/

































































































































































































































































































































Text
БИОЛОГИЯ
I
1
А К А Д E M И Я Н А У К
С-0 ЮЗА СОВЕТСКИХ СОЦИАЛИСТИЧЕСКИХ РЕСПУБЛИК
ТРУДЫ ЗООЛОГИЧЕСКОГО ИНСТИТУТА т. XXXIX
БИОЛОГИЯ
ПТИЦ
И 3 Д А Т Е Л Ь С Т В О «И А У К А»
МОСКВА 1966 ТЕ1ГПНГРАД
УДК 598.2
Главный редактор
Директор Зоологического института АН СССР
академик Б. Е. Быховский
Редакционная коллегия:
М, Н. Дубинина, А. И. Иванов (редактор тома), И. М. Лихарев,
Г. С. Медведев, А, Н. Световидов
2-10-6
900-66
Р. Л. Потапов
ПТИЦЫ ПАМИРА
R. L. Potapov. The birds of the Pamirs
СОДЕРЖАНИЕ
Стр.
Предисловие 3
Природные условия Памирского нагорья 9
Систематический обзор гнездящихся на Памире птиц 16
Сезонная жизнь птиц Памирского нагорья 81
Орнитогеография Памирского нагорья 99
ПРЕДИСЛОВИЕ
Памирское нагорье — самый высокий горный массив в нашей стране.
Географическое положение его на границе Передней и Центральной Азии,
своеобразные ландшафты, напоминающие Тибет, исключительно суровые
для этих широт природные условия — все это привлекало на Памир много-
численных исследователей, в том числе и зоологов.
Первые сведения о птицах Памира доставил натуралист Ф. Столичка,
посетивший южную часть нагорья в 1874 г. Путевые заметки Столичкп
вместе с описаниями птиц, собранных его спутником Биддульфом, были
опубликованы Б. Шарпом (Sharpe, 1891).
В 1877 г. на Памир через перевал Кызыларт поднялся знаменитый
русский зоолог Н. А. Северцов. Плохая ноябрьская погода не позволила
ему проникнуть в глубь нагорья, и, не дойдя до оз. Каракуль, он повернул
обратно. В следующем, 1878 г. Северцов совершил новое путешествие на
Памир, пройдя на этот раз через значительную часть нагорья, до озер
Яшилькуль и Сассыккуль на юг. Несмотря на то, что Северцов был на
Памире во второй половине августа и сентябре, он собрал значительный
материал о гнездящихся и пролетных птицах Памира. В ряде работ,
опубликованных в 1879, 1880 и 1883 гг., Н. А. Северцов дал хорошее
описание авифауны Памира и ее особенностей. Сборы Н. А. Северцова
хранятся в коллекциях Зоологического института АН СССР (ЗИН
АН СССР).
В 1884 г. на Памире работал геолог Д. Л. Иванов, посетивший почти
все районы нагорья. Не будучи зоологом, он, однако, дал очень интерес-
ное описание фауны Памира (1885). *
В 1887 г. Г. Е. и М. Е. Грум-Гржимайло, процдя в июле через перевал
Кызыларт, спустились в котловину qp,. Каракуль, затем прошли в котло-
вину оз. Рангкуль и через хр. Сарыкол перевалили в китайскую часть
Памира — Тагдумбаш-Памир. Дойдя до истоков р. Дандим-Дарьи, экс-
— 3 -
1*
педиция перевалила через южную часть хр. Сарыкол и по рр. Бейку,
Аксу и Акбайталу вернулась обратно. Небольшие коллекции, очень плохо
датированные, хранятся в ЗИН АН СССР.
В 1890 г. на северных склонах Куньлуня, близ юго-восточной окра-
ины Памира (уроч. Тохтахон), работала экспедиция М. В. Певцова.
Участник экспедиции П. К. Козлов собрал небольшую коллекцию птиц,
хранящуюся в ЗИН АН СССР; результаты наблюдений опубликованы
(Козлов, 1899).
В 1892 г. на Памире собирал птиц Маурер; его сборы хранятся в Зо-
ологическом музее Московского государственного университета.
В 1894 г. через Тагдумбаш-Памир прошел В. Л. Эббот, сборы которого
обработал Ч. Ричмонд (Richmond, 1896).
В 1895 г. в работах русско-английской разграничительной комиссии
принимал участие английский натуралист А. Олкок. Материалы, собран-
ные Олкоком на юго-восточной окраине Памира, опубликованы им
в 1897 г. (Alcock, 1897).
В 1901 г. небольшую коллекцию птиц собрал на Памире зоолог
Н. В. Богоявленский. Его коллекция хранится в Зоологическом музее
Московского государственного университета. В это же время на Памире
работал Д. Н. Головнин, опубликовавший свои наблюдения (Головнин,
1901).
В 1909 г. в Тагдумбаш-Памире (уроч. Тагарма и оз. Малый Каракуль)
коллектировал для М. А. Мензбира М. Н. Дивногорский. Его сборы
хранятся в коллекции ЗИН АН СССР. В 1936 г. А. М. Судиловская
обработала эти сборы и опубликовала результаты обработки.
Летом 1913 г. Памир посетил Л. А. Молчанов. Сборы были им обра-
ботаны совместно с Н. А. Зарудным и результаты обработки опубликованы
в 1915 г.
После установления на Памире Советской власти, с конца 20-х годов,
началось интенсивное исследование территории нагорья. В 1928 г. была
организована Советско-германская экспедиция, которой был собран и не-
большой орнитологический материал, обработанный впоследствии
А. Я. Тугариновым (1930). Немецкий энтомолог и зоогеограф В. Райниг,
принимавший участие в этой экспедиции, опубликовал интересную ра-
боту по зоогеографии Памира (Reinig, 1930). В 1929 г. небольшую коллек-
цию птиц в Заалайском хребте собрал препаратор Зоологического инсти-
тута АН СССР В. И. Шель.
В 1930 г. английский орнитолог Ф. Ладлоу прошел из Кашгара в Каш-
мир через китайскую часть Памира. Результаты исследований он опуб-
ликовал совместно с Н. Кинниром (Ludlow, Kinnear, 1933—1934).
В 1932 г. Зоологический отряд Таджикской комплексной экспедиции
под руководством М. П. Розанова пересек Памир с севера на юг по обыч-
ному маршруту. Коллекция птиц, собранных отрядом, обработана
Г. П. Дементьевым (1935) и хранится в ЗИН АН СССР.
Все эти исследования проводились маршрутным порядком и не могли
дать достаточно полного представления о фауне Памира. Только с 1934 г.
началось стационарное изучение птиц Памира. В этом году Р. Н. Меклен-
бурцев работал на западной границе Памира в верховьях р. Шахдара и
на оз. Яшилькуль. В 1936 г. эти работы были продолжены. Зиму 1936/37 г.
он зимовал в центре нагорья (уроч. Башгумбез). В ряде работ, опубли-
кованных Р. Н. Мекленбурцевым, содержится много интересных сведений
о гнездовой и зимней жизни птиц Памира.
В 1940 г. была опубликована работа А. И. Иванова «Птицы Таджики-
стана», в которой были сведены все данные о птицах Памира, известные
к 1937 г., когда была закончен? рукопись книги.
— 4 —
В 1950 г. появилась работа А. Б. Кистяковского, основанная на мате-
риалах, собранных в пределах Бадахшана в 1937 г. Коллекции погибли
во время войны.
С 1955 по 1960 г. на Памире работал орнитолог И. А. Абдусалямов,
который провел стационарные исследования орнитофауны в районе Па-
мирской биостанции, у оз. Рангкуль и в районе Кызылрабата. Им опубли-
кован ряд работ, где впервые освещается биология многих птиц нагорья.
Большой коллекционный материал, собранный автором, а также
А. В. Поповым, который коллектировал здесь в 1955 г., хранится в Ин-
ституте зоологии и паразитологии им. акад. Е. Н. Павловского АН Тад-
жикской ССР (ИЗИП) и частично был использован при написании настоя-
щей работы.
Зимой 1960/61 г. на Памире работал зоолог того же института
Л. А. Арутюнов. Некоторые устные сведения, любезно сообщенные мне
Л. А. Арутюновым, также были использованы в этой работе.
Мне удалось посетить Памир в летние сезоны 1954, 1956, 1958, 1960,
1961 и 1962 гг. Исследования проводились как на маршрутах, охвативших
основные районы нагорья, так и на стационарах. Стационары, числом 11,
были расположены в наиболее характерных, «модельных», местах Памир-
ского нагорья. Основным стационаром все годы являлась Памир-
ская биологическая станция АН Таджикской ССР.1 Станция располо-
жена в уроч. Чечекты, на конусе выноса речки того же названия, у юж-
ного подножия западной части хр. Музкол, на высоте 3860 м над ур. м.
В окрестностях станции представлены все характерные биотопы нагорья,
кроме озер и приозерных лугов. К самой станции подходят полупустынные
ровные пространства, занимающие большую часть долины р. Акбай-
тала и нижние части склонов окружающих гор. Субальпийские прибрежные
луга тянутся вдоль речек Мукора и Чечекты. Альпийские луга встре-
чаются в верхнем течении этих речек, по склонам гор, в местах с избыточ-
ным увлажнением и в ледниковых цирках Мукор и Чечекты, располо-
женных на высоте 4600 и 4710 м над ур. м. На склонах гор, покрытых скуд-
ной полупустынной растительностью, широко представлены осыпи и
скалы. При выходе из цирков встречается типичный моренный ландшафт.
Небольшой висячий ледник спускается к цирку Зор-Чечекты, сползая
с северного склона пика Караджилга (он же Зор), высотой 5680 м. На хреб-
тах, окружающих пик, хорошо выражены субнивальный и нивальный
пояса. Линия вечных снегов проходит на высоте 5300 м на южном склоне
и 5100 м на северном.
Стационар Аличур-1 был расположен в среднем течении р. Аличура,
у оз. Акбалык (3800 м), среди обширных болотистых лугов с небольшими
озерками, тянущихся по обоим берегам реки. С лугами граничат полу-
пустыни, которые занимают более возвышенные части долины и пологие
склоны окружающих гор. Долины правых притоков Акджилги и Базар-
дара в нижних своих частях представляют ровные поверхности со слабым
наклоном, а в верховьях резко сужаются и приобретают типичный аль-
пийский характер. Работа на стационаре проводилась в июне—июле
1954 г.
Сходные местообитания представлены и на стационаре Аличур-2,
но по причине большей высоты (4000 м) луга здесь меньше развиты. Над
правым берегом долины возвышается крупный километровой высоты
скальный массив. Работа здесь велась в августе 1954 и в июне 1961 гг.
Стационар Яшилькуль располагался у места впадения в озеро р. Али-
чура (3734 м). При впадении река образует род дельты, где хорошо раз-
виты луга, как и по всей нижней части долины. У устья Аличура находятся 1
1 Далее в тексте сокращенно — Биостанция.
— 5 -
оошпрпые, хорошо прогреваемые солнцем мелководья озера, кое-где
растет тростник. Остальные берега озера почти лишены растительности,
и большая глубина начинается сразу же у берегов. Работа здесь проводи-
лась в июле и сентябре 1954 г. и в июле 1962 г.
Стационар Зоркуль находился на восточном берегу этого второго по
величине озера Памира, среди нагромождений древних морен. Озеро
расположено на высоте 4125 м в котловине, занимающей обширное пони-
жение между Ваханским и Южноаличурским хребтами. Озеро по проис-
хождению моренно-подпрудное и образовалось после того, как морены
максимальной фазы оледенения перегородили западную часть котловины.
Рис. 1. Долина р. Аксу у скального массива Акташ.
Кое-где по берегам озера тянутся узкие полоски лугов, которые больше
всего развиты на восточном берегу, в месте впадения в озеро проток, сое-
диняющих его с небольшими озерами Кукджигит и Чаканкуль. На об-
ширных плоских поверхностях, занимающих большую часть котловины,
а также на моренах развиты пустынно-степные растительные ассоциации.
В восточной части озера, более мелкой, есть несколько островов. Работа
на этом стационаре проводилась в сентябре 1956 г.
Стационар Каракуль был организован на юго-западном берегу озера,
у устья Музкола. Каракуль — самое крупное озеро Памира: площадь
водного зеркала 369 км2, а максимальная глубина 238 м. Вода в озере
горько-соленая. Берега лишены растительности, только в нескольких
местах имеются небольшие пятна сильно засоленных лугов. Погребен-
ные льды залегают во многих местах по берегам и частично покрывают
дно озера. Большая часть котловины занята пустынными растительными
формациями, местами имеются пески. Работа здесь проводилась в июне
1961 г. и в июне—июле 1962 г.
Стационар Кызылрабат помещался в верхнем течении Аксу, на вы-
соте 4000 м. Вниз по реке от стационара простираются обширные площади
лугов, занимающих почти всю пойму, где имеется целый ряд мелких
озер. На надпойменных террасах развиты обычные полупустыни. Ближе
- 6 —
к окружающим хребтам залегает широкая полоса древних морен, по-
крытых местами степной растительностью. Впадающие в Аксу реки Беик,
Ханюлы, Кашкасу и Бахмыр сравнительно многоводны и текут по узким
долинам, отличающимся довольно теплым климатом, так как сюда
в меньшей степени проникают холодные ветры. Долина р. Кызылрабата
завалена древними моренами; она более холодная и меньше увлажнена.
Здесь господствует полупустынная растительность. По островкам на
Рис. 2. Скалы в альпийском поясе.
р. Аксу, так же как и в ущелье р. Бейка, кое-где имеются заросли ивня-
ков, иногда достигающие 1 м в высоту. Над правым берегом поймы воз-
вышается огромной стеной полуторакилометровой высоты скальный мас-
сив Акташ (рис. 1). По левому берегу тянутся пологие горы с выходами
крутых скал — массив Аюджолу. Работа здесь велась в июле и августе
1954 г., в августе 1956 г. и в июле 1961 г.
Стационар грот Шахты располагался на конусе выноса у подножия
южной оконечности хр. Найзаташ, с восточной, подветренной стороны,
на высоте 4100 м. Под конусом простирается обширная долина Куртек-
сай, совершенно безводная. На склонах гор вокруг стационара много
скал и осыпей, с верхней части хребта стекают небольшие ручейки, не
достигающие его подножия (рис. 2). Хребет в этом месте немногим выше
— 7 —
5000 м, ледников здесь нет, и только у вершинного гребня залегают не-
большие снежные поля. Работа здесь велась в июле—августе 1960 г.
Стационар Кокуйбель, в верхнем течении этой реки (3800 м), поме-
щался на надпойменной террасе, почти совершенно лишенной раститель-
ности. Значительная часть поймы занята лугами, сильно засоленными.
Здесь проходит верхняя граница ивы, которая на своем верхнем пределе
представлена небольшими угнетенными растениями около 20 см высотой.
Окружающие склоны гор и обширные ровные пространства долины пред-
ставляют собой типичную полупустыню. Наблюдения здесь проводились
в июне—июле 1958 г.
Рис. 3. Котловина оз. Рангкуль и хр. Музкол.
Стационар Рангкуль был расположен на озере того же названия.
Проточное, пресное озеро Рангкуль соединяется протокой с бессточным,
соленым озером Шоркуль. По берегам Рангкуля расположены значитель-
ные площади лугов, у берегов сильно заболоченных. Выше, над лугами,
располагаются ровные пространства котловины, покрытые полупустын-
ной растительностью (рис. 3). Кое-где встречаются небольшие участки,
занятые степными ассоциациями. С южной стороны котловины над озе-
рами возвышаются высокие отвесные скалы. На это озеро совершались
регулярные выезды в течение всей работы автора на Памире.
Стационар Шитхарв помещался в кишлаке того же названия в южном
Бадахшане, на высоте 2650 м. Долина Пянджа здесь почти сплошь за-
нята садами и посевами. В ущельях Шитхарв и Сарышитхарв пред-
ставлены типично бадахшанские ландшафты — необычайно крутые склоны
гор, почти лишенные растительности, гигантские осыпи и стремительные
потоки, которые несутся по дну ущелий, окруженные узкой лентой ту-
гайных зарослей из ивы, тополя, березы, облепихи, смородины, шипов-
ника и других пород. Автор работал здесь в июле 1956 г.
Все стационары располагались в субальпийском поясе. Однако всюду
предпринимались многочисленные экскурсии в выше расположенные
пояса — альпийский и субнивальный. Лучше всего во время таких экс-
— 8 —
курсий был изучен альпийский пояс хребтов Базардара, Музкол и Най-
заташ.
Вертикальное распределение птиц, гнездящихся на нагорье, выясня-
лось только путем находок возможно большего числа гнезд. Одновре-
менно с визуальными наблюдениями изучался микроклимат гнездовых
убежищ с помощью минимальных, максимальных и срочных термометров^
а в некоторых случаях — температурный режим насиживания. Последний
выяснялся при помощи микроэлектротермометров: датчик-термистор
помещался в подстилке гнезда, среди яиц, а от него за какое-либо укрытие
шел провод, к которому в нужное время можно было, не вспугивая птицы,
подключать сам прибор.
Помимо изучения гнездовой жизни, собирались материалы и о других
сторонах птичьей жизни — питании, сезонной периодичности явлений,
численности и ее колебаниях. Было найдено и изучено свыше 400 гнезд
с птенцами или яйцами 27 видов птиц, кроме того, были собраны много-
численные сведения о размножении еще 21 вида. Во время полевых работ
удалось собрать свыше 600 экз. тушек птиц 74 видов, большая часть кото-
рых поступила в коллекции ЗИН АН СССР.
Кроме собственных коллекционных сборов и дневников, был использо-
ван богатый коллекционный материал Зоологического института АН СССР,
собранный в свое время в пределах Памира Н. А. Северцовым, Г. Е. и
М. Е. Грум-Гржимайло, Л. А. Молчановым и другими, всего около
700 экземпляров птиц.
Во время проведения полевых работ автору была оказана неоценимая
помощь директором Памирской биологической станции К. В. Станюко-
вичем и сотрудником этой станции Л. Ф. Сидоровым, а в последние годы
также директором Хорогского ботанического сада А. В. Гурским. При на-
писании настоящей работы большую помощь оказали автору советы и
указания сотрудников Орнитологической лаборатории Зоологического
института АН СССР — А. И. Иванова, Е. В. Козловой, И. А. Нейфельдт
и К. А. Юдина. Немало способствовали выполнению настоящей работы
ценные консультации, данные Б. К. Штегманом. Автор пользуется слу-
чаем выразить всем этим товарищам свою глубокую благодарность. Осо-
бенно благодарен автор профессору Л. А. Портенко за ценные советы,,
критические замечания и то большое внимание, которое он уделил автору.
Фотографии, иллюстрирующие текст, выполнены автором.
ПРИРОДНЫЕ УСЛОВИЯ ПАМИРСКОГО НАГОРЬЯ
Физико-географическое положение, границы.
Памир входит в систему горных поднятий, обрамляющих Центральную
Азию с запада. Своеобразие его природы в значительной степени обуслов-
лено тем, что он расположен на стыке высочайших хребтов Азии:
Тянь-Шаня, Куньлуня, Каракорума и Гиндукуша, причем гребни этих
хребтов и смежных поднятий изолируют Памир от равнин северной
Индии, Средней и Центральной Азии.
Границы Памирского нагорья четко выражены горными хребтами.
Северной границей служит Заалайский хребет с главной вершиной —
пиком Ленина (7134 м над ур. м.). Особенно величественно выглядит этот
хребет с севера, из Алайской долины, над которой он возвышается ледя-
ной стеной четырехкилометровой высоты. Снеговая линия проходит здесь
на высоте 4300—4500 м, а многочисленные ледники спускаются к самому
подножию хребта. Заалайский хребет служит мощным барьером для воз-
душных масс, несущих осадки с севера и северо-запада. По этой причине
южные склоны хребта, обращенные к нагорью, осадков почти не получают
и являются одним из самых сухих мест Памира. Это обстоятельство вместе
- 9 -
с южной экспозицией склонов сказалось на высоте снеговой линии, кото-
рая проходит здесь выше, чем где-либо в СССР, на уровне 5200 м (Забиров,
1955), а по некоторым данным — даже на высоте 5500 м (Станюкович,
1960), т. е. на целый километр выше, чем на северных склонах хребта.
Фактической природной границей нагорья служит снеговая зона хребта,
так как обильно увлажненные северные склоны ниже снеговой линии
имеют совершенно иной, «алайский» природный облик.
На востоке границу Памира составляет целая система хребтов (Кинг-
тау, Кашгарский, южная часть Сарыкольского), простирающаяся от
долины р. Маркансу на юг до долины р. Ташкургана. Наиболее высок
из них Кашгарский хребет с вершинами Конгур (7719 м) и Музтагата
(7545 м). К югу от этого хребта граница, огибая с севера долину р. Ташкур-
гана, переходит на хр. Сарыкол и продолжается по нему к югу вплоть
до долины р. Вахджирджилги. Снеговая линия на западных склонах
Сарыкола проходит на высоте 5000 м (Забиров, 1955). О высоте ее на во-
сточных склонах сведений нет, но некоторые данные (Юнатов, 1961) го-
ворят о значительно большем увлажнении восточных склонов, а следо-
вательно, и о более низком положении снеговой линии. Считать восточной
границей Памира весь хребет Сарыкол, как это делают некоторые (Ки-
стяковский, 1950; Забиров, 1955), на наш взгляд, неправильно. Все
долинные понижения между северной частью названного хребта и хреб-
тами Кингтау и Кашгарским имеют вполне памирский облик. Эта часть
Памира нередко именуется в различных источниках Тагдумбаш-Памиром.
Южная граница Памира проходит по хр. Гиндукуш от перевала Баро-
гпль (3804 м) до перевала Килик (4755 м) и далее на восток по широтной части
хр. Музтаг, обрамляющей с юга долину Вахджирджилги. Верховья этой
реки, вплоть до слияния ее с р. Хунджеробом, по своей природе, вероятно,
также являются частью Памирского нагорья. Грандиозные гребни Гин-
дукуша и Каракорума, со средними высотами 5500—6500 м, почти нацело
перехватывают осадки, несомые индийскими муссонами. Львиная их доля
выпадает на южных склонах вышеназванных хребтов. Северные склоны,
обращенные к Памиру, получают осадков намного меньше. Только бла-
годаря северной экспозиции и местным особенностям горно-долинной
циркуляции воздушных масс здесь развито значительное оледенение и
снеговая линия проходит на высоте всего 4800 м.
Западная граница Памира не столь резка, как предыдущие, но тем не
менее достаточно отчетлива. Разными авторами она проводится примерно
одинаково от хр. Академии Наук до устья р. Маца, впадающей в р. Памир.
Положение этой границы в значительной степени обусловлено наличием
ряда крупных меридиональных поднятий: хр. Академии Наук, хр. Зу-
лумарт, группа пика Советских Офицеров в хр. Музкол, массив Бакчи-
гир и другие вершины (Сидоров, 1960). Вдоль этой границы между Пами-
ром и Бадахшаном залегает неширокая переходная полоса, имеющая
смешанные природные черты как Памира, так и Бадахшана. В некоторых
местах этой переходной полосы, среди глубоких ущелий бадахшанского
типа расположены своеобразные островки памирской природы на высо-
ких (4100—4300 м) перевальных плоскостях в верховьях некоторых рек
(Рамаиф, Марджанай и др.). К западу от указанной границы долины рек,
впадающих в Пяндж, приобретают каньонообразный характер и создают
резко расчлененный ландшафт, сильно отличающийся от ровных, широких
долин Памира. Обращает на себя внимание определенная высота, на ко-
торой проходит западная граница Памира при пересечении поперечных
долин (Баляндкиик, Кокуйбель, Западный Пшарт, Мургаб, Памир,
Вахан-Дарья и др.). Эта высота колеблется в небольших пределах (3500—
3700 м) и в конечном итоге совпадает с верхней границей древесно-кустар-
никовой растительности в указанных долинах. Напрашивается вывод,
- 10 -
что там, где нет четких границ в виде окраинных хребтов, граница Памира
определяется именно этим признаком. Автор имел возможность посетить
пограничные области между Памиром и Алаем, между Памиром и Бадах-
шаном, и везде природная граница нагорья являлась также и границей
распространения характерного высокогорного орнитокомплекса Памира,
сильно отличающегося от орнитофауны Алая и Бадахшана. Судя по ли-
тературным данным, эта граница также хорошо прослеживается на во-
стоке и юге нагорья.
Таким образом, Памир в настоящей работе принимается в тех границах,
в которых понимает его большинство исследователей, начиная с Н. А. Се-
верцова, причем считается невозможным объединение его с Бадахшаном,
что делают некоторые авторы, прежде всего по фаунистическим сообра-
жениям. Принципиальная разница между Памиром и Бадахшаном уже
достаточно подробно освещена в литературе (Кистяковский, 1950; Станю-
кович, 1952; Сидоров, 1960, и др.), поэтому мы считаем необходимым на
данном этапе наших знаний отказаться от неправильной терминологии,
по которой Бадахшан именуется Западным Памиром, а собственно Памир
Восточным Памиром.
Орография. Окраинные хребты Памира по высоте значительно
превосходят горы, расположенные во внутренних частях нагорья, и Памир
в целом можно представить себе как своеобразную чашу с приподнятыми
краями (Гвоздецкий, 1959). Такое чашеобразное или ячеистое строение
отдельных горных систем вообще характерно для Центральной Азии (Си-
ницын, 1959). Само Памирское нагорье представляет собой систему широ-
ких озерных котловин и речных долин, поднятых на высоту 3600—4200 м
над ур. м. и разделенных хребтами небольшой, до 1—2 км, относительной
высоты, сильно разрушенными денудационными процессами. Большин-
ство хребтов вытянуто с востока на запад и наибольшую высоту имеет
в западных частях. Между ними залегают широкие долины. Для централь-
ной части нагорья характерны крупные бессточные озерные котловины
(Каракульская, Рангкульская), имеющие в диаметре 20—40 км.
К югу от Рангкульской котловины вплоть до Ваханского хребта тя-
нутся системы небольших, ниже 5000 м, хребтов, разделенных широкими
•безводными долинами. Весь этот район прорезает с юго-востока на се-
веро-запад широкая долина самой крупной реки Памира — Аксу. После
слияния с р. Акбайталом Аксу круто поворачивает на запад, уже под
названием Мургаб. Еще одна долина меридиональной ориентации зале-
гает между Сарыкольским и Кашгарским хребтами, к востоку от Рангкуль-
ской котловины.
Реки и озера. С ледников Памира берет начало целый ряд рек,
являющихся основными истоками Аму-Дарьи. Это прежде всего р. Аксу,
с притоками Беик, Истык и Западный Пшарт, которая при выходе из
Сарезского озера получает название Бартанг, затем Вахан-Дарья, Памир,
Аличур, Кокуйбель и Баляндкиик. Это самые крупные реки Памира,
вода в которых есть круглый год. Другие реки, нередко текущие по очень
широким и длинным долинам (Восточный Пшарт, Акбайтал), существуют
только летом, во время таяния снегов. Целый ряд речек стекает с гребней
снежных хребтов по боковым долинам, причем большинство из них даже
в период максимального таяния снегов не достигает основных водных арте-
рий или же достигает их на очень короткое время в отдельные часы суток.
Такие потоки берут начало у ледников и снежников на вершинах хребтов
и протекают по довольно узким долинкам с крутым падением и обрыви-
стыми берегами. Особенно много таких речек в высоких хребтах, как на-
пример Музкол, Базардара, Южноаличурский. В отличие от этих бурных
речек крупные реки Памира, протекающие по пологим долинам, имеют
более или менее спокойное течение, меандрируют в рыхлых наносах и
— 11
в июле, во время бурного таяния снегов местами выходят из берегов,
затопляя прибрежные луга. Все реки Памира — ледникового питания,
и пик паводка бывает в июле — самом теплом месяце лета. Весенний паво-
док, вызываемый таянием снегов субальпийского пояса, выпавших за
зиму, невелик, однако воды этого паводка успевают проделать большую
эрозионную работу.
Памир богат озерами, в этом отношении он напоминает Тибет. Здесь
имеется 20 сравнительно крупных озер и около трех десятков мелких.
Самое крупное озеро Памира — Каракуль. Второе по величине озеро —
Рис. 4 Оз. Рангкуль.
Зоркуль, немногим уступает ему по величине Яшилькуль. Затем идет
моренно-подпрудное пресное озеро Чакматинкуль, откуда берет начало
р. Аксу, пресное озеро Рангкуль (рис. 4) и соленое Шоркуль, расположен-
ные в одной котловине, соленое озеро Сассыккуль к юго-востоку от Яшиль-
куля, проточные озера Чаканкуль и Кукджигит в Зоркульской котловине
и проточное озеро Малый Каракуль в долине р. Кешкиберсу (китайская
часть Памира). Мелкие озера, площадью менее 1 км2, которых на Памире
более 30, по своему происхождению и расположению делятся на две
группы. Одни из них располагаются в обширных речных долинах и озер-
ных котловинах среди заболоченных лугов и образовались в результате
таяния погребенных под почвой льдов. Таких озер много в верховьях
долины Аксу и в Аличурской долине. Озера другой группы разбросаны
по перевальным плоскостям и или образовались при заполнении впадин
талыми водами, или же имеют моренно-подпрудное происхождение. В не-
которых местах есть озера, пересыхающие в засушливые годы. Многие
озера Памира, так же как и в Тибете, носят явные следы усыхания. Озера
Каракуль, Рангкуль, Сассыккуль и многие другие в отдаленном прошлом
имели гораздо' большие размеры. Воды Каракуля, например, заполняли
всю его огромную котловину и, как доказывает ихтиофауна озера (Берг.
1931), имели сток в долину р. Кокуйбеля.
— 12 —
Во многих местах нагорья, главным образом в верховьях Аксу и в до
лине Аличура, имеются теплые и горячие источники. Нередко такие ис-
точники выходят на земную поверхность на дне озер и рек, благодаря
чему целые участки водной поверхности в таких местах не замерзают
в течение всей зимы.
Климат. Факторы, определяющие основные черты памирского
климата, следующие:2 1) географическое положение, близкое к субтро-
пическим шпротам (37—39° с. ш.); 2) значительная удаленность от океа-
нов; 3) положение в области высочайших хребтов континента и огромная
высота над уровнем океана; 4) большая высота окружающих хребтов
по сравнению с уровнем остальных пространств нагорья; 5) широкое
распространение ровных долинных пространств — основных геоморфо-
логических элементов нагорья — и небольшая относительная приподня-
тость над ними горных массивов, имеющих к тому же пологие, слабо
изрезанные склоны; 6) разреженный растительный покров, использую-
щий незначительную часть солнечной энергии и не препятствующий
излучению ее с поверхности; 7) почвенный покров, способный благодаря
скелетности своего строения к внутрипочвенной конденсации влаги,
что имеет существенное значение для микроклимата; 8) слабое развитие
зимой снегового покрова — только в южной части нагорья и на короткий
срок.
Формирующийся под влиянием перечисленных факторов климат Па-
мира является прежде всего резко континентальным. Для Мургаба пока-
затель континентальности равен 248%, тогда как максимальная конти-
нентальность, известная па земле, равна 263% (Хотан). Следует заметить,
что климатические условия в Мургабе гораздо мягче, чем во многих рай-
онах нагорья, и в некоторых из них величина континентальности должна
быть еще более высокой, приближаясь к максимальным, существующим
на земле. Основная особенность континентального климата — резкие
колебания температур — выражена на Памире в полной мере. Самая
низкая температура, отмеченная в январе на высоте 4000 м, —50° С, самая
высокая в июле, на высоте 3700 м 31 ° С. Годовая амплитуда достигает,
следовательно, 81°. Максимальные суточные колебания: в будке 31 °,
на почве 60°.
Осадков в целом по нагорью очень мало, не более, чем в пустынях
Средней Азии (50—300 мм в год). Высокая сухость нагорья обусловлена
тем, что окружающие горные хребты перехватывают почти все осадки.
Южная часть нагорья получает несколько больше осадков, чем северная.
Летом сюда докатываются влажные массы индийского муссона, а зимой
повышенную влажность создают местные особенности горно-долинной
циркуляции воздуха.
Инсоляция на Памире приближается к величинам, максимальным для
земного шара (1.62 кал. на 1 см2 в сентябре, в полдень). Этому способ-
ствует почти всегда ясное небо. Среднее количество облачных дней для
Мургаба — 3.9 в месяц.
Температура, влажность, годовое количество осадков весьма различны
в разных частях нагорья. Каждому поясу свойствен свой климат, причем
климатические различия между поясами весьма значительны.
В субальпийском поясе только 4 месяца имеют средние температуры
выше 0°; однако ночные заморозки продолжаются все лето п безморозный
период практически отсутствует. При ночных заморозках в мае и июне
температура падает иногда до —10°, а в августе и сентябре до —18°.
Особенно сильны ночные заморозки в обширных понижениях — озерных
котловинах пли речных долинах, куда ночью с окружающих гор стекает
2 По Л. Ф, Сидорову (1960).
— 13 —
холодный воздух и где образуются так называемые «озера холода». На
склонах окружающих гор ночные температуры выше. Климат субальпий-
ского пояса неодинаков: южные части заметно теплее, чем северные.
Здесь сказывается разница в широте в полтора земных градуса. Заметна
разница и в количестве осадков: чем ближе к окраинам нагорья, тем их
выпадает больше. Поэтому наиболее увлажнены районы, примыкающие
к горному обрамлению Памира, за исключением Каракульской котловины,
а самые сухие расположены в центральных частях нагорья. Здесь есть
места, где осадков иной раз выпадает не более 20 мм в год. Но и в наиболее
влажных местах субальпийского пояса количество осадков не превышает
300 мм. Особенности горно-долинной циркуляции воздушных масс в юж-
ных частях нагорья (Сидоров, 1960) обусловливают возможность ежегод-
ного, хотя и кратковременного существования снегового покрова, отсут-
ствующего в остальных районах нагорья. Малоснежность — одна из
характерных черт зимы субальпийского пояса Памира. Следует заметить,
что для Памира характерен весенне-летний максимум осадков, что свой-
ственно всей Центральной Азии. Воздух субальпийского пояса очень сух;
среднемесячная влажность особенно низка летом. Среднегодовая влаж-
ность 32.3%, гораздо ниже, чем в пустынях Средней Азии, хотя в июле—
августе она примерно такая же. Летом в течение суток она может падать
до абсолютных минимумов в 5—10%. В соответствии с количеством выпа-
дающих осадков влажность воздуха выше в окраинных частях нагорья.
Альпийский пояс имеет более холодный и более влажный климат.
Средняя годовая температура в альпийском поясе хр. Музкол равна
—8° С, средняя температура трех летних месяцев колеблется от 5 до 8° С.
Верхнюю часть альпийского пояса захватывает полоса максимальных
осадков, расположенная на Памире в пределах 4600—5000 м над ур. м.
(Станюкович, 1960): здесь в год выпадает 350—400 мм осадков. Распределе-
ние осадков по альпийскому поясу нагорья подчинено тем же закономер-
ностям, что и в субальпийском. На окраинных хребтах Памира, особенно
на внешних их склонах, осадков выпадает столько, что здесь существует
мощное оледенение.
Субнивальный пояс, который можно назвать поясом тающих снежни-
ков (от 4800 м и до снеговой линии), довольно узок. Высота и ширина
его колеблется в зависимости от экспозиции склона и высоты снеговой
линии. Этот пояс характеризуется еще более низкими температурами
(средняя температура июля 2 ° С) и большим количеством осадков (да
500 мм в год). Субстрат здесь летом всегда сырой, пропитан снеговой
водой. Ночью почва покрывается ледяной коркой. В этом поясе вообще
не бывает безморозных дней.
Нивальный пояс, или пояс вечных снегов, характеризуется почти
полным отсутствием жизни. Снег и лед покрывают всю поверхность гор,
и круглый год здесь царит мороз. Этот пояс простирается от линии веч-
ных снегов до самых высоких точек хребтов и местами занимает большие
пространства, образуя ледяную пустыню.
Положение линии вечных снегов в разных районах нагорья весьма
варьирует, находясь в тесной зависимости от экспозиции склонов, коли-
чества осадков и географической широты местности. В целом же для Па-
мира характерно необычайно высокое, выше, чем где-либо в СССР, поло-
жение снеговой линии, в среднем на высоте 5100 м.
Для всех поясов Памира характерны сильные ветры, преимущественно
западных румбов, дующие в течение почти всего года. Наибольшей силы
они достигают в широких долинах, ориентированных с запада на восток
(Зоркуль—Памир, Аличур, Маркансу) и представляющих собой как бы
гигантские аэродинамические трубы. Ветры свирепствуют здесь непре-
рывно, иногда утихая только на рассвете. Низкие температуры,, господ-
— 14 —
ствующие на открытых пространствах нагорья, в значительной степени
обусловлены охлаждающим действием ветров. В этом легко убедиться,
сравнив для примера климат просторной долины верхней Аксу и одной
из узких боковых долинок в массиве Акташ. Небольшие извилистые ска-
листые ущелья в этом массиве, каких много на Памире, хорошо защищены
от холодных ветров, что сразу сказывается как на растительности, так
и на животном мире. Во многих закрытых от ветра ущельях ива идет
вверх до 4100 м, тогда как в открытой ветрам долине Аксу низкорослые
кустики ивы едва достигают высоты 3900 м.
Необходимо подчеркнуть, что климат каждой долины, каждой котло-
вины и каждого склона в значительной степени зависит от местных усло-
вий (орография, экспозиция, глубина долинного вреза и т. д.). Поэтому
при общем господстве сурового континентального климата в различных
местах нагорья условия сильно варьируют и холодные места чередуются
со своеобразными климатическими оазисами. В этом отношении средние
показатели, характеризующие как климат нагорья в целом, так и климат
отдельных поясов, недостаточны для понимания особенностей распреде-
ления животного мира. В таких случаях важен микроклимат, имеющий
решающее влияние во многие ответственные моменты жизни организма.
Напомним, что солнечный нагрев на Памире очень силен, и не будь по-
стоянного охлаждающего влияния сильных ветров, здесь летом царила бы
настоящая жара. Поэтому в защищенных от ветра местах на южных и
восточных склонах температура бывает довольно высокой даже в альпий-
ском и субнивальном поясах. В щелях под камнями осыпи, например,
на высоте 4700 м температура воздуха в середине августа достигала 39°,
а ночью не опускалась ниже 6° С. Подобных температурных убежищ осо-
бенно много в скалах и осыпях на самой различной высоте, и их наличие
позволяет многим видам птиц не только гнездиться по всему нагорью, но
и подниматься на гнездовье вверх, до наибольших в пределах СССР высот.
Растительность. Растительный покров Памира очень беден
и носит ярко выраженный пустынный характер. Примечательно полное
отсутствие древесной растительности и почти полное кустарниковой. По-
следняя представлена в основном ивой и встречается или в самых низких
частях нагорья, или же в хорошо защищенных от ветра теплых ущельях,
как например Бахмальджилга и Беик. В ущелье Бахмальджилги заросли
ивы особенно густы и достигают высоты 2 м.
Сомкнутый растительный покров наблюдается только на субальпий-
ских и альпийских лугах, которые занимают ничтожную часть площади
нагорья. Луга эти с весьма скудным видовым составом растительности
приурочены к местам с хорошим почвенным увлажнением и существуют
только за счет грунтовых вод. В субальпийском поясе луга занимают
пониженные пространства у рек и озер. Благодаря мерзлотным процессам,
происходящим в почве, она вся покрыта буграми и ямами, поэтому боль-
шинство лугов Памира по виду напоминает кочковатое болото. Низко-
травность — характерная черта как альпийских, так и субальпийских
лугов. Средняя высота травостоя 11—12 см (Сидоров, 1960). Правда,
в некоторых местах по долинам Аксу и Аличура он может достигать зна-
чительной высоты, до 30, а то и до 60 см, но таких мест очень мало. Наибо-
лее крупные массивы субальпийских лугов расположены в долинах Аксу,
Аличура, Кокуйбеля и в озерных котловинах Каракуля и Рангкуля.
Альпийские луга никогда не занимают больших площадей и представлены
небольшими лужайками по берегам альпийских потоков, на днищах лед-
никовых цирков или на склонах гор, в местах с хорошим почвенным
увлажнением.
Большую часть площади нагорья занимают полупустынные ассоциации
с сильно разреженным растительным покровом. Наиболее характерны
— 15 —
терескенно-полынные формации, занимающие обширные пространства
^межгорных понижений и нижние части склонов гор. Эти формации состоят
из редко разбросанных кустиков полыни или терескена. Низкие, призе-
мистые кустики последнего редко бывают выше 20—25 см, так как их не-
прерывно объедает скот. В местах, где регулярного выпаса нет, терескен-
ники развиты хорошо, кустики его достигают большей высоты и сильно
ветвятся. Терескеновые полупустыни распространены только в суб-
альпийском поясе; выше 4300 м терескен, как правило, в горах не растет.
Кое-где на окраинах нагорья, особенно в южной его части, где зимой
снеговой покров предохраняет растения от вымерзания, встречаются
степные ассоциации, но площади, занимаемые ими, ничтожны.
В альпийском поясе тоже господствуют полупустынные ассоциации,
но уже с другим видовым составом растительности. Во многих местах
преобладают подушечники. Они покрывают разреженными группами
•склоны гор, поднимаясь вверх до субнивальной полосы. Полупустыня,
граничащая с вечными снегами, является характерной для Памира чертой,
свойственной также и Тибету. Местами в нижних частях альпийского
пояса встречаются густые заросли полукустарничков.
Растительность субнивального пояса имеет довольно скудный видовой
состав; она представлена мелкими травянистыми растениями, ютящимися
в щелях между камнями. Их низкие, прижатые к земле побеги вегетируют
в очень сжатые сроки, в конце июля—августе.
Такова в целом краткая характеристика природных условий Памир-
ского нагорья. Основные черты ее — суровый, резко континентальный
климат и скудная растительность полупустынного типа, произрастающая
на обширных равнинах межгорных понижений и пологих склонах гор.
СИСТЕМАТИЧЕСКИЙ ОБЗОР ГНЕЗДЯЩИХСЯ НА ПАМИРЕ ПТИЦ
В настоящее время мы располагаем сведениями о гнездовании 50 видов
птиц в пределах Памирского нагорья. В опубликованных до настоящего
времени списках гнездящихся птиц Памира (Мекленбурцев, 1949; Кистяков-
ский, 1950; Абдусалямов, 1961) содержится соответственно 53, 55 и 42 вида.
Помимо птиц, гнездование которых на Памире доказано, в эти списки
включено еще 13 видов, прямых сведений о гнездовании которых на на-
горье нет, причем для 12 из них гнездование на Памире вообще вряд ли
возможно. Все эти виды будут рассмотрены в настоящем разделе, в таких
случаях названия птиц приводятся без порядкового номера.
Большой баклан — Phalacrocorax carbo sinensis (Schaw et Nodd.).
Регулярно встречается в летнее время на многих озерах и реках нагорья:
на Каракуле, Яшилькуле, на Аксу. Основываясь на этом, Р. Н. Меклен-
бурцев (1949) и И. А. Абдусалямов (1961) посчитали его гнездящимся,
но в действительности у нас нет тому никаких доказательств. Никто еще
не находил на Памире ни гнезд, ни птенцов бакланов. Указание И. А. Аб-
дусалямова (1961) на встречу выводков бакланов на Сарезском озере,
т. е. уже, собственно, в Бадахшане, ошибочно. Мне пришлось быть в это же
время на озере. Там держались не выводки, а группы взрослых птиц,
промышлявших рыбу в устьях рек. Уместно напомнить, что весной первые
закраины на озере появляются только в конце апреля; до этого времени
существование бакланов на скованном льдом озере немыслимо. Следо-
вательно, птицы могут приступить к гнездованию не ранее начала мая,
но тогда у них никак не могут быть летные птенцы в середине июля, когда
вел наблюдения И. А. Абдусалямов.
В конце июня 1961 г. большие скопища бакланов наблюдались в устье
р. Музкола на Каракуле. Сотни птиц собирались сюда утром, к вечеру
улетая ночевать на острова. В 1962 г. одиночки и группы тоже регулярно
— 16 —
кунор, они гнездятся почти на километр ниже, а еще севернее, в бассейне
р. Толы (48—49° с. ш.) гнездящихся гусей находили на высоте всего 1000 м
(Козлова, 1930). На Памире в настоящее время все гнездовья гусей рас-
положены в субальпийском поясе, причем наиболее высоко они гнездятся
на оз. Зоркуль — 4125 м над ур. м. Выше расположенных подходящих
озер на Памире нет, что, видимо, и определяет здесь верхнюю границу
гнездования. В Тибете, где многие озера расположены гораздо выше, гуси
гнездятся до 5000 м.
Численность когда-то была очень высокой. Сейчас она крайне низка.
По моим данным, в 1962 г. на Каракуле обитало 25—30 пар, на Зоркуле —
50—60 пар и на Рангкуле — не более 5 пар. Кроме гнездящихся, на ука-
занных озерах проводит лето некоторое количество холостых птиц, общее
число которых в 1962 г. не превышало 100.
В пределах нагорья гуси гнездятся только на островах крупных прес-
ных и соленых озер. Гнездование на скалах в настоящее время не наблю-
далось, за исключением нескольких сомнительных случаев на Рангкуле.
В конце прошлого века Д. Л. Иванов встречал выводки нелетных горных
гусей по всем мелким озеркам в долинах Аличура и Аксу, на которых
не было островов. Поэтому приходится предполагать, что птенцы были
в данных случаях выведены в скалах и потом приведены к воде. Как из-
вестно, в Тибете и Ладаке при наличии крупных озер с островами гуси
гнездятся именно там, но при отсутствии последних они переходят на
гнездование в скалах (Бианки, 1907; Osmaston, 1928; Schafer, 1938).
После окончания сезона размножения горные гуси появляются и на дру-
гих озерах нагорья.
Горный гусь обладает сравнительно легким, сильным полетом и спо-
собен к довольно сложным маневрам в воздухе, что особенно необходимо
при гнездовании в скалах. Некоторые наблюдатели (Пржевальский, 1876)
считают, что они при спуске кувыркаются через голову, но у меня сложи-
лось впечатление, что они просто падают вниз, переваливаясь с крыла на
крыло. Когда стая снижается таким образом с большой высоты, то слы-
шится сильный шум, наподобие того, какой издает на ухабистой дороге
автомашина, груженная порожними ящиками. Если смотреть на снижа-
ющуюся стаю сзади, то видно только, как птицы беспорядочно, в разных
направлениях, все время меняющихся, стремительно падают вниз. Все
это происходит настолько быстро, что подробностей разглядеть не удается.
На земле горные гуси ходят и даже бегают легко, почти не перевали-
ваясь; хорошо ныряют, особенно во время линьки. В прозрачных водах
памирских озер бывает хорошо видно, как спасающиеся под водой от пре-
следования линные гуси энергично гребут как ногами, так и лишенными
перьев крыльями. Голос горного гуся низкий, гораздо ниже, чем у гу-
менника. На полете он звучит, как гнусавое, низкое «гак. гак» или
«хок. хок». Беспокоясь у гнезд, они издают низкие, стонущие звуки,
вроде «ааа. аааа» или «уо. уо». Вообще же горный гусь не шумлив,
и даже большие сборища осенью производят сравнительно мало шума.
Памирские горные гуси славятся своей осторожностью и обычно не
подпускают человека ближе 200 м.
Следует отметить еще одну особенность горных гусей — их колони-
альное гнездование. Среди остальных гусей колониальное гнездование
известно только у белого гуся — Chen hyperboreus (Pall.).
Р. Н. Мекленбурцев наблюдал прилет гусей на озерах Рангкуль и
Яшилькуль в начале апреля. С. Гедин 18 апреля встретил у Рангкуля
большие скопления гусей. В это время на Памире стоит еще настоящая
зима, и температура по ночам нередко падает до —30°. Озера покрыты
льдом, и только у берегов появляются небольшие закраины, но места
гнездования — плоские островки — уже свободны от снега и льда. Вскоре
— 18 —
встречались по всему озеру, не образовывая больших скоплений. Тем же
летом мной были осмотрены все острова озера, и нигде не было замечено
признаков гнездования бакланов. Местные жители, хорошо знающие
озеро, ни разу не встречали ни гнезд, ни птенцов бакланов.
Черный аист — Ciconia nigra (L.). Р. Н. Мекленбурцев (1937) встретил
выводок из 2 старых и 3 молодых птиц на лугах Аличура у Башгумбеза
24 июля 1934 г. Добыть аистов не удалось, но Р. Н. Мекленбурцев пред-
положил, что это — местные птицы. Мне также один раз встретилась
группа из 3 аистов в низовьях Аличура (Акджар) 2 июля 1954 г. Это были
явно холостые птицы, которых вообще много летом в южном Таджикистане.
Виденные Р. Н. Мекленбурцевым и мной птицы были во всяком случае
уже летными и по оставлении гнезда могли прикочевать на Памир. Если же
этих птиц считать местными, гнездовыми, то пришлось бы предположить,
что черные аисты начинают откладку яиц на нагорье в первой половине
апреля, когда по ночам температура падает до —30°, стоит еще настоящая
зима, реки скованы льдом, а луга покрыты наледями и снегом. В таких
условиях аисты вряд ли могут вообще существовать, не говоря уже о гне-
здовании.
1. Горный гусь — Eulabeia indica (Latham). В настоящее время горный
гусь гнездится только на четырех озерах нагорья: Каракуле, Зоркуле,
Рангкуле и Малом Каракуле. На оз. Рангкуль еще в 50-х годах гуси оби-
тали в большом числе, но из-за постоянного разорения колоний на легко-
доступных островах они почти перестали здесь гнездиться. Исчезли гуси
из долины Аличура, с оз. Сассыккуль, с озер перевала Кызылрабат, с вер-
ховьев Аксу, хотя еще в конце прошлого века они гнездились там во мно-
жестве (Д. Л. Иванов, 1885). Сокращение мест гнездования происходило
повсеместно и привело к тому, что ареал горного гуся в настоящее время
из сплошного превратился в прерывистый. Как известно, основная часть
ареала охватывает нагорье Тибета, восточный Цайдам, Кукунор и систему
Наныпаня.’На западной окраине Центральной Азии он населяет Ладак
и горами Каракорума и западного Куньлуня проникает на Памир, кото-
рый составляет самую крайнюю северо-западную часть сплошного аре-
ала. Далее к северо-востоку по северной части горного обрамления Цент-
ральной Азии распространение горного гуся становится прерывистым.
На Тянь-Шане в настоящее время он гнездится достоверно только в трех
местах: на озерах Сонкуль и Чатыркуль и на мелких озерках Покровских
сыртов (Янушевич и др., 1959). Дальше к северо-востоку изолированная
северная часть ареала охватывает юго-восток Алтая, Монгольский Алтай,
Хангай, Хэнтэй и бассейн оз. Хубсугул. В область низменных центрально-
азиатских пустынь горный гусь не идет, нет его и в Гобийском Алтае.
Горные гуси зимуют в теплых долинах северной Индии, северной
Бирмы и Юньнани (Китай), т. е. в ближайших к местам гнездования теп-
лых странах (Козлова, 1952). Экземпляр горного гуся, добытый зимой
в низовьях Пянджа (А. И. Иванов, 1940), видимо, является случайно за-
летным. За три года работы па зимовках птиц в низовьях Пянджа и Вахша
автору ни разу не пришлось его встретить или хотя бы слышать о нем.
Данных кольцевания почти нет. Горный гусь, окольцованный птен-
цом па оз. Сонкуль 13 июля 1959 г., был добыт 25 марта 1960 г. в Гилгите
(Индия).3
Высота обитания горного гуся над уровнем моря, как и у многих дру-
гих горных птиц, зависит от широты местности, в конечном итоге опреде-
ляющей климат и высоту снеговой линии. В юго-восточном Тибете на
33° с. ш. эти птицы не гнездятся ниже 4400 м. На 4° севернее, на оз. Ку-
3 См. Journ. Bombay Nat. Hist. Soc., v. 58(1), 1961.
2 Биология птиц — 17 —
после прилета гуси, не дожидаясь окончательного вскрытия озер, присту-
пают к гнездованию.
По описанию Н. М. Пржевальского (1876), брачные игры происходят
в воздухе. При этом самец гоняется за самкой, кувыркаясь на лету. Осо-
бые игры происходят и на земле (Тристан и Звескин, 1960).
Сроки гнездования, видимо, мало зависят от конкретных климатиче-
ских условий на каждом озере. Главное — чтобы были свободны от снега
места гнездования. На оз. Каракуль, например, стаивание льда происхо-
дит почти на месяц позже, чем на Рангкуле, а сроки размножения гусей
на этих озерах одни и те же. Вообще на всем протяжении ареала горного
гуся разница в сроках вывода птенцов невелика и не превышает 15—
20 дней.
Гусыни приступают к откладке яиц во второй половине мая. На остров-
ках памирских озер гуси гнездятся небольшими колониями от 2 до 15 пар,
часто вместе с буроголовыми чайками и крачками. Величина колонии за-
висит от количества удобных для гнездования мест и от общей численности
гусей. На оз. Каракуль, где численность гусей непрерывно уменьшается,
в 1961 г. колонии насчитывали по 3—9, а в 1962 г. — по 2—6 гнезд в каж-
дой. В то же время известно, что на озерах южного Тибета (оз. Рамцо),
где численность гусей высока, гнездовые колонии насчитывают до несколь-
ких тысяч птиц (Baker, 1935). Небольшие островки гуси предпочитают
большим островам; некоторые из них, имея 15—20 м в поперечнике, воз-
вышаются над водой не более чем на 0.5 м. Обычно гуси избирают островки,
лишенные растительности, совершенно ровные или заваленные камнями,
но на Рангкуле они гнездились на островах, заросших осокой.
Гнезда на островах Каракуля представляли собой небольшие углубле-
ния на голой почве, выстланные небольшим количеством пуха, перемешан-
ного с землей. Ямки были окружены плотными пуховыми валиками, со-
ставлявшими края гнезда. Диаметр гнезда в среднем 31 см (20 гнезд).
Гнезда располагались в 4—10 м одно от другого. На островах Рангкуля,
заросших травой, гуси по наблюдениям И. А. Абдусалямова, ямок не
рыли, и гнездо представляло собой просто небольшую площадку, окру-
женную пуховым валиком. На голых островках тот же автор наблюдал
в гнездах выстилку из веточек терескена и других растений, чего на Ка-
ракуле не было. Неизвестен такой факт и для Ладака, но в юго-восточ-
ном Тибете, по наблюдениям Шефера (Schfifer, 1938), гуси, гнездившиеся
на скалах, выстилали гнезда веточками карликовой березы.
Нормальная величина кладки для Памира — 3—4 яйца: по 3 яйца
в 8 найденных гнездах, по 4—в 13, по 5 — в 3, по 6 — в 2 и по 7 —
в 2 гнездах. При гибели кладки гусыня вторично откладывает 3 яйца.
В таких случаях гнезда делаются небрежно, наспех, из стеблей осоки,
и в них совсем отсутствует пух. Известно, что кладки горных гусей могут
заключать до 12 яиц (Янушевич, 1955), но в таких случаях возникает пред-
положение, что в одно гнездо несутся две или три гусыни. Подобного мне-
ния придерживается и Бекер (Baker, 1935). В колониях на оз. Каракуль
в некоторых случаях были обнаружены слегка насиженные, покинутые
кладки, засыпанные землей. Эти гнезда не имели вокруг характерного
пухового валика. И. А. Абдусалямов находил на Рангкуле под некоторыми
гнездами с яйцами закопанные в землю другие кладки. Подобные случаи
описаны и для Тянь-Шаня, где гусыни закапывают кладку в водоросли
и над ней строят новое гнездо (Тристан и Звескин, 1960). В таких случаях
можно предположить захват гусыней чужого гнезда, но точных наблюде-
ний, объясняющих это явление, нет.
Яйца горных гусей чисто белого цвета, с очень толстой скорлупой.
Толщина скорлупы яиц с Памира 0.47—0.57, в среднем 0.51 мм (в Тибете,
- 19 —
2*
по измерениям Шефера, 0.52—0.58 мм). Размеры 20 яиц с Каракуля:
75.5-90.0x53:9-60.0 мм.
В колониях на оз. Каракуль в июне 1961 г. во всех осмотренных гнез-
дах были яйца с птенцами накануне вылупления и только в одном нахо-
дились однодневные птенцы. Срок насиживания — 29—30 дней (Тристан
и Звескин, 1960), следовательно, в данном случае гуси приступили к от-
кладке яиц в конце второй декады мая. Это крайний срок для Памира,
особенно для Каракуля, где в конце мая свободны ото льда только неболь-
шие закраины. В 1962 г. первый выводок появился тоже 27 июня.
После вылупления пуховые птенцы долго держатся у островков, где
они вывелись. Гусята, вышедшие из яиц на скалах, добираются до воды
сами, следуя на крик родителей. При этом они сваливаются с отвесных
скал, иногда сильно ударяясь о камни, но без всякого вреда для себя
(Schafer, 1938). Пуховички в возрасте до одной недели способны прыгать
через препятствия, правда, не в такой степени, как птенцы огари, да и
способность эта быстро исчезает. Во всяком случае наличие такого реф-
лекса указывает на сравнительную древность гнездования горных гусей
на скалах.
Только что вылупившиеся птенцы имеют сернисто-желтую окраску,
особенно интенсивную на боках и брюшке; темя серое; спина и крылышки
тоже серые, с оливковым оттенком и незначительной примесью желтых
тонов. В этом возрасте они очень походят на пуховичков серого гуся
(Anser anser L.), но хорошо отличаются наличием двух светлых полос,
заходящих с боков на спину позади крыльев. Желтый цвет постепенно
бледнеет и через месяц исчезает, дольше всего сохраняясь на голове и шее.
Окраска верхней стороны тела становится буровато-серой, а нижней —
серовато-белой. На 30—33-й день распускаются широкие плечевые перья
серого цвета, с охристыми каймами, и начинают распускаться кроющие
перья крыла; перья заменяют пух на спине. Маховые перья выходят из
чехлов на 47—50-й день. В возрасте 55 дней молодые надевают полный
осенний наряд, отличаясь от взрослых птиц окраской головы и шеи.
Линька головы и шеи начинается у молодых в первых числах октября,
в возрасте около 100 дней; в течение следующих 20 дней молодежь приобре-
тает характерную для взрослых окраску этих частей тела, но черные по-
лосы еще испещрены мелкими белыми перышками. Окончательный взрос-
лый наряд они надевают зимой, к февралю.
Пуховички в возрасте от 2 до 25 дней, содержавшиеся в неволе, боль-
шую часть времени проводили на суше, кормясь свежей травой или отды-
хая. К воде они подходили в среднем один раз за 3 часа. При этом иногда
по почину одного из них они бросались в воду и начинали стремительно
нырять, то выскакивая па поверхность, то вновь исчезая под водой, уси-
ленно работая при этом крыльями. Подобное оживление продолжалось
не более 10—15 сек., после чего птенцы выходили на берег, сушились и
продолжали кормиться. Все время пребывания в воде не превышало обычно
5 мин. В природе пуховички проводят в воде гораздо больше времени.
Видимо, это имеет большое значение для нормального развития и функ-
ционирования копчиковой железы. У пуховичков, содержавшихся в не-
воле и проводивших большую часть времени на суше, этого, вероятно,
не произошло. Впоследствии оперение у подросших молодых при купании
сильно намокало. Они плавали, глубоко погрузившись в воду, хотя
привычки стремительно нырять и резвиться под водой не потеряли.
Попытки к полету у содержавшихся в домашних условиях птенцов
наблюдались с момента появления маховых перьев. Птицы время от вре-
мени принимались стремительно бежать вперед, усиленно взмахивая
крыльями. Пробежав 20—30 м, они останавливались и возобновляли
прерванную кормежку. Птенцы начали взлетать во время таких пробежек
— 20 —
в возрасте 70 дней и в течение следующих 5—7 дней вполне приобрели
способность к полету. В естественных условиях молодые птицы подни-
маются на крыло, видимо, раньше. В этот период иногда можно было
наблюдать, как молодые птицы, слегка приподняв крылья, начинали
бежать друг за другом по небольшому кругу, радиусом около полутора
метров. Такая беготня длилась обычно 10—15 сек.
Птенцы горного гуся очень легко приручаются, что уже было отмечено
раньше (Козлова, 1930). Нам удалось воспитать без особых затруднений
4 птенцов, взятых на следующий день после вылупления. Они всегда бе-
жали на голос воспитателя, даже будучи уже взрослыми, и ничем в своем
поведении, кроме склонности и способности к полету, не отличались от
домашних гусей. Прирученные гусята очень долго сохраняли птенцовый
писк, вплоть до ноября.
Как только молодежь поднимается на крыло, у гусей устанавливается
определенный ритм суточной жизни, сохраняющийся вплоть до отлета.
День птицы проводят на безопасных островах и отмелях озер или же на
воде вдали от берега, а перед закатом летят на луга и болота в поймах
речек, впадающих в озера, или на мелкие озерки. Птицы кормятся там
всю ночь и с рассветом возвращаются на дневки.
В сентябре на озерах наблюдается заметное увеличение количества
гусей. Они появляются и там, где летом отсутствовали, например на Сас-
сыккуле, Яшилькуле и других озерах. В первых числах сентября на-
чинается движение гусей на зимовки. Над Памиром стаи летят к юго-
западу, пересекая хребты нагорья на большой высоте (Alcock, 1897;
Мекленбурцев, 1946). По моим наблюдениям, пролетные стаи летят иногда
на высоте около 7000 м над ур. м. Первая стая в 1956 г. появилась над
Рангкулем 1 сентября. Основная масса гусей, скопляющаяся на этом
озере осенью, отлетает в середине октября, и только небольшие группы
в случае благоприятной погоды иногда задерживаются до первых чисел
ноября. В середине ноября озеро покрывается льдом; Каракуль и Зор-
куль замерзают раньше, и гуси соответственно раньше оттуда уле-
тают.
Интенсивная линька взрослых происходит в первой половине июля.
На Рангкуле И. А. Абдусалямов находил маховые перья по берегам
с 10 июля. Два экземпляра, добытые 14 июля 1956 г. на Каракуле, со-
вершенно не могли летать. У самки маховые перья были в чехликах,
у самца же только появились кисточки. 24 июля 1962 г. на островах
в восточной части Каракуля держались небольшие стаи нелетных гусей.
У добытого самца маховые перья только начали отрастать. Экземпляр
в коллекции ЗИН АН СССР, добытый на этом озере в середине августа
1890 г., уже перелинял. В то же время у двух птиц, добытых М. П. Роза-
новым 14 августа 1932 г. на Зоркуле, линька была в разгаре. У самца
маховые перья отросли наполовину, у самки — на 3/4 нормальной длины.
В целом линька гусей памирской популяции заканчивается к концу ав-
густа. Частичная осенняя линька происходит только у молодых.
Весной в рационе гусей преобладает растительная пища, главным
образом осоки по прибрежным лугам, а также различные водные расте-
ния, преимущественно рдесты. Летом, в период размножения, большую
роль в питании играют различные беспозвоночные, прежде всего бокоплавы
(Gammarus lacustris), в огромном числе населяющие многие озера, а также
взрослые тендипедиды и их личинки, которые в июле—августе в отдель-
ные дни образуют на поверхности воды сплошные пленки по нескольку
квадратных метров. Осенью, перед отлетом, наряду с водными растениями
и различными беспозвоночными в пищу идут осоки. Планктон и бентос
памирских озер весьма различен в отдельных водоемах, соответственно
и рацион гусей на различных озерах неодинаков.
— 21 —
Врагов у горного гуся, немного. Гнездование на островах исключает
разорение гнезд хищными млекопитающими, а из птиц некоторый урон
гусиным колониям наносит только ворон. И. А. Абдусалямов (1958а)
уже указывал на тот вред, который наносят вороны колониям на оз. Ранг-
куль. Мне также приходилось находить расклеванные воронами яйца по-
близости от гнездовых колоний на Каракуле. Только там, где гусиные
гнезда расположены среди колоний буроголовых чаек, они не подвер-
гаются нападениям ворона.
При сравнительной бедности Памира дичью правильная промысло-
вая эксплуатация горных гусей может дать экономический эффект. Необ-
ходимые для этого мероприятия не требуют особых материальных затрат
и сводятся исключительно к упорядочению сроков охоты и охране гнездо-
вых колоний на озерах. Даже эти примитивные меры в короткий срок помо-
гут восстановить прежнее обилие горного гуся на озерах нагорья и до-
биться того, что ежегодно осенью общее число гусей на Памире будет до-
стигать примерно 15—17 тысяч птиц. Из этого числа без особого вреда
для популяции можно будет отстреливать примерно треть, т. е. 5 тысяч
гусей. Принимая во внимание добычливость охоты на этих птиц при их
обилии, возможность отстрела такого количества гусей при указанной
численности вполне реальна.
Использовать горных гусей для нужд человека можно и путем одо-
машнивания. Птенцы этого гуся в первые дни жизни легко приручаются
и становятся совершенно ручными. Важно, что горные гуси свободно
переносят низкие температуры, по крайней мере до —30°, и в условиях
Памира могли бы стать хорошим исходным материалом при создании
опытных гусиных ферм. При зимнем содержании горных гусей вполне
можно использовать теплые и горячие источники, каких на Памире не-
мало. Во всяком случае в КНР уже сделаны первые успешные опыты по
полувольно му содержанию горного гуся на оз. Кукунор, на высоте 3206 м
над ур. м. (Chi, 1961). Следует иметь в виду, что хозяйственное исполь-
зование горных гусей в настоящее время является единственным способом
для эксплуатации богатых кормовых запасов памирских озер.
2. Огарь — Casarca ferruginea (Pall.). Обычная птица Памирского
нагорья. Распространена шире горного гуся, так как гнездится одиноч-
ными парами, менее прихотлива в выборе водоемов и меньше страдает
от преследования человека, гнездясь в недоступных скалах. Вертикаль-
ное распространение на нагорье столь же широкое. Главная масса ога-
рей гнездится в пределах субальпийского пояса, но отдельные пары не-
редко гнездятся и выше — до высот 4500 —4600 м.
Численность довольно высокая, но трудно поддается учету. На озерах
Рангкульской котловины, по данным И. А. Абдусалямова, гнездится
около 70 пар, что, по-моему, сильно преувеличено. На оз. Каракуль,
во много раз большем по размерам и имеющем неплохие кормовые условия,
регулярно гнездится не более 25—30 пар. В долине р. Аличура, на про-
тяжении 30 км, гнездится 15—20 пар. Там, где огарей мало беспокоят,
они более многочисленны даже на маленьких озерках. Так, в первых чис-
лах июля 1958 г. на двух маленьких озерках на перевале Карабулак
(хр. Музкол) мы обнаружили около двух десятков взрослых птиц и 30
только начавших оперяться молодых. Помимо гнездящихся, на Памире
держится большое число холостых птиц, которые образуют в верховьях
Аксу, на Каракуле, Зоркуле и особенно на Яшилькуле большие скопления
(на восточном берегу Яшилькуля— до нескольких сотен особей).
Огарь обитает на озерах любых размеров и различной степени соле-
ности. Предпочтение отдается мелким водоемам, хорошо прогреваемым
солнцем и богатым органической жизнью. Необходимое условие — на-
— 22 —
личие в радиусе до 3 км скал или пойменных обрывов, где устраивается
гнездо.
Полет легкий, сильный и отличается высокой маневренностью. На
Памире во время размножения огарь ведет себя как истинно горная птица.
Ее часто можно видеть летящей среди скал или сидящей на маленьких
карнизах на головокружительной высоте в окружении ряда высокогор-
ных памирских птиц. Огари свободно летают вдоль скальных стен как
вверх, так и вниз, часто пользуясь скользяще-пикирующим полетом.
Голос — протяжное, гнусавое «аанг. . аааннннгр. .», откуда и местное
название птицы — ангыр. Этот крик издается при тревоге или крайнем
возбуждении. В спокойном состоянии они покрякивают хриплым голосом
явно утиного, а не гусиного тембра. Самка, когда ведет птенцов, часто
крякает низким голосом. В больших скоплениях птицы иногда издают
звуки, напоминающие трели лягушек. По земле передвигается легко,
хорошо плавает и прекрасно ныряет.
Прилет ранний, в конце марта—начале апреля (Мекленбурцев, 1946).
С момента прилета держатся на оттаявших лужках и отмелях озер или
у краев тающих ледяных полей, покрывающих в это время большие уча-
стки долин. Птицы появляются одиночками и парами. Численность ста-
билизируется в конце апреля, когда прилет и, видимо, пролет заканчи-
ваются.
Брачное оживление начинается в конце апреля—начале мая. Пары
с криками кружат над долинами, то и дело улетая в скалы и высматривая
ниши для устройства гнезд. Плотность населения огарей в известной
степени зависит от наличия удобных для гнездования мест, но и при на-
личии таковых особых скоплений на гнездовьях не образуется. В верхо-
вьях р. Аличура на участке скальной стены размером 500x200 м гнезди-
лось 4 пары; правда, шум от них был как от нескольких десятков птиц.
Огарь гнездится на Памире исключительно в скалах или пойменных об-
рывах, выбирая расщелины, пещерки и глубокие ниши. При осмотре
пещер в скалах мне часто встречались следы огарей. Наиболее высоко
расположенное гнездо было найдено мной в верховьях р. Аличура, в скаль-
ной нише на высоте 4550 м над ур. м. Геоботаник Л. Ф. Сидоров встретил
выводок птенцов на озерах перевала Кызылрабат (4420 м) и на оз. Чу-
куркуль (4425 м). Птицы гнездились в скалах у этих озер на высоте 4500—
4600 м. В то же время они не гнездятся на озерах перевальной плоскости
в верховьях р. Рамаифа (4400 м), где слишком холодно, снеговая линия
проходит на высоте 4800 м и по берегам озер лежат снежники.
Гнезда в скалах обычно бывают совершенно недоступны, и за гнездо-
выми нишами удавалось наблюдать только в бинокль. В верховьях
р. Аличура две гнездовые ниши помещались в отвесной двухсотметровой
стене южной экспозиции, в 150 м над ее основанием и в 4350 м над ур. м.
Вход в каждую из них имел в диаметре около полуметра, а глубина была
столь значительной, что насиживающую птицу не было видно. В много-
численных трещинах и нишах вокруг гнездились клушицы, скалистые
голуби, темнобрюхие улары и пара пустельг. В период наблюдений, с 5
по 15 июня, самки насиживали, улетая по утрам и вечерам кормиться
на болотистые луга, расположенные под горой. Перед вылетом на кор-
межку птицы делали несколько кругов вокруг гнезда, причем самец все
время кричал, а потом уходили вниз. По возвращении самка, сделав не-
сколько кругов, садилась в нишу, а самец, крича, делал еще один круг и
улетал на луга, где и кормился весь день. На ночевки самцы возвраща-
лись в скалы и ночевали где-нибудь поблизости от гнезда.
В 1961 г. первый выводок в этих местах был обнаружен 11 июня —
наиболее ранняя дата для Памира, а для Аличура тем более, так как эта
долина—одна из самых холодных на нагорье. Выводок был встречен на
— 23 —
крутой осыпи, на высоте 4500 м, в 500 м над долиной. Над осыпью возвы-
шалась отвесная скальная стена с гнездовой нишей в 50 м над основанием.
Экспозиция тоже была южной. Оказавшийся поблизости самец с большой
скоростью пикировал на людей раз за разом, едва не задевая крыльями.
Самка в это время поспешно спускалась вниз по осыпи, призывно покря-
кивая и ведя за собой выводок пуховичков, которые непрерывно пищали.
При нашем приближении самка взлетела и присоединилась к самцу.
Птенцы попрятались под камнями и непрерывно подавали голос, благо-
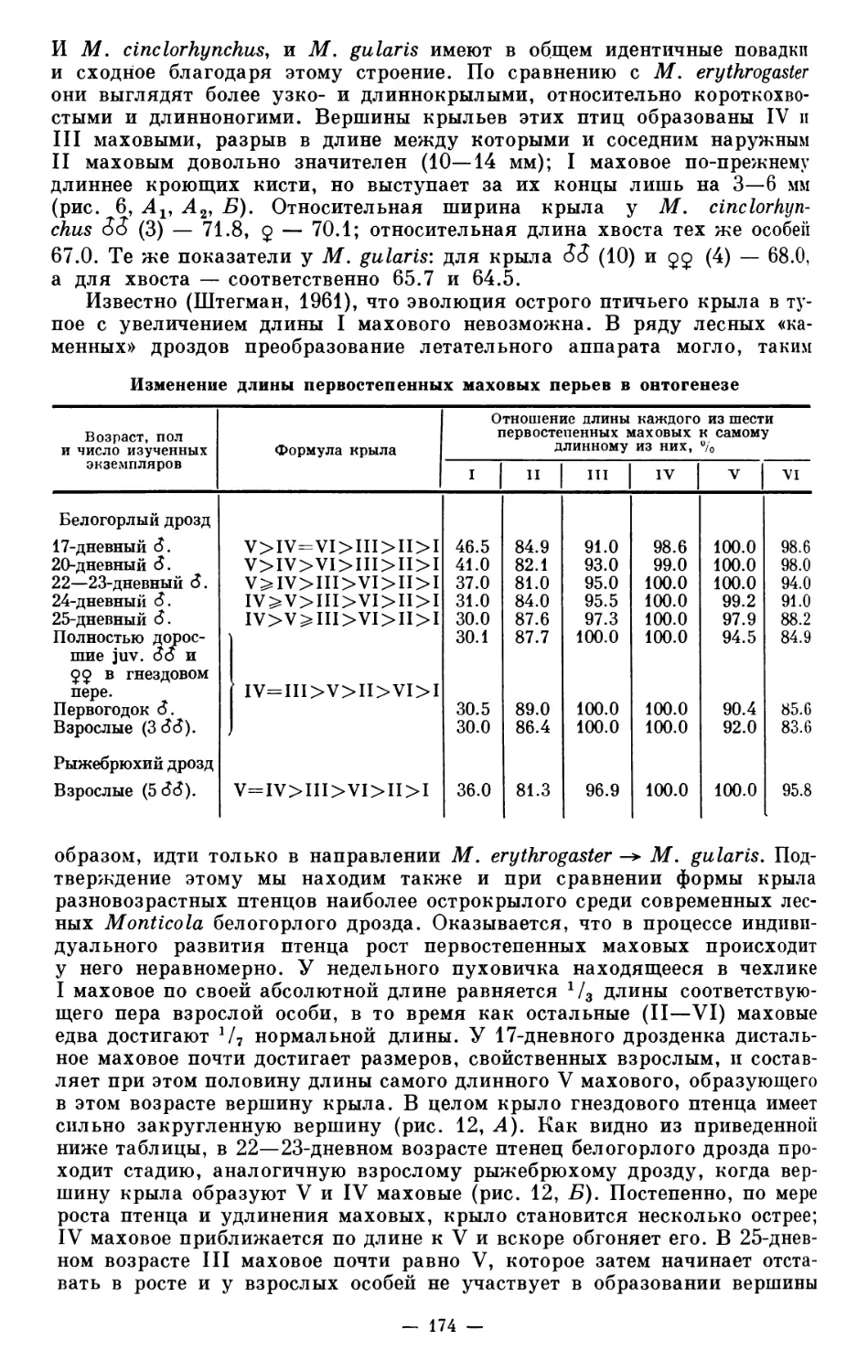
даря чему двоих удалось быстро разыскать (рис. 5). У одного из них при
падении из гнезда был слегка разбит клюв, но сам птенец от этого не по
Рис. 5. Однодневные птенцы огари — Casarca jerruginea (PalL).
страдал. Гнездовая ниша находилась в отвесной скале, и пуховички пры-
гали с высоты 50 м, падая на каменную осыпь. Местное население убе-
ждено, однако, что огарь переносит своих птенцов на воду в клюве. В связи
с этим интересно сравнить, что рассказывали тибетцы по этому поводу
орнитологу Ладлоу (Baker, 1935). По их словам, огари не переносят птен-
цов. После вылупления самка просто выталкивает птенцов из гнезда,
а стоящий внизу самец принимает их на распростертые крылья, чем
предохраняет от ушибов. Однако такая сложная процедура вряд ли су-
ществует на самом деле. Скорее всего птенцы сами выскакивают на зов
матери. А. Г. Банников (Банников и Тарасов, 1957) наблюдал переход
пуховичков от скал к озеру ночью, тогда как в нашем случае это происхо-
дило днем. То же пишут Шефер (Schafer, 1938) и Янушевич с соавторами
(1959). Правда, было несколько случаев, когда на озерках поблизости от
наших стоянок пуховички появлялись ранним утром, и было очевидно,
что они пришли сюда ночью. Необходимо учитывать время, которое тре-
буется выводку для преодоления подчас больших расстояний от гнезда
до воды. В наблюдавшемся нами случае птенцам предстояло спуститься
по крутым осыпям и скалам по вертикали на 500 м и потом добираться еще
2 км по изрытым оврагами конусам выноса и кочковатым лугам до ближай-
шего озерка. Для такого путешествия может мало оказаться и суток.
— 24 —
Учитывая, что продолжительность насиживания у огари равняется
28 дням, можно высчитать, что огари в данном случае начали откладку
яиц в начале мая. Это очень ранняя дата; судя по наблюдениям других
лет и в других местах, большинство огарей начинает кладку во второй
половине мая, как и горные гуси. В той же долине Аличура первые вы-
водки появились: в 1937 г. 26 июня (Мекленбурцев, 1946), в 1954 г.
27 июня. На Каракуле выводки пуховичков Л. А. Молчанов (Молчанов
и Зарудный, 1915) наблюдал 25 июня. Таким образом, огарь на Памире
гнездится на 2.5 месяца позже, чем на равнинах южного Таджикистана.
В Ладаке и Тибете сроки размножения те же, что и на Памире.
Судя по количеству птенцов в выводках, огари откладывают 6—9 яиц.
Обычно в выводке бывает 5—6 птенцов, и чем старше выводок, тем он
меньше, так как гибель их от различных хищников велика, особенно на
мелких озерках. Правда, и здесь птенцы довольно энергично скрываются
от преследователей, прекрасно ныряя и незаметно затаиваясь под нависаю-
щими берегами.
У птенцов до недельного возраста сильно развит инстинкт прыгания
через препятствия. Натыкаясь на какую-либо преграду, птенец начинает
упорно прыгать на нее, пытаясь преодолеть, даже если это будет высокая
стена. В возрасте 3 дней птенцы после нескольких попыток вскакивали
на преграду 30 см высоты. Они прыгают, отталкиваясь сразу обеими ла-
пами и пытаясь зацепиться за преграду острыми, цепкими коготками.
Наткнувшись на непреодолимое препятствие, птенец может прыгать на
него до полного изнеможения. У птенцов, содержавшихся в неволе, такое
поведение продолжалось до недельного возраста, но на свободе оно сохра-
няется дольше, до 10—11 дней. В таком возрасте птенцы настолько
прибавляют в весе, что при неудачных прыжках сверху вниз могут раз-
биться насмерть. В условиях неволи стремление прыгать совершенно уга-
сало на 9-й день и на смену приходила явная боязнь высоты. Такие птенцы,
например, уже боялись спускаться по ступенькам крыльца. О силе ног
птенцов и о способности их с момента рождения преодолевать значитель-
ные расстояния пешком сообщалось ранее (Банников и Тарасов, 1957).
Выводки огарей, до подъема молодых на крыло, держатся все время
на озерах, где кормятся в основном животным кормом у берегов. В воде
птенцы, содержавшиеся в неволе, оставались подолгу и более охотно, чем
птенцы горных гусей. На озерах Памира птенцы огарей начинают подле-
тывать уже в начале августа и в середине месяца поднимаются на крыло.
Содержавшиеся в неволе молодые огари приобретали способность к полету
в возрасте 70 дней.
После того как молодые поднимутся на крыло, выводки присоеди-
няются к скоплениям холостых птиц на больших озерах и держатся
вместе большими стаями вплоть до отлета. Кормятся они большей частью
на мелких местах, где вода хорошо прогревается. Ночуют здесь же,
тогда как во время гнездования многие летают на ночевку в скалы. Пра-
вильных суточных перелетов, как у горных гусей, у огарей не наблюда-
лось.
В сентябре число огарей заметно возрастает за счет прилетных особей.
На крупных озерах скапливаются большие стаи. Большая часть отлетает
в середине сентября, задолго до ледостава. Иногда, при теплой погоде,
одиночки и небольшие группы могут задерживаться до начала ноября.
Складывается впечатление, что процесс линьки маховых перьев проте-
кает очень быстро, и птица теряет способность к полету только на короткий
срок. Несмотря на обилие огарей на Памире, ни мне, ни другим исследова-
телям, работавшим здесь, не удалось ни разу встретить линных, нелетав-
ших птиц, даже на местах их скоплений, где, судя по кучам перьев на
берегах, собирались линяющие птицы. Такого же мнения придерживаются
— 25 -
А. Г. Банников и П. П. Тарасов (1957). Только в одном случае (Януше-
вич и др., 1959) удалось поймать пару линных птиц на оз. Сонкуль
17 июля. На Памире, по наблюдениям И. А. Абдусалямова (1961), линька
начинается 5—7 июля. В 1962 г. при обследовании большого скопления
огарей в восточной части оз. Яшилькуль в середине июля абсолютно все
птицы были летными.
Питание изучено плохо. Можно только сказать, что в отличие от гор-
ных гусей животная пища играет у них главную роль. Благодаря быстрой
усвояемости различных беспозвоночных желудки у большинства добытых
огарей оказывались пустыми или содержали неопределимые остатки. Пу-
ховички огарей, как и других уток, часто склевывают различных беспо-
звоночных с поверхности воды.
На Памире огарь является объектом спортивной охоты, но из-за низ-
кого качества мяса добывается мало. Местное население нередко ловит
птенцов и откармливает их до двухмесячного возраста, тогда мясо их
бывает довольно вкусно. Огарь, как и горного гуся, можно использовать
для домашнего разведения. Птенцы, взятые в возрасте 1—2 дней, хорошо
выживают.
3. Кряква — Anas platyrhynchos L. (?). По сообщениям местных жите-
лей, какая-то утка, по описаниям напоминающая самку кряквы, регулярно
гнездится на разливах р. Аксу у Тохтамыша. По их словам, в 20-х числах
июля 1961 г. здесь наблюдался выводок из 6 пуховичков. Киргизы назы-
вали эту утку «эрдек», как обычно именуются различные утки рода Anas.
Сообщают также, что эта утка попадается на ключах у Акташа зимой.
Как любезно сообщил нам Л. А. Арутюнов, в указанном месте зимой
ему попадались из уток только кряквы.
4. Большой крохаль — Mergus merganser orientalis Gould. На Памире
крохаль населяет субальпийский пояс, изредка поднимаясь в нижние
части альпийского. Численность невысокая. По всему Аличуру гнездится
не более 5—6 пар; в верховьях Аксу пара от пары гнездится на расстоя-
нии 2—3 км.
Распределение по местам гнездования происходит в конце апреля.
Гнездятся крохали в скалах и в наибольшем числе встречаются там, где
имеется сочетание скал и водоемов, богатых рыбой, например в верхо-
вьях р. Аксу у Акташа. Гнезда устраивают в нишах отвесных скал.
Гнездовая ниша, обнаруженная в верховьях Аксу, находилась в отвесной
скальной стене южной экспозиции, в 60 м над ее основанием. Столь же
недоступными были гнезда, найденные Р. Н. Мекленбурцевым на р. Али-
чуре. Сроки гнездования более поздние, чем у других водоплавающих
нагорья. У скал Акташа в середине мая 1961 г. крохали держались па-
рами и целыми днями кормились на ручьях, вытекающих из-под скал и
изобилующих рыбой; 18 мая одна пара крохалей упорно кружилась
у стенки скалы, время от времени присаживаясь на края многочисленных
ниш, очевидно, выбирая место для гнезда. В середине июля здесь же
попадались исключительно одиночные самцы; самки, видимо, сидели
на гнездах; 11 июля была встречена самка в скалах правого берега р. Ха-
нюлы, летевшая по ущелью вверх; это было в 400 м над долиной, на высоте
4300 м; самка, вероятно, возвращалась на гнездо после кормежки. Пер-
вый выводок на реке появился через 5 дней, 16 июля: 7 пуховичков в со-
провождении самки быстро спускались вниз по стремнинам. В 1954 г.
первые выводки в этих местах были встречены 15 июля. В 1956 г. в двух
выводках, встреченных здесь же 18 августа, было по 5 почти выросших
молодых.
На р. Аличуре в 1962 г. первый выводок, состоявший из 6 пуховичков
в возрасте 2—3 дней, был встречен 12 июля. Выводок сопровождали обе
старые птицы, которые при приближении лодки взлетели. Предоставлен-
- 26 —
ные самим себе птенцы разбились на две группы. Они поплыли в разные
стороны, вверх и вниз по течению, придерживаясь круто нависших бере-
гов и непрерывно ныряя. Сообщение Р. Н. Мекленбурцева (1946) о появ-
лении первых выводков в июне относится, вероятно, к году с очень теплым,
ранним летом.
В пределах нагорья существует определенная зависимость сроков
размножения крохалей от высоты местности. Так, на Сарезском озере
(3230 м) выводки появляются в середине июня, в нижнем течении р. Мур-
габа (3500 м) — 30 июня (Молчанов и Зарудный, 1915), в верховьях
Аксу и в низовьях Аличура (3800—3900 м) — в середине июля. Следова-
тельно, основная масса крохалей на нагорье начинает гнездиться в сере-
дине-конце мая, несколько позднее, чем на сыртах Тянь-Шаня.
По мере подрастания птенцов выводки постепенно спускаются по рекам
вниз, к озерам. Молодые поднимаются на крыло поздно. Выводки нелет-
ных молодых, размером уже со взрослую птицу, встречались до 10 сен-
тября.
Осенью некоторая часть популяции покидает Памир, спускаясь по
системам Пянджа и Тарима вниз, но часть регулярно зимует по незамер-
зающим участкам рек и озер у теплых ключей, в частности на оз. Булун-
куль, на Аксу и на Аличуре.
Интенсивная линька у гнездящихся крохалей происходит в конце
июля. Холостые птицы линяют раньше. На оз. Каракуль 2 июля 1962 г.
на одном из островов была встречена группа из 30—40 птиц, сидевших
с подветренной стороны на берегу. Линька у них была в разгаре, маховые
перья отросли лишь наполовину, и только несколько старых самцов
могло летать. Часть линных птиц бросилась убегать по берегу, прячась
за камнями, а часть спасалась по воде, то ныряя, то безуспешно пытаясь
взлететь.
Пищей крохалей на Памире служит рыбья молодь представителей
родов Schizothorax и Nemachilus, которой изобилуют многие реки и озера.
5. Беркут — Aquila chrysaetos daphanea Sev. На Памире гнездится
в пределах субальпийского пояса, охотится же во всех поясах. Числен-
ность невысока. Редкие пары гнездятся в крупных скалистых массивах
нагорья, несколько чаще встречаясь на окраинных хребтах, более изре-
занных и изобилующих скалами. В центральных частях нагорья среди
широких долин и пологих хребтов беркут почти не встречается. Заметных
колебаний численности не наблюдалось. Во время охотничьих вылетов
орлы часто поднимаются очень высоко, до линии вечных снегов. Нередко
их приходилось видеть парящими над самыми гребнями хребтов. На
одном из гребней близ Биостанции у беркутов имелось постоянное место
отдыха на высоте 5200 м, где птицы подолгу отдыхали и чистились.
В связи с большим разнообразием объектов питания способы охоты
у беркутов довольно разнообразны. За пищухами эти орлы охотятся,
расхаживая по осыпям ранним утром, когда зверьки наиболее активны.
При этом они постепенно поднимаются вверх, прыжками, помогая себе
взмахами крыльев. Охота на зайцев производится парящим полетом на
высоте всего 1.5—2 м над склоном. При этом используются сильные
восходящие токи воздуха, появляющиеся после 10—11 час. утра. Низко
парящие беркуты тщательно осматривают крупные камни, в тени которых
зайцы любят прятаться днем. Эта охота происходит в пределах субальпий-
ского пояса, так как зайцы редко поднимаются выше 4300 м. Примерно
так же охотятся беркуты и на уларов, поднимаясь при этом до 5000 м.
Сурков они добывают, внезапно появляясь из-за перегиба склона или
из-за гребня и хватая зверьков, далеко отбежавших от своих нор.
Р. Н. Мекленбурцев (1946) наблюдал воздушные игры беркутов с де-
кабря и весь январь. Единственное гнездо было найдено 4 июля 1958 г.
— 27 —
у Сарезского озера, в 800 м над уровнем последнего, и помещалось в не-
большой пещерке южной экспозиции, у основания скальной стенки;
в гнезде находилась скорлупа разбитого кем-то яйца, отложенного явно
весной этого года. По одной находке трудно судить, какие высоты пред-
почитает беркут для гнездования, но на сыртах Тянь-Шаня гнезда не на-
ходили выше 3600 м (Янушевич, и др., 1959). В юго-восточном Тибете
Шефер (Schafer, 1938) нашел, что эти орлы наиболее многочисленны на вы-
сотах 2400—4000 м, хотя сами гнезда не были найдены. Беркут гнездится
рано, поэтому на Памире он вряд ли может гнездиться в альпийском
поясе, который к началу гнездования (конец марта) находится под снегом,
а морозы достигают —40°.
Молодые беркуты становятся особенно многочисленными на нагорье
с конца июля, когда, вероятно, и происходит их вылет из гнезд. Число
беркутов, как и остальных крупных хищных птиц, повышается к октябрю,
но затем резко падает. Видимо, большая часть популяции на зиму покидает
Памир, откочевывая вниз. На зимовку остаются только редкие пары и
одиночки, очевидно старые птицы (Мекленбурцев, 1946). Обращает на себя
внимание густое зимнее опушение птиц. У экземпляров с Памира длина
пуха в нижней части груди равна в среднем 74—75 мм, тогда как у зимних
A. ch. julva (НаЫ.) из Предкавказья и Закавказья она равна
42 мм.
Главной пищей служат сурки, зайцы, улары, хотя при случае беркут
не брезгует и мелкой добычей. В июле, в разгар гнездового сезона, орлы
время от времени навещают луга субальпийского пояса, где ищут по болот-
цам птенцов куликов или огарей, вызывая переполох в птичьем населении
лугов. По наблюдениям геоботаников К. В. Станюковича и Л. Ф. Сидо-
рова, беркуты летом нападают на ягнят архаров. Козлят козерогов, судя
по нашим наблюдениям, они не трогают. Летом 1958 г. я вел непрерывное
недельное наблюдение за жизнью стада козерогов в 50 голов, где было
около 20 маленьких козлят. Орлы регулярно посещали козерожьи паст-
бища, но на козерогов и козлят не обращали внимания, причем козероги
тоже не реагировали на появление беркутов. В зимнее время, по данным
Р. Н. Мекленбурцева (1946), пищу беркута составляют главным образом
зайцы и улары. На падали этот орел наблюдался только раз.
Орлан-долгохвост — Haltaeetus leucoryphus (Pall.). Обычен на рыб-
ных озерах и реках субальпийского пояса, в альпийском же поясе из-за
отсутствия таковых не встречается.
Первые птицы появляются в конце апреля, с началом вскрытия рек и
озер, становясь многочисленными в конце мая (Мекленбурцев, 1946).
Одиночные долгохвосты нередки летом на рр. Аксу, Мургабе и А личу ре,
на озерах Зоркуль, Яшилькуль. На оз. Булункуль в конце июля 1954 г.
наблюдалось целое скопление этих птиц, которые сидели по болотистым
берегам, добывая изобиловавшую здесь рыбу. У двух добытых особей же-
лудки, пищеводы и рты были до отказа набиты ею.
Гнездование на Памире не доказано; ни гнезд, ни птенцов никто на
нагорье не находил. Имеется только наблюдение над брачными играми
(воздушные гонки) с мая по июнь, когда орланы держались у скал (Мек-
ленбурцев, 1946). В коллекции ЗИН АН СССР хранятся 3 молодые
птицы в гнездовом наряде, добытые у Мургаба 20 июля и 13 ноября
и на Аличуре 6 сентября. Взрослые же птицы, как и молодые в возрасте
2—3 лет, добывались на Памире неоднократно. Сходная картина наблю-
дается в Тибете, где орланы появляются в мае и все лето встречаются,
часто в большом количестве, по берегам рыбных водоемов или на склонах
гор, где ловят пищух. Несмотря на то что орлан-долгохвост включен
в список гнездящихся в Тибете птиц (Козлова, 1952), гнезда его там не
найдены.
— 28 —
Необходимо иметь в виду, что на равнинах Индии, где орланы гнез-
дятся в большом числе, цикл размножения очень ранний и к маю, когда
здесь начинается сезон дождей, молодые уже покидают гнезда. Возможно,
что именно дожди являются причиной широкого разлета как старых, так
и молодых птиц из мест гнездования в аридные области Средней и Цент-
ральной Азии. С другой стороны, широкий разлет молодых после гнездо-
вания к северу, видимо, вообще характерен для орланов. Во всяком слу-
чае у американского белоголового орлана — Н. I. leucocephalus (L.). —
путем кольцевания удалось установить, что после вылета молодых из
гнезд в апреле они разлетаются из Флориды далеко к северу, достигая
Канады. Одна из птиц, покинув гнездо 15 апреля, за месяц покрыла по
прямой 1600 км к северу (Broley, 1947). Одногодки и холостые птицы могут
начинать кочевки гораздо раньше. Очевидно, придется вообще пересмот-
реть существующее представление о гнездовом ареале орлана-долго-
хвоста, так как во многих случаях он заносится в списки гнездящихся
птиц на основе нескольких встреч в летнее время разновозрастных осо-
бей. Отлет начинается с конца сентября.
Основную пищу орланов на нагорье составляет рыба, иногда они
поедают и падаль.
Бурый гриф — Aegypius monachus (L.). Мне неоднократно приходилось
встречать летом одиночных птиц в разных районах нагорья, но никаких
данных о гнездовании бурого грифа на Памире нет.
6. Снежный сип — Gyps htmalayensts Hume. Снежный сип на Памире
сравнительно малочислен и гнездится только в субальпийском поясе.
Судя по литературным данным, этот сип встречается в Тянь-Шане гораздо
чаще (Шнитников, 1949; Штегман, 1954; Янушевич и др., 1959). По-види-
мому, он не столь привержен к высокогорью, как принято считать: в Тянь-
Шане гнезда находили главным образом в верхнем поясе леса, в зоне
арчи (Спангенберг и Дементьев, 1935). Самые крупные колонии в Гима-
лаях расположены в пределах 1200—2400 м, хотя в Гилгите сип гнез-
дится до 3000 м, а в Тибете — до 4200 м (Baker, 1935). Только в юго-
восточном Тибете, по наблюдениям Шефера, снежные сипы поднимаются
до 4400 м, становясь выше очень редкими.
Вертикальное распространение снежного сипа на Памире тоже под-
тверждает его стремление гнездиться возможно ниже, а горизонтальное —
приверженность к местам с сильно пересеченным скалистым рельефом,
которых на нагорье не так уж много. В наибольшем числе сипы встре-
чаются в самых низких частях нагорья, причем только там, где имеются
крупные скалистые массивы (верховья рр. Баляндкиика и Аксу, озерные
котловины Рангкуля и Яшилькуля). В то же время в поисках корма сипы
могут подниматься очень высоко, и нередко приходилось встречать птиц,
паривших на высоте около 6000 м над ур. м. В Гималаях кормившихся
на падали сипов наблюдали на перевале Тангла, на высоте 5700 м над
УР- м.
Численность низкая. Это объясняется большой высотой нагорья и
сравнительно слабо расчлененным рельефом. Иногда, впрочем, число
снежных сипов на нагорье может заметно возрастать. Н. А. Северцов
в 1878 г. нашел его на Памире редким, М. П. Розанов, проходивший
через Памир в 1932 г., пишет об очень малом числе крупных хищных
птиц, в том числе и сипов, указывая, что им часто попадались трупы
павших животных, совершенно не тронутые хищниками. В 1954 г. снеж-
ных сипов на нагорье было довольно много и они регулярно встречались
в различных районах. В 1956, 1958, 1960 гг. их было значительно меньше,
в 1961 г. мне за все лето только трижды встретились пары сипов, все
в районе Кызылрабата, а летом 1962 г. в Каракульской котловине и в до-
лине р. Аличура сипы не встречались.
— 29 —
Гнездование начинается, видимо, в марте. В пределах нагорья найдено
только 2 гнезда. Первое было обнаружено в пещере Мататаш (4000 м),
в скалах южного берега оз. Рангкуль. Пещера расположена в совершенно
отвесной стене северной экспозиции, в 120 м над ее основанием. Дно ее
было покрыто толстым слоем птичьего помета. Группа альпинистов, под-
нявшаяся сюда в начале апреля 1958 г. в поисках легендарных сокровищ,
обнаружила в глубине пещеры гнездо, представлявшее собой груду му-
сора, на которой лежало яйцо. Оно было свежим или слабо насиженным.
Окраска яйца чисто белая, размеры 101.4x72.8 мм. Средние размеры
яиц с Тянь-Шаня 99.0x71.4 мм, с Гималаев — 94.8x70.1 мм. Во время
посещения пещеры людьми сипы кружили над ними, но близко не подле-
тали. По данным И. А. Абдусалямова (1961), в скалах у Рангкуля гнезди-
лось несколько пар сипов, причем каждая из них занимала отдельное
ущелье.
Второе гнездо было найдено геоботаником Л. Ф. Сидоровым в июле
1902 г. Добраться до него не удалось, но в бинокль оно было отчетливо
видно. Гнездо помещалось в глубокой нише отвесной скалы, в 100 м над
ее основанием (4100 м над ур. м.). Оно было явно брошено, так как сипы
у гнезда не появлялись, а в самом гнезде в середине лета находилось
большое белое яйцо. Эта скала восточной экспозиции возвышается над
левым берегом заболоченной поймы р. Аксу, против массива Акташ, где
пары сипов наблюдаются регулярно.
Первых подлетков у Рангкуля И. А. Абдусалямов наблюдал в начале
августа.
Число снежных сипов осенью резко возрастает, нередко они встре-
чаются целыми скоплениями (Мекленбурцев, 1946). Большинство их
покидает Памир в начале зимы, и на зимовку остаются только немногие
одиночки, которые кормятся отбросами и падалью у населенных пунктов.
В это время они теряют свою осторожность и нередко становятся добычей
охотников. Длинный, густой пух снежных сипов помогает им переносить
очень низкие температуры — до —50°. Но и они иногда гибнут зимой,
вероятно от голода, так как нам трижды приходилось находить весной
мумифицированные морозами и сухостью трупы в прекрасном зимнем опе-
рении.
Местные жители высоко ценят мягкий теплый пух сипов и шкуры его
используют для изготовления шапок и т. п. Желчь также ценится, как
лекарство от многих заболеваний.
7. Бородач — Gypaetus barbatus hemachalanus (Hutton). Как и снежный
сип, бородач придерживается на гнездовье субальпийского пояса, предпо-
читая места с сильно расчлененным рельефом. Как и в Гиссаро-Карате-
гине (Попов, 1959) и на Тянь-Шане, в альпийском поясе не гнездится.
Только в Тибете, по наблюдениям Шефера, они достигают больших высот:
гнезда были обнаружены на высоте 4700 м. На Памире бородач гнез-
дится в крупных скалистых массивах и каньонах. Охотничьи угодья разно-
образны, но большей частью это склоны гор альпийского и верхней части
субальпийского поясов. Далеко от склонов не удаляется и над широкими
пространствами межгорных впадин появляется редко.
По численности бородач заметно превосходит снежного сипа. Более
многочислен по окраинным хребтам нагорья, где пара от пары селится
на расстоянии 15—20 км.
По способам охоты эти птицы напоминают беркутов. Чаще всего боро-
дачи парят вдоль крутых склонов на небольшой высоте, высматривая
сурков, или расхаживают пешком у сурочьих колоний, подкарауливая
зверьков у нор. От других крупных хищников отличаются меньшей осто-
рожностью.
— 30 —
Увеличение численности происходит в марте за счет прилета откоче-
вывавших на зиму особей. Старое гнездо было найдено в каньоне р. За-
падного Пшарта (4000 м). Оно располагалось на карнизе скалы южной
экспозиции, в 15 м над основанием и представляло кучу набросанных
в беспорядке веток терескена, мирикарии, ивы, клочков шерсти и облом-
ков костей. Под гнездом валялись кости зайца и сурка, сравнительно
свежие. Вероятно, незадолго до этого гнездо было обитаемо и разорено
вследствие его сравнительной доступности.
Гнездование начинается в марте, так как в начале августа появляются
молодые птицы, покинувшие гнезда.
Каждая пара имеет в период гнездования обширный охотничий уча-
сток, около 100 км2, который регулярно облетает утром и вечером. Опре-
деленных маршрутов при этом нет. В районе Биостанции охотничий уча-
сток пары птиц занимал южный отрог хр. Музкол в районе пика Карад-
жилга. В мае—начале июня приходилось видеть главным образом
одиночных бородачей; самки в это время, вероятно, насиживали. Позднее,
после вылупления птенцов, в июне—июле бородачи охотились парами.
В поисках добычи они медленно парили над самыми склонами гор, ис-
пользуя сильные восходящие токи воздуха, то поднимаясь вдоль гребней,
то спускаясь до нижних частей склонов. Заметное предпочтение оказы-
валось склонам, сильно изрезанным оврагами, с хорошо развитой луговой
растительностью и колониями сурков.
В сентябре количество бородачей заметно увеличивается. Осенью и в
первую половину зимы они положительно многочисленны. Р. Н. Меклен-
бурцев (1946) насчитывал в это время не менее 10 встреч в день. Такая
концентрация объясняется бесснежностью памирской зимы; сюда соби-
раются птицы, изгнанные глубокими снегами из соседних горных районов;
во второй половине зимы число их сильно сокращается.
Летом основной пищей служат сурки, большей частью молодые,
а также зайцы и, видимо, улары. В желудке взрослой самки, добытой
9 июля 1962 г. у оз. Яшилькуль, обнаружены остатки молодого сурка
(Скалли наблюдал у перевала Санжу, в западном Куньлуне, как бородачи
охотились на изобиловавших здесь сурков, не обращая внимания на раз-
бросанные повсюду трупы лошадей). Зимой, по опросным сведениям,
бородачи кормятся на свалках у населенных пунктов и всегда слетаются на
падаль. Летом на падали бородачей встречать не приходилось, но в сен-
тябре 1954 г. бородач был пойман руками на перевале Койтезек, где
группа этих птиц разделывала лежавший у дороги труп ишака; одна из
них так наелась, что при приближении людей не могла взлететь. Есть
наблюдения геоботаников (Л. Ф. Сидоров), подтверждающие, что боро-
дачи разбивают крупные кости, сбрасывая их сверху на камни.
Балобан — Falco cherrug milvipes Jerd. Впервые был добыт на Памире
Н. А. Северцовым у входа в ущелье Кызыларт осенью 1879 г. Он же тогда
наблюдал балобана в глубине нагорья и предполагал его гнездование.
Р. Н. Мекленбурцев во время зимовки встречал одиночных птиц 4 раза,
причем последнего балобана наблюдал 8 мая. И. А. Абдусалямов (1961)
несколько сезонов наблюдал одиночных, вероятно бродячих, соколов на
скалах у Рангкуля. Добытый им 2 июля самец интенсивно линял, семен-
ники не были развиты. Вероятно, Памир следует исключить из области
гнездования балобана из-за отсутствия прямых доказательств гнездова-
ния последнего на нагорье.
8. Пустельга — Cerchneis tinnunculus tinnunculus (L.). Пустельга
населяет все главные хребты нагорья, гнездясь исключительно в субаль-
пийском поясе и не поднимаясь на гнездовье выше 4300 м. Численность
всюду низка. В верховьях р. Аличура 2 пары гнездились на расстоянии
6 км друг от друга. В южных оконечностях хребтов Найзаташ и Музкол
— 31 —
гнездилось по одной паре. На гнездовье связаны со скалами и далеко от
них не удаляются.
В связи со скудостью растительного покрова и сильными ветрами
способы охоты пустельги на нагорье несколько иные, чем на равнинах.
Птицы в поисках добычи летают низко над землей или лавируют среди уте-
сов, внезапно появляясь из-за укрытий и нападая врасплох на кормя-
щихся птиц или грызунов. Иногда пустельга охотится в открытых местах,
бросаясь сверху на стайки воробьиных птиц, кормящихся на земле. За
шесть полевых сезонов только раз удалось видеть, как пустельга охоти-
лась над полями Биостанции, летая характерным для нее трепещущим
полетом.
Р. Н. Мекленбурцев (1946) впервые упомянул пустельгу как гнездя-
щуюся в скалах Аличура, но подробности при этом не указывались, и
факт гнездования ее на нагорье остался под некоторым сомнением. Сейчас,
после находок мной 3 гнезд, нескольких только что вылетевших из гнезд
выводков и многочисленных наблюдений за явно гнездовыми парами,
можно безусловно утверждать, что пустельга регулярно гнездится на
нагорье.
К размножению памирская популяция приступает в начале июня,
почти на 2.5 месяца позднее, чем в низинном Таджикистане, и на месяц
позднее, чем на сыртах Тянь-Шаня. Гнезда помещаются в нишах скал
обычно южной экспозиции. Единственное гнездо, до которого удалось
добраться, помещалось в узкой щели под крышей высокого (6 м) камина,
расположенного в скальной стене южной экспозиции (хр. Найзаташ,
центральный Памир). Солнце в щель не попадало, здесь всегда царил
полумрак. Гнездо — простая ямка в субстрате на небольшом карнизе —
не имело специальной подстилки. Птенцы сидели на слое объедков и
помета; высота места гнезда 4150 м над ур. м. Другое гнездо, найденное
в верховьях Аличура, располагалось в глубокой нише огромной скалы,
также южной экспозиции, в 100 м над основанием, на высоте 4250 м над
ур. м. Еще одно гнездо было устроено в нише скалистого останца Чатыр-
таш, с северной его стороны, в 40 м над основанием скалы, на высоте
4100 м над ур. м.
В гнездах на Аличуре в период наблюдений с 5 по 20 июня самки
насиживали. В гнезде, найденном в хр. Найзаташ 26 июля, находилось
4 птенца, готовых к вылету; два из них покинули гнездо на следующий
же день, а два других — 28 июля. После вылета они еще 2 дня сидели
у гнезда, выпрашивая корм у взрослых птиц громким отрывистым криком.
Такой же выводок, только что покинувший гнездо, был встречен 6 августа
в скалах южной части Рангкульской котловины, на высоте 3900 м над
ур. м.
Число соколков в августе заметно увеличивается, в сентябре идет
слабый пролет. В августе и сентябре пустельги при охоте нередко наве-
щают альпийский пояс, где встречались до высот 4700 м. В Тибете во
время охотничьих вылетов они поднимаются до 5500 м (Schafer, 1938).
Р. Н. Мекленбурцев (1946) изредка встречал пустельгу в зимнее время.
Вероятно, в поисках корма отдельные особи залетают на нагорье в ясную
солнечную погоду из соседних районов. Предполагать регулярную зи-
мовку здесь вряд ли возможно.
В питании пустельги на нагорье преобладают птицы. У гнезда, най-
денного в хр. Найзаташ, в большом количестве валялись остатки каменок,
горихвосток, снежных воробьев, преимущественно подлетков. Нередко
наблюдались нападения на горных вьюрков, рогатых жаворонков, удодов
и др. Обычные места охоты — пойменные обрывы и скалы — имеют на
Памире наибольшую плотность птичьего населения, но почти лишены
грызунов. В питании пустельги последние играют явно подчиненную
— 32 —
роль. В том же гнезде в хр. Найзаташ были найдены остатки двух круп-
ных, до 18 см в длину, пищух Ochotona macrotis.
В связи с этим представляют интерес находки, сделанные при археоло-
гических раскопках в гроте Шахты. Этот грот, известный своими рисун-
ками неолитического времени, расположен в 50 м ниже камина, где было
найдено гнездо пустельги (Найзаташ). В гроте под крышей располагалось
старое гнездо пустельги, устроенное аналогичным образом и, судя по на-
текам помета, занимавшееся много лет подряд. При раскопках прямо
под гнездом в слое, датированном ранней бронзой (2000—2500 лет до н. э.),
были найдены нижняя челюсть пустельги, а также ряд костей мелких
грызунов (Ochotona macrotis, Lagurus luteus, Microtus sp.), которые в отли-
чие от костей козерогов, встречавшихся в наибольшем числе, никак не
могли быть добычей древних обитателей грота и скорее всего являются
остатками добычи пустельги. Можно предположить, что в отдаленном
прошлом, когда климат Памирского нагорья был мягче, а луговая расти-
тельность была распространена шире, грызунов соответственно было
значительно больше и в питании пустельги они играли главную роль.
Вместе с постепенным иссушением Памира и ухудшением климата грызу-
нов становилось все меньше и пустельга была вынуждена постепенно
переключаться на другую добычу, главным образом на птиц, так как
рептилий на Памире нет, а крупных насекомых очень мало. Этот процесс
превращения пустельги в орнитофага, возможно, продолжается на Па-
мире и по сей день.
9. Темнобрюхий улар — Tetraogallus himalayensis himalayensis Gray.
Памир населяет номинальный подвид, распространенный также в Каш-
мире и западных Гималаях. При этом памирские птицы заметно отли-
чаются от гималайских более светлой окраской верхней стороны тела,
обусловленной сухим климатом нагорья. По окраске спины некоторые
памирские экземпляры приближаются к форме Т. h. grombczewskii Bianchi,
еще более светлой, населяющей Куньлунь к западу от хр. Русского,
включая последний.
Гималайский улар, как известно, обитает в хребтах Саура и Тарба-
гатая, в Джунгарском Алатау, на всем Тянь-Шане, в хребтах Памиро-
Алая (за исключением хребтов на юго-западе страны — Кугитанг, Баба-
таг, Актау) и Гиндукуша, в хр. Спингар, в системах Куньлуня и Нань-
шаня, в западных Гималаях, где проникает к югу до 31° с. ш. (Lamprey,
1954). Весьма характерно, что субальпийский и альпийский пояса наз-
ванных хребтов отличаются высокой аридностью. Гималайского улара
нет в Гималаях к югу от 31 ° с. ш., где благодаря прямому воздействию
муссона влажность резко возрастает (Mani, 1962). Не выяснено, как далеко
заходит темнобрюхий улар в Тибет. Вероятно, он населяет только горное
обрамление Тибета с запада, севера и частично востока, но не проникает
в глубь нагорья.
На Памире улары населяют все хребты, за исключением небольшой
группы низких безводных гор к югу от Рангкульской котловины. Диапа-
зон высот, в которых гнездится улар на Памире, простирается от 3800 до
4800 м. Высота гнездования темнобрюхого улара зависит не только от на-
личия определенных местообитаний, но и от преследования человеком.
Наиболее высоко улары гнездятся там, где их все время тревожат люди.
В районе Биостанции частые экскурсии сотрудников и местных жителей
в горы загоняют уларов на самые высокие гребни хребтов, где они гнез-
дятся до высот 4800 м (цирк Зор), вместе с тибетским уларом. Выше, по
нашим наблюдениям, улары не гнездятся. В местах, где нет населения,
эти птицы гнездятся всего в 100—150 м над днищами долин, там, где
склоны становятся более крутыми и появляются осыпи и скалы. Гнездовья
в таких случаях располагаются на высотах 4100 —4300 м, что можно
3 Биология птиц — 33 —
считать нормальным для памирской популяции. В Бадахшане, где ложа
долин расположены ниже, понижаются и места гнездования уларов.
В Гиссаро-Каратегине они гнездятся на высотах всего 2500—3800 м (По-
пов, 1959). На севере ареала, в низких кульджинских горах, улары гнез-
дятся начиная с 900 м, а в южной части, в Гарвале, — с 2400 м (Baker,
1935), т. е. намного ниже, чем на Памире. Следовательно, в целом темно-
брюхого улара нельзя отнести к высокогорным птицам.
На Памире улар населяет крутые, пустынные склоны гор, каменники,
скалы с наличием вблизи ручьев или речек и небольших пятен лугов.
Только на Памире вследствие специфических условий нагорья улары
гнездятся много выше верхней границы леса, так как все нагорье распо-
ложено выше этой границы. В остальных частях ареала они предпочитают
гнездиться в верхней части лесного пояса, будь то разреженные ельники,
как на Тянь-Шане (Кузьмина и Савинов, 1953; Кузьмина, 1955), пояс
арчи в Гиссаро-Каратегине (Попов, 1959) или заросли дуба в Гиндукуше
(Paludan, 1959). Явно избегают только густых сомкнутых лесов. Там
улары гнездятся выше границы леса, но все же в поясе кустарников.
Места гнездования уларов на Куньлуне и в восточном Тибете неизвестны.
Следовательно, повсюду, где есть подходящие условия, улар предпочи-
тает гнездиться в субальпийском и верхней части лесного пояса.
Численность уларов на Памире высока, но птицы расселены по хреб-
там крайне неравномерно. По моим наблюдениям, нормальной можно
считать плотность, когда на 1 км маршрута вдоль склона на оптимальной
высоте встречаются 3 пары. Есть места (Южноаличурский хребет, хр.
Базардара), где на 1 км маршрута встречается 5—6 пар, и, наоборот,
в других местах можно встретить 1—2 пары только на 4—5 км пути.
Крылья улара во время полета работают энергично только при взлете
и посадке. Улары взлетают обычно с крутых склонов гор или со скал,
бросаясь вниз и планируя с потерей высоты на противоположный склон
долины. Во время такого полета слышен резкий свист, производимый
жесткими перьями распростертых крыльев. Улар, как правило, не машет
крыльями почти до самой посадки. Во время полета развивается большая
скорость. Получаемую при этом инерцию птицы используют, чтобы перед
посадкой взлететь вверх по склону на 80—100 м. Это значительно компен-
сирует первоначальную потерю высоты. Приближаясь к склону, улары
меняют угол атаки крыла, ставя его плоскость под тупым углом к направле-
нию полета, и птицу несет вверх инерция и сила сопротивления воздуха.
Для одного такого полета, который улары совершали каждый вечер,
перемещаясь с дневок на вершине горы Мукор у Биостанции на противо-
положный склон долины, было высчитано, что за 1.5 км перелета птицы
теряли в высоте только 400 м. На ровной плоскости улары, даже застиг-
нутые опасностью, взлететь не могут и бегут до ближайшего склона, где
переходят на скользяще-пикирующий полет. Подобную картину наблю-
дал и Б. К. Штегман (1954). Кажется совершенно невероятным, чтобы
улары могли «парить совершенно свободно в потоках восходящего воз-
духа», как утверждает М. А. Кузьмина (1955, стр. 186). Приводимый ею
случай, когда улар взлетел на 70-метровую скалу вдоль вертикального
ее края на неподвижных крыльях объясняется совершенно так же — инер-
цией, развитой при скользяще-пикирующем полете. Вообще полет улара
назвать парящим невозможно, достаточно услышать тот резкий свист,
который бывает при этом слышен. Правильнее называть такой полет
скользяще-пикирующим.
На кормежках, обычно поднимаясь вверх, улары передвигаются нето-
ропливым шагом. Нельзя, на наш взгляд, согласиться с М. А. Кузьминой
(1955) в том, что лапы уларов не приспособлены к лазанию. Улары лазают
по скалам не хуже кекликов. Несколько раз я наблюдал, как улар сво-
- 34 —
бодно поднимался по отвесной скале, используя мельчайшие выступы
и карнизы. При отсутствии такой способности улары вряд ли стали бы
гнездиться в скалах, где всюду преобладают вертикальные плоскости.
На Памире в таких местах улары столь же многочисленны, как и на поло-
гих склонах. Следует заметить, что при лазании по маленьким выступам
отвесных стен как раз не нужно иметь широко расставленных пальцев
ног. Наоборот, чем уже лапа, тем удобнее помещать ее на узких карнизах
скал. Ширина лапы играет роль при передвижении по сыпучему или вяз-
кому субстрату, а не на скалах.
Бег улара не слишком быстр, особенно на ровной плоскости. Но на
высотах свыше 4000 м скорость, с которой улар передвигается вверх*
человеку не под силу даже на пологих склонах.
Наши наблюдения показывают, что у уларов нет особого брачного
крика. Обычная позывка как самцов, так и самок — громкий свистг
начинающийся на низкой ноте, быстро переходящий на высокие ноты..
К свисту прибавляются различные концовки: он или вновь заканчивается
низкой нотой, или после него раздается отрывистое квохтанье, либо свист
заканчивается низким урчаньем, переходящим в отрывистое «ко-ко-ко-ко»,
тоже с понижением в конце. Этот свист, который многие считают брачным
криком, улары чаще всего издают при тревоге. Не раз приходилось наблю-
дать, как улары принимались свистеть, едва завидя человека. Такого же
мнения придерживается и Д. Н. Головнин (1901). Уходя от опасности,
улары издают хриплый звук «кх. .кх», напоминающий тревожное цо-
канье фазанов. При полете издается звонкое непрерывное клохтанье,
что-то вроде «кли-кли-кли-кли-кулигалу. .кли-кли-кли-кулигалу-кули-
галу». Все эти голоса особенно часто слышны весной, во время брачного
оживления.
Улары славятся своей осторожностью. Подойти к ним на дробовой
выстрел удается очень редко. Заметив внизу охотника, улары быстро
идут вверх, все время пересвистываясь. Если охотник внезапно появ-
ляется над ними сверху, улары бросаются вниз и планируют на другое
место.
Разбивка на пары, по И. А. Абдусалямову, начинается в конце апреля
и сопровождается частыми драками самцов. В середины мая мы заставали
уларов уже разбившимися на пары. Одиночки встречались редко. Драк
между самцами мне наблюдать не приходилось. Разгар токования прихо-
дится на середину—конец мая. С утра и до вечера, слегка затихая днем,
в горах звучат крики уларов. На утренних и вечерних кормежках самцы,
сопровождая самок, то и дело поднимают вертикально хвост и с низким
квохтаньем, приспустив крылья, обхаживают самок. Во время брачного
оживления суточный ритм жизни не нарушается. На рассвете птицы
парами спускаются с ночевок в скалах на кормежки, часто выходя по
глубоким саям к самому основанию склонов. Отдельные пары кормятся
на расстоянии 50—100 м друг от друга. В полдень птицы, закончив кор-
межку, поднимаются назад, на дневной отдых, вновь спускаясь на кор-
межку вечером, но уже не так низко. Если уларов не тревожат, то самки
начинают гнездиться в местах дневного отдыха у первых скал на склонах.
Осмотр 20 старых гнезд показал, что улары охотнее всего гнездятся на
скалистых обрывах по соседству с подвижными осыпями, в маленьких
пещерках и каминах южной экспозиции. Гнездо обычно скрыто под нави-
сающим камнем или скалой. Оно представляет собой ямку в щебне,
скудно выстланную ветками терескена и перьями наседки (рис. 6, 7).
Кладка начинается в последней декаде мая или начале июня. Наиболее
рано размножение наблюдалось в котловине Рангкуля, где самка с послед-
ним яйцом, готовым к сносу, была добыта 22 мая (Абдусалямов, 19576).
Самые поздние выводки встречены в скалах Найзаташ в 1960 г., когда
- 35 —
3*
пуховички в возрасте 4 дней были найдены 26 июля. Улары на Памире
гнездятся позже, чем в других частях ареала: на пол месяца позже, чем
на сыртах Тянь-Шаня, и на месяц, чем в Ладаке.
Рис. 6 Самка темнобрюхого улара — Tetraogallus himalayensis
Gray — на гнезде
Судя по числу птенцов и по находкам гнезд, сделанным местными жи-
телями, число яиц в кладке обычно равно 6. Судя по литературным дан-
Рис. 7. Гнездо темнобрюхого улара — Tetraogallus himalayensis Gray.
ным, величина кладки варьирует в различных частях ареала: в Тянь-
Шане — от 5 до 9 яиц (Кузьмина, 1955), в Гималаях — 4—6, редко больше
(Osmaston, 1928; Baker, 1935).
36
В найденной мной кладке было 6 свежих яиц, серых, слегка желтова-
тых, с большим количеством темно-коричневых пятен и точек разной
величины, разбросанных по всей поверхности; одно яйцо было почти без
пятен. Размеры: 65.8x43.7, 66.8x46.0, 67.6x47.2, 67.6x47.3, 68.0x47.0,
68.0x48.1 мм. Гнездо было найдено 7 июня 1961 г. в верховьях р. Аличура,
в скалах правого борта долины, на высоте 4250 м. Оно помещалось под
нависающим выступом на широком балконе скалы южной экспозиции.
Насиживавшая самка подпускала человека на 5—6 м, после чего сры-
валась и стремительно планировала вниз. Возвращалась она к гнезду
пешком, незаметно и осторожно, не ранее чем через полчаса. Самец уча-
стия в насиживании не принимал, но все время держался поблизости.
Рис. 8. Самка темнобрюхого улара — Tetraogallus himalayensis Gray, —
отводящая от выводка.
Самка слетала с гнезда кормиться только днем, когда сильный солнечный
нагрев не давал кладке слишком остывать. Температура на подстилке во
время насиживания колебалась в пределах 22—28° и заметно зависела от
температуры воздуха. Наблюдения над гнездом велись в течение 12 дней;
19 июня кладка была взята в коллекцию. К этому времени в яйцах только
сформировались зародыши. Температура внутри модели гнездового убе-
жища колебалась в пределах от —6° до 33°. Вокруг гнезда нередко выпа-
дал снег, но хорошая защита от ветра заметно сказывалась. Здесь было
гораздо теплее, чем в долине на 220 м ниже, где все время дули холодные
ветры. Близ найденного гнезда, на той же высоте, держались еще 2 пары
уларов. Гнезда их находились на недоступных карнизах, в 300—400 м от
найденного гнезда.
После вылупления птенцов самка, часто вместе с самцом, выводит их
на альпийские лужайки, где они питаются только растительным кормом—
свежими травами. В недельном возрасте птенцы уже употребляют
такой грубый корм, как побеги терескена. При опасности они затаиваются
в камнях, где благодаря своей окраске совершенно незаметны. Самка
усиленно отводит человека от птенцов. Она то падает, чуть ли не кувыр-
каясь вниз по камням, то бежит, волоча крылья с тревожным квохтаньем
(рис. 8). Один из встреченных выводков, в котором птенцы уже перепар-
хивали, сопровождали самка и самец. Они пришли в крайнее волнение,
— 37 —
не улетали при нашем приближении, а уводили птенцов, издавая ранее
не слышанный мною звук — глухой, низкий стон, отдаленно напоми-
нающий вой собаки.
Выводки держатся близ водопоев, но временами удаляются от них,
спускаясь в нижние части склонов и иногда по конусам выноса выходят
почти на днища долин. В районе Сарезского озера самки с птенцами пас-
лись в боковых долинках на высоте 3800 м, среди зарослей карликового
тополя у русла потока. В конце июля—августе, когда птенцы подрастают,
выводки поднимаются в верхнюю часть альпийского пояса, где вегетация
в разгаре. Месяцем раньше сюда поднимаются холостые птицы. В литера-
туре высказывалось мнение, что самцы, когда самки заняты насижива-
нием, сбиваются в стаи и держатся отдельно, не проявляя интереса
к гнезду, а впоследствии и к выводку. Наблюдения на Памире не под-
тверждают этого: самец все же находится близ гнезда, но держится очень
скрытно и выдает себя только голосом на утренних кормежках и днем,
когда сопровождает кормящуюся самку. Наряду с ними в стороне от
гнездовий держатся небольшие группы холостых уларов по 5—15 птиц.
Число птиц в этих группах нередко значительно больше, чем гнездящихся
вокруг самок. Из таких групп в июле добывались как самцы, так и самки.
Количество не занятых гнездованием уларов бывает большим и в 1960 г.
в окрестностях Биостанции составляло 40% всей популяции. Вероятно,
улары становятся половозрелыми в возрасте не менее полутора лет.
В августе у подросших птенцов выводка устанавливается суточный
ритм, который затем соблюдается всю жизнь: спуск утром на кормежки,
дневной отдых на местах, расположенных немного ниже ночевок, вечер-
няя кормежка и уход вечером на ночевки в скалы. Суточные вертикальные
перемещения в летне-осеннее время не превышают 400—500 м. Спустив-
шись на кормежки утром, улары медленно поднимаются наверх, насы-
щаясь по дороге. Сезонные вертикальные миграции более значительны.
В августе—сентябре улары живут на высотах 4800—5000 м, в безлюдных
местностях — на полкилометра ниже. Не приходилось слышать, чтобы
кто-нибудь встречал на Памире уларов выше 5200 м. В субнивальный
пояс (4800 —5000 м) они поднимаются редко, а линию вечных снегов не
пересекают никогда. Утверждение И. А. Абдусалямова (1957а) о том, что
улары в послегнездовой период поднимаются до высоты 6000 м, основано
на ошибке в определении высоты. В местах, где работал упомянутый автор,
ни одна вершина не достигает такой высоты.
В октябре, когда снеговая линия начинает опускаться ниже, улары
тоже спускаются вниз и зимуют на нижних частях склонов. Зимой во
время суточных перемещений они не поднимаются выше субальпийского
пояса и иногда спускаются даже на плоскости долин. Гонимые бескорми-
цей, улары теряют осторожность, нередко подходя вплотную к населен-
ным пунктам. Известны случаи, когда улары в поисках корма заходили
в конюшни и жилые помещения. Образ жизни зимой тот же — утренние
и вечерние кормежки и дневной отдых. Отдыхают и ночуют улары на вы-
соте 3900—4000 м, в нишах и пещерках скал южной экспозиции.
Самцы в мае не имели следов линьки. Самки, добытые в середине и
конце июля у выводков, меняли мелкое перо. Перья крыльев и хвоста
были сильно изношены. У самки, добытой из стаи холостых птиц 2 августа,
интенсивно линяло контурное перо, маховые и рулевые перья. Часть ма-
ховых была в чехлах, а другая выросла незначительно. Поэтому самка
планировала с трудом, неуверенно и легко была сбита брошенным камнем.
Пища почти исключительно растительная. Сильный крючковатый клюв
уларов, как заметил Б. К. Штегман (1954), хорошо приспособлен для копа-
ния в твердом грунте, откуда они извлекают корни и клубни растений.
Насекомых на Памире мало, и улары только случайно захватывают их
— 38 —
вместе с травой. Этим они отличаются от тянь-шанских птиц, в летнем
рационе которых преобладают насекомые. Зимнее питание не изучено.
Судя по зимнему помету, пища состоит из стеблей и корешков отмерших
растений, собранных на бесснежных склонах. Летом кормятся исключи-
тельно свежей зеленью, поедая листья, почки, стебли и цветки Festuca
coelestis, Роа litwinowiana, Oxytropis immersa, Potentilla sericea, Primula
sp. и многих других трав (Абдусалямов, 1957а).
Мясо уларов превосходного качества, но охота на них весьма трудна.
Поэтому, несмотря на сравнительное обилие, улары добываются в незна-
чительном числе. Птенцы уларов, взятые в возрасте 10—12 дней и старше,
хорошо приручаются, но попыток разведения уларов еще не предпринима-
лось.
10. Тибетский улар — Tetraogallus tibetanus tibetanus Gould. На Памир-
ском нагорье проходит северо-западная граница ареала тибетского улара.
Помимо Памира, этот улар населяет, как известно, Тибетское нагорье и
окружающие его горные системы. Распространение на Памире до сих пор
точно не выяснено. По Р. Н. Мекленбурцеву (1946), он населяет восточную
часть Южноаличурского хребта до перевала Харгуш, горы у Мур га ба,
у р. Аксу в районе Тохтамыша и по среднему течению р. Акбайтала. Мной
найден в южной части хр. Музкол (Биостанция), а И. А. Абдусалямов на-
шел этого улара в скалах у оз. Рангкуль. Много тибетских уларов в Са-
рыкольском хребте, где у перевала Сарыташ они встречены несколькими
исследователями. Тибетские улары чаще встречаются в хребтах восточной
окраины Памира, входящих в систему Куньлуня. Отсюда на севере он
проник и в хр. Музкол, непосредственно соединяющийся с Сарыколом,
а на юге — в Южноаличурский хребет и хр. Базардара, по которым
распространился на запад по крайней мере до перевала Xapryni, а, может
быть, и до Рушанского хребта.4 Граница ареала, проходящая по нагорью,
незначительно пульсирует. Во многих местах тибетские улары то появ-
ляются, то исчезают. В центральной части Памира, например, есть долина
под названием Куртек-Сай («куртек» — киргизское название тибетского
улара), что говорит о том, что когда-то эти птицы здесь водились. Сейчас
здесь они не встречаются совершенно.
Местообитания те же, что у темнобрюхого улара, только на большей
высоте. Находки гнезд, птенцов, наблюдения в гнездовой период свиде-
тельствуют о том, что тибетский улар гнездится в пределах альпийского
пояса (4300 —4800 м). На такой большой высоте эти улары обитают и
в других частях ареала, но на более влажных окраинах высокой Централь-
ной Азии они гнездятся ниже: в Ладаке — до 3900 м, в южном Тибете —
до 3600 м. Складывается впечатление, что тибетские улары более влаго-
любивы, чем темнобрюхие, и в засушливых высокогорьях Центральной
Азии выбирают большие высоты из-за большей влажности.
Численность невысокая и подвержена резким колебаниям. Например,
в цирке Зор, в окрестностях Биостанции, в 1954—1955 гг. обитало не-
сколько выводков тибетского улара, в 1956 г. там был найден только
1 выводок, а в 1960 и 1962 гг. не было найдено ни одного.
Полет тибетских уларов мне наблюдать не удалось; при каждой из
немногих встреч они уходили вверх по склону, не взлетая даже при выст-
релах. Судя по описаниям Шефера, полет тибетского улара такого же
скользяще-планирующего типа, как у темнобрюхого, но еще более стреми-
телен и ловок. По словам Шефера, они пикируют вниз наподобие хищной
птицы, развивая большую скорость и проделывая при этом такие фигуры
4 В коллекции ЗИН АН СССР имеется 4 экземпляра, датированные 24 VIII,
1 и 6 IX, на этикетках которых значится «Рушан».
— 39 —
высшего пилотажа, что приходится удивляться, как они при этом не рас-
шибаются о скалы.
Голос заметно отличается от голоса предыдущего вида, особенно свист,
более тихий и высокий.
Единственное найденное и описанное И. А. Абдусалямовым гнездо
(1957а) помещалось на участке глинистой почвы среди обломков скал,
на высоте 4500 м. Экспозиция склона не указана. 26 мая кладка содержала
6 насиженных яиц. Для Памира это очень ранняя кладка, но лето 1956 г.,
когда было найдено гнездо, было самым теплым за последние 30 лет.
Нормально же кладка происходит в конце мая—начале июня, как и
в Ладаке (Baker, 1935) и в северо-восточном Тибете (Бианки, 1907).
В цирке Зор мы наблюдали тем же летом молодых птиц, немного уступав-
ших в размерах взрослым, 22 июля. Холодным летом 1960 г. Л. А. Ару-
тюнов встретил выводок пуховичков в середине июля на перевале Сары-
таш (Сарыкол).
Суточные вертикальные кочевки у тибетских уларов еще меньше,
чем у темнобрюхих, так как они в большей степени привязаны к опреде-
ленным частям склонов. Сезонные миграции могут быть значительными и
зависят от количества снега, выпадающего в горах. Следует иметь в виду,
что миграции тибетского улара совершаются только пешком, так как
птицы неспособны к активному полету. По сообщениям местных охотни-
ков, в снежные зимы улары спускаются до подножия хребтов, когда,
очевидно, становится возможным переход через долины, с одного хребта
на другой. Существуют места, где эти птицы скапливаются на зимовках
в большом числе, разбредаясь по окрестным хребтам с наступлением
весны. Р. Н. Мекленбурцев (1946) наблюдал их всю зиму у своей зимовки,
но летом не встречал ни одного. На этом основании он решил, что тибет-
ские улары ушли вверх и не гнездятся ниже 5000 м. Между тем снеговая
линия в районе зимовки проходит на высоте 5100 м, и маловероятно,
чтобы улары гнездились у вечных снегов, в узкой стометровой полосе
субнивального пояса. Проще предположить, что с наступлением весны
улары откочевали с мест зимовки в другие места.
Добытые 20 октября птицы усиленно линяли. Линькой было охвачено
мелкое оперение, рулевые и маховые перья уже отросли (Абдусалямов,
1961).
Питание сходно с питанием темнобрюхого улара. У добытых в летнее
время птиц в зобах обнаружены цветки и побеги Oxytropis immersa,
Sausurea pamirica, Chorispora songorica; в зобах октябрьских птиц — кор-
невища осок и луковицы лилейных (Абдусалямов, 1957а). В зимнее время,
по Р. Н. Мекленбурцеву (1946), улары кормятся почками и вегетативными
побегами Stellaria humifisa, Роа Litwinowiana, Festuca coelestis, Smelow-
skia calycina, Nepeta kokanica, Oxytropis immersa. Saxifraga hirculus,
Potentilla sericea, Alopecurus mucronatus, Primula sp., Eurotia cera-
toides.
11. Кеклик — Alectoris kakelik subsp. (?). Кеклик встречен в двух
местах нагорья. Р. Н. Мекленбурцев (1946) нашел его в конгломератовых
обрывах каньона Карасу, а пара пуховичков была добыта экспедицией
Грум-Гржимайло в июне на р. Бейке. Мне кеклик не встретился ни разу,
несмотря на неоднократное посещение этих мест. В каньон Карасу кеклик
попадает из долины Мургаба, где встречается регулярно в 40—50 км ниже
устья Карасу Кеклики поднимаются сюда, вероятно, только в особенно
теплые годы, которым сопутствует высокая численность этих птиц в со-
седних районах Бадахшана. Можно с уверенностью утверждать, что
в 1954, 1960 и 1961 гг. они на Карасу не гнездились. На р. Бейке его сей-
час тоже нет. Местные жители, хорошо знающие кеклика, утверждают,
что здесь он не водится. Имеется подозрение, что пуховички, доставлен-
— 40 —
ные Грум-Гржимайло, были добыты на одноименной речке, но стекающей
по другую сторону перевала Беик с южного склона Сарыкола, где значи-
тельно теплее и растительность богаче. Вероятно, гнездование кеклика на
окраинах нагорья носит эпизодический характер. По ущельям внешнпх
склонов пограничных хребтов Памира кеклики гнездятся в болыпомчисле,
поднимаясь вверх до 3600 м.
Взрослых птиц с Памира я не видел. На Карасу может заходить
А. к. kakelik Falk, на Беик — обитающий рядом А. к. pallescens Hellmayr.
12. Лысуха—Fulica atra atra L. Регулярно появляется на озерах
Памира на весеннем и осеннем пролетах, причем нередко часть птиц
остается на все лето. Летом лысухи наблюдались и добывались на всех
крупных памирских озерах. В особенно теплые годы отдельные пары мо-
гут даже гнездиться. Гнездо лысухи на оз. Зоркуль нашел в конце июля
зоолог Е. Н. Яковлев. Оно было устроено на одном из островов в восточной
части озера среди гнезд горных гусей. В гнезде находилось несколько
яиц, и насиживавшая птица подпустила человека почти вплотную.
В 1961 г. на озерке в верховьях р. Бахмыра (4070 м) старое гнездо
лысухи было найдено в густых зарослях высокой осоки. Видимо, отдель-
ные попытки лысух гнездиться на озерах Памира кончаются неудачно,
так как до сих пор никто не видел и не добывал здесь птенцов этой птицы.
Находка гнезда лысухи на оз. Зоркуль является рекордной по высоте
над уровнем океана. До сих пор наиболее высоким местом, где они гнез-
дятся, было оз. Сонкуль на Тянь-Шане — 3016 м (Янушевич и др., 1959).
В Кашмире они не гнездятся выше 2400 м (Baker, 1935).
И. А. Абдусалямов (1961) наблюдал в 1956 г., как несколько стай лысух
прилетело на оз. Шоркуль в конце июля на линьку. В начале августа боль-
шинство их уже бурно линяло. Линька проходила в очень сжатые сроки.
У добытых 3 августа птиц не было ни маховых, ни рулевых перьев, а в се-
редине августа они уже могли пролетать 15—20 м. Птицы покинули
озеро в середине сентября.
13. Короткоклювый зуёк — Charadrius mongolus pamirensis Richm.
На Памире распространен повсеместно по межгорным понижениям
субальпийского пояса, не поднимаясь на гнездовье выше 4300 м.
Любимые места гнездования — открытые, плоские пространства с щеб-
нистым, глинистым или песчаным грунтом, вблизи лугов или воды.
Иногда зуйки гнездятся очень далеко от воды, за 10—15 км. Нередки
они и в долинах, заваленных древними моренами, т. е. в местах с весьма
неровным рельефом.
Численность местами довольно высокая, достигает на некоторых участ-
ках максимальной плотности — 1 пара на 1 км2 (котловина озер Рангкуль,
Яшилькуль, побережье Каракуля, долина среднего течения Аличура,
долина р. Карасу). В нешироких, заваленных древними моренами долин-
ках южного Памира пара от пары гнездится на расстоянии 2—3 км. Реже
заселены безводные долины, где 1 пара встречается на 10 км маршрута.
Прилет наблюдался Р. Н. Мекленбурцевым 6 мая. В Рангкульской
котловине они прилетают в конце апреля. Брачные игры начинаются
вскоре после прилета. Птицы гоняются друг за другом низко над землей,
со своеобразным скрипучим криком «чи-у-рри. чи-у-рри». Этот крик,
судя по описаниям, свойствен ладакским и восточнотибетским зуйкам
(Schafer, 1938; Ali, 1946). Приходилось слышать голоса токующих зуйков
в мае в середине ночи. О токовании самцов по ночам пишет и Р. Н. Меклен-
бурцев (1946).
Зуйки гнездятся на открытых площадках, причем гнездо помещается
либо под кустиком терескена, либо под прикрытием камня, либо совер-
шенно открыто. Иногда гнезда устраиваются среди отмерших стеблей
куста терескена, засыпанного песком. Гнезда не имеют подстилки, как и
— 41 —
в Ладаке, но гнезда, найденные в северо-западных Гималаях, содержали
тонкую выстилку из травы (Whistler, 1925).
Верхний предел гнездования определяется на Памире наличием под-
ходящих биотопов, которые представлены только до высоты 4300 м. Наи-
более высоко зуек гнездится в долине р. Кызылрабата, где выводок нелет-
ных птенцов был встречен на высоте 4250 м. В Ладаке они гнездятся не-
сколько выше, до 4500 м (Osmaston, 1928), а в юго-восточном Тибете—
даже до 5100 м (Schafer, 1938).
Кладка состоит из 3 яиц. Самки несутся в последней декаде мая—
начале июня. Гнезда зуйков часто разоряют, и птицы вынуждены делать
Рис. 9. Птенец короткоклювого зуйка — Charadrius mongolus
pamirensis Richm.
повторные кладки. Поэтому гнездовой сезон растянут и пуховичков можно
встречать вплоть до конца июля.
Кроме гнездовых, летом встречаются стайки холостых, годовалых
птиц, отличающихся от половозрелых тусклой окраской. Из такой стайки
в середине июля были добыты 2 самки с совершенно неразвитыми фолли-
кулами. Очевидно, короткоклювые зуйки становятся половозрелыми
только на втором году жизни, как и большеклювые зуйки — Ch. lesche-
naultii Less. (Козлова, 1961). Стайки холостых птиц кочуют летом очень
широко, заходя даже на склоны гор до 4350 м.
Колониального гнездования, о котором пишет Р. Н. Мекленбурцев
(1946), не наблюдалось, однако в удобных местах пары гнездились близко
друг от друга (не ближе 100 м одна от другой).
Встреченные выводки птенцов всегда сопровождали две взрослые
птицы. Особенно сильное беспокойство проявляли родители тех выводков,
где находились пуховички в возрасте до 7 дней (рис. 9). Самка при этом
усиленно отводила — падала на землю и билась на ней, продвигаясь
вперед ударами крыльев, или бежала в сторону, волоча крыло и падая на
бок. Время от времени птицы бросались на людей, налетая почти в упор
с резким криком «увит. (тувит)». Беспокойство начиналось, когда
человек приближался к выводку ближе 100 м. Позднее, когда птенцы уже
хорошо бегали, самки не беспокоились так сильно и не отводили. До
— 42 —
подъема на крыло выводки кормятся на ровных полупустынных участках
долин или на склонах, иногда по окраинам лугов.
В конце июля молодые и старые птицы сбиваются в стаи по 40—50
птиц, которые держатся до отлета в озерных котловинах. Наряду с этими
стаями встречаются еще выводки с пуховичками. В пустынных долинах
в конце лета встречаются только небольшие стайки по 5—10 птиц.
Отлет происходит в августе, и основная масса исчезает в первой поло-
вине этого месяца. Запоздалые выводки задерживаются до сентября.
Н. А. Северцов добыл молодую самку на Аличуре 7 сентября.
Пища состоит главным образом из различных жуков, остатки которых
встречены во всех осмотренных желудках. В поисках жуков зуйки охотно
посещают места стоянок скота — большие площадки, покрытые слоем
помета. Зуйки часто кормятся по берегам озер и рек, где поедают различ-
ных беспозвоночных, бродя почти по брюшко в воде. На пойменных лугах
они не кормятся, явно избегая сомкнутой растительности.
14. Перевозчик — Actitis hypoleucos (L.). Населяет самые низкие части
нагорья — низовья рр. Аксу—Мургаба, Аличура, Памира, Кокуйбеля
и котловины озер Рангкуль и Яшилькуль, местами поднимаясь до 3800 м.
Таким образом, на нагорье Памира перевозчик гнездится выше, чем
где-либо на территории ареала. На сыртах Тянь-Шаня он не гнездится
выше 3600 м (Янушевич и др., 1959), В Ладаке его нет, а в Кашмире он
поднимается только до 2700 м (Baker, 1935). Гнездование в Тибете не дока-
зано.
Численность невелика, не более 1 пары на 1 км побережья. Холостые
птицы встречаются чаще.
Прилетает на озера Памира в конце апреля. Единственное гнездо,
найденное здесь И. А. Абдусалямовым на оз. Рангкуль, было устроено
в куче полусгнивших трав, выброшенных водой, на одном из островков
и содержало 9 июля 4 яйца.
С конца июля на озерах и речках субальпийского и альпийского поя-
сов появляются одиночные кочующие птицы, старые и молодые. В сен-
тябре на некоторых озерах (Рангкуль, Яшилькуль) их скапливается до-
вольно много, но к концу месяца они все отлетают. Последняя встреча
отмечена 1 октября на р. Памире.
15. Травник — Tringa totanus eurhinus Oberh. Большинство птиц
гнездится на кочковатых, болотистых участках обширных субальпийских
лугов поблизости от воды, в пределах высот 3700—4000 м. Верхний предел
распространения определяется наличием больших луговых массивов
с высокой травой; выше всех расположены такие луга у оз. Зоркуль
(4150 м), здесь и проходит верхняя граница гнездования травника. В Ла-
даке и Тибете при наличии подходящих местообитаний травники гнез-
дятся и выше, до 4700 м (Osmaston, 1928; Baker, 1935; Schafer, 1938).
Эти кулики на нагорье предпочитают равнинные ландшафты и избегают
сильно расчлененного рельефа.
В наибольшем числе травники гнездятся у оз. Рангкуль и по рр. Аксу
и Аличуру, образуя в подходящих местах на кочковатых болотах и ост-
ровках небольших озер колонии по 3—10 пар; редки на р. Кокуйбеле
(1 пара на 10 км маршрута), у озер Яшилькуль и Зоркуль и очень редки на
Каракуле.
Р. Н. Мекленбурцев (1946) застал травников в большом числе на Бу-
лункуле 5 мая. Теплой весной 1961 г. травники появились на Рангкуле
в конце апреля. На местах гнездования, расположенных на высоте
4000 м и более (р. Аличур), они появляются позднее, в середине или конце
мая.
С момента прилета и до середины июня на болотистых лугах держатся
небольшие крикливые стайки, по 6—15 птиц. Они кормятся, с протяж-
— 4.3 —
ными криками гоняются друг за другом или, собравшись тесной кучкойг
начинают громко кричать, вытягивая шеи. В середине мая 1961 г. на лугах
у Акташа (3900 м) они уже держались парами, хотя гнездование началось
здесь значительно позже — в конце июня. Такая задержка объясняется
медленным ростом трав. Травники устраивают свои гнезда только в гу-
стых зарослях осок не менее 20 см высоты. В долине Аличура тем же ле-
том откладка яиц также началась в конце июня. В среднем течении реки
(оз. Акбалык, 3800 м), где вегетация начинается раньше, некоторые пары
приступают к размножению в начале июня, хотя самки, не закончившие
кладку, добывались и здесь до 22 июня. В целом же сроки размножения
на Памире поздние, даже на сыртах Тянь-Шаня травники гнездятся почти
на месяц раньше.
Найденные гнезда были устроены в густой траве на кочках и малень-
ких островках. В колонии, найденной у Акташа, на кочках-островках
небольшого озерка гнездилось 6 пар травников, причем каждое гнезде
располагалось на отдельном островке. Гнезда представляли собой ямки,
выстланные травой и скрытые сверху густой осокой. Число яиц в гнездах
3—4. 20 июля в некоторых гнездах птенцы уже вылупились, в других
шло вылупление, в двух гнездах находились сильно насиженные кладки.
В колонии на озерке Бахмыр (4070 м), обнаруженной 18 июля 1961 г.,
птенцы уже вылупились. Вылупление в гнезде шло не одновременно,
растягиваясь на сутки и более; поэтому птенцы покидали гнездо пооди-
ночке, разбредаясь в траве поблизости. Только раз, в долине Аличура
у Акбалыка, было найдено гнездо, в котором находилось 3 недавно вы-
лупившихся птенца, обогреваемых старой птицей. Недавно вылупившиеся
птенцы искусно прячутся в траве, и, несмотря на издаваемый ими писк,
даже кропотливые поиски обычно были безуспешными; взрослые птицы
при этом проявляли сильное беспокойство, с криками летая над теми ме-
стами, где прятались птенцы, задерживаясь над ними на несколько мгно-
вений трепещущим полетом. Беспокойство охватывало всех членов коло-
нии, но особенно сильно беспокоилась та пара, чье гнездо было ближе
всех к наблюдателю. Птицы из дальних гнезд только тревожно кричали,
тогда как травники с ближних гнезд налетали на человека в упор. Отвода
от птенцов ни разу не наблюдалось.
Летные молодые появляются в конце августа и вскоре после этого
вместе со взрослыми покидают нагорье, так как уже в начале сентября
сильные ночные заморозки на большую часть суток сковывают льдом
главные места кормежек — мелкие водоемы и болотца. Одиночные трав-
ники задерживаются дольше. В сентябре 1956 г. слабый пролет шел
в юго-западном направлении, дата последней встречи — 21 сентября.
В желудках добытых птиц находились трудноопределимые остатки
мелких беспозвоночных. Основные места кормежек — илистые берега
водоемов, где травники часто бродят по брюшко в воде.
16. Черноголовый хохотун — Larus ichtyaetus Pall. Гнездясь на озе-
рах зоны степей и полупустынь, черноголовый хохотун, кроме того, под-
нимается довольно высоко в горы, где в настоящее время достоверно най-
ден гнездящимся только на Кукуноре (3200 м). Сведения для Памира не
столь определенны. Считалось, что в пределах нагорья хохотуны гнез-
дятся на Каракуле, Зоркуле и Яшилькуле. На последнем из них
Р. Н. Мекленбурцев в начале сентября 1935 г. наблюдал даже плохо
летавших молодых, что весьма сомнительно: здесь нет островов, где могли
бы гнездиться эти чайки. В настоящее время в гнездовой сезон на озерах
Каракуль и Яшилькуль хохотуны не встречаются совершенно (наблюде-
ния 1954, 1961 и 1962 гг.). Только на Зоркуле они довольно обычны. Это
озеро богато рыбой, которую сравнительно легко добывать на мелководьях
у восточного берега.
- 44 —
Экспедиция Ф. Столички добыла неполовозрелую особь 14 апреля
1874 г. в верховьях р. Вахан-Дарьи, южнее Зоркуля. Сам Зоркуль в это
время бывает подо льдом.
В летнее время часто встречали этих чаек в восточной части озера, но
гнезд их на островах не находили.
Осенью 1956 г., 15—28 сентября, мы наблюдали хохотунов на Зоркуле
ежедневно. Группы их держались на отмелях восточного берега. Старые
и молодые птицы отдыхали днем в обществе буроголовых чаек и огарей,
вылетая по зорям на ловлю лжеосманов, изобиловавших на мелководье.
Охотились они парами и реже одиночками. Молодая самка, добытая
20 сентября, хорошо летала. Отлет происходит с началом ледостава,
в октябре.
17. Буроголовая чайка — Larus brunnicephalus Jerd. Собранные мной
материалы позволяют сделать некоторые замечания о таксономическом
положении буроголовой чайки. Существуют разногласия относительно
того, считать ли ее самостоятельным видом или только подвидом L. ri-
dibundus L. Отличия у взрослых птиц буроголовых и обыкновенных чаек
сводятся в основном к окраске крыльев: большему распространению бе-
лого поля на маховых перьях обыкновенной чайки и меньшему — у бу-
роголовой. В бассейне Тарима добыт ряд экземпляров, которые можно
было бы считать переходными по окраске крыльев. Не касаясь спорного
таксономического положения таримской популяции, можно отметить ряд
важных систематических признаков, отличающих буроголовую чайку
от обыкновенной. Во-первых, у буроголовых чаек крыло несколько более
широкое и тупое. Во-вторых, гнездовой наряд буроголовой чайки серой
окраской верха тела резко отличается от такового обыкновенной чайки.
У молодых обыкновенных чаек коричневый затылок отделен от спины
белым зашейком, тогда как у буроголовых чаек серая окраска головы
и спины сливается. Различна окраска и пуховых птенцов: при сходном
расположении пятен птенцы буроголовой чайки окрашены в светло-серый
цвет в отличие от окрашенных в желто-коричневые тона птенцов обыкно-
венной чайки. В-третьих, голос взрослых птиц очень похож па голос
черноголового хохотуна, заметно отличаясь от голоса обыкновенной
чайки.
Кроме того, несмотря на широкую изменчивость размеров и окра-
ски яиц у обеих форм, яйца буроголовых чаек в целом заметно крупнее
и бледнее окрашены, причем в окраске их совершенно отсутствуют олив-
ковые тона, столь характерные для яиц обыкновенных чаек. Следует
особо подчеркнуть, что по распространению и экологии буроголовые
чайки являются высокогорными птицами, и ареал их нигде не перекры-
вается с ареалом обыкновенной чайки (гнездование последней на оз. Ку-
кунор требует подтверждения). Следовательно, возможность гибридиза-
ции исключена. Наличие в бассейне Тарима особей с широким развитием
черного цвета на крыльях может являться следствием индивидуальной
изменчивости и не связано с гибридизацией. Наши данные, таким обра-
зом, подтверждают заключение Е. В. Козловой (1952) о весьма далеком
родстве буроголовой и обыкновенной чаек, являющихся несомненно
двумя различными видами.
Памирское нагорье — единственное место в нашей стране, где буро-
головая чайка найдена на гнездовье. Ареал этого вида охватывает Тибет,
Ладак, Цайдам и бассейн оз. Кукунор. Зимовки расположены в Китае
(в бассейне Тарима и на юго-востоке), по восточному побережью Индии,
в Бирме. В пределах Памирского нагорья буроголовая чайка гнездится
в субальпийском поясе, на оз. Каракуль. На оз. Зоркуль. при обследо-
вании озера в июле 1962 г., зоолог Г. Сапожников не нашел гнезд этого
вида.
— 45 —
На Каракуле чайки селятся на небольших островах близ берегов
восточной и юго-западной частей озера (рис. 10). Островки эти — затоп-
ленные древние морены или остатки коренного берега — покоятся на
постепенно тающих погребенных льдах.
Голосовые реакции довольно однообразны и сводятся к вариациям
низкого, хриплого крика «аа. . .аа», который может быть то более отры-
вистым, то протяжным и иногда звучит, как грачиное «краа». Изредка изда-
ется отрывистый звонкий крик «ау. .» и еще реже — трескучее «ка-ка-ра».
Иногда в колониях слышались труднопередаваемые звуки, слегка напо-
Рис. 10. Колония буроголовых чаек —Larus brunnicephalus Jerd. —на оз. Каракуль.
минавшие весеннее бормотание тетерева. Обычно в колониях стоит тишина
и голоса чаек слышны редко, оглушительный шум поднимается только
во время тревоги.
Чайки появляются па Каракуле в конце апреля, когда поверхность
озера еще покрыта льдом и свободны только небольшие участки у бере-
гов, близ устьев впадающих речек. Размножение начинается через месяц
после прилета. Самки часто строят свои гнезда среди колоний гусей,
которые в конце мая заняты насиживанием. Гнездовые постройки разно-
образны. Встречаются массивные, хорошо построенные гнезда, но иногда
это лишь небрежно набросанные кучки стеблей. Диаметр гнезд 20—28 см,
высота 3—8 см. Сооружаются они из стеблей злаков (Hordeum) и рдестов
(Potamogeton), целые валы которых лежат на прибойной линии. Гнезда
расположены в колониях тесно друг к другу, иногда вплотную, прямо
на голой земле, среди разбросанных всюду камней. Самки начинают
нестись в начале июня, но даже в одной колонии это происходит неодно-
временно: в то время как некоторые птицы сидят на гнездах, другие еще
только их строят. Очень поздние кладки были обнаружены в маленькой
колонии из 4 гнезд 26 июня 1961 г., на маленьком островке в заливе
Музкол. Одно гнездо еще пустовало, в другом было 2 свежих, а в двух
других — по 3 слегка насиженных яйца (рис. 11). Видимо, небольшие
колонии подвержены разорению чаще и чайки вынуждены делать по-
вторные кладки. Средний размер 13 яиц 60.7x41.0 мм. В целом они
— 46 -
заметно меньше яиц птиц из Ладака (средний размер 61.3x42.6; Baker,
1935). Количество яиц в кладке — 1—3. Из 52 осмотренных гнезд в 7
было по 1 яйцу, в 22 — по 2 и в 23 — по 3 яйца.
В колонии на маленьком островке у о. Южного 28 июня 1962 г. во
многих гнездах из яиц вылуплялись птенцы. Кое-где встречались 2—3-днев-
ные птенцы, но большинство гнезд содержало сильно насиженные кладки.
Колония занимала вершину островка, возвышающегося на 2—3 м над
водой. Ближе к воде располагались гнезда горных гусей и крачек. Сам
островок находится в полосе «ветровой тени», закрытый от леденящих
Рис. 11. Гнездо буроголовой чайки — Larus brunnicephalus Jerd.
Оз. Каракуль.
северных ветров громадой о. Южного. При появлении людей чайки
взлетели с тревожными криками и несколько минут кружились над
островом, но вскоре успокоились и расселись по гнездам, хотя люди
находились в 10 м от них. Только иногда отдельные птицы налетали
на меня в упор, как бы собираясь ударить.
Вторая крупная колония, не менее 50 пар, помещалась на одном
из островков у восточного берега оз. Каракуль. Сам остров, такая же
древняя морена, как и первый, и таких же размеров, отделился от бе-
рега, видимо, совсем недавно. 24 июля в колонии находились довольно
крупные птенцы. При нашем приближении взрослые птицы, громко
крича, согнали молодежь в 2 большие стаи и погнали к берегу, бук-
вально насильно сталкивая птенцов в прибой. Многих птенцов крупные
волны вкупе с ураганным ветром сразу же выбрасывали обратно, и они
затаивались, забиваясь в щели среди прибрежных камней. Но большин-
ству удалось отплыть благополучно, и они качались на крупных волнах
с наветренной стороны острова, окруженные эскортом взрослых птиц.
Птенцы были разновозрастными, но все в гнездовом пере; длина крыла
у добытых колебалась от 171 до 210 мм.
Размножение буроголовых чаек на Памире происходит раньше, чем
в южном Тибете и Ладаке, где массовая откладка яиц начинается с се-
редины июня (Baker, 1935; Ali, 1946).
— 47 —
В сентябре 1956 г. на оз. Зоркуль эти птицы в большом числе дер-
жались вместе с черноголовыми хохотунами и обыкновенными чайками
по отмелям восточного берега. Самка в гнездовом наряде была добыта
19 сентября. Отлетают в октябре.
На Зоркуле пищей служат главным образом лжеосманы (Schysopy-
gopsis sp.). У самки, добытой 19 сентября, в зобу и желудке были только
эти рыбы, до 10 см длиной. На Каракуле, где рыбы очень мало, чайки
питаются гаммаридами (Молчанов и Зарудный, 1915) и ручейниками.
Желудки взрослых и молодых птиц с о. Чаячьего 24 июля 1962 г. были
набиты ручейниками (Astratus alaicus Mart.). Желудок взрослой птицы
содержал 100—150 экз. этих насекомых.
Как редкая птица, нигде более в СССР не встречающаяся, заслужи-
вает всемерной охраны.
18. Речная крачка — Sterna hirundo tibetana Saund. Тибетский подвид
речной крачки населяет высокогорные водоемы Центральной Азии, в том
числе и Памирского нагорья, где гнездится на озерах субальпийского
пояса. Кроме озер Каракуль и Зоркуль, крачки обитают на Малом Ка-
ракуле и на мелких озерках в поймах Аксу и Аличура. На речных остро-
вах они не гнездятся, как и на мелких озерках, расположенных на пере-
вальных плоскостях;-видимо, нет их и на лишенном рыбы оз. Рангкуль.
Гнездовья крачек расположены в пределах высот 3750—4150 м. В Тибете
крачки гнездятся вплоть до 4800 м (Schafer, 1938).
Численность на Памире невелика. В 1962 г. на Каракуле гнездилось
не более 40 пар. На островках озер поймы Аксу уАкташа в 1961 г. гнез-
дилось 6—8 пар.
Полет, голос и другие черты поведения тибетского подвида такие же,
как и у номинального.
Первых крачек на оз. Рангкуль И. А. Абдусалямов наблюдал 7 мая.
В долине Аличура появление отмечено 14 мая (Мекленбурцев, 1946). Наи-
более раннее гнездование наблюдалось на оз. Каракуль, где откладка
яиц происходила в начале июня. Несколько позже они гнездятся в вер-
ховьях Аксу и на Зоркуле. Позднее всех гнездятся крачки в среднем
течении Аличура.
Гнезда крачек отличаются друг от друга на разных озерах и даже
на разных островах. Так, в заливе Музкол на Каракуле, на маленьких
островках с темным грунтом гнездовая подстилка нередко совсем отсут-
ствовала, гнездо представляло собой ямку, и темные яйца своей окраской
сливались с грунтом. Поскольку крачки на таких островах гнездятся
поодиночке, то маскировка гнезда имеет для них большое значение. Там,
где крачки гнездятся в колониях чаек и гусей, надобность в маскировке
отпадает и птицы строят довольно большие гнезда из стеблей водных
трав и терескена. В колониях чаек и гусей гнезда крачек располагаются
по краям островов, ближе к воде. Острова на Каракуле почти лишены
растительности, и гнезда строятся на голой почве. В пойме Аксу на
островках небольших озер, заросших осокой, гнезда помещались в густой
траве и имели травяную подстилку. Диаметр лотка 9.0—11.5 см, гнезда —
13.5—20 см. Из 28 осмотренных гнезд 18 содержали по 3 и 10 — по 2 яйца.
В южном Тибете в кладках бывает только 2 яйца (Schafer, 1938). Сред-
ние размеры 20 яиц 43.4x30.6 мм, т. е. более крупные, чем у номиналь-
ного подвида, и даже крупнее, чем у тибетских крачек из Ладака (средний
размер 41.9x30.5 мм; Baker, 1935).
27 июня 1961 г. в гнездах крачек на Каракуле шло вылупление птен-
цов. 28—29 июня 1962 г. большинство гнезд содержало сильно насижен-
ные кладки и только в двух находились 1—2-дневные птенцы. В вер-
ховьях Аксу у Акташа на небольших озерках 20 июля 1961 г. крачки
были заняты выкармливанием птенцов, прятавшихся в густых зарослях
— 48 —
осоки. Птенцов найти не удалось. Было найдено только 1 гнездо с 3
сильно насиженными яйцами на островке-кочке, густо заросшей травой.
Это была явно повторная кладка. Птенцы тибетских крачек по окраске
не отличимы от птенцов номинального подвида.
Отлет не прослежен. В начале сентября наблюдались большие стаи
у оз. Яшилькуль, а в конце сентября — на Зоркуле.
На Аличуре и Аксу главный объект питания — рыба, на Каракуле —
гаммариды, ручейники, личинки тендипедид и рыба.
19. Скалистый голубь — Columba rupestris turkestanica But. Туркестан-
ский скалистый голубь, населяющий горы Средней и западные части
Центральной Азии, является обычной гнездящейся птицей Памира. Он
населяет субальпийский и нижнюю часть альпийского пояса, поднима-
ясь на гнездовье до 4500 м. Примерно до такой высоты гнездятся голуби
и в Тибете (Schafer, 1938). На таких высотах Памира, правда, голуби
редки; основная масса популяции гнездится в пределах субальпийского
пояса.
Численность наиболее высока там, где скалы непосредственно гра-
ничат с лугами или посевами (Кызылрабат, Шадпут, Рангкуль, Мургаб,
Биостанция). В других местах нагорья редок, но встречается регулярно.
Численность его в земледельческих районах Бадахшана заметно выше.
Скалистый голубь обладает довольно быстрым и маневренным полетом.
Нередко можно наблюдать пары и стайки этих птиц, стремительно ле-
тящих вдоль скалистых склонов на большой абсолютной высоте, невзи-
рая на ураганный ветер. Наиболее активно голуби кормятся по утрам
и вечерам. Днем они чаще всего отдыхают в тени на карнизах скал.
Голос — частое, низкое урчанье «уррр. . УРРРР- уррр». Очень осто-
рожны и редко подпускают на дробовой выстрел.
Р. Н. Мекленбурцев наблюдал прилет первых птиц 23 марта, когда
на нагорье еще стоит зима. С момента прилета стайки голубей кормятся
в широких долинах субальпийского пояса, на проталинах. Брачное
оживление начинается в конце апреля. С этого времени и до конца июля
воркование слышится постоянно. Сезон размножения растянут и длится
с мая по август. Там, где голубей много, они гнездятся колониями и
стаями летают на кормежки и водопой. При низкой численности они
гнездятся далеко друг от друга и стай летом не образуют.
Гнезда строят в нишах скал южной экспозиции. Самка с готовым
к сносу яйцом добыта у Биостанции 28 мая 1960 г. В 1961 г. в конце мая
птицы были заняты насиживанием. У оз. Рангкуль на высоте 3900 м
в скале южной экспозиции была обнаружена небольшая колония. 9 июля
из ниш здесь было выпугнуто 15 пар, но многие птицы не покинули ниш,
несмотря на стрельбу. В гнездах, вероятно, находились маленькие птенцы.
В том же году молодые птицы, недавно покинувшие гнездо, были встре-
чены на р. Бейке и в районе Биостанции 15 июля.
После вылета из гнезд голуби кочуют небольшими стайками вплоть
до глубокой осени. В августе—сентябре на кормежках они поднимаются
до верхних частей альпийского пояса (4700 м), ночуя там в скалах.
Осенние стаи голубей на Памире никогда не достигают таких размеров,
как в Бадахшане. К концу октября большинство голубей покидает Памир,
но в крупных населенных пунктах (Мургаб) отдельные стайки могут за-
держиваться до декабря, а возможно, и зимовать (Мекленбурцев, 1946).
Пищу составляют семена дикорастущих и культурных растений.
Во время сева они нередко кормятся на полях, подбирая семена, плохо
заделанные в землю. Во вторую половину лета у Биостанции голуби
кормились на альпийских лугах исключительно семенами Polygonum
viviparum, массовое плодоношение которого происходит с конца июля.
Водопои посещаются регулярно, обычно днем, после утренней кормежки.
Биология птиц
— 49 —
Скалистый голубь является объектом спортивной охоты и добывается
осенью в немалом количестве.
Белогрудый голубь — Columba leuconota Vig. До сих пор в орнитоло-
гической литературе появляются указания о гнездовании белогрудого,
или снежного, голубя на Памире. За 6 полевых сезонов мне не пришлось
встречать этой птицы, так же как и И. А. Абдусалямову. Говорить
о гнездовании этой птицы на Памире нет никаких оснований. Единствен-
ный раз, правда, местные жители сообщили нам, что встретили странного
голубя, по описаниям — явно снежного, в середине июля в верховьях
р. Шинды, на высоте около 4500 м. Стая из 4 птиц сидела на берегу по-
тока и пила воду. Вспугнутые, они ушли вверх по ущелью. По словам
местных жителей, раньше такого голубя они не встречали.
20. Тибетская саджа — Тchangtangia tibetana (Gould). Ареал вида охва-
тывает Тибет, Куньлунь, степи близ Кукуиора, Ладак и Памир (Коз-
лова, 1952). Наиболее низко саджи гнездятся у оз. Кукунор (3200 м),
хотя обычно они не селятся ниже 3600 м, поднимаясь в Тибете до 5700 м
(Schafer, 1938). В настоящее время на Памире гнездится только в не-
скольких местах: в котловинах озер Рангкуль и Каракуль, в среднем
течении р. Акбайтала и р. Аксу, а также в долине Тузджилга, в восточ-
ной оконечности Южноаличурского хребта, и у оз. Салангур. Все пе-
речисленные места гнездования расположены в субальпийском поясе.
Тибетские саджи гнездятся недалеко от воды, на широких плоско-
стях межгорных равнин, покрытых полупустынной растительностью и
иногда всхолмленных конусами выносов и древними моренами. Вне гнез-
дового сезона они также придерживаются плоских равнин и пологих
склонов гор, поднимаясь осенью в нижние части альпийского пояса.
Пересеченного рельефа явно избегают.
Численность в конце прошлого века была высокой, и Д. Л. Иванов
(1885) встречал многочисленные стайки по всему нагорью. В настоящее
время по всему нагорью гнездится не более полусотни пар. Как облик,
так и поведение тибетской саджи весьма своеобразны и только подчерки-
вают правильность выделения этой птицы в отдельный род. Полет ее
напоминает полет чернобрюхого рябка, только более замедлен. Он на-
столько характерен, что по нему сразу можно узнать птицу. Голос ти-
бетской саджи — громкий крик «ав-ва» или «ув-ва», откуда и местное
название — «увак». Стайка самцов при приближении опасности сначала
уходит пешком, переговариваясь гнусавым, отрывистым звуком «а. »,
затем перелетает на другое место, издавая при полете такие же отрыви-
стые звуки, но разных тонов, отчего получается разноголосица, несколько
напоминающая перекличку летящих лебедей. Самка при тревоге хрипло
квохчет «ка-ка-ка-ка-ка-ка-ра!». По земле саджи передвигаются легко и
быстро.
Весеннее оживление начинается в конце мая, когда из табунков вы-
деляются отдельные пары. Наряду с гнездящимися все лето встречаются
стайки холостых, неполовозрелых птиц. Такая же картина наблюдалась
в южном Таджикистане у чернобрюхого рябка. Кладка происходит
в конце мая—начале июня. Для гнездования выбираются плоские участки
долин, до 1.5 км в поперечнике. Гнезда устраиваются на голой почве,
под защитой кустика терескена или совершенно открыто. И. А. Абду-
салямов (19596) нашел в гнездах слабую выстилку из перьев и стеблей.
Вполне очевидно, что обильная выстилка демаскировала бы гнездо, кото-
рое благодаря покровительственной окраске яиц и насиживающей птицы
незаметно даже вблизи. На Тибете и в Ладаке, за малым исключением,
гнезда саджи также лишены подстилки (Baker, 1935).
У самцов и самок, добытых И. А. Абдусалямовым 6 июня 1957 г.
в среднем течении р. Акбайтала, были хорошо развиты наседные пятна;
- 50 -
у одной из самок в яйцеводе находилось готовое яйцо; тогда же было
найдено гнездо с 3 яйцами. Этим же автором насиженная кладка была
найдена в Рангкульской котловине 21 июня. Размеры яиц 33.0x50.0 мм.
В среднем течении р. Акбайтала саджи гнездятся на плоском про-
странстве между берегом и отрогом хребта.Ровная глинистая поверхность
кое-где усыпана щебенкой и совершенно лишена растительности; только
ближе к склону торчат чахлые кустики терескена. 24 июня 1961 г. здесь
были встречены 3 пары птиц и выводок, состоявший из 2 пуховичков
в возрасте 2—3 дней. Выводок сопровождала группа взрослых птиц —
3 самца и самка. Самка стала усиленно отводить. Самцы держались поо-
Рис 12. Птенец тибетской саджи — Tchangtangia tibetana (Gould).
даль, хотя один из них тоже пытался отводить. Пуховички были обна-
ружены только после часа упорных поисков. Оба сидели, прижавшись
к земле и настолько сливались с грунтом, что разглядеть их можно
было только на расстоянии 1 м (рис. 12). Обнаруженные птенцы начали
громко пищать; голос их напоминал писк новорожденных щенят. Слыша
их, самка подходила к человеку на 8—10 м и потом с криком взлетала.
Пуховички ходили с трудом, неуклюже и быстро остывали на холодном
ветру.
Окраска птенцов такова: от конька клюва по бокам головы идет белая
полоска. Такого же цвета полоски идут от ноздри к глазу и от угла рта
под глаз, к ушной области. Основной тон верхней стороны тела светло-
коричневый, с рыжеватым оттенком. На спине видны 3 узкие белые
полосы и неясные полосы из белых пятнышек по бокам и на крыльях.
Шея сверху покрыта более длинным пухом, и полосы здесь неясны.
Нижняя сторона тела серовато-белая, за исключением рыжеватых лап
и подбородка. От птенцов обыкновенной саджи они отличаются, помимо
строения лап, отсутствием срединной белой полосы на голове и Попе-
речных белых полос на спине, меньшей пятнистостью верхней стороны
тела и рыжеватым подбородком, который у пуховичков обыкновенной
саджи чисто белый. У птенцов тибетской саджи толще клюв: высота его
у 3-дневного птенца обыкновенной саджи 3.7 мм, у тибетской — 5.4 мм.
В 1954 г. 12 августа у оз. Рангкуль, на пустынном пространстве
у склонов гор был встречен выводок из 3 птенцов, уже начавших опе-
ряться и хорошо бегавших. Это был явно запоздалый выводок. В осталь-
- 51
4*
ной части ареала сроки размножения приходятся на май—июнь. Ранние
кладки на юге Тибета бывают уже в апреле.
В июле 1962 г. на озерах Яшилькуль и Каракуль наблюдалось слабое
движение саджей к западу и северо-западу. Летели стайки взрослых
птиц по 3—12 в каждой, с перерывами в 2—3 дня. Птицы были необы-
чайно осторожны, и ни одной добыть не удалось.
Зимний период в жизни саджей хорошо описан Р. Н. Мекленбурце-
вым (1946). Всю зиму птицы пасутся на бесснежных южных склонах,
на высоте около 4500 м. Во вторую половину зимы, во время сильных
снегопадов, они спускаются в широкие долины и появляются близ че-
ловеческих поселений. По сообщениям зимовщиков Биостанции, саджи
в последние зимы держались небольшими табунками в старых моренах
у оз. Каракуль, в долине Акбайтала у Биостанции и в ряде других долин,
где обитали в пределах 3700—4200 м. По наблюдениям Р. Н. Меклен-
бурцева, птицы целыми днями кормились по бесснежным пятнам на скло-
нах гор, нередко вместе с тибетскими уларами.
Некоторые наблюдения показывают, что тибетские саджи оседлы не
в такой степени, как это принято считать. Выше упоминалось о слабом
движении саджей на запад в июле. Р. Н. Мекленбурцев указывает, что
саджи появились на зимовке у Башгумбеза в середине ноября, а до этого
наблюдался слабый лёт птиц на запад на большой высоте, т. е. явно
птицы летели далеко. Весной саджи с зимовки исчезли. Вероятно, позд-
ней осенью птицы улетают из центральных районов Тибета, т. е. из
Джангтанга, где зимние условия слишком суровы, и зимуют в местах
с более мягким климатом. Любопытно, что Р. Н. Мекленбурцев наблю-
дал лёт саджей к западу, находясь на западной границе ареала. Куда
детели саджи — остается загадкой, так как западнее Памира никто сад-
жей не находил ни зимой, ни летом.
Можно полагать, что Памир был заселен саджами в геологическом
прошлом именно во время таких осенних кочевок.
Пища целиком растительная: зимой, по Р. Н. Мекленбурцеву, —
почки и побеги Oxytropis immersa, Smelowskia calycina, Stellaria humi-
fisa; летом зобы добытых птиц были набиты побегами Oxytropis hisiticula
(Абдусалямов, 19596).
Сравнительно недавно саджа являлась объектом спортивной охоты,
но сейчас всюду редка и попадает под выстрелы охотников случайно.
Но. несмотря на отсутствие специальной охоты, эти птицы в силу своей
доверчивости находятся на Памире, а следовательно и в СССР, на
грани истребления. Необходимо строгое запрещение охоты на эту
птицу.
21. Филин — Bubo bubo hemachalanus Hume. Этот подвид филина, на-
селяющий высокогорья Центральной Азии, гнездится и на Памире, где
найден летом в следующих местах: в скалах у Рангкуля, Акташа и вер-
ховьев р. Аличура, в скалистых долинках юго-восточных отрогов хребта
Базардара, у озер Яшилькуль и Булункуль и в конгломератовых об-
рывах по р. Мургабу. Все места гнездования расположены в пределах
субальпийского пояса. Следов пребывания его выше 4300 м встречать
не приходилось.
Брачные крики достигают наибольшей интенсивности в феврале, когда
птицы кричат и днем (Мекленбурцев, 1946). Гнезда с яйцами не найдены.
В Тибете такие находки делались в марте и в июне (Schafer, 1938). Сле-
довательно, гнездовой сезон в высокогорьях сильно растянут. На Памире
я слышал брачные крики в верховьях Аличура утром 10 июня.
Зимой филины держатся в тех же местах, но появляются и у насе-
ленных пунктов. Так, геоботаник Л. Ф. Сидоров добыл филина в пос. Кы-
зыл рабат.
- 52 -
В погадках, собранных у оз. Рангкуль, были обнаружены кости
следующих животных: огарь (2 случая), чирок (2), рогатый жаворонок (4),
дрозд (1), темнобрюхий улар (1), тибетская саджа (2), камышница (1),
лысуха (1), ржанка (5), зуёк короткоклювый (4), перевозчик (2), крачка (1),
горный гусь (1); млекопитающие — заяц (32), пищуха большеухая (6),
хомячок серый (4), памирская полевка (3), сурок длиннохвостый (1) и
желтая пеструшка — один случай (Громов и Егоров, 1953). В верховьях
Аличура в погадках были обнаружены кости: полевки (10), пищухи (4),
зайца (4), сурка (1), пустельги (2), мелких воробьиных (3). Некоторые
погадки состояли из костей рыб (Мекленбурцев, 1946). Таким образом,
главными объектами охоты являются зайцы, мелкие грызуны, рыба и раз-
личные птицы. Малый процент сурков объясняется тем, что они выходят
из нор поздно, после восхода солнца, и райо уходят на ночевку.
22. Домовый сыч — Athene noctua subsp.(?). В коллекции ЗИН АН СССР
хранятся 2 экземпляра молодых сычей с недоросшими маховыми перьями;
добытые экспедицией Грум-Гржимайло на р. Бейке в конце июня. Однако,
как и в случае с кекликом, на этикетках мог подразумеваться другой
Беик, стекающий с хребта от одноименного перевала к югу. В Михман-
Юлы добыл сыча Олкок (Alcock, 1897), а И. А. Абдусалямов наблюдал
его в середине августа на Зоркуле. Если сыч и гнездится на Памире,
то только по южной его окраине.
Сизоворонка — Coracias garrulus semenovi Loud, et Tschusi. Внесена
в список гнездящихся птиц Р. Н. Мекленбурцевым (1949) и А. Б. Ки-
стяковским (1950) после того, как экспедиция М. П. Розанова добыла
2 взрослых самцов у Кызылрабата в июле—августе. Сизоворонки в лет-
нее время неоднократно встречались и мне, но всегда это были одиноч-
ные, бродячие птицы без каких-либо признаков гнездования. Вряд ли
вообще возможно размножение этих теплолюбивых птиц на холодном
нагорье, где даже простое существование их затруднено. Об этом сви-
детельствуют остатки погибших птиц, иногда находимые на Памире.
Удод — Upupa epops epops L. Сказанное о предыдущем виде целиком
можно отнести и к удоду, с той только разницей, что одиночные бродя-
чие птицы летом встречаются довольно часто.
23. Полевой жаворонок — Alauda gulgula subsp.(?). Поющие полевые
жаворонки были встречены только в среднем течении Аличура (Акбалык,
3850 м) в июне—июле 1954 г. и в июле 1956 г. Они начинали петь с вос-
ходом солнца и до полудня, несмотря на сильный ветер. При ветре токо-
вые полеты продолжались недолго, 1—2 мин., и не выше 30 м над землей.
Большую часть времени птицы проводили в густой траве лугов. Найти
гнездо и добыть взрослых птиц не удалось, поэтому вопрос о подвиде
остается открытым. Вероятнее всего здесь обитает A. gulgula lhamarum Mei-
nertz., который в большом числе гнездится по рр. Гунту и Шотбулаку
до высоты по крайней мере 3600 м (Мекленбурцев, 1946). Там жаворонки
вьют гнезда среди густой осоки, приступая к размножению в конце мая.
Однако здесь может гнездиться и другой жаворонок, A. arvensis dulcivox
Brooks, который населяет Алайскую долину и часто добывается на Па-
мире на пролетах.
Летом 1961 г. жаворонков у Акбалыка не удалось обнаружить.
24. Тонкоклювый жаворонок — Calandrella acutirostris acutirostris Hume.
Гнездится спорадично в субальпийском поясе нагорья. Обычен в котло-
винах озер Рангкуль, Яшилькуль и Булункуль, в долине Аксу—Мур-
габа вверх до Кызылрабата, в долине Аличура от Акбалыка до устья
и в долинах Сарыкола, Тагармы, Памира и Вахан-Дарьи.
Судя по литературным данным, тонкоклювые жаворонки гнездятся
в довольно сухих местах, однако крайняя сухость нагорья слишком вы-
сока и для них. Поэтому они селятся здесь в наиболее влажных местах,
— 53 -
получающих около 300 мм осадков в год, чем и объясняется их спорадич-
ное гнездование. Индикатором повышенного количества осадков для суб-
альпийского пояса Памира является степная растительность, которая
есть везде, где гнездятся эти жаворонки. Помимо влажности, важен еще
рельеф. На пологих склонах древних морен они еще гнездятся, но на
склоны гор не идут. Одно из важных условий — близость водоемов.
В выборе характера растительного покрова эти жаворонки не так строги
и гнездятся как на голой почве, так и в густой траве лугов.
Численность наиболее высока у озер Рангкуль и Яшилькуль.
В остальных местах жаворонков меньше, на 1 км маршрута встречается
7—10 пар.
Тонкоклювые жаворонки особенно активны в безветренные часы, но
поющие птицы встречаются и во время сильных ветров. Токовые полеты
прекращаются, когда скорость ветра превышает 5—6 м сек. Голос до-
вольно однообразен и звучит как «цирр» или «циа». При тревоге эта по-
зывка сдваивается. В отличие от рогатого жаворонка тонкоклювые жа-
воронки очень пугливы.
В котловине Рангкуля И. А. Абдусалямов наблюдал прилет 17 мая.
Сроки размножения поздние. Пение начинается с июня, причем жаво-
ронки одинаково поют как на земле, так и в воздухе. Разгар пения падает
на июль. Песня состоит из несложных фраз, повторяемых несколько раз
с небольшими интервалами.
В районе Кызылрабата жаворонки гнездились в полосе пологих, ста-
рых морен между горами и в пойме реки. Большинство селилось па
плоских поверхностях древних террас и в сухих руслах, усыпанных
галькой, где растительность была представлена только редкими кусти-
ками терескена. В среднем течении р. Мургаба они гнездились на ма-
леньких площадках среди кустиков мирикарии, часто у самого русла.
В устье Аличура жаворонки вили гнезда как на ровных, пустынных
поверхностях террас, так и на лугах, в густой траве, но при этом избе-
гали сырых участков. В среднем течении реки они совсем не гнездились
в засушливой долине, а предпочитали лучше увлажненные северные
склоны морен по левому ее краю.
Гнездо — плотная, аккуратно свитая из старых стеблей постройка,
более компактная, чем у рогатого жаворонка. Размеры гнезд (3): наруж-
ный диаметр 74—120 мм, диаметр лотка 51—70 мм, глубина лотка 35 —
40 мм. Не было замечено и признаков сооружений из камешков для
защиты от ветра, о чем писал Хингстон (Hingston, 1927). Только гнезда,
устроенные в полупустыне, имели с подветренной стороны выкладку
из кусочков почвенной корки, как и у рогатого жаворонка (см. ниже).
Яиц в кладке 3, размеры их (6) 19.8x14.4 мм.
Раньше всего гнездятся тонкоклювые жаворонки в среднем течении
р. Мургаба (3500 м). Здесь 5 августа 1961 г. молодые птицы уже держа-
лись стайками вместе со старыми; вывод был давно закончен. Была най-
дена только одна насиженная кладка, возможно повторная. Позднее
гнездятся жаворонки в Рангкульской котловине (3780 м), где И. А. Аб-
дусалямов наблюдал вылетевших из гнезд подлетков 18 июля. В это же
время выводят жаворонки в устье Аличура (3760 м). В 1962 г. вылупле-
ние началось здесь 10 июля. Позднее всего гнездятся птицы в верховьях
Аксу (4000 м), где в середине июля в найденных гнездах были слегка
насижеиные кладки.
Эмбриональный пух у птенцов расположен на надглазничных, заты-
лочном, спинном, плечевых, локтевых и бедренных участках кожи. Он
густой, серого цвета. Кожа на верхней стороне тела темно-шиферного
цвета, резко отличается от светлой нижней стороны тела. Валик в углах
рта слегка желтоватый.
— 54 —
Тонкоклювые жаворонки держатся на местах гнездовий до самого от-
лета. Осенью они появляются и в тех местах, где отсутствовали летом.
В сентябре стайки тонкоклювых жаворонков встречаются всюду. Отлет
происходит в октябре.
25. Рогатый жаворонок — Eremophila alpestris diluta (Sharpe). Одна
из обычнейших птиц нагорья. Основная масса популяции гнездится в пре-
делах субальпийского пояса, и только редкие пары поднимаются до вы-
соты 4400 м. В теплые годы возможно гнездование на больших высотах,
до 4500 м, где нередко наблюдались взрослые птицы. В юго-восточном
Тибете эти жаворонки гнездятся до 5300 м (Schafer, 1938), в Ладаке и
южном Тибете — до 4500 м (Baker, 1934).
Излюбленные места гнездования — пологие склоны, усыпанные ще-
бенкой, ровные площадки на террасах, плоские вершины старых морен,
сухие окраины кочковатых лугов и возделанные поля. Рогатые жаво-
ронки заселяют также обширные плоскости межгорных понижений, но
очень редки в совершенно безводных местностях. Гнездятся они и на
крутых склонах гор, и на лугах, и даже в узких альпийских долинках
у бурных потоков. Определенно избегают только высокотравных боло-
тистых лугов и песков. На кормежках старых и молодых птиц можно
встретить решительно всюду.
По численности рогатые жаворонки уступают лишь горным вьюркам.
В подходящих местообитаниях пара от пары гнездится на расстоянии
70—100 м. В безводных долинах центрального Памира, где жаворонков
меньше всего, расстояние между отдельными парами нередко достигает
1—3 км.
Рогатые жаворонки — сравнительно малоподвижные птицы, более
оживленные только в брачный период. Голос — меланхоличное «тси-ли»
или'«тсп-ли-ли», которое как самцы, так и самки повторяют на разные
лады. Летают неохотно и большую часть дня проводят на земле, кормясь
или отдыхая среди камней. Ночуют здесь же, забиваясь под камни или
кустики терескена.
Весеннее оживление начинается в апреле. Хорошо выраженной песни
у самцов нет, просто несколько обычных позывов быстро и громко по-
вторяется в унылой минорной гамме. Эта нехитрая песенка распевается
обычно на земле или при погоне за самкой или соперником. В воздухе
в отличие от равнинных популяций Средней Азии и Казахстана не поют.
Пары занимают определенные гнездовые участки, откуда изгоняют
посторонних особей. Первые пары приступают к размножению в начале
мая, но таких немного. Большинство птиц начинает гнездиться в конце
мая. Гнездовой период растянут, и гнезда с яйцами попадаются до на-
чала августа. Н. А. Северцов добыл подлетка даже 18 сентября. Есть
основания полагать, что на Памире часть популяции, начиная гнез-
диться очень рано, делает 2 вывода в лето. Иначе невозможно объяснить
частую встречаемость гнезд с кладками в июле—августе. Сильно растянут
сезон размножения у рогатых жаворонков и в других высокогорьях
Центральной Азии, в частности в Ладаке, где они также, видимо, выво-
дят дважды в лето (Baker, 1934).
Гнездо строит в основном самка. Сначала вырывается ямка, в которой
и свивается гнездо таким образом, что верхний край его находится вро-
вень с землей. Ямка выкапывается с подветренной стороны укрытия
(камень, куст терескена). В большинстве мест Памира летом преобла-
дают ветры юго-западных румбов, поэтому большинство гнезд довольно
точно ориентировано по сторонам света, помещаясь с северо-восточной
стороны от укрытия. Во время рытья субстрат отбрасывается назад, об-
разуя по подветренному краю гнезда валик. Поскольку свежие земляные
выбросы, резко выделяясь на светлой, выгоревшей на солнце поверх-
— 55 —
ности, сильно демаскируют гнездо, птицы вынуждены маскировать их,
закрывая кусочками поверхностной корки или камешками, покрытыми
пустынным загаром. В результате получается впечатление искусственной
стенки, сооруженной у края гнезда, которой Хингстон (Kingston, 1927)
ошибочно приписал функцию защиты от ветра. На самом деле никакой
защиты от ветра такое сооружение не оказывает, о чем убедительно го-
ворит его положение всегда с подветренной стороны.
Гнезда построены примерно по одному типу. Варьируют размеры и
прочность самой постройки, что зависит от индивидуального опыта
птицы. Чашечка вьется из отмерших стеблей злаков, обычно весьма не-
Рис. 13. Гнездо рогатого жаворонка — Eremophila alpestris
dilute, (Sharpe).
брежно, хотя встречаются и аккуратные постройки. Гнездо выстилается
исключительно растительным пухом, чаще всего сильно опушенными се-
менами терескена. В южном Тибете выстилка бывает иногда из перьев
или из шерсти. Размеры гнезд таковы (15): наружный диаметр 90—130 мм,
диаметр лотка 60—80 мм, глубина его 30—50 мм и толщина дна 15—20 мм.
В полной кладке 1—4 яйца. Из 30 найденных гнезд с полными кладками
3 содержали по 1 яйцу, 16 — по 2, 10 — по 3 и 1 — 4 яйца. В последнем
случае гнездо было найдено под перевалом Кызыларт, на высоте 4200 м
и 14 июля содержало 3 птенцов и 1 яйцо-болтун. Гнездо с кладкой из
4 яиц нашел на Рангкуле И. А. Абдусалямов, но такие случаи — исклю-
чение. Наиболее обычны кладки из 2—3 яиц, как и на Тибете (рис. 13).
В одни годы (1961, 1962) встречаются преимущественно кладки из 2 яиц,
в другие (1954, 1956) — из 3. Связь величины кладок с погодными усло-
виями не прослеживается. Видимо, в данном случае имеет значение воз-
раст птиц. В то же время малые размеры кладок рогатых жаворонков
на Памире безусловно являются приспособлением к нехватке корма.
На меньших высотах, как в Алайской долине, число яиц в кладках воз-
растает до 4—6 (Янушевич и др., 1959).
Окраска яиц однообразна. Белый фон равномерно покрыт неясными
серовато-фиолетовыми пятнами, поверх которых разбросаны более резкие
светло-коричневые пятнышки. В зависимости от густоты этих пятен
общий фон, нередко даже в одной кладке, может колебаться от светло-
— 56 —
Температура насиживания в гнездах рогатых
жаворонков
Гнездо № 1 Гнездо № 2
время темпера- тура поч- вы, °C темпера- тура в гнез- де, °C время Д o' а я q СУ Е СС со S Г £
3 ч. 30 м. 28.2 3 ч. 30 м. 19.2
9 20 — 30.0 9 30* 24.8
10 0 — 30.0 10 00* 24.8
10 25 — 33.8 11 00 30.3
10 32 27.0 27.6 И 30 27.3
И 00 29.0 30.3 12 00 28.2
12 00 . 35.5 30.9 13 00 29.7
12 30 35.0 32.2 16 00 27.8
13 00 36.0 30.9 17 00* 29.8
18 00 — 31.2 18 00* 28.8
18 20 — 31.7 19 00 30.3
19 00 — 31.6 19 30* 27.3
20 00 — 31.4 20 00 28.5
20 30 — 30.4 20 30 26.5
21 30 — 26.8 21 00* 32.1
21 45 — 29.2 21 30* 24.3
Примечание. Звездочкой отмечены случаи, ког-
да птица в гнезде отсутствовала.
и вечером, около 17 час. Температура
в 2 гнездах микроэлектротермометром.
Таблица 1
серого до темно-коричневого, почти шоколадного цвета. Средние размеры
6 яиц 24.4x16.7 мм.
Насиживает самка. Самцы в это время держатся близ гнезда, вступая
в драки с другими жаворонками, если те подлетают близко. На кор-
межках самцы сопровождали самок, которые во время насиживания кор-
мились обычно дважды в день: утром, когда солнце поднималось доста-
точно высоко (11 —12 час.),
насиживания определялась
Результаты приведены в
табл. 1.
Срок насиживания, про-
слеженный в 1 гнезде, равен
И дням. Развитие птенцов
шло следующим образом.
1-й день. Вес птенца
2.71 г. Он покрыт густым се-
рым пухом, расположенным
на надглазничных, затылоч-
ном, спинном, плечевых, кис-
тевых, бедренных и брюш-
ных участках кожи. Валик
в углах рта желтого, зев —
оранжевого цвета. На языке
3 черных пятна и 1 впереди
языка, в вершине нижней че-
люсти. Ушные отверстия от-
крыты, птенцы реагируют на
циканье.
2-й день. Пух распу-
шился, глаза намечены бо-
роздами. На затенение момен-
тальной реакции не было, при
тактильном раздражении пте-
нец сжимался. Кожа на спи-
не стала более пигментиро-
ванной, серо-черной.
3-й день. Глаза приоткрылись, птенцы реагируют на голос наблю-
дателя, вскидывая голову и выпрашивая корм. Издают слабое циканье.
Как и накануне, стараются держаться на солнечной стороне гнезда.
4-й день. Вес 4.5 г. Глаза открылись совсем. Появились пеньки
перьев на крыльях, спине, шее и по бокам брюшка. Глаза большей частью
закрыты, но и в таком положении птенцы реагируют на изменение све-
товой обстановки. Самка, как и ранее, продолжает обогревать птенцов.
5-й день. Вес 5.25 г. Наметились все птерилии. Перестали вскиды
вать голову на голос наблюдателя, на лапах стоять еще не могут.
6-й день. Почти все перья, кроме маховых, вышли из чехликов.
7-й день. Вышли из чехликов маховые перья, первые маховые едва
проклюнулись. Верх приобретает пеструю окраску.
8-й день. Первые маховые распускаются медленнее остальных,
вышли из чехликов рулевые перья, птенцы сидят молча, при испуге
издают тонкий писк, стараются держаться головой к ветру.
9-й день. Птенцы покидают гнездо.
От этой схемы иногда бывают отклонения. Маховые перья иногда по-
являются на 6-й день, а птенцы уходят из гнезда на 8-й. Нередко с 3—4-го
дня развитие птенцов становится неравномерным и один из них сильно
отстает в росте, что приводит к задержке в гнезде на 1—2 дня. В 1962 г.
— 57 —
было проведено несколько измерений, выясняющих приобретение птен-
цами теплокровности. 2-дневный птенец, обогреваемый самкой, имел тем-
пературу тела 31.5° при наружной температуре воздуха 16.5°. У остав-
ленного без обогрева птенца через минуту температура понизилась
до 28.0°, еще через минуту — до 27.5°, и еще через минуту — до 25.5°.
Птенцы покидают гнездо нелетными, способность к полету появляется
через 5—7 дней.
Процент разорения гнезд выше, чем у других птиц Памира. Из 30
найденных гнезд было разорено 12. Большинство гнезд разоряют собаки,
лисицы и клушицы.
Поднявшись на крыло, молодые птицы держатся небольшими стай-
ками и одиночками, и только в августе начинают сбиваться в большие
стаи, достигающие максимальных размеров в конце сентября. Состав та-
ких стай непостоянен, они то рассыпаются, то собираются вновь. К ноя-
брю значительная часть популяции откочевывает с нагорья и здесь
остаются преимущественно самцы (Мекленбурцев, 1946), которые кочуют
небольшими стайками, обычно близ населенных пунктов.
Весной жаворонки часто кормятся на посевах, собирая семена, плохо
заделанные в землю. В гнездовой период корм в основном животный.
Осенью, с прекращением таяния снегов, когда ледниковые потоки не до-
стигают нижних частей долин, жаворонки регулярно поднимаются в верх-
ние части субальпийского пояса на водопой.
26. Горная ласточка — Ptyonoprogne rupestris rupestris (Scop.). Гнез-
дится в субальпийском поясе Памира. Здесь она всюду редка, но распро-
странена шире, чем другие ласточки. Отдельные пары найдены на гнез-
довье в следующих местах: р. Мургаб (3600 м), р. Аксу (3700 м), р. Ка-
расу (Джамантал, 3600 м), оз. Рангкуль (3900 м), приток Кокуйбеля
Карабулак (3900 м), Западный Пшарт (4000 м), хр. Найзаташ (4200 м).
На Алпчуре не найдены, они гнездятся только на одном из притоков,
в теплом каньоне р. Бахмальджилги (3800 м). Выше 4200 м на гнездовье
не найдены. В Ладаке они гнездятся выше, до 4500 м (Osmaston, 1927;
Ali, 1946).
Гнездится на крупных скалах, реже пойменных обрывах. У верхнего
предела распространения заселяет только скалы. Кормиться залетают
в нижние части альпийского пояса. Численность низкая. Одиночные пары
гнездятся далеко друг от друга, колоний не образуют.
В отличие от городских горные ласточки охотятся главным образом
вдоль вертикальных стен и обрывов, причем на лету почти вплотную
прижимаются к поверхности скал. Полет при этом гораздо менее стре-
мительный, но зато более маневренный, чем у городских ласточек. Каждая
пара имеет свой участок, который облетает по определенным маршрутам.
Приверженность горных ласточек к вертикальным плоскостям, обычно
южной экспозиции, объясняется тем, что днем скалы сильно нагреваются
солнцем. Вдоль них образуются восходящие токи теплого воздуха, за-
хватывающие насекомых, в большом числе ютящихся в густой раститель-
ности у подножия скал. Часть насекомых ласточки, вероятно, снимают
прямо с поверхности скал.
Первых ласточек у Рангкуля И. А. Абдусалямов наблюдал 5 мая.
Пролетные одиночки встречаются до середины июня. К гнездованию при-
ступают поздно, в июле. Сообщение И. А. Абдусалямова о кладке у Ранг-
куля в конце мая ошибочно. Даже в Алайской долине, где на высоте
3000 м сроки размножения птиц более ранние, горные ласточки начинают
гнездиться в конце июня (Янушевич и др., 1959).
Гнезда сооружаются на стенах скал и береговых обрывах, всегда под
нависшим козырьком. Они примечательны необычайно густой выстилкой,
состоящей в основном из пуха бородачей и сипов. Б. Гейликман, между
— 58 —
прочим, не раз наблюдал в горах Армении, как горные ласточки брали
пух из гнезд белоголовых сипов (устное сообщение). В меньшей степени
используется шерсть козерогов. Гнезда устраиваются на высоте несколь-
ких десятков метров над основанием скалы и совершенно недоступны.
Нередко они помещаются в колониях городских ласточек.
На р. Карабулаке эти ласточки вили гнезда 4—5 июля (1958 г.).
В ущелье Бахмальджилги 11 июля 1962 г. ласточки насиживали. Два
гнезда, найденные в скалистых долинках хр. Найзаташ на высоте 4200 м,
26—28 июля 1960 г. содержали слегка насиженные кладки. В ущелье
Западного Пшарта в одном из гнезд 24 августа 1954 г. находилось 4
вполне оперившихся птенца, в другом 28 августа было 3 птенца накануне
вылета и 1 яйцо-болтун. Таким образом, в разные годы сроки размноже-
ния примерно одинаковы и выкармливание птенцов падает на август —
месяц наиболее спокойный в отношении ветров. Отлет происходит сразу же
после вылета молодых, так что в сентябре их встречать уже не прихо-
дилось.
27. Деревенская ласточка — Htrundo rusttca rustica L. Встречается на
Памцре только по долине Аксу—Мургаба, вверх до Кызылрабата, но
гнездится, видимо, нерегулярно. В 1932 г. М. П. Розанов добыл в Кы-
зылрабате молодую ласточку, которая хотя и имела недоросшие руле-
вые, вполне могла залететь сюда из долины верхнего Пянджа. В июле
1961 г. несколько ласточек все лето держалось в Кызылрабате, но не
гнездилось. В 1957 г. И. А. Абдусалямов наблюдал, как в Мургабе эти
ласточки носили глину для гнезд 21 мая. Мне они в Мургабе вообще
не встречались. Кроме Кызылрабата, бродячие особи встречены летом
у Рангкуля, у Каракуля и у Биостанции.
28. Городская ласточка — Delichon urbica meridionalis Hart. Эта лас-
точка, будучи многочисленной в Бадахшане, на Памире редка и встре-
чается только в нижней части субальпийского пояса. До высоты 4000 м
она поднимается в немногих местах. Городская ласточка найдена на
гнездовье: на рр. Аксу—Мургабе с притоками, вверх до долины р. Каш-
касу, у оз. Рангкуль, у оз. Яшилькуль и по Аличуру вверх до 3850 м,
в верховьях Западного Пшарта до 4050 м.
Прилет на оз. Рангкуль наблюдался 12 мая (Абдусалямов, 1961).
Пролетные ласточки встречались до середины июня.
К гнездованию приступают поздно, в июне. Около 10 осмотренных
колоний состояли из групп тесно прилепленных друг к другу гнезд и
располагались под нависшими карнизами скал южной экспозиции. В са-
мом центре находились старые гнезда, 2—3-летней давности, на них были
налеплены новые, в количестве, составлявшем примерно треть общего
числа гнезд. Встречались колонии, состоявшие из нескольких новых
гнезд, или одни старые, брошенные птицами. Иногда попадались одиноч-
ные гнезда; все они были встречены на верхнем пределе распростране-
ния, что свидетельствует о тенденции в теплые годы к расширению вер-
тикальных пределов гнездования. Гнезда обычно устраиваются в нишах
и щелях скал, закрываемых искусственной глиняной стенкой с летком.
Судя по нескольким слоям выстилки в гнездах и учитывая, что ласточки
на Памире делают только 1 вывод за лето, можно думать, что гнезда
используются несколько лет подряд.
Число яиц в кладке 2—3, меньше, чем в Ладаке, где в нормальной
кладке 4 яйца. На равнинах, как известно, городские ласточки откла-
дывают до 7 яиц. Размеры 2 яиц из одной кладки: 20.8x13.2,
20.5X13.9 мм.
В верховьях Западного Пшарта (4000 м) 2 июля 1961 г. ласточки
еще лепили гнезда. В ущелье Бахмальджилги (3800 м) 11 —12 июля
1962 г. в большинстве гнезд птицы насиживали, в остальных были све-
— 59 —
жие кладки. В долине р. Бейка (4100 м) 15 июля 1961 г. в гнездах нахо-
дились слегка насиженные кладки. На Западном Пшарте 24 августа
1954 г. в большей части гнезд птенцы были накануне вылета, некоторые
уже покинули гнезда. В целом сроки размножения на Памире более
поздние, чем в Ладаке, где птенцы вылетают из гнезд в июле (Osmaston,
1927). Иногда наблюдается массовая гибель птенцов накануне вылета
во время внезапных похолоданий. При обследовании гнезд в 1961 г. во
многих из них в слоях, соответствовавших 1959 г., находились муми-
фицированные трупики вполне оперившихся птенцов, по 2—3 в каждом
гнезде. Такие же случаи наблюдал И. А. Абдусалямов на Рангкуле.
Птенцы гибнут незадолго до вылета, когда им нужно максимальное ко-
личество корма. В этом отношении горные ласточки, которые селятся
разрозненными парами и более приспособлены к добыче корма в горах,
во время похолоданий гораздо лучше переносят бескормицу и в отличие
от городских ласточек успешно выкармливают 4 птенцов.
После вылета молодых городские ласточки на нагорье не задержи-
ваются и исчезают в начале сентября. Спустя некоторое время на Памире
начинается пролет более северных популяций. Последняя стайка из
3 птиц была встречена на Зоркуле 19 сентября 1956 г.
29. Ворон — Corvus corax tibetanus Hodgs. Гнездится по всему Памир-
скому нагорью в субальпийском поясе, навещая остальные пояса во время
поисков пищи. Всюду редок. Скопления молодых и старых птиц наблю-
дались только на свалках у Мургаба, у Кызылрабата и у оз. Булункуль.
По наблюдениям Р. Н. Мекленбурцева, брачные игры начинаются
в феврале и продолжаются около месяца. В середине апреля вороны
держатся скрытно, в глухих местах у скал. Первые хорошо летавшие
молодые были встречены 12 июня, но они могли подняться на Памир
с меньших высот, где весна наступает раньше. Иначе пришлось бы пред-
положить, что вороны на Памире приступают к откладке яиц в начале
апреля, когда стоит еще настоящая зима. Столичка наблюдал гнездо-
строение у Акташа 4 мая (Sharpe, 1891). По некоторым наблюдениям,
вороны могут гнездиться на нагорье еще позже, в середине мая. В 1961 г.
на стационаре Аличур-2 пара воронов встречалась регулярно до 20 июня,
когда наблюдения были прерваны. Пара эта держалась в гигантских ска-
лах над стационаром в одном и том же месте, нападая на появлявшихся
поблизости крупных хищных птиц. Судя по поведению воронов, гнездо
их располагалось на высоте 4200—4300 м, в одной из многочисленных
ниш.
По-видимому, сроки размножения растянуты, как и в других частях
ареала. Шефер (Schafer, 1938) добыл в юго-восточном Тибете насижи-
вавшую самку 8 июня, а 24 августа наблюдал подлетков.
Помимо гнездовых пар, летом встречаются группы холостых птиц. Их
всегда много у Мургаба на свалках, так же как и в Кызылрабате. Боль-
шие скопления воронов наблюдались в июле 1954 г. на оз. Булункуль,
где птицы кормились у берегов рыбой.
Молодой самец с недоразвитым контурным оперением добыт у Мургаба
14 июля (Дементьев, 1935).
Зимой, по наблюдениям Р. Н. Мекленбурцева, вороны держатся.у на-
селенных пунктов, в то же время регулярно облетая большие территории
в поисках корма.
Ворон кормится отбросами на свалках, разоряет гнезда птиц, особенно
гусей, нападает на птенцов огарей и, видимо, уларов, поедает раститель-
ную пищу на лугах и мелких беспозвоночных по берегам водоемов,
охотно питается рыбой, которую ловит на мелководьях, часто кормится
на падали. Один из работников Биостанции наблюдал в январе успеш-
ное нападение ворона на зайца.
— со -
30. Клушица — Pyrrhocorax pyrrhocorax centralis Stres. Клушицы гнез-
дятся по всему нагорью, даже в самых безводных местах. Основная масса
обитает в пределах субальпийского пояса, отдельные пары гнездятся и
выше, до 4500 м. Они предпочитают гнездиться в скалах, конгломерато-
вых обрывах и изредка в развалинах могил поблизости от лугов или
посевов. В небольшом числе населяют и безводные долины.
Численность наиболее высока близ больших массивов лугов у озер
Рангкуль, Яшилькуль, Булункуль и по долине Аксу.
Прекрасные летные качества клушиц отмечались многими наблюда-
телями. Часто приходилось видеть, как стаи клушиц, закончив кор-
межку на полях Биостанции, низко над землей летели к ближайшему
склону, где старались поймать восходящий ток воздуха. В таком потоке
птицы начинали описывать широкие круги, не удаляясь от склона и
быстро поднимаясь вверх. Иногда стая теряла восходящий поток; тогда
птицы снова переходили на активный полет, пока не ловили новый.
Нередко за каких-нибудь 5 мин. стая поднималась подобным образом
на такую высоту, что переставала быть видимой в бинокль. И все это
проделывалось подчас без единого взмаха крыльев. Достигнув нужной
высоты, клушицы слегка сгибали крылья и начинали плавно скользить
вниз, направляясь к местам отдыха или гнездовьям. Птицы спускались
при этом довольно быстро и с малой потерей высоты, так как по време-
нам, используя инерцию, меняли угол атаки крыла и взлетали вверх.
При этом полет замедлялся, но клушицы складывали крылья, и скорость
полета опять быстро возрастала.
Голос — звонкое, слегка гнусавое, визгливое «ийок-ки» или «кьяааа»,
по общему звучанию похож на крик обыкновенных галок.
Брачное оживление начинается в апреле. В конце этого месяца
Р. Н. Мекленбурцев наблюдал гнездостроение. Оно сильно затягивается,
судя по тому, что подлетки появляются не ранее конца июня. Клушицы
гнездятся как группами, так и отдельными парами в нишах скал и бере-
говых обрывов. Чаще всего гнезда помещаются в неглубоких нишах скал.
Основание гнезда строится из веток терескена, а лоток густо выстилается
шерстью. Судя по количеству птенцов в выводках, число яиц в кладке 3,
очень редко 4.
8 июня 1962 г. в гнезде, расположенном в глубокой щели скалы юж-
ной экспозиции, на высоте 4300 м (Биостанция), находились довольно
большие птенцы. 10 июня 1961 г. у останца Чатырташ, в верховьях
Аличура, под гнездовой нишей у основания скалы было найдено 3 не-
давно выброшенных кем-то разновозрастных птенца. У старшего махо-
вые перья наполовину вышли из чехликов. 7—8 июля 1962 г. происходил
массовый вылет из гнезд у устья Аличура. В гнезде, найденном у Кызыл-
рабата 10 июля 1961 г., были 3 птенца, выскочившие наружу, как только
до него добрались. Здесь же, в долине р. Бейка, 2 выводка, только что
покинувшие гнездо, были встречены через 5 дней.
Птенцы незадолго до вылета из гнезда встречают взрослых, прино-
сящих корм, особыми криками, напоминающими протяжный стон. Такие
крики издают и подлетки еще 2—3 дня после вылета из гнезда. Взрослые
птицы кормят птенцов 3—4 раза в час. Спустя несколько дней после
вылета выводки объединяются в крупные стаи, кочующие по лугам и
склонам гор. Осенние стаи клушиц на Памире никогда не достигают
таких огромных размеров, как в долинах Бадахшана, но иногда на-
считывают до ста птиц. К зиме большая часть популяции откочевывает
с нагорья, а на зиму остаются небольшие группы, концентрирующиеся
у населенных пунктов.
Птенцов выкармливают главным образом беспозвоночными, которых
старые птицы добывают на лугах, в прибрежной грязи, на склонах гор
— 61
под камнями и даже на диких и домашних животных. Не раз удавалось
наблюдать, как клушицы садились на спины отдыхающих сибирских ко-
зерогов и склевывали с них паразитов. Нередко клушицы разоряют
гнезда мелких птиц, в частности бледной завирушки, пустынной каменки
и рогатого жаворонка. Большинство мелких воробьиных тревожно реаги-
рует на появление клушиц у гнезд.
Осенью, зимой и весной основной корм растительный. Во время ве-
сеннего сева стаи клушиц наносили некоторый урон опытным посевам*
Биостанции, собирая не только плохо заделанные в почву семена, но>
и выкапывая их из земли. Осенью в окрестностях зимовки Р. Н. Меклен-
бурцева клушицы кормились на склонах гор, выкапывая клубни и корни
растений.
31. Альпийская галка — Graculus graculus jorsythi (Stol.). Гнездится
в основном по окраинам нагорья. Она обитает в Заалайском хребте,
в скалах Акташа, в долине р. Аличура, в верховьях р. Памира и
у оз. Рангкуль, т. е. в наиболее увлажненных местах нагорья с боль-
шим количеством скал и лугов. Вертикальное распределение у альпий-
ской галки такое же, как и у клушицы, но выше 4200 м она не подни-
мается. Везде, где гнездится альпийская галка, гнездится также и клу-
шица. В наибольшем числе альпийская галка обитает в долинах
Бадахшана, где местами она гораздо многочисленнее клушиц.
Местообитания те же, что и у клушиц, но в несколько более увлаж-
ненных местах. В еще большей степени, чем клушица, альпийская галка
связана с лугами, что было уже объяснено Б. К. Штегманом (1955).
Приверженность альпийских галок к влажным скалистым местообита-
ниям и земледельческим районам хорошо прослеживается и в других
частях Центральной Азии. Альпийские галки не гнездятся в пустынных
внутренних частях Ладака и Тибета, заселяя только хорошо увлажнен-
ные окраинные хребты.
Большие колонии в пределах нагорья находятся только в верховьях
р. Памира, несколько меньше их в верховьях Западного Пшарта и у Ак-
таша. Численность подвержена сильным колебаниям. Например, альпий-
ские галки встречались у Акташа в 1954 г. в большом числе, а в 1961 г.
там вообще не были обнаружены.
В скалах Аличура первые пары появляются в начале марта (Меклен-
бурцев, 1946). Биддульф наблюдал их в большом числе у Акташа в на-
чале мая 1874 г. (Sharpe, 1891). В середине июля 1954 г. у Акташа дер-
жались большие стаи по 30—40 птиц, среди которых было много молодых.
Альпийские галки в небольшом числе зимуют на нагорье (Мекленбур-
цев, 1946).
32. Стенолаз — Tichodroma muraria L. Будучи обычной птицей окру-
жающих горных хребтов, стенолаз гнездится на Памире в очень малом
числе. Гнездовые пары были встречены в низовьях р. Бейка, у оз. Ранг-
куль, в ущельях восточных склонов хр. Найзаташ, в скалах Акташа
и у Биостанции, в диапазоне высот 3800—4600 м.
По всему нагорью стенолазы обитают в одних и тех же биотопах —
хорошо прогреваемых солнцем отвесных скалах, защищенных от ветров,
обычно поблизости от воды, особенно там, где вода течет прямо по стен-
кам скал. Только в таких местах эта строго насекомоядная птица может
найти пищу в достаточном количестве. Именно поэтому стенолаз гнез-
дится так высоко в центральном Памире (4600 м), где субальпийский
пояс очень сух, и только в альпийском поясе имеются описанные выше
биотопы. На более увлажненных окраинах нагорья он гнездится ниже,
на высоте 3900—4100 м. Стенолаз вообще может подниматься на гнездовье
очень высоко. Мейнертцхаген, например, встретил выводок этих птиц на
перевале Каракорум (6300 м) (Meinertzhagen, 1927). Наблюдения индий-
— 62 -
ских энтомологов (Mani, 1962) подтверждают, что и на таких больших
высотах в скалах все же имеется достаточно нагреваемых солнцем убе-
жищ с теплым микроклиматом, где обитает ряд «нивальных» насекомых.
Вероятно, именно недостаточная влажность обусловливает отсутствие
этой птицы во внутренних частях Ладака и Тибета.
Прилет происходит в мае. Гнездование позднее. В районе Биостанции,,
на высоте около 4700 м, 28 июля 1961 г. самец гонялся за самкой и пел.
Судя по поведению самки, она была занята насиживанием. Выкармли-
вавшие стенолазы были встречены в низовьях р. Бейка 18 июля 1961 г.
Откочевка большинства особей с Памира происходит в конце августа.
Одновременно через нагорье идет пролет стенолазов в юго-западном и
восточном направлениях. Добытые в это время птицы интенсивно линяли,
шла смена контурных, рулевых и маховых перьев. Часть птиц спускается
на зимовку в теплые предгорья Гиндукуша и Бадахшана, а часть —
в долины западной Кашгарип, где зимой стенолазы обычны. Во время
ясной, солнечной погоды стенолазы залетают на нагорье и зимой, так как
Р. Н. Мекленбурцев встречал одиночных птиц 4 декабря и 7 марта. Та-
кие находки показывают, что стенолазы способны переносить довольно
низкие температуры.
33. Белобрюхая оляпка — Cinclus cinclus leucogaster Вр. Населяет все
окраинные хребты Памира, а также хребты Музкол, Базардара и Южно-
аличурский. Она селится по берегам бурных потоков, текущих среди
скал, а таких мест на нагорье немного. Ее нет в безводных горах цент-
ральной части нагорья, а также на больших реках со спокойным течением.
На Сарезском озере, в пограничной области между Памиром и Бадах-
шаном, восточную часть бассейна — устье Мургаба — населяет белобрю-
хая оляпка, а западную — бурая (С. pallasi Temm.). Последняя под-
нимается там на высоту 3850 м, но в пределы нагорья не заходит.
Вертикальное распределение также определяется наличием подходя-
щих местообитаний. На р. Западном Пшарте оляпки гнездятся от устья
(3300 м) до верховьев (4200 м). В хребтах Музкол и Базардара они гнез-
дятся в пределах высот 4000—4200 м, поднимаясь в нижнюю часть аль-
пийского пояса. Распространение их вверх ограничивает отсутствие
крупных потоков. В южном Памире (рр. Беик, Ханюлы) нижний пре-
дел гнездования находится на высоте 4000 м, так как дальше реки вы-
ходят из ущелий на широкий простор долины Аксу.
Численность невысокая. Пара от пары гнездится не ближе 1 км.
Обычно в долине протяженностью 7—8 км гнездится 2—3 пары. На реках
южного Памира, которые достаточно многоводны с ранней весны, оляпки
появляются с середины мая. В долинах хр. Музкол оляпки прилетают
в начале июня, когда речки перестают пересыхать на ночь.
Места гнездования однотипны. Ими служат отвесные скалы, под кото-
рые бьет вода. Гнезда строятся на маленьких площадках в 0.2—3 м над
водой, в начале—середине июня. Новое гнездо сооружается недалеко
от прошлогоднего, и нередко на стенах скал встречается рядом не-
сколько старых гнезд. Размеры гнезд (3): наружная длина 20.5—24.0 см,
ширина 22—27 см, высота 14—16 см. Размеры летка 7—8x3 см. Раз-
меры гнездовой камеры: высота 8, ширина 6.5 см.
В верховьях Западного Пшарта 12 июля 1961 г. найдено свежее
гнездо с яйцом-болтуном. В гнезде, найденном 12 июля 1962 г. в ущелье
Бахмальджилги, находилось 2 насиженных яйца. В низовьях р. Бейка
гнездо, найденное 11 июля, содержало 1 крупного птенца, у которого
маховые и рулевые перья начали выходить из чехликов. Цвет их черный,
со светло-коричневыми концами. Спина была покрыта серыми перьями,
имевшими слегка охристое основание. Перья на брюшных и бедренных
птерилиях только что вышли из чехликов и были светло-желтого цвета.
— 63 —
Зев светло-оранжевый, клюв оранжевого цвета с желтым концом. Ширина
валиков в углах рта 5.1 мм, цвет их белый, у концов клюва переходящий
в желтый.
Самка, выкармливавшая птенцов, была добыта у Биостанции 19 июля
1962 г. Подлеток, которого еще кормили взрослые, был добыт в вер-
ховьях Западного Пшарта (4200 м) 24 августа 1954 г. Сроки гнездования,
следовательно, сравнительно поздние.
Оляпки покидают Памир вскоре после вывода птенцов, в сентябре,
спускаясь в долины Бадахшана, Кашмира и Кашгарии. Ранний отлет
их объясняется тем, что в сентябре таяние снегов почти прекращается,
бурные потоки превращаются в небольшие ручейки, промерзающие ночью,
а крупные реки начинают покрываться льдом. Возможна зимовка оди-
ночных птиц на теплых ключах в верховьях Аксу.
34. Каменка-плясунья — Oenanthe isabellina (Temm.). На Памире свя-
зана с местами, где выпадает около 300 мм осадков в год. Поэтому рас-
пространение ее на нагорье спорадично и не выходит за пределы нижней
части субальпийского пояса. Гнездится по долине р. Аличура вверх
до 4000 м и в верховьях Аксу (3900—4000 м). Н. А. Северцов указывает
на гнездование плясуньи на Каракуле; мной она здесь не найдена. Воз-
можно, гнездование в Зоркульской котловине, где имеются подходящие
места и где ее добыла экспедиция Столички.
На Памире плясунья населяет выровненные пространства, пологие
склоны морен и террас в непосредственной близости от лугов.
Гнездование позднее. На стационаре Аличур-2, где гнездилась одна
пара, в период наблюдений с 5 по 20 июня 1961 г. самка, вероятно, на-
сиживала, так как появлялась редко и ненадолго. Самец же целыми
днями держался на окраинах лугов, где все время пел, совершая иногда
короткие брачные полеты. При взлете слышалось легкое щебетанье, на-
поминающее голос жаворонка. В песне воспроизводились голоса травни-
ков и особенно часто повторялся протяжный свист, которым местные
пастухи сгоняют скот. То же самое наблюдалось на стационаре Аличур-1
в конце июня—начале июля 1954 г. У Кызылрабата плясуньи гнездились
в моренных холмах, примыкающих к лугам. В середине июля 1961 г.
самцы уже не пели. 9 июля наблюдался вылет выводка из гнезда, устро-
енного в заброшенной сурчине. Тревожный крик самки у выводка —
громкий непрерывный крик «так-так-так-так-так». Через день подлетки
откочевали от гнезда на несколько десятков метров, а еще через день
вообще исчезли из района наших работ. В 1962 г. у устья Аличура вылет
подлетков из сурочьей норы наблюдался 8 июля.
Отлет ранний. Плясуньи покидают нагорье сразу же после вывода
птенцов. Пролет более северных популяций идет с августа по октябрь.
35. Пустынная каменка — Oenanthe deserti oreophila (Oberh.). Один
из наиболее характерных обитателей Памира. Пустынная каменка насе-
ляет все долины и овраги и во многих безводных местностях является
единственной гнездящейся птицей. Основная масса популяции гнездится
в пределах 3600—4300 м, но в благоприятные годы отдельные пары под-
нимаются по теплым, сухим склонам до 4550 м. В Ладаке и Тибете они
гнездятся немного выше, до 4950 м (Baker, 1933).
Пустынные каменки населяют как скалы и речные террасы, так и
склоны различной крутизны, заваленные камнями и осыпями. Нередко
гнездятся в скалах у самых лугов, вместе с плясуньей, но в отличие от
нее никогда не селятся на ровных местах. Чаще всего они встречались
в нижней части склонов гор на границе с ровными пространствами меж-
горных впадин.
По численности занимают третье место после горного вьюрка и рога-
того жаворонка. В пустынных долинах центрального Памира пара от
- 64 -
пары гнездится на расстоянии 1 км, в скалах и обрывах вдоль лугов
пары гнездятся через 100—200 м. Нормальной можно считать плотность
каменок по долине р. Чечекты у Биостанции, где на протяжении 9 км
по склонам вокруг потока гнездится 12—15 пар.
Охотятся каменки главным образом на склонах гор, нередко вылетая
на пустынные плоскости и на луга. Часто взлетают на отдельные круп-
ные камни и с них высматривают добычу. Во время охоты нередко бро-
саются вниз по склону навстречу восходящим потокам воздуха и на
трепещущих, а иногда и совсем неподвижных крыльях задерживаются
на несколько секунд на одном месте. Высмотрев добычу, птица стреми-
тельно бросается вниз. По земле передвигается сильными прыжками.
Р. Н. Мекленбурцев наблюдал прилет пустынных каменок в середине
апреля. Первыми появляются самцы. В течение всего мая через нагорье
идет слабый пролет самцов, самок летит меньше. У местных птиц пары
образуются в начале мая, что сопровождается непрерывными драками сам-
цов, которые во время размножения очень подвижны, часто совершают
токовые полеты и изгоняют с гнездового участка других каменок и даже
горихвосток. Позывка самца — меланхоличная трелька «тилили-тюлюлю»,
наиболее часто слышна во время драк и при опасности. Некоторые счи-
тают эту позывку песней. Гнусавые трели, перемежающиеся с короткими
и длинными свистами, иногда бывают слышны при токовых полетах и
у гнезд. Пение слышно до конца августа.
Гнездо сооружается главным образом самкой. Из 27 найденных гнезд
11 помещалось в щелях и нишах скал, 7 — в норах и щелях пойменных
обрывов, 6 — под камнями на склонах и 3 — в каменных стенах загонов
для скота. Они устроены так же, как гнезда пустынных каменок на рав-
нинах Средней Азии и Ирана (Зарудный, 1900). Это громоздкие постройки,
состоящие из 3 слоев разнородного материала. Рыхлую основу состав-
ляют веточки терескена, корешки и стебли травянистых растений. Часто
эта основа заполняет всю гнездовую полость. На рыхлом основании
вьется чашечка, аккуратно свитая из более тонкого растительного ма-
териала, перемешанного с шерстью. Выстилка состоит из толстого слоя
шерсти козерогов и домашних животных. Постройка гнезда длится не
менее 10 дней. Нормальное число яиц в кладке 4, реже 3, еще реже 5.
Повторные кладки содержат 2—3 яйца. Размеры яиц одной кладки:
21.2x16.0, 21.0x15.8, 22.4x15.8 мм.
Большая часть каменок приступает к откладке яиц в начале июня,
хотя И. А. Абдусалямов находил полные кладки уже 23 мая. Длитель-
ность насиживания, прослеженная в 2 гнездах, равна 13 дням. Одно-
дневный птенец весит 2.8 г. Длинный птенцовый пух расположен на над-
глазничных, затылочном, спинном, плечевых и бедренных участках кожи.
Корм для птенцов обычно собирается на склонах и лужайках, располо-
женных нередко на 300 м по вертикали ниже гнезда. В гнезде, найденном
4 июля 1962 г. в стенке каменной изгороди, находилось 5 явно голодаю-
щих птенцов, так как взрослые птицы не справлялись с выкармливанием
такого большого выводка. Птенцы были очень тощими и, несмотря на
большие размеры, имели маховые перья еще в пеньках. В гнездах же
с нормальной для Памира кладкой в 3—4 яйца каменки всегда успешно
выкармливают выводок (рис. 14) и никаких задержек в росте и развитии
птенцов не замечалось. Вылет птенцов происходит на 14-й день. Позывка
подлетков — отрывистый писк; подлетающих родителей они встречают
легким трещаньем. Температура только что покинувшего гнездо подлетка
36° при температуре воздуха 15°. Взрослые кормят подлетков еще 7—8,
иногда до 10 дней. Первые подлетки появились в 1961 г. 25 июня, мас-
совый вылет шел в начале июля. В 1960 и 1962 гг. сильное похолодание
в конце мая задержало гнездование и первые подлетки появились
5 Биология птиц
— 65 —
30 июня. Гнезда пустынных каменок из-за плохой маскировки часто ра-
зоряются; повторные кладки нередки, и подлетки встречаются до начала
августа.
С середины июля, когда выводки разлетаются. Памир буквально на-
водняет масса одиночных молодых каменок, которые встречаются всюду,
поднимаясь до высоты 4700 м. С середины августа появляются пролет-
ные старые птицы, у которых линька идет полным ходом. Пролет закап-
чивается к октябрю.
Каменка-плешанка — Oenanthe hispanica pleschanka (Lepech.). Вклю-
чена в списки гнездящихся птиц Памира А. Б. Кистяковским (1950).
Рис. 14. Пустынная каменка — Oenanthe deserti oreophila
(Obcrh.) — с кормом у гнезда
На наш взгляд, никаких оснований для этого нет. На Памире каменка-
плешанка встречается только на пролетах.
36. Черноголовый чекан — Saxicola torquata тайга (Pall.). В. Эббот
добыл насиживавшую самку на восточной окраине Памира, видимо в до-
лине Сарыкола, 11 июня 1894 г. В коллекцию было взято также гнездо
с яйцами, принадлежавшее добытому экземпляру. Местность указана
неточно (Тагдумбаш-Памир). Высота находки 3900 м.
Самка этого вида была добыта экспедицией Грум-Гржимайло в июле
1885 г. на р. Бейке. Мной на Бейке не найден. Интенсивный пролет через
нагорье идет в августе—сентябре.
37. Горихвостка-чернушка — Phoenicurus ochruros phoenicuroides (Moore).
Туркестанская горихвостка-чернушка, населяющая гористые части Сред-
ней и Центральной Азии, является обычной птицей нагорья. Основная
часть популяции гнездится в пределах субальпийского пояса, и только
отдельные пары поднимаются до 4350 м.
Местообитания схожи с таковыми пустынной каменки, но обязательно
наличие поблизости хотя бы небольших лужаек. Эта горихвостка яв-
ляется обычной птицей долин и покрытых гигантскими осыпями склонов
переходной зовы между Памиром и Бадахшаном, на высотах 3200—
3700 м.
Особенно много чернушек в местах с кустарниковой растительностью,
хотя сама чернушка в кустарниках не гнездится и редко в них кормится,
— 66 —
явно предпочитая окружающие скалы. В долине р. Бейка вдоль скал,
нависших над зарослями ивняка, в середине июля 1961 г. встречено 3 вы-
водка на протяжении 1 км. Численность чернушек довольно высока по
всем узким, теплым ущельям субальпийского пояса. На открытых про-
странствах межгорных впадин они не встречаются.
Прилетают в конце апреля (Абдусалямов, 1961). В середине мая пары
занимают гнездовые участки. Самцы в это время очень агрессивны и всту-
пают в драки не только друг с другом, но и с самцами краснобрюхих
горихвосток. По сравнению с чернушками, обитающими на меньших
высотах (Алайская долина, Бадахшан), самцы памирской популяции
отличаются меньшей интенсивностью и звучностью пения. Громкая песня
чернушки в щели Бордобо в Заалайском хребте (4200 м) звучит, как
«чи-ко. чи-чи-чи-чи-чи-ко». Песня памирских самцов варьирует. Пер-
вый вариант можно передать слогами «ви-те-ре-ре. ви-те», а второй —
«рри-чи-чи-чи-чи-тиу (далее следует оригинальный звук, в точности вос-
производящий шорох осыпающихся камней). чри-чи-чпу». Концовка
бывает громкой и резкой. Иногда песня заканчивается тихой трелькой.
Среди поющих самцов несколько раз встречались особи в наряде, свой-
ственном самкам.
В холодное лето 1960 г. птицы приступили к постройке гнезд в конце
мая, а в еще более холодном 1962 году — в начале июня. Из найденных
гнезд 10 помещалось под камнями на склонах и 4 — в трещинах и поло-
стях скал, обычно южной экспозиции. Гнездо представляет собой довольно
рыхлую постройку. Основание состоит из рыхлой кучи стеблей и кореш-
ков, иногда с примесью мха и растительного пуха. Чашечка свивается
из тонких стебельков, шерстинок и пуха. Выстилка довольно толстая и
делается из шерсти, волоса, перьев и растительного пуха. Размеры 5 гнезд:
наружный диаметр 125—140 мм, высота 45—73 мм, диаметр лотка 53—
70 мм и глубина его 32—38 мм. Нормальное число яиц в кладке — 4.
Яйца светло-голубоватые, почти белые, без пестрин. Средний размер
И яиц 20.60 X 15.32 мм, вес свежих яиц 1.85—2.00 г. Самки несутся
сначала июня, в случае затяжной весны — с середины месяца. Насижива-
ние длится 13 дней. Однодневный птенец одет густым пухом, достигающим
на спине И мм и расположенным на тех же участках кожи, что и у обыкно-
венной горихвостки. Температура насиживания незадолго перед вылупле-
нием равна в среднем 31°; температура тела 12-дневного птенца в гнезде
39.3° В этом возрасте птенцов кормят с интервалами в 5—7 мин. Птенцы
покидают гнезда на 13-й день и держатся вблизи еще неделю. Подлетки
сначала сидят под камнями, ожидая корма, а под конец уже сами следуют
за взрослыми. Позывка подлетков звучит, как «ци-кре. ци-кре-кре»,
напоминая тревожную позывку взрослых птиц — «ци-тек. . ци-тек-тек».
Массовый вылет происходит в первой половине июля. С конца этого
месяца одиночные молодые, начавшие кочевку, встречаются по всему
нагорью. Запоздалые кладки, вызванные, вероятно, разорением гнезд,
нередки. Свежие кладки встречались до 1 июля.
По окончании гнездового периода начинаются широкие кочевки как
старых, так и молодых птиц. В сентябре через нагорье идет заметный
пролет чернушек в юго-западном направлении. Дата последней встречи
в 1956 г. — 10 октября.
Линька начинается в конце июля и продолжается до конца августа.
Еще 28 августа попадались старые птицы, у которых маховые перья только
начинали выходить из чехликов.
38. Краснобрюхая горихвостка — Phoenicurus erythrogaster grandis
(Gould). Обычная гнездящаяся птица Памира, где населяет верхнюю часть
субальпийского пояса и альпийский пояс от 3900 до 4700 м. Из насекомо-
ядных птиц нагорья этот вид гнездится наиболее высоко. Нет его в без-
— 67 —
5*
водных хребтах и долинах центрального Памира. В бассейне Аличура
они населяют берега только самых верхних частей ледниковых потоков,
такая же картина наблюдается и в других местах.
Гнездовья располагаются преимущественно у потоков в отвесных ска-
лах и на крутых склонах гор или морен, усыпанных камнями и осыпями,
обычно поблизости от лугов. Своей привязанностью к горным потокам
эти горихвостки напоминают Chaimarornis leucocephala (Vig.), о чем уже
упоминалось ранее (Osmaston, 1927). Вероятно, этой привязанностью
к влажным местообитаниям и объясняется то обстоятельство, что они не
гнездятся в нижних частях субальпийского пояса, где влажность очень
низка.
В наибольшем числе краснобрюхие горихвостки селятся вдоль пото-
ков, берега которых покрыты узкими полосками лугов. Здесь пара от
Рис. 15. Самец краснобрюхой горихвостки — Phoentcurus
erythrogaster grandis (Gould).
пары гнездится на расстоянии 300—400 м. Более редки они на скалистых
склонах у водораздельных гребней, 1 пара на 1.5 км маршрута. В таких
местах птицы выбирают наиболее влажные участки, обычно у выходов
грунтовых вод, иногда среди снежников. Нередко горихвостки гнездятся
в крупнообломочных осыпях, под которыми текут ручейки.
Краснобрюхие горихвостки — молчаливые и довольно подвижные
птицы. Охотятся, передвигаясь сильными прыжками и время от времени
перелетая на другое место. Добычу берут на земле, но нередко и в воздухе.
Птицы, зимующие в нижних поясах гор, окружающих Памир, начинают
подниматься вверх в марте. Последние зимовавшие горихвостки наблю-
дались в предгорьях Гиссарского хребта 1 апреля (А. И. Иванов, 1949).
В 1961 г. первые горихвостки появились у Биостанции 23 марта.
Р. Н. Мекленбурцев встречал их на Аличуре с 4 апреля. 4 апреля добыл
эту птицу у Акташа Биддульф (Sharpe, 1891). Весенний пролет длится
весь апрель и начало мая. На пролете краснобрюхие горихвостки придер-
живаются луговых массивов широких долин, избегаемых летом. На места
гнездовья первыми поднимаются самцы, которые нередко вынуждены
держаться еще среди сплошных снегов (рис. 15). Распределение по гнез-
довым участкам сопровождается ожесточенными драками самцов как друг
с другом, так и самцами чернушек и пустынных каменок. Поют самцы
очень редко. За 2 летних сезона удалось слышать эту песню только 6 раз.
- 68 -
Не упоминает о песне краснобрюхих горихвосток и А. Кыдыралпев (1959).
Песня состоит из меланхолических, громких свистов, по тону несколько
напоминающих свист черного дрозда. Свисты прерываются щебетаньем
или циканьем. Во время пения также воспроизводится тот особый звук,
напоминающий шорох осыпающихся камней, который характерен для
пения чернушки.
С середины мая самки начинали искать места для гнезд, а в конце
этого месяца гнездостроение шло полным ходом. Строит гнездо преимуще-
ственно самка, это же наблюдается и в Тянь-Шане. Гнездо представляет
собой большую, громоздкую постройку, весом свыше 100 г. Основанием
гнезда служит беспорядочная куча корешков и крупных стеблей, иногда
наполовину заполняющая гнездовое убежище. На этом основании соору-
жается чашечка, свитая или из тонких стеблей Stipa glariosa, или почти
целиком из шерсти козерогов и домашних овец с примесью перьев и мха.
Выстилка чаще всего делается из перьев птиц — голубя, улара, синей
птицы и др., реже из шерсти. Размеры гнезд: наружный диаметр 130—
140 мм, высота 80 мм, диаметр лотка 65—78 мм, глубина 36—47 мм. Судя
по размерам, гнезда краснобрюхпх горихвосток па Памире более мас-
сивны, чем на Тянь-Шане. Из 13 найденных гнезд 8 были устроены в ни-
шах и трещинах скал, 4 — под камнями в осыпях п 1 — в нише берегового
обрыва.
Полная кладка содержит 4 яйца, так же как и в Ладаке, тогда как
в Тянь-Шане полная кладка состоит из 5 яиц. Средний размер 5 яиц
22.7 X 15.2 мм. Цвет отнюдь не голубоватый, как пишет И. А. Абдуса-
лямов (1961), а белый с красноватыми крапинками, сосредоточенными на
тупом конце. Самки начинают кладку в конце мая, хотя похолодания
могут ее сильно задержать. Запоздалые кладки, вызванные, видимо,
разорением гнезд, встречаются до середины июля. Одна из них, найденная
15 июля 1961 г. на р. Бейке, содержала 3 свежих яйца. Гнездо было сде-
лано явно наспех, и громоздкое основание, обычное в других гнездах,
здесь отсутствовало, но шерстяная чашечка толщиной 9 мм была свита
тщательно. Самец участия в насиживании не принимает, хотя держится
у гнезда и сопровождает самку на кормежках. Насиживание в одном из
гнезд продолжалось 14 дней. Опушение только что вылупившегося птенца
типично для рода Phoenicurus. Эмбриональный пух темно-коричневого
цвета располагается на надглазничных, затылочном, спинном и плечевых
участках кожи. На голове пух достигает в длину 12 мм.
Гнезда нередко строятся очень высоко над лужайками, откуда берется
корм для птенцов, и птицам при выкармливании приходится далеко и
высоко летать. В одном случае горихвостки носили корм с лужайки
в гнездо, расположенное на 250 м по вертикали выше, на высоту 4700 м.
Частота прилетов при этом была не менее 1 раза в 10 мин. В середине дня
интенсивность кормления резко падала, снижаясь до 2 прилетов в час.
Температура тела птенцов накануне вылета равнялась 40°, птенцы поки-
дали гнездо на 14-й день. Более длительного пребывания в гнездах, как
на сыртах Тянь-Шаня (Кыдыралиев, 1959), не наблюдалось.
У гнезд с птенцами горихвостки теряют свою обычную осторожность,
особенно самцы, которые кормят птенцов в 4—5 м от наблюдателя. В другое
время взрослые птицы редко подпускают человека ближе 30 м.
Подлетки остаются у гнезда еще 1—2 недели, старые птицы кормят
молодых не менее 7 дней после вылета. Голос подлетков такой же, как
у подлетков чернушки. Массовый вылет происходит в середине июля.
После распада выводков горихвостки по-прежнему держатся на тех же
высотах и только во время снегопадов спускаются ниже.
Откочевка птиц с нагорья начинается в сентябре. В 1956 г. наблю-
дался оживленный пролет этих горихвосток в верховьях р. Памира. Птицы
— 69 -
двигались с перевала Башгумбез в Южноаличурском хребте вниз по до-
лине, временами собираясь в небольшие группы. Отлет заканчивается
в середине октября.
Старые птицы энергично линяют с конца июля, заканчивая линьку
к середине сентября. Молодые перелинивают в первый, взрослый наряд
позднее — к октябрю.
Варакушка — Су anosy Ivia svecica tianschanica Tug. Указана А. Б. Ки-
стяковским (1950) как гнездящаяся по всему Памиру, в том числе и в Ка-
ракульской котловине. Это явная ошибка. Нигде на территории нагорья
варакушки не гнездятся, но бывают в большом числе на пролете с сере-
дины августа п до ноября.
Каменный дрозд — М on tic о la saxatilis turkestanicus Zar. А. Б. Кистя-
ковский (1950) приводит этого дрозда в качестве гнездящейся птицы юж-
ного Памира. Я не располагаю никакими данными о гнездовании камен-
ного дрозда на Памире. Он обычен здесь только на осеннем пролете с ав-
густа по октябрь.
39. Гималайская завирушка — Laiscopus himalayanus (Blyth). Населяя
горные системы, с севера и запада окружающие Центральную Азию,
гималайские завирушки не проникают в глубь этой страны. Гнездясь
в поясе хорошо развитых и обильно увлажненных альпийских лугов, они
не находят необходимых условий в засушливых высокогорьях Централь-
ной Азии. По этой же причине гималайские завирушки не гнездятся во
внутренних частях Памирского нагорья, а населяют только более влаж-
ные окраинные хребты. Здесь они найдены в двух местах: в Заалайском
хребте и на западных отрогах Южноаличурского хребта.
По Р. Н. Мекленбурцеву (1946), завирушки населяют днища ущелий
у подножия перевалов, где разбросаны снежные и ледяные поля. Здесь
завирушки держатся на россыпях камней, чередующихся с мокрыми
лугами, покрытыми низкорослой осокой.
В верховьях Шахдары гималайские завирушки появляются в первой
половине апреля. 11 июня найдено гнездо с 5 сильно насиженными яй-
цами. Первые летные молодые встречены 20 июля (Мекленбурцев, 1946).
В Заалайском хребте Л. А. Молчанов добыл молодую птицу у перевала
Кызыларт 6 августа. Здесь же добывал старых и молодых птиц Н. А. Се-
верцов.
40. Бледная завирушка — Prunella fulvescens fulvescens (Sev.). В отличие
от предыдущего вида бледная завирушка селится в более ксерофильных
биотопах и широко распространена в горах Передней, Средней и Цент-
ральной Азии, причем диапазон высот, населяемых ею, очень широк.
В горных странах с обильным атмосферным увлажнением эта завирушка
выбирает более сухие участки на склонах гор (Сушкин, 1938; Штегман,
1954). Примечательно, что даже для такого ксерофильного вида, как
бледная завирушка, памирский климат слишком сух, и в своем распро-
странении по нагорью бледные завирушки связаны с местообитаниями,
имеющими несколько более влажный микроклимат.
Вертикальное распространение на нагорье довольно широкое. Бледные
завирушки гнездятся в субальпийском и в нижней части альпийского
пояса до высоты 4600 м.
Эта завирушка наиболее многочисленна в узких долинах с кустарни-
ковой растительностью, где на 1 км маршрута встречается 3 пары, а также
близ ручьев на склонах, особенно там, где скалы и осыпи чередуются
с небольшими лужайками. Численность заметно колеблется. Например,
они изобиловали в районе Биостанции в 1960 г., но в 1962 г. численность
их резко снизилась.
Большую часть времени бледные завирушки проводят на склонах,
среди камней. Корм разыскивают, передвигаясь быстрыми прыжками
— 70 —
п изредка перелетая с места на место. Позывка — тихая мелодичная
трель, несколько более звонкая, чем у Prunella modularis (L.).
Завирушки появляются на нагорье в середине апреля (Sharpe, 1891;
Мекленбурцев, 1946). До начала гнездования они кочуют по долинам
парами и одиночками и постепенно, с таянием снегов, поднимаются на-
верх. Размножение начинается в конце мая—начале июня, как и на Тянь-
Шане. Распределение по гнездовым участкам сопровождается частыми
драками самцов. Песня самца состоит из 2—3 трелей, следующих одна
за другой в различных тональностях. Самец в гнездостроении не участ-
вует, а только сопровождает самку, приносящую гнездовой материал.
Гнездо строится около 10 дней, причем половина времени уходит на со-
оружение шерстяной выстилки. Маскируются гнезда плохо, часто видны
снаружи и нередко разоряются, что вызывает повторные кладки. Все
10 найденных гнезд помещались в нишах и щелях скал или крупных
камней. Предпочитается южная экспозиция, хотя иногда гнезда поме-
щаются в глубоких щелях, куда не проникает солнце. Самая низкая тем-
пература, отмеченная в модели гнездового убежища в период насижива-
ния, равнялась —0.5° (при наружных минимумах до —7°). Гнездовая
постройка довольно громоздкая. На рыхлом основании из веточек и
корешков терескена, Stipa glariosa, Hordeum и других растений вьется
аккуратная чашечка из тонких стеблей осок и злаков. Чашечка высти-
лается толстым слоем шерсти, иногда с небольшим количеством перьев.
Размеры гнезд: наружный диаметр 155—170 мм, высота 70 мм, диаметр
лотка 50—65 мм, глубина 35—65 мм. Яиц обычно 3, очень редко 4, тогда
как на Тянь-Шане полная кладка содержит 5 яиц (Кыдыралиев, 1962).
Цвет яиц чисто голубой; размеры их из одной кладки: 20.0 X 14.0,
20.0 X 14.0, 20.1 X 13.8 мм. В свежем яйце желток отличается необыч-
ным красным цветом.
Кладка начинается в начале июня. Самая ранняя из найденных мной
кладок была начата 6 июня (1960 г.); И. А. Абдусалямов в 1956 г. нашел
кладку, начатую 31 мая. Самец в насиживании не участвует, по крайней
мере днем. Все светлое время суток он держится близ гнезда, распевая
на вершинах скал. Птенцы вылупляются на 13-й день. Вес однодневного
птенца 1.6 г. Птенцы хорошо опушены. Кожа их очень темная, почти
черная. В возрасте 3 дней птенцы уже довольно громко пищат. Темпера-
тура тела у 4-дневных птенцов при обогревании взрослой птицей 35°,
через полчаса после прекращения обогрева она падает до 31 ° При тревоге
птенцы завирушки могут выскочить из гнезда уже на 11-й день. Массовый
вылет из гнезд происходит в первой половине июля, хотя едва летающие
подлетки встречались в 1961 г. до 3 августа, а Н. А. Северцов добывал их
даже в конце этого месяца. Растянутость периода гнездования бледной
завирушки наблюдается и в других высокогорьях. На Тянь-Шане
Б. К. Штегман наблюдал подлетков 14 августа, в Тибете гнезда с кладками
п птенцами можно находить в течение всего августа (Baker, 1933).
После вылета птенцы держатся у гнезда еще 4—5 дней. Далее выводки
сбиваются в небольшие стайки, которые кочуют по лужкам на склонах
гор до отлета. Р. Н. Мекленбурцев наблюдал последних завирушек
30 ноября.
41. Белая трясогузка — Motacilla alba personata Gould. Гнездится
у оз. Яшилькуль и по долине Мургаба до высоты 3600 м, в отдельные
теплые годы поднимаясь до 4000 м. В Ладаке эта трясогузка также не
поднимается выше 3600 м (Osmaston, 1928).
Первые трясогузки появляются на нагорье очень рано, в конце марта.
Гнездование позднее. Единственное гнездо, найденное И. А. Абдусалямо-
вым, было устроено у теплого источника в сарае, на высоте 4000 м и
28 июля 1960 г. содержало 2 насиженных яйца. Гнездование трясогузки
— 71 —
здесь было явно случайным, и она отсутствовала в этом месте в 1954,
1956 и 1961 гг.
Помимо гнездовых птиц, на нагорье нередко встречаются одиночные
бродячие особи. Осенью через Памир идет заметный пролет белых трясо-
гузок. Пролет начинается с первых чисел августа и длится до ноября.
Горная трясогузка — Motactlla cinerea melanope Pall. Горная трясо-
гузка включена в список гнездящихся птиц Памира Р. Н. Мекленбурце-
вым (1949). Насколько мне известно, гнездовых находок на нагорье нет.
Эти трясогузки встречаются только в осеннее время, начиная с августа.
42. Желтоголовая трясогузка — Motactlla citreola calcaratus Hodgs.
Этот подвид, населяющий высокогорья Средней и Центральной Азии,
является обычной птицей Памира. Большая часть популяции гнездится
в пределах 3700—4000 м, но отдельные пары поднимаются до 4400 м.
Немногим выше гнездятся они в Ладаке и Тибете — до 4600 м, причем
в юго-восточном Тибете основная масса селится в тех же пределах, что
и на Памире (Schafer, 1938).
Желтоголовые трясогузки в большом числе населяют только обширные
болотистые луга субальпийского и альпийского поясов. На небольших
лужках по краям альпийских потоков они не встречаются. Там, где их
много, пара от пары гнездится нередко на расстоянии 50 м.
Первыми появляются в начале апреля самцы (Мекленбурцев, 1940).
Пролет одиночных особей идет до начала июня. На лугах у Акташа 18 мая
1961 г. встречались исключительно самцы. Число их в это время бывает
невелико и достигает нормального уровня к началу июня. В середине
этого месяца начинается гнездостроение. Гнезда помещаются под навис-
шими кочками и скрыты сверху или выступом кочки, или нависающей
осокой. В одном случае гнездо было свито у входа в сурчиную нору,
в другом — в выемке берегового обрыва. У верхнего предела гнездова-
ния, на высотах свыше 4100 м, гнезда трясогузок устраиваются исключи-
тельно в закрытых убежищах. Гнездо аккуратно свивается из стеблей
старых осок и кобрезий, чашечка обычно выстлана толстым слоем черной
шерсти. Размеры гнезд (5): наружный диаметр 110—120 мм, высота 53 —
80 мм, диаметр лотка 63—70 мм, глубина 25—48 мм. В полной кладке
4—5 яиц. Размеры их из одной кладки 19.9—20.6 14.5—14.95 мм.
Самец принимает участие в насиживании и выкармливании птенцов.
Сроки размножения сильно колеблются. Наиболее ранняя из найден-
ных кладок была начата 7 июня (Акбалык, 3850 м), самая поздняя —
7 июля (Шадпут, 4000 м). И. А. Абдусалямов нашел гнездо с 2 свежими
яйцами даже 17 июля (Акташ, 3850 м). Столь поздние кладки являются
явно повторными, взамен утраченных. Гнезда трясогузок, помимо разо-
рения различными хищниками, нередко гибнут во время летних павод-
ков. Так, в 1961 г. в верховьях р. Аксу 17 июля во время паводка было
затоплено находившееся под наблюдением гнездо с птенцами в возрасте
7 дней.
Только что вылупившиеся птенцы покрыты густым эмбриональным
пухом, расположенным на надглазничных, затылочном, спинном, плече-
вых и локтевых участках кожи. Клюв темно-желтый, зев оранжевый.
У 5-дневного птенца, взятого из гнезда, температура тела падала за 5 мин.
с 39.1° до 32.1°, при наружной температуре воздуха 13.2е. Оставшиеся
в гнезде птенцы охлаждались за это время только на 1°.
После вылета из гнезда молодые трясогузки кочуют по нагорью не-
большими стайками и в одиночку, нередко образуя скопления на местах
кормежек. Отлетают в конце августа—начале сентября. Линька происхо-
дила с конца июля. В конце августа птицы были в свежем оперении.
43. Снежный воробей — Montifringilla nivalis alpicola (Pall.). Являясь
оседлой птицей Памира, гнездится по всему нагорью, но не выходит за
пределы субальпийского пояса, не встречаясь в альпийском поясе даже
на кормежках. Большая часть популяции вообще не поднимается выше
4000 м. Наибольшая высота находок гнезд — 4150 м.
Снежный воробей определенно связан с наиболее влажными местами
субальпийского пояса нагорья, где гнездится в скалах, обрывах и осыпях
близ лугов или возделанных полей. Гнездится и в населенных пунктах.
В пустынных безводных долинах не встречается. Основными местами
кормежек являются субальпийские луга, небольшие лужайки у подножия
скал и окрестности загонов для скота. В наибольшем числе встречен в са-
мых низких местах нагорья, в частности в скалах у покрытой лугами
долины р. Мургаба (3600 м).
Сроки размножения те же, что на Тянь-Шане и в Ладаке. Брачное
оживление начинается в конце мая. Самцы проделывают особые токовые
полеты, описание которых было дано Б. Осместоном (Osmaston, 1927).
Птица взлетает вверх с какого-нибудь возвышения и, взяв разгон, проле-
тает по инерции несколько метров с неподвижными крыльями, поднятыми
над спиной. При этом на расправленных крыльях бросаются в глаза
белые поля. Полет сопровождается звонкими, с металлическим оттенком
выкриками «чинь. .чинь. .чинь». Особой брачной песни самец не имеет.
Пары снежных воробьев селятся сравнительно далеко друг от друга.
Ничего похожего на колониальное гнездование не наблюдалось. Гнезда
устраиваются в норах под камнями осыпей или в щелях скал, нередко
на расстоянии до 2 м от поверхности. Реже они попадаются в нишах конгло-
мератовых обрывов или под крышами домов. Иногда в одной и той же щели
птицы гнездятся несколько лет подряд, надстраивая старое гнездо или
сооружая новое рядом со старым. Гнездо представляет собой грубую
постройку из стеблей злаков, только внутренняя часть чашечки свита
более тщательно и почти целиком состоит из шерсти. Размеры гнезда:
наружный диаметр 140 мм, высота 80 мм, диаметр лотка 75 мм и глубина
45 мм. В кладке 4 яйца белого цвета. Размеры яиц из одной кладки
32.1—32.6 X 17.0—17.1 мм. Кладка происходит в первой половине июня.
Случаи разорения гнезд редки по причине их труднодоступности, и за-
поздалые кладки почти не встречаются. Насиживает только самка. Одно-
дневный птенец хорошо опушен. Эмбриональный пух расположен на
надглазничных, затылочном, плечевых, локтевых и спинном участках
кожи. Длина его на спине 10 мм. Через сутки после вылупления птенец
весит 2.45 г. Птенцы выкармливаются исключительно животным кормом.
Взрослые птицы собирают насекомых и пауков с земли, передвигаясь
шагом или прыжками. В поисках корма они иногда улетают на расстояние
до 1 км от гнезда. У гнезд взрослые птицы держатся открыто и близко
подпускают наблюдателя. Сильное беспокойство проявляется только
у гнезд с птенцами накануне вылета. При появлении опасности в таких
случаях птицы почти непрерывно кричат «так-так-так-так-так».
По вылете из гнезда подлетки держатся около него еще 2—3 дня,
а потом начинают кочевать вслед за взрослыми, которые подкармливают
пх еще не менее 3 дней. Осенью снежные воробьи сбиваются в небольшие
стайки и кочуют по окраинам лугов, скапливаясь у населенных пунктов.
Здесь они особенно многочисленны зимой, но и в это время нередко уле-
тают кормиться на склоны гор (Мекленбурцев, 1946).
Большую часть года снежные воробьи питаются семенами диких и
культурных растений. Часто кормятся на вспаханных полях, поедая плохо
заделанные в почву семена овса и ячменя. Летом важную роль в питании
играют насекомые, которыми выкармливают птенцов.
44. Полевой воробей — Passer montanus pallidus Zar. И. А. Абду-
салямов (1961) наблюдал 25 мая 1957 г., как эти воробьи строили гнезда
в кишлаке Шаймак. Нам эти воробьи встречались только на пролете.
— 73 —
Гнездование на Памире, если оно и имеет место, явно случайно. В Шай-
маке воробьи не гнездились ни в-1954, ни в 1961 гг.
45. Горный вьюрок — Leucosticte brandti pamirensis Sev. Подробные
сведения о гнездовании горного вьюрка на Памире были опубликованы
нами ранее (Потапов, 1963). В этой работе памирские горные вьюрки рас-
сматривались как подвид вида L. arctoa, в том объеме, в каком он прини-
мался Л. А. Портенко (1960), т. е. включавшего группы центрально-
азиатских, алтайских, саянских, монгольских, сибирских, курильских
и американских форм. В настоящей работе признается более удобным
каждой из этих групп подвидов придавать видовое значение.
Ареал памирского подвида охватывает весь Памир, Бадахшан, хребты
Зеравшанский, Туркестанский, Гиссарский и Каратегинский, Гиндукуш.
Этот же подвид обитает и в Гил гите (Вакег, 1926).
Гнездовья располагаются на крутых склонах гор в альпийском и суб-
нивальном поясах, в пределах высот 4400—5000 м. На кормежках регу-
лярно встречаются на всех лугах нагорья, независимо от высоты местности.
Горный вьюрок — самая многочисленная птица нагорья. Несколько
реже встречается в пустынных горах центрального Памира. Колебаний
численности по годам не замечалось.
Горные вьюрки — стайные птицы. Даже во время размножения они
продолжают кормиться и собирать корм для птенцов стаями, хотя состав
таких стай непостоянен и все время меняется.
Полет горных вьюрков быстрый и сильный, с частыми взмахами длин-
ных, острых крыльев. Вьюрки способны преодолевать сопротивление
сильнейших памирских ветров. Во время кочевок они одинаково свободно
летят как вниз, так и вверх, легко набирая высоту. По земле передвигаются
как скачками, так и шагом. Голос — грубая позывка, звучащая как «чррр»
или «чря-чря». Она характерна для обоих полов. Кроме нее, самцы издают
иногда особый писк, сходный с тем, какой издают самцы зябликов на
пролете.
Размножение, как и у других вьюрковых птиц нагорья, позднее —
начинается оно в июле.
Брачной песни у самцов нет. Перед спариванием самец, прыгая вокруг
самки, издает частые, 2—3 в секунду, чирикающие звуки «чив. чив.
чив. чив». При этом плечи у него приподняты над спиной, концы кры-
льев опущены, а хвост вздернут кверху. Эта церемония несколько отлична
от таковой, описанной у американского L. tephrocotts atrata (French,
1959). В период брачных игр стаи не распадаются, но отдельные пары то
и дело отделяются от кормящихся птиц и носятся друг за другом. Во время
брачных игр самка нередко симулирует голодного подлетка — раскры-
вает клюв и трясет крыльями, а самец ее кормит.
Гнезда устраиваются в щелях отвесных скал и осыпях на большой
высоте. К началу гнездования эти места освобождаются из-под снега,
хотя кое-где еще встречаются снежники. Самое верхнее из найденных
гнезд помещалось на высоте 4810 м, но приходилось встречать птиц,
таскавших корм птенцам на осыпи, расположенные на высоте 5000 м,
почти под снеговой линией. Очевидно, снеговая линия в условиях Памира
является верхним пределом гнездования вьюрков, так как выше нее
встречать этих птиц не приходилось. Нижнюю границу гнездования на
нагорье определяет, видимо, влажность атмосферы, которая ниже границы
гнездования этих птиц опускается ниже необходимого для них минимума.
Следует отметить, что полоса гнездования горных вьюрков на Памире
совпадает с полосой максимальных осадков. Здесь их выпадает около
400 мм, в два с лишним раза больше, чем в субальпийском поясе (Станю-
кович, 1960). Вероятно, именно этим следует объяснить «высокогорность»
горных вьюрков в пустынной Центральной Азии, где только самые
— 74 —
верхние пояса гор получают необходимое для этих птиц количество
влаги.
На Памире горные вьюрки не образуют гнездовых колоний, а гнездятся
рассеянными группами по 5—10 пар. Для гнездования выбирается
какой-либо небольшой скалистый цирк, склон или кулуар с подходящим
субстратом и микроклиматом. Места таких поселений непостоянны и
меняются из года в год. Явное предпочтение отдается склонам западных
и северных экспозиций, по-
лучающим больше влаги.
Гнезда строятся в ска-
лах и осыпях. Все 5 най-
денных гнезд были устро-
ены под камнями в осыпях
различной крутизны. Ак-
куратная чашечка сви-
вается из стеблей злака
Calamagrostis anthoxant
hoides, на рыхлом осно-
вании, состоящем из об-
ломков веточек терескена
п мха. Выстилка или со
всем отсутствовала (2 слу-
чая), или состояла из тон-
кого слоя шерсти козерога.
По верхнему краю гнезда
обычно вплетались круп-
ные перья птиц (рис. 16).
Самка откладывает 4,
реже 3 белых яйца. Размер
яиц из одной кладки:
21.8 X 16.3, 22.2 X 16.3,
22.35 X 16.2 и 22.5 X 16.7 мм.
Насиживают только сам-
ки. В начале насижива-
ния они слетают вниз
на луга кормиться, где их
нередко подкармливают
самцы; в конце насижи-
вания самки сидят плотно
и почти не слетают с гнезда,
Рис. 16. Гнездо горного вьюрка — Leucosticte
brandti pamirensis Sev
а самцы кормят их более
или менее регулярно.
Как и у всех предста-
вителей рода Leucosticte,
у самцов и самок во
время выкармливания птенцов развиваются особые подъязычные мешки —
приспособление для переноса корма с мест кормежек к далеко отстоящим
гнездам. Сначала мешки развиваются у самцов, которым приходится
кормить насиживающих самок. Эти мешки образуются впячиванием вниз
ротовой полости с обеих сторон языка. Такие впячивания сходятся под
языком и тесно прилегают друг к другу, но не сообщаются. По мере на-
полнения они сильно выдаются вниз. Под их давлением mm. mylo-hyoideus
расходятся в стороны, а тканевая оболочка мускулов сильно растяги-
вается и окружает оба мешка наружной оболочкой. Наполненные кормом
мешки сильно выдаются за дно ротовой полости и настолько выпячивают
кожу подбородка, что перья на нем встают торчком.
Однодневный птенец горного вьюрка хорошо опушен. Пух расположен
на надглазничных, затылочном, плечевых, локтевых, бедренных, голенных
и спинном участках кожи. Наибольшую густоту и длину (13 мм) пух имеет
на спине и плечах. Яйцевой зуб белый, валик в углах рта светло-желтый,
потом он становится желтым. Зев без пятен, красного цвета. Первые
3—4 дня после вылупления птенцов самка почти не покидает гнезда;
корм ей и птенцам носит самец. На 4-й день устанавливается нормальный
режим кормления — 1 прилет в 40 мин. Слуховые отверстия открываются
у птенцов уже через 2 дня, глаза прорезаются на 5-й день, на 6-й по-
являются пеньки маховых и рулевых перьев, обозначаются грудная и
спинная птерилии. Маховые перья выходят из чехликов на 8-й день,
перья на грудной и спинной птерилиях появляются днем раньше. Птенец
полностью оперяется на 13—14-й день, только на голове еще остается
эмбриональный пух. Вылет в одном из гнезд произошел на 16-й день.
Птенцы выкармливались исключительно семенами трав молочной и
восковой спелости, тогда как американские вьюрки кормят птенцов, как
правило, насекомыми. Пищевод птенцов сильно растяжим, как у птен-
цов клеста, и вмещает сразу большое количество семян. Сам пищевод
резко сдвинут вправо, как у птенцов пустынного снегиря.
На другой день после вылета птенцы свободно перелетали па 100—
200 м, но большую часть времени сидели на скалах и подавали голос,
весьма напоминающий позывку подлетков воробьев. Массовый вылет из
гнезд происходит с середины августа, в особенно теплые годы он может
происходить на неделю раньше.
Наряду с гнездящимися летом встречаются холостые птицы, состав-
ляющие до 30% популяции и не имеющие никаких признаков размноже-
ния. Размеры их несколько меньше, чем размножающихся самцов и самок.
Это говорит о том, что горные вьюрки становятся половозрелыми только
на втором году жизни.
После вывода птенцов вьюрки сбиваются в огромные стаи, по 100—
500 птиц, которые в сентябре кормятся у самой снеговой линии, спу-
скаясь ниже только во время снегопадов.
Горные вьюрки зимуют на нагорье в сравнительно большом числе,
кормясь на бесснежных склонах гор. К маю значительная часть популя-
ции откочевывает с Памира и здесь остаются главным образом небольшие
стайки старых птиц. В конце мая численность вьюрков вновь начинает
возрастать и стабилизируется к концу июня.
Памирские горные вьюрки — исключительно растительноядные
птицы. Всю зиму и весну вплоть до июня они питаются семенами различ-
ных альпийских растений, разыскивая их на лужайках, на склонах гор
среди камней и особенно на влажных участках почвы, недавно освобо-
дившихся из-под снега. Под снегом и в снегу сохраняется больше семян,
а кроме того, во влажной почве они быстрее трогаются в рост и питатель-
ное значение их для вьюрков значительно выше, чем совершенно сухих
семян, разбросанных ветром по пустынным склонам.
С середины июля, когда начинается массовое созревание семян травя-
нистых растений, вьюрки питаются исключительно ими. Семена восковой
спелости они вылущивают из зеленых оболочек, и клювы вьюрков начиная
с этого времени и до конца августа всегда бывают выпачканы в зелени.
Этими же семенами вьюрки выкармливают и птенцов. Насекомые в пище-
водах и желудках как старых птиц, так и птенцов попадались редко и,
видимо, захватывались случайно, вместе с растительным кормом.
На Памире считают, что горные вьюрки вредят посевам, выкапывая
из земли посаженные семена. На самом деле горные вьюрки никогда не
выкапывают семена из земли к поедают только те, которые остались на
поверхности и все равно обречены на гибель. Большие стаи горных вьюр-
— 76 —
ков на свежевспахапных полях питаются главным образом семенами сор-
ных растений, чем безусловно приносят пользу.
46. Горная коноплянка — Cannabina flavirostris pamtrensis (Zar. et
Harms). Темный и мелкий подвид горной коноплянки, населяющий
горные хребты Памиро-Алая, гнездится главным образом в субальпий-
ском поясе Памира, хотя отдельные пары в теплые годы поднимаются
до высот 4500—4600 м. Распространена на нагорье спорадично. Обычна
в котловинах озер Яшилькуль, Булункуль, Рангкуль, в долинах Аличура
и его притоков, Мургаба, Карасу (в нижней части), Кокуйбеля, Шадпута,
в районе Биостанции. Нерегулярно гнездится в верховьях Аксу (много
в 1954 и 1956 гг., нет в 1961 г.). Не найдена в Каракульской котловине
и в сухих горах центрального Памира.
В низовьях Аличура, Бахмальджилги и у оз. Рангкуль, где конопля-
нок особенно много, пары селятся в 100—150 м друг от друга. В остальных
местах более редки.
Большую часть времени горные коноплянки проводят на лугах, где
собирают корм в густой траве. Значительно реже они кормятся на горных
склонах, покрытых полупустынной растительностью. Ночуют в щелях
скал. Весь сезон размножения на местах кормежек держатся стаями.
Состав этих стай непостоянен и все время меняется. Правильнее их рас-
сматривать как сообщества на кормных местах.
В. Эббот застал этих птиц на оз. Малый Каракуль (3500 м) уже 3 апреля.
На Рангкуле и Аличуре прилетают в начале мая (Мекленбурцев, 1946;
Абдусалямов, 1961). К размножению приступают поздно — в июле, когда
начинается созревание семян травянистых растений. Первую половину
лета птицы живут небольшими стайками. Брачное оживление наступает
в конце июня. Стайки при этом не распадаются, но отдельные пары на-
чинают встречаться в местах гнездования. Гнезда строятся в первых
числах июля. В 1962 г. с 6 по 15 июля гнездостроение наблюдалось в ни-
зовьях Аличура. Гнездо строит самка, самец только сопровождает ее.
Материалом служат старые стебли трав, которые птицы собирают на лугах
л затем тащат наверх, на склоны гор. Гнездо представляет собой довольно
плотную постройку с глубоким лотком, тщательно выстланным шерстью
и пухом. Оно помещается или среди камней, или в больших кустиках
терескена, кузиний, а также в ивняках, на кустах по окраинам зарослей.
И. А. Абдусалямов находил гнезда в каменных оградах у полей. Гнезда
строятся среди камней даже при наличии кустарников. Например,
в ущелье Бахмальджилги в начале июля коноплянки строили гнезда как
в ивняках, так и в каменных осыпях на окружающих склонах. В полной
кладке 4—5 яиц размером 17.2 X 11.2 мм (Абдусалямов, 1961), они
заметно мельче, чем у тибетского и ладакского подвидов.
Наиболее ранняя кладка, состоявшая из 5 яиц, была найдена 6 июля
1958 г. под перевалом Карабулак (3800 м). Большинство птиц начинает
кладку в середине июля. В период насиживания самцы нередко кормят
самок, прилетающих на кормежки на луга, при этом самки трясут крыль-
ями и пищат, как подлетки. Птенцов выкармливают главным образом
очищенными семенами, в частности семенами Lepidium ruderale. Насе-
комых в вскрытых зобах встречалось мало, вероятно, птицы захватывали
их случайно, вместе с растительным кормом.
Первые подлетки в 1960 г. появились 12 августа, массовый вылет
обычно происходит в середине этого месяца. После вылета птенцов горные
коноплянки держатся шумными стайками на местах гнездования вплоть
до отлета, который происходит в конце октября—начале ноября.
47. Обыкновенная чечевица — Carpodacus erythrinus ferghanensis
(KozL). Чечевица — единственная древесно-кустарниковая птица, оби-
тающая на Памире. В пределах нагорья чечевицы гнездятся только в одном
— 77 —
месте — в ущелье Бахмальджилги (3800 м). Вероятно, это самое высокое
место гнездования чечевицы в СССР. В окружающих Памир горных
странах чечевицы обитают во всех ущельях с древесно-кустарниковыми
зарослями, поднимаясь до 3500 м, лишь немного не доходя до верхней
границы кустарниковой растительности. В ущелье Бахмальджилги чече-
вицы гнездятся в густых и высоких — до 2 м — зарослях ивняка. Числен-
ность небольшая. На всем 3-километровом протяжении ивняков в середине
июля 1962 г. насчитывалось 6—7 поющих самцов. В середине июля встре-
чались в основном только они; самки, видимо, сидели на гнездах. Гнездо
со свежей кладкой из 5 яиц я нашел 9 июля 1958 г. в устье Мургаба
(3270 м), а только что вылетевших из гнезда подлетков — 5 августа 1961 г.
в среднем течении реки (3400 м) — все в пределах Бадахшана. В конце
августа на нагорье появляются многочисленные стайки пролетных чече-
виц, которые скапливаются у населенных пунктов. Пролет заканчивается
к октябрю.
48. Большая чечевица — Carpodacus rubicilla severtzovi Sharpe. Этот
подвид, населяющий горы Средней Азии, Кашмир и северо-западные
Гималаи, принадлежит к оседлым птицам Памира, где спорадично гнез-
дится в субальпийском поясе нагорья. Выше 4200 м не подни-
мается.
Большие чечевицы предпочитают гнездиться там, где имеется сочетание
скал, лугов и воды, хотя отдельные пары гнездятся иногда в 8—10 км от
ближайших лугов. В большом числе обитают также в ущельях с кустар-
никовой растительностью, например по рр. Бахмальджилге и Бейку,
но в кустарниках не гнездятся. Нередко селятся на конгломератовых
обрывах древних террас. Наибольшая плотность — 1 пара на 300 м марш-
рута вдоль пойменных обрывов и подножий скал — отмечена в районе
Биостанции, в долинах верхних притоков Аксу, у оз. Рангкуль, по
рр. Мургабу, Кокуйбелю, Западному Пшарту, Бахмальджилге, в ска-
листых долинах гор Найзаташ, в Аличурской долине, в котловине Акджар.
В остальных местах редка и совсем не найдена у Каракуля.
Полет большой чечевицы тяжелый и мало маневренный. Кормится она
на лугах или пустынных склонах среди камней и часто перелетает с места
на место. Днем любит отдыхать на карнизах скал, в тени. Держится
одиночками или небольшими группами по 3—4 птицы. Голос — резкое,
гнусавое «кви» или «ввиии». Самцы нередко издают особую позывку,
звучащую как медленная, звонкая трель «вви. юв-юв-юв», с пониже-
нием тона в конце. Очень осторожны.
Гнездование позднее. Всю весну и первую половину лета большие
чечевицы кормятся на лугах и склонах гор субальпийского пояса парами
или группами по 3 особи. Эти группы состоят обычно из самки, старого
и молодого самцов. Брачное оживление наступает в конце июня. Самцы
вступают в драки друг с другом и поют. Специально брачной песни они
не имеют, а только часто и с большим азартом повторяют свою обычную
позывку. Гнезда вьют в начале июля. Они помещаются в нишах выветрен-
ных скал или обрывов или в глубоких трещинах под нависшими косырь-
ками скал, причем вход в гнездо в таких случаях расположен снизу.
Единственное гнездо, найденное И. А. Абдусалямовым, помещалось именно
в таком месте. Оно представляло собой довольно рыхлую постройку сле-
дующих размеров: наружный диаметр 180 мм, диаметр лотка 90 мм,
глубина 37 мм. По размерам оно существенно не отличалось от гнезд
из Ладака. То же самое можно сказать о размерах и окраске яиц. Боль-
шие чечевицы гнездятся в скалах и каменных осыпях, видимо, только
на нагорьях Тибета и Памира. В остальных же частях ареала, в частности
в южном Тибете и Ладаке, эти птицы вьют гнезда на кустах и даже на
деревьях, на высоте 2 м от земли (Baker, 1934).
— 78 —
Вылет птенцов из гнезд происходит во второй половине августа.
На зиму остается только часть популяции, преимущественно старые
самцы, которые держатся близ населенных пунктов (Мекленбурцев, 1946).
49. Красный вьюрок — Pyrrhospiza punicea kilianensis Vaurie. Крас-
ный вьюрок относится к той группе альпийских птиц, которые обитают
в хорошо увлажненных окраинных хребтах Центральной Азии, не про-
никая в глубь засушливых высокогорий. Этот вьюрок найден на гнездовье
в Шугнанском и Шахдаринском хребтах, недалеко от границ Памира.
А. В. Попов добыл здесь подлетка на высоте 3700 м. М. Н. Дивногорский
добыл 2 старых птиц в июле и августе 1909 г. на восточных склонах Каш-
гарского хребта. И, наконец, И. А. Абдусалямов добыл пару гнездовых
птиц на наиболее влажной окраине Памира — в верховьях Аксу у Ак-
таша — в июле 1960 г. на высоте 4100 м.
Местообитания — сочетание скал, каменников, воды и луговой расти-
тельности в местах, получающих не менее 350 мм осадков в год. В одном
из ущелий Акташа встречен вместе с большой чечевицей и горным вьюрком.
Исключительно редкая птица. За 6 лет не встречена мной ни разу.
В тех местах, где она добыта И. А. Абдусалямовым, в 1961 г. не была
найдена.
Заслуживает внимания почти полное сходство пения красного вьюрка
и большой чечевицы, о чем уже упоминалось в литературе (А. А. Куз-
нецов, 1960).
Сроки размножения на Памире такие же, как и у других вьюрковых
птиц. У добытой 9 июля самки в яйцеводе находилось сформировавшееся
яйцо, но без скорлупы; у добытого тогда же самца гонады были сильно
увеличены (Абдусалямов, 1962). Таким образом, откладка яиц происходит
в середине июля, вылет из гнезд — в середине августа.
50. Пустынный снегирь — Bucanetes mongo Ileus (Swinh.). Одна из
обычных птиц Памира. Эта птица, вероятно, единственная, населяющая
субальпийский и альпийский пояса от самых нижних частей нагорья
вплоть до высоты 4750 м. Большая часть популяции тем не менее гнездится
в пределах субальпийского пояса.
Пустынные снегири населяют самые различные местообитания с раз-
личными режимами температуры и влажности. Они гнездятся в сухих,
пустынных оврагах центральной части Памира, на склонах гор, приле-
гающих к поймам рек, в скалах верхней части альпийского пояса, не-
вдалеке от первых снежников. Обязательное условие — наличие воды
не далее чем в 1—2 км.
Численность подвержена заметным колебаниям. У Биостанции, на-
пример, пустынные снегири гнездились в большом числе в 1954 и 1961 гг.
и почти совершенно отсутствовали в 1958 и 1960 гг. Средними по числен-
ности были 1956 и 1962 годы. В годы сравнительного обилия пара от пары
гнездится на расстоянии 300—400 м, тогда как в годы с малой численно-
стью в районе Биостанции гнездились всего 2—3 пары.
Стайки пустынных снегирей часто кормятся на окраинах лугов или
на пустынных склонах рядом с горными коноплянками и очень похожи
на них своим поведением. Но в отличие от них снегири кормятся преиму-
щественно на участках с разреженной растительностью полупустынного
типа и, как правило, не заходят в густую траву лугов или посевов. По-
зывка взрослых птиц двух типов: это слегка гнусавое «вжинь», или звон-
кое «ви-тю», с несколько меланхолическим оттенком.
Первые птицы появляются на нагорье в начале мая. Весь май и июнь
они малочисленны, держатся парами или небольшими стайками на пу-
стынных участках склонов, невдалеке от воды. Основная масса появляется
сравнительно позднее, в середине лета. В 1954 г. пустынные снегири по-
явились в большом числе в начале июля, в 1961 и 1962 гг. — с середины
— 79 —
июля. Вскоре вслед за этим птицы приступают к размножению. Разбивка
на пары происходит в течение нескольких дней и сопровождается драками
и азартным пением самцов. Специальной брачной песни у пустынного
снегиря нет, просто он громко повторяет подряд свои позывки, разно-
образя их несколькими гнусавыми звуками. Почти сразу же начинается
гнездостроение. Самец участия в нем не принимает, только сопровождает
самку, собирающую гнездовой материал. В это время нередко происходит
кормление самцом самки: самка захватывает своим клювом клюв самца
и трясет крыльями, как подлеток.
Гнезда помещаются в укрытиях среди камней или скал (рис. 17). Одно
пз найденных гнезд помещалось в полости разрушенного выветриванием
Рис. 17. Пустынный снегирь — Bucanetes mongolicus (Swinh ) —
на гнезде.
камня, лежавшего на пологом склоне морены (4150 м), другое — под на-
висшим камнем на крутом склоне оврага и было открыто сбоку (3900 м),
третье было устроено в неглубокой нише отвесной скалы (4000 м). Экспо-
зиция всюду была южная. Гнезда устроены по одному типу. На рыхлом
основании из обломков стеблей, корешков полыни и терескена вьется
сравнительно аккуратная чашечка из тонких стеблей злаков. Выстилка
состоит из тонкого слоя шерсти. Размеры одного из гнезд: наружный
диаметр 135 мм, высота 60 мм, диаметр лотка 60 мм, глубина 45 мм. В пол-
ной кладке 5 яиц, размеры их из одной кладки: 17.7—18.7 X 14.2—14.9 мм.
Птенец суточного возраста весил 2.3 г и был хорошо опушен. Эмбриональ-
ный пух расположен на надглазничных, затылочном, спинном, плечевых,
локтевых и бедренных участках кожи. Глаза открываются на 3-й день,
тогда же появляются пеньки маховых, которые выходят из чехликов на
7-й день. Самка обогревала птенцов по крайней мере до 5-дневного воз-
раста. Самка сидит на гнезде очень плотно, подпуская наблюдателя до
2 м. Вылетающие птенцы окрашены в бежево-коричневые тона, только
основания маховых перьев темные. Птенцы выкармливаются исключи-
тельно семенами трав восковой спелости. Старые птицы приносят корм
— 80 —
сразу большими порциями, помещая его в сильно растяжимых пищеводах.
Пищеводы птенцов также способны сильно растягиваться и вмещать боль-
шие порции корма. У птенцов пустынного снегиря, как и у птенцов гор-
ного вьюрка, пищеводы сильно смещены вправо.
В 1958 г. у Сарезского озера (3270 м) гнездостроение наблюдалось
15 июля. В 3 гнездах, найденных в 1961 г., кладки были начаты 15, 19
п 26 июля. Массовый вылет в том же году происходил в середине августа.
Первое время после вылета выводки держатся у гнезд, затем сбиваются
в большие стаи и до отлета кочуют по нагорью. Отлетают в конце сентября.
На зимовки пустынные снегири спускаются в равнины Таджикистана и
Кашгарии. Экспедиция Форсайта встретила их зимой в большом числе
у Кашгара (Sharpe, 1891).
Краснокрылый чечевпчник — Rhodopechys sanguinea Gould. Л. А. Мол-
чанов (Молчанов и Зарудный, 1915) наблюдал стайку этих птиц в июне
1913 г. в низовьях р. Памира, на самой границе нагорья. Видимо, на этом
основании чечевичник включен Р. Н. Мекленбурцевым (1949) в список
гнездящихся птиц Памира. Гнезд его на территории нагорья, насколько
нам известно, никто не находил. Ни мне, ни И. А. Абдусалямову вообще
не пришлось его встретить на нагорье ни разу. В своем распространении
этот вид, вероятно, обходит Памирское нагорье, гнездясь на меньших
высотах.
СЕЗОННАЯ ЖИЗНЬ ПТИЦ ПАМИРСКОГО НАГОРЬЯ
1. 3 и м н и й период. Зимний период в жизни птиц Памира
наименее изучен. При написании этого раздела была использована работа
Р. Н. Мекленбурцева (1946), зимовавшего в верховьях Аличура в 1936—
1937 гг., некоторые наблюдения сотрудников Биостанции и ряд сведений,
любезно предоставленных Л. А. Арутюновым, зимовавшим на нагорье
в 1960/61 г.
Несмотря на очень суровые климатические условия, на Памире зимует
сравнительно большое количество птиц. Зимовка некоторых видов воз-
можна благодаря ряду благоприятных факторов. Во-первых, на большей
части субальпийского пояса снежный покров отсутствует. Только в южной
части Памира, в верховьях Аксу и котловине оз. Зоркуль, снег выпадает
регулярно и образует сплошной покров, сохраняющийся 2—3 месяца.
Тем не менее и здесь значительные участки южных склонов всю зиму
свободны от снега. Отсутствие снегового покрова значительно расширяет
кормовые возможности для птиц и позволяет им питаться осыпавшимися
семенами растений. Во-вторых, благодаря действию теплых источников
целые участки рек и озер не замерзают, что дает возможность зимовать
водоплавающим птицам. В-третьих, весьма благоприятен для птиц длин-
ный световой день — 9 ч. 40 м. в середине декабря. И, наконец, не менее
благоприятен для птиц сильный солнечный нагрев в дневные часы, когда
даже в самый разгар зимы обнаженный субстрат нагревается до плюсовых
температур. Резкое уменьшение теплоотдачи в дневное время и длитель-
ный срок кормежки позволяют птицам накопить некоторый запас энерге-
тических ресурсов, необходимый для переживания морозной ночи.
Неблагоприятные условия памирской зимы проявляются в виде очень
низких температур, главным образом ночных. Средняя температура
января в субальпийском поясе —19.5°, в отдельные ночи температура
падает до —50° Зимних метеорологических наблюдений для альпийского
пояса нет, но, очевидно, температуры там еще ниже. Относительная влаж-
ность воздуха в зимние месяцы заметно возрастает и составляет в январе
54%. Сильные ветры, которые не прекращаются на нагорье и зимой, только
усугубляют отрицательное действие низких температур. Отрицательным
0 Биология птиц
- 81 —
фактором является большая продолжительность зимы. В течение 7 меся-
цев, с октября по апрель, среднесуточная температура воздуха держится
ниже 0° и становится положительной только со второй декады мая.
На Памире в зимнее время зарегистрировано 23 вида. В это число не
входят птицы, встреченные на нагорье в конце зимы (март—начало
апреля), т. е. явно пролетные, сбившиеся с пути весенние мигранты (чи-
бис, белая трясогузка, сорокопут-жулан и многие другие). Залет их на
холодное нагорье во время весеннего пролета возможен благодаря срав-
нительно высоким дневным температурам, которые в марте достигают
иногда 11°.
Таблица 2
Состав зимней орнитофауны Памира
Вид Примечани
Anas platyrhynchos L. A. strepera L. Bucephala clangula (L.) Mergus merganser orientalis Gould. Aquila chrysaetos daphanea Sev. Gyps himalayensis Hume. Gypaetus barbatus hemachalanus Hutt. Falco cherrug milvipes Jerd. Cerchneis tinnunculus tinnunculus L. Tetraogallus tibetanus tibetanus Gould. T. himalayensis himalayensis Gray. Tchangtangia tibetana (Gould). Bubo bubo hemachalanus Hume. Alauda arvensis dulcivox Brooks. Eremophila alpestris diluta (Sharpe). Corvus corax tibetanus Hodgs. C. corone orientalis (Eversm.). Pyrrhocorax pyrrhocorax centralis Stres. Graculus graculus jorsythi (Stol.). Tichodroma muraria L. Montifringilla nivalis alpicola (Pall.). Leucosticte brandti pamirensis Sev. Carpodacus rubicilla severtzovi Sharpe. Обычны на теплых ключах. Редкие одиночки на теплых ключах. Редкие стайки на теплых ключах. Одиночкп на теплых ключах. Зимует часть популяции. То же. » » Бродячие одиночки. Залеты одиночек. Зимует вся популяция. То же. » » » » Небольшие группы у населенных пунк- тов. Зимует часть популяции. Зимует вся популяция. Залеты одиночек. Зимует часть популяции. То же. Залеты одиночек. Зимует вся популяция. Зимует часть популяции. То же.
Основное ядро зимующих птиц составляют виды оседлые. Таковых 18
(табл. 2). Среди них имеется только 5 видов, популяции которых совер-
шенно оседлы и не выходят во время зимних кочевок за пределы нагорья.
У остальных 13 видов большая часть популяции покидает зимой нагорье,
спускаясь в долины Синьцзян-Уйгурского автономного района Китая,
Кашмира и Таджикистана. На зимовку остаются преимущественно старые
птицы, в основном самцы. Из других птиц, гнездящихся на нагорье, поле-
вой жаворонок и стенолаз тоже встречаются зимой, но местные популяции
их скорее всего откочевывают южнее, а на нагорье зимуют более северные
популяции. Один вид, черная ворона, несомненно залетный. Из всех
птиц, зимующих на нагорье, только 2 вида — гоголь и черная ворона —
не встречены здесь летом.
Все зимующие птицы проводят зиму в пределах субальпийского пояса.
Только улары и саджи поднимаются регулярно в нижние части альпийского
пояса, где кормятся по бесснежным пятнам пли на копанках архаров.
Кроме них, в альпийский пояс изредка залетают вороны и крупные хищ-
ники. Горные склоны в пределах альпийского пояса обычно покрыты
снегом, который исчезает лишь в мае. Таким образом, вертикальные
кочевки на нагорье в течение зимы выражены слабо и происходят почти
82 -
целиком в пределах субальпийского пояса. Они проявляются в том, что
во время сильных снегопадов, весьма редких в зимнее время, птицы спу-
скаются со склонов гор в долины. Зимой почти вся птичья жизнь сосредо-
точивается у населенных пунктов. Непосредственно связаны с ними 6 ви-
дов и еще 6 регулярно посещают окраины поселков.
Среди зимующих птиц больше всего зерноядных — 8 видов. Падалью
регулярно питаются 5 видов, 4 вида являются всеядными, 2 вида питаются
исключительно живой добычей, 3 вида обитают на водоемах и питаются
водной растительностью, беспозвоночными и рыбой. Целиком насекомо-
ядных птиц нет, за исключением стенолаза, факт зимовки которого
остается под сомнением.
Суточная жизнь птиц в зимнее время почти не изучена. Судя по наблю-
дениям Р. Н. Мекленбурцева и Л. А. Арутюнова, птицы кормятся весь
день, в середине дня нередко греясь на солнце в защищенных от ветра
местах. Ночевки расположены в скалах. В качестве ночных убежищ
выбираются небольшие, но глубокие пещеры или ниши с узким входом.
Во многих из них, судя по толстым слоям зимнего помета, на ночевки регу-
лярно собирается большое число мелких птиц. Они ночуют на карнизах
под нависшими сводами, сбиваясь в большие, плотные кучи, взаимно со-
гревая друг друга. Пещеры с большим количеством зимнего помета птиц
я находил преимущественно на высотах 3900—4300 м, т. е. на высотах,
где ночные температуры зимой падают до —50°. В таких условиях окучи-
вание птиц на ночевках и взаимное их согревание является важным при-
способлением, помогающим лучше переносить ночные морозы. Подобное
явление наблюдалось и в Андах, где на высокогорных плато птицы ночуют
в пещерах и расщелинах скал и даже ледников большими скоплениями.
Более 20 видов, главным образом вьюрковых, сбиваются при этом в плот-
ные кучи до 200 экземпляров и более (Pearson, 1953; Dorst, 1957).
2. Весенний пролет. Весенний пролет на Памире очень
слабый и является только отголоском мощной пролетной волны, охваты-
вающей горы и равнины Средней Азии с февраля по май. Большую часть
этого времени на Памире сохраняются зимние условия, и птицы, проле-
тающие в марте и апреле, обходят нагорье стороной. На Памире в это
время появляются только одиночки и небольшие стайки сбившихся с пути
мигрантов, которые нередко гибнут здесь от холода и бескормицы. С ап-
реля, когда начинается памирская весна и вскрываются озера и реки,
количество пролетных птиц заметно увеличивается, главным образом
за счет водоплавающих, но все же остается небольшим. В табл. 3 сведены
все данные, существующие по весеннему пролету на Памире. Большинство
приводимых дат заимствовано из работ Р. Н. Мекленбурцева (1946) и
И. А. Абдусалямова (1961).
Как можно судить по приведенным данным, весенний пролет большин-
ства птиц через нагорье носит случайный характер. Следует иметь в виду,
что в таблицу включены все встречи редких пролетных, возможно даже
залетных, видов за много лет. Ежегодно через нагорье весной летит не
более 20—25 видов. Обращает на себя внимание отсутствие на весеннем
пролете почти всех куликов, журавлей, гусей и многих других. Завален-
ные снегами мощные хребты, окружающие Памир, являются серьезным
препятствием для большинства весенних мигрантов.
3. Гнездовой период. Сроки появления отдельных видов
весной еще недостаточно выяснены. В конце марта заметно увеличивается
количество крупных хищников. Одновременно начинается разлет этих
птиц, а также клушиц и воронов от населенных пунктов по местам гнездо-
вания. В конце марта появляются скалистые голуби, которые в первое
время кормятся в широких субальпийских долинах. В начале апреля
прилетают бледные и гималайские завирушки, которые тоже держатся
- 83 -
6*
Таблица 3
Весенний пролет птиц на Памире
Вид Время пролета Характер пролета
Ardea cinerea L . V Редкие одиночки.
Anas platyrhynchos L. III—V Небольшие стайки.
A. strepera L. IV—V Стайки и одиночки.
A. acuta L. III—V । Редкие стайки.
A. penelope L. . V » »
*4. querquedula L. V » »
A. crecca L. III—V » »
*Л. clypeata L. . IV—V Большие стаи.
Aythya ferina (L.) III—V Небольшие стайки.
★Cygnus sp. . . V Редкие одиночки.
★Aquila heliaca Sav. V » »
Accipiter nisus nisus (L.) . . V—VI » »
Aesalon columbarius lymani (Bangs) III—IV » »
Crex crex (L.) . ... V » »
Porzana pusilia pusilia (Pall.) . IV—V » »
Gallinula chloropus chloropus (L.) . IV » »
Fulica atra atra L. IV Одиночки, группы.
Burhinus oedicnemus ssp. V Редкие одиночки.
★Vanellus vanellus (L.) . . III—IV » »
★Numenius arquatus arquatus (L.). ? » »
★Numenius phaeopus ssp. . . . ? » »
★Gelochelidon nilotica nilotica (Gm.) V » »
★Columba eversmanni Bp. . . . IV » »
Streptopelia turtur arenicola (Hart.) V—VI Небольшие стайки.
*£. orientalis meena (Sykes) V » »
★Alcedo atthis ssp. V Редкие пары и одиночки.
Upupa epops epops L. IV—V Редкие одиночки.
Alauda arvensis ssp III—V Стайки и одиночки.
★Melanocorypha bimaculata (Menetr.) IV Редкие одиночки.
Hirundo rustica rustica L. .... V—VI » »
★Riparia riparia diluta Sharpe et Wyatt. V » »
★Muscicapa parva albicilla Pall. V » »
Saxicola torquata maura (Pall.) V » »
★Oenanthe hispanica pleschanka (Lep.) . . VI » »
★Phoenicurus phoenicurus phoenicurus (L.) V Редкие одиночки.
Myophonus caeruleus turkestanicus Zar. V » »
Turdus ruficollis atrogularis Jar. III—V » »
★Sylvia nisoria merzbacheri Schal. V » »
★Laiscopus collaris rufilatus (Sev.) IV » »
Motacilla alba personata Gould III—V » »
M. a. dukhunensis Sykes . . IV—V » »
★Anthus trivialis trivialis (L.) V » »
Lanius schach erythronotus Vig. . . . V » »
L. collurio isabellinus Hempr. et Ehr. V » »
★L. minor Gm. . V » »
Pastor roseus (L.) VI » »
★Sturnus vulgaris porphyronotus Sharpe V » »
Л*. v. poltaratskyi Finsch. . ... II—III Редкие стайки.
Passer domesticus bactrianus Zar. et Kudasch. V Редкие одиночки.
P. montanus ssp. . ... V » »
P. hispaniolensis transcaspicus Tschusi V » »
★P. ammodendri ssp. V » »
Fringilla coelebs L V—VI » »
★Fringillauda nemoricola altaica Ev. V » »
★Emberiza pusilia Pall. V » »
Примечание.
Виды, которые бывают на пролете нерегулярно,
отмечены звездочкой.
в долинах. Озера в это время покрыты льдом, но у берегов образуются
многочисленные закраины, где держатся горные гуси, прилетающие в на-
чале апреля. Позднее, в середине апреля, появляются огари, а в конце —
буроголовые чайки, крачки, желтоголовые трясогузки, краснобрюхие
- 84 —
горихвостки и горихвостки-чернушки. Все они обитают первое время
в нижней части субальпийского пояса. Движение птиц вверх, на места
гнездовий, начинается в первой декаде мая, когда прилетает большинство
гнездящихся птиц. Последними, в середине мая, прибывают тонкоклювые
жаворонки. Движение птиц вверх продолжается до начала июля, когда
освобождаются из-под снега верхние части альпийского пояса и гнездя-
щиеся там горные вьюрки и пустынные снегири занимают гнездовые тер-
ритории. Сюда же поднимаются холостые улары, а несколько позднее —
и выводки. Кроме того, виды, гнездившиеся первоначально в нижних
поясах гор, в случае потери кладки приступают к повторной, как правило,
на большей высоте. В конце июня и в июле гнезда с запоздалыми клад-
ками отдельных пар встречаются нередко на самых верхних границах
вертикального распространения вида.
Необходимо упомянуть, что все сроки прилета и начала гнездования
птиц, относящиеся к марту, апрелю и началу мая, могут резко меняться
в ту или иную сторону в зависимости от характера весны.
Для многих певчих птиц Памира характерна слабая активность пения
самцов. Азартно распевающие самцы — явление довольно редкое. Самцы
большинства тех видов, которые, обитая на меньших высотах, поют много
и хорошо, на Памире поют слабее, как бы вполголоса. Это характерно
для рогатого жаворонка, горихвостки-чернушки, бледной завирушки,
каменки-плясуньи, пустынной каменки. Очень редко поют самцы красно-
брюхих горихвосток и стенолазов, почти не поют оляпки. С одной стороны,
подобное явление объясняется угнетающим действием холодных ветров.
Пение птиц можно слышать в редкие часы или даже минуты безветрия.
Сильными ветрами объясняется и то обстоятельство, что рогатые жаво-
ронки на Памире никогда не поют в воздухе, как самцы равнинных попу-
ляций. С другой стороны, причиной малой песенной активности самцов
может являться необычайно бедная звуковая среда нагорья. Большей
частью здесь царит безмолвие, нарушаемое только свистом ветра или сла-
бым шумом горного потока. На большое влияние звуковой среды на пение
птиц указывал в свое время А. С. Мальчевский (1959). Можно сказать, что
на Памирском нагорье нет хорошо поющих птиц. Единственное исключе-
ние составляют малые жаворонки, которые подолгу поют в воздухе, не-
взирая на сильные ветры.
Ряду птиц (горный гусь, огарь, беркут, врановые) свойственны особые
воздушные игры, которые благодаря изменчивой ветровой обстановке
отличаются подчас очень сложными эволюциями на большой высоте.
Весеннее оживление у некоторых оседлых птиц начинается очень рано
(беркут, филин, ворон). Однако низкие температуры воздуха еще долго
не дают птицам возможности приступить к размножению. Еще в конце
марта температура воздуха по ночам опускается до —40°. Крупные хищ-
ные птицы и ворон начинают гнездиться только месяц спустя, в апреле
и даже мае. Таким образом, размножение этих рано гнездящихся на рав-
нинах птиц на нагорье сильно запаздывает. По сравнению с равнинами
юго-западного Таджикистана на той же широте гнездование указанных
видов запаздывает на полтора месяца.
В начале мая, при отсутствии похолоданий, приступает к размноже-
нию часть клушиц и рогатых жаворонков. Такое раннее гнездование бы-
вает не каждый год и целиком зависит от погоды. Возможность его обу-
словлена тем, что солнечный нагрев в начале мая довольно высок и на
поверхности почвы и скал температура поднимается иногда до 30°. Но
основная масса клушиц и рогатых жаворонков гнездится позднее, в по-
следней трети мая, когда средняя температура воздуха поднимается до
2.6°. В конце мая приступают к размножению и другие птицы, гнездя-
щиеся на голой поверхности почвы: горный гусь, тибетская саджа, ко-
— 85 —
роткоклювый зуёк и др. Горные гуси нередко начинают кладку уже во
второй трети мая; в это же время гнездятся и огари. В конце мая также
начинается гнездование ряда видов, гнездящихся в скалах (пустельги,
скалистого голубя, уларов, пустынной каменки, горихвосток). Несколько
позднее, в первой трети июня (средняя температура декады 3.9°), начи-
нают гнездиться завирушки и снежный воробей. Примерно в это же
время, нередко даже со второй трети июня (средняя температура декады
5.2°) приступают к размножению птицы, гнездящиеся на субальпийских
лугах: травники, перевозчики, трясогузки. Сроки начала гнездования
этих птиц определяются временем вегетации луговой растительности, что,
в конечном счете зависит от высоты местности над уровнем моря. В сере-
дине июня начинают размножение каменки-плясуньи. В конце июня
(средняя температура декады 6.5°) приступают к строительству гнезд
городские ласточки. Горные ласточки гнездятся еще позднее, в начале
июля (средняя температура декады 7.8°). И, наконец, в середине июля
(средняя температура декады 8.8°), с началом массового созревания семян
альпийских и субальпийских трав, начинают гнездиться вьюрковые птицы.
Вылет птенцов у вьюрковых птиц и у ласточек затягивается до конца ав-
густа. Позднее всех заканчивают вывод птенцов рогатые жаворонки,
нередко в середине сентября. Это время можно считать концом гнездового
сезона на Памире. Таким образом, в субальпийском поясе нагорья сезон
размножения длится 5.5 месяца. В альпийском поясе сезон более сжат
и продолжается 3.5 месяца. При этом нужно учитывать, что гнездо-
вание одних и тех же видов в различных частях нагорья проходит в раз-
ные сроки и в целом запаздывает по направлению снизу вверх, а также
в открытых широких долинах. Раньше всех гнездятся птицы в долине
р. Мургаба, в наиболее низкой части нагорья (3600 м); несколько позже
размножение начинается в котловинах озер Рангкуль и Яшилькуль
(3700—3800 м); позднее всех гнездятся птицы в холодных долинах Аличура
и Зоркуля (4000—4300 м).
Из птиц, гнездящихся на Памире, только рогатый жаворонок регулярно
имеет вторую кладку, поэтому и период гнездования у него соответственно
растянут. У всех остальных птиц имеется только одна нормальная кладка.
Растянутость сроков гнездования у некоторых видов (травник, пустын-
ная каменка и др.) объясняется различными сроками гнездования на раз-
ных высотах, а также частой гибелью гнезд.
В условиях сурового, резко континентального климата Памира выбор
места для гнезда является важным условием успешного размножения
птиц. Обилие разнообразных убежищ в скалах и осыпях естественно
приводит к тому, что большинство видов (25) устраивает свои гнезда
исключительно здесь, а еще 7 видов гнездятся более или менее регулярно.
Огромное разнообразие щелей, трещин, полостей различной величины
и с различным микроклиматом дает возможность широкого выбора. На
Памире к гнездованию в скалах перешли многие древесно-кустарниковые
птицы. Этот процесс шел, видимо, постепенно, с поднятием Памирского
нагорья, ухудшением климата и деградацией древесно-кустарниковой
растительности. Главной причиной являются, видимо, низкие темпера-
туры. Даже в тех местах, где имеются небольшие заросли ивняков
(рр. Беик, Шинды, Карасу), птицы в них совершенно не гнездятся, пред-
почитая скалы и осыпи, где гнезда гораздо лучше защищены от ночных
заморозков. Такие виды, как большая чечевица, красный вьюрок, бледная
и гималайская завирушки, в более теплых Ладаке и южном Тибете, при
наличии соответствующей растительности, гнездятся на ветвях деревьев
и кустарников. Подобное явление указывает на пластичность гнездо-
строительных инстинктов. Например, горная коноплянка гнездится на
нагорье как на кустиках растений, так и в камнях, в зависимости от кли-
- 86 -
магических условий местности. Кроме того, эти явления могут указывать
на исторически недавний переход указанных видов на гнездование в ска-
лах. На сравнительную легкость перехода древесно-кустарниковых птиц
к жизни в скалах указывал еще А. Б. Кистяковский (1950). На Памире
к гнездованию в скалах переходят не только древесно-кустарниковые
птицы, но и луговые (желтоголовая трясогузка) и водоплавающие. Инте-
ресно отметить, что все обычные на нагорье водоплавающие птицы (горный
гусь, огарь, большой крохаль) гнездятся в скалах или по крайней мере
способны там гнездиться. Эта тенденция к переходу на гнездование в скалы
птиц, первоначально с ними не связанных, по сути дела свойственна орни-
тофаунам всех крупных горных поднятий.
У ряда видов гнезда располагаются прямо на поверхности голой почвы.
Четыре из них гнездится только на островах больших озер, обычно ли-
шенных растительности: горный гусь, черноголовый хохотун, буроголо-
вая чайка, речная крачка. На островах гнезда строятся с подветренной
стороны гребня или под укрытием больших камней. Нередко выбираются
островки, расположенные в ветровой тени более крупных островов или
береговых мысов. Два вида — тонкоклювый и рогатый жаворонки —
гнездятся на плоских пространствах долин, где выбирают укрытые от
ветра места и обычно помещают гнезда с подветренной стороны кустика
или камня. И только два вида — тибетская саджа и короткоклювый
зуёк — нередко устраивают гнезда на совершенно голой поверхности, но
при этом выбирают для гнездования теплые, низкие участки долин и озер-
ных котловин. Никто из открыто гнездящихся видов, за одним исключе-
нием, не поднимается на гнездовье выше 4200 м, обитая только в субаль-
пийском поясе. Только редкие пары рогатых жаворонков иногда подни-
маются до высот 4500 м.
Четыре вида помещают свои гнезда в густой траве субальпийских лугов,
так как в большинстве случаев только в субальпийском поясе трава до-
стигает высоты, достаточной для маскировки гнезда. Два вида — пере-
возчик и полевой жаворонок — очень редки на нагорье и гнездятся в огра-
ниченном числе мест. Два других вида — травник и желтоголовая трясо-
гузка — являются массовыми, причем наибольшее число особей гнездится
в пределах 3700—4100 м. Вероятно, верхнюю границу гнездования трав-
ника определяет именно высота травостоя на лугах. Желтоголовая трясо-
гузка, гнездящаяся на больших высотах, в верхней части субальпийского
пояса и в нижней части альпийского, где ночные заморозки особенно
сильны, переходит на гнездование в закрытых убежищах.
Небольшая группа птпц гнездится в постройках человека. Настоящих
синантропов на Памире почти нет, только полевой воробей и деревенская
ласточка изредка гнездятся в поселках долины Аксу. В постройках чело-
века, чаще всего в развалинах могил и каменных оградах, гнездятся
клушицы, горихвостки-чернушки, пустынные каменки, снежные воробьи,
горные коноплянки — т. е. птицы, обычно гнездящиеся в скалах.
Как видно из вышеизложенного, большая часть памирских птиц
гнездится в закрытых убежищах. Открыто гнездящихся видов только 12,
причем некоторые из них (горный гусь, желтоголовая трясогузка) нередко
устраивают гнезда в закрытых убежищах. Тибетский и темнобрюхий улары
иногда помещают свои гнезда открыто на площадках среди скал, но это
бывает редко. Большинство найденных мной гнезд уларов помещалось
в нишах и было закрыто сверху.
Температура в гнездовых убежищах, расположенных в скалах и осы-
пях, как показали измерения, заметно выше температуры окружающего
воздуха. Температура измерялась непосредственно в месте расположения
гнезда или в сооруженных поблизости моделях гнездовых убежищ. На-
пример, в гнездовом убежище бледной завирушки в щели скалы южной
— 87 —
экспозиции на высоте 4080 м минимальная температура за период с 9 по
18 июня была равна 4°, тогда как на открытом воздухе она опускалась
до —4.5°. В другом гнездовом убежище того же вида на высоте 4100 м в скале
восточной экспозиции с 19 по 21 июня минимальная температура была
— 0.5°. В гнездовом убежище пустынной каменки под камнем на склоне
южной экспозиции, на высоте 3900 м, с 4 по 17 июня максимальная тем-
пература равнялась 23.0°, а минимальная 1.5°, при наружном максимуме
17.0° и минимуме —4.6°. В гнездовых убежищах горных вьюрков на высотах
4600—4800 м минимальная температура была на 4—5°, а максимальная —
на 10—12° выше наружной. Это вызывается сильным нагревом камней,
скрывающих гнездо, лучами солнца. Такому нагреву способствует южная
экспозиция склонов и скальных стен, которую предпочитает большинство
птиц — 84 из 121 найденного гнезда располагались на склонах южной
экспозиции. Исключение составляют только горные вьюрки, выбирающие
для гнездования более холодные, но зато и лучше увлажненные склоны
северных и западных экспозиций. Важным условием, часто сильно влияю-
щим на выбор экспозиции, является защита от ветра. Наветренная сто-
рона скал, имеющих даже южную экспозицию, птицами обычно не за-
селяется. Особенно заметно стремление птиц гнездиться на склонах юго-
восточных экспозиций в широких межгорных долинах, где дуют холодные
ветры преимущественно западных румбов. В глубоких извилистых уще-
льях, хорошо укрытых от ветров, приверженность птиц к склонам южной
экспозиции не так сильна. На склонах южной экспозиции охотно селятся
многие открыто гнездящиеся птицы.
Как уже упоминалось, тонкоклювый и рогатый жаворонки нередко
прибегают к маскировке своих гнезд, закрывая свежие выбросы земли
камешками или кусочками почвенной корки, покрытыми пустынным
загаром. Упомянутые виды жаворонка являются единственными среди
птиц Памира, специально маскирующими свои гнезда. Для некото-
рых других открыто гнездящихся видов — тибетской саджи и коротко-
клювого зуйка — хорошей маскировкой служит покровительственная
окраска яиц и насиживающих птиц. Отсутствие гнездовой постройки
в таких случаях вызвано тем, что постройка сильно демаскировала бы
гнездо. Даже у крачек, гнездящихся отдельными парами на голых остро-
вах, постройка обычно отсутствует, яйца бывают темноокрашенными и
цвет их совершенно сливается с темным фоном грунта. Такое гнездо трудно
заметить даже на малом расстоянии.
Мелкие птицы, гнездящиеся в скалах и осыпях, помещают свои гнезда
в хорошо закрытых убежищах, где специальная маскировка не нужна,
но нередко их гнезда бывают укрыты плохо, видны снаружи и часто ра-
зоряются.
Многие птицы, гнездящиеся на склонах гор, устраивают гнезда зна-
чительно выше тех мест, где они кормятся и берут корм для птенцов.
Наибольший разрыв между местами гнездования и кормежек бывает у гор-
ного вьюрка, до 1000 м по вертикали. У огари и пустынного снегиря такой
разрыв достигает 500 м, у пустынной каменки и краснобрюхой гори-
хвостки 300—400 м, но в большинстве случаев он не превышает 50—100 м.
Это вызывается большей частью стремлением птиц помещать свои гнезда
в наиболее безопасных местах, какими являются верхние части склонов,
мало посещаемые различными хищниками. Иногда это объясняется кли-
матическими условиями, различными на разных высотах. Так обстоит
дело у горного вьюрка и, по-видимому, у тибетского улара.
Подобная закономерность не является всеобщей и наблюдается главным
образом у певчих птиц. У уларов, например, в начале июня, когда проис-
ходит откладка яиц, места кормежек расположены значительно ниже
гнезд. Впоследствии, с началом вегетации растительности на больших
- 88 —
высотах в альпийском поясе, птицы поднимаются выше и гнезда оказы-
ваются внизу. Поэтому после вылупления птенцов выводки сразу же
поднимаются наверх. Крупные хищные птицы и вороны, гнездясь пре-
имущественно в субальпийском поясе, в поисках корма часто посещают
вышерасположенные склоны, вплоть до вечных снегов.
Верхний предел гнездования большинства видов непостоянен и зависит
от погодных условий лета. В теплые годы многие птицы поднимаются на
гнездовье значительно выше, чем в холодные. В табл. 4 приведены данные
о верхнем пределе гнездования некоторых видов в окрестностях Биостан-
ции в теплый 1956 и в холодный 1960 годы.
Для певчих птиц, регулярно гнездящихся в скалах, характерны очень
крупные, громоздкие гнезда, свитые довольно небрежно, так как строи-
тельство гнезд в полостях среди кам-
ней не требует большого искусства.
Большинство гнезд устроено по одно-
му типу и состоит из 3 слоев. Первый
слой, наиболее грубый, из крупных
обломков веток и корней, составляет
рыхлое основание или платформу,
на которой далее сооружается ча-
шечка гнезда. Это рыхлое основание
иногда очень массивно и у красно-
брюхих горихвосток и пустынных
каменок, например, заполняет всю
полость гнездового убежища, что в не-
малой степени способствует утепле-
нию гнезда. Чашка гнезда бывает сви-
та более аккуратно из тонких стеблей
злаков с небольшой примесью шерсти и перьев. Выстилка
исключительно из шерсти и перьев. В большинстве гнезд она бывает сде-
лана очень тщательно и имеет значительную толщину. У некоторых видов,
Таблица 4
Верхний предел гнездования
(в м над ур. м.)
некоторых птиц в разные годы
Вид 1956 г. I960 г.
Темнобрюхий улар 4800 4500
Горная коноплянка 4700 4300
Рогатый жаворонок Краснобрюхая гори- 4500 4300
хвостка 4700 4400
Бледная завирушка 4400 4200
почти
состоит
например у клушицы и снежного воробья, чашечка и выстилка пред-
ставляют собой единое целое, сооруженное из шерсти животных. Только
у горного вьюрка выстилка бывает очень тонкой и нередко совсем отсут-
ствует, что не совсем понятно. Гнездясь выше, чем другие птицы, в осо-
бенно суровых условиях, вьюрки, казалось бы, должны иметь в гнезде
хороший утепляющий слой. Толстую выстилку из шерсти делают желто-
головая трясогузка и оляпка. В гнездах жаворонков шерсти обычно нет
п выстилка целиком состоит из растительного пуха, главным образом
пз опушения семян терескена. В гнездах городских ласточек, хорошо
закрытых от ветра, выстилка очень скудна, тогда как в более открытых
гнездах горных ласточек она обильна и состоит из теплого пуха крупных
хищных птиц.
Для многих видов птиц Памира характерно уменьшение числа яиц
в кладке, нередко весьма значительное, по сравнению с величиной кладок
у популяций, гнездящихся на меньших высотах или на хребтах, грани-
чащих с обширными равнинами (Тянь-Шань, Гималаи) (табл. 5). Нетрудно
заметить, что все перечисленные виды являются насекомоядными или по
крайней мере выкармливают насекомыми птенцов.
Численность насекомоядных птиц на Памире вообще очень мала и вме-
сте с фактом уменьшения величины кладки у них ясно свидетельствует
о нехватке корма. Действительно, насекомых на Памире очень мало.
В большом количестве они встречаются только на заболоченных лугах
у субальпийских озер и в меньшей степени — по берегам рек и леднико-
вых потоков, а также в осыпях у тающих снежных полей. Как показали
исследования индийских энтомологов (Mani, 1962), высокогорные насе-
— 89 —
Таблица 5 Величина кладки у некоторых видов на Памире и на меньших высотах
Количество яиц в кладке
Вид на низких горах и равнинах на Пами- ре
Рогатый жаворонок 4—5 1—3
Городская ласточка 3—7 3
Скалистая ласточка 4—5 3
Клушица .... 3—9 3
Белобрюхая оляпка 4—5 1—2
Пустынная каменка 4—6 3—4
Г орихвостка-чернушка 5-6 4
Бледная завирушка 3-5 3
Белая трясогузка 4—5 2
Снежный воробей 4—6 4
комые Центральной Азии, за малым исключением, являются гигрофилами,
обитают в местах с влажным микроклиматом и просто не могут сущест-
вовать на обширных ровных пространствах и склонах гор Памира, со-
вершенно лишенных влаги и снегового покрова зимой. А снеговой покров
особенно важен в суровой обстановке высокогорий, так как предохра-
няет насекомых от вымерзания.
Помимо неблагоприятных климатических условий, сравнительная
бедность энтомофауны Памира объясняется тем обстоятельством, что он
находится в центре сплетения высочайших хребтов, отгораживающих
его от равнинных пространств Средней и Центральной Азии. Дело в том,
что в горных системах, непосред-
ственно граничащих с равнинами, на
склонах гор, особенно в верхних поя-
сах, непрерывно выпадают органи-
ческие осадки в виде спор, пыльцы,
семян, мертвых и живых насекомых,
увлеченных воздушными потоками на
большую высоту и осаждающихся
на стоящих на их пути вершинах
хребтов. Эти органические осадки,
интенсивность которых может быть
очень большой, служат источником
обильной пищи высокогорным насе-
комым, паукам и птицам (Штегман,
1954; Mani, 1962). Например, на юж-
ном склоне хребта Пир-Панджал,
на участке снежного поля площадью
в 10 м2 на высоте около 4000 м за
20 мин. в мае—июне выпадало не ме-
нее 400 экз. различных мертвых на-
секомых. Замерзая, они аккумулиру-
ются на поверхности ледников и снеж-
ных полей, образуя целые слои, которые к зиме заносятся снегом, а в даль-
нейшем, при таянии снегов, вытаивают наружу. Кроме того, на склоны
хребтов, возвышающихся над равнинами, нередко вторгаются активно
мигрирующие массы насекомых, большая часть которых при этом обычно
гибнет на снегу и ледниках. Такие случаи хорошо известны для Гималаев,
хребтов, окружающих Ферганскую долину, и для некоторых гор Север-
ной Америки (Mani, 1962). На Памире ничего подобного не наблюдается.
Иными словами, мы наблюдаем в данном случае полную аналогию с ат-
мосферными осадками. Высочайшие горные хребты, окружающие Памир,
стоят на пути воздушных масс, перехватывают как атмосферные осадки,
так и «воздушный планктон» и к тому же препятствуют проникновению
на нагорье масс мигрирующих насекомых.
О том, что на уменьшение плодовитости птиц на больших высотах
влияет именно недостаток пищи, а не специфические физические условия
высокогорья, говорит тот факт, что у видов, кормовая база которых
обильна, величина кладки не уменьшается. Например, желтоголовые
трясогузки, гнездящиеся на болотистых лугах, изобилующих насекомыми,
откладывают такое же количество яиц, как и на равнине. Откладывают
нормальное число яиц вьюрковые птицы, выкармливающие птенцов расти-
тельным кормом, имеющимся в достаточном количестве, и т. д.
По характеру режима насиживания птицы Памира делятся на две
группы. Открыто гнездящиеся виды кормятся, прерывая насиживание,
по утрам и вечерам. Утренняя кормежка начинается не ранее того, как взо-
шедшее солнце хорошо прогреет воздух п почву, что гарантирует оставляе-
— 90 —
мую кладку от переохлаждения. В дневные часы птица насиживает, не-
смотря на то, что эти часы наиболее теплые. Днем накал почвы бывает
нередко очень велик, до 60°, и насиживающая птица предохраняет кладку
от перегрева. В пасмурную погоду открыто гнездящиеся птицы кормятся
и днем, но такие дни на Памире редки. Вторая кормежка бывает незадолго
до захода солнца.
Закрыто гнездящиеся птицы в период насиживания встречаются на
кормежках в любое время дня, но большая часть их предпочитает кор-
миться в середине дня, в наиболее теплые часы.
Температура насиживания измерялась в гнездах как крупных (темно-
брюхий улар), так и мелких птиц. У всех находившихся под наблюдением
видов наблюдалась явная зависимость температуры насиживания от на-
ружной температуры воздуха — она повышалась днем и понижалась
ночью. У рогатых жаворонков температура насиживания колебалась
в пределах 24.3—33.8°, у желтоголовой трясогузки — 26.0—27.0°, у го-
рихвостки-чернушки — 28.8—31.5°, у пустынной каменки — 30.0—31.5°,
у темнобрюхого улара — 22.0—28.0°.
Птенцы большинства видов воробьиных птиц Памира имеют хорошо
развитый эмбриональный пух, независимо от того, являются ли они от-
крыто или закрыто гнездящимися. В одних случаях, например у пред-
ставителей родов Phoenicurus и Prunella, имеется более длинный пух, чем
у представителей тех же родов на равнинах, но у многих видов (рогатый
и тонкоклювый жаворонки, пустынная каменка и др.) эмбриональный
пух развит также, как и у равнинных популяций. Это только подтверждает
предположение, высказанное ранее А. С. Мальчевским (1959), что тепло-
изолирующее значение эмбрионального пуха ничтожно по сравнению с тем
теплом, которое получает пойкилотермный птенец от обогревающей его
старой птицы. Следует подчеркнуть только, что все открыто гнездя-
щиеся птицы имеют хорошо опушенных птенцов. Но на нагорье, видимо,
вообще отсутствуют виды, у которых птенцы не имеют эмбрионального
пуха.
Большой интерес представляет вопрос о том, каким образом птенцы
открыто гнездящихся видов избегают губительного действия на развиваю-
щийся организм сильного ультрафиолетового излучения, свойственного
высокогорью. Пока только удалось заметить, что у всех открыто гнездя-
щихся птиц кожа птенцов на верхней стороне тела сильно пигментирована.
Вероятно, темная пигментация играет роль предохранительного экрана.
Она особенно сильна у птенцов тибетской саджи, рогатого и малого
жаворонков и в меньшей степени — у птенцов травника, буроголовой
чайки, речной крачки и желтоголовой трясогузки. Сильная пигментация
птенцов бледной завирушки определенно указывает на сравнительно
недавний переход к закрытому гнездованию. Отсутствие пигментации
у птенцов темнобрюхого улара может показаться на первый взгляд про-
тиворечием сказанному, так как птенцы этого вида с момента вылупления
находятся на открытых склонах гор. На самом деле выводки уларов пер-
вое время держатся среди каменников или крупнообломочных осыпей,
где птенцы большей частью скрываются в тени камней.
В развитии птенцов особых отклонений от нормы не наблюдалось.
Счучаи долгого, свыше 20 дней, пребывания в гнездах птенцов некоторых
видов, как например снежного воробья, краснобрюхой горихвостки, ги-
малайской и бледной завирушек, отмеченные А. Кыдыралиевым для
Тянь-Шаня, на Памире не наблюдались. Причиной подобных задержек
на Тянь-Шане была длительная бескормица, вызываемая внезапными
похолоданиями и снегопадами. На Памире же длительных снегопадов
в летнее время почти не бывает, так как на нагорье выпадает вообще очень
мало осадков, не более, чем в пустынях Средней и Центральной Азии.
— 91 —
По той же причине на Памире не наблюдается массовой гибели гнезд
во время снегопадов, что нередко случается в горах Тянь-Шаня. За 6 лет-
них сезонов только один раз, в середине июня 1960 г., глубокий снег,
выпавший в котловине оз. Каракуль и пролежавший три дня, погубил
многие гнезда рогатых жаворонков. Иногда во время похолоданий в ав-
густе, гибнут птенцы городских ласточек.
В отличие от птиц высокогорий Тянь-Шаня большинство насекомо-
ядных видов Памира компенсирует хронический недостаток корма умень-
шением величины выводка. В тех редких случаях, когда на Памире кладка
у какого-либо насекомоядного вида бывает больше нормальной, здесь
также наблюдается голодание птенцов и развитие их сильно замедляется.
После того, как молодые птицы поднимутся на крыло, что у большин-
ства видов происходит в конце июля—августе, начинаются массовые пе-
ремещения. Ласточки и каменка-плясунья покидают нагорье сразу после
вывода птенцов, плясунья — в конце июля и ласточки — в конце августа.
Другие виды остаются на местах гнездования вплоть до отлета, сбиваясь
здесь в большие стаи (горный гусь, чайки, горная коноплянка, пустын-
ный снегирь) или стайки (травник). Долго остаются на местах гнездования
краснобрюхие горихвостки. У многих видов происходят горизонтальные
перемещения в пределах субальпийского пояса. Многие виды концентри-
руются на лугах у больших озер (клушицы, альпийские галки), на озерах
(огари, крохали), или у населенных пунктов (рогатый жаворонок, снеж-
ный воробей).
Большинство птиц, закончив сезон размножения, начинает широко
кочевать по нагорью. Многие из них (перевозчик, тонкоклювый жаворонок,
белая трясогузка) и в это время не поднимаются выше субальпийского
пояса, тогда как другие в равной степени встречаются и в альпийском
поясе (хищные птицы, ворон, стенолаз, пустынная каменка, желтоголовая
трясогузка и др.). Эти птицы не образуют стай, выводки их распадаются
вскоре после вылета птенцов из гнезд. Для некоторых видов в конце ав-
густа—сентябре отмечено движение вверх (саджа, скалистый голубь,
рогатый жаворонок, горный вьюрок). Стайки их кормятся на альпийских
лужайках, где в это время происходит массовое созревание семян, в ча-
стности Polygonum viviparum, семенами которого кормится большое
число птиц. Массовое перемещение вниз начинается в конце сентября,
когда температура воздуха в альпийском поясе резко падает, начинаются
сильные снегопады и склоны гор постепенно скрывает снег.
Заканчивая обзор летней орнитофауны Памира, необходимо добавить,
что, помимо гнездящихся, в летнее время встречается довольно много
птиц, как правило, здесь не размножающихся. Состав и количество этих
птиц бывает различным в разные годы. Здесь кочуют и не достигшие поло-
вой зрелости молодые (кулики, хищники), и уже кончившие размножение
на меньших высотах, и поднявшиеся на нагорье в поисках корма (орлан-
долгохвост) или на линьку (кряква, чирок-свистунок, лысуха), и отстав-
шие во время весеннего пролета больные птицы, и случайно залетевшие
стаи (испанский воробей), и наконец, взрослые птицы, по каким-либо
причинам не размножавшиеся в данном году (большой баклан, хищники).
Испанские воробьи, например, раньше не оставались на лето, но в 1962 г.
целая стая провела все лето у Биостанции. Бакланы, изобиловавшие на
оз. Каракуль в 1961 г., в 1962 г. были здесь малочисленны.
4. Период осенних миграций. Время отлета местных
популяций с нагорья часто трудно установить, так как с середины августа
до сентября на Памире идет пролет птиц, движущихся с Тянь-Шаня, из
Казахстана и Сибири на индийские зимовки. Перерыв между отлетом
местной популяции и появлением пролетных особей замечен только у ка-
менки-плясуньи. В табл. 6 указаны даты последних встреч, которые,
— 92 —
Таблица 6
Осенний пролет птиц на Памире
Вид Время пролета Конец отлета гнездящихся птиц Характер пролета
Phalacrocorax carbo sinensis (Shaw et Nodd.) — Конец IX Пролета нет.
Ardea cinerea L VIII—IX — Стаи, одиночки.
★Ixobrychus minutus (L.) IX — Редкие одиночки.
★Ciconia nigra (L.) . . VII—VIII — Только залетные.
Eulabeia indica (Lath.) IX 10 X Большие стаи.
★Tadorna tadorna (L.) . VIII — Редкие одиночки.
Casarca ferruginea (Pall.) IX 10 X Стаи.
Anas platy rhynchos L. V 111—X — Стайки.
A. strepera L. VIII—IX — »
A. acuta L IX—X — »
*4. angustirostris (Menetr.) V11—X — Редкие стайки.
A. querquedula L. VIII—X — Стайки.
A. crecca L. . VIII—X — »
*Л. clypeata L. ... IX — Редкие стайки.
* Ay thy a nyroca (Giild.) VIII—IX — » »
*Л. juligula (L.) . . . VIII — » »
★Bucephala clangula (L.) IX — Стайки.
★Mergus albellus (L.) IX — Одиночки.
Neophron percnopterus percnop- terus (L.) . . . VIII-IX Редкие одиночки.
Aegypius monachus (L.) . . VIII—X — Одиночки и группы.
Circus cyaneus cyaneus (L.) IX — Редкие одиночки.
C. macrourus (Gm.) . . VIII—IX — Одиночки.
C. aeruginosus aeruginosus (L.) VIII—IX — Одиночки и группы.
★Haliaeetus albicilla (L.) IX — Редкие одиночки.
H. leucoryphus (Pall.) . . . — X Пролета нет.
★Milvuskorschun korschun (Gm.) VIII — Одиночки.
★Buteo rufinus rufinus Cretzhm. VIII — »
★Accipiter nisus nisus (L.) VIII—X — Редкие одиночки.
Cerchneis tinnunculus tinnuncu- lus (L.) .... VIII—IX » »
★Aesalon columbarius lymani (Bangs) . ... IX — » »
★Falco peregrinus babylonicus Irby X » »
F. cherrug milvipes Jerd. VIII—IX — Одиночки.
★Coturnix coturnix (L.) IX—X — Редкие одиночки.
Crex crex (L.) X — » »
Porzana pusilia pusilia (Pall.) IX — » »
Gallinula chloropus chloropus (L.) IX — » »
Fulica atra atra L VII—IX — Редкие стайки.
★Tetrax tetrax orientalis (Hart.) IX — Редкие одиночки.
★Chlamydotis undulata macque- eni Gray. IX — » »
★Squatarola squatarola (L.) . . IX — » »
★Pluvialis dominica fulva (Gm.) VIII—IX — » »
★Charadrius dubius curonicus Gm. . VIII—IX » »
Ch. mongolus pamirensis Richm. — 7 IX Пролета нет.
★Ch. alexandrinus alexandrinus L. . .... VIII — Редкие одиночки.
★Chettusia gregaria (Pall.) X — » »
★Vanellus vanellus (L.) . . VIII—IX — » »
★Himantopus himantopus hi- mantopus (L.) . . IX — » »
Tringa ochropus (L.) VII—IX — Одиночки и группы.
T. glareola L. . VII—IX — Редкие одиночки.
T. nebularia (Gunn.) VIII—IX — » »
*T. totanus totanus L. IX — » »
— 93 —
Таблица 6 (продолжение)
Вид Время пролета Конец отлета гнездящихся птиц Характер пролета
Tringa t. eurhinus (Oberh.) IX 21 IX Небольшие группы.
♦T. erythropus (Pall.) IX — Редкие одиночки.
*7. stagnatilis (Bechst.) IX — » »
Actitis hypoleucos (L.) VII—IX 1 X » »
★Xenus cinereus (Giild.) VIII—IX — » »
★Phalaropus lobatus (L.) VIII—IX — » »
★Ph. fulicarius (L.) . . VIII — Случайный залет.
★Arenaria interpres interpres (L.) VIII — Редкие одиночки.
Philomachus pugnax (L.) VIII—IX — Небольшие стайки.
Calidris minuta (Leisl.) VII—IX — » »
C. temminckii (Leisl.) VII—IX — » »
♦C. testacea (Vieill.) VIII-IX — » я »
*C. alpina ssp. IX — Редкие одиночки.
★Limicola falcinellus (Pont.) . . VIII — » »
Gallinago gallinago gallinago (L.) VIII—X — » »
★G. solitaria solitaria Hoags. IX — » »
★Numenius arquatus arquatus (L.) IX — » »
★Limosa limosa limosa (L.) IX — » »
Larus canus heinei Hom. VIII—IX — Одиночки и группы.
L. ichtyaetus Pall. — X Пролета нет.
L. brunnicephalus Jerd. — X » »
L. ridibundus L. VIII—IX — Редкие одиночки.
★Chlidonias leucoptera (Temm.) IX — » »
Sterna hirundo tibetana (Saund.) — X Пролета нет.
★Columba livia neglecta Hume IX — Редкие одиночки.
C. rupestris turkestanica But. — X Пролета нет.
★C. eversmanni Bp. X — Редкие одиночки.
Streptopelia turtur arenicola
(Hart.) . ... VIII—X — Одиночки и стайки.
S. orientalis meena (Sykes). VIII-X — » »
★Cuculus canorus subtelephonus
Zar. VIII—IX — Редкие одиночки.
★Otus scops pulchellus Pall. IX — » »
★0. brucei (Hume). IX — » »
★Alcedo atthis ssp. . . IX — » »
Coracias garrulus semenovi Loud.
et Tshusi VII—VIII — » »
Upupa epops epops L. VII—IX — Одиночки.
★Apus apus ssp. IX — Редкие одиночки.
★Jynx torquilla torquilla L. . IX — » »
Alauda arvensis dulcivox Brooks. IX—XII — Стайки и одиночки.
Л. gulgula lhamarum Mei-
nertzh. IX—X — Редкие стайки.
Calandrella acutirostris acuti-
rostris Hume IX—X X Стайки.
Ptyonoprogne rupestris rupest-
ris (Scop.) . VIII—IX IX Редкие одиночки.
Hirundo rustica rustica L. . . VIII—IX — » »
Delichon urbica meridionalis
Hart. VIII—IX IX Одиночки и стайки.
Oriolus oriolus ssp. . . . VIII—IX — Редкие одиночки.
★Corvus corone sharpei Oates . X — » »
★C. c. orientalis Ev. . . VIII — » »
Tichodroma muraria (L.) . . VIII—IX IX » »
Cinclus cinclus leucogaster Bp. — X Пролета нет.
★Muscicapa striata neumanni
Poche . . .... IX — Редкие одиночки.
Saxicola torquata maura (Pall.) VIII-IX — Обычен.
Oenanthe isabellina (Temm.). VIII—IX — Обычна.
Oe. deserti oreophila (Oberh.). . VIII—IX X »
Oe. hispanica pleschanka (Lep.). IX—XI Редкие одиночки.
— 94 —
Таблица 6 (продолжение)
Вид Время пролета Конец отлета гнездящихся птиц Характер пролета
Monticola saxatilis turkestani- cus Zar. . ... VIII—IX Редкие одиночки.
★M. solitaria pandoo Sykes . IX — » »
Phoenicurus ochfuros phoenicu- roides (Moore) VIII-X X Обычна.
Ph. erythrogaster grandis (Gould) IX—X X »
★Ph. caeruleocephalus Vig. IX — Редкие одиночки.
Cyanosylvia siecica pallidogu- laris (Zar.) . . . VIII-X Обычна.
C. s. tianschanica Tug VIII—XI — »
★Calliope pectoralis ballioni Sev. . . . VIII — Редкие одиночки.
Myophonus caeruleus turkesta- nicus Zar. . . ... VIII—IX — » »
★Turdus rujicollis atrogularis Jar. ... XI — » »
Phylloscopus trochiloides v tri- ll anus Blyth . VIII-IX — Обычна.
Ph. inornatus humei Brooks VIII-IX — Редкие одиночки.
★Hippolais caligata caligata (Licht.) .... VIII — » »
*H. c. rama (Sykes). . VIII—IX — » »
★Acrocephalus dumetorum Blyth. . . .... VIII—IX — » »
♦A. agricola agricola (Jerd.). VIII — » »
★Tribura major (Brooks). IX — » »
★Locustella naevia straminea Seebohm. . ... VIII—IX » »
★Sylvia nisoria merzbacheri Schal. . . VIII — » »
*5. communis rubicola Stres IX — » »
*5. curruca althaea Hume VIII-IX — » »
*5. c. margelanica Stolzm. VIII-IX — » »
*£. mystacea Menetr. . . . IX — » »
Laiscopus himalayanus (Blyth) — ? Пролета нет.
Prunella fulvescens fulvescens (Sev.) . .... IX—XI XI Небольшие стайки.
Motacilla alba personata Gould VIH-XI — Редкие одиночки.
M. a. dukhunensis Sykes VIII—X — » »
M. cinerea melanope Pall. VIII-IX — » »
Budytes citreola calcarata (Hodgs.) . ... VIII—X X » »
★B. flava beema (Sykes). IX — » »
★B. f. melanogrisea Hom. . . VIII — » »
Anthus trivialis trivialis (L.) VIII—IX — Одиночки и группы.
A. t. harringtoni With. . . VIII—IX — » »
Lanius schach erythronotus Vig. VIII-IX — Редкие одиночки.
L. collurio isabellinus Hemp, et Ehrrenb VIII—IX » »
L. c. phoenicuroides Schal.. VIII—IX — » »
Pastor roseus (L.). VIII-IX — » »
Sturnus vulgaris porphyronotus Sharpe IX — »
S. v. poltaratskyi Finsch . IX—X — »
Passer domesticus bactrianus Zar. et Kud IX—XI » »
★P. hispaniolensis transcaspi- cus Tschusi . . X Большие стаи.
★Fringilla coelebs L. . X — Редкие одиночки.
Cannabina flavirostris pami- rensis Zar. et Harms. . . XI Пролета нет.
C. cannabina fringillirostris Bp. et Schleg. VIII — Редкие одиночки.
— 95 —
Таблица 6 (продолжение)
Вид Время пролета Конец отлета гнездящихся птиц Характер пролета
Carpodacus erythrinus fergha-
nensis (Kozl.) VIII—IX — Небольшие стайки.
Bucanetes mongolicus (Swinli.) IX—X X » »
Emberiza leucocephalos Gm. X — » »
E. bruniceps Brandt. VIII—IX — Одиночки, стайки.
E. buchanani Blyth. VIII—IX — » »
Примечание. Виды, которые встречаются на пролете нерегулярно, отмечены звездочкой.
вероятно, относятся к пролетным популяциям. Очевидно, у большинства
перелетных видов местные популяции успевают покинуть нагорье до от-
лета последних мигрантов. Исключение составляют только горные гуси,
которые остаются на озерах вплоть до ледостава и покидают Памир вместе
с последними пролетными стаями.
Осенний пролет на Памире, как и всюду, несравненно богаче и интен-
сивнее весеннего. Тем не менее половина летящих через Памир видов
(68 из 140) бывает здесь на пролетах далеко не каждый год и в столь
малом числе, что, может быть, правильнее говорить об осенних залетах
птиц, сбившихся с пути. Большое их число объясняется тем, что Памир
расположен поблизости от важного пролетного пути, который ведет из
Сибири, Казахстана и Тянь-Шаня в Индию, в обход Центральной Азии,
вдоль подножий ее горного обрамления. По пролету в Центральной Азии,
особенно в Тибете, сведений очень мало, но существующие данные позво-
ляют судить о том, что большинство мигрантов обходит Тибет стороной,
вдоль его западных и восточных окраин. Следуя таким путем из Восточного
Казахстана и Сибири, птицы обходят Памирское нагорье с запада, при
общем направлении пролета на юго-запад. По наблюдениям В. Н. Шнит-
никова (1949), общее направление осеннего пролета в Семиречье тоже
юго-западное. Птицы летят здесь вдоль подножия и по западным окраинам
Тянь-Шаня к верховьям Сыр-Дарьи и Аму-Дарьи. Дальше их путь лежит
через западные отроги Памиро-Алая к перевалам Гиндукуша. Через
хребты Памиро-Алая летит свыше 150 видов птиц (А. И. Иванов, 1950),
но подавляющее большинство их пролетает западнее Памирского на-
горья. Сравнительное обилие уток и куликов на Памире объясняется на-
личием многочисленных озер, богатых животным и растительным кормом,
весьма малочисленных на остальной территории Памиро-Алая. Кроме
того, часть водоплавающих птиц и куликов летит на зимовки в южную
Азию через системы Лобнора и Тарима и пересекает стык хребтов Памиро-
Алая и Куньлуня в самом узком месте (Пржевальский, 1883). При этом
значительная их часть попадает на озера Памира.
Попав на Памир, большинство мигрантов двигается далее в том же
юго-западном направлении. В районе Памирской биостанции и озер Ранг-
кульской котловины наблюдался пролет ряда видов (горный гусь, белая
и желтоголовая трясогузки, лесной конек, краснобрюхая горихвостка,
варакушка и др.) по горным долинам через перевалы южных гребней
хр. Музкол к югу и юго-западу, в долину р. Мургаба, изобилующую бо-
лотистыми лугами. Дальше птицы летят к юго-западу в долину р. Али-
чура, а затем в долину р. Памира, через перевалы Южноаличурского
хребта. Через один из них, перевал Башгумбез, в последних числах сен-
тября 1956 г. шло движение вниз, к р. Памиру, скворцов, белых трясогу-
зок, лесных коньков, серых славок, пустынных каменок, плешанок
— 96 —
п краснобрюхих горихвосток. Дальше они следовали по р. Памиру вниз,
где шел пролет орланов-долгохвостов, зеленых пеночек, черноголовых
чеканов и варакушек в теплую, изобилующую садами и полянами до-
лину верхнего Пянджа, в обход мощных ледяных узлов Каракорума
и восточного Гиндукуша. Дальше птицы следуют через долины афганского
Бадахшана и восточного Афганистана. По сообщению Ахтара (Akhtar,
1955), птицы пересекают Гиндукуш по ряду перевалов, в частности через
перевал Перван (2890 м). Одновременно он отмечает, что через Гималаи
от Кашмира до Сиккима видимого пролета нет. Впрочем, такие крупные
птицы, как горный гусь, могут лететь прямо через Каракорум.
Небольшая часть птиц, например удод, стенолаз, летят в восточном
направлении на зимовки в теплые долины южной Кашгарии. И, наоборот,
наблюдается движение птиц в западном направлении, например испан-
ского воробья (Абдусалямов, 1961).
Пролет птиц через Памир начинается в конце июля, когда на озерах
нагорья появляются небольшие группы различных куликов. Одновре-
менно увеличивается количество уток. В августе число пролетных птиц
растет с каждым днем, достигая максимума в сентябре. В октябре количе-
ство их резко падает, так как нередко с первых чисел этого месяца уста-
навливается зимняя погода и снега закрывают перевалы, т. е. как раз
в то время, когда осенний пролет в Средней Азии наиболее интенсивен.
Все же в октябре и даже в ноябре выпадают временные улучшения погоды,
которыми пользуются одиночки и стайки сбившихся с пути мигрантов
для перелета через нагорье. Р. Н. Мекленбурцев (1946) наблюдал в 1936 г.
в верховьях р. Аличура стайки скворцов, полевых жаворонков и одиноч-
ных белых трясогузок, каменок-плешанок и варакушек до середины ноября.
При внезапных ухудшениях погоды такие поздние мигранты попадают
в тяжелые условия и ищут спасения в домах и постройках человека.
Гибель птиц на нагорье во время осеннего перелета — явление довольно
обычное. Трупы погибших от истощения и замерзших птиц — лесных
коньков, длиннохвостых сорокопутов, зеленых пеночек и других — при-
ходилось находить часто. Подобные случаи вообще нередки в высоко-
горьях Центральной Азии. Непрерывно меняющиеся условия погоды,
снежные бураны, резкие длительные похолодания, сопровождающиеся
ураганными ветрами, — внезапно делают горные системы совершенно
непреодолимыми для птиц. Исследователи в горах Центральной Азии не-
редко и в большом числе находили трупы пролетных пернатых (Козлов,
1899; Shipton, 1938; Мекленбурцев, 1946; Кистяковский, 1950; Singh,
1961, и др.).
Относительная и абсолютная высота хребтов сама по себе не служит
сдерживающим фактором. Не раз наблюдалось, как при благоприятной
погоде различные птицы, в том числе и мелкие воробьиные, без труда пре-
одолевали перевалы высотой до 6700 м (Meinertzhagen, 1954). Высота,
которой достигают птицы при перелете через хребты Центральной Азии,
бывает очень большой. По сообщению Мейнертцхагена (Meinertzhagen,
1954), на фотографии солнца, сделанной в Дехра-Дуне, у южного подно-
жия центральных Гималаев, в полдень 19 сентября 1919 г. была ясно видна
стая гусей, пересекающих солнечный диск и летящих к югу. Высчитано,
что они летели на высоте около 9 км. Вероятно, на этой высоте стая
пересекла главный гребень Каракорума, ледяную стену 7-километровой
высоты, нередко скрытую в это время высокой шапкой снеговых туч.
На Памире не раз стаи гусей наблюдались на высотах свыше 6000 м
над ур. м.
Подавляющее большинство пролетных птиц, двигаясь через Памир,
придерживается субальпийских и альпийских лугов вдоль рек и озер.
На них, в прибрежной грязи и на мелководьях к концу лета развивается
7 Биология птиц
— 97 -
богатая органическая жизнь. Многие реки и озера изобилуют рыбой,
а богатые пищевые запасы окрестных лугов привлекают немало зернояд-
ных птиц.
Большинство мелких воробьиных летит через нагорье в одиночку,
придерживаясь наиболее благоприятных местообитаний. Одиночных ва-
ракушек, каменок, черноголовых чеканов приходилось встречать в совер-
шенно пустынных долинах на высоте около 4000 м над ур. м. Они быстро
продвигались в юго-западном направлении, придерживаясь мелких обрыв-
чиков, подножий морен и пересохших русел ручьев. В холодной котло-
вине оз. Зоркуль, которую вообще избегали пролетные воробьиные,
во второй половине сентября лесные коньки, зеленые пеночки, славкп-
завирушки, различные каменки, варакушки и другие редкими одиночками
летели к западу по северному берегу озера. Все они двигались вдоль
береговых обрывов, защищавших их от холодных ветров. С подветренной
стороны крутые берега хорошо прогревались солнцем. Там создавался
настолько теплый микроклимат, что человек, замерзший на холодном
ветре, в таком месте быстро отогревался. На берегах озер Рангкуль
и Шоркуль птицы использовали для защиты от ветров и для кормежки
глубокие щели и трещины в береговых обрывах.
Большинство водоплавающих местами кормежек и отдыха избирали
хорошо прогреваемые солнцем мелководья, где держались с подветрен-
ной стороны береговых мысов.
От низких ночных температур (до —19° в сентябре) птицы искали на
ночь различные убежища. Ряд наблюдений в верховьях высокогорных
долин и по берегам озер и рек показал, что трясогузки, коньки, пеночки,
славки и другие мелкие воробьиные забивались на ночь в мелкие ямки,
в щели береговых обрывов и скал и особенно часто — под нависшие дерно-
вины береговых обрывчиков у самой воды. Эти убежища покидались
только на восходе солнца.
По ночному пролету данных почти нет, только в сентябре наблюдался
ночной пролет камышниц. Как в августе, так и в сентябре был сделан ряд
наблюдений в разные часы ночи и в разную погоду, но ни разу не удалось
слышать голоса летящих птиц.
Изменения погоды сильно влияли на интенсивность пролета, что осо-
бенно хорошо наблюдалось в окрестностях Памирской биостанции.
Обширные опытные посевы и луга всегда привлекали большое количе-
ство пролетных птиц. Во время снегопадов, когда бушующие на перевалах
метели закрывали дальнейший путь, число птиц на полях резко увеличи-
валось. В густых посевах и среди построек Биостанции можно было
встретить камышниц, иволг, пестрых каменных дроздов, синих птиц
и многих других, причем в поисках корма птицы нередко залетали в жи-
лые помещения. Пролет возобновлялся с малейшим улучшением погоды,
когда птицы, поднимаясь к истокам речек, стремились как можно быстрее
проскочить через заваленные снегом перевалы. В ясные дни количество
пролетных птиц в окрестностях Биостанции заметно уменьшалось. Неза-
долго до сильных похолоданий пролет заметно усиливался. Например,
за день до резкого похолодания, начавшегося 1 октября 1956 г., вниз
по р. Памиру шел интенсивный по сравнению с предыдущими днями
пролет. Но все же не все успели проскочить до снегопадов вниз, в теплую
долину Пянджа. 2 октября, уже на второй день непрерывной снежной
метели, вниз по долине двигались массы откочевывавших с верховьев
реки рогатых жаворонков и горных вьюрков, среди которых попадались
стайки бледных завирушек, одиночные белые трясогузки, лесные коньки,
краснобрюхие горихвостки и варакушки. Несомненно, что и на больших
высотах птицы заблаговременно реагируют на изменения погоды, но все же
в меньшей степени, чем на равнинах. Слишком уж непостоянна погода
— 98 —
в горах, и смена одного ее режима другим совершается^подчас совершенно
внезапно, за несколько часов и по нескольку раз в течение суток.
Среди пролетных воробьиных нередко встречаются интенсивно линяю-
щие особи. Вероятно, у горных популяций ряда видов (зеленая пеночка,
стенолаз, пустынная каменка, горихвостка-чернушка) не все птицы успе-
вают закончить линьку к моменту отлета.
Одной из причин бедного осеннего пролета через нагорье является
отсутствие древесно-кустарниковой растительности. Многие птицы, свя-
занные с ней, чувствуют себя на нагорье весьма неуютно. Чечевицы,
сорокопуты, славки, пеночки, темнозобые дрозды, вертишейки обычно
придерживаются густых посевов, каменных и проволочных оград, нагро-
мождений крупных камней, крутых склонов и скал, избегая ровных по-
верхностей. Одновременно приходилось наблюдать, как резко менялось
поведение таких птиц при пролете через безлесное нагорье. Пеночки,
например, двигались вдоль крутых склонов, лазая под нависающими
камнями, как бледные завирушки, и перелетая от одного укрытия к дру-
гому. Славки-завирушки, которым всеь время приходилось держаться
на земле, довольно ловко охотились на насекомых, напоминая движениями
в воздухе мухоловок, а на земле — зарянок. Ястребиные и серые славки
были менее подвижны и охотились, слетая с какого-либо возвышения на
землю, где заметили добычу. По земле они передвигались тяжеловато,
напоминая вьюрков.
Регулярность осеннего пролета ряда видов через нагорье сильно ко-
леблется. Птицы то отсутствуют в одном году, то изобилуют в другом.
Например, в 1956 г. был сильный пролет славок-завирушек {Sylvia сиг-
ruca althaea Hume), осенью 1954 и 1960 гг. они не появлялись совершенно.
Болотные луни летели в большом числе в 1956 г. и были редки в остальные
годы. В 1954 г. проходил сильный пролет турухтанов, осенью в остальные
годы их было мало и т. д.
ОРНИТОГЕОГРАФИЯ ПАМИРСКОГО НАГОРЬЯ
1. Вертикальное распределение птиц Памира
Своеобразие природных условий Памира обусловило весьма сложное
распределение птиц на нагорье. Прежде всего оно крайне неравномерно.
Исключительная сухость климата приводит к тому, что при выборе гнездо-
вых местообитаний влажность часто играет большую роль, чем темпера-
тура. Из 50 видов 40 совершенно ясно показывают приверженность к бо-
лее влажным местообитаниям, а из остальных десяти восемь имеют в та-
ких местах наиболее высокую численность. Сухость памирского климата
оказывается слишком высокой даже для тех птиц, которые в более влажных
областях считаются ксерофильными (каменка-плясунья, тонкоклювый
жаворонок, бледная завирушка и др.). Поэтому существует резкий кон-
траст между пустынными холодными долинами или озерными котлови-
нами, с одной стороны, и пойменными лугами или хорошо увлажненными
ущельями, с другой. Места, хорошо прогреваемые солнцем, достаточно
увлажненные и защищенные от холодных ветров, представляют собой
своеобразные оазисы высокогорной пустыни, и большинство птиц гнездится
именно здесь. Эти оазисы, поглощавшие основное внимание орнитологов,
в значительной степени затушевывают истинную картину вертикального
распределения птиц на нагорье. Сам облик памирских ландшафтов спо-
собствует тому, что у многих создается впечатление о Памире как о слабо
расчлененном плато, расположенном целиком в альпийском поясе. На-
личие вертикальной поясности в пределах нагорья до последнего времени
оставалось незамеченным орнитологами или даже вообще отрицалось
7*
— 99 —
(Кистяковский, 1950). Таким образом, в орнитологической литературе
сложилось мнение о Памире как о высокогорной альпийской пустыне,
не имеющей высотных поясов. Однако такое мнение противоречит всему
тому, что известно о природе гор. Тем более странным было бы это по от-
ношению к Памиру, где перепад высот достигает 4000 м. Поскольку геобо-
таники уже давно выделили на Памире несколько вертикальных поясов,
природные условия которых заметно отличаются друг от друга, логично
было бы предположить неодинаковый характер орнитофауны на разных вы-
сотах.
Следует особо остановиться на следующем моменте. На Памире, как
и на других высокогорьях Центральной Азии, благодаря сильному ни-
Рис. 18. Схема распределения гнездящихся птиц Памира по высотам.
I — субальпийский пояс; II — альпийский пояс; III — субнивальный пояс. По
оси абсцисс — высота над уровнем моря (в м); по оси ординат — число видов.
велирующему влиянию резко континентального, пустынного климата
границы между поясами значительно менее резки, чем на окраинных хреб-
тах этой страны, например на Тянь-Шане или Гималаях.
Большинство видов птиц, гнездящихся на Памире, не поднимается
выше субальпийского пояса. Как можно видеть на рис. 18, число гнездя-
щихся видов особенно резко падает между 4200 и 4300 м над ур. м., т. е.
как раз на границе субальпийского и альпийского поясов. Выше в аль-
пийском поясе количество видов постепенно убывает с высотой, незна-
чительно в нижней части пояса и более заметно в верхней его части,
выше 4500 м. В субальпийском поясе картина совершенно иная. Количество
гнездящихся птиц на «дне» Памирского нагорья, в нижней части субаль-
пийского пояса, сравнительно невелико и с высотой начинает быстро
увеличиваться, достигая максимума на высоте 4000 м. Затем количество
видов постепенно начинает убывать, но все же на верхней границе субаль-
пийского пояса гнездится не меньше птиц, чем на нижней, хотя видовой
состав здесь иной.
Подобная картина объясняется тем, что высоты 4000 м достигает боль-
шая часть птиц, поднимающихся на Памир из соседних, более низких
горных систем, и в то же время до этой высоты спускаются многие высо-
когорные виды, ниже не гнездящиеся. Образно говоря, на высоте около
4000 м происходит как бы встреча высокогорного орнитокомплекса с низ-
— 100 —
когорным. Кроме того, благодаря особенностям ландшафта главные
озера Памира расположены в пределах высот 3700—4100 м, и целая группа
птиц, связанных с ними, как высокогорных, так и широко распространен-
ных, естественно, гнездится именно в этих высотных пределах. Такие,
отнюдь не высокогорные виды, как травник, огарь, кряква, лысуха,
находят для себя подходящие условия в субальпийском, «озерном» поясе
Памирского нагорья и вновь появляются на гнездовье только в пред-
горьях Памиро-Алая, минуя окружающие Памир горные системы с их
резко расчлененным рельефом.
Таким образом, вместе с температурным фактором, играющим решаю-
щую роль в определении верхней границы распространения птиц в горах,
немаловажное значение на Памире имеет и рельеф. Это особенно хорошо
видно при рассмотрении верхних пределов гнездования птиц на Памире:
Высота над уровнем
моря, м
Перевозчик . . . . 3780
Кряква и обыкновенная чечевица 3800
Полевой жаворонок 3850
Тонкоклювый жаворонок, городская ласточка, ка-
менка-плясунья и белая трясогузка 4000
Беркут и снежный воробей . . . 4050
Снежный сип, гималайская завирушка и красный
вьюрок . . . 4100
Бородач, лысуха, травник, буроголовая чайка, реч-
ная крачка и большая чечевица . 4150
Тибетская саджа, филин, скалистая ласточка, аль-
пийская галка и белобрюхая оляпка 4200
Пустельга и короткоклювый зуёк 4250
Ворон . 4300
Большой крохаль и горихвостка-чернушка 4350
Горный гусь и желтоголовая трясогузка . 4400
Скалистый голубь, рогатый жаворонок и клушица 4500
Огарь и пустынная каменка 4550
Стенолаз, бледная завирушка и горная коноплянка 4600
Краснобрюхая горихвостка 4700
Пустынный снегирь . 4750
Темнобрюхий и тибетский улары 4800
Горный вьюрок 5000
То же можно сказать и о факторах, определяющих нижнюю границу
гнездования. Только к температурному и ландшафтному факторам здесь
прибавляется еще и фактор влажности. Вероятно, определенный минимум
влажности определяет нижнюю границу гнездования тибетского улара
и горного вьюрка.
В целом, вертикальное распределение птиц на Памирском нагорье
отличается следующими особенностями.
Прежде всего субтропические широты, в которых расположено на-
горье, в сочетании с высокой сухостью климата обусловили чрезвычайно
высокое положение снеговой линии в горах Памира, выше, чем где-либо
в СССР. Вслед за ней столь же высоко поднимается растительный и жи-
вотный мир. Не удивительно поэтому, что подавляющее большинство
птиц, населяющих нагорье, гнездится здесь выше, чем в других горных
системах нашей страны.
Широкое развитие пустыни в субальпийском и альпийском поясах
нагорья, так же как и в ниже расположенных поясах окружающих гор-
ных систем, вызвало весьма широкое вертикальное распространение
многих видов. Особенно примечателен в этом отношении пустынный сне-
гирь, гнездящийся от самых предгорий Памиро-Алая, от высот около 1000 м
над ур. м., до верхней границы альпийского пояса Памира. Короче, все
— 101
ксерофильные виды в условиях крайне континентального, аридного па-
мирского климата отличаются широким вертикальным распространением.
Наоборот, влаголюбивые виды, которых, кстати, очень немного, отли-
чаются сравнительно узким вертикальным и спорадическим горизонталь-
ным распространением. Типичным примером такого вида является горный
вьюрок. И если ксерофильные виды в большинстве своем гнездятся в ниж-
ней части нагорья, будучи все же весьма теплолюбивыми, то мезофильные
связаны с более холодным альпийским поясом, где влажность в 2—3 раза
выше, чем в субальпийском поясе.
2. Зоогеографический анализ памирской орнитофауны
Из 50 гнездящихся на Памире видов птиц обычны только 36 видов.
Остальные 14 встречаются на лучше увлажненных окраинах или в самых
низких частях нагорья, где обитают в небольшом числе, а некоторые из
них вообще гнездятся нерегулярно. Поэтому при анализе мы их рассмат-
ривать не будем.
Среди видов, характерных для нагорья, 5 являются убиквистами и ши-
роко распространены в самых различных ландшафтах Палеарктики (пу-
стельга, беркут, филин, ворон, городская ласточка); 3 из них — беркут,
филин и ворон — представлены высокогорными центральноазиатскими
формами.
Довольно большая группа птиц, 9 видов, связана в своем распростра-
нении на нагорье с хорошо увлажненными лугами, озерами и ре-
ками. Из них 2 вида — горный гусь и буроголовая чайка — являются
эндемиками Центральной Азии. 3 вида: большой крохаль, речная крачка
и желтоголовая трясогузка, широко заселяющие соответствующие биотопы
Палеарктики, — представлены особыми центральноазиатскими формами.
Группа пустынно-степных видов представлена хорошо. Причем 2 вида
из 7 — тибетская саджа и тонкоклювый жаворонок — являются энде-
миками центральноазиатских высокогорий. Как и буроголовая чайка,
в прошлом они были связаны с ровными пространствами, которые в про-
цессе тектонических движений были подняты на большую высоту. То же
можно сказать и о короткоклювом зуйке. Остальные 4 вида — рогатый
жаворонок, пустынная каменка, каменка-плясунья и пустынный снегирь—
широко распространены в пустынно-степной зоне Палеарктики, причем
рогатый жаворонок и пустынная каменка представлены высокогорными
центральноазиатскими подвидами.
Наиболее многочисленна (12 видов) группа, характерная для горных,
большей частью аридных областей юга Палеарктики. Многие из этих ви-
дов (бородач, темнобрюхий улар, клушица, стенолаз) гнездятся иногда
очень высоко, но ни один из них не может быть отнесен к исключительно
высокогорным видам. Для всех характерно обитание в сильно пересе-
ченных ландшафтах гор средней высоты, причем многие нередко гнездятся
на высотах, немногим превышающих уровень океана (клушица, горная
ласточка, горихвостка-чернушка и др.). Вообще только 3 вида из 36,
обычных на нагорье, являются действительно высокогорными не только
на Памире, но и в других частях своего ареала. Это тибетский улар, красно-
брюхая горихвостка и горный вьюрок. Все они являются весьма специали-
зированными и, вероятно, довольно древними обитателями альпийского
пояса Центральной Азии.
В целом получается весьма любопытная картина. На Памире, в самых
высоких горах нашей страны, высокогорные птицы составляют незна-
чительную часть орнитофауны, основной состав которой образуют низко-
горные или пустынно-степные виды, характерные для аридных областей
южной Палеарктики. Это объясняется тем, что большинство настоящих
102 —
альпийцев приспособлено к обитанию в сравнительно узких экологиче-
ских нишах, отличающихся прежде всего повышенной влажностью и опре-
деленным минимумом положительных температур лета. Поэтому они
не находят для себя подходящих условий на пустынных нагорьях Цен-
тральной Азии и гнездятся только на окраинных хребтах этой страны,
получающих достаточно влаги (Columba leuconota, Laiscopus himalayanus,
L. collaris, Grandala coelicolor, Pyrrhospiza punicea и др.). На это впервые
указала Е. В. Козлова (1952) при анализе происхождения тибетской ор-
нитофауны. На Памире подобная закономерность повторяется как бы
в миниатюре. Во внутренние части нагорья проникают только 3 альпий-
ских вида, а остальные гнездятся по окраинным хребтам, и ареал их коль-
цом охватывает Памир (Columba leuconota, Laiscopus himalayanus, Pyrrhos-
piza punicea и др.). Три вида альпийцев, гнездящихся на Памире: тибет-
ский улар, краснобрюхая горихвостка и горный вьюрок, — вероятно,
лучше других приспособлены к низким летним температурам и поэтому
способны существовать на тех высотах нагорья, которые получают до-
статочное для них количество влаги.
Необходимо обратить внимание на тот факт, что настоящих альпий-
ских видов в природе вообще очень мало. Кажущаяся их многочислен-
ность происходит от того, что нередко обитание какого-либо вида в аль-
пийском или даже субальпийском поясе одной из горных систем бывает
достаточным для причисления его к альпийцам, что, конечно, неверно.
В целом из 36 видов, обычных для Памира, более одной трети (13 видов)
имеет несомненно центральноазиатское происхождение. Среди них на-
ходится 5 видов, являющихся эндемиками Центральной Азии. Из осталь-
ных 23 видов около половины (9) представлено хорошо обособленными
центральноазиатскими подвидами.
3. Зоогеографическое районирование Памирского нагорья
Вопросы зоогеографического районирования Памира в свое время были
весьма запутаны благодаря объединению в одно целое двух совершенно
различных природных областей — Памира и Бадахшана. М. А. Мензбир
(1914) выделил Памир в самостоятельный участок и, приняв во внимание
значительную примесь тибетских элементов, объединил Памир с Ладаком
в Западно-Тибетский округ. Этим было подчеркнуто отличие орнито-
фауны Памира от других зоогеографических участков Средней Азии,
тяготеющих к Средиземноморью. Правильно определив, таким образом,
место Памира в зоогеографической схеме деления Палеарктики, М. А. Менз-
бир тем не менее допустил ошибку, обусловленную малой изученностью
области. Он объединил в Памирский участок как Памирское нагорье,
так и Бадахшан — страны, совершенно разные по природным условиям.
Впервые граница между Памиром и Бадахшаном была проведена
Б. С. Виноградовым и др. (1935). При этом Бадахшан (Западный Памир,
по терминологии авторов) соединялся с Восточно-Бухарским участком,
а собственно Памир (Восточный Памир) рассматривался как нерезко
выраженная переходная область.
Г. П. Дементьев в 1935 г., обработав сборы М. П. Розанова, дал крат-
кий зоогеографический анализ Памира, в котором тоже рассматривал
как отдельные области «Западный Памир» (Бадахшан) и «Восточный
Памир» (Памир), но тем не менее дал для них общий список видов. В этом
списке среди обилия средиземноморцев, населяющих ущелья Бадахшана,
совершенно тонут немногие центральноазиатские виды, свойственные
Памирскому нагорью. На основании этого списка автор сблизил Памир
не с Тибетом, а с Тянь-Шанем. Подтверждающий правомочность такого
сближения список видов, якобы общий этим странам, почти не содержит
— 103 —
видов, гнездящихся на Памире (Pica pica hemileucopter a, Dryobates leucop-
terus leptorhynchus. Alectoris kakelik falki и др.). Г. П. Дементьев писал,
что многие «общие Памиру и Тянь-Шаню формы в Тибете вовсе отсутст-
вуют» (Дементьев, 1935 : 94). На это следует возразить, что таких форм
всего две и, кроме того, они не являются характерными для Памира,
обитая в самых низких частях или на окраинах нагорья (пустельга и пе-
ревозчик).
В 1936 г. опубликовал свои материалы по млекопитающим и птицам
Памира Р. Н. Мекленбурцев. Он справедливо указал на то, что М. А. Менз-
бир не делал различий между «Западным» и «Восточным» Памиром, ко-
торые, по словам автора, представляют собой совершенно различные об-
ласти. Р. Н. Мекленбурцев соединил с Горно-Бухарским участком фауну
долины Пянджа и его притоков — крупных рек Бадахшана. Высокогор-
ные области водоразделов между этими реками он предложил отнести
к «Восточному Памиру». При этом была дана карта, где примерно на-
мечена граница между «Западным» и «Восточным» Памиром. Высотный
принцип ее построения в общем верен, но альпийский пояс хребтов Ба-
дахшана исследован еще слишком недостаточно, чтобы делать окончатель-
ный вывод.
Б. К. Штегман (1938) сближал Памир с Тибетом, причем под Памиром
он понимал именно нагорье, а не территорию Горно-Бадахшанской АО.
А. И. Иванов (1940) также подчеркнул разницу между Бадахшаном
и Памиром, отказавшись при этом от неправильной терминологии «За-
падный» и «Восточный» Памир. Он показал, что на выровненных простран-
ствах нагорья находит свой западный предел ряд видов тибетских степей
и полупустынь.
В 1949 г. Р. Н. Мекленбурцев опубликовал специальную работу,
посвященную орнитогеографии «Западного» и «Восточного» Памира,
Уже из названия работы видно, что автор рассматривает Бадахшан и Па-
мирское нагорье как одну страну, хотя в тексте основное внимание уделено
тому, чтобы показать резкие фаунистические различия между этими об-
ластями. Несмотря на ряд фактических ошибок, вызванных слабой изу-
ченностью гнездовой орнитофауны Памира в то время, автор дал правиль-
ную в принципе картину распространения птиц на Памире и в Бадахшане,
но объяснил ее, на наш взгляд, неверно. Прежде всего Р. Н. Мекленбур-
цев считал, что фаунистические различия между «Восточным и Западным
Памиром. объясняются современными экологическими, а не истори-
ческими зоогеографическими причинами» (Мекленбурцев, 1949 175).
Однако у нас нет никаких оснований считать экологические причины
не зоогеографическими. Кроме того, существующие резкие экологические
различия между Памиром и Бадахшаном возникли не сразу, поскольку
эти области прошли длительные, на первых этапах, возможно, сходные,
а потом все более расходящиеся пути развития. Следовательно, есть здесь
и исторические причины. Если автор под историческими зоогеографи-
ческими причинами понимал формирование и развитие фаун, то с этим
тем более нельзя согласиться. Памир и Бадахшан не являлись сколько-
нибудь заметными центрами формообразования у птиц, а заселялись из
соседних областей. На месте же представители того или иного типа
фауны претерпевали весьма незначительные изменения, не выходящие
за рамки вида. При этом процессы заселения Памира и Бадахшана шли
по-разному и орнитофауны их имеют, следовательно, исторически сло-
жившиеся отличия. Нельзя также согласиться с Р. Н. Мекленбурцевым,
когда он трактует «Восточный» и «Западный» Памир частями Средней
Азии. Это правомочно только для последнего.
В 1950 г. вышла в свет работа А. Б. Кистяковского, посвященная зоо-
географии Памира. Отдавая дань традициям 30-х годов нашего века, автор
— 104 —
все еще именует Бадахшан и Памир «Западным» и «Восточным» Памиром,
хотя впоследствии доказывает, что эти области принадлежат различным
зоогеографическим округам. Таким образом, названный автор сам себе
противоречит, объединяя все же эти области под названием Памир и опре-
деляя его границы административными границами Горно-Бадахшанской
АО, считая их естественными. Однако естественной границей между при-
родными областями никак не может быть река, протекающая в узком
ущелье и имеющая совершенно одинаковые берега. А именно по такой
реке, по Пянджу, проходит у А. Б. Кистяковского западная граница
Памира. Неправ А. Б. Кистяковский и тогда, когда утверждает, что эти
границы общеприняты. Крупнейшие исследователи Памира, начиная
с Н. А. Северцова, всегда различали Памир и Бадахшан и никогда не
делали попыток объединить их в нечто целое. При этом анализ орнито-
фауны «Восточного Памира», как, между прочим, и в работах Р. Н. Мек-
ленбурцева, делается без учета особенностей вертикального распределе-
ния птиц. По сути дела, здесь вообще отрицается поясность «Восточного
Памира», подчеркивается однообразие условий по всему нагорью, что,
как мы видим, далеко не верно. В списки гнездящихся птиц «Восточного
Памира» включены многие виды, которые в действительности здесь не
гнездятся. Эти ошибки объяснялись слабой изученностью гнездовой орни-
тофауны нагорья, но они заметно снизили, а в некоторых случаях п свели
на нет ценность теоретических выводов автора. Например, тянь-шань-
ская варакушка рассматривается А. Б. Кистяковским как доледниковый
реликт Каракульской котловины и делается вывод, что эта котловина
в период максимального оледенения была свободна ото льда. Однако
в действительности варакушка здесь, как и везде на Памире, не гнездится.
Но, несмотря на эти ошибки, со многими выводами можно согласиться.
Совершенно правильно А. Б. Кистяковский сблизил Памир с Ладаком
и отнес его фаунистически к Центральной Азии, а Бадахшан, «Западный
Памир» — к Средней Азии. Не вызывает возражений и хорошо аргу-
ментированное автором значение Памира как зоогеографического участка.
Только при этом не оговорено его островное положение и оторванность
от высокогорий Тибета и Ладака.
Нельзя не упомянуть о том, что в последнее время некоторые авторы
выступили против дробного зоогеографического деления для горных си-
стем, вместо чего предлагают проводить анализ орнитофауны только по
методу «типов фауны» Б. К. Штегмана. В частности, в последнее время
такую попытку предприняли Л. С. Степанян (1960) и Р. Л. Бёме (1963).
Еще раньше Б. К. Штегман (1946) справедливо указывал на разнородность
тянь-шаньской фауны и на невозможность выделять здесь зоогеографи-
ческие участки, проектируя их на одну плоскость карты. В противном
случае вместо зоогеографического, фаунистически однородного участка
мы получили бы нагромождение из различных высотных поясов со столь же
различными типами фауны. Исходя из этих соображений, Л. С. Степанян
впал в другую крайность, утверждая, что «зависимость современного раз-
мещения птиц от конкретных экологических условий и обнаруживающаяся
при этом мозаичность фауны Тянь-Шаня, по нашему мнению, исключают
необходимость зоогеографического деления этой страны» (Степанян,
1960 : 374). Таким образом, отрицается необходимость зоогеографического
деления крупных горных систем. На наш взгляд, такое деление все же
вполне возможно и необходимо, так как в конечном итоге дает конкретные
пространственные границы распространения тех же типов фауны. Только
при этом необходимо учитывать не только вертикальные, но и горизон-
тальные поясные границы. Определенный пояс какого-нибудь горного
хребта вполне может быть единицей зоогеографического районирования,
как и ландшафтная зона.
— 105 —
В целом зоогеографическое положение Памира можно характеризо-
вать следующим образом. Памир является пустынным нагорьем, подня-
тым выше границы древесной растительности и окруженным горными си-
стемами с резко расчлененным рельефом, с глубокими теплыми долинами,
покрытыми зарослями деревьев и кустарников. Орнитофауна этих глу-
боких долин резко отличается от памирской, и ареал многих видов, гнез-
дящихся здесь, охватывает Памир сплошным или почти сплошным
кольцом (Alectoris kakelik, Corvus corone, Pica pica, Oriolus oriolus, Serinus
pusillus, Emberiza cia, Oenanthe hispanica, Phylloscopus collybita, Ph. tro-
chiloidesK мн. др.), на что обратил еще внимание Р. Н. Мекленбурцев (1949).
Таким образом, орнитофауна Памирского нагорья, имеющая ярко вы-
раженный центральноазиатский высокогорный облик, занимает изолиро-
ванное, островное положение по отношению к Тибету. Непосредственно
с высокогорьями Тибета Памир соединяет только хр. Каракорум с его
северным отрогом — хр. Музтаг. Альпийский и субнивальный пояса
этих хребтов являются как бы узким каналом, по которому возможно
взаимное проникновение тибетских и памирских птиц. Но следует учи-
тывать, что по середине этого канала располагается гигантская ледяная
пустыня массива Хиспар, которая делает такой обмен фаунистическими
элементами если не невозможным, то во всяком случае весьма затрудни-
тельным.
На Тибетском нагорье, по последним сведениям (Козлова, 1952),
гнездится 31 вид птиц, из них 21 вид следует считать общим с Памиром
(отмечены звездочкой).
★Eulabeia indica (Lath.)
★Casarca ferruginea (Pall.)
★Gypaetus barbatus (L.)
★Gyps himalayensis Hume
★Haliaeetus leucoryphus (Pall.)
★Tetraogallus tibetanus Gould
Grus nigricollis Przew.
★Charadrius mongolus Pall.
★Tringa totanus L.
★Larus brunnicephalus Jerd.
★Sterna hirundo L.
* Tchangtangia tibetana (Gould)
★Columba rupestris Pall.
★Athene noctua (Scop.)
★Bubo bubo (L.)
* Corvus corax L.
*Pyrrhocorax pyrrhocorax (L.)
Pseudopodoces humilis (Hume)
Monti/ringilia adamsi Adams
M. henrici (Oust.)
M. ruficollis Blanf.
M. blanjordi Hume
M. taczanowskii Przew.
★Leucosticte brandti (Bp.)
Kozlowia roborowskii (Przew.)
★Carpodacus rubicilla (Giild.)
★Eremophila alpestris (L.)
E. teleschowi Przew.
Melanocorypha maxima Gould
★Calandrella acutirostris Hume
★Phoenicurus erythrogaster (Giild.)
Основное отличие памирской орнитофауны от тибетской состоит
в том, что на Памире отсутствуют такие характерные тибетские птицы,
как земляные вьюрки, ложносойка и черношейный журавль. В этом
отношении орнитофауна Памира более сходна с орнитофауной Ладака,
где тоже отсутствуют земляные вьюрки и ложносойка, но там все же гнез-
дится черношейный журавль.
Многие исследователи, начиная с М. А. Мензбира, сравнивали орнито-
фауны Памира и Ладака и в результате сближали их или признавали
совершенно различными. Сравнение орнитофауны этих стран вполне пра-
вомочно. Памир и Ладак — единственные, значительные как по площади,
так п по высоте нагорья, расположенные к западу от Тибетского плато,
но в отличие от Памира Ладак непосредственно примыкает к Тибету и рас-
положен на тех же широтах. ЕслиМ. А. Мензбир сблизил орнитофауны
Памира и Ладака и объединил их в одну Западно-Тибетскую подпровин-
цпю, то Г. П. Дементьев, наоборот, считал их совершенно различными
(1935). В доказательство Г. П. Дементьев привел список птиц, представ-
ленных в Ладаке и на Памире разными подвидами. С одной стороны, в этом
— 106 —
списке немало бадахшанских птиц, на Памире не гнездящихся. Кроме
того, судя по списку, под термином «Ладак» автор понимал территорию,
входящую в административные границы Ладака. Но в этих границах
страна столь же неоднородна, как и Горно-Бадахшанская АО. Северная
ее часть, изрезанная глубокими лесистыми ущельями, резко отличается
от южной, где расположены высокогорные плато, подобные тибетским.
Это — плато Рупшу, поднятое на высоту около 4500 м, с рядом больших
и малых озер и расположенное к востоку от него плато с оз. Пангонг,
непосредственно примыкающее к Тибету. Между ними залегает глубокая
долина р. Инда, покрытая древесно-кустарниковой растительностью
и имеющая фауну, совершенно отличную от фауны нагорий. Долины Инда
и Шайока прорезают всю территорию Ладака с юго-востока на северо-за-
пад, и по ним древесно-кустарниковая фауна теплых долин проникает
почти к самому подножию Тибета. Таким образом, в Ладаке мы имеем
два типа орнитофауны — орнитофауну глубоких долин и орнитофауну
высокогорных плато, и смешивать их в одну вряд ли целесообразно.
Кажется очевидным, что при сравнении орнитофаун Памира и Ладака
необходимо брать только орнитофауну высокогорий Рупшу и Пангонга,
т. е. сравнивать сравнимые вещи. Ниже приводится список птиц (47 видов),
обитающих на высокогорьях Ладака
с Памиром).
Podiceps cristatus cristatus (L.)
*Eulabeia indica (Lath.)
*Casarca ferruginea (Pall.)
*Mergus merganser orientalis Gould
* Aquila chrysaetus daphanea Sev.
*Gypaetus barbatus hemachalanus Hutton
*Gyps himalayensis Hume
*Cerchneis tinnunculus interstinctus Horsf.
Perdix hodgsoniae hodgsoniae (Hodgs.)
*Alectoris kakelik pallescens Hume
*Tetraogallus himalayensis himalayensis
Gray
*T. tibetanus tibetanus Gould
Grus nigricollis Przew.
*Charadrius mongolus atrifrons Wagl.
*Tringa totanus eurhinus (Oberh.)
*Actitis hypoleucos (L.)
*Larus brunnicephalus Jerd.
*Sterna hirundo tibetana Saund.
*Columba rupestris turkestanica But.
*Tchangtangia tibetana (Gould)
*Bubo bubo hemachalanus Hume
Apus apus pekinensis Swinh.
*Calandrella acutirostris tibetana Brooks
*Eremophila alpestris elwesi Blanf.
*Ptyonoprogne rupestris rupestris (Scop.)
*Delichon urbica cashmeriensis Gould
(звездочкой отмечены виды, общие
*Corvus corax tibetanus Hodgs.
^Pyrrhocorax pyrrhocorax himalayensis
Gould
*Graculus graculus forsythi (Stol.)
* Tichodroma muraria muraria L.
*Cinclus cinclus cashmeriensis Gould
★Oenanthe deserti oreophila (Oberh.)
*Phoenicurus ochruros phoenicuroides (Mo-
ore)
★Ph. erythrogaster grandis (Gould)
★Calliope pectoralis pectoralis
Gould
Phylloscopus affinis Tick.
Sylvia curruca althaea Hume
Laiscopus collaris nipalensis Blyth
Prunella rubeculoides (Moore)
* P. fulvescens fulvescens (Sev.)
* Motacilla alba personata Gould
M. a. alboides (Hodgs.)
★ M. citreola calcaratus Hodgs.
★ Monti f ringilia nivalis adamsi (Adams)
Passer domesticus parkini (Whistl.)
★Leucosticte brandti haematopygia Gould
* Cannabina flavirostris ladacensis (Mei-
nertzh.)
★Carpodacus rubicilla rubicilloides Przew.
Следовательно, в Ладаке гнездится почти такое же количество видов.
Из них 38 видов являются общими с Памиром, причем 26, более половины
всех птиц, представлено теми же самыми подвидами. Многие виды предста-
влены формами, очень близкими между собой (Charadrtus mongolus ра-
mirensis и Ch. т. atrifrons, Cannabina flavirostris pamirensis к C. f. ladacen-
sis и др.). Вообще, в Ладаке количество характерных тибетских птиц
по сравнению с Памиром увеличивается только на 1 вид, что очень немного,
если принять во внимание непосредственную близость Ладака и Тибета.
Гнездование остальных 9 видов, отсутствующих на Памире, объясняется
несколько более благоприятными условиями среды. Более южное, на 4°,
положение и повышенная по сравнению с Памиром влажность (снеговая
— 107 —
линия проходит в среднем на высоте 5100 м) дают возможность проникать
кустарниковой растительности местами до высоты 4800 м. Эти кустарники
(Caragana sp. и др.), образуя местами целые заросли, придают особый
колорит стране и позволяют многим птицам (Prunella rubeculoides, Carpo-
dacus rubicilla rubicilloides, Phylloscopus af finis) подниматься высоко в горы.
В то же время влажность высокогорий Ладака недостаточна для того,
чтобы здесь могли гнездиться влаголюбивые альпийские виды, населяю-
щие окраинные хребты Центральной Азии (Columba leuconota, Laiscopus
himalayensis, Grandala coelicolor и др.), — закономерность, опять-таки
общая с Памиром. По причине малой влажности здесь очень редки сте-
нолаз и альпийская галка, a Calliope pectoralis найден на гнездовье только
в одном месте, под перевалом Тзака, па высоте 4350 м, отличающемся
теплым местным климатом и обильным грунтовым увлажнением (Osmas-
ton, 1927).
В конечном итоге, на наш взгляд, орнитофауны высокогорий Памира
и Ладака очень близки к тибетской и фаунистически несомненно отно-
сятся к Тибетской провинции. Общая орнитофаунам Памира и Ладака
черта — отсутствие тибетских земляных вьюрков и ложносойки — явля-
ется наиболее существенным отличием от орнитофауны Тибета. В то же
время отличия авифаун Памира и Ладака позволяют рассматривать их
как отдельные зоогеографические участки Тибетской провинции.
Рассмотрим теперь взаимоотношения орнитофаун Тянь-Шаня и Памира.
Поскольку Тянь-Шань — страна неизмеримо более обширная, чем Памир,
с большим разнообразием условий и с гетерогенной фауной, с памирской
орнитофауной правильным будет сравнивать только орнитофауну сыртов
Тянь-Шаня, т. е. выровненных высокогорий субальпийского и альпий-
ского поясов страны, расположенных в центральных ее частях. Орнито-
фауна тянь-шаньских сыртов изучена в настоящее время довольно хорошо.
Ниже приводится список птиц, обычных на сыртах (звездочкой отмечены
виды, общие с Памиром).
*Eulabeia indica (Lath.)
*Casarca jerruginea (Pall.)
* Aquila chrysaetos daphanea Sev.
Aegypius monachus (L.)
*Gyps himalayensis Hume
*Сураё1и5 barbatus hemachalanus
Hutto Q
*Cerchneis tinnunculus tinnunculus (L.)
* Tetraogallus himalayensis severtzowi Zar.
*Charadrius mongolus pamirensis Richm.
*Tringa totanus eurhinus (Oberh.)
*Actitis hypoleucos (L.)
*Columba rupestris turkestanica But.
* Athene noctua plumipes Swinh.
*Alauda arvensis dulcivox Brooks
*Eremophila alpestris albigula (Bp.)
*Ptyonoprogne rupestris rupestris (Scop.)
* Deli chon urbica meridionalis (Hart.)
*Corvus corax tibetanus Hodgs.
* Pyrrhocorax pyrrhocorax centralis Stres.
* Graculus graculus jorsythi Stol.
Oenanthe oenanthe argenthea (Lonnb.)
* Oe. isabellina (Temm.)
* Phoenicurus ochruros phoenicuroides (Mo-
ore)
* Ph. erythrogaster grandis (Gould)
* Tichodroma muraria L.
*Laiscopus himalayanus (Blyth)
*Prunella julvescens julvescens (Sev.)
*Motacilla alba personata Gould
*Budytes citreola calcaratus (Hodgs.)
Anthus spinoletta blakistoni Swinh.
* Mo ntif ringilia nivalis alpicola (Pall.)
*Leucosticte brandti brandti Bp.
*Cannabina jlavirostris korejevi (Zar. et
Harms.)
*Carpodacus rubicilla severtzovi Sharpe
Как видно из этого списка, виды, гнездящиеся на тянь-шаньских
сыртах, почти все гнездятся и на Памире, причем большинство из них
представлено теми же подвидами. Но на сыртах Тянь-Шаня отсутствуют
такие характерные тибетские виды, как тибетский улар, тибетская саджа,
буроголовая чайка, тибетская форма речной крачки, тонкоклювый жаво-
ронок. Это объясняется большей удаленностью сыртов от Тибета, малой
их площадью и сравнительно высокой влажностью климата, что, по-ви-
димому, не подходит для ксерофильных тибетских птиц. Весьма показа-
— 108 —
тельно в этом отношении отсутствие на сыртах такого широко распро-
страненного в Центральной Азии ксерофильного вида, как пустынная
каменка, которую здесь заменяет обыкновенная каменка. Если Памир яв-
ляется как бы островом высокогорной центральноазиатской орнитофауны,
то сырты Тянь-Шаня — это маленькие островки той же фауны. Орнито-
фауна альпийского пояса хребтов Тянь-Шаня, окружающих сырты, весьма
близка к альпийской фауне горного обрамления Центральной Азии и пред-
ставлена тем же набором видов, что и на окружающих Памир хребтах,
и даже несколько богаче. Здесь появляется, в частности, альпийская
завирушка. Следовательно, сырты Тянь-Шаня, населенные характерной
центральноазиатской фауной, имеющей, правда, ряд местных особенно-
стей, вызванных более северным положением, изоляцией и большим
количеством осадков, вероятно, заслуживают выделения в особый уча-
сток Центральноазиатской, а не Среднеазиатской провинции.
4. К истории развития и формирования памирской орнитофауны
Тема эта неоднократно развивалась различными исследователями,
начиная от Н. А. Северцова (1880). Вначале эволюция ландшафтов Памира
рассматривалась только в общем плане, при построении схем развития
ландшафтов Средней и Центральной Азии (Мензбир, 1914; Сушкин,
1925; Дементьев, 1938). Первая попытка конкретно воссоздать историю
формирования памирской орнитофауны была предпринята Р. Н. Меклен-
бурцевым (1949), а вслед за ним — А. Б. Кистяковским (1950). Схемы
возникновения и формирования орнитофауны Памирского нагорья, пред-
ставленные этими авторами, в общем были согласованы с данными геологии
того времени. Несмотря на некоторые существенные разногласия,5 6 оба
автора исходили из того, что море исчезло с территории Памира к неогену,
когда началось интенсивное горообразование, и к началу четвертичного
периода памирская система гор была уже сформирована. Оба автора спра-
ведливо полагали, что Памир центром видообразования не был, а заселялся
из соседних областей, вместе с развитием определенных ландшафтов.
В пользу такого предположения говорят отсутствие на нагорье эндемич-
ных видов и очень малое количество подвидов. С тех пор вопрос этот более
не обсуждался. В то же время за последние десятилетия на территории
нагорья получили большой размах геологические, гляциологические, ар-
хеологические и геоботанические исследования, которые дали много новых
материалов, позволяющих в настоящее время по-новому и с большей
достоверностью воссоздать историю развития памирских ландшафтов,
а вместе с ними и историю формирования памирской орнитофауны.
Прежде всего, данные геологических исследований заставили совер-
шенно по-иному взглянуть на время образования и темпы поднятия Па-
мирской горной системы. Приходится теперь считать, что процесс подня-
тия Памира проходил почти целиком в плейстоцене. По данным неотекто-
нпки, он не закончился и до сих пор. В палеогене территория нагорья
была целиком под водой. Море постепенно мелело, и в олигоцене появи-
лась суша. В миоцене, по О. К. Чедия (1962), площадь Памира пред-
ставляла в целом сушу с полого-холмистым рельефом. На месте современ-
ных хребтов Заалайского и Академии Наук существовали островные
останцы доолигоценового рельефа. Они могли быть остатками островов
мелкого моря, покрывавшего территорию Памира в палеогене. Красно-
бурая окраска миоценовых отложений Памира косвенно указывает на
жаркий климат. Смена в плиоцене этой окраски на серую, по мнению мно-
5 Р. Н Мекленбурцев признавал покровное оледенение, а А. Б. Кистяковский
не признавал.
— 109 —
гих исследователей, указывает на похолодание климата. Оно могло быть
вызвано усилением континентальности, обусловленной продолжавшимся
отступанием моря и начавшимся поднятием страны. Первые данные пыль-
цевого анализа с Памира (Пахомов, 1962), которые как раз относятся
ко времени, переходному от третичного к четвертичному периоду (видимо,
верхний плиоцен), показали следующую картину. Сильно холмистую
местность, носившую уже, по-видимому, горный характер, покрывала
темнохвойная тайга. Среди пыльцы древесных пород преобладает пыльца
ели (Picea) — 59%. Далее следуют сосна (Pinus) — 7%, пихта (Abies) —
6%, кедр (Cedrus) — 5%, тсуга (Tsuga) — 5%, ликвидамбр (Liquidam-
Ьаг) — 2%, береза (Betula) — 1%, ольха (Alnus) — 1% и лещина (Согу-
lus) — 1%. Такое разнообразие пород, сочетание представителей родов,
произрастающих в разной экологической обстановке, может косвенно ука-
зать на существовавшую уже тогда разницу условий, вызванную поясно-
стью страны, т. е. на горный характер местности. На это же указывает
большое разнообразие хвойных пород, характерное в настоящее время
только для гор теплых стран северного полушария. О фауне, населяющей
эти горы, мы можем только гадать. Вполне вероятно, что здесь, на стыке
гималайских и сибирских таежных формаций, фауна носила смешанный
характер, но была явно таежного типа. Наличие пустынных и даже степных
форм при таком характере растительности исключается. Возможно, что
выше лесного пояса существовал уже субальпийский, а, может быть,
и альпийский пояс, которые в таком случае должны были отличаться
большой влажностью и напоминать современную альпику Гималаев.
Однако отсутствие в пробах пыльцы характерных субальпийских растений,
например рододендрона или арчи, что, правда, может быть и случайным,
наличие этих поясов не подтверждает.
Нижний плейстоцен. Продолжавшееся поднятие гор Па-
мира при высокой влажности климата привело к возникновению и быст-
рому развитию в конце описываемого периода мощного первого оледе-
нения, которое многие авторы (Марков, 1935; Чедия, 1962; Васильев,1962)
считают полупокровным или даже покровным. В последнем случае вся
жизнь на нагорье была бы уничтожена. Однако большинство современ-
ных данных говорит о том, что покровного оледенения все-таки не было
и в период максимального развития ледников оставались значительные
площади, свободные от снега и льда. Даже на юго-восточном Памире,
куда стекали гигантские ледники из крупного центра оледенения — тек-
тонического узла Шаарунг, разделяющего Памир и Тибет (толщина
ледника у Акташа была не менее 1 км), оледенение было полупокровного
типа (Рейман и Сидоров, 1962). В центральном Памире большая область
была свободна от ледников. Ото льда была свободна котловина оз. Ранг-
куль и окружающие ее горы, к югу вплоть до Аксу, горный массив между
нижним течением Аксу и верховьями Карасу и Истыка, а также значи-
тельные участки к югу от оз. Каракуль и в районе перевала Найзаташ.
В этих районах хребты имеют незначительную относительную высоту
и в настоящее время почти все расположены ниже снеговой линии. При-
нимая во внимание более или менее равномерный темп поднятия Памира
и окружающих его хребтов, можно полагать, что и в те времена централь-
ные части нагорья были ниже окраин, представляя собой сравнительно
ровные участки, напоминавшие современные тянь-шаньские сырты. Эти сво-
бодные от ледников центральные участки нагорья были в период первого
оледенения окружены гигантским ледяным кольцом, которое, по-видимому,
было сплошным. Непосредственная связь лесных поясов Тянь-Шаня
и Гималаев в районе Памира была, вероятно, нарушена, но лесная расти-
тельность могла оставаться кое-где в центре нагорья по днищам глубоких
долин и была более развита на внешних склонах нагорья. Альпийский пояс
- 110 —
должен был быть развит лучше в центральных частях Памира, а по окраи-
нам, вероятно, тянулся узкой полосой, выклиниваясь в тех местах, где
ледники вторгались прямо в лес. Подобные условия вряд ли способство-
вали продвижению тибетско-гималайских альпийских видов на и через
нагорье, это движение началось значительно позднее.
Средний плейстоцен. Отступание ледников, начавшееся
в конце нижнего плейстоцена, привело к тому, что большая часть нагорья
оказалась свободной ото льда. Судя по отложениям ленточных глин
в долине Аличура, межледниковый период, предшествовавший последнему,
по счету второму, оледенению, длился не менее 80 000 лет. В конечном
счете от начала межледникового периода нас отделяет примерно 120000 лет.
Судя по остаткам ископаемой пыльцы в ленточных глинах в долине ниж-
него течения Аличура (Акджар), климатические условия на всем протя-
жении межледниковой эпохи не оставались неизменными, а менялись до-
вольно значительно. Ландшафты нагорья заметно изменились. Продукты
разрушения гор и огромные морены погребли под собой большое число
долин и значительно выровняли поверхность. Во многих районах (Али-
чур, Салангур, Рангкуль и др.) морены, перегораживая долины, играли
роль запруд и создавали многочисленные ледниковые озера. Образование
большого количества озер на нагорье — наиболее характерная черта
межледниковой эпохи Памира. Некоторые из озер имели очень крупные
размеры. В частности, водная площадь Каракуля была по крайней мере
вдвое больше современной, а глубина на 100 м больше. Озеро было про-
точным и имело сток в долину р. Кокуйбеля. Немногим уступал ему
по размерам древний Рангкуль. Древний Чакыл-Агыркуль имел в длину
50 км, а глубину 60 м. Большое озеро покрывало значительную часть до-
лины Тагарма (Н. Т. Кузнецов, 1962).
Судя по многочисленным отпечаткам водных и околоводных растений
(Myriophyllum boamicum, Monocotyledonia gen. sp., Potamogeton sp.,
P. natanellus, Butomus sp., Butomus junceus, Bolbaschenus cempactus,
Equisetum sp. и др.), обнаруженным в ленточных глинах, озера имели
довольно богатую органическую жизнь, а следовательно и птичье насе-
ление.
Одновременно с отступанием ледников лесная растительность вновь
стала продвигаться внутрь Памира, и уже самые нижние слои глин межлед-
никового периода из долины р. Аличура дают большой процент (43—90%)
содержания пыльцы древесных пород. Судя по остаткам пыльцы, видовой
состав памирских лесов по сравнению с доледниковым периодом сильно
изменился. Преобладающими породами на все время межледниковья
становятся кедр, предположительно гималайский (Cedrus deodara), и сосна,
что свидетельствует о более умеренном и сухом климате. Об этом же
говорит малый процент пыльцы ели и почти полное отсутствие пыльцы
ликвидамбра и тсуги. Постепенное нарастание сухости, связанное с про-
должавшимся поднятием окраинных хребтов страны, очевидно, продолжа-
лось до середины межледниковья и привело к резкому сокращению лесного
пояса Памира. Возможно, это было вызвано значительным похолода-
нием, на что косвенно указывает заметно больший процент ели в средних
слоях глин межледникового периода. В этих слоях преобладает пыльца
кустарниковых и травянистых пород, от 62 до 87 %. Альпийский и субаль-
пийский пояса в это время уже должны были занимать значительные
площади и по-прежнему отличались влажным климатом, хотя местами
в субальпике уже могли появляться участки с небольшим увлажнением,
где развивалась растительность типа степной, а, может быть, и полупу-
стынной. Можно предполагать, что в период межледниковья и особенно
в последние его стадии, предшествовавшие второму оледенению, альпий-
ская фауна Центральной Азии была представлена на Памире гораздо
— 111
полнее, чем в настоящее время, и ряд ее представителей проник на Тянь-
Шань именно через Памир.
В конце межледникового периода влажность климата вновь начинает
расти, а параллельно с этим вновь большое развитие получает древесная
растительность (30—59% всей ископаемой пыльцы). Преобладающей
породой по-прежнему являлся кедр.
Верхний плейстоцен. Повышение атмосферной влажности
и прогрессивное похолодание климата Памира, вызванное непрекращаю-
щимся поднятием нагорья, привело к тому, что около 30 000 лет назад
значительная часть страны вновь оказалась покрытой мощными ледни-
ками. Тем не менее второе оледенение было слабее первого, и площадь,
свободная от фирна и льда, была по крайней мере в два раза больше той,
которая оставалась во время первого оледенения. Второе оледенение
имело несколько стадий наступления ледников, причем первая стадия
была самой мощной. Последняя, самая слабая стадия закончилась при-
мерно 6000 лет тому назад. Последовательное уменьшение размеров
стадий оледенений указывает на все возраставшую сухость климата.
Погребенные горизонты торфяников, располагавшихся на месте древних
озер и болот долины Аличура и хронологически относящихся к этому же
времени, свидетельствуют в свою очередь о резком похолодании климата.
Можно предположить, что в результате продолжавшегося поднятия
Памир примерно в это время оказался поднятым выше границ леса,
и лесной пояс совсем исчез с территории нагорья. Только в глубоких до-
линах должны были существовать леса тугайного типа, а на склонах гор
оставались арчевники.
Антропоген. Последний межстадиальный период второго оле-
денения не закончился до сих пор. Подавляющее большинство ледников
Памира отступает и в настоящее время, что вызвано непрекращающимся
иссушением климата. Можно определенно утверждать, что современные
климатические условия Памира гораздо суровее, чем в начале последнего
меж стадиального периода. В пользу этого свидетельствуют прежде всего
археологические находки (Ранов, 1961а, 19616; Бутомо и др., 1962).
Человек появился на высотах Памира вскоре после начала отступания
ледников последней стадии оледенения, в V—VI тысячелетиях до н. э.
Судя по остаткам стоянок, неолитические люди не жили здесь постоянно,
а приходили, вероятно, только в летнее время, привлеченные обилием
диких животных, главным образом архаров и козерогов, кости которых
в большом числе содержатся во всех культурных горизонтах неолита
и бронзы. На зиму охотники покидали Памир, спускаясь, по-видимому,
в теплые долины Кашгарии. Во всяком случае на неолитических и мезо-
литических стоянках Памира не было обнаружено и признака каких-
либо жилищ, а раскопанные в верховьях р. Маркансу очаги (стоянка
Ошхона) были открытого типа (Ранов и Сидоров, 1960). Это также сви-
детельствует о более теплом летнем климате нагорья, так как вряд ли
в настоящих условиях даже хорошо приспособленные к невзгодам неоли-
тические люди смогли бы жить здесь на открытых стоянках.
Древесная растительность тугайного типа из ив, берез, тополей и дру-
гих пород, а в ряде мест по склонам гор и арча просуществовали на Памире
вплоть до нашей эры. Об этом также говорят некоторые находки археоло-
гов. При раскопках на стоянке Ошхона, расположенной на высоте 4200 м,
были извлечены из кострищ угли березы, арчи и терескена, растений, ко-
торые и сейчас произрастают бок о бок в некоторых ущельях Бадахшана.
Абсолютный возраст угля 9530 + 130 лет. Логично предположить, что эти
растения произрастали в районе стоянки, а не транспортировались изда-
лека. В таком случае, учитывая, что верхний предел произрастания дре-
весных пород на определенной широте подвержен очень незначительным
— 112 —
колебаниям и что на данной широте он проходит сейчас на высоте 3500—
3600 м над ур. м., вполне вероятно, что эта местность поднялась за 9.5 ты-
сяч лет на 600—700 м (Бутомо и др., 1962).
Древесная растительность окончательно исчезла с территории нагорья,
по-видимому, на рубеже нашей эры, когда даже днища долин поднялись
выше верхней границы леса. Кустарниковая растительность сохранилась
в отдельных рефугиумах и до нашего времени, но при этом везде, где она
есть, наблюдаются ясные следы ее постепенной деградации, что указывает
на продолжающееся ухудшение климата.
Произрастание в прошлом хвойных лесов на Памире и соседних хреб-
тах, их постепенная деградация и исчезновение в связи с поднятием страны
п ухудшением климатической обстановки, более долгое существование
пояса арчи и его исчезновение по тем же причинам уже в историческое
время — все это хорошо подтверждает современное распространение
птиц, связанных с этими типами растительности. Прежде всего в этом отно-
шении показательно распространение кедровок. Обитающая в Кашмире
кедровка, Nucvjraga caryocatactes multipunctata Gould, могла попасть сюда
только из Тянь-Шаня, двигаясь по хвойным лесам Памиро-Алая. Это
станет ясным, если мы рассмотрим систематическое взаимоотношение
евразийских кедровок. Все подвиды N. caryocatactes L. отчетливо делятся
на две группы: более примитивную, с коротким, толстым клювом южную
группу (N. с. hemispila Vig., N. с. macella Thayer et Bangs, N. c. yunnanen-
sis Ingr., N. c. owstoni Ingr.) и более измененную северную (N. с. caryoca-
tactes L., N. c. macrorhynchos Brehm, N. c. wolfi Jord., N. c. relicta Reich.,
N. c. kamtschatkensis Bar., N. c. japonicus Hart., N. c. rothschildi Hart.).
Кашмирские кедровки явно относятся к северной группе и могли попасть
в Кашмир только с севера. После того как прямая связь хвойных лесов
Гималаев и Тянь-Шаня прервалась, в Кашмире успела образоваться
особая, хорошо обособленная форма. Распространяясь постепенно дальше
к юго-востоку, по хвойному поясу Гималаев, кашмирские кедровки встре-
тились с южными N. с. hemispila. В районе совместного обитания в цент-
ральных Гималаях эти кедровки обитают на разных высотах. Логично
предположить, что кашмирские кедровки, как более северные, будут
занимать большие высоты, а гималайские, как более южные, — меньшие.
Так и наблюдается на самом деле (Baker, 1934). При этом нижняя граница
кашмирского и верхняя гималайского подвидов перекрывается. На опре-
деленных высотах эти подвиды обитают совместно, помесей, насколько
известно, не образуют и, по существу, должны рассматриваться как
подвиды разных видов кедровок. Центральные Гималаи — единствен-
ное место, где перекрываются ареалы северной и южной групп кедро-
вок.
Как уже говорилось выше, северные кедровки могли проникнуть
в Кашмир только через хвойные леса Памиро-Алая, несомненно сущест-
вовавшие в те времена. Иначе трудно допустить, чтобы эти узко специали-
зированные птицы, вся жизнь которых связана с хвойными лесами,
отважились бы на длительное путешествие через обширные безлесные вы-
сокогорья, совершенно чуждые им экологически. Маловероятность та-
кого предположения подтверждает тот факт, что за почти столетнюю исто-
рию изучения орнитофауны Памиро-Алая кедровки здесь ни разу не на-
блюдались и не добывались.
Распространение птиц через хвойные леса Памиро-Алая шло не только
с севера на юг, но и в обратном направлении. Есть основания предполагать,
что королек — Regulus regulus tristis Pleske — попал на Тянь-Шань из
лесов Гималаев (Штегман, 1946); с такой же долей вероятности это можно
предполагать и для черной синицы — Parus ater rujipectus Sev., и для
клеста-ел овика — Loxia curvirostra tianschanica Laubm.
8 Биология птиц
— ИЗ —
На восточных склонах Памирского нагорья, в хребтах Кингтау, Каш-
гарском и в бассейне верховьев р. Тизнафа, и сейчас имеются массивы
хвойного леса, которые образуют ясно выраженный пояс только в горах
Кингтау, а дальше к югу встречаются небольшими изолированными уча-
стками. Наиболее изолирован самый южный участок — ельники Тизнафа.
Обитающая здесь рыжешейная синица — Parus rubidiventris rufonuchalis
Blyth — могла попасть сюда только из лесов Тянь-Шаня и только в том
случае, если связь хвойных лесов Тизнафа и Тянь-Шаня была прямой.
Несмотря на значительную изоляцию в настоящее время, тизнафская
популяция рыжешейной синицы ничем не отличается от номинального
подвида, населяющего Тянь-Шань. Это говорит о том, что изоляция не
столь давняя и что сплошной пояс хвойных лесов по восточным склонам
Памира перестал существовать также не очень давно.
Ряд птиц, связанных с арчовыми зарослями, проник из Кашмира
в Ал ай и Тянь-Шань [Mycerobas carnipes speculigerus (Brandt), C er thia
himalayana taeniura Sev., Parus rubidiventris rujonuchalis (Blyth), Phoeni-
curus coeruleocephalus Vig., Calliope pectoralis ballioni Sev.] и наоборот
(Leptopoecile sophiae sophiae Sev.). Это опять же могло иметь место только
в том случае, если бы арчовые насаждения образовывали непрерывный
пояс от Гималаев до Тянь-Шаня. Сейчас этот пояс имеет значительный
разрыв на территории Памира и Бадахшана. Поскольку в данном случае
мы имеем разрыв ареалов одних и тех же подвидов или же между подви-
дами, едва различимыми между собой, ясно, что разрыв этот недавнего
происхождения.
Параллельно с исчезновением древесно-кустарниковой растительности
шли и другие важные изменения в растительном покрове нагорья. Вплоть
до начала нашей эры на Памире широко была распространена раститель-
ность степного и лугового типа, но в результате дальнейшего усыхания,
за последние два тысячелетия степи почти исчезли с высот Памира, а пло-
щади лугов сильно сократились. Небольшие участки степной раститель-
ности, сохранившиеся в ограниченном числе мест, сейчас всюду являются
индикаторами большей увлажненности и наличия зимой снегового по-
крова, без которого такая растительность в условиях Памира вымер-
зала бы зимой.
Большой интерес представляет находка неолитических рисунков
в гроте Шахты (Ранов. 19616), о чем уже упоминалось (см. стр. 33). В со-
хранившейся части рисунка, изображавшего облавную охоту, ясно раз-
личаются изображения медведя и двух кабанов, поражаемых стрелами.
Ближайшие места, где водятся кабаны, отстоят от грота Шахты на сотни
километров. Вряд ли в те времена племена могли кочевать так широко,
и еще менее вероятно, чтобы они столь живо изображали сценки охоты на
зверей, которых уже давно не видели. Вероятнее предположить, что кабаны
в те времена водились в непосредственной близости от грота Шахты, про-
никая сюда из южного Таджикистана по тугайным зарослям долины Али-
чура—Гунта. В сцене охоты обращает на себя внимание странная фигурка
охотника, которую можно рассматривать как изображение человека, за-
маскированного под страуса. Такая трактовка сугубо предположительна,
но вполне вероятно, что неолитические охотники, зимовавшие на равни-
нах Кашгарии, знали эту птицу, которая могла дожить в пустынях Цент-
ральной Азии до исторических времен (Lowe, 1931).
Ископаемых остатков птиц на Памире до сих пор обнаружено очень
мало. Это, во-первых, плечевые кости какой-то крановой птицы и утки,
а также бедренная кость перепела и плечевая — стрепета, найденные
среди костных остатков неолитической стоянки Ошхона в долине Мар-
кансу; во-вторых, — бедренная кость перепела и нижняя челюсть пустель-
ги, извлеченные из слоя бронзового века в гроте Шахты.
- 114 —
В свете изложенных фактов и предположений общий ход изменений
орнитофауны Памира в антропогене можно представить следующим об-
разом. В период, последовавший за вторым оледенением, климат которого
был мягче и влажнее, чем сейчас, орнитофауна нагорья была гораздо
богаче и существенно отличалась от современной. Здесь, с одной стороны,
гнездился целый ряд птиц, связанных с древесно-кустарниковой расти-
тельностью, в частности с арчой. Гораздо богаче должна была быть пред-
ставлена группа альпийских видов, обитающих ныне только по окраинным
хребтам нагорья или вообще здесь не встречающихся (белогрудый голубь,
красный вьюрок, альпийская и гималайская завирушки и др.). Можно
считать, что эти птицы вместе с горной коноплянкой, горным вьюрком,
большой чечевицей и некоторыми другими видами были наиболее древними
обитателями нагорья. Вообще орнитофауна Памира в период, следовавший
непосредственно за последним оледенением, весьма напоминала современ-
ную орнитофауну Тянь-Шаня, только примесь гималайских форм должна
была быть более значительной. Позднее всех Памир должны были засе-
лить такие ксерофильные формы, как тибетская саджа, пустынная ка-
менка или пустынный снегирь. Параллельно с этими изменениями, вместе
с деградацией и исчезновением сначала древесной, а потом и кустарниковой
растительности постепенно исчезали древесно-кустарниковые птицы, за
исключением тех, которые сумели приспособиться к новым условиям и
перейти на гнездование в камнях и скалах. С ростом сухости климата,
с уменьшением осадков исчезли многие влаголюбивые виды, в частности
ряд альпийских птиц. Можно предполагать, что в связи с продолжаю-
щимся поднятием нагорья условия существования здесь продолжают
ухудшаться, а вместе с этим становится беднее и орнитофауна.
Резюмируя вышеизложенное, можно сказать, что Памир превратился
в холодную высокогорную пустыню сравнительно недавно. Изменения
климата, вызвавшие этот процесс, были обусловлены не прекратившимся
до сих пор поднятием как самого нагорья, так и окружающих его горных
систем, перехватывавших большую часть осадков. О молодости современ-
ной памирской орнитофауны, являющейся дериватом тибетской и занимаю-
щей, как уже показывалось, островное, изолированное от Тибета поло-
жение, свидетельствует полное отсутствие видовых эндемиков и ничтожно
малый процент эндемичных подвидов, что вряд ли могло иметь место,
если бы изоляция была более длительной.
ЛИТЕРАТУРА
Абдусалямов И. А. 1957а. Новые данные по птицам Восточного Памира.
Докл. АН Тадж. ССР, 22 35—37.
Абдусалямов И. А. 19576. Материалы по биологии гималайского и тибетского
уларов на Памире. Тр. Тадж. унив., сер естеств. наук, 2 179—187.
Абдусалямов И. А. 1958а. К экологии птиц Ранг-Кульской долины. Изв. Отд,
естеств. наук АН Тадж. ССР, 1 115—119.
Абдусалямов И. А. 19586. Некоторые данные по экологии горного гуся Eula-
beia indica Latham на Памире. Тр. АН Тадж. ССР, LXXX1X : 233—239.
Абдусалямов И. А. 1959а. Распределение птиц Памира по биотопам. Тр.
Узбекск. унпв., нов. сер., 88 73—81.
Абдусалямов И. А 19596. К биологии тибетской саджи. Орнитология, 2 218—
220.
Абдусалямов И. А. 1961. Птицы долины озера Ранг-Куль на Памире. Тр.
Инет. зоол. и наразитол. АН Тадж. ССР, 21 : 1—151.
Абдусалямов И. А. 1962. О гнездовании красного вьюрка на Памире. Орнито-
логия, 5 249—250.
Банников А. Г. п П. П. Т а р а с о в. 1957. К биологии огаря (Tadorna ferru-
ginea Pall.). Тр. Бюро кольцев., 9 208—214.
Бёме Р. Л. 1963. Опыт орнитогеографнческого районирования высокогорий Палеарк-
тики. Орнитология, 6 227—234.
Берг Л. С. 1931. Рыбы. Тр. Памирск. экснед. 1928 г., VIII, Зоология 23—28.
- 115 -
8*
Бианки В. Л 1907. Материалы для авифауны Монголии и восточного Тибета.
Тр экспед. Русск. геогр. общ., соверш. в 1899—1901 гг., V : 1—251.
Б у т о м о С. В., В. А. Ранов, Л. Ф. С и д о р о в и И. А. Ш и л к и н а. 1962.
Палеогеографические результаты изучения высокогорной стоянки каменного
века на Памире. Докл. АН СССР, 146, № 6 : 1380—1382.
Васильев В. А. 1962. Стратиграфия четвертичных отложений Таджикистана.
В сб.: Новейший этап геологического развития территории Таджикистана,
Душанбе : 5—34.
Виноградов Б. С., Е. Н. Павловский и К. К. Флеров 1935. Звери
Таджикистана. Тр. Тадж. базы АН СССР, I : 1—276.
Гвоздецкий Н. А. 1959. О географическом понятии «Памиро-Алай» и расчле-
нении гор Средней Азии на горные системы В сб.: Вопросы физической геогра-
фии СССР, изд. Моск, унив.: 138—142.
Г е д и н С. 1899. В сердце Азии. СПб.
Головнин Д. Н. 1901. Очерки охоты на Памирах. VI Охота на Памирах по перу.
Природа и охота, IX 1—16.
Громов И. М. и О. В. Егоров. 1953. Материалы по питанию и хозяйственному
значению филина Восточного Памира и Копет-Дэга. Зоол. журн., XXXII,
5 : 964—978.
Гурвич В. Ф 1957. Фауна озера Каракуль и ее формирование. Дисс., Ташкентск.
унив.
Дементьев Г. П. 1935. Материалы по авифауне Памира. Тр Таджикско-Памирск.
экспед. АН СССР, XXXII, Л. : 61—95.
Дементьев Г. П. 1938. К вопросу о взаимоотношениях Палеарктической и Восточ-
ной областей на примере туркестанской и индийской авифауны. Бюлл. Моск,
общ. испыт. природы, Отд. биол., XLVII, 5—6 : 343—356.
Забиров Р. Д. 1955. Оледенение Памира. М.
Зарудный Н. А. 1900. Экскурсия по северо-восточной Персии и птицы этой
страны. Зап. АН, сер. VIII, X, 1 1—262.
Иванов А. И. 1940. Птицы Таджикистана. Тр. Тадж. базы АН СССР, X : 1—300.
Иванов А. И. 1949. Зимняя орнитофауна окрестностей Душанбе. Тр. Тадж. фил
АН СССР, XIX : 49—72.
Иванов А. И. 1950. Зимовки и пролет птиц на территории Памиро-Алая. в сб.:
Памяти акад. П. П. Сушкина, Л. : 266—289.
Иванов Д. Л. 1885. Охота на Памире. Природа и охота, 1 1—48.
Капитонов В. И. и Ф. Б. Чернявский. 1960. Воробьиные птицы низовий
Лены. Орнитология, 3 : 80—97.
Кистяковский А. Б. 1950. Материалы по зоогеографии Памира. Научн. зап.
Киевск. унив., IX, 6 5—58.
Козлов П. К. 1899. Маршрут Тибетской экспедиции М. В. Певцова (1889—1891 гг.)
п географическое распределение млекопитающих и птиц в местностях, исследо-
ванных ею. Ежегодн. Зоол. музея Акад, наук, 4 257—287.
Козлова Е. В. 1930. Птицы юго-западного Забайкалья, северной Монголии и
Центральной Гоби. Материалы Комисе, по исслед. Монгольск. и Тувинск. Нар.
Республ. АН СССР, 12 1—396.
Козлова Е. В. 1952 Авифауна Тибетского нагорья, ее родственные связи и исто-
рия. Тр. Зоол. инет. АН СССР, IX, 4 : 964—1028.
Козлова Е. В. 1961. Кулики, ч. 1. Фауна СССР, нов. сер., 80 (Птицы, II, 1, ч. 2) :
1—500.
Кузнецов А. А. 1960. О кашмирском красном вьюрке. Орнитология, 3 351—361.
Кузнецов А. А. 1962. Состав, численность и размещение авифауны высокогорья
Киргизского хребта. Орнитология, 4 : 351—361.
Кузнецов Н. Т. 1962. Древние озера Центральной Азии. Природа, 1 : 108—
110.
Кузьмина М. А. 1955. Материалы по экологии и морфологии темнобрюхого улара
и кеклика. Зоол. журн., XXXIV, 1 : 175—190.
Кузьмина М. и £. Ф. С а в ино в. 1953. Материалы по экологии темнобрю-
хого улара (Tetraogallus himalayensis sewertzowi Zar.) в Заилийском Алатау.
Зоол. журн., XXXII, 6 : 1234—1240.
Кыдыралиев А. 1959. Размножение краснобрюхой горихвостки на Централь-
ном Тянь-Шане. Орнитология, 2 : 209—213.
Кыдыралиев А. 1962. Бледная завирушка в Центральном Тянь-Шане. Орнито-
логия, 4 : 256—259.
Мальчевский А. С. 1959. Гнездовая жизнь певчих птиц. Л.
Марков К. К. 1935. История рельефа и оледенения Памира. В сб.: Пять лет по
Памиру, М.—Л. : 251—324.
Мекленбурцев Р. Н. 1936. Материалы по млекопитающим и птицам Памира.
Тр. Среднеазиатск. гос. унив., сер. Villa, Зоология, 22 : 1—40.
Мекленбурцев Р. Н. 1937. Заметки о новых и редких для Памира видах птиц.
Бюлл. Среднеазиатск. гос. унив., Биология, 22 : 285—290.
— 116 —
Мекленбурцев Р. Н. 1946. Зимующие и пролетные птицы Восточного Памира.
Бюлл. Моск. общ. испыт. природы, Отд. биол., LI, 1 87—110.
Мекленбурцев Р. Н. 1949. Об орнитологических взаимоотношениях между
Восточным и Западным Памиром. Бюлл. Среднеазиатск. гос. унив., Биология,
28 : 157—177.
Мензбир М. А. 1914. Зоологические участки Туркестанского края и вероятное
происхождение фауны последнего. Прилож. 4-е к Временнику Общ. содейств.
успехам опытн. наук им. X. С. Леденцова, М., 1 144.
Молчанов Л. А. и Н. А. Зарудный. 1915 К авифауне Памира. Ежегодн.
Зоол. музея Акад, наук, 19 : 439—477.
Пахомов М. М. 1962. Первые результаты палинологических исследований кайно-
зойских отложений Памира. В сб.: Новейший этап геологического развития
территории Таджикистана, Душанбе : 59—68.
Попов А. В. 1958. Питание птиц в Бадахшане плохо заделанными в почву семенами
пшеницы. Тр. АН Тадж. ССР, LXXXIX : 209—215.
Попов А. В. 1959. Птицы Гиссаро-Каратегина. Душанбе.
По ртенк о Л. А. 1960. Воробьиные. Птицы СССР, ч. IV, Опред. по фауне СССР,
изд. Зоол. инет. АН СССР, 69 : 1—419.
Потапов Р Л. 1956. К орнитофауне Памира. Докл. АН Тадж. ССР, 15 57—62.
Потапов Р. Л. 1957. Новые орнитологические находки в Таджикистане. Докл.
АН Тадж. ССР, 22 39—41.
Потапов Р. Л. 1958. О пролете птиц на Памире осенью 1956 г. Тр. АН Тадж. ССР,
LXXXIX : 195—208.
Потапов Р. Л. 1962а. Гнездование пустельги (Cerchneis tinnunculus tinnunculus L.)
на Памире. Зоол. журн., XLI, 8 : 1265—1266.
Потапов Р. Л. 19626. Особенности гнездования птиц на Памире. Матер.
III Всесоюзн. орнитол. конфер., Львов, 2 : 147—148 (тезисы).
Потапов Р. Л. 1963. О биологии горного вьюрка (Leucosticte arctoa pamirensis
Sev.) на Памире. Зоол. журн., XLII, 5 : 716—726.
Пржевальский Н. М. 1876. Монголия и страна тангутов, т. II. СПб.
Пржевальский Н. М. 1883. Из Зайсана через Хами в Тибет и на верховья
Желтой реки. СПб.
Ран о в В А. 1961а. Изучение памятников каменного века на Восточном Памире
в 1958 г. Тр. Инет, истории АН Тадж. ССР, XXVII : 19—35.
Ран о в В. А. 19616. Рисунки каменного века в гроте Шахты. Сов. этнография,
6 : 70—81.
Ранов В. А. иЛ. Ф. Сидоров. 1960. К вопросу об изменениях природных усло-
вий Памира в голоцене. Докл. АН Тадж. ССР, III, 1 : 21—24.
Рейман В. М. иЛ. Ф. Сидоров. 1962. О древнем оледенении юго-восточного
Памира. Докл. АН СССР, 147, 2 : 452—453.
Северцов Н. А. 1873. Вертикальное и горизонтальное распределение Туркестан-
ских животных. Изв. Общ. любит, естествозн., антропол. и этногр., VIII,
2 : 1—155.
Северцов Н. А. 1877. О зоологических (преимущественно орнитологических)
областях внетропических частей нашего материка. Изв. Русск. геогр. общ.,
XIII, 3 : 125-155.
Северцов Н. А. 1879. Заметки о фауне позвоночных Памира. Зап. Турк. отд.
Общ. любит, естествозн., антропол. и этногр., I, 1 : 58—89.
Северцов Н. А. 1880а. Об орографическом образовании Высокой Азии п его
значении для распространения животных. Речи и протоколы VI Съезда русск.
естествоиспыт. и врачей, СПб. : 94—118.
(Северцов Н.) Severtzow N. 18806. Etudes sur le passage des oiseaux dans
1’Asie Centrale, particulierement par le Ferghana et le Pamir. Bull. Soc. natur.
Moscou, LV, 1 : 234—287.
(Северцов H.) Severtzow N. 1883. On the birds of the Pamir range.
Ibis : 48—83.
Сидоров Л. Ф. 1959. К вопросу о древнем оледенении Памира. ДАН АН СССР,
127, 4 : 860—861.
Сидоров Л. Ф. 1960. Луга Памира. Дисс. Л., Ботанич. инет. АН СССР.
Синицын В. М. 1959. Центральная Азия. М.
(Спангенберг Е. иДементьев T.)Spangenberg Е. et G. Demen-
ti е f f 1935. Contribution a 1’avifaune du Tian-Chan occidental (Monts Ale-
xandrowski). Alauda, 7 : 336—366.
Станюкович К. В. 1952. Еще раз о том, что называть Памиром. Изв. Всесоюзн.
геогр. общ., 84 : 407—409.
Станюкович К. В. 1560. Растительность высокогорий СССР, т. I. Душанбе.
Степанян Л. С. 1959. Некоторые особенности размножения птиц в высокогорье
Тянь-Шаня. Бюлл. Моск. общ. испыт. природы, Отд. биол., LXIV, 2 : 126—128.
Степанян Л. С. 1960. Размещение авифаунпстических комплексов в Центральном
Тянь-Шане. Орнитология, 3 366—376
— 117 —
С у д и л о в с к а я А. М. 1936. Птицы Кашгарии (преимущественно по сборам
М. Н. Дивногорского). М.
Сушкин П. П. 1925. Зоологические области Средней Сибири и ближайших частей
Нагорной Азии и опыт истории современной фауны Палеарктической Азии.
Бюлл. Моск. общ. испыт. природы, Отд. биол., XXXIV : 7—86.
Сушкин П. П. 1938. Птицы Советского Алтая. Л.
Тристан Д. Ф. и А. Г. 3 в е с к и н. 1960. О биологии горного гуся (Eulabeia
indica Latham) в Тянь-Шане. Зоол. журн., XXXIX, 1 : 145—146.
Трофимов А. К. 1962. О возрасте и истории развития древних оледенений За-
падного и юго-восточного Памира. В сб.: Новейший этап геологического раз-
вития территории Таджикистана, Душанбе : 253—282.
Тугаринов А. Я. 1930. Птицы. Тр. Памирск. экспед. 1928 г., II, Зоология 83—
86.
Ч е д и я О. К. 1962 История геологического развития территории Таджикистана
в кайнозое. В сб.: Новейший этап геологического развития территории Таджи-
кистана, Душанбе : 283—309.
Шест оперов Е. Л. 1929. Материалы для орнитофауны Илийского края. Бюлл.
Моск, общ испыт. природы, Отд. биол., XXXVIII, 1—2 154—248.
Шнитников В. Н. 1949. Птицы Семиречья. М.
Штегман Б. К. 1938. Основы орнитогеографического деления Палеарктики.
Фауна СССР, нов. сер., 19 : 1—157
Штегман Б. К. 1946. Основные типы авифауны Тянь-Шаня и их генезис. Вести.
АН Казахск. ССР, 7—8 : 36—43.
Штегман Б. К. 1954. О птицах высокогорной зоны Заилийского Алатау. Тр. Лен
общ. естествоиспыт., СХХП, 4:255—276.
Штегман Б. К. 1955. К вопросу об адаптивных особенностях и филогенетических
соотношениях вороновых птиц. Зоол. журн., XXXIV, 6 : 1357—1378.
Ш те г м а н Б. К. 1957. Жизнь в субнивальной зоне. Природа, 10 : 93—95.
Юнатов А А. 1961. К познанию растительного покрова западного Куньлуня и
прилегающей части Таримской впадины. В кн.: Куньлунь и Тарим. Очерки
природных условий. М. : 133—153.
Янушевич А. И. 1955. Горный гусь. Природа, 3 112—ИЗ.
Янушевич А. И., П. С. Тюрин, И. Д. Яковлева, А. Кыдыралиев
иН. Н.Семенова. 1959—1961. Птицы Киргизии. Фрунзе, I, 1959; II, 1960;
III, 1961.
Akhtar S. А. 1955. Bird migration and fowling in Afghanistan. Journ. Bomb. Nat.
Hist. Soc., 53, 1 : 49—53.
A 1 с о c k A. W. 1897. Report upon the natural hisotry of the Pamir Boundary Comis-
sion Birds. Report on the proc, of the Pamir Boundary Comission, VIII : 55—99.
A 1 i S. 1946. An ornithological pilgrimage to lake Manosarovar and mount Kailas.
Journ. Bomb. Nat. Hist. Soc., 46, 2 : 286—308.
Baker S. 1922—1930. The fauna of British India. Birds. 2-d ed. London, I, 1922;
II, 1923; III, 1924; IV, 1925; V, 1926; VI, 1927; VII, 1928; VIII, 1930.
Baker S. 1932—1935. The nifidication of birds of the Indian Empire. London, I, 1932;
II, 1933; III, 1934; IV, 1935.
В г о 1 e у C. L. 1947. Migration and nesting of the Florida Bald Eagles. Wilson Bull.,
59 2 : 3_______20
C h i Y. L. 1961. Bird island. OrnithoL Mitteil., 13, 12 : 22—224.
D о r s t J. 1957. La vie sur les hauts plateaux andins de Perou. Terre et Vie, 104 : 3—50.
D о r s t J. 1962. Aspects of bird ecology in the Peruvian high Andes. Abstracts of pa-
pers, XIII Intern. Ornith. Congress, Ithaca : 15—16.
French N. R. 1959. Life history of the Rosy Black Finch Auk, 76 : 159—180.
Kingston R. W. 1927; Bird notes from the Mount Everest expedition of 1924.
Journ. Bomb. Nat. Hist. Soc., 32, 1—2 : 320—329
L a m p г e у H. F. 1954. Birds seen above the tree-line in Tehri-Garwal in the Central
Himalayas. Journ. Bomb. Nat. Hist. Soc., 52, 2—3 : 610—615.
Lowe P. R. 1931. Struthious remains from China and Mongolia, with descriptions
of Struthio wimani, Struthio anderssoni, and Struthio mongolicus spp. nov. Pa-
laeontol. Sinica, ser. С, VI, 4 : 1—40.
Ludlow F., N. В. К i n n e a r. 1933—1934. A contribution to the ornihtology of
Chinese Turkestan. Ibis : 240—259, 440—473, 658—694, 95—125.
Maclaren P. I. R. 1948. Notes on the birds of the Gyantse road, southern Tibet,
May 1946. Journ. Bomb. Nat. Hist. Soc., 47, 2 : 301—308.
Mani M. S. 1962. Introduction to high altitude entomology. Insect life above the tim-
ber line in the North-West Himalaya. London.
Meinertzhagen R. 1927. Ladakh, with special reference to its natural history.
Geograph. Journ., LXX, 2 : 129—163.
Meinertzhagen R. 1954. Birds of Arabia. London.
Osmas ton B. R. 1927—1928. Notes on the birds of Kashmir. Journ. Bomb. Nat.
Hist. Soc., 31, 4, 1927 975—999; 32, 1, 1928 134—153.
— 118 —
Paludan К 1959. On the birds of Afghanistan. Kobenhavn.
Pearson О. P. 1953. Use of caves by hummingbirds and other species at high alti-
tudes in Peru. Condor, 55 : 17—20
R e i n i g W. F. 1930. Versuch einer zoogeographischen Gliederung des Pamir Gebietes.
Zool. Jahrb., Abt. System., Okol , Geograph, d. Tiere, 59, 4 : 447—504.
Richmond C. W. 1896. Catalogue of a collection of birds, made by Dr. Abbott in
Eastern Turkestan, the Tian-Shan mountains and Tagdumbash Pamir, Central
Asia, with notes on some of the species. Proc. U. S. Nat. Mus., XVIII : 569—591.
Schafer E. 1938. Ornithologische Ergebnisse zweier Forschungsreisen nacb Tibet.
Journ. f. Ornithol. Sondrheft, 86 : 1—349.
Sharpe R. B. 1891. Scientific results of the Second Yarkand mission. Aves. London :
1-153.
Sh i p t о n E. 1938. The Shaksgam Expedition, 1937. Geograhp Journ., XCI, 4 : 313—
339.
Singh G 1961. The eastern steppe Eagle [Aquila nepalensis nepalensis (Hodgs)j
on the south col of Everest. Journ. Bomb. Nat. Hist. Soc., 58, 1 : 270.
Whistler H. 1925. The birds of Lahul, N -W. Himalaya. Ibis : 152—208.
И. А. Нейфельдт
ЛЕСНЫЕ «КАМЕННЫЕ» ДРОЗДЫ
I. A. N е u f е 1 d t. The Wood «Rock»-Thrushes
СОДЕРЖАНИЕ
Стр.
Введение 120
Современное распространение лесных «каменных» дроздов 124
Повадки и образ жизни лесных «каменных» дроздов 135
Основные черты внешней морфологии лесных «каменных» дроздов 148
О родовой самостоятельности лесных «каменных» дроздов 175
Вероятная история рода Orocetes 176
ВВЕДЕНИЕ
Лесные «каменные» дрозды — это Monticola erythrogaster (рыжебрю-
хий дрозд), М. cinclorhynchus (синегорлый дрозд) и М. gularis (белогорлый
Дрозд).
Два первых вида описаны в 1831 г. по экземплярам из Гималаев Ви-
горсом (Vigors) соответственно как Turdus erythrogastra и Petrocincla,
cinclorhyncha. Через два года после этого (Jardine a. Selby, 1833) появилось
первое изображение рыжебрюхого дрозда, переименованного авторами
в Petrocincla rufwentris. Оба лесных вида оказались, таким образом, объ-
единенными вместе с описанными еще Линнеем скальными формами sa-
xatilis и solitarius. В 1837 г. Свайнсон (Swainson) выделил синегорлого
дрозда в особый род Petrophila, а спустя три года Грей (Gray, 1840) пред-
ложил для той же формы родовое название Orocetes, которое на протяжении
последующих сорока лет весьма широко использовалось при обозначении
лесных видов и только однажды (Horsfield a. Moore, 1854) было применено
к пестрому каменному дрозду (М. saxatilis).
Третий лесной вид — белогорлый дрозд — стал известен в науке зна-
чительно позднее. Первоначальное его описание принадлежит Свайно
(Swinhoe, 1862), присвоившему птице, добытой в окрестностях Пекина,
название Orocetes gularis и тем самым подчеркнувшему ее близость не
к скальным, а к лесным формам. В работе 1863 г. (Swinhoe, 1863а), снаб-
женной, кстати, очень удачно выполненным цветным рисунком самца
белогорлого дрозда, Свайно уже вполне определенно говорит о тесном род-
стве О. gularis и О. cinclorhynchus, проявляющемся не только в сходстве
окраски их оперения, но и в особенностях образа жизни этих птиц.
Необходимо подчеркнуть, что в 40 —80-х годах прошлого столетия
лишь немногие исследователи понимали род «Каменные» дрозды очень
широко или же, наоборот (Blyth, 1849), совершенно произвольно, осно-
— 120 —
вываясь на каких-то формальных признаках, группировали виды в так-
сономические единицы более высокого ранга. Большинство же системати-
ков этого периода без специальной аргументации и в значительной сте-
пени интуитивно совершенно справедливо выделяло восточноазиатские
лесные формы в самостоятельный род. К сожалению, их точка зрения не
получила своего дальнейшего развития и была забыта с выходом в свет
пятого тома «Каталога птиц Британского музея», в котором Сибом (See-
bohm, 1881) признал существование только одного обширного рода Mon-
ticola Boie, 1822. В орнитологии утвердилось представление о «каменных»
дроздах как о единой монолитной группе, включающей наряду с действи-
тельно скальными ксерофильными формами, живущими на юге Евразии
п в Африке, три восточноазиатских лесных вида. Относительно же поло-
жения М. gularis, М. cinclorhynchus и М. erythrogaster в системе, степени
пх родства между собой и генетических связей с остальными Monticola
мнения специалистов неоднократно менялись.
Сибом, например, так представлял себе структуру этого рода: М. sa-
xatilis, М. cyanus, М. solitaria, М. cinclorhyncha, М. rupestris. М. exp-
loratory М. breripes, М. erythrogaster, М. gularis, М. rujocinerea.
Интересующие нас лесные азиатские формы оказались в его схеме изоли-
рованными друг от друга, вклинившимися между ними африканскими
эндемиками.
Вариант системы, опубликованный Шарпом (Sharpe, 1903), в общем
очень близок к предыдущему, с той лишь разницей, что пестрый камен-
ный дрозд рассматривается в нем в качестве самостоятельного рода.
Хартерт (Hartert, 1910) отводит М. gularis место между М. saxatilis
и М. solitarius. Сходным образом сгруппированы виды и в работах ряда
советских орнитологов (Дементьев, 1937; Портенко, 1954).
Сравнивая М. gularis и М. cinclorhynchus, Мейзе (Meise, 1934) пришел
к заключению, что различия между ними лишь подвидового ранга. Не
углубляясь в вопросы филогении и руководствуясь только тем, что сине-
горлый дрозд был описан раньше, чем белогорлый, Мейзе впервые трак-
тует М. cinclorhynchus как политппический вид, с чем вряд ли можно
согласиться.
Во время своих экспедиций в Приморский край Л. М. Шульпин собрал
большой материал по биологии белогорлого дрозда — вида, до того вре-
мени совершенно не изученного. Ему, правда, не удалось найти гнезда и
описать еще неизвестные яйца и пуховичков этой птицы, но он был первым
исследователем, детально познакомившим нас с ее образом жизни и повад-
ками на гнездовье, а также, что особенно ценно, попытавшимся привлечь
все известные в ту пору фактические данные по распространению, эколо-
гии и внешней морфологии к рассмотрению вопроса об истории группы
«каменных» дроздов восточной Азии. Нет надобности излагать здесь под-
робно содержание статьи Шульпина (1931). Достаточно будет напомнить
лишь те основные выводы, которые смогут понадобиться нам в дальней-
шем. Сравнивая особенности окраски и рисунка оперения молодых в гнез-
довом наряде, а также взрослых самок трех лесных Monticola, Шульпин
пришел к верному заключению о близком родстве этих форм между собой.
Самцы восточноазиатских подвидов синих каменных дроздов, как известно,
внешне похожи на самцов рыжебрюхого дрозда. Не усмотрев в этом сход-
стве конвергенции, Шульпин использовал его в качестве основы, на ко-
торой якобы возможно объединение скальных и лесных форм. «М. gula-
ris— cinelorhyncha, М. solitaria и М. erythrogastra разошлись все из одного
фокуса; при этом первые далее уклонились, оставшись в лесной среде,
М, solitaria сильно отошли и видоизменились в иной экологический тип,
и лишь М. erythrogastra недалеко ушел от исходного типа» (Шульпин,
1931 : 283). Цитируемый автор, таким образом, считает исходной, претер-
- 121 -
певшей в процессе эволюции наименьшие изменения окраску рыжебрю-
хого дрозда, а филогенетически наиболее молодой — окраску типа
Л/. gularis—М. cinclorhynchus. Как будет показано ниже, анализ возраст-
ных и сезонных нарядов лесных «каменных» дроздов убеждает нас в обрат-
ном. Что же до остальных представителей принимаемого ныне рода Моп-
ticola, то Шульпин, специально ими не занимавшийся, высказал только
предположение о том, что все африканцы стоят ближе к пестрому камен-
ному дрозду.
В противоположность старым схемам, подразделявшим род Monti-
cola на 10 монотипических видов или на 8 видов, из коих 2 [solitarius и
cinclorhynchus) образовывали подвиды, система Р. Мейнерцхагена (R. Mei-
nertzhagen, 1951), базирующаяся на широкой концепции вида, признает
существование только 4 видов, включающих 14 форм. Это М. saxatilis
saxatilis, М. s. rufocinerea, М. s. sclateri, M. s. brevipes, M. s. explorator,
M. rupestris; M. gularis gularis, M. g. cinclorhyncha,
M. g. angolensis; M. solitarius solitarius, M. s. longirostris, M. s. pandoo,
M. s. philippensis и M. s. erythroyastra. Как видно, группа лесных
«каменных» дроздов оказалась расчлененной; ее отдельные представители
истолковываются как подвиды двух разных видов. Белогорлый и сине-
горлый дрозды — две близкие формы вида gularis, а не cinclorhynchus,
и в этом отношении система Мейнерцхагена представляет собой некоторый
шаг вперед по сравнению, например, со схемой Мейзе. Однако в тот же
вид gularis ошибочно переведен и имеющий с ним очень мало общего южно-
африканский angolensis. Что же касается остальных африканских камен-
ных дроздов, то всем им, кроме М. rupestris, отведено место подвидов
М. saxatilis и расположены они в порядке, вряд ли отражающем их фило-
гению. Рыжебрюхого дрозда Мейнерцхаген довольно смело сближает
с синими каменными дроздами и, считая его, по-видимому, наиболее моло-
дым среди М. saxatilis, перемещает в самый конец списка. Классификация,
предложенная Мейнерцхагеном, в известной степени прагматична, по-
скольку основана она только на некоторых признаках самого общего
внешнего сходства между взрослыми самцами без учета истинного род-
ства. Подобное искусственное объединение под одним видовым названием
на самом деле неблизкородственных форм создавало большие трудности
для всех, кто пытался, используя подобную систему, объяснять современ-
ное распространение отдельных представителей рода Monticola или же
реконструировать его вероятную историю. Сам Мейнерцхаген выдвинул
следующую гипотезу возникновения разорванного ареала вида gularis
в принимаемом им объеме. Этот исследователь считал, что М. g. angolen-
sis, в окраске оперения старых самцов которого он усмотрел якобы черты
примитивности, следует считать формой, исходной для всех gularis.1
Возможно, что группа возникла где-то на севере и попала на юг (в Аф-
рику?) в ледниковую эпоху. С отходом ледника часть птиц, по мнению на-
званного специалиста, вновь распространилась к северу и обосновала
колонии в Гималаях (М. cinclorhynchus) и в северо-восточной Азии (М. gu-
laris). Если даже принять взгляды Мейнерцхагена в той их части, которая
касается оценки таксономического ранга отдельных компонентов и гене-
тических связей между ними, то и при этом трудно себе представить воз-
можность быстрого обособления столь далеко разошедшихся форм,
как синегорлый и белогорлый дрозды. Не понятны в свете этой гипотезы
причины, побудившие их предков предпринять в голоцене очень дальнее
переселение через обширные необлесенные пространства из южной Аф-
1 По непонятным причинам в работе Мейнерцхагена эта архаичная, с точки зре-
ния самого автора, форма стоит в конце системы и расценивается лишь как подвид
М gularis.
— 122 —
рики в чуждую этим птицам Азию, а также происхождение миграций двух
азиатских лесных Monticola и размещение мест их зимовок (полная потеря
связей с африканским континентом). Что же до вида solitarius, то Мейнерц-
хаген считает, что он не пошел в плейстоцене на юг до Африки и имел ре-
фугиум в юго-восточной Азии. Когда же условия позволили, синие ка-
менные дрозды двинулись на север, распространились на запад и восток,
образовав в лесистой части южных Гималаев особую популяцию (М. eryth-
rogaster).
Взгляды Мейнерцхагена на структуру рода Monticola не завоевали
широкого признания. Из современных орнитологов их некоторое время
разделял Бисвас (Biswas, 1953), который уже в 1961 г. нашел необходимым
присоединиться в этом вопросе к Вори (Vaurie, 1959).
Иную точку зрения высказал Рипли (Ripley, 1952а). Взаимоотношения
внутри рода Monticola, тесно связанного с чеканами и каменками и слу-
жащего как бы вводным к трибе настоящих дроздов, представляются этому
ученому следующим образом: М. rupestris', М. explorator explorator,
М. е. tenebriformis', М. brevipes brevipes, М. b. pretoriae; М. rufocinereus
rufocinereus, М. г. sclateri, М. г. tenuis, М. г. angolensis; М. saxatilis', М. gu-
laris cinclorhynchus, М. д. gularis; М. rufiventris (erythrogas-
ter)', М. solitarius solitarius, М. s. longirostris, M. s. scorteccii, M. s. behnkei,
M. s. magnus, M. s. philippensis, M. s. pandoo, M. s. madoci. Как видно,
все азиатские лесные «каменные» дрозды рассматриваются им в одном ряду
и занимают то же место, что и в схеме Хартерта (между пестрыми и си-
ними дроздами). Белогорлый и синегорлый дрозды по-прежнему трак-
туются Рипли лишь как подвиды, порядок расстановки которых, призван-
ный отражать предполагаемое направление эволюции в пределах группы,
обратный тому, что мы видели у Мейнерцхагена. Важным положительным
моментом системы Рипли, придающим ей более естественный вид, является
объединение вместе всех эндемичных африканских видов и помещение
их в самом начале, до М. saxatilis. Обращает на себя внимание еще одна
деталь: рыжебрюхие восточноазиатские формы М. solitarius поставлены
не перед синими, т. е. не сразу после М. erythrogaster, а в конце списка.
Этим как бы оттеняется обособленное положение всей группы синих ка-
менных дроздов и независимое параллельное развитие у некоторых ее
представителей окраски, похожей на окраску М. erythrogaster.
Из новейших систем следует остановиться на схеме, принятой Вори
(Vaurie, 1959). Еще в 1955 г., занимаясь ревизией палеарктических Тиг-
didae, он подверг критике взгляды тех систематиков, которые вслед за
Мейзе расценивали белогорлого и синегорлого дроздов лишь как подвиды
одного вида. Вори тогда совершенно справедливо подметил, что кажуща-
яся на первых порах весьма незначительной разница во внешнем виде
между двумя упомянутыми формами, разорванными в гнездовой период
расстоянием порядка 2500 км, на самом деле гораздо большая, чем между
симпатричными в Гималаях синегорлым и рыжебрюхим дроздами, видовая
самостоятельность которых ни у кого не вызывает сомнения. М. gularis
и М. cinclorhynchus, по его мнению, не что иное, как аллопатрические
виды. Интересно, что в работе 1961 г. Рипли (Ripley, 1961) отходит от
своих прежних взглядов и, следуя за Вори, рассматривает синегорлого
дрозда в качестве самостоятельного вида, а через три года после этого он
(Ripley, 1964) вновь возвращается в вопросе о таксономическом ранге сине-
горлого и белогорлого дроздов к трактовке Мейзе. Вторым важным положи-
тельным моментом, выгодно отличающим схему Вори от всех рассмотренных
ранее, является перемещение лесных «каменных» дроздов как группы
относительно молодой в конец системы и придание им тем самым большей
самостоятельности. Предложенная этим систематиком классификация
палеарктических Monticola выглядит так: М. saxatilis', М. solitarius solita-
— 123 —
rius. M. s. longirostris. M. s. pandoo. M. s. philippensis; M. rufiventris I
(erythrogaster). M- cinclorhynchus, M. gularis. Некоторые поло- I
жения этой системы по-прежнему спорны или малоубедительны. Вряд ли
можно, например, считать обоснованной принятую в ней последователь-
ность размещения видов азиатских лесных Monticola. при которой рыже-
брюхий дрозд оказался формой наиболее примитивной, а белогорлый —
сильно изменившейся.
Приведенного довольно беглого обзора, однако, вполне достаточно для
того, чтобы представить себе, сколь различно интерпретируются лесные
«каменные» дрозды восточной Азии даже современными орнитологами.
Это обстоятельство, с которым я столкнулась во время обработки материала
по белогорлому дрозду, и побудило меня заняться более углубленным ис-
следованием всей группы. В результате сравнительного изучения внешней
морфологии различных возрастов, оологических признаков, особенностей
распространения, экологии и образа жизни каменных дроздов я пришла
к заключению, что действительно три вышеупомянутых лесных вида объ-
единяются в естественную группу и должны быть противопоставлены
истинным каменным дроздам как отдельный род. В дальнейшем тексте
приводится подробное описание особенностей, позволяющих сгруппиро-
вать каменных дроздов в два рода, а также определить систематическое
положение и филогению одного из них — лесных «каменных» дроздов.
Основой для изучения лесных Monticola послужили собственные сборы
и наблюдения над М. gularis в Амурской обл., коллекции Зоологического
института АН СССР (ЗИН АН СССР), Зоомузеев Киевского (ЗМ КГУ),
Московского (ЗМ МГУ), Дальневосточного (ДВ ГУ) университетов
и Дальневосточного филиала Сибирского отделения АН СССР (ДВ ФАН),
а также литературные источники, откуда почерпнуты основные данные
о распространении и биологии М. cinclorhynchus и М. erythrogaster.
Всем коллегам-орнитологам, оказывавшим помощь в получении пе-
речисленных материалов, особенно А. Б. Кистяковскому и Л. А. Смо-
горжевскому, предоставившим в мое распоряжение серию добытых ими
в Приамурье белогорлых дроздов, я приношу искреннюю благодарность.
СОВРЕМЕННОЕ РАСПРОСТРАНЕНИЕ ЛЕСНЫХ «КАМЕННЫХ» ДРОЗДОВ
Ареалы 2
Рыжебрюхий дрозд—М. erythrogaster (рис. 1, 1) —спорадически
гнездится к югу от р. Янцзы, в лесах средневысотных гор Наньлпна.
где, по данным Лятуша (La Touche, 1925), прослежен к северу почти до
границы между провинциями Фуцзянь и Чжэцзян. Обычен также в под-
ходящих биотопах Юньнань-Гуйчжоуского нагорья. Далее на запад об-
ласть распространения этого вида охватывает Сино-Тибетские горы, юго-
восточную оконечность Тибетского нагорья и простирается в Гималаи.
Там М. erythrogaster живет южнее Главного хребта от юго-западной Сы-
чуани и юго-востока Тибетского автономного района в Китае, через Бу-
тан, север Западного Бенгала, Сикким, Непал (хр. Махабхарат), северо-
западный Уттар-Прадеш, Химачал-Прадеш, до южного Джамму и Каш-
мира (Малые Гималаи, хребты Наг-Тибба, Дхаоладхар, Пир-Панджал).
В очень небольшом числе доходит до Северо-Западной пограничной про-
винции Пакистана. Известен из Ассама, где обитает южнее Брахмапутры
в горах Шиллонгского массива, а также из северной Бирмы, откуда про-
никает до округа Чин и княжества Каренни. Возможно, что какое-то
2 Общие сведения о географическом распространении рыжебрюхого и синегор-
лого дроздов заимствованы из сводок Чжена (Cheng, 1958), Вори (Vaurie, 1959) и Рипли
(Ripley, 1961).
124 -
количество птиц населяет горные области северного Лаоса и Вьетнама
(Delacour et Jabouille, 1931).
В гнездовой период рыжебрюхий дрозд приурочен преимущественно
к высотам 1219—2286 м над ур. м. Здесь, ниже линии устойчивого зимнего
снегового покрова, в зоне произрастания субтропических влажных горных
О ' X 2 <ШП) 3 // ♦ > 5 ® « // 7 = «
Рис. 1. Современные ареалы лесных «каменных» дроздов:
1 — ареал рыжебрюхого дрозда; 2 — зимние находки рыжебрюхого дрозда за пределами гнездовой
области; з — область гнездовий синегорлого дрозда; 4 — места пролета синегорлого дрозда; 5 —
область зимовок синегорлого дрозда; 6 — гнездовая область белогорлого дрозда; 7 — места пролета
белогорлого дрозда (вне гнездовой области); 8 — область зимовок белогорлого дрозда; знак вопроса —
места возможного гнездования рыжебрюхого дрозда.
лесов он находит оптимальные защитные и кормовые условия. Небольшая
часть птиц проникает выше, достигая в восточных Гималаях высоты
3353 м над ур. м. В отличие от предыдущей, практически оседлой популя-
ции, сезонные перемещения которой почти не выходят за пределы указан-
ных высот, эта группа М. erythrogaster с наступлением холодов вынуждена
предпринимать вертикальные кочевки значительно большей протяжен-
ности. Известно, например, что в Химачал-Прадеше (Индия) рыжебрюхий
- 125 -
дрозд спускается во второй половине октября с хр. Дхаоладхар и дер-
жится до марта на высотах 1800—365 м над ур. м., а летом вновь заби-
рается почти до 3048 м (Whistler, 1926). В Шимле, по данным Джонза
(Jones, 1926, 1947), зимой живет у подножий гор в окрестностях Калка,
а иногда попадается и на равнине близ Амбалы (500 м над ур. м.). Не-
сколько восточнее, около Найни-Тал, пара птиц была обнаружена в фев-
рале на высоте 2043 м (R. Meinertzhagen, 1927). В Непале много экземпля-
ров добыли в районе Тансинга (1400 м над ур. м.) в декабре—январе,
а близ Катманду (1982—2377 м над ур. м.) — в феврале (Rand a. Flem-
ming, 1957). Стивенс (Stevens, 1925) и Али (АН, 1962) пишут, что в Сик-
киме в холодный сезон эти дрозды откочевывают обычно до 1000—600 м.
Примерно в той же зоне добывали их в середине ноября в Бутане, хотя
местами еще в начале сентября они бывали на высоте 2438 м над ур. м.
(Ludlow a. Kinnear, 1937). В более поздней своей работе Ледлоу и Киннир
(Ludlow a. Kinnear, 1944) упоминают о птицах, собранных в юго-восточном
Тибете на высотах 600—365 м во второй половине февраля; одиночки,
однако, и там оставались зимовать значительно выше, примерно на 1982 м
(Ludlow, 1951). Со второй половины марта и в течение всего лета М. eryth-
rogaster занимали в той же местности высотную зону 1982—3200 м над ур. м.
Рипли (Ripley, 1952b) во время своего путешествия по Ассаму видел их
в ноябре в горах Нага на высоте 1219—2133 м. В Китае, в провинциях
Юньнань, Гуандун, Фуцзянь, Хубэй, Сычуань и др., встречаются круглый
год (Bangs a. Phillips, 1914; Rensch, 1923; La Touche, 1925; Rothschild.
1926, 1927; H. Caldwell a. J. Caldwell, 1931) и, судя по хранящемуся
в коллекции ЗИН АН СССР самцу от 31 января из Мынны (Юньнань),
зимой также бывают значительно ниже тысячеметровой отметки. Изве-
стно, правда, сообщение Райли (Riley, 1926) об одном, по-видимому за-
летном, экземпляре, который был добыт около Лицзяна (4572 м над ур.
м.) почти у границы зоны вечных снегов. В северо-восточной Бирме,
к востоку от р. Инмай, рыжебрюхие дрозды найдены в феврале на абсо-
лютной высоте 1525 м (Smythies, 1949), а южнее, в автономном госу-
дарстве Шан, несколько особей было добыто в том же месяце на горе
Лоймве, 1700 м над ур. м. (Schaunensee, 1946а). За пределами очерченного
ареала рассматриваемый вид обнаружен только в северо-западном Таи-
ланде: Мехонгсон, Чиангмай, Лампун (рис. 1, 2). Дейгнан (Deignan,
1945, 1946), например, сообщает, что с начала ноября и по крайней мере
до 6 февраля немного птиц регулярно держалось там в годы его иссле-
дований в субтропическом вечнозеленом лесу по вырубкам и вдоль
заросших дорог в хребтах Кун-Тан, Танентаунджи и на горе Донсутеп
(1200—1830 м над ур. м.).
Область гнездовий синегорлого дрозда — М. cinclorhynchus (рис. 1.
3) — охватывает южные отроги Гималаев с примыкающими к ним хребтами
Махабхарат, Дхаоладхар и Пир-Панджал от Бутана до Джамму и Каш-
мира и в этой своей части совпадает с ареалом рыжебрюхого дрозда. Од-
нако далее она заходит на труднодоступное Читральское нагорье в Се-
веро-Западной пограничной провинции Пакистана и простирается еще
западнее на горные массивы Нуристана и хр. Сафедкох в Афганистане.
Изолированная колония этих птиц находится в Ассаме, в горах Кхасп
и Качар, где они также симпатричны с М. erythrogaster. Бекер (Baker,
1924), а вслед за ним и Али (Ali, 1953) называют в качестве мест обитания
М. cinclorhynchus еще и горы северной Бирмы. Большинство же совре-
менных авторов считает бирманские сообщения неточными, нуждающи-
мися в проверке.
В пору размножения синегорлый дрозд предпочтительно занимает
высоты в 1219—2743 м над ур. м., так что часть его популяции системати-
чески гнездится несколько выше предыдущего вида в зоне, лежащей над
— 126 -
линией устойчивого зимнего снегового покрова. Какое-то число птиц
поднимается летом и до 3048 м.
В противоположность М. erythrogaster М. cinclorhynchus на зиму не
остается в пределах гнездовой области. На севере западного Пенджаба,
в окрестностях Равалпинди (1830 м над ур. м.), его наблюдали только
до 23 сентября (Whistler, 1930). С хр. Дхаоладхар исчезает, как правило,
в сентябре, но случается, что одиночки задерживаются там до начала
октября. И октября у Кангры обнаружены дрозды, спустившиеся уже
до 750 м (Whistler, 1926). В Сиккиме, по свидетельству Али (АИ, 1962),
отсутствуют с ноября по апрель. На границе с Бутаном, близ Ташигонга,
один экземпляр, вероятно из начавших миграцию, был добыт 12 октября
на высоте 914 м над ур. м. (Ludlow a. Kinnear, 1937). На зимовке на юге
штата Майсур (Индия) первые появлялись 27 ноября (Betts, 1929). Вес-
ной на местах гнездовий в разных частях ареала наиболее ранние отмечены
во второй половине марта—апреле (Stevens, 1925; Whistler, 1926; Ripley,
1950; Rand a. Flemming, 1957; Paludan, 1959), хотя некоторые запоздав-
шие особи попадались на пролете в штате Махараштра еще до 23 марта
(Ali a. Abdulali, 1937).
Основная масса птиц летит через крутые, хорошо облесенные горы
Сивалик и окружающие их болотистые джунгли тераев. Далее, миновав
почти сплошь возделанную Гангскую равнину с разбросанными среди
полей рощами — остатками некогда бывшего здесь леса, следуют на юг
п юго-запад через одетые субтропическими и тропическими лесами цепи
окраинных хребтов Деканского плоскогорья, захватывая иногда и более
равнинные участки в штатах Западный Бенгал и Мадрас (рис. 1, 4). Не-
безынтересно отметить, что на вершинах гор Сатпуры и хр. Майка л а
(штат Мадхья-Прадеш) пролетные синегорлые дрозды оказываются в об-
становке, весьма сходной, с точки зрения условий обитания, с гималай-
ской. Небольшая часть мигрантов идет западнее, через засушливые,
непустынные области Раджастхана (горы Аравалли) и штат Гуджарат
(Сурат). Тайсхерсту (Ticehurst, 1922) в свое время сообщили об одной
птице, которая в течение нескольких дней держалась близ Карачи. Пере-
секая, таким образом, почти всю континентальную Индию, М. cinclor-
hynchus достигают основных мест зимовок в Западных Гатах и лежащих
к югу от них массивах Анамалаи, Нил гири и Пал ни, от Пуны до штата
Керала (рис. 1,5). Изредка попадаются в холодный сезон в северо-запад-
ном Пакистане (Whitehead a. Magrath, 1909). Некоторые особи, возможно
из ассамской популяции, проводят зиму в западной и центральной Бирме,
проникая на юг до провинции Ракхайн. Повсюду в негнездовой области
живут ниже 1525 м над ур. м.
Белогорлый дрозд — М. gularis — единственный представитель
группы лесных Monticola, обитающий на территории нашей страны. Имею-
щиеся в литературе сведения позволяют следующим образом охарактери-
зовать его современный ареал (рис. 1, 6).
В СССР живет от Забайкалья до Приморья. По свидетельству Т. Н. Га-
гиной (1960), М. gularis был даже добыт восточнее г. Улан-Удэ, в густой
тайге по р. Гурульбе. В небольшом числе гнездится на юге Читинской обл..
где обнаружен на склоне Тасейской сопки и в районе гольца Саранакача
(Павлов, 1948), у горы Алханай и в верховьях р. Ингоды (Гладков, 1954),
а также около Бальзино (самка в коллекции ЗИН АН СССР, добытая
Б. С. Виноградовым). Очень характерен для покрытых негустым лесом
плато и низких горных кряжей Приамурья от линии Большой Невер —
Джалинда на западе до устья Амура на востоке (Штегман, 1931; Кистя-
ковский, 1959: Нейфельдт и Соколов, 1960; Кистяковский, Лоскот и
Смогоржевский, 1962; Колонии, 1963). Наиболее северные находки в этой
части ареала известны из Бомнака на Зее (экземпляр В. Дорогостайского
- 127 -
в ЗИН АН СССР), окрестностей Новороссийки и Усть-Норска на р. Се-
лемдже (сборы Л. А. Смогоржевского и Ю. А. Волненко в ЗМ КГУ),
устья р. Верхнего Мельгина на Бурее (Кистяковский и Смогоржевский,
1964) и с оз. Эворон (Воробьев, 1954). Далее к югу, в Хабаровском и При-
морском краях, по наблюдениям Л. М. Шульпина (1931) и Е. П. Спан-
генберга (1960, 1965), белогорлый дрозд обычен в хребтах Сихотэ-Алиня
от Софийска до Сучана. Нигде, однако, не живет на морском побережье,
а вблизи обширных низменностей и широких долин крупных рек ввиду
редкости там подходящих биотопов распространен спорадично (только
по холмам) или же вовсе отсутствует на гнездовье.
За пределами Советского Союза рассматриваемый вид свойствен се-
веро-восточным провинциям Китая, где селится в типичных для него
стациях — на севере и в центральной части Большого Хингана, а также
в невысоких горах и на плоскогорьях Малого Хингана; найден в подсту-
пающих к Харбину сопках и средневысотных горах восточной Маньчжу-
рии; нередок еще в горах Ляосп — в районе Тунлинского леса к северу
от Великой китайской стены (Ingram, 1909; La Touche, 1920; Meise, 1934;
Shaw, 1936; Yamashina, 1939; Piechocki, 1958; материалы ЗИН АН СССР).
По покрытым лесом хребтам проникает из Китая в северную часть п-ова Ко-
рея, в пограничные провинции КНДР — Хамгён-Пукдо и Пхёнан-Пукдо
(Kuroda, 1918; Austin, 1948; коллекция ЗИН АН СССР). Имеются указа-
ния, что какое-то число птиц нерегулярно гнездится в уцелевших местами
лесах гористых окрестностей Пекина (Swinhoe, 1861, 1862, 1863b, 1871;
Wilder a. Hubbard, 1938). Совсем недавно, летом 1962 г., во время фауни-
стического обследования юга провинции Шаньси группа китайских зо-
ологов впервые обнаружила, что М. gularis живет там на облесенных се-
верных склонах хр. Чжунтяошань (Тан Сань-цзу и др., 1965). В свете
этой находки выглядит теперь гораздо более обоснованным мнение
Л. М. Шульпина, считавшего, что добытый М. Березовским в первой де-
каде октября на юге провинции Ганьсу молодой самец является не про-
летным, а местным.
Относительно вертикального размещения этого дрозда известно сле-
дующее. В Приамурье он занимает местность, возвышающуюся на 290—
500 м над ур. м. В Сихотэ-Алине, по А. М. Шульпину (1931), не живет
ниже 700 м; А. А. Назаренко (устное сообщение) наблюдал его в горах
южного Приморья от 300 м до верхней границы леса (1600 м над ур. м.
на горе Облачной). В Большом Хингане белогорлого дрозда добывали на
высоте 1158 м над ур. м. (Ingram, 1909), в северной части п-ова Корея най-
ден летом на плоскогорье Киндзадор, на высоте 914 м (Янковский, 1898),
а у самого южного предела распространения — в китайской провинции
Шаньси — живет в поясе лесов у 1500-метровой отметки. При нанесении
на физическую карту всех пунктов, откуда достоверно известны летние
находки М. gularis, становится очевидным, что он обитает, в общем, ниже
двух предыдущих видов. По мере продвижения с юга на север белогорлый
дрозд занимает все меньшие высоты, нигде, однако, не опускаясь в пору
размножения ниже 200 м.
Так же как и М. cinclorhynchus, М. gularis — типичный перелетный
вид, гнездовая область и места зимовок которого удалены друг от друга
на значительное расстояние.
Осенью с родины отлетает довольно поздно. В Забайкалье, близ Кяхты,
по-видимому, мигрирующие особи были добыты в несвойственных им био-
топах 15 сентября, а около Кайластуя, на р. Аргуни — 10 сентября (Га-
гина, 1960). А. Я. Тугаринов (1932) сообщает о самце, добытом 7 сентября,
во время пролета в восточной Монголии на р. Халхин-Голе. В коллекциях
ЗИН АН СССР и ДВ ФАН хранятся самцы от 15 сентября, доставленные
К. А. Воробьевым из южного Приморья, а в материалах ДВ ГУ — три
— 128 —
птицы, собранные на пролете В. И. Лабзюком 10, 18 и даже 28 сентября
на о. Большой Пелис, в заливе Петра Великого. По данным Ямасины
(Yamashina, 1939), в Маньчжурии этих птиц встречали еще 12—14 сен-
тября. Южнее, в китайской провинции Хэбэй, близ Пекина, в Ганьсу и
на севере п-ова Корея они задерживаются до конца сентября и даже до
октября (Swinhoe, 1861; Березовский и Бианки, 1891; Shaw, 1936; Wilder
a. Hubbard, 1938; Austin, 1948).
Мигрируют через весь Восточный Китай (рис. 1, 7). 27 сентября и
5 октября были добыты близ Чжэньцзяна (La Touche, 1906), в сентябре —
около Шанхая (Styan, 1894), 20 и 27 октября — в провинции Фуцзянь
(Mell, 1922).
Зимовки располагаются в юго-восточных провинциях Китая и в Индо-
китае (рис. 1, 8). В южной части Вьетнама, например, эти дрозды обна-
ружены в горах Чыонгшон, близ Джиринга и в окрестностях Тайнина;
очень обычны зимой в Камбодже, особенно в поросших лесом горах Кра-
вань, близ Бокара (914 м над ур. м.); в Таиланде зимуют в восточной, при-
мыкающей к Меконгу, части плато (Кеммарат, У бон), на севере — на
хр. Кун-Тан и в районе горы Донсутеп — на высотах порядка 450—
465 м над ур. м., на юго-востоке страны (Чантабури и Трат), а также к югу
от перешейка Кра; в небольшом числе бывают в западной и центральной
Бирме, главным образом на нагорье в автономном государстве Шань и
в хр. Ракхайн (Robinson, 1915; Robinson a. Kloss, 1919; Baker, 1924;
Delacour, 1929; Deignan, 1945, 1963; Schaunensee, 1946b; Smythies, 1953).
Случается, что в холодный сезон посещают п-ов Малакка (Robinson, 1928).
Свои зимовки покидают, по-видимому, не ранее конца апреля, так
как известно (Macdonald, 1904), что еще в этом месяце их добывали в Бирме
угоры Попа. Весной пролетные замечены в Китае, в провинции Фуцзянь,
9 мая; в мае же найдены в провинции Цзянси, близ Цзюцзяна, и около
Циндао, в провинции Шаньдун; во второй половине мая появлялись в про-
винциях Хэбэй (Пекин и Циньхуандао) и Ляонин (Инкоу, Нючжуан,
Аныпань), а также у Харбина и в Большом Хингане (La Touche, 1899,
1920; Бианки, 1902; Ingram, 1909; Kuroda, 1932; Meise, 1934; Wilder
a. Hubbard, 1938; Shaw a. Liu, 1940). В КНДР во время миграции следуют
преимущественно вдоль северной границы страны, но иногда летят и юж-
нее. Наиболее ранние добыты в разные годы на юге п-ова Корея, в про-
винции Чолла-Намдо 20 мая, а на севере — в Пхёнан-Намдо 23 мая,
в Пхёнан-Пукдо И мая (Austin, 1948). В СССР, в окрестностях Владиво-
стока, К. А. Воробьевым (1954) замечен пролет 11 мая. В лишенной дре-
весной растительности местности на южном берегу оз. Ханка и на п-ове Де-
Фриза первые встречены 26 мая (Шульпин, 1931; Омелько, 1956). В кол-
лекции ЗМ КГУ есть самка, добытая В. М. Лоскотом 20 мая на нижнем
Амуре, а в ЗИН АН СССР хранится самец, убитый Б. К. Штегманом
24 мая у южной оконечности Буреинского хребта. На юге Амуро-Зейского
плато в течение четырех лет, когда велись мои наблюдения, передовые
появлялись 19—26 мая.
Бпотопическое размещение
Современные ареалы лесных «каменных» дроздов определяются исклю-
чительной приуроченностью этих птиц к определенным растительным груп-
пировкам, очертания которых они в значительной степени повторяют.
Рыжебрюхий дрозд (М. erythrogaster) ведет лесной образ жизни. По
данным Рипли (Ripley, 1961), гнездится предпочтительно в сырых субтро-
пических горных лесах и в субтропических влажных сосновых лесах. Пер-
вый тип насаждений (рис. 2, /), свойственный наиболее влажной восточной
части Гималаев, а также горам Ассама, западной Бирмы и Южного Ки-
j 9 Биология птиц — 129 —
тая, произрастает на высотах 915—1830 м. Он характеризуется высоким
(до 30 м) несомкнутым пологом древесной растительности, верхний ярус
в котором образован вечнозелеными дубами и каштанами с примесью
ясеня, бука, ольхи и берез; местами на более возвышенных участках
встречаются массивы сосны и других хвойных. Средний ярус и подлесок
в таком лесу хорошо выражены; травяной покров беден или совсем отсут-
Рис. 2. Вертикальное размещение лесных «каменных»
дроздов в Гималаях (схематизировано).
I — субтропический горный лес; II — субтропический влажный
сосновый лес; III — влажный умеренный лес гималайского типа;
IV — сырой умеренный лес; V — зона обитания рыжебрюхого
дрозда: а — спорадично, только летом, б — только зимой, в —
круглый год; VI — зона летнего обитания синегорлого дрозда:
а — регулярно, б — редко. По оси ординат — высота над уровнем
моря (в м).
ствует; изредка разрастаются бамбуки; широко представлены лианы и
эпифиты. Масса вырубок, зарастающих порослевым дубом и кустарниками.
Второй тип насаждений (рис. 2, II) как бы замещает предыдущий в Гима-
лаях к западу от Сиккима. Его западная (южный Кашмир) и верхняя
(2286 м над ур. м.) границы полностью совпадают с соответствующими
границами распространения М. erythrogaster. В виде пятен субтропические
влажные сосновые леса найдены также в подходящих условиях в Индии,
в Ассаме (горы Кхаси и Нага), в Китае (в юго-восточной части Тибетского
автономного района, в Юньнани и западной Сычуани). Это почти чистые
сосновые насаждения, преимущественно из Pinus longifolia (чир) и
Р. khasia, сильно пострадавшие от пожаров. Видное место принадлежит
— 130 —
в них дубам и рододендронам. Травяной покров обычно хорошо развит;
подлесок же выражен плохо, за исключением низкорослых вечнозеленых
дубов, разрастающихся в районах с повышенным увлажнением. На ров-
ных, слабонаклонных площадях чировые леса уже в значительной сте-
пени сведены. Птицы селятся не в густых высокоствольных лесах обоих
упомянутых типов, а в их разреженных, более светлых участках вблизи
опушек, зарастающих вырубок и гарей с сохранившимися одиночными,
старыми, высокими деревьями. Охотно гнездятся и на облесенных склонах
гор. Рыжебрюхие дрозды очень консервативны в выборе мест обитания,
свидетельством этого является их исключительная привязанность к древ-
ним лесным биотопам влажной субтропической зоны. С вырубанием этих
лесов в районах интенсивной хозяйственной деятельности человека со-
кращается и ареал М. erythrogaster. По-видимому, именно этим следует объ-
яснять спорадичность распространения вида в горах Южного Китая, где
он явно обречен на полное исчезновение.
Все исследователи, наблюдавшие рыжебрюхого дрозда, подчеркивали,
что и зимой он продолжает оставаться птицей негустых субтропических
лесов, определенно избегающей пространств, лишенных древесной расти-
тельности, и не проявляющей какой-либо привязанности к скалам. Ранее
уже говорилось, что какая-то небольшая часть популяции этих дроздов
поднимается выше зоны субтропических лесов почти до 3353 м (рис. 2,
7, а). Оставаясь лесными обитателями, они занимают там насаждения
умеренного пояса, защитные и кормовые условия в которых летом мало
отличимы от субтропических. Чемпионом (Champion, 1936) описаны два
тппа подобных лесов.3 По всей длине Гималаев (от Читрала до юго-восточ-
ного Тибета) на высотах 1525—3353 м над ур. м. произрастают так назы-
ваемые влажные умеренные леса гималайского типа (рис. 2, III). Это
довольно редкие высокоствольные насаждения, первый ярус в которых
образуют на южных склонах сосна и кедр, а на северных — пихта, ели
и местами лиственница. Во втором ярусе представлены такие листопадные
породы, как орех, платан, клен, ясень и березы. Хорошо развитый под-
лесок включает как вечнозеленые (дубы, рододендроны, лавры), так и
листопадные породы (лещина, боярышник, сирень, рябина, черемуха,
шиповник и др.). По мере движения на запад широколиственные вечнозе-
леные виды, смешиваясь с доминирующими хвойными, становятся все
более редкими. В лесу довольно много мхов и мало древесных эпифитов.
Древостой во многих местах сильно пострадал от пожогов и обрубки су-
чьев при расчистке земель под пастбища. В лесу обилие поваленных об-
горевших деревьев и сухостоя. Сходного типа насаждения занимают свой-
ственные им высоты в горных ущельях юго-западной и северной Сычуани,
а также на северо-западе Юньнани. Здесь, в Китае, они уцелели в своем
первобытном состоянии только в наиболее труднодоступных районах,
на крутых обрывах, а также в заповедных «священных рощах». Более
узкую и низкую зону (1830—2895 м) занимает сырой умеренный лес
(рис. 2, IV), растущий в лучше увлажняемых областях Гималаев, лежащих
восточнее 88е в. д. Основными лесообразующими породами в нем являются
зашедшие из субтропического пояса вечнозеленые дубы, лавры и каштаны,
достигающие 15—18 м высоты, а также листопадные клены и вязы. Под-
лесок хорошо развит, много эпифитов, мхов и папоротников.
По всей вероятности, субтропические горные леса (главным образом
первого типа), в которых в настоящее время гнездится и зимует большин-
ство М. erythrogaster, следует считать исконными биотопами вида. Вселе-
ние же его в леса умеренного пояса и проникновение на высоты, где
в холодный сезон года подолгу лежит снег и создаются, таким образом, усло-
? -------------
3 В дальнейшем названия типов лесов приводятся по Чемпиону (Champion, 1936).
- 131 —
9*
вия, неблагоприятные для существования насекомоядных птиц, произо-
шло позднее. Предки рыжебрюхих дроздов должны были первоначально,
по-видимому, освоить участки умеренных горных лесов, лежащие по со-
седству с субтропическими, в том же высотном поясе, и лишь после этого
смогли беспрепятственно распространиться по ним выше.
Синегорлый дрозд (М. cinclorhynchus). так же как и предыдущий вид,
экологически связан с лесом. В пору размножения он предпочитает
сильно изреженные и, по свидетельству Али (АН, 1962), более светлые
и открытые, чем избираемые М. erythrogaster. сосновые и смешанные дре-
весные насаждения с хорошо развитым травяным покровом или кустар-
никовым ярусом. Прежде всего это уже знакомые нам влажные сосновые
леса субтропического пояса гор, к которым приурочено распространение
М. cinclorhynchus. живущих на Шиллонгском плато и на южных, отли-
чающихся сравнительной сухостью климата склонах западных Гималаев.
Выше 1830 м над ур. м. сплошные чировые леса не идут и уступают свое
место влажным умеренным лесам гималайского типа, в которых синегор-
лый дрозд тяготеет все же больше к растительным формациям с преоблада-
нием сосны, дубов и лиственных пород умеренного пояса. По этим лесам
он проникает в относительно засушливые пограничные области восточного
Афганистана. Палудан (Paludan, 1959) нашел, например, что М. cinclor-
hynchus живет там в хвойных насаждениях около местечка Пашки и в низ-
корослых дубравах близ Вамы рядом с другими гималайскими лесными
видами, такими, как Psittacula himalayana. Lophophorus impejanus. Puc-
rasia macrolopha. Dendrocopos himalayensis. D. auriceps. Pericrocotus brevi-
rostris. Phylloscopus tytleri. Garrulus lanceolatus и др. Массивы Нуристана
и Спингара, до которых еще доходят индийские муссоны, являются одно-
временно и северо-западным форпостом влажных умеренных лесов ги-
малайского типа, и западным пределом распространения всех перечислен-
ных птиц.
Во время миграций синегорлый дрозд сохраняет свою привязанность
к лесным биотопам. В ряде мест, где на пути его следования встречаются
горные леса, подобные тем, что растут на вершинах Сатпуры и Майкала,
он оказывается в обстановке, весьма сходной с гималайской. Вынужден-
ные, однако, пересекать на пролете по существу весь полуостров Индо-
стан, птицы, естественно, попадают в совершенно безлесные аридные
районы.
На зимовках живут преимущественно в зоне субтропических горных
лесов, пятна которых обнаружены ботаниками (Champion, 1936) даже
в Западных Гатах на высоте 1067—1525 м над ур. м. и в горах Нилгири
и Пални. Напомню, что на родине (в Гималаях) М. cinclorhynchus никогда
не селится в подобных насаждениях, которые, однако, охотно использует
для гнездовий М. erythrogaster. Помимо негустых, светлых лесных участ-
ков, синегорлые дрозды нередко посещают в холодный сезон плантации
кофе и кардамона, расположенные по склонам гор и на равнине.
Белогорлый дрозд (М. gularis) — третий представитель рассматривае-
мой группы птиц — тоже определенно лесной житель. Еще никому не
приходилось видеть его в пору размножения в низменной местности, ли-
шенной древесной растительности. На юге Амуро-Зейского плато этот
весьма обычный там вид занимает дубово-сосновые и сосново-листвен-
ничные леса на пологих, хорошо дренируемых склонах сопок, предпочти-
тельно северной или северо-западной экспозиции. Изредка гнездится на
восточных и северо-восточных склонах и вовсе не живет на сухих, сильно
инсолируемых южных. Первый ярус в подобных лесах состоит из сосны
и лиственницы, сильно разреженных в результате рубки и пожаров. Оди-
ночные уцелевшие в нем старые деревья достигают большой высоты. Вто-
рой ярус представлен обычно монгольским дубом и даурской березой.
— 132 —
Разрастающиеся в условиях затененности и повышенного увлажнения
(дожди, туманы) кустарники (рододендрон и леспедеца) и порослевой дуб
образуют густой подлесок. Иногда шлейфы склонов бывают заняты трудно-
проходимым мелколесьем из молодых осин, плосколистных берез и лист-
венниц. Одиночные пары селятся во вторичных дубовых насаждениях на
склонах плато вблизи сосняков или лиственничников, напоминающих
в какой-то мере биотопы синегорлого дрозда в Нуристане.
Насколько удается судить по имеющимся в литературе весьма отрывоч-
ным сведениям, в общем, в аналогичной обстановке живут М. gularis
и в остальных частях своей гнездовой области.4 Так, К. А. Воробьев (1954)
пишет, что нашел гнездо и встретил выводки на р. Горин и близ оз. Эво-
рон на склонах, одетых лесом из осины, берез, дуба и лиственницы.
Е. П. Спангенберг (1960) сообщает о двух известных ему поселениях этих
дроздов, располагавшихся в старых, светлых смешанных насаждениях,
главным образом из осин, берез, лиственниц и елей, покрывавших сопки
в среднем течении р. Хунгари. В Сихотэ-Алине, по Л. М. Шульпину
(1931), наиболее излюбленные стации белогорлого дрозда — гребни гор
и склоны, покрытые гарью или смешанным захламленным лесом из ильма,
ясеня, клена с значительной примесью в древостое хвойных (лиственница,
ель, кедр), густым кустарниковым и травяным покровом. У северной
границы распространения, в верховьях р. Зеи, В. Дорогостайский (1915)
добыл в мае самца, державшегося среди сухостоя в лиственничнике,
окруженном марями. На плоскогорье Киндзадор в северной части п-ова Ко-
рея встречен в лесу из хвойных и лиственных пород деревьев в конце июня
(Янковский, 1898), а южнее — в китайской провинции Шаньси — пред-
почитает северные склоны, заросшие сосной, березами, дубами, туйей,
шиповником и спиреей (Тан Сань-цзу и др., 1965).
Занимая более разнообразные, нежели М. erythrogaster и даже М. cin-
clorhynchus, лесные биотопы, белогорлые дрозды, однако, всюду предъ-
являют к ним одни и те же обязательные требования: приподнятость рель-
ефа, определенная крутизна и экспозиция склонов с характерным микро-
климатом, многоярусность и разреженность древостоя, смешанный его
состав с непременным присутствием одиночных старых сосен, лиственниц
или других больших деревьев, возвышающихся над лесом. Применительно
к Сихотэ-Алиню Л. М. Шульпин первоначально считал также необходи-
мым наличие каменистого или скалистого грунта в лесу, хотя позднее
он писал (Шульпин, 1931 271), что «решительно не мог заметить какой-
либо привязанности птицы к скале как таковой». Многолетние наблюдения
в Приамурье убеждают меня в том, что характер субстрата не играет ка-
кой-либо существенной роли при выборе этим видом гнездового участка.
На родине по окончании периода размножения, а также в поисках
корма белогорлые дрозды нередко посещают и другие типы насаждений,
спускаются с холмов к их подножию, поросшему непроходимым мелко-
лесьем, встречаются в чистых, светлых, сырых лиственничниках, березня-
ках и дубравах, держатся на плато и на облесенных террасах падей. На
пролете бывают и в местности, лишенной древесной растительности.
На р. Аргуни, например, обнаружены осенью на степных сопках, а в восточ-
ной Монголии — в ильмовых рощах. В Китае мигранты попадались в су-
хих сосняках, «священных» заповедных рощах, парках и садах. Так же
как и синегорлые дрозды, зимой предпочитают гнездовые стации рыжебрю-
хого дрозда — субтропические горные леса, занимая особенно охотно
участки с нарушенным растительным покровом. Штреземан и Хейнрих
4 Большинство приводимых ниже данных относится к летним встречам взрослых
особей или (в лучшем случае) слетков, а не к находкам гнезд и потому не всегда может
точно характеризовать гнездовые стации изучаемого вида.
— 133 —
(Stresemann u. Heinrich, 1940) отмечали, что в Бирме, на горе Виктория,
М. gularis держались в сильно захламленном и ежегодно горевшем лесу
на высоте 1400 м над ур. м.
Заканчивая обзор современного географического и экологического
распространения лесных «каменных» дроздов, можно сказать следующее.
В противоположность остальным Monticola все представители этой
группы не обнаруживают никакой связи с африканским континентом или
Европой. Они являются эндемиками юго-восточной
Азии, даже во внегнездовое время не выходящими за ее пределы.
Судя по их нынешнему распространению, размещению зимовок и путей
пролета, лесные Monticola также и автохтоны этой части Азии;
здесь они возникли и развились.
Рыжебрюхий и синегорлый дрозды симпатричны на значительном про-
странстве Гималаев, причем первый вид имеет явную тен-
денцию к проникновению далее на восток, в бо-
лее влажные районы Бирмы и Южного Китая, тогда как вто-
рой там вовсе не бывает иопределе н но тяготеет к обла-
стям с относительно сухим климатом, лежащим
северо-западнее Сиккима. В Гималаях экологически они в какой-то сте-
пени замещают друг друга.
Белогорлый дрозд живет сейчас обособленно от двух упомянутых форм.
Однако изолированные гнездовья этого вида, обнаруженные в Китае
южнее области его сплошного летнего распространения, свидетельствуют
о более обширном сплошном ареале М. gularis
в прошлом, южная граница которого, по-видимому, не была столь
удалена от ареала М. erythrogaster.
Среди лесных «каменных» дроздов есть формы практически оседлые
и настоящие мигранты. Так, живущие бок о бок в ряде районов Гималаев
синегорлый и рыжебрюхий дрозды ведут себя с наступлением холодов
по-разному: одни предпринимают дальние перелеты, другие остаются на
родине и лишь несколько спускаются с высоких гор вниз.
Областью совместного обитания трех видов
является западная Бирма, где они встречаются
во время зимовок. Это обстоятельство лишний раз подтверждает
высказанное ранее предположение о том, что корни происхожде-
ния изучаемой группы находятся где-то на юге
Азии.
Ни один представитель лесных Monticola не гнез-
дится ниже двухсотметровой отметки. Интересно
при этом, что меньшая по сравнению с М. cinclorhynchus и М. erythrogas-
ter высота местности, в которой живут М. gularis. компенсируется большей
ее широтой, благодаря чему достигаются в общем
весьма сходные условия обитания трех рассмат-
риваемых видов в пору размножения.
На всем пространстве своего ареала М. erythrogaster. М. cinclorhynchus
и М. gularis связаны с древесной растительностью и исчезают вместе с ее
уничтожением. Скалистых и безлесных ландшафтов
определенно избегают. Наибольшим консерватизмом в вы-
боре мест гнездования отличается рыжебрюхий дрозд, тогда как ежегодно
мигрирующие через несвойственные им низинные ксерофильные биотопы
синегорлый и белогорлый дрозды приобрели определенный диапазон эко-
логической пластичности, позволивший им расселиться далее к западу
и северу.
Строгая приуроченность лесных Monticola к лесам субтропического
и умеренного поясов и исключительное однообразие занимаемых этими
птицами в течение года стаций говорят о том, что леса — исконные
— 134 —
биотопы группы. Наиболее древними из них
следует признать, вероятно, субтропические влаж-
ные горные леса, в которых и по сей день во все сезоны живут
М. erythrogaster. Два других вида сохраняют связь с первобытными био-
топами лишь зимой.
ПОВАДКИ И ОБРАЗ ЖИЗНИ ЛЕСНЫХ «КАМЕННЫХ» ДРОЗДОВ
Почти все, кому приходилось видеть рыжебрюхого дрозда (М. erythro-
gaster), называли главную особенность его поведения в гнездовой период —
удивительную скрытность, осторожность и даже пугливость. Джонз
(Jones, 1947) пишет, что в лесах близ Шимлы присутствие этой птицы
удается установить только по ее пению или беспокойной позывке. Дер-
жатся М. erythrogaster всегда уединенно, стай или незначительных скопле-
ний не образуют не только летом, но и в холодный сезон года. По данным
Бекера (Baker, 1924) и Али (АН, 1962), многими своими повадками напо-
минают горихвосток и настоящих дроздов одновременно. Поющие самцы
сидят на сухих макушках или высоко расположенных ветвях старых со-
сен, пихт и других хвойных, растущих в лесу близ открытых, разрежен-
ных участков или дорог. Временами спускаются на землю, где добывают
корм. Поднятые оттуда, тотчас скрываются среди ветвей деревьев. Зимой
становятся доверчивее — бывают около человеческих поселений, отыски-
вают пищу в садах, имеют обыкновение садиться на фруктовые деревья,
телеграфные провода, а иногда и на крыши домов. Очень знаменательно,
что в местах, где в лесу попадаются участки с выходами скал, птицы ни-
когда на них не присаживаются.
Самец имеет приятную трелевую песню дроздового типа. При испуге
и беспокойстве оба пола издают звуки, которые одни орнитологи (Stan-
ford а. Мауг, 1941) сравнивают с тревожным криком белки из рода Тошей-
tes, а другие (Jones, 1926; АН, 1962) характеризуют как отрывистое низ-
кое «тиканье», повторяемое через определенные интервалы и перемежаю-
щееся с резкими, неприятными звуками, напоминающими тревожный
крик сойки. Наиболее точное описание голосовых реакций рыжебрюхого
дрозда принадлежит Рипли (Ripley, 1952b), который наблюдал зимой за
потревоженной парой в горах Нага (Ассам). Он пишет, что обеспокоен-
ные птицы перелетали по вершинам высоких сосен с грубой трескотней,
прерываемой иногда высоким, пронзительным свистом, присаживались
чаще вдоль, а не поперек толстых ветвей и задирали вверх хвосты. Кстати,
о последней особенности — подергивании хвостом, подмеченном не только
этим автором. У М. erythrogaster эти движения не имеют ничего общего
с известным частым «дрожанием» хвоста у горихвосток и пестрых камен-
ных дроздов. Они напоминают скорее аналогичную реакцию представи-
телей рода Turdusn отличаются своей медлительностью и размеренностью.
Гнездо вьют оба члена пары. Для помещения его используют прикор-
невые дупла, ниши в почве или между корнями деревьев, а при каменистом
грунте еще и расщелины скал, прикрытые нависающими пучками травы
(Hume a. Oates, 1890; Baker, 1933). Конструкция, наружные размеры
и материал гнезд варьируют, что, как и у большинства других птиц, за-
висит не только от индивидуальных способностей строителей, но и от ха-
рактера укрытий, в которых предполагается разместить гнезда, а также
от наличного материала, пригодного для их возведения. Из Шимлы Джонз
описывает гнезда, стенки которых состояли из сухой травы и прутиков,
а выстилкой служили мелкие корешки и травинки. В коллекции Бекера
и Юма были такие же постройки из травы, корешков и веточек и гнезда
другого типа, свитые из листьев деревьев, папоротников и лишайников;
в качестве внутренней отделки в них использовался тоже растительный
— 135 —
материал (тонкие корешки, мох, мелкие соломинки диких злаков), но
иногда попадались шерсть и волос.
Кладка яиц у основной массы начинается в апреле—мае. Так, 20 и
21 мая в гнездах, найденных в Китае, были уже птенцы (La Touche, 1899).
В Сиккиме Али (АИ, 1962) датирует гнездовой период рыжебрюхого дрозда
маем—июнем. Почти все кладки из числа доставленных Бекеру из Ассама
были собраны в мае—июне, однако птенцов там находили и в первую не-
делю мая. Хранящиеся в ЗИН АН СССР полные свежие кладки добыты
в Гималаях 16 апреля и 1 мая. В Непале, по Бисвасу (Biswas, 1961),
начало размножения приходится, вероятно, на апрель, поскольку у до-
бытых там 9, 14 и 21 апреля самца и самок гонады были сильно увеличены.
Далее в Гималаях, близ Кумаона, самые ранние яйца обнаружены5апреля,
а у Найни-Тал птенцы на вылете оказались в гнездах слишком рано —
29 апреля и 7 мая (Baker, 1933). В Шимле размножение протекает в конце
апреля—середине мая (Jones, 1947). У Дхармсалы свежие кладки были
20 мая (Hume a. Oates, 1890). Уистлер (Whistler, 1930) сообщает, что
гнезда с сильно насиженными яйцами были найдены близ Марри 30 мая,
а в районе Равалпинди — еще четыре гнезда в мае—июле. Самые поздние
кладки от 23 июля известны из Ассама. На этом основании Бекер выска-
зал предположение о существовании у М. erythrogaster двух нормальных
циклов размножения в году, хотя материал, которым оперирует этот
автор, вряд ли может быть сравним, так как он происходит из удаленных
друг от друга и экологически весьма различных районов. Более вероятно,
что здесь, как и в случаях с другими лесными «каменными» дроздами,
имело место лишь повторное или запоздалое гнездование. Как правило,
рыжебрюхие дрозды успевают вывести и вырастить птенцов только один
раз — в самое сухое и теплое время года, до наступления продолжитель-
ного дождливого муссона.
Нормальное число яиц в полной кладке — 4 (Hume a. Oates, 1890;
La Touche, 1899; Whistler, 1930; Jones, 1947; материалы ЗИН АН СССР).
Правда, Бекер, а с его слов и Али, называют 4—6 яиц, чаще 5—6, а иногда 3.
Нередко в кладках бывают неоплодотворенные яйца. Латушу, например,
были известны два гнезда, в одном из которых оказалось 2 птенца и 2 бол-
туна, а в другом — 3 птенца и 1 болтун.
Обогревом кладки и птенцов занята только самка (Jones, 1947). Све-
дения Бекера о том, что насиживают оба родителя, вероятно, основаны
на каком-то недоразумении; они не подтверждаются позднейшими ис-
следователями. Выкармливают молодых самец и самка.
Пищу отыскивают преимущественно в лесу на поверхности почвы,
а зимой также и в садах близ населенных пунктов. Добывают главным
образом насекомых (преобладают прямокрылые), иногда собирают дожде-
вых червей и мелких моллюсков. У экземпляров, добытых в декабре
в Бирме, в виде незначительной примеси к основным кормам обнаружены
остатки ягод и семена (Stanford а. Маут, 1941). Бекер, а за ним и Али
пишут о змеях, ящерицах, лягушках и даже о рыбе как о пищевых объек-
тах рыжебрюхого дрозда. Чрезвычайная скудность сведений об этом виде
не позволяет пока составить ясного представления о всех потребляемых
им кормах; однако уже сейчас очевидно, что в пищевом рационе такой
сравнительно небольшой и слабой насекомоядной птицы позвоночные
животные, хотя бы и очень мелкие, могут играть только подчиненную
роль. Приемы охоты М. erythrogaster, по данным Бекера, напоминают
повадки горихвостки, в моменты, когда, подкараулив насекомое с какой-
нибудь точки дерева, птица бросается за ним вслед и ловит на лету. В про-
тивоположность двум другим видам лесных Monticola рыжебрюхий дрозд
любит рыться в гниющей листве деревьев на земле, проделывая это та-
ким же образом, как и все представители рода Turdus.
- 136 —
Одним из очень интересных моментов биологии рассматриваемого
впда является воспитание им птенцов обыкновенной кукушки. В пре-
делах его гнездовой области живут две близкие формы — Cuculus canorus
bakeri и С. с. telephonus.3 Первая из них приурочена в своем распростране-
нии к субтропическим горным лесам, произрастающим на высотах 1219—
1830 м над ур. м. в Юньнани и юго-восточном Тибете (Китай), Бирме
и в Индии на Шиллонгском плато (Ассам). Она занимает, таким образом,
центральную и наиболее древнюю часть ареала не только рыжебрюхого
дрозда, но, по-видимому, и всей группы лесных Monticola. В богатой
биологической коллекции Бекера (Baker, 1942) находится 22 кладки
М. erythrogaster, содержащие яйца С. с. bakeri, окраска которых очень
близка к окраске яиц вида-воспитателя. Сказанное позволяет предпола-
гать давность связей между этим паразитом и хозяином; возможно, что
их истоки восходят почти ко времени обособления ветви лесных «камен-
ных» дроздов. Гнездовья второй азиатской формы обыкновенной кукушки,
являющейся близким родственником bakeri, простираются от Западной
Сибири до Камчатки, а на юг — через п-ов Корея, Северный Китай и
Центральную Азию до Гималаев. С М. erythrogaster С. с. telephonus встре-
чается на сравнительно небольшой территории Гималаев, поскольку там
она предпочитает высоты (2000—3657 м над ур. м.), редко посещаемые
этим дроздом. Среди просмотренных Бекером кладок с яйцами азиатской
кукушки только одна принадлежала М. erythrogaster. Собрана она была
в окрестностях Марри — у западного предела распространения обоих
видов. Как и в предыдущих случаях, кукушечье яйцо имело окраску
мимикрирующего типа.
Биология синегорлого дрозда (М. cinclorhynchus) изучена к настоящему
времени значительно лучше, чем образ жизни рыжебрюхого дрозда.
Исследователи, имевшие возможность наблюдать ту и другую птицу
в природе, отмечали, что повадки М. cinclorhynchus в пору размножения
очень сходны с повадками его «larger cousin» (М. erythrogaster). Подобное
замечание полевых орнитологов представляет особую ценность, так как
далеко не все систематики, как мы помним, усматривали у этих видов
признаки близкого родства.
Бейтс и Лоутер (Bates a. Lowther, 1952) пишут, что в Кашмире М. cin-
clorhynchus — обитатели древесного яруса светлых сосновых лесов, на
землю спускаются редко, только чтобы схватить там какое-нибудь насеко-
мое, после чего тотчас возвращаются на прежнее место. В гнездовой
период скрытны и пугливы. Живут уединенно и даже зимой попада-
ются в одиночку или парами (Betts, 1929; Ali, 1953, 1954). Джердон (Jer-
don, 1862), правда, видел однажды группу в 4—5 особей. Некоторые
авторы особо подчеркивают чрезвычайную молчаливость этих птиц
в негнездовое время. Вблизи скал и на скалах их никто не отмечал. Исклю-
чение представляют только данные Бекера, которыми следует пользо-
ваться с большой осторожностью, так как туземные коллекторы, достав-
лявшие материал названному ученому, могли ошибаться в своих определе-
ниях.
О пении синегорлого дрозда написано довольно много. Все специа-
листы сходятся на одном — песня громкая, приятная («прелестная»),
мелодичная, но унылая. Блайс (Blyth), державший самца в неволе, уловил
некоторое сходство его песни с песней зарянки (Jerdon, 1862). Поет больше
по утрам и вечером, сидя на макушке или ветви высокого хвойного дерева.
В момент присаживания на ветку птица медленно поводит хвостом вверх
5 Изучение некоторых черт внешней морфологии обыкновенных кукушек, особен-
ностей их распространения и образа жизни позволяет мне присоединиться к тем авто-
рам. которые признают азиатский подвид telephonus.
— 137 —
и вниз, как и при сильном беспокойстве, однако не трясет им, как гори-
хвостка. Листер (Lister, 1953, 1954), которому посчастливилось наблюдать
М. cinclorhynchus близ Дарджилинга в разгар брачного периода, так
характеризует его пение: песня явно различимого дроздового типа, но от-
дельные звуки не очень ясные и трудно передаваемые; это как бы основа,
на которой могут быть исполнены различные вариации, чередующиеся
друг с другом. Между строфами нормальной громкой песни птица встав-
ляет другие более грубые звуки, слышимые и различимые только на не-
большом расстоянии. В общем они похожи на отдельные разрозненные
ноты из обычной песни, произносимые нерегулярно, в иной тональности
и очень невнятно. В свете существующих представлений о структуре
птичьей песни (Мальчевский, 1959) это пение синегорлого дрозда следует,
по-видимому, относить к категории подпесни, хотя сам Листер расцени-
вает ее как песню, имеющую вторичное происхождение.
О каких-нибудь токовых полетах, совершаемых самцом в воздухе,
в литературе не упоминается. Есть только ряд наблюдений, что в моменты
сильного возбуждения, перелетая с дерева вниз или на другую присадоч-
ную точку, он продолжает петь на лету.
По свидетельству Бекера, принципиальная разница между синегорлым
и рыжебрюхим дроздами при выборе ими гнездовых участков заключается
в том, что первый вид предпочитает занимать в лесу валы и насыпи, тогда
как второй тяготеет больше к обрывистым склонам, покрытым древесной
растительностью. Гнездование обоих видов тесно связано с почвой. Наи-
более распространенными, а следовательно типичными для М. cinclorhyn-
chus являются постройки, размещенные в ямках среди корней, в прикор-
невых дуплах, в естественных нишах и углублениях в субстрате или
в пустотах, образовавшихся на пологих склонах в результате выпадения
камня, комка земли или выгнившего корня. Такие гнезда обычно хорошо
замаскированы свисающей травой или кустарниками. Бекер (Baker, 1933)
приводит также случаи гнездования синегорлого дрозда на выступах
скал в лесу и даже в щелях полузаросшей, старой, заброшенной стены.
Возможно, что при каких-то исключительных обстоятельствах этот вид
поселяется в несвойственной для него обстановке. Не исключено, однако,
и другое — неточность определения птиц лицами, коллектировавшими
для Бекера. Так или иначе, общим для всех гнезд рассматриваемого лес-
ного «каменного» дрозда было помещение их в укрытиях, обязательно
защищенных сверху, сзади и с боков.
Гнезда довольно рыхлые, очень похожи на постройки М. erythrogaster,
только несколько аккуратнее и, сообразно размерам самих птиц, меньшей
величины. Их материал и конструкция варьируют в зависимости от места
расположения и индивидуальных способностей возводивших их дроздов.
Все известные Бекеру гнезда синегорлого дрозда имели внутреннюю
выстилку из нежных корешков и стебельков трав. В чировых лесах Каш-
мира Бейтс (Bates, 1935) нашел 4 гнезда, из которых два состояли целиком
из сосновой хвои с незначительной примесью в наружной облицовке
тоненьких веточек, одно было обильно выстлано такой же хвоей, одно
свито из хвои сосны, веточек и моха. Кроме того, Бейтс и Лоутер видели
постройки, в наружные края которых птицы вплели кусочки коры, а к обыч-
ному материалу, выстилающему лоток, добавили немного шерсти. Юм
упоминает еще гнезда, сделанные из сухих листьев.
Жизнь в несколько более сухом климате на западе Гималаев, где
влияние муссона сказывается меньше, чем на востоке, а также позднее
возвращение в районы гнездовий с зимовок сдвигают сроки размножения
этого вида по сравнению с рыжебрюхим дроздом больше на июнь—начало
июля. Основная масса самок несется в мае—июне (Hume a. Oates, 1890;
Oates, 1905; Osmaston, 1926; Whistler, 1930; Jones, 1947; Ali, 1962; кол-
- 138 —
лекция ЗИН АН СССР), апрельские кладки — исключительная редкость.
Из Кашмира, где синегорлый дрозд приступает к размножению, как
правило, в мае, известна одна очень ранняя кладка — от 13 апреля.
Одиночки откладывают яйца и в июле, а случается, что и в начале августа.
Нет, однако, никаких оснований относить подобную растянутость гнездо-
вого периода за счет нормальных вторых кладок.
Законченная кладка содержит 3—5, обычно 3—4 яйца. Насиживает
их одна самка. Бейтс и Лоутер, подолгу наблюдавшие и фотографиро-
вавшие этих птиц в Кашмире, ни разу не застали самца за подобным
занятием. Основой же для широко распространенного в литературе
мнения о том, что все заботы о потомстве, в том числе и обогрев кладки
и птенцов, в равной степени ложатся на обоих партнеров, послужило,
вероятно, сообщение Маршалла( Hume a. Oates, 1890), видевшего самца
в гнезде с большими птенцами. Однако самец мог оказаться там и в момент
кормления молодых, что еще никак не говорит о его участии в насижива-
нии. Сидит самка очень плотно, даже может быть поймана на гнезде.
Бейтс и Лоутер (Bates a. Lowther, 1952 : 92) так описывают ее поведение:
«Согнанная с гнезда, самка молча улетала прочь, вниз по склону, на кото-
ром жила; обратно возвращалась неохотно и только после того, как убе-
ждалась, что за ней не следят; ввиду своей чрезвычайной осторожности
является очень трудным объектом для съемок».
Сведения о кормах и приемах их добычи весьма отрывочны. Али,
например, сообщает, что в Сиккиме свою основную пищу — насекомых —
синегорлые дрозды собирают на ветвях и со стволов деревьев. На зимовках
тоже преимущественно насекомоядны, изредка ловят ящериц и мелких
лягушек. У Джердона (Jerdon, 1862 516) находим такую фразу: «По-
видимому, часто поедает различные плоды и ягоды». Мне же думается,
что корма растительного происхождения играют в пищевом рационе
самую второстепенную роль. Охотятся двумя способами: выслеживают
добычу, сидя на ветке дерева, и затем преследуют ее в воздухе, как боль-
шинство мелких Turdus, или же роются в лесной подстилке подобно
настоящим дроздам (АИ, 1953).
О поведении самок вне гнезда практически ничего неизвестно, из-за
их чрезвычайной скрытности и молчаливости.
Как и М. erythrogaster, М. cinclorhynchus воспитывает кукушат. В пре-
делах гнездовий С. с. bakeri живет только небольшая ассамская популяция
синегорлых дроздов, остальные же обитают совместно с С. с. telephonus.
Несмотря на это, большинство обнаруженных у этого вида кукушечьих
яиц [по Бекеру (Baker, 1942), 10 из 11] принадлежало первой форме.
Окраска яиц вида-паразита всегда имитировала окраску яиц лесных
«каменных» дроздов.
Осторожность белогорлого дрозда (М. gularis), по словам
Л. М. Шульпина, может быть сравниваема только с осторожностью Огео-
cincla sibirica. Мне не приходилось видеть сибирских дроздов в достаточ-
ном числе и потому трудно судить о их повадках. На основании же много-
летних продолжительных наблюдений над М. gularis я могу совершенно
определенно признать его наиболее осторожным и пугливым среди всех
известных мне Passeres. Только тщательная маскировка и бесшумное
передвижение дают наблюдателю возможность приблизиться к этой птице
на ружейный выстрел. Даже в разгар брачного периода, когда кажется,
что самец целиком поглощен своим пением, достаточно одного неосторож-
ного движения, чтобы белогорлый дрозд обнаружил подходящего человека
и тотчас исчез в подлеске. В нижнем ярусе леса, где, кстати, проходит
большая часть жизни самки, обоих партнеров удается видеть с гораздо
меньшего расстояния. Создается впечатление, что там они чувствуют
себя в родной стихии. Умело маскируясь среди ветвей и листвы,
— 139 —
птицы нередко сами подлетают к наблюдателю, проявляя при этом ско-
рее любопытство, нежели страх.
Наиболее заметны М. gularis весной и в начале лета, пока поют самцы.
С вылетом молодых родители становятся особенно молчаливыми и почти
ничем не выдают своего присутствия. Так же ведут себя они и на пролете.
В Приморье только что появившиеся весной дрозды этого вида держались
тихо, не издавая никаких звуков; новые пришельцы стремительно броса-
лись сверху в гущу невысокого деревца или кустов и присаживались
там спокойно на нижних ветвях (Шульпин, 1931).
Как и два других лесных «каменных» дрозда, белогорлый — типичный
лесной житель. Даже в горах Сихотэ-Алиня, где в лесу нередки обнаже-
ния каменистого грунта и выходы скал, никому из исследователей не при-
ходилось видеть его сидящим на камне или скале, подобно тому как это
делают постоянно многие африканские виды Monticola. а также наши
синие и пестрые каменные дрозды.
Почти все орнитологи, писавшие о М. gularis. совершенно справедливо
называли его лучшим певцом среди птиц Сибири и Дальнего Востока.
Петь самцы начинают весной, как только достигнут гнездовых мест. На
юге Амуро-Зейского плато интенсивное регулярное пение было слышно
с конца мая до середины июня. С появлением молодых наступал заметный
спад активности, хотя они продолжали петь и при выводках до 13—17
июля, но не в полную силу и с большими перерывами.
Белогорлый дрозд принадлежит к числу дневных певцов. Сидя на ма-
кушке самого высокого дерева, растущего на вершине сопки, он запевает,
как только первый луч восходящего солнца позолотит вершины холмов,
и продолжает петь в течение всего светлого времени суток, интенсивнее
все же на зорях. Вечером умолкает после того, как солнце скроется за
видимой с высоты линией горизонта. Внизу, в падях, в эту пору уже
воцаряется полумрак, становится прохладно и сыро, вступают в права
такие ночные и сумеречные певцы, как большой козодой, уссурийская
совка, большой погоныш, соловей-красношейка, различные сверчки и др.
В первой декаде июня в Амурской обл. М. gularis запевали в 3 ч. 30 м.—
3 ч. 45 м. по местному времени. Особенно охотно и подолгу пели они в яс-
ную, тихую погоду, при сильном ветре иногда умолкали. В разгар размно-
жения продолжали петь при любом ветре и в дождь. Майские утренние
заморозки не оказывали заметного влияния на деятельность этих птиц.
Песня белогорлого дрозда состоит из двух хорошо отличимых частей.
Первая из них (основная) — это собственно песня, громкая, регулярно
повторяемая, заслужившая столько похвал за свою красоту, сложность
и многообразие переходов. Состоит она из протяжных флейтовых нот,
звучащих торжественно и печально. Двух абсолютно одинаковых песен
нет; каждая птица поет по-своему. В концевые строфы своей песни отдель-
ные (далеко не все) самцы включают свистовые отрывки из песен пли позы-
вок других видов птиц. Обследованные мной популяции верхнеамурских
белогорлых дроздов особенно охотно заимствовали строфы из песен дуб-
ровника и уссурийского черноголового дубоноса. Вообще же пересмеш-
ничество занимает очень небольшой отрезок песни дрозда, ядро которой
всегда сохраняет специфику, присущую именно этому виду.
Для пения избирается макушка или сухая ветвь высокой, старой
сосны пли лиственницы, растущей на вершине холма. На участке, заня-
том парой, бывает несколько таких излюбленных присадочных мест. Если
самца не беспокоить, то в пору, когда самки сядут на яйца, он может
пробыть на одном дереве, почти в одной точке, не меняя своего положения,
более получаса и вернется сюда же после некоторого перерыва. Во время
пения птица иногда слегка взъерошивает перья на темени и затылке, так
что снизу удается рассмотреть голубую «шапочку»; всегда сильно разду-
— 140 —
вает горло, словно демонстрируя имеющееся на нем ослепительно белое
пятно; немного приспускает крылья, обнажая рыжую поясницу и белые
пятна, образующие «зеркальце» на второстепенных маховых. Сидит она
при этом почти неподвижно, не делая резких поворотов и движений.
Здесь же по соседству ловит на лету промелькнувшее насекомое и тотчас
возвращается обратно.
Вторая часть песни белогорлого дрозда обычно ускользает от слуха
орнитологов, занятых маршрутными исследованиями и интенсивно кол-
лектирующих. Это неприметная, едва слышимая с расстояния в не-
сколько метров, невнятная и торопливая скрипучая-трель, почти лишен-
ная свистовых нот и перегруженная обрывками голосовых реакций бли-
жайших соседей по биотопу. Такое пение правильнее будет, вероятно,
как и в случае с синегорлым дроздом, расценивать как подпесню, ибо оно
не столь демонстративно, как основная песня, воспроизводится нерегу-
лярно и лишь при определенных обстоятельствах. Самец исполняет эту
часть своей песни не «для всех» и не с вершины самого высокого в округе
дерева, а только в моменты особенно сильного возбуждения, находясь
в подлеске поблизости от самки. В период разбивки на пары и откладки
яиц, когда самки более подвижны, сильно возбужденные самцы часто
меняют место пения пли же, прервав свою песню, исчезают внизу в за-
рослях мелколесья и кустарников. Со стороны при этом кажется, что они
вовсе умолкают, хотя на самом деле, оказавшись около своей партнерши,
самец исполняет вполголоса «только для нее» подпесню. Весной к одной
самке нередко слетаются одновременно 2—3 самца, занимающие соседние
высотные рубежи. Они щебечут, свистят, как будто стараются переще-
голять друг друга, напоминая в такой момент шумную стаю рассевшихся
по деревьям молодых скворцов. Однако скоро самец — член пары отго-
няет пришельцев.
Каких-либо своеобразных токовых полетов, подобных тем, которые
известны для скальных форм каменных дроздов (Jahn, 1942; Bates а.
Lowther, 1952; Koffan a. oth., 1956; Farkas, 1962a, 1962b, 1963; Юдин,
устное сообщение), M. gularis не совершает. Иногда в разгар брачного
периода, заметив самку, неожиданно вылетевшую на какое-то мгновенье
из нижнего яруса леса и усевшуюся где-нибудь довольно открыто на ниж-
нюю ветвь деревца или на макушку куста, самец устремляется к ней,
продолжая петь на лету. Оказавшись поблизости от самки, ее партнер
принимает характерную позу «ухаживания»: вытягивает шею, приподни-
мается на ногах и вдруг, изогнув круто шею, резким движением опускает
голову, словно кланяясь. Все это время самец исполняет подпесню, пере-
межающуюся иногда отрывками из основной песни. В большинстве слу-
чаев подобное поведение предшествует спариванию, которое происходит
тут же, в чаще кустарников, на небольшом расстоянии от поверхности
земли. При перемещении самки с места на место самец следует за ней по
низу и, догнав ее, вновь повторяет свой «ритуальный танец».
Самка в догнездовое время отличается исключительной скрытностью
п молчаливостью, и, хотя она и присутствует постоянно где-то поблизости
от самца, заметить ее почти невозможно. Лишь изредка, преследуемая
партнером, она ненадолго покидает нижний ярус леса и внезапно появ-
ляется высоко в кроне старой сосны или лиственницы для того, чтобы
вскоре снова исчезнуть в тенистом подлеске.
Все известные мне гнезда белогорлого дрозда по характеру их разме-
щения могут быть разделены на три группы. В первую входят постройки,
свитые в пустотах, образовавшихся в почве на склонах после выпадания
комка земли или камня, выгорания или выгнивания корня дерева или же
на месте обвалившегося входа в заброшенную нору грызуна (рис. 5, Л).
С особой охотой птицы селились в таких укрытиях, расположенных под
— 141
лежащей толстой ветвью или поваленным деревом, среди корней, около
пня или кочки. Подобный тип чисто наземного гнездования, по-видимому,
наиболее свойствен рассматриваемому виду; в нишах были устроены 10 из
18 изученных мной гнезд, а также оба описанные в литературе (Воробьев,
1954; Спангенберг, 1960). Вторую, значительно меньшую, но все же весьма
распространенную группу составляют постройки, помещенные в прикор-
невых дуплах (5 случаев из 18). Укрытия этого рода возникли в резуль-
тате выгнивания и частого обгорания оснований стволов толстых сосен,
лиственниц, реже дубов (рис. 3). Для гнезда дрозды использовали
только те прикорневые дупла, входные отверстия которых были обращены
в сторону, противоположную склону, т. е. вниз, к ложбине. Защитные
условия в укрытиях второй группы в принципе ничем не отличались от
условий, существующих в укрытиях первой группы. В тех случаях,
когда дно дупла оказывалось очень неровным, воздвигнутые на его ос-
нове гнездовые постройки были несколько приподнятыми над поверх-
ностью земли. Гнезда третьей группы (их мне известно только 3) распола-
гались в полудуплах, удаленных от субстрата на 20, 60 и даже на 100 см,
и принадлежали тем парам, которые поселялись по краю сырых, затапли-
ваемых во время муссонных дождей падей. Подобный тип гнездования, по
всей вероятности, нехарактерен для вида в целом и, как мы видели, не
встречается у двух других представителей группы, живущих высоко в го-
рах. Можно думать, что он развился как приспособление для защиты по-
томства от чрезмерного увлажнения в тех случаях, когда белогорлые
дрозды по каким-то причинам занимали участки слабо наклонные или
вовсе лишенные уклона, на которых обычные наземные постройки неми-
нуемо погибли бы от воды (Нейфельдт и Соколов, 1960). Общим для всех
типов укрытий было обязательное присутствие у них, кроме дна, трех сте-
нок и крыши.
Несмотря на кажущееся на первый взгляд обилие в лесу подобных
ниш, прикорневых дупел и полудупел, птицы предъявляют к ним еще
какие-то, одним им известные требования и потому в ряде случаев испы-
тывают явную нехватку в подходящих укрытиях. Мне не раз приходилось
видеть, как тщательно и подолгу выбирает самка место для своего буду-
щего гнезда. Случалось, что, уже начав строительство, она вскоре убеж-
далась в непригодности занятого дупла или ниши (из-за размеров, неудач-
ного расположения на слишком пологом или крутом склоне, подверженном
к тому же чрезмерному воздействию солнечных лучей или господ-
ствующих ветров, приносящих осадки, или же по причине плохой замаски-
рованности) и оставляла его, принимаясь вскоре за поиски нового укры-
тия. Интересно, что птицы охотно занимают однажды использованные
гнездовые места, если из них извлечь прошлогоднюю постройку. Так,
в 1961 г. одна пара белогорлых дроздов, вероятно, не нашедшая для
себя подходящего укрытия, загнездилась только 23 июня — в день, когда
из обнаруженного на ее участке прикорневого дупла был выброшен мате-
риал развалившегося старого гнезда, принадлежавшего тому же виду.
Форма, размеры и конструкция гнезд М. gularis варьируют в зависи-
мости от места их расположения. Наиболее примитивно были устроены
гнезда, помещенные на почве в неглубоких, блюдцеобразных ямках. Они
состояли из тонкого слоя сосновой хвои и сухих травинок — материала,
образующего в постройках более сложной конструкции только выстилку
лотка. Некоторые дрозды ограничивались вообще несколькими хвоин-
ками, положенными на дно естественного углубления в земле, ближе
к наружному краю ниши (рис. 3, 4). Значительно распространеннее,
однако, гнезда, свитые в укрытиях с покатым и неровным дном. Они отли-
чаются большими наружными размерами, рыхлостью и неряшливостью.
В них хорошо различимы три слоя: наружный — из листьев, одиночных
— 142 —
Рис. 3. Расположение гнезд белогорлого дрозда.
Вверху — в нише на почве; внизу — в прикорневом дупле лиственницы.
тонких веточек с примесью лишайников, моха, кусочков лиственничной
или сосновой коры; внутренний — из хвои сосны; выстилка из сосновой
хвои, мелких травинок и корешков. Самыми же громоздкими оказались
постройки, помещенные в прикорневых дуплах и полудуплах, дно которых
имело уклон в сторону входного отверстия. Здесь, прежде чем приступать
к возведению собственно гнезда, птицы создавали для него прочную гори-
зонтальную платформу из беспорядочно набросанных веточек дуба, лист-
венницы, сосны, рододендрона, грубых стеблей травянистых растений,
кусочков коры и сухих дубовых листьев. Этот же. материал шел обычно
и на строительство наружных стенок гнезда, что придавало всему соору-
жению весьма неряшливый вид (рис. 3, Б). Два других слоя в таком
гнезде состояли из сосновой хвои, мелких соломинок диких злаков и ко-
решков. В противоположность очень примитивным постройкам, занимав-
шим пустоты в почве, гнезда последнего типа имели стенки неодинаковой
толщины. Те из них, которые соприкасались со стенками дупла, были
очень тонкими, а обращенные к входу — массивными, утолщенными.
В одном и том же гнезде передняя стенка бывала в 60—170 мм толщиной,
а задняя соответственно 10—3 мм. Изменчива и высота таких сооружений:
100—125 мм у наружного края укрытия и 60—75 мм около задней его
стенки. Размеры же лотка всегда оставались почти стабильными: диаметр
лотка 90—100 мм, его глубина 33—47 мм.
К гнездованию белогорлые дрозды в массе приступают вскоре после
возвращения с зимовок. Главная роль в этом процессе принадлежит
самке. Она не только собирает поблизости строительный материал, но
и вьет гнездо. Делает все молча и незаметно, а потому, как, впрочем, и
в остальное время, увидеть ее не так-то просто. Самец в известных мне
случаях лишь подносил прутики и травинки, укладыванием их и формов-
кой лотка не занимался. Зато он первый замечал приближение опасности
и подавал сигнал тревоги. На устройство гнезда средней сложности птицы
тратят три-четыре дня, а на самую примитивную постройку — почти
в два раза меньше. В день завершения строительства самка лишь слегка
поправляет лоток и выстилает его нежным материалом. Это занятие отни-
мает немного времени, и потому вторую половину дня оба партнера часто
бывают свободны и около гнезда не появляются, хотя и держатся где-то
рядом. На следующее утро самка приступает к кладке. По наблюдениям
только за одной особью, в гнездо садится около 8—9 час. утра и к 11 час.—
11 ч. 30 м. сносит яйцо. В каждом отдельном случае самка остается после
этого еще некоторое время в гнезде, а затем улетает и возвращается только
утром следующего дня, чтобы отложить очередное яйцо. Пока несущаяся
самка находилась в гнезде, самец держался поблизости в кронах деревьев;
потревоженный, он начинал издавать звуки, очень напоминающие про-
тяжный писк испуганной зарянки (Erithacus rubecula). Самка подпускала
к себе очень близко, а затем молча оставляла гнездо и скрывалась в под-
леске.
В полной кладке бывает 5—8 яиц, обычно (в 44% известных мне слу-
чаев) 6 яиц, реже (в 22% случаев) 5. Два осмотренных в Амурской обл.
гнезда содержали по 7 яиц и одно — 8. В поздних кладках оказалось 3
и 2 яйца, а в описанной К. А. Воробьевым (1954) — 7 (6 птенцов и болтун).
На юге Амуро-Зейского плато к размножению большинство пар при-
ступало в первой половине июня. 1 июня замечено спаривание белогорлых
дроздов. Первое яйцо появилось 1 июня в одном гнезде, 3 июня — в двух,
6 июня — в одном, 7 июня — в четырех гнездах, а также 11, 14 и 19 июня.
Часть самок начала нестись во второй половине этого месяца: 21 июня
(один случай), 23 июня (два случая) и по одному случаю 24 и 27 июня.
Бывают, правда очень редко, поздние кладки. В 1959 г., например, об-
наружено гнездо, в котором первое яйцо было отложено 4 июля. Белогор-
— 144 —
лые дрозды, как правило, размножаются один раз в году и только при по-
тере кладки гнездятся повторно. Встречающаяся у этого вида растяну-
тость гнездового периода в одном районе обусловлена рядом причин,
среди которых наиболее существенны, по-видимому, следующие: 1) не-
достаток своевременно обнаруживаемых укрытий, пригодных для помеще-
ния в них гнезд; 2) первоначально неудачный выбор места гнездования и
вызванная этим необходимость оставлять незавершенные или готовые
постройки, а порой и гнезда с яйцами и отыскивать новые убежища;
3) неодновременное начало гнездостроения и неодинаковые его темпы у раз-
ных особей; 4) повторное размножение, обусловленное разорением гнезд.
Исключительная бедность сведений об изучаемом виде позволяет лишь
в самых общих чертах наметить сроки его размножения в других частях
гнездовой области. В коллекции ЗИН АН СССР есть, например, самка
с большими наседнымп пятнами, добытая 6 июня в Тунлинском лесу
(Китай). Пихоцкий (Piechocki, 1958) сообщает о молодых, которых добы-
вали в Малом Хингане 1—4 августа. В Приморье Л. М. Шульпин (1931)
наблюдал 10 июня спаривание белогорлых дроздов и застрелил в тот же
день самца, семенники которого достигали максимальных размеров. В та-
ком же примерно состоянии были гонады у самцов от 2 и 5 июня с Хунгари,
а 14 июня там же найдено недостроенное гнездо (Спангенберг, 1960).
К. А. Воробьев, впервые описавший гнездо М. gularis. найденное им на
р. Горин, отметил, что 9 июля в нем были птенцы на вылете, а 29 июля
там же встречен выводок. В материалах ЗМ МГУ хранятся слетки от
27 июня, доставленные Н. А. Гладковым из Читинской обл. Г. В. Колонии
(1963) считает, что встреченные им 10 июля близ Большого Невера беспо-
коившиеся птицы держались, по-видимому, у гнезда.
На основании приведенного обзора создается впечатление, что дрозды
этого вида, живущие на различных широтах, гнездятся приблизительно
в одни и те же сроки, очень близкие к срокам размножения синегорлых
дроздов.
Насиживание кладки начинается после снесения предпоследнего,
а чаще последнего яйца. Обогревом яиц, а позднее и птенцов занята только
самкаг которая сидит очень плотно, позволяя иногда приблизиться
к гнезду почти на расстояние вытянутой руки. В противоположность
самцу, который при появлении человека на занятом парой участке начи-
нает сильно беспокоиться, самка в первый период насиживания по-преж-
нему скрытна и молчалива. Вспугнутая, слетает с гнезда, не издавая ни-
каких звуков, и кажется, что исчезает среди густого подлеска. На самом же
деле, отлетев на несколько метров, самка тихо возвращается в район
гнезда и усаживается сбоку или позади от наблюдателя, искусно маски-
руясь среди пестрых от солнечных бликов ветвей и листвы деревьев. При-
мерно за 5—6 дней до появления птенцов, будучи согнанной с гнезда, она
начинает проявлять беспокойство, выражая его протяжным звуком, вроде
«ци-и-и», несколько более резким и низким по тональности, нежели писк
самца.
Инкубационный период длится 14—15 суток, но в случае необходи-
мости самка может просидеть на яйцах и дольше. Из 96 яиц в обследо-
ванных гнездах вывелось 92 птенца и только 4 яйца (по одному в четырех
гнездах) оказались неоплод отворенными. Один болтун оказался также
п в гнезде, найденном К. А. Воробьевым. Вылупление птенцов в пределах
одного гнезда занимает от одних до полутора суток. У большинства пар
оно приходилось на последние числа июня—начало июля и только при
позднем размножении растягивалось до конца июля.
Первые дни после появления молодых самка ведет себя так же, как и
в конце периода насиживания яиц: подолгу находится в гнезде, не летает
за кормом, неохотно взлетает. С подрастанием дроздят она становится
Ю Биология птиц
— 145 —
более беспокойной и самоотверженно защищает выводок, подлетая иногда
очень близко к потревожившему ее человеку. Инстинкт охраны потомства
берет верх над врожденной осторожностью и пугливостью. Теперь уже
оба партнера встречают пришельца на некотором расстоянии от гнезда
характерным заряночьим свистом. В моменты сильного раздражения они
отрывисто трещат и чекают, слегка приседают и подергивают в такт хвос-
тами. Как и оба рассмотренные ранее вида лесных «каменных» дроздов.
М. gularis не трясет часто хвостом, а лишь размеренно поводит им вверх
и вниз подобно настоящим дроздам. Л. М. Шульпин заметил также, что
и «кланяется» он не так сильно, как скальные формы.
Во время охоты за насекомыми, которые являются их главным источ-
ником питания, белогорлые дрозды ведут себя как зарянки или гори-
хвостки. Они то склевывают добычу с травы или ветвей и листьев деревьев
и кустарников, то схватывают ее на лету, преследуя в воздухе или у самой
поверхности земли. По характеру посадки и сильному, ловкому маневрен-
ному полету эти птицы — нечто среднее между зарянками и представите-
лями рода Turdus. Роющимися в лесной подстилке или разыскивающими
корм на почве мне их наблюдать не приходилось, хотя такая возмож-
ность не исключена, например, во время миграции, когда белогорлые
дрозды оказываются порой в местности, лишенной древесной раститель-
ности. Забота о кормлении птенцов ложится на обоих родителей, однако
самка прилетает к гнезду все же чаще самца. В принесенной порции бы-
вает 1—2 насекомых, достаточных для того, чтобы утолить голод 1 — 2 птен-
цов.
Как и его ближайшие родичи, белогорлый дрозд — типичная насеко-
моядная птица. У трех экземпляров, добытых в мае у Циндао, гусеницы,
жуки и другие насекомые составляли, по данным Шоу и Лю (Shaw a. Liu.
1940), 99?о от общего числа кормовых объектов. Л. М. Шульпин (1931)
обнаружил в желудках собранных им особей остатки одних жуков, среди
которых попался очень крупный, длиной почти 13 мм. От Е. П. Спанген-
берга узнаем, что в обследованных им желудках оказались хитиновые
фрагменты насекомых и мелкие моллюски. Анализ собственных материа-
лов и визуальные наблюдения у гнезд за кормящими птицами также убеж-
дают меня в том, что круг объектов их питания, по крайней мере на ро-
дине, ограничивается насекомыми. Состав кормов, приносимых птенцам,
в общем мало чем отличается от летнего рациона взрослых особей и само-
стоятельно охотящихся молодых. Наибольшее значение для тех и для дру-
гих имеют, вероятно, прямокрылые — наиболее массовые и легкодоступ-
ные насекомые в гнездовых биотопах М. gularis. Они встречены в 12%
просмотренных желудков и в 75% пищевых проб, полученных от гнездо-
вых птенцов. Нередко попадались также жуки-щелкуны (от 25 до 31 %
встреч), бабочки (10—20%) и гусеницы (25—37%) совок.6 В июле бело-
горлые дрозды, как, впрочем, и многие другие насекомоядные птицы,
обитающие на юге Амуро-Зейского плато, систематически поедали круп-
ных клопов-щитников (45°о встреч). Молодежь, кроме того, получала
бабочек-медведиц (1О?о), гусениц тополевой ленточницы (15%) и пядениц
(15%). Растительные остатки в пище этого вида не найдены.
Совсем недавно мне посчастливилось обнаружить в одном из гнезд
белогорлого дрозда яйца двух самок азиатской кукушки (С. с. telephonus\
которые были окрашены, хотя и не одинаково, но все же по общему для
яиц всех лесных Monticola типу (рис. 4). Среди дальневосточных воробьи-
ных никто не несет яйца, сходные по своему цвету с яйцами М. gularis.
6 Первая цифра в скобках означает частоту встреч кормов в 30 исследованных
желудках взрослых птиц и слетков, а вторая — встречаемость тех же объектов в 30 про-
бах, полученных от гнездовых птенцов методом перевязок.
— 146 —
Нп в одном из гнезд, принадлежащих другим Passeres, не были также
найдены кукушечьи яйца с подобной пигментацией скорлупы. Все это дает
основание предполагать, что у северного предела распространения группы
лесных «каменных» дроздов живет особая географическая раса азиатских
кукушек, издавна на них паразитирующая (Нейфельдт, 1965).
Из предыдущего изложения следует, что поведение и образ жизни
М. erythrogaster, М. cinclorhynchus и М. gularis характеризуют их тоже
как представителей единой лесной группы птиц, прекрасно отличаю-
щейся от всех остальных Monticola. По ряду черт биологии лесные «ка-
менные» дрозды представляют параллель зарянкам и настоящим дроздам.
Напомню, что Лэк (Lack, 1954), специально изучавший голосовые реакции
Erithacus rubecula, среди которых он совершенно справедливо считал
наиболее ценными для таксономических целей не пение, а разного рода
позывки, пришел к заключению, что этот вид (зарянка) является скорее
миниатюрным дроздом, нежели соловьем, варакушкой пли горихвосткой,
с которыми ее сближают обычно систематики.
Связь ныне вымерших представителей лесных Monticola, от которых
ведут свое начало рассматриваемые формы, с лесом возникла, по-види-
мому, очень давно, еще в пору становления группы. В противном случае
невозможно себе представить, каким путем эти птицы смогли стать воспи-
тателями большой популяции кукушек (предков нынешних С. с. bakeri),
являющихся исконными обитателями горных лесов Бирмы и Юго-Запад-
ного Китая.
В противоположность типичным дендрофилам, живущим высоко
в кронах деревьев, таким, например, как личинкоеды пли свиристели,
лесные «каменные» дрозды, вероятно, сравни-
тельно недавно освоили верхний ярус леса и
ведут свое начало от каких-то форм, населяв-
ших подлесок в негустых низинных насажде-
ниях субтропического типа. Об этом свидетельствуют прежде всего
чрезвычайная пугливость и осторожность самцов во время пребывания
на макушках деревьев и постоянное их стремление в случае опасности
укрыться внизу, в кустарниках. С нижним же ярусом леса, где эти птицы,
кстати, ведут себя более доверчиво, нежели наверху, связаны такие важ-
ные моменты их биологического цикла, как брачные игры и спаривание,
размножение и кормодобывание; здесь же протекает вся жизнь самки,
птенцов-слетков, а по окончании гнездовых дел также и самца.
Живя в подлеске, предки лесных Monticola никогда,
видимо, не гнездились открыто на деревьях.
В пользу их изначального гнездования в естественных углублениях и
неровностях почвы говорит отсутствие у всех современных представителей
группы необходимых для обитателей древесного яруса гнездостроительных
навыков. Дальнейшая эволюция этих дроздов шла в направлении приспо-
собления к условиям горных лесов, где, по-видимому, и осуществился
впервые их переход от открытого наземного гнездования к жизни в укры-
тиях, способных защитить примитивные постройки и находящиеся в них
кладки или птенцов не только от избыточного увлажнения, но и от пря-
мого смыва с наклонной поверхности сильным водным потоком. Сперва
это могли быть небольшие пустоты в мягком или скалистом грунте обле-
сенных склонов (тип гнездования, свойственный и поныне М. erythro-
gaster, М. cinclorhynchus и большинству М. gularis), а потом и прикорне-
вые дупла деревьев.
Судя по всему, предки лесных Monticola вели образ
жизни, близкий к тому, котор ы й известен в на-
стоящее время для рыжебрюхого дрозда — вида,
сохранившего в своей экологии много архап-
— 147 —
10*
ческих черт. Белогорлый же дрозд по этим при-
знакам кажется наиболее измененным, тогда как
синегорлый занимает как бы промежуточное положение между двумя
названными формами.
ОСНОВНЫЕ ЧЕРТЫ ВНЕШНЕЙ МОРФОЛОГИИ ЛЕСНЫХ
«КАМЕННЫХ» ДРОЗДОВ
Ввиду ограниченности и неравноценности имевшихся в моем распоря-
жении коллекционных материалов по экзотическим Monticola, а также
невозможности восполнить этот пробел сведениями, почерпнутыми в ли-
тературе, пришлось ограничить изучение морфологии лесных «каменных»
дроздов параллельным рассмотрением лишь некоторых признаков внеш-
него строения с привлечением данных по их изменению в онтогенезе.
Принятая в настоящем разделе, не такая, как в предыдущих, после-
довательность размещения видов не случайна, а практически наиболее
удобна для дальнейшего анализа и понимания разбираемого вопроса.
Яйца
В ряду морфологических критериев, обычно привлекаемых специали-
стами к решению эволюционных проблем, оологические признаки Pas-
seres незаслуженно оставляют без должного внимания, хотя они, «как
исходные (в онтогенезе) признаки, могут указывать на более древние
связи, проявление которых у взрослых форм может быть и не столь от-
четливым» (Кузякин, 1954 27). Неправильно было бы, конечно, думать,
что, объединяя совершенно разных птиц только по окраске или струк-
туре.скорлупы их яиц, можно построить систему, в какой-то мере отра-
жающую филогению группы. Это так же нереально, как и попытки систе-
матизировать, базируясь на любых других (географических, экологических,
этологических, сравнительноанатомических) признаках, рассмат-
риваемых в отрыве друг от друга. В комплексе же с остальными оологи-
ческие признаки могут немало способствовать выяснению генетических
связей между близкими видами.
В кладках белогорлого дрозда (М. gularis) обнаружены яйца следую-
щих двух хорошо различимых типов: 1) основной бледно-розовый или
нежно-палевый фон скорлупы густо и равномерно усеян очень мелкими,
видимыми только при внимательном рассмотрении, глубокими крапин-
ками светло-терракотового цвета, которых на тупом конце яйца бывает
иногда значительно больше, так что, сливаясь они образуют на нем по-
темнение; 2) такой же, как у яиц первой группы, основной фон испещрен
частыми, мелкими, глубокими пятнами бледно-терракотового цвета,
а поверх них в общем довольно равномерно разбросаны отчетливые круп-
ные пестрины темно-терракотового или ржавчато-бурого цвета. Между
этими двумя типами есть ряд переходов, однако в пределах одного гнезда
встречались всегда одинаково окрашенные яйца (рис. 4). Форма их и
размеры варьируют незначительно. Преобладают кладки с яйцами обыч-
ной овоидной формы; попадались и заметно вытянутые вдоль главной оси
пли же относительно короткие и округлые яйца. Размеры (29): 21.3—
25.3x16.0—18.0 мм, средние —23.2x17.4 мм.
Яйца синегорлого дрозда (М. cinclorhynchus), насколько можно судить
по его кладкам, хранящимся в ЗИН АН СССР, а также по хорошим опи-
саниям, имеющимся в литературе (Hume a. Oates, 1890; Baker, 1933;
Bates a. Lowther, 1952), очень похожи на яйца белогорлого дрозда. Ок-
раска их варьирует в общем в тех же пределах. На больших сериях удается
обнаружить иногда почти чисто белые яйца с венчиком из красновато-
148 —
бурых пятен на тупом конце. По форме преобладают более округлые,
нежели у предыдущего вида, яйца. Размеры (6): 23.0—24.0 Х17.9—18.0 мм,
средние — 23.1 Х18.0 мм.
У рыжебрюхого дрозда (М. erythrogaster) основная окраска яиц варьи-
рует от палево-желтовато-кремовой, почти белой, до темной буровато-
палевой. Такой фон бывает покрыт мелкими пли красноватыми, или тер-
ракотовыми пятнами и мазками, которые распределяются или равномерно
по всей поверхности яичной скорлупы, или же (главным образом на свет-
лых яйцах) концентрируются у тупого конца, образуя подобие венчика.
Рис. 4. Гнездо с полной кладкой белогорлого дрозда и двумя яйцами
обыкновенной кукушки.
Эти яйца походят на яйца двух предыдущих видов, однако у М. erythrogas-
ter встречается еще третий тип окраски яиц, не отмеченный пока для бело-
горлого и синегорлого дроздов. В коллекции ЗИН АН СССР он представ-
лен кладкой, собранной 1 мая 1917 г. в Шимле. Поверхность таких яиц
настолько часто усеяна темно-терракотовыми и бурыми пятнами, что
сгущаясь, они полностью закрывают основной фон и придают скорлупе
однотонную темную окраску. Интересно, что сходным образом было пиг-
ментировано одно из кукушечьих яиц, найденных мной в гнезде М. gula-
ris. По своей величине яйца рыжебрюхого дрозда несколько крупнее, что
вполне отвечает большим размерам самой птицы. Форма их изменчива,
однако в пределах, описанных выше; преобладает овоидный тип. Про-
меры 75 яиц из коллекции Бекера (Baker, 1933), 8 из них в настоящее
время находятся в фондах ЗИН АН СССР: 24.3—29.5x19.0—21.1 мм,
средние — 26.8x19.9 мм.
Как видно из приведенного обзора, на стадии яйца три
азиатских лесных вида Monticola обнаруживают
удивительное сходство. Некоторые вариации окраски яиц
практически не отличимы у разных видов, а все известные индивидуаль-
ные и видовые отклонения в их пигментации укладываются в один общий
для группы «розовый» тип окраски, близкий к заряночьему. Известно,
— 149 —
что остальные Monticola, в том числе все африканские эндемики, несут
гораздо более сложно пигментированные яйца, совершенно не похожие
на яйца лесных «каменных» дроздов, — голубые или зеленовато-голубые
с бурыми пятнышками или без них (Гладков, 1954; Archer a. Godman,
1961; Mackworth-Praed a. Grant, 1963). Ни у первой, ни у второй группы
птиц ни разу, даже в виде аномалии, не встречены яйца другого., не свой-
ственного ей типа окраски. Диморфность яиц у представителей принимае-
мого ныне рода Monticola, включающего азиатских лесных и скальных
«каменных» дроздов, доставившая столько неприятностей орнитологам,
пытавшимся, как например Лэк (Lack, 1958), найти ее объяснение, ста-
новится, таким образом, вполне понятной. Дело в том, что Monticola,
по-видидимому, сборная группа.
Если даже считать, что все представители нынешнего обширного рода
Monticola произошли от какого-то общего корня, то при этом наблюдае-
мая очень четкая разница между скальными и лесными формами на самой
ранней стадии индивидуального развития, на стадии яйца, свидетель-
ствует об очень давнем обособлении обеих ветвей от основного ствола и об
их длительном самостоятельном существовании.
Возрастные и сезонные наряды
Пуховой наряд
Птенцы белогорлого дрозда (М. gularis) появляются на свет слепыми,
с закрытыми слуховыми отверстиями, голыми, с редким, но довольно
длинным (12—13 мм) золотисто-рыжим пухом над глазами, на затылке,
плечах и передней части спины (рис. 5). Кожа у недавно вылупившегося
птенца желтовато-оранжевого цвета, ротовая полость и язык желтые,
складки в углах рта светло-желтые, почти белые.
По данным Тайсхерста (Ticehurst, 1926), у птенцов синегорлого дрозда
(М. cinclorhynchus) в этом возрасте на тех же самых участках тела разви-
вается точно такое же рыжее опушение.
Пуховой наряд рыжебрюхого дрозда (М. erythrogaster) пока, к сожа-
лению, никем не описан. Оологические признаки, а также особенности
окраски и структуры ювенального оперения этого вида позволяют, од-
нако, предполагать, что пуховички М. erythrogaster ничем принципиально
не отличаются от рассмотренных выше.
На следующем, более позднем этапе своего
развития три лесных вида, таким образом, тоже, вероятно,
очень близки между собой и прекрасно отли-
чаются от скальных форм, эмбриональное опушение кото-
рых хотя и располагается на тех же птерилиях, но имеет совершенно
иную — серую окраску (Witherby a. oth., 1958). Вместе с тем по отсут-
ствию локтевой пуховой птерилии все Monticola не могут быть спутаны
с настоящими дроздами (род Turdus) и очень близки по этому признаку
к зарянкам и соловьям.
Пуховой наряд носится птенцами недолго. У белогорлого дрозда,
например, уже на 2-е сутки становятся хорошо заметными пеньки расту-
щего мелкого пера, а на 4-й день жизни молодых над поверхностью кожи
появляются «иголочки» — вершины пеньков маховых. Если дроздят не
беспокоят, то в нормальных условиях они покидают гнездо, достигнув
15—16-дневного возраста (рис. 5). Такие птенцы хорошо оперены,
хотя далеко еще не все перья, особенно на хвосте и крыльях, закончили
свой рост. На голове и спине у них сохраняются отдельные пушинки —
последние следы предыдущего наряда. К 3-недельному возрасту формиро-
вание первого перьевого (гнездового) наряда в общем заканчивается и
— 150 —
молодежь приступает к частичной линьке в зимнее перо. Необходимо
заметить, что у лесных «каменных» дроздов ювенальные маховые и руле-
вые прекращают свой рост позднее. К тому времени уже успевают сме-
ниться большие партии мелкого гнездового пера на зимнее.
Рис. Птенцы белогорлого дрозда.
Вверху — 2-дневный; внизу — 15-дневный.
Гнездовой наряд
Птенцовые признаки строения и поведения являются, как известно,
более примитивными, нежели признаки взрослых птиц, причем на ранних
стадиях развития могут проявляться некоторые черты, утраченные
— 151 —
взрослыми особями. Структура, окраска и рисунок перьевого покрова
птицы в онтогенезе изменяются обычно в сторону усложнения, что, ве-
роятно, отражает их филогению. Изучение возрастных нарядов представ-
ляет поэтому для нас особый интерес, как могущее дать очень много для
выяснения родства между отдельными видами и для выявления степени
их измененности по сравнению с исходными формами. Все это в свою
очередь способно пролить свет на общее направление эволюции группы.
Гнездовой наряд — самый первый перьевой наряд, надеваемый птен-
цами всех воробьиных. Прежде чем приступать к его характеристике,
рассмотрим кратко некоторые структурные особенности ювенального
контурного пера лесных «каменных» дроздов.
В опахале дефинитивного пера всех Monticola (в современном понима-
нии) хорошо различимы следующие три отдела: 1) видимая (контурная)
часть опахала, которая образована бородками первого порядка (rami)
и короткими жесткими бородками второго порядка или лучами (radii)
с отходящими от них шипиками или крючочками, обеспечивающими сцеп-
ление всего опахала; 2) пуховая часть, структурно очень напоминающая
пух, лежит ниже; в этом отделе отходящие от стержня пера бородки пер-
вого порядка несут хорошо развитые, очень длинные и мягкие radii,
лишенные каких-либо сцепляющих приспособлении; 3) самая нижняя
шелковистая часть опахала, которой свойственно недоразвитие лучен.
Оба последних отдела обычно скрыты на теле птицы под контурными ча-
стями соседних перьев и практически не участвуют в формировании
окраски покровов. 7 Очень существенным моментом является то, что ба-
зальная (пуховая) часть опахала пера развивается в онтогенезе по тем же
принципам, что и первичный пух, являющийся наиболее примитивным
структурным образованием покрова птицы, тогда как формирование апи-
кальной (контурной) части пера подчиняется совсем иным закономерно-
стям (Войткевич, 1962).
Перо ювенального наряда каменных дроздов даже при самом поверх-
ностном рассмотрении кажется более рыхлым и рассученным, чем дефини-
тивное. При 50—60-кратном увеличении удается увидеть, что в опахале
их гнездового пера представлены только два верхних отдела, из коих
пуховой занимает гораздо большую площадь, нежели контурный. У лес-
ных форм контурный отдел пера все же всегда очень хорошо выражен,
в то время как у скальных его размеры минимальны. М. saxatilis и М. so-
litarius имеют гнездовое перо, у которого почти вся видимая часть опахала
пуховая и только некоторые radii, отходящие у самой вершины, имеют
небольшую длину и сцеплены друг с другом посредством крючочков. Таким
образом, уже по наличию или отсутствию в опахалах ювенальных перьев
контурной части представители современного рода Monticola могут быть
без труда разделены на две группы.
Оказывается, что соотношение пуховой и контурной частей на опаха-
лах одноименных перьев неодинаково и у разных видов лесных «каменных»
дроздов. Относительно примитивным среди них по этому признаку следует
признать белогорлого дрозда — вид, у которого наряду с ювенальным
оперением, имеющим довольно большую, плотную контурную часть, на
груди и брюшке развиваются перья с сильно рассученными опахалами,
бородки которых не сцеплены друг с другом. Рыжебрюхого же дрозда,
гнездовое перо которого по своей структуре и рисунку приближается во
многом к оперению взрослой самки, можно расценивать как более моло-
дую форму.
При дальнейшей характеристике нарядов и при описании окраски, рисунка
и структурных особенностей пигментации перьев лесных «каменных» дроздов всюду
имеется в виду только дистальная (видимая) часть опахал.
— 152 —
Еще в гнездовом пере у трех рассматриваемых видов лесных «камен-
ных» дроздов четко выступают половые различия, проявляющиеся глав-
ным образом в иной пигментации маховых и рулевых, а также в отличной
тональности окраски всего мелкого оперения. У белогорлого дрозда
самцы и самки легко различаются с 9—10-дневного возраста, т. е. еще
в период пребывания в гнезде.
Ниже приводятся описания гнездовых нарядов лесных Monticola.
Самки в гнездовом пере
Белогорлый дрозд (М. gularis). Экземпляры коллекции ЗИН АН
СССР (22) 11 июля 1927 г., Приморье; 28 июля 1959 г., Амурская обл.
Все мелкое перо верха желтоватое или рыжевато-желтое (на мантии п
боках шеи светлее, почти палевое), с широкими черными вершинными
каймами. На пояснице к основному цвету примешиваются ржавые тона.
Верхние кроющие хвоста (надхвостье) густо-ржавые, без черных вершин-
ных оторочек. Оперение низа нежно-палевого цвета, с узкими, черными
вершинными каемками. Налегая друг на друга, такие перья придают
«чешуйчатость» рисунку зоба и груди. На горле и брюшке каймы на перьях
очень узки или вовсе отсутствуют, и потому эти участки выделяются своей
однотонной, почти лишенной поперечных полос окраской. В принципе
рисунок всего мелкого оперения одинаковый — палевое поле и черная
вершинная кайма, ширина которой варьирует в зависимости от топогра-
фии пера.
Маховые и рулевые бурые. Три последние пары второстепенных махо-
вых, а также все кроющие второстепенных маховых несут па внешних
краях опахал узкие желтовато-ржавые каймы. У части молодых самок
в основании наружных опахал большинства второстепенных маховых
бывает хорошо заметно круглое желтоватое пятно, образующее на раскры-
том крыле «зеркальце».
Синегорлый дрозд (М. cinclorhynchus). В моем распоряжении не было
гнездовых птенцов этого вида.
По данным Сибома (Seebohm, 1881), половые различия в окраске
покровов птенцов синегорлого дрозда проявляются в неодинаковой
окраске маховых и рулевых, цветность которых совпадает с их окраской
у взрослых особей. Этот автор не приводит подробного описания юве-
нального наряда и только указывает, что молодые своей пятнистой ок-
раской напоминают старых самок. Учитывая большое сходство взрослых
самок белогорлого и синегорлого дроздов, можно думать, что и в птенцо-
вом возрасте они мало отличимы. Быть может, у самок последнего вида
не бывает светлых пятен на второстепенных маховых и не столь отчетливо
заметно горловое пятно.
Рыжебрюхий дрозд (М. erythrogaster). Я не имела возможности держать
в своих руках самку этого вида в гнездовом наряде.
Джердон (.Terdon, 1862) пишет, что молодые самцы и самки М. erythro-
gaster отличаются друг от друга все той же окраской перьев хвоста и
крыльев; в остальном же полы окрашены сходно.
Более подробная характеристика гнездового наряда приведена ниже,
при описании покровов молодого самца.
Самцы в гнездовом пере
Белогорлый дрозд (М. gularis). Экземпляры коллекции ЗИН АН СССР
из Амурской обл. (d>0): 7 июля 1959 г., 17 июля 1957 г., 19 (2), 21 и
31 июля 1959 г., 9 августа 1959 г.
— 153 —
Окраска и рисунок мелкого контурного пера в общем повторяют
наряд молодой самки. Однако на сериях самцы в гнездовом пере выглядят
более темно и контрастно окрашенными, с меньшей примесью рыжеватых
тонов, которые заменяются палевыми.
Маховые молодых самцов буровато-черные. В основании наружных
опахал большинства второстепенных маховых (кроме 1-го, реже и 2-го)
у всех без исключения особей отчетливые чисто белые пятна, образую-
щие «зеркальце».8 Последние три пары второстепенных маховых, а также
все кроющие второстепенных маховых, как и у самок, с хорошо заметными
желтовато-ржавыми вершинными каймами. Рулевые перья черные,
с голубовато-аспидным налетом на верхней стороне.
Синегорлый дрозд (М. cinclorhynchus). В коллекциях наших зоологи-
ческих учреждений нет самцов этого вида в гнездовом пере.
У Бекера (Baker, 1924) находим, что молодой самец сходен с самкой,
только у него уже в раннем возрасте на крыльях и хвосте появляется голу-
бизна. Мне думается, что самцы этого вида в ювенальном пере должны
быть очень похожими на одновозрастных самцов белогорлого дрозда и
иметь те же «зеркальца», рыжее надхвостье и горловое пятно, возможно
не столь контрастное, как у М. gularis. Основные различия между самцами
этих несомненно близких видов сводятся в гнездовом наряде, по-видимому,
только к наличию у синегорлого дрозда заметной примеси голубого на
внешних опахалах маховых перьев.
Рыжебрюхий дрозд (М. erythrogaster). Экземпляр (<5), частично выли-
нявший из гнездового пера, в коллекции ЗИН АН СССР от 26 августа
1886 г. из Найни-Тал (Индия).
Мелкое оперение верха нежно-палевого цвета, с черными вершинными
каймами, от ширины которых зависит в общем тональность окраски
покровов на разных участках тела. Так, голова и шея выглядят более
светлыми, нежели спина; несколько темнее их оказывается ушная область,
а по краям век и на боках шеи вырастают перышки, почти вовсе лишенные
черных оторочек. Верхние кроющие хвоста такого же, как у молодых
белогорлых дроздов, темно-ржавого цвета, причем каждое перо имеет
черную кайму — свидетельство того, что для М. erythrogaster это прими-
тивный, постепенно утрачивающийся признак. В окраске низа преобла-
дают не палевые, а желтовато-бурые тона. Все перья, в том числе и на
брюшке, с черными концами; только в области горлового пятна оторочки
на опахалах сильно сужаются.
Маховые и рулевые буровато-черные, с интенсивным ультрамарино-
вым налетом, правда не столь блестящим, как у взрослых самцов. Послед-
ние партии второстепенных маховых и все верхние кроющие крыла не-
сут на концах палевые каемки.
В процессе изучения ювенальных нарядов трех азиатских лесных
Monticola удалось установить, что эти виды близки между собой не только
по строению, но и по рисунку и окраске гнездового пера и вместе с тем
прекрасно отличимы от скальных форм по следующим основным призна-
кам.
Во-первых, как уже упоминалось выше, л е с н ы е «к а м е н н ы е»
дрозды имеют ювенальные перья с довольно
хорошо выраженной контурной частью опахал,
у скальных же преобладает пуховая.
Во-вторых, у всех азиатских лесных видов четко
проявляется половой диморфизм, пол птенцов может
быть определен в последние дни их пребывания в гнезде; у М. solitarius
8 Счет всех маховых ведется в проксимальном направлении.
— 154 —
молодые самцы и самки различаются с большим трудом, а у М. saxatilis
и большинства африканских эндемиков они практически не отли-
чимы.
В-третьих, принципиально иная расцветка покровов у представителей
этих двух групп: отчетливое светлое горловое пятно,
«зеркальце» или его следы на крыльях, рыжее
надхвостье у нерыжехвостых форм, вершинные каймы
только на последних партиях маховых — у лесных Monticola; отсутствие
горлового пятна, «зеркальца», а также рыжего цвета на верхних кроющих
хвоста (у нерыжехвостых видов) и широкие светлые оторочки по краю
опахал всех без исключения маховых перьев — у скальных Monticola.
В целом все перечисленные особенности ювенальных покровов харак-
теризуют лесных «каменных» дроздов как группу более молодую, нежели
скальные каменные дрозды.
Обнаружение у птенцов рыжебрюхого дрозда — вида, не имеющего
во взрослом состоянии никаких следов рыжего цвета на дорсальной сто-
роне тела, — таких же, как у белогорлого дрозда, ржаво-рыжих перьев
на пояснице и надхвостье говорит о несомненной древности этого приз-
нака и о происхождении всех лесных Monticola от ка-
кого-то общего предка, верхние кроющие
хвоста у которого были рыжими.
Из современных представителей лесной группы М. gularis на этом
основании может быть признан наиболее близким к исход-
ной форме.
Линька из гнездового пера, как уже говорилось, начинается у лесных
«каменных» дроздов по достижении птенцами примерно трехнедельного
возраста. Протекает она очень бурно и охватывает только мелкое контур-
ное оперение. Первыми сменяются плечевые и мелкие кроющие крыла,
а также перья на спине и боках груди (экземпляры ЗИН АН СССР —
Я. gularis, S3, 19 и 20 июля 1959 г., Амурская обл.). Постепенно процесс
линьки захватывает перья на пояснице, горле и подмышечные (экземпляры
в коллекции ЗИН АН СССР из Амурской обл. — М. gularis, 8, 19 июля
1959 г., J» 28 июля 1959 г.). Находящиеся в материалах ЗИН АН СССР
молодые самцы М. gularis из Приамурья от 31 июля 1959 г. и М. erythro-
gaster из Индии от 26 августа 1886 г. с доросшими до нормы ювенальными
маховыми и рулевыми сменили уже большие партии перьев на брюшке,
горле, боках тела, спине, пояснице, а также мелкие кроющие маховых.
Гнездовое перо сохранилось у них преимущественно на голове, шее, груди
п надхвостье. В ЗИН АН СССР есть также молодая самка белогорлого
дрозда, добытая в Приморье 21 августа 1921 г., которая вступила в завер-
шающую фазу этой линьки. В конце августа—начале сентября боль-
шинство лесных «каменных» дроздов надевает наряд первогодков. У птен-
цов позднего вывода (к ним следует отнести самца М. gularis, доставлен-
ного М. Березовским из южной Ганьсу) смена перьев прекращается лишь
в октябре.
Первая в жизни настоящая линька является у лесных Monticola,
таким образом, частичной. Маховые и рулевые гнездового наряда сме-
няются только следующей осенью. По цветности оперения зимний наряд
первогодков такой же, как и зимний наряд взрослых птиц, и в этой связи
его можно считать окончательным. К весне в результате обнашивания
светлых каемок на вершинах перьев оба зимних наряда превращаются
в абсолютно не отличимые друг от друга по окраске брачные наряды
первогодков и старых особей. Качественно же они различны, ибо наряд
первых комбинированный (он образован новым мелким оперением и юве-
нальными маховыми и рулевыми), тогда как у вторых все перо новое,
сменившееся во время полной годовой линьки.
— 155 —
Подобная последовательность смены возрастных нарядов существует,
однако, не у всех представителей современного рода Monticola. Так же
как и азиатские лесные «каменные» дрозды, линяют синие каменные
дрозды (М. solitarius) и, по данным Фаркаша (Farkas, 1962а, 1962b, 1963),
некоторые южноафриканские формы (М. brevipes, М. rupestris, М. explo-
rator). Зато молодые пестрые каменные дрозды (М. saxatilis) обладают
двукратной линькой. Осенью, претерпев такую же, как у лесных «камен-
ных» дроздов, неполную линьку из гнездового пера, они надевают первый
зимний наряд первогодков, в котором полы едва различимы, но зато по
отсутствию голубого цвета на голове и шее прекрасно узнаются среди
зимующих птиц молодые самцы. Простой износ пера этого наряда еще не
может дать яркого и контрастного брачного наряда самцов и потому
в феврале—марте первогодки М. saxatilis частично линяют второй раз.
В коллекции ЗИН АН СССР есть два самца из Эфиопии от 18 и 20 марта
со следами еще незакончившейся весенней линьки. Новый наряд перво-
годков, назовем его предбрачным, несет теперь все признаки зимнего
наряда взрослых пестрых каменных дроздов, с той лишь разницей, что
у молодых светлые каемки на свежем весеннем оперении гораздо уже,
чем у старых осенью и зимой. Возможно, что в этом есть определенный
смысл, так как в распоряжении первогодков на формирование брачного
наряда остается очень мало времени, за которое не успели бы стереться
более широкие каймы. К началу размножения популяция пестрых ка-
менных дроздов внешне выглядит однородной, молодежь не отличима от
старых птиц. Очень вероятно, что дважды линяют и некоторые другие
скальные формы.
Выпадение у лесных «каменных» дроздов дополнительной предбрачной
линьки молодых иформирование у них окончательного
наряда сразу после гнездового — несомненный
шаг вперед по сравнению с М. saxatilis, имеющим примитивный
первый зимний наряд.
Наряды первогодков и взрослых птиц
На вторую осень своей жизни лесные Мо/г^со/а-первогодки одновре-
менно со взрослыми птицами проходят полную годовую линьку, после
которой обновляется все оперение. Я располагаю очень небольшим
материалом, характеризующим этот процесс только у одного вида —
М. gularis. Сроки линьки старых белогорлых дроздов находятся в прямой
связи со сроками их размножения и потому сильно растянуты. Так,
в Амурской обл. ни у одного экземпляра из числа добытых от выводков
с маленькими птенцами, даже в августе, не обнаружены признаки смены
пера. Не имела следов линьки и самка от 1 августа 1932 г., добытая
К. А. Воробьевым на р. Горин (материалы ЗМ МГУ). У пар, загнездив-
шихся раньше, обновление покровов происходило довольно интенсивно
уже во второй половине июля. Ниже приведен перечень известных мне
линных белогорлых дроздов.
17 июля 1957 г., Амурская обл. (ЗИН АН СССР) — мелкое опере-
ние очень обтерто, частью выпало; включились в линьку VIII первостепен-
ные маховые, IX и X маховые этого разряда — в виде пеньков.
3, 24 июля 1928 г., Приморье (ЗИН АН СССР) — очень обношенное
старое перо, многие мелкие перья выпали; VIII первостепенные маховые
отсутствуют, IX — развернувшиеся пеньки, X — нормальной длины.
5, 24 июля 1959 г., Амурская обл. (ЗИН АН СССР) — линяет мелкое
перо на горле, зобу, груди, плечах, а также мелкие кроющие крыла;
156 —
VIII первостепенные маховые и их кроющие развились примерно наполо-
вину, IX и X еще в чехлпках.
(5, 28 июля 1932 г., Приморье (ЗМ МГУ) — меняется мелкое перо, глав-
ным образом на надхвостье, плечах, спине и крыльях; VI первостепенные
2
маховые в виде пеньков, VII — развернувшиеся пеньки, VIII — у,
а IX и X — нормальной длины.
5?, 1 августа 1925 г., Читинская обл. (ЗИН АН СССР) — происходит
бурная смена мелкого пера, заметное количество свежих перьев па зобу,
боках тела и среди кроющих крыла; VI первостепенные маховые —лоп-
I 2 3
нувшие пеньки, VII — -, VIII — —, IX — нормы, а X —доросли
полностью.
Ежегодная линька белогорлых дроздов протекает в основном на ро-
дине этих птиц. Сентябрьские экземпляры, как правило, одеты в свежее
перо. Некоторые особи включаются в миграцию, не успев окончательно
перелинять, с недоросшими до нормы перьями хвоста.
Определить возраст лесных «каменных» дроздов во время их пребыва-
ния на зимовках практически невозможно, так как молодежь и взрослые
птицы в свежем пере окрашены идентично. Поздним летом, когда на местах
гнездовий тоже могут быть встречены полностью вылинявшие первогодки
и дрозды старше года, сменившие почти все мелкое оперение, «старики»
легко узнаются по сильно обтрепанным или линяющим маховым и руле-
вым.
Надеваемый осенью зимний наряд несет уже все признаки окраски
следующего (брачного) наряда, однако отличается от него тусклыми крас-
ками благодаря имеющимся на всех мелких перьях светлым каймам,
которые приглушают основные цвета. Часть оперения имеет иногда до-
полнительные, снашивающиеся к весне, предвершинные полосы или пятна,
которые являются рудиментами рисунка, существовавшего некогда у пред-
ков современных лесных Monticola. Удивительное сходство окраски
свежего пера старых птиц с нарядом первогодков дает основание предпо-
лагать, что у лесных «каменных» дроздов, как, впрочем, и у других пер-
натых, зимний наряд примитивнее брачного.
Преобразование рисунка пера, появление новых красок
на его опахале начинается обычно с вершинной
части, лежащей ниже зоны обнашивания. Весной,
в брачную пору, после того как сотрутся каймы зимнего наряда, эти
иначе окрашенные, ранее неприметные периферийные участки выступают
наружу и при случае могут послужить материалом для отбора.
Дегенерирующие, исчезающие цвета сохра-
няются в основании или пристержневом отделе
контурной части опахала и, как правило, бывают скрыты
под соседними перьями.
Яркое самцовое оперение знаменует собой конечную ступень развития
окраски определенного вида. Оно бывает настолько изменено, что уста-
новить по нему родство даже близких форм бывает зачастую очень сложно.
Вместе с тем сходная окраска может развиться параллельно у самцов
неродственных форм на совершенно разной основе. В этой связи, помимо
изучения возрастных и сезонных нарядов особей обоего пола, весьма
ценным оказывается выявление всякого рода ненормальностей, прояв-
ляющихся в окраске оперения. Представляют также интерес и признаки,
подверженные большой индивидуальной изменчивости (свидетельство
молодости или исчезновения признака). Сами по себе они не могут быть
использованы в диагностических целях, но зато могут дать очень много
— 157 —
для определения диапазона и направления возможных преобразований
окраски покровов птицы в процессе эволюции.
После этих некоторых предварительных замечаний, без которых
невозможен анализ фактических данных, содержащихся в последующих
разделах работы, перейду к описанию нарядов. Во избежание повторений
наряды первогодков и взрослых лесных «каменных» дроздов характери-
зуются одновременно. Не следует, однако, забывать, что при своем боль-
шом внешнем сходстве наряды эти все же представляют собой разные
этапы возрастных изменений покровов.
Самки в первом годовом и окончательном нарядах
Белогорлый дрозд (М. gularis) осенью и зимой в свежем пере. Экзем-
пляры ЗИН АН СССР: j juv. — 21 августа 1920 г. из Приморья; j ad. -
1 августа 1925 г. из Читинской обл.
Основная тональность окраски верха оливково- или желтовато-бурая.
Каждое перо с черной предвершинной поперечной полосой и золотистой
каймой. Оперение низа имеет тот же рисунок, но черные полосы на нем
значительно уже, а фон светлее, почти желтовато-белый, благодаря
чему отчетливо видна поперечная исчерченность зоба и груди. На горле и
брюшке перья вовсе лишены черных полос.
Маховые и рулевые, как и в ювенальном наряде, бурые, с значитель-
ной примесью охристо-бурого по краю наружных опахал. Три последние
пары второстепенных маховых и все большие кроющие перья крыла
с желтовато-ржавой оторочкой. Так же как и у молодых, у части взрослых
самок на наружных опахалах большинства второстепенных маховых есть
желтовато-белое округлое пятно, образующее крыловое «зеркальце»
(рис. 6, ах, AJ, которое у особей, его не имеющих, заменяется едва замет-
ной рыжевато-бурой полоской (рис. 6, я2, А2).
Брачный наряд самок всех видов лесных Monticola достигается за счет
простого стирания светлых вершинных каемок на перьях предыдущего
наряда, а также частичного износа и выцветания наиболее экспонирован-
ных участков опахал, без какой-либо дополнительной линьки. Благодаря
этому к весне меняется не только общая тональность окраски оперения, но
и его рисунок.
Белогорлый дрозд весной и летом в брачном наряде. Экземпляры:
1) с отчетливым крыловым «зеркальцем», — 29 мая 1962 г. и 6 июня
1962 г. из Хабаровского края (ЗМ КГУ); 4 июля 1961 г., 20 и 28 июля
1959 г. из Амурской обл. (ЗИН АН СССР); 2) с редуцированным «зеркаль-
цем», j2 — 20 мая 1959 г. из Хабаровского края (ЗМ КГУ); 6 июня 1930 г.
из Китая; 13 июля и 9 августа 1959 г. из Амурской обл. (ЗИН АН СССР).
У самки этого вида в брачном наряде почти целиком исчезает харак-
терный для зимнего пера желтовато-бурый налет и становятся преобла-
дающими на дорсальной стороне оливково-серые тона, а на вентральной —
чистые белые, палевые и черные. В общем рисунке покровов отчетливо
выступают участки, соответствующие разно окрашенным участкам опе-
рения самца: это верх головы, спина, поясница и надхвостье, уздечка,
«уши», бока шеи, горловое пятно, грудь и брюшко (рис. 7, А).
Верх головы (лоб, темя и затылок), дорсальная сторона шеи, плечи и
передний отдел спины (мантия) оливково-серые. На голове среди одно-
цветных перьев попадаются и с черными наствольями. На плечах и мантии
все оперение с черными концевыми пятнами. Поясница, надхвостье, бока
шеи и головы, а также весь низ одеты перьями, основная окраска которых
беловато-палевая или палевая. Различное соотношение фона и черных
полос обеспечивает различный рисунок опахал этих перьев, что в свою
очередь создает неодинаковую окраску определенных участков тела
158 —
Рис. 6. Крылья самок лесных «каменных» дроздов.
Рис. В. С. Рождественской.
4 _ белогорлого дрозда с четким крыловым «зеркальцем», — второстепенное махо-
вое такого крыла; Д2 — белогорлого дрозда с рудиментарным «зеркальцем», а2 — вто-
1 остспенное маховое такого крыла; Б — синегорлого дрозда; В — рыжебрюхого дрозда.
Рис. 7. Самки лесных «каменных» дроздов в брачном наряде
(схематизировано). Рис. В. С. Рождественской.
А — белогорлый дрозд; Б — синегорлый дрозд; В — рыжебрюхий дрозд.
птицы. Так, оперение на границе лба с надклювьем, уздечка, перья во-
круг глаз, на щеках, брюхе и подхвостье вовсе лишены каких-либо пятен
пи полос (случаются только отдельные перышки с пестринами); отсут-
ствует темный рисунок почти на всем оперении, покрывающем бока шеи.
лроющие ушей, перья на груди и боках тела с неширокими вершинными
займами, придающими чешуйчатость окраске этих областей. Зоб и бока
орла выглядят темнее, так как, кроме обычной черной оторочки, каждое
IX перо имеет широкую полосу того же цвета, проходящую поперек опа-
хала. Оперение поясницы и надхвостья наряду с вершинной несет еще
одну или две широкие черные полосы. В центре горла чисто белое поле
( виде неправильного треугольника.
Рис. 8. Самка белогорлого дрозда около гнезда с птенцами.
Рулевые оливково-серые. Маховые буровато-серые, с значительной
примесью охристо-бурого по краю наружных опахал. Светлые оторочки
на последних партиях второстепенных маховых отсутствуют, зато отчет-
ливо видна отмеченная ранее диморфность окраски оснований наружных
опахал этих перьев. На сериях удалось обнаружить, что изменение рас-
цветки опахал второстепенных маховых находится у самок белогорлого
дрозда в коррелятивной зависимости от других варьирующих признаков
в окраске их перьевого покрова. Например, у группы особей, имеющих
ярко выраженное крыловое «зеркальце», всегда хорошо представлены
горловое пятно, лишенная пестрин область на границе лба и надклювья,
а также светлые участки на боках шеи. У экземпляров же, у которых
присутствуют только следы «зеркальца», все перечисленные особенности
окраски несколько затушеваны, границы между темными и светлыми
участками сглажены.
Наблюдаемые индивидуальные отклонения в окраске мелкого опере-
ния самок, выражающиеся в общем в большем или меньшем развитии
темных или светлых (фоновых) красок, обусловлены разной степенью
износа опахал у птиц, живущих в различной экологической обстановке,
а также у экземпляров, добытых в разное время.
В принципе покровы взрослой самки обладают
цветностью, близкой к рисунку ювенального
И Биология птиц
— 161 —
оперения (рис. 8), что позволяет считать их относительно более при-
митивными, чем наряды самцов.
В виде отклонений от нормы в окраске самок белогорлого дрозда
иногда встречаются элементы яркого самцового оперения — признаки
так называемой «петухоперости». Мне известны два таких экземпляра:
1 августа 1932 г. из Приморского края (коллекция ЗМ МГУ), у кото-
рой почти все мелкие перья темени и затылка с примесью кобальтово-
голубого цвета; 6 июня 1930 г. из Китая (ЗИН АН СССР), с отдельными
голубыми перышками на голове.
Синегорлый дрозд (М. cinclorhynchus) осенью и зимой в свежем пере.
В СССР в коллекциях самки в этом наряде отсутствуют. Не удалось также
найти их описание в доступной мне литературе. По-видимому, осенью и
зимой оперение этого вида отличается от брачного наличием таких же.
как у самок белогорлого дрозда, дополнительных желтоватых каемок на
вершинах опахал, которые придают общий оливковый оттенок окраске
всех покровов.
Синегорлый дрозд весной и летом в брачном наряде. Экземпляр (j) —
от 8 мая 1875 г. из Кумаона (Индия) в коллекции ЗИН АН СССР.
Весь верх, в том числе и задняя часть спины с надхвостьем, оливково-
серый. В отличие от белогорлого дрозда у самок этого вида плечевые и
перья спины, так же как лоб, мантия, темя и затылок, без черных вершин-
ных пятен, которые развиваются только на пояснице и верхних кроющих
хвоста (рис. 7, Б). Перья низа беловато-палевые, с некоторой примесью
охристых тонов. На горле и брюшке они не имеют темных каемок; на бо-
ках головы и шеи перья несут узкие серовато-бурые оторочки; на зобу
и груди, а также га боках тела каймы на вершинах опахал становятся
несколько шире, однако не достигают размеров полос, имеющихся на
соответствующих мелких перьях у белогорлого дрозда. Горловое пятно
присутствует, но очертания его ре столь определенны и границы размы-
тые (рис. 7, Б).
Перья хвоста оливково-серые, маховые такие же или слегка буровато-
серые. В основании наружных опахал второстепенных маховых нет свет-
лого пятна, но на месте, соответствующем его положению, как и у части
самок М. gularis, имеется рыжевато-бурая полоска (рис. 6, Б), которая,
хотя и не бросается сразу в глаза, все же не может остаться незамеченной.
Интересно, что Сибом (Seebohm, 1881 321), располагавший гораздо
большим материалом по синегорлому дрозду, включил этот признак
даже в описание наряда взрослой самки.
Как и у предыдущего вида, у самок М. cinclorhynchus бывают случаи
частичной «петухоперости», выражающиеся в появлении среди перьев
низа отдельных ржаво-рыжих перышек, свойственных наряду взрослого
самца.
В целом самки синегорлого дрозда окрашены менее пестро и светлее
самок белогорлого дрозда.
Рыжебрюхий дрозд (М. erythrogaster) осенью и зимой в свежем пере.
Я располагала лишь одним экземпляром (р), добытым в 1862 г. в Непале
(фонды ЗИН АН СССР).
Как и у ранее описанных видов, у самок М, erythrogaster свежее опере-
ние выглядит довольно равномерно и однотонно окрашенным благодаря
дополнительным золотисто-серым оторочкам, развивающимся на его
опахалах и затушевывающим основные цвета.
Верх темно-оливково-серого или буровато-серого цвета. Голова и дор-
сальная сторона шеи без пестрин; спина, плечи, поясница и надхвостье
с чешуйчатым рисунком, образованным узкими, черными предвершинными
каймами на опахалах перьев. Низ тела имеет основную окраску охристо-
или желтовато-палевую. На каждом пере значительное развитие черного
— 162
цвета; на уздечке, вокруг глаз, на подбородке, горле и боках шеи — это
сравнительно узкие вершинные каймы, которые на ушных перьях и боках
горла становятся шире, оставляя свободной только пристержневую часть
опахал; остальные перья низа имеют одинаковый рисунок — каждое
с двумя широкими, черными поперечными полосами и золотистой (та-
кой же, как и на всех остальных мелких перьях) оторочкой.
Маховые и рулевые оливково-бурые. Все второстепенные маховые
без заметных вершинных каемок. В базальной части их наружных опахал
ржаво-бурые поля, почти невидимые в свежем пере.
Терминальные каймы зимнего наряда у рыжебрюхого дрозда отли-
чаются очень незначительной шириной и потому снашиваются, как пра-
вило, еще в середине зимы. Весной в так называемом брачном наряде
самки имеют оперение, опахала которого стираются настолько, что на
поверхность выступают многие особенности окраски, скрытые обычно
под черными предвершинными полосами налегающих друг на друга
перьев.
Рыжебрюхий дрозд весной и летом в брачном наряде. Экзем-
ляр (j) — от 17 мая 1875 г. из Индии (коллекция ЗИН АН СССР).
Весь верх, включая поясницу и надхвостье, такого же, как у синегор-
лого дрозда, оливково-серого цвета, с той лишь разницей, что у рассматри-
ваемого вида все перья на плечах, спине, пояснице и надхвостье имеют
черные настволья и неширокие каймы (рис. 7, В). Мелкое оперение на
границе лба с надклювьем, на уздечке, вокруг глаз, на подбородке, горле
п боках шеи после обнашивания концевых полос становится одноцветным
палево-белым. Бока горла, кроющие ушей и весь остальной низ выглядят
темно- и пестроокрашенными. Наличие на опахалах перьев, одевающих
эти участки тела, не одной, а двух широких черных полос обеспечивает,
даже при снашивании пера наполовину, присутствие на его конце чер-
ного цвета (рис. 7, В). Светлое горловое пятно и сильно исчерченный чер-
ным низ у самок М. erythrogaster бывают обязательно (Smythies, 1948).
Рулевые оливково-серые. Маховые буровато-серые, с рыжеватой ото-
рочкой по краю наружных опахал. В базальном отделе внешних опахал
большинства второстепенных маховых небольшие охристо-рыжие пятна,
соответствующие по своему положению белым пятнам, образующим кры-
ловое «зеркальце» у самок белогорлого дрозда (рис. 6, В).
У находящейся в ЗИН АН СССР самки рыжебрюхого дрозда среди
нормальных перьев спины и надхвостья попадаются ультрамариновые,
такие же, как у самцов. Вероятно, как и у ранее описанных двух видов
лесных «каменных» дроздов, «петухоперость» самок — не столь уж редкое
явление. Латуш (La Touche, 1925), например, заметил, что у особей, добы-
тых в Китае, примесь синего цвета бывает у самок на голове, хвосте и не-
которых других участках.
Из приведенных описаний видно, что, как и в ювенальном наряде,
взрослые самки М. gularis, М. cinclorhynchus и М. erythrogaster весьма
сходны по своей окраске. В противоположность самкам остальных видов
Monticola все они прекрасно отличаются от самцов в окончательном на-
ряде полным отсутствием в покровах яркого рыжего или
синего оперения (имеется в виду нормальная окраска взрослых
особей). Напомню, что у М. solitarius на маховых и рулевых перьях ста-
рых самок всегда заметен налет синего цвета; у М. saxatilis взрослые
самки, как и самцы, имеют совершенно одинаковые рыжие рулевые и верх-
ние кроющие хвоста; у трех африканских видов — М. angolensis, М. brevi-
pes и М. rupestris — рыжие не только хвосты, но и зоб, грудь и брюшко.
У самого маленького африканца — М. rufocinerea, — также относимого
современными систематиками к каменным дроздам, даже в брачном наряде
половой диморфизм сводится к незначительной разнице в тональности
— 163 —
И*
окраски мелкого оперения: у самок к голубовато-серому цвету на спине,
голове, горле и зобу примешиваются буроватые краски.
У самок лесных «каменных» дроздов в брачном наряде всегда есть
светлое горловое пятно и «зеркальца» или их следы на
крыльях, — признаки, не встречающиеся у самок других видов Monticola.
Большое сходство самок трех лесных азиатских
видов в окончательном наряде, так же как и почти
одинаковая окраска их в гнездовом пере, гово-
рит не только о несомненном близком родстве
этих птиц, но и об их относительной молодости
и недавней дифференциации.
Крыловое «зеркальце» в ряду лесных Monticola — дегенерирующий
признак. У самок рыжебрюхого дрозда, самцы которого лишены светлых
пятен на крыльях, в основаниях наружных опахал второстепенных
маховых имеются участки, идентичные по своему положению и окраске
тем буроватым полоскам, которые развиваются на месте «зеркальца»
у некоторых самок белогорлого дрозда. Это обстоятельство дает основание
рассматривать такие участки как следы некогда бывшего
у предков М. erythrogaster крылового «зеркальца».
Наряду с упомянутой тенденцией к исчезновению пятен на второсте-
пенных маховых, тоже от М. gularis к М. erythrogaster, постепенно просле-
живается параллельное изменение окраски мелкого оперения взрослых
самок. Оно сводится в общем к уменьшению горлового
пятна, общему посерению покровов дорсальной сто-
роны тела, распространению пятнистости на все
перья низа и усложнению их рисунка.
Самцы в первом годовом и окончательном нарядах
Л. М. Шульпин, собравший уникальную для своего времени коллек-
цию белогорлых дроздов, впервые обратил внимание на то, что в окраске
взрослых самцов этого вида довольно часто встречаются отклонения,
выражающиеся в варьировании размеров и конфигурации белого горло-
вого пятна и «зеркальца» на крыльях, в большем или меньшем развитии
черного цвета на перьях боков горла, шеи и груди, в разном соотношении
черного и рыжего на боках головы и спине, а также в появлении у некото-
рых особей голубого налета на перьях спины. Сам он не придал этому
серьезного значения и посчитал все наблюдаемые им случаи отклонениями
в окраске, являющимися «проявлением в той или иной степени рисунка
самки» (Шульпин, 1931 274).
На просмотренной мной большой (около 70 экз.) серии самцов М. gu-
laris удалось установить, что изменчивость этих признаков коррелиро-
вана и что в природе существуют два географически никак не локализо-
ванных типа окраски самцового оперения белогорлых дроздов. Очень
вероятно, что им соответствуют и наметившиеся ранее две группы самок.
В пределах ареала вида, в одних биотопах друг с другом живут, таким
образом, пары, состоящие из разно окрашенных самцов и самок.
Первую группу самцов белогорлого дрозда, у которых, как увидим
ниже, в окраске покровов получают большое развитие рыжие тона, на-
зовем условно «рыжей», а вторую — «черной». Между ними есть ряд
переходов, как бы отражающих разные стадии изменения пигментации
оперения на пути от группы № 1 к группе № 2.
«Рыжий» белогорлый дрозд (М. gularis) осенью и зимой в свежем пере.
Я располагала только одним экземпляром (3) от 24 сентября 1930 г. из
Китая (материалы ЗИН АН СССР).
~ 164 —
Рис. 9. Пигментация бородок первого и второго порядка в контурной части опахал оперения
самцов лесных «каменных» дроздов (схема).
Прерывистая линия — граница зоны износа бородок свежего пера при образовании брачного наряда»
Обозначения см. в тексте.
Верх головы и шеи («шапочка»), а также мелкие верхние кроющие
крыла грязно-кобальтово-голубые, без характерного блеска, благодаря
тому, что, помимо золотисто-бурой оторочки, свойственной большинству
свежих перьев, каждое голубое перо имеет в зимнем наряде неширокое
предвершинное черное пятно или кайму — рудиментарный признак
(рис. 9, A-IX). Оперение лба на границе с надклювьем, уздечка, перья
вокруг глаз, бока горла и шеи («зализы»), зоб, грудь, бока, задняя часть
спины, поясница и надхвостье ржаво-рыжие; брюшко и подхвостье
несколько светлее, почти рыжие. Все перья с узкими желтыми каемками,
придающими «чешуйчатость» окраске низа. «Зализы» вплотную подходят
к голубой «шапочке». Кроющие ушей в основании ржавые, однако большая
видимая часть их опахал черного цвета, с такими же, как и на черных
перьях дорсальной стороны шеи, плечевых и мантии, оливково-желтыми
или белесыми каймами. Среди черных перьев дорсальной стороны попа-
дается много ржавых с черными вершинами или целиком ржаво-рыжих
со светлой оторочкой. У некоторых «рыжих» самцов черные перья мантии
в зимнем наряде имеют двуцветную оторочку: по наружному краю
опахала проходит присутствующая обычно на всем свежем оперении
желтоватая кайма, а базальнее — ржавая — свидетельство того, что
у предковых форм рыжий (ржавый) цвет получал большее развитие
(рис. 9, A-VIII, в).9 Посредине горла — пятно, образованное белыми
перьями, часть из которых со слабым рыжеватым налетом.
Рулевые черные с голубовато-аспидным налетом на верхней стороне
опахал. Маховые буровато-черные с хорошо заметным светло-голубым
налетом по внешнему краю опахал. Три последние пары второстепенных
маховых, а также все кроющие маховых с палевыми каемками на верши-
нах. В основании наружных опахал всех, кроме I или I и II второстепен-
ных маховых, большие, четко ограниченные белые пятна, образующие
«зеркальце».
Брачный наряд самцов формируется в результате изменения окраски
и рисунка покровов, происходящего, как и в случае с самками, вне пе-
риода линьки, за счет обнашивания периферических отделов опахал.
Первыми обычно стираются светлые оторочки зимнего пера, затушевы-
вавшие основные яркие краски, а за ними — дополнительные предвер-
шинные пятна и каймы (рис. 9). Все цвета становятся в брачном наряде
чистыми, насыщенными. Голубые перья (у самцов М. erythrogaster —
ультрамариновые) приобретают к весне шелковистый отлив, который
обусловлен отражением лучей света от гладкой поверхности лишенных
radii дистальных участков бородок первого порядка (Войткевич, 1962).
Дело в том, что в голубых и синих перьях лесных «каменных» дроздов
меланин распределяется только в вершинах radii — апикально, по терми-
нологии Франка (Frank, 1939). Основания же бородок второго порядка
в месте их отхождения от ramus прозрачны, так как вовсе лишены пиг-
мента. При трении находящиеся на концах бородок первого порядка
лучи поэтому легко обламываются в своей депигментированной части,
освобождая, таким образом, более прочные голубые (или синие) ramus.
Необходимо отметить, что Франк — первый исследователь, имевший воз-
можность изучать окраску и структуру птичьего пера с помощью электрон-
ного микроскопа, — также обратил внимание на случаи частичной потери
radii на вершинах опахал. Из-за ограниченности материала (Франк имел
дело только с перьями брачного наряда) он не смог, однако, понять при-
чину возникновения подобных структур.
9 Ранее (Нейфельдт и Соколов/I960) эта особенность пигментации покровов сам-
цов белогорлого дрозда была ошибочно принята мной за признак возрастного димор-
физма в зимнем наряде.
- 165 —
«Рыжий» белогорлый дрозд весной и летом в брачном наряде (рис. 10).
Экземпляры (55): 26 мая 1961 г., 30 мая 1957 г., 3 июля 1961 г., 11 июля
1959 г., 17 июля 1957 г., 24 июля 1959 г. из Амурской обл. (ЗИН
АН СССР); 26, 27, 28 мая 1961 г., 27 и 29 мая. 1 июня 1962 г. из Хабаров-
ского края; 29 мая 1963 г. из Амурской обл. (ЗМ КГУ).
«Шапочка» и мелкие верхние кроющие крыла кобальтово-голубые
с атласным блеском. Оперение лба на границе с надклювьем, уздечка,
Рис. 10. «Рыжий» самец белогорлого дрозда
с кормом для птенцов.
перья вокруг глаз, на
боках горла и шеи, на зо-
бу, груди, боках тела,
задней части спины, пояс-
нице и надхвостье густо-
ржавые. «Зализы» обяза-
тельно распространяются
до голубой «шапочки»
(рис. 11, Ах). Брюшко и
подхвостье несколько свет-
лее — ржаво-рыжие. Бока
головы (уши) почти цели-
ком черные, однако скры
тая под широкими черны-
ми каймами часть опахал
ушных перьев сохранила
следы их первоначальной
(древней) ржавой окраски
(рис. 9, A-VII). В этой
связи особый интерес пред-
ставляет самец, добытый
в Северном Китае 5 июня,
у которого все перья
ушной области были ры-
жими, а черный цвет рас-
полагался у них только
по самому краю опахал
(Rensch, 1923). Верхняя
сторона шеи, плечи и
передняя часть спины
черные. Среди черных
перьев могут попадаться
и такие, пристержневая или базальная часть которых рыжего цвета, и даже
целиком ржавые перья, что говорит о некогда более широком распро-
странении рыжины на дорсальной стороне тела белогорлых дроздов
(рис. 9, A-VIII, а, б). На горле чисто белое пятно в форме несколько
растянутого треугольника.
Рулевые почти черные, сверху с голубовато-аспидным налетом. Махо-
вые окрашены так же, как и в предыдущем наряде, с той лишь разницей,
что последние партии этих перьев и их кроющие не имеют палевых кае
мок.
При переходе от группы «рыжих» к группе «черных» в окраске самцов
М. gularis хорошо прослеживается тенденция к постепенному замеще-
нию ржаво-рыжего цвета на боках головы, шеи и задней
части спины черным, а также к распространению чер-
ноты на рыжие участки боков горла, зоба и бо-
ков тела. Намечается также уменьшение размеров гор-
лового пятна, зарастание его рыжими и черными перьями, и в ка-
кой-то степени сокращение крылового «зеркальца».
- 166 —
В коллекциях оказалось довольно много самцов, которые по харак-
теру пигментации оперения могут быть отнесены к переходной группе.
Это следующие экземпляры: 24 мая 1928 г., 6 июня 1958 г., 10 июня 1958 г.,
И июня 1957 г., 25 июня 1957 г., 13 июля 1959 г. из Амурской обл.;
8 июня 1926 г., 10 июня 1926 г., 20 июня 1926 г. и 24 июля 1926 г. из При-
морья (материалы ЗИН АН СССР); 27 мая 1961 г., 27 мая 1962 г., 31 мая
1959 г., 31 мая 1962 г. и 2 июня 1962 г. из Хабаровского края; 29 мая (2)
и 2 июня 1963 г. из Амурской обл. (фонды ЗМ КГУ).
У всех этих птиц рыжие «зализы» на боках шеи уже не контактируют
непосредственно с голубой «шапочкой»; их разъединяет пока еще очень
узкая зона из черных перышек. На лбу и боках головы значительно увели-
чилось количество черного. На боках тела, пояснице, горле и зобу черные
перья еще единичны.
«Черный» белогорлый дрозд осенью и зимой в свежем пере. Экземпляры
коллекции ЗИН АН СССР: juv. <5 — 31 июля 1959 г. из Амурской обл.;
ad. 3 — 15 сентября 1948 г. из Приморья. К этой же группе следует от-
нести молодого самца, добытого М. Березовским в южной Ганьсу в начале
октября (материалы ЗМ МГУ), а также экземпляр от 7 сентября, добы-
тый А. И. Ивановым в восточной Монголии (Тугаринов, 1932), который, по
устному сообщению Б. К. Штегмана, был в свежем оперении и имел почти
заросшее горловое пятно при очень пестрой окраске верха.
Верх головы и шеи, а также верхние мелкие кроющие крыла такого же,
как у «рыжих» самцов, цвета. Оперение лба по границе с надклювьем и
уздечка черные, в виде незначительной примеси сохраняются ржавые
перышки. Вокруг глаз, на боках шеи, всей спине и плечах черные перья
с характерными для зимнего наряда оливково-желтыми каймами. Окраска
низа, поясницы и надхвостья густо-ржавая, на брюшке и подхвостье,
как и у особей первой группы, светлее. На пояснице, боках горла и зобу
многие рыжие перья имеют на концах черные пятна, расположенные вне
зоны обнашивания. Размеры горлового пятна сильно сокращены по срав-
нению с тем, что мы видели у «рыжих» самцов; масса рыжих перьев захо-
дит на белое поле и разбивает его на изолированные участки. Цветность
мелкого оперения не столь контрастна, как в брачном наряде, благодаря
золотистым оторочкам на вершинах опахал.
Маховые и рулевые такие же, как и в соответствующем наряде «рыжих»
птиц. Возможны вариации в величине и числе белых пятен, образующих
«зеркальце».
«Черный» белогорлый дрозд весной и летом в брачном наряде. Эк-
земпляры: 33 — 29 мая 1961 г., 31 мая 1961 г., 7 июня 1914 г., 6 июля
1928 г. из Амурской обл.; 26 мая 1927 г. и 20 июня 1926 г. из Приморья;
20 июня с п-ова Корея (материалы ЗИН АН СССР); 28 мая и 31 мая (2)
1962 г. из Хабаровского края; 29 мая 1963 г. из Амурской обл. (ЗМ КГУ).
«Шапочка» и мелкие кроющие крыла такие же, как и у представите-
лей предыдущей группы, — кобальтово-голубые с атласным блеском.
Оперение лба по границе с надклювьем, уздечка, перья вокруг глаз,
кроющие ушей, перья на боках и верхней стороне шеи, всей спине и
плечевые черные. Голубая «шапочка» нигде не соприкасается с рыжим
оперением (рис. 11, А2). У некоторых особей на уздечке и вокруг глаз
вырастают отдельные рыжие перышки — остатки бывшего здесь некогда
сплошь рыжего оперения (рис. 9, Б-V, а), а на черных перьях мантии
в качестве нового признака появляется сизый налет. В общем, у «черных»
самцов существует как бы сплошная черная маска, сливающаяся с чер-
ным же оперением спины. Бока горла у птиц этой группы по-прежнему
ржавые, но много перьев, у которых ржавой остается лишь базальная
пли пристержневая часть опахал, тогда как их вершины уже черного
цвета, бывают и целиком черные перья, которые образуют по обе стороны
— 167 —
горлового пятна (на месте находившихся там рыжих перьев) черные
«усы» (рис. 9, Б IV, б, в; 11, А2). Черный цвет распространяется и
на некоторые ржавые перья зоба, боков тела, поясницы и надхвостья, где
пока занимает только предвершинную, не обнашиваемую часть опахал
(рис. 9, Б-I, б, Б-П, б). Весь остальной низ остается ржавым, более
светлым на брюшке и подхвостье. Горловое пятно белое, но оно сильно
сужено и деформировано по сравнению с тем, что мы видели у «рыжих»
самцов. Часть его поля заросла белыми перьями, концы которых имеют
рыжие пятна, много целиком рыжих перьев и даже рыжих с черными
вершинами; попадаются на горле и совершенно черные перья. Подобная
полиморфность окраски перьев на одном небольшом участке как бы от-
ражает возможный путь перехода от «рыжих» форм к «черным»
(рис. 9, Б-Ill, а—д).
Рулевые и маховые окрашены так же, как и у других самцов этого
вида. Разница лишь в том, что у «черных» особей на I и II, а иногда
на I—IV второстепенных маховых отсутствуют белые пятна, образующие
крыловое «зеркальце».
Довольно широкий диапазон вариаций окраски взрослых самцов бе
логорлых дроздов в пределах каждой группы обусловлен неравномерным
стиранием и неодинаковым выцветанием оперения у разных особен.
Синегорлый дрозд (М. cinclorhynchus) осенью и зимой в свежем пере.
В коллекции ЗИН АН СССР только один экземпляр (5), добытый
в 1877 г. в Индии.
Верх головы и шеи, мелкие верхние кроющие крыла и все горло ко
бальтово-голубые с оливково-желтым налетом благодаря каймам на всех
перьях; на «шапочке» некоторые перья, как и у белогорлого дрозда,
зимой имеют еще дополнительные предвершинные черные пятнышки
(рис. 9, B-III, IV, IX). Уздечка, бока головы и шеи, спина и плечи чер-
ные, с чешуйчатым (зимним) рисунком, образованным желтоватыми кай-
мами на концах перьев. Поясница, надхвостье и весь низ рыжие, с та-
ким же скобчатым рисунком.
Перья крыльев буровато-черные с голубоватым налетом не только на
наружных опахалах первостепенных маховых, но и на второстепенных
и многих кроющих крыла. «Зеркальце» хорошо выражено. Последние три
второстепенных маховых и все кроющие маховых, как и у предыдущего
вида, с рыжевато-желтыми оторочками, снашиваемыми к весне.
Синегорлый дрозд весной и летом в брачном наряде. Экземпляры
ЗИН АН СССР (SS): 1863 г., Непал; 8 мая 1875 г., Алмора (Индия).
«Шапочка», мелкие верхние кроющие крыла и все горло кобальтово
голубые с атласным блеском. Оперение лба на границе с надклювьем,
уздечка, перья вокруг глаз, кроющие ушей, бока шеи, плечи и спина
такие же черные, как и у «черных» самцов М. gularis. Зоб, грудь и весь
остальной низ, а также поясница и надхвостье однотонные ржаво-рыжие
(рис. И, Б).
Рулевые буровато-черные, на них, как и на перьях мантии, бывает
заметный голубой налет. Маховые такого же цвета, причем все второ-
степенные маховые, кроме I, с белыми пятнами в основании наружных
опахал.
Как видно, самец синегорлого дрозда очень похож на самцов «черных»
белогорлых дроздов. Главное отличие между ними заключается в том,
Рис. И. Самцы лесных «каменных» дроздов в брачном наряде (схематизировано).
Рис. В. С. Рождественской.
Ai — белогорлый дрозд («рыжий» тип); А2 — белогорлый дрозд («черный» тип); Б — синегорлый
дрозд; В — рыжебрюхий дрозд. 1 — белый цвет; 2 — ржаво-рыжий; з — ржавый; 4 — каштаново-
красный; 5 — черный; 6 — кобальтово-голубой с атласным блеском; 7 — ультрамариновый с атлас-
ным блеском; 8 — черновато-синий без блеска.
— 168 —
что у М. cinclorhynchus горло имеет голубую окраску, занимающую об-
ласть, соответствующую горловому пятну М. gularis и лежащим по бокам
от него участкам рыжего цвета.
Рыжебрюхий дрозд (М. erythrogaster) осенью и зимой в свежем пере.
Экземпляры коллекции ЗИН АН СССР (33): 31 января 1959 г. из Юнь-
нани, зима 1869/70 г. из Дарджилинга и две птицы без дат из Гималаев.
Общий тон окраски верха ультрамариновый. На пояснице, надхвостье
и мелких кроющих крыла краски чистые, так как даже в зимнем наряде
перья на этих участках не имеют светлых оторочек (рис. 9, Г, II). Опе-
рение верхней стороны головы («шапочка») и шеи с менее заметными,
нежели у двух предыдущих видов, предвершинными черными пятныш-
ками и оливково-палевыми или палевыми каймами, придающими ему
грязный оттенок (рис. 9, Г-IX). На спине и плечах, окрашенных в черно-
вато-синий цвет, каждое перо на вершине имеет такую же, обнашивае-
мую к весне двуцветную кайму, создающую чешуйчатый рисунок мантии.
Горло матово-синего цвета, концы перьев с типичными зимними каем-
ками. Остальной низ каштаново-красный; вскоре после осенней линьки
на многих перьях бывают видны узенькие палевые оторочки, которые
стираются еще зимой. Оперение вдоль границы лба с надклювьем, уз-
дечка, перья вокруг глаз и кроющие уха черные, с едва заметными
каемками. Среди ушных перьев бывают и такие, у которых присутствует
синий цвет в окраске краев опахал. Такой же легкий синеватый налет
имеют, как правило, черные перья на боках шеи.
Рулевые с вентральной стороны буровато-черные, а с дорсальной
ультрамариново-синие. Маховые буровато-черные. Видимые при сложен-
ном крыле участки опахал этих перьев ультрамаринового цвета. Послед-
ние партии второстепенных маховых и все кроющие маховых перьев
в зимнем наряде с палевой вершинной каймой. «Зеркальце» или его
следы отсутствуют.
Первогодки и полностью вылинявшие старые птицы у рыжебрюхого
дрозда в свежем пере практически не различимы. И те, и другие имеют
на вершинах второстепенных маховых и их кроющих неширокие пале-
вые каймы, которые очень скоро стираются и в середине зимы уже не-
заметны. Сибом (Seebohm, 1881), не имевший, по-видимому, возможности
сравнивать наверняка разновозрастных самцов в свежем осеннем наряде,
ошибочно считал, что этот признак (каймы на второстепенных маховых)
может быть использован для определения молодых. Единственное, что
привлекло мое внимание при просмотре экземпляров М. erythrogaster
коллекции ЗИН АН СССР, это то, что тональность оперения верхней
стороны тела у упомянутого самца из Юньнани не ультрамариновая,
а кобальтово-синяя, приближающаяся к таковой М. gularis и М. cinclor-
hynchus. Мне неизвестен возраст этой птицы. Если бы она была действи-
тельно первогодком, то такая особенность ее окраски представляла бы
определенный интерес, как возможная переходная ступень в эволюции
нарядов самцов лесных «каменных» дроздов. Нельзя не заметить, что и
при рассмотрении в 50—60-кратную лупу бородки первого порядка опа-
хала синего пера юньнаньского экземпляра оказались по своей окраске
почти не отличимыми от rami перьев «шапочки» М. gularis и вместе с тем
они были совсем непохожи на соответствующие бородки покровов взрослых
М. erythrogaster.
Рыжебрюхий дрозд весной и летом в брачном наряде. В коллек-
ции ЗИН АН СССР хранятся три экземпляра (33) в этом наряде: без
даты из Дарджилинга, 1862 г. из Непала и 1877 г. из Сиккима.
После обнашивания концевых каемок на перьях зимнего наряда, за-
тушевывавших основные краски, к весне у рыжебрюхих дроздов стано-
вятся отчетливо видными все те основные отделы, которые соответствуют
— 170 —
разно окрашенным участкам перьевого покрова самцов М. gularis и
М. cinclorhynchus, хотя цветность их иная (рис. 11, В).
В брачном наряде верх головы и шеи, мелкие кроющие крыла, по-
ясница и надхвостье ультрамариновые с атласным блеском. Горло тускло-
синее, так как в опахалах одевающих его перьев у бородок первого по-
рядка не обламываются наполненные меланинами лучи — radii
(рис. 9, Г-Ш, IV). Оперение лба, вокруг глаз и на уздечке, кроющие
ушей и перья на боках шеи черные. Каудальная часть шеи, плечи и спина
черновато-синие, поскольку к цвету синих rami примешивается чернота
заполненных меланинами и потому не снашиваемых radii (рис. 9, Г-VI,
VII). Вообще «посинение» этих перьев произошло, видимо, относительно
недавно и процесс этот далеко не завершен, так как во многих бород-
ках первого порядка еще чередуются черные и структурно иные синие
участки. Зоб, грудь, брюшко, бока тела и нижние кроющих хвоста каш-
таново-красные .
У ныне живущих рыжебрюхих дроздов в нарядах взрослых самцов
и первогодков белый цвет полностью отсутствует. У самок мы ранее об-
наружили горловое пятно и следы «зеркальца», говорящие о том, что
этот вид происходит от форм, обладавших названными признаками.
Очень ценным в этой связи является оставшееся почему-то незамеченным
небольшое примечание, сделанное Джердоном (Jerdon, 1862 515) к опи-
санию М. erythrogaster, в котором он говорит, что попадаются взрослые
самцы, у которых на горле заметен белый участок, образованный свет-
лыми основаниями некоторых перьев. Позднее Давид и Устале (David
et Oustalet, 1877) подтвердили, что в отдельных случаях, в виде аномалии,
у самцов рассматриваемого вида в центре синего горла бывает белое
пятно. Этот несомненно атавистический признак, изредка появляющийся
у рецентных рыжебрюхих дроздов, является еще одним фактом, подтвер-
ждающим, что такие на первый взгляд внешне различные птицы, как
М. gularis и М. erythrogaster, на самом деле очень близки филогенети-
чески.
Во введении к настоящей работе уже говорилось, что некоторые авторы
сближают М. erythrogaster с рыжебрюхими азиатскими формами синих
каменных дроздов, придавая решающее значение пестрой (каштаново-
рыжей с синим) окраске тех и других. Изучение нарядов этих птиц и их
изменений в онтогенезе лишний раз убеждает меня в том, что лесные
рыжебрюхие дрозды и М. solitarius philippensis имеют весьма поверхно-
стное сходство и что их окраска развилась независимо на совершенно
разной основе.
Окраска мелкого оперения рыжебрюхих дроздов варьирует довольно
значительно, что, как и в случае с М. gularis, следует связывать с разной
степенью изношенности и выцветания опахал. Среди дроздов этого вида,
добытых в одной местности на сравнительно небольшой территории, можно
найти экземпляры, прекрасно отличающиеся друг от друга большей или
меньшей насыщенностью каштанового цвета в покровах низа или разной
степенью развития синего на горле. Наверно, на основании этих при-
знаков, обусловленных, как выяснилось, разным состоянием пера и
варьирующих независимо от географического распространения отдельных
особей (Rand a. Flemming, 1957), был в свое время описан особый под-
вид М. erythrogaster sinensis (A. Meinertzhagen, 1927), признанный немно-
гими специалистами.
Заканчивая описание нарядов взрослых самцов лесных «каменных»
дроздов, необходимо подвести некоторые итоги.
При всем кажущемся на первых порах различии в окраске их брачного
оперения самцы трех рассмотренных лесных видов имеют много черт
внешнего сходства, отличающих их одновременно от самцов скальных
— 171
форм. В качестве таких основных признаков могут быть названы: 1) обя-
зательное развитие у лесных Monticola черного цвета на
боках головы и шеи; 2) полное отсутствие в окра-
ске рулевых перьев рыжих тонов; 3) наличие изолирован
ного горлового пятна (белого или синего), не контак-
тирующего непосредственно с синим оперением других
частей тела птицы; 4) присутствие на дорсальной стороне обособлен-
ных участков (верх головы и предплечья), покровы которых пре-
красно выделяются среди остального оперения, даже при сходной
окраске, своим атласным блеском (рис. 11).
В ряду лесных «каменных» дроздов по окраске и рисунку перьевого
покрова наиболее примитивными следует считать
«рыжих» самцов белогорлого дрозда, а более всего
уклонившимися от исходной формы — самцов рыжебрюхого дрозда. Из-
менение окраски самцового оперения в группе лесных Monticola обна-
руживает тенденцию к распространению пигмента в депигментированпые
перья горлового пятна и «зеркальца» с последующим усилением мелани-
новой пигментации (от рыжей до черной) и переходом к синей окраске
этих участков, к редукции рыжего цвета на перьях дорсальной стороны
и боков головы и замене его сперва черным, а затем синим цветом раз-
личной тональности, к потемнению окраски низа (от рыжей до каштаново-
рыжей).
Ржавый, ржаво-рыжий, каштаново-рыжий (или каштаново-красный)
и черный цвета, преобладающие в окраске оперения самцов лесных
Monticola, представляют собой последовательные стадии окисления одного
пропигмента — меланогена. В противоположность этим, так называемым
химическим цветам синяя окраска перьев достигается за счет перестройки
структуры клеток в бородках первого порядка контурной части опахал.
Меланиновая пигментная подкладка в комбинации с такими клетками
и создает различные варианты голубой и синей окраски.
Известно, что темпы пигментообразования в перьях находятся в за-
висимости от уровня происходящих в организме птицы окислительных
процессов, характер и интенсивность которых меняются с изменением
условий окружающей среды, в первую очередь температуры, влажности,
освещенности и кормов (Войткевич, 1962). В обстановке влажного суб-
тропического или тропического климата южной Азии, в которой, по-ви-
димому, обитали предки лесных «каменных» дроздов и в которой до сих пор
постоянно живут оседлые М. erythrogaster, меланогенез должен был, та-
ким образом, протекать значительно быстрее, чем в гораздо более суро-
вых, подверженных сезонным колебаниям климата условиях современного
ареала М. gularis. Совершенно ясно, однако, что переход в процессе
эволюции группы от одной пигментной окраски оперения к другой занял
намного меньше времени, чем весьма сложный и трудный этап превра-
щения такой окраски в структурную. В этой связи становится очевидным,
что при кажущемся на первых порах весьма незначительном различии
в окраске белогорлого и синегорлого дроздов на самом деле эти виды
отстоят друг от друга по этому признаку дальше, чем синегорлый и
рыжебрюхий дрозды.
При рассмотрении пигментации не целых перьев, а лишь отдельных
структурных элементов (ramus и radii) контурной части их опахал у сам-
цов трех видов лесных Monticola удается составить почти непрерывный
ряд постепенного перехода от чисто белой окраски через рыжую и черную
к структурным голубой и синей, отражающий в общих чертах эволюцию
окраски оперения от типа «рыжих» М. gularis до М. erythrogaster
(рис. 9, A-III, а, б; Б-Ш, а-д\ Б-IV, а-в\ Б-V, а, б; Б-VI-
VIII-, B-V-VIII-, Г-V, VII, а, б; Г-VI, а, б; Г-VIII, III, IV, II, /А).
— 172 —
Анализ нарядов самцов и самок трех современных азиатских лесных
видов, а также их возрастных и сезонных изменений позволяет сделать
следующие весьма важные для понимания эволюции группы выводы:
1) б е л ы е «зеркальца» на крыльях и такого же цвета горло-
вое пятно, атласно-голубые мелкие кроющие перья
крыла и «шапочка», р ж а в о - р ы ж и е брюшко, пояс-
ница и надхвостье с одновременным значитель-
ным распространением этого цвета на заднюю часть
спины, бока головы и шеи — признаки филогенетически
наиболее древние, в той или иной степени свойственные общей
предковой форме лесных Monticola', 2) из ныне живущих представителей
группы только некоторые («рыжие») взрослые самцы
М. gularis сохранили в своем внешнем облике все
эти архаические черты. У самцов остальных («черных»)
М. gularis, всех М. cinclorhynchus и М. erythrogaster большинство пере-
численных признаков в дефинитивном пере не столь отчетливо выражено
пли вовсе утрачено, однако следы их хорошо прослеживаются в нарядах
самок и гнездовых птенцов.
Сказанное заставляет считать окраску типа
«рыжих» самцов белогорлого дрозда относи-
тельно наиболее примитивной, близкой к ис-
ходной. Предки синегорлого дрозда произошли,
по всей вероятности, от форм типа «черных»
М. gularis и в свою очередь дали начало рыжебрю-
хим дроздам, самцы которых в настоящее время
несут черты наибольшей измененности в окра-
ске перьевого покрова.
Некоторые особенности внешнего строения
По сравнению с типичными скальными формами, такими, например,
как М. saxatilis и М. solitarius, все лесные «каменные» дрозды имеют
довольно закругленное крыло с первым маховым, обязательно далеко вы-
дающимся за кроющие кисти, относительно длинный, слегка ступенча-
тый хвост и укороченные плюсны. Все это — признаки, характеризую-
щие внешние морфологические адаптации к жизни в лесных условиях.
Группа лесных Monticola развивалась в довольно однородной обста-
новке лесов субтропической и умеренной зоны и в этой связи слабо ди-
вергировала. Некоторые наблюдаемые различия в строении отдельных
видов этой группы находят свое объяснение в особенностях современного
образа жизни птиц.
Так. среди них рыжебрюхий дрозд, обитающий оседло в исконных
лесных биотопах, обладает самыми закругленными и широкими крыльями,
длинным хвостом и укороченными цевками. Вершину его крыла обра-
зуют V и IV маховые и не доходящее до их концов всего на 2—3 мм
III маховое; I маховое на 10—16 мм выдается за концы кроющих
кисти и практически достигает половины длины крыла (рис. 6, В). От-
носительная ширина такого крыла (отношение длины I второстепенного
махового к длине крыла, в °о) у семи самцов — 77.9, у двух самок —
74.7. Размеры хвоста коррелированы с шириной крыла; относительная
длина хвоста (отношение длины хвоста к длине крыла, в ?й) у тех же
самцов — 85.2, а у самок — 82.5.
Два других вида тоже тесно связаны с лесом, однако с наступлением
холодов они предпринимают перелеты, во время которых преодолевают
по воздуху большие расстояния, а на отдыхе и кормежках нередко ока-
зываются в несвойственных для представителей группы стациях.
— 173 —
И М, cinclorhynchus, и М. gularis имеют в общем идентичные повадкп
и сходное благодаря этому строение. По сравнению с М. erythrogaster
они выглядят более узко- и длиннокрылыми, относительно короткохво-
стыми и длинноногими. Вершины крыльев этих птиц образованы IV и
III маховыми, разрыв в длине между которыми и соседним наружным
II маховым довольно значителен (10—14 мм); I маховое по-прежнему
длиннее кроющих кисти, но выступает за их концы лишь на 3—6 мм
(рис. 6, А19 Л2, Б). Относительная ширина крыла у М. cinclorhyn-
chus $$ (3) — 71.8, $ — 70.1 ; относительная длина хвоста тех же особей
67.0. Те же показатели у М. gularis: для крыла 5S (10) и (4) — 68.0,
а для хвоста — соответственно 65.7 и 64.5.
Известно (Штегман, 1961), что эволюция острого птичьего крыла в ту-
пое с увеличением длины I махового невозможна. В ряду лесных «ка-
менных» дроздов преобразование летательного аппарата могло, таким
Изменение длины первостепенных маховых перьев в онтогенезе
Возраст, пол и число изученных экземпляров Формула крыла Отношение длины каждого из шести первостепенных маховых к самому длинному из них, %
I п ш IV V VI
Белогорлый дрозд 17-дневный 3. V>IV=VI>III>II>I 46.5 84.9 91.0 98.6 100.0 98.6
20-дневный <5. V>IV>VI>III>II>I 41.0 82.1 93.0 99.0 100.0 98.0
22—23-дневный <5. V^IV>III>VI>II>I 37.0 81.0 95.0 100.0 100.0 94.0
24-дневный d. IV^V>III>VI>II>I 31.0 84.0 95.5 100.0 99.2 91.0
25-дневный d. IV>V^III>VI>II>I 30.0 87.6 97.3 100.0 97.9 88.2
Полностью дорос- шие juv. do и 30.1 87.7 100.0 100.0 94.5 84.9
2$ в гнездовом пере. IV=III>V>II>VI>I
Первогодок d. 30.5 89.0 100.0 100.0 90.4 85.6
Взрослые (3 dd). 30.0 86.4 100.0 100.0 92.0 83.6
Рыжебрюхий дрозд Взрослые (5 dd). V=IV>III>VI>II>I 36.0 81.3 96.9 100.0 100.0 95.8
образом, идти только в направлении М. erythrogaster -> М. gularis. Под-
тверждение этому мы находим также и при сравнении формы крыла
разновозрастных птенцов наиболее острокрылого среди современных лес-
ных Monticola белогорлого дрозда. Оказывается, что в процессе индиви-
дуального развития птенца рост первостепенных маховых происходит
у него неравномерно. У недельного пуховичка находящееся в чехлике
I маховое по своей абсолютной длине равняется 1/3 длины соответствую-
щего пера взрослой особи, в то время как остальные (II—VI) маховые
едва достигают 1 /7 нормальной длины. У 17-дневного дрозденка дисталь-
ное маховое почти достигает размеров, свойственных взрослым, и состав-
ляет при этом половину длины самого длинного V махового, образующего
в этом возрасте вершину крыла. В целом крыло гнездового птенца имеет
сильно закругленную вершину (рис. 12, Л). Как видно из приведенной
ниже таблицы, в 22—23-дневном возрасте птенец белогорлого дрозда про-
ходит стадию, аналогичную взрослому рыжебрюхому дрозду, когда вер-
шину крыла образуют V и IV маховые (рис. 12, Б). Постепенно, по мере
роста птенца и удлинения маховых, крыло становится несколько острее;
IV маховое приближается по длине к V и вскоре обгоняет его. В 25-днев-
ном возрасте III маховое почти равно V, которое затем начинает отста-
вать в росте и у взрослых особей не участвует в образовании вершины
— 174 —
крыла (рис. 12, В, Г). Сказанное позволяет предполагать, что заост-
ренное крыло М. gularis эволюционировало из типа
крыла М. erythrogaster и что тип лесной оседлой на-
секомоядной птицы являлся, таким образом,
первичной жизненной формой для всех лесных
«каменных» дроздов.
Среди ныне живущих представителей группы рыжебрюхий дрозд,
сильно уклонившийся от исходной формы по своей окраске, сохранил
образ жизни и общий тип строения, близкие к предковым, тогда как
синегорлый и белогорлый дрозды — виды, обнаруживающие примитив-
ность в структуре и пигментации покровов — в
этом отношении более изменились.
Что же касается размеров лесных Monticola,
то самым крупным из них является М, erythrogaster.
Вес добытых в Ассаме двух самцов, по данным
Рипли (Ripley, 1952b), 56.0 и 59.0 г, а самок — 48.0
и 56.0 г. Длина крыла взрослых особей: 88 (7) —
118.0—125.0 мм, 25 (2) — 116.0 и 118.0 мм (мате-
риалы ЗИН АН СССР); по Бисвасу (Biswas, 1961),
38 (4) - 121.5-129.0 мм, 25 (5) - 117.0-122.0 мм.
Длина хвоста тех же экземпляров соответственно:
88 — 100.0—110.0 мм, 29 — 95.0—96.0 мм; 33 —
100.0—105.0 мм, 52 — 96.0—100.0 мм. Абсолютная
длина клюва (от оперения лба) у хранящихся в ЗИН
АН СССР рыжебрюхих дроздов: 88 (7) — 18.6—
20.0 мм, 22 (2) — 18.0 мм, а длина цевки у них же
соответственно 24.6—27.0 и 27.0 мм.
Два других вида — М. cinclorhynchus и М. gu-
laris — несколько мельче. Палудан (Paludan, 1959)
пишет, что пять самцов синегорлого дрозда из Афгани-
стана весили 29.0—33.0 г, в среднем 31.5 г. Вес взрос-
лых белогорлых дроздов из Амурской обл.: 88 (12) — 32.0—37.3 г,
средний — 34.3 г; 2$ (5) — 32.2—40.2 г, средний — 36.4 г; из Китая
(Shaw, 1936): 88 (41) — 29.0—41.0 г, средний — 34.0 г; 22 (12) —
33.0—43.0 г, средний — 36.0 г. Главнейшие промеры этих птиц: М. cin-
clorhynchus — длина крыла 88 (3) 102.0—104.0 мм, 9 — 97.0 мм (мате-
риалы ЗИН АН СССР); по Бисвасу (Biswas, 1961), 8 — 105.0 мм,
22 (4) — 97.0—101.0 мм; абсолютная длина хвоста тех же особей соот-
ветственно: 88 — 69.0 мм, 5 — 70.0 мм; о — 64.0 мм, 22 — 65.0—66.5 мм;
длина клюва у хранящихся в ЗИН экземпляров: 38 (3) — 17.0—18.0 мм,
2 — 18.0 мм, а цевки соответственно 25.0—25.7 и 24.7 мм; М. gularis
(материалы ЗИН АН СССР) — длина крыла 88 (10) 96.0—102.5 мм,
29 (4) — 95.0—97.0 мм; длина хвоста соответственно 62.0—69.0 и 62.0—
65.0 мм; длина клюва 15.5—17.5 и 15.5—17.0 мм, а длина цевки 22.5—
24.5 и 23.0-24.4 мм.
Половой диморфизм в размерах очень незначителен; самцы несколько
крупнее самок, хотя вес последних, особенно в период кладки яиц, может
немного превышать вес самцов.
О РОДОВОЙ САМОСТОЯТЕЛЬНОСТИ ЛЕСНЫХ «КАМЕННЫХ» ДРОЗДОВ
Изучение всех перечисленных материалов не оставляет сомнения
в том, что в современный род Monticola искусственно (главным образом
на основании некоторых черт самого общего внешнего сходства между
самцами в брачном наряде) объединены птицы, весьма различные по об-
Рис. 12. Изменение
формы крыла бело-
горлого дрозда в
онтогенезе (схема).
А — 17-дневный птенец;
Б — 22-дневный птенец;
В — 25-дневный птенец;
Г — взрослая птица.
- 175 —
разу жизни, повадкам, особенностям индивидуального развития, морфо-
логии и своему происхождению и что поэтому его необходимо разбить
по крайней мере на два самостоятельных рода.
К первому из них — роду Каменных дроздов (Monticola) — относятся
африканские эндемики — М. rupestris (Vieillot), М. explorator (Vieillot),
М. brevipes (Waterhouse), M. rufocinerea (Ruppell), M. angolensis Sousa,
а также распространенные в Северной Африке и довольно широко на
юге Евразии М. saxatilis (L.) и М. solitarius (L.). Поскольку в доступных
мне коллекциях африканские виды были представлены неполностью и
зачастую единичными экземплярами, то судить о филогенетических свя-
зях и путях эволюции внутри этой группы не представляется возможным.
Однако на основании того, что удалось узнать о каменных дроздах по
имеющимся материалам и литературе, создалось впечатление, что и в та-
ком объеме Monticola гетерогенны по своему составу. Кажется, например,
очень сомнительной принадлежность к каменным дроздам таких птиц,
как М. rufocinerea и, вероятно, близких к ним африканских же М. ango-
lensis, которых некоторые авторы (Ripley, 1952а) рассматривают в ка-
честве подвидов одного вида. Это некрупные лесные птицы, гнездящиеся
в дуплах деревьев, над поверхностью земли, совершенно не связанные
в своей жизни со скалами, морфологически и многими своими повадками
напоминающие горихвосток (Mackworth-Praed a. Grant, 1960, 1963;
Archer a. Godman, 1961). Весьма близки между собой экологически и
по своему строению, а также и по окраске М. rupestris, М. explorator,
М. saxatilis и, по-видимому, М. brevipes. Все они — типичные обитатели
сухих каменистых и обрывистых безлесных склонов гор пли высоких
плато, покрытых редкими кустарниками; гнездятся в трещинах и рас-
щелинах скал (Koffan a. oth., 1956; Farkas, 1962а, 1962b, 1963; Mack-
worth-Praed a. Grant, 1963; Broekhuysen, 1965). От представителей этой,
вероятно африканской по происхождению, группы хорошо отличаются
синие каменные дрозды (М. solitarius), центр возникновения которых
находится, по-видимому, где-то в южной Азии.
Ко второму роду — Лесных горных дроздов, за которым по закону
приоритета следует сохранить старое название Orocetes Gray, 1840 —
принадлежат три монотипических вида — О. gularis Swinhoe, О. cinclo-
rhynchus (Vigors) и О. erythrogaster (Vigors). Представители этой группы —
эндемики и автохтоны юго-восточной Азии, эволюционировавшие само-
стоятельно, вне связи со скальными формами. Предки Orocetes более
всего, вероятно, походили по окраске на современных «рыжих» О. gularis,
а по образу жизни и строению — на О. erythrogaster, но имели еще более
закругленные крылья и большее развитие рыжего цвета на спине и боках
головы.
ВЕРОЯТНАЯ ИСТОРИЯ РОДА OROCETES
Накопившиеся к настоящему времени сведения о распространении,
экологии и морфологии рецентных видов лесных горных дроздов,
а так же данные по палеогеографии Азии дают возможность высказать
некоторые соображения о вероятной истории рода Orocetes. Понятно, ко-
нечно, что с помощью таких материалов, без палеонтологических данных,
могут быть приблизительно реконструированы лишь самые основные вехи
того длинного и сложного пути, который был пройден этой группой птиц
за многие миллионы лет.
Можно предположить, что какой-то гипотетический предок рода Oro-
cetes, носивший уже все признаки этого рода и ведущий свое происхож-
дение от наземногнездящихся птиц, населявших участки негустых суб-
тропических лесов приподнятых равнин и плато юго-восточной Азии,
— 176 —
развился в позднем олигоцене—миоцене в горных лесах субтропического
облика, произраставших на незначительных высотах в Юго-Западном
Китае и северной Бирме. Это была типичная насекомоядная лесная форма,
обитавшая в нижнем ярусе, которая с уходом в горы стала постепенно
переходить от открытого гнездования на почве к жизни на склонах
в естественных укрытиях, способных защитить гнезда и потомство от
сильного намокания или смыва во время ливней. В зависимости от ха-
рактера субстрата это могли быть или небольшие пустоты и ниши в мяг-
ком грунте, или расщелины в скалах. Иногда птицы, по-видимому,
помещали свои рыхлые постройки под прикрытием лежащего на склоне
дерева или толстой ветви, образовывавших подобие навеса. Первичные
Orocetes вели оседлый образ жизни и имели свойственные лесным обита-
телям закругленные и относительно короткие крылья, длинный хвост и
умеренной длины цевки. По внешнему облику они более всего походили
на «рыжих» белогорлых дроздов, хотя очень возможно, что рыжий цвет
у их самцов распространялся и на ушные перья, и на перья мантии.
Окраска яиц у предков лесных горных дроздов, вероятно, уже прибли-
жалась к таковой у современных форм. Быть может, к этому времени
следует относить установление связи между древними Orocetes и обыкно-
венными кукушками — предками нынешних С. с. bakeri, жившими
в тех же биотопах.
Освоив пояс субтропических лесов в средневысотных горах Юньнани
и северной Бирмы, предки Orocetes проникли оттуда и в примыкавшие
участки лесов умеренного пояса, занимавшие те же высоты. Найдя там
вполне благоприятную обстановку, птицы легко приспособились к новым,
несколько более суровым и в какой-то мере подверженным сезонным
климатическим колебаниям условиям. Жизнь в пограничных с субтропи-
ческими районах лесов умеренного пояса позволила им впоследствии не
только освоить весь этот пояс в древних хребтах Юго-Западного Китая,
но и беспрепятственно расселиться в неогене через покрытую редкой
древесной растительностью равнинную территорию Восточного Китая
к северу, в Маньчжурию, Приморье и Приамурье. В этих областях,
имевших в миоцене—плиоцене умеренный, но более теплый, влажный
и менее сезонный, чем сейчас, климат, предки Orocetes заняли в первую
очередь, по-видимому, покрытые лесом склоны гор и плато Большого
Хингана и Сихотэ-Алиня, а затем распространились по сходным биото-
пам в другие гористые районы. В крайних южных и северных частях
своего теперь довольно обширного сплошного ареала первичные Orocetes
оказались в среде, абиотические и биотические факторы которой были
довольно различными; разрыв между ними усиливался в связи с про-
грессировавшим на севере в неогене общим похолоданием. Надо думать,
что у южных особей, меланогенез в оперении которых должен был про-
текать в субтропиках весьма интенсивно, уже тогда довольно часто стали
появляться индивидуальные отклонения окраски самцового оперения
в сторону типа «черных» О, gularis. Малозаметные в свежем пере, эти
новые признаки окраски в брачную пору выступали вполне отчетливо
и, подхваченные отбором, могли закрепиться в популяции. У северных
птиц этот процесс протекал очень медленно и не привел пока к заметным
сдвигам по сравнению с исходной окраской.
С поднятием Гималаев, примерно в начале плиоцена, для особей южной
популяции, к тому времени сильно изменивших свой внешний облик,
представилась возможность миграции по горным лесам субтропического
и отчасти умеренного пояса на запад, почти до Кашмира. Одновременно
с предками Orocetes в молодые Гималаи вселились и другие птицы, обра-
зовавшие там впоследствии обширный комплекс эндемичных гималайских
видов западнокитайского и бирманского происхождения. Колонизация
12 Биология птиц
— 177 —
лесных Гималаев животными, так же как и растениями, происходила,
таким образом, с востока, преимущественно из Юньнани (Meinertzhagen,
1928; Вульф, 1944). Поскольку к моменту появления предков южных
Orocetes в Гималаях в горные леса еще не успели проникнуть их возмож-
ные конкуренты со стороны Индостана (между древними материками
Гондваны и Ангарским еще долго существовал рукав моря Тетис, пре-
пятствовавший вселению с юга), птицам не пришлось осваивать новые
экологические пиши и менять свой образ жизни. Попав высоко в горы,
Orocetes сохранили привязанность к своим коренным биотопам, что в свою
очередь не повлекло за собой каких-либо существенных преобразований
их летательного аппарата, ног и клюва.
Проявлявшийся уже и в ту далекую от нас пору известный консер-
ватизм лесных горных дроздов в выборе мест обитания послужил, ве-
роятно, тем основным препятствием, которое пе позволило им (в проти-
воположность, например, предкам синих каменных дроздов) выйти на
морское побережье и затем распространиться на Японские острова, не-
однократно восстанавливавшие свою сухопутную связь с материком на
протяжении третичного и четвертичного периодов. В силу тех же причин
распространение древних Orocetes в низменных восточных частях Север-
ного и Среднего Китая, занятых в плиоцене ландшафтами степного и
саванного характера с лесами, росшими главным образом вдоль русел
рек и в ключевых урочищах предгорий, должно было быть спорадиче-
ским. Птицы лишь использовали эту плохо облесенную территорию при
своем первоначальном движении к северу, жить же на ней остались
очень немногие, основавшие в подходящих местах небольшие изоли-
рованные колонии. В конце плиоцена—плейстоцене именно здесь, в наи-
более слабом звене ареала лесных горных дроздов, могла произойти его
дизъюнкция. Возможно, что первым толчком к этому явилось образова-
ние широких, совершенно лишенных древесной растительности долин
крупных китайских рек и последовавшее затем интенсивное образование
лёсса. Неоднократные морские трансгрессии, имевшие место в четвертич-
ный период, при которых, по данным Г. У Линдберга (1955), воды океана
покрывали не только морское побережье, но и заливали равнинные
участки внутри азиатского континента (при предпредпоследней транс-
грессии до изогипсы 150—180 м), усугубили этот разрыв и привели
к окончательной изоляции северных и южных форм. Небезынтересно на-
помнить в этой связи, что ни один вид современных Orocetes не живет
в местности, лежащей ниже двухсотметровой отметки.
Уже в верхнем плиоцене различия в окраске северных Orocetes и
южных, занимавших площадь, соответствующую по своему положению
центральной части современного ареала синегорлого и рыжебрюхого
дроздов, носили характер хороших подвидовых. Первые (предки О. gula-
ris) приобрели, вероятно, черноту на перьях ушной области и мантии,
уклонившись, таким образом, очень незначительно от исходной формы.
Зато вторые (предки нынешних О. cinclorhynchus и О. erythrogaster) могли
иметь структурно иную расцветку покровов на горле и интенсивную чер-
ную пигментацию дорсальной стороны, боков головы и шеи. Те и другие
продолжали вести образ жизни лесных насекомоядных птиц и сохраняли
соответствующие ему закругленные крылья, относительно длинный хвост
и короткие плюсны. Северная часть популяции была оседлой, а южная
в той своей части, которая проникла с поднятием гор на высоту 2500 м
и более, вынуждена была, по-видимому, предпринимать сезонные верти-
кальные кочевки. Это обстоятельство не привело, однако, к заметным
конструктивным преобразованиям их органов передвижения.
После окончательной изоляции северная популяции лесных горных
дроздов дала начало отдельному виду, который в ходе своей дальнейшей
— 178 —
истории по окраске мало изменился. Предки О. gularis обособились, та-
ким образом, от общего ствола ранее двух других форм.
В силу каких-то пока неясных причин еще в доледниковую эпоху
в гималайской части южной популяции Orocetes наметился разрыв, при-
мерно в той области (Сикким—Бутан), в которой сейчас проходит кли-
матическая граница между более влажными восточными и относительно
сухими западными Гималаями. На существование здесь какого-то эколо-
гического барьера, повлиявшего на распространение и последующее
формообразование флоры в умеренном поясе Гималаев, обратил в свое
время внимание Е. В. Вульф (1944).
Наступившее в плейстоцене похолодание, сопровождавшееся в Гима-
лаях и некоторых горах Дальнего Востока ледниковыми явлениями, ока-
зало решающее влияние на окончательное обособление гималайской по-
пуляции от южнокитайской и на независимое формирование сезонных
перелетов у предков белогорлого и синегорлого дроздов.
На интересующей нас территории плейстоценовые ледники достигали
наибольшего развития в Гималаях, где в ряде мест они спускались до
1200—1500 м над ур. м., достигая зоны пышной растительности (Сини-
цын, 1962). Вызванное оледенением понижение постоянной снеговой ли-
нии повлекло за собой сдвиг всех вертикальных поясов и изменение
в них условий существования. Предки О. cinclorhynchus, гонимые холо-
дами и наступавшей после выпадения снега бескормицей, откочевывали
зимой к югу, в богатые лесами горные области западной части Индостана.
В теплое время года они возвращались в гнездовую область, где занимали
свои исконные биотопы. Из оседлых птицы превращались в дальних
мигрантов.
На Дальнем Востоке и в Северном Китае ледниковый период, по
данным В. М. Синицына, отозвался общим похолоданием, появлением
резкой сезонности смены времен года с отрицательными температурами
и снегопадами зимой, расширением пояса вечной мерзлоты и образова-
нием небольших ледников в самых высоких частях Сихотэ-Алиня, Боль-
шого Хингана и Маньчжуро-Корейских гор. Хотя эти события не носили
характера катастроф, они вызвали большие изменения в условиях оби-
тания предков О. gularis, вынудив их искать в холодный сезон года но-
вые места, более подходящие по своей экологической обстановке. Птицы
страдали зимой на родине не столько от холода, сколько от отсутствия
необходимых кормов. Вертикальные сезонные перемещения в случае
с О. gularis не могли уже спасти их, так как во время максимального оле-
денения физико-географическая обстановка внеледниковых областей Даль-
него Востока была довольно суровой — ландшафт холодной лесостепи
(Никольская, 1961), а во время трансгрессий моря воды его затапливали
всю низменную территорию. Будучи довольно консервативными лесными
обитателями, предки О. gularis не могли также пережить ледниковое
время в лежащих южнее безлесных областях Среднего Китая. Оставался
один выход — миграции дорогой предков, некогда распространившихся
к северу из южной Азии, в обратном направлении. В поисках оптимальных
условий для своих зимовок белогорлые дрозды пересекли весь Китай и
на юге страны достигли границ ареала китайской части южной популя-
ции Orocetes (предков рыжебрюхих дроздов). В районы, занятые абори-
генами, стали на длительное время вселяться пришельцы с севера; однако
не выдержав конкуренции со стороны будущих О. erythrogaster, белогор-
лые дрозды вынуждены были мигрировать южнее, до мест своих нынешних
зимовок, лежащих в основном в Индокитае. Эти длительные сезонные
выселения, как и в случае с О. cinclorhynchus послужили началом воз-
никновения регулярных сезонных миграций О. gularis. В ходе дальней-
шей истории приспособление к диареальному существованию привело
12аК-
— 179 —
к преобразованию летательного аппарата обеих форм: произошло неко-
торое удлинение и заострение их крыльев и укорочение хвостов.
На месте первичного очага возникновения группы (в Юго-Западном
Китае и северной Бирме), территория которого с третичного периода не
испытывала серьезных изменений климата и ландшафта, развивались
О. erythrogaster. Оседлая жизнь в довольно стабильных влажных и теп-
лых условиях горных субтропических лесов позволила им сохранить
почти без изменений повадки, образ жизни и строение (в частности, ле-
тательный аппарат), унаследованные от предков, и вместе с тем повлекла
за собой значительное изменение их окраски, а возможно, и размеров.
В конце плейстоцена на юго-востоке Азии оформились, таким образом,
три хорошо отличимых друг от друга по морфологии, экологии и по-
ведению аллопатрических вида: О. gularis, О. cinclorhynchus и О. erythro-
gaster.
В голоцене ареалы лесных горных дроздов приобрели современные
очертания. С отступанием ледника, по мере нормализации условий су-
ществования на родине, О. gularis и О. cinclorhynchus не только возвра-
тились на оставленную ими в период похолодания территорию, но и за-
воевали новые области для гнездования. Так, «ксерофилизация» на путях
пролета и приобретенный тогда же некоторый диапазон экологической
пластичности помогли синегорлым дроздам освоить относительно сухие,
светлые сосновые леса и распространиться по ним через все Гималаи
на запад до горных массивов Спингара и Нуристана. К послеледнико-
вому же времени относится, по-видимому, и вселение оседлого рыжебрю-
хого дрозда в восточные Гималаи, где ныне он симпатричен с перелетным
синегорлым дроздом.
В историческое время ареалы лесных горных дроздов (особенно О. gu-
laris и О. erythrogaster) претерпели большие изменения. Под влиянием
интенсивной хозяйственной деятельности человека, продолжительность
которой исчисляется несколькими тысячелетиями (при наличии огромного
по численности населения), первичные леса на огромной территории
Центрального и Южного Китая, бывшей Маньчжурии и нашего Даль-
него Востока, а также во многих районах Гималаев полностью сведены,
что привело к резкому уменьшению численности Orocetes и к значитель-
ному сокращению их гнездовой области. Если на первых порах выборочная
рубка и возникавшие ранней весной небольшие пожары оказывали даже
какое-то благоприятное влияние на численность этих птиц (появлялось
больше разреженных участков, пригодных для поселения в них Orocetes,
а также прикорневых дупел, выгоравших в основаниях стволов деревьев),
то последующее освоение всех сколько-нибудь доступных земель под паст-
бища и посевы с полным вырубанием древесной растительности стало
для них катастрофой.
ЛИТЕРАТУРА
Березовский М. и В. Бианки. 1891. Птицы Ганьсуйского путешествия
Г. Н. Потанина, 1884—1887. Материалы по орнитологии Китая, главным обра-
зом южной части провинции Ганьсу. СПб. : 99.
Бианки В. 1902. К орнитофауне Маньчжурии. Ежегодн. Зоол. музея Акад, наук,
7 14—16.
В о й т к е в и ч А. А. 1962. Перо птицы, М.
Воробьев К. А. 1954. Птицы Уссурийского края. М. : 237—240.
Вульф Е В. 1944. Историческая география растений. М.—Л.
Гагина Т. Н. 1960. Новые данные о распространении некоторых птиц в Восточной
Сибири. Тр. пробл. и тем. совещ. ЗИН АН СССР, IX : 95.
Гладков Н. А. 1954. Род Каменные дрозды — Monticola Boie, 1822. В кн.: Птицы
Советского Союза, 6. М. : 479—492.
Дементьев Г. П. 1937. Воробьиные. Полный определитель птиц СССР, IV.
М.-Л. : 259-261.
— 180 —
Дорогостайский В. 1915. Предварительное сообщение о поездке в Яблоно-
вый хребет по поручению Академии наук в 1914 г. Зап. Акад, наук : 407.
Кистяковский А. Б. 1959. Новые данные о северных границах распростране-
ния маньчжурского орнитофаунистического комплекса на Амуре. Тез. докл.
Второй всесоюзн. орнитол. конфер., Ill, М. : 80—81.
Кистяковский А. Б., В. И. Лоскот и Л. А. Смогоржевский.
1962. Новые данные о северной границе маньчжурской фауны. Матер. Третьей
всесоюзн. орнитол. конфер., 2. Львов : 25—26.
Кистяковский А. Б. и Л. А. Смогоржевский. 1963. Новые дан-
ные о границе китайского орнитофаунистического комплекса на р. Бурее.
Зоогеография суши, Тез. докл. III Всесоюзн. совещ. по зоогеогр., Таш-
кент : 131—132.
Кистяковский А. Б. и Л. А. Смогоржевский. 1964. О границе ки-
тайского орнитофаунистического комплекса на реке Бурее. Научн. докл. Высш
школы, биол. науки, 3 26—29.
Колонии Г. В. 1963. К орнитофауне верхнего Амура. Орнитология, 6 : 472.
Кузякин А. П. 1954. Учет оологических признаков и особенностей гнездовья
в классификации птиц. Бюлл. Моск. общ. испыт. природы, Отд. биол., LIX,
6 : 27—35.
Линдберг Г. У. 1955. Четвертичный период в свете биогеографических данных.
М,—Л.
Мальчевский А. С. 1959. Гнездовая жизнь певчих птиц. Изд. ЛГУ.
Нейфельдт И. А. 1965. Об одном, еще не известном виде — воспитателе обыкно-
венной кукушки. Зоол. журн., XLIV, 1 : 140—142.
Нейфельдт И. А. и Б. В. Соколов. 1960. Гнездование лесного каменного
дрозда в Верхнем Приамурье. Орнитология, 3 : 236—250.
Никольская В. В. 1961. Палеогеография ледникового времени. В сб.: Даль-
ний Восток, М. : 73—79.
ОмелькоМ. А. 1956. О перелетах птиц на полуострове Де-Фриза Тр. Дальневост,
фил. АН СССР, III (6) : 354—355.
Павлов Е. 1948. Птицы и звери Читинской области. Читиздат : 66, 142.
Портенко Л. А. 1954. Птицы СССР, ч. III. Опред. по фауне СССР, изд. ЗИН
АН СССР, 54 : 212-218.
Синицын В. М. 1962. Палеогеография Азии. М.—Л.
Спангенберг Е. П. 1960. О редких и интересных птицах среднего течения Хун-
гари и верхней Удоми. Охр. прир. и озеленение, 4 : 141—142.
Спангенберг Е. П. 1965. Птицы бассейна реки Имана. Сб. трудов Зоол. музея
МГУ, IX : 193—194.
Тан Сань-цзу, Ma-Юн, Ван Тя-цюн, ВанЦзы-юй и Чжоу
Н э - у. 1965. Фауна птиц и млекопитающих хр. Чжунтяо в пров. Шаньси.
Acta Zool. Sinica, 17, 1 : 95.
Тугаринов А. Я. 1932. Птицы восточной Монголии по наблюдениям экспе-
диции 1928 г. Тр. Монг, комиссии АН СССР, I : 235—236.
Штегман Б. (Stegmann В.) 1931. Die Vogel dauro-mandschurischen Ueber-
gangsgebietes. Journ. f. Ornithol., LXXXIX, 2 : 207—209.
Штегман Б К. 1961. Рудимент дистального махового в крыле птицы. Тр. Зоол.
инет. АН СССР, 29 : 227—256.
Шульпин Л. М. 1931. Лесной «каменный» дрозд (Monticola gularis Sw.) восточной
Азии. К истории группы каменных дроздов. Ежегодн. Зоол. музея Акад, наук,
32 269-288.
Янковский А. 1898. Орнитологический дневник с 5 мая по 5 ноября 1898 года
(экспедиция Русского географического общества в Корею и Маньчжурию).
Зап. Приамурск. отд. Русек, геогр. общ., III, 3 : 121—122.
А 1 i S. 1953. The birds of Travancore and Cochin. Bombay : 67—68.
Ali S. 1954. The birds of Gujarat, pt. II Journ. Bomb. Nat. Hist. Soc., 52, 4 : 471.
A 1 i S. 1962. The birds of Sikkim. Madras : 279—280.
Ali S. a. H. A b d u 1 a 1 i. 1937 The birds of Bombay and Selsette. Journ. Bomb.
Nat. Hist. Soc., 39, 3 : 523.
Archer G. a. E. M. God man. 1961. The birds of British Somaliland and the
gulf Aden. Edinburgh—London : 1148—1150.
Austin O. L. 1948. The birds of Korea. Bull. Mus. Compar. Zool., 101, 1 : 207—208.
Baker E. St. 1924. The fauna of British India including Ceylon and Burma. Birds,
II, London : 169—178.
Baker E. St. 1933. Nidification of birds of Indian Empire, II (Turdidae—Sturnidae).
London : 147—153.
Baker E. St. 1942. Cuckoo problems. London.
Bangs O. a. J. C. Phillips. 1914. Notes on a collection of birds from Yunnan.
Bull. Mus. Compar. ZooL, 58 : 292.
Bates K. S. P. 1935. Notes on the habits of some Indian birds. Journ. Bomb. Nat.
Hist. Soc., 37, 4 904—906.
— 181 —
Bates R. S. P. a. E. H. N. Lo w ther 1952. Breeding birds of Kashmir. Oxford
Univ. Press : 91—94.
Betts F. N. 1929. Notes on the birds of Coorg. Journ. Bomb. Nat. Hist. Soc., 33,
3 544.
Biswas B. 1953. A checklist of genera of Indian birds. Records of Ind. Mus., Cal-
cutta, 50, 1 : 47.
Biswas B. 1961. The birds of Nepal, pt. 5. Journ. Bomb. Nat. Hist. Soc., 58,
3 668—670.
В 1 у I h E. 1849. Catalogue of the birds in the Museum Asiatic Society. Calcutta : 164—
165.
Broekhuysen G. J. 1965. Nesting behaviour of the Sentinel Thrush Monticola
explorator (Vieillot). Ostrich, XXXVI, 1 : 41—42.
Caldwell H. R. a. J. C. Caldwell. 1931. South China birds. Shanghai : 66—69.
Champion H. G. 1936. A. preliminary survey of the forest types of India and Burma.
Ind. Forest Records, I, 1 : 1—286.
Cheng Tso-Hsin. 1958. A distributional list of Chinese birds, II. Passeriformes, Pe-
king : 152—153.
David A. et E. О u s t a 1 e t. 1877. Les oiseaux de la Chine. Paris : 159—160.
Deignan H. G 1945 The birds of Northern Thailand. U. S. Nat. Mus. Bull.,
186 423—424.
Deignan H. G. 1946. Notes on birds of Northern Siam. Not. Nat. Acad. Nat. Sc. Phi-
ladelphia, 173 : 10.
Deignan H. G. 1963. Checklist of the birds of Thailand. U. S. Nat. Mus. Bull.,
226 165.
Delacour J. 1929. On the birds collected during the fourth expedition to French
Indo-China, p. II. Ibis : 417.
Delacour J. etP. J abouille. 1931. Les Oiseaux de 1’Indochine Francaise, III.
Paris : 122—124
Farkas T. 1962a. Zur Biologie und Ethologie der siidafrikanischen Arten der Gattung
Monticola (Boie). I Teil. Monticola brevipes (Waterhouse). Vogelwelt, 83, IV : 97—
116.
Farkas T. 1962b. Zur Biologie und Ethologie der siidafrikanischen Arten der Gattung
Monticola (Boie). II Teil. Monticola rupestris L. Vogelwelt, 83, VI : 161—173.
Farkas T. 1963. Zur Biologie und Ethologie der siidafrikanischen Arten der Gattung
Monticola (Boie). Ill Teil. Monticola explorator L. Vogelwelt, 84, I : 11—22.
Frank F. 1939. Die Farbung der Fogelfeder dutch Pigment und Struktur. Journ.
f. Ornith., 87, 3 : 426—523.
Gray G. R. 1840. A list of the genera of birds. London : 21.
Hartert E. 1910. Die Vogel der palaarktischen Fauna, I. Berlin : 671—676.
Horsfield T. a. F. Moore. 1854. A catalogue of birds in the Museum of the Hon.
East-India Company, 1. London : 188—189.
Hume А. О. a E. W. О a t e s. 1890. The nests and eggs of Indian birds, II. London :
102—106.
Ingram C. 1909. The birds of Manchuria. Ibis : 426.
Jahn H. 1942. Zur Oekologie und Biologie der Vogel Japans. Journ. f. Ornithol., 90,
1—2 : 177—179.
Jardine W. a. F. S e 1 b y. 1833. Illustrations of ornithology, I, III. Edinburgh,
I 167; III : 129, pl. CXXIX.
J e r d о n T. C. 1862. The birds of India, I. Calcutta : 514—516.
Jones A. E. 1926. Notes on birds of Ambala district Journ. Bomb. Nat. Hist. Soc.,
31, 4 : 1002.
Jones A. E. 1947. The birds of the Simla and adjacent hills, p. II. Journ. Bomb. Nat.
Hist. Soc., 47, 2 : 239—240.
К of fan К., T. Farkas a. I. J. Ferguson-Lees. 1956. Photographic
studies of some less familiar birds. LXXI. Rock-Thrush. British Birds, 49 : 268—
271.
Kuroda N. 1918. Notes on Korean and Manchurian birds. Annot. Zool. Jap., IX,
4 : 539—540.
Kuroda N. 1932. A fifth lot of bird-skins from Manchuria. Tori, 7, 35 : 422.
Lack D. 1954. Call-notes Erithacus and convergence. Ibis, 96, 2 : 312—314
Lack D. 1958. The singinificance of the colour of Turdinae eggs. Ibis, 100, 2 145—
166.
La Touche J. 1899. Notes on the birds of North-west Fohkien. Ibis 179—180.
La Touche J. 1906. Fild-notes on the birds of Chinkiang, Lower Yangtse basin,
p. II. Ibis 627.
La Touche J. 1920. Notes on the birds North-east Chihli, in North. China.Ibis : 884.
La Touche J. 1925. A handbook of the birds of Eastern China, I, 2. London : 123—
125.
Lister M. D. 1953. Secondary song of some Indian birds. Journ. Bomb. Nat. Hist.
Soc., 51, 3 702.
— 182 —
Lister M. D. 1954. A contribution to the ornithology of the Darjeeling area. Journ.
Bomb. Nat. Hist. Soc., 52, 1 40.
Ludlow F. 1951. The birds of Kongbo and Pome, south-east Tibet. Ibis, 93, 4 : 561.
Ludlow F. a. N. В Kinnear. 1937. The birds of Bhutan and adjacent territories
of Sikkim and Tibet, p. II. Ibis, : 276—277.
Ludlow F. a. N. B. Kinnear. 1944. The birds of south-eastern Tibet. Ibis,
86 191
Macdonald I. C. 1904. Occurence of the White-Throated Rock-Thrush, Petro-
phila (Monticola) gularis in Burma. Journ. Bomb. Nat. Hist. Soc., 15, 4 : 727.
M а с к w о r t h - P r a e d C. W. a. G. H. B. Grant. 1960. Birds of Eastern and
Nordth Eastern Africa. Afr. Handb. of birds, London, 2 : 258—259.
M а с к w о r t h - P r a e d C. W. a. G. H. B. Grant. 1963. Birds of the Southern
third of Africa. Afr. Handb. of birds, London, 2 : 138—141.
M e i n e r t z h a g e n A. C. 1927. On a new race of Rock-Thrush (Monticola). Bull.
Brit. Orn. Club., 47 : 148—149.
Meinertzhagen R. 1927. Systematic results of birds collected at high altitudes
in Ladak and Sikkim. Ibis, III, 4 : 580.
Meinertzhagen R. 1928. Some biological problems connected with the Himalaya.
Ibis, IV, 3 : 480—533.
Meinertzhagen R. 1951. Some relationships between African, Oriental, and
Palaearctic genera and species, with review of the genus Monticola. Ibis, 93,
3 443—459.
M e i s e W. 1934. Die Vogelwelt der Mandschurei. Abhandl. u. Ber. Mus. Tierkunde
Dresden, 18, 2 : 43.
Mell R. 1922. Beitrage zur Fauna Sinica. Arch. f. Naturgesch., A, 10 : 65—66.
Oates E. W. 1905. Catalogue of the collection of birds eggs in the British Museum,
IV. London : 130—131.
Osmaston В. B. 1926. Notes on the birds of Kashmir. Journ. Bomb. Nat. Hist.
Soc., 31, 4 : 985.
P a 1 u d a n K. 1959. On the birds of Afghanistan. Vedensk. Medd. Dansk. natur. For.,
122 196.
Piechocki R. 1958. Beitrage zur Avifauna Nord- und Nordost-Chinas (Mandschu-
rei). Abhandl. u. Ber. Mus. Tierkunde Dresden, 24 : 153.
Rand A. L. a. R. L Flemming. 1957. Birds from Nepal. Fieldiana, Zool., 41,
1 : 158.
R e n s c h B. 1923. Zoologische Ergebnisse der Walter Stotznerschen Expeditionen
nach Szetschwan, Osttibet und Tschili. Turdinae, II. Teil. Abhandl. u. Ber. Mus.
Tierkunde Dresden, 16, 2 : 48.
Riley J. H. 1926. A collection of birds from the provinces of Yunnan and Szechwan,
China, made for the National Geographic Society by Dr. Joseph F. Rock. Proc.
U. S. Nat. Mus., 70, 5 : 1—70.
Ripley S. D. 1950. Birds from the Nepal. Journ. Bomb. Nat. Hist. Soc., 49, 3 : 388—
389.
Ripley S. D. 1952a. The Thrushes. Postilia. Yale Peabody Mus., 13 : 1—48.
Ripley S. D. 1952b. A collection of birds from the Naga Hills. Journ. Bomb. Nat.
Hist. Soc., 50, 3 490.
Ripley S. D. 1961. A synopsis of the birds of India and Pakistan, together with these
of Nepal, Sikkim, Bhutan and Ceylon. Madras.
R i p 1 e у S. D. 1964. Genus Monticola Boie. Check-listof birds of the World. Cambridge —
Massachusetts, X : 135—140.
Robinson H. G. 1915. On the birds collected by Mr. G. Boden Kloss on the coast
and islands of South-easterm Siam. Ibis : 752.
Robinson H. G. 1928. The birds of the Malay Peninsula. London : 232—233.
Robinson H. а. С. В. К 1 о s s. 1919. On the birds from South Annam and Cochin
China. Ibis : 594.
Rothschild L. 1926. On the avifauna of Yunnan, with critical notes. Novit. Zool.,
33 : 256.
Rothschild L. 1927. On the avifauna of Yunnan, with critical notes. Suppl., Novit.
Zool., 34 : 40.
Schaunensee R. M. 1946a. On two collections of birds from the southern Shan
States, Burma. Proc. Acad. Nat. Sci. Philadelphia, XCVIII : 117.
Schaunensee R. M. 1946b. On the Siamese birds. Proc. Acad. Nat. Sci. Phila-
delphia, XCVIII : 70.
Seebohm H. 1881. Catalogue of the Passeriformes, or perching birds in the collection
of the British Museum. Turdidae. Catalogue of the Birds in the British Mus.,
V : 312—327.
Sharpe R. B. 1903. A hand-list of the genera and species of birds, IV. London : 144—
Shaw T. H. 1936. The birds of Hopei province. Zool. Sinica, ser. «В», The vertebrates
of China, XV, 1, Peking : 686—688.
— 183 —
Shaw T. H. a. С. T. L i u. 1940. The food of some wild birds. Bull. Fan. Mem. Inst.,
Biol. Zool. ser., X : 144.
Smythies В. E. 1948. Some birds of the Gandak-Kosi watershead, including the
pilgrim trail to the sacred lake of Gosainkund. Journ. Bomb. Nat. Hist. Soc.,
47, 3 : 441.
Smythies В. E. 1949. A reconnaisance of the N’Mai HKA Drainage, Northern
Burma. Ibis., 91, 4 : 641.
Smythies В. E. 1953. The birds of Burma. London—Edinburgh : 132—133.
Stanford J. К. a. E. Mayr. 1941. The Vernay-C utting expeditionen to Northern
Burma, p. III. Ibis : 228—229.
Stevens H. 1925. Notes on the birds of the Sikkim Himalayas, p. V. Journ. Bomb.
Nat. Hist. Soc., 30, 2 : 360—361.
Stresemann E. u. G. Heinrich. 1940. Die Vogel des Mount Victoria. Mitt
Zool. Mus. Berlin, 24, 2 : 222—223.
S t у a n F. W. 1894. Notes on the ornithology of China. Ibis : 333.
Swainson W. 1837. On the natural history and classification of birds, II. London:
232.
Swinhoe R. 1861. Notes on ornithology taken betveen Takoo and Peking in the
neighbourhood of the Reiho River, Province of Chelee, North China, from August
to December, 1860. Ibis : 332—333.
Swinhoe R. 1862. On some Tientisin birds, collected by Mr. Fleming R. A., in the
possession of Mr. Whiteley. Proc. Zool. Soc. London : 318.
Swinhoe R. 1863a. Additions and corrections to the ornithology of Northern China.
Ibis : 93—94.
Swinhoe R. 1863b. Catalogue of the birds of China, with remarks principally on
their geographical distribution. Proc. Zool. Soc. London : 282.
Swinhoe R. 1871. A revised catalogue of the birds of China and its islands,
with description of new species, references to former notes, and occasional
remarks. Proc. Zool. Soc. London : 368.
Ticehurst С. B. 1922. The birds of Sind, p. II. Ibis : 642.
Ticehurst G. B. 1926. On the down-plumage of some Indian birds. Journ. Bomb.
Nat. Hist. Soc., 31, 2 : 368—378.
V a u r i e Ch. 1955. Systematic notes on Palearctic birds. № 14. Turdinae: the genera
Erithacus, Luscinia, Tarsiger, Phoenicurus, Monticola, Erythropygia and Oenanthe.
Amer. Mus. Novitates, 1731 : 20—24.
V a u r i e. Ch. 1959. The birds of the Palearctic fauna. Passeres. London : 357—361.
Vigors N. A. 1831. Observations on a collection of birds from the Himalayan Moun-
tains with characters of new Genera and Species. Proc. Zool. Soc. London: 171—172.
Whistler H. 1926. The birds of Kangra District, Punjab. Ibis : 560.
Whistler H. 1930. The birds of the Rawal Pindi District, N.-W. India. Ibis : 92.
Whitehead С. H. T. a. H. A. F. M a g r a t h. 1909. On the birds of Kohat and
Kurram, Northern India, p. II. Ibis : 223.
W i 1 d e r G. D a. H. W. Hubbard 1938 Birds of Northeastern China.
Peking : 297.
W i t h e r b у H. F., F. C. R. J о u r d a i n, N. E. T i c e h u r s t a. B. W. T u-
c к e r. 1958. The handbook of British birds, II. London : 141—144.
Yamashina Y. 1939. Note on the specimens of Manchurian birds chiefly made by
Mr. H Orii in 1935 Tori, X, 49 : 498.
А. В. Кречмар
ПТИЦЫ ЗАПАДНОГО ТАЙМЫРА
А. V. Krechmar. The birds of the western parts of the Taimyr Peninsula
СОДЕРЖАНИЕ
Стр.
Введение . 185
Краткая характеристика природных условий западного Таймыра 190
Систематический обзор птиц западного Таймыра 197
Сезонные явления в жизни птиц западного Таймыра 299
ВВЕДЕНИЕ
Территория западною Таймыра, охваченная нашими исследованиями,
почти целиком относится к бассейну р. Пясины (рис. 1). Лишь крайний
юг исследованного участка, окрестности Хантайского озера и р. Хан-
тайки, принадлежит непосредственно системе Енисея. Восточная граница
исследованной территории лишена каких-либо достаточно ярко выражен-
ных естественных рубежей и поэтому весьма условна. Нами она прово-
дится от восточного конца Хантайского озера к устью Авама, а оттуда,
пересекая самые верховья правых притоков Пясины, выходит к восточ-
ному побережью Пясинского залива. Таким образом, исследованный
участок достаточно велик и простирается с севера на юг на 600—700 км,
а с запада на восток — на 250—350 км. Последовательная смена ланд-
шафтов от арктических тундр в низовьях Пясины до северотаежных
участков в долинах Хантайки и Рыбной и у Норильских озер, а также
наличие своеобразных высокогорных комплексов плоскогорья Путорана
создают особые условия для распределения фауны западного Таймыра.
Крайняя суровость климата территории, лежащей у северной оконеч-
ности азиатского материка, между 68° и 74° с. ш., способствует проявле-
нию целого ряда особенностей экологии обитающих там наземных по-
звоночных. Не меньший интерес представляют и зоогеографические осо-
бенности местной фауны западного Таймыра, характеризующейся
взаимопроникновением приблизительно одинакового количества видов и
подвидов как западного, так и восточного происхождения.
Изучение и освоение Таймырского полуострова и прилежащей к нему
части Сибири благодаря значительной удаленности и крайне суровому
климату до последних десятилетий шло очень медленно.
Первым натуралистом, уделившим значительное внимание флоре и
фауне Таймыра, был А. Ф. Миддендорф (1869), посетивший крайний север
Сибири во время своего знаменитого путешествия 1843—1844 гг. В этой
поездке Миддендорф уделил основное внимание изучению центрального
— 185 -
и северного Таймыра, побывав в верховьях Пясины и на р. Дудыпте
лишь проездом ранней весной и поздней осенью 1843 г.
Весной и летом 1877 г. низовья Енисея посетил английский натура-
лист Генри Сибом (Seebohm, 1901).
Г. Попгем, посетивший низовья Енисея летом и осенью 1895 г., опу-
бликовал большую часть своих орнитологических наблюдений в журнале
«The Ibis» (Popham, 1897, 1898, 1901).
В 1900—1901 гг. морское побережье западного Таймыра посетило
судно Русской полярной экспедиции «Заря». Многочисленные остановки
«Зари» у побережья западного Таймыра и ее вынужденная зимовка дали
возможность натуралистам экспедиции А. А. Бируле и Г. Э. Вальтеру
собрать интересные фаунистические и биологические материалы (Вальтер,
1902; Бируля, 1907).
Летом 1907 и 1908 гг. в низовьях Енисея работал А. Я. Тугаринов,
базировавшийся в окрестностях Луковой протоки на Бреховских остро-
вах, у пос. Гольчихи и в устье Глубокой, на берегу Енисейского залива.
Результаты, полученные по обработке собранных там материалов, содер-
жатся в работе «Материалы по птицам Енисейской губернии», опубли-
кованной А. Я. Тугариновым и С. А. Бутурлиным в 1911 г.
Летом и осенью 1914 г. в окрестностях Гольчихи работала английская
натуралистка Мод Хэвиленд. Результаты ее орнитологических исследо-
ваний напечатаны в нескольких небольших статьях (Haviland, 1915).
В связи с неотложными задачами освоения Крайнего Севера в первые
годы Советской власти на Таймыре работали самые разнообразные эк-
спедиции, по исследованию фауны они уделяли мало внимания.
Заслуживает упоминания опубликованная в 1930 г. статья Е. О. Яков-
лева «К границам распространения промысловых зверей и птиц в Ту-
руханском крае», написанная по материалам, собранным им в 1920—
1924 гг.
В 1929—1930 гг. в устье Пясины, с зимовкой, работал И. И. Колю-
шев — участник Пясинской экспедиции Сибторга. Собранные им сведе-
ния по фауне наземных позвоночных этого участка морского побережья
опубликованы в сборнике «Материалы по изучению Сибири» (1933).
В июле—октябре 1932 г. южную часть западного Таймыра проездом
посетил В. Н. Скалой, опубликовавший результаты своих орнитологи-
ческих наблюдений в журнале «Le Gerfaut» за 1935—1939 гг.
А. В. Михеев, побывавший летом 1941 г. в районе Дудинки, пос. Голь-
чихи, фактории «Сопочная Карга» и рр. Глубокой и Каменки, в 1948 г.
опубликовал статью «Охотничьи богатства Енисейско-Пясинской тундры».
Весьма отрывочные данные, содержащиеся в этой работе, относятся
только к прибрежной зоне Енисея и Енисейского залива.
В самые последние годы, вплоть до настоящего времени, в низовьях
Енисея работали экспедиции Института географии АН СССР под руко-
водством Е. Е. Сыроечковского, выпустившего по наземной фауне целый
ряд небольших статей. Одна из этих статей, «Птицы Хантайского озера
и прилежащих гор Путорана», изданная в 1961 г., относится непосред-
ственно к исследованной нами территории. Недостаток фактических ма-
териалов значительно снижает научное значение статьи.
Наряду с исследованиями, касающимися фауны западного Таймыра,
нельзя не упомянуть основных работ, посвященных смежным террито-
риям.
На западе, на Гыданском полуострове, в 1927 и 1928 гг. работал
С. П. Наумов, впоследствии опубликовавший работу «Млекопитающие
и птицы Гыданского полуострова».
На восточном Таймыре ранней весной и летом 1928 г. работал А. И. Тол-
мачев. Материалы его исследований были обработаны совместно с А. Я. Ту-
- 186 —
Рис. 1 Картосхема западного Таймыра.
гариновым и опубликованы в работе «Материалы для авифауны восточ-
ного Таймыра».
Наконец, на северном Таймыре, базируясь на метеостанции «Озеро
Таймыр», в 1947—1949 гг. работал В. М. Сдобников. Результаты иссле-
дований по фауне млекопитающих и птиц он опубликовал в целом ряде
статей.
Исследованиями зоологов на западном Таймыре были охвачены
в основном только наиболее легко доступные участки суши, прилегаю-
щие к морскому побережью и долине Енисея. Внутренние водораздель-
ные пространства, как и долина р. Пясины, берущей свое начало к северу
от полярного круга, остались практически не изученными. Побывавшие
здесь проездом А. Ф. Миддендорф и В. Н. Скалой не смогли собрать
значительных материалов по сезонным условиям их маршрутов, приуро-
ченных к ранней весне или к осени. Между тем именно эти наименее
изученные водораздельные пространства представляют для зоологов наи-
больший интерес благодаря отсутствию интразональных элементов фауны,
характерных для долин больших рек, текущих в меридиональном на-
правлении. Из-за почти полного отсутствия постоянных селений человека
животный мир этих внутренних пространств почти полностью сохранил
свой первобытный характер.
Большая часть наших исследований на западном Таймыре, проводив-
шихся непрерывно с 1957 по 1964 г. и охвативших все сезоны года, от-
носится к бассейну р. Пясины и окрестностям так называемых Нориль-
ских озер в западной части плоскогорья Путорана. Сравнительно хорошо
изученная другими исследователями фауна морского побережья и долины
Енисея, почти не посещавшихся во время наших экспедиционных поез-
док, в настоящую работу не включается. Наиболее длительные исследо-
вания производились у истоков Рыбной, в окрестностях Пуринскпх
озер, в устье Дудыпты и близ устья Амундакты.
Стационар близ истоков р. Рыбной, в окрестностях которого мы ра-
ботали в 1958, 1959 и 1964 гг. в общей сложности более 11 месяцев, был
устроен на правом берегу реки, в 3 км от западного берега оз. Кета.
Рельеф местности слабо холмистый, высота над уровнем моря около
120 м, огромное количество мелких озер. Преобладающий ландшафт —
лиственничная и елово-лиственничная тайга с подлеском из багульника
и карликовой березки. По берегам оз. Кета есть участки лиственничного
редколесья и заболоченных открытых пространств. На вершинах сухих
холмов и на небольших островках оз. Кета встречаются сухие камени-
стые и поросшие дриадой участки, напоминающие арктическую тундру.
От края плоскогорья Путорана избушка стационара удалена на 18—20 км.
Таким образом, в непосредственной близости от стационара на р. Рыбной
находились все ландшафты, наиболее характерные для окрестностей Но-
рильских озер.
Стационары на Пуринских озерах и близ устья Большой Дюрюсы,
около которых в 1960—1963 гг. исследования производились свыше 6 ме-
сяцев, расположены близко друг от друга и в очень сходных ландшафт-
ных условиях. Слабо холмистая равнина покрыта необозримыми кочкарни-
ковыми мохово-осоковыми, мохово-пушицевыми и пятнистыми тундрами.
В низинах и по берегам местами довольно многочисленных озер обычны
заросли полярных ив, немногим более метра в высоту, на склонах хол-
мов и на некоторых участках водоразделов есть карликовая березка и
голубика, а на хорошо дренированных местах обычны участки дриадово-
кассиопеевой тундры. В поймах Агапы, Пуры и Большой Дюрюсы масса
озер, перемежающихся с хорошо дренированными песчаными гривами.
Обычны там и участки низинных полигональных болот. Следовательно,
- 188 -
близ этих стационаров были сосредоточены почти все элементы ландшаф-
тов, обычных для типичной тундры.
Близ полярной станции «Кресты», в устье Дудыпты, где мы базиро-
вались в 1958, 1960, 1961 и 1962 гг. около 5 месяцев, тундры носят
несколько более южный характер. Здесь произрастают отдельные
чахлые деревья — лиственницы, а заросли ольховых кустарников зани-
мают значительные площади. Есть и участки равнинных плоско-бугри-
стых тундр, а также сухие каменистые холмы. Положение стационара
на берегу р. Пясины благоприятствовало наблюдениям над сезонными
миграциями многих тундровых птиц.
Наконец, стационар на берегу оз. Кета против устья Амундакты,
близ которого мы работали в июле и августе 1964 г., был устроен у самого
подножия плоскогорья Путорана, в месте, где было сравнительно легко
на него подниматься. Поэтому там была возможность совершать даже
однодневные экскурсии в горы как до границы древесной растительности,
так и в гольцовый пояс. В непосредственной близости от стационара
было много поросших высокоствольным елово-лиственничным лесом тер-
рас и крупнокаменистых осыпей. Многочисленные мелкие острова, обра-
зованные выносами р. Амундакты, были также легко доступны для ис-
следования.
Помимо этих четырех основных стационаров, во время длительных
лодочных или пеших маршрутов вдоль рр. Пясины, Пуры, Агапы, Но-
рилки, Талой, Глубокой, Рыбной, Хантайки, Могокты, а также озер Пя-
сино, Мелкого, Глубокого и некоторых других были сделаны десятки
временных стоянок длительностью от 1 до 10 дней. Некоторые из них
посещались нами повторно на протяжении ряда лет. Много наблюдений
было сделано и во время переездов.
Во время полевых исследований основное внимание уделялось изуче-
нию деталей размножения, характера и сроков миграций, а также рас-
пределения видов по элементам ландшафта. Во всех случаях, когда это
было возможно, проводился сбор материалов по питанию, динамике чис-
ленности, особенностям сезонной жизни и другим сторонам экологии.
За весь период полевых работ было найдено и исследовано около
500 гнезд, принадлежащих 66 видам, и собраны сведения по размноже-
нию еще 28 видов. Во время наших исследований на западном Таймыре
было собрано более 450 экземпляров птиц, принадлежащих 98 видам.
Подавляющая часть этих сборов находится в коллекциях Зоологического
института АН СССР.
Во время полевых исследований значительное содействие мне было
оказано сотрудниками Научно-исследовательского института Крайнего Се-
вера М. X. Геллером, В. А. Забродиным, М. И. Климантовым, В. А. Герке,
В. Куксовым и Б. Б. Боржоновым, а также участниками экспедиции
Гельминтологической лаборатории АН СССР В. Л. Контримавичусом,
Е. Сыроечковским и В. Трофименко.
В. В. Леонович, участник двух наших экспедиций в бассейне р. Пя-
сины, любезно предоставил в мое распоряжение свои дневники. Энто-
молог Ю. И. Чернов, также участник нашей поездки летом 1960 г., ока-
зал мне неоценимую помощь, взяв на себя определение содержимого
желудков птиц. При систематической обработке собранных материалов
я пользовался любезным содействием многих сотрудников Лаборатории
наземных позвоночных Зоологического института. Всем этим товарищам
считаю своим долгом выразить искреннюю благодарность.
Особенно я благодарен профессору Л. А. Портенко, который своим
исключительно внимательным отношением и ценными советами содейст-
вовал завершению работы.
Все фотографии в тексте работы выполнены автором.
— 189 —
КРАТКАЯ ХАРАКТЕРИСТИКА ПРИРОДНЫХ УСЛОВИЙ ЗАПАДНОГО ТАЙМЫРА
Исследованная нами территория включает западную и юго-западную
часть собственно Таймырского полуострова и прилегающую к ней с юга
северо-западную область плоскогорья Путорана. Находящиеся у запад-
ных отрогов этого плоскогорья озерные и речные долины также охвачены
нашими исследованиями. Таким образом, не вся нами изученная терри-
тория является Таймыром в буквальном смысле слова.
Р е л ь е ф. Более трех четвертей площади западного Таймыра занято
западной частью так называемой Таймырской депрессии — обширной низ-
менности, лежащей между устьями Енисея и Хатанги (Сакс, 1945).
На юге исследованной территории эта низменность граничит с северным
краем Средне-Сибирского плоскогорья, а в нижнем течении Пясины в нее
включаются западные отроги хр. Бырранга. Близ своей центральной
части Таймырская депрессия представляет собой слабо холмистые или
почти плоские равнины водоразделов, перемежающихся с обширными
ровными пространствами древних морских и речных террас. Абсолютная
высота наиболее возвышенных холмов изредка достигает 150—180 м,
обычно же они не превосходят 50—100 м (Сакс, 1945).
Более пересеченный рельеф, сопровождаемый и большими абсолют-
ными высотами, характерен для участков, расположенных по краям Тай-
мырской депрессии, близ отрогов хр. Бырранга и Средне-Сибирского пло-
скогорья. Здесь наблюдается типичный моренный ландшафт, характери-
зующийся обилием холмов высотой 25—50 м. Все сколько-нибудь зна-
чительные понижения между холмами обычно бывают заняты многочис-
ленными озерами. Многие озера соединены между собой, образуя системы,
но в ряде случаев лежат в глубоких котловинах, замкнутых высокими
холмами. Плоские низины, «лайды», иногда сменяющие типичный морен-
ный ландшафт по краям депрессии, обязаны своим происхождением ис-
чезнувшим более значительным озерам (Сакс, 1945).
Современные речные долины в пределах Таймырской депрессии не
превышают 5—7 км в ширину, лишь в редких случаях достигая 12—15 км.
Во многих местах реки вообще не имеют поймы и текут просто среди
холмов водораздельных плато или возвышенных морских террас. Поймы
характеризуются наличием многочисленных песчаных грив и сухих буг-
ров, перемежающихся с бесчисленными озерами.
Вблизи гор Бырранга на водоразделах начинают появляться выходы
трапповых пород. В предгорьях Бырранга высота отдельных холмов до-
стигает 300 и даже 400 м над ур. м. Здесь слабо расчлененные горные
массивы перемежаются с широкими заболоченными долинами. В бассейне
р. Тареи преобладает мелкосопочный ландшафт с редкими более высо-
кими холмами (Сакс, 1945).
На юге исследованной территории большие площади занимают горы
Путорана, являющиеся северо-западной возвышенной частью Енисейско-
Ленской платформы. Плоскогорье Путорана в своей центральной части
возвышается до 1800 —2000 м над ур. м., по периферии расчленено глу-
бокими долинами, отделяющими столовые возвышенности. Такие возвы-
шенности и отдельные горы имеют очень характерные очертания и до-
стигают 500—600 м над уровнем долин (Суслов, 1935). Более или менее
обширные возвышенности обычно имеют плоские или слабо холмистые
вершины и обрывистые склоны, иногда круто спускающиеся к долине
почти вертикальной стеной. Вершины отдельно стоящих гор часто имеют
форму острого ребра. Крутые края плато нередко прорезаются неболь-
шими эрозионными рытвинами, которые, однако, не нарушают его мас-
сивности. В верхних частях склонов можно увидеть глубокие ледниковые
кары (Суслов, 1935). Один довольно обширный горный массив —Лан-
— 190 —
такайский камень — полностью отделен от основного плоскогорья доли-
нами рр. Рыбной и Хантайки и имеет более сглаженные края и меньшие
абсолютные высоты.
Прорывающие массивный край основного плато узкие, но глубокие
долины, имеющие смешанное, тектонически-ледниковое происхождение
(Суслов, 1935), заняты рядом больших озер, обычно вытянутых в ши-
ротном направлении. Озера эти, по своему облику очень напоминающие
фьорды Скандинавии, имеют протяженность 40—120 км и с запада под-
гружены конечными ледниковыми моренами. Обрывающиеся к берегам
озер склоны плоскогорья часто очень круты, изобилуют осыпями и вы-
ходами скальных пород. Горизонтальное залегание и неоднородность
слагающих плоскогорье пластов сильно способствуют образованию ха-
рактерной ступенчатости горных склонов. При этом горизонтальные
террасы перемежаются с крутыми скалистыми обрывами или осыпями
из крупных глыб диабазов (Суслов, 1935; Михайлов, 1947). Некоторые
склоны, сложенные из более рыхлых пород, характеризуются наличием
мелкощебнистых осыпей. Гребни верхнего края обрывов под влиянием
выветривания часто приобретают сложную зубчатость. Поверхность плато
по большей части покрыта россыпью диабазов. Здесь много пятен мелких
камней и земли, окруженных более крупными камнями.
Многочисленные реки, впадающие в большие Норильские озера, текут
по дну крупных ледниковых долин, иногда соединяющих впадины двух
разных озер.
Особо важную роль среди ландшафтов юга исследованной территории
пграет так называемая «Норильская долина», имеющая протяженность
с севера на юг около 140 км, а с запада на восток до 60—65 км. В наибо-
лее пониженной своей части, близ устья Рыбной, эта долина лежит над
уровнем моря около 40 м, а на водоразделе рр. Рыбной и Хантайки ее
отметки достигают 110—120 м (Михайлов, 1947). При приближении к пе-
риферии уровень дна долины постепенно повышается. Рельеф дна Но-
рильской долины слабо холмистый, с исключительным обилием мелких
озер. Озера эти обычно лежат на разных уровнях и соединены друг
с другом целой системой узких ручейков, иногда с каменистым ложем.
Отдельные возвышенности здесь обычно не превышают 20—30 м над
уровнем ближайших озер. Благодаря обилию моренных отложений, вы-
стилающих дно Норильской долины, некоторые из протекающих здесь
рек имеют порожистый характер. Таковы Талая, Глубокая, Рыбная
в своем верхнем течении, Хантайка и некоторые из их притоков. Кое-где,
даже в центральной части долины, имеются выходы коренных пород.
Прорезая их, реки образуют характерные пороги и даже водопады, на-
пример Орон на р. Рыбной, Хантайские пороги и др. Ближе к централь-
ной части долины холмистый рельеф местами сглаживается и уступает
место довольно значительным, ровным заболоченным низинам — «лайдам».
На водоразделе рр. Рыбной и Хантайки отмечены узкие и длинные
(до 30 км) гряды коренных пород, относительная высота которых дости-
гает 100 м (Михайлов, 1947). Наконец, особого типа рельеф образуют
большие конечные морены, подпирающие с запада многие из больших
Норильских озер. Озеро Кета, например, подпружено мореной, вал ко-
торой достигает 50—60 м высоты и 5 км ширины.
Климат и растительность. Исследованная территория
находится между 68 и 74° с. ш., поэтому климат ее чрезвычайно суров.
Почти на всей площади западного Таймыра господствуют среднегодовые
температуры ниже —10° С. Полярный день, длящийся 1.5—2.5 месяца,
и несколько менее продолжительная полярная ночь оказывают значи-
тельное влияние па жизнь растительного и животного мира. Однако
в связи со значительным протяжением с севера на юг и некоторыми
— 191
особенностями рельефа территория эта хараклеризуется значительным
разнообразием ландшафтно-климатических условий.
Наибольшую площадь на западном Таймыре занимает типичная тундра,
простирающаяся от побережья Карского моря до широты устья Черной.
Она совпадает с полосой наиболее холодного лета, и среднеиюльская
температура там обычно не превышает 11 ° С. Восемь месяцев в году ха-
рактеризуются отрицательными средними температурами. Из остальных
четырех месяцев собственно летними являются только июль и август,
так как в сентябре, а особенно в июне много дней также бывает мороз-
ными. Легкая доступность пясинских тундр для северных ветров бла-
годаря небольшим высотам северных отрогов хребта Бырранга способст-
вует почти регулярным весенним похолоданиям, часто сопровождаемым
сильными пургами. Так, очень сильная пурга, вызвавшая гибель зна-
чительного числа пролетных куликов и воробьиных, бушевала 11 и
12 июня 1961 г. В следующем, 1962 г. значительное похолодание (до —7°,
—10°C) было нами отмечено 12—14 июня. Не оказывает смягчающего
влияния на климат тундры и такая сравнительно крупная река, как
Пясина, потому что ее истоки берут начало значительно севернее поляр-
ного круга. Пясина вскрывается обычно только в третьей декаде июня,
в то время как Енисей на той же широте — в начале этого месяца. За-
мерзает Пясина также на месяц раньше Енисея, в конце сентября или
в первых числах октября. Господствующие в тундре сильные ветры в со-
четании с пересеченным рельефом способствуют крайне неравномерному
распределению снегового покрова. С возвышенных мест снег зачастую
сдувается совсем, а в разного рода понижениях и распадках он скапли-
вается в громадных количествах. Такое неравномерное распределение
снега накладывает свой отпечаток на характер растительности. В местах
не очень пониженных, например в неглубоких приозерных котловинах,
в речных поймах и на верхней части оврагов, скопившийся за зиму снег
полностью успевает растаять к началу лета. В таких случаях он является
хорошей защитой для идущих далеко на север кустарниковых форм ив-
няка и зарослей карликовой березки. На дне глубоких оврагов и у кру-
тых склонов северной экспозиции снег залеживается до конца лета,
а в некоторые годы не успевает растаять и до наступления зимы. В таких
местах происходит образование снежников, характерных и для более
южных районов западного Таймыра (Суслов, 1935). На месте растаявших
снежников растительность или совсем не успевает развиться, или остается
весьма разреженной.
Из типов растительных сообществ, характерных для этой безлесной
зоны, следует прежде всего отметить осоково-моховой и пушицево-мохо-
вой кочкарниковый тип тундры (Виноградова, 1937), покрывающей поло-
гие склоны и вершины холмов на пространстве от устья Дудыпты до
устья Пуры и даже несколько севернее.
При этом пушицево-моховая кочкарниковая тундра, часто с кустарни-
ками и лишайниками, обычно встречается в условиях ухудшенного дренажа
на участках незначительных понижений, на нижних частях склонов,
около высыхающих водоемов. Кочки, обязанные своим образованием пу-
шице, имеют 10—30 см в высоту. Помимо пушицы и лишайников, на них
произрастают камнеломки, мытники, гречиха, а также некоторые осоки.
Пространства между кочками покрыты в основном мхами. Кустарники
ивы и карликовой березки обычно бывают расположены по ложбинам
между кочками, что обеспечивает им защиту в зимнее время.
Осоково-моховая с кустарниками тундра большей частью бывает при-
урочена к возвышенным, слабо увлажненным и недостаточно дренирован-
ным местам. Микрорельеф такой тундры характеризуется наличием лож-
бин разных размеров и сухих кочек и бугорков между ними. На кочках
— 192 —
преобладают осока и дриада. Иногда на кочках встречаются кустики
голубики и брусники, а также некоторые виды мхов и лишайников.
В ложбинах между кочками растут различные мхи, по которым стелются
кустарники ив и карликовой березки. Среди этой формации часто встре-
чаются различного размера пятна голого грунта.
Дриадово-моховая пятнистая тундра встречается в местах с хорошим
дренажем на прибрежных буграх у обрывов, на сухих холмах и на ка-
менистых склонах морен и распространена как на севере, так и на юге
исследованной территории. На выпуклостях и кочках в такой тундре
основным компонентом являются дриада и некоторые представители раз-
нотравья; мхов мало. Зато в ложбинках преобладают мхи. характерна
Кассиопея, а в более южных тундрах также толокнянка, голубика и
мелкий багульник. Дриадово-моховая тундра часто характеризуется на-
личием пятен голого грунта самых разнообразных размеров и форм.
Участки моховой тундры с кустарниками приурочены к пойменным
плоским островам и некоторым возвышенным террасам, особенно по бе-
регам рек у склонов коренного берега. Микрорельеф такой тундры очень
слабо кочковатый. Сплошной моховой ковер покрыт редкими кустиками
ивняка, осок и других растений. Пятнистость обычно не выражена.
Арктическая полигональная тундра занимает вершины и верхние части
склонов холмов на севере тундровой зоны. Значительные (до 80%) пло-
щади голого субстрата в такой тундре перемежаются с трещинами, порос-
шими дриадой, Кассиопеей, камнеломками и некоторыми другими видами
растений; мхов и кустарников ничтожно мало (Виноградова, 1937).
На многих склонах в низовьях Пуры можно было наблюдать постепенный
переход от типичной арктической тундры (на вершине холма) к кочкар-
никовой осоково-моховой (внизу).
Плоско-бугристые тундровые болота встречаются на плоских, не-
сколько пониженных участках приречных террас и водоразделов, главным
образом в южной части тундровой зоны. Этот ландшафт представляет
собой плоские бугры, в поперечнике от 3 до 15 м и высотой 0.3—0.5 м
(Виноградова, 1937), разделенные гипново-осоковыми мочажинами в виде
канав или округлых ям. Как бугры, так и мочажины характеризуются
торфянистыми почвами. На буграх обычны ерниково-моховые заросли,
иногда растут багульник и голубика. Ива и разнотравье встречаются
значительно реже, очень обычны лишайники. В мочажинах наиболее
обильны осока и пушица.
Низинные полигональные болота распространены главным образом
на участках плоских пойменных террас. Они представляют собой забо-
лоченные мохово-осоковые мочажины, разделенные довольно правильной
сетью перекрещивающихся узких мохово-кустарниковых гряд. В некото-
рых случаях мочажины переходят в небольшие мелкие озера с зарослями
у берегов. Такого типа болота отмечены нами как на юге тундровой зоны,
так и в низовьях Пясины.
Полоса кустарниковой тундры достигает своего наибольшего развития
южнее устья Дудыпты и тянется до устья Черной, где постепенно пере-
ходит в лесотундру. К югу от устья Дудыпты вдоль Пясины обычны
ольшаники, местами достигающие 2 м в высоту. Заросли низкорослых
ивняков и ерника здесь достигают большого развития и местами покры-
вают почти всю поверхность тундры. Ниже устья Дудыпты ольха встре-
чается только небольшими куртинками на склонах холмов. Самые север-
ные местонахождения ольхи близ Пясины отмечены в 30 км ниже устья
Агапы, а в водораздельной тундре отдельные кустики были встречены
мной даже в верховьях Пуры. Ивняки проникают па север, по-видимому,
вплоть до устья Пясины. Во всяком случае в 50 км ниже устья Пуры,
в защищенных от ветра распадках близ реки Пясины нам случалось ви-
13 Биология птиц — 193 —
деть кусты высотой более метра. Повсюду в пределах типичной тундры
ивняки занимают понижения рельефа, заносимые зимой толстым слоем
снега.
Проходящая южнее кустарниковых тундр зона лесотундры выражена
на западном Таймыре слабо. Она окаймляет северные и северо-западные
склоны плоскогорья Путорана, значительно расширяясь лишь у берега
Енисея. В климатическом отношении лесотундра характеризуется более
теплым и сухим летом. Метеорологических данных из припясинской ле-
сотундры нет, поэтому приходится ссылаться на сведения метеостанции
в Дудинке, находящейся близ северной границы приенисейской лесо-
тундры. Средняя температура июля там достигает 12° С, а среднегодовая
поднимается до —10.7Q С. В связи с более благоприятными метеорологи-
ческими условиями в лесотундре произрастает хотя и угнетенная, но
все же настоящая древесная растительность из лиственницы. Особенно
хорошо развиты лиственничные лесонасаждения по берегам рек и много-
численных озер. Отдельные угнетенные деревца проникают далеко
в тундру и были встречены нами даже в 30 км севернее устья Дудыпты.
В пясинской лесотундре наблюдается сочетание лиственничных куртин
с подлеском из ольхи, ивняков, белой березы, карликовой березки, ба-
гульника и некоторых других кустарников и кустарничков с участками
открытой тундры. Последние приурочены по большей части к водоразде-
лам и вершинам многочисленных холмов. Особенно обычны участки мо-
хово-кустарниковых, пятнистых и каменистых тундр (Шумилова, 1933).
Наконец, в южной части исследованной территории, на участке, за-
щищенном с севера и востока плоскогорьем Путорана, а с запада — мас-
сивом Лантакайский камень, находятся значительные площади северота-
ежного ландшафта, непосредственно переходящие в зону туруханской
и эвенкийской тайги. Климат этого защищенного горами участка приоб-
ретает заметные черты континентальности. По данным метеостанции аэ-
ропорта «Надежда», находящегося на крайнем северо-западе этой зоны,
на высоте 225 м над ур. м., средняя температура июля поднимается до
13.6° С. Многолетняя среднемесячная июля у пос. Валек, находящегося
на 25 км севернее, но на высоте 80 м над ур. м., достигает 14.6° С. Число
дней с температурами выше 0, 5, 10 и 15° (за три года, по данным метео-
станции в Надежде) составляет соответственно 105, 82, 49, 20 дней.
Восточнее и юго-восточнее, в окрестностях Норильских озер, климат еще
более континентальный. Количество осадков невелико — в районе На-
дежды оно составляет 386 мм, из которых 236 мм выпадает летом. Однако
благодаря особенностям рельефа и господствующим зимой сильным вет-
рам в долины Норильских озер, а также Рыбной и Хантайки много снега
наносится с окружающих участков плоскогорья Путорана. По этой при-
чине глубина снегового покрова в хорошо облесенной местности к весне
достигает 80, 100 см и более. Средняя годовая скорость ветра в окрест-
ностях Норильска достигает 5.7 м/сек. Наиболее бурные периоды — на-
чало и конец зимы, когда скорость ветра в отдельные дни достигает
38 м/сек. и более. Основное направление бурных ветров южное, юго-
восточное и юго-западное.
В связи с более теплым летом снежников здесь немного, но зато
некоторые из них достигают исключительного развития. Это связано,
видимо, с частными особенностями рельефа, способствующими накопле-
нию огромных масс снега. Длина снежников в долинах рр. Амундакты
и Камюстяха измеряется километрами.
К сожалению, геоботанических описаний этой территории в литера-
туре пока нет, но даже при беглом взгляде на местность в долинах
рр. Рыбной и Хантайки с какой-нибудь возвышенной точки бросается
в глаза преобладание лесного ландшафта в отличие от типичного лесо-
— 194 —
тундрового облика берегов Енисея на соответствующей широте. Таежный
характер растительности близ оз. Лама подчеркивает и С. П. Суслов
(1935).
Высокоствольный смешанный лес чаще всего тяготеет к склонам гор
южной экспозиции, где растет иногда полосой в несколько километров
шириной. По террасам и не очень крутым склонам такой лес иногда под-
нимается до высоты 200—250 м над ур. м. Особенно хорошо развита
древесная растительность в каньонах некоторых горных речек и по се-
верным берегам таких озер, как Лама, Глубокое, Кета, Хантайское.
Несколько реже попадаются участки такого леса близ центра речных
Рис. 2 Лиственничный лес в долине р. Рыбной.
долин; там они чаще приурочены к берегам рек и более или менее круп-
ных озер. Наконец, в южной части исследованной территории, в вер-
ховьях Хантайки, высокоствольные елово-лиственничные леса местами
вытесняют остальные типы древесных насаждений.
Преобладающей древесной породой в таких лесах являются сибирская
лиственница, в меньшем количестве встречаются ель и белая береза.
Количество ели и березы варьирует в зависимости от местности. Например,
процент ели значительно выше на берегах восточной оконечности оз. Кета
а в лесах, удаленных от краев речных долин, наблюдается наибольшее
преобладание лиственницы. Высота древостоя обычно колеблется от 12
до 15 м, отдельные деревья бывают несколько выше. Подлесок и подрост
в таких лесах выражены слабо, поверхность земли большей частью по-
крыта ковром из мхов и лишайников. Лишь кое-где, чаще всего на склонах
плоскогорья, существует обильный подлесок из ольхи, крупных ивняков,
карликовой березки, багульника и голубики. Изредка встречаются также
рябина и малина.
Лиственничный лес, во многих местах довольно разреженный (рис. 2),
является, пожалуй, самым типичным элементом местного ландшафта в ок-
рестностях Норильских озер. Он занимает обширные площади в долинах
— 195 —
13*
рр. Рыбной, Глубокой, Норилки, Хантайки и типичен для долин речек,
впадающих в большие Норильские озера с южной стороны.
Сибирская лиственница, составляющая более 90древесной расти-
тельности такого леса, достигает высоты 8—12 м, древостой разреженный,
бросается в глаза суховершинность многих деревьев и обилие сухостоя,
В подавляющем большинстве случаев лишайниково-моховая подстилка
в лесу покрыта густым подлеском из карликовой березки, багульника,
голубики и разных видов ив. Местами же мы видим только ягельный
и мохово-ягельный покров почвы.
Березово-лиственничный смешанный лес с примесью ели, ольхи и
разных видов ив чаще всего растет куртинами, вкрапленными в другие
растительные ландшафты, и приурочен большей частью к берегам мелких
озер, ручьев и склонам небольших холмов. Состоит такой лес в основном
из сибирской лиственницы с древостоем в 8—12 м, обычно значительного
количества белой березы, незначительной примеси ели и подлеска из ив,
ольхи, красной смородины, можжевельника, шиповника, карликовой бе-
резки, багульника, голубики и других кустарников и кустарничков.
Деревья в таком лесу растут густо, местами есть обильный подрост.
Тундроподобные открытые пространства, «лайды», кое-где поросшие
чахлым лиственничным лесом, занимают значительные площади в долинах
рр. Рыбной и Хантайки. Расположены они почти всегда в плоских низи-
нах, на водоразделах мелких речек. Эти унылые открытые пространства,
по своему облику несколько сходные с обширными торфяными болотами
европейского Севера, по характеру своей растительности очень напоми-
нают южные плоско-бугристые тундры. Их растительность состоит из
мхов, лишайников, осок, карликовой березки, багульника, местами встре-
чаются отдельные чахлые лиственницы или небольшие их поросли, в не-
которых местах — более или менее обширные заросли низкорослых по-
лярных ив. Довольно сильно развит кочкарник, на увлажненных участ-
ках много болот и озер.
Если рассматривать ландшафты окрестностей Норильских озер, нельзя
не упомянуть о многочисленных небольших островках и каменистых при-
брежных грядах, по своему облику очень сходных с участками аркти-
ческой тундры. Каменисто-щебнистый субстрат здесь покрыт зарослями
дриады, мхов и лишайников. Местами много площадок голого грунта,
кустарники развиты слабо. Своим возникновением подобные ландшафты
обязаны как доступности ветрам, так и положению у крупных водоемов,
освобождающихся от льда только в середине или конце июля.
Горные элементы ландшафта плоскогорья Путорана находятся в усло-
виях гораздо более сурового климата, что связано как со сравнительно
большими высотами, так и с доступностью для ветров любых направлений.
Метеорологических данных для северной части Путорана, к сожалению,
нет. По нашим наблюдениям, весна там наступает недели на 2—3 позднее,
чем в долинах, а осень — соответственно раньше. Поэтому в местах с от-
рицательным рельеформ, особенно у склонов северной экспозиции, в пло-
скогорье отмечено много снежников; некоторые из них сохраняются до
следующей зимы.
Как уже сказано выше, елово-лиственничные высокоствольные леса
на склонах южной экспозиции поднимаются до высоты 200—250 м над ур. м.
На террасах и пологих склонах, расположенных на высоте 300—350 м,
преобладают уже лиственничные леса, часто со значительной примесью
белой березы и ольховых кустарников. В долинах ручьев, стекающих
с плоскогорья, на этой высоте отмечены хорошо развитые заросли высокого
тальника. Некоторый интерес представляет нахождение разреженного
березового леса на одной из террас гор Тонель, на высоте около 300 м.
Низкорослые искривленные березы там растут в 4—7 м друг от друга;
— 196 —
почва покрыта мхами и лишайниками с редкими кустиками карликовой
березки и черники. На высоте 350—450 м, в зависимости от характера
рельефа, находится лиственничная или ольховая лесотундра. В сравни-
тельно защищенных от .ветров местах куртинки лиственничного редко-
лесья поднимаются до высоты 500—550 м, а ольховники даже несколько
выше. Отдельные искривленные лиственницы, как и заросли низкорос-
лого ивняка по логам на склонах южной экспозиции, достигают высот
800 м над ур. м. и более.
На ровных, не защищенных с севера плато уже на высоте 400—450 м
преобладает пятнистая мохово-лишайниковая тундра с редкими кустиками
карликовой березки. На склонах с выходами камней и других хорошо
дренированных местах на этой и большей высоте нередко встречаются
участки дриадовой тундры. В логах и приозерных понижениях есть не-
которое подобие лугов, где растет купальница и другие влаголюбивые
растения. На плоских вершинах, на высоте 600—800 м, находятся по-
крытые лишайниками россыпи камней с редкими кустиками дриады между
ними. Низкорослые кустики карликовой березки встречаются здесь лишь
изредка.
СИСТЕМАТИЧЕСКИЙ ОБЗОР ПТИЦ ЗАПАДНОГО ТАЙМЫРА
1. Краснозобая гагара — Gavia stellata stellata (Pontopp.). При наличии
подходящих водоемов очень обыкновенна на гнездовье всюду в пределах
западного Таймыра, выходя на побережье Карского моря. Она одинаково
многочисленна как на мелких лесных озерах, так и на припойменных тунд-
ровых водоемах, где плотность ее гнездования достигает в среднем 1 —
3 пары на каждые 10 озер. В числе немногих птиц изредка гнездится на
озерах плоскогорья Путорана, на высоте 450—500 м.
Прилетает краснозобая гагара сравнительно поздно, сразу после
появления талой воды поверх льда на озерах и образования промоин на
реках. В истоках Рыбной она была впервые отмечена в 1958 г. 13 июня,
в 1959 г. 4 июня и в 1964 г. 8 июня. Валовой ее пролет в эти годы проходил
там соответственно 14—20, 9—15 и 10—21 июня. Весной 1962 г. в устье
Дудыпты краснозобая гагара была отмечена 12 июня, ее пролет проходил
там 15—25 июня. На Пуринских озерах краснозобая гагара появилась
в 1961 г. 17 июня. Летят весной краснозобые гагары в одиночку, парами
или небольшими группами, по 3—5 птиц. Большие скопления они обра-
зуют в рано освобождающихся от льда истоках рек, вытекающих из боль-
ших озер (р. Рыбная), или в заберегах таких крупных озер, как Пурин-
ские.
В противоположность большинству видов птиц, связанных с водой,
краснозобые гагары приступают к размножению не сразу после прилета,
а только дней через 10—15. За это время уровень воды в озерах, особенно
пойменных, спадает после весеннего паводка и гнездящиеся у самой
кромки берега гагары к концу насиживания уже не могут оказаться в де-
сятках или даже в сотнях метров от воды. В предшествующее гнездова-
нию время пары гагар держатся поблизости от своих будущих гнездовий,
в заберегах или просто на участках залитой половодьем тундры.
В первой половине июля краснозобые гагары приступают к гнездо-
ванию. В противоположность другим видам гагар они избирают самые
маленькие водоемы, многие из которых даже нельзя назвать озерами.
Зачастую это просто заполненные водой ямы или канавы метров 5 шириной
и с низкими заболоченными берегами. Рыбы в таких водоемах не бывает,
поэтому гагары приносят корм птенцам с близлежащих рыбных озер
(Тугаринов и Бутурлин, 1911). Три найденных нами гнезда были устроены
на берегах луж, причем два из них располагались в непосредственной
- 197 —
близости от колонии серебристых чаек и бургомистров. Лоток диаметром
30—40 см во всех трех случаях представлял просто умятую во мху пло-
щадку со скудной выстилкой из стеблей осок и других болотных растений.
Гнезда находились в 20—60 см от воды, к которой всегда вели два хорошо
заметных лаза. По короткой крутой дорожке гагара сходит с гнезда в воду,
а по более длинной и пологой поднимается на гнездо.
Полная, слегка насиженная кладка из двух яиц найдена в пойме Пуры
8 июля 1963 г. Две наполовину насиженные кладки осмотрены в том же
году 14 июля на острове Пуринского озера. Яйца грязно-оливкового
цвета с темно-бурыми редкими пятнами. Размеры 6 яиц: 67 х47, 69.9 X45.8,
71.8x46, 72.6x46.7, 67.5x45.2, 68.8x46.2 мм.
Вылупление птенцов происходит в конце июля—начале августа.
Так в гнезде, найденном в пойме р. Агапы 26 июля, было два только что
выклюнувшихся птенца. Обе старые птицы находятся у выводка до тех
пор, пока молодые не научатся летать, т. е. до последних чисел августа
или первых чисел сентября.
В конце августа и в начале сентября можно было наблюдать неболь-
шие группы гагар, с гоготанием летающих над тундрой или лесом. Хо-
рошо выраженного осеннего пролета краснозобых гагар ни разу наблю-
дать не случалось; обычно они незаметно исчезают к моменту замерзания
мелких озер в конце сентября.
В желудках и пищеводах 5 добытых экземпляров найдены рыбьи
кости и чешуя. Более крупные рыбы достигали 15 см длины. Во время
весеннего пролета случалось наблюдать гагар, кормившихся в лужах
талой воды среди болотистой тундры, где они отыскивали, по-видимому,
беспозвоночных.
2. Чернозобая гагара — Gavia arctica arctica (L.). Как и предыдущий
вид, чернозобая гагара является обычной гнездящейся птицей всюду
на западном Таймыре, доходя, вероятно, до морского побережья (Колю-
шев, 1933). Во время нашей экспедиции в низовьях Пясины летом 1961 г.
чернозобые гагары были там довольно обыкновенны. Что касается числен-
ности, то этот вид не только не уступает краснозобой гагаре, но местами
встречается даже гораздо чаще. Так, на каждые 10 озер, обследованных
в тундре, лесотундре или северной тайге, приходилось от 2 до 5 пар черно-
зобых гагар.
На юге западного Таймыра, в истоках Рыбной, чернозобые гагары впер-
вые отмечены в 1958 г. 10 июня, в 1959 г. 4 июня, а в 1964 г. 7 июня. Ва-
ловой их пролет там наблюдался в 1958 г. 14—26 июня, в 1959 г. 6—19 июня,
в 1964 г. 10—20 июня. В устье Дудыпты в 1962 г. чернозобые гагары появи-
лись 6 июня, а закончился их пролет там 19—20 июня. В районе Пурин-
ских озер первые гагары в 1961 и 1963 гг. отмечены 18 и 11 июня, а их
пролет там проходил соответственно с 19 по 25 и с 16 по 22 июня. Сроки
появления и пролета гагар весной зависят прежде всего от наличия за-
берег, промоин и даже больших луж талой воды. Во время поздней весны
1961 г. случалось наблюдать одиночных гагар на мелких тундровых
озерах, плававших в воде, выступившей поверх льда, и нырявших в пои-
сках корма под лед, в проталины не шире метра. Во время валового про-
лета, который совпадает по времени с интенсивным образованием заберег
на реках и озерах, гагары летят чаще одиночно или небольшими группами,
хотя нередки стайки до 10 и более птиц. В районе Пуринских озер, где
направление пролета было более заметно, птицы летели в основном с за-
пада на восток.
В этот период в удобных для кормления полыньях и заберегах наблюда-
лись значительные скопления чернозобых гагар. В истоках Рыбной
и в устье Дудыпты нам случалось видеть более чем 15—20 птиц одновре-
менно. В. В. Леонович (устное сообщение) 16 июня 1961 г. встретил на за-
- 198 -
берегах р. Агапы более 70 чернозобых гагар. Весенний пролет заканчи-
вается в третьей декаде июня, после чего отдельные пары распределяются
по многочисленным уже оттаявшим озерам.
Как и другие виды гагар, чернозобая приступает к размножению только
после того, как озера войдут в берега после весеннего паводка. В одной
и той же местности сроки гнездования гагар бывают иногда довольно
сильно растянуты в зависимости от гидрологического режима водоемов
различного типа. На пойменных озерах гагары гнездятся несколько позже,
чем в водораздельной тундре. В противоположность краснозобой гагаре
чернозобая чаще всего избирает для гнездования озера средних размеров,
Рис. 3. Насиживающая чернозобая гагара — Gavia arctica (L.).
диаметром 150 —800 м и более, избегая самых мелких заболоченных водое-
мов. Чтобы избежать разрушительного действия прибоя, на больших
озерах она обычно устраивает гнезда в небольших заливах или около
выдающихся мысов, чаще всего на моховом или мохово-осоковом берегу,
возвышающемся над уровнем воды на 10—20 см; иногда используется
сфагновая сплавина. Лоток гнезда представляет собой умятую во мху
и осоке площадку диаметром 30—50 см, удаленную от воды на 20—60 см.
Яйца по форме и окраске сходны с яйцами краснозобой гагары, их размеры:
77.5x51.5, 76.5x48.3, 73.3x58.1 мм. Во всех 3 найденных полных клад-
ках было по 2 яйца, в выводках также отмечалось чаще всего по 2 пуховых
птенца.
В очень раннюю весну 1960 г. две кладки ненасиженных яиц (одна
из них неполная) были найдены в верховьях Пясины 24—25 июня, на Пу-
ринских озерах в 1961 г. 9 июля было обнаружено гнездо с неоконченной
кладкой, а в 1962 г. слабо насиженные яйца найдены 1 июля. Таким обра-
зом, в годы с нормальным ходом весны гагары заканчивают кладку в пер-
вой декаде июля. Исключение составляют птицы, гнездящиеся на поймен-
ных водоемах. Пока одна из птиц насиживает (рис. 3), вторая обычно
199 —
держится в непосредственной близости от гнезда. Вылупление птенцов
наблюдалось в 1963 г. на Пуринских озерах 22—24 июля. В первые двое
суток после вылупления птенцы, покрытые темно-серым пухом, еще
довольно беспомощны, плохо плавают и совсем не могут нырять. В конце
июля и в начале августа на многочисленных тундровых и лесных озерах
почти ежедневно наблюдались пары старых птиц с пуховыми птенцами.
В последних числах августа и в начале сентября молодые гагары подни-
маются на крыло, после чего выводки ведут кочевой образ жизни до самого
отлета, соединяясь иногда по нескольку особей вместе. Начиная со второй
декады августа на некоторых рыбных тундровых озерах были заметны
небольшие скопления чернозобых гагар, насчитывавшие иногда до де-
сятка и более особей, по-видимому, это были холостые птицы.
Улетает чернозобая гагара перед третьей декадой сентября, когда
нормально замерзают мелкие тундровые и лесные озера. На реках, за-
мерзающих несколько позже, чернозобые гагары в противоположность
крохалям и нырковым уткам, как правило, не задерживаются.
Чернозобая гагара — типичный ихтиофаг. Ныряя за рыбой, она спо-
собна пробыть под водой более минуты и проплыть 80—100 м, хотя во время
обычной кормежки гагары выслеживают свою добычу с поверхности воды
и во время погружений проплывают не более 15—20 м. Как показали на-
блюдения с высокого берега, гагара под водой гребет только ногами,
не прибегая к помощи крыльев.
3. Белоклювая гагара — Gavia immer adamsi (Gray). Белоклювая га-
гара в противоположность мнению некоторых авторов, которые считают
ее птицей морских побережий, на западном Таймыре довольно обычный
гнездящийся вид внутренних водоемов; на юг она доходит почти до се-
верной границы таежной зоны. Судя по нескольким встречам белоклю-
вых гагар летом 1964 г. на оз. Кета, они, возможно, изредка гнездятся
и в северотаежных местообитаниях. На всей этой территории, в зоне тундры
и лесотундры, она гнездится спорадично, не будучи приурочена к какому-
нибудь определенному ландшафту. Как показали подсчеты В. В. Леоно-
вича (устное сообщение) во время весеннего пролета гагар на р. Агапе
в 1961 г., численность пролетных белоклювых гагар уступает численности
весьма обыкновенных чернозобых всего в 4 раза. Следовательно, для внут-
ренних районов Таймыра эта гагара не является редким видом.
Сроки весеннего прилета белоклювых и чернозобых гагар почти совпа-
дают. Весной 1961 г. первые птицы наблюдались в низовьях Агапы 14 июня,
их валовой пролет отмечен 14—20 июня. В 1962 г. в устье Дудыпты весен-
ний пролет белоклювых гагар наблюдался 5—20 июня. Наконец, в 1963 г.
на Пуринских озерах слабо выраженный пролет проходил 11—18 июня.
Явно пролетный экземпляр белоклювой гагары был добыт нами в верховьях
Рыбной 19 июня 1964 г. Интересно заметить, что наиболее интенсивный
весенний пролет этих гагар был отмечен на р. Агапе, в месте, где она
течет с запада на восток. Летят белоклювые гагары парами или в одиночку,
только изредка группами до 3—5 птиц. Как и другие виды гагар, они
образуют на местах отдыха и кормежки временные скопления. К последней
декаде июня весенний пролет обычно заканчивается.
Сразу после прилета пары белоклювых гагар держатся в заберегах
озер около мест гнездования. Вечером и ночью можно было ежедневно
в течение двух недель слышать стоны и вопли этих птиц, пока уровень
воды в водоемах не установился.
Для своего гнездования белоклювые гагары избирают средней величины
и довольно крупные озера, до 2—3 км в длину, с прозрачной водой и пес-
чаным или каменистым дном. Такие водоемы, как правило, удалены от
речных пойм, но богаты рыбой. Оба найденных нами гнезда были устроены
таким образом, что прибойная волна никак не могла повредить насижи-
— 200 —
вавшим птицам. В одном случае оно было расположено на изрезанном
заливами берегу небольшого островка рядом с двумя гнездами серебристых
чаек, в другом — на небольшом озерке, отделенном от основного озера
узкой, но глубокой канавкой. В обоих случаях гнезда были устроены
на довольно сухом берегу, выше уровня озера сантиметров на 20—40,
и маскировались кустиками шерстистой ивы и карликовой березки.
Оба гнезда находились в 30—40 см от воды, к которой вели два хорошо
различимых лаза: пологий, для подъема на гнездо, и крутой, для спуска
в воду. Сами гнезда представляли собой слегка выложенные мхом и осокой
Рис. 4. Белоклювая гагара — Gavia immer adamsi (Gray) — на гнезде.
площадки диаметром около 50—60 см, постепенно переходящие в лазы
к воде. Две найденные кладки содержали по 2 яйца, в одной было единст-
венное яйцо. Интересно отметить, что две кладки были найдены в одной
и той же гнездовой ямке в 1962 и в 1963 гг. По окраске и форме яйца не
отличимы от яиц чернозобой гагары, но несколько крупнее. Размер 3 из-
меренных яиц: 88.5x55.5, 87x55.8, 87.3x55 мм.
В исключительно раннюю весну 1960 г. в гнезде, найденном у истоков
Пясины, 27 июня были немного насиженные яйца. В 1962 г. в окрестно-
стях Пуринских озер 7 июля найдены почти свежие яйца, в 1963 г. в этом же
гнезде отложенное яйцо было обнаружено 1 июля. Озера в это время еще
почти полностью были покрыты льдом. Таким образом, белоклювые
гагары, очевидно, нормально заканчивают кладку в конце июня—на-
чале июля. Этим более ранним по сравнению с другими гагарами срокам
гнездования несомненно способствует скорее устанавливающийся уровень
внепойменных озер.
В середине июля поражало исключительное обилие комаров, сплош-
ным слоем покрывавших яйца белоклювой гагары. Очевидно, они очень
сильно беспокоили насиживающую птицу, о чем свидетельствовали хорошо
заметные на скорлупе яиц кровавые пятна от раздавленных гагарой
насосавшихся насекомых. Насиживает кладку в основном самка (рис. 4).
— 201 —
Самец принимает участие в насиживании, сменяя самку на 2—3 дневных
часа, пока она кормится. В 1962 г. удалось сделать довольно интересные
наблюдения в период насиживания. Самка вела себя у гнезда осторожно,
соскальзывала в воду на расстоянии 100—150 м от приближающегося
наблюдателя и выплывала на поверхность уже метрах в 80 от берега.
Пока я осматривал гнездо, птица обычно молча плавала на середине
озера. Возвращалась она на гнездо только когда люди скрывались за
холмом. Если я заставал на гнезде самца, он подпускал к себе метров
на 15, затем нырял и выплывал шагах в 20 от берега. Здесь он с громким
криком, напоминающим ржание, выпрыгивал из воды на всю длину своего
тела и делал угрожающие выпады в мою сторону, приближаясь иногда
метров на 10. После 5—10 мин. такого поведения птица успокаивалась
и спокойно плавала метрах в 30 —40 от берега. Гагара проявляла замет-
ное беспокойство при появлении в окрестностях ее гнезда серебристых
чаек или поморников и в таких случаях даже в моем присутствии прибли-
жалась к гнезду, опасаясь за его благополучие. Взлетают белоклювые
гагары неохотно. Если их не тревожили, то они отправлялись на кормежку
и возвращались обратно к гнезду, проплывая по узкому, шириной 20—
25 см, ручейку, соединявшему озеро, где находилось гнездо, с другим,
более крупным озером, в котором гагары обычно кормились.
Насиживание длится не менее 25 дней. Вылупление птенца в гнезде,
найденном в 1962 г., происходило 31 июля. Во втором яйце можно было
услышать писк еще не выклюнувшегося птенца. Пуховой птенец белоклю-
вой гагары покрыт темно-серым пухом и так же беспомощен, как одноднев-
ные птенцы чернозобой гагары. Нырять он не может, быстро намокает
в воде и, попав в нее, стремится как можно скорее выбраться на берег.
Обе старые птицы проявляли около новорожденного птенца крайнее
беспокойство.
Ясно выраженного осеннего пролета у белоклювой гагары во внутрен-
них областях Таймыра не наблюдалось. Одиночные холостые птицы и мел-
кие их группы случалось встречать на реках и озерах со второй декады
августа до конца этого месяца.
По своей осторожности белоклювая гагара значительно превосходит
гагар других видов. Держится она обычно на крупных водоемах, далеко
от берега и взлетает неохотно. Как показали наблюдения с лодки, бело-
клювая гагара при передвижении под водой пользуется ногами; крылья
при этом у нее плотно прижаты к телу.
4. Лебедь-кликун — Cygnus cygnus (L.). Кликун — обыкновенная гнез-
дящаяся птица у Норильских озер и на р. Рыбной. У оз. Пясино граница
его распространения смыкается с гнездовой областью тундрового лебедя.
Гнездящиеся пары кликунов распределены сравнительно редко — одна
встреча на 30—50 км маршрута. Они всюду придерживаются наиболее
глухих лесных озер.
Среди водоплавающих северной тайги в предгорьях Путорана кликун
является первым вестником весны. Весной 1958 г. пары этих лебедей
были встречены в истоках Рыбной 19 мая, в день нашего приезда. Судя
по многочисленным следам на льду реки, они держались там уже давно.
В 1959 г. первая пара была отмечена в полынье истоков Рыбной уже 8 мая,
а в 1964 г. — вечером 6 мая. В это время стояла еще настоящая зима
и температура в ночное время иногда опускалась ниже —30 °C. В течение
мая прилетевшие кликуны держались на незамерзающих почти круглый
год полыньях по рекам, вытекавшим из Норильских озер. За время дневной
экскурсии случалось наблюдать одновременно по десятку и более лебедей.
В середине и в двадцатых числах мая обычно заканчивается прилет старых
кликунов, которые появляются парами. В конце мая и начале июня чаще
встречались мелкие группы неполовозрелых особей, кочевавших по озе
— 202 —
рам весь июнь. Стай кликунов ни разу наблюдать не случалось. Пролет
шел только днем. В ночные часы нам неоднократно попадались лебеди,
спавшие в значительном удалении от берега на льду реки и озер.
В конце мая и в первой половине июня кликуны уже встречались
на проталинах недалеко от мест гнездовий. В конце июня и в первых числах
июля в 1958 г. на 80 км маршрута в верховьях и среднем течении Рыбной
мы три раза встречали беспокоившиеся пары. Лебеди с криком летали
около небольших заросших лесных озер с сильно изрезанными берегами,
поросшими труднопроходимыми зарослями карликовой березки и ивняка.
Вылупление птенцов, очевидно, происходит в середине июля. Выводок
из трех пуховых птенцов размером с шилохвость был обнаружен на до-
вольно большом лесном озере близ истоков Рыбной 24 июля 1959 г. Ста-
рые птицы держались у выводка и подпустили лодку метров на 20.
Птенцы не могли нырять и были легко пойманы на воде. Первого летного
молодого в сопровождении двух старых птиц удалось наблюдать в истоках
Рыбной 26 сентября 1959 г.
Находящиеся у выводков кликуны теряют маховые вскоре после
вылупления птенцов, в третьей декаде июля. Так, оба старых лебедя,
наблюдавшихся у выводка 24 июля, при попытке взлететь потеряли сразу
все первостепенные маховые. У старого самца, добытого на оз. Конон
7 августа, недоросшие маховые выступали из чехлов на 30 —40 мм. Лии-
ных холостых кликунов мы ни разу не видели.
Осенний отлет происходит у Норильских озер в конце сентября и в на-
чале октября, сразу после замерзания мелких водоемов. В 1959 г. в послед-
ний раз кликуны наблюдались у истоков Рыбной 6 октября. Отлетают
они семьями, пролетных стай осенью не бывает. Летят кликуны на зи-
мовки в западном направлении и довольно медленно, как показала добыча
в Пермской обл. 16 ноября 1959 г. молодого лебедя, окольцованного нами
24 июля 1959 г. близ оз. Кета.
5. Тундровый лебедь — Cygnus bewickii Yarrell. Тундровые лебеди
гнездятся всюду в тундре и лесотундре, в бассейне р. Пясины. Южнее,
у Норильских озер, они встречались только на пролете. Численность
их на гнездовье не очень велика. Во время авиамаршрута над богатой
озерами поймой р. Агапы в начале августа 1963 г. на протяжении 100 км
было отмечено всего два выводка. Приблизительно такая же плотность заре-
гистрирована у р. Пуры. Во время плавания по ней в июле 1961 г. отдель-
ные лебеди и пары встречались не чаще чем через 50—80 км. В особенно
благоприятных, богатых озерами местах гнездящихся лебедей бывает
больше, и тогда пары встречаются в 3.5—5 км друг от друга (например,
в некоторых частях поймы Агапы, по сообщению В. В. Леоновича).
Весенний пролет тундровых лебедей наблюдался в истоках Рыбной
в 1958 г. 19 мая—3 июня, в 1959 г. 23—30 мая. а в 1964 г. 23 мая—22 июня.
В последнем случае, по-видимому, имел место и пролет холостых птиц
на места линьки. В устье Дудыпты в 1962 г. первые лебеди появились
3 июня, пролет небольших групп наблюдался 6—15 июня. На местах гнездо-
вий у р. Агапы и Пуринских озер хорошо выраженного прилета замечено
не было. На Агапе появление лебедей весной 1961 г. зарегистриро-
вано 31 мая; 8—10 июня там шел их массовый прилет (сообщение В. В. Лео-
новича). На гораздо позже вскрывающемся 2-м Пуринском озере, распо-
ложенном на 40 км севернее, лебеди появились только 17—20 июня.
В 1963 г. они наблюдались там с 4 июня. Таким образом в среднем тунд-
ровые лебеди прибывают на места гнездовий в конце мая—начале июня,
т. е. дней на 20 позже кликунов. В северной полосе тайги их весенний про-
лет проходит лишь немногим раньше. При этом отдельные группы в за-
тянувшиеся весны, например в 1958 г., задерживаются на несколько дней
на открытой воде в истоках рек. На пролете у Норильских озер и в устье
— 203 —
Дудыпты часто случалось наблюдать стайки из 5—20 птиц, а у Пуринских
озер обычно летели пары и одиночные лебеди.
Единственное найденное нами в 1961 г. гнездо было устроено на не-
затопляемой паводками сухой гриве, в пойме близ устья Малой Пуры.
До ближайшего тундрового озера было 600—700 м, а до реки метров 500.
Гнездо представляло собой кучу мха и осок около 1.5 м в диаметре и 25—
30 см высотой, с глубоким лотком, выстланным злаками. Насиживающую
птицу на громоздкой постройке было видно более чем за километр. 21 июля
в гнезде было 4 яйца, внутри которых уже пищали птенцы. При нашем
приближении насиживавшая самка прижалась ко дну глубокого лотка
так, что остались видны только S-образно изогнутая шея и верх спины.
Подпустив людей шагов на 70, она слетела и долгое время кружилась
вблизи, с угрожающим шипением приближаясь метров на 15. Иногда она
садилась метрах в 70—100 и пыталась отводить. Летом 1960 г. на озерах
в пойме р. Агапы встречены 2 выводка, состоявшие из 2—3 пуховых птен-
цов. 9 сентября на небольшом озере близ устья Дудыпты наблюдался
выводок из 4 уже подлетывавших молодых. Очевидно, к середине сентября
все птенцы поднимаются на крыло. Что касается линьки старых птиц
при выводках, то она начинается не раньше начала августа, так как
в конце июля мы еще наблюдали лебедей, летавших над тундрой.
Осенью 1959 г. осенний пролет тундровых лебедей в истоках Рыбной
длился с 27 сентября по 4 октября. Лебеди летели группами по 4—15 птиц
прямо на юг.
6. Белолобый гусь — Anser albifrons Scop. Белолобый гусь гнездится
в тундрах севернее устья Дудыпты, но не повсеместно. Его гнездовой
ареал состоит из отдельных участков, на некоторых из них численность
белолобого гуся не уступает численности гуменника.
Весенний пролет начинается несколькими днями позже, чем у гумен-
ников. В 1961 г. в тундре у 2-го Пуринского озера белолобые гуси впервые
замечены 9 июня. С этого же времени начался и их валовой прилети пролет,
который длился до 22 июня. Весной 1962 г. несколько южнее, в устье*
Дудыпты, белолобые гуси появились 28 мая, а их валовой пролет проис-
ходил 5—10 июня. В 1963 г. у Пуринских озер они были впервые отме-
чены 5 июня, а 6 и 7 июня наблюдался довольно интенсивный пролет
в северном направлении. У Норильских озер пролетные белолобые гуси
встречались лишь изредка. Во время пролета они летят обособленными
от других гусей стаями по 10—25 птиц. Только иногда мелкие группы
и одиночки присоединяются к гуменникам. Направление пролета белоло-
бых гусей северное, реже северо-западное или северо-восточное. Числен-
ность их на пролете в среднем течении Пясины не уступает численности
гуменников. Во всяком случае белолобые гуси составляют половину про-
дукции весенней охоты у местных охотников. Кормятся и отдыхают эти
гуси там же, где и гуменники, и подобно им тяготеют к первым появив-
шимся в тундре лужам талой воды.
В бассейне Пясины белолобый гусь гнездится отдельными очагами.
В таких местах численность его иногда превышает численность гуменника.
Участки повышенной численности белолобого гуся обнаружены нами
у р. Большой Дюрюсы, в окрестностях 2-го Пуринского озера и в низовьях
Пуры. Как и гуменники, белолобые гуси устраивают свои гнезда в раз-
личных типах тундры. Из 9 осмотренных гнезд 4 были расположены
в холмистой мохово-осоковой тундре, два —в кочкарниково-ивовой кустар-
никовой тундре, одно — в типичном полигональном болоте, на моховой
перемычке между заполненными водой ямами, одно — на мохово-лишай-
никовой с кустарником сухой гриве и одно — в дриадовой тундре у Пу-
ринского озера. Гнезда чаще всего представляют собой ямки, умятые
среди кочек и мха и довольно скудно выстланные смесью надерганных
— 204 -
в непосредственной близости прошлогодних растений с пухом и мелким
пером гусыни. От гнезд гуменника гнезда белолобых гусей обычно отли-
чаются несколько меньшим содержанием мха и травы в подстилке лотка,
которая состоит в основном из пуха и пера. Осмотренные нами полные
Рис. 5. Пара белолобых гусей — Anser albifrons Scop. — у гнезда.
кладки содержали 3, 3, 3, 4, 5, 6, 6 и 6 яиц. Их окраска несколько свет-
лее, чем у яиц гуменника. Размеры 16 яиц: макс. 85.2x55, мин. 73.5X
Х50.5, в среднем 79.2x51.6 мм. Как и гуменники, белолобые гуси в боль-
шинстве приступают к гнездованию сразу после прилета, во второй по-
ловине июня. Пары, гнездящиеся в южной части ареала, уже начинают
кладку, когда еще продолжается интенсивный пролет гусей на север.
Судя по состоянию яичника у самки, добытой на Пуринских озерах 22 июня
1961 г., 5 яиц у нее было отложено, а шестое, уже одетое в скорлупу,
находилось в яйцеводе. Только что оконченная кладка из 6 яиц в 1963 г.
— 205 —
была найдена 19 июня. В 1961 г. слегка насиженные кладки обнаружены
4 и 6 июля. Следовательно, сроки начала насиживания у белолобых
гусей, как и у гуменников, в значительной степени зависят от метеоро-
логических условий и падают на вторую половину июня—начало июля.
Во время насиживания самец почти все время держится рядом с сидя-
щей на яйцах самкой и первый замечает приближение опасности (рис. 5).
Вылупление птенцов у белолобого гуся происходит с первой декады
июля до конца этого месяца. Летом 1960 г. пуховые птенцы более чем
недельного возраста были пойманы уже 18 июля. С другой стороны,
в 1961 г. в низовьях Пясины птенцы такого же возраста наблюдались только
4 августа. В 1963 г. выводки из недавно вылупившихся птенцов в массе
появились 13—15 июля.
Рис. 6. Выводок белолобых гусей — Anser albifrons Scop.
Самцы всегда держатся около выводков вместе с самками (рис. 6),
и предположение Е. С. Птушенко (1952), что они присоединяются к холо-
стым особям и начинают линьку вместе с ними, безусловно ошибочно.
У выводка старые птицы ведут себя смелее, чем гуменники, подпускают
лодку или человека на 10—15 м, а потом самоотверженно отводят. В этот
период белолобые гуси держатся преимущественно на мелких тундровых
реках и озерах, где часто собираются по нескольку выводков вместе.
20 июля 1961 г. во время нашего трехкилометрового маршрута вдоль
р. Большой Дюрюсы наблюдалось 8 выводков белолобых гусей, в то время
как выводков гуменника на этом же участке было всего пять. В 1963 г.
на том же участке реки было не менее 10 выводков. Позднее, в десятых
числах августа, выводки белолобых гусей, как и выводки гуменников,
собираются на более крупных реках до подъема на крыло, т. е. до конца
августа. В это время выводки белолобых гусей и выводки гуменников
часто держатся вместе и образуют общие стайки по 20—40 птиц.
Линька старых гусей у выводков начинается в первых числах августа
и заканчивается в конце этого месяца, к тому времени, когда молодые
становятся летными. Хорошо выраженный пролет холостых гусей на
места линьки наблюдался у Пуринских озер в 1961 и 1963 гг. в конце
— 206 —
июня и начале июля. Стаи в два—три десятка особей, выстроившись
треугольником, летели на довольно большой высоте в северном и северо-
западном направлениях. Но в противоположность наблюдениям
В. М. Сдобникова (1959а) на Таймырском озере количество гусей, летящих
на места линьки, на западном Таймыре всегда во много раз меньше, чем
на весеннем пролете. Собираются на линьку холостые белолобые гуси
(рис. 7) часто на тех же водоемах, где и гуменники, и нередко образуют
общие с ними стаи. Чаще всего встречаются стаи из 26—50 гусей, но не-
редки и скопления до 200—300 особей. Как и у гуменников, у белолобых
гусей соотношение между количеством холостых и размножающихся
особей в разные годы резко меняется. В 1961 г., например, было много-
Рис. 7. Линные белолобые гуси — Anser albifrons Scop. — в низовьях р. Пуры.
холостых линных гусей и мало выводков, а в 1963 г. — наоборот. Линька
маховых у белолобых гусей начинается несколько позднее, чем у гумен-
ников, в двадцатых числах июля. По нашим наблюдениям, сделанным
в низовьях Пуры в 1961 г., белолобые гуси теряли маховые 20—29 июля.
У шести линных гусей, добытых на 2-м Пуринском озере 24 июля 1962 г.,
маховые еще не выступали из пеньков. К третьей декаде августа холо-
стые белолобые гуси уже заканчивают линьку.
Литературные указания на осенний отлет белолобых гусей с 20 по
30 августа (Наумов, 1931) относятся, очевидно, только к холостым осо-
бям. Довольно ясно выраженный осенний пролет наблюдался нами в вер-
ховьях Пясины 3—13 сентября 1958 г. Отдельные стаи белолобых гусей
были замечены во время валового пролета гуменников 30 сентября—
2 октября 1959 г. у истоков Рыбной и 27—29 сентября 1964 г. под Нориль-
ском.
7. Пискулька — Anser erythropus (Brunn.) (рис. 8). Распространение
пискульки на западном Таймыре очень своеобразно. Лишь изредка
встречаясь в северной тайге у Норильских озер, она является очень обыч-
ной гнездящейся птицей лесотундры у оз. Пясино и в верховьях Пясины.
Затем она в небольшом количестве гнездится в кустарниковой тундре
— 207 —
вплоть до устья Дудыпты. Скопления линных пискулек отмечены
В. В Леоновичем в арктической тундре в бассейне р. Тареи и даже у Тай-
мырского озера В. М. Сдобниковым (1959а).
Слабо выраженный весенний пролет пискулек в 1958 г. наблюдался
6—8 июня, а в 1959 г. — 27 мая и 1 июня в истоках Рыбной. Птицы летели
стаями по 15—20 особей, изредка в одиночку, в северном направлении.
Гнездо, найденное нами 8 июля 1964 г. у западного берега оз. Кета,
было устроено на сухой кустарниковой кочке среди болота, метрах в 70
от берега озера. Располагалось оно совсем открыто, и насиживающую
птицу совсем не скрывали росшие рядом кустики карликовой березки
Рис. 8 Пискулька — Anser erythropus (Brunn.) — у гнезда.
и багульника. Лоток гнезда был обильно выстлан смесью пуха с мелкими
фрагментами сухих растений. Лежавшие в гнезде 5 яиц находились на
последней стадии насиженности, вылупление гусят происходило на сле-
дующий день — 9 июля. У гнезда держались обе старые птицы. В ле-
сотундре из 4 осмотренных гнезд два были расположены на возвышенном,
поросшем смешанным мелколесьем полуострове у северо-восточного берега
оз. Пясино, а 2 других — на небольшом островке посредине озера, близ
истоков Пясины, в лесотундре. В последнем случае гнезда находились
в ближайшем соседстве с колонией полярных крачек, с многочисленными
гнездами морянок и средних крохалей и даже двумя парами серебристых
чаек. Впрочем, несколько пар полярных крачек гнездилось и побли-
зости от гнезд пискулек, найденных на оз. Пясино. Все гнезда были уст-
роены среди густой поросли ольхи, лиственницы и карликовой березки.
Насиживавшие птицы затаивались и подпускали человека на несколько
шагов. В 1960 г. кладки из 5 и 6 яиц, осмотренные на оз. Пясино 24 июня,
были очень сильно насижены, а в гнездах, найденных в истоках Пясины
27 июня и заключавших 4 и 5 яиц, птенцы должны были на днях выклю-
нуться. Весна в 1960 г. была очень ранняя, и сроки гнездования у боль-
шинства птиц были сдвинуты дней на 15 раньше. Поэтому нормальным вре-
— 208 —
менем вылупления птенцов у пискульки на западном Таймыре следует
считать, очевидно, 10—15 июля.
В конце июня 1960 г. в истоках Пясины часто наблюдались стаи по
20—30 пискулек, летавших над лесотундрой в разных направлениях.
Небольшие группы наблюдались в первых числах июля близ устья Ду-
дыпты. Интересно, что в период линьки маховых значительные скопления
ппскулек встречены В. В. Леоновичем в арктической тундре у р. Тареи,
километров на 300—400 севернее ближайших гнездовий этого вида.
Довольно оживленный осенний пролет в 1959 г. наблюдался в истоке
Рыбной с 27 сентября по 1 октября. Стаи летели в южном направлении
как днем, так и ночью.
8, 9. Гуменник — Anser fabalis fabalis (Lath.), A. f. sibiricus Alph.
Из всех гусей, гнездящихся на западном Таймыре, гуменник является
самым обыкновенным. Только на некоторых северных участках исследо-
ванной территории он несколько уступает в численности белолобому
гусю. Будучи редкой гнездящейся птицей северной тайги, гуменник обычен
в лесотундре и достигает максимальной численности в среднем течении
Пясины, где в некоторых местах благоприятной для гнездования тундры
приходится на 1 км2 3—5 гнезд. На многочисленных совершенно не посе-
щаемых человеком реках и озерах западного Таймыра очень обычны ты-
сячные скопления линных гуменников.
В окрестностях Норильских озер гуменники появляются в середине
мая. В 1958 г. в истоках Рыбной 19 мая шел уже довольно интенсивный
пролет. В 1959 г. первый гусь там был замечен только 20 мая, а валовой
пролет проходил 24 мая—3 июня. В 1964 г. первая группа гусей в истоках
Рыбной наблюдалась 17 мая, но наиболее интенсивный пролет проходил
31 мая—1 июня, когда и закончился. Весной 1961 г. в тундре у Пурин-
ских озер три гуменника наблюдались 22 мая, в день нашего прибытия
на место полевых работ, однако валовой пролет в условиях сильно за-
тянувшейся весны проходил 7—21 июня. До этого времени рано приле-
тевшие гуси держались на сравнительно сильно оттаявших отмелях рек,
например на Агапе, и почти совершенно отсутствовали в глубине тундры,
еще сплошь покрытой снегом. В 1962 г., тоже с несколько затянувшейся
весной, первые гуси появились в устье Дудыпты 22 мая, а их валовой про-
лет проходил 1—8 июня. В 1963 г. первый гусь наблюдался во время силь-
ной южной пурги 21 мая, а массовое появление гуменников отмечено
в первых числах июня. Весеннего пролета гуменников в 1963 г. не удалось
проследить. Очевидно, они появились во время сильных туманов, на-
блюдавшихся там в начале июня.
Время валового пролета гуменников обычно совпадает со временем
интенсивного образования проталин. Как уже отмечалось (Сдобников,
1959а), гуменники в период весеннего пролета очень охотно пользуются
попутным ветром, даже во время сильного снегопада. Более того, по на-
шим наблюдениям на западном Таймыре, самые первые гуси почти всегда
появляются вслед за южными пургами. В случае резких похолоданий
пролет гуменников не только прерывается, но и происходит частичное
возвращение уже прилетевших гусиных стай к югу. Так, после сильной
северной пурги 13 июня 1961 г., в течение трех дней при резком северном
ветре шел довольно интенсивный пролет гусиных стай на юг. Возврат
гусей был замечен и в устье Дудыпты после похолодания 7 июня 1962 г.
Во время валового весеннего пролета гуменники летят как стаями
по 20—30 птиц, так и группами из 3—6 особей. В ясную, тихую погоду
они чаще летят на высоте 30—70 м, а в ветреную и снежную — над са-
мой землей. Очень охотно гусиные стаи придерживаются богатых прота-
линами берегов и отмелей рек, текущих в меридиональном направлении.
С появлением открытой воды на болотах в таких местах собирается на от-
14 Биология птиц
— 209 —
дых огромное количество всех видов водоплавающих. Очень привлекают
их также рано освобождающиеся от снега, поросшие мелким хвощом
островки и отмели рек и больших озер.
В тайге у Норильских озер численность пролетных гусей невелика,
и за день с одного места редко можно увидеть более двух десятков летящих
на север стай. Это, очевидно, объясняется удаленностью Норильских
озер от основных пролетных путей. Зато в бассейне р. Пясины в лучшие
дни весеннего пролета стаи и мелкие группы гуменников пролетают через
одно и то же место каждые 10—15 мин. в течение почти всего дня. На юге
обследованной территории конец пролета определить довольно легко,
в то время как севернее, на местах массового гнездования гуменников,
его заметить труднее вследствие интенсивных кочевок уже осевших гусей.
Часть гуменников образует пары еще до прилета на места гнездования.
Во всяком случае в пролетных стаях, наблюдавшихся весной в истоках
Рыбной, можно было различить отдельные пары. Сразу после прилета
гусей на места гнездования ежедневно можно наблюдать брачные полеты,
когда сразу два самца летают за одной самкой. В это время гуси очень
заметны, повсюду слышится их гоготание и непрерывно происходят
местные передвижения пар и небольших групп во всех направлениях.
Такое поведение гусей характерно для второй половины июня.
Гнездятся гуменники не далее 500—600 м от какого-нибудь водоема,
чаще всего в кочкарниковой мохово-пушицевой или мохово-осоковой
тундре. В лесотундре они устраивают гнезда в смешанном мелколесье
по берегам небольших озер. Иногда они избирают места с хорошим об-
зором, поселяясь на обрывистых берегах озер, а также на вершинах и скло-
нах холмов. Из 47 найденных гнезд гуменников 31 было устроено в типич-
ной для окрестностей Пуринских озер мохово-осоковой и мохово-пуши-
цевой тундре, большей частью на пологих склонах увалов, 11 помещалось
на обрывистых берегах озер, 4 — в пятнистой тундре и 1 — на мохово-
лишайниковом бугре среди болота. Гуменники не теряют своего «стадного»
инстинкта даже в период гнездования, и отдельные пары нередко устраи-
вают свои гнезда в непосредственной близости друг от друга. Два гнезда,
найденные в холмистой кочкарниковой тундре 7 июля 1962 г., были уст-
роены в 7—8 м одно от другого. 20 июня 1963 г. на обрывистом берегу
острова 1-го Пуринского озера было найдено два гнезда, расположенных
метрах в 15 друг от друга. Нередко гуменники гнездятся и в непосредст-
венной близости от гнезд хищников — мохноногого канюка и сокола-сап-
сана. Около двух из четырех осмотренных в 1963 г. гнезд сапсанов гнезди-
лось по одной паре гуменников. В четырех случаях отдельные гнезда
гуменников были найдены около гнезд мохноногих канюков и один раз —
среди колонии серебристых чаек и бургомистров. Однако такого тесного
сожительства с хищниками, какое наблюдается у краснозобой казарки,
у гуменника нет.
Гнезда устраиваются обычно в углублениях между кочками, иногда
на верхушках крупных кочек или просто на голой земле по уступам бе-
реговых обрывов. Кустики карликовой березки и полярных ив в кочкар-
никовой тундре не могут создать для гнезд надежного укрытия, поэтому
насиживающую гусыню почти всегда можно заметить издали. Материалом
для выстилки гнезда служит смесь мелких перьев и пуха, выщипанных
на груди, с травой и мхом, надерганными в непосредственной близости от
гнезда. Из растительных материалов построено и основание гнезда.
Чаще всего это мхи, прикорневые части осок, кустики дриады, выдернутые
вместе с корнями, и стебли злаков. Соотношение между количеством рас-
тительного материала и перьев очень сильно варьирует в разных гнездах.
Количество пуха в лотке гнезда заметно возрастает к концу насиживания.
Размеры гнезда весьма изменчивы и находятся в зависимости от характера
— 210 —
места гнездования. Гнезда, устроенные в сухих углублениях между вы-
сокими кочками, гораздо менее массивны, чем расположенные в сырой
равнинной тундре. Наиболее мощные постройки из злаков и осок попада-
лись на земляных береговых уступах.
К строительству гнезд и откладке яиц гуменники приступают сразу
после прилета. В яйцеводе самки, добытой у истоков Рыбной 21 мая 1958 г.,
было готовое к сносу яйцо. В тундре гуменники начинают гнездиться
несколько позднее, в первых числах июня. В 1963 г. с нормальным ходом
весны первое гнездо с только что оконченной кладкой из 5 яиц было най-
дено у Пуринских озер 15 июня. Из 11 гнезд, найденных в этом году между
18 и 21 июня, 8 содержали уже законченные кладки. В холодную весну
Рис. 9. Насиживающий гуменник — Anser jabalis Lath.
1961 г. первая оконченная кладка из 4 яиц зарегистрирована 3 июля.
В 4 гнездах, осмотренных 5—7 июля 1962 г., были обнаружены кладки
недельной насиженности. Исключительно ранней весной 1960 г. гуменники,
судя по появлению птенцов в начале июля, приступили к кладке в мае.
Величина кладки у гуменника, как и у других видов гусей, подвержена
значительным колебаниям, зависящим от хода весны. 4 полные кладки,
найденные в 1961 г., содержали 2, 3, 3, 4 яйца. Весна в том году была хо-
лодная, пролет гусей затянулся, прилетевшие в тундру птицы не имели
достаточных запасов подкожного жира. В большинстве выводков, наблю-
давшихся летом того же года, было по 2—3 птенца. Гнезда гуменников
в 1961 г. встречались значительно реже, чем в другие годы. В 1962 г.,
также отличавшемся довольно холодной весной, 8 найденных у Пурин-
ских озер полных кладок содержали 1,2,2, 2, 3, 3, 3 и 5 яиц. В условиях же
хорошей весны 1963 г. среднее число яиц в 29 оконченных кладках почти
достигало 5. Не особенно редки были кладки из 7 яиц, а кладка с 1 яйцом
была найдена только один раз. Размеры яиц гуменника (146): макс. 92.5 X
Х57.5, мин. 73.2x48.3, в среднем 81.6x53.7 мм.
О степени заселения парами гусей одних и тех же участков тундры
в разные годы можно судить по количеству находимых гнезд. В 1961 г.
на Пуринских озерах и р. Пуре найдено 4 гнезда, в 1962 г. в районе Пу-
ринских озер — 8 гнезд, а в 1963 г. там же было осмотрено 34 гнезда.
— 211 —
14*
Насиживает только самка (рис. 9). Сходя с гнезда, что чаще всего бывает
в полуденные часы, она прикрывает яйца выстилкой с краев лотка. Са-
мец чаще всего сидит рядом с гнездом или кормится поблизости. При при-
ближении человека он первым настораживается и часто становится замет-
ным уже издали.
Во время наших маршрутов по заселенной гусями тундре в конце июня
и в первой половине июля самцы, а также присоединяющиеся к ним самки
и холостые гуси взлетали и с тревожным гоготанием кружились около
человека. К концу периода насиживания гуси становятся более скрытными.
Сроки вылупления птенцов у гуменника на западном Таймыре также
сильно колеблются в зависимости от хода весны. В 1960 г. новорожден-
ные гусята были встречены в устье Дудыпты уже в первых числах июля.
Летом 1961 г. первый выводок птенцов в возрасте 1—2 дней отмечен на
р. Пуре 18 июня, в 1962 г. 2 птенца такого же возраста наблюдались
17 июня. В сравнительно благоприятных условиях 1963 г. гнездо с только
что выклюнувшимися птенцами впервые найдено 12 июля. Массовое вы-
лупление гуменника в тундре западного Таймыра в годы с нормальной
весной происходит, очевидно, 13—15 июля.
На второй день после вылупления последнего птенца взрослые гуси
ведут выводок к ближайшему водоему. В это время выводки чаще всего
можно встретить на более крупных реках, таких, как Пясина, Агапа,
Пура, и на больших озерах. Пользуясь этими крупными водоемами как
путями расселения, они собираются на немногих благоприятных в кормо-
вом отношении озерах и небольших равнинных речках. Держатся выводки
гуменников чаще всего по нескольку на одном, нередко небольшом во-
доеме, иногда в ближайшем соседстве с холостыми линными гусями.
На богатых озерами низинах все гусиные выводки из года в год держатся
на одних и тех же немногих озерах. На их берегах, как правило, много
осок, хвощей, местами бросаются в глаза гладко вытоптанные гусями
участки чистого мха. Очень охотно гуси живут и на небольших тундровых
речках, например на Большой Дюрюсе, а также в низовьях речек, впа-
дающих в Пуринские озера. Держатся они там довольно оседло до середины
или до последних чисел августа, пока гусята не достигнут размера красно-
зобой казарки. Тогда многие выводки снова переходят на крупные реки,
где собираются в небольшие стаи, по 15—30 птиц. Во время маршрута
по р. Агапе 18 августа 1960 г. на протяжении 60 км нами было встречено
5 таких групп молодых гуменников, иногда вместе с белолобыми
гусями. Некоторые из молодых гусей уже пробовали летать. В годы с нор-
мальными метеорологическими условиями массовый подъем на крыло
молодых, а также и старых размножавшихся гуменников происходит
в конце августа и в начале сентября.
Линька гуменников при выводках и холостых птиц протекает в разные
сроки. Гуси, у которых интенсивно линяло мелкое перо, неоднократно
добывались в первых числах июля. Обычно это были размножавшиеся
птицы. Линька маховых у них начиналась в первой декаде августа. Ста-
рые птицы у 10 выводков, встреченных на р. Дюрюсе 31 июля 1962 г.,
еще могли летать. У встреченных на р. Агапе 10 августа 1960 г. гуменни-
ков первостепенные маховые находились в состоянии интенсивной линьки.
Старые гуси заканчивают линьку, а молодые становятся Летными почти
одновременно, в конце августа или в начале сентября.
Линька холостых гуменников начинается и заканчивается гораздо
раньше. В последних числах июня и в начале июля у Пуринских озер
ежедневно наблюдались стаи, насчитывавшие десятки птиц, летевших
на места линьки в северном и северо-западном направлениях. 7—9 июля
1961 г. на низинных припойменных участках в верховьях Пуры нами были
встречены многочисленные стаи гусей, прилетевших на линьку.
— 212 —
Линяют гуменники на реках и озерах, в самых глухих, редко посе-
щаемых человеком участках тундры. К таким водоемам можно причислить
все притоки Пясины от устья Агапы до мыса Входного и озера в бассейне
всех этих рек. Лишь сама Пясина, да озера, находящиеся близ рыбачьих
избушек, редко посещаются линяющими гусями. Численность их на не-
которых водоемах западного Таймыра бывает очень велика. Трудно или
почти невозможно произвести более или менее точный учет гуменников
даже на отдельных водоемах, так как во время линьки они часто сме-
шиваются в общие стаи с белолобыми гусями. Численность линных гусей
можно проиллюстрировать следующими наблюдениями, сделанными во
время нашего маршрута по р. Пуре летом 1961 г. Стаи, содержавшие в сред-
нем по 150 гусей, были встречены через каждые 3—4 км. В местах, где
холмистая тундра подходила вплотную к реке, гуси попадались через
каждые 2—2.5 км, на участках с широкими песчаными отмелями — че-
рез 6—7 км. Если учесть, что общая протяженность нашего маршрута
по Пуре составляла не менее 350 км, то получается, что на этой реке
линяет не менее 15 000 гусей. Почти на всех мелких притоках Пуры,
по крайней мере на их приустьевых участках, держатся стаи от несколь-
ких десятков до сотен линных гуменников и белолобых гусей.
В некоторые годы, например в 1963 г., численность холостых линяю-
щих гусей бывает во много раз меньшей, чем обычно. Зато количество
размножавшихся пар в 1963 г. было чрезвычайно велико. Это интересное
явление, отмеченное в 1946 г. на Таймырском озере (Сдобников, 1959а),
нельзя объяснить особенно благоприятными метеорологическими усло-
виями на местах гнездования. В 1960 г. в условиях исключительно ранней
и теплой весны и хорошего лета было очень много линных холостых гусей,
а количество выводков было обычным. Это отклонение от нормы, возможно,
зависит от условий, сложившихся еще на зимовках и во время пролета
на север.
Начало массовой линьки холостых гуменников происходит обычно
в середине июля. Приблизительно половина всего количества гуменников,
встреченных на р. Агапе 13 июля 1960 г., уже не могла летать, остальные
взлетали неохотно. Многие, взлетев, тут же падали в воду, потеряв на
наших глазах почти все маховые. В 1961 г. на р. Пуре первый линный
гуменник был замечен 10 июля, начало массовой линьки наблюдалось
15 июля. В 1962 г. на 1-м Пуринском озере 16 июля мы видели стаю не
менее чем из 300 гуменников, причем только 50—70 гусей сохраняло спо-
собность к полету. В 1963 г. начало линьки у гуменников отмечено 12 июля,
массовая линька проходила в середине месяца. Линька маховых проходит
очень быстро. У гусей, добытых 28 июля 1961 г. в низовьях Пуры, маховые
уже выступали из пеньков на 20—30 мм. Первых гуменников, поднявшихся
после линьки на крыло, мы отметили на р. Агапе 2 августа, а стаи хорошо
летавших гуменников наблюдались 7 августа 1960 г. В первой декаде
августа большинство холостых гусей уже оканчивало линьку.
Только что перелинявшие гуси еще держатся около мест линьки не-
большими группами, с криком летают над тундрой, как бы пробуя силы
своих еще не окрепших крыльев. К середине августа они собираются
в крупные стаи, которые кочуют по богатой голубикой тундре, поросшим
хвощом отмелям и другим кормным местам до самого отлета. В конца
августа к ним присоединяются начавшие летать молодые с выведшими
их старыми птицами. После этого гуменники держатся в богатых кормами
местах уже значительными стаями, насчитывающими иногда сотни гусей.
С другими видами в этот период они, как правило, уже не смешиваются.
Осенний отлет гусей происходит всегда в сентябре, в очень сжатые
сроки. Обычно гуменники держатся в тундре до тех пор, пока не ударят
морозы и не выпадет снег, после чего они сразу исчезают. В 20-х числах
— 213 —
сентября 1961 г. у устья Дудыпты проходил массовый отлет гуменников
во время первой пурги. Гуси стаями и небольшими группами весь день
летели низко над тундрой, пользуясь попутным северным ветром. В про-
тивоположность весеннему пролету очень хорошо выражен осенний пролет
гуменников в районе Норильских озер. В 1959 г. он наблюдался нами
в истоках Рыбной 30 сентября и 1 октября. Гуси летели стаями до 50—
70 птиц на высоте 100—150 м. Всю вторую половину дня 1 октября такие
стаи следовали друг за другом через короткие промежутки времени.
Потом, через несколько минут, следующие стаи появлялись выше или
ниже по реке. Все гуси летели строго на юго-восток. Пролет продолжался
и ночью. 2 октября гусей уже не было видно. Не менее оживленный про-
лет наблюдался и осенью 1964 г. под Норильском. В период с 27 по 29 сен-
тября, сразу после резкого похолодания, сопровождавшегося северным
ветром и снегопадом, многочисленные стаи гусей летели в юго-восточном
направлении. В некоторые моменты наиболее интенсивного пролета в поле
зрения было более чем по 10 стай. К 30 сентября пролет закончился.
Судя по направлению летящих осенью птиц и многочисленным наход-
кам в теле добытых весной гуменников чугунной дроби китайского про-
исхождения, гнездящиеся на западном Таймыре гуменники пролетают
через территорию Китайской Народной Республики, а, возможно, и зи-
муют там.
Пути пролета гусей осенью, видимо, отличаются от весенних, которые
приурочены в основном к долинам Енисея и Пясины. Но часть пясинской
популяции гуменников и во время своего осеннего пролета придерживается
Енисея, поскольку молодой гуменник, окольцованный нами 19 августа
1960 г. на р. Агапе, был добыт осенью того же года в устье Ангары.
В питании гуменника, как и других видов гусей, в весеннее время
основную роль играют хвощи. Их прошлогодние побеги поедаются гу-
сями на ранее всего оттаивающих отмелях рек и некоторых озер. Прошло-
годние пушицу и осоки и их корневища гуменники выщипывают на немно-
гочисленных в это время проталинах. Летом, особенно в период скоплений
гусей на линьку, основную их пищу составляют свежевыросшие пушица
и осоки, очень обильные на низинах с озерами. Хвощи поедаются в это
время в меньшем количестве, преимущественно по берегам более крупных
рек. Осенью стаи гусей охотно кормятся голубикой, в изобилии растущей
на склонах холмов в кустарниковой тундре и лесотундре. Во время осен-
него пролета в лесотундре и тайге гуменники кормятся голубикой, а оста-
навливаясь на берегах рек и озер, — также хвощом.
Для определения подвида гуменников, гнездящихся на западном
Таймыре, в нашем распоряжении была серия в 21 экземпляр, добытый
близ западного берега оз. Кета весной и в начале лета 1964 г. При сравнении
этого материала с сериями гусей с европейского Севера и северо-востока
Сибири, хранящимися в коллекции Зоологического института, стано-
вится очевидным, что примерно половина наших экземпляров принадле-
жит к номинальной форме A. f. fabalis (Lath.). Этот подвид является, видимо,
единственным для всего материкового Севера к западу от низовьев Лены
(Портенко, 1939), хотя достоверно гнездящихся экземпляров из самых
западных частей ареала явно недостаточно. Некоторые из наших экземпля-
ров по общей форме клюва очень напоминают подвид A. f. sibiricus Alph.,
северо-западная граница ареала которого проходит где-то по южному
Таймыру (Johansen, 1945). Произведя измерения длины надклювья у тех
19 экземпляров, пол которых был точно определен вскрытием, мы полу-
чили: 1) для 11 самцов — минимум 58, максимум 72, а в среднем 66.6 мм;
2) для 8 самок — минимум 51, максимум 66 и в среднем 61.1 мм. 3 самых
длинноклювых самца (надклювья в 72, 71 и 69 мм) и 4 самки (надклювья
66, 66, 64 и 64 мм) как по размерам, так и по форме клюва относятся
— 214 —
к A, j. sibiricus Alph. 5 экземпляров (3 самца и 2 самки) обладают пере-
ходными признаками. Таким образом, на нашем материале хорошо про-
слеживается стык ареалов А. /. fabalis и A. f. sibiricus, которые на запад-
ном Таймыре дают ряд переходных форм.
10. Краснозобая казарка — Rufibrenta ruficollis (Pall.). Всюду в бас-
сейне р. Пясины, к северу от устья Агапы, краснозобая казарка является
обычной гнездящейся птицей. По сообщению Л. Н. Мичурина, отдельные
выводки краснозобых казарок встречаются и на побережье Карского моря,
например на р. Убойной. Гнездование казарки в верховьях Дудинки
(Рогачева, 1959) крайне сомнительно и никакими фактическими данными
не подтверждалось. Во время наших многолетних исследований красно-
зобая казарка ни разу не была обнаружена не только в лесотундре, но
н в южной части кустарниковой тундры. Будучи гораздо менее много-
численной, чем гуменник и белолобый гусь, краснозобая казарка на своих
гнездовьях встречается все же довольно часто.
В тундру она прилетает позже других видов гусей. В устье Дудыпты
весной 1962 г. казарки впервые наблюдались 4 июня, на 2-м Пуринском
озере в 1961 г. они появились только 10 июня. В 1963 г. первая пара там
была отмечена 15 июня. Между тем пролет других видов гусей начался
уже давно. Заметно выраженного весеннего пролета казарок мы не на-
блюдали ни на Пуринских озерах, ни в устье Дудыпты, а у Норильских
озер не встречали их совсем.
Начиная с середины июня в окрестностях Пуринских озер пары или
небольшие группы казарок попадались уже почти ежедневно. Большая
часть тундры в это время была еще покрыта снегом, а на некоторых озе-
рах еще только появлялись забереги. В конце июня, со вскрытием тундро-
вых рек и появлением воды у берегов крупных озер, почти все казарки
собираются непосредственно у мест гнездования.
На обследованной территории можно выделить четыре типа местооби-
таний краснозобых казарок на гнездовье.
Первый тип гнездовий — гнезда устраиваются на подмытых крутых
берегах рек и больших озер на земляных уступах или трещинах оседаю-
щих глыб земли у верхнего края обрыва. Гнездовья такого типа обнару-
жены на р. Агапе и у Пуринских озер.
Вторым типом гнездовий казарки следует считать сравнительно по-
логие склоны на границе речных долин с верхней террасой. В таких
местах казарки гнездятся на склонах поперечных глинистых гребней,
отлого спускающихся в долину. В некоторые годы в ложбинах между
этими гребнями снег задерживается почти до конца лета. Местообитания
такого типа характерны для берегов р. Пуры и ее притоков.
Третьим типом гнездовий казарки являются выходы скал на берегах
рек или даже скалистые острова. Здесь птицы устраивают свои гнезда
в дриадовой тундре у края обрыва. В низовьях Пуры гнезда краснозобой
казарки были найдены именно в таких местах.
Наконец, в более редких случаях эти гуси гнездятся на довольно низ-
ких островах с моховым и мохово-кустарниковым покровом. Такого типа
гнездовья обнаружены нами на 2-м Пуринском озере и в среднем течении
Пуры.
Замечательно, что все зарегистрированные нами гнезда были устроены
в непосредственной близости от гнезд хищных птиц, в подавляющем
большинстве соколов-сапсанов. Вблизи 18 осмотренных гнезд сапсанов
были либо обнаружены гнезда казарок, либо казарки держались здесь
небольшими стайками. В трех случаях, на скалистом острове и на низмен-
ных островах, наблюдалось гнездование казарок среди колоний серебри-
стых чаек и бургомистров, и только один раз два прошлогодних гнезда
были найдены рядом с гнездом мохноногого канюка. Факты сожительства
— 215 —
краснозобой казарки с соколом-сапсаном описаны для гыданской тундры
С. П. Наумовым (1931), для п-ова Ямал Б. М. Житковым (1912). Гнезд
казарок, устроенных на значительном расстоянии от гнезд хищников,
мы не встречали ни разу. Обычно гнезда казарок находятся в радиусе
50—100 м от гнезда сапсана, но иногда и в непосредственной близости
от него. Гнездо казарки, найденное в низовьях одного из притоков Пуры
16 июля 1961 г., было устроено на площадке глинистого гребня, в 1.5 м
от находившегося на том же гребне гнезда хищника. Обычно около каж-
дой пары сапсанов гнездится 2—6 пар казарок. Соколы, по нашим наблюде-
ниям, никогда не трогают краснозобых казарок и не разоряют их гнезд.
Среди найденных у гнезд остатков пищи сапсанов ни разу не было обна-
ружено костей или перьев казарок или других видов гусей. Основную
пищу соколов составляют в этот период ржанки и другие кулики, а также
белые и тундряные куропатки. Более загадочны случаи гнездования ка-
зарок среди колоний известных разорителей птичьих гнезд — серебри-
стых чаек и бургомистров. По наблюдениям, сделанным в одной из ко-
лоний на 2-м Пуринском озере в июле 1963 г., вспугнутые с гнезд чайки
после своего возвращения никогда не разоряли устроенных прямо среди
колонии и ничем не защищенных гнезд казарок, гуменника, морянок
и краснозобых гагар. Все эти птицы, вспугнутые человеком, возвращались
к гнездам позднее чаек. Не трогали чайки даже бегавших среди колонии
новорожденных птенцов казарок. С другой стороны, именно в колониях
чаек насиживающие казарки подпускали человека вплотную и взлетали
только при попытке дотронуться до них. Эти факты совместного гнездо-
вания пока еще неясны и требуют специальных исследований. Во всех
случаях сожительства с хищными птицами казарки надежно защищены
от главного разорителя птичьих гнезд — песца.
Гнезда свои краснозобые казарки устраивают в углублениях под
прикрытием кустиков полярных ив, карликовой березки или прошло-
годних злаков, а иногда в ямках и трещинах обрывов вообще без всякого
прикрытия. В самом начале кладки гнездо представляет собой ямку
глубиной 5—8 см около 15—20 см в поперечнике. Оно очень скудно вы-
стлано стеблями и листьями прошлогодних злаков и других растений.
Со временем же лоток обильно выстилается темно-серым пухом с брюха
самки. Количество яиц в законченных кладках из 30 гнезд, найденных
в 1961, 1962 и 1963 гг., колебалось от 1 до 8, в годы с нормальной весной
равно в среднем 6—7. Попгем (Popham, 1898) находил и 9 яиц в кладке.
Яйца одноцветные беловатые, с легким оливково-желтоватым оттенком.
Их размеры (45 яиц): макс. 73.9x47, мин. 65.5x43.3, в среднем 69.6X
Х45.2 мм.
В годы с нормальным ходом весны краснозобые казарки начинают
нестись почти сразу после прилета, во второй декаде июня. В 1963 г.
в гнезде, осмотренном 20 июня на острове 1-го Пуринского озера, была
неполная кладка из 3 яиц. Законченная кладка из 8 еще не насиженных
яиц была найдена в том же году уже 25 июня. В годы с более суровой
весной, например в 1961 г., казарки с прилета держались парами и не-
большими группами неподалеку от гнезд сапсанов, а к кладке приступили
только в конце июня или даже в начале июля. На освободившихся от снега
поросших хвощом отмелях озер, где часто кормились краснозобые ка-
зарки, в 1961 г. мы дважды находили их яйца, лежавшие прямо на земле.
Первое гнездо с 1 только что отложенным яйцом было обнаружено тогда
28 июня. Неоконченную кладку из 3 яиц нам удалось найти 4 июля 1961 г.
В верхнем течении Пуры 7 июля 1961 г. было осмотрено 5 гнезд со слегка
насиженными кладками. Весной 1961 г. около каждого гнезда сапсана
в конце июня наблюдалось до 10 и более казарок, но в первых числах
июля почти все они куда-то исчезли, а гнездились только 1—2 пары.
— 216 —
Пока самки насиживают, самцы не сидят непосредственно около гнезд,
как гуменники и белолобые гуси, а держатся в 20—100 м от гнезда на снеж-
никах, на берегу или на воде ближайшего водоема. Если, как это обычно
бывает, на небольшом участке гнездится несколько пар казарок, то все
самцы соединяются в общую стайку.
Вылупление птенцов в годы с нормальным ходом весны происходит
в середине июля. В 1963 г. 14 июля в одном из гнезд уже проклюнулись
птенцы. После несколько запоздавшей весны 1962 г. 22 июля в одном
из 4 найденных гнезд находилось 5 только что вылупившихся птенцов,
а во втором гнезде птенцы уже проклюнули скорлупу. На следующий
день, 23 июля, птенцы выклюнулись и в двух других гнездах. 24 июля
на 2-м Пуринском озере было найдено еще одно гнездо казарки с 6 только
что выклюнувшимися птенцами. Таким образом, в 1962 г. вылупление
птенцов краснозобой казарки проходило очень дружно 21—25 июля.
В 4 гнездах, осмотренных в низовьях Пуры 24 июля 1961 г., вылупление
птенцов не начиналось. В 1960 году, отличавшемся очень ранней весной,
на одном из озер у р. Агапы 22 июля было обнаружено 2 выводка с птен-
цами более чем недельного возраста.
В первые сутки птенцы остаются в гнезде и отогреваются самкой.
Только после этого они способны следовать за взрослыми до ближайшего
водоема, который обычно находится не далее 200—300 м. Держатся вы-
водки на небольших и средних речках или на озерах, недалеко от мест
гнездования (рис. 10). Пуховые птенцы 2—3-дневного возраста прекрасно
плавают и ныряют, почти не уступая в этом отношении старым птицам.
Самка с самцом всегда находятся с птенцами. Дважды нам случалось
наблюдать соединенные выводки, когда 7 и 12 гусят плавали в сопро-
вождении 4 старых казарок. При приближении к выводку лодки или че-
ловека самец самоотверженно отводит, в то время как самка старается
как можно быстрее увести гусят подальше от опасности.
Молодые казарки, как и другие виды гусей на западном Таймыре,
начинают летать только во второй половине августа или даже в начале
сентября.
В это же время старые казарки у выводков заканчивают линьку ма-
ховых. Начинают линять они не раньше начала августа.
— 217 —
Холостые краснозобые казарки линяют на наиболее удаленных от
рыбачьих избушек реках и озерах. Значительных скоплений во время
линьки они не образуют. В самой большой из встреченных нами стай
было не более 150 птиц. Группы по 10—30 особей были встречены на
участке р. Пуры длиной около 60 км не менее 10 раз. Это была наивысшая
численность линных казарок, какую случалось отметить за весь период
экспедиций. С другими видами линных гусей краснозобые казарки обычно
не смешиваются. Зато, как это заметил и в тундре Гыданского полуострова
С. П. Наумов (1931), стайки линных казарок на р. Пуре очень часто дер-
жатся также в ближайших окрестностях гнезд сапсанов.
Начинают линьку холостые краснозобые казарки одновременно с гу-
менниками, в середине июля. 16 июля 1962 г. на Пуринском озере наблю-
Рис. 11. Линные краснозобые казарки—Rujibrenta rujicollis (Pall.).
далась стайка десятка в полтора только что потерявших маховые красно-
зобых казарок (рис. 11). В 1961 г. 15 июля было много птиц, уже начавших
линьку. У казарки, добытой 22 июля 1960 г. на р. Большой Дюрюсе,
маховые выступали из чехлов на 35—50 мм. 7 августа 1960 г. впервые
наблюдались холостые краснозобые казарки, полностью закончившие
линьку.
Хорошо выраженного осеннего пролета мы не наблюдали, хотя и встре-
чали на р. Пясине в конце августа отдельные стаи по 2—3 десятка казарок
в каждой.
11. Шилохвость — Anas acuta L. Шилохвость довольно обыкновенна
на гнездовье в северной тайге у Норильских озер и в лесотундре близ
истоков Пясины. В кустарниковой тундре у устья Дудыпты она встре-
чается уже гораздо реже, в верховьях Пуры немногочисленна, а севернее
уже не гнездится совсем. Небольшие скопления линных холостых шило-
хвостей наблюдались на тундровых речках у Пуринских озер и в бас-
сейне р. Агапы.
У истоков Рыбной первое появление шилохвостей в 1958 г. отмечено
1 июня, а 6—15 июня шел массовый прилет и пролет. В 1959 г. они на-
блюдались там уже 27 мая; их пролет начался 30 мая и закончился к 4—
5 июня. В 1964 г. первые шилохвости появились в истоках Рыбной 20 мая,
а их пролет и массовый прилет наблюдался 2—8 июня.
В тундру шилохвости прилетают несколько позже, когда появляются
лужи на болотах. В очень позднюю весну 1961 г. у Пуринских озер шило-
— 218 —
хвости впервые наблюдались 21 июня, в 1963 г. они были там уже 6 июня.
В 1962 г. в устье Дудыпты первая пара появилась 5 июня. В пределах
тундры хорошо выраженного пролета шилохвостей не бывает. Во время
весеннего пролета в южной части исследованной территории во многих
стаях насчитываются десятки птиц. Самцы и самки прилетают одновре-
менно, хотя держатся зачастую отдельными группами. В истоках Рыбной
можно было видеть многие десятки шилохвостей одновременно. Они от-
дыхали на кромке льда у речных промоин. Пролетные стаи чаще всего
наблюдались в утренние и вечерние часы. Все они летели в северном на-
правлении.
Сразу после прилета, в первых числах июня, случалось встречать
отдельные пары. В течение почти всего июня ежедневно можно было ви-
деть самцов (одного или даже двух), следовавших за самкой, летевшей
над рекой или озерами. Во второй половине июня самки замечались уже
значительно реже. Самцы начинали собираться в стайки. В последних
числах июня и в начале июля шилохвости в северной тайге попадались
довольно редко: гнездящиеся самки начали насиживание, а холостые
вместе с самцами откочевали на места линьки.
Гнездятся шилохвости чаще всего в участках северной тайги, очень
богатых мелкими озерами, и в лесотундре. В тундре они обычно селятся
возле припойменных водоемов или по заросшим берегам озер на плоской
равнине. Найденные у р. Рыбной 5 гнезд были расположены на рас-
стоянии 1, 60, 60, 100, 200 м от берегов ближайших водоемов в густых
зарослях карликовой березки. Лишь одно гнездо было устроено среди
камней на моренной гряде. Единственное найденное нами в тундре у Пу-
ринских озер гнездо помещалось у колонии серебристых чаек и также
было хорошо защищено густым ерником. Все гнезда представляли собой
просто ямки во мху, выстланные серым пухом или смесью пуха и в незна-
чительном количестве — растительного материала.
В северной тайге у Норильских озер шилохвости приступали к на-
сиживанию 18—25 июня. В гнезде, найденном 26 июня 1958 г. на берегу
оз. Кета, было 10 свежих яиц. В гнезде, найденном в моренной гряде
у западного берега оз. Кета 19 июня 1964 г., была незаконченная кладка
из 4 яиц. В дальнейшем самка снесла еще 2 яйца. Во втором гнезде, най-
денном в тот же день на поросшем багульником торфяном бугре, было
8 ненасиженных яиц, эта кладка была окончена 21 июня, когда было сне-
сено последнее, 9-е яйцо. В двух гнездах, осмотренных 3 июля 1958 г.
по соседству с водопадом Орон на р. Рыбной, было 7 и 8 яиц более чем
недельной насиженности. Как и другие утки, шилохвость, уходя, закры-
вает яйца пухом с краев гнезда.
Вылупление птенцов на западном Таймыре происходило 10—20 июля.
В 1958 г. первый выводок только что вылупившихся птенцов наблю-
дался на р. Норилке 20 июля, в 1959 г. выводки в 7 и 8 однодневных птен-
цов отмечены близ истоков Рыбной 12 и 14 июля. 18 июля снова встретился
выводок. Один из пяти птенцов в нем весил 25.5 г. В 1963 г. в пойме Пуры
уже 8 июля отмечен выводок из 8 утят, которых самка вела к воде.
В гнезде шилохвости, осмотренном на 2-м Пуринском озере 14 июля,
все 6 яиц были надклюнуты. Выводки шилохвостей чаще всего держались
на мелких илистых заливах таежных рек и крупных озер, а также на не-
больших заросших таежных и тундровых водоемах. Повсюду замечались
большие колебания сроков развития птенцов в разных выводках. 6 ав-
густа 1958 г., например, в одном из мелких илистых заливов оз. Глубокого
я одновременно наблюдал птенцов величиной с чирка и совсем маленьких.
У птенца, добытого на притоке Хантайки 5 августа 1959 г., на брюхе
и спине уже появились мелкие перья, хвост имел в длину 40 мм, а на
крыльях только пробивались пеньки маховых. Первых способных к по-
— 219 —
лету молодых шилохвостей случалось видеть у Норильска в двадцатых
числах августа. В истоках Рыбной 29 августа 1959 г. наблюдался выводок
из 4 очень хорошо летавших молодых, а 3 сентября на одном из озер
в истоках Пясины были отмечены 2 хорошо летавшие молодые, но был до-
быт и птенец с еще не вполне выросшими маховыми. К началу сентября
подавляющее большинство молодых шилохвостей на западном Таймыре
уже хорошо летает.
Скопления холостых шилохвостей на линьку в пределах исследован-
ной территории нами отмечены в бассейне р. Агапы и на Пуринских озе-
рах. Здесь шилохвости выбирали тихие извилистые речки, текущие по
равнинной тундре и соединенные с многочисленными озерами сложной
системой проток. Берега таких речек и проток местами покрыты густыми
зарослями полярных ив, заливаемых половодьем в конце июня и начале
июля. Сходные участки есть в верховьях Большой Дюрюсы и в низинной
тундре между 1-м и 2-м Пуринскими озерами. В первых числах июля
здесь ежедневно случалось наблюдать стаи, прилетавшие на линьку:
10 июля 1962 г. на Большой Дюрюсе была отмечена первая шилохвость,
потерявшая маховые; 12 июля в верховьях этой речки наблюдалась стайка
из 12 самцов, только что начавших линять. В этот и следующие дни в за-
топленной тундре окрестностей Пуринских озер было уже много линных
шилохвостей и чирков. Как и гуси, шилохвости в начале линьки очень
упитаны. Заканчивают линьку они в конце июля и начале августа.
Во время поездки по Большой Дюрюсе 31 июля 1962 г. мы убедились, что
приблизительно половина всех встреченных шилохвостей уже летала.
Хорошо выраженный осенний пролет шилохвостей наблюдался в 1959 г.
у истоков Рыбной. Утки летели в ночные часы. Весь сентябрь стаи каж-
дую ночь садились кормиться на илистые, поросшие хвощом отмели
р. Рыбной. Наибольшее количество утиных стай было учтено в середине
и в двадцатых числах сентября. Многие из них насчитывали десятки
и даже сотни особей. В последний раз шилохвости были отмечены 4 сен-
тября. Осенью 1964 г. в связи с высоким уровнем воды в р. Рыбной, за-
топившей все илистые отмели, шилохвостей не наблюдалось совсем.
Несколько желудков шилохвостей, добытых во время весеннего про-
лета в истоках Рыбной, содержали в большом количестве бокоплавов
(Gammaridae) и вегетативные части придонных растений.
12. Свиязь — Anas penelope L. На северной окраине тайги у Нориль-
ских озер свиязь на гнездовье очень обыкновенна и по численности не
уступает шилохвости. В лесотундре в истоках Пясины она встречается
уже довольно редко, а в тундре у р. Агапы не найдена вовсе.
Весной свиязь прилетает раньше всех других уток. В 1958 г. у истоков
Рыбной первая пара отмечена 21 мая, массовый прилет происходил 6—
12 июня. В 1959 г. первые свиязи появились на р. Рыбной также 21 мая,
однако многочисленные стаи наблюдались только 30 мая и 4 июня. В 1964 г.
валовой пролет свиязей происходил в истоках Рыбной 8—11 июня. В де-
сятых числах июня повсюду на речках и озерах был слышен брачный
крик самцов и крякание самок: самцы гонялись за самками. В конце июня
свиязи, как и шилохвости, держатся очень скрытно.
Кладки начинаются рано. Самка, добытая в истоках Рыбной 6 июня
1959 г., уже отложила первое яйцо. Судя по появлению выводков, свиязи
начинают насиживать одновременно с шилохвостями.
В районе оз. Кета 12 июля 1959 г. был добыт пуховой птенец весом
207 г; 23 июля 1958 г. птенец в возрасте нескольких дней попался в кап-
кан, поставленный на ондатру; 26 июля 1959 г. на притоке Хантайки
мы видели птенцов величиной почти с чирка; 18 августа 1958 г. на р. Глу-
бокой наблюдалось около 20 утят приблизительно одинакового возраста
с двумя или тремя самками. Птенцы еще не летали, так как их маховые
- 220 —
выступали из чехлов всего на 2—3 см. Такого же возраста были и утята,
добытые 28 и 29 августа 1964 г. в одном из заливов оз. Кета, хотя в неко-
торых наблюдавшихся там же выводках они уже начали летать. Утенок,
добытый 24 августа 1959 г. на одном из озер близ истоков Рыбной, весил
590 г, маховые у него выступали из чехлов на 5—6 см. Летных молодых
свиязей случалось наблюдать в конце августа и в начале сентября, но
некоторые выводки очень запаздывают: 12 сентября 1959 г. у истоков
Рыбной была добыта молодая птица, еще не умевшая летать.
В первой половине сентября летные выводки ежедневно наблюдались
в илистых протоках Рыбной и на мелких лесных озерах. Пролетные
стаи, насчитывавшие зачастую сотни свиязей, наблюдались на отмелях
р. Рыбной каждую ночь с 16 по 27 сентября 1959 г. В последний раз мы
встретили свиязь 4 октября.
13. Чирок-клоктун — Anas formosa Georgi. Чирок-клоктун на запад-
ном Таймыре — очень редкая гнездящаяся птица. 28 мая 1960 г. нам
удалось добыть одиночного селезня в окрестностях Норильска, другой
самец был добыт из пары 29 июня 1963 г. у р. Большой Дюрюсы; у самки
там явно где-то было гнездо. Чаще встречались клоктуны близ истоков
Рыбной весной 1964 г., когда мы видели парочки и даже стайки этих чир-
ков 14, 16, 20, 27 июня. В последнем случае добыли селезня от самки,
видимо гнездящейся. Несколько сомнительны сведения Е. Е. Сыроечков-
ского (1961), который во время весеннего пролета уток у Малого Хантай-
ского озера видел будто бы до 7—8 клоктунов в день. Весной 1958, 1959
и 1964 гг. в истоках Рыбной, всего в 60 км от Малого Хантайского озера
и в совершенно сходных ландшафтных условиях, нами были добыты мно-
гие сотни уток и среди них только 3 клоктуна. В затопленной половодьем
тундре у р. Большой Дюрюсы 12 июля 1962 г. было добыто несколько клок-
тунов, только что потерявших маховые. Очевидно, одиночные особи
иногда линяют в тундре западного Таймыра вместе с шилохвостями и чир-
ками-свистунками.
14. Чирок-свистунок — Anas crecca crecca L. Распростренение чирка-
свистунка на западном Таймыре полностью совпадает с областью гнездо-
вания шилохвости, но всюду он встречается значительно реже. Во время
линьки маховых холостые чирки встречаются в тундре вместе с шилохво-
стями.
На места гнездования чирки прилетают значительно позже других
речных уток. В истоках Рыбной в 1958 г. они появились 8 июня, в 1959 г.
5 июня, а в 1964 г. 10 июня. Их массовый прилет и пролет в эти годы там
наблюдался соответственно 14—20, 7—9 и 11—13 июня. В устье Дудыпты
весной 1962 г. чирки появились 6 июня, а у Пуринских озер в 1963 г.
13 июня. Во время весеннего пролета у Норильских озер были нередки
стайки более чем по 10 чирков; в тундре они чаще встречались парами
или небольшими группами.
К размножению чирки приступают почти сразу после прилета. В се-
редине июня всюду на небольших лесных озерах и ручьях по ночам слы-
шалось их оживленное крякание. Они очень охотно селятся около мел-
ких луж и ям, заполненных водой. Гнездо, найденное у р. Рыбной, на краю
кочковатого болота, среди кустиков мелкого тальника и карликовой бе-
резки, помещалось около едва заметной лужи. 4 яйца лежали в углубле-
нии между кочками, обильно выстланном пухом. 2 июля 1958 г. яйца
были уже значительно насижены. У самки, добытой у оз. Кета 18 июня
1958 г., было отложено 5 или 6 яиц.
Вылупление птенцов у чирков, гнездящихся в окрестностях Нориль-
ских озер, происходит во второй декаде июля. На одном из озер у устья
Талой 27 июля 1958 г. был отмечен выводок птенцов недельного возраста.
17 августа 1959 г. на одном из сильно заболоченных озер у правого берега
— 221 —
Рыбной наблюдались 5 птенцов величиной почти со старую утку, но с не-
развитыми маховыми. Добытый 23 августа молодой чирок весил 220 г,
его маховые выступали из чехлов на 2 см. Пух в очень небольшом коли-
честве сохранился у него только на надхвостье. В одном из встреченных
26 августа 1959 г. выводков птенцы уже летали. В конце августа всюду
можно было наблюдать хорошо летавших молодых чирков. В первых чи-
слах сентября случалось встречать стаи, в которых насчитывалось 20—
30 чирков, кормившихся на илистых отмелях р. Рыбной.
Линька холостых чирков-свистунков протекает в тех же местах и в те же
сроки, что и у шилохвостей. В верховьях Большой Дюрюсы 12 июля
1962 г. мы добыли около десятка чирков, менявших весенний наряд на
летний. Птицы взлетали очень неохотно, очевидно, маховые у них ослабли
накануне линьки. В затопленной кустарниковой тундре попалась тогда же
и стайка чирков, только что утративших маховые. Все добытые в этот
период свистунки были очень жирными. Больших скоплений линных чир-
ков нам видеть не случалось. В первых числах августа у большинства
чирков линька заканчивалась. Осенний пролет в истоках Рыбной наблю-
дался 5—15 сентября 1959 г. 20 сентября отмечены только оди-
ночки. Самые последние чирки отлетели из истоков Рыбной 26 сен-
тября.
15. Гага-гребенушка — Somateria spectabilis (L.). Гага-гребенушка —
хотя и немногочисленная, но определенно гнездящаяся птица в нижнем
и среднем течении Пясины. Южным пределом ее гнездового ареала яв-
ляются низовья Агапы. Близ устья Дудыпты, гребенушек в период гнез-
дования нам встречать уже не случалось.
В 1961 г. отдельные стаи и пары гаг наблюдались у 2-го Пуринского
озера 20—25 июня. Весной 1962 г. единственный экземпляр гребенушки
был отмечен и добыт близ устья Дудыпты 10 июня. По заберегам Пурин-
ских озер и по всей реке Пуре небольшие группы самок встречались
в июле 1961, 1962 и 1963 гг. В конце июля они еще хорошо летали. Летом
самцы гребенушек попадались очень редко.
Гнездо с 4 свежеотложенными яйцами было найдено В. В. Леоновичем
7 июля 1961 г. в пойме Агапы, в осоке среди низинной тундры. Выводок
из 6 или 7 пуховых птенцов встречен нами на маленьком озере в пойме
Агапы в начале августа 1960 г.
16. Хохлатая чернеть—Aythya fuligula (L.). У Норильских озер,
а также в бассейне рр. Рыбной и Хантайки хохлатая чернеть — обыч-
ная гнездящаяся птица. В лесотундре близ истоков Пясины мы ее уже не
встречали.
Весной 1959 г. в верховьях Рыбной хохлатая чернеть впервые отмечена
4 июня. Довольно заметный прилет наблюдался 6—13 июня 1959 г. и
10—11 июня 1964 г.
Как и морские чернети, хохлатые расселяются по лесным озерам сразу
после появления на льду талой воды. В это время они встречаются
преимущественно парами.
У самки, добытой у истоков Рыбной 24 июня 1958 г., диаметр наиболь-
шего фолликула равнялся 22 мм, яйцевод, по-видимому, был активен.
У другой самки, добытой в тот же день, насколько можно было судить по
состоянию яичника, было уже отложено 4 яйца.
Выводок из пуховых птенцов в возрасте нескольких дней наблюдался
на одном из озер у оз. Глубокого 6 августа 1958 г. Птенец, добытый близ
истоков Рыбной 4 сентября 1959 г., весил 660 г, его маховые выступали из
чехлов на 52 мм. Осенью 1964 г. массовый подъем на крыло молодых
хохлатых чернетей на озерах близ истоков Рыбной происходил 5—10 сен-
тября; двух хорошо летавших молодых чернетей удалось добыть на р. Рыб-
ной 22 сентября 1959 г.
— 222 —
Как и морская чернеть, хохлатая отлетает очень поздно. В 1958 г.
последняя птица была добыта 14 октября, во время ледостава на р. Рыб-
ной.
17. Морская чернеть — Aythya marila marila (L.). В северной полосе
тайги у Норильских озер и в лесотундре у верховьев Пясины морская чер-
неть очень обыкновенна, но в зону тундры она заходит недалеко и север-
нее истоков Пуры нам не встречалась.
В тайге у истоков Рыбной весной 1959 г. морские чернети появились
1 июня, 4—9 июня проходил их валовой пролет и прилет. В 1964 г. массо-
вый прилет чернетей наблюдался там 7—11 июня. В устье Дудыпты в
в 1962 г. первые чернети отмечены 7 июня, их пролет продолжался 15—
19 июня. Весной 1963 г. первых чернетей мы наблюдали у Пуринских озер
16 июня.
Морские чернети приступают к размножению во второй декаде июня.
В это время пары чернетей очень подвижны и их можно часто наблюдать,
особенно в ночные часы, не только в тайге, но и в богатой озерами тундре.
Особенно охотно держатся пары чернетей у небольших озер с заросшими
берегами, на лужах, нередко в соседстве с синьгами, турпанами и хохла-
тыми чернетями.
В яйцеводе самки, добытой близ истоков Рыбной 19 июня 1959 г., на-
ходилось готовое к откладке яйцо. Здесь же 28 июня 1959 г. на берегу не-
большого лесного озера было найдено гнездо, помещавшееся на кочке у са-
мой воды, под прикрытием ольхового куста. 8 яиц лежало просто на
примятой прошлогодней траве. К концу насиживания в лотке появилось
небольшое количество пуха. 30 июня я видел рядом с гнездом самца мор-
ской чернети; птенцы в этом гнезде выклюнулись 19 июля; самка при этом
сидела на гнезде. 22 июля 1960 г. в тундре у р. Агапы наблюдался выводок
из 7 недавно вылупившихся птенцов. Для морской чернети, как и для дру-
гих нырковых уток, очень характерен разнобой в сроках размножения
у разных пар. 17 августа 1959 г. у истоков Рыбной был добыт пуховой пте-
нец весом в 160 г. Другой утенок, добытый 26 августа, весил 410 г, его ма-
ховые уже начали выступать из лопнувших пеньков. Птенцы из разных
выводков у оз. Кета 31 августа 1959 г. весили 570 и 490 г, маховые у них
выступали на 3 и 5 мм. Приблизительно такие же птенцы были добыты
там и 4 сентября. 11 сентября 1959 г. впервые наблюдались едва летавшие
молодые чернети, но регулярно стали встречаться летавшие молодые
только во второй половине сентября.
Линных самок при выводках случалось встречать во второй половине
августа. 26 августа 1959 г. на одном из озер у истоков Рыбной была добыта
линная самка, у которой маховые едва выступали из пеньков.
Отлетают морские чернети поздно, в конце сентября и в начале октября.
Больших стай они не образуют; держатся выводками или небольшими
стайками.
18. Синьга — Melanitta nigra nigra (L.). Среди гнездящихся водо-
плавающих птиц в северной полосе тайги у Норильских озер синьга, по-
жалуй, самый многочисленный вид. В лесотундре у истоков Пясины синьга
встречается в меньшем числе, а в кустарниковой тундре у устья Дудыпты
вообще очень редка. Севернее, в типичной тундре, выводки синьги нам ни
разу не попадались. Численность синьги на гнездовье местами очень высо-
кая. Отдельные выводки встречены на большинстве лесных озер у истоков
Рыбной. Южнее, у р. Хантайки, синьга опять встречается реже.
Весной 1958 г. в истоках р. Рыбной первая пара наблюдалась 31 мая,
массовый прилет происходил 8—18 июня. В 1959 г. передовые синьги
появились 4 июня, массовый прилет наблюдался 7—20 июня. Весной
1964 г. валовой прилет происходил 8—12 июня. Птицы прибывали обычно
стаями в 10—30 особей, реже парами и небольшими группами. Первое
— 223 —
время после прилета они держались на многочисленных полыньях вер-
ховьев Рыбной и только после вскрытия лесных озер, во второй половине
июня, количество синьги на реке заметно сокращалось.
Образование пар происходит еще до прилета на места гнездования.
Расселившись по только что вскрывшимся мелким лесным водоемам, синьги
держатся только парами. Во второй половине июня на заберегах боль-
шинства озер ежедневно наблюдались пары, между отдельными самцами
происходили драки. При этом один из них, разбежавшись на поверхности
воды и громко хлопая крыльями, наскакивал на другого. Некоторые самцы
синьги нападали не только на самцов своего вида, но и на турпанов.
К откладке яиц большинство самок приступает во второй половине
июня. У самки, добытой близ истоков Рыбной 13 июня 1959 г., два яйца
были уже отложены, однако из 8 самок, добытых 14—17 июня, только
одна снесла первое яйцо. Самка, добытая 20 июня, снесла уже 3 яйца,
а диаметр трех наибольших фолликулов у нее равнялся 30, 26, 16 мм.
Судя по состоянию яичника самки, добытой 27 июня 1959 г., можно было
заключить, что она отложила 6 яиц и закончила кладку.
Все 3 найденных нами гнезда синьги помещались среди довольно гу-
стых кустов ольхи, багульника и карликовой березки. Одно из них было
устроено у конца сильно выдающегося мыса на берегу Рыбной, второе —
на островке в дельте Кета-Ирбо, а третье — просто в кустарниковой кур-
тинке близ восточного склона моренной гряды западного берега оз. Кета.
Первые два гнезда находились на расстоянии 5—10 м от воды, а третье —
в 200—300 м. Устройство самих гнезд такое же, как у других нырковых
уток. Выстилка лотков состояла из сухих листьев и злаков в смеси с серым
пухом. В первом гнезде, осмотренном 26 июня 1964 г., была полная кладка
из 8 недавно отложенных светло-оливковых яиц (рис. 12). Во втором
гнезде 8 июля была кладка из 7 яиц более чем недельной степени насижен-
ности. Наконец, в третьем гнезде, найденном в этот же день, было 5 только
что отложенных яиц.
Выводки синьги чаще всего встречаются на небольших таежных озе-
рах с торфянистыми или заросшими осокой берегами, но нередко они по-
— 224 —
падались и на спокойных плесах крупных лесных рек, например на Глу-
бокой, Рыбной и Талой.
7 совсем маленьких пуховых утят, которых самка вела к воде, наблю-
дались на берегу оз. Кета 21 июля 1959 г. Другой выводок был обнаружен
на одном из лесных озер 24 июля. Только что вылупившиеся птенцы были
замечены у оз. Конон 7 августа 1959 г. В 1964 г. 2 выводка в 4 и 6 пуховых
птенцов не более чем недельного возраста наблюдались близ устья Амун-
дакты 8 августа. Вес одного из птенцов, пойманных 17 августа 1959 г.
в истоках Рыбной, равнялся 470 г, маховые выступали из пеньков на 2—
3 мм, пух только частично сменился пером. У птенцов, добытых 20 августа,
маховые показались на 5—8 мм. Другой утенок 23 августа весил 510 г,
маховые выступали из чехлов на 10 мм. Молодая птица 2 сентября весила
800 г, маховые у нее выступали из пеньков на 33 мм, пух сохранился
только кое-где на спине и надхвостье. 14 сентября 1959 г. мы видели на
оз. Кета 2 выводка еще не умевших летать молодых. Из 3 птиц, наблюдав-
шихся на р. Рыбной 20 сентября 1959 г., одна еще не летала, и две другие
едва отрывались от воды. Во второй декаде сентября большинство молодых
поднимается на крыло, но отдельные выводки сильно запаздывают в своем
развитии и, возможно, погибают. Так, 2 сентября 1959 г. на одном из озер
близ истоков Рыбной был добыт пуховой птенец весом всего 340 г, у него
не было даже пеньков маховых.
С середины июля до середины августа на р. Рыбной и на больших озе-
рах постоянно встречались стаи из 10—30 холостых самок. У некоторых
интенсивно линяло мелкое перо. Более значительные стаи самцов неодно-
кратно наблюдались на оз. Кета и на р. Глубокой с июля по сентябрь,
однако ни разу не случалось наблюдать синьг, меняющих маховые.
Исчезает синьга одновременно с замерзанием озер и небольших рек,
в конце сентября или даже в начале октября. На крупных реках и озерах
одиночные птицы и даже стаи задерживаются дольше. Хорошо выражен-
ного осеннего пролета синьги на западном Таймыре нет.
19. Турпан — Melanitta fusca fusca (L.). Границы гнездового ареала
турпана на западном Таймыре те же, что у синьги. Он наиболее обычен
в тайге окрестностей Норильских озер и совершенно отсутствует в типич-
ной тундре. Численность его в некоторых местах у Норильских озер не-
много уступает численности синьги, но все же турпан является одной
из самых обыкновенных гнездящихся уток. Как и синьга, турпан очень
характерен для средних и мелких северотаежных озер.
Весной 1959 г. первый турпан отмечен в истоках Рыбной 2 июня, од-
нако в массе эти утки появились только 9—15 июня. В 1958 г. валовой
прилет наблюдался 12—16 июня. В 1964 г. первая пара появилась в исто-
ках Рыбной 31 мая, а массовый их прилет происходил 3—11 июня. Основ-
ная масса самок появилась позднее самцов, 9—12 июня.
С появлением заберег на мелких лесных озерах пары турпанов поки-
дают плесы рек, на которых держались после прилета. В окрестностях
истоков Рыбной 20—23 июня 1959 г. на очень многих водоемах попадались
отдельные пары. В яйцеводе добытой самки найдено готовое к откладке
яйцо. Как видно было по состоянию яичника, самка, добытая на р. Рыб-
ной 23 июня 1959 г., снесла уже 6 яиц. Самка, добытая 28 июня, уже
окончила кладку из 6 или 7 яиц.
Гнездо турпана, найденное в окрестностях водопада Орон на р. Рыбной
3 июля 1958 г., было устроено в зарослях прошлогодней осоки на малень-
ком островке среди озера. В лотке, выстланном сухими стеблями осок
и злаков, лежало 8 слегка насиженных яиц.
Выводки встречались как на мелких лесных озерах, так и на плесах
рек с тихим течением, часто вместе с выводками синьги. При птенцах
наблюдались только самки.
15 Биология птиц
— 225 —
Темпы развития молодых турпанов на озерах близ истоков Рыбной
могут быть проиллюстрированы данными, приведенными в табл. 1. 11 и
13 сентября 1959 г. я наблюдал 3 выводка еще не летавших молодых.
Вполне самостоятельные молодые турпаны наблюдались на р. Рыбной
22 сентября 1959 г. Приблизительно в те же сроки происходило развитие
молодых и в 1964 г.
Линька самок при выводках происходит в конце августа и в первой
половине сентября. У двух самок, застреленных на одном из озер у исто-
ков Рыбной 4 сентября 1959 г., маховые выступали из чехлов на 68 и 78 мм.
Скоплений линных самцов нам встречать не случалось. Значительные
стаи хорошо летавших старых птиц временами встречались на оз. Кета
с начала июля до конца сентября.
Отлетают турпаны в конце сентября и в начале октября, накануне
замерзания рек и озер. Исчезают незаметно, массового отлета не наблю-
далось. Таблица 1 20. Морянка — Сlangula hye- Развитие птенцов турпана осенью 1959 г. malts (L.). Морянка — самый
Дата Вес птенца, г mhui ичиилиплып ЙИД Jo и выступающих лесотундре бассейна р. Пясины. Имаховых У Норильских озер она гнездится перьев, мм спорадично во вкраплениях тундры
17 августа 23 августа 26 августа { 31 августа 4 сентября 730 870 900 970 1100 1000 1030 960 1060 1150 и лесотундры у берегов озер Мел- 10—15 кого, Глубокого, Кета, Хантай- 25 ского и др., а также на некоторых 20 озерах тундроподобных мохово- 25 кустарниковых участков водораз- 45 делов. 36 В 1958 г. морянки отмечены 35 в истоках Рыбной 6 июня, а мас- 37 совый прилет и пролет наблюдался 8—15 июня. Весной 1959 г. морян- ки появились там уже 1 июня; явно выраженный пролет про-
до л жался до середины июня.
В тундру морянки прилетают несколько позже, одновременно с по-
явлением первых луж талой воды. Весной 1962 г. передовые стаи
наблюдались в устье Дудыпты 7 июня, массовый прилет и пролет их отме-
чен 9—20 июня. У Пуринских озер в 1961 г. первая стая наблюдалась
11 июня, а массовое появление морянок отмечено только 25 июня—4 июля,
в период интенсивного образования заберег. В 1963 г. первые морянки
наблюдались там 14 июня, 17—20 июня их стало уже довольно много.
Весной морянки чаще летят стаями по 10—15 птиц, но нередки и большие
стаи, в которых насчитывается иногда более сотни особей. Над участками
открытой воды стаи летят обычно очень низко, но над сушей чаще всего
на высоте 100—300 м. Прилетев на места гнездований, морянки держатся
первое время в заберегах рек и озер и на мелких лужах среди тундры.
В конце июня и в начале июля у кромки льда, плавающего на больших
озерах, попадались многие десятки морянок одновременно. В это время
они ведут себя очень оживленно, кричат, гоняются друг за другом и стай-
ками летают во всех направлениях. Погоня за самками и драки двух или
более самцов наблюдаются в этот период в любой час дня, но в ночное
время морянки в подавляющем большинстве спят на краю льдин. Оживлен-
ное поведение самцов морянок продолжается всю первую декаду июля,
когда большая часть самок насиживает.
Гнездятся морянки в различного типа тундрах на большем или меньшем
расстоянии от воды. Некоторые из осмотренных гнезд были устроены у са-
- 226 -
мой воды, но было немало и таких, которые находились на расстоянии
десятков или даже сотен метров от ближайшего озера. Очень охотно
гнездятся морянки на небольших островах, особенно если на них есть
колонии чаек или крачек. В начале лета 1964 г. на нескольких маленьких
островках в устье Кета-Ирбо и у залива Онган, близ северного берега
оз. Кета, нами было осмотрено 4 гнезда морянок. На одном островке среди
лесотундрового озера близ истоков Пясины 27 июня 1960 г. было найдено
8 гнезд морянок. На острове Чаячьем на 2-м Пуринском озере 14 июля
1963 г. нами было осмотрено 3 гнезда и 1 гнездо было найдено на малень-
ком моховом островке среди озера в пойме Агапы 18 июля 1960 г. Во всех
случаях неподалеку гнездились отдельные пары крачек. Чаще всего мо-
рянки гнездятся среди редких кустиков карликовой березки и полярных
ив, не играющих почти никакой роли в укрытии гнезда или насиживающей
птицы. Из 25 найденных нами гнезд 16 помещалось именно в разреженном
кустарнике, 6 было расположено на совершенно открытом месте и только
3 — среди густых кустиков полярной ивы. Для устройства гнезда морянки
часто пользуются естественными углублениями подходящего размера
или просто уминают ямку среди мха и прошлогодней травы. Первые
из отложенных яиц лежали на скудной подстилке из стебельков злаков
и осок, а также кусочков мха, но к концу кладки лоток обильно высти-
лался темно-серым пухом. Количество яиц в 17 осмотренных нами полных
кладках колебалось от 1 до 9, чаще всего их было 5—8.
К кладке яиц морянки приступают сразу после прилета. Самка, добы-
тая у истоков Рыбной 9 июня 1959 г., уже снесла 2 яйца. В гнезде, найден-
ном у западного берега оз. Кета 26 июня 1959 г., было 8 свежеотложенных
яиц. Во всех 8 гнездах, осмотренных в верховьях Пясины 27 июня 1960 г.,
кладки были уже закончены, а некоторые из них даже значительно наси-
жены. В тундре у Пуринских озер 2 гнезда, содержавших по 6 свежеот-
ложенных яиц, были найдены 28 июня 1963 г. В условиях поздней весны
1961 г. морянки начали кладку в июле. Яйца в 3 гнездах, осмотренных
у устья Кета-Ирбо 7—10 июля, были более чем недельной насиженности.
2 неполные кладки, с 1 и 3 яйцами, были осмотрены в окрестностях 2-го Пу-
ринского озера только 5 июля.
Насиживающая самка благодаря своей покровительственной окраске
почти незаметна и обычно подпускает человека вплотную. Уходя на
кормежку, что птица делает как в дневные, так и в ночные часы, она всегда
закрывает яйца пухом с краев гнезда. Замечено, что в теплые июльские
дни самки покидают свои тщательно укрытые пухом насиженные кладки
на 8—10 дневных часов. Самцы не принимают никакого участия в наси-
живании, не проявляют заботы о выводке, хотя и держатся первое время
недалеко от гнезд.
Вылупление птенцов в пойме Агапы летом 1960 г. происходило 18 июля.
Во внепойменных водоемах оно, вероятно, происходит несколько раньше.
В 1961 году, отличавшемся холодной весной, выводок из 7 пуховых птен-
цов в возрасте нескольких дней, наблюдался в низовьях Пясины 7 авгу-
ста. У молодых морянок, добытых в пойме Агапы 9 августа, маховые уже
выступали из чехлов на 25—30 мм. Птенцы, отстреленные на одном из
озер в устье Агапы 19 августа 1961 г., весили еще меньше 400 г, а их махо-
вые выступали из чехлов на 10 мм.
В годы с ранней весной молодые поднимаются на крыло в конце авгу-
ста, но обычно в первой половине сентября. В некоторых запоздавших
выводках птенцы не успевают вполне вырасти до наступления холодов
и погибают.
Сотенные стаи самцов, очевидно, приготовившихся к отлету на места
линьки, наблюдались на Пуринских озерах в середине июля. Больших
— 227 -
15*
скоплений линных морянок нам наблюдать не случалось. Чаще всего
стайки заключали по 10—30 птиц, в основном холостых самок.
Линные морянки, с выводками и без них, держались на небольших
тундровых озерах и реках. Все птицы, наблюдавшиеся в последних числах
июля, еще хорошо летали. Первая начавшая линьку морянка была добыта
3 августа 1960 г., много линных морянок наблюдалось в середине августа.
К концу месяца все холостые самки поднимаются на крыло.
Линька самок при выводках начинается в середине и в 20-х числах
августа, а заканчивается в начале сентября. 19 августа 1961 г. на одном
из озер в устье Агапы были добыты самки, недавно потерявшие маховые.
У некоторых птиц началась и линька рулевых.
Отлетают морянки перед самым ледоставом, во второй половине сен-
тября. Заметно выраженного осеннего пролета морянок на западном
Таймыре нет, они исчезают исподволь. Впрочем, 20 сентября 1959 г.
на оз. Кета наблюдались стаи, в которых насчитывались многие сотни
морянок. Осенью 1959 г. последние морянки близ истоков Рыбной были
отмечены 6 октября.
21. Гоголь — Bucephala clangula (L.). У Норильских озер, а также
в верховьях Рыбной и Хантайки гоголь довольно редок, но гнездится
регулярно. В типичной лесотундре в верховьях Пясины он уже не встре-
чается.
Весной 1958 г. первые гоголи были отмечены в истоках Рыбной 1 июня,
чаще они стали встречаться только 6—12 июня. В 1959 г. появление
гоголей зарегистрировано 30 мая, их прилет продолжался до 15—19 июня.
С появлением заберег на мелких лесных озерах, во второй половине
или в конце июня, случалось видеть пары гоголей почти ежедневно. На
реке в это время наблюдались только самцы.
Достаточное количество подходящих для гнездования дупел находят
гоголи в высокоствольных елово-лиственничных лесах, где стволы де-
ревьев нередко достигают 50—70 см в диаметре. Правда, из-за приурочен-
ности таких лесов к склонам гор южной экспозиции некоторые гнезда,
видимо, бывают удалены от водоемов на значительные расстояния. Так
22 июля 1964 г. в высокоствольном смешанном лесу на склоне гор Тонель,
более чем в километре от воды, был найден мертвый однодневный птенец
гоголя. Судя по этой находке и недельному возрасту 5—6 пуховых птен-
цов, наблюдавшихся в верховьях Хантайки 31 июля 1959 г., период
насиживания у гоголя кончается в начале третьей декады июля.
22. Луток — Mergus albellus (L.). Луток — редкая гнездящаяся птица
района Норильских озер. В лесотундре в верховьях Пясины за время
наших экспедиций он не был встречен ни разу.
10 июня 1959 г. и 19 июня 1964 г. пары лутков наблюдались на про-
моинах в устьях ручейков, вытекающих из лесных озер близ истоков Рыб-
ной. Одиночный самец был встречен на озере в устье Норилки 18 июня
1960 г. На одном из таежных озер близ истоков Рыбной 8 сентября 1964 г.
был найден выводок поднявшихся на крыло молодых лутков. Наконец,
11 сентября 1959 г. в илистом заливе р. Рыбной наблюдался выводок
хорошо летавших молодых.
23. Большой крохаль — Mergus merganser merganser L. На севере
таежной зоны, у Норильских озер, большой крохаль довольно обыкно-
венен. В лесотундре у верховьев Пясины гнездование его сомнительно,
хотя небольшие стайки встречались там летом 1960 г. В 1964 г. в связи
с длительной оттепелью, державшейся с 13 по 25 мая, большие крохали
прилетели уже 23 мая.
В отличие от средних крохалей они более разборчивы и держатся
далеко не на всяких водоемах. В пределах исследованной территории
эти птицы встречались исключительно на наиболее крупных реках, как
— 22S -
например Рыбной, и на озерах только с прозрачной водой и каменистыми
берегами. Весной 1959 г. первый крохаль отмечен у истоков Рыбной 30 мая,
4—6 июня пары и небольшие группы наблюдались ежедневно.
18 июля 1959 г. на лесистом острове в верховьях Рыбной было найдено
гнездо, устроенное под сдвоенным стволом упавшей сухой ели, в 10—15 м
от реки. В лотке, обильно выстланном пухом, лежало 11 значительно
насиженных яиц.
16 сентября в верховьях Рыбной наблюдался выводок из еще не умев-
ших летать птенцов. Во время осеннего отлета в последний раз мы видели
стаю больших крохалей 4 октября 1959 г.
24. Средний крохаль — Mergus serrator serrator L. Средний крохаль
довольно многочислен в тайге у Норильских озер и в лесотундре у вер-
ховьев Пясины. В тундре этот крохаль гнездится редко и самой северной
точкой его распространения по Пясине следует считать — «Машкины
яры», километров на 30 ниже устья Агапы, где летом 1960 г. наблюдался
выводок.
В 1958 г. первые крохали появились в истоках Рыбной 6 июня, их
массовый прилет начался двумя днями позже. Весной 1959 г. первые
особи средних крохалей прилетели 1 июня, а 7 июня они наблюдались
уже в значительном количестве. В 1964 г. их прилет происходил 7—9 июня.
Прилетают крохали большей частью парами, реже небольшими стайками
и в одиночку. Первое время они держатся на полыньях, например на
р. Рыбной, но после вскрытия озер их можно было столь же часто встре-
чать на любых водоемах.
3 из 5 найденных гнезд были устроены на небольшом островке, посреди
лесотундрового озера близ истоков Пясины. Они были скрыты в густых
зарослях ивняка, ольхи и мелкорослой лиственницы. 2 других гнезда были
устроены на небольшом облесенном островке в верховьях Рыбной. Одно
из них помещалось между несколькими стволами березы и было защищено
густейшим переплетением ветвей багульника. Второе гнездо находилось
в чаще леса под прикрытием наклонного ствола лиственницы. Гнезда
представляли собой умятые во мху и прошлогодней траве ямки, обильно
выстланные пухом. В первых 3 гнездах 27 июня 1960 г. находилось 7,
12 и 14 совершенно ненасиженных яиц. Одно из найденных в верховьях
Рыбной гнезд 3 июля 1964 г. содержало 7 яиц более чем недельной стадии
насиженности; во втором гнезде в этот же день было 8 совершенно свежих
яиц. Возможно, кладка была неполная.
Птенцы у средних крохалей появляются в конце июля и даже в начале
августа. Выводки обычно держатся на крупных и мелких таежных озерах,
на маленьких равнинных и горных речках и ручьях и на более крупных
реках, таких, как Рыбная, Глубокая, Хантайка, Пясина. 29 июля 1959 г.
на одном из притоков Хантайки наблюдался выводок из совсем маленьких
птенцов. 7 августа 1959 г. на той же речке мы встретили 2 выводка: в одном
из них птенцы были уже величиной с чирка, а в другом — в возрасте
2—3 дней. Птенец, добытый в одном из заливов оз. Глубокого 7 августа
1958 г., весил 92 г. 15 августа 1958 г. в протоке Глубокой наблюдался
выводок из 16 птенцов, ловивших мальков. Добытый утенок весил 200 г.
Маховые у молодого крохаля, пойманного 30 августа 1959 г., не превы-
шали 3 см. В большом выводке, наблюдавшемся на р. Рыбной 2 сентября
1959 г., все 19 птенцов еще не могли летать. Спасаясь, они только бежали
по воде. У добытых 10 сентября 1959 г. молодых птиц маховые выступали
из чехлов тоже только на 3 см. 21 сентября на одном из озер близ истоков
Рыбной молодые крохали еще только пытались подняться на крыло.
В третьей декаде сентября, однако, в большинстве выводков молодые
летали.
— 229 —
Средние крохали отлетают очень поздно, задерживаясь на речных
полыньях до ледостава. Близ истоков Рыбной одиночные птицы еще
встречались 6 октября 1959 г., а на небольшой полынье у устья Ду-
дыпты один крохаль наблюдался даже в десятых числах октября
1961 г.
25. Полевой лунь — Circus cyaneus (L.). По сведениям Е. Е. Сыроеч-
ковского (1961), самка полевого луня была добыта в окрестностях Хан-
тайского озера 27 мая 1960 г. Нами этот хищник наблюдался лишь од-
нажды, 23 мая 1964 г., в верховьях Рыбной. Очевидно, это залетный вид,
отдельные особи которого в период весеннего прилета изредка попадают
значительно севернее своего гнездового ареала.
26. Орлан-белохвост — Haliaetus albicilla albicilla (L.). В тайге
у Норильских озер и по рр. Хантайке и Рыбной орланы гнездятся регу-
лярно.
В тундре и лесотундре по р. Пясине нами наблюдались только
кочующие холостые особи. Во всяком случае гнезда и выводки орланов
во время наших разъездов там ни разу не попадались. Один из орланов,
добытых нами в тундре у р. Агапы, оказался годовалой молодой птицей,
а второй — очень старой холостой самкой.
Прилетают орланы рано, в первых числах мая. В 1959 г. они были
замечены на оз. Кета уже 4 мая. Весной 1964 г. орланы прилетели 5 мая;
в середине этого месяца в истоках Рыбной они наблюдались уже в значи-
тельном количестве. В самые первые дни после прилета орланы держались
по соседству с избушками рыбаков и поедали брошенную около прорубей
рыбу. Несколько позже, во время прилета водоплавающих, они собира-
лись у промоин в истоках рек, например р. Рыбной, где охотились на уток
и гусей. В таких местах в конце мая и в начале июня можно было одно-
временно видеть трех-четырех орланов, сидевших на сухих вершинах
лиственниц. В третьей декаде июня, по окончании пролета водоплаваю-
щих и с вскрытием водоемов, орланы распределяются по окрестной тайге
более равномерно. Холостые птицы начинают свои летние кочевки, а раз-
множающиеся орланы в летнее время держатся близ гнезд.
Судя по встречаемости отдельных орланов и их выводков, пары гнез-
дятся на расстоянии 10—20 км друг от друга. Одно из осмотренных нами
гнезд помещалось на самой вершине лиственницы, на высоте 12—15 м
от земли. Сама постройка гнезда была очень громоздкой — не менее
1.3—1.5 м в диаметре и около 2 м высоты. Лиственница с гнездом находи-
лась на расстоянии около 50 м от берега одного из заливов оз. Кета,
среди довольно густого смешанного леса. Уже 7 мая 1964 г. это гнездо
было занято парой орланов. Во время посещения гнезда 7 июля обе старые
птицы проявляли заметное беспокойство. 27 августа это гнездо было уже
покинуто выводком. Другое гнездо орланов было осмотрено нами 25 июля
1959 г. в высокоствольной елово-лиственничной роще на водоразделе
рр. Рыбной и Хантайки. Оно лежало на земле, у основания высокой лист-
венницы, с вершины которой рухнуло после сильного ветра. Рядом
с гнездом, прямо на земле, затаился вполне оперившийся птенец, у кото-
рого маховые выступали из чехлов на 10—15 см. Около него лежали
остатки окуней, щук, налимов, тайменей, ондатр и молодых средних
крохалей. Неподалеку от этого гнезда находилось два других, пустых,
устроенных на лиственницах вполдерева, на высоте 8—10 м. Очевидно,
что орланы гнездились в этой роще из года в год. Старые орланы вели
себя у гнезда довольно осторожно. При посещении этого места 8 августа
я наблюдал молодого белохвоста, парившего в сопровождении старых
птиц высоко над лесом. Другая семья, состоявшая из одного молодого
и двух старых орланов, наблюдалась в верховьях Глубокой ежедневно
с 11 по 20 августа 1958 г.
— 230 —
До самого отлета выводки кочуют недалеко от места гнездования.
Отлетают они в конце сентября—начале октября. Близ истоков Рыбной
осенью 1959 г. орланы в последний раз были отмечены 3 октября.
Из всех гнездящихся на западном Таймыре птиц орлан-белохвост
наиболее осторожен. Человека, идущего на лыжах или пешком, он редко
подпускает на 300 —400 шагов. Во время весенней охоты на водоплаваю-
щую дичь белохвосты, сидя на вершинах высоких деревьев, издали зорко
следят за людьми, а после их ухода отыскивают и поедают подранков.
Орланы часто едят падаль. В тундре их постоянно случалось видеть у мест
разделки оленьих туш и у куч выброшенной испорченной рыбы. В мае
и в начале июня, когда реки еще подо льдом и орланы лишены возможно-
сти ловить рыбу, в истоках Рыбной нам очень часто случалось видеть их
неуклюжие и всегда неудачные попытки нападать на уток и гусей. Од-
нажды, 19 мая 1958 г., мы наблюдали нападение старой самки на сидевшую
у края промоины выдру. Зверь вовремя заметил угрожавшую опасность
и скрылся под лед. Орлан сел на льдину и стал выжидать появления
скрывшейся выдры, но был застрелен. Иногда эти хищники успешно
ловят молодых зайцев. 14 августа 1964 г. на песчаном берегу оз. Кета,
близ устья Амундакты были найдены свежие остатки сильно подросшего
зайчонка, а рядом на песке виднелись отчетливые отпечатки ног орлана.
Но основную летнюю пищу орланов составляет рыба. Как было уже давно
отмечено некоторыми наблюдателями, орланы иногда нападают на добычу,
которую они не в силах вытащить из воды. В 1959 г., в сухой жаркий
июльский день, я видел в верховьях Рыбной орлана, сидевшего на берегу
реки, мокрого и неспособного к полету. Может быть, он был затащен в воду
крупным тайменем. Один такой таймень, весом более 20 кг, пойманный
нами на притоке Хантайки 29 июля 1959 г., носил на спине около жиро-
вого плавника свежий след лапы белохвоста.
27. Мохноногий канюк (зимняк) — Buteo lagopus kamtschatkensis De-
ment. Всюду в бассейне р. Пясины мохноногий канюк очень обыкновенен
на гнездовье. У Норильских озер он встречается несравненно реже,
гнездится на скалистых обрывах отрогов плоскогорья Путорана, где
изредка встречается также в альпийском поясе.
Прилетает в тундру канюк довольно рано. Весной 1963 г. у Пуринских
озер первый канюк был отмечен после сильной южной пурги 21 мая,
одновременно с первым гуменником. В 1961 г. наша экспедиция прибыла
в тундру 22 мая, а в 1962 г. — 19 мая и оба раза застала уже прилетевших
канюков. Однако массовое появление канюков, а иногда даже хорошо
выраженный их пролет, в тундре наблюдались обычно в конце мая и в са-
мых первых числах июня. Значительные скопления пролетных зимняков
отмечены нами у крутых обрывистых берегов Пуринских озер и р. Агапы.
В таких местах, изобилующих укромными, хорошо защищенными от про-
низывающих ветров уголками, иногда случалось наблюдать до десяти и
более особей одновременно. В середине июня 1961 г. мохноногие канюки
совершенно исчезли из своих весенних убежищ и, очевидно, откочевали
в более богатые кормами участки тундры. Хорошо выраженный весенний
пролет наблюдался близ устья Дудыпты весной 1962 г. Особенно охотно
они придерживались невысоких грив дриадовой тундры, протянувшихся
параллельно р. Пясине. С 23 мая по 6 июня здесь все время можно было
видеть небольшие группы и одиночек, летевших низко над землей в се-
верном направлении. Многие отдыхали на высоких кочках или пастях.
В десятых числах июня пролет закончился. Южнее, близ западного берега
оз. Кета, массовый пролет канюков на север был отмечен в период с 12
по 19 мая 1964 г., во время сильной оттепели.
С появлением в тундре многочисленных проталин канюки приступают
к гнездованию. Гнезда свои они устраивают в самых разнообразных ме-
— 231 —
стах. На юге исследованной нами области зимняки гнездятся в отрогах
Путорана. Найденное здесь в 1958 г. гнездо было расположено на недо-
ступном для человека скалистом уступе обрыва на краю плоскогорья,
в 150 м от подножия горы. Случаев гнездования мохноногих канюков на
деревьях мы не отметили. В тундре и лесотундре гнезда устроены прямо
на земле, на склонах пологих холмов или просто на равнинных водо-
разделах. Чаще канюки гнездятся на уступах обрывов по берегам озер
и рек или на крутых склонах распадков, наконец, на небольших обо-
собленных холмиках или гребнях в долинах многочисленных тундровых
ручьев. Из 18 осмотренных нами гнезд 3 были расположены в кочкарни-
ковой тундре на очень пологих склонах, 8 — на уступах крутых береговых
обрывов, 6 — на холмиках и гребнях по краям долин, и 1 — на скалах
плоскогорья Путорана.
Основания гнезд обычно представляли собой плоские кучи небрежно
сложенных сухих веток полярных ив и других кустарников диаметром
40—80 см и всего 5—10 см высотой. При недостатке сухих прутьев ранней
весной, когда тундра еще покрыта снегом, канюки используют для по-
стройки гнезд не совсем обычный материал. В основаниях гнезд мы не-
сколько раз находили недавно сброшенные небольшие рога северного
оленя. Лоток гнезда диаметром около 25 см обильно выстилается тонкими
прутиками, а также сухими стеблями злаков и осок. На краю гнезд мохно-
ногих канюков нам часто случалось находить зеленые ветви Кассиопеи,
свежие побеги полярных ив, багульника, пучки зеленого мха и дриады
и даже крупные зеленые стебли хвоща. Эта особенность зимняка совер-
шенно аналогична с таковой у обыкновенного канюка, который приносит
в свои гнезда зеленые ветви лиственницы, березы или других де-
ревьев.
В случаях гнездования на одном месте в продолжение нескольких лет
подряд канюки обычно не надстраивают старые гнезда, а делают вблизи
новые. Поэтому очень часто поблизости, в нескольких десятках метров
от жилого гнезда, можно встретить одно или несколько старых. Если
уступ с гнездом невелик и удобных мест близко нет, то канюки строят
новое гнездо рядом со старым, как бы продолжая старую постройку.
Такие сложные гнезда были найдены нами в низовьях Пясины 2 и 8 ав-
густа 1961 г.
Заселенность тундры мохноногими канюками зависит от количества
мышевидных грызунов. В годы высокой численности леммингов и полевок,
например в 1960 и 1963 гг., гнезда канюков у Пуринских озер отстояли
на расстояние 3—6, а иногда даже 1.5—2 км одно от другого. В бедный
мышевидными грызунами 1961 год прилетевшие в большом количестве
канюки ко времени размножения исчезли, а оставшиеся пары гнездились
в 10—20 км друг от друга. В низовьях Пясины, где количество грызунов
тогда было средним, гнезда канюков встречались через каждые 6—8 км
маршрута.
Количество яиц в кладках также завпсит от кормовых условий птиц,
что подтверждалось рядом наблюдений. В 2 гнездах, осмотренных в 1960 г.
у Пуринских озер, было по 5 птенцов, а в 3-м — 4 птенца и 1 яйцо-болтун.
В 1963 году, отличавшемся несколько меньшей численностью грызунов,
в 5 гнездах оказалось по 4 яйца и в 1 — 3 птенца. В 1961 г. в 3 гнездах,
найденных в низовьях Пуры и Пясины, где численность леммингов была
средней, обнаружены 3 птенца и болтун, 2 яйца и птенец и 2 яйца. Нако-
нец, 3 из 4 гнезд, исследованных в среднем течении Пясины и у Пурин-
ских озер в 1961 и 1962 гг., когда леммингов и полевок было очень мало,
содержали по 2 яйца, а 1 гнездо — всего лишь единственное яйцо.
Размеры яиц (20): макс. 61.7 X 47.2, мин. 53.2 X 42.1, в среднем
56 X 44.1 мм.
— 232 —
Приступают к насиживанию канюки довольно рано. В южной части
исследованного района, у оз. Кета, уже 27 мая наблюдалось гнездо с на-
сиживавшей птицей. В тундре канюки приступают к гнездованию не-
сколько позже, когда места для устройства гнезд полностью освобождаются
от снега и появляются многочисленные проталины, на которых можно
собрать строительный материал.
В нормальную весну 1963 г., в одном из гнезд, найденных у Пуринских
озер, кладка из 4 яиц была окончена 15 июня. Вылупление последнего
птенца в этом гнезде происходило 17 июля, следовательно, эмбриональное
развитие длилось 32 дня. В раннюю весну 1960 г. в гнезде у устья Ду-
дыпты первый птенец выклюнулся уже 6 июля. В 1961 г. вылупление
первого птенца в одном из гнезд в долине р. Пуры происходило 19 июля.
Насиживает всегда самка, а самец приносит ей пищу, состоящую из лем-
мингов и полевок. В годы обилия мышевидных грызунов, например
в 1963 г., на краях гнезд даже в период насиживания часто случалось на-
ходить остатки копытных леммингов, реже обских леммингов и полевок-
экономок. В 1961 г., когда грызунов в тундре было очень мало, около
одного гнезда у Пуринских озер были найдены остатки нескольких
тундряных куропаток.
Птенцы растут довольно быстро и дней через 35—40 после вылупления
начинают летать. 12 августа 1960 г. в 2 гнездах у Пуринского озера стар-
шие птенцы при нашем приближении улетели. В гнезде, найденном в ни-
зовьях Пясины 8 августа 1961 г., выклюнулись еще не все птенцы. Однако
этот случай нехарактерен даже для холодного 1961 года, так как 2 августа
в другом гнезде у старшего птенца маховые уже выступали из чехлов
на 6—12 мм. В годы с нормальной весной у гнездящихся в тундре канюков
молодые покидают гнезда в третьей декаде августа.
Хорошо выраженного отлета, какой наблюдал А. Я. Тугаринов (Ту-
гаринов и Бутурлин, 1911) на Енисее, нам обычно наблюдать не случалось.
Лишь осенью 1960 г., в период обилия леммингов, в устье Дудыпты были
отмечены местные скопления канюков, державшихся там вплоть до пер-
вого снегопада. Ко времени выпадения снега, в конце сентября или в на-
чале октября, канюков в тундре уже почти нет. Летят они на юго-запад,
как показал случай добычи на Украине молодой птицы, окольцованной
нами в августе 1960 г. у Пуринских озер.
Общеизвестно, что мохноногий канюк является типичным миофагом.
Это подтверждается остатками пищи, найденными почти у всех осмотрен-
ных нами гнезд, и вскрытием желудков нескольких добытых летом эк-
земпляров. Однако особенно в годы низкой численности мышевидных
грызунов мохноногие канюки очень охотно едят падаль. В конце мая и
в начале июня 1961 г. нам ежедневно случалось наблюдать группы
в 3—5 канюков, поедавших либо внутренности убитых на охоте оленей,
либо зайцев, положенных в качестве приманки для песцов. Как показали
довольно многочисленные наблюдения, в этот период канюки очень охотно
ловят также белых и тундряных куропаток.
28. Ястреб-тетеревятник — Accipiter genttlis buteoides (Menzb.).
Ястреб-тетеревятник — очень редкая гнездящаяся птица северотаежных
ландшафтов исследованной территории.
Выводок этих ястребов и одиночные молодые неоднократно наблюда-
лись нами на северном берегу оз. Кета в период с 20 августа по 15 сен-
тября 1964 г. Птицы держались у каменистых осыпей, где охотились за
пищухами. 30 мая 1960 г. самец тетеревятника был добыт в окрестностях
Хантайского озера (Сыроечковский, 1961).
Судя по полному отсутствию встреч тетеревятников в 1958 и 1959 гг.,
можно предположить, что эти ястреба гнездятся в окрестностях Нориль-
— 233 —
ских озер не всегда, а только в годы, наиболее благоприятные в климати-
ческом (1960 г.) или пищевом (1964 г.) отношении.
В желудке молодой птицы, добытой 19 августа 1964 г., были найдены
остатки пищухи.
29. Дербник — Aesalon columbarius regulus (Pall.). Дербник на запад-
ном Таймыре гнездится всюду в тайге и лесотундре, заходя на север до
крайней границы древесной растительности. В кустарниковой тундре
близ устья Дудыпты гнездование его сомнительно. Хотя одиночные
птицы и пары регулярно встречались весной и осенью даже в низовьях
Агапы, однако летом эти мелкие соколы там никогда не наблюдались.
Дербники появляются весной одновременно с мелкими куличками и
воробьиными, в конце мая и в начале июня. В окрестностях оз. Кета
первый дербник отмечен в 1958 г. 1 июня, в 1959 г. — 27 мая. Весной
1964 г., во время сильной оттепели, вызвавшей преждевременный прилет
целого ряда видов мелких птиц, первый дербник был отмечен близ истоков
р. Рыбной уже 21 мая. В погоне за многочисленными лапландскими по-
дорожниками и другими мелкими птицами дербники весной нередко
залетают в тундру. 10 июня 1961 г. одиночный дербник наблюдался даже
в окрестностях 2-го Пуринского озера. Охотившиеся на мелких птичек
дербники весной 1962 г. ежедневно наблюдались около построек полярной
станции Кресты, в устье Дудыпты. В середине июня они исчезли.
Летом дербники ведут довольно скрытный образ жизни и поэтому даже
на местах гнездования в тайге и лесотундре встречаются сравнительно
редко. Однако, как можно было судить по распределению выводков
в осеннее время, отдельные пары гнездились не дальше чем в 8—10 км
одна от другой, а иногда и значительно ближе. Дербники обычно гнездятся
на земле, хотя в отдельных случаях селятся и среди выступов скал. Так,
при посещении 22 июля 1964 г. одного ледникового кара близ верхней
границы древесной растительности в плоскогорье Путорана, мы наблю-
дали, как у отвесных скал с тревожным криком летал дербник. Добраться
до гнезда, к сожалению, не удалось. Летом 1964 г. два гнезда дербников,
находившихся на расстоянии не более 1.5—2 км одно от другого, были
найдены в устье Амундакты, близ южного берега оз. Кета. Оба гнезда
были устроены в разреженном лиственничном лесу, перемежающемся
небольшими участками заросших кустарником открытых пространств.
Несмотря на наличие большого количества подходящих деревьев, гнезда
помещались на земле. Одно из них было устроено на мохово-лишайнико-
вой кочке с северной стороны большого ольхового куста, росшего на
совершенно открытом месте. Второе гнездо было расположено в густейших
ивняково-ерниковых зарослях, также на высокой сухой кочке. Устрой-
ство гнезд было весьма примитивным — птенцы лежали просто на пло-
щадках, хорошо умятых среди мхов и лишайников. В первом гнезде
27 июля было 5 покрытых белым пухом птенцов, маховые у которых уже
выступали из чехлов на 12—15 мм. 29 июля птенцы перебрались на со-
седнюю кочку, в метре от гнезда. 4 августа у старшего птенца маховые
достигали уже 2/3 нормальной длины, а пух сохранился только в виде
торчащих в разных местах пучков. 14 августа молодые дербники уже почти
не отличались от взрослых, хотя и держались вместе со старыми птицами
в ближайших окрестностях гнезда. Четыре птенца, обнаруженных во
втором гнезде, были несколько моложе и приобрели способность к полету
только после 20 августа.
8 августа 1958 г. выводок из 3 только что покинувших гнездо птенцов
был обнаружен в редком лиственничном лесу на берегу небольшого озера
в окрестностях оз. Глубокого. Двое из них, помоложе, сидели на корнях
у основания дерева, а самый старший — на его наклонном стволе не выше
метра от земли. Никаких признаков гнезда на ближайших деревьях обна-
- 234 —
ружить не удалось. Очевидно, птицы, как и в первых двух случаях, гнез-
дились на земле, на торфяных буграх или в ямке у основания дерева. При
нашем приближении старые соколки с громким криком летали рядом,
присаживаясь на вершины ближайших лиственниц. Старший птенец мог
уже довольно хорошо летать, но самый молодой был еще довольно беспо-
мощен. Средний птенец весил 185.5 г, длина его крыла 143 мм, хвоста —
87 мм. 14 августа 1958 г. в редком лиственничном лесу на западном берегу
оз. Кета наблюдался выводок из 3 хорошо летавших молодых. Они сидели
на вершинах лиственниц на расстоянии 50—100 м друг от друга. Само-
стоятельно ловить птиц молодые еще не умели и громкими криками встре-
чали старых дербников, подлетавших к ним с добычей.
Судя по нашим наблюдениям у гнезд, самка во время выкармливания
птенцов все время находится поблизости, в то время как самец летает
за добычей часто за много километров. Так, от гнезд, расположенных
в устье Амундакты в 400 —500 м от берега оз. Кета, самцы летали на охоту
на противоположный берег озера, каждый раз пересекая 3—4 км откры-
той воды. 2 сентября 1958 г. в верховьях Пясины мы встретили выводок
из 2 молодых дербников при двух взрослых. Молодые были уже совер-
шенно самостоятельными, хотя выводок еще держался вместе.
Перед осенним отлетом, который происходил во второй половине сен-
тября, дербникп встречались поодиночке. Очевидно, вскоре после того,
как птенцы станут вполне самостоятельными, выводки разбиваются.
В это время они ведут кочевой образ жизни и снова появляются в тундре
на местах скоплений мелких птиц. Около рыбачьих избушек в устье
Агапы, где осенью держится масса трясогузок, каменок, рогатых жаво-
ронков и других воробьиных, в конце августа и начале сентября 1960
и 1961 гг. нам ежедневно случалось наблюдать одного или двух дербников.
Исчезают дербники осенью довольно рано, после отлета основной
массы мелких птиц. Осенью 1959 г. в верховьях Рыбной они в последний
раз наблюдались 17 сентября.
В тайге, лесотундре и тундре западного Таймыра дербник питается
исключительно мелкими птицами, которых он очень ловко преследует как
на открытом месте, так и среди зарослей. Нам случалось наблюдать
успешную охоту этих соколков на чечеток, белых трясогузок, подорожни-
ков, дроздов, бекасов и других куликов. В 6 вскрытых желудках были
обнаружены остатки мелких воробьиных, обычно от 2—3 экземпляров.
Среди них удалось установить наличие чечеток, варакушек и трясогузок.
30. Сокол-сапсан — Falco peregrinus leucogenys Brehm. Сапсан на за-
падном Таймыре почти всюду — обычная гнездящаяся птица. В тайге
у Норильских озер и в бассейне рр. Рыбной и Хантайки он сравнительно
редок, что объясняется недостатком в речных и озерных долинах скал,
удобных для гнездования. Зато в тундре и лесотундре сапсанов очень много,
и по рекам с обрывистыми берегами гнездящиеся пары встречаются через
каждые 15—20 км.
Прилетают сапсаны в третьей декаде мая. В истоках Рыбной в 1959 г.
первый из них отмечен 28 мая, однако вследствие редкости там соколов
первые появившиеся особи могли быть пропущены. Весной 1964 г. ло-
вивший пуночек сапсан наблюдался там уже 14 мая. Гораздо более до-
стоверны сроки появления сапсанов на местах их гнездований в тундре.
Весной 1961 г. первая пара была зарегистрирована у Пуринских озер
25 мая, в 1963 г. первый сокол там отмечен 21 мая, а в устье Дудыпты
летевшая на север птица наблюдалась 24 мая 1962 г. Массовое появление
сапсанов в тундре обычно происходит в последних числах мая.
После прилета пары соколов держатся у обрывов в местах своих гнездо-
ваний. 1—7 июня 1963 г. на крутых берегах у Пуринского озера неодно-
кратно случалось наблюдать брачные полеты самца. Сокол поднимался
— 235 —
на высоту 100—150 м и оттуда с громким, хриплым криком, планируя,,
спускался по спирали вниз, иногда присаживаясь на оголенный от снега
уступ берегового обрыва.
В южной части обследованной территории, у Норильских озер, соколы
избирают для гнездования уступы береговых скал; иногда, хотя и редко,
они гнездятся и среди утесов плоскогорья. В подобных местах беспокоив-
шиеся пары наблюдались 30 июня 1958 г. в верховьях Макуса и 1 августа
1959 г. у порогов р. Хантайки. В тундре и лесотундре сапсаны чаще всего
гнездятся на уступах глинистых или песчаных обрывов рек и крупных
озер или на площадках глинистых гребней на склоне между основной тер-
расой и долиной реки или озера. Там, где тундровые реки прорезают вы-
ходы траппов, как например в низовьях Пуры, сапсаны гнездятся на вы-
ступах скал.
Гнезда бывают расположены на площадках ближе к верхнему краю
обрыва; обычно это ямки, устроенные на голой земле (13 гнезд), и лишь
изредка лоток уминается в прошлогодней траве (3 гнезда). Чаще всего
гнезда помещались на склонах южной экспозиции и были хорошо защи-
щены от ветров земляными уступами или крупными кочками. Лоток обычно
бывает в диаметре около 25 см, глубина его 3—7 см. Яйца лежат просто
на земле или умятой прошлогодней траве, и только края лотка обрам-
ляются сухими палочками, соломинками, птичьими костями, обломками
камней и прочим мусором, который птица собирает в радиусе менее
метра. Иногда более мелкий растительный мусор выстилает и дно лотка.
В непосредственной близости от жилых гнезд всегда можно найти не-
сколько старых, как это заметил С. П. Наумов (1931) на Гыданском полу-
острове. Привязанность сапсанов к излюбленным обрывам и даже от-
дельным уступам очень велика, хотя соколы, по-видимому, не используют
одну гнездовую ямку два года подряд. Во время наших маршрутов по тунд-
ровым речкам некоторые из этих излюбленных обрывов посещались нами
ежегодно в течение трех лет, и в большинстве случаев каждый раз сапсаны
там гнездились. Остатки пищи и помет, накапливающиеся у обрывов за
многие годы, иногда сильно способствуют пышному развитию там ра-
стительности. В середине и конце июля, в период массового цветения
тундровых растений, гнездовья соколов заметны уже издали, как и не-
которые песцовые норы. Величина кладки у соколов-сапсанов колеблется
в различные годы в зависимости от характера весны, хотя числен-
ность гнездящихся пар из года в год остается приблизительно одина-
ковой.
Гнезда, осмотренные в 1960 году, отличавшемся очень ранней весной,
содержали 2, 2, 3 и 3 яйца или птенца. В условиях очень поздней весны
1961 г. в найденных у Пуринских озер и на р. Пуре гнездах было 1, 2,
2, 3, 3 яйца или птенца. В 1962 году, также отличавшемся несколько за-
поздавшей весной, количество яиц и птенцов в осмотренных гнездах
было равно 1, 2, 3 и 3. Нормально протекавшей весной 1963 г. условия
для размножения сапсанов были, очевидно, оптимальными и все 4 осмот-
ренных гнезда содержали по 4 яйца.
Яйца сапсанов грязно-кирпичного цвета, интенсивность их окраски
сильно варьирует. Их размеры (13 яиц): макс. 55x43.1, мин. 50.5x40.3,
в среднем 52.7x41.3 мм.
К устройству гнезд и к откладке яиц сапсаны приступают во второй
или третьей декаде июня. Гнездо с полной кладкой из 4 совершенно
ненасиженных яиц найдено у Пуринских озер 20 июня 1963 г. Другая
кладка из 4 яиц, осмотренная там же 25 июня, была уже слегка насижена.
Слабо насиженная кладка из 3 яиц в 1961 г. была обнаружена 29 июня.
В 1962 г. 3 несколько насиженных яйца были найдены в гнезде блиа
устья Агапы 2 июля.
— 236 —
Насиживает только самка, а самец либо охотится в тундре, либо сидит
на излюбленном уступе обрыва или скалы, метрах в 20—50. При прибли-
жении к гнезду человека самец ведет себя очень агрессивно и зачастую
проносится в 2—3 м от головы наблюдателя. Самка гораздо осторожнее
самца и так близко к человеку не подлетает. Агрессивное по отношению
к хищникам поведение соколов прочно обеспечивает безопасность их
гнезд. За весь период полевых работ нам неизвестно ни одного случая
разорения четвероногими хищниками гнезд сапсанов. Этим пользуются
некоторые виды гусей, в особенности краснозобые казарки, которые очень
охотно гнездятся в непосредственной близости от соколиных гнезд.
Рис. 13. Сапсан — Falco operegrinus Tunst. — у гнезда.
Вылупление птенцов сапсанов в тундре западного Таймыра обычно
происходит в третьей декаде июля и лишь изредка — в середине этого
месяца. В гнезде, осмотренном на р. Агапе 16 июля 1960 г., было 2 ново-
рожденных птенца. В другом гнезде 19 июля 1960 г. яйца были сильно
насижены. В гнезде, осмотренном в низовьях Пуры 28 июля 1961 г.,
вылупился первый птенец (рис. 13). Только что выклюнувшиеся пухо-
вые птенцы были обнаружены в 2 гнездах в верховьях Пуры 22 и 24 июля
1962 г. В холодную погоду самка отогревает новорожденных птенцов,
а в жаркую и солнечную защищает их от теплового удара. Птенцы разви-
ваются быстро: в 2 гнездах, осмотренных 10 августа 1960 г., они уже
полностью сменили пух на перо и* перебрались на расположенные в не-
скольких метрах от гнезд глинистые уступы.
В третьей декаде августа молодые сапсаны поднимаются на крыло.
21 августа 1960 г. в одном из распадков на берегу Пясины наблюдался
выводок из 3 хорошо летавших молодых, но ловить добычу они еще не
умели и их кормили старые птицы. Выводки из 2 хорошо летавших, но
еще сопровождавшихся взрослыми птицами молодых сапсанов, были
дважды отмечены в верховьях Пясины 31 августа 1958 г. Они еще держа-
лись на обрывах в непосредственной близости от своих гнездовий.
Слабо выраженный пролет наблюдался 5—20 сентября 1959 г. в вер-
ховьях Рыбной. В этот период охотящиеся па пролетных уток сапсаны
встречались почти ежедневно. В 1959 г. последний сапсан был отмечен
там 27 сентября.
— 237 —
В условиях ранней весны недавно прилетевшие в тундру сапсаны
питаются в основном белыми и тундряными куропатками. 9 июня 1961 г.
преследуемая соколом куропатка спаслась, сев на снег прямо у наших
ног. В это время года нередко случалось видеть сапсанов, ловивших
пуночек. В тайге у Норильских озер сапсаны весной п осенью обычно
охотятся на водоплавающих. 16 июня 1958 г. в истоках Рыбной была
добыта самка, преследовавшая шилохвость. Весной и осенью изредка
случалось наблюдать также нападения сапсанов (правда, неудачные)
на гусей и гагар.
Летом, в период гнездования, нам ни разу, однако, не случалось ви-
деть сапсанов, преследовавших птиц крупнее куропатки. Возможно, это
объясняется тем, что хищник не может принести крупную птицу к своему
гнезду. Беглый просмотр остатков пищи около двух десятков гнезд
и вскрытие нескольких желудков сапсанов, добытых летом, показали,
что основную добычу составляли кулики и куропатки. В большом коли-
честве найдены остатки (крылья) золотистых и бурокрылых ржанок,
тулесов, турухтанов, чернозобиков, мелких воробьиных, чирков, белых
и тундряных куропаток и один раз — крылья длиннохвостого поморника.
Остатков гусей и казарок около гнезд не было обнаружено ни разу.
31. Кречет—Falco gyrfalco L. В пределах исследованной нами тер-
ритории кречет нигде не был нами обнаружен на гнездовье. Не был отме-
чен он также на восточном Таймыре А. И. Толмачевым (Тугаринов и Тол-
мачев, 1934), а к западу, в Гыданской тундре, — С. П. Наумовым (1931).
Однако в коллекциях Зоологического института АН СССР имеется
самка, добытая у гнезда А. Я. Тугариновым на р. Боганиде 7 июля 1916 г.
По-видимому, кречет на западном Таймыре, как и в других местах своего
ареала, является редкой гнездящейся птицей.
Осенью же и зимой этот крупный сокол регулярно наблюдался нами
в тайге и лесотундре западного Таймыра. 2 сентября 1958 г. мы наблю-
дали в верховьях Пясины пару кречетов, сидевших на вершинах чахлых
лиственниц. Регулярно отмечались кречеты, ловившие белых куропаток,
в октябре—ноябре 1958 г. у пос. Потапово на Енисее, а в декабре 1958 г.
в тайге у Хантайского озера. В декабре 1960 г. кречет был встречен под
Норильском. Во всех случаях зимних встреч кречеты были отмечены
в ближайшем соседстве с белыми куропатками, в период их наивысшей
численности в тайге и лесотундре. В конце зимы и весной мы этих со-
колов ни разу не встречали.
32. Белая куропатка — Lagopus lagopus koreni Thayer et Bangs. Белая
куропатка — довольно многочисленная гнездящаяся птица в северной
полосе тайги, в лесотундре и тундре к северу до среднего течения Пуры.
В низовьях Пясины и Пуры она уже не попадалась.
Из тундры белые куропатки на зиму откочевывают в лесотундру и тайгу.
На места гнездований у Пуринских озер они в массе возвращаются в мае
и начале июня, с появлением первых проталин. Ко времени нашего самого
раннего приезда в тундру 10 мая 1963 г. там уже держались немногочислен-
ные стаи куропаток. Однако массовое продвижение куропаток происхо-
дит одновременно с весенней кочевкой самок диких северных оленей на
места отела с середины мая до начала июня. 7 июня 1961 г. в окрестностях
2-го Пуринского озера наблюдался особенно оживленный пролет куро-
паток. Многочисленные стада диких оленей двигались в это время в северо-
западном направлении, кое-где останавливаясь, чтобы попастись. Ку-
ропатки повсюду кормились непосредственно среди раскапывавших снег
оленей. Отдельные их стаи и небольшие группы перелетали с места на место
вслед за медленно идущими животными. Чаще всего встречались стайки
по 20—25 птиц. Надо льдом довольно широкого Пуринского озера стаи
и одиночные куропатки летели, не присаживаясь, на запад и северо-за-
— 238 —
над. Когда было много оленей, наблюдалось и много куропаток, и наобо-
рот. Весной 1962 г. в устье Дудыпты ярко выраженный пролет небольших
стай был отмечен в последних числах мая. Куропатки двигались вслед
за стадами диких оленей в северном и северо-западном направлениях.
Таким образом, совместные кочевки куропаток и диких оленей носят
характер хорошо выраженной закономерности. Весной 1963 г. в связи
с общим уменьшением численности куропаток их весенний пролет был вы-
ражен слабо.
В мае и в начале июня куропатки держатся в тундре на немногочи-
сленных проталинах, местах, где снег выдут ветрами, а также на копани-
цах оленей и зайцев-беляков. Заросли ивняка, приуроченные к берегам
рек, распадкам и низинам между холмами, в это время еще покрыты
толстым слоем снега. Очень многочисленны куропатки на прибрежных
участках речных долин, поросших мелкими кустиками полярных ив,
где нанесенный с речных отмелей песок способствует более быстрому
таянию снега. В одном таком месте на берегу Агапы 3—5 июня 1961 г.
каждый день наблюдалась стая более чем в 150 куропаток. Будучи спуг-
нуты, они затем снова возвращались на прежнее место. В конце мая
и в начале июня куропатки по-прежнему держатся небольшими группами,
по 5—10 особей, хотя не очень редки также большие стаи по 50 и даже
200—300 особей. Такие стаи состоят в основном из самцов, хорошо за-
метных благодаря темной окраске оперения на шее. Иногда, в более или
менее теплую погоду и преимущественно в ночные часы, в мае и в начале
июня слышалось токование отдельных самцов. В тайге у Норильских
озер больших стай куропаток весной наблюдать не случалось, чаще
всего там встречались стайки по 5—7 птиц, державшихся в зарослях ив-
няка по берегам рек и озер.
Во время интенсивного таяния снега, в середине июня, происходит
распределение куропаток по гнездовым участкам. В тундре они избирают
плоские кочкарниковые пространства, изобилующие зарослями карли-
ковой березки и полярных ив. Местами, например у Пуринских озер
и в низовьях Агапы, это припойменные или лежащие близ берегов озер
низины, реже плоские водораздельные тундры. Южнее, близ устья Ду-
дыпты, куропатки гнездятся на больших пространствах слабо всхолмлен-
ной кочкарниковой с кустарниками тундры. В таких местах численность
гнездящихся пар, по подсчетам, сделанным в 1960, 1961 и 1962 гг., как
самцов, так, позднее, и выводков, достигает 12—14 на 1 км2. В некоторые
годы, например в 1963 г , численность куропаток снижалась до 2—3 пар
на 1 км2. В северной полосе тайги у Норильских озер белые куропатки
особенно охотно гнездятся в разреженном лиственничном лесу с густым
кустарниковым подлеском, у берегов больших озер, на лесных опушках
и обширных тундроподобных болотах. Численность гнездящихся пар
здесь несколько меньшая, чем в тундре, и достигает, по наблюдениям
1958, 1959 и 1964 гг. у озер Кета и Глубокое, 4—8 пар на 1 км2. Во время
образования пар токование самцов бывает особенно интенсивным. В се-
редине или конце июня в заселенной куропатками тундре повсюду можно
наблюдать самцов, неподвижно сидящих на высоких кочках или переле-
тающих с места на место. В северной полосе тайги самцы, охраняющие
свои гнездовые участки, очень часто сидят на вершинах обособленно стоя-
щих лиственниц. Драки между самцами наблюдались в 1961 г. у Пурин-
ских озер 25 июня—10 июля. Громкие и непрерывные крики токующих
куропаток в этот период можно было слышать в любой час дня и ночи.
В середине июля начавшие летнюю линьку самцы даже в годы с затянув-
шейся весной (1958 и 1961 гг.) полностью прекращали токование.
10 найденных гнезд представляли собой ямки, умятые во мху и прошло-
годней злаково-осоковой растительности между кочками или на кочках..
— 239 —
Скудная выстилка из сухих листьев ив и нескольких перьев с брюха
линяющей самки находилась далеко не во всех гнездах. Все гнезда были
устроены под прикрытием довольно редких кустиков карликовой березки
или полярных ив. В густых зарослях кустарника, так же как и на совер-
шенно открытых местах, куропатка гнезд не устраивает. В 5 осмотренных
нами законченных кладках содержалось 9, 9, 10, 11 и 12 яиц. Размеры
(20 яиц): макс. 43.3x32, мин. 39.8x29.5, в среднем 41.8x30.5 мм.
К гнездованию самки приступают вскоре после образования пар.
В северной полосе тайги, у оз. Кета, неполная кладка из 10 яиц была
обнаружена 24 июня 1959 г. Через день птица закончила кладку, отло-
Рис. 14. Насиживающая белая куропатка — Lagopus lagopus кот ent
Thayer et Bangs.
жив 11-е яйцо. Неоконченная кладка из 5 яиц была найдена в тундре
у устья Дудыпты 25 июня 1962 г. 2 неполные кладки, содержавшие 3
и 6 яиц, были осмотрены в пойме Большой Дюрюсы 5 июля того же года.
В 1961 г. в окрестностях 2-го Пуринского озера 2 гнезда, в каждом из
которых было по 9 совершенно ненасиженных яиц, были найдены 3
и 8 июля.
Насиживавшие самки подпускали человека очень близко (рис. 14),
но, сорвавшись с гнезда, летели далеко. Самцы почти всегда держались
поблизости и в некоторых случаях пытались отводить. Вылупление птенцов
происходит во второй половине июля. Однако в 1960 году, отличавшемся
исключительно ранней и теплой весной, выводок с птенцами в возрасте
нескольких дней был обнаружен у устья Дудыпты уже 8 июля, тогда
как в 1961 и 1962 гг. в начале июля куропатки только начинали насижи-
вание. В тот же день, 8 июля 1960 г., там были найдены еще 2 выводка
с только что вылупившимися птенцами. В позднюю весну 1962 г. первый
выводок, состоявший из 10 птенцов с маховыми длиной 8—10 мм, заре-
гистрирован только 27 июля. В гнезде, осмотренном в истоках Пуры
13 июля 1963 г., вылупление птенцов должно было произойти не раньше,
чем через неделю. В северотаежных ландшафтах близ берега оз. Кета
— 240 —
летом 1964 г. выводки с птенцами в возрасте 2—5 дней наблюдались
17 и 21 июля. Выводки куропаток держатся поблизости от мест гнездо-
вании на участках равнинной кустарниковой тундры, а в тайге — на лесных
опушках и болотах, особенно на склонах, поросших редким лиственнич-
ным лесом, у берегов больших озер. В августе куропатки перебираются
на богатые ягодами голубики склоны холмов или в заросли морошки на
болотистых низинах. У выводков почти всегда встречались как самка,
так и самец. По наблюдениям, сделанным в тундре у р. Агапы летом 1960 г.,
выводки куропаток с начала августа зачастую объединяются по нескольку
вместе; 1 августа, на поросшем ивняком берегу тундряного озера наблю-
далась стайка из 15—20 молодых с 3 или 4 взрослыми куропатками;
4 августа в низинной кустарниковой тундре наблюдалась стая из более
чем 50 куропаток, в подавляющем большинстве это были хорошо летавшие
птенцы.
Растут птенцы быстро. Выводок из 8 или 9 хорошо летавших молодых
был обнаружен в пойме Дюрюсы 17 июля 1960 г. Вес птенца, добытого
21 июля 1960 г., равнялся 105 г. 2 птенца, добытые там 4 августа 1960 г.,
весили 242 и 282.5 г, а отстрелянные 7 августа — 320 и 415 г. Молодой
самец, добытый в пойме Агапы 14 августа 1960 г., весил 430 г, а на груди
у него уже появились перья, характерные для взрослого самца. В 1958 году,
отличавшемся довольно холодной весной, птенец, добытый у оз. Глубокого
7 августа, весил 200 г, а молодая птица, отстрелянная 16 августа, — 300 г.
К концу августа молодые всегда почти достигают размера взрослых и ста-
новятся совершенно самостоятельными. В этот период выводки и стаи
по 30—50 птиц очень любят держаться в верховьях небольших тундро-
вых распадков, заросших карликовой березкой и ивняком.
Начало осенних кочевок белых куропаток отмечено в начале октября,
когда выводки и даже небольшие стаи осенью 1959 г. появились в север-
ной полосе тайги близ истоков Рыбной. Много белых куропаток, стаями
п небольшими группами медленно двигавшихся к югу, наблюдалось в устье
Дудыпты в конце октября 1960 и 1961 гг. после обильных снегопадов,
засыпавших тундровые кустарники. Под Норильском зимой 1960 г. мно-
гочисленные стаи куропаток появились в начале и в середине декабря.
В богатой зарослями ивняков пойме Енисея, километров на 100 южнее
Дудинки, осенью 1957 г. много куропаток наблюдалось в конце октября
и в начале ноября. В некоторые дни их кочевки носили характер хорошо
выраженного пролета, когда отдельные стаи летели на юг на высоте 70—
100 м. В пойме Енисея куропатки многочисленны в продолжение зимы
и являются одним из главных объектов охотничьего промысла.
Линька белых куропаток на западном Таймыре происходит в следующие
сроки. Весенняя линька самцов начинается в середине апреля или в на-
чале мая. 22 мая 1961 г. у Пуринских озер уже наблюдались птицы, почти
закончившие линьку. Первые перья летнего наряда появляются у птиц
в начале (у Норильских озер) или даже в середине июля (у р. Пуры).
В конце июля и в начале августа спинка и грудь у них полностью наде-
вают летний наряд, брюхо обычно остается белым все лето. В третьей
декаде августа начинают появляться заметные признаки зимней линьки:
обозначаются пеньки опушений ног и довольно многочисленные белые
перья на боках и шее. В конце сентября и первых числах октября боль-
шая часть особей уже полностью надевает зимний наряд.
У самок первые летние перья на голове и шее появляются только в конце
мая. Верхняя сторона тела у них быстро надевает летний наряд, и начав-
шие насиживание птицы в конце июня и в первых числах июля полностью
сливаются с окружающим фоном тундры. Зимние перья на брюхе появ-
ляются одновременно с осенними в третьей декаде июля. К последним
числам сентября самки становятся полностью белыми.
16 Биология птиц
— 241 —
Молодые куропатки в середине августа на нижней стороне тела при-
обретают уже много белых перьев, рост которых идет одновременно
с первыми осенними перьями. Молодые птицы становятся полностью
белыми одновременно со старыми, в конце сентября или в начале октября.
На протяжении большей части года белые куропатки питаются почти
исключительно концевыми побегами и почками кустарников и деревьев.
Зимой 1957/58 г. в пойме Енисея нам ежедневно случалось наблюдать
Рис 15 Самец тундряной куропатки — Lagopus
mutus pleskei Serebr
стаи кормящихся куропаток.
Птицы быстро бегали по
снегу между ивовыми куста-
ми, с характерным треском
обламывая замерзшие веточ-
ки. При этом вся стая обычно
находилась в движении, пере-
мещаясь вдоль густых кус-
тарниковых зарослей. Ха-
рактерный треск, производи-
мый при обламывании веток,
был слышен в тихую погоду
на расстоянии 150—200 ша-
гов. Особенно интенсивно
кормились куропатки на рас-
свете. Отдельные птицы, а
иногда и стаи нередко корми-
лись ветками и почками,
сидя на высоких ивовых кус-
тах пли даже на березах.
Особенно часто случалось
наблюдать кормящихся на
деревьях куропаток в конце
зимы, в марте и апреле.
33. Тундряная куропат-
ка — Lagopus mutus pleskei
Serebr. (рис. 15). На западном
Таймыре тундряная куропат-
ка гнездится от арктической
тундры на морском побережье
(Бируля, 1907; Колюшев,
1933) до холмистых мохово-
осоковых и мохово-пушице-
вых кочкарниковых тундр на
широте устья Агапы. Кроме того, она в незначительном количестве встре-
чается в альпийском поясе плоскогорья Путорана. Южная граница ее
гнездового ареала в тундре в разные годы может значительно смещаться
в зависимости от характера весны. В 1960 году, отличавшемся ранней
и очень теплой весной, выводки тундряных куропаток ни разу не были
обнаружены южнее Машкиных яров на р. Пясине, в 30 км ниже устья
р. Агапы. Летом 1961 года, известного своей очень холодной, затянув-
шейся весной, тундряные куропатки были очень многочисленны в устье
Агапы, в окрестностях Пуринских озер и, можно полагать, значительно
южнее. В 1962 г. после довольно холодной весны тундряные куропатки
отмечены нами в холмистой тундре километров на 30 севернее устья
Дудыпты, а отдельные пары — даже у Крестов. Наконец, в условиях нор-
мальной весны 1963 г. тундряные куропатки почти не гнездились южнее
верховьев Пуры. Как и белые куропатки, тундряные зимой в массе отко-
чевывают к югу, в лесотундру и северную тайгу. В тундре остаются лишь
— 242 —
очень немногочисленные особи (Колюшев, 1933). Весной на юге гнездового
ареала они появляются в апреле—июне одновременно с белыми куропат-
ками. Оба вида куропаток кочуют вместе с многочисленными стадами ди-
ких оленей. Встречать больших стай нам ни разу не случалось, весной
чаще всего наблюдались группы из 3—5, реже 7—10 особей. Очень часто
эти куропатки встречались среди стай белых куропаток. Весной они дер-
жались преимущественно на оголенных от снега участках возвышенной
тундры и на местах кормежки диких северных оленей или зайцев.
Во второй декаде июня 1961 г. пары куропаток занимали свои гнездо-
вые участки, избирая холмистые кочкарниковые и пятнистые тундры.
Тундряные куропатки очень часто селятся на сильно пересеченных скло-
нах многочисленных тундровых распадков. В конце июня и в начале июля
можно было ежедневно наблюдать самцов, сидящих на кочках, и слышать
их токование. Судя по встречам самцов в июне 1961 г. в холмистой тундре
у 2-го Пуринского озера, численность гнездящихся пар достигала там
3—4 на 1 км2. Приблизительно столь же густо была заселена тундра
и в верхнем течении Пуры. В середине и в двадцатых числах июня рядом
с самцами обычно случалось поднимать и самок, однако в конце июня
и в начале июля самки держались более скрытно и затаивались среди ко-
чек. В 1961 г. самки начали откладку яиц в середине июня, а в годы с более
ранней весной они неслись уже в начале месяца. Куропатка, добытая
20 июня 1961 г., снесла 3 или 4 яйца. Выводки из 7 и 9 уже летавших
молодых были встречены в низовьях Пясины 7 августа 1961 г. Возле
одного выводка держалось 3 старых птицы. Добытый птенец весил 73 г,
его маховые выступали из чехлов на 50—55 мм. Птенец, добытый из вы-
водка в альпийском поясе плоскогорья Путорана на высоте около 800 м
над ур. м. 24 июля 1964 г., весил 45 г и мог довольно хорошо летать. В пер-
вых числах сентября 1961 г. наблюдались выводки у устья Агапы. Моло-
дые достигали 2/3 размера старых и держались в общих стаях с моло-
дыми белыми куропатками.
Откочевывать к югу тундряные куропатки начинают в конце октября
и 'начале ноября, часто вместе с белыми куропатками. В эти месяцы
они наблюдались нами в 1960 и 1961 гг. в устье Дудыпты. В приенисейской
лесотундре, километров на 100 южнее Дудинки, в 1957 г. они появились
в ноябре. Под Норильском тундряные куропатки в некоторые годы встре-
чались всю зиму. Осенью 1959 г. близ истоков Рыбной довольно много
их появилось уже в середине октября. Очевидно, птицы спустились
с плоскогорья. На зимовках, как и во время весенних кочевок, обычно
в стаях не бывает более 30 особей. Чаще тундряные куропатки держались
группами по 6—10 птиц. Как и белые куропатки, тундряные очень охотно
придерживаются кустарниковых зарослей, где кормятся теми же побегами
и почками. Общества белых куропаток они не избегают: напротив, неболь-
шие группы и одиночки часто к ним присоединяются. Численность встре-
ченных нами зимой в тайге и лесотундре тундряных куропаток была обычно
более чем в 10 раз ниже численности белых.
В зобах 6 птиц, добытых весной 1961 и 1962 гг.у Пуринских озер
находились почти исключительно листья дриады и только в очень незна-
чительном количестве — почки полярных ив, голубики и карликовой
березки. Зобы двух самцов, добытых 2 августа 1961 г. в низовьях Пясины,
также были наполнены листочками дриады.
34. Каменный глухарь — Tetrao parvirostris parvirostris Вопар. В тайге
у Норильских озер каменный глухарь очень редок, хотя несомненно гнез-
дится. 20 октября 1958 г. в смешанном лесу под Норильском были обна-
ружены его следы, очень хорошо отпечатанные на выпавшем снегу. В дру-
гой раз следы глухаря были отмечены на берегу таежного озера близ
истоков Рыбной 17 мая 1959 г. Старый самец, пролетевший над Рыбной,
— 243 —
16*
наблюдался 19 июня 1964 г. В редком лиственничном лесу у западного
конца оз. Кета 21 июня 1959 г. была добыта петухоперая глухарка со слабо
развитым яичником, птица весила всего 1440 г. Зоб у нее был набит яго-
дами брусники. Норильские охотники глухаря не знают, но эвенки, про-
мышляющие у Хантайского озера, добывают его ежегодно. По сведениям
местного населения, какие-то глухари не очень редко встречаются близ
берега Енисея на 100 километров южнее Дудинки.
35. Рябчик — Tetrastes bonasia sibiricus But. Несмотря на то, что в тайге
в окрестностях Норильских озер по ландшафтным условиям есть много
мест, подходящих для рябчиков, мы их там ни разу не встречали. У Хан-
тайского озера и р. Хантайки рябчики, по словам жителей, изредка
гнездятся. В лесотундре вдоль берегов Енисея на тех же самых широтах
они довольно обыкновенны. В окрестностях пос. Потапово в октябре
1957 г. и в марте 1958 г. нам случалось видеть и добывать рябчиков и даже
целые их выводки неоднократно.
36. Тулес — Squatarola squatarola (L.). На западном Таймыре тулес
весьма обыкновенен в тундре от устья Дудыпты до побережья Карского
моря (Вальтер, 1902; Колюшев, 1933). Типичным местообитанием его
являются очень сухие, песчаные, дриадово-кассиопеевые, невысокие бугры
и гривы в речной пойме или среди плоской низинной тундры. В таких
местах численность тулеса может быть сравнительно высокой: 28 июля
1960 г., пройдя 4.5—5 км вдоль поймы Агапы, мы встретили 3 пары.
Прилетают тулесы в тундру одновременно с бурокрылыми ржанками.
Весной 1962 г. первые тулесы, летевшие вдоль Пясины на север, были
отмечены близ устья Дудыпты 4 июня.
Сразу после прилета птицы занимают удобные для гнездования участки.
Гнездо с 4 не насиженными яйцами было найдено на сухой гриве, порос-
шей дриадой, в пойме Пуры 22 июля 1962 г. Оно было расположено на от-
крытой песчаной площадке и представляло собой неглубокую ямку,
очень скудно выстланную кусочками сухих прутиков полярной ивы
и лишайниками. Яйца имели следующие размеры: 53.5x35.3, 50.2x34.3,
52x36, 52.3x35 мм.
Самец и самка при приближении человека проявляли крайнее беспо-
койство и еще издали встречали его тревожным криком. Прежде чем взле-
теть, насиживавший тулес отбегал в сторону шагов на 50 или 100. Столь же
опасливо он возвращался к кладке. Подлетавших к гнездовому участку
серебристых чаек и поморников тулесы преследовали с очень большим
ожесточением. 14 июля 1963 г. гнездящаяся пара была, однако, обнаружена
на сухой части острова 2-го Пуринского озера, метрах в 150 от довольно
большой колонии серебристых чаек и бургомистров. В 1960 году, отли-
чавшемся очень ранней весной, выводок пуховых птенцов наблюдался
в пойме Большой Дюрюсы уже 22 июля. Один из них весил 40 г, на крыльях
у него едва появились пеньки с кисточками. Обе старые птицы держались
при выводке.
Основным кормом тулеса служат наземные насекомые, прежде всего
жужелицы и живущие в почве личинки двукрылых. Почти во всех желуд-
ках были найдены взрослые жужелицы Pterostichus (Cryobius) sp., Р. ver-
miculosus, P. costatus, Amara alpina, Carabus{\Hpe4^p и Чернов, 1963). В 6 из
13 исследованных желудков обнаружены личинки Tipula. Эти две группы
насекомых совершенно разнокачественные по экологии, и составляют
основу питания тулесов. В единичном числе встречены летающие формы:
Chironomidae, Muscidae, Tortricidae, Culicidae.
37. Золотистая ржанка — Pluvialis apricarius altifrons (Brehm). Зо-
лотистая ржанка очень обыкновенна в северной полосе тайги у Нориль-
ских озер, в лесотундре у верховьев Пясины и в тундре на север до устья
Дудыпты. Севернее она встречается значительно реже, а на широте устья
— 244 —
Агапы и Пуринских озер уже просто редка. В тайге и лесотундре она
населяет открытые тундроподобные участки и торфяные болота, иногда
гнездится на покрытых кочкарником склонах моренных гряд. В тундре
у устья Дудыпты пары ржанок были наиболее многочисленны в кочкарни-
ках и на плоско-бугристой равнине. Летом 1960 г. их численность в таких
местах достигала 3—4 пар на 1 км маршрута.
В северной тайге у западного берега оз. Кета ржанки появляются
сравнительно поздно. В 1958 г. они прилетели 13 июня, а в 1959 г. —
между 10 и 16 июня. На торфяных болотах у оз. Глубокого беспокоив-
шихся птиц случалось наблюдать 8—10 августа 1958 г. В конце августа
1959 г. ржанки близ истоков Рыбной уже не встречались.
Хорошо выраженного осеннего пролета у них подметить не удалось.
38. Бурокрылая ржанка — Pluvialis dominicus julvus (Gm.). Бурокры-
лая ржанка в бассейне р. Пясины обнаружена нами начиная с устья
Дудыпты, где она мало уступает по численности золотистой ржанке, до
тундр близ устья Пуры, а возможно и севернее. В лесотундре у верховьев
Пясины она не найдена. В отличие от золотистой ржанки бурокрылая
предпочитает более сухие кочкарники и очень обычна на склонах и даже
на вершинах бесчисленных в Таймырской тундре увалов. Численность
гнездящихся пар падает по мере продвижения к северу. Если у Пурин-
ских озер и в верховьях Пуры пары ржанок встречались через каждые
200—400 м маршрута, то в низовьях Пясины в начале августа 1961 г.
они наблюдались только через 1—2 км пути.
В тундру ржанки прилетают в начале июня, сразу после образования
обширных проталин на холмах и возвышенных берегах рек и озер. Весной
1962 г. стаи появились в устье Дудыпты 4 июня и до середины этого месяца
шел их интенсивный пролет. В 1961 г. первая одиночная птица отмечена
у Пуринских озер 6 июня, однако большие стаи появились только 12 июня
и исчезли они уже в середине месяца. В 1963 г. небольшие группы ржа-
нок впервые наблюдались там 4 июня, а к 12 июня их пролет уже закон-
чился. Как в устье Дудыпты, так и у Пуринских озер птицы летели с юга
на север. Более значительные стаи, в которых насчитывалось около трех
десятков особей, отмечены в устье Дудыпты; у Пуринских озер встречались
преимущественно группы из 3—7 ржанок. Чаще всего пролетные стаи
останавливались на многочисленных протянувшихся с севера на юг гривах
сухой дриадовой тундры.
В первые же дни пролета ежедневно случалось наблюдать токование
самцов. Издаваемый ими резкий брачный посвист очень напоминает токо-
вой крик ту леса. Его можно было слышать почти до конца июня,
а в 1961 г. — даже в первой декаде июля. Многочисленные прочные
пары, занявшие свои гнездовые участки, отмечены с 15—20 июня. У самки,
добытой 20 июня 1962 г. у устья Дудыпты, яйцевод был развит хорошо,
а диаметр наибольшего фолликула равнялся 12 мм. В условиях более ран-
ней весны 1963 г. добытая в окрестностях Пуринских озер самка уже
15 июня снесла первое яйцо. Гнездо с полной кладкой из 4 ненасиженных
яиц было найдено там 17 июня. Другое гнездо было обнаружено в сухой
лишайниковой дриадово-кассиопеевой тундре у 1-го Пуринского озера
12 июля 1962 г. 4 значительно насиженных яйца лежали в ямке, выстлан-
ной исключительно кусочками лишайников. Яйца имели следующие раз-
меры: 47x33.5, 45.6x33.5, 45x32, 46.7x33.2 мм. У гнезда держатся
обе птицы и при приближении человека отводят, хотя и не подпускают
на близкое расстояние. Вылупление молодых происходит приблизительно
во второй декаде июля, хотя некоторые выводки довольно сильно запазды-
вают. Судя по поведению многочисленных пар, наблюдавшихся в вер-
ховьях Пуры 12 июля 1963 г., у них уже появились пуховые птенцы.
Ржанки вели себя исключительно смело, близко подпускали людей,
- 245 -
а длиннохвостых поморников преследовали с большим ожесточением.
Птенец, пойманный 14 августа 1960 г. в холмистой тундре у 1-го Пурин-
ского озера, уже вполне оперился, осталось только немного пуха на го-
лове, шее и боках. Маховые выступали из чехлов на половину своей длины.
Старые птицы держатся вместе с молодыми, пока они не станут вполне
самостоятельными. К концу августа обычно все бурокрылые ржанки ис-
чезают из тундры. Сколько-нибудь выраженного осеннего пролета за-
метить нельзя. Состав пищи соответствует характеру местообитаний птицы.
сухих участках тундры Carabidae
(табл. 2).
39. Галстучник — Charadrius
hiaticula hiaticula L. Повсюду
на исследованной территории
галстучник очень обыкновенен.
На юге западного Таймыра мы
В основном преобладают обитающие на
Таблиц
Летние корма бурокрылой ржанки
(низовья р. Агапы, июль 1960 г.,
12 желудков)
а
2
Пищевые объекты
Всего Carabidae (im.)
В том числе:
Pterostichus (Cryobius)
sp.
Р. vermiculosus
Р. costatus
Carabus sp.
A mar a alpina
Silphidae (im.) . .
S taphylinidae (im.) .
Chrysomela septentrionalis
Diptera
Bibionidae
Листья растений .
Семена.
18
И
1
3
2
1
1
1
1
1
10
?
4
1
1
1
1
1
1
1
1
1
2
1
встречали выводки на отмелях
притоков Хантайки, а на севере
он отмечен И. И. Колюшевым
(1933) как обычная гнездящаяся
птица прибрежной полосы и
островов Пясинского залива.
При наличии соответствующих
местообитаний численность гал-
стучников может достигать 2—
3 пар на 1 км маршрута. Имен-
но такая частота встреч зареги-
стрирована в начале августа
1959 г. на отмелях р. Могокты
и в июле 1960 г. на берегах
Агапы и некоторых крупных
озер, расположенных непода-
леку от нее.
Весной 1958 г. у истоков
Рыбной галстучники впервые
были отмечены 13 июня, в 1959 г.
первая пара наблюдалась
2 июня, но массовое появле-
ние зарегистрировано 12 июня. В 1964 г. первые галстучники там наблю-
дались 31 мая. В тундре близ устья Дудыпты весной 1962 г. стаи, летевшие
к северу, ежедневно наблюдались с 6 по 15 июня. В окрестностях Пурин-
ских озер в 1961 г. они появились 12 июня, а в 1963 г. — 6 июня. Более
позднее появление галстучников в южной части исследованной террито-
рии может быть объяснено тем, что отмели и песчаные возвышенности осво-
бождаются от снегового покрова в тундре скорее, чем в северной полосе
тайги, где снег не в такой степени выдувается ветрами.
Вскоре после прилета, 12—15 июня, галстучники уже токовали.
В это время пары занимают гнездовые участки. На западном Таймыре
замечено два основных типа местообитаний галстучников. Во-первых, это
плоские песчаные или мелкогалечные берега и отмели рек и крупных
озер.
В таких условиях галстучники встречались на западном Таймыре
повсюду как в тундре, так ина севере таежной зоны. Вторым типом гнез-
довий являются пятна сухой земли, песка или гальки около изрезанных
склонов многочисленных в тундре распадков и ручьев. Местообитания
второго типа особо характерны для тундры. На крупных Норильских
озерах галстучники охотно заселяют высокие каменистые острова.
— 246 —
Гнездо галстучник устраивает на совершенно открытом месте. Это
просто ямка в песке, земле или мелкой гальке, окруженная валиком из
камешков или кусочков дерева. На рано освободившейся от снега вершине
моренной гряды близ западного берега оз. Кета 21 июня 1964 г. было
найдено гнездо с неполной кладкой из 3 яиц. 4-е яйцо, уже полностью
одетое скорлупой, было обнаружено в яйцеводе, добытой у гнезда самки.
3 яйца в гнезде, найденном на галечниковой косе близ устья Кета-Ирбо
8 июля 1964 г., были уже значительно насижены. В условиях очень ран-
ней весны 1960 г. гнездо с 4 значительно насиженными яйцами было
найдено 24 июня на гальке близ истоков Пясины. В типичной тундре
гнездо с 4 ненасиженными яйцами было обнаружено 2 июля 1962 г. на
склоне одного из распадков берега Пясины, на песчаном выбросе нежи-
лой песцовой норы. В период откладки яиц и в начале насиживания
самцы ожесточенно прогоняют со своего гнездового участка чужих птиц
своего вида. При приближении человека насиживающая птица подобно
ржанкам и тулесу отбегает в сторону, после чего начинает отводить, про-
делывая очень характерные телодвижения.
Вылупление птенцов у галстучников обычно происходит в конце вто-
рой декады июля. В гнезде, найденном на каменистом островке оз. Кета
17 июля 1964 г., 2 птенца уже вылупились, причем один был значительно
крупнее другого. Последний птенец полностью освободился от скорлупы
только к вечеру 18 июля. Таким образом птенцы вылуплялись не одно-
временно, а в течение 2.5—3 суток, из чего можно заключить, что галстуч-
ники начинают насиживание сразу после сноса первого яйца. Во время
вылупления птенцов старые птицы немедленно уносят яичную скорлупу.
Выводок пуховых птенцов недельного возраста наблюдался на берегу
Агапы 17 июля 1960 г. Другой выводок из 4 однодневных птенцов был
найден на берегу оз. Кета 21 июля 1959 г. Птенцы были еще совсем бес-
помощными, вес одного из них равнялся 10.5 г. Обе старые птицы обогре-
вали птенцов. При этом одна из них проделывала это охотно и сидела на
птенцах значительное время, а другая, хотя и садилась на пищавших
от холода птенцов, но в ту же минуту бросала их и с беспокойным писком
бегала рядом. В 1964 г. на галечниковой косе у оз. Кета новорожденные
птенцы галстучников также были обнаружены 21 июля. В 1961 году, отли-
чавшемся очень холодной весной, пуховой птенец был пойман в низовьях
Пясины 7 августа. У него маховые выступали из чехлов на 5—10 мм. Вы-
водок еще не летавших птенцов наблюдался в низовьях Могокты 2 ав-
густа 1959 г., начавший летать молодой там был пойман 3 августа. Птенцы
в выводке, замеченные у западного берега оз. Кета еще 21 июля 1959 г.,
11 августа уже хорошо летали и по величине почти не отличались от дер-
жавшихся при выводке обеих старых птиц. Таким образом, птенцы гал-
стучников достигают полного роста менее чем в 3 недели. В середине
августа выводки кочуют по песчаным и илистым отмелям недалеко от мест
гнездования. К концу этого месяца они постепенно откочевывают.
17—19 августа на р. Глубокой наблюдались стаи галстучников, летевших
на запад. Отдельные группы и одиночки задерживались в тундре и север-
ной тайге до последних чисел августа и даже до начала сентября. Самой
поздней была встреча 3 птиц 3 сентября 1958 г. в верховьях Пясины.
Как видно из табл. 3, галстучники в период гнездования питаются
самыми различными насекомыми.
40. Хрустан — Eudromias morinellus (L.). Хрустан постоянно гнез-
дится на самом севере исследованной территории, в отрогах хр. Бырранга
и достигает даже побережья Карского моря (Бируля, 1907; Колюшев,
1933). Южнее он ежегодно регулярно встречался нам во время весеннего
пролета. В 1963 г., однако, отдельные пары гнездились в холмистой дриа-
довой тундре в окрестностях Пуринских озер, на 200 км южнее своих
— 247 —
Таблица 3
Летние корма галстучника
(верховья р. Пясины и низовья р. Агапы,
1—5 августа 1960 г., 9 желудков)
3
х
ф
X з
Д) г4
Ч
О 2
2 х
Пищевые объекты
ч ч 2 2 S 2
2 х ЛЮ
К ж в о я о
Всего Carabidae (im.)
В том числе:
Pterostichus vermicu-
losus ....
Pt. (Cryobius) sp.
Bembidion sp. .
Dytiscidae (im., 1.)
Всего Diptera
В том числе:
Fungivoridae (1.)
Rhagionidae (1.).
Limoniidae (1.)
Muscidae (im.) .
Culicidae (im.) .
Chironomidae (1.) .
Cordiluridae (im.)
обычных гнездовий на севере исследованной территории. Возможно,
гнездились они там и в 1961 г. Гнездование хрустана в альпийском поясе
плоскогорья Путорана не доказано прямыми наблюдениями. Однако
в начале августа 1964 г. там наблюдались большие стаи, состоявшие как
из старых, так и молодых особей, которые вряд ли успели откочевать
с побережья Карского моря. Изредка хрустаны гнездятся на напоминаю-
щих арктическую тундру каменистых гривах близ берегов Норильских
озер, на высоте не более 100—120 м над ур. м.
Весной 1958 г. небольшие группы хрустанов наблюдались 4—13 июня
на каменистой моренной гряде у западной оконечности оз. Кета. В 1961 г.
у Пуринского озера пролетные
пары впервые отмечены 16 июня.
Затем они постоянно встреча-
лись там вплоть до 29 июня.
В 1963 г. первая стайка наблю-
далась 7 июня, 9—14 июня
ежедневно встречались стайки и
отдельные пары. Все хрустаны
держались на участках дриадо-
вой или пятнистой возвышенной
тундры.
14 июня 1963 г. дважды слу-
чалось наблюдать токование
хрустанов на земле. В 1961 г.
оно отмечено 22 июня. Самец
расхаживал перед самкой, плав-
но поднимая и опуская расправ-
ленные крылья. Токование сам-
цов несколько раз случалось
видеть и 21 июня 1964 г. на мо-
ренной гряде близ западного
берега оз. Кета. Одной из до-
бытых там в этот же день са-
мок, судя по состоянию ее яич-
ника, было уже отложено
3 яйца. Две самки, добытые
у Пуринских озер 14 июня
1963 г., также уже отложили
1962 г. никаких признаков хру-
39
5
11
25
3
10
67
2
2
1
2
9
30
10
3
1
1
12
9
2
1
1
1
1
3
3
отметить, что летом
по 3 яйца. Интересно
станов на этом же месте обнаружено не было. Стайки, состоявшие из холо-
стых хрустанов, наблюдались 5—8 августа 1961 г. в каменистых россыпях
в низовьях Пясины.
Выводок хорошо летавших молодых наблюдался в сухом русле ручья
близ р. Агапы 9 августа 1960 г. Вероятно, это были птицы, уже откоче-
вавшие с мест гнездования. 6 августа 1964 г. в альпийском поясе плоско-
горья Путорана был добыт хорошо летавший молодой хрустан.
41. Фифи — Tringa glareola (L.). Фифи — самый многочисленный
кулик у Норильских озер. Он очень обыкновенен и в лесотундре верховьев
Пясины, заходит даже в тундру до широты устья р. Агапы. Всюду фифи
очень характерен для берегов мелких заболоченных водоемов и заросших
болот близ более крупных озер. В богатых болотами и озерами окрестно-
стях р. Рыбной на каждом километре маршрута обычно встречалось не-
сколько пар, особо заметных в период появления выводков.
В 1958 г. первые фифи в истоках Рыбной были отмечены 8 июня, од-
нако массовое появление их зарегистрировано только в первые по-настоя-
щему теплые дни. В следующем, 1959 г. фифи прилетели уже 1 июня,
— 248 —
a 4 июня происходил их валовой прилет. Весной 1964 г. во время сильной
оттепели токовой крик фифи впервые отмечен 18 и 19 мая, но полной уве-
ренности в столь раннем появлении этих куликов все-таки нет. Совер-
шенно достоверно токующая птица наблюдалась тогда 31 мая. Как и
многие другие птицы, фифи в начале весны преимущественно держались
на кромке льда р. Рыбной и в устьях ручьев. Стай весной ни разу видеть
не случалось. Фифи появлялись парами или поодиночке. Как уже отмеча-
лось, токовавших фифи можно было слышать уже в день их появления
на гнездовье, но особенно интенсивно токование проходило в середине и
в 20-х числах июня. В это время кулики держались на небольших заросших
осокой болотах, около заболоченных берегов озер и ручьев или просто
на разреженных увлажненных участках тайги.
Найденные гнезда были устроены в самых разнообразных условиях.
Одно из них было расположено на ягельной подстилке сухого листвен-
ничного леса, 5— на сухих осоковых кочках по краям болот, 1 — среди
кустиков багульника на сухой лесной опушке и 4 — на деревьях, в прош-
логодних гнездах темнозобых дроздов, на высоте 2—4 м. Около гнезд,
расположенных на земле, всегда было небольшое прикрытие, состоявшее
из редких кустиков карликовой березки, багульника или просто пучков
прошлогодних осок. Лотки гнезд представляли собой ямки, умятые среди
осок, мхов и лишайников без всякой выстилки. В случаях гнездования
фифи в старых гнездах дроздов в них не было замечено никаких призна-
ков перестройки.
В большинстве осмотренных гнезд содержалось по 4 яйца, но в двух
гнездах были полные кладки по 3 яйца, а в одном — 2 яйца. Окраска
общего фона яиц варьировала от светло-оливкового до буро-песочного.
В годы с нормальным ходом весны птицы приступают к насиживанию
в конце второй декады июня. Свежая кладка из 3 яиц была найдена в исто-
ках Рыбной 22 июня 1958 г. 3 гнезда, яйца которых содержали эмбрионы
длиной около 10 мм, были осмотрены в окрестностях водопада Орон на
р. Рыбной 2 и 3 июля 1958 г. В очень раннюю весну 1960 г. кладки из 2 и
3 яиц, отложенные в гнездах дроздов, были обнаружены В. В. Леоновичем
под Норильском уже 12 июня.
В период насиживания фифи очень доверчивы и обычно подпускают
человека вплотную. Судя по наличию у обоих полов наседных пятен,
насиживают и самец, и самка.
Вылупление птенцов нормально происходит во второй декаде июля.
Летом 1964 г. первые пары фифи, своим беспокойным поведением указы-
вавшие на присутствие птенцов, отмечены 15 июля. Гнездо с 4 только
что выклюнувшимися птенцами было найдено В. Герке близ западного
берега оз. Кета 11 июля 1959 г. Около новорожденных птенцов беспокои-
лись обе старые птицы. Когда один из птенцов, совсем еще беспомощный,
был вынут из гнезда и положен на кочку в метре от него, одна из птиц
сразу села его отогревать, в то время как вторая быстро вернулась на
гнездо с остальными 3 птенцами. Все телодвижения отогревавших птенцов
фифи полностью аналогичны таковым у пепельного улита. Как и у по-
следних, птенцы фифи в первые дни своей жизни питаются в основном
комарами, которых ловят с большой ловкостью. Взрослые фифи у вывод-
ков, по нашим наблюдениям, также все время ловят кишащих на траве и
в воздухе комаров. У выводка из 4 птенцов примерно двухдневного воз-
раста, найденного там 14 июля, была всего одна птица. У выводков,
наблюдавшихся в дальнейшем, также обычно беспокоились только оди-
ночные фифи, судя по нескольким вскрытиям — самцы. Очевидно, самки
вскоре покидают выводок. В отличие от пепельных улитов самцы далеко
не во всех случаях остаются при выводках до отлета молодых, а обйчно
покидают последних через 5—8 дней после их вылупления. Летом 1959 г.
— 249 -
нам случалось ловить птенцов такого возраста, в одиночку отыскивавших
пищу на лесных болотах, причем старых фифи нигде поблизости не было.
Замечены довольно дальние передвижения выводков. Двухдневный
птенец, окольцованный 14 июля, был пойман 17 июля на расстоянии около
1 км от места кольцевания. У птенца еще только начали появляться пеньки
маховых. На берегу одного из лесных озер у западного берега оз. Кета
23 июля 1959 г. был пойман другой птенец, с пеньками маховых не более
2—3 мм. 26 июля 1958 г. близ устья Рыбной был найден мертвый птенец
фифи, подавившийся крупным дождевым червем. У него уже значительно
развились перья, и маховые выступали из чехлов на 10 мм. 1 августа
1959 г. на берегу Могокты были пойманы два уже перепархивавших
молодых фифи. У этого выводка беспокоилась одиночная старая птица.
Хорошо летавшие молодые наблюдались на берегу Норилкп 2 августа
1958 г. В том же году много летных молодых было встречено в одном из
заливов оз. Глубокого 6 августа. Таким образом, в первых числах
августа подавляющее большинство молодых становится вполне самостоя-
тельными. С этого времени фифи встречаются во множестве на илистых
берегах ручьев и проток, на осоковых и травянистых болотах и просто
на кромке воды многочисленных небольших озер. Держатся они как вы-
водками, так и одиночно. Во второй декаде августа большая часть из них
откочевывает с мест гнездования, и с середины августа фифи встречались
редко. Отдельные птицы попадались, однако, даже в первых числах
сентября. В последний раз одиночные молодые фифи отмечены в вер-
ховьях Пясины 3 сентября 1958 г., а у истоков Рыбной — 4 сентября
1959 г.
42. Щеголь — Tringa erythropus (Pall.). Щеголь очень обыкновенен на
гнездовье как в окрестностях Норильских озер, так и в лесотундре близ
истоков Пясины, но гнездится спорадично. В местах наивысшей числен-
ности, например близ истоков Глубокой и в лесотундре около пос. Курья
на р. Пясине, гнездящиеся пары встречались через каждые 200—250 м
маршрута. Типичные местообитания — захламленные лесные опушки
с отдельными болотистыми лужами и окраины кочкарниковых болот, ок-
руженных лесом. В лесотундре щеголь придерживается открытых боло-
тистых низин, в зону тундры заходит недалеко и севернее устья Дудыпты
вообще не был встречен.
Весной 1959 г. первая пара наблюдалась в истоках Рыбной 1 июня.
Щеголи держались на отмелях вместе с пепельными улитами. В середине
июня пары наблюдались в типичных для них местообитаниях и, по всей
вероятности, начали насиживание. В 1964 г. слабо выраженный про-
лет щеголей наблюдался там 31 мая—11 июня, потом все птицы ис-
чезли.
Гнездо, найденное недалеко от водопада Орон 2 июля 1958 г., было рас-
положено на большой кочке, поросшей карликовой березкой и багульни-
ком, на краю довольно сухого болота. 4 яйца лежали в плоском лотке,
выстланном прошлогодними листьями карликовой березки. В 1960 году,
отличавшемся необычайно ранней весной, недавно вылупившиеся пуховые
птенцы были найдены В. В. Леоновичем в верховьях Пясины уже 1 июля.
У молодого щеголя, пойманного на болотистом берегу озера близ истоков
Рыбной 13 июля 1959 г., уже начали появляться маховые. Хорошо ле-
тавшие молодые наблюдались около оз. Глубокого 5 и 6 августа 1958 г.
Отдельные беспокойно кричавшие щеголи, сидевшие на вершинах де-
ревьев, и даже пары наблюдались там еще 10 августа. В большинстве
случаев у выводков беспокоилась одна птица. Две из них, добытые 10 ав-
густа 1958 г., оказались самцами. Молодые птицы встречались большей
частью поодиночке на илистых берегах заливов оз. Глубокого и р. Глу-
бокой. В середине августа почти все щеголи исчезли. В 1958 г. они в по-
— 250 —
следний раз были отмечены 16 августа (р. Глубокая), а в 1959 г., в вер-
ховьях р. Рыбной — 17 августа.
43. Пепельный улит — Heteroscelus incanus brevipes (Vieill.). Пепель-
ный улит — обычная, а местами даже многочисленная птица в северотаеж-
ных ландшафтах у Норильских озер и в бассейнах рр. Рыбной и Хан-
тайки.
В пределах лесотундры пепельные улиты встречены только близ
истоков Пясины, хотя не исключено их гнездование у рек и ручьев у се-
верных отрогов плоскогорья Путорана. На подобных, иногда даже без-
лесных горных речках они были обнаружены В. И. Капитоновым (1962)
в северной Якутии. Значительно реже гнездится пепельный улит в аль-
пийском и субальпийском поясах плоскогорья Путорана, где летом
1964 г. отдельные выводки наблюдались на высотах до 750 м над ур. м.
Численность пепельного улита в некоторых местах довольно велика: во
время экскурсии 30 июня 1958 г. вдоль берега Камюстяха (приток р. Рыб-
ной) отдельные пары встречались через каждые 200—300 м береговой
линии. Несколько меньшая численность этих улитов отмечена на галечном
берегу в верховьях Рыбной и Хантайки летом 1959 г.
Прилетают пепельные улиты раньше большинства других куликов.
В 1958 и 1964 гг. у истоков Рыбной первые пары были замечены 31 мая,
а в 1959 г. — 1 июня. Пепельные улиты появляются парами или в оди-
ночку, стай весной ни разу встречать не случалось. Ко времени их прилета
русла многочисленных каменистых ручьев, на которых селятся эти кулики,
еще погребены под толстым слоем снега. Поэтому только что прилетев-
ших улитов обычно случалось наблюдать в рано вскрывающихся вер-
ховьях Рыбной, где они кормились на кромке льда или на немногочислен-
ных обнажившихся песчаных либо каменистых отмелях. Основу их пищи
составляли, судя по визуальным наблюдениям, веснянки, которые
в массе появлялись в конце мая и начале июня. Несколько позднее, когда
под снегом начинали пробиваться многочисленные ручейки талой воды,
пепельные улиты очень охотно кормились в их устьях. И только ко второй
половине июня, по мере интенсивного стаивания снега, пары пепельных
улитов распределялись по местам гнездований.
Непременным условием гнездовых местообитаний пепельных улитов
на западном Таймыре является наличие галечниковых или моренных
русел или берегов озер. В некоторых случаях это берега довольно крупных
рек, прорезающих моренные гряды, таких, как Рыбная, Талая, Хантайка,
а иногда небольшие ручейки, соединяющие лежащие на разных высотах
мелкие озера. На каменистых берегах таких ручьев пепельные улиты
обнаружены нами повсюду на водоразделе рр. Рыбной и Хантайки и близ
истоков Пясины. В случаях гнездования пепельных улитов на берегах
больших Норильских озер они явно предпочитают устья впадающие в них
ручьев или ближайшие окрестности снежников. Абсолютная высота
местности особого значения для гнездования, по-видимому, не имеет. По
нашим наблюдениям, улиты одинаково охотно гнездятся как на высоте
50—100, так и 250—400 м над ур. м. На больших высотах просто реже
встречаются подходящие местообитания. Берега рек и ручьев, около кото-
рых гнездятся улиты, в ряде случаев были покрыты хорошим елово-лист-
венничным лесом, часто с труднопроходимым подлеском из ивняков,
ольхи, белой и карликовой березы, или завалены буреломом. Такие ха-
рактерные местообитания часто встречались на рр. Рыбной и Хантайке,
на ручьях по водоразделам и вдоль берегов оз. Кета. В истоках Пясины и
на некоторых реках плоскогорья Путорана улиты встречались в листвен-
ничном редколесье, а также на совсем открытых тундроподобных прост-
ранствах. Часто селятся они в непосредственной близости к снежникам,
прорезаемым реками.
— 251 —
Токование пепельного улита можно было услышать с первых дней
после прилета и до двадцатых чисел июня. Токовая песня очень напоми-
нает брачный крик фифи, но гораздо звонче. Как и фифи, токующий пе-
пельный улит часто присаживается на вершины деревьев. Особенно ожив-
ленно токовали улиты в середине июня, когда их можно было услышать
во всякое время суток. В конце месяца они стали более скрытными,
и чаще попадались кормившиеся одиночки. В 1964 г. улиты токовали до
первых чисел июля.
Начало насиживания нормально падает на третью декаду июня.
Самка, добытая у истоков Рыбной 28 июня 1958 г., уже закончила кладку,
отложив 4 яйца. 19 июля 1958 г. геолог Ю. Я. Лифшиц (личное сообще-
Рис. 16 Насиживающий пепельный улит — Heteroscelus incanus brevipes (Vieill.) —
в гнезде темнозобого дрозда — Turdus naumanni eunomus.
ние) в верховьях Макуса, на высоте около 320 м над ур. м., нашел гнездо.
Оно помещалось в ямке между валунами, на участке поросшей карлико-
вой березкой горной лесотундры и содержало 4 яйца. Второе гнездо,
найденное В. В. Леоновичем в истоках Пясины, было осмотрено нами
27 июня 1960 г. Оно находилось в лиственничном редколесье с кустами
ольхи и ивняков и с нижним ярусом из мхов, лишайников, осок, голубики
и карликовой березки и было расположено метрах в 100 от галечникового
берега Пясины и в 200—250 м от каменистого русла, вытекающего из
озера ручья. Кладка из 4 яиц лежала в прошлогоднем гнезде темнозобого
дрозда, устроенном на лиственнице на высоте 2—2.5 м от земли. Яйца
пепельного улита ярко-голубые, усеянные темно-бурыми пятнами сред-
ней величины, образующими венчик на тупом конце. В очень раннюю
весну 1960 г. яйца были уже наполовину насиженные. Их вес: 18.41,
18.83, 19.3, 19.7 г. Размеры: 43.5x30, 43.7x30, 43.6x30.5, 44.5x30 мм.
Насиживала самка (рис. 16), а самец при нашем приближении проявлял
крайнее беспокойство. В моменты наибольшего возбуждения его беспо-
койные крики переходили в токовую песню. Так как у добытого самца
- 252 —
были обнаружены наседные пятна, он, очевидно, тоже принимает участие
в насиживании.
Первый выводок из 4 только что вылупившихся птенцов наблюдался
нами в верховьях Рыбной 16 июля 1959 г.; взятый птенец весил 12 г.
Птенцы в сопровождении одной взрослой птицы держались в сильно
увлажненном устье каменистого ручейка, впадавшего в р. Рыбную, среди
кустиков полярных ив, осоковых кочек и разного растительного мусора,
принесенного половодьем. Шныряя среди растений, птенцы тихо пописки-
вали; старая птица отвечала им негромкой мелодичной трелью. Намокнув
и простыв, они начинали особенно громко пищать. В ответ находившийся
Рис 17 Пепельный улит — Heteroscelus incanus brevipes (Vieill.) —
обогревающий птенцов
поблизости старый улит издавал трель и становился в очень характерную
позу. Он немного приседал и вытягивал шею вперед по направлению
к птенцам. Замерзшие малыши прятались к нему под крылья, по двое
с каждой стороны. Старый улит, распушив оперение, обогревал их
(рис. 17). Обсохнув и отогревшись, птенцы затем вылезали и начинали
энергично искать насекомых, в основном очень многочисленных в то время
комаров, которых они не только склевывали с травы, но и хватали на лету.
Точно так же ловила комаров и старая птица. Даже во время согревания
птенцов она схватывала сидевших или даже пролетавших поблизости
насекомых. Комары, пик численности которых падает на середину июля,
видимо, составляют основную пищу птенцов многих видов куликов в пер-
вую неделю их жизни.
Массовое вылупление птенцов пепельного улита в верховьях Рыбной
отмечено 18 июля 1959 г., когда на протяжении 2 км береговой линии было
встречено 4 выводка. Все они держались на заросших ивняком берегах,
недавно освободившихся от полых вод. 19 июля там было поймано
3 птенца. Пеньки маховых у них уже выступали на 2—2.5 мм. Их вес:
15.5, 12.5, 12.2 г. В 1964 г. выводки с совсем маленькими птенцами были
отмечены на северном берегу оз. Кета 22 и 25 июля. Около большинства
этих выводков держалось по две старых птицы, вопреки тому, что отметил
— 253 —
К. А. Воробьев (1959) на хр. Черского. По нашим наблюдениям с расстоя-
ния всего нескольких метров, в отогревании птенцов почти одинаково при-
нимают участие оба старых улита. Возле выводков, встреченных на р. Хан-
тайке 31 июля и 1 августа 1959 г., было как по одному, так и по два бес-
покоившихся старых улита. Путем тщательных наблюдений, сделанных
за целым рядом выводков пепельных улитов на берегу оз. Кета летом
1964 г., удалось установить, что обе старые птицы обычно встречаются
лишь у самых маленьких птенцов, возраст которых не превышает 1—2
дней. Позднее самки бросают молодых, которые с этого времени полностью
остаются на попечении самцов. Нередко самки покидают птенцов сразу
после их вылупления, и тогда у выводков с самого начала наблюдается
только одна взрослая птица. Во всяком случае все 6 улитов, добытых от
выводков с подросшими птенцами в период с 29 июля по 12 августа 1964 г.,
оказались самцами. Все отстреленные в это же время одиночные птицы,
державшиеся на галечниковых берегах особняком, были хорошо упитан-
ными самками. По мере роста птенцов инстинкт заботы о потомстве за-
метно слабеет и у самцов, но все же они обычно держались вместе с моло-
дыми до самого их отлета.
Птенцы растут быстро. У молодого улита, найденного мертвым 24 июля
1959 г. в сухом каменистом русле ручья, маховые уже немного выступали
из лопнувших пеньков. Молодые, наблюдавшиеся на водоразделе рр. Рыб-
ной и Хантайки 7 и 9 августа 1959 г., уже отлично летали, хотя их еще
сопровождали старые птицы, беспокоившиеся при приближении человека.
Некоторые выводки заметно запаздывают. 31 июля 1959 г. в верховьях
Хантайки птенцы из выводков были в возрасте всего 3—4 дней. Несколько
сдвинуты сроки размножения и у улитов, гнездящихся в альпийском
поясе плоскогорья Путорана. На берегу одного небольшого озерка на
высоте более 400 м 13 августа 1964 г. был пойман птенец не старше 8—
*9 дней. Однако к концу первой декады августа почти все молодые пепель-
ные улиты становятся совершенно самостоятельными и сразу начинают
откочевывать с мест гнездований.
На каменистом берегу западной оконечности оз. Кета 21 июля 1959 г.
случалось видеть одиночных улитов. На 10 км маршрута их было отме-
чено 8. По-видимому, это были покинувшие выводки самки.
В середине августа количество пепельных улитов резко упало, хотя
отдельные птицы и выводки регулярно встречались на протяжении всего
месяца. В это время они наблюдались исключительно на кромке галечни-
ковых или валунных берегов рек и ручьев. Осенью 1959 г. небольшая
стайка пепельных улитов в верховьях Рыбной в последний раз была
отмечена 8 сентября.
44. Перевозчик — Actitis hypoleucos (L.). Перевозчик оказался обычным
гнездящимся куликом на берегах таежных речек и ручьев в окрестностях
Норильских озер. В лесотундре близ истоков Пясины он уже отсутство-
вал. Особенно много перевозчиков было встречено в июле—августе 1959 г.
на р. Могокте — одном из притоков Хантайки. Там пары и выводки по-
падались через каждые 200—300 м береговой линии. Сходная численность
была отмечена в том же году и в верховьях Рыбной. На р. Норилке пере-
возчики встречались гораздо реже.
Весной 1958 г. первые перевозчики появились в истоках Рыбной
11 июня, а их массовый прилет происходил 19 июня, после освобождения
от снега и льда берегов реки. В более теплую весну 1959 г. перевозчики
появились там уже 3 июня, а 6—13 июня наблюдались во множестве.
В 1964 г. первая птица была отмечена 13 июня. С момента прилета эти
кулички уже держались парами. Их излюбленными местообитаниями
были песчаные или галечниковые берега рек и небольших речек, особенно
в местах впадения в них ручейков.
— 254 —
Гнездо с 4 ненасиженными яйцами было найдено в верховьях Рыбной
1 июля 1964 г. Оно было устроено в лиственничном с примесью белой
березы лесу, метрах в 50 от берега реки. Само гнездо помещалось под
защитой редких кустиков карликовой березки и багульника, а выстилка
его лотка состояла из сухих листьев белой березы.
Вылупление птенцов у перевозчиков происходит во второй декаде
июля. Во время экскурсии по берегу Рыбной 18 июля 1959 г. наблюда-
лось несколько пар беспокоившихся птиц. 19 июля был пойман пуховой
птенец в возрасте нескольких дней. Весил он 14 г, на крыльях уже появи-
лись пеньки маховых длиной 12 мм, наметились и пеньки кроющих крыла.
28 июля 1959 г. в верховьях Могокты был найден мертвый молодой пере-
возчик с хорошо развитыми маховыми. 31 июля 1959 г. в верховьях Хан-
тайки наблюдался хорошо летавший молодой. В 1959 г. первый летный
выводок в устье Талой также отмечен 31 июля. В большинстве выводков,
встреченных в низовьях р. Могокты 2 августа 1959 г., молодые хорошо
летали. Вскоре после подъема на крыло молодых перевозчики начинают
отлетать с гнездовий. 12—19 августа 1959 г. в верховьях Рыбной они встре-
чались лишь изредка, а последняя птица была замечена 23 августа.
Осенью 1964 г. перевозчики наблюдались 2 сентября.
45. Мородунка — Xenus cinereus (Guld.). Повсюду в окрестностях
Норильских озер мородункЯ — обычная гнездящаяся птица. Типичные
местообитания ее — песчано-илистые и галечниковые берега рек и боль-
ших озер. На мелких таежных водоемах мородунки встречались редко.
В пределах исследованной нами территории она ни разу не была встре-
чена севернее устья Норилки.
Весной 1958 г. в истоках Рыбной мородунки появились 16 июня. Их
массовый прилет отмечен 18 июня, после оттаивания значительных пло-
щадей галечниковых и илистых отмелей. В 1959 г. токование нескольких
мородунок было слышно уже 2 и 3 июня, а 4 июня отмечено их массовое
появление. Весной 1964 г. первые мородунки прилетели 2 июня, но в зна-
чительном количестве наблюдались только с 10 июня.
В первые дни прилета мородунки держались вместе с пепельными
улитами и фифи на кромке льда и в устьях ручьев. Позднее они распреде-
лились по береговой линии рек и озер более равномерно, и с этого времени
пары явно придерживались определенных мест. Спаривание мородунок
случалось наблюдать в истоках Рыбной 20 июня 1959 г. В 1959 г. токова-
ние этих куличков стало гораздо менее интенсивным 24—25 июня, но
в более холодную весну 1958 г. их брачное оживление длилось дней на
10 дольше.
Гнезда свои мородунки устраивают в самых разнообразных условиях.
Одно из них, содержавшее неполную кладку из 3 яиц, было найдено на
берегу заболоченного таежного озера 24 июня 1964 г. Оно помещалось
на опушке мелкого березняка с отдельными лиственницами, на совер-
шенно сухом месте, метрах в 5 от воды. Лоток гнезда был умят во мху п
окружен редкими кустиками злаков и плауна. Его выстилка состояла из
сухих березовых листьев и разного растительного мусора. При посещении
гнезда 27 июня в нем было обнаружено уже 4 яйца. Гнездо со значительно
насиженной кладкой из 4 яиц было найдено близ западного берега оз. Кета
2 июля 1959 г. Оно было расположено на идущей вдоль берега озера мо-
ренной гряде, на совершенно открытом месте среди небольших камней и
кусочков дерева. Яйца лежали в неглубокой выстланной всяким мусором
ямке и благодаря своей окраске поразительно сливались с окружающим
фоном. Гнездо было удалено на 50—60 м от ближайшего заболоченного
озерка; 11 пюля птица еще продолжала насиживание. Третье гнездо было
устроено на небольшом низинном островке близ устья Кета-Ирбо, в по-
лосе растительного мусора, идущей вдоль песчаной косы. Выстилка лотка
— 255 —
состояла из сухих палочек и щепок, с боков гнездо было хорошо защи-
щено двумя довольно толстыми, сухими палками, лежавшими на земле
параллельно. 10 июля 1964 г. в этом же гнезде наблюдалось вылупление
птенцов. Судя по сделанным у гнезд наблюдениям, в насиживании прини-
мают участие обе птицы. Во время вылупления птенцов как самка, так и
самец проявляли крайнее беспокойство, но в более поздний период у вы-
водков обычно наблюдалась только одна птица (рис. 18). Птенцы более
чем недельного возраста часто ведут уже совершенно самостоятельный
образ жизни.
Несколько пар мородунок, своим беспокойным поведением указы-
вавших на присутствие выводков, наблюдались на берегу оз. Кета 18 июля
Рис. 18. Мородунка — Xenus cinereus (Giild.) — с однодневными
птенцами
1959 г. 3 птенца, весившие 16.5, 17.0 и 19.5 г, с пеньками маховых в 12—
13 мм, были пойманы там 19 июля. Выводок из недавно вылупившихся
птенцов был найден в устье Рыбной 20 июля 1958 г. Таким образом, вы-
лупление птенцов мородунки чаще всего происходит во второй декаде
июля.
Как и у других куликов, птенцы мородунки развиваются быстро.
В выводке, наблюдавшемся 21 июля 1964 г. у оз. Кета, птенцы уже в зна-
чительной мере сменили пух на перо. 31 июля 1958 г. в устье Талой был
пойман пытавшийся летать молодой куличок. Птенец из другого выводка,
добытый там 2 августа, имел уже наполовину освободившиеся от чехлов
маховые. В этот же день наблюдался и хорошо летавший молодой. Неко-
торые выводки, однако, несколько запаздывают в своем развитии: 7 ав-
густа был пойман еще не умевший летать молодой, маховые которого
выступали из чехлов еще на половину своей длины.
Как и другие кулики, начавшие вести самостоятельный образ жизни,
молодые мородунки вместе со старыми птицами начинают откочевку
к югу. Начиная с 10—12 августа они уже встречались лишь изредка,
хотя полностью исчезли только к началу сентября.
46. Круглоносый плавунчик — Phalaropus lobatus (L.). Круглоносый
плавунчик довольно обыкновенен в северной полосе тайги, лесотундре и
— 256 —
тундре до низовьев Пуры на севере. Однако севернее Пуринских озер
круглоносые плавунчики гнездятся значительно реже, чем на юге иссле-
дованной нами территории, и в низовьях Пясины полностью сменяются
плосконосыми.
Весной 1958 г. в верховьях Рыбной круглоносые плавунчики появи-
лись 10 июня, а в 1959 г. — 1 июня. В тундре близ устья Дудыпты
в 1962 г. первые плавунчики были зарегистрированы 7 июня, а на местах
гнездования у Пуринских озер они были отмечены в 1961 г. 21 июня,
а в 1963 г. — 8 июня. В первые дни после прилета наблюдались небольшие
стайки, но дней через 5—6 плавунчики встречались исключительно парами.
На севере тайги и в лесотундре плавунчики гнездятся на небольших
озерах и просто лужах с заболоченными или заросшими осокой берегами.
В тундре они чаще всего встречаются на водоемах в речной пойме и на
полигональных болотах, значительно реже на небольших лужах по влаж-
ным водоразделам.
Гнезда, найденные В. В. Леоновичем в пойме Агапы 6—10 июля 1961 г.,
были устроены среди травы на берегу заболоченного озера и содержали
совершенно ненасиженные кладки.
Плавунчиков, выказывавших сильное беспокойство, случалось наблю-
дать на берегу маленького лесного озера близ истоков Рыбной 13 и 14
июля 1959 г.; 14 июля там был найден мертвый птенец в возрасте 2—3 дней.
Летом 1958 г. молодой плавунчик, весивший 14 г, был пойман в устье
Талой 30 июля. Его маховые выступали из чехлов на 8—9 мм. Взрослых
плавунчиков поблизости не было, и птенец вполне самостоятельно разы-
скивал корм на берегу протоки. Сильно подросший птенец, наблюдавшийся
на одном из озер близ р. Глубокой 12 августа 1958 г., также держался со-
вершенно самостоятельно. Видимо, самцы часто покидают птенцов уже
вскоре после их вылупления. Хорошо летавшие молодые наблюдались
у р. Глубокой 15 августа 1958 г.
Десятки кочевавших круглоносых плавунчиков наблюдались в ни-
зовьях Агапы 30 июля п 1 августа 1960 г. К десятым числам августа пла-
вунчики полностью исчезли из тундры. В 1961 году, отличавшемся очень
поздней весной, эти кулички встречались там еще в конце августа.
47. Плосконосый плавунчик — Phalaropus julicarius (L.). Плосконо-
сые плавунчики, обыкновенные в устье Пясины (Колюшев, 1933), дохо-
дят на юге своего гнездового ареала до широты устья Агапы. Весной 1961 г.
они впервые были отмечены 29 июня, хотя возможно, что прилетели и на
несколько дней раньше. Небольшие группы кормились на полузатоплен-
ном участке берега 2-го Пуринского озера. В условиях нормальной весны
1963 г. прилет зарегистрирован там 11 июня.
Как и круглоносые плавунчики, плосконосые гнездятся на заросших
осокой берегах небольших тундровых озер. 6 июля 1961 г. сильно беспокоив-
шийся самец наблюдался на берегу болотистой лужи близ северного бе-
рега Пуринского озера. В одной из мохово-осоковых кочек была обнару-
жена умятая птицей пустая гнездовая ямка, хорошо замаскированная
нависавшим сверху пучком сухой осоки. Гнездо с 4 яйцами, находивши-
мися на последней стадии насиженности, было найдено в окрестностях
2-го Пуринского озера 13 июля 1963 г. Оно было устроено среди осоки
в травянистой слабо кочкарниковой низине, метрах в 5 от небольшой
болотистой лужи. У гнезда беспокоился только самец.
48. Камнешарка — Arenaria interpres oahuensis (Blox.). Камнешарка,
очень обыкновенная на Таймырском побережье (Бируля, 1907), в пределах
исследованной нами территории встречается только на пролете и в не-
большом числе.
16 июня 1958 г. одиночная камнешарка была добыта В. Герке близ
истоков Рыбной. Весной 1961 г. В. В. Леонович наблюдал в пойме Агапы
17 Биология птиц
— 257 —
пары и небольшие группы с 9 по 18 июня. Некоторые камнешарки держа-
лись вместе с бурокрылыми ржанками. Мы наблюдали их 6 и 19 августа,
в окрестностях Пуринских озер и у устья Агапы. В обоих случаях оди-
ночные птицы кормились на захламленном берегу блпз построек чело-
века.
49. Турухтан — Philomachus pugnax (L.). Турухтан, хотя и в неболь-
шом количестве, но несомненно доходит до морского побережья (Бируля,
1907; Колюшев, 1933).
Нами летом 1961 г. турухтаны наблюдались в низовьях Пуры,
а В. В. Леоновичем — близ устья Тареи. Типичные местообитания турух-
танов — равнинные заболоченные участки, обычные на водоразделах
притоков Рыбной и Хантайки, а в тундре — в пойме рек. В лесотундре
близ истоков Пясины турухтаны очень обыкновенны на болотистых низи-
нах, примыкающих к многочисленным озерам.
В тайге у Норильских озер, у истоков Рыбной, турухтаны весной
1958 г. впервые были отмечены 14 июня, а в 1959 г. — 9 июня. Сколько-
нибудь выраженного весеннего пролета там не наблюдалось. Очевидно,
пролетный путь, как и у большинства птиц, проходит в стороне. В тундре
у устья Дудыпты весной 1962 г. первые турухтаны наблюдались 10 июня,
а 12 июня шел их массовый пролет на север. В условиях поздней весны
1961 г. на местах гнездования у Пуринских озер турухтаны были отмечены
только 22 июня, а в 1963 г. — 8 июня. Кроме того, хорошо выраженный
пролет (исключительно самцов) наблюдался у Пуринских озер с 18 июня
по 4 июля 1963 г. Птицы небольшими стаями летели в северном и северо-
западном направлениях.
После прилета, в середине июня, турухтаны вели себя очень ожив-
ленно, и на многочисленных проталинах ежедневно случалось наблюдать
бои самцов. К началу июля турухтаны становятся малозаметными. Самцы,
по-видимому, откочевывают с мест гнездований, а самки начинают насижи-
вание. Все 8 найденных нами гнезд были устроены на кочках среди кочкар-
никовых мохово-осоковых болот. Их устройство было самое примитивное:
простые ямки, умятые птицами среди стеблей осок и злаков, и моховая
подстилка. Обычно они хорошо маскировались нависающими пучками
прошлогодних осок.
Очевидно, турухтаны начинают кладку в двадцатых числах июня.
Кладка из 4 яиц, найденная близ западного берега оз. Кета 27 июня
1958 г., была очень слабо насижена. В 4 кладках, осмотренных в долине
р. Рыбной 2 июля 1958 г., яйца были значительно насижены и содержали
эмбрионы длиной 1.5—2 см. В условиях ранней весны 1960 г. в гнезде,
найденном в устье Норилки 21 июня, яйца были насижены слабо. В позд-
нюю весну 1961 г. слабо насиженные яйца турухтана были обнаружены
в гнезде, найденном в верховьях Пуры 8 июля. В условиях нормальной
весны 1963 г. первые выводки с птенцами в возрасте 2—4 дней были обна-
ружены в пойме Пуры 10 и 11 июля. В 1960 г. маленькие пуховые птенцы
были найдены в верховьях Пясины 30 июня. Летом 1960 г. хорошо летав-
шие молодые турухтаны ежедневно наблюдались в пойме Агапы, начиная
с 30 июля. В 1958 г. у оз. Глубокого вполне самостоятельный молодой был
добыт 10 августа.
Сразу после подъема на крыло молодых турухтаны начинают свои
кочевки к югу. 14 августа 1959 г. на болоте блпз западного берега оз. Кета
наблюдалась стая, в которой насчитывалось не менее 25 особей. В другой
раз такая же стая была отмечена на берегу р. Глубокой 16 августа 1958 г.
В последней декаде августа турухтаны встречались только изредка.
Как видно из приведенной табл. 4, состав пищи турухтанов, добытых
осенью, довольно разнообразен. Интересно, что в ней значительный про-
цент составили растительные остатки. С значительным потреблением ту-
— 258 -
рухтанами растительной пищи согласуется и большее, чем у других
куликов, количество гальки в их желудках.
50. Кулик-воробей — Calidris minuta (Leisl.). Кулик-воробей — много-
численная гнездящаяся птица в тундрах севернее истоков Пуры вплоть
до побережья Карского моря (Колюшев, 1933). Южная граница гнездова-
ния в разные годы испытывает некоторые смещения, но в общем проходит
немного южнее Пуринских озер. Летом 1960 г. в низовьях Агапы были от-
мечены только одиночные особи; в 1961 г., по сообщению В. В. Леоновича,,
они там не гнездились совсем, хотя в 30 км севернее, у 2-го Пуринского
озера, нами были встречены от-
дельные пары, а в верховьях
Пуры найдены гнезда. Летом
1962 г. эти кулички у Пурин-
ских озер не гнездились, но
в 1963 г. там было найдено
5 гнезд и наблюдалось множе-
ство выводков.
На южной границе гнездо-
вий у Пуринских озер, весной
1961 г. кулики-воробьи появи-
лись 16 июня, а в 1963 г. —
10 июня. На весеннем пролете
у устья Дудыпты стайки, ле-
тевшие в северном направле-
нии, ежедневно встречались 5—
12 июня. В первое время после
прилета, в середине или в двад-
цатых числых июня, ежедневно
случалось наблюдать оживлен-
ное токование. При этом сразу
2—4 куличка с характерным
чириканьем низко летали над
берегами тундровых водоемов.
К 22—25 июня 1963 г. токо-
Таблица 4
Осенняя пища турухтана
(р. Агапа, август 1960 г., 10 желудков)
Пищевые объекты
Trichoptera (1.)
Trichoptera (im.)
Carabidae (im.)
Dytiscidae (im.).
Haliplidae (im.)
Tipulidae (1.)
Arctiidae (1.)
Notostraca
Amphipoda
Copepoda
Gastropoda
Sphaeridae ....
«Луковички» горца живо-
родящего
7
2
4
10
1
3
5
6
4
?
4
7
35
2
1
4
5
1
1
1
2
3
5
1
2
4
вание куликов-воробьев почти полностью прекратилось.
Гнездятся кулики-воробьи в разных местах, но всегда достаточно
сухих. Непременным условием является только наличие на расстоянии
15—100 м участков увлажненной низменной тундры или берегов водое-
мов. 5 из 10 осмотренных гнезд были расположены в мохово-осоковой
тундре с кустарниками, 2 — в моховой, с редкими тальниковыми кустами,
2 — в дриадовой тундре поблизости от поймы и 1 — на песчаном, порос-
шем редкими кустиками ив островке р. Пуры. Гнезда всегда представляли
собой просто ямку, умятую во мху и сухой осоке, иногда выстланную
очень незначительным количеством сухих листочков карликовой березки,
полярных ив, осок и злаков. Гнезда обычно лишь слегка маскируются
с боков кустиками карликовой березки и полярных ив. Во всех гнездах,
найденных в 1963 г., полные кладки содержали по 4 яйца. В условиях
суровой весны 1961 г. из 4 найденных гнезд только в одном было 4 птенцаг
в трех же других — 3, 2, 2 яйца. По своей окраске яйца кулика-воробья
заметно отличаются от яиц белохвостого песочника более теплыми, желто-
ватыми тонами.
К устройству гнезд кулики-воробьи приступают через несколько дней
после прилета. В яйцеводе у самки, добытой у Пуринских озер 14 июня
1963 г., было готовое к сносу первое яйцо. Гнездо с 4 ненасиженными
яйцами было найдено 18 июня. Летом 1961 г. гнездо с 2 слабо насиженными
яйцами было осмотрено в верховьях Пуры 7 июля. Насиживающие птицы
— 259 —
17*
вели себя у гнезда очень доверчиво, некоторые особи позволяли трогать
себя коротким прутиком. Вспугнутый с гнезда куличок обычно отводит,
а если наблюдатель долго не уходит, то прямо у него на глазах возвра-
щается к кладке и продолжает насиживание. В 1963 г. 10—14 июня в ок-
рестностях Пуринских озер вылупление птенцов в гнездах у куликов-
воробьев происходило очень дружно. 10 июня 1963 г. на границе влажной,
богатой водоемами поймы Пуры с сухой береговой террасой было обнару-
жено сразу 2 выводка однодневных пуховых птенцов. Один из птенцов ве-
сил 4.8 г. На следующий день выводков было уже много повсюду. Кладка
из 4 яиц накануне вылупления молодых была отмечена 14 июля 1963 г.
В последний раз однодневные птенцы наблюдались в 1963 г. 18 июля.
В 1961 г. новорожденные пуховые птенцы в среднем течении Пуры впервые
появились 18 июля.
Выводки обычно придерживались более увлажненных низин и берегов
водоемов, около них всегда находилась только одна старая птица. У 1-го
Пуринского озера в 1963 г. летные молодые были впервые отмечены
26 июля. 7 августа 1961 г. в низовьях Пясины был пойман вполне взрос-
лый, но еще не летавший птенец. Летные выводки держатся на местах,
где они вывелись, всего несколько дней, после чего начинают кочевку
к югу. В 1960 г. в низовьях Агапы появилось много молодых куликов-
воробьев. 19 августа 1958 г. небольшие стайки наблюдались и в северной
тайге у р. Глубокой. Больших стай отлетающих куликов, подобных
тем, которые наблюдал В. М. Сдобников (19596) в устье Нижней Тай-
мыры, мы не встречали.
51. Белохвостый песочник — Calidris temminckii (Leisl.). В Нориль-
ской долине белохвостый песочник гнездится отдельными, изолирован-
ными друг от друга очагами. В лесотундре верховьев Пясины он повсюду
очень обыкновенен и достигает наибольшей численности в тундре между
устьем Дудыпты и Пуринскими озерами. На берегу Пясины на 30 км
выше устья Агапы 2 июля 1962 г. пары встречались через каждые 100—
150 м. Несколько меньшая численность была отмечена в том же году в ок-
рестностях Пуринских озер: пары на берегу попадались через 200—300 м.
К северу от Пуринских озер количество этих куличков падает, а в нижнем
течении Пясины и Пуры во время нашей поездки летом 1961 г. белохво-
стые песочники совсем не встречались. Не были они найдены и на побе-
режье Карского моря (Бируля, 1907; Колюшев, 1933).
Весной 1958 г. первые особи были отмечены в истоках Рыбной 6 июня;
массовый пролет наблюдался до двадцатых чисел июня. В 1959 г. первая
стайка появилась там 30 мая и пролет длился до середины июня. В тундре
у устья Дудыпты первые стайки наблюдались в 1962 г. 3 июня, а их пролет
продолжался до середины июня. В 1961 г. белохвостые песочники появи-
лись у Пуринских озер 11 июня, а в 1963 г. — 6 июня. Массовый их при-
лет в 1963 г. происходил 7—9 июня.
Уже во время пролета всюду наблюдались токующие самцы. Но осо-
бенно интенсивным становится токование во второй половине июня. В это
время повсюду можно было видеть куличков, как бы висящих в воздухе,
на высоте 3—10 м, и слышать их характерную трель. По наблюдениям
27—30 июня, на берегу р. Пясины каждый самец токовал на одном и
том же месте в течение круглых суток с очень небольшими перерывами.
Типичными гнездовыми местообитаниями белохвостого песочника
является моховая тундра со слабо выраженным кочкарником и с ред-
кими кустиками полярных ив и карликовой березки. Такие места чаще
всего находятся на пологих, обычно южных склонах холмов и на берегах
водоемов. У Норильских озер песочники гнездятся на поросшей кустар-
ником прибрежной полосе рек, а особенно охотно на покрытых мхом участ-
ках мелких островов крупных озер. На одном таком островке, расположен-
— 260 —
ном на оз. Кета, близ устья Кета-Ирбо, 8 июля 1964 г. было найдено сразу
5 гнезд белохвостых песочников. Очень охотно гнездятся белохвостые
песочники, как это заметил Н. А. Гладков в бухте Тикси (Гладков, 1958),
около немногочисленных в тундре поселений человека. Около каждой
построенной в тундре рыбачьей избушки гнездятся 1—3 пары. Здесь их
привлекают насекомые, в изобилии размножающиеся на отбросах. Все
19 осмотренных гнезд представляли собой ямки, умятые во мху или
прошлогодней траве без специальной выстилки. Они маскировались ред-
кими кустиками карликовой березки и полярных ив, а также сухими стеб-
лями и листьями злаков и осок. 15 гнезд содержали по 4 яйца и 4 — по
3 яйца.
В 1963 г. первое гнездо с неоконченной кладкой из 2 яиц было
найдено у Пуринских озер 18 июня. На следующий день в нем оказалось
3-е, последнее яйцо, и птица приступила к насиживанию. Второе гнездо,
с кладкой из 4 яиц, было найдено 22 июня, и третье — 30 июня 1963 г.,
с наполовину насиженными яйцами. В 1960 г. 2 гнезда со значительно
насиженными кладками были отмечены в верховьях Пясины 24 июня и
5 июля. В условиях 1961 г. слегка насиженные яйца были обнаружены
в окрестностях истоков Пуры 3 и 7 июля, а близ устья Агапы летом
1962 г. — 2 гнезда с кладками такой же степени насиженности — 2 июля.
В 5 гнездах, осмотренных на оз. Кета 8 июля 1964 г., кладки были недавно
закончены. Кладка с 4 значительно насиженными яйцами отмечена там
18 августа.
Обычно у гнезда попадалась только одна самка; в начале насиживания
самцы токовали поблизости. В нормальную весну массовое вылупление
птенцов происходило после 10 июля. В верховьях Пуры 10 июля 1963 г.
были найдены как гнездо с сильно насиженными яйцами, так и выводок
однодневных птенцов. С этого времени ежедневно встречалась масса вывод-
ков, которые держались на более увлажненных участках низин. В 1962 г.
вылупление происходило позже. 29 июля в верховьях Пуры в гнезде
оказалось 4 надклюнутых яйца. Выводки попадались в конце месяца.
В 1960 г. 22 июля в пойме Агапы был пойман птенец, у которого маховые
выступали из чехлов на 18—20 мм, а 26 июля молодые уже хорошо летали.
Стайку хорошо летавших молодых птиц в 1963 г. случилось встретить
в низовьях Агапы 28 июля. В первой декаде августа подавляющее боль-
шинство молодых уже летает, хотя отдельные выводки запаздывают
даже в благоприятные для гнездования годы. 7 августа 1960 г. на берегу
одного из озер в пойме Агапы был пойман птенец с маховыми, выступав-
шими из чехлов на 25—30 мм.
Небольшие стайки, в которых насчитывалось не более 15—20 особей,
ежедневно встречались на отмелях рек и озер во второй декаде августа.
В 1958 г. на отмелях р. Глубокой 16—20 августа наблюдались скопления
стай песочников, очевидно, начавших пролет. В конце августа песочники
попадались гораздо реже, хотя отдельные птицы и даже стайки были
отмечены в верховьях Рыбной 4 сентября 1959 г.
52. Краснозобик — Calidris testacea (Pall.). Типичный обитатель
арктических тундр, краснозобик не встречался южнее низовьев Пуры.
В. В. Леонович, спускаясь в 1960 и 1961 гг. по Пясине до устья Тареи,
также не обнаружил его южнее. На побережье Карского моря краснозо-
бик многочислен (Бируля, 1907).
Весенний пролет был отмечен у 2-го Пуринского озера 15—18 июня
1961 г., в устье Дудыпты — 10—15 июня 1962 г. В 1963 г. в окрестностях
Пуринских озер мы видели стайки лишь дважды: 31 мая и 14 июня.
У Норильских озер краснозобики ни разу не были встречены даже на
пролете. Во многих случаях эти песочники летели в общих стаях с черно-
зобиками и бурокрылыми ржанками и вместе с ржанками кормились на
— 261
освободившихся от снега возвышенных участках тундры. Все краснозо-
бики летели в северном направлении.
На местах гнездования, в низовьях Пуры, нам случалось наблюдать
этих песочников только в конце июля 1961 г., когда у всех куликов уже
вывелись птенцы. Отдельные пары, своим беспокойным поведением ука-
зывавшие на присутствие выводков, чаще всего встречались в сильно ув-
лажненных распадках, в местах, где еще сравнительно недавно лежал
снег. Здесь, на участках моховой тундры со слабо развитым осоковым
кочкарником, отдельные пары встречались приблизительно через каждые
200—300 м маршрута. В одном случае с одного места наблюдались сразу
две беспокоившиеся пары этих куликов. Пары проявлявших сильное беспо-
койство краснозобиков в 1960 и 1961 гг. наблюдались В. В. Леоновичем
близ устья Тареи 24 и 22 июля. В. М. Сдобников (19596) также наблюдал
у выводков обеих старых птиц. Возможно, что самцы краснозобиков,
которые, по наблюдениям А. А. Бирули (1907), принимают участие в наси-
живании, держатся у выводков только в первые дни после вылупления
птенцов. В пользу такого предположения говорят наблюдения Л. А. Пор-
тенко (1959), который видел у выводков только самок.
53. Чернозобик — Calidris alpina centralis (But.). В южной части
тундры, близ устья Дудыпты, у Пуринских озер и в пойме Агапы, черно-
зобик очень обыкновенен, а местами и многочислен. Севернее, в низовьях
Пуры, он встречался нам лишь изредка, а на морском побережье, судя
по сведениям целого ряда исследователей (Вальтер, 1902; Бируля, 1907;
Колюшев, 1933), он не гнездится совсем. Типичные местообитания черно-
зобиков — кочкарниковые увлажненные, в особенности припойменные
участки. Гнездятся они и в слабо холмистой мохово-осоковой кочкарнико-
вой и плоско-бугристой тундрах. В подходящих местах численность черно-
зобиков довольно велика: на 7 км маршрута в равнинной кочкарниковой
тундре в окрестностях устья Дудыпты 5 июля 1960 г. было найдено
2 гнезда и 1 выводок.
На весеннем пролете близ истоков Рыбной чернозобики наблюдались
13—18 июня 1958 г. и 4 июня 1959 г. В устье Дудыпты весной 1962 г. они
впервые были отмечены 3 июня, а 4—10 июня шел массовый пролет черно-
зобиков на север. На местах гнездований у Пуринских озер в 1961 г. чер-
нозобики появились 11 июня, а в 1963 г. — 9 июня. Во время весеннего
пролета чернозобики неоднократно наблюдались в общих стаях с красно-
зобпками. Сразу после прилета повсюду в болотистых местах слышалось
токование чернозобиков. В 1961 г. оно прекратилось только 4 июля,
а в 1962 г. — 28 июня. В этот период птицы держались парами около бо-
лотистых луж и на проталинах.
Гнезда чернозобики устраивают в довольно разнообразных условиях
рельефа, но всегда в кочкарниковой тундре. Из 6 найденных нами гнезд
три были расположены на равнинной мохово-осоковой кочкарниковой
тундре, а три — в холмистой кочкарниковой тундре. Все гнезда представ-
ляли собой простые ямки, умятые птицей в кочках и защищенные сверху
или с боков пучками прошлогодних осок и злаков. В лотке некоторых
гнезд обнаружена скудная выстилка из сухих листьев карликовой бе-
резки, полярных ив и злаков. В 4 гнездах было по 4 яйца и только в од-
ном— 2. Размеры измеренных яиц: 36.5x25, 37.9x25.4, 35.7x25.3 и
34x25.2 мм. Чернозобики приступают к насиживанию в третьей, а
в годы с более ранней весной — во второй декаде июня. У самки, добытой
близ устья Дудыпты 19 июня 1962 г., диаметр наибольшего фолликула
равнялся 8 мм, а яйцевод был развит очень сильно. Неполная кладка из
2 яиц была найдена в окрестностях 2-го Пуринского озера 27 июня 1961 г.
2 оконченные и даже значительно насиженные кладки из 4 яиц были
найдены у Пуринских озер 5 июля 1962 г. В третьем гнезде, осмотренном
— 262 —
там 12 июля, в яйцах обнаружены наполовину сформировавшиеся птенцы.
С другой стороны, летом 1960 г. массовое вылупление птенцов чернозобика
происходило 5 июля. В этот же день за одну экскурсию было найдено
3 выводка однодневных птенцов, которые в двух случаях еще находились
в гнездовых ямках.
Выводки хорошо летавших молодых чернозобиков летом 1960 г. наблю-
дались уже 1 августа, а в 1961 г. — в середине августа. Как и другие
кулики, чернозобики сразу после подъема на крыло молодых начинали
откочевки к югу. Небольшие стайки, кормившиеся вместе с белохвостыми
песочниками и галстучниками, наблюдались на отмелях тундровых озер
в продолжение всей первой декады августа 1960 г. Небольшую стайку
чернозобиков, державшихся вместе с галстучниками, случилось наблю-
дать на западном берегу оз. Кета 14 августа 1959 г. К началу сентября
в тундре отмечались только одиночные птицы.
54. Дутыш — Calidris melanotos (Viell.). В тундре у Пуринских озер
и в низовьях Агапы дутыш является хотя и относительно редким, но бе-
зусловно гнездящимся видом. Сопоставляя эти данные с находками
А. И. Толмачева на восточном Таймыре (Тугаринов и Толмачев, 1934),
к югу от Таймырского озера и В. М. Сдобникова (19596) в окрестностях
бухты Ожидания, на северном берегу этого озера, мы приходим к выводу,
что дутыш распространен в тундре почти всего Таймырского полуострова,
за исключением его северных частей.
Во время наших работ в тундре у Пуринских озер в 1961—1963 гг.
характерное токование самцов нередко случалось слышать на участках
низкой заболоченной тундры во второй половине июня. Слышал дудение
и В. В. Леонович, работавший в пойме Агапы весной 1961 г. Самих кули-
ков случалось видеть только изредка.
29 июня 1963 г. на участке мохово-пушицевой кочкарниковой боло-
тистой тундры с кустами багульника, близ поймы Большой Дюрюсы, было
найдено гнездо дутыша. Лоток — ямка во мху — был слегка выстлан
сухими стебельками осоки и листьями карликовой березки; 4 слабо наси-
женных яйца имели светло-оливковую окраску с темно-бурыми пят-
нышками. Их размеры: 38.8x26.8, 37.8x26.8, 37.5x27.2 и 38x27 мм.
Самец держался недалеко от гнезда, а самка при нашем приближении
отводила.
55. Песчанка — Calidris alba (Pall.). В пределах исследованной тер-
ритории песчанка изредка встречается на весеннем пролете.
В 1958 г. небольшая стайка держалась на обнажившейся от снега косе
в истоках Рыбной 30 мая. Весной 1959 г. одиночная песчанка наблюдалась
там 28 мая. Небольшие группы и одиночки были встречены В. В. Леоно-
вичем в пойме Агапы 10—20 июня 1961 г.
56. Обыкновенный бекас — Gallinago gallinago gallinago (L.). В преде-
лах западного Таймыра обыкновенный бекас гнездится всюду к югу от
верховьев Пуры. Численность его невелика, а в тундре он положи-
тельно редок. Придерживается бекас наиболее увлажненных мест. На
севере таежной зоны он довольно обыкновенен на заросших осокой бе-
регах небольших озер и в прилегающих к этим озерам болотах, в тундре
обычен на припойменных участках.
Весной бекасы прилетают сравнительно поздно, после образования
значительного количества проталин. В 1958 г. в истоках Рыбной мы в пер-
вый раз видели бекасов и слышали их токование 13 июня, в 1959 г. —
8 июня, в 1964 г. — 18 июня. Ранней весной 1960 г. они появились под
Норильском уже в первых числах июня. В устье Дудыпты, где проталины
образуются раньше, чем в тайге, в 1962 г. бекасы отмечены 5 июня. На
северном пределе своего ареала, у 2-го Пуринского озера, в условиях
исключительно холодной весны 1961 г. бекасы появились только 21 июня.
— 263 —
С момента прилета и до десятых чисел июля можно было наблюдать токо-
вание. Особенно охотно бекасы токовали в утренние и вечерние часы. В ме-
стах их наивысшей численности, в заболоченном лесу под Норильском
весной 1960 г. случалось видеть одновременно 2—4 токующих бекасов.
Выводок из 4 хорошо летавших молодых наблюдался в пойме Агапы
2 августа 1960 г. В середине августа бекасов было уже очень мало. С а мы е
поздние встречи у устья Агапы в 1960 г. были зарегистрированы 21 ав-
густа, а в 1961 г. — 20 августа. На болотах близ истоков Рыбной в по-
следней декаде августа 1964 г. наблюдался слабо выраженный пролет
бекасов.
57. Азиатский бекас — Gallinago stenura (Вопар.). Распространение
азиатского бекаса на западном Таймыре полностью совпадает с областью
гнездования обыкновенного, однако он всюду гораздо многочисленнее
и придерживается более сухих местообитаний. В тайге азиатские бекасы
наиболее обыкновенны по соседству с покрытыми кочкарником морен-
ными грядами и другими холмами, на гарях и сухих болотах. В тундре
они также встречаются на холмистых кочкарниковых и даже дриадовых
участках. Одним из условий, необходимых для гнездования азиатского
бекаса, является наличие в непосредственной близости от сухих возвы-
шенностей более увлажненных и даже заболоченных низин, пригодных
для кормежки выводков. Поэтому токование обоих видов бекасов на запад-
ном Таймыре зачастую можно наблюдать одновременно.
Прилетают азиатские бекасы несколько раньше, чем обыкновенные.
Близ истоков Рыбной весной 1958 г. их появление было зарегистрировано
8 июня, в 1959 г. — 6 июня, а в 1964 г. — 13 июня. В тундре у Пуринских
озер в 1961 и 1963 гг. они были отмечены 7 и 8 июня.
Токование наблюдалось с первых дней их прилета, но особенно ин-
тенсивно проходило во второй половине июня, после того как бекасы
заняли места гнездований. В этот период случалось видеть одновременно
2—3 бекасов, с характерными криками кружившихся над занятым участ-
ком. Токовые полеты азиатских бекасов такие же, как у обыкновенных,
только издаваемый крик напоминает короткий сиплый свист, а вместо
характерного для нашего бекаса «блеяния» слышится резкий звук рас-
секаемого птицей воздуха при ее стремительном снижении. Токование
чаще слышалось в утренние и вечерние часы, однако в теплую весеннюю
погоду эти бекасы токовали весь день. Весеннее оживление заканчивается
только к десятым числам июля.
За все время полевых исследований нами осмотрено 3 гнезда азиатских
бекасов. Первое было найдено на краю сухого кочкарникового болота
в бассейне р. Рыбной 30 июня 1958 г. Оно было устроено на сухой кочке
под прикрытием сухих пучков осок и злаков. Лоток представлял собой
ямку, умятую в кочке и очень скудно выстланную стебельками сухой
травы. В гнезде было 2 оливково-зеленоватых с темно-бурыми пятнами
яйца. Размеры одного из них 43.5x30.5 мм. Яйца были значительно наси-
жены и содержали эмбрионы длиной около 10 мм. Второе гнездо, найден-
ное в устье Норилки В. В. Леоновичем 22 июня 1960 г., было устроено
недалеко от склона холма, покрытого горелым лесом, среди кустов ольхи
и карликовой березки; 4 яйца были уже несколько насижены. Наконец,
третье гнездо было найдено нами в тундре в верховьях Пуры 21 июля
1962 г. Оно было расположено в сухой мохово-кустарниковой тундре,
на основной террасе, метрах в 30 от склона, спускающегося к болотистой
пойме Пуры. Лоток гнезда был выстлан сухими листьями ив и карликовой
березки и защищен редкими кустиками последней. Кладка состояла из
4 яиц в последней стадии насижен пости у эмбрионов был хорошо развит
пух. Размеры яиц: 48.1 Х31, 46.3x30.4, 45.7 хЗО.5 и 47 хЗО.1 мм. Насижи-
вавшая птица при приближении человека отводила. 14 июля 1959 г. на
- 264 —
небольшом болоте у западного берега оз. Кета мы наблюдали азиатского
бекаса, проявлявшего крайнее беспокойство. Очевидно, там был выводок.
1 августа 1958 г. мы видели выводок прекрасно летавших молодых в устье
Талой. Хорошо летавших молодых случилось встретить 3 августа 1959 г.
на залуженном берегу р. Могокты. В конце августа 1964 г. азиатские бе-
касы ежедневно наблюдались вместе с обыкновенными бекасами и гаршне-
пами на болотах близ истоков Рыбной. Видимо, это был слабо выражен-
ный пролет.
Во второй половине августа подавляющая часть азиатских бекасов на
западном Таймыре с мест гнездования отлетает.
58. Гаршнеп — Lymnocryptes minima (Brunn.). На севере таежной
зоны у Норильских озер гаршнеп редок на гнездовье. На гнездование его
в лесотундре западного Таймыра указывал В. Н. Скалой (1938). Север-
нее, в низовьях Агапы, гаршнеп, возможно, изредка гнездится, так как
В. В. Леонович в июне 1961 г. несколько раз слышал там токование.
Нами гаршнепы наблюдались только в верховьях Рыбной. 30 июня
1958 г., пройдя 30 км по редким лиственничным лесам и открытым заболо-
ченным пространствам, мы нашли гнездо, а в другом месте добыли самку
с почти готовым к откладке яйцом в яйцеводе. Гнездо было устроено на
кочкарниковом осоковом болоте по соседству с несколькими лужами и не-
большими озерками и было расположено на сухой осоковой кочке. Это
была просто ямка, умятая птицей среди стеблей прошлогодних трав.
Кладка из 4 совершенно не насиженных яиц хорошо маскировалась навис-
шими над ней пучками сухой травы. Птица у гнезда была добыта. Кроме
этих случаев, гаршнеп наблюдался и был добыт еще на заболоченном
озере в истоках Рыбной 24 августа 1959 г. В это же самое время гаршнепы
наблюдались там и в 1964 г., когда они появились и исчезли вместе с про-
летными бекасами.
59. Средний кроншнеп — Numenius phaeopus phaeopus (L.). Средний
кроншнеп является довольно обыкновенной гнездящейся птицей в окрест-
ностях Норильских озер и в бассейне р. Хантайки. Севернее, в лесотундре
верховьев Пясины, кроншнепы уже не гнездятся. Типичные местообитания
средних кроншнепов — кочкарниковые, поросшие редкими чахлыми
лиственницами, открытые участки на водоразделах в Норильской долине.
Здесь птицы гнездились группами по 2—3 пары и встречались через 10—
15 км маршрута.
Весной 1958 г. у западного берега оз. Кета кроншнепы были впервые
отмечены 13 июня. Возможно, что они прилетели на несколько дней
раньше, так как участок, где они постоянно встречались, не посещался
нами несколько дней. В 1959 г. прилет кроншнепов был отмечен 3 июня.
К гнездованию кроншнепы, по-видимому, приступают во второй поло-
вине июня. У самки, добытой близ истоков Рыбной 17 июня 1959 г.,
диаметр наибольшего фолликула равнялся 15 мм, а яйцевод был очень
сильно развит. Гнездо с 4 значительно насиженными яйцами (эмбрионы
длиной около 2 см) было осмотрено в долине р. Камюстяха 1 июля 1958 г.
Оно было расположено на кочке, среди редких кустиков багульника и
карликовой березки, на сухом, поросшем редкими лиственницами, кочкар-
никовом болоте. Около гнезда беспокоились обе старые птицы. Метрах
в 300 от этого места наблюдался третий кроншнеп, сидевший на сухой
вершине лиственницы и тревожно кричавший. Видимо, поблизости было
второе гнездо.
2 августа 1958 г. в устье Талой мы встретили кроншнепов, беспокоив-
шихся у выводка. 27 июля и 6 августа на сухом болоте в верховьях
Могокты наблюдалась беспокоившаяся птица, оказавшаяся самцом. Судя
по его поведению, птенцы еще не летали. Хорошо летавший молодой крон
шнеп попался на болотистом берегу оз. Глубокого 7 августа 1958 г.
— 265 —
60. Малый веретенник — Limosa lapponica lapponica (L.). Малый вере-
тенник найден на гнездовье в тундре от устья Дудыпты на юге до верхнего
течения Пуры на севере. На этой территории он гнездится только отдель-
ными очагами, месторасположение которых меняется по годам. В ни-
зовьях Пясины и Пуры летом 1961 г. веретенники нам ни разу не попада-
лись.
На побережье Карского моря А. А. Бируля (1907) встречал только
стаи кочующих птиц, а работавший в устье Пясины И. И. Колюшев (1933)
вообще не упоминает этого кулика. Таким образом, малый веретенник на
западном Таймыре заходит только в самую южную часть арктической
тундры. Что касается гнездования его в тайге и лесотундре, то за все
время полевых работ в верховьях Рыбной нами только однажды, в сере-
дине июня 1959 г., наблюдалась пара, державшаяся на одном месте не-
сколько дней, но, к сожалению, из нее был добыт только самец.
Весной 1959 г. в истоках Рыбной веретенники наблюдались 12 июня;
в тундре близ устья Дудыпты в 1962 г. они были отмечены 4 июня. Север-
нее, у Пуринских озер, первая пара в 1961 г. была встречена 7 июня.
Токование самцов в устье Дудыпты отмечено 15—17 июня 1962 г.
Местообитания всех встреченных в гнездовой период малых веретен-
ников очень характерны. Иногда это были сильно увлажненные, а иногда
и более сухие, усеянные крупными осоково-пушицевыми кочками участки
тундры. Такие участки обычно приурочены к руслам ручейков, текущих
на дне довольно обширных распадков.
Пары беспокойно кричавших веретенников, судя по их поведению —
у выводков, наблюдались близ устья Дудыпты 6 июля 1960 г. Сильно
тревожившаяся самка была добыта близ истоков Пуры 6 июля 1963 г.
Как было видно по состоянию яичника, она снесла 4 яйца. Во время четы-
рехкилометрового маршрута по руслу одного из ручьев в верховьях
Пуры 10 июля 1963 г. наблюдались три беспокоившиеся пары. Все птицы
держались в самой низкой, недавно освободившейся от снега части долины.
В желудке веретенника, добытого около устья Дудыпты 6 июля 1960 г.,
была обнаружена целая «коллекция» довольно редких жуков: Carabus
truncaticollis polaris, Pterostichus costatus, Pterostichus sp., P. vermiculosus,
Elaphrus latipennis (Carabidae), Chrysomela tolli (Chrysomelidae), Lepyrus
sp. (Curculionidae).
61. Длиннохвостый поморник — Stercorarius longicaudus longicaudus
Vieill. (рис. 19). Из всех видов поморников, гнездящихся в тундре бас-
сейна р. Пясины, длиннохвостый является самым многочисленным. Бу-
дучи обычной птицей морского побережья (Бируля, 1907), длиннохвостый
поморник на гнездовье доходит к югу вплоть до границы древесной расти-
тельности; в устье Дудыпты он уже довольно многочислен. В годы высокой
численности мышевидных грызунов, например в 1960 г., отдельные пары
длиннохвостого поморника регулярно встречались даже в лесотундре
у северного берега оз. Пясино, где, судя по поведению отдельных пар,
возможно, и гнездились. У Норильских озер только один раз, 6 июня
1959 г., наблюдалась залетная пара.
Длиннохвостые поморники прилетают в тундру позднее других видов
поморников: весной 1962 г. в устье Дудыпты они появились только
16 июня и сразу в большом количестве. В окрестностях Пуринских озер
весной 1961 г. эти поморники также были впервые отмечены 16 июня,
а в 1963 г. — 7 июня. Весной поморники появляются с юга и летят в се-
верном направлении, как это очень четко наблюдалось весной 1961 г.
Тогда птицы летели небольшими группами или отдельными парами, сле-
довавшими друг за другом через короткие промежутки времени. По-ви-
димому, гнездящиеся на западном Таймыре длиннохвостые поморники,
летящие с мест зимовок вдоль морского побережья, поворачивают к югу
— 266 —
вдоль русла Енисея, на котором раньше всего появляются забереги и
полыньи. И только долетев до южной границы тундр, птицы сворачивают
в сторону и распределяются по местам гнездований. В 1961 г. в южных
тундрах наблюдалась депрессия численности мышевидных грызунов.
Это и вызвало массовое передвижение поморников к северу, где коли-
чество леммингов было средним. Вообще, в тех случаях, когда грызунов
в тундре мало, поморники совершают внезапные и довольно далекие пере-
мещения. Так, с 16 по 20 июня 1962 г. в ближайших окрестностях поляр-
ной станции Кресты в устье Дудыпты держались многие сотни поморни-
ков. Однако 21—22 июня они внезапно исчезли, а одна из окольцованных
Рис. 19. Пара длиннохвостых поморников — Stercorarius longicaudus Vieill.
птиц была поймана 22 июня в г. Дудинке на Енисее, более чем за 200 км от
места кольцевания.
Длиннохвостые поморники, обитающие в любых типах тундры, в раз-
ные годы испытывают значительные колебания численности. Если летом
1960 г., в период высокой численности леммингов и полевок, пары помор-
ников встречались в пойме Агапы 4—5 раз на 10 км маршрута и многие из
них гнездились, то в 1961 г. они там почти не наблюдались, хотя были
обыкновенны в низовьях Пуры и Пясины, где количество грызунов было
средним. То же наблюдалось и в 1962 г., когда мышевидных было по-преж-
нему мало. Летом 1963 г., одновременно со вспышкой размножения лем-
мингов и полевок, количество поморников в пойме Агапы было таким же,
как в 1960 г.
Гнездо длиннохвостого поморника было найдено на участке дриадовой
тундры в среднем теченип Пуры 18 июля 1961 г. Единственное сильно
насиженное яйцо лежало на примятых птицей лишайниках безо всякой
подстилки. Обе птицы при нашем приближении вели себя довольно спо-
койно. В первой декаде августа 1961 г. летных молодых поморников в ни-
зовьях Пясины еще не было видно. В условиях ранней весны 1960 г. по-
морники приступили к размножению значительно раньше. 10 августа
1960 г. наблюдались 2 выводка с 2 хорошо летавшими молодыми птицами
— 267
Таблица 5
Пища длиннохвостого поморника
(устье р. Дудыпты и р. Агапы, 1960 г.)
° a
о
Q.
К
R
2 s
Пищевые объекты
V io го
A S ИО ИО
И ®
Обский лемминг
Полевка Миддендорфа
Завязи голубики
Agriotidae . . .
Amara alpina (im.)
Pterostichus {Cryobius) im.
P. costatus (im.)
Carabus (im.)
Dytiscidae (im.). .
Curculionidae (im.)
A raneina
Перезимовавшие ягоды
голубики
Обский лемминг
Ягоды голубики
Ягоды морошки
Водяной жук
в каждом. Выводки поморников встречались ежедневно до двадцатых
чисел августа. В последних числах августа и в начале сентября длинно-
хвостые поморники почти полностью отлетают. Осенних скоплений, по-
добных наблюдавшимся А. А. Бирулей (1907) в заливе Вальтера и столь
характерных для серебристых чаек, нам ни разу наблюдать не слу-
чалось.
Как показала В. И. Осмоловская (1948) для южного Ямала, длинно-
хвостый поморник чаще других поморников использует растительную
пищу и насекомых. Как видно из табл. 5, питание поморников, добытых
нами на западном Таймыре, до-
вольно разнообразно. Однако,
как показали многочисленные
визуальные наблюдения, длин-
нохвостые поморники, при их
высокой численности, являются
основным врагом чечеток, тря-
согузок, подорожников и дру-
гих мелких воробьиных. Осо-
бенно часто поморники пресле-
дуют птиц осенью и весной,
причем одинаково успешно ло-
вят их в воздухе, и на земле.
В годы низкой численности мы-
шевидных, особенно весной, в
питании длиннохвостых помор-
ников значительную роль играет
и падаль. Очень часто помор-
ники держатся около рыбачьих
избушек, где поедают выбро-
шенные отбросы.
62. Короткохвостый помор-
ник — Stercorarius parasiticus
(L.). Короткохвостый поморник
довольно обыкновенен в тундре
севернее устья Дудыпты. В го-
ды вспышек размножения по-
левок, например в 1964 г., от-
наблюдались даже в окрестностях
Июль, 8 желудков
5
1
7
4
2
13
1
3
1
1
1
5
1
1
2
2
5
1
2
1
1
1
2
40
Август, 9 желудков
3
2
5
1
3
7
7
1
дельные птицы время от времени
оз. Кета. Обычно он встречается реже длиннохвостого поморника. Числен-
ность его колеблется в зависимости от количества мышевидных грызунов,
так же как у длиннохвостого поморника.
Прилетают короткохвостые поморники довольно рано. Весной 1962 г.
в устье Дудыпты они появились 6 июня, но в массе наблюдались только
18 июня вместе с длиннохвостыми поморниками. В тундре у Пуринских
озер первый короткохвостый поморник весной 1961 г. замечен 1 июня,
однако в значительном количестве они появились только 19—25 июня.
Весной 1963 г. эти поморники прилетели сразу в большом количестве ве-
чером 6 июня. Наблюдавшиеся стаи летели с северо-запада на юго-восток.
Такое же точно направление пролета этих птиц было отмечено В. В. Лео-
новичем в пойме Агапы весной 1961 г.
Повсюду и чаще всего короткохвостые поморники встречались на
припойменных участках или в равнинной кочкарниковой тундре. Гнездо
с двумя сильно насиженными яйцами было найдено В. В. Леоновичем
близ устья Дудыпты 5 июля 1960 г. Гнезда, содержавшие по 1 ненаси-
женному яйцу, были найдены в пойме Агапы 8 и 10 июля 1961 г. Помор-
ники от гнезд отводили.
268 -
По своему питанию, как указывает ряд авторов для поморников юж-
ного Ямала (Дунаева и Кучерук, 1941; Осмоловская, 1948), короткохво-
стый поморник является типичным хищником. Остатки мышевидных были
встречены в 4 из 6 желудков птиц, обследованных весной 1963 г. Остатки
рыбы были найдены в двух желудках, а хитин жуков Carabus — в трех
желудках. У поморников, добытых в низовьях Агапы летом 1960 г., помимо
остатков грызунов и мелких птиц, в желудках были найдены такие мел-
кие насекомые, как комары Aedes. У них же встречены крупные жуки
Amar a alpina.
63. Средний поморник — Stercorarius pomarinus (Temm.). Область рас-
пространения среднего поморника на западном Таймыре совпадает с аре-
алами гнездования двух других видов поморников, однако всюду он
встречается значительно реже. Изредка залетает и в северотаежные ланд-
шафты близ Норильских озер. Так, 27 июня 1958 г. одиночный поморник
наблюдался в истоках Рыбной. В годы с низкой численностью мышевид-
ных грызунов, например в 1961 и 1962 гг., средние поморники, вероятно,
совсем не гнездились.
Весной 1961 г. средние поморники появились у 2-го Пуринского озера
12 июня. Они держались в небольшом числе около рыбачьей избушки
в течение нескольких дней. Весной 1962 г. в устье Дудыпты, а позднее
и на Пуринских озерах средние поморники ни разу не были встречены.
В 1963 г. средние поморники появились у Пуринских озер 9 июня и затем
ежедневно наблюдались все лето. Судя по состоянию яичника самки, до-
бытой 15 июня, она уже снесла первое яйцо.
В желудках двух особей, добытых весной 1963 г., были найдены только
остатки копытных леммингов.
64— 66. Серебристая чайка — Larus argentatus heuglini Вгее, L. a. ante-
lius Ired., L. a. vegae Palmen. Серебристая чайка очень обыкновенна как
в северной полосе тайги у Норильских озер, так и в открытой тундре
до самого морского побережья (Бируля, 1907). В северной полосе тайги
она довольно многочисленна по берегам и островам крупных Норильских
озер, таких, как Мелкое, Глубокое, Кета и Хантайское. На небольших
лесных озерах она не гнездится. В лесотундре у истоков Пясины и
в тундре серебристые чайки селятся на берегах любых водоемов.
На берегу оз. Кета весной 1958 г. первые особи были замечены 31 мая,
в 1959 г. — 23 мая, а в 1964 г. — 25 мая. В устье Дудыпты весной 1962 г.
серебристые чайки были впервые отмечены 25 мая, однако в большом
количестве они появились только 5 июня. Старые птицы прилетели во
множестве 10 июня. В тундре у Пуринских озер в 1961 г. серебристые
чайки были отмечены 28 мая, а их массовый прилет происходил 1—5 июня.
Молодые чайки наблюдались там только 21 июня. В окрестностях 1-го
Пуринского озера весной 1963 г. передовые чайки были отмечены
31 мая. Очень интенсивный прилет происходил 5 и 6 июня. Все птицы
летели с юга. 7 июня прилет закончился.
В первое время чайки держатся у заберег рек и озер, недалеко от мест
гнездования, а чаще около рыбачьих избушек и других жилых построек,
где они вместе с поморниками собирают различные отбросы.
Чайки приступают к гнездованию только после оттаивания мелких
тундровых водоемов и примыкающих к ним болот. На юге исследованной
нами территории чайки обычно устраивают гнезда на каменистых или
песчаных островках больших озер. В таких местах, например на острове
близ истоков Пясины, гнездятся сразу многие десятки пар. В тундре
они одинаково охотно поселяются как на мелких островках и торчащих
из воды кочках и камнях на небольших озерах (Бируля, 1907), так и на
низинных островах среди больших водоемов. Здесь они образуют иногда
значительные колонии, насчитывающие более 40—50 пар, в то время как
— 269 -
на небольших озерах обычно гнездятся группами по 2—5, редко до
10—15 пар. Известны случаи гнездования серебристых чаек на скалистых
выступах высоких берегов и островов р. Пуры и просто на кочках среди
полузатопленных полигональных болот. В составе их колоний иногда
находятся бургомистры в количестве нескольких пар и полярные крачки.
Гнезда представляют собой кучки из стеблей осок, злаков и пучков
мха с примесью прутиков тальника и карликовой березки и бывают са-
мых различных размеров. Когда они помещаются на кочках среди низин-
ной или даже затопленной тундры, их высота достигает 10—15, а по-
перечник в основном 40—50 см. Гнезда, устроенные на скалах или на
очень сухих моховых буграх, требуют гораздо меньше материала. Очень
часто пары, гнездящиеся в одной колонии в совершенно одинаковых
условиях, используют для постройки гнезд разные строительные мате-
риалы. Так, среди более чем трех десятков гнезд, осмотренных в июле
1963 г. на острове 2-го Пуринского озера, некоторые представляли собой
смесь стеблей осок и злаков, основа других состояла из мха, третьи
были сложены как из мха, так и из осок и злаков и т. п. Лоток обычно
выстилается сухими стебельками и корнями злаков и осок. Многие гнезда
используются из года в год или через несколько лет снова, в зависимо-
сти от наличия или отсутствия корма в окрестностях колонии. В результате
многолетнего накопления погадок и действия помета почва около гнезд
зачастую резко выделяется среди окружающего фона. Среди крупных
колоний, в которых гнезда расположены на расстоянии 2—3 м друг от
друга, земля бывает покрыта хорошо утоптанным зеленым мхом, густо
усеянным погадками. На ней развиваются довольно пышные кусты карли-
ковой березки и даже заросли цветковых растений. Иногда встречаются
участки с хорошим злаковым задернением, но с сильно вытоптанной и
выщипанной травой.
Численность чаек, гнездящихся в колониях, меняется в разные годы.
В 1963 году, сравнительно богатом мышевидными грызунами, на о. Чая-
чьем среди 2-го Пуринского озера было осмотрено более 30 гнезд сереб-
ристых чаек, а в 1962 г., когда леммингов п полевок было очень мало,
чайки там почти не гнездились. Они просто держались в окрестностях
колонии. В богатой озерами пойме Агапы летом 1960 г. небольшие посе-
ления серебристых чаек были отмечены на болотистых островках и кочках
среди многих озер, однако в августе 1961 г., после резкого снижения
численности грызунов, чайки там почти совсем исчезли. Следовательно,
численность серебристых чаек, так же как количество поморников и мох-
ноногих канюков, в значительной мере зависит от наличия леммингов
и полевок.
Из 48 гнезд, осмотренных в 1960—1963 гг., 31 содержало по 3 яйца,
14 — по 2 и 3 — по 1 яйцу. При этом большая часть кладок, содержавших
менее 3 яиц, найдена в 1961 и 1962 годах, более бедных мышевидными
грызунами.
К откладке яиц серебристые чайки приступают начиная с середины
июня и до первых чисел июля, в зависимости от метеорологических усло-
вий и месторасположения гнезд. Чайки, гнездящиеся в поймах, гнездятся
позднее поселяющихся на незатопляемых островах. Вероятно, в поймах
много кладок гибнет при особенно высоких подъемах воды.
В 2 гнездах, осмотренных в верховьях Пясины 27 июня 1960 г., яйца
были уже значительно насижены. У чаек, гнездящихся на заливаемом
полигональном болоте между 1-м и 2-м Пуринскими озерами, 9 июля
1962 г. яйца были очень слабо насижены. Массовое вылупление птенцов
у чаек на о. Чаячьем, на 2-м Пуринском озере, в 1963 г. происходило
14—16 июля. В небольшой колонии, исследованной в среднем течении
Пуры в 1961 г., птенцы выклевывались 18 и 19 июля. В одном гнезде.
— 270 —
однако, были обнаружены совершенно не насиженные яйца. По-види-
мому, это была повторная кладка. В гнезде, осмотренном 31 июля 1964 г.
на небольшом островке среди оз. Кета, яйца находились еще на середине
срока насиженности. Эта кладка несомненно является повторной, тем бо-
лее, что после многоснежной зимы весной 1964 г. наблюдалась массовая
гибель от затопления гнезд целого ряда видов поселившихся на островках
птиц. У серебристых чаек, гнездившихся на скалистом острове в низовьях
Пуры, 24 июля 1961 г. оказались сильно насиженные кладки. Птенцы
в возрасте нескольких дней были пойманы в пойме Агапы 18 июля
1960 г.
Молодые едва летавшие чайки наблюдались на одном из озер близ
устья Агапы 24 августа 1960 г. 2 плохо летавших птенца отмечено в вер-
ховьях Рыбной 13 августа 1959 f. В массе молодые чайки становятся
самостоятельными только в конце августа. В это время они вместе со
старыми откочевывают из глубины тундры на берега наиболее крупных
рек и озер, где образуют значительные скопления у рыбачьих избушек
и у мест постановки сетей. Здесь иногда собираются многие десятки осо-
бей. В таких местах стаи чаек держатся до самого отлета, который про-
исходит одновременно с ледоставом на более крупных реках в конце
сентября и в начале октября. Около поселений человека, где много мяс-
ных и рыбных отбросов, небольшие стайки молодых чаек, по нашим на-
блюдениям, задерживаются до конца октября и впоследствии замерзают.
Судя по добыче в верховьях Лены 28 сентября 1960 г. молодой птицы,
окольцованной нами в августе того же года в низовьях Агапы, серебристые
чайки отлетают на зимовки в юго-восточном направлении.
Анализ содержимого более 30 желудков чаек, добытых в тундре за-
падного Таймыра летом 1960 г., показал почти полное отсутствие в их
пище насекомых. В подавляющем большинстве желудки содержали
остатки леммингов и полевок, а значительно реже — рыб и птиц. Беглый
просмотр громадного количества разбросанных около гнезд погадок
также показал преобладание в пищевом рационе чаек мышевидных гры-
зунов. Остатки рыбы были встречены в меньшем количестве, а остатки
птиц, в том числе уток и куропаток, попадались лишь изредка. Скорее
всего чайки поедают уснувшую рыбу, так же как и расклевывают погиб-
ших или ослабевших птиц, хотя они активно ловят и птенцов; 22 июля
1959 г. мы наблюдали как серебристая чайка пыталась поймать утят синьги.
Очень часто, особенно весной и осенью, серебристые чайки кормятся на
трупах северных оленей, причем около одного павшего животного иногда
их собираются многие десятки. Рыбаки ненавидят серебристых чаек за то,
что они расклевывают рыбу, попавшую в сети на небольшой глубине.
Подавляющее большинство добытых нами серебристых чаек по ок-
раске своей мантии очень похожи на экземпляры А. Я. Тугаринова, со-
бранные им в Енисейском заливе. Все эти чайки относятся к подвиду
L. a. heuglini Вгее. Среди них имеются отдельные особи со значительно
более светлой мантией, по своим признакам уклоняющиеся в сторону
L. a. vegae Palmen., и даже типичные представители этого подвида. Нако-
нец, две чайки, добытые на гнездовье в южной части исследованной тер-
ритории, близ оз. Кета, являются типичными L. a. antelius Ired. и почти
не отличимы от ямальских.
67. Сизая чайка — Larus canus heinei Hom. Сизая чайка изредка гне-
здится у Норильских озер; севернее же, в лесотундре и тундре в бассейне
р. Пясины, она ни разу не была встречена.
Весной 1959 г. близ истоков Рыбной сизые чайки появились 1 июня.
В течение всего лета они изредка наблюдались также на водоразделе
рр. Рыбной и Хантайки. 19 июля 1964 г. близ северного берега оз. Кета
была добыта гнездившаяся где-то поблизости самка. В 1958 г. сизые
271
чайки несколько раз были замечены на рр. Глубокой и Талой. Хорошо
летавшая молодая птица была добыта из выводка на западном берегу
оз. Кета 25 августа 1959 г.
68. Бургомистр — Larus hyperboreus pallidissimus Port. Бургомистр ре-
гулярно гнездится на тундровых реках и озерах и распространен на юг
до низовьев Дудыпты. По численности он в 8—10 раз уступает серебристой
чайке. Судя по наблюдениям А. А. Бирули (1907), в тундре он встречается
чаще, чем на морском побережье. Количество бургомистров, как и сереб-
ристых чаек и поморников, в значительной мере зависит от численности
мышевидных грызунов.
Прилетают бургомистры в третьей декаде мая, раньше большинства
других птиц. В 1962 г. близ устья Дудыпты они появились 21 мая и дер-
жались приблизительно неделю, после чего отлетели к северу. Весной
1961 г. у Пуринских озер эти крупные чайки были отмечены 26 мая,
а в 1963 г. — 24 мая.
После прилета бургомистры бывают довольно обыкновенны до сере-
дины июня, так как в это время они очень охотно посещают окрестности
поселений человека и собирают отбросы. Во второй половине июня они
уже встречаются почти исключительно около гнездовий. Селятся они
обычно совместно с серебристыми чайками как на низменных островах
и даже кочках среди озер, так и на скалах. Три гнезда, осмотренные
в колонии серебристых чаек на о. Чаячьем 15 июля 1963 г., представляли
собой очень массивные, диаметром более 60 см, постройки из мха и корне-
вищ осоки. Только одно гнездо, найденное там же, помещалось на сухой
дерновинной кочке и отличалось небольшими размерами. В общей коло-
нии чаек гнезда бургомистров располагались несколько особняком, вдоль
кромки берега. Гнезда, осмотренные на скалистом острове р. Пуры
24 июля 1961 г., ничем не отличались от расположенных рядом с ними
гнезд серебристых чаек. Яйца бургомистров, обычно в числе 3 и только
изредка 2, по своей окраске не отличимы от яиц серебристых чаек, а по
размерам разнятся весьма незначительно. 4 найденных в 1963 г. яйца
имели размеры: 79x56, 75x53.3, 74x54.5 и 82.3x52.5 мм.
Сроки размножения бургомистров те же, что и у серебристых чаек.
Благодаря такому сходству гнездовых местообитании и одинаковым сро-
кам размножения эти два вида на западном Таймыре иногда образуют
помеси. Такой птицей является экземпляр, добытый нами близ Пурин-
ского озера 4 июня 1963 г. Эта чайка, в общем очень похожая на бурго-
мистра, по окраске первостепенных маховых напоминает серебристую
чайку. Птица была добыта из пары и, судя по хорошо развитым семен-
никам, принимала участие в размножении.
В гнездах, осмотренных в низовьях Пуры 24 июля 1961 г., были сильно
насиженные яйца. В 1963 г. на о. Чаячьем вылупление птенцов происхо-
дило 15 июля. Сильно подросшие птенцы, начавшие надевать перья,
наблюдались на озерах в пойме Агапы 28 июля 1960 г.
В конце августа и в сентябре среди скоплений серебристых чаек бурго-
мистры не наблюдались. Видимо, они исподволь кочуют к морскому по-
бережью.
Погадки у гнезд бургомистров состояли в основном из костей и шерсти
леммингов и полевок. В незначительном количестве встречались кости
и чешуя рыб и оленья шерсть. Ранней весной бургомистры очень часто
едят падаль.
69. Малая чайка — Larus minutus Pall. Залеты малых чаек на иссле-
дованную территорию были отмечены дважды. Один раз одиночная старая
птица наблюдалась в верховьях Пясины в начале августа 1963 г. В по-
следней декаде августа 1964 г. большая стая молодых малых чаек дер-
жалась близ истоков Рыбной в течение нескольких дней.
- 272 —
70. Полярная крачка — Sterna paradisaea Pontopp. Полярная
крачка — обычная гнездящаяся птица как в тайге у Норильских озер,
так и в тундре до морского побережья (Бируля, 1907). Гнездится на бе-
регах и островах больших и малых водоемов, а также на вершинах хол-
мов, расположенных недалеко от воды.
Весной полярные крачки прилетают сравнительно поздно, с появле-
нием первых заберег на озерах и значительных талых луж. В 1958 и
1964 гг. в верховьях Рыбной первые крачки были отмечены 8 июня,
в 1959 г. — 3 июня. В устье Дудыпты весной 1962 г. крачки впервые
наблюдались 10 июня. В окрестностях Пуринских озер они появились
в 1961 г. 21 июня, а в 1963 г. — 8 июня.
В южной части исследованной территории, у Норильских озер, крачки
гнездятся на торфяных буграх и небольших болотистых островках среди
небольших таежных озер, а также на выходах мелкой гальки и песка
у гребней гряд, расположенных недалеко от водоемов. Только на боль-
ших озерах, как Хантайское, Кета, Мелкое и др., крачки селятся на
низких песчаных или галечных островах и косах. В тундре они гнездятся
исключительно на низких берегах и островах водоемов всех типов, а на
возвышенностях мы их гнезд ни разу не находили. Довольно часто крачки
селятся поблизости от колоний серебристых чаек, хотя всегда распола-
гают свои гнезда на противоположном конце островка. Больших ко-
лоний на западном Таймыре они не образуют, таковых не наблюдали
ни А. А. Бируля (1907), ни И. И. Колюшев (1933) на побережье Карского
моря. В наибольших осмотренных нами колониях насчитывалось не более
15—20 пар. Чаще крачки гнездятся мелкими поселениями, по 3—5 пар,
или даже отдельными парами.
Все найденные нами гнезда крачек были крайне примитивны. Если
они помещались на сухих холмах или отмелях, то это были неглубокие
ямки, окруженные очень небольшим валиком из камешков или сухого
растительного мусора. Гнезда, расположенные на торфяных буграх, коч-
ках или просто на мху, также представляли собой сделанные птицами
углубления, обрамленные небольшим количеством сухих стеблей злаков
и осок. 2 гнезда, найденные 15 июля 1963 г. на о. Чаячьем, представляли
собой ямки на широкой полосе растительного мусора, тянувшейся вдоль
берега острова и отложенной во время половодья. В большинстве осмот-
ренных гнезд было 2 яйца и только изредка — 1. В насиживании прини-
мает участие и самец, который ненадолго сменяет улетающую на кор-
межку самку. Крачки очень активно отгоняют от гнезда крупных птиц,
в особенности поморников и чаек. В большинстве случаев крачки на-
падают даже на приближающихся к гнездам людей. Крачки около гнезда,
найденного на западном берегу оз. Кета летом 1959 г., были настолько
агрессивны, что без шапки невозможно было фотографировать их гнездо.
К размножению крачки приступают вскоре после прилета. Погоня
самцов за самками и спаривание наблюдались в верховьях Рыбной
13 июня 1958 г. 2 слегка насиженных яйца были найдены 25 июня. В более
раннюю весну 1959 г. гнездо с несколько насиженными яйцами было
обнаружено у западного берега оз. Кета уже 18 июня, а 5 июля в нем
вылупился первый птенец. Там же в 1964 г. вылупление птенцов крачки
было отмечено только с 10 июля; в 2 других гнездах, осмотренных
в тот же день, яйца были сильно насижены. Гнездо с 2 яйцами, находив-
шимися в последней стадии насиженности, было найдено 18 июля 1964 г.
Такие сдвиги в сроках размножения крачек в 1964 г. легко объясняются
гибелью многих гнезд в половодье, вызвавшей повторное гнездование.
В гнездах, осмотренных в верховьях Пясины 27 июня 1960 г., яйца были
значительно насижены и содержали эмбрионы длиной около 1.5 см.
Кладка из 2 сильно насиженных яиц была найдена в среднем течении
18 Биология птиц
— 273 —
Пуры 19 июля 1961 г. Летом 1963 г. на о. Чаячьем на 2-м Пуринском
озере вылупление птенцов происходило 13—15 июля. Первые два дня
птенцы находятся в гнезде и их обогревает старая птица, впоследствии
они держатся в зарослях осоки на берегу водоема недалеко от гнезда.
У молодых крачек, наблюдавшихся в пойме Агапы 18 июля 1960 г., ма-
ховые выступали из чехлов на 20 —30 мм. 2 августа молодые в этой
колонии поднялись на крыло. Летом 1958 г. на одном из озер близ р. Глу-
бокой хорошо летавшие молодые крачки наблюдались 7 августа. Само-
стоятельно добывать пищу они еще не умели, и родители продолжали
их кормить. После того как молодые приобретают полную самостоятель-
Таблица 6
Пища полярной крачки
(р. Агапа, июль 1960 г., 30 желудков)
ность, во второй декаде августа,
полярные крачки откочевывают
на берега более крупных рек и
озер, где образуются временные
Пищевые объекты
Рыба
Notostraca
Amphipoda
Dytiscidae (1.) .
Dytiscidae (im.)
Trichoptera (im.)
Lepidoptera (im.) .
Cordiluridae (im.)
Muscidae (im.) .
Empididae (im.)
Culicidae (im.) . . .
Chironomidae (im., p.)
11
19
?
20
2
41
1
1
1
1
?
?
9
2
4
3
2
6
1
1
1
1
9
8
массового выплода мошки. Как видно
скопления, иногда из многих
десятков птиц. Отлетают крач-
ки в конце августа. Осенью
1959 г. близ истоков Рыбной
они не встречались уже после
20 августа. Выводок летных мо-
лодых еще наблюдался на оз.
Кета 26 августа 1964 г.
Для полярных крачек очень
характерно питание насекомы-
ми, плавающими на поверхности
воды или неглубоко в ней. Во
время массовых вылетов весня-
нок в первых числах июля
1958 г. большие скопления кра-
чек наблюдались на р. Рыбной
около водопада Орон. Много
крачек было на р. Глубокой
15—19 августа 1958 г., в дни
из табл. 6, в желудках гнездя-
щихся в тундре крачек постоянно встречаются и наземные насекомые,
снесенные ветром в воду.
71. Обыкновенная кукушка — Cuculus canorus canorus L. В северной
полосе тайги у Норильских озер кукушки, как правило, очень обыкно-
венны, а местами даже многочисленны. Но в некоторые годы, например
в 1964 г., северная граница ее ареала временно отступает к югу, тогда
кукушка становится положительно редкой. В лесотундре близ истоков
Пясины она уже не встречается совсем. Типичные местообитания обыкно-
венных кукушек — куртинки смешанного леса на берегах рек и не-
больших лесных озер. Довольно многочисленны они также в высоко-
ствольных лесах, окаймляющих горные склоны южной экспозиции. Во вто-
рой половине июня, в теплые дни, здесь можно было слышать с одного
места кукование 3—4 самцов.
Весной 1958 г. кукушки появились близ истоков Рыбной 8 июня,
в первый по-настоящему теплый день. В более теплом 1959 году они были
отмечены 4 июня, а в исключительно теплую весну 1960 г. — в конце
мая.
Кукование самцов начинается сразу после прилета, обычно в утрен-
ние и вечерние часы. Позже, в двадцатых числах июня, они кукуют
в продолжение круглых суток. В начале июля кукушки кукуют уже
более вяло, а к середине месяца, а в некоторые годы и раньше, кукование
совсем прекращается.
— 274 —
По нашим наблюдениям на западном Таймыре, кукушки обычно под-
кладывают яйца в гнезда пеночек-весничек и таловок. 11 июня 1959 г.
нами было обнаружено яйцо какой-то кукушки в гнезде овсянки-крошки.
Яйцо было раза в полтора крупнее четырех лежавших в этом же гнезде
яиц овсянки и отличалось более холодным, фиолетовым оттенком.
Хорошо летавшие молодые наблюдались в окрестностях оз. Конон
6—7 августа 1959 г. и у р. Глубокой — 10 августа 1958 г., в это время
их еще кормили пеночки, в большинстве случаев — таловки. Кукушо-
нок, добытый 10 августа 1958 г., весил 77 г, его рулевые имели в длину
88 мм.
12 августа 1958 г. мы наблюдали вполне самостоятельную молодую
кукушку. После 20 августа кукушки больше не встречались.
72. Глухая кукушка — Cuculus optatus Gould. Глухая кукушка —
обычная гнездящаяся птица северной полосы тайги. По-видимому, она
лучше приспособлена к неблагоприятным условиям высоких широт, чем
обыкновенная кукушка. В 1964 г., когда количество обыкновенных ку-
кушек в Норильской долине резко снизилось, численность глухой ку-
кушки осталась на прежнем уровне. В типичную лесотундру близ исто-
ков Пясины, она на западном Таймыре не проникает, что не совпадает
с наблюдениями В. И. Капитонова (1958) в низовьях Лены. По числен-
ности глухая кукушка обычно уступает обыкновенной только у р. Но-
рилки, а несколько южнее, близ истоков Рыбной и в бассейне Хантайки,
местами даже превосходит последнюю. Никакой разницы в характере
местообитаний у обоих видов кукушек не было нами замечено.
В северной полосе тайги, близ западного берега оз. Кета, крик глухой
кукушки впервые был отмечен в 1958 г. 11 июня, в 1959 г. — 12 июня,
а в 1964 г. — 13 июня.
Особенно интенсивным было токование глухих кукушек в связи с по-
теплением: в 1958 и 1964 гг. 15 июня и в 1959 г. 13 июня. Их крик слы-
шался преимущественно в ночные часы. 16 июня 1959 г. на лиственнице
у берега р. Рыбной мы наблюдали пару. Почти ежедневно, особенно
во второй половине июня, случалось наблюдать самцов, прогонявших
других кукушек со своих участков. С другой стороны, сами кукушки
зачастую подвергались нападениям мелких воробьиных, в особенности
темнозобых дроздов. Голоса глухих кукушек полностью замолкли
в 1958 г. 10 июля, в 1959 г. — в конце июня, а в 1964 г. — 14 июля.
Летом 1959 г. хорошо летавших, но еще короткохвостых молодых
случалось наблюдать в устье Могокты и блйз оз. Конон 1—7 августа.
В 1958 г. 6 августа покинувший гнездо птенец был добыт в островке
смешанного леса в окрестностях оз. Глубокого. Он весил 68 г и имел
рулевые длиной 44 мм. Во всех случаях молодых кукушек выкармливали
пеночки-таловки.
Вполне самостоятельных молодых случалось видеть на западном бе-
регу оз. Кета 14 августа 1959 г. Одиночные птицы через некоторые про-
межутки времени летели берегом озера в южном направлении. В третьей
декаде августа нам уже не случалось наблюдать глухих кукушек.
73. Белая сова — Nyctea scandiaca (L.). Белая сова в пределах иссле-
дованной территории, по-видимому, нормально не гнездится, хотя и встре-
чается в гнездовое время. Очевидно, ее гнездовой ареал приурочен
к морскому побережью и островам Северного Ледовитого океана (Бируля,
1907; Тугаринов и Бутурлин, 1911; Колюшев, 1933). Проверка много-
численных сообщений рыбаков о гнездовании совы всегда давала отрица-
тельные результаты. В годы низкой численности леммингов и полевок
нам нередко случалось наблюдать белых сов, кочующих по тундре в июле
и августе, в то время как в годы с большим количеством мышевидных,
например в 1960 и 1963 гг., они летом не встречались совсем. Это, по-
— 275 —
18*
видимому, объясняется тем, что в годы с хорошими кормовыми услови-
ями большая часть белых сов гнездится в прибрежных арктических
тундрах, а в менее благоприятные годы они кочуют по всему Таймыру.
Как и куропатки, совы откочевывают на зиму в полосу северной тайги
и в лесотундру. Осенью 1958 г. близ устья Дудыпты нескольких сов
мы наблюдали в середине сентября, а в 1960 и 1961 гг. они там появи-
лись только в середине октября, вслед за многочисленными стаями белых
и тундряных куропаток. Осенью 1957 г. приблизительно в такое же
время появились совы и в приенисейской лесотундре, на 100 км южнее
Дудинки. Весной совы возвращаются в тундру вместе с куропатками,
но часто ненадолго задерживаются в местах, изобилующих перелетной
птицей. Близ западного берега оз. Кета пролетные особи наблюдались
в 1958 г. в конце мая, а в 1959 г. с 19 по 25 мая. Весной 1962 г. близ
устья Дудыпты несколько белых сов держалось с 19 мая по 15 июня.
В тундре у Пуринских озер совы регулярно встречались в 1961 г. 3—5 июня,
а в 1963 г. — 10 мая—10 июня. По нашим наблюдениям, поздней осенью
белые совы питаются почти исключительно куропатками, которых с оди-
наковым успехом ловят как на открытом месте, так и среди кустов ив-
няка. При этом нельзя было не удивиться стремительности полета хищ-
ника, нападавшего на добычу. Иногда, в годы низкой численности лем-
мингов, совы нападают на довольно крупных птиц. 21 июля 1962 г.
в верховьях Пуры мы спугнули сову с гуменника, наполовину съеден-
ного ею. В конце мая 1961 г. у обрывов берега 2-го Пуринского озера
были найдены остатки растерзанного канюка. Судя по свежим следам
на снегу, сова напала на зимняка, спавшего на береговом уступе. В одной
из найденных нами погадок совы был обнаружен целый череп горностая.
74. Ястребиная сова — Surnia ulula ulula (L.) (рис. 20). В пределах
исследованной нами территории ястребиная сова гнездится только на
юге, у Норильских озер, да и то не ежегодно. Весной и летом 1958 и
1959 гг. она не встречалась там совсем. В 1964 г., во время обилия полевок,
ястребиные совы неоднократно наблюдались близ истоков Рыбной в мае
и начале июня. Но особенно много их стало в конце августа и в сентябре,
когда за дневную экскурсию случалось видеть 3—4 совы. Птицы сидели
на сухих вершинах лиственниц и поэтому были хорошо заметны издали.
Возможно, привлеченные обильной пищей совы прикочевали сюда и из
расположенных южнее участков тайги.
Подобная же картина была отмечена и зимой 1957/58 г., когда на
севере таежной зоны и в лесотундре было много мышевидных грызунов.
Ястребиные совы наблюдались тогда во время каждой экскурсии в при-
енисейской лесотундре и в тайге у оз. Хантайского. Ближе к весне,
когда грызуны стали менее доступны, совы нападали на попавшихся
в петли белых и тундряных куропаток и поедали их.
75. Болотная сова — Asio flammeus flammeus (Pontopp.). Болотная
сова изредка гнездится в окрестностях Норильских озер и даже заходит
в тундру на север до устья Агапы. 30 июня 1958 г. она наблюдалась на
тундроподобном болоте в бассейне р. Рыбной. Найденное близ истоков
Рыбной гнездо помещалось на довольно большой кочке среди поросшего
карликовой березкой мохового болота. Это была площадка, умятая среди
мха и лишайников на вершине кочки и хорошо защищенная кустиками
багульника и карликовой березки; рядом с гнездом лежало довольно
много перьев, утерянных линяющей птицей; 8 июля 1964 г. в гнезде
было 7 сильно насиженных яиц. 19 июня 1962 г. мы видели сову, летев-
шую вдоль берега р. Пясины в северном направлении. Выводок, состо-
явший из 4 хорошо летавших молодых и 2 старых птиц, наблюдался
в ивняковых зарослях близ устья Агапы 21 августа 1960 г. Он постоянно
встречался в окрестностях рыбачьих избушек до первых чисел сентября.
— 276 —
На этом же месте наблюдался выводок болотных сов и в конце августа
1961 г., а 5 августа 1963 г. там была отмечена одиночная птица. Таким
образом, болотные совы гнездятся на западном Таймыре ежегодно, не-
зависимо от численности мышевидных грызунов, хотя летом 1964 г.,
во время обилия полевок, их было заметно больше. В отличие от белой
и ястребиной болотная сова деятельна только в сумерки и в ночные часы.
76. Трехпалый дятел — Picoides tridactylus tridactylus (L.). В преде-
лах исследованной территории трехпалый дятел изредка встречается
Рис. 20. Ястребиная сова — Surnia ulula ulula (L.).
в тайге у Норильских озер. Здесь его местообитания приурочены к высо-
коствольному елово-лиственничному лесу у обращенных к югу обрывов
плоскогорья Путорана. 2 мая 1959 г. и 27 мая 1958 г. в таких лесах
у склонов гор Кета неоднократно случалось наблюдать трехпалых дят-
лов, издававших типичную для них весной «дробь». Южнее, в бассейне
р. Хантайки, трехпалые дятлы гнездятся и в смешанных рощах на водо-
разделах. 5 августа 1959 г. близ берега р. Могокты мы наблюдали пару
дятлов, долбивших сухую лиственницу. В третьей декаде августа трех-
палые дятлы стали встречаться гораздо чаще и во всех типах леса. Среди
них наблюдались также молодые. Отдельные птицы на зиму не откочевы-
вают из окрестностей Норильских озер. Один трехпалый дятел был
добыт в окрестностях Норильска в феврале 1958 г.
77. Рогатый жаворонок — Eremophila alpestris flava (Gm.). Рогатый
жаворонок — обычная, хотя и не очень многочисленная птица в тундре
— 277 —
севернее р. Дудыпты. На этой территории они характерны для дриадо-
вых и сухих пятнистых тундр на склонах и вершинах грив и холмов.
У Норильских озер рогатый жаворонок в незначительном количестве
встречается на весеннем и осеннем пролете. В альпийском поясе плоско-
горья Путорана хотя и гнездится, но определенно редок.
Первые пролетные жаворонки в многоснежную весну 1958 г. были
отмечены близ западного берега оз. Кета 2 июня. Слабо выраженный
пролет наблюдался до 12 июня. В следующем, 1959 г. пролет проходил
в более сжатые сроки, 28—30 мая. Стайки держались на снегу у воды
или на галечниковых отмелях, где жаворонки поедали веснянок и бо-
коплавов. В 1964 г. стайки летевших к северу рогатых жаворонков на-
блюдались там 20—31 мая. Столь раннее появление этих птиц, видимо,
было связано с сильной оттепелью, устойчиво державшейся почти всю
вторую половину мая.
В тундре около устья Дудыпты летевшие на север стаи впервые на-
блюдались 4 июня 1962 г. Пролет закончился к концу первой декады
июня. Весной 1961 г. первые жаворонки наблюдались 5 июня, а их мас-
совый прилет и пролет происходил 8 и 9 июня. В более теплом 1963 году
первая птица была там отмечена 28 мая, но только 6—8 июня жаворонки
появились во множестве. Сразу после прилета, во второй декаде июня,
у рогатых жаворонков происходит образование пар и расселение по удоб-
ным для гнездования местам. В хорошую погоду на таких местах в сере-
дине и даже в конце июня ежедневно случалось слышать пение самцов.
12—15 июня 1963 г. на участке сухой тундры близ вершины холма не-
однократно наблюдались пары, ожесточенно преследовавшие одиночек,
залетевших на их гнездовые участки.
Все 5 найденных нами гнезд были расположены на участках дриадово-
кассиопеевой или сухой лишайниково-моховой пятнистой тундр. 3 гнезда
помещались в довольно глубоких ямках на совершенно открытом месте,
а 2 других были устроены сбоку сравнительно больших кочек. Гнезда
представляли собой довольно аккуратные постройки из стеблей и листьев
сухих злаков, осок и Кассиопеи. Одно из гнезд, устроенное на совершенно
открытом месте, было окружено валиком из кусочков мха, благодаря
чему оно полностью сливалось с окружающим фоном. Во всех гнездах
лоток был выстлан растительным пухом пушицы, ивы и других растений,
без малейшей примеси перьев. В гнездах было 2, 2, 4, 4 и 5 яиц или птен-
цов. Размеры яиц в одной из осмотренных мной кладок были следующие:
22.8x16.4, 21.9x16.5, 22.8x16.9, 22.4x16.5 и 23.2x16.9 мм. Окраска
яиц светло-оливковая, с расплывчатыми мелкими крапинками.
В нормально протекавшую весну 1963 г. гнездо с 5 ненасиженными
яйцами было найдено в окрестностях Пуринских озер 21 июня. В 1961 г.
кладки из 2 и 4 незначительно насиженных яиц отмечены 27 июня и
3 июля. Гнездо с птенцами в возрасте 2—3 дней в верховьях Пуры было
осмотрено 11 июня 1961 г. Птенцы в гнезде, найденном 8 июля 1963 г.,
были одеты светло-желтым пухом по краям головы, на спине и плечах;
по всему телу сквозь пух проглядывали пеньки.
В сухой тундре у Пуринских озер много хорошо летавших молодых
рогатых жаворонков наблюдалось 20—25 июля 1963 г. Некоторых из них
еще выкармливали старые птицы. В 1961 г. летные молодые были от-
мечены в низовьях Пясины только в начале августа. Выводок хорошо
летавших молодых жаворонков наблюдался в альпийском поясе плоско-
горья Путорана (около 800 м над ур. м.) 6 августа 1964 г.
В тундре бассейна Пясины рогатые жаворонки отлетают в конце сен-
тября и первой декаде октября. Только небольшие группы запоздавших
птиц несколько задерживаются около жилищ человека. В долине Енисея
их отлет несколько запаздывает, и осенью 1957 г. около пос. Потапово
— 278 —
нам случалось наблюдать жаворонков даже в конце октября. В полосе
северной тайги, у западного берега оз. Кета, стаи жаворонков наблюда-
лись 2—6 октября.
78. Береговая ласточка — Riparia riparia kolymensis But. Береговая
ласточка является очень обычной гнездящейся птицей Норильской до-
лины. В лесотундре близ истоков Пясины она уже не встречается, хотя
в июне 1962 г. залетные одиночки наблюдались несколько раз севернее
устья Дудыпты. Весной 1958 г. близ западного берега оз. Кета берего-
вые ласточки были впервые отмечены 26 июня, в день появления первых
комаров. В 1959 г. они прилетели уже 16 июня, а в 1964 г. —
20 июня.
К гнездованию они приступают сразу после прилета. 29 июня 1958 г.
мы наблюдали ласточек, выкапывавших норки в торфяных буграх на бе-
регу одного из таежных озер, расположенных близ истоков Рыбной.
В круто обрывающихся пластах торфа было около 15 наполовину закон-
ченных норок. При нашем приближении из них вылетело несколько птиц,
но большая часть ласточек летала низко над озером, несмотря на то, что
был час ночи и температура воздуха опустилась ниже нуля. 20 июня
1959 г. в подобных же торфяных буграх близ устья Кета-Ирбо мы осмот-
рели другую небольшую колонию береговых ласточек. Птицы доканчи-
вали строительство гнезд. 21 июля 1958 г. в земляном обрыве берега
р. Рыбной около ее устья была осмотрена более значительная колония.
В ней насчитывалось более трех десятков жилых норок. В большинстве
из них уже появились птенцы, так как ласточки беспрерывно подлетали
с кормом. Вылет птенцов происходит в конце июля или в начале августа.
В тихий вечер 7 августа 1959 г. мы видели массу ласточек, ловивших
насекомых над водной поверхностью оз. Ко нон. Среди добытых птиц
оказались и молодые. После вылета молодых береговые ласточки значи-
тельными стаями держатся близ мест гнездований весь август. В середине
и даже в конце августа, в период выплода мошки, местные скопления
береговых ласточек, летавших над водой, наблюдались на рр. Глубокой
и Рыбной в 1958 и 1959 гг. В это время они иногда встречаются даже
севернее гнездового ареала: 29 августа 1958 г., во время массового появ-
ления мошек, десятки береговых ласточек наблюдались даже близ истоков
Пясины. В первых числах сентября эти птицы исчезают.
79. Городская ласточка — Delichon urbica lagopoda (Pall.). В северной
полосе тайги у Норильских озер ласточки гнездятся отдельными очагами.
Обе найденные нами в природных условиях колонии были расположены
в скалистых каньонах рек — у водопада Орон на р. Рыбной и около
верхнего порога на р. Хантайке. Кроме того, городские ласточки регу-
лярно гнездятся на каменных строениях г. Норильска. 2 июля 1958 г.
в колонии у водопада Орон ласточки только начали строительство гнезд.
В колонии на р. Хантайке, осмотренной 1 августа 1959 г., возле гнезд
носились стаи. Можно предполагать, что в стаях было много молодых,
уже покинувших гнездо.
80. Кукша — Perisoreus infaustus yakutensis But. Кукша (рис. 21)
довольно обыкновенна в тайге у Норильских озер. В лесотундре близ
истоков Пясины она уже не найдена.
Типичные местообитания кукш — высокоствольный елово-лиственнич-
ный лес с примесью белой березы, занимающий значительные площади
у обращенных к югу горных склонов и вдоль берегов рек и озер. Осо-
бенно часто встречались кукши по берегам оз. Кета и на р. Глубокой.
Некоторые особи там, видимо, зимуют, прокармливаясь поблизости от
избушек рыбаков. Мы наблюдали кукш с 21 апреля 1959 г., сразу после
прибытия на место полевых работ, и до середины октября. В ноябре
кукш стало мало.
— 279 —
К гнездованию кукши приступают очень рано. Выводок вполне само-
стоятельных молодых кукш наблюдался в верховьях Рыбной уже
24 июня 1964 г., при этом птенцы были почти не отличимы от старых птиц.
Во второй половине лета кукши обычно держатся одиночно. Местное
население не любит кукш за то, что они регулярно поедают подвешенную
для сушки рыбу, а в период промысла спускают капканы и пасти, по-
ставленные на песцов и соболей.
81. Ворон — Corvus corax corax L. В отрогах плоскогорья Путорана,
особенно в окрестностях Норильских озер, ворон гнездится регулярно.
В период осенних кочевок, в конце сентября и в начале октября 1960
Рис. 21. Кукша — Perisoreus injaustus yakutensis But.
и 1961 гг., выводки воронов наблюдались в тундре несколько севернее
устья Дудыпты, куда они прилетали подбирать отбросы от убитых север-
ных оленей. В северной полосе тайги ворон — одна из немногих регу-
лярно зимующих птиц. Довольно много их ежегодно держалось на окраи-
нах г. Норильска, а во время поездки в декабре 1957 г. на оз. Хантайском
мы ежедневно наблюдали воронов около трупов загрызенных волками
домашних оленей.
Брачные игры одной пары воронов мы наблюдали у южного склона
скалистых Вальковских гор 1 мая 1958 г. В мае мы видели воронов близ
истоков Рыбной, но почти всегда одиночек. 7 мая 1964 г. мы наблюдали
пару воронов в ледниковом каре в верховьях одного из притоков Кета-
Ирбо. Судя по беспокойному поведению птиц, у них здесь было гнездо.
В это время у них, вероятно, уже началось насиживание. В июне и июле
вороны встречались лишь изредка. Молодой ворон в сопровождении
двух старых наблюдался на р. Глубокой 5—18 августа. Все это время
птицы держались на одном месте, не удаляясь более чем на 3—4 км. Два
выводка, состоявшие из 3 и 6 воронов, постоянно держались у западного
берега оз. Кета с начала августа до начала октября 1959 г. Только с се-
редины октября вороны стали встречаться поодиночке.
В ранневесенний период, осенью и зимой вороны питаются почти
исключительно падалью. 28 апреля—4 мая 1959 г. они регулярно поедали
брошенную рыбаками гнилую рыбу. В августе—сентябре выводки обычно
— 280 —
наблюдались у берегов озер Кета и Глубокого, где они также подбирали
мертвую рыбу или отбирали добычу у серебристых чаек. 7 сентября
1958 г. в лесотундре близ истоков Пясины три ворона кормились на
внутренностях убитых нами оленей, отогнав предварительно стаю более
двух десятков чаек.
У старого самца, добытого 30 августа 1959 г. близ истоков Рыбной,
мелкое перо находилось в состоянии интенсивной линьки.
82. Серая ворона — Corvus corone sharpei Oates. Регулярно залетает,
а в отдельные годы, возможно, и гнездится в некоторых пунктах Но-
рильской долины. Близ стационара у истоков Рыбной одиночки наблю-
дались 22 июня 1958 г. и 9 июня 1959 г. Весной 1964 г. здесь наблюдалась
уже пара ворон. Птицы, впервые отмеченные 17 мая, наблюдались почти
ежедневно вплоть до 24 июня, пока одна из них не была подстре-
лена.
83. Черная ворона — Corvus corone qrientalis Eversm. Черная ворона,
не очень редкая в пойме Енисея (Тугаринов и Бутурлин, 1911), в бас-
сейне Пясины наблюдалась только один раз — 21 июня 1962 г., близ
устья Дудыпты. Пара весь день держалась на льду р. Пясины, но к ве-
черу исчезла.
84. Кедровка — Nucifraga caryocatactes macrorhynchos Brehm. Нигде
в пределах исследованной территории кедровка не гнездится. Имеющийся
в нашей коллекции залетный экземпляр привезен сотрудником Научно-
исследовательского института сельского хозяйства Крайнего Севера
М. И. Климантовым из пос. Потапово, расположенного на берегу Енисея
на 100 км южнее Дудинки. Зимой 1959 г. эта птица прилетела к зданию
конторы, во дворе которой и погибла от истощения.
85. Пухляк — Parus montanus borealis Selys. В тайге у Норильских
озер пухляк в небольшом количестве встречается регулярно. В лесо-
тундре у истоков Пясины пухляки нами ни разу не наблюдались. 17 июня
1962 г. парочка залетных пухляков перепархивала в пучках старой про-
волоки и среди прошлогодней травы около полярной станции Кресты
в устье Дудыпты. На следующий день они исчезли.
27 мая 1958 г., во время экскурсии по высокоствольному смешанному
лесу близ южных склонов гор Кета, мы несколько раз встречали неболь-
шие стайки и пары пухляков. Выводки вполне самостоятельных молодых
наблюдались в смешанном лесу на берегу р. Глубокой 16—19 августа
1958 г. Пухляки, по-видимому, в небольшом количестве остаются на
всю зиму в тайге западного Таймыра. Во время экскурсий в окрестно-
стях пос. Потапово на Енисее и близ истоков Хантайки в ноябре—де-
кабре 1957 г. небольшие стайки пухляков встречались почти ежедневно.
86. Светлоголовая гаичка — Parus cinctus cinctus Bodd. Светлоголо-
вая гаичка — довольно обыкновенная гнездящаяся птица в тайге близ
Норильских озер и в бассейне рр. Рыбной и Хантайки, где придержи-
вается как высокоствольного елово-лиственничного леса, в изобилии про-
израстающего вдоль южных склонов гор, так и смешанных лиственнично-
березовых рощ, обычных по берегам рек и озер. В лесотундре близ исто-
ков Пясины светлоголовые гаички, видимо, не гнездятся и встречаются
только во время кочевок во внегнездовое время.
В весенний период они иногда встречаются в совместных стайках
с пухляками. Пары постоянно наблюдались на берегу оз. Кета уже
в середине апреля 1959 г. Однако в это время они встречались почти
исключительно у избушки, где подбирали отбросы. Небольшие стайки
светлоголовых гаичек регулярно наблюдались зимой 1957/58 г. близ
пос. Потапово на Енисее и около истоков Хантайки.
87. Поползень — Sitta europaea asiatica Gould. Поползни, по-видимому,
изредка гнездятся в северной полосе тайги у Норильских озер. Они ре-
— 281 —
гулярно встречались нам в островках смешанного леса близ истоков
Рыбной только в сентябре 1959 и 1964 гг. и в начале октября 1959 г.
Один раз, 27 сентября 1959 г., поползень наблюдался в обществе светло-
головых гаичек. Зимой 1957/58 г. они регулярно встречались в приени-
сейской лесотундре около пос. Потапово.
88. Черноголовый чекан — Saxicola torquata stejnegeri (Рагг.). Черно-
головый чекан в небольшом количестве регулярно гнездится в северной
полосе тайги у Норильских озер и в лесотундре близ истоков Пясины.
Отдельные пары гнездятся и в тундре, проникая на север до широты Пу-
ринских озер. В тайге и лесотундре черноголовый чекан придерживается
участков угнетенного лиственничного леса с густой порослью багульника,
карликовой березки и голубики. Именно в таком месте 3 июля 1958 г.
было найдено гнездо близ водопада Орон. Оно было устроено у основания
большой кочки, заросшей осокой, багульником и карликовой березкой.
Стенки и основание гнезда сплетены из сухих стебельков злаков и осок,
а лоток выстлан перьями. В гнезде лежали 4 слабо насиженных яйца.
89. Каменка — Oenanthe oenanthe oenanthoides (Vig.). ;B пределах ис-
следованной территории каменка гнездится повсеместно. Не была встре-
чена она только в высокогорных ландшафтах плоскогорья Путорана.
В северной полосе тайги у Норильских озер она характерна для изо-
билующих выходами валунов моренных гряд, а также для берегов рек
и крупных озер. В лесотундре и тундре бассейна р. Пясины каменка
более многочисленна и придерживается крутых, обрывистых берегов или
сильно пересеченных участков водораздельной тундры. Очень часто она
селится в непосредственной близости от избушек рыбаков. Численность
каменки в нижнем течении Агапы довольна велика: на 1 км береговой
линии в июле в среднем встречалось 2 выводка.
Весной каменки появляются довольно рано. В 1958 г. близ истоков
Рыбной их пролет происходил 9 и 10 июня, а в 1959 г. с 30 мая по 5 июня.
Весной 1964 г. первая пара там была отмечена 5 июня. Как и другие
мелкие птицы, каменки в это время кормились многочисленными веснян-
ками на кромке речного льда. Весной 1962 г. в тундру близ устья Дудыпты
каменки прилетели 5 июня. В окрестностях Пуринских озер в 1961 и
1964 гг. каменки появились в один и тот же день — 7 июня.
На западном Таймыре каменки гнездятся в щелях и трещинах в земле
на береговых обрывах, между камнями и в подходящих нишах в забро-
шенных постройках человека. В бедной подходящими для гнездования
местами тундре мы находили гнезда в нежилых норах леммингов и пес-
цов. 18 июня 1959 г. в моренной гряде на берегу оз. Кета было осмотрено
гнездо с неполной кладкой из 2 яиц. 23 июня птица окончила кладку,
отложив 6-е яйцо. 8 июля в гнезде были уже птенцы в возрасте 2—3 дней.
Гнездо с 5 слегка насиженными яйцами было найдено в тундре близ
устья Агапы 2 июля 1962 г. В гнезде, устроенном в норке копытного
лемминга близ 1-го Пуринского озера, 18 июля 1963 г. был обнаружен
вполне оперившийся и хорошо летавший птенец. Его рулевые выступали
из чехлов приблизительно на 10 мм, а на маховых чехлы сохранились
только в их основании, выступая на 6—8 мм. Неподалеку от гнезда
в зарослях полярных ив размещались другие молодые, уже покинувшие
гнездо. Летом 1961 г. в низовьях Пясины хорошо летавшие птенцы держа-
лись у входа в покинутую песцовую нору, где, как было видно по боль-
шому количеству помета, они вывелись. Массовое появление выводков
на берегу р. Агапы было отмечено 20—21 июня 1960 г.
В некоторых случаях выводки каменок довольно долго держатся в бли-
жайших окрестностях гнезда. В конце августа каменки начинают отле-
тать к югу, временно задерживаясь около поселений человека. В десятых
числах сентября они полностью исчезают.
— 282 —
90. Варакушка — Cyanosylvia svecica svecica (L.). Варакушка является
обычной гнездящейся птицей на всей обследованной нами территории
от северной полосы тайги у Норильских озер до арктических тундр в ни-
зовьях Пясины.
В тайге и лесотундре варакушки гнездятся на лесных опушках и на
поросших кустарником берегах рек и озер. Их основные местопребы-
вания в тундре — поросшие тальником низины и пересеченные берега
водоемов. Там, где особенности рельефа способствуют произрастанию
хотя бы небольших кустиков полярных ив, варакушки гнездятся и в арк-
тических тундрах недалеко от морского побережья. В таких условиях
они поселяются и в непосредственной близости к постройкам человека.
Местами численность варакушек довольно высока — 10 июля 1961 г. на
подходящих участках берега р. Пуры, в 100—120 км от ее истоков, от-
дельные пары встречались через каждые 100 м.
В северной полосе тайги, близ истоков Рыбной, весной 1958 г. ва-
ракушки появились 13 июня, а в 1959 г. — 4 июня. В тундре у устья
Дудыпты в 1962 г. они были впервые отмечены 10 июня. В окрестностях
Пуринских озер сильно затянувшейся весной 1961 г. первая птица
наблюдалась 25 июня, а в нормальных условиях 1963 г. —
8 июня.
В те же дни, когда наблюдались первые прилетевшие особи, случа-
лось слышать и пение самцов. Оно становилось наиболее интенсивным
уже через неделю, когда варакушки приступали к постройке гнезд.
В тундре у Пуринских озер птица со строительным материалом в клюве
была замечена 15 июня 1963 г. Вполне законченное, но еще пустое гнездо
было обнаружено там 20 июня.
Все 14 найденных нами гнезд были устроены однотипно — сбоку более
или менее крупных злаково-осоковых или моховых кочек, также близ
береговых склонов или у ям и распадков. Постройка обычно помещалась
с южной или юго-восточной стороны кочки и была прикрыта нависав-
шими сверху пучками прошлогодней травы или кустиком карликовой
березки. Гнездо обычно было полузакрыто сверху и свито из сухих сте-
бельков злаков и осок. Поперечник лотка 60—65 мм, глубина
45—50 мм. Его выстилка всегда состояла из тонких стеблей и
корешков злаков и никогда не содержала птичьих перьев или
оленьих волос. В 9 из 12 гнезд с полными кладками, осмотренных в раз-
ные годы, было 5, в двух — 4 и в одном — 6 яиц или птенцов. Насижи-
вает только самка, самец носит ей корм. Первое яйцо в гнезде, найден-
ном у западного берега оз. Кета в 1958 г., было отложено 19 июня;
22 июня самка снесла последнее, 4-е, яйцо и приступила к насиживанию.
Полная, несколько насиженная кладка из 5 яиц была отмечена 2 июля
1958 г. В следующем, 1959 г. 2 гнезда со свежеотложенными кладками
из 5 и 6 яиц были найдены на берегу р. Кета-Ирбо 20 июня. В 1964 г.
гнездо с ненасиженной кладкой из 5 яиц было обнаружено там 27 июня.
В тундре у Пуринских озер слегка насиженная кладка осмотрена 10 июля
1961 г. Летом 1962 г. 2 наполовину насиженные кладки были обнару-
жены там 7 и 8 июля. В гнезде, найденном в тундре у Пуринских озер
19 июля 1962 г., было 5 птенцов, у которых маховые начали выступать
из чехлов. Гнездо с 5 птенцами в возрасте 3—4 дней зарегистрировано
у западного берега оз. Кета 10 июля 1959 г. В другом гнезде, осмотрен-
ном нами 13 июля, птенцы уже вполне оперились. В третьем гнезде,
разысканном там же 14 июля, часть птенцов уже вылетела. Выводки
хорошо летавших молодых варакушек ежедневно наблюдались у Пурин-
ских озер после 25 июля 1963 г. и в северной полосе тайги у рр. Глубо-
кой и Хантайки — в конце июля и первых числах августа 1958, 1959
и 1964 гг. К началу сентября почти все варакушки отлетают.
— 283 —
91. Дрозд-белобровик — Turdus iliacus iliacus L. Белобровик изредка
гнездится в северной полосе тайги у Норильских озер, а еще реже —
в лесотундре близ истоков Рыбной.
Весной 1958 г. близ истоков Рыбной белобровики появились 8 июня,
в 1964 г. они наблюдались там 6 июня. Все лето белобровики изредка
встречались в высокоствольном смешанном лесу вдоль склонов гор Кета
и у берегов Рыбной и Хантайки. Гнездо с 4 немного насиженными яйцами
было найдено под Норильском 12 июня 1960 г. В другом гнезде, осмот-
ренном там 17 июня 1960 г., было 5 только что вылупившихся птенцов.
6 сильно насиженных яиц было обнаружено в третьем гнезде, найденном
в низовьях Норилки 18 июня 1960 г. Наконец, в четвертом гнезде, осмот-
ренном в этот же день, было 4 сильно насиженных яйца и 2 новорожден-
ных птенца. Естественно, что в годы с более нормальным ходом весны
сроки размножения белобровиков бывают не такими ранними. Осенью
белобровики исчезали одновременно с темнозобыми дроздами в конце
сентября.
92. Дрозд-рябинник — Turdus pilaris zarudnyi Loud. Рябинник, как и
белобровик, гнездится в тайге у Норильских озер и в лесотундре близ
истоков Пясины. По численности он несколько превосходит белобровика,
но все же встречается сравнительно редко. Его местообитания — опушки
небольших куртин смешанного леса, особенно близ склонов холмов, бо-
гатых зарослями голубики.
Три гнезда, содержавшие по 6 свежих или слегка насиженных яиц,
были найдены в устье Норилки 17 июня 1960 г.
Выводок покинувших гнездо молодых рябинников летом 1958 г.
впервые наблюдался нами 23 июля близ устья Рыбной. Весь август 1959 г.
эти дрозды держались на богатой ягодами моренной гряде у западного
берега оз. Кета. В последний раз выводок рябинников был отмечен
29 августа 1958 г. около истоков Пясины.
93, 94. Темнозобый дрозд — Turdus naumanni eunomus Temm. и
T. п. naumanni Temm. Темнозобый дрозд гнездится до северной гра-
ницы древесной растительности и в этих пределах довольно многочислен.
В плоскогорье Путорана темнозобый дрозд поднимается до самой гра-
ницы древесной растительности (400—600 м над ур. м.), где еще очень
обыкновенен. В кустарниковой тундре он уже не встречается, хотя за-
леты одиночных птиц и зарегистрированы осенью 1960 и 1961 гг. близ
устья Агапы. Гнездится темнозобый дрозд во всех типах лиственничного
и смешанного леса.
Весной 1958 г. близ истоков Рыбной эти дрозды впервые были отме-
чены 4 июня, а в 1959 г. первая птица наблюдалась уже 24 мая, когда
лиственничная тайга имела еще зимний облик. В 1964 г. первые дрозды
прилетели 31 мая, 8—10 июня 1958 и 1964 гг. происходил довольно ин-
тенсивный пролет небольших стаек и одиночек в северном направлении.
Птицы придерживались хорошего леса на берегу р. Рыбной.
С первого дня появления ежедневно слышалось пение самцов, сидев-
ших на вершинах деревьев. Особенно интенсивно пели дрозды в ночные
часы, несмотря на мороз. Летом 1958 г. дрозды пели до последних чисел
июля. 4 июня 1959 г. впервые замечены пары, беспокоившиеся на за-
нятых участках. С этого времени в рощах смешанного леса на берегах
озер и р. Рыбной пары попадались через каждые 200—300 м маршрута.
Гнезда дрозды устраивали на лиственницах или елях, у ствола, на вы-
соте 1—7 м.
Некоторые были расположены на пнях у самой земли. Обычно
гнезда были аккуратно свиты из тонких прутиков и сухих стебельков
злаков и осок, обмазанных грязью.
— 284 —
Количество яиц в 7 гнездах, осмотренных в верховьях Рыбной летом
1958 и 1959 гг., было 4—5. В более теплом 1960 году нх обычно было 5—6.
Яйца зеленовато-голубые, с мелкими бурыми крапинками.
Весной 1959 г. блпз западного берега оз. Кета первое гнездо с не
оконченной еще кладкой было найдено 16 июня; последнее, 5-е, яйцо
было отложено 18 июня, и с этого дня началось насиживание. В другом
гнезде, обнаруженном там же 24 июня, оказалась неполная кладка
из 3 яиц; 26 июня было снесено 4-е яйцо, и птица начала насиживать.
Весной 1960 г. первые гнезда, с неоконченными кладками из 2 и 3 яиц,
были осмотрены под Норильском 12 июня. В 1964 г. полностью закоп-
ченное гнездо, в котором кладка еще не была начата, найдено 18 июня.
Гнезда с насиживающими птицами были тогда отмечены 21—26 июня и
даже 10 июля. 2 гнезда, содержавшие птенцов двухдневного и четырех-
дневного возраста, были найдены в окрестностях водопада Орон 3 июля
1958 г. Летом 1960 г. первое гнездо с недавно вылупившимися птенцами
отмечено 22 июня.
Первый покинувший гнездо птенец был пойман 12 июля 1959 г. Он
уже мог пролететь большое расстояние, но на основаниях маховых еще
сохранились чехлы длиной 15 мм. С 13 июля 1959 г. выводки дроздов
стали встречаться ежедневно. Особенно много дроздов, покинувших
гнезда, наблюдалось близ западного берега оз. Кета 21 июля. Летом
1958 и 1964 гг. в окрестностях оз. Кета массовое появление выводков
темнозобого дрозда также было отмечено в начале третьей декады июля.
Интересны массовые перекочевки выводков темнозобых дроздов в субаль-
пийский пояс плоскогорья Путорана, наблюдавшиеся в конце июля—
начале августа 1964 г. Выводки дроздов встречались даже на высоте
свыше 700 м над ур. м. в жалких кустиках полярных ив на склонах
южной экспозиции, а несколько ниже, у границы древесной раститель-
ности, их было огромное количество. Скорее всего такое вертикальное
перемещение дроздов было вызвано массовым выплодом насекомых, ко-
торый в субальпийском и альпийском поясах значительно запаздывает.
После первого заморозка, случившегося 11 августа 1964 г., все дрозды
спустились в долину оз. Кета.
В августе выводки переходят на питание ягодами и соответственно
держатся в лиственничном редколесье с подлеском из голубики, особенно
на склонах холмов, а также по берегам рек и больших озер. В подобных
местах выводки и небольшие стайки дроздов ежедневно наблюдались
в августе и сентябре 1958—1959 гг., на берегах озер Кета, Глубокого
и близ истоков Пясины. В начале сентября 1959 г. было особенно отчет-
ливо заметно объединение выводков в стаи. В третьей декаде сентября
дроздов близ истоков Рыбной стало заметно меньше, а 1 октября 1959 г.
наблюдалась последняя одиночная птица.
Для определения подвидов в нашем распоряжении имелась серия
в 20 экземпляров добытых в окрестностях оз. Кета темнозобых дроздов,
в том числе 18 взрослых птиц. Подавляющее большинство из них отно-
сится к подвиду Т, п. eunomus Temm. Однако при сравнении этих дроздов
с сериями, хранящимися в коллекциях Зоологического института,
Т. п. naumanni Temm. из Якутии, становится очевидным наличие у не-
которых из них целого ряда промежуточных между этими двумя подви-
дами признаков. Два или три из наших экземпляров благодаря сильному
развитию рыжих тонов на боках тела настолько приближаются к форме
Т. п, naumanni, что могут быть даже причислены к последней.
95. Пеночка-весничка — Phylloscopus trochilus septentionalts (Brehm).
Весничка довольно обыкновенна в северной полосе тайги у Норильских
озер, многочисленна в лесотундре близ истоков Пясины и изредка встре-
чается в тундре вплоть до окрестностей Пуринских озер, где на блюда -
- 285 -
лась в начале августа 1960 г. и в конце июня 1961 г. Численность весни-
чек в некоторые годы резко падает. Например, летом 1964 г., даже в се-
веротаежных ландшафтах у оз. Кета она встречалась лишь изредка.
В тайге и лесотундре веснички держатся в зарослях кустарников
на лесных опушках, а в тундре — среди ивняков в логах и приозерных
низинах, особенно близ речных пойм.
Прилетает весничка раньше других пеночек. Весной 1958 г. близ
истоков Рыбной она появилась 6 июня, в 1959 г. — 30 мая, а в 1964 г. —
10 июня.
Пение весничек можно было слышать с самого дня их прилета. К се-
редине июля оно почти прекращалось, но в августе возобновлялось и
изредка слышалось до самого отлета.
2 июля 1958 г. в редком лиственничном лесу с примесью белой березы
в окрестностях водопада Орон было найдено гнездо. Оно было устроено
сбоку моховой кочки, среди кустиков карликовой березки и имело ти-
пичный для пеночек облик, со входом сбоку. Лоток был обильно выложен
зимними перьями белых куропаток. В гнезде было 4 ненасиженных яйца.
Выводки покинувших гнездо молодых весничек были отмечены на водо-
разделе рр. Рыбной и Хантайки 25 июля 1959 г. Отлетают веснички
в первой декаде сентября.
96. Пеночка-теньковка — Phylloscopus collybita tristis Blyth. Распро-
странение теньковки на западном Таймыре обычно полностью совпадает
с распространением веснички. По устному сообщению В. В. Леоновича,
изолированный очаг гнездования теньковок летом 1961 г. был обнаружен
им в низовьях Агапы. В некоторые годы, например в 1964 г., теньковка,
наоборот, не гнездится даже в окрестностях Норильских озер. В отли-
чие от веснички теньковка чаще встречается в островках высокоствольного
смешанного леса со значительной примесью белой березы. По численности
она в несколько раз уступает гораздо более обыкновенным весничке и
таловке.
Весной теньковка прилетает одновременно с весничкой. В 1959 г.
в тайге близ истоков Рыбной она впервые была отмечена 30 мая. С этого
момента ее пение было слышно в течение всего июня и до середины июля,
а отдельные самцы возобновили его в конце августа. В начале сентября
теньковки исчезли.
97. Пеночка-таловка — Phylloscopus borealis talovka (Port.). В северной
полосе тайги у Норильских озер таловка многочисленнее веснички. В ле-
сотундре близ истоков Пясины она встречается уже гораздо реже, а
в тундру не проникает совсем. В плоскогорье Путорана, у границы
древесной растительности, таловки также не наблюдались. Придержи-
ваются таловки всех типов смешанного леса с подлеском из ив, ольхи
и карликовой березки. Не избегают и зарослей кустов по берегам
рек.
Прилетают таловки поздно. Весной 1958 г. они появились в верховьях
Рыбной 22 июня, в 1959 г. — 18 июня, а в 1964 г. — 20 июня. Интенсив-
ное пение самцов слышалось ежедневно с прилета до последних чисел
июля. Некоторые самцы пели и до самого отлета в августе.
Гнездо таловки было найдено 19 июля 1959 г. на склоне моренной
гряды близ берега р. Рыбной. Оно помещалось в углублении под моховой
кочкой и содержало 5 птенцов в возрасте 5—6 дней. Выводки таловок
в массе появились в последних числах августа. Летом 1959 г. близ за-
падного берега оз. Кета эти пеночки были особенно многочисленны 12—
19 августа. Исчезли они в первых числах сентября. Осенью 1959 г. по-
следняя таловка была отмечена 4 сентября.
98. Пеночка-зарничка — Phylloscopus inornatus inornatus (Blyth).
В тайге у Норильских озер и бассейнов рр. Рыбной и Хантайки зарничка
— 286 -
гнездится регулярно. В лесотундре близ истоков Пясины ее уже нет.
Чаще всего зарничка встречается в смешанном лесу разного возраста
с преобладанием белой березы. По численности она значительно усту-
пает весничке и таловке, но в некоторых подходящих местообитаниях
довольно обыкновенна.
Весной 1959 г. близ истоков Рыбной зарнички прилетели 10 июня,
а в 1964 г. — 15 июня. Пение самцов можно было слышать с первого
дня их появления, но особенно оживленно они пели в конце июня. Самка
с готовым к сносу яйцом была добыта под Норильском 16 июня 1960 г.
Выводок хорошо летавших молодых птичек наблюдался близ оз. Конон
19 июля 1959 г. Осенью зарнички отлетают незаметно. В 1959 г. они
в последний раз наблюдались 19 августа.
99. Малый мухолов — Siphia parva albicilla (Pall.). В пределах ис-
следованной территории северная граница распространения малого му-
холова проходит в окрестностях оа. Кета и истоков Рыбной. Здесь из-
редка встречались отдельные пары. Южнее, на водоразделе рр. Хантайки
и Рыбной, малый мухолов гнездится более регулярно, избирая светлый
и сухой смешанный лес среднего возраста. В подобном лесу неподалеку
от оз. Конон 25 июля 1959 г. наблюдался выводок вполне самостоятель-
ных молодых мухоловов.
100. Сибирская завирушка — Prunella montanella montanella (Pall.).
Сибирская завирушка найдена на гнездовье под Норильском и в лесо-
тундре у истоков Пясины. В окрестностях оз. Кета, Мелкого, Глубокого
и по рр. Хантайке, Рыбной, Глубокой и Талой летом 1958 и 1959 гг.
сибирские завирушки отмечены не были. Осенью 1964 г. они там, однако,
появились. Близ оз. Кета мы наблюдали тогда завирушек как у границы
древесной растительности в плоскогорье Путорана, так и в долинных
смешанных лесах.
Весной 1960 г. пение самцов слышалось ежедневно 7—30 июня.
В. В. Леоновичем под Норильском 12 июня было найдено гнездо, устро-
енное на ели, на высоте около 1 м от земли. Основной строительный ма-
териал — мох, снаружи гнездо было укреплено сухими травинками и
мелкими прутиками. Яиц еще не было. Второе гнездо, с неоконченной
кладкой из 4 яиц, осмотренное в тот же день, было устроено на березовом
кусте, на высоте 40 см, и содержало неполную кладку из 4 яиц; 13 июня
самка снесла 5-е яйцо и начала насиживание.
101, 102. Белая трясогузка — Мо tact Ila alba ocularis Swinh. и
M. a. dukhunensis Sykes. Белая трясогузка гнездится как у Норильских
озер, так и в лесотундре близ верховьев Пясины. В тундре она довольно
многочисленна до истоков Пуры, севернее обыкновенна только близ ры-
бачьих избушек и других жилых строений. По характеру местообитаний
белая трясогузка имеет много общего с каменкой, хотя более привязана
к берегам водоемов и к жилищам человека. Почти у каждой расположен-
ной в тундре рыбачьей избушки гнездится 2—3 пары.
Прилетают весной белые трясогузки в конце мая или в первых числах
июня. Близ истоков р. Рыбной первые особи были замечены в 1958 г.
31 мая, в 1959 г. — 28 мая, а в 1964 г. — 21 мая. Массовый прилет и
пролет происходил соответственно 6—10, 1—5 июня и 25 мая—4 июня.
Птички стайками и в одиночку летели вдоль р. Рыбной, присаживаясь
на снег для ловли веснянок. Около полярной станции Кресты, в устье Ду-
дыпты, весной 1962 г. первые белые трясогузки были отмечены 4 июня.
В тундре у Пуринских озер в 1961 г. они появились 12 июня, а в 1963 г. —
5 июня. Довольно интенсивный прилет и пролет наблюдался там весной
1963 г. 7 и 8 июня, когда отдельные пары или одиночки через небольшие
промежутки времени летели в северном направлении, вдоль узкого пе-
решейка между двумя тундровыми озерами.
— 287 —
Сразу же после прилета пары белых трясогузок встречаются близ
мест своих будущих гнездований — у обрывистых берегов рек и озер,
в местах нагромождения плавника или камней и в поселках.Трясогузки,
гнездящиеся в щелях береговых обрывов, часто селятся в непосредствен-
ной близости от гнезд мохноногих канюков. Пары, гнездящиеся у из-
бушек, устраивают свои гнезда либо в щелях и нишах строений, либо
среди пустых бочек и разного другого хлама в непосредственной бли-
зости от них. Одно из гнезд, найденное нами весной 1961 г. близ избушки
на 2-м Пуринском озере, было расположено прямо на земле рядом с боль-
шой кочкой и хорошо замаскировано нависавшим пучком сухой травы.
Довольно массивное основание гнезда состоит из мха и большого коли-
чества сухих стеблей злаков и осок. Лоток обычно обильно выстилается
оленьим волосом. Из 10 осмотренных нами полных кладок 7 заключали
по 5 яиц, 2 — по 6 и в одной было только 1 яйцо; судя по незначительной
степени насиженности 26 июля, несомненно это была повторная кладка.
В 1963 г. первое гнездо с неполной кладкой из 2 яиц было найдено
в окрестностях Пуринских озер 20 июня. В гнезде, найденном 23 июня,
было 5 уже несколько насиженных яиц; вылупление птенцов в этом
гнезде происходило 30 июня. В третьем гнезде, найденном там же, птенцы
выклюнулись 3 июля 1963 г. В 1962 г. в 2 гнездах выклевывание птен-
цов было отмечено 9 июля. В значительно более холодную весну 1961 г.
2 гнезда с совершенно ненасиженными кладками, содержавшими по 5 яиц,
были осмотрены у 2-го Пуринского озера 4 июля. В одном из гнезд у Пу-
ринских озер 19 июля 1962 г. птенцы уже полностью оперились и их
маховые наполовину освободились от чехлов. Недавно покинувшие гнездо
молодые трясогузки в течение нескольких дней держатся скрытно, и
только 22—26 июля выводки становятся очень заметными. В это время
они уже очень хорошо летают и начинают вести вполне самостоятельный
образ жизни. Ближе к осени, во второй половине августа, трясогузки
становятся особенно многочисленными в окрестностях избушек, а в на-
чале сентября происходит их отлет. В северной тайге у Норильских
озер осенью 1959 г. последние белые трясогузки наблюдались 14 сен-
тября.
Судя по нашим сборам, на исследованной территории обитает два под-
вида белой трясогузки: в тундре бассейна Пясины распространена
М. a. ocularis, а в северотаежных ландшафтах Норильской долины
гнездится М. a. dukhunensis.
103. Горная трясогузка — Motacilla cinerea melanope Pall. Горная
трясогузка изредка гнездится среди скал верхней границы древесной
растительности в плоскогорье Путорана. 3 августа 1964 г. в горах То-
нель на высоте около 500 м над ур. м. наблюдался выводок. Рулевые
у добытого из этого выводка птенца достигали приблизительно половины
нормальной длины. У выводка беспокоились обе старые птицы.
104. Желтоголовая трясогузка — Motacilla citreola citreola Pall.
Желтоголовая трясогузка на гнездовье довольно обыкновенна в тайге
у Норильских озер и в лесотундре близ истоков Пясины. Отдельные
пары регулярно гнездятся и в тундре, проникая на север до широты
Пуринских озер. Наиболее типичные местообитания желтоголовых тря-
согузок — ерниковые сухие болота и заболоченные лесные опушки, осо-
бенно у берегов небольших озер. В таких местообитаниях отдельные
пары летом 1958 и 1959 гг. встречались близ истоков Рыбной через каж-
дые 300 —500 м маршрута.
Весной 1958 г. близ истоков Рыбной первая желтоголовая трясогузка
была отмечена 8 июня, в 1959 г. — 2 июня, а в 1964 г. — 31 мая.
В тундре у Пуринских озер весной 1963 г. гнездившаяся пара появилась
только 21 июня.
— 288 —
Первое гнездо было найдено 3 июля 1958 г. на болотистой гари близ
берега р. Рыбной, недалеко от водопада Орон. Оно помещалось сбоку
кочки под прикрытием куста багульника и было построено из сухих
стеблей злаков и осок; выстилка лотка совсем не содержала перьев.
В лотке лежало 5 беловатых, слабо насиженных яиц. Во втором гнезде,
найденном на болоте, поросшем редкими лиственницами, близ истоков
р. Рыбной, 13 июля 1959 г. было 5 вполне оперившихся птенцов. Летом
1958 г. плохо летавший, но уже покинувший гнездо птенец был пойман
близ устья Рыбной 26 июля. В следующем, 1959 г. выводки хорошо ле-
тавших молодых наблюдались во множестве уже 23 июля. В тундре
у Пуринских озер молодые покинули гнездо также 22—25 июля. Особенно
много летных молодых птиц было отмечено в верховьях Хантайки и
на р. Могокте в конце июля и в начале августа 1959 г. У западного берега
оз. Кета желтоголовые трясогузки полностью исчезли к первым числам
сентября 1959 г.
105. Желтая трясогузка — Motacilla flava plexus Thayer et Bangs.
У Норильских озер желтая трясогузка очень обыкновенна и по числен-
ности не уступает желтоголовой, однако в лесотундре близ истоков Пя-
сины мы ее уже не встречали.
В окрестностях западного берега оз. Кета желтые трясогузки весной
1958 г. прилетели 12 июня, а в 1959 г. — 2 июня. Как и другие виды
трясогузок, они первое время держались на берегах реки, где ловили
веснянок. 2 июля 1958 г. на небольшом болоте в распадке среди леса
близ берега р. Рыбной было найдено гнездо. Оно было устроено сбоку
большой кочки под кустом багульника и содержало слегка насиженную
кладку из 5 сероватых яиц. Второе гнездо было найдено на тундропо-
добном ерниково-лишайниковом участке западного берега оз. Кета 8 июля
1964 г. Оно помещалось с южной стороны большой кочки среди мха и
лишайников. Материалом для его постройки служили стебельки хвощей
и злаков, а также кусочки мха; выстилка лотка состояла из сухой хвои
лиственницы и небольшого количества оленьего волоса. В гнезде содер-
жалось 5 яиц более чем недельной насиженности.
21 июля 1958 г. в устье Рыбной мы наблюдали самку с кормом
в клюве; 27 июля на этом же месте был отмечен выводок хорошо летав-
ших молодых. Много молодых трясогузок наблюдалось в начале августа
1958 г. близ берега оз. Глубокого и в конце июля 1959 г. у р. Могокты.
Отлетают они одновременно с желтоголовыми трясогузками. Осенью
1959 г. у истоков Рыбной последняя трясогузка наблюдалась 31 августа.
106. Краснозобый конек — Anthus cervina rufogularis Brehm. Красно-
зобый конек довольно обыкновенен в лесотундре близ истоков Пясины
и в тундре до среднего течения Пуры. В низовьях Пясины, так же как и
у Норильских озер, краснозобый конек гнездится несколько реже.
Очень обычен он в высокогорных ландшафтах плоскогорья Путорана,
где одинаково часто встречается в горной тундре и лесотундре. В тундре
он селится преимущественно в поросших мелким ивняком логах и на
увлажненных низинах, особенно в богатых кустарниками поймах рек.
В тайге и в лесотундре конек характерен для открытых участков тундро-
подобных болот. В местах его наивысшей численности, в пойме Агапы
и в верховьях Пуры, летом 1960 и 1963 гг. отдельные пары встречались
через каждые 200—300 м маршрута.
Довольно интенсивный весенний пролет наблюдался нами близ запад-
ного берега оз. Кета 13—15 июня 1958 г. Весной 1961 г. в тундре у Пу-
ринских озер коньки появились 10 июня, а в 1963 г. они были там от-
мечены 11 июня.
С первых дней после прилета и даже во время пролета случалось
видеть поющих самцов. Оживленное пение слышалось и в десятых числах
19 Биология птиц
— 289 —
июля. 3 из 5 найденных нами гнезд были устроены сбоку кочек, одно —
прямо в углублении без всякого прикрытия и одно — в ямке у камня.
Материалом служили сухие стебельки осок и злаков, лоточек в двух
случаях состоял исключительно из сухих стебельков и корней, а в осталь-
ных случаях для выстилки гнезда были использованы олений волос,
перья или хвоя лиственниц. 4 измеренных нами яйца имели следующие
размеры: 19.7x15, 19.2x15.4, 20x15.7 и 20.7x15 мм. Сроки гнездова-
ния коньков варьируют в зависимости от месторасположения гнезд.
У самки, добытой у западного берега оз. Кета 21 июня 1964 г., в яйце-
воде было найдено готовое к откладке яйцо. В гнезде, найденном в вер-
ховьях Пуры 11 июля 1963 г. и устроенном на склоне тундрового рас-
падка, недавно освободившемся от снега, было 4 только что отложенных
яйца. В другом гнезде, расположенном на участке возвышенной дерно-
винной тундры, 12 июля 1962 г. было 4 болтуна и 2 птенца в возрасте
5—6 дней. В 2 гнездах, обнаруженных у западного берега оз. Кета 8
и 9 июля 1964 г., было 4 и 5 значительно насиженных яиц. Наконец,
гнездо, осмотренное на участке пятнистой мохово-дриадовой тундры
в среднем течении Пуры, 14 июля 1961 г. содержало 3 немного насижен-
ных яйца. Молодых, покинувших гнездо и ведущих вполне самостоя-
тельный образ жизни, случалось встречать в начале и середине августа.
Осенний пролет краснозобых коньков мы наблюдали в верховьях
Рыбной в 1959 и 1964 гг. в последнюю декаду августа. В это время еже-
дневно встречались стайки по 10—20 птиц, часто вместе с желтыми и
желтоголовыми трясогузками. К началу сентября все коньки исчезли.
В желудках коньков, добытых в начале июня 1960 г. под Нориль-
ском, во множестве обнаружены Lithobiidae, личинки листоедов и жуки
Aphodius sp. В желудках птиц, убитых в тундре у р. Агапы, встречены
самые разнообразные насекомые различных экологических групп: жуже-
лицы (Pterostichus, Bembidion, Elaphrus), листоеды, личинки мясных мух
(Protophonnia terrae-novae), мелкие перепончатокрылые и водяные клещи.
Преобладали насекомые нижней части травостоя и поверхности почвы.
107. Горный конек — Anthus spinoletta harmsi Zar. Горный конек до-
вольно обычен в плоскогорье Путорана. Здесь он гнездится у верхней
границы древесной растительности и в различных типах горных тундр,
поднимаясь вверх почти до гольцового пояса. Любимые его местообита-
ния — нагромождения каменных глыб у выветрившихся скал. В таких
местах 22 июля 1964 г. мы неоднократно наблюдали беспокоившихся
птиц с кормом в клюве. Выводки хорошо летавших молодых горных
коньков регулярно наблюдались в конце июля и в начале августа 1964 г.
Как старые, так и молодые птицы были исключительно осторожны.
108. Свиристель — Bombycilla garrulus (L.). Под Норильском свири-
стель встречается очень редко. Несколько южнее, в верховьях Рыбной,
он гораздо более обыкновенен, а в бассейне р. Хантайки местами даже
многочислен. В типичной лесотундре у северной оконечности оз. Пясино
и в верховьях Пясины свиристель не был найден.
Весной эти птицы появляются рано. Около избушки стационара близ
истоков Рыбной первая пара наблюдалась в 1959 г. 20 мая, а в 1964 г. —
24 мая. Но ежедневно встречать свиристелей нам случалось только на
экскурсии в бассейне р. Хантайки, в июле—августе 1959 г. Здесь они
придерживались небольших рощ елово-лиственничного леса с примесью
березы по берегам рек. В 1964 г. свиристели встречались близ истоков
Рыбной почти ежедневно. Выводок из 5 птиц впервые отмечен в верховьях
Могокты 28 июля 1959 г. Через несколько дней, 31 июля, в верховьях
Хантайки были добыты из выводка вполне самостоятельные молодые
птицы. В 1964 г. летный выводок свиристелей наблюдался у северного
берега оз. Кета уже 17 июля. Особенно часто стали встречаться выводки
— 290 -
и даже небольшие стайки в первой декаде августа. С середины августа
до конца сентября 1959 г. они регулярно встречались и в верховьях
Рыбной.
У добытых в последних числах августа птиц проходила интенсивная
линька мелкого пера.
Осенью 1959 г. свиристели последний раз наблюдались у истоков
Рыбной 27 сентября.
109. Серый сорокопут — Lanius excubitor sibiricus Bogd. Серый сороко-
пут гнездится в Норильской долине, особенно там, где много участков
горелого леса. Не исключено, что он гнездится и в лесотундре у берегов
оз. Пясино, хотя близ истоков р. Пясины мы серых сорокопутов ни разу
не наблюдали. Южнее, в верховьях Рыбной и в бассейне Хантайки,
серый сорокопут был отмечен как на гнездовье, так и на пролете.
Весной 1964 г. первый сорокопут у оз. Кета был отмечен уже 17 мая.
В июле 1958 г. и в июне 1960 г. одиночные птицы регулярно наблюдались
как близ устья Рыбной, так и в низовьях Норилки. Летом 1964 г. серые
сорокопуты регулярно наблюдались в верховьях Рыбной и у устья Амун-
дакты. Выводок из 5 молодых в сопровождении старых птиц был отмечен
на небольшом островке в устье Талой 1 августа 1958 г., он держался в сме-
шанном мелколесье, выросшем на месте пожара. Добытый молодой сороко-
пут весил 72 г, его хвост имел в длину 108 мм.
Судя по многочисленным остаткам мелких птиц, разбросанным по
островку, сорокопуты держались здесь уже давно, а возможно, и гнез-
дились. Среди остатков пищи удалось определить перья чечеток и ноги
светлоголовых гаичек.
Осенний пролет серых сорокопутов удалось хорошо проследить осенью
1959 и 1964 гг. у западного берега оз. Кета. Здесь вдоль берега озера с се-
вера на юг протянулась довольно высокая моренная гряда, местами по-
росшая редкими лиственницами с густым подлеском из ольхи, ив, багуль-
ника и голубики. Эти кусты осенью буквально кишели чечетками, пеноч-
ками и другими мелкими воробьиными. Здесь старые и молодые серые
сорокопуты ежедневно наблюдались в период с 14 августа по 20 сентября
1959 г. Обычно они неподвижно сидели на вершинах лиственниц, откуда
стремительно бросались вниз за добычей. Нередко в это время сорокопуты
встречались и на берегу р. Рыбной. 20 сентября мы наблюдали сороко-
пута в последний раз, как он ловил чечетку. Ей удалось набрать высоту,
п обе птицы, поднимаясь по спирали, скрылись из виду. В желудках трех
добытых тогда сорокопутов обнаружены остатки двух мелких воробьи-
ных, мышевидного грызуна, жука-плавунца и двух кобылок.
НО. Сибирский жулан — Lanius cristatus cristatus L. В окрестностях
Норильска, близ озер Мелкого, Глубокого, Кета, а также во всем бассейне
р. Рыбной сибирский жулан гнездится очень редко. В лесотундру он не
проникает совсем. Однако южнее оз. Конон, лежащего на водоразделе
рр. Рыбной и Хантайки, отдельные пары и выводки нам встречались ре-
гулярно в июле и августе 1959 г.
Гнездо жулана было найдено нами у края поросшего ерником болота
на опушке смешанного леса близ северного берега оз. Кета. Оно было
устроено на небольшой ели, на высоте 2—2.5 м от земли. Основание гнезда
представляло собой массивную постройку из толстых стеблей злаков
и тонких веточек лиственниц, его лоток был выстлан тонкими стебельками
злаков и осок в смеси с перьями белых куропаток. 22 июля 1964 г. в гнезде
находилось 6 птенцов, маховые у которых выступали из чехлов на 6—8 мм.
Выводки жуланов держались на границе куртин смешанного леса с тундро-
подобными открытыми пространствами. В одном из таких мест в верховьях
Могокты выводок хорошо летавших молодых впервые наблюдался 27 июля
1959 г. С этого дня выводки жуланов встречались ежедневно вплоть
1/а 19 Биология птиц
- 291 —
до 8 августа, когда мы снова вернулись в бассейн р. Рыбной. 10 августа
1958 г. выводок, состоявший из 5 птиц, был встречен в редком листвен^
ничном лесу близ юго-западного берега оз. Глубокого. Во всех случаях
как старые, так и молодые жуланы были очень осторожны.
111. Полевой воробей — Passer montanus montanus (L.). В южной
части исследованной территории полевые воробьи гнездятся регулярно,
хотя на зиму частично отлетают к югу. Нескольких воробьев мы видели
в декабре 1957 г. в пос. Таймыр на Хантайском озере, а в г. Дудинке
они гнездятся в довольно большом количестве. Отлетавших к югу пролет-
пых одиночек несколько раз случалось видеть в начале ноября 1957 г.
в пос. Потапово на Енисее.
Особый интерес представляют случаи проникновения воробьев далеко
в тундру, притом через места, совершенно не населенные человеком.
В конце июня 1960 г. пара воробьев несколько раз наблюдалась около
нашей палатки в верховьях Пясины. В июле этого же года В. В. Леонович
(устное сообщение) нашел воробьев на гнездовье близ устья Тареи. 19 июля
птенцы у них покинули гнездо. В июне 1963 г. мы добыли воробья из пары,
державшейся в течение нескольких дней около избушки на берегу 2-го
Пуринского озера. Еще интереснее случай добычи воробья близ устья
Пясины (Колюшев, 1933). Воробьи, проникшие так далеко на север,
с наступлением зимы несомненно либо ' откочевывают к югу, либо поги-
бают.
112. Юрок—Fringilla montijringilla montifringilla L. В северной
полосе тайги у Норильских озер и в бассейне рр. Хантайки и Рыбной
юрок довольно обыкновенен. По склонам гор Путорана юрки поднимаются
до верхней границы древесной растительности. Севернее, в лесотундре
близ истоков Пясины, они уже не были найдены. Типичные местооби-
тания юрков — сухой смешанный лес в холмистой местности на берегах
рек и озер.
Весной 1959 г. первый юрок наблюдался в верховьях Рыбной уже
1 июня. Массовое появление этих птиц было отмечено только 7—8 июня.
В более суровых условиях 1958 г. юрки прилетели значительно позднее —
19 июня. В 1964 г. первые юрки появились там 4 июня, а их прилет про-
исходил 10—13 июня.
Весь июнь и до середины июля ежедневно случалось слышать их ха-
рактерный крик.
Гнездо с неполной кладкой из 4 яиц было найдено под Норильском
В. В. Леоновичем 12 июня 1960 г. Оно помещалось на лиственнице в 3.5 м
от поверхности земли. Другое гнездо, содержавшее 3 не насиженных яйца,
было осмотрено близ устья Норилки 22 июня 1960 г. Птенец с еще корот-
ким хвостом, покинувший гнездо, был впервые отмечен в верховьях Рыб-
ной 18 июля 1959 г. Летом 1958 г. в устье Талой первый выводок был
встречен 29 июля. Особенно много выводков наблюдалось в бассейне
Могокты и в верховьях Хантайки в конце июля и в начале августа 1959 г.
Юрки держались выводками вплоть до двадцатых чисел августа. Неболь-
шие стайки встречались близ западного берега оз. Кета после 21 августа.
Осенью 1959 г. юрки окончательно исчезли к началу сентября.
113, 114. Чечетки — Acanthis flammea flammea (L.) и A. f. exilipes
(Coues). Чечетка — обычная гнездящаяся птица всюду от тайги у Но-
рильских озер до тундр в низовьях Пясины. В плоскогорье Путорана она
найдена на высоте свыше 800 м над ур. м., где гнездится в распадках
и на склонах южной экспозиции. В таежных ландшафтах численность
чечеток в различные годы меняется очень сильно, что связано, по-видимому,
с урожаями лиственничных шишек. В начале мая 1958 и 1964 гг., после
обильного плодоношения лиственницы, чечеток в Норильской долине
наблюдалось очень много. Однако в 1959 г. на тех же самых местах, за
— 292 —
длительную дневную экскурсию удавалось встретить не больше 1—2 пар
чечеток. В тундровой части ареала колебания численности гнездящихся
там чечеток значительно менее резки.
В годы обилия лиственничных шишек чечетки держатся в северотаеж-
ных ландшафтах в течение всей зимы. В тундре они появляются в разное
время. В 1961 г. в окрестностях 2-го Пуринского озера они были впервые
отмечены 12 июня, а 14—20 июня встречались довольно многочисленные
стаи. Весной 1963 г. чечетки изредка наблюдались с 15 июня, но в большом
количестве они появились только в первой декаде июля, когда основные
заросли ивняка полностью освободились от снега.
В южной части исследованной территории чечетки гнездятся во всех
типах лиственничного и смешанного леса, но чаще всего в небольших
лесных островках, расположенных в богатой озерами местности. В тундре
чечетки чаще всего гнездятся в зарослях кустарников, обычно приурочен-
ных к склонам распадков и приозерным низинам. Гнезда свои они устраи-
вают в самых разнообразных условиях. В северотаежных ландшафтах
мы наблюдали их на лиственницах у самого ствола или на концах густых
ветвей, на торцах обломанных трухлявых пней, в извилинах корявых
березовых стволов, в развилках кустов ольхи и т. п. Одно гнездо, найден-
ное близ западного берега оз. Кета 5 июля 1964 г., помещалось в полу-
дупле обломка лиственничного ствола. В тундре чечетки располагают
свои гнезда в ветвях полярных ив или ольхи. В одном случае гнездо
было расположено просто на кочке под прикрытием нависавшей сверху
ветки тальника. Поскольку высота растущих в тундре кустарников
редко превышает 1 м, то и гнезда располагались низко. В северотаежных
ландшафтах, несмотря на наличие высоких деревьев, птицы также лишь
изредка гнездятся на высоте более 3—4 м.
Гнезда сплетаются из сухих стебельков злаков, снаружи иногда обли-
цовываются кусочками лишайников и бересты. Лотки всех осмотренных
гнезд были обильно выстланы растительным пухом и перьями куропаток.
Количество яиц в осмотренных нами кладках колебалось от 2 до 5,
чаще их было 4.
Чечетки, гнездящиеся в северотаежных ландшафтах, приступают
к гнездованию рано. Даже в довольно холодную весну 1958 г. гнездо
со слегка наспженпой кладкой было найдено близ западного берега оз. Кета
уже 16 июня. Вылупление птенцов в нем происходило 24 или 25 июня.
В гнезде, осмотренном там же в 1959 г., вылупление птенцов происходило
26 июня, а 8 или 9 июля они уже вылетели. В условиях сильно затянув-
шейся весны 1964 г. вылупление птенцов в большинстве найденных гнезд
происходило в начале июля. 12 июля 1959 г. в верховьях Рыбной был
отмечен выводок подросших молодых чечеток с рулевыми достаточно
полной длины. Особенно много выводков наблюдалось в середине июля
1959 г. и 21—29 июля 1958 г.
Чечетки, гнездящиеся в тундре, приступают к гнездованию значительно
позднее, часто во второй декаде или даже в конце июля. Такие же сроки
приводит С. П. Наумов (1931) и для тундры Гыданского полуострова.
Поздние находки кладок в 1960 г., в первый год нашей работы в тундре,
навели на мысль о наличии вторых выводков. Но в дальнейшем эта мысль
была оставлена, так как за все время полевых работ мы ни разу не видели
летных молодых раньше первых чисел августа. Столь позднее гнездова-
ние, видимо, связано со сроками освобождения от снега и зазеленения
кустарниковых зарослей, в ветвях которых чечетки строят гнезда. Как
показали наблюдения, они обычно не гнездятся в еще не одетых листьями
кустах, очевидно из-за активной хищнической деятельности поморников.
Летом 1960 г. гнездо с 4 ненасиженными яйцами было найдено в пойме
Агапы 26 июля. Вылупление птенцов в другом гнезде, осмотренном
— 293 —
в том же году, происходило 3 августа. 5 гнезд, найденных летом 1963 г.,
в период с 26 по 30 июля, содержали как неоконченные кладки, так и зна-
чительно насиженные яйца и даже птенцов в возрасте нескольких дней»
В гнезде, осмотренном в низовьях Пясины 2 августа 1961 г., было 5 почти
готовых к вылету птенцов.
Осенью 1958 г. стаи чечеток впервые наблюдались близ оз. Глубокого
16 августа, а в 1959 г. у западного берега оз. Кета 22 августа. Особенно
много стай, иногда состоявших из десятков птиц, было там отмечено в конце
августа и в начале сентября. Это безусловно связано с массовой откочев-
кой чечеток с мест гнездований в тундре. Птицы держались в лиственнич-
ном редколесье и в кустах ольхи. С середины сентября чечеток ста-
новится значительно меньше.
Для определения подвидов гнездящихся на западном Таймыре чече-
ток нами собрана серия в 38 экземпляров. Все эти птицы были сравнены
с сериями чечеток из разных частей их ареала, хранящимися в коллекциях
Зоологического института АН СССР. Более чем половина западнотаймыр-
ских чечеток, судя по сильному развитию белого цвета в оперении над-
хвостья, безусловно относится к подвиду A. f. exilipes (Coues). Остальные
экземпляры несколько темнее, чем A. /. exilipes, по общий тон их окраски
все-таки несколько более светлый, чем у типичных A. f. flammea (L.).
Это относится даже к экземплярам var. holboellii, наиболее близким
кА./. flammea (L.). Таким образом, на западном Таймыре преобладают
светлоокрашенные чечетки А. /. exilipes (Coues) и формы, обладающие
признаками, промежуточными между этими двумя подвидами.
Строгой приуроченности подвидов к определенным типам местооби-
таний мы подметить не смогли. Среди экземпляров, добытых у гнезд в ти-
пичной тундре, есть очень близкие кА./. flammea, в то время как в северо-
таежных ландшафтах у Норильских озер нормально гнездятся А. /. exi-
lipes (Coues).
Что касается var. holboellii, то эта большеклювая форма является,
по нашему мнению, экологической расой, приспособившейся к питанию
семенами лиственницы. Во всяком случае в годы обильного плодоношения
лиственницы (1958, 1964 гг.) таких чечеток всегда бывает много, тогда
как в остальные сезоны мы их не встречали совсем.
115. Белокрылый клест — Loxia leucoptera bifasciata (Brehm). Бело-
крылый клест в небольшом количестве регулярно гнездится в Норильской
долине. В годы урожая лиственничных и еловых шишек, например в 1958
и 1964 гг., эти клесты на западном Таймыре были очень многочисленны,
регулярно встречались всю зиму и проникали даже в лесотундру.
Зимой и весной 1958 г. пение самцов мы слышали регулярно с марта
до первых чисел июня. Гнездятся клесты в различные сроки. Иногда они
приступают к размножению уже в марте. Выводок еще короткохвостых
молодых клестов наблюдался в среднем течении р. Фокиной 11 апреля
1958 г. Другой выводок, в котором старые птицы еще кормили молодых,
наблюдался там 12 апреля 1958 г. При этом следует отметить, что средняя
температура марта в том году приближалась к —30° С.
Огромное количество клестов, в том числе и молодых, было встречено
1 мая 1958 г. близ склонов Хараелахских гор, недалеко от Норильска.
Но более нормальным временем начала гнездования белокрылых клестов
на западном Таймыре является май или даже начало июня. В яйцеводе
самки, добытой близ истоков Рыбной 12 мая 1964 г., было найдено
почти готовое к сносу яйцо. 23 мая 1964 г. в редком смешанном лесу близ
истоков Рыбной было найдено гнездо. Оно было устроено на суховершин-
ной лиственнице на высоте 6—7 м от земли, у самого ствола, с западной
его стороны. Внешний диаметр постройки около 15 см, высота 8 см, лото-
чек был около 5.5 см в поперечнике и 4 см глубиной. Основной строитель-
- 294 —
ный материал — злаково-осоковые волокна, перемешанные с древесными
лишайниками, растительным пухом и перьями куропаток. Специальной
выстилки в лотке не было. Снаружи гнездо было облицовано тонкими
веточками лиственниц и полностью сливалось со стволом и основаниями
ближайших сучьев. У гнезда наблюдалась пара клестов, выказывавших
сильное беспокойство. В яйцеводе добытой самки было найдено готовое
к откладке первое яйцо. 1 июня 1958 г. близ западного берега оз. Кета
мы наблюдали самку со строительным материалом в клюве. 24 июня 1964 г.
в верховьях Рыбной наблюдались покинувшие гнездо молодые, которых
еще кормили старые птицы.
Весной и летом 1959 г. в тех же самых местах клесты попадались
лишь изредка.
116. Клест-еловик — Loxia curvirostra ermakt Sushk. Клест-еловик
встречается в северной полосе тайги близ Норильских озер, очень редок,
и то только в годы урожая еловых шишек. Мы наблюдали небольшие
стайки 1 мая 1958 г. в смешанном, высокоствольном лесу в 25 км к северу
от Норильска.
117. Обыкновенная чечевица — Carpodacus erythrtnus erythrtnus (Pall.).
Обыкновенная чечевица — немногочисленная, но регулярно гнездящаяся
птица в окрестностях Норильских озер. Самым северным ее местонахожде-
нием в пределах исследованной нами территории является устье Норилки,
где весной 1960 г. наблюдались поющие самцы, а В. В. Леоновичем было
найдено прошлогоднее гнездо. Особенно много чечевиц мы наблюдали
в смешанном мелколесье на берегу Могокты в конце июля и в начале ав-
густа 1959 г. Весной 1958 г. близ истоков р. Рыбной пение самцов было
впервые отмечено 15 июня, в 1959 г. оно не прекращалось вплоть до де-
сятых чисел августа.
118. Щур — Pinicola enucleator pacata Bangs. В тайге у Норильских
озер щур довольно обыкновенен. В лесотундре близ истоков Пясины
он уже не встречается. В плоскогорье Путорана щуры изредка встречаются
у верхней границы древесной растительности.
Весной 1958 г., во время хорошего урожая лиственничных и еловых
шишек, щуры стали встречаться уже в начале апреля. Они держались
парами или группами по 3—5 особен в островках лиственничного и сме-
шанного леса на берегу р. Фокиной. Особенно много щуров наблюдалось
во время экскурсии 1 мая 1958 г. у склонов гор Хараелах. Многие самцы
оживленно пели.
Летом 1958, 1959 и 1964 гг. щуры регулярно встречались повсюду
в окрестностях оз. Кета и вдоль рр. Хантайки, Рыбной, Талой и Глубокой.
У самки, добытой близ склонов гор Кета 20 июня 1959 г., диаметр наиболь-
шего фолликула равнялся 10 мм, яйцевод был деятельным. В яйцеводе
самки, добытой близ истоков Рыбной 18 июня 1964 г., было обнаружено
готовое к сносу яйцо. Наседное пятно у нее было развито хорошо. 7 июля
1959 г. в верховьях Рыбной мы наблюдали самку, проявлявшую сильное
беспокойство: по-видимому, где-то вблизи находилось ее гнездо. Осенью
1959 г. щуры в последний раз были отмечены 1 сентября.
По нашим наблюдениям, щуры ранней весной питаются семенами
хвойных и особенно набухающими почками лиственницы. Клювы и опере-
ние двух птиц, добытых близ истоков Рыбной 26 мая 1958 г., были вы-
мазаны ягодами в фиолетовый цвет. Видимо, щуры долгое время питались
прошлогодней голубикой, собирая ее на оттаявших склонах холмов.
119. Полярная овсянка — Embertza pallasi polaris (Midd.). Полярная
овсянка изредка гнездится по берегам больших Норильских озер и в суб-
альпийском поясе плоскогорья Путорана. В лесотундре на р. Пясине
нам она ни разу не встречалась. Ее характерные местообитания — откры-
тые ландшафты и лесные опушки, поросшие кустиками ольхи и ивняка
— 295 —
Весной 1959 г. на западном берегу оз. Кета полярная овсянка была
впервые отмечена 10 июня, а в 1964 г. — 7 июня. Всего нами было осмот-
рено 3 гнезда полярных овсянок. Одно из них, устроенное на багульни-
ковом болоте с редкими кустиками ольхи, было найдено у берега оз. Кета
27 июня 1958 г. Оно было расположено на кочке под кустиком карликовой
березки и по виду очень напоминало гнездо овсянки-крошки. В лотке
лежало 5 свежеотложенных яиц, окраска которых также очень напоминала
окраску яиц овсянки-крошки. Второе гнездо было обнаружено на опушке
смешанного леса, на узком перешейке между озерами близ западного берега
оз. Кета. Оно помещалось на поросшей кустиками багульника и карли-
ковой березки моховой площадке, окруженной более высокими кустиками
ольхи. Само гнездо было сплетено из тонких стеблей и листьев злаков,
выстилка лотка состояла из сухой хвои лиственницы. 29 июня 1964 г.
в гнезде была полная кладка из 5 слегка насиженных яиц. Третье гнездо,
найденное на островке близ устья Кета-Ирбо, поросшем кустами ольхи
и ивняка, было устроено на сухой моховой кочке и отличалось от преды-
дущего выстилкой лотка, которая состояла просто из более тонких сте-
бельков злаков; 7 июля 1964 г. в этом гнезде находилось 5 яиц, а 8 июля
там происходило вылупление птенцов.
Насиживала только самка, хотя самец держался рядом с гнездом и при
приближении человека проявлял сильное беспокойство.
4 августа 1964 г. в устье Амундакты наблюдался выводок полярных
овсянок. 12 августа 1964 г. вполне самостоятельные молодые птицы были
встречены в зарослях ольхи в плоскогорье Путорана, на высоте 400—
500 м над ур. м.
Выводок хорошо летавших молодых полярных овсянок наблюдался
на берегу оз. Глубокого 7 августа 1958 г.
120. Овсянка-крошка — Emberiza pusilia (Pall.). В северной полосе
тайги близ Норильских озер овсянка-крошка является самой многочис-
ленной из всех гнездящихся там птиц. В лесотундре у истоков Пясины
она также очень обыкновенна, но в тундру заходит в очень незначительном
числе и севернее низовьев Агапы не была встречена ни разу. Численность
этих овсянок может быть очень высокой: летом 1958 и 1959 гг. в разных
типах лиственничного леса у верховьев Рыбной пары селились в 50—100 м
одна от другой.
Прилетают овсянки-крошки в конце мая или в начале июня. Весной
1958 г. близ истоков Рыбной первая птица была отмечена 2 июня, но мас-
совое их появление было зарегистрировано только 9 и 10 июня. В 1959 г.
они прилетели 30 мая, а 6 июня повсюду можно было наблюдать пары.
В 1964 г. они впервые отмечены 2 июня, а массовый прилет происходил
3—8 июня.
Гнездятся эти овсянки во всех типах лиственничного и смешанного
леса, чаще всего на участках с разреженным древостоем и подлеском из
багульника и карликовой березки. Все 38 осмотренных нами гнезд были
устроены весьма однотипно в углублениях сбоку кочек или просто в под-
ходящих ямках. Сами постройки были сплетены из тонких сухих стеблей
злаков и осок, лоток выстилался более тонкими стеблями и корешками.
Прикрытием для гнезд обычно служили составлявшие нижний ярус леса
кустики карликовой березки, багульника, голубики и черники. Число
яиц в найденных гнездах обычно колебалось от 3 до 6, чаще их было 5.
Кладка из единственного насиженного яйца, осмотренная 14 июля 1959 г.,
несомненно была повторной. Яйца имеют слегка охристую окраску с бу-
рыми точечками и черточками.
К размножению овсянки-крошки приступают во второй декаде июня,
когда появляется большое количество проталин. В это время повсюду
слышалось пение самцов. Овсянка, строившая гнездо, весной 1958 г.
— 296 —
впервые наблюдалась 18 июня; на следующий день, 19 июня, было найдено
гнездо с 1-м яйцом; 22 июня овсянка снесла последнее, 4-е, яйцо. В сере-
дине третьей декады июня 1958 г., судя по нескольким найденным в это
время гнездам, почти все овсянки закончили кладки и приступили к на-
сиживанию. В 1964 г. из-за исключительного обилия снега и связанного
с этим позднего образования проталин большинство овсянок-крошек
закончило кладки только в самых последних числах июня или в начале
июля.
В 3 гнездах, найденных близ западного берега оз. Кета 30 июня и 2 июля
1959 г., были сильно насиженные кладки. 7 июля 1959 г. в 2 гнездах ока-
залось 4 и 5 птенцов приблизительно трехдневного возраста. 10 июля
птенцы уже начали выскакивать из гнезд и затаиваться рядом, несмотря
на то, что маховые перья у них еще только начинали выступать из чехлов;
12 июля оба гнезда уже пустовали.
Хорошо летавшие молодые овсянки-крошки летом 1958 и 1959 гг.
наблюдались после 21—23 июля. Особенно заметны были они в конце июля
и в начале августа, так как в это время составляли основу пернатого на-
селения в нижнем ярусе леса. С середины августа количество овсянок
заметно уменьшалось, а окончательно отлетали они к первым числам
сентября. Осенью 1959 г. близ истоков Рыбной овсянки-крошки в послед-
ний раз наблюдались 4 сентября.
В желудках 7 овсянок, добытых в июне 1960 г. близ устья Норилки
и в верховьях Пясины, были обнаружены только насекомые, причем
преобладали влаголюбивые формы, обитающие в моховой дернине (Ti-
pulidae, Limoniidae, Rhagionidae). Встречены также взрослые долгоножки,
хирономиды, листоеды, жужелицы.
121. Лапландский подорожник — Calcarius lapponicus lapponicus (L.).
Лапландский подорожник является самой характерной и многочисленной
птицей тундры. В лесотундре он уже не столь обыкновенен и заселяет
лишь более обширные открытые участки, а в полосу северной тайги не
проникает совсем. Самое южное местонахождение гнездящихся подорож-
ников было отмечено весной 1960 г. у юго-восточного берега оз. Пясино.
В небольшом количестве лапландский подорожник гнездится и в альпий-
ском поясе плоскогорья Путорана, где встречается на поросших ерником
открытых пространствах на высоте 500—600 м над ур. м. В тундре подо-
рожник населяет самые разнообразные ландшафты, но чаще всего участки
кочкарниковой мохово-осоковой и мохово-пушицевой тундры. Не избегает
он и порослей карликовой березки и низкорослых полярных ив. В таких
местах гнездящиеся пары подорожников встречаются через каждые 50—
100 м.
Прилетают лапландские подорожники в конце мая или в начале июня.
В 1961 г. первая одиночная птица была отмечена у Пуринских озер 29 мая,
в 1963 г. небольшая стайка наблюдалась 31 мая. В тундре близ устья
Дудыпты весной 1962 г. их пролет начался 2 июня. Наиболее интенсивный
пролет на север наблюдался в эти годы 3—18 июня. Подорожники по боль-
шей части придерживались проталин на холмах, где земля лучше прогре-
валась. В период весеннего пролета нередко встречались стаи из многих
десятков птиц, но чаще наблюдались небольшие группы.
Через несколько дней после прилета, в теплые солнечные дни, самцы
уже начинали петь. С этого времени подорожники разбились на пары
и занимали гнездовые участки. При расселении подорожников 7—8 июня
1963 г. нередко случалось наблюдать довольно ожесточенные драки между
отдельными самцами.
Гнезда подорожники устраивают очень однотипно в углублениях сбоку
осоковых, осоково-пушицевых или моховых кочек с кустарниками. Именно
таким образом было расположено подавляющее большинство из 34 осмот-
— 297 —
ренных нами гнезд. Только немногие гнезда были устроены между двумя
кочками или просто в углублении под прикрытием кустика карликовой
березки.
Одно гнездо, найденное на моховом бугре среди болотистой низины
27 июня 1963 г., по своему месторасположению резко выделялось среди
остальных. Оно было устроено на дне ямки глубиной около 15 см и около
8 см в поперечнике. Большинство осмотренных гнезд находилось с юж-
ной, юго-западной или юго-восточной стороны кочек. Они были сплетены
из сухих стеблей и листьев осок или злаков. Наружная сторона гнезда
обычно скреплялась кусочками мха. Лоточек с обильной выстилкой
из перьев и оленьей шерсти. В гнездах, осмотренных у Пуринских озер
летом 1963 г., были кладки из 5—6 яиц. В более суровых условиях
1961 и 1962 гг. подорожники насиживали 4—6, а в одном случае —
только 3 яйца.
Весной 1963 г. постройка гнезда самкой впервые наблюдалось 12 июня.
Неполная кладка из 4 яиц была найдена 17 июня 1963 г., на следующий
день самка снесла последнее, 5-е, яйцо и начала насиживание. 19—23 июня
1963 г. во всех найденных гнездах были уже полные кладки. В холодную
весну 1961 г. первое гнездо с неоконченной кладкой из 2 яиц было найдено
в окрестностях 2-го Пуринского озера 25 июня. В некоторых найденных
тогда гнездах даже в начале июля были неполные кладки. Только что
начатая кладка из 1 яйца была отмечена 2 июля и весной 1962 г. Вылупле-
ние птенцов летом 1963 г. происходило очень дружно 27—30 июня. Лишь
в немногих осмотренных нами гнездах оно задерживалось па 3—4 дня.
Слабо насиженная кладка из 4 яиц, найденная 13 июля 1963 г., видимо,
была повторной. В 1961 г. первое гнездо с 2 только что вылупившимися
птенцами и 1 яйцом было найдено в долине р. Пуры 13 июля. В 1962 г.
вылупление птенцов подорожника впервые отмечено 8 июля.
Летом 1963 г. первый покинувший гнездо молодой подорожник был
пойман в пойме Пуры 10 июля. Его рулевые только на 8—10 мм освобо-
дились от чехлов. Много хорошо летавших молодых подорожников было
отмечено в тундре у Пуринских озер 16—20 июня 1963 г. В 1961 г. летные
молодые регулярно встречались в бассейне р. Пуры только после 25 июля,
а в 1962 г. — после 20 июля. Летом 1963 г. старые птицы кормили хорошо
летавших молодых вплоть до двадцатых чисел июля.
Образование стай начинается во второй декаде августа. После 20 ав-
густа, а особенно в конце месяца наблюдался хорошо выраженный отлет
к югу. 27—29 августа 1960 г. на берегу р. Пясины близ устья Агапы по-
стоянно встречались стаи по 50—100 особей, летевших вверх по реке.
В первых числах сентября подорожников в тундре уже оставалось мало,
а к двадцатым числам они полностью исчезли. Довольно хорошо выражен-
ный пролет подорожников наблюдался в верховьях Рыбной 6—7 сентября
1964 г.
122. Пуночка — Рlectrophenax nivalis pallidior Salom. Во время наших
поездок мы ни разу не встречали пуночек, гнездящихся южнее низовьев
Пуры. Здесь 23 июля 1961 г. в недоступных утесах наблюдались много-
численные пары. Хорошо летавшие молодые, но еще с короткими хвостами
летом 1961 г. были впервые встречены в низовьях Пясины 3 августа.
Изредка, например холодным летом 1961 г., отдельные пары гнездились
в постройках человека значительно южнее, в частности близ устья Агапы.
В скалистых ландшафтах плоскогорья Путорана пуночка не гнездится.
На пролете пуночка очень обыкновенна в южной части исследованной
территории. Она характерна тем, что из всех мелких птиц первой прилетает
весной, а осенью последней отлетает к югу.
В долине Енисея, на 100 км южнее удинки, весной 1958 г. первые
луночки были отмечены 17 апреля, а в 1959 г. у западного берега оз. Кета —
— 298 —
27 апреля. В тундре у Пуринских озер и близ устья Дудыпты пуночки
появляются только в начале мая. Даже во время самого раннего приезда
в тундру, 10 мая 1963 г., мы их уже заставали на берегу 1-го Пуринского
озера. Хорошо выраженный пролет наблюдался весь май, а в тундре обычно
длился до середины июня. В это время пуночки особенно охотно дер-
жались около жилищ человека.
Отлет к югу начинается в первых числах или в середине сентября.
В окрестностях Норильских озер пуночки появляются только в первых
числах октября. Окончательно исчезают они к середине октября, хотя
вдоль берега Енисея осенью 1958 г. встречались даже в ноябре.
СЕЗОННЫЕ ЯВЛЕНИЯ В ЖИЗНИ ПТИЦ ЗАПАДНОГО ТАЙМЫРА
Зима. Как видно из характеристики природных условий западного
Таймыра, зима там очень длинная и холодная. Даже на юге исследован-
ной территории, у Норильска, среднесуточная отрицательная темпера-
тура воздуха держится около 8.5 месяцев в году. При этом средние темпе-
ратуры двух-трех самых холодных месяцев часто опускаются ниже —30 ° С,
с абсолютными минимумами, достигающими —50° и более. Многодневные
пурги со скоростью ветра до 40 м/сек. и полярная ночь, длящаяся в за-
висимости от широты местности 1.5—2 месяца, являются неотъемлемой
частью и без того крайне холодной таймырской зимы. Метеорологические
условия зимних месяцев на плоскогорье Путорана, на высоте 800—1500 м
над ур. м., видимо, еще более суровы.
Поэтому вполне естественно, что типичная тундра и альпийский пояс
плоскогорья Путорана в середине зимы практически совсем лишены орни-
тофауны. Исключения могут составлять лишь немногочисленные тундря-
ные куропатки, а в годы с высокой численностью леммингов — белые
совы.
Лесотундра благодаря наличию островков древесной растительности
и не заносимых снегом кустарниковых зарослей более благоприятна для
зимовки птиц. Немало способствует этому обстоятельству и несравненно
большая освоенность лесотундры человеком. В ней во множестве зимуют
как белые, так и тундряные куропатки. Птицы эти держатся в основном
в тальниковых зарослях речных пойм, по ручьям и на опушках лесных
куртин. Обычны здесь зимой хищники — белая сова и кречет, связанные
в своем питании с куропатками. В годы высокой численности мышевидных
грызунов обычной зимней птицей лесотундры является ястребиная сова.
На юге лесотундровой полосы изредка встречается трехпалый дятел.
Близ поселков и в особенности в местах забоя оленей иногда зимуют
вороны. Из мелких воробьиных очень редко, в начале и конце зимы,
встречаются стайки светлоголовых гаичек или пухляков, а в годы хоро-
шего урожая лиственничных шишек можно встретить стайки чечеток,
белокрылых клестов и щуров. Вообще же следует отметить, что одной
из самых характерных особенностей лесотундровой орнитофауны запад-
ного Таймыра является редкость мелких воробьиных зимой.
В северотаежном ландшафте окрестностей Норильских озер этот спи-
сок дополняется кукшей и каменным глухарем. Ворон, светлоголовая
гаичка, пухляк и трехпалый дятел здесь гораздо более обыкновенны,
чем в лесотундре. Возможно, здесь зимует и поползень, неоднократно
встречавшийся нам в конце октября 1959 г. В годы высокого урожая лист-
венничных и еловых шишек белокрылые клесты, щуры и чечетки бывают
иногда даже многочисленными. В другие же зимы северотаежные ланд-
шафты почти так же бедны мелкими воробьиными, как и лесотундровые.
Вообще из зимующих на западном Таймыре птиц только белая и тунд-
ряная куропатки являются видами, приспособленными к ночевкам в снегу,
- 299 —
и с почти неограниченной кормовой базой. Поэтому местами они бывают
многочисленными. Трехпалый дятел, кукша и синицы к середине зимы
значительно уменьшаются в числе, очевидно откочевывая в более южные
участки тайги. Возможно, что поползни к середине зимы откочевывают
все без исключения. Оставшиеся кукши, синицы и даже вороны придержи-
ваются ближайших окрестностей жилищ человека.
Прилет и пролет птиц весной. Первыми прилетными
птицами на западном Таймыре являются пуночки. Их стайки появляются
в Норильской долине в третьей декаде апреля. Приблизительно в одно
время с пуночками начинают продвигаться на север в тундру белые и тунд-
ряные куропатки^Эти перемещения куропаток в южных тундрах западного
Таймыра носят характер хорошо выраженного перелета. Основная волна
пролета куропаток приходится на конец мая и начало июня и по времени
и направлению совпадает с массовыми перекочевками диких северных оле-
ней. Пролет куропаток обычно полностью заканчивается к концу первой
декады июня.
Соколы-сапсаны прилетают в тундру одновременно с массовым появле-
нием куропаток, в третьей декаде мая. Из рано прилетающих птиц следует
отметить орлана-белохвоста, прилет которого в северной полосе тайги
происходит в начале и середине мая. Рано прилетают также передовые
лебеди обоих видов и некоторые виды уток. В незамерзающих даже в зим-
ние месяцы истоках Рыбной пары лебедей-кликунов в 1959 г. были впер-
вые отмечены 9 мая, а в 1964 г. — даже 6 мая, в полной зимней обстановке;
в двадцатых числах мая там наблюдались и первые утки. В последней
декаде мая, а иногда и в середине этого месяца появляются на западном
Таймыре передовые гуменники. Заметной разницы между сроками по-
явления передовых гусей в тундре и в северной полосе тайги Норильской
долины нам подметить не удалось. Прилетают в третьей декаде мая также
мохноногие канюки и бургомистры. Таков ранневесенний состав орнито-
фауны.
Только после резкого перелома в ходе весны, который обычно проис-
ходит в первых числах июня, начинается массовый прилет и пролет по-
давляющего большинства птиц. Начало настоящей весны, непосредственно
предшествующее валовому прилету, характеризуется резким потеплением
воздуха и обычно выпадением мокрого снега, переходящего в дождь.
При этом в тундре сразу появляется довольно много проталин, а в се-
верной полосе тайги образуются полыньи на реках и происходит интенсив-
ное таяние снега. На второй день после такого резкого потепления обычно
появляются первые белые трясогузки, а в последующие дни прилетают
и другие виды птиц. Эта основная волна прилета и пролета, падающая
на первую половину июня, очень характерна для западного Таймыра.
За этот короткий промежуток времени, как показывают многолетние
наблюдения, происходит прилет и пролет основной массы особей более
чем 50 видов. Сначала прилетают почти все кулики и водоплавающие,
крачки, кукушки, многие воробьиные. У большинства этих птиц отдель-
ные особи появляются значительно раньше, но пролет основной массы
популяции обычно происходит в начале июня. К таким птицам относятся
гуменники, оба вида лебедей, ряд уток, некоторые кулики, дрозды, лап-
ландские подорожники.
Вторая, менее мощная волна весеннего пролета, охватывает такие виды,
как гагары и некоторые поздно прилетающие воробьиные, например бе-
реговая и городская ласточки, пеночки-таловки, чечетки (в тундре)*
Эти птицы обычно прилетают во второй и даже третьей декаде июня.
К началу июля прилет всех видов птиц нормально заканчивается. Сроки
появления ряда птиц и характер их пролета испытывают значительные
колебания, зависящие от погодных условий. В случае ранних потеплений
— 300 —
прилет передовых особей, а иногда даже значительной части популяций
некоторых видов происходит раньше. Например, в условиях исключи-
тельно ранней весны 1960 г. валовой прилет и даже пролет большинства
видов водоплавающих, куликов и многих воробьиных в окрестностях
Норильска наблюдался уже в конце мая. Подобное явление имело место
и весной 1964 г., когда в результате значительного потепления (до 10 ° С),
наступившего после 13 мая и устойчиво державшегося более 10 дней,
белые трясогузки прилетели 24 мая, а серый сорокопут был отмечен даже
17 мая. Как показали многочисленные наблюдения, даты появления пере-
довых птиц многих видов в различные годы гораздо более постоянны, чем
сроки их массового пролета. Это особенно хорошо видно на примере
лебедя-кликуна, гуменника, некоторых куликов и лапландского подо-
рожника. Сроки пролета у подавляющего большинства видов как в годы
с нормальной (1959, 1963 гг.), так и с поздней (1958, 1962 гг.) весной были
крайне сжаты. Только в условиях сильно затянувшейся холодной весны
1961 г. прилет и пролет многих видов происходил гораздо дольше. По-
явление уток в значительной степени зависит от наличия открытой воды.
Немногочисленные полыньи в верховьях некоторых северотаежных рек
появляются в конце мая и в первых числах июня. На них скапливаются
почти все гнездящиеся в ближайших окрестностях водоплавающие.
На такой полынье в верховьях Рыбной весной 1958, 1959 и 1964 гг. мы
ежедневно наблюдали тысячные стаи разных видов пластинчатоклювых,
державшихся там до середины и даже до конца июня, т. е. до времени
вскрытия многочисленных таежных озер. Подобные же местные скопления
уток и гагар были отмечены весной 1961 и 1962 гг. в заберегах рр. Агапы
и Дудыпты. Первые талые лужи очень привлекают и гусей. На совсем
небольшом, но уже оттаявшем болоте близ устья Дудыпты 6—12 июня
1962 г. мы ежедневно наблюдали сотни гуменников, белолобых гусей,
а иногда и краснозобых казарок. Ранние полыньи и незамерзающие
верховья северотаежных рек в начале июня, в период выплода веснянок,
привлекают некоторых воробьиных, таких, как белая, желтая и желто-
головая трясогузки, рогатые жаворонки, дрозды, пеночки, а кроме того,
и некоторых куликов. В тундре многие из рано прилетающих птиц на-
ходят убежище и пищу у крутых береговых обрывов. Во время холодной
и поздней весны 1961 г. именно в таких хорошо защищенных от ветра
и пурги местах чаще всего попадались мохноногие канюки, соколы-сап-
саны, пуночки, лапландские подорожники, рогатые жаворонки, ка-
менки.
Большое значение для птиц, рано прилетающих в тундру, имеют об-
ширные песчаные отмели медленно текущих рек и озер. Господствующие
в тундре сильные ветры зачастую сдувают снег с таких отмелей и рассеи-
вают мелкий песок по поверхности снега на сравнительно больших пло-
щадях. Под действием солнечных лучей такой смешанный с песком снег тает
при гораздо более низкой температуре, чем чистый снег. Благодаря этому
в прирусловых частях пойм некоторых рек образуются своеобразные
оазисы весны, где в конце мая и в начале июня струятся ручейки, а зна-
чительные пространства отмелей и даже берегов полностью освобо-
ждаются от снега. Именно в таких местах собираются рано прилетевшие
кулики, воробьиные и особенно гуси, так как многие освободившиеся
от снега отмели бывают покрыты свежей порослью хвощей, являющихся
для всех видов гусей лакомой пищей. Весной 1961 г. в окрестностях
2-го Пуринского озера в начале июня стояла настоящая зима и первые
перелетные птицы наблюдались в ничтожном количестве, но на песчаных
отмелях р. Агапы, всего в 40 км южнее, происходил оживленный прилет
гусей, лапландских подорожников, рогатых жаворонков и некоторых
куликов.
20 Биология птиц — 301 —
Такое неравномерное распределение недавно прилетевших птиц,
образующих местами значительные скопления, очень типично для запад-
ного Таймыра. Более равномерно распределяются птицы только во вто-
рой половине июня, когда всюду образуется много проталин, а на мел-
ких водоемах появляется вода у берегов. У подавляющего большинства
видов вслед за расселением обычно начинается размножение.
Что касается направления пролета, то все виды воробьиных, кулики,
кукушки, многие водоплавающие, серебристые чайки и некоторые дру-
гие птицы, над которыми имеются конкретные наблюдения, появляются
с юга. Основной пролетной магистралью для большинства из них яв-
ляется р. Пясина. Очень хорошо выраженный пролет вдоль ее берегов
наблюдался нами близ устья Дудыпты весной 1962 г. При этом большая
часть птиц летела прямо вдоль Пясины с юга, но некоторые виды, напри-
мер гуси, частично прилетали к руслу реки с юго-запада, из долины
Енисея, и даже с юго-востока. Севернее, у Пуринских озер, основная
масса птиц летела на северо-запад и па север. Видимо, большая часть
гнездящихся на западном Таймыре видов сворачивает от русла Енисея
на северо-восток несколько южнее Дудинки, пролетая широким фронтом
над лесотундрой прямо к верховьям Пясины. От русла этой реки по ее
притокам и прямо через водоразделы они уже растекаются по всей окре-
стной тундре, а частично даже сворачивают на восток, вдоль северной
кромки плоскогорья Путорана, в авамскую лесотундру. В защищенной
почти со всех сторон горами Норильской долине весенний пролет гнез-
дящихся в тундре птиц обычно бывает выражен слабо. Здесь в основном
наблюдается прилет местных видов, т. е. гнездящихся в северной полосе
тайги. В тех редких случаях, когда пролет здесь бывает выражен силь-
нее, например весной 1964 г., основное его направление—с юго-востока
на северо-запад, к истокам Пясины.
Интересно отметить, что некоторые виды птиц, пролетные пути кото-
рых нормально проходят вдоль морского побережья, во многих случаях
также появляются с юго-запада. К таким птицам принадлежат гагары,
морянки, поморники. Очевидно, значительная часть этих видов под-
нимается вверх по Енисею, где забереги появляются уже в конце мая,
а оттуда вместе с основной массой перелетных птиц проникает на запад-
ный Таймыр в восточном и северо-восточном направлениях. Хорошо
выраженный пролет всех трех видов гагар в восточном направлении был
отмечен В. В. Леоновичем в долине р. Агапы весной 1961 г. Многочис-
ленные средние поморники, массовый прилет которых у Пуринских озер
наблюдался нами 6 июня 1963 г., появились с севера и северо-запада.
С этой же стороны обычно появляются и бургомистры.
Период размножения. В годы обильного плодоношения
лиственницы первыми приступают к гнездованию белокрылые клесты.
Недавно покинувшие гнездо молодые клесты были нами добыты в сред-
нем течении р. Фокиной И апреля 1958 г. Однако столь ранние сроки
характерны не для всей популяции, так как весной 1964 г. в верховьях
Рыбной 23 мая нами было найдено только что построенное гнездо птицы
этого вида. Очень рано, в мае или даже в апреле, приступают к размно-
жению гнездящиеся на юге исследованной нами территории врановые —
ворон и кукша. Мохноногий канюк, гнездящийся в окрестностях Но-
рильских озер на скалистых уступах обращенных к югу склонов гор,
также приступает к насиживанию в мае. Но таких птиц, которые при-
ступают к гнездованию до июня, т. е. еще зимой, на западном Таймыре
очень мало. Подавляющее большинство видов, гнездящихся в южной
части исследованной территории, начинает нестись, как правило, во вто-
рой декаде июня, а к насиживанию приступает в третьей декаде или
даже в конце этого месяца, когда более или менее устанавливается средне*
— 302 -
ъуютнъя толъжителънля начала насиживания
у птиц, таеадцшцхъя я тундре и я северной, полосе таити, почти не раз-
личаются. Это можно объяснить более медленным таянием снега в обле-
сенной местности. В открытой тундре, наоборот, снеговой покров быстро
разрушается под действием ветра и солнца, особенно на склонах и вер-
шинах холмов. Некоторые различия в сроках начала насиживания у раз-
личных видов и даже у особей одного вида часто зависят от особенностей
расположения гнезд. Гораздо большими бывают различия в сроках раз-
множения у птиц по годам. В условиях исключительно ранней весны
1960 г. такие птицы, как гуменник, шилохвость, морянка, мохноногий
канюк, сапсан, турухтан, чернозобик и некоторые другие, гнездились
на 10—25 дней раньше, чем в позднюю весну 1961 г. Судя по находкам
нескольких подросших птенцов в первой декаде июля 1960 г., белые
куропатки приступили к размножению почти на месяц раньше, чем
в 1961 или 1962 гг. При этом у многих видов, в особенности позднее при-
летающих, разница в сроках насиживания меньше в годы с ранней и
нормальной (1963 г.) весной, чем в годы с нормальной и поздней весной.
Большинство видов птиц, гнездящихся на западном Таймыре, устраи-
вает свои гнезда на земле. Из 61 вида, найденных на гнездовье в тундре,
к таковым относится 57. Даже в северной полосе тайги близ Норильских
озер из 82 видов гнездящихся птиц устраивают свои гнезда на земле
больше половины — 49 видов. Что касается общих закономерностей
размещения устроенных на земле гнезд, то в большинстве случаев они
бывают расположены на возвышенных точках рельефа, особенно по скло-
нам холмов южной экспозиции и по северным берегам водоемов. В наи-
большей степени эта закономерность выражена в многоснежные годы,
когда большую часть июня почти вся почва еще покрыта снегом, на фоне
которого резко выделяются немногие оттаявшие бугры и небольшие
участки склонов. Именно на таких рано оттаивающих бугорках в усло-
виях очень многоснежной весны 1964 г. были найдены кладки шило-
хвостей, пискульки, галстучника, варакушки и некоторых других птиц,
которые, судя по датам начала насиживания, приступили к гнездованию
в середине и даже в начале июня, при почти сплошном снеговом покрове.
Подобная же картина наблюдалась и в. 1961 г. в окрестностях Пуринских
озер, когда вследствие сильно затянувшейся и многоснежной весны очень
многие из найденных гнезд также были устроены на местах наиболее
ранних проталин. В годы с ранней и особенно малоснежной весной при-
уроченность гнезд к буграм и южным склонам выражена заметно слабее.
Поэтому в 1963 г. свыше 100 гнезд, найденных в слабо холмистой тундре
близ тех же самых Пуринских озер, были распределены сравнительно
равномерно. От мощности снегового покрова сроки размножения многих
гнездящихся на земле птиц зависят даже в большей степени, чем от тем-
пературного режима весны. Именно этим обстоятельством, по-видимому,
и объяснялась небольшая разница в сроках начала насиживания у мно-
гих видов в условиях холодной, но малоснежной весны 1958 г. и сравни-
тельно теплой, но очень многоснежной весны 1959 г. Помимо водораз-
дельных пространств и возвышенных берегов, очень характерными ме-
стами гнездования некоторых птиц являются небольшие пойменные
гривы и террасы, не затопляемые половодьем. Поскольку в речных пой-
мах и по соседству с ними обычно орнитофауна бывает богаче, особенно
в зоне тундр, а таких незаливаемых мест не очень много, то количество
гнезд здесь иногда бывает очень велико. Так, на небольших песчаных
гривах, с разными типами тундр наверху, осмотренных нами летом
1963 г. в верховьях Пуры, гнезда встречались нам через каждые 20—
50 м, причем чаще всего у краев этих возвышенностей. Здесь гнездились
следующие виды: краснозобая гагара, белолобый гусь, морянка, белая
— 303 —
20*
куропатка, тулес, белохвостый песочник, кулик-воробей, рогатый жаво-
ронок, лапландский подорожник и, возможно, еще многие другие. При
заселении пойм многие птицы, однако, подвергаются угрозе затопления
чрезмерно поднявшимися полыми водами. Паводки в среднем течении
и в низовьях Пясины бывают очень сильными: весной 1962 г. уровень
реки у метеостанции Кресты, в устье Дудыпты, поднялся более чем
на 10 м. Именно по причине весенних паводков сроки гнездования чаек,
гагар, гусей и уток, гнездящихся на берегах и кочках пойменных озер
и болот, значительно запаздывают. Как показали наблюдения в окрест-
ностях Пуринских озер, эта особенность касается и гагар, гнездя-
щихся на многих водораздельных водоемах, так как даже незначитель-
ный подъем воды ведет к гибели их гнезд, устроенных у самой кромки.
В годы с обильными снегопадами, например в 1964 г., наблюдался до-
вольно значительный подъем воды даже на больших Норильских озе-
рах, которые благодаря своей громадной площади в нормальные годы
сохраняют довольно стабильный уровень. В начале июля 1964 г. у запад-
ного конца оз. Кета мы наблюдали массовое затопление гнезд галстуч-
ников, белохвостых песочников, крачек, морянок и других видов, гнез-
дящихся в дельте Кета-Ирбо.
Вообще острова и сильно выдающиеся мысы являются, пожалуй,
самым излюбленным местом гнездования ряда видов, особенно некото-
рых куликов, чаек и водоплавающих. Стремление этих птиц гнездиться
на островах, особенно если они расположены на озерах со сравнительно
постоянным уровнем воды, одинаково характерно для птиц тундры,
лесотундры и северной тайги.
Например, на о. Чаячьем, на 2-ом Пуринском озере, не более
200—250 м в длину и около 80 м в самом широком месте, помимо 32 гнезд
серебристых чаек и 4 гнезд бургомистров, было найдено 4 гнезда поляр-
ных крачек, 3 гнезда морянок, 3 гнезда краснозобых казарок, 2 гнезда
куликов-воробьев, 2 гнезда краснозобых гагар, 1 гнездо гуменника и
1 — шилохвости. Кроме того, там же наблюдалась пара сильно беспо-
коившихся тулесов, у которых несомненно тоже было гнездо. Почти
все найденные на острове гнезда были расположены на восточной его
половине, рядом с колонией чаек и даже внутри нее. Судя по наличию
старых гнездовых ямок рядом с новыми гнездами, морянки гнездятся
на одних и тех же местах из года в год.
Несомненно, столь большое скопление гнезд разных видов птиц свя-
зано с присутствием чаек и крачек, умеющих прогонять хищников и
в данном случае — песца. Во второй половине лета остров соединяется
с берегом болотистым перешейком и становится доступным для проник-
новения песца.
На вышеупомянутых островках в устье Кета-Ирбо и на других остро-
вах оз. Кета, близ входа в залив Онган, вместе с полярными крачками
гнездились морянки, синьга, средний крохаль, мородунка, галстучник,
белохвостый песочник, полярная овсянка и овсянка-крошка.
Островки с менее разнообразным населением нам неоднократно встре-
чались и на озерах в пойме Агапы и Пуры. Но почти во всех случаях
там гнездились серебристые чайки и полярные крачки. Более подробного
рассмотрения заслуживает небольшой островок, осмотренный нами
27 июня 1960 г. на одном из озер близ истоков Пясины, в типичной лесо-
тундре. Вдоль южного берега поросшего кустами ольхи, полярных ив,
лиственничным мелколесьем и карликовой березкой, на нем была обна-
ружена колония полярных крачек из 10—15 пар, 2 гнезда серебристых
чаек, 9 гнезд морянок, 3 гнезда средних крохалей, 2 — пискулек, 1 —
белоклювой гагары и 1 гнездо белохвостого песочника (рис. 22). Еще
некоторые гнезда могли остаться незамеченными. Во всех случаях сов-
— 304 —
местного гнездования фактов разорения чайками гнезд других птиц не было
замечено. Очевидно, что в данном случае при совместном гнездовании
проявляются какие-то более сложные инстинкты, которые в непосредст-
венной близости гнездовий заглушают более простые инстинкты питания.
Не менее интересны факты гнездования птиц рядом с хищниками,
давно отмеченные в литературе (Бируля, 1907; Тугаринов и Бутурлин,
1911; Наумов, 1931; Сдобников, 1959а; Леонович и Залетаев, 1963).
На западном Таймыре классическим примером такого рода является со-
жительство краснозобой казарки с соколами и чайками.
Птиц, гнездящихся исключительно в скалах, на западном Таймыре
почти нет.
Из 25 видов, гнездящихся на деревьях и кустарниках, 19 устраивают
свои гнезда самостоятельно и только 6 большей частью гнездятся в дуплах
© / @ 2 О 3 ® * © 5 ° е € 7 04
Рис. 22. Схема размещения птичьих гнезд на острове одного из озер близ истоков
р. Пясины лето 1960 г.
1 — заросли кустарников; 2 — колония полярных крачек; з— гнезда серебристых чаек; 4 — гнездо
белоклювой гагары; 5 — гнезда пискулек; 6 — гнезда морянок; 7 — гнезда средних крохалей;
8 — гнездо белохвостого песочника.
или гнездах других птиц, почти их не видоизменяя. Отсутствие дупел
в значительной степени ограничивает продвижение к северу ряда видов.
Среди них следует упомянуть малого мухолова, обитающего только на
юге Норильской долины, где чаще попадаются покинутые дупла дятлов
и синиц. Из птиц второй группы довольно редкие здесь гоголь и луток
чаще всего гнездятся в высокоствольных смешанных лесах на склонах
южной экспозиции, где на месте выгнивших суков в сухих лиственницах
попадается довольно много крупных дупел.
Довольно большая группа птиц (15 видов), как правило, гнездится
открыто на деревьях и кустарниках. Сюда относятся орлан-белохвост,
кречет, кукша, 3 вида дроздов, сибирская завирушка, свиристель, оба
вида сорокопутов, юрок, чечетки, обыкновенная чечевица, клесты и щур.
Все эти виды характерны для северной полосы тайги и в значительно
меньшей степени — для лесотундры. В типичной тундре гнездятся только
чечетки. При устройстве своих гнезд на деревьях птицы этой группы
чаще всего используют лиственницу, густые, покрытые многочисленными
побегами ветви которой благоприятствуют укреплению и сохранению
гнезд. Высота их над землей обычно не превышает 5—6 м, чаще ниже —
3—4 м. Исключение представляет орлан-белохвост, который обычно воз-
двигает свою мощную постройку на высоте 8—10 м или даже на самой
вершине большого дерева.
Сроки гнездования у птиц, устраивающих свои гнезда на деревьях
и кустах, как правило, дней на 5—10 опережают те же сроки у видов,
гнездящихся на земле, и испытывают меньшие колебания из года в год.
— 305 —
Исключение составляют гнездящиеся в тундре чечетки, приступающие
к гнездованию только во второй декаде или даже в конце июля.
Если говорить о птицах, гнездящихся на деревьях, нельзя обойти
молчанием факты гнездования фифи и пепельного улита в старых гнездах
темнозобых дроздов. Весной 1960 г. В. В. Леоновичем и нами было най-
дено 4 кладки фифи и 1 кладка пепельного улита в прошлогодних дроздо-
вых гнездах, устроенных на высоте 2—5 м от земли. Возможно, это ин-
тересное явление, зарегистрированное именно в 1960 г., объясняется
исключительно теплой весной, вследствие чего оба вида куликов при-
ступили к размножению несколько раньше обычного, в то время как
на земле снег еще не успел растаять.
Величина кладки птиц, гнездящихся на западном Таймыре, также
подвержена значительным колебаниям по годам, зависящим как от ме-
теорологических особенностей на местах гнездования, так и от общих
кормовых условий и от физического состояния птиц. Последнее связано
с условиями зимовок и пролета. Позднее гнездование во время холодных
весен при недостатке мест, удобных для устройства гнезд, ведет к сильному
истощению птиц, прилетевших в нормальные сроки. Иногда наблюдались
случаи «выкидывания» готовых к откладке яиц просто па местах кор-
межки или на снегу. Интересно отметить, что в условиях суровой весны
1961 г. снизилась величина кладок даже у таких птиц, как кулик-воро-
бей, обычно отличающихся большим постоянством величины кладки.
Вылупление птенцов у большинства видов птиц на западном Таймыре
проходит в менее сжатые сроки, чем начало насиживания, что, естест-
венно, связано со значительной разницей в длительности эмбрионального
развития птенцов. Первыми начинают вылупление птенцы дроздов и
гнездящихся в таежных местообитаниях чечеток, у которых длительность
насиживания не превышает 11—12 дней; у этих птиц мы находили птен-
цов начиная с 25 июня вплоть до первых чисел июля. Влияние, оказы-
ваемое на сроки появления птенцов у этих птиц особенностями весны,
незначительно. В годы с нормальной весной примерно в такие же сроки
или в среднем на 2—3 дня позднее происходит появление птенцов у лап-
ландского подорожника, отмеченное нами в 1963 г. 27 июня—5 июля
(7 гнезд). В связи с гнездованием на земле в условиях тундры сроки эти
сдвигаются в зависимости от характера весны, на 15—20 дней. Примерно
в те же самые сроки появляются птенцы также у некоторых других
воробьиных — рогатого жаворонка, каменки, варакушки, белой трясо-
гузки, полярной овсянки и овсянки-крошки. У всех этих птиц в годы
с нормальными метеорологическими условиями массовое вылупление
птенцов происходит в первой декаде июля, реже в конце июня. Несколько
позднее, обыкновенно во второй декаде июля, появляются птенцы и
у черноголового чекана, пеночек, желтой и желтоголовой трясогузок,
краснозобого и горного коньков. В общем, у всех воробьиных, за исклю-
чением гнездящихся в тундре чечеток, вылупление птенцов происходит
с последней декады июня до середины июля.
Вылупление птенцов у большинства куликов и водоплавающих про-
исходит гораздо дружнее и в годы с нормальной весной (1959, 1963 гг.)
было приурочено ко второй декаде июля. Это время обычно совпадает
с резким потеплением и связанным с ним массовым вылетом комаров.
О крайне сжатых сроках появления пуховых птенцов у некоторых видов
водоплавающих и куликов можно судить по следующим примерам.
Во всех 5 гнездах краснозобой казарки, осмотренных в период с 22
по 24 июля в верховьях Пуры и на Пуринских озерах, происходило
вылупление птенцов.
Летом 1963 г. первое гнездо гуменника, в котором птенцы проклю-
нули скорлупку яиц, было найдено 10 июля. Второе гнездо, с 4 только что
- 306 —
вылупившимися, но уже обсохшими птенцами, было найдено 12 июля.
13—14 июля на р. Пуре и Пуринских озерах мы наблюдали уже около
десятка выводков с птенцами в возрасте 1—3 дней. 15 июля началось
вылупление гусят и в последнем из найденных в 1963 г. гнезде гуменника.
С 12 по 18 июля 1959 г. на озерах близ западного берега оз. Кета
почти ежедневно наблюдались выводки однодневных птенцов шилохвости.
10 июля 1963 г. было отмечено сразу 4 или 5 выводков однодневных
пуховых птенцов куликов-воробьев и белохвостых песочников. До этого
дня мы находили только их кладки. В последующие дни, 13. 14 и 15 июля,
выводки наблюдались ежедневно.
В 1959 г. гнездо с только что вылупившимися птенцами фифи было
найдено у западного берега оз. Кета 11 июля, 12—17 июля ежедневно
встречались новые выводки и отводившие старые птицы. Так же дружно
происходило тогда появление птенцов и у пепельного улита, первый
выводок которого нами обнаружен 16 июля 1959 г.
В условиях исключительно ранней весны 1960 г. на протяжении
дневного маршрута по тундре 5 июля было найдено сразу 3 выводка
однодневных птенцов чернозобика; в двух случаях они находились еще
в гнездах.
Приблизительно в это же время происходит массовое вылупление и
в гнездах чаек и крачек. Внутри отдельных колоний птенцы выклевы-
ваются в очень сжатые сроки, исключения составляют лишь пары, за-
гнездившиеся вторично после гибели первой кладки. У Норильских озер,
где полярная крачка гнездптся на незатопляемых островах и просто на
сухих гривах, вылупление птенцов у нее происходит несколько раньше,
в конце первой декады июля (1959, 1963 гг.). Значительно позднее,
в третьей декаде июля или даже в августе, происходит вылупление птен-
цов у многих нырковых уток, и особенно у крохалей. Последнее связано,
по-видимому, как с более поздними сроками размножения, так и с огром-
ной величиной кладки, которая у среднего крохаля нередко достигает
15—16 яиц. В зависимости от метеорологических условий сроки вылуп-
ления птенцов белой куропатки колеблются почти в пределах месяца,
падая то на начало (1960 г.), то на конец июля (1961 г.). Почти в такой же
степени непостоянно и время появления птенцов у дневных хищников,
например у мохноногого канюка и сокола-сапсана.
В общем, можно заключить, что у большинства видов гнездящихся
на западном Таймыре птиц вылупление птенцов нормально происходит
во второй декаде июля. Только у видов, строящих теплые гнезда, наси-
живание заканчивается в последней трети июня или в начале июля,
а у некоторых даже раньше.
Ко времени появления птенцов у большинства воробьиных, в конце
июня или в начале июля, как правило, наступает довольно резкое по-
тепление погоды и среднесуточные температуры поднимаются значительно
выше 0°, а иногда даже достигают 9°, 10° С и более. Это способствует
повышению активности насекомых, столь необходимых почти всем во-
робьиным для выкармливания птенцов. По мере их роста к концу первой
декады июля температура воздуха неуклонно повышается, количество
насекомых возрастает. Все это вместе с круглосуточным солнечным осве-
щением в значительной степени способствует удовлетворению возрастаю-
щих с каждым днем потребностей птенцов в пище. Однако в связи с на-
личием ночных заморозков и кратковременных похолоданий некоторые
взрослые птицы часто бывают заняты отогреванием даже подросших
птенцов. Возможно, именно поэтому сколько-нибудь заметной разницы
между темпами роста птенцов воробьиных на западном Таймыре и
в условиях умеренного климата подметить не удалось. Подавляющее
большинство видов воробьиных в годы с нормальной весной полностью
- 307 —
заканчивает гнездовую жизнь с 15 по 25 июля. Случаев вторичного их
гнездования на западном Таймыре даже в условиях ранней весны мной
ни разу не было отмечено. Значительная группа куликов полностью за-
канчивает цикл размножения лишь немного позднее большинства во-
робьиных. Хотя вылупление в гнездах большинства куликов и происхо-
дит во второй декаде июля, рост птенцов идет настолько быстро, что
в первой декаде августа большинство из них уже может летать. Этому
безусловно способствуют оптимальные температурные условия, обычно
достигающие своего максимума во второй половине июля. Благодаря
многочисленности кровососущих насекомых обычно начиная с десятых
чисел, а реже с начала июля быстро растущие птенцы многих куликов
всюду находят обильный корм.
В первый и второй день после вылупления птенцов у выводков мно-
гих видов куликов (фифи, пепельный улит, щеголь, мородунка и некото-
рые другие) наблюдались сразу самец и самка. У птенцов 3—4-дневного
возраста у большей части видов куликов обычно находились только
самцы. Однако и у них инстинкт заботы о потомстве со временем заметно
угасает. Поэтому у многих видов, например у фифи, мородунки и кругло-
носого плавунчика, подросшие птенцы уже ведут самостоятельный образ
жизни. У других, например у пепельного улита, самцы держатся вместе
с уже поднявшимися на крыло молодыми. У галстучников обе старые
птицы держатся с выводком до отлета.
Во второй декаде августа обычно приобретают самостоятельность
птенцы полярных крачек, серебристых чаек и бургомистров.
Птенцы большинства водоплавающих, вылупление которых происхо-
дит приблизительно одновременно с птенцами куликов, требуют для сво-
его развития значительно больше времени. Быстрее всего приобретают
самостоятельность птенцы свиязи, шилохвости и чирков. Молодые этих
видов в наиболее ранних выводках поднимаются на крыло еще до сере-
дины августа. Обычно же они начинают летать только к первым числам
сентября. В самом конце августа и в первой декаде сентября заканчивают
свое развитие молодые лебеди-клпкуны, тундровые лебеди, пискульки,
белолобые гуси, гуменники и краснозобые казарки. Особенно медленно
идет рост птенцов у нырковых уток. Даже в условиях необыкновенно
теплой осени 1959 г. в первых числах сентября подавляющее большинство
молодых чернетей, турпанов, синьг, средних крохалей и других нырко-
вых уток, гнездящихся в окрестностях оз. Кета, еще не могли летать.
Массовый подъем на крыло молодняка нырковых уток начинается только
во второй декаде сентября. При наступлении ранних холодов, возможно,
запоздавшие выводки погибают. Приблизительно к концу первой декады
сентября, т. е. тоже незадолго до ледостава, приобретают способность
к полету и молодые гагары всех трех видов.
Говоря о размножении птиц, нельзя умолчать об его особенностях
в разные годы, не связанных с температурными условиями. Влияние
численности мышевидных грызунов на размножение птиц-миофагов на
западном Таймыре обычно бывает очень сильным. Если летом 1960 г.
в пойме Агапы и у Пуринских озер серебристые чайки и бургомистры
гнездились почти на каждом подходящем островке, то в 1961 и в 1962 гг.
их здесь почти совсем не было видно. Так же интенсивно там размножа-
лись и длиннохвостые поморники, выводки которых во второй половине
августа 1960 г. встречались через каждые 4—5 км маршрута. В 1961
и в 1962 гг. в тех же местах наблюдались только редкие одиночные по-
морники. Не в меньшей степени зависит от численности мышевидных и
размножение мохноногого канюка. В 1960 и 1963 гг. в окрестностях
Пуринских озер гнезда канюков находились на расстоянии 3—6 или
даже 1.5—2 км одно от другого и содержали по 4—5 яиц или птенцов.
— 308 —
Летом 1961 и 1962 гг. канюки там гнездились в 10—20 км пара от пары,
а количество яиц в кладках не превышало двух. В том же 1961 году
в низовьях Пясины, где количество грызунов было средним, гнезда ка-
нюков были расположены в 6—8 км одно от другого и содержали по-
2—4 яйца или птенца.
Об интенсивности размножения некоторых водоплавающих в значи-
тельной степени можно судить по соотношению количества птиц при
выводках и числа холостых особей, собирающихся на линьку. После
исключительно ранней и теплой весны 1960 г. в пойме Агапы и у Пурин-
ских озер количество как выводков, так и линных холостых особей гу-
менников и белолобых гусей летом было нормальным. В условиях же
1963 г., когда весна по сравнению с 1960 г. была более поздней и холод-
ной, нами в тех же самых местах было найдено свыше трех десятков
гнезд гуменников, а количество наблюдавшихся выводков было значи-
тельно большим, чем в 1960 г. Линных гусей летом 1963 г., напротив,
наблюдалось очень мало. Сопоставляя эти данные, можно вывести за-
ключение, что в массовом размножении гусей летом 1963 г. приняла
участие почти вся популяция этих птиц. Одной из возможных причин
таких неожиданных вспышек размножения гусей может являться хорошая
физиологическая подготовленность в благоприятных условиях, сложив-
шихся на местах зимовок и путях пролета.
Послегнездовая жизнь и кочевки птиц перед
отлетом. Большинство таймырских птиц приступает к отлету вскоре
после завершения размножения, но есть виды и даже целые группы
(воробьиные, дневные хищники), которые улетают не сразу. У таких ви-
дов как молодые, так и старые особи ведут в этот период кочевой образ
жизни. Период этот интересен в том отношении, что очень многие птицы
в поисках пищи совершают довольно дальние перекочевки, появляясь
даже в чуждых им ландшафтных условиях. Так, молодые темнозобые
дрозды даже до образования стай, в конце июля и в начале августа,
в массе кочуют в альпийский и субальпийский пояса плоскогорья Путо-
рана. Сюда их привлекают многочисленные в это время насекомые, пик
численности которых в горах по сравнению с долинами запаздывает на
10—15 дней. Поползни, которые в условиях северной тайги западного
Таймыра очень редки на гнездовье, в конце августа и в начале сентября
регулярно встречаются как в окрестностях Норильских озер, так и не-
сколько севернее, в лесотундре. Во время осенних кочевок становится
повсюду обыкновенной такая спорадично гнездящаяся птица, как сибир-
ская завирушка. В тундре в этот период встречается целый ряд видов,
гнездящихся значительно южнее. В лесотундре близ истоков Пясины
в последних числах августа 1958 г. наблюдались, например, большие
стаи береговых ласточек. Около поселков человека и у кустарников по
речным берегам, где собиралось много мелких птиц, в первых числах
сентября мы регулярно встречали дербников. Около мест охоты на диких
северных оленей в устье Дудыпты и даже севернее каждую осень дер-
жатся выводки воронов. Именно во вторую половину лета чаще встре-
чаются залетные виды, гнездящиеся значительно южнее. Так, 20—25 ав-
густа 1964 г. близ западного берега оз. Кета регулярно встречалась стая
более чем в сотню молодых малых чаек.
Очень типичны для тундрового ландшафта многочисленные холостые
поморники и чайки, в течение всего лета ведущие бродячий образ жизни.
Эти птицы в глубине тундры обычно попадаются в одиночку или неболь-
шими группами, собираются в большие стаи у становищ рыбаков, в местах
разделки мяса и других подобных местах. Поморники иногда даже со-
вершают сравнительно дальние залеты на север таежной полосы. Напри-
мер, летом 1964 г. в окрестностях оз. Кета мы неоднократно наблюдали
— 309 -
короткохвостых поморников, привлеченных многочисленными полевками.
Изредка встречающиеся летом в тундре одиночные орланы-белохвосты,
видимо, также принадлежат к негнездящимся особям. Во всяком случае
такими являются оба добытых нами экземпляра: очень старая самка и
второгодок.
Осенний пролет и отлет. В отличие от прилета отлет
гнездящихся на западном Таймыре птиц растянут и длится с начала
августа до конца сентября или даже первых чисел октября. Первыми
отлетают многие виды куликов; их выводки, поднявшись на крыло, почти
никогда не задерживаются у мест гнездования больше, чем на несколько
дней. Во второй декаде августа повсеместно замечается почти полное
исчезновение лесотундровых и северотаежных куликов: перевозчиков,
фифи, пепельных улитов, мородунок. Поскольку гнездовья всех этих
видов на западном Таймыре находятся у северного предела, то отлетают
они незаметно, исподволь, не образуя таких стай, какие наблюдаются
у пролетных типично тундровых куликов, которые в это же время на-
блюдаются там на пролете.
Почти одновременно с куликами, в двадцатых числах августа, закан-
чивают отлет и полярные крачки. В середине августа улетают молодые
птицы обоих видов кукушек, пролет которых наблюдался нами у запад-
ного берега оз. Кета 14 августа 1959 г. Гнездящиеся в северной полосе
тайги воробьиные, как и кулики, исчезают незаметно; их местные ко-
чевки переходят в отлет постепенно. Во второй половине августа больше
всего мелких птиц держится в кустарниковых зарослях по берегам рек
и озер, в мелколесье на лесных опушках. Здесь во множестве встречаются
разные виды пеночек, овсянки-крошки, полярные овсянки, варакушки,
желтые и желтоголовые трясогузки. Количество их к концу августа
о каждым днем уменьшается, и в начале сентября встречаются уже только
одиночные птицы. Темнозобые дрозды и чечетки в конце августа и в на-
чале сентября собираются в значительные стаи и задерживаются дольше
большинства других видов. В годы с теплой осенью, особенно при обиль-
ном урожае ягод, дрозды встречаются вплоть до конца сентября и даже
первых чисел октября (1959 г.). Одновременно с дроздами улетают и
свиристели. Сравнительно поздно проходит отлет у больших сорокопу-
тов, которые наблюдались нами близ западного берега оз. Кета вплоть
до 20 сентября 1959 г. Полное исчезновение их в 1959 г. совпало с резким
уменьшением числа чечеток и некоторых других мелких птиц, служивших
им пищей. В годы обильного плодоношения лиственницы чечетки задер-
живаются гораздо дольше, а некоторые из них даже зимуют.
Из птиц, гнездящихся в тундре, раньше других начинают отлетать
краснозобые коньки. Небольшие стайки их мы наблюдали 7—8 августа
1963 г. в истоках Пясины. Птицы летели вдоль берега реки к югу. У за-
падного берега оз. Кета краснозобые коньки в довольно значительных
количествах наблюдались в последней декаде августа 1959 и 1964 гг.
Лапландские подорожники окончательно исчезают из тундры только
к 20—25 сентября. Дольше всех задерживаются в тундре рогатые жа-
воронки, которые в устье Дудыпты были встречены еще 8 октября, и
особенно пуночки, окончательно отлетающие только к середине октября.
Пролет этих видов в лесотундре у Норильских озер выражен
слабо.
Из водоплавающих раньше других начинают свое продвижение к югу
стаи шилохвостей и чирков; хорошо выраженный их осенний пролет
в верховьях Рыбной наблюдался нами в течение почти всего сентября
1959 г. В это же самое время там наблюдался мало оживленный отлет
перелинявших холостых гуменников и белолобых гусей и хорошо замет-
ный осенний пролет соколов-сапсанов.
— 310 —
Массовый отлет нырковых уток, составляющих подавляющее большин-
ство гнездящихся на западном Таймыре водоплавающих, начинается
только с наступлением ледостава на мелких таежных или тундровых
озерах, во второй половине или в конце сентября. Так, 20—25 сентября
1960 г., накануне ледостава на Пясине, у устья р. Дудыпты, когда все
тундровые озера уже замерзли, на реке наблюдались многочисленные
стаи синьги, турпанов, чернетей, морянок, крохалей. Такая же картина
была отмечена и в верховьях Рыбной в конце сентября—начале октября
1959 г. Все нырковые летели вдоль рек к югу. Отдельные отставшие
утки задерживались на полыньях до середины октября и, судя по неко-
торым наблюдениям, погибали.
В исключительно сжатые сроки, во второй половине сентября или
в начале октября, проходит отлет гнездящихся на западном Таймыре
гусей. Основная масса их задерживается на местах гнездования и линьки
до первых более или менее значительных морозов, часто сопровождаю-
щихся северными пургами.
Основная часть популяции серебристых чаек также отлетает к югу
в конце сентября или начале октября, в период ледостава или первых
обильных снегопадов.
Таким образом, отлет гнездящихся на западном Таймыре птиц факти-
чески заканчивается к началу октября. В тундре к этому моменту остаются
только белые и тундряные куропатки и полярные совы, которые окон-
чательно откочевывают оттуда только в ноябре, когда обильные снего-
пады полностью заносят кустарники.
В лесотундровом и северотаежном ландшафте остаются зимующие
птицы, часть которых, впрочем, на середину зимы все же отлетает за
пределы западного Таймыра.
ЛИТЕРАТУРА
Б и р у л я А. А. 1907. Очерки из жизни птиц полярного побережья Сибири. Зап.
Акад, наук, 8-я сер., XVIII, 2 : 1—156.
(Вальтер Г.) Walter Н. 1902. Ornithologische Beobachtungen an der westlichen
Taimyrhalbinsel vom September 1900 bis August 1901. Ежегодн. Зоол. муз.
Акад, наук, 7 : 152—160.
Виноградова А. Н. 1937. Геоботанический очерк оленьих пастбищ района реки
Пясины. Тр. Арктич. инет., LXIII : 5—46.
Воробьев К. А. 1959. Результаты орнитологических исследований хребта Чер-
ского. Орнитология, 2 : 115—121.
Гладков Н. А. 1958. Птицы заполярной Якутии (бухта Тикси). Проблемы Се-
вера, 2 : 169—193.
Дунаева Т. Н. и В. В. К у ч е р у к. 1941. Материалы по экологии наземных
позвоночных тундры южного Ямала. Матер, к познанию фауны и флоры СССР,
изд. Моск. общ. испыт. природы, нов. сер., отд. зоол., 4 : 1—80.
Житков Б. М. 1912. Птицы полуострова Ямал. Ежегодн. Зоол. муз. Акад, наук,
17 : 311-370.
Капитонов В. И. 1958. О распространении кукушек. Природа, 4 : 104.
Капитонов В. И. 1962. Орнитологические наблюдения в низовьях Лены. Орни-
тология, 4 : 40—61.
Колюшев И. И. 1933. Материалы по фауне и промыслу западного угла Таймыр-
ского полуострова. Матер, по изуч. Сибири, IV, Томск : 68—106.
Кречмар А. В. и Ю. И. Чернов. 1963. Материалы по трофическим связям
некоторых птиц-энтомофагов в тундрах западного Таймыра. Учен. зап. Моск,
обл. пед. инет., CXXVI, Зоол., 6 : 93—100.
Леонович В, В. и В. С. Залетав в. 1963. Под защитой сильных. Природа,
3 : 91—96.
Миддендорф А. Ф. 1869. Путешествие на север и восток Сибири, ч. II, отд. 5.
Сибирская фауна : 1—311.
Михайлов Н. Н. 1947. Геоморфологические наблюдения в западной части гор
Путорана. Вопр. географии, 3 : 7—26.
Михеев А. В. 1948. Охотничьи богатства Енисейско-Пясинской тундры. Охрана
природы, 4 108—119.
— 311 —
Наумов С. П. 1931. Млекопитающие и птицы Гыданского полуострова (северо-
западная Сибирь). Тр. Полярн. комиссии АН СССР, 4 : 1—106.
Осмоловская В. И. 1948. Экология хищных птиц полуострова Ямала. Тр.
Инет, географии АН СССР, XLI : 5—77.
Портенко Л. А. 1939. Фауна Анадырского края. Птицы, чч. 1 и 2. Тр. Научно-
исслед. инет, полярн. землед., животное, и промысл, хоз., 5 : 1—209; 6 : 1—198.
(Портенко Л. A.) Portenko L. А. 1959. Der Siechelstrandlaufer, Erolia fer-
ruginea (Pontopp.). Journ. f. Ornithol., 100, 2 : 141—175.
Птушенко E. C. 1952. Отряд гусеобразные. В кн.: Птицы Советского Союза, 4.
М. : 247-343.
Рогачева Э. В. 1959. Очерки фауны млекопитающих и птиц енисейской тайги и
лесотундры. В сб.: География населен, паземи. животн. и методы его изучен.f
М.—Л. : 197—207.
Сакс В. Н. 1945. Новые данные о геологическом строении р. Пясины. Тр. Горно-
геол. упр., 16 : 3—54.
Сдобников В. М. 1959а. Гуси и утки Северного Таймыра. Тр. научно-исслед.
инет, сельск. хоз. Крайнего Севера, 9, Норильск : 154—183.
Сдобников В. М. 19596. Кулики Северного Таймыра. Тр. научно-исслед. инет,
сельск. хоз. Крайнего Севера, 9, Норильск : 184—207.
(Скалой В. Н.) S с а 1 о n W. N. 1935, 1937—1939. Les oiseaux du Sud de Taimir.
Gerfaut, 25, 1935 : 9—20, 201—217; 27, 1937 : 108—120, 181—195, 220 —225;
28, 1938 : 23—32, 91—105, 165—175; 29, 1939 : 32—48.
Суслов С. П. 1935. К геоморфологии района Норильских озер (оз. Лама). Тр. Инет,
физич, геогр., 14 : 119—176.
Сыроечковский Е. Е. 1961. Птицы Хантайского озера и прилежащих гор
Путорана (Средняя Сибирь). Учен. зап. Красноярск, пед. инет., 20, 2 : 89—119.
Тугаринов А. Я. и С. А. Бутурлин. 1911. Материалы по птицам Енисей-
ской губернии. Зап. Красноярск, отд. Русск. геогр общ., 1 1—440.
Тугаринов А. Я. и А. И. Толмачев. 1934. Материалы для авифауны вос-
точного Таймыра. Тр. Полярн. комиссии АН СССР, 16 : 1—47.
Шумилова Л. В. 1933. Материалы по изучению оленьих пастбищ в районе озера
Пясино и Норильских гор в Туруханском крае. Матер, по изуч. Сибири, IV,
Томск : 1—24.
Яковлев Е. О. 1930. К границам распространения промысловых зверей и птиц
в Туруханском крае. Тр. Зоол. секции Средне-Сибирск. гос. геогр. общ., Зоол.
сборн., 1, Красноярск : 51—61.
Haviland М. D. 1915. A summer on the Yenisei (1914). London : 1—328.
Johansen H. 1945. Om racer af Saegdgaas. Dansk. Ornithol. Forenings. Tidskr.,
38 : 106—127.
P о p h a m H. L. 1897. Notes on birds observed on the Yenisei River, Siberia, in 1895.
Ibis : 89—108.
P о p h a m H. L. 1898. Further notes on birds observed on the Yenisei River, Siberia.
Ibis : 489—520.
P о p h a m H. L. 1901. Supplementary notes on the birds of the Yenisei River.
Ibis : 449—458.
Seebohm H. 1901. The birds of Siberia. London 1—512.
СОДЕРЖАНИЕ
Стр.
Р. Л. П о т а п о в. Птицы Памира ........................... 3
И. А. Н е й ф е л ь д т. Лесные «каменные» дрозды.......... 120
А В. Кречмар. Птицы западного Таймыра...................... 185
CONTENTS
Page
R. L. Potapov. The birds of the Pamirs ...................... 3
I. A. N e и f e 1 d t. The Wood «Rock»-Thrushes............ 120
A V. К г e c h m a r. The birds of the western parts of the Taimyr Peninsula 185
БИОЛОГИЯ ПТИЦ
Труды Зоологического института
том 39
Утверждено к печати
Зоологическим институтом
Академии наук СССР
Редактор издательства Л. Ф. Пухальская
Художник Д. А. Андреев
Технический редактор Л. М. Семенова
Корректоры О. М. Лаушкина и М. А. Судакова
Сдано в набор 2/VI 1966 г. Подписано к печати 5/Х 1966 г.
РИСО АН СССР Ke 55-96В. Формат бумаги 70х108,/1<. Бум. л. 10.
Печ. л. 193/4 + 1 вкл. (’/< печ. л.) = 28 усл. печ. л. Уч.-изд. л. 27.45.
Изд. Кв 2970. Тип. зак. Кв 922. М-27827. Тираж 2000. Бумага типо-
графская Кв 1
Цена 1 р. *94 к. + переплет 10 к.
Ленинградское отделение издательства «Наука»
Ленинград, В-164, Менделеевская лин., д. 1
1-я тип. издательства «Наука». Ленинград, В-34, 9 линия, д. 12
ИЗДАТЕЛЬСТВО «НАУКА»
ИМЕЮТСЯ В ПРОДАЖЕ КНИГИ
Войткевич А. А. Перо птицы. (Морфология, разви-
тие, линька и нейро-гормональная регуляция). 1962.
288 стр. Цена 1 р. 43 к.
Новик И. Е. Биология размножения и искусствен-
ное осеменение сельскохозяйственной птицы. 1964.
140 стр. Цена 83 к.
Организация и методы учета птиц и вредных
грызунов. 1963. 256 стр. Цена 1 р. 69 к.
Портенко Л. А. Птицы СССР. Часть 4. Отряд
Воробьиных (Определители по (фауне СССР. Вып. 69).
1960. 415 стр. Цена 2 р. 49 к.
Работы по экологической морфологии птиц и
млекопитающих. Труды Института морфологии
животных. Вып. 32. 1960. 259 стр. Цена 1 р. 60 к.
Труды Второй Прибалтийской орнитологической
конференции. 1957. 427 стр. Цена 2 р. 55 к.
Фауна СССР. Птицы. Том II, вып. 3. Ржанкооб-
разные. Подотряд: чистиковые. 1957. 144 стр. Цена 40 к
Якоби В. Э. и др. Функциональная морфология птиц.
1964. 95 стр. Цена 42 к.
Заславский М. А. Изготовление чучел птиц, скеле-
тов и музейных препаратов. Таксидермия птиц.
1966. 252 стр. Цена 2 р. 07 к.
Книга представляет собой подробное руководство но изго-
товлению, монтировке и хранению чучел различных птиц, ске-
летов и анатомических музейных препаратов. В ней системати-
зированы современные методы и технические приемы препари-
рования птиц, изготовление научных коллекционных тушек,
художественных чучел, скелетов и всевозможных препаратов
с описанием новейших способов применения наиболее доступ-
ных пластмасс и других химических соединений.
Особенное внимание уделяется изготовлению чучел птиц
в различных динамических позах, что является существенным
при построении биологических групп и диорам. Подробно разби-
раются способы изготовления искусственных частей чучел: греб-
ней, глаз, муляжей голов для комбинированных чучел.
1"- ' - -- - ------------------------ |
В книге приведены обширные материалы по уходу за кол-
лекциями (хранение, дезинсекция) и даны современные методы
построения экспозиции естественно-исторического музея.
Текст сопровождается большим количеством оригинальных
иллюстраций, детально поясняющих различные рабочие про-
цессы.
ЗАКАЗЫ НА КНИГИ НАПРАВЛЯЙТЕ ПО АДРЕСУ:
Москва, В-463, Мичуринский пр., 12. Магазин «Книга—почтой».
Ленинград, Д-120, Литейный пр., 57. Магазин «Книга—почтой».
или обращайтесь в ближайший к месту вашего жительства
магазин «Академкнига»
АДРЕСА МАГАЗИНОВ «АКАДЕМКНИГА»:
Москва, ул. Горького, 8 (магазин Кг 1)
Москва, ул. Вавилова, 55/5 (магазин № 2)
Ленинград, Д-120, Литейный пр., 57
Свердловск, ул. Белинского, 71-в
Новосибирск, Красный пр., 51
Киев, ул. Ленина, 42
Харьков, Уфимский пер., 4/6
Алма-Ата, ул. Фурманова, 139
Ташкент, ул. К. Маркса, 29
Ташкент, ул. Шота Руставелли, 43
Баку, ул. Джапаридзе, 13
Уфа, пр. Октября, 129
Уфа, Коммунистическая ул., 49
Фрунзе, пр. Дзержинского, 41 I