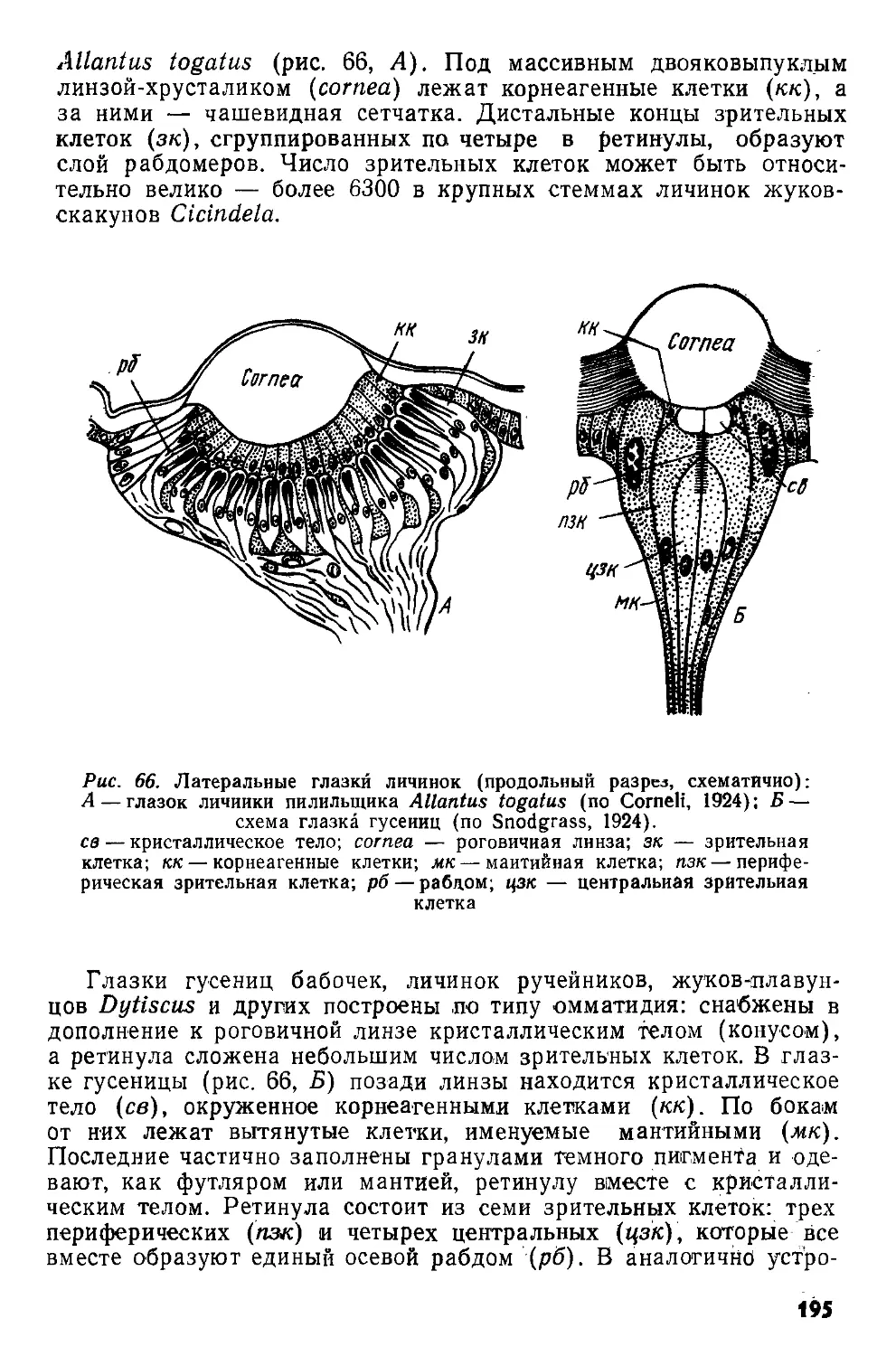
/










































































































































































































Author: Мазохин-Поршняков Г.А.
Tags: insecta hexapoda насекомые энтомология физиология насекомых
Year: 1977




Text
РУКОВОДСТВО ПО ФИЗИОЛОГИИ ОРГАНОВ ЧУВСТВ НАСЕКОМЫХ
Под редакцией профессора Г. А. МА30ХИНА-П0РШНЯК0ВА
ДОПУЩЕНО МИНИСТЕРСТВОМ ВЫСШЕГО И СРЕДНЕГО СПЕЦИАЛЬНОГО ОБРАЗОВАНИЯ СССР в качестве учебного пособия для студентов биологических специальностей ВЫСШИХ УЧЕБНЫХ ЗАВЕДЕНИЙ
ИЗДАТЕЛЬСТВО
МОСКОВСКОГО УНИВЕРСИТЕТА 1977
УДК 595.7,—11.
Рецензенты:
акад. М. С. Гиляров-, кафедра зоологии и дарвинизма Московского педагогического института им. В. И. Ленина
Руководство по физиологии органов чувств насекомых. Под ред. проф. Г. А. Мазохина-Поршня-кова. М., Изд-во Моск, ун-та, 1977.
224 с., с ил. Библиогр. 36 назв., 11 табл.
Руководство содержит описания строения и функций органов чувств насекомых (механо-, слуховых и обонятельных рецептов, термо*, гигро- и фоторецепторов). Часть разделов книги посвящена общим принципам строения и работы нервной системы, механизмам пространственной ориентации и реакциям насекомых на магнитные и электрические поля. Авторы руководства, опираясь на многолет* ний опыт преподавания на кафедре энтомологии МГУ, создали сводку современных знаний по этим важнейшим с точки зрения теории и практики биологии вопросам.
Авторы: Рустем Девлетович Жантиев, Юрий Алексаи-дрович Елизаров, Георгий Александрович Мазохии-Поршияков, Владимир Борисович Чернышов.
21008 — 079 , Р -------------84—77
077(02) —77
(J) Издательство Московского университета, 1977 г.
ПРЕДИСЛОВИЕ
В настоящее время энтомологи разного профиля, включая и специалистов по прикладной энтомологии, вынуждены широко опираться на достижения сенсорной физиологии насекомых. В этой бурно развивающейся области исследования на стыке биологии, физики и химии накоплено много ключевых фактов и сделано немало важных обобщений, но все они в значительной мере остаются достоянием узких специалистов. По крайней мере две причины привели к относительно малой доступности современных сведений об устройстве и работе органов чувств насекомых. Во-первых, сенсорная физиология как наука, опирающаяся на непривычные биологу физико-математические подходы к явлениям, ограждена барьером из специальных понятий и терминов, непонятных непосвященному. Во-вторых, большинство новых сведений по сенсорной физиологии рассеяно в многочисленных, зачастую труднодоступных зарубежных журналах. Что касается обзорной физиологической литературы, изданной на русском языке, то она или безнадежно устарела, как, например, переведенная в 1953 г. «Физиология насекомых» Р. Шовена, или написана в форме, рассчитанной только на специалистов. Иными словами, ни в отечественной, ни в мировой литературе сейчас нет современной сводки по сенсорной физиологии насекомых, пригодной для усвоения как основ, так и последних достижений в этой достаточно сложной отрасли знаний.
Предлагаемое «Руководство» ставит своей целью хотя бы частичное устранение причин, препятствующих широкому освоению мировых достижений физиологии органов чувств насекомых. Требование доступности изложения по возможности увязано в нем с задачей описания всех основных сведений и новейших данных в рассматриваемой области.
Читатель обратит внимание на определенное различие в уровнях изложения материалов по механорецепции, хеморецепции и фоторецеп-
3
ции, но оно не связано с индивидуальными вкусами или интересами авторов соответствующих глав, а отражает степень изученности разных видов чувствительности животных, В частности, о зрении человека и животных, в изучении которого участвовали такие великие исследователи, как Максвелл и Гельмгольц, мы знаем гораздо больше, чем, например, о хеморецепции. Поэтому если в первом случае можно было сосредоточить внимание на механизмах процессов и подходах к их изучению, то во втором случае пришлось ограничиться главным образом описательным изложением.
Авторство «Руководства» распределено следующим образом. Предисловие, введение, главы об общей характеристике нервной системы и органов чувств насекомых, о фоторецепции и механизмах пространственной ориентации насекомых, заключение, а также общая редакция текста принадлежат Г. А. Мазохину-Поршня-кову; главы о механорецепции и слухе написаны Р. Д. Жантиевым; главы о хеморецепции, восприятии температуры и влажности — Ю. А. Елизаровым; глава о реакции насекомых на электрические и магнитные поля — В. Б. Чернышовым. Создавая руководство, авторы опирались на многолетний опыт чтения лекций и ведения практических занятий по физиологии и биофизике органов чувств насекомых для студентов кафедры энтомологии Московского государственного университета. Но оно рассчитано не только на студентов и аспирантов университетов: книга может служить справочным пособием и для научных работников, так как содержит значительно больше сведений, чем обычный учебник по физиологии.
Введение
РОЛЬ ОРГАНОВ ЧУВСТВ В ЖИЗНИ НАСЕКОМЫХ
Органы чувств неотделимы от центральной нервной системы организма. Если последняя несет функцию управления, координируя физиологические процессы и поведенческие реакции организма, то органы чувств через посредство своих сигналов связывают центральную нервную систему как с внешним миром, так и с внутренней средой организма. Чувствующие, или рецепторные, клетки, рассеянные по телу или объединенные в сложные рецепторные органы, служат своеобразными «окнами» во внешний мир и внутреннюю среду организма. Информация, поступающая через них в центральную нервную систему, чрезвычайно разнообразна и, как мы увидим ниже, совершенно необходима для организации целесообразного поведения, а также для биологически оправданного и согласованного функционирования физиологических систем организма.
Выполнение всех трех непременных жизненных задач организма: питания, размножения и расселения,— обеспечивающих сохранение вида, возможно только благодаря непрерывному контролю со стороны разнообразных чувствующих органов. Рецепторы, совместно с их мозговыми центрами, называемые вместе анализаторами, не только выделяют из фона те или иные предметы и явления, т. е. отвечают на вопрос «что?», но и устанавливают положение предмета в пространстве, т. е. отвечают на вопрос «где?».
Рассмотрим на примерах, как органы чувств позволяют выполнить названные выше жизненные задачи и какие вопросы возникают у исследователя при наблюдении за сенсорным поведением насекомого.
Питание. Перед большинством появляющихся на свет личинок и взрослых насекомых встает необходимость поиска пищи для собственного пропитания, а в случае имаго — и для будущего потомства. Для монофагов задача особенно сложна: требуется найти, например, строго определенное растение. Точно так же самка одно-ядного паразита должна разыскать среди множества видов один определенный да еще обнаружить его в толще ствола дерева (например некоторые наездники). В поисках пищи для себя или для потомства насекомым помогают все органы чувств, обращенные во внешнюю среду, но в особенности обонятельные, вкусовые и зрительные. Среди обонятельных и вкусовых стимулов известны разнообразные пищевые аттрактанты, которые избирательно при
5
влекают различных насекомых. Ими могут быть сложные химические соединения растительного или животного происхождения, а чаще —простые соединения типа глюкозидов и эфирных масел, содержащиеся в листьях или плодах растений. Для кровососов ат-трактийны пот животных и выдыхаемая ими углекислота. К числу аттрактантов относится даже дым, привлекающий некоторых златок, откладывающих яйца на опаленные огнем ослабленные деревья. Описаны курьезные случаи, когда на стадион опускались мириады этих жуков, привлеченные папиросным дымом неистовых болельщиков. Ряд пищевых аттрактантов уже синтезирован и используется в качестве приманки для ловушек в борьбе с вредными видами. К числу зрительных пищевых стимулов относятся окраска, размер и форма съедобного объекта, иногда — характер его движения. При кажущейся самоочевидности этих стимулов как потенциальных признаков пищи нельзя забывать об их относительности и фактической сложности использования ими. Так, один и тот же объект, например белый цветок, в разных условиях освещения отражает свет разного спектрального состава; видимая форма предмета зависит от ракурса; видимый размер предмета зависит от расстояния до него; для того чтобы оценить характер движения объекта, нужно прежде всего отличить вызванное им смещение изображения на сетчатке от смещения этого же изображения вследствие собственного движения наблюдателя. Поэтому только сложный механизм константности восприятия позволяет зрительно опознавать предметы по их абсолютным признакам, инвариантным к перечисленным выше и многим другим преобразованиям. В поиске и выборе пищи участвуют и другие рецепторы: влажности, температуры, тактильные, слуховые. Например, жужжащий звук летящей пчелы привлекает ее врагов: ос-филантов Philanthus triangulum и шершней Vespa crabro. Канадская паразитическая муха-тахина Calcondamyia auditrix находит своего хозяина— цикаду —почти исключительно по ее песне, поэтому заражает только самцов.
Размножение. Наиболее характерная форма поведения, связанная с размножением, — поиск полового партнера. Причастность органов чувств к обслуживанию полового поведения совершенно очевидна, и, пожалуй, именно в этой сфере проявляются удивительные возможности, заложенные в устройстве рецепторных систем насекомых. Главную роль в поиске и опознании полового партнера у большинства насекомых играет чувство обоняния, узко настроенное на восприятие полового аттрактанта. Среди неподдаю-щегося перечислению множества запахов самец безошибочно выделяет один, именно тот, который принадлежит самке его вида, хотя может реагировать и на запахи близких видов. Половой аттрактант самки возбуждает хеморецепторы самца при ничтожной концентрации молекул в воздухе, что позволяет ему найти самку с расстояния (в рекордном случае) до 12 км. Самец, в свою очередь, нередко имеет органы «очарования», пахучий секрет кото
6
рых — афродизиак — предрасполагает самку к копуляции. Иначе говоря, видоспецифичными пахучими сигналами обмениваются оба половых партнера, что обеспечивает надежность их встречи.
Недавно показано на дубовой листовертке Tortrix viridana, что половой феромон попадает в организм самки из личиночного кормового растения и определяется химизмом последнего. Поэтому самки, воспитавшиеся на диете А, не привлекают самцов, выросших на диете Б. Это обстоятельство ведет к репродуктивной изоляции популяций и может быть причиной возникновения временных (обратимых) внутривидовых форм.
У дневных видов и у светящихся насекомых особенно значительна в половом поведении роль зрения. Окраска крыльев и всего тела, характер полета и некоторые другие визуальные признаки служат для дневных бабочек, стрекоз, многих мух и других насекомых специфическими сигналами самца и самки, легко улавливаемыми их фасеточными глазами. Иногда эти признаки настолько специфичны для насекомых, что об их существовании мы можем судить только с помощью специальных приборов. Например, мы не видим невооруженным глазом различия в отражении крыльями ультрафиолетовых лучей, которое является у некоторых бабочек действенным вторичнополовым признаком. В ряде случаев в зрительной системе насекомых удалось выявить специальные детекторы цвета, узко настроенные на восприятие окраски полового партнера. Общеизвестна оптическая сигнализация у жуков-светляков, но не все подозревают, насколько сложно она организована. Каждый вид имеет свои опознавательные огни — светящиеся пятна, отличающиеся конфигурацией и временными параметрами. На вспышку видоспецифичного сигнала самца его избранница отвечает через строго определенный интервал времени призывным свечением. Строгая видовая специфика набора сигналов и ответов обеспечивает надежную связь и в то же время служит этологическим барьером, если несколько видов обитают совместно.
Вызывает удивление своей сложностью в половом поведении и акустическая сигнализация. На фоне разнообразных шумов (даже очень громких) кузнечики, сверчки и некоторые другие насекомые за десятки метров выделяют призывную песню полового партнера и пеленгуют направление источника звука. Кроме призывной песни существуют и другие сигналы: копуляционные, угрожающие и территориальные. Способность слухового анализатора к тонкой видоспецифичной настройке порождает, в частности, возникновение местных диалектов территориальных песен, хорошо изученных у саранчовых Британских островов.
Расселение. Расселение требует в первую очередь надежной ориентации в пространстве, иначе животное будет перемещаться хаотично и не сможет покинуть исходную территорию. Расселение, сопряженное с ориентацией, может быть как активным — разлет, расползание, так и пассивным — перенос ветром или водой. При активном расселении насекомые ориентируются в основном зри
7
тельно по наземным вехам и небесному компасу в виде солнца, поляризации света голубого неба и луны. При этом наведение на цель становится возможным благодаря механизму одного из таксисов, позволяющему на основе сигналов от рецепторов удерживать локомоторную ось в избранном направлении. «Штурманское искусство» насекомых, способных вносить поправку к избранному курсу на суточное смещение небесных ориентиров, почти не уступает искусству птиц использовать небесный компас. Возможно, насекомые, как и птицы, ориентируются и по магнитному полю Земли. При пассивном переносе, например ветром, насекомые выбирают определенную позу, способствующую направленному переносу тела по воздуху, на основе информации от ветрочувствительных волосков и других рецепторов.
Все названные формы активности связаны либо с локомоцией, либо с удерживанием определенного положения тела в пространстве, а также отдельных частей тела относительно друг друга. И то и другое возможно только на основе информации, поступающей от специальных датчиков. К ним относятся в первую очередь различные механорецепторы, чувствительные к растяжению, сжатию или крутящему моменту — стимулам, прилагаемым к кутикуле, соединительной ткани и мышцам в результате или внешнего воздействия, или внутреннего усилия, или только веса данной части тела. Сигналы механорецепторов обеспечивают контроль позы, координацию движений частей тела при беге, плавании, завивании кокона, копуляции и т. п., а также сигнализируют о разрыве контакта с субстратом, направлении и скорости смещения тела при движении.
О роли сенсорных сигналов в осуществлении двигательных реакций насекомых дает хорошее представление анализ броска богомола Mantis religiosa на добычу. Богомол, поворачивая голову, выслеживает добычу зрительно и может схватить ее даже тогда, когда она находится сбоку от его продольной оси. Следовательно, центр, управляющий броском, должен располагать информацией как о направлении на жертву относительно головы богомола, так и о положении головы относительно переднегруди с ее хватательными ногами. Информацию первого рода дают глаза, информацию второго рода дают механорецепторы—две пары так называемых волосковых пластинок в шейной области. Если перерезать нервы от всех шейных волосковых пластинок (деафферентировать управляющий центр), то надежность броска падает до 20—30% против; 85% в норме. При деафферентации только одной левой стороны промахи учащаются, причем замечается тенденция богомола направлять бросок правее цели. Сигналы, поступающие только от правых шейных пластинок, истолковываются управляющим центром как поворот головы вправо.
Афферентный контроль ходьбы осуществляется исключительно большим набором механорецепторов: в частности, за стимуляцию тех или иных ножных мышц леваторов и депрессоров отвечают
8
определенные рецепторы лапки, голени, бедра. Некоторые из них, например колоколовидные сенсиллы, расположены так, что их возбуждают силы натяжения, возникающие в ноге, когда насекомое нормально стоит. Поэтому если разрушить механорецепторы ноги, то у насекомого нарушается механический аспект ходьбы: аллюр, скорость и т. п. Поза при ходьбе часто регулируется за счет обратной связи с волосковыми пластинками, которые контролируют угол между коксой и трохантером (вместе с бедром). Палочник Caraussius morosus в норме свободно удерживает тело над грунтом. Зазор между ними сохраняется и тогда, когда насекомое несет на себе груз вчетверо тяжелее тела. Если же волосковые пластинки повреждены, то палочник начинает касаться субстрата даже под тяжестью собственного тела.
Из всех форм локомоции наиболее требователен в отношении сенсорной информации полет. Афферентные сигналы не только вызывают полет, они же необходимы и для его поддержания и регулирования. Хорошо известен так называемый тарзальный рефлекс: отрыв ног от опоры у многих насекомых вызывает полет или плавательные движения (например у водяных клопов — белостома-тид), сразу же прекращающиеся при возобновлении контакта с субстратом. Датчиками тарзального рефлекса служат несколько типов механорецепторных сенсилл в ногах. К числу рецепторов, поддерживающих полет, относятся ветрочувствительные волоски на голове и крыльях. Их фазово-тонические сигналы зависят от скорости и направления воздушного потока и могут не только поддерживать и регулировать полет, но и запускать его. У пчел, мух, тлей к автоматической стабилизации полета причастен также Джонстонов орган антенн. Его сигналы наряду с другими датчиками регулируют работу крыльев: чем больше давление воздуха на жгут антенны, тем меньше амплитуда взмахов ипсилатерального крыла. Легко представить, что на основе такой петли отрицательной обратной связи автоматически удерживается прямолинейное направление полета.
Рецепторы участвуют в регуляции не только локомоторной системы, но и практически всех других физиологических систем и органов. Их участие в управлении процессом пищеварения, например, очень демонстративно у кровососущих комаров. Самки комаров Anopheles питаются не только кровью позвоночных, но и пьют так называемые «свободные жидкости»: выступающий из растений сок, росу и т. п. При этом только кровь поступает непосредственно в кишечник, а другие жидкости первоначально хранятся в слепом ответвлении пищевода — в объемистом пищевом резервуаре. Но если в эксперименте комар выпьет открыто лежащую каплю крови, без прокола покрова жертвы, то кровь поступает не в кишечник, а также в пищевой резервуар, и насекомое вскоре погибает. Дело в том, что направлением тока всасываемой насекомым жидкости управляют находящиеся на хоботке и в глотке рецепторы.
9
Примером рецепторной активации желез внутренней секреции может служить зависимость линьки кровососущего клопа Rhodnius от объема выпитой крови: личинка линяет только после того, как выпьет определенную порцию крови, причем единовременно. Если ту же порцию крови личинка получает в несколько приемов, с перерывами между отдельными актами кровососания, то она не линяет. Эксперименты крупнейшего английского энтомофизиолога В. Вигглесворса показали, что зависимость между линькой и кро-вососанием довольно сложна. Линька наступает под действием гормона экдизона, выделяемого проторакальной железой, стимуляцию которой обеспечивают сигналы нейросекреторных клеток мозга. Мозговой центр, в свою очередь, активируется сигналами определенных рецепторов, в том числе рецепторов растяжения, которые расположены в стенках брюшка клопа. Эти рецепторы срабатывают только тогда, когда кишечник расширяется до некоторого порогового объема, что наступает при поступлении в него определенной порции крови. Точно так же сигналы о растяжении прямой кишки, например, запускают акт дефекации, сигналы о растяжении протоков половых желез самки информируют центральную нервную систему о готовности организма к яйцекладке и т. п. Приведенные примеры убедительно показывают, что согласованная работа внутренних органов зависит от информации, поступающей с интерорецепторов.
Приведенных выше сведений достаточно, чтобы объяснить причину нарастающего из года в год интереса к сенсорной физиологии насекомых. Всем, кто изучает взаимоотношение насекомых и других животных с внешней средой или пытается управлять поведением полезных и вредных видов, необходимы сведения об устройстве и работе органов чувств. У каждого исследователя, которого с научной или прикладной целью интересуют поиск кормового растения фитофагом или хозяина паразитом, привлечение полового партнера, сезонные или суточные миграции и другие подобные реакции организма, всегда возникает вопрос: на основе каких внешних стимулов и как животное ведет направленный поиск? Иначе говоря, задача объяснения мотивации выбора и механизмов наведения на цель неизбежно возникает во всех исследованиях, прямо или косвенно связанных с анализом поведения целого организма или работы отдельных его органов. В равной мере без учета рецепторного контроля внутренних органов нельзя плодотворно изучать динамику развития организма.
Есть и другая причина, способствовавшая бурному развитию физиологии органов чувств насекомых и животных в целом,— это бионический аспект проблемы рецепции. Рецепторы животных обычно превосходят по многим параметрам аналогичные по назначению датчики, конструируемые в настоящее время человеком. Поэтому понятно стремление изучить ту или иную живую систему, чтобы создать сходное по принципу работы техническое устройство. Физиология органов чувств в сравнении с большинством дру
10
гих биологических дисциплин продвинулась далеко вперед в результате включения в свой арсенал подходов, привнесенных на пути бионического поиска физиками, кибернетиками, математиками. Для бионики недостаточно только качественных характеристик, а необходимы количественные параметры живой системы, переводимые на язык математики.
Если говорить более конкретно, то инженеров интересуют органы чувств насекомых как потенциальные прототипы технических устройств с исключительно высокой чувствительностью, помехоустойчивостью, избыточностью конструкции, сочетающейся с миниатюрностью и низкой затратой энергии на работу. Чувствительность рецепторных клеток насекомых практически доведена до физического предела. Так, чтобы возбудить обонятельную клетку на антенне самца тутового шелкопряда, настроенную на восприятие полового аттрактанта самки, достаточно контакта с одной молекулой этого вещества. Зрительную клетку фасеточного глаза может возбудить единичный фотон. Механорецепторная клетка так называемого подколенного органа улавливает колебания субстрата, амплитуда которых меньше диаметра атома водорода. При этом рецепторы отличаются от известных технических датчиков информации поразительной помехоустойчивостью. Мы уже отмечали, что кузнечик выделяет (опознает) видоспецифичную песню на фоне •самых разнообразных звуков. Пчела издалека зрительно опознает известный ей цветок среди множества других предметов, сходных по размеру, окраске и форме. Избыточность конструкции живых систем проявляется в том, что разрушение части органа не выводит его из строя, причем у насекомых это свойство сочетается с крайней миниатюрностью всех органов.
Во всех без исключения рецепторных системах бионики особенно стремятся расшифровать высокоэффективные биологические способы выделения сигнала из шума. Наряду с этим в обонятельном анализаторе главный объект поиска — способы организации исключительно высокой и избирательной чувствительности к запахам, в слуховом анализаторе — способы пеленгации источника звука и опознания его сигналов, в зрительном анализаторе — механизмы анализа поляризации света и восприятия невидимых человеком лучей.
Достижения сенсорной бионики, насколько можно судить по доступным публикациям ’, пока скромнее успеха, достигнутого собственно сенсорной физиологией, обогащенной физическим подходом, заимствованным из бионики. Как пример успеха назовем создание прибора для измерения скорости самолетов относительно Земли, работающего на принципе восприятия движения фасеточным глазом, обнаруженном у жука-долгоносика Chlorophanus. Неоднократно сообщалось о создании акустических приборов, при-
1 Известно, что за рубежом бионические исследования широко финансируются военным ведомством и многие из них имеют соответствующую направленность, не подлежащую широкой огласке.
11
влекающих (и уничтожающих) кровососущих комаров, и ультразвуковых излучателей, имитирующих крик летучих мышей и тем отпугивающих вредных ночных бабочек, которые слышат эти звуки. В борьбе с непарным шелкопрядом и родственными видами удачно используются ловушки с половым аттрактантом (например синтетическим диспарлуром). Усовершенствованы световые ловушки с излучателем ультрафиолетовых лучей, особенно привлекательных для ночных насекомых.
Как для биоников, так и для биологов разных специальностей представляет большой интерес связанная с изучением рецепторов проблема опознания образов, кратким изложением которой мы и закончим обзор роли органов чувств в жизни насекомых.
Поиск того или иного объекта всегда опирается на различение (дискриминацию) внешних стимулов и их модальностей, за что целиком ответственны рецепторы, поскольку они стоят на «входе» организма. Но целенаправленный выбор возможен только при условии совпадения рецепторных сигналов от объекта с его описанием или признаками, заложенными в центральной нервной системе организма. Поэтому выбор объекта определяется не только поступающей извне сенсорной информацией, но и той, что содержится в генетической или индивидуальной памяти организма. Выбору предшествует опознание объекта как такового, сличение с эталонным представлением о нем, уже существующим в центральной нервной системе.
В связи с этим встает фундаментальный вопрос: в какой форме хранится в памяти насекомых описание объектов —в виде конкретных признаков каждого из них в отдельности или обобщенного представления? Следующий пример пояснит нашу мысль. Когда пчела безошибочно находит свой улей по окраске (пчеловоды давно заметили, что окраска облегчает поиск, и поэтому рядом стоящие ульи окрашивают в разные цвета), то неискушенному наблюдателю может показаться, что дело обстоит совсем просто. Пчела, как известно, может различать цвета, поэтому она и опознает свой улей по окраске. Но в действительности она опознает улей как таковой, не путает его с другими предметами, идентично окрашенными. Задачу для пчелы можно усложнить, поставив на улей какой-либо предмет, искажающий вид улья. Формально, на языке описания данной ситуации рецепторами глаза, здесь объект иной, тем не менее натренированная пчела и в этих условиях опознает его как улей. Значит, пчела хранит в памяти образ улья — некоторое обобщенное представление о нем, которое, как легко догадаться, может возникнуть только в результате личного опыта, многократного возвращения в улей в разных ситуациях и выделения в процессе формирования образа главных оптических признаков улья.
Способность медоносной пчелы к зрительному обобщению недавно подтверждена в специальных экспериментах, в которых обучение насекомого вели на разных объектах, но относящихся по од
12
ному общему всем им признаку к тому же самому классу подкрепляемых (пищей) объектов, которым был противопоставлен класс неподкрепляемых объектов. Прежде эта логическая операция считалась привилегией исключительно высших животных с объемистым мозгом, в поведении которых некоторые исследователи усматривали признаки «элементарного рассудка».
Проблема опознания образов оказалась в центре внимания не только биологов, но и конструкторов «думающих» машин. Дело в том, что визуальное опознание у человека и животных инвариантно ко многим преобразованиям узнаваемого объекта. Знакомое лицо мы опознаем анфас и в профиль, на фотографии, по контурному рисунку и даже на карикатуре. Опознанию предшествует выделение каких-то узловых признаков, и на их базе следует логическая операция обобщения и формирования образа. Но какие признаки и как их обобщает мозг — далеко не всегда известно, и в этом трудность создания алгоритмов и программ для компьютеров, например, читающих тексты, набранные разным шрифтом. Не все требуемые здесь эксперименты возможны на человеке, а отдельные из них, в особенности с хирургическим вмешательством, выполнимы только на животных. Отсюда понятна актуальность изучения сложных форм поведения насекомых, в данном случае визуального поведения пчел. Сравнительно малое число нейронов в сетчатке глаза и в особенности в головном ганглии делает пчел, по сравнению с высшими позвоночными, более доступным объектом для изучения периферических и центральных механизмов обобщения и опознания образов.
Г лава I
ОБЩАЯ ХАРАКТЕРИСТИКА НЕРВНОЙ СИСТЕМЫ И ОРГАНОВ ЧУВСТВ НАСЕКОМЫХ
1. КРАТКИЕ СВЕДЕНИЯ ОБ АНАТОМИИ И ГИСТОЛОГИИ НЕРВНОЙ СИСТЕМЫ
По особенностям организации и функции нервная система насекомых может быть подразделена на центральную, симпатическую и периферическую.
Центральная нервная система имеет метамерное строение: она состоит из серии нервных узлов, или ганглиев, соединенных в нервную цепочку межсегментными стволами — коннективами. Каждый ганглий парный, его левая и правая половины соединены более или менее выраженными комиссурами. Передний отдел нервной цепочки из надглоточного и подглоточного ганглиев называют головным мозгом, а остальные ганглии — брюшной нервной цепочкой. Число ганглиев в брюшной цепочке у разных филогенетических групп насекомых и даже разных стадий их индивидуального развития различно, однако степень интеграции ганглиев, как правило, не коррелирует с уровнем поведения насекомого.
Надглоточный ганглий — самый крупный и наиболее сложный нервный узел, сходный по функциям с головным мозгом позвоночных. Его образуют, по крайней мере, три слившихся сегментальных ганглия: прото-, дейто- и тритоцеребрум. Более крупный протоце-ребрум (рис. 1, PC) тесно связан с фасеточными глазами (или стеммами у личинок Holometabola) и дорсальными глазками, а его грибовидные тела (гт) и примыкающие к ним мозговые ядра служат высшим ассоциативным центром, где замыкаются условные связи. Грибовидные тела наиболее развиты у общественных насекомых, ведущих, в сравнении с остальными, более сложный образ жизни и обладающих высокой способностью к научению. Двустороннее разрушение грибовидных тел влечет выпадение приобретенных навыков, но безусловные реакции на те же стимулы у насекомого сохраняются. Дейтоцеребрум (DC) связан с антеннами, их сенсорным аппаратом и мышцами, поэтому считается обонятельным центром. Миниатюрный тритоцеребрум (ТС) иннервирует верхнюю губу, а также связывает надглоточный ганглий с симпатической нервной системой.
14
Подглоточный ганглий, также состоящий из трех сегментальных нервных масс, обслуживает сенсиллы и мышцы ротовых частей — соответственно мандибул, максилл и лабиума, а также рецепторы и эффекторы шейной области.
Брюшная нервная цепочка начинается тремя грудными ганглиями, которые у одних насекомых лежат раздельно, у других —
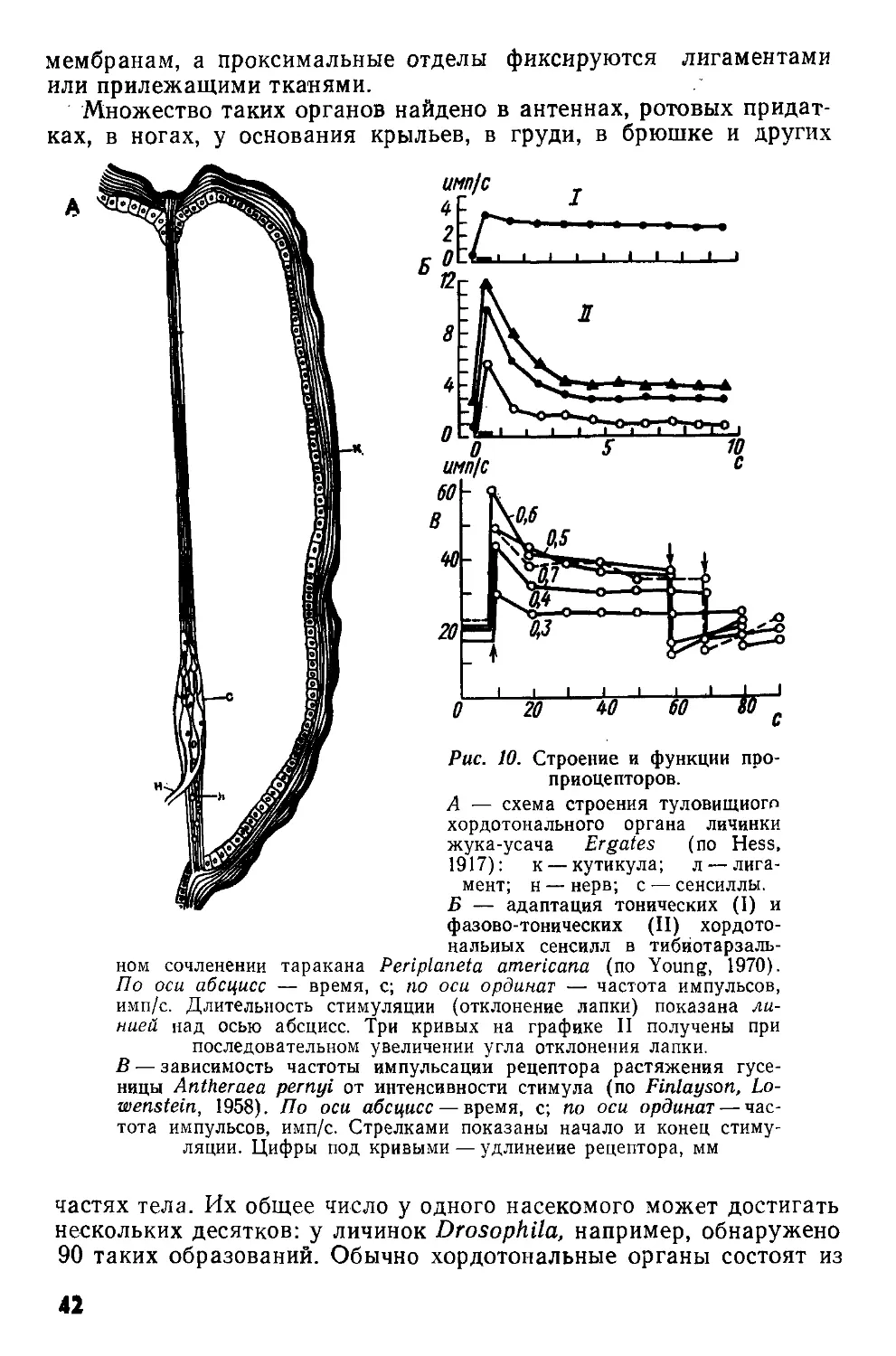
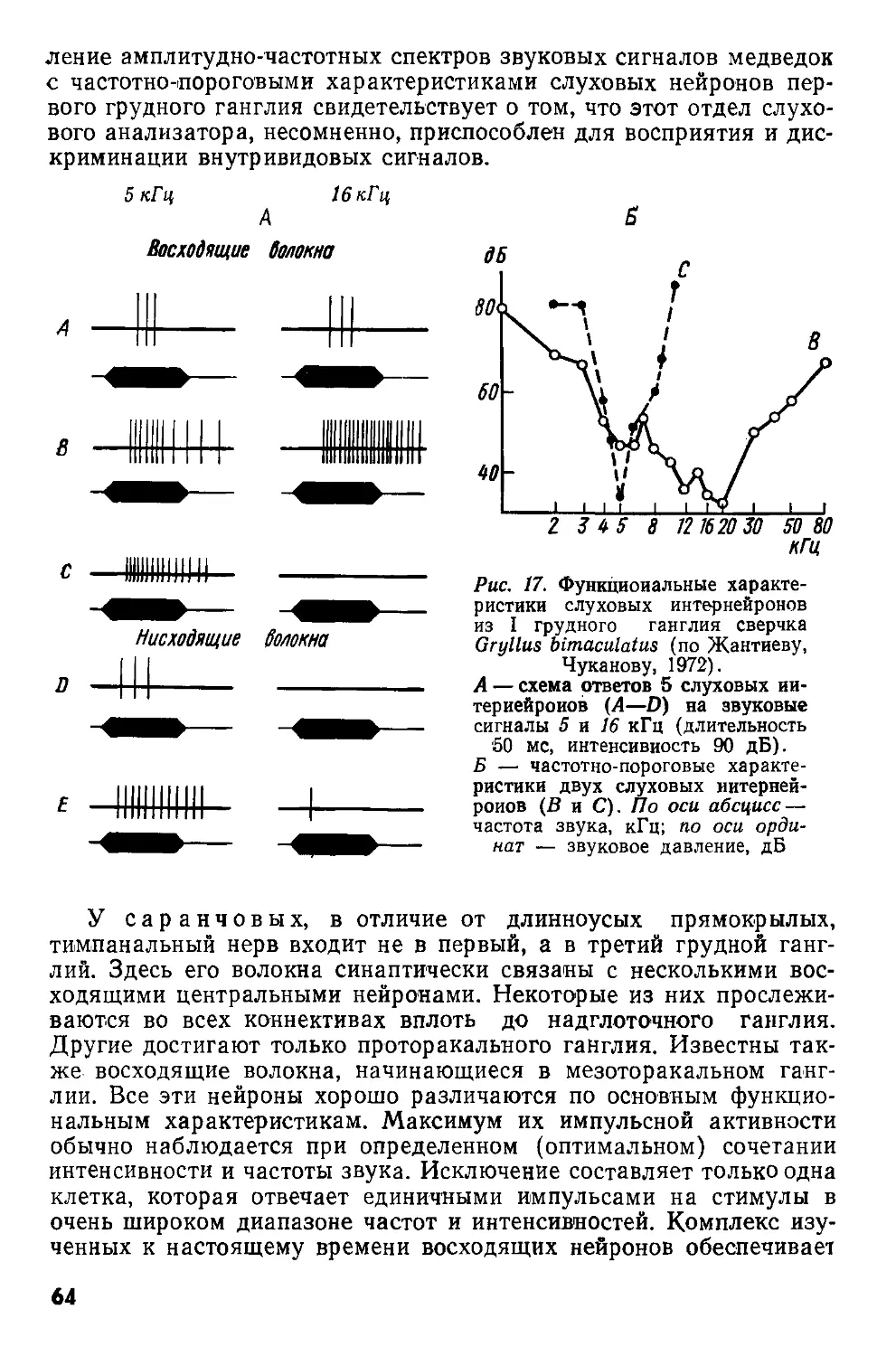
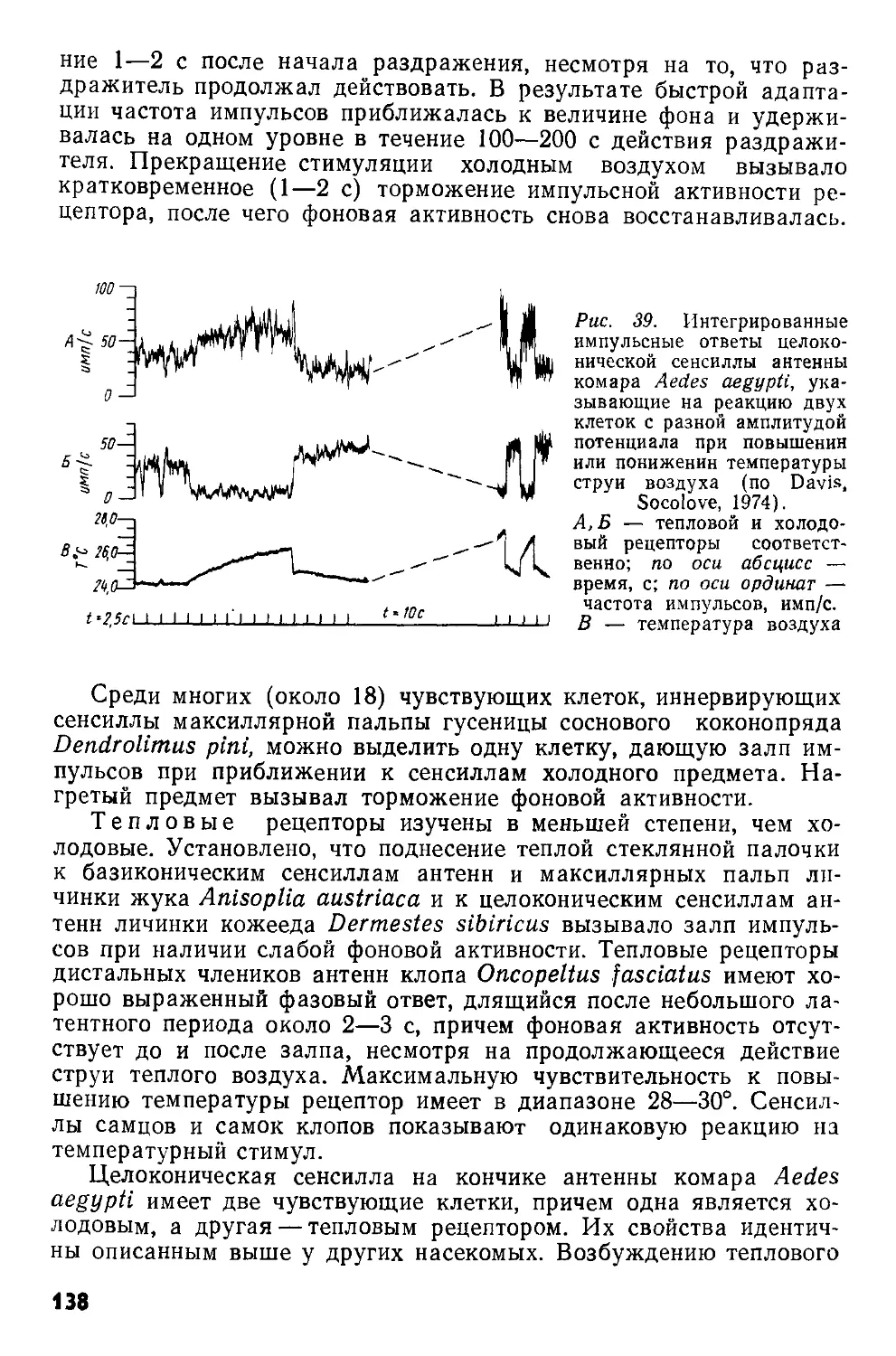
Рис. 1. Схематизированная картина фронтального среза через надглоточный ганглий сверчка Gryllus campestris (по Huber, I960).
Границы нейропиля прото-(PC), дейто-(ДС) и трн-тоцеребрума (ГС) обведены прерывистой линией. Ассоциативные ядра протоцере-брума обведены сплошными линиями.
ан — антеннальный нерв; вп — возвратный придаток, ножки грибовидного тела: гт — грибовидное тело; дн — двигательные нейроны дейтоцеребрума; клн+фк — клнпео-лабральный нерв н фронтальный коннектив; кЗ — комиссура тритоцере-
брума; мп — медиальный придаток ножки грибовидного тела; мц — межцеребральная часть; нет — ножка грибовидного тела; опт — оптические доли; оцн — оцеллярный нерв; цт — центральное тело; птм — протоцеребральный мост. Нервные пути: 1—3 — оптический тракт; 4 — оптическая комиссура; 5 — оцеллярный тракт; 6 — комиссура дейтоцеребрума; 7 — ольфакторно-глобулярный тракт; 8 — афферентные нервы антенн; 9 — эфферентные аксоны, идущие к мышцам антенн; 10 — связь между прото- и дейтоцеребрумом; 11—моторные аксоны, идущие к верхней губе; 12—нервные пути, связывающие надглоточный ганглий с подглоточным
сливаются вместе (иногда только мезо- и метаторакальный ганглии) или даже объединяются с несколькими (и изредка со всеми) брюшными ганглиями. Грудные ганглии ответственны в первую очередь за управление движениями ног и крыльев. В них могут также замыкаться условные локомоторные рефлексы.
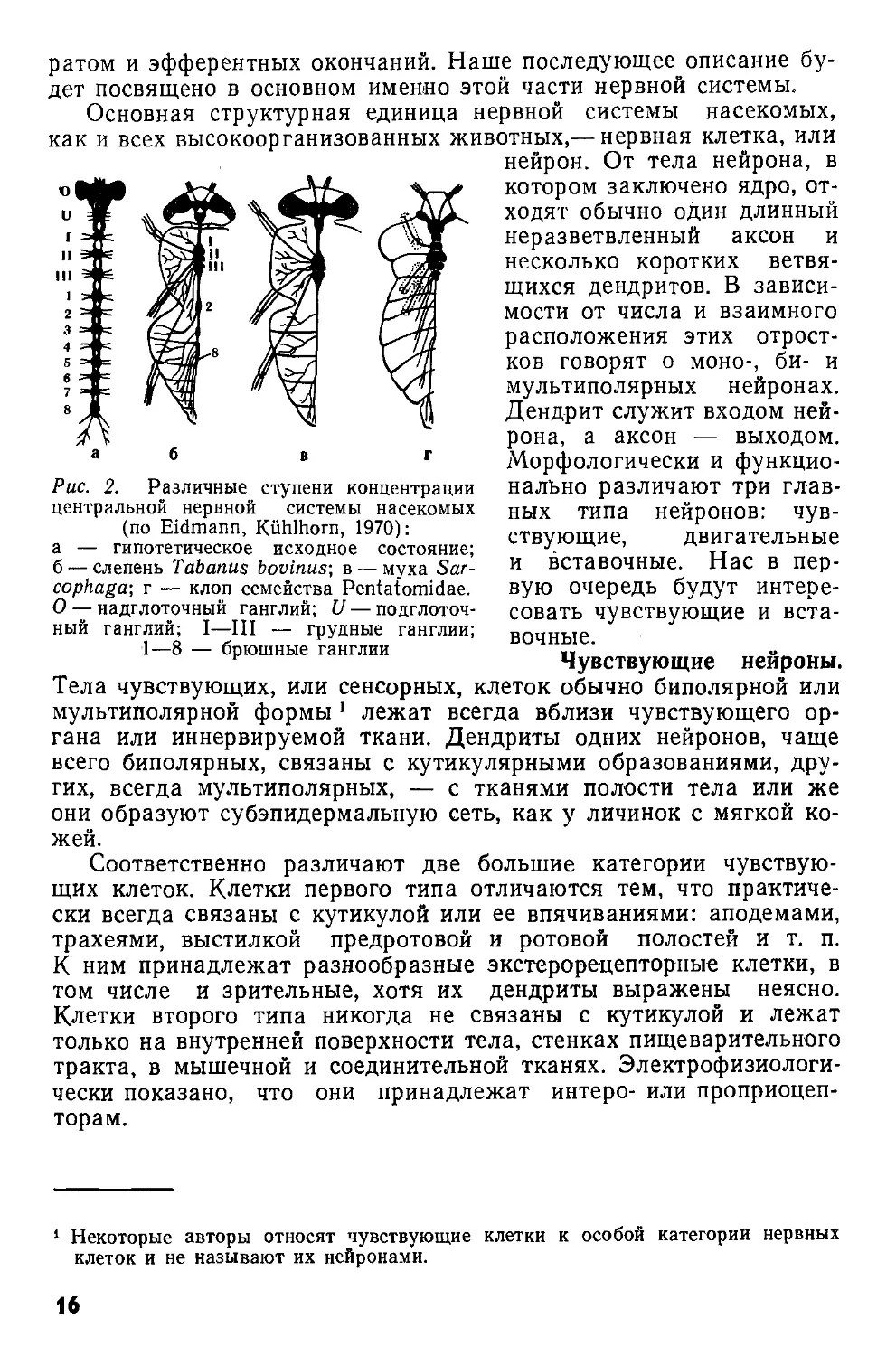
Менее сложно устроены брюшные. ганглии, число которых не превышает 8, иннервируют мышцы брюшных сегментов и их придатков и получают от них сенсорные нервы. Представление о том, в каких комбинациях могут сливаться ганглии брюшной нервной цепочки, дает рис. 2.
Симпатической, или вегетативной, нервной системы, регулирующей работу внутренних органов, и связанного с ней нейросекреторного аппарата насекомых мы практически не будем касаться в дальнейшем изложении.
Периферическая нервная система состоит из афферентных и эфферентных нервов, чувствующих клеток со вспомогательным аппа-
15
ратом и эфферентных окончаний. Наше последующее описание будет посвящено в основном именно этой части нервной системы.
Основная структурная единица нервной системы насекомых,
как и всех высокоорганизованных
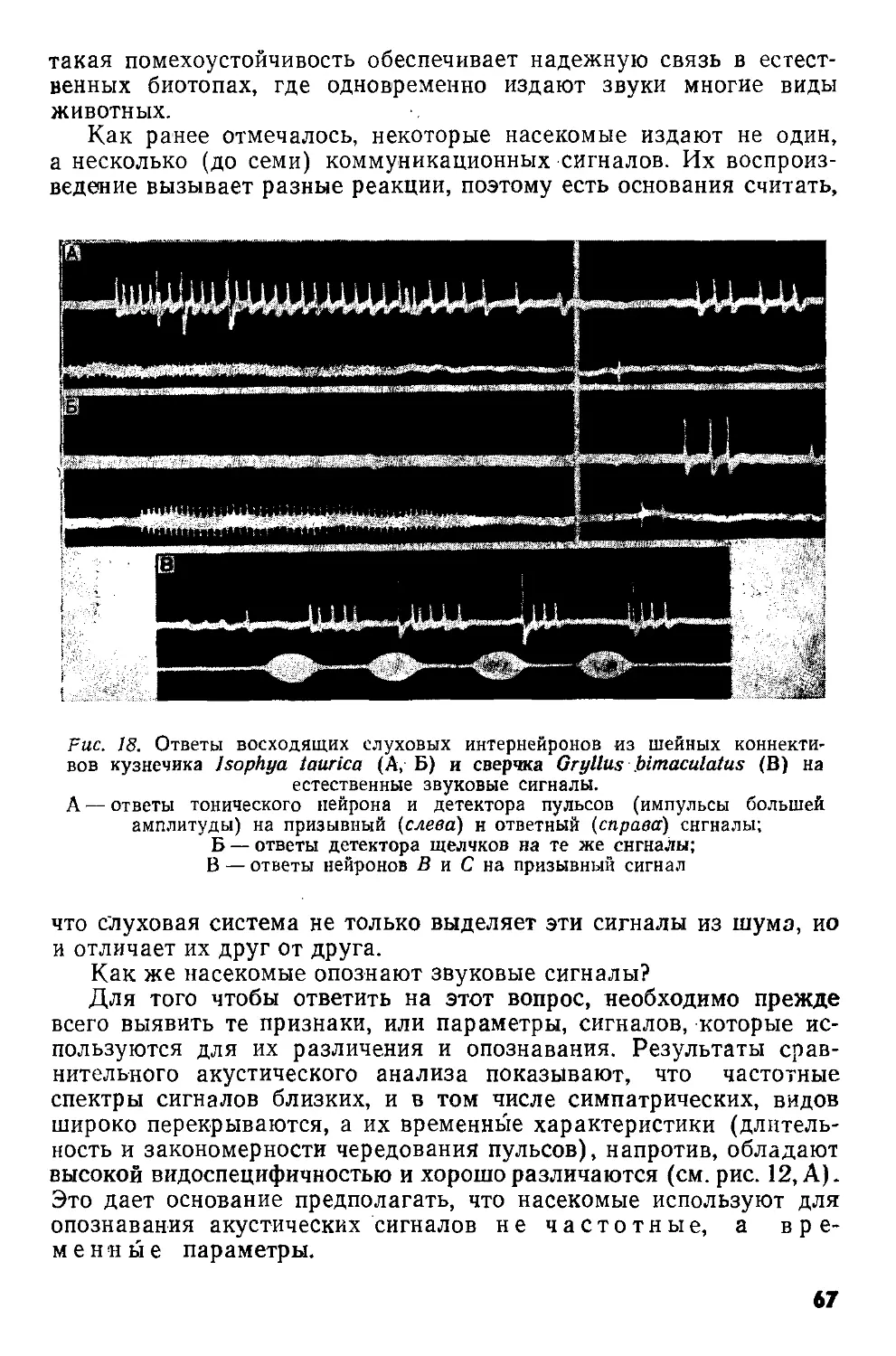
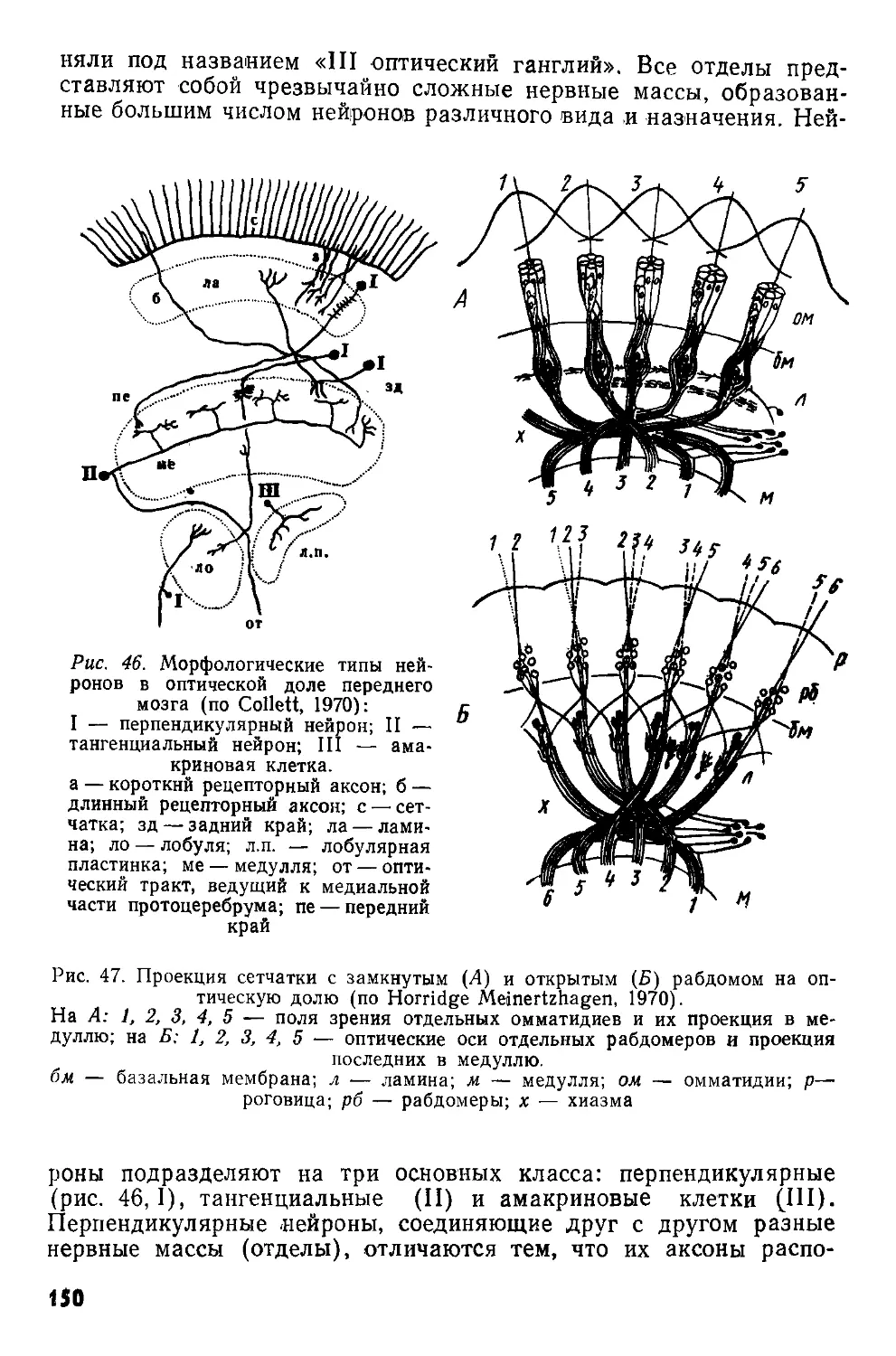
а б в г
Рис. 2. Различные ступени концентрации центральной нервной системы насекомых (по Eidmann, Kiihlhorn, 1970):
а — гипотетическое исходное состояние; б — слепень Tabanus bovinus-, в — муха Sarcophagus г — клоп семейства Pentatomidae. О — надглоточный ганглий; U — подглоточный ганглий; I—III — грудные ганглии;
1—8 — брюшные ганглии
животных,— нервная клетка, или нейрон. От тела нейрона, в котором заключено ядро, отходят обычно один длинный неразветвленный аксон и несколько коротких ветвящихся дендритов. В зависимости от числа и взаимного расположения этих отростков говорят о моно-, би- и мультиполярных нейронах. Дендрит служит входом нейрона, а аксон — выходом. Морфологически и функцио-
нально различают три главных типа нейронов: чувствующие, двигательные и вставочные. Нас в первую очередь будут интересовать чувствующие и вставочные.
Чувствующие нейроны.
Тела чувствующих, или сенсорных, клеток обычно биполярной или мультиполярной формы 1 лежат всегда вблизи чувствующего органа или иннервируемой ткани. Дендриты одних нейронов, чаще
всего биполярных, связаны с кутикулярными образованиями, дру-
гих, всегда мультиполярных, — с тканями полости тела или же они образуют субэпидермальную сеть, как у личинок с мягкой кожей.
Соответственно различают две большие категории чувствующих клеток. Клетки первого типа отличаются тем, что практически всегда связаны с кутикулой или ее впячиваниями: аподемами, трахеями, выстилкой предротовой и ротовой полостей и т. п. К ним принадлежат разнообразные экстерорецепторные клетки, в том числе и зрительные, хотя их дендриты выражены неясно. Клетки второго типа никогда не связаны с кутикулой и лежат только на внутренней поверхности тела, стенках пищеварительного тракта, в мышечной и соединительной тканях. Электрофизиологически показано, что они принадлежат интеро- или проприоцепторам.
1 Некоторые авторы относят чувствующие клетки к особой категории нервных клеток и не называют их нейронами.
16
Аксоны чувствующих клеток идут непосредственно в соответствующие ганглии ЦНС, иногда находящиеся непосредственно в головном мозгу, например оптические или обонятельные центры. Вопрос о каналах связи рецепторных клеток с нервным центром чрезвычайно важен для правильной интерпретации работы анализатора и механизма управления поведением насекомого. Теперь, по-видимому, все признают несостоятельным прежнее мнение о том, что в некоторых рецепторных системах, например в антеннах клопа Rhodnius, имеет место срастание аксонов нескольких чувствующих клеток в единое волокно. Но замыкание группы рецепторов на один периферический нейрон второго порядка, т. е. потеря «адреса» входного сигнала, характерно для первого оптического ганглия насекомых. Смысл такого способа связи с центром, ведущего к частичной потере информации от совокупности датчиков, пока не всегда ясен (см. ниже).
Нервная ткань, в том числе и сенсорные клетки, происходят из эктодермы. Их принадлежность покрову тела выражается и в том, что связь чувствующего органа с ЦНС устанавливается центростремительно. Так, В. Вигглесворс показал на клопе Rhodnius, что перерезанный афферентный нерв регенерирует в направлении к ЦНС. Точно так же во время каждой линьки, когда образуются дополнительные рецепторы, чтобы обслуживать увеличивающуюся поверхность тела, их чувствующие клетки посылают аксоны центростремительно.
Выявленный на гистологических препаратах факт центростремительного развития аксона может стать одним из оснований для важного заключения о том, что путь от чувствующей клетки до ЦНС прямой, без синаптического переключения. Вблизи рецепторных клеток и афферентных нервов встречаются другие, например, нейроглиальные (питающие) клетки, но они не имеют отношения к передаче рецепторного сигнала.
Двигательные (моторные) нейроны. Тела этих обычно униполярных нейронов лежат всегда в сегментальном ганглии, чаще с его вентральной стороны и с боков. Их дендриты густо ветвятся в нейропиле ганглия, вступая в синаптические контакты с вставочными нейронами или даже непосредственно с разветвлениями аксонов чувствующих клеток. Аксоны двигательных нейронов уходят за пределы ганглия к мышцам, где заканчиваются множеством моторных концевых пластинок, или бляшек. Одну и ту же мышцу иннервируют так называемые быстрые и медленные двигательные волокна: по первым идут импульсы, вызывающие быстрые сокращения мышцы, по вторым — вызывающие тоническое сокращение. При этом тело иннервирующего двигательного нейрона может лежать в ганглии как того же сегмента, которому принадлежит мышца, так и соседнего. Часто встречается контралатеральное положение тела двигательного нейрона по отношению к эффектору.
Вставочные нейроны, или интернейроны, выполняют функцию передачи возбуждения с одного нейрона на другой. Они связы
17
вают чувствующие клетки с моторными, причем в последовательную цепь может быть включено несколько клеток, и тогда говорят об интернейронах 2-го, 3-го и более высокого порядка. Их обычно униполярные небольшие (от 2—3 мкм и меньше) тела лежат всегда по периферии ганглия, а отростки, в числе которых аксон, нередко т-образный, вступают внутрь ганглия, в его нейропиль. Интернейроны могут быть сегментальными или мультисегментальны-ми, с восходящими и нисходящими волокнами. Последние участвуют, например, в управлении локомоторными реакциями. Благодаря множеству связей интернейроны образуют коммутационные сети для выработки нервных команд, направляемых по специальным трактам в составе комиссур и коннектив.
Среди восходящих волокон у насекомых обращают на себя внимание аксоны так называемых гигантских интернейронов, в частности, берущие начало в последнем брюшном ганглии и идущие по цепочке до грудных и даже до головных центров. Они отличаются большой длиной и толщиной (до 30 мкм и более), а главное, высокой скоростью проведения сигналов. Так, если в обычных афферентных двигательных нервах импульсы распространяются со скоростью 1,5—3,0 м/с, то в гигантском волокне, например таракана Periplaneta, скорость проведения может достигать 5—7 м/с. Считалось, что с помощью таких «быстрых» интернейронов осуществляется реакция убегания, когда опасность грозит насекомому сзади, но теперь этот взгляд признан несостоятельным.
Очень сложна область ганглиозного нейропиля, в котором генерируются нервные команды, управляющие поведением насекомого. Ее образует переплетение отростков дендритов моторных нейронов с разветвлениями ассоциативных и афферентных волокон, связанных друг с другом множеством синаптических контактов. Волокна здесь особенно тонкие: от 0,1 мкм по сравнению 1—3 мкм на выходе из дорсальных (двигательных) и вентральных (чувствующих) корешков. Они содержат протонейрофибриллы диаметром 100— 200 А, митохондрии, гранулярный эндоплазматический ретикулум, нейросекреторные включения и прочие элементы. Синаптическая передача основана на поперечных и продольных контактах волокон или на контакте типа терминальной пуговки. Ультраструктура синаптических образований напоминает таковую синапсов у позвоночных: пресинаптическое волокно в районе контакта содержит синаптические пузырьки, наполненные химическим медиатором. Между трехслойными пре- и постсинаптическими мембранами находится синаптическая щель шириной 100—300 А. Как полагают, у насекомых имеется не только холинэнергический, но и адренэнер-гический механизм синаптической передачи. Не исключается также возможность электрической передачи возбуждения с помощью соответствующих синапсов, в которых импульсы передаются в результате электрического воздействия пресинаптического тока на мембрану постсинаптического волокна. Такие электрические си
18
напсы недавно обнаружены между зрительными интернейронами— детекторами движения, где они обеспечивают максимально высокую скорость проведения сигналов.
2. ОСОБЕННОСТИ СЕНСОРНОЙ СИСТЕМЫ
Сенсорные органы представляют собой специализированные образования, предназначенные улавливать энергию внешних или внутренних стимулов для активации их нервных клеток, чьи сигналы информируют ЦНС о характере стимуляции. К таким стимулам, посредством которых организм регулирует свои отношения с окружающей средой, относится прежде всего энергия частиц и молекул, слагающих мир. Так, зрительные клетки поглощают фотоны определенной энергии, отраженные окружающими предметами или исходящие непосредственно от их источника. Лучистое тепло улавливают другие клетки путем абсорбции фотонов более широкого энергетического уровня. Вкус и запах улавливаются путем утилизации потенциальной энергии, заложенной во взаимном притягивании и отталкивании частиц, образующих атом. Звук обнаруживается посредством использования энергии движения частиц молекулярного уровня.
Все клетки организма работают за счет внешней энергии, получаемой в конечном счете в виде молекул или частиц. Но чувствующие клетки отличайтся от остальных, по крайней мере, в двух отношениях. Во-первых, энергия внешнего воздействия служит для чувствующей клетки только стимулом, или триггером, для запуска процессов, совершаемых за счет потенциальной, накопленной через механизм обмена энергии самой клетки. Поэтому энергия, необходимая для возбуждения рецептора, всегда меньше той, которую несет его выходной сигнал. Во-вторых, чувствующая клетка имеет на выходе электрическую энергию, обязательно передаваемую другим клеткам, которые сами не способны воспринимать энергию данного внешнего воздействия.
Обратимость изменения состояния в ответ на внешнее воздействие позволяет чувствующей клетке служить датчиком информации, однако в этой роли к ней предъявляются очень жесткие требования. Рецептор не сможет дать много полезной организму информации, если он так груб, что не чувствителен даже к большим изменениям энергии на входе, или, наоборот, так «совершенен», что реагирует на незначительные флюктуации входного сигнала. Кроме того, польза рецептора была бы небольшой при одинаковой реакции его на разные виды энергии или простой сигнализации о наличии или отсутствии стимула. С точки зрения запросов организма у рецептора должны быть: 1) некоторый оптимум чувствительности; 2) способность выделять определенные виды стимулов; 3) способность улавливать не только появление и исчезновение стимула, но и величину, направление и скорость его
19
изменения. Природа позаботилась об удовлетворении этих запросов. Хотя для клеточной протоплазмы характерна умеренная чувствительность ко многим видам энергии и реакция на многие параметры стимулов, рецепторы специализировались на восприятие определенного вида энергии. Специализация рецептора выражается в повышенной чувствительности каждого к «своему» виду стимула и его определенным параметрам. Это оказалось возможным, например, за счет приобретения рецепторными клетками специальных свойств на молекулярном уровне. Так, приобретение зрительными клетками фотолабильного пигмента позволило поглощать лучистую энергию в определенном диапазоне длин волн и отличать свет от теплового воздействия лучей.
В дополнение к специализированной чувствительности как таковой рецепторные клетки приобрели окружение в виде вспомогательных структур, предопределяющих род воспринимаемой энергии. В простейшем случае это — изгибающийся под действием внешних сил кутикулярный волосок, в более сложном — хрусталик, ирисовые клетки и другие элементы глаза, обслуживающие фоторецепторную часть — сетчатку, или тимпанальная мембрана органа слуха. Названные структуры могут не только влиять на чувствительность такой комбинированной рецепторной сенсиллы (например удаление хрусталика ухудшает зрение, а прободение тимпанальной мембраны ухудшает слух), но и предопределяют, какие параметры стимула (величина, скорость изменения и т. п.) могут быть использованы. Можно сказать вслед за известным американским физиологом В. Дж. Детье, что «очищение» рецепторных органов от генерализованных признаков и неизбирательности ответа прослеживается на всем эволюционном пути как их сенсорных клеток, так и дополнительных структур.
В чем же специфика сенсорной системы насекомых? Два типичнейших признака насекомых — малая величина тела и наличие экзоскелета — накладывают на их сенсорную систему и весь нервный аппарат определенные ограничения. Прослеживаемая в филогенезе насекомых миниатюризация тела ведет к укорочению нервных путей и сокращению времени поступления нервных импульсов от рецептора к нервному центру и от последнего к эффектору. Это может повлечь за собой, при прочих равных условиях, более высокую реактивность и подвижность малого по размерам организма по сравнению с крупным. И действительно, среди класса насекомых у мелких форм тараканов, прямокрылых, жуков, перепончатокрылых замечена тенденция реагировать быстрее по сравнению с крупными формами. Кроме того, малая величина тела неизбежно вызывает уменьшение числа нейронов, возможных в данном объеме. Если мозг млекопитающих содержит более 1010 нейронов, то во всей нервной системе таракана Periplaneta число нейронов— порядка 105, а в каждом отдельном ганглии его брюшной цепочки — от 102 До 103 нейронов. При небольшом числе нейронов уменьшается информационная емкость нервной системы, но это об
20
стоятельство, как мы увидим ниже, не ограничивает процветания насекомых. Более того, редукции числа нейронов у них способствует образование специальных гигантских волокон, так характерных для этих членистоногих. Например, гигантские волокна у Ре-riplaneta занимают 12% площади поперечного сечения брюшной нервной цепочки.
По мнению физиолога Редера (1959), относительное преимущество нервной системы из малого числа крупных единиц перед нервной системой, образованной многими небольшими единицами, легко понять, если допустить, что скорость реакции для ее обладателя важнее, чем детали информации. Так, чтобы обнаружить и успеть избежать врага, высокая скорость реагирования дает насекомому большую выгоду, чем подробная информация о враге. Для насекомого-хищника скорость броска также важнее подробного представления о жертве. С этой точки зрения вполне понятно развитие системы гигантских волокон, проводящих нервные команды быстрее, хотя бы и ценой потери потенциальной информации, которую мог бы передать нерв того же сечения, но составленный из большого числа обычных (тонких) волокон.
Пример экономности нервных элементов мы находим и в мб-торной системе насекомых. Мышцы насекомых иннервируются не сотнями; двигательных волокон, а единицами: их может быть четыре, два и даже одно на целую мышцу. Всю мускулатуру второй пары ног личинки стрекозы Aeschna обслуживают только 6 мотонейронов. У пустынной саранчи такую сложную мышечную систему, как крыловая, обслуживают не более чем по 80 мотонейронов в мезо- и метаторакальном ганглиях. Однако миниатюрный двигательный аппарат'насекомых работает не менее прецизионно, чем даже у позвоночных, а по скорости сокращения мышц насекомые намного превосходят их.
Очевидно и ограничение числа нейронов в собственно сенсорной системе насекомых. Оно связано не только с миниатюрностью тела, но и с тем, что тело заключено в жесткую, лишенную чувствительности кутикулу. Если у млекопитающих с их мягкими покровами кожные рецепторы исчисляются миллионами, то покров насекомых относительно малочувствителен, поскольку энергия внешних стимулов может достичь чувствующих клеток только в отдельных точках тела, специально предусмотренных природой. Например, на вентральной поверхности каждого брюшного кольца личинки четвертого возраста клопа Rhodnius находится до 420 рецепторов; на всей ноге мухи Phormia — менее 500 рецепторов; общее число тактильных рецепторов на теле трутня медоносной пчелы не превышает 3000. И только на высокоспециализированных сенсорных областях — в глазах и на антеннах — число рецепторов велико, хотя и здесь их все же меньше, чем в органах зрения и обоняния позвоночных. В фасеточном глазу максимальное число рецепторных (зрительных) клеток обнаружено у стрекоз — приблизительно до 210 000. Обычно же их всего несколько тысяч
21
или даже сотен, тогда как в сетчатке позвоночных — миллионы. На каждой антенне трутня медоносной пчелы до 500 тыс. сенсорных клеток.
С экономностью числа нервных клеток связана интересная особенность нервной системы насекомых, заключающаяся в том, что для совершения рефлекторного ответа организма достаточно возбудить одну-единственную чувствующую клетку. Так, стимуляция одного чувствующего нейрона волоска на оральном диске мух вызывает движение хоботка, т. е. запускает один из начальных этапов пищевой реакции. Сходным образом, раздражение одного тактильного нейрона может вызвать в ряде случаев последовательность поведенческих ответов — от простого отдергивания придатка до убегания насекомого.
Экономность сенсорной системы насекомых выражается, наконец, и в том, что, по-видимому, подавляющее большинство их рецепторных клеток, независимо от модальности или расположения на теле,— первичночувствующие. Так называемые «первичные» (первичночувствующие) рецепторные клетки выполняют двойную функцию: они не только воспринимают энергию внешнего стимула, но и генерируют и сами же передают импульсы в ЦНС. «Вторичными» (вторичночувствующими) рецепторами называют такие, у которых чувствующий нейрон возбуждается не внешним стимулом, а через посредство неразрывно связанной с ним одной или нескольких собственно воспринимающих клеток. К вторичным рецепторам у высокоразвитых животных относят рецепторы органов слуха, зрения, вкуса, боковой линии, а к первичным — обонятельные и тактильные клетки, мышечные веретена.
Чувствующий нейрон (или группа нейронов) у насекомых, как правило, объединен с дополнительными обслуживающими клетками в элементарный орган, называемый сенсиллой. Как нейрон, так и функционально связанные с ним клетки, дающие выход к кутикулярным структурам или внутренним стенкам тела, происходят путем деления одной материнской эктодермальной клетки. Полагают, что за возможным исключением фоторецепторов все сенсиллы, столь характерные насекомым, представляют собой гомологи кожного волоска, или хеты. Они состоят минимум из трех клеток: волоскообразующей (трихогенной), гнездообразующей (тррмогенцой) и чувствующего нейрона. Сенсиллы рассеяны по телу поодиночке или образуют поля, либо объединены в сложные органы, такие, как, например, фасеточные глаза и тимпанальные (слуховые) органы.
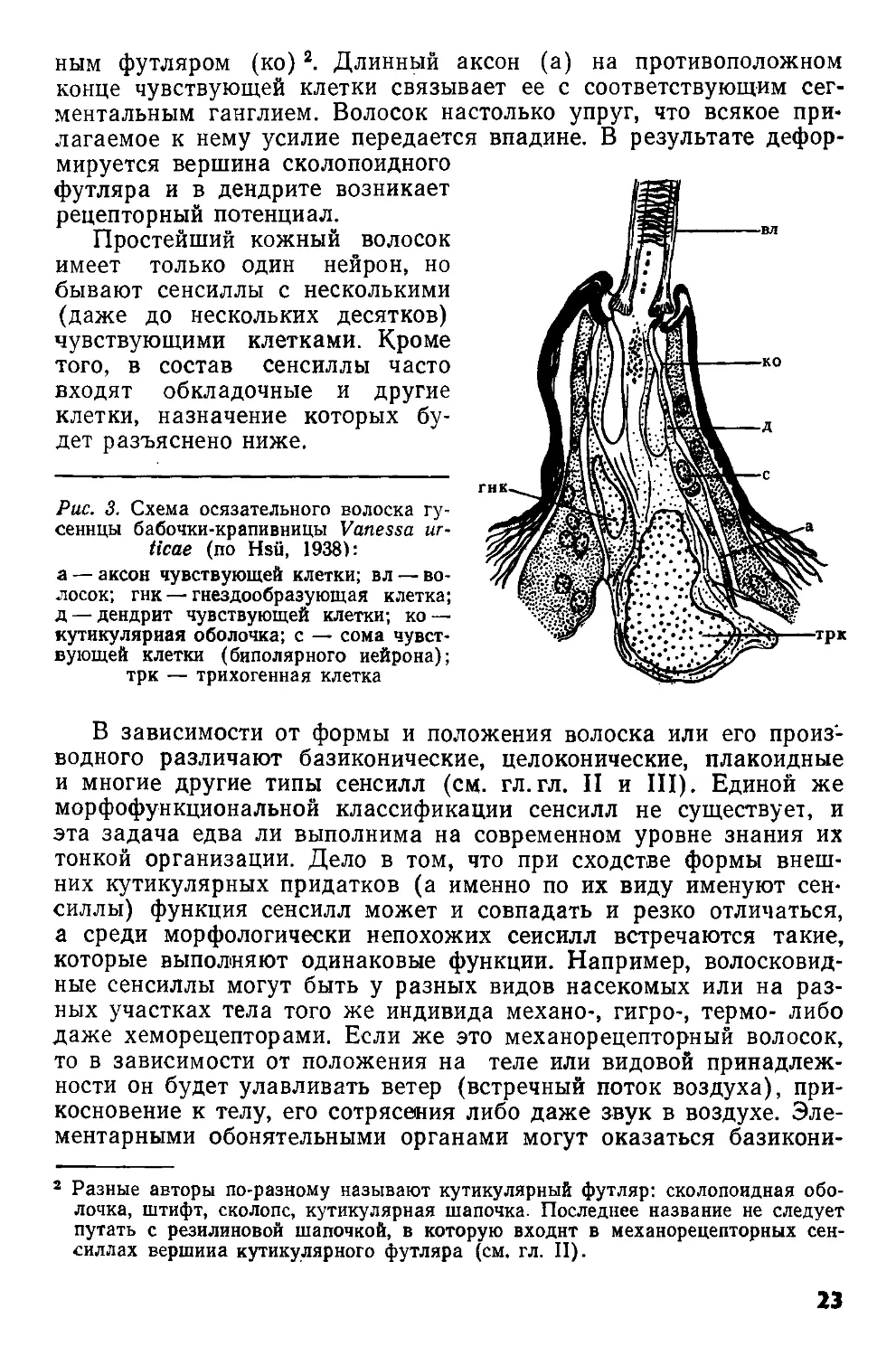
Общий план строения сенсилл рассмотрим на примере наиболее простой — трихоидной — сенсиллы, или кожного волоска (рис. 3). Его функция полимодальна: осязание и восприятие колебания воздуха. Полый кутикулярный волосок (вл) сидит в гибкой мембранозной впадине. С утолщенным основанием волоска контактирует вершина короткого неразветвленного дендрита (д) чувствующей клетки, одетого в своей терминальной части кутикуляр
22
ным футляром (ко)2. Длинный аксон (а) на противоположном конце чувствующей клетки связывает ее с соответствующим сегментальным ганглием. Волосок настолько упруг, что всякое прилагаемое к нему усилие передается впадине. В результате дефор
мируется вершина сколопоидного футляра и в дендрите возникает рецепторный потенциал.
Простейший кожный волосок имеет только один нейрон, но бывают сенсиллы с несколькими (даже до нескольких десятков) чувствующими клетками. Кроме того, в состав сенсиллы часто входят обкладочные и другие клетки, назначение которых будет разъяснено ниже.
Рис. 3. Схема осязательного волоска гусеницы бабочки-крапивницы Vanessa иг-ticae (по Hsu, 1938):
а — аксон чувствующей клетки; вл — волосок; гнк — гнездообразующая клетка; д — дендрит чувствующей клетки; ко — кутикулярная оболочка; с — сома чувствующей клетки (биполярного нейрона);
трк — трихогенная клетка
В зависимости от формы и положения волоска или его производного различают базиконические, целоконические, плакоидные и многие другие типы сенсилл (см. гл. гл. II и III). Единой же морфофункциональной классификации сенсилл не существует, и эта задача едва ли выполнима на современном уровне знания их тонкой организации. Дело в том, что при сходстве формы внешних кутикулярных придатков (а именно по их виду именуют сенсиллы) функция сенсилл может и совпадать и резко отличаться, а среди морфологически непохожих сеисилл встречаются такие, которые выполняют одинаковые функции. Например, волосковидные сенсиллы могут быть у разных видов насекомых или на разных участках тела того же индивида механо-, гигро-, термо- либо даже хеморецепторами. Если же это механорецепторный волосок, то в зависимости от положения на теле или видовой принадлежности он будет улавливать ветер (встречный поток воздуха), прикосновение к телу, его сотрясения либо даже звук в воздухе. Элементарными обонятельными органами могут оказаться базикони-
2 Разные авторы по-разному называют кутикулярный футляр; сколопоидная оболочка, штифт, сколопс, кутикулярная шапочка. Последнее название не следует путать с резилиновой шапочкой, в которую входит в механорецепторных сенсиллах вершина кутикулярного футляра (см. гл. II).
23
ческие и плакоидные, целоконические и некоторые другие сенсиллы. Даже такие специализированные сенсиллы, как хордотональные, реагируют на звук, вибрацию, растяжение частей тела. Наконец, известны и такие сенсиллы, которые одновременно служат двум целям. Например, волоски на хоботке мухи Calliphora— одновременно механо- и хеморецепторы. Одни чувствующие клетки таких сложных сенсилл возбуждаются при сдавливании, а другие — исключительно химическими стимулами.
Представление о многообразии видов чувствительности насекомых дает следующая классификация, в которой, повторяем, пока только отдельным модальностям стимулов удается указать морфологически строго определенный тип сенсиллы, например восприятию фотонов света — светочувствительную клетку с рабдомом, содержащим фотопигмент.
I. Механорецепция
1. Тактильная чувствительность (осяза-
ние) 2. Интеро- и проприорецепторная чувст-
вительность (оценка механических состояний внутри тела и взаимного поло- Контактное
жения его частей) 3. Статическое чувство (чувство равнове- восприятие
‘ сия) 4. Сейсмическая чувствительность (вос-
приятие сотрясений) 5. Слух. Дистантное восприятие
//. Термо- и гигрорецепция
Восприятие конвекционной и лучистой Контактное
тепловой энергии, а также парообраз- и дистантное
ной и капельно-жидкой влаги восприятие
III. Хеморецепция
1. Вкус Контактное восприятие
2. Обоняние Дистантное восприятие
IV. Фоторецепция
1. «Кожная» светочувствительность Только дистантное
2. Зрение восприятие
Помимо этого насекомые реагируют также на силу гравитации, на магнитные и электрические поля, но специфических рецепторов, предназначенных исключительно для восприятия названных стимулов, у них пока не обнаружено. Реакция обеспечи
24
вается прежде всего возбуждением разнообразных механорецепторов, локализованных в местах сочленений тела: головы с грудью, переднегруди со среднегрудью и, конечно, отдельных частей ног. Что касается чувствительности к магнитному полю и умеренному по силе полю статического электричества, то скорее всего и она связана с возбуждением проприорецепторов, контролирующих растяжение и взаимное положение частей тела.
Таким образом, большинство типов рецепторов насекомых «обращено» во внешнюю среду. Это — экстерорецепторы, воспринимающие внешние стимулы изначально контактным способом, а по мере специализации — и на расстоянии, т. е. .дистантно. Но столь же важны для жизни насекомых интеро- и проприорецеп-торы, сигнализирующие о раздражителях внутренней среды и двигательной системы организма. Интеро- и проприорецепторы, хотя и «обращены внутрь» организма, но при определенных условиях, как, например, в случае восприятия гравитации или магнитного поля, могут давать информацию и о внешней среде. В последнем случае ЦНС отличает (опознает) реакцию, например, проприорецепторов при активном сгибании суставов от реакции на гравитацию. Совпадение сигналов от рецепторов в разных частях тела истолковывается как действие силы тяжести, а несовпадение— как следствие локомоции.
3. ФИЗИОЛОГИЧЕСКИЕ МЕХАНИЗМЫ СЕНСОРНОЙ СИСТЕМЫ
И МЕТОДЫ ИХ ИЗУЧЕНИЯ
Последовательность процессов, протекающих в возбужденных рецепторах насекомых, сводится к следующей схеме. Адекватный раздражитель, взаимодействуя с рецептирующими элементами дендрита чувствующей клетки, вызывает такие локальные процессы, которые изменяют проницаемость их поверхностной мембраны. В результате возникают ионный ток и, как следствие, деполяризация клеточной мембраны, регистрируемая в виде медленного рецепторного потенциала (рис. 4, А, Б). Когда деполяризация достигает некоторого критического уровня, в определенной зоне клетки, как полагают, в аксонном холмике, возникают импульсы, или спайки, распространяющиеся по аксону в направлении ганглия. Мембрана хёморецепторных клеток под действием одних веществ деполяризуется, под влиянием других — гиперпо-ляризуется. В последнем случае импульсный ответ в аксоне клетки затормаживается. Градуальный деполяризационный потенциал клетки называют также генераторным3, поскольку он приводит к генерации быстрого ответа клетки в виде импульсов, возникаю
3 Во вторичночувствующих рецепторах позвоночных рецепторный потенциал отличают от генераторного.
25
щих, как известно, по закону «все или ничего» (рис. 4, В). Импульсы бывают двухфазными или многофазными, как, например, в аксоне вкусовых и многих механорецепторных клеток. Моно-фазные импульсы механорецепторных нейронов отличаются увеличением их амплитуды по мере нарастания генераторного потенциала.
Рис. 4. Ответы рецепторных клеток.
А, Б — рецепторный потенциал зрительной клетки мухи Luci-На, зарегистрированный внутриклеточно; запись на А сделана при освещении в тысячу раз выше, чем запись Б (по Naka, 1961).
В — рецепторный потенциал с наложенными на нем импульсами, зарегистрированный вне-клеточно от вкусовой сенсиллы мухи Phormia regina-, стимуляция 0,5 М раствором глюкозой (по Uehara, Morita, 1972)
Процессы трансформации энергии стимула, приводящие к изменению проницаемости рецепторной мембраны, пока изучены не настолько, чтобы можно было говорить о них с уверенностью и подробно (в какой-то мере исключение составляют фоторецепторы позвоночных животных).
Итак, если мы будем считать, что рецепторы насекомых являются первичными, то, следовательно, у них одна и та же сенсорная клетка и рецептирует стимул, и генерирует залп импульсов. Нет сомнений, что именно так обстоит дело в хеморецептор-ных или механорецепторных сенсиллах. Но зрительные клетки насекомых следует отнести скорее всего ко вторичным рецепторам, как и у высших животных. При их возбуждении, в период увеличения деполяризации, наблюдаются так называемые миниатюрные потенциалы, происхождение которых объясняют выделением порций медиатора из рабдомера, представляющего собой рецептор, объединенный с сенсорным нейроном — зрительной клеткой.
Деполяризационный ответ рецепторной клетки, как и частота импульсации в ее аксоне, в определенных пределах пропорционален логарифму интенсивности стимула. Максимальная амплитуда медленного ответа близка к величине потенциала покоя клетки — 30—70 мВ. В аксонах разных рецепторов импульсы могут следовать как в течение всего периода действия раздражителя, так и только после его включения (on-ответ) или выключения (off-ответ); помимо on-off-ответов в волокнах часто регистрируется спонтанная (фоновая) импульсация. Соответственно разли
26
чают тонические рецепторы, медленно адаптирующиеся к воздействию или неадаптирующиеся совсем, и фазовые, реагирующие только на изменение стимула. Фазовые рецепторы обычно быстро адаптируются — на повторные воздействия отвечают постепенно уменьшающимся ответом.
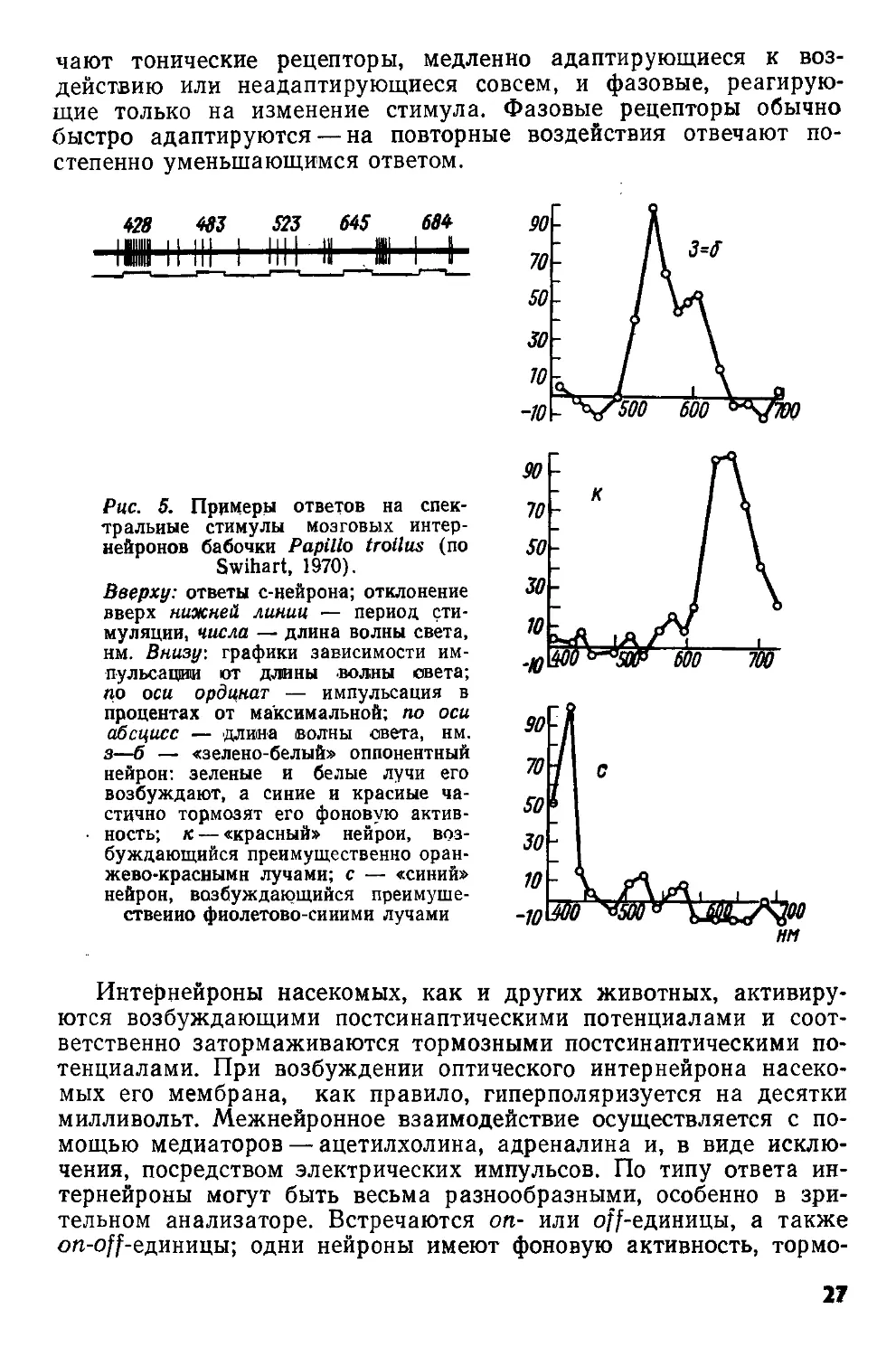
428 483 523 645 684
Рис. 5. Примеры ответов на спектральные стимулы мозговых интернейронов бабочки Papilla troilus (по
Swihart, 1970).
Вверху: ответы с-нейрона; отклонение вверх нижней линии — период стимуляции, числа — длина волны света, нм. Внизу: графики зависимости им-пульсации ют длины волны света; по оси ординат — импульсация в процентах от максимальной; по оси абсцисс — длина волны света, нм. з—б — «зелено-белый» оппонентный нейрон: зеленые и белые лучи его возбуждают, а синие и красные частично тормозят его фоновую активность; к — «красный» нейрон, возбуждающийся преимущественно оранжево-красными лучами; с — «синий» нейрон, возбуждающийся преимущественно фиолетово-синими лучами
Интернейроны насекомых, как и других животных, активируются возбуждающими постсинаптическими потенциалами и соответственно затормаживаются тормозными постсинаптическими потенциалами. При возбуждении оптического интернейрона насекомых его мембрана, как правило, гиперполяризуется на десятки милливольт. Межнейронное взаимодействие осуществляется с помощью медиаторов — ацетилхолина, адреналина и, в виде исключения, посредством электрических импульсов. По типу ответа интернейроны могут быть весьма разнообразными, особенно в зрительном анализаторе. Встречаются on- или off-единицы, а также on-off-единицы; одни нейроны имеют фоновую активность, тормо
27
зящуюся при стимуляции рецепторов, у других она отсутствует; бывают адаптирующиеся к непрерывной стимуляции и неадап-тирующиеся нейроны (рис. 5).
О сложном межнейронном взаимодействии в мозге насекомых при различных входных (рецепторных) сигналах можно судить по такому примеру. Замечено, что у мухи Boethcherisca ре-regrina с закрашенными фасеточными глазами на активность одиночных интернейронов переднего, среднего и заднего мозга, связанных с обонятельными сенсиллами и механорецепторами антенн, влияют сигналы от простых глазков. При освещении глазков мухи спонтанная активность одних единиц усиливается, а других тормозится. Выключение света по-разному сказывается и на ответах разных интернейронов, вызванных, например, пахучим стимулом: одни из них усиливаются, другие ослабляются. Первые встречаются преимущественно в среднем мозгу, вторые же приурочены к дорсальной области переднего и вентральной части среднего и заднего мозга.
Для того чтобы проникнуть в механизмы работы сенсорной системы, необходим разносторонний подход к ее изучений). Физиологию в широком смысле интересуют следующие важнейшие моменты: 1) способы, которыми рецептор детектирует адекватные стимулы, т. ё. выделяет и поглощает энергию адекватного раздражителя; 2) молекулярные и ионно-обменные процессы в рецепторной клетке, приводящие в конечном итоге ,к генерации импульсов; 3) способы кодирования рецепторной информации; 4) механизм межнейронного взаимодействия и связи в ассоциативных центрах, а также способы обработки ими рецепторной информации. Завершает эту цепь поиска вопрос о соответствии между видами поведения целого организма и детектируемой им информацией 4.
Коротко остановимся на подходах и методах, которыми располагает современная физиология органов чувств.
Морфофункциональный подход, традиционный для биолога, приобрел теперь более широкие и даже новые возможности благодаря использованию не только разных вариантов световой, но и электронной, в том числе сканирующей, микроскопии. Возможность различать внутриклеточные и межклеточные структуры, измеряемые ангстремами, служит хорошей основой для суждения о механизмах поглощения и трансформации внешней энергии в рецепторе и передачи нервного сигнала по аксону. Здесь велика роль гистохимии, особенно много давшей для расшифровки молекулярных процессов в возбужденных фоторецепторах.
Электрофизиологический подход. Успех в понимании работы сенсорной и всей нервной системы насекомых, достигнутый в последние 10 лет, обязан, пбжалуй, в первую очередь именно электрофизиологическому подходу, в особенности микроэлектродному
4 В последнее время этот вопрос относят к области этологии животных.
28
методу регистрации активности нервных клеток.. Для внутриклеточного отведения электрической активности применяют тончайшие стеклянные микропипетки (диаметр кончика 0,6—0,3 мкм), заполненные 2М КС1. Такой микроэлектрод вводят в сенсиллу или нервную ткань до тех пор, пока скачком не изменится потенциал покоя на 40—(60 мВ, что свидетельствует о проколе клеточной мембраны 5. Внутриклеточным микроэлектродом можно регистрировать как генераторный потенциал клетки, так и ее импульсную активность. Активность нейронов может быть зарегистрирована и внеклеточным электродом. В зависимости от положения его кончика и точек приложения второго (индифферентного) электрода отводят либо медленный суммарный потенциал возбужденных клеток в виде, например, электроретинограмм (ЭРГ) или электроантеннограмм (ЭАГ), либо быстрый потенциал — экстра-клеточные импульсы. Экстраклеточные электроды используют не только для регистрации ответа нейрона, но и для его стимуляции, например, током или химическим агентом, как при излучении вкусовых волосков. Через регистрирующий электрод специальной краской можно пометить место отведения, что особенно важно при внутриклеточной регистрации.
Моделирование. Моделирование как способ изучения сенсорных органов и нейронных сетей все шире входит в арсенал физиолога. Исходя из параметров системы, создают либо физическую (например электронную), либо математическую модель ее. В последнем случае используют ЭВМ, для которой разрабатывают последовательность специальных команд —программу. Так, известны физические модели отдельных нейронов и целых систем, например цветового зрения некоторых животных. На ЭВМ многие исследователи пытаются моделировать механизм периферического кодирования признаков и центрального опознавания образов.
Поведенческий подход. Наблюдение за поведением животного следует считать, строго говоря, единственным адекватным способом для суждения о назначении и различительных возможностях его органов чувств. Биолога интересуют прежде всего те свойства сенсорных систем, которые реализуются в поведении, а точнее, те рецепторные сигналы, которые контролируют реакцию животного. Поэтому наблюдение за врожденными или приобретенными в ходе научения реакциями в норме и после нарушения целостности рецепторных органов или ассоциативных центров дает очень много для понимания их функции. Изменение поведения при частичной искусственно вызванной деафферентации позволяет выявить роль тех или иных рецепторов в управлении реакциями. Регистрируют как реакции отдельных частей тела, например выдвижение хоботка в ответ на стимуляцию вкусовых сенсилл или отдергивание ноги при ее механической стимуляции, так
s Поверхность клеточной мембраны имеет отрицательный знак по отношению к содержимому клетки. Поэтому введенный внутрь клетки микроэлектрод будет положительным относительно индифферентного электрода.
29
и моторные ответы целого организма — приближение к стимулу или уход от него. Реакция может быть как безусловнорефлекторной, так и на основе условного рефлекса, замыкающегося в простейшем случае, как показано известным физиологом Дж. Хор-риджем (1962), на уровне грудных ганглиев нервной цепочки. Подключив последовательно к источнику тока симметричные лапки у двух обезглавленных тараканов Periplaneta atnericana, он заметил, что то насекомое, которое в процессе обучения получало удар тока лишь при опускании ноги, замыкавшей цепь «наказания», в контрольных опытах чаще держало ее приподнятой, чем партнер, получавший удар вне зависимости от положения своей ноги.
Особенно легко вырабатывается условный рефлекс у общественных перепончатокрылых пчел, муравьев, ос, чем исследователи широко пользуются для изучения механизмов рецепторного управления поведением. Например, кардинальные сведения о цветовом зрении и обонянии медоносной пчелы были получены именно в опытах с обучением насекомого.
Глава II
МЕХАНОРЕЦЕПЦИЯ
Насекомые обладают очень разнообразными и совершенными рецепторами для восприятия механических стимулов. Их развитие и специализация протекали под влиянием трех основных факторов: приспособления к жизни в чрезвычайно разнообразных условиях среды, совершенствования способов передвижения (в том числе полета) и усложнения поведения. Все эти факторы содействовали развитию механизмов ориентации в пространстве и координации движений, что, в свою очередь, способствовало повышению уровня организации механорецепторов. С их помощью насекомые воспринимают почти любые внешние и внутренние механические стимулы: прикосновение, движение воздуха или воды, силу гравитации, угловые ускорения, вибрации субстрата, звуковые и ультразвуковые колебания, смещения различных частей тела и внутренних органов и др.
Сравнительные исследования механорецепторов насекомых свидетельствуют о том, что оии существенно различаются по уровню функциональной специализации, в результате чего некоторые из них могут реагировать на разные стимулы, а одно и то же механическое воздействие воспринимается несколькими рецепторами. Это обстоятельство, а также условность подразделения некоторых стимулов препятствуют разработке единой классификации механорецепторов. В настоящем руководстве мы будем придерживаться сравнительно простой системы, принятой большинством современных исследователей.
1. КРАТКИЕ СВЕДЕНИЯ
О МОРФОЛОГИИ МЕХАНОРЕЦЕПТОРОВ
Насекомые воспринимают разнообразные механические стимулы с помощью сенсилл трех основных типов: трихоидных, кампаниформных и хордотональных, а также «рецепторов растяжения». Каждый из этих элементов может функционировать самостоятельно, но чаще всего они концентрируются в определенных местах тела или объединяются со вспомогательными структурами, образуя более или менее сложные органы (тимпанальные, хордотональные и т. п.).
Прежде чем приступить к описанию физиологии перечисленных элементов, целесообразно рассмотреть некоторые особенности их ультраструктурной организации.
31
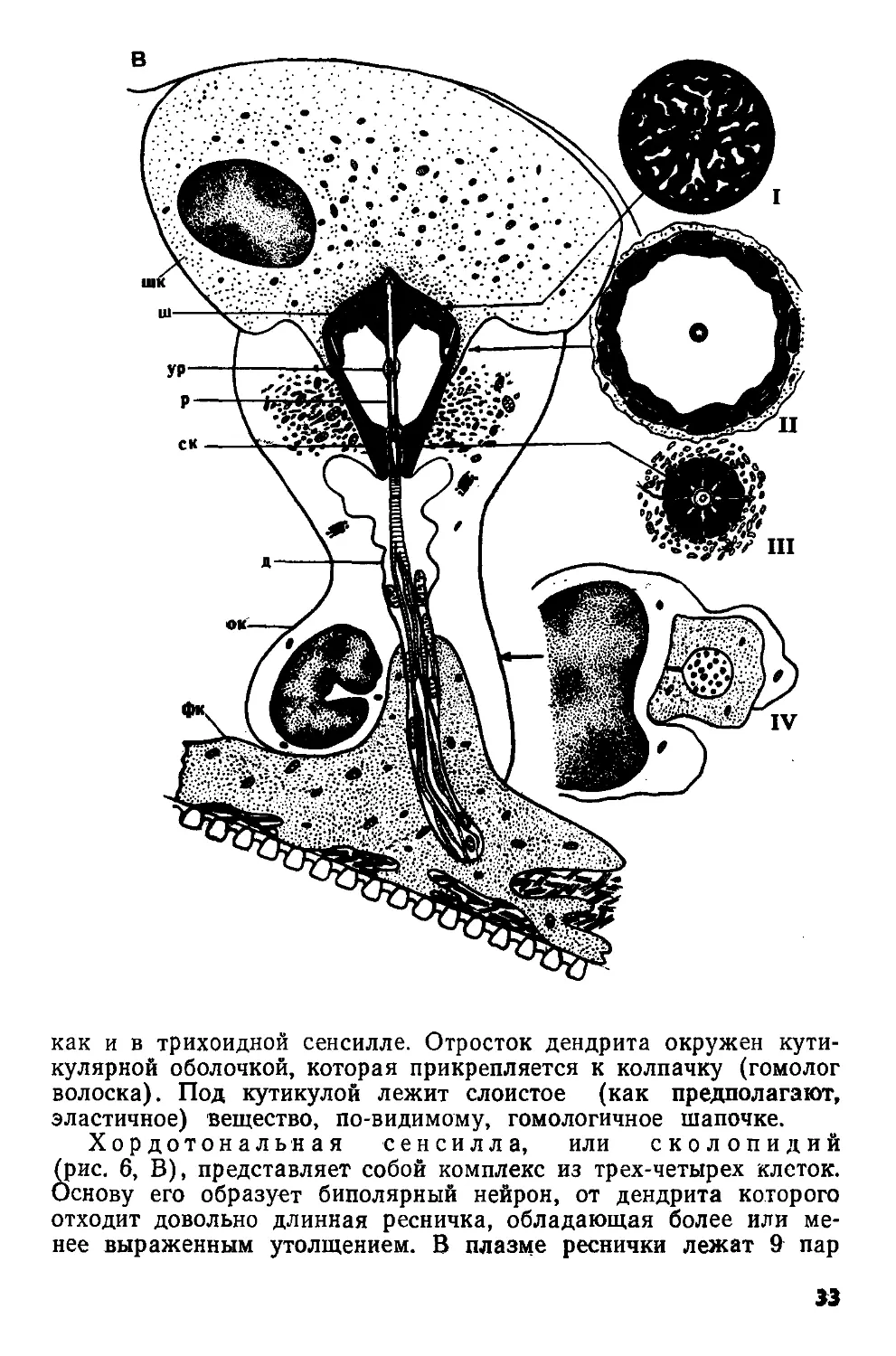
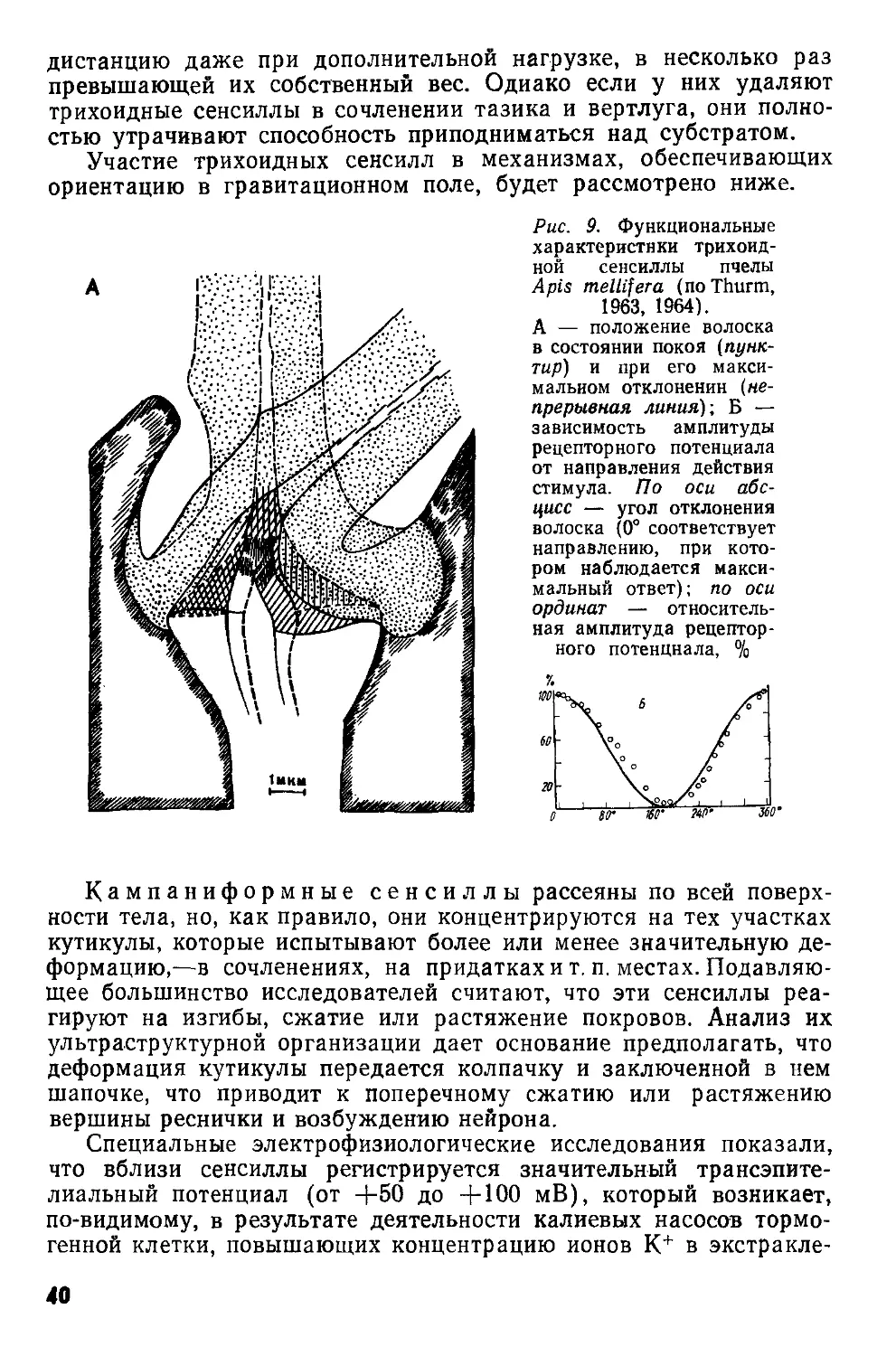
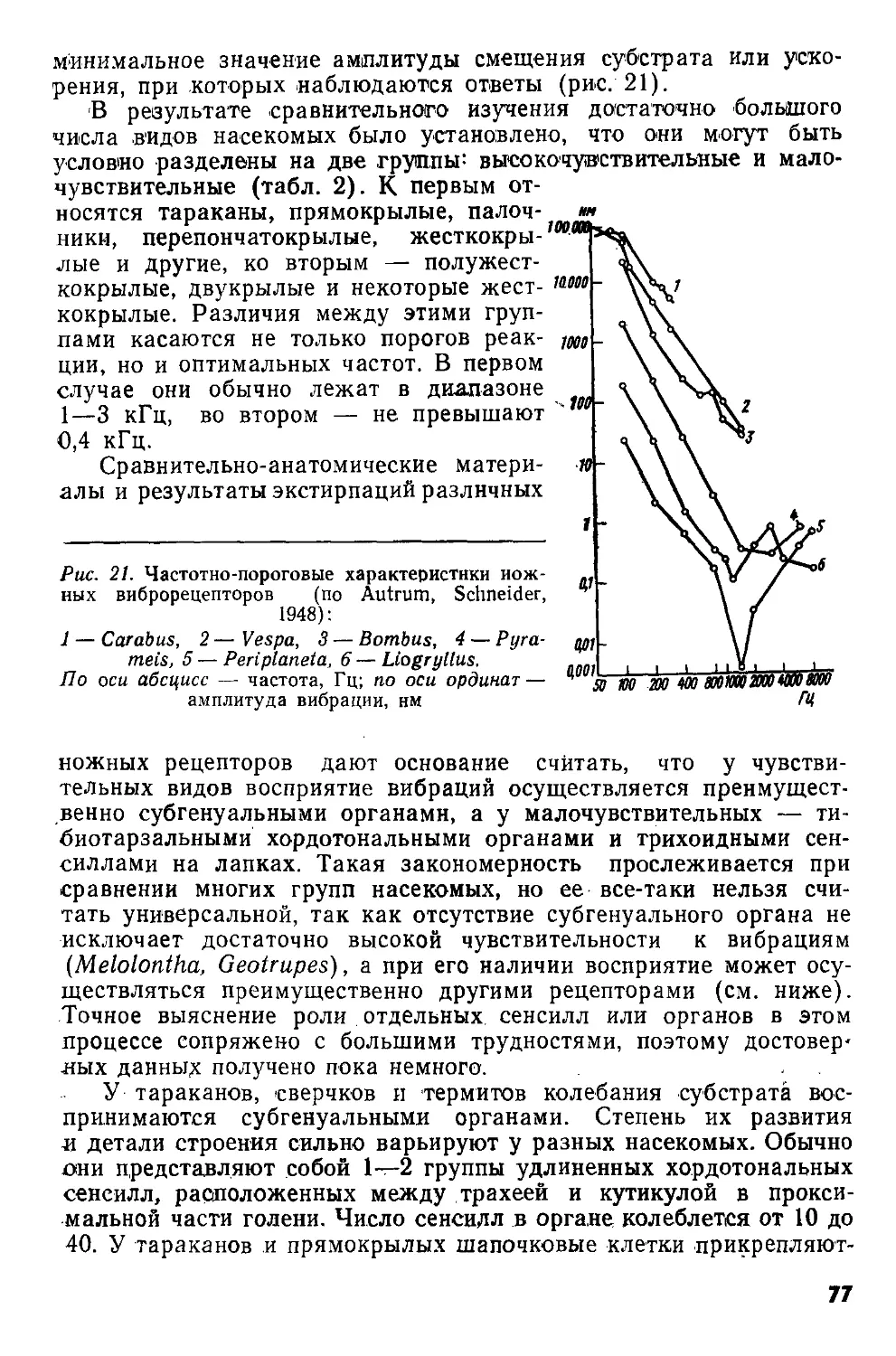
Рис. 6. Схемы строения механорецепторных сенсилл: А — трихоидной, Б — кампаниформной сенсилл пчелы Apis mellifera (по Thurm, 1964); В — хордотональной сенсиллы из тимпанального органа кузнечика Decticus verrucivorus (по Жантиеву, 1969).
вл — волосок; д — дендрит; ко — кутикулярная оболочка; к — кутикула; м — мембрана; ок — обкладочная клетка; р — ресничка; ск — сколопс; тт — трубчатое тело; трк — трихогенная клетка; ур — утолщение реснички; фк — фиброзная клетка; ш — шапочка; шк — шапоч-кован клетка.
I—IV — поперечные срезы
Общий план строения трихоидной сенсиллы (рис. 6,А) был изложен ранее. Исследование деталей ее организации показало, что сочленовная мембрана, обеспечивающая подвижность волоска, состоит из эластичного белка — резилина. Изнутри к ней примыкает губчатое образование, получившее название шапочки. Дендрит биполярного нейрона заканчивается веретеновидным отростком, представляющим собой модифицированную ресничку. В его цитоплазме обнаружено 9 пар фибрилл, связанных с корешковым аппаратом дендрита. Дистальная часть отростка, входящая в канал шапочки, содержит множество микротрубочек (трубчатое тело). Снаружи отросток окружен тонкой кутикулярной оболочкой, соединяющейся с основанием волоска. Большая часть дендрита заключена в складки двух вспомогательных клеток (трихогенной и тормогенной), к которым примыкают глиальные клетки.
У кампаниформной сенсиллы (рис. 6, Б) биполярный нейрон и вспомогательные клетки имеют такое же строение,
32
как и в трихоидной сенсилле. Отросток дендрита окружен кутикулярной оболочкой, которая прикрепляется к колпачку (гомолог волоска). Под кутикулой лежит слоистое (как предполагают, эластичное) вещество, по-видимому, гомологичное шапочке.
Хордотональная сенсилла, или сколопидий (рис. 6, В), представляет собой комплекс из трех-четырех клеток. Основу его образует биполярный нейрон, от дендрита которого отходит довольно длинная ресничка, обладающая более или менее выраженным утолщением. В плазме реснички лежат 9 пар
13
периферических фибрилл, связанных с корешковым аппаратом дендрита, а ее утолщенная часть обычно содержит три извитые нити. Снаружи дистальная часть дендрита и ресничка окружены ребристым сколопсом—внутриклеточным продуктом обкладочной клетки. Дистальные концы ребрышек сколопса примыкают к конусовидной или колоколовидной шапочке — экстраклеточному образованию, пронизанному многочисленными полостями и каналь-
Рис. 7. Схемы строения рецепторов растяжения (по Osborne, Finlayson, 1962) личинки ручейника Phryganea (А) и уховертки Forficula (Б)
цами. В центральной части шапочки расположен канал, в который входит дистальная часть реснички. Снаружи шапочку окружает шапочковая, или прикрепительная, клетка, обычно содержащая многочисленные тонофибриллы, а проксимальная часть дендрита заключена в складки фиброзной клетки и тяжи волокнистого материала, обеспечивающего прикрепление нейрона к близлежащим структурам.
Сенсиллы такого типа входят в состав тимпанальных, субге-нуальных и хордотональных органов. В некоторых антеннальных и ножных хордотональных органах найдены модифицированные сенсиллы, отличающиеся от описанных главным образом наличием двух нейронов и соответственно двух ресничек в одном ско-лопсе. Джонстоновы органы состоят из сенсилл, включающих по два или три нейрона. В последнем случае один из нейронов обладает недоразвитой ресничкой, а у двух других ее дистальная часть утолщена. Шапочка в этих сенсиллах превращается в длинный тонкостенный футляр, вершина которого обычно входит в сочленовную мембрану, соединяющую 2-й и 3-й членики антенны.
Рецепторы растяжения в отличие от описанных выше сенсилл представляют собой одиночные мультиполярные нейроны, расположенные в полости тела. Их дендриты ветвятся в соединительнотканных тяжах ил,и мышцах, в связи с чем различают два основных типа таких рецепторов. г
34
Примером первого из них могут служить органы, найденные Б брюшке у личинок стрекоз. Каждый сегмент брюшка содержит три пары клеток, связанных с соединительнотканными тяжами, занимающими продольное, дорсовентральное или косое положение. Подобные рецепторы обнаружены у поденок, тараканов, уховерток (рис. 7, Б), жуков и др. Специальные гистологические исследования показали, что окончания дендритов лишены глиальной оболочки (Шванновских клеток) и заключены в капсулу из соединительной ткани.
У прямокрылых в брюшке расположены рецепторы второго типа, связанные с продольными мышцами. Каждый из них состоит из крупного, окруженного капсулой мультиполярного нейрона и модифицированного мышечного волокна. Аналогичные рецепторы чешуекрылых и ручейников отличаются тем, что их мульти-полярные нейроны контактируют со специализированными мышечными волокнами, обладающими слабой поперечной исчерчен-ностью и очень крупными ядрами. К каждому такому волокну подходит отдельный мотонейрон (рис. 7,А).
2. ФУНКЦИИ ТАКТИЛЬНЫХ РЕЦЕПТОРОВ
При соприкосновении насекомых с твердым субстратом или при их перемещении в воздушной или водной среде возбуждаются механорецепторы, но наиболее важную роль в восприятии контактных стимулов играют рассеянные по всему телу трихоидные сенсиллы. Обычно они концентрируются на тех склеритах и придатках, которые чаще всего соприкасаются с окружающими предметами или интенсивней обдуваются во время полета.
Морфологические данные свидетельствуют о том, что, несмотря на сравнительно простую организацию и однотипность строения, эти сенсиллы часто обладают признаками достаточно выраженной специализации, указывающими на возможность избирательной рецепции определенных механических стимулов. Результаты физиологических и этологических экспериментов позволяют разделить трихоидные сенсиллы наземных насекомых на две группы. Представители одной из них возобуждаются при контакте с твердыми телами и могут быть названы собственно осязательными, в то время как другие приспособлены для восприятия движения воздуха и получили название ветрочувствительных. У водных форм аналогичные рецепторы должны возбуждаться во время плавания, но они изучены еще недостаточно, поэтому мы ограничимся рассмотрением двух упомянутых выше групп.
Детали строения осязательных рецепторов варьируют в значительных пределах, но почти все сенсиллы этого типа обла-, дают сравнительно жесткими толстостенными волосками и мало-эластичнЫми сочленовными мембранами. Помимо простых механо
35
рецепторов у насекомых встречаются сенсиллы со смешанными функциями, реагирующие на механические и химические стимулы. В полость их волосков входят отростки хеморецепторных нейронов (см. стр. 83).
Согласно существующим представлениям, при отклонении волоска из положения равновесия его основание или сочленовная
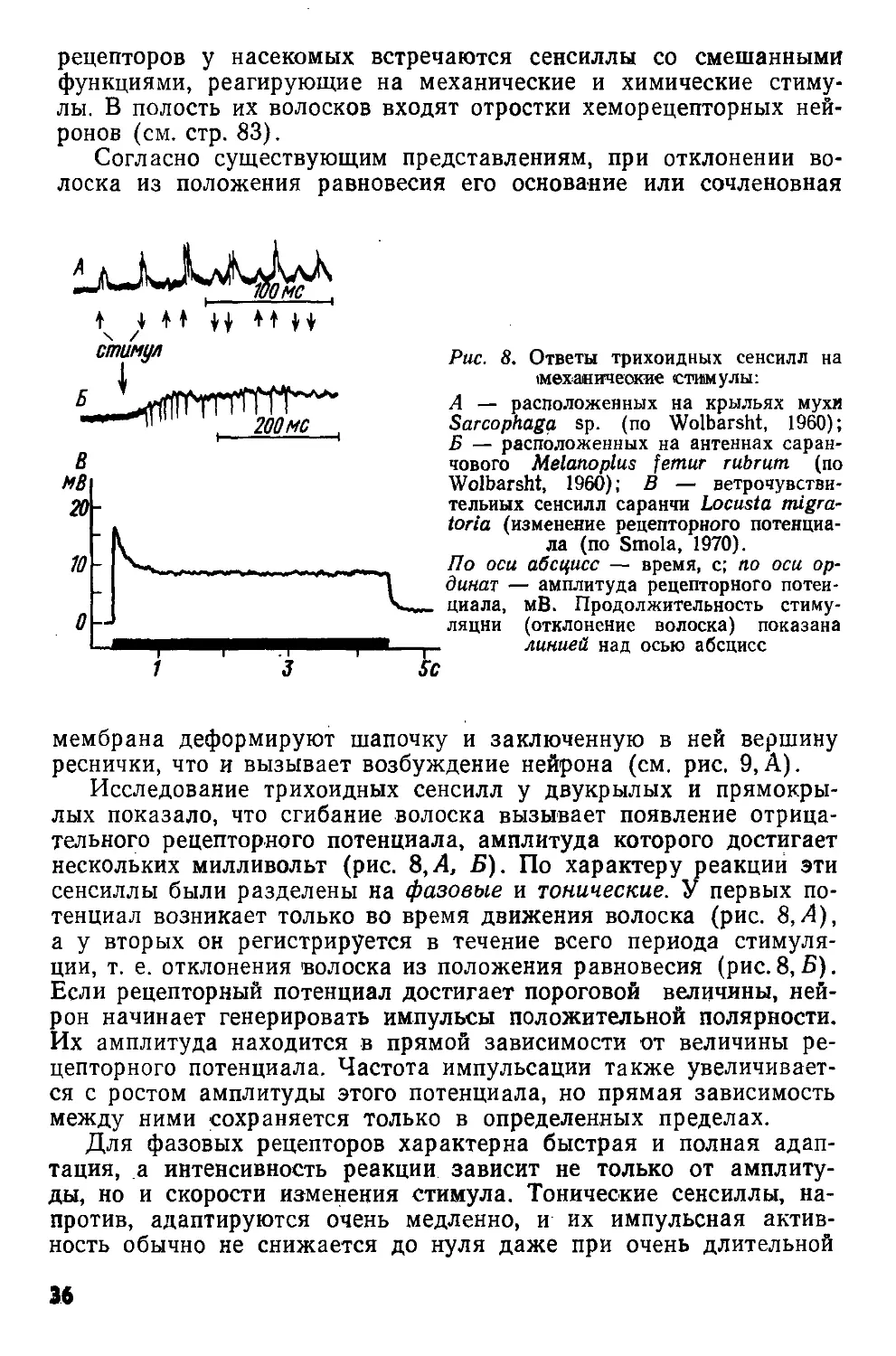
Рис. 8. Ответы трихоидных сенсилл на механические стимулы:
А — расположенных на крыльях мухи Sarcophaga sp. (по Wolbarsht, 1960); Б — расположенных на антеннах саранчового Melanoplus femur rubrum (по Wolbarsht, 1960); В — ветрочувстви-тельиых сенсилл саранчи Locusta migra-toria (изменение рецепторного потенциала (по Smola, 1970).
По оси абсцисс — время, с; по оси ординат — амплитуда рецепторного потенциала, мВ. Продолжительность стимуляции (отклонение волоска) показана линией над осью абсцисс
Л
мембрана деформируют шапочку и заключенную в ней вершину реснички, что и вызывает возбуждение нейрона (см. рис. 9,А).
Исследование трихоидных сенсилл у двукрылых и прямокрылых показало, что сгибание волоска вызывает появление отрицательного рецепторного потенциала, амплитуда которого достигает нескольких милливольт (рис. 8,А, Б). По характеру реакции эти сенсиллы были разделены на фазовые и тонические. У первых потенциал возникает только во время движения волоска (рис. 8,А), а у вторых он регистрируется в течение всего периода стимуляции, т. е. отклонения волоска из положения равновесия (рис.8,В). Если рецепторный потенциал достигает пороговой величины, нейрон начинает генерировать импульсы положительной полярности. Их амплитуда находится в прямой зависимости от величины рецепторного потенциала. Частота импульсации также увеличивается с ростом амплитуды этого потенциала, но прямая зависимость между ними сохраняется только в определенных пределах.
Для фазовых рецепторов характерна быстрая и полная адаптация, а интенсивность реакции зависит не только от амплитуды, но и скорости изменения стимула. Тонические сенсиллы, напротив, адаптируются очень медленно, и их импульсная активность обычно не снижается до нуля даже при очень длительной
36
стимуляции. Эти различия дают основание предполагать, что сенсиллы первого типа играют существенную роль при кратковременных соприкосновениях с субстратом—при движении, ощупывании и т. п., а тонические элементы функционируют главным образом во время длительных контактов — при копуляции, яйцекладке и др.
Используя информацию от рассеянных по всему телу трихоид-пых сенсилл, насекомые получают представление о субстрате и окружающих предметах практически при всех видах жизнедеятельности. Без тактильной рецепции невозможны ни простые двигательные акты, ни сложные формы поведения. Особенно велико значение осязания у насекомых, активных в ночное время или живущих в темноте — в норах, скважинах, пещерах, в толще субстрата и других подобных местах.
Ветрочувствительные сенсиллы в отличие от осязательных обладают более тонкими волосками и чрезвычайно эластичной сочленовной мембраной, обеспечивающей исключительно высокую чувствительность этих рецепторов к перемещениям окружающего воздуха. Такие сенсиллы встречаются на многих частях тела насекомых. Лучше других в настоящее время изучены рецепторы, расположенные на голове у саранчи и на церках многих ортоптероидных насекомых, поэтому, говоря о ветрочувствительных сенсиллах, мы будем иметь в виду прежде всего эти рецепторы.
Морфологические исследования показали, что основание волоска заканчивается двумя небольшими отростками, которые вхо-. дят в соответствующие ямки в кутикуле, в результате чего образуется сочленение, облегчающее качание волоска в определенной плоскости. Отклонение волоска в одном из двух возможных направлений вызывает деполяризацию мембраны нейрона и появление медленного рецепторного потенциала. Его амплитуда в течение нескольких миллисекунд достигает максимума, а затем (если волосок не возвращается в исходное положение) падает до определенного уровня, на котором остается до конца стимуляции (рис. 8, В). Такой фазово-тонический характер изменения потенциала позволяет сенсилле реагировать как на колебания, так и на стационарные отклонения волоска. У каждой сенсиллы деполяризация мембраны наступает только при смещении волоска в определенном направлении. Отклонение его в противоположную сторону (от положения равновесия) вызывает гиперполяризацию. Эти различия можно объяснить тем, что вершина отростка дендрита не совпадает с продольной осью волоска или (в случае цер-кальных сенсилл) окружена кутикулярными складками только с трех сторон, поэтому качание волоска оказывает на отросток неэквивалентное воздействие. По-видимому, одно из движений приводит к сжатию, а противоположное — к растяжению его дистальной части. При увеличении угла отклонения волоска до 12— 25° происходит возрастание амплитуды как фазового, так и тони
37
ческого компонентов рецепторного потенциала, но в первом случае наблюдается логарифмическая, а во втором — линейная зависимость. Эти закономерности нарушаются, если волосок отклоняют за пределы, ограниченные сочленовным валиком.
При достижении пороговой величины рецепторный потенциал вызывает появление в аксоне импульсных разрядов. Их частота пропорциональна амплитуде потенциала в области плато. Интенсивность ответа рецептора зависит не только от величины силы, отклоняющей волосок, но и от скорости его смещения. Так, в сенсиллах на голове саранчи максимальная частота импульсации и наибольшая скорость ее нарастания наблюдаются при очень быстром отклонении волоска, если же смещение волоска замедлено, то импульсация нарастает постепенно, и фазовая часть ответа может вообще отсутствовать.
Хотя каждая сенсилла обладает направленной чувствительностью, ее возможности в этом отношении очень ограничены. Сравнительные данные свидетельствуют о том, что плоскости качания близлежащих сенсилл ориентированы по разному, поэтому при изменении направления стимула возбуждаются разные группы рецепторов. Совокупность таких элементов, очевидно, представляет собой рецептивное поле, передающее информацию о направлении потока воздуха, воздействующего на данный участок кутикулы.
Тщательные исследования трихоидных сенсилл на голове саранчи показали, что эти рецепторы участвуют в запуске и поддержании полета. Аксоны их биполярных нейронов входят в головные ганглии, где они синаптически связаны с нисходящими фазовыми и тоническими (фазово-тоническими) интернейронами, влияющими на работу нейронов крылового аппарата.
Возбуждение трихоидных сенсилл на церках тараканов и сверчков, происходящее при обдувании насекомого или быстром приближении к нему какого-нибудь предмета, обычно вызывает реакцию убегания. В течение длительного времени считалось, что афферентные импульсы возбуждают в последнем брюшном ганглии несколько гигантских нейронов, идущих через все брюшко в метаторакальный ганглий, где они запускают реакцию мотонейронов третьей пары ног. Новейшие исследования показали, однако, что мотонейроны связаны синаптически не с гигантскими волокнами, а с идущими параллельно небольшими нейронами, обладающими меньшей скоростью проведения. Функции гигантских волокон, продолжающихся до надглоточного ганглия, пока остаются не совсем ясными.
3. ФИЗИОЛОГИЧЕСКИЕ ОСОБЕННОСТИ ПРОПРИОЦЕПТОРОВ
Проприоцепторами называют механорецепторы, посылающие в ЦНС информацию о положении, деформации и смещениях раз
38
личных частей тела *. Их функционирование обеспечивает координацию всех подвижных органов и тканей насекомого в состоянии покоя и во время любых двигательных актов. При экспериментальном выключении проприоцепторов насекомые теряют способность поддерживать естественные позы, двигаться и целесообразно реагировать на внешние воздействия.
Функции проприоцепторов у насекомых выполняют чувствительные элементы всех описанных ранее типов (стр. 31): трихоид-ные, кампаниформные, хордотональные сенсиллы и рецепторы растяжения.
Трихоидные сенсиллы, выполняющие функции проприоцепторов, обычно образуют очень густые скопления (так называемые волосковые пластинки) в местах контакта подвижно сочлененных склеритов — на переднем крае груди, на щупиках, тазиках, вертлугах и т. п. Возбуждаются они при соприкосновении с межсегментной мембраной или поверхностью близлежащего склерита. Так, волоски, покрывающие передний край груди, реагируют на движения головной капсулы, а аналогичные сенсиллы тазика стимулируются при его перемещении относительно плей-рита.
Электрофизиологические эксперименты свидетельствуют о том, что уровень импульсной активности этих рецепторов пропорционален степени изгиба волосков. Асимметрия сочленовной области волосков (рис. 9, А) оказывает значительное влияние на амплитуду ответа. Зависимость реакции сенсиллы от направления отклонения волоска представлена на рис. 9, Б. Рецепторный потенциал одиночной сенсиллы после отклонения волоска быстро достигает максимума, затем падает до определенного уровня и сохраняется почти без изменений вплоть до конца стимуляции. Такой характер адаптации позволяет отнести эти сенсиллы к фазово-тоническим рецепторам.
Трихоидные сенсиллы играют очень важную роль в процессе локомоции и ориентации насекомых. Ранее (см. гл. I) были рассмотрены функции шейных волосков у богомолов. Другие примеры участия этих рецепторов в сложных двигательных реакциях дают некоторые летающие насекомые. Так, у стрекозы Апах голова обладает определенным моментом инерции, поэтому в полете при вращении туловища вокруг продольной оси она может смещаться относительно груди и возбуждать проторакальные сенсиллы. Поступающая от них информация используется для коррекции работы крыльев, компенсирующих вращательные движения.
Аналогичным образом контролируется работа ног. Многие насекомые, передвигаясь по твердому субстрату, удерживают тело на некотором расстоянии от его поверхности и сохраняют эту
1 Некоторые проприоцепторы, воспринимающие механическое состояние внутренних органов (пищеварительной, кровеносной систем и др.), называют интеро-рецепторами.
39
дистанцию даже при дополнительной нагрузке, в несколько раз превышающей их собственный вес. Одиако если у них удаляют трихоидные сенсиллы в сочленении тазика и вертлуга, они полностью утрачивают способность приподниматься над субстратом.
Участие трихоидных сенсилл в механизмах, обеспечивающих ориентацию в гравитационном поле, будет рассмотрено ниже.
Рис. 9. Функциональные характеристики трихоид-ной сенсиллы пчелы Apis mellifera (noThurm, 1963, 1964).
А — положение волоска в состоянии покоя (пунктир) и при его максимальном отклонении (непрерывная линия)-, Б — зависимость амплитуды рецепторного потенциала от направления действия стимула. По оси абсцисс — угол отклонения волоска (0° соответствует направлению, при котором наблюдается максимальный ответ); по оси ординат — относительная амплитуда рецепторного потенциала, %
Кампаниформные сенсиллы рассеяны по всей поверхности тела, но, как правило, они концентрируются на тех участках кутикулы, которые испытывают более или менее значительную деформацию,—в сочленениях, на придатках и т. п. местах. Подавляющее большинство исследователей считают, что эти сенсиллы реагируют на изгибы, сжатие или растяжение покровов. Анализ их ультраструктурной организации дает основание предполагать, что деформация кутикулы передается колпачку и заключенной в нем шапочке, что приводит к поперечному сжатию или растяжению вершины реснички и возбуждению нейрона.
Специальные электрофизиологические исследования показали, что вблизи сенсиллы регистрируется значительный трансэпителиальный потенциал (от +50 до +100 мВ), который возникает, по-видимому, в результате деятельности калиевых насосов тормо-генной клетки, повышающих концентрацию ионов К+ в экстракле-
40
точной полости вокруг дистальной части дендрита и его отростка. При механической стимуляции (сгибании кутикулы) происходит уменьшение величины этого потенциала, сходное с фазово-тоническим изменением рецепторного потенциала других механорецепторов. Предполагается, что амплитуда трансэпителиального потенциала контролируется сопротивлением мембраны реснички, изменяющимся при деформации ее дистальной части.
На различных участках тела насекомого наряду с одиночными круглыми сенсиллами обычно встречаются группы сенсилл с эллиптическими колпачками. Их продольные оси в пределах группы параллельны друг другу и направлению основных сил, деформирующих данный участок кутикулы. На крыльях эти сенсиллы обычно располагаются вдоль жилок, а на ногах — параллельно их продольной оси. Благодаря такой конструкции и положению эллиптические сенсиллы избирательно реагируют на механические воздействия, в результате чего несколько групп по-разному ориентированных сенсилл могут передавать информацию в ЦНС не только об интенсивности стимула, но и о его направленности. Электрофизиологические данные указывают на то, что сенсиллы одной группы функционируют как единое целое и представляют собой более или менее изолированный проприоцептивный орган.
Многочисленные наблюдения и эксперименты свидетельствуют о том, что кампаниформиые сенсиллы участвуют в регуляции разных двигательных актов. Обычно количество и степень развития этих рецепторов находятся в прямой зависимости от подвижности и маневренности насекомых. Так, например, у летающего долгоносика Sitophilus orizae на крыльях гораздо больше таких сенсилл, чем у нелетающего Sitophilus granarium.
У тараканов стимуляция кампаниформных сенсилл на вертлугах вызывает рефлекторное опускание ног. Сенсиллы, расположенные на вертлуге и в проксимальной части бедра, принимают участие в тарзальном рефлексе, т. е. запускают полет при потере контакта конечностей с субстратом.
Очень важную роль играют кампаниформиые сенсиллы в регуляции полета. Поступающая от них информация влияет на работу мышц, обеспечивающих изменение подъемной силы крыльев, как это показано на пустынной саранче. При выжигании этих рецепторов на крыльях существенно нарушается способность насекомого маневрировать в полете. Изучение импульсной активности сенсилл на задних крыльях саранчи показало, что при периодической деформации крыла ответ сенсилл синхронизируется с ритмом раздражения вплоть до частоты 65—70 Гц, причем уровень реакции зависит от ускорения деформации.
Хордотональные органы состоят из нескольких (как исключение, одиночных) сенсилл, натянутых между подвижно сочлененными сегментами или частями тела (рис. 10, Л). Их шапоч-ковые клетки обычно прикрепляются к кутикуле или сочленовным
41
мембранам, а проксимальные отделы фиксируются лигаментами или прилежащими тканями.
Множество таких органов найдено в антеннах, ротовых придатках, в ногах, у основания крыльев, в груди, в брюшке и других
Рис. 10. Строение и функции проприоцепторов.
А — схема строения туловищного хордотонального органа личинки жука-усача Ergates (по Hess, 1917): к — кутикула; л — лига-мент; н — нерв; с — сенсиллы.
Б — адаптация тонических (I) и фазово-тонических (II) хордото-нальиых сенсилл в тибиотарзаль-
ном сочленении таракана Periplaneta americana (по Young, 1970). По оси абсцисс — время, с; по оси ординат — частота импульсов, имп/с. Длительность стимуляции (отклонение лапки) показана линией над осью абсцисс. Три кривых на графике II получены при
последовательном увеличении угла отклонения лапки.
В — зависимость частоты импульсации рецептора растяжения гусеницы Antheraea pernyi от интенсивности стимула (по Finlayson, Lowenstein, 1958). По оси абсцисс — время, с; по оси ординат — частота импульсов, имп/с. Стрелками показаны начало и конец стимуляции. Цифры под кривыми — удлинение рецептора, мм
частях тела. Их общее число у одного насекомого может достигать нескольких десятков: у личинок Drosophila, например, обнаружено 90 таких образований. Обычно хордотональные органы состоят из
42
однотипных сенсилл, но в некоторых из них встречаются сенсиллы с одним и двумя нейронами (см. стр. 34). В настоящее время можно считать доказанным, что эти органы фукционируют как проприоцепторы, воспринимающие смещение между органами и тканями под действием внешних и внутренних сил. Механизм работы отдельных сенсилл не вполне ясен, но большинство исследователей считает, что возбуждение их нейронов происходит при деформации сенсилл, приводящей к взаимодействию шапочки и дистальной (возможно, и расширенной) части реснички. Не исключена возможность, что характер этого взаимодействия различен в разных органах.
Электрофизиологические эксперименты показали, что у ракообразных и насекомых в ножных хордотональных органах имеются мелкие тонические и крупные фазовые (или фазово-тонические) элементы (рис. 10, Б). Следовательно, в пределах одного органа существует определенная функциональная специализация сенсилл. Одни из них передают информацию преимущественно о положении сочлененных участков ноги, а другие —об их движении.
Тщательное морфологическое и физиологическое исследование тибиотарзальных хордотональных органов у таракана позволило получить некоторые представления о механизме их работы. В частности, было установлено, что они реагируют на опускание и отведение лапки назад, но не проявляют активности при ее возвращении в исходное положение. Учитывая конструктивные особенности сочленения, можно с достаточной уверенностью предполагать, что возбуждение сенсилл происходит при их растяжении, а не при расслаблении или сжатии. На основании структурных различий сенсиллы, входящие в состав этого органа, были разделены на три группы. Одну из них, состоящую из сравнительно крупных, проксимально расположенных сенсилл, удалось идентифицировать с фазово-тоническими элементами, а другую, дистально лежащую группу мелких сенсилл — с тоническими.
Один из наиболее сложных хордотональных органов — Джонстонов орган, помещающийся во втором членике антенн. После первых морфологических работ стало очевидным, что его радиально расположенные сенсиллы должны реагировать на смещение третьего членика (и связанного с ним жгутика) относительно второго. Последующие физиологические исследования подтвердили это предположение. Стимуляция Джонстонова органа происходит как при активном ощупывании антеннами различных предметов, так и во время движения насекомого в воздушной или водной среде. Доказано также, что этот орган воспринимает звуковые колебания и силу тяжести.
Электрофизиологические исследования показали, что у мух Calliphora отклонение аристы и ее возвращение в исходное положение вызывают появление в электроантеннограмме двух значительных по амплитуде волн (до +5 мВ), возникающих, по-видимому, в результате суммации ответов множества синхронно реаги
43
рующих сенсилл Джонстонова органа. Амплитуда первой волны зависит от интенсивности стимула. Анализ электрической активности дает основание предполагать, что некоторые сенсиллы отвечают на любые колебания аристы, в то время как другие возбуждаются только при определенном направлении стимула. Подобная избирательность была обнаружена и при микроэлектродной регистрации ответов одиночных сенсилл в Джонстоновом органе саранчи. Помимо фазовых реакций Джонстонова органа в антеннах наблюдаются и тонические ответы каких-то иных рецепторов (возможно, трихоидных сенсилл), передающих информацию о положении жгутика относительно второго членика.
Рецепторы растяжения, несмотря на их морфологическое разнообразие, обладают сходными физиологическими характеристиками. В состоянии покоя (при минимальном натяжении) спонтанная импульсная активность обычно поддерживается на уровне 5—10 имп/с. При растяжении рецептора импульсация быстро возрастает до максимума, а затем снижается до определенного уровня, на котором она может оставаться в течение длительного времени (рис. 10,В). В момент расслабления импульсная активность резко падает (иногда до 0), после чего наблюдается постепенное восстановление исходного уровня импульсации. Благодаря такой фазово-тонической реакции рецептор растяжения передает информацию как о переходных процессах, так и о стационарных смещениях близлежащих структур. Частота импульсов в начальной фазе разряда возрастает пропорционально увеличению интенсивности стимула, но при экстремальном растяжении рецептора эта зависимость нарушается. На частоту импульсации оказывает влияние также скорость, с которой изменяется длина соединительнотканного тяжа или мышцы. Так, в рецепторах, лежащих у основания крыльев саранчи, частота разрядов и темп ее нарастания максимальны при быстрых перемещениях крыла. Во время ритмического качания крыловой пластинки рецептор отвечает синхронными залпами импульсов только на ее подъем, причем увеличение частоты колебаний приводит к уменьшению числа импульсов в разрядах. Специальные эксперименты показали, что удаление рецепторов растяжения или искусственная стимуляция идущих от них аксонов оказывают существенное влияние на ритм работы крылового аппарата саранчи. Анализ полученных результатов свидетельствует о том, что эти органы выполняют функцию обратной связи, стабилизируя уровень активности центральных нейронов, управляющих работой крыловых мышц.
По-видимому, не менее важную роль рецепторы растяжения играют в регуляции других локомоторных реакций, и в частности червеобразных движений личинок. Кроме того, есть основания думать, что они участвуют в контроле ритмических движений брюшка в процессе дыхания. Известно также, что у клопа Rhodnius растяжение абдоминальных рецепторов тормозит кровососание и стимулирует выделение гормона линьки.
44
4. ФИЗИОЛОГИЧЕСКИЕ ХАРАКТЕРИСТИКИ ОРГАНОВ РАВНОВЕСИЯ
Наземные формы. Простейшие наблюдения и эксперименты убеждают нас в том, что насекомые уверенно ориентируются в гравитационном поле. Если поместить насекомое с закрашенными глазами на вертикальную пластинку, то оно, как правило, будет двигаться вверх или вниз, демонстрируя положительный или отрицательный геотропизм. Отклоняя пластинку в ту или иную сторону, можно заметить, что насекомое сохраняет направление движения (относительно Земли). Аналогичные наблюдения могут быть проведены в естественных условиях. Так, живущие в песке жуки (Bledius, Dyschirius) роют норки, выдерживая вертикальное направление с точностью до 3°. Пчелы также определяют это направление в затемненных ульях и используют его во время танцев для передачи информации о положении источника пищи. Многие насекомые обладают способностью не только двигаться вдоль направления силы тяжести, но и сохранять в процессе ориентации определенный угол по отношению к вертикали.
Эти и многие другие факты свидетельствуют о том, что наземные насекомые воспринимают силу тяжести, однако никаких специализированных органов для ее рецепции у них не обнаружено. Разрешить это противоречие удалось сравнительно недавно после серии исследований, выполненных на пчелах, муравьях и некоторых других насекомых. Результаты многочисленных экспериментов показали, что некоторые части тела (голова, антенны, брюшко), несмотря на малые размеры, обладают достаточным весом, чтобы оказывать ощутимое воздействие на контактирующие с ними проприоцепторы. А так как сила этого воздействия зависит от положения тела в пространстве, то, сравнивая информацию, поступающую от разных рецепторов, насекомое может ориентироваться по отношению к направлению силы тяжести. Последовательное выключение разных механорецепторов позволило выделить те органы, которые играют первостепенную роль в этом процессе. Ими оказались скопления трихоидных сенсилл (волосковые пластинки), расположенные на антеннах, шее, тазиках, у основания брюшка и некоторых других сочленениях (рис. 11,А). Сравнительное изучение их функций показало, что селективное отключение рецепторов по-разному влияет на поведение насекомых. Так, у пчел и муравьев блокирование шейных и петиолярных (брюшных) сенсилл снижало точность ориентации в большей степени, чем отключение антеннальных или тазиковых рецепторов. В результате других экспериментов было установлено, что с помощью петиолярных и ножных сенсилл пчела определяет только общее направление оси тела (вверх или вниз), а при перемещении в горизонтальной плоскости или под углом к вертикали она использует главным образом шейные рецепторы.
45
Функционирование описанного механизма ориентаций сопряжено с двумя значительными трудностями. Одна из них определяется необходимостью одновременной оценки постоянно изменяющейся информации, получаемой от многих рецепторов, а другая состоит в полифункциональности этих рецепторов — их способности реагировать не только на изменение положения тела в пространстве, но и на смещение его частей в процессе локомоции. Точность, с которой насекомое ориентируется в гравитационном поле, свидетельствует о том, что его ЦНС успешно справляется с решением этих задач.
Водные формы. Многие водные насекомые в отличие от наземных обладают специальными органами равновесия.
У личинок двукрылых сем. Limnobiidae они напоминают статоцисты моллюсков и ракообразных (рис. И,Б). На последнем сегменте тела у них имеется пара открывающихся наружу мешочков, содержащих мелкие песчинки (статолиты). Отверстие органа прикрыто несколькими щетинками, на его внутренней поверхности (в двух точках) расположены короткие чувствительные волоски. Изнутри к нему подходит мышца. При ее периодических сокращениях в полость поступает вода, приводящая в движение статолиты. Их перемещение, по-видимому, воспринимается чувствительными волосками. Удаление этих образований приводит к нарушению положительного геотропизма у личинок, поэтому есть основания считать их органами равновесия.
Органы другого типа, называемые иногда «внешними статоцистами», найдены у многих водных полужесткокрылых. Принцип их работы основан на том, что сила, действующая на воздушные запасы (в трахеях или на вентральной стороне тела), всегда направлена к поверхности воды. Поэтому при изменении положения насекомого относительно вертикали на его теле происходит закономерное перераспределение воздуха, которое может восприниматься специальными механорецепторами.
У гладышей (Notonectidae) и гребляков (Corixidae) пузырьки воздуха, заключенные между головой и переднегрудью, соприкасаются с короткими четырехчлениковыми антеннами. Изменение объема этих пузырьков или их смещение воспринимаются, по-видимому, Джонстоновыми органами и волосками на третьем членике. Удаление антенн приводит к нарушению нормальной ориентации насекомых во время плавания.
У нимф водяных скорпионов (Nepa) на вентральной стороне брюшка имеется пара дыхательных каналов, покрытых длинными гидрофобными волосками. На четырех сегментах эти каналы слегка расширены и снабжены короткими сенсорными хетами, воспринимающими изменения объема выступающих здесь пузырьков воздуха. В имагинальной фазе у водяных скорпионов рядом с дыхальцами на трех брюшных сегментах появляются довольно сложные органы, состоящие из соприкасающихся грибовидных хет и мелких папилл (рис. 11, В). К каждому из этих элементов подхо-
46
Рис. 11. Органы равновесия.
А — положение на теле муравья Formica polyctena скоплений трихоид-ных сенсилл (волосковых пластинок) (по Mark!, 1965);
Б — схема строения статоциста личинок двукрылых сем. Limnobiidae (по Wolff, 1922);
В — схема строения органа равновесия водяного скорпиона Nepa rubra (по Thorpe, Crisp, 1947).
гс — грибовидные сенсиллы; н — нерв; сп — сенсорные папиллы; т — трахея
дит биполярный нейрон. Под «шляпками» грибовидных хет лежит слой воздуха, который сообщается через дыхальце с полостью трахеи. При перемещении насекомого в пространстве происходит изменение гидростатического давления и соответствующее изменение объема полости органа, что приводит к смещению грибовидных хет. Электрофизиологические опыты показали, что каждая такая хета и связанный с ней нейрон представляют собой фазово-тонический рецептор. Оценивая различия в гидростатическом давлении, воздействующем на три пары органов, насекомое может с высокой точностью определять свое положение относительно направления силы тяжести. В экспериментальных условиях водяные скорпионы отличают горизонтальную поверхность от наклонной, если угол между ними достигает 2°, что соответствует перепаду давлений 0,24 мм водяного столба. Разрушение рецепторов полностью лишает насекомое способности ориентироваться в гравитационном поле.
Глава III
СЛУХ
Многие насекомые обладают более или менее специализированными органами, воспринимающими звуковые волны в воздушной, водной и твердой средах. Хотя между этими явлениями нет принципиальных различий, рецепторы, реагирующие на звуки в воздухе и воде, принято называть слуховыми, а воспринимающие колебания субстрата — виброрецепторами. Следуя традиции, мы рассмотрим их независимо в двух разделах данной главы.
В естественных условиях слуховые органы насекомых могут реагировать на разнообразные звуки как биотического, так и абиотического происхождения, но в подавляющем большинстве случаев их основная функция состоит в восприятии внутривидовых коммуникационных сигналов. Акустическая сигнализация играет в жизнедеятельности многих насекомых очень важную роль. Достаточно сказать, что с ее помощью осуществляется встреча особей противоположного пола, поддерживается репродуктивная изоляция у близких видов и регулируется размещение насекомых в биотопе. Несмотря на большое разнообразие конкретных проявлений акустического поведения, насекомые чаще всего используют слуховые анализаторы для решения двух основных задач: опознавания видоспецифических или биологически важных сигналов и локализации источника звука. Механизмы, обеспечивающие успешное внутривидовое общение и акустическую ориентацию, представляют значительный интерес, поэтому их рассмотрению будет уделено несколько больше внимания, чем некоторым другим вопросам. Начать же эту главу целесообразно с краткой характеристики звуковых сигналов насекомых.
1. КОММУНИКАЦИОННЫЕ
ЗВУКОВЫЕ СИГНАЛЫ
Общее число сигналов, вызывающих или сопровождающих специфические поведенческие реакции у насекомых, достигает тринадцати, но число сигналов, издаваемых особями одного вида, не превышает семи.
К сожалению, до настоящего времени не удалось выработать ни единой классификации, ни системы терминов для обозначения этих звуков. Однако большинство исследователей так или иначе признают существование сигналов следующих типов ’.
1 Так как звуки общественных насекомых распространяются преимущественно через субстрат, мы рассмотрим их в специальном разделе (см. стр. 79).
49
Рис. 12. Физические характеристики звуковых сигналов прямокрылых насекомых.
А — осциллограммы призывных сигналов симпатрических видов сверчков (по Жаитиеву, Дубровину, 1974): 1 —Oecanthus turanicus Uv., 2—Gryl-lodinus macropterus Fuen., 3 — Tartarogryllus tartarus Sauss., .4 — T. burdi-galensis Latr., 5 — Mela.nogryllus desertus Pall., 6 — Modicogryllus pallipal-pis Tarb., 7 — Gryllus bimaculatus Deg., 8 — Pteronemoblus heydeni Fisch. Б—Г — амплитудно-частотные спектры: Б — призывного сигнала сверчка Gryllus bimaculatus; В — прекопуляционного сигнала его же (по Жантие-ву, Чуканову, 1972); Г — призывного сигнала кузнечика Tettigonia cantans.
Отметка времени — 100 мс
Призывные сигналы издаются «спонтанно» половозрелыми самцами очень многих видов насекомых. Как правило, они предназначены для привлечения самок, но неоднократно отмечалось, что эти звуки выполняют и некоторые дополнительные функции. Так, например, у сверчков в зависимости от обстоятельств они могут влиять на акустическую активность других самцов и их распределение в колонии.
Для большинства сигналов этого типа характерна относительная стабильность физических параметров, и прежде всего временной организации, определяющей их видоспецифический «рисунок» (рис. 12,А). Помимо прямокрылых насекомых призывные звуки издают цикады (Auchenorrhyncha), некоторые клопы (Pentatomi-dae, Corixidae), жуки-долгоносики (Curculionidae) и двукрылые (Trypetidae).
Помимо сигналов, издаваемых с помощью специальных органов, значительной привлекательностью для насекомых иногда обладают звуки полета. Особенно велико значение этих звуков в жизнедеятельности многих двукрылых (Culicidae, Ceratopogonidae, Tabani-dae) и, возможно, некоторых перепончатокрылых (Megachile, An-
50
thophora, Apanteles и др.) - Тщательное исследование поведения нескольких видов комаров показало, что самцы отыскивают самок преимущественно по звуку и могут привлекаться искусственными сигналами, если их частота соответствует частоте биения крыльев самок данного вида.
Ответные сигналы издаются самками саранчовых, некоторых кузнечиков, клопов, цикадок в ответ на призывное пение самцов. У саранчовых самки издают звуки, постепенно приближаясь к поющему самцу. Их сигналы могут закономерно чередоваться. Самец при этом либо остается неподвижным, либо двигается навстречу самке. У многих листовых кузнечиков (Phaneropterinae) самки отвечают на сигналы самцов с определенной задержкой, причем ее величина специфична для каждого вида.
Копуляционные сигналы подробно изучены у прямокрылых насекомых (саранчовых и сверчков), и, кроме того, имеются сведения об аналогичных звуках у цикад, клопов, жуков и некоторых двукрылых.
51
У прямокрылых обычно выделяют три типа сигналов, так или иначе связанных непосредственно со спариванием: прекопуляцион-ные, копуляционные и посткопуляционные. Первые из них выполняют три основные функции: поддерживают репродуктивную изоляцию у симпатрических видов, свидетельствуют о готовности самцов к спариванию и побуждают самок к копуляции. Как правило, сигналы этого типа хорошо отличаются по интенсивности, частотным и временным параметрам от призывных звуков и, кроме того, обладают более или менее явственными видоспецифическими свойствами. Звуки второго типа издаются самцами некоторых саранчовых во время копуляции и, по-видимому, способствуют удержанию самки. Посткопуляционные сигналы характерны для тех видов сверчков, у которых самка может оставаться с самцом в период между копуляциями. Предполагается также, что эти звуки удерживают самку от преждевременного удаления сперматофора.
Территориальные сигналы изучены еще недостаточно, но есть основания думать, что они выполняют очень важные функции, регулируя размещение насекомых в биотопах. Трудности их изучения определяются главным образом тем, что у многих видов аналогичные реакции вызывают обычные призывные сигналы самцов.
Сравнивая акустическое поведение некоторых насекомых, можно выделить две разновидности этих звуков: аггрегационные и предупреждающие. Первые из них описаны у саранчовых, цикад, клопов и жуков (Passalidae). Обычно они оказывают привлекающее воздействие на особей обоего пола (в этом их отличие от призывных сигналов), в результате чего образуются более или менее крупные скопления насекомых, самцы которых иногда осуществляют «хоровое пение». Сигналы этого типа могут издаваться не только самцами и самками, но и личинками. Предупреждающие сигналы вызывают противоположные реакции, удерживая животных на определенном расстоянии друг от друга. Они регистрируются у насекомых, ведущих оседлый образ жизни,— у многих саранчовых и кузнечиков.
Если в результате конкурентных отношений самцы сближаются на недопустимую дистанцию, они издают сигналы агрессии. Конфликты обычно возникают при вторжении самца на чужую территорию или появлении двух самцов около самки. У сверчков эти звуки хорошо отличаются от других внутривидовых сигналов по интенсивности и временным параметрам и обычно сопровождаются активной борьбой соперничающих самцов.
Аналогичные сигналы описаны у многих кузнечиков, саранчовых и некоторых жуков.
Сигналы протеста издают очень многие насекомые в момент опасности или нападения. Примерами таких сигналов могут служить резкие «вскрикивания» цикад, подвергшихся нападению птиц, или характерное «скрипение» взятого в руки жука-усача. Несмотря на то что эти звуки описаны у представителей почти
52
всех отрядов насекомых, обладающих звуковыми органами, их адаптивное значение остается неясным. Неоднократно высказывалось предположение, что они оказывают отпугивающее влияние на хищников, однако экспериментальных подтверждений этой гипотезы получить не удалось.
2. ФИЗИЧЕСКИЕ ХАРАКТЕРИСТИКИ ЗВУКОВЫХ СИГНАЛОВ
Как известно, насекомые издают звуки с помощью пяти основных механизмов:
1) трением более или менее специализированных частей подвижно сочлененных склеритов (фрикционные органы); .
2) вибрацией специализированных мембран (тимбальные органы);
3) вибрацией крыльев и связанных с ними структур;
4) пропусканием воздуха через отверстия;
5) ударением различными частями тела по субстрату.
При изучении физических характеристик звуковых сигналов основное внимание уделяется обычно анализу временных и частотных параметров. Измерение их уровня звукового давления сопряжено со значительными трудностями, поэтому сравнимых данных получено пока немного. Другие физические параметры остаются почти совершенно неизученными.
В настоящее время в биоакустике не существует общепринятой терминологии, поэтому используемые в дальнейшем термины требуют некоторых пояснений.
В большинстве акустических сигналов насекомых можно выделить элементарные звуковые посылки — пульсы, возникающие в результате однократного срабатывания фрикционного или тим-бального органа. Иногда в их заполнении различают щелчки — амплитудные всплески, соответствующие удару острого канта {plectrum) по единичному выступу зазубренной пластинки (pars stridens). Периодически повторяющиеся группы пульсов образуют серии (рис. 12, А, 3—7). Сигналы неопределенной длительности, состоящие из непрерывно чередующихся с одним и тем же интервалом пульсов, называют трелями (рис. 12,А, 2, 8).
Временные параметры. Только звуки, возникающие во время полета насекомых, представляют собой непрерывно длящиеся гармонические колебания. Во всех других сигналах обычно заметна более или менее отчетливо выраженная амплитудная модуляция. Если ее глубина достигает 100%, то звуки распадаются на отдельные посылки (импульсно-модулированный сигнал). Для большинства звуковых сигналов насекомых характерны быстрые переходные процессы и довольно крутые фронты пульсов, играющие существенную роль в восприятии и распознавании видоспецифических сигналов.
53
Длительность пульса колеблется у насекомых от 0,8 мс до 1 с, но в большинстве случаев она составляет 10—30 мс. Сравнительно простые сигналы состоят из трелей или стабильно повторяющихся серий, образованных более или менее стандартными пульсами. Частота чередования пульсов в трелях и сериях лежит в пределах 5—800 в секунду. Серии у изученных видов состоят из 2—70 пульсов. Их длительность колеблется от 15 до 1500 мс. Частота чередования серий варьирует у разных видов от 0,5 до 20 в секунду.
Сравнивая большое количество амплитудно-модулированных звуковых сигналов насекомых, можно выделить три основных способа усложнения их ритмической организации:
1) закономерное изменение амплитуды пульсов в сериях (возрастание от начала к концу, чередование пульсов разной амплитуды и др.);
2) изменение длительности и частоты чередования пульсов в сериях;
3) чередование серий и отдельных пульсов или разных серий.
У большинства прямокрылых насекомых при повышении температуры частота повторения пульсов и серий возрастает (обычно линейно), а их длительность сокращается.
Звуки, издаваемые при помощи ударных механизмов, обычно представляют собой короткие щелчки, повторяющиеся у некоторых видов с определенной частотой (до 100 в секунду). Для вибраций, возникающих при царапании субстрата (например у голодных личинок шершней), характерны большая продолжительность и очень изменчивая амплитуда.
Частотные параметры. Все звуки насекомых могут быть условно разделены на «тональные» и «шумовые». Первые из них представляют собой гармонические колебания, дающие линейчатые спектры (рис. 12, Б), вторые — сложные шумоподобные сигналы, для которых характерны сплошные спектры, занимающие более или менее широкую полосу частот (рис. 12, Г).
Тональные звуки возникают при полете многих насекомых. Их основная частота обычно соответствует частоте биения крыльев, но, кроме того, их спектры содержат высокочастотные гармоники. Результаты анализа этих звуков у некоторых видов насекомых свидетельствуют о том, что наиболее низкочастотные звуки издают стрекозы, бабочки и некоторые прямокрылые (5—50 Гц). У многих двукрылых и перепончатокрылых основная частота составляет несколько сотен герц, а у некоторых мелких двукрылых она достигает 2 кГц.
С помощью фрикционных органов тональные сигналы издают почти все сверчкообразные и некоторые кузнечики (Нотогосогу-phus, Drepanoxiphus). Их пульсы имеют почти синусоидальное заполнение. В призывных сигналах сверчков и медведок основная частота лежит в диапазоне 1,5—8 кГц (рис. 12,Б), а у кузнечиков она смещена в область ультразвука (16—30 кГц).
54
Шумовые сигналы возникают при работе всех перечисленных ранее механизмов звукоизлучения. Их частотные характеристики разнообразны и с трудом поддаются классификации. Сравнение полученных к настоящему времени спектрограмм свидетельствует о том, что полоса излучаемых частот варьирует от нескольких сотен герц до десятков килогерц. В некоторых спектрах легко выделяются одна или несколько областей доминирующих частот, в других — амплитуда составляющих бывает приблизительно одинаковой. Относительная величина спектральных компонентов может заметно меняться не только у особей одного вида, но и у одного животного при разных условиях (в частности, при разных положениях микрофона). Тем не менее уже в первых сравнительных биоакустических работах было отмечено, что сигналы представителей крупных таксономических групп насекомых (до надсемейства включительно) занимают определенные диапазоны частот. В дальнейшем были получены материалы, указывающие на то, что специфическими частотными характеристиками могут обладать не только сигналы видов, относящихся к одному семейству, но и роду. Так, например, у всех изученных видов рода Tettigonia имеются максимум на доминирующих частотах 8—10 кГц и два других максимума: 18—22 и 34—44 кГц. Огибающая их спектров в целом очень специфична, и ее можно легко отличить от огибающих спектров других кузнечиков сем. Tettigoniidae.
Зависимость частоты звуков от температуры изучена еще недостаточно. В некоторых случаях повышение температуры приводит к повышению частоты (Oecanthus), в других такая закономерность не наблюдается (Grt/llus).
Определить минимальную интенсивность акустических сигналов насекомых чрезвычайно трудно, так как для этого необходима очень чувствительная аппаратура. Максимальные уровни звукового давления (УЗД), приближающиеся к ПО—115 дБ2, зарегистрированы у кузнечиков и цикад. Некоторое представление о сравнительной интенсивности звуков разных групп насекомых дает табл. 1.
Вполне понятно, что перечисленные нами свойства звуковых сигналов связаны между собой и целиком определяются работой звуковых органов.
В 1963 г. Дюмортье предложил разделить основные параметры звуков членистоногих на «физические» и «биологические». Под первыми понимались параметры сигналов, определяющиеся механическими свойствами звукоизлучающих структур, а под вторыми — те особенности сигналов, которые контролируются работой нервно-мышечного аппарата. Такую классификацию нельзя считать достаточно строгой, но она все же отражает существенную закономерность, состоящую в том, что частотные характеристики звуков зависят главным образом от механических свойств звуковых органов,
2 УЗД измеряется в децибелах относительно 0,00002 Н/м2.
55
Таблица I
УЗД акустических сигналов некоторых насекомых, дБ
Насекомоа УЗД
Orthoptera Listrocaelidae Psedophyllidae Conocephalidae Tettigoniidae Bradyporidae (Ephippigerinae) Gryllidae Oecanthidae Gryllotalpidae Acrididae Homop tera Cicadidae Coleop tera Hymenoptera Formicidae Lepidoptera Sphingidae 100—110(8—10) 98—102(10) 100—110(10) 100—115(10) 90— 95(100) 90(10) 60(10) 90(100) 70 90—100(10) 30— 60 74(0,5) 65(5)
Примечания: 1. В таблице, за небольшим исключением, даны максимальные значения УЗД призывных сигналов.
2. В скобках указано расстояние от микрофона до объекта, см,
а временной рисунок сигнала определяется преимущественно работой некоторых элементов ЦНС и связанных с ними мышц, приводящих в движение эти органы.
3. СТРОЕНИЕ
СЛУХОВЫХ ОРГАНОВ
Насекомые воспринимают звуковые колебания с помощью Джонстоновых и тимпанальных органов. В экспериментальных условиях на низкочастотные звуки реагируют также некоторые трихоидные сенсиллы (особенно ветрочувствительные волоски), однако служит ли для них звук адекватным раздражителем и воспринимают ли они его в естественных условиях, остается пока неясным.
Джонстоновы органы используются для рецепции звука двукрылыми (Culicidae, Chironomidae, Drosophilidae и др.) и, возможно, цикадками (Auchenorrhyncha). У большинства из них эти органы имеют такое же строение, как и у других насекомых, но у комаров наблюдаются гипертрофия второго членика антенн и значительное увеличение числа сенсилл (до нескольких тысяч)
56
Рис. 13. Слуховые органы насекомых.
А — схема строения Джонстонова органа комара сем. Culicidae (по Risler, 1955): I, II — первый и второй членики антенны; ан — антеннальный нерв; бп — базальная пластинка (отросток III членика антенны) ; ж — жгутик антенны с волосками; н — нерв Джонстонова органа; с — хордотональные сенсиллы.
Б — схема строения тимпанального органа кузнечика Decticus verrucivo-rus, вид с дорсальной стороны.
В — то же, поперечный срез (по Schwabe, 1906).
к — кутикула; н — нейроны; его — субгенуальный орган; по — промежуточный орган; тм —• тимпанальная мембрана; с — хордотональные сенсиллы; сг — слуховой гребень; т —• трахея; тн — тимпанальный нерв
(рис. 13, А). Их шапочковые клетки прикрепляются к специальным отросткам третьего членика, вдающимся глубоко в полость второго. Жгутик антенн снабжен длинными, радиально расположенными волосками.
Рис. 14. Слуховые органы насекомых. А — схема строения тимпанального органа совки (Noctuidae) (по Eggers, 1919 с изменениями по Roeder, Treat, 1961): вм — воздушные мешки; л — ли-гамент; тм — тимпанальная мембрана; тн — тимпанальный нерв; с —• хордотональные сенсиллы.
Б — схема строения крылового тимпанального органа Chrysopa sp. (по Miller, 1970): глм — гладкая часть тимпанальной мембраны; гом — гофрированная часть тимпанальной мембраны; т — трахея; с — хордотональные сенсиллы. В — схема строения слухового органа бражника Celerio lineata (по Roeder et al., 1970): г — глаз; па — пальпы; пи —
пилифер; х — хоботок
Тимпанальные органы — наиболее сложные слуховые органы насекомых. Они имеются у многих прямокрылых, цикад, полужесткокрылых и бабочек. Детали их строения довольно сильно варьируют не только в пределах отрядов, но и семейств, поэтому мы ограничимся рассмотрением наиболее типичных примеров.
У длинноусых прямокрылых (Dolichocera) тимпанальные органы находятся в голенях передних ног. У некоторых видов они открываются наружу двумя овальными мембранами, у других мембраны прикрыты складками кутикулы так, что снаружи заметны только одна или две узкие щели.
Полость между барабанными перепонками выполнена двумя отростками ножной трахеи (рис. 13,Б,В). На дорсальной стороне одного из них расположено несколько групп хордотоиальных сенсилл. Проксимальная группа представляет собой субген у ал ь-ный орган. Она состоит из множества длинных сенсилл, распо
58
ложенных веерообразно; их шапочковые клетки сходятся в одной точке и прикрепляются к стенке голени. Промежуточный отдел образован немногими сенсиллами, натянутыми между кутикулой и трахеей. Слуховой гребень состоит нз субинтегу-ментных сенсилл (их шапочковые клетки не прикреплены к кутикуле). Они располагаются в один ряд и постепенно уменьшаются в размерах к дистальному концу голени.
В отличие от кузнечиков, сверчков и медведок у саранчовых, бабочек и цикад тимпанальные органы находятся в груди или первом членике брюшка. Так, например, у бабочек из семейства совок (Noctuidae) в сочленовной мембране между грудью и брюшком имеются два очень глубоких вдавления (рис. 14,А). Их передние стенки образованы тимпанальными мембранами (по две с каждой стороны). К наружной мембране изнутри подходит тонкий нерв, заканчивающийся двумя хордотональными сенсиллами. Приблизительно так же устроены тимпанальные органы у саранчовых и цикад, но там число хордотональных сенсилл достигает нескольких десятков и даже сотен.
У некоторых чешуекрылых (Satyridae) и сетчатокрылых сравнительно простые тимпанальные органы расположены в крыльях. У златоглазки (Chrysopa) они представляют собой вздутия радиальных жилок, заполненные гемолимфой (рис. 14, Б). С вентральной стороны эти органы затянуты тонкой гофрированной мембраной. К ее проксимальной части прикрепляется 25 хордотональных сенсилл.
Очень своеобразные слуховые органы были найдены недавно у некоторых бражников (Sphingidae, Chaerocampinae) (рис. 14, В). Они состоят из двух компонентов: заполненного трахейными мешками второго членика нижнегубных щупиков и соприкасающегося с ним пилифера 3. Звуковые волны вызывают колебания щупика, которые передаются рецепторам, расположенным в пилифере.
4. ФУНКЦИИ
СЛУХОВЫХ ОРГАНОВ
Как известно, приборы для регистрации звука реагируют на един из трех его параметров: звуковое давление, градиент давления или смещение частиц среды.
Прибор, регистрирующий звуковое давление, представляет собой толстостенную камеру, одна из стенок которой замещена туго натянутой мембраной. Заключенный в этом резервуаре воздух сообщается с наружной средой через небольшое отверстие, благодаря чему внутри камеры поддерживается атмосферное давление. В звуковом поле на диафрагму прибора действует сила, пропор
3 Palpi peliferi — небольшие подвижно сочлененные придатки, представляющие, по-видимому, части редуцированной верхней губы.
59
циональная разности между давлением звука и атмосферным давлением. Если длина волны значительно превышает размеры такого приемника, то его чувствительность не зависит от направления прихода звука.
Приемники градиента давления представляют собой натянутую на жесткую рамку и открытую с двух сторон мембрану (например
Рис. 15. Адаптация двух а-клеток из тимпанального органа саранчи Schi-stocerca gregaria к звуковому раздражителю (3 кГц) интенсивностью 80 дБ (I) и 90 дБ (II) (по Michel-sen, 1966).
По оси абсцисс — время, с; по оси ординат — число импульсов за 96 мс; толстые линии — длительность стимуляции
ленточный микрофон). Сила, действующая на мембрану в звуковом поле, зависит от угла, под которым звуковые волны падают на ее поверхность, поэтому такой приемник является направленным.
Колебания частиц воздуха можно наблюдать под микроскопом, если ввести в звуковое поле аэрозоль или какие-нибудь тела с минимальной массой (например очень тонкую кварцевую нить).
В течение длительного времени предполагалось, что все слуховые органы насекомых воспринимают смещение или градиент давления. Однако в последние годы были получены данные, свидетельствующие о том, что, по крайней мере, тимпанальные органы саранчовых и кузнечиков представляют собой сложные приемники, реагирующие как на давление, так и на градиент давления.
Анализ импульсной активности одиночных нейронов показал, что в разных слуховых органах и даже в пределах одного некоторые сенсиллы могут адаптироваться по-разному. Чаще всего через несколько миллисекунд после включения звука частота импульсов быстро возрастает до максимума, а затем более или менее быстро падает до определенной величины, после чего либо остается в течение длительного времени на этом уровне, либо постепенно снижается (рис. 15). Реже наблюдаются медленное возрастание и быстрый спад частоты импульсации. Первые несколько импульсов; обычно генерируются в сенсиллах тимпанальных органов синхронно, в результате чего в их суммарных ответах регистрируется так называемая синхронная фаза, состоящая из нескольких волн большой амплитуды. При длительной стимуляции она сменяется асинхронной фазой, а после выключения звука наблюдается небольшое последействие.
При стимуляции Джонстоновых органов и церкальных трихоидных сенсилл низкочастотными звуками регистрируются суммарные
60
потенциалы, представляющие собой гармонические колебания, частота которых равна или кратна частоте звука.
Абсолютная чувствительность большинства слуховых рецепторов насекомых невелика: пороги реакции, регистрируемые обычными методами, лежат в области 30—40 дБ. Использование некоторых специальных методов позволяет обнаруживать слабые ответы при УЗД, не превышающем 7—10 дБ. Максимальной чувствительностью обладают Джонстоновы органы (порог около 0 дБ).
Частота импульсации отдельных сенсилл тимпанальных органов возрастает при увеличении интенсивности стимула до 30— 40 дБ над порогом реакции. Наличие в сложных слуховых органах сенсилл с разной чувствительностью значительно расширяет их динамический диапазон.
Важной особенностью тимпанальных органов насекомых следует считать чрезвычайно малую временную константу. У большинства изученных видов она составляет всего 2—3 мс. Это позволяет насекомым различать отдельные звуковые пульсы, следующие с частотой 100—300 в секунду.
При изучении функций слуховых органов насекомых большое внимание всегда уделялось их частотным характеристикам. На первых этапах этих исследований была выдвинута гипотеза, отрицавшая способность слуховых рецепторов дискриминировать звуки по частоте. Иначе говоря, считалось, что в отличие от позвоночных у насекомых все сенсиллы слухового органа «настроены» на один диапазон частот. В ходе дальнейших работ, проводившихся на представителях разных отрядов насекомых, были получены многочисленные подтверждения этой гипотезы. Однако в последние годы удалось установить, что тимпанальные органы прямокрылых имеют более сложную функциональную организацию, чем предполагалось ранее, и, в частности, содержат группы сенсилл, по-разному реагирующих на звуки разных частот.
У саранчовых в изолированных тимпанальных органах выделены четыре группы сенсилл, обладающих разными функциональными свойствами (рис. 16). Три из них (а, Ь, с) реагируют преимущественно на звуки низких частот — от 1 до 10 кГц (оптимум чувствительности— 2—4 кГц), а одна (d) воспринимает высокочастотные колебания в диапазоне 6—40 кГц (оптимум чувствительности—12 кГц). Исследование механических свойств тимпанальной мембраны с помощью метода лазерной голографии показало, что частотные характеристики четырех групп сенсилл определяются резонансными свойствами тех специализированных участков тимпанальной мембраны, к которым они прикрепляются.
У кузнечиков и сверчков изучение тимпанальных органов затрудняется тем, что к ним примыкают сенсиллы подколенного органа, часть из которых посылает волокна в тимпанальный нерв. Тем не менее, сочетая электрофизиологические методы с перерезками веточек тимпанального нерва, удалось получить частотные характеристики этих органов. Так, было показано, что у кузнечи-
61
ков подколенный орган не реагирует на звуковые колебания, превышающие 1 кГц. Промежуточный орган отвечает на звуки в диапазоне от 1 до 15 кГц (оптимум на 5—7 кГц), а слуховой гребень воспринимает высокочастотные колебания от 1 до 100 кГц (оптимум на 12—30 кГц).
5(?Г\С V
Рис. 16. Строение и функциональные характеристики тимпанального органа саранчи.
А —схема строения тимпанального органа: а—d — четыре группы хордотональных сенсилл; тн — тимпанальный нерв; тм — тимпанальная мембрана (по Gray, 1960).
Б — частотно-пороговые характеристики четырех групп хордотональных сенсилл в изолированном тимпанальном органе (по Michelsen, 1969). По оси абсцисс — частота звука, кГц; по оси ординат — его интеисивиость, дБ
05
75
70-
65-
60-
55-
50-
45-
40-
। | । > । । I 1 । I I Г 1.1 1_[ I—
7 1,5 2 2,53 4 56 8 1012 1620253040
КГц
а d
tf /\
Аналогичные результаты были получены при изучении функциональной организации тимпанальных органов сверчков. Низкочастотные звуки (1 —15 кГц) вызывают ответы проксимальной части трахеального органа, гомологичной промежуточному органу кузнечиков. Оптимум чувствительности этой группы сенсилл лежит в области 4—5 кГц. Дистальная часть трахеального органа воспринимает звуки более высоких частот — от 10 до 50 кГц (оптимум чувствительности— 16 кГц).
У медведок, как и у сверчков, на частотно-пороговых кривых тимпанальных органов выделяются две зоны повышенной чувствительности: на 1,5—2 кГц и на 16—18 кГц,— поэтому можно предполагать, что сенсиллы у этих насекомых также образуют два функциональных отдела, по-разному реагирующих на звуки высоких и низких частот.
В отличие от прямокрылых в тимпанальных органах цикад и чешуекрылых пока не удается обнаружить сенсилл с разными частотными характеристиками. Оптимум их чувствительности у цикад лежит в области 3—7 кГц, а у чешуекрылых он смещен в ультразвуковой диапазон— 15—60 кГц. Джонстоновы органы и трихоид-ные сенсиллы реагируют на звуки сравнительно низких частот (до 1—3 кГц).
62
5. ЦЕНТРАЛЬНЫЕ ОТДЕЛЫ СЛУХОВЫХ АНАЛИЗАТОРОВ
Афферентные волокна, идущие от тимпанальных органов, входят в грудные ганглии, а волокна Джонстонова органа поступают непосредственно в головной мозг (надглоточный ганглий).
Исследование функциональной организации высших отделов слуховых анализаторов проводилось до настоящего времени только на прямокрылых и чешуекрылых насекомых.
У кузнечиков в первом грудном ганглии сначала были найдены 2 крупных Т-образных нейрона, посылающих свои ветви как в надглоточный ганглий, так и во II и III грудные ганглии. Чаще всего эти клетки дают фазовые ответы, т. е. реагируют на включение или быстрое изменение амплитуды звука. Контралатеральные рецепторы оказывают на них слабое возбуждающее и сравнительно сильное тормозное влияние. Последующие эксперименты показали, что в грудном отделе имеется несколько Т-образных слуховых волокон, а также восходящие и нисходящие волокна. Среди них были найдены фазовые, тонические, фазово-тонические нейроны, а также клетки, характер ответов которых определялся условиями стимуляции. Уровень импульсной активности почти всех нейронов зависит от интенсивности и частоты звука, достигая максимума только при определенном их сочетании. На реакции большинства изученных элементов влияет также направление прихода звука, причем форма полученных диаграмм изменяется при изменении интенсивности и частоты сигнала.
У сверчков в первом грудном ганглии лучше других изучены 5 центральных слуховых нейронов (рис. 17): 3 восходящих (А, В, С) и 2 нисходящих (D, Е). Все они возбуждаются низкочастотным отделом тимпанального органа (оптимум на 5 кГц), а клетки А, В и Е, кроме того, связаны с высокочастотными сенсиллами (оптимум на 16 кГц). Вследствие этого на частотно-пороговых кривых нейронов С и D имеется только один оптимум, а у нейронов А, В и Е достаточно отчетливо выражены две зоны повышенной чувствительности. По характеру реакций на длительные сигналы эти клетки могут быть разделены на фазовые (A, D) и тонические (В, Е). При определенных условиях стимуляции почти все они испытывают тормозное влияние со стороны других нервных элементов. Контралатеральные рецепторы оказывают на некоторые нейроны только тормозное (С), а на другие — тормозное и возбуждающее воздействие (В).
Слуховые анализаторы медведок изучены еще недостаточно, но имеющиеся данные позволяют думать, что они имеют много общего с гомологичными анализаторами сверчков. В коннективах, отходящих от I грудного ганглия, у двух видов медведок удалось проанализировать ответы пяти клеток. По функциональным характеристикам некоторые из них близки к слуховым нейронам сверчков (А и С), но другие существенно отличаются от них. Сопостав-
63
ление амплитудно-частотных спектров звуковых сигналов медведок с частотно-пороговыми характеристиками слуховых нейронов первого грудного ганглия свидетельствует о том, что этот отдел слухового анализатора, несомненно, приспособлен для восприятия и дискриминации внутривидовых сигналов.
Рис. 17. Функциональные характеристики слуховых интернейронов из I грудного ганглия сверчка Gryllus bimaculatus (по Жантиеву, Чуканову, 1972).
А — схема ответов 5 слуховых интернейронов (4—D) на звуковые сигналы 5 и 16 кГц (длительность 50 мс, интенсивность 90 дБ).
Б — частотно-пороговые характеристики двух слуховых интернейронов (В и С). По оси абсцисс — частота звука, кГц; по оси ординат — звуковое давление, дБ
5 кГц 16 кГц
А
Восходящие Волокна
с —НИЩИ || -------
У саранчовых, в отличие от длинноусых прямокрылых, тимпанальный нерв входит не в первый, а в третий грудной ганглий. Здесь его волокна синаптически связаны с несколькими восходящими центральными нейронами. Некоторые из них прослеживаются во всех коннективах вплоть до надглоточного ганглия. Другие достигают только проторакального ганглия. Известны также восходящие волокна, начинающиеся в мезоторакальном ганглии. Все эти нейроны хорошо различаются по основным функциональным характеристикам. Максимум их импульсной активности обычно наблюдается при определенном (оптимальном) сочетании интенсивности и частоты звука. Исключение составляет только одна клетка, которая отвечает единичными импульсами на стимулы в очень широком диапазоне частот и интенсивностей. Комплекс изученных к настоящему времени восходящих нейронов обеспечивает
64
передачу информации об акустических сигналах в диапазоне от 3 до 40 кГц при уровнях звукового давления от 40 до ПО дБ..Приблизительно половина этих клеток испытывает влияние со стороны контралатеральных рецепторов. Кроме того, на реакции некоторых слуховых интернейронов значительное воздействие оказывают афферентные волокна, не связанные с тимпанальными органами, но реагирующие на низкочастотные колебания.
У чешуекрылых тимпанальный нерв входит в третий грудной ганглий, но его волокна прослеживаются и в других грудных ганглиях. В мезоторакальном ганглии найдены две группы восходящих нейронов второго порядка. Ипсилатеральные волокна идут отсюда в подглоточный ганглий, а контралатеральные достигают только проторакального ганглия. Ответы этих клеток, получивших название «релейных», мало чем отличаются от реакций афферентных нейронов. Более существенное преобразование информации производят нейроны другого типа, обнаруженные в мезоторакальном ганглии. Они отвечают на звуковые посылки любой длительности только одним импульсом. В грудном отделе ЦНС чешуекрылых регистрируются ответы и некоторых других слуховых волокон, но они изучены еще недостаточно. Несмотря на сравнительную простоту организации, восходящие слуховые пути могут передавать значительный объем информации об основных параметрах звуковых сигналов (интенсивности, частоте повторения, направленности и др.).
Высшие отделы слуховых анализаторов, расположенные в надглоточном ганглии, исследованы у саранчи, кузнечиков и некоторых чешуекрылых (Noctuidae).
В первом случае нейроны, реагирующие на звук, были обнаружены в задней латеральной части протоцеребрума и в передней латеральной части дейтоцеребрума. Тщательное изучение их функциональных характеристик показало, что некоторые клетки возбуждаются сигналами, обладающими . определенным сочетанием интенсивности, частоты, длительности и частоты повторения. Иначе говоря, эти нейроны ведут себя хак фильтры, выделяющие звуки с определенными параметрами. Сопоставляя активность некоторых клеток до и после отключения одного из тимпанальных органов, удалось установить, что контралатеральные рецепторы оказывают на них преимущественно тормозное воздействие; На основании этого было высказано достаточно обоснованное предположение об участии нейронов надглоточного ганглия не только в распознавании, но и локализации звуковых сигналов.
У кузнечиков слуховые нейроны также расположены преимущественно между прото- и дейтоцеребрумом. По большинству характеристик они оказались сходными с интернейронами грудных ганглиев. Различия между ними проявляются главным образом в характере изменений активности во время стимуляции, длительности ответов и зависимости их реакций от ориентации насекомого в звуковом поле.
65
Исследование слухового центра в надглоточном ганглии чешуекрылых прежде всего показало, что и высший отдел слухового анализатора представлен у этих насекомых сравнительно небольшим количеством элементов. С каждой стороны протоцеребрума удалось обнаружить по одному фазовому и по два тонических нейрона. Активность одного из них подавлялась звуком больших интенсивностей, что указывает на его возможную связь с низкочувствительной клеткой тимпанального органа и позволяет рассматривать этот нейрон как важный элемент механизма, обеспечивающего изменение поведения бабочек при приближении летучей мыши.
6. МЕХАНИЗМЫ ОПОЗНАВАНИЯ КОММУНИКАЦИОННЫХ ЗВУКОВЫХ СИГНАЛОВ
О том, что насекомые слышат и опознают коммуникационные звуковые сигналы, свидетельствуют реакции двух основных типов: изменение акустической активности (фонореакция) и двигательные реакции (фонотаксис). В первом случае наблюдаются активация, торможение, альтернация (чередование) или синхронизация пения. Во втором — приближение, удаление или иные перемещения насекомого относительно источника звука.
Избирательность акустических реакций можно легко наблюдать у видов, издающих призывные и ответные звуки. Так, самки кузнечиков рода Isophya отвечают на каждый пульс сигнала самца одним—тремя короткими щелчками (рис. 18, А, Б). Помещая перед садком с половозрелыми самками поющих самцов разных видов прямокрылых, в том числе и близких симпатрических видов Isophya, нетрудно заметить, что ответные сигналы издаются только на призывные звуки своего вида.
Многочисленные исследования положительного фонотаксиса у разных групп прямокрылых дают убедительные доказательства того, что самок привлекают видоспецифические призывные сигналы. Реакции обоих типов сохраняются и при воспроизведении сигналов, записанных на магнитную ленту, поэтому нет оснований сомневаться в том, что насекомые реагируют именно на звук, а не на какие-то иные факторы.
Лабораторные эксперименты показывают, что насекомые не только отличают видоспецифические сигналы от других звуков, предъявляемых последовательно, но и опознают их при одновременном звучании, т. е. выделяют полезный сигнал из шума. В результате специального изучения помехоустойчивости акустической системы связи у кузнечиков рода Metrioptera было установлено, что самки проявляют положительный фонотаксис даже в том случае, если шум (звук симпатрического вида) превышает по интенсивности сигналы «своих» самцов в 3 раза. Совершенно очевидно, что
66
такая помехоустойчивость обеспечивает надежную связь в естественных биотопах, где одновременно издают звуки многие виды животных.
Как ранее отмечалось, некоторые насекомые издают не один, а несколько (до семи) коммуникационных сигналов. Их воспроизведение вызывает разные реакции, поэтому есть основания считать,
Fuc. 18. Ответы восходящих слуховых интернейронов из шейных коннекти-вов кузнечика Isophya taurica (А, Б) и сверчка Gryllus bimaculatus (В) на естественные звуковые сигналы.
А — ответы тонического нейрона и детектора пульсов (импульсы большей амплитуды) на призывный (слева) н ответный (справа) сигналы;
Б — ответы детектора щелчков на те же сигналы;
В — ответы нейронов В и С на призывный сигнал
что слуховая система не только выделяет эти сигналы из шума, ио и отличает их друг от друга.
Как же насекомые опознают звуковые сигналы?
Для того чтобы ответить на этот вопрос, необходимо прежде всего выявить те признаки, или параметры, сигналов, которые используются для их различения и опознавания. Результаты сравнительного акустического анализа показывают, что частотные спектры сигналов близких, и в том числе симпатрических, видов широко перекрываются, а их временные характеристики (длительность и закономерности чередования пульсов), напротив, обладают высокой видоспецифичностью и хорошо различаются (см. рис. 12, А). Это дает основание предполагать, что насекомые используют для опознавания акустических сигналов не частотные, а временные параметры.
67
Тщательный анализ сигналов, зарегистрированных при разных условиях или полученных от разных особей одного вида, свидетельствует о том, что временные параметры существенно различаются по степени изменчивости. Поэтому для опознавания сигналов, по-видимому, должны использоваться не все, а только некоторые, наиболее стабильные элементы.
Для выяснения реальной роли тех или иных акустических компонентов в процессе опознавания проводят специальные этологические эксперименты, в которых сравнивают реакцию насекомых на естественные и искусственные звуки. Обычно для этого используют одну из описанных выше реакций — звуковые ответы или фонотаксис. Сначала активным насекомым воспроизводят естественные (чаще всего призывные) сигналы, а затем их определенным ббразом трансформируют или заменяют имитациями (моделями) с изменяемыми параметрами. Так, например, сверчкам, издающим призывные звуки, которые состоят из четырехпульсовых серий, предъявляют модельные сигналы; в них произвольно изменяются длительность пульсов, частота их повторения, число в серии, длительность серий и т. д.
Результаты таких экспериментов, проведенных на разных группах прямокрылых, показали, что временная организация сигнала действительно играет решающую роль в опознавании, но разные виды насекомых могут использовать для этого разные компоненты. Так, самки симпатрических видов рода Hotnorocoryphus различают призывные трели самцов только по частоте повторения пульсов. У некоторых стеблевых сверчков (Oecanthus) сигналы опознаются главным образом по частоте повторения серий. Сверчки рода Gryllus используют для этой цели частоту повторения пульсов и длительность серий, а некоторые виды кузнечиков рода Isophya — длительность пульсов.
В течение длительного времени принято было считать, что насекомые не могут распознавать звуки по частоте. Однако результаты новейших исследований свидетельствуют о том, что частотные параметры все же используются для опознавания некоторых коммуникационных сигналов. При рассмотрении этого вопроса необходимо прежде всего помнить, что слуховые органы обладают определенной частотной избирательностью, поэтому диапазон воспринимаемых ими частот может не перекрываться со спектрами сигналов некоторых групп насекомых. Так, например, многие кузнечики, по-видимому, почти не слышат призывных сигналов сверчков, что, несомненно, облегчает их звуковую коммуникацию в естественных условиях. Некоторые листовые кузнечики {Isophya) издают звуки в сравнительно узкой полосе частот и обладают соответствующим образом «настроенными» рецепторами, позволяющими использовать частоту для опознавания видоспецифических призывных звуков. Их самки отвечают на модельные сигналы только в том случае, если их частотные характеристики приближаются к естественным.
68
Этологические опыты, проводившиеся на сверчках из родов Oecanthus и Scapsipedus, показали, что изменение частоты в диапазоне, доступном для слуховых рецепторов, не оказывает существенного влияния на привлекательность искусственных призывных сигналов для самок, но аналогичные опыты, поставленные на представителях других родов Grylloidea (Acheta, Gryllus, Teleogryllus, Scapteriscus), дали противоположные результаты. Причем в некоторых случаях повышение частоты призывного звука до 12—15 кГц приводило к смене положительного фонотаксиса на отрицательный.. По-видимому, для устранения этих противоречий потребуются до-: полнительные исследования, но результаты последних экспериментов и электрофизиологические данные (см. далее) указывают на то, что у сверчков и медведок эти параметры играют существенную роль в процессе опознавания.
Для двукрылых частота, по-видимому, служит единственным критерием, по которому они могут различать звуки, возникающие в полете. Хорошо известно, например, что самцов комаров привлекают тональные сигналы, если их частота приближается к частоте биения крыльев летящей самки.
Физиологические механизмы, обеспечивающие опознавание звуковых сигналов, находятся в стадии интенсивного изучения. Накопленные к настоящему времени факты пока не позволяют составить цельного представления о работе слухового анализатора, но они отражают существенные закономерности функционирования некоторых его отделов.
Обработка акустической информации проходит в слуховых анализаторах три основных этапа. Первый из них осуществляется в слуховых органах4, второй — в интернейронах грудных ганглиев, третий —в слуховых центрах надглоточного ганглия.
Рецепторы отвечают на естественные сигналы синхронными залпами импульсов, передающими информацию о длительности звуковых посылок и закономерностях их повторения, т. е. с большей или меньшей четкостью отражают временной рисунок («pattern») сигнала. Частота импульсов в разряде зависит от интенсивности звука. У прямокрылых характер ответа определяется не только временными, но и частотными параметрами сигнала. Так, у сверчков призывные и агрессивные сигналы возбуждают преимущественно низкочастотный отдел слухового органа, оптимум активности которого совпадает с основной частотой этих звуков, а прекопуля-ционные сигналы, содержащие ультразвуковые компоненты, воспринимаются в основном высокочастотным отделом. Следовательно, в данном случае первый этап различения внутривидовых сигналов осуществляется уже на уровне рецепторов.
Слуховые интернейроны, расположенные в грудных ганглиях, обладают разными функциональными характеристиками и в соответствии с этим по-разному отвечают на одни и те же сигналы.
4 Здесь и далее речь идет только о тимпанальных органах.
69
Регистрируя активность восходящих волокон в шейных коннекти-вах, можно отметить следующие основные закономерности.
Интенсивность сигнала влияет не только на частоту импульса-ции в разрядах, но и на число работающих нейронов, причем ее увеличение приводит либо к возрастанию, либо к подавлению активности некоторых клеток.
У прямокрылых насекомых отдельные интернейроны связаны только с высокочастотными или низкочастотными сенсиллами, поэтому информация о частоте звука может передаваться по разным каналам. Наряду с этим существуют клетки, имеющие двухвершинные характеристики, указывающие на то, что они возбуждаются обоими отделами тимпанального органа.
Быстро адаптирующиеся клетки реагируют только на начало звука или отвечают на каждый пульс одним-двумя импульсами, подчеркивая тем самым ритмическую организацию сигнала. Тонические нейроны, напротив, отражают не только частоту повторения, но и длительность звуковых посылок (рис. 18, В).
Сравнение реакций комплекса восходящих интернейронов на разные призывные сигналы свидетельствует о том, что в грудных ганглиях акустическая информация претерпевает заметные преобразования, приводящие к выделению и контрастированию многих параметров сигнала. Такая предварительная обработка, по видимому, необходима для успешного функционирования высших отделов анализатора. Однако на этом этапе еще не происходит детектирования видоспецифического призывного сигнала.
Сопоставляя далее ответы грудных интернейронов на внутривидовые сигналы, можно заметить, что каждый из них вызывает специфическую реакцию определенных клеток. Так, у сверчков на призывные сигналы отвечают нейроны С. или (при больших интенсивностях) В и С, на прекопуляционные — преимущественно ВиВ1, ана звуки агрессии — только В. Еще большая селективность наблюдается у некоторых кузнечиков (Isophyd), издающих длительные пульсы и короткие щелчки. В грудном отделе их слухового анализатора обнаружены «детекторы пульсов» и «детекторы щелчков» — нейроны, реагирующие только на один из этих сигналов (рис. 18, А, Б).
Приведенные, а также некоторые другие факты дают основание предполагать, что в грудных ганглиях осуществляется важный этап различения и, возможно, опознавания внутривидовых сигналов.
Изучение высших отделов слухового анализатора насекомых только начинается. Судя по предварительным данным, в надглоточном ганглии не удается обнаружить детекторы, отвечающие только на видоспецифические призывные сигналы, но у саранчи все же найдены клетки, обладающие достаточно высокой избирательностью. Некоторые из них реагируют на сравнительно длинные (20 мс) низкочастотные посылки, повторяющиеся с частотой 20 в секунду. Другие возбуждаются очень короткими (0,5—5 мс) высокочастотными сигналами. Сравнение ответов этих нейронов на
70
искусственные и естественные звуки показйло, что они приспособлены для выделения стридуляционных (призывных) и мандибулярных (возникающих при щелкании мандибул) звуков. Таким образом, в надглоточном ганглии продолжается процесс различения внутривидовых сигналов. Механизмы выделения и опознавания видоспецифических призывных сигналов пока остаются неизученными, но есть все основания предполагать, что они будут исследованы в ближайшем будущем.
7. ПРОСТРАНСТВЕННЫЙ СЛУХ
Слуховые рецепторы. В 1940 г. Памфри высказал предположение, что все слуховые рецепторы насекомых реагируют не на звуковое давление (как у позвоночных животных), а на смещение, или градиент давления. Приемники такого типа обладают направленной чувствительностью (направленностью), что создает теоретическую возможность монауральной локализации источника звука. Если звуковые колебания воспринимаются более или менее открытой с двух сторон мембраной (тимпанальные органы), то слуховой орган должен обладать независимой от частоты звука двусторонней характеристикой направленности (при длинах волн, не сравнимых с его величиной). В тех случаях, когда звуковые колебания приводят в движение не мембрану, а легкий подвижный стержень (трихоидные сенсиллы, Джонстоновы органы), характеристики направленности приемника должны существенно зависеть от его конструктивных особенностей (формы, способов прикрепления подвижных структур и др.).
При экспериментальной проверке этой гипотезы вначале были получены положительные результаты. Так, оказалось, что комары могут находить источник звука с помощью одного слухового органа. Ранее аналогичные опыты были поставлены на сверчках. Электрофизиологические исследования показали, что изолированный тимпанальный орган саранчи обладает двусторонней направленностью (рис. 19, А) и, следовательно, может рассматриваться как приемник градиента давления. Такие же результаты были получены при изучении слуховых органов кузнечиков. Правда, изменение их чувствительности было незначительным, но ее уменьшение при фронтальной стимуляции препарата позволяло думать, что и эти слуховые органы реагируют на градиент давления.
|В дальнейшем, однако, стали накапливаться данные, противоречащие упомянутой гипотезе Памфри или ставящие под сомнение ее универсальность. Так, в поведенческих экспериментах не подтвердилась способность некоторых сверчков и кузнечиков к монауральной локализации источника звука. Исследования зависимости направленности тимпанальных органов саранчи от частоты звука дали противоречивые результаты. В некоторых экспериментах диаграммы направленности изменялись с изменением
74
частоты, в других такой закономерности не наблюдалось. Специальное изучение этого вопроса показало, что тело саранчи оказывает существенное влияние на прохождение звуковых волн, в результате чего тимпанальные органы на высоких частотах (10—20 кГц) работают как приемники давления, а на низких (около 3,5 кГц) в какой-то мере реагируют и на градиент давления.
Рис. 19. Характеристики направленности слуховых органов:
90- А—изолированного тимпанального органа саранчи (по Pumphrey, 1940); Б — тимпанального органа сверчка Gryllus Ыта-culatus (по Жатнневу и др., 1975).
а, б — до и после удаления контралатеральной
передней ноги.
Частота звука 5 кГц, интенсивность 84 дБ. По радиусам отложена амплитуда ответов в дБ относительно максимума
Некоторые особенности строения и положения тимпанальных органов кузнечиков давали основания предполагать, что они в значительно большей степени соответствуют требованиям, предъявляемым к идеальным приемникам градиента давления. Однако исследования зависимости характеристик их направленности от частоты звука показали, что в диапазоне 5—8 кГц они имеют почти круговые диаграммы и, следовательно, ведут себя как приемники давления, а на частотах 10—20 кГц диаграммы приобретают форму эллипса, что указывает на возможность восприятия градиента давления.
Характеристики направленности тимпанальных органов сверчков изучали только на оптимальных частотах — в области 5 и 16 кГц. При вращении препарата в звуковом поле перепады чувствительности достигали 6—11 дБ. Полученные диаграммы приближались по форме к кардиоиде (5 кГц) или эллипсу (16 кГц). После удаления контралатеральной ноги и закрывания отверстия трахеи в трохантере чувствительность ипсилатерального органа на 5 кГц возрастала и асимметрия диаграмм уменьшалась (рис. 19, Б). Отсюда возникло предположение, что звук, приходящий по трахее с контралатеральной стороны, ослабляет амплитуду колебаний тимпанальной мембраны.
Для снятия характеристик направленности тимпанальных органов чешуекрылых (Noctuidae) использовали короткие щелчки. Полученные диаграммы показали, что пороги реакции снижались при стимуляции рецепторов под углом 90° к плоскости мембраны
72
и значительно возрастали при переносе источника звука на противоположную сторону тела. Учитывая частотные характеристики тимпанальных органов, можно предполагать, что их направленность возникает в результате экранирующего влияния тела бабочки. В других -опытах было установлено, что ответы этих рецепторов зависят не только от угла, под которым приходит звук, но и от положения крыльев.
Суммируя результаты описанных выше экспериментов, можно прийти к выводу, что тимпанальные органы насекомых не подчиняются закономерностям, характерным для простых приемников -градиента давления.
В отлнчие от тимпанальных органов трихоидные сенсиллы и Джонстоновы органы в силу своих конструктивных особенностей могут реагировать только на смещение. Действительно, звуковые колебания воспринимаются в этих органах тонкими волосками -или жгутиками антенн, подвижно сочлененными с кутикулой с помощью эластичной мембраны. В звуковом поле переменный ток воздуха захватывает эти образования внутренним трением и увлекает их в направлении колебания частиц. Если бы такие органы (были снабжены рецепторами, реакция которых отражала направление колебаний подвижных элементов, то с их помощью -можно было бы успешно локализовать источник звука. Как ранее отмечалось, в трихоидных сенсиллах на церках тараканов имеются структуры, ограничивающие колебания волоска в определенной плоскости, а возбуждение нейрона наступает только в том случае, если волосок отклоняется в одном из двух возможных направлений. Имея набор по-разному ориентированных сенсилл такого типа, можно довольно точно определить направление на источник стимуляции (колебания воздуха и, возможно, низкочастотные звуки).
Значительно больший интерес в этом отношении представляют Джонстоновы органы, в состав которых входит несколько групп хордотональных сенсилл. Результаты их исследования у комаров электрофизиологическими методами в основном сводятся к следующему.
При помещении электрода во второй членик антенны и стимуляции препарата звуками низких частот (50—500 Гц) удается зарегистрировать суммарный синхронный ответ. Амплитуда регистрируемого потенциала в определенных пределах зависит от интенсивности звука и его частоты. Максимальная амплитуда ответа наблюдается при частотах, близких к резонансной частоте антенны. Регистрируемый потенциал иногда помимо основной частоты содержит еще и первую гармонику. Специально проведенные эксперименты показали, что гармоника появляется в том случае, когда звук падает на антенну под углом. Если же фронт звуковой волны перпендикулярен к антенне, то ответ содержит только основную частоту. При стимуляции антенн комара иглой, соединенной с прибором, сообщавшим ей синусоидальные колебания, было установ
73
лено, что амплитуда первой гармоники и ее отношение к амплитуде основной частоты зависят от угла, под которым совершаются движения иглы.
На основании этих данных была предложена следующая теория работы Джонстонова органа. 'При движении антенны вдоль ее продольной оси базальная пластинка (и отходящие от нее отростки) стимулирует сенсиллы радиальной серии, в результате чего возникает суммарный потенциал, частота которого соответствует частоте стимулирующего звука. Если фронт звуковой волны параллелен оси антенны, то она совершает качательные движения, а базальная пластинка попеременно стимулирует две группы хордотональных сенсилл, что приводит к удвоению частоты регистрируемого потенциала. В промежуточных случаях, когда звук падает на антенну под углом меньше 90°, в этом потенциале наблюдаются колебания двух частот (основной частоты и первой гармоники), причем соотношение их амплитуд пропорционально углу, под /которым действует стимул.
В процессе дальнейших исследований было установлено, что соотношение амплитуд основной частоты и первой гармоники зависит не только от направления стимуляции, но и от частоты звука, а также от положения электрода во втором членике антенн. Поэтому данная теория в настоящее время нуждается в серьезных поправках.
Центральные слуховые нейроны. Тимпанальные органы у насекомых разделены расстоянием от нескольких миллиметров до 3—4 см, поэтому существует возможность восприятия и оценки различий в их стимуляции. При этом теоретически могут быть использованы фазовые, временные и амплитудные различия.
Исследование функциональной организации тимпанальных органов показало, что они не имеют механизмов, необходимых для кодирования и передачи информации о фазовых изменениях звука, поэтому для предположений об использовании этого параметра в процессе локализации нет никаких оснований.
Интерауральные различия по времени могут достигать у наиболее крупных насекомых 0,1—0,15 мс. В большинстве же случаев они, потвидимому, не превышают 10—30 мкс. Возможность использования столь малых временных различий5 в пространственном слухе насекомых по ряду причин представляется маловероятной и требует специального доказательства.
Различия в интенсивности стимуляции симметричных тимпанальных органов возникают по двум причинам: из-за их направленной чувствительности и в результате экранирующего влияния тела насекомого. Влияние последнего фактора становится замет
5 Значения иитерауральиых временных различий даны для случая, когда звук падает под углом 90° к продольной оси тела насекомого. При локализации источника звука отклонения от курса обычно не достигают 30—40°, поэтому эти различия будут еще меньшими.
74
ным при длинах волн, сравнимых с телом насекомого, и возрастает с ростом частоты.
Электрофизиологические исследования, выполненные в течение (последнего десятилетия, позволили установить наличие перекрестных тормозных и возбуждающих связей между симметричными отделами слуховых анализаторов в грудных и головных ганглиях (рис. 20). Впервые такие связи были обнаружены при изучении
Рис. 20. Изменения ответов слуховых интернейронов в грудном отделе ЦНС при вращении насекомого в звуковом поле.
А — ответы нейрона С сверчка Gryllus Ы-maculatus (по Жан-тневу и др., 1976): а, б — до и после перерезки контралатерального тимпанального нерва. Частота 5 кГц, интенсив-
ность 57 дБ. По радиусам отложена интенсивность ответа в дБ относительно
максимума.
Б — ответы нейрона F саранчи Locusta migratoria (по Kalmring et al., 1972): a, 6 — до и после разрушения контралатерального тимпанального органа. Частота звука 12 кГц, иитенсивиость 63 дБ. По радиусам — число импульсов в ответе на звуковую посылку длительностью 20 мс
слуховой системы кузнечиков, впоследствии их удалось найти у саранчовых, сверчков, медведок и чешуекрылых.
Сравнение полученных к настоящему времени фактов позволяет наметить некоторые общие закономерности в реакциях центральных слуховых нейронов при изменении интерауральных различий в стимуляции.
Прежде всего следует отметить, что ответы многих интернейронов зависят от положения источника звука в пространстве и закономерно изменяются при его перемещении6 (или вращении насекомого), в результате чего могут быть получены соответствующие диаграммы (см. рис. 20). Наблюдающиеся при этом различия в ответах касаются как частоты импульсации, так и величины скрытых периодов реакции. Различия в порогах реакции симметричных нейронов колеблются в значительных пределах и могут достигать 40—60 дБ. Наряду с этим встречаются клетки, совершенно не реагирующие на вращение насекомого в звуковом поле.
Контралатеральный тимпанальный орган оказывает на активность нейронов как тормозное, так и возбуждающее воздействие.
s Здесь и далее речь идет о перемещениях в горизонтальной плоскости.
75
Обычно одно из них преобладает, а их соотношение зависит не только от интерауральных различий в стимуляции, но и от интенсивности, а иногда и частоты звука. В некоторых случаях наблюдаются «парадоксальные» реакции, когда усиление стимуляции ипсилатерального органа вызывает повышение импульсацни контралатеральных интернейронов.
Значительный интерес представляют клетки, ответы которых изменяются только при изменении интерауральных различий стимуляции и не зависят (в широких пределах) от интенсивности звука.
Использование ритмично повторяющихся звуковых посылок, особенно следующих с частотой повторения пульсов в видоспецифических сигналах, существенно усиливает различия в ответах симметричных нейронов.
|Сравнение реакций слуховых интернейронов грудных и головных ганглиев свидетельствует о том, что между ними нет принципиальных различий, но асимметрия в активности протоцеребраль-пых нейронов может быть выражена в меньшей степени, чем у элементов из грудных ганглиев.
Исследование центральных отделов слуховых анализаторов насекомых началось сравнительно недавно, поэтому многие особенности их функционирования при локализации источника звука остаются еще неизученными. Тем не менее проведенные эксперименты позволили выявить два важных факта: существование перекрестных связей на нескольких уровнях слуховых анализаторов и участие в них многих нейронов с разными характеристиками. Сопоставление этих данных с результатами этологических опытов дает основание предполагать, что реципрокные влияния (особенно тормозное, см. рис. 20) повышают контрастность в реакциях симметричных нейронов, в результате чего, по-видимому, увеличиваются разрешающая способность системы и точность локализации, а использование нескольких каналов с разными перекрестными связями повышает пластичность этого механизма и помехоустойчивость при изменяющихся условиях ориентации.
8. ВИБРОРЕЦЕПЦИЯ
Многочисленные наблюдения и электрофизиологические данные свидетельствуют о способности насекомых воспринимать вибрации субстрата. Рецепторы, реагирующие на эти колебания, локализуются в ногах7. Регистрируя суммарную активность сенсорных нейронов в ножных нервах, можно получить представление о чувствительности этих рецепторов. За порог реакции принимается то
7 Исключение составляют только трихоидные сенсиллы на стернитах саранчовых и, возможно, булавовидные хеты на церках сверчковых.
76
минимальное значение амплитуды смещения субстрата или ускорения, при которых наблюдаются ответы (рис. 21).
В результате сравнительного изучения достаточно большого числа видов насекомых было установлено, что они могут условно разделены на две группы: высокочувствительные и чувствительные (табл. 2). К первым относятся тараканы, прямокрылые, палоч-ники, перепончатокрылые, жесткокры-100 лые и другие, ко вторым — полужесткокрылые, двукрылые и некоторые жест-кокрылые. Различия между этими группами касаются не только порогов реак-ции, но и оптимальных частот. В первом случае они обычно лежат в диапазоне 1—3 кГц, во втором — не превышают 0,4 кГц.
Сравнительно-анатомические материалы и результаты экстирпаций различных
быть мало-
Puc. 21. Частотно-пороговые характеристики ножных виброрецепторов (по Autrum, Schneider, 1948):
1 — Carabus, 2 — Vespa, 3 — Bombus, 4 — Pyra-meis, 5 — Periplaneia, 6 — Liogryllus.
По оси абсцисс — частота, Гц; по оси ординат — амплитуда вибрации, нм
- юо -
in-
ly
Ц01
«о 200 w
гц
ножных рецепторов дают основание считать, что у чувствительных видов восприятие вибраций осуществляется преимущественно субгенуальными органами, а у малочувствительных — ти-биотарзальными хордотональными органами и трихоидными сенсиллами на лапках. Такая закономерность прослеживается при сравнении многих групп насекомых, но ее все-таки нельзя считать универсальной, так как отсутствие субгенуального органа не исключает достаточно высокой чувствительности к вибрациям (Melolontha, Geotrupes), а при его наличии восприятие может осуществляться преимущественно другими рецепторами (см. ниже). Точное выяснение роли отдельных сенсилл или органов в этом процессе сопряжено с большими трудностями, поэтому достовер-пых данных получено пока немного.
У тараканов, сверчков и термитов колебания субстрата воспринимаются субгенуальными органами. Степень их развития и детали строения сильно варьируют у разных насекомых. Обычно они представляют собой 1—2 группы удлиненных хордотональных сенсилл, расположенных между трахеей и кутикулой в проксимальной части голени. Число сенсилл в органе колеблется от 10 до 40. У тараканов и прямокрылых шапочковые клетки прикрепляют-
77
Чувствительность насекомых к вибрациям (по Schneider, 1950)
Таблица 2
Группа Виды Оптимальная частота, кГц Пороговая амплитуда, мкм
I Periplaneta americana 1,4 0,000004
Decticus verrucivorus 2,0 0,000036
Tachycines asynamorus 1,0 0,000068
Liogryllus campestris 1,5 0,0001
Melolontha melolontha 1,8 0,0001
Vanessa atalanta 3,0 0,00029
Carausius sp 1,5 0,00035
Satyri dae 2,0 0,0004
Geotrupes sylvaiicus 1,5 0,00048
Stenobothrus sp 3.0 0,00062
Vanessa io 2,0 0,00068
Aeschna cyanea 3,0 0,0009
Apis mellifera 2,5 0,013
Andrend nitida 1,0 0,018
Camponotus sp. 2,0 0,019
Forficula auricularia 1,2 0,019
Bombus soroensis 1,5 0,032
Vespa crab'o 1,5 0,036
Pterostichus sp 0,8 0,040
Silpha obscura 1,0 0,045
Agrotis sp 1,2 0,057
Pseudophonus sp 0,75 0,061
п Reduvius personaius 0,25 1,0
Rhodntus prolixus 0,4 1,3
Eristalis sp 0,2 6,59
Calliphora erithrocephala 0,3 17,4
ся к кутикуле в одной точке, а тела биполярных нейронов образуют дугу, поэтому в целом орган имеет веерообразную форму (см. рис. 13,Б). Несколько наружных, более или менее обособленных сенсилл часто рассматриваются как самостоятельный прилежащий орган.
У тараканов (Periplaneta) ответы субгенуального органа регистрируются в диапазоне 0,025—6 кГц. Оптимум чувствительности лежит в области 1—5 кГц. Минимальные пороги реакции достигают 10-7—10“10 см. Прилежащий орган воспринимает только низкочастотные колебания в пределах 30—700 Гц и отличается меньшей чувствительностью (пороги—10-в см). Помимо хордотональ-ных органов на вибрации субстрата отвечают также кампани-формные сенсиллы, расположенные в проксимальной части голени, но они возбуждаются достаточно сильными колебаниями с частотой, не превышающей 100 Гц.
78
Сходные результаты были получены при изучении ножных рецепторов сверчков (Gryllus)&. Оптимум чувствительности субгенуальных органов передних ног лежит в диапазоне 0,8—4 кГц, а задних — 0,4—0,5 кГц. Минимальные пороги (по амплитуде смещения) во всех трех случаях различались мало (1,4Х10-8—2,2Х Х10-8 см), но передние рецепторы оказались менее чувствительными к ускорению. Искусственное утяжеление задних ног не влияло на функции рецепторов, поэтому можно считать, что их характеристики определяются только механическими свойствами самих субгенуальных органов. Сравнение органов трех пар ног показывает, что их оптимальные частоты находятся в обратной зависимости от собственной массы.
Эти данные находятся в соответствии с модельными представлениями, согласно которым субгенуальный орган рассматривается как эластичная мембрана с массой в центре, натянутая в сосуде с жидкостью. Сотрясение такой системы вызывает смещение массы и колебания мембраны, регистрируемые измерителем (сенсиллами) .
У муравьев-листорезов (Atta) ножные рецепторы реагируют на звуки в диапазоне 0,05—4(7) кГц. Оптимум чувствительности по амплитуде смещения лежит в области 1—3 кГц (минимальный порог— 1,ЗХ10~7 см), а по ускорению — 0,1—2 кГц (порог — 2,5 см/с2). Наиболее чувствительные сенсиллы находятся в передних йогах, причем солдаты менее восприимчивы к вибрациям, чем рабочие. Селективное разрушение разных рецепторов показало, что на колебания субстрата реагируют главным образом кампани-формные сенсиллы, расположенные в области сочленения вертлуга и бедра. Роль субгенуальных органов в этом случае остается неясной.
Вибрации, воспринимаемые насекомыми в естественных условиях, возникают под действием абиотических факторов (ветер, дождь и т. п.) и в результате активности животных (в том числе и особей своего вида). Обычно сотрясения субстрата происходят при передвижении, питании, прокладывании ходов, строительстве и других видах жизнедеятельности. Но кроме того, многие насекомые специально стучат по окружающим предметам различными частями тела или царапают их мандибулами8 9. В некоторых случаях субстрат (например сухие листья) служит только для усиления звука, в других же он используется как канал Связи, а распространяющиеся в нем колебания служат сигналами, несущими определенную информацию. Такая коммуникация хорошо развита у общественных насекомых (термитов, муравьев, пчел, ос), а так
8 Помимо реакции субгенуальных органов в передних ногах регистрировали также ответы каких-то других низкочастотных мехаиорецепторов, расположенных проксимальнее тибиотарзального сочленения.
9 В некоторых случаях (например у муравьев) через субстрат передаются также колебания, издаваемые фрикционными органами.
79
же у веснянок (Plecoptera), ceHoejno3(Psocoptera), жуков-точильщиков (Anobiidae) и др. У одиночно живущих насекомых эти сигналы используются для привлечения особей противоположного пола. Термиты стучат по стенкам гнезда в случае опасности. Воспроизведение этих колебаний вызывает у них серию защитных реакций. Голодные личинки ос получают пищу, если они царапают стенки ячеек мандибулами. Некоторые муравьи (Atta, Megaponera), оказавшись под слоем почвы, издают «сигналы бедствия», привлекающие внимание особей, находящихся на поверхности. Виды, живущие в древесине (Camponotus), стучат по субстрату при появлении опасности или повреждении гнезда. Очень большую роль подобные сигналы играют в жизни пчел. Малейшие сотрясения улья вызывают у них активную защитную реакцию. Доказано также, что матки медоносной пчелы отвечают на «кваканье» других маток или его имитацию только в том случае, если сигнал передается через субстрат. У безжалых пчел (Melipona) разведчица стимулирует фуРажиров на вылет за пищей, издавая специальные звуки. Их восприятие осуществляется виброрецепторами, так как покрытие пола улья резиной полностью исключает реакцию.
Анализ функциональной организации виброрецепторов свидетельствует о том, что они адаптированы для восприятия коммуникационных сигналов. Так, у термитов (Zootermopsis angusticollis) оптимальная частота сенсилл субгенуальных органов (1,15 кГц) почти совладает с доминирующей частотой сигнала тревоги (1,14 кГц). Аналогичное соответствие имеется между частотным оптимумом рецепторов муравьев (Atta) и спектром прошедших через почву «сигналов бедствия».
Помимо вибраций твердого субстрата насекомые могут воспринимать колебания поверхностной пленки воды. Водомерки (Gerri-dae) используют такие волны не только для обнаружения добычи (насекомых, плавающих на поверхности), но и для внутривидовой коммуникации в период размножения. Восприятие колебаний осуществляется различными механорецепторами ног. Минимальные пороги их реакций достигают 1 мкм на частоте 200-—300 Гц.
У жуков-вертячек (Gyrinidae) эти функции выполняет Джои-стонов орган. Второй членик их антенн скользит по воде, а подвижно сочлененный с ним утолщенный жгутик располагается над поверхностью. Колебания второго членика вызывают смещение его относительно жгутика, что приводит к возбуждению хордотональных сенсилл. В целом антенна представляет собой вибрационную систему с резонансной частотой, близкой к 250 Гц. В области частот, не превышающих 150 Гц, она работает как измеритель ускорений. Согласно электрофизиологическим данным, рецептор реагирует на колебания, амплитуда которых достигает нескольких микрон, причем его чувствительность возрастает с увеличением частоты. Во время движения жуки могут определять искривления поверхностной пленки (мениск) вблизи преград с расстояния 0,5— 1,5 см и избегать столкновений.
80
Глава IV
ХЕМОРЕЦЕПЦИЯ
Хеморецепция— восприятие химических стимулов —считается одним из древнейших видов чувствительности. Биологическое значение запахов и веществ контактного действия для насекомых может рассматриваться в двух аспектах. С одной стороны, химические стимулы служат ориентирами, по которым насекомые могут отыскивать и дифференцировать пищевые субстраты, определять оптимальные места для яйцекладки, избегать опасности. С другой стороны, они могут обеспечивать связь между особями различных полов, распознавание особей своего вида или своей семьи у общественных насекомых, передачу сигналов тревоги или меток, ограничивающих территорию, а также следовых меток.
Спектр запахов и веществ контактного действия, воспринимаемых хеморецепторными органами насекомых, чрезвычайно широк. Он включает в себя цветочные и другие запахи, выделяемые растениями, на которых питаются насекомые, а также запахи разлагающихся растительных и животных субстратов, привлекающих насекомых для откладки яиц. Кровососущие насекомые крайне чувствительны к запахам, выделяемым человеком и теплокровными животными. Хищные насекомые и насекомые-паразиты могут отыскивать свои жертвы по пахучему следу. Большое значение в жизни насекомых имеют вещества, выделяемые ими посредством специальных желез в окружающую среду и воздействующие на других особей того же вида, вызывая изменение поведения или действуя иа процессы развития. Для обозначения таких веществ немецкими энтомологами П. Карлсоном и М. Люшером был предложен термин «феромоны».
Различают феромоны, действующие через обонятельные рецепторы (половые аттрактанты, вещества тревоги, маркирующие запахи) и через контактные хеморецепторы (маточное вещество пчел, вещества, влияющие на развитие половых особей термитов, и т.п.). Феромоны первой группы, запускающие определенную поведенческую реакцию животного, американский физиолог Е. Вильсон назвал «релизерами». Феромоны второй группы оказывают «первичный эффект»; они замедляют или ускоряют половое созревание других особей, действуя на метаболизм через эндокринные органы. Советский энтомолог Киршенблат предложил называть подобные биологически активные вещества «телергонами» и разработал для них довольно сложную классификацию, учитывающую действие этих соединений как между особями одного вида, так и между представителями разных видов.
81
До настоящего времени не потеряла своего значения и широко используется в работах по хеморецепции насекомых терминология, предложенная В. Детье, Л. Брауном и К. Смитом в 1960 г., определяющая химические стимулы по характеру их действия на объект. Вещества, обеспечивающие направленное движение к источнику стимуляции или от него, были названы аттрактантами и репеллентами; вещества, вызывающие резкое замедление движения или остановку,— аррестантами; вещества, действие которых приводит к началу движения или какого-либо рефлекторного акта (питание, яйцекладка),—стимулянтами; вещества, тормозящие реакцию,— детеррентами.
Обычно различают две категории хеморецепции: обоняние и вкус. Обонятельные, или ольфакторные, органы воспринимают запахи — пары летучих соединений, присутствующих в воздухе в очень малой концентрации (сотни и тысячи молекул на 1 см3). В отличие от них на вкусовые органы действуют вещества, растворенные в воде в значительно больших концентрациях. Каких-либо иных принципиальных различий в первичных механизмах обонятельной и вкусовой рецепции до настоящего времени не обнаружено. Однако возможные различия, по-видимому, должны существовать, поскольку у многих насекомых или. их личинок, обитающих в водной среде, вкусовые и обонятельные органы морфологически разобщены и имеют разные физиологические характеристики (по результатам поведенческих опытов).
Следует также отметить, что вкусовые рецепторы насекомых имеют более широкий диапазон функций, чем у других представителей животного мира. Если, например, у позвоночных животных вкусовые чувствительные клетки локализованы в ротовой полости и функционируют только в связи с актом питания, то у насекомых аналогичные рецепторы можно встретить также и на конечностях, яйцекладе и других придатках тела. Весь комплекс хеморецепторов, реагирующих при контакте с субстратом, у насекомых называют органами контактной хеморецепции, выделяя из него собственно вкусовые органы, расположенные на челюстях, околорото-вых придатках и в периферических отделах передней кишки.
В отдельных работах можно встретить упоминание о третьей категории хеморецепции — «общем химическом чувстве», иногда наблюдаемом у насекомых. Однако на основании результатов экспериментальных исследований в настоящее время считается, что в явлениях, приписываемых «общему химическому чувству», имеет место неспецифическое раздражение клеток нервной цепочки насекомого парами веществ, проникающих сквозь кутикулу покровов или выстилку трахей, либо аналогичное воздействие реагентов на любые другие чувствительные органы, расположенные на поверхности тела насекомого.
Несмотря на широкое многообразие молекулярной природы запахов и веществ контактного действия, служащих адекватными раздражителями обонятельных органов и органов контактной хе
82
морецепции, а также на различное развитие этих органов у насекомых, отличающихся по своей биологии и экологии, следует отметить общность принципов строения и значительное сходство физиологических свойств отдельных хеморецепторов.
1. ОБЩИЕ ОСОБЕННОСТИ СТРОЕНИЯ
И КЛАССИФИКАЦИЯ ХЕМОРЕЦЕПТОРНЫХ ОРГАНОВ
Клеточный аппарат сенсилл. Выше отмечалось, что хеморецеп-торные органы, функционирующие как экстерорецепторы, построены по типу сенсилл. В состав каждой сенсиллы входит «эпидермальный клеточный комплекс», включающий в себя чувствующие, обкладочные (трихогенную и тормогенную) и Шванновские клетки, а также «кутикулярный отдел», имеющий различную форму и закрывающий отверстие в кутикуле, куда подходят отростки чувствующих клеток.
Чувствующие клетки хеморецепторных сенсилл — биполяры. Периферический отросток клетки, часто называемый дендритом, отходит от апикального полюса клетки и заканчивается в кутикулярном отделе сенсиллы. В большинстве хеморецепторных сенсилл, как обонятельных, так и контактных, значительная часть периферического отростка чувствующей клетки обычно заключена в плотный чехол — сколопоидную оболочку, которая может быть либо общей для всех чувствующих клеток сенсиллы, либо окружать каждый периферический отросток в отдельности. Сколопоидная оболочка обычно заканчивается у основания волоска сенсиллы, хотя в ряде случаев, что свойственно контактным хемо-рецепторным сенсиллам, продолжается в кутикулярный отдел, доходя до его вершины, либо, сливаясь со стенками волоска, делит его полость на два отсека. Сколопоидная оболочка представляет собой продукт секрета трихогенной, или внутренней обкладочной, клетки.
Физиологическое значение сколопоидных оболочек в хеморе-щепторных сенсиллах можно рассматривать с двух сторон. Во-первых, стенки Оболочек образуют внутри сенсиллы систему полостей, которые служат проводниками, участвующими в распределении электрических токов при возбуждении чувствующих клеток. Во-вторых, сколопоидная оболочка важна при линьке, когда на месте старой сенсиллы формируется новая. Вместе с заключенными периферическими отростками она связывает старую сенсиллу с но-вообразующейся почти до самого конца линьки (возможно, для поддержания функции органа либо для преграждения доступа ли-ночной жидкости к чувствующим клеткам). Новый волосок оказывается сформированным за день до линьки, но содержит старую сколопоидную оболочку с нефункционирующими дендритами. Новая сколопоидная оболочка откладывается вокруг старой. Когда
83
наступает линька, старая оболочка вытаскивается из новой, в которую затем подрастают периферические отростки чувствующих клеток.
Периферические отростки чувствующих клеток в хеморецептор-ных сенсиллах имеют цилиарное строение (рис. 22). Проксимальная часть отростка расширена, содержит большое число митохондрий, вакуоли и пластинчатые тела. В дистальном отделе периферический отросток сужается и приобретает структуру реснички, содержащей девять пар периферических и иногда одну пару центральных фибрилл, которые затем замещаются микротрубочками, диффузно распределенными по всей ширине периферического отростка. Между микротрубочками могут встречаться скопления мелких пузырьков. В настоящее время реснички, обнаруженные в чувствительных клетках ольфакторных, вкусовых, а также механорецепторных клетках насекомых, рассматриваются Я. А. Винниковым (1971) как общий признак строения рецепторных клеток беспозвоночных и позвоночных животных.
IB основании реснички в цитоплазме периферического отростка хорошо заметно электрон неплотное базальное тело, от которого й направлении перикариона отходят поперечно исчерченные корешки. Проксимальная часть периферического отростка чувствующей клетки содержит митохондрии, пузырьки, мультивезикуляр-ные тела и отдельные части эндоплазматического ретикулюма. Значительную часть тела клетки занимает ядро. Цитоплазма содержит обычный набор органелл. Центральные отростки чувствующих клеток сенсиллы вместе с выростами Шванновских клеток образуют нервный ствол. Нервные стволы, отходящие от отдельных сенсилл, сливаясь, образуют общий нерв придатка: хоботка, антенны, пальцы и т.д.
Периферические отростки в пределах одной сенсиллы могут тесно сближаться друг с другом, причем расстояние между мембранами соседних дендритов достигает 180—200 А. На их поверхности могут возникать специализированные структуры — десмосомы. Обнаружены два типа десмосом: обычные и септальные. Обычные десмосомы, или пластинки прикрепления, представляют собой локальные электронно-плотные расширения подмембранных областей контактирующих дендритов. Пластинки прикрепления образуются также при контакте мембран периферического отростка и трихогенной клетки. Часто десмосомы локализованы на выростах, которые образованы трихогенной клеткой. Ширина десмосом 0,1—0,15 мкм, а межмембранное расстояние составляет 170—200 А. Септальная десмосома характеризуется наличием поперечных мостиков, или септ, которые соединяют плазматические мембраны соседних клеток. Иногда дендрит образует пальцеобразный вырост, который охватывается выростами трихогенной клетки.
Трихогенная и тормогенная клетки, окружая пучок периферических отростков чувствующих клеток, образуют межклеточную «4
Рис. 22. Ультраструктура чувствующих клеток хе-морецепторных сенсилл насекомых.
А — поперечный срез контактной хеморецеп-торной базиконической сенсиллы мокреца Culi-coides fascipennis на уровне реснички периферического отростка чувствующей клетки; Б — поперечный срез обонятельной трихоидной сенсиллы комара Culex pi-piens на уровне микротрубочек периферических отростков; В — продольный срез периферических отростков чувствующих клеток базиконической обонятельной сенсиллы слепня Haematopota plu-vicdis на уровне реснички; Г — поверхность трихогенной клетки контактной хеморецепторной сенсиллы слепня Chry-sops caecutiens, граничащая с внутренней полостью сенсиллы.
бт — базальное тело; в — внутренняя полость сенсиллы; м — митохондрии; мв — микровиллы; мт — микротрубочки; пз — пузырьки; по — периферический отросток чувствующей клетки; со— сколопоидная оболочка; трк — трихогенная клетка
полость сенсиллы, которая продолжается в просвет кутикулярного отдела. Поверхность этих клеток, образующая дно и боковые стенки полости, покрыта многочисленными микроворсинками, в основании которых расположены митохондрии (рис. 22, Г). Для обкладочных клеток характерен хорошо развитый аппарат Гольджи, что говорит об их секреторной функции и о важной роли в формировании жидкого содержимого полости сенсиллы и просвета ее кутикулярного отдела, т.е. среды, обеспечивающей нормальное функционирование периферических отростков чувствующих клеток в момент генерации возбуждения. Весь эпидермальный клеточный комплекс подстилается базальной мембраной гиподермы, переходящей в соединительнотканную оболочку нервного ствола сенсиллы.
Классификация хеморецепторных сенсилл. В настоящее время еще не выработан единый принцип классификации хеморецепторных сенсилл, однако можно проследить две тенденции в разделении их по типам.
Первая связана с функциональными характеристиками сенсилл, прежде всего с проницаемостью кутикулярного отдела для красителей. В 1954 г. американский энтомолог Е. Слайфер показала, что после помещения лапок комаров и мух в раствор красителя прокрашиваются только кончики хем'очувствителыных волосков, причем глубина проникновения краски возрастает с увеличением времени окрашивания. Если же такой операции подвергнуть чувствительные волоски на антенне саранчи, то произойдет равномерное окрашивание кутикулярного отдела сенсиллы. Е. Слайфер связала этот эффект с толщиной стенок кутикулярного отдела и предположила, что краситель проходит через поры, локализованные в первом случае лишь на кончике волоска, а во втором — по всей его поверхности. Это предположение в дальнейшем подтвердилось в электронно-микроскопических исследованиях. Первая группа волосков была отнесена к вкусовым, вторая—к обонятельным сенсиллам.
По Е. Слайфер (1970), все хеморецепторы насекомых делятся на две группы: толстостенные и тонкостенные (табл. 3). К первой группе органов с контактной хеморецепторной функцией относятся многочисленные волоски и колышки (pegs), встречающиеся на поверхности кутикулы, колышки в углублениях кутикулы и папиллы— небольшие кутикулярные бугорки, к апикальным порам которых подходят чувствительные окончания. Вторая группа хеморецепторов— тонкостенные обонятельные органы, куда также входят волоски и колышки, расположенные на поверхности тела насекомых или в кутикулярных ямках. Кроме того, к тонкостенным хеморецепторам следует относить бороздчатые волоски, плакоид-ные органы, а также своеобразные петлеобразные органы, обнаруженные на антеннах галлиц.
Удачная по своему замыслу система классификации хеморецепторных органов, предложенная Е. Слайфер, имеет основной
86
Таблица 3
Классификация хеморецепторных сенсилл
По Слайфер, 1971 По Снодграссу, 1935 и Шнейдеру, 1969
ТИП модальность ТИП модальность
А. Толстостенные хеморецепторы 1. Волоски на поверхности 2. Колышки на поверхности 3. Колышки в ямках 4. Папиллы Б. Тонкостенные хеморецепторы 5. Волоски на поверхности 6. Колышки на поверхности 7. Колышки в ямках 8. Бороздчатые волоски 9. Плакоидные органы 10. Петлеобразные сенсиллы 11. Сложные чувствительные органы вкус обоняние 1. Хетоидные сенсиллы: а) бороздчатые б) ветвящиеся 2. Трихоидные сенсиллы: а) с острым кончиком б) с тупым кончиком 3. Базиконические сенсиллы: а) длинные б) короткие 4. Стилоконические сенсиллы 5. Булавовидные » . . 6. Плакоидные » . . 7. Целоконические » . . 8. Ампуловидные » . . 9. Лопастевидные » . . 10. Целосферические » . . 11. Со складчатой булавой 12. Сложные чувствительные органы вкус то же вкус обоняние вкус, обоняние то же вкус обоняние то же » » » » обоняние? то же » » обоняние
недостаток, заключающийся в том, что ори ее использовании невозможно отразить все разнообразие строения хеморецепторных органов. Поэтому наибольшее распространение получила терминология, в которой типы хеморецепторных органов обозначаются соответственно морфологии кутикулярного отдела (рис. 23). Этот подход, введенный в 1935 г. Р. Снодграссом и получивший свое развитие в работах известных специалистов по хеморецепции насекомых— американского энтомолога В. Детьё и немецкого физиолога Д. Шнейдера,—в настоящее время опирается на данные сканирующей электронной микроскопии, позволяющей выявлять мельчайшие особенности ультра морфслоги и кутикулярного отдела хеморецепторных сенсилл.
Соответственно наружной морфологии хеморецепторные волоски разделяются на хетоидные сенсиллы — длинные прямые волоски с толстой кутикулой и острым кончиком и трихоидные сенсиллы— длинные, обычно слегка изогнутые волоски. Поверхность хетоидных сенсилл несет продольные борозды либо, как, например, на антеннах короедов, имеет мельчайшие ответвления. Бороздчатые хетоидные сенсиллы обычно контактные хеморецепторы, а ветвящиеся, видимо, имеют механорецепторную функцию.
87
Расположенные на хоботке мух трихоидные сенсиллы с бороздчатой поверхностью и острым кончиком относятся к вкусовым сенсиллам. Гладкая поверхность трихоидных сенсилл обычно сочетается с тонкой кутикулой и закругленной вершиной волоска. Таковы многочисленные обонятельные сенсиллы на антеннах самцов сатурний и волнянок.
Рис. 23. Типы хеморецепторных сенсилл насекомых: а — хетоидная; б — трихо-идная с острым кончиком; в — трихоидная с тупым кончиком; г — базикониче-ская; д—стилоконичеокая; е — булавовидная; ж — плакоидная; з — целоконическая; и — ампуловидная; к — лопастевидная; л — целосферическая; м — со складчатой булавой
Как хеморецепторы наиболее широко распространены базико-нические сенсиллы. Это — короткие волоски или конусы с закругленным кончиком, встречающиеся на антеннах, челюстях, пальцах, яйцекладе и других частях тела насекомых. Базиконические сенсиллы функционируют либо как обонятельные, либо как контактные хеморецепторы в зависимости от их внутреннего строения.
На антеннах бабочек и челюстях гусениц встречаются стило-кони веские сенсиллы, представляющие собой маленький чувствительный конус на вершине кутикулярного выроста. Булавовидные сенсиллы, расширенные у вершины и соединяющиеся ножкой с кутикулой, обнаружены на максиллярных пальпах мокрецов и комаров. У коллембол аналогичные органы на антеннах имеют крупные складки и выпячивания.
К обонятельным органам относятся небольшие базиконические сенсиллы, скрытые в углублении кутикулы, имеющем вид капсулы с отверстием наружу. Такие образования называются целоконическими, когда капсула находится у поверхности кутикулы, и ампуловидными—когда капсула углублена в полость антенны и соединена с ее поверхностью тонким каналом. На антеннах некоторых бабочек из семейства совок найдены лопастевидные сенсиллы, погруженные в кутикулу таким образом, что снаружи видна лишь вершина кутикулярного отдела. Целосферические сенсиллы, недавно обнаруженные на антеннах жуков,—заключенные в кутикулярную капсулу сферические образования со сложной перфорацией стенок. Обонятельная функция сенсилл двух последних типов еще экспериментально не подтверждена.
88
Для перепончатокрылых и многих других насекомых характерны обонятельные плакоидные сенсиллы, встречающиеся главным образом на антеннах. Кутикулярный отдел сенсиллы имеет вид овальной или круглой пластинки, иногда слегка выступающей над поверхностью кутикулы.
Как и в первом случае, предложенная классификация хеморе-цепторных органов насекомых не может полностью отразить их разнообразие. Типы сенсилл могут перекрываться. Так, например, длинные базиконические сенсиллы практически неотличимы от коротких трихоидных сенсилл с тупым кончиком. Существует много переходных форм. Отдельные сложные обонятельные органы, образовавшиеся в результате слияния многих сенсилл, вообще не поддаются классификации и рассматриваются индивидуально.
К тому же существующее деление сенсилл по типам не отражает их функциональной принадлежности. Так, базиконические сенсиллы могут быть или вкусовыми, или обонятельными, или воспринимающими изменение влажности. Большинство трихоидных. сенсилл — механорецепторы, некоторые целоконические сенсиллы на антеннах комаров выполняют функцию температурных рецепторов. Окончательная классификация сенсилл может быть произведена лишь при комплексном использовании электронно-микроскопического и электрофизиологического методов.
2. МОРФОЛОГИЧЕСКАЯ ХАРАКТЕРИСТИКА КОНТАКТНЫХ ХЕМОРЕЦЕПТОРНЫХ СЕНСИЛЛ
Локализация. Основное назначение контактных хеморецептор-ных органов насекомых — определение пригодности субстрата для питания и яйцекладки. Поэтому они встречаются на тех участках тела насекомого или его придатках, которые используются для отгрызания и размельчения пищи, а также для ощупывания поверхности предметов, т. е. на челюстях, околоротовых придатках, яйцекладах и дистальных члениках ног. Для того чтобы определить локализацию контактных хеморецепторов, растворы различных вкусовых веществ наносили на различные участки тела насекомого и регистрировали его ответную реакцию, которая проявлялась в характерных рефлекторных движениях ротовых частей, хоботка, околоротовых придатков. В случае, если отдельные сенсиллы были хорошо различимы при малых увеличениях микроскопа (лабеллярные волооки высших мух), производили раздражение отдельных чувствительных волосков растворами соли или сахара, заключенными в микропипетку. Окончательная идентификация чувствительных волосков как контактных хеморецепторов может быть осуществлена по методике, предложенной в 1954 г. американскими исследователями Е. Ходжсоном и К. Редером, когда микропипетка с раздражающим раствором одевается на вершину
89
волоска и одновременно служит электродом, регистрирующим биоэлектрическую) активность чувствующих клеток сенсиллы.
Многочисленные исследования показали, что контактные хеморецепторы у насекомых из разных отрядов в основном локализованы на лабиальных и максиллярных пальцах, челюстях, лигуле и гипофарингое. У насекомых, имеющих хоботок, контактные хеморецепторы хорошо развиты на дистальных члениках ног, преимущественно на лапках. Они обнаружены также на антеннах пчел, муравьев, кузнечиков, термитов, бабочек из семейства совок, на яйцекладах мух, орехотворок и наездников-ихневмонид.
У насекомых с ортоптероидным типом ротового аппарата (тараканы, прямокрылые, многие жуки) все его части — мандибулы, макоиллы, верхняя и нижняя губы — участвуют в откусывании и размельчении пищи. Хеморецепторы в этом случае диффузно и равномерно распределены по внутренней поверхности ротовых придатков. У саранчи Schistocerca gregarina мелкие трихоидные сенсиллы размером 35 мкм присутствуют на всех частях ротового аппарата, а базиконические сенсиллы найдены на клипеолабраль-ном комплексе и гипофарингсе. На эпифаринпсе личинок стрекоз обнаружено множество сенсилл, представляющих собой выпуклые участки мезокутикулы с центральной порой, куда подходят периферические отростки чувствующих клеток. У имаго стрекоз поверхность эпифарингса покрыта многочисленными базиконически-ми сенсиллами.
У гусениц бабочек контактные хеморецепторы сконцентрированы на дистальных участках мамиллярных и лабиальных пальп и представлены базиконическими и стилоконическими сенсиллами. Аналогичное распределение вкусовых сенсилл было обнаружено у личинок щелкунов рода Agriotes и личинок хлебного жука Ani-soplia austriaca. В последнем случае стилоконические сенсиллы присутствуют также на максиллах и на внутренней поверхности верхней губы личинки жука. У имаго бабочек контактные хеморецепторы найдены на лабеллах и лапках ног и отсутствуют на лабиальных и максиллярных пальпах. Сходная картина наблюдается у Thysanura и Diptera.
У высших двукрылых вкусовые хеморецепторы расположены на вентролатеральной поверхности лабелл и представлены трихо-идными сенсиллами разного размера. Самые длинные волоски у мух Calliphora erythrocephala достигают 300 мкм и расположены по периферии орального диска. Волоски внутренних рядов имеют меньшие размеры, а на вентральной поверхности хоботка встречаются маленькие, около 10 мкм, «интерпсевдотрахеальные папиллы». Такое различие в длине волосков рассматривается как приспособление для увеличения числа контактов рецепторов с поверхностью во время приема пищи, когда лабеллы находятся в развернутом состоянии. Общее количество лабеллярных трихоидных сенсилл у мух колеблется от 220 у Calliphora vomitoria до 28—30 у Stomoxys calcitrans.
90
Еще в 1926 г. Д. Минничем было показано, что мухи вытягивают хоботок при контакте дистальных сегментов йог с раствором сахара. Позже тарзальная чувствительность была обнаружена у комаров, слепней, клопов, бабочек, саранчовых и многих других насекомых. По своей морфологии тарзальные хеморецепторы мало отличаются от трихоидных лабеллярных сенсилл. Однако число контактных хеморецепторов на лапках трех пар ног неодинаково. Так, тарзусы передних ног мухи Phormia regina несут 303 сенсиллы, а тарзусы задних ног—147 сенсилл. Такая же особенность распределения тарзальных сенсилл установлена для осенней жигалки Stomoxys calcitrans, кровососущих комаров Aedes aegyp-ti, некоторых видов слепней. Увеличение числа сенсилл на лапках передних ног свидетельствует об их большей чувствительности к контактным раздражителям, что также подтверждается физиологическими тестами.
У насекомых с колюще-сосущим ротовым аппаратом (комары, тли, клопы) контактные хеморецепторные органы можно разделить на две группы соответственно выполняемым функциям. Это, во-первых, трихоидные и базиконические сенсиллы внешней поверхности нижней губы, используемые при ощупывании поверхности субстрата и при питании свободными жидкостями, и, во-вторых, сенсиллы колющих стилетов и стенок пищевого канала, функционирующие при проколе тканей.
Контактные хеморецепторные органы насекомых представлены четырьмя типами сенсилл: трихоидными, базиконическими и сти-локоническими, а также чувствительными папиллами.
Ультраструктура контактных хеморецепторных сенсилл. Наиболее полно строение и функциональные свойства трихоидных хеморецепторных сенсилл исследованы на примере лабеллярных хеморецепторов двукрылых (рис. 24, 25).
Кутикулярный отдел сенсиллы представляет собой толстостенный волосок, который разделен продольной перегородкой на две полости, названные по толщине ограничивающей их кутикулы: толстостенной и тонкостенной. Перегородка волоска, по-видимому, сколопоидная оболочка, окружает периферические отростки чувствующих клеток и одной стороной спаяна с внутренней поверхностью стенки волоска. В толстостенной полости к вершине волоска проходят периферические отростки клеток. Тонкостенная полость волоска сообщается с межклеточной полостью сенсиллы. Основание волоска связано с кутикулой лабеллума тонкими тяжами, имеющими, по-видимому, кутикулярную природу. Эти тяжи обеспечивают отклонение волоска при контакте с субстратом, а также приведение его в исходное 1состояние.
На вершине волоска имеется одна или несколько пор диаметром около 500 А, соединяющих толстостенную полость с внешней средой. Иногда поры смещены несколько ниже вершины волоска, Имеющей в этом случае форму папиллы (рис. 25,Б). Тонкостенная
91
полость заканчивается слепо под вершиной волоска на расстоянии нескольких микрон.
При электронномикроскопическом исследовании вершины хемо-рецепторной сенсиллы у мух Phormia regina и Calliphora vicina
Рис. 24. Схема строения контактной хеморецеп-торной трихоидной сенсиллы мошки Boophthora erythrocephala (по Елизарову, Чайке, 1975):
по — периферический отросток; рк — рецепторная клетка; см — сочленовная мембрана; со — сколопоидная оболочка; тлп — толстостенная полость; тнп — тонкостенная полость; ток — тор-могенная клетка; трк — трихогенная клетка; цо — центральный отросток; швк — Шванновская клетка
обнаружены два состояния поры: открытое и закрытое. Водные растворы и влажный воздух вызывают открывание поры, тогда как действие сухого воздуха, фиксаторов и прикосновение к волоску приводят к закрыванию поры. Механизмы, осуществляющие от-
92
Рис. 25. Контактные хеморецепторные сенсиллы.
А — трихоидные лабеллярные сенсиллы на краю орального диска хоботка мухи Protophormia terraenovae; Б — пора на кончике трихоидной сенсиллы; В — вязкое содержимое, выступающее из поры; Г — поперечный срез волоска на лабеллуме комара Culex pipiens; Д — поперечный срез лабеллярной трихоидной сенсиллы мошки Boophthora erythrocephala на уровне трубчатого тела (тт) механорецептора.
Обозначения, как на рис. 24
крывание и закрывание пор в трихоидных сенсиллах, пока остаются невыясненными. Видимо, они связаны с изменением давления внутри волоска, так как сократительных волокон или каких-либо тяжей в полости волоска не обнаружено. Предполагается, что в кончике волоска в состав кутикулы входит структурный белок «резилин», обеспечивающий подвижность складок, закрывающих пору.
На вершине волоска могут наблюдаться капли вязкой жидкости, выступающей из поры (рис. 25, В). Вязкий субстрат служит посредником между растворами вкусовых веществ и чувствительными мембранами периферических отростков. В отдельных трихоидных сенсиллах отмечаются пучки филаментов из электронно
93
плотного материала, соединяющиеся с мембраной периферического отростка, что говорит об определенном сходстве с поротрубчатой системой обонятельных сенсилл.
Число рецепторных клеток в контактных хеморецепторных трй-хоидных сенсиллах не превышает пяти, хотя в отдельных сенсиллах их может быть и меньше. Периферические отростки доходят до вершины волоска, кроме отростка одной из клеток, который заканчивается у основания волоска характерным образованием, называемым «трубчатым телом» (рис. 25, Д). Как показали электрофизиологические исследования, этот отросток принадлежит ме-ханорецепторной клетке. Трубочки, содержащиеся в дистальной части периферического отростка механорецептора, имеют диаметр 150 А и состоят из электронноплотного материала. В центре трубчатого тела они расположены гексагонально, на периферии — палисадно. Между собой трубочки связаны перпендикулярными слоями электронноплотного вещества. Стимуляция механорецепторной клетки происходит в момент сжатия трубчатого тела, возникающего при движении волоска.
Кутикулярный отдел базиконических сенсилл с контактной хеморецепторной функцией представляет собой полый толстостенный конус, погруженный в углубление кутикулы и соединенный с верхним краем расположенного под ним канала сочленовной мембраной. Между основанием конуса и стенками канала может быть натянута тонкая дополнительная мембрана, состоящая из отдельных тяжей.
Отличительной особенностью строения кутикулярного отдела базиконических сенсилл служит отсутствие продольной перегородки в его полости. Периферические отростки чувствующих клеток, окруженные сколопоидной оболочкой, проходят в центре кутикулярного отдела к терминальной поре, диаметр которой около 300 А. Сколопоидная оболочка прикреплена к стенкам конуса только в области его вершины. Внутри футляра, образованного сколопоидной оболочкой, отходят продольные выросты, частично разделяя периферические отростки клеток.
Базиконические сенсиллы иннервируются тремя-пятью чувствующими клетками. В отдельных сенсиллах, например в терминальных сенсиллах максиллярных пальп саранчи Schistocerca gre-garina, их число может доходить до 11 на одну сенсиллу.
Вкусовые папиллы. Интерпсевдотрахеальные папиллы высших мух представляют собой небольшие конусы, не превышающие по высоте 7 мкм. Они находятся в кутикулярных ямках на такой глубине, что вершина конуса слегка выступает над поверхностью кутикулы. Папиллы встречаются у многих мух на вентральной поверхности хоботка в промежутках между псевдотрахеями.
Исследование ультраструктуры папилл мух Boettcherisca pereg-rina показало большое сходство в строении папилл и контактных хеморецепторных базиконических сенсилл. Эпидермальный клеточ
94
ный комплекс папиллы содержит четыре чувствующие клетки, две трихогенные и одну тормогенную. Одна из чувствующих клеток служит механорецептором и образует трубчатое тело в основании папиллы. Периферические отростки клеток имеют структуру ресничек. Как и в предыдущих типах сенсилл, обкладочные клетки образуют две полости: базальную, где расположены основания ресничек, и межклеточную полость сенсиллы, доходящую до основания кутикулярного конуса. Дистальные части периферических отростков чувствующих клеток заключены в сколопоидные оболочки, заканчивающиеся у основания папиллы. В узком ее просвете сколопоидных оболочек не обнаружено, дендриты лежат свободно в жидком содержимом кутикулярного отдела. Характер возникновения возбуждения при раздражении клеток папиллы поэтому должен несколько отличаться от процессов, происходящих при возбуждении клеток в двуполостной трихоидной контактной хемо-рецепторной сенсилле.
Другие типы контактных хем о рецепторных сенсилл. Контактная хеморецепторная функция свойственна стило-коническим сенсиллам личинок чешуекрылых и жуков, мелким чувствительным шипам и порам, обнаруженным на клипео-лабру-ме саранчи и в цибариальной полости мухи комаров, а также чувствительным бугоркам на антеннах бабочек-совок, куполообразным сенсиллам на яйцекладах наездников и т.п. Все эти органы имеют поры на вершине кутикулярного отдела, 3—5 чувствующих клеток с цилиарной структурой периферических отростков, обкладочные клетки, образующие межклеточные полости сенсиллы, сколопоидные оболочки, окружающие периферические отростки целиком или в отдельности. Механо рецепторов в таких сенсиллах не обнаружено.
3. МОРФОЛОГИЧЕСКАЯ ХАРАКТЕРИСТИКА ОБОНЯТЕЛЬНЫХ СЕНСИЛЛ
Локализация. У большинства насекомых обонятельные сенсиллы расположены на антеннах. Экспериментально это впервые было показано в 1921 г. Е. Фишером, который наблюдал постепенное уменьшение интенсивности ольфакторной реакции вплоть до ее полного исчезновения в результате последовательной ампутации члеников антенны пчелы. Однако у некоторых мух и комаров обонятельные сенсиллы могут встречаться также и на околоротовых придатках. Регистрация биопотенциалов от антеннального нерва и нервной цепочки сверчка показала, что обонятельные сенсиллы у этого насекомого находятся на околоротовых придатках вместе с контактными хеморецепторами.
Рассматривая распределение ольфакторных сенсилл на антеннах насекомых, можно увидеть различную степень их концентра-
95
ции на поверхности кутикулы. Диффузное распределение сенсилл наблюдается у тараканов, некоторых прямокрылых и перепончатокрылых насекомых, в частности у ос, пчел и наездников.
Для комаров характерна группировка сенсилл на различных участках флагеллума антенны. Обонятельную функцию у этих на-' секомых выполняют тонкостенные волоски, разделяемые на три типа по своей величине. Один тип сенсилл доминирует на дистальных члениках флагеллума антенны комаров. Сенсиллы другого типа сконцентрированы на проксимальных члениках, а третьего типа равномерно распределены по флагеллуму антенны.
Следующую ступень концентрации обонятельных сенсилл можно встретить у дневных бабочек. На большей части члеников антенны, начиная с дистального конца флагеллума, базиконические тонкостенные сенсиллы собраны в тесные группы, локализованные на медиальной стороне члеников и ограниченные от других участков кутикулы. Скопление обонятельных сенсилл, образующих рецепторное поле, окружено гребневидными утолщениями кутикулы. Такой тип обонятельного органа свойствен антеннам бабочек из семейств нимфалмд и бархатниц. У бабочек из семейства белянок чувствительные поля имеют меньшие размеры и углублены по отношению к поверхности кутикулы членика антенны.
Дальнейшее развитие обонятельных органов наблюдается у двукрылых насекомых из подотряда короткоусых мух. Здесь на третьем членике антенны встречаются так называемые обонятельные ямки, представляющие собой глубокие впячивания кутикулы членика, открывающиеся наружу небольшими отверстиями. Дно этих впячиваний покрыто бавиконическими сенсиллами, воспринимающими запахи. В отдельных случаях, связанных с олигомеризацией антенн, например, у личинок комаров и некоторых жуков, обонятельные сенсиллы могут сливаться, образуя массивные органы с единым кутикулярным отделом. Наиболее ярко такие органы выражены у цикад, где второй членик антенны плотно покрыт розетками уплощенных пальцеобразных выростов, выходящих из углубления кутикулы и представляющих собой результат слияния многих обонятельных сенсилл.
Таким образом, на примере наездников, комаров, дневных чешуекрылых, мух и цикад мы видим процесс постепенного усложнения обонятельного органа насекомых — от равномерно распределенных по антенне сенсилл до специализированных многоклеточных образований.
(Самым характерным структурным признаком обонятельных сенсилл насекомых следует считать многочисленные поры их кутикулярного отдела. Впервые поры были обнаружены в базиконических сенсиллах кузнечика Romalea microptera, когда было показано, что кутикула базиконических сенсилл проницаема для водных растворов кристаллического фиолетового и что краситель проникает через поры, имеющиеся в кутикулярном отделе. В последние
96
Рис. 26. Кутикулярные отделы обонятельных сенсилл.
А — трихоидные обонятельные сенсиллы на поверхности флагеллума антенны бабочки волнянки Orgyia prisca-, Б — булавовидные сенсиллы максиллярных пальп комара Culex pipiens; В — плакоидные сенсиллы на поверхности флагеллума антенны самца пчелы Eucera cinerea
10 лет поры широко исследовались с помощью электронного мик-
роскопа.
Ультраструктура обонятельных сенсилл. Кутикулярный отдел плакоидной сенсиллы имеет вид тонкой кутикулярной пластин
ки овальной или продолговатой формы (рис. 26,(В). В настоящее
время плакоидныё сенсиллы обнаружены на антеннах жуков, равнокрылых хоботных и перепончатокрылых.
Плакоидные сенсиллы пчелы были первыми хемо-рецепторными сенсиллами насекомых, исследованными с помощью электронного микроскопа. Очень тонкая сочленовная мембрана сенсиллы пересечена 120—150 радиально идущими утолщениями, между которыми
Рис. 27. Схема строения плакоид-ной сенсиллы жука Acilius sulca-tus (по Иванову, 1969): бм — базальная мембрана; дк--диск; дс — дистальный сегмент; ит — наружное тельце; пс — проксимальный сегмент периферического отростка; пр — проток железы; ст — стенка наружного скелета антенны; трк — трихоген-ная клетка; швк — Шванновская клетка.
Остальные обозначения, как на рис. 22.
расположены поры. Дальнейшие электронно-микроскопические исследования показали, что поверхность пластинки и сочленовной мембраны представляет собой слой эпикутикулы, под которым расположены зубцы, состоящие из менее электронноплотной кутикулы.
В промежутках между зубцами обнаруживаются поры, диаметр которых составляет около 100 А. В состав плакоидной сенсиллы пчелы входят 12—18 чувствующих клеток, периферические отростки которых заканчиваются под сочленовной мембраной, т. е. в области локализации пор.
Кроме сочленовной мембраны поры могут располагаться также по всей поверхности кутикулярной пластинки плакоидной сенсиллы. У осы Nasonia vitripennis вся пластинка перфорирована небольшими неправильными отверстиями, через которые проходит
98
раствор кристаллического фиолетового при нанесении его на наружную поверхность сенсиллы.
Иное строение кутикулярного отдела плакоидных сенсилл у жука Acilias silvaticus (рис. 27). Здесь полость кутикулярного диска, погруженного в чашеобразное углубление кутикулы, связана с каналом, пронизывающим кутикулу антенны. Через этот канал проходят два периферических отростка чувствующих клеток, разветвляющихся в полости диска. Верхняя пластинка снабжена порами, диаметр которых увеличивается в направлении наружной поверхности.
Уникальная плакоидная сенсилла описана у наездников- Coleoi-des brunneri. В ней находятся две продолговатые куполообразные пластинки, подвешенные изнутри к кутикулярному отделу сенсиллы. Этими пластинками внутренняя полость сенсиллы разделяется на три канала. Ветви периферических отростков заполняют две трети срединного канала, а два боковых канала и треть срединного заняты тормогенной клеткой. Авторы приписывают этим сенсиллам восприятие инфракрасного излучения. К разновидности плакоидных сенсилл могут быть отнесены также сложные чувствительные органы — ринарии, локализованные на антеннах тлей.
Трихоидные обонятельные сенсиллы с тонкими стенками представляют собой один из наиболее распространенных типов обонятельных сенсилл насекомых (см. рис. 26,А). Они характеризуются небольшой толщиной кутикулы и наличием пор. Число пор на одном волоске может достигать 1500—2500. Показано, что поры трихоидных сенсилл шелкопряда связаны с периферическими отростками чувствующих клеток поровыми трубками.
Трихоидные обонятельные^ сенсиллы имеют различное число чувствующих клеток. Так, в сенсиллах короеда Scolytis multistria-tus обнаружены 2 клетки, в сенсиллах муравья Lasius fuligino-sus—20—30, а в сенсиллах клопа Lygaeus kalmii — 50—60 клеток.
Периферические отростки чувствующих клеток обонятельных сенсилл обычно начинают ветвиться в основании волоска. Однако в некоторых трихоидных сенсиллах, например на антеннах шелкопряда Bombyx mori, периферические отростки не ветвятся.
Своеобразно устроены трихоидные сенсиллы ногохвостки Onychiurus sp. Поры и поровые трубки кутикулярного отдела подстилаются аморфным электронноплотным материалом, секретируемым оберточными клетками, служащим, по всей вероятности, стимулопроводящим элементом сенсиллы.
Разновидностью трихоидных сенсилл считаются сенсиллы антенн эмбий Ptilocerembia sp. и тараканов Arenivaga sp. Это толстостенные волоски, кутикула которых имеет поры. Такого рода сенсиллы сочетают в себе признаки толстостенных и тонкостенных хеморецепторов.
Тонкостенные базиконические сенсиллы широко распространены на антеннах насекомых и, согласно электрофизиологи-
99
Рис. 28. Строение обонятельных сенсилл.
А — базиконические сенсиллы на антенне мухи Protophormia terraenovae; Б — поперечный срез кутикулярного отдела базиконической сенсиллы москита Phlebotomus papatasi-, В — поперечный срез антеннальной трихоидной сенсиллы бабочки павлиноглазки Actias artemis.
п — пора; пт — поровые трубочки; по — периферические отростки
ческим данным, могут функционировать как типичные обонятельные рецепторы.
Наружный кутикулярный отдел базиконической сенсиллы имеет вид небольшого шипа, стенки которого перфорированы многочисленными порами (рис. 28,А,Б). Базиконические сенсиллы, сохраняя общий план строения, различаются у разных видов насекомых по числу чувствующих клеток, величине кутикулярного отдела, а также по числу и размеру пор.
100
Кутикулярный отдел базиконических сенсилл обычно имеет толщину стенок 0,05—0,3 мкм с гексагонально размещенными порами. Такое размещение пор наиболее экономично с точки зрения эффективного использования площади. Кутикулярный отдел базиконических сенсилл мухи Phormia regina имеет 180—360 пор, а в базиконических сенсиллах жука Necrophorus sp. число пор достигает 13 000—17 000. Наружное отверстие поры не превышает 50 А.
В полости кутикулярного отдела базиконических сенсилл периферические отростки рецепторных клеток интенсивно ветвятся. В 1967 г. в первых электронномикроскопических работах Е. Слай-фер установила, что к порам подходят тонкие ветви, диаметр которых равен 200 А. Позже было показано, что в порах находятся не кончики периферических отростков, а пучки тонких нитей, которые образуются путем выпячивания стенки периферического отростка. Эти тончайшие нити диаметром 200 А были названы «поровыми филаментами», или поровыми нитями. Им приписывается функция первичного взаимодействия с молекулами пахучих веществ. Однако в 1969 г. немецким морфологом К- Эрнстом было показано, что в базиконических сенсиллах жука Necrophorus трубочки, сходные с поровыми нитями, не являются выростами мембран периферических отростков, а имеют экстраклеточное происхождение. В период онтогенеза сенсилл трубочки, связанные с порами, встречаются в просвете шипа еще до вхождения периферических отростков в полость последнего. В пользу экстраклеточной природы трубок говорит также их небольшой диаметр, равный 150—200 А, в то время как диаметр тончайших ветвей периферических отростков — около 500 А. Кроме того, толщина стенки трубочек составляет около 30 А, что не соответствует толщине клеточной мембраны. Таким образом, поры и поровые трубки образуют единую по происхождению поро-трубчатую систему, которая формируется в процессе онтогенеза оберточными клетками и, видимо, служит для проведения молекул пахучих веществ к периферическим отросткам.
На максиллярных пальпах комаров и москитов встречаются булавовидные органы, имеющие тонкую кутикулярную стенку (см. рис. 26,Б). Изучение поведения комаров и электрофизиологические исследования показали, что булавовидные органы имеют чувствующие клетки, воспринимающие углекислоту, и клетки с ольфакторной функцией.
Длина булавовидных органов составляет 16—17 мкм, а их диаметр — 5—6 мкм. Они иннервируются тремя биполярными нейронами. Толщина кутикулы булавовидного органа равна в среднем 0,1 мкм. Через весь слой кутикулы булавовидных органов проходят поровые каналы. С каждым поровым каналом связаны 2—3 поровые трубки диаметром 100—140 А и толщиной стенок 30 А. Поровые трубки представляют собой кутикулярные образования, и их сердцевина электроннопрозрачна. Начинаясь от прок
101
симальной части порового канала, они простираются в полость волоска на расстояние до 0,1 мкм. Иногда периферические отростки образуют спиралеобразную пластинчатую структуру, состоящую из складок их мембран.
Целоконическая сенсилла представляет собой конусообразный или волосковидный орган, погруженный в кутикулу. Це,-локонические сенсиллы широко распространены среди насекомых н встречаются у представителей многих отрядов.
В связи с тем, что число целоконических сенсилл на единицу площади невелико, исследование их ультраструктуры затруднено, и в настоящее время нет полных сведений о строении этих сенсилл.
По данным световой и электронной микроскопии, в целоконических сенсиллах прямокрылых имеется несколько чувствующих клеток, периферические отростки которых подходят к вершине конуса. Периферические отростки не ветвятся и заключены в сколопоидную оболочку. Вершина конуса имеет отверстие, которое обнаруживается при нанесении растворов красителей. Сходное строение имеют целоконические сенсиллы антенн уховерток.
В отличие от вышеописанного типа стенки целоконических сенсилл бабочки Danaus gillipus перфорированы многими порами, однако поровые трубки отсутствуют.
У некоторых видов стрекоз Lygoptera и Anisoptera имеются как простые, так и сложные целоконические сенсиллы. Кутикулярный конус сенсилл пронизан порами. Сложные сенсиллы представляют собой несколько кутикулярных отделов, отличающихся по внешнему виду, которые находятся в углублении кутикулы.
Целоконические сенсиллы присутствуют на антеннах слепней в разном количестве в зависимости от вида насекомого и обнаруживают поразительную общность ультраморфологии. Наибольшее число целоконических сенсилл (300—400) насчитывается у слепней родов Tabanus и Hybomitra, среднее (150—180)—у видов рода Haematopota и наименьшее (40—120) —у видов рода Chrysops.
Сложные органы обоняния. Кроме сенсилл с четко выраженной ольфакторной функцией в последнее время в литературе появились сообщения о новых типах чувствительных органов, ультра-структурная организация которых свидетельствует об их обонятельной функции.
Опоясывающие нити. Ультраструктура тонкостенных кутикулярных нитей, окружающих многими петлями антенны самцов и самок видов семейства Cecidomyidae, исследована у галли-цы Contarinis sorghicola. Опоясывающие нити по своему строению подобны тонкостенным обонятельным сенсиллам; их диаметр свыше 1 мкм, а стенки пронизаны порами. На поперечных срезах через нити виден просвет, заполненный ветвями периферических отростков чувствующих клеток.
Чувствительные придатки, или конусы. Антеннальный придаток личинки жука Ctenicera destructor — сложный чув
102
ствительный орган, образованный соединением 12 базиконических сенсилл. В придатке имеются 36 биполярных чувствующих клеток, распределенных в 12 групп по три в каждой. Кроме них в состав чувствительного придатка входят по 12 трихогенных, тормо-генных, трофических и нейриллемных клеток. Все клетки, принадлежащие одной элементарной сенсилле, отделены от соседних мембрановидной перегородкой. В конусе периферические отростки чувствующих клеток ветвятся и заполняют все его пространство. Кутикула конуса пронизана небольшими трубочками, диаметр которых равен 300 А. Непосредственной связи периферических отростков с трубочками не обнаружено.
Кутикулярные выросты, напоминающие базиконические сенсиллы и связанные с 30 чувствующими клетками, описаны на антенне муравья Camponotus vagus. Поры и поровые трубки имеются только в апикальной части выростов. Кольцевидное утолщение стенок выростов делит внутреннюю полость сенсиллы на две камеры. В нижней камере находится специфическая жидкость, через которую должны пройти молекулы пахучих веществ, прежде чем достигнут периферических отростков чувствующих клеток.
Постантеннальный орган ногохвосток по своему строению отличается от обычной схемы строения сенсиллы. В постантеннальном органе Onychiurus sp., имеющем предположительно хемо- или гигрорецепторную функцию, перикарионы чувствующих клеток локализованы в головном мозгу. Имеются и другие ульт-раструктурные особенности, выделяющие этот орган среди остальных сенсилл: наличие двух зон ресничек в периферических отростках и отсутствие корешковых структур. Сходный орган описан также и у бессяжковых из семейства Acerentomidae.
Целосферические сенсиллы обнаружены только на антеннах жуков Necrophorus sp. Кутикулярный отдел целосферических сенсилл, имеющий вид сферы, полностью погружен в углубление кутикулы. Углубление открывается наружным отверстием диаметром 1—2 мкм. Стенка кутикулярного отдела пронизана порами, с которыми связаны мешковидные образования из волокнистого материала. В сенсиллах первого подтипа эти образования оканчиваются во впячиваниях третьей обкладочной клетки, которая выстилает внутреннюю сторону кутикулярного отдела сенсиллы. В сенсиллах второго подтипа мешковидные структуры окаймляют наружную полость сенсиллы. В сенсилле найдены две чувствующие клетки, периферические отростки которых окружены;со всех сторон сколопоидной оболочкой. По-видимому, проникновение молекул пахучих веществ к ветвям периферических отростков осуществляется путем диффузии через сколопоидную оболочку.
Сложные чувствительные органы сконцентрированы на флагеллуме антенн цикады Pyrops candelarla. Они имеют вид округлых участков сильно складчатой перфорированной кутикулы. Общее число пор на пластинке достигает 40000—160000. В СО
ЮЗ
став каждой сенсиллы, считаемой сложным обонятельным органом, возникшим из нескольких базиконических сенсилл, входит большое число чувствующих клеток, собранных в 25—35 групп по 6—10 клеток в каждой. Периферические отростки каждой группы окружены чехлом, который образован базальной клеткой. В терминальной части периферические отростки ветвятся, причем каждая ветвь связана с несколькими порами.
Стимулопроводящая система обонятельных сенсилл. За исключением отдельных примеров, все перечисленные типы обонятельных сенсилл характеризуются пористостью стенок кутикулярного отдела. Для того чтобы вступить во взаимодействие с рецепторными зонами мембран чувствительных сенсилл, молекулы раздражителя должны проникнуть во внутреннюю полость сенсиллы, чему способствует особая стимулопроводящая система, включающая в себя поры и соединенные с ними поровые трубочки или филаменты в стенке сенсиллы.
В обонятельных волосках обнаружено несколько типов организаций стимулопроводящей системы. В толстостенных сенсиллах на антенне таракана Arenivaga sp. описаны поровые каналы диаметром 100—200 А и длиной до 1 мкм, соединяющие с внешней средой полость волоска, где находятся периферические отростки. Проксимальные части каналов часто бывают расширенными, образуя капсулы, заполненные электроннопрозрачным содержимым. Внутренняя поверхность стенок каналов выстлана электронноплотной пленкой. Использованием различных режимов фиксации препаратов, действием органических растворителей и гидрофильных красителей была показана неоднородность содержимого каналов. Оказалось, что капсулы заполнены липидами, а электронноплотная пленка представляет собой липопротеиновую мембрану, которая выстилает и внутреннюю полость волоска.
Большинство обонятельных сенсилл насекомых имеет тонкие стенки кутикулярного отдела. В обонятельных сенсиллах кровососущих двукрылых и самцов бабочек сатурний наиболее типичное строение поры — коническая полость, пронизывающая стейку сенсиллы и открывающаяся наружу узким отверстием (около 100 А). Полость содержит 4—8 поровых трубочек, обычно выходящих из нее в просвет сенсиллы и во многих случаях вступающих в контакт с мембранами разветвлений периферических отростков. Отверстие поры может находиться в углублении, в отдельных случаях принимающем вид капсулы. При этом дно капсулы образуется периферическими отростками поровых трубочек.
Проводящая роль поровых трубочек в антеннальных сенсиллах жуков Necrophorus vespillo была показана Эрнстом в 1969 г. Он наблюдал проникновение частиц коллоидного серебра (протаргола) в просвет трубочки, хотя выхода частиц в полость сенсиллы не происходило. Аналогичная картина была описана для поротрубчатой системы дорсального органа личинки мухи Musca do-mestica, а также для поровых каналов толстостенных обонятель-
104
них сенсилл клеща Amblyomma americanum. Вместе с тем обонятельное сенсиллы органа Галлера этого клеща, видимо, имеют гидрофобное содержимое пор, так как гранулы серебра скапливаются лишь в начале поры. Длительная экспозиция в растворе протаргола приводила к возникновению электронноплотных филаментов из мелкодиспергированного материала в порах, по размерам и расположению очень похожих на поровые трубочки насекомых, но без центральной светлой части. Эти филаменты могли достигать мембран разветвлений периферического отростка чувствительной клетки сенсиллы.
Присутствие тяжей, или филаментов, в порах тонкостенных обонятельных сенсилл было показано у таракана Arenivaga sp. Тяжи имеют липидную природу, так как растворяются в ацетоне, исчезают при изменении режима фиксации, ведущему к вымыванию липидов, не пропускают в полость волоска через поры водорастворимые красители.
Существуют две точки зрения на связь поровых трубочек, или филаментов, с кутикулой. С одной стороны, считают, что трубочки представляют собой вещество непрерывной мембраны, подстилающей кутикулу волоска сенсиллы изнутри. С другой стороны, вещество, из которого состоят трубочки, отождествляют с эпикутикулой волоска. Так, обонятельные сенсиллы тутового шелкопряда имеют трехслойную эпикутикулу. Наружный электронноплотный слой не заходит в пору. Средний слой, прозрачный на электронных микрофотографиях, заполняет сердцевину микротрубочки, а внутренний слой эпикутикулы, по плотности равный наружному, формирует стенки микротрубочки. Поскольку материал, составляющий содержимое н стенки микротрубочек, у этого насекомого не растворяется в органических растворителях, ему приписывают свойства «стабилизированных липидов», состоящих из полимерных единиц. Если учесть, что значительная часть поровых трубочек (на некоторых срезах волоска до 4О°/о) вступает в контакт с мембранами периферических отростков чувствующих клеток, то можно говорить о прямой связи в виде липидного мостика поверхности кутикулы волоска, куда попадают молекулы пахучих веществ, с рецепторной мембраной. Установлено, что сенсиллы на антеннах самца тутового шелкопряда адсорбируют 80—100% бомбикола. попавшего на антенну, хотя их поверхность составляет лишь около 13% площади антенны. Количественные измерения порога чувствительности самца тутового или непарного шелкопрядов к феромонам самки, полученные при использовании как поведенческого, так и электрофизиологического критериев, показали, что возбуждение самца наступает, когда на антенны действует струя воздуха, содержащая около 100 молекул аттрактанта в 1 см3 воздуха. Если допустить, что все молекулы адсорбируются и попадают на поверхность волосков и учесть площадь поперечного сечения поровых каналов, которая составляет 0,03% от площади волосков, то за 30 с (максимальное время латентного периода реакции) при
105
скорости потока 1 см/с вероятность прямого попадания хотя бы одной молекулы в пору равна 0,1. Расчеты приводят к выводу, что стимулопроводящая система должна активно участвовать в транспортировке молекул раздражителя не только внутри порового канала и поровых трубочек, но и на поверхности волоска.
Несомненная общность строения стимулопроводящей системы' в обонятельных сенсиллах у большого числа исследованных представителей различных отрядов насекомых говорит об ее важной роли в первичных процессах обонятельной рецепции членистоногих. Очевидно, что функциональная роль поро-трубчатой системы у насекомых тесно связана с явлением сверхвысокой чувствительности к специфическим запахам, поскольку в обонятельных органах позвоночных аналогичных морфологических образований не обнаружено.
4. ФИЗИОЛОГИЯ КОНТАКТНЫХ ХЕМОРЕЦЕПТОРНЫХ СЕНСИЛЛ
Качественный диапазон каналов, по которым поступает информация через хеморецепторы у насекомых, крайне узок. Так, если в обонятельном эпителии кролика содержится около 100 млн. чувствительных клеток, то у гусеницы бражника всего 48. Вместе с тем в способности распознавать пищевые субстраты по запаху насекомые не уступают высшим животным. Выбор пищи у мухи Phormia regina обеспечивается функционированием лишь 250 ла-беллярных волосков и 65 папилл на хоботке.
Малочисленность чувствующих клеток в воспринимающей системе неизбежно приводит к увеличению специализации отдельных ее элементов. Хеморецепторные органы насекомых отличаются от позвоночных более высокой специфичностью рецепторных клеток. Вместе с тем нельзя предполагать, что насекомые живут в мире альтернативных химических сигналов. Поскольку синаптические контакты у насекомых не найдены ни на периферии, ни в афферентных путях, у них, по-видимому, отсутствует предварительная интеграция сигнала, а первичный анализ импульсной активности осуществляют нервные центры, и они же формируют ответ на предъявленный химический стимул.
Пороги и чувствительность
Первые результаты по определению чувствительности контактных хеморецепторов, процессов суммации, адаптации, зависимости ответов от качества раздражающего стимула и т. д. были получены путем изучения поведения насекомых или простейших движений их конечностей, хоботка, околоротовых придатков при раздражении сенсилл растворами и чистыми веществами.
106
Результаты сопоставления вкусовой чувствительности насекомых и человека показывают, что на широкий круг химических соединений она измеряется примерно одними и теми же величинами. Так, человек ощущает вкус сахара при концентрации 0,02 М, а пчела отличает раствор этого вещества от дистиллированной воды при концентрации 0,06 М. Слепень Tabanus положительно реагирует на растворы сахарозы в концентрациях 0,005—0,11 М. Однако многие бабочки вытягивают хоботок при контакте раствора сахарозы с тарзусом, когда концентрация раствора равна 0,00001 М.
Хеморецепторы, расположенные на тарзусе и лабеллах одного и того же вида насекомых, имеют разную величину порога реакции на растворы вкусовых веществ. Пороговая концентрация сахарозы, необходимая для возникновения рефлекторной пищевой реакции мухи Phormia regina, равна 0,0098 М, если раствор наносить на тарзальные волоски. При раздражении лабеллярных волосков положительный ответ возникает на 0,0001 М концентрацию сахарозы.
У мух Calliphora наблюдаются обратные отношения: тарзальные рецепторы в шесть раз более чувствительны к раствору сахара, чем лабеллярные. У пчел, собирающих сладкий нектар, вкусо^ вые рецепторы хоботка малочувствительны к сахару. Пороговая концентрация для лабеллярных рецепторов равна 0,125—0,062 М, в то время как контактные хеморецепторы антенн воспринимают раствор сахарозы в концентрации 0,07—0,01 М. Малая чувствительность лабеллярных рецепторов у насекомых, питающихся растворами углеводов, биологически оправдана, так как насекомые должны поглощать при питании более концентрированные растворы сахаров.
Таблица 4
Пороговые концентрации вкусовых веществ для человека и пчелы, М (по Редеру, 1963)
Вещество Человек Пчела
Хлористый натрий Соляная кислота Сахароза Хинин 0,09 0,001 0,02 1 Х10-7 0,24 0,001 0,06 1 X10-5
Для веществ, которые вызывают у насекомых отрицательную пищевую реакцию, или реакцию отвергания (хлористый натрий, соляная кислота, хинин), установлены пороговые значения концентраций, близкие к тем, которые наблюдаются у человека. Так, человек начинает отличать раствор хлористого натрия от дистиллированной воды при концентрации 0,09 М, пчела — при 0,24 М,
107
слепни Tabanus sp.—при 0,8 M, мухи Phormia regina — при 0,58 М. Пороговые концентрации кислоты, хинина и сахарозы для пчел и человека имеют величину одного и того же порядка (табл. 4).
Однако к этим данным следует относиться с определенной осторожностью, поскольку в качестве критериев использовались не ответы хеморецепторов насекомых, а рефлекторные реакции всего организма или отдельного органа в ответ на химическую стимуляцию. Даже в случае раздражения отдельной хеморецепторной сенсиллы на хоботке или лапке насекомого следует учитывать физиологическое состояние насекомого, определяющее порог рефлекторной реакции.
Электрофизиологические исследования
В 1954 г. американскими физиологами Е. Ходжсоном и К. Редером была предложена методика регистрации биоэлектрической активности контактных хеморецепторных сенсилл, используемая и в настоящее время (рис. 29). В опыте тонкая пипетка (диаметром 10—20 мкм) заполнялась раствором электролита и служила активным электродом. Если в ткани тела или головы насекомого ввести другой, индифферентный электрод и привести пипетку в контакт с кончиком волоска, то можно зарегистрировать импульсы, отражающие активность чувствующих клеток сенсиллы. Контактным химическим раздражителем в данном случае будет электролит, заполняющий пипетку. Подобный метод не применим для неэлектролитов. Для раздражения сенсиллы неэлектролитами, растворимыми в воде, например различными сахарами, в качестве растворителя для раздражающего вещества используют либо крайне слабый раствор хлористого натрия, либо раствор электролита, нейтрального для хеморецепторных клеток. Иногда регистрируют потенциалы через отверстие в стенке волоска, из полости сенсиллы, куда микроэлектрод вводят через сочленовную мембрану (рис. 29). Во всех случаях потенциалы чувствующих клеток соседних сенсилл обычно не регистрируются, так как клетки данной сенсиллы отделены базальной мембраной от полости тела и от чувствительных клеток других сенсилл. Импульсы в ответах контактной хеморецепторной сенсиллы можно разделить на несколько типов по величине их амплитуды. Каждая из амплитуд соответствует активности одной из чувствующих клеток.
При раздражении контактных хеморецепторных сенсилл растворами хлористого натрия или других солей в концентрации 0,1—0,5 М регистрируется залп одинаковых по амплитуде (около 1 мВ) и форме импульсов, отражающих активность чувствующей клетки, специфичной к растворам электролитов. Эту клетку обозначают как солевой, или L-рецептор. Другая клетка в сенсилле возбуждается при раздражениях растворами глюкозы, сахарозы
108
и других углеводов, потребляемых насекомыми в пищу, и называется сахарным, или S-рецептором. Его импульсы несколько меньше по амплитуде, чем импульсы L-рецептора, когда они регистри-
руются одновременно. В кончике волоска дендрита третьей клетки, которую часто lj^-рецептором. Ее возбуждение происходит при действии на сенсиллу чистой воды.
Результаты последних исследований говорят о том, что солевые рецепторы (L) можно разделить на две группы: анионные (£]) и катионные (L2). Они вызывают реакцию отвергания у мух, неиспытывающих чувства жажды. Когда же мухи находятся долгое время без воды, раздражение катионного рецептора вызывает положительную пищевую реакцию, в то время как раздражение анионного соответствует реакции отвергания.
Если во время регистрации импуль-
расположено окончание называют водным, или
Рис. 29. Способы регистрации биопотенциалов хеморецепторных сенсилл насекомых (по Hodgson, 1964):
А — регистрация микропипеткой от кончика волоска контактной хеморецептор-ной сенсиллы; Б — регистрация через отверстие в стенке волоска; В — реги-
сов в ответ на раздражение растворами сахара или соли согнуть волосок при помощи раздражающей пипетки, то можно зарегистрировать импульсы небольшой амплитуды. Они отражают активность М-рецептора — механо-рецепторной клетки, которая присутствует в сенсилле совместно с хемо-рецепторными (Lb L2, 5, UP) клетка-
ми.
Одна из особенностей импульсов, регистрируемых по методике Е. Ходжсона и К. Редера при помощи микропипетки, заключается в их положительной полярности. Причину этого явления объяснили результаты одновременной регистрации биопотенциалов
страция микроэлектродом через кутикулярную стенку придатка; Г — регистрация суммарного потенциала от целого придатка — антенны. S — раздражитель; R — активный электрод; I — индифферентный электрод
на различных уровнях длинного (100—200 мкм) лабеллярного волоска мухи Phormia regina. Дендритный холмик, где возникает импульс, отстоит достаточно далеко от места регистрации. В мо-
мент появления импульса в основании дендрита, где мембрана
становится негативной, в кончике волоска возникает положитель-
ное отклонение потенциала. Импульс распространяется по двум направлениям: к центральной нервной системе по аксону клетки и к кончику волоска по ее дендриту. Когда импульс достигает верхушки волоска, положительная фаза переходит в отрицательную фазу.
109
Регистрация активности хеморецепторных клеток во вкусовой папилле у мухи Phormia regina показала несколько иную картину. В то время как импульсы сахарного рецептора имеют положительную полярность, импульсы солевого рецептора, зарегистриро-В|анные в тех же условиях, имеют отрицательную полярность. Подобные результаты были получены и в опытах при отведении биопотенциалов от маленьких (30—40 мкм) хеморецепторных воло-
Рис. 30. Ответы контактной хеморецепторной сенсиллы на лабеллуме мухи Calliphora (по Morita, Yamashita, 1959):
А — на раздражение 0,25-молярным раствором сахарозы до (а) и после (б) отравления; Б — на раздражение 0,25-молярным раствором сахарозы (а), смесью 0,25-молярного раствора сахарозы и 0,05-молярного раствора СаС12 (б) и 0,05-молярным раствором СаС12 (в).
Отметка времени — 20 мс
сков на хоботке комара Aedes aegypti, где импульсы L-рецептора характеризуются отрицательным знаком отклонения потенциала, а импульсы S-рецептора — положительным.
Генераторный потенциал хеморецепторной клетки представляет собой медленное отрицательное колебание с амплитудой в несколько милливольт, быстро возрастающее в момент начала действия раздражителя и поддерживающееся на постоянном уровне в течение всего времени его действия. На фоне генераторного потенциала регистрируются импульсы положительной полярности с частотой, снижающейся по мере действия раздражителя (рис. 30,Л). При раздражении хеморецептора хлористым калием и гуанином генераторный потенциал приобретает обратный знак, рецепторная мембрана гиперполяризуется, и импульсы исчезают (рис. 30,5). При раздражении сенсиллы смесью веществ, возбуждающей несколько рецепторных клеток, регистрируется суммарный генераторный потенциал.
110
Свойства солевого рецептора
Е. Ходжсоном и К. Редером в 1954 г. на лабеллярных хеморецепторах мухи Phormia regina было установлено, что интенсивность стимула зависит в основном от вида катиона и в меньшей степени от аниона молекулы. Эффективность раздражающего раствора пропорциональна ионной подвижности химических веществ. Видимо, на рецепторной поверхности солевого рецептора существуют отрицательные (анионные) заряды, вступающие в связь с катионами раздражителя. Связь устанавливается чрезвычайно быстро, так как латентный период возбуждения рецептора очень мал и составляет несколько миллисекунд. Отсутствие температурного эффекта и эффекта pH (в диапазоне pH 3—11) в реакции солевого рецептора на раздражитель позволяет предположить, что взаимодействие между катионами раздражителя и мембранными акцепторами солевого рецептора носит физический характер и не включает энзимохимические реакции.
Сравнение функций солевого рецептора мухи с реагирующей на соль рецепторной клеткой крысы позволило сделать вывод, что катионы реагируют с некоторым акцептором мембраны и величина ответа прямо пропорциональна числу ионов, вступающих с ним в реакцию. Как и у позвоночных животных, ответы солевого рецептора в сенсиллах насекомых подчиняются теории Л. Бейдлера (1954), основное уравнение которой имеет следующий вид:
с с 1 И Rmax K-Rmax
где С — концентрация стимула;
7? —величина ответа клетки при данной концентрации; /?тах — величина ответа, когда заняты все участки акцептора;
К — константа равновесия.
Все параметры этого уравнения, кроме константы равновесия (К), были получены экспериментально. При этом графическое вы-С
ражение зависимости — от С представляет собой прямую линию. R
Константа равновесия служит мерой сродства раздражителя с рецептором. Небольшие величины найденных констант равновесия подтвердили предположение об адсорбции раздражителя на мембране чувствительной клетки и неферментативной природе реакции.
Л. Бейдлер, а вслед за ним Д. Эванс и Д. Меллон, проверявшие его теорию в 1962 г. на вкусовых рецепторах мухи, считают, что акцепторами, или активными участками мембраны, являются полиэлектролиты, имеющие заряженные боковые цепи, и предполагают, что в основе комплекса «стимул — акцептор» лежит образование водородных связей между катионами стимула и функциональными группами акцептора с отрицательным зарядом.
111
Несколько иных представлений о механизме восприятия раздражителя солевым рецептором насекомых придерживается английский физиолог К. Рис (1968). В основе его теории лежит предположение о наличии в мембране хеморецептора пор, внутренняя поверхность которых имеет фиксированные отрицательные заряды, в результате чего мембрана имеет относительную проницаемость к различным ионам.
Экспериментальное исследование зависимости между электрофизиологическим ответом солевого рецептора и физико-химическими параметрами молекул раздражителя—ионной активностью и ионным радиусом катионов и анионов солей — дало положительную корреляцию. Однако она наблюдается только в том случае, когда величина диаметра ионов рассчитывается без учета их гидратации. Теория К. Риса, в отличие от теории Л. Бейдлера, дает объяснение процессам адаптации, наступающей в рецепторе при длительном действии раздражающего стимула. В момент раздражения происходит быстрое возрастание концентрации ионов в жидкости, омывающей окончания периферических отростков чувствующих клеток. Это приводит к адсорбции положительно заряженных ионов на порах мембраны, что повышает ее сопротивление и приводит к уменьшению деполяризационного потенциала. Последнее вызывает падение частоты импульсов, соответствующее процессу адаптации рецептора.
Другая теория возбуждения хеморецепторов, в которой возникновение возбуждения солевого рецептора обусловливается изменением проницаемости клеточной мембраны под действием стимула, была выдвинута в 1972 г. К- Оттером. Клеточная мембрана рецептора рассматривается как каркас из бимолекулярного слоя фосфолипидов, в котором липидные молекулы расположены перпендикулярно к плоскости мембраны гидрофильными группами наружу. Кроме того, в мембране могут присутствовать глобулярные и спиральные молекулы протеинов, и изменение проницаемости мембраны вкусовых клеток для ионов, наступающее во время стимуляции, происходит в результате открытия и запирания «организованных» и «статистических» пор. «Организованные» поры могут быть образованы спиральными сегментами пептидов, стабилизированных гидрофобными посредниками с липидами мембран. Знак заряда внутреннего пространства пор находится в зависимости от последовательности аминокислот в цепочках пептидов. «Статистические» поры появляются в результате теплового движения молекул мембраны, плотность «упаковки» которых определяет число пор. Ионы Са++ связывают молекулы мембраны путем формирования звеньев между анионными участками.
При раздражении рецептора хлоридами щелочных металлов ионы С1~ связывают ионы Са++ и тем самым ослабляют «упаковку» молекул мембраны, что ведет к ослаблению проницаемости. Этому процессу противодействует фиксация щелочных катионов на фосфатных группах молекул мембраны, которая определяется
112
сродством катионов к фосфатным группам. Подобными механизмами К. Оттер объясняет ответы вкусовой клетки на раздражение неорганическими солями аммония, солями щелочных и щелочноземельных металлов. Действие растворенных в воде органических раздражителей зависит от адсорбционных свойств молекул раздражающего вещества и от их распределения в системе вода — липидный слой и внутри двухслойной мембраны.
Свойства сахарного рецептора
Экспериментальные данные, полученные Е. Ходжсоном в электрофизиологических опытах на мухе и при изучении поведенческих реакций, показали, что изомерия молекул сахаров является одним из основных качеств молекул, определяющих стимулирующий эффект. Так, Р-арабиноза оказалась более сильным раздражителем, чем L-арабиноза. Однако стимулирующая эффективность сахаров, спиртов, кислот и других органических соединений зависит не только от стереоизомерии, но и от степени растворимости, атомного веса, разветвленности молекулярной цепочки, пространственной структуры молекулы.
В настоящее время считают, что на рецепторной мембране даже одной чувствующей клетки — сахарного рецептора — есть несколько типов так называемых «рецепторных участков», которые вступают в связь с молекулами раздражителя. «Рецепторные участки» для глюкозы соединяются с гидроксильными группами молекулы. Другие соединения также будут действовать на эти участки, если наклон их гидроксильных групп к основной плоскости молекулы колеблется в пределах ±19°.
В водных растворах сахара обычно присутствуют в виде 5—6-членных колец — фуранозных или пиранозных группировок. Голландский энтомолог Л. Шоонховен в 1974 г., после проведения испытаний большой группы полигидроксильных соединений, образующих 6-членные кольца, предположил, что наибольшим стимулирующим действием для контактных рецепторов клеток, локализованных в медиальных стилоконических сенсиллах гусеницы Den-drolimus pint, обладают вещества с определенным расположением гидроксильных групп:
Три экваториальные гидроксильные группы пиранозного кольца (1, 2, 3) не должны быть стерически изолированы другими группами или радикалами соседних атомов углерода (V). Стери-
113
ческая доступность данных трех функциональных групп обеспечивает их взаимодействие с соответствующими акцепторами на мембране сахарного рецептора. Конформационные изменения формы молекулы в конфигурации «кресла» (С1—1С) не влияют на эффективность действия раздражителя на рецепторную клетку-
Регистрация биоэлектрической активности рецепторов медиальной стилоконической сенсиллы гусеницы Denarolimus pini показала правильность такого предположения (табл. 5).
Таблица 5
Эффективность стимулирующего действия циклических шестичленных углеводов для контактных хеморецепторов гусеницы Dendrolimus pini
(по Шоонховену, 1974)
Эффективность Вещество
Высокая (90—130) pD-ксилоза > pD-галактоза >• р Д-глюкоза > мезоинозиол > Р£)-глюкоза > аО-арабиноза >• рД-фукоза
Средняя (60—20) сахароза > 2-диокси-рО-глюкоза > рО-метилглюкозид > аД-арабиноза > aD-ликсоза > глиоксалевая кислота
Малая илн отсутствие ответа аД-сорбоза; pD-фруктоза; NaCl; аа-трегалоза; Р£)-рнбоза; D-сорбитол; aD-манноза
П р имечание. Эффективность оценивается по частоте афферентной импульсации (указана в скобках), имп/с.
Рассматривая сахарный рецептор как систему, продуцирующую потенциалы действия, и используя кумулятивные графики, отражающие процесс накопления импульсов во времени, Л. Шоон-ховен обнаружил определенную аналогию между кумулятивными графиками, отражающими процессы адаптации рецептора, и кривыми, отражающими зависимость скорости ферментативной реакции от концентрации субстрата. Для обработки материала было использовано уравнение Михаэлиса-Ментен:
1 _ Лм _L 1
1-4 4max S + 4max
где S — концентрация стимула;
A — ответ рецептора на данную концентрацию;
Ап.™ — ответ на концентрацию, вызывающую максимальную реакцию рецептора;
Км— константа Михаэлиса.
114
Как и в случае уравнения Л. Бейдлера, наблюдается линейная зависимость импульсного ответа рецептора от молярной концентрации раздражителя. Константа Михаэлиса в данном случае отражает сродство молекул раздражителя к акцепторам рецепторной мембраны сахарного рецептора. Эффективность веществ, раздражающих контактные хеморецепторы медиальной стилоконической сенсиллы гусеницы Dendrolimus pini, вычисленная по константе Михаэлиса, уменьшается в следующей последовательности: мезо-инозитол > р D-глюкоза > ₽ D-ксилоза > д-лактон D-глюконовой кислоты.
Многочисленные поведенческие и электрофизиологические исследования контактных хеморецепторов мух показали, что на стимулирующую активность молекул сахаров большое влияние оказывает их оптическая изомерия. В 1972 г. японские физиологи X. Морита и другие, сопоставляя экспериментальные данные с теоретическими расчетами, детально изучили сродство молекул стимула к рецепторным мембранам, влияние положения отдельных активных центров в молекулах а- и р-глюкозы и их производных на стимулирующую активность: положение ОН-группы, наличие и положение полярных и неполярных групп — метильной, этильной, п-нитрофенильной.
Еще в ранних работах Э. Эванс (1963) предположил, что хемо-рецепторная мембрана сахарного рецептора имеет два акцепторных участка различного типа, один из которых вступает во взаимодействие с молекулой глюкозы, другой — с молекулой фруктозы. Дисахарид сахароза соединяется с «фруктозным» участком. Дальнейшее развитие представлений о возбуждении сахарного рецептора было проделано X. Морита и А. Шираиши в 1968 г. Эти авторы теоретически проанализировали предположение Эванса и пришли к заключению, что оно верно при действии типа сахарозы, когда образуется комплекс 1:1 — «рецепторный участок»: дисахарид. В случае моносахаридов необходимо образование другого комплекса — 2 :1, когда две молекулы стимула одновременно связываются одним рецепторным участком. Поэтому авторы предложили модель «рецепторного участка», состоящего из двух субъединиц. Возбуждение возникает, если один «рецепторный участок» одновременно занимают две молекулы моносахаридов или одна молекула дисахарида. В наибольшей степени для объяснения функционирования такого «рецепторного участка», состоящего из двух субъединиц, соответствует аллостерическая макромолекула.
Это объясняется двумя причинами. Во-первых, только макромолекулярная структура может обеспечить специфическое восприятие сахаров; во-вторых, незаряженные молекулы сахара могут вызвать электрические изменения в рецепторной мембране только посредством конформационных изменений рецепторного участка, и эти изменения могут быть связаны с аллостерическим переходом.
115
Первым экспериментальным подтверждением гипотезы о существовании вкусового рецепторного белка было выделение Ф. Да-столи и С. Прайсом в 1966 г. «сладкочувствительного» и «горько-чувствительного» белков из языка быка. В последние годы был сделан ряд попыток выделить аналогичный белок из сахарного рецептора насекомых. На основании некоторого параллелизма констант Михаэлиса для а-глюкозидазы и величины поведенческого порога для сахарного рецептора мух было сделано предположение, что этот фермент может быть рецепторным протеином для сахаров. Позже многие исследователи обнаружили независимо друг от друга а-глюкозидазу в экстрактах лабеллума и ног мух. Было установлено, что этот фермент имеет три изозима: Pl, Р2, РЗ, причем Р2 обладает наибольшим сродством к ряду сахаров. Недавно а-глюкозидазная активность была обнаружена на вершине хеморецепторного волоска мухи. Константа Михаэлиса этого фермента находилась в соответствии с таковой изозима Р2 в препаратах лабеллума и ног мух.
Таким образом, имеются данные, позволяющие рассматривать изозим Р2 а-глюкозидазы как наиболее вероятный акцептор для сахарного рецепторного комплекса в хеморецепторных сенсиллах мух.
Факторы, влияющие на чувствительность
Эндогенные факторы. Работа хеморецепторных органов существенно зависит от физиологического состояния организма насекомого. Под термином «физиологическое состояние» обычно обозначают весь сложный комплекс изменений внутренней среды организма, соответствующий стадиям развития, возрасту, функционированию пищеварительного тракта, репродуктивным процессам и т. д.
Давно известно, что чувствительность насекомых к таким химическим раздражителям, как сахара, изменяется в зависимости от возраста или степени насыщенности насекомого пищей или водой. Ответы контактных хеморецепторных клеток мухи Phormia regina зависят от возраста насекомого, степени его насыщения и некоторых генетических факторов. Е. Ходжсон утверждает, что некоторые нейросекреторные гормоны повышают чувствительность хеморецепторов насекомых.
При отведении биопотенциалов от лабеллярных хеморецепторных сенсилл у самок комаров Aedes aegypti в ответ на раздражение 0,2 М раствором хлористого калия и сахарозы оказалось, что максимальная частота импульсов в ответе как при раздражении хлористым калием, так и при раздражении сахарозой наблюдается в течение первых суток после выхода комаров из куколки. Затем интенсивность ответов снижается н становится минимальной
116
на шестые-седьмые сутки, т. е. сразу же после первого кровососа-ния. Постепенно незначительное увеличение ответов сменяется еще более глубоким снижением их интенсивности на десятые— тринадцатые сутки, во время их первой яйцекладки. Через некоторое время интенсивность ответов вновь возрастает.
Кровососание, развитие яиц и последующая яйцекладка сопровождаются значительными перестройками в гормональном составе крови самок комаров. Все эти данные говорят о том, что в колебаниях чувствительности организма насекомого к химическим стимулам при разных физиологических состояниях немалую роль играют изменения чувствительности самих рецепторных клеток. Очевидно, наряду с прямой нервной регуляцией чувствительности рецепторов важное значение здесь играет состояние внутренней среды организма, проявляющееся через гормональную регуляцию рецепторов.
Экзогенные факторы. В настоящее время внимание многих исследователей обращено на решение вопроса о взаимодействии между специфическими чувствующими клетками в хеморецепторных сенсиллах насекомых при раздражении их несколькими раздражителями одновременно. Наряду с установлением факта специфической чувствительности L-, S-, W- и ЛЬрецепторов к раздражению соответственно моновалентными солями, сахарами, водой и механической стимуляцией имеются данные, которые указывают, что эта специфичность относительна.
Во-первых, на ответы хеморецепторных клеток действует такой фактор, как температура. Сами хеморецепторы не отвечают на непосредственное действие тепла или холода, но частота импульсов в афферентном залпе L- и S-рецепторов в ответ на раздражение солью или сахаром резко изменяется при изменении температуры всего препарата. Вместе с тем изменение температуры раздражающего раствора, охлаждение или согревание кончика волоска не влияют на ответ. Так, колебания температуры раствора от 2 до 41° не изменяют величины пороговой концентрации раздражителя при регистрации рефлекторных реакций и частоту импульсов в афферентном залпе сенсиллы. Проведенные опыты показали, что температурный эффект проявляется на уровне тел хеморецепторных клеток, где генерируются импульсы.
Во-вторых, механические раздражители, например качание волоска, не только возбуждают Af-рецептор, но и в определенной степени влияют на S- и L-рецепторы. Среди лабеллярных волосков мухи Lucilia sericata встречаются такие, где приложение механического стимула в течение первой минуты действия химического раздражителя вызывает увеличение частоты S- и L-рецепто-ров. Адаптированные к химическим стимулам S- и L-рецепторы не отвечают на механическое раздражение. Однако пока еще не представлено прямых доказательств того, что механические раздражения, вызывающие ответ S- и L-рецепторов, не являются для них неспецифическим раздражением.
117
Действие смеси раздражителей
Давно известно, что смесь веществ, например хлористого натрия и глюкозы, вызывает в специфических рецепторах ответ меньшей интенсивности, чем раздражение этими же веществами в тех же концентрациях в отдельности. Показано, что добавление к раствору хлористого натрия таких веществ, как СаС12, MgCl2, Уо2(ЫОз)2, HgCl2, FeCl2, ВаС12, этилового спирта, маннозы и сахарозы, тормозит реакцию Л-рецептора, а добавление к раствору -сахарозы NaCl, СаС12, FeCl2, HgCl2, холинхлорида, уксусной кислоты и октиламина тормозит реакцию S-рецептора.
Среди этих веществ можно выделить октиламин, который не оказывает раздражающего влияния ни на какую из рецепторных клеток сенсиллы, а тормозит как S-, так и L-рецепторы. Степень торможения зависит от концентрации октиламина. Раздражение раствором СаС12 лабеллярных трихоидных хеморецепторов у мухи Calliphora vomitporia также вызывает торможение S- и Л-рецепто-ров. Знак генераторного потенциала при этом становится положительным, т. е. рецепторная мембрана гиперполяризуется. Однако, если раздражать раствором СаС12 интерпсевдотрахеальную папиллу— вкусовые органы на оральном диске мухи Phormia regina, то можно зарегистрировать афферентные залпы импульсов. Очевидно, реактивные свойства возбуждаемых рецепторных мембран в чувствующих клетках, специфических к раздражению солями, различны у этих двух органов.
Интересны процессы торможения в клетке, специфичной к раздражению водой (^-рецептор). Ответ рецептора максимален при действии дистиллированной воды. Добавление любого вещества, как имеющего тормозные свойства (октиламин), так и раздражителя для других рецепторов (сахароза, хлористый натрий), вызывает торможение водного рецептора, выражающееся в резком уменьшении частоты импульсов. Постепенное снижение концентрации водного раствора соли или сахара сопровождается повышением частоты импульсов водного рецептора.
Смеси растворов солей и сахаров могут оказывать на контактные хеморецепторы и соответственно на пищевую реакцию либо возбуждающее, либо тормозящее действие в зависимости от соотношения компонентов в смеси. Тормозящее действие смеси растворов сахара и соли проявляется при любой концентрации взятых веществ в диапазоне от 0,01 до 1,0 М. Это хорошо заметно на примере контактных хеморецепторов комара Aedes aegypti. Если сравнить серии адаптационных кривых ответов S- и Т-рецепторов при раздражении растворами сахарозы и хлористого калия, взятых раздельно, то обнаруживается зависимость ответа от их концентрации. Зависимость более четко выражена для L-рецептора и слабее для S-рецептора. Обращает на себя внимание то, что во
118
всех случаях при раздражениях растворами эквимолярных концентраций сахарозы и хлористого калия частота импульсов 5-рецептора превышает частоту импульсов L-рецептора.
Раздражение лабеллярных сенсилл комара смесью растворов хлористого калия и сахарозы обычно вызывает уменьшение частоты импульсов для большинства концентраций. Это хорошо заметно для растворов хлористого калия всех концентраций в диапазоне от 0,05 до 1,0 М при сравнении частоты импульсов соответствующего рецептора в каждой концентрации с опытами, где раздражители использовались раздельно. Интересно, что смесь растворов сахарозы и хлористого калия эквимолярных (0,5 М) концентраций вызывает в хеморецепторных сенсиллах комаров афферентный залп, состоящий преимущественно из импульсов 5-рецептора. Приблизительно одинаковое число импульсов 5- н Л-рецепто-ров в первые 200 мс после начала раздражения наблюдается при действии на сенсиллу смеси растворов 0,1 М сахарозы и 0,9 М хлористого калия.
Можно предположить три возможных объяснения эффекта торможения в смеси двух растворов. Во-первых, физико-химическое взаимодействие между молекулами соли и сахара перед их воздействием на рецепторную мембрану. Во-вторых, взаимодействие молекул сахара и соли на рецепторной мембране — «конкурентное» торможение. В-третьих, взаимодействие между 5- и Л-рецеп-торными клетками.
Статистической обработкой афферентных залпов импульсов было установлено, что возбуждение Л-рецептора возникает независимо от раздражения 5-рецептора и наоборот. Некоторые вещества не вызывают возбуждения этих рецепторов, но оказывают тормозящий эффект в смеси с раздражителем. Поэтому было высказано мнение, что тормозящий эффект в смеси раздражителей вызван изменениями коэффициентов диффузии и термодинамической активности компонентов в растворе.
Однако последние работы в этом направлении говорят о том, что тормозные эффекты при действии специфических ингибиторов (СаС12, уксусная кислота, октиламин и др.) и ослабление реакции в ответ на раздражение смесью двух или более раздражающих компонентов связаны с процессами, происходящими в рецепторной мембране. Так как в рецепторной мембране предполагается наличие специфических участков, избирательно взаимодействующих с молекулами определенной формы, то в простейшем случае имеются два типа таких участков. Первый реагирует с молекулами, вызывающими возбуждение, что приводит к деполяризации мембраны, а второй — с молекулами, оказывающими тормозной эффект и приводящими к гиперполяризации мембраны. Взаимодействие возбудимых и тормозных специфических участков мембраны при действии многокомпонентных раздражителей, очевидно, обеспечивает тот или иной характер ответа, выраженный в виде афферентного залпа импульсов.
119
Специфичность чувствующих клеток у фитофагов
Своеобразие питания фитофагов определяет свойства рецепторных клеток. Среди жуков-листоедов рода Chrysolina один из видов считается монофагом зверобоя, богатого гликозидом гиперазидом. Исследование набора рецепторных клеток тарзальных сенсилл этого жука позволило выделить одну клетку, которая возбуждается только этим алкалоидом. Другие виды жуков Chrysolina, не являющиеся монофагами зверобоя, не имеют этого рецептора, хотя характеристики остальных чувствующих клеток в тарзальных сен силлах совпадают.
Возбуждение тех или иных специфических клеток в контактных и обонятельных сенсиллах часто связывают со способностью насекомых дифференцировать пищевые субстраты, определять качество пищи. Так, активность сахарного и водного рецепторов мух обычно связывают с положительными пищевыми реакциями, а солевого— с отрицательными. Эта зависимость хорошо проявляется при действии на сенсиллы больших концентраций (0,5—1,0 М) сахара или соли. Специфичность рецепторов, однако, не абсолютна, и уменьшение концентрации раздражителя обычно приводит к тому, что в ответе участвуют две или три чувствующие клетки сенсиллы. К тому же отмечается значительная вариабельность контактных хеморецепторных сенсилл по набору рецепторов и возбудимости чувствующих клеток как при сравнении сенсилл, расположенных на различных участках лабеллума или тарзуса, так и при повторных раздражениях одной и той же сенсиллы каким-либо раздражителем. Анализируя ответы отдельных сенсилл, в которых возбуждены две или более клетки, некоторые авторы даже приходят к выводу, что специфичности индивидуальных клеток к конкретным химическим соединениям не существует, а информация кодируется по изменению частоты импульсов, поступающих по афферентным волокнам в центральную нервную систему.
Аналогичная картина наблюдается и при исследовании вкусовых сенснлл гусениц, где исследовались ответы максиллярных сенсилл монофагов и олигофагов на соки предпочитаемых и не-предпочитаемых растений. В этих опытах не было обнаружено достоверной разницы в электрофизиологических ответах на соки растений, поедаемых или отвергаемых гусеницами.
Исследования характера специфичности вкусовых сенсилл гусениц чешуекрылых показали большое разнообразие как числа специфических чувствующих клеток в максиллярных стилокониче-ских сенсиллах, так и химических соединений, многие из которых имеют сигнальное значение при узнавании гусеницами кормового растения (табл. 6).
Широкий набор специфических рецепторных клеток у гусениц отражает разнообразие пищевых субстратов и способность насекомых к переходу с одного растения на другое при питании. Это
120
Таблица б
Специфичность рецепторных клеток в максиллярных стилоконических сенсиллах у личинок различных насекомых
(по Шоонховену, 1974)
Насекомое Сенсилла
медиальная латеральная
1 2 3 4 1 2 3 4
Manduca sexta соль СП ИНОЗ СОЛЬ С/Г сал ИНОЗ
Sphinx ligustri кон ИНОЗ с кон иноз
Laothoe populi кон по/сал амк с по/сал
Pieris brassicas соль соль Дет г. г. амк с/г Г. г. ант
Limantria dispar соль Дет сорб СОЛЬ с/г
Euproctis chrysorrhoca соль ИНОЗ СОЛЬ с ИНОЗ
Dendrolimus pint кон иноз амк с
Adoxophyes orana соль соль амк с фло иноз
Phylosamia cynthia соль кон Г иноз амк с/г Г иноз
Bombyx mori соль соль Дет вода соль с/г г ИНОЗ
Обозначения: соль — избирательный ответ на соли; кон — на коннесин; сп — на спирт; по—на популии; сал — на салицил; дет — на детерренты; г — на глюкозу; иноз — на инозитол; г. г. — иа горчичные глюкозиды; сорб — на сорбитол; амк — иа аминокислоты; с — на сахарозу; ант — на антоциан.
подтверждается экспериментально. Так, перевод гусениц бражника Manduca sexto, на искусственную диету привел к изменению пищевых реакций, которое выражалось в том, что гусеницы стали поедать растения, ранее отвергаемые ими. Добавление салицила в диету этой гусеницы, питающейся на томате, позволило направленным воспитанием повысить чувствительность рецепторов к новому кормовому растению — капусте. Направленный сдвиг чувствительности рецепторов у гусениц можно получить в течение одной генерации. Однако чтобы путем направленного воспитания получить изменения в реактивности контактных рецепторов к различным концентрациям хлористого натрия у мух и комаров, требуется 8—10 поколений.
Личинки хлебного жука Anisoplia austriaca, обитающие в почве и питающиеся корнями злаков, ограничены в возможностях передвижения и выборе пищи по сравнению с гусеницами. Видимо, поэтому у них более четко проявляется разница в ответах контактных хеморецепторных сенсилл на предпочитаемые и отвергаемые растения (табл. 7).
В стилоконической сенсилле хлебного жука на раздражение глюкозой отвечают три чувствующие клетки, а на раздражение хлористым натрием — одна. Спектры ответов на водные экстракты корней различных растений напоминают ответы на соль и сахар, хотя частота импульсов имеет меньшие значения.
Исследование индивидуальных характеристик нескольких де-
121
Таблица 7
Реакция стилоконической сенсиллы личинки хлебного жука на контактное раздражение соками растений
(по Елизарову, Куэн, 1969)
Тип растения Предпочитав’ мость Число клеток Частота афферентной нмпульсации, имп/с
I тип II тип ш тип сумма
Кукуруза + 3 21,7 19,2 14,2 55,1
Пшеница + 3 25,2 20,5 8,1 51,8
Огурец ± 2 13,5 26,1 — 39,5
Люпин ± 2 8,7 22,4 —— 31,1
Чеснок — 1 58,0 — — 58,0
Табак — 1 66,2 — —— 66,2
Хлористый натрий, 0,1 М 1 82,2 — —— 82,2
Глюкоза 3 54,4 17,1 7,2 78,7
сятков хеморецепторных волосков на лабеллуме мухи Musca do-mestica по типам ответов на стандартный набор раздражителей (сахара и соли) позволило выделить солевой, сахарный и два промежуточных рецептора (рис. 31). Следует отметить, что набор ре-
Рис. 31. Физиологические характеристики солевого (£1,2,3), сахарного (S) и промежуточных (W,J) рецепторов в лабеллярной сенсилле мухи Musca domestica.
По оси ординат — импульсная активность чувствующих клеток сенсилл на стандартный набор раздражителей, имп/с: 0,2-молярнын раствор глюкозы (а); 0,01-, 0,1- и 1-молярные растворы NaCl (соответственно б, в, г).
1 — сенсиллы адорального ряда, 2 — аборального. Каждая кривая — среднее из 20 повторностей
цепторов в сенсиллах хоботка одной и той же мухи варьирует в значительной степени. Например, из четырех хеморецепторных клеток сенсиллы три могут иметь характеристики солевого рецептора, а одна — сахарного. Могут встречаться также волоски, где солевых рецепторов вообще нет.
Специфичность рецепторов в лабеллярных сенсиллах мухи Musca domestica подтверждают результаты экспериментов, где определялся эффект действия оубаина — ингибитора мембранной Ма+-К+-активируемой АТФ-азы. Система АТФ — На+-К+-активи-руемая АТФ-аза — одна из основных систем в комплексе процессов, обеспечивающих активный транспорт ионов через клеточные
122
мембраны. Оказалось, что высокие концентрации оубаина (10-2М) вызывают необратимое подавление импульсной активности всех чувствующих клеток лабеллярной сенсиллы за 18—20 мин. Если же регистрировать ответы рецепторных клеток в первые 200 мс после нанесения раздражителя в смеси с различными концентрациями оубаина, то тормозной эффект оубаина проявляется лишь в концентрации 10-5М и неодинаково для различных рецепторных клеток в сенсилле. Сахарный рецептор снижает частоту
Рис. 32. Тормозной эффект действия оубаина в смеси с 0,1-молярным раствором хлористого натрия (по Грицай, 1975).
По оси абсцисс — концентрация оубаина, М; по оси ординат — импульсная активность лабеллярной сенсиллы мухи Musca domestica в процентах от частоты импульсов прн контрольном раздражении. Пунктир — ответ сенсиллы на контрольные раздражения 0,1 М раствором NaCl в каждой концентрации оубанна.
1 — интенсивность ответов солевого рецептора при раздражении смесью растворов оубаина и хлористого натрия; 2, 3 — то же для промежуточного и сахарного рецепторов соответственно
импульсов в ответе на 75%, а промежуточный — на 55%. Действие оубаина на солевой рецептор не сопровождается тормозным эффектом (рис. 32).
Перфорация хеморецепторной сенсиллы непосредственно на кончике волоска позволяет подвергать воздействию оубаина в первые доли секунды только хеморецепторную мембрану периферического отростка чувствующей клетки — участка, где возникает генераторный потенциал. Место возникновения потенциалов действия— дендритный холмик — удалено от вершины волоска на 500 мкм. Раздражающий раствор может проникнуть к этому участку по внутренней полости волоска, но его концентрация в содержащейся в просветах жидкости должна сильно снизиться.
Именно этой особенностью строения хеморецепторной сенсиллы можно объяснить довольно высокие концентрации оубаина и длительное время, необходимое для подавления АТФ-азной активности в месте возникновения потенциала действия. Обращает на себя внимание тот факт, что при длительном действии оубаина все три рецептора прекращают работу приблизительно одновременно в зависимости от концентрации ингибитора. Этот факт позволяет предположить одинаковую АТФ-азную активность в зонах возникновения потенциалов действия для трех рецепторных клеток сенсиллы.
Использование микропипетки для регистрации импульсной активности лабеллярных хеморецепторов мухи позволило обнаружить тормозной эффект действия оубаина в первые 200 мс адекватного раздражения. Кратковременность этого процесса позво
123
ляет сказать, что полученный эффект связан с действием оубаина на хеморецепторную мембрану, находящуюся непосредственно под перфорацией волоска. Очевидно, хеморецепторные мембраны солевого, сахарного и промежуточного рецепторов обладают различной чувствительностью к оубаину и, следовательно, различной ферментативной активностью.
5. ФИЗИОЛОГИЯ
ОБОНЯТЕЛЬНЫХ СЕНСИЛЛ
Пороги и чувствительность
Обонятельные органы насекомых изучены в меньшей степени, чем сенсиллы контактной хеморецепции. До недавнего времени основным методом изучения ольфакторных органов служило наблюдение за характером поведенческих реакций при действии на насекомого раздражающих запахов. Этот метод не потерял своего значения и в настоящее время, особенно при изучении характера влияний химических соединений с репеллентными и аттрактивными свойствами.
Принято считать, что насекомые обладают очень высокой чувствительностью к запахам. Однако, как и для вкусовой чувствительности, экспериментально было показано, что пороги возбуждения для большинства запахов у насекомых близки к тем, которые наблюдаются у позвоночных животных и человека. Так, если пороговая концентрация запаха аммиака для человека равна 1,7-10“6 М/л, то для мухи Phormia regina ее величина — 2,0-Ю'6 М/л. Человек более чувствителен к запаху бензола (поро-
Таблица 8
Значения пороговых концентраций обонятельных стимулов, М/л
(по Prosser, Brown. 1962; Boeckhetal, 1966; Елизарову, 1971)
Объект Ионон Фенилпропиловый спирт Скатол Масляиан кислота Капроновая кислота
Человек . . . 5,0хЮ-13 1,1Х10-11 3,0X10-4 1,1X10-4 3,3X10-!°
Пчела .... 2,5х10-и 3,8хЮ'12 2,ЗХ1О'8 1,7X10’» 3,6x10-!°
Собака .... 5,0х10-1в — —— 1,5x10-4 6,0 ХЮ’18
Угорь .... 9,7 ХЮ'1» 3,7Х1О-18 — — —
говые концентрации соответственно равны 6,8-10~8 М/л и 6,4-Ю-6 М/л) и к запаху этанола (5,4-10_6 М/л и 7,2-Ю-3 М/л). Ко многим сильно пахнущим соединениям чувствительность насекомых меньше, чем у других животных (табл. 8).
124
Определение чувствительности насекомых к запахам при последовательном удалении члеников антенн показало, что пороги на запах возрастают соответственно числу удаленных члеников. Такие результаты были получены при исследовании реакций пчел на запах кумарина и комаров — на запахи некоторых репеллентов. Отпугивающее действие репеллентов сохранялось, если у комара оставался хотя бы один членик флагеллума, несущий ольфакторные сенсиллы. Знак реакции при этом не изменялся. Величина ольфакторного порога зависела от числа возбужденных сенсилл.
Здесь интересно провести аналогию с тем, что у насекомых с высокоразвитым обонянием количество обонятельных сенсилл увеличено. Известно, что у самцов многих насекомых обоняние развито в большей степени, чем у самок (комары, бабочки, некоторые жуки). Антенны самцов, на которых локализованы обонятельные сенсиллы, гораздо сложнее, чем у самок, и содержат большее количество сенсилл. У самца пчелы на одной антенне содержится до 30 тыс. плакоидных сенсилл, а у рабочей пчелы — 6 тыс. Показано, что мухи, питающиеся на навозе и пользующиеся обонянием при отыскании пищи, имеют большее количество обонятельных сенсилл, чем мухи, питающиеся на цветах, у которых главную роль в поисках пищи играет зрение. Паразитирующие формы с редуцированной обонятельной функцией имеют малое количество обонятельных сенсилл. Так, на антенне вши их всего 9—10.
Зависимость величины ольфакторного порога от количества возбуждаемых сенсилл можно назвать пространственной суммацией. Однако уменьшение порогов не говорит об увеличении чувствительности хеморецепторов. Пороги возбуждения в данных опытах— это не истинные пороги хеморецептора, а пороги рефлекторного ответа организма или отдельного органа (в зависимости от методики опыта) на обонятельный стимул. С увеличением числа рецепторов наблюдается увеличение числа афферентных импульсов в центральную нервную систему, где и происходят процессы суммации, искажающие истинную величину поведенческого ответа.
Электрофизиологические исследования
Электроантеннограмма. Значительно более удачными оказались исследования потенциалов антенн насекомых. Если к дистальному и проксимальному концам антенн присоединить электроды, то при раздражении струей запаха регистрируется медленный потенциал— электроантеннограмма (ЭАГ), причем вершина антенны становится электроотрицательной по отношению к ее основанию. Амплитуда ЭАГ определяется концентрацией пахучего вещества в струе воздуха, форма — видом вещества. Наркотизация приводит
125
к исчезновению ответа. Запах полового аттрактанта вызывает ЭАГ только у самцов Bombyx mori. Медленные потенциалы позже были зарегистрированы от антенн других чешуекрылых, а также от антенн термитов, тараканов и жуков при раздражении их запахами пищевых субстратов.
Так же, как и в чувствительных клетках контактных хеморецепторных сенсилл, в момент действия химического стимула в периферических отростках обонятельных клеток развивается деполяризация— генераторный потенциал. На антеннах насекомых, особенно самцов чешуекрылых, воспринимающих половые аттрактанты, локализованы тысячи сенсилл одного типа с рецепторными клетками, воспринимающими данный запах. Суммирование генераторных потенциалов, одновременно возникающих в множестве чувствующих клеток, формирует электроантеннограмму. Когда ольфакторные сенсиллы равномерно распределены по поверхности антенны, форма ЭАГ соответствует форме генераторного потенциала отдельной клетки.
С помощью ЭАГ удалось получить данные по специфичности полового аттрактанта у различных видов бабочек. Оказалось, что у представителей семейства сатурний половые аттрактанты самок не имеют видовой специфичности. Антенны самцов Hyalophora prometea, Н. galleta, Philosamia Cynthia реагировали на половой аттрактант, взятый от любой самки этих трех видов. Однако половые аттрактанты самок данных видов бабочек не вызывали ответа в антеннах самцов Bombyx mori, и, в свою очередь, половой аттрактант самки Bombyx mori не действовал на антенны самцов сатурний.
Оригинальное исследование по определению специфичности реакций самцов бабочек-сатурний на половые аттрактанты самок было проведено Е. Преснером в 1968 г., который использовал в качестве критерия ЭАГ самцов бабочек. У каждого из самцов 104 видов сатурний были зарегистрированы ЭАГ в ответ на действие запахов половых аттрактантов, полученных из самок тех же 104 видов. У большинства видов максимальный ответ ЭАГ наблюдался на запах самки своего вида и нескольких близких видов, т. е. реакция была неспецифической. Лишь 5 видов из 104 имели максимальный ответ антенн самцов только на запах самки своего вида. Специфический запах имели самки Aglia tau из Средней Европы, Hamileuca eglanteruna — из США, Antheraea yamamai — из Японии и с Дальнего Востока, Gunanisa maja — из Кении и Loepa katinka — из Индии. Их половой аттрактант вызывал максимальную ЭАГ только у самцов своего вида. Однако самцы двух первых видов реагировали максимальной ЭАГ не только на запахи самок своего вида, но и на запахи самок Citheronia brissotii и Eacles magnifica.
Хотя запах полового аттрактанта самки может стимулировать самцов своего и других видов, он никогда не действует на самок своего и любого другого вида.
1Э6
Известны случаи использования ЭАГ в испытаниях репеллентов против тараканов и при анализе некоторых природных запахов, являющихся для насекомых пищевыми аттрактантами. Так, в процессе регистрации ЭАГ у 50 видов пчел при раздражении запахами экстрактов 11 видов орхидей оказалось, что максимальная амплитуда ЭАГ соответствовала запаху того вида орхидей, для которого данная пчела была опылителем.
Методика регистрации ЭАГ широко используется при тестировании половых аттрактантов насекомых. Определение эффективности фракций экстрактов пахучих желез, промежуточных и окон-
а 5 I
Ч ю т
cis tram ЛАЛЛ/VW”"
inn cis V
'"'«АЛЛАЛАЛЛ"'
ЛАААААЛЛ’" нтр
Рис. 34. Реакции ольфакторных рецепторов антенн самца тутового шелкопряда на запах синтетического полового аттрактанта самки «бом-бикола» (по Schneider, 1967).
Рис. 33. Чувствительность самцов тутового шелкопряда Bombyx mori к запаху синтетического полового аттрактанта самки «бомбнкола» (гексадекадиен-10 транс-12-цис-ол) и его гомологов (по Kasang, Kaissling, 1972).
а — изомерия; б — формула; в — порог ЭАГ на источник стимула, мкг
По оси абсцисс — число молекул бомбнкола на 1 см3 воздуха; по оси ординат — величина потенциала, мВ.
ГП — генераторный потенциал обонятельной сенсиллы (максимальное и минимальное значение); ЭАГ — электроантеннограмма. Стрелками обозначены пороговые концентрации поведенческого ответа
чательных продуктов синтеза половых аттрактантов у многих совок и листоверток, некоторых жуков и клопов проводилось с использованием ЭАГ. В последние годы препараты антенн или целых насекомых соединяют с выводной системой газожидкостных хроматографов, на которых производится разделение экстрактов половых аттрактантов. Параллельно с записью результатов фракционирования производится запись ЭАГ, что позволяет уже в процессе анализа дифференцировать активные фракции экстракта.
По амплитуде ЭАГ удалось количественно обозначить разницу в реакциях самцов тутового шелкопряда Bombyx mori на запахи полового аттрактанта и его гомологов. Максимальная чувстви-
127
тельность наблюдалась только при раздражении антенн самца синтетическим половым аттрактантом — бомбиколом (гексадекадиен- 10-транс- 12-цис-ол-1) . Изменение геометрии молекулы — использование 10-цис-12-трансизомер а — снижает чувствительность в 100 раз. Спирт с одной двойной связью еще в меньшей степени раздражает рецепторы антенны, а чувствительность к полностью насыщенному гексадеканолу-1 в миллион раз ниже, чем к половому аттрактанту (рис. 33).
Несмотря на определенные удобства использования ЭАГ в качестве биотеста в исследованиях половых аттрактантов, следует
а д 6 г
1В 6'7 /Vw\M/W\ /,5 40° И2± A/V\yW\AA у‘/о5 № t-s A/WWVW\
W 7-? /vV\_JwW\ i,s*id5
11 7-S 2,1 ’ID1
Puc. 35. Чувствительность самцов непарного шелкопряда Lymantria dispar к запаху синтетического полового аттрактанта самки «диспарлура» (цис-7,8-эпокси-2-мети-локтадекан) и его гомологов.
а — число атомов С в цепочке; б — место эпоксигруппы; в — формула; г — порог реакции ЕДао, мкг
отметить, что чувствительность этого метода во много раз ниже, чем использование в качестве критерия специфической реакции самца на запах самки характерного поведения: трепетание крыльев, изгибание брюшка и т. д. Д. Шнейдер в своих работах указывал, что пороговая концентрация бомбикола в поведенческих опытах равна 102 молекул на 1 см3 воздуха, тогда как изменения амплитуды ЭАГ наступают только при концентрации 107 моле-кул/см3 (рис. 34).
Аналогичные соотношения, указывающие на резко выраженную специфическую чувствительность к запаху, были получены на самцах непарного шелкопряда Lymantria dispar в отношении синтетического полового аттрактанта — диспарлура. Диспарлур (Ч«с-7,8-эпокси-2-метилоктадекан)—углеводород с эпоксигруппой во втором положении 18-углеродной цепочки. Изменение ее длины на один атом С, перемещение эпоксигруппы на одно положение вправо или влево, использование трансизомера — все эти незначительные изменения структуры молекулы приводили к снижению его эффективности на 5—7 порядков (рис. 35). Обращает на себя внимание, что пороговые значения для гомологов из разных рядов довольно близки друг другу и резко отличаются от величин, полученных для полового аттрактанта. В данной серии экспериментов в качестве критерия специфической реакции было использовано характерное трепетание крыльев самца бабочки. Когда же ответ регистрировали в виде ЭАГ самца, то разница между теми же гомологами составляла лишь 2—3 порядка.
128
Регистрация биопотенциалов от одиночных обонятельных сенсилл насекомых в отличие от записи ЭАГ позволяет на клеточном уровне судить о механизмах различения специфических запахов и выделения их из сопутствующих летучих соединений. На антеннах жуков Necrophorus vespillo обонятельные сенсиллы содержат только по одной чувствующей клетке. Регистрация биоэлектрической активности этих сенсилл микроэлектродом, введенным в осно-
Рис. 36. Электрическая активность обонятельной
хеморецепторной сенсиллы на антенне жука Nec- °' ---—•—1—и
rophorus vespillo (по Boeckh, 1962). ,_____,_______tw
Раздражитель — запах гнилого мяса; ai — начало, аг — конец раздражения
вание сенсиллы, при раздражении антенны каким-либо запахом позволила выявить медленный генераторный потенциал, на фоне которого наблюдался залп импульсов, причем наибольшая частота импульсов соответствовала восходящей фазе медленного потенциала (рис. 36). Ответы на раздражение различными запахами варьировали по амплитуде и крутизне нарастания медленного потенциала, что сопровождалось изменением частотного спектра импульсов. Некоторые вещества, например циклопентан, вызывали медленное положительное колебание, которое соответствовало торможению чувствительной клетки. Импульсы при этом исчезали. Оказалось, что возбуждающий или тормозящий эффект в ряду жирных кислот зависит от числа атомов углерода в цепочке (табл. 9).
Таблица 9
Зависимость ответов обонятельной сенсиллы Necrophorus vespillo от длины углеродной цепочки в ряду жирных кислот
(по Беку, 1962)
Кислота Формула Число атомов Реакция
Пропионовая СН3СН,СООН 3
Масляная CHsCHgCHgCOOH 4
Капроновая СН3(СН2)3СН2СООН 6 4“
Гептановая СН3(СН2)4СН2СООН 7 +
Каприловая СНз (СН2)5СН2СООН 8 +
Пеларгоновая СН3 (СН2)0СН2СООН 9 +
Каприновая СНз (СН2)7СН2СООН 10 0
Стеариновая • СНз(СН2)15СН2СООН 18 0
Олеиновая СНз (СН2)7СН=СН (СН2)7СООН 18 +
Примечания: (—) — гиперполяризация мембраны, торможение; (-]-) — деполяризация мембраны, возбуждение; (0) — ответа нет.
Генераторный потенциал и частота афферентных импульсов в залпе были минимальными при раздражении хеморецептора парами кислоты, имеющей в цепочке четыре атома углерода, и мак-
129
симальными, когда число достигало семи-восьми. Дальнейшее уве-
личение числа атомов углерода вызывало снижение ответа, который исчезал при действии каприновой кислоты, имеющей 10 атомов углерода. Кислоты, имеющие менее четырех атомов углерода в молекуле, вызывали торможение чувствительной клетки в обонятельной сенсилле Necrophorus vespillo, сопровождающееся гиперполяризацией мембраны. У жуков Necrophorus humator подобный эффект вызывали молекулы жирных а_______I__________, кислот, имеющие менее шести атомов
углерода в молекуле. Подобные наб--------- людения ответов обонятельных сен-
Рис. 37. Импульсная активность специфичной к бом-биколу рецепторной клетки в антеннальной сенсилле самца тутового шелкопряда при раздражении различными концентрациями запаха бомбнкола (по Schneider, 1974):
а, б, в — 10~4; г — 10~3; д — 10~2; е — 10~‘ мкг;
ж — 1 мкг
силл на антенне саранчи на запахи жирных кислот показали такую же зависимость ответа от длины углеродной цепочки. Интенсивность импульсного ответа сенсиллы увеличивалась при возрастании числа атомов углерода от трех до шести и уменьшалась при дальнейшем увеличении длины цепочки до восьми атомов. Запахи жирных кислот с числом атомов углерода меньше трех и больше восьми были неэффективными.
Многие запаховые вещества, вызывающие привлечение или отпугивание насекомых, имеют в составе молекулы функциональную группу (гидроксильную, карбоксильную, эпоксигруппу и т. д.) и алкильный радикал. Использование разнообразных тестов: полевые и ольфактометрические опыты, изучение рефлекторных реакций, регист
рация потенциалов антенн или отдельных сенсилл — приводит к общему заключению о том, что' максимальный раздражающий эффект наблюдается при строгом сочетании положения функциональной группы с параметрами алкильного гидрофобного радикала. Изменение положения функциональной группы либо увеличение или укорочение алкильного радикала хотя бы на одну группу СН2 ведет к резкому изменению чувствительности обонятельного рецептора на данный химический
стимул.
Отведение биопотенциалов от обонятельных сенсилл на антенне самца бабочки Antheraea pernyi, имеющих три чувствующие клетки, показало, что две клетки в сенсилле отвечали на раздражение запахами фенилацетата, терпинеола, изосафрола, гераниола и других веществ, генерируя залпы импульсов разной интенсивности. Третья клетка, генерирующая импульсы большой амплитуды, возбуждалась только при раздражении запахом полового аттрак
130
танта, не отвечая ни на какие другие раздражители. Ответ этой клетки был фазным и характеризовался быстрой адаптацией. Из двух чувствующих клеток, входящих в состав трихоидной сенсиллы антенны самца Bombyx mori, одна была высокочувствительной к запаху полового аттрактанта — бомбикола. Регистрируя ответы этой клетки, Д. Шнейдеру с сотрудниками удалось установить, что изменение частоты следования импульсов наступает, когда на сенсиллу попадает хотя бы одна молекула бомбикола (рис. 37).
Электрофизиологические исследования показали, что в сенсиллах на антенне пчелы Apis mellifera есть отдельные клетки, настроенные на восприятие СОг- В плакоидных сенсиллах трутня и рабочей пчелы были обнаружены специфические клетки двух типов: одна — реагирующая на запах полового аттрактанта самки, другая — на запах секрета железы Насонова, выделяющей следовой феромон. Клетки, реагирующие на углекислый газ, присутствуют также в булавовидных сенсиллах на пальпах комаров Aedes aegypti. Ни по своей морфологии, ни функционально они не отличаются от ольфакторных клеток, иннервирующих ту же сенсиллу.
Рассматривая спектры чувствующих клеток в обонятельных сенсиллах, Шнейдер выделяет специфические чувствующие клетки (odour specialists), реагирующие на узкий круг химических соединений, например на половые аттрактанты, и клетки генерализованного типа (odour generalists), отвечающие на раздражение широким кругом веществ.
В последнее время появились работы, указывающие на то, что чувствующие клетки в обонятельных сенсиллах насекомых могут иметь очень узкий спектр и реагировать на одно-два соединения, не относящихся к феромонам, а связанных с пищевым поведением данного вида. Так, в трихоидных антеннальных сенсиллах долгоносика Hylobius abietes выявлена чувствующая клетка, приходящая в возбуждение только при раздражении ненасыщенным спиртом анетолом, тогда как другие запахи либо не возбуждают эту клетку, либо вызывают эффект торможения (рис. 38). Кроме «анетолового» рецептора в сенсиллах на антенне этого долгоносика обнаружены «трансвербенольная» и «циклогексановая» клетки.
Исследования спектров обонятельных чувствующих клеток генерализованного типа в сенсиллах антенн различных насекомых при действии стимулов широкого круга разнообразных химических соединений позволили выявить типы рецепторов, с неперекрываю-щимся набором запахов. Так, у саранчи Locusta migratoria обнаружены четыре типа клеток, включающих рецепторы жирных кислот, аминогрупп, и рецепторы, реагирующие на влажный и.сухой воздух. На антеннах таракана Periplaneta americana обнаружены четыре типа рецепторов: пентаноловый, октаноловый, масляной и муравьиной кислот. У водного жука Dytiscus marginalis описаны
131
Рис. 38. Импульсная активность специфических рецепторных клеток в обонятельных сенсиллах антенн жука Hylabius abietus (по Mustaparta, 1973):
а — ответ на запах анетола; б — ментола; в — а-пинена; г — 5-гексен-2-она
клетки с четырьмя реакционными спектрами, у муравья Lasius fuliginosus — с одиннадцатью, у пчелы Apis mellifera — с десятью. Следует отметить, что усложнению поведения насекомого соответствует увеличение числа специализированных чувствующих клеток в обонятельном аппарате антенн.
Роль смесей в восприятии половых аттрактантов
Продолжающиеся исследования половых аттрактантов насекомых накопили ряд фактов, указывающих на то, что феромонные железы насекомых совместно с веществами, имеющими сигнальное значение для самца или самки, выделяют либо вещества-синергисты, либо ингибиторы. Использование в феромонной связи двухкомпонентных запахов дает возможность понять механизм изоляции родственных видов, для которых половым аттрактантом служит одно и то же вещество. В настоящее время определено молекулярное строение аттрактантов более чем для десяти видов листоверток. Анализ полученных соединений показал их большое
132
сходство внутри таксономических групп семейства. Для подсемейства Tortricinae характерны 12-углеродные ненасыщенные спирты или ацетаты с двойной связью в 7—10-м положениях. Аттрактанты бабочек подсемейства Olethreinae имеют 14-углеродную цепочку с двойной связью в 11-м положении. Аттрактантом для нескольких видов может быть одно и то же вещество. Так, установлен одинаковый аттрактант для листоверток Argyrotaenia volutinana и Ostrinia nubialis— цис-\ 1-тетрадеценилацетат. Однако в естественных условиях самцы этих бабочек, вылетая одновременно, не ошибаются в выборе самок. Оказалось, что добавление к аттрактанту его геометрического изомера — транс-11-тетрадеценилацетата — вызывает различные ответы самцов этих видов в зависимости от величины добавки. Смесь, в которой трансизомер присутствует в количестве 4%, привлекает в основном листоверток Ostrinia nubialis, а увеличение концентрации второго компонента до 8% вызывает массовое привлечение самцов листовертки Argyrotaenia volutinana.
Дне-11-тетрадеценилацетат также относится к необходимым компонентам в смеси аттрактантов для листоверток Adoxophyes огапа и Clepsis spectrana, хотя в чистом виде и не привлекает самцов. Добавление второго компонента — цнс-9-тетрадеценилацетата в отношении 2:3 вызывает положительную реакцию у Adoxophyes огапа, а в отношении 3:1 —у Clepsis spectrana.
Если трансизомер тормозит привлекающее действие цисЛ 1-тетрадеценилацетата для Argyrotaenia volutinana, то додецилацетат, будучи его синергистом, увеличивает привлечение в полевых опытах до 170%. Регистрация биопотенциалов ольфакторных сенсилл на антеннах этой листовертки позволила выявить две клетки, одна из которых специфична к цасизомеру, а вторая реагирует на оба изомера. Смесь цис- и транс-11-тетрадеценилацетата вызывает торможение первой клетки, а смесь цасизомера и додецилацетата— вещества, не действующего на рецепторы,— напротив, резкое возрастание частоты ее импульсов. Очевидно, процессы, обеспечивающие синергизм и торможение аттрактанта происходят на уровне специфической рецепторной мембраны первой клетки. Вторая клетка генерализованного типа. Ее воздействие на реакцию опосредовано через нервные центры.
Специфичность в нервных центрах
Высокая избирательность насекомых к определенным запахам связана с присутствием в хеморецепторных антеннальных сенсиллах специфических чувствующих клеток. Их аксоны в составе антеннального нерва входят в дейтоцеребрум (обонятельный центр мозга) насекомого и образуют синаптические контакты в его нейропиле. Регистрация биоэлектрической активности дейтоцеребру
133
ма, проведенная у тараканов Blatta orientalis и Periplaneta ате-ricana, самцов тутового и непарного шелкопрядов при раздражении антенн запахом соответствующего аттрактанта самки, показала, что в залпе импульсов дейтоцеребрума можно выделить ответы отдельных нейронов, реагирующих на половой аттрактант и не возбуждающихся при действии неспецифических запахов. Так, ненасыщенные жирные кислоты, этанол, ацетон и другие химические соединения обычно либо вообще не вызывали ответа, либо ответ был иным как по частоте, так и по набору импульсов.
Специфичность в реакциях отдельных нервных клеток проявляется у чешуекрылых не только на уровне дейтоцеребрума, но и прослеживается на уровне ганглиев грудного отдела нервной цепочки. При раздражении антенн самца непарного шелкопряда запахом диспарлура в птероторакальном ганглии регистрируется импульсная активность нейронов, разряжающихся в характерном ритме трепетания крыльев, вызванном действием полового аттрактанта самки. Соответственно и электрофизиологический ответ крыловых мышц при раздражении самца шелкопряда аттрактантом самки по своим частотным характеристикам резко отличается от реакции на другие запахи.
Общую схему формирования стереотипного поведения насекомого при восприятии специфических запахов можно представить как наследственно закрепленную систему нейрональных связей от центральных отделов обонятельного анализатора до мотонейронов грудного отдела нервной цепочки. Специализированный канал проведения возбуждения начинается отдельными чувствующими клетками в сенсиллах и прослеживается на уровне дейтоцеребраль-ных нейронов, где происходит конвергенция возбуждения от нескольких чувствующих клеток на один ассоциативный нейрон. В обонятельном центре мозга насекомого, а в отдельных случаях и в проторакальном ганглии происходит формирование импульсного сигнала, запускающего мотонейроны в определенном ритме возбуждения. Такая схема приемлема для объяснения восприятия половых аттрактантов и некоторых пищевых стимуляторов.
Как считает Г. А. Мазохин-Поршняков (1975), генетически закрепленная система восприятия химических стимулов, организованная по типу «детектора», не исключает параллельного функционирования у того же насекомого генерализованного канала восприятия широкого круга химических раздражителей. Такой канал связан с активностью многих чувствующих клеток в сенсиллах. В этом случае химический раздражитель вызывает неспецифический ответ, обычно связанный с общим возбуждением насекомого. У насекомых, которым свойственны сложные формы поведения (пчелы, муравьи), генерализованный вход сигнала доминирует, и возникновение отдельных актов поведения при химической стимуляции определяется прежде всего характером обработки информации на различных уровнях хорошо развитых ассоциативных центров мозга.
134
Принимая во внимание определяющее значение возбуждения специфических рецепторных клеток в сенсиллах, следует заметить, что мы еще очень мало знаем о тех процессах, которые обусловливают их возбуждение. Экспериментальные данные свидетельствуют о преобладающей роли физико-химических, скорее всего адсорбционных, процессов на молекулярном уровне, протекающих на мембране хеморецептора. На это же указывает и зависимость эффекта от геометрии молекул, ццс-трансконфигурации и даже от оптической изомерии молекул запахового раздражителя. Углубление исследований по физиологии хеморецепции насекомых приводит, однако, к появлению ряда фактов, которые трудно объяснить с общепринятых позиций понимания процессов возбуждения обонятельных и вкусовых рецепторов. Это прежде всего сверхвысокая чувствительность обонятельных органов насекомых к запахам половых аттрактантов, когда для появления специфической реакции самцов тутового шелкопряда требуется менее одной молекулы в секунду на обе антенны бабочки. Пока еще трудно объяснить факты прилета бабочек с расстояния 6—10 км на запах аттрактанта. Некоторые авторы считают, что вследствие диффузии в воздухе запаховый стимул становится подпороговым уже на расстоянии около 100 м от неоплодотворенной самки. Неясны механизмы изменения знака реакции насекомых на запах и контактное раздражение в зависимости от концентрации вещества, а также особенности восприятия запаховых смесей и сложных растворов. Работы в этом направлении, несомненно, будут иметь значение для решения общих вопросов обонятельной и вкусовой рецепции.
Глава V
ТЕРМОРЕЦЕПЦИЯ
И ГИГРОРЕЦЕПЦИЯ
1. ТЕМПЕРАТУРНЫЕ РЕЦЕПТОРЫ
Локализация и строение сенсилл. Восприятие температуры и способность ориентироваться по температурному градиенту—неотъемлемое свойство всех насекомых. Температура окружающей среды — важный экологический фактор, обеспечивающий нормальную жизнедеятельность организма насекомого. Для насекомых характерна реакция термопреферендума — способность собираться в зоне оптимальной температуры. Тепло становится одним из стимулов, по которому кровососущие насекомые — комары и клопы — осуществляют ориентацию на хозяина-прокормителя. Многие стволовые вредители, например жуки-усачи, отличают ослабленные деревья от здоровых по разнице температур их поверхности.
Чувствительными органами насекомых, реагирующими на изменение температуры, служат трихоидные, базиконические и целоконические сенсиллы, расположенные главным образом на антеннах. Так, у таракана Periplaneta americana крупные волоски, чувствительные к изменению температуры, составляют 0,1% от всех сейсилл на антенне. Температурные рецепторы в целоконических сенсиллах найдены на антеннах саранчи, пчелы, различных видов комаров рода Aed.es, личинок кожеедов Dermestes sibiricus. Многие насекомые и их личинки имеют температурные рецепторы в базиконических сенсиллах на максиллярных и лабиальных пальцах. Это — гусеницы коконопрядов и сатурний, почвообитающие личинки жуков (табл. 10). Удаление антенн и пальп в эксперименте не снимает температурную чувствительность у сверчков и тараканов. Предполагают, что терморецепторы у этих насекомых могут быть также и на лапках ног, на брюшке, церках и т. д.
Ультраструктура сенсилл, чувствительных к изменению температуры окружающего воздуха, до настоящего времени не изучена.
Известные температурные рецепторы насекомых реагируют на конвекционное тепло — энергию молекул нагретого воздуха. Предположение об ориентации насекомых по инфракрасному излучению нагретых тел пока не нашло экспериментального подтверждения. Вместе с тем высокая чувствительность к незначительным изменениям температуры позволяет говорить о способности насекомых чувствовать температурный градиент при удалении от источника тепла. Так, чувствительные к понижению температуры клет-
136
Таблица 10
Распределение обонятельных, вкусовых и температурных рецепторов на придатках головы личинок жуков
(по Елизарову и др., 1968; Елизарову, Куэн, 1971)
Сенсиллы Модальность рецепторов
Anisoplia austriaca Agriotes sp. Dertnestes sihirica Tribolium destructor
Базиконические
концевого членика антенн обоняние вкус вкус вкус
предпоследнего членика антенн тепло — -— —
Антеннального органа — (?) обоняние —
Целоконическая антенны — тепло ——
Стилэконические
верхней губы вкус — — —
максилл ВКУС вкус — —
Базиконические
максиллярных пальп тепло вкус вкус вкус
лабиальных пальп вкус вкус вкус вкус
ки в трихоидных антеннальных сенсиллах Periplaneta americana реагируют пятикратным увеличением частоты импульсов по сравнению с фоном при понижении температуры окружающего воздуха всего на 1°. Аналогичные рецепторы гусениц Dendrolimus pint и Philosamia cynthia увеличивают частоту импульсов на 150— 300% при изменении температуры на Г в течение 0,5 с. Сходная картина наблюдается в температурных рецепторах пчел и комаров. Температурный градиент над поверхностью кожи человека изменяется от 32 до 34° на расстоянии более 40 см. Конвекционные потоки теплого воздуха, идущие от человека, могут иметь локальные температурные различия на 1—2°, что используется комарами при ориентации на небольших расстояниях.
Свойства холодовых и тепловых рецепторов. Температурные рецепторы насекомых можно разделить на два типа: одни из них увеличивают частоту импульсов при снижении температуры, другие— при увеличении температуры. Холодовые рецепторы были впервые обнаружены на антенне пчелы Apis mellifera. Введение вольфрамового электрода в целоконическую сенсиллу позволило зарегистрировать фазово-тонический ответ чувствующей клетки на резкое понижение температуры. Чувствительность изменения частоты импульсов в тонической части ответа была равна 2 имп/с/град. Холодовой рецептор с фазово-тоническим ответом был обнаружен также в целоконической сенсилле антенны саранчи. Импульсный ответ холодового рецептора трихоидной сенсиллы антенны таракана Periplaneta americana имел хорошо выраженный фазовый характер. Уменьшение температуры на 0,5° вызывало увеличение фоновой активности с 25 до 50 имп/с. В любом случае высокая частота импульсов поддерживалась лишь в тече-
137
ние 1—2 с после начала раздражения, несмотря на то, что раздражитель продолжал действовать. В результате быстрой адаптации частота импульсов приближалась к величине фона и удерживалась на одном уровне в течение 100—200 с действия раздражителя. Прекращение стимуляции холодным воздухом вызывало кратковременное (1—2 с) торможение импульсной активности рецептора, после чего фоновая активность снова восстанавливалась.
Рис. 39. Интегрированные импульсные ответы целоконической сенсиллы антенны комара Aedes aegypti, указывающие на реакцию двух клеток с разной амплитудой потенциала при повышении или понижении температуры струи воздуха (по Davis, Socolove, 1974).
А, Б — тепловой и холодовый рецепторы соответственно; по оси абсцисс — время, с; по оси ординат — частота импульсов, имп/с. В — температура воздуха
Среди многих (около 18) чувствующих клеток, иннервирующих сенсиллы максиллярной пальпы гусеницы соснового коконопряда Dendrolimus pint, можно выделить одну клетку, дающую залп импульсов при приближении к сенсиллам холодного предмета. Нагретый предмет вызывал торможение фоновой активности.
Тепловые рецепторы изучены в меньшей степени, чем холодовые. Установлено, что поднесение теплой стеклянной палочки к базиконическим сенсиллам антенн и максиллярных пальп личинки жука Anisoplia austriaca и к целоконическим сенсиллам антенн личинки кожееда Dermestes sibiricus вызывало залп импульсов при наличии слабой фоновой активности. Тепловые рецепторы дистальных члеников антенн клопа Oncopeltus fasciatus имеют хорошо выраженный фазовый ответ, длящийся после небольшого латентного периода около 2—3 с, причем фоновая активность отсутствует до и после залпа, несмотря на продолжающееся действие струи теплого воздуха. Максимальную чувствительность к повышению температуры рецептор имеет в диапазоне 28—30°. Сенсиллы самцов и самок клопов показывают одинаковую реакцию на температурный стимул.
Целоконическая сенсилла на кончике антенны комара Aedes aegypti имеет две чувствующие клетки, причем одна является холодовым, а другая — тепловым рецептором. Их свойства идентичны описанным выше у других насекомых. Возбуждению теплового
138
рецептора соответствует торможение холодового и наоборот (рис. 39). Ответ рецепторов фазово-тонический. Быстрое возрастание частоты импульсов над величиной фоновой активности в начале залпа проявляется при изменении температуры на 0,05° и составляет 4 имп/с для холодового рецептора и 3,2 имп/с для теплового. Частота импульсации в тонической части залпа теплового рецептора увеличивается при достижении температуры около 24° и достигает максимума при 28,5° (35 имп/с), а в случае холодового рецептора имеет пик при 26° (30 имп/с). Чувствительность тонической части залпа теплового рецептора равна 5—7 имп/с/град, а холодового — 5—6 имп/с/град. Действие на целоконическую сенсиллу инфракрасного излучения с длиной волны 10 мкм, двуокиси углерода, влажного воздуха, запахов аттрактантов и репеллентов не вызывало изменений фоновой активности.
2. РЕЦЕПТОРЫ ВЛАЖНОСТИ
Локализация и строение. Исследования восприятия влажности окружающего воздуха показали, что многие насекомые хорошо различают степень увлажнения воздуха и могут ориентироваться по градиенту концентраций водяного пара. Так, комары избегают
Рис. 40. Реакции мух Drosophila melanogaster на влажность в альтернативной камере прн перепаде влажности от 77 до 100% (по Perttunen, Syrjamaki, 1958):
1 — нормальные особи; 2 — мухи, экспонированные в сухом воздухе; 3,4 — мухи с удаленными антеннами.
По оси абсцисс — время, мин; по оси ординат — интенсивность распределения во влажную камеру, %
очень высокой влажности воздуха и в диапазоне 95—85% относительной влажности различают ее с точностью до 1%. В диапазоне 85—30% относительной влажности чувствительность комаров падает до 40%.
Многочисленные опыты с экстирпацией антенн, пальп и других придатков показали, что рецепторы влажности тараканов, муравьев, жуков, вшей, пчел, многих двукрылых и других насекомых локализованы на антеннах. Большинство таких результатов было получено в альтернативных камерах или в камерах с градиентом влажности, позволяющих насекомым либо осуществить выбор из двух значений влажности, либо продемонстрировать гиг-ропреферендум — предпочтение оптимального диапазона влажности воздуха.
Мухи Drosophila melanogaster в альтернативной камере с относительной влажностью 100 и 77% предпочитают низкую влажность (рис. 40). Если же мух продержать в сухом воздухе 5 часов
139
при 25°, то они будут предпочитать влажный воздух. Удаление антенн вызывает исчезновение реакции у нормальных мух и значительное ослабление ее у мух, экспонированных перед опытом в сухом воздухе. Вероятно, рецепторы влажности локализованы не только на антеннах, но и на других участках тела насекомых.
Последовательное удаление члеников антенн у прямокрылых и жуков сочетается с уменьшением интенсивности реакции выбора в альтернативной камере, причем величина реакции коррелирует с числом сенсилл определен-ного типа на оставшихся
i I t
Puc. 41. Импульсная активность чувствительных клеток третьего антеннального сегмента гусениц Manduca sexta (по
Dethier, Shoonhoven, 1968):
а — при действии на антенну струи воздуха с относительной влажностью 65%; б — при действии на аитеииу воздуха с относительной влажностью 5%. Стрелки — начало и конец раздражения
члениках. Это — целоконические тонкостенные сенсиллы у саранчи Melanopus bu-vittatus и базиконические сенсиллы жука Oryza.eph.ilis surinamensis. Электронномикроскопическое изучение базиконических сенсилл, чувствующие клетки которых реагируют на изменение влажности, показало их большое сходство с базико-ническими обонятельными сенсиллами. Это тонкостен-
ные конусы с толщиной кутикулы около 0,1 мкм. Кутикула перфорирована порами с наружным отверстием 0,01 мкм и внутренним — 0,1 мкм. Полость сенсиллы заполняют ветвящиеся периферические отростки двух чувствующих клеток. Обкладочные клетки снабжены микроворсинками с многочисленными митохондриями. Стенки волоска сенсиллы тесно спаяны с кутикулой антеннального членика, что исключает возможность колебаний сенсиллы. Поро-трубчатой системы в рецепторах влажности не обнаружено.
Электрофизиологические исследования. В. Детье и Л. Шоон-ховеном в 1968 г. было установлено, что на антеннах гусениц Manduca sexta, Pieris гарае и Heliotis zea кроме холодовых рецепторов имеются чувствующие клетки, реагирующие на изменение влажности воздуха, т. е. специализированные рецепторы влажности. Введение вольфрамового микроэлектрода в третий членик антенны гусеницы позволило зарегистрировать активность чувствующих клеток. Оказалось, что в ответ на струю воздуха с относительной влажностью 65% наблюдается залп импульсов с несколькими амплитудами (рис. 41, а). Действие на антенну сухого воздуха приводит к торможению импульсной активности (рис. 41,6). Воздушный поток в обоих случаях имел одинаковую температуру— 30°. Струя холодного воздуха также вызывает постепенное возрастание частоты импульсов. Этот эффект объясняется тем, что снижение температуры воздуха всегда сопровождается повыше
но
нием относительной влажности воздуха в замкнутой системе, что и наблюдалось в условиях эксперимента.
На поперечных срезах у вершины максиллярных стилетов клопов родов Triatoma и Rhodnius обнаружены периферические отростки трех чувствующих клеток. Электрофизиологическое исследование показало, что эти клетки не относятся к хеморецепторам, но отвечают на механическую стимуляцию. При раздражении стилетов воздухом, имеющим высокую влажность, было выяснено, что две клетки реагируют не только на механическое раздражение, но и на влажность воздуха, т. е. представляют собой гигрорецепторы. Частота импульсов за 400 мс после начала раздражения достигала 10—12 имп/с при влажности 50% и 18—20 имп/с при влажности воздушной струи 100% и температуре 30°. Далее частота падала, достигая фонового уровня (1—3 имп/с) через 1,5—2 с, несмотря на продолжающееся действие раздражителя. Интенсивность ответа уменьшалась при понижении температуры струи воздуха. Вычисление внутренней энергии воздушных потоков с различной влажностью и разной температурой позволило обнаружить логарифмическую зависимость между термодинамической функцией — энтальпией и ответом рецепторов. Зависимость прослеживалась в диапазонах температур от 20 до 42° и относительной влажности от 0 до 100%.
Большинство исследователей температурных рецепторов и рецепторов влажности насекомых считают маловероятным существование специфических для данных категорий рецепции мембран в чувствующих клетках. Охлаждение или нагревание, а также действие влажного воздуха, вероятно, приводят к механическим деформациям кутикулярного отдела сенсилл либо к изменению химического состояния субстрата, прилежащего к мембранам чувствующих клеток. Таким образом, термо- и гигрорецепция могут быть частными случаями механо- или хеморецепции. Об этом свидетельствуют значительное сходство ультраструктуры сенсилл, а также полученные в электрофизиологических экспериментах данные, указывающие на равенство внутренней (термодинамической) энергии стимула и энергии механического раздражения, вызывающих одинаковый ответ рецептора.
Глава VI
ФОТОРЕЦЕПЦИЯ
Среди сенсорных систем животных, в их числе и насекомых, зрительный анализатор занимает особое место. По сложности организации и многообразию даваемой информации он далеко превосходит другие органы чувств, а рецепторные элементы глаза настолько специализированы, что их трудно отнести к первично-чувствующим клеткам. У многих насекомых параллельно с развитием совершенного полета и подвижного образа жизни органы зрения достигли удивительного совершенства, став основным датчиком информации для дальней и ближней ориентации.
Морфологически у насекомых различают три типа глаз: ^фасеточные, или сложные, 2) личиночные глазки, или с т е м м ы, замещающие фасеточные у видов с неполным превращением, и 3) дорсальные (теменные) глазки, или о ц е л л и, свойственные имаго и экзоптериготным личинкам и функционально дополняющие сложные глаза. Наиболее изучены фасеточные глаза, отличающиеся особой сложностью строения,— главные органы зрения у имаго. Поэтому именно им будет посвящено основное содержание главы.
1. СОВРЕМЕННОЕ ПРЕДСТАВЛЕНИЕ О ФУНКЦИОНАЛЬНОЙ ОРГАНИЗАЦИИ ФАСЕТОЧНОГО ГЛАЗА
Фасеточные, или сложные, глаза имеют почти все взрослые насекомые подкласса Pterygota (кроме блох и некоторых других групп) и их личинки у форм с неполным превращением. В подклассе бескрылых насекомых только у Machilidae встречаются глаза, близкие по своей организации к фасеточным. У остальных представителей насекомых место фасеточных глаз занимают простые латеральные глазки.
Строение омматидиев
Омматидии—структурные единицы фасеточного глаза — имеют свой собственный светопреломляющий (диоптрический), светоизолирующий (экранирующий) и светочувствительный (фоторецепторный) аппарат. У разных насекомых число омматидиев различно. Чаще всего оно достигает нескольких тысяч, вплоть до 28 тыс. у некоторых стрекоз и бабочек.
142
Светопреломляющий аппарат омматидиев образован прозрачными роговичной линзой и кристаллическим конусом (рис. 42, 1, 2). С внешней стороны роговица омматидия имеет вид более или менее выпуклой шестигранной фасетки. При электронно-микроскопических исследованиях было обнаружено, что фасетки дневных и ночных бабочек, двукры
лых, ручейников покрыты сеткой из прозрачных кутикулярных бугорков высотой около 200 нм и на столько же отстоящих друг от друга. На модели глаза, а также путем соответствующих расчетов показано, что наружная сетка из бугорков снижает отражение света и тем повышает (на 5%) прозрачность роговицы. Подобно просветляющему, или ан-тирефлексному, покрытию у современной оптики, сетка облегчает проникновение лучей с 1=320— 700 нм из воздуха в более плотную среду глаза. Ее биологиче-
Рис. 42. Схема омматидия оптикосупер-познциоииого глаза в состоянии световой (б) и темиовой (а) адаптации (по Ног-ridge, 1972; Meyer-Rochow, 1972, видоизменено) :
1 — роговичная линза; 2 — роговичный коиус; 3 — кристаллические клетки; 4 — кристаллический конус; 5 — гранулы экранирующего пигмента; 6 — придаточные пигментные клетки; 7 — кристаллический тракт; 8 — ирисовые пигментные клетки; 9 — тела ретинулярных клеток; 10 — прозрачная зона; 11 — рабдом; 12 — базальная зрительная клетка; 13— трахейный тапетум; 14 — базальная мембрана; 15 — аксоны зрительных кле-
ток
ская роль дополняется также камуфлированием глаза за счет устранения блеска его поверхности.
Роговичная линза как собирательный светопреломляющий аппарат функционирует совместно с непосредственно примыкающим к ней кристаллическим конусом. Последний представляет собой прозрачное плотное или жидкое тело, образованное четырьмя Зем-перовыми (кристаллическими) клетками — их телами или экстра-клеточным выделением (рис. 42, 3). Сравнительно недавно австралийский исследователь Г. А. Хорридж обнаружил, что кристал
143
лические клетки продолжаются в виде тонких кристаллических нитей (тракта) (рис. 42, 7) в глубь омматидия. Это особенно заметно в оптикосуперпозиционных глазах, где им приписывают роль световода.
Светоизолирующий аппарат омматидия представлен набором пигментных клеток, содержащих гранулы экранирующего пигмента. Этот аппарат необходим для защиты рабдома от бокового за-света и повышения направленной чувствительности омматидия. В дистальной части омматидия всегда присутствуют две корне-агенные клетки и не менее четырех ирисовых пигментных клеток (рис. 42, 8), а в аппозиционных глазах имеется еще несколько ретинальных пигментных клеток, лежащих в проксимальной половине омматидия. Кроме того, гранулы защитного пигмента присутствуют в теле зрительных клеток и в межклеточной среде. Различают оммохромы — более темные (красные, коричневые) пигменты и птерины — более светлые. Первые поглощают лучи в широком диапазоне спектра, кроме красных лучей, вторые — главным образом ультрафиолетовые (УФ) лучи. Иногда защитные пигменты частично или полностью отсутствуют. Такие светлоглазые мутанты известны, например, среди мух, и их широко используют для исследования различных зрительных функций. В глазах опти-косуперпозиционного типа гранулы защитного пигмента периодически переходят из дистального положения в проксимальное и наоборот в зависимости от интенсивности освещения, времени суток и других причин (рис. 42, а, б).
Светочувствительный аппарат омматидия образуют чаще всего 8—9 зрительных, или ретикулярных, клеток с четко выраженными аксонами, связывающими их с оптическим центром. Зрительные клетки (рис. 42, 9) лежат обычно в 2—3 яруса, а одна или две из них, так называемые базальные клетки (12), асимметрично смещены к базальной мембране, подстилающей дно глаза. Аксон базальной клетки идет непосредственно во второй оптический ганглий, а не оканчивается в первом, как у остальных зрительных клеток. Разные зрительные клетки омматидия отличаются различной спектральной н поляризационной чувствительностью.
Вдоль оптической оси омматидия (иногда ярусами) расположены рабдомеры зрительных клеток, образующие в совокупности рабдом (рис. 42, 11). Различают замкнутый (спаянный) раб-дом (рис. 43, А) — когда отдельные рабдомеры тесно объединены друг с другом, и открытый (рис. 43, Б) —если отдельные рабдомеры (кроме двух) разобщены и оптически изолированы друг от друга. Каждый рабдомер сложен большим количеством микроскопических трубочек — микровилл — диаметром около 500 А, лежащих перпендикулярно оптической оси омматидия (рис. 43, В, Г). Микровиллы, образованные двойной мембраной толщиной около 50 А, содержат зрительный пигмент, концентрация которого достигает 107 молекул на рабдомер. Поэтому рабдомер является собственно фоторецепторным элементом зрительной
144
Рис. 43. Электронные микрофотографии поперечного среза раб-дома.
А — медоносная пчела Apis mel-lifera (по Goldsmith, 1962); 1—8 — зрительные клетки; d — скопление десмосом; ег — эндоплазматический ретикулум; f — нити кристаллических клеток; Б, В, Г — муха Drosophila melanogaster (по Wolken, 1961): Ri—Ri— семь раб-домеров; Г—реконструкция участка рабдомера, рассеченного вдоль
клетки, функциональным аналогом наружного членика палочек и колбочек сетчатки позвоночных.
Замечательная особенность структуры рабдома заключается в упорядоченном расположении его микровилл. Например, у медоносной пчелы микровиллы восьми наиболее крупных клеток расположены в двух взаимно перпендикулярных направлениях, при
145
этом ориентация трубочек в рабдомерах соседних омматидиев остается одинаковой на больших участках глаза. У других насекомых картина ориентации микровилл может быть иной, но они всегда по-разному направлены в несимметричных рабдомерах. Такая структура рабдома при дихроизме поглощения света молекулами зрительного пигмента обеспечивает анализ поляризации лучей (см. ниже).
Проекция внешнего мира на растр зрительных клеток
Вершины рабдомеров, по крайней мере в глазу мух, лежат в фокальной плоскости оптики омматидия. Они одеты бесструктурными «шапочками», облегчающими вхождение внутрь лучей за счет более низкого коэффициента преломления, чем у собственно рабдомера (рис. 44, А, д). Что касается качества изображения, создаваемого оптикой омматидия, то оно малосущественно для зрения, поскольку его детали не могут быть разрешены ретикулярными клетками. Объяснение этому следует искать в особенностях проекции сетчатки в зрительном ганглии, при которой сигналы разных зрительных клеток теряют свой «адрес» уже на первом интернейроне.
Отдельные рабдомеры или весь рабдом замкнутого типа обладают свойствами волновода, что обеспечивает проведение лучей и их поглощение зрительным пигментом по всей длине рабдома. Дело в том, что к рабдомеру со стороны тела зрительной клетки примыкают структуры, имеющие более низкий показатель преломления. Поэтому часть лучей на границе раздела претерпевает полное внутреннее отражение. Так, у мухи Calliphora erythroce-phala показатель преломления (п) рабдомеров равен 1,349, а у окружающих его структур— 1,344—1,340; угол полного внутреннего отражения для такого рабдомера-волновода составляет 83° (рис. 44, А).
Поле зрения отдельного рабдомера Calliphora erythrocephala в среднем около 7°. Но имеющая особо важное функциональное значение полуширина поля зрения — угол, в пределах которого интенсивность истечения света снижается наполовину от максимального,— отдельного рабдомера этой мухи равна всего 1,2°. Судя по электрофизиологическим измерениям, поле зрения замкнутого рабдома аппозиционных глаз тоже не превышает 8°. Однако в опти-косуперпозиционных глазах, например жука-плавунца, оно существенно шире — практически охватывает поле зрения целого глаза. Поэтому поля зрения смежных омматидиев (а их оси расходятся под углом обычно от 1 до 3—4°) даже в аппозиционных глазах заметно перекрываются. Впрочем, это обстоятельство, как полагают (см. ниже), благодаря механизму латерального торможения не ухудшает остроту зрения насекомого.
146
Со времени классических исследований фасеточного глаза членистоногих венским офтальмологом А. Экснером (1891) единодушно принималось, что диоптрический аппарат омматидия представляет собой преломляющий цилиндр — такую линзу, которая сложена слоями с показателем преломления, возрастающим к центру. Теперь всеобщность этого утверждения подвергли сомнению. Толь-
Рис. 44. Схема аппозиционного глаза мухи Calliphora erythrocephala (по Seitz, 1968):
А — клеточные элементы омматидия и их показатели
преломления: а — роговица, б — кристаллический конус, в — пигментная клетка, г — Земперова клетка, д — заключенный в «шапочку» вырост рабдомера, е — рабдомер, ж — зрительная клетка;
Б — группа из семи (I—VII) омматидиев. Черным кружком изображен один из семи (1—7) рабдомеров каждого омматидия, смотрящий в одну и ту же точку пространства
ко у нескольких видов насекомых, в частности у светляка Phausis splendidula, показатель преломления действительно возрастает к центру кристаллического конуса, а у остальных насекомых диоптрический аппарат аналогичен обычной собирательной линзе (рис. 44, А). В аппозиционных и оптикосуперпозиционных глазах эта линза рисует в каждом омматидии обособленное прямое или перевернутое (у разных насекомых по-разному) изображение приблизительно в плоскости шапочек рабдомеров. Мы уже знаем, что качество этого изображения, иногда (у мух, например) довольно высокое, не определяет степени детализации воспринимаемой картины. Видеть отдельные детали изображения препятствует своеобразие проекции сетчатки в зрительном ганглии, а у насекомых с единым замкнутым рабдомом к тому же все зрительные клетки,
147
независимо от геометрии изображения стимула на рабдоме, получают равное количество света. В результате внешний мир анализируется насекомыми с точностью до растра омматидиев, а не отдельных зрительных клеток, как у позвоночных. В этом смысле сохраняется прежнее — мюллеровское — представление о мозаичности зрения насекомых.
Рис. 45. Упрощенная схема возникновения сетчаточного изображения в аппозиционном (а), оптико- (б) и нейросуперпозиционном (в) глазу (по Kirschenfeld, 1970).
1 — диоптрический аппарат омматидия; 2 — зрительные клетки с замкнутым (а и б) или открытым (в) раб-домом; 3—проекция аксонов зрительных клеток на оптические патроны ламины.
Заштрихованы те рабдомеры, на которые попадают параллельно идущие лучи света (показаны стрелками)
Неожиданной оказалась проекция внешнего мира На растр зрительных клеток у мух. Углы (2—4°) между оптическими осями семи рабдомеров каждого омматидия в глазу мухи находятся в таком соответствии с углами разобщения соседних омматидиев, что в семи смежных омматидиях имеется по одному рабдомеру, смотрящему в одну и ту же точку пространства (рис. 44, Б). Иначе говоря, семь рабдомеров одного омматидия смотрят в разные точки пространства, но те же точки видят по одному рабдомеру в шести соседних омматидиях. Аксоны шести зрительных клеток, рабдомеры которых направлены в одну и ту же точку, сходятся в одном и том же оптическом патроне ламины, где их сигналы суммируются. Поэтому между числом светящихся точек на сетчатке мухи и числом оптических патронов существует соотношение 1:1, т. е. число оптических патронов равно числу омматидиев. При такой нейрооптической конструкции за счет увеличения эффективности входного зрачка из многих фасеток светосила глаза повышается в 7 раз по сравнению с классическим аппозиционным глазом, как, например, у медоносной пчелы. Поэтому глаза мух теперь выделяют в особый тип нейросуперпозиционных глаз.
Соответственно сказанному мы будем различать у насекомых три морфологических типа фасеточных глаза: аппозиционный, оп-тикосуперпозиционный и нейросуперпозиционный. Аппозиционные глаза свойственны большинству дневных насекомых, кроме двукрылых и клопов. Глаза этого типа приспособлены для работы при избытке света. Анатомически для них характерны удлиненный рабдом, достигающий вершины кристаллического конуса, и постоянная пигментная изоляция смежных омматидиев (рис. 44, А; 45, а). Оптикосуперпозиционные глаза ветре-
148
чаются у подавляющего большинства ночных насекомых. Здесь рабдом укорочен, между ним и кристаллическим конусом имеется отчетливо выраженный кристаллический тракт (см. рис. 42, а). Защитный пигмент в удлиненных ирисовых пигментных клетках мигрирует в зависимости от состояния адаптации глаза. При пониженном освещении, в том числе вечером и ночью, гранулы пигмента сосредоточиваются в дистальном положении, в результате исчезает экранизация пространства между кристаллическими конусами и вершинами рабдомеров, и в один и тот же омматидий могут попадать лучи, прошедшие через группу смежных фасеток (рис. 45, б). Еще не ясно до конца, как работают оптикосуперпо-зиционные глаза. Но у некоторых ночных жуков и бабочек, а также у толстоголовок Hesperiidae при дистальном положении ирисового защитного пигмента безусловно имеет место суперпозиция лучей на рабдомах, чем достигается повышенная чувствительность к свету, хотя и ценой потери качества изображения. При дневном положении защитного пигмента эти глаза функционируют, по-видимому, аналогично аппозиционным; ретинальное изображение образуется за счет аппозиции лучей, прошедших через разные фасетки, т. е. без наложения друг на друга. Нейросуперпози-ц и о н н ы е глаза известны только у двукрылых — насекомых с открытым рабдомом. В них нет суперпозиции лучей на сетчатке, но имеет место суперпозиция сигналов, исходящих от зрительных клеток разных омматидиев, в первом оптическом ганглии (рис. 45, в). Глаза этого типа, как и аппозиционные, обеспечивают высокое разрешение точек пространства, но отличаются от вторых повышенной светосилой. Недаром мухи активны при такой низкой освещенности, при которой не летают другие дневные насекомые.
2. ОПТИЧЕСКИЕ ГАНГЛИИ
И МЕХАНИЗМЫ
ПРЕДВАРИТЕЛЬНОЙ ОБРАБОТКИ
ЗРИТЕЛЬНОЙ ИНФОРМАЦИИ
Оптические ганглии и типы их интернейронов
Зрительный центр фасеточных глаз находится в парных оптических (зрительных) долях, представляющих собой боковые части переднего мозга. В каждой оптической доле различают четыре отдела: 1) ламину (I оптический ганглий), обычно непосредственно подстилающую дно глаза; 2) медуллю (II оптический ганглий), связанную с ламиной внешней хиазмой; 3) лобулю и 4) лобулярную пластинку, или лобулюс (рис. 46). Два последних отдела, связанных с медуллей внутренней хиазмой, ранее объеди-
149
няли под названием «III оптический ганглий». Все отделы представляют собой чрезвычайно сложные нервные массы, образованные большим числом нейронов различного вида .и назначения. Ней
Рис. 46. Морфологические типы нейронов в оптической доле переднего мозга (по Collett, 1970):
I — перпендикулярный нейрон; II — тангенциальный нейрон; III — амакриновая клетка.
а — короткий рецепторный аксон; б — длинный рецепторный аксон; с — сетчатка; зд — задний край; ла — ламина; ло — лобуля; л.п. — лобулярная пластинка; ме — медулля; от — оптический тракт, ведущий к медиальной части протоцеребрума; пе — передний край
Рис. 47. Проекция сетчатки с замкнутым (А) и открытым (5) рабдомом на оптическую долю (по Horridge Meinertzhagen, 1970).
На А: 1, 2, 3, 4, 5 — поля зрения отдельных омматидиев и их проекция в ме-дуллю; на Б; I, 2, 3, 4, 5 — оптические оси отдельных рабдомеров и проекция последних в медуллю.
бм — базальная мембрана; л — ламина; м — медулля; ом — омматидии; р— роговица; рб — рабдомеры; х — хиазма
роны подразделяют на три основных класса: перпендикулярные (рис. 46,1), тангенциальные (II) и амакриновые клетки (П1). Перпендикулярные нейроны, соединяющие друг с другом разные нервные массы (отделы), отличаются тем, что их аксоны распо
150
лагаются более или менее перпендикулярно к поверхности ганглия, в котором они лежат. Тангенциальные нейроны связывают по горизонтали параллельные ряды перпендикулярных клеток, а также образуют прямую связь между ипси- и контралатеральными оптическими долями. Амакриновые клетки — это нейроны, отростки которых не выходят за пределы одного отдела оптической доли. Они встречаются в разных ганглиях и имеют окончания на тех же уровнях, что и нейроны остальных классов,
В дистальной части ламины плотно лежат друг возле Друга так называемые «оптические патроны». В каждом оптическом патроне сходятся в виде пучка обычно шесть коротких волокон зрительных клеток, синаптически контактирующих здесь с двумя-тремя перпендикулярными клетками — нейронами второго порядка, а также с несколькими центробежными волокнами. Число оптических патронов в ламине равно числу омматидиев.
Соответственно двум типам рабдома описаны два типа проекции растра сетчатки на ламину. У насекомых с замкнутым раб-домом в каждом оптическом .патроне сходятся аксоны одного и того же омматидия, т. е. наблюдается полная и прямая проекция короткоаксонных ретинальных клеток каждого омматидия на I оптический ганглий (рис. 47, А). У мух и, вероятно, у других насекомых с открытым рабдомом в каждом омматидии сходятся аксоны группы смежных омматидиев, именно тех ретинальных клеток;, рабдомеры которых смотрят в одну и ту же точку пространства (рис. 47, Б).
Длинные волокна, принадлежащие базальным клеткам омматидия, проходят ламину насквозь, затем вместе с перпендикулярными волокнами вступают в наружную хиазму и оканчиваются в виде пучков в медулле. За счет этих пучков возникает столбча-тость медулли. Проекция оптических патронов ламины на медул-лю полная, но зеркально повернутая в вертикальной плоскости.
Для наглядности можно записать, что некоторой последовательности омматидиев — 1, 2, 3, 4, 5 соответствует в ламине тот же ряд оптических патронов, а в медулле он переходит в преобразованный ряд — 5, 4, 3, 2, 1 (рис. 47, А, Б).
Связь первых двух оптических ганглиев с третьим еще сложнее и пока недостаточно изучена. Полагают, что имеется не менее шести раздельных путей проекции сетчатки на третий оптический ганглий. Что касается более высоких связей, то оптические волокна, судя по данным электрофизиологических опытов, прослеживаются в средней части протоцеребрума и в его грибовидных телах.
В настоящее время точная картина нервных связей в зрительном центре усиленно изучается многими исследователями-морфологами совместно с электрофизиологами. Раскрытие ее даст ключ к пониманию механизмов обработки зрительной информации, к чему так стремятся конструкторы «думающих» машин.
151
Электрическая реакция глаза и передача сигналов зрительной клеткой
Под действием света в зрительной клетке возникает харак-
терный рецепторный потенциал, имеющий положительный знак
при внутриклеточном отведении (см. рис. 4,Л,Б). Совокупность
nnnniMitinniiiiiiMiiiiiiiiiiiuiniuiniiJinii___
1/155 с
этих рецепторных потенциалов возбужденной сетчатки образует основную часть ЭРГ, регистрируемой на роговице в виде медленного столообразного отрицательного потенциала, сохраняющегося в течение всего периода за-света глаза (рис. 48, а). Другой (быстрый) компонент двухфазной ЭРГ, наблюдаемый у мух, бабочек, стрекоз и некоторых других насекомых, имеет ганглиозное происхождение (рис. 48, б). После удаления оптических ганглиев или выключения их стрихнином и другими агентами двухфазная ЭРГ с быстрыми on- и off-пиками превращается в медленную монофазную отрицательную ЭРГ обычного вида.
Рис. 48. Электроретииограммы:
а — комара Anopheles maculipennis (по Мазохину-Пор-шнякову, 1965); б, в, г — мухи Calliphora erythrocephala (по Autrum, 1958, видоизменено).
ЭРГ иа в, г получены при мелькающем освещении. Нижние кривые — отметки раздражения; частота стимуляции обозначена на рисунке; с — свет; г — темнота
Возбуждение зрительной клетки передается по ее аксону, по-видимому, электротонически, а не в форме дискретных спайков, как у хеморецепторных и большинства других чувствующих клеток насекомых. Разные авторы при тщательном специальном поиске не нашли внутриклеточных импульсов в фоторецепторах стрекоз, прямокрылых, мух, рабочих медоносных пчел. Только у трутня медоносной пчелы видели одиночный спайк, накладывающийся на фазу нарастания медленного деполяризационного потенциала зрительной клетки.
У мечехвоста Litnulus эксцентрическая клетка разряжается залпом импульсов, легко отводимым внутри- и внеклеточно. Зрительный центр у него удален от глаза, и связь омматидиев с ним осуществляет пучок длинных рецепторных аксонов. У насекомых оптические гаНглии сближены с рецепторами глаза, аксоны ретикулярных клеток короткие (например у саранчи перелетной — 200 мкм), поэтому неудивительно, что им свойственна безымпуль-сная передача сигналов сетчатки. Пока остается неизвестным, по аксонам каких клеток омматидия и каким образом передается ин
152
формация одновременно о яркости, цвете и поляризации стимула. Ясно одно: ^визуальная информация о внешнем мире поступает в мозг с точностью до растра омматидиев. Она может передаваться по двум разным каналам: коротким аксонам зрительных клеток и более длинному волокну базальной клетки. Если сигналы всех зрительных клеток одного омматидия суммируются на первом интернейроне в ламине, то базальная клетка может передавать свой сигнал непосредственно в медуллю.
Детектирование признаков и переработка информации в зрительном центре
В оптических ганглиях происходит сложная обработка зрительной информации, поступающей от сетчатки глаза. Некоторые механизмы этой обработки уже выявлены, другие — интенсивно изучаются.
Сначала у позвоночных, затем у насекомых были открыты тангенциальные ганглиозные нейроны, избирательно реагирующие на определенные параметры зрительного стимула: размер, характер движения, цвет и т. п. (рис. 49). Такие, иногда чрезвычайно узкоспециализированные интернейроны получили название детекторов. Детекторы возникли у животных как способ экономного описания внешнего мира и передачи в высшие ассоциативные центры только существенной для опознания объектов информации, отфильтрованной от ее избытка. Нет сомнения в том, что у низших животных, у насекомых в особенности, сигналов некоторых видов детекторов достаточно для запуска врожденных стереотипных реакций.
У мух, бабочек, жуков во II и III оптических ганглиях, а также в средней части переднего мозга при электрофизиологических исследованиях обнаружено много видов зрительных детекторов с разнообразными свойствами. Их рецептивные поля варьируют от 1—6° до размера поля зрения целого глаза и могут быть как монокулярными, так и бинокулярными. В последнем случае одновременная стимуляция левого и правого глаз нередко вызывает в нейронах реципрокно-тормозной ответ. Одни детекторы длительно реагируют на продолжительную стимуляцию, другие обладают «привыканием», но наиболее замечательное свойство большинства их — дирекциональная (направленная) чувствительность к движению: реакция в основном или исключительно на те стимулы, которые движутся в строго определенном направлении (рис. 49, а, б, в). Движение в «предпочитаемом» направлении возбуждает нейрон, а в прямо противоположном подавляет его фоновую активность.
Простейшими среди зрительных детекторов являются нейроны «тревоги», реагирующие только на затемнение части глаза или на начало движения объекта в любом направлении. Импульсации
153
клеток такого рода, сигнализирующих о всяких изменениях в поле зрения, в принципе достаточно для своевременного запуска реакции убегания (рис. 49, г, д, е).
-фф.
Рис. 49. Импульсная активность нейронов—детекторов направления движения в зрительной системе стрекозы Aeschna viridis (по Зенкину, Писареву, 1969):
а, б — реакция нейрона на движение соответственно черной или белой полосы снизу вверх; в — реакция того же нейрона на выключение света; г — реакция нейрона «с привыканием» на движение темного пятна диаметром 3° (вторая стимуляция следовала через 20 с после первой); д— реакция off-элемента (детектора затемнения) на выключение света; е, ж — реакции предыдущего нейрона на движение темной полосы к центру и от центра рецептивного поля глаза; з — реакция off-элемента на изменение освещения.
Стрелками показано направление движения тестовых объектов; калибровка—100 мкв; отметка времени — 0,5 с
Более сложны и интересны детекторы направления движения. Они избирательно разряжаются импульсами при движении объекта сверху вниз или снизу вверх, слева направо или справа налево, в ростральном или каудальном направлениях и т. д. Понятно, что на основе набора подобных нейронов с их бинокулярными перекрестно-тормозными связями легко организовать детектирование и автоматический контроль продольного и поперечного крена в полете и при плавании. Благодаря им же тело насекомого легко
154
удерживается в определенной точке пространства, например при сносе течением, а также создается механизм отличения собственного движения от смещения фонового окружения. Бинокулярные нейроны, чувствительные либо к ростральному, либо к каудальному движениию, лежат в основе управления оптомоторной реакцией, служащей для компенсации отклонений от прямолинейного курса движения. Свойства этих нейронов у пластинчатоусых жуков хорошо совпадают с характеристиками их оптомоторной реакции, как например, у навозных жуков Geotrupes.
Из визуальных детекторов пищи примечательны детекторы небольшого движущегося пятна, первоначально описанные у лягушек, а затем у стрекоз. У имаго Aeschna — это клетки с «привыканием», реагирующие на движение только маленьких (диаметром 1—6°) темных или светлых пятен. Детекторы движущегося пятна имеют прямое отношение к организации пищевого поведения: с их помощью насекомые опознают мелкую добычу, и, кроме того, видимо, осуществляется запуск реакции преследования у охотящейся стрекозы.
Зрительные детекторы позволяют опознавать по окраске особей своего вида. Одним из примеров могут служить мозговые интернейроны тропической Morpho amathonte, возбуждаемые движущимися стимулами голубого цвета и тормозимые стимулами с большей длиной волны. Вся зрительная система этой атласно-голубой бабочки хорошо приспособлена для обслуживания брачного поведения, причем названные детекторы специально настроены на опознавание полового партнера. В какой мере зрение управляет поведением бабочек-морфид, можно судить по распространенному в тропиках способу их привлечения и лова: берут голубую блестящую пластинку или зеркало и этим предметом размахивают в воздухе.
Таким образом, ганглиозные нейроны-детекторы обеспечивают прежде всего механизм выделения признаков, необходимых, а иногда и достаточных для опознавания определенных объектов. Кроме того, они являются датчиками информации, достаточной для формирования целесообразной моторной команды, т. е. для запуска поведенческого ответа и наведения организма на цель на основе алгоритма одного из таксисов (см. ниже).
Электрофизиологические исследования позволили выявить в глазах животных другой очень интересный механизм предварительной обработки зрительных сигналов. Речь идет о так называемом «латеральном торможении» нейронов, первоначально описанном и подробно изученном в фасеточном глазу мечехвоста Limulus Нобелевским лауреатом X. Хартляйном, а затем обнаруженном в сетчатке позвоночных и оптическом ганглии насекомых.
Регистрируя импульсную реакцию пучка аксонов от одиночного омматидия мечехвоста, идентичную электрическому ответу аксона отдельной зрительной клетки, X. Хартляйн обратил внимание
155
на то, что ответ омматидия зависит не только от возбуждающего стимула, но и от активности соседних омматидиев. Рецептивные единицы мечехвоста оказались взаимозависимыми, при этом взаимодействие между ними было строго тормозным: каждый возбужденный омматидий тормозил соседние омматидии и в то же время сам тормозился ими. Торможение было тем больше, чем сильнее
Рис. 50. Частота импульсов в пучке волокон отдельного омматидия при освещении ступенчатым яркостным узором глаза мечехвоста (по Ratliff, Hartline, 1959).
На разные участки глаза последовательно проецировали с помощью полупрозрачной маски прямоугольную яркостную ступеньку с соотношением интенсивности левого и правого полей, равным 4: 1 (вставка в верхнем правом углу; по оси ординат — оптическая плотность маски).
1 — изменение возбуждения исследуемого омматидия при различных положениях яркостной ступеньки, когда освещен только этот омматидий, а все остальные затемнены непрозрачным экраном. Кривая возбуждения полностью воспроизводит (в логарифмическом масштабе) ступенчатый перепад в иитенсивностн освещения, т. е. в отсутствие торможения. Отдельные точки — частота импульсов (цифры справа) тестового рецептора при освещении яркостной ступенькой.
2 — изменение возбуждения (частота импульсов — цифры слева) того же омматидия и для тех же положений яркостной ступеньки после снятия экрана. В этих условиях был освещен весь глаз, и в результате торможения рецепторов кривая возбуждения тестового омматидия резко отличается от той, которая получена в предыдущем опыте. Масштабная черта по абсциссе равняется 0,5 мм на поверхности глаза. По оси ординат справа и слева — частота импульсов в секунду
возбуждался омматидий. Оно передавалось через сплетение тончайших боковых веточек аксонов рецепторных клеток — нейропиль— проксимальнее базальной мембраны. У насекомых, как полагают, взаимотормозные отношения существуют между оптическими патронами ламины.
Благодаря механизму латерального торможения, как видно на рис. 50, усиливается различие в возбуждении по-разному освещенных соседних омматидиев. Единицы сильнее освещенной левой половины глаза, примыкающие к зоне слабого освещения (левая половина кривой 2), испытывают со стороны этой зоны меньшее торможение, поэтому реагируют более частыми импульсами, чем те омматидии, которые окружены зоной одинакового возбуждения. И наоборот, омматидии правой половины глаза, примыкающие к зоне более сильного засвета (правая половина кривой 2), тормозятся ею сильнее, чем друг другом, и поэтому они реагируют более редкими импульсами. В результате подчеркивается краевой контраст на границе перепада освещенности на сетчатке, что в конечном счете способствует выявлению контура или границы по-разному освещенных участков. Расчеты показали также, что механизм латерального торможения, устраняя последствия перекрыва
156
ния полей зрения смежных омматидиев, улучшает разрешающую способность глаза. Идея латерального взаимодействия между соседними рецепторами положена в основу некоторых технических декодирующих устройств, предназначенных, например, для расшифровки нечетких фотоснимков.
3. КОНТРАСТНАЯ ЧУВСТВИТЕЛЬНОСТЬ, АДАПТАЦИЯ И РЕТИНОМОТОРНЫЕ ЯВЛЕНИЯ
Для нормального видения, позволяющего различать многообразие окружающих предметов, глаз должен иметь высокую контрастную чувствительность. Контрастную чувствительность сетчатки измеряют как А///-100%, где Д/ — минимальное визуально контролируемое изменение интенсивности света, а I — исходная интенсивность света. У хорошо видящих животных, в том числе у насекомых, она достигает 5% и даже 1%. Это значит, что, например, пчелы или мухи замечают разницу в интенсивности света на 5% и меньше. Общая же освещенность в естественных условиях изменяется в громадном диапазоне: от 100 тыс. лк в летний полдень до 0,2 лк при полной луне и 0,0005 лк безлунной ночью. Как же обеспечивается в зрительной системе высокая контрастная чувствительность при работе в условиях непостоянной освещенности, изменяющейся в миллионы раз?
Прежде всего нужно учесть, что в природе животных окружают почти всегда несамосветящиеся объекты, которые только отражают свет, падающий извне. Но такие объекты отличаются по интенсивности отражения не более, чем в 20—30 раз, и именно такой сравнительно узкий перепад яркостей должен замечать их глаз. Последнее достигается тем, что амплитудные характеристики реакции фоторецепторов приходятся наиболее крутой частью как раз на такой узкий перепад интенсивностей света. Для этого необходимо, чтобы при изменении общего уровня освещения количество поглощаемых рецепторами фотонов изменялось бы по возможности мало. В таком случае амплитудные характеристики будут как бы сдвигаться, подстраиваясь своей крутой частью к данному уровню освещения. Разнообразные опыты показали, что именно так обстоит дело и в глазу камерного типа позвоночных, и в совершенно иначе устроенном фасеточном глазу насекомых.
Регуляция количества света, поглощаемого рецепторами, достигается благодаря согласованной работе нескольких механизмов зрительной адаптации, или приспособления глаза для функционирования при широко меняющейся освещенности. О состоянии адаптации обычно судят по пороговой энергии света, достаточной для появления электрического ответа глаза (ЭРГ) или моторной реакции целого организма. Образец кривых изменения
157
пороговой чувствительности глаза в процессе темновой адаптации представлен на рис. 51. У дневных насекомых с аппозиционными (Argynnis) и нейросуперпозиционными (Calliphora) глазами чувствительность глаза в темноте возрастает только в десятки раз, а у ночных, например у бабочек совок или пядениц, она увеличивается на 4—5 десятичных логарифмических порядков. У пер-
Рис. 51. Кривые изменения чувствительности глаза ночной бабочки Cerapterix graminis (/, 2) и дневной оа бочки Argynnis adippe (3) в ходе темновой адаптации (по Bernhard, Ottoson, 1960):
1 — случай, когда ирисовый пигмент не перемещался;
2 — при перемещении пигмента.
По оси ординат — логарифмы пороговой интенсивности света; по оси абс-
вых — это однофазный и короткий процесс, у вторых — двухфазный и продолжительный. Начальные 8—10 мин темновая адаптация идет и у тех и у других практически одинаково, но затем чувствительность ночных видов с оптикосуперпозиционными глазами круто повышается: наступает вторая фаза адаптации, а у дневных видов светочувствительность сохраняется на прежнем уровне. Гистологический контроль положения ирисового защитного пигмента в омматидиях показал, что перегиб адаптационной кривой ночных бабочек хорошо совпадает по времени с началом перехода этого
пигмента в положение, характерное для ночи. То и другое наступает в среднем на 6—8 мин темновой адаптации, и оба эти процесса заканчиваются примерно за 30—50 мин.
Световая адаптация глаза, подстройка его чувствительности для работы при ярком освещении, идет гораздо быстрее.
Таким образом, к адаптации глаза причастны ретиномоторные явления, одним из примеров которых служит перемещение гранул защитного пиг-
цисс — время, мин мента в ирисовых пигментных клетках оптикосуперпозиционного глаза. Регуляция доступа света к фоторецепторам в данном случае целиком механическая. Под действием света или утром, в силу внутреннего физиологического ритма, или «часов», насекомого, гранулы пиг-
мента мигрируют проксимально, что по результату аналогично сужению входного зрачка. Темнота или наступление вечера стимулируют обратный процесс: гранулы пигмента собираются у дистального конца клеток, благодаря чему больше лучей света достигает рабдома каждого омматидия.
Миграции экранирующего пигмента в ирисовых и других пигментных клетках нередко сопутствует изменение длины рабдома. Под действием избытка света он укорачивается, что ухудшает
158
условия его освещения. В темноте и при низкой освещенности рабдом медленно (часами) удлиняется, как бы тянется к свету. Адаптационные изменения длины рабдома замечены в секомых.
Но как объяснить быстрые изменения чувствительности глаза дневных насекомых? Известны, по крайней мере, два механизма быстрой адаптации глаза. Первый из них связан с фотохимическими превращениями зрительного пигмента в рецепторах. Под действием света концентрация исходного зрительного пигмента падает, и, следовательно, уменьшается вероятность встречи фотонов с. неразложеннымн молекулами исходного фотопигмента. В темноте идет ресинтез фотопигмента, что и обеспечивает повыше ние светочувствительности рецептора. Первая (быстрая и короткая) фаза темновой адаптации оптикосуперпозиционного глаза, возможно, отражает именно этот фотохимический процесс в сетчатке.
Второй механизм адаптационной подстройки чувствительности глаза основан на изменении волноводных свойств рабдома, регулирующих в конечном счете угловое поле зрения рецептора. Так, у саранчи Locusta migratoria при световой адаптации палисад из вакуолей эндоплазматического ретикулюма вокруг рабдома распадается, и вместо него здесь скапливаются митохондрии, коэффициент преломления которых ближе к таковому рабдома, чем у палисада. В результате ухудшаются условия светопроведения, меньше света удерживается рабдомом при многократном отражении лучей на границе с митохондриями, а поле зрения рабдома при этом сужается. В итоге падает уровень светочувствительности глаза. И наоборот, образование палисада в темноте повышает светочувствительность глаза, поскольку уменьшается угол полного внутреннего отражения рабдома, что приводит к расширению его поля зрения.
Сходное явление описано у мухи Musca domestica. Оно наблюдалось за счет миграции гранул экранирующего пигмента в цитоплазме зрительных клеток. При освещении глаза гранулы экранирующего пигмента быстро, в течение нескольких секунд, мигрируют к рабдомерам, вследствие чего в последних нарушаются условия для полного внутреннего отражения и свет частично рассеивается, частично поглощается экранирующим пигментом. Это влечет за собой сужение поля зрения рецепторов, что показано по уменьшению угла, в пределах которого свет вызывает электрический ответ зрительной клетки. Таким способом фоторецепторы мухи защищаются от избытка света. Регуляция осуществляется, по-видимому, на основе петли обратной связи между интенсивностью возбуждения зрительной клетки и количеством мигрирующего в ней защитного пигмента, т. е. она похожа по своей сущности на так называемый «зрачковый рефлекс» камерного глаза позвоночных.
159
4. ОСТРОТА ЗРЕНИЯ, ВОСПРИЯТИЕ ФОРМЫ И ОБЪЕМА
Разрешающая способность
Острота зрения, или разрешающая способность, характеризует способность глаза различать степень расчлененности объектов, в том числе отличать две точки от одной более крупной. Измеряют ее тем минимальным расстоянием между двумя объектами, при котором они видны как два раздельных объекта. При прочих равных условиях острота зрения животных определяется в основном степенью расчлененности: 1) зрительного растра, или растра рецептивных единиц, имеющих самостоятельные поля зрения, и 2) оптического изображения на этом растре сетчатки. У позвоночных с их глазами камерного типа рецептивной единицей сетчатки может служить каждая зрительная клетка в отдельности, при этом условии качество ретинального изображения приобретает большое значение для восприятия деталей внешнего мира. У насекомых, судя по особенностям проекции зрительных клеток омматидиев в первый оптический ганглий, такой рецептивной единицей, воспринимающей расчлененность, служит целый омматидий, а не его отдельные фоторецепторы. Поэтому качество ретинального изображения внутри омматидия несущественно для различения деталей внешнего мира — зрительный центр декодирует поступающую информацию с точностью до числа омматидиев, а не их зрительных клеток. Таким образом, разрешающую способность фасеточного глаза характеризует в первую очередь угловая плотность омматидиев, т. е. число омматидиев в пределах некоторого угла. Чем больше омматидиев приходится на один и тот же угол, т. е. чем ближе сведены их оптические оси, тем выше острота зрения.
Зависимость между угловой плотностью омматидиев и остротой зрения легко понять из следующего рассуждения. Допустим, что на темном фоне имеются две светлые точки. То, что мы видим их как два светлых пятна, разделенных темным промежутком, а не слившихся в одно сплошное большое пятно,— это результат прежде всего различного возбуждения соседних рецепторов сетчатки. Поэтому, чтобы судить, разрешает или не разрешает глаз какие-то объекты, достаточно сравнить интенсивность возбуждения смежных рецепторов этими объектами. Две светлые точки насекомое увидит как две — заметит между ними темный промежуток — только при условии, что ретинальное изображение этих точек, вместе с промежутками между ними, захватит по меньшей мере три омматидия. В таком случае промежуточный омматидий может оказаться возбужденным слабее, чем краевые омматидии, а это и есть условие для отличия двух светлых точек от одной более крупной. Но двух смежных омматидиев недостаточно, чтобы всегда отличать две точки от одной, равной им по суммарной площади.
160
Соотношение между разрешающей способностью глаза и величиной межомматидийных углов или угловой плотностью омматидиев поясняет рис. 52. Глаз А позволяет разрешить светлые точ
ки 1 и 2 потому, что они возбуждают краевые омматидии а и ч сильнее, чем промежуточный омматидий б. Дело в том, что угло-
вое расстояние между точками 1 и 2 вдвое больше угла а между осями смежных омматидиев. Две светлые точки, разобщенные углом 2 а и больше, очевидно, всегда будут возбуждать краевые омматидии не так, как промежуточный, т. е. в принципе всегда могут быть различимы насекомым. Комбинируя положение теста относительно осей омматидиев, легко себе представить, что в некоторых условиях возможно различение точек, разделенных углами менее 2 а, но более а. Глаз Б, у которого углы между осями омматидиев существенно больше, не позволит разрешить те же точки 1 и 2, т. е. не даст возможности насекомому отличить их от одной более крупной точки 3.
Глаза насекомых не обладают аккомодацией: радиус кривизны роговичной линзы у них постоянен. Поэтому для фасеточного глаза существует простая зависимость между расстоянием до объекта и числом различимых деталей: чем ближе объект, тем больше деталей видит насекомое.
Не следует думать, что острота зрения фасеточного глаза может неограниченно возрастать при увеличении угловой плотности омматидиев, т. е. за счет уменьшения диаметра каждого из них. Известный английский ис-
Рис. 52. Зависимость остроты зрения фасеточного глаза от угловой плотности омматидиев (по Мазохину-Поршиякову, 1965).
Глаз А разрешает точки I и 2, так как они разделены углом, вдвое большим угла а между оптическими осями омматидиев.
Глаз Б с меньшей угловой плотностью омматидиев те же точки 1 и 2 не разрешает. Здесь угловое расстояние между точками равно а, поэтому глаз их не отличит от одной более крупной точки. 3.
а, б, в — отдельные омматидии; О — оптическая ось омматидиев; одинаково возбужденные омматидии выде-
лены светлым
следователь Г. Барлоу (1952) показал, что при малых диаметрах фасеток предел увеличению разрешающей силы глаза кладет дифрак-
ция на входном зрачке омматидия и что у перепончатокрылых, в частности у медоносной пчелы, угловая плотность омматидиев в средней части глаза оказалась оптимальной с этой точки зрения. Иначе говоря, в глазу пчелы выдержана оптимальная конструкция: оси омматидиев сближены ровно Настолько, чтобы обеспечить максимальную остроту зрения, допускаемую дифракцией лучей с длиной волны более 300 нм.
<61
Рис. 53. Гусеница бражника Sphinx ligustri (А) и растровое изображение ее (Б) с теми деталями, которые разрешает глаз стрекозы Aeschna с расстояния 9,5 см (по Мазохину-Поршнякову, 1958)
Разрешающую способность аппозиционного глаза насекомых, рассчитанную по угловой плотности омматидиев, промоделировал Мазохин-Поршняков (1958) фотографическим путем. В основу модели положены три предпосылки: 1) каждый омматидий является рецептивной единицей, посылающей нервный сигнал, независимый от сигналов своих соседей; 2) величина сигнала определяется общим количеством света, попадающего во входное отверстие омматидия; 3) свет, исходящий от каждой точки поля зрения глаза, воздействует только на один омматидий. Некоторая схематичность допущений сделана намеренно ради упрощения технического решения задачи. В действительности поля зрения смежных омматидиев частично перекрываются, и между омматидиями существуют взаимо-тормозные отношения, хотя эти два эффекта в значительной мере, как мы уже знаем, компенсируют друг друга.
Задача исследователя заключав лась в том, чтобы получить на фотобумаге изображение предмета, разбитое на столько отдельных элементов, сколько омматидиев охватывает ретинальное изображение этого предмета. В пределах каждого элемента участок изображения должен иметь равномерную плотность, имитирующую реакцию ом-в него свет.
матидйя на весь попадающий
Для этого негативное изображение предмета проецировали на покрытую молочным стеклом перфорированную пластинку — растр «омматидиев», разбивающую изображение на заданное число элементов. Под перфорированную пластинку помещали фотобумагу, на которой печаталась искомая картина, воспроизводящая степень подробности различения насекомым деталей этого или иного объекта. На рис. 53 изображена гусеница бражника Sphinx ligustri с теми подробностями, которые может заметить насекомое, имеющее глаза аппозиционного и нейросуперпозиционного типов. Это изображение в длину покрывает 45 отверстий растра. Для таких насекомых, как стрекозы Aeschna или мухи Eristalis, у которых углы между осями омматидиев равны примерно 1°, эта фотография воспроизводит гусеницу при рассматривании на расстоянии 9,5 см. Для бабочек Zygaena с вдвое меньшей угловой плотностью омматидиев эта же фотография будет соответствовать случаю, когда
1<И
они смотрят на гусеницу с вдвое меньшего расстояния, т. е. когда угловой размер гусеницы достигнет 90°.
Ряд исследователей пытались измерить остроту зрения насекомых по их поведению. Например, судя по «критическому» периоду полос вертикального черно-белого узора, нанесенного внутри цилиндра, при котором вращение этого цилиндра перестает вызывать у сидящего внутри насекомого оптомоторную реакцию — компенсационное движение головы или всего тела при смещении поля зрения,— максимальная острота зрения действительно хорошо совпадает с величиной 2а. Именно для медоносной пчелы, мухи Calliphora erythrocephala, жука Chlorophanus критический период узора равен соответственно 1,96°; 4°; 14°, а удвоенный угол-между осями смежных омматидиев равен у этих насекомых соот-' ветственно 1,8°; 4,8°; 14°.
Личинки палочника Dixippus morosus стремятся «вползти» на нарисованные на бумаге вертикальные черные полосы, признавая их за стебли растений. Если полосы нарисованы слишком часто, то насекомые перестают на них реагировать. Оказалось, что минимальный угловой период полос, при котором они привлекают палочников, приблизительно вдвое больше межомматидийного угла в глазу насекомого, т. е. тоже равен около 2а.
Нередко даже опытные исследователи оценивают максимальную остроту зрения по минимальному размеру видимых предметов, незаконно подменяя одно понятие другим. Чтобы увидеть маленький предмет, т. е. отличить присутствие предмета в поле зрения от его отсутствия, совсем не требуется высокая разрешающая способность глаза, но важно, чтобы он имел достаточно высокую контрастную чувствительность. Поясним эту мысль на примере охотящейся стрекозы. Она замечает муху с расстояния в несколько метров, когда угловой размер добычи в несколько раз меньше межомматидийного угла а. Иными словами, стрекоза видит муху с такого расстояния, с которого она уже не может отличить двух близко расположенных мух от одного более крупного насекомого. Появление темной точки в поле зрения единственного омматидия уменьшает количество падающего на него света, и этого достаточно, чтобы отличить случай присутствия точки от случая ее отсутствия. При этом точка будет видна тем дальше, чем выше контрастная чувствительность фоторецепторов и чем больше контраст в яркости между точкой и фоном, на котором она находится. Таким образом, чтобы увидеть сколь угодно маленький предмет, достаточно одного рецептора, но чтобы отличить его от другого маленького предмета, требуется минимум три рецептора.
Разрешающая способность глаза в принципе зависит также от контрастной чувствительности, величины поля зрения и характера нервного взаимодействия его рецептивных единиц. Чем выше контрастная чувствительность зрительного растра, тем лучше острота зрения, так как тем больше перепадов освещенности заметит насекомое. Мы уже отмечали, что поля зрения соседних оммати
163
диев частично перекрываются. Это может снизить потенциальную остроту зрения при данной угловой плотности омматидиев, поскольку наложение друг на друга полей зрения ведет к сглаживанию различий в состоянии возбуждения соседних омматидиев. Однако мы знаем, что в оптическом ганглии насекомого имеется механизм подчеркивания контуров, который функционально компенсирует отрицательный эффект наложения полей зрения у соседних омматидиев.
До сих пор мы подразумевали, что глаз замечает расчлененность окружающей картины в результате неодинакового возбуждения разных омматидиев одновременно, т. е. зрительный анализатор оценивает расчлененность по различию сигналов от разных клеток сетчатки. Возможен и другой способ восприятия расчлененности, когда последовательно, в разные моменты времени, сравнивается возбуждение одних и тех же зрительных единиц. Глаз, построенный на таком принципе, смог бы различать на движущейся картине Даже больше деталей, чем это обеспечивает угловая плотность зрительных единиц. В зрительном анализаторе человека и, видимо, всех позвоночных животных этот принцип не реализован, но у насекомых, некоторых паукообразных и ракообразных скорее всего есть зрительный механизм, основанный на трансформации пространственной последовательности стимулов во временную последовательность возбуждения одних и тех же рецепторных единиц. Известный немецкий физиолог Г. Аутрум показал, что фасеточный глаз быстро летающих насекомых, для которых особенно важно различать предметы в движении, отвечает тем требованиям, которым должен удовлетворять обсуждаемый гипотетический зрительный механизм. Именно восприятие расчлененности на движущейся картине как последовательности мельканий возможно при условии малой инерционности сетчатки и ее высокой контрастной чувствительности. Как показали электрофизиологические измерения критической частоты мельканий и прямое наблюдение за поведением насекомых в оптомоторном барабане с периодическим узором полос, для хороших летунов — мух, пчелиных, стрекоз— мелькания не сливаются до тех пор, пока их частота станет равной 250—300 в секунду (см. рис. 48, в, г). Вспомним, что человек не видит смены кадров на киноэкране, следующих с частотой уже 16 в секунду. Инерционность нашего глаза намного ниже, чем у быстро летающих насекомых. О высокой контрастной чувствительности фасеточного глаза уже говорилось — она достигает 1 %, что достаточно для видения при движении.
Еще одна особенность фасеточного глаза явно возникла как приспособление для видения в полете. У ряда подвижных насекомых, таких, как мухи и пчелиные, межомматидийные углы по горизонтальному диаметру глаза крупнее, чем по вертикальному. Дело в том, что для различения деталей во время движения глазу достаточно небольшого числа омматидиев, вплоть до одного, и явно невыгодна большая угловая плотность зрительных единиц, так
164
как при этом освещенность соседних омматидиев будет изменяться (мелькать) слишком быстро для того, чтобы рецепторы успевали реагировать на такие мелькания. Поэтому в направлении смены объектов в поле зрения при движении (по горизонтали) меж-омматидийные углы у хороших летунов в два-три раза шире, чем по вертикали (рис. 54).
Рис. 54. Величина углов между оптическими осями омматидиев рабочей пчелы Apis mellifera в вертикальном (а, б, г) и горизонтальном (в. д) направлениях.
а, б, в — результаты измерения del Portillo (1936);
г, д — результаты измерения Baumgarten (1928). Измерения получены разными способами.
По оси ординат — порядковые номера омматидиев сверху вниз (4) и спереди назад (Б); по оси абсцисс — величина углов
Прямым свидетельством реализации у животных принципа зрения в движении можно считать необычное устройство глаз у некоторых паукообразных и ракообразных. Например, в глазу морского рачка Copilia позади сравнительно крупной линзы лежит всего четыре зрительные клетки, но они подвижны за счет ритмических сокращений специальных мышц. Такая сетчатка из четырех элементов непрерывно колеблется, сканируя ретинальное изображение с частотой 5—6 раз в секунду. Малое число рецепторов здесь компенсируется их подвижностью, что, очевидно, позволяет рачку различать детали и форму предметов. Сканирующая сетчатка известна также в глазах некоторых пауков и клещей. Недавно описана особая мышца в глазах мух, периодически подергивающая проксимальную часть слоя рецепторов.
Человек не видит предметы, если их изображение неподвижно относительно сетчатки. Но стабилизация ретинального изображения осуществима только в эксперименте, а в реальных условиях глаз непрерывно совершает движения: произвольные скачки — смену точек фиксации, непроизвольные скачки, дрейф, тремор и другие, препятствующие возникновению, как говорят, «пустого» поля. Не исключено, что мышца в фасеточном глазу мух служит именно тому, чтобы обеспечить восприятие, периодически двигая рецепторы относительно сетчаточного изображения, поскольку сами глаза неподвижны относительно головы.
Существует любопытная и, возможно, глубокая по своему физиологическому смыслу аналогия между движениями глаз человека при рассматривании предметов и движениями головы у хищных мух-ктырей во время визуального выслеживания добычи. Человек не может плавно следовать взором от одной точки предмета к другой — глаза непроизвольно и очень быстро, за сотые до
165
ли секунды, скачком переходят из одного положения в другое. Киносъемка охотящегося ктыря показала, что и его голова очень быстрыми скачками переходит из одного положения в другое, когда муха обращает внимание на новые объекты. Но и человек и ктырь плавным движением (первый — глаз, второй — всей головы) прослеживают движущийся предмет; скачки в этом случае никогда не наблюдаются.
Различение формы и восприятие объема
До последнего времени исследователи связывали восприятие формы насекомыми исключительно с движением ретинального изображения относительно рецепторов, когда графические (пространственные) свойства предметов могут восприниматься как последовательность мельканий или последовательность возбуждения во времени одних и тех же омматидиев. Эта гипотеза возникла на основе следующих наблюдений. Пчелиных, бабочек, мух и других актофильных насекомых расчлененные фигуры спонтанно привлекают сильнее фигур с ровным сплошным контуром или однородным (нерасчлененным) внутренним полем. Колеблющиеся на ветру цветки насекомые посещают чаще, чем неподвижные цветки. Но самое главное доказательство состоит в том, что никому не удавалось обучить пчел различать фигуры с простым контуром, в частности отличать треугольник от квадрата или круга равной площади. Однако в тех же опытах пчелы различали, например, лучистые фигуры: 8- и 16-лучевые звезды, а также отличали крест и звезду от круга. «Решают» пчелы и более сложные задачи на опознавание, явно руководствуясь статистическими признаками рисунка фигур — повторяемостью одних и тех же сочетаний, например светлого и темного,— если это черный рисунок на белом фоне. После соответствующего научения пчелы в опытах Л. И. Францевича (1968) различали стандартного размера черно-белые случайные рисунки, предлагаемые по специальной программе компьютером, если они относились к разным статистическим классам в указанном выше смысле. Поэтому многие авторы утверждали, что насекомые могут различать фигуры только по степени расчлененности их контура или внутреннего поля, т. е. путем сравнения частоты мельканий, возникающих в глазу при движении насекомого относительно объекта или самого объекта относительно глаза. В действительности дело обстоит иначе. Пчелы при длительном обучении все же способны отличать, например, треугольник от квадрата, особенно если контур фигур многократно повторен, что позволяет насекомому оценивать форму как издалека, так и вблизи, когда в поле зрения помещается только часть ее. Опознать эти фигуры по частоте и последовательности мельканий практически невозможно. Следовательно- пчелы могут воспринимать пространственные отношения предметов или элементов рисунка (видеть
166
форму), в том числе и по картине возбуждения разных рецепторов одновременно. Из этого следует, что насекомым свойственны оба рассмотренных выше механизма анализа расчлененности поля зрения.^
Насекомые различают предметы по размеру. Например, пчелы различают два круга при условии различия их площадей в 2 раза и больше; но осы и мухи делают это более тонко: различие становится возможным уже при условии отношения площадей фигур не менее чем 1 : 1,5.
Насекомые, отличающиеся быстрым полетом или хорошо прыгающие, а также большинство хищников воспринимают также объемность предметов и хорошо оценивают глубину пространства. Стереоскопичность зрения достигается тем, что поля зрения обоих глаз впереди и внизу частично перекрываются: в правом и левом глазах есть омматидии, оптические оси которых сведены или хотя бы параллельны друг другу. Такие (корреспондирующие) пары омматидиев «видят» предмет в одной и той же точке пространства. Для разных точек пространства по мере удаления от глаза Корреспондирующими окажутся разные пары омматидиев, что в принципе достаточно для улавливания изменения глубины. Корреспондирующие омматидии, например, у богомолов лежат в специальной зоне глаза причудливой формы, одностороннее ослепление которой ведет к утрате способности насекомого безошибочно схатывать добычу. В норме же богомол, как и охотящаяся личинка стрекозы, никогда не выбрасывает ловчий аппарат, когда расстояние до жертвы больше последнего. Стереоскопичность зрения важна не только охотникам за добычей, но и всем быстро летающим и быстро бегающим формам для обеспечения посадки, своевременного маневра при облете препятствий или преследования полового партнера. В случае посадки сигнал для своевременного вытягивания ног посылают особые детекторы, описанные на примере зрительной системы мух.
В заключение этого раздела обратим внимание на функциональную неоднородность фасеточного глаза, четко проявляющуюся в ряде отношений в поведении насекомого, но не всегда морфологически ясно выраженную. Так, взрослая стрекоза Libellula quadrimaculata различает цвета только с помощью вентральной половины глаза, сложенной более мелкими зеленоватыми фасетками. Дорсальная половина глаза цветослепа, но обладает высокой чувствительностью к коротковолновым лучам: ультрафиолетовым, фиолетовым и синим, что облегчает стрекозе замечать мелкую добычу на фоне неба. В глазу богомола Stogmatoptera описана особая зона с повышенной остротой зрения. Эта так называемая фо-веа, как уже говорилось выше, необходима и достаточна для точной оценки дистанции броска на добычу. У водомерок Gerris только дорсальная половина глаза с особым расположением микровилл в рабдомерах пригодна для анализа направления поляризации света и астрономической ориентации. У медоносной пчелы
167
не замечено резких морфологических различий между верхней и нижней половинами глаза, но функционально они не равноценны. Пчела различает цвет обеими половинами глаза; но для опознава-
ния объектов по форме и размеру
Рис. 55. Ориентация тела медоносной пчелы, летящей над горизонтальной поверхностью (по Мазохииу-Поршнякову, Таимовой, 1973):
А — зона глаза, которой пчела смотрит на близлежащие горизонтальные предметы; Oi—Oi — медиальная ось тела; Ог—Ог — продольная ось головы; Г—Г — линия горизонта
пригодна в основном нижняя половина, а для анализа поляризации служит исключительно верхняя половина глаза.
Во время полета над горизонтальной поверхностью пчела удерживает медиальную ось тела примерно на 10° выше линии горизонта. Благодаря этому, а также
постоянному наклону головы ротовыми частями несколько вперед она смотрит вниз вентральной третью поля зрения (рис. 55, А), как раз приспособленной для опознавания предметов по форме, размеру и цвету. Что же касается медиальной
и дорсальной третей глаза, направленных в полете в основном вверх, то они предназначены для измерения и удерживания ме-нотаксических углов ориентации по солнцу и поляризации света голубого неба и, конечно, для общего обзора.
5. ВОСПРИЯТИЕ ПОЛЯРИЗАЦИИ СВЕТА
Изучая сигнализацию у медоносных пчел, К- Фриш сделал в 1948 г. интересное открытие. Он уже знал, что возвратившаяся з улей пчела-разведчица ориентирует свой «танец» по солнцу: прямолинейная часть «танца» на горизонтальной поверхности направляется к солнцу под тем же углом и с той же стороны, под которым по отношению к солнцу нужно искать место взятка. Но оказалось, что разведчица правильно ориентирует свой «танец» и в отсутствие солнца, если только видит хотя бы небольшой участок голубого неба. Стоило облакам закрыть небо над смотровым отверстием в экспериментальной камере, и «танец» пчелы становился беспорядочным. К. Фриш заключил, что пчела видит на голубом небе какой-то компас, по которому ориентируется так же легко, как и по солнцу. Физики, к которым обратился исследователь, сказали, что этим компасом может служить поляризация света. Рассеиваемые молекулами воздуха солнечные лучи оказываются плоско (линейно) поляризованными, при этом поляриза
168
ция голубого неба под углом 90° к солнцу может достигать 70%. Электрический вектор поляризованного луча так связан с положением солнца, что оказывается перпендикулярен плоскости, образованной следующими тремя точками: солнце, созерцаемая точка неба и центр глаза. Поэтому плоскость поляризации света в каждой точке голубого неба будет параллельна обозначенной выше плоскости.
Для проверки значения поляризации света К. Фриш поместил над пчелой специальную поляроидную пленку, плоско поляризующую проходящий сквозь нее свет. Оказалось, что таким поляроидом можно управлять направлением «танца» пчелы: при повороте пленки на некоторый угол на такой же угол изменяется направление «танца». Следовательно, пчела отличает поляризованные лучи (свет голубого неба) от деполяризованных (свет, рассеиваемый облаками) и может ориентироваться как по солнцу, так и по картине поляризации света неба, свободно переходя от одного из этих ориентиров к другому.
В дальнейшем оказалось, что не только пчелы, но и все насекомые и их свободноживущие личинки, а также паукообразные и ракообразные воспринимают поляризацию света и способны ориентироваться по плоскости поляризации лучей. Сведения об этой способности членистоногих получены двумя методами: наблюдением за поведением животного при повороте плоскости поляризации падающего на него света и регистрацией электрического ответа при освещении поляризованными лучами. Жук-навозник Geot-rupes, посаженный в цилиндр с однородно окрашенными стенками, удерживает определенное направление движения, если видит над собой голубое небо или непосредственно солнце. При появлении над цилиндром облаков путь жука становится хаотичным. Если медленно поворачивать над цилиндром пленочный поляроид, то и ползущий жук поворачивается в ту же сторону. Аналогично ведут себя под поляроидом жук-кравчик Lethrus apterus, когда он возвращается в норку, и многие другие насекомые и их личинки. Водные ракообразные, клещи, мухи Drosophila предпочитают при этом даже в покое сохранять постоянную ориентацию тела к плоскости поляризации света: одни виды — параллельно этой плоскости, другие — перпендикулярно к ней.
Электрическую реакцию глаза на характер поляризации света удается заметить при регистрации как ЭРГ, так и ответа отдельной зрительной клетки. При изменении высоты ЭРГ на вспышки поляризованного света и такой же интенсивности деполяризованного света было замечено, что в первом случае она больше приблизительно на 30%. Если же один омматидий или их группу освещали вспышками света, поляризуемого последовательно в разных направлениях, амплитуда ЭРГ не изменялась. При внутриклеточном отведении ответа рецепторов мух на вспышки поляризованного света показано, что амплитуда реакции изменяется на 15—20% при повороте плоскости поляризации на 90°. Такое из
169
менение соответствует тому, которое можно получить, вдвое изменяя интенсивность света. Важно, что эффект действия поляризации света выявляется при освещении глаза лучами только коротковолновой половины спектра.
Вскоре после открытия поляризационной чувствительности глаза пчелы была высказана гипотеза о ее механизме. Этот механизм может быть основан на поляризационных свойствах рабдо-
Рис. 56.
Упрощенная схема механизма восприятия поляризации света.
а — модель К. Фриша: розетка из восьми секторов поляроидной пленки, поляризующих свет в направлении стрелок; б — многообразие направлений колебания электрического вектора в неполяризованном луче; в — направление колебаний электрического вектора в дан-г Й е ном плоскополяризованном луче; г — схема рабдома медоносной пчелы, образованного зрительными клетками № 1—8; д — картина возбуждения рабдома пчелы неполяризованным светом; е — картина возбуждения рабдома пчелы светом, поляризованным по схеме в. Степень зачернения рабдомеров на д, е пропорциональна степени поглощения
ими света
меров зрительных клеток, если их розетка (на поперечном сечении) подобна радиальному поляроиду, в котором каждый рабдо-мер поляризует свет в плоскости, например, параллельной ориентации своих микровилл. Чтобы сделать эту мысль наглядной, К. Фриш сконструировал модель рабдома отдельного омматидия. Восемь (по числу известных в то время зрительных клеток в омматидии пчелы) треугольников из поляроидной пленки были сложены вершинами внутрь так, что каждый сектор поляризовал свет параллельно своему основанию (рис. 56, а). Если смотреть через эту модель на белое облако — источник деполяризованного света, то все восемь секторов выглядят одинаковыми. При рассматривании же голубого неба на модели виден крестообразный узор из светлых и темных секторов, неодинаковый для разных участков небосвода.
В соответствии с таким представлением о механизме анализа поляризованного света оказались приведенные выше, а также многие другие результаты электрофизиологических экспериментов. Поэтому исследователи пришли к заключению, что анализатором поляризации является каждый рабдомер зрительных клеток. При освещении ретинулы омматидия пчелы неполяризованным светом каждый рабдомер, как теперь установлено, общий у двух смежных зрительных клеток, поглощает лишь часть лучей (рис. 56, д). При освещении светом, поляризованным, как показано на рис. 56, в, два диаметрально противоположных рабдомера клеток 7+8 и 3+4 поглощают максимальное количество лучей. Два других, принадлежащих клеткам 1+2 и 5 + 6, реагируют минимально, так как их микровиллы перпендикулярны плоскости колебаний падающих лучей (рис. 56, е). Различия в возбуждении со
170
седних симметричных пар клеток омматидия могут быть сигналом о поляризации для оптического центра, поскольку каждая зрительная клетка имеет свое собственное нервное волокно.
Впоследствии это представление было уточнено тем, что реакция на поляризацию была замечена не у всех клеток, а только у тех, которые были чувствительны к коротковолновым лучам. Теперь стало понятным, почему пчелы не ориентируются по плоскости поляризации длинноволновых (%>500 нм) лучей спектра, и можно себе представить, как совокупность фоторецепторов омматидия, наряду с анализом поляризации, осуществляет спектральный анализ света, необходимый для цветоразличения.
Почему же отдельная зрительная клетка обладает свойствами поляроида? Выше мы уже обращали внимание на периодическую ультраструктуру рабдомеров и ее закономерное построение у группы соседних омматидиев. В мембранах, покрывающих уложенные определенным образом трубочки-микровиллы рабдомеров, содержатся молекулы зрительного пигмента, а органические молекулы поглощают преимущественно те лучи, которые поляризованы в направлении их продольной оси (явление так называемого дихроизма). Экспериментально показано, что линейно поляризованный свет поглощается сильнее, когда электрический вектор лучей параллелен микровиллам, а не перпендикулярен им. Поэтому совокупность одинаково ориентированных микровилл с параллельно лежащими на них молекулами фотопигмента может служить искомым анализатором поляризованного света.
Развитое здесь представление о механизме анализа поляризации нельзя считать окончательным. Недавно появились высказывания о фильтрующей роли дистальных рабдомеров по отношению к проксимальным, получающим благодаря такому расположению максимально поляризованный свет. Другие исследователи обращают внимание на то, что свет распространяется по рабдомеру-волноводу в виде узоров, известных под названием «электромагнитных мод», форма и интенсивность которых зависят от характера поляризации падающего на глаз света. В зависимости от направления поляризации лучей, за счет особенностей этих мод, разные зрительные клетки омматидия могут быть возбуждены по-разному.
К поляризации света чувствительны не только фасеточные глаза, но и стеммы имаго, и латеральные личиночные глазки, у которых слой рабдомеров не всегда имеет упорядоченную ориентацию. Для таких простых глазков сигналом о поляризации света может служить появление на сетчатке некоторого яркостного узора, зависящего от направления электрического вектора поляризованных лучей. Дело в том, что не все косо падающие на роговицу поляризованные лучи неба будут проникать внутрь: лучи, плоскость колебания которых параллельна отражающей поверхности, будут преимущественно отражаться, тогда как лучи, колеблющиеся в плоскости, перпендикулярной первым, войдут внутрь с меньшими
17.1
потерями, в результате на сетчатке образуется неравномерное распределение света, связанное с поляризацией падающих лучей.
Чувствительность глаза к поляризации обеспечивает членистоногим не только широкие возможности астрономической ориентации, но и помогает опознавать некоторые земные объекты. Если посмотреть сквозь поляроидную пленку, постепенно поворачивая ее вокруг зрительной оси, то легко заметить непостоянство бликов на различных предметах: они то гаснут, то возникают вновь, причем по мере исчезновения блика на одном предмете он может появиться на другом. Это объясняется тем, что свет, отраженный даже едва блестящими поверхностями диэлектриков, в той или иной степени поляризован. Поэтому одинаковые для невооруженного человеческого глаза блики на разных предметах могут быть различными для глаза насекомых, и эта скрытая особенность бликов может служить для насекомого дополнительным признаком для распознавания объектов и их взаимного расположения. Плоско поляризуют свет водная поверхность водоемов и влажных предметов, покрытые воскоподобным налетом листья, особенно молодые, венчики многих цветков, покровы тела у многих насекомых и прочие природные объекты.
И все же способность реагировать на поляризацию света нельзя считать уникальной для глаза членистоногих — в слабой степени она свойственна и человеческому глазу. Так, при рассматривании через поляроидную пленку равномерно освещенной светлой поверхности примерно каждый третий-четвертый человек отчетливо видит на ней желтоватую сноповидную фигурку, поворачивающуюся при повороте пленки. Этот так называемый «эффект Гай-дингера» некоторые люди замечают, когда пристально смотрят на безоблачное небо под углом около 90° к солнцу, где поляризация максимальна. Механизм его, как полагают, связан с нарушением радиального расположения нервных структур в центральной ямке сетчатки — в фовеа. Однако эффект Гайдингера в наших обычных зрительных оценках не играет никакой роли, тогда как способность членистоногих визуально реагировать на поляризацию света для них жизненно важна.
6. ЦВЕТОВОЕ ЗРЕНИЕ
Способность различать цвета — одна из важнейших функций зрения, кажущаяся самоочевидной, но далеко не всеми правильно понимаемая, каждый светочувствительный аппарат, будь то фотографическая эмульсия, фотоэлемент или глаз животного, может неодинаково реагировать на свет, отличающийся по спектральному составу. Но о цветовом зрении говорят только тогда, когда разная реакция на излучения различного спектрального состава не зависит от их интенсивности. Отсутствие взаимозаменяемости видимых излучений произвольного спектрального состава — необходимый и формально достаточный критерий цветового зрения.
172
Для человека и высокоорганизованных животных, таких, как позвоночные и насекомые, в определение цветового зрения следует включить способность опознавать *окраску предметов независимо от цвета освещения. Так, пчелы, как и человек, могут отличать, например, белые предметы от желтых даже тогда, когда первые освещены желтым светом, а вторые — белым, т. е. и те и другие предметы отражают свет одного и того же спектрального состава. Это называется константным восприятием окраски, без которого операция различения отраженного света по спектральному составу была бы недостаточной для надежной ориентации в многообразии окружающих предметов. В природе все живые организмы встречаются в основном с такими объектами, которые не светятся сами, а только отражают свет, падающий извне. Поэтому константность — чрезвычайно важное свойство зрительного анализатора, делающее его совершенным прибором для распознавания несамосветящихся предметов, спектральная характеристика которых зависит от цвета освещения. Механизм константности еще не выяснен до конца, но ясно, что он основан на сопоставлении спектральной информации от разных частей глаза и к нему причастны интернейроны с рецептивным полем, «оппонентным» по реакции на коротко- и длинноволновые спектральные лучи (см. ниже).
Изучение цветового зрения животных доступно путем регистрации их разнообразных реакций, зависящих от спектрального состава света. Это могут быть реакции целого организма (его поведение) или реакции отдельных частей зрительной системы (вызванные светом электрические потенциалы сетчатки и зрительных центров, фотохимические превращения и морфологические изменения в рецепторах под действием света и т. п.). 'Следует, однако, отметить, что если для понимания механизма цветоразличения может оказаться достаточным знание принципа кодирования в рецепторах информации о цвете и картины передачи соответствующих сигналов на разных этажах зрительного анализатора, то для окончательного суждения о цветоразличительных возможностях животного необходим только поведенческий эксперимент. Известны примеры (кошка), когда потенциальные цветоразличительные возможности сетчатки глаза почти не реализуются в поведении, и животное ведет себя в эксперименте и в обычной обстановке практически цветослепым. Это важно помнить биологу, когда он, опираясь на выводы, например электрофизиологических опытов, объясняет физиологический механизм той или иной реакции животного на цветные стимулы, особенно при этологических исследованиях.
Периферический аппарат цветоразличения
Понятие о светоприемниках. В колориметрии — науке об измерении цвета — вопрос о многообразии различимых глазом цветов решается путем нахождения числа светоприемнйков в сетчатке и
173
спектральной чувствительности каждого из них в отдельности. Под светоприемниками понимают рецепторы с различной спектральной чувствительностью, передающие в оптический центр независимые и невзаимозаменяемые сигналы. Морфологически им соответствуют рецепторы сетчатки, как правило, с различным пигментом и обособленным каналом связи — аксоном. Светоприемни-ки находятся на входе зрительной системы, поэтому излучения, неразличимые — тождественные — для светоприемников, неразличимы и для зрительного центра. Светоприемники обладают свойством аддитивности, которое выражается в том, что излучения А и В, приводящие светоприемник в одно и то же состояние, обычно остаются для него неразличимыми и после прибавления к каждому любого третьего излучения С, т. е. если А = В, то и Л + С=В + С. Аддитивность светоприемников позволяет характеризовать их спектральной чувствительностью в строго физическом смысле.
Необходимым условием для спектрального анализа света сетчаткой считается присутствие в ней минимум двух типов светоприемников с различной спектральной характеристикой. При одном работающем светоприемнике любые излучения могут привести его в тождественное состояние, в результате чего декодирующее центральное устройство по величине сигналов светоприемника сможет судить одновременно только об одном параметре света — о его интенсивности, а не о цвете. При двух и большем числе одновременно работающих светоприемников с несовпадающей спектральной чувствительностью по совокупности их сигналов возможен анализ не только интенсивности, но и спектрального состава света. Например, большинство пар монохроматических излучений, равновозбуждающих светоприемник А, окажется нетождественным для светоприемника В. Поэтому в величине и соотношении ее у сигналов двух светоприемников А и В будет закодирована информация как об интенсивности, так и спектральном составе света. Три типа светоприемников обеспечат различение большего числа излучений. В центральной ямке сетчатки человека имеется, как известно, три типа светоприемников. Минимум три типа светоприемников существует и в глазу пчелиных и дневных бабочек, образ жизни которых связан с необходимостью опознавать по окраске цветущие растения.
Спектральная чувствительность светоприемника определяется в первую очередь тем зрительным пигментом, который заключен в фоторецепторе. В ряде случаев на пути лучей к фоторецептору стоят цветные светофильтры в виде, например, экранирующего пигмента, селективно пропускающего свет. Поэтому спектральная чувствительность светоприемника может зависеть также и от оптических свойств тех сред, которые фильтруют свет на пути к рецепторам.
Измерение спектральной чувствительности светоприемников. Из предыдущего изложения ясно, что изучение механизма цвето
174
вого зрения необходимо начинать с измерения числа типов свето-приемников глаза и относительной спектральной чувствительности каждого в отдельности. Существует несколько методов изучения светоприемников.
Чаще всего прибегают к наиболее простому методу равных ответов. Он заключается в подборе и измерении энергии различных монохроматических лучей, вызывающих равную по амплитуде ЭРГ глаза или равную поведенческую (обычно фототропиче-скую) реакцию насекомого. Результат эксперимента представляют обычно в виде кривой величин, обратных числу квантов (1/Q) или энергии (1/£) спектральных стимулов, вызывающих равный ответ.
Получаемая «кривая равных спектральных ответов» темноадаптированного глаза сама по себе почти не несет информации ни о числе светоприемников, ни о спектральной чувствительности каждого из них. Но если дополнительно измерить кривую равных ответов при фоновом цветном подсвете, то по ее изменению можно с большой вероятностью судить о числе зрительных пигментов в сетчатке и на основе этого косвенно предполагать число светоприемников и их спектральную характеристику. Идея заключается в том, что фоновое цветное излучение, например синее, сильнее разлагает («утомляет») прежде всего тот зрительный пигмент, который более чувствителен к нему. Поэтому кривая равных ответов, измеренная при адаптации к синему, будет отличаться от кривой темноадаптированного глаза только в том случае, если сетчатка содержит различные зрительные пигменты. Остается неясным, оДНа’кб,: находятся ли разные пигменты в разных фоторецепторах или они содержатся во всех рецепторах в виде смеси. Между тем о разных типах светоприемников можно было бы говорить только в первом случае.
Надежнее в этом отношении метод измерения спектральной эффективности отдельных зрительных клеток. Глаз возбуждают с помощью специального стимулятора обычно равноквантовыми и равными по продолжительности вспышками различных монохроматических лучей, регистрируя при этом внутриклеточным микроэлектродом амплитуду деполяризационного потенциала фоторецептора (рис. 57, Д). Непосредственно в эксперименте получают так называемую «кривую спектральной эффективности» — кривую, огибающую амплитуды ответов клетки на равные спектральные стимулы (рис. 57, 5). Светоприемники целесообразнее характеризовать относительной спектральной чувствительностью— приведенной к максимуму кривой, обратной отношению энергии света различных длин волн, одинаково возбуждающего фоторецептор. Ее легко рассчитать из кривой спектральной эффективности, если известна зависимость амплитуды потенциала фоторецепторной клетки (в мВ) от интенсивности света.
Для расчета кривой спектральной чувствительности фоторецептора достаточно иметь амплитудные характеристики данной клет
175
ки для различных монохроматических лучей, но такой эксперимент более длителен и поэтому трудновыполним, так как клетка с введенным в нее микроэлектродом «живет» недолго — несколько минут.
Рис. 57. Кривая спектральной эффективности зрительной клетки с л=540 нм кузнечика Tetligonia cantans (по Вишневской, Мазохину-Поршиякову, 1972). А — снимок с экрана осциллографа ответов клетки на . 19 равноэнергетических спектральных стимулов с Х=390—669 нм. Б — кривая спектральной эффективности (1) и рассчитанная по ней кривая спектральной чувствительности (2). Вертикальные черточки — среднее квадратичное отклонение. По оси ординат в одном и том же масштабе в относительных единицах отложены амплитуды ответов клетки (для 1) и величин, обратных энергии света, вызывающего равные по амплитуде спектральные ответы (2); по абсциссе — длина волны света, нм
. Метод микроспектрофотометрии — изящный прием определения спектрального поглощения пигментов in situ. Именно этим методом недавно впервые были определены кривые погло-: щения колбочковых зрительных пигментов человека. Он заключа-. ется в том, что с помощью микроспектрофотометра измеряют спектральное поглощение отдельного фоторецептора непосредственно в живой сетчатке. Диаметр светочувствительного элемента фоторецептора составляет всего несколько микрометров, у насекомых диаметр рабдомера равен 0,5—2,0 мкм — отсюда трудность использования метода, так как требуется сфокусировать свет строго на одном рабдомере, а не на нескольких.
Наиболее надежен в отношении изучения спектральной чувствительности сетчатки метод мелькающей колориметрии, или метод нуль-установки, посредством которого судят о числе светоприемников по числу базисных (основных) излучений, необходимых и достаточных для воспроизведения в смеси друг с другом всех спектральных цветов. Он основан на подборе визуальных цветовых равенств (тождеств) и измерении энергии отдельных их компонентов: базисных и монохроматических излучений. В разработке его большую роль сыграл выдающийся отечественный специалист в области физиологической оптики проф. Н. Д. Нюберг. Эксперимент ведется на колориметре замещения, позволяющем быстро заменить монохроматическое излучение
176
на смесь базисных цветов и наоборот. Критерием тождественности для насекомого каждого монохроматического излучения подобранной смеси базисных цветов принимают отсутствие ЭРГ в момент их замены друг на друга. Сама возможность такой подмены, остающейся незамеченной глазом, служит доказательством объективности и надежности метода.
Величину отношения энергии (в процентах от максимума) каждого базисного цвета (£\_ ) в их смесях друг с другом к энер-
гии каждого визуально тождественного этим смесям монохроматического цвета (£%) изображают в виде так называемых кривых сложения базисных цветов. Кривые сложения, т. е. функции
Ек
- характеризуют с точностью до их линейной комбина-
л~п
ции спектральную чувствительность каждого светоприемника. Если видимый глазом спектр имеет одномерные — монохроматические — области, где работает по одному светоприемнику, то кривые сложения базисных цветов легко пересчитать непосредственно в
кривые спектральной чувствительности светоприемников.
Колориметрический метод рассчитан не на определение зрительных пигментов сетчатки, а только на то, чтобы непосредственно найти число фактически работающих в глазу светоприемников и определить их спектральную чувствительность. Иными словами, он более других методов адекватен задачам описания светоприемников сетчатки и измерения многообразия цветов, различимых глазом. Именно колориметрические измерения послужили в свое время математику Грассману экспериментальной основой для выведения трех известных законов оптического смешения цветов для
человеческого глаза.
Зрительные пигменты. Зрительный акт начинается с момента поглощения фотонов зрительным пигментом фоторецептора. Молекула фотопигмента состоит из двух непременных частей: прозрачной белковой — о пси на и красящего хромофора — ретин а л я, представляющего собой альдегид витамина Ai или Аг. Под действием поглощенного кванта света молекула пигмента фотоизоме-ризуется: переход из цис- в трансконфигурацию. В этом и состоит единственная роль поглощаемого фотона, так как все дальнейшие реакции в фоторецепторе уже не зависят от наличия света. Чтобы вызвать посредством фотоизомеризации пигмента реакцию зрительной клетки, достаточно хотя.бы одного кванта света.
Зрительных пигментов насекомых известно пока немного, что связано с трудностями их выделения и идентификации. Например, чтобы извлечь зрительный пигмент у медоносной пчелы, американскому исследователю Т. Голдсмису (1958) пришлось измельчить 20 тыс. голов этого насекомого. Экстрагируют фотопигменты насекомых, обычно дигитаниновым .раствором в нейтральном фосфатном буфере. Чаще всего у насекомых встречаются фртопигменты типа родопсина (палочкового пигмента позвоночных) с Хтах меж-
177
ду 440—510 нм и вторым пиком поглощения в ультрафиолетовых лучах. Например, для мух характерен родопсин с Хтах = 495 нм. Фотохимический цикл его превращения может быть описан как /495«гМ580: нативный пигмент /495 под действием света переходит в термостабильный фотопродукт — метародопсин АТ580, который регенерирует в исходный пигмент не только в темноте, но и под действием света, в особенности желтого. В глазу, адаптированном к обычному «белому» свету, между /495 и количеством
Рис. 58. Кривые спектральной чувствительности светоприемников комнатной мухи Musca domestica (4) и стрекозы Libellula quadrimaculata (Б) (по Мазохину-Поршнякову, 1965).
а, б — коротко- и длинноволновые светоприемники мухи; в — единственный светоприемник верхней половины глаза стрекозы; г, д — коротко- и длинноволновые светоприемники нижней половины глаза стрекозы.
По оси ординат — относительная чувствительность к равиоэиергетическому спектру в процентах от максимума; по оси абсцисс — длина волны света, нм
Л1580 устанавливается фотохимическое равновесие. Видимо, особый зрительный пигмент, чувствительный в основном к ультрафиолету, свойствен только насекомым. Такой фотопигмент с \пах =345 нм извлечен из головы булавоуски Ascalaphus maca-ronius и некоторых других насекомых.
Влияние фильтрующего аппарата на спектральную чувствительность глаза. Аппарат светоизоляции фоторецепторов подробнее изучен у мух, у которых он особенно сильно влияет на спектральную чувствительность и другие параметры глаза. В норме в глазах Calliphdra и других мух есть два типа экранирующих пигментов; красные оммохромы и желтоватые птерины. Первые задерживают все лучи, кроме красных, вторые задерживают в основном ультрафиолетовые лучи. Поэтому все спектральные лучи могут проникать к рабдому отдельного омматидия только через его собственную фасетку, а красные — и через соседние фасетки, так как экранирующий пигмент для них прозрачен. В результате чувствительность глаза мух к красным лучам повышена: кривая равных ответов имеет характерный пик у X—620 нм, а сетчатка оказалась наделенной своеобразным дополнительным светоприемни-ком. У мух нет специального красночувствительного зрительного пигмента, но есть красночувствительный (%шах =620 нм) светоприемник «фильтровой» природы. Он представляет собой, по-видимому, совокупность из шести рабдомеров каждого омматидия, которые, по сравнению со сдвоенным центральным (седьмым и вось
178
мым) рабдомером, находятся в преимущественных условиях освещения красными лучами за счет красных рефлексов от экранирующего пигмента и проникновения красных лучей со стороны смежных омматидиев. Вместе с основным зеленочувствительным (Хтах ~500 нм) светоприемником этот дополнительный «фильтровой» светоприемник, в основе которого лежит тот же самый или близкий родопсиноподобный зрительный пигмент, позволяет мухам различать цвета (рис. 58, .4).
Системы цветового зрения. Видимый насекомыми спектр шире, чем у человека: он включает ультрафиолетовые лучи с длиной волны вплоть до 240 нм, хотя в естестенной обстановке лучи с Z<300 нм отсутствуют (см. ниже, рис. 60, Д). О наборе свето-приемников у разных насекомых дает представление табл. 11.
Таблица 11
Примеры светоприемников у взрослых насекомых
Насекомое ^max светопРием‘ иика, нм Способ определения
Пчела Apis mellifera (рабочая) 340; 430; 460; 530 внутриклеточно, равноквантовая стимуляция
Шмель Bombus distinguendus (самка) 365; 440—450; 540—550 по ЭРГ, колориметрически
Оса Paravespula germanica (самка) 374; 455; 530 и подъем у 389 по ЭРГ, методом равных ответов
Муравей Formica cunicularia (рабочий) 470; 520 по ЭРГ, колориметрически
Бабочка Heliconius erato 440; 490; 600 по ЭРГ, равноквантовая стимуляция
Бабочка Papilio troilus Клоп Notonecta glauca: 483; 587; 645 по ЭРГ, равнокваитовая стимуляция
иижияя часть глаза 350; 420; 567 внутриклеточно, равноквантовая стимуляция
Жук Cetonia autata 365; 500 по ЭРГ, колориметрически
Кузнечик Troglophilus neglectus 500 внутриклеточно, равнокваитовая стимуляция
Кузнечик Tettigonia cantans 400; 540 и подъем у 390 внутриклеточно, равиоквантовая стимуляция
Таракан Periplaneta americana 365; 507 внутриклеточно, равноквантовая стимуляция
Термит Anacanthotermes ahnge-rianus Стрекоза Libellula pulchella-. 500 по ЭРГ, колориметрически
верхняя часть глаза нижняя » » 420 380; 518; 550 по ЭРГ, методом равных ответов
Цветослепые формы, по крайней мере среди взрослых насекомых, встречаются крайне редко. К ним относятся скрытоживущие виды: пещерный кузнечик Troglophilus neglectus, термит Anacantho
179
termes ahngerianus, мучной хрущак Tenebrio molitor, которые имеют по одному-единственному светоприемнику.
В основном приходится иметь дело с ди- и трихроматами, периферический цветоразличающий аппарат которых образован двумя или тремя типами светоприемников. Иногда, как у медоносной пчелы, в глазу бывает четыре спектральных типа рецепторов, но поведенческие опыты показывают, что цветовое зрение пчелы трихроматическое и, значит, сигналы двух близких рецепторов с Хтах, равной 430 и 460 нм, взаимозависимы. Трихроматами среди насекомых бесспорно являются пчелиные и, возможно, дневные бабочки.
Для всех насекомых характерна обычно высокая чувствительность к ультрафиолету, определяющаяся специальным «ультрафиолетовым» светоприемником, как у булавоуски Ascalaphus та-caronius или жука-бронзовки, а также частично (иногда целиком, как у мух) вторым пиком поглощения (в ультрафиолетовых лучах) у родопсиноподобного зрительного пигмента. Поэтому видимый насекомыми спектр простирается от ультрафиолетовых лучей (%=300 нм) до красных включительно (Х=650 нм). Судя по колориметрическим измерениям в электрофизиологическом эксперименте и результатам поведенческих опытов, близкие монохроматические цвета насекомые дифференцируют наиболее четко в средней части спектра (зеленые и сине-фиолетовые цвета), где резко изменяется спектральная чувствительность у разных светоприемников. Цвета, лежащие на краях спектра, а именно: оранжевокрасные (с одной стороны) и далекие ультрафиолетовые (с другой стороны),— насекомые различают слабо.
У трихроматов (пчелиных) для цветоразличения, видимо, особенно велика роль рецепторов ультрафиолета. Например, медоносные пчелы легко отличают смесь из 99% желтых лучей с Х=588 нм +1 % УФ с %=360 нм от 100% желтых лучей с %—588 нм.
У некоторых насекомых, в особенности у хищников, охотящихся в воздухе, замечено различие в спектральной чувствительности верхней и нижней частей глаза. Так, у стрекоз Libellula (рис. 58, Б), булавоуски Ascalaphus macaronius верхняя половина глаза цветослепа, но обладает высокой чувствительностью к лучам, рассеиваемым голубым небом; нижняя половина глаза способна к цветоразличению.
Кодирование и передача спектральной информации в мозг
Судя по физиологическим и морфологическим признакам, отдельный омматидий представляет собой единицу, достаточную для спектрального, а также поляризационного анализа света. Различными способами показано, что слагающие его зрительные клет
180
ки неодинаковы по спектральной чувствительности. Так, у медоносной пчелы в каждом омматидии имеется три типа светоприемников; у американского таракана Periplaneta americana из восьми зрительных клеток омматидия пять являются рецепторами с Хтах =507 нм и три клетки — рецепторами с лшах = 365 нм.
О способе передачи с сетчатки сигналов о цвете и их последующей обработке пока известно немного. Считается, что элект-
Рис. 59. Модель передачи информации о цвете в зрительной системе бабочки Epargireus clarus (по Swihart, 1969):
1 — «сииий» рецептор; 2 — «зеленый» рецептор; 3 — цветное ганглиозное волокно; 4 — вставочный интернейрон с «задержкой»
внутри омматидия
рическая связь между разными рецепторами практически отсутствует. В таком случае можно думать, что постретинальные сигналы отдельного омматидия поступают в оптические патроны ламины по многим, обычно по шести независимым, каналам, а в медуллю поступает информация, по крайней мере, по двум разным каналам: аксонам базальных клеток и интернейронам оптического патрона. К этому нужно добавить, что в действительности в каждом омматидии есть обычно две базальные клетки, а в оптическом патроне — два-три перпендикулярных интернейрона плюс несколько центробежных волокон.
В оптических ганглиях и в особенности в грибовидных телах переднего мозга удается электрофизиологически зарегистрировать волокна, отличающиеся специфической реакцией на свет различных длин волн, что говорит об их причастности к обработке спектральной информации (см. рис. 5). У бабочки Epargyreus clarus замечен особенно интересный тип цветного «оппонентного» волокна. Это волокно интенсивно разряжается импульсами при освещении глаза синим светом, очень коротко отвечает на зеленые лучи, но не возбуждается «белыми» лучами. Здесь синий свет тормозит ответ волокна на зеленые лучи и наоборот; при этом ответ на синий свет возникает без задержки, а на зеленый — с задержкой. В сетчатке этой бабочки только два спектральных типа рецепторов: с Хтах =440 нм (синий светоприемник) и Хтах =540 нм (зеленый светоприемник), — поэтому легко представить себе модель связанного с ними цветного оппонентного волокна (рис. 59).
У другой бабочки — Papilio troilus — среди разнообразных цветных интернейронов мозга наиболее интересны волокна с двумя пиками чувствительности — в синем и оранжевом участках спектра. Эта бабочка спонтанно предпочитает синие и оранжевые цветки растений, поэтому вполне вероятно, что именно названные интернейроны управляют ее пищевым поведением.
181
Поведенческие реакции на цветные стимулы
В начале раздела о цветовом зрении мы уже говорили о том, что знания периферических и даже центральных механизмов зрения еще недостаточно для правильного предвидения визуальных реакций целого организма. Дело в том, что с помощью зрительного и любого другого анализатора животное не только различает стимулы или объекты, но и опознает их как таковые — выбирает определенные объекты среди многих. Целесообразные поведенческие реакции всегда развиваются с участием врожденной или приобретенной памяти. Именно память позволяет насекомому опознать требуемый объект как таковой, например, как съедобный, но о содержании памяти мы можем пока судить исключительно по поведению животного. В этом заключается незаменимая роль поведенческого эксперимента.
Опыты с регистрацией поведения в ответ на те или иные стимулы— технически более простой способ изучения цветового зрения, и они были поставлены на насекомых раньше электрофизиологических измерений чувствительности фасеточного глаза. Мы остановимся на двух видах поведенческих реакций на цвет: спонтанном (врожденном) предпочтении и выборе на основе индивидуального научения.
Замечено, что многие антофильные насекомые: бабочки, мухи, различные пчелы — избирательно посещают цветущие растения, предпочитая те из них, которые имеют определенную окраску. Эксперименты с цветными макетами подтвердили этот вывод. Так, если поместить в обширный садок с недавно вышедшими из куколок голодными парусниками Papilio troilus макеты цветков из пурпурной, синей, зеленой, желтой, оранжевой, красной бумаги, а также из серой бумаги разной светлоты, то бабочки будут садиться преимущественно на синие и пурпурные, отчасти на оранжевые макеты. Тот факт, что преимущественный выбор синих, пурпурных и оранжевых макетов был сделан в присутствии серых разной светлоты, позволяет заключить об опознавании предлагаемых макетов именно по цвету, а не по светлоте. В этом опыте бабочки демонстрируют разную реакцию на объекты, окрашенные по-разному, независимо от яркости (светлоты) последних, а это отвечает сделанному нами выше определению цветового зрения.
Мухи Eristalis, жуки Meligaethes, а также тли в период питания предпочитают желтые окраски. Стремление к желтому положено в основу привлекающего действия липких или наполненных водой ловушек для тлей и других вредных насекомых. В разные периоды жизни насекомого его цветовое представление может измениться, в частности тлей во время расселительных полетов привлекают преимущественно синие поверхности — цвет безоблачного неба как признак открытого пространства.
182
Врожденное стремление насекомого к излучениям определенного спектрального состава непосредственно свидетельствует о его способности к различению цвета. Этой особенностью поведения можно пользоваться как способом быстрой проверки способности насекомого к цветоразличению.
Но более гибок и информативен другой способ доказательства и подробного изучения цветового зрения, основанный на выработке условного рефлекса на определенный цвет. Чаще всего вырабатывают пищевой рефлекс на различение цветных образов. К. Фриш еще в 1914 г. первым поставил такой эксперимент на пчелах. На специальном столике он разложил в виде шахматного поля 16 бумажных квадратиков, среди которых 15 были изготовлены из «ахроматической» бумаги разной светлоты: от белой до черной, а 16-й квадратик был вырезан из голубой бумаги. На голубом образце была сладкая приманка, а на остальных в аналогичных сосудах — вода. После нескольких посещений приманки пчелы находили голубой образец в любом месте шахматного поля и садились на него даже тогда, когда приманку снимали с него и все образцы покрывали новым чистым стеклом. В условиях такого эксперимента пчелы могли опознать голубой образец только по цвету, а не по его светлоте. Если бы они ориентировались по светлоте голубого квадратика, то среди ахроматических образцов обязательно нашелся бы равносветлый голубому, и пчелы садились бы на него так же часто, как на голубой. В действительности же обученные пчелы практически не ошибались в выборе голубого, что исчерпывающим образом доказывает их способность к различению цвета.
Впоследствии также путем обучения Г. А. Мазохиным-Поршня-ковым была показана способность пчел дифференцировать близкие окраски, например желтую и оранжевую. В этом опыте шахматное поле было сложено из оранжевых квадратиков разной светлоты, среди которых присутствовал один желтый квадратик с приманкой. После обучения в контрольном эксперименте желтый образец был заменен другим, желтым, но более темным, и, несмотря на это, пчелы садились только на желтый квадратик.
В широкоизвестных опытах с обучением пчел К. Даумер доказал, что для этих насекомых солнечный белый свет неотличим от смеси трех спектральных цветов: 55% желтого с А,=588 нм плюс 30% сине-фиолетового с Х=440 нм и плюс 15% ультрафиолетового с А.=360 нм. Тем самым оказалось, что ультрафиолетовые лучи оказываются для глаза пчел дополнительным цветом к смеси желтых и сине-фиолетовых лучей, а смесь желтых и ультрафиолетовых лучей дает «пчелиный пурпурный» цвет, отсутствующий в спектре'. Как известно, пурпурный цвет, воспринимаемый челове-
1 Дополнительными цветами называют такую пару цветов, которые при смешении друг с другом дают белый (серый) цвет, т. е. дополняют друг друга до белого. Здесь имеется в виду оптическое смешение излучений, а не механическое смешение красок на палитре.
183
ком и тоже отсутствующий в спектре, представляет собой смесь синих лучей с красными и считается дополнительным к зеленому цвету (рис, 60, Б). Результаты этих выдающихся опытов наряду с последующими электрофизиологическими экспериментами послу-
жили основанием для утверждения
УФ | »T f И 3 РI Т~~ збо 150 Ш 45о гоо iso sx iwfeem
Рис. 60. Границы видимого спектра (Д) для человека (/), медоносной пчелы Apis mellifera (2), мухи Drosophilla melanogaster (3) и цветовой круг (Б) для медоносной пчелы (4) и человека (5) (рис. 4,5 — по Daumer, 1956).
Заштрихованы (Б) области основных цветов, которые при оптическом смещении друг с другом дают все остальные цвета. Между ними лежат промежуточные цвета, возникающие при оптическом смешении двух соседних основных цветов. Цвета противоположных секторов являются дополнительными. Числа— длина волны света, нм. Буквы в кавычках — условные названия пчелиных цветов.
ж — желтый, з — зеленый, к — красный, о — оранжевый, п — пурпурный, с — синий, с-з — сине-зеленый, уф — ультрафиолетовый, ф — фиолетовый, ф-с—фиолетово-синий
о трихромазии зрения пчел..
Пчелы очень быстро и прочно запоминают окраску пищевых объектов. Достаточно двух-трех подкреплений выбора, чтобы пчела на всю жизнь (летом — две-три недели) запомнила цвет кормушки, в которой она нашла сладкую приманку. Показано, что пчелы способны запоминать комбинацию из двух-трех окрасок в качестве отличительного признака пестро раскрашенных предметов. Например, они легко научаются безошибочно выбирать тестовую карту с приманкой, составленную из оранжевых и голубых квадратиков, среди четырех других карт без приманки: оранжевой с желтым, голубой с желтым, желтой с зеленым и зеленой с оранжевым. Среди такого набора карт подкрепляемую комбинацию красок можно запомнить только по двум признакам: «есть и оранжевый и голубой» или «нет ни желтого, ни зеленого». Несомненно, что пест-
рая раскраска венчиков — двухцветная у нивяника (Leucanthemum vulgare) или трехцветная у пикульника (Galeopsis speciosa) — помогает пчелам опознать эти растения среди красочного разнообразия других. При скучен-
ном расположении ульев пчеловоды иногда красят их также в два цвета, чтобы облегчить пчелам поиск своей семьи.
Другие насекомые, например осы Vespidae, не имеющие такой тесной связи с цветущими растениями, как пчелиные, хотя и легко научаются отличать одну окраску от другой, но совершенно не способны запоминать парные и тем более тройственные сочета
ния цветов.
184
Удивительную сложность визуального поведения пчел, связанного с распознаванием окрасок, может иллюстрировать еще один пример. В ходе тренировки по специальной схеме у пчелы удается выработать условный рефлекс на обобщение типа окраски по признаку «двухцветность», а именно: пчела научается выбирать всегда двухцветную тестовую карту независимо от конкретных цветов, составляющих ее. Например, обученная пчела выбирает только двухцветную карту среди ранее незнакомого ей набора из трех карт: 1) двухцветная —голубой-(-желтый цвета, 2) одноцветная голубая и 3) одноцветная желтая. В другом наборе: 1) оранже-вый-|-синий, 2) сплошь синяя и 3) сплошь оранжевая карты — она избирает также двухцветную. Такой сложный выбор основан на обобщенном представлении о двухцветности, на противопоставлении всех двухцветных образцов всем одноцветным образцам.
В заключение этого раздела, в котором особенно часто употреблялись названия воспринимаемых человеком цветов, следует предостеречь читателя о незаконности переноса наших ощущений на восприятие животных. Ощущение цвета, запаха и прочего стимула всегда субъективно, т. е. принципиально невозможно ощутить ощущения другого субъекта, чтобы убедиться в их тождественности. Но можно (и именно так поступают исследователи) проверить сходство реакций у разных индивидов на физически тождественные стимулы, и в этом смысле можно говорить о равной способности различать или отождествлять разные раздражители. Поэтому, когда говорилось, что пчела отличает «желтый цвет от зеленого», имелось в виду, что те излучения, которые называем желтыми мы, и пчелы могут отличать от излучений, вызывающих у нас ощущение зеленого цвета. Но как выглядят для пчел, как ощущаются.ими эти излучения—мы не знаем.
Окраски растений и покрова тела насекомых, различимые фасеточным глазом
Разнообразие окрасок несамосветящихся объектов — растений, большинства животных, минералов и т. п„ — которое мы привыкли видеть в естественной обстановке, порождено тем, что, во-первых, разные предметы по-разному отражают спектральные лучи, и, во-вторых, солнечный свет есть смесь монохроматических лучей широкого непрерывного диапазона спектра. Так, оранжевые окраски отличаются от желтых тем, что первые сильнее поглощают синефиолетовые и зеленые лучи. Если же смотреть на эти окраски при искусственном освещении натриевой лампой, то большинство образцов оранжевой окраски мы не отличим от желтых. Свет натриевой лампы монохроматический, желтый, а желтые и оранжевые окраски отражают эти лучи практически одинаково сильно, поэтому под натриевой лампой названные образцы визуально неразличимы. Из этого примера ясно, как зависит возможность разли
185
чения окрасок от их отражательной характеристики и спектрального состава освещения.
Экспериментально доказано, что зрительный анализатор человека и, видимо, всех цветозрячих животных приспособлен для опознавания предметов главным образом по их спектральной отражательной характеристике, а не спектральному составу тех лучей, которые отражают данные предметы в данный момент времени. Один и тот же предмет в зависимости от цвета освещения может отражать свет разного спектрального состава, и задача зрительного анализатора заключается в том, чтобы выделить ту инварианту, которая характеризует предмет. Последнее достигается благодаря автоматическому (у человека — подсознательному) введению «поправки на освещение» (Гельмгольц) к цвету рассматриваемого предмета за счет сравнения отражаемого им спектра со спектром других предметов в поле зрения. Мы уже говорили, что это свойство зрения называют константностью восприятия окраски. Константность цветовосприятия позволяет опознать предмет в разных условиях освещения, когда он отражает свет разного спектрального состава. Например, пчела зрительно замечает цветок одуванчика не только на солнце, но и в тени, где он объективно зеленый, поскольку в спектре голубого неба, создающего освещение в тени, очень мало желтых и тем более оранжевых лучей. Точно так же белый цветок в тени — объективно зеленовато-голубой за счет освещения голубым небом и зелеными рефлексами (отраженными лучами) от листьев растений, но это обстоятельство не затрудняет поиск насекомым, привыкшим кормиться на белых цветах.
Для человека многообразие окрасок природных объектов порождается избирательным отражением ими света в диапазоне спектра от фиолетовых лучей до красных включительно. Но насекомые чувствительны к ультрафиолетовым лучам, поэтому характер отражения этих лучей позволяет им распознавать окраску предметов в той же мере, как на наши цветовые оценки влияют сине-фиолетовые лучи. Прежде всего для глаза насекомых не идентичны по цвету все те окраски, которые мы называем белыми. Например, давно замечено, что пчелы легко отличают кажущиеся нам одинаково белыми две поверхности, одна из которых выкрашена цинковыми белилами, а другая—свинцовыми. Свинцовые белила гораздо сильнее отражают ультрафиолетовые лучи; они «действительно белые» для насекомых в отличие от цинковых белил, которые сильно поглощают ультрафиолетовые лучи и поэтому имеют для насекомых цвет, дополнительный к ультрафиолетовому. В частности, для пчел дополнительным к ультрафиолетовому является сине-зеленый цвет.
Измерение коэффициента отражения спектральных лучей цветками растений показало, что белые венчики, как правило, слабо отражают ультрафиолетовые лучи и, следовательно, выглядят для насекомых окрашенными в дополнительный к ультрафиолетовому цвет. Желтые, оранжевые и красные цветки отражают ультра
186
фиолетовые лучи иногда очень сильно. Из них цветок красного мака Papaver rhoeas выглядит для насекомых, не чувствительных к красным лучам, например для пчел, чисто ультрафиолетовым, поскольку венчик мака практически не отражает иных лучей, кроме красных и ультрафиолетовых. Неожиданной оказалась раскраска для глаза насекомых некоторых цветков, кажущихся человеку
Рис. 61. Цветки и листья лапчатки Potentilla anserina (а) и лютика Ranunculus aces (б) (по Мазохину-Поршнякову, 1965).
Снимок А сделан в ультрафиолетовых лучах (внизу — шкала стандартов с коэффициентами отражения: 22, 11, 6 и 3%). Снимок Б сделан в видимом человеком свете
однотонножелтыми. Всем знакомы очень похожие друг на друга желтые цветки гусиной лапки Potentilla anserina и лютика едкого Ranunculus асег, но на снимке в ультрафиолетовых лучах они выглядят резко различными (рис. 61). Цветок гусиной лапки неравномерно отражает ультрафиолет — периферия венчика отражает сильнее (около 20%), чем его внутренняя часть вместе с генеративными элементами (около 2%) 2. Поэтому цветок гусиной лапки в отличие от цветка лютика выглядит насекомым двухцветным за счет «скрытого» (от взора человека) рисунка. Скрытые рисунки помогают насекомым не только различать разные цветки, но и быстрее находить нектар. Узор, за счет более сильного поглощения ультрафиолетовых лучей внутренней частью цветка, служит
2 Для оценки интенсивности отражения обычно фотографируют вместе с растением шкалу с известными коэффициентами отражения. Тогда, сравнивая на негативе или позитиве плотность почернения элементов растения с элементами шкалы, можно судить о степени отражения света растением.
187
пчелам визуальным указателем нектара и пыльцы. Как только пчела подползает к указателю нектара, даже на искусственно раскрашенной модели цветка, она опускает голову и выбрасывает хоботок, готовясь к приему пищи. Указатели нектара могут иметь различную окраску, не обязательно ультрафиолетовую.
Скрытые рисунки несут на крыльях некоторые дневные бабочки, у которых особенности отражения телом ультрафиолетовых
Рис. 62. Фотоснимки самца (1) и самки (2) бабочки Gonepteryx rhamni, сделанные в ультрафиолетовых лучах с Х=Э65 нм (по Мазо-хину-Поршнякову, 1965)
лучей следует считать проявлением скрытого для глаза человека полового диморфизма. Так, на снимке в ультрафиолетовых лучах самец бабочки-лимонницы Gonepteryx rhamni имеет скрытый рисунок за счет более сильного отражения этих лучей основной частью поля переднего крыла (рис. 62). Самка лимонницы слабо и равномерно по всему полю переднего и заднего крыла отражает ультрафиолетовые лучи. Таким образом, самец лимонницы выглядит для самки двухцветным, а не однотонножелтым, каким мы воспринимаем его. Еще более интересен факт существования резкого полового диморфизма в окраске у таких видов, у которых человеческий глаз его практически не замечает. Например, белые и издалека (в полете) неразличимые нами самец и самка капустной белянки Pieris brassicae или боярышницы Aporia crataegi выглядят для насекомых по-разному за счет неодинакового отражения ультрафиолета. Самцы почти не отражают эти лучи, поэтому имеют для глаза насекомых цвет, дополнительный к ультрафиолетовому, тогда как самки за счет более сильного отражения лучей по всему спектру выглядят для насекомых иначе. Различие в отражении ультрафиолетовых лучей разными участками крыла, па наш глаз одинаково окрашенными, зависит в одних случаях от содержания особых пигментов, например птерина, в других — от оптических свойств и ориентации самих чешуек. Аналогичные различия у разных участков лепестка или венчика в целом, как полагают, только пигментной природы.
188
7. УСТРОЙСТВО И РАБОТА
ПРОСТЫХ ГЛАЗКОВ
Дорсальные глазки и их назначение
Дорсальные, или теменные, глазки свойственны взрослым насекомым, а также личинкам — нимфам — видов с неполным превращением. В типичном случае имеются три глазка, которые расположены в виде треугольника на лобно-теменной части головы: ближе к ротовому аппарату лежит непарный медиальный глазок, а выше него, по бокам, находятся два латеральных глазка. Медиальный глазок, парный по своему происхождению, нередко крупнее остальных, но он может отсутст'вовать. Реже встречаются случаи, когда редуцированы только боковые глазки.
Морфологи утверждают, что у ряда представителей разных семейств и разных отрядов глазки вовсе отсутствуют. Однако совсем недавно у дневных бабочек (Rhopalocera), бражников, медведиц и представителей других семейств чешуекрылых, ранее считавшихся лишенными глазков, на темени вблизи антенн обнаружено по паре небольших (26 мкм) глазков. У бражников в глубине головы под каждым внешним глазком лежит связанный с ним внутренний глазок. Строение и электрическая реакция на свет этих внутренних глазков характерны для обычных глазков других насекомых.
Типичный внешний глазок имеет все три непременных для настоящего органа зрения морфофункциональных элемента: светопреломляющий, светочувствительный и светоизолирующий. Светопреломляющей линзой чаще всего служит прозрачное двояковыпуклое утолщение кутикулярного покрова (рис. 63, A, cornea). Под ним находится тонкий слой прозрачных корнеатенных клеток, образовавших линзу. К слою корнеагенных клеток обычно непосредственно примыкает вогнутая по форме линзы сетчатка, сложенная из отдельных ретинул, в которые сгруппированы по две-три удлиненные зрительные клетки (рис. 63, А, Б). Число ретинул варьирует от нескольких десятков до нескольких сотен. Внутри ретинулы, вдоль ее оси, лежит рабдом (рис. 63, А, Б, рб), сложенный из рабдомеров отдельных зрительных клеток. Зрительная клетка имеет богатый эндоплазматический ретикулюм, многочисленные митохондрии и обычно гранулы темного экранирующего пигмента. Проксимальный конец клетки переходит в аксон (Б, ак), уходящий в оцеллярный нерв. Между ретинулами иногда вклиниваются узкие ретинальные пигментные клетки (А, рп) либо клетки трахеального тапетума (Б, до), хорошо отражающего свет. Светоизоляция сетчатки достигается в одних случаях тем, что ее кольцом охватывают столбчатые гиподермальные клетки, заполненные темным пигментом (А, гк). В других случаях, когда отсутствуют ретинальные пигментные клетки, сетчатка целиком заклю
189
чена в непрозрачный бокал, сложенный пигментными клетками (Б, гк).
Нередко встречаются отступления от описанного выше плана строения глазка. Например, у таракана Periplaneta ретинулы расположены хаотично, многослойной массой, так что их рабдомеры направлены в разные стороны. У первичнобескрылых насекомых,
Рис. 63. Строение дорсальных глазков насекомых (продольный разрез, схематично): А — схема типичного глазка (по Weber, 1949); Б — устройство бокового дорсального глазка стрекоз (роговичная линза удалена) (по Ruck, Edwards, 1964).
ак — аксоны зрительных клеток в области их синаптического контакта с ин-
тернейронами; гк — эпителиальные (ги-
подермальные) пигментные клетки; <3 —
дендриты интернейронов; до — дистальные окончания клеток тапетума; зк —
зрительные клетки, сгруппированные в ретинулы; кк — корнеагенные клетки; рб — рабдом; нк — иепигментированные клетки оцеллярной ножки, часть которых футляром одевает нервные волокна; гл— расширенная часть глазка;
рп — ретинальные пигментные клетки
в частности у Machilis, роговичная линза едва намечается, а ретинулы и корнеагенные клетки лежат вперемешку друг с другом, не образуя обособленных слоев.
Оптический ганглий каждого глазка лежит вблизи его сетчатки в утолщенной части так называемой оцеллярной ножки, представляющей собой заключенную в футляр из непигментированных клеток (Б, нк) совокупность нервных волокон, уходящих в межцеребральную часть переднего мозга. В этом ганглии аксоны зрительных клеток вступают в многочисленные синаптические контакты с дендритами нейронов второго порядка (5, <Э), тела которых
190
лежат, видимо, в межцеребральной части мозга. Существенно, что число этих постсинаптических волокон значительно меньше числа пресинаптических волокон и что часть их отличается большой толщиной. В ножке медиального глазка саранчи Locusta migratoria содержится только шесть постсинаптических волокон: два более толстых, диаметром около 8 мкм, и четыре меньшей толщины — около 4 мкм. На обработанных хлоридом кобальта препаратах медиального глазка саранчи Schistocerca vaga обнаружено 17 больших билатерально симметричных глазковых интернейронов. В каждой ножке имеется по семь больших интернейронов, из которых четыре связаны сразу с двумя глазками. Один из интернейронов медиального глазка имеет в мозгу билатеральную связь, остальные ограничены каждый своей частью мозга. Обнаружена также связь ганглиозных интернейронов с оптическими ганглиями фасеточных глаз, в первую очередь с комиссурой между левым и правым оптическими ганглиями.
Судя по электрофизиологическим данным, сигналы с глазков влияют на активность нервных клеток не только переднего, но и среднего и заднего мозга головы. При освещении глазков спонтанная активность одних нервных клеток усиливается, других — тормозится. Замечена также импульсация в шейных коннективах нервной цепочки в ответ на возбуждение глазков: включение света или движение в их поле зрения предметов, контрастирующих с фоном.
Таким образом, дорсальные глазки связаны с мозгом чрезвычайно малым числом анатомически обособленных нервных каналов, и трудно ожидать, чтобы они передавали много информации. Но влияние сигналов с глазков прослеживается на многих мозговых центрах и даже на «исполнительной» части центральной нервной системы за пределами головы.
Электрическая активность глазка зависит от реакции его зрительных клеток и интернейронов оцеллярной ножки. Зрительные клетки реагируют на увеличение интенсивности света нарастающим деполяризационным потенциалом до 40 мВ. Крупные интернейроны ножки отвечают на свет гиперполяризационным потенциалом, максимальная амплитуда которого достигает 30 мВ. Для того чтобы вызвать ответ такого интернейрона, достаточно одной десятой интенсивности света, вызывающего реакцию отдельной зрительной клетки. Благодаря множеству контактов с разными рецепторами сетчатки динамический диапазон ответов интернейронов глазка, по сравнению с его зрительными клетками, сдвинут на более низкие интенсивности света.
Экстраклеточно легко отвести от глазка двухфазную ЭРГ с отрицательными и положительными волнами on-эффекта, а от его ножки—спонтанную импульсацию, подавляемую светом. ЭРГ, например у саранчи-шистоцерки, состоит из трех компонентов, .генерируемых соответственно деполяризационным потенциалом рецепторов, гиперполяризацией интернейронов и их off-активностью.
191
нп
Из этих компонентов наиболее лабилен электрический ток, связанный с деполяризацией зрительных клеток. Судя по этому главному компоненту ЭРГ, мелькания сливаются для глазка стрекозы Pachydiplax только тогда, когда частота вспышек превышает 200 раз в секунду. Для глазка медоносной пчелы критическая частота слияния мельканий еще выше — 265 в секунду.
Рис. 64. Кривая равных ответов медиального дорсального глазка пчелы Apis mellifera (по Goldsmith, Ruck, 1958):
1 — значения, полученные при уравнивании высоты отрицательной волны on-эффекта ЭРГ; 2 — значения, полученные при уравнивании высоты положительной волны on-эффекта ЭРГ.
По оси ординат — величины, обратные числу квантов света, вызывающего ЭРГ равной высоты; за 1,0 принято значение для Х=490 нм; по оси абсцисс — длина волны света, нм
В темноте интернейроны оцеллярной ножки мух, пчелиных, саранчовых и других насекомых спонтанно генерируют 40—70 импульсов в секунду. Освещение сначала сильно подавляет частоту импульсации, а при достаточно ярком свете активность интернейрона совсем прекращается. Но по истечении двух-трех минут устанавливается новый уровень импульсации, который тем ниже, чем выше освещенность. При выключении света частота импульсов возрастает, но через две-три минуты возвращается к стационарному темновому уровню. Оба переходных процесса, видимо, отражают адаптационные изменения, протекающие в глазке достаточно быстро.
Следовательно, по данным электрофизиологии, глазок служит не только для регистрации изменения освещения, но и для того, чтобы отличать неизменное освещение от темноты. Поскольку частота постоянной импульсации глазков градуально зависит от силы света, они могли бы давать информацию и об абсолютной освещенности. Наконец, благодаря глазкам центральная нервная система насекомого получает в темноте высокое тоническое возбуждение.
О спектральной чувствительности глазков дает представление Кривая равных ответов (ЭРГ) медиального глазка медоносной пчелы (рис. 64). Она двухвершинная, причем пик ультрафиолета (%=340 нм), если кривую рассчитать по положительной волне on-ответа ЭРГ, превышает подъем в сине-зеленой (%=490 им) части спектра. Зависимость формы ЭРГ от спектрального состава стимулирующего света можно расценивать как одно из указаний на цветоразличительную способность глазка.
Понять назначение дорсальных глазков до конца можно было бы при наличии их у всех насекомых, ведущих, открытый активный образ жизни и лишенных признаков редукции других
192
органов или упрощения общего уровня организации. Но морфологи утверждают, что нет глазков, например, у слепней. Зрительное поведение слепней приниципиально не отличается от поведения других мух с глазками, и в таком случае нужно думать, что функции глазков все же могут компенсироваться фасеточными глазами. С другой стороны, глазки в качестве дополнительного органа зрения появились у членистоногих очень рано. Они были уже у трилобитов и есть у мечехвостов в дополнение к их латеральным глазам, напоминающим фасеточные глаза насекомых.
Если судить ,по поведению насекомых и результатам электрофизиологических экспериментов, глазки физиологически не равноценны фасеточным глазам и не заменяют последние, а только дополняют их. Мухи, пчелы, муравьи, саранчовые и другие насекомые с закрашенными фасеточными глазами и интактными дорсальными глазками, ведут себя как слепые, не ориентируясь даже по обычному источнику света. Вместе с тем закрашивание глазков, хотя бы одного из трех, более или менее существенно нарушает фотокйнетические реакции: они замедляются, теряют четкость и требуют для своего осуществления более интенсивного освещения. Поэтому уже давно сложилось мнение, согласно которому глазки регулируют фотокинетические реакции организма, обычно усиливая их при оптимальном или пониженном освещении и тормозя их при чрезмерном освещении. Предполагается, что сигналы с глазков влияют в первую очередь на чувствительность интернейронов фасеточного глаза, хотя стимулирующий эффект глазков замечен даже на сердечной активности насекомого: их освещение вызывает тахикардию у неподвижно сидящей мухи.
Впоследствии выяснились другие функции глазков. И прежде предполагали, что глазки имеют отношение к осуществлению „полета. По крайней мере, была замечена корреляция между бескрылостью и отсутствием глазков в пределах одного и того же таксона. Так, глазков нет у бескрылых форм трипсов, тлей, муравьев, но они имеются у крылатых. У хороших летунов они, как правило, крупнее, чем у слабых летунов. Например, у неравнокрылых стрекоз Anisoptera, в сравнении с медлительными Zygop-tera, глазки крупнее, с более сложной сетчаткой, причем медиальный глазок ориентирован вперед, а не вверх, как у равнокрылых стрекоз. Но только теперь электрофизиологическим методом—по импульсной активности в нервной цепочке — обнаружили, что глазки могут служить анализаторами движения крупных объектов, учитывающими направление их движения относительно стрекозы, в частности приближение или удаление. Так, медиальный глазок Sympetrum flaveolutn сигнализирует о движении крупного предмета в основном сверху вниз, а латеральные глазки — о движении вверх-вниз.
Причастность глазков к ориентации непосредственно замечена по поведению мух Sarcophaga и нескольких видов шмелей. Эти насекомые с закрашенными фасеточными глазами, но не тронуты
193
ми глазками реагируют на поворот плоскости поляризации света, Глазки отличаются от фасеточных глаз намного большей светосилой и могут функционировать даже в сумерках, когда света недостаточно для работы фасеточных глаз. В результате суточный период активности насекомого удлиняется благодаря глазкам. По крайней мере, на перепончатокрылых замечено, что при увеличении размера глазков расширяется диапазон освещенностей, в которых насекомое активно. Различные виды пчел и ос прекращают фуражировку при определенной пороговой интенсивности освещения, которая оказывается тем ниже, чем крупнее глазки, причем виды с глазками более 0,37 мм в диаметре летают даже ночью. Если у медоносной пчелы закрасить дорсальные глазки, то она утром вылетает позднее, а вечером возвращается в улей раньше, чем контрольные интактные пчелы. Следовательно, глазки действительно позволяют насекомому оценивать абсолютную освещенность, что предполагалось исходя из результатов описанных выше электрофизиологических опытов.
Латеральные глазки
5
Рис. 65. Расположение стемм на голове гусеницы Isia Isabella (по Dethier, 1942) (вид сбоку):
1,2,6 — тройные глазки; 3,4,5 — одинарные глазки; 7 — ротовой аппарат
Латеральные глазки встречаются у личинок насекомых с полным превращением и отдельных представителей имаго, например у ногохвосток, чешуйниц, блох, самцов червецов и некоторых других. У первых — это временные органы зрения, именуемые с т е м м а -м и. В процессе метаморфоза стеммы, как правило, дегенерируют, не превращаясь в сложные глаза, и редко переходят имаго в неизменном виде.
Латеральные глазки располагаются по бокам головы в количестве от одного до шести (обычный случай), редко их больше. Если глазков несколько, то не все они развиты в равной мере: обычно два бывают крупнее остальных (рис. 65). Диаметр глазков варьирует у разных насекомых от 50 мкм
до 0,5 мм, неодинаково и их устройство. Более примитивные из них близки к дорсальным глазкам, а более сложные напоминают своей организацией отдельный омматидий фасеточного глаза. Примером первого типа может служить глазок личинки пилильщика
194
Allantus togatus (рис. 66, Д). Под массивным двояковыпуклым линзой-хрусталиком {cornea) лежат корнеагенные клетки (кк), а за ними — чашевидная сетчатка. Дистальные концы зрительных клеток (эк), сгруппированных noi четыре в ретинулы, образуют слой рабдомеров. Число зрительных клеток может быть относительно велико — более 6300 в крупных стеммах личинок жуков-скакунов Cicindela.
Рис. 66. Латеральные глазки личинок (продольный разрез, схематично): А — глазок личинки пилильщика Allantus togatus (по Corneli, 1924); Б — схема глазка гусениц (по Snodgrass, 1924).
св—кристаллическое тело; cornea — роговичная лннза; зк — зрительная клетка; кк — корнеагенные клетки; мк—мантийная клетка; пзк—периферическая зрительная клетка; рб —рабдом; цзк — центральная зрительная клетка
Глазки гусениц бабочек, личинок ручейников, жуков-плавунцов Dytiscus и других построены по типу омматидия: снабжены в дополнение к роговичной линзе кристаллическим телом (конусом), а ретинула сложена небольшим числом зрительных клеток. В глазке гусеницы (рис. 66, Б) позади линзы находится кристаллическое тело (св), окруженное корнеагенными клетками (кк). По бокам от них лежат вытянутые клетки, именуемые мантийными (мк). Последние частично заполнены гранулами темного пигмента и одевают, как футляром или мантией, ретинулу вместе с кристаллическим телом. Ретинула состоит из семи зрительных клеток: трех периферических (пж) и четырех центральных (цзк), которые все вместе образуют единый осевой рабдом (рб). В аналогично уст'ро-
195
енных латеральных глазках личинки муравьиного льва насчитывается до 40—50 удлиненных зрительных клеток.
Оптические свойства и предполагаемая функция глазков гусеницы своеобразны. Роговичная линза и кристаллическое тело функционируют как единая оптическая система. Эта система чрезвычайно короткофокусна (/«20 мкм), поэтому ее относительное отверстие превышает 1 :1,0 и, следовательно, светосила глазка очень велика. Благодаря малой величине фокусного расстояния создаваемое оптической системой изображение незначительно перемещается по глубине, когда объект приближается к глазку или удаляется от него. Поэтому резкое изображение объекта всегда остается в плоскости рабдома, вытянутого вдоль оптической оси. Поле зрения отдельного глазка такого типа, измеренное у личинки муравьиного льва электрофизиологическим способом, равно примерно 47°, а его чувствительность даже под углом 8° к оптической оси не превышает 50% от максимальной.
В глазках с трехраздельной роговицей (рис. 65, 1, 2, 6) образуются соответственно три изображения одной и той же точки. Последнее обстоятельство, а также узкое поле зрения и вытянутый «двухэтажный» рабдом свидетельствуют в пользу того, что отдельный глазок гусеницы не предназначен для восприятия деталей предметов и функционирует подобно одиночному омматидию сложного глаза. Шесть глазков расположены так, что каждый имеет обособленное, практически независимое поле зрения. Поэтому гусеница одновременно видит с каждой стороны головы, очевидно, только шесть независимых точек, каждая из которых есть совокупность всех лучей, попадающих в один глазок. Мозаика этих точек слишком груба, чтобы различать детали и форму предмета. Но малочисленность зрительных единиц, возможно, компенсируется их движением: «привычкой» большинства гусениц раскачивать переднюю часть тела из стороны в сторону и таким способом сканировать окружающие предметы. В разделе о восприятии формы мы уже отмечали, что сканирования поля зрения даже единичным чувствующим элементом в принципе достаточно для оценки визуальной расчлененности окружающей картины.
Стеммы личинок, построенные по типу дорсальных глазков, обладают, вероятно, теми же оптическими свойствами, что и последние. К сожалению, мы вынуждены это только предполагать, так как латеральные глазки личинок и с анатомической и с функциональной стороны изучены незаслуженно мало.
Электрическую реакцию глазка—ЭРГ — отводили только у гусениц бабочек, личинок комара Aedes aegypti и муравьиного льва Euroleon nostras. У первых двух ЭРГ монофазная, роговично-отрицательная, но у муравьиного льва двухфазная: с on- и о//-пика-ми, направленными в противоположные стороны. В ходе темновой адаптации ЭРГ глазка гусеницы тутового шелкопряда увеличивается примерно в течение одного часа. На свету ее амплитуда стабилизируется быстрее: за 8—10 мин. Слияние мельканий наступает
196
Рис. 67. Кривые дифференциального спектра родопсина (/) и спектральной чувствительности белоглазой (2) и нормальной (3) личинок комара Aedes aegypti (по Seldin, 1972).
По оси ординат — логарифмы спектрального поглощения (для /) или относительной чувствительности (для 2 и 3); по оси абсцисс — длина волны света, нм
уже при 25—30 вспышках в секунду света умеренной яркости, но для личинки муравьиного льва критическая частота мельканий выше — более 40 мельканий в секунду.
В глазках личинки комара Aedes aegypti обнаружен микро-спектрофотометрическим .способом зрительный пигмент родопсин с Хтах=515 нм и для этих же глазков была измерена по ЭРГ кривая равных спектральных ответов (рис. 67). У белоглазых личинок-мутантов, лишенных экранирующего пигмента, спектральная чувствительность зрительного пигмента хорошо совпадает с кривой равных ответов глазка, из чего можно заключить, что личинка комара цветослеПа. Противоположный вывод следует из электрофизиологических измерений спектральной чувствительности глазков гусеницы тутового шелкопряда. Ее кривая равных ответов с пиками в ультрафиолетовом и зеленом участках спектра изменяет свою форму при цветовой адаптации. Глазки гусеницы, по-видимому, имеют два светочувствительных пигмента, что обеспечивает ей возможность цветоразличения.
Этот вывод хорошо согласуется с наблюдением за поведением гусениц дневных бабочек Vanessa и Pieris. В период питания зеленая, желтая и оранжевая окраски привлекают гусениц, а синяя отталкивает; причем обе группы окрасок гусеницы отличают от серых поверхностей разной свет
лоты. Ко Времени окукливания привлекающими становятся темные тона: коричневый и черный. Скорее всего, цветовое зрение имеют и личинки колорадского жука Leptinotarsa decemlineata, глазки которых отличаются высокой контрастной чувствительностью. Зеленую бумагу личинки жука предпочитают серым бумагам, а эти последние с коэффициентами отражения 87 и 93% или 54 и 41% отличают друг от друга.
Показано, что гусеницы бабочек могут различать размер и форму предметов. Из двух равных по ширине вытянутых прямоугольных фигур гусеницы монашенки Lymantria monacha предпочитают более высокую, причем с расстояния 30 см они различают фигуры высотой 9 и 10 см. Сужающаяся кверху фигура привлекает их сильнее фигуры той же высоты и площади, но сужающейся книзу или имеющей параллельные края.
Гусеницы бабочек Malacosoma, Choristoneura, личинка пилильщика Neodiprion реагируют на поворот плоскости поляризации света и могут пользоваться как ориентиром поляризацией света голубого неба.
1*7
Г лава VII
РЕАКЦИИ НАСЕКОМЫХ НА ЭЛЕКТРИЧЕСКИЕ И МАГНИТНЫЕ ПОЛЯ
Любой организм постоянно находится в электрических и магнитных полях. Однако влияние этих факторов никогда не бывает столь очевидным, как, например, света и температуры. Реакция на электрические и магнитные поля в большинстве случаев очень изменчива, что зависит как от индивидуальных различий объектов, так и геофизического фона, на котором проводится опыт. Малая выраженность реакции иногда приводит к тому, что за реакцию на электромагнитное поле иногда принимают другие эффекты: температурные, акустические или вибрационные.
Несмотря на большие трудности исследования реакций насеко-мых на электромагнитные поля, результаты экспериментов в ряде случаев удается повторять. Часть из них выстраивается в единую линию, подтверждая друг друга. Именно о таких результатах, свидетельствующих о перспективности подобных исследований, и пойдет речь в данной главе.
Вопрос о том, какие рецепторы воспринимают действие электрических или магнитных полей, еще менее ясен. В одних случаях наблюдаемые реакции можно объяснить раздражением механорецепторов под влиянием механических сил, возникающих в электромагнитном поле, в других — возможными изменениями каких-то физико-химических показателей тканей тела и воды, содержащейся в них, в третьих — остается предположить существование гипотетического специализированного рецептора, хотя существование этого рецептора никогда и никем не было доказано.
Рассмотрим реакции насекомых на факторы, воздействующие на них в естественной обстановке, а также искусственно создаваемые в экспериментах.
1. Электрические факторы:
а) электростатический заряд поверхностей окружающих предметов и градиенты потенциала;
б) переменное низкочастотное электрическое поле;
в) ионизация и электропроводность воздуха.
2. Электромагнитные колебания.
3. Магнитные факторы:
а) постоянное магнитное поле;
б) переменное низкочастотное магнитное поле.
198
Строго говоря, любые изменения как электрического, так и магнитного полей представляют собой электромагнитные колебания. Тем не менее есть физические основания называть переменным электрическим и переменным магнитным полями такие, которые создаются искусственно, например в конденсаторе и соответственно в соленоиде или около его при подаче на них относительно низкочастотного потенциала (не выше 1 —10 кГц).Электромагнитными колебаниями мы будем называть все природные колебания, от низких до высоких частот, а также искусственные колебания высоких частот (десятки кГц и выше). Следует подчеркнуть также, что между многими из этих факторов в природе наблюдаются сложные взаимодействия.
1. РЕАКЦИИ НА ЭЛЕКТРИЧЕСКИЕ ФАКТОРЫ
Электростатический заряд поверхности и градиенты потенциала. Электростатические заряды возникают на поверхности различных предметов в результате их трения друг о друга или о воздух. Эти местные заряды могут достигать сотен вольт. Кроме того, значительная разность потенциала постоянно имеет место между поверхностью земли с находящимися на ней предметами и верхними слоями атмосферы и облаками. В ясную устойчивую погоду возникающий градиент потенциала у поверхности земли равен 100—130 В/м (вольт на метр), а при приближении грозы он увеличивается до десятков тысяч.
Тело движущегося насекомого в результате трения о субстрат и воздух приобретает электрический заряд, подобно любому движущемуся предмету. Этот заряд зависит скорее от подвижности насекомого, чем от размеров его тела, и варьирует как по знаку, так и по абсолютной величине. Измерения зарядов тела рабочих муравьев дали очень изменчивую величину в пределах от ±10-11 до 5-10-10 К. Не исключено, что различные острые выросты на теле насекомых (щетинки, шипы и рога) способствуют стоку избыточного заряда в воздух.
При движении насекомого (особенно при его полете) благодаря такому электрическому заряду возникают также и слабые переменные электромагнитные поля (аурополя), частота которых равна частоте взмахов крыльев.
Заряд тела насекомого взаимодействует с зарядами окружающих предметов. Возникающие механические силы прямо пропорциональны произведению взаимодействующих зарядов и в некоторых случаях могут быть достаточными для того, чтобы их восприняло насекомое. Эти силы препятствуют движению насекомых вплоть до их прилипания к заряженной поверхности, а также приводят к налипанию на тело мелких пылевых частиц. Можно видеть, как у дрозофил в искусственном поле слегка расходятся
199
крылья, подобно листочкам электроскопа. В очень больших по напряженности полях бабочки платяной моли буквально прилипают крыльями к заряженным поверхностям. Если заряжены только определенные участки пластиковой поверхности, то мелкие осы-наездники при движении по ней, подходя к заряженной зоне, замедляют бег и усиленно ощупывают антеннамй дорогу перед собой. По той же причине ловушки из полиэтилена, процеживающие воздух, собирают значительно меньше насекомых, если эти ловушки по каким-то причинам оказываются заряженными.
Заряд поверхности можно снять, если предмет поместить в садок, обтянутый металлической заземленной сеткой (клетка Фарадея). Кроме того, клетка Фарадея будет препятствовать наведению потенциала на поверхность предметов верхними слоями атмосферы. Можно ожидать, что если заряды поверхностей как-то мешают насекомым, то условия внутри клетки Фарадея окажутся для них более благоприятными. Действительно, внутри клетки Фарадея наездники откладывают больше яиц. Накрывание кустов можжевельника металлической сеткой приводило к заметному возрастанию численности насекомых на этих кустах. Аналогичным образом, если накрывать поверхность почвы клетками Фарадея, то в поверхностном слое почвы оказывается в 1,5 раза больше мелких почвенных членистоногих, чем на участках, покрытых сеткой из изолятора. Особенно заметно возрастает численность некоторых ногохвосток и клещей.
Поведение насекомых между пластинами заряженного конденсатора также показывает тормозящее влияние электрического поля. При подаче на пластины потенциала, создающего поле, близкое или несколько большее по напряженности, чем атмосферное поле в грозовую погоду, мухи-дрозофилы перестают бегать. Сходным образом, но менее отчетливо реагируют на поле некоторые жуки-кожееды. Бабочки же платяной моли при включении поля нередко приходят в возбуждение, а потом останавливаются. Реакция насекомых совершенно не зависит от того, контактируют ли они непосредственно с токопроводящей поверхностью. По-видимо-му, чем меньше насекомое, тем резче выражена его реакция на поле. Интересно, что дрозофилы заметно реагируют даже на относительно слабое электрическое поле, вплоть до 200 В/м (примерно такое поле возникает при прохождении облака над головой), хотя и очень быстро адаптируются к такому полю.
Отметим, что, несмотря на очевидность реакции дрофозил и других насекомых на электрическое поле, она отличается плохой воспроизводимостью. Зимой дрозофилы, как правило, вообще не реагируют на поле. Летом же уровень их реакции при, казалось бы, строго одинаковых условиях варьирует от 13 до 93%. Реакцию на поле редко удается наблюдать в городской лаборатории и значительно чаще в сельской местности. Есть основания предполагать, что чувствительность дрозофил, пчел, а возможно, и других насекомых к электрическому полю уменьшается с повышением влаж
200
ности, а также инфразвукового и вибрационного фона. Инфразвуки же возникают по многим причинам: при приближении атмосферного фронта, от гроз, магнитных бурь, ветра; транспорт и промышленные установки также нередко генерируют инфразвуки.
Почти не изучена ориентация насекомых в электрическом поле. В конденсаторе с вертикальным направлением силовых линий большинство дрозофил ориентирует ось тела параллельно силовым линиям поля. Сообщалось, что майские жуки предпочитают определенные направления по отношению к горизонтально идущим силовым линиям. Однако эти предпочитаемые направления сложным образом менялись во времени.
Итак, электростатическое поле влияет на поведение насекомых, явно препятствуя их движению, нарушая их «комфорт», а также приводит к отталкиванию друг от друга одноименно заряжающихся конечностей их тела. Эффект варьирует, по-видимому, в зависимости от нескольких факторов: от величины заряда тела насекомого, его физиологического состояния и уровня возбужденности, а также от положения оси тела по отношению к силовым линиям поля. Однако было бы преждевременным объяснять все реакции насекомых на электрическое поле только механическими взаимодействиями зарядов.
Переменное низкочастотное электрическое поле. Поле, создаваемое синусоидально меняющимся во времени током промышленной частоты (50 Гц), влияет на поведение насекомых аналогично постоянному полю. Однако при одинаковой напряженности поля реакция дрозофил на переменное поле в 1,5—2 раза выше, чем на постоянное.
Своеобразно влияет переменное поле на общественных пчел и ос. У одиночных особей видимая реакция может отсутствовать, но при помещении целого улья или гнезда между пластинами конденсатора она резко выражена. Внутри такого улья и гнезда быстро повышается температура, насекомые приходят в возбуждение, и часто вся семья вылетает из гнезда. Осы при этом нередко сцепляются в клубок.
Интересно, что этот эффект наблюдается только в том случае, когда пластины конденсатора параллельны сотам и, следовательно, бегающие насекомые пересекают силовые линии поля. Соответственно пчелы реагируют только при вертикальном положении, пластин, а осы — горизонтальном. Заметим, что дрозофилы как в постоянном, так и переменном поле стремятся расположить ось своего тела по возможности параллельно силовым линиям поля.
Итак, реакция на переменное поле (50 Гц), по-видимому, сходна с реакцией на постоянное, но резче выражена. Возможно, что в переменном поле медленнее наступает адаптация или на насекомых влияет частое перераспределение зарядов внутри тела.
Ионизация и электропроводность воздуха. Ионизация воздуха, влияя на его электропроводность, играет значительную роль в регулировании поверхностных зарядов. Кроме того, возможно воз
201
действие ионов вдыхаемого воздуха на физиологические процессы. В воздухе присутствуют ионы двух типов: легкие и тяжелые. Легкие ионы обладают положительным или отрицательным зарядом. В природе уровень ионизации варьирует в зависимости от местности, растительного покрова, загрязненности воздуха, сезона и погоды. Как правило, количество ионов в воздухе относительно очень мало, причем чаще всего число отрицательных ионов несколько превышает число положительных. Соответственно изменения электропроводности воздуха также очень незначительны.
Легкие ионы оседают на взвешенных в воздухе твердых частицах и капельках жидкости, образуя электрически малоактивные тяжелые ионы.
Неоднократно предполагалось, что уровень ионизации воздуха влияет на процессы жизнедеятельности. Считается, что это влияние связано с концентрацией положительных и отрицательных ионов, а также с соотношением между ними.
Влияние ионов на насекомых относительно мало изучено. Отмечалось возбуждающее действие отрицательных ионов на пчел, а положительных ионов — на мух, а также влияние соотношения их концентраций на некоторых бабочек. Повышение концентрации положительных ионов в эксперименте явно способствовало линьке тлей. Однако поведение дрозофил совершенно не зависело от уровня ионизации воздуха как при колебаниях ионизации в естественных диапазонах, так и при повышении уровня ионизации на несколько порядков при помощи радиоактивного коллектора. Аналогичным образом не наблюдалось никакой корреляции между поведением дрозофил и естественными колебаниями электропроводности воздуха, как положительной, так и отрицательной.
Рецепторы, воспринимающие уровень ионизации воздуха, не известны.
2. РЕАКЦИИ
НА ЭЛЕКТРОМАГНИТНЫЕ КОЛЕБАНИЯ
Частотный спектр электромагнитных колебаний, возникающих в атмосфере (атмосферики), очень широк — от 0,01 Гц (и ниже) и достигает десятков мегагерц. В быту хорошо известны атмосферики радиочастотного диапазона, создающие радиопомехи. Уровень их меняется в зависимости от близости атмосферных фронтов, отражательной способности ионосферы, а также от состояния геомагнитного поля, например от наличия геомагнитных бурь.
Широко известно, что поведение насекомых зависит от погоды, даже при наблюдениях в лаборатории, а следовательно, может коррелировать с атмосфериками. Именно такая корреляция наблюдалась при изучении поведения пчел, а также интенсивности дыхания термитов. Однако влияние самих атмосфериков никем доказано не было. В частности, электромагнитные колебания, создава
202
емые на близком расстоянии искрящимися контактами и имитирующие атмосферики, совершенно не влияли на поведение дрозофил.
Еще сложнее вопрос о влиянии на поведение насекомых магнитных бурь. Предполагают, что на организм влияют атмосферики очень низких частот. Неоднократно наблюдалась достоверная корреляция между уровнем возмущенности геомагнитного поля и численностью сборов насекомых на свет кварцевой лампы (но не лампы накаливания). Корреляция была положительной для жуков и бабочек яблоневой плодожорки и отрицательной для совок и длинноусых двукрылых. В дни магнитных бурь вероятность искажения суточного ритма некоторых жуков-кожеедов при наблюдении даже в стабильных лабораторных условиях возрастала вчетверо.
Геомагнитная буря — сложный комплекс геофизических явлений. Корреляция между показателями поведения насекомых и индексами геомагнитной возмущенности далеко не всегда достоверна. Действующее на насекомых начало геомагнитной бури пока неизвестно, однако не исключено, что здесь большую роль играют инфразвуки, генерируемые бурей.
Итак, влияние на насекомых электромагнитных колебаний естественного происхождения остается недоказанным.
3. РЕАКЦИИ
НА МАГНИТНЫЕ ФАКТОРЫ
Постоянное магнитное поле. Геомагнитное поле характеризуется очень низкой напряженностью по сравнению с полями, обычно применяемыми в технике. Его вариации невелики и составляют, как правило, доли процента от абсолютной напряженности.
Неоднократно сообщалось, что насекомые ориентируются определенным образом в естественном или искусственном магнитных полях. Однако, как мы покажем далее, большинство из этих работ может быть подвергнуто сомнению.
Еще со времен Фабра энтомологи, наблюдая удивительно точную ориентацию насекомых в пространстве, предположили, что последние пользуются магнитным компасом. Однако ни одна попытка изменить курс движения насекомого искусственным магнитным полем не была успешной.
Приобрели широкую известность работы, согласно которым насекомые (мухи, жуки, прямокрылые, термиты) останавливаясь, располагают ось своего тела преимущественно параллельно или перпендикулярно силовым линиям магнитного поля. Еще ранее сообщалось, что царицы термитов ориентируют ось своего тела по линии север--юг.
Ориентация оси тела параллельно или перпендикулярно силовым линиям подтверждалась как визуальными наблюдениями, так и фоторегистрацией направлений осей тела. Однако в дальнейшем
203
выяснилось, что в результате неизбежной неточности измерений в таких наблюдениях имеет место своеобразная психологическая ошибка. Наблюдатель непроизвольно смещает величины измеряемых углов к основным осям координат, вследствие чего и формируется ложная крестообразная картина предпочитаемых направлений. Следовательно, нельзя утверждать, что предпочитаемая ориентация действительно имеет место хотя бы в некоторых случаях.
По ряду причин некоторое сомнение вызывает интерпретация наблюдений, показывающих скопление насекомых (пчел, дрозофил) у полюсов искусственного магнита. В одном из таких случаев удалось убедительно показать, что скопление у полюсов магнита возникло из-за большего нагрева последнего при освещении, по сравнению с окружающими поверхностями. По-видимому, в подобных опытах необходим строжайший контроль за температурой и освещенностью.
Для некоторых явлений магнитная ориентация остается, единственно допустимым объяснением. Так, недавно было показано/ что жужелицы :при их движении на местности. явно предпочитают определенные направления, казалось бы, связанные с положением солнца. Они закономерно изменяются «по кругу» соответственно движению солнца по небосводу, но изменения курса продолжаются как в дневное, так и в ночное время, когда ориентация по солнцу невозможна.
Более того, эти же суточные изменения ориентации сохраняются в закрытом помещении при диффузном освещении сверху и даже в пещере. Скорее всего их можно объяснить постоянно меняющейся магнитной ориентацией. .
Аналогичное явление наблюдалось у майских жуков. Подобно жужелицам, они предпочитали определенные направления по отношению к силовым линиям магнитного поля, но их ориентация изменялась в течение суток более сложным образом^ Ориентация майских жуков по магнитному полю, по-видимому, сочетается с ориентацией по другим факторам. Во всяком случае очень небольшое изменение (всего на 1,4°) направленности искусственного магнитного поля по отношению к странам света приводило к резким и достоверным изменениям ориентации.
Отмечалось, что майские жуки каким-то образом «запоминают» направление, в котором они летят от мест выплода в долинах к горным лесам, где питаются имаго. В дальнейшем самки возвращаются по тому же маршруту в долины для откладки яиц. В лабораторных опытах только что вышедших из куколок жуков жестко закрепляли в определенном направлении. Спустя некоторое время жуки явно стремились сохранить то же направление движения или прямо ему противоположное. Однако поле искусственного магнита закономерно изменяло эту ориентацию.
Не исключено, что именно таким образом, не имея каких-либо видимых ориентиров, находят точное направление предварительно выдрессированные соответствующим образом общественные осы.
204
Возможно, что по этой же причине сохраняют направление подземного хода термиты после предварительной зрительной ориентации по наземному ориентиру.
Совсем недавно было обнаружено, что жуки — мучные хрущаки, находившиеся в течение длительного времени в контейнере, освещенном сбоку, «запоминают» направление бегства от света и сохраняют это же направление в камере с однородным рассеянным освещением. Компенсация геомагнитного поля кольцами Гельмгольца полностью устраняет эту ориентацию. Интересно, что если направление световых лучей регулярно меняется в течение суток, жуки способны запоминать эти изменения и затем «проигрывать» их без каких-либо световых ориентиров. Кажется очевидной аналогия явно магнитной ориентации этих жуков с тем, что наблюдается у других насекомых.
Итак, хотя постоянная ориентация насекомых в магнитном поле маловероятна, вполне возможно, что это поле используется насекомыми при отсутствии других, более очевидных ориентиров.
В ряде работ показано воздействие магнитного поля на ориентацию насекомых по отношению к силе тяжести. Так, курс "поднимающихся по наклонной плоскости дрозофил, отклоняется в зависимости от ориентации этой плоскости по странам света. Отклонение бывает максимальным, если мухи при своем движении пересекают силовые линии геомагнитного поля. Искусственное магнитное поле еще более увеличивает это отклонение.
Аналогичным образом пчелы допускают максимальную ошибку в своем танце на вертикальной плоскости сот, если улей ориентирован, так, что танцующие пчелы пересекают, силовые линии магнитного поля. Величина этих ошибок закономерно изменяется на протяжении суток, причем если пчелы танцуют на двух сторонах вертикальной площадки, суточные кривые этих ошибок зеркальны по отношению друг к другу. Напомним, что направленность силовых линий геомагнитного поля закономерно изменяется в течение суток.
Компенсация геомагнитного поля кольцами Гельмгольца значительно снижает ошибку ориентации пчел по отношению к силе тяжести. Если же производить регулярную подстройку колец к изменяющемуся направлению силовых линий геомагнитного поля, то ориентация пчел становится практически безошибочной.
Рецепторы, обеспечивающие направленное восприятие магнитного поля, неизвестны. Живой организм в норме не имеет никаких ферромагнитных включений, аналогичных стрелке компаса. Однако недавно сообщалось, что вещество, из которого состоит тело насекомого обладает парамагнитными свойствами, т. е. способно приобретать магнитные свойства при достаточно длительном воздействии внешнего магнитного поля. Наблюдения за поведением насекомых в какой-то мере подтверждают этот вывод. Так, курс
1 Сухое насекомое, растертое в порошок.
205
жужелицы достоверно становится более точным после ее прохождения в поле искусственного магнита. С другой стороны, компенсация геомагнитного поля снижает ошибку ориентации пчел к силе тяжести не сразу, а спустя некоторое время.
Итак, имеется ряд фактов, показывающих, что магнитное поле воспринимается насекомыми и играет определенную роль в их пространственной ориентации особенно при отсутствии других ориентиров. Один из возможных механизмов ориентации в магнитном поле — восприятие механических сил, возникающих при пересечении насекомым силовых линий поля.
Переменное магнитное поле инфразвуковых частот, создаваемое при помощи электромагнитов, возбуждает некоторых насекомых, однако наблюдавшиеся эффекты оказались невоспроизводимыми. Здесь не исключалась возможность воздействия на насекомых вибрационных и акустических помех.
* *
*
Влияние на насекомых электрических и магнитных полей остается почти не исследованным. Только сейчас имеющиеся твердо установленные факты начинают укладываться в какую-то систему. Становятся все более очевидными сложность и взаимозависимость реакций насекомых на естественные абиотические факторы.
Можно надеяться, что дальнейшее изучение этой труднейшей проблемы приведет к открытиям, значение которых выйдет далеко за пределы энтомологии. В самом деле, именно во влиянии магнитных и электрических полей или факторов, связанных с ними в геофизическом комплексе, предполагается причина широкоизвестных патологических реакций человека и животных на погоду и магнитные бури. Не исключено, что именно этот комплекс геофизических факторов — одна из причин удивительной способности насекомых и других живых организмов «воспринимать» время суток и года, находясь, казалось бы, в полной изоляции от естественной среды.
Распутывание клубка влияний геофизических факторов может привести и к обнаружению новых средств воздействия на поведение, размножение и развитие вредных насекомых. Наконец, исследование реакций насекомых на электрические и магнитные поля, несомненно, раскроет способы их восприятия, а возможно, будут найдены и новые рецепторы.
Глава VIII
МЕХАНИЗМЫ
ПРОСТРАНСТВЕННОЙ
ОРИЕНТАЦИИ
1. ПЕРВИЧНАЯ И ВТОРИЧНАЯ ОРИЕНТАЦИЯ И КОНТРОЛИРУЮЩИЕ ЕЕ СИСТЕМЫ
Тело насекомого и любого другого животного всегда ориентировано по отношению к какому-либо внешнему фактору: гравитации, свету, ветру и т. п. Различают первичную ориентацию — удерживание типичного, или исходного, положения тела в пространстве и вторичную — нацеливание локомоторной оси организма на определенный стимул, выбор которого определяется внутренней мотивацией. Внутренней мотивацией, или потребностью определенного действия, — понятием, необходимым для корректного описания поведения животных, — мы будет называть сменяющие друг друга стремления: отыскать пищу, встретить полового партнера, вырыть норку и иную потребность целенаправленного действия, вытекающую из общего физиологического состояния организма и генетической детерминации проявления и последовательности биологических процессов.
Первичная ориентация, или сохранение основного положения тела в пространстве, достигается у насекомых различными способами: за счет сохранения контакта конечностей с субстратом, путем удерживания тела под определенным углом к направлению силы тяжести, световых лучей и другими факторами, — т. е. на основе тактильной, проприоцептивной, зрительной и другой информации. Так, ползущее насекомое стремится сохранить контакт всех ног с субстратом, в противном случае оно взлетает или совершает плавательные движения, если это обитатель воды. Летающие насекомые, за исключением стрекоз, двукрылых и некоторых других, падают вниз и начинают кружиться, когда гасят свет. Такая реакция на темноту возникает, возможно, потому, что в норме эти насекомые удерживают тело спиной к свету — в сторону неба. Если, например, личинку жука-плавунца осветить снизу, то она переворачивается и начинает плавать брюшком кверху. У тех насекомых (мух, стрекоз), которые способны летать в темноте, к стабилизации положения тела причастны различные механорецепторы. О их роли в сохранении позы и координации частей тела достаточно подробно говорилось в главе о роли органов чувств в поведении и разделах, посвященных механорецепции.
207
Вторичная ориентация тела всегда накладывается на его исходное положение, но она не обязательно сопровождается поступательным движением. Например, заметив движущийся объект, личинка стрекозы-коромысла принимает позу слежения, зрительно фиксируя свою потенциальную добычу. Сидящая на месте муха-ктырь синхронно поворачивает голову, следя за пролетающими мимо насекомыми. В более простом случае насекомое выбирает положение тела под определенным углом к направлению действия какого-либо фактора. Мухи-дрозофилы, например, усаживаются либо параллельно, либо перпендикулярно плоскости поляризации света (в зависимости от его интенсивности). Прежде чем разбирать способы, с помощью которых насекомое задает и длительно сохраняет неизменным направление своей локомоторной оси, остановимся на некоторых общих схемах управления, рассматриваемых в биологической кибернетике.
Процессы управления могут быть либо такими, когда успех действия предопределяется до его начала, либо такими, когда достижение цели дополнительно корректируется но ходу действия. Примерами их могут служить соответственно выстрел из пушки, прицельность которого предопределена, в момент наводки, и пуск самонаводящегося снаряда, который в полете дополнительно корректирует свое направление на цель. Первые процессы осуществляются по схеме так называемой открытой контролирующей системы, вторые — по схеме замкнутой контролирующей системы, т. е. схемы с петлей обратной связи. В основе ориентации насекомых лежат обе схемы управления. Так, хватательный бросок богомола Mantis religiosa управляется открытой контролирующей системой: его направление заранее задается на основе сигналов от глаз и проприоцепторов шейной области и не корректируется по ходу развития. Ориентация по источнику света или направлению гравитации — пример управления на основе замкнутой петли обратной связи, когда отклонение от однажды выбранного угла движения по отношению к ориентиру влечет за собой компенсирующий его поворот. Сходным образом достигается, например, удерживание исходного направления полета в темноте, а также поддерживание его скорости. Здесь Джонстонов орган антенн, ветрочувствительные волоски на голове и крыльях служат датчиками сигналов для управления системы с обратной связью.
Другой пример замкнутой контролирующей системы, постоянно работающей у двигающегося насекомого, дает управление оптомоторной реакцией. Здесь петля отрицательной обратной связи замыкается через посредство сигналов от глаз, и благодаря ей насекомое автоматически (непроизвольно) сводит к минимуму сдвиг на сетчатке изображения окружающей картины, что корректирует первичную и вторичную ориентацию тела.
Итак, однажды заданное на основе первичной или вторичной ориентации положение всех трех осей тела прочно удерживается насекомым с помощью разнообразных корректирующих механиз
208
мов. Они стабилизируют положение тела, предотвращая пассивное смещение под действием внешних сил: ветра, колебания субстрата и т. п. Но вместе с тем насекомое может активно принять любое положение в пространстве. Представляет интерес выяснить, выключаются ли при таких активных движениях названные контролирующие механизмы или же они работают постоянно? Если верно последнее предположение, то позиционное изменение должно было бы соответствовать ожидаемому значению стимула в контролирующей системе. На примере мухи Eristalis показано, что в ходе активного движения не выключаются, по крайней мере, оптомоторные контролирующие механизмы. Так, если изменить пространственное соотношение между фоторецепторами и эффекторами путем поворота головы на 180°, то муха станет неспособной активно двигаться. Способность двигаться утрачивается потому, что следующий за каждым изменением положения тела в пространстве стимулирующий эффект оказывается противоположным тому, который ожидается. Последующие «корректирующие» действия еще более увеличивают ошибку. Из результатов этого и подобных опытов вытекает принцип реафферентации, состоящий в том, что для каждого активного движения сохраняется некоторая копия эфферентных сигналов, посланных мышцам, которая сравнивается с реафферентными, сигналами, вызванными движением. Благодаря тому же самому корректирующему механизму, что и при удерживании устойчивого положения тела (что важно как защита от внешних воздействий), эфферентные сигналы продолжают поступать к мышцам, пока эфферентная копия и реафферентные сигналы не «скомпенсируют» друг друга. Такая схема управления известна в технике под названием петли обратной связи с ожидаемым значением активно регулируемой величины.
2. ТАКСИСЫ И ТРОПИЗМЫ
Общепринятой классификации форм двигательной активности и способов ориентации насекомых до сих пор нет. Отчасти повинны в этом исследователи, описавшие под одним и тем же названием разные явления, но главная причина кроется в многообразии и сложности самих изучаемых явлений.
Чтобы составить верное представление о проблеме ориентации, условимся о следующем. Таксисом будем называть только принцип ориентации, или алгоритм наведения на цель, который по идее не зависит от рода ориентирующего стимула и управляет не обязательно поступательным движением организма. Таксисов известно пока всего пять: фобо-, клино-, тропо-, тело- и менотаксис. Таксису будем противопоставлять понятие «тропизм» — целенаправленный двигательный акт, мотивированный внутренним состоянием организма. В основе тропизма лежит концентрация внимания живот-
209
лого на определенном сигнальном стимуле, выбор которого определяет внутренняя мотивация действий.
Обычно термины «таксис» и «тропизм» употребляют как синонимы, а многие авторы их вообще не различают. Однако в таком случае невозможно отличить принцип ориентации от биологической мотивировки выбора привлекающего стимула. Таксис позволяет движению быть направленным, а тропизм делает его целенаправленным. Тропизмы указывают на признаки (сигналы), по которым животное находит пищу, укрытие или полового партнера, а таксисы объясняют способ, каким животное локализует в пространстве требуемый объект и удерживает путь к нему. Таксисы целесообразно классифицировать и именовать по алгоритму ориентации, а тропизмы— по ориентирующему фактору (фото-, хемо-, геотропизм и т. д.). Из сказанного понятно, что тропизм всегда имеет знак — положительный или отрицательный и что один и тот же тропизм может осуществляться на основе разных таксисов. Например, положительный фототропизм может осуществляться с помощью механизма тело- или тропотаксиса, в таком случае соответственно через фототело- или фототропотаксис. Тропизмы включаются, видимо, при действии «ключевых стимулов» эталогов, и к их запуску могут быть причастны узконастроенные чувствующие или вставочные нейроны-детекторы, о которых упоминалось в предыдущих главах. Сам выбор насекомым того или иного таксиса зависит от окружающей обстановки, расстояния до цели, а также от индивидуальных особенностей организма. Ниже мы будем рассматривать только таксисы и не коснемся тропизмов. Механизм последних почти совсем не расшифрован, да и относятся тропизмы скорее к области этологии, чем к физиологии органов чувств.
Фоботаксис — простейший по принципиальной схеме и исполнению способ ориентации, который следует рассматривать филогенетически самым древним. В его основе лежат кинезы, которыми принято именовать изменения локомоторной активности организма в ответ на изменение внешнего воздействия. Такие изменения активности можно назвать вынужденными: замедление или ускорение движения — ортокинез и изменение частоты смены направления движения — клинокинез. Например, у многих насекомых и их личинок наблюдается фотоортокинез, выражающийся в ускорении движения при увеличении интенсивности освещения. Столь же обычен термоортокинез — ускорение движения с ростом температуры. Клинокинетические реакции известны в отношении температуры, влажности, запаха и других факторов. Например, в зоне угрожающих температур насекомое чаще изменяет направление движения, чем в зоне оптимума. Таким образом, алгоритм фобо-таксиса предельно прост: чем дальше условия от оптимума, тем быстрее двигайся и чаще изменяй направления движения. Такой алгоритм случайного самонаведения позволяет насекомому в конце концов найти оптимальную зону и дольше удерживаться в ней, чем где-то в стороне. Фоботаксисом легко объяснить происхожде
210
ние скоплений насекомых в местах с предпочитаемыми температурой, влажностью, освещенностью. Насекомые прибегают к фобо-таксису обычно в обстановке с малым градиентом стимулов, в диффузном поле, где нет четко выраженных ориентиров, либо когда неожиданно попадают в неблагоприятные условия или бывают резко возбуждены.
Все остальные таксисы отличаются от фоботаксиса тем, что их алгоритмы, во-первых, балансные, предусматривающие выравнивание последовательных или одновременных возбуждений следящих рецепторов, во-вторых, они обеспечивают направленное наведение на объект поиска.
Клинотаксис — ориентация путем сравнения и достижения постоянного уровня возбуждения одних и тех же рецепторов в процессе активного перемещения их в разные точки стимулирующего поля. Наведение локомоторной оси организма на стимул (или от него, в зависимости от знака тропизма) достигается путем последовательного маятникообразного движения антеннами или всей головой. В положении максимального (минимального) возбуждения рецепторов насекомое поворачивает тело так, чтобы оно приняло нормальное относительно этих рецепторов положение. Ориентация на основе клинотаксиса напоминает способ локализации источника звука человеком, глухим на одно ухо: чтобы точнее определить направление, откуда исходит звук, он вынужден поворачивать голову влево и вправо, сравнивая при этом ощущения громкости.
Фотоклинотаксис легко наблюдать у личинок комнатной мухи Musca domestica, избегающих свет вплоть до окукливания. Благодаря последовательным движениям передней частью тела они выбирают такое положение, в котором наименее возбуждены фоторецепторы, лежащие в складке головоглоточного скелета. Клино-таксическая ориентация по градиенту запаха описана у медоносной пчелы. Когда пчелу с одной антенной (вторая антенна ампутирована) помещали в V-образную камеру, одно колено которой .содержало пахучую приманку, то в ходе поиска пчела поворачивала свою единственную антенну из стороны в сторону. Если же антенну приклеивали к голове, то у пчелы наблюдались маятникообразные колебания всего тела. При относительно больших расстояниях до источника запаха насекомое прибегает к клино-таксической ориентации, если даже у него функционируют обе антенны. Так, например, поступает самец тутового шелкопряда Bombyx mori на расстоянии свыше 20 см от самки. Характерный «рыскающий» полет типичен для крупных навозных и падальных жуков, когда они ищут пищу. Но здесь клинотаксис, вероятно, комбинируется с типичным фоботаксисом, так как по мере приближения к источнику запаха — зоне оптимума — скорость полета и число смен направления резко уменьшаются.
Если алгоритм клинотаксиса предусматривает баланс последо-. вательных возбуждений одних и тех же рецепторов, то в основе
241
остальных таксисов лежит уравнивание синхронных возбуждений разных рецепторов.
Тропотаксис — наведение локомоторной оси тела на стимул (или от него) путем достижения равновесия в возбуждении симметричных рецепторов (глаз, антенн и др.). Тропотаксическая ориентация известна по отношению к источнику света, тепловых лучей, звука, гравитации и других стимулов. Примером такой ориентации может служить поведение медоносной пчелы в упомянутой выше У-образ-ной камере, в одном из колен которой заключена пахучая приманка. В норме пчела, расставив антенны в стороны, безошибочно находит приманку. Если антенны закрепить на лбу в перекрещенном положении, то пчела всякий раз поворачивает в непахнущее колено. Насекомое удаляется от источника запаха потому, что более сильные сенсорные импульсы вызывают соответствующие моторные команды не на той стороне тела, которой пчела должна повернуться для достижения баланса возбуждения рецепторов правой и левой антенн. Когда же антенны лежат параллельно друг другу или расставлены под некоторым углом, то насекомое ориентируется тропотаксически. Если антенны сведены так, что их вершины почти соприкасаются, то пчела ищет приманку только клинотакси-чески.
Ось, вокруг которой лежат антенны и другие парные рецепторные органы, хотя и совпадает с морфологической серединой, основа ее в конечном счете остается физиологической. Это доказывается, в частности, сравнением поведения интактной и ослепленной на один глаз личинки жука-плавунца Acilius, которая тропотакси-чески ориентирует свое тело по источнику света как в горизонтальной, так и в вертикальной плоскостях. В норме она удерживает положение спиной к свету, предупреждая не только вращательные движения по горизонтали (путем сохранения равенства возбуждения правой и левой групп стемм), но и килевую качку, что достигается за счет баланса между возбуждением передних и задних стемм. Односторонне ослепленная личинка непрерывно поворачивается.
Тропотаксис отличается от во многом сходно проявляющегося в поведении организма телотаксиса (см. ниже) следующими цри-знаками. 1. Насекомое, помещенное перед двумя одинаковыми источниками раздражения, например лампами, движется по средней линии между ними и только в непосредственной близости сворачивает к одному из них, т. е. переходит к телотаксису. 2. При нарушении симметрии рецепторов, например путем закрашивания одного глаза, насекомое совершает даже в параллельном пучке света манежные (круговые) движения, оставаясь зрячей стороной к стимулу.
Телотаксис — наведение на стимул путем достижения такого положения тела, когда особая зона осевых, направленных прямо вперед чувствующих элементов, например зона фиксации глаза, обращена строго к стимулу. Каждое смещение стимула из зоны
212
фиксации вызывает компенсационный поворот тела, «возвращающий» стимул в положение фиксации. Телотаксис позволяет избирать направление исключительно в сторону интересующего стимула, а не от него. Телотаксис сохраняется у насекомого после закрашивания одного из двух симметричных глаз. Поэтому можно думать, что в основе его лежит сложный монолатеральный баланс возбуждения рецепторных клеток, симметричных относительно других рецепторных клеток, ориентированных строго вперед.
Чаще всего телотаксис направляет зрительные поведенческие реакции насекомых, такие, как ориентация самца бабочки или мухи, преследующего самку; ориентация стрекозы, догоняющей комара; ориентация пчелы, когда она садится на цветок. Другим хорошо проанализированным примером телотаксического наведения может служить уже не раз упоминавшийся нами хватательный бросок богомола. Однако не всегда легко решить, ориентируется ли насекомое в данный момент на основе телотаксиса или тропотаксиса. Известны случаи, когда у одного и того же насекомого, например пчелы, в разных условиях и в разное время жизни проявляется либо телотаксис, либо тропотаксис. Изменяется способ ориентации и в зависимости от того, какой участок глаза возбужден, поскольку для телотаксиса пригодна только его фовеальная часть.
Менотаксис— наведение на цель путем выбора и сохранения некоторого постоянного угла между продольной осью тела и направлением на какой-либо посторонний (случайный) стимул-ориентир, например источник света или звука. Менотаксическая ориентация 'по существу аналогична ориентации по компасу или движению по азимуту, изобретенному человеком, и, строго говоря, включает в себя телотаксис, который можно рассматривать частным случаем менотаксиса, когда угол между направлением на ориентирующий стимул и продольной осью тела равен нулю.
Менотаксис открывает насекомому принципиально иные, отличные от других таксисов возможности ориентации в пути или в покое. При этом нет необходимости ориентироваться непосредственно по привлекающему или отталкивающему стимулу, а можно выбрать любой посторонний ориентир или веху. В качестве последней служат источник света (солнце, небо, луна) или броский, выделяющийся среди других предмет, а также источник звука, направление ветра или течения, направление силы гравитации.
По-видимому, особенно широко прибегают насекомые к мено-таксической ориентации по отношению к оптическим вехам. Их фасеточный глаз является удобным угломерным инструментом, позволяющим, например медоносной пчеле, вести отсчет углов с точностью вплоть до Г. Ориентация по солнцу впервые была замечена у муравьев, затем астрономическую ориентацию подробно изучали у медоносной пчелы, клопа Vella и многих других насекомых, способных избирать в качестве вехи не только солнце, но и направление поляризации света голубого неба.
213
Большой интерес представляет способность насекомых транспонировать (переносить) менотаксический угол ориентации с одного ориентира на другой, например переносить угол относительно источника света к направлению силы тяжести и наоборот. Когда насекомое попадает в темноту, то на наклонной поверхности оно ползет под определенным углом к направлению силы гравитации, зависящем от предшествовавшего светового угла ориентации. Ме-
Рис. 68. Указание направления медоносной пчелой Apis mellifera, «танцующей» на вертикальных сотах (по Frisch von, 1956—1957).
1 — улей; 2 — место кормления; 3 — схема виляющего «танца» пчелы, прямолинейный пробег которого отклонен на 40е слева от вертикали и тем указывает на цель под углом 40° слева от солнца
Экспериментально установлено,
доносная пчела, как это показано К- Фришем, исполняет так называемый виляющий «танец» на вертикальных сотах улья под тем же углом к гравитации, под которым она видела солнце во время полета к источнику пищи (рис. 68). Полет в направлении к солнцу здесь отмечается направлением «танца» прямо вверх. Отклонение в ту или другую сторону от направления на солнце передается равным угловым отклонением влево
или вправо от направления вверх. Присутствующие при «танце» разведчицы пчелы-фуражиры, покидая улей, поступают наоборот: транспонируют увиденный гео-менотаксический угол ориентации к углу фотомено-таксическому.
что геоменотаксические углы
пчела отсчитывает главным образом по степени наклона головы относительно груди, регистрируемого проприоцепторами в обла-
сти шеи: дорсальными и вентральными волосковыми пластинками. Когда пчела стоит на горизонтальной поверхности, те и другие волосковые пластинки равномерно прижаты головой. Когда пчела ползет вверх, то голова наклоняется вентрально вследствие ее низкого центра тяжести. В этом положении вентральные волосковые пластинки сдавливаются сильнее дорсальных, причем тем больше, чем круче подъем. Когда же пчела ползет вниз, то максимально возбуждаются дорсальные пластинки. Причастность этого механизма к оценке гравитационных углов доказывают опыты с перерезанием нервов, идущих от волосковых пластинок, или изменением положения центра тяжести головы (дополнительным грузом), после чего ухудшалась способность пчелы ориентировать свой «танец» относительно направления силы тяжести.
214
Поворачивая в вертикальное положение поверхность, по которой ползет муравей Myrmica, ориентирующийся фотоменотаксичес-ки по лампе, можно испытать его способность к транспонированию угла ориентации, если выключить свет в помещении. Оказалось, что для муравья световой угол равняется четырем зеркально-симметричным гравитационным углам (рис. 69, б). В аналогичных
Рис. 69. Перенос менотаксического светового угла (а) к гравитационному муравьем Myrmi- а са (б), жуком Geotrupes (в), жуком Cocci-nella (г) и ручейником Limnophilus (д) (по
Jander, 1957; Frisch, Lindauer, 1963).
С — направление светового луча; Г—направление силы тяжести. Тонкими стрелками показано направление движения насекомых под соответствующими углами к С и Г
опытах у жука Coccinella (г) обнаружены два зеркально-симметричных гравитационных угла, а у другого жука — Geotrupes (в) — один, причем при ориентации головой вниз. Совсем иным оказался перенос углов ручейником Limnophilus (д), который относит световой угол к гравитационному как 3:2, т. е. н? соответственно, как другие насекомые, а пропорционально.
Исходный биологический смысл переноса менотаксических углов с одного ориентира на другой состоит в том, чтобы в неожиданно изменившейся обстановке удержать прямолинейное движение, которое дает больше шансов найти «выход», чем неориентированное хаотичное движение. Последнее в силу всегда существующей незначительной асимметрии локомоторного аппарата неизбежно выведет насекомое на круговую траекторию. Известно, например, что заблудившийся человек описывает круги именно по этой причине. Но у медоносной пчелы транспонирование ориентационных углов приобрело сигнальное значение: направлением «танца» разведчица сообщает другим членам общины о направлении полета за взятком или к новому улью (в период роения).
Способность переносить угол световой ориентации к гравитационному углу и наоборот прямо указывает на существование в центральной нервной системе насекомого единого центра менотак-сиса, являющегося, возможно, общим для всех балансных таксисов центром управления. Топография и морфология этого центра пока точно неизвестны. Предположительно его отождествляют с грибовидными телами переднего мозга, где прослеживаются волокна нейронов — детекторов движения, безусловно входящие в состав физиологического механизма, таксисов. Хорошим примером внешнего выражения сложности такого управляющего центра может служить, например, внесение поправки к курсу на смещение солнца, что необходимо насекомым при астрономической ориентации.
215
3. АСТРОНОМИЧЕСКАЯ ОРИЕНТАЦИЯ
Легко себе представить, что для ползущего или летящего насекомого солнце или направление поляризации плоскости голубого света неба удобно в качестве менотаксического ориентира в той же мере, что и броские наземные вехи. Вся сложность состоит в том, что ни солнце, ни зависящая от его положения картина поляризации небосвода не остаются на месте, а сами перемещаются с угловой скоростью 15° в час. И если для кочующего насекомого: мухи, жука, стрекозы, — не столь важно, что утром солнце стояло на востоке, а днем оказалось на юге (в течение их коротких перелетов с одного поля на другое солнце смещается незначительно и не препятствует удерживать прямолинейность пути), то для насекомых, совершающих дальние перелеты или регулярно летающих из гнезда туда и обратно, движение солнца по орбите приобретает принципиальное значение. Чтобы безошибочно на протяжении дня пользоваться небесным компасом, нужно учитывать суточное движение солнца и уметь вносить поправку к своему курсу, а для этого необходимо располагать часами.
Бабочки-чертополоховки Vanessa cardui в отдельные годы устремляются весной со Средиземноморского побережья Африки на север, пересекая Средиземное море, и достигают южной Европы. Потомки этих мигрантов, перевалив Альпы, летят в центральную и даже северную Европу. Ежегодно стаи бабочек-монархов Danaus plexippus весной летят из Флориды и южной Калифорнии до Канады — за 3 тыс. км, а их потомство осенью возвращается на юг зимовать. Путь бабочек пролегает частично вдоль побережья, частично — над однообразными, лишенными ориентиров прериями. Медоносная пчела летает на фуражировку в радиусе до 5 км от улья. За сотни метров уходят от своих гнезд пустынные муравьи Cataglyphis, вынужденные находить путь назад исключительно по небесному компасу.
Какие же непосредственные наблюдения и факты доказывают «штурманское искусство» насекомых, их умение вносить поправку к курсу на движение солнца по небосводу?
В одном из опытов К. Фриша наблюдаемый улей сначала стоял у озера и группа меченых пчел несколько дней подряд кормилась на противоположном берегу на столике, стоящем на 200 м западнее улья. Однажды утром перевезли улей в другое незнакомое пчелам место, где по четырем сторонам горизонта на расстоянии 200 м от улья были выставлены такие же столики с приманкой, возле которых дежурили наблюдатели. Оказалось, что пчелы полетели только к западному столику, не взирая на то, что солнце стояло на юго-востоке, а не на западе, как во время последней кормежки у озера.
М. Линдауер провел еще более эффектный опыт. В тропической зоне, в т. Перадении (Шри Ланка), он приучил пчел находить
216
приманку в полуденное время к северу от улья (рис. 70). В ночь с 23-го на 24-е апреля он вместе с ульем перелетел на самолете севернее, в г. Пуну (Индия). В полдень 24-го апреля солнце проходит через зенит на широте как раз между названными городами, но в Перадении оно видно на севере, а в Пуне — под тем же уг-
Рис. 70. Годичный путь полуденного положения солнца на небосводе (по Мазохину-Порш-някову, 1965).
Слева наверху — положение солнца относительно городов Перадеиия и Пуна 24-го апреля в 12 ч по местному солнечному времени. Стрелкой вверх показано направление, в котором пчелы искали приманку в Перадении. Стрелкой вниз обозначено направление, в котором они полетели на поиск приманки в Пуне (по- данным опытов Lindauer, 1957)
лом на юге. Как же это отразилось на ориентации пчел, привыкших в полдень летать за пищей на север, т. е. в направлении на солнце? В Пуне они также устремились прямо .на солнце, но это соответствовало направлению на юг. Опыт, как и предыдущий, убедительно доказывает умение пчел ориентироваться по солнцу.
Возвращаясь из Перадении на родину, М. Линдауер захватил с собой пчел в Мюнхен. Тем самым пчелы оказались неожиданно переброшенными по долготе на 69° западнее, и их внутренние «часы» показывали на 4,5 часа больше местного мюнхенского времени, т. е. пчелиные «часы» не были согласованы с положением солнца на небосводе для долготы Мюнхена. Соответствующие опыты показали, что первое время перевезенные пчелы ориентировались только по наземным вехам, а не по солнцу. Из других опытов также следует, что воспитанные при искусственном освещении пчелы, попав в естественную обстановку, сразу не могут пользоваться солнцем как компасом. Для этого они должны в течение нескольких дней наблюдать суточный ход светила, чтобы «поставить» свои «часы» по местному солнечному времени: синхронизировать их с положением солнца на небосводе в данном пункте.
Неудивительно, что пчелы северного полушария, перевезенные в Бразилию более четырех веков тому назад, дали начало местным расам, которые привыкли компенсировать движение солнца против часовой стрелки. Они приспособились к то-му, что в южном полушарии солнце движется справа налево и днем стоит на севере. Более того, те семьи пчел, которые обитают в пределах тропического пояса — между тропиками Рака и Козерога, — должны два раза в год перестраиваться в этом отношении, так как в течение года полуденное положение солнца дважды пересекает небесный экватор (см. рис. 70).
Насколько полно синхронизированы «часы» пчел с движением солнца, видно из следующих опытов М. Линдауера. Известно, что
217
пчела-разведчица, вернувшись в улей, указывает направление к обнаруженному дуплу для поселения роя с помощью виляющего «танца», строго ориентированного по отношению к солнцу через геоменотаксический угол. Однажды экспериментатор побудил пчелу-разведчицу, которая накануне нашла место для поселения роя, танцевать ночью... и она указала правильное направление. Знакомства с дневной частью солнечной орбиты оказалось достаточным, чтобы пчела экстраполировала ночной путь светила под горизонтом по своим внутренним «часам» (рис. 71).
Рис. 71. Схема синхронизации «часов» медоносной пчелы Apis mellifera (слева) и клопа Vella caprai (справа) с движением солнца (по Мазо-хину-Поршнякову, 1961).
Схема дана для периодов равноденствия, когда солнце восходит в 6 ч и заходит в 18 ч по местному солнечному времени.
1,2 — направление хода «часов» животного днем и ночью
В других экспериментах того же автора пчелы, наблюдавшие в своей жизни только нисходящую четверть солнечной орбиты (до полудня улей содержали в темном помещении), утром искали приманку на незнакомой местности, куда их перевезли накануне, в том же направлении, что и в период послеполуденных тренировок.
Среди одиночных насекомых астрономическая ориентация подробно изучена у водяного клопа Velia caprai. Было замечено, что клопы одного из водоемов г. Фрайбурга (ФРГ), если их потревожить на берегу, убегают всегда на юг, что соответствует направлению на водное пространство. Точно к югу убегают они в любое время дня, ориентируясь, как оказалось, по солнцу и поляризации света неба и учитывая при этом суточный ход светила. В комнате потревоженные клопы убегают тоже в определенном направлении по отношению к лампе, которую они, очевидно, принимают за солнце. Аналогично ведут себя они и ночью, если горит лампа. В ходе этих наблюдений было установлено, что величина угла ориентации по отношению к солнцу (лампе) изменяется в течение суток, но не так, как у медоносной пчелы. В полдень и в полночь этот угол равен нулю—насекомые бегут прямо к лампе, от полудня до заката он увеличивается с правой стороны насекомого, а после заката и до полуночи уменьшается. Наконец, от полуночи до восхода угол ориентации увеличивается с левой стороны насекомого, а затем вплоть до полудня уменьшается. Из этого следует, что «часы» водяных клопов, управляющие изменением ориентационного угла, ночью идут в направлении, противоположном дневному. Ночью по «часам» клопов солнце движется вспять: с запада через юг на восток. Подобные «часы» обнаружены у других членистоногих: пауков, ракообразных, которые также могут ориентироваться по небесному компасу и компенсировать суточный ход солнца (см. рис. 71).
218
ЗАКЛЮЧЕНИЕ
Физиология органов чувств животных, в том числе насекомых, за два последних десятилетия продвинулась далеко вперед. Только за этот период оригинальных экспериментальных работ опубликовано не меньше, чем за всю предшествовавшую историю ее существования. Наряду с общим научно-техническим прогрессом важное значение для бурного и разностороннего накопления знаний об устройстве и работе рецепторных органов имело преодоление философско-методологического барьера, ограждавшего биологию от широкого использования для изучения живых организмов физико-математических подходов и методов так называемых точных наук. На стыке сенсорной физиологии и этих наук возникли новые направления исследований, относимые теперь к биофизике органов чувств, биологической кибернетике и бионике. В дополнение к наблюдательно-описательному подходу и преимущественному изучению работы сенсорного органа в целом, по поведению животного, физиологи стали широко использовать методы электрофизиологии, электронной микроскопии, физического и математического моделирования, позволившие углубиться в клеточные механизмы и расшифровать межнейронные взаимодействия в отдельных сенсорных аппаратах животных, в том числе насекомых.
Первоначально усилия были направлены на изучение рецепторных входов, передающих информацию более высоким отделам соответствующих анализаторов. В результате мы располагаем достаточно подробными и глубокими знаниями о функциональных параметрах многих рецепторов, первичных физико-химических процессах их возбуждения, кодировании информации о стимулах и передаче сигналов рецепторными клетками. Фото- и механорецепторы изучены с этой стороны детальнее и глубже других. Впрочем, в самые последние годы появились работы, подробно описывающие процессы, происходящие при восприятии насекомыми запаха: адсорбцию молекул пахучего вещества на поверхности сенсиллы, перенос их к рецепторным мембранам, динамику возбуждения чувствующей клетки, ферментативное расщепление молекул раздражителя. Но вопрос о роли специфических рецепторных белков в процессе восприятия запахов еще остается нерешенным.
Сведения о .рецепторных входах, даже самые подробные, все же не позволяют составить верное представление о тех сложных и совершенных механизмах, которые обеспечивают насекомому ориентацию в пространстве и координацию движений. Поэтому в последнее время интерес исследователей заметно переместился на изучение организации нервных сетей и ганглиозных механизмов, посредством которых передается и обрабатывается информация, поступающая от рецепторов. Изучение тонкого строения межнейронного взаимодействия н принципов обработки и выделения существенной информации в оптических, акустических, хеморецеп-
219
торных центрах надглоточного ганглия насекомых — это, пожалуй, сейчас основное направление концентрации усилий физиологов, работающих в области сенсорной физиологии насекомых. Такие исследования дают материал для специалистов, конструирующих сложные автоматические системы, в том числе роботы, которые должны «уметь» распознавать разнообразные внешние стимулы и на основе последних самостоятельно вырабатывать программу действий.
Перспектива дальнейшего развития физиологии органов чувств насекомых связывается также с задачей регуляции численности вредных и полезных видов без глубого вмешательства в природу, без загрязнения среды. Глубокое знание периферических и центральных рецепторных механизмов позволит воздействовать на насекомых путем сигнального управления их поведением. Уже сейчас синтезированные половые феромоны используются в практике борьбы с некоторыми насекомыми-вредителями, позволяющей строго избирательно воздействовать на фауну защищаемых угодий. Синтезирован ряд высокоэффективных репеллентов для кровососущих насекомых и клещей. Нанесение таких репеллентов на кожу тела, пропитка ими одежды или защитных сеток остаются пока единственным надежным способом защиты человека от «гнуса» в тайге или тундре. Известны также примеры успешного использования в практических целях оптических аттрактантов для насекомых.
ЛИТЕРАТУРА
ОБЩИЕ РУКОВОДСТВА
Проссер Л. П., Браун Ф. Сравнительная физиология животных. М., «Мир», 1967.
Шовен Р. Физиология насекомых. М., ИЛ, 1953.
Шовен Р. Поведение животных. М., «Мир», 1972.
В u 11 о с k Т. Н., HorridgeG. A. Structure and function in the nervous system of invertebrates. Vol. I, II. San Francisco, 1965.
D e t h i e г V. G. The physiology of insect sences. 2 Ed. London — N. Y., Methuen, 1972.
Rockstein M. (Ed.). The physiology of Insecta. Vol. II, 2 Ed. London — N.Y., Academic Press, 1974.
Roeder K. D. (Ed.). Insect physiology. N. Y., Wiley, 1953.
Wigglesworth V. B. The principles of insect physiology. 6 Ed. London — N.Y., E. Dutton, 1965.
МЕХАНОРЕЦЕПЦИЯ И СЛУХ
Биоакустика. Под ред. В. Д. Ильичева. М., «Высшая школа», 1975.
Жантиев Р. Д. Ультрастоуктуриая организация механорецепторных сенсилл насекомых. — «Жури. общ. биол.», 1969, т. 30, № 2.
Жантиев Р. Д. Звуковая сигнализация членистоногих. — В сб.: Звуковая сигнализация животных. Пущино, Изд-во Науч, центра биол. исслед. АН СССР, 1974.
220
Ж антиев Р. Д. Акустическая ориентация насекомых. — В сб.: Чтения памяти В. А. Холодковского. Докл. на 26-м ежегодном чтении. Л., «Наука», 1975.
Попов А. В. Сравнительное исследование коммуникационных звуковых сигналов и некоторых принципов организации слуховой системы цикад и прямокрылых. — «Тр. ВЭО», 1969, т. 53.
Свидерский В. Л. Нейрофизиология полета насекомых. Л., «Наука», 1973.
Alexander R. D. Acoustical communication in Arthropods. — «Ann. Rev.
Entom.», 1967, vol. 12.
Haskell P. T. Insect sounds. London, Witherby, 1961.
Michelsen A. Hearing in invertebrates. — In: Handbook of sensory physiology. Ed. board: H. Autrum, vol. V/I, Berlin — Heidelberg — N. Y., Springer-Verl., 1974.
XEMO-, ТЕРМО- И ГИГРОРЕЦЕПЦИЯ
Елизаров Ю. А. Хеморецепцня насекомых и репелленты. — «Успехи совр. бнол.», 1962, т. 54, вып. 4.
Елизаров Ю. А. Хеморецепцня кровососущих членистоногих. — «Успехи совр. биол.», 1965, т. 59, вып. 2.
Елизаров Ю. А., Синицына Е. Е. Физиологические особенности хеморецепторных органов насекомых. — «Тр. ВЭО», 1969, т. 53.
Миняйло В. А., Ковалев Б. Г. Половые феромоны и их применение в борьбе с вредителями. — В кн.: Энтомология, т. 2. М., Изд-во ВИНИТИ, 1973.
Хеморецепция насекомых. Вильнюс, т. 1, 1971; т. 2, 1975.
Jacobson М. Insect sex pheromones. London — N.Y., Academic Press, 1972.
К a i s s 1 i n g К. E. Insect olfaction. — In: Handbook of sensory physiology. Ed. board: H. Autrum, vol. IV/I, Berlin — Heidelberg—N. Y., Springer-Verl., 1971.
P i n e t J. M., Bernard J. Essai d’interpretation du mode daction de la va-reur d’eau et de la temperature sur un recepteur d’insecte. — «Ann. zool. ecol. anim.», 1972, vol. 4, No. 4.
S 1 i f e r E. H. The structure of arthropod chemoreceptors. — «Ann. Rev. Entom.», 1970, vol. 15.
ФОТОРЕЦЕПЦИЯ
Грибакин Ф. Г. Ультраструктурная организация фоторецепторов насекомых. — «Тр. ВЭО», 1969, т. 53.
Мазохин-Поршняков Г. А. Зрение насекомых. М., «Наука», 1965.
Мазохин-Поршняков Г. А. Современное состояние изучения зрения насекомых.— «Успехи совр. биол.», 1971, т. 72, вып. 2.
Мазохин-Поршняков Г. А. Механизмы цветоразличения у насекомых. — В кн.: Итоги науки и техники. Бионика, т. I. М., ВИНИТИ, 1973.
Францевич Л. И. Функциональная морфология зрительного анализатора насекомых. — «Журн. эвол. биох. и физиол.», 1971, т. 7, № 2.
Bernhard С. G. (Ed.). The functional organisation of the compound eye. London, Pergamon Press, 1966.
Horridge G. A. (Ed.). The compound eye and vision of insects. Oxford, Clarendon Press, 1974.
Snyder A. W., Menzel R. (Ed.). Photoreceptor optics. Berlin — Heidelberg — N. Y., Springer-Verl., 1975.
Wehner R. (Ed.). Information processing in the visual systems of Arthropods. Berlin, Springer-Verl., 1972.
Yagi N., Koyama N. The compound eye of Lepidoptera. Tokyo, 1963.
ПРОСТРАНСТВЕННАЯ ОРИЕНТАЦИЯ, РЕАКЦИЯ НА ЭЛЕКТРОМАГНИТНЫЕ ПОЛЯ
Frisch von К. Tanzsprache und Orientierung der Bienen. Berlin, Springer-Verl., 1965.
Lindauer M. (Red.). Orientierung der Tiere im Raum. Teil 1: Sinnes- und neu-rophysiologische Grundlagen. Stuttgart, Fischer-Verl., 1973.
221
ОГЛАВЛЕНИЕ
Предисловие ......................................................... 3
Введение. РОЛЬ ОРГАНОВ ЧУВСТВ В ЖИЗНИ НАСЕКОМЫХ ’ 5
Глава!.
ОБЩАЯ ХАРАКТЕРИСТИКА НЕРВНОЙ СИСТЕМЫ И ОРГАНОВ
ЧУВСТВ НАСЕКОМЫХ.....................................................14
1. Краткие сведения об анатомии н гистологии нервной системы ............................................14
2. Особенности сенсорной системы................................19
3. Физиологические механизмы сенсорной системы и методы их изучения.......................................25
Глава II.
МЕХАНОРЕЦЕПЦИЯ.................................................31
1. Краткие сведения о морфологии механорецепторов . 31
2. Функции тактильных рецепторов.........................35
3. Физиологические особенности проприоцепторов .... 38
4. Физиологические характеристики органов равновесия . . 45
Глава III.
СЛУХ...........................................................49
1. Коммуникационные звуковые сигналы.........49
2. Физические характеристики звуковых сигналов ... 53
3. Строение слуховых органов.................56
4. Функции слуховых органов..................59
5. Центральные отделы слуховых анализаторов .... 63
6. Механизм опознавания коммуникационных звуковых сигналов ............................................66
7. Пространственный слух.....................71
8. Виброрецепция.............................76
Глава IV.
ХЕМОРЕЦЕПЦИЯ.............................................81
1. Общие особенности строения и классификация хеморецепторных органов....................................83
2. Морфологическая характеристика контактных хеморецепторных сенсилл.................................89
3. Морфологическая характеристика обонятельных сенсилл . 95
4. Физиология контактных хеморецепторных сенсилл . . . 106
5. Физиология обонятельных сенсилл.........................124
Глава V.
ТЕРМОРЕЦЕПЦИЯ И ГИГРОРЕЦЕПЦИЯ...................................136
1. Температурные рецепторы.................................136
2. Рецепторы влажности.....................................139
222
Глава VI.
ФОТОРЕЦЕПЦИЯ.............................................142
1. Современное представление о функциональной организации сложного глаза.....................................142
2. Оптические ганглии и механизмы предварительной обработки зрительной информации . . ....... 149
3. Контрастная чувствительность, адаптация и ретиномоторные явления........................................157
4. Острота зрения, восприятие формы и объема .... 160
5. Восприятие поляризации света....................168
6. Цветовое зрение.................................172
7. Устройство и работа простых глазков.............189
Глава VII.
РЕАКЦИИ НАСЕКОМЫХ НА ЭЛЕКТРИЧЕСКИЕ И МАГНИТНЫЕ ПОЛЯ.....................................................198
1. Реакции на электрические факторы...............199
2. Реакции на электромагнитные колебания..........202
3. Реакции на магнитные факторы...................203
Глава VIII.
МЕХАНИЗМЫ ПРОСТРАНСТВЕННОЙ ОРИЕНТАЦИИ ... 207
1. Первичная и вторичная ориентация и контролирующие ее системы '..........................................207
2. Таксисы и тропизмы..............................209
3. Астрономическая ориентация .....................216
Заключение...............................................219
Л ит е рат у ра..........................................220