выполняют ориентирую-
ориентирующую функцию для гидрофобных частиц, самый же контакт реализу-
реализуется за счет межмолекулярных взаимодействий. Возникновение
электрического заряда на поверхности характерно прежде всего для
золей и суспензий, дисперсная фаза которых представлена тверды-
твердыми частицами. Заряд на поверхности образуется при адсорбции ио-
ионов, диссоциации и переходе ионов из одной фазы в другую. Фосфат
железа, каолин всегда, даже в кислой среде, заряжаются отрицатель-
отрицательно, а гидроксид железа и целлюлоза - всегда положительно. В ре-
результате возникает (р - электрический потенциал поверхности. За-
63
ряд частиц определяется потенциалобразующими ионами, которые
уравновешиваются противоионами из адсорбционного и диффузно-
диффузного слоев. Образуется двойной электрический слой, удерживаемый у
частицы вместе с слоем жидкости до границы скольжения. Адсорб-
Адсорбционный слой имеет размеры < 1 нм. Диффузный слой в зависимо-
зависимости от концентрации ионов имеет толщину 1-1000 нм, но меньше,
чем граница скольжения, и при движении возникает электрокине-
электрокинетический дзета-потенциал (Q. Эту величину можно достаточно
точно измерить. Добавление в среду одновалентных противоионов
приводит к компенсации потенциалобразующего слоя, и С, = 0.
Многовалентные катионы, как Fe3*, могут вызывать перезарядку по-
поверхности. Это явление в крупных технологических масштабах ис-
используется для коагуляции микроорганизмов при очистке воды.
Микроорганизмы для прикрепления образуют особые вещества,
находящиеся под генетическим контролем.
Коллоидные явления обусловливают устойчивость суспензий
планктонных организмов, нарушение которой приводит к их оседа-
оседанию, например в конце цветения. Устойчивость дисперсных систем
означает способность сохранять свой состав. Различают седимента-
ционную и агрегативную устойчивость. Седиментационная устойчи-
устойчивость определяется способностью частиц противостоять оседанию
под действием силы тяжести и зависит от дисперсности. Противо-
Противодействует оседанию сила трения в дисперсионной среде и броунов-
броуновское движение. Агрегативная устойчивость подразумевает сохране-
сохранение межфазовой поверхности и поверхностной энергии. Она опреде-
определяется способностью противостоять слипанию частиц с образовани-
образованием агрегатов. Для твердых частиц слипание называется коагуляци-
коагуляцией. Процесс этот имеет определяющее значение в микробиологии,
например при очистке воду, когда коагулировавшие хлопья оседа-
оседают. Изменение агрегативной устойчивости обусловливается присут-
присутствием электролитов, влияющих на двойной слой. Специфическая
адсорбция ионов на поверхности приводит к ослаблению электриче-
электрического потенциала и слипанию частиц. Повышение заряда потенциал-
образующего слоя приводит к увеличению диффузного слоя. При
концентрационной коагуляции происходит сжатие диффузного
слоя. Такие изменения происходят в почве, когда избыток Na приво-
приводит к вымыванию Са и Mg, нарушению структуры двойного слоя и
концентрационной коагуляции. Порог коагуляции - это наимень-
наименьшая концентрация электролита, при которой начинается коагуля-
коагуляция. Есть эмпирическое правило Шульце-Гарди, описывающее ко-
коагуляцию. Чем выше валентность коагулирующего иона, тем
меньше его нужно для коагуляции: порог коагуляции обратно про-
пропорционален шестой степени валентности иона электролита.
Однако многовалентные ионы могут перезаряжать поверхность,
меняя величину и знак ^-потенциала. Эти процессы важны для
сорбции на слизях коллоидов тяжелых металлов, например Fe и А1.
64
По этой причине слизистые циано-бактериальные маты обра-
образуют сорбционный барьер для тяжелых металлов, действующий
иногда в геологическом масштабе при образовании отложений
редких элементов. Очень наглядно такой процесс происходит для
коллоидного раствора золота, оседающего на поверхности циано-
бактерий.
При увеличении числа частиц коллоидов образуется каркас из
дисперсной фазы, пространство между которым заполнено диспер-
дисперсионной средой. Образование структуры обусловлено контактами
между частицами. Такие структуры могут быть упорядоченными и
неупорядоченными. Они относятся к связнодисперсным и обладают
прочностью. Подобно твердому телу, они противодействуют внеш-
внешним деформирующим силам. Высокомолекулярные соединения -
белки и полисахариды - образуют связнодисперсные системы. Это
студни, сохраняющие свою форму под действием внешней нагрузки
и стабилизирующие погруженные в них дисперсные частицы, как
бактериальные клетки. Те парагистологические структуры микроб-
микробных сообществ, которые были рассмотрены выше, отражают жизнь
микроорганизмов в коллоидной среде со всеми присущими ей элек-
электрокинетическими явлениями. Поверхностный заряд живой клетки
играет при этом первостепенную роль.
Коллоидная среда обитания создает каркас, по которому могут
передвигаться крупные скользящие организмы с трихомным строе-
строением. Напротив, мицелиальный план строения приспособлен к раз-
развитию в почве с поровым пространством между твердыми частицами.
Пелофильная микрофлора в илах содержит очень большое число
нитчатых и трихомных скользящих бактерий. Выделение их счита-
считается трудной задачей. Среди этих бактерий по включениям серы
опознаются бесцветные серобактерии, образующие шелковистые
белые слои и покровы "серного мата". Большинство этих организ-
организмов зависят от ацетата как дополнительного или основного субстра-
субстрата. Например, Ф. Виддель с сотрудниками1' нашли, что в эти маты в
тесной ассоциации с серобактериями включаются нитчатые сульфат-
редукторы рода Desulfonema, использующие широкий набор
жирных кислот от ацетата до капроата. Таким образом, в иле при-
присутствует ассоциация сульфатредукторов, окисляющих продукты
брожения, с серобактериями, окисляющими H2S и удаляющими О2 с
продукцией сульфата, в тесном контакте друг с другом. Особенно
наглядно такое сообщество проявляется на примере взаимодействия
довольно тонких трихомов Desulfonema с гигантскими нитями
Thioploca, пересекающими кислородный градиент и создающими
малый серный цикл.
11 См.: Fukui M., Teske A., Assmus В. et al. Physiology, phylogenetic relationships, and
ecology of filamentous sulfate-reducing bacteria (genus Desulfonema). Arch.
Microbiol. 1999. Vol. 172. P. 193-203.
3. Г. А. Заварзин
65
Рекомендуемая литература
Х.Вавилин В.А., Васильев В.Б., Рытое СВ. Моделирование деструкции орга-
органического вещества сообществом микроорганизмов. М.: Наука, 1993. 208 с.
2. Громов Б.В., Павленко Г.В. Экология бактерий. Л.: Изд-во ЛГУ, 1989.
Ъ.Заварзин Г.А. Литотрофные микроорганизмы. М.: Наука, 1972. 376 с.
4. Заварзин Г.А. Недарвиновская область эволюции //Вести. РАН 2000 Т 70
№ 5. С. 403-411. '
5. Заварзин Г.А., Колотилова Н.Н. Введение в природоведческую микробио-
микробиологию М.: Книжный дом "Университет", 2001. 256 с.
6. Звягинцев Д.Г, Почва и микроорганизмы. М.: Изд-во МГУ, 1987. 256 с.
Т.Клюйвер А., Ван Ниль К. Вклад микробов в биологию. М.: Изд-во иностр
лит., 1959.
8. Кожевин П.А. Микробные популяции в природе. М.: Изд-во МГУ, 1989.
9. Стейниер Р., Эйдельберг Э., Ингрэм Дж. Мир микробов: В 3 т.' М.: Мир,
10. Экология микроорганизмов / Ред. А.И. Нетрусов. М.: Академия, 2003.
11. Alexander M. Microbial communities and interactions: A prelude // Manual of envi-
environmental microbiology. Wash. (D.C.), 1997. P. 5-13.
12. Biology of prokaryotes / Ed. J.W. Lengeler et al.; Stuttgart; N.Y.: Thieme-
Blackwell, 1999.
13.Biology of anaerobic microorganisms / Ed. A.J.B. Zehnder. NY etc- Wilev
1988.872 р. •• *. ¦
14. Brock biology of microorganisms / Ed. M.T. Madigan et al. 8th ed. Upper Saddle '
River (N.J.): Prentice Hall, 1997.
15. Marshall K.C. Interfaces in microbial ecology. Cambridge: L.: Harward Univ. press, '
1976. 156 p. ' j
16. Perry J., Staley J.T. Microbiology dynamics and diversity. Fort Wort (NYV
Sounders College Publ., 1997.
17. The prokaryotes: A handbook of habitats, isolation and identification of bacteria/ Ed
A. Balows et al. etc. Springer, 1992. Vol. 1-4.
18. Tolker-Nielsen Т., Molin S. Spatial organization of microbial biofilm communuties // '
Microbiol. Ecol. 2000. Vol. 40. P. 75-84. i
Лекция 3
ЦИАНО-БАКТЕРИАЛЬНЫЕ МАТЫ
3.1. ЦИАНО-БАКТЕРИАЛЬНЫЕ МАТЫ
КАК МОРФОЛОГИЧЕСКИ ОФОРМЛЕННОЕ
АВТОНОМНОЕ СООБЩЕСТВО
Слово "мат" стало широко применяться в английской литерату-
литературе для разнообразных бентосных микробных сообществ и биопле-
биопленок, в том числе и рыхлых. Общим для матов считается плотная упа-
упаковка биомассы в продолжающиеся слои с четким и вместе с тем
флуктуирующим вертикальным градиентом химических и физиче-
физических факторов. Биопленка представляет более общее понятие, каса-
касающееся протяженного развития любого микробного сообщества на
поверхности. В наиболее "чистом" виде биопленка на поверхности
воды называется гидробиологами нейстоном.
В русской литературе Б.Л. Исаченко1 A927 г.) четко различал
два типа строения бентосных фотосинтезирующих сообществ. Он
использовал предложенный Палласом A773 г.) термин "кожа" для
бентосных сообществ Microcoleus chthonoplastes в виде плотной эла-
эластичной массы и термин "войлок" для переплетенной массы нитча-
нитчатых зеленых водорослей Vaucheria или иных. Оба термина хорошо
соответствуют разным условиям развития сообществ: в альго-бак-
териальном "войлоке" остается свободное пространство с водой, до-
доступной даже для беспозвоночных, в то время как циано-бактери-
альная "кожа" представляет плотный гель, напоминающий 1-2%
агар.
Циано-бактериальные маты составили особый предмет в раз-
развитии представлений о микробном сообществе. Есть ряд причин к
этому.
1) Циано-бактериальные маты представляют автономное сооб-
сообщество, где присутствуют и продукционная ветвь углеродного цикла,
осуществляемая цианобактериями, и деструкционная ветвь, осуще-
осуществляемая разными бактериями. Чтобы обозначить гетерогенность
этого сообщества, его было предложено писать через дефис циано-
1 Исаченко БЛ. Микробиологические исследования над грязевыми озерами //
Тр. Геол. ком. Н.С. 1927. Вып. 148. См.: БЛ. Исаченко. Избр. тр. М.; Л.:
Изд-во АН СССР, 1951. Т. 2. С. 26-143.
3*
67
бактериальное, в отличие от сообщества цианобактерий, состав-
составленного одной группой организмов.
2) Циано-бактериальные маты — в высокой степени консерва-
консервативные сообщества, архитектура которых похожа в разных местах
обитания; прежде всего это относится к матам морских лагун, сход-
сходным по всему земному шару.
3) Циано-бактериальные маты представляют строго морфоло-
морфологически оформленные сообщества разнородных организмов. Эди-
фикатором, или формообразующим компонентом, этих сообществ
послужили образующие гликокаликс цианобактерий.
4) Циано-бактериальные маты заведомо представляют древней-
древнейшее сообщество на Земле, имеющее четкую геологическую лето-
летопись в качестве строматолитов - литифицированных слоистых
матов. Ведутся дискуссии о наибольшей древности первых строма-
строматолитов, которые одни авторы относят к первым проявлениям жиз-
жизни на Земле, другие - датируют несколько позже, но все равно очень
давно, около 3,5 млрд лет назад.
5) Циано-бактериальные маты стали излюбленным объектом
изучения взаимодействия разных трофических групп микроорганиз-
микроорганизмов. Благодаря применению микроэлектродной техники доказано
существование в них резких окислительно-восстановительных гра-
градиентов в диапазоне менее 100 мкм.
6) Тесное взаимодействие и контакт трофических групп микро-
микроорганизмов в циано-бактериальном сообществе предполагает про-
происхождение из этого сообщества ветвей наиболее массовых видов
бактерий, прежде всего протеобактерий. Здесь особенно высока ве-
вероятность латерального генетического обмена.
Сходство в морфологии разных матов очень велико, несмотря
на их широкое климатическое и географическое распространение.
Сходство распространяется на морские, лагунные (гиперсоленые),
внутриконтинентальные маты высокоминерализованных озер и в
меньшей степени - на маты термальных источников, которые сле-
следует разобрать отдельно. Маты эфемерных пресных водоемов на
грунте (в лужах) не имеют развитой деструкционной ветви. Маты
встречаются в разных природных условиях, но общим для всех их
местообитаний является отсутствие беспозвоночных с "пастбищным"
питанием. Отрицательная корреляция матов с беспозвоночными яв-
является четко установленным фактом, хотя причины отсутствия
хищников могут быть различны: высокие соленость, температура,
концентрация сероводорода. Поэтому сейчас маты, за исключением
эфемерных на грунте, обычны лишь в экстремальных условиях.
Экстремальные условия, собственно говоря, и определяются по от-
отсутствию высшей растительности и сопутствующих ей животных.
Очевидно, что этого ограничения для матов не было в докембрии,
пока на Земле не появились зоотрофные животные. Труднее отве-
ответить на вопрос, почему цианобактерий не оказались вытесненными
68
из матов эволюционно более поздними эукариотными водорослями.
Альго-бактериальные маты пока не привлекли такого внимания из-
за менее четкой структуры.
В настоящее время наиболее подробно изучены: 1) маты гипер-
гиперсоленых морских лагун, 2) маты континентальных водоемов, 3) ма-
маты термальных источников. Все эти места обитания резко отличают-
отличаются друг от друга, и очевидно, что организмы одного места обитания
совершенно не приспособлены для развития в другом. Их видовой
состав крайне различен. Тем большее значение имеет сходство мор-
морфологической и трофической структуры, которое выступает как
закономерность, лежащая над филогенетической таксономией и оп-
определяющая функциональное положение и функциональные харак-
характеристики каждого организма.
Особо следовало бы рассмотреть субаэральные маты на грунте
или скопления почвенных водорослей, среди которых особенно при-
примечательны содержащие гетероцисты азотфиксирующие цианобак-
цианобактерий Nostoc. Такой циано-бактериальный покров свойственен пус-
пустынным областям с эфемерным увлажнением.
Маты имеют строго последовательное слоистое строение. По-
Поскольку слои мата ярко окрашенны, они получили, например, для
песчаных побережий Балтики, немецкое название Farbstreifensand-
watt - "полосато-окрашенные песчаные болота".
3.2. МАКРОСТРУКТУРА
ЦИАНО-БАКТЕРИАЛЬНЫХ МАТОВ
Циано-бактериальный мат представляет прокариотное бентос-
ное сообщество, объединенное в физическую структуру слизью
(гликокаликсом) как формообразующим средством, превращаю-
превращающим сообщество в морфологически единое образование - донную
кожу.
В циано-бактериальном мате взаимодействуют представители
разных трофических групп, осуществляя полный цикл биогенных
элементов. Макроструктура мата отражает последовательность
трофических взаимодействий между группами организмов. По тер-
терминологии экологов мат представляет собой консорциум с синузием
цианобактерий и аноксигенных фототрофов и гильдиями деструк-
деструкторов. Трофические взаимодействия в мате определяются двумя до-
доминирующими циклами: углерода и серы. Структура мата вне зави-
зависимости от его топического положения - прибрежного морского,
лагунного гиперсоленого, озерного аталассофильного, термального
вулканического - определяется последовательностью трофических
взаимодействий и транспортными процессами.
Энергетика циано-бактериального мата определяется светом и
его проникновением в мат. Смена дня и ночи определяет два его со-
69
стояния: на свету и в темноте. На свету доминируют продукционные
процессы, в темноте - деструкционные. В связи с тем, что время сме-
смены дня и ночи меньше времени размножения бактерий, они должны
приспосабливать свой обмен к существованию в переменном режи-
режиме. Это приспособление осуществляется как за счет внутриклеточ-
внутриклеточных запасных веществ, так и за счет пулов внеклеточного вещества
в сообществе.
Следующим определяющим фактором является диффузия кисло-
кислорода и его обмен с атмосферой. Днем существует избыток кислорода,
пересыщение им, поскольку пузырьки кислорода улавливаются струк-
структурой мата. Ночью кислород полностью потребляется, и его пополне-
пополнение диффузией из атмосферы оказывается совершенно недостаточным
для поддержания аэробных условий под слоем оксигенных фотоавто-
трофов. Поэтому организмы в верхних слоях мата вынуждены перехо-
переходить от аэробиоза, включая условия гипероксии, к анаэробиозу.
Под слоем оксигенных фототрофов располагается слой бесцвет-
бесцветных нитчатых серобактерий, работающих в режиме тиобиоса на
У//////////////////////
Д<8 V <€ Д<8 V
Ъ> V В>Д Ъ> V <&
///////////////////777/
//7/////77///777//777//
///У///////////////////
П
ш
А с8 V
цианобактерии
слои карбоната
кальция
слой развития
пурпурных бактерий
с отложением гипса
сульфатредуцирую-
щие бактерии
IV
Рис. 3.1. Строение циано-бактериального мата [по: Бактериальная..., 2002]
I - зона оксигенного фотосинтеза с развитием цианобактерии; II - зона аноксигенного фо-
фотосинтеза с развитием пурпурных бактерий; III - зона захоронения цианобактерии с просло-
прослоями карбоната кальция; IV - аноксигенная зона развития сульфатредуцирующих бактерий
70
Рис 3.2. Поперечные срезы матов: алкалофильного (а) и галофильного (б) [по:
Герасименко и др., 2003]
а - зона оксигенного фотосинтеза с минеральными слоями; б - зона аноксигенного фото-
фотосинтеза; в - зона захоронения цианобактерии; г - зона сульфатредукции
границе сероводородной и оксигенной зон (рис. 3.1, 3.2). Здесь же
обильны нитчатые скользящие флексибактерии, морфологически
сходные с витреосциллами. Этим организмам приписывают способ-
способность к хищничеству путем лизиса жертвы, но на самом деле она не
подтверждена выделением чистых культур. На границе проникновения
света располагаются пурпурные бактерии, особенно с поглощением
в ближней инфракрасной области, поскольку длинноволновые лучи
менее рассеиваются. Спектральные характеристики аноксигенных
71
Рис. 3.2. (окончание)
фототрофных бактерий хорошо согласуются с законами хроматиче-
хроматической адаптации: максимумы их поглощения располагаются в окнах
пропускания хлорофилла цианобактерий. Они определяют анаэроб-
анаэробное окисление сероводорода в сульфат на свету, а ночью подвижные
формы мигрируют к поверхности и существуют за счет окисления
соединений серы. Под ними располагается слой анаэробных органо-
трофных бактерий, которые осуществляют разложение мортмассы
продуцентов. Слой этот не характеризуется морфологически и
обычно находится под воздействием поднимающегося снизу серово-
сероводорода.
Глубже располагается зона сульфатредукции, в которой завер-
завершается разложение органических веществ и начинается цикл серы.
Черные сульфиды железа окрашивают эту зону и служат ее надеж-
надежным индикатором.
72
3.3. ФОТОТРОФНЫЙ СЛОЙ
Физическая структура мата диктуется особенностями развития
первичных продуцентов и эдификаторов мата - цианобактерий. Они
образуют псевдоткань, в которой развиваются остальные группы
организмов. Развитие цианобактерий в мате определяется в первую
очередь освещенностью. Маты развиваются в мелководных услови-
условиях, и для них важна прозрачность воды над матом, обеспечиваемая
удалением мути как неорганической, так и планктонной.
Состав оксигенных фототрофов в матах изучен относительно
хорошо благодаря тому, что идентификация цианобактерий и водо-
водорослей основана на морфологических критериях. В матах доминиру-
доминируют либо осцилляториевые, либо энтофизалисовые цианобактерий.
В гиперсоленых водоемах характерным эдификатором является
Microcoleus - организм, образующий пучок трихомов внутри общего
слизистого чехла. Эти чехлы оттесняют бактерии в пространство
между ними и создают вокруг пучка трихомов световые колодцы.
Microcoleus образует особенно плотную кожу на дне соленых мор-
морских лагун с соленостью вплоть до примерно 17% и представляю-
представляющую за счет внеклеточной слизи водоупор.
Распространены эти маты повсеместно и очень сходны в разных
местах океанического побережья. Сейчас маты с Microcoleus явля-
являются наиболее распространенными в прибрежных частях морских
водоемов. У нас в стране они были подробно изучены в лагунах Ара-
батской стрелки на Сиваше (Заварзин и др., 1993). Аналогичные ма-
маты известны для берегов Персидского залива, на берегах залива
Акаба в Красном море, на Пиренейском полуострове по побережью
Средиземного моря, в Калифорнийском заливе. В общем они при-
приурочены к семиаридному и аридному климату. В гумидном климате
они не столь заметны. При повышенном содержании H2S распро-
распространение получают различные виды Phormidum, Lyngbya, образую-
образующие маты за счет переплетения трихомов в ткань. Маты нитчатых
организмов, получающие механическую прочность за счет перепле-
переплетения нитей в ткань или войлок, не служат водоупорами.
Верхний слой мата занимают оксигенные фототрофы, плот-
плотность которых достаточна для полного самозатенения хлорофил-
хлорофиллом. Это достигается в слое около 1 мм и менее. Таким образом,
продукционная активность мата находится на пределе, обусловлен-
обусловленном проникновением света. На глубине 0,1-0,3 мм освещенность
возрастает по сравнению с поверхностью. Это обусловлено рассея-
рассеянием света. В кварцевом песке с частицами 125-250 мкм вертикаль-
вертикальная составляющая освещенности падает логарифмически от 100%
на поверхности до 0% на глубине 2 мм. Вертикальная составляющая
отраженного света возрастает к поверхности до 20% от падающего
света. Наконец, скалярная составляющая возрастает к поверхности
до 200% от падающего света (рис. 3.3, а).
73
Интенсивность света, % от падающей радиации
О 50 100 150 200
а
Рис. 3.3. Профили освещенности и фотосинтеза в циано-бактериальных
матах
а - профиль интенсивности света [по: Kuhl et al., 1994]; Еи - отраженная, Ed - проникаю-
проникающая, Eq - скалярная составляющие в кварцевом песке с размером частиц 125—250 мкм. Ве-
Величины даны в % от радиации на поверхности осадка;
б - профиль фотосинтеза, концентрации О2 и интенсивности света [по: Lassen et al.,
1992]
Фотосинтез, нмоль Оз • см 3 • с '
О 10 20 30
-0,5
« 0,5
I
40
С>2, мкМ
500
1000
200 400 600
Скалярная радиация, мкЭйншт • м~2 • с
Рис. 3.3. (окончание)
При избыточной освещенности цианобактерии уходят в глубину,
и нередко под затеняющим предметом поверхность осадка становит-
становится зеленой из-за выползающих на поверхность цианобактерии. Эта
вертикальная миграция особенно хорошо видна в течение суток. Для
фототрофов мата в полной мере действует правило хроматической
адаптации - приспособленности к поглощению света, проходящего
через вышележащий слой. Аноксигенные фототрофы, из которых
особенно характерна пурпурная Thiocapsa, иногда Chromatium, соста-
составляют отчетливо видимый вишневый слой на глубине около 1 мм, от-
отделенный от верхнего зеленого слоя бесцветным промежутком.
Проникновение света в мат было измерено Б. Иоргенсеном с по-
помощью микросветоводов (рис. 3.3, б). В верхнем 0,0-1,5 мм слое мата
74
доминирует поглощение хлорофилла (Хл) а (пики при 440 нм и
675 нм) и каротиноидов D50-550 нм), обусловленное диатомовыми.
На глубине 1,5-2,0 мм в области развития Microcoleus и Phormidium
помимо сигнала Хл а появляется сигнал фикоцианина F20-625 нм).
Ниже 2,5 мм появляется сигнал бактериохлорофилла (Бхл) а (807 нм и
865 нм) и Бхл с G46 нм) за счет присутствия пурпурных и зеленых ано-
ксигенных фототрофов. Близкий инфракрасный свет, используемый
комплексом бактериохлорофилл-белок, рассеивается в 10—100 раз
меньше, чем видимый свет. Поэтому наблюдается четкое распределе-
распределение групп фототрофов соответственно хроматической кооперации
(рис. 3.4). Зеленые серные бактерии имеют Бхл c,d,e с поглощением
700-800 нм; пурпурные с Бхл а - 800-900 нм; с Бхл Ъ - 1000-1050 нм.
75
400 500 600 700 800 900 1000 1100
нм
Рис. 3.4. Хроматическая кооперация фототрофов [по: Perry, Staley, 1997]
а - цианобактерий; б - пурпурные бактерии; в - зеленые бактерии; / - сцитонемин; 2 - ка-
ротиноиды; 3 - хлорофиллы (Хл), бактериохлорофиллы (Бхл); 4 - фикобилины
Особо следует рассмотреть действие на маты ультрафиолета.
Известно, что UV-B B80-315 нм) оказывает стерилизующее дейст-
действие и способствует образованию "озоновых дыр". Тем не менее, ко-
когда не было кислородной атмосферы и соответственно защитного
слоя озона, циано-бактериальные сообщества процветали. Распре-
Распределение энергии в Вт/м2 примерно таково: видимый + инфракрас-
инфракрасный D00-1000 нм) - 1200-800; UV-A C15-400 нм) - 40-50; UV-B
B80-315 нм) - 3-4. Фотосинтез Microcoleus угнетался на 60% приме-
примесью 8% UV-B к фотосинтетически активной радиации (ФАР), при-
причем ингибирующий эффект простирался на глубину фотической зо-
зоны 0,7 мм в песке. UV-A угнетает Calothrix thermalis, действуя на син-
синтез тетрапирролов. Суточная миграция цианобактерий обусловлена
в основном реакцией на UV-A с порогом около 1,3 Вт/м2.
Особенности распределения ФАР в рассеивающих средах приво-
приводят к тому, что максимум фотосинтеза, измеренный по выделению
кислорода в матах, находится примерно на глубине 1 мм. Впрочем,
76
эта глубина сильно зависит от освещенности, и при слабой интенсив-
интенсивности максимум мигрирует вверх. Благодаря микроэлектродам с
очень быстрым откликом на изменения О2 было показано, что
градиент О2 мгновенно, за доли секунды, реагирует на изменение ос-
освещенности. Таким образом, и в пространстве, и во времени в мате
происходят быстрые изменения химических градиентов, в том числе
важнейшего из них - градиента О2. Дыхание фототрофов представ-
представляется постоянным в течение 4 сек. Динамический баланс между
продукцией О2 и его потреблением на дыхание в мате приводит к то-
тому, что при освещении концентрация О2 превышает насыщающую,
и это отлично видно по выделению пузырьков газа. При затемнении
О2 полностью используется в течение нескольких минут. Избыточ-
Избыточный кислород токсичен для цианобактерий. В этом одна из причин
их развития с неспецифическими аэробными спутниками. Развитие
Microcoleus зависит от бактерий-спутников: если штамм организма
не обладает каталазой, то в чистой культуре на свету он быстро де-
деградирует, в то время как рост его с галофильными аэробными спут-
спутниками происходит нормально.
Цикл азота в мате связан в основном с анаболическими процес-
процессами. По элементному составу маты очень близки к отношению
Редфильда С : N : Р = 106 : 16 : 1.
В них могут идти микробные процессы.
Состав биомассы [(СН2ОI0б(МН3I6Н3РО4] = [А].
Аэробная деградация:
[А] + 138О2 = 106СО2 + 16HNO3 + H3PO4 + 122Н2О;
Д G'q = -3190 кДж • моль глюкозы.
Нитратное дыхание:
[А] + 94,4HNO3 = 106СО2 + 55,2N2 + 1Н3РО4 + 177,2Н2О;
A Gq = -3030 кДж • моль-1 глюкозы.
Восстановление железа:
[А] + 212Fe2O3 + 848H+ = 106СО2 + 16NH3 + 1Н3РО4 + 424Fe2+ + 530H2O;
424FeOOH 742Fe2+
Д G6 = -1410 кДж • моль-1 глюкозы.
Сульфатное дыхание (сульфидогенез):
[А] + 53 SO2." = Ю6СО2 + 16NH3 + Ш3РО4 + 53S2- + Ю6Н2О;
Д G6 = -380 кДж • моль глюкозы.
Метаногенез:
[А] = 53СО2 + 53СН4 + 16NH3 + 1Н3РО4;
Д G6 = -350 кДж • моль глюкозы.
Фиксация азота, измеряемая по редукции ацетилена, обнаружи-
обнаруживает четкую зависимость от ФАР, находясь в противофазе к ней, но
зависит и от доступной энергии. Максимум азотфиксации имеет ме-
место ранним утром на рассвете и на закате, когда ФАР снижается
примерно до 1/10 дневного максимума. Величина фиксации зависит
77
от доминирующей формы: Lyngbya фиксирует многократно больше,
чем Microcoleus, но суммарная фиксация достаточна, чтобы обеспе-
обеспечить лишь 15-20% от дневного потребления углерода. Днем проис-
происходит энергичное потребление СО2 и синтез запасных органических
веществ, а вечером и ночью - дыхание и фиксация азота с образова-
образованием запасного азотистого вещества цианофицина, полимера аспа-
рагиновой кислоты и аргинина, который может служить и энергети-
энергетическим источником, сбраживаясь в орнитиновом цикле. Дополни-
Дополнительным источником азота служит NH4, поступающий из анаэробной
зоны разложения мортмассы под фотическим слоем. Концент-
Концентрация NHj резко падает в фотической зоне. Для медленно растуще-
растущего мата Microcoleus рецикл азота в системе оказывается достаточ-
достаточным.
В ночное время мат становится анаэробным, и цианобактерии
должны получать энергию хемотрофным путем. На темновой мета-
метаболизм цианобактерии обращали мало внимания, между тем он
очень важен для сообщества. Цианобактерии в качестве основного
запасного вещества на свету накапливают полиглюкозу. Ночью в
аэробных условиях она расходуется на дыхание. Если резервуар рас-
растворенного кислорода мал, как это имеет место в матах, то быстро
наступают аноксические условия и цианобактерии вынуждены пере-
переходить к энергетически менее выгодному брожению. Запас углево-
углеводов у цианобактерии бывает настолько велик, что может обеспечи-
обеспечивать даже темновую азотфиксацию в восстановленных условиях.
Цианобактерии способны осуществлять разные типы брожения:
часть из них осуществляет гомо- или гетероферментативное молоч-
молочнокислое брожение, часть — смешанное и даже гомоацетатное.
Ферменты этих брожений конститутивны у цианобактерии. Продук-
Продуктами брожений являются СО2, Н2, формиат, лактат, ацетат, этанол2.
Из списка продуктов видно, что к их использованию приспособ-
приспособлены сульфатредукторы, и в сообществе вероятно образование
H2S, который может окисляться в серу как хемотрофными аэроба-
аэробами, так и аноксигенными фотосинтетиками. То и другое возможно
днем. Многие тиофильные цианобактерии обладают гидрогеназой и
способны к сероредукции с образованием H2S, при этом не происхо-
происходит выделения Н2. Таким образом, для сообщества выгодно дневное
накопление элементной серы, которое действительно наблюдается
в мате не только для аноксигенных фототрофов, но и тионовых ба-
бактерий.
Microcoleus сбраживает гликоген, но не свое осмопротекторное
вещество гликозилглицерол, с образованием ацетата, этанола, фор-
миата, Н2. В присутствии серы образуется вдвое больше ацетата,
2 Stal L. Cyanobacterial mats and stromatolites // The ecology of cyanobacteria / Ed.
B.A. Whitton, M. Potts. Kluwer academic, 2000. P. 61-120.
78
резко снижается содержание остальных продуктов, и вместо Н2 про-
происходит образование H2S. Таким образом, происходит типичный
ацетогенный сдвиг:
Глюкоза = 2ацетат + 2СО2 + 8[Н]; 8[Н] + 4S = 4H2S.
В результате можно утверждать, что эдификатор циано-бактери-
альных матов, Microcoleus chthonoplastes, отлично приспособлен к
суточным изменениям режима.
Внутриклеточная сера как запасное вещество, пригодное слу-
служить резервом катаболизма как донор электрона в аэробных и как
акцептор в анаэробных условиях, на самом деле представляет капли,
заключенные в белковую мембрану и располагающиеся в периплаз-
периплазме. Образование серы по Трюперу3 начинается с окисления H2S или
сульфанной группы тиосульфата мембраносвязаной сульфид:хинон
редуктазой или периплазматическим флавоцитохромом, причем в
случае тиосульфата сульфит окисляется и выбрасывается. Актива-
Активация цепей молекул внутриклеточной серы происходит путем ее вос-
восстановления с последующим окислением в сульфит сирогем сульфит-
редуктазой. Окисление сульфита в сульфат происходит либо на
наружной стороне мембраны в периплазме сульфит:оксидоредукта-
зой, либо в цитоплазме через аденозинфосфосульфат (АФС) с вы-
выбросом сульфата АТФ-сульфурилазой. Обмен такого рода наблюда-
наблюдается у представителей Ectothiorhodospiraceae, Chlorobiaceae, пурпур-
пурпурных несерных бактерий, зеленых нитчатых фототрофов, цианобак-
цианобактерии, а также у многих хемолитотрофных серных бактерий, но не
у эукариот.
В ночное время окисление соединений серы оказывается основ-
основной статьей расхода кислорода, и, следовательно, тионовые и серо-
серобактерии играют важную роль в поведении системы в целом. Спо-
Способность бесцветных серобактерий окислять и восстаналивать внут-
внутриклеточную серу, возможно, унаследована ими от аналогичных по
строению трихомных цианобактерии. Цикл серы в циано-бактери-
альном мате описал ван Гемерден4. В трофических группировках
циано-бактериального мата наблюдается тесная взаимозависимость
между несколькими основными группами бактерий, располагающи-
располагающимися последовательно: 1) оксигенными цианобактериями; 2) сероба-
серобактериями; 3) аноксигенными фототрофами; 4) сульфатредуцирую-
щими бактериями (СРВ). Группы имеют противоположные требо-
требования: сульфид токсичен для цианобактерии, кислород токсичен для
L
3 Truper H.G. Dissimilatory sulfur metabolism and prokaryotes // X Intern, symp. on
phototrophic prokaryotes, Barcelona, Spain, Aug. 26-31,2000: Program and abstracts.
Barcelona, 2000. P. 40.
4 Ende F.P. den, Gemerden H. van. Relationships between functional groups of organ-
organisms in microbial mats // Microbial mats: Structure, development and environmental
significance / Ed. L.G. Stal, P. Caumette. B. etc.: Springer, 1994. P. 339-354.
79
СРБ и аноксигенных фототрофов. Спектр фотосинтетически актив-
активной радиации (ФАР) для аноксигенных фототрофов частично комп-
комплементарен ФАР для цианобактерий. Благодаря светорассеянию на
минеральных частицах пурпурные бактерии располагаются под циано-
бактериями на 1-2 мм ниже. Серобактерии защищают цианобактерий
от сероводорода, а пурпурных и СРБ - от кислорода. Скорость роста
Microcoleus стимулируется сероводородом до 0,15 мМ и полностью
угнетается выше 0,2 мМ. Для аноксигенных фототрофов максимум
скорости роста лежит при 0,2 мМ и медленно снижается к 1 мМ, при-
причем при высокой концентрации накапливается гликоген на ночь.
Типичным аноксигенным фотосинтетиком в мате является
Thiocapsa roseopersicina. Организм способен к фототрофному и хе-
мотрофному росту, использованию полисульфидов, ассимиляции
низкомолекулярных органических веществ. Кислород подавляет
фотосинтез, но не рост, аэробно организм растет, как тиобациллы.
Днем в аэробных условиях пигменты не синтезируются, а ночью в
анаэробных - пигменты быстро синтезируются за счет резерва
гликогена.
Окисление соединений серы осуществляют тионовые бактерии,
близкие к Thiobacillus thioparus. При снижении в газовой смеси воз-
воздуха от 70% до 30% образование сульфата линейно уменьшается,
причем содержание растворенного кислорода удерживается ниже
детектируемого уровня, и при этом линейно возрастает образование
серы как конечного продукта. Сродство тиобацилл к сульфиду
больше, чем у тиокапсы, но зато тиокапса может расти при мень-
меньшем содержании кислорода, и в результате в бинарной культуре при
70% воздуха численность тиобацилл начинает снижаться, а тио-
капс - возрастать. Бинарная культура оказывается устойчивой,
поскольку тиобациллы снижают содержание кислорода ниже детек-
детектируемого уровня, и становится возможным синтез пигмента фото-
трофами.
Наибольшая численность СРБ наблюдается в верхнем оксигени-
рованном слое. Утверждается, что нитрат может ими восстанавли-
восстанавливаться в аммоний, углекислота - в ацетат, восстанавливаются селе-
нат, уран, железо и (!) даже О2. Устойчивость СРБ к О2 различна,
причем токсичен не сам О2, а продукты его неполного окисления;
особенно возрастает токсичность О2 при окислении сероводорода и
тиолов, что ведет к образованию супероксида и перекиси. Оказа-
Оказалось, что обычные субстраты СРБ могут служить донорами элект-
электрона при восстановлении О2 и при этом идет транспорт протона и
образование АТФ. Многие СРБ а) восстанавливают сульфат в тио-
тиосульфат или тритионат; б) осуществляют диспропорционирование;
в) аналогично тиобациллам окисляют соединения серы: сульфит,
тиосульфат, полисульфиды. Отсюда делается заключение, что СРБ
отнюдь не так специфичны, как рисовалось, начиная с работ Бейе-
ринка и до Постгейта. Это касается широкого круга соединений
80
серы и их превращений. Далее, отдельные виды ведут себя по-разно-
по-разному по отношению к разным соединениям, далеко не всегда следуя
термодинамике. Все СРБ относятся к типичным вторичным анаэро-
анаэробам и используют продукты брожения, а не углеводы или белки.
Поэтому они нуждаются в цепочке обычных анаэробов как трофи-
трофических предшественников.
3.4. БИОСТАБИЛИЗАЦИЯ ОСАДКОВ
Галофильные маты развиваются на побережьях, где движение
осадков вдоль берега приводит к образованию кос и лагун. Движе-
Движение осадков создает физически неустойчивую среду, обитание в ко-
которой затруднено. Интерстициальное пространство в песчаных
осадках заселяется группой мелких организмов - псаммоном. Од-
Одновременно происходит заселение водорослями, чаще всего диато-
диатомовыми, которым движение осадка приносит меньше всего вреда.
Среди первопоселенцев присутствуют и цианобактерий. Развитие их
приводит к сукцессионным изменениям и закреплению осадка бла-
благодаря склеиванию частиц слизью, выделяемой цианобактериями.
Процесс получил название биостабилизации осадков и был исследо-
исследован для Северного моря.
Внеклеточные полимерные вещества, или экзополимеры, харак-
характерны для многих прокариот. Они синтезируются в ответ на стрес-
стрессы и создают микроусловия для заключенных в них микроорганиз-
микроорганизмов, обеспечивают фиксацию микроорганизмов в определенном
месте. Экзополимеры состоят из очень больших (> 100000 дальтон)
молекул, как правило, анионной природы, обеспечивающих (из-за
присутствия уроновых кислот) их катионобменные свойства. Экзо-
Экзополимеры по химической природе бывают либо полисахаридами,
либо протеинами. Гидрофильные экзополимеры представлены пре-
преимущественно гетерополисахаридами, сильно гидратированы и
склонны к диспергированию, в то время как гидрофобные обязаны
своими свойствами метальным и ацетильным группам или белковой
природе; гидрофобность обеспечивает связывание клетки с твер-
твердым субстратом. Состав и состояние экзополимеров в большой сте-
степени зависит от природных условий развития организмов и часто на-
нарушается в лабораторных культурах.
Третичная структура экзополимеров обусловливает три состоя-
состояния: 1) конденсированного геля, 2) рыхлой слизи, 3) коллоидного
раствора. Между этими состояниями есть плавные переходы. Экзо-
Экзополимеры обозначают как слизь и капсулу. Капсула тесно связана с
индивидуальной клеткой, слизь распространяется без прямого кон-
контакта с ней. На состояние экзополимеров очень влияет температура,
и при ее понижении они переходят в более конденсированное и упо-
упорядоченное состояние (см.: 2.2.4. "Коллоидная среда обитания
микроорганизмов").
81
Упорядочение экзополимеров обусловлено двумя различными
механизмами. Полимерные цепочки гетерополисахаридов упорядо-
упорядочиваются благодаря катионным мостикам щелочноземельных метал-
металлов Са и Mg; гликопротеины сшиваются прочными дисульфидными
связями -S=S-. Щелочноземельные металлы склонны к обмену и мо-
могут быть удалены хелирующими агентами. В зависимости от степени
конденсации экзополимеров они могут представлять препятствие для
проникновения крупных частиц, например бактериальных клеток, в
чехлы цианобактерий, а с другой стороны, служить сетью для удер-
удерживания макромолекул, таких, как экзоферменты. Наличие слизи во-
вокруг клетки обусловливает характер диффузии веществ в полимер-
полимерной матрице. Гелеобразная слизь придает сообществу прочность и
характер "кожи". Такая "кожа" обладает, как уже отмечалось, водо-
водонепроницаемыми свойствами наподобие агарового геля.
Альтернативой слизи может служить "ткань", образуемая пере-
переплетением нитевидных организмов. В этом случае прочность струк-
структуры обусловливается трением нитей или их склеиванием и прочно-
прочностью нитей на разрыв. Пространство между нитями может быть
заполнено слизью, но может быть и свободно. В этом случае меха-
механическую прочность обеспечивает сравнительно тонкий чехол, как
у Phormidium, и прочность самого трихома. "Ткань" не менее проч-
прочна, чем "кожа", но не обладает водоудерживающими свойствами и
напоминает рыхлую бумагу. Этот тип строения матов у крупных
нитчатых эукариотных водорослей получил название "войлока".
По своим механическим свойствам оба русские названия гораздо более
точны, чем английский "мат". Последнее слово совсем не следует
применять к рыхлым, легко взмучиваемым пушистым бентосным
сообществам. Такие сообщества удерживаются в гидродинамически
спокойной среде в виде наилка на поверхности уплотненного осадка.
Следующим шагом к устойчивости служит цементация мине-
минеральными частицами. Цементация осуществляется чаще всего кар-
карбонатами с включением силикатов, но может происходить и за счет
фосфатизации. Минерализация мата приводит к его превращению в
органо-седиментационную породу и вместе с тем вынуждает бакте-
бактерии передвигаться на поверхность мата. В структуру мата включа-
включаются минеральные частицы, принесенные извне течением. Преиму-
Преимущественной областью развития матов служат приливные отмели
(tidal flats). В зависимости от величины частиц они разделяются на
песчаные (> 63 нм), илистые B-63 нм), глинистые (< 2 нм).
Физическая устойчивость мата в среде зависит от свойств экзо-
экзополимеров, объединяющих разнородные организмы в оформленное
сообщество с закономерной физической структурой. Устойчивость
определяется способностью противостоять движению воды, вызван-
вызванному течениями или ветром. Оценить ее можно по взмучиванию ми-
минеральных частиц под действием гидравлического удара. Для этого
используется воздух или вода под давлением и отмечается давление,
82
при котором поверхность мата нарушается. При увеличении скоро-
скорости течения происходит привнос частиц и их оседание на поверхно-
поверхности, седиментация. Микробный мат служит ловушкой для оседаю-
оседающих минеральных частиц. При дальнейшем увеличении скорости те-
течения выше некоторой критической величины начинается размыва-
размывание осадка, эрозия. Очевидно, что в природных условиях периоды
седиментации и взмучивания меняются в зависимости от условий, и
на профиле осадков можно отмечать периоды штормов.
Частицы осадка скрепляются органическим веществом, которое
адсорбировано на поверхности мата или же находится в интерстици-
альном (поровом) пространстве. Когда гидравлическое воздействие
переходит некоторый порог, происходит вынос интерстициального
органического вещества, но адсорбированное вещество сохраняет-
сохраняется, пока не будет стерто абразией. Отсюда возникает зависимость
устойчивости осадка к гидравлическому удару от содержания орга-
органического вещества, составляющего процент-доли процента от
осадка. Тем не менее оно оказывает склеивающее действие на оса-
осадок. Слизистый слой цианобактериальной кожи на дне водоема пол-
полностью предотвращает взмучивание осадка и поэтому ценится тех-
технологами. Водопроницаемость его крайне низка. Таким образом, в
осадке следует рассматривать несколько размерных интервалов:
микрониши, соответствующие биопленке в десятки-сотню микрон;
средний уровень, соответствующий миллиметрам; макрошкалу, оп-
определяющую застойные и подвижные участки на побережье в мет-
метровом-километровом диапазоне.
3.5. ГАЛОФИЛЬНЫЙ МАТ
Галофильные маты образуются в условиях, когда испарение
преобладает над поступлением влаги, дождевой или же при промывке
течениями. Такие условия преобладают в семиаридной зоне и тропи-
тропиках. В результате движения осадка вдоль побережья образуются
сначала песчаные косы, составляя цепь островов, например Фриз-
Фризские острова на побережье Северного моря, затем они замыкаются,
образуя замкнутые или полузамкнутые лагуны, как Сиваш на бере-
берегу Азовского моря. Моделью образования солеродного или эвапо-
ритового бассейна считается Кара-Богаз-Гол на Каспии. В лагунах
происходит испарение морской воды, приводящее к засолению. При
концентрации морской воды последовательно выпадают соли, сна-
сначала карбонаты, затем гипс, затем при примерно 25% солености вы-
выпадает галит - поваренная соль. Этот процесс используется в соле-
солеварнях, которые служат крупномасштабными моделями естествен-
естественных процессов образования эвапоритов - осадочных минералов и
пород хемогенного происхождения. ;
Физические условия существования галофильного сообщества,
помимо обусловленных высокой соленостью, характеризуются ма-
83
.
лой гидродинамической подвижностью среды в мелководных лагу-
лагунах и эвапоритовой тягой за счет испарения, направленной снизу
вверх. Повышение солености предотвращает развитие животных и
водорослей, за исключением рачка Anemia (Decapoda), живущего в
условиях солености примерно до 10%, и галофильной зеленой водо-
водоросли Dunaliella salina, образующей глицерин в качестве осмопротек-
тора и при высокой солености придающей красный цвет осадку за
счет каротиноидов. Таким образом, в соленых лагунах развивается
прокариотное сообщество практически без вмешательства эукари-
отных организмов. Здесь нет даже протестов. Это галофильное со-
сообщество составляет целостную систему, которая может служить
аналогом тех систем, которые существовали на Земле до появления
эукариот. Большинство результатов по циано-бактериальным ма-
матам получено именно на основе галофильных матов.
Биологические особенности организмов, входящих в галофиль-
галофильное циано-бактериальное сообщество, обусловливаются их способ-
способностью переносить высокую концентрацию солей вне клетки. Спо-
Способность противостоять осмотическому стрессу предполагает повы-
повышение осмотического давления внутри клетки.
3.5.1. Галофильные микроорганизмы
Жизнедеятельность клетки в соленой среде может обусловли-
обусловливаться двумя различными механизмами: 1) устойчивостью всех фер-
ферментов и компонентов клетки к высокому содержанию солей, "вы-
"высаливающих" белки; 2) удалению солей из клетки и замене их на ор-
органические осморегуляторные вещества. Известно, что оба меха-
механизма действуют. Первый механизм действует у экстремально гало-
галофильных аэробных архей Halobactenaceae и анаэробных бактерий
Halanaerobiales. При этом доминирующий во внешней среде ион на-
натрия внутри клетки уравновешивается ионом калия. Второй меха-
механизм включает большое количество органических веществ, кото-
которые должны удовлетворять нескольким требованиям: быть низко-
низкомолекулярными, не вымываться из клетки через мембрану, проти-
противостоять обычному метаболизму клетки, их концентрация должна
поддаваться регулирующему механизму в соответствии с меняю-
меняющейся соленостью среды, они должны быть совместимы с обменом
клетки.
К осморегуляторным веществам относятся следующие классы
соединений: 1) сахара и сахароспирты, прежде всего сахароза и тре-
галоза, и полиолы глицерин и арабит, а также гетерозиды (глико-
зилглицерол); 2) глицинбетаин; 3) тетрагидропиримидины (эктоин);
4) альфа-аминокислоты (пролин, глутамин); 5) N-ацетилирован-
ные диаминокислоты (iV-дельта-ацетилорнитин) и карбоксамиды
глутамина; 6) метилированные серные соединения, как диметил-
сульфониопропионат (рис. 3.5). Концентрация осмолитов в клетке
84
СН2ОН
CH2OH
' -о.
но
НО
О
носнР
Сахароза
:н2он
Гликозилглицерол
СН
сн3сн3
Бетаин
НО
Трегалоза
СНз
СН3—S-v^~v.
3 coo
Диметилсульфониопропионат
or
н2
Пролин
Н
о
coo3
СН3 g н
R = Н Эктоин
R = ОН Гидроксиэктоин
Н Н СС
Ncc-ацетилорнитин
NH2 I
I NH
Л
NH,
4
ш
NS-карбамоилглутаминамид ^-ацетилглутамилглутаминамид
Рис 3.5. Осморегуляторные соединения галофильных и галотолерантных бак-
бактерий [по: Galinski, Triiper, 1994]
85
Разные
галофилы
Microcoleus
chtho'nopltastes
(гетерозид)
Cladophora
.(некромасса)
Hqkmaerobiwn
Sporokdobacter
1 2 3
5 6
7 8
Proteobacteria
^„Сульфатре- у
дукторы., J
*¦ Тионовые -'
"(S-цикл)
Рис. 3.6. Трофические связи между микроорганизмами в галофильном циано-
бактериальном сообществе при анаэробном разложении осморегуляторных ве-
веществ
превышает 0,5 М^. Цианобактерии в качестве осмопротекторов ча-
чаще всего имеют трегалозу или гетерозиды. Концентрация Сахаров
редко превосходит 500 мМ, но у Microcoleus при предельной концен-
концентрации солей может достигать 30% от веса клетки.
Аэробные галофильные бактерии относятся преимущественно
к альфа- и гаммапротеобактериям. У них основными осмопротек-
торными веществами служат глицинбетаин и эктоин (Deleya,
Halomonas, Vibrio alginolyticus, а также Ectothiorhodospira,
Rhodospirillum). Глицинбетаин служит "обобществленным" осморе-
гулятором сообщества, и большинство организмов охотно транс-
транспортируют его извне внутрь клетки, приобретая таким способом ус-
устойчивость к осмотическому стрессу. Эктоин был открыт сравни-
сравнительно недавно, но оказался доминирующим у эубактерий.
Разложение осморегуляторных веществ в галофильном сообще-
сообществе осуществляет характерная группа галоанаэробов. Среди них
Halanaerobium, Halobacteroides специализированы на сбраживании
дисахаридов и гликозилглицерола, a Acetohalobium сбраживает гли-
глицинбетаин. Образующийся при этом триметиламин используется ха-
характерной группой метилотрофных галофильных метаногенов
5 Galinski E.D., Truper H.G. Microbial behaviour in salt stressed ecosystems // FEMS !*
Microbiol Rev, 1994. Vol. 15. P. 95-108. f'
86
3,5
15
20
25
30 % NaCl
Рис. З.7. Адаптивная динамика метилотрофных галофильных метаногенов
(Т.Н.Жилина, неопубл.)
1 - Methanosarcina; 2 - Methanococcus methylutens; 3 - Methanococcus tindarius;
4 — Methanohalophilus halophilus Z-7982T; 5 — Methanohalophilus halophilus Z-7601; 6 -
Methanohalopilus portugalensis Z-7401; 7 - Methanohalopilus portugalensis Z-7302; 8 -
Methanohalobium evestigatum Z-7408; 9 - Methanohalobium evestigatum Z-7303T
Methanohalobium. В результате в сообществах высокоминерализо-
высокоминерализованных водоемов создаются условия для специфического метаболи-
метаболического пути, обусловленного разложением органических осмопро-
текторных веществ, которые по мере увеличения минерализации
отвлекают на себя все ббльшую часть продукции (рис. 3.6, 3.7).
3.6.АЛКАЛОФИЛЫЮЕ СООБЩЕСТВО СОДОВЫХ ОЗЕР
Микробные процессы в природной обстановке обусловлены их
непосредственной связью с первичными процессами в геосфере. Со-
Содовые озера, образующиеся в результате испарения продуктов угле-
кислотного выщелачивания во внутриконтинентальных бессточных
областях, в принципе могут быть противопоставлены морским та-
лассофильным местообитаниям. При высокой минерализации такие
водоемы оказываются неподходящими местами для высших орга-
организмов, включая эукариот. Вместе с тем известно, что они служат
местами необычайно пышного развития микробной жизни. Эта
пышность поражала всех естествоиспытателей, начиная с Б.Л. Иса-
Исаченко, который был пионером исследования микробиологии содо-
содовых озер Кулундинской степи, а также исследователей африканских
озер.
Содовые озера представляют собой лишь маленькие модели,
что-то вроде природного мезокосма. В зависимости от условий они
очень разнообразны по гидрохимии. Их малые размеры обусловли-
обусловливают большую нестабильность в зависимости от колебаний клима-
климата. В геологическом масштабе времени это явно эфемерные образо-
87
¦к
5,1
¦I,
вания. Будучи экстремальными местами обитания, они, очевидно,
своеобразны и населены сообществом экстремальных алкалофи-
лов. Такое сообщество должно быть приспособлено к жизни в высо-
высокоминерализованной среде с избытком Na+ и очень низким содержа-
содержанием Н+, что должно влиять на энергетику.
При рН > 9 в осадок уходит большинство металлов, оставляя до-
доминирующим катионом Na и до некоторой степени Mg, раствори-
растворимость которого контролируется образованием магнезита и доломи-
доломитов. Отсутствие Са и Fe в растворе приводит к тому, что удаляются
основные катионы, связывающие фосфат в нерастворимые минера-
минералы, и в щелочной среде он перестает быть лимитирующим элемен-
элементом, если достигает водоема. В роли лимитирующего элемента мог
бы выступать азот, но для цианобактерий и анаэробов его отсутст-
отсутствие не служит препятствием, поскольку эти организмы обладают
способностью к азотфиксации. В мелких водоемах с постоянным по-
поступлением эолового материала в виде пыли вряд ли могут быть
лимитирующими другие элементы, хотя в сообществе для обеспече-
обеспечения их доступности должны быть развиты способы комплексообра-
зования и транспорта. Моделью для изучения химии и биологии со-
содовых водоемов служат Великие Африканские озера и экстремаль-
экстремально минерализованные водоемы Восточно-Африканского рифта,
прежде всего экваториальное озеро Магади, находящееся в услови-
условиях недавней вулканической деятельности, цепь водоемов Вади-эль-
Натрун к западу от Александрии и озера Центральной Азии (Завар-
зин, 1993; Заварзин и др., 1999). Переходную форму составляют так
называемые степные озера с обильным развитием высшей расти-
растительности по берегам.
Для выявления трофической структуры сообщества прежде все-
всего необходимо выявить источник органического вещества. В содо-
содовых водоемах, особенно Африки, первичными продуцентами слу-
служат цианобактерий. Причина заключается в высоком содержании
фосфатов, достигающем десятков миллиграмм в литре. В результа-
результате развития цианобактерий водоемы евтрофицируются, подобно
тому как это происходило в равнинных водохранилищах за счет
смыва удобрений с полей. Алкалофильные цианобактерий очень
разнообразны и включают различных синехококков, афанокапсу,
рабдодерму, формидии. Поскольку их систематика основана на мор-
морфологии, флористические описания были составлены гидробиоло-
гидробиологами на основании микроскопии природного материала. Выделение
культур цианобактерий на искусственных средах позволило опреде-
определить их поля развития в координатах минерализация-рН и соле-
соленость-щелочность. Области развития цианобактерий указывают,
что цианобактерий вообще приспособлены к щелочным условиям
наземных углекислых вод, где их разнообразие очень велико. Не слу-
служит ограничивающим фактором рН и потому, что при развитии фо-
фотосинтеза в плотных массах водорослей показатель поднимается до
рНП. Можно без преувеличения утверждать, что для цианобакте-
цианобактерий содовые водоемы служат наиболее характерным местообитани-
местообитанием, из которого их не вытеснили эукариоты, в противоположность
морю, где разнообразие цианобактерий не так велико. Доминирую-
Доминирующей планктонной формой цианобактерий в африканских озерах
служат виды спирулины, являющейся основным продуцентом и эди-
фикатором в этих водоемах. Развитие планктонных цианобактерий
характерно для африканских содовых озер, и малая прозрачность
воды вытесняет бентосные формы на отмели.
Циано-бактериальные маты в содовых водоемах были описаны
для внутренних областей Пиренейского п-ва. В общем они анало-
аналогичны галофильным матам по строению. При этом ведущее значе-
значение имеет минерализация. Структура алкалофильного циано-бакте-
риального мата была подробно изучена для бессточного содового
озера Хилганта в Бурятии. Здесь также доминировал Microroleus.
Архитектура алкалофильного мата оказалась очень сходной с архи-
архитектурой галофильного. Таким образом, общие закономерности,
обусловленные проникновением света и последовательностью хи-
химических реакций, оказываются определяющими строение - или па-
рагистологию, по терминологии В. Крумбайна, - бентосного циано-
бактериального сообщества. Эти закономерности универсальны.
В высокоминерализованных водоемах характерны такие вто-
вторичные вселенцы, как зеленая одноклеточная водоросль Dunaliella,
для содовых водоемов - D. viridis, выделенная нами из оз. Магади в
рассоле с 260 г/л соды. Но для происхождения наземной биоты бо-
более важны почвенные водоросли. В пробах из разных содовых озер
постоянно обнаруживался в виде небольшой примеси очень мелкий
зеленый кокк с хлорофиллами аиЬ, далеко уступающий по разме-
размерам присутствующим здесь цианобактериям. Он был выделен в
культуру и оказался представителем рода почвенных водорослей
Chlorella - С. minutissima - вид, который ранее в таких условиях не
регистрировался. Это можно считать указанием на связь микрофлоры
содовых озер с микрофлорой окружающей почвы. При понижении
минерализации разнообразие эукариот возрастает, и происходит пе-
переход к "нормальному" сообществу с участием протистов, беспозво-
беспозвоночных и развитием водорослей.
Даже в экстремальных условиях оз. Магади при насыщении во-
воды троной (NaHCO3 • Na2CO3 ¦ 2Н2О) над 30-метровым слоем содовых
отложений при минерализации 260 г/л разнообразие микробной
жизни очень велико. Под микроскопом видны самые разнообразные
формы бактериопланктона, и можно утверждать, что морфологиче-
морфологическое разнообразие в алкалофильном сообществе покрывает все из-
известное разнообразие прокариот: здесь есть и многочисленные спи-
спирохеты, и простекобактерии с выростами, множество изогнутых ми-
кроциклусов, спирилл, палочек, кокков, угловатые плоские клетки
архей. Менее разнообразны сообщества озер Центральной Азии.
89
Типовым процессом сульфидогенеза в алкалофильном сообще-
сообществе оказалась водородная сульфатредукция. Хотя сульфатредукция
как ведущий процесс в содовых водоемах была описана еще в
1930-х годах Б.Л. Исаченко, возбудитель процесса в культуру выделить
не удавалось до 1995 г., когда это было сделано у нас в лаборатории.
Возбудителем процесса оказались вибрионы, использующие в каче-
качестве единственного субстрата Н2 или формиат. Они восстанавлива-
восстанавливают сульфат в сульфид. Сначала был выделен Desulfonatronovibrio
hydrogenovorans, оказавшийся свойственным как центрально-афри-
центрально-африканскому озеру Магади, так и центрально-азиатским озерам с мень-
меньшей минерализацией и совсем другим климатом. Тем не менее
штаммы имели практически полную гомологию ДНК. Отсюда воз-
возникает вопрос в связи с конвенцией по биоразнообразию, объявля-
объявляющей государственную собственность на свои генетические ресур-
ресурсы: приложима ли она к микробам, распространяемым с пылью по
всему свету? Не следует ли более внимательно отнестись к микроб-
микробным сообществам в природе, для которых необходимы "заповедни-
"заповедники для микробов"? Вслед за Desulfonatronovibrio был выделен еще
один сульфатредуктор - толстый вибрион Desulfonatronum. Клетки
этого типа отчетливо наблюдаются в сообществе с целлюлозораз-
лагающими микроорганизмами на ранних стадиях разложения цел-
целлюлозы. На поздних стадиях их сменяют крупные палочки того же
рода, использующие ацетат синтрофно (рис. 3.8).
Образование ацетата в среде осуществляет широкий спектр аце-
тогенных бактерий, использующих разнообразные субстраты.
Ацетогены имеют специальный метаболический путь, характе-
характеризуемый ферментом СО-дегидрогеназой. Из экстремально алкало-
фильных гомоацетогенов биохимически пока изучена Natroniella
acetogena, выделенная нами из содовых рассолов оз. Магади. Она
стала объектом биохимических исследований относительно новой
группы гомоацетогенов, экологическая роль которых в трофиче-
трофической структуре микробных сообществ была неясна, но вырисовыва-
вырисовывается как все более и более значительная. Natroniella принадлежит
новой филогенетической ветви галоанаэробов, недавно выделенных
в новый порядок Halanaerobiales. Галоанаэробы приурочены к мор-
морским гиперсоленым условиям. Natroniella развивается при высокой
концентрации NaCl и предельно высокой щелочности. Устойчи-
Устойчивость клетки к этим условиям зависит от необходимости поддержи-
поддерживать энергизацию мембран, при остановке обмена она быстро лизи-
руется.
Следующую группу ацетогенов составляют организмы, сбражи-
сбраживающие аминокислоты, в том числе аргинин, входящий в такой ха-
характерный запасной продукт цианобактерий, как цианофицин.
К ним относятся описанные нами в 1998 г. Tindallia и Natronincola.
Основными продуктами их обмена являются ацетат и аммиак. Они
используют крайне ограниченное число соединений как субстрат
90
Cyanobacteria
(^Мортмасса J)
Гидролитики
-NH3) I : '—>( Мономеры
Метилотрофные
метаногёны' •
Methanosalsus
H2S;) - Ectothiorhodospira
Рис. 3.8. Трофические связи в алкалофильном сообществе
брожения, но в их числе есть и орнитин, являющийся продуктом
темнового обмена цианобактерий при разложении цианофицина.
Наконец, среди первичных анаэробов особое место занимают
спирохеты. Их регулярное появление в содовых озерах легко реги-
регистрируется микроскопически, но выделение в чистую культуру тра-
традиционно считается трудным. Из содовых озер были выделены три
вида спирохет: Spirochaeta alkalica, S.africana, S.asiatica. Эти организ-
организмы сбраживают моно- и дисахариды и относятся к группе диссипо-
91
трофов - организмов, использующих рассеиваемые низкомолеку-
низкомолекулярные вещества, например из мест гидролиза полисахаридов. Про-
Продуктами обмена брожения оказываются ацетат, водород и некото-
некоторые другие соединения, как этанол. В совместной культуре с суль-
фатредукторами на сахарах образуется сероводород, но ни водород,
ни ацетат не появляются. Интересно, что спирохеты оказались
очень устойчивыми по отношению к сероводороду, в то время как
целлюлозоразрушающие бактерии - чувствительными.
В высокоминерализованных местообитаниях доминирует С-1
путь метаногенеза. Из С-1 соединений Methanosalsum zhilinae исполь-
использует для образования метана метанол или метиламины аналогич-
аналогично тому, как это имеет место для галофильных сообществ лагун
Сиваша.
Образование восстановленных соединений донными анаэроба-
анаэробами, в то время как первичные продуценты цианобактерии образуют
кислород, пузырьками поднимающийся к поверхности, позволяет
сформироваться в сообществе окислительному аэробному фильтру.
Его исследовал на тех же образцах Д.Ю. Сорокин. Он обнаружил
экстремально алкалофильные тионовые бактерии, которые окисля-
окисляют сероводород в сульфат. Далее были найдены алкалофильные
водородные бактерии. Алкалофильные нитрификаторы, вообще
говоря, были известны, но озадачивающей находкой явилось откры-
открытие алкалофильного организма, способного окислять и метан в уг-
углекислоту, и аммиак в нитрит. Нитрификаторы и метанотрофы
очень похожи по цитологии - их клетки набиты слоями мембран, но
аммиак служит сильным ингибитором окисления метана у всех дру-
других метанотрофов. Это первый случай такого совмещения функций.
Другую группу алкалофильных метано- и метилотрофов выделили
из центрально-азиатских озер Ю.А. Троценко и его сотрудники.
Из щелочных озер Африки британские исследователи выдели-
выделили разнообразных аэробных органотрофов. Все эти организмы от-
относятся к протеобактериям. Под слоем троны на поверхности рассо-
рассола развиваются вишнево-красные архебактерии Natronobacter и
Natronococcus, открытые В.Д. Грантом и Б. Тиндаллом - пионерами
исследования оз. Магади. Эти организмы используют эвапоритовую
тягу для концентрации органических веществ, подобно галобактери-
ям. Наличие бактериородопсина позволяет им иметь дополнитель-
дополнительный источник энергии - свет.
Особое значение в содовых водоемах приобретает серный цикл
и участвующие в нем аноксигенные пурпурные бактерии. Нередко
вода содовых озер приобретает красную окраску. Пурпурные бакте-
бактерии представляют анаэробный окислительный фильтр, работаю-
работающий на свету в отсутствие О2. Типичным обитателем содовых водо-
водоемов служат пурпурные бактерии рода Ectothiorhodospira, способ-
способные окислять сероводород, откладывая капли серы вне клеток, до
сульфата. Они могут также использовать водород и ацетат. Это массо-
92
вые обитатели содовых водоемов. Однако в последние годы
В.М. Горленко открыл еще целый ряд представителей аноксиген-
ных алкалофильных бактерий. В их числе оказались очень изящные
тонкие пружинки Heliorestis - представителя гелиобактерий, образу-
образующих совершенно иную филогенетическую ветвь. Массовое разви-
развитие пурпурных бактерий делает их наиболее значительными во вто-
вторичной продукции. В аэробных условиях в цилиндрах Виноградско-
го развитие пурнурных было жестко связано с развитием сульфат-
редуцирующих бактерий и было очень мощным. Приходится при-
признать, что пары пурпурные фототрофы и сульфатредукторы пред-
представляют звенья одного цикла. Интересно, что и разложение целлю-
целлюлозы резко ускорялось на свету при участии пурпурных бактерий,
при этом не появлялись промежуточные продукты.
Итак, можно резюмировать, что трофическая сеть в алкало-
фильном сообществе полна, и большую роль в нем в высокоминера-
высокоминерализованных озерах играет серный цикл. Вопрос о функциональной
полноте алкалофильного сообщества получает положительный от-
ответ. Второй вопрос касается филогенетического положения. Если
суммировать полученные результаты и показать на филогенетиче-
филогенетическом дереве положение выделенных и описанных видов, то оказыва-
оказывается, что фактически все главные ветви имеют своих представите-
представителей среди алкалофилов. В особенности это характерно для протео-
бактерий, цианобактерии, галоанаэробов, грамположительных
"клостридий" и "бацилл", спирохет, архей Euryarchaeota. Ограниче-
Ограничений здесь нет даже для высокоминерализованных водоемов. Вместе
с тем алкалофилы оказываются филогенетически отличными от
аналогичных нейтрофилов на уровне рода-вида, но не представляют
самостоятельной филогенетической ветви, которая исключила бы
возможность рассматривать это сообщество при выявлении источ-
источника разнообразия наземной биоты.
3.7. ТЕРМОФИЛЬНЫЙ МАТ
Термофильные маты развиваются в термальных источниках с
нейтральным рН и СО2 как доминирующим газом. Повсеместно, от
Йеллоустона в Америке до Узона на Камчатке и от Исландии до Но-
Новой Зеландии, они имеют характерный внешний вид, который для
водотоков был назван "V-типом". Он обусловлен распределением
температуры: на стрежне потока она выше, чем у берегов, и это оп-
определяет верхнюю границу роста цианобактерии фототрофов. Она
лежит несколько выше 60°. После работ Т. Брока в 1960-х годах на
источнике Октопус-Спринг в Йеллоустоне его сообщество стало
моделью для изучения термофилов (Brock, 1986).
У выхода термального источника - грифона - при температуре
выше 70° развивается тионовая бактерия Thermothrix thiopara и со-
93
путствующие ей серовосстанавливающие бактерии. Организм окис-
окисляет сероводород в серу при очень низком содержании кислорода.
У выхода, в самой горячей зоне, нити термотрикса розоватые от
гема, затем они покрыты белым слоем серы. Рост термотрикса
прекращается ниже 55-57°. Ярко белые космы его достигают де-
десятков сантиметров в длину и располагаются по стрежню потока.
Это типичный кренофил. Цепи слегка изогнутых длинных клеток
термотрикса переплетаются между собой и поэтому удерживают-
удерживаются в потоке в виде цепи. Организм отличается очень большой ско-
скоростью роста.
Вслед за термотриксом идет рыжеватый слой фотоорганотроф-
ной бактерии Chloroflexus. Это длинная, нитевидная бактерия с чех-
чехлом опять-таки образует космы в горячей зоне около 70°. Надо ска-
сказать, что хлорофлексусов, по-видимому, довольно много, но в куль-
культуру выделены лишь отдельные представители. Chloroflexus разви-
развивается вместе с одноклеточной цианобактерией Synechococcus lividus
и составляет фронт сообщества, обращенный к выходу горячей во-
воды. Почему фотоорганотрофный организм оказался перед сообще-
сообществом, а не сзади? Причина заключается в конвекционном потоке:
горячая вода экстрагирует органические вещества из фронта циано-
бактерий и охлаждаясь опускается вниз, образуя придонный проти-
противоток, который очень просто наблюдать в стеклянных проточных
камерах с точечным вводом горячей воды.
Фронт густо-зеленого развития циано-бактериального сообще-
сообщества располагается на изотерме около 60°, которую следует считать
нормой для верхнего предела развития термофильных цианобакте-
рий. Фронт составляет ветвящаяся цианобактерия Mastigocladus
laminosus {Fischerella laminosa). Организм этот имеет очень изменчи-
изменчивую морфологию. Он служит эдификатором для термофильного ци-
циано-бактериального сообщества. За ним следуют разные виды
Phormidium, прежде всего P. laminosum. Виды формидиум образуют
микроколонии на поверхности мата, которая представляется своего
рода мозаикой из сплетений нитей разного диаметра. Видовое назва-
название "laminosum" отчетливо указывает на способность организма об-
образовывать слои. Действительно, термофильное циано-бактериаль-
ное сообщество отличается очень четким слоистым строением. При
высокой скорости роста термофильных фототрофов на образова-
образовании слоев сказывается суточный цикл: в источнике на Узоне с выно-
выносом коллоидной серы образовывались отчетливые слои серы ночью,
а цианобактерий днем, образуя слоеный мат толщиной в сантиметр.
В плотном термофильном мате образование слизи имеет мень-
меньшее значение, чем в галофильном. Быстрый ток горячей воды за-
заставляет организмы опускаться вниз, образуя плотную ткань из вет-
ветвящихся трихомов Mastigocladus или переплетающихся трихомов
Phormidium. При этом организмы образуют отчетливые мозаичные
группировки. В горячей воде, даже обдаваемые кипящей водой из
94
к
к
II
гейзеров, как в Долине Гейзеров, Камчатка, циано-бактериальные
маты плотно связаны с субстратом. Определяющим здесь служит
кренофильный образ жизни сообщества. Термофильный цианобак-
териальный мат отличается очень высокой скоростью роста. Мак-
Максимальная плотность хлорофилла достигается очень быстро. Струк-
Структура мата позволяет удерживаться одноклеточным Synechococcus,
которые подробно изучены в Октопус-Спринг в Йеллоустоуне.
В России аналогичной моделью послужили маты кальдеры Узон,
Камчатка (Кальдерные микроорганизмы, 1989).
Аноксигенные фототрофы не характерны для термофильного
мата, и в этом его отличие от мезофильных матов минерализован-
минерализованных водоемов. Они появляются при температуре около 50°. К ним
относятся Chlorobium tepidum.
Термофильный мат содержит полный набор термофильных де-
деструкторов, развивающихся при 70-80°. Они очень разнообразны и
интересны как перспективные объекты биотехнологии. Наиболь-
Наибольшую известность приобрел аэробный органотроф Thermus - проду-
продуцент широко применяемой термофильной ДНК-полимеразы. Анаэ-
Анаэробные процессы осуществляются крайне разнообразными организ-
организмами, способными до температуры в 80 °С использовать очень
широкий круг веществ; выше этой температуры происходит резкое
снижение разнообразия. Аллохтонная целлюлоза разлагается под-
подробно изученным Caldocellulosoruptor (=Anaerocellum) и многочис-
многочисленными и разнообразными термофильными клостридиями. Инте-
Интересен Dictioglomus, образующий крупные шары из объединенной
поверхностной мембраны, внутри которых в виде распирающей
шар спирали располагаются упругие нити организма. Известны и
термофильные спирохеты: морская Spirochaeta thermophila и кон-
континентальная S.caldaria. Гомоацетатные бактерии представлены
Clostridium thermoautotrophicum — умеренным термофилом, способ-
способным использовать водород и широкий набор углеводов.
Вторичные анаэробы представлены и метаногенной, и сульфидо-
генной ветвями. К серной ветви относится термофильный сульфатре-
дуктор Thermodesulfobacterium commune, описанный учеником Т. Бро-
ка Г. Зейкусом, с более или менее обычным для сульфатредукторов
набором доноров электрона, но представляющий самостоятельную
филогенетическую линию высокого ранга. Восстановление молеку-
молекулярной серы, столь характерной для термальных мест обитания, осу-
осуществляет ацетат-использующая Desulfurella, открытая Е.А. Бонч-
Осмоловской и представляющая самостоятельный подкласс среди
протеобактерий. Термофильный метаногенез осуществляется водо-
род-использующей Methanobacterium thermoautotrophicum, открытой
тем же Г. Зейкусом и очень подробно исследованной как модель во-
водородных метановых архебактерий. Ацетокластический путь пред-
представлен Methanothrix thermoacetophila, открытой Т.Г. Ягодиной и
А.Н. Ножевниковой в термах Узона. В то же время следует заметить,
95
что кокковидные термофильные метаногены пока не найдены, и ме-
таносарцины для этих условий не характерны.
Совершенно неожиданной, но широко распространенной груп-
группой организмов оказались открытые В.А.Светличным анаэробные
карбоксидотрофы. Эти организмы осуществляют реакцию СО +
+ Н2О = СО2 + Н2, аналогичную парогазовой конверсии СО в химиче-
химической промышленности, и полностью превращают 100% СО в водо-
водород без ингибирования конечным продуктом. Кроме СО, другие со-
соединения не используются. Ключевой фермент - СО-дегидрогеназа -
на редкость термоустойчивая. Организм, названный Carboxydothermus
hydrogenoformans, был обнаружен во многих гидротермах. Загадку
представляет источник СО, необходимый для этой узкоспециализи-
узкоспециализированной группы.
Для термального сообщества нами четко установлено существо-
существование термофильных железо-восстанавливающих бактерий, кото-
которые образуют магнетит. Организмы способны развиваться синт-
рофно и восстанавливать железо за счет ацетата совместно с други-
другими факультативно анаэробными бактериями. Термофильные магне-
магнетит-образующие бактерии были найдены в Долине Гейзеров, где от-
отсутствует серный цикл, в отличие от Узона, а затем и в ряде терм по
всему земному шару. Вскоре аналогичные организмы были описаны
австралийскими учеными в подземных термах. Аэробные организ-
организмы бактериального окислительного фильтра изучены значительно
хуже. Интерес представляет водородная бактерия Calderobacterium
(= Hydrogenobacter), которая растет автотрофно с использованием
ацетатного пути усвоения СО2 и относится к самым древним эубак-
териям. О тионовой бактерии Thermothrix thiopara уже упоминалось.
Что очень важно, - это отсутствие метанокисляющих бактерий и,
следовательно, незамкнутость цикла Зёнгена в термофильном сооб-
сообществе. Несмотря на все усилия К. Штеттера и у нас, метанотрофы
в термальных условиях выше 60 °С не были найдены и окисление
метана сообществом не регистрировалось. Лишь недавно появилось
сообщение о термофильном метанотрофе с оптимумом выше 60 °С.
Термофильный мат отличается способностью перерабатывать вос-
восстановленные газы вулканических эксгаляций
3.8. ЦИАНО-БАКТЕРИАЛЬНЫЕ МАТЫ
В СРАВНИТЕЛЬНОМ АСНЕКТЕ
Сопоставление типичных представителей функциональных
групп в мате позволяет судить об инвариантности трофических
взаимоотношений в этих сообществах. О составе циано-бактери-
альных сообществ, развивающихся в разных экстремальных усло-
условиях, можно судить по табл. 3.1, составленной по нашим данным,
где приведены типичные представители функциональных групп.
96
се
ё 8
и
3
ш
о
X
а 2
?.!
s §•
о. 5
s
X
S
I
4. Г. А. Заварзин
•е-
о
¦е-
о
¦§•
I
О.
g
I
о —
II
О
О.
I
IS
I
111
К S «w
I
5 ^ §• ? S
S ¦= S :5 -2
И
о.
3
i!
Il
5 «
;S 3
1^
111
S ^ -a
'5 J3 <s
~ -c vo
S - о
з а о
i.
[
л
ю
о
с
т
в
¦е-
о
f
К
х
Э
1
ё
2
f
I
о.
•в* с
II
I
00
I
I
1
11
р
« 3
ж
g
I
2
с
3
о
5
1
с
I I
4>
-a
I
I
§
s s
II
a a
it
alls
^ ^ .!^ -J.
•Si
I
I
V
3
|
|
I
I
Ill
^11
в а ь
>• e-
•8-
S
*1l I ?
и tfl i
Q Q ^ 3
a с С
^?5
•II
3 § -5
III
3t
I
5"
-0
&
о
8.
о
a.
6
3
I
a.
I
3
CQ
к
3.9. МАТЫ СУЛЬФУРЕТЫ И ТИОБИОС
Циано-бактериальный мат представляет полную систему, вклю-
включающую первичных продуцентов. Однако возможны условия, при
которых мат образуется за счет разложения аллохтонного органи-
органического вещества в донных осадках, завершающегося в море суль-
фидогенезом. В этом случае возникает резкий перепад между при-
придонной водой, в той или иной степени содержащей поступающий
сверху кислород, и донными отложениями, откуда поступает H2S.
В этих условиях развивается микробное сообщество, получившее
название сулъфуреты. В этом сообществе осуществляется серный
цикл, инициируемый сульфатредукторами, разлагающими оседаю-
оседающее сверху органическое вещество. Если приток оксигенированной
воды сверху ограничен, например в стратифицированных водоемах,
то здесь развиваются окисляющие серу организмы: в фотической
зоне - серные пурпурные или зеленые аноксигенные фототрофы, в
темноте - хемосинтетики.
Примером стратифицированного водоема служит Черное море,
и, моделируя условия в нем, Егунов обнаружил образование в ци-
цилиндрах на некоторой глубине от поверхности зоны развития окис-
окисляющих серу микроорганизмов, получившей название "пленки Егу-
нова". Некоторые аналоги такой пленки действительно существуют
в природе, образуя узкие зоны развития в области хемоклина - рез-
резкого изменения химических условий в стратифицированном водо-
водоеме, но это, конечно, не пленки, а зоны скопления организмов. Это
зона развития "градиентных" организмов, типичными представите-
представителями которых служат бесцветные серобактерии, как тиоспириллы,
Macromonas, Thiovulum, трудно поддающиеся культивированию, но
хорошо различимые под микроскопом. Одноклеточный, очень под-
подвижный Thiovulum диаметром 10-20 мкм образует висячую бахрому.
Клетки соединяются тонкими слизистыми нитями и образуют неж-
нежную сеть, отделяющую оксигенированную воду от обогащенной се-
сероводородом. Они создают свой собственный диффузионный слой в
воде и контролируют диффузионные потоки кислорода и сульфида.
Донные осадки имеют более резкий градиент H2S и О2, и поэто-
поэтому здесь возникает возможность развития другого сообщества, по-
получившего название тиобиос. В этом случае один организм может
пересекать границу между двумя зонами. Для этого он должен обла-
обладать достаточными размерами. Название это было предложено для
крупных беспозвоночных, червей и моллюсков. Часто эти организ-
организмы обладают симбиотическим питанием за счет хемосинтезирую-
щих симбионтов. Лучшим примером этого типа питания может слу-
служить гигантская, метровых размеров рифтия, развивающаяся у гид-
гидротермальных сероводородных выходов на дне океана и имеющая
мощную систему транспорта кислорода к расположенной внутри ор-
организма трофосоме, содержащей тионовые бактерии.
99
Аналогично О2 изменяются градиенты H2S. В результате оказы-
оказывается, что в морских осадках восстановление сульфатов с образова-
образованием H2S происходит только ниже зоны с кислородом и нитратом,
на глубине 1,1 мм, и прекращается на глубине 2,1 мм вследствие ис-
исчерпания сульфата. Полное окисление H2S происходило в слое
0,3 мм с временем оборота в несколько секунд. Из этого примера
можно вывести по меньшей мере два следствия:
1) серный цикл сульфатредукции-окисления H2S может служить
дополнением к окислительной системе донных осадков с временем
оборота в секунды-десяток секунд;
2) градиент в осадке достаточно резкий, чтобы мог существо-
существовать микробный тиобиос, т.е. организмы, пересекающие градиент
от H2S-3OHbi до кислородной зоны, так как для этого достаточно
иметь длину в сотню мкм, которой достигают нитчатые серобакте-
серобактерии, как Thioploca. Аналогичная по строению микроколеусу трихом-
ная серобактерия Thiploca образует маты с массой 1 кг/м2 сырого ве-
веса в зоне апвеллинга у побережья Чили. Бактерия имеет диаметр
15-40 мкм и длину до 7 см. Пучки по сотни трихомов заключены в об-
общий слизистый чехол, внутри которого они скользят со скоростью
нескольких мкм/с. Чехлы образуют слой, толщиной в сантиметры, и
торчат над поверхностью дна, как газон. Внутри каждой клетки
имеется вакуоль, занимающая 80% ее объема. Бактерии поглощают
из воды нитрат, концентрация которого в воде составляет
20-30 ммоль, и концентрируют его в вакуоли до 0,5 М. Затем они
скользят вниз в зону сульфидогенеза и окисляют сероводород до
элементарной серы с восстановлением NOJ в NH4. Сера затем
может быть окислена до сульфата. Тиоплоки поддерживают кон-
концентрацию сульфида в поровой воде на микромолярном уровне. Не-
Некоторые гигантские Beggiatoa диаметром в 100 мкм имеют такие же
вакуоли и окружают гидротермальные выходы матами в 1-3 см или
рыхлыми массами в 10-20 см. Слой цитоплазмы у этих гигантских
бактерий имеет толщину лишь в 2-3 мкм.
Другим примером бактериального тиобиоса, основанного на
пересечении слоя градиента между сульфидогенной и оксигениро-
ванной зонами, может служить описанный Б.В. Перфильевым
"Thiodendron", развивающийся в виде синевато-белых медузопо-
добных масс на дне сероводородных источников (оз. Чокрак на
Керченском п-ве) или в море. На разрезе колоний тиодендрона от-
отчетливо видно его ритмическое слоистое строение. Тиодендрон
оказался симбиотическим сочетанием спирохет и сероокисляющих
бактерий.
Из приведенных примеров видно, что существование очень рез-
резких градиентов в структурированном сообществе позволяет взаимо-
взаимодействовать противоположно направленным процессам и замыкать
циклы внутри одного сообщества.
Рекомендуемая литература
1. Гусев М.В., Никитина К.А. Цианобактерии. М.: Наука, 1979. 228 с.
2. Еленкин А.А. Синезеленые водоросли СССР. М; Л: Изд-во АН СССР, 1936.
679 с.
3. Заварзин Г.А. Эпиконтинентальные содовые водоемы как предполагаемые
реликтовые биотопы формирования наземной биоты // Микробиология.
1993. Т. 62, № 5. С. 789-800.
4. Заварзин Г.А., Герасименко Л.М., Жилина Т.Н. Цианобактериальные сооб-
сообщества гиперсоленых лагун Сиваша // Там же. С. 1113-1127.
5. Заварзин ГА., Жилина Т.Н., Кевбрин В.В. Алкалофильное микробное сооб-
сообщество и его функциональное разнообразие // Там же. 1999. Т. 68, № 5.
С. 579-599.
6. Кальдерные микроорганизмы. М.: Наука, 1989.
7. Определитель пресноводных водорослей СССР / Ред. М.М. Голлербах. М.:
Сов. наука, 1951.
8. Пиневич А.В., Аверина С.Г. Оксигенная фототрофия. СПб., Изд-во СПбГУ
2002. 236 с.
9. Функциональная структура цианобактерии / Ред. Б.В. Громов. Л.:
Изд-во ЛГУ, 1986.
10. Biostabilization of sediments / Ed. W.E. Krumbein et al. Oldenburg: Bill, and
Inform. System Univer., 1994. 528 p.
11. Brock T. Thermophiles: General molecular and applied microbiology. N.Y.; Toronto.
Wiley, 1986.
12. Microbial mats: Structure, development and environmental significance / Ed.
L.J. Stal, P. Caumette. B. etc.: Springer, 1994. 466 p.
13. Microbial mats: Physiological ecology of benthic microbial communities / Ed.
Y. Cohen, E. Rosenderg. Wash. (D.C.): Amer. Soc. for Microbiol., 1989. 494 p.
14. The molecular biology of cyanobacteria / Ed. D.A. Bryant. Dordrecht: Kluwer, 1994.
881 p.
15. The ecology of cyanobacteria: Their diversity in time and space / Ed. B.A. Whitton,
M. Potts. Dordrecht: Kluwer, 2000.
100
L
Лекция 4
АТМОСФЕРА И БАКТЕРИИ
4.1. АТМОСФЕРА И БИОТА
Среди других оболочек Земли атмосфера представляет неболь-
небольшой динамичный, хорошо перемешанный резервуар с малым време-
временем пребывания в нем компонентов и быстрыми транспортными
процессами. Собственные химические процессы в атмосфере обу-
обусловлены в первую очередь солнечной энергией, приводящей на ос-
освещенной поверхности к серии взаимосвязанных фотохимических
реакций. Интенсивность реакций обусловлена меридиональным рас-
распределением получаемой энергии с максимумом на экваторе
(рис. 1.4). Сопоставление качественного состава атмосферы и организ-
организмов с газовыми функциями приводит к выводу, что все компоненты ат-
атмосферы, за исключением инертных газов, находятся под контро-
контролем организмов, часто узкоспециализированных по одному газу.
Справедливо и обратное, все биогенные макроэлементы, за исклю-
исключением фосфора, имеют воздушную форму миграции. История фор-
формирования атмосферы состоит в постепенном превращении под воз-
воздействием биоты продуктов дегазации Земли в воздух. Современная
жизнь развивается на дне воздушного океана и активно взаимодей-
взаимодействует с ним, подвергаясь его воздействию и, в свою очередь, влияя
на него.
Определяющими при формировании состава атмосферы послу-
послужили два процесса: 1) удаление из атмосферы углекислоты дегаза-
дегазации, связанной в конечном счете в карбонаты Са и Mg в результате
химического выветривания, и 2) образование О2, диоксигена, за счет
деятельности оксигенных фототрофов с эквивалентным захороне-
захоронением органического углерода (рис. 1.2). Связывание СО2 возможно в
абиотических реакциях углекислотного выщелачивания, создающих
маршрут от кристаллических изверженных пород к глинам и урав-
уравновешивающим их карбонатным породам. Углекислотное выщела-
выщелачивание идет с достаточной скоростью только при повышенных
температуре и давлении. Оно значительно ускоряется в биологиче-
биологически опосредованных процессах. Образование кислорода представля-
представляет исключительно биогенный процесс с циклом оксигенного фото-
102
синтеза и аэробного дыхания. Аэробное разложение мортмассы
обусловливает создание повышенной концентрации СО2 в местах
разложения и, соответственно, ускорение выщелачивания. Итак,
подлинным источником эволюции атмосферы является биотическая
трансформация СО2.
Газовый состав атмосферы первоначально образовался в ре-
результате дегазации Земли, основной этап которой быстро прошел
на самых ранних стадиях формирования планеты, но дегазация про-
продолжается до сих пор при вулканизме и по разломам. Современный
поток эндогенных газов идет через формирующуюся океаническую
кору в океан и достигает поверхности только после установления
равновесия с массой вод глубокого океана. Дополнительным источ-
источником служит вулканическая регазация, прежде всего на островных
дугах. В целом этот газовый поток называют "дыханием планеты".
В нем следует различать две составляющие: магматическую (дегаза-
(дегазация) и метаморфическую (регазация), связанную с рециклом осадоч-
осадочных пород. При высокой температуре и давлении реакция воды с на-
нагретыми породами приводит к образованию восстановленных со-
соединений. Этот газовый поток может перерабатываться подземной
микробиотой, часто гидрогенотрофной и термофильной. При выхо-
выходе газов на поверхность в низкотемпературных газогидротермах
преимущественно углекислотного состава (мофетах) развивается
сообщество термофильных микроорганизмов, которое можно рас-
рассматривать как аналог сообщества раннего докембрия, трансформи-
трансформировавшего газы дегазации в воздух.
Переработка восстановленных газов эндогенного происхожде-
происхождения в подземном пространстве осуществляется "глубинной микро-
микробиотой" (deep subterranean biosphere) с преимущественным участием
анаэробных гидрогенотрофных термофильных и гипертермофиль-
гипертермофильных микроорганизмов, распространяющихся в области высокого
термического градиента до глубины примерно 3 км. Эти организмы
развиваются в трещинном пространстве пород и подземной гидро-
гидросфере. Конечными продуктами их обмена служат H2S и СН4. Гипер-
Гипертермофилы обнаружены при глубоком бурении нефтяных скважин
и в общем аналогичны тем, которые развиваются в естественных га-
газогидротермах.
Атмосфера взаимодействует с океаном, который служит для нее
основным источником водяных паров. С поверхностными водами
океана атмосфера находится в физико-химическом взаимодействии,
обусловленном растворением газов пропорционально их парциаль-
парциальному давлению (закон Генри). Океан представляет большой резер-
резервуар растворенных газов, которые могут подвергаться в нем транс-
трансформации с участием биоты. Взаимодействие океана с атмосферой
в первую очередь физико-химическое, поскольку в пелагиали океа-
океана циклы биотических элементов замкнуты за счет аэробной дегра-
деградации оседающей мортмассы планктона. С атмосферой взаимодей-
103
ствуют поверхностный фотический слой, где развивается фито-
фитопланктон, и находящийся под ним слой регенерации биотических
элементов. Если фотический слой составляет около 30±10 м, то вме-
вместе со слоем регенерации он составляет 100-200 м.
Океан служит областью седиментации карбонатов. В осадках
развивается микрофлора донных отложений, образующая восстано-
восстановленные газы (газогенерирующий этап), прежде всего H2S с домини-
доминированием сульфатредукции как заключительного этапа анаэробной
деградации мортмассы и развитием на поверхности ила организмов,
окисляющих соединения серы (сообщество сульфуреты), и даже
придонные воды обычно оксигенированы. Благодаря высокому
слою оксигенированной воды донные выделения из океана не дости-
достигают поверхности, даже в случае Черного моря - модели состояния
древнего стратифицированного океана. Особый случай представля-
представляет локальное просачивание метана, наподобие грязевого вулканиз-
вулканизма на суше (холодные метановые "сипы"). Источником этого мета-
метана, помимо деятельности метаногенов, может служить разложение
газогидратов метана. На дне океана в области спрединга на выходе
эндогенных газов, образующихся при контакте морской воды с пе-
перегретыми породами базальтов океанической коры, развиваются
особые микробные сообщества подводных гидротерм (термальные
глубоководные "оазисы"), в которых продукция органического ве-
вещества осуществляется за счет хемосинтеза и окисления газов кис-
кислородом фотосинтетического происхождения, приносимого в глуби-
глубину холодными океаническими водами.
Твердая поверхность суши прямо контактирует с атмосферой.
Находящаяся в субаэральных условиях биота непосредственно взаи-
взаимодействует с атмосферой, и поэтому ее влияние на состав атмосфе-
атмосферы гораздо значительнее и, главное, быстрее, чем воздействие мор-
морской биоты. Взаимодействие атмосферы с сушей происходит через
слой "почвенного воздуха" - газов, находящихся в пористом про-
пространстве почвы и горных пород. Состав почвенного воздуха замет-
заметно отличается от состава атмосферы в связи с тем, что здесь идут ре-
реакции как с грунтовыми водами, так и с минеральными компонента-
компонентами. Почвенный воздух находится под влиянием жизнедеятельности
биоты, и в первую очередь микробиоты. Над поверхностью почвы и
до вершины растительного покрова располагается воздушное про-
пространство, получившее название "аэротоп". Дыхание почвы пред-
представляет наиболее быстрый механизм газообмена с атмосферой.
Среди природных объектов с особой интенсивностью газообме-
газообмена следует выделить амфибиальные ландшафты - такие природные
системы, в которых есть наиболее тесное взаимодействие между
твердой, жидкой, газообразной фазами. Под амфибиальными ланд-
ландшафтами (см. Лекцию 6) можно понимать болота, гидроморфные
почвы, мелководные внутренние водоемы, приливные отмели, лагу-
лагуны. Здесь создаются условия для тесного взаимодействия генериру-
104
ющего газы анаэробного сообщества и окисляющего их аэробного
сообщества. Быстрый газообмен с атмосферой обусловливает уси-
усиленное влияние этих сообществ на ее состав.
В каждой из рассмотренных сред развивается микробное сооб-
сообщество с сопряженными трофическими связями. Среди организмов,
входящих в эти сообщества, необходимо выделить организмы с га-
газовой функцией.
4.2. ПАРНИКОВЫЙ ЭФФЕКТ И МИКРООРГАНИЗМЫ
Состояние земной поверхности обусловлено энергией, получае-
получаемой от Солнца, т.е. радиационным балансом. Эта энергия обуслов-
обусловливает, во-первых, тепловой климатический режим и, во-вторых,
после поглощения фотосинтетически активной радиации (ФАР) де-
деятельность биосферной трофической системы. ФАР изменялась в
течение геологического времени, но считается постоянной, с не-
небольшими колебаниями, в течение исторического времени, хотя ее
величина зависит, например, от облачности, не говоря о суточных и
сезонных колебаниях.
Радиационный баланс атмосферы определяется:
1) радиацией, полученной извне атмосферы от Солнца, соответ-
соответственно солнечной постоянной Е = 1380 Вт • м-2, эта энергия распре-
распределяется в солнечном спектре так:
10% X < 400 нм - ультрафиолет, включая UV-B,
45% X 400-750 нм - видимый свет, включая ФАР,
45% X > 750 нм - инфракрасные тепловые лучи;
2) радиацией, профильтрованной через атмосферу с поглощени-
поглощением и доходящей до поверхности Земли, где часть ее поглощается и
переходит в тепловую энергию, а часть отражается:
02 поглощает радиацию с X < 210 нм и не допускает радиацию с
X < 50 нм,
03 поглощает 210 < Х< 310 нм (UV-B),
Н2О и СО2 поглощают инфракрасную радиацию с X > 750 нм
особенно эффективно в диапазоне 8000-20000 нм.
Радиация, дошедшая до Земли, отражается от облачного покро-
покрова, затем от поверхности льдов, океана, континентов. Отраженная
радиация сдвигается в длинноволновую область. Отраженная радиа-
радиация определяется уравнением:
Е - A - а)/4 = sT4,
где Е = 1380 Вт • м-2; а - альбедо; s - постоянная Больцмана
5,67 • 10~8 Вт • м~2 • К-4; 4 - отношение поверхности к сечению диска
Земли; Т - температура в градусах Кельвина. В присутствии атмо-
атмосферы отраженная радиация поглощается соответственно
Е ¦ A - а)/4 = fsT<>,
105
2,5
ffl 2,0
1,5
0,5
0,2 0,6 1,0 1,4 1,8 2,2 2,6
Длина волны, мкм
3,0
Рис. 4.1. "Окна пропускания". Интенсивность солнечного излучения за предела-
пределами атмосферы (I) и на уровне моря B) в зависимости от длины света [по: Иси-
Исидоров, 2001]
Заштрихованы участки, соответствующие линиям поглощения указанных компонентов
где f = 0,61 - поглощение атмосферой. Отраженная радиация может
уходить обратно в окружающее пространство через "окна" пропус-
пропускания. Они располагаются в области 8000-12000 нм (рис. 4.1). Не-
Небольшое увеличение поглощения в окнах пропускания резко сказы-
сказывается на радиационном балансе: инфракрасная отраженная радиа-
радиация попадает в ловушку и не рассеивается в окружающем простран-
пространстве. Это явление и называется "парниковым эффектом", а газы,
поглощающие в указанном диапазоне, - "парниковыми". Явление
это было описано в начале XX в. Сванте Аррениусом, который
предсказал повышение температуры Земли при увеличении содер-
содержания СО2 от сжигания топлив. Одним из первых, кто обратил вни-
внимание на предстоящее изменение климата, был М.И. Будыко.
Парниковый эффект обусловливает среднюю температуру у по-
поверхности Земли 15 °С, в то время как без этого поглощения она бы-
была бы -18 °С, как на Луне. Отсюда ясно, что наряду с прямым полу-
получением энергии Солнца важно удержание тепла атмосферой. Пар-
Парниковый эффект обеспечивает нахождение Земли между двумя
крайними устойчивыми состояниями: оледенения, как на Марсе, или
разогрева, как на Венере. К парниковым газам относят: пары воды,
углекислоту, метан, закись азота, окислы азота, озон, окись углеро-
углерода, хлорфторуглеводороды. Пары воды в наибольшей степени обу-
обусловливают парниковый эффект и его сильную положительную
обратную связь: с повышением температуры их содержание в атмо-
атмосфере увеличивается.
106
В окнах пропускания помимо СО2 радиацию поглощают СН4 и
N2O, но особенно сильно - инертные в тропосфере галогенуглеводо-
роды, которые в дополнение реагируют в стратосфере под воздей-
воздействием жесткого ультрафиолета с разрушением стратосферного
озона. СН4 поглощает примерно в 20 раз сильнее, чем СО2 (в пере-
пересчете на молекулу), a N2O - в 320 раз. Фреоны поглощают еще силь-
сильнее. На основе поглощения и времени пребывания в атмосфере был
рассчитан "парниковый потенциал":
Газ
"Парниковый потенциал"
по IPCC1 1992, 1994
со2
сн4
N2O
CFC-11
CFC-12
HCFC-22
HFC-134а
1
11
320
4000
8500
1700
1300
Моделирование показывает, что атмосфера очень чувствитель-
чувствительно откликается на изменение своего состава прежде всего радиаци-
радиационным балансом и соответственно изменением климата.
Содержание углекислоты возросло с XVII столетия на 25%, с
290 ppmv до 355 ppmv, прирост ее содержания за этот период по
данным на 1991 г. составлял 1,5 ppmv/год, что соответствует
1,6 Гт С/год. Прирост за последнее столетие коррелировал с числен-
численностью человечества. Углекислота атмосферы (рис. 4.2, 4.3) пред-
представляет малый динамичный резервуар, отражающий равновесие с
растворенной углекислотой океана, дисбаланс между стоком и
продукцией в биотическом цикле, захоронением в виде карбона-
карбонатов при седиментации и регазацией в геологическом цикле. По-
Пополнение резервуара углекислоты в прошлом шло за счет эндоген-
эндогенных источников. Ее содержание в воздухе коррелирует с темпера-
температурой.
Метан образуется микроорганизмами и разрушается в тропо-
тропосфере в результате фотохимических реакций. С 1850 г. концентра-
концентрация возросла с 0,8 ppmv до 1,7 ppmv. Начиная с 1990 г. прирост мета-
метана снизился с 20 ppmv/год до 10 ppmv/год.
Закись азота образуется микроорганизмами при нитрификации
и денитрификации как промежуточный продукт. Считается, что
увеличение ее образования обусловлено применением азотистых
удобрений.
Окись углерода не является парниковым газом, но влияет на
окислительное состояние атмосферы, снижая окисление метана.
Источником служат пожары, фотохимические реакции окисления
углеродистых веществ.
IPCC. Radiative forcing and climate change: Report of the Scientific Assessment
group (WGI) on the Intergovernmental Panel on Climate Change (IPCC), 1994.
107
ГтС/год
Рис. 4.2. Среднемесячная концентрация СО2 по меридиональным станциям
Тихого океана (Australian Bureau Meteorology)
Станции: / - Alert; 2 - Barrow; 3 - Mauna Loa; 4 - Cape Grim; 5 - South Pole
Сезонные колебания, связанные с ассимиляцией СО2 в вегетационный период, сильнее всего
в континентальном бореальном поясе G,2 - Аляска) и влияют на все Северное полушарие
вплоть до тропиков C - Гаваи). В океаническом южном полушарии колебания сглажены
D - Тасмания, 5 - Южный полюс). Наземные экосистемы имеют наиболее быстрый обмен с
атмосферой. Перенос воздушных масс через тропическую зону конвергенции замедлен
Окислы азота образуются в результате высокотемпературных
реакций на воздухе, особенно в двигателях внутреннего сгорания, но
также при грозовых разрядах, и служат важными катализаторами
фотохимических реакций.
Двуокись серы образуется при сжигании серосодержащих топ-
лив, особенно сернистого угля, а также и при окислении биогенного
диметилсульфида (ДМС).
Следует обратить внимание, что техногенные источники парни-
парниковых газов совпадают по составу с вулканическими источниками.
108
1980
1985
1990
1995
2000
Рис. 4.3. Межгодовое варьирование глобального наземного потока по монито-
мониторингу 513С в Тасмании [по: Francey et al., 2001]
/ - поступление от топлив; 2 - СО2 в атмосфере; 3 - океан; 4 - наземный сток, оценка по
|3С; 5 - разность потока в океан
Причина заключается в высокотемпературных реакциях. Отсюда
следует, что в природе вероятно существование естественных меха-
механизмов стока этих газов, и моделью их могут служить микроорга-
микроорганизмы вулканических мест обитания - кальдерная микрофлора.
Важнейшим свойством климатической системы является нали-
наличие положительных обратных связей. При повышении температу-
температуры воздуха увеличивается содержание паров воды в атмосфере, в
большой степени отвечающих за поглощение теплового излучения
и ведущих к потеплению. При понижении температуры происходит
образование льда, повышающего сухость атмосферы и увеличиваю-
увеличивающего отражение солнечной радиации, что влечет за собой похолода-
похолодание. При повышении температуры воды океана уменьшается рас-
растворимость углекислоты и соответственно происходит ее выброс в
атмосферу, что опять-таки ведет к потеплению. Все эти связи ведут
к большой неустойчивости климатической системы.
Критическим для проблемы антропогенного влияния является
корреляция газового состава атмосферы и температуры в прошлом.
Газовый состав атмосферы и температуру прошлого научились оп-
определять с большой точностью по составу газовых включений, а
температуру - по изотопии кислорода. В результате было установ-
установлено, что климат резко менялся не только в течение геологических
эпох, но и десятилетий. Первым примером таких изменений послу-
послужило резкое потепление в Северной Атлантике - климатическое
109
явление Дансгаарда-Оешгера. Причины этих резких колебаний па-
леоклимата неясны. Неясно, что предшествует чему: повышение
температуры увеличению концентрации парниковых газов или
же наоборот.
Кроме мониторинга химического состава атмосферы важней-
важнейшим фактом стало исследование состава газов, уловленных в ледя-
ледяных кернах Антарктиды. Керны были получены на станции "Восток"
в буровой установке по разработке Института Арктики и Антаркти-
Антарктики в Ленинграде и послужили для анализа пузырьков газа. Захват га-
газов атмосферы происходит при последовательном спрессовывании
снега в фирн и затем в лед, что происходит на глубине первых мет-
метров; только на этом последнем этапе прекращается миграция газов
по ходам. Таким образом, состав газа в пузырьках соответствует со-
составу атмосферы в ту или иную эпоху с обобщением по меньшей ме-
мере в десятки лет. Ледяные керны были исследованы хроматографи-
чески и масс-спектрометрически и дали корреляцию температуры,
определяемой по изотопу 18О, с содержанием СО2 и СН4. Удалось
проследить состав атмосферы на протяжении 160 тыс. лет. Анало-
Аналогичная работа была проведена в Гренландии и привела к заклю-
заключению о резких колебаниях климата в доантропогенную эпоху с
изменением около 5°С за десятилетие. Информация об эпизодах
внезапного потепления климата нашла свое подтверждение в дан-
данных палеоклиматологии, по слоям роста кораллов, индикаторным
организмам (фораминиферам) в донных отложениях океанов.
Основной вывод заключается в том, что температура и содер-
содержание парниковых газов в атмосфере положительно корре-
коррелируют.
Климат зависит не только от парниковых газов. Значение имеет
и ряд иных факторов эндогенной и экзогенной природы. К ним от-
относятся:
1) астрономические факторы: изменение солнечной постоянной,
эксцентриситета земной орбиты с периодом 93 000 лет, наклона зем-
земной оси с периодом 41 000 лет, прецессия оси с периодом 21 000 лет
(циклы Миланковича). В масштабе десятков тысяч лет эти факторы
являются ведущими и получили хорошее подтверждение для не-
нескольких сот тысяч лет (Graedel, Crutzen, 1992). Астрономические
факторы хорошо согласуются с ледниковыми периодами2;
2) геологические факторы: тектонические процессы, вулкани-
вулканические извержения, эрозия и выветривание горных пород, положе-
положение континентов;
3) океанические факторы: Эль Ниньо (теплое течение у запад-
западных берегов Южной Америки, обусловленное атмосферными ци-
циклоническими системами над Тихим океаном) и менее выраженная
2 Muller R.A., MacDonald GJ. Ice ages and astronomical causes: Data spectra] analysis
and mechanisms. В.: Springer; Chicherster; L.: Praxis, 2000. 318 p.
110
Северо-Атлантическая осцилляция, изменение термогалинной цир-
циркуляции вод океана, изменение уровня океана (эвстатическая кри-
кривая), образование льда;
4) биологические факторы: влияние растительности на альбедо
и эвапотранспирацию. Они эффектно иллюстрированы Дж. Ловело-
ком на примере "мира фиалок" (Daisy World): пусть часть поверхности
Земли покрыта белыми фиалками с высоким альбедо. При повыше-
повышении температуры фиалки размножаются и увеличивают отражаю-
отражающую поверхность, снижая температуру и обусловливая отрицатель-
отрицательную связь. Это пример саморегуляции "Земли как живого организма",
или модной гипотезы Геи. К той же категории относится выделение
морским фитопланктоном диметилсульфида, который в воздухе
превращается в конечном итоге в сульфат, служащий центрами кон-
конденсации для воды облачного покрова Земли.
Однако главным фактором влияния биоты на климат оказыва-
оказывается ее участие в образовании и поглощении парниковых газов, по-
поскольку основным стоком для СО2 является поглощение фотоавто-
трофами, а источником разных парниковых газов служит аэробная
и анаэробная деструкция мортмассы. Эмиссия парниковых газов на
90% обусловлена деятельностью микроорганизмов. Только 10%
эмиссии СО2 с суши связано с сжиганием топлив. Роль животных су-
сущи количественно незначительна. В антропогенной продукции ме-
метана, за исключением источников, связанных с добычей ископаемых
топлив, в первую очередь с открытыми разработками угольных ме-
месторождений, образование метана обусловлено изменениями усло-
условий обитания метаногенного сообщества, например на рисовых по-
полях. Закись азота образуется при сжигании биомассы, но основным
ее источником служат микробные процессы цикла азота. Цикл ле-
летучих соединений серы связан с гниением и мощным процессом
сульфатредукции. Так называемая "антропогенная продукция" пар-
парниковых газов обусловлена в значительной степени изменением ус-
условий обитания микробного сообщества в антропогенно-модифици-
антропогенно-модифицированном ландшафте.
Сопоставление газов, эмитируемых с поверхности Земли в атмо-
атмосферу, с разнообразием специализированных физиологических
групп бактерий показывает, что для отдельных газов можно указать
циклы с бактериями-генераторами газов и бактериями-окислителя-
бактериями-окислителями. Эти циклы представляют важнейший глобальный биотический
механизм и рассматриваются подробно ниже.
Побудительным мотивом к действиям в связи с изменениями
климата послужили антропоцентрические соображения. Сначала
панику вызвало появление "озоновых дыр" над Антарктидой, а по-
потом и в Северном полушарии. Уменьшение стратосферного озона
ведет к увеличению UV-B, достигающего обитаемой поверхности
Земли, и угрожает здоровью. Образование "дыр" приписывается
фреонам, хотя есть и другие участники. Последовал "Монреальский
Ш
протокол", запрещающий выпуск фреонов. Для микробиологов это
обернулось повышенным интересом к организмам, способным ис-
использовать хлорированные соединения, особенно в анаэробных ус-
условиях, проводя реакцию дегалогенизации.
Когда американец Килинг в результате многолетних наблюде-
наблюдений, начиная с 1958 г., за содержанием газов в меридиональной сети
морских обсерваторий нашел увеличение концентрации СО2 в пос-
последние десятилетия, то это свидетельствовало об изменении балан-
баланса в биосфере в сторону преобладания источников над стоками, или
деструкции над продукцией. Неприродным источником СО2 служит
сжигание углеродных топлив, которое суммируется с предположи-
предположительно находящимся в равновесии балансом продукции и деструк-
деструкции. Концентрация СО2 в значительной степени определяет парни-
парниковый эффект. Возможность глобального изменения климата вы-
вызвала необходимость превентивных действий вплоть до заключения
"Рамочной Конвенции по климату" в 1992 г., согласно которой все
страны должны воздерживаться от увеличения эмиссии парниковых
газов и должны:
- способствовать уменьшению источников парниковых газов,
- поддерживать существующие стоки парниковых газов,
- расширять стоки парниковых газов,
- сохранять резервуары предшественников парниковых газов,
прежде всего органического углерода.
Химия атмосферы стала одной из наиболее актуальных проблем
естествознания. Обыватель думает, что источник проблемы - толь-
только в промышленности и сжигании топлив. На самом деле, антропо-
антропогенная деятельность лишь усиливает дисбаланс в атмосфере на
5-10%, и этого оказывается достаточно, чтобы климатическая сис-
система вышла из равновесия. Основным источником парниковых га-
газов на Земле является деятельность микроорганизмов.
4.3. ПАРАДОКС КИСЛОРОДА
Основным биогенным газом атмосферы стал кислород. Он из-
изменил состояние поверхностных оболочек Земли, как гидросферы,
так и литосферы. Парадокс кислорода - диоксигена, О2, - заключа-
заключается в том, что термодинамически он должен был бы исключить су-
существование неокисленных соединений, но этого не происходит по
кинетическим причинам. Пожалуй, это наиболее наглядный пример
ограниченности термодинамического подхода к природоведению.
Свободный диоксиген малореакционноспособен, и это обуслов-
обусловливает возможность его продукции оксигенными фототрофами и
существования жизни в кислородной атмосфере. Если бы реакции
О2 определялись термодинамикой, а не кинетикой, то в оксигени-
рованной атмосфере органические вещества быстро бы окислились в
112
Таблица 4.1
Стандартные окислительно-восстановительные потенциалы для одно-
и двуэлектронных реакций диоксигена в воде
[по: Active oxygen in biochemistry, 1995]
Реакция
O2 + e~ = Q~2
Ог + е- + 2Н+ = Н2О2
Н2О2 + е" + Н+ = Н2О + ОН
ОН + е" + Н+ = Н2О
О2 + 2е~ + 2Н+ = Н2О2
Н2О2 + 2е~ + 2Н+ = 2Н2О
Продукт
Супероксид
Пероксид
Гидроксил
Вода
Пероксид
Вода
Е, В (рН 7,25)
-0,33
+0,89
+0,38
+2,31
+0,281
+1,349
СО2 и Н2О. Поэтому реакции О2 служат лучшим примером зависи-
зависимости биосферы от каталитических функций организмов. Оксиге-
нирующие ферменты не только обусловливают скорость реакций,
но и определяют их специфичность, связывая субстраты окисления
в контролируемом положении и снижая до минимума спонтанное
окисление промежуточными продуктами. Восстановление диоксиге-
диоксигена требует одновременного переноса четырех электронов, что огра-
ограничивает возможность такой реакции:
О2 + 4Н+ + 4е- = 4Н2О, Е° = +0,815 В.
Ступенчатое восстановление О2 ведет к появлению его реакци-
онноспособных форм (табл. 4.1). Относительно низкий потенциал
одноэлектронного восстановления диокисгена служит одним из фак-
факторов, ограничивающих его реакционноспособность, но вместе с
тем диоксиген не может реагировать, пока не произошло его восста-
восстановление. Поэтому химическое восстановление диоксигена проис-
происходит лишь с сильными восстановителями, такими, как металлы, се-
сероводород, или же при высокой энергии активации, например при
повышении температуры, вызывающей цепную реакцию сгорания.
Промежуточные продукты восстановления кислорода в высокой
степени реакционноспособны и поэтому токсичны. Ферменты, обес-
обеспечивающие использование окислительной способности диоксиге-
диоксигена, либо стабилизируют метаболиты, содержащие супероксид, либо
осуществляют двухэлектронный перенос. Для одноэлектронного
восстановления требуется сильный восстанавливающий агент,
такой, как восстановленные флавины и гидрохиноны. Активация О2
флавиновыми монооксигеназами происходит без участия металла и
включает промежуточное образование супероксида, быстро реаги-
реагирующего с окисленным флавином с образованием гидропероксида
как промежуточного продукта.
ИЗ
Нетоксичные реакции оксигенирования включают образование
комплексов с металлами, обычно Fe(II) и Cu(I). Соединения, образу-
образующиеся при восстановлении О2, т.е. супероксид (О2), пероксид
(О2~) или оксид (О2-), могут быть стабилизированы координацией
с металлом. Такие реакции позволяют преодолеть кинетические и
термодинамические барьеры.
Диоксиген появился на Земле в результате оксигенного фото-
фотосинтеза3. Абиогенная продукция О2 в результате фотолиза паров во-
воды количественно невелика и происходит в области, где возможно
быстрое расходование окислителя в фотохимических реакциях, на-
например для образования окислов азота из крайне инертного динитро-
гена N2. При биогенном образовании О2 возникают те же трудности
с возможностью появления токсичных промежуточных продуктов.
Оксигенный фотосинтез должен провести реакцию выделения О2
так, чтобы не возникали промежуточные продукты, а из сферы ре-
реакции уходил бы сразу относительно инертный диоксиген О2. Эта за-
задача решается в фотосистеме II (ФСП) таким образом, что молеку-
молекула воды оказывается в окружении четырех атомов Мп, взаимодейст-
взаимодействующих с фотохимическим окислителем хлорофиллом а с
Ет =+600-800 мВ, а восстановитель сбрасывается на резервуар
пластохинона. Фотосистема II представляет универсальный меха-
механизм для всех цианобактерий, водорослей и растении. Поэтому про-
продукция О2 в глобальном масштабе определяется плотностью хлоро-
хлорофилла на единицу освещенной поверхности вне зависимости от того,
в каких оксигенных организмах находится хлорофилл. Биологиче-
Биологические механизмы оксигенного фотосинтетика должны обеспечить
быстрое удаление О2 внутри клетки, что обеспечивается фотоды-
фотодыханием, и из клетки, иначе неизбежен окислительный стресс. Про-
Проще всего задача решается для одноклеточного планктонного орга-
организма в аноксической среде, куда О2 просто диффундирует. Если в
этой среде есть такие восстановители, как Fe2+ или H2S, то угроза на-
накопления О2 снимается. Однако при повышении плотности клеток
возникают ограничения, достигающие своего предела в биопленках,
где отток О2 замедлен, и он может образовывать пузырьки, создавая
вокруг них условия насыщения 100% О2. В результате физическая
форма сообщества, определяющая транспортные процессы в нем,
оказывается критической. Высокая плотность требует создания эф-
эффективных механизмов детоксикации в клетке. Вторая сторона ки-
кислородного парадокса заключается в образовании токсических
форм при дыхании и восстановлении О2.
Доступность О2 для аэробных организмов определяется его транс-
транспортом. Поэтому простейшим механизмом для снижения токсичности
3 Пингвин А.В., Аверина С.Г. Оксигенная фототрофия. СПб.: Изд-во СПбУ,
2002. 236 с.
114
О2 служит усиление диффузионного барьера для газа с помощью ма-
малопроницаемых оболочек, как у аэробных азотфиксирующих орга-
организмов, например гетероцист цианобактерий, и переход клетки в мик-
микроаэробные условия или же за счет очень быстрого использования О2
при дыхании, как у азотобактера. Толстые слизистые капсулы ограни-
ограничивают диффузию О2 к клеткам в биопленках, но это действие приоб-
приобретает значение при толщине слизистого слоя более 0,1 мм вследствие
исключения всех форм переноса, кроме молекулярной диффузии. Еще
более эффективным способом снижения концентрации О2 служит ис-
использование его для дыхания в плотных колониях клеток: внутри та-
таких образований создается аноксическая зона, так как скорость погло-
поглощения О2 превышает скорость его диффузии извне. Особое место за-
занимают аэротолерантные анаэробы, обладающие механизмами дето-
детоксикации, но не использующие О2 для дыхания.
Диоксиген О2 может быть восстановлен в воду переносом четырех
электронов под действием фермента цитохромоксидазы. Окисление
может осуществляться также присоединением одного атома кислоро-
кислорода монооксигеназами или двух атомов диоксигеназами. Обычно осу-
осуществляются обратимые реакции диоксигена с металлами, причем
участвующие в них медь и железо окружены гидрофобным белком,
защищающим комплекс металла с кислородом от дальнейших реак-
реакций, которые привели бы к необратимому окислению металла. Хотя
начальные реакции сходны, последующие превращения крайне разно-
разнообразны - от переноса кислорода гемоглобином до реакции с оксиге-
назами или переноса четырех электронов с синтезом воды цитохромо-
ксидазой.
Цитохромоксидаза катализирует перенос электронов от цито-
хрома с на кислород с образованием воды. Она содержит два атома
меди (Си и Сив) и два гема (а и а3).
Перенос электронов сопровождается образованием промежу-
промежуточных продуктов восстановления кислорода. Промежуточные про-
продукты связаны с ферментом и в свободном состоянии не обнаружи-
обнаруживаются.
В дыхательной цепи, однако, имеется ряд одноэлектронных пе-
переносчиков, которые неизбежно образуют токсические формы не-
неполного восстановления кислорода. Поэтому аэробные организмы
помимо способности использовать О2 как конечный акцептор элек-
электрона должны обладать эффективной защитой от активных форм
кислорода. К таким ферментам относятся супероксиддисмутаза,
каталаза, пероксидазы.
Супероксиддисмутаза (СОД) переводит О2 в менее токсичный
пероксид Н2О2, который в свою очередь разлагается каталазой на
воду и О2. Супероксиддисмутаза присутствует практически во всех
аэробных организмах и содержит либо Fe2+, либо Мп3+. Синтез ката-
лазы происходит в ответ на окислительный стресс. Неэнзиматиче-
115
ским механизмом снижения токсичности служит обилие восстанов-
восстановленных соединений, например глутатиона, концентрация которого в
клетке может достигать 10 мМ.
Ряд субстратов, таких, как, например, углеводороды, не могут
быть использованы без предварительного оксигенирования. Активи-
Активирующие диоксигенирующие ферменты, как Р-450, метанмонооксиге-
наза, тирозиназа, имеют сходные центры для связывания О2, но рядом
с ними расположены центры связывания окисляемого субстрата.
Монооксигеназы включают один из атомов О2 в окисляемый субстрат,
а второй атом окисляют в воду. За исключением флавиновых фермен-
ферментов, образующих пероксидную активную форму для окисления, боль-
большинство оксигеназ содержит металл. Р-450 осуществляет окисление
углеводородов за счет супероксидного промежуточного состояния.
Другую возможность для оксигенирования представляет активация
субстрата до такого состояния, в котором он может реагировать с О2,
как, например, для катехинов и липооксигеназы.
Растворимость О2 в воде невелика и составляет в равновесии с
современной атмосферой 250 мкмоль/л при 25°С. Низкая раствори-
мость кислорода обусловливает зависимость аэробных организмов
не столько от его резервуара в среде, сколько от масс-переноса.
Даже при небольшой концентрации окисляемых веществ О2 быстро
используется аэробами, которые служат главными агентами созда-
создания анаэробных условий. Практически доступность О2 для аэробов
определется по "биохимическому потреблению кислорода" (БПК),
когда определяется убыль растворенного О2 в замкнутом сосуде,
обычно за 5 суток (БПК5). Возможность развития аэробов опреде-
определяется в первую очередь обменом с атмосферой. Микроорганизмы
различаются по способности использовать О2. Некоторые организ-
организмы развиваются только при пониженном содержании О2; они полу-
получили название микроаэрофилов или микроаэробов.
Для многоклеточных организмов скорость диффузии О2 недос-
недостаточна, и потребовался механизм связывания и транспорта кисло-
кислорода в тканях. Такими переносчиками служат гемоглобин и сходные
с ним соединения.
У растений для транспорта О2 в корни имеется проводящая сис-
система аэренхимы, развитая у водных растений.
4.3.1. Роль микроорганизмов в формировании
кислородной атмосферы
Формирование состава современной атмосферы Земли, в пер-
первую очередь характеризующейся высоким содержанием О2, обусло-
обусловлено деятельностью одной-единственной группы микроорганиз-
микроорганизмов - оксигенных цианобактерий, обладающих уникальной систе-
системой расщепления воды для получения восстановителя с выделением
О2 как отбрасываемого продукта реакции. Устройство фотосинте-
116
тического центра - фотосистемы II - было рассмотрено ранее. По-
Поскольку хлоропласты всех протестов, водорослей, растений счита-
считаются в конечном итоге производными от симбиотических цианобак-
цианобактерий, то получается вполне прямолинейная картина: биосфера
есть результат развития цианобактерий и переработки продуктов
их жизнедеятельности в биотических и абиогенных реакциях. При
всей утрированности этой формулировки в ней содержится суть
проблемы.
Универсальность ФС II определяет условия генерации кислоро-
кислорода. К ним относятся доступность фотосинтетически активной ради-
радиации и, следовательно, ограничение области продукции дневной по-
поверхностью, которая автоматически совпадает с поверхностями раз-
раздела суши и атмосферы и океана и атмосферы. Время продукции за-
зависит от суточных циклов. Поэтому фотосинтетики приспособлены
к импульсному характеру фототрофного обмена и должны в виде
альтернативы ему для темного периода суток иметь иной тип обме-
обмена для обеспечения энергии поддержания. Особенностью обмена
фотоавтотрофов служит то, что они не ограничены углеродистыми
веществами и могут запасать их в подвижные резервы и даже ис-
использовать для построения оболочек, например в виде целлюлозы.
Зависимость ФС II от хлорофилла позволяет точно определять
область развития первичных продуцентов О2 по плотности хлоро-
хлорофилла, оцениваемой в диапазоне 600-700 нм, делая возможным при-
применение дистанционных аэрокосмических методов4. Между содер-
содержанием хлорофилла и продукцией имеется линейная зависимость,
четко установленная для водоемов с фотосинтетиками, имеющими
короткий жизненный цикл. Для растительного покрова зависимость
несколько сложнее, потому что приходится вносить поправку на све-
светорассеяние, обусловленное архитектурой фитоценоза, а также дос-
доступностью воды в разных растительных сообществах. По А.Т. Мок-
роносову (Круговорот углерода..., 1999), каждый килограмм хлоро-
хлорофилла обеспечивает в среднем за период вегетации связывание
145±29 кг атмосферного углерода.
Несмотря на то что О2 при действии ФС II происходит из воды,
суммарная реакция оксигенной фототрофии уравновешивается свя-
связыванием СО2 в биомассу [СН2О] по уравнению:
СО2 + Н2О = [СН2О] + О2.
Схема вовлечения СО2 в обмен фотоавтотрофа представлена на
рис. 4.4.
Географическую область продукции О2 сообществом организ-
организмов нельзя определить по плотности хлорофилла, поскольку его
выделение уравновешивается дыханием. В масштабе экосистем
ААндрузДж., Бримблекумб П., Джикелз Т., Лисе П. Введение в химию окружа-
окружающей среды. М.: Мир, 1999. 271 с
117
I Пограничный слой
1НСОз
Цитоплазматическая
мембрана
РБФ
ФГА
¦ Карбоангидраза
Фермент с
карбоангидразной
функцией
Рис. 4.4. Механизм концентрации Снеорг у цианобактерий [по: Kaplan, 1999]
Схематически представлены компоненты цианобактериальной клетки, участвующие в
Слсорг концентрирующем механизме: пограничный слой, цитоплазматическая мембрана и
цитоплазма с областью тилакоида и карбоксисомой. Отмечены три возможных перенос-
переносчика HCOJ : СМР - первичный HCOJ транспортер ABC типа, let В - предположитель-
предположительно осуществляющий симпорт Ыа+/НСОз или антипорт Na+/H+, и транспортер
неизвестной природы с низким сродством к HCOJ (на схеме отмечен вопросительным
знаком). Вынос HCOJ из клетки осуществляется с помощью этих переносчиков (пока-
(показано только для СМР) и путем диффузии.
Предполагается и наличие активного транспорта для СО2. Накопленный HCOJ
проникает в карбоксисому, где за счет активности карбоангидразы создается
высокая концентрация СО2 вблизи Рубиско. Часть СО2 фиксируется, а часть диф-
диффундирует наружу. Регенерация рибулозо-1,5-бисфосфата осуществляется в карбок-
118
L
область продукции О2 определяется дисбалансом продукции и дест-
деструкции и прямо соответствует количеству неразложенного органи-
органического вещества в почве или донных осадках.
Разнообразие организмов, занимающих функциональную нишу
первичных продуцентов - оксигенных фотоавтотрофов, составляет
предмет ботаники. Цианобактерий, микроводоросли-протисты, ма-
макроводоросли, бессосудистые и сосудистые растения в общем обра-
образуют монофилетическое дерево, хотя на уровне эукариот-протист
оно возникает комбинаторным путем с разными нефотосинтетиче-
скими предшественниками. Разные ветви дерева водорослей обнару-
обнаруживают параллелизм постепенно усложняющихся форм, указывая
на общие законы дифференциации, не зависящие от общности про-
происхождения. Параллелизм форм водорослей явился одним из наибо-
наиболее крупных эмпирических обобщений описательной ботаники
1920-х годов.
Экофизиология разных водорослей позволяет им занимать все
биотопы, где есть свет и влага. Вместе с тем определенные группы
доминируют в определенных условиях. Цианобактерий предпочита-
предпочитают щелочные среды. Они составляют большинство организмов в
экстремальных условиях - в термальных источниках, при высокой
солености, в содовых озерах. Существенно, что цианобактерий со-
составляют важную группу среди почвенных водорослей, развиваю-
развивающихся на увлажненном грунте и, следовательно, могут быть отнесе-
отнесены к континентальным формам. Они относительно немногочислен-
немногочисленны в море. Строение циано-бактериального сообщества (мата) бы-
было разобрано подробно (см. Лекцию 3), альго-бактериальные сооб-
сообщества сложнее из-за присутствия в них животных.
Лабораторные культуры микроводорослей, начиная с выделен-
выделенной М. Бейеринком Chlorella, постепенно становятся все более при-
привлекательным объектом микробиологии и физиологии растений.
Идентификация водорослей и цианобактерий основывается на мик-
микроскопии и морфологических критериях, причем некоторые из
идентификационных признаков, например агрегаты, образование
поверхностных оболочек, нередко теряются в лабораторных куль-
культурах. Для цианобактерий расхождение морфологической класси-
классификации и генетической на основании молекулярно-биологических
признаков значительно. Поэтому пока физиологические исследова-
исследования микроводорослей в культурах служат лишь важным дополнени-
дополнением к тому, что получили альгологи в работах in situ.
Разнообразие оксигенных фототрофов очень велико и рассмат-
рассматривается применительно к водорослям альгологами, создавшими
для их идентификации внушительный набор определителей. В Рос-
России используется многотомный "Определитель пресноводных водо-
водорослей", составленный Ботаническим институтом в Санкт-Петер-
Санкт-Петербурге. Систематика цианобактерий (синезеленых водорослей) была
составлена в начале 1920-х годов Л. Гейтлером, а в России -
119
А.А. Еленкиным на основе микроскопических наблюдений природ-
природного материала. Она была изменена на основе изучения чистых
культур в соответствии с Бактериологическим кодексом после ра-
работ Р. Стениера, показавшего, в частности, большую зависимость
морфологии цианобактерий от условий культивирования. Коллек-
Коллекция культур цианобактерий в России создана Б.В. Громовым. Одна-
Однако чистые культуры цианобактерий оказались очень неустойчивы-
неустойчивыми, и обычно они поддерживаются в смешанной культуре. Следую-
Следующим шагом явилась молекулярно-биологическая классификация на
основе 16S рДНК, которая еще более сократила число генородов.
Вместе с тем СВ. Шестаковым и его сотрудниками создана полная
карта генома одного из видов Synechocystis, позволяющая выделить
функционально значимые гены. Несмотря на все морфологическое
разнообразие цианобактерий, их обмен довольно сходен и различа-
различается на уровне адаптации к топическим условиям. Возникновение
этой группы представляет "бутылочное горлышко" эволюции, ко-
когда из сравнительно немногих организмов возникло разнообразие.
Цианобактерий располагаются у корня ствола филогенетического
дерева разнообразия живых организмов и в основании всех биогеохи-
биогеохимических превращений, послуживших "коридором обитаемости" для
высших организмов. Физиология их описана довольно подробно5.
Экофизиология водорослей пока выходит за пределы микро-
микробиологии и служит предметом работы альгологов и физиологов рас-
растений. Для субаэральных условий прямое определение суммарной
деятельности оксигенных организмов затруднено высокой концент-
концентрацией О2 в воздухе. В субаквальных условиях растворимость О2
низка и составляет при 20 °С 8 мг/л, т.е. 0,25 мМ, что обеспечивает
быстрый отток газа от продуцента. В воде фотосинтез удобнее все-
всего определять по продукции О2 либо химически по Винклеру, как
был получен основной массив данных, либо электрохимически. Для
этого сравнивается содержание О2 в освещенной и затемненной
склянках за определенное время.
Определяющим фактором для фотосинтеза служит доступность
света, и, следовательно, идет конкуренция за освещенную поверх-
поверхность. Развитие сообщества ограничивается максимальной плотно-
плотностью хлорофилла на единицу поверхности, около 1000 мг/м2 для
умеренного пояса. При такой плотности начинается самозатенение,
которое и служит ограничивающим фактором, определяя верхний
предел для численности фотосинтезирующих организмов. С другой
стороны, свет доступен лишь в течение дня, а ночью фототрофные
организмы должны переходить на поддерживающий обмен за счет
внутриклеточных запасов.
5 См., например: Гусев М.В., Никитина К.А. Цианобактерий. М.: Наука, 1979;
Stal L. Cyanobacterial mats and stromatolites // The ecology of cyanobacteria / Ed.
B.A. Whitton, M. Potts. Kluwer academic, 2000. P. 61-120.
120
Следующим по значимости фактором служит приток биогенных
элементов. Он определяется содержанием доступных форм элемен-
элемента в среде и его транспортом к месту обитания организмов. Следует
различать регенерационный поток, образующийся вследствие авто-
автолиза и разложения мортмассы деструкторами в сообществе, экс-
экспортный поток, обусловленный выносом, привнос извне течениями
или же за счет извлечения из горной породы. Эти потоки четко раз-
различаются для сообщества фитопланктона. Лимитирующие факторы
различны для разных групп фототрофов. Для цианобактерий, среди
которых много азотфиксаторов, наибольшее значение имеет дос-
доступность фосфатов, для водорослей на первом месте обычно быва-
бывает азот, а фосфат следует за ним. Далее следует серия микроэлемен-
микроэлементов, начиная с железа, обычно рассматриваемая физиологами расте-
растений применительно к культивируемому виду.
4.4. ДЫХАНИЕ ОРГАНОТРОФОВ
Биогенным стоком для кислорода атмосферы служит его аэроб-
аэробное восстановление в СО2 органотрофами, которое называют дыха-
дыханием:
[СН2О] + О2 = СО2 + Н2О.
Среди бактерий дыхание свойственно наиболее многочислен-
многочисленным и разнообразным в современных условиях группам протеобак-
терий и грамположительных организмов. Поскольку в течение поч-
почти столетия микробиологическая техника была приспособлена к ис-
исследованию аэробных органотрофов, именно здесь накоплены ос-
основные сведения о микробном разнообразии. Аэробные органотро-
фы, вероятно, сформировались в рамках циано-бактериального со-
сообщества, где было изобилие органического вещества и с суточной
периодичностью возникал избыток кислорода вплоть до гиперок-
сии. Ситуация была разной в планктонном сообществе, где раство-
растворенный О2 распределялся в массе воды, и в бентосном сообществе,
где его пузырьки могли механически удерживаться. Иную ситуацию
представляли разреженные циано-бактериальные пленки на влаж-
влажном грунте со свободным обменом с атмосферой.
Вся совокупность аэробных органотрофных организмов суще-
существует за счет дыхания. Традиционно их называют гетеротрофами,
хотя, строго говоря, этот термин относится к анаболизму. Дыхание
представляет наиболее выгодный энергетически процесс, но зави-
зависит от доступности О2, резервуар которого мал вследствие малой
растворимости, и доступность целиком определяется потоком к ме-
месту развития организма или колонии. Поэтому деятельность аэроб-
аэробных органотрофов определяется конкуренцией и за доступный Сорг,
и за О2.
121
Развитие аэробных органотрофов лимитируется, следователь- ¦
но, двумя факторами: доступным органическим веществом и прито- ;
ком О2. Если резервуар доступного органического вещества, напри- |
мер в виде растительных остатков, может быть значительным, то ¦
доступность кислорода зависит только от его притока и определяет- '
ся скоростью переноса. Для органотрофов скорость переноса О2 !
представляет ограничивающий фактор для микроместообитания со- ¦
общества. Это хорошо прослеживается для биопленок, иногда с же- г
стким послойным распределением микроорганизмов. i
Конкуренция за кислород, служащий универсальным акцепто- f
ром для аэробов, ведет к общей конкуренции в сообществе аэроб-
аэробных организмов и может быть преодолена за счет более высокого
сродства к кислороду у одних организмов, чем у других. По этому
признаку различаются аэробы и микроаэрофилы. Такое разделе-
разделение оправдано для водных организмов с более или менее устойчи-
устойчивыми концентрациями О2 в определенных слоях воды. Для поч-
почвенных организмов вероятны резкие колебания от полной доступ-
доступности О2 в почвенном воздухе до задержек в его поступлении при
дождях. Поэтому здесь большое значение приобретают адаптив-
адаптивные способности организма. Вместе с тем в почве благодаря ее
пористости микроорганизмы находятся в условиях обмена с атмо-
атмосферой через почвенный воздух, причем обеспечивается и отток
СО2. В еще большей степени адаптация к колебаниям в содержании
кислорода требуется для обитателей циано- и альгобактериальных
сообществ, которые должны противостоять гипероксии.
В сумме органотрофы осуществляют реакцию, обратную фо-
фотосинтезу: разлагают [СН2О], поглощают кислород, выделяют
СО2. Об успехе деятельности органотрофов можно судить по оста-
остаточному органическому веществу в местообитании: отсутствие
его указывает на полноту действия сообщества органотрофных
аэробов.
Переход от аэробного к анаэробному существованию обуслов-
обусловливает у факультативных анаэробов прежде всего смену типа ката-
катаболизма. Для этого существуют две возможности: A) переход к бро-
брожению или B) использование иного, чем О2, окислителя. Брожение
при исчерпании О2 осуществляют многие органотрофы, в их числе
наиболее подробно изученные энтеробактерии. Любопытным при-
примером могут служить цианобактерии, сбраживающие ночью накоп-
накопленную за день полиглюкозу. Число факультативных анаэробов-ор-
ганотрофов очень велико, и можно думать, что этот тип метаболиз-
метаболизма возник в циано- и альго-бактериальных сообществах, где обиль-
обильное содержание доступного органического вещества сочетается с
периодической аноксией по ночам.
Факультативные анаэробы с окислительным обменом могут ис-
использовать окислители, образовавшиеся при избытке О2 в сообще-
сообществе. Прежде всего это нитраты, служащие субстратом для денитри-
122
фикаторов, кроме того, окислы железа и марганца, а также молеку-
молекулярная сера для некоторых сероредукторов.
В деятельности биоты следует различать три крупных этапа:
первый состоит в фотоавтотрофном кислород-углекислотном цикле
с образованием Сорг и О2 в эквимолекулярных количествах; второй
обусловлен газогенерацией под действием преимущественно анаэ-
анаэробных организмов с выделением восстановленных газов; третий
представлен бактериальным окислительным фильтром, который
предотвращает выход этих газов в атмосферу.
В дополнение к углеродному циклу следует добавить циклы азо-
азота, серы, железа. Эти циклы действуют как в субаэральных, так и в
субаквальных условиях.
С прямым действием цикла сопряжены биотически опосредо-
опосредованные реакции, к которым в первую очередь следует отнести реак-
реакции выветривания горных пород. Эти реакции термодинамически
детерминированы и могут идти самопроизвольно, однако их кинети-
кинетика будет иной, чем под воздействием биоты. Важнейшей из реакций
выветривания является реакция углекислоты с магматическими си-
силикатами, ведущая к связыванию углекислоты в форме карбонатов.
4.5. ОКИСЛИТЕЛЬНЫЙ БАКТЕРИАЛЬНЫЙ ФИЛЬТР
Другую область конкуренции за О2 представляет окислитель-
окислительный бактериальный фильтр (рис. 4.5), в котором происходит окис-
окисление восстановленных газов и летучих органических веществ, про-
продуцируемых тазогенерирующими анаэробами или же происходящих
из геосферных источников, как углеводороды над нефтегазовыми
месторождениями.
Аэробное окисление восстановленных газов осуществляется га-
зотрофами - водородными бактериями, серными и тионовыми бак-
бактериями, нитрификаторами, карбоксидобактериями, метано- и ме-
тилотрофами. Они составляют сообщество "окислительного бакте-
бактериального фильтра". Следует заметить, что газотрофами являются
исключительно бактерии. Эукариоты не сохранили таких функций.
Суммарным результатом действия анаэробного сообщества и бакте-
бактерий окислительного фильтра является полное окисление органиче-
органического вещества по последовательности:
[СН2О] -» Брожение -» Вторичные анаэробы -» Газотрофы + О2 =
= СО2 + Н2О + Окисленные неорганические соединения.
Формирование состава атмосферы представляет геологический
процесс и соответственно обусловлено резервуарами с большими вре-
временами пребывания. Эти резервуары образуются в результате дисба-
дисбаланса в циклах с гораздо меньшими временными характеристиками -
вплоть до суточных. Короткие циклы более понятны биологам.
123
¦е-
1
8
?
A
X
ев
s
о.
я
О
ft.
Вместе с тем образование больших резервуаров обусловлено неспо-
неспособностью микроорганизмов полностью замкнуть циклы. Биогеохи-
Биогеохимическая сукцессия обусловлена накоплением неиспользуемого веще-
вещества, побочного или остаточного продукта жизнедеятельности.
Общие условия деятельности микроорганизмов при эмиссии
парниковых газов в атмосферу могут быть представлены схемой:
N,0
ФОТОХИМИЯ В
ТРОПОСФЕРЕ
ТРАНСПОРТ В
ТРОПОСФЕРЕ
ДЕСТРУКЦИЯ
МОРТМАССЫ,
НИТРИФИКАЦИЯ
Метанотрофы, метилотрофы,
карбоксидобактерии
водородные, тионовые,
серобактерии
Уровень воды
БАКТЕРИАЛЬНЫЙ
АЭРОБНЫЙ
ОКИСЛИТЕЛЬНЫЙ
ФИЛЬТР
ТРАНСПОРТ
В ПОЧВЕННОМ ВОЗДУХЕ
ИЛИ ВОДНОЙ ТОЛЩЕ
Метаногены Сульфидогены
АНАЭРОБНЫЙ
БАКТЕРИАЛЬНЫЙ
ГАЗОГЕНЕРАТОР
Образно говоря, систему можно сравнить с котлом на огне: па-
паром из котла занимаются химики атмосферы, крышку представляет
бактериальный фильтр, а варится в котле поступающий в нее орга-
органический углерод главным образом в виде нерастворимых полиме-
полимеров первичных продуцентов.
Выход газов в атмосферу определяется характером транспорта
через аэрируемые поверхностные слои, где развивается окислитель-
окислительный бактериальный фильтр газотрофов (рис. 4.5). Если этот транс-
транспорт происходит в виде пузырьков, как это бывает с метаном на бо-
болотах, то микроорганизмы не успевают его окислить, и газ проска-
проскакивает в атмосферу. То же происходит, если имеются некие каналы
проводимости - трещины, трубы. Если поток диффузионный, то
обычно бактериальный фильтр развивается до такой степени, что
полностью перехватывает поток газов. Поток в виде пузырьков
(эбулиционный) для микроорганизмов недоступен.
Рыхлая почва оказывается для организмов носителем окисли-
окислительного газового фильтра (рис. 4.6). В последнее время получили
развитие точные методы исследования газообмена почвы с помо-
125
2S3I i 2 . -. •. з
Рис. 4.6. Схема развития микробных процессов в почве и обмена с приземным
воздухом
/ - водные пробки; 2 - аэробные организмы; 3 - анаэробные организмы
Схема газообмена микробиоты почвы с приземным воздухом. Обмен происходит че-
через поры аэрации. Микроорганизмы развиваются в виде биопленок на поверхности поч-
почвенных частиц. При этом создаются два типа условий для развития анаэробов и генерации
восстановленных газов: 1) внутри комочков почвы с разлагаемым органическим вещест-
веществом; 2) внутри полостей, закрытых водными пробками. Аэробы (изображены палочками)
развиваются на поверхности ходов аэрации, например, по корневым ходам, используя ор-
органическое вещество и обогащая почвенный воздух СОг со снижением содержания СЬ.
Анаэробы внутри комочков почвы (Л) защищены от проникновения От слоем аэробов на
поверхности, использующих продукты брожения внутри комочка и составляющих газо-
газовый фильтр. Анаэробы во временно замкнутых полостях (В) находятся в условиях непо-
непостоянного режима и представлены факультативными анаэробами, продуцирующими, на-
например, N2O. Увлажнение и испарение создают непостоянный солевой режим и вынужда-
вынуждают почвенные организмы иметь механизмы осмоадаптации. В результате дыхания поч-
почвенный воздух отличается от приземного. Такая же система образуется в бактериальном
окислительном фильтре, в том числе, создаваемом искусственно для предотвращения вы-
выхода окисляемых и дурно пахнущих газов
щью высокочувствительных газохроматографических и спектраль-
спектральных методов. Здесь особенно важно исследование кинетики процес-
процесса при концентрации газов, близкой к атмосферной. Такие методы
были использованы Р. Конрадом при изучении потребления газов
почвенным сообществом.
Для диффузионного потока из донных отложений озер обычно
достаточно 10 м, как это было показано в оз. Плен для окисления
метана. В почве для окисления дурно пахнущих продуктов необходи-
необходимо более 1 м рыхлого грунта, что подтверждено кладбищенской
практикой.
Однако действие бактериального фильтра зависит не только от
характера транспорта газов из генерирующих слоев и мощности по-
126
L
тока, но и от встречного потока кислорода воздуха. В почве этот по-
поток осуществляется по порам аэрации, образованным на месте раз-
разрушенных корней в первую очередь. Поток газов в почве идет пре-
прежде всего диффузионным путем, и это - медленный перенос по
извилистым ходам пор аэрации. Перенос воздуха усиливается коле-
колебаниями температуры, ветром у поверхности и резко ослабляется
образованием водяных пробок в порах аэрации. В результате ниже-
нижележащие слои оказываются изолированными от атмосферы и в них
быстро происходит накопление углекислоты и снижение содержа-
содержания кислорода. Концентрация углекислоты в почвенном воздухе
возрастает вплоть до 10%, соответственно уменьшается количество
кислорода.
4.6. УГЛЕКИСЛОТА
Наиболее важным для биоты газом атмосферы служит углекис-
углекислота, в итоге дающая начало кислороду атмосферы как побочному
продукту обмена, хотя по биохимической реакции О2 образуется из
воды. Поступающая эндогенная СО2 имела два пути превращения:
1) реакция углекислотного выщелачивания пород в субаэральных
условиях вела к образованию и накоплению масс осадочных карбо-
карбонатов, преимущественно доломитов CaMg(CO3J; 2) реакция фотоав-
тотрофного образования Сорг с характерным изотопным составом
513С около -25%с при 813С ювенильной углекислоты около -5%с. На-
Накопление Сорг происходило в осадочных породах, причем в древних
породах его примерно столько же, сколько в современных, и вело к
освобождению эквивалентного количества О2. Масса Сорг в осадоч-
осадочных породах указывает на нижний предел суммарного образования
О2 за историю Земли.
Поглощение О2 происходило в реакциях с эндогенными восста-
восстановленными веществами, наиболее наглядным результатом этого
стали геологические отложения окислов железа в железистых квар-
кварцитах, соответствующих примерно 40% Сорг осадочных горных
пород, остальное приходится на сульфаты океана. Накопление О2 в
атмосфере по исчерпании резервуара восстановленных веществ ли-
литосферы привело к переходу около 2 млрд лет назад к кислородной
атмосфере, датируемой по появлению красноцветных пород. Уста-
Установление аналогичного современному кислород-углекислотного ци-
цикла на основе деятельности циано-бактериального сообщества про-
произошло в период от 2 до 1 млрд лет назад, когда начали распростра-
распространяться эукариоты-протисты. Геологическим свидетельством этой
эпохи служат массы строматолитов, представляющих литифициро-
ванные остатки циано-бактериальных сообществ, в которых проис-
происходит накопление биогенных карбонатов. Грубая ошибка состоит в
предположении, что воздействие кислорода на биоту началось с воз-
возникновения кислородной атмосферы. Чтобы появиться в атмосфере,
127
02 должен был до того локально синтезироваться биотой в массо-
массовом количестве, чтобы заполнить громадные резервуары раство-
растворенного О2 гидросферы и затем приходящей в равновесие с водой
атмосферы. Кислород атмосферы не представляет причину разви-
развития аэробной биоты. Поэтому циано-бактериальное сообщество с
аэробными бактериями следует рассматривать как предшествую-
предшествующее формированию современной атмосферы. Другие предположе-
предположения, например о гидрогенотрофной подземной биосфере или об ос-
основанном на фотосинтетическом аноксигенном серном цикле "пур-
"пурпурном океане" с образованием сульфата как конечного продукта,
приходится рассматривать как чисто гипотетические. Около 2 млрд
лет назад сформировался и серный цикл с участием сульфатредук-
торов, как об этом можно судить по изотопным соотношениям, хо-
хотя сами сульфатредукторы появились много раньше. Серный цикл
был особенно важен для формирования состава вод океана.
Углекислота, как уже отмечалось, представляет важнейший
парниковый газ. Основной ее источник в атмосфере - дыхание мик-
микроорганизмов. Например, для России эмиссия СО2 из почвы по
меньшей мере в 8 раз превосходит эмиссию из индустриальных ис-
источников. Суммарный поток углекислоты из местообитания
или ландшафта есть мера деятельности деструкторов в этом сооб-
сообществе.
Для наземных систем оценивается дыхание почвы. Дыхание
почвы измеряется камерным методом. Для этого на поверхности
почвы устанавливают газонепроницаемый колпак и измеряют выде-
выделение углекислоты доступным методом: поглощением Ва(ОНJ с по-
последующим титрованием, с помощью инфракрасного анализатора,
газохроматографически. Сейчас разработаны очень сложные и
очень дорогие приборы для автоматического определения дыхания
почвы. Однако методические трудности связаны не с измерением
химических компонентов, а с биологическими особенностями. Что-
Чтобы получить величину дыхания, нужно подавить поглощение угле-
углекислоты при фотосинтезе. Это достигается затенением камеры.
Но дыхание зависит от экссудатов растений, которые выделяются
при фотосинтезе. Следовательно, надолго затенять нельзя. Далее,
дыхание зависит от температуры, а под колпаком развивается заме-
замечательный парниковый эффект. При короткой экспозиции за дыха-
дыхание легко принять обмен резервуара почвенного воздуха с призем-
приземным слоем. Концентрацию газов в почвенном воздухе можно изме-
измерять с помощью диффузионных камер Паникова: в почву на разной
глубине закладывают патроны с мембраной, не пропускающей воду,
но проницаемой для газов, и измеряют концентрацию газов газохро-
матографическим методом. Метод довольно инерционен. Сущест-
Существенно лучше определение потоков СО2 с помощью инфракрасных
анализаторов. Следующее затруднение возникает с мозаичностью
условий даже на небольших площадях.
128
L
со2
V
ФОТОСИНТЕЗ
ДЫХАНИЕ i БИОМАССА
листьев корней i (первичная продукция)
GPP
"Дыхание"
почвы
корни и
ризо-
ризосфера
Мортмасса +
Вторичная продукция
НПП (NPP)
1 год
~ 10 лет
гумус
торф
НЭП
(NEP)
НБП
(NBP)
Рис. 4.7. Баланс углерода в наземных системах
Обратите внимание на возможность двойного счета. Дыхание корней, входящих в биомас-
биомассу, учитывается два раза. СО2, например, по 14СО2, может быть учтена несколько раз.
Долгоживущая биомасса + мортмасса + вторичная продукция входят в нетто-экосистем-
ную продукцию. NBP - нетто^биомная продукция с накоплением устойчивого органиче-
органического вещества (торф, гумус) за время более 10 лет
При интерпретации дыхания микроорганизмов по эмиссии СО2
(рис. 4.7) следует точно знать, что именно измеряется. Критическим
является время. Полное поглощение СО2 при автотрофной ассими-
ассимиляции - полная первичная продукция (GPP) - определяется по вклю-
включению 14С в биомассу или по коррелирующему признаку — поверх-
поверхностной плотности хлорофилла. Нетто-первичная продукция (NPP)
учитывается по приросту биомассы и включает потери прЬ дыхании
растения. Экосистемная нетто-продукция (NEP) исключает потери
Сорг на дыхание органотрофов за время вегетационного сезона. Ме-
Между тем эмиссия СО2 на поверхность почвы включает зависящие от
идущего фотосинтеза корневое дыхание, разложение экссудатов ри-
зосферной микрофлорой и микоризой, разложение короткоживу-
щего корневого опада. Поэтому эмиссию необходимо сопоставлять
с полной первичной продукцией, которая вдвое больше нетто-пер-
нетто-первичной продукции.
Чтобы избавиться от недостатков камерного метода, пользуются
микрометеорологическим способом: устанавливают мачту с распо-
расположенными на разной высоте датчиками и по градиенту компонен-
компонентов вычисляют разность между поглощением и эмиссией углекислоты.
5. Г. А. Заварзин
129
Для оценки градиента важна турбулентность и соответственно изме-
измерение скорости ветра. Чтобы коррелировать эти показатели, разра-
разработана достаточно сложная математическая модель. Микрометео-
Микрометеорологический подход дает интегрированную оценку потока в зави-
зависимости от рельефа местности. Хорошо он действует в аэротопе
леса или над открытым полем, где потоки воздуха более или менее
стабильны. Вместо мачты можно применять аэростат или самолет,
измеряющий концентрацию над большой поверхностью. Но все эти
методы очень дорогие и относятся скорее к области физики и химии
атмосферы, чем микробиологии.
Для данного типа почвы варьирование выделения углекислоты не
слишком велико, так как она уравновешивается с жидкой фазой, но
вместе с тем для неоднородного растительного покрова пространст-
пространственную неоднородность следует учитывать. Еще важнее временная
неоднородность, обусловленная суточным ходом, погодными услови-
условиями, сезонностью. Обычно дыхание почвы ограничивают вегетаци-
вегетационным сезоном. Однако дыхание продолжается и под снегом, а газо-
газообмен с атмосферой зависит от того, образовался ли на поверхности
почвы ледяной покров или же снег лег на влажную почву. В целом не
следует гнаться за чрезмерной точностью в определении суммарного
дыхания, не учитывая всех этих побочных факторов.
Тем не менее удалось составить карту почвенного дыхания для
России. Оказалось, что величина почвенного дыхания варьирует в
пределах от 1 до 18 т СО2/га за вегетационный период. Наименьшая
эмиссия установлена для глеевых почв тундры и северной тайги,
наибольшая - для пахотных перегнойных торфянистых почв и чер-
черноземов. В северных почвах особенно четко выражена корреляция
дыхания почвы с температурой с резким переходом около + 15°С.
Суммарный годовой поток углекислоты в большой степени зависит
от: 1) продолжительности вегетационного сезона; 2) площади, занятой
данным типом почвы. Ограничивающим фактором в полученных дан-
данных почвенного дыхания служит процесс переноса газов между поч-
почвой и атмосферой. Для таежной зоны средние потоки СО2 из поч-
почвы в течение вегетационного сезона находятся в диапазоне
0,5-3,3 г/(м2 • сут) под естественными ценозами и примерно в два раза
больше в агроценозах. Для территории России приблизительная вели-
величина ежегодной эмиссии углекислоты, полученная суммированием
почвенного дыхания по площадям почвенных группировок, составила
4,29 млрд т углерода. Эта величина близко согласуется с оценкой нет-
нетто-первичной продукции в 4,4 млрд т углерода (±20%) для той же тер-
территории и недоучитывает ряд естественных источников, например
разложение древесных остатков. Для баланса по углероду дыхание
почвы должно согласовываться с общей продукцией (GPP) подземной
части растительного покрова (Круговорот углерода..., 1999).
Каков вклад в почвенное дыхание разных групп организмов?
Решают эту задачу по-разному. Один способ заключается в том,
130
чтобы из образца почвы выбрать все корешки и измерить дыхание.
Другой способ состоит в том, чтобы ингибировать дыхание разных
групп организмов, например эукариот-грибов, антибиотиками, или
бактерий другими антибиотиками и судить о вкладе разных групп
организмов на основе ингибиторного анализа. Если нарушить почву
или донные отложения перемешиванием, то нарушается микростру-
микроструктура сообщества, от которой и зависит газообмен. Особенно легко
повреждается мицелий. Отделить корневое дыхание растений от
дыхания микроорганизмов ризосферы довольно трудно. Тем не ме-
менее предполагается, что корневое дыхание растений составляет при-
примерно 1/3 от дыхания почвы, а дыхание грибов составляет около 2/3
от дыхания микроорганизмов.
В водных системах выделение углекислоты нельзя измерять по
приросту углекислоты из-за того, что концентрация ее в бикарбо-
натной системе вод велика, и трудно уловить малые изменения, ко-
которые к тому же зависят от изменений растворимости. Деструкцию
удобнее измерять изотопным методом по выделению радиоактив-
радиоактивной углекислоты из меченого субстрата. Для гетерогенных систем
это делать трудно, так как трудно добиться равномерного внесения
материала. Чаще всего применяют болтушки донных осадков, в ко-
которых нарушена микростратификация. Чтобы избежать этого, в ко-
колонки ила изотоп вносят на проволоке, покрытой слоем клея
(например, ПВА). Общим ограничением здесь служит необходи-
необходимость использовать определенные субстраты от индивидуального
химического соединения, вроде ацетата, до суммы растворимых со-
соединений в виде меченой биомассы.
Оценка деструкции для океана изотопным методом была про-
проведена Ю.И. Сорокиным в 1960-х годах. А в 1950-х годах величину
первичной продукции радиоактивным изотопом углерода опреде-
определил Стиман Нильсен. Величина деструкции оказалась выше про-
продукции, и возникла проблема согласования баланса для океаничес-
океанических систем.
Между массой организмов и продукцией нет прямЪй зависимо-
зависимости: организмы с коротким жизненным циклом могут давать значи-
значительную продукцию при малой биомассе за счет быстрого оборота.
А организмы с длительным циклом накапливают биомассу. Именно
таково соотношение между фитопланктоном и остальными форма-
формами жизни в океане. Для годового цикла в продуктивной зоне океана
дыхание биоты в 10 раз превышает прирост ее массы, составляя со-
соответственно 3 Гт массы углерода в биоте и 36 Гт в выделяемой уг-
углекислоте.
Для того чтобы составить баланс по углероду, необходимо опре-
определить потребление СО2 при продукции и выделение СО2 при дест-
деструкции в определенном месте и затем решить, на какую площадь
можно распространить эти значения. Возникает классификацион-
классификационная задача, предшествующая картированию и определению площа-
5* ' ш
дей. Наиболее удобными для определения границ ландшафтов ока- f
зались почвенные карты. Следующий вопрос состоит в возможно- f
сти отнести полученные данные к годовому циклу, найдя величины, -1
соответствующие периоду биологической активности. Обычно fe
пользуются величиной безморозного периода, приравнивая его к ве- **
гетационному периоду, приводимому в агрофизических справочни- f
ках. Но в природе деятельность микроорганизмов продолжается и *
после сбора урожая.
При оценке роли кислород-углекислотного цикла для формиро-
формирования состава атмосферы приходится обращаться к геологическим
временным масштабам. Тогда резервуары с длительностью пребы-
пребывания в годы, такие, как при учете годичного цикла, и даже в сотни
лет для максимальных сроков пребывания в биомассе, можно счи-
считать постоянными и малыми. Наибольшее значение приобретают
резервуары устойчивого органического углерода почвы (торфа, гу-
гумуса, каустобиолитов) и рассеянного вещества осадочных горных
пород - керогена, соответствующие дисбалансу в углеродном цикле
и оставшемуся неиспользованным эквимолекулярному количеству
кислорода атмосферы и окисленных соединений литосферы. Угле-
Углекислота регулируется карбонат-силикатным процессом химическо-
химического выветривания, компенсирующим поступление эндогенной угле-
углекислоты.
Кислород-углекислотный цикл с участием продуцентов-фотоав-
тотрофов и деструкторов-органотрофов дал наиболее широкие ни-
ниши для разнообразия живых существ, вместив помимо исходных
прокариот все растения, грибы, животные.
Анаэробный цикл СО2 прежде всего связан с метаном, полно-
полностью зависящим от микроорганизмов, образующих "цикл Зёнгена";
метан наиболее важен для микробиологов и рассматривается от-
отдельно.
4.7. ЛЕТУЧИЕ СОЕДИНЕНИЯ АЗОТА
Основную массу атмосферы составляет инертный динитроген
N2. По-видимому, он имеет эндогенное происхождение, хотя опреде-
определение его содержания в магматических газах особенно ненадежно
из-за загрязнения подсасываемым воздухом. Вместе с инертными га-
газами его можно рассматривать как фон. Небольшая часть динитро-
гена усваивается в процессе синтеза биомассы сообществом и посту-
поступает в нее в стехиометрическом соотношении, определяемом содер-
содержанием белка и нуклеиновых кислот. Поэтому цикл азота и его ле-
летучих соединений жестко связан с циклом органического углерода
отношением < 1/6. Единственным входом в цикл азота служит азот-
фиксация, осуществляемая только прокариотами (рис. 2.3). Она слу-
служит для перевода азота из атмосферного резервуара в преходящий
132
резервуар биомассы и мортмассы. В органических соединениях
азот в восстановленной форме присутствует в виде аминогрупп в
белке и в героциклах азотистых оснований. Естественные источни-
источники связанного азота, атмосферные и вулканические, количественно
второстепенны. Важные изменения в азотный цикл внесло произ-
производство минеральных азотистых удобрений, приобретшее широкие
масштабы в XX в. Таким образом, на входе в цикл азота располага-
располагается азотсодержащая мортмасса. Обсуждая состав атмосферы, мы
не будем полностью рассматривать цикл азота, в котором органиче-
органический азот и растворенный азот минеральных соединений количест-
количественно играет подчиненную роль. Основной резервуар азота нахо-
находится в атмосфере. Вовлечение его в биогеохимические реакции в
подавляющей степени обусловлено биотическими факторами. Вме-
Вместе с тем летучие соединения окислов азота играют определяющую
роль в фотохимии атмосферы, инициируя окислительный цикл, как
об этом сказано выше.
Летучие соединения азота удобнее начать рассматривать с амми-
аммиака. Образование аммиака происходит при аммонификации органи-
органических азотистых соединений. Этот процесс идет в анаэробных ус-
условиях с образованием аммиака как основного или единственного
продукта. В аэробных условиях часть аммиака перехватывается на
нитрификацию, а часть - на конструктивные процессы растениями -
и возвращается в рецикл. В анаэробных условиях источником аммо-
аммония может служить аммонифицирующая нитратредукция, при кото-
которой, в отличие от денитрификации, образуются не N2 и окислы азота,
а аммиак.
Улетучивание аммиака происходит в щелочных условиях, когда
из иона аммония образуется аммиак. Такие условия довольно редки.
Характерным источником аммиака атмосферы служат пастбища, где
локально создается высокая концентрация аммиака за счет разложе-
разложения карбамида, а в просторечии - мочевины. Мочевина поступает при
разложении мочевой кислоты в орнитиновом цикле. Разложение мо-
мочевины осуществляет группа уробактерий, сейчас практически вы-
выпавших из внимания. Уробактерии, как, например Sporosarcinaxurea,
обладают уреазой, под действием которой карбамид распадается на
аммиак и углекислоту. В водной среде образуется бикарбонат аммо-
аммония - великолепное средство для подщелачивания среды, из-за чего
мочевину употребляют в импортной зубной пасте для борьбы с кари-
кариесом. Уробактерии в высокой степени алкалотолерантны и создают
среду с рН > 10, при которой аммиак и улетучивается.
В атмосфере аммиак как единственная летучая щелочь играет
очень важную роль, нейтрализуя SO2, превращающуюся в серную
кислоту кислотных дождей. Аммиак соединяется с ней, образуя кис-
кислый сернокислый аммоний NH4HSO4. Выпадение сернокислого ам-
аммония служит важным фактором удобрения омбротрофных, т.е.
133
имеющих дождевое питание, мест обитания, например лесо-болот-
ных экосистем и верховых болот.
В аэробных условиях, когда источником азота является ион ам-
аммония, происходит нитрификация. Она ограничена с кислой сторо-
стороны диссоциаций азотистой кислоты: в недиссоциированном виде
азотистая кислота представляет сильный яд и мутаген. Первая ком-
комбинированная культура аммонификатора и возбудителей первой и
второй фаз нитрификации, давшая нитрификацию органического
азота, была получена В.Л. Омелянским, но его учитель С.Н. Вино-
градский проворчал, что такой результат можно было предвидеть, и
дело дальше не пошло. На самом деле отношения здесь очень непро-
непростые. Впоследствии такие примитивные трофические цепи были ис-
использованы для обезнитрачивания, важнейшего процесса в очист-
очистных сооружениях, предотвращающего евтрофикацию сточными во-
водами. Здесь вслед за ступенями аммонификации и нитрификации
была включена ступень денитрификации.
Нитрификация служит мощным окислительным процессом.
При разложении органического вещества в очистных сооружениях
по потреблению кислорода она занимает второе место после образова-
образования углекислоты. Расход кислорода на нитрификацию в аэротенках
приходится учитывать как существенную часть затрат. Нитрифика-
Нитрификация осуществляется высокоспецифичными организмами-монотро-
фами в две фазы: I - окисление аммиака в высокореакционоспособ-
ный нитрит, который появляется лишь в ничтожных концентрациях
как промежуточный продукт; II - окисление нитрита в нитрат, кото-
который является термодинамически стабильной формой в окислитель-
окислительной атмосфере Земли. Если следовать термодинамическим равнове-
равновесиям, то доминирующей формой азота в биогенной кислородной ат-
атмосфере должен был бы быть нитрат. Однако нитрат накапливает-
накапливается лишь в условиях невозможности его использования растениями, в
геохимических ловушках. Такими ловушками служат селитряницы,
где разложение азотистых органических веществ не сопровождает-
сопровождается вымыванием нитрата или включением в биотический круговорот.
Высолы нитратов появляются при эвапоритовой тяге.
Деятельность нитрификаторов в целом ведет к переводу соеди-
соединений азота в растворимые формы. Однако по пути нитрификации
в качестве промежуточного продукта образуется N2O, которая мо-
может выделяться в атмосферу, составляя один из важных источников
этого газа в связи с легкостью обмена с атмосферой. Накопление
нитритов как при окислении аммиака, так и при нитратредукции мо-
может приводить к образованию в побочных реакциях весьма токсич-
токсичных нитрозаминов.
Денитрификация и образование окислов азота. Нитраты и ни-
нитриты становятся исходным материалом для образования окислов
134
азота N0 и N02, получивших общее обозначение N0^. Эти окислы
образуются и при высокотемпературных реакциях с участием воз-
воздуха - пожарах, электрических разрядах в атмосфере, но особенно
в двигателях внутреннего сгорания. Их участие в фотохимических
реакциях в тропосфере делает их исключительно важными загряз-
загрязнителями. Высшие окислы азота легко вымываются из атмо-
атмосферы.
Ключевая роль окислов азота в тропосферном фотохимическом
цикле как катализаторов окислительных процессов упоминалась вы-
выше. Образование N0 имеет существенное значение в химии тропо-
тропосферы. Измерение потоков N0 часто обнаруживает не только образо-
образование, но и поглощение, причем потоки очень варьируют в зависи-
зависимости от условий и времени. Обычная величина потока составляет
от -12 до +250 нг N/(m2 ¦ с). Образование N0, потребление, диффу-
диффузия регулируются независимо.
Главным источником N0 считается денитрификация. Образова-
Образование окислов азота было обнаружено еще в XIX в. при открытии де-
денитрификации. Дополнительными источниками служат нитратное
дыхание, свойственное очень широкому кругу организмов; гетеро-
гетеротрофная денитрификация; химическая дисмутация нитрита. Денитри-
фикаторы образуют N0 при восстановлении нитрита. Продукция
N0 при денитрификации на порядки превышает продукцию в аэроб-
аэробной зоне, однако требует строго анаэробных условий, при которых
обмен с воздухом затруднен. Сродство денитрификаторов к N0
очень высоко, и Км < 6нМ. Регуляция обмена N0 зависит не только
от регуляции на уровне клетки, но и от смены микробных популя-
популяций. Одним из определяющих факторов для состава популяций слу-
служит температурная адаптация. Повышение рН кислых почв сильно
увеличивает разнообразие NO-образующих организмов.
Закись азота N20 относится к числу постоянных микроприме-
микропримесей атмосферы с содержанием 0,3 ррт, причем в Северном полу-
полушарии ее содержание выше. Концентрация ее быстро возрастает за
последнее время, и это приписывают возрастающему употреблению
азотных удобрений. По своему парниковому эффекту N20 превосхо-
превосходит метан и углекислоту, однако наибольший интерес она представ-
представляет в связи с реакциями в стратосфере. N20 представляет химиче-
химически устойчивый газ со сравнительно высокой растворимостью.
Однако через тропосферу она проходит без больших потерь, так
как совершенно не участвует в фотохимических реакциях в тропо-
тропосфере. Вымывание атмосферными осадками для нее незначительно
по сравнению с высшими окислами азота. Для возбуждения N20 не-
необходим УФ с длиной волны меньше 340 нм. Тогда N20 реагирует с
активным кислородом, образующимся при фотолизе озона, и дает
две молекулы N0, которые участвуют в фотохимическом цикле, ве-
ведущем к понижению концентрации озона.
135
Концентрация N2O остается постоянной на протяжении тропо-
тропосферы и убывает лишь с переходом в стратосферу. Это иллюстри-
иллюстрирует отсутствие тропосферного стока. Перенос N2O через границу
тропосферы оценивается в 10-15 Тг/год, а суммарная мощность ре-
реакции образования N0 - 1-1,6 Тг/год, и соответственно время пре-
пребывания N2O в атмосфере оценивается в 100-150 лет.
Источники N2O находятся на поверхности Земли, и ими счита-
считаются наземная и океаническая денитрификация и нитрификация.
Для океанической эмиссии N2O поток рассчитывается по разности
концентрации в воде и атмосфере и оценивается величиной порядка
40 Тг/год. Дисбаланс между поглощением и выделением N2O соста-
составляет для океана около 5 Тг/год. В воде океана концентрация N2O от
0,2 до 1,2 мкг/л. Наиболее велика она на глубине 350-1000 м под фо-
тическим слоем в зоне температурного скачка, а наименее - у по-
поверхности. Отсюда следует, что N2O образуется на некоторой глуби-
глубине и поднимается к поверхности. Соотношение форм азота в его неор-
неорганических соединениях составляет в Атлантике N2O : NO2 : NOJ =
= 1 : 30 : 1000, а в Перуанском апвеллинге -1:1: 140.
Растворимость N2O в морской воде в зависимости от температу-
температуры при атмосферном давлении составляет:
Температура 0 5 10 15 20 25 30
Растворимость, 1,077 0,887 0,741 0,627 0,537 0,466 0,407
см3 газа/см3 воды
Источником N2O служит исключительно деятельность микроор-
микроорганизмов. Она образуется при нитрификации и денитрификации как
промежуточный продукт, причем при денитрификации возможно ее
потребление. Существует довольно много организмов, способных
использовать растворенную N2O как терминальный окислитель.
В аэробных условиях N2O окисляется перекисями в побочных реак-
реакциях, катализируемых микроорганизмами, например под воздейст-
воздействием каталазы.
В агросистемах около 20% азота может превращаться в N2O, и из
50 Тг/год удобрений может образовываться около 6 Тг/год N2O с еже-
ежегодным приростом концентрации за счет антропогенного источника в
0,4%. Уже давно было установлено, что количество N20,
выделяемой при денитрификации, зависит от нагрузки нитратом, при-
причем при повышении содержания NO3 выход N2O возрастает вслед-
вследствие предпочтительного восстановления нитрата. В почве концен-
концентрация нитрата может варьировать очень сильно по профилю. Избы-
Избыточная концентрация нитрата при применении азотных удобрений
свыше 300 кг азота/га применялась под хлопчатником и привела к
обогащению подземных вод нитратом. Другим фактором является
создание анаэробиоза. После дождя, когда поры аэрации блокиру-
блокируются водяными пробками, N2O выделяется в атмосферу.
136
Ключевым ферментом, определяющим обмен N2O, служит ре-
дуктаза закиси азота, которая встраивается в электронтранспорт-
ную цепь у нитрификаторов I фазы и у денитрификаторов. Это синий
Cu-содержащий белок. Свойства редуктазы закиси азота изучены
очень подробно6. Важнейшей ее особенностью является зависи-
зависимость от доступности меди как микроэлемента. Недостаток меди
сильно влияет на ход денитрификации.
N2O, подобно N2, рассматривается как соединение с тройной свя-
связью. Редуктаза закиси азота имеет открытый Р. Петровой специфи-
специфический газообразный ингибитор - ацетилен, и этот прием одно вре-
время широко использовался для оценки денитрификации: по образова-
образованию N2O при ингибировании ацетиленом можно судить о ходе
процесса денитрификации даже на фоне большой концентрации N2.
4.8. ВОДОРОДНЫЕ БАКТЕРИИ
Водорода в атмосфере очень мало, хотя он является постоянным
продуктом реакций анаэробного разложения органического вещест-
вещества и довольно устойчив фотохимически. Малое количество Н2, по-
поступающего в атмосферу, обусловлено A) его потреблением анаэ-
анаэробными гидрогенотрофными организмами и B) эффективным ба-
бактериальным фильтром, в котором участвуют аэробные гидрогено-
трофы, способные снижать концентрацию Н2 до уровня, на котором
он и наблюдается в атмосфере.
Источником Н2 служат газы, образуемые первичными анаэро-
анаэробами в кислотогенной (иначе - водородной) фазе анаэробного раз-
разложения органических веществ и диффундирующие из анаэробной
зоны в аэробную, где оптимальные условия для газотрофов созда-
создаются в области хемоклина и оксиклина. Это местообитание обычно
коррелирует с микроаэробными условиями. Другим источником Н2
могут служить фототрофные организмы, в том числе и цианобакте-
рии; здесь содержание О2 высоко. Выделение Н2 цианобактериями
может служить причиной того, что среди водородных бактерий мно-
много организмов, происхождение которых связано, скорее всего, с циа-
но-бактериальными сообществами.
Под водородными бактериями понимают организмы, способные
использовать реакцию 2Н2 + О2 = Н2О в качестве энергодающей.
Водородные бактерии были открыты в начале XX в., вскоре вслед
за открытием хемосинтеза, и подробное описание их газообмена бы-
было дано в 1910 г. А.Ф. Лебедевым, который сформулировал вывод о
том, что "энергетический процесс идет независимо от ассимиляции
углекислоты". Почти все водородные бактерии способны использо-
6Berks B.C., Ferguson SJ., MoirJ.W.B., Richardson D.Y. Enzymes and associated elec-
electron transport systems that catalyze the respiratory reduction of nitrogen oxides and
oxyanions // Biochim. et biophys. acta. 1995. Vol. 1232. N 3. P. 97-173.
137
вать помимо водорода разнообразные органические вещества, и
первый описанный организм не случайно был назван "Bacillus рап-
totrophus" - "всеядный", что резко отличало его от строго специали-
специализированных хемосинтетиков, описанных С.Н. Виноградским. Водо-
Водородные бактерии получили таксономическое название
"Hydrogenomonas", и в качестве тривиального термин "гидрогеномо-
нады" вполне употребим и сейчас. С начала 1960-х годов Х.Г. Шле-
гель избрал один штамм водородных бактерий, Alcaligenes
("Hydrogenomonas") eutrophus H-16, в качестве универсальной моде-
модели для исследования биохимии хемосинтетиков, своего рода Е. coli, в
этой физиологической группе. В результате этой работы, проведен-
проведенной с участием многих студентов Института микробиологии в
Гёттингене, водородные бактерии стали одними из наиболее полно
и всесторонне изученных. В СССР для аналогичной цели был ис-
использован штамм A. eutrophus Z-1 как модель возможного производ-
производства белка на водороде за счет электролиза. Широкому изучению
водородных бактерий предшествовала разработка методов культи-
культивирования микроорганизмов в определенной газовой среде, потре-
потребовавшая изменения многих навыков микробиологов. Водород-кис-
Водород-кислородная смесь очень взрывоопасна и недаром заслужила название
"гремучего газа". Работа даже с немногими литрами ее требует спе-
специальных мер предосторожности. При обеспечении газообмена
газ-жидкость водородные бактерии растут исключительно быстро
и достигают высоких плотностей, что сделало их интересными био-
биотехнологическими объектами. Угроза взрыва газовой смеси может
быть сильно сокращена применением электролиза 2Н2О = 2Н2 + О2
вплоть до культивирования в электролизере. Технология культиви-
культивирования водородных бактерий в массовом масштабе была разрабо-
разработана в Институте биофизики АН СССР в Красноярске.
В общих чертах обмен гидрогеномонад включает реакцию гид-
рогеназы, сопряженную с мембранной электрон-транспортной це-
цепью и синтезом АТФ с О2 как терминальным акцептором, и в анабо-
анаболизме - автотрофную ассимиляцию СО2 через цикл Кальвина как у
фотоавтотрофов. Таким образом, это самый простой тип обмена,
поскольку газы не требуют транспортных механизмов и продукт ре-
реакции нетоксичен. Все гидрогеномонады способны расти органо-
трофно, причем излюбленными веществами служат органические
кислоты и другие продукты первичных анаэробов.
Впоследствии оказалось, что такой тип обмена, как у гидрогено-
гидрогеномонад, довольно широко распространен среди аэробных микроорга-
микроорганизмов, принадлежащих к разным родам. Необходимым условием
служит наличие "потребляющей" гидрогеназы, которая есть у мно-
многих азотфиксирующих организмов, например у ризобиев. Большин-
Большинство водородных бактерий относится к грамотрицательным псевдо-
псевдомонадам, часть из которых была переклассифицирована, один из
родов носит характерное название Acidovorax. Относительно мало
138
водородных бактерий среди грамположительных организмов, хотя
они есть среди нокардий, артробактеров и термофильных бацилл,
как Bacillus schlegelii. Сейчас проверка на способность к автотроф-
ному росту с Н2 должна входить в список тестов, используемых для
характеристики каждого организма. Органотрофный рост боль-
большинства водородных бактерий не дает ясной картины, в какой мере
Н2 может быть использован как дополнительный источник энергии
организмами, не обладающими автотрофной ассимиляцией СО2.
Водородные бактерии способны существовать в широком диапа-
диапазоне физико-химических условий, но внимание привлекали мезо-
фильные нейтрофилы, к которым относится большинство описанных
организмов. Был найден ряд термофильных форм, от умеренно тер-
термофильных до бацилл, растущих при 70 °С. Открытые совсем недав-
недавно психроактивные водородные бактерии, которые важны для биоце-
биоценозов тундры России, принадлежат к артробактерам и Acidovorax.
Самостоятельную группу составили экстремально термофиль-
термофильные водородные бактерии, обнаруженные в гидротермах. Одна из
бактерий, выделенная из термофильного циано-бактериального ма-
мата на Камчатке, получила название Calderobacterium. Это длинная
палочка, растущая при 80 °С, масса которой окрашена в красный
цвет цитохромом с. При высокой температуре растворимость кисло-
кислорода резко падает и организм вынужден иметь эффективный меха-
механизм его использования. Организм оказался строгим литоавтотро-
фом и, в отличие от всех других водородных бактерий, не использу-
использует органические вещества. Одновременно в Японии был выделен
сходный организм, получивший название Hydrogenobacter. Необыч-
Необычным оказался у него способ ассимиляции СО2 посредством восстано-
восстановительного цикла трикарбоновых кислот. Третий организм, относя-
относящийся к той же группе экстремально термофильных водородных ба-
бактерий, получил название Aquifex и оказался принадлежащим к самой
глубокой ветви эубактерий на 16S рДНК филогенетическом дереве.
Способность к окислению № имеется у ацидофильных термо-
термофильных архей из Sulfolobaceae. О способности к окислению Н2 у аци-
ацидофильных протеобактерий группы Acidiphilium пока не сообщалось.
Алкалофильные водородные протеобактерий недавно выделены из
содовых озер с рНЮ и высокой минерализацией. Таким образом, во-
водородные бактерии по своим экофизиологическим характеристикам
способны покрыть почти все поле физико-химических условий.
4.9. КАРБОКСИДОБАКТЕРИИ
Содержание СО в атмосфере имеет зимний максимум и летний,
минимум, объясняемые фотохимическими процессами. Содержание
СО колеблется в пределах 0,1-0,3 ppmv. При концентрации
1000 ррт СО летален для человека, а при 100 ppmv вызывает отра-
отравление. Допустимой считается концентрация ниже 9 ppmv.
139
CO - в высокой степени токсичный бесцветный газ без запаха,
имеющий порог воспламенения 15-71% в воздухе и 16-93% в кисло-
кислороде. В зависимости от температуры его растворимость составляет:
Температура, °С 0 24 30 60
Растворимость, мкл/мл; 35,37 21,42 19,98 14,88
Важнейшие реакции СО и изменение их свободной энергии Гиб-
бса AG/0 (кДж) следующие:
2СО + О2 = 2СО2 -514,21
2СО + О2 + 2Н2О = 2Н2СО3 -497,42
СО + Н2О = НСООН +2,08
СО + NOJ = СО2 + NO2 -183,0
4СО + SO2." = S2- + 4СО2 -198,3
СО + Fe3+ + H2O = СО2 + 2Fe2+ + 2Н+ -168,46
НСООН + NOJ = СО2 + NO2 + Н2О -185,10
СО + Н2О = СО2 + Н2 -19,93
СО + 2NO = СО2 + N2O -326,06
4НСООН = 4СО2 + 4Н2 -88,03
Почвы обычно представляют сток для атмосферной СО, причем
в них идут одновременно процессы поглощения и выделения газа.
Наблюдаемая концентрация СО - равновесная для этих процессов.
Выделение СО зависит от содержания органического вещества, и
считается, что оно обусловлено абиогенной химической реакцией, в
то время как поглощение идет за счет биологических процессов.
Выделению СО способствуют повышенная температура и сухость
почвы. Содержание СО в почвенном воздухе коррелирует с суточ-
суточным ходом температуры, увеличиваясь днем и уменьшаясь ночью.
В плохо аэрируемых горизонтах происходит образование СО, и со-
содержание ее возрастает до 3-6 ppmv. Известно, далее, образование
СО растениями при освещении, которое, однако, имеет очень не-
небольшую величину. Рассматривая проблему СО, нужно признать,
что естественные концентрированные источники этого газа неиз-
неизвестны. Часто СО рассматривают как чисто антропогенную примесь
в атмосфере. Основным источником служит сжигание топлив, ответ-
ответственное за 3/4 эмиссии, пожары. Организмы, окисляющие СО, назы-
называют карбоксидобактериями, или карбоксидотрофами. Они предста-
представлены хемолитоавтотрофами, способными катализировать окисле-
окисление СО в СО2, использовать энергию этой реакции, ассимилировать
углекислоту, быть устойчивыми к высокой концентрации СО.
Предположение, что СО служит источником энергии для микро-
микроорганизмов, было высказано в начале XX в. в лаборатории Бейе-
ринка в связи с изучением воздушного питания и олигокарбофилии.
Карбоксидобактерии были описаны Кистнером в 1953 г. под назва-
названием Hydrogenomonas carboxydovorans, но их культура утеряна.
140
Потом карбоксидобактерии были изолированы и описаны Г.А. Завар-
зиным, Э.У. Санжиевой и А.Н. Ножевниковой как Seliberia carboxy-
dohydrogena, Pseudomoans carboxydoflava, Pseudomonas gazotropha,
Comamonas compransoris, Achromobacter carboxydus, позднее к ним
добавили Arthrobacter sp.ll/х, термофильный Bacillus schlegeliP. За-
Затем последовало новое уточнение родовой принадлежности, и они
получили новые родовые названия. Вследствие сходной процедуры
выделения все они более или менее одинаковы физиологически, хо-
хотя принадлежат к разным родам. Все выделенные штаммы подавля-
подавлялись 2,5% NaCl, что указывает на их принадлежность к наземным
организмам. Но попытки выделения из моря, насколько автору из-
известно, и не делались.
Все известные карбоксидобактерии были выделены в культуру в
атмосфере, содержащей десятки процентов СО. Такая концентрация
создается в исключительных случаях. В атмосфере СО относительно
мало, и эти организмы не обладают достаточным сродством к ней, что-
чтобы обеспечить биологический сток из атмосферы, в отличие, напри-
например, от метанотрофов или водородных бактерий. Более того, метанол
служит сильным ингибитором реакции фермента карбоксидобактерии,
реагируя с пятивалентным молибденом реакционного центра.
Аэробные карбоксидобактерии могут расти и как водородные
бактерии, это факультативные автотрофы, способные использо-
использовать широкий круг органических соединений. Можно сказать, что
это водородные бактерии, обладающие дополнительной способно-
способностью к использованию СО. Эта способность обусловлена фермен-
ферментом СО-дегидрогеназой, сейчас подробно исследованной В. Свет-
Светличным и О. Мейером. Он определяет возможность использования
СО в энергетическом обмене аэробных карбоксидобактерии. Этот
фермент совершенно отличен от анаболической СО-дегидрогеназы
анаэробов. У большинства аэробов СО является мощным ингибито-
ингибитором терминального участка дыхательной цепи. Устойчивость к СО
карбоксидобактерии определяется разветвленной цепью переноса
электронов. Свободные водород или формиат не являются проме-
промежуточными продуктами окисления СО как можно было бы думать
по реакциями
СО + Н2О = СО2 + Н2 или СО + Н2О = НСООН.
Аэробные организмы, окисляющие СО, характеризуются очень
медленным ростом, обычно инкубация требует 1 месяц. Часто они
угнетаются высокой концентрацией фосфата, но тем не менее сре-
среда, рекомендованная для их культивирования, содержала 10 г/л фос-
фосфатных солей!
Карбоксидобактерии используют СО как наихудший субстрат,
предпочитая органические вещества и водород. Они способны к мик-
7 Заварзин Г.А. Водородные бактерии и карбоксидобактерии. М.: Наука. 1978.
141
сотрофному росту, при котором анаболизм осуществляется за счет
органических веществ, а окисление СО дает дополнительную энер-
энергию. При этом ни СО, ни СО2 не дают радиоактивной метки в био-
биомассе.
Ферментом СО:акцептор оксидоредуктазой занялись очень де-
детально, т.к. этот фермент использовали для удаления СО, например
в фильтрах сигарет. Источником кислорода при окислении СО слу-
служит вода. Фермент работает с искусственными акцепторами элект-
электрона, такими, как метиленовый синий. Он очень специфичен по суб-
субстрату и способен лишь медленно окислять НАДН. Кажущаяся Км
составляла 53 мкМ СО для P. carboxydovorans и 63 мкМ СО для
P. carboxydohydrogena. Молекула фермента содержит две молекулы
ФАД, две - молибдена, 8 железо-серных групп. Фермент активиру-
активируется селенитом. Локализован фермент на внутренней стороне цито-
плазматической мембраны.
Устойчивость к СО составляет иную часть проблемы и связа-
связана прежде всего с терминальными оксидазами. Имеется разветв-
разветвленная цепь цитохромов на уровне цитохромов b и о. В разных
условиях культивирования поток электронов идет по разным мар-
маршрутам.
Исследование возможности деятельности карбоксидобактерий в
удалении СО из атмосферы показало, что их сродство к СО слиш-
слишком низко и эти организмы приспособлены к токсичной концентра-
концентрации СО. Такие концентрации СО образуются при парогазовой кон-
конверсии углеродистых топлив и метана или же при подземных пожа-
пожарах.
Биологическое окисление СО приписывается неспецифическо-
неспецифическому окислению. Неспецифическое окисление СО идет под действием
метанмоноксигеназы (ММО) у метилотрофов Methylococcus capsula-
tus, Methylosinus trichosporium. У этих организмов окисление требует
НАДН, генерируемого в обмене.
Окисление СО в анаэробных условиях по реакции СО + Н2О =
= СО2 + Н2 с очень маленьким выходом энергии в -19,93 кДж/моль
было открыто в 1990 г. в лаборатории Заварзина В. Светличным8.
Организм был обнаружен в гидротермах Курильских островов и
оказался способен проводить эту реакцию при 70 °С. Он получил
название Carboxydothermus и сейчас стал одним из наиболее изу-
изученных организмов в связи с полным превращением СО в Н2 и
возможностью замены обратимой реакции парогазовой конвер-
конверсии при производстве водорода биотехнологическим процессом.
Т.Г. Соколовой удалось выделить из гидротерм целый ряд орга-
организмов.
% Светличный В.А., Соколова ТТ., Герхардт И., Заварзин Г.А. Новая группа
анаэробных термофильных карбоксидобактерий // ДАН СССР. 1990. Т. 314.
С. 742-744.
142
4.10. ЛЕТУЧИЕ УГЛЕВОДОРОДЫ
И БАКТЕРИАЛЬНЫЙ ФИЛЬТР
Вынос на поверхность газов глубинного происхождения
В.И. Вернадский назвал "газовым дыханием Земли". Понятие "бак-
"бактериальный фильтр" по отношению к горючим углеводородным га-
газам миграционного потока из подпочвенных осадочных пород было
введено Г.А. Могилевским9, в 1937-1939 гг. установившим окисле-
окисление этих газов в почвенном слое. Впоследствии это явление было
использовано им для поиска нефтегазовых месторождений, над ко-
которыми особенно активно развивались бактерии, способные исполь-
использовать высшие гомологи метана. Окисление метана метанотрофами
связано с циклом Зе'нгена, идущим в местах разложения органиче-
органического вещества, при котором высшие гомологи метана не образуют-
образуются. В этом отношении окисление метана не является процессом, при-
приуроченным к газовым аномалиям. Иное дело представляет окисле-
окисление летучих высших гомологов метана углеводородоокисляющими
микроорганизмами, которое оказалось приурочено к глубинным ис-
источникам этих газов. В газовых месторождениях с содержанием ме-
метана 80-90% углеводороды Q-Cs составляют 1—15%, причем их
концентрация возрастает с глубиной. В попутном нефтяном газе
сумма тяжелых углеводородов составляет 25^10%. Над газовыми и
нефтяными месторождениями образуются аномалии в содержании
углеводородов в газовой фазе пород и почвы. Аномалии приуроче-
приурочены к потокам газов из глубины. Массоперенос из глубин на дневную
поверхность осуществляется по зонам трещиноватости пород за
счет фильтрационного и диффузионного процессов. Необходимым
условием развития окислительного бактериального фильтра служит
доступ кислорода. В почве и рыхлых породах обеспечивается доступ
атмосферного кислорода из почвенного воздуха или же переносимо-
переносимого подземными водами. В этой зоне смешения встречных газовых
потоков и формируется микробное сообщество окислительного ба-
бактериального фильтра из микроорганизмов, использующих летучие
углеводороды. Наиболее благоприятными для жизнедеятельности
организмов, окисляющих неметановые летучие углеводороды, слу-
служат подпочвенные аэрируемые горизонты до уровня грунтовых вод
и зоны неотектонической трещиноватости. Обычные пластовые
температуры для нефтегазоносных бассейнов не превышают 100 °С,
но область развития окисляющих углеводороды организмов бакте-
бактериального фильтра находится у нас в стране в зоне температур ме-
менее 10 °С, а в подземных водах 4 °С.
Метанокисляющие организмы способны соокислять большое
число летучих веществ в неростовом процессе. Продукты неполно-
9 Могилевский Г.А. Микробиологический метод поисков газовых и нефтяных
залежей. М.; Л.: Госпромтехиздат, 1953. 56 с.
143
го окисления этих соединений могут быть токсичными для метило-
трофов и обусловливают развитие в сообществе с ними организмов-
спутников, завершающих окисление и устраняющих токсическое
действие. Поэтому сообщество оказывается более устойчивым, чем
чистые культуры метанотрофов, к примесям в природном газе.
Ю.Р. Малашенко особое внимание обратил на соокисление этана с
попутным образованием ацетальдегида и других продуктов непол-
неполного окисления, представляющих заведомо токсичные вещества.
Окисление летучих гомологов метана осуществляет широкий
круг микроорганизмов, для которых нет необходимости в синтезе
С-С связи и которые могут совмещать продукты окисления росто-
ростового субстрата с центральным метаболизмом; они относятся к груп-
группе аэробных органогетеротрофов, характеризуемых возможностью
использования углеводородных субстратов. Организмы, использую-
использующие летучие углеводороды, довольно разнообразны и включают
ряд грамположительных аэробов. Способность к окислению углево-
углеводородов у них распределена неравномерно: часть штаммов довольно
специфична по отношению к субстрату, окисляя один-два н-алкана,
часть обладает более широкими возможностями. Ключевой реакци-
реакцией служит окисление концевой метильной группы. Наибольшее вни-
внимание было уделено организмам, окисляющим пропан и бутан. Ти-
Типичными, но не единственными организмами этой функциональной
группы являются родококки.
Организмы, окисляющие углеводороды, способны использовать
их крайне низкие концентрации. Минимальная концентрация пропа-
пропана оценивается в 0,001%, и для летучих углеводородов минимум на-
находится на уровне ниже десятых частей на миллион. Растворимость
газообразных н-алканов в воде мала, составляя до 60 мг/л.
Многочисленные бактериальные съемки, проведенные на тер-
территории СССР, показали, что пропан- и бутанокисляющие бактерии
практически не обнаруживаются в местах разложения органическо-
органического вещества. Вместе с тем они приурочены к нефтеносным районам
как в почве, так и подземных водах по всей территории страны вне
зависимости от климатических поясов. Развитие организмов, ис-
использующих неметановые м-алканы, в природных биоценозах воз-
возможно в основном за счет газов, мигрирующих из нефтегазоносных
залежей. В природоведческом отношении существенно, что весь ми-
миграционный поток углеводородных газов может быть полностью
окислен бактериями, образующими мощный биологический окисли-
окислительный фильтр в зоне нефтяных и газовых месторождений. Описа-
Описание мест распространения пропан- и бутанокисляющих организмов
на территории СССР дано в обзоре10. В качестве индикаторных ор-
организмов используют культуры Rhodococcus rhodochrous и R. ruber.
10 Оборин А.А., Стадник Е.В. Нефтегазопоисковая геомикробиология. Екате-
Екатеринбург, 1996.
144
Первоначально оценивали рост этих организмов при посеве в ста-
стаканчики, которые ставили в эксикаторы с газовой смесью. Затем
стали применять посев шприцем в пенициллиновые флаконы на
элективные среды с пропаном, что дает лишь качественный ответ,
вполне достаточный в большинстве случаев. Наконец, в последнее
время разработаны иммунофлуоресцентные методы учета родокок-
ков в природных средах.
4.11. МИКРООРГАНИЗМЫ
КАК АЭРОЗОЛЬНОЕ ЗАГРЯЗНЕНИЕ АТМОСФЕРЫ
Микроорганизмы, в особенности их споры, специально предна-
предназначенные для распространения воздушным путем, представляют
типичное аэрозольное загрязнение атмосферы. Бактериальные аэ-
аэрозоли атмосферы рассматриваются как взвешенные мигрирующие
частицы или капли диаметром менее 20 мкм. В воздушной среде ми-
микроорганизмы не развиваются, и нет доказательств их размножения,
например, в дождевых каплях, хотя одно-два деления допускаются.
Однако перенесенные ветром микроорганизмы могут служить ис-
источником инфекции - эпифитотии или эпизоотии. Именно этот эпи-
эпидемиологический аспект привлекал особое внимание в микробиоло-
' гии атмосферы. Поэтому методом определения числа взвешенных
бактерий было проращивание их на чашках Петри, экспонирован-
экспонированных для оседания частиц на их поверхности. Однако большинство
бактерий в атмосфере либо мертвы, либо относятся к организмам в
так называемом "некультивируемом состоянии", т.е. не прорастаю-
прорастающим на средах. Численность бактериального аэрозоля составляет в
лесу и над полем 300-1500 колониеобразующих единиц на кубометр,
возрастая до десятков тысяч и миллионов при сельскохозяйствен-
сельскохозяйственных работах. Среди колоний бактерий, собранных днем, больше
пигментированных форм. Большинство бактерий аэрозолей грам-
положительные. Средний размер частиц бактериального аэрозоля
составляет 3,6 мкм вследствие того, что бактерии прикрепляются к
частицам пыли, часто растительного происхождения. Считается,
что большинство бактерий смывается в воздух с листьев и в мень-
меньшей степени - с поверхности почвы. Выживание бактерий в аэрозо-
аэрозоле является функцией размеров частиц. Оно определяется также
воздействием повышенной температуры, солнечного света, дегидра-
дегидратации. Одним из механизмов снижения повреждений является нали-
наличие трегалозы, способствующей устойчивости мембран. Числен-
Численность бактерий в воздухе варьирует в течение дня, что определяется
микрометеорологическими условиями. Имеется минимум численно-
численности ночью, пик - при восходе, обусловленный подъемом воздуха от
нагреваемой поверхности, полуденный максимум и вечерний спад к
ночному минимуму, указывающий на роль термодиффузии. Годовой
145
цикл для умеренных широт, прослеженный для Москвы и Монреаля,
имеет четкий зимний минимум и летний максимум, обусловленные
снеговым покровом и летней пылью. Естественным источником для
культивируемых микроорганизмов служат прежде всего раститель-
растительность и почва.
Удаление частиц из атмосферы зависит от их размеров. Сухое
отложение имеет порог около 1 мкм: частицы меньшего размера ве-
ведут себя как газы, частицы большего размера оседают вследствие
тяжести, а частицы в 1 мкм имеют наименьшую скорость осажде-
осаждения. Еще более отчетливо это вырисовывается для влажного осаж-
осаждения. Коэффициент вымывания (доля частиц, вымываемых за 1 с)
составляет до 10-2 для частиц меньше 0,1 мкм, которые быстро коа-
коагулируют, и для частиц более 10 мкм, оседающих под действием си-
силы тяжести, в обоих случаях вымывание происходит за часы. Части-
Частицы около 1 мкм имеют коэффициент вымывания 10~5, и их время
пребывания в атмосфере составляет недели. Микроорганизмы попа-
попадают в размерную категорию с наибольшим временем пребывания
в состоянии аэрозоля.
4.12. Эволюция состава атмосферы
Эволюция состава атмосферы и океана тесно взаимосвязаны и проходили,
главным образом, до начала геологической истории - 4,6-3,8 млрд лет назад.
Холодная аккреция Земли предполагает дегазацию после достижения ею кри-
критических размеров вследствие разогревания. Основным аргументом служит
малое содержание инертных газов, прежде всего неона, которые не могли быть
захвачены из космоса. Образование Земли при ударе планетезималей предпо-
предполагает немедленную дегазацию вследствие разогрева при ударе с выделением
воды и газов. На период аккреции и дегазации отводится 10-100 млн лет с разо-
разогревом до 1500 К (близь температуры плавления силикатных пород). В этот пе-
период атмосфера могла содержать пары воды и терять водород, как это про-
произошло на Венере. Есть предположение, что часть воды поступила с падением
комет. Краткий обзор гипотез показывает, что происхождение атмосферы и
океана до конца метеоритной бомбардировки остается в области гаданий.
После окончания бомбардировки произошла конденсация пара в океан, а в
атмосфере остались N2, CO2, СО и частично Н2 при отсутствии в приземном
слое О2, следы которого могли быть в стратосфере за счет фотолиза; модель
дает для О2 концентрацию 10~14 современного уровня. Водород мог диссипиро-
вать в космос. Особенно интересна судьба СО2, которая считается основным
продуктом дегазации на основании аналогии с современным вулканизмом. По
меньшей мере 1/3 СО2 из имеющихся сейчас 1023 г углерода карбонатов (Буды-
ко и др., 1985), составляющих существенную часть осадочных пород в
2,4 • 1024 г, могла быть в первоначальной атмосфере и постепенно удалилась из
нее реакцией с силикатами в субаэральных условиях на континентах. Если же
Земля была покрыта водой, то процесс мог затянуться на сотни миллионов лет.
Моделирование термического режима поверхности Земли с учетом на 1/3 мень-
меньшей светимости раннего Солнца дает исходную температуру для 1 атм около
40 °С и 85 °С для 10 атм давления. Существование углекислотной атмосферы не
146
может быть ни отвергнуто, ни подтверждено геологическими свидетельствами
из-за отсутствия геологической летописи.
При меньшей светимости Солнца температура (остывшей!) поверхности
Земли должна была обеспечиваться в сотни раз более высоким содержанием
СО2, в десятые доли атмосферы в позднем архее. Роль другого парникового га-
газа - метана, который мог продуцироваться из геологических источников Н2 и
СО2 метаногенными гипертермофильными археями или абиогенно, пока не об-
обсуждается.
Для формирования сульфатов Кастинг1 предположил серию фотохимиче-
фотохимических реакций с участием О и ОН радикалов, источники которых в аноксической
атмосфере неясны, но вполне могут быть получены по аналогии с атмосферой
Венеры, содержащей 96% СО2, 3,4% N2, и в ррт: 69 О2, 18,6 Аг, 186 SO2. Объем
воды океана предполагается близким к современному с самого начала. При ма-
максимально эффективном фотолизе содержание сульфатов в первичном океане
должно было быть в 30 раз меньше.
При высоком давлении СО2 океан можно считать "содовым" в том смысле,
что основными анионами были HCOJ иС1 , но, в противоположность мнению
Кемпе и Дегенса2, не из-за высокого рН, который привел бы к немедленному
удалению СО2 из атмосферы с образованием хемогенных карбонатов и, следо-
следовательно, охлаждению в связи с уменьшившимся парниковым эффектом, а по
доминирующему аниону. Для углекислотной атмосферы рН океана предполага-
предполагается несколько ниже рН6.
Предложенная Кастингом картина ранней эволюции океана и гидросферы
не является единственной. Ключевым событием для выбора сценария служит
прекращение бомбардировки Земли. По данным для Луны пик бомбардировки
приходился на 4,2-3,9 млрд лет назад, а древнейшие породы Исуа и Акилеа не
несут следов бомбардировки и представляют слоистые железистые кварциты
без следов падения астероидов. Аккреция Земли в разных вариантах гипотез
предполагает разогрев ее поверхности от полного расплавления с последующей
послойной дифференциацией до более растянутых во времени столкновений,
которые вели лишь к частичному и временному разогреву и дегазации в горя-
горячих точках. Начало накопления воды могло иметь место при достижении про-
то-Землей половины радиуса от современной. Различие в исходных моделях
еще раз подтверждает необходимость двигаться от известного вглубь, а не на-
наоборот. Предположение Г. Аррениуса, что Земля могла стать обитаемой мик-
микроорганизмами во время или вскоре ее образования, основывается на гипотезе
гетерогенной аккреции3.
Представление о том, что в архей 3,8 млрд лет назад Земля вступила с уже
сформированным биогеохимическим циклом, аргументируется Щидловским на
основе изотопии карбонатов и керогена. Сорг составляет 0,5-0,6% из 2,4 • 1024 г
осадочных пород, что составляет 1,3 • 1022 г, а карбонатов в 40 раз больше. В те-
течение геологического времени Сорг колебался относительно среднего 0,5% не-
незначительно (со значительным превышением в сланцах), что указывает на по-
1
1 Kasting J.F. Evolution of the Earth's atmosphere and hydrosphere. Hadean to recent //
Organic geochemistry: Principles and applications / Ed. M.H. Engel and S.A. Marko.
N.Y., L.: Plenum press, 1993. P. 611-621.
2 Kempe S., Degens E.T. An early soda ocean? // Chem. Geol. 1985. Vol. 53. P. 95.
3 Arrhenis C, Upland A. Accretion of Moon and Earth and the emergence of life // Ibid.
2000. Vol. 169. P. 69-82.
147
стоянство отношения между фототрофной ассимиляцией СО2 и скоростью за-
захоронения Сорг в седиментационном процессе. Это относится и к Исуа с содер-
содержанием Сорг 0,6%. Продуктивность бентосных систем в циано-бактериальных
матах составляет 8-12 г Сорг/(м2 • сут). Отсюда можно полагать продуктивность
земной системы примерно постоянной в пределах зависимости от влажной по-
поверхности и находящейся близко к насыщению.
Автотрофная ассимиляция ведет к облегчению изотопии Сорг первичными
продуцентами в зависимости от первой реакции используемого пути
на 20-30 %о. Сопоставление изотопного состава Сорг на протяжении всей истории
Земли свидетельствует об основном пути его происхождения от обладающих
рибулозобисфосфатным путем автотрофов, что было установлено уже в 1975 г.
Отклонения от среднего предполагают, по мнению Шидловского, включение
метанотрофии, ведущей к значительному облегчению изотопного состава Сорг,
однако по меньшей мере одно событие (Фортескью 2,7 млрд лет назад) имело
место до накопления кислорода в атмосфере. Тяжелый Сорг Исуа приписывали
метаморфизации, но последующие определения (Mojzsis et. al., 1999) показали
присутствие обычного для биогенного материала изотопного облегчения.
Относительно изотопии серы Шидловский убежден, что фототрофное
окисление серы предшествовало оксигенному фотосинтезу. Поскольку сульфат
присутствовал в отложениях архея до развития окисленной атмосферы, то био-
биогенным источником его могли бы быть только аноксигенные фототрофы.
Рекомедуемая литература
1. Будыко М.В., Ронов А.Б., Яншин АЛ. История атмосферы. Л.: Гидрометео-
издат, 1985. 208 с.
2. Грегори Ф. Микробиология атмосферы. М.: Мир, 1964. 376 с. '
З.Заварзин Г.А. Литотрофные микроорганизмы. М.: Наука, 1972. 324 с.
А.Заварзин Г.А. Водородные бактерии и карбоксидобактерии. М.: Наука,
1978.
5.3аварзин Г.А. Бактерии и состав атмосферы. М.: Наука, 1984. 199 с.
6. Исидоров В.А. Органическая химия атмосферы. СПб.: Химия, 2001. 352 с.
1. Исидоров В.А. Экологическая химия: (Учебник для вузов). СПб.: Химия,
2001.304 с.
8. Круговорот газов в природе. М.: Наука, 1979.
9. Круговорот углерода на территории России /НТП "Глобальные изменения
природной среды и климата". М., 1999.326 с. (Избр. науч. тр. по пробл. "Гло-
"Глобальная эволюция биосферы. Антропогенный вклад").
10. Graedel T.A., Crutzen P.J. Atmospheric change: An Earth system perspective. N.Y.:
Freeman, 1992.
W.Lighthart B. The ecology of bacteria in the alfresco atmosphere // FEMS Microbiol.
Rev. 1997. Vol. 23. P. 263-274.
12. Mojzsis S.J., Arrhenius G., McKeegan K.D. et al. Evidence for life on Earth before
3800 millions years ago // Nature. 1996. Vol. 384. P. 55-59.
13. Schidlowski M. The initiation of biological processes on Earth: Summary of empiri-
empirical evidence // Organic geochemistry: Principles and applications / Ed. M.H. Engel,
S.A. Marko. N.Y.; L.: Plenum press, 1993.
14. Schidlowski M. Sedimentary carbon isotope archives as recorders of early life:
Implications for extraterrestrial scenarios // Fundamentals of life. 2002. P. 308-329.
Лекция 5
МИКРОБИОЛОГИЯ ВОДНЫХ СРЕД
ОБИТАНИЯ
L
5.1. ВОДОЕМЫ И ГИДРОЛОГИЧЕСКИЙ ЦИКЛ
Бактерии не способны к жизнедеятельности вне жидкой воды.
Этот основной постулат бактериологии определяет существование
бактерий в определенных пределах температуры и окислительно-
восстановительного потенциала. Реакции, несовместимые с водой,
т.е. находящиеся за пределами термодинамического поля устойчи-
устойчивости воды, не могут служить бактериям в качестве источника энер-
энергии для жизнедеятельности. Это исключает из рассмотрения множе-
множество реакций, связанных с твердой фазой; здесь реакции могут идти
только на поверхности и обусловлены переходом вещества в раство-
растворенное состояние. Это важно для разложения гидрофобных соеди-
соединений (из которых внимание сконцентрировалось на углеводородах,
но в природе гораздо важнее липиды) или минералов вроде серы.
В почве, в горных породах бактерии развиваются в поровом про-
пространстве, заполненном водой. В воздухе бактерии только пережива-
переживают в виде аэрозоля как стадия расселения. Таким образом, вся эколо-
экология микроорганизмов - это деятельность микробов в жидкой воде и
вызываемые ими в воде химические изменения. Поэтому гидросферу
можно считать истинной средой обитания микроорганизмов.
Поведение воды на земной поверхности определяется гидроло-
гидрологическим циклом (рис. 5.1). Движущей силой цикла служит атмо-
атмосферный перенос влаги за счет испарения с поверхности океана;
атмосферный гидрологический цикл оказывается определяющей
частью климатической системы и ее зональности; он служит связу-
связующим звеном океанической и континентальной систем. Гидрологи-
Гидрологический цикл работает как перколятор: испаряющаяся вода приходит
в равновесие с газами атмосферы, перераспределяется с потоками
воздуха в атмосфере и, "почти дистиллированная", выпадает на по-
поверхность суши; здесь она выщелачивает горные породы, создавая
кору выветривания и приходя в неполное равновесие с минералами
поверхностных пород, возвращается в океан в виде речного стока,
обогащенного растворенными веществами и взвесью минералов
("твердым стоком"), перерабатываемыми в бассейнах седиментации -
основной зоне формирования осадочных пород. Под действием
149
Циркуляция
атмосферы
'0,013 Мкм3
Звапотранспирация
71 ккм3
Газы
дегазации
Океан
1350 Мкм3
ц Гидротермальный
фг.юнч
Под земные воды '<
8" Мкм3
1 Подводный
ккм3/год - потоки
Мкм3 -резервуары
Рис. 5.1. Гидрологический цикл
силы тяжести вода стекает вниз по поверхности земли, занимая де-
депрессии и обусловливая этим транспорт растворенных веществ.
Основные резервуары воды: 1) океан', 2) наземные водоемы, 3) во-
водотоки-реки, 4) амфибиальные ландшафты, 5) подземная гидросфе-
гидросфера, 6) криосфера. Океан оказывается ловушкой для стока (вплоть до
времен геологического рецикла) и источником атмосферной влаги.
Распределение воды по резервуарам приведено в табл. 5.1.
Меньшее внимание привлекали подземные воды, хотя они име-
имеют первостепенное значение для установления химизма поверхност-
поверхностных вод. Кроме того, подземная гидросфера рассматривается как
местообитание организмов, взаимодействующих с потоками эндо-
эндогенного вещества из глубин Земли. В особенности интересно пове-
поведение микробиоты в осадочной оболочке.
Обмен с подземными водами имеет первостепенное значение
для установления химического состава вод. Здесь выделяются воды
подземного стока и гидротермальные воды зоны рифтогенеза. Дви-
Движущим механизмом служит атмосферный перенос влаги, считаю-
считающийся почти равным по масштабам речному стоку.
Таким образом, водоемы как среда обитания микроорганизмов,
разделяются на две большие группы: во-первых, океаны как конеч-
конечные аккумулирующие водоемы стока с постоянством химического
150
Таблица 5.1
Распределение воды по резервуарам
А [по: Алекин, Ляхин, 1984] Б [по: Graedel, Crutzen, 1993]
А. Резервуар
Объем
тыс. км;
Б. Резервуар
Объем,
% от общего
Мировой океан
Подземные воды
из них пресные
Ледники
Криосфера
Озера пресные
соленые
Болота
Реки
Атмосфера
Всего
1338000
23400
10530
24064
300
91
85
11
2
13
1385984
Океаны
Подземные воды
Криосфера
Озера и реки
Почвенная влага
Атмосфера
Биота
Всего
1350000
8000
29000
100
100
13
1
1387000
96,5
0,6
1,7
0,01
0,0000001
состава, который мог меняться лишь в течение миллиардов лет; во-
вторых, постоянно обновляющиеся континентальные воды с резер-
резервуарами в 0,2 млн км3 и ежегодными осадками в 0,1 млн км3. Воды
криосферы и подземной гидросферы выведены из этого круговоро-
круговорота как застойные карманы, и их следует рассматривать отдельно.
Сток воды с континентов в океан составляет 46 тыс. км3, из них реч-
речной - 40,5; ледовой - 3,3; подземной - 2,2. Средняя минерализация
речного стока - 76,8 мг/л, в то время как соленость океана составля-
составляет 35000 мг/л (Алекин, Ляхин, 1984).
Сравнение среднего состава вод речного стока и состава вод оке-
океана показывает их различие (табл. 5.2). Ионный состав речных вод
обусловлен процессом химического выветривания на континентах.
Состав воды океана не может быть произведен из речной воды
простым испарением. Здесь участвует комбинация более сложных
механизмов.
Важнейшим является карбонатное равновесие океана. Ведущим
для удаления карбонатов считается биологическое образование скеле-
скелетов рифостроителей, например кораллов в тропических водах, моллю-
моллюсков, а в толще воды - планктонных кокколитофорид и фораминифер.
В прошлом роль осадителей карбонатов выполняли микробные сооб-
сообщества предшественников строматолитов, причем вместе с кальцием
в толщи доломитов уходил и магний. Кремний удаляется в виде опала
диатомовыми и радиоляриями. Морская вода не насыщена относи-
относительно кремния, и это означает, что механизм удаления эффективнее
выщелачивания и выноса из поровой воды осадков. Образование суль-
151
Таблица 5.2
Средняя концентрация химических элементов в гидросфере
[по: Корж, 1989]
Элемент
Li
В
С
N
О
F
Na
Mg
Концентрация, г-моль/л
Реки
3,6 -10-7
1,6 ¦ Ю-6
-
-
-
5,3 ¦ Ю-6
2,3 ¦ 10
1,4 • 10
Океан
2,5 • Ю-5
4,0 • 10
2,3 • 10~3
3,0 • Ю-5
62,5
6,8 • Ю-5
4,7 • 10
5,3 • Ю-2
Элемент
А1
Si
Р
S
С1
к
Са
Вг
Sr
Концентрация, г-моль/л
Реки
1,8 • 10
3,7 • 10
1,3 • Ю
1,1 • КГ4
1,8 • 1(Н
3,8 • Ю-5
3,4 • 1(И
2,5 • 10
6,8 • 10-7
Океан
4,2 • 10"8
7,5 • Ю-5
1,9 • 10-6
2,8 ¦ Ю-2
5,5 • 10
1,0 • Ю-2
1,0 • Ю-2
8,3 • 10
8,7 • Ю-5
фидов обязано бактериальному сульфидогенезу как главному меха-
механизму анаэробной деструкции в осадках.
В последние годы привлекает внимание конвекционный гидро-
гидротермальный механизм в зонах спрединга. Он рассматривается в ка-
качестве важного механизма выноса ионов из морской воды, но коли-
количественные оценки потоков очень неопределенны. Холодная насы-
насыщенная вода порового пространства осадков просачивается через
трещины базальтов на дне, достигает термального потока с глуби-
глубинами около 2 км, что требует тысячелетий, реагирует с флюидами и
выносится на поверхность на флангах поднимающихся базальтов.
Для магния реакция описывается следующим уравнением:
11 Fe2Si04+2 Н2О+ 2 Mg2++2 SO2." =
фиалит морская вода
= Mg2Si3O6(OHL + 7Fe3O4 + FeS2+ 8SiO2.
сепиолит магнетит пирит опал
Выходящие термальные воды, как известно, служат местом обиль-
обильного развития микробной жизни в океане на основе хемосинтеза и
окисления восстановленных эндогенных веществ кислородом фото-
фотосинтеза, насыщающим холодную глубинную воду.
Время пребывания элемента в воде служит важнейшим показате-
показателем его круговорота (табл. 5.3). Наибольшей скоростью круговорота
обладает бикарбонат, находящийся в обмене с атмосферой, и каль-
кальций с временем пребывания около 1 млн лет. Основным источником
кальция служит речной сток, а дополнительным - циркуляция воды
в срединно-океанических хребтах. Удаление кальция из современно-
152
Таблица 5.3
Время пребывания элементов в воде океана
[по: Holland et al., 1986]
Компонент
Концентрация,
ммоль/кг
Время пребывания, годы
Na+
Mg2+
Са2+
К+
Br
468,0
53,2
10,2
10,2
545,0
28,2
2,4
0,84
«0,01
4,8 • 107
1,0 • 107
8,5 • 105
5,9 • 106
7,3 • 107
7,9 • 106
8,0 • 104
1,0 • 108
« 7 ¦ 104
го океана идет, главным образом, в виде СаСО3, меньшую, но каче-
качественно важную роль в балансе играет осаждение гипса
CaSO4 • 2Н2О, ангидрита CaSO4 и доломита CaMg(CO3J. Практиче-
Практически весь СаСО3 осаждается биологически в поверхностных водах эу-
кариотными организмами. Осадки докембрия представлены преи-
преимущественно доломитом. Принимается, что состав воды океана не
менялся существенно в течение 0,9 млрд лет, судя по составу эвапо-
ритов1. Однако концентрация солей в воде может варьировать в пре-
пределах 2-3 раз, не влияя на порядок образования солей.
Водоемы резко различаются по своей гидрохимии. Морские во-
водоемы и производные от них талассофилъные отличаются постоян-
постоянством состава и доминированием хлорида натрия и сульфата магния
как основных составляющих. Они представляют собой океаниче-
океаническую воду разной степени концентрации вплоть до эвапоритовых
бассейнов, состав воды которых обусловливает химическую садку
солей и отложение из рассолов водорастворимых минералов. Ата-
лассофилъные водоемы не связаны по своему минеральному составу
ни с нынешней морской водой, ни с погребенными отложениями
морских солей. Разнообразие гидрохимии таких озер приводит к
развитию в них своеобразных микробных сообществ.
Континентальные воды очень разнообразны по своему составу:
от пресных вод горных потоков, вытекающих из-под ледников, до
минерализованных озер аридного климата. Гидрохимия и классифи-
I
1 Borchert H., Muir R.O. Salt deposits: The origin metamorphism, and deformation of
evaporites; N.Y., Van Nostrand, 1984; Zharkov.M.A. History of Paleozoic salt accu-
accumulation. N.Y.: Springer, 1984.
153
кация континентальных вод изложена в монографии О.А. Алекина
и Ю.И. Ляхина A984).
Континентальные воды представляют малые резервуары с не-
небольшим временем пребывания воды в них. Химический состав их
варьирует в широких пределах в зависимости от времени контакта с
твердой минеральной фазой, промывного режима, и поэтому сред-
средние значения имеют мало смысла. Следует отметить, что континен-
континентальные воды содержат мало NaCl, если только они не образовались
путем размывания древних морских осадков, как соленые озера,
вроде Мертвого моря или ряда озер к северу от Каспия. Микроорга-
Микроорганизмы пресных вод чувствительны к 3,5% NaCl, что служит тестом
на морское или пресноводное происхождение. В пресных водах до-
доминирует бикарбонат-ион, поступающий из атмосферы и из почвен-
почвенных вод области водосбора. В формировании ионного состава пресных
вод главную роль играет процесс химического выветривания как из-
изверженных пород, так и осадочных, вынесенных на поверхность в
результате тектонических процессов. Плащ осадочных пород образо-
образовался уже на ранних этапах развития Земли, и поэтому повторное вы-
выщелачивание их играет большую роль в круговороте минеральных ве-
веществ. Остаточным продуктом выщелачивания служит кора выветри-
выветривания. В современных условиях на формирование состава континен-
континентальных вод большое влияние оказывают почвенные воды района
водосбора, находящиеся под влиянием растительного покрова. В про-
прошлом, до силура, растительный покров отсутствовал.
Критическим фактором в формировании состава континенталь-
континентальных вод является время их контакта с твердой фазой. Подобно тому
как на формирование состава атмосферы влияют биогенные факторы
почвенного воздуха и аэротопа, так на формирование состава вод влия-
влияют интерстициальные (находящиеся между твердыми частицами) во-
воды, одним из примеров которых может быть почвенная влага и грунто-
грунтовые воды. В равновесие с минералами вмещающих пород могут прий-
прийти только подземные воды при очень больших временах пребывания.
Интересной моделью конечного результата химического вывет-
выветривания горных пород на площади водосбора могут служить мине-
минерализованные бессточные озера, представляющие своего рода лизи-
лизиметры. Они очень разнообразны по гидрохимии, особенно в услови-
условиях подземного питания. Состав их вод нужно корректировать на оса-
осаждение минералов в пределах водосбора.
5.2. ВОДОЕМ КАК МОДЕЛЬ ЭКОСИСТЕМЫ
В экологии водоем является моделью экосистемы с геохимиче-
геохимическим круговоротом веществ. Баланс веществ в водоеме поддается
более легкому определению, чем в иных экосистемах. Приток и вы-
вынос можно оценить по речному стоку. Растворенные вещества более
или менее равномерно распределяются в объеме воды, и для них су-
существуют хорошо разработанные и унифицированные методы
154
"мокрой химии". Это связано с решающим значением воды в жизни
человечества и всего живого мира. Особенно удобны в качестве мо-
моделей экосистем озера. Роль микроорганизмов в них была подробно
изучена у нас в стране школой СИ. Кузнецова, которым был разра-
разработан подход к выяснению микробиологических процессов, послу-
послуживший неким шаблоном в течение уже полустолетия. Работы этой
школы опубликованы в серии монографий (Кузнецов, 1952; Кузне-
Кузнецов, 1970; Горленко и др., 1977 и т.д.), которые могут быть использо-
использованы как дополнительное чтение. Моделью в исследованиях
СИ. Кузнецова служило озеро Глубокое недалеко от Звенигорода.
Экосистема водоемов была исследована гидробиологом Г.Г. Винбер-
гом на основе цикла Сорг/О2 как ведущего процесса для водной толщи.
В водоеме рассматриваются биогеохимические круговороты, в
первую очередь углерода и кислорода, затем азота, серы, железа.
Внимание микробиологов сосредоточивается на скоростях процес-
процессов, катализируемых специфическими группами микроорганизмов,
например железобактерий или серобактерий. Далее, существенно
распределение процессов по профилю в водном столбе. Необходи-
Необходимым условием является зависимость от фотического слоя первич-
первичной продукции. В зависимости от стратификации водоема происхо-
происходит послойное распределение доминирующих групп организмов.
В особенности проявляется оно при достаточно выраженном влия-
влиянии донных отложений, в которых доминируют анаэробные процес-
процессы, служащие источником восстановленных веществ и газов; такое
влияние проявляется в относительно мелководных водоемах, к ко-
которым принадлежит большинство озер. Отсюда толщу водоема
можно рассматривать как пространство, заключенное между двумя
слоями: фотическим слоем первичной продукции и донными отложе-
отложениями с источником восстановленных соединений. Значительно мень-
меньше внимания микробиологи уделяли изменениям в водоеме во време-
времени, которые хорошо известны гидробиологам и обусловлены перио-
периодичностью развития фитопланктона в виде сменяющих друг друга
волн цветения, обусловленных сезонностью и исчерпанием биогенных
элементов и затем их регенерацией при деструкции мортмассы.
Взаимосвязь процессов в водоеме позволяет рассматривать населя-
населяющих его микроорганизмов как единое сообщество, связанное матери-
материальным переносом вещества, но для анализа удобнее разделить его на
три подсистемы: фртическую зону, донные отложения, водную массу.
5.2.1. Лимнология
В водоеме создается стратификация водного столба за счет на-
нагрева верхней части воды. Граница между теплой и холодной водой
может быть очень резкой и называется термоклином. Перенос вод-
водных масс через термоклин ограничен, и вещества проходят зону
температурного скачка за счет медленного диффузионного процес-
процесса. Над зоной температурного скачка располагается эпилимнион, а
155
под ним гиполимнион. В эпилимнионе большую роль играет турбу-
турбулентное перемешивание воды ветром, а также суточные колебания,
обусловленные охлаждением воды ночью и конвекцией в затишных
местах, например между стеблями водных растений. С термоклином
часто согласуется хемоклин - резкое изменение химического соста-
состава воды в связи с доминированием в гиполимнионе восстановленных
соединений, а в эпилимнионе - окисленных. Главным окислителем в
эпилимнионе служит кислород, находящийся в равновесии с кисло-
кислородом воздуха по закону Генри. В часы фотосинтеза содержание ки-
кислорода может быть избыточным, а ночью снижаться. Хемоклин
часто совпадает с оксиклином, обусловленным растворенным кис-
кислородом. В гиполимнионе содержание кислорода обусловливается
переносом его через зону температурного скачка и балансом между
процессами потребления и поступления. Проникновение света опре-
определяет фотическую зону, где возможен фотосинтез. Проникнове-
Проникновение света зависит от мутности воды, обусловленной минеральными
компонентами и развитием планктона. Для потребления кислорода
в гиполимнионе важен баланс между поступлением кислорода свер-
сверху, образованием его в фотическом слое, потреблением на окисле-
окисление растворенного органического вещества (РОВ), органического
вещества частиц - "дождя трупов"и пеллетов-фекалий взвешенного
органического вещества (ВОВ), и поднимающихся со дна восстанов-
восстановленных соединений - продуктов брожений. К ним в первую очередь
относятся газы метан и сероводород, летучие жирные кислоты.
Таково состояние водоема в период стагнации летом. Осенью
наступает осеннее перемешивание: вода охлаждается сверху, и начи-
начинается конвекционное выравнивание химических показателей с про-
проникновением окисленной воды к восстановленным донным отложени-
отложениям. Зимой образование ледового покрова останавливает и ветровое
перемешивание, и обмен газов с атмосферой. Поэтому происходит
использование резервуара растворенного кислорода. Растворимость
кислорода невелика - 0,25 ммоль/л, поэтому постепенно происходит
восстановление водной массы и могут возникать заморные явления
в зависимости от поступления окисляемых веществ. Весной при
вскрытии водоема вода оказывается обогащенной биогенами и на-
начинается весенняя вспышка цветения, связанная с весенним пере-
перемешиванием. Описанная картина относится к озерам, располо-
расположенным в климатическом поясе с выраженными сезонными коле-
колебаниями.
Из этой картины выпадают меромиктические водоемы, сохра-
сохраняющие постоянную стратификацию за счет того, что нижний слой
воды по тем или иным причинам, обычно вследствие более высоко-
высокого содержания солей, обладает большей плотностью, чем вышеле-
вышележащие, и имеется пикноклин. Чаще всего такая стратификация обу-
обусловлена наслоением пресной воды на соленую морскую. Примером
служит оз. Могильное на о-ве Кильдин, впервые исследованное
Б.Л. Исаченко в начале XX века.
156
Если перепад плотности захватывает фотическую зону, то на гра-
границе происходит развитие пурпурных бактерий, а нижние слои охва-
охвачены сероводородной зоной. Стратификация дает возможность обра-
образования переходной зоны, которую составляют градиентные организ-
организмы, развивающиеся в виде узкой зоны в области перепада концентра-
концентраций, если нет физического перемешивания. Легко показать, что их
развитие приводит к сужению зоны перепада и в отсутствие мешаю-
мешающих влияний создает резкий барьер. Крупным примером такого водо-
водоема служит Черное море, где в зоне хемоклина наблюдаются слои с
повышенной концентрацией микроорганизмов, но, конечно, не плен-
пленка. Очевидно, что баланс процессов в водоеме в сильнейшей степени
определяется его масштабами и будет разным для лесного водо-
водоема, как оз. Глубокое, или для оз. Байкал с огромной массой воды,
или для морей, где глубины, течения находятся в ином масштабе
величин. Различия в плотности воды вследствие различий в темпе-
температуре и содержании солей обусловливают термогалинную цирку-
циркуляцию, имеющую первостепенное значение для течений на глуби-
глубине, в особенности в океане.
Толща воды служит средой обитания бактериопланктона,
который зависит от первичной продукции органических веществ фито-
фитопланктоном и переработки его зоопланктоном. Развитие в водоеме ор-
ганизмов-фильтраторов может привести к выеданию бактериопланк-
бактериопланктона до уровня примерно 105 клеток/мл. Считается, что планктонные
микроорганизмы гомогенно распределены в толще воды. На самом
деле часть из них ассоциирована со взвешенными частицами.
Водоем ограничен снизу донными отложениями ила, на которых
развивается бентос и в которых идут преимущественно анаэробные
микробные процессы. Условия обитания в иловых отложениях и
донных осадках совершенно отличны от условий обитания в водной
толще. Для донных отложений характерна слоистость, переходящая
в микрозональность. Такие микрозоны были продемонстрированы
Б.В. Перфильевым для донных отложений озер Карелии, где четко
выявлялись зоны отложения окислов железа и марганца. В илах раз-
развивается пелофильная микрофлора, среди которой много организ-
организмов, приспособленных к ползанию в рыхлой массе осадков.
Сбоку водоем ограничен прибрежной зоной, составляющей со-
совершенно особое местообитание - экотон, которым называют по-
пограничную зону между двумя ландшафтами (см. Лекцию 6). Нередко
экотон представлен амфибиальным ландшафтом: низовым болотом,
зарослями тростника (маршем), приливной отмелью, мангровыми за-
зарослями, лагунами, сабхами. В экотоне, как правило, располагаются
геохимические барьеры, представляющие фундаментальное понятие
геохимии. Вследствие резкого скачка кислотно-щелочных, окисли-
окислительно-восстановительных и других условий на геохимическом барь-
барьере многие соединения переходят в другую форму и часто осаждают-
осаждаются. Геохимические барьеры поддерживаются не только транспортом
вещества из контрастных зон навстречу друг другу, но и деятельно-
157
стью микробных сообществ переходной зоны, которые создают рез-
резкие градиенты вследствие быстрой трансформации веществ.
В амфибиальных ландшафтах вся водная масса находится под
влиянием процессов в донных отложениях. Двумя экосистемами, ог-
ограничивающими понятие амфибиальных ландшафтов, служат мел-
мелководные окраинные моря и заливы океана, с одной стороны, и за-
заболоченные земли, с другой. Между ними располагается весь спектр
от лагун и приливных отмелей или мангровых зарослей в морских
условиях до равнинных озер и болот в континентальных. Для амфи-
амфибиальных ландшафтов трудно сказать, имеем ли мы дело с водо-
водоемом или же затопленными грунтами или почвой. Поскольку в мел-
мелководье контакт между водной, газовой, твердой фазами наиболее
тесный, то здесь и разыгрываются наиболее интенсивные биотиче-
биотические события с участием микроорганизмов, обусловленные близким
пространственным контактом аэробных и анаэробных сообществ.
Помимо экотонов к амфибиальным ландшафтам принадлежат
крайне мелководные водоемы и болота, включая затопленные поч-
почвы, с преобладанием процессов, происходящих в насыщенных водой
отложениях. Взаимодействие их с водной толщей в большой степе-
степени зависит от размеров водоема.
Разнообразие гидрохимии озер ведет к тому, что часто они рассма-
рассматриваются как уникальные местообитания, и особенности, выявлен-
выявленные для одного из них, лишь в ограниченной степени можно перенести
на другие. Для бактериолога важно, что рассматривать деятельность
бактерий в водоеме вне тесной связи с фитопланктоном и зоопланкто-
зоопланктоном, - особенно представленных микроорганизмами-протистами и
фильтраторами с бактериотрофным питанием, - можно лишь в огра-
ограниченных пределах. Эти пределы сводятся в первую очередь к геохи-
геохимическим циклам элементов, которые контролируются бактериями.
Помимо минерального состава вод, степени их минерализации,
определяющим для водоемов служит цикл углерода в них. В зависи-
зависимости от источника поступления и концентрации органического ве-
вещества водоемы различаются по степени трофии: олиготрофные,
мезотрофные, евтрофные. В олиготрофных водоемах содержание
и продукция органического вещества невелики, и резервуар кисло-
кислорода достаточен для поддержания аэробных условий в течение всего
времени, примером их могут служить горные озера, оз. Байкал,
оз. Ладожское, оз. Онежское в отсутствие антропогенного загрязне-
загрязнения. Евтрофными называют водоемы с обильной продукцией
органического вещества и, соответственно, обогащенной им водой.
К евтрофикации ведет усиленная продукция автохтонного органи-
органического вещества, например за счет избыточного поступления
биогенов и развития первичной продукции, или же аллохтонного
органического вещества, поступающего в экосистему водоема из-
извне. Поступление трудно разлагаемых гумусовых веществ ведет к
образованию болотных дистрофных водоемов гумидной зоны с
темноокрашенной коричневой водой. Общее содержание Сорг лишь
158
отчасти характеризует состояние водоема. Значительно точнее от-
отражает состояние биотических процессов в водоеме биологическое
потребление кислорода (ВПК), выражающее в мг О2 потребление
кислорода на окисление веществ биотой.
Численность бактерий в озерах коррелирует с трофностью во-
водоема:
Тип водоема
Олиготрофный
Мезотрофный (переходный)
Евтрофный
Дистрофный
Евтрофный искусственный
Численность
бактерий,
• 1С? клеток/мл
50
1000
2000-10000
1000-2000
10000
Процессы физические, химические, биологические тесно связа-
связаны в водоеме в единую систему с множеством обратных связей.
5,2.2. Экология водных микроорганизмов
Вода как объект исследования микроорганизмов отличается от
почвы прежде всего легкостью прямого микроскопического наблю-
наблюдения. Отсюда множество знаний о таких формах организмов, кото-
которые, являясь массовыми, не поддаются культивированию. Микроско-
Микроскопическое исследование водных микроорганизмов началось в середине
XIX в., прежде всего Ф. Коном, до начала развития методов культи-
культивирования бактерий, и продолжалось независимо как побочный
продукт наблюдений гидробиологов за водорослями и протистами.
Первыми в поле зрения микроскопистов попали крупные нитчатые
бактерии, как Beggiatoa, Thioploca, Crenothrix, Sphaerotilus, Gallionella,
Leptothrix, Peloploca. Их развитие носит характер массовых обраста-
обрастаний, они формируют дерновинки, маты и составляют бактериальный
бентос, хотя такой термин микробиологи редко употребляют, а гово-
говорят скорее о биопленках. До сих пор такие организмы с трудом под-
поддаются лабораторному культивированию. Они легко идентифициру-
идентифицируются по характерной морфологии под микроскопом наряду с синезе-
леными водорослями (цианобактериями) и служат индикаторными
формами для характеристики состояния водоема. Описание этих ор-
организмов составило большой массив знаний о водной микрофлоре,
накапливавшийся примерно с 1880-х до 1920-х годов. После примене-
применения С.Н. Виноградским микроскопического подсчета бактерий в поч-
почве наступил период количественного учета бактерий в воде. Сначала
водупросто испаряли в цилиндре на поверхности предметного стекла.
Следующим этапом в развитии прямых методов было использование
мембранных фильтров из нитроцеллюлозы. Они дали возможность
изучать бактериопланктон при очень низкой концентрации клеток в
воде. Через мембранный фильтр пропускали определенный объем
159
воды, бактерии окрашивали эритрозином, фильтр подсушивали, про-
просветляли иммерсионным маслом и под микроскопом подсчитывали
бактерии. Метод этот в практику контроля водоснабжения ввел
А.С. Разумов, а в очень широком масштабе для морей применил
А.Е. Крисе. Сейчас предпочитают окрашивать бактерии на фильтре
люминесцентными красителями и считать в эпилюминесцентном ми-
микроскопе. Прямой метод дал, во-первых, точное количественное
представление о численности водных организмов, во-вторых, ясное
представление об их морфологическом разнообразии.
Прямому методу противостояли методы культивирования Р. Ко-
Коха с подсчетом числа колоний на агаризованных средах и идентифи-
идентификацией индикаторных организмов во главе с Escherichia coli для
установления "коли-титра", разработанные санитарными бактерио-
бактериологами в 1880-е годы. Численность бактерий, определенная методом
высева, оказывается в 100-10 000 раз ниже результатов прямого
счета. Применяются всякие ухищрения, чтобы уменьшить этот раз-
разрыв. Наименьшее различие получается при применении разбавленных
и голодных сред и подсчете микроколоний, но и в этом случае разни-
разница составляет десятки и сотни раз. Различие в численности водных ба-
бактерий при сравнении данных прямого метода и высева тем больше,
чем чище вода: для сточных вод оно составляет десятки и сотни раз,
для чистых олиготрофных вод - десятки тысяч. Но дело не только в
численности: микроскоп с очевидностью убеждал, что культивируют-
культивируются не те организмы, которые наблюдаются в природе.
Противоречие между прямыми и культуральными методами
представляет важнейшее противоречие природоведческой микро-
микробиологии, наиболее отчетливо иллюстрируемое на примере водной
микробиологии. При доминировании медицинских микробиологов
был принят Кодекс номенклатуры бактерий, по которому узаконен-
узаконенным мог быть только тот таксон бактерий, который представлен
"чистой культурой живого штамма в общепризнанной коллекции
культур". В результате уцелели только те "морфовиды", которые
были закреплены традицией, как Thioploca, Gallionella и т.д., а об ос-
остальных постарались забыть. С другой стороны, применение пря-
прямых молекулярных методов идентификации дало множество "кло-
"клонов" последовательностей, о которых ничего нельзя сказать, кроме
как об их филогенетическом положении и местонахождении отбора
пробы. Молекулярные методы расцениваются как "современные", а
микроскопические как "архаические". Исследователь роли микро-
микроорганизмов в природе - наблюдатель - может иметь иное мнение.
Разработка молекулярных методов идентификации и филогенети-
филогенетического положения "некультивируемых" организмов привела к по-
пониманию того, что микрофлора океана, например, представлена со-
совсем другими организмами: фотосинтезирующим пикопланктоном,
массовыми планктонными археями. Фактически это подтверждение
другими методами вывода, который был сделан на заре бактериоло-
бактериологии водными микробиологами.
160
5.2.3. Бактериопланктон
Планктон классифицируется по размеру микроорганизмов
(в мкм):
Фемто-
0,02-0,2
Вирусы
Пико-
0,2-2,0
Бактерии
Нано-
2,0-20
Микро-
20-200
Фитопланктон
Мезо-
200-2000
Протозоо-
планктон
Некто?
Вплоть
до кита
Водная толща служит местообитанием бактериопланктона, тес-
тесно связанного в своей жизнедеятельности с другими планктонными
организмами и создающего классический экологический треуголь-
треугольник продуценты-консументы-редуценты. Первые представлены
фитопланктоном от известных планктонных водорослей до мель-
мельчайших пикопланктонных форм, консументы - зоопланктоном, а
все бактерии-деструкторы отнесены к редуцентам.
Метод подсчета на мембранных фильтрах дал количественные
представления о численности микроорганизмов безотносительно к
их специфическим функциям. Метод особенно надежен для исследо-
исследования планктона, который предположительно представлен отдель-
отдельными организмами, свободно парящими в объеме воды. Развитие
бактериопланктона определяется доступностью питательных ве-
веществ, находящихся в очень сильном разбавлении и, следовательно,
требующих для их использования высокого сродства к субстрату.
Концентрации используемого органического вещества в океане нахо-
находятся в пределах десятка-тысячи мкг/л, т.е. 10~9, ppb (partes pro billion).
Вертикальное распределение бактериопланктона очень харак-
характерно, и получено множество кривых численности в зависимости от
глубины. Как правило, в олиготрофных водоемах численность бак-
бактериопланктона имеет максимум в гиполимнионе, чаще всего сразу
под максимумом фитопланктона. В евтрофных водоемах максимум
чаще наблюдается в металимнионе в зоне хемоклина, иногда очень
резкий, хотя из-за небольшой глубины таких водоемов численность
бактерий может быть высокой по всему объему. В общем, числен-
численность бактерий следует за продуцентами фитопланктона и их отми-
отмиранием. Та же закономерность наблюдается и в сезонной численно-
численности. Максимум численности бывает в период летней стагнации в ию-
июне-июле, минимум - зимой.
Численность бактерий соответствует не всему органическому
веществу, определяемому методом химического окисления (ХПК -
химическое потребление кислорода, определяемое по перманганат-
ной или бихроматной окисляемости), а лишь легко усвояемой его
части. Особенно наглядно такое различие между ХПК и ВПК для
дистрофных гуматных вод. Взвешенное органическое вещество по-
полимеров либо оседает на дно в мелководных водоемах, либо исполь-
используется в микроагрегатах-пеллетах по мере оседания. Экосистемная
(нетто-) продукция не может служить критерием трофности: она
6. Г. А. Заварзин
161
одинакова в арктических и тропических озерах вследствие очень вы-
высокой деструкции в тропиках. Широко используемое понятие
"евтрофикация" становится далеко не таким ясным, когда пытают-
пытаются установить простые и универсальные критерии.
Численность бактериопланктона контролируется животными-
фильтраторами, иногда крупными, которые очищают воду от взве-
взвеси. Нередко чистота воды зависит именно от эффективности удале-
удаления бактериопланктона, а не от возможностей его развития. Это
справедливо и для экосистемы коралловых рифов, и для многих во-
водоемов. Зоопланктон, однако, ориентирован преимущественно на
фитопланктон как свою пищевую базу. Роль микроскопических жи-
животных с бактериотрофным питанием в водных экосистемах,
особенно протистов, очень велика. Нижним пределом для эффек-
эффективного питания фильтраторов считается плотность 105±1 клеток ба-
бактерий в 1 мл воды. Наличие и сильное воздействие консументов
отличает водные микробные системы от наземных, где микробы
могут найти убежище в физической нише.
Фитопланктон, как правило, имеет свободную от бактериальных
обрастаний поверхность клеток, пока не началось отмирание. Есть
специальные группы бактерий-хищников, сходных с бделловибриона-
ми, которые способны нападать на микроводоросли; они были иссле-
исследованы Б.В. Громовым и дали пример любопытной группы
организмов. Однако, как правило, живые клетки фитопланктона сво-
свободны от эпифитов. Вместе с тем получить бактериально чистую
культуру водорослей крайне трудно. Проблема взаимоотношения во-
водорослей и бактерий особенно настойчиво разбиралась в связи с раз-
разработкой систем жизнеобеспечения. Отмирающие клетки водорос-
водорослей сразу же становятся объектом заселения их бактериями, среди
которых многочисленны организмы с прикрепительными дисками,
как у каулобактеров. Микроскопия таких отмирающих клеток пока-
показывает, что они .становятся похожими на ершики для мытья пробирок
из-за множества палочковидных бактерий, прикрепляющихся к ним.
Внутри более крупных клеток отмерших водорослей можно иногда
наблюдать микроколонии бактерий, но в основном процессы разложе-
разложения идут снаружи. Взвешенное органическое вещество служит пита-
питательным субстратом зоопланктона, в первую очередь, фильтраторов.
Бактерии служат первичными деструкторами того, что называ-
называется растворенным органическим веществом, но для выхода ве-
веществ в раствор необходимы либо автолиз, либо разрушение под
действием внешних факторов. Важным путем провоцирования лизи-
лизиса у водорослей служат фаги и вирусы, которые рассматриваются
как агенты, определяющие конец цветения при высоких плотностях
водорослей. Альгофаг хлореллы был открыт Н.Б. Заварзиной2, а
2 Заварзина Н.Б., Проценко А.Е. О лизисе культур Chlorellapyrenoidosa Pringh//
Докл. АН СССР. 1958. Т. 122, № 5. С. 936-939; Заварзина Н.Б. Литический
агент в культурах Chlorella pyrenoidosa Pringh // Там же. 1961. Т. 137, № 2.
162
через пять лет были открыты цианофаги3. Затем они стали объек-
объектом интенсивного изучения как важный экологический фактор. Фа-
голизис цианобактерий и водорослей приводит к резкому выбросу
растворимых легкоусваиваемых веществ гибнущей популяцией во-
водорослей. В 1990-х годах обнаружение вирусов цианобактерий пока-
показало, что численность частиц составляет до миллиона на мл, 5%
цианобактерий в море содержали фаги, принадлежащие ко всем
группам пресноводных фагов. Особый интерес вызвали фаги циано-
цианобактерий пикопланктона. В целом можно полагать, что не только
зоопланктон, но и вирусы могут служить фактором регуляции чис-
численности фитопланктона, особенно в конце цветения при высоких
плотностях. Минимальная плотность популяции хозяина для фаго-
фаговой инфекции >104 клеток/мл.
5.2.4. Круговорот углерода
Цикл углерода в водоеме включает следующие стадии:
' 1) первичная продукция, лимитируемая биогенами (N, P, Si, Fe...);
2) регенерационная продукция, обеспечиваемая биогенами, ос-
освобождаемыми при деструкции в фотической зоне и непосредствен-
непосредственно под ней;
3) экспортная продукция - вынос вещества из фотической зоны;
4) вторичная продукция - синтез биомассы органотрофами-дест-
рукторами;
5) синтез биомассы цепью консументов;
6) синтез биомассы в фекальной цепи деструкции (пеллеты);
7) синтез биомассы деструкторов в осадках (пелофильная мик-
робиота).
Заведомая ограниченность применения культуральных методов к
водной микробиологии из-за большого вклада некультивируемых ор-
организмов породила вариант "бескультурной микробиологии", где оце-
оцениваются процессы, а не организмы. Для таких работ не обязательна
специальная подготовка в области микробиологии, и они эффектив-
эффективны в руках экологов с широким образованием. Наибольших успехов
в этом отношении достигли гидробиологи, которым была важна сум-
суммарная групповая оценка баланса процессов. Лидером здесь был
Г.Г. Винберг, исследовавший цикл СО2/О2 в озерах, за которым пос-
последовали различные разработчики тех же принципов в школе микро-
микробиологов СИ. Кузнецова. В результате накоплены обширные коли-
количественные данные об интенсивностях процессов, но обычно на еди-
единичных станциях и очень редко при мониторинге.
Поскольку в водоеме проходят последовательные волны цвете-
цветения и отмирания водорослей, обусловленные использованием био-
биогенных элементов и их последующей регенерацией при деструкции,
3 Safferman R.S., Morris M.F. Algal viruses: Isolation // Science. 1963. Vol. 140.
P. 679-680.
6* 163
г
одиночные измерения очевидно недостаточны и могут создать лож-
ложное впечатление. Это относится даже к водоемам с экстремальными
условиями. Волны цветения накладываются на сезонные изменения.
Волны цветения обусловлены прежде всего внутренними причина-
причинами в системе первичных продуцентов.
Для оценки скорости процессов круговорота углерода исполь-
используется определение баланса кислорода, концентрацию которого
можно измерить очень точным иодометрическим титрованием, ме-
методом Винклера. Берут темную и светлую склянки с одинаковыми
пробами воды. Продукция оценивается по выделению кислорода
при фотосинтезе в светлой склянке, считая, что выделение кисло-
кислорода эквивалентно синтезу органического вещества. Деструкцию
определяют в темной склянке вариантом "биохимического потреб-
потребления кислорода" БПК5, широко применяемого для сточных вод.
По методу Винберга баланс между продукцией и деструкцией оп-
определяют в пробах воды в светлой и темной склянках, которые
подвешивают на тросе в водоеме при естественной освещенности и
температуре. Разность между светлой и темной склянками дает чи-
чистую продукцию за рассматриваемый промежуток времени, обыч-
обычно сутки. Определение кислорода титрованием или электрохими-
электрохимически можно заменить быстрыми и гораздо более чувствительны-
чувствительными радиоизотопными методами.
Радиоизотопный метод дал возможность определить продукцию
аноксигенных фотосинтетиков. В результате были получены очень
четкие кривые, указывающие на приуроченность процесса к хемо-
клину и границе сероводородных и кислородных вод. Этим же мето-
методом оценивается хемосинтез. Опять-таки развитие хемосинтетиков
и ассимиляция ими углекислоты приурочены к зоне хемоклина -
нижней границе проникновения кислорода и верхней границе посту-
поступления восстановленных веществ, прежде всего сероводорода и
метана из глубины. До какой-то степени можно судить и об орга-
организмах, вызывающих процесс. Надежно это делается в отношении
аноксигенных фототрофов, идентифицируемых микроскопически,
как и фитопланктонные организмы. Менее надежно - для хемо-
хемосинтетиков, где прямые методы отсутствуют, а культуральные го-
говорят скорее о корреляции. Вторичная продукция биомассы оцени-
оценивается по гетеротрофной фиксации углекислоты, составляющей
около 6% от биомассы. Вторичная продукция может быть велика
в водоемах с большим поступлением аллохтонного органического
вещества.
Гидробиологический метод оценки процессов круговорота угле-
углерода и серы в водоемах поставил эту область микробиологии на
прочную количественную основу. Бактериопланктон может быть
отделен от более крупных планктонных организмов фильтрованием
через крупнопористый фильтр. При интерпретации результатов,
полученных таким способом, необходимо постоянно иметь в виду,
что агрегаты бактерий и бактерии, связанные с взвешенными части-
164
Г цами, остаются на фильтре, и результат может быть ошибочным.
г Этого не происходит при обычном скляночном методе.
f Концентрация растворенных органических веществ в воде очень
i невелика и, за исключением евтрофных водоемов, не превышает не-
Г скольких мг/л. Поэтому организмы бактериопланктона в значитель-
значительной части представлены олиготрофами. Изучение олиготрофов -
один из редких и блестящих случаев, когда за микроскопическим
обнаружением организмов последовало их выделение в культуру и
выяснение физиологии. Доминирование олиготрофов в водных сис-
системах было установлено в 1940-х годах А.С. Разумовым, который
отнес сюда организмы, растущие только на "голодном агаре", "вод-
"водном агаре" и очень медленно образующие на нем мелкие колонии.
Именно эта линия продолжается до сих пор при исследовании вод
океана. В начале 1960-х годов Д.И. Никитин4, применив электрон-
электронный микроскоп для прямого наблюдения микроорганизмов, обнару-
обнаружил "новые и необычные формы микроорганизмов" со своеобраз-
своеобразными выростами и придатками, которые совершенно разрушили
представление о формах бактерий как "биллиардной шаре, кие и
пробочнике". Организмы, развивавшиеся в озерах и дистиллирован-
дистиллированной воде, были идентифицированы микроскопически. Они имели
длинные клеточные стебельки и получили название Caulobacter. Не-
Несколько раньше А.С. Разумов описал массовую форму "Gallionella
kljasmiensis", где клетки были объединены в розетки неклеточными
стебельками и размножались почкованием. Впоследствии организм
был переклассифицирован и по номенклатурным правилам получил
название Planctomyces. Несколько культур таких организмов были
выделены Шлесснером в лаборатории П. Хирша в Киле. Планкто-
мицеты составили отдельный класс организмов по 16S рДНК гену, и
прямые молекулярно-генетические методы показали, что это массо-
массовые формы в природе, хотя в культуре получены лишь немногие и
они очень отличаются от обычных бактерий. Из других олиготро-
олиготрофов Л.В. Васильевой и Д. Сталей удалось выделить в культуру ряд
олиготрофных простекобактерий с характерными клеточными при-
придатками. Стебельки и выросты сильно увеличивают периплазмати-
ческое пространство клетки, в котором расположены системы
транспорта, обеспечивающие рост при низкой концентрации суб-
субстрата. Развитие олиготрофных организмов определяет нижний
предел органического вещества в воде. Так, высокое сродство к уг-
углеводам приводит к тому, что в воде растворенные углеводы прак-
практически отсутствуют.
К олиготрофам не следует относить организмы, развивающиеся
на взвешенных частицах органического вещества. Как уже упомина-
упоминалось, отмершие планктонные организмы облеплены бактериями, и
времени оседания на немногие десятки метров достаточно для полно-
4 Никитин Д.И., Васильева Л.В., Лохмачева. Редкие и необычные формы поч-
почвенных микроорганизмов. М.: Наука, 1966. 71 с.
165
го разложения доступной части. Эти организмы плохо поддаются
количественному учету, так как каждое микросообщество представ-
представляет лишь одну "колониеобразующую единицу". Прямое микроско-
пирование дает указание на сильное развитие бактерий в микроаг-
микроагрегатах.
Особую задачу представляет так называемый "водный гумус".
Это стойкие органические вещества, по-видимому, аналогичные
настоящему гумусу в биологическом смысле. Они представляют
громадный резервуар органического углерода, растворенного в
очень низкой концентрации в воде океана. До какой степени они
могут служить субстратом для развития бактерий, например при
так называемом "скляночном эффекте" - резком возрастании чис-
численности бактерий при выдерживании проб воды в склянках, - не-
непонятно.
В водоемах со значительным содержанием сульфатов деструк-
деструкция в иле ведет к образованию сероводорода при сульфатредукции.
Если в водоеме поступление железа недостаточно для связывания
сероводорода в сульфиды, то он выходит из донных отложений.
Его окисление при доступе кислорода происходит на поверхности
ила серобактериями и тионовыми, способными вести процесс при
минимуме притока кислорода, когда концентрация его находится
за пределами обнаружения методом Винклера. Сероводород мо-
может окисляться без потребления кислорода аноксигенными фото-
трофами, и в этом случае в толще воды на границе проникновения
света происходит массовое развитие пурпурных или зеленых серо-
серобактерий.
Рассматривая цикл углерода в водоеме в целом, заключал
СИ. Кузнецов свой анализ (Кузнецов, 1970), необходимо признать,
что основная масса органических веществ не только синтезируется,
но и успевает вновь минерализоваться в водной массе озера за счет
аэробных процессов. Часть прежде всего полимерного органическо-
органического вещества в виде детрита попадает в иловые осадки. Эта часть
совершенно незначительна в олиготрофных и глубоководных водо-
водоемах и может быть значительной в евтрофных мелководных водо-
водоемах или таких их участках, как заливы. В иловых осадках развива-
развиваются преимущественно анаэробные процессы. Продукты анаэроб-
анаэробного разложения, прежде всего метан, поднимаются в водную толщу
озера и там окисляются аэробной микрофлорой. Процессы, проис-
происходящие в толще ила и на его поверхности, следует рассмотреть от-
отдельно.
С циклом углерода тесно связан цикл кислорода. Растворимость
кислорода в воде заключается между 11 мг/л при 4 °С и 8 мг/л при
25 °С и резко снижается при повышении температуры. Для боль-
большинства рыб замор наступает при 2 мг/л. Дефицит кислорода обу-
обусловлен наличием окисляемых органических веществ, когда в воде
появляются сероводород и закисное железо. В зоне температурного
скачка может наблюдаться максимум кислорода в связи с фотосин-
166
тезом и затруднением обмена с воздухом и фотической зоной. Про-
Профиль кислорода указывает на характер процессов в водоеме и очень
показателен. Важнейшим процессом является дыхание бактерий,
которое составляет 0,2 -*¦ 100 • 10~12 мгДклетка • час) при весе клетки
10-12 мг. Сопоставление этих величин с численностью бактерий убе-
убедило, что их дыхание и окисление органических веществ составляет
главный вклад в баланс кислорода в гиполимнионе в соотношении
1 моль кислорода на 1 грамм-атом органического углерода. Ско-
Скорость потребления кислорода подвержена очень резким колебаниям
в зависимости от поступления окисляемых веществ. Важнейшим из
этих окисляемых веществ является метан, поднимающийся из дон-
донных отложений как конечный продукт распада в анаэробных усло-
условиях. Окисление метана требует трех молекул кислорода на молеку-
молекулу газа и поэтому сильно сказывается на балансе кислорода. В опре-
определенные сезоны оно может составлять 70% от общего потребления
кислорода. За метаном по интенсивности потребления кислорода
следует нитрификация.
Круговорот углерода в водной толще водоема можно предста-
представить в виде следующей схемы:
Первичная продукция
...Фитопланктон,
планктрн
Гидролиз
Вторичная продукция
Бактерио-
планктон
I
Бен гас
i Анаэробный ил
Захоронение
I
167
Оседающее взвешенное органическое вещество поступает в дон-
донные отложения, характер которых сильнейшим образом зависит от
придонных течений и привноса минеральной взвеси в процессе седи-
ментогенеза. В олиготрофных водоемах донные отложения обычно
сложены озерными песками и имеют хороший обмен с придонной
водой. Здесь идут процессы выщелачивания минеральных компонен-
компонентов донных отложений. Оседающее органическое вещество образует
на поверхности наилок, который легко взмучивается и подвергается
разложению в аэробных условиях. Глинистые грунты связаны с муть-
евыми потоками, и в них легко происходит захоронение органическо-
органического вещества в связи с его физическим запечатыванием; такое фи-
физически иммобилизованное вещество становится недоступным для
бактерий и дает начало глинистым сланцам с высоким содержанием
органического углерода. При этом развиваются восстановительные
процессы, приводящие к мобилизации железа и марганца (оглеению).
В евтрофных водоемах обильное поступление взвешенного органиче-
органического вещества (ВОВ) приводит к образованию рыхлых органиче-
органических илов - сапропеля. Он сносится в котловины и углубления.
Процессы распада органического вещества в иловых отложени-
отложениях обусловлены тем, что в них поступает относительно устойчивая
взвесь, сохранившаяся при оседании в водной толще. Для озер более
половины осадков составляет лигнино-гумусовый комплекс, до Г0%
приходится на битумы. В иловом слое создаются анаэробные усло-
условия уже в нескольких миллиметрах от поверхности. Разложение
клетчатки, гемицеллюлоз идет преимущественно по пути метаноге-
неза с образованием в качестве промежуточных продуктов летучих
жирных кислот и значительного количества ацетата.
На поверхности ила развивается своеобразная микрофлора, со-
состоящая из нитчатых скользящих бактерий. Характерным предста-
представителем ее служит, например, Peloploca, которую до сих пор не уда-
удалось культивировать.
Анаэробные процессы ведут к выделению из донных отложений
углекислоты, метана, аммиака, восстановленного железа и марган-
марганца. Последние поступают в придонные слои при отсутствии выделе-
выделения сероводорода, который начинает серный цикл и характеризует-
характеризуется в талассофильных водоемах черными илами, обогащенными
сульфидом, - сначала аморфным гидротроилитом FeS • пН2О, кото-
который затем реакцией с серой превращается в устойчивый пирит FeS2.
В отсутствие сероводорода Fe2+ диффундирует в придонную воду.
5.2.5. Цикл железа в континентальных водоемах
Цикл железа парагенетически связан с метаногенезом и идет в
аталассофильных олиготрофных водоемах. В результате образует-
образуется озерная железная руда в виде конкреций. Благодаря своей свое-
своеобразной морфологии и естественной окраске окислами железа же-
168
лезобактерии сразу же стали объектами изучения микроскопистов.
С ними связано начало геологической микробиологии, когда в 1830-е
годы Г.Х. Эренберг предположил, что образование осадочных руд
связано с деятельностью железобактерий. В целом, эта группа пред-
представляет свидетельство безуспешных усилий традиционной микро-
микробиологии и основывается на прямых наблюдениях в природе
Н.Г. Холодным.
Исследования образования железной руды в озерах Карелии по-
позволили Б.В. Перфильеву5 установить микрозональное строение
иловых отложений. Опыты были основаны на формировании вто-
вторичного диагенетического профиля. Пробы донных отложений
отбирали в 200 мл граненые стаканы, заполненные на 2/з осадком, и
оставляли на очень длительные сроки. В осадке происходило восста-
восстановление металлов, и под поверхностью осадка образовывались
тонкие миллиметровые зоны темно-коричневых окислов марганца
и охристые прослойки гидроокислов железа. Окислы металлов от-
отлагались в виде оформленных структур, принадлежащих организ-
организмам группы Metallogenium для марганца и Siderococcus для железа.
Обе эти формы рассматриваются сейчас американскими исследова-
исследователями как продукты жизнедеятельности микроорганизмов. Однако
их удается культивировать в чистой культуре, пересевать и менять
хозяина. Процесс основан на продукции перекиси водорода и разло-
разложении ее другим организмом в симбиотической паре. Он был найден
для пары псевдомонад и затем оказался универсальным перекисным
механизмом окисления. Доказательство перекисного механизма
было впервые получено В.В. Балашовой6 с Mycoplasma laidlawii, ко-
которая образует перекись при окислении органических веществ в ми-
микроаэробных условиях. Если в среде присутствует железо, то оно
катализирует разложение перекиси, но само окисляется в охристый
осадок. Процесс окисления железа функционально заменяет реак-
реакцию каталазы.
Образование микрозон железобактериями в лаборатории легче
всего наблюдать в культуре Gallionella по ван Нилю. Культуру ведут
в пробирке с 1 см слоем осадка FeS ¦ пН2О. Гидротроилит готовят
смешением растворов соли железа и сульфида натрия с последую-
последующей промывкой осадка кипящей водой. Gallionella развивается на
стенках пробирки в виде белых пушистых колец, последовательно
спускающихся вниз. Организм, подобно другим железобактериям,
микроаэробный. Как правило, Gallionella развивается в холодной
воде, обогащенной бикарбонатом железа. Характерная морфология
5 Перфильев Б.В., ГабеД.Р. Изучение методом микробного пейзажа бактерий,
накопляющих марганец и железо в донных отложениях // Роль микроорганиз-
микроорганизмов в образовании железомарганцевых озерных руд. М.; Л.: Наука, 1964.
С. 16-53.
6 Балашова В.В. Микоплазмы и железобактерии. М: Наука, 1964.
169
Gallionella с витыми стебельками, сложенными множеством нитей,
служила предметом многочисленных не очень убедительных иссле-
исследований.
Железо мобилизуется в затопленной почве в результате глеево-
го процесса и часто поступает в озеро в виде твердого стока взвеси
с окружающих болот. Образование охры и болотной руды чаще все-
всего обусловлено развитием Leptothrix ochraceae. Организм представ-
представлен цепочкой псевдомонад, заключенной в трубчатый чехол, как у
Sphaerotilus, но ожелезненный. Цепочка клеток выходит из чехла,
оставляя пустую охристую трубку. Массы чехлов Leptothrix образу-
образуют плотные отложения охры. В анаэробных условиях происходит
быстрое восстановление железа.
Классическим местом развития железобактерий являются выхо-
выходы углекислых подземных вод в источники. Помимо упомянутых
здесь иногда развивается нитчатая бактерия Toxothrix, оставляющая
за собой нити гидроокислов железа и образующая плотные слои-
слоистые массы охры в источниках.
Чтобы закончить с железобактериями, следует упомянуть мно-
многочисленные и разнообразные организмы, у которых гидроокислы
железа откладываются в слизистой капсуле по реакциям катионно-
го обмена с кислыми полисахаридами. Отложения придают харак-
характерный вид клеткам, и такие организмы описаны под собственными
названиями, например среди "Siderocapsaceae". Процесс усиливается
при разложении железных солей органических кислот, например
цитратов, оксалатов. В природе он особенно важен для гуматов же-
железа, образующихся в гумидном поясе.
Образование железных и железо-марганцевых конкреций, пред-
представляющих собой лепешки размером от боба до чашки Петри, обу-
обусловлено восстановлением железа в донных отложениях, миграцией
его на поверхность, преимущественным окислением на топографи-
топографических высотах в токе придонной воды, физико-химическим накоп-
накоплением на поверхности окислов и минералов. Образование конкре-
конкреций представляет автокаталитический процесс с участием микроор-
микроорганизмов. По-видимому, аналогично образованию "бобовой руды" в
озерах идет образование представляющих рудные скопления полей
железо-марганцевых конкреций в глубоководных частях океана.
Очень наглядной иллюстрацией бактериальных процессов мо-
может служить разделение железа и марганца при образовании оса-
осадочных месторождений. Железо и марганец сходны по своим физи-
физико-химическим характеристикам, но у марганца более высокий
окислительный потенциал. Поэтому сначала при образовании оса-
осадочных месторождений происходит отложение окислов железа, а
потом - марганца. Такое распределение хорошо наблюдается в мес-
месторождениях Кривого Рога, Керчи.
Общая картина круговорота железа в континентальных водо-
водоемах представлена на схеме:
170
Fe + СО2
Порода + Вода
рН<5
Анаэробиоз
Мобилизация в
подземных водах
Мобилизация
в болотах¦
Fe2+ + 2НСО3-
Gallionella Анаэробное^»
Toxothrix окисление
аноксигенными
фототрофами
Fe-гуматы
ДИСТРОФНЫЕ
ВОДОЕМЫ
Leptothrix
"Siderocapsaceae"
Взвесь
ЕВТРОФНЫЕ
ВОДОЕМЫ
Leptothrix
Blastocaulis
Сток
Siderococcus
' Gallionella
-Болотная руда-"
ОЛИГОТРОФНЫЕ
ВОДОЕМЫ
Озерная руда
_J
Metallogenium
t
Fe2+ + 2HCO3-
Донные отложения
Захоронение Восстановление
в аэробных железа бактериями
условиях I
Geobacter и другие магнетит-
образующие бактерии
Fe3O4
-Desulfovibrio -
i Leptospirillum
Fe2* pH<4
i Acidithiobacillus
FeS • nH2O
\
FeS2.
Сульфиды
5.2.6. Водотоки и водопользование
Микробиология рек связана в основном с их использованием в
качестве источников чистой воды. Микробное население водотоков
разделяется на часть, увлекаемую потоком воды с большим количе-
количеством взвешенных частиц, на микробные сообщества обрастаний,
способные противостоять смыву, на сообщества затишных мест, где
происходит осаждение иловых частиц. Особое место в водотоках
имеет сообщество обрастаний, которое получило название крено-
фильной микрофлоры. Организмы обрастаний находятся под меха-
механическим воздействием потока и поэтому имеют ряд приспособле-
приспособлений, обеспечивающих плотную связь с субстратом. С другой сторо-
стороны, при высокой скорости потока диффузионный пограничный слой
становится меньше и питание растворенными веществами облегча-
облегчается. Прежде всего это относится к кислороду, который насыщает-
171
ся воздухом за счет турбулентного перемешивания. Для развития в
потоке возможны два способа приспособления. В одном случае соз-
создается плотный контакт с твердой поверхностью на дне. Микроор-
Микроорганизмы прикрепляются слизью или даже минеральными осадками,
как Toxothrix охристым осадком железа или осаждающие известняк
цианобактерии. Другой вариант осуществляется в более медленно
текущих водах, когда к твердой поверхности прикрепляются нити,
развевающиеся в потоке в виде косм. Эту стратегию осуществляет
Sphaerotilus, Thiothrix, Crenothrix, а в термальных источниках -
Thermothrix thiopara, изогнутые клетки которого цепляются друг за
друга. Для нитчатых кренофильных бактерий обычно наличие вла-
влагалища, в которое заключена цепь клеток или трихом. Кренофилы
не были объектами физиолого-биохимических исследований.
Водотоки рассматриваются как основной источник чистой воды.
По течению воды можно различать зоны самоочищения. Эти зоны
были исследованы для альпийских ручьев и речек и получили назва-
название зон сапробности. С гор стекает чистая олигосапробная вода, по
мере продвижения потока по равнине в нее поступают органические
вещества и она становится мезосапробной. Загрязнение стоками пе-
переводит воду в разряд полисапробной. Каждая зона характеризуется
своими индикаторными организмами, и, в частности, Sphaerotilis яв-
является индикатором полисапробности. Самоочищение проходит в
обратном порядке. Основным фактором самоочищения водотоков
служит их усиленная аэрация по сравнению с другими водными сре-
средами при постоянном обновлении среды около биопленок. В очи-
очищенной воде БПК5 не должно превышать 3 мг/мл.
Проблема чистой воды - одна из важнейших для человечества.
При обсуждении формирования вод следует учитывать ряд аспектов.
1. Водосборную площадь. Она представляет катену - стоковую
серию ландшафтов от автономных к подчиненным. Автономные
ландшафты в нашей равнинной стране представлены лесо-болотны-
ми системами. В них нужно выделить: а) верховые болота с омбро-
трофным (дождевым) питанием, играющие роль водонапорных ба-
башен с ультрапресной водой и создающие обильные гумусовыми ве-
веществами, дистрофные воды; б) леса с эвапотранспирационным ци-
циклом и с подстилкой, где происходит интенсивное разложение орга-
органических веществ; в) агроэкоценозы, служащие источником диф-
диффузного загрязнения со смывом биогенов; г) индустриальные и се-
селитебные площади, служащие локальными источниками сточных
вод. По существу, воды поверхностного стока образуются из поч-
почвенного раствора, и время их формирования составляет менее 1 года.
2. Первичные водотоки - малые реки. Именно к ним относятся
уровни сапробности.
3. Микробное сообщество гидроморфных луговых почв речных
долин и амфибиальных ландшафтов низовых болот. Оно составляет
главный действующий фактор водоохранной зоны. Прибрежные *
172
леса резко сокращают сток мутной воды и снижают бактериальные
загрязнения. Для удержания твердого стока достаточно 30 м лесной
полосы. Низовые болота могут служить источником вторичного за-
загрязнения.
4. Крупные реки с разработанными речными долинами. Они
служат для оценки стока с крупных водосборных площадей. Водое-
Водоемы - проточные озера и водохранилища наиболее интенсивно изу-
изучались в водной микробиологии в плане круговорота веществ в озе-
озерах. Вследствие евтрофикации водоемы могут служить источником
вторичного загрязнения воды.
Основной функцией кооперативного микробного сообщества
здесь является разложение аллохтонного органического вещества.
Наилучшие условия для самоочищения достигаются при гравий-
гравийном или песчаном дне и отсутствии заиливания рыхлыми органиче-
органическими осадками. Самоочищение усиливается с прогреванием воды
летом и затухает зимой. С завершением самоочищения вода стано-
становится лишенной микроорганизмов и чрезвычайно уязвимой для
загрязнения.
При рассмотрении стокового пути вод следует отметить два уз-
узловых участка интенсивных микробных процессов, ведущих к очистке
воды: 1) переход почвенных стоков в воды водотоков; 2) первые эта-
этапы фильтрации через донные отложения и верхние горизонты водо-
вмещающего слоя. Эти процессы проходят в поровом пространстве
и поэтому обусловлены действием микробных сообществ, развива-
развивающихся в биопленках.
5.3. ПОДЗЕМНЫЕ ВОДЫ
Из общего объема вод суши в 90 млн км3 на подземные воды
приходится 60; криолитозону - 29; озера - 0,75; почву - 0,075; реки -
0,0012 (млн км3). Воды подземной гидросферы, находящиеся под
дном морей и океанов, не оцениваются. Питание речного стока на
45% в лесной зоне осуществляется подземными водами, в лесостеп-
лесостепной - на 35%, в степной - на 20%. Питание воды верхних зон интен-
интенсивного водообмена связано с дневной поверхностью. Выделяют
верхнее почвенно-грунтовое, глубоко грунтовое, подземное пита-
питание. Подземные воды разделяются на горизонты: верхний с минера-
минерализацией до 1 г/л, промежуточный солоноватый от 1 г/л, соленый
выше 10 г/л, нижний рассольный выше 35 г/л. Эти воды генетически
связаны с морскими: при перемещении суши относительно базиса
они остаются погребенными в породах. Подземные воды располага-
располагаются в водоносном горизонте, выходы которого на дневную поверх-
поверхность связаны с областью питания и областью разгрузки7.
7 Толстой МЛ., Малыгин В.А. Геология и гидрогеология. М.: Недра, 1988.
173
Подземные воды привлекают внимание исследователей по ряду
причин, как естественнонаучных, так и прагматических. Тесный и
длительный контакт подземных вод с вмещающими породами ведет
к установлению равновесия с минералами этих пород, и таким обра-
образом их состав отражает направление выщелачивания. При длитель-
длительном пребывании в резервуаре процессы затухают и возобновляются
лишь при притоке свежей воды, например при закачке в нефтяные
месторождения, или же на изливе этих вод на дневную поверхность
в источниках и при подземном стоке в моря. Подземные воды слу-
служат источником чистой воды, иногда и ценных минеральных вод.
С другой стороны, в подземные резервуары осуществляется закачка
отходов. Все эти действия идут в очень широких масшатабах, и ан-
антропогенное изменение подземной гидросферы, особенно в насе-
населенных районах, очень существенно. Подземные воды представля-
представляют важнейший ресурс и являются объектом гидрогеологии.
Представление о подземной гидросфере сильно расширилось в свя-
связи с бурением на нефть на глубину нескольких километров. Вопрос
о "нижней границе биосферы" был поставлен Б.Л. Исаченко, когда
на нефтяных скважинах Закавказья был обнаружен излив розовой
воды, обусловленный развитием пурпурных серобактерий. Пара-
Парадокс развития фотосинтетиков в глубине сейчас объясняют их спо-
способностью к хемосинтезу. Общее мнение склоняется сейчас к тому,
что подземная гидросфера населена микроорганизмами, по мень-
меньшей мере, на глубину нескольких километров, но в ней главную
роль играют гидрогенотрофы, связанные с эндогенными источника-
источниками химических веществ.
В подземных водах особую роль играет взаимодействие с твер-
твердыми частицами; собственно важнейшие процессы идут в поровом
пространстве породы с ее растворением в случае образования кар-
карста до цементации в местах развития подщелачивания.
Просачивание воды через пористые породы ведет к их освобож-
освобождению от микроорганизмов, которые можно рассматривать как ин-
индикаторы загрязнения и поступления воды с поверхности; их появ-
появление указывает на наличие подземных водотоков. Фильтрация во-
воды через породы есть путь получения чистой воды. Вместе с тем в
подземных водах, как и в океане, распространены формы "ультра-
"ультрамикробов" размером около 0,3 мкм, возникающие в результате го-
голодания и находящиеся в анабиотическом состоянии. Эти формы
фильтруются через породы с проницаемостью 1,2-18 дарси на рас-
расстояния по меньшей мере 200 м. Поры более 1 мкм доступны для их
проникновения. При нормальном питании ультрамикробы восстана-
восстанавливают размеры нормальных бактерий с образованием обильных
экзополисахаридов.
Обычно коллекторами для подземных вод служат пористые оса-
осадочные породы, например песчаники. В гранитных породах вода на-
находится в трещинах и разломах. Большинство микробов в подзем-
174
ной гидросфере находится в прикрепленном состоянии, и плотность
их составляет 103-107 клеток/мл воды, что сопоставимо с поверхно-
поверхностными водами. Судя по молекулярной идентификации и культураль-
ным методам, наиболее распространен Acinetobacter, за ним следуют
Bacillus, Desulfovibrio, Desulfomicrobium, Pseudomonas, Rhodocyclus,
Shewanella, Thiomicrospira, Zoogloea. Важнейшими процессами в грани-
гранитах являются мобилизация железа и образование сероводорода. Когда
эти воды приходят в соприкосновение с водотоками по трещиновато-
сти, создаются условия для развития аэробных литотрофов.
Подземные воды широко варьируют по гидрохимическому со-
составу от артезианской питьевой воды до насыщенных рассолов со-
солей. Однако все эти воды отличаются недостатком кислорода вплоть
до полного анаэробиоза и возрастанием температуры с глубиной
вследствие геотермического градиента. В этом отношении они смы-
смыкаются с гидротермами. Поэтому можно ожидать развития в них
своеобразной микрофлоры, в том числе и гипертермофильной. Раз-
Развитие подземной микрофлоры происходит над зоной катагенеза, где
доминируют процессы термического превращения веществ.
Развитие микроорганизмов в подземных водах ведет к образова-
образованию газов. Доступное органическое вещество разлагается с удале-
удалением следов растворенного кислорода и насыщением воды углекис-
углекислотой, в том числе и от деятельности первичных анаэробов, проду-
продуцирующих органические кислоты. Продукты брожения используются
вторичными анаэробами. Метаногенез представляется важнейшим
процессом, ведущим к образованию газовых месторождений. За ним
следует сульфидогенез с образованием сероводородных вод и це-
цементацией порового пространства карбонатами и сульфидами. Осо-
Особый интерес представляет образование высших углеводородов,
которые не являются продуктами катаболизма микроорганизмов, а
возникают в результате преобразования этих продуктов в химиче-
химических реакциях. Однако с развитием знаний об экстремофильных ор-
организмах область возможных биологических реакций расширяется,
хотя для доказательства этих реакций требуется работа в конкрет-
конкретных местообитаниях.
Среди экстремальных микроорганизмов особо следует отметить
анаэробных гипертермофилов, которые представляют собой наибо-
наиболее вероятных обитателей подземной гидросферы, в некоторых ус-
условиях оказавшихся вблизи поверхности. Среди них важны лито-
трофные организмы, использующие водород эндогенных реакций в
нижележащих слоях на контакте подземной гидросферы с нагреты-
нагретыми изверженными породами. В соленых водах нефтяных месторож-
месторождений Африки, по данным французских исследователей, важную
группу составили галоанаэробы и галофильные сульфатредукторы.
В нефтяных водах глубинных скважин Северного моря широко рас-
распространенными оказались термофильные железо-, марганец- и ни-
тратвосстанавливающие организмы. В породах распространены ме-
175
таногены, составляющие значительную часть микробного населения
подземных вод, причем метилотрофные и ацетокластические метано-
гены доминируют в неглубоких подземных водах, а литотрофные рас-
распространены вне зависимости от органического вещества, причем
источником энергии предполагается водород геологического проис-
происхождения. Силикаты железа изверженных пород реагируют с анокси-
генными водами с продукцией водорода, что ведет к серпентинизации.
В результате оказывается возможным выращивать в лаборатории
анаэробов с базальтом, выщелачивание которого служит единствен-
единственным источником энергии. Нерастворимые акцепторы электрона, как
окислы железа и марганца или гипс, оказываются особенно характер-
характерными окислителями в условиях подземной гидросферы.
Одним из важных последствий деятельности железовосстанавли-
вающих бактерий в подземной гидросфере является ее насыщение
Fe(II), которое может представлять значительные затруднения в во-
водоочистке. К этим бактериям относятся Geobacter, Geothrix, Bacillus,
Shewanella. В культуре необходим прямой контакт клеток с окислами,
но в природе возможны и комплексы, и коллоиды. Добавление хели-
рующих соединений, в том числе гуматов, усиливает восстановление
железа в подземных водах in situ. Гуматы работают как переносчик-
шаттл между клеткой и окислом. Это действие в лаборатории симу-
симулируют хиноны: микробы восстанавливают хиноны в гидрохинон, ко-
который реагирует с Fe(III), восстанавливая его и окисляясь в хинон.
В области просачивания поверхностных вод действует окисли-
окислительный бактериальный фильтр, создающий условия для окисления
углеводородов и восстановленных неорганических соединений.
Многие подземные хранилища, особенно для радиоактивных отхо-
отходов, предполагают соприкосновение с цементом, которое создаст
высокощелочные условия вплоть до рН 13,5. В Иордании существу-
существуют источники подземных вод в богатом органическим веществом
мергеле с рН 12,9. Исследование 16S рРНК выявило присутствие в
этих источниках протеобактерий, в том числе Acinetobacter, демон-
демонстрирующее развитие в гиперщелочных водоемах широкого разно-
разнообразия бактерий.
5.4. ОКЕАН
5.4.1. Зональность океана
Открытый океан не следует рассматривать как однообразную
морскую массу. Он разделяется на несколько регионов, между кото-
которыми происходит медленный обмен благодаря системе течений,
поверхностных и глубинных (рис. 5.2). В океане можно выделить
Рис. 5.2. Обмен поверхностных и глубинных вод в океане
[по: Me Cartney, 1994]
176
11
is a
полярные зоны (Арктика и Антарктика), зоны циркуляции в сред-
средних широтах, апвеллинга в экваториальных водах.
Океан разделяется на пелагиаль с большими глубинами и нери-
тическую область, находящуюся вблизи континентов, подвержен-
подверженную большому влиянию процессов, происходящих на дне, и деятель-
деятельности бентоса.
Для биолога пелагиаль океана не представляет однородной мас-
массы перемешанной воды, а разделяется на географические области,
определяемые климатом, течениями, влиянием континентов
(рис. 5.3). Они были предложены Свердрупом и, соответственно,
включают: полярный биом, область западных ветров (westerlies),
пассатную область (trades), береговую пограничную зону и внезо-
нальные области: переходные слои и бедные хлорофиллом области
в центральных частях океана. Дистанционное зондирование хлоро-
хлорофилла в океане отчетливо выделяет эти области.
Полярный биом характеризуется сезонностью и наличием ледо-
ледового покрова. При таянии его весной образуется поверхностный оп-
опресненный слой богатый нитратами, обеспечивающими цветение.
Устойчивый градиент плотности около отступающего весной ледо-
ледового покрова создает условия для максимального развития биоты.
Цветение продолжается обычно дней 10 и завершается исчерпанием
нитрата на севере. Вокруг Антарктиды исчерпания нитрата не про-
происходит. Образующийся при похолодании лед всплывает и включа-
включает в себя водоросли, в то время как термогалинная стратификация
обеспечивает устойчивость. Проникновение света сквозь лед созда-
создает возможность развития на рыхлой нижней стороне льда биоцено-
биоценоза, включающего водоросли и всплывающие бактерии с газовыми
везикулами, исследованные Д. Сталей.
В зоне западных ветров имеется сильное ветровое перемешива-
перемешивание, изменение освещенности из-за сезонности и облачности. По атмо-
атмосферному полярному фронту распространяются штормы. Пикноклин
устанавливается весной, и за ним следует цветение фитопланктона,
особенно в Северной Атлантике, где оно представляет исключи-
исключительное явление среди наблюдаемых дистанционными методами яв-
явлений. Оно было исследовано Свердрупом и послужило моделью
для объяснения цветения. Причина заключается в обогащении нит-
нитратами подповерхностного слоя. Цветение начинается, когда пере-
перемешивание выводит этот слой на фотическую поверхность. За ним
следует олиготрофный период с преимущественно регенерацион-
ным циклом и NHj как ведущей формой азотистого питания.
Иногда лимитация вызывается силикатами, поскольку основное
трофическое звено составляют диатомея Chaetoceros и копепода
Calanus. В более низких широтах цветение вызывается ветровым
сгоном и подъемом вод с нитратом. Здесь располагается переходная
зона к олиготрофному "голубому" океану. Весной стратификация
178
60
120
180
120
Рис. 53. Зональность океана [по: Кафаров, Кудряшов, 2000]
Северное полушарие: А — полярная (арктическая), Б - субполярная (субарктическая), В —
умеренная (бореальная), Г - субтропическая, Д - тропическая (пассатная), Е - экватори-
экваториальная
Южное полушарие: Ж -тропическая, 3 - субтропическая, И - умеренная, К - субполярная
(субантарктическая), Л — полярная (антарктическая) (Арабскими цифрами A—13) обозна-
обозначены соответствующие природные зоны суши)
воды поддерживает на плаву крупные диатомовые и фитофлагелля-
ты, располагающиеся на пикноклине, а летом доминирование пере-
переходит к неоседающему пикопланктону и мелким эукариотам.
Пассатная область обычно называется тропическим океаном, за-
занимающим 45% площади. В центральной части океанов располагают-
располагаются олиготрофные халистазы, обусловленные круговыми течениями.
Солнечная радиация поддерживает термическую стратификацию, и
пикноклин очень устойчив. Пелагический биом здесь очень разнооб-
разнообразен и оценивается как климаксное состояние экосистемы. Вместе с
тем малая продуктивность совпадает с областью развития пикопланк-
тона. Основное внимание привлекло развитие азотфиксирующей
цианобактерии Trichodesmium, хотя азотфиксацию осуществляют и
симбиотические организмы с диатомовыми, и сообщества в Саргассо-
вом море. Развитие Trichodesmium прослеживается в зонах выпадения
эоловой пыли, обеспечивающей сообщество другими элементами.
Прибрежные области очень различаются и друг от друга и вну-
внутри их самих. Основным фактором служит взаимодействие с сушей.
Речной сток особенно важен в экваториальной области, где с суши
179
выносится очень много материала, в особенности в северную часть
Индийского океана реками Восточной Азии, а в Атлантический оке-
океан - Нигером, Конго, Амазонкой. Границей области следует считать
континентальный склон, который служит препятствием для глубоких
течений и создает фронт. На западных берегах континентов создает-
создается апвеллинг, обеспечивающий высокую продуктивность, особенно у
побережья Перу и Калифорнии, вблизи течения Гумбольдта, у Канар-
Канарских островов, вблизи течения Бенгуела у юга Африки.
Островные дуги Курильских и Алеутских островов, в которые
упираются течения, создают условия для почти постоянного цвете-
цветения фитопланктона, в то время как центральная часть Охотского
моря олиготрофна. В зоопланктоне прибрежной области большую
роль играют личинки многоклеточных бентосных организмов.
Особые области создаются в случае образования опресненного
пикноклина вследствие сильных дождей, как в западной части тро-
тропического Тихого океана, где возможны минимумы кислорода в глу-
глубине фотической области и развитие пурпурных бактерий; в облас-
областях ограничения роста не азотом и фосфором, а другими элементами,
например железом, как в восточной Пацифике; там, где раститель-
растительноядный зоопланктон не в состоянии сдерживать вспышки развития
фитопланктона. Перечисленные области океана разделяются на
"провинции". Они требуют рассмотрения тех особых ситуаций,
которые здесь создаются8.
Биогеохимические циклы С, N, Р в океане обусловлены энерги-
энергией Солнца, поступающей на поверхность океана, где может разви-
развиваться фитопланктон. Фотическая зона простирается менее, чем на
200 м. Ниже этого слоя идут процессы деструкции. В океане строит-
строится трофическая система, дополняемая транспортными процессами.
C-N-P циклы в океане имеют и глобальные и региональные состав-
составляющие с характерным временем от часов, суток и сезонов (для ло-
локальных процессов), до еще более длительных периодов, когда во-
вовлекается глубинная циркуляция вод.
Утверждается, что олиготрофный океан в качестве продуктив-
продуктивного начала имеет бактериальное звено, в противоположность про-
продуктивным зонам, где доминирует эукариотный фитопланктон диа-
диатомовых, кокколитофорид, жгутиковых. Биологическая помпа в
океане извлекает 5-7,4 Гт/год углерода, примерно 10% от глобальной
продукции. Вместе с тем пелагиаль океана имеет замкнутый цикл
органического вещества, в котором происходит полное разложение
оседающей первичной продукции, не достигающей абиссали. Отсю-
Отсюда следует, что биотическое звено пелагиали океана мало влияет на
глобальные процессы, не имея дисбаланса по элементам. Масса во-
воды глубокого океана работает прежде всего как физическая и физи-
физико-химическая система с определяющими процессами растворения
8 Longhurst A. Ecological geography of the sea. San Diego: Acad. press, 1998.
180
веществ в зависимости от температуры и давления. Она же осущест-
осуществляет медленные транспортные процессы соответственно конвейе-
конвейеру глубинных течений.
Органический углерод в море оценивается операционно: к рас-
растворимому относят проходящий через фильтр 0,2-1 мкм:
Растворенный органический углерод
Коллоидный (< 1 мкм) органический углерод
Мертвый органический углерод (РОС)
Биомасса
В том числе:
бактерии
протисты
планктон (фито- и зоо-)
Всего органического углерода
По меньшей мере половина органического углерода в океане
имеет возраст более 6000 лет, и, следовательно, доля доступного
для органотрофов углерода много меньше. Взвешенный органиче-
органический углерод (детрит) по массе превосходит живой на один-два
порядка.
Микроорганизмы в море распределены не равномерно или хао-
хаотически, а сгруппированы в сгущения разного размера (Sorokin,
i Таблица 5.4
Распределение микробной биомассы в океане
[по: Sorokin, 1999]
ГтС
100
100
20
1,6
0,4
0,1
1,2
222
%
45
45
9
0,7
0,2
0,1
0,5
Область океана
Глубина, м
Площадь, | Объем,
• 108км2 I • 106км3
Биомасса,
• 1012гС
Неритическая(над
континентальным
шельфом)
Эпипелагическая
Мезопелагическая
Батипелагическая
Абиссопелагическая
Эуфотическая
Афотическая
Всего
0-100
100-200
0,36
0,36
Провинции океана
0-100
100-200
200-1000
1000-4000
>4000
3,26
3,26
3,26
3,19
2,1
Океан в целом
0-100
100-200
>200
3,62
3,26
3,62
3,6
3,6
32,6
32,6
316
805
183
36
1304
1376
135^04
36-108
204-611
82-245
158^74
20-60
2,3-6,9
339-1016
118-353
180-541
637-1910
21,2
5,7
32,0
12,8
24,8
3,1
0,4
53
19
28
100
181
1999), что представлено в табл. 5.4. В ней принимаются следующие
оценочные значения:
плотность бактерий 1,035-1,093 г/см3;
сухой вес/упакованная биомасса обычно оценивается в 20%;
углерод/объем обычно принимается 0,2 ¦ 10~12 г/мкм3;
углерод/клетка бактерии 10 • 10~15 г/клетка.
Воды океана отличаются большим постоянством химического
состава. Они характеризуются соленостью, варьирующей в узких
пределах. Соленость есть суммарное содержание в граммах всех
твердых минеральных растворимых веществ, содержащихся в 1 кг
морской воды, при условии, что бром и йод замещены эквивалент-
эквивалентным количеством хлора, все углекислые соли переведены в окислы,
а все органические вещества сожжены при температуре 480 °С
(г. Стокгольм, 1899). Между содержанием хлор-иона в морской воде
и общей концентрацией солей существует жесткое соотношение.
В 1966 г. ЮНЕСКО рекомендовала соотношение между соленостью
и хлорностью для морской воды Ъ%о = 1,81578 С1%о, где знак %о
"промилле"(рго mille) означает весовые части на тысячу. За стан-
стандартную морскую воду принята вода из центральной части Бискай-
Бискайского залива.
Искусственная морская вода имеет следующий химический со-
состав (в промилле по массе, или г/кг): NaCl - 23,926; Na2SO4 - 4,008;
КС1 - 0,677; NaHCO3 - 0,196; KBr - 0,098; H3BO3 - 0,026; NaF - 0,003;
MgCl2 - 53,27 мл 1М раствора; СаС12 - 10,33 мл 1М раствора; SrCl2 -
0,9 мл 0,1 M раствора.
Отсюда фоновая среда для культивирования морских микроор-
микроорганизмов (в г/л): NaCl - 24; Na2SO4 - 4; КС1 - 0,7; NaHCO3 - 0,2; KBr-
0,1 и 1М растворы (в мл): MgCl2 - 53; СаС12 - 10. Далее добавляются
растворы микроэлементов. Следует особо заметить, что в связи с
постоянством состава воды океана морские микроорганизмы очень
чувствительны к его изменениям и многие не растут на искусствен-
искусственных растворах, требуя натуральную морскую воду. Далее следует
заметить, что повышение солености при испарении морской воды
оказывается неблагоприятным для морских организмов, и их сменя-
сменяют галофилы.
5.4.2. Фитопланктон и первичная продукция
Начало углеродного цикла в открытом океане определяется
развитием фитопланктона. Величину продукции можно оценить либо
прямыми методами по ассимиляции 14С или продукции О2 в склян-
склянках, либо по коррелирующему признаку - содержанию хлорофилла,
который можно определять в толще воды по флуоресценции, а для
поверхностных вод - дистанционными методами. Типичное соотно-
182
шение хлорофилла и продукции (по Lally, Parsons, 1997) приведено
ниже:
Индекс ассимиляции,
мг С / (мг Хлорофилла а ¦ час)
2-14
2-3,5
6-10
0,2-1,0
9-17
Условия ограничения
Общий предел
Низкая температура B-4 °С)
Высокая температура (8-18 °С)
Недостаток биогенов
Избыток биогенов
Дистанционное зондирование подтвердило полученное ранее су-
судовыми методами распределение областей продукции в океане и
связь ее с глубинными течениями, выносящими на поверхность пи-
питательные вещества (рис. 5.4). Океан оказался весьма неоднороден,
и в нем имеются области высокой продуктивности в прибрежных зо-
зонах и в зонах апвеллинга и очень низкой продуктивности "голубо-
"голубого", или олиготрофного, океана.
Продукция (первичная продукция) - количество органического
вещества в граммах, создаваемого организмами в единице объема
воды (или под единицей площади акватории) за единицу времени.
В высокопродуктивной зоне фотосинтез фитопланктона идет до
проникновения 10% падающей ФАР, в малопродуктивной - до 0,1%
ФАР, что и считается нижней границей зоны фотосинтеза. Диато-
Диатомовые приурочены к высокопродуктивным зонам, повсеместны
празинофиты, динофлагелляты (особенно в теплых водах), кокко-
литофориды, силикофлагелляты в холодных. Зеленые водоросли
отсутствуют. Продукция заключается в пределах 100-500 мг
С/(м2 • сут). 77% первичной продукции дают диатомовые, 22% - пе-
ридинеи, 1% - цианобактерии (сейчас оценка вклада цианобактерий
существенно возросла за счет пикопланктона). Оценка продукции в
разных зонах океанов дает следующие значения:
Средняя годовая продукция г С/(м2 • год)
Континентальный апвеллинг (Перу, , 500-600
Бенгуела)
Континентальный шельф (Европа) 300-500
Субарктический океан (Северные Атлантика, 150-300
Пацифика)
Антициклонические вихри (Саргассы) 50-150
Планктонная экосистема ограничена прежде всего поступлени-
поступлением биогенов к первичным продуцентам с очень коротким жизнен-
жизненным циклом. Лимитация роста фитопланктона определяется в пер-
первую очередь поступлением нитратов и фосфатов, но также зависит
и от микроэлементов, в числе которых оказалось железо. Фото-
трофные продуценты имеют малую биомассу и очень высокую
183
Рис. 5.4. Первичная продукция в океане [по: Кобленц-Мишке, 1985]
а - по результатам определения радиоуглеродным методом; б - по косвенным данным; 1 -
< 100; 2 - 100-150; 3 - 150-250; 4 - 250-500; 5 - >500 мгС/м2; б - границы круговоротов.
Круговороты: Сб Ар - субарктический; Сб Тс - северный субтропический; Сб Тю -
южный субтропический; Тс - северный тропический; Тю - южный тропический; Тар -
тропический (Аравийского моря); Тб - тропический (Бенгальского залива); Сб Ан - суб-
субантарктический; Ан — антарктический
оборачиваемость, обеспечивая развитие гораздо большего по массе
зоопланктона. Поэтому планктонная система определяется рецик-
рециклом биогенов:
Новая
продукция
NO3
Апвеллинг
Регенерированная
продукция
-Аммонификация —
Рецикл—
Нитрификация
Фотическая
зона
Зона
регенерации
Осаждение
N,
орг
Нижняя граница продуктивной зоны определяется скачком в со-
содержании биогенов ("нутриклин") и находится на глубине несколь-
нескольких десятков метров. Освобождение биогенов в фотической зоне со-
создает малый цикл "регенерационной продукции".
Основную роль в фитопланктоне продуктивных областей океана
играют микроскопические водоросли и протисты, дающие заведомо
более 50% первичной продукции и представлявшиеся единственными
продуцентами до относительно недавнего времени. Важнейшими
группами фитопланктона являются диатомовые водоросли и фитоф-
лагелляты: динофлагелляты, хризомонады, криптомонады. Эукари-
отный фитопланктон появился во время неопротерозойской револю-
революции около 1 млрд лет назад, регистрируемой по массовому распро-
распространению акритарх.
Хризомонады имеют скелетные структуры, состоящие из кар-
карбоната кальция, как у Emiliana, Coccolithus, Pontosphaera, и вызыва-
вызывают цветения в теплых окраинных морях, внося значительный вклад
в известковые осадки на дне океана. К силикофлагеллятам относят-
относятся наиболее обычные Distephanus и Dictiocha. В теплых морях цвете-
цветение вызывает Phaeocystis с крупными слизистыми агрегатами кле-
клеток. Динофлагелляты, как Gymnodinium, имеют крупные размеры и
хитиноидные панцири, часть из них способна к фаготрофии. Диноф-
Динофлагелляты развиваются в конце вегетационного сезона и, обладая
185
способностью к вертикальной миграции, ночью опускаются к пик-
ноклину и питательным веществам, а днем поднимаются на свет,
перемещаясь на 20 м по вертикали. Основную массу фитопланктона
в умеренных и холодных водах составляют планктонные диатомо-
диатомовые водоросли. Часто они имеют выросты для парения, как наибо-
наиболее распространенные Chaetoceros, Asterionella, Thalassiosira. С ними
связаны кремнистые осадки умеренных широт.
Важную часть автотрофного планктона составляют инфузории
с включенными в клетку и функционирующими хлоропластами
жертв или симбиотическими водорослями. Наиболее известен
Mesodinium как массовая форма с очень широким пределом приспо-
приспособления к внешним условиям и вызывающая цветения, особенно в
водах с неустойчивым режимом. Мезодиниум обладает очень высо-
высокой скоростью миграции - 2,5 мм/с и способен проходить через пик-
ноклин за питательными веществами.
Для развития фитопланктона характерны ритмические явления,
связанные с массовым развитием одной из форм - цветением, ее от-
отмиранием и сменой другой формой, приспособленные к сезонным
событиям. В морях умеренного климата максимумы фитопланктона
наблюдаются весной и осенью с разделяющими их минимумами в се-
середине лета и зимой. Весной при таянии льда диатомовые развива-
развиваются на образующемся на поверхности пикноклине, как на жидком
дне. Образованное в периоды цветения органическое вещество ис-
используется в органотрофную фазу минимума хлорофилла.
В состав морского фитопланктона входят цианобактерии и пи-
коэукариоты, участие которых недоучитывалось в прошлом из-за
потерь при сборе и фиксации материала. Цианобактерии в планкто-
планктоне привлекли особое внимание, во-первых, в связи с азотфиксацией
Trichodesmium и гетероцистных цианобактерии Richelia inracellularis,
симбионтов диатомей Rhizosolenia, Hemialus, Chaetoceros, во-вторых,
в связи с присутствием в пикопланктоне, представленном
Synechocystis, Synechococcus, Prochlorococcus. Иную группу представ-
представляют цианобактерии прибрежной зоны, которые следует рассмат-
рассматривать как представителей бентоса9.
Планктонным цианобактериям сейчас уделяют все больше вни-
внимания (Paerl, 2000). Допускается, что в современном океане пико-
планктон поставляет до 30-50% первичной продукции. Распределе-
Распределение хлорофилла между фракциями по размерности обнаружило
существование в олиготрофном малопродуктивном океане пико-
планктона как основной части продуцентов. В нем в 1979 г. были об-
обнаружены по оранжевой флуоресценции фикоэритрина при возбуж-
возбуждении 540 нм мелкие медленнорастущие виды цианобактерии, а за-
9 Golubii S., Le Campion-Alsumard Т., Campbell S.E. Diversity of marine cyanobacte-
ria // Bull, de Inst. oceanogr. Monaco. 1994. Special number 19: Marine cyanobacte-
ria. P. 53-76.
186
тем и прохлорофиты. Им приписывалось 10% первичной продукции,
но в малопродуктивном олиготрофном океане. Численность их дос-
достигает 106 клеток/мл. Они приурочены к относительно теплым во-
водам и не встречаются в полярных. Все они облигатные фототрофы,
хотя возможность ассимиляции органических веществ имеется.
К азотфиксации они неспособны, и nif ген у них не обнаружен, они ис-
используют аммоний и нитрат, но могут ассимилировать и аминокисло-
аминокислоты. В условиях лимитации фосфатом они синтезируют периплазмати-
ческий фосфат-связывающий белок, передающий фосфат к системе
высокой аффинности, сходной с системами пресноводных цианобакте-
цианобактерии. Имеется и система высокой аффинности к железу. Причины доми-
доминирования этого типа цианобактерии в олиготрофном океане неясны.
Ниже приводится список и некоторые свойства морских планк-
планктонных цианобактерии (по Paerl, 2000).
Группа
Одноклеточные
одиночные
колониальные
Нитчатые
В том числе
образующие
гетероцисты
Род
Synechococcus
Prochlorococcus
Synechocystis
Aphanothece
Merismopedia
Lyngbya
Oscillatoria
Phormidium
Spirulina
Trichodesmium
Anabaena
Aphanizomenon
Nodularia
Richelia
^-фиксация Характеристики
- Обычны в олиготроф-
олиготрофном океане
- Цветение в гиперсоле-
гиперсоленых лагунах
- Цветение в лагунах
+ В богатых водах
- В богатых водах
+ Агрегаты, часто
с протестами
+ С планктонными
водорослями
- Одиночно
и с протестами
- В гиперсоленых водах
+ + + Агрегаты при цвете-
цветении в тропическом
океане
+ Одиночно в лагунах
+ Цветение в богатых
водах
+ Цветение
в прибрежных водах
+ Эндосимбионт пелаги-
пелагических диатомовых
Азотфиксирующие цианобактерии распространены в разных
условиях в прибрежной зоне, но для открытого океана интерес пред-
представляет массовая колониальная форма красного цвета
Trichodesmium в тропическом океане, лимитированном по азоту, спо-
способность которого к азотфиксации была установлена только в
1961 г. Другой потенциальный азотфиксатор - осциляториевая циа-
нобактерия Katagnymene - пока описана лишь в одиночных работах.
187
Далее были обнаружены коккоидные диазотрофные цианобакте-
рии, обнаружен и ген nifli. Возможно, что Trichodesmium не исчер-
исчерпывает фототрофных диазотрофов океана. Развитие его лимитиру-
лимитируется температурой, доступностью фосфата и Fe. Современная оцен-
оценка азотфиксации в море составляет 80-125 Тг N/год. Trichodesmium
может быть ответственен за 65 Тг N/год10.
Многоклеточные водоросли развиваются в прибрежной зоне и
на мелководье. Макроводоросли в океане принадлежат трем глав-
главным группам: зеленым, красным, бурым. Красные и бурые почти не
имеют пресноводных представителей и развиваются в море. Макро-
Макроводоросли развиваются в прибрежной литоральной зоне с большой
интенсивностью процессов биологического круговорота, взаимо-
взаимодействием с сушей, приливно-отливным и волновым режимом. Зеле-
Зеленые водоросли разделились на две большие ветви, из которых пра-
зинофиты, по-видимому, представлены микрофоссилиями акритар-
хами в геологической летописи.
Из-за повторных явлений эндосимбиоза многие группы водорос-
водорослей возникли как химеры. Так, Chlorella, Chlamydomonas оказались
филогенетически сборными группами. Зеленых водорослей в план-
планктоне моря мало11.
Вертикальное распределение фитопланктона зависит от двух фа-
факторов: проникновения света сверху и поступления биогенных эле-
элементов снизу, из зоны деструкции. Поэтому максимум развития при-
приходится не на поверхностные воды, а располагается ниже на глубине
десятка-двух метров с четкими максимумами хлорофилла и активно-
активности ассимиляции. Виды фитопланктеров обнаруживают четкую стра-
стратификацию. На эту общую картину накладываются локальные влия-
влияния, например мутность воды в прибрежной зоне. При оценке авто-
трофной ассимиляции и первичной продукции по хлорофиллу прихо-
приходится учитывать их не строгую корреляцию (см. выше Lally, Parsons,
1997). Ассимиляционная величина хлорофилла Ахл определяется
отношением ассимилированного углерода в мг С/(м3 • час) к содер-
содержанию хлорофилла в мг/м3. Первичная продукция варьирует от
0,2-0,4 г С/(м2 • сут) в олиготрофном океане до 4-6 г С/(м2 • сут) в зонах
апвеллинга. Самозатенение фитопланктона приводит к верхнему пре-
пределу плотности 20-40 г/м3. Скорость ассимиляции следует за свето-
световым режимом суток с максимумом в полдень (Sorokin, 1999).
В олиготрофом океане основная часть хлорофилла находится в
клетках очень мелких прохлорофит Prochlorococcus и цианобакте-
10 Саропе D.G., Carpenter E.A. Nitrogen fixation by marine cyanobacteria: Historical
and global prespectives // Bull. Inst. oceanogr. Monaco. 1994. Special number 19:
Marine cyanobacteria. P. 235-256.
1' Medlin L., and Simon N. Phylogenetic analysis of marine phytoplankton // Molecular
approaches to the study of the ocean / Ed. K.E. Cooksey. L.: Chapman and Hall, 1998.
P. 162-186.
188
рий Synechococcus, которые с трудом, но поддаются культивированию.
Их скорости оседания очень малы, плотности ниже выедания зооплан-
зоопланктоном. В олиготрофном океане продукция зависит только от регене-
регенерации, и отношение новой продукции к общей составляет около ОД.
Напротив, в области апвеллинга это отношение может быть 0,8.
При высоком содержании биогенов в прибрежных водах иногда
происходит массовое развитие водорослей, особенно перидиней, от-
ответственных за токсические "красные приливы". Высокая плот-
плотность фитопланктона создает благоприятные условия для лизиса во-
водорослей фагами и вирусами фемтопланктона, которые оказались
не только многочисленны в водах морей, но и важным фактором ли-
лизиса продуцентов.
Численность вирусов в море составляет от 104 до 108 частиц в
1 мл воды и коррелирует с плотностью микробного населения. Мор-
Морфологически они разнообразны. Частицы вирусов не представляют
устойчивого компонента и после выхода в открытую воду теряются
из нее за относительно короткое время, причем численность частиц
резко варьирует. Они могут исчезать при фильтрации зоопланкто-
зоопланктоном. Вирусы рассматриваются как факторы, контролирующие чис-
численность фитопланктона. Оценочная величина лизиса бактерио-
планктона, обусловленного фагами, составляет 10% в сутки.
Synechococcus лизируется на 1-30% в сутки. Вирусы определенно иг-
играют роль ограничивающего фактора при цветении, поскольку цве-
цветение вызывается генетически однородной популяцией организмов,
например Emiliania huxlei. Вместе с тем цианобактерии быстро соз-
создают устойчивые к фаголизису популяции12.
5.4.3. Регенерационный цикл азота в океане
Определяющим для фитопланктона служит цикл азота, поэтому
особое значение приобретает роль бактерий в цикле азота в море.
Согласно гипотезе К. Брандта A899) азотфиксация на суше уравно-
уравновешивается денитрификацией в море. Работы Б.Л. Исаченко13
A951 г.) в Баренцевом море продемонстрировали присутствие в оке-
океане всех групп бактерий цикла азота, чем было доказано, что цикл
может быть замкнут.
Развитие фитопланктона в современном океане определяется в
первую очередь доступностью связанного азота. Согласно гипотезе
Свердрупа весеннее развитие фитопланктона обусловливается по-
12 Proctor M. Marine virus ecology // Molecular approaches to the study of the ocean /
Ed. K.E. Cooksey. L.: Chapman and Hall, 1998. P. 113-130. {"virus is a noncellular
genetic element that enlists a living cell for its own reproduction"... The extracellular
form - the phage or virus particle - is metabolically inert.).
13 Исаченко БЛ. Исследования над бактериями Северного Ледовитого океана //
Тр. Мурм. науч.-пром. эксп. 1906 г. Пг, 1914.
189
ступлением нитрата из глубоких вод. После исчерпания нитрата на-
наступает фаза деструкции с освобождением аммония, который ис-
используется для регенерационной продукции, поскольку нитрифика-
торы не могут конкурировать с ассимиляцией азота. Тем не менее, в
море установлены специфические для морских условий формы нит-
рификаторов, Nitrosococcus, которые и отвечают за образование ни-
нитратов как основной формы азота в глубинных водах. Однако ис-
источник нитрата в придонных водах неясен. Денитрификация воз-
возможна только в аноксигенных условиях либо в осадках, либо
локально. Тем не менее ей приписывается регулирующая роль факто-
фактора, не допускающего избыточной продукции.
Баланс элементов и регуляция на глобальном уровне
Биотические процессы в массе вод открытого океана определя-
определяются развитием фитопланктона и последующим его потреблением в
цепи консументов и при бактериальной деградации. Масса фито-
фитопланктона в море относительно невелика, и первичная продукция
поддерживается быстрой оборачиваемостью в регенерационном ци-
цикле. Развитие фитопланктона происходит в фотическом слое и в
океане ограничено доступностью в первую очередь азота и фосфо-
фосфора. Остальные лимитирующие факторы, как кремний или железо,
проявляются в региональных условиях. Состав биомассы фитоплан-
фитопланктона определен отношением Редфильда C:N:P= 106: 16: 1, а с
учетом расхода на дыхание растворенного О2 - 138. По уточненным
данным, это отношение колеблется в пределах
Р : N : С : (С + Сдых) = 1 : 16 (±1) : 117 (±14) : 170 (±10).
Отсюда следует, что фитопланктон определяет соотношение
биогенных элементов в современном океане. В большинстве случа-
случаев лимитирующим компонентом служит нитрат, образуемый при
нитрификации аммонийного азота, освобождаемого при разложе-
разложении мортмассы фитопланктона, при потреблении зоопланктоном
(продуктом служит мочевина), при бактериальной аммонификации
или же при лизисе. Этот азот используется в регенерационном цик-
цикле в фотическом слое и под ним. Экспортная продукция уходит в
глубокий океан и создает там отношение N : Р = 15. Пополнение ци-
цикла идет за счет азотфиксации, которая хорошо известна для сооб-
сообществ Trichodesmium, развивающихся только в теплых водах и лими-
лимитированных фосфатом. Цианобактериальный пикопланктон
способностью к азотфиксации не обладает. Для областей высокой
продуктивности, как в Южном океане вокруг Антарктиды, следует
искать иные пути азотфиксации, например симбиотической фикса-
фиксации с диатомовыми водорослями. Потеря азота из системы может
идти за счет денитрификации в аноксических условиях.
Разложение органического вещества сбалансировано с потреб-
потреблением О2 при дыхании и поэтому может вести к образованию ано-
190
ксических условий при избыточном поступлении экспортной про-
продукции. Сейчас доля аноксических условий в океане оценивается в
14%, и их распределение совпадает с известными областями денит-
денитрификации и избытка новой продукции фитопланктона над имею-
имеющимся запасом растворенного О2 в 331 мкмоль/кг при 2° и 35%о со-
солености.
Источником фосфора в океане служит выветривание континен-
континентов и вынос фосфатов в океан, где их концентрация 2,2 мкмоль/кг
при общем резервуаре 3,1 • 1015 моль. Фосфат экспортной продукции
может захораниваться в осадках и выходить из цикла в трех основ-
основных процессах захоронения: 1) органического фосфата; 2) Са-фос-
фатов; 3) Fe-фосфатов. В аноксических условиях происходит осво-
освобождение фосфата в водную толщу и вынос его при апвеллинге. В
зоне апвеллинга происходит полное использование лимитирующего
компонента.
Отсюда может быть построена модель круговорота в современ-
современном океане с взаимосвязанными резервуарами кислорода, фосфора,
нитрата, обусловленными жесткими соотношениями элементов в
продукции фитопланктона. Резервуар кислорода определяется захо-
захоронением Сорг в осадках. Таким образом, скорость седиментации в
сочетании с новой продукцией фитопланктона является главным ре-
регулирующим фактором. Денитрификация в аноксических условиях
ограничивает образование новой продукции и, следовательно, ведет
к уменьшению аноксических условий. С другой стороны, критиче-
критическим для развития фитопланктона, и особенно цианобактерий, слу-
служит выветривание на континентах с поступлением фосфатов с реч-
речным стоком. В аноксических условиях происходит освобождение
фосфатов из осадков под действием анаэробных процессов.
В аноксических условиях происходит, с одной стороны, усиление
захоронения органического вещества из-за меньшей скорости разло-
разложения, а с другой - ускорение выделения фосфатов из аноксических
осадков. Тогда работают положительные обратные связи, и система
производит больше захороненного органического вещества и соот-
соответственно больше О2, и усиление первичной продукции обусловли-
обусловливает положительную обратную связь. Избыточный кислород, если он
не употреблен на окисление выходящих из системы восстановителей,
таких как окислы железа или сульфат, усиливает разложение и оста-
останавливает освобождение фосфатов из осадков. О2 осуществляет от-
отрицательную обратную связь в биотической системе океана.
Циано-бактериальное сообщество протерозоя, лимитированное
фосфором, создавая избыточную новую продукцию, способствова-
способствовало образованию аноксических условий на глубине и освобождению
фосфатов (и железа) из осадков. Удаление фосфора из цикла, на-
например при образовании месторождений фосфатов, связанных по-
повсеместно с циано-бактериальными матами, приводит к снижению
продукции и, соответственно, потребления О2 на дыхание и появле-
191
нию избыточного окислительного потенциала (например для закис-
ного железа, мигрировавшего при аноксии). Цианобактериальный
океан протерозоя мог не иметь лимитации по азоту в отличие от со-
современного эукариотного, и тогда только поступление фосфатов из
областей субаэрального углекислотного выщелачивания и субак-
вальных гидротерм определяло циклы. Отсюда углекислотное вы-
выщелачивание в субаэральных условиях на континентах могло играть
определяющую роль не только в удалении СО2, но и в создании усло-
условий "содового континента", обеспечивающего преимущественную
миграцию фосфата. Отсутствие растительного покрова освобожда-
освобождало фосфор от улавливания в резервуаре наземной биомассы/мор-
тмассы. Роль биогенного выщелачивания субаэральными матами и
формирования протерозойских палеопочв (Retallack, 1990) ока-
оказывается определяющей также и для понимания круговорота в
океане14.
5.4.4. Деструкция
Растворенное и взвешенное органическое вещество
Система планктона океана находится под контролем физиче-
физических факторов, из которых на первом месте стоит освещенность и
суточные ритмы с вертикальным движением планктона. Сле-
Следующие ритмы задает сезонность. Пики весенного развития
фитопланктона и зоопланктона несколько разнесены по вре-
времени, и зоопланктон иногда запаздывает. Тогда обычная для моря
цепь продуцент-консумент-редуцент меняется на цепь проду-
продуцент-редуцент с разложением органического вещества в донных
осадках.
В горизонтальном направлении развитие планктона определя-
определяется поверхностными течениями и областями циркуляции, из кото-
которых важны крупные вихри, которые перемещают массы воды. Ци-
Циклонические вихри с низким давлением в середине приводят к ап-
веллингу с подъемом холодной воды. Апвеллинг возникает и при
сгоне воды от берега ветром, как у берегов Чили, Перу или Запад-
Западной Африки.
Органические вещества поступают воду в виде экссудатов - рас-
растворенного органическое вещества (РОВ) и оседающего взвешенно-
взвешенного органического вещества (ВОВ) отмершего фитопланктона. Дест-
Деструкция и регенерация биогенов в продуктивной зоне обеспечивают-
обеспечиваются зоопланктоном, поедающим ВОВ и выделяющим растворенные
продукты обмена, и бактериопланктоном, использующим РОВ. Зона
14 Retallack GJ. How to find a Precambrian paleosol // Early organic evolution:
Implications for mineral and energy resources / Ed. M. Schidlowski et al. N.Y.:
Springer, 1990.
192
деструкции располагается под фотической зоной и часто опирается
на пикноклин.
Разложение РОВ определяется свободным бактериопланкто-
бактериопланктоном, который в большинстве относится к олиготрофным органо-
трофам с широким пищевым спектром. Легко растворимые низ-
низкомолекулярные вещества представляют преимущественный суб-
субстрат для бактерий, развивающихся дисперсно. Концентрация
РОВ лежит в пределах нанно- и микромолей на литр, составляя
30-100 мкг С/л в поверхностных водах океана и 50-150 мкг С/л в
морях. Они распределены между десятками разнообразных
компонентов, каждый из которых требует своей транспортной си-
системы в клетке, т.е. составляет нанномоли/л, близко к нижнему
порогу усвоения бактериями. Важнейшим источником служит вы-
выделение фитопланктоном низкомолекулярных веществ, как экссу-
экссудатов. Зоопланктон выделяет растворимые продукты обмена, в
частности мочевину.
Взвешенное органическое вещество ВОВ потребляется зоо-
зоопланктоном и агрегатами бактерий, обрастающих оседающие час-
частицы с превращением 1/4 углерода в биомассу бактериальных кле-
клеток. При этом скорость процесса деструкции лимитируется разло-
разложением полимеров оболочек, в особенности гидролизом хитина.
Потребление О2 и деструкция органического вещества являет-
является основной функцией бактериопланктона в экосистеме водоемов.
Дыхание бактериопланктона обычно составляет 70% от потребле-
потребления О2 в пробе воды. Развитие бактериопланктона определяет раз-
разность между поступлением органического вещества и его стацио-
стационарной концентрацией, которая при высокой плотности бактерио-
бактериопланктона может приближаться к нулю даже при значительном
притоке.
Деструкцию (по Sorokin, 1999) определяют на основании отно-
отношения продукции бактериопланктона к дыханию, которое предста-
' вляет коэффициент К2 - эффективность использования энергии, со-
1 ставляющую в природных условиях 0,25-0,35 со средним значени-
' ем 0,32. В лабораторных условиях обычны величины 0,4-0,5. Для
* расчета используется бактериальная продукция Р мг/л сырой массы,
; итогда дыхание МЬ = Р{\-К2)/К2 = Р ¦ 0,68/0,32 = Р ¦ 2,125. Для пере-
« счета этой величины в углерод она умножается на 0,2; для пересче-
* та в потребление О2 мг/(л • сут.) - на 2,67; для пересчета дыха-
' ния в доступный органический углерод употребляется коэффици-
; ент 0,375.
| Другая оценка получается из общего дыхания микропланктона,
* составляющего 0,9-0,95 от разложения органического вещества.
' Бактериопланктон отделяется фильтрованием через 1-2 мкм
¦ фильтр, разливается по склянкам, инкубируется 10—20 час при тем-
1 пературе in situ. Начальная и конечная концентрация О2 определяет-
определяется по Винклеру.
7. Г. А. Заварзин
193
Деструкция в планктоне
Подвижный зоопланктон поедает ВОВ, в том числе бактерио-
планктон, и превращает его в фекальные пеллеты, обогащенные
бактериями. Пеллеты могут служить пищевым субстратом для сле-
следующего ряда фильтраторов, в том числе подвижно мигрирующих в
вертикальном направлении. Зоопланктон переваривает водоросли и
выбрасывает избыточный азот в виде мочевины или аммония, кото-
которые служат субстратом нитрификаторов. В состав зоопланктона
входят крупные формы, как копеподы, поедающие диатомовые во-
водоросли и фитофлагеллят, а также микрозоопланктон. Зоопланк-
Зоопланктон зависит в своем развитии от фито- и бактериопланктона и рас-
располагается над термоклином в фотической зоне.
Наннозоопланктон с размерами 2-20 мкм представлен протиста-
ми, преимущественно нанногетеротрофами, инфузориями, личинка-
личинками многоклеточных. Нанногетеротрофы составляют 90% биомассы
этой размерной категории и представлены фаготрофными жгутико-
жгутиковыми: Bodonidae, Protomonadinae, Choanoflagellida.
Микрозоопланктон относится к размерной категории
20-200 мкм. Наиболее харакерны для морского микрозоопланкто-
микрозоопланктона радиолярии, фораминиферы, тинтинниды, личинки многокле-
многоклеточных. Саркодовые протисты формально разделены на супер-
суперклассы Rhizopodea с классами амеб Lobosea и фораминифер
Granuloreticulosea (карбонатный скелет) и суперкласс Actinopodeac
классами Acanthorea (акантарии с стронциевым скелетом),
Polycystenea (радиолярии с кремневым), Phaeodaria (радиолярии с
кремневым), Heliozoa (солнечники). Радиолярии и фораминиферы
играют значительную роль в биогеохимических циклах благодаря
образованию скелетов. Многие протисты образуют ассоциации с
фототрофными организмами в виде экто- и эндосимбиоза. В та-
такие ассоциации обычно входят симбиотические фототрофы, как у
радиолярий и фораминифер. Хотя питание для этих протистов по-
поступает от улавливаемого планктона, вместе с тем у них развито
симбиотическое питание. Планктонные фораминиферы распро-
распространены в поверхностных водах, богатых планктоном с числен-
численностью до 105 особей/м3. Симбиотрофные формы ограничены фо-
фотической зоной на глубине 10-50 м. Им свойственна суточная ми-
миграция от репродуктивной зоны 200 м до верха фотической зоны.
В качестве симбионтов у них существуют динофлагеллаты. Бак-
Бактериальные компоненты системы не описаны. Бентосные фора-
фораминиферы достигают плотности 103 особей/см2 при миллиметро-
миллиметровых размерах и имеют симбионтов из числа динофлагеллат и
красных водорослей. Симбионт ассимилирует метаболические
продукты с азотом и фосфором и продуцирует фотосинтетаты,
получая возможность вертикальной миграции и защиту от фильт-
фильтраторов.
194
Микрозоопланктон располагается в нижней части фотической
зоны над термоклином. Значительная часть планктонных протистов
собирается в агрегаты "морского снега" на мортмассе зоопланктона
и фекальных пеллетах. Вне фотической зоны микрозоопланктон
очень редок. Микрозоопланктон представлен в основном Protozoa -
сборной группой организмов с органотрофным или фаготрофным
питанием. Именно наличие последней группы служит важным фак-
фактором, ограничивающим плотность бактериопланктона. Пищевая
пирамида зоопланктона отчасти следует размерным классам, но не-
некоторые крупные формы оказываются обильными, в то время как
пищевая база поддерживается благодаря высокой оборачиваемости.
Трофическая структура зоопланктона по потребляемой пище согла-
согласуется с классификацией по размерам, хотя некоторые представите-
представители протистов способны питаться добычей, крупнее себя с помощью
ловчего аппарата - щупалец, которыми крупные фораминиферы и
радиолярии способны захватывать даже многоклеточную добычу.
По питанию среди органотрофных протистов различают 1) питаю-
питающихся бактериями, 2) питающимися водорослями, 3) хищных. В ка-
каждую из трофических групп попадают представители разных круп-
крупных таксонов. Вместе с те(м в состав таксонов родового ранга входят
организмы разных размеров, и поэтому существенна видовая харак-
характеристика. Из-за большого разнообразия зоопланктона и обычной
ограниченности знаний таксономистов одной группой трудно соста-
составить общую картину.
Скелетные остатки фито- и зоопланктона обусловливают харак-
характерные зоны осадков в пелагиали океана: в областях высокой проду-
продуктивности осадки преимущественно кремнистые из-за развития
диатомовых и кремнежгутиковых, до глубины лизоклина (уровня
карбонатной компенсации) распространены карбонатные осадки
("глобигериновые илы"), наконец, в пелагических глубоких частях
океана дно покрывают красные глины, свидетельствующие о том,
что разложение органического вещества и растворение тонкодис-
тонкодисперсных минеральных осадков в процессе оседания прошло полно-
полностью. Осадки планктона существенны для циклов кальция и кремния.
Сезонное распределение бактериопланктона следует за разви-
развитием фитопланктона, создавая максимум в гетеротрофную фазу.
Вертикальное распределение бактериопланктона (табл. 5.5) имеет
небольшой максимум в поверхностном слое воды, в зоне испарения,
и основной - на глубине 20-50 м под зоной развития фитопланктона
на сезонном термоклине или же на поверхностях пикноклина при
смешении вод, например при таянии льдов в полярных морях. В ге-
гетеротрофную фазу сезонного развития бактериопланктон может
ь достигать плотности 15-400 мг/м3 A0б клеток/мл) и превышать мас-
массу фитопланктона, становясь основной пищевой базой для зооплан-
зоопланктона. Зоопланктон способен снижать плотность бактериопланкто-
бактериопланктона до 10М05 клеток/мл.
Т* 195
Таблица 5.5
Обмен и плотность бактериопланктона
[по: Sorokin, 1999]
Бассейн
Состоя-
Состояние
Тип
Плотность
N.-106
кл/мл
В, мг/м3
влажной
био-
биомассы
Продуктивность
Р,
мг/(м3 х
хсут)
Ц(Р/В)
Дыхание, мкг
Ог/(л • сут)
Mb
TMR
Гипер- Загрязненная Min 10
евтороф- лагуна
ный Мах 40
2000
500
10000 3000
0,2 600 2000
0,5 5000 8000
Евтроф-
ный
Лагуны,
заливы
Океанский
апвеллинг
Озера
Мезо-
троф-
ный
Олиго-
троф-
ный
Тихий
океан
5
10
2
5
3
8
Слои анокси- 10
генного
фотосинтеза 40
Внутренние 1,5
моря
Озера 3
Умеренный 1
океан 2
Слои хемо- 1
синтеза,
хемоклин 2,5
Озера 0,5
0,8
Промежуточ- 0,1
ные
Антарктиче- 0,2
ские воды
Глубокие 0,01
воды > 800
0,02
2000
3000
400
2000
600
2000
2000
8000
200
400
100
300
200
500
40
70
10
20
1
2
500
1500
200
5000
300
800
20
150
100
300
50
200
40
100
15
50
3
5
0,02
0,06
0,2
0,5
0,4
0,8
0,3
0,7
0,01
600
1700
250
600
380
900
0,03
0,5 150
0,2
0,4
20
1,5 60
0,15 4
1500
3000
300
1000
500
1500
200
1,5
0,5
0,8
0,05
400
100
300
600
150
400
30
80
5
0,30 6 7
0,02 0,03 0,04
0,03 0,08 0,15
f
I
* N - общее число клеток, В - биомасса, Р - продукция, ц(Р/В) - коэффициент про-
продукции за сутки, Mb - дыхание бактериопланктона, TMR - общее дыхание микро-
микропланктона.
В условиях, когда деятельность зоопланктона подавлена, напри-
например ночной аноксией, дыхание бактериопланктона может приво-
приводить к коллапсу экосистемы. В этих случаях даже при малой продук-
продуктивности плотность бактериопланктона может достигать значи-
значительных величин. По величинам потребления О2 определяется
трофность водоемов.
Развитие бактерий на взвешенных частицах органического ве-
вещества приводит к экспорту бактерий из продуктивной зоны вместе
с "дождем трупов" без возможности вернуться обратно. Это экс-
экспортная продукция. Создается подобная хемостату система выноса
из продуктивной зоны и необходимость постоянной инокуляции.
Особую возможность для бактерий избежать выноса в глубины и
приобрести способность к активной вертикальной миграции пред-
представляет образование ассоциаций с зоопланктоном.
Оседающее органическое вещество в виде "морского снега" из
довольно крупных агрегатов ВОВ с разнообразным сообществом
бактерий на них выходит из продуктивной зоны и вместе с тем обо-
обогащает холодные глубокие воды биогенами. Считается, что 75-95%
органического вещества разлагается в зоне 500-1000 м; 5-10% орга-
органического веществ'а достигают глубины 2000-3000 м. Основную
часть осадка составляют минеральные скелеты планктона, хорошо
прослеживаемые по характеру ила на дне. Таким образом, область
регенерации находится сразу под фотической зоной и не простира-
простирается в глубину. Перенос вещества в сильнейшей степени зависит,
как от горизонтальных течений, так и от скольжения масс воды по
областям с разной плотностью. Экспорт питательных веществ из
фотической зоны в глубокие воды определяется даунвеллингом хо-
холодных вод. Подъем холодных вод на поверхность происходит толь-
только в местах апвеллинга, и здесь создаются условия для развития фи-
фитопланктона за счет импорта биогенов из глубины океана. Области
высокой продуктивности в прибрежной зоне имеют своим источни-
источником биогены терригенного материала.
Бактерии океана
Исследование функционального разнообразия бактерий океана
было начато в 1907 г. Б.Л. Исаченко15, который в своих работах до-
доказал получением культур, что в полярном океане присутствуют все
основные физиологические группы бактерий и, соответственно,
имеется возможность замкнуть все циклы. Эта работа была продол-
продолжена Зо Беллом в 1940 г. (Zo Bell, 1946) в связи с исследованием
нефтяных месторождений. В 1950-х годах А.Е. Крисе и его немного-
15 Исаченко Б Л. Исследования над бактериями Северного Ледовитого океана //
Тр. Мурм. науч.-пром. эксп. 1906 г. Пг., 1914. См.: Исаченко БЛ. Избранные
труды: В 2 т. М.; Л.: Наука, 1951.
196
197
численные сотрудники провели гигантскую работу по подсчету ор-
организмов в глубоководных частях океана на мембранных фильтрах
со стандартной окраской эритрозином (Крисе, 1959). Сейчас эта ра-
работа повторяется с помощью флуоресцентных красителей, а в пос-
последнее время и с помощью олигонуклеотидных флуоресцентных
проб (FISH).
Молекулярные данные по последовательностям нуклеотидов
указывают на существование 9 филогенетических ветвей морских
прокариот, из которых только две имеют культивируемых предста-
представителей. Как показал анализ их филогенетического положения,
выделенные из моря культуры бактерий преимущественно были
представлены грамотрицательными подвижными палочками, аэ-
аэробными копиотрофами, легко вырастающими на чашках Петри.
Большинство культивируемых морских бактерий относится к гам-
гамма-ветви Proteobacteria. Для таксономии морских псевдомонад ис-
поль-зовали род Alteromonas, в котором потом выделили роды
Alteromonas, Pseudoalteromonas, Marinomonas, Shewanella, Glaciecola.
Морских спирилл относили к роду Oceanospirillum, среди которых есть
представители как гамма-, так и альфа-протеобактерий. Филогенети-
Филогенетически родственная группа Vibrio близка по обмену энтеробактериям и
способна к брожению, что позволяет ей быть совместимой с пребыва-
пребыванием в животных. Среди выделенных культур барофилов преоблада-
преобладает факультативно анаэробная Shewanella. Большое число штаммов
протеобактерий изолированы из прибрежных условий Тихого океана
В. Михайловым16 и относятся к разным родам протеобактерий.
Альфа-протеобактерии выделяются значительно реже, но по
данным определения прямым методом они многочисленны в море.
Как и следовало ожидать, альфа-протеобактерии гораздо более спе-
специализированы и плохо адаптируются к росту в лаборатории. К чис-
числу культивируемых относятся группировки Roseobacter и
Sphingomonas. Группировка Roseobacter представлена фенотипически
разными организмами с исключительно морским местообитанием.
Первые представители этого рода были выделены с поверхности во-
водорослей и были отнесены к эритробактериям, поскольку синтезиро-
синтезировали фотосинтетически активный бактериохлорофилл а и могли
использовать его для синтеза АТФ в аэробных условиях, но не для ро-
роста. Другие виды Roseobacter не синтезируют бактериохлорофилл.
Многие из них метаболизируют серные соединения, в частности, ди-
метилсульфопропионат, предшественник диметилсульфида (ДМС).
Erythrobacter, переименованный в Sphingomonas, в отличие от
других пресноводных эритробактерий, обитает в море. Это аэробные
подвижные палочки с полярным жгутиком, образующие желтые ко-
16 Михайлов В.В., Кузнецова Т.А., Еляков Г.Б. Морские микроорганизмы и их
вторичные биологически активные метаболиты. Владивосток: Дальнаука,
1999. 132 с.
198
1
лонии, из-за чего они часто попадали в число флавобактерий. Один
из штаммов Sphingomonas оказался "ультрамикробом" из-за очень
маленьких размеров и способности к олиготрофному росту.
Морские метанотрофы и метилотрофы разделены между аль-
альфа- и гамма-подклассами протеобактерий. Их представители были
выделены из моря В.Ф. Гальченко17 и классифицированы в числе
обычных родов. Для моря они представляют функционально важ-
важную группу, предотвращая возможность выноса из осадков метана,
в том числе метана кристаллогидратов.
Вторую крупную филогенетическую группу в море составляет
не получившая таксономического названия ветвь Cytophaga-
Flavobacterium-Bacteroides ("CFB-ветвь" эубактерий). В целом CFB-
группа очень разнообразна и морфологически, и филогенетически.
Культуры их представлены аэробными или факультативно анаэроб-
анаэробными хемоорганотрофами, многие обладают скользящим движени-
движением и приурочены к разложению таких макромолекул, как хитин, агар,
ДНК, целлюлоза. Большинство культур морских CFB отнесено к се-
семейству Flavobacteriaceae. Они в большинстве ассоциированы с по-
поверхностями как "морской снег", микро- и макроводоросли, обладают
скольжением и способностью гидролизовать макромолекулы.
Планктомицеты представляют крупную самостоятельную ветвь
бактерий и в начале были описаны как пресноводные организмы.
В море установлены клоны ДНК этих пока в большинстве некуль-
тивируемых организмов, преимущественно ассоциированных с
"морским снегом" и разлагающимися остатками водорослей. Они
обнаруживаются в этих агрегатах по флуоресцентным олигонукле-
отидным пробам в виде обычных палочек.
Среди бактериопланктона открытого океана под эуфотической
зоной пелагические кренархеоты составляют большую часть пико-
планктона, и численность их достигает 39% на глубинах более
1000 м. Численность архей Мирового океана составляет
1,3 • 1028 клеток и эубактерий 3,1 ¦ 1028. Большинство архей предста-
представлено планктонными пелагическими кренархеотами с неизвестной
функцией, составляющими 20% всего морского пикопланктона. В про-
продуктивной зоне преобладают эубактерий. И архей, и бактерии обна-
обнаруживают увеличение численности в придонных водах18.
Глубоководные микроорганизмы
Глубоководный бактериопланктон находится в условиях посто-
постоянно низкой температуры с очень небольшими колебаниями и
может состоять из психрофилов с крайне ограниченными возмож-
возможностями адаптации из-за медленности синтетических процессов. Со-
11 Гальченко В.Ф. Метанотрофные бактерии. М.: ГЕОС, 2001. 500 с.
18 Kanter M.B., De Long E.F., Karl DM. Archaeal dominance in the mesopelagic zone
of the Pacific ocean //Nature. 2001. Vol. 409. P. 507-510.
199
держание органического вещества здесь крайне низкое, и поэтому
бактерии представлены карликовыми формами. Глубокие воды
океана отличаются большим постоянством условий солености, тем-
температуры, давления. Поэтому можно ожидать, что большинство
организмов здесь относится к медленно растущим психрофилам и
барофилам, приспособленным к условиям среды обитания. Приме-
Применение методов, не связанных с культивированием, пока служит наи-
наиболее важным источником информации. Как уже отмечалось, моле-
молекулярные методы идентификации выявили присутствие в планктоне
океана значительного числа архебактерий, относящихся к Сге-
narchaeota, в то время как в продуктивной зоне преобладают
Eubacteria.
Глубокие воды океана не имеют первичных продуцентов. Бак-
Бактериальное население зависит от продукции на поверхности и тех
остатков, которые не успевают разложиться при оседании. Большая
вероятность достичь дна есть у остатков крупных организмов. Харак-
Характерным указанием на такой процесс служат конодонты - зубы ис-
ископаемых акул. Дна могут достигать фекальные пеллеты, например,
с высоким содержанием хитина членистоногих. Они обычно покры-
покрыты колонизовавшими их бактериями. Но скорости деградации орга-
органического вещества в холодных водах очень невелики.
Исключение представляют гидротермальные оазисы с эндогенным
источником восстановителя и области просачивания восстановленных
углеводородов, прежде всего метана, в том числе и из кристаллогидра-
кристаллогидратов. В этой области большой интерес представляет анаэробное окисле-
окисление метана консорциумами метаногенов с сульфатредукторами, уста-
установленное в области просачивания метана из кристаллогидратов.
Прямое наблюдение метановых сипов в Черном море обнаружи-
обнаружило выходы, покрытые красноватой коркой изотопно легких карбо-
карбонатов с массой нитчатых бактерий, напоминающих Methanosaeta
(MethanothrixY9. Попытки культивирования биопленки оказались
неудачными, и о действующих агентах можно было судить только
по микроскопии.
Исследователями из Бременского института морской микробио-
микробиологии в местах развития мата Beggiatoa над метановыми сипами с
80 мМ СН4 из разлагающихся кристаллогидратов была найдена не-
необычайно мощная сульфатредукция, достигавшая рекордной вели-
величины 140 ммоль/(м2 • сут), в то время как вокруг она вообще не об-
обнаруживалась20. Было обнаружено исчерпание сульфата на глубине
19 Пименов Н.В., Русанов И.И., Поглазова М.Н. и др. Бактериальные обраста-
обрастания на коралловидных постройках в местах выхода метановых газовыделе-
газовыделений в Черном море // Микробиология. 1997. Т. 66, К» 3. С. 421-428.
20 Velantine D.L., Reeburgh W.S. New perspectives on anaerobic methane oxidation //
Environ. Microbiol. 2000. Vol. 2, № 5. P. 477-484; Boetius A., Raveschlag K.,
Schubert CJ. et al. A marine mibrobial consortium apparently mediating anaerobic
oxidation of methane // Nature. 2000. Vol. 407, № 5. P. 623-626.
200
4
12 см осадка с соответствующим образованием сероводорода. Мик-
Микроскопия с применением специфической флуоресцентной молеку-
лярно-биологической пробы для in situ гибридизации на новую ветвь
архей из Methanosarcinales обнаружила их массовое развитие в мета-
метановых сипах и отсутствие вокруг; они не окрашивались DAPI. Ана-
Аналогичная специфическая окраска на протеобактерии показала при-
присутствие кокков группы DesulfosarcinalDesulfococcus. Консорциум
состоял из внутреннего агрегата размером в 2-3 мкм из сотни кле-
клеток архей по 0,5 мкм, окруженных слоем в 1-2 клетки из примерно
200 сульфатредукторов. Консорциумы располагались слоем на глу-
глубине 1-2 см и достигали численности в 70 млн/мл. Выход энергии на
моль метана при давлении около 8 МПа составлял около —40 кДж, и
разрешенные концентрации промежуточных веществ для обратного
метаногенеза составляли от 10-10 до 10-9 М для водорода и от 3 ¦ 10-'2
до 3 • 10-8 М для ацетата. Высокое давление метана на глубине ука-
указывает на участие барофильных организмов в окислении метана из
кристаллогидратов. Таким образом, факт существования консорци-
консорциумов метаногенов и сульфатредукторов, способных к окислению
метана за счет сульфата, можно считать установленным, хотя пока
они не поддаются культивированию.
5.4.5. Донные осадки
Развитие процессов в морских осадках в большой степени зави-
зависит от характера этих осадков, распределяющихся по поверхности
дна океана закономерным образом: 1) глубоководные красные гли-
глины с очень медленной жизнью в них из-за недостатка органическо-
органического вещества и низкой температуры; 2) карбонатные осадки, образу-
образующиеся в теплых морях, обрезанные снизу уровнем лизоклина и
обусловленные оседанием "глобигеринового ила"; 3) кремнистые
осадки в областях повышенной продуктивности с развитием диато-
диатомовых; 4) осадки в зоне континентального сноса в окраинных морях.
Прибрежная область с взаимным влиянием терригенного сноса
и приливных течений представляет особый ландшафт. Примером
исследований приливных отмелей послужила бухта Яде на западе
Германии, представляющая эстуарии рек Эмса, Везера, Эльбы (так
называемое Wadden See). Приливы здесь превосходят 3 м и обнажа-
обнажают на два-три десятка километров обширную топкую илистую пло-
плоскость, прорезанную каналами по руслам рек и стекающей прилив-
приливной воды. Серая поверхность ила покрыта коричневатой пленкой
диатомовых. Ил уплотняется с глубиной десятка сантиметров и пе-
переходит в черные сульфидные глины с FeS как основной формой се-
серы и морским содержанием сульфата 28 мМ. Верхний слой состоит
из слоистых силикатов размерами < 63 мкм с поровым пространст-
пространством 67% на поверхности и 28% на глубине 17 см. Пленка диатомо-
диатомовых создает пересыщение О2 на глубине 0,3 мм, откуда О2 не успе-
201
вает обмениваться с вышележащей водой, задерживаясь в поровом
пространстве. Ниже он быстро используется, что ведет к с повы-
повышенному содержанию растворенного неорганического углерода
вследствие дыхания. Еще ниже располагается зона с Fe и Мп, где ма-
мало сульфидов, но наиболее высока скорость их образования. Осадки
содержат почти 3% органического углерода при положительной
корреляции с глинами. Ниже 8 см повышается содержание органи-
органических кислот до 50 мкМ ацетата, 30 мкМ лактата, присутствует
формиат. В Wadden See лимитирующим фактором для разложения
органического вещества служила температура21.
Осадки очень богаты микрофлорой и содержат 1-6 млрд клеток
в 1 см3 осадка. Из клеток, красящихся DAPI, 17-44% удалось припи-
приписать на основе FISH-проб определенным филогенетическим группам.
Цитофаги-флавобактерии A5-25%) с численностью 6-108 кле-
клеток/см3 оказались важной морской группой микроорганизмов; пред-
предполагается, что они, располагаясь в начале трофической цепи дест-
деструкции ВОВ, предопределяют численность остальных групп бакте-
бактерий. Развитие "цитофаг"в анаэробной зоне связано с возможностью
развития этих организмов как анаэробных гидролитиков. Сульфат-
редукторы Desulfovibrio B,9 • 108) и Desulfococcus с Desulfosarcina
B,8 ¦ 108) составляли следующую по численности за цитофагами
группу. Протеобактерии а-, C-, у-(наполовину S-окисляющие эндо-
симбионты), ?-ветвей, планктомицеты, веррукомикробы, актиноба-
ктерии с высоким Г + Ц составили по 1-6% каждой группы. Планк-
Планктомицеты и веррукомикробы не определяются пробой на бактерии.
На поверхности было много аэробных Arcobacter из группы условно
патогенных кампилобактеров22.
Рекомендуемая литература
1. Алехин О.А.,Ляхин Ю.И. Химия океана. Л.: Гидрометеоиздат, 1984. 343 с.
2. Биология гидротермальных систем / Отв. ред. А.В. Гебрук. М.: КМК Press,
2002. 543 с.
Ъ.Гаррелс P.M., Крайст ЧЛ. Растворы, минералы, равновесия. М.: Мир, 1961,
4. Горленко В.М., Дубинина Г.А., Кузнецов СИ. Экология водных микроорга-
микроорганизмов. М.: Наука, 1977. 289 с.
5.Дривер Дж. Геохимия природных вод. М.: Мир, 1985. 440 с.
6. Исаченко БЛ. Избранные труды. В 2 т. М.; Л.: Наука, 1951.Т. 1: 408 с; т. 2:
430 с.
7. Кузнецов СИ. Роль микроорганизмов в круговороте веществ в озерах. М.:
Изд-во АН СССР, 1952. 300 с.
21
Biostabilization of sediments / Ed. W.E. Krumbein, D.M. Paterson, L.J. Stal.
Oldenburg, 1994.
22 Llobet-Brossa E., Roselo-Mora R., Amann R. Microbial community composition of
Wadden sea sediments as revealel by fluorescence in situ hybridisation // Appl.
Environ. Microbiol. 1998. Vol. 64. P. 2691-2696.
8. Кузнецов СИ. Микрофлора озер и ее геохимическая деятельность. Л.: Нау-
Наука, 1970. 440 с.
9. Крисе А.Е. Морская микробиология (глубоководная). М.: Изд-во АН СССР
1959.
Ю.Лисицын АЛ. Процессы океанской седиментации. М.: Наука, 1978. Уточ-
Уточненное издание: Lisitzin А.Р. Oceanic sedimentation: Lithology and geochemistry
Wash.: (D.C.), 1996.
11. Сорокин Ю.И. Черное море. М.: Наука, 1982. 216 с
12. Физическая география Мирового океана / Отв. ред. К.К. Марков. Л.- Наука
1980.364 с.
13. Химия морей и океанов. М.: Наука, 1997. 415 с.
14. KarlDM., Dobbs F.C Molecular approaches to microbial biomass estimation in the
sea // Molecular approaches to the study of the ocean / Ed. K.E. Cooksey. L •
Chapman and Hall, 1998. P. 29-90.
15. Lally СМ., Parsons T.R. Biological oceanography: An introduction. L : Butterworth
1997.
\6.Sorokin Yu. Aquatic microbial ecology. Leiden: Backhuys, 1999.
I8.Z0 Bell С Marine microbiology. Waltham, (Mass.), 1946.
202
1
Лекция 6
АМФИБИАЛЬНЫЕ ЛАНДШАФТЫ, ЭКОТОНЫ
И ГЕОХИМИЧЕСКИЕ БАРЬЕРЫ
6.1. СРЕДА ОБИТАНИЯ МИКРОБНЫХ СООБЩЕСТВ
Сообщество организмов развивается в определенной среде оби-
обитания. Для отдельных видов микроорганизмов среда обитания мо-
может быть ограничена сообществом, располагающимся в микрони-
микронише. Мозаика местообитаний слагается в экосистему более высокого
уровня, и при этом необходим иерархический пространственный
подход. Для решения многих задач следует рассматривать инте-
интегральную деятельность микробных сообществ на больших про-
пространствах, например при определении дыхания почвы или эмиссии
газов в пределах корней одного растения, одного поля, ландшафта, (
региона, страны. С другой стороны, взаимодействуя с геосферой и!
друг с другом, микробные сообщества меняют условия своего обита
ния. Обратная связь от микробного сообщества к среде обитания
очень велика, и особенно велика связь его с геосферой. В наиболее t
общей форме можно утверждать - и в этом состоит цель книги - что'
микробные сообщества создали биосферу. В этом утверждении нет *
преувеличения, поскольку вся система биогеохимических циклов
контролируется бактериями, первыми обитателями Земли. В наибо-
наиболее концентрированной форме взаимодействие микроорганизмов с
геосферой происходит в контрастных системах, где велико взаимо- (
действие трех сред - воздушной, водной, минеральной - и существу-'
ет латеральный перенос вещества из соседних ландшафтов.
6.2. МИГРАЦИЯ ЭЛЕМЕНТОВ
Миграция элементов осуществляется в разных формах, и здесь-
следует отметить вклад в нее живых организмов. Перенос вещества'
осуществляется в водной или воздушной среде. Здесь мы не обсуж-1
даем геологическую миграцию в виде расплавов, хотя она составляв
ет исходный пункт для трансформации веществ на поверхности Зем-
Земли. Излияния лавы дают изверженные породы, переработка кото-"
рых на поверхности Земли и составляет исходный пункт в геохимии
ландшафтов. Особый случай составляют гидротермы и дегазация*
204
при вулканизме. В гидротермальных условиях на глубине при контак-
контакте паров воды с нагретыми породами происходит образование
флюидов, из которых наиболее характерен флюид NaCl. Термаль-
Термальные воды в вулканических поясах и при подводном вулканизме в
океане выносят на поверхность химические элементы. В гидротер-
гидротермальном процессе и при дегазации происходит химическая диффе-
дифференциация.
Механическая миграция начинается с выветривания горных по-
пород, их измельчения главным образом под действием физических
факторов: перепадов температуры, воздействия воды, ветра. Обра-
Образующиеся мелкие частицы с неизмененным минеральным составом
переносятся водой в виде твердого стока (денудация), пылевых час-
частиц (дефляция). Сортирование их происходит под действием силы
тяжести, и при этом тяжелые частицы сортируются на месте, обога-
обогащая песчаную фракцию Ti, Zr. Они накапливаются в седиментаци-
онных ловушках, обычно образованных понижениями рельефа.
В результате создаются россыпи. Перенос взвешенного вещества
- водотоками осуществляется в виде твердого стока. Ветровой пере-
- нос пыли обусловливает формирование залежей лёсса. Механиче-
' екая миграция приводит к эрозии поверхности и образованию раз-
1 дробленной породы с большой поверхностью.
* Химическая миграция начинается с химического выветривания
1 (выщелачивания), которое переводит химические соединения в рас-
¦ твор. Порядок перевода соединений в раствор определяется их рас-
' творимостью в воде. Отношение содержания элемента в воде к его
i содержанию в дренируемой породе дает коэфициент водной мигра-
' ции. Он может быть низким для элемента с высоким кларком, на-
'- пример кремния, и высоким для элемента с низким кларком. Выще-
* лачивание сильно ускоряется под действием кислот, в первую оче-
* редь углекислоты, растворенной в дождевой и подземных водах.
* Континентальные воды характеризуются ионным стоком. Этот
¦¦ сток хорошо оценивается по "шестикомпонентному" анализу реч-
речной воды и площади водосбора. Он составляет десятки и сотни тонн
¦» на 1 км2 водосбора в год. Выщелачивание многократно усиливается
под действием микроорганизмов, локально образующих углекисло-
углекислоту, органические кислоты, комплексообразователи. Биогенное вы-
выщелачивание составляет важнейший раздел природоведческой мик-
'робиологии. К наиболее яркому примеру биогенного выщелачива-
выщелачивания следует отнести сернокислотное выщелачивание в зоне выхода
в окислительную зону сульфидных руд. Окисление сульфидов идет
под действием тионовых бактерий и оставляет лишь кремневый ске-
скелет, при этом выносится даже алюминий. Вынос осуществляется с
топографических высот. Элювиальные ландшафты относятся к
автономным, а аллювиальные - к подчиненным.
- Биогенная миграция в наземных ландшафтах обусловлена мощ-
мощными процессами извлечения элементов проводящей системой рас-
205
тений и мицелием микроорганизмов. Наиболее наглядным приме-
примером может служить эвапотранспирация растений, дающая сущест-
существенный вклад в атмосферный гидрологический цикл и выносящая
на поверхность почвенный раствор. В море биогенная миграция свя-
связана с деятельностью образующего скелет планктона и является оп-
определяющей для миграции кремния у диатомовых, кальция - у кок-
колитофорид и фораминифер, стронция - у акантарий, фосфора - у
скелетных животных.
Миграция веществ в водной среде осуществляется в виде ионно-
ионного стока, твердой взвеси. Коллоиды представляют доминирующую
форму миграции для кремнезема, который коагулируя дает гель, в
частности запечатывающий микрофоссилии и затем превращаю-
превращающийся в аморфный опал. Коллоидные растворы образуют и гумусо-
гумусовые вещества. В целом это важнейшая форма миграции веществ в
природе, по меньшей мере равноценная ионной форме миграции.
Выпадение коллоидов из раствора происходит вследствие измене-
изменения их заряда, потери гидратационной оболочки. В коллоидном со-
состоянии мигрируют глинистые минералы с сорбированными на их
поверхности элементами. Миграция железа в водотоках идет тоже в
виде коллоидов.
Газовая миграция имеет особое значение, потому что этой фор-
формой миграции обладают биогенные элементы, и состав атмосферы
находится под контролем биоты, перерабатывающей продукты де-
дегазации Земли. Этот раздел подробно рассматривался в связи с ат-
атмосферой. Миграция газов в хорошо перемешанной атмосфере соз-
создает одинаковые условия для развития жизни на Земле.
Миграция органических веществ и их комплексов с минераль-
минеральными имеет первостепенное значение для гумидного пояса. Тропи-
Тропические реки Рио Негро, Нигер, Амазонка, Меконг несут огромное
количество растворенного и взвешенного органического вещества.
Некоторые из них получили даже название "черных". Значительна
миграция растворимых форм гуминовых веществ в бореальной зоне
Северной Евразии, где многие реки имеют коричневую окраску.
Органические вещества речного стока отлагаются в эстуариях, об-
образуя обширные конусы выноса.
Транспорт вещества обусловлен приложением внешней энергии,
источником которой для поверхности Земли является поглощаемая
радиация Солнца. Для миграции воздушных масс разработана модель
глобальной циркуляции атмосферы, для водных масс - модель цирку-
циркуляции в океане. Другим источником энергии для миграции вещества в
ландшафте является гравитационная энергия. Под действием силы
тяжести вещество мигрирует вниз, и именно эта энергия определяет
стоковые серии, заканчиваясь подчиненными ландшафтами. Нако-
Наконец, в другом временнбм масштабе действуют тектонические процес-
процессы, обусловленные внутренней энергией Земли, с тектоникой плит,
горообразованием, обменом вещества в литосфере.
206
i *
* <*
В соответствии с гравитационной миграцией вещества разрабо-
разработана классификация ландшафтов: 1) элювиальный на водоразделах
с глубоким залеганием подземных вод и выносом вещества; 2) суб-
аквальный, получающий вещества с твердым и жидким стоком и
представляющий седиментационную ловушку, как бессточное озе-
озеро; 3) супераквальные с близким залеганием грунтовых вод и воз-
возможностью переноса вещества из них при испарении, как, например,
при формировании солончаков. В общем смысле следует различать
субаэральные наземные ландшафты и субаквальные - водные.
6.3. ЭКОТОН И ГЕОХИМИЧЕСКИЙ БАРЬЕР
Мозаичность ландшафтов и элементов внутри ландшафтов
(patches) предопределяет наличие многочисленных границ между
разными компонентами природной среды. Этот пограничный ланд-
ландшафт получил название экотон. Примером экотонов может слу-
служить литораль моря, прибрежные болота - марши, опушка леса.
Собственно говоря, 'экотон представляет ландшафт особого рода,
внутри которого происходит взаимодействие между двумя средами.
Условия обитания в экотонах отличны от условий существова-
существования в смежных ландшафтах. Они обусловлены транспортом вещест-
вещества через границу между двумя различными средами обитания. В ка-
каждом из ландшафтов существует равновесное состояние вещества.
При переходе через границу вещество попадает в иную среду, где со-
создаются иные условия для равновесия. Вещество оказывается неста-
нестабильным, и происходит его трансформация. Переход из одного
состояния в другое сопровождается отмиранием организмов и, сле-
следовательно, появлением доступного органического вещества для
обитателей экотона. Экотон рассматривается как особое местооби-
местообитание на границе двух ландшафтов. В нем формируется специфиче-
специфическое сообщество микроорганизмов, связанное с превращением ве-
веществ, мигрирующих из одного ландшафта в другой. Для такой
ситуации очень продуктивным оказалось понятие геохимического
барьера - переходной зоны, где происходит резкое изменение усло-
условий. Оно подробно разработано А.И. Перельманом1. Геохимиче-
Геохимический барьер и экотон - близко коррелирующие понятия. В экотоне
обычно создается геохимический барьер. Микробиологи редко
пользуются понятием экотона как слишком крупномасштабным для
них. Геохимический барьер - понятие другого масштаба, чем эко-
экотон, в котором может быть множество геохимических барьеров,
как, впрочем, они есть и в любом ландшафте. Геохимический барь-
барьер может быть сжат до размеров микрозоны.
Чтобы понять, что такое геохимический барьер, можно пере-
перечислить разные типы барьеров: окислительный, кислородный,
1 См.: Перелъман А.И. Геохимия ландшафта. М: Высш. шк., 1975. 496 с.
207
щелочной, сероводородный, сорбционный и др. Часть веществ про-
проходит через барьер, часть осаждается. Например хлор-ион не задер-
задерживается ни на одном из перечисленных барьеров, а металлы с пе-
переменной валентностью, как железо, на всех этих барьерах перехо-
переходят в иное состояние. Масштабы геохимического барьера могут
быть различными - от литорали моря до микрозоны в осадках. Раз-
Различным оказывается и способ транспорта через барьер - от разгруз-
разгрузки континентальных подземных вод в море (залив Куронг в Австра-
Австралии или Восточный Балхаш с доломитообразованием) до молеку-
молекулярной диффузии в поровом пространстве. Итак, геохимическим
барьером для какого-нибудь элемента или его соединения называет-
называется такая (пограничная) среда, где происходит резкое снижение его
миграционной способности.
Каждый барьер специфичен по отношению к веществам, участ-
участвующим в реакции. Например, порядок осаждения гидроокислов оп-
определяется произведением растворимости соответствующих осад-
осадков и рН осаждения. Поэтому геохимический барьер представляет
механизм разделения элементов в осадочном процессе и их накопле-
накопления. При этом образуются парагенетические ассоциации минералов.
За осаждением основного вещества следует соосаждение с ним, на-
например, микроэлементов. Образование барьера обусловлено диффу-
диффузией веществ навстречу друг другу в пористой среде, скоростью ре-
реакции осаждения, цементацией или кольматацией порового про-
пространства с образованием водоупора. Особенно характерно это для
почв, где на горизонтах вмывания образуются так называемые пан-
панцири, например железные. Барьер представляет собой динамиче-
динамическое образование: например, если фронт восстановительной среды
продвинется за барьер, то осажденные окисленные соединения пос-
после восстановления будут перенесены к новому положению фронта.
Так образуются роллы урановых руд на фронте окисленных вод.
Естественно, что мощность барьера определяется резервуаром ве-
веществ и буферной емкостью среды, в которую барьер продвигается.
Пространство, занятое барьером, зависит от скорости реакции, на-
например осаждения. При низкой температуре кинетика реакций
нередко позволяет химическому соединению длительное время на-
находиться в метастабильном состоянии. Барьер тогда получается раз-
размытым.
Микроорганизмы обладают способностью использовать энер- *
гию окислительно-восстановительных реакций веществ, находя-
находящихся в метастабильном состоянии в области термодинамической
устойчивости продукта. При этом они значительно ускоряют дости-
достижение термодинамического равновесия, включая иной путь для
обычной химической реакции. В применении к геохимическому
реноса к барьеру вследствие создания резкого градиента. В то же
время плотность организмов, участвующих в реакции, достигает
здесь максимума. Типичными примерами геохимических барьеров в
водной среде служат оксиклин и хемоклин, к которым приурочены
максимумы развития определенных организмов. Организмы такого
типа, называемые градиентными, должны обладать как рядом ки-
кинетических характеристик, так и биологической способностью удер-
удерживаться в зоне смешения.
Серию геохимических барьеров можно наглядно воспроизвести
в "колонке Виноградского" - цилиндре, на дне которого идет разло-
разложение органического вещества с образованием сероводорода, а
выше в виде ряда полос располагаются слои развития групп микро-
микроорганизмов. Принцип "колонки Виноградского" осуществляется в
разных экосистемах. Для градиентных организмов образование гео-
геохимического барьера обусловлено последовательностью реакций и
концентрацией реагирующих веществ. Слоистая структура сообще-
сообщества поддерживается либо закреплением в слизистом матриксе, либо
активной миграцией в зону оптимума. Условием сохранения служит
спокойная гидродинамика с преобладанием диффузионных процес-
процессов переноса. В воде пленки нет, но есть узкие слои повышенной
концентрации микрооргнизмов в области хемоклина. Очень многие
организмы образуют пленки в цилиндрах, удерживаясь за стенки.
Такие пленки очень интенсивно поглощают кислород, и под ними
может создаваться микроаэробное пространство. Это наглядный
пример того, как создается геохимический барьер для кислорода,
хотя, конечно, утрированный из-за очень высоких концентраций ор-
органического вещества. Строго говоря, хемоклин в водоемах в пери-
период стагнации и есть геохимический барьер. Такой хемоклин может
быть размытым, особенно для медленно реагирующих веществ.
Образование геохимического барьера в виде пленки на границе
двух сред хорошо известно для поверхностного слоя ила с пелофиль-
ными микроорганизмами или же нейстона - пленки на поверхности
воды, удерживаемой силами поверхностного натяжения. Если нет
механических помех из-за ветра и волнения, например между стеб-
стеблей болотных растений, то такая пленка составляет окислительный
газовый фильтр и содержит разнообразные организмы, в том числе,
например, метанотофов, окисляющих болотный газ. В пористой
среде диффузия приводит к образованию микрозон, продемонстри-
продемонстрированных Б.В. Перфильевым для железо- и марганец-окисляющих
организмов: в стакане с песком из осадков карельских озер на не-
некотором расстоянии от поверхности образуются четкие полоски
окислов железа и марганца с связанными с ними организмами. Вы-
Высокая плотность организмов в биопленках или матах обусловлена
барьеру это означает ускорение реакции и соответственно сжима- ' * высокой интенсивностью процессов химического преобразования,
ние барьера в полоску, где с каждой стороны реагирующие вещест- *
ва находятся в минимуме. Это влечет ускорение диффузионного пе-
208
1
и они представляют биологическую ипостась геохимического
барьера.
209
Для природоведческой микробиологии формирование геохими-
геохимического барьера представляет фундаментальное понятие, и поэтому
необходимо рассмотреть формы деятельности в разных барьерах.
В барьере происходит образование наиболее характерных аутиген-
ных сингенетических минералов. Они маркируют положение плен-
пленки в осадочных породах после литификации и цементации.
Испарительный барьер. Он формируется в аридном и семиарид-
ном климате с превышением испарения над притоком. Испарительный
барьер для морской воды образуется в лагунах с просачивающейся
водой и имеет строго характерную последовательность выпадения
солей, начиная с карбонатов, гипса, кончая галитом. В континен-
континентальных условиях испарительный барьер свойственен солончакам в
почве и бессточным содовым озерам. Образующиеся минералы и
породы носят название эвапоритов и иногда дают месторождения
солей. В пористой среде воды разделяются на гравитационные и по-
ровые. Гравитационные воды уходят в водотоки, а поровые прихо-
приходят в равновесие с вмещающей породой. При испарении образуется
эвапоритовая тяга, соли подтягиваются к поверхности, и возникает
засоление. В таких условиях развиваются галофильные организмы,
характерные группировки которых следуют уровню солености и хи-
химическому составу доминирующих солей. Галофильные организмы
адаптированы к NaCl, их сообщество, исследованное на примере
морских лагун, было рассмотрено отдельно (см. Лекцию 3). В ата-
лассофильных условиях испарительный барьер приводит к форми-
формированию содовых озер и содовых солончаков - местообитанию ал-
калофильного микробного сообщества.
Щелочной барьер зависит от рН среды и ее щелочности. Щелоч-
Щелочной барьер может возникать, например, при выходе на поверхность
насыщенных углекислотой подземных вод и их быстрой дегазации.
В растворе остаются щелочные и щелочно-земельные металлы, ко-
которые создают щелочной резерв и выпадают в осадок. Щелочной
барьер особенно важен для осаждения щелочноземельных металлов
и отложения карбонатов. На нем осаждаются Fe, Ca, Mg, Ba, Sr, Cr,
Zn, Cu, Ni, Co, Pb, Cd в порядке убывания кларков. На осаждение
влияет реакция с анионами, образование комплексов, например кар-
карбонатных, как для комплекса уранила, обеспечивающего его хоро-
хорошую миграцию. Микроорганизмы создают щелочной барьер в ре-
результате удаления сильных кислот, доминирующий процесс при
этом - сульфатредукция, ответственная за создание щелочности в
подземных водах, солончаках. Локальное подщелочение может воз-
возникать при аммонификации. Удаление СО2 при фотосинтетической
ассимиляции также повышает рН и создает в дневное время щелоч-
щелочной барьер, сочетающийся с окислительным. В анаэробных услови-
условиях гораздо более характерно возникновение кислой обстановки за
счет образования органических кислот и углекислоты. Разложение
же ацетата в метаногенном сообществе приводит к подщелочению
210
среды. Очень характерный щелочной барьер возникает при смеше-
смешении кислых "купоросных" вод, образуемых при сернокислотном вы-
выщелачивании, с пресными. Такие кислые воды мобилизуют катио-
катионы, выщелачивая породу, но при смешении с нейтральными водами
и повышении рН происходит образование щелочного барьера, и, к
примеру, в случае впадения кислого ручья с железом в пресную ре-
реку наблюдается выпадение лимонита. Щелочной барьер особенно
важен при карбонатной цементации: образуемый карбонатный це-
цемент скрепляет обломки в монолитную породу, например брекчию.
Кислородный барьер. Кислородный барьер - это граница между
аэробной и анаэробной зонами, и она имеет первостепенное значе-
значение для всех микробных процессов. Кислородный барьер создается
ниже оксиклина интенсивным поглощением кислорода при дыхании
микроорганизмов. Металлы с переменной валентностью в окислен-
окисленном состоянии обычно обладают меньшей растворимостью, чем в
восстановленном, и при окислении выпадают в осадок. Каждое со-
соединение окисляется в соответствии определенным окислительно-
восстановительным потенциалом (Eh). Например, железо окисляется
раньше марганца, и это приводит к тому, что отложения марганца
образуются ниже по течению или выше по профилю, чем отложе-
отложения железа. Окислительно-восстановительный барьер представляет
специальные условия для литотрофных организмов, получающих
свою энергию от такой реакции. К ним нужно отнести железобакте-
железобактерий, серных и тионовых бактерий, нитрификаторов. Это удобный
пример для того, чтобы объяснить роль бактерий в барьере. Ско-
Скорость ферментативной реакции многократно превышает скорость
химической реакции, и поэтому литотрофные микроорганизмы по-
получают преимущество. В зоне их развития мигрирующее восстанов-
восстановленное вещество метастабильно, и, если бы не было бактерий, оно
мигрировало бы дальше и барьер получался бы размазанным, а не в
виде строго локальной зоны.
Окислительно-восстановительная реакция двусторонняя. В фор-
формировании кислородного барьера участвует, с одной стороны, окси-
генный фотосинтез, создающий насыщение кислородом при осве-
освещении, а с другой - удаление кислорода при аэробном дыхании.
Дыхание создает кислородный барьер за счет веществ, которые са-
сами по себе электрохимически неактивны. Поднимающиеся из анаэ-
анаэробной зоны вещества, например метан, служат субстратом окисле-
окисления микроорганизмами. Деятельность микроорганизмов является
основной причиной формирования кислородного барьера, препятст-
препятствующего проникновению кислорода вниз из атмосферы или фоти-
ческой зоны. Существенно, что кислородный барьер зависит от
дневного света.
Минералы, образующиеся в окислительном барьере, указывают
на обстановку в момент их формирования. Особенно хорошим индика-
индикатором служат минералы железа. В окислительной обстановке обра-
211
зуются рыжие и бурые охры, обусловленные трехвалентным желе-
железом. В отсутствие сульфата в заболоченных почвах зеленоватую
окраску дает восстановленное железо. Наконец, в присутствии серо-
сероводорода образуется черный гидротроилит. Эти минералы хорошо
маркируют барьеры. Окислительные барьеры образуют четкую пос-
последовательность в морских донных осадках: сначала в узкой поверх-
поверхностной пленке ила происходит окисление доступного органического
вещества и отложение окисленного железа, затем начинается черная
зона сульфатредукции, под ней располагается зона метаногенеза.
Глеевый барьер в глинистой почве образуется в отсутствие суль-
сульфата при избытке органического вещества. Как правило, он сочетает-
сочетается с анаэробным метаногенным сообществом. Первоначально, однако,
происходит восстановление железа, придающее почве зеленоватый
цвет. Глеевый процесс типичен для гидроморфных почв гумидного
климата, для болот, где развивается характерное микробное сообще-
сообщество. Оно особенно интересно для областей с периодическим затопле-
затоплением, как, например, на рисовых полях с резким переходом от восста-
восстановления железа к его окислению в периоды осушения. Застойный
характер вод блокирует вынос вещества и способствует оглеению.
Сероводородный барьер связан не только с удалением кислоро-
кислорода, но и со связыванием большинства металлов в нерастворимые
сульфиды вплоть до цементации породы ими. Доминирует образова-
образование черного гидротроилита FeS-/z(H2O). Осадочные сульфидные
месторождения в большинстве случаев связаны с деятельностью
сульфидогенного сообщества в прошлом. Развитие этого сообщест-
сообщества, как правило, определяется поступлением доступного органиче-
органического вещества. Образование сероводородного барьера обусловле-
обусловлено в первую очередь, если не исключительно, деятельностью суль-
фатвосстанавливающих организмов. Сообщество организмов при
доминировании серного цикла носит название сульфуреты, для него
характерны интенсивно идущие превращения серы с зональным рас-
расположением участвующих в них группировок организмов.
Сорбционный барьер. На поверхности минеральных частиц про-
происходят катионообменные реакции. В этом отношении особенно ха-
характерно поведение глинистых минералов, необратимо сорбирую-
сорбирующих калий в первую очередь. Большой поверхностью обладают
коллоиды, преимущественно отрицательно заряженные, как гель
кремнекислоты, глинистые минералы, гумусовые вещества. На от-
отрицательно заряженных коллоидах сорбируются микроэлементы, и
это приводит к их концентрации. Ионообменные свойства пород оп-
определяют обменный комплекс почвы. Ионообменными свойствами
обладает и гликокаликс организмов, связывающих металлы. Уроно-
вые кислоты особенно хороши для связывания металлов благодаря
карбоксильным группам и сульфогруппам полисахаридов. В резуль-
результате пленка цианобактерий с их развитыми чехлами оказывается
областью сорбции ряда металлов. Можно наблюдать даже образо-
212
вание "позолоченных" цианобактерий и в лаборатории, и в микро-
фоссилиях. Такому сорбционному барьеру приписывают образова-
образование золотых месторождений Витватерсранда в Южной Африке.
Из перечисленных разных типов геохимических барьеров ясно,
что деятельность микроорганизмов может приводить к образова-
образованию их разных комбинаций.
В природоведческой микробиологии понимание закономерно-
закономерностей физической миграции веществ в ландшафтах, особенно напра-
направление водотока и формирование барьера, играет определяющую
роль. В тех масштабах, с которыми имеет дело микробиолог, каж-
каждый элемент ландшафта представляет местообитание для своего ми-
микробного сообщества. Большая трудность связана с переходом от
микроусловий к усредненной характеристике пространства. При
этом разумно оперировать не средней пробой почвы или осадков, а
строго определять структурный элемент, для которого характерно
данное сообщество, - микрозональность развития.
В целом экотоны представляют своеобразные линейно вытяну-
вытянутые ландшафты на границе двух ландшафтов. Одним из планетарно
важных экотонов и геохимических барьеров служит граница между
морем и континентом. Здесь происходит смешение вод двух разных
систем: морской хлоридно-сульфатной и континентальной бикарбо-
натной. На границе развивается сульфатредукция и создается серо-
сероводородный барьер, а в дополнение к кислотно-щелочному - иногда
сорбционный, иногда испарительный барьеры. Они служат местом
осаждения разных элементов вплоть до образования рудных осадоч-
осадочных месторождений, например меди. Поэтому для реконструкции
палеообстановок первостепенное значение имеет фациальный ана-
анализ. Вопрос состоит в том, какие воды служат источником питания
для образующихся осадков: морские или континентальные? Осаж-
Осаждение карбонатов в экотоне на границе моря и суши ведет к образо-
образованию рифогенных структур, укрепляющих цепи береговых отме-
отмелей и создающих за ними пространство лагун. В лагунах происходят
интенсивные биологические процессы седиментации и диагенеза.
В докембрии образование лагун и карбонатных платформ было
обусловлено деятельностью циано-бактериального сообщества, со-
здавашего после литификации пояса строматолитов. В результате
микробное сообщество формировало ландшафт экотона.
Важнейшим примером расположения геохимического барьера в
экотоне служат некоторые амфибиальные ландшафты.
6.4. АМФИБИАЛЬНЫЕ ЛАНДШАФТЫ
Амфибиальные ландшафты - термин, взятый из канадской тер-
терминологии. Амфибиальные ландшафты представляют более широ-
широкое понятие, чем болота, включая и морские системы, например
приливные отмели (англ. tidal flats), мангры, марши (англ. marsh),
213
ватты (нем. Watts). Такое расширенное понимание амфибиальных
ландшафтов позволяет сравнивать талассофильные экосистемы с
хлоридным и сульфатным морским составом вод, с аталассофиль-
ными континентальными экосистемами с преимущественно бикар-
бонатным составом. В США рассматривают 5 типов увлажненных
земель: 1) морские, 2) в эстуариях, 3) речные, 4) озерные, располага-
располагающиеся в депрессиях или реках, перегороженных дамбами, 5) собст-
собственно болотные, к которым отнесены все неприливные увлажнен-
увлажненные земли с древесной растительностью, кустарниками, мхами и ли-
лишайниками. Для России преимущественное значение имеют собст-
собственно болотные экосистемы. Краевые морские условия включают
дельты, береговые и системы барьерных островов, лагуны и эстуа-
эстуарии (дельты рек в приливной зоне, от лат. aestus - прилив), прилив-
приливные отмели. В приморских системах источником воды служат при-
приливы, заливающие морской водой прибрежные равнины с пологим
рельефом. Они служат ярким примером обширных экотонов - по-
пограничных ландшафтов между морем и сушей. Амфибиальные
ландшафты пограничной зоны имеют два характерных типа перехо-
перехода: лагуны, располагающиеся параллельно берегу и отделенные от
моря цепью островов или пересыпью вследствие движения осадков
вдоль берега, и эстуарии, располагающиеся в понижениях релье-
рельефа суши, часто в местах впадения рек перпендикулярно береговой
линии. Эта зона имеет определяющее значение для отложения
осадков.
Для микробиологии амфибиальные ландшафты представляют
первоочередной интерес, потому что здесь возрастает биогеохими-
биогеохимическая активность из-за резкого окислительно-восстановительного
градиента в вертикальном профиле с участием анаэробных и аэроб-
аэробных организмов в составе тесно взаимодействующего микробного
сообщества с минимальными ограничениями для транспорта ве-
веществ. Постоянное затопление обусловливает ведущую роль анаэ-
анаэробных процессов в донных отложениях, а застойный режим огра-
ограничивает вынос микроорганизмов. В профиле амфибиальных ланд-
ландшафтов формируется геохимический барьер, восстановительный и
часто сероводородный внизу и окислительный наверху в фотиче-
ской зоне и при контакте с воздухом. Микробное сообщество в ам-
амфибиальных ландшафтах резко разделяется на анаэробный генера-
генератор восстановленных веществ в донных отложениях и окислитель-
окислительный фильтр в контакте с атмосферой (см. также Лекцию 4). Отно-
Отношения здесь аналогичны тем, которые имеют место между иловыми
осадками водоемов и прилегающим придонным слоем воды. Анаэ-
Анаэробный генератор образует восстановленные газы по пути сульфи-
догенеза (в случае высокой минерализации) или метаногенеза (при
лимитации сульфатом). Накапливаются медленно метаболизируе-
мые вещества, которые могут захораниваться в осадках, образуя
торф или, после диагенеза, горючие сланцы и иные каустобиолиты.
214
Морские амфибиальные ландшафты служат важной областью от-
отложения минералов, вплоть до рудообразования. В амфибиальных
ландшафтах понижена гидродинамика, и поэтому происходит осаж-
осаждение взвешенных веществ. Этим они отличаются от береговых
экосистем, обращенных в сторону открытого водоема с волновым
прибоем и интенсивным перемещением веществ. Пониженная гид-
гидродинамика служит причиной оседания веществ и создания области
седиментогенеза с последующими диагенетическими процессами. К
амфибиальным ландшафтам можно отнести и эпиконтинентальные
мелководья, где отношение водной массы к донным отложениям не-
невелико и состояние водной толщи определяется процессами, идущи-
идущими на дне. В рассматриваемую категорию попадают также ланд-
ландшафты аридного и семиаридного климата с солончаками, сорами,
сабхами. До известной степени к амфибиальным ландшафтам мож-
можно причислить и озерные системы, если рассматривать их в совокуп-
- ности с водами гидроморфных почв, а небольшие озера - как свое-
своеобразный аналог лизиметров.
Амфибиальные ландшафты могут представлять либо автоном-
автономные системы, как верховые болота, либо минеротрофные подчинен-
подчиненные ландшафты с высокой первичной продукцией, как низовые бо-
болота, либо пограничные зоны между двумя ландшафтными зонами,
составляя экотоны. В экотонах геохимический барьер создается за
счет латерального переноса вещества. Последнее очень характерно
для морских побережий. Такие амфибиальные ландшафты оказыва-
оказываются зависящими от привноса аллохтонного вещества. Интенсив-
Интенсивность микробных процессов определяется наличием доступного ор-
органического вещества, автохтонного, или же аллохтонного.
Амфибиальные ландшафты еще более разнообразны, чем озе-
озера. Среди них можно выделить такие однотипные и распространен-
распространенные ландшафты, как соленые прибрежные лагуны, заросли трост-
тростника, сфагновые болота, содовые озера и солончаки. Состав воды в
этих ландшафтах зависит от ее морского или континентального
происхождения. По отношению к солености (содержанию СГ или
других солей) различают воды:
Наземные болотные
Морские (талассофильные) (минерализованные)
Гипергалинные Гиперсоленые
Соленость, %
Эугалинные (euhaline)
Миксогалинные (brackish)
Полигалинные
Мезогалинные
Олигогалинные
Опресненные
> 40 (рассол)
Эусалинные (eusaline) 30-40 (рассол)
Миксосалинные 0,5-30 (рассол)
Полисалинные 18—30 (рассол)
Мезосалинные 5,0-18
Олигосалинные 0,5-5
Пресные < 0,5
Ультрапресные < 0,01
Приведенная выше классификация вод содержит выходящие за
пределы нормы экстремальные условия, например гиперсоленые,
215
щелочные, и она удобна для топической характеристики местооби-
местообитаний микробных сообществ.
В переходной зоне от суши к морю особенно развиты процессы
седиментогенеза. Они простираются по континентальному склону,
но особенно интенсивно идут за экотоном. В зоне накопления осад-
осадков развиты процессы микробного диагенеза, ключевые для геоло-
геологической микробиологии.
6.5. СЕДИМЕНТОГЕНЕЗ
В упрощенном виде образование бассейна седиментации в гео-
геологическом масштабе связано с расхождением континентальной,
коры (рифтингом) и выходом океанических базальтовых подстила-
подстилающих пород на краю континента. Время процесса составляет пер-
первые сотни миллионов лет. В образовавшейся депрессии происходит
развитие глубокого осадочного бассейна с последовательно распо-
расположенными нефтематеринскими, пористыми нефтегазосодержащи-
ми, запечатывающими осадочными толщами.
В континентальных седиментационных системах различают че-
четыре типа отложений: флювиальные (аллювиальные конусы и ре-
реки, меандрирующие, или блуждающие), эоловые (пустыни), озерные,
гляциальные. Эоловые отложения ведут к формированию толщ
тонкодисперсного лёсса и грубодисперсного песка и относятся к вну-
триконтинентальным областям.
Озерные системы составляют около 1% поверхности континен-
континентов при их высоком нынешнем стоянии. Депрессии, в которых рас-
располагаются озера, имеют различное происхождение: рифты, кра-
терные явления, гляциальные процессы, запруды разного происхо-
происхождения. Крупнейшим озером является Каспий. Открытые озера
имеют сток и постоянную береговую линию, закрытые озера - не-
неустойчивые уровень и береговую линию. Седиментация в озерах от-
отличается от морской: 1) меньшим значением ветрового перемешива-
перемешивания; 2) намного более быстрым накоплением осадков из-за малого
отношения размеров озерной депрессии к площади водосбора; 3) от-
отсутствием приливных явлений.
На морском побережье образуются длинные узкие полосы на-
накопления песка, параллельные береговой линии и пересекаемые
скалистыми выступами берега и приливными каналами, эстуариями
рек, заливами (рис. 6.1). Барьерные острова аналогичны основной
суше, но отделены от нее лагунами или маршами. Условия осажде-
осаждения в них определяются приливами: микроприливная зона 0-2 м;
мезоприливная - 2-4 м; макроприливная - > 4 м. Барьерные условия
создаются в микроприливной зоне, где энергия движения воды недос-
недостаточна для их разрушения. Подъем и опускание уровня моря созда-
создает прибрежные равнины. Основным источником материала для
216
Дюны
Лагуна
Марш
Приливная
отмель
Эстуарий —
Приливные
каналы
Рис. 6.1. Общая картина барьерных островов в приливной зоне [по: Boggs,
1995]
Острова образуются за счет перемещения осадков вдоль берега волнами. Прилив промы-
промывает каналы, перпендикулярно береговой линии, особенно в эстуариях. Отмели образу-
образуются за счет турбуленции. Дюны преимущественно эоловые. Лагуна параллельна берегу.
В ней образуются глинистые приливные отмели из тонкодисперсных осадков. Песчаный
пляж располагается в области действия волн. Выше прилива располагается марш
построения барьерных островов служит движение быстро оседаю-
оседающих морских осадков (песка) под действием волн.
За барьерными островами создается система лагун - относи-
относительно небольших, полузамкнутых морских водоемов, определяе-
определяемых как полоса морской воды вблизи моря или же сообщающаяся с
ним, но отделенная барьерной полосой суши в виде рифа или песча-
песчаного бара. Лагуны простираются параллельно береговой линии в от-
отличие от эстуариев, представляющих собой перпендикулярные ей
заливаемые речные долины. Часто они ассоциированы с речными
дельтами, барьерными островами, приливными отмелями. Эстуарии
относятся к наиболее продуктивным системам и рассматриваются
как нефтематеринские области.
Приливные отмели, или ватты, развиваются в области высоких
приливов при отсутствии действия волн на пологом рельефе. Это
очень пологая равнина, достигающая в глубину десятков и сотен ки-
километров, пересеченная множеством каналов (нем. Wattsee) с боло-
болотистыми илистыми отложениями, разделенная на три зоны:
subtidal - ниже уровня самого глубокого отлива, подверженная са-
самым сильным приливным течениям; intertidal - между уровнями
217
средних приливов и отливов, один или два раза в сутки находящаяся
в субаэральном состоянии; supratidal - лежащая выше уровня прили-
приливов и покрытая водой только в случае нагонов. Во влажном клима-
климате она покрыта маршем, в аридном - сабхой. Источником наносов
служит море, и осадки поступают на сушу в своего рода "дельте на-
наоборот", по приливным каналам, и откладываются между ними,
Маты цианобактерий особенно важны в супра- и интерприливной
зонах как улавливающие и связывающие агенты. Здесь описаны си-
силикатные отложения, но аналогичная картина имеет место и с кар-
карбонатными осадками.
Илы (англ. muds) преобладают в супра- и верхней интерприлив-
интерприливной зонах, выше начинаются обильные отложения растительного
материала. В аридном климате слои ила могут прерываться отложе-
отложениями гипса, галита, трещинами усыхания, а в высоких широтах -
клиньями льда. Карбонатные отложения на приливных отмелях в
общем те же, что и на песчаных, но более пологие. На приливных
отмелях особенно отчетливо сказываются трансгрессия и регрессия
моря.
Карбонатные отложения в условиях в виде платформ (не рифов)
образуются в условиях теплого климата на мелководном шельфе,
занимавшем ранее обширные площади. Они возникали в геологиче-
геологическом прошлом на краю кратонов, в интракратонных бассейнах, на
открытых в море банках. Они создаются также в маргинальных ус-
условиях на пляжах, в лагунах, приливных отмелях. Окаймленные
платформы, края которых имеют подъем, резко обрываются в глу-
глубину, как Большой Барьерный риф Австралии. У неокаймленных
платформ такого края нет; в современных условиях они образуются
в умеренных водах. Примером изолированной платформы могут
служить Багамы, неокаймленной - западные берега Флориды, Пер-
Персидский залив.
Во всех перечисленных типах ландшафтов идут очень активные
микробные процессы, обусловленные как высокой собственной
продуктивностью этих экосистем, так и привносом осадочного ал-
лохтонного материала.
Силикокластические морские условия осадконакопления харак-
характерны для шельфов. Континентальный шельф со средней шириной
75 км имеет типичный уклон 4° и обрывается в среднем на глубине
130 м континентальным склоном, который на пассивном крае кон-
континента сглажен отложениями и круто уходит на глубины
1500-4000 м. 20-25% всех осадочных пород составляют песчаники,
сложенные зернами песка от 0,06 до 2 мм, цементированные матри-
матрицей. Основой является кварц SiO2, составляющий 50-60%; мелко-
мелкокристаллический кварц носит название кремень. Происходит кварц
из изверженных пород или переработанного старого песчаника.
Далее идут глинистые минералы, составляющие в песчаниках при-
примерно 5%. Акцессорными служат тяжелые минералы весом > 2,9.
218
1
Цементирующими служат либо новообразованные силикаты, чаще
всего опал, либо карбонаты, как известняк или доломит. Переотло-
Переотложение кремния в виде опала приводит к тому, что органические ос-
остатки, например циано-бактериальные маты, окремняются и захо-
захораниваются в том состоянии, каком они были в момент начала обра-
образования кремневого желвака. На петрографических срезах в таких
синевато-черных желваках обнаруживаются микрофоссилии превос-
превосходной сохранности. Они выглядят темно-коричневыми на про-
прозрачном фоне. При растворении кремней микрофоссилии дисперги-
диспергируются без следа.
Сланцы - это силикокластические породы, сложенные тонко-
тонкодисперсными глинами или суглинками, часто тонкослоистыми; их
относят к аргиллитам. Они обогащены органическим веществом.
Сланцы составляют примерно 50% всех осадочных пород. 20-50%
минералов в них представлены глинистыми, примерно столько же
кварцем, 10-20% шпатами, 5-10% карбонатами, постоянно присут-
присутствуют пирит и сидерит при довольно высоком содержании орга-
органического углерода 1-10%. Кварц, шпаты, глинистые минералы
имеют терригенное происхождение. Сланцы отлагаются всюду, где
энергия штормовых волн недостаточна для взмучивания осадка.
Особенно характерны они для морских условий в приконтиненталь-
ной зоне. Именно в этой зоне происходит захоронение остатков рас-
растений и снесенной наземной и бентосной микрофлоры, в том числе
цианобактерий. Они запечатываются в глинах и в результате силь-
сильного уплотнения превращаются в сплющенные плоские пластинки
органостенных микрофоссилии. Органическое вещество в них в
процессе диагенеза переходит в кероген.
Различают три основные провинции осаждения:
- континентальный блок на кратоне, имеющем с одной стороны
пассивный край континента, а с другой - орогенный пояс, или зону
конвергенции плит. Материнские породы состоят из изверженных
плутонических пород, метаморфических, осадочных пород, но со-
содержат мало вулканического материала. Поступающие осадки
обычно состоят из кварцевого песка, полевых шпатов с высоким
отношением калиевых шпатов к плагиоклазам, а также обломков
метаморфических и осадочных пород. Осадки отлагаются в краевых
морских бассейнах или внутри континентов;
- магматические (вулканические) дуги располагаются в зонах
столкновения плит и имеют вулканический материал, часто с плагио-
плагиоклазом. Отложение происходит перед дугой или же за ней;
- рецикл орогенных провинций осуществляется в зоне кон-
конвергенции плит, где происходит воздымание плит с располагаю-
располагающимися на них осадочными породами, и глубже, вплоть до области
базальтов.
Важнейшим процессом в осадках является диагенез. Его можно
разделить на две стадии. В первой стадии, чаще всего биогенной,
219
идут интенсивные процессы переработки аллохтонного органиче-
органического вещества. Наиболее типичен для морских условий сульфидо-
генез, который в пористых осадках идет до исчерпания сульфата в
интерстициальной воде. После исчерпания сульфата и глубже идет
метаногенез. Продуктами бактериального диагенеза служат аморф-
аморфные минеральные осадки, которые с разной скоростью для разных
веществ кристаллизуются сначала в метастабильные минералы, а
потом, по мере созревания, приобретают окончательные стабиль-
стабильные формы. Дальнейшие этапы диагенеза связаны с геологическим
формированием осадков при их погружении. Этот этап диагенеза
происходит при температуре выше, чем температура выветривания,
но ниже требуемой для метаморфизма. Основными действующими
факторами служат геотермальный градиент и возрастающая соле-
соленость поровых вод.
6.6. ОСАДКИ И ЗАХОРОНЕНИЕ УГЛЕРОДА.
КЕРОГЕН.
НЕФТЕМАТЕРИНСКИЕ ПОРОДЫ
В глобальном цикле углерода основным резервуаром Сорг,
рый ответственен за эквивалентное образование О2 атмосферы, яв-
является рассеянный углерод осадочных пород, получивший название
керогена. В отличие от биологии, рассматривающей продукцию ор-
органического вещества и его участие в современном круговороте, в
геологическом масштабе времени важно захороненное органиче-
органическое вещество, т.е. та его часть, которая не подверглась деструкции.
Оно выходит из круговорота и определяет биогеохимическую сук-
сукцессию, в данном случае вследствие сопряжения циклов углерода и
кислорода оксигенными фототрофами с образованием резервуара
О2, эквивалентного захороненному органическому веществу. В ре-
результате оказывается, что захоронение Сорг вследствие дисбаланса
между продукцией и деструкцией является главной движущей силой '
перехода поверхностных оболочек Земли в окисленное состояние. '
Важнейшей причиной образования органического углерода осадоч-'
ных пород и, соответственно, избыточного кислорода атмосферы со'
всей цепью последующих событий служит неспособность микробно- '
го сообщества в определенных условиях полностью разложить орга-
органический углерод мортмассы продуцентов.
Образование керогена подробно изучено в связи с генезисом
нефтематеринских пород, поскольку он считается предшественни-
предшественником нефти и газа. Кероген определяется как "осадочное органиче-
органическое вещество, не растворимое в обычных органических раствори-
растворителях" или органическое вещество осадочных пород, не растворимое
ни в щелочах, ни в кислотах, ни в обычных органических раствори-
растворителях. По другому определению кероген - это "тонкорассеянный
220
органический материал, освобождаемый из осадочных пород после
обработки кислотами (НС1 + HF)". Кероген - это заведомая смесь
веществ и остатков всегда в виде частиц. Палинологи утверждают,
что кероген состоит главным образом из остатков организмов и от-
отсюда дают ему определение "палинологическое органическое веще-
вещество": «то, что видно в неизмененных термально осадках, главным
образом состоит из морфологически распознаваемых частиц биоло-
биологического происхождения, таких, как "древесина", кутикула, водо-
водоросли, цисты динофлагеллят, споры. Отсюда следует, что кероген
возник вследствие избирательного сохранения биологически проду-
продуцированного устойчивого материала» (Tyson, 1995). Остальное, как
пленки гумуса, относится к аморфной фракции.
Захоронение Сорг связано с образованием осадков, и поэтому
значение имеет то количество органического вещества, которое до-
достигает поверхности ила. При разложении органического вещества,
например фитопланктона, выделяются три фазы. Сначала происхо-
происходит переход биомассы в мортмассу. В первой фазе (фракция Ml)
происходит автолиз клетки с освобождением растворимого органи-
органического вещества, и считается, что этот этап не контролируется ба-
бактериями, а обусловлен внутренними причинами. При этом теряет-
кото- ' ся 25-40% Сорг и 40-60% органического азота, поступающего в реге-
-'орг
нерационный цикл продукции. Эта фаза протекает в течение не-
нескольких дней. Вторая фаза (М2) начинается после бактериальной
колонизации мортмассы и продолжается от 3/4 года до 2 лет. За год де-
деградирует 75-80% фитопланктона. Фаза зависит от гидролитической
деятельности бактерий, которая определяется доступной для колониза-
колонизации поверхностью и температурой. В этой фазе резко возрастает по-
потребление О2. В третьей фазе (МЗ) происходит накопление устойчиво-
устойчивого Сорг, и оно составляет несколько процентов за год. С длительностью
разложения 1000 лет в осадки поступает до 12% Сорг. Изменение орга-
органического вещества в этой фазе мало зависит от температуры и био-
турбации. Вещество, уходящее ниже 1-2 м от поверхности осадка, за-
захоранивается надолго, и его содержание по профилю мало изменяется.
Захоронение углерода зависит от продуктивности, и его содер-
содержание в осадках увеличивается примерно в два раза с возрастанием
продуктивности в 1,6-1,7 раза. Однако прямой зависимости между
продукцией и захоронением нет, поскольку для захоронения нужно
осаждение большой массы осадков. Было сформулировано предста-
представление, что "степень бактериальной деградации критически зависит
от скорости седиментации независимо от природы и количества от-
отлагаемого органического вещества" (Tyson, 1995). Установлена за-
зависимость между накоплением органического вещества и скоро-
скоростью осаждения: логарифм Сорг прямо пропорционален логарифму
общего накопления осадков в г/(м2 • год).
Сейчас это представление качественно модифицировано в
функцию метаболизируемого органического вещества, достигаю-
221
щего определенной зоны диагенеза. Критическим является переход
захораниваемого органического вещества через оксический/анокси-
ческий барьер в зону сульфатредукции. Между концентрацией О2 в
придонной воде в 1 м от поверхности ила и содержанием Сорг имеет-
имеется четкая зависимость: при концентрации О2 около 1,5 мг/л содержа-
ние QPr B осадках начинает резко расти от 1% до 6% в полностью
аноксических осадках. "Междисциплинарные геологические свиде-
свидетельства в пользу корреляции между качеством и количеством органи-
органического вещества, с одной стороны, и дизоксической (аноксиче-
ской) - оксической палеобстановкой - с другой, остаются неоспори-
неоспоримыми" (Tyson, 1995). Современное захоронение для оседающего
Сорг, проходящего границу аноксии в осадках, составляет 3-29% при
среднем значении 9,8%.
Граница доступности кислорода в современных осадках обычно
определяется на глаз по окисленному железу; как правило, проник-
проникновение О2 в осадки ограничивается 5 мм в тонкодисперсном осадке
и менее 20 мм в песке. В отсутствие биотурбации проникновение 02
в осадки шельфа составляет 1-2 мм, в то время как в пелагических
осадках - 20-40 мм. Переходная зона находится в области потенциа-
потенциала +250 ± 50 мВ. Аэробное разложение доминирует до 0,3 мг/л О2.
B.C. Рибур (W.S. Reeburgh) предложил следующие зоны для
обычно принимаемой последовательности бактериального разло-
разложения органического вещества:
Окис-
Окислитель
О2
NOJ
Мп
Группа
Зона
Конц.,
мМ
Е,
кДж/моль
Название i
Аэробы Ох
Денитрификаторы NR
Mn-редукторы Мп
Fe-редукторы Fe
Сульфатредукторы SR
Метаногены Me
Fe
СО,
<0,09
<0,04
<0,3g
<0,7g
30
730-100
710-100
600-50
-50
-200
-250
-^75
-448
-349
-114
-77
-58
Oxic
Suboxic
Suboxic (
Suboxic
Anoxic
Anoxic
Разница между оксическим и аноксическим диагенетическиш
режимами для бактерий состоит в возможности или невозможности
реакций оксигенирования, необходимых для деструкции восстанов-
восстановленных органических соединений. Окислительный барьер создается
использованием доступного органического вещества.
Таким образом, через аноксический барьер проходят в основном
частицы, подвергшиеся в той или иной степени гумификации. По со-
составу геохимики условно классифицируют кероген как бедный во-
водородом, гуминовый - преимущественно предшественник газа - и
как богатый водородом, сапропелевый - предшественник нефти.
Поскольку захораниваемое органическое вещество в виде керогена
состоит в основном из частичек биологического происхождения, то
можно утверждать, что кислородная атмосфера образовалась за
222
счет мортмассы, перешедшей в анаэробную зону и запечатанной
там процессами седиментации.
В неопротерозое происходило перестроение процесса седимен-
седиментации органического вещества в морских бассейнах. Углеводороды
осадков протерозоя происходят в основном от бактерий и других ор-
ганотрофов, и это указывает на полную переработку мортмассы
первичных продуцентов. С началом кембрия сохранность водорос-
водорослевых липидов, таких, как изопреноиды - продукты деградации
хлорофилла, возрастает в связи с их транспортом в осадки в быстро
погружающихся фекальных пеллетах зоотрофных организмов.
Полная переработка медленно оседающей мортмассы фитопланк-
фитопланктона в протерозое предполагает участие сульфатредукторов и соот-
соответственно формирование восстановительных условий в водной
толще, как это происходит сейчас в некоторых районах Индийского
океана.
Примером условий образования потенциально нефтематерин-
ских пород венда может служить реконструкция палеоэкологиче-
палеоэкологических условий тех центральных районов Русской плиты, которые рас-
рассматриваются как потенциальный источник для формирования
древних нефтей и битуминоидов2 (рис. 6.2). Критическим является
бассейн седиментации. В глубоких бассейнах происходит почти пол-
полное разложение оседающего органического вещества и накопление
фрамбоидного пирита, свидетельствующего о переходе осадков в
аноксические условия. В мелководных бассейнах главную роль иг-
играет поступление терригенно-глинистых осадков, приводящее к бы-
быстрому захоронению органических остатков практически без дест-
деструкции.
В какой мере рассматриваемые закономерности могут быть
приложены к раннепротерозойским углеродистым породам? Чер-
Черные сланцы Курской магнитной аномалии (КМА) были образованы
микробным матом с доминантой, органостенные микрофоссилии
которого подобны современной цианобактерии Microcoleus. В обра-
образовании шунгитового осадка Карелии принимали участие коккоид-
ные цианобактерии3. В обоих случаях наблюдается ассоциация угле-
углеродистых пород с кремнистыми. Время ведет к графитизации орга-
* нического углерода древних пород.
f На основании широкого распространения строматолитов в
' Сибири было предположено, что типичным для образования оса-
' дочных пород на этой территории в рифее был обширный эпикон-
2 Афанасьева М.С., Бурзин М.Б., Михайлова М.В. Кузьменко Ю.Т. Условия об-
образования потенциально нефтематеринских пород. // Геология нефти и газа.
1995. N 4. С. 42-45.
j 3 Жмур СИ., Горленко В.М., Розанов А.Ю. и др. Цианобактериальная бентос-
ная система - продуцент углеродистого вещества шунгитов нижнего протеро-
протерозоя Карелии//Литология и полезн. ископаемые. 1993. N 2. С. 122-124.
223
Относительно глубоководная часть бассейна
Одиночный и колониальный планктон Плавающие массы эохолыний
О
Мелководная часть
бассейна
Вынос терригенного
материала с суши
Некромасса планктона
2^*ЙЙЙЙ1» \ /^"Распресненные
Планктонные серные eferrmi^ воды эохолыний
бактерии\
\ Полная деструкция планктонного органического вещества
\ / I Соленая \ '
Некромасса серных | морская
Песчано-алевритистые
осадки
бактерий
вода с
| сероводородом
Редкие
остатки
планктона
/Немногочисленные нитчатые
/ бентосные цианобактерии
Донные илистые осадки, обогащенные органическим веществом,
переполненные фрамбондальным пиритом
Планктон в наиболее
открытых частях Пятна цветения
бассейна Нитч;
Предположительно
гидродинамически
активные части
бассейна
.атыхцианобактерий Заросли макрофитов
Крайне мелководные части бассейна
Бентосные нитчатые цианобактерии
В результате сезонных поступлений в бассейн
больших масс терригенно-глинистых осадков
происходит быстрое захоронение некромассы ,
планктона и бентоса практически без деструкции
Рис. 6.2. Реконструкция палеоэкологических условий в бассейне центральных
районов Русской плиты во время накопления осадков в начале Редкинского (а)
и в Поваровское (б) время позднего венда [по: Афанасьева и др., 1995]
тинентальныи водоем с относительно малыми глубинами, позволяв-
позволявшими развиваться бентосным циано-бактериальным сообществам4.
Впоследствии это представление нашло свое подтверждение не только
относительно карбонатных пород, но и других осадков. Развитие
эпиконтинентальных водоемов имело место в талассократические
эпохи, связанные с глобальными потеплениями, и обусловлено из-
изменениями климата для больших промежутков времени.
Поскольку условием образования богатых органическим веще-
веществом пород служит быстрый переход массы отмирающих первич-
первичных продуцентов через границу оксической и аноксической зон,
бентосные циано-бактериальные сообщества с резкой границей в
4 Крылов 14.Н., Заварзин Г.А. 1988. Условия образования карбонатных толщ в
верхнем рифее Южного Урала // Докл. АН СССР. Т. 300. N 5. С. 1223-1225.
224
миллиметры между оксическими и аноксическими слоями должны
были бы хорошо сохраняться. Такой путь образования нефтемате-
ринских пород описывает М.Б. Бурзин для отложений вендского до-
маника.
Трудно объяснить, почему заведомо образовавшиеся из циано-
бактериальных матов карбонатные строматолиты содержат так ма-
мало органики. В карбонатах органическое вещество не достигает та-
такой степени полимеризации, как в глинистых породах. Причиной
может служить формирование карбонатных отложений в оксиче-
оксической зоне с полным разложением органического вещества.
Оценивая итоги палеонтологических исследований, А.Ю. Роза-
Розанов5 пришел к заключению, что "согласно всей совокупности дан-
данных по высокоуглеродистым породам, многие бассейны прошлого,
в которых отлагались черносланцевые толщи, не были глубоковод-
глубоководными, как обычно полагали, а представляли собой огромные эпи-
континентальные бассейны с природной аноксией". Основу Сорг чер-
черных сланцев представляет кероген.
Кероген состоит как из биогенных остатков, так и из продуктов
их диагенетического превращения. Основные количественно опре-
определяемые химические компоненты керогена представлены нор-
нормальными, разветвленными и циклическими углеводородами, серо-
серосодержащими алкилированными углеводородами, алкилфенолами,
произошедшими из липидов, лигнина. Они составляют меньшую
часть Сорг керогена. В основном он представлен органическими
остатками, которые претерпели изменения во время и после захоро-
захоронения. В отличие от битумов и нефтей кероген не мигрирует и оста-
остается на месте захоронения. Кероген разделяется на структурирован-
структурированную (частично углефицированную) и аморфную (гумифицирован-
ную) фракции. В противоположность Р.В. Тайсону, другие авторы
считают основной по массе частью керогена аморфный кероген, об-
образующий пленки во вмещающей породе. Образование нефти из ке-
керогена представляет реакцию диспропорционирования, дающую бо-
богатую водородом подвижную фазу и лишенный водорода углистый
остаток, который может переходить в графит. Образование нефтей
и углей представляет альтернативные пути захоронения Сорг, при-
причем угли раньше рассматривались как материнские породы для газа.
Битумы представляют часть органического вещества, которая мо-
может быть экстрагирована органическими растворителями и состав-
составляет обычно 30-50 мг/г Сорг керогена.
Превращение керогена в нефть прослеживается по ряду параме-
параметров созревания Сорг содержащих его соединений. Одним из подхо-
подходов является представление о "мертвом углероде" с разделением ис-
исходного керогена на A) инертную часть, превращающуюся в графит
3 Розанов А.Ю., Заварзин ГЛ. Бактериальная палеонтология // Вестн. РАН.
1997. Т. 3. С. 241-245.
8. Г. А. Заварзин
225
при высоком давлении и температуре, B) реактивную, которая дает:
а) лабильный Сорг с последующим превращением в нефть и газ,
б) устойчивую часть, которая превращается в газ в зоне катагенеза
при высоком давлении и температуре. Моделью процесса может
быть образование следов нефтей из относительно свежего органи-
органического вещества в гидротермальном процессе, как это наблюда-
наблюдалось на Узоне, термальных площадках Калифорнии. Газ и нефти
представляют части захороненного Сорг, которые способны к мигра-
миграции в поровом пространстве пород и скапливаются в отдалении от
места их образования. Выход нефти и газа в окислительную зону с
доступом О2 и в меньшей степени SO^" обусловливает возможность
их микробной деградации после длительного периода геологическо-
геологического захоронения.
Образование1 нефтяных и газовых месторождений из нефтема-
теринской осадочной породы, обогащенной органическим вещест-
веществом в виде керогена, происходит в процессе геологического катаге-
катагенеза с характерным временным периодом в сотни миллионов лет,
Миграция углеводородов в пористой породе приводит к отсутствию
прямой связи с нефтематеринской породой и накоплением в ловуш-
ловушках. Образовавшиеся в восстановительной зоне при высокой темпе-
температуре и давлении углеводороды всплывают, занимая максимально
высокое положение в проницаемом слое породы. Дальнейший их
подъем ограничивает газоупорная плотная порода либо в виде сво-
свода (антиклиналь), либо в виде запирающего пласт сброса. Углеводо-
Углеводороды могут скапливаться и в меньших запечатанных вторичными
барьерами объемах нефтепроницаемых пород, часто на глубинах
3-4 км. Присутствие смеси жидких и газообразных углеводородов
приводит к образованию газоконденсатных месторождений.
6.7. БОЛОТА
Наземные амфибиальные экосистемы часто стали называть
американским термином ветленды (англ. wetlands). Согласно анг-
английскому определению "ветленды представляют площади, в кото-
которых (а) уровень воды находится на или вблизи поверхности почвы
значительную часть вегетационного периода и (б) почвы покрыты
активной растительностью в период насыщения водой". Это опреде-
определение соответствует русскому понятию болот. Болотные системы
представляют важнейший ландшафт для обеспечения устойчивости
в биосфере. В глобальном отношении они служат стоком СО2 и ис-
источником СН4 - важнейших парниковых газов. В региональном мас-
масштабе они определяют гидрологический цикл, располагаясь, как
правило, в области питания рек. Болота относятся к амфибиальным
ландшафтам с доминированием высшей растительности. Этим они
отличаются от морских лагун или от гидротерм, где развиваются ци-
226
анобактериальные маты. Болота относятся к внутриконтиненталь-
ным ландшафтам с пресной или ультрапресной водой.
Все амфибиальные ландшафты характеризуются тесным взаи-
взаимодействием доминирующей анаэробной зоны с мелкой аэробной
зоной. Масса воды, большей частью застойной, относительно неве-
невелика. Отсюда характерной чертой биоценозов амфибиальных ланд-
ландшафтов является тесное взаимодействие аэробных и анаэробных
организмов. Среди анаэробов безусловно доминируют бактерии.
Восстановительные процессы в анаэробной зоне оказывают опреде-
определяющее влияние на химический состав воды и ход минералообразо-
вания. Типичным выражением этих процессов служит образование
болотной железной руды.
В соответствии с употреблением слова "болото" в русской лите-
литературе с этим ландшафтом связано образование торфа - слабораз-
ложившейся гумифицированной массы растительных остатков.
Таким образом, болото может быть охарактеризовано как ланд-
ландшафт с нарушенным процессом минерализации органического ве-
вещества. Неполноценность анаэробного микробного сообщества в
разложении растительных остатков ведет к накоплению в болотах
Сорг, которому эквивалентно накопление О2, образовавшегося при
фотосинтезе. Таким образом, в глобальном масштабе болота слу-
служат не только стоком СО2, но и источником О2, хотя их продуктив-
продуктивность ниже, чем во многих других экосистемах. Дефектность про-
процесса минерализации растительной мортмассы, преимущественно
лигноцеллюлозы, сообществом микроорганизмов болота обуслов-
обусловливает дисбаланс в углеродном цикле наземных экосистем. Точно
так же дефектность анаэробного сообщества в циано-бактериаль-
ных матах приводит к накоплению липидного материала мембран,
служащего предшественником керогена при образовании нефтей.
Пресный и ультрапресный состав болотных вод определяет направ-
направленность процесса разложения органического вещества метаноген-
ным сообществом. В противоположность заболоченным землям с
повышенной соленостью сульфидогенез в болотах не доминирует.
Тонкий слой аэрированной почвы или воды ограничивает возмож-
возможности окисляющих газы организмов, прежде всего метанотрофов
"бактериального фильтра", и потому болота служат важнейшим ис-
источником восстановленных газов, в первую очередь метана, в атмо-
атмосфере. Болота и лесо-болотные экосистемы служат важнейшим ис-
источником образования углей.
По определению, сформулированному Всесоюзным ботаниче-
ботаническим обществом в 1966 г., "...болото - тип земной поверхности, посто-
постоянно или длительное время увлажненной и характеризующейся со-
соответственным почвообразовательным процессом".
Заболоченные земли (то же wetlands) формально определяют
как "земли, переходные между наземными и водными системами,
где вода подходит к поверхности или покрывает ее тонким слоем".
8» ' 227
Им свойственно: 1) наличие гидрофитов; 2) наличие твердого суб-
субстрата, классифицируемого как гидроморфная почва, 3) насыщение
субстрата (почвы или торфа) водой или его затопление водой каж-
каждый год во время вегетационного сезона. Внутренние заболоченные
земли характеризуются наличием торфа, мелководьем, раститель-
растительностью, включающей Typha, Saggitaria, Pontederia, Phragmites,
Glyceria, Carex. Северные торфяные болота характеризуются мощ-
мощным торфонакоплением и подразделяются на два типа: bog (верхо-
(верховое, преимущественно моховое) и fen (низовое, преимущественно
осоковое). Имеется ряд юридических определений "увлажненных
земель:
Охрана болот. Заболоченные земли привлекли внимание преж-
прежде всего как местообитания перелетных птиц и стали охраняемыми
территориями согласно Раамсарской конвенции. Но по существу
они составляют важнейший компонент гидрологической сети. Если
ранее единственно правильной политикой относительно болот при-
признавалось их осушение, то теперь это время прошло, хотя продол-
продолжается нарушение состояния болот из-за изменения гидрологиче-
гидрологических условий. Болота представляют малую хозяйственную ценность
для владельцев земель, но они важны как общественное достояние,
прежде всего для регулирования качества воды.
Это понимание биосферной функции болот нашло отражение в законода-
законодательстве США. Охрана болот в США регулируется двумя указами президента
Картера 1977 г.: Executive Order 11990 Protection of Wetlands требует рассматри-
рассматривать охрану болот как важную часть политики: "Каждое агенство должно руко-
6 The term "wetlands" ...refers to lowland cowered with shallow and sometimes tempo-
temporary or intermitten waters. They are refered to by such names as marshes, swamps,
bogs, wet meadows, potholes, sloughs, and riveroverflow lands. (U.S.Fish and Wildlife
Serv. Circular. 1956. N 39), т.е. увлажненные земли - это низины, покрытые
мелкой и иногда временной водой.
Канадское определение: "...области, где преобладают влажные почвы,
имеющие уровень грунтовых вод вблизи или над поверхностью минеральной
почвы большую часть талого сезона, поддерживая гидрофильную раститель-
растительность". ("Wetland is defined as a land having the water table at, near, or above the land
surface or which is saturated for a long enough period to promote wetland or aquatic
processes as indicated by hydric soils, hydrophylic vegetation, and various kinds of
biological activity which are adapted to the wet environment" - Canadian Wetland
Registry).
Английское определение: «.. .wetland is "an area in which: (a) water table is at
or near the soil surface for significant part of the growing season; and (b) soils are cov-
covered by active vegetation during the period of water saturation"».
Официальное определение в США дано в разделе 404 Clean Water Act
Amendments, 1977: "The term wetlands means those areas that are inundated or satu-
saturated by surface or ground water at a frequency and duration sufficient to support, and
that under normal cirumstances do support, a prevalence of vegetation typically adapt-
adapted for life in saturated soil conditions. Wetlands generally include swamps, marshes,
bogs, and similar areas".
228
водить и предпринимать действия к минимизации деструкции, потере или дегра-
деградации болот и сохранять и усиливать естественные и полезные свойства бо-
болот...". Второй указ касался заливных площадей: Executive Order 11988.
Floodplain Management устанавливает аналогичную федеральную политику для
защиты заливных земель, требуя от агенств избегать, где это возможно, актив-
активности на заливных землях. Раздел 404 Federal Water Pollution Control Act с допол-
дополнениями 1977 г. стал основой для широкой программы защиты увлажненных зе-
земель. Сначала этот акт воспринимали только относительно водотоков, но ре-
решение суда в пользу Совета защиты природных ресурсов включило в понима-
понимание "вод США" также болотные воды: "Как жизненно важные для окружаю-
окружающей среды области (болота) составляют продуктивный и ценный обществен-
общественный ресурс, ненужное изменение или деструкция которого следует отклонять
как противоречащее общественным интересам". Важные с общественной точ-
точки зрения переувлажненные земли не представляют ценности для владельца.
Поэтому фермеры в США подняли яростную кампанию против применения ак-
актов охраны этих земель, утверждая, что они теряют возможность продать их
для развития и им труднее получить займы. Увлажненные земли, даже под паш-
пашней, сохраняют способность удалять загрязнения, но мелиорированные теряют
способность улучшать качество поверхностных и грунтовых вод ниже по тече-
течению. История чрезмерной мелиорации в СССР с тяжелыми экологическими по-
последствиями широко известна.
Среди континентальных болот по типу питания различают авто-
автономные ландшафты верховых болот с дождевым питанием и подчи-
подчиненные ландшафты низовых болот, аккумулирующих сток. Конт-
Контраст между верховыми и низовыми болотами определяется недос-
недостатком минеральных компонентов в первых и их избытком во вто-
вторых. Для болот разработана детальная классификация, приведенная
В.В. Мазингом в многоязычном словаре болотоведения ("тельмато-
логии"O. За исходные взяты англоязычные термины.
- Swamp - болото - увлажненные земли с доминированием деревьев или
кустарников; в Европе покрытое лесом сфагновое болото могут называть бо-
болотом (swamp), а иногда так называют и земли, покрытые травянистой расти-
растительностью. У нас в эту категорию попадают черноольшанники с особым ре-
режимом азотного питания, обусловленным симбиотической азотфиксацией
Frankia.
- Marsh - маршевое болото (иногда у нас этому соответствуют тростнико-
тростниковые заросли), часто или постоянно заливаемое пространство, приспособленное
к водонасыщенной почве, характеризующееся травянистой растительностью;
по европейской терминологии, марш имеет минеральную почву и не накаплива-
накапливает торфа. Марши часто связаны с приливными отмелями (tidal flats), представ-
представляющими обширную илистую поверхность, обнажаемую при отливе моря. Под-
Подпор приливной волны вызывает застой воды на вышележащей континенталь-
континентальной равнине, прорезанной множеством рек, и образование гидроморфных почв
в условиях пресной воды. В немецком языке для этих систем употребляется
обобщающий термин die Watt, включающий и отмели без растительности
(Sandwatt). Здесь микробы создают своеобразное сообщество, напоминающее
колонку Виноградского, сжатую до сантиметра - Streiffarbsandwatt.
7 Мазинг В.В. Немецко-английско-шведско-финско-эстонско-русский термино-
терминологический словарь по болотоведению. Тарту, 1960. 188 с.
229
- Bog - верховое сфагновое болото (Hochmoor), накапливающее торф, ко-
которое не имеет значительных притоков или стоков, кислое, дистрофное, с дож-
дождевым питанием (омбротрофное). Для нашей страны это принципиально важ-
важная экосистема.
- Moor - верховое болото, в европейской терминологии синоним торфяни-
торфяника, a highmoor - это приподнятое болото, в то время как lowmoor - торфяник в
депрессии, не поднимающийся над своим периметром.
- Muskeg - сфагновое болото, большие пространства торфяников обычно
в Канаде и на Аляске.
- Fen - низовое болото с накоплением торфа, которое питается дренажны-
дренажными водами из окружающей минеральной почвы и обычно имеет травянистую
растительность, преимущественно осоковую. Это минеротрофные болота, бо-
богатые кальцием с нейтральной реакцией среды, замыкающие катену.
- Peatland - торфяник, общее название для любой увлажненной почвы, на-
накапливающей частично разложенный растительный материал.
- Mire - топь, в европейской терминологии синоним торфяника.
- Bottomland - пойма, низинные земли по берегам водотоков, обычно аллю-
аллювиальной равнины периодически заливаемые; они часто покрыты лесом, и их
называют заливаемыми лесами.
- Floodplains - заливные земли, периодически заливаемые территории на
берегах рек и озер.
- Wet meadow - болотистый луг, травяная земля с увлажненной почвой, но
без стоячей воды.
- Reedswamp - тростниковое болото, марш с доминированием тростника
Phragmites, термин, особенно распространенный в Восточной Европе.
- Slough - болотистая или мелководная озерная система с медленно проте-
протекающей водой.
- Pothole - мелкие болотистые пруды.
- Playa - термин, употребляемый в США для болотистых прудов, но гораз-
гораздо чаще для приморских низин. В аридном климате это сабхи (себхи, sabha -
арабское название), у нас в Прикаспии - соры (шоры), солончаки.
- Tidal flat - приливные отмели, плоские пространства у низких берегов
океана, периодически заливаемые во время прилива и состоящие из mud flat в
нижней части и заросших высшей растительностью торфяников на морских от-
отложениях в верхней части. На побережье Северного моря этот ландшафт назы-
называют Wattensee, часто он прорезан эстуариями.
- Mangroe - заросли мангровых деревьев в приливной зоне тропически
морей.
- Полигональное болото - болото, образуемое в тундре на вечной мерзло-
мерзлоте и разделенное ледовыми клиньями на ряд блюдец-полигонов; оно наиболее
важно для севера России.
.- Аапа - сфагновые болота Финляндии и Карелии в виде валов.
Все эти термины имеют не вполне точные переводы в других
языках, например, в русском отсутствует эквивалент swamp, вероят-
вероятно, точнее всего относящийся к болотам Полесья; англичане отно-
относят swamp к тростниковым зарослям. Разнообразие болот предпола-
предполагает соответствие каждому типу болота не только растительного, но
и микробного сообщества. В особенности велико различие между
морскими болотами с достаточным количеством сульфата для доми-
доминирования сульфидогенеза и серного цикла и пресными болотами с
доминированием метаногенеза. Для северной Евразии наиболее
важны верховые сфагновые болота на водораздельных пространст-
пространствах, низовые болота у водотоков, влажная тундра.
Главным признаком собственно болот служит накопление тор-
торфа - не полностью разложенных остатков растений, пропитанных
гуминовыми веществами. Таким образом, функциональным призна-
признаком этой экосистемы служит дисбаланс между продукцией и дест-
деструкцией, ведущий к накоплению органического углерода. Ключе-
Ключевым процессом оказывается замедленное разложение растительных
остатков органотрофным микробным сообществом. Замедленное
разложение обусловливается в первую очередь двумя ловушками:
1) анаэробной ловушкой, блокирующей разложение лигноцеллюло-
зы, 2) холодной ловушкой, тормозящей все процессы микробного
разложения. В дополнение действуют такие факторы, как повы-
повышенная кислотность, накопление трудно доступных гуматов. Мик-
Микробное разложение оказывается заторможенным на ранних фазах.
Большее значение анаэробиоза иллюстрируется быстрым торфона-
коплением в тропических болотах, где высокая температура. Анаэ-
Анаэробиоз обусловлен избыточным увлажнением при отсутствии под-
подвижности вод и ограничением обмена с воздухом. Избыточное
увлажнение возникает вследствие того, что болото возникает на во-
доупоре. Этот водоупор может образовываться не только на таких
тонкодисперсных минеральных грунтах, как глины, но и из-за фор-
формирования торфа, и в результате возникает положительная обрат-
обратная связь. На севере водоупор создается на вечной мерзлоте. Осуше-
Осушение болот ведет к стремительному биологическому выгоранию тор-
торфа из-за аэрации. Таким образом, определяющим свойством всех
торфяников служат подавленная микробная деструкция и избыточ-
избыточное образование устойчивых органических веществ гуминовой при-
природы.
Гидрология является определяющим фактором болотной экоси-
экосистемы. Баланс воды в болоте складывается из суммы атмосферных
осадков, поверхностного притока (включая наводнения) и подземно-
подземного притока (для низовых болот), потерь в виде эвапотранспира-
ции, поверхностного и подземного стоков. Болота характеризу-
характеризуются застойным режимом воды. Время пребывания воды в болоте
определяется отношением общего притока к объему воды. Струк-
Структурные и функциональные характеристики зависят прежде всего от
гидрологического режима, который влияет на доступность пита-
питательных веществ. Гидравлическая проницаемость болот составляет
(в см/с • 10-5): верховое болото слабой степени разложения - 500;
средней степени разложения - 80; сильной степени разложения - 1;
осоковое болото: слой 0-50 см - 310; слой 100-150 см - 6. Отсюда
видно, что положительная обратная связь ведет к экспансии болота
за счет уплотнения торфянистых остатков.
Географическое распределение болот строго соответствует
климатическим поясам избыточного увлажнения. Общая площадь
230
231
х103км2
1500 г
1000-
0
N 90 80 70 60 50 40 30 20 10 0 10 20 30 40 50° S
Рис. 6.3. Меридиональное распределение болот на Земном шаре
/ - по Aselman, Crutren, 1989,2 - по Matthews, Fung, 1987
болот оценивается от 5,3 • 1012 до 8,6 • 1012 м2 в зависимости от уче-
учета сезонно заливаемых равнин Южной Америки. Оценки разных ав-
авторов сильно различаются. Болота проходят поясом избыточного
увлажнения между 50-70° С.Ш. (рис. 6.3.) и распределяются на тер-
территории стран следующим образом (в млн км2): Канада - 1,27;
США - 0,23; Аляска - 0,325; СССР (97% Россия) - 1,5; Европа -
0,0067; Юго-Восточная Азия - 0,244; Китай - 0,031; Ближний Вос-
Восток - 0,008; Южная Америка - 1,5 (временно заливаемые равнины);
Центральная Америка - 0,0175; Африка - 0,356; Австралия -
0,00166. По примененным 6 классам болот они распределяются так:
bogs - 1,9; fens - 1,5; swamps - 1,1; floodplains - 0,8; marsh - 0,4. Тро-
Тропические болота гумидного пояса Экваториальной Африки, Индо-
Индонезии, Южной Америки (Амазония) отличаются очень высокой про-
продуктивностью, и вытекающие из них реки часто имеют темную во-
воду из-за обилия гуминовых веществ. Амазонка служит одним го
главных источников сноса растворенного органического вещества в
океан. Анализ распределения болот на территории Северной Евра-
Евразии, проведенный С.Э. Вомперским A994) в связи с определением
естественного углеродного баланса России, показал, что оторфо-
ванные и заболоченные земли занимают 369 млн га., из них 3/4 - в
зоне вечной мерзлоты с общим запасом Сорг 113,5 Гт.
232
Продуктивность болота зависит от того, покрыто ли оно стоя-
стоячей водой или же имеется проток. Во втором случае продуктивность
существенно выше. Деструкция оказывается наименьшей в постоян-
постоянно анаэробных условиях и наиболее высокой при чередовании аэ-
аэробных и анаэробных условий.
Почвы увлажненных земель классифицируются как 1) мине-
минеральные почвы, 2) органические торфянистые, или гистосоли, когда
они содержат более 20% и до 35% органического вещества по сухо-
сухому весу:
Характеристики почв
Почвы
минеральные
органические
Органические вещества, %
рН
Плотность, г/см^
Пористость, %
Проницаемость
Водоудержание
Питательные вещества (биогены)
Катионобменная способность
обусловлена полимерами
Менее 20 или 35
Близь нейтральные
1-2
45-55
Высокая (кроме глин)
Низкое
Много
Низкая, металлы
Более 20 или 35
Кислые, рН 4
0,2-0,3
80
Варьирует
Высокое
Мало
Высокая, водород
Уроновых кислот
Минеральным называют дно с содержанием органического ве-
вещества менее 20%. Вода считается кислой при значении рН < 5,5 и
щелочной при рН > 7,4, а примерно нейтральной при рН 5,5-7,4.
Территория России почти полностью, за исключением юга, относит-
относится к кислой зоне.
Гидрохимия вод амфибиальных ландшафтов зависит от типа пи-
питания. Для морских экосистем - это разной степени разбавления
морская вода или, в лагунах, концентрированная вследствие испаре-
испарения. Для наземных ландшафтов тоже существуют очень большие
вариации - от ультрапресных вод моховых верховых болот вплоть
до насыщения солями, выпадающими на сорах или себхах в виде га-
лита, а на содовых солончаках - в виде карбонатов натрия. Вода болот
России обнаруживает сезонные изменения состава. Наименьшее
значение минерализации приходится на паводковый период, летом
вдет постепенное нарастание, осенью минерализация снова снижается
за счет дождей. Зимой происходит концентрация вследствие вымора-
вымораживания с максимумом при наибольшем промерзании. На низинных
болотах Приуралья минерализация менялась от 25,8 до 2350,8 мг/л,
а на верховых (омбротрофных) - от 9,8 до 146,3 мг/л8. Состав вод ни-
низовых болот резко выраженный гидрокарбонатно-кальциевый.
8 Черняев A.M., Черняева Л.Е., Еремеева М.Н. Гидрохимия болот: Урал и Пря-
уралье. Л.: Гидрометеоиздат, 1989. 429 с.
233
17
Рис. 6.4. Распределение болот в России, [по: Stolbovoi, McCallum, 2002]
Классы: 1 - сфагновые (> 50 см); 2 - мелкооторфованные C0-50 см); 3 - облесенные
A0-30 см); 4 - кустарниковые A0-30 см); 5 - заторфованные леса (< 10 см); 6-травяные
болота
Кальций служит показателем евтрофности и хорошо коррелирует с
рН. При содержании Са2+ > 20 мг/л рН 7 ± 0,5, а при Са2+ < 10 мг/л
растительность олиготрофная и рН может снижаться. Аммоний со-
содержится в воде верховых болот на уровне 1-3 мг/л, а в низовых -
около 5 мг/л. Фосфатов очень мало: от 0,045 до 0,2 мг/л, и максимум
их приходится на зимний период. Существенную часть минеральных
веществ составляет кремнекислота с сильными сезонными колеба-
колебаниями и максимумом в апреле. Железо болотных вод имеет четкий
зимний максимум и летний минимум. С железом связано образова-
образование зеленоватого глеевого горизонта в болотистых почвах. Источ-
Источником растворенного железа является его восстановление железо-
редуцирующими психрофильными органотрофными бактериями с
образованием сидерита, но не магнетита. Весной при таянии снега
железо поступает в воду, и происходит развитие железобактерий,
чаще всего психрофильной Gallionella, с образованием обильных ох-
охристых осадков (рис. 6.4).
6.8. ВЕРХОВЫЕ БОЛОТА -
АВТОНОМНЫЕ РЕЛИКТОВЫЕ БИОЦЕНОЗЫ
Верховые болота являются наиболее типичными представите-
представителями автономных ландшафтов. Они развиваются на водоразделах и
в своем питании ограничены тем, что поступает к ним из воздуха с
осадками и пылью. От минерального грунта они отделены слоем
234 "'
торфа. Вода отличается низкой минерализацией, составляющей де-
сятки-сотню мг/л, низкой электропроводностью, низким значением
рН 3-5, обусловленным органическими кислотами, углекислотой,
ионным обменом на поверхности растений. Фотосинтез может по-
повышать рН. Цикл серы здесь минимизирован. Таким образом, вер-
верховое болото представляет прежде всего углеродный цикл при ми-
минимуме сочетания с другими циклами. Вместе с тем цикл углерода
замкнут далеко не полностью, и углерод уходит в резервуар дли-
длительного времени пребывания, несмотря на медленный процесс ас-
ассимиляции по сравнению с другими растительными сообществами.
Господствующей растительностью на верховых болотах явля-
являются сфагновые мхи. Они считаются древнейшими растениями на
суше. Персистентность мхов связывают с особенностями их биоло-
биологии. Они устойчивы к изменению условий. Отсутствие покровных
тканей позволяет им очень быстро набирать влагу. Вместе с тем они
устойчивы к высушиванию, гербарные мхи оживали через 50 лет.
Сфагны растут и в тундре, и в горячих источниках Южных Курил.
Развиваясь на поверхности скал, мхи вместе с лишайниками создают
торфяной и гумусовый остаток, удерживающий влагу. Таким обра-
образом, мхи - и пионеры сущи, и очень древние растения, создающие
реликтовые сообщества. Верховые болота с доминированием сфаг-
нов представляют чрезвычайно своеобразные экосистемы. Они
прослеживаются с пермского периода, когда обнаружены остатки
Protosphagnales, открытые М.Ф. Нейбург в пермских угольных отло-
1, жениях Евразии. Листья Protosphagnales, в отличие от современных
' сфагнов, имеют срединную жилку, но в остальном представляют
такую же сеть хлорофиллоносных клеток и окружающих их водо-
водоносных. Образование современных болот на территории Евразии
началось после оледенения. Мхи создают очень агрессивное сооб-
сообщество, образуя изолирующий слой, на котором не прорастают се-
семена других растений. У них развиты и антибиотические свойства.
Мхи, живущие во влажных условиях в гумидном климате, оказыва-
оказывают важное влияние на регулирование водного режима. В этом отно-
отношении они до известной степени противопоставляются сообществам
["' лишайников, обитателям сухих мест и топографических высот.
Растительность верховых сфагновых болот отличается крайне
низким биоразнообразием, и в состав сообщества помимо сфагнов
входит четко ограниченный круг растений-сфагнофилов, преиму-
преимущественно вересковых кустарничков. Сфагны представляют край-
крайний пример социальных растений, развиваясь в виде сообщества -
сфагнеты. В России встречается 42 из известных 300 видов рода
Sphagnum, с наиболее распространенными S.angustifolium, S.balticum,
Sfuscum, S.magellanicum. Доминирующие виды сфагнов варьируют в
зависимости от 1) омбротрофного или минеротрофного питания;
2) степени увлажнения; 3) расположения на краю или в середине
болота. Наиболее важными факторами оказались климат и хими-
235
ческий состав воды. Но сфагны и сами по себе служат мощным сре-
дообразующим фактором. Sphagnum развивается в условиях низкого
содержания катионов, особенно кальция, так как его ткань, содер-
содержащая много уроновых кислот, служит хорошим катионообменни-
ком, приводя к подкислению среды. В случае притока минеральных
веществ сфагны оказываются неконкурентоспособными по сравне-
сравнению с цветковыми растениями. Поэтому они представляют экстре-
мофилов-олиготрофов по отношению к минеральному питанию.
Сфагны создают специфическую среду и в рамках ландшафта, и
на уровне дерновины. Тонкий и длинный стебелек сфагна не имеет
ризоидов и покрыт веточками. На вершине веточки образуют плот-
плотные головки с звездчатыми листочками, занимающими площадь
2-4 см2. Стебелек окружен эпидермисом из бесцветных клеток с по-
порами. Под ними идет буроватый древесинный цилиндр, а централь-
центральная часть стебелька занята токостенными клетками сердцевины.
Ббльшая часть листочков состоит из крупных гиалиновых клеток с
порами и поперечными утолщениями. Между ними зажаты тонкие
хлорофиллоносные клетки, сеть которых напоминает зеленую во-
водоросль - водяную сеточку Hydrodictyon. Форма хлорофиллоносных
клеток служит определительным признаком сфагнов. Гиалиновые
клетки образуют капиллярную сеть, позволяющую сфагнам удер-
удерживать стократные количества воды. Сфагны относятся к типич-
типичным кальциефобам.
Структура сфагнового покрова вполне закономерна. До 5 см в
глубину располагаются прямостоячие стебли "зеленого мха", далее
до 15-20 см идет слой "белого мха" (очес) из потерявших хлорофилл
вследствие самозатенения растеньиц. На глубине 20 см в зависимо-
зависимости от уровня воды обычно начинается переход в "бурый мох" и на-
начало торфообразования. Растеньица сохраняют все листья выше са-
самого высокого уровня воды и теряют все листья ниже самого низко-
низкого. Благодаря особенностям сфагнов вода в болоте претерпевает
*
Отличительной особенностью сфагнов служит их способность
удерживать влагу гиалиновыми клетками листьев. Вода и углекис-
углекислота поступают в сфагны без проводящей системы и устьиц. Уро-
Уровень воды служит определяющим фактором в развитии сфагнов.
Вода поднимается в капиллярном пространстве между стеблями и
листьями мха. Рост мха зависит от влажности головок. Пассивная
зависимость от влаги и колебаний уровня воды компенсируется ус-
устойчивостью к высушиванию и быстрым откликом на увлажнение,
причем быстрее восстанавливается дыхание и медленнее - фотосин-
фотосинтез. Фотосинтез зависит и от влажности, и от содержания воды. Для
сфагнов естественное содержание влаги составляет 660-1200% и оп-
определяет соотношение проникновения воды снизу и света и углеки-
углекислоты сверху. В насыщенном водой мхе фотосинтез возрастает с
увеличением СО2 до 8000 ррт. Ассимиляция СО2 идет как у обыч-
обычных С-3 растений со скоростью 100-300 мг СО2 / (м2 • час). Жизненная
стратегия сфагнов представляет компромисс между водным режи-
режимом, доступностью света, поступлением СО2 и минеральным пита-
питанием. Малая зависимость сфагнов от минеральных веществ опреде-
определяет выигрыш сфагнов при их конкуренции с сосудистыми растениями
в ультрапресных водах. Они представляют выраженных кальциефо-
бов. Сфагны отличаются эффективной системой ассимиляции свя-
связанного азота, которая позволяет им выигрывать не только у сосу-
сосудистых растений, но и у микроорганизмов-деструкторов. Сфагнум
содержит мало азота с отношением С : N = 72. Прирост сфагнов за-
зависит от рецикла азота в сообществе. Легко деградируемые азоти-
азотистые соединения разлагаются микробами, масса которых невелика,
и соответственно невелика и масса азотистых соединений, иммо-
иммобилизуемых в микробной биомассе, где относительно велик вклад
глюкозаминов стенок грибов. Повышение содержания азота приво-
приводит к проигрышу сфагнов по отношению к сосудистым растениям.
f Часть азота остается в трудноразлагаемых азотистых соединениях,
резкие изменения. Обилие уроновых кислот, служащих мощным ка- * * которые переходят в гумусовые вещества торфа в затопленной ана-
тионобменником, приводит к повышению кислотности до рН 3,0-4,5
и созданию среды, ограничивающей развитие нейтрофильных рас- * ? ко в последнее время С.Н. Дедыш удалось установить, что ацидо-
тений. Кроме этого, сфагны выделяют антибиотическое вещество •
сфагнол, подавляющее бактерий. Особенности структуры сфагно %
вого покрова приводят к удержанию взвешенных частиц и очистке *
воды от механических примесей. Гиалиновые клетки с порами слу-
служат ловушками для бактерий, которых легко наблюдать внутри и
которые не образуют микроколоний, указывающих на размноже-
эробной зоне; при осушении они могут снова использоваться. Толь-
"# фильные метанотрофы сфагновых болот Methylocella, Methylocystis
* обладают нитрогеназой, сходной с нитрогеназой ацидофильного
** азотфиксатора Beijerinkia. Для этих организмов характерно подав-
* ление роста связанным азотом, и они представляют "нитрофобов".
"* Болота представляют собой глобальную ловушку для органиче-
* ского углерода,- обусловленную подавлением микробного разложе-
разложение, как это происходит на волокнах растительных остатков. Оби- * ' ния в анаэробной среде. В результате образуется торф, представля-
лие продуктов разложения сфагнов в виде фульвокислот придает* ** ющий спресованные, не полностью разложенные остатки растений,
высокую цветность воде, которую относят к категории "дистроф-^;
ных вод". Содержание минеральных компонентов в ней минимально
и составляет величину порядка
электропроводность.
236
|* пропитанные гумусовыми веществами. Моховые болота - достаточ-
достаточно простые системы, где место цианобактерий в циано-бактериаль-
100 мг/л. Соответственно низка ^W* ном мате или водорослей в альго-бактериальном сообществе зани-
• мают в качестве первичных продуцентов мхи.
237
Продукция сфагновых болот, растительного сообщества сфаг-
неты составляет важную часть ежегодной продукции растительно-
растительности бореальной зоны. Сфагны располагаются в виде сплошного
покрова, имея до 6000 листьев/см2. Число растений Sphagnum соста-
составляет около 46 200 головок/м2 в пределах 25 000-60 000 головок/м2.
Сухой вес сфагнов увеличивается с глубиной вследствие уплотнения
и сворачивания стебельков, составляя в верхних 10 см 10,6 г/дм2; на
уровне 10-20 см - 21,4 г/дм2; на уровне 20-30 см - 32,6 г/дм2;
40-50 см - 36,8 г/дм2. Годовой прирост составляет 1,1-2,5 см/год.
Прирост торфа на верховом болоте оценивается в 1-0,8 мм/год. Ис-
Исходя из плотности сфагнов 104 головок растеньиц на м2 и площади
сфагновых болот России в 2 • 1012 м2, легко оценить численность
сфагнов в 1016-1017 головок растеньиц. Вес головки сфагна в 2 см,
соответствующий годовому приросту, составляет около 20 мг, из
них половина приходится на углерод. Отсюда годовой сток углерода
в сфагны - нетто-первичная продукция (НПП) составляет для Рос-
России около 0,1 Тг С/год, но нетто-экосистемная продукция (НЭП)
значительно меньше из-за деструкции. Прирост торфа - нетто-био-
мная продукция (НБП) для России составляет 0,017 Гт С/год (Вом-
перский, 1994).
''¦ Обмен углерода в сфагнете обусловлен деструкцией доступного
вещества, которое поступает преимущественно в виде углеводов, и
превращением его в гуминовые вещества, прежде всего в фульво-
кислоты. Таким образом, обмен углерода в сфагнете представлен в
значительной степени прямым маршрутом от первичных продуцен-
продуцентов к устойчивым органическим веществам в виде фульватов и гу-
мифицированных остатков растений. Бактерии сфагнеты, несмотря
на невысокую общую численность, очень разнообразны и относят-
ся к разным филогенетическим линиям. Функционально они пред-
ставлены преимущественно олиготрофными сахаролитическими аэ- *
робами, приспособленными к существованию в ультрапресной х<й'
лодной кислой воде. Анаэробное разложение органических веществ*
торфа в значительной степени подавлено. Аэрация, возникающая*
при осушении болот, устраняет существование анаэробной ловушки*
для углерода, и торф начинает быстро использоваться различными*
органотрофами. . ':
Избыточный углерод сбрасывается в виде торфа и служит твер-^
дым субстратом для выхода ближе к свету: подобно циано-бактери-+
альным матам сфагны поднимаются к свету "на трупах" предшест-*
венников. Рыхлый верхний слой сфагна служит хорошим теплоизо-*'
лятором. Он может нагреваться выше температуры окружающего*
воздуха, в то время как вода на глубине одного-двух десятков санти-*
метров остается холодной; моховой покров служит аккумулятором*
холода. Растительный покров сфагнового болота развивается в виде*
системы кочек и мочажин. В мочажинах происходит более быстрой
разложение мха, несмотря на более высокую продукцию, чем на*
238
кочках. Макроструктура кочек и депрессий между ними очень напо-
напоминает картину строматолитов, где нетто-прирост на возвышениях
создает положительную обратную связь для фотосинтетиков, а уси-
усиленная деструкция в депрессиях создает каналы между ними. Далее,
все верховое болото развивается в виде щита с увеличенным приро-
f стом торфа в середине в результате дисбаланса между продукцией и
1 разложением. Общая аналогия между сфагновым болотом и полем
строматолитового циано-бактериального сообщества достаточно
велика, несмотря на принципиальные различия в минеральных ком-
компонентах, что указывает на общие закономерности развития фото-
синтезирующего покрова в амфибиальных ландшафтах.
1 Сообщество сфагнеты включает и ряд сосудистых растений, в
; особенности вересковых кустарничков с их специфической микори-
\ зой. Для них характерен физиологический ксерофитизм. Сосуди-
| стые растения образуют транспортный путь для. газов, пересекаю-
пересекающий границу аэробной и анаэробной зоны. По этому пути происхо-
^ дит поступление воздуха в бескислородную зону и выход болотного
* газа из анаэробной зоны в атмосферу. В особенности характерны
для эмиссии газа кочки осок, прежде всего пушицы Eriophorum. Xa-
; рактерной особенностью этих растений является способность к раз-
развитию корневой системы в затопленном торфе в бескислородных
'* условиях. Чтобы избежать аноксии корней, почти все сосудистые
( растения выработали структурные приспособления в виде аэренхи-
аэренхимы, служащей для транспорта газов. В наземных растениях порис-
пористость составляет 2-7% объема, а у болотных - до 60%. Аноксия сти-
_г мулирует переключение с клеточного дыхания на образование эта-
нола, который диффундирует из корней наружу. Формирование
~ аэренхимы регулируется этиленом, продукция которого стимулиру-
стимулируется при затоплении. Этилен, в свою очередь, стимулирует целлюла-
зу кортекса, приводя к дезинтеграции клеточной стенки и создавая
структуру сот. Главным фактором в поступлении О2 служит порис-
' тость корней, причем важна радиальная потеря О2, при которой на
поверхности корней отлагается окисное железо, представляющее
^естественный индикатор поступления воздуха. Часто эти отложения
¦ содержат массу эпифитных железобактерий группы сидерокапс.
Недостаток минерального питания на кислых и торфянистых
'почвах привел к образованию у вересковых кустарничков, таких,
'как Vaccinum (брусника), Oxicoccus (клюква), эндотрофной арбуску-
*лярной микоризы. Под слоем отмершего мха кустарнички образуют
^плотную переплетенную сеть, которая связана отходящими от эпи-
эпидермиса корней гифами микоризы, пронизывающими торф. Ниже
*они отмирают, оставляя в торфе горизонтальные слои. Микоризы
вересковых представляют мантию до 85 мкм. Массу микоризы
[ обычно оценивают по содержанию глюкозамина. Микориза прони-
проникает только снаружи в клетки кортекса, где формируются клубки, об-
обновляющиеся каждые 10-15 дней. С корнями вересковых ассоцииро-
239
ван медленно растущий аскомицет, заполняющий до 40% клеток
корня. Микориза усиленно потребляет органические вещества в
присутствии аммония. Симбиоз определяется поступлением легко
используемых органических веществ от растения и минеральных ве-
веществ, в первую очередь иммобилизованного железом фосфора, из
разлагаемого торфа от микоризы. Транспорт кислорода от аэренхи-
аэренхимы растения в мицелий гриба не обсуждается. Симбиоз грибов с ве-
вересковыми исследовал в 1950-х годах Г. Бургеф9 и выделил грибы-
симбионты на безазотистой среде с крахмалом при рН4. Удался и
ресинтез консорциума. Сфагны проращивали из спор на стерильном
торфе при высокой влажности, и для прорастания они нуждались в
присутствии видов грибов Mortirella, Trichoderma. Для ресинтеза си-
системы требуются четыре компонента: мох, зеленое растение, два
гриба. Грибы-симбионты обладали целлюлозоразлагающей актив-
активностью, использовали пектин, дубильные вещества, прежде всего
катехины, и обильно образовывали меланины, служившие исход-
исходным материалом для синтеза гумусовых веществ. Мох, зеленое рас-
растение, грибы образовывали взаимообусловленную трофическую си-
систему.
Деструкция в верховых болотах происходит в кислой среде, бла-
благоприятной для компостирования и сохранения полуразложившихся
растительных остатков в виде торфа. Деструкцию в болотах изуча-
изучали главным образом in situ по разложению растительной массы в ме-
мешочках и потере веса. Определяющим для скорости разложения
служит содержание лигнина. Сфагны содержат 1-2% лигнина и со-
состоят преимущественно из полисахаридов, которые поставляют в
воды смесь различных олигосахаридов. Напротив, вересковые со-
содержат до 30% лигнина. Деструкция начинается с вымывания легко- *
растворимых веществ, прежде всего из листьев, где их содержится
до 30%. Несмотря на малое содержание лигнина, сфагны устойчивы'
к разложению не только из-за кислой среды, но и вследствие специ-
специфического ингибирования микроорганизмов, хорошо известного по'
применению сфагнов как стерилизующих повязок. Хлопковая нить'
по испытанию на разрыв в присутствии сфагнов разлагалась гораз-'
до медленнее. Отсюда следует, что в присутствии сфагнов специфи-
специфически подавляется разложение целлюлозы, а не азотистых веществ.'1
Данные по разложению болотных растений отчетливо показывают,11
что в первые два года происходит мобилизация примерно половины
азота опада из 12% разложенного органического вещества для сфаг-"
на и 35% - для пушицы с отношением С : N, достигающим 110 на4
второй год. На третий год это соотношение составляет 40, указывая1"
на вторичное закрепление азота деструкторами. Разложение такв
растений низовых болот, как тростник и осока, идет более равно-'
9 Burgeff H. Mikrobiologie des Hochmoores mit besonderer Berucksichtigung def
Erikazeen - Pilz - Symbiose. Stuttgart: Fischer, 1961. 178 S. *
240
мерно. В результате запас N и Р в торфе верховых болот выше, чем
в осоковых. Вместе с тем и содержание экстрагируемого фосфора в
сфагновых болотах выше, чем в минеротрофных, где фосфор свя-
связывается кальцием, железом, алюминием. Разложение оценивается
у сфагнов 0,5% сутки, вдвое меньше, чем у осок, и идет быстрее все-
всего на глубине 5 см.
Медленное отмирание микроорганизмов в сфагновом торфе мо-
может быть проиллюстрировано встречаемостью по профилю торфа
дрожжей. В сфагновом болоте основная масса дрожжей представле-
представлена фитобионтами. В отличие от почвенных горизонтов, где числен-
численность дрожжей резко снижается при переходе к минеральным сло-
слоям, в торфе численность остается постоянной на глубину разреза.
Доминируют эпифиты Rhodotorula mucilaginosa, Sporobolomyces
roseus и эврибионт Cryptococcus albidus, составляющие на мхах и
подстилке 80%. В торфяно-болотистых почвах доминирует
Cryptococcus gilvescens, не встречающийся в других почвах. Дрожжи
используют здесь растворимые сахара, и их стратегия оценивается
как стратегия патиентов, приспособленных к низкой температуре и
осолодению растительной массы (как при низовом 6°С брожении в
пиве), но росту в аэробных условиях, а не спиртовому брожению.
Критическим для разложения оказывается уровень грунтовых
вод, определяющий границу между аэробной и анаэробной зонами.
Разложение интенсивнее всего идет в тонком слое на поверхности
воды, приближающемся по условиям к нейстонной пленке, и в воде,
поднимающейся по смоченной поверхности стеблей. В анаэробной
зоне сфагновых болот происходит подавление разложения расти-
растительных бстатков, в отличие от минеральных низовых болот, где
осуществляется интенсивный биологический цикл. В нормальных
условиях сфагновое болото служит долгосрочной ловушкой для уг-
углерода, несмотря на малую первичную продукцию. При уровне
воды 15 см от поверхности болото служит стоком СО2, при уровне
> 30 см - источникам. Основной причиной высокой нетто-экоси-
стемной продукции служит подавление деструкции. При устранении
анаэробной ловушки начинается быстрое биологическое выгорание
торфа вплоть до минерального грунта. Это подтверждается опытом
осушения болот как в Белоруссии, так и Индонезии.
Замедление деструкции в болотах хорошо иллюстрируется вы-
высокой сохранностью органических веществ в торфе. Прежде всего
это древесные остатки с лигнином. К ним следует добавить и остат-
остатки животных. В болотах хорошо сохраняются трупы людей неолита
(Moorleiche на побережье Северного моря) и археологические пред-
предметы, включая ткани. Болота служат не только глобальным стоком
СО2 в органический углерод, но и локальным источником накопле-
накопления угля. Образование угля в процессе длительного диагенеза обу-
обусловлено потерей кислорода и водорода. Часто угли захватывают
при своем образовании метан, и он является причиной взрывов в
241
угольных шахтах, а при открытой добыче - эмиссии метана в воз-
воздух. Таким образом, наземные болота, в противоположность мор-
морским, служат источником образования угля, а не нефти.
Микроорганизмы-деструкторы разных болот должны обладать
различными экофизиологическими характеристиками. Очень при-
приблизительно можно составить матрицу для доминирующих условий:
Продукция/
деструкция
Продуценты
Деструкция
Трофическая
цепь блоки-
ровна
Деструкторы-
аэробы
Бактерии-
анаэробы
Тундра влажная
Баланс
Мохово-кустар-
ничковая
растительность
Неполная
В егетационным
сезоном
Грибы и бактерии-
аэробы, психро-
филы и эфемеры
Психрофилы,
маслянокислые,
гомоацетатные
Верховое болото
Накопление
торфа
Сфагны
Неполная
Кислотностью,
анаэробиозом
Грибы, ацидо-
филы.олиго-
трофы
В торфе слабый
метаногенез
Низовое болото
Накопление
торфа
Сосудистые расте-
растения
Полная
Анаэробиозом,
продукция преоб-
преобладает над
деструкцией
Бактерии, ней-
трофилы, газо-
трофы
Метаногенное и
отчасти сульфи-
догенное сооб-
сообщества
В аэробной зоне мохового болота сочетаются условия психро-
филии и олиготрофии, и для организмов этой зоны можно предло-
предложить название психроолиготрофов.
Сфагновые болота, несмотря на замедленность процессов в них,
служат одним из наиболее значимых источников биогенной эмиссии
метана, особенно в России. В сфагновых болотах работает ацидо-
ацидофильный цикл Зёнгена. Метаногенное сообщество в торфе развива-
развивается. Установлено, что метаногенез в сфагновом торфе стимулиру-
стимулируется добавлением органических кислот, прежде всего ацетата, но не
метанолом или метиламинами, что составляет резкий контраст с
солеными водами. Можно полагать, что здесь доминирует С-2 путь '
метаногенеза, хотя возможно и использование Н2 + СО2. В этом слу-
случае, однако, отчетливую конкуренцию метаногенам составляют го-
гомоацетатные бактерии, роль которых подробнее разбирается в свя- 1
зи с психрофильным сообществом. Вообще говоря, развитие мета- '*
ногенов сильно подавляется при рН < 6, но в очистных сооружениях *"'
удалось зарегистрировать развитие метаносарцины и при рН < 4, '
однако культура была крайне неустойчива. Тем больший интересу
представляет тот факт, что именно сфагновые болота служат важ-
важнейшим для России источником метана. При всей слабой продукции^'
метана в сфагновом болоте иногда оказывается, что слой торфа
плавает на газовой подушке болотного газа. Такие "сплавины" харак-
242
*
терны для краев зарастающих водоемов. Эмиссия метана в атмосфе-
атмосферу определяется дисбалансом между продукцией метаногенами и
окислением метанотрофами. Потребление метана образцами торфа
из болот подавлялось даже небольшой концентрацией нитрата. Ме-
танотрофное сообщество сфагнеты оказалось представленным
азотфиксирующими ацидофильными метанотрофами Methylocapsa,
составившими новую группу среди С-1 использующих организмов.
Ацидофильные метанотрофы были выделены С.Н. Дедыш и сейчас
представлены видами нового рода Methylocella, разными для разных
мест обитания - болота, леса, тундры10. Организм доминирует в сла-
слабокислой среде до рН 4,5. При повышении рН > 6 метилоцеллы вы-
вытесняются обычными метанотрофами, свойственными низовым бо-
болотам.
Рекомендуемая литература
1. Боч М.С., Мазинг В.В. Экосистемы болот СССР. Л.: Наука, 1979. 188 с.
2. Вомперский С.Э. Роль болот в круговороте углерода / Биогеоценотические
особенности болот и их рациональное использование. М.: Наука, 1994.
С. 5-37. (Чтения памяти академика В.Н. Сукачева; XI).
3. Мазинг В.В. Структурная организация болот // Там же. С. 38-60.
4. Перельман A.M., Касимов Н.С. Геохимия ландшафта. М.: Астрея-2000, 1999.
768 с.
5. Сидоренко А.В. Новое в учении о биосфере. М.: Наука. 1984. 16 с.
6. Смоляницкий Л.Я. Некоторые закономерности формирования дернины
сфагновых мхов // Ботанич. журн. 1977. Т. 15. С. 21—31.
7. Boggs S. Principles of sedimentology and stratigraphy. 2nd ed. Englewood Cliffs.
(N.J.): Prentice-Hall, 1995.
8. Tyson R.V. Sedimentary organic matter: Organic facies and palynofacies. L., etc.:
Chapman and Hall, 1995.
ч ¦*¦
10Дедыш С.Н. Метанотрофные бактерии кислых сфагновых болот // Микро-
'' биология. 2002. Т. 71. N 6. С. 741-754.
Лекция 7
МИКРОБИОЛОГИЯ ПОЧВЫ
7.1. ИСТОРИЯ
Почвоведение
Русское почвоведение в отличие от французской школы агрохи-
агрохимиков и физиологов растений середины XIX в., послужившей от-
отправной точкой для создателя почвенной микробиологии С.Н. Ви-
ноградского, зародилось в 1871 г. на основе наук о Земле на кафед-
кафедре геологии и минералогии Санкт-Петербургского Университета.
Его создателем стал В.В. Докучаев, опубликовавший в 1883 г. "Рус-
"Русский чернозем". "В России почвоведение возглавлялось геологами,
и его развивали не как раздел геологии, а как самостоятельную на-
научную дисциплину" (Зонн, 1999, с. 23). Следуя геологической тради-
традиции, русское почвоведение основывается на генезисе почв и "про-
"профильном подходе", имеющем в виду прежде всего формирование
почвенного профиля. Практическая значимость новой науки опре-
определялась запросами агрономии с понятием "плодородия почвы". •
В результате описания шурфов создавалась важная для землеполь- i
зователей почвенная карта на исходно визуально-морфологической '
основе с привязкой к ней ряда химических показателей. Для перене- <
сения результатов исследования шурфов на площади требовалось *
создание классификации почв и ее воплощение на картографиче- *
ской географической основе. Комплекса идей о почве, выдвинутых |
в короткое время последней четверти XIX в., совпавшей с эпохой
"золотого века" общей микробиологии, оказалось достаточно для *
их разработки огромной армией почвоведов-докучаевцев в течение *
более столетия и для их неустанной борьбы с инакомыслием. *
Когда после работ В.В. Докучаева предмет изучения возник пе- •
ред глазами исследователей, создалась необходимость его всесто-5*
роннего изучения. Именно эта работа и была проделана первым по-%
колением после Докучаева, несшим новое, в том числе и вне основ- *
ного русла. Неполнота генетического подхода была довольно быст-к
ро отмечена и восполнена уже первым поколением почвоведов-уче-ч-
ников Докучаева. Наиболее точно свой скептицизм выразил»
В.И. Вернадский, сказав, что докучаевское понимание почвы "не яв- *
ляется единственным и окончательным" (Зонн, 1999, с. 22-28). Для;
244
ряда проблем докучаевское "профильное" почвоведение составляет
необходимую, но недостаточную часть, поскольку не ориентирова-
ориентировано на процессы динамического равновесия.
Связь между почвой и ландшафтом, учитывая значение релье-
рельефа, растительности, поверхностных вод, выраженное прежде всего
в биогеохимическом понимании проблемы миграции элементов, в
особенности в связи с формированием коры выветривания, была
разработана в 1940-х годах. Б.Б. Полыновым1 и его последователями
как геохимия ландшафта. Полынов выступил с идеей о почвообра-
зующей роли растений "петрофитов", т.е. первичных поселенцев на
скальных породах и рухляке. При этом несколько в стороне остает-
остается покров пенеплена с тонкодисперсными продуктами выветривания,
очевидно представляющими иную линию эволюции поверхности.
В крупном масштабе геологического времени специальный ин-
интерес представляют взгляды Ю.Ф. Готшалка A941 г.), который выде-
выделил педосферу, охватывающую всю толщу собственно почвы и
коры выветривания. Педосфера включает в себя: ризосферу - от
поверхности Земли до предела проникновения корней с верхней ча-
частью, собственно почвой, содержащей наиболее измененные мате-
материнские породы, и подпочвой - со слабоизмененными остатками;
грунт - нижнюю часть педосферы, лежащую на коре выветривания
(по: Зонн, 1999, с. 255).
В настоящее время сформировался острый интерес к тому аспе-
аспекту почвоведения, который отражает экологические функции почв
в ландшафте (биогеоценозе), понимаемом как единство раститель-
растительного покрова и почвы как на локальном, так и на региональном (зо-
1 нальном) и, в меньшей степени осознанном, планетарном биосфер-
s ном уровне. В этом отношении наиболее значимы почвы, находящи-
* еся вне сельскохозяйственного использования, например для России
1 это прежде всего почвы лесного фонда, составляющие 68% почв
\ страны. При этом "горизонтальные" ландшафтные, часто стоковые
' (катены, дренаж), серии представляют иногда больший интерес, чем
1 "вертикальные" генетические. Здесь выявляется четыре основных
аспекта:
* 1) взаимодействие почвы с растительным покровом в первую
* очередь через корневую систему с микоризой и ризосферой; при
* этом почва упрощенно понимается как корнеобитаемый слой зем-
lt ной поверхности;
" 2) взаимодействие почвы с атмосферой через почвенный воздух;
3) участие в формировании состава континентальных вод как
'' поверхностного стока, так и глубинных; особое значение эта функ-
* ция почв приобретает на равнинных территориях гумидного клима-
3 та, где время контакта метеорных вод с жизнедеятельным слоем
1 Полынов Б.Б. Кора выветривания. Л., 1934; Полынов Б.Б. Геохимические
ландшафты // Вопросы минералогии, геохимии и петрографии. М.; Л., 1946.
245
земной поверхности больше и протекает даже под покровом сезон-
но-промерзающего слоя круглогодично;
4) для больших временных промежутков приобретает значение
взаимодействие почвы с грунтом и корой выветривания, где основ-
основную роль играет биогенное выщелачивание исходной горной поро-
породы (педолиз) и новообразование почвенных минералов, с одной сто-
стороны, и гумуса - с другой.
7.2. ПОЧВЕННАЯ МИКРОБИОЛОГИЯ
Почвенная микробиология традиционно является областью ин-
интересов отечественной микробиологии. Начало ее лежит в трудах
С.Н. Виноградского, особенно относящихся ко второму этапу его
деятельности, основанному на применении прямых методов. Затем
последовали работы ряда исследователей, из которых особенно
нужно отметить Н.А. Красильникова, рассматривавшего почвенную
микробиологию как синтетическую дисциплину "биология почв" с
особенным вниманием к взаимодействию микроорганизмов с корне-
корневой системой растений, но так же учитывавшего роль грибов,
почвенных водорослей, простейших. Однако впоследствии домини-
доминирование получила почвенная бактериология, особенно распростра-
распространившаяся благодаря работам Е.Н. Мишустина. Почвенные микро-
микробиологи кафедры биологии почв, возглавляемой Д.Г. Звягинцевым,
также работали с акцентом на бактериологию в соответствии с за-
запросами времени. Для работы в области почвенной микробиологии
студенты, окончившие биологический факультет, должны ознако-
ознакомиться прежде всего с химией почвы и только при анализе более :
крупных объектов переходить к изучению ландшафтов. '
Почва представляет освещаемую твердую поверхность Земли, >
покрытую растительным покровом, и, начиная с силура, примерно !
350-400 млн лет назад, обеспечивает выход растений в аэротоп - i
слой атмосферы между верхушками растений и поверхностью зем- ¦
ли. Как операционное биологическое понятие почву удобнее всего *
определить как "корнеобитаемый слой земной поверхности". При г
этом за пределами определения окажутся лишенные высшей расти- ¦
тельности участки с водорослевой или моховой растительностью и !
лишенные органического вещества области первичного почво-'
образовательного процесса: обнаженные грунты, пески, скалы. Для
последних удобным обозначением служит "обнаженная кора вывет-'
ривания", обычно заселенная микробными биопленками. В коре вы-
выветривания доминируют элювиальные процессы выноса. Как "био- *
косное тело" почва отличается от минеральной фазы и грунта обо- *'
гащением органическим веществом. Органическая почва растет из*1
воздуха. В корнеобитаемом слое появляется структура, обусловлен-'
ная корневой сетью, и меняется гидрологический режим. Растение5
пересекает две среды обитания и становится мощным транспорт-
ным агентом для потоков вещества. Растение создает своей корне-
корневой системой особую область для развития микроорганизмов. В пер-
первую очередь это обусловлено эвапотранспирацией растения, обеспе-
обеспечивающей тягу почвенного раствора корнями. Следующим фактором
служат корневые выделения, обеспечивающие рост органотрофных
организмов ризосферы. Далее, растение вступает в симбиотические
отношения прежде всего с грибами-микоризниками, но также и с
симбиотическими бактериями, из которых наибольшее внимание за-
заслужили симбиотические фиксаторы азота. Таким образом, задаю-
задающим параметром становится экофизиология растительного покро-
покрова, характеризующего ландшафт.
Примитивные почвы, лишенные растительного покрова, гос-
господствовали на поверхности Земли в докембрии и были представле-
представлены наземными циано- и альго-бактериальными матами с вероятным
ранним появлением грибов, предварительным условием для кото-
которых служило появление кислорода в атмосфере. Переходный пери-
период к покрову сосудистых растений могли составлять лишайники и
мхи. Вынос фотосинтезирующей поверхности в аэротоп мог про-
произойти лишь при создании жестких структур углеродного скелета,
представляющего композитную конструкцию из лигноцеллюлозы,
включающей фибриллы целлюлозы, склеенные сетью гемицеллюло-
зы и армированной лигнином. Лигноцеллюлоза у древесных растений
на 1/3 состоит из лигнина и в общем синтезе органического вещества в
наземных экосистемах составляет не менее 20 Гт/год. Лигноцеллюлоза
не подвергается автолизу при отмирании растения и составляет основ-
основную массу поступающего в почву органического вещества.
В почвенно-растительном покрове следует различать фотоав-
тотрофную наземную часть, дополняемую гетеротрофной под-
подземной корневой частью растения, и органотрофную группировку
грибов и бактерий. Фотоавтотрофы осуществляют фотосинтез
только в дневные часы и зависят от света. Гетеротрофное дыхание
| идет круглосуточно. Органотрофы действуют круглосуточно, а так-
также вне вегетационного сезона, разлагая листьевой и корневой опад.
" Разложение лигноцеллюлозы осуществляется в первую очередь
грибами. Отсюда вытекают три трофических маршрута:
Миколитический
Лигнолитический (вторичная биомасса
грибов)
Хитиназы
Целлюлозолитический
Целлюлозы
Целлодекстрины
Олигосахариды
Лигниназы, оксидазы
Фенолы
N-глюкозамин
246
1
Разложение мохового покрова на поверхности почвы, например
в бору-зеленомошнике со сплошным покровом гипновых мхов, вклю-
включает стадии: а) отмирающего белесого мха (очёс) обычно вследст-
вследствие самозатенения и автолиза; б) торфа, представляющего гумифи-
247
цированные, не полностью разложенные растительные остатки, со-
сохраняющиеся в анаэробных условиях как на сфагновых болотах;
в) разложения торфа на границе с минеральной почвой.
Физические условия существования в почве с пористым про-
пространством между минеральными частицами, недостатком влаги
обусловливают преимущества для мицелиального строения микро-
микроорганизмов. Мицелий может обволакивать твердые растительные
остатки и осуществлять гидролиз в непосредственном контакте с ни-
ними. Поэтому почву можно рассматривать как растительно-грибную
систему. Бактерии развиваются в почвенной влаге и образуют био-
биопленки на поверхности минеральных и органических частиц.
В почвенной системе можно выделить три подраздела с разны-
разными условиями жизни для микробиоты: а) первичные продуценты
растения с их корневой системой, населенной консорциумом микро-
микроорганизмов, включающим микоризу, бактерии ризосферы; б) мик-
микробное сообщество, осуществляющее деструкцию мортмассы,
конечным продуктом которого является гумус; в) микробное сооб-
сообщество, взаимодействующее с минеральной частью, конечным
результатом чего является превращение минералов материнской
породы в педогенные минералы с наиболее характерной группой
глинистых минералов. Между этими подразделами системы сущест-
существует тесное взаимодействие, обусловленное, например, действием
корневых выделений и продуктов разложения мортмассы на мине-
минералы, но тем не менее каждый из разделов удобнее рассматривать
отдельно.
Совокупная деятельность почвенной биоты оценивается по га-
газообмену - выделению углекислоты. Для проведения таких исследо-
исследований не требуется специально микробиологических знаний: дается
интегральная оценка с употреблением характерного термина "био-
та". В продукционной части цикл углерода оценивается интегрально
во времени по нетто-продукции, т.е. накопленной массе органиче-
органического вещества. Деградационную ветвь можно оценивать по разно-
разности между первичной продукцией и нетто-продукцией. Прямыми
способами являются определение скорости фотосинтеза по погло-
поглощению СО2, содержание которой в воздухе мало, (а не по выделе- *
нию О2, как в водных системах), и деструкции - по выделению СО2-
"дыханию почвы". '
<
7.3. ПОЧВЕННАЯ БИОЛОГИЯ '
Почва как среда обитания организмов представляет гетероген-
ную трехфазную систему, включающую почвенный воздух, почвен-' t
ную влагу, минеральные частицы (рис. 7.1). В результате деятельно-*'
сти живых организмов в почве добавляется мертвое органическое
вещество - гумус - и живая масса корней и микроорганизмов. '* ¦
Эти компоненты превращают выветрелые и выщелоченные горныей^
Консорциум растения
Гумусовый ряд
деструкции
Минеральный ряд
выветривания
Осадки
Грибы сапрофиты | —"" <¦
•Микофильные-. • : _
^.бактерии . V' "f1 Зелоты
^^^ I Гумус
Песок
Глина
Грунт
Рис. 7.1. Микробиологические аспекты почвоведения
породы в почву. Выщелачивание представляет ключевой процесс
воздействия микроорганизмов на горные породы. Но превращение
их в почву зависит от накопления не разложенного микроорганизма-
микроорганизмами органического углерода - процесса гумификации, наиболее важ-
важного, магистрального пути в почве как экосистеме. Образование гу-
гумуса есть побочный результат незамкнутости цикла углерода в на-
наземных экосистемах, возникающего на основе дисбаланса между
продукционной ветвью фотосинтезирующих наземных организмов,
способных выносить фотосинтезирующий аппарат в воздух, в аэро-
* топ (от лишайников и мхов до деревьев и трав) и деградационной
1 ветвью, обусловленной деятельностью микроорганизмов. Наличие
ч газовой фазы отличает почву от донных отложений в бассейнах се-
* диментации, где происходит, например, образование богатых орга-
1 ническим веществом сланцев.
Почва как среда обитания микроорганизмов в отличие от вод-
водных систем имеет ряд характерных особенностей:
1) путь углерода ведет к образованию резервуара гумуса как
стойкого органического вещества,
2) доминирующей группой являются мицелиальные организмы -
грибы и актиномицеты,
3) продукционная ветвь в толще почвы обусловлена корневой
системой растений.
Главным органическим компонентом, поступающим в почву, яв-
является лигноцеллюлоза. Лигнин вообще не свойственен водным ор-
248
249
ганизмам. Несмотря на возможность рассматривать его как основ-
основной предшественник гумуса и несомненную роль в его образован™,
предположение о прямой последовательности лигнин-гумус явно
недостаточно, т.к. гумус образуется и из предшественников, содер-
содержащих мало лигнина, например в торфяных сфагновых болотах или
же при первичном почвообразовательном процессе под лишайника-
лишайниками на выветрелых скалах, и содержит довольно много азота. В этих
случаях и роль корневой системы отсутствует. Являются ли гумусо-
гумусовые вещества продуктом аэробной переработки органических ос-
остатков грибами или же они образуются и анаэробно в затопленном
слое болот из торфа?
Все эти вопросы получают неопределенные ответы, универ-
универсальное решение отсутствует.
Гетерогенность почвы обусловлена наличием разных по разме-
размерам частиц и эта иерархия размеров должна постоянно учитывать-
учитываться. Она приведена ниже (по: Paul, Clark, 1989):
Категория
Макро агрегат
Микроагрегат (песок)
(разлагаеамя органика)
Корневые волоски (устой-
(устойчивая органика)
Крупные поры (устойчи-
(устойчивая органика)
Поры (устойчивая мине-
минеральная часть)
Размеры, мкм
>2000
200
20
2
0,2
компонент
Корни и гифы
Растительные и
грибные остат-
остатки
Органические
остатки (гумус)
Алюмосилика-
Алюмосиликаты, окислы, по-
полимеры, сорби-
сорбированные на
глинах
Компоненты
Частицы,
макропоры
Корешки, ги-
гифы, минераль-
минеральные частицы
Гифы, бакте-
бактерии, глина
Бактерии, гу-
гумус, глинистые
минералы
Глинистые
минералы
Доступным для развития микроорганизмов является поровое
пространство почвы порядка микрон, заполненное водой и почвен-*
ным воздухом. Микроорганизмы занимают менее 0,01 порового1
пространства. Положение внутри пор до известной степени защищав
ет микроорганизмы от хищников. Микропоры менее 0,2 мкм, напри-И
мер чехол из глинистых минералов, физически защищают органиче-*'
ские вещества от воздействия экзоферментов. ^
Разные почвы имеют агрегаты разного размера, меняющиеся в*']
зависимости от состояния почвы и, например, ее обработки. Воз-''
душно сухие агрегаты обычно состоят из агломератов водопрочных'
агрегатов меньшего размера. Характерный масштаб агрегатов с^
ставляет от долей до десятка миллиметров. Формирование агрега-^ '¦
тов происходит при участии микроорганизмов. Оно обусловлено^
склеивающим действием бактериальных слизей на мелкие мине-
минеральные компоненты почвы, прежде всего глинистые частицы. Гли-
Глинистые частицы адсорбируют бактерии или при высокой дисперс-
дисперсности способны коагулировать бактериальные клетки из коллоид-
коллоидных растворов. Агрегаты скрепляются и армируются мицелием
микромицетов, оплетающим частицы снаружи. Органические час-
частицы и коллоиды гуминовых веществ взаимодействуют с мине-
минеральными компонентами, создавая органо-минеральную структуру
агрегатов.
При микроскопическом анализе агрегатов в их центральной
части обнаруживаются комки бактерий, а снаружи облекающий
войлок мицелия. Бактерии проникают в поровое пространство аг-
агрегата с порами не менее 0,6 мкм, если поры не заняты почвенной
влагой. Внутренняя часть агрегата остается влажной, в то время
как поверхность подсушивается и находится в равновесном состоя-
состоянии с влагой почвенного воздуха. Агрегат почвы оказывается мес-
местообитанием микробного сообщества, созданным при участии вза-
взаимодействующих в нем функционально различных групп микроор-
микроорганизмов.
Распределение влаги в агрегате ведет к тому, что его централь-
центральная часть оказывается восстановленной из-за медленной диффузии
02 по капиллярам и поглощения О2 в наружном слое аэробными
органотрофами. Установление анаэробиоза в центральной части аг-
агрегата зависит от диффузионных процессов и наличия легкодоступ-
легкодоступного органического вещества. Математические модели агрегата по-
показали, что при размерах агрегата 10 мм внутри должна возникать
анаэробная зона. Прямые измерения с помощью микроэлектродной
техники установили, что анаэробная зона возникает в агрегатах раз-
размером 4 мм. Эксперименты по выделению газообразных продуктов
обмена микроорганизмов показали, что СН4 может продуцировать-
продуцироваться при размерах агрегатов менее 2 мм, но его продукция возрастает
с увеличением размеров агрегата. Такая же картина наблюдается
для N2O, продуцируемой факультативными анаэробами денитрифи-
каторами. Напротив, продукция СО2 в результате дыхания значи-
значительно усиливается с уменьшением размеров агрегатов и увеличени-
увеличением поверхности. Продукция восстановленных газов увеличивается с
г увеличением влажности, достигая максимума по достижении
^"капиллярной влагоемкости", а затем падает с появлением водных
пробок2.
Почвенный воздух располагается в порах аэрации, образующих
'либо каналы проводимости, например по сгнившим корням, либо
лабиринт между агрегатами, и от этого зависит длина пути газа до
Степанов АЛ., Манучарова Н.А., Полянская Л.М, Продуцирование закиси
азота бактериями в почвенных агрегатах // Почвоведение. 1997. № 8.
С. 973-976.
250
251
атмосферы. Поровое пространство почвы зависит от ее характера и
обычно составляет 50-60% объема, что вместе с растворенным кис-
кислородом создает резервуар доступного для почвенной биоты окис-
окислителя, находящийся в динамическом равновесии со слоем атмосфе-
атмосферы в растительном покрове (аэротопом).
Скорость диффузии q (г/см2 • с) зависит от коэффициента диф-
диффузии в почве D (см2/с), концентрации С (г/см3), длины пути z (см):
q = DdCldz.
Газ
Коэффициент
диффузии, см2 • с
Воздух
Вода • 10
Раствори-
Растворимость, см3/л
20 °С
Водный потенциал
МПа
-1,5
-10
-25
-65
Бар
-15
-100
-250
-650
Д
0,99
0,93
0,83
0,62
Осмолиты, вес/объем
NaCl
2,0
12,3
25,3
Сахароза
17
52
70
83
Организмы
Rhizobium, Nitrosomonas
Clostridium, Mucor
Micrococcus, Penicillium
Xeromyces
i
co2
o2
N2
0,161
0,205
0,205
0,177
0,180
0,164
8,878
0,031
0,015
Отсюда важнейшим показателем служит аэрация почвы, опре-
определяющая переход от аэробиоза к анаэробиозу. Она осуществляет-
осуществляется через поры аэрации в основном под действием диффузии, обусло-
обусловленной градиентом концентрации. Достаточно небольшой разницы
в парциальном давлении, чтобы кислород двинулся в почву, а угле-
углекислота стала выходить из нее.
Поры блокируются водой с коэффициентом диффузии в
10 000 раз меньше, чем на воздухе. Частицы почвы размером более
3 мм не имеют кислорода в центральной части. Возможность суще-
существования в них анаэробов зависит, однако, от скорости поглощения
кислорода аэробами, а она, в свою очередь, определяется доступным
для окисления веществом. В отсутствие дыхания корней B0-40% от
общего дыхания почвы) и микроорганизмов F0-80%), например в
промерзшей почве, состав почвенного воздуха быстро уравновеши-
уравновешивается с атмосферным.
Почвенная влага, в том числе пленочная, является местом оби-
обитания бактерий и отчасти грибов, мицелий которых может выхо-
выходить в газовую фазу. Почвенная вода имеет матричный потенциал,
зависящий от ее связывания с поверхностью частиц, и осмотический
потенциал, зависящий от растворенных веществ. Почвенная вода
рассматривается как гравитационная, капиллярная, гигроскопиче-
гигроскопическая. После удаления гравитационной воды остается вода, определя-
определяемая почвенной влагоемкостью. Капиллярная вода имеет водный
потенциал от -0,01 до -0,03 МПа и доступна для роста микробов, ко-
когда наблюдается их наибольшая активность. Ниже -1,5 МПа начи-
начинается завядание растений. При -30 МПа организмы сохраняют'
лишь 10% активности, измеряемой по скорости разложения органи-
органического вещества.
252
I
I
1
Щ Ш
Вода в почве входит в состав почвенного раствора, служащего
местообитанием бактерий. Состав почвенного раствора определяет-
определяется прежде всего карбонат-бикарбонатным равновесием, от которо-
которого зависят его рН, а также и растворимость минералов как почво-
образующей породы, так и педогенных. Важнее всего поведение ти-
поморфных элементов: Са, Fe, Al, Si. Почвенный раствор подвержен
испарению при засухе и промывному режиму при дождях, и состав
его непостоянен. Поэтому почвенная биота должна приспосабли-
приспосабливаться к переменному режиму в отношении осмотического давления.
Кроме минеральных компонентов, в почвенный раствор поступают
органические вещества из корневой системы растений и продукты
микробного разложения. Устойчивую часть составляют коллоиды
гуминовых веществ, обладающие комплексообразующим действи-
действием. Физико-химическая система почвенного раствора зависит от
многих переменных и очень сложна. От нее зависит состояние поч-
почвенной биоты, и прежде всего бактерий.
- 7.4. РАСПРЕДЕЛЕНИЕ МИКРООРГАНИЗМОВ
В ПОЧВЕ
Очень много усилий было уделено изучению распределения ми-
миЗ
кроорганизмов в почве и подсчету их численности. Задача решалась
* учетом на средах с идентификацией колоний, прямым счетом с по-
мощью разных способов микроскопии, определением химическими
методами по характерным компонентам, определением фермента-
ферментативной активности, молекулярно-биологическими методами.
Критическим является распределение организмов по почвенно-
почвенному профилю: 1) на поверхности почвы в области подстилки и опада;
2) в аэрируемом слое с развитой корневой системой; 3) ниже уровня
почвенных вод.
Разлагающийся растительный опад традиционно является обла-
областью развития гидролитических аэробных организмов. В связи с тем
что опад представлен в первую очередь лигноцеллюлозой, он пред-
представляется оптимальным местом для развития сапрофитных грибов.
^Ежегодный опад составляет 100-300 г/и2 для травянистых систем и
к200—800 г/м2 для леса. К характерным сапрофитным грибам в под-
'стилке относят Alternaria, Cladosporium, "Mycelia sterilia", в то время
[""как в нижележащем гумусовом слое доминируют Penicillium,
253
Trichoderma, Fusarium. Разложение опада ведет к характерной сук-
сукцессии грибов. Прямой учет живого и мертвого мицелия в почве по-
показывает, что он составляет главную часть микробной биомассы,
особенно в лесных почвах.
Очень важным показателем является так называемый "подсти-
"подстилочный коэффициент" - отношение ежегодного опада к подстилке,
который показывает скорость разложения растительных остатков.
На севере в тундре он составляет 1 : 200, а на юге в субтропиках
200 : 1 (Перельман, Касимов, 1999). Продукты разложения грибами
опада и ветоши служат субстратом для микофильных бактерий, при-
причем особое значение приобретает разложение фенольных соедине-
соединений. Отмерший мицелий грибов также служит субстратом для бак-
бактерий с актиномицетами как характерной группой, идущей по от-
отмершему мицелию. Разложение остатков в хвойных лесах приводит
к формированию темноокрашенных гумусовых вод.
Наибольший интерес представляет ассоциация микроорганиз-
микроорганизмов с корневой системой живых растений. Корни располагаются в
почвенном горизонте, наиболее богатом органическим веществом.
Взаимодействие с корневой системой включает три области: 1) об-
область почвы с непосредственным воздействие корней - ризосфера;
2) поверхность корня - ризоплана; 3) ткань корня.
В ризосфере наблюдается действие корневых экссудатов, содер-
содержащих разнообразные органические вещества, и корневого опада.
Экссудаты содержат широкий набор углеводов, аминокислот, орга-
органических кислот. Корневой опад дает лигноцеллюлозу и полисаха-
полисахариды слизи (mucigel), составляющие 80% потери углерода корнем.
Из суточной ассимиляции 11% уходит на микробное дыхание и 2% -
в органическое вещество почвы.
В ризосфере происходит падение численности микроорганизмов
от поверхности корня:
Расстояние, мм 0-1 1-5 5-10 10-15 15-20
Численность, млрд/см3 120 96 41 34 13
Разнообразие микроорганизмов вблизи корня определяется раз-
разнообразием поступающих веществ, разнообразием трофических
взаимодействий микроорганизмов между собой (гидролитики и дис-
сипотрофы, бактериолитическая петля) и влиянием специфических
веществ растений. Здесь можно ожидать очень широкий круг орга-
нотрофных аэробных организмов, пищевые потребности которых
ориентированы на экссудаты. Как правило, эта область развития
микроорганизмов характеризуется избытком органического углеро-
углерода и лимитирующим содержанием азота и фосфора.
На поверхности листьев в филлосфере развиваются организмы,
специфически взаимодействующие с растением, а также паразиты -
бактериальные и грибные. Сапрофиты используют гуттацию, сли-
слизетечение.
254
7.5. ДЕГРАДАЦИЯ ОРГАНИЧЕСКОГО ВЕЩЕСТВА
В ПОЧВЕ
7.5.1. Разложение растительных остатков
Органическое вещество почвы прирастает за счет фотоавто-
трофной ассимиляции углекислоты из воздуха растительным по-
покровом. При этом работает циклический механизм, при котором
углекислота, образуемая при дыхании гетеротрофных корней и
органотрофном дыхании биоты почвы в неосвещенной части
почвенного профиля, возвращается в аэротоп - приземный
слой атмосферы в пределах растительного покрова. Дисбаланс в
цикле приводит к выводу части Сорг в соединения с большим
временем пребывания углерода, составляющие малую часть по-
потока углерода, но за счет необратимости процесса создающие
главные резервуары Сорг на Земле. Устойчивые органические ве-
вещества составляют собственно почвенный углерод. Пополнение
цикла происходит за счет углекислоты атмосферы. Таким об-
образом, почва в своей органической компоненте растет за счет воз-
Духа.
Необходимо оценить основные потоки Сорг в почве, чтобы по-
понять роль специфических групп микроорганизмов, катализирующих
превращения вещества на определенных этапах их преобразования.
Вход в систему представляет СО2, ассимилированная растением, вы-
выход — устойчивые гумусовые вещества почвы. Пути углерода в сис-
системе для разных ландшафтных зон различны: в черноземах под тра-
травянистым покровом происходит накопление гумуса, в гидроморф-
ных почвах избыточного увлажнения накапливается торф, при дос-
достаточной аэрации происходит полное сгорание органического веще-
вещества, как в подзолах и глеевых почвах, противопоставляемых в ка-
качестве "минеральных почв" почвам "органическим". Различие меж-
между этими почвами определяется полнотой деструкции растительной
мортмассы.
Органическое вещество почвы отличается по своему составу от
характеризуемого соотношением Редфильда органического вещест-
вещества морских водорослей.
С : N : Р соотношение в наземных системах:
Морские водоросли
Почвенные бактерии
Деревья
Почвенная органика
Целина
Пахота с удобрением
Сильно выветрелая почва
С
106
31
500
191
119
200
N
16
5
16
6
9
10
Р
1
1
1
1
1
1
255
Главным маршрутом деградации для растительного покрова со-
современного типа, утвердившегося на Земле около 350 млн лет назад,
служит разложение скелетного материала наземных растений, пред-
представленного лигноцеллюлозой. Она составляет около 1/2 всего уг-
углерода, ассимилированного растениями. Лигноцеллюлоза представ-
представляет медленно метаболизируемый углерод растительной мортмас-
сы. Для России поступление лигноцеллюлозы составляет величину
порядка 2 Гт С/год. Из них не менее 250 Мт С/год поступает в виде
лесной древесины.
Древесина появилась на Земле поздно, через 200 млн лет после
появления скелетных животных и начала фанерозоя. В это время на
Земле уже давно была полностью оксигенированная атмосфера. Ра-
Растительный покров, которому было суждено существенно модифи-
модифицировать атмосферный гидрологический цикл, в своем существова-
существовании зависел от атмосферной влаги. Поэтому с растительным покро-
покровом связаны аэробные ксерофильные мицелиальные эукариотные
органотрофные организмы - грибы, являющиеся основными дест-
деструкторами и в своей главной трофической роли в биосфере ответст-
ответственные за разложение растительной мортмассы. В палеонтологиче-
палеонтологической летописи растения и грибы появляются одновременно, грибы,
возможно, несколько раньше.
Древесина представляет собой композитный скелетный матери-
материал, состоящий из волокон целлюлозы, армирующих сеть из геми-
целлюлоз, связанных, в свою очередь, с лигнином, физически обле-
облекающим волокна целлюлозы. По химическому составу все эти без-
безазотистые соединения различаются. Композитный состав древеси-
древесины предопределяет необходимость ее разложения сообществом спе-
специализированных организмов, способных разлагать отдельные ком-
компоненты. Ключевым этапом служит дезинтеграция скелетных стру-
структур, требующая деполимеризации макромолекул. При этом про-
процесс должен происходить в субаэральных условиях, при достаточ-
достаточной, но не избыточной влажности.
Лигноцеллюлоза поступает в почвенно-растительную систему в
виде стоячей мортмассы погибших деревьев; опада - валежника и
подстилки; корневого опада в глубоких слоях почвы с образующи-
образующимися по корневым ходам порами аэрации. Поступление нераствори-
нерастворимого органического вещества в виде твердой фазы предопределяет
значение физического контакта с ним организмов-деструкторов в
виде скользящих и слизистых форм бактерий, мицелиальных микро-
микроорганизмов. Опосредованную роль играют зоотрофные организмы,
способные захватывать и измельчать твердые частицы и осуществ-
осуществлять биотурбацию, но и у них ключевой процесс деполимеризации
обусловлен деятельностью микрофлоры пищеварительного тракта.
Химический состав растений включает 2-15% белка и 10% рас-
растворимых соединений, 10-60% целлюлозы, 10-30% гемицеллюлозы,
5-30% лигнина. Итак, основная проблема для микроорганизмов на-
256
земных систем - гидролиз органических полимеров. Разложение
азотистых соединений - белка и нуклеиновых кислот - не представ-
представляет значительной проблемы. Его и аэробно, и анаэробно осуществ-
осуществляют разнообразные организмы пептолитического пути, деятель-
деятельность которых оценивается по образованию аммиака и которые по-
поэтому давно получили название аммонификаторов. Деградация до-
доминирующих в растительных остатках нерастворимых безазоти-
безазотистых соединений осуществляется в сахаролитическом пути.
Качественно пути деградации выглядят, как представлено на
схеме:
Белки и нукле-
нуклеиновые кислоты
A)
РАСТИТЕЛЬНАЯ МОРТМАССА
Полисахариды Лигнин
B)
C)
РО43
пептолитическии
путь
сахаролшпический
путь
окислительная
деградация
микроболитическая
петля
D) G)—
амино-
аминокислоты
ванилиновая,
феруловая кислоты
I
удаление боковых цепей
—~ ароматические ядра
СО2
U1D1 ¦
I I
E) поликонденсация
F) ГУМУС
Разложение 1000 мкг органического углерода в грамме почвы по
приведенной выше схеме дает следующие величины (Paul, Clark, 1989):
Категория органического вещества
1 - легко разлагаемое
2 - медленно разлагаемое
3 - лигнин
4 - продукты микроорганизмов
5 - защищенное органическое вещество
6 - старое органическое вещество
7 - устойчивые микробные продукты
9. Г. А. Заварзин
Резервуар,
мкг/г почвы
150
650
200
6
5000
7000
4
Константа
к, сут~'
0,2
0,08
0,01
0,8
3-КГ4
10
-7
0,3
Эффектив-
Эффективность Y, %
60
40
10
40
20
20
25
257
Скорость разложения широко варьирует в зависимости от кли-
климатических условий. Так, для соломы получены данные (Paul, Clark,
1989):
Условия
Лабораторные
Нигерия
Англия
Канада
Время полу-
полураспада, сут
9
17
75
160
к, 1/сут
0,08
0,04
0,01
0,003
Относительная
скорость разло-
разложения
1
0,5
0,125
0,05
Из приведенных значений видно, что зависимость скорости раз-
разложения от климата очень высока. Более того, скорость разложе-
разложения в условиях Канады, близких к нашей стране, в 10 раз ниже, чем
в тропической Нигерии. Отсюда из-за замедленной скорости разло-
разложения Россия имеет положительный баланс углерода, хотя далеко
отстает от тропического пояса по первичной продукции (см.
рис. 1.2-1 А, табл. 7.1).
Растительные остатки представляют сложную смесь быстро и
медленно деградируемых соединений. Растительный опад с мягкими
тканями быстро подвергается разложению группировкой микроор-
Таблица 7.1
Запасы органического углерода в слое 0-100 см почв и торфов России
[по: Орлов и др., 1996]
Зона
Полярно-тундровая
Лесотундровая
Среднетаежная
Южно-таежная
Лесостепная
Степная
Сухостепная
Полупустынная
Всего по равнинам
Кавказ
Южная Сибирь
Северная Сибирь
Курило-Камчатская обл.
Всего по горным областям
Всего по России
Площадь,
млн га
180,7
233,6
237,8
236,6
126,4
79,9
28,2
15,0
138,2
8,2
248,0
238,3
26,3
575,8
1714,0
Сорг, млн т
Всего
19225,6
39357,8
51988,4
61952,2
38378,1
21347,9
2824,9
1092,3
236167,2
1026,6
29061,6
27674,9
2216,6
59979,7
296146,5
В том числе
сельскохозяйственные
Пахота
0
36
81,1
3617,8
11334,2
9562,7
1378,4
161,4
26171,6
97,9
1024,7
0
0
1122,6
27294J
Прочие
0
254,7
820,0
6269,7
5349,9
4802,3
668,7
515,0
18022,2
416,1
1698,3
0
0
2124,4
20136,7
258
**¦/*'•
*>
2
1
V
i
О
о
о
с
?
о
S
о
х
X
о.
а>
о
л
В
§
I
U
s
ь
$
700-1000
1000-1300
1300-1700
1700-2200
El 3000-4400
¦Ц 4400-6000
¦M 6000-9000
ШШ -'«ИМ)
Я^^ непочвенные объекты
^^"l терри1Ч>рия республик
вывшего СССР
——~> границы административных
областей, округов
Авторы: В. Н. Кудеяров, И. Н. Курганова
Рис. 7.3. Годовая эмиссия углекислого газа из почв России [по: Кудеяров,
Рис. 7.4. Картограмма запасов гумуса в почвах России, т/га [по: Орлов, 1999]
/ - < 90; 2 - 90-120; 3 - 120-150; 4 - 150-180; 5 - 210-240; б - 240-270; 7 - 270-300; 8 - 320-350; 9 - 420-450; 10 - 450-480; //-510-540; 12 - 570-600-
/.?-> 1000
Рис. 7.5. Гидролиз целлюлозы [по: Tomme et al., 1995]
/ - ГАК; 2 - ксилогликан; 3 - поперечные сшивки; 4 - фенольные сшивки
Модель фрагмента первичной клеточной стенки травянистых растений иллюстриру-
иллюстрирует трудности, с которыми сталкиваются микроорганизмы при гидролизе растительных
остатков. Модель основания на анализе колеоптиля кукурузы и др. однодольных. Пока-
Показан один слой микрофибрилл целлюлозы непосредственно после деления клеток; множе-
множество таких слоев срастаются, образуя клеточную стенку. Микрофибриллы связываются
глюкуроноарабиноксиланами (ГАК), скрепляющимися с микрофибриллами фенольным
сшивками. В стенках многих других высших растений во взаимосвязях участвуют молеку-
молекулы ксилогликана, присоединяющиеся водородными мостиками к поверхности микрофиб-
микрофибрилл
ганизмов, участвующих в компостировании, включающей как
аэробные, так и анаэробные формы, например гетероферментатив-
ные молочнокислые бактерии, маслянокислые бактерии, в основ-
основном использующие углеводы с образованием консервирующих орга-
органических кислот. Скорость процесса разложения приводит к тому,
что развитие этой группировки зимогенных бактерий носит эфемер-
эфемерный характер. Из легкоразлагаемых углеводов первое место зани-
занимает крахмал как запасное вещество с группировкой аэробных и
анаэробных амилолитических бактерий. Более устойчивы к разло-
разложению структурные компоненты растительной ткани, как
262
пектин. Имеется специализированная группа бактерий, осуществля-
осуществляющая мацерацию растительной ткани и представленная бактериями
мочки льна. Защитные слизи и камеди разлагаются медленнее, но и
здесь действует специфическая группировка пуллуланолитических
бактерий с гидролитическим ферментом пуллуланазой. В слизете-
слизетечениях развивается Leuconostoc, в свою очередь образующий слизь.
Растительные оболочки (рис. 7.5) содержат структуры, состав-
составленные из: а) микрофибрилл целлюлозы с кристаллическими и
аморфными участками, последние легче гидролизуются; б) гемицел-
люлозы с ксиланами и глюкоманнанами, разделяющими целлюлоз-
целлюлозные фибриллы; в) ксилана из ксилозной цепи с глюкуроновой кис-
кислотой и арабинозой, этерифицированной феноловыми кислотами, в
свою очередь связанной с лигнином ковалентными связями; г) лиг-
лигнина, представляющего нерегулярную сеть из ароматических ком-
компонентов, как фенилпропан, и устойчивого к микробному разложе-
разложению; д) пектина, представляющего слизи срединной пластинки, из
полиуроновых кислот, например галактуроновой кислоты, этери-
фицированных метанолом. Метанол отщепляется метилэстеразой.
Метальные группы пектина и лигнина служат источником С-1 со-
соединений для метилотрофов. В водорослях аналогом пектина явля-
является альгиновая кислота. Водоросли не содержат лигнина, являюще-
являющегося характерным веществом для устойчивого углеродного скелета
наземной растительности.
В море целлюлоза не является доминирующим структурным по-
полисахаридом. Здесь ее заменяет хитин скелета морских беспозво-
беспозвоночных, по обилию на Земле представляющий второй или третий
полисахарид. На суше помимо беспозвоночных источником хитина
служит клеточная стенка грибов. Гидролизуется хитин экзофермен-
том хитиназой. Хитин гидролизуют микофильные грибы, а также
широкий набор бактерий как аэробных, в том числе актиномицеты,
так и анаэробных. Среди бактерий следует отметить помимо стреп-
томицетов аэробные псевдомонады, цитофаги, флавобактерии, арт-
робактеры, бациллы, Lysobacter (лизирующий структурно сходный с
хитином муреин бактерий) и анаэробные клостридии,
Halanaerobacter. Характерными потребителями ацетилглюкозамина
служат планктомицеты.
Как уже отмечалось, полиглюкоза представлена растворимыми
легкодоступными соединениями, например крахмалом, и труднодос-
труднодоступными нерастворимыми компонентами оболочек, прежде всего
целлюлозой. Крахмал содержит неразветвленную амилозу с а-1,4- и
разветвленный амилопектин с ос-1,6-связями. Это легкодоступное
соединение, гидролизуемое многими микроорганизмами, образую-
образующими амилазу. Довольно легко используется и пектин, для которо-
которого характерными являются Erwinia, Pectinatus и организмы мочки
льна "Bacillus felsineus". Осуществляют гидролиз пектина пектина-
зы с характерным побочным продуктом метанолом.
263
Целлюлоза имеет молекулярный вес около миллиона, Р-1,4- связь
в цепочке и водородные связи между параллельными цепочками.
Гидролизуется целлюлоза целлюлазным комплексом, который осу-
осуществляет сначала разрушение кристаллической структуры, затем
деполимеризацию цепочки на дисахарид целлобиозу экзоглюкана-
зой или целлодекстрины эндоглюканазой. Целлобиоза гидролизуется
целлобиазой с образованием глюкозы, которая может транспорти-
транспортироваться в клетку. Целлюлозолитические организмы представляют
важнейшую группу, открывающую основной маршрут деструкции
органического углерода на Земле. Разложение целлюлозы оказыва-
оказывается лимитирующим звеном в круговороте углерода в наземных
экосистемах и подробнее рассмотрено ниже.
Гемицеллюлозы представляют сборную группу полимеров гек-
соз, пентоз, уроновых кислот. Обычно доминируют ксилоза и ман-
ноза. Гемицеллюлозы сами по себе легко гидролизуются, но в при-
природе они обычно связаны с другими полимерами, и их разложение
затруднено.
Ксилан представляет гетерополимер из цепи ксилозы с арабино-
зой, глюкозой, галактозой, и другими сахарами как боковыми цепя-
цепями. Типичной группировкой бактерий, деградирующих ксилан, явля-
являются клостридии с низким Г + Ц, как Clostridium polysaccharolyticum,
С. cellulovorans, С. xylanolyticum, С. xylanovorans, а также представите-
представители родов Eubacterium, Fibrobacter, Butyrivibrio, Selenomonas,
Thermotoga. Он относительно легко сбраживается анаэробными ор-
организмами, которые используют широкий набор олигосахаридов,
образующихся после гидролиза ксиланазами.
Гидролиз полиглюкозы и вообще полисахаридов микроорганиз-
микроорганизмами обычно подавляется при высоком содержании продуктов гид-
гидролиза в среде. Отсюда этот маршрут специально обусловлен взаи-
взаимодействием гидролитиков с диссипотрофами.
Хитин состоит из полимера N-ацетил-Б-глюкозамина. Как уже
отмечалось, это основной компонент клеточной стенки грибов.
Хитин ассоциируется с другими полимерами. Его разложение в
большой степени зависит от физического состояния и доступности
поверхности для действия хитиназы и других гидролаз. Темноокра-
шенные пигменты меланины обусловлены связью Сахаров с феноль-
ными соединениями. Аминосахара, включая ацетилглюкозамин и
ацетилмурамовую кислоту, присутствуют и в стенке бактерий. Раз-
Разложение стенок грибов идет относительно быстро и легко под дей-
действием как бактерий, так и микофильных грибов, которые домини-
доминируют в более кислых условиях. Однако в целом стенки и мембраны
представляют относительно устойчивый компонент микробной
мортмассы. Считается, что аминосоединения микробных стенок
служат предшественниками азота й гумусе.
Лигнин представляет условие появления наземной растительно-
растительности. Это разветвленный полимер фенил-пропаноида. Лигнин обра-
264
зуется химической поликонденсацией фенола под действием свобод-
свободного радикала и не имеет закономерного строения. Он связывается
с целлюлозой и гемицеллюлозами, инкрустируя их. В результате
получается устойчивое, вследствие своей гетерогенности и неупоря-
неупорядоченности, соединение, которое не поддается прямому энзиматиче-
скому гидролизу. Деградация лигнина осуществляется почти исклю-
исключительно аэробными микроорганизмами. Исключение составляет
микробное сообщество пищеварительного тракта термитов. Дегра-
Деградация лигнина почти недоступна прокариотам, и о возможности их
участия в деградации меченого по углероду лигнина идут споры. Во
всяком случае, процесс если и есть, то очень слабый.
Деградацию лигнина осуществляют грибы "белой гнили"
Polyporus, Pleurotus, Phanerochaete. Обычно для начала деградации
необходимо наличие легкодоступного субстрата дыхания. Напро-
Напротив, грибы "бурой гнили" разлагают углеводные компоненты и ос-
оставляют фенольную часть, которая окисляется и темнеет. Продук-
Продукты деградации лигнина, в том числе ароматические соединения,
доступны как аэробным, так и анаэробным бактериям. Проведено
очень большое число работ по разложению модельных компонен-
компонентов лигнина, таких, как кофеиновая кислота. Особенно быстро раз-
разложение идет в аэробно/анаэробном сообществе.
Частицы органического вещества представлены в основном
оболочками отмерших организмов и состоят из нерастворимых по-
полимеров. Они входят в относительно медленно метаболизируемую
органику, которая становится доступной для использования осмо-
трофными организмами только после гидролиза до олигомеров.
Гидролиз осуществляется вне клетки, и поэтому гидролитические
экзоферменты должны быть устойчивы и активны в условиях сре-
среды. Для экстремофильных гидролитических организмов эта необхо-
необходимость накладывает дополнительное ограничение: кроме поддер-
поддержания внутриклеточного гомеостаза клетки должны вырабатывать
устойчивые и активные белки-экзоферменты. Гидролитические эк-
экзоферменты необходимы группам гидролитических организмов, ис-
использующим целлюлозу и другие полисахариды, хитин, нераствори-
нерастворимые белки, липиды.
Гидролитические экзоферменты бактерий могут либо распола-
располагаться на внешней поверхности клетки, либо секретироваться в среду.
Во втором случае продукты гидролиза могут стать доступными не
только продуценту, но и случайным организмам в окружении. Типич-
Типичную группировку такого типа и составляют диссипотрофы. Располо-
' жение гидролитических ферментов на клеточной поверхности пред-
¦' полагает их заякоривание в липидной части поверхностных струк-
структур, что, в свою очередь, предполагает необходимость тесного
физического контакта клетки с гидролизуемым материалом. Отсю-
Отсюда физическая организация взаимодействия клетки с гидролизуе-
*" мым материалом приобретает первостепенное значение. С одной
стороны, организм должен выработать механизмы адгезии к гидро-
лизуемому субстрату, а с другой — сохранить способность переме-
перемещаться с уже выработанных участков на свежие. Это требование
может быть выполнено для организмов, образующих подвижные
клетки для расселения, или же, что еще выгоднее, для организмов со
скользящим типом движения, расползающихся по поверхности суб-
субстрата. Образование слизистой капсулы на поверхности гидролизу-
емого сложного субстрата создает возможности для кооперативно-
кооперативного действия сообщества разнородных организмов, воздействующих
на разные компоненты сложного нерастворимого субстрата. Регу-
Регуляция синтеза экзоферментов подчиняется общим правилам катабо-
литной репрессии и индукции. Считается, что обычно организмы не
способны синтезировать экзоферменты в присутствии растворимых
легкоусваиваемых веществ, в том числе продуктов гидролиза.
Альтернативой гидролитическому пути разложения полимеров
служит расщепление взвешенного органического вещества с нерегу-
нерегулярным строением под действием активного кислорода. Примером
такого способа деградации служит процесс разложения лигнина эк-
зоферментами, использующими как источник активного кислорода
Н2О2, образующуюся при дыхании клетки, он носит общее название
соокисления труднодоступных субстратов. Очевидно, что этот спо-
способ деградации доступен только аэробам. Он распространен преиму-
преимущественно у грибов. Разложение негидролизуемого полимерного
вещества анаэробами происходит медленно и в водных экосистемах
часто требует участия сообщества микроорганизмов с сульфатреду-
кторами как терминальной группировкой. В наземных условиях
сульфидогенез ограничен, и основным путем разложения в анокси-
ческой обстановке служит метаногенез.
Суммарный процесс разложения целлюлозы в почве оценивает-
оценивается аппликационным методом по И. Вострову: в почвенный профиль
закладывается ткань и после инкубации выявляется область разру-
разрушения, связанная с определенным слоем почвы.
7.5.2. Разложение целлюлозы
Эффективный гидролиз целлюлозы требует взаимодействия не-
нескольких гидролаз, и поэтому организмы секретируют разные эн-
доглкжаназы и экзоглюканазы (целлобиогидролазы). Целлюлазная
система целлюлолитических организмов всегда ассоциирована с
другими гидролазами полисахаридов, особенно ксиланазами.
Целлюлазы содержат два домена: каталитический и связываю-
связывающий. Такой принцип организации свойственен и другим гидролазам
полисахаридов. Связывающие домены могут входить в S-слой бак-
бактерий. Целлюлазы и ксиланазы секвенированы для многих организ-
организмов, и у бактерий их строение широко варьирует. Для каждого се-
семейства ферментов классификация, основанная на их последова-
266
тельности, находится в противоречии с принятой филогенией бакте-
бактерий, снова подтверждая, что эти гены распространялись путем ин-
интенсивного латерального переноса. Это особенно свойственно се-
семействам ферментов, содержащих гены и из грамотрицательных и
из грамположительных организмов: например, сходны целлюлазы у
Erwinia и Cellulomonas. Вместе с тем активный центр каталитическо-
каталитического домена гидролаз отличается стереоспецифичностью и консерва-
консервативностью. Он включает в разных типах реакции либо глутамино-
вую, либо аспарагиновую кислоты для нуклеофильной атаки через
образование промежуточного оксокарбониевого продукта.
Целлюлозосвязывающий домен обеспечивает связывание с цел-
целлюлозой. У грибов он содержит 33-36 аминокислот и очень консер-
консервативен. У бактерий этот домен варьирует и обнаруживается в
целлюлазах, ксиланазах, хитиназах; он содержит около сотни ами-
аминокислот, у бацилл и клостридий - 130-170 аминокислот. От этого
домена зависит специфичность энзима по отношению к форме поли-
полисахарида. Другим компонентом целлюлаз служит участок, обеспе-
обеспечивающий связь с клеткой целлюлозолитического организма, обычно
богатый пролином.
Целлюлозолитическая система микроорганизмов обычно вклю-
включает набор гидролаз полисахаридов. Это наблюдается для разложе-
разложения не только природного материала, но и очищенной целлюлозы.
В результате организмы содержат гены нескольких целлюлаз. На-
Например, у штамма Clostridium thermocellum их было обнаружено 15.
Некоторые целлюлозолитические организмы имеют набор фермен-
ферментов, достаточный для разложения нативной растительной целлюлозы.
Такие наборы есть у грибов бурой древесной гнили, у клостридиев,
организмов рубца. У фитопатогенных и симбиотических организ-
организмов имеется специфический неполный набор целлюлаз для внедре-
внедрения через растительную стенку в корни. Неполный набор целлюлаз
возможен и для организмов, являющихся членами целлюлозолити-
целлюлозолитического консорциума; такие организмы не способны расти в чистой
культуре на целлюлозе, но активно размножаются в сообществе.
Целлюлозолитические энзимные системы разделяются на
"не-комплексирующие" (или "не-агрегирующие", растворимые) и
комплексирующие системы. Первые свойственны преимуществен-
преимущественно аэробным грибам и бактериям. Они выделяются в среду как
экзогидролазы в виде нескольких растворимых целлюлаз и деполи-
мераз полисахаридов. Комплексирующие системы типичны преиму-
преимущественно для анаэробов.
Синергическое действие растворимых целлюлаз приводит к то-
тому, что смесь ферментов имеет более высокую активность, чем сумма
их активностей. Предложено следующее объяснение синергическо-
му усилению действия. Эндоглюканазы разрывают целлюлозную
цепочку в местах нарушения кристаллической структуры. В этих
разрывах появляются точки приложения для экзоглюканаз (или
267
целлобиогидролаз). Гидролиз концов кристаллических участков
обнажает новые места нарушения кристаллической структуры. Гид-
Гидролиз целлобиозы Р-глюкозидазой приводит к появлению глюкозы
как конечного продукта, ингибирующего гидролазы. В этой схеме
независимо действующие ферменты без взаимодействия белок-бе-
белок-белок усиливают кооперативное действие.
Комплексирующие системы целлюлаз представляют собой по-
полиферментные комплексы, получившие название целлюлосомы.
Они характерны для клостридиев и в качестве модели подробно изу-
изучены у Clostridium thermocellum. Целлюлосомы образуют протубе-
протуберанцы на поверхности клетки и содействуют адсорбции клетки на
нерастворимой целлюлозе. Индивидуальные целлюлосомы устой-
устойчивы в воде и обладают высокой целлюлолитической активностью
вне зависимости от клетки. Они содержат 14-50 полипептидов,
большинство из которых обладает а-1,4-глюканазной или ос-1,4-кси-
ланазной активностью. Клостридии содержат многочисленные гены
глюканаз, представляющие ферменты различных семейств. Цел-
Целлюлосомы под электронным микроскопом выглядят как несколь-
несколько (более четырех) цепочек из 5-8 глобул. Предполагается, что
это индивидуальные эндоглюканазы на общем основании. Такая
система способна координированно атаковать целлюлозу с образо-
образованием в качестве продукта декстрина. Белок основания связыва-
связывается с целлюлозой и заякоривает фермент на ней. Он предположи-
предположительно обеспечивает перемещение ферментного комплекса по
гидролизуемому субстрату. Белки целлюлосомы и соответствую-
соответствующие гены идентифицированы. Для высокой активности целлюло-
целлюлосомы критическим оказывается координированное действие фер-
ферментов.
Аналогичные целлюлосомам системы ферментов установлены
у Ruminococcus, Bacteroides cellulosolvens, Bacteroides (=Fibrobacter)
succinogenes, Acetivibrio cellulolyticus - анаэробных организмов, кото-
которые плотно прикрепляются к гидролизуемой целлюлозе. Прикреп-
Прикрепление дает определенное преимущество в смешанных культурах. На
поверхности клеток были обнаружены аналогичные целлюлосомам
протуберанцы. Часть эндоглюканаз обнаруживается в везикулах
капсулы руминококков. Однако генетического подтверждения сход-
сходства с целлюлосомами клостридиев пока нет, хотя они реагируют с
антителами для целлюлосом.
У разных организмов ферменты целлюлозолитической системы
синтезируются конституитивно или же индуцируются. Индуктором
синтеза целлюлаз и ксиланаз может служить целлобиоза, продукт
экзоглюканаз, и целлодекстрины как продукты эндоглюканаз. Ин-
Индукторами могут служить и иные олигосахариды. Особенно эффек-
эффективным индуктором считается сефароза. С другой стороны, эти же
соединения могут ингибировать синтез. Поэтому существенна их
концентрация в среде и транспорт в клетку.
268
*
i
t
i
I
I
Из рассмотрения действия целлюлозолитических организмов и
ферментов можно вывести необходимость по меньшей мере трех
уровней кооперативного действия. Первый уровень состоит в дейст-
действии различных ферментов на субстрат. При этом ферменты либо
могут действовать индивидуально и при этом иметь синергический
эффект, либо быть организованы в структуру, обладающую коопе-
кооперативным действием, как в целлюлосоме. На уровне организма воз-
возможен синтез полного набора ферментов одним организмом, спо-
способным расти на целлюлозе в чистой культуре и осуществлять ее
полный гидролиз. При неполном наборе ферментов используется
только часть полимерного материала, хотя рост еще возможен; ли-
либо рост отсутствует, несмотря на наличие целлюлаз. Наконец, на
уровне кооперативно действующего сообщества разные организмы,
синтезирующие разные гидролазы, оказывают совместное действие
и развиваются в виде сообщества, быстро гидролизуя субстрат, хотя
поодиночке они на целлюлозе не растут или же рост их ограничен.
Синтрофия обусловлена не анаболической зависимостью от факто-
факторов роста в сообществе, а кооперативным воздействием на сложный
субстрат.
Следующий шаг состоит в разложении сложного взвешенного
вещества, где присутствуют и разные полисахариды, и белки, и ли-
пиды, как это имеет место при разложении мортмассы. Здесь воз-
возможно не только синергическое действие, но и реакции торможе-
торможения, например за счет протеолиза незащищенных экзоферментов.
Физическая организация на микроуровне приобретает первостепен-
первостепенное значение, включая защиту ферментных комплексов от угрозы
извне.
Вся обширная группировка наземных целлюлозолитических ми-
микроорганизмов и их сателлитов сформировалась после развития на
Земле растительного покрова. Широкое распространение этой тро-
трофической группировки могло иметь место в конце силура 0,35 млрд
лет назад, хотя водные целлюлозолитические бактерии могли раз-
развиваться с неопротерозоя около 1 млрд лет назад, особенно в связи
с распространением нитчатых водорослей в водоемах. В период гос-
господства цианобактерий целлюлозолитические организмы имели ог-
ограниченный источник субстрата. У бактерий процесс освоения но-
нового субстрата шел за счет латерального переноса генов на основе
самых разнообразных филогенетических ветвей. Гидролитическая
активность есть типичный пример подготовительного метаболизма.
Вопрос состоит в совместимости с морфо-физиологическими осо-
особенностями. Особенно подходящими оказались организмы со сколь-
скользящим движением, способные двигаться по поверхности твердого
субстрата, как цитофаги, и с мицелиальным строением, как актино-
мицеты.
Основной группировкой наземных организмов, ориентирован-
ориентированной на разложение твердой фазы растительного вещества - оболоч-
269
ки клетки, являются эукариотные мицелиальные организмы. Грибы
в целом представляют группировку, трофически ориентированную
на разложение твердой фазы растительного органического вещест-
вещества в субаэральных условиях. Подавляющая масса грибов так или
иначе связана с разложением растительного материала либо как са-
профиты, либо как паразиты. К этому следует добавить симбиоти-
ческие отношения грибов микоризы с корневой системой растений.
Итак, в целлюлозолитический маршрут входят следующие груп-
группировки:
1) гидролиз целлюлозы грибами "бурой гнили" осуществляется
очень широким кругом аэробных организмов, составляющих важ-
важнейшую часть группировки сапрофитных грибов и некоторых фито-
патогенных;
2) аэробные бактерии с характерной группировкой цитофаг;
3) анаэробные бактерии с характерной группировкой клостри-
дий; гидролиз целлюлозы археями пока неизвестен в противопо-
противоположность участию термофильных архей в протеолитическом пути.
4) аэробные сахаролитические диссипотрофы, как, например,
простекобактерии, но также многочисленные органотрофные орга-
организмы;
5) анаэробные сахаролитические диссипотрофы, как, например,
спирохеты, но также очень многочисленные сахаролитические
анаэробы, образующие трофическую систему в метаногенном сооб-
сообществе с последовательным использованием продуктов обмена тро-
трофических предшественников.
7.5.3. Разложение лигнина
Лигноцеллюлоза составляет почти половину годичной продук-
продукции наземной растительности. Лигноцеллюлоза составляет 90-98%
древесины. По обилию лигнин стоит на втором месте после целлю-
целлюлозы. Лигнин, составляя углеродный скелет растений, трудно разла-
разлагается и поэтому представляет один из важных компонентов резер-
резервуаров углерода с длительным временем пребывания, и его разло-
разложение представляет узкое место в круговороте углерода в наземных
системах. В лигноцеллюлозе лигнин связан с гемицеллюлозой, в то
время как его связь с целлюлозными фибриллами скорее физиче-
физическая, чем химическая. Поэтому пути деградации лигноцеллюлозы
могут идти по двум независимым маршрутам: разложение целлюло-
целлюлозы группировкой "бурой гнили" или же разложение гемицеллюлоз-
но-лигнинового комплекса организмами "белой гнили", наиболее
известным примером которых может служить опенок Armillariella, a
наиболее частым лабораторным объектом - Phanerochaete. Грибы,
растущие на лигноцеллюлозе представляют наиболее разнообраз-
разнообразную и многочисленную группировку сапрофитных грибов, макро- и
микромицетов. Позднее появление лигноцеллюлозы в истории био-
270
ты в фанерозое, связанное с появлением наземного растительного
покрова, предполагает существование в условиях ранее созданной
аэробной атмосферы и развитие грибов как наземной группировки
органотрофных ксерофильных аэробных организмов, развиваю-
развивающихся в виде мицелия на твердом субстрате. Разложение лигнина
происходит за счет аэробных процессов с участием активного кисло-
кислорода, необходимого для разрыва связей между фенилпропановыми
частями молекулы лигнина. Предполагается, что ароматические со-
соединения, образующиеся при разложении лигнина, лишь замедлен-
замедленно разлагаются в аэробных условиях и являются исходным матери-
материалом для конденсации в молекулы гуминовых соединений, возмож-
возможно, с участием микробных внеклеточных полимераз в биологически
опосредованных реакциях.
Обычно считается, что в анаэробных условиях лигнин инертен,
но это не относится к олигомерным продуктам его разложения, ко-
которые могут метаболизироваться бактериями, атакующими в анаэ-
анаэробных условиях фенолы (Schink et al., 2000).
Анаэробное разложение полученных из лигнина соединений мо-
может осуществляться в условиях денитрификации, сульфидогенеза,
метаногенеза и подробно исследовано в последние десятилетия
Г. Фуксом, Б.Шинком после пионерских работ Г. Зейкуса по разло-
разложению лигнина бактериями. Эти исследования особенно усилились
в связи с изучением деградации галогенизированной ароматики, по-
поступающей от ксенобиотиков в виде загрязнения.
Общая схема разложения древесины включает разделение ее на
компоненты, которые разлагаются разными группировками микро-
микроорганизмов.
Древесина
Воска, смолы Белок
Лигноцеллюлоза
Лигниназа
Лигнин
Олиголигнолы
растворимые
Целлюлозные компоненты
Гемицеллюлозы Целлюлоза
Гумус
Метилированные
фенилпропановые
компоненты
Ароматические
соединения
- Ксилан
Ксиланаза
- Пентозы
- Гексозы
Целлюлозы
Целлодекстрин
Целлобиоза
Глюкоза
Уроновые кислоты
Дезинтеграция лигнина включает несколько этапов. На первом
из них происходит образование лигнолов, растворимых остатков
лигнина, представляющих случайное собрание гидрофильных моле-
271
кул умеренного молекулярного веса. Образуются эти молекулы в
результате действия "лигниназ" - ферментов, продуцирующих при
соокислении органического вещества продукты неполного восста-
восстановления О2 - перекиси и даже радикалы. Перекисный механизм
разложения лигнина приводит к хаотическому разрыву связей, но
образующиеся осколки оказываются более доступны для микроор-
микроорганизмов, и поэтому их концентрация в системе определяется дина-
динамическим равновесием между продукцией и разложением. Следую-
Следующий этап разложения приводит к образованию крупных молекул
ароматических кислот: феруловой, сирингиловой, циннамовой, ва-
ванилиновой. В дополнение к ним имеется множество осколков стру-
структуры в виде циклических соединений, особенно фенолов, катехи-
нов. В результате деметилирования поступают и С-1 соединения.
Размеры молекул этих субстратов позволяют осуществлять их осмо-
трофное использование с поступлением в клетку. Широкий набор
субстратов допускает две возможности: действия микроорганизмов-
универсалистов, использующих все эти соединения внутри клетки,
или же действия микробов-специалистов в составе сообщества. Обе
эти возможности осуществляются. Многие грибы, растущие на дре-
древесине, оказываются в достаточной степени универсалистами.
Разложение циклических соединений аэробными микроорганиз-
микроорганизмами внутри физической структуры разлагаемых растительных
остатков неизбежно приводит к дефициту кислорода, которому спо-
способствует влажность. Аноксические условия способствуют разви-
развитию бактериальной микрофлоры, способной к разложению цикли-
циклических соединений без участия кислорода и без доступа внешних
окислителей, как нитрат или сульфат. В этих условиях, помимо спо-
способности к брожению, оказывается важным использование органи-
органических веществ как акцепторов электрона. В качестве примера та-
таких процессов можно назвать фумаровое дыхание и, что особенно
интересно, открытое недавно хиноновое дыхание. В какой мере хи-
ноны могут служить диффундирующим переносчиком между вос-
восстановленной частью структуры и поверхностью, где они могут
окислиться и вернуться обратно, а не быть съеденными аэробами,
неясно.
Разложение лигнина, как было упомянуто выше, ведет к образо-
образованию веществ разной молекулярной массы. Среди них доминируют
циклические соединения. Предполагается, что эти соединения, об-
обладающие реакционно-способными функциональными группами,
могут реагировать между собой, объединяясь в крупные макромоле-
кулярные соединения путем химической конденсации, возможно,
под действием неспецифически активирующих процесс ферментов.
Происходит так называемая поликонденсация с синтезом гумусовых
веществ. В нее вовлекается ряд веществ микробного происхожде-
происхождения. Эта схема описывает представления о происхождении гуми-
новых соединений из продуктов разложения лигнина. При этом ис-
272
<
1
ходные химические структуры лигнина в гумусе уже перестают
быть опознаваемыми вследствие перестройки на уровне относи-
относительно мелких молекул, вплоть до фенолов.
Описываемая последовательность событий в главном для назем-
наземных биоценозов маршруте превращений органического вещества
вряд ли может быть прослежена по судьбе химических структур вви-
ввиду их разнообразия. В общих чертах предполагается первоначальная
деполимеризация лигнина от молекулярных масс порядка мегадаль-
тон к их осколкам размером в тысячи-сотни дальтон с последую-
последующим укрупнением в макромолекулы порядка десятков-сотен тысяч
дальтон. Поскольку сам лигнин подвергается при своем биосинтезе
процессу ферментативной конденсации из фенольных предшествен-
предшественников, то представляется вероятной иммобилизация аналогичным
образом органического углерода путем образования гумусовых со-
соединений, но из гораздо большего разнообразия предшественников
и не под таким строгим контролем, как в клетке растения. При этом
прямое участие биоты уже ограничено, и это лишает процесс специ-
специфичности.
В масштабах России процесс вероятного синтеза гумуса из лиг-
лигнина как основного предшественника количественно согласуется с
поступлением лигноцеллюлозы, которое с избытком перекрывает
оценку синтеза гумуса порядка 200 Мт С/год, если считать время
пребывания углерода в гумусе около тысячи лет. Дополнительным
источником вещества в гумусе, и особенно азотистых компонентов,
следует считать деградацию мицелия грибов как вторичных проду-
продуцентов мортмассы в трофической системе. Отсюда следует, что в
образовании гуминовых соединений лигнин в первую очередь игра-
играет роль как поставщик циклических соединений, но он является не
единственным предшественником, а лишь важным составным ком-
компонентом.
Лигноцеллюлоза представляет важную составную часть торфа -
не полностью разложенных гумифицированных растительных ос-
остатков. Микробные группировки, участвующие в процессе деграда-
деградации лигноцеллюлозы, следующие:
1. Лигнолитический маршрут:
Группировка микроорганизмов, разлагающих лигнин
- грибы "белой гнили": крайне разнообразные аэробы с пере-
кисным механизмом разрушения лигнина, включая микромицеты и
шляпочные грибы с длительным жизненным циклом; десятки тысяч
видов гидролитических организмов;
- сопутствующие гидролитикам аэробные диссипотрофы, ис-
использующие продукты гидролиза лигнина, в первую очередь цикли-
циклические, фенольные соединения;
- анаэробы с лигнолитической функцией отсутствуют! (на са-
самом деле это очень медленный процесс, осуществляемый сообщест-
сообществом);
273
- метилотрофные организмы, использующие продукты демети-
лирования лигнина, но прежде всего пектина, и включающие аэроб-
аэробные и анаэробные формы;
Микофилъная группировка бактерий
- аэробные гидролитики, разлагающие мортмассу грибов с ха-
характерной группировкой хитинолитических организмов;
- следующие за ними диссипотрофы, в том числе использующие
глюкозамин;
- анаэробные диссипотрофы, способные сбраживать низкомо-
низкомолекулярные органические соединения, в том числе посредством хи-
нонового дыхания.
2. Ксиланолитический маршрут, названный так из-за доминиро-
доминирования в нем разложения ксилана как важнейшей полимерной частя
гемицеллюлоз. Он приводит к освобождению лигнина и целлюлозы
из гемицеллюлозной матрицы. В нем участвуют грибы, дрожжи, ба-
бактерии, осуществляющие двухэтапное разложение субстрата:
- гидролитические аэробные и анаэробные организмы с фер-
ферментом ксиланазой;
- диссипотрофы, использующие преимущественно пентозы как
аэробно, так и анаэробно для брожения.
Целлюлозолитический маршрут описан выше.
7.5.4. Разложение древесины ксилофагами
Разложение древесины с ее последующей гумификацией пред-
представляет длительный процесс с участием разных групп организмов и
их сменой по ходу разложения. Ведущую роль в разложении играют
грибы-ксилотрофы, которым приписывается 90% разлагаемой дре-
древесины. Поэтому лес можно обозначить как древесно-грибную сис-
систему. В распространении грибов большую роль играют насекомые-
ксилофаги, осуществляющие механический перенос зародышей
грибов. Мицелиальное строение грибов дает им большое преимуще-
преимущество перед бактериями, которые не могут перемещаться в мертвой
древесине и обычно следуют за гифами грибов, образуя микофиль-
ную группировку, в которой специальную роль играют азотфикса-
торы.
Апикальный рост грибов обеспечивает их перемещение внутри
плотного субстрата и позволяет им образовывать мицелиальную
сеть с проникновением в полости сосудов и трахеид. Транспорт ве-
веществ осуществляется внутри гиф гриба благодаря порам между
клетками гифы и активному перемещению внутри клеток. У неко-
некоторых грибов, как у опенка, есть "транспортные гифы" в ризоморфах.
Вокруг гиф имеются слизистые чехлы, удерживающие внеклеточ-
внеклеточные гидролазы в непосредственной близости от гиф и ограничиваю-
ограничивающие рассеяние продуктов гидролиза. Грибная зона лигнолитических
грибов окружается черной каймой (ксилостромой) отложения поли-
274
*
I
I
4
меризованных полифенолов и МпО2. В грибной зоне стабилизиру-
стабилизируются влажность и газообмен.
Группировки грибов, связанных с деревьями, разделяют на:
Микогоризонты
фотосинтезирующего слоя
стволового слоя
мортмассы
Преобладают
паразиты
эпифиты (+ лишайники)
факультативные сапротрофы
и паразиты
сапротрофы
подземного слоя
симбионты и микориза
Практическая значимость грибов-ксилотрофов вызвала боль-
большой интерес к их изучению как в отношении систематики, так и
функциональном. Грибы, населяющие древесину, крайне разнооб-
разнообразны и специализированы по своим функциям. Часть из них явля-
является симбионтами, не приносящими видимого вреда древесине или,
как "микоризные грибы, заведомо полезными. Ксилотрофы имеют-
имеются среди базидиомицетов, аскомицетов, анаморфных (несовершен-
(несовершенных) грибов и некоторых хитридиевых. Многие из них идентифици-
идентифицируются по плодовым телам как трутовики.
В функциональном отношении следует провести границу между
паразитическими грибами, развивающимися в живом растении и со-
составляющими предмет фитопатологии, и сапрофитами, развиваю-
развивающимися на погибших деревьях, в опаде или в мертвой древесине
стволов. Впрочем, многие ксилопаразиты, как опенок, способны
как развиваться на мертвой древесине, так и поражать живые дере-
деревья. Их главное отличие от сапротрофов - способность преодоле-
преодолевать защитные барьеры живого дерева.
Жизненный режим грибных сообществ, развивающихся на сухо-
сухостое, за счет погребенной в земле древесины корней, в опаде и под-
подстилке, существенно различается в первую очередь из-за режима
влажности. Поэтому здесь доминируют разные виды. Взаимоотно-
Взаимоотношения в сообществах грибов в сильной степени обуславливаются об-
образованием ими физиологически активных веществ, антибиотиков и
фунгицидов в том числе. Общими условиями развития ксилотрофов
следует считать избыток углерода по отношению к азоту и фосфо-
фосфору. В древесине С : N : Р = 500 : 16 : 1, а в биомассе грибов С : N = 6.
Конкуренция грибов за питательные компоненты связана, во-пер-
во-первых, с колонизацией первичных ресурсов, во-вторых, сукцессией при
освоении уже колонизованного субстрата. Конкуренция идет в
очень жесткой форме, включая упомянутые антибиотические реак-
реакции, микопаразитизм, физическое вытеснение мицелием, перехват
i продуктов гидролиза. По ходу разложения наблюдается смена доми-
доминирующих видов. Сначала через раневую поверхность живого дере-
дерева проникают грибы-паразиты, вызывающие заболонную гниль
275
водопроводящей ткани и изменяющие ее цвет. После вымывания
экстрактивных веществ для развития гриба требуется вторичное ув-
увлажнение. Срубленная древесина подвергается побурению, связан-
связанному с автолитическими процессами (прелость). На следующей ста-
стадии (подпар) проникают грибы, осваивающие экстрактивные вещест-
вещества паренхимных клеток. Затем начинается развитие собственно кси-
лотрофов в виде белых пятен на буром фоне (мраморная гниль). Эта
стадия сменяется мягкой гнилью, когда древесина теряет прочность.
Далее развиваются типичные сапротрофы валежника. Стадии эти
протекают несколько лет. За ними следует гумификация уже разло-
разложенной древесины. Колонизация пней достигает максимума в 2-5 лет.
Развитие грибов в древесине приводит к закреплению азота и
фосфора в мицелии, причем грибная масса замещает древесную: при
разложении на 20% масса грибов составляет 43 мг/г древесины, при
разложении на 32% - 200,8 мг/г и при глубоком разложении на 61%
снижается до 192 мг/г. Животные не играют большой роли в разло-
разложении древесины3.
Колонизация древесины грибами отличается от разложения от-
отдельных компонентов ее тем, что осуществляется в ткани растения
и требует гистологического подхода. Разные типы грибов различа-
различаются по стратегии колонизации. Из мест инфекции грибы проходят
по сердцевинным лучам и только оттуда проникают в остальную
древесину. Переход от одной древесной клетки к другой происходит
у грибов через простые, полуокаймленные и окаймленные поры
клеток ксилемы с образованием канала для микрогиф механически
или с помощью ферментов (грибы бурой гнили), или же путем рас-
распространения в срединной пластинке (грибы белой гнили). Возбуди-
Возбудители деструктивных гнилей, разлагающие целлюлозу, проникают в
клетки древесины из сердцевинных лучей отдельными разветвлен-
разветвленными гифами, а лигнинразрушающие грибы - пучком гиф, который
затем обильно разветвляется. Затем такой гриб заполняет полость
клетки густым переплетением гиф, причем мицелий иногда собира-
собирается в клубки. Поскольку разложение субстрата достигается дейст-
действием экзоферментов, дающих усваиваемые грибом низкомолеку-
низкомолекулярные продукты, стратегия гриба состоит в том, чтобы не допус-
допустить их рассеяния из ближайшего окружения гриба. Крупные моле-
молекулы экзоферментов сорбируются на деградируемом субстрате и в
рыхлой слизи; мелкие молекулы должны свободно диффундировать
от места их образования к грибу, и поэтому локализация микрооби-
микрообитания внутри клетки и участка ткани оказывается выгодной, чтобы
свести к минимуму рассеяние вовне. Стратегия сводится к созданию
микроусловий на расстоянии быстрой молекулярной диффузии, т.е.
в сотни микрон. Вторая задача связана с транспортом веществ
3 Покаржевский А.Д. Геохимическая экология наземных животных. М: Наука, '
1985. С. 250. |
276
1
вдоль мицелия гриба, представляющего целостный организм, а не
отдельную клетку. Части мицелия несут разные задачи (Рабинович
и др., 2001).
По характеру действия грибы делят на:
- недеструктивные, окрашивающие древесину за счет разложе-
разложения белков (синяя гниль);
- вызывающие мягкую гомогенную гниль с образованием тем-
ноокрашенных гумусовых веществ шоколадного цвета; большей ча-
частью это аскомицеты и несовершенные грибы, к которым относят-
относятся виды Chaetomium, Stachybotris, Trichoderma; при этом потребляет-
потребляется преимущественно целлюлоза;
- вызывающие бурую гниль, преимущественно базидиомицеты;
побурение связано с накоплением ароматических хромофоров - хи-
нонов, феноксисоединений;
- вызывающие белую гниль базидиомицеты; они сохраняют
структуру древесины за счет удаления хромофоров типа хинонов;
считается, что они разрушают лигнин, но при этом в сопряженном
процессе идет потребление и целлюлозы.
Различие между грибами белой и бурой гнили устанавливают по
реакции Бавендамма - способности к окислению и конденсации гал-
галловой кислоты в агаризованной среде как критерию способности к
разрушению лигнина. Реакция эта зависит от полифенолоксидаз
(лакказы). На самом деле грибы белой и бурой гнили используют
оба компонента, но с разными скоростями и могут быть охарактери-
охарактеризованы показателем Y = С/(С + L), отнесенным к разложенной морт-
массе, где С - целлюлоза, L - лигнин.
Грибы мягкой гнили относятся к гумификаторам, способным и к
разложению лигнина, и к образованию гумуса. Гифы их распростра-
распространяются в древесине преимущественно вдоль целлюлозных волокон,
секретируя фермент лишь в определенных участках гиф, создавая
каверны с коническими вершинами 0,6-13 х 5-140 мкм. Предполага-
Предполагают, что целлюлазы распространяются вдоль волокон целлюлозы.
Набор гидролаз, катализирующих разложение полисахаридов,
включает целлюлазно-гемицеллюлазный комплекс и вспомогатель-
вспомогательные эстеразы. Кроме того, присутствуют оксидоредуктазы, прежде
всего полифенолоксидазы, описанные у микромицетов-гумифика-
торов. Лакказы ксилотрофов имеют оптимум при рН 4-5, а у мик-
ромицетов он близок к нейтральному. Лакказы мягкой гнили связа-
связаны с полимеризацией низкомолекулярных фенолов и детоксика-
цией катехинов, таннинов, стильбенов с вовлечением свободных
углеводов и аминокислот. Фермент микромицетов целлобиозоде-
гидрогеназа окисляет олигосахариды, но не моносахариды с восста-
восстановлением хинонов, фенокси-радикалов или Fe(III) и Cu(II).
Возбудители бурой гнили избирательно разрушают полисаха-
ридную часть, оставляя лигниновый скелет. Сначала (до 10% поте-
потери веса) идет использование гемицеллюлоз, маннана, затем (до 50%
277
потери веса) используется целлюлоза. При этом растет раствори-
растворимость в 1 %-ной щелочи. Считается, что бурая гниль является типич-
типичным процессом гумификации путем накопления предшественников
гумуса из остаточного лигнина. Возможны два пути: конденсация в
присутствии азотистых веществ оксикоричной, ванилиновой, окси-
бензойной, феруловой, протокатеховых кислот как продуктов де-
деградации лигнина или же синтез предшественника гумуса через ши-
киматный путь в самом грибе.
Для гидролиза упорядоченной целлюлозы в древесине особую
роль играет неферментативный гидролиз ("химия Фентона"). Окси-
дазный свободнорадикальный механизм химии Фентона (Fe2+ +
+ Н2О2) основан на окислении олигосахаридов в присутствии О2 глю-
козо-, галактозо-, целлобиозодегидрогеназами, образующими пере-
перекись. Последняя в присутствии Fe2+ разлагается с образованием ра-
радикала ОН', способного неспецифически окислять как целлюлозу,
так и лигнин. Fe2+ регенерирует в реакции целлобиозодегидрогена-
зы (флавоцитохрома):
Fe2+ + Н2О2 + Н+ = Fe3+ + Н2О + ОН",
Целлобиоза + 2Fe3+ = Целлобионолактон + 2Fe2+ + 2Н+.
Реакции радикалов ОН' идут неспецифически, и при этом мо-
могут участвовать различные формы активного кислорода. Преиму-
Преимущественно Р-расщеплению подвергаются алифатические С-С свя-
связи. Начальным этапом аэробной генерации одноэлектронного
окислителя является активация О2, восстановитель для кото-
которого должны поставлять внутриклеточные системы гриба. Расти-
Растительные полифенолы - таннин и галловая кислота - являются
мощными ингибиторами фентоновских процессов, связывающими
Fe2+.
Грибы бурой гнили разрушают лигнин по окислительному меха-
механизму при действии пероксидаз и активного кислорода. Процессы
эти в принципе аэробные и зависят от поступления молекулярного
кислорода. Эти реакции не дают энергии для гриба, но, подобно вне-
внеклеточным гидролазам, участвуют в многоступенчатом превраще-
превращении нерастворимого материала в диффундирующие низкомолеку-
низкомолекулярные соединения, доступные для осмотрофа. Продуктами окисле-
окисления являются разнообразные циклические соединения, для усвоения
которых в универсальных клеточных процессах требуется подгото-
подготовительный метаболизм.
Разложение лигнина в древесине происходит без непосредствен-
непосредственного контакта с гифами на диффузионном расстоянии. Грибы обла-
обладают набором ферментов, трансформирующих нефенольные аро-
ароматические структуры в легко метаболизируемые ациклические со-
соединения. Основное действие связано с лакказами и Мп-пероксида-
зами.
278
1
i
V
У
Характерным метаболитом лигнинразлагающих грибов являет-
является оксалат, в количественном отношении превышающий другие ор-
органические кислоты, как янтарная, фумаровая. Предполагается, что
лигнинразрушающие грибы разлагают также грубый гумус в под-
подстилке. "Для России суммарную деятельность лигнинразрушающих
грибов можно оценить в 1/4-1/3 от продукции лесов, что составит
примерно 80 млн т С/год. К этой величине следует добавить суммар-
суммарный вклад грибов в разложение гумуса в лесных почвах, особенно на
подзолах, что составит приблизительную величину порядка
100 млн т С/год.
7.6. ГУМУС
Органическое вещество почвы на 10-20% состоит из углеводов,
на 20% из азотистых соединений, 10-20% алифатических соедине-
соединений, остальное представлено ароматикой. Вещество, остающееся
после экстракции щелочью, пирофосфатом, называется гумином.
Вещество, осаждаемое из щелочного экстракта при подкислении на-
называется гумусовыми кислотами. Вещество, остающееся в подкис-
подкисленном экстракте, называется фулъвокислотами. Все эти вещества
темноокрашенны благодаря присутствию ароматики.
Считается, что компоненты лигнина являются предшественни-
предшественниками почвенных гуминовых соединений, для которых тоже харак-
характерно присутствие фенольных групп и поликонденсация в гетеропо-
лимерное соединение с высоким молекулярным весом. В системном
плане гумус представляет устойчивый остаточный продукт разло-
разложения растительных остатков микробным сообществом, не поддаю-
поддающийся дальнейшему прямому разложению. Химически же он явля-
является продуктом трансформации этих остатков в устойчивое соедине-
соединение. Гумус следует отличать от торфянистых остатков - покры-
покрытых гумусовыми пленками не полностью разложенных остатков
растений, как правило, накапливающихся при избыточном увлажне-
увлажнении. Наличие гумуса отличает почву от обнаженного грунта или
горной породы. Гуминовые соединения ответственны за водный ре-
режим вследствие гигроскопичности, за структуру почвы как склеива-
склеивающий агент, за состав почвы вследствие комплексообразования.
Разложение гуминовых соединений осуществляется медленно и
идет, главным образом, путем соокисления.
Гумусовые вещества не имеют регулярной структуры, и молеку-
молекулы их не имеют одинакового строения, хотя имеют сходные молеку-
молекулярные веса и набор функциональных химических групп. По эле-
элементному составу (вес %) они содержат:
С Н N S О Зола
Фульвокислоты 49,5 4,5 0,8 0,3 44,9 2,4
Гуминовые кислоты 56,4 5,5 4,1 1,1 32,9 0,9
279
По функциональным группам (в мг-экв/г):
-ОСН3 -СООН Фенольные-ОН Кислотность
Фульвокислоты 0,5 9,1 3,3 12,4
Гуминовые кислоты 1,0 4,5 2,1 6,6
Гумус разлагается микробным сообществом и в зависимости от
условий представляет динамический резервуар устойчивого органи-
органического углерода почвы. Время пребывания углерода в этом резер-
резервуаре оценивается по радиоактивному углероду и составляет до
1000 лет. Размеры этого резервуара в глобальном масштабе оцени-
оцениваются величиной 1500 • 1015 г и в 3 раза превосходят содержание уг-
углерода в биомассе. Географически гумус накапливается в гумидном
поясе при равнинном ландшафте. С уменьшением поступления рас-
растительных остатков, например при распашке целины, содержание
гумуса падает примерно на 1/3. Это служит прямым указанием на
его деградацию, а не только на вынос при обработке почвы.
Разложение гумуса микроорганизмами представляет особый ин-
интерес. Для разлагающей гумус микрофлоры было предложено Ви-
ноградским название автохтонная. С этой микрофлорой бились до-
довольно много. В общем типичными организмами, развивающимися
на гуминовых веществах, оказались артробактеры и нокардии.
Это олиготрофные организмы в том смысле, что они растут при
крайне низком содержании доступной органики, но совершенно от-
отличны от диссипотрофов. Олиготрофные нокардии медленно разла-
разлагают труднодоступное органическое вещество. Диссипотрофы ис-
используют легко доступную органику в очень низкой концентрации.
В прибрежных водах водорастворимые гуминовые вещества
обычно составляют 20-60% РОВ. Они представляют собой органи-
органические кислоты с карбоксильными и фенольными группами, возни-
возникающие различным путем как остаточные продукты деградации,
так и путем конденсации4. Ранее их рассматривали как очень устой-
устойчивые, но сейчас по разложению синтетического 14С-лигнина (поли-
меризат кониферилового спирта) как единственного источника
энергии установлено, что они могут поддерживать рост микроорга-
микроорганизмов.
7.6.1. Анаэробное разложение фенольных соединений
В кислородной атмосфере микробная деградация ароматиче-
ароматических соединений осуществляется грибами и бактериями, которые
используют О2 для активации субстрата в реакциях оксгенирования.
В результате действия моно- и диоксигеназ вследствие введения гид-
роксигрупп исходные вещества преобразуются в небольшое число
ключевых соединений, из которых наибольшее внимание уделяется
4 Thurman EM. Organic geochemistry of natural waters. Dordrecht: Nijhoff/Junk, 1985.
280
катехину, протокатехиновой кислоте, гентизату. У них, в свою оче-
очередь, под действием оксигеназ подвергается разрыву ароматиче-
ароматическое кольцо вблизи гидроксигрупп. Образовавшиеся ненасыщенные
органические кислоты подвергаются дальнейшему разложению
обычно через ацетильные или сукцинильные производные. Каждая
реакция оксигенирования в высокой степени экзотермична и обу-
обусловливает выделение в виде тепла около 300 кДж/моль реактанта.
Эта энергия не может быть использована организмом для синтеза
АТФ. Подготовительные реакции оксигенирования составляют це-
цену возможности дальнейшего использования продуктов окисления
как субстрата энергетического обмена.
Очевидно, что, подобно гидролитикам, оксигенирующие орга-
организмы образуют широкий спектр усвояемых растворимых ве-
веществ, и это создает условия для обильного шлейфа диссипотро-
диссипотрофов, сопровождающих их деятельность. Нерастворимые полимер-
полимерные ароматические вещества обусловливают структурную органи-
организацию сообщества с доминирующей группировкой мицелиальных
организмов, инициирующих распад твердых лигнифицированных
остатков в опаде или мертвых растительных тканях. Вымываемые
ароматические соединения и продукты их дальнейшего преобразо-
преобразования в виде растворимого "водного гумуса", фульвокислот, фено-
фенолов поступают в почвенный раствор или осадки водоемов. Высо-
Высокая потребность в О2 для оксигенирования и его использование
приводят к ограниченной доступности кислорода в плотных струк-
структурах растительного опада.
Ароматические продукты распада могут поступать в анаэроб-
анаэробные зоны и там разлагаться за счет восстановительных процессов5.
Образование СН4 при разложении ароматических соединений было
обнаружено еще в 1930-х годах на примере анаэробных очистных
сооружений. Установленным фактом является способность ана-
анаэробных организмов деградировать ароматические соединения. Био-
Биохимические пути были исследованы лишь в 1980-х годах с образова-
образованием в качестве метаболитов бензоил-КоА, резорцина, флорглюцина.
Путь с бензоил-КоА считается наиболее широким, вовлекающим
фенолы, гидроксибензоат, анилин, крезолы. Анаэробные сообщест-
сообщества разлагают эти вещества как по метаногенному пути с участием
бродилыциков и метаногенов, так и в условиях сульфидогенеза, при-
причем выделена чистая культура сульфатредуктора, использующего
нафталин. Модельным объектом служат денитрификаторы. Воз-
Возможным акцептором служит Fe(III), имеющее промежуточный
окислительно-восстановительный (О-В) потенциал. Катехин, быст-
быстро разрушаемый аэробами, медленнее разлагается в анаэробных
условиях.
1
5 Schink В., Philipp В., Midler J. Anaerobic degradation of phenolic compounds //
Naturwissenschaften, 2000. Bd. 87. P. 12-23.
281
Анаэробное разложение фенола и гидроксибензоатов было ис-
исследовано Г. Фуксом на примере денитрифицирующей бактерии
Thauera aromatica. Начальным этапом служит фосфорилирование
оксигруппы неизвестным донором. Далее происходит дефосфорили-
рование с карбоксилированием в пара-положении и затем присоеди-
присоединение КоА. После образования бензоил-КоА ароматическое кольцо
разрывается путем восстановления с переносом 4-х электронов, воз-
возможно, с промежуточным образованием радикала. Реакция требует
затраты двух молекул АТФ на каждый электрон, участвующий в ре-
реакции восстановления. Затраты компенсируются при последующем
окислении образующихся карбоновых кислот, например при денит-
рификации.
Гидрохинон деградируется сульфатвосстанавливающими бакте-
бактериями Desulfococcus и бродилыциками Syntrophus gentiane через об-
образование гентизата в результате карбоксилирования ароматиче-
ароматического кольца и затем этерификации карбоксила КоА, за которой
следует бензоил-КоА путь. Промежуточным соединением при дру-
другом пути разложения ароматических соединений служит гидрокси-
гидрохинон, образующийся путем гидроксилирования и декарбокси-
лирования исходных веществ.
В целом можно сказать, что анаэробное разложение ароматиче-
ароматических соединений происходит по-разному и включает разные ступени
подготовительного метаболизма, связанные с затратой энергии.
Критическим является О-В потенциал и термодинамика общего
процесса, зависящая от акцептора электрона для завершающих ре-
реакций катаболизма.
7.6.2. Гуматы как субстраты микробного метаболизма
Гуминовые соединения, несмотря на свое обилие в природных
условиях и очевидное участие в процессах трансформации органи-
органического вещества, о чем легко было заключить по исчезновению
их бурой окраски при оглеении и подзолообразовательном процес-
процессе, долгое время рассматривали как инертные конечные продукты
обмена с медленным разложением в аэробных условиях под дейст-
действием "автохтонной микрофлоры Виноградского", в основном в ре-
результате соокисления преимущественно артробактерами, корине-
формами, нокардиями. Эти представления резко изменились в ре-
результате работ Д. Ловли по обмену железа. Он и его последовате-
последователи установили, что гуматы могут участвовать в обмене в трех вари-
вариантах:
- в качестве переносчиков-комплексообразователей между не-
нерастворимыми окислами металлов, прежде всего железа, и бактери-
бактериями-восстановителями;
- в качестве акцепторов электрона при анаэробном "хиноно-
вом" дыхании;
282
- в качестве доноров электрона для анаэробного дыхания6.
Все эти возможности осуществляются довольно широким кру-
кругом организмов, из которых следует выделить две группы: во-пер-
во-первых, организмов, близких к сульфатредукторам, как Geobacter, и,
во-вторых, организмов, близких к энтеробактериям и осуществля-
осуществляющих смешанное брожение. Универсальным механизмом, опреде-
определяющим возможность использования гуматов как электрон-
обменивающих компонентов, служит наличие в их структуре хи-
нонов, которые могут реагировать с экскретируемыми клеткой
хинонами, а те, в свою очередь, с хинон-содержащим участком це-
цепи переноса электрона клетки. Восстановление гуматов согласу-
согласуется с содержанием в них хинонов и происходит с образованием
радикалов. Цитохромы не принимают прямого участия в обмене с
окислами железа, и клетки нуждаются в гидрофобном взаимодей-
взаимодействии с частицами нерастворимых окислов. В результате оказыва-
оказывается, что способности восстанавливать гуматы и соединения Fe(III)
жестко коррелируют. Восстановленные гуматы реагируют с
Fe(III), и меру их окисления можно определить по образованию
Fe(II).
Вместо гуматов стал широко использоваться их плохо раствори-
растворимый химический аналог, антрахинон-2,6-дисульфонат (AQDS), кото-
который при восстановлении в AHQDS приобретает красно-коричневую
окраску, быстро исчезающую при окислении на воздухе; окраску
можно регистрировать спектрофотометрически при 450 нм. Работу
с AHQDS приходится вести в строго анаэробных условиях.
AQDS/AHQDS стала рутинной парой для имитации участия гумино-
вых соединений в окислительно-восстановительных реакциях. В хи-
ноновом дыхании промежуточным соединением при восстановлении
в гидрохинон служит семихинон. Показано, что в гуминовых соеди-
соединениях переход хинон/гидрохинон аналогичен переходу в модельном
соединении AQDS. Во всех случаях микроорганизмы, реагирующие
с AQDS, могли использовать и гуматы.
6 Lovley D.R., Phillips EJ. Novel mode of microbial energy metabolism: Organic carbon
oxidation coupled to dissimilatory reduction of iron and manganese // Appl. Environ.
Microbiol. 1988. Vol. 54. P. 1472-1480; Lovley D.R., Coates J.D., Blunt-Harris E.L.
et al. Humic substances as electron acceptors for microbial respiration // Nature. 1996.
Vol. 382. P. 445-448; Lovley D.R., FragaJL., Coates J.D., Blunt-Harris E.L. Humics
as an electron donor for anaerobic respiration // Environ. Microbiol. 1999. Vol. 1.
P. 89-98; Benz M., Schink В., Brune A. Humic acid reduction by Propionibacterium
freudenreichii and other fermentating bacteria // Appl. Environ. Microbiol., 1998.
Vol. 64. P. 4507^4512; Coates J.D. et al. Recovery of humic-reducing bacteria from a
diversity of environments // Ibid. P. 1504-1509; Scott D., McKnight DM., Blunt-
Harris E.L. et al. Quinone moities act as electron acceptors in the reduction of humic
substances by humic-reducing microorganisms // Environ Sci. and Technol. 1998.
Vol. 32. P. 2984-2989; Newmann D.K., Kolter R. A role of excreted quinones in the
extracellular electron transfer // Nature. 2000. Vol. 405. P. 94-97.
283
Восстановленный AHQDS может биологически окисляться в
анаэробных условиях нитратом или фумаратом аммоний-образую-
аммоний-образующими организмами, как Geobacter или Wolinella:
4 AHQDS + NOJ + 2Н+ = 4 AQDS + NHj + 3H2O.
Способность окислять AHQDS нитратом с продукцией N2 свой-
свойственна не всем денитрификаторам, хотя некоторые типичные де-
нитрификаторы осуществляют эту реакцию. В качестве акцепторов
могут служить также арсенат и селенат. В концентрации 0,1 мМ
AHQDS может служить переносчиком восстановителя между орга-
организмами, окисляющими ацетат с ним как акцептором (Geobacter), и
организмами, использующими его как донор электронов (Wolinella).
Способность восстанавливать внеклеточные хиноны была обнару-
обнаружена у всех гипертермофилов7. Само по себе восстановление AQDS
значит немногим более, чем восстановление метиленовой синей или
тетразолия в культурах микроорганизмов. Это в большей или мень-
меньшей степени универсальная реакция микроорганизмов. Важна воз-
возможность перенести эти наблюдения на природные условия, т.е.
проследить за динамикой гумусовых соединений8.
Тем не менее работы Д. Ловли установили существование окис-
окислительно-восстановительного цикла гуматов, в котором они служат
не единственным источником энергии для микроорганизмов, но ско-
скорее посредниками между ними и нерастворимыми восстановителями
и окислителями в почве. Благодаря быстрому окислению на воздухе
они становятся хорошим посредником между анаэробной микро-
микрофлорой затопленных почв и аэрируемыми слоями почвенного про-
профиля. Кроме гуматов в природе на роль окислительно-восстанови-
окислительно-восстановительных переносчиков могут претендовать и другие фенольные со-
соединения. При этом имеют значение не только собственно биологи-
биологические реакции, но и чисто химические взаимодействия, особенно с
нерастворимыми компонентами.
Хиноны в гуматах могут служить окислительно-восстановитель-
окислительно-восстановительными переносчиками и в химических реакциях при выделении Н2
гидрогеназой и, таким образом, снимать ингибирующий эффект при
синтрофном окислении несбраживаемых соединений. Внеклеточ-
Внеклеточные хиноны могут также служить и защитным механизмом для ана-
анаэробов против О2. Работа по окислительно-восстановительному ци-
циклу гуматов дает ключ к универсальному биогеохимическому меха-
механизму в богатых органическим веществом условиях переходной зо-
зоны увлажненных почв.
7 Lovley D.L., Kashefi К., Vargas M. et al. Reduction of humic substances and Fe(III)
by hyperthermophilic microorganisms // Chem. Geol. 2000. Vol. 169. P. 289-298.
8 Cervantes F.J., Van der Velde S., Lettinga C, Field J.A. Competition between
methanogenesis and quinone respiration for ecologically important substances //
FEMS Microbiol. Ecol. 2000. Vol. 34, N 2. P. 161-172.
284
1
Отсюда оказывается возможным выявить роль анаэробных "ав-
"автохтонных" организмов в сложной системе микробного сообщества,
устанавливающих химический шунт с первичными анаэробами и,
возможно, с синтрофной группировкой.
7.7. ОРГАНО-МИНЕРАЛЬНЫЕ СОЕДИНЕНИЯ В ПОЧВЕ
На поверхности глинистых частиц почвы происходит связыва-
связывание органических веществ. Особое значение имеет прочное взаимо-
взаимодействие гуминовых веществ с глинистыми минералами (рис. 7.6).
Во взаимодействии с отрицательно заряженными минералами уча-
участвуют аминогруппы, образующие ионную связь, металл-органиче-
металл-органические комплексы многовалентных металлов, как Fe, образующие
мостик между отрицательно заряженной поверхностью глинистого
минерала и карбоксильной группой гуминовой кислоты. Водород-
Водородные связи определяют связывание полимеров, например реакцион-
реакционно-способной свежеобразованной бактериальной слизи, с поверхно-
поверхностью глинистых частиц, и поэтому биологическая активность в почве
приводит к образованию стойких органо-минеральных соединений,
чего не происходит, например, при смешении торфа с глиной. Орга-
Органические соединения на поверхности глин защищены как от химиче-
химического, так и биотического окисления. Попавшие в межслойное про-
пространство даже такие легкодоступные вещества, как пептиды, ока-
оказываются недоступными для окисления. В крайнем выражении это
ведет к образованию черных глинистых сланцев, вплоть до горю-
горючих. С другой стороны, образование органического покрова на по-
поверхности минерала предотвращает его дальнейшее разрушение
при выветривании. Органические вещества, связываясь с Fe или А1
на поверхности кристалла, могут блокировать точки роста и предот-
предотвращать укрупнение педогенных минералов.
Отсюда возникает связь между образованием глинистых мине-
минералов при выветривании и геологическим захоронением Сорг, в свою
очередь, ведущим к повышению окислительного статуса атмосфе-
атмосферы. В результате за О2 атмосферы отвечает выветривание с образо-
образованием в конечном итоге глин, связывающих восстановленный Сорг
с эквивалентным освобождением О2.
7.8. МИНЕРАЛЬНЫЙ СОСТАВ ПОЧВЫ
Минеральный состав почвы формируется из унаследованных из
материнской породы литогенных минералов и вновь образованных
педогенных минералов, важнейшими из которых являются глины.
Физическое выветривание ведет к раздроблению исходной породы и
образованию песка, состоящего из кварца и полевых шпатов преи-
преимущественно. Кварц частично литогенный, а частично - образован
285
й
CM
00
СТ
I
I
в
о
с
I
S
«
X
К
S
I
1
о*
ю
о
о
В"
1
в
i
а.
в результате выветривания как его конечный продукт. Песчаные
почвы образуются при отложении песка из водной среды как мор-
морские донные осадки, речные и озерные, флювиогляциальные и со-
состоят почти на 90% из кварца. Основные породы, известковые, гли-
глинистые, лёсс содержат мало кварца. Полевые шпаты состоят на
80-90% из щелочных шпатов, и их содержание уменьшается с увели-
увеличением времени выветривания и переходом к мелким фракциям.
Слюды оказываются в средней фракции, предшествующей глинам.
Минеральный состав почвы формируется в результате выветри-
выветривания, под которым понимается трансформация минералов и пород
на поверхности Земли. В выветривании участвуют физические, хи-
химические, биологические процессы.
Физическое выветривание ведет к измельчению породы при ме-
механическом воздействии. Давление ведет к растрескиванию породы;
ветер с песком приводит к истиранию; сезонные и суточные перепа-
¦' ды температуры - к разному расширению зерен минералов и воз-
возникновению мономинеральной смеси, а прогрев поверхности - к от-
отслаиванию поверхностных слоев; замерзание льда в трещинах ведет
к давлению до 200 МПа; кристаллизация, например, гипса в трещинах
известняка разрывает породу; давление корней и гиф расширяет
трещины. При этом в ранее монолитной породе образуется поровое
пространство, заполняемое влагой и представляющее микрообита-
микрообитание для митофильных микроорганизмов.
Химическое выветривание ведет к трансформации минералов
вследствие реакций на их поверхности, и поэтому величина поверх-
поверхности имеет первостепенное значение. Ключевую роль играет пере-
переход элементов из кристаллической решетки в раствор с последующим
возможным образованием педогенных минералов, из которых важ-
важнейшими являются глины, карбонаты, окислы. Химическое выветри-
выветривание обусловлено гидролизом, при котором происходит замена кати-
катионов металлов породообразующих минералов на протоны; реакция
ускоряется в растворах минеральных и органических кислот:
_Si_O-Me + Н+ = -Si-0-Н + Ме+,
-С-О-Ме + Н+ = -С-О-Н + Ме+.
Поскольку кислоты являются важнейшим продуктом обмена
микроорганизмов, их жизнедеятельность приводит к сильнейшему
ускорению выветривания. Конечным продуктом разложения орга-
органического вещества является углекислота, и при повышении ее кон-
концентрации особенно возрастает растворимость таких карбонатных
пород, как известняки и труднорастворимые доломиты. Поэтому
все органотрофы являются мощными агентами выветривания. Суб-
Субстратом кислотного выветривания служат полевые шпаты (рис. 7.7),
которые передают в раствор щелочные металлы. Порошки поле-
полевых шпатов вследствие обмена Na+ или К+ на Н+ дают щелочную
287
Ортоклаз
KAlSi3Og
К
Na "
NaAlSi3Og
Альбит
Ca
[H2O + CO2]
CaAl2Si208
— Анортит
Na+
+
(CO2 + H2O)
NaHCO3 + H+
Глины
Al2Si205@HL
Каолинит
Al2Si4O10(OHJ
Монтморилонит
Ca2+
+
BCO2 + 2H2O)
Ca(HCO3J + 2H+
I
CaCO3 + CO2 + H20
2NaAlSi3Og + 2H2O + 2CO2 = Al2Si4O10(OHJ + 2Na+ + 2HCO3-+ 2SiO3
2NaAlSi3O8 + 2H+ + 9H2O = Al2Si505@HL + 2Na+ + 4H4Si04
Рис. 7.7. Углекислотное выветривание полевых шпатов
В качестве выщелачивающего агента вслед за углекислотой из
разлагаемой мортмассы могут выступать соединения азота. После
аммонификации они подвергаются бактериальной нитрификации, и
образующаяся азотная кислота нейтрализуется катионами материн-
материнской породы. Особенно сильно такое выщелачивание наблюдается
на известняках в условиях избыточного поступления азота в виде
продуктов жизнедеятельности животных. Однако усвоение нитра-
нитратов растениями или разложение в анаэробной зоне денитрификато-
рами ведет к отложению кальцита.
Специальную роль играет химическое выветривание в присутст-
присутствии сульфидных минералов, служащих субстратом питания для
группы тионовых бактерий. В результате прямого биологического
воздействия на сульфидные минералы образуется серная кислота,
которая ведет к сернокислотному выщелачиванию с образованием
обеленных пород, из которых вынесены все металлы и остается
лишь силикатный скелет или формируются глины. Одновременно с
окислением Fe и S здесь происходит и кислотный гидролиз. Реакция
окисления для пироксена имеет вид:
4CaFeSi2Q6 + 6Н2О + 8Н2СО3 + О2 =
= 4 Са(НСО3J + 4 FeOOH + 8 H2Si03.
Значительно реже наблюдается алкалинолиз, при котором вы-
выщелачивающим агентом служит ион ОН-, связывающийся с алюми-
алюминием:
К-шпат + НОН = Н-шпат + К+ + ОН-,
KAlSi3O8 + 8Н2О = А1(ОНK + 3H4Si04 + КОН.
Тенденция выветривания минералов следует из диаграммы их
' устойчивости. Эмпирически установлена следующая устойчивость к
выветриванию: оливин <гранат <пироксен омфибол <биотит пла-
плагиоклаз <ортоклаз <мусковит <кварц. Типичные минералы указы-
указывают на гумидность климата: для аридного - это гипс, кальцит, оли-
оливин; семиаридного - биотит, иллит, смектит; гумидного - хлорит и
каолинит; очень влажного - гиббсит, гематит, гетит, анатаз. Быст-
Быстрее всего выносятся Na, Ca, Mg, а К остается связанным в калиевых
шпатах и глинах. Из анионов выносятся хлориды и сульфаты, а
фосфаты связываются кальцием, алюминием, железом. Кремний
выносится с самого начала, но вынос его идет ступенчато с образо-
образованием сначала иллита и смектита, а затем обедненного Si каолини-
каолинита; в результате накапливаются окислы Al, Fe, Ti. Оценивается вы-
выветривание9 по выносу Na, К, Ca, Mg и составляет в умеренно влаж-
влажном климате 0,2-0,8 квал/(га • год). Крупнокристаллический гранит
подвергается глубокому физическому выветриванию, а базальты
более устойчивы, но по минеральному составу продукты разруше-
разрушения гранитов изменяются меньше, чем глубокоизмененные при вы-
выветривании минералы базальтов. Вулканические породы со стекла-
стеклами выветриваются легче.
7.9. КОРА ВЫВЕТРИВАНИЯ
При обсуждении микробных процессов в педосфере микробио-
микробиолог сталкивается с необходимостью понять, что такое педосфера
как среда обитания микроорганизмов. В нее включаются два разде-
раздела поверхности суши: 1) собственно почва и 2) кора выветривания,
где осуществляется взаимодействие с твердой минеральной фазой
воды, почвенного воздуха и газов, микроорганизмов. Обычно преи-
'Scheffer / Schachtschabel Lehrbuch der Bodenkunde 13 durchgesehene Auflage von
P. Schachschabel et al. Stuttgart: Enke, 1992. S. 19.
288
10. Г. А. Заварзин
289
мущественное внимание придается границе почвы и коры выветри-
выветривания (или подпочвы, грунта) в профиле, в то время как особенно-
особенности латеральных потоков почвенного раствора и твердого вещества
представляются менее важными.
Главным свойством собственно почвы как среды обитания мож-
можно считать наличие процессов первичной продукции, обусловлен-
обусловленных деятельностью фотоавтотрофных организмов, представленных
начиная с девона растительным покровом сосудистых растений.
Этот покров не только обеспечивает доминирование в почве цикла
органического углерода как ведущего процесса, но и особенности
водного цикла с эвапотранспирацией и роль корневой системы как
структурного фактора. В педосфере, предшествовавшей развитию
растений и почвы в ее современном понимании, ту же роль выпол-
выполняли мхи, лишайники, альго- и циано-бактериальные сообщества.
Продукционная функция почвы обозначается словом "плодородие",
к сожалению, слишком ассоциирующимся с сельскохозяйственными
аспектами. Фотоавтотрофия определяет расположение почвы на ос-
освещаемой "дневной поверхности".
Кора выветривания для микробиолога представляет область
преимущественно деструкционных процессов, поскольку автотроф-
ный синтез органического вещества играет несущественную роль.
Геологи здесь придают преимущественное значение абиотическим
процессам химического выветривания, обусловленного выщелачи-
выщелачиванием и переосаждением минералов циркулирующими растворами
с главной ролью растворенной СО2 как летучей кислоты. Против
такого представления энергично протестовал Б.Б. Полынов считав-
считавший, что "формирование остаточной коры выветривания (элювия)
нельзя представлять себе как процесс абиотический" (Полынов,
1956, с. 455). Кора выветривания располагается под почвой как ис-
источником органических веществ, в особенности устойчивых типа гу-
гумуса, и поэтому Полынов в 1950 г. заявил: "Итак, я утверждаю, что
элювий, как и другие формы коры выветривания, не является ре-
результатом только абиотических процессов и в своем формировании
находится в зависимости от процессов почвообразования" (Там же).
Формирование педосферы слагается из двух процессов: с одной
стороны, новообразование органического вещества продуцентами и
преобразование его с сопутствующей трансформацией минераль-
минеральных компонентов, а с другой стороны, биогенное выветривание, ко-
которое влечет за собой образование остаточных и преобразованных
минеральных компонентов материнской породы. Почва составляет
тонкий каталитический слой на поверхности, от которого действие
распространяется вглубь.
Является ли почва новообразованием или преобразованием по-
породы? Крайним примером новообразования может служить накоп-
накопление торфа в автономных ландшафтах, каковы верховые болота,
или органогенного горизонта в тундре, или мелкозема под лишайни-
290
ками, где роль исходных минеральных компонентов минимальна.
Другую крайность представляют песчаные почвы, где основу соста-
составляет мелкораздробленная материнская порода. Еще более вырази-
выразительным примером могут служить подзолы, где на первый план вы-
выходит преобразование материнской породы. Между этими крайними
примерами располагаются различные варианты почв. В общем сле-
следует помнить, что "почва растет вверх", и в этом смысле является
новообразованием. В росте вверх участвует не только образование
органического вещества, но и латеральный перенос, в первую оче-
очередь ветровой. Вынос приводит к выравниванию рельефа за счет де-
денудации, эрозии, но не за счет почвообразования.
Конечным результатом выветривания в пределах педосферы
можно считать преобразование каркасных силикатов изверженных
пород в слоистые силикаты глин, с одной стороны, а с другой - обра-
образование осадочных карбонатов не столько в профиле выветривания,
сколько в бассейне седиментации:
Изверженные породы —
Глины
Карбонаты
Общепринято объяснение происхождения глин как продукта хи-
химического выветривания минералов типа полевых шпатов и слюд.
При этом глубокие части коры выветривания сохраняют структуру
и строение коренных пород, хотя первичные минералы полностью
превращаются в глинистые. Особенно наглядно это преобразование
в вулканических областях с серно-кислотным или гидротермальным
выветриванием. Однако на самом деле основную трудность предста-
представляет перестройка кристаллической структуры алюмосиликатов.
Уже в 1934 г. в книге "Кора выветривания" Б.Б. Полынов свел
все случаи образования в зоне выветривания и гидросфере углекис-
углекислых солей в две общие категории: 1) образование карбонатов путем
непосредственного действия СО2 на горные породы и минералы по
реакции RSiO3 + СО2 + Н2О = RCO3 + SiO2 + Н2О и 2) образование
карбонатов в реакции солей СаС12 + Na2CO3 = СаСО3 + 2NaCl. На са-
самом деле процесс A) не так прямолинеен и включает микробное
разложение мортмассы с образованием СО2 и органических кислот,
которые служат мощным выщелачивающим агентом; разложение
солей Са и кислот ведет к образованию карбонатов. Точно так же
процесс B) включает транспорт и смешение потоков Са2+ и СО3
или же дегазацию раствора с бикарбонатом.
Основное внимание уделялось тому, что Б.Б. Полынов называл
ортоэлювиальным процессом с образованием элювия на месте об-
образования на водоразделе в автономном ландшафте из изверженных
пород, что легко наблюдать в горных областях. Для территории
291
России гораздо более значимы процессы параэлювиалъные, идущие
при преобразовании в почвы осадочных пород большей частью мор-
морского происхождения, в первую очередь глин, а также известняков.
Для европейской части особое значение имеет преобразование
флювиогляциальных отложений в виде песков, а для аридной зоны -
лёссов как эоловых отложений. Поскольку осадочные породы уже
подверглись глубокой химической переработке, то закономерности
их выветривания иные. Для известняков это повторный цикл выще-
выщелачивания с карстовыми явлениями, но и с нарастанием защищаю-
защищающего карбонаты от вымывания слоя почвы с восходящим потоком
эвапотранспирации. Для глин - взаимодействие с устойчивым орга-
органическим веществом гумуса и образование способных к миграции
комплексов с возрастающей ролью коллоидных процессов.
В связи с развитыми Б.Б. Полыновым и его учениками, в пер-
первую очередь А.И. Перельманом, М.А. Глазовской, представлениями
о геохимии ландшафтов, следует сказать, что главным в них послу-
послужила геохимия элементов на основе валового анализа образцов;
минералогия оказалась совсем в тени, а органическая геохимия прак-
практически не используется. Поэтому для микробиолога, заинтересо-
заинтересованного как в преобразовании минералов, так и в превращении
органического вещества, эти представления могут служить лишь от-
отправной точкой исследования. <
7.10. ПАЛЕОПОЧВЫ
Термин "почва" в русской литературе не соответствует термину
"soil" в англоязычной, куда попадают и грунт, и так называемые
"коры выветривания". Тем не менее и в английской литературе гео-
геологи подразумевают под нижней границей почвы область обильно-
обильного распространения жизни, исключая из этого "подземную биосфе-
биосферу" пористых горных пород. Для абиогенных продуктов деградации
пород употребляется термин "реголит", как для лунных минералов.
В привычном смысле почва - это корнеобитаемый слой наземной
поверхности, но такое представление может рассматриваться толь-
только с конца силура. До этого времени идут процессы выветривания и
выщелачивания вне воздействия растений, но под воздействием
микробиоты. Можно полагать, что биологические процессы на по-
поверхности суши были обусловлены альго-бактериальными сообще-
сообществами или же, позднее, включали и сообщества грибов и водорос-
водорослей. Почва в этом смысле отличается от грунта, который может
образовываться из осадков в депрессиях. Поэтому изучение погре-
погребенных поверхностных слоев суши докембрия, точнее "досилура",
может дать представление о преимущественном вкладе атмосфер-
но-абиогенных процессов в формирование поверхностных слоев су-
суши. При этом не следует, однако, забывать, что циано-бактериаль-
ные маты существовали на суше по меньшей мере уже 2 млрд лет
292
I
I
Щ
*
%
назад. Аналогами таких сообществ могут служить современные со-
сообщества пустынь, такыров, соров, где доминируют сообщества
низших растений.
Почвы, не имеющие растительного покрова, рассматриваются
как области первичного почвообразовательного процесса.
Поскольку корнеобитаемый слой появился на Земле лишь
~ 300 млн лет назад, а 3 млрд лет развития биоты прошло без сосу-
сосудистых растений, то для рассмотрения докембрийских (точнее, до-
силурийских) "палеопочв" удобнее пользоваться термином педо-
сфера для обозначения поверхностного слоя земной поверхности,
находящегося под интенсивным воздействием биоты и приведше-
приведшего к формированию древних кор выветривания. Не следует вос-
воспринимать растительный покров как нечто "напочвенное" с ли-
листьевым опадом, без мощной транспортной системы сосудистых
растений. Корневая система создала не только транспорт органи-
органики вглубь почвы за счет корневых выделений и мортмассы корне-
корневых остатков, но и сформировала ток почвенного раствора за счет
эвапотранспирации из глубины почвы, в отличие от испарения с
поверхности в отсутствие корней. Для предшествующих этапов
развития приходится принимать как аналог эфемерный покров на
грунте современных почвенных водорослей и циано-бактериаль-
ных матов.
В геологическом масштабе времени необходимо думать о после-
последовательном возникновении в истории Земли накладывавшихся
друг на друга наземных биоценозов и общих для них биогеохимиче-
биогеохимических процессах:
- циано-бактериальном - с начала появления наземной биоты;
- альго-бактериальном - не позднее 1 млрд лет назад;
- альго-грибном (возможно, также лихенизированном) с первы-
первыми гифоподобными микрофоссилиями - около 1 млрд лет назад
(Мейен, 1987);
-растительно-грибном, начавшемся примерно 300 млн лет назад
с образованием почвы в ее современном понимании с корневой сис-
системой растительного покрова, причем, возможно, ранее началась
биотурбация, обусловленная почвенными животными в альго-бак-
териальных матах.
В этих сообществах представлены как продукционная, так и де-
струкционная ветви углеродного цикла. Перечисленные этапы мо-
могут быть соотнесены с этапами эволюции наземного мира от прока-
риотных обитателей увлажненной поверхности Земли до появления
Paleotrachites как древесно-микоризной системы в позднем силуре с
началом "макро-палеоботаники" и эволюцией сосудистых растений.
Сукцессия на минеральной педосфере при заселении ее живыми
организмами представляет приблизительный аналог биогенетиче-
биогенетического закона К. Бэра, конечно понимаемый не слишком буквально,
поскольку сосудистые растения успешно могут быть первопоселен-
293
цами, развиваясь на минеральных породах и не нуждаясь в этом от-
отношении в предшественниках.
В интерпретации процессов на суше следует различать два пери-
периода: 1) примерно до 2 млрд лет назад в атмосфере, лишенной кисло-
кислорода, и 2) после начала накопления кислорода в атмосфере и распро-
распространения красноцветных пород. Корни растений появляются с де-
девона, в силуре есть возможные следы ризоморф, в позднем ордови-
ордовике есть ходы животных, вероятно, питавшихся обильной раститель-
растительной пищей.
Палеопочвы докембрия были идентифицированы Г. Реталла-
ком10. Докембрийские палеопочвы имеют такую же последователь-
последовательность горизонтов в цвете, текстуре, минералогии, химическом соста-
составе, как и современные. Есть примеры накопления глин, карбонатов,
гипса, аморфного кремния, марганца. Источником ошибок при ин-
интерпретации погребенных горизонтов как палеопочв может быть их
сходство с гидротермическими изменениями пород. Горизонты па-
палеопочв могут быть сопоставлены с современной номенклатурой:
органогенный О-горизонт на поверхности, обогащенный торфом;
элювиальный Е-горизонт, обогащенный кварцем; Bt-горизонт, обо-
обогащенный кварцем или биотитом; или Bs-горизонт с полуторными
окислами железа; или Bk-горизонт. Наиболее характерным призна-
признаком палеопочв служат педогенные глины. Криогенные структуры
просматриваются в палеопочвах 2,3 млрд лет в виде клиньев, запол-
заполненных песком.
Таким образом, профиль палеопочв или кор выветривания
очень сходен с тем, что наблюдается для минеральных компонент
современных почв. Под вопросом остается степень участия в преоб-
преобразовании выветрелой породы в почву выщелачивания органиче-
органическими веществами, продуцируемыми биотой.
Первичный почвообразовательный процесс привлекал внима-
внимание почвоведов, и его легко можно наблюдать на обнажениях скал,
где развивается классическая сукцессия от первопоселенцев мик-
микробного сообщества с водорослями и цианобактериями как проду-
продуцентами к лишайникам, накапливающим грубый гумус или торф в
трещинах и под коркой, и затем к высшим растениям. Эту последо-
последовательность можно проследить во всех климатических поясах от Ка-
Карелии до Монголии. Следует заметить, что заселение рыхлой вулка-
вулканической пирокластики сейчас идет иначе, начинаясь с высших рас-
растений. В то же время на пенеплене - выровненом снесенном матери-
материале в Закаспии - развиваются сообщества такыров или соров (сабх-
в английской литературе), более всего напоминающие циано-бакте-
риальные маты.
10 Retallack GJ. How to find a Precambrian paleosol // Early organic evolution:
Impications for mineral and energy resources / Ed. M. Schidlowski et al. N.Y.:
Springer, 1990.
294
Рекомендуемая литература
1. Бабьева И.П., Зенова Г.М. Биология почв. М.: Изд-во МГУ, 1989. 336 с.
2. Виноградский С.Н, Микробиология почвы. М.: Изд-во АН СССР, 1952.
З.Добровольская Т.Г. Структура бактериальных сообществ почв. М.: ИКЦ
"Академкнига", 2002. 282 с.
4. Дыхание почвы: Сб. науч. тр. Пущино, 1993. 144 с.
5. Звягинцев Д.Г. Почва и микроорганизмы. М.: Изд-во МГУ, 1987. 256 с.
6. Зонн СВ. История почвоведения России в XX в.: Неизвестные и забытые
страницы: В 2 т. М.: Ин-т географии РАН, 1999.
7. Ммшенецкий А.А. Микробиология целлюлозы. М.: Изд-во АН СССР, 1953.
440с.
8. Красильников И. А. Микроорганизмы почвы и высшие растения. М.: Изд-во
АН СССР, 1958. 464 с.
9. Мейен СВ. Основы палеоботаники, 1987.
10. Научные основы устойчивости лесов к дереворазрушающим грибам. М.:
Наука, 1992. 222 с.
11. Оллиер К. Выветривание. М.: Недра, 1987.
12. Орлов Д.С Химия почв. М.: Изд-во МГУ, 1985.
13. Орлов Д.С. Запасы, поступление и круговорот органического углерода в поч-
почвах России // Круговорот углерода на территории России. М., 1999. С. 271-299.
14. Орлов Д.С, Бирюкова О.Н., Суханова Н.И. Органическое вещество почв
Российской Федерации. М.: Наука, 1996. 254 с.
15. Паников Н.С Кинетика роста микроорганизмов. М.: Наука, 1991.
16. Перельман А.И., Касимов Н.С. Геохимия ландшафта. М.: Астрея-2000, 1999.
768 с.
17. Печуркин Н.С. Популяционная микробиология. М.: Наука, 1978.
18. Полынов Б.Б. Избранные труды. М.; Л.: Изд-во АН СССР, 1956. 751 с.
19. Рабинович МЛ., Болобова А.В., Кондращенко В.И. Теоретические основы
биотехнологии древесных композитов / Отв. ред. A.M. Безбородов. Кн. 1.
Древесина и разрушающие ее грибы. М.: Наука, 2001. 264 с.
20. Решетникова И.А. Деструкция лигнина ксилотрофными макромицетами.
М.: Изд-во МГУ, 1997. 197 с.
21. Рипачек В. Биология дереворазрушающих грибов. М.: Лес. пром-ость, 1967.
258 с.
22. Структурно-функциональная роль почвы в биосфере. М.: ГЕОС, 1999.
278 с.
23. Barker W.W., Welch S.A., Banfield J. Biogeochemical weathering of silicate mine-
minerals // Rev. in Miner. Geomicrobiology. 1997. Vol. 35. P. 391-^428.
24. Colberg PJ. Anaerobic microbial degradation of cellulose, lignin, oligolignols, and
monoaromatic lignin derivatives // Biology of anaerobic organisms / Ed.
A.J. Zehnder. N.Y.: Wiley, 1988. P. 333-372.
25. Coates J.D. et al. Recovery of humic-reducing bacteria from a diversity of environ-
environments // Appl. Environ. Microbiol. 1998. Vol. 64. P. 1504-1509.
26. Hattori T. Soil aggregates as microhabitats for microorganisms // Rep. Inst Agr. Res.
Tohoku Univ. 1988.Vol. 37. P. 23-26.
27. Holmen K. The global carbon cycle // Global biogeochemical cycles / Ed.
S.S. Butcher et al. Acad. Press. 1992. P. 239-263. Repr., 1994: 1998.
28. Krauskopf K.B., Bird D.K. Introduction to geochemistry. 3rd ed., N.Y.: McGraw-
Hill, 1995.
29. Lovley D.R., Phillips EJ. Novel mode of microbial energy metabolism: Organic car-
carbon oxidation coupled to dissimilatory reduction of iron and manganese // Appl.
Environ. Microbiol. 1988. Vol. 54. P. 1472-1480.
295
30. Lovley DM., Coates J.D., Blunt-Harris E.L. et al. Humic substances as electron
acceptors for microbial respiration // Nature. 1996. Vol. 382. P. 445^48.
31. Lovley D.R., Fraga J.L., Coates J.D., Blunt-Harris E.L. Humics as an electron donor
for anaerobic respiration // Environ. Microbiol. 1999. Vol. 1. P. 89-98.
32. Paul E.A., Clark F.E. Soil microbiology and biochemistry. San Diego, etc.: Acad
press, 1989.274 р.
33. Schink В., Philipp В., Muller J. Anaerobic degradation of phenolic compounds //
Naturwissenschaften. 2000. Vol. 87. P. 12-23. •
34. Schwamman D.W. Life.temperature, and the Earth: The self organizing biosphere.
N.Y.: Columbia Univ. press, 1999.
35. Schwamman D.W., Volk T. Biotic enhancement of weathering and the habitability
of the Earth // Nature. 1989. Vol. 340. P. 457-459.
36. The global carbon cycle / Eds. B. Bolin et al. N.Y.: J. Wiley, (SCOPE-13). 1979.
37. White A.F., Brantley S.L. Chemical weathering of silicate minerals: An overview//
Rev. Miner. 1995. Vol. 31. P. 11-22.
Лекция 8
ИСТОРИЧЕСКАЯ МИКРОБИОЛОГИЯ
8.1. РОЛЬ ПРОКАРИОТНОГО МИРА
В СТАНОВЛЕНИИ БИОСФЕРЫ
Для оценки роли разных групп организмов в биосфере природа
поставила гигантский эксперимент. Современная Земля с ее расти-
растительным покровом и животным миром представляет заключитель-
заключительный этап развития, который сложился только после длительной,
предшествовавшей истории биосферы. Формирование мира назем-
наземных растений относится к силуру примерно 420 млн лет назад, а ми-
мира многоклеточных животных - к венду около 600 млн лет назад.
Все эти сложные и разнообразные живые существа смогли сущест-
существовать лишь в уже сложившейся биосфере, обязанной своим проис-
происхождением микроскопическим организмам. Отсюда история био-
биосферы - это в подавляющей степени история микробов и того, как
они создали биосферу, в которой появились более сложно организо-
организованные существа. Последние трансформировали течение биотиче-
биотических процессов на Земле, но не изменили суть системы. Поэтому, ре-
реконструируя жизнь докембрия, можно получить представление об
основе жизненной машины планеты, еще не заслоненной красками
цветов и движением мягких и пушистых животных, т.е. перейти от
видимости к сути. Базовая система процессов - скрытая основа су-
существа биосферы - сложена деятельностью "невидимых", и ее мож-
можно понять и реконструировать из знания прошлого, когда на нее не
накладывались и ее не изменяли последующие события, к которым
приспосабливался мир микробов.
Микропалеонтология дает последовательную картину развития
мира микроскопических существ примерно с 1,5 млрд лет назад
(рис. 8.1), когда появились относительно крупные протисты и био-
биосфера была в общем сходна с современной, хотя еще не было ни рас-
растительного покрова на суше, ни организмов со скелетом, сменив-
сменившим мягкотелых животных венда в кембрии.
Еще более ранние этапы относятся к эпохе существования толь-
только прокариот, которые одни ответственны за становление биосферы
как первое сообщество живых организмов. С начала появления жиз-
жизни на Земле прокариоты действовали в основном как катализаторы
297
Первичные
продуценты Fe Почва
Древнейшие Metazoa
Древнейшие Protista
Строматолиты
Автотрофы по 812С/13С
Рис. 8.1. Важнейшие биотические события в истории Земли
геохимических реакций, сопряженных в первую очередь с циклом
органического углерода, начинающегося деятельностью фотоавто-
трофных оксигенных цианобактерий. Именно этот первый доку-
документирующий этап и позволяет обнаженно выяснить пути формиро-
формирования биосферы. Здесь состояние геосферы задает первоначальные
условия, и биота лишь трансформирует протекание процессов, ма-
материальным источником для которых служит геосфера. Поэтому
геосфера первична, а биота вторична.
Роль прокариотного мира в становлении биосферы основывает-
основывается на том простом факте, что прокариоты были первыми обитате-
обитателями Земли. Их функциональное разнообразие сложило систему от-
отношений биоты и геосферы, в более узком смысле - литосферы.
Все остальные организмы вписывались в эту ранее сложившуюся
систему отношений. Они не могли противоречить ей и удержива-
298
лись только в том случае, если согласовывались с биогеохимической
системой прокариот, взаимодействующих с геосферой. На следую-
следующем этапе было возможно изменение ранее сложившейся системы
за счет ее новых компонентов, с преимущественным влиянием на
некоторые количественные потоки, но не принципиальным измене-
изменением существенных связей. Вопреки принятому представлению,
частное не может изменить общее, если оно первоначально не сог-
согласуется с ним, поскольку оно не может реализоваться. Поэтому
эволюция высших организмов вписывается в прокариотную био-
биосферу как среду своего обитания как в предшествующую данность.
Историю биоты на Земле можно представить в виде схемы ад-
аддитивной эволюции с этапами.
I. Прокариоты.
И. Прокариоты + протесты.
III. Прокариоты + протисты + многоклеточные.
Определяющими на каждом этапе являются первичные проду-
продуценты - оксигенные фотоавтотрофы:
1) цианобактерий,
2) цианобактерий + водоросли,
3) цианобактерий + водоросли + растения.
Поскольку они занимают одну и ту же нишу по отношению к
свету, которая ограничивается плотностью хлорофилла на единицу
обитаемой поверхности с самозатенением как верхним пределом, то
величина первичной продукции приблизительно постоянна в преде-
пределах пЮ2 Гт С/год, где п, судя по современной продукции, равно 1-2.
Вклад каждой группы фотоавтотрофов менялся в течение эволю-
эволюции и в зависимости от талассо- или теократических эпох и климата.
Выход растений в аэротоп - воздушную среду обитания - увеличил
обитаемую поверхность (рис. 8.2).
Цель этой лекции заключается в том, чтобы проследить на при-
примере реликтовых микробных сообществ, как менялось взаимодейст-
взаимодействие биогеохимических циклов в преимущественно прокариотный
период развития биосферы. Этот период начинается в архее с пер-
первым появлением биоты одновременно с началом геологической
%
ВоДа
Активный
микробный
мат
Осадок
Пластовые
Куполообразные
и желваковые
Столбчатые
Рис. 8.2. Предполагаемое чередование форм строматолитов по экотону
299
истории, но нет оснований полагать, что биота играла сколько-ни-
сколько-нибудь значительную роль в становлении природы Земли в этот преи-
преимущественно геологический период и тем более в период ее догео-
догеологической истории. Подлинное развитие прокариотной биосферы
начинается с позднего архея и в течение протерозоя привело к ста-
становлению всех биогеохимических циклов современного типа. Клю-
Ключевым событием следует признать переход к кислородной атмосфе-
атмосфере, происшедший в протерозое в период 2,2-2,0 млрд лет назад в ви-
виде анаэробно/аэробной революции и обусловленный сопряжением
биотических циклов Сорг и О2. В конце протерозоя появляются про-
тисты-эукариоты, заменяющие продуцентов-прокариот в ведущем
цикле органического углерода, но не изменяющие системы циклов.
Микробная биосфера занимает 3/4 истории Земли, а исключительно
прокариотная - больше половины. Поэтому бессмысленно пытать-
пытаться понять движущие силы геосферно-биосферной эволюции, исходя
из конечного состояния системы.
Из приведенной в начале книги схемы сопряжения биогеохими-
биогеохимических циклов следует, что СО2 участвует в двух циклах: органиче-
органического и неорганического углерода. Первый привел к формированию
оксической (кислородной) атмосферы за счет дисбаланса фотоавто-
трофной продукции и деструкции, второй явился основным резерву-
резервуаром депонирования углекислоты в виде карбонатов с эквивалент-
эквивалентным образованием выщелоченных пород главным образом в виде
глин. Фотоавтотрофы способны развиваться только на освещенной
("дневной") поверхности, и конечным результатом их деятельности
является замена части СО2 на О2. Неполное окисление Сорг ведет к
сохранению эквивалентной части О2 и переходу поверхностных обо-
оболочек в окисленное состояние. Углекислотное выветривание пород
может идти и под поверхностью с выносом прежде всего Na, Ca, Mg
подземными водами, и связыванием СО2 в карбонаты. Движущей
силой углекислотного выветривания является атмосферный гидро-
гидрологический цикл с испарением воды из океана, служащего конеч-
конечным резервуаром для континентального стока. Быстрое углекис-
углекислотное выветривание идет в гидротермальных условиях с трансфор-
трансформацией изверженных пород в глины. Отложившиеся в неглубоких
частях океана карбонаты становятся объектом геологического ре-
рецикла, обусловленного тектоникой. Современный рецикл карбонатов,
обусловленный эукариотной биотой океана, оценивается по времени
пребывания кальция в океане в 1 млн лет. Отложение карбонатов обу-
обусловливает нейтральную реакцию среды на Земле. В результате опре-
определяется господствующий ?h-pH режим планеты. Циклы органическо-
органического и неорганического углерода не полностью замкнуты, и это ведет к
биогеохимической сукцессии, являющейся основным механизмом эво-
эволюции геосферно-биосферной системы (Заварзин, 2000).
В эволюции геосферно-биосферной системы было несколько
переходных периодов. Важнейшим из них было формирование кис-
300
дородной атмосферы, отмечаемое по появлению красноцветных по-
пород субаэрального происхождения около 2 млрд лет назад и сокра-
сокращению образования окисленных железистых кварцитов. К этому же
переходному времени относится и время распространения серного
цикла в океане, датируемое по изотопам серы и развитию сульфат-
редукции, начиная с 2,7 млрд лет назад, предварительным условием
для которого служит достаточная концентрация сульфатов в мор-
морской воде и установление близкого к современному состава вод оке-
океана. В этот период на Земле биота была представлена прокариота-
прокариотами, включавшими цианобактерий и бактерий, или кратко - циано-
бактериальными сообществами.
Около 1 млрд лет назад произошло распространение эукариот-
ных организмов, прежде всего протистов, уже в оксической атмо-
атмосфере. Они сменили цианобактерий прежде всего как основная
группа фитопланктона океана. Затем с началом кембрия 0,6 млрд
лет назад у эукариот возник формируемый в компартментализован-
ной клетке скелет и произошла перестройка цикла кальция с обра-
образованием карбонатов скелетными рифостроителями. Около 0,3 млрд
лет назад появился растительный покров суши и возникла почва как
корнеобитаемый слой, сменив на суше кору выветривания.
Изложенная история событий показывает, что при обсуждении
эволюции следует исходить из большой системы прокариотной био-
биосферы, взаимодействующей с геосферой, в то время как попыт-
попытки понять эволюцию исходя из биологии позвоночных заведомо
неполны.
8.1.1. Границы приложимости актуалистического метода
Актуалистический подход ограничен сходством современной
обстановки с реконструируемой обстановкой прошлого. Для этого
могут быть использованы локальные явления и объекты, которые
мы считаем аналогичными имевшимся в прошлом. По современным
животным можно с определенной вероятностью реконструировать
вымершие организмы, относящиеся к тем же таксономическим
группам. Актуалистический подход перестает работать, когда усло-
условия совершенно отличны от современных. По-видимому, границу
актуалистического подхода приходится поставить в раннем архее в
"догеологической" истории Земли. Здесь действуют логические
конструкции и аналогии сравнительной планетологии.
Предполагается, что планеты земной группы Марс, Земля, Ве-
Венера образовались из одинакового исходного материала, но вследствие
разного положения относительно Солнца история их была различ-
различна. Марс утратил атмосферу, Венера сохранила плотную атмосферу
и утратила гидросферу, Земля сохранила и атмосферу и гидросферу.
В дегазации планет происходила потеря летучих компонентов, из
которых важны вода, углекислота, азот, инертные газы. Диссипация
301
водорода на Венере привела к потери ею воды и созданию углекис-
лотной атмосферы с мощным парниковым эффектом. Почему Зем-
Земля изменила углекислотную атмосферу и избежала "убегающего
парникового эффекта"?
Углекислота на Земле обнаруживается в осадочной оболочке в
виде карбонатов щелочноземельных металлов. Для их образования
необходимо освобождение металлов из изверженных пород основ-
основного состава. Такой процесс называется химическим выветривани-
выветриванием. Утратив металлы, эти породы превращаются в глины. Следова-
Следовательно, должен существовать баланс карбонатов и глин, причем
часть элементов, прежде всего Na и С1, оказывается в океане. Уда-
Удаление СО2 из атмосферы может происходить при контакте извер-
изверженных пород с атмосферой, т.е. субаэрально. Для реакции гидро-
гидролиза необходима вода. Поэтому местом реакции служит увлажнен-
увлажненная твердая поверхность в контакте с газовой фазой. Скорость реак-
реакции увеличивается с повышением температуры, давления, концент-
концентрации СО2. На современных вулканах пропариваемые породы в низ-
низкотемпературных гидротермах (мофетах) превращаются в глины. В
древнейших (более 3 млрд лет) сильно метаморфизированных поро-
породах Алданского щита присутствуют карбонаты в виде мраморов, и,
следовательно, процесс выветривания был интенсивным.
Отсюда ключевым явлением в ранней истории Земли оказыва-
оказывается вынос кальция и магния из изверженных пород. Образование
карбонатов определяет уход атмосферы из зоны "убегающего" пар-
парникового эффекта и близьнейтральную обстановку на поверхности
Земли. Трудность состоит в том, что металлы находятся в твердой
фазе, в то время как углекислота и хлор могут находиться и в газо-
газовой, и водной фазах, создавая исходно кислые условия. В какой ме-
мере химические процессы срединно-океанических хребтов с конвек-
конвекцией воды современного океана через нагретые породы могут вос-
воспроизводить обстановку ранней Земли? Во всяком случае, хемо-
трофные сообщества гипертермофильных микроорганизмов здесь
развиваются.
В современных условиях формирование карбонатов, главным об-
образом в виде СаСО3, в подавляющей степени идет биогенным путем
через образование скелетов морских организмов. Этот этап стал воз-
возможен только после неопротерозойской революции с началом кемб-
кембрия. Как происходило образование карбонатов в период господства
прокариот, когда не могло быть внутриклеточного концентрацион-
концентрационного механизма? Свидетельством биогенного образования карбона-
карбонатов Са и Mg служат доломитизированные толщи строматолитов. До-
Доломитизация указывает на повышенный рН и карбонатную щелоч-
щелочность, что не согласуется с высоким рСО2 в атмосфере. Происходил
ли этот процесс в океане или же в субаэральных условиях?
Рассматриваемые явления можно изобразить следующей схе-
схемой:
302
f
*
i
j
¦
f
t
1
%
i
f
i
%
i
i
Вода
CO2 атмосферы
Изверженные породы
Глины-
Карбонаты
Соденая вода моря
8.1.2. Геосфера и историческая геология
История Земли начинается с раннего после аккреции периода,
когда происходила дегазация планеты с образованием атмосферы,
гидросферы в виде протоокеана, первичной суши. За появлением
этих трех сред можно следить по осадочной летописи, которая начи-
начинается 3,8-3,9 млрд лет назад метаморфизированными осадочными
отложениями Исуа на юго-западе Гренландии, продолжается
3,5 млрд лет назад на северо-западе Австралии, а также сильно ме-
метаморфизированными породами Алданского щита. Все, что проис-
происходило ранее, находится в области реконструкции.
Биологи склонны придавать излишнее значение абсолютным
датировкам, не учитывая того, что эти датировки относятся к от-
отдельным образцам, а не ко всей поверхности Земли в целом. Корре-
Корреляция геологических событий по времени представляет специаль-
специальную задачу стратиграфии. Попытка оперировать "точными" датами
несостоятельна не только из-за неоднородности географической по-
поверхности в прошлом, как и в настоящем, но и из-за того, что син-
синхронизация событий ранней геологической истории пока возможна
лишь для интервалов во многие десятки и сотни миллионов лет.
Приводимая ниже очень беглая история становления геосферы
имеет своей целью прежде всего ограничить полет фантазии биоло-
биологов-экспериментаторов.
По сложившимся сейчас на основе сравнительной планетологии
представлениям Земля возникла 4,5-^,6 млрд лет назад, как это сле-
следует из изотопного возраста метеоритов, путем аккреции из раз-
раздробленного вещества. Свидетельства метеоритной бомбардировки
сохранились на Марсе, Меркурии и на Луне в виде кольцевых удар-
ударных кратеров. Судя по поверхности Луны, наиболее интенсивная ме-
метеоритная бомбардировка приходилась на период 4,2 млрд лет назад
и закончилась 3,9 млрд лет назад. Принимается, что планеты земной
группы - Меркурий, Земля, Марс - имеют холодное происхождение,
но в конце формирования они проходили стадию "океана магмы" с
расплавленным наружным слоем, перемешанным падающими пла-
нетезималями. На Земле следов метеоритной бомбардировки не со-
сохранилось. Период от 4,5 до 3,8 млрд лет реконструируется по изо-
303
топным соотношениям в магме и газах. Эти данные указывают на
дифференцировку 4,4-4,3 млрд лет назад с выделением основной
массы "летучих": паров воды и газов. Теплопотери в начале' были
многократно выше современных, и эндогенные процессы играли
первостепенную роль. Светимость Солнца была на 25-30% меньше
и возрастала примерно линейно. В тепловом режиме Земли боль-
большую роль мог играть парниковый эффект за счет паров воды и уг-
углекислоты1.
Предполагаемая последовательность термических событий на
Земле ограничивает время, предшествовавшее началу осадочной
геологической летописи, 3,8-3,9 млрд лет назад, вероятным разогре-
разогревом при метеоритной бомбардировке, несовместимым с обитаемо-
обитаемостью, хотя возможность холодных "островов" не исключена. С дру-
другой стороны, утверждения в находках бактериоморфных остатков в
некоторых метеоритах группы углистых хондритов с возрастом
4,5-^,6 млрд лет переводят вопрос о появлении жизни на Земле в об-
область космических пределов. Логические реконструкции биологов
с исходным организмом "прогенотом" следует оценивать скепти-
скептически.
При дегазации происходило быстрое, судя по моделям - в тече-
течение сотни миллионов лет, выделение основной массы "летучих", к
которым прежде всего относятся пары воды, азот, углекислота. От-
Относительно других компонентов, особенно восстановленных газов
аммиака, метана, сероводорода, твердого суждения нет. Считается,
что в первичной атмосфере отсутствовал кислород и она была ано-
ксической, хотя кислород был необходим уже для формирования
древнейших окислов железа в формации Исуа 3,8 млрд лет назад.
Дегазация с формированием гидросферы и первичной атмосферы
представляет главный механизм, который определил дальнейшее
взаимодействие биосферы и геосферы. Главным компонентом ат-
атмосферы, на который воздействовали биота и литосферные процес-
процессы, была углекислота. Ее удаление из атмосферы при возрастании
светимости Солнца обеспечило дальнейшее нахождение поверхно-
поверхности Земли в тепловом режиме обитаемости.
При таком масштабе времени необходимо учитывать независи-
независимую от биосферы геологическую эволюцию Земли, обусловленную
прежде всего ее энергетическим балансом, направленным на необ-
необратимую потерю внутренней энергии вследствие рассеяния в косми-
космическое пространство и приобретения энергии извне от Солнца.
Внешняя энергия обусловливает тепловую климатическую машину
и вызываемое ей выветривание и латеральное перемещение масс.
Необратимая геологическая эволюция Земли составляет ту среду, в
Ч \
1 Впервые на него указали Л.М. Мухин и В.И. Мороз. См.: Письма в "Астрон.
журн." 1977. Т. 3. С. 78; а также см.: Сорохтин ОТ., Ушаков С.А. Развитие
Земли. М.: Изд-во МГУ, 2002. 560 с.
304
которой происходит биологическая эволюция с ее обратными связя-
связями на геосферу в ответ на прямые воздействия извне.
Историческая геология сейчас может дать только некую общую
(с существенными пробелами) картину событий протерозоя и лишь
в очень гадательной форме - раннего архея (Хаин и др., 1997; Schopf,
1983; 1992; Schopf, Klein, 1992). Общая картина геологической эво-
эволюции оперирует крупными эпохами (сотни миллионов лет) и стре-
стремится дать планетарную обстановку рассматриваемого периода.
И временной, и пространственный масштабы подхода не соответст-
соответствуют запросам биолога. Если пользоваться аналогией с современ-
современной биосферой, то можно было бы утверждать, исходя из общих
соображений, что жизнь на Земле либо была невозможна, либо
должна быть совершенно иной. Средняя температура современной
Земли в 15° предполагает преобладание организмов, чувствитель-
чувствительных к повышению температуры. Все соображения, исходящие из
средних значений, очевидно неприемлемых при анализе современ-
современной биосферы, как "средняя температура по больнице", возникают в
сравнительной планетологии и по отношению к прошлому Земли.
Поэтому необходимо опираться на эмпирические данные, которые
исходно локальны. Для функционирующей биосферы в принципе
важна ее неоднородность, предполагающая перенос вещества между
регионами.
В отличие от геологии, рассматривающей последовательность
образования комплексов пород на основании разрезов, для биолога
важнее площадное распределение физико-географических условий
"географической оболочки Земли", входящих в понятие "палеогео-
"палеогеографии", реконструирующей ландшафтные обстановки геологиче-
геологического прошлого, включая климатические условия. Основой для ре-
реконструкции служит прежде всего фациальный анализ. При этом ус-
устанавливаются области сноса, континентального осадконакопления,
химический состав палеоводоемов и их осадков. О климате прошло-
прошлого можно судить по типам кор выветривания, зависящим от тепла и
влаги. В аридном климате образуются соли, гипс, седиментацион-
ные доломиты, а в гумидном - горючие сланцы. В тропической и
субтропической зоне создаются литологические формации: карбо-
карбонатные и гипсовые, эвапоритовые, карбонатно-сульфатные. В уме-
умеренно теплой зоне континентальные красноцветы теряют карбонат-
ность. Для реконструкции климата важен минералогический состав
аутигенных минералов в осадочных породах.
Обсуждение роли микробиоты в прошлом Земли, в том числе в
архее и протерозое, должно строиться на фациальной, а не плане-
планетарной основе. Фации представляют отражение ландшафтов в седи-
ментогенезе. По более точному определению, это комплекс отло-
отложений, отличающийся составом и физико-географическими услови-
условиями образования от соседних отложений того же времени. Какие
фации доминировали в ту или иную эпоху, насколько они сходны с
305
современными, или, точнее, насколько им аналогичны современ-
современные, составляет геологический аспект непрерывности развития био-
биосферы, поскольку организмы приспосабливаются к существенным
для них факторам ландшафта и лишь во вторую очередь, например,
к петрографическим особенностям пород. Тесная связь микробиоты
с геологической средой и ее участие в трансформации минералов от-
отличают палеомикробиологию от палеоэкологии фанерозоя, осно-
основанной на индикаторных организмах и их остатках. Микробная
жизнь находит наиболее богатые условия для себя на границе двух
сред, в области термодинамической метастабильности соединений,
транспортируемых из одной среды в другую, и формирования геохи-
геохимического барьера, часто совпадающего с экотоном. Гетероген-
Гетерогенность планетарной среды обитания есть непременное условие разви-
развития жизни.
Вместе с тем фации формировались под воздействием двух хо-
хорошо перемешанных и поэтому однородных сред: атмосферы и оке-
океана. Здесь не случайно вместо слова "гидросфера" употреблено сло-
слово "океан" как единая водная масса - Панталасса. Континентальные
воды не перемешаны, а очень разнообразны и локальны, географи-
географически изолированы. Отсюда они могут быть и источником разно-
разнообразия жизни, и рефугиумами для сообществ прошлого. Атмосфе-
Атмосфера и океан в течение истории Земли эволюционировали, меняя свой
состав с развитием вулканического и осадочного процессов, но глав-
главным образом под воздействием взаимосвязанных биогенных циклов
органического углерода-кислорода-углекислоты. Поэтому, напри-
например, прибрежные фации зоны седиментации в начале протерозоя и
в конце протерозоя оказываются в совершенно иных геохимических
обстановках.
Как уже упоминалось, микроорганизмы развиваются в области
термодинамической стабильности продукта реакции и метастабиль-
метастабильности субстратов-реактантов. В качестве контрпримера приводят
фотосинтез океана. Но фотосинтез обусловлен десятками-сотней
метров фотической зоны, поверхностным слоем, в котором быстро
происходит исчерпание питательных веществ и жизнь поддержива-
поддерживается за счет регенерационного цикла. "Новая" продукция возможна
лишь за счет поступления веществ в продукционную зону извне за
счет выноса апвеллингом из глубокого океана, что опять-таки озна-
означает регенерационный цикл, но в большем масштабе. Для океана в
целом "новую" продукцию обеспечивают континентальный снос за
счет химического субаэрального выветривания и зона спрединга за
счет гидротермального выноса. Зависимость деструкции от метаста-
бильных органических веществ очевидна. Таким образом, для раз-
развития микробных сообществ необходима неоднородная среда, и об-
областью "сгущения" жизни служит геохимический барьер или экотон
в широком смысле. В общей форме можно сказать, что важнейшей
областью для биоты была промежуточная зона между континентом
306
I
*
I
I
f
*
1
i
i
*
i
?
i
t
4
I
«
«
I
i
*
*
4
№
f
*
t
и океаном, вплоть до континентального склона. Размеры этой мел-
мелководной области колебались в зависимости от эвстатических коле-
колебаний уровня океана, обусловленных тектоникой и климатом. Но
жизнь, особенно микробная, определяется локальными условиями
малых экосистем. Поэтому попытки на основе локальных находок
привязать явления жизни к масштабу всей планеты должны воспри-
восприниматься с большими ограничениями: жизнь процветала в некоторых
очагах, которые могли сохраниться локально и в общей изменив-
изменившейся обстановке планеты в целом. На этом и основывается рекон-
реконструкция жизни прошлого с помощью реликтовых сообществ - оби-
обитателей необычных современных ландшафтов, которые могут быть
сопоставлены с фациями прошлого.
То, что такие сообщества сохранились в экстремальных местах
обитания и содержат преимущественно или исключительно бакте-
бактерии, показывает, что эволюция ведет к сужению базиса жизни и ее
ограничению "нормальными" условиями, где процветает раститель-
растительный покров, хотя потенциал к занятию экстремальной ниши у выс-
высших организмов имеется, что доказывается вселением отдельных
массовых видов в экстремальные местообитания, например зеленой
водоросли Dunaliella в гиперсоленые лагуны или красной водоросли
Cyanidium в область высокотемпературного сернокислотного выве-
выветривания. Альтернативой представлению о сужении базиса жизни
может служить представление о смещении его вследствие деятель-
деятельности биоты и поддержания устойчивости новой среды обитания.
Примером может служить формирование почвы как корнеобитае-
мого слоя на поверхности суши, служащей вместе с растительным
покровом основой ландшафта. Поверхность коры выветривания в
досилурийское время могла быть средой развития циано-бактери-
альных или альго-грибных сообществ со многими сходными функ-
функциями, которые позволили вселиться в нее первым растениям, но не
была почвой с сосудистыми растениями, решившими задачу транс-
транспорта через границу двух сред с атмосферой. "Нормальные" условия
современности есть результат биосферной сукцессии.
С другой стороны, представление о том, что архейский или ран-
непротерозойский океан был местом обитания цианобактерий, явно
ложно, если считать этот океан аналогичным современному. В сов-
современном океане цианобактерий, несмотря на выгодную лимитиро-
лимитированную азотом нишу олиготрофного океана, появляются как массо-
массовые вторичные вселенцы в условиях достаточного фосфорного
питания. Именно в таких условиях происходит цветение немногих
видов планктонных цианобактерий, как Trichodesmium. Отсюда сле-
следует прямолинейное заключение, что цианобактерий первоначально
процветали в водах суши или аналогичных им. Сильным контраргу-
контраргументом является развитие морского фототрофного прокариотного
пикопланктона с Synechococcus, Prochlorococcus, который еще недо-
недостаточно изучен с физиологических позиций, но отвечает за сущест-
307
венную часть первичной продукции и может рассматриваться как
реликт биоты первичного океана, доминирующий в олиготрофных
халистатических областях. Тем не менее для типичных цианобакте-
рий современный океан представляет чужеродную среду обитания,
где процветают эукариотные протисты в планктоне и многоклеточ-
многоклеточные водоросли на шельфе.
Прямое сопоставление современных сообществ с теми, кото-
которые развивались в условиях архея и протерозоя, может быть сдела-
сделано лишь в общей форме. По-видимому, наименее измененными в
истории Земли оказались условия жизни глубокой подземной про-
кариотной биосферы, точнее, обитателей подземной гидросферы,
развивающихся в пористых породах за счет продуктов химических
реакций воды с нагретыми породами и летучих выделений магмы.
Такое сообщество хемосинтетиков может быть и персистентным,
и реликтовым в полном смысле слова. Донором электронов для не-
него служит водород от серпентинизации, а акцепторами - сера, как
у серозависимых архей, или углекислота, как у метанобразующих.
Принципиально важна была бы находка литотрофных гомоаце-
татных обитателей подземной биосферы. Среди хемосинтези-
рующих организмов способностью использовать воду как окисли-
окислитель обладают СО-окисляющие анаэробы Carboxydothermus, обра-
образующие водород из воды в катаболической реакции СО + Н20 =
= СО2 + Н2.
Какими были условия образования фаций в геологической исто-
истории Земли? На этот вопрос отвечают геохронология и историческая
геология, к краткому обзору которых следует перейти. Несмотря на
явную связь микробиоты с "маленьким теплым прудом" как местом
формирования экосистемы и сообщества, биологу для реконструк-
реконструкции нужно четко представлять, о какой эпохе идет речь и какие ус-
условия доминировали в эту эпоху.
8.2. ГЕОХРОНОЛОГИЯ И ИСТОРИЧЕСКАЯ ГЕОЛОГИЯ
В исторической геологии биолога интересуют аспекты, связан-
связанные с поверхностью Земли, с общим составом атмосферы и океана,
фациальной обстановкой прежде всего в среде осадконакопления
(рис. 1.3). Седиментогенез включает две стороны: источники сноса и
условия накопления. На суше развивается процесс выветривания
(денудации) с образованием остаточных кор выветривания и накоп-
накоплением осадков в понижениях. Растворимые соли и тонкие взвеси
выносятся в море. Источники отложений исследованы гораздо сла-
слабее, чем осадки, отражающие свою историю. Отложения определя-
определяются климатической зональностью, подробно разработанной
Н.М. Страховым в теории литогенеза. Важнейшими аспектами кон-
континентального литогенеза служит развитие его в условиях гумидно-
го или же аридного климата, определяющее глубину химического
308
ч
f
выветривания, тесно связанного с деятельностью микробиоты.
В понимании климатов прошлого существенно эмпирическое поня-
понятие климатических циклов с максимумами и минимумами увлажне-
увлажнения. В цикл входят четыре стадии: тепло-сухо, тепло-влажно, хо-
холодно-влажно, холодно-сухо. Наиболее четко климатические цик-
циклы проявляются в континентальных и мелководно-осадочных обра-
образованиях при спокойном тектоническом режиме. К сожалению, об
условиях на поверхности древней суши геологи могут сказать очень
немного из-за уничтожения поверхности выветриванием.
Стратиграфы выработали достаточно сложную и обильную
терминами описательную классификацию истории Земли, с которой
сопоставляется история биоты. Эта классификация включает иерар-
иерархическое подразделение времени и соответствующих ему отложе-
отложений. Геохронологическая шкала включает следующие подразде-
подразделения:
- зон (архей, протерозой, фанерозой; часто архей и протерозой
объединяют общим названием докембрий);
- эра составляет первые сотни миллионов лет и соответствует
крупным изменениям в органическом мире; в фанерозое - это па-
палеозойская, мезозойская, кайнозойская; в стратиграфии эре соот-
соответствует группа;
- период составляет десятки миллионов лет, и в фанерозое раз-
различают 12 периодов, начиная с кембрия и кончая четвертичным; в
стратиграфии - это система с характерными для нее крупными био-
биологическими таксонами;
- эпоха имеет стратиграфический эквивалент - отдел - и обыч-
обычно составляет первые десятки миллионов лет, обозначаемые как
ранняя, средняя, поздняя часть периода и соответственно нижний,
средний, верхний отдел;
- век соответствует стратиграфическому ярусу и включает от-
отложения нескольких миллионов лет, обычно обозначаемые по мест-
местности стратотипического разреза;
- хронозона в 1-3 млн лет обозначается по типичному виду зо-
зонального комплекса.
Временные датировки устанавливают прежде всего по данным
распада естественно-радиоактивных элементов в составе минералов
горных пород. В них первоначальное содержание изотопа убывает
по экспоненте и соответственно накапливаются продукты распада.
Осадочные породы часто датируются по возрасту подстилающих и
перекрывающих изверженных пород. Метод "абсолютной" геохро-
геохронологии особенно важен для докембрия, где отсутствуют индика-
индикаторные ископаемые. Условия на Земле до трансформации ее по-
поверхностных оболочек биотой имели в качестве независимого фак-
фактора геологические события, прежде всего поступление материала
из глубин Земли. Для докембрия международная классификация и
классификация Северной Евразии различаются (рис. 8.3).
309
Эра
Период
2500
Основание кембрия
-1600-
а
Неопротерозой-Ш]
650 -
Криогений
850 -
Тоний
1000
Стений
1300
Эктазий
1400 -
Калиммий
Статерий
- 1800
Орозирий
- 2050
Рясий
- 2300
Сидерий
Акротем;
о.
2500±50
I
Эонотема | Эратема
)снование кембрия
Систе
1650 ±50
Верхний
3150 ±50-|
Нижний
•650 ±20
Верхний
(каратавий)
¦1000 ±50-
Средний
(юрматиний)
-1350 ±20-
Нижний
(бурзяний)
Верхний
Карелии
1900±50-
Нижний
Карелии
Венд
Рис. 8.3. Соотношение международной (А) и российской стратиграфической
шкал (Б) периодизации основных событий в докембрии. Даты в млн лет [по:
Семихатов, 1993]
Доархейский (гадейский) период - от начала формирования
Земли до появления первых пород с возрастом менее 3,9 млрд лет -
описывается на основании общих соображений и сопоставления с
данными для Луны, относящимися примерно к этому интервалу.
Представляется, что в этот период шла бомбардировка Земли пла-
нетезималями с локальным разогревом и проплавлением коры,
которую В.Е. Хаин назвал призрачной - фантомной. Призрачны и
суждения биологов о начале жизни в такой обстановке. Можно ска-
сказать, что геологическая среда в этот период так отличалась от ар-
архейской, что фантомная биота этого времени испытывала бы боль-
большие затруднения, приспосабливаясь к последующей среде с атмо-
атмосферой и гидросферой. В этот период шла интенсивная дегазация
Земли с удалением летучих, прежде всего воды, так что в следую-
следующий, "геологический" этап своего существования - архей - Земля
вступает с гидросферой и атмосферой. Этот ранний этап характери-
310
ц
i
V
i
I
i
зуется однонаправленной эволюцией примерно до 3,5-2,5 млрд лет,
когда начинают доминировать циклические процессы.
"Конец раннеархейского времени, т.е. около 3,5 млрд лет назад,
ознаменовался формированием океанической и континентальной
коры, мантии, гидросферы и атмосферы, делающей ее сходной с со-
современным обликом Земли..." (Хаин и др., 1997, с. 98). В раннеар-
хейское время началось отложение железистых кварцитов из
водной среды в Исуа, и по сумме данных существование биоты в вы-
высшей степени вероятно. Таким образом, появлению жизни на Земле
предшествует этап формирования геологической среды с атмосфе-
атмосферой, гидросферой, литосферой, и можно говорить о первоначальной
биоте, правда, не двигаясь дальше признания факта ее существова-
существования. Вместе с тем важно сделать оговорку о различиях в понимании
терминов геологами и биологами: геологи, говоря об океане и кон-
континенте, обычно имеют в виду океаническую или континентальную
кору, а биологи подразумевают под океаном субаквальные, а под
континентом (сушей) - субаэральные условия жизни.
Основанный на датировании по изверженным породам, анализ
архея не дает ясного представления о процессах выщелачивания на
континентах и химической дифференциации при седиментогенезе.
Представляется вероятным господство физического выветривания
и углекислотного выщелачивания, ведущего в первую очередь к вы-
выносу щелочных металлов во многих местонахождениях строматоли-
строматолитов и наиболее древних микрофоссилий, обычно интерпретируемых
как сходные с цианобактериями.
В это время происходило образование железистых кварцитов.
Для него характерны также строматолиты и формирование карбо-
карбонатных платформ. Развиты гидротермальные процессы. Гидрохи-
Гидрохимия архейского океана неясна, однако она должна допускать образо-
образование кварцитов из кремния. Возможной моделью для нее служит
взаимодействие воды с магматическими породами в современных
вулканах и образование хлоридно-натрового флюида. В водоемах
архея, хотя бы локально, уже присутствуют сульфаты и происходит
отложение гипса. Химическое выветривание на суше связано в пер-
первую очередь с углекислотным выщелачиванием основных и ультра-
ультраосновных изверженных пород и связыванием углекислоты с щелоч-
щелочноземельными металлами в карбонаты. Испарение воды в аридной
зоне по исчерпании резерва кальция делает вероятным появление на
суше крупных водоемов содового типа.
Окислительные процессы в субаэральной обстановке отсутству-
отсутствуют, но в мелководных строматолитах Карелии с возрастом
2,02 млрд лет откладывается рассеянный гематит, придающий им
"красноцветный" облик и свидетельствующий о локальной окисли-
окислительной обстановке. Таким образом, конец архея-начало протеро-
протерозоя, по-видимому, имеет уже достаточно развитую прокариотную
биосферу, взаимодействующую с поступлением свежего извержен-
311
ного материала из глубин в условиях бескислородной атмосферы.
Места обитания микроорганизмов вполне разнообразны, и имеются
типичные фации. Вместе с тем обитаемая поверхность суши - слой
в несколько миллиметров, куда проникал свет, - должна была бы
дать место сообществу, аналога которого в современных условиях
нет: аноксическому циано-бактериальному сообществу, способному
отдавать кислород в бескислородную атмосферу (рис. 8.2).
Архей кончается хорошо выраженным рубежом гранитоидного
магматизма на уровне 2,6-2,5 млрд лет. Предполагают, что в раннем
протерозое (нижнем Карелии) единый материк Пангея 0 раздробился
на протоплатформы, в основании которых лежат отложения нижнего
протерозоя, с подвижными поясами между ними. Раздробление прояв-
проявлялось в появлении рифтов в континентальной коре. Вероятно дейст-
действие тектоники плит с соответствующими ей зонами спрединга и субдук-
ции. Легкие материки перемещались на поверхности как пенка на
молоке, или, что точнее, как корка на поверхности жидкой лавы.
В протерозое около 2,0-1,9 млрд лет назад происходит сдавлива-
сдавливание подвижных поясов и возникают новый гигантский материк Пан-
Пангея I и его антагонист океан Панталасса. На эволюцию состава мор-
морской воды указывает появление растворенных в ней карбонатов.
К раннему протерозою относится и переход состава атмосферы от
бескислородной к кислородной. Наиболее характерными членами
разреза протоплатформенных чехлов были пестроцветные, конти-
континентальные и прибрежно-морские мелководные толщи терриген-
ных осадков, углеродистых сланцев. На платформах образуется и
частично сохраняется осадочный чехол. В океане он постоянно по-
поступал в рецикл. Осадки составлены преимущественно карбонатами
и глинами мелководных бассейнов. Толщи известняков, а чаще до-
доломитов, со строматолитами и железистые кварциты типичны для
раннепротерозойских подвижных зон.
На территории Северной Евразии наиболее изучены нижнепро-
нижнепротерозойские отложения Карелии и Украинского щита. В Карелии
нижнепротерозойские образования подразделяются на комплексы:
сумий, сариолий, ятулий, суйсарий. По В.В. Мележику, здесь господ-
господствовала палеогеографическая обстановка относительно неболь-
небольших водоемов, сменившаяся морскими условиями. В них формиро-
формировались доломитизированные строматолиты. Очень характерны для
этой зоны шунгиты - отложения восстановленного углерода, иначе
высокоуглеродистые негорючие сланцы. На Украинском щите в уз-
узкой меридиональной полосе Курско-Криворожской подвижной сис-
системы с возрастом 2,6-2,3 млрд лет образовалась мощная железоруд-
железорудная курская магнитная аномалия - КМА. Снизу она представлена
терригенными породами, а вверху - толщей железистых кварцитов
с мощностью серии 2,5 км. Ранний протерозой является временем
самого крупного железорудного накопления в планетарном масшта-
масштабе, не повторившегося впоследствии.
312
4
4
¦и
i
Ранний протерозой - время радикальной геохимической пере-
перестройки. К причинам ее относят превышение уровня срединно-
океанических хребтов уровнем вод; рост железного ядра Земли; из-
изменение состава атмосферы. В эту эру происходят: обогащение
кремнеземом выплавляющихся из мантии магматических пород; по-
поступление в воды океана при гидратации кремнезема и карбонатов с
широким распространением осадочных карбонатов и кварцитов.
Кварциты встречаются практически во всех толщах нижнего проте-
протерозоя. Особенно характерно они сочетаются с отложениями железа.
Морская обстановка разнообразна, и наблюдаются следы усыхания
и появление эвапоритов, включающих гипсы, что свидетельствует о
наличии в морских водах сульфатов. В континентальной обстановке
сформировались красноцветные осадочные отложения конгломера-
конгломератов, гравелитов, аркозов. Они окрашены в красный цвет, свидетель-
свидетельствуя о присутствии в атмосфере свободного кислорода.
Физико-химическая обстановка протерозойского перехода была
смоделирована А. Дроздовской, исходя из термодинамических рав-
равновесий минералов. Переход восстановительной обстановки в окис-
окисленную получился очень резким. Предложенная ею схема химиче-
химической эволюции основана на "титровании" системы океан-атмосфе-
океан-атмосфера-осадочные породы возрастающими к прошлому количествами
элементного углерода, что и задавало исходно восстановительные
условия. Ряд термодинамически обусловленных равновесий рисует
картину химической эволюции. Особенно наглядно в модели полу-
получается период образования железорудных формаций, отмечающих
исключительно резкий переход от восстановленного состояния к
другому, окисленному.
Ранний протерозой послужил временем первой биогенной рево-
революции в геохимической истории Земли вследствие биогеохимиче-
биогеохимической сукцессии, вызванной накоплением в атмосфере кислорода.
Именно к концу этого времени на основе цикла органического угле-
углерода произошло формирование системы биогеохимических циклов
современного типа. Прекратилось позднеархейское накопление оса-
осадочного золота. Ранний протерозой занят переходным железным
циклом, приведшим к железорудному накоплению.
Поздний протерозой начинается 1650 млн лет тому назад ка-
карельской тектономагматической эпохой и в русской литературе но-
носит название рифея, соответствующего большей части мезопроте-
розоя и неопротерозою по международной шкале. Важной границей
служит гренвильская тектономагматическая эпоха 1000 млн лет на-
назад, отделяющая мезопротерозой от неопротерозоя (каратавия с ка-
ратауской и лахандинской сериями). Заканчивается рифей 600 млн
лет назад байкальской тектономагматической эпохой, после кото-
которой начинается венд. На основе палеомагнитных данных считается,
что в начале рифея существовал суперконтинент Пангея I, куда вхо-
входили 1/2-2/3 современной континентальной коры. Палеоклиматиче-
313
екая обстановка реконструируется по осадочным породам и корам
выветривания - палеоиндикаторам климата. Большие объемы вы-
высокомагнезиальных известняков в раннем и среднем рифее свиде-
свидетельствуют об очень теплом азональном климате с температурой в
пределах до 40°, судя по изотопии кислорода. Существовали и обла-
области аридного климата. В конце среднего рифея произошло пониже-
понижение температуры, снизились карбонатообразование, отложение
кремней. Сформировались резкие климатические зоны. В позднем
рифее и раннем венде образовались мощные ледниковые отложе-
отложения как континентальные, так и морские. С ними коррелируют же-
железорудные залежи. С началом неопротерозоя согласуется форми-
формирование современного состава воды океана, устанавливаемое по со-
соотношению солей в эвапоритах.
Неопротерозой связан с распространением эукариотных микро-
микроорганизмов с фаготрофным питанием, развитием многоклеточных
водорослей. С биологической точки зрения этот период представля-
представляет неопротерозойскую революцию - конец доминирования только
прокариот и наслоение на созданную ими глобальную биогеохими-
биогеохимическую систему эукариот. Примером этой эпохи служит лахандин-
ская серия, где Б.В. Тимофеевым на основе органостенных микро-
фоссилий впервые были описаны смешанные танатоценозы прока-
прокариот с эукариотами. Стремительно происходит формирование разных
линий протистов - очень разнообразных одноклеточных и колони-
колониальных эукариот. Среди первичных продуцентов распространение
получают разные линии макроскопических водорослей, включая
красные и бурые, которые занимают литораль, где ранее доминиро-
доминировали строматолиты.
Венд завершает протерозой, от 600±10 млн лет назад, и продол-
продолжается до 535-540 млн лет. Представление о венде было разработано
Б.С. Соколовым, назвавшим этот период вендом (от имени балтий-
балтийского племени вендов, или венедов). В это время происходит разви-
развитие многоклеточных животных, дифференцированных, но лишен-
лишенных скелета; они известны под названием "эдиакарской фауны из
Австралии". Фауна венда представлена бесскелетными гигантскими
животными, либо фильтраторами, либо симбиотрофами, как совре-
современные кораллы. Присутствие хищников-консументов сомнитель-
сомнительно. В составе продуцентов распространены макроскопические водо-
водоросли - вендотениды, описанные М.Б. Гниловской. Венд начинается
с мощнейшего варангерского оледенения, приведшего к снижению
уровня Мирового океана. Льдами покрыты континентальные участки
и часть моря, о чем свидетельствуют перенесенные льдом породы и
характерные следы движения ледников по поверхности. Вместе с
тем имеется прихотливая климатическая зональность, обусловлен-
обусловленная иным расположением платформ. Так, Сибирь, занятая огром-
огромным Сибирским морем с повышенной соленостью, располагалась
между 10° и 30°, в то время как Европа находилась вблизи Южного
314
ц
«I
щ
щ
11
полюса. Антарктика, Китай, Индостан находились в экваториаль-
экваториальных широтах. Во второй половине венда произошло сильное потеп-
потепление, распространились карбонатные формации, в связи с таянием
льдов поднялся уровень Мирового океана. Началось развитие мас-
массивов биогермов. Температура мест образования строматолитов со-
составляла тогда 35^5°.
В кембрии, начинающем палеозой, распространение получает
скелетная фауна, но не происходит резкого изменения флоры. Кем-
Кембрий явно связан с появлением зоотрофного питания: хищников,
способных заглатывать крупную добычу, и растительноядных жи-
животных, обладающих пастбищным типом питания. Внезапное появ-
появление разнообразных типов животных в течение нескольких милли-
миллионов лет не нашло своего объяснения. Г.П. Леонов2 предположил,
что заселение океана скелетными организмами произошло из внут-
риконтинентальных бассейнов, разнообразных по своей химии, пос-
после вендского цикла регрессии и трансгрессии океана. Гипотеза вну-
триконтинентального происхождения скелетной фауны явно не
имеет отношения к прокариотам, которые если и мигрировали из
эпиконтинентальных водоемов в океан, то сделали это гораздо
раньше. Скелет кембрийских животных был известковый, кремние-
кремниевый или хитиновый с фосфатами. Наиболее известны членистоно-
членистоногие трилобиты. Строматолиты перестают быть единственными ри-
фообразователями, а вместо них появляются изящные археоциаты и
кишечнополостные. Имеются моллюски. Таким образом, кембрий
функционально вводит в пищевые цепи зоотрофов, а в геохимиче-
геохимические циклы - концентрационные (а не каталитические, как ранее)
процессы на основе внутриклеточной биоминерализации, позволяю-
позволяющей строить наружные скелеты. Вместе с тем сохраняются ранее
возникшие продуценты: флора морского фитопланктона, бентос-
ных водорослей макрофитов. В этом отношении современный оке-
океан представляет реликтовую систему, сформировавшуюся в проте-
розое-рифее.
В начале палеозоя формируются суперконтиненты Гондвана и
Лавразия. Крупные изменения происходят в конце силура-девоне,
т.е. среднем палеозое. Революция заключается в выходе расти-
растительности на сушу с мохообразными как первопоселенцами. В кон-
конце силура появляются первые наземные растения, в девоне они
расселяются по суше. В континентальных глинах ("сухарных као-
линитовых глинах") силура, подстилающих угленосные слои, обна-
обнаруживается разнообразная микофлора, примечательно сходная с
современной. В карбоне начинается обильное угленакопление в
амфибиальных ландшафтах, подробно рассмотренное палеобота-
палеоботаниками (Мейен, 1987), и создается растительный покров с корне -
обитаемой почвой.
2 Леонов Г.П. Историческая геология. М.: Изд-во МГУ, 1980. 344 с.
315
10
о
-10
-20
-30
-40
-50
Аномалия
ломагунди -ятулий
- B,0 - 2,3 млрд лет)
Терминальная Позднепермская
неопротерозойская аномалия
аномалия (около 0,26 млрд лет)
(около 0,6 млрд лет) / Современные
морские
карбонаты
''Франсвильская ^
аномалия орг
(около 2 млрд лет)
5|Аномалия Фортескью
(около 2,7 млрд лет)
Архей
3,5
Современные
морские
осадки
2,5
Протерозой | Ф анерозой]
"Г
1,5
Г
0,5
Рис. 8.4. Изотопный состав органического и неорганического углерода на про-
протяжении истории Земли [по: Schidlowski, 2002]
а - сводные кривые распределения 5С в карбонатах (Скар6) и в захороненном органиче-
органическом веществе (Сорг) на протяжении 3,8 млрд лет геологической истории Земли; данные
формации Исуа пересмотрены из-за возможного позднего загрязнения;
б- фракционирование изотопов углерода основными группами растений и микроорганиз-
микроорганизмов и основные примеры величин изотопного фракционирования углерода в неорганиче-
неорганических соединениях. Треугольниками отмечены средние значения; PDB - карбонаты белем-
белемнитов, принятые за стандарт для 13С/|2С
Исключительно важным объективным показателем для рекон-
реконструкции деятельности биоты в геологическом прошлом представ-
представляется изотопный состав осадочных пород, содержащих биогенные
элементы, в первую очередь углерод и серу. Деятельность микроор-
микроорганизмов приводит к фракционированию изотопов, причем в био-
биомассу поступают преимущественно легкие изотопы. Изотопное
фракционирование углерода определяется отношением изотопов
13Q12C в частях на тысячу (промилле, %о). За стандарт (PDB) приня-
принято отношение 13С/12С = 88,99 в раковине белемнитов. Отсюда
5>ЗС = [(>зС/>2Спроба - 13С/'2Сстандарт) - 1] • 1000.
Снижение показателя 813С характеризует все формы биогенно-
биогенного углерода (с поправкой на дискриминацию тяжелого изотопа при
диффузии), образованного из углекислоты атмосферы или бикарбо-
бикарбонатов океана, и обычно заключается между -20 и -30%о. Постоянст-
Постоянство изотопного отношения органического углерода для почти 4 млрд
лет истории доказывает вовлечение его через рибулозобисфосфат-
ный цикл. С этим геохимическим фактом следует согласовывать все
реконструкции предполагаемой ранней биоты, например ряда ано-
316
3
I
\
ш
m
m
ч
m
щ
m
*
I I I I I
-40
-30
613С[%с, PDB]
-20
С3 - растения
i i i I I i i i
-10 0
I I I 1 1 I I I I I I I I I I I I I I I I I I I I
Морские карбонаты |
Морские бикарбонаты ¦
I СС>2 атмосферы ¦
Растворенная СО2 ¦
- растения С
Толстянковые С
Водоросли I
Цианобактерии г
(культивируемые)
Цианобактерии
(из природных местообитаний) С
I
Аноксигенные
фототрофные
бактерии
Метаногены с
-85%о <1
I Chromatiaceae
"з Rhodospirillaceae
I
'
'
I Chlorobiaceae
Chloroflexaceae
D Метанотрофные бактерии
i i i i i i I i i i i i i i i i I i i i i i i i i i I i i i i i i i i i 1 i i i i i i i i i i i i i i i
-40 -30 -20 -10 0 5
Рис. 8.4. (окончание)
ксигенных фототрофов, лишенных рибулозобисфосфатного цикла.
Локальные или временные отклонения от отношения - 26±7%с тре-
требуют специального объяснения в каждом случае. Резкое облегчение
происходит при окислении метана. Стандартная картина распреде-
распределения изотопов углерода в истории Земли дана М. Шидловским
(Schidlowski, 2002) с указанием главных групп организмов (рис. 8.4).
В этой картине следует обратить внимание на необычно резкое
облегчение углерода в немногих пробах Исуа, соответствующее ме-
танотрофии, для которой, однако, не было мыслимого источника
окислителя. Поэтому М. Шидловский предполагает последующее
изменение соотношения изотопов вследствие метаморфизма, кото-
который претерпели эти породы. Краткое и резкое утяжеление углеро-
углерода карбонатов имеет место в ятулий, для которого имеется хорошая
летопись строматолитов в Прионежье.
Аналогичная ситуация имеет место с изотопами серы. Широкое
распространение сульфатредукции, ведущей к облегчению биоген-
биогенных сульфидов, имеет место ранее 2 млрд лет назад, хотя отдельные
пробы указывают на наличие этого процесса и ранее 3 млрд лет.
317
8.3. БИОЛОГИЧЕСКАЯ ПЕРИОДИЗАЦИЯ
Биологическая периодизация с биосферных позиций выглядит
несколько иначе, чем геологическая. Тем не менее общие подходы
совпадают не только для фанерозоя, где периодизация шла на осно-
основе смены характерных групп живых организмов, но и для докемб-
докембрия, если рассматривать биосферно-геосферную систему. Между пе-
периодами нет резких границ, такая граница, как появление скелетной
фауны в кембрии, представляет скорее исключение. Как правило,
имеется период более или менее длительного перехода, когда новые
явления накладываются на существующую систему и изменяют ее.
Далее внутри системы происходят изменения, которые носят харак-
характер биосферной сукцессии и определяются исчерпанием или запол-
заполнением резервуаров. Старое остается основой возникшей системы,
и лишь некоторая часть старого отмирает. Другая часть старого
приспосабливается к новой надстройке. Поэтому вся система адди-
аддитивна, и "несовершенные", примитивные организмы продолжают
составлять ее основу, образуя связи с новыми компонентами. В от-
отличие от постепенных изменений возможны катастрофические пе-
перестройки, связываемые чаще всего с изменениями климата. В этом
случае идет заселение в освободившиеся экологические ниши и про-
пространства, и первые поселенцы соответствуют уже новой обстанов-
обстановке. Впоследствии они могут сохраняться в изменяющихся условиях в
силу толерантности и способности к адаптации, как сохраняются
растительные формации с длинным жизненным циклом.
Обращает внимание совпадение появления некоторых новых
групп в протерозое с оледенениями. Оледенения согласуются с эв-
статическими колебаниями уровня океана и соответственно с рег-
регрессиями и трансгрессиями моря. Отсюда следует предположение о
том, что нестабильная равнинная часть континента с разнообразны-
разнообразными амфибиальными ландшафтами и эпиконтинентальными водо-
водоемами была источником биоразнообразия, в то время как относи-
относительно устойчивый глубокий океан оставался консервативным. Для
биологической эволюции можно предложить несколько крупных
этапов, в которых новые группировки накладываются на предыду-
предыдущие, как на фундамент.
Первый этап составляет добиосферная эволюция геосферной
системы в архее, которая приводит к формированию основных ти-
типов местообитания организмов: протоконтинентальной суши, пер-
первичных водоемов протоокеана с неясным гидрохимическим соста-
составом, лишенной кислорода атмосферы. Вместе с тем сформированы
главные геологические механизмы физического и химического
выветривания, вулканизма, тектоники, формирования осадочных
пород. Геологический этап, охватывающий миллиард лет истории
Земли, накладывается на "догеологическую" космическую историю
планеты. Важнейшей границей между ними служит завершение де-
318
газации летучих, создающей первоначальные атмосферу и водоемы.
Дегазация продолжается, затухая, и в дальнейшем, но основная мас-
масса летучих уже выделилась в доархейский период. Таким образом,
геологическая система уже включила в себя важнейшие среды жиз-
жизни, нашедшие свое отражение в фациях. Этап этот приходится рас-
рассматривать как добиосферный, не потому что там нет жизни, мик-
робиота присутствует, но она, по-видимому, не является фактором,
меняющим лицо Земли.
При отсутствии фактических данных возможно сопоставление
предполагаемых условий на Земле с возможностями ныне существу-
существующего разнообразия прокариот. В частности, гидрогенотрофные
гипертермофильные обитатели подземной биосферы служат канди-
кандидатами на роль первых обитателей планеты. Примером таких сооб-
сообществ могут быть сообщества термальных выходов на поверхности
суши и в подводных гидротермах. Для последних древнейшей наход-
находкой микрофоссилий считаются нитевидные формы диаметром 0,5-2
и длиной до 300 микрон и почти полностью сложенные кристаллами
пирита, из Пилбара (Австралия) в отложениях, интерпретируемых
как глубинная гидротерма с возрастом более 3,2 млрд лет3. Факти-
Фактический материал о возможных древних формах жизни появился с ис-
исследованием метеоритов, возраст которых равен возрасту Земли.
В нескольких углистых хондритах с высоким содержанием углерода
были найдены остатки, которые можно сопоставить с биотическими
формами. Исследование сколов метеоритов в сканирующем элек-
электронном микроскопе обнаружило формы, подобные бактериям. Ин-
Интерпретация их как микрофоссилий вызвала бурную дискуссию в
основном критического характера, как это было и при находках
первых микрофоссилий в отложениях докембрия. Действительно,
возможность имитации минералами морфологии микроорганизмов
очень велика. Тем не менее одновременное присутствие органиче-
органических веществ вместе с "бактериоморфами" в метеоритах ставит под
сомнение их интерпретацию как свидетельство абиотической пред-
эволюции - "биопоэза".
В раннем архее господствуют однонаправленные изменения и
сравнительно невелика роль циклических процессов. В условиях
продолжающейся дегазации можно предполагать развитие микро-
микроорганизмов, способных участвовать в прямых трофических маршру-
маршрутах, основанных на эндогенных источниках. Наиболее вероятным
источником электронов для катаболизма может служить водород.
Возможными акцепторами электрона служат сера, углекислота, во-
вода. Выходы эндогенных газов, очевидно, сочетаются с повышенным
3 Rasmussen В. Filamentous microfossils in a 3,235-million-year-old volcanogenic mas-
massive sulphide deposit // Nature. 2000. Vol. 405, № 6787. P. 676-679. Westall F. et al.
Early Archean fossil bacteria and biofilms in hydrothermally-influenced sediments
from the Barberton belt, South Africa // Precamb. Res. 2001. Vol. 106. P. 93-116.
319
тепловым потоком и требуют способности жить в зоне конденсации
паров воды. Среди современных микроорганизмов таким условиям
соответствует экофизиология гипертермофильных архей
Crenarcheota. Продуктом серозависимых архей является сероводо-
сероводород, термофильных метаногенов - метан. В геосферном масштабе
важна судьба этих веществ. Метан служит важным парниковым га-
газом, и обусловленный им парниковый эффект мог компенсировать
меньшую на 25-30% светимость Солнца. Сероводород должен был
сильнейшим образом вызывать осаждение сульфидов. Могла ли
служить стоком для метана и сероводорода фотохимия в бескисло-
бескислородной атмосфере? Могли ли микроорганизмы составить одно зве-
звено в абиогенном геосферном цикле? С первичной биотой Земли,
скорее всего, можно сопоставить современную подземную биоту
гидротермальных областей.
Следующий этап господства прокариотной жизни, "прокарио-
тий", ведет к формированию биосферы, способной изменить лицо
Земли. Он начинается циано-бактериальными сообществами в бес-
бескислородной обстановке, а достигает максимума около 2 млрд лет
назад, когда была сформирована система катализируемых микроорга-
микроорганизмами биогеохимических циклов с ведущим циклом органического
углерода и сопряженными с ним циклами углекислоты и кислорода.
К 2 млрд лет назад имеется уже полноценная система оксигенных
цианобактерий и соответственно бактериальная система деструк-
деструкции. Дисбаланс Сорг между этими двумя системами, НГП (нетто-гео-
сферная продукция), служит движущей силой обогащения атмо-
атмосферы кислородом. Карбонатный маршрут ведет к созданию основ-
основного резервуара углекислоты в виде карбонатов щелочноземельных
металлов, выводящего ее из цикла выщелачивания на суше. В конце
этого этапа в полную силу начинает работать серный цикл с биоген-
биогенным сульфидогенезом и обусловленным либо фототрофными анаэ-
анаэробными бактериями, либо аэробными серными бактериями биоген-
биогенным сульфатогенезом; дисбаланс в цикле ведет к накоплению
сульфатов в океане. Особенностью этапа является сопряжение био-
биологического цикла кислорода с геологическим резервуаром железа,
создающее накопление окисных форм железа в кремнистых осадках.
Собственно говоря, в течение "прокариотия" полностью формирует-
формируется биогеохимическая машина планеты, которая затем поддерживает-
поддерживается бактериями-катализаторами геохимических превращений.
Когда система биогеохимических циклов вполне сформирована,
накоплен кислород в атмосфере и нет нужды в связанном азоте, на
прокариотную систему в течение неопротерозойской революции на-
накладываются эукариоты. Этот этап, окончательно сложившийся
около 1 млрд лет назад, можно назвать "протистий", так как именно
распространение протистов, прежде всего в планктоне, приводит к
тому, что эукариоты становятся господствующими продуцентами.
Возникновение многих параллельных рядов протистов происходит
320
. на основе способности эукариот к комбинаторному соединению с
« другими организмами. Протесты вводят в живой мир фаготрофию,
i т.е. способность захватывать внутрь клетки-организма твердые час-
] тицы, отделяя их от цитоплазмы в пищеварительной вакуоли. Такой
j тип питания не совместим со строением прокариотной клетки,
j Он предполагает переход к ббльшим размерам, чем у прокариот,
j которые служат добычей протестам, как хищникам, так и фильтра-
j торам. В биогеохимическом плане лимитация фототрофной продукции
органических веществ фосфатом переходит к лимитации связанным
азотом, который прокариоты с известными энергетическими затра-
затратами могли получать из атмосферы, а эукариоты - не могут. Олиго-
трофный океан до известной степени представляет реликт этого
этапа.
Развитие протистов-эукариот ведет к появлению многоклеточ-
ности у водорослей и занятию ими фотической зоны в мелководьи.
Формирование тканей на основе дифференциации функций клеток
в колонии идет по параллельным последовательностям в разных ря-
рядах протистов. Путь к дифференцированному многоклеточному
тканевому организму через колонию одноклеточных довольно хо-
хорошо прослежен. Этот этап можно назвать "метазоем" с указанием
на формирование многоклеточности. Он начинается 1 млрд млн лет
назад. Характерными его представителями являются как мягкие ор-
организмы венда, так и скелетные организмы кембрия, создающего
все разнообразие животных. В биогеохимическом смысле появле-
появление скелетов у губок и кишечнополостных на основе трансбиомине-
рализации с переходом к ним функции рифообразователей предста-
представляет крупное событие. Вместе с тем "метазой" вводит в систему
консументов и хищников с зоотрофным питанием, начиная гонку
размеров. У зоотрофов появляется пищеварительный канал, в ко-
котором развивается сообщество симбиотических микроорганизмов.
Крупные Metazoa создают для анаэробного микробного сообщества
новую среду обитания - свой пищеварительный тракт.
Что предшествовало появлению на суше наземной растительно-
j сти? По-видимому, уже ранее здесь были мицелиальные грибы,
I поскольку само происхождение сосудистых растений связывают с
, симбиозом их вероятных предшественников - харовых водорослей с
j грибами. Водные грибы известны с венда или ранее. Девонские рас-
I тения уже имеют микоризу, дополняющую корневую систему. На воз-
j никшую лесную систему продуцентов и деструкторов, которую
i можно назвать "древо-гриб", накладываются консументы-животные.
Возможно, что еще ранее существовали альго-грибные сообще-
сообщества как обитатели суши, сменившие циано-бактериальные маты
амфибиальных ландшафтов и почвенные водоросли. Конечным эта-
этапом взаимодействия с грибом служит лихенизация гриба. Примеча-
Примечательна имитация лишайниками кустистой и листоватой формы рас-
растений. Не были ли первоначальными обитателями суши, в том числе
П. Г. А. Заварзин
321
каменных осыпей, сообщества, аналогичные литофильным лишай-
лишайникам? Трофическая система растений и грибов создает основу
ландшафта и приводит к появлению собственно почвы вместо коры
выветривания, или педосферы, обогащенной органическим вещест-
веществом, или торфом, как это наблюдается сейчас на первых стадиях
сукцессии в горах в "первичном почвообразовательном процессе".
Переходу к растительному покрову на суше предшествовал пе-
период, в котором наземная биота была представлена грибами и, по-
видимому, почвенными водорослями или цианобактериями как пер-
первичными продуцентами. Эта ассоциация естественна: можно наблю-
наблюдать, как быстро грибы заселяют влажную поверхность погибаю-
погибающего циано-бактериального мата. Для появления наземной расти-
растительной биоты имеются палеонтологические свидетельства. Камен-
Каменноугольному периоду на суше около 380 млн лет назад предшеству-
предшествуют каолинитовые огнеупорные ненабухающие "сухарные глины"
(flint clays), нередко подстилающие каменноугольные пласты. Хроно-
стратиграфически они коррелируют с первыми углями и бокситами.
Эти глины образовывались в песчано-глинистых и озерно-болотных
отложениях на окислительно-восстановительном барьере и генети-
генетически связаны с гумидными корами выветривания и продуктами их
переотложения потоками. Сухарные глины почти полностью утра-
утрачивают щелочные и щелочноземельные элементы и обогащаются
алюминием. Они четко приурочены к среднему девону - раннему
карбону. Предполагается, что в раннем карбоне сухарные глины по-
покрывали едва ли не половину суши и их образование прекратилось с
трансгрессией моря и сменой терригенного осадконакопления отло-
отложением морских карбонатов. Впоследствии эти отложения были
размыты эрозией. Предполагается, что они формируются в услови-
условиях усиленного промывного режима гумидного климата за счет кор
выветривания пород докембрийских кристаллических щитов в тео-
теократические эпохи.
Исследование этих глин в сканирующем электронном микро-
микроскопе обнаружило очень хорошо сохранившиеся остатки мицелия и
спороношения грибов, настолько сходных с современными, что их
сочли современным загрязнением. Углерод в составе каолинитизи-
рованных микрофоссилий не обнаружен4. Однако еще в 1916 г.
М.Д. Залесский сообщал о "новых ископаемых микроорганизмах"
как сингенетичных углефикации водорослевых осатков. Процесс
очень быстрого окремнения и образования микрофоссилий сейчас
хорошо документирован. Он идет под влиянием выпадения из алло-
фановых гелей монокристаллического каолинита и быстрого погру-
4 Лукин А.Е. Генезис сухарных глин и проблемы экологической минерало-
минералогии //Минерал, журн., 1993. N 3. С. 20-26; Лукин А.Е. Природа сухарных глин
и общие проблемы эволюции биосферы / Биокосные взаимодействия: жизнь
и камень. Материалы 1 Международного симпозиума. СПб., 2002. С. 89-91.
322
жения в зону затрудненного водообмена. Новообразованные цемен-
цементирующие железисто-глиноземистые каолинитовые агрегаты пред-
представлены сростками фоссилизированного мицелия. Аэробность и
ксерофитность грибов обусловливает их становление в раннепалео-
зойскую эпоху, по-видимому, как доминирующих деструкторов на
суше. Нижней границей их возможного распространения служит об-
образование оксической атмосферы. Таким образом, развитие этого
грибного мира на суше предшествовало появлению растительного
покрова с его хорошо известными ранними микоризами на корнях
древесных растений.
Следующим этапом становится появление растительного покро-
покрова на суше с выходом фотосинтезирующей поверхности в атмосфе-
атмосферу. Этот этап можно назвать "плантий", и он начинается с среднего
палеозоя. Цикл органического углерода существенно перестраива-
перестраивается, и глобальная фотосинтезирующая поверхность возрастает и
становится более устойчивой по сравнению с мелководными эпи-
континентальными водоемами и амфибиальными ландшафтами, где
могли развиваться циано-бактериальные, а затем альго-бактериаль-
ные сообщества. Выход в аэротоп растений становится возможен с
формированием ими твердой основы стебля, сложенной лигноцел-
люлозой, и созданием транспортной сосудистой системы. По своей
функции органический скелет растения должен быть построен из
трудноразлагаемого вещества. Следовательно, возникает задача
развития соответствующей группы деструкторов. Если прокариоты
справляются с разложением целлюлозы как компонента клеточных
стенок водорослей, то разложение лигнина оказалось для них непо-
непосильной задачей. Лигнин разлагается грибами на основе перекисно-
го механизма, продуцирующего реакционноспособные не полно-
полностью восстановленные формы кислорода. Отсюда процесс разложе-
разложения лигнина становится облигатно аэробным. Ограничение доступа
кислорода приводит к захоронению органики в каустобиолитах с со-
соответствующей экосистемной нетто-продукцией кислорода.
Если оставить в стороне принципиально важное образование
скелетов, развитие мира зоотрофных животных-консументов в виде
пищевой пирамиды относится скорее к вопросам биоразнообразия,
чем изменению биогеохимических циклов. Они важны для регуля-
регуляции состояния сообщества. Животные в значительной степени зави-
зависят от микробного сообщества своего пищеварительного тракта.
Микробное сообщество продуцирует всасываемые эпителием низ-
низкомолекулярные восстановленные продукты, которые затем окис-
окисляются в аэробном метаболизме животного. В отличие от протестов
захват твердых частиц эпителием кишечника животных не имеет
первостепенного значения, питание осуществляется всасыванием.
Животные в большинстве не имеют собственной целлюлазы. Поэ-
Поэтому они облигатно зависят от бактериального сообщества. После-
Последующую эволюцию в фанерозое можно рассматривать как быстрое
и*.
323
увеличение биоразнообразия, возникновение очень сложной и спе-
специфической взаимозависимости в системе, например растений от на-
насекомых-опылителей.
В избранном масштабе времени увеличение сложности не ведет
к устойчивости системы, напротив, ее сложность и специализация
повышают вероятность отказов в отдельных звеньях, и соответст-
соответственно эволюция живого мира и увеличение нарастающей сложности
ведут к неустойчивости и легкоиндуцируемым внешними причинами
катастрофам.
8.4. ДИВЕРСИФИКАЦИЯ ПРОКАРИОТ
ВО ВРЕМЯ ПАЛЕОПРОТЕРОЗОЯ
Для решения задачи о возможном участии биоты в ранней эво-
эволюции биосферы — примерно до 2 млрд лет назад - нужно ответить
на следующие связанные между собой вопросы о поведении соеди-
соединений типоморфных элементов.
Сорг осадочных пород определяет количество О2, ушедшего в ат-
атмосферу в результате деятельности циано-бактериального сообще-
сообщества и захоронения Сорг в виде керогена. Количественная оценка
массы содержавших Сорг осадочных пород архея-палеопротерозоя
невозможна из-за геологического рецикла. Поэтому приходится
опираться на концентрацию Сорг в анализированных образцах, и
вывод зависит от выборки. В некоторых образцах осадков архея со-
содержание Сорг не уступает более поздним породам. Первичными
продуцентами могли служить оксигенные цианобактерии. История
строматолитов свидетельствует о том, что циано-бактериальные со-
сообщества, вероятно, существовали с 3,5 млрд лет тому назад, хотя
для древнейших строматолитов есть вероятность ошибочной интер-
интерпретации слоистых осадочных структур, а нитчатые микрофоссилии
могут принадлежать и иным, чем цианобактерии, трихомным орга-
организмам. Однако большинство исследователей склоняется к интер-
интерпретации этих микрофоссилии как цианобактерии.
О2 в атмосфере до рубежа 2 млрд лет, вероятно, отсутствовал,
хотя об этом надежно можно судить только по субаэральным отло-
отложениям. О наличии или отсутствии О2 в атмосфере вряд ли можно
судить по субаквальным осадкам, поскольку в них возможны анок-
сические условия. Раннее появление О2, необходимое для метано-
трофии 2,7 млрд лет назад (рис. 8.4), возможно.
Fe в виде окислов четко регистрируется в период до 2 млрд лет
назад в виде крупномасштабных отложений железистых кварцитов.
Источником Fe в них могли служить гидротермальные процессы.
Для кислорода окислов железа есть два варианта: оксигенный фото-
фотосинтез цианобактерии с продукцией О2 с последующим окислением
Fe(II) в Fe(III) и отложением смеси Fe3O4 и Fe2O3 или же аноксиген-
ный "железный" фотосинтез. В обоих случаях необходимым услови-
324
ем является отсутствие H2S в местах отложения. Процесс образова-
образования месторождений железистых кварцитов закончился около 2 млрд
лет назад.
SC>4~ океана, причем концентрация сульфатов стала близкой к
современной ранее 2 млрд лет назад, имеет следующие возможности
происхождения:
- фотохимическое окисление соединений серы в атмосфере при
фотолизе воды аналогично Венере с последующим вымыванием в
океан; сейчас в стратосфере Земли содержатся сульфаты в "слое
Юнга";
- биотическое окисление восстановленных соединений серы за
счет О2, продуцированного цианобактериями в оксигенном фото-
фотосинтезе;
- биотическое окисление за счет аноксигенного фотосинтеза
серных бактерий.
Расчет М. Шидловского вероятной продукции О2 по Сорг осадоч-
осадочных пород приводит к заключению о фотосинтезе цианобактерии
как источнике кислорода и в сульфатах, и в окислах железа. Но та-
такой процесс обязательно предполагает уравновешивание растворен-
растворенного О2 водоемов, в которых идет фотосинтез, с атмосферой и, сле-
следовательно, существование оксигенированной атмосферы во все
время существования железистых кварцитов, т.е. с 3,8 млрд лет назад.
Это "зеленый океан" с обильной продукцией органического вещест-
вещества. Если же расчет Сорг осадочных пород архея-палеопротерозоя
преувеличен, то возможность аноксигенного образования окислов
железа и сульфатов становится вероятной. В этом случае аноксиген-
ный фотосинтез в состоянии обеспечить образование и окислов же-
железа, и растворенных сульфатов океана, который можно было бы
обозначить как "пурпурный океан". Локально такие условия суще-
существуют сейчас в некоторых стратифицированных местах Индийского
океана. При аноксигенном фотосинтезе продукция органического ве-
вещества существенно меньше и лимитирована поступлением H2S
и Fe(II) из эндогенных источников. По изотопии серы в составе
барита в Фиг-Три C,2 млрд лет назад) уже давно было высказано
предположение, что источником сульфата служило окисление серо-
сероводорода пурпурными бактериями, а не окисление в оксической ат-
атмосфере5.
Палеонтологи, исследующие древнейшую летопись Земли,
встречаются с многочисленными трудностями. Древнейшие осадоч-
осадочные породы метаморфизированы, псевдофоссилии, возникшие в
5 Thode H.G., Goodwin AM. Further sulfur and carbon isotope studies of late Archaean
iron-formations of the Canadian Shield and the rise of sulfate-reducing bacteria //
Developments and interactions of the Precambrian atmosphere, lithosphere and bios-
biosphere/Ed. B. Nagy, at al. Amsterdam: Elsevier, 1983. P. 229-248. (Developments in
Precambrian Geology; Vol. 7).
325
геологических процессах, легко могут одурачить исследователя, но
тем не менее установлено, что осадочные породы с возрастом
3,5 млрд лет содержат микрофоссилии, строматолиты; имеются гео-
геохимические свидетельства существования сложных микробных со-
сообществ, содержавших в качестве первичных продуцентов авто-
трофные, скорее всего, фототрофные организмы с характерным
фракционированием изотопов углерода. Эти геологические наблю-
наблюдения согласуются со сравнительно-биологическими представления-
представлениями, что первичные экосистемы были анаэробными и состояли из
прокариотных организмов (Schopf, 1983; Schopf and Klein, 1992). Тем
не менее прямые данные по ископаемым свидетельствам жизни в те-
течение первого миллиарда лет остаются слабодокументированными,
требуя совместной работы представителей наук о Земле и микро-
микробиологов, чтобы осветить первоначальную вспышку эволюции.
Следует принять, что только одна группа прокариот - цианобакте-
рии - вследствие своих размеров и разработанной морфологической
систематики пока поддается идентификации как микрофоссилии.
Об остальных организмах приходится судить главным образом по
геохимическим свидетельствам их деятельности. Вместе с тем толь-
только микропалеонтология дает не реконструкцию, а прямое (с поправ-
поправкой на возможность ошибочной интерпретации) свидетельство. Не-
Невольная концентрация внимания на первичных продуцентах кисло-
кислорода и Сорг создает одностороннюю картину, а, как было показано
ранее, биогеохимия определяется деятельностью сообщества. В дан-
данном случае необходимо представлять и деятельность деструкторов-
органотрофов.
Палеонтологическая летопись палеопротерозоя известна гораз-
гораздо лучше, чем архея. Цианобактерии возникли в архее и/или раннем
протерозое и могли быть представлены рассеянными популяциями
кокков и нитей, находимыми в древнейших породах. Кремни поздне-
позднего архея-раннего протерозоя содержат фоссилии, вероятно, циано-
цианобактерии, но уже в несколько более молодых породах возрастом
примерно 2,2-2,0 млрд лет были найдены древнейшие бесспорные
цианобактерии. Примечательной особенностью этих древнейших
микрофоссилии, точно так же как самых молодых протерозойских
цианобактерии, является то, что они практически неотличимы от
современных видов, живущих в сходных условиях (Сергеев и др.,
2003). Например, Eoentophysalis belcherensis, известный из многих па-
лео- и мезопротерозойских отложений приливной зоны, практиче-
практически идентичен современной цианобактерии Entophysalis major. Точ-
Точно так же встречающиеся на приливных отмелях эллипсовидные
фоссилии, относимые к роду Archaeoellipsoides, можно интерпрети-
интерпретировать как сохранившиеся репродуктивные стадии (акинеты) циано-
цианобактерии, сходных с современными представителями широко рас-
распространенного рода АпаЪаепа. Открытие акинет в отложениях па-
палеопротерозоя особенно интересно в связи с данными молекулярной
326
f
«f
биологии: в филогении, основанной на последовательности 16S
рРНК, организмы, образующие акинеты, относят к самым молодым
ветвям филогенетического дерева цианобактерии6. Находка ископа-
ископаемых акинет показывает, что диверсификация цианобактерии в
принципе была завершена к тому времени, когда мы видим их пер-
первые бесспорные микрофоссилии. Из уст биохимиков постоянно слы-
слышен вопрос: "Как на основании сходства формы можно судить о
функции?". Если я вижу человека с ушами, то могу предполагать с
большой вероятностью, что он слышит, хотя заключение о том, что
он понимает то, что слышит, часто ошибочно. К счастью, цианоба-
цианобактерии имеют лишь ограниченное число морфологических паралле-
параллелей среди прокариот, хотя мощные бентосные маты трихомных се-
серобактерий, как Thioploca, развиваются глубже фотической зоны.
Вывод об идентичности палеопротерозойских цианобактерии
современным представляет один из принципиальных эмпирических
фактов биологии. Эволюции этих организмов на фоне меняющейся
геосферы и биосферы не происходило. Отсюда следует, что их
бактерии-спутники, развивающиеся в совместном циано-бактери-
альном сообществе, тоже не изменились, тем более что их среда
обитания - внутреннее пространство циано-бактериального сооб-
сообщества - заведомо устойчивее, чем геосферные изменения от бески-
бескислородной к кислородной атмосфере. Отсюда можно сделать вывод,
что эволюции прокариот, начиная по меньшей мере с палеопротеро-
палеопротерозоя, вообще не было. Этот вывод чрезмерен, так как бактерии при-
приспосабливаются к новым условиям, включая антропогенные, доста-
достаточно быстро.
Для возникновения биоразнообразия прокариот, отражающего-
отражающегося прежде всего в разнообразии биохимических функций, удобнее
всего обратиться к представлению о латеральном переносе блоков
генов. Такого рода комбинаторика предполагает возможность при-
приспособления и к новым экологическим нишам, возникающим в ре-
результате геосферной эволюции, и, что значительно более ясно, к
эволюции самого сообщества, вызывающего сукцессионные изме-
изменения среды обитания - биосферы. Представление о комбинатор-
комбинаторном генетическом возникновении биоразнообразия прокариот допу-
допускает большое несоответствие между функцией и филогенией. Оно
и наблюдается в действительности. Наилучшие условия для обмена
генетической информацией существуют в плотных сообществах.
Таким сообществом является циано-бактериальный мат. Комбина-
Комбинаторная генетическая система прокариот позволяет возникать функ-
функционально возможным организмам, например использующим новые
продукты обмена. Если такие организмы не возникнут, то в сооб-
сообществе произойдет накопление неиспользуемого продукта, произой-
6 Giovannoni SJ., Turner S., Olsen GJ. at al. Evolutionary relationships among
cyanobacteria and green chloroplasts //J. Bacteriol. 1988. Vol. 170. P. 3584-3592.
327
дет быстрая сукцессия и сообщество придет в тупик. Было показа-
показано, что функциональная (фенотипическая) система бактерий пред-
представляет собой комбинаторное разнообразие (Заварзин, 1974). Во-
Вопрос состоит в том, откуда взялось первоначальное разнообразие
генов? Ответа нет, можно предполагать, что оно "свалилось с не-
неба", но в раннем протерозое уже существует функционально пол-
полноценное микробное сообщество, а не разрозненные виды. Авто-
Автономное микробное сообщество, по определению, такое, которое
может существовать вследствие замкнутых циклов биогенных эле-
элементов самостоятельно в течение длительного времени. Циано-
бактериальное сообщество явно представляет пример такого сооб-
сообщества.
Предположение о том, что первоначальное разнообразие орга-
организмов "свалилось с неба" имеет свою историю. Во-первых, биосфе-
биосфера основывается на ряде сопряженных циклов, отдельные этапы ко-
которых катализируются разными функционально организмами. Поэ-
Поэтому В.И. Вернадский сформулировал тезис, согласно которому
один вид не может существовать на Земле длительно, а должно бы-
было существовать сообщество видов. Это положение исключает воз-
возможность монофилии в экосистемном контексте. Элементарная ло-
логика говорит о том, что единичный организм невозможен так же,
как вечный двигатель: он не может быть одновременно и продуцен-
продуцентом, и деструктором. Ближе всего к такому состоянию подходят фо-
тоавтотрофы. Днем они продуценты, а ночью деструкторы. Но для
замыкания цикла им пришлось бы съесть и свои структурные ком-
компоненты и самоуничтожиться. Во-вторых, ряд авторов обсуждали
гипотезу панспермии, вынося происхождение жизни за пределы Зе-
Земли и ее геологической истории. Сейчас гипотеза существования
жизни вне Земли и, главное, до формирования Земли получила не-
некоторое подтверждение. Одна ограниченная группа метеоритов, уг-
углистых хондритов, содержит не только обильное углеродистое ве-
вещество, но в нем обнаруживаются и фоссилизированные структуры,
морфологически очень сходные с фоссилизированными цианобак-
териями. Результаты не говорят о том, что жизнь на Землю была
занесена метеоритами, поскольку микрофоссилии достоверно мерт-
мертвы, но допускают, что сходная с земной жизнь существовала вне Зе-
Земли, имея возраст 4,6 млрд лет, т.е. "метеоритный возраст Земли".
Поэтому умозрительный тезис Вернадского о "вечности жизни" по-
получает подтверждение.
Важность вопроса о бактериоморфных остатках в метеоритах
заставляет несколько подробнее остановиться на них. По существу-
существующим представлениям метеориты имеют астероидное или кометное
происхождение. Возраст большинства метеоритов близок к возрас-
возрасту Земли. Они разделяются на каменные, железные, железокамен-
ные в зависимости от состава. Для бактериальной палеонтологии
наибольший интерес представляют углистые хондриты.
328
I
i
i
i
1
«
<
i
i
ч
v
«
I
I
I
t
V
4
i
%¦
i
i
i
#
В последние годы в углистых метеоритах Мурчисон, Ефремов-
ка, Алленде, Мигей, Оргей были выявлены литифицированные кок-
коидные и нитчатые формы, аналогичные по морфологии и разме-
размерам современным бактериальным формам и их литифицированным
древним аналогам. В метеоритах Ефремовка и Мурчисон обнаруже-
обнаружены бактериоморфные минерализованные образования, внешне име-
имеющие морфологию типа современной цианобактерии Gloeocapsa.
Здесь же найдены и коккоидные формы типа синехококков. В них
встречаются и литифицированные нитчатые формы, аналогичные
многим современным трихомным цианобактериям. Иногда эти нити
объединены в агрегаты в виде пучков. Основным современным ме-
методом исследования бактериоморфных образований в метеоритах
явилась сканирующая электронная микроскопия сколов каменного
материала.
Значительная часть того, что известно сейчас о бактериоморф-
бактериоморфных структурах в метеоритах, была получена на основании светооп-
тических исследований в 1970-х годах7. Однако превозмогла крити-
критическая оценка. Такая же ситуация имеет место и сейчас.
А.Ю. Розанов суммировал возражения в группы.
1. Все "биоморфные структуры" - результат засорения при па-
падении метеорита на Землю.
2. Наннометровый уровень объектов ("наннобактерий") невоз-
невозможен для живого организма.
3. Морфология абиогенных материалов на наннометровом уров-
уровне плохо изучена, и, возможно, они сходны с бактериями морфоло-
морфологически.
4. Заведомо абиогенные полимерные кристаллы неотличимы от
фоссилизированных микробных и даже точнее - цианобактериаль-
ных форм.
Все эти возражения имеют свои основания. Прежде всего "био-
"биоморфы" метеоритов - не живые бактерии, а минеральные псевдо-
псевдоморфозы. Свежее засорение имеет место, но оно хорошо отличимо
от минеральных структур. Обсуждение наннометровых объектов
вряд ли продуктивно, поскольку нет достаточного опыта в изучении
минеральных образований на этом уровне, и бактериологи еще дале-
далеко не освоились с "наннобактериями". К объектам микронных раз-
размеров эти возражения явно не относятся. Сходство минералов, на-
например железа, с бактериальными формами очень велико, и ошиб-
ошибки такого рода в высшей степени вероятны. Особенного внимания
заслуживают аморфные минералы и ранние стадии их кристаллиза-
кристаллизации. Здесь требуется крайняя осторожность в интерпретации.
Однако в целом А.Ю. Розанов заключает: "Принципиальное
морфологическое единство земных микробных организмов - как со-
современных, так и древних - с псевдоморфозами по микроорганиз-
^f 7 Л/agy B. Carbonaceous meteorites. N.Y.: Elsevier, 1975.
J
329
мам из углистых хондритов дает основание для утверждений о един-
единстве микробиологического мира Земли и космических объектов"8.
Сходство минеральных образований с живыми организмами за-
заставляет меня задать себе вопрос об ограниченности пространства
логических возможностей для геометрических фигур. В кристалло-
кристаллографии оно установлено математически точно. Для живых организ-
организмов и аморфных сгустков оно может быть перекрывающимся.
Сходство нередко ставит в тупик наблюдателя-микроскописта и для
современных объектов, а тем более для ископаемых.
Эволюция цианобактерий представляется критической для ис-
истории Земли, поскольку только они приобрели способность исполь-
использовать для фотосинтеза воду как восстановитель. В цианобактери-
альном фотосинтезе молекулярный кислород образуется как бросо-
бросовый продукт, и аккумуляция его в атмосфере и океане привела к бы-
быстрым изменениям в биосфере. В бентосном циано-бактериальном
сообществе и сейчас основными деструкторами являются анаэробы.
Аэробные организмы скорее играют роль стока для того малого ре-
резервуара О2, который остается в мате, и для детоксикации продуктов
неполного окисления. Поэтому можно ожидать в Карелии большего
дисбаланса фотосинтез/дыхание, чем это наблюдается в современ-
современных сообществах, и, следовательно, большей экосистемной продук-
продукции кислорода. Стоком для кислорода были физические резервуары
океана и атмосферы, в свою очередь находившиеся во взаимодейст-
взаимодействии с восстановленными веществами эндогенного происхождения.
В глобальном масштабе значение имеет НГП - нетто-геосферная
продукция Сорг, который захоранивается.
Для конца раннего протерозоя особое значение имеет ятулий
(около 2 млрд лет назад), следующий за первым значительным гу-
ронским оледенением. В этот период происходит резкое утяжеление
углерода карбонатов до +0,8% и даже +1,2%, что связывается с мас-
массовым развитием строматолитов и выносом облегченного углеро-
углерода -2,5% в Сорг (Семихатов и др., 1999). Картина развития стромато-
строматолитов в ятулий карельского щита представлена В.В. Макарихиным
и В.А. Мележиком (Melezhik et al., 1997). Земля в палеопротерозое
испытала ряд тектонических изменений, в результате которых воз-
возникли стабильные континенты, создав возможности для долгосроч-
долгосрочного захоронения органического вещества на шельфе и платформах
осадочных пород. Высокая скорость захоронения органического уг-
углерода, отмечаемая по аномально высокому изотопному отноше-
отношению углерода в палеопротерозойских карбонатах и органическом
веществе, сочетающаяся с уменьшающимся потоком восстановлен-
восстановленных газов и ионов из вулканических и гидротермальных источников,
привела к возрастанию атмосферного кислорода от менее чем 1%
1
I
I
*
щ
4
8 А.Ю. Розанов. Бактериально-палеонтологический подход к изучению метео-
метеоритов // Веста. РАН. 2000. Т. 70, № 3. С. 214-223.
330
#
4
Щ
4
¦
¦
4
(от нынешнего содержания) до 10-15% уже 2,2-2,0 млрд лет назад.
Это изменение отмечено в распределении чувствительных к кисло-
кислороду минералов в осадочных породах и в профилях выщелачивания
древних почвенных горизонтов.
Результатом было изменение состояния биосферы. Возникли
новые условия на суше и на поверхности океана, откуда растворен-
растворенный кислород уходил в атмосферу. Другим возможным источником
кислорода является наземный циано-бактериальный мат, в своем
обмене с атмосферой не имевший мощного буфера растворенного
О2 океана. Однако для морских условий известны литифицирован-
ные циано-бактериальные сообщества строматолитов, а на суше не
было условий для литификации и неясны условия захоронения Сорг,
являющиеся обязательным условием экосистемной продукции кис-
кислорода. В геологическом масштабе как эквивалент кислорода важ-
важны богатые органическим веществом сланцы, а не быстро оборачи-
оборачивающиеся динамические резервуары. В океане для развития аэро-
аэробов или анаэробов первостепенное значение имела термогалинная
циркуляция: в случае формирования устойчивого пикноклина анаэ-
анаэробные сообщества продолжали процветать в глубоком бескисло-
бескислородном океане и осадках.
Аэробные бактерии обладают способностью использовать
молекулярный кислород для окисления органического вещества,
фундаментально изменив биогеохимический цикл Сорг. Аэробные
органотрофы, представленные сейчас самой массовой группой
протеобактерий, филогенетически имеют точку ветвления, близ-
близкую к цианобактериям (Zavarzin et al., 1990). Функционально они
могли получить условия для своего развития не в момент появле-
появления кислородной атмосферы, а с самого начала выделения О2 ок-
сигенными фототрофами. В сообществе сразу могла образоваться
трофическая связь с аэробными организмами, использовавшими
его как наиболее выгодный сток электрона. Более того, если в
планктонных условиях О2 мог удаляться из сферы реакции со ско-
скоростью турбулентной диффузии, то в условиях плотной популяции
бентосных организмов его концентрация могла повышаться до по-
появления пузырьков и создания 100%-ного насыщения и кислород-
кислородного стресса. Поэтому локальные места обитания с постоянным
источником О2 днем возникли задолго в геологическом смысле,
чуть ли не за миллиард лет до появления красноцветов как индика-
индикаторов окислительной атмосферы. Все рассуждения о "точке Пас-
тера", "точке Юри" совершенно бессмысленны в приложении к ба-
бактериальному сообществу из-за гетерогенности условий в масшта-
масштабе обитания микромира.
Вместе с тем предварительным условием для неопротерозой-
неопротерозойской революции с появлением крупных в десятки микрон эукариот-
ных протистов следует считать возникновение кислородной среды
обитания для окислительного по существу или аэробного метабо-
331
лизма эукариот. Суть заключается в транспортных ограничениях
внутри крупной клетки и необходимой вследствие этого компар-
тментализации. Принципиальным поворотом в биологии стала
теория симбиогенетического происхождения эукариот Л. Маргу-
лис, предполагающая системное, а не точечное изменение на этом
этапе. Сейчас ясно, из кого произошли органеллы эукариотной
клетки, но остается неясным, что представлял собой хозяин. Соз-
Сознавая, что путь шел от разнородного микробного сообщества с ци-
анобактериями-прохлорофитами как предшественниками хлоро-
пластов и протеобактериями, предшественниками митохондрий
через эндосимбиоз, мы плохо представляем организм, настолько
лишенный гетерофобии, что он мог стать матрицей для той комби-
комбинаторной игры геномов целых организмов, которая послужила ос-
основой неопротерозойской революции. Функционально предшест-
предшественник эукариот представляется фаготрофным прокариотом.
Здесь приходится на время отвлечься от функционального био-
сферно-геосферного подхода и обратиться к возникновению био-
биоразнообразия прокариот. Для описания разнообразия прокариот
мощным орудием стало установление родства организмов по гену
16S рРНК, ставшее основой в новых определителях бактерий и
учебных курсах.
Филогенетическое дерево прокариот, построенное по этому
единственному гену, относящемуся к внутренней сфере организма,
не дает представления о его функции в биосферно-геосферной сис-
системе. Считается, что точка ветвления указывает на последователь-
последовательность возникновения. Вместе с тем неясно, идет ли "молекулярный
хронометр" мутаций с одинаковой скоростью во всех условиях. Есть
сравнительно немного корреляций крупных физиологических групп
микроорганизмов с филогенетической системой. Для малочислен-
малочисленных групп говорить о корреляции физиологии и филогении не сле-
следует. Одну ветвь составляют оксигенные фототрофы - цианобакте-
рии и с ними прохлорофиты. Целостную ветвь составляют мицелит
альные прокариоты актиномицеты. В одной ветви сосредоточены
спирохеты. Не продолжая этого перечисления, следует заметить,
что в любой биологической классификации, как правило, складыва-
складывается такая ситуация, в которой некоторые группы невысокого так-
таксономического ранга очень многочисленны, в то время как другие,
резкое отличие которых заставляет придать им значение таксона
высокого ранга, представлены немногими видами. Далее, современ-
современная система бактерий, основанная на выделенных в чистую культу-
культуру и сохраняемых в коллекциях видах, заведомо отличается от того,
что есть в природе. Филогенетическая система в принципе ориенти-
ориентирована на выяснение генеалогии отдельного вида на основе сходст-
сходства генома.
Тем не менее можно сделать некоторые обобщения. Первое из
них было сделано относительно гипертермофилов и экстремальных
332
t
4
4
i
4
4
i
Щ
Щ
4
ц
#
«I
4*
термофилов9. И среди бактерий, и среди архей эти организмы име-
имеют наиболее глубокие точки ветвления на филогенетическом дере-
дереве. Строго говоря, из этого следует, что их сходство с другими орга-
организмами минимально. Однако возможен вывод, что это наиболее
, древние организмы. Вывод этот очень соблазнителен тем, что сог-
согласуется с представлением о первичных гидротермальных условиях
обитания в архее, сохранившихся как реликт в современной обста-
обстановке. Альтернативным объяснением может служить повышенная
скорость изменения генома в экстремальных условиях из-за необхо-
необходимости репарации. Второе обобщение было сделано относительно
наиболее разнообразной группы грамотрицательных бактерий, по-
получившей название Proteobacteria (от гр. Proteus - изменчивый),
которые составляют основную массу культивируемых организмов.
Они коррелируют с условиями циано-бактериального сообщества,
причем можно усмотреть некоторые трофические тенденции в под-
подклассах протеобактерий (Zavarzin et al., 1990). Сюда входит большин-
большинство аэробов и вообще организмов с окислительным метаболизмом.
Следующее обобщение состоит в том, что и анаэробы-клостридии
(в широком смысле) имеют точку ветвления вблизи цианобактерий,
указывая на некоторый комплекс общности. Его можно характери-
характеризовать как комплекс организмов водной среды обитания. Однако
эта тенденция далека от того, чтобы можно было говорить о моно-
филетическом происхождении рассматриваемой группы, например
от пурпурных бактерий. Линия актиномицетов явно связана с назем-
наземными условиями.
Возвращаясь к рассмотрению сообщества микроорганизмов,
следует еще раз напомнить, что для существования в геологическом
масштабе времени это сообщество должно включать организмы,
осуществляющие все трофические функции, т.е. в него должны вхо-
входить продуценты и деструкторы для всех компонентов продуктов и
мортмассы продуцентов. В сообществе, имеющем длительное суще-
существование, должен осуществляться закон сохранения масс. Фактиче-
Фактически даже в реликтовых сообществах не наблюдается трофической
системы монофилетического происхождения. Это заключение при-
приходится принять как эмпирическое обобщение.
9 Kandler О. Early life on Earth. Columbia, N.Y.: Bengtsons, 1994. P. 152-160. (Nobel
symposium 84).
333
8.5. ВОЗНИКНОВЕНИЕ
ЭУКАРИОТНЫХ МИКРООРГАНИЗМОВ
И НЕОПРОТЕРОЗОЙСКАЯ РЕВОЛЮЦИЯ
Выяснение истории происхождения по "молекулярным часам"
имеет основой спорное логическое допущение. На самом деле все
ныне живущие организмы с их предками имеют одинаковый воз-
возраст. Почему считается, что часы остановились для Chloroflexus, но
продолжают идти для Zea mats'] Превращение деревьев сходства по-
последовательностей в генеалогию представляет не бесспорную логи-
логическую операцию. Сравнение молекулярных последовательностей
предполагает, что эукариоты имели историю почти такую же дли-
длительную, как прокариоты; однако самые древние эукариоты могли
быть простыми анаэробными гетеротрофами, которые имели огра-
ограниченное значение для древнейших экосистем. Поскольку развитие
богатых кислородом экосистем было предварительным условием
для эволюции аэробов с дыхательным метаболизмом вне непосред-
непосредственного контакта с источником кислорода, можно предполагать,
что первое появление распознаваемых эукариотных микрофосси-
лий следует ожидать в породах, датируемых после геохимических
свидетельств возрастания кислорода в атмосфере. На практике
идентификация палеопротерозойских ископаемых как эукариот
сталкивается с большими трудностями. В ранних исследованиях
крупные размеры клеток и наличие небольших органических вклю-
включений в них интерпретировали как ископаемые ядра и органеллы.
Однако эти включения представляют, вероятно, сжавшуюся прото-
протоплазму, не имеющую никакого отношения к первоначальной струк-
структуре; такие включения дают ненадежное средство для отличия иско-
ископаемых эукариот от прокариот. Размеры могут быть полезным кри-
критерием, потому что большинство прокариотных клеток меньше
60 мкм в максимальном измерении. Однако аномально крупные ба-
бактерии могут быть ошибочно классифицированы по этому призна-
признаку, в то время как многие эукариотные клетки много меньше
60 мкм. Более того, хотя большинство бактерий меньше 60 мкм,
многие цианобактерии могут образовывать внеклеточные обертки,
достигая значительно больших размеров, и хорошо сохраняются как
микрофоссилии в ископаемом виде. В результате лучшим средством
для признания микрофоссилии как эукариот оказывается распозна-
распознавание морфологии, свойственной только клеткам с ядром в совре-
современной биосфере. Первые одноклеточные микрофоссилии с дейст-
действительно сложной морфологией были найдены в мезопротерозое
A,5-1,1 млрд лет назад). Однако в породах с возрастом 1,7-1,9 млрд
лет найдены крупные, до нескольких сантиметров, отпечатки в виде
сплющенных или вдавленных остатков, возможно эукариотного
происхождения. Присутствие эукариот в сообществах палеопроте-
розоя подкрепляется также присутствием в палеопротерозойском
334
*
ti
t
I
битуме стеранов - биомаркеров, которые считаются свойственными
преимущественно эукариотам, но на самом деле есть и у бактерий.
Таким образом, хотя индивидуальные линии доказательства потен-
потенциально открыты для альтернативных интерпретаций, сумма свиде-
свидетельств предполагает, что эукариоты были широко распростране-
распространены 1,7-1,9 млрд лет назад (Knoll, 1992), хотя есть мнение, что их ми-
микрофоссилии бесспорны лишь с 850 млн лет10. Биологическая при-
принадлежность этих организмов в целом неясна, и ни одна из их мор-
морфологических особенностей не связывает эти ископаемые с ныне
живущими линиями водорослей или протистов. По крайней мере,
некоторые могли принадлежать к вымершим группам ранних
эукариот. Тем не менее мелкие, подобные Chlorella эукариоты
практически неотличимы от прокариот, и нельзя исключить воз-
возможность того, что мелкие коккоидные микрофоссилии из древ-
древних отложений окажутся остатками эукариотных, а не прокариот-
прокариотных клеток11.
Принципиальный вывод из данных палеонтологии заключается
в том, что функциональная организация биосферы изменяется ред-
редко, с длинными периодами относительной стабильности, перемежа-
перемежающимися короткими периодами адаптивной радиации или вымира-
вымирания12. Вслед за важными биологическими и геосферными изменени-
изменениями в палеопротерозое длительная эра мезопротерозоя
A600-1000 млн лет назад) представляется сравнительно спокойной.
Ассоциации микрофоссилии типа Ганфлинта исчезают из летописи
вместе с железными формациями, в которых они обнаруживают-
обнаруживаются, но циано-бактериальные ассоциации из окремненных карбона-
карбонатов приливной зоны сходны и с палеопротерозойскими, и с совре-
современными.
Принципиальным биологическим событием во время перехода
от мезо- к неопротерозою (от среднего к позднему рифею) была ра-
радиация "высших" эукариот, отмеченная разнообразными новыми
формами в палеонтологической летописи и подкрепленная сравне-
сравнением молекулярных последовательностей ныне живущих протистов
(Knoll, 1992). Это изменение не было внезапным; оно началось в
позднем мезопротерозое и продолжалось в течение неопротерозой-
неопротерозойской эры. Его причина неясна, хотя возможными кандидатами явля-
являются эволюция сложности цитоскелета и происхождение классиче-
классически мейотических форм и полового размножения. В результате в те-
Ю Cavalier-Smith T. The phagotrophic origin of eukaryotes and phylogenetic classifica-
classification of protozoa // Intern. J. Syst. and Evol. Microbiol. 2002. Vol. 52. P. 297-354.
11 Kazmierczak J. The biology and evolutionary significance of Devonian volvocaceans
and their Precambrian relatives // Acta Palaeontol. Pol. 1981. Vol. 26, N 3/4.
P. 299-337.
12 Knoll A.H., Sergeev V.N. Taphonomic and evolutionary changes across the
Mezoproterozoic-Neoproterozoic transition //Neues. Jb. Geol. Palaontol. Abh. 1995.
Vol. 195, N 1/3. P. 289-302.
335
чение относительно короткого времени дифференцировались все
основные группы протестов и водорослей, а также предки живот-
животных, грибов, растений, изменив природу и сложность биотических
сообществ. Наглядно эта диверсификация просматривается среди
органостенных микрофоссилий Лаханды и Мироедихи, описанных
Б.В. Тимофеевым и Т. Герман (Герман, 1990; Тимофеев и др.,
1976).
Остатки эукариотных организмов, которые могут быть с уве-
уверенностью отнесены к ныне живущим крупным группам, это банги-
офиты - многоклеточные красные водоросли, сохранившиеся в ок-
ремненных мелководных карбонатах формации Хантинг в Северной
Америке. Предполагаемые зеленые и, возможно, хромофитовые
водоросли известны из мезопротерозоя, и эта диверсификация под-
подкрепляется как молекулярными биомаркерами, так и морфологией
ископаемых. Появление и распространение крупных многоклеточ-
многоклеточных водорослей в местах, ранее занятых циано-бактериальными со-
сообществами, совпадает с уменьшением строматолитов. В этом же
интервале времени морфологически сложные микрофоссилий мор-
морского фитопланктона, обычно относимые к акритархам, начинают
диверсифицироваться и обнаруживают более быстрые скорости
эволюции; практическим следствием оказывается, что в стратигра-
стратиграфии неопротерозоя эукариотные микрофоссилий оказываются
очень полезными для биостратиграфической корреляции. Остатки
грибов найдены в отложениях верхнего неопротерозоя, в венде13,
где они встречаются с обильными остатками Metaphyta и Metazoa
(Соколов, 1997). Палеонтологические свидетельства взрыва разно-
разнообразия эукариот в отложениях неопротерозоя поддерживаются не-
независимыми данными палеобиохимии и молекулярной биологии.
"Большой взрыв" эволюции эукариот дал предков трех групп мно-
многоклеточных организмов, доминирующих в современных ландшаф-
ландшафтах: животных, растений и грибов. Итак, палеонтологические и
молекулярные данные свидетельствуют, что основная радиация эу-
эукариотных организмов была около 1000 миллионов лет назад при
начале неопротерозойской эры.
В биогеохимическом плане неопротерозойская революция ведет
к существенному ограничению: утрате способности фитопланктона
к азотфиксации. Отсюда ограничение первичной продукции рецик-
рециклом азота. "Новая" продукция возможна лишь в условиях взаимо-
взаимодействия с прокариотным миром, сохранившим способность к азот-
фиксации. Взаимодействие идет в первую очередь с деструкторами,
для которых нужен избыток углеродного питания. Животные-кон-
сументы, помимо своей роли как регуляторов плотности продуцен-
13 Бурзин М.Б. Древнейший хитридиомицет (Mycota, Chytridiomycetes incertae
sedis) из верхнего венда Европейской платформы // Фауна и экосистемы гео-
геологического прошлого. М.: Наука, 1993. С. 21-33.
336
тов по модели хищник-жертва, вносят специфический вклад в био-
биогеохимию биоминерализацией скелета.
В истории Земли наблюдалось несколько периодов резкого из-
изменения биогеохимических циклов и связанной с ними перестройки
биосферы. Побудительной причиной для этих перестроек служили,
по-видимому, изменения в геосфере, связанные с большим или мень-
меньшим поступлением материала в зону деятельности живых существ.
Причины времени перестроек в геосферно-биосферной системе не
выяснены, но их существование не вызывает сомнения. В цикличе-
циклических механизмах биосферы основную роль играет неполное замыка-
замыкание биогеохимических циклов, в первую очередь неполнота дест-
деструкции (минерализации) органического углерода, приведшая к са-
самой крупной перестройке - переходу к окислительной атмосфере.
Ниже приведены некоторые из периодов изменения.
I. Около 2 млрд лет назад заканчивается железорудная эпоха,
обусловленная формированием гигантских осадочных отложений
окислов железа магнетита и гематита; эти окислы поглотили
значительную долю всего кислорода, образованного за историю
Земли.
П. Ранее 2 млрд лет назад начинается и продолжается доныне
эпоха серного цикла с развитием сульфатредукции, причем сульфа-
сульфаты моря накапливают другую половину кислорода; на этом же рубе-
рубеже 2 млрд лет начинает расти кислород в атмосфере, составляя до
5% всего О2, образованного за историю Земли.
III. Около 0,8 млрд лет назад что-то происходит с циклами ще-
щелочноземельных элементов: прекращается внеклеточное осаждение
карбонатов кальция в строматолитах и вывод углекислоты в геоло-
геологический резервуар карбонатов, сокращается образование доломи-
доломитов, и только 0,54 млрд лет назад начинается внутриклеточное обра-
образование известковых скелетов. Значение концентрирующей функ-
функции животных столь очевидно представлено биогенными породами,
что не может быть преуменьшено.
Все эти перестройки связаны с историей микробных экосистем,
и их следует рассмотреть отдельно на основе ранее разобранных ци-
циклов и катализирующих их сообществ микроорганизмов.
8.6. ПЕРЕХОД К МИРУ ТКАНЕВЫХ ОРГАНИЗМОВ -
ФАНЕРОЗОЮ
С биогеохимической позиции переход к фанерозою определяет-
определяется началом формирования скелета животными. Начиная с этого мо-
момента концентрационная функция высших организмов оказывает
принципиальное влияние на циклы кальция, фосфора, кремния и не-
некоторых других элементов. Концентрация элементов происходит
внутриклеточно и внутри организма и поэтому в меньшей степени
337
зависит от условий окружающей среды. Растворение скелетов пос-
после отмирания организмов определяется не столько термодинами-
термодинамическими условиями, сколько кинетикой. Поэтому скелеты орга-
организмов составляют структурную основу рифогенных формаций
фанерозоя.
Образованию скелетных структур, по которым отмечается кем-
кембрий и начинает в полной мере действовать биостратиграфия с ин-
индикаторными организмами, предшествует венд - эпоха возникнове-
возникновения бесскелетных организмов (Соколов, 1997). Они известны как
эдиакарская фауна главным образом по отпечаткам в песчано-гли-
нистых осадках. Существовали они относительно короткое время и
не оставили потомков. Одновременно с ними существовали разнооб-
разнообразные прокариоты, протисты, макроводоросли, микрофоссилии,
которых находят в тех же породах. Эдиакарские организмы были
лишены покровов, поддающихся литификации. Они представлены
преимущественно неподвижными бентосными организмами. Спосо-
Способы питания этих лишенных пищеварительного тракта и ротового
отверстия организмов остаются неясными. Допускается даже осмо-
трофное питание14, что для существ таких размеров маловероятно.
Предполагается, что многие из них были способны к симбиотроф-
ному питанию совместно с низшими организмами. Идее симбиотро-
физма очень способствовало открытие симбиотрофного питания у
гигантских погонофор вестиминтифер - обитателей глубоководных
гидротермальных оазисов. Эти организмы принадлежат к тиобиосу
и развиваются за счет окисления сероводорода симбиотическими ти-
оновыми бактериями, располагающимися в специальном органе -
трофосоме. Погонофора осуществляет транспортные функции, дос-
доставляя с помощью гемоглобиновой системы транспорта кислород,
захватываемый венчиком ярко-красных лопастей в оксигенирован-
ной зоне, и сероводород из восстановительной области. Тиобиос со-
составляет заметную часть обитателей морского дна. Аналогичное
симбиотрофное питание возможно за счет окисления метана15.
Обширная группа наиболее примитивных многоклеточных ор-
организмов относится к "Parazoa", среди которых губки обладают спо-
способностью к образованию скелета, и одни из наиболее древних по-
подобных губкам организмов - археоциаты - образуют удивительно
красивые ажурные скелеты. Межклеточное пространство в теле гу-
губок заполнено разнообразными, иногда очень своеобразными бак-
бактериями и археями. Функционирование этих симбиотических комп-
комплексов еще находится на ранних стадиях расшифровки. Однако
можно утверждать, что симбиотический тип существования живот-
14 Burzin M.B., Grazhdankin D.V., Bronnikov А.А. The mystery of the Ediacaran orga-
organisms //J. Journals. 1998. Vol. 2, № 1. P. 47-53.
15 Малахов В.В., Галкин СВ. Вестиментиферы - бескишечные беспозвоночные
морских глубин. М.: КМК sci. press, 1998. 205 с.
338
ных продвинулся очень далеко, как можно судить по экосистеме ко-
коралловых рифов, возникшей много позже.
Роющие организмы с пищеварительным трактом оставили следы
в столь древних осадках, что интерпретация этих следов часто вызы-
вызывает сомнения, не имитируют ли их газовые ходы. Детритофагия
представляет один из древнейших способов зоотрофного питания. Ре-
Результатом явился факт формирования внутри пищеварительного тра-
тракта микробного сообщества, обеспечивающего пищеварение живот-
животного, как универсальной для зоотрофов системы питания16. Многие
компоненты пищи не могут быть превращены ферментами животно-
животного в усвояемые молекулы, и поэтому участие в питании анаэробного
микробного сообщества необходимо.
В биогеохимическом плане следующие революционные собы-
события связаны, однако не столько с концентрирующей функцией жи-
животных, сколько с эволюцией первичных продуцентов. В позднем
силуре происходит выход на сушу зеленых растений, способных вы-
вынести в воздушную среду фотосинтетический аппарат и при этом не
погибнуть от высыхания. Задача была решена с помощью развития
проводящей системы для воды, ненужной водорослям, и формирова-
формирования углеродного скелета из целлюлозы и лигнина, способного под-
поддерживать фотосинтетический аппарат в аэротопе. Неясно, не была
ли эта задача решена ранее лишайниками как симбиотическими ор-
организмами, способными развиваться в воздушной среде. Существу-
Существуют предположения, что наземные растения были тесно связаны с
микоризоподобными грибами, участвовавшими в создании транс-
транспортной системы. Так или иначе в течение геологически короткого
времени произошла биологическая революция, связанная с появле-
появлением наземного растительного покрова, резко изменившего условия
жизни на континентах. Сюда относятся избыток органического уг-
углерода в форме лигноцеллюлозы, изменение атмосферного гидро-
гидрологического цикла за счет эвапотранспирации, изменение денудации
за счет образования корневого войлока.
Природоведческая микробиология этого этапа развития биоты
на планете в своих существенных чертах описывает современное со-
состояние. Важно отметить, что этот этап основывался на уже сло-
сложившейся системе биогеохимических циклов, что не было нужды
изобретать что-то принципиально новое, и могли быть использова-
использованы генетические ресурсы прошлого. Вместе с тем возникли новые
возможности для взаимодействия с растением в ризосфере, ризопла-
не, филлосфере, в проникновении в проводящую систему, парази-
паразитизме. Гораздо менее осознаваемым является тот факт, что на этом
этапе прокариоты взаимодействовали с уже сложившейся системой
продуценты + деструкторы в виде растения + грибы.
16 Уголев A.M. Эволюция пищеварения и принцип эволюции функций: Элемен-
Элементы современного функционализма. Л.: Наука, 1985. 544 с.
339
Итак, с 2 млрд лет тому назад на Земле существует микробная
система современного типа, на которую постепенно налагались все
более усложнявшиеся последующие формы. Детализация общей
системы и оценка участия в ней разных функциональных групп сос-
составляет суть аддитивной эволюции, происходящей по законам био-
биосферной сукцессии.
Рекомендуемая литература
1. Бактериальная палеонтология / Ред. А.Ю. Розанов. М.: ПИН РАН, 2002.
188 с.
:. Герман Т.Н. Органический мир миллиард лет назад. Л.: Наука, 1990.
i. Заварзин Г.А. Фенотипическая систематика бактерий: Пространство логи-
логических возможностей. М.: Наука, 1974.
Заварзин Г.А. Бактерии и состав атмосферы. М: Наука, 1984. 199 с.
Заварзин Г.А. Развитие микробных сообществ в истории Земли // Пробле-
Проблемы доантропогенной эволюции биосферы. М.: Наука, 1993. С. 212-222.
Заварзин Г.А. Становление биосферы // Микробиология. 1997. Т. 66, № 6.
С. 725-734.
Заварзин Г.А. Недарвиновская область эволюции // Вестн. РАН. 2000. Т. 70,
№5. С. 403^111.
Заварзин ГА. Роль микробиоты в образовании пород // Изв. РАН. Секция
наук о Земле. 2001. № 7. С. 131-137.
Заварзин Г.А. Роль комбинаторных событий в развитии биоразнообразия//
Природа. 2002. № 1. С. 12-19.
Заварзин Г.А. Микробный геохимический цикл кальция // Микробиология.
2002. Т. 71, № 1.С. 5-22.
МаргелисЛ. Роль симбиоза в эволюции клетки. М.: Мир, 1983. 352 с.
Мейен СВ. Основы палеоботаники. М., 1987.
Палеонтологический журнал. 2003. Специальный выпуск.
Сергеев В.Н. Окремненные микрофоссилии докембрия и кембрия Урала и
Средней Азии. М.: Наука, 1992. 139 с.
Сергеев В.Н., Герасименко Л.М., Заварзин Г.А. Протерозойская история
цианобактерий и их современное состояние // Микробиология. 2003. Т. 71,
№6.
Семихатов М.А., Раабен М.Е., Сергеев В.Н. и др. Биотические события и
положительная изотопная аномалия карбонатного углерода 2,3-2,06 млрд
лет тому назад // Стратиграфия. Геохим. корреляция. 1999. Т. 7, № 5.
С. 3-27.
Соколов Б.С. Очерки становления венда. М.: КМК Лтд, 1997.
Сорохтин ОТ., Ушаков С.А. Развитие Земли. М.: Изд-во МГУ, 2002. 560 с.
Тимофеев Б.В., Герман Т.Н., Михайлова Н.С. Микрофоссилии докембрия,
кембрия и ордовика. Л.: Наука, 1976. 106 с.
Федонкин М.А. Бесскелетная фауна венда и ее место в эволюции метазоа.
М.: Наука, 1987. 176 с.
Хаин В.Е., Короновский Н.В., Ясаманов Н.А. Историческая геология. М.:
Изд-во МГУ, 1997. 448 с.
Янкаускас Т.В., Герман Т.Н., Михайлова Н.С. и др. Микрофоссилии докем-
докембрия СССР. Л.: Наука, 1989. 190 с.
Knoll A.H The early evolution of eukaryotes: a global perspective // Science. 1992.
Vol. 256. P. 622-627.
ч
9.
10.
11.
12.
13.
14.
15.
16.
17.
18.
19.
20.
21.
22.
23.
24. Melezhik V.A., Fallick A.E., Makarikhin V.V., Lyubtsov V.V. Links between
Palaeoproterozoic palaeogeography and rise and decline of stromatolites:
. FennoScandian Shield // Precambrian Res. 1997. Vol. 82. P. 311 -348.
25. SchopfJ.W. (ed.) Earth's earliest biosphere, its origin and evolution. Princeton; N.Y.:
Princeton Univ. press, 1983. 543 p.
26. SchopfJ.W. (ed.) Major events in the history of life. Boston: Jones and Barlet, 1992.
27. SchopfJ.W., Klein C. (eds.) The Proterozoic biosphere: A multidisciplinary study.
Cambridge: Cambridge Univ. press, 1992. 1348 p.
28. Schidlowski M. Search for morphological and biogeochemical vestiges of fossil life
in extraterrestrial settings: Utility of terrestrial evidence // Astrobiology: The quest
for the conditions of life. В.; Heidelberg: Springer, 2002. P. 373-386.
29. Zavarzin G.A., Stackebrandt E., Murray R.G.E. A correlation of phylogenetic diver-
diversity of the Proteobacteria with the influences of ecological forces // Canad. J.
Microbiol. 1990. Vol. 31. P. 1-6.
340
СПИСОК ЗАИМСТВОВАННЫХ РИСУНКОВ
И ТАБЛИЦ
Рисунки
Рис 1.4.
Sellers W.D. Physical climatology. Chicago: Univ. of Chicago press.
Рис 1.5.
Graedel T.A., Crutzen P.J. Atmospheric change. An Earth System Perspective. N.Y.:
W.H. Freeman and Company. 1992. 448 p.
Рис 1.6.
Ibid.
Рис. 2.6.
J0rgensen B.B., Revsbech N.P. Diffusive boundary layers and the oxygen uptake of sed-
sediments and detritus // Limnol. and Oceanogr. 1985. Vol. 30. P. 111-122.
Рис. 3.1. Бактериальная палеонтология. / Ред. А.Ю. Розанов. М.: ПИН РАН,
2002. 188 с.
Рис. 3.2.
Герасименко Л.М., МитюшинаЛЛ., Иамсараев Б.Б. Маты Microcoleus из алкало-
фильных и галофильных сообществ // Микробиология. 2003. Т. 72, № 1. С. 84-92.
Рис. 3.3.
A. Kuhl M., Lassen С, Jorgensen В.В. Optical properties in microbial mats: Light meas-
measurements with fiber-optic microprobes // Microbial mats: Structure, development and
environmental significance. B. etc., 1994. (Далее: Microbial mats).
B. Lassen C, Ploug H., Kuhl M. et al. Oxygenic photosynthesis and light distribution in
marine microbial mats // Microbial mats; Lassen C, Ploug H., J0gensen B.B. Microalgal
photosynthesis and special scalar irradiance in coastal marine segiments of Limfiorden,
Denmark // Limnol. and Oceanogr. 1992. Vol. 37. P. 760-772.
Рис З.4.
Perry J., Staley J.T. Microbiology dynamics and diversity. Wort (N.Y.): Sounders
College Publ., 1997.
Рис. З.5.
Calinski E.D., Truper H.G. Microbial behaviour in salt stressed ecosystems // FEMS
Microbiol. Rev. 1994. Vol. 15. P. 95-108.
Рис. 4.1.
Исидоров В.А. Органическая химия атмосферы. СПб.: Химиздат, 2001. 352 с.
342
Рис 4.2.
Australian Bureau Meteorology
Рис 4 3.
Francey R.L., Allison C.E., Trudinger CM. et al. The anterannual variation in global
atmosphere 813C and its link to net terrestrial exchange // VI Intern, carbon dioxide conf.
2001. Vol. 1. P. 43-46.
Рис. 4.4.
Kaplan A., Reinhold L. CO2-concentrating mechanisms in photosynthetic
microorganisms // Annu. Rev. Plant Physiol. and Plant Mol. Biol. 1999. Vol. 50.
P. 539-570.
Рис 5.2.
McCartney M.S. Oceanus. 1994. Vol. 37. P. 5-8.
Рис. 53.
Богданов Д.В. по: Кафаров AM., Кудряшов В.А. Морская биогеография. М.:
Наука, 2000. С. 17.
Рис 5.4.
Кобленц-Мишке О.И. Фотосинтетическая первичная продукция // Биологиче-
Биологические ресурсы океана. М.: Агропромиздат, 1985. С. 48-62.
Рис. 6.1.
Boggs S. Principles of sedimentology and stratigraphy. Englewood Cliffs: Prentice-Hall,
1995.
Рис. 6.2.
Афанасьева М.С., Бурзин М.Б., Михайлова М.В., Кузьменко Ю.Т. Условия об-
образования потенциально нефтематеринских пород // Геология нефти и газа.
1995. № 4. С. 42-45.
Рис 63.
Aselmann /., Crutzen P. Global distribution of natural freshwater wetlands and rice pad-
paddies, their net primary productivity and possible methane emssions // J. Atmosph. Chem.
1989. Vol. 8. P. 307-358.
Mathews E., Fung F. Methane emission from natural wetlands: global distribution area
and environmental characteristics of sources // Global Biogeochem. Cycles, 1987.
Vol. 1. P. 61-86.
Рис. 6.4.
Stolbovoi V., McCallum. Land resources of Russia, Intern. Inst. for Appl. Systems
Analysis and the Russ. Acad. of ScL, Laxenburg (Austria), 2002. CD-ROM.
Рис. 7.2.
Ibid.
Рис. 7.З.
Кудеяров В.Н. Почвенные источники углекислого газа на территории России //
Круговорот углерода в России. 1999.
Рис. 7.4.
Орлов Д.С. Запасы, поступление и круговорот органического углерода в поч-
почвах России // Круговорот углерода в России. М., 1999.
Рис. 7.5.
Tomme P., Warren R.A., Gilkes N.R. Cellulose hydrolysis by bacteria and fungi // Adv.
Microbiol. Physiol. 1995. Vol. 35. P. 1-81.
Рис. 7.6.
Stevenson FJ. Humus chemistry: genes, composition, reactions. N.Y.: Wiley, 1982.
343
Рис. 8.4.
Семихатов М.А. Новейшие шкалы общего расчленения докембрия: Сравне-
Сравнение // Стратиграфия. Геол. корреляция. 1993. Т. 1, № 1. С. 6-20.
Рис. 8.5.
Schidlowski M. Sedimentary carbon isotope archives as recorders of early life:
Implications for extraterrestrial scenarios // Fundamentals of life // Ed. G. Palyi et al.
2002. P. 307-329.
Таблицы
Таблица 4.1.
Active oxygen in biochemistry, 1995.
Таблица 5.1.
А. Алекин О.А., Ляхин Ю.И. Химия океана. Л.: Гидрометеоиздат, 1984. 343 с.
Б. Graedel ТА., Crutzen PJ. Atmospheric change. An Earth System Perspective. N.Y.:
W.H. Freeman and Company. 1992. 448 p.
Таблица 5.2.
Корж В Д. Геохимия элементного состава океанской воды // Химия морей и
океанов. М.: Наука, 1997. 415 с.
Таблица 5.3.
Holland H.D., Lazer В., McCaffrey M. Evolution of the atmosphere and oceans // Nature.
1986. Vol. 320. P. 27-39.
Таблица 5.4.
Sorokin Yu. Aquatic microbial ecology. Leiden: Backhuys, 1999.
Таблица 5.5.
To же. :
Таблица 7.1.
Орлов Д.С., Бирюкова О.Н., Суханова Н.И. Органическое вещество почв
Российской Федерации. М.: Наука, 1996. 254 с.
СОДЕРЖАНИЕ
Предисловие.
Лекция 1
БИОСФЕРА И МИКРОБЫ
1.1. Биосферный подход 5
1.2. Система биогеохимических циклов 10
1.2.1. Цикл органического углерода 10
1.2.2. Сопряжение циклов элементов с циклом Сорг 12
1.3. Геобиофизика 17
1.4. Ландшафт 19
1.5. Глобальная циркуляция и ландшафты 23
Рекомендуемая литература 29
Лекция 2
МИКРОБНОЕ СООБЩЕСТВО
2.1 Трофическая структура сообщества 30
2.1.1. Кооперация в сообществе 30
2.1.2. Энергетика сообщества 40
2.1.3. Межвидовой перенос электрона. Синтрофия 41
2.1.4. С-2 путь и ацетогенез 44
2.1.5. Экологические ниши 46
2.1.6. Терминология 50
2.2. Физическая организация сообщества 52
2.2.1. Парагистология сообществ 52
2.2.2. Межвидовой транспорт в сообществе 55
2.2.3. Градиенты и транспорт в сообществе 59
2.2.4. Коллоидная среда обитания микроорганизмов 61
Рекомендуемая литература 66
345
Лекция 3
ЦИАНО-БАКТЕРИАЛЬНЫЕ МАТЫ
3.1. Циано-бактериальные маты как морфологически оформлен-
оформленное автономное сообщество 67
3.2. Макроструктура циано-бактериальных матов 69
3.3. Фототрофный слой 73
3.4. Биостабилизация осадков 81
3.5. Галофильный мат 83
3.5.1. Галофильные микроорганизмы 84
3.6. Алкалофильное сообщество содовых озер 87
3.7. Термофильный мат 93
3.8. Циано-бактериальные маты в сравнительном аспекте 96
3.9. Мат сульфуреты и тиобиос 99
Рекомендуемая литература 101
Лекция 4
АТМОСФЕРА И БАКТЕРИИ
Атмосфера и биота 102
Парниковый эффект и микроорганизмы 105
Парадокс кислорода 112
4.3.1. Роль микроорганизмов в формировании кислородной
атмосферы 116
Дыхание органотрофов 121
Окислительный бактериальный фильтр 123
Углекислота 127
Летучие соединения азота 132
Водородные бактерии 137
Карбоксидобактерии 139
Летучие углеводороды и бактериальный фильтр 143
Микроорганизмы как аэрозольное загрязнение атмосферы 145
Эволюция состава атмосферы 146
Рекомендуемая литература 148
4.1.
4.2.
4.3.
4.4.
4.5
4.6.
4.7.
4.8.
4.9.
4.10,
4.11.
4.12,
Лекция 5
МИКРОБИОЛОГИЯ ВОДНЫХ СРЕД ОБИТАНИЯ
5.1. Водоемы и гидрологический цикл 149
5.2. Водоем как модель экосистемы 154
5.2.1. Лимнология 155
5.2.2. Экология водных микроорганизмов 159
5.2.3. Бактериопланктон 161
5.2.4. Круговорот углерода 163
5.2.5. Цикл железа в континентальных водоемах 168
5.2.6. Водотоки и водопользование , 171
346
5.3. Подземные воды 173
5.4. Океан 176
5.4.1. Зональность океана 176
5.4.2. Фитопланктон и первичная продукция 182
5.4.3. Регенерационный цикл азота в океане 189
5.4.4. Деструкция 192
5.4.5. Донные осадки 201
Рекомендуемая литература 202
Лекция 6
АМФИБИАЛЬНЫЕ ЛАНДШАФТЫ, ЭКОТОНЫ
И ГЕОХИМИЧЕСКИЕ БАРЬЕРЫ
6.1. Среда обитания микробных сообществ 204
6.2. Миграция элементов 204
6.3. Экотон и геохимический барьер 207
6.4. Амфибиальные ландшафты 213
6.5. Седиментогенез 216
6.6. Осадки и захоронение углерода. Кероген. Нефтематеринские
породы , 220
6.7. Болота 226
6.8. Верховые болота - автономные реликтовые биоценозы 234
Рекомендуемая литература 243
Лекция 7
МИКРОБИОЛОГИЯ ПОЧВЫ
7.1. История 244
7.2. Почвенная микробиология 246
7.3. Почвенная биология 248
7.4. Распределение микроорганизмов в почве 253
7.5. Деградация органического вещества в почве 255
7.5.1. Разложение растительных остатков 255
7.5.2. Разложение целлюлозы 266
7.5.3. Разложение лигнина 270
7.5.4. Разложение древесины ксилофагами 274
7.6. Гумус 279
7.6.1. Анаэробное разложение фенольных соединений 280
7.6.2. Гуматы как субстраты микробного метаболизма 282
7.7. Органо-минеральные соединения в почве 285
7.8. Минеральный состав почвы 285
7.9. Кора выветривания 289
7.10.Палеопочвы 292
Рекомендуемая литература 295
347
Лекция 8
ИСТОРИЧЕСКАЯ МИКРОБИОЛОГИЯ
8.1. Роль прокариотного мира в становлении биосферы 297
8.1.1. Границы приложимости актуалистического метода 301
8.1.2. Геосфера и историческая геология 303
8.2. Геохронология и историческая геология 308
8.3. Биологическая периодизация 318
8.4. Диверсификация прокариот во время палеопротерозоя 324
8.5. Возникновение эукариотных микроорганизмов и неопротеро-
неопротерозойская революция 334
8.6. Переход к миру тканевых организмов - фанерозою 337
Рекомендуемая литература 340
Список заимствованных рисунков и таблиц 342
Научное издание
Заварзин Георгий Александрович
ЛЕКЦИИ
ПО
ПРИРОДОВЕДЧЕСКОЙ
МИКРОБИОЛОГИИ
Утверждено к печати
Ученым советом
Института микробиологии
Российской академии наук
Зав. редакцией Н.А. Степанова
Редактор М.М. Гусева
Художник Ю.С. Шлепер
Художественный редактор Ю.И. Духовицкая
Технический редактор В.В. Лебедева
Корректоры ТА. Печко, Т.Н. Шеповалова
Подписано к печати 06.08.2003
Формат 60x90 Vi6. Гарнитура Тайме
Печать офсетная
Усл.печ.л. 22,0. Усл.кр.-отт. 22,0. Уч.-изд.л. 23,6
Тираж 510 экз. Тип. зак. 4483
Издательство "Наука"
117997, Москва, Профсоюзная ул., 90
E-mail: secret@naukaran.ru
Internet: www.naukaran.ru
Санкт-Петербургская типография "Наука"
199034, Санкт-Петербург, 9-я линия, 12
Магазины "Книга-почтой"
121099 Москва, Шубинский пер., 6; 241-02-52
197345 Санкт-Петербург, ул. Петрозаводская, 7Б; (код 812) 235-05-67
Магазины "Академкнига" с указанием отделов "Книга-почтой"
690088 Владивосток, Океанский пр-т, 140 ("Книга-почтой"); (код 4232) 45-27-91
620151 Екатеринбург, ул. МгСмина-Сибиряка, 137 ("Книга-почтой"); (код 3432)
55-10-03
664033 Иркутск, ул. Лермонтова, 298 ("Книга-почтой"); (код 3952) 46-56-20
660049 Красноярск, ул. Сурикова, 45; (код 3912) 27-03-90
220012 Минск, проспект Ф. Скорины, 72; (код 10375-17) 232-00-52, 232-46-52
117312 Москва, ул. Вавилова, 55/7; 124-55-00
113105 Москва, Варшавское ш., 9, строение 1Б (книжная ярмарка "Централь-
"Центральная", 5 этаж); 737-03-33 (доб. 50-10)
117192 Москва, Мичуринский пр-т, 12; 932-74-79
103054 Москва, Цветной бульвар, 21, строение 2; 921-55-96
630091 Новосибирск, Красный пр-т, 51; (код 3832) 21-15-60
630090 Новосибирск, Морской пр-т, 22 ("Книга-почтой"); (код 3832) 35-09-22
142290 Пущино Московской обл., МКР "В", 1 ("Книга-почтой"); A3) 3-38-80
443022 Самара, проспект Ленина, 2 ("Книга-почтой"); (код 8462) 37-10-60
199034 Санкт-Петербург, В.О., 9-я линия, 16; (код 812) 323-34-62
191104 Санкт-Петербург, Литейный пр-т, 57; (код 812) 272-36-65
199164 Санкт-Петербург, Таможенный пер., 2; (код 812) 328-32-11
194064 Санкт-Петербург, Тихорецкий пр-т, 4; (код 812) 247-70-39
634050 Томск, Набережная р. Ушайки, 18; (код 3822) 51-60-36
450059 Уфа, ул. Р. Зорге, 10 ("Книга-почтой"); (код 3472) 24-47-74, факс
(код 3472) 24-46-94
450025 Уфа, ул. Коммунистическая, 49; (код 3472) 22-91-85
Коммерческий отдел, г. Москва
Телефон 241-03-09
E-mail: akadem.kniga@g 23.relcom.ru
Склад, телефон 291-58-87
Факс 241-02-77
WWW. AK-Book. naukaran.ru
По вопросам приобретения книг
просим обращаться также
в Издательство по адресу:
117997 Москва, ул. Профсоюзная, 90
тел. факс @95) 334-98-59
E-mail: initsiat @ naukaran.ru
Internet: www.naukaran.ru