/




























































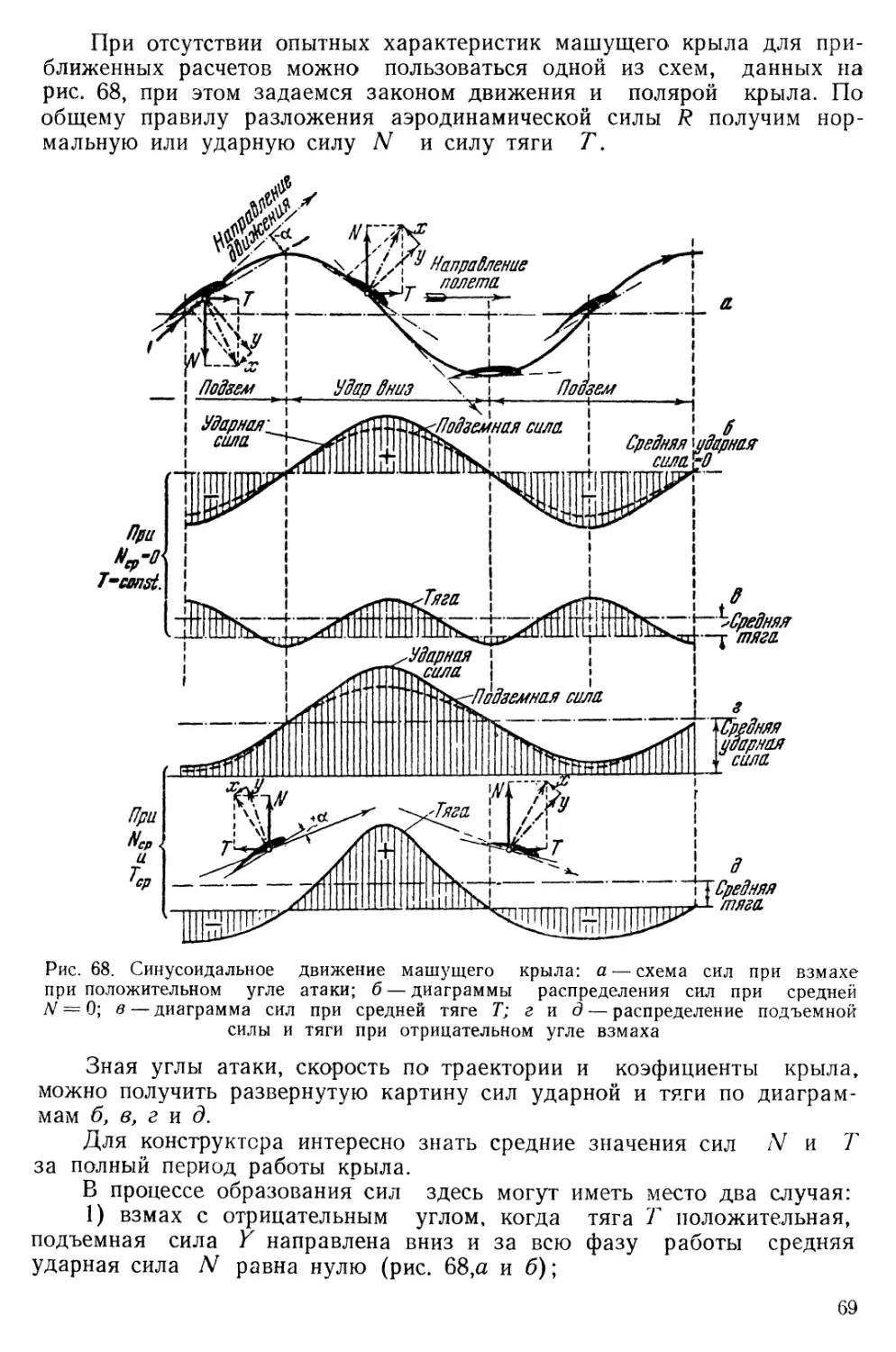

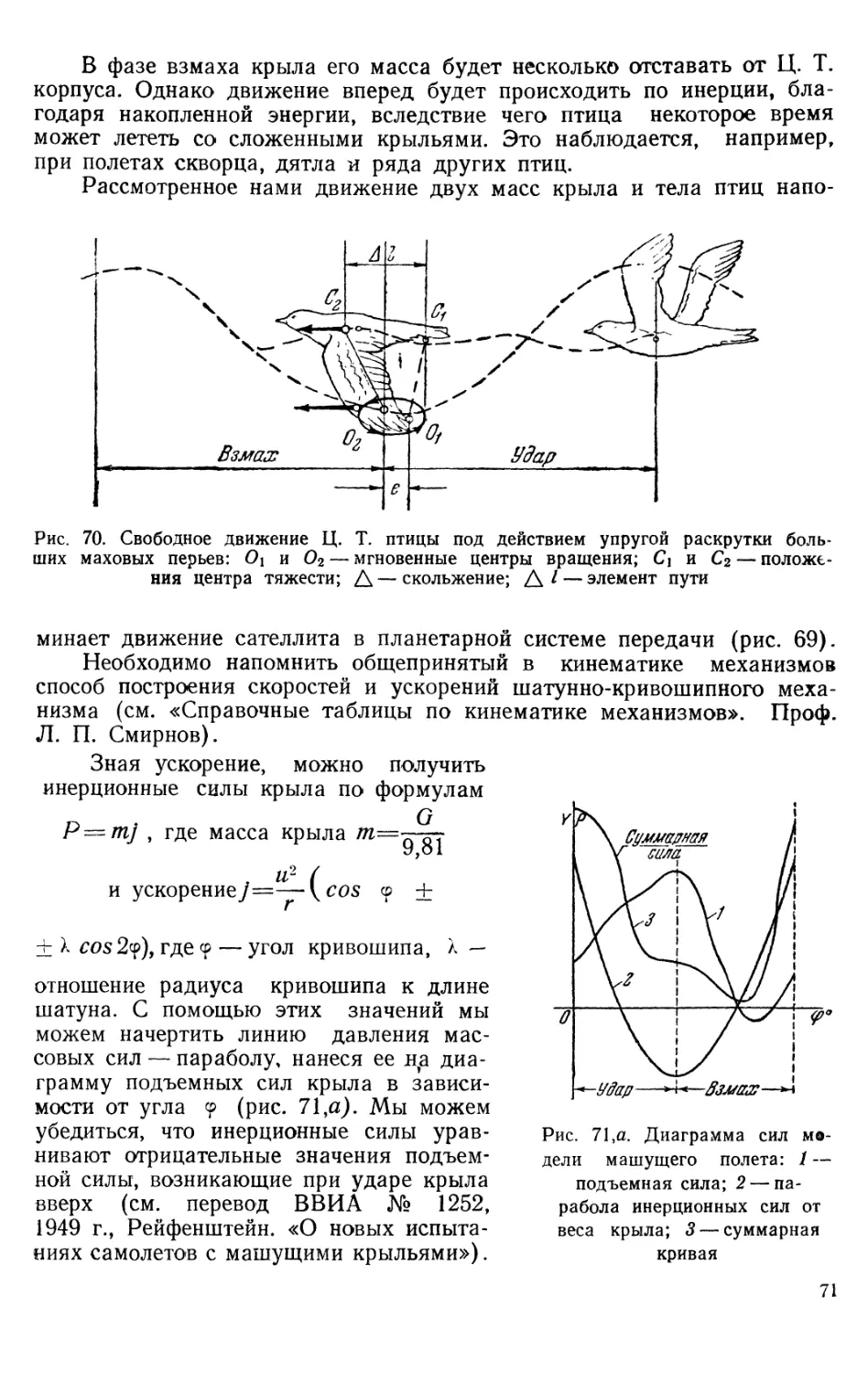






























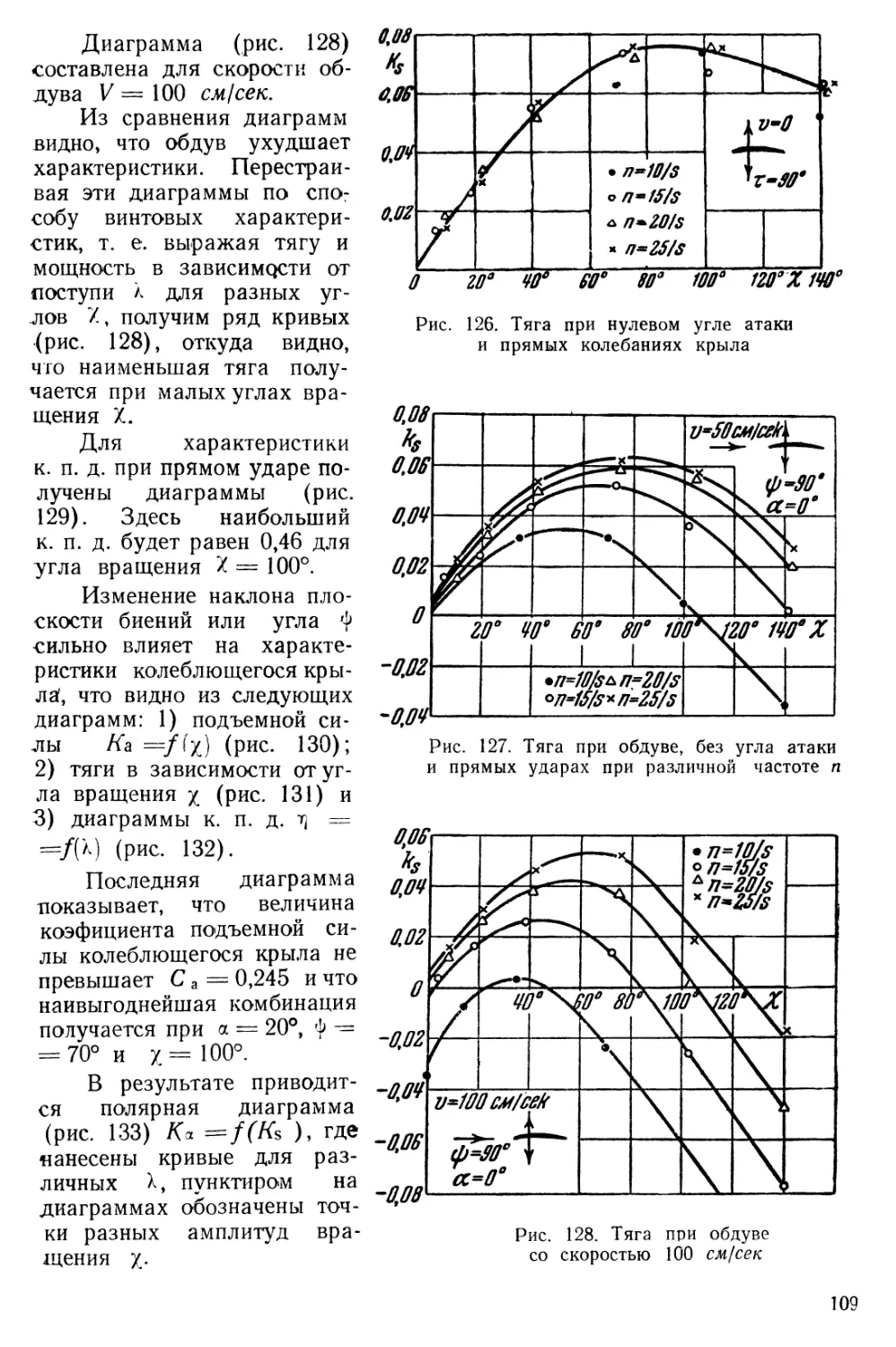

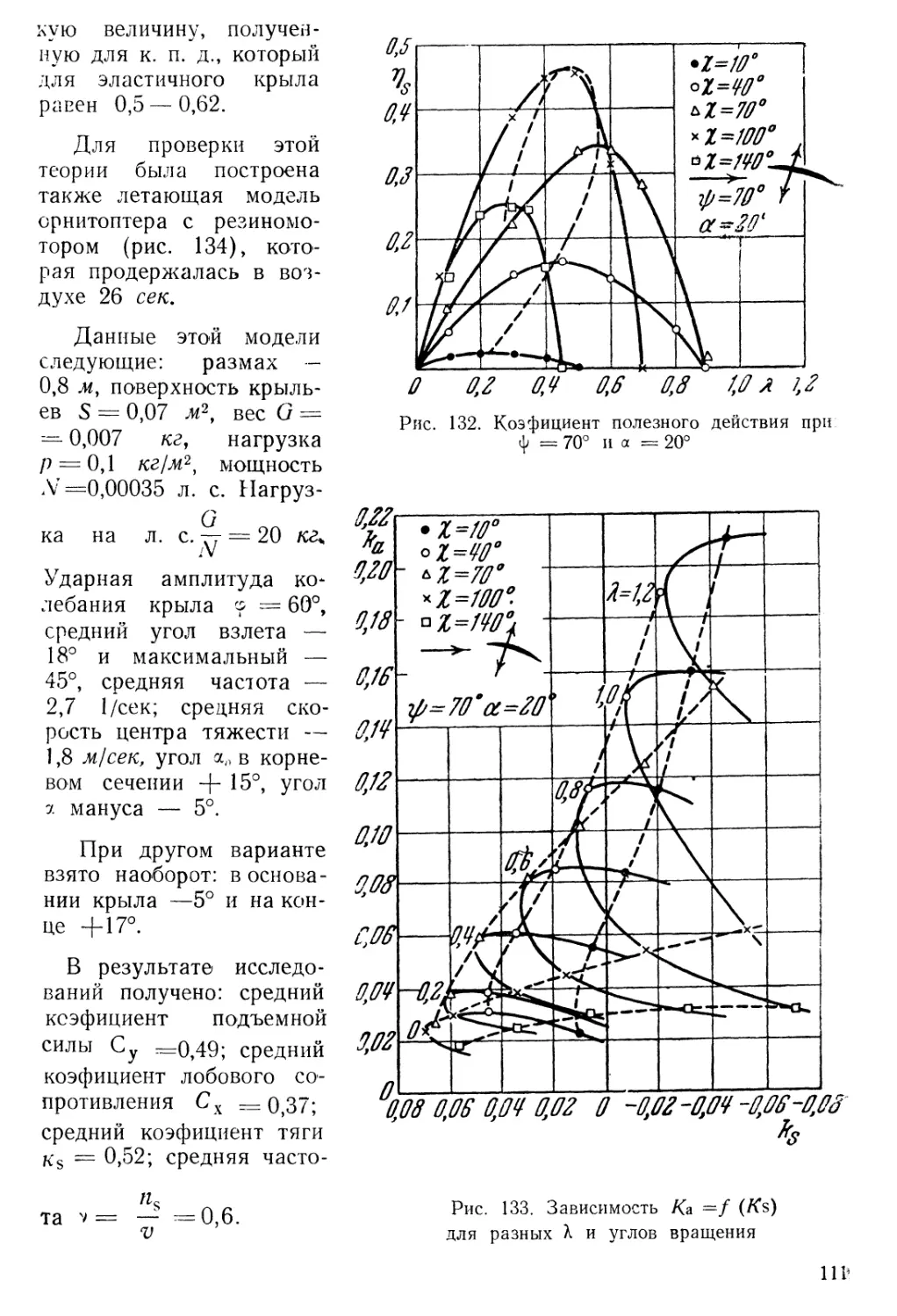





















Text
И. Н. ВИНОГРАДОВ
АЭРОДИНАМИКА ПТИЦ-ПАРИТЕЛЕЙ
ИЗДАТЕЛЬСТВО ДОСАРМ * МОСКВА-1951
ВСЕСОЮЗНОЕ ДОБРОВОЛЬНОЕ ОБЩЕСТВО
СОДЕЙСТВИЯ АВИАЦИИ
И. Н. ВИНОГРАДОВ
АЭРОДИНАМИКА ПТИЦ-ПАРИТЕЛЕЙ
В книге инженера И. Н. Виноградова рассмотрены полет и устройство крыла птиц-парителей с точки зрения современной аэродинамики.
Автор объясняет высокие полетные качества птиц эффектами проницания крыла воздухом и особой его кинематикой.
В книге приведены основные положения теории тяги в машущем полете и некоторые результаты по лабораторным исследованиям моделей птиц и элементов колеблющегося крыла на орнитоптерных приборах.
Книга рассчитана на общественность Досава и всех интересующихся проблемой машущего полета.
ПРЕДИСЛОВИЕ
Замечательные исследования ученых по прикладной аэродинамике, по теории крыла и винта дали прочную базу для теоретического расчета современных самолетов. Классические исследования Н. Е. Жуковского, С. А. Чаплыгина, замечательные работы других советских ученых, а также многочисленные теоретические и экспериментальные работы зарубежных исследователей дали достаточно законченную теорию тех механических процессов, которые вызывают образование сил, действующих на самолет в установившемся полете.
Гораздо менее подробно изучены те процессы, которые протекают в воздухе при неустановившемся полете, в частности, при машущем полете. Если колебания крыльев с малой амплитудой, т. е. различного рода вибрации крыльев, лопастей пропеллера и т. п., имеющих весьма важное значение в современной авиационной технике, вызвали появление многочисленных теоретических и экспериментальных работ, то работа машущих крыльев при больших амплитудах во многих частях еще совершенно не изучена; в самое последнее время появились работы, представляющие, несомненно, только первые шаги в этой области.
Причиной такого положения является недостаточный экспериментальный материал, который мог бы служить базою для построения полной теории работы машущих крыльев и возможности расчета сил, возникающих при этом.
За последние годы прйвился ряд интересных исследований в этой области. Работа инженера М. К. Тихонравова «Полет птиц и машины с машущими крыльями» (1-е изд. 1937 г.; 2-е изд. 1949 г.), обширное и весьма важное исследование профессора Н. А. Гладкова «Биологические основы полета птиц» (1949 г.), работы профессора Г. С. Шестаковой — все они дают обширный и важный материал по динамике машущего полета.
К этой же области относится и предлагаемая работа инженера И. Н. Виноградова «Аэродинамика птиц-парителей». Автор собрал
большой материал, во многих вопросах дал ему интересное освещение и сделал заслуживающие большого внимания выводы. Несомненно, что чтение этой книги принесет много интересного для всех работающих в этой области.
За последние годы интерес к исследованиям машущего полета чрезвычайно возрос и среди инженеров, и специалистов механиков, и среди биологов. Показателем этого может служить факт образования при Досаве секции машущего полета, которая объединяет и инженеров и биологов. Автор этой книги является одним из организаторов и деятельных работников секции. Можно пожелать, чтобы книга наряду с другими работами привлекла внимание инженеров, биологов и теоретиков к этой интересной и многообещающей для механики области.
Профессор Вл. Голубев.
ВВЕДЕНИЕ
В истории авиации и воздухоплавания можно установить несколько периодов, когда среди ученых и изобретателей наблюдался повышенный интерес к идее машущего полета. Ввиду неудач и сопряженных с ними разочарований интерес к проблеме полета птиц временами ослабевал, но затем появлялся вновь.
В начале XX столетия, в связи с бурным развитием моторной авиации, наступил перерыв в исследованиях машущего полета(. И только в начале тридцатых годов, после первых успехов безмоторного летания, снова пробуждается интерес к изучению полета птиц. В некоторых конструкциях планеров были воспроизведены внешние очертания и профили птичьих крыльев.
В последнее время развитие реактивной техники снова отодвинуло на второй план исследование полета птиц. Эта проблема остается нерешенной и в наши дни.
Итак, в течение 140 лет со времени постройки первой летающей модели все попытки воспроизвести машущий полет птиц оказались безрезультатными. Причинами такого положения являются: во-первых, неясность самой схемы работы крыла птицы и, во-вторых, механический перекос формул и методов аэродинамических исследований к колебательному движению крыла птицы. В этом, отчасти есть вина орнитологов, выводами которых пользовались изобретатели приборов с машущими крыльями.
Орнитологи совершенно не обращали внимания на процессы, проходящие внутри крыла, и на самую структуру его туннельных образований. До сего времени существовало представление о непроницаемости крыльев, что автоматически перешло в нашу техническую литературу. Между тем фотосъемка телеобъективом помогла обнаружить проницание крыла потоком.
Вполне естественно, что аэродинамические испытания мертвых птиц в трубе не соответствовали условиям работы живого организма, вследствие чего были получены очень низкие числовые показатели по сравнению с самолетом и планером. Автор считает, что настало время заново пересмотреть весь накопленный материал наблюдений, пополнить его и дать оценку работе отделов крыла птицы с точки зрения известных в самолетостроении аналогичных устройств и создать новую
методику эксперимента в целях установления физической картины машущего полета.
Вопросы аэродинамики птиц интересовали многих русских ученых.
Родоначальник моторной авиации русский офицер А. Ф. Можайский после долгого изучения полета птиц осуществил постройку первого в мире самолета с паровым двигателем.
Знаменитый аэродинамик профессор Н. Е. Жуковский в своем первом докладе «О воздухоплавании» в 1898 г. высказывал идеи, которые легли в основу динамики орнитоптера. Он говорил: «Совершенная летательная машина отбрасывает вниз возможно большее количество воздуха с возможно меньшей скоростью».
К славной плеяде русских изобретателей — пионеров машущего полета — следует отнести кандидата С.-Петербургского университета В. М. Михневича, который в 1871 г., за 19 лет до появления книги «Полет птиц» французского ученого Марэя, представил в ученое отделение Морского технического комитета подробную записку «О летании птиц и устройстве воздухоплавательного снаряда». В его машущем аппарате крылья шарнирно крепились к перекладине, а на них устанавливались специальные рычаги, концы которых были стянуты пружиной.
Технический комитет, исходя из того, что вопрос о летании по воздуху на практике недостаточно исследован, решил «оставить предложение г. Михневича без последствий».
Лейтенант В. Л. Спицын в 1880 г. на основе изучения полета птиц предложил аппарат, у которого четыре машущих крыла поднимались вверх ребром и опускались плашмя при частоте двух ударов в секунду.
Ряд других русских изобретателей занимался изучением машущего полета птиц для применения его принципов в конструкциях орнитоптеров.
Весьма поучительный эксперимент по изучению полета птиц производил в 1885— 1890 гг. доктор Арендт в г. Симферополе.
Он замораживал живых птиц при различных положениях крыльев и хвоста, затем поднимал их на змее и, сбрасывая с высоты 50 м, наблюдал их планирующие и даже парящие полеты.
В 1908 г. летчик А. В. Шиуков производил заклеивание крыльев живых птиц, лакировал их поверхности, причем в той или иной степени птица теряла способность к полетам.
В 1908—1910 гг. доктор Е. П. Смирнов создал оригинальную теорию динамического парения птиц, основанную на свойствах крыла. По его заключению летательный аппарат птицы представляет весьма совершенный парашют, эластичность элементов которого и щелевидные образования имеют весьма большое влияние на качество полетов.
Знаменитый русский летчик, автор мертвой петли П. Н. Нестеров в 1913 г. построил самолет на основе изучения полета птиц. Главнейшую особенность его аппарата составляло отсутствие вертикального оперения, функции которого выполнялись горизонтальным оперением подобно птицам. Кроме того, в своем изобретении П. Н. Нестеров впер-
ьые осуществил перемену угла атаки крыла от 0° до 9° движением особого эксцентрика.
Работа инж. М. К. Тихонравова «Полет птиц и машины с машущими крыльями», изданная в 1937 г., основана на изучении полета птиц. Она дает богатый статистический материал по орнитологии, обработанный на основании аэродинамических представлений того времени. Вопросам .построения научной теории машущего полета посвящены труды (10 и 119 члена корреспондента Академии наук СССР профессора В. В. Голубева, в которых он впервые изложил стройную вихревую теорию возникновения тяги машущего крыла и указал на большое значение вязкости воздуха.
Из последних наших отечественных работ должен быть отмечен труд доктора биологических наук Н. А. Гладкова — «Полеты в природе». Он углубляет связь между орнитологией и аэродинамикой в целях объяснения вопросов, связанных с парением птиц, кинематикой движения их крыльев и приспособлениями для затягивания срывов потока.
Большой интерес представляет работа доктора биологических наук Г. С. Шестаковой «Полет птиц» и ее исследование в области структуры поверхностного слоя птицы.
В настоящее время работу птичьего крыла мы можем рассматривать с точки зрения современной аэродинамики, на основе, результатов изучения пограничного слоя потока, обтекающего крыло.
В предлагаемой книге анализ работы элементов крыла был произведен на основании данных многолетних наблюдений и изучения фото-и киносъемок различных птиц в полете. Выводы автора об особенностях работы проницаемого крыла подтвердились при испытании моделей птиц в аэродинамической трубе в ноябре 1950 г.
Новые данные о структуре крыла птиц-парителей, полученные при участии профессора Г. С. Шестаковой, были доложены автором в институте эволюционной морфологии Академии наук СССР в 1948 г., а также в Министерстве авиационной промышленности и 24 октября 1948 г. на конференции в Центральном аэроклубе Досава.
В книге приведены некоторые материалы для приближенного аэродинамического расчета и материалы по волновой структуре отходящего за крылом птицы потока
В связи с новыми понятиями о свойствах птичьего крыла в книге встречаются новые термины, как-то: «проницание крыла», «приспособи-тельность обтекания», «ударная амплитуда», «удар крыла», «увязание крыла в воздухе» и др. При пользовании графиками из иереводной литературы следует руководствоваться переходными формулами, помещенными в тексте.
Всесоюзное добровольное общество содействия Авиации ведет большую работу среди широких масс населения. Среди других секций в Досаве существует секция машущего полета, члены которой изучают полеты в природе и работают над созданием летательных аппаратов с движущимися или машущими крыльями.
1 В конце книги приведен список использованной автором литературы. Цифры в скобках, встречающиеся в тексте (например, (1) или (10 и 11) и т. д.), обозначают порядковый номер труда, приведенного в этом списке.
Новая проблема, столь увлекательная по своему содержанию, несомненно, заинтересует молодежь, перед которой открываются новые перспективы летать на планерах, превращенных в орнитоптеры.
В нашей стране благодаря заботам партии, правительства и лично, товарища Сталина созданы все условия для развития науки и техники.
Разрешение проблемы машущего полета — это одна из задач авиационной техники, открывающая новые широкие горизонты.
Автор приносит глубокую благодарность профессору Г. С. Шестаковой за консультации в вопросах анатомического строения птиц и предоставленные материалы для изучения их крыльев и кандидату технических наук инженеру И. П. Толстых за ценные указания и советы по существу содержания этого труда. Автор считает своим долгом отметить, что большую помощь в его работе оказал ему покойный академик заслуженный деятель науки В. П. Ветчинкин.
ГЛАВА 1
КАК УСТРОЕН ЛЕТАТЕЛЬНЫЙ АППАРАТ ПТИЦ
ОСНОВНЫЕ ПОНЯТИЯ О СТРУКТУРЕ КРЫЛА
Строение крыла птицы изучено в настоящее время очень подробно, и мы воспользуемся основными терминами орнитологии, чтобы сохранить единство понятий в определении частей крыла и выполняемых ими функций.
Рассматривая внешний наряд крыла птицы (рис. 1 и 2), мы отмечаем:
1. Большие маховые перья, называемые также первостепенными (рис. 2; I—X).
2. Малые маховые, или второстепенные (рис. 2; 1 —12).
3. Плечевые перья L и подмышечные М.
4. Кроющие, наложенные черепицеобразно на ряды основных перьев (рис. 1,а и Ь).
5. Крылышко К, или лучевой предкрылок.
Костная структура крыла (рис. 2) состоит:
1. Из плечевой кости А.
2. Предплечья В, состоящего из двух костей — лучевой С и локтевой D.
3. Мануса FH, состоящего из костей запястья F, пястья Е и фаланг пальцев Н, G, J.
Эти кости представляют связанную между собой суставами систему костных рычагов; все предплечье работает, как шарнирный параллелограмм.
Плечо соединено с корпусом через плечевой шарнир А и приводится в движение грудным мускулом при опускании плеча и подъемным мускулом при его подъеме.
Движение в шарнире ограничено. Вращение около поперечной оси известно в анатомии под названием пронации и супинации, происходит в очень узких пределах (от —5° до +50°) (у человека кости предплечья могут поворачиваться около продольной оси почти до полного перекрещивания). При полностью раскрытом крыле кости предплечья образуют с плечом угол от 90° до 140° в зависимости от вида птиц и применяемого ими способа полета. В устройстве крыла обращает на себя внимание треугольная кожная складка ДА (потаги-ум)—она выравнивает передний край крыла до прямой линии и создает при парении глубокую вогнутость нижней поверхности крыла.
ПЕРЬЕВОЙ АППАРАТ
Рассмотренный нами костный скелет крыла птицы (рис. 2) покрыт малыми и большими веерно-расходящимися перьями.
Группа больших маховых перьев в количестве от 10 до 12 прикреплена к запястью Е и фалангам пальцев Я, Л эта группа с самостоя-
тельными, разнообразными движениями образует треугольную поверхность мануса, занимающую от 30 до 40% всей площади крыла.
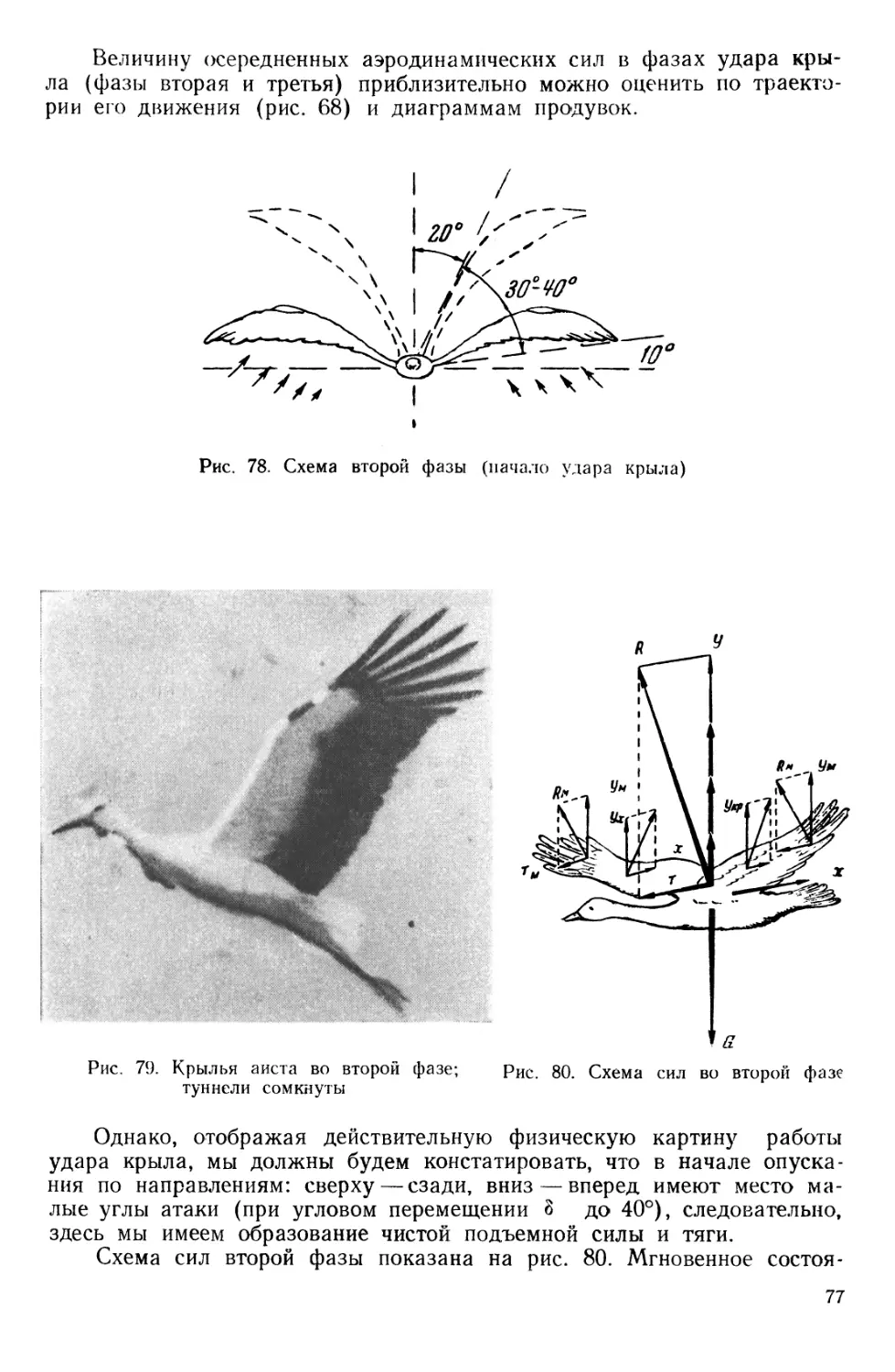
Первое короткое перо I очень напоминает лопасть пропеллера и отличается наибольшей жесткостью, угол установки основания пера 35°, а конца около 15°. Это перо вытягивает крыло при подъеме.
Второе, более длинное II имеет несколько меньшую закрученность и является как бы контрпропеллером, выравнивающим поток от перво
го пера. Остальные перья (рис. 3) имеют широкое желобчатое или S -образное основание и длинный узкий конец, саблевидно изогнутый и легко деформируемый потоком. Углы установки этих перьев у коршуна в свободном распущенном состоянии в сечении 4 (рис. 11) даны в таблице 1.
Эти перья иногда называют пропеллирующими, так как они дают тягу в машущем полете.
Задняя
Рассматривая детально большие маховые перья, мы заметим, что переднее перо является гораздо более узким и жестким, чем задние, на поверхности последних имеется широкая волнистая складчатость, которая расходится при движении крыла. Подобная структура пера
Таблица 1
№ пера
I II III IV V VI
Угол в основании пера 35° 20° 15° 10° 10° 8°
и упругий очин наилучшим образом обеспечивают пружинный эффект отдачи в начале фазы подъема крыла.
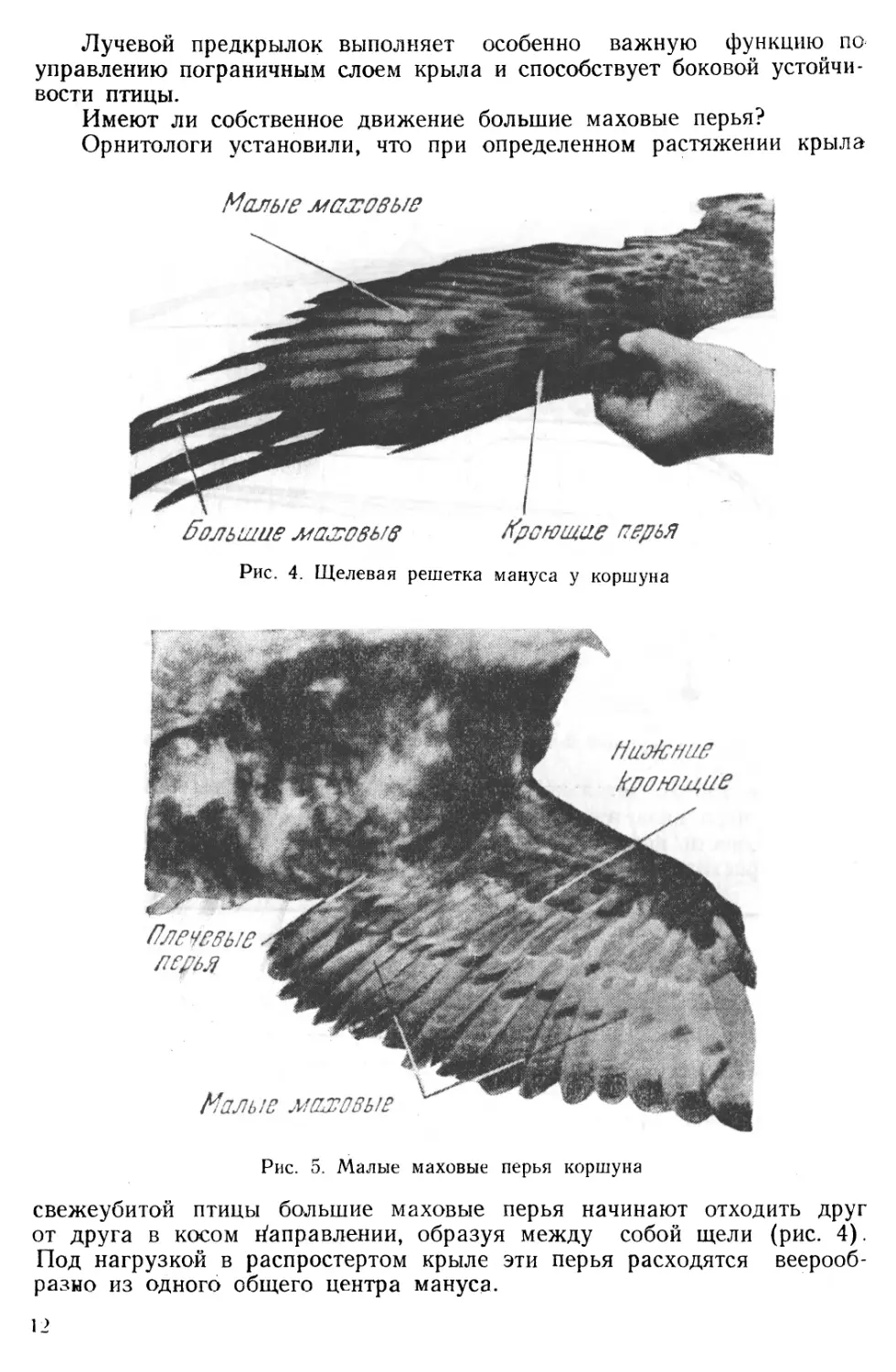
В зоне шарнира мануса и непосредственно над основанием больших перьев (рис. 2) находится лучевой предкрылок К, известный под латинским названием «алюля».
Перышки этого образования прикреплены к первому пальцу мануса (рис. 4) и через особую систему мускульных связей имеют самостоятельное движение.
Лучевой предкрылок выполняет особенно важную функцию по управлению пограничным слоем крыла и способствует боковой устойчивости птицы.
Имеют ли собственное движение большие маховые перья?
Орнитологи установили, что при определенном растяжении крыла
Рис. 4. Щелевая решетка мануса у коршуна
Рис. 5. Малые маховые перья коршуна
свежеубитой птицы большие маховые перья начинают отходить друг от друга в косом Направлении, образуя между собой щели (рис. 4). Под нагрузкой в распростертом крыле эти перья расходятся веерообразно из одного общего центра мануса.
При дальнейшем растяжении крыла каждое перо разворачивается около продольной оси своего стержня, пбд определенным углом к горизонту.
Кроме того, в полете под действием аэродинамических сил концы больших маховых перьев быстро колеблются в вертикальном направлении. Такого рода колебательные движения перьев играют большую роль в образовании тяги для поступательного движения.
Наблюдая птицу на светлом фоне неба, можно заметить, что концевая область мануса, вследствие указанного явления, иногда вырисовывается несколько расплывчатой или размытой.
Поперечное щелеобразов ание и эластичность перьев составляют весьма существенное отличие природного крыла от самолетного.
МАЛЫЕ МАХОВЫЕ ПЕРЬЯ
Эта группа перьев пТиц в количестве до 20 у сухопутных и до 37 у морских парителей прикреплена к костям предплечья параллельно друг другу.- Длина всех маховых перьев одинакова, опахала их симметричны относительно оси стержня (рис. 5).
Имея S-образное сечение опахал, малые маховые перья образуют туннели (рис. 6), сообщающиеся через нижние кроющие перья с подкрыльной областью.
При виде сзади перья имеют определенные углы поворота по отношению к плоскости плечевой области, постепенно возрастающие к шарниру плеча.
Рис. 6. Туннели перьев в средней области
В таблице 2 приведены углы поворота перьев при полном раскрытии крыла.
Та б
№ пера
1-4 5—6 7-8 9—10 10-12
40°—22° 20'’—15° 13° 10°—5° 5°
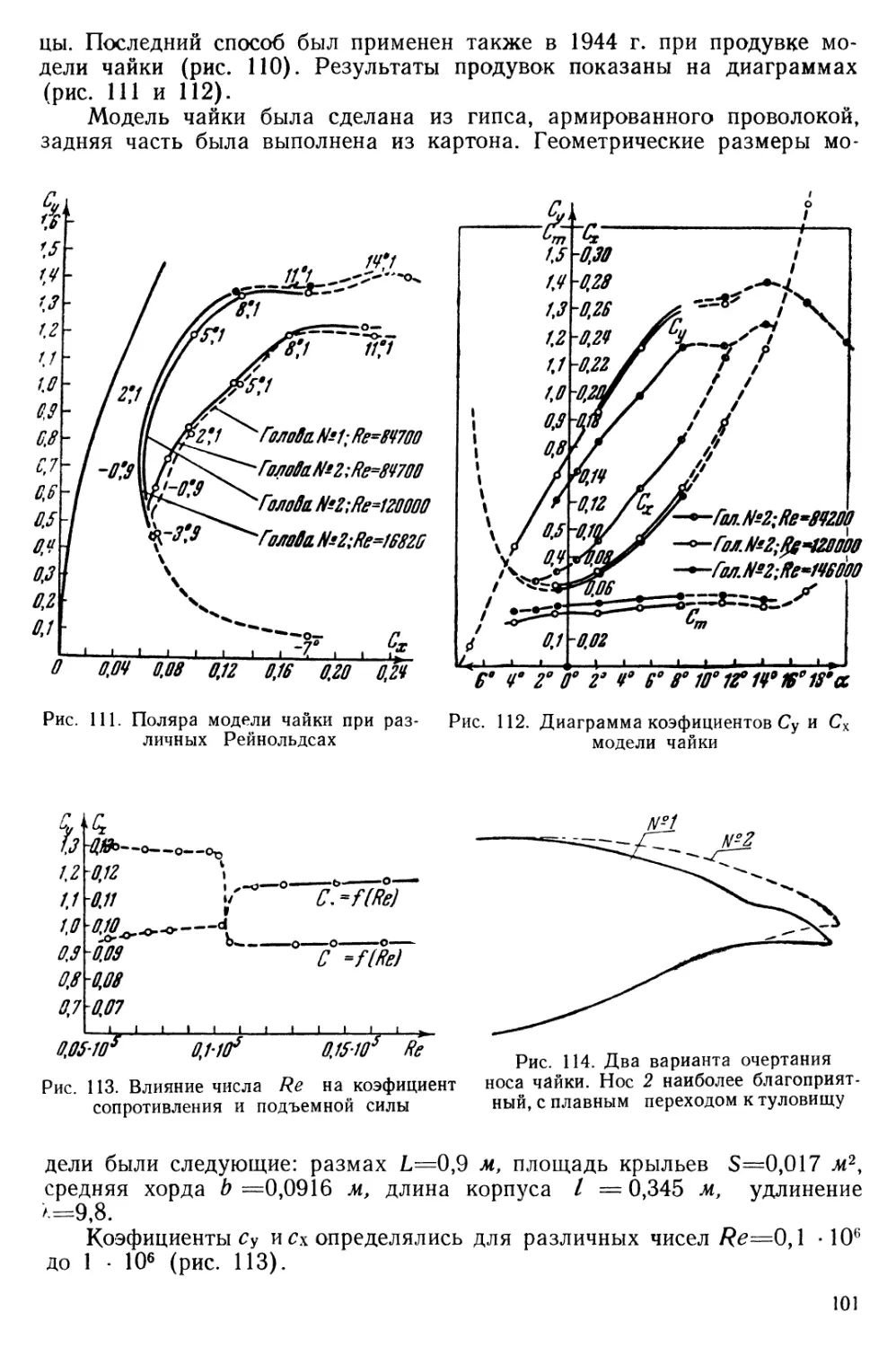
Наибольший поворот этих перьев, согласно фото, имеет место в фазе подъема крыла, очевидно, для пропуска воздуха. При обратном движении перья накладываются, образуя туннели (рис. 7).
Рис 7. Наполнение туннелей левого крыла чайки воздухом при взмахе
Туннели плечевой области имеют переменный объем и выполняют весьма важное назначение (см. главу IV).
ОБЛАСТЬ ПАРАШЮТНЫХ ОТВЕРСТИЙ КРЫЛА
Обособленную группу перьев составляют подмышечные у основания крыла М (рис. 2).
При парящем полете они выполняют роль зализа крыла самолета, при взмахе же они частично перепускают воздух из подкрыльной области в ряды верхних кроющих перьев для сдувания пограничного слоя. В момент крутого парашютирования через них прорывается поток для устойчивого спуска, как у парашютов через отверстие в зенитной точке.
Плечевые группы перьев с наружной стороны покрыты тройным рядом покровных перышек, а с внутренней — одним или двумя рядами легко отходящих и менее плотных перьев.
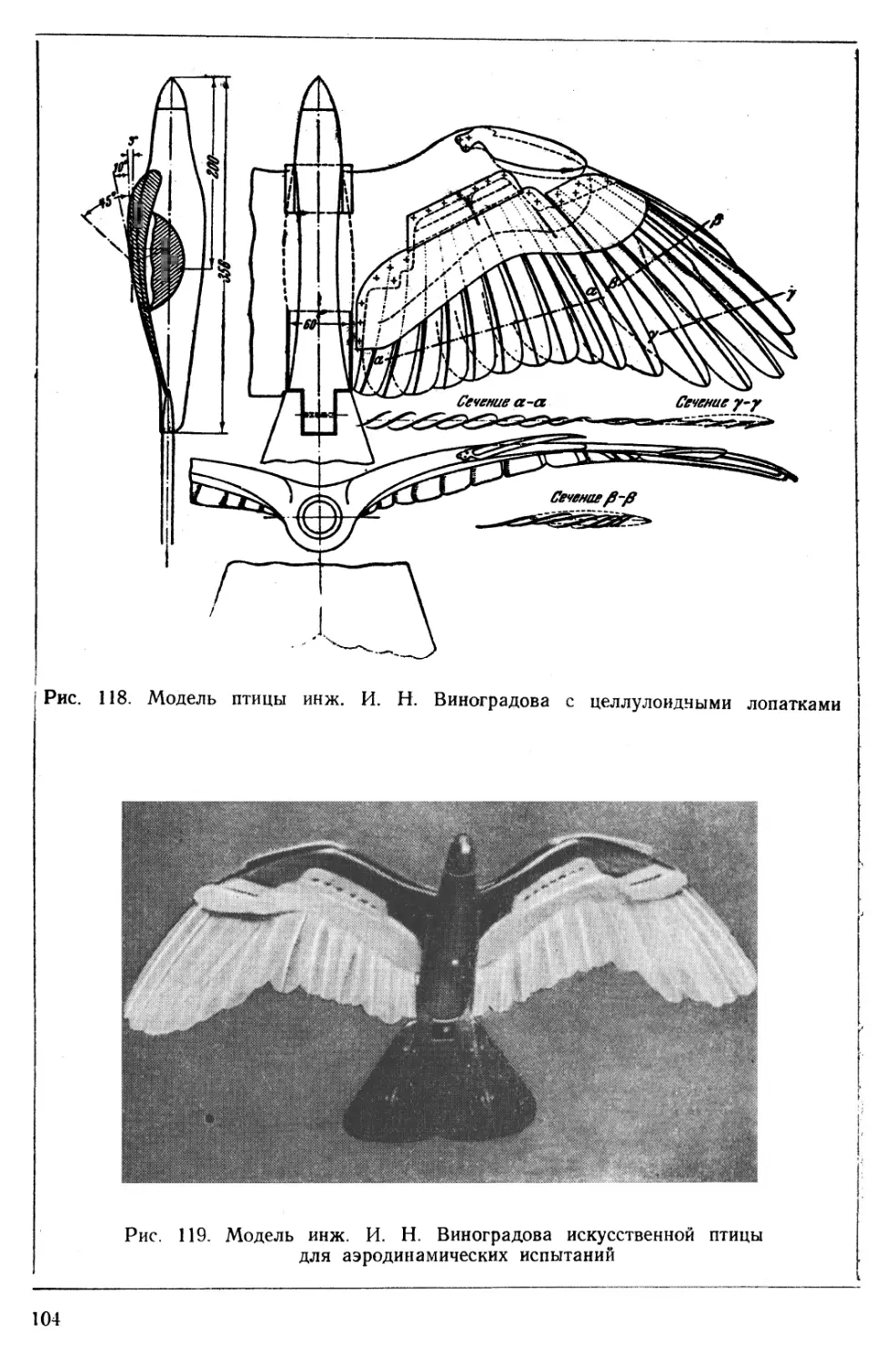
ПРИСПОСОБЛЕНИЯ, ПОДОБНЫЕ МЕХАНИЗАЦИИ КРЫЛЬЕВ САМОЛЕТОВ
Путем тщательного изучения устройства крыла птицы были обнаружены в нем хорошо известные в самолетостроении элементы механизации (рис. 8).
На схеме а показана область решетчатых крыльев системы^ С. А. Чаплыгина, соответствующая вееру мануса у птицы.
На схеме б мы видим аналогичный алюле предкрылок самолета и известное многощелевое устройство.
Морская, птица фрегат в дает пример своеобразного применения закрылка фаулера в природе — мощную посадочную механизацию, уве-
Рис. 8. Устройства крыла птицы, напоминающие механизацию самолета
личивающую подъемную силу крыла самолета. Крыло сокола в сочетании с хвостовым оперением дает пример посадочного щитка на самолете. (В правом углу (рис. 8) на схеме д показаны средства механизации крыла самолета).
Проницаемость крыла птицы, кроме видимых щелей, обнаруживается также в подслойном прорыве потока в рядах кроющих перьев. Отсюда следует один вывод, что крылья птиц не обладают герметичностью, кроме внешнего обтекания по контуру профиля, они проницаются потоком, воздействующим на пограничный с л о й, а в итоге на лобовую и Подъемную силы крыл.а.
Это — главное отличие природного крыла от самолетного, влияющее на пограничный слой.
КОНТУРНЫЕ И ПРОФИЛЬНЫЕ ОЧЕРТАНИЯ КРЫЛА ПТИЦЫ
Крыло птицы является в высшей степени подвижным. Птица легко изменяет в полете очертания контура профиля, а также размах, глубину и хорду.
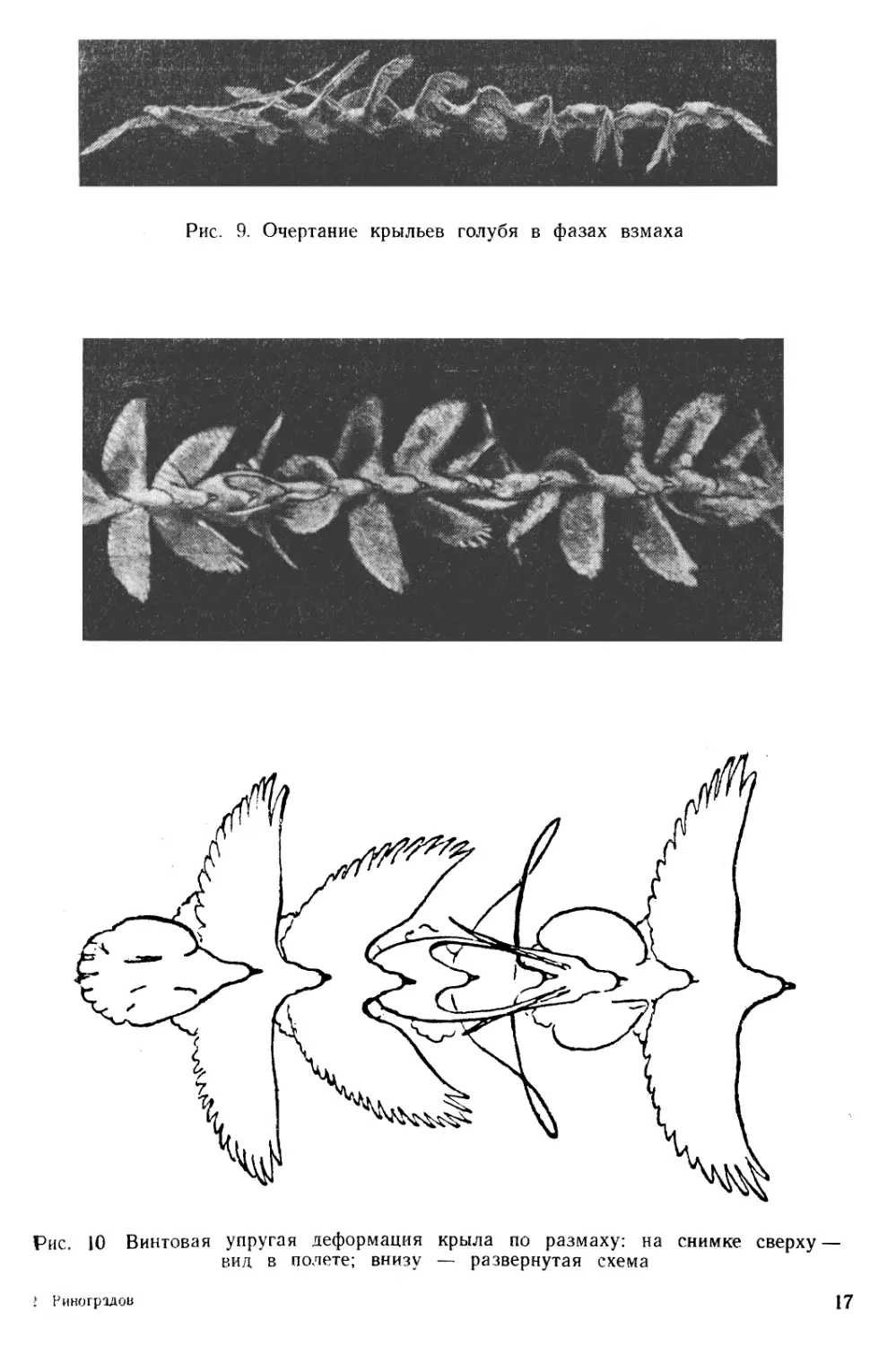
При киносъемке птиц в полете установлено, что их крылья в различные фазы взмаха отличаются большим разнообразием контурных очертаний (рис. 9).
При взмахе элементы крыла движутся с различными окружными скоростями, причем его внешний отдел, или манус, обгоняет в своем движении область предплечья, благодаря чему крыло принимает форму опрокинутой вниз чаши или форму ладьи с веерообразными и загнутыми вверх концами.
У некоторых птиц, например, у голубя и болотного луня, в верхней мертвой точке мы видим предельное сближение крыльев; для уток характерной особенностью положения крыльев в нижней точке являются отвесно опущенные концы мануса.
Площадь проекции крыла при взмахах то увеличивается до полных габаритов распластанного крыла, то уменьшается, при этом манус движется с различными ускорениями.
Одновременно изменяется стреловидность в плане и претерпевает упругие изменения опорная поверхность опахал крыла, деформирующихся по винтовой или, вернее, гиперболоидной поверхности (рис. 10), с ясно выраженным переходом в зоне мануса.
Не менее замечательны изменения профиля крыла в полете.
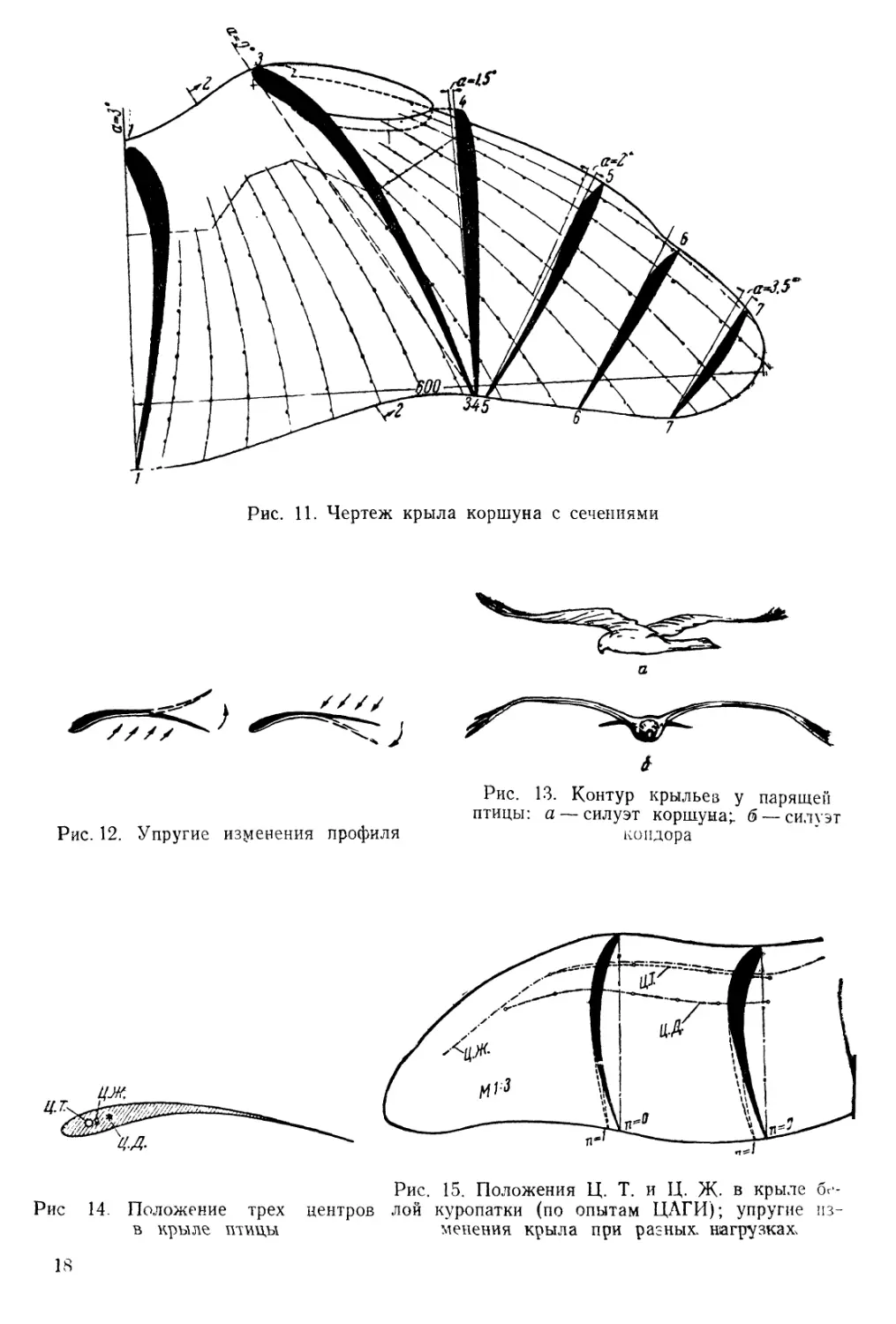
При измерении полностью раскрытого крыла коршуна установлено, что очертания его профиля изменяются от глубоковогнутого при основании до двояковыпуклого (рис. 11). Контур профиля в плечевой области у орла и коршуна приближается к профилю Г461, в средней зоне у лучевого предкрылка — к ЦАГИ 692 и на конце — к двояковыпуклому профилю. Следует отметить, что в зоне мануса носовая часть профиля клинообразная благодаря двум первым пропеллерным сильно изогнутым перьям.
Самый профиль крыла птиц под нагрузкой принимает S-образный вид (рис. 12). Контуры крыльев парящей птицы показаны на рис. 13.
Ввиду эластичности перьев многие исследователи утверждают, что при парении передняя или носовая кромка крыла ниже заднего ребра (рис. 13), так что положительного угла атаки по отношению к горизонту у парящей птицы может и не быть1.
1 Имея в виду, что в плечевой области у птиц профиль крыла сильно вогнутый и имеет подъемную силу при отрицательных углах атаки до — 5°, это явление вполне возможно.
Кроме того, птица обычно парит в восходящем потоке, так что положительного угла атаки по отношению к горизонту у птиц может и не быть.
Рис. 9. Очертание крыльев голубя в фазах взмаха
Рис. 10 Винтовая упругая деформация крыла по размаху: на снимке сверху — вид в полете; внизу — развернутая схема
> Виноградов
17
Рис. 11. Чертеж крыла коршуна с сечениями
Рис. 12. Упругие изменения профиля
Рис. 13. Контур крыльев у парящей птицы: а — силуэт коршуна;, б — силуэт кондора
Рис. 15. Положения Ц. Т. и Ц. Ж. в крыле бе-
Рис 14. Положение трех центров лой куропатки (по опытам ЦАГИ); упругие пз-в крыле птицы менения крыла при разных, нагрузках.
СОПРОТИВЛЯЕМОСТЬ КРЫЛЬЕВ НАГРУЗКАМ
Расположение плечевых костей птицы является наиболее благоприятным для работы крыла на прочность. Здесь наблюдается тот случай, известный из практики самолетостроения, когда центр тяжести (Ц.Т.) совпадает с его центром жесткости (Ц.Ж-), т. е. с той точкой, около которой крыло поворачивается под действием крутящего момента. Аэродинамическая сила приложена в центре давления (Ц.Д.) (рис. 14).
Если же все три центра совмещаются в одной точке (Ц.Ж.), то кручения не будет и крыло только изгибается. Эти условия являются необходимыми для устранения вибрации крыла, и птицы в совершенстве этим пользуются, избегая возникновения больших силовых напряжений в структуре крыла. На рис. 15 показано положение линий всех трех центров в крыле белой куропатки при испытании его на прочность (16) под нагрузкой, вес которой изменялся от единичного, равного половине веса птицы (п=1), до 13-кратного, при котором крыло сломалось. Пунктиром обозначено смещение профиля при нагрузке однократным весом. Расчетом на прочность было установлено, что материал костей крыла по своей крепости приближается к мягкой стали.
УГЛЫ УСТАНОВКИ И УГЛЫ АТАКИ КРЫЛЬЕВ У БОЛЬШИХ ПТИЦ
Угол установки крыла или угол между хордой корневого сечения и осью корпуса птицы-парителя находится в пределах от с? = 12° до 26° при сложенном состоянии крыльев и может меняться в полете до -J-3501.
Непосредственным измерением крыльев коршуна установлено, что они имеют геометрическую крутку.
Если принять в крыле коршуна (рис. И) за базу сечения 3, то получим углы закрутки 3 ненагруженного крыла и углы установки р-г? (таблица 3).
Таблица 3
Сечения
1 2 3 4 1 1 5 6
3° 3° Г 40' оэ 1°30' 2° 3°36'
3+? 15° 13°40' 12° 13°30' 1 14° 14°36'
1 Это изменение угла установки происходит периодически под действием мускулов.
Углы установок перьев мануса и малых маховых перьев коршуна в среднем сечении, считая от конца крыла, даны в таблице 4.
Таблица 4
№ пера
17 1 16 15 14 13 12 11 10 9
?ш 30° 25° 20° 15° 10° 8° С о 10° 15’
+ Ф 50° 45° 40° 35° 30° 28° 25“ 30й 35°
Повидимому, в машущем полете перья мануса работают в закри-тической области углов атаки при отсутствии срыва потока вследствие
Рис. 16. Упругие изменения чаши мануса в полете: / — плечевая область; II — манус
Рис. 17. Чашевидность и закрученность крыла коршуна в свободном состоянии
упругих колебаний. Крылья крупных птиц можно рассматривать состоящими из двух чашевидных углублений (рис. 16), где I— плечевая, или пазушная область с глубокой вогнутостью; II — область мануса, повернутая на угол А® . При опускании крыло вследствие упругости перьев распрямляется и концевые части перьев мануса дают обратный саблевидный изгиб (рис. 16).
На рис. 17 можно проследить чашевидность крыла коршуна по натянутой нити.
По сравнению с крылом самолета парашютообразная структура крыла птицы является одним из отличительных его свойств.
Рис. 18. Винтовой изгиб по размаху крыла чайки
Рис. 19. Очертание раскрытых крыльев коршуна
При посадке с парашютированием (рис. 31) были определены: установочный угол 26°, средние углы атаки для плечевой области крыла 58—60° и для мануса 62—70°.
РАЗМАХ И УДЛИНЕНИЕ КРЫЛЬЕВ ПТИЦ
В результате изучения крыльев сухопутных и морских парителей было установлено различие в удлинении, очертаниях и углах закрутки.
Альбатросы, осуществляющие динамическое парение над волнами, имеют размах до L = 3,4 м и удлинение Z = 16. Серебристая чайка имеет удлинение 9,8; крылья закручены по размаху и походят на пропеллер (рис. 18).
Крыло коршуна и орла одинаково хорошо приспособлено для парения как в слабых термических потоках, так и в сильных восходящих течениях около кучевых облаков и в потоках обтекания. Удлинение их крыльев порядка X = 5—6, очертания близки к трапецевидным с закругленными концами (рис. 19). Из наблюдений установлено, что короткие и широкие крылья хорошо приспособлены к подъему, длинные и узкие — к быстрому полету.
ГЛАВА II
ВИДЫ И ПРИЕМЫ ПОЛЕТОВ ПТИЦ
Исследование полета птиц имеет своею целью практическое использование вновь открытых эффектов крыла для летательных аппаратов.
Для конструкторов орнитоптера весьма важно отделить существенное и главное в механизме птицы, знание кинематики движения крыла и параметров, необходимых для проектирования аппаратов с машущими крыльями.
С этой стороны знакомство с формой крыла и его структурой, начиная с представителей летающих животных юрской эпохи земли, может представить известный интерес для строителей орнитоптера.
ПТИЦЫ ПЕРВИЧНОЙ ЭПОХИ ЗЕМЛИ
При раскопках в сланцевых отложениях были найдены: первые птицы — археорникс, археоптерикс и летающие животные: птеронадон, птеродактиль и др.
М. К. Тихонравов (2) утверждает, что по' одному из отпечатков он определил площадь крыльев птеродактиля, равную 3 м2, и нашел удлинение их, равное л — 16,6, а другого —л = 20, размах же крыльев находится в пределах от 6 м до 8,2 мх.
Птеронадон — разновидность летающего животного, он своими складчатыми крыльями (рис. 20) напоминает летучую мышь. Летательная перепонка его располагалась между чрезвычайно развитыми «мизинцами», брюшной стороной туловища и иногда маленькими ногами. Предполагают, что его вес достигал 20 кг, т. е. по весу и размаху крыльев он вдвое превосходил наибольшую из существующих птиц.
Археоптерикс имел перьевое оперение современных птиц (рис. 21) и своеобразный хвост. В меловую эпоху существовало до 24 разновидностей птиц, мало отличавшихся от современных. О летных качествах
‘Полет птиц и машины с машущими крыльями» Оборонгиз, 1949 г.
птеронадона и других крылатых гигантов первичной эпохи земли1 существуют различные мнения. Одни из ученых предполагают, что птеро-надон представлял собой своеобразный «мускуллет», способный к ограниченным планирующим спускам; другие допускают, что они были совершенные летуны.
Исходя из удельной нагрузки на крыло птеродактиля (6—10 кгДч2).
Рис. 20. Птеронадон
можно предполагать, что вертикальная скорость снижения при планировании могла быть не больше, чем у современных парителей; в таком случае летающие животные были приспособлены к парению в термических потоках и в потоках воздуха, обтекающего склоны гор.
Конвекция воздуха и восходящие потоки в ту эпоху были более мощные, чем в наше время, благодаря поверхностному нагреву земли и более сильной солнечной радиации; вполне возможно, что птеродактиль хорошо ими пользовался и отлично парил. Обладая длинными крыльями, подобно современному альбатросу, он мог подниматься с прибрежных скал и с гребней морских волн.
На основании изучения даже этого скудного материала о летающих животных мы приходим к выводу, что птерозавров следовало бы изучать подробнее. Для этого необходим, конечно, фундаментальный материал и привлечение специалистов палеонтологов.
1 Проф. И. Вальтер. «Земля до появления человека». СПБ, 1912 г.
ВИДЫ ПОЛЕТОВ птиц
Орнитологи считают, что число видов пгиц, существующих на земле, может доходить до 9 000— 10 000.
Разные птицы применяют различные приемы полета, ввиду большого разнообразия орнитология их не классифицирует. Наиболее часто
Рис. 21. Археоптерикс
встречающиеся приемы полета: 1) машущий; 2) пропеллерный; 3) планирующий; 4) парящий; 5) парашютирующий. Разновидностями машущего полета являются: а) трепещущий, б) вибрационный, в) ныряющий, г) ракетообразный, д) резонансный.
Машущий полет наиболее распространен в природе. При машущем полете широко раскрытые крылья движутся, опускаясь сверху— сзади, вниз—вперед, а затем в обратном направлении, но уже с большей скоростью1. При этом полете мускульное усилие трансформируется в тягу и подъемную силу крыла. Во время опускания крыла вниз опахала перьев прижимаются друг к другу и создают поверхность различ
1 Следует отметить, что неравномерность взмахов и опускание крыла наблюдается не у всех птиц; у крупных тиц отношение времени взмахов вверх и вниз равно 2:3.
ной плотности и проницаемости, крыло поворачивается на отрицательные углы. При обратном движении угол атаки возрастает; при полном взмахе раскрываются решетки плечевой области; туннельные перья несколько расходятся в средней области, так что крыло становится силь-
Рис. 22. Чайка в парящем полете но проницаемым для воздуха, вследствие чего уменьшается вредное сопротивление при подъеме.
В зависимости от скорости взмаха крыльев птица может летать быстрее или медленнее; имеет значение и то, как высоко поднимает крылья птица и как низко их опускает.
Во время скоростного полета птица уменьшает несущую поверхность крыльев, при медленном, наоборот, увеличивает. Особо важное значение имеют в машущем полете и при парении пальцеобразно расставленные пропеллерные перья.
Пропеллерный вид полета встречается при горизонтальном установившемся движении, когда функции крыла разделены; средняя область является несущей, а колеблющийся манус пропеллирующим, создающим тягу для поступательной скорости.
Планирующий и парящий виды полетов достаточно хорошо изучены.
Конфигурация крыльев птиц-парителей нередко служит образцом для конструкторов планера. На рис. 22 заснято парение чайки. Здесь
псГгок проходит через туннели. На рис. 23 концы крыльев чайки опущены вниз, слегка загнуты на конце и своим очертанием напоминают опрокинутую букву дубль-вэ (U7). При парении чайка делает крен благодаря выдвиганию крылышка; ее хвост раскрыт на полный веер,
Рис. 23. Образование W при парении чайки
вследствие чего создается эффект, подобный эффекту крыла с выдвижным закрылком.
Аист при посадке (рис. 24,Л и 24,Б) имеет сильно загнутые вверх концы крыльев. Такое же очертание крыльев имеют при парении и другие птицы с большой удельной нагрузкой: орлы, коршуны, пеликаны и т. д.
Парашютирующий полет, т. е. крутой спуск под углом 40 — 60 ° Ъри неподвижных крыльях, практикуют чайки, луни, вороны, голуби и другие крупные птицы; среди небольших птиц, также пользующихся этим приемом для точной посадки, можно отметить жаворонка.
Трепещущий полет при сильно поднятом корпусе и частых взмахах мануса применяется некоторыми птицами для «стояния на месте», например, им пользуется кобчик, северная крачка, пустельга и некоторые виды воробьиных птиц (серая мухоловка, трясогузка и др.).
Вибрационный полет представляет собой разновидность машущего полета, во время которого взмахи крыльев происходят чрезвычайно быстро.
Этот вид полета позволяет птице также держаться на месте в спокойном воздухе; этим пользуются колибри.
Самые маленькие колибри, весящие 1,8 г, делают около 60 взмахов в секунду. В момент «стояния» около цветка работа крыльев
Рис. 24,Л. Удар крылом вниз при стоянии ча месте. Концы кроющих и малых перьев повернуты к корпусу; алюля поднят; перья мануса сильно изогнуты
Рис. 24,Б. Посадка аиста в гнездо, видны приподнятые перышки благодаря проницанию потоком и концевая решетка мануса
колибри была заснята на киноленту, причем оказалось, что конец крыла описывает вытянутую восьмерку и совершает вращательное движение, резко меняя угол атаки и профиль (рис. 25). Подобно многим насекомым колибри могут летать также назад.
Маленькие птицы, например, скворцы, трясогузки, дятлы, пользуются волнообразным или ныряющим видом полета, во время которого
Рис. 26. Взлет фазана
при периодическом складывании крыльев они летят подобно стреле, теряя при этом высоту с последующим подъемом.
Подобный же вид полета встречается и у воробья. При крутых взлетах он делает крыльями до 13 ударов в секунду, поднимаясь под углом более 60°.
Наиболее крутые взлеты совершают алтайские горные индейки, или улары. Этот вид полета называется ракетообразным; так как он сопровождается большим шумом, то его называют также хлопающим. Подобным же способом летает фазан. Его полет весьма маневренный,
он может даже останавливаться в воздухе и совершать разворот до 180° (рис. 26).
Рис. 27. Положение кистей крыла: а — у хлопающих птиц; б — у парителей
Не обладая сильными грудными мускулами для такого вида полета, хлопающая птица быстро утомляется и переходит на планирование. Особенности полета этих птиц связаны со структурой их крыльев.
Наиболее характерной особенностью внешней архитектуры крыла хлопающих птиц является сильная чашевидность в плечевой зоне и перевернутое W при виде спереди. Благодаря этому птица взлетает без разбега буквально «с места», что мы и наблюдаем у куропаток, тетеревов и фазанов.
Анатомия крыла у этих птиц показывает^, что локтевой сустав (рис. 27,а) соединен с более коротким, плечом, нежели у парителей (рис. 27,6) и, кроме того, угол между костями, плечевой и локтевой, у первых ближе к Э0°, а у вторых тупой, что, повидимому, влияет на частоту взмаха крыла и на необходимые мускульные усилия.
В таблице 5 приведены углы сопряжения костей крыла различных видов птиц (рис. 28) в свободном состоянии и отношение длин локтевой /л и плечевой 1ГТЛ костей.
Резонансный полет наблюдается при групповых полетах птиц, когда они располагаются в строю «клином» или по треугольнику.
Таблица 5
п т и ц а i 7 /л /пл
Аист 120 150 0,42
Степной орел . . 120 130 1,14
Коршун 70 140 1,4
Сорока 80 145 1,38
Кобчик 80 160 1,68
Сарыч ...... 50 ПО 1,34
При одновременных взмахах птицы заставляют колебаться отходящие потоки так, что сзади летящие, обычно более слабые, получают дополнительную поддерживающую силу. Эта своеобразная кооперация в приро-
Рис. 28. Схема расположения костей крыла
де чрезвычайно' экономит энергию птиц, особенно при дальних перелетах.
Данные птиц в зависимости от наиболее часто применяемого ими вида полета приведены в таблице 6.
Таблица 6
I Вид полета Название птиц Скорость полета (км/час) Площадь крыла (ж2) Размах крыла («) Удлинение Нагрузка на м2 поверхности Число колебаний! (—) Вес птицы (кг) Вес мускулов в 9о % от общего веса
Ракетообраз- Тетерев 35 0,165 0,9 4,9 14,4 3,5 2,38 —
ный Фазан 50 0,181 0,75 3,1 7,47 — 1,35 34,5
Куропатка . . . 40 0,0548 0,62 7,34 7,85 3 0,43 —
Машущий и Степной орел 80 0,5 1,650 5,5 10,6 1,0 5,3 18,3
парящий Коршун .... 60 0,215 1,26 7,6 4,7 1,5 1,0 18,6
Альбатрос . . . 72 0,720 3,4 16 13,2 — 9,5 —
Серая чайка** . 38 0,334 1,48 7,65 2,66 — 0,89 —
Трепещущий Жаворонок . . . [ 0,020 0,32 5,06 1,83 — 0,037 —
Вибрационный Ласточка** . . . 112 ' 0,013 0,336 8,96 1,59 — 0,02 —
Стриж** .... 114 j 0,012 0,38 11,8 3,77 — 0,046 —
** По данным М. К- Тихонравова.
ПРИЕМЫ ПИЛОТАЖА У ПТИЦ
Комбинируя различные виды полета и изменяя очертание профиля, размах и площадь крыла при разных частотах и амплитудах колебаний, птица достигает большого разнообразия в пилотаже.
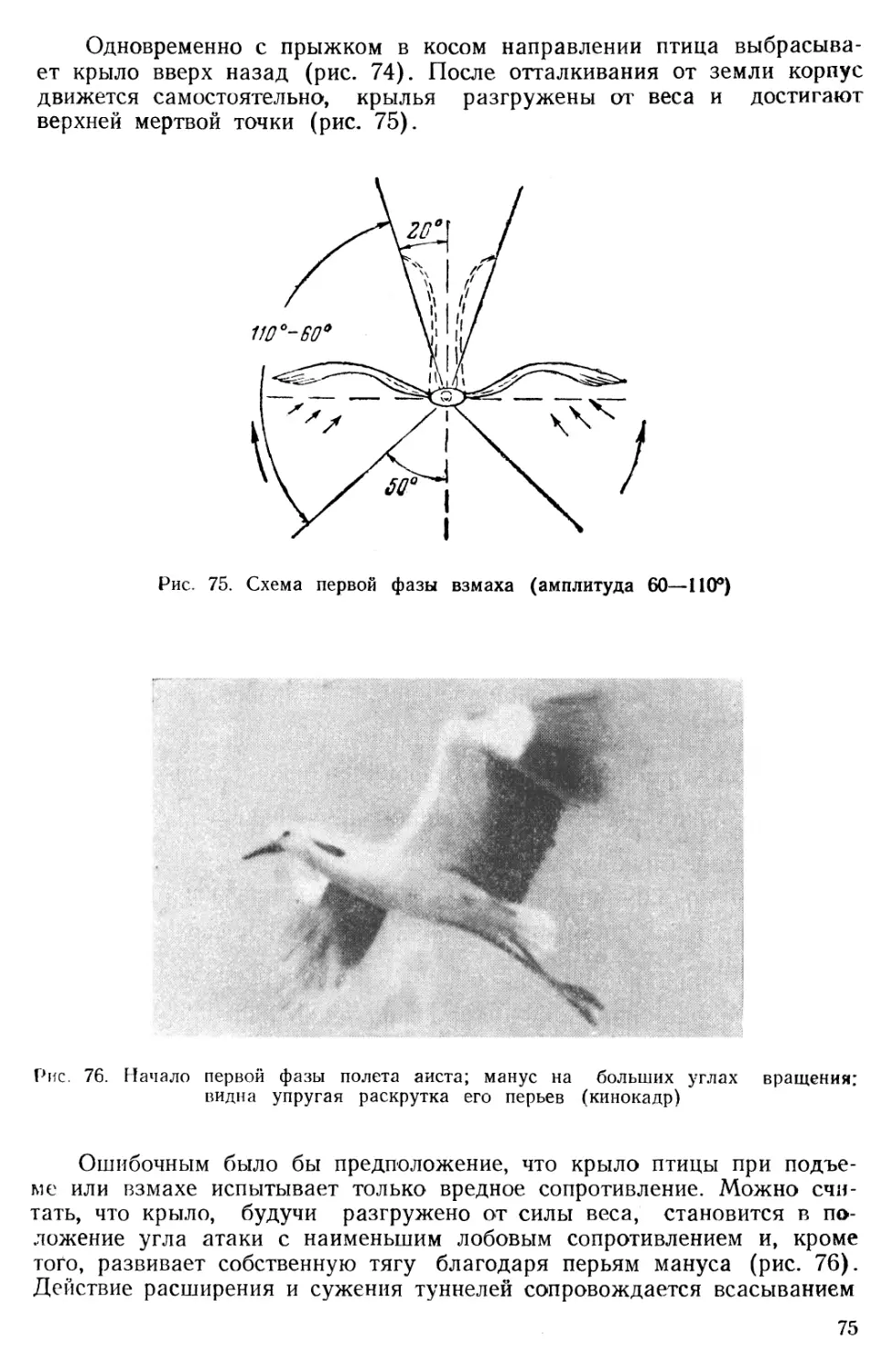
В природе можно видеть почти все виды пилотажного искусства, освоенного человеком на самолете, включая штопор и мертвые петли. Вместе с тем следует считать исключительной принадлежностью птиц такие маневры, как стояние на месте, крутое парашютирование с любой высоты и торможение крыльями под прямым углом к направлению движения.
Приведем описание наблюдений за полетами птиц. На рис. 29 заснят прием посадки в гнезде болотного луня.
Спуск происходит с вертикально поднятыми крыльями, невидимому, птица использует здесь парашютный эффект благодаря сопротивлению открытых плечевых решеток и опущенному хвосту.
На рис. 30 показан спуск чайки в гнездо парашютированием, а на схеме рис. 31—углы поворота крыла по отношению к оси корпуса, достигающие в корневом сечении 20° и в концевом 38°.
Интересен маневр, применяемый пустельгой при ее охоте за полевой мышью (рис. 32). Перед броском вниз птица стоит на месте, используя трепещущий вид полета, и затем, нацелившись, падает со сложенными вверх крыльями в намеченную точку, не изменяя положения корпуса. Захват добычи сопровождается одновременным распусканием крыльев и переходом на подъем.
Подобный прием вертикального спуска условимся называть «пикированием плашмя», так как здесь при сложенных вверх крыльях туловище имеет малый наклон к горизонту и хвост поднят почти вертикально.
Этот маневр птицы, несомненно, представляет интерес для конструкторов самолета.
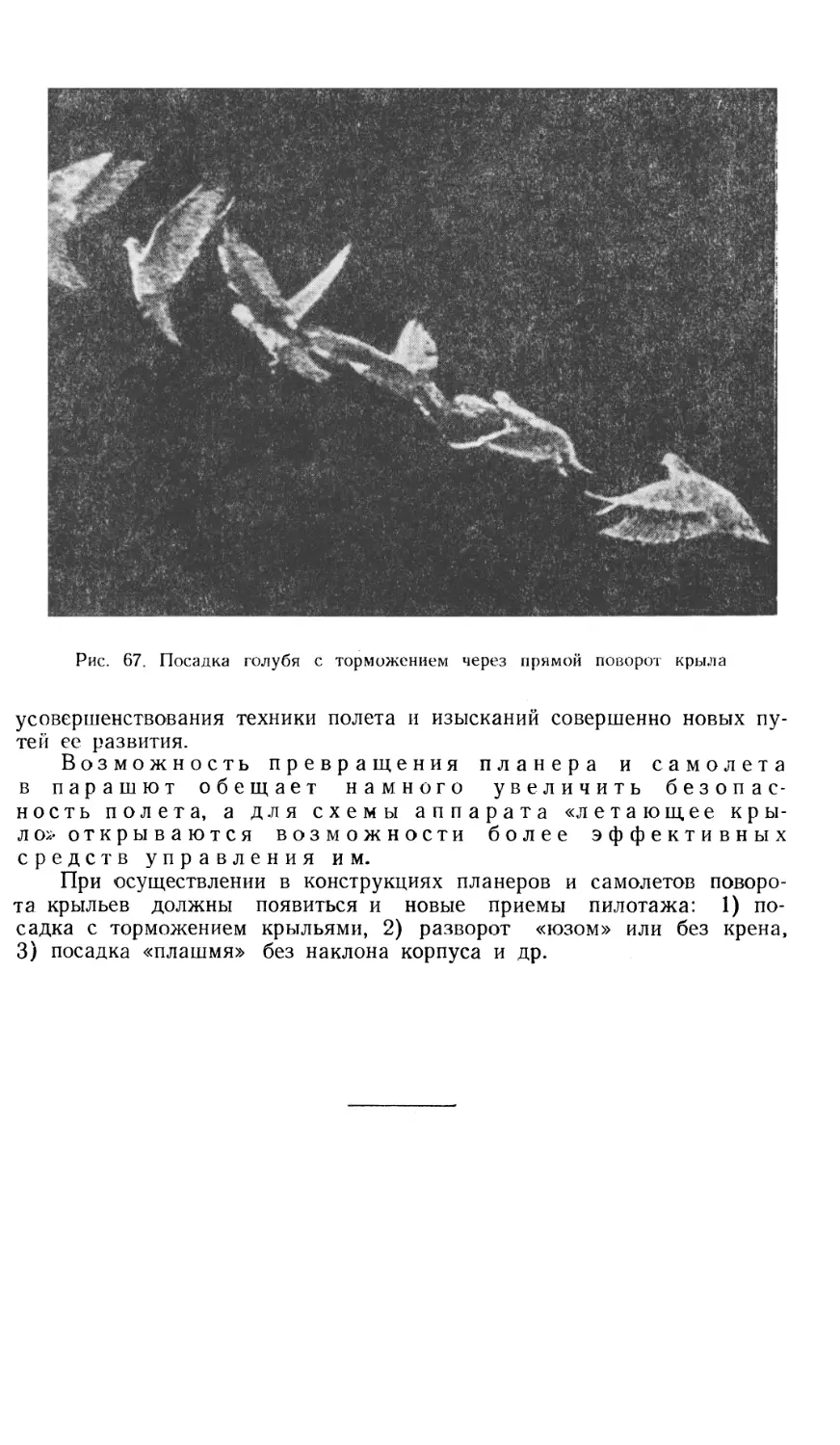
На рис. 33 мы видим посадку олуши (баклана) на отвесную скалу у своего гнезда, при этом корпус ставится в вертикальное положение и птица делает несколько ударов крыльев, направленных против движения, чем смягчает посадку; этот же прием выполняет и голубь (рис. 67).
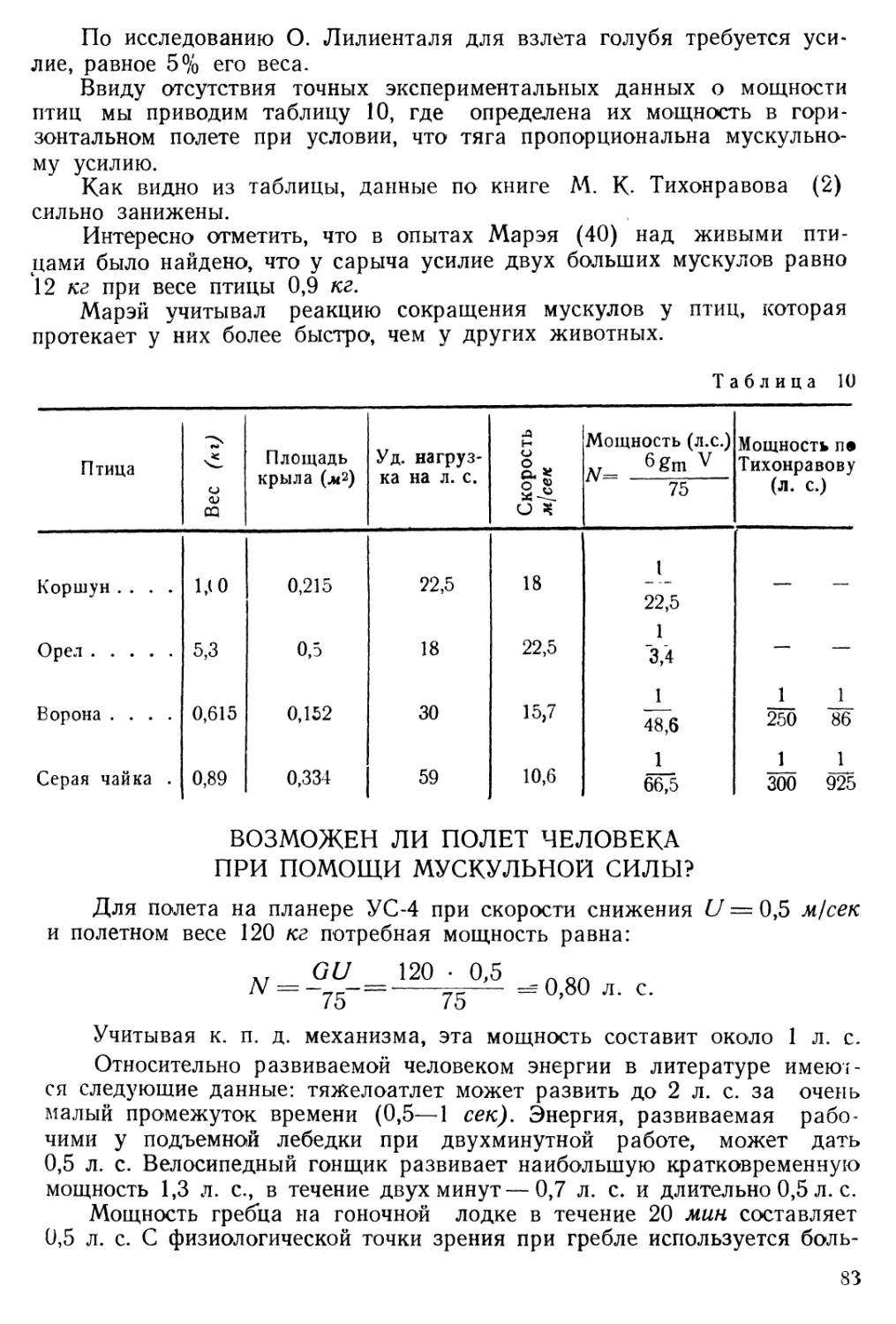
ЛЕТНЫЕ ХАРАКТЕРИСТИКИ ПТИЦ
Собственная скорость птицы относительно воздуха определена для многих видов птиц с большой степенью точности.
Наибольшая скорость отмечена у стрижа — до 144 км/час.
Почтовый голубь в тихую погоду при продолжительном полете делает 60 км/час.
В условиях преследования добычи скорость хищной птицы значительно возрастает. Так, сокол-сапсан при пикировании на добычу развивает скорость до 360 км/час вместо 95 км/час в горизонтальном полете.
Рис. 29. Лунь парашютирует в гнездо
Н. Виноградов
33
Рис. 30. Чайка садится в гнездо под углом атаки 30—50°
Рис. 31. Схема углов парашютирования чайки
Рис. 32. Пустельга охотится за полевой мышью
Рис. 33. Олуша тормозит крылом при посадке на скалу
Дальность перелета без спуска установлена для окольцованных чибисов. При попутном ветре из Англии в Ньюфаундленд через Атлантику они покрывают весь путь протяжением 3 500 км за 24 часа.
Наблюдениями с самолета установлена следующая наибольшая высота полета: для ворон и галок — 2 000 м, для жаворонка — 1 900 м, для аиста и сарыча — 900 м, для лебедей — 300—500 м, для журавлей — 800—1 200 м.
Используя восходящие потоки, гималайские грифы, поднимаясь с Эвереста в Гималаях, достигают высоты более 8 000 м над уровнем мор*я. На Кавказе наблюдаются полеты горных орлов выше Эльбруса и Казбека. В связи с этим интересно сравнить достижения птиц и рекордные достижения планеристов.
На таблице 7 приведены горизонтальные и вертикальные скорости, полученные при наблюдениях за парением птиц (42).
Таблица
Птица Скорость при парении (м/сек) Наименьшая скорость сниже-ния (м/сек) Угол планирования
Альбатрос 14 0,42 1 :33
Красный коршун Ю,5 0,67 1:16
Стервятник 9,2 0,55 1 :17
Ястреб 8.1 0,5 1 :16
Таблица
Название планера Горизонтальная скорость (м/сек) Скорость снижения (м/сек) Угол планирования
КАИ-3 • . 15,2 0,55 1 :27,7
Ш-5 13,3 0,77 1 :17,3
ДК-3 • ... 18 0,55 1 :32.8
КИМ-2 15,6 0,64 1:24.2
Другими словами, с высоты 10 м альбатрос может планировать на расстояние до 200 м, коршун — 90—170 ж, ястреб — 90—160 м.
По данным таблицы 8 следует, что планеры обладают несколько большими горизонтальными скоростями, чем птицы, и почти одинаковой с ними скоростью снижения.
Из небольшого перечня примеров пилотажа птиц видно, что возможности птичьего крыла весьма значительны и приходится признать, что птицы более «приспособлены» к воздуху, чем самолеты, обладают большей устойчивостью и управляемостью на всех режимах. В дальнейших главах мы постараемся выявить, в чем именно выражается «приспособленность» летательного аппарата птиц в условиях воздушной среды.
ГЛАВА 111
РЕЗУЛЬТАТЫ ИЗУЧЕНИЯ ПОЛЕТА ПТИЦ
ИССЛЕДОВАНИЕ ЖИВЫХ ПТИЦ
Произведенные киносъемки полета голубей в конце прошлого столетия показали большую ценность этого способа изучения полета птиц. Они позволили разделить полет на отдельные фазы, проследить изменение положения крыльев и траекторию их движения (рис. 34). Хотя
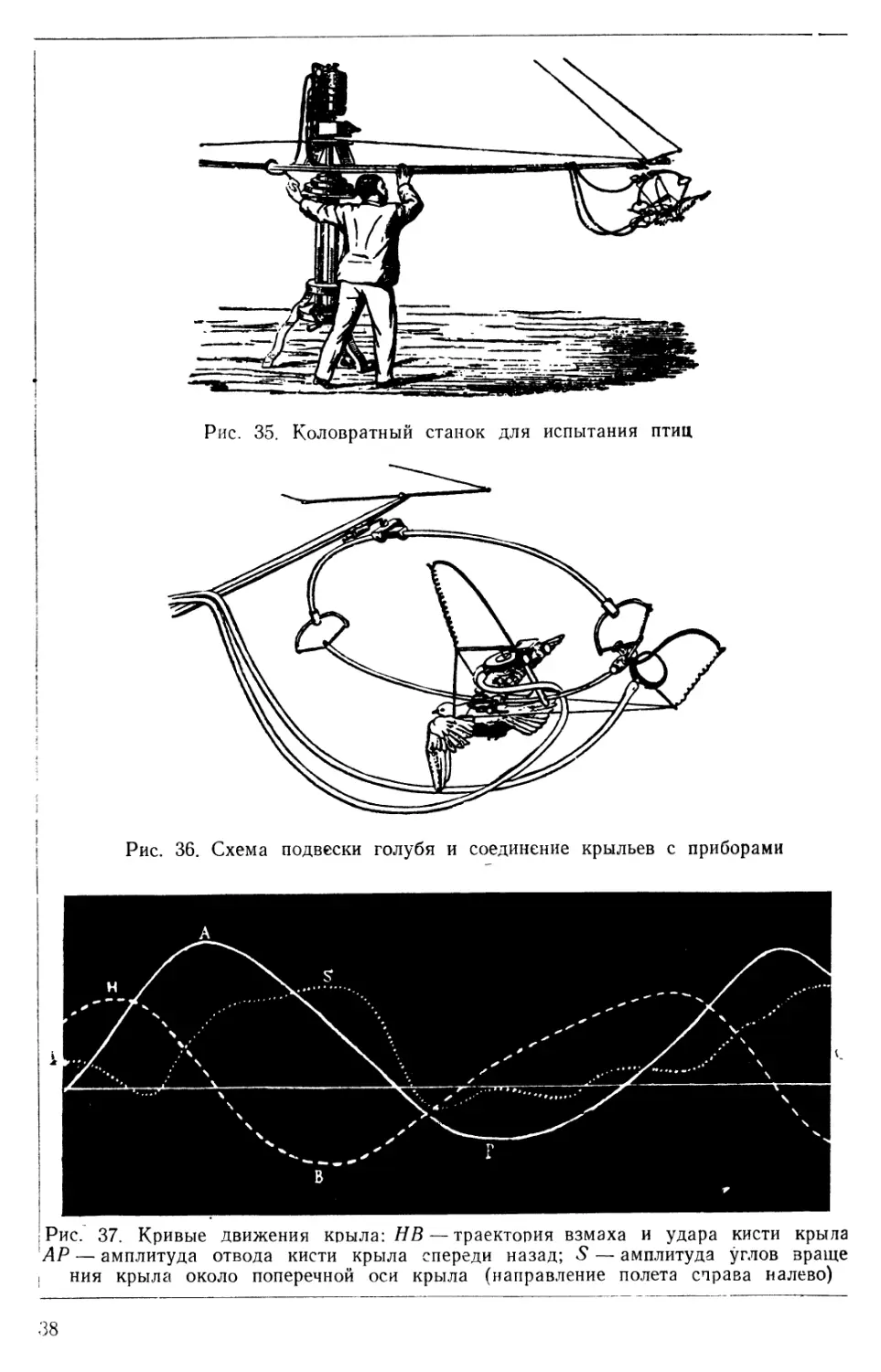
наибольшая скорость съемки не превышала Vso в секунду, результаты ее помогли создать методику проведения лабораторных опытов с живыми птицами. Прибор Марэя (рис. 35) представлял собой коловратную машину переносного типа, которая устанавливалась на открытом воздухе. На длинном плече рычага, перпендикулярно к нему, на легком эллиптическом кронштейне (рис. 36) подвешивалась испытуемая птица, корпус которой охватывал особый бандаж. Плечевая область крыла шелковыми шнурами соединялась с рычагами пневматического манипулятора для записи вертикальных движений и для записи движений крыла по горизонтали к мембранному цилиндру (40).
Воздух, нагнетаемый через ручную резиновую грушу, подводился в мембранные коробки по резиновым трубкам.
Движения крыльев птиц записывались на вращающемся барабане с миллиметровой бумагой.
На рис. 37 представлены кривые движений крыла голубя. АР — запись движения при отводе кисти крыла, НВ — запись траектории
1
Рис. 36. Схема подвески голубя и соединение крыльев с приборами
Рис. 37. Кривые движения крыла: НВ — траектория взмаха и удара кисти крыла АР — амплитуда отвода кисти крыла спереди назад; 5* — амплитуда углов враще । ния крыла около поперечной оси крыла (направление полета справа налево)
крыла и кривая S — изменение угла атаки при вращении. Из диаграммы следует, что 1) продолжительность движения плеча в вертикальном направлении при подъеме много меньше, чем при опускании; 2) когда крыло поднимается, углы атаки возрастают, а при опускании наоборот; 3) нуль кривой S соответствует моменту, когда крыло горизонтально.
Рис 38. Эллиптическая кривая вершины плеча (направление движения слева направо)
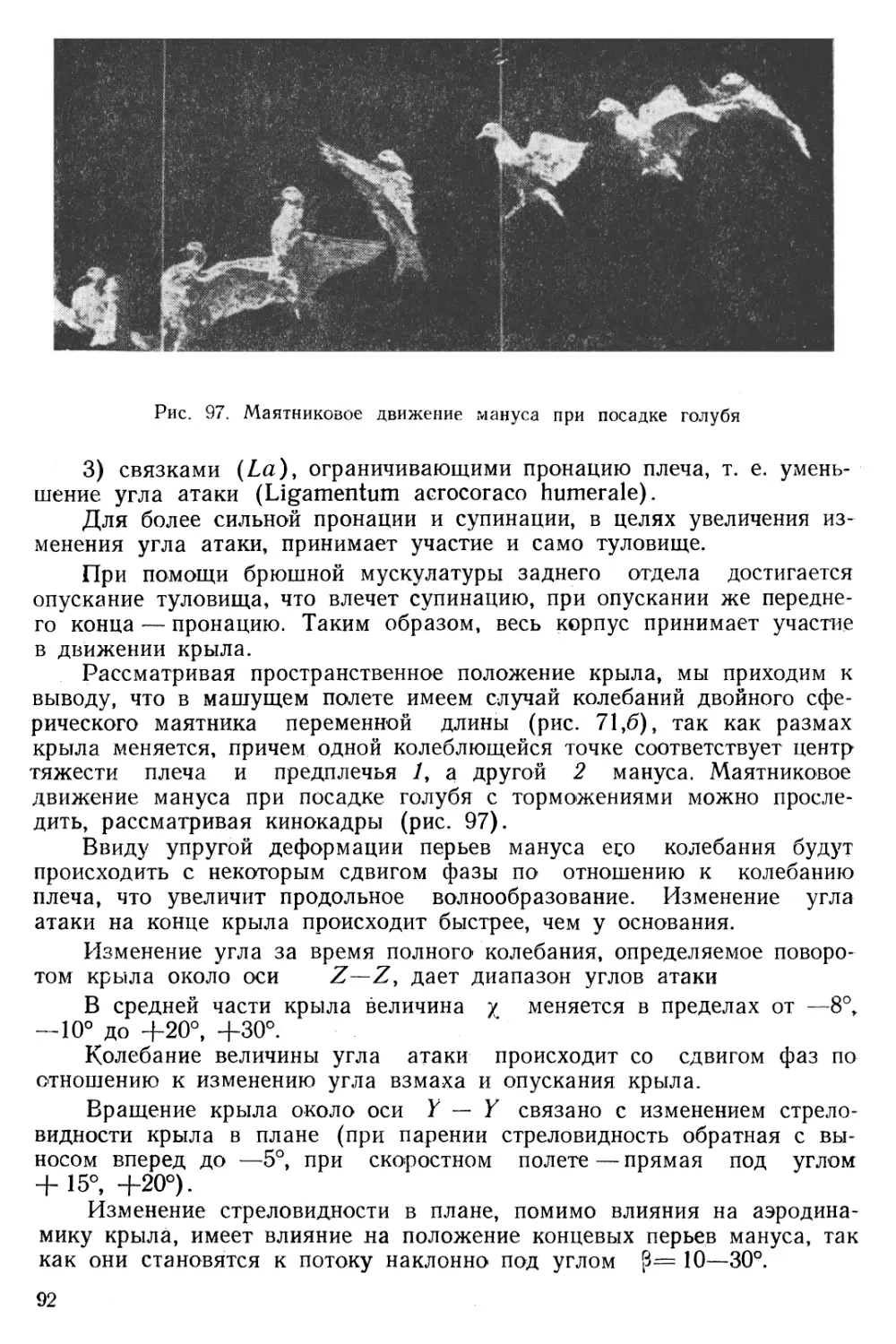
Весьма важным построением, на основе этих двух кривых, явилась траектория локтевой конечности плеча или вершины его во время полного колебания.
На рис. 38 показана эта траектория, согласно которой плечо голубя при махании на месте идет вниз и вперед, потом возвращается назад, описывая кривую, близкую к эллипсу.
В этих опытах впервые была установлена неравномерность взмаха и удара крыла. Возвратное движение при взмахе крыла есть акт более быстрый и зависит от изгиба локтя. Неравенство времени подъема и опускания всегда наблюдалось у птиц с большой поверхностью крыла и с малой частотой колебаний. Продолжительность подъема и опускания крыла в шестидесятых долях секунды указана в таблице 9.
Таблица 9
Птица Время одного колебания Подъем (взмах) Опускание (удар крыла)
Утка 6,66 3 3,66
Голубь „ . 7,5 3 4,5
Сарыч 21,5 8,5 1 13
Эти результаты были получены методом электрического хроногра-фирования на особой установке.
ОПЫТЫ ПО ЗАКЛЕИВАНИЮ КРЫЛЬЕВ ПТИЦ
В России еще в 1908 г. целая серия опытов над воронами была проделана в Тифлисе одним из пионеров авиации летчиком А. В. Шиу-ковым. Он установил, что при заклеивании мануса птица полностью теряет способность к полету, заклеивание же средней части крыла вынуждает ее к хлопающему виду полета с более частыми взмахами, при этом птица скоро утомляется. А. В. Шиуковым тогда же было замечено, что покрытие лаком перьев птицы лишало ее способности к полету,, чем подтверждается наша мысль о роли пограничного слоя у птиц.
В те же годы подобные эксперименты были произведены над голубями доктором Е. П. Смирновым, который так описывает свои наблюдения (6):
«Взяв тонкий, тюлевый, шелковый дамский вуаль, я оклеил его с обеих сторон тончайшей папиросной бумагой. В итоге получилась своеобразная чрезвычайно легкая ткань, в то же время хорошо сопротивляющаяся разрыву при давлении. Затем приобрел на Щукином у знакомого голубятника хорошего, способного к парению турмана. Захлороформировав его, я разложил его на столе грудью вверх и, растянув его крылья, оклеил их с нижней стороны так, чтобы крылья могли беспрепятственно растягиваться и складываться наподобие японского складного веера. При этом, вполне понятно, просветы щелей, образующиеся при растяжении крыла, между большими маховыми перьями уже не зияли, а были затянуты легкой тканью.
Когда эта хлопотливая и требующая некоторой сноровки операция была закончена, я привел голубя в чувство и после того, как он достаточно отдохнул и оправился в тесной клетке, пустил его на дворе на волю, чтобы посмотреть, как он будет себя вести.
Почувствовав себя на свободе, мой турман сделал несколько сильных взмахов своими крыльями и поднялся на воздух. Но полет его (как я и ожидал заранее) скорее напоминал характерное порхание или даже, если так можно выразиться, мерцание бабочки-капустницы в-воздухе, или в лучшем случае — летучей мыши.
Он все время мелькал в воздушной среде, тяжело и непрерывно хлопая своими крыльями. Такой полет стоил ему значительных усилий,, он, повидимому, скоро утомился и сел для отдыха на край крыши сеновала. Через некоторое время, достаточно отдохнув, он распустил крылья и смело ринулся вниз головой, очевидно, намереваясь использовать парящий полет. Однако это ему совершенно не удалось и, перевернувшись в воздухе через голову, он удержался в воздухе лишь посредством учащенных ударов крыльями. Видимо, ошеломленный такой непредвиденной неудачей, он быстро вернулся на прежнее место. Несколько раз он возобновлял эти попытки парения, но всякий раз неудачно. Наконец, отлетев от крыши слишком далеко, но не успев вовремя вернуться к ней для отдыха и не рассчитав своих сил, при одном неловком движении грузно упал с высоты на землю и сломал одно из своих крыльев, так что я из сострадания должен был его тут же прикончить».
Из этого опыта доктор Е. П. Смирнов делает следующие выводы: «Если парение птиц происходит именно так, как учит теория Лилиенталя, и птица при своем парении ставит крылья под определенным углом к горизонту, то с этой точки зрения затягивание щелей между большими маховыми перьями тонкой и легкой тканью для птицы было бы не только совершенно безразлично, но представляло бы даже некоторые выгоды.
Однако опыт с достаточной убедительностью показал мне, что пти.-иа при таких условиях совершенно утрачивает способность к парению и что эти щели должны играть немаловажную роль в общем механизме парения»1.«
Выводы этого замечательного русского орнитолога подтвердились спустя 27 лет в аналогичных опытах В. Ф. Савельева и при наблюдениях автора, описанных в главах III и IV.
Этими опытами установлено следующее:
1. Устранение щелей и разрезного устройства больших маховых перьев лишало крыло его эффективности, так что птица вынуждена была применить иной режим колебаний с затратой чрезмерных и утомительных усилий.
Выключение концевой тяги мануса заставляло возмещать ее более частым колебанием всей поверхности крыла.
2, Заклеивание с нижней стороны изменяло характер обтекания крыльев птицы, в результате подъемная сила оказывалась недостаточной для горизонтального полета.
3. При заклеивании крыльев птицы ее летательный аппарат походил на простое крыло планера и самолета, которое для машущего полета явно невыгодно.
Очевидно эти факты следует учесть конструкторам орнитоптеров, сообщая крыльям необходимую проницаемость.
ОДЫМЛЕНИЕ КРЫЛЬЕВ
В целях изучения протекания воздуха через крыло птицы в 1947 г. был поставлен следующий простой опыт. Под крыло коршуна в зону плечевой области и туннелей мануса была направлена струя воздуха из шланга воздуходувного агрегата пылесоса с широким прямоугольным насадком. Вентилятор засасывал воздух и дым от горящей пакли, облитой керосином. При этом было обнаружено:
1) отклонение потока за крылом до 30° к горизонту;
2) просачивание воздуха через верхние кроющие ряды перьев;
3) сильный ток струек из туннелей мануса;
4) вибрирующее состояние его перьев.
Для большей наглядности при повторных опытах на верхнюю поверхность крыла насыпался серпантин, который позволял проследить по движению его прохождение потока в зоне кроющих и второстепенных перьев.
1 Эти выводы подтверждены в опытах с живыми птицами в 1949 г. проф. Г. С. Шестаковой на биологической станции Академии наук СССР
При этом случайно было обнаружено, что крыло, свободно опираясь на две точки, примерно в шарнире и под лучевым предкрылком, находясь под действием струи из шланга, держалось совершенно свободно и не имело никаких стремлений к повороту около вертикальной оси или к запрокидыванию около поперечной. Это устойчивое положение сохранялось и при одной опоре.
Отсюда можно сделать вывод, что крыло с- проницанием, имеющее глубоковогнутый профиль у основания и двояковыпуклый на конце, при отрицательной крутке обладает хорошей устойчивостью.
Скорость обдува во время эксперимента составляла около 6 м/сек при направлении потока под углом в 40°. Точка приложения потока переносилась как в глубину, так и по размаху крыла.
При опытах с искусственными моделями птиц в аэродинамической трубе, произведенных по заданию ЦК Совета Досава, в 1950 году был получен спектр, подтвердивший дымовую обдувку.
Крыло модели (рис. 119) было, оснащено сверху и снизу шелковинками и заснято в потоке при скорости 15 м/сек. Полученные фотографии показывают вход воздуха в туннель на всем диапазоне летных углов птицы, от 0° до 20°, а также прорыв потока на верхней поверхности вдоль хорды крыла. Спектр из шелковинок обнаружил значительный скос потока за минусом, а также вихревой шнур, отходящий в средней области мануса, а не с конца крыла, как у обычных жестких крыльев. Замечено также, что у лопаточных крыльев концевые срывы отсутствуют даже при углах 18 — 20°.
ГЛАВА IV
ФОТО-АЭРОДИНАМИЧЕСКИЙ МЕТОД ИЗУЧЕНИЯ КРЫЛЬЕВ
ВЫВОД ИЗ ОДНОГО ПОВЕРОЧНОГО РАСЧЕТА
В целях объяснения полета птиц в различных трудах исследователей работа птичьего крыла рассматривается как работа особого аэродинамического профиля, движущегося в потоке с переменным углом атаки. При восходящем движении крыла с положительным
Рис 39 Силовые схемы: А — при опускании и В — при подъеме крыла
углом атаки возникает положительная подъемная сила N и отрицательная тяга Т по схеме В рис. 39. При опускании крыла угол атаки, уменьшается до отрицательного и крыло дает положительную тягу и подъемную силу по схеме А рис. 39.
Если провести приближенный поверочный расчет взлета орла’ по формулам аэродинамики, то при самых оптимальных коэфициентах подъемной силы, указанных на диаграмме (рис. 40), получается, что •он отрывается при скорости 36 км/час, тогда как прямым наблюдение »ем установлено, что птица подымается, нередко с добычей, при скорости 15—18 км/час.
1 Данные орла приведены в таблице 7, коэфициенты приведены на рисунке для «соответствующей области
На диаграмме (рис. 40) показан этот результат расчета, где пересечение вычисленной кривой для числа колебаний п = 2,5 х/сек с линией, соответствующей весу 5,4 кг, наступает при скорости около 10 м/сек. Чтобы получить действительное совпадение, надо для ско-
Рис. 40. Диаграмма поверочного расчета орла: Y— подъемная сила при л=2,5 1/сек: К — поправочный коэфициент
рости 4 м/сек найденную подъемную силу увеличить в 5,5 раза, что показывает кривая К.
О. Лилиенталь на основе своих опытов (13) считает, что эффект крыла в машущем состоянии в 9 раз больше, чем при простом обтекании крыла в режиме планирования. В нашем примере среднее значение коэфициента Су для скорости отрыва 4 м/сек будет равно 6.
Объяснить количественную сторону явления на основе простых аэродинамических уравнений оказалось невозможным, продувки же чучел птиц в трубе показали очень низкое качество.
На основе продувок вороны в ЦАГИ в 1928 г. было найдено качество порядка — 4,5 и угол наиболее пологого планирования — 12,5°^ т. е. это полностью расходится с тем, что мы наблюдаем в природе.
Ясно, что расхождение следует искать в неправильной трактовке физической схемы работы крыла. В живой птице различные отделы крыла исполняют какие-то неизвестные нам функции, и сочетание их дает замечательные эффекты в полете.
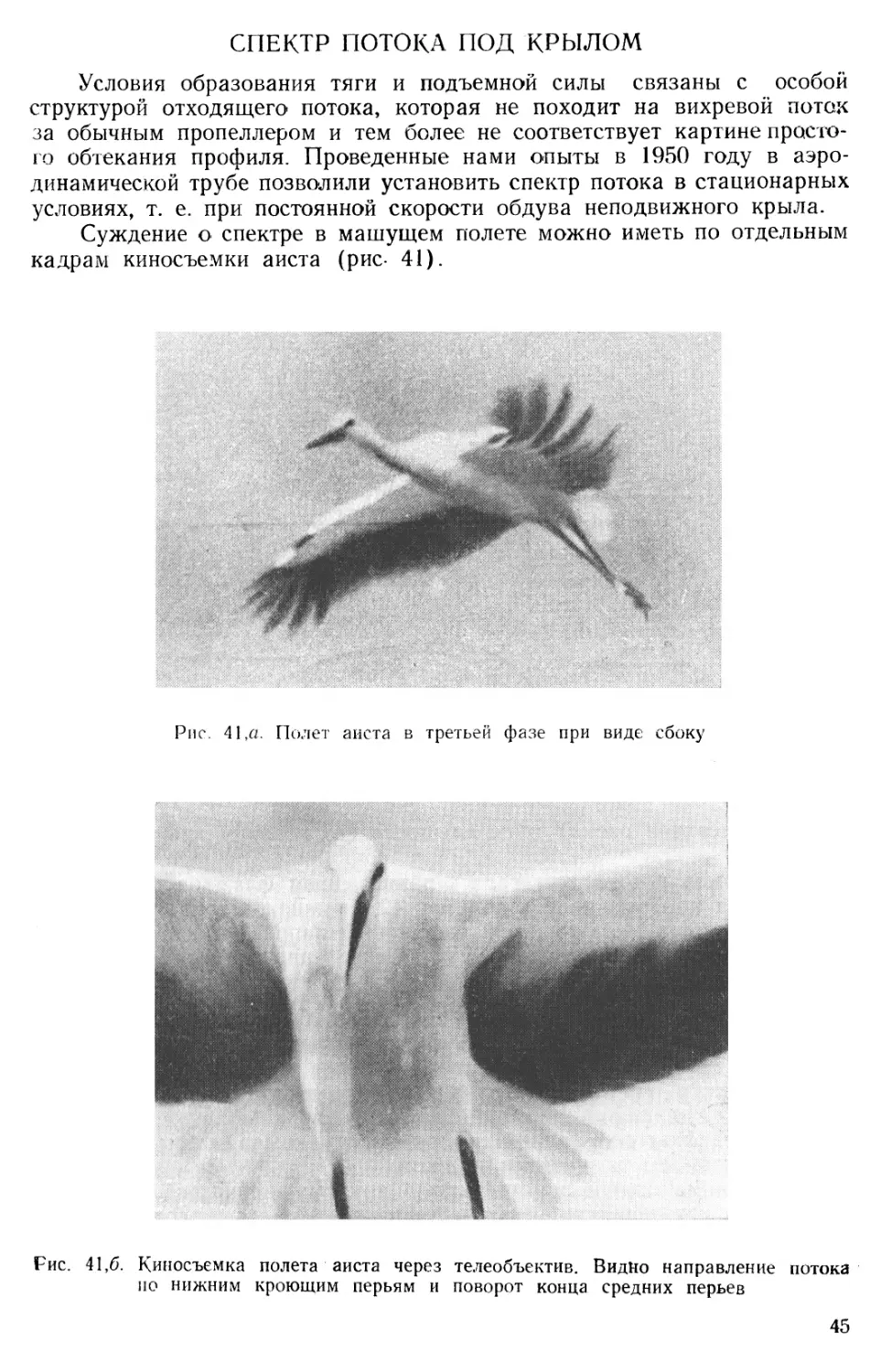
СПЕКТР ПОТОКА ПОД КРЫЛОМ
Условия образования тяги и подъемной силы связаны с особой структурой отходящего потока, которая не походит на вихревой поток за обычным пропеллером и тем более не соответствует картине простого обтекания профиля. Проведенные нами опыты в 1950 году в аэродинамической трубе позволили установить спектр потока в стационарных условиях, т. е. при постоянной скорости обдува неподвижного крыла.
Суждение о спектре в машущем полете можно иметь по отдельным кадрам киносъемки аиста (рис- 41).
Рис. 41,а. Полет аиста в третьей фазе при виде сбоку
Рис. 41,6. Киносъемка полета аиста через телеобъектив. Видйо направление потока по нижним кроющим перьям и поворот конца средних перьев
На основании отклонения концевых перьев (рис. 41,6) можно заметить вращательное движение частиц воздуха в плечевой области.
Эти фотографии подтверждают ранее произведенные опыты на коловратной машине (41) с крыльями птицы фрегат, где флажками было установлено вращательное движение потока в средней части крыла
Рис. 42. Радиальное течение воздуха под крылом фрегата
(рис. 42, верхняя схема) и перемещение его к концу крыла (средняя схема).
Видимость потока достигалась размещением флажков по косо натянутой под крылом проволоке (рис. 42, нижняя схема); движение флажков показывало, что поток растекается направо и налево по размаху крыла и, кроме того, имеет и обычное циркуляционное течение (верхняя схема). Это обстоятельство является весьма важным для волнообразования при упругих колебаниях крыла, что рассматривается в главе IX.
ЭФФЕКТ ПРОНИЦАНИЯ КРЫЛА ПОТОКОМ ВОЗДУХА
Вторым фактором, влияющим на работу крыла птицы, является его проницаемость воздушным потоком.
При внимательном осмотре крыла крупных хищников легко проследить по изношенности перьев течение воздуха на поверхности крыла и внутри его. На фотографии крыла коршуна (рис. 4 и 5) видны рельефно выступающие образования с обеих сторон крыла, направлен
ные к корпусу в области второстепенных машущих перьев и наружу в области мануса.
Поверхность крыла птицы образована наложением S-образных перьев (рис. 6), внутри которых находятся конические туннели желобчатой формы. На снимке пикирующего ястреба (рис. 43) они проекты -
Рис. 43. Темные полосы туннелей у пикирующего ястреба руются в виде отдельных черных полос, перемежающихся с более светлыми. Туннельность наиболее сильно выражена в средней зоне крыла.
При рассмотрении крыльев у мертвого коршуна удалось установить восемь различно спрофилированных туннелей в области второстепенных перьев. Наше предположение, что воздушный поток протекает здесь в фазе взмаха из подкрыльной зоны через слабо прилегающий нижний ряд кроющих перьев, подтвердилось при опытах в трубе с моделями птиц в 1950 г.
На фотографии двух чаек в полете видно, что туннели наполнены воздухом (рис. 7). Здесь же, у основания крыла, видна решетка плечевой области, своего рода жалюзи, перепускающие поток в момент подъема крыла. Каждое отдельное перо при этом обдувается потоком, создавая элементарную подъемную силу.
Установлено, что ренТетки образуют различные углы наклона, возрастающие к корпусу; у некоторых птиц они открываются при парашютировании со сложенными крыльями, как, например, это видно на рис. 29.
Работу туннелей впервые автору пришлось наблюдать в 1921 г. у гнезда ястреба. При ветре силой в 5—6 м/сек птица висела в воздухе на расстоянии не больше одного метра, защищая свой выводок. При этом заметно было раздвижение туннелей, изменение площади хвостового веера и небольшое колебание носовой части крыла при угле атаки около 45 — 50°. Птица держалась против ветра очень устойчиво в течение 10—15 секунд, не меняя своей позиции, пока порывы ветра
Рис. 44,а. Блокировка перьев чайки
не сбивали ее, но она тотчас снова становилась в положение «висения на месте».
Естественно1 было предположить, что изменение объема туннелей должно сопровождаться явлениями всасывания и сжатия воздуха. Однако роль туннелей была выяснена значительно позднее, когда были получены фотоснимки чаек в различных фазах полета. Результаты исследований методом заклеивания крыльев, примененные нашими исследователями, окончательно убеждают в особо важном значении туннельных образований.
Туннели в крыле напоминают собой своеобразные мехи. При подъеме крыла они засасывают воздух из носовой подкрыльной зоны через легкоотходящий нижний ряд кроющих1. Когда же крыло движется вниз, происходит блокировка эластичного конца перьев
Рис. 44,6. Раскрытие туннелей вдоль образующей создает отсос пограничного слоя: А — схема протока через туннель; Б — схема сдувания вдоль пера
1 Среди орнитологов существует мнение, что прорыв потока легче может произойти на нижней поверхности, где стык туннелей менее плотный. Однако в таком случае нельзя объяснить образование большой подъемной силы крыла.
Другое возражение состоит в том, что сжатый в туннелях воздух скорее откроет более слабые кроющие перья. При этом нужно иметь в виду, во-первых, пуховый уплотнитель в основании очина пера и, во-вторых, направление бороздок пера, которые будут препятствовать обратному движению воздуха из туннелей.
(рис. 44,а). А так как при этом площадь крыльев сокращается, туннели уменьшаются в объеме. В результате сжатый воздух прорывает долевой стык туннелей (рис. 44,6) и вызывает сдувание пограничного слоя, а следовательно, увеличение подъемной силы крыла.
Прорыв воздуха в нисходящей фазе движения крыла происходит в
Рис 45 Чайка на режиме крутого парашютирования
зоне верхних кроющих перьев, о чем свидетельствует сильная изношенность их концов.
Надо полагать, что в этом случае прорывается воздух из аванкамер туннелей, особенно в средней части плечевой области. Близ шарнира крыла, где находится парашютный клапан или плечевые перья, образующие зализ, воздух прорывается из пазушной зоны под верхние ряды кроющих перьев (подслойный прорыв). Замечательно то, что кроющие перья области мануса не носят следов прорыва, так как здесь туннели концевой блокировки не имеют, они оканчиваются в виде сопла.
Второстепенные перья у сарыча имеют на конце следы износа о г вихревых струек, вытекающих из туннелей.
Вся картина проницания крыла парителя становится ясной при рассмотрении фотографии чайки на режиме крутого парашютирования
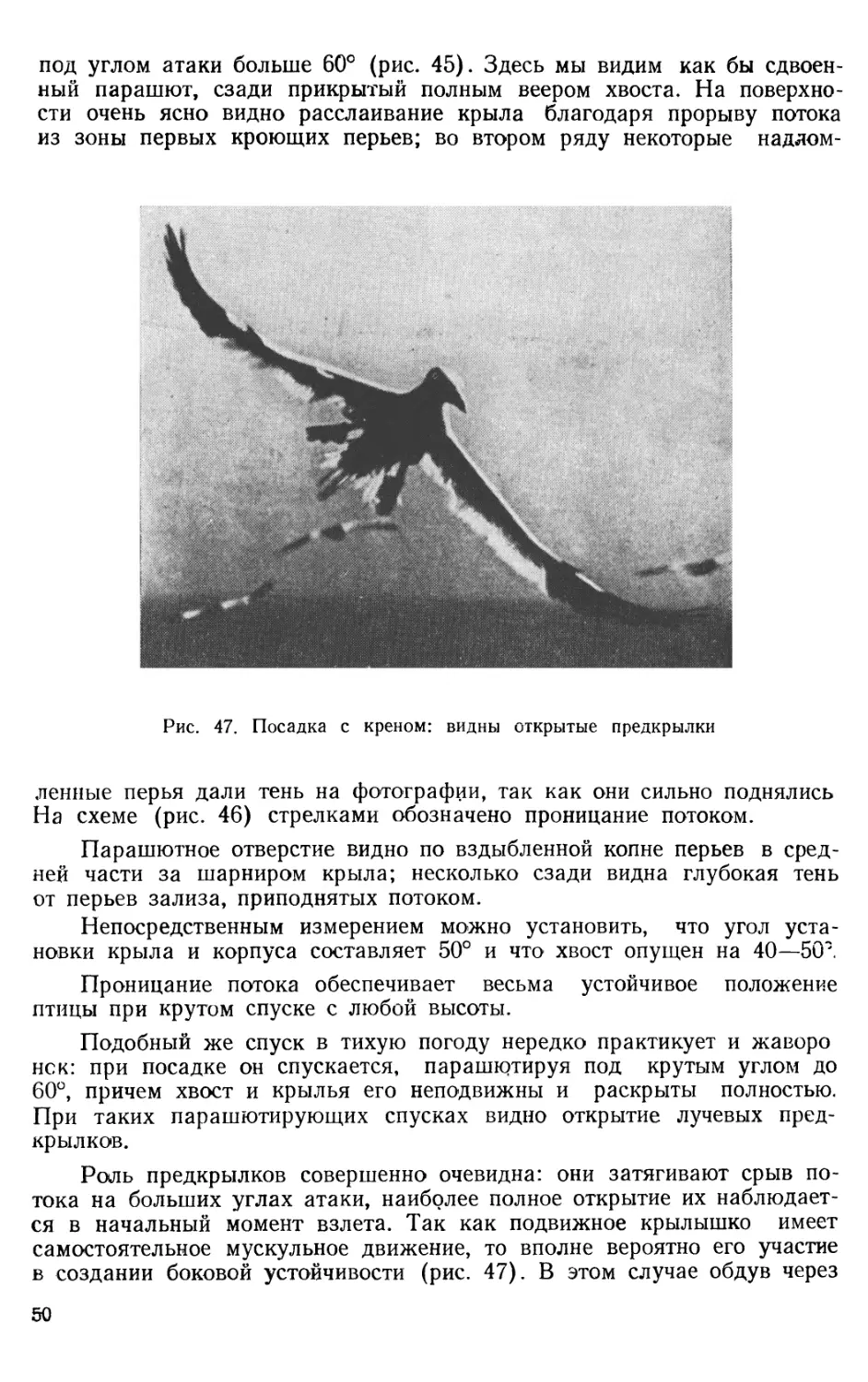
под углом атаки больше 60° (рис. 45). Здесь мы видим как бы сдвоенный парашют, сзади прикрытый полным веером хвоста. На поверхности очень ясно видно расслаивание крыла благодаря прорыву потока из зоны первых кроющих перьев; во втором ряду некоторые надлом-
Рис. 47. Посадка с креном: видны открытые предкрылки ленные перья дали тень на фотографии, так как они сильно поднялись На схеме (рис. 46) стрелками обозначено проницание потоком.
Парашютное отверстие видно по вздыбленной копне перьев в средней части за шарниром крыла; несколько сзади видна глубокая тень от перьев зализа, приподнятых потоком.
Непосредственным измерением можно установить, что угол установки крыла и корпуса составляет 50° и что хвост опущен на 40—5(Г.
Проницание потока обеспечивает весьма устойчивое положение птицы при крутом спуске с любой высоты.
Подобный же спуск в тихую погоду нередко практикует и жаворо нск: при посадке он спускается, парашютируя под крутым углом до 60°, причем хвост и крылья его неподвижны и раскрыты полностью. При таких парашютирующих спусках видно открытие лучевых предкрылков.
Роль предкрылков совершенно очевидна: они затягивают срыв потока на больших углах атаки, наиболее полное открытие их наблюдается в начальный момент взлета. Так как подвижное крылышко имеет самостоятельное мускульное движение, то вполне вероятно его участие в создании боковой устойчивости (рис. 47). В этом случае обдув через
него идет вдоль передней кромки мануса, вызывая увеличение подъемной силы на конце крыла или возникновение кренящих моментов при раздельном действии предкрылков.
ПРИСПОСОБИТЕЛЬНОЕ НАРУШЕНИЕ ОБТЕКАЕМОСТИ КРЫЛА
К процессам, присущим крылу птицы, относится также приспособительное нарушение обтекаемости.
При первом поверхностном знакомстве с устройством перьевого аппарата обращает на себя внимание елочно-гофровая структура перьев, состоящая из бороздок, идущих под углом к очину пера в 40—45° в широкой части и до 10° в узкой (рис. 48). Шаг бороздчатых углублений составляет: в основании пера в среднем 1 мм, а ближе к концу — 1,5—2 мм. При наложении перьев в образовавшихся туннелях бороздки идут в противоположных направлениях. Такое их расположение вызывает вихреобразование уже внутри туннелей, следовательно, скорость протекания воздуха в них уменьшается.
Сильная пушистость в носовой части под кроющими перьями рассматривается в орнитологии как защитное утепляющее приспособление, однако слой пуха перекрывает основание пера до аванкамер. У некоторых пород птиц, как, например, у кобчика, под пуховой прослойкой находятся нитяные кроющие перья. Эта пушистость напоминает собой род сетки или шарик семени одуванчика, плавающего в воздухе и пе-
реносимого воздушными потоками на далекое расстояние.
Над хвостом той же птицы и в задней части спины имеется целая копна из нитяных перьев с прослойкой из пуха (многослойная сетка) (рис. 49). Образ жизни кобчика объясняет назначение такого приспособления. Кобчик применяет «стояние на месте» для охоты за мышами, в это время его крылья совершают вибрационные движения, при которых перья мануса отбрасывают поток в плечевую область, продувая его через парашютный клапан и копнистое образование нитяных перьев, что необходимо для устойчивости1.
Из аэродинамики известно, что сопротивление трения, зависящее от вязкости, может быть в 3— 4 раза больше, чем сопротивление формы, и что оно больше на малых скоростях. Таким образом, гофровая структура пера, пушистость, нитяные перья являются приспособительным нарушением обтекаемости крыла.
Рис. 48. Направление бороздок в опахале пера: 1—передний край; 2-— опахало пера
1 Другими наглядными примерами использования вязкости воздуха крылом птицы является маневр зависания на одном крыле, часто практикуемый воронами, и крутое парашютирование, подобное изображенному на рис. 45.
До сего времени остается необъясненным тот факт, что внутри крыльев птиц содержатся значительные массы воздуха, например, в перьях утки имеется 670 см3 при весе всех перьев в 65 г. Какое влия-
Рис. 49. Копна из нитяных перьев кобчика
ние оказывает на работу крыла периодическое проницание хотя бы 50% этой массы воздуха? Возможно, что картина обтекания будет существенно отличаться от принятой в аэродинамике.
ИСПОЛЬЗОВАНИЕ ВЯЗКОСТИ ВОЗДУХА
Сопротивление трения воздуха, зависящее от вязкости, в современных самолетах представляет наибольший процент (около 30%) в балансе всех сопротивлений. Его вредное влияние стараются уменьшить полировкой поверхности, потайной клепкой и другими способами.
Поверхность птичьего крыла далеко не совершенна. Как показывает микроструктура маховых перьев, нижняя поверхность опахал большей частью сильно ребристая, верхняя — слабо ребристая с выступающими на поверхности рядами ворсинок, образующих значительную шероховатость верхней поверхности крыла1. Наряду с этим проток воздуха через крыло птицы проходит через внутренние сетчатые камеры, образованные особым пухом и нитяными перышками в основании очина перьев. Воздух выходит наружу через верхние кроющие черепицеобразные перья (рис. 1 и 24,Л). Наличие протоков обнаружено фотографией на режимах больших углов атаки и в третьей фазе удара крыла (гл. VII).
Чтобы понять эти явления, необходимо вспомнить, что такое вязкость (19).
1 Наблюдения производились в микроскоп в орнитологическом кабинете Института эволюционной морфологии Академии наук СССР.
Под вязкостью понимают силы внутреннего трения. Силы вязкости стремятся затормозить быстро движущиеся частицы и ускорить более медленные.
Вязкость стремится уничтожить вихри, которые в вязкой жидкости постепенно рассеиваются и исчезают. С другой стороны, вязкость может создавать вихри. Особенно сильные вихреобразования наблюдаются возле самой поверхности тел, обтекаемых потоком. Благодаря тре
Рис. 50. Молекулярное движение частиц воздуха как причина вязкости
Рис. 51. Коэфициент сопротивления шара в зависимости от числа Рейнольдса (Re): I- - при больших Re, II—при малых Re
нию о поверхность происходит некоторое отставание скорости потока на поверхности (рис. 50).
Слой, где происходит это явление, называется пограничным слоем, его толщина около 1 мм при существующих габаритах крыла. Физическая сущность вязкости объясняется молекулярным строением вещества.
Молекулы газов и капельных жидкостей обладают по кинетической теории вполне определенными средними скоростями беспорядочного движения, зависящими лишь от их температуры. Движение молекул производит перемешивание слоев текущей жидкости. На рис. 50 приведены следующие обозначения:
I — слой быстро движущихся молекул.
II — слой медленно движущихся молекул.
Медленные молекулы, попадая в более быстрые слои, будут их затормаживать.
Вязкость обычно определяют при помощи волосных сосудов или трубок. Ее же определяют особым коэфициентом рь размерность которого кг1 м~2 сек1.
В практике для целей расчета и измерений пользуются кинематическим коэфициентом вязкости:
— • кг° м2 сек -1. Р
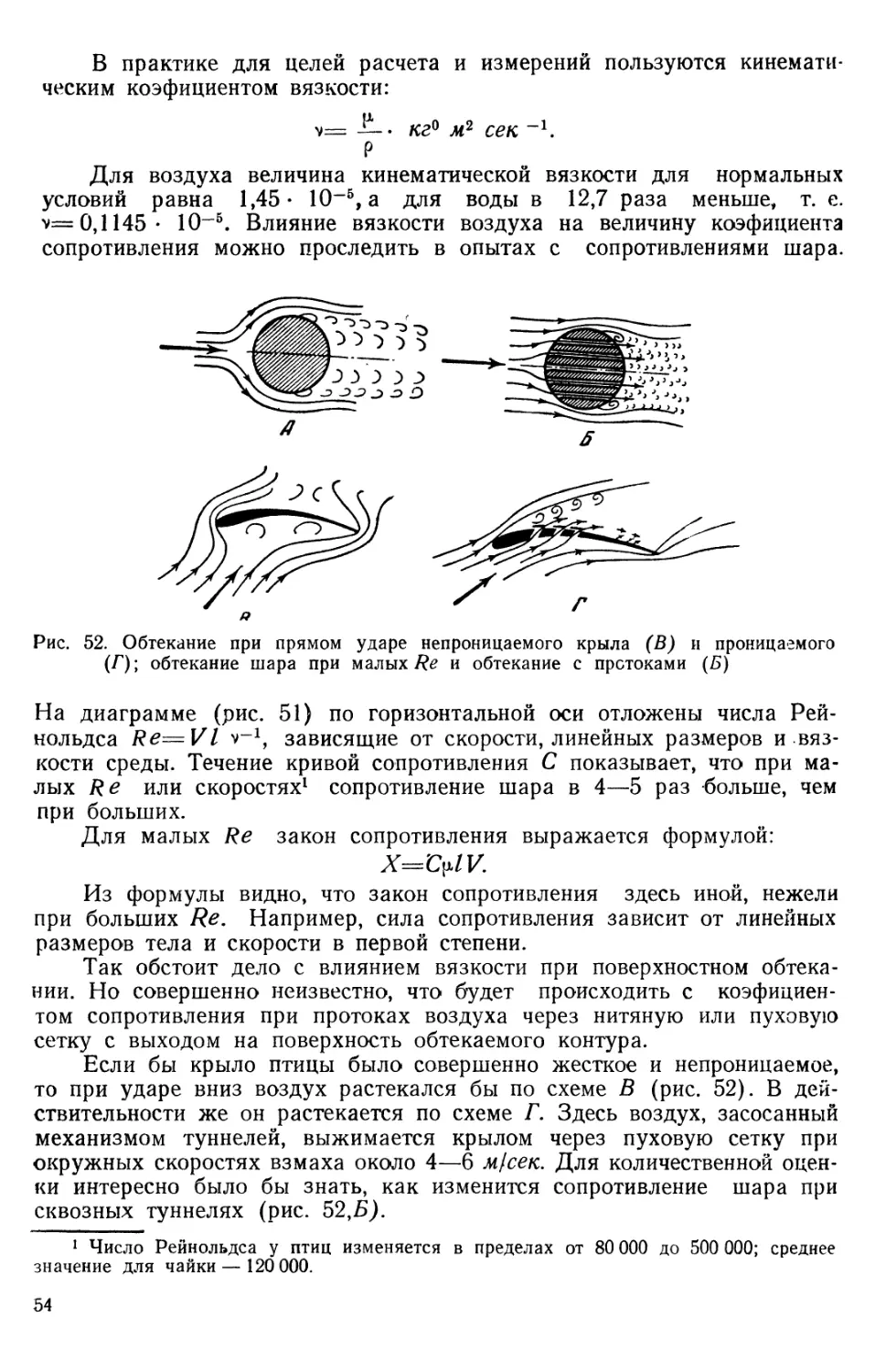
Для воздуха величина кинематической вязкости для нормальных условий равна 1,45- 10~5, а для воды в 12,7 раза меньше, т. е. v=0,1145- 10-5. Влияние вязкости воздуха на величину коэфициента сопротивления можно проследить в опытах с сопротивлениями шара.
Рис. 52. Обтекание при прямом ударе непроницаемого крыла (В) и проницаемого (Г); обтекание шара при малых Re и обтекание с протоками (Б)
На диаграмме (рис. 51) по горизонтальной оси отложены числа Рейнольдса Re=Vl v-1, зависящие от скорости, линейных размеров и вязкости среды. Течение кривой сопротивления С показывает, что при малых Re или скоростях1 сопротивление шара в 4—5 раз больше, чем при больших.
Для малых Re закон сопротивления выражается формулой: X~C^IV.
Из формулы видно, что закон сопротивления здесь иной, нежели при больших Re, Например, сила сопротивления зависит от линейных размеров тела и скорости в первой степени.
Так обстоит дело с влиянием вязкости при поверхностном обтекании. Но совершенно неизвестно, что будет происходить с коэфициентом сопротивления при протоках воздуха через нитяную или пуховую сетку с выходом на поверхность обтекаемого контура.
Если бы крыло птицы было совершенно жесткое и непроницаемое, то при ударе вниз воздух растекался бы по схеме В (рис. 52). В действительности же он растекается по схеме Г. Здесь воздух, засосанный механизмом туннелей, выжимается крылом через пуховую сетку при окружных скоростях взмаха около 4—6 м{сек. Для количественной оценки интересно было бы знать, как изменится сопротивление шара при сквозных туннелях (рис. 52,Б).
1 Число Рейнольдса у птиц изменяется в пределах от 80 000 до 500 000; среднее значение для чайки — 120 000.
Качественную оценку этого явления на малых скоростях можно получить из приведенных кривых на рис. 53 сопротивлений сеток туннельного радиатора.
Более мелкая сетка за № 6 при скорости V— 6 м/сек, дает сопротивление в 12 раз большее, нежели сетка № 1, а при скорости У = = 4 м/сек — почти в 15 раз.
Так как крыло птицы при парении и при парашютировании имеет куполообразную форму парашюта, то сопротивление его с проницанием будет весьма значительным.
С другой стороны, сама; направленность потока, проницающего крыло, будет содействовать его устойчивому положению.
Рис. 53. Кривые сопротивления сеток в туннельном радиаторе
Это и объясняет нам новый термин «увязание крыла в воздухе», которым мы и будем пользоваться при дальнейшем изложении процессов машущего полета.
В дополнение можно привести и другие примеры использования вязкости в природе. Сибирская белка-летяга совершает планирующие спуски до 25 м, так же планируют индийская белка тагуан и обезьяна-пропитекус (из Явы) (8). В данных случаях пушистый хвост и меховой покров спины заменяют им перьевой аппарат вязкости птиц1.
Влиянием вязкости воздуха можно объяснить, например, поворот птиц на месте при зависании на одном крыле, а также устойчивость при маневрах и весьма малые радиусы их виражей.
Физическая картина работы крыла в машущем полете дополняется новым видом сопротивления: сопротивлением проницанию потокам воздуха. Влияние вязкости воздуха наблюдается при малых скоростях протока воздуха через крыло. Однако в фазах машущего полета это
1 Отсутствие оперения у перепончатокрылых (типа летучей мыши) ограничивает их возможность к парению и стоянию на месте, их основным видом полета по «необходимости является вибрационный и притом на короткие расстояния.
состояние сменяется принудительным сокращением туннелей; начинает ся сдувание пограничного слоя, крыло приобретает высокие подъемные свойства.
В этом переменном состоянии потока около крыла и внутри его и заключается «приспособительность» обтекания у птиц, которое связа-но с приемами полета и режимами.
Замечательно то, что птица вторых режимов (т. е. при критических углах атаки) не боится, тогда как для самолета они опасны. Маленькие ястреб и ворона, только что оперившиеся, при первых же уроках полета дают прекрасный объект для понимания этого явления. Их неуверенный полет при больших углах с полным открытием поверхности допускает прямое наблюдение работы крыла на взлете.
Эта картина физического состояния резко отличается от простого обтекания жестких крыльев самолета или планера и должна явиться предметом специальных лабораторных исследований.
УПРУГИЙ ЭФФЕКТ БОЛЬШИХ МАХОВЫХ ПЕРЬЕВ
На кинокадрах полета чаек, заснятых в Баренцовом море Академией наук СССР, видно следующее: чайки попарно движутся в резонансном полете, причем крылья их несколько сдвинуты в фазах движения.
В этом состоянии большие маховые перья на фото оказываются смазанными в один и тот же момент или в одной и той же позиции. Это явление обнаруживает также фотоснимок полета чаек, сделанный телеобъективом при скорости 1 : 500 в секунду (рис. 54), на котором концы мануса расплывчаты.
Рис. 54. Раскрутка больших перьев в конце 4-й фазы
При рассмотрении снимков горных орлов (при броске вниз и при парении) оказалось, что большие маховые крылья отходят вперед и вверх, образуя веер и многопланное расположение лопаток. Схема этих двух деформаций по вертикали и горизонтали представлена на рис. 55. Сопоставляя эти факты, необходимо- сделать сле-
Рис. 55. Крутильная и изгибная деформация больших перьев: zis —отвод вперед;. A h — прогиб; М — скручивающий момент
дующий вывод: в момент подъема крыла перо, как сильно изогнутая пружина, распрямляется и быстро возвращается в исходное положение, отбрасывая поток назад, и тем самым создает толчок, преодолевающий инерцию крыла1.
Этим действием больших маховых перьев и объясняется расплывчатость снимка мануса в начале фазы подъема.
Так как манус крыла имеет большую скорость, чем плечевой отдел, то в сочетании с указанным эффектом он ограничивает растекание потока, загоняя часть его в среднюю зону, где образуется повышенное давление. Надо полагать, что деформированные упругие второстепенные перья произведут такой же эффект отграничения потока вдоль ребра схода крыла. Действие упругого возврата их напоминает первые пружинные самолетные посадочные щитки, применявшиеся в 1917—1918 гг. С точки зрения прочности такое крыло обладает антивибрационными свойствами.
Из содержания данной главы мы приходим к следующим выводам:
1. Машущее крыло создает особые условия обтекания, подсчет силы сопротивления которого не может быть выполнен по формулам аэродинамики.
2. При установлении схемы работы крыла птицы должны быть приняты во внимание:
а) всасывающее действие его туннелей;
б) прорыв потока в сопряжении крыла с корпусом и долевой, в стыке туннелей;
в) пружинный эффект отдачи больших маховых перьев.
1 Этот эффект больших маховых перьев изучал Марэй. ,,Le vol des oiseaux". Paris, J 898 (см. стр. 120).
ГЛАВА V
КАК ОСУЩЕСТВЛЯЕТСЯ УПРАВЛЕНИЕ ПОГРАНИЧНЫМ СЛОЕМ В КРЫЛЕ ПТИЦЫ
ПРЕДСТАВЛЕНИЕ О ПОГРАНИЧНОМ СЛОЕ
Чтобы установить состояние потока около движущегося контура, надо знать состояние пограничного слоя, который в случае сильной турбулентности может в несколько раз увеличить лобовое сопротивление крыла и значительно снизить подъемную силу. Интересно было бы знать, принадлежат ли крылья птиц к числу ламинарных или умеренно турбулизированных.
Как известно из аэродинамики, на поверхности крыла, обтекаемого потоком воздуха, возникает трение, величина которого зависит от степени шероховатости и от скорости потока. Благодаря трению скорости частиц в тонком слое, прилегающем к поверхности, изменяются от местной скорости до нуля, при этом происходит расслаивание потока (рис. 56). На границе двух слоев, имеющих разные скорости, происходит вращение частиц или, как говорят, турбулизация потока, которая может увлечь и вышележащие слои.
Элементарные вихри, возникающие по обе стороны профиля, на конце его соединяются, образуя вихревой след или спутную струю. Толщина пограничного слоя, где происходит это явление, вообще может изменяться в широких пределах, однако при небольших габаритах крыла его величина составляет от 0,5 до 1 мм. Интересно отметить, что интенсивность турбулентности в нем может значительно превышать интенсивность турбулентности в набегающем потоке1.
При обтекании тел рассматривают три вида пограничного слоя (рис. 57)—турбулентный, ламинарный и смешанный (26).
Экспериментально установлено, что турбулентное сопротивление в 4—5 раз больше ламинарного, т. е. того слоя, где поток движется без завихрений. Отсюда вполне понятно стремление уменьшить сопротивление трения. Кроме того, для крыльев самолета найдены особые ламинарные профили, отрыв потока у которых начинается в расстоянии 75% хорды от носка ее.
1 Инж. Дородницын. «Переход ламинарного пограничного слоя в турбулентный и ламинарный профили». Труды ЦАГИ № 563, 1945 г.
Обычно более ранний отрыв потока и нарушение ламинарного обтекания наблюдаются при увеличении угла атаки, так что на критических углах, близких к 16—20°, вся поверхность обтекается турбулентным потоком.
Воздействие на пограничный слой (33) можно осуществить сдува
Рис. 56. Схема скоростей в пограничном слое крыла и области потока: 1 — пограничный слой;
2 —вихревой след или спутная струя; 3 — потенциальный слой; Pi, Р%, Рз — давление в потоке
Рис. 57. Схемы пограничного слоя: а — ламинарного; б — турбулентного; в — смешанного
нием его через особые щели и отсосом путем особых механических устройств; в таких случаях значительно возрастет подъемная сила крыла. Особенно эффективным методом для увеличения подъемной силы является создание принудительной циркуляции скорости, не зависящей от угла атаки, при которой возможно достигнуть увеличения подъемной силы крыла в 3—4 раза1.
МЕТОДЫ, ПРИМЕНЯЕМЫЕ ПТИЦАМИ
Из некоторых наблюдений можно предположить, что птицы используют первый и последний способы управления пограничным слоем. Во второй и третьей фазах возникает сдувание пограничного слоя при помощи подслойного прорыва потока из. области парашютного клапана и долевого прорыва стыка туннелей. В результате подъемная сила крыла сильно возрастает. Схема обдува показана на рис. 58.
Здесь первая щель в области лучевого предкрылка, вторая и третья щели находятся в зоне кроющих перьев 2.
1 В. В. Столбов. «Самолетостроение последних лет». Оборонгиз, 1940 г.
2 Следует обратить внимание на то, что щели в крыльях птиц идут не вдоль крыла, а поперек, т. е. по хорде, так что крыло расчленяется на ряд узких и длинных пластин с удлинением от !/ю До Уго, а в некоторых случаях и до Vso- Известно, что крылья такого типа сохраняют поток без отрыва до углов атаки 50°. В этом случае щелевое устройство крыла увеличивает критический угол, а не Сд . Проф. Г. Бураго. «Приближенная теория крыльев малых удлинений». (См. Труды научно-технической конференции. 1944 г., т. III. ВВИА).
Сдувание наблюдается и в фазе подъема крыла, когда в плечевой области второстепенные перья поворачиваются и пропускают поток сверху вниз (рис. 59).
Так как сами решетки имеют несущий S-образный профиль, то они также создают местную подъемную силу.
Управление пограничным слоем по способу создания циркуляции.
Рис 58. Схема отсоса и сдувания пограничного слоя: 1 — действие луп; 2 и 3— отсос через кроющие ряды перьев; 4 — сдувание через туннельный прорыв потока;
5 — туннель; 6 — нижние клапанные перья
Рис. 59 Силовая схема крыла при подъеме: 1 — область плечевых решеток, создающих подъемную силу; 2 — область мануса с тягой
не зависящей от угла атаки, применяется птицами при вибрационном действии мануса в трепещущем полете и при стоянии на месте.
Обычно при наступлении критического угла атаки нарушается обтекание профиля, и крыло теряет свою подъемную силу (рис. 60,6/).
Однако птица может стоять на месте под углами свыше 40°, при этом колеблющийся манус напоминает вентилятор1, отсасывающий по
1 Механизм образования тяги для стояния на месте создается теми же пропел лирующими перьями мануса.
граничный слой средней несущей части крыла (рис. 60,6), ввиду этого завихрения (рис. 60, в) исчезают, поток становится плавным и на аэродинамических весах установлено возрастание подъемной силы в 4 раза. Аналогично этому полет совы также подтверждает нашу мысль:
Рис. 60. Возникновение циркуляции, не з»ависящей от угла атаки, при управлении пограничным слоем: а — срыв обтекания на критических углах; б — спектр крыла при управлении пограничным слоем; в — схема обтекания на закритических углах
несмотря на сильную бахромистость передней кромки перьев она летает при больших углах атаки.
Рассматривая основные способы управления пограничным слоем, мы наблюдаем замечательное свойство птичьего крыла: приспособительное изменение обтекаемости потоком, при котором происходят различные процессы как внутри, так и вне крыла. Летательный аппарат птицы осваивает воздух, пропуская его через толщу нитяной пуховой набивки, и как бы «выжимает» его в определенные фазы взмаха, создавая сдувание пограничного слоя.
Открытие указанных свойств птичьего крыла позволяет объяснить подъем с добычей при двукратном полетном весе орла или сапсана.
При первых же своих взмахах птица автоматически приводит в действие весь аппарат управления пограничным слоем, который столь значительно увеличивает подъемную способность крыла.
ГЛАВА VI
УСТОЙЧИВОСТЬ и управляемость птиц
УСЛОВИЯ РАВНОВЕСИЯ
Так как крыло птицы движется по траектории, близкой к синусоиде^ при непрерывно изменяющихся углах атаки, то естественно предположить, что условия равновесия и устойчивости здесь должны быть иные по сравнению с самолетом или планером.
Из аэродинамики известно, что состояние равновесия в полете зависит от положения центра тяжести1 и центра давления воздушных сил.
Рис. 61. Равновесие сил в горизонтальном полете
Возникающие продольные моменты птица компенсирует действием; хвостового оперения или изменением стреловидности крыльев и кривизной профиля. Из наблюдений установлено, что колебания корпуса/ при этом весьма незначительны. В установившемся горизонтальном движении (диаграммы а и в, рис. 68) могут быть приняты схемы (рис. 61), когда центр давления занимает переменное положение по отношению к центру тяжести. Произвольным изменением угла атаки крыльев, благодаря повороту в шарнире, птица создает моменты, которые, повиди-мому, уравновешиваются моментом хвостового оперения.
В случае полета с добычей центр тяжести птицы переходит назад
1 В орнитологии положение центра тяжести определяется позади плечевого шарнира птицы.
и может достигнуть предельного положения 30—35% корневой хорды крыла (нормальная центровка для птиц — около 15—20% хорды).
Для случая планирования или парения схема сил, действующих на птицу, дана на рис. 62. Здесь центр давления совпадает с центром тяжести птицы и слагающие веса G и подъемной силы Y зависят от угла наклона траектории 0 и равны соответственно Y = О2 и X = Gv
Рис. 62. Схема сил при планировании
СПОСОБЫ СОХРАНЕНИЯ УСТОЙЧИВОСТИ1
Особый интерес для авиационных конструкторов представляет вопрос об осуществлении птицами устойчивости относительно трех пространственных осей Z—Z, X — X, Y соответственно различают устойчивость продольную, поперечную и путевую (рис. 63).
Постоянно действующими факторами устойчивости у птиц являются: эластичность перьев, переменная стреловидность крыла в плане и низкое положение центра тяжести.
Профиль крыла, образованный эластичными перьями, в полете де
Рис. 63. Положение трех пространственных осей крыла
формируется и принимает S-образное очертание (рис. 64), что приближает его к устойчивым профилям, для которых положение Ц. Д. не меняется при режимах летных углов атаки.
1 И. Виноградов. Статья «Проектирование легкого самолета». Журнал «Самолет», №№ 8, 9, 10, 1937 г.
Необходимым изменением стреловидности в плане птица достигает, повидимому, целесообразной центровки, чем весьма эффективно сохраняет продольную устойчивость.
Как показывают продувки моделей, большие птицы (орлы, коршуны, аисты и др.) обладают большей устойчивостью в полете, не
жели мелкие воробьиные, но которые ввиду этого являются более маневренными, нежели первые.
Устойчивое равновесие в случае
'Рис. 64 Профили сечений крыла сокола: корневое и среднее при нагружении однократным п = 1 и двукратным весом п — 2
потери хвоста у больших птиц вообще исключено, у малых—возможно. Для сохранения продольной устойчивости при различных Эволюциях птица пользуется возможностью изменения угла атаки, переменной площадью хвостового оперения и отклонениями его вверх или вниз.
Орнитологи, занимавшиеся воспитанием молодых птиц, описывают первые неуверенные их учебные полеты, при которых можно проследить отмеченные действия крыльев и хвоста. В большинстве случаев при полете с рук, как наблюдал автор, молодые ворона и ястреб совершали взмахи с полной амплитудой, хвост был полностью раскрыт и траектория полета была волнистой благодаря грубым движениям хвоста. Замечательно, что при этом путевая устойчивость сохранялась вполне, а случаев безразличного равновесия никогда не наблюдалось.
ПУТЕВАЯ УСТОЙЧИВОСТЬ ПТИЦ
Путевую устойчивость птиц необходимо рассматривать вместе с боковой устойчивостью1.
У птиц роль вертикального оперения выполняет корытообразное искривление (например, у голубя и чайки) боковых перьев хвоста или раздвижной V-образный хвост (у ласточек и стрижей).
Кроме того, крыло птицы-парителя имеет отрицательную концевую закрутку, сообщающую крылу значительную путевую устойчивость.
Весьма сильным средством воздействия на путевую устойчивость является раздельное поднятие алюли и искривление хвоста. Изменение кривизны мануса крыла также является одним из средств восстановления путевой устойчивости.
На основании продувок моделей можно утверждать, чтЬ проницание крыла потоком и управляемый пограничный слой крыла усиливают
1 Д. Кузьменко и Б. Висленев «Теория авиации». Военгиз. 1936 г.
стабилизирующие действия всех выше отмеченных способов сохранения путевой устойчивости.
Низкое положение центра тяжести, поперечное V крыла создают моменты, выравнивающие боковое скольжение.
Рис. 65 Искривление, крыла чайки на вираже
В некоторых случаях пологого планирования крылья парителей образуют перевернутое V с отгибом вверх больших маховых перьев (рис. 65).
Моменты скольжения птица регулирует в широких пределах, благодаря приспособительному нарушению обтекаемости.
КАК ПТИЦА УПРАВЛЯЕТ ПОЛЕТОМ
Как видно из фотоснимков (рис. 32 и 45) диапазоны углов отклонения крыла и хвоста птиц достигают широких пределов: хвостовое оперение может быть опущено вниз до 45° и поднято вверх до 60°, а углы поворота крыла могут изменяться от —5° до -|-50о. Переход на пикирование или на крутой подъем птица осуществляет вращением крыла около поперечной оси, в нормальном же полете необходимые углы атаки крыла достигаются действием хвостового оперения.
Хвост птиц можно рассматривать как стабилизатор с переменными площадью и углом атаки, а также как орган, создающий моменты около поперечной и вертикальной осей.
Рулевые перья укреплены посредством специальной мускулатуры к заднему отделу позвоночника (пигостилю). Особенностью их действия являются:
1) винтовой поворот около продольной оси, усиливающий мрмент крена;
2) изменение площади, которая при парении и посадке является несущей.
Площадь хвостового оперения у коршуна в скоростном полете составляет 29% от площади крыльев, а при парении и посадке полностью раскрытый веер хвоста увеличивает площадь еще на 5—6%.
По своему устройству рулевые перья напоминают малые машущие
Рис. 66. Парашютирование чайки с приподнятым корпусом
перья. Они отличаются лишь тем, что имеют меньшее туннельное образование и меньший саблевидный изгиб.
Птица может различным образом ориентировать положение своего* корпуса относительно направления полета. Например, может осуществить крутое планирование «плашмя» (рис. 29 и 46); перейти на парашютирование с приподнятым или опущенным корпусом (рис. 30 и 66} и, наконец, подобно самолету, может пикировать в направлении оси своего корпуса.
Осуществляя посадку с торможением, птица может поставить свои крылья почти под прямым углом к движению.
На рис. 67 засняты последовательные положения крыльев голубяг при посадке на крышу, с торможением, вертикальным поворотом корпуса и ударами крыла навстречу потоку.
На рис. 97 показан тот же маневр при снижении с тем отличием^ что корпус в последний момент отклонен назад, хвост сильно отжаг вперед, чтобы избежать опрокидывания через голову при посадке.
Столь разнообразные приемы полета достигаются у птиц комбинацией поворотного движения крыла с отклонением хвостового оперения, которое может действовать как руль высоты и руль направления (рис. 24).
Управление кренами достигается сокращением площади одного крыла и открытием алюли на стороне поднимающегося крыла (рис. 47). Из главы V известно, какое это имеет значение для создания подъемной силы, а тем самым и для боковой устойчивости.
Длинная шея уток и гусей также облегчает управление и центровку птиц в полете.
Из этого краткого обзора методов управления, осуществляемых птицами, видно, какие перспективы открываются у конструкторов для 66
Рис. 67. Посадка голубя с торможением через прямой поворот крыла
усовершенствования техники полета и изысканий совершенно новых путей ее развития.
Возможность превращения планера и самолета в парашют обещает намного увеличить безопасность полета, а для схемы аппарата «летающее крыло» открываются возможности более эффективных средств управления им.
При осуществлении в конструкциях планеров и самолетов поворота крыльев должны появиться и новые приемы пилотажа: 1) посадка с торможением крыльями, 2) разворот «юзом» или без крена, 3) посадка «плашмя» без наклона корпуса и др.
ГЛАВА VII
СИЛОВАЯ СХЕМА КРЫЛА ПТИЦЫ
В этой главе приведены факторы работы машущего крыла, от которых зависит, по нашему мнению, образование тяги и подъемной силы крыла и представление о которых необходимо иметь, чтобы правильно судить о кинематике крыла и об условиях, влияющих на прочность его конструкции. Допущенная схематизация в изложении процессов машущего крыла оправдывается отсутствием разработанных методов аэродинамического расчета.
ОСОБЕННОСТИ ДВИЖЕНИЯ КРЫЛА
Для уяснения процесса образования тяги и подъемной силы машущим крылом птицы надо иметь в виду следующее: 1) шарнирное соединение с корпусом, 2) переменное положение оси вращения крыла, 3) синусоидальный характер движения, 4) колебание центра тяжести птицы.
Плечевой шарнир у больших птиц отличается своеобразной работой мышц, которые приводят в движение крыло1.
При взмахах и ударах крыла появляются инерционные силы движущихся масс плечевого и концевого разделов крыла.
Оси поворота крыла, относительно которых происходят взмах и изменения установочного угла, автоматически принимают наивыгоднейшее положение для создания подъемной силы и тяги. Как мы увидим далее в фазе взмаха и при подпрыгивании, во время отрыва от земли крылья имеют небольшую подъемную силу.
СИНУСОИДАЛЬНЫЙ ЗАКОН ДВИЖЕНИЯ
Возникновение аэродинамических сил связано со скоростью движения машущего крыла по траектории, которая при установившемся горизонтальном полете является синусоидой.
1 См. главу IX.
При отсутствии опытных характеристик машущего крыла для приближенных расчетов можно пользоваться одной из схем, данных на рис. 68, при этом задаемся законом движения и полярой крыла. По общему правилу разложения аэродинамической силы /? получим нормальную или ударную силу Af и силу тяги Т.
Рис. 68. Синусоидальное движение машущего крыла: а — схема сил при взмахе при положительном угле атаки; б — диаграммы распределения сил при средней N — 0; в — диаграмма сил при средней тяге Т; г и д — распределение подъемной силы и тяги при отрицательном угле взмаха
Зная углы атаки, скорость по траектории и коэфициенты крыла, можно получить развернутую картину сил ударной и тяги по диаграммам б, в, г и д.
Для конструктора интересно знать средние значения сил N и Т за полный период работы крыла.
В процессе образования сил здесь могут иметь место два случая:
1) взмах с отрицательным углом, когда тяга Т положительная, подъемная сила Y направлена вниз и за всю фазу работы средняя ударная сила N равна нулю (рис. 68,а и б);
2) взмах с положительным углом атаки (рис. 68,д), когда тяга имеет отрицательное значение, а подъемная сила положительная и за весь период крыло развивает среднюю положительную ударную силу jVcp (рис. 68,г).
Первый случай дает наибольшую скорость, а второй наибольшую подъемную силу, следовательно, первым случаем птица будет пользоваться в горизонтальном полете, а вторым — при подъеме1.
ИНЕРЦИОННЫЕ КОЛЕБАНИЯ В МАШУЩЕМ ПОЛЕТЕ
Кроме аэродинамических сил, в машущем полете большую роль имеют инерционные колебания крыла и тела птицы, которые возникают от изменения знака ускорения центра тяжести машущего крыла и от упругой раскрутки мануса.
Этот случай становится явным из схемы траекторий движения центра тяжести корпуса и крыла (рис. 69), полученных на основе анализа
Рис. 69. Движение центров тяжести крыла и корпуса
киносъемки машущего полета. Как видно из схемы, масса тела птицы движется, повидимому, по параболе, а вершина крыла — по синусоиде.
При ударе Ц. Т. крыла обгоняет Ц. Т. корпуса, а при взмахе отстает. Это явление напоминает известный эффект раскачивания на качелях, когда каждый раз, благодаря подталкиванию качелей «в такт», их движения могут достигнуть больших размахов колебания.
Это явление в механике объясняется теоремой о движении центра тяжести.
В конечной фазе опускания крыла, когда меняется направление его движения, упругая раскрутка мануса усиливает общий эффект подталкивания «в такт» (рис. 70).
Очевидно, чем длиннее крыло, тем сильнее будет бросок вперед. Следовательно, длиннокрылые птицы — стрижи, альбатросы, кулики— должны обладать большим ускорением центра тяжести, нежели ширококрылые голуби, коршуны, куропатки и др.
1 В опытах с эластичными машущими крыльями установлено, что они дают под емную силу при взмахе, если скорость полета больше 8 м/сек (см. гл. IX).
В фазе взмаха крыла его масса будет несколько отставать от Ц. Т. корпуса. Однако движение вперед будет происходить по инерции, благодаря накопленной энергии, вследствие чего птица некоторое время может лететь со сложенными крыльями. Это наблюдается, например, при полетах скворца, дятла и ряда других птиц.
Рассмотренное нами движение двух масс крыла и тела птиц напо-
е
Рис. 70. Свободное движение Ц. Т. птицы под действием упругой раскрутки больших маховых перьев: О\ и О2 — мгновенные центры вращения; Cj и С2 — положения центра тяжести; Л — скольжение; Д/— элемент пути
минает движение сателлита в планетарной системе передачи (рис. 69).
Необходимо напомнить общепринятый в кинематике механизмов способ построения скоростей и ускорений шатунно-кривошипного механизма (см. «Справочные таблицы по кинематике механизмов». Проф. JI. П. Смирнов).
Зная ускорение, можно получить инерционные силы крыла по формулам п • G
P = mj , где масса крыла
9,о1
и ускорение/=— \ cos ср +
± X cos 2ср), где ср — угол кривошипа, X — отношение радиуса кривошипа к длине шатуна. С помощью этих значений мы можем начертить линию давления массовых сил — параболу, нанеся ее на диаграмму подъемных сил крыла в зависимости от угла ср (рис. 71,а). Мы можем убедиться, что инерционные силы уравнивают отрицательные значения подъемной силы, возникающие при ударе крыла вверх (см. перевод ВВИА № 1252, 1949 г., Рейфенштейн. «О новых испытаниях самолетов с машущими крыльями»).
Рис. 71,а. Диаграмма сил модели машущего полета: 1 — подъемная сила; 2 — парабола инерционных сил от веса крыла; 3 — суммарная кривая
Это явление заслуживает всестороннего исследования, поскольку от него зависит плавный ход механизма в машущем полете и проч ность колеблющихся частей, подвергающихся инерционным нагрузкам1.
МАЯТНИКОВОЕ ДВИЖЕНИЕ КРЫЛА
Применяя к крылу птицы термин «маятниковое движение», мы лишь упрощаем схему поперечных колебаний крыла в целях наглядного представления.
В технической литературе этот термин применяется там, где восстанавливающей силой является не только сила тяжести, но центробежные силы вращающихся масс (см. журнал «Техника воздушного флота» № 3 за 1939 г.). Аналогия с двойным сферическим маятником оправдывается тем, что крыло совершает колебания около трех пространственных осей.
Мы рассмотрели движение масс в продольном направлении, однако нельзя оставить без внимания и упругое поперечное колебание крыла. На самом деле крыло состоит из двух колеблющихся частей: плечевой и мануса. Манус имеет самостоятельное сложное движение с центром тяжести, далеко выдвинутым по размаху крыла.
Поэтому крыло можно представить в виде двух материальных то-
Рис. 71,6. Схема двойного маятника: 1 — Ц. Т. средней области; 2 — Ц. Т. мануса
чек, совершающих движение по типу двойного сферического маятника около продольной оси (рис. 71,6).
В процессе махания крыло приобретает волнообразный изгиб, подобный тому, который наблюдается в опыте с резиновой трубкой: если один конец ее закрепить, а другому сообщить резкий толчок, то трубка изгибается волнообразно.
Это обстоятельство, повидимому, является весьма важным в образовании силы тяги крыла.
СХЕМА РАБОТЫ МАШУЩЕГО КРЫЛА
Для более наглядного представления физической схемы работы крыла парителей мы расчленим процесс образования подъемной силы и тяги в машущем полете на четыре отдельные фазы согласно кино-
1 При расчете орнитоптера необходимо соблюдать определенное соотношение между массой крыла и корпуса при движении крыла вверх и вниз. В среднем вес крыла по отношению к весу прибора должен находиться от ]Л до 7s, как это установлено при пролете моделей.
съемкам (рис. 34). Первая фаза—подъем крыла или взмах (6—8); вторая фаза — опускание крыла до 40° от верхней мертвой точки (9—10).
при малых углах атаки; условимся называть эту фазу «началом удара крыла». Третья фаза—парашютный удар вниз или опускание (1—2) в пределах до 30°; четвертая фаза — упругое действие мануса и отталкивание (4—5) в пределах до 50° (рис. 54).
Те же фазы хорошо заметны на кинокад-
рах полетов аиста (рис. 72,а), отдельные увеличения которых представлены на рис. 41а 76, 79 и 82.
Полная амплитуда колебания 6 от верхней мертвой точки крыла до нижней равна ПО—130°, а в некоторых случаях может доходить до 180°.
Принятое нами положение крыльев при взмахе и ударе можно проследить на фотоснимке рис. 72,6, где заснят подъем стаи чаек и видны первая фаза (средняя часть рисунка), вторая (на левой стороне) и четвертая (в правом верхнем углу рисунка).
Скорость движения крыла V слагается из поступательной v и окружной и и при опускании и подъеме крыла (рис. 39) па соответствующем радиусе в зависимости от частоты колебаний
\ сек /
V = У + t? (I),
2 s ‘ п
гдеи = —, s — длина дуги, опи
санной крылом.
Средняя окружная скорость на конце крыла составляет у птиц 6,4— 8,2 м!сек.
Рис. 72,а. Кинокадр полета аиста при виде сбоку
1
Рис. 72,6. Фазы крыльев чаек в
момент взлета при виде сзади
ПЕРВАЯ ФАЗА
Отрыв тяжелых птиц от земли происходит после разбега (аисты, журавли), а средних по весу — в большинстве случаев с места, причем
Рис. 73. Трехсуставные сочленения костей ног птицы
Tnj~RT
Рис. 74. Схема сил при отрыве от земли
птицы осуществляют прием подпрыгивания. Благодаря тройному сочленению костей ног (рис. 73) и растяжению мускульных связей прыжок птицы позволяет ей получить начальную скорость порядка 4—5 м/сек.
Одновременно с прыжком в косом направлении птица выбрасывает крыло вверх назад (рис. 74). После отталкивания от земли корпус движется самостоятельно, крылья разгружены от веса и достигают верхней мертвой точки (рис. 75).
Рис. 75. Схема первой фазы взмаха (амплитуда 60—ПО0)
Рис. 76. Начало первой фазы полета аиста; манус на больших углах вращения; видна упругая раскрутка его перьев (кинокадр)
Ошибочным было бы предположение, что крыло птицы при подъеме или взмахе испытывает только вредное сопротивление. Можно считать, что крыло, будучи разгружено от силы веса, становится в положение угла атаки с наименьшим лобовым сопротивлением и, кроме того, развивает собственную тягу благодаря перьям мануса (рис. 76). Действие расширения и сужения туннелей сопровождается всасыванием
Рис 77 Открытие плечевых решеток аисга в первой фазе (кинокадр)
воздуха и сдуванием пограничного слоя через решетки плечевой зоны (рис. 77), так что в результате аэродинамическое сопротивление крыла полностью компенсируется собственной тягой на взмахе.
Схема сил, действующих на птицу в момент отрыва, дана на схеме рис. 74, где через J обозначено ускорение, m — масса тела тицы (т = )>^и —составляющие аэродинамической силы по вертикали
х У,о1 /
и горизонтали. Сила толчка Re равна инерции веса mj , Состояние равновесия при отрыве можно выразить через уравнения:
= ХТ = 0, ГЛ4 = 0 (II)
ВТОРАЯ ФАЗА
Положение крыла во второй фазе характеризуется схемой рис. 78, где дано среднее угловое перемещение на 35—40° вниз от верхней точки подъема крыла.
С момента начала опускания крыла туннели сглаживаются и крыло птицы принимает нагрузку от веса ее корпуса. На рис. 79 показана вторая фаза полета аиста1.
1 Все снимки полетов аиста заимствованы из научно-популярного фильма «История одного кольца»
Величину осередненных аэродинамических сил в фазах удара крыла (фазы вторая и третья) приблизительно можно оценить по траектории его движения (рис. 68) и диаграммам продувок.
(начало удара крыла)
Рис. 78. Схема второй фазы
Однако, отображая действительную физическую картину работы удара крыла, мы должны будем констатировать, что в начале опускания по направлениям: сверху — сзади, вниз — вперед имеют место малые углы атаки (при угловом перемещении 3 до 40°), следовательно, здесь мы имеем образование чистой подъемной силы и тяги.
Схема сил второй фазы показана на рис. 80. Мгновенное состоя
ние равновесия выражается через уравнение (II). В развернутом виде это уравнение будет для подъемной силы:
2ГКР + 2ГМ —0 = 0 (III),
для тяги:
2ТМ + 2Ткр - X = О (IV),
А4М А4кр — О,
где О —вес птицы;
X — лобовое сопротивление;
Ум —подъемная сила мануса;
/М —тяга мануса;
Укр — подъемная сила крыла и
ГКр—тяга средней части крыла.
ТРЕТЬЯ ФАЗА
Переход к третьей фазе (рис. 41,о) достигается вращением крыла в шарнире на наивыгоднейшие углы атаки, при этом движение крыла будет косо направленным вниз под углом порядка 30° к горизонту (установлено для аиста, для других птиц значительно меньше) (рис. 81).
Картина обтекания для средней и концевой части будет различна* а именно: система крыльев птицы воспроизводит схему работы двух
Рис. 81. Схема положения корпуса аиста и направление удара крыла в первой и третьей фазах
спаренных парашютов при маневре скольжения, а манусохМ создается тяга при обтекании решетчатого крыла на больших углах.
Как нам известно из главы III, в этом случае происходит мгновенный и сильный выход воздуха из кроющих рядов перьев и одновременно подслойный и долевой прорыв из туннелей. Все это влияет на со
стояние пограничного слоя, уменьшая сопротивление крыла и увеличивая его подъемную силу. Вместе с тем под крылом возникает интенсивное поперечное волнообразование вследствие поперечных колебаний крыла. В результате происходит сложение продольных и поперечных колебаний \ повышающих общую тягу всех частей крыла Трудность учета всех факторов в работе столь необычного в инженерной практике способа пропеллирования заставляет нас, при количественной оценке, объединить работу этой фазы со второй, тогда в вышеприведенных уравнениях (I и III) удобно принять, что тяга и подъемная сила являются слагающими общей аэродинамической силы /?, т. е.
pl/2
где 7
Т2,3 = ^?cosa
— /?sina R~CTqS
(V), (VI), (VII),
a —угол атаки, Сг на наивыгоднейшем режиме
равен 1,0—1,2 и в скоростном — 0,55 — 0,6. Указанные величины получены в результате лабораторных опытов. На взлетном режиме’ можно принять значение Сг =5 — 6, как это показывает поверочный расчет орла (рис. 40).
ЧЕТВЕРТАЯ ФАЗА
В заключительной четвертой фазе крыло имеет вид опрокинутой вниз чаши (рис. 82 и 83). Благодаря упругой раскрутке больших маховых перьев движение центра тяжести ускоряется (рис. 70) как по горизонтали, так и по вертикали и последнее движение совпадает с
Рис. 82. Кинокадр полета аиста в четвертой фазе
1 Эго явление рассматривается в гл. IX.
&
Рис. 83. Крылья олуши в четвертой фазе
Рис 84. Взлет серой вороны: заметно проницание потока через кроющие перья в плечевой области; перья мануса. сильно изогнуты
началом взмаха. Прямым наблюдением над полетом вороны установлено, что она взмывает вверх на 1,2—1,5 м в конце первого же удара крыла вниз (рис. 84).
В двух последних фазах удара крыльев птица находит в центре давления ту опору, при которой возможно мускульное подтягивание всего корпуса птицы к крылу и некоторый подъем плечевого шарнира.
Рассмотренная четырехфазная схема работы крыла птицы наблюдается при взлете на полных взмахах (амплитуда до 140°) и в скоростном полете.
В горизонтальном полете при неполной амплитуде мануса в 40° или 60° ясно выраженными являются первая и вторая фазы. Для подсчета усилий здесь удобно привести схему раздельных функций крыла, считая, что средняя часть является несущей или подъемной, концевая — пропеллирующей.
<6 Виноградов
ГЛАВА VIII
МУСКУЛЬНЫЕ УСИЛИЯ птиц
КОСВЕННЫЕ МЕТОДЫ ОЦЕНКИ УСИЛИЙ
Приближенная оценка усилий, затрачиваемых птицей в полете, была произведена О. Лилиенталем, однако прямых измерений в полете сил тяги птиц до сего времени не производилось1.
В биологии имеются косвенные методы оценки мускульного усилия? по весу мускулов: грудного, опускающего крыло (musculus pectoralis) „ и действующего при подъеме крыла (musculus supracoracoideus).
Вес грудных мускулов птиц qm в процентах от полного веса следую-
щий:
утки 18% орла 18,2%
кулика 18,25% коршуна 18,6%
луня 18,19% вороны 16%
воробья 20% чайки 12%
фазана 16,2%
Принимая затрачиваемую мощность пропорциональной весу муску-
лов, профессор В. С. Пышнов дает формулу коэфициента мощности^
птиц в случае висения на месте:
\ 2з \Отр /
G и для полета по горизонталям: —j-------------
к 3 (^тр 0 3 где к — качество,
Отр — вес грудных мускулов,
I — размах в м.
По данным инж. М. К. Тихонравова средняя мощность больших: птиц составляет 0,00124 л. с.
Указанные величины, повидимому, представляют максимальные усилия, развиваемые птицей при полном напряжении ее сил. Трансформация мускульного усилия птицы в тягу и подъемную силу осуществляется крылом со всеми присущими ему качествами.
Доктор биологических наук Н. А. Гладков среднее мускульное усилие птиц считает равным 10% от веса.
1 В опытах Марэя тяга птиц измерялась на ротативной установке.
По исследованию О. Лилиенталя для взлета голубя требуется усилие, равное 5% его веса.
Ввиду отсутствия точных экспериментальных данных о мощности птиц мы приводим таблицу 10, где определена их мощность в горизонтальном полете при условии, что тяга пропорциональна мускульному усилию.
Как видно из таблицы, данные по книге М. К- Тихонравова (2) сильно занижены.
Интересно отметить, что в опытах Марэя (40) над живыми птицами было найдено, что у сарыча усилие двух больших мускулов равно 12 кг при весе птицы 0,9 кг.
Марэй учитывал реакцию сокращения мускулов у птиц, которая протекает у них более быстро, чем у других животных.
Таблица 10
Птица Вес (кг) Площадь крыла (л«2) Уд. нагрузка на л. с. Скорость м/сек Мощность (л.с.) Мощность пв Тихонравову (л. с.)
6gm V 75
Коршун .... 1,(0 0,215 22,5 18 1 22,5 — —
Орел 5,3 0,5 18 22,5 1 3,4 — —
Ворона .... 0,615 0,152 30 15,7 1 48,6 1 250 1 86
Серая чайка . 0,89 0,334 59 10,6 1 66,5 1 300 1 925
ВОЗМОЖЕН ЛИ ПОЛЕТ ЧЕЛОВЕКА ПРИ ПОМОЩИ МУСКУЛЬНОЙ СИЛЫ?
Для полета на планере УС-4 при скорости снижения U = 0,5 м/сек и полетном весе 120 кг потребная мощность равна:
кт GU 120 • 0,5 nQn
N = = 0,80 л. с.
75 75
Учитывая к. п. д. механизма, эта мощность составит около 1 л. с.
Относительно развиваемой человеком энергии в литературе имеют -ся следующие данные: тяжелоатлет может развить до 2 л. с. за очень малый промежуток времени (0,5—1 сек). Энергия, развиваемая рабочими у подъемной лебедки при двухминутной работе, может дать 0,5 л. с. Велосипедный гонщик развивает наибольшую кратковременную мощность 1,3 л. с., в течение двух минут—0,7 л. с. и длительно 0,5 л. с.
Мощность гребца на гоночной лодке в течение 20 мин составляет 0,5 л. с. С физиологической точки зрения при гребле используется боль
шая часть мускулатуры, чего нельзя достигнуть педалями ножного управления.
В силу биологических особенностей организма человек неспособен к длительному напряжению своей сердечной деятельности и дыхательных органов. Так, например, установлено, что количество сокращений сердечных мышц в минуту у голубя — 192, утки — 212, коршуна — 258, тогда как у человека — 64. Далее, дыхательная поверхность у человека 13 см2 на 1 г веса, у голубя — 173 см2, у колибри — 666 см2. Частота дыхания у голубя в спокойном состоянии — 29, в полете — 450; у ворон изменяется от 31 до 390, у человека—от 17 до 20 в минуту.
Физическая мощность зависит от процента участия всей мускулатуры в процессе движения.
При педальной системе, как на велосипеде, используется около 3% мускульного веса, а при движении сгибания и выпрямления (например, при гребле) —свыше 22%.
Следовательно, этот способ наиболее выгоден для орнитоптеров, так как движения сгибания и выпрямления должны производиться в умеренном темпе, в ритме ударов машущих крыльев.
Равномерное движение корпуса не оказывает влияния на спокойное управление аппаратом, чего нельзя сказать о быстрых и постоянно изменяющихся движениях на педалях. Кратковременное выполнение горизонтального полета (при U = 0) или получение более пологого угла планирования зависит от конструкции аппарата и степени тренированности пилота.
Использование мускульных усилий в процессе полета планеров может уменьшить скорость их снижения, например, при переходе от одного термического потока к другому и для парения в вертикальных воздушных потоках небольшой скорости.
Применение машущего крыла для планера даст новое содержание технике безмоторного полета и новый вид увлекательного спорта. Технически вполне осуществимой явится постройка моторного мускуллета с двигателем 1,5—2 л. с. в помощь мускульным усилиям человека, т. е. летательного прибора личного пользования, доступного для всех.
ГЛАВА IX
ФИЗИЧЕСКОЕ СОСТОЯНИЕ ПОТОКА ВОЗДУХА ЗА КРЫЛОМ ПТИЦЫ
Для того, чтобы объяснить явления, происходящие как вокруг крыла птицы, так и позади его, необходимо знать структуру отходящего потока.
Некоторое представление о характере потока за крылом можно иметь, наблюдая: 1) парение птиц над морскими волнами, 2) групповые полеты чаек и 3) журавлиный строй при осенних перелетах.
Рис. 85. Парение над волнами
В первом случае мы имеет пример динамического парения, когда энергия колебания волны сообщается воздуху. Птица при этом подставляет действию потока свои крылья, которые в силу упругости то отгибаются вверх, то возвращаются в исходное положение (рис. 85). Наибольшее давление на крыло будет соответствовать гребню волны, когда птица взмывает, и наименьшее — углублению волны, когда птица опускается. При этом эластичные концы перьев автома4-тически увеличивают кривизну крыла и сохраняют подъемную силу. Известно, что альбатросы часами могут парить над волнами и иногда встречаются в открытом море за сотни километров от берега. Траектория их парения повторяет траекторию волны или очень близка к ней.
При групповом полете чаек отдельные их пары совершают ритмические колебания с определенным сдвигом фаз (рис. 86). По-видимому, первая птица отбрасывает поток, несущий кинетическую энергию, которая воспринимается второй чайкой.
Рис. 86. Кинокадры полета стаи чаек: видны отдельные пары в одном ритме колебаний
Чайки могут сбиваться и лететь в противоположных фазах, при большой скорости, но в установившемся движении их колебания одинаковы. Птицы как бы связаны невидимой нитью (рис. 87). Тог же эффект можно наблюдать при полете журавлей в строю «клином».
Рис. 87. Полет звена в одном ритме
РЕЗОНАНСНЫЙ ПОЛЕТ
С помощью киносъемок полета журавлей удалось установить, что птицы машут с одним и тем же периодом, частотой и амплитудой взма-
Рис. 88. Полет стаи лебедей ха, сохраняя между собой определенное расстояние; такого рода машущий полет мы условились называть резонансным (рис. 88).
Причиной полетов в виде строя треугольником или клином является особая структура потока за крылом птицы. Можно предполагать, что последовательные сгущения и разряжения воздуха, производимые
машущим крылом, распространяются вслед за волнообразной траекторией крыла (рис. 89). Летящая уступом сзади птица использует энергию волны, подставляя ее действию свои крылья. Возможно и другое предположение, что поток за крылом находится в синхронном или свя-
Рис. 89. Траектория конца крыла птицы
занном движении с двумя вихревыми дорожками по теории проф. В. В. Голубева.
Как показывает спектр этих дорожек за машущим крылом, заснятых в гидроканале инж. Я. Е. Полонским, они имеют волновой характер (рис. 106 и 107).
Весьма вероятно, что эти сбегающие дорожки вызывают колебательное состояние потока за крылом соответствующей частоты и периода, что в конечном итоге даст поток волнового типа сзади машущего крыла.
На самом деле, если бы поток был просто вихревой, то птица, летящая сзади, испытывала бы затруднения в полете подобно самолету, попавшему в струю от винта впереди идущей машины.
Между тем птицы в резонансном полете строго соблюдают свое построение и отбившиеся от стаи быстро отстают. Только при волновом характере потока за машущим крылом и можно объяснить использование энергии его в резонансном полете птиц.
ВОЛНООБРАЗОВАНИЕ ЗА МАШУЩИМ КРЫЛОМ
Суммируя все три приведенных щей картине физического состояния
случая, мы приходим к следую-потока за машущим крылом
тгицы.
«Рис. 90. Горизонтальная проекция крыла в машущем полете
1. Последовательное сближение и распластывание крыльев при взмахах (рис. 90) вызывает поперечное волнообразование, которое усиливается благодаря радиальному течению воздуха (41) по размаху крыла (рис. 42).
2. Вслед за движением крыла распространяется продольная волна.
Повидимому, сбегающие вихри распадаются вблизи крыла вследствие эластичности его перьев (рис. 91). В крайних положениях мануса по траектории движения сбегает двойная
Рис. 91. Распад вихрей за средней областью крыла
вихревая дорожка, также быстро распадающаяся вблизи крыла.
3. В результате этого происходит сложение поперечных и продольных волн в виде результирующей волны, отходящей из средней зоны крыла, на границе обдува лучевым предкрылком (рис. 92).
Манус крыла дает собственное волнообразование, усиливающее действие средней зоны крыла. Это представление об изолированном действии концевого отдела крыльев имеет подтверждение в устройстве лентовидных крыльев вымпелевого козодоя (рис. 93). Далеко отходящие за крыло ленты облегчают сход вихрей из средней зоны крыла; перья мануса находятся за его пределами.
Наши соображения о значимости волнообразования за крылом птицы находят авторитетные подтверждения в работе «О крылатых пропеллерах» Н. Е. Жуковского1. Он говорит следующее:
«Перемещение птиц в воздухе под действием внутренних сил будет нами только тогда вполне выяснено, когда будет обстоятельно изучен фундаментальный фактор этого перемещения — воздушная волна».
Подобием волнообразования за крылом птиц является хорошо изученное проф. В. В. Шулейкиным движение угрей и хвоста рыб 2. По их телу от головы к хвосту пробегает волна подобно опыту с резиновой трубкой, если один конец закрепить, а другой встряхнуть. Эта волна вызывается не отдельными взмахами, а непрерывной работой мышц, которые ритмически то сокращаются, то распластываются, благодаря этому каждая точка тела рыбы или угря совершает колебание
влево и вправо. При этом, чем ближе к хвосту лежит соответствующая точка, тем больше запаздывает она в своих колебаниях.
В результате от головы к хвосту бежит постоянная «твердая волна» (рис. 94) и тело благодаря сопротивлению трения будет отталкиваться в воде и двигаться в направлении, обратном этому движению. Самая волна оказывает давление на тело, скорость же распространения ее должна превышать скорость поступательного движения тела.
У длиннокрылых птиц имеется аналогичный аппарат принудительного воздействия для волнообразования — это двойное маятниковое колебание крыла, переменное наполнение туннелей воздухом, разглаживание их и радиальное течение воздуха. Весьма важным сходственным
1 Полное собрание сочинений Н. Е. Жуковского, том VI, 1937 г.
2 В. В Шулейкин. «Физика моря». Изд. Академии наук СССР, 1949 г.
Рис. 95. Характер нарастания волн при разных амплитудах колебания
фактором с движением хвоста рыб является амплитуда мануса, которая превышает амплитуду плечевой области. О характере нарастания волн вдоль крыла можно судить по диаграмме (рис. 95) В. В. Шулейкина, вычисленной им для различных амплитуд с интервалом в 0,1 периода колебаний (время, в течение которого хвост, в нашем случае ма-нус, сделает полный размах вверх и вниз и обратно, пройдя через оба крайних положения). В машущем полете птиц необходимо учесть также непрерывно действующее продольное волнообразование от взмахов и ударов крыла.
Сложение этих волн и дает тот удивительный эффект тянущей силы, которая особенно хорошо просматривается в движениях стрижей, морских чаек и альбатросов.
Теперь становится понятным, по чему при опытах с заклеиванием крыльев у птиц так резко понижалась
их летная способность. Ухудшение полета наступало не только вслед ствие потери управления пограничным слоем, но и в результате того, что исчезал аппарат поперечного волнообразования, поэтому сильно падала тяга, и полет птиц напоминал полет бабочки.
Рассматриваемая нами картина волнового потока за крылом птицы показывает, что теория машущего крыла, осуществляющего тягу и подъемную силу, ставит задачу отыскания связи между упругими колебаниями системы и аэродинамикой крыла, проницаемого потоком.
КИНЕМАТИКА КРЫЛА ПТИЦЫ
Движение крыла птицы полностью зависит от особенностей анатомического устройства.
Рассматривая связи плечевого шарнира, биология устанавливает три вида движения крыла относительно пространственных осей (рис. 63):
1) ротационное движение — около поперечной оси Z—Z (на угол х);
2) вертикальное — с амплитудой о (взмах) (около оси Х—Х );
3) горизонтальное — относительно оси Y—Y (на угол ф).
Ротационное движение плечевой кости осуществляет изменение угла атаки, переводя его из положительного значения на отрицательное. Наи больший угол поворота у чайки составляет: /=40°, у голубя/-=30° у воробьиных птиц значительно меньше. Этот вид вращения крыла
в орнитологии называется пронацией и супинацией соответственно убыванию или возрастанию угла атаки.
Изменение же угла может быть плавным и происходить в одном направлении (у парителей) или же это изменение будет знакопеременным с колебаниями (при вибрационном и трепещущем полете).
Рис 96 Занос крыла чайки вперед на угол 30°
Изменение угла атаки является очень важным фактором возникновения тяги и подъемной силы, как это установлено в лабораторных опытах над колеблющимися крыльями насекомых.
Вертикальное перемещение — это колебания крыла в пределах — 110 — 140° при полном взмахе всей поверхности.
Третий вид вращения в плечевом шарнире осуществляется приведением и отведением плеча в горизонтальной плоскости на угол ф, когда птица складывает крыло, прижимая его к корпусу или распрямляет его, изменяя стреловидность в плане. На рис. 96 виден занос крыла чайки при посадке в гнездо. По нашим измерениям этот угол составляет 30°.
Ограничение движений плеча достигается особыми связками или сухожилиями:
1) связками, ограничивающими отведение плеча вперед (Ligamen-tum scapulo humerale);
2) связками (Li), ограничивающими подъем плеча и супинацию его, т. е. увеличение угла атаки (Ligamentum coraco humerale);
Рис. 97. Маятниковое движение мануса при посадке голубя
3) связками (La), ограничивающими пронацию плеча, т. е. уменьшение угла атаки (Ligamentum acrocoraco humerale).
Для более сильной пронации и супинации, в целях увеличения изменения угла атаки, принимает участие и само туловище.
При помощи брюшной мускулатуры заднего отдела достигается опускание туловища, что влечет супинацию, при опускании же переднего конца — пронацию. Таким образом, весь корпус принимает участие в движении крыла.
Рассматривая пространственное положение крыла, мы приходим к выводу, что в машущем полете имеем случай колебаний двойного сферического маятника переменной длины (рис. 71,6), так как размах крыла меняется, причем одной колеблющейся точке соответствует центр тяжести плеча и предплечья /, а другой 2 мануса. Маятниковое движение мануса при посадке голубя с торможениями можно проследить, рассматривая кинокадры (рис. 97).
Ввиду упругой деформации перьев мануса ецо колебания будут происходить с некоторым сдвигом фазы по отношению к колебанию плеча, что увеличит продольное волнообразование. Изменение угла атаки на конце крыла происходит быстрее, чем у основания.
Изменение угла за время полного колебания, определяемое поворотом крыла около оси Z—Z, дает диапазон углов атаки
В средней части крыла величина / меняется в пределах от —8°, — 10° до +20°, +30°.
Колебание величины угла атаки происходит со сдвигом фаз по отношению к изменению угла взмаха и опускания крыла.
Вращение крыла около оси Y — Y связано с изменением стреловидности крыла в плане (при парении стреловидность обратная с выносом вперед до —5°, при скоростном полете—прямая под углом + 15°, +20°).
Изменение стреловидности в плане, помимо влияния на аэродинамику крыла, имеет влияние на положение концевых перьев мануса, так как они становятся к потоку наклонно под углом 10—30°.
Для нас представляет интерес тот случай, когда в установившемся горизонтальном полете работает концевой отдел, выполняя пропеллиру-ющие функции, а средняя часть является несущей.
Непосредственным измерением крыла коршуна (крыло было ампутировано и защемлено в плечевой области) было установлено, что при неподвижном плече (humerus) амплитуда мануса ом составляет — 5° +35°, отрицательное значение соответствует полному раскрытию крыла и положительное — сокращению его на Ф = 20°.
Для мануса найдено' отклонение вверх от горизонтали на 18° и вниз на 27°, причем первому положению соответствует складывание веера назад на угол Ф =20° и второму — полное раскрытие (рис. 98).
Рис. 98. Углы отклонения мануса
Случай колебания мануса можно уподобить движению сферического маятника, совершающего квазигармонические колебания
Рассмотренные нами положения могут дать некоторый материал для построения теории машущего полета и экспериментов с летающими моделями.
ТЕОРИЯ ПРОПУЛЬСИВНЫХ ВИХРЕЙ ПРОФ. В. В. ГОЛУБЕВА
Как известно, на основании знаменитой теоремы Н. Е. Жуковского (рис. 99) подъемная сила крыла выражается в виде уравнения:
r=JPJ7Z,
где J — циркуляция скорости по контуру профиля (j, р - плотность (кгм - А/сек?), V —скорость полета (м/сек), I — размах (м).
По теории Н. Е. Жуковского крыло самолета может быть заменено системой вихревой пелены (рис. 100). Эта пелена неустойчива, недалеко за крылом она разрывается и свертывается в два мощных вихревых шнура по концам крыла, имеющих ту же циркуляцию скорости (рис. 101). Действие этих вихрей выражается в отклонении потока вниз со скоростью U, называемой индуктивной скоростью (рис. 102).
1 Так называются колеблющиеся системы, у которых коэфициент упругости зависит от времени.
Причина подъемной силы может быть объяснена как следствие отброса потока вниз со скоростью индукции U, По теореме количества движения величина ее равна массе набегающего потока, помноженной
Рис. 99. Циркуляционное течение скорости по контору крыла
Рис. 100. Свертывание вихревой пелены
Рис. 101. Схема П-образных вихрей Рис. 102. Образование силы индук-
тивного сопротивления
на скорость отклонения его вниз. Так как крыло воздействует на струю в диаметре размаха, то секундная масса I2, V р и подъ-
емная Y = U • ms .
Для определения сил, действующих на крыло, весьма важно найти схему движения воздуха за крылом. У крыла обычного самолета такой схемой является схема с вихревыми усами.
Если крыло, имеющее малые толщину и угол атаки, совершает очень малые колебания, как это имеет место при флаттере, то принимают, что от крыла отходит плоская вихревая пелена.
Однако амплитуда колебания и угол атаки крыла птицы, как мы уже установили, не являются весьма малыми и поэтому схема вихревой пелены не пригодна для построения теории машущего крыла.
Для построения этой теории при любых предположениях о величинах амплитуды колебания, угла атаки и толщины профиля советский ученый В. В. Голубев1 принял схему вихревой дорожки. Эта схема предполагает, что в крайних положениях крыла, производящего возвратно-поступательные колебания с одинаковой скоростью опускания и поднятия, от него отделяются, дискретные вихри, образующие вихревую дорожку. В период перехода крыла из одного крайнего положения в другое в пограничном слое происходит образование и «созревание» этих вихрей. На рис. 103 помещена схема такой дорожки.
1 В. В. Голубев. «Механизм образования тяги машущего крыла». ВВИЛ им. Н. Е. Жуковского, 1946 г. Труды ’научно-технической конференции 1944 г., т. III
Если в канале на плохо обтекаемое тело набегает поток жидкости, то от тела периодически отрываются вихри, образующие за ним хорошо видимую (если поверхность жидкости посыпана порошком талька) дорожку, называемую дорожкой Кармана. На рис. 104 помещена схема такой дорожки.
Мы видим, что вихревая дорожка за машущим крылом, принятая в схеме В. В. Голубева, и вихревая дорожка Кармана за плохо обтекае-
Рис. 103. Схема пропульсивных вихрей В. В. Голубева
Рис. 104. Схема вихрей Кармана
мым телом отличаются лишь направлением вращения вихрей. Карма-новская вихревая дорожка создает за телом течение U жидкости, направленное в сторону движения тела, вследствие чего на тело будет действовать реактивная сила в противоположном направлении- Эта сила называется лобовым сопротивлением тела (сопротивление формы).
Вихревая дорожка за машущим крылом (рис. 103) создает течение жидкости (воздуха) в направлении, противоположном направлению поступательного движения крыла, и приводит к реактивной силе, действующей на крыло в направлении этого движения, т. е. дает тягу крыла.
Следовательно, энергия, затрачиваемая на создание обычным крылом вихрей дорожки Кармана, создает лобовое сопротивление (сопротивление формы). В случае машущего крыла эта энергия идет на создание тяги.
В свою теорию В. В. Голубев вводит упрощающее положение, что непрерывная вихревая пелена заменяется действием пограничного слоя крыла или влиянием вязкости.
Рассмотрение вихревых дорожек В. В. Голубева, отходящих в крайних положениях крыла, позволяет решить задачи о машущем крыле при любых размерах его, различных величинах взмаха и удара.
В результате можно определить подъемную силу Y и силу тяги или сопротивления X по формулам: рУГ, здесь буквой Г обозна-
чена циркуляция:
Г=—izb Vcosa cosO—sin В— , L
сила тяги
Х=0,94 pfecos о I/ (Hsin 8 -4 wcosS) [ V + 1,2 w], где b — средняя хорда (в м), а и В- —углы атаки, приведенные на схеме (рис. 39), и и w —скорости опускания и подъема, а и 8 — полусумма и полуразность углов:
Р1+и2 И1— 1Х2
2 ’ 2 ’
?i— ~2 и р-2= Н2.
В последней формуле выбран определенный режим колебания, зависящий от скорости полета:
ширина дорожек h n OQ1 период колебания I ’
При этом условии вихревые дорожки будут устойчивыми.
Уравнение сопротивления или тяги позволяет объяснить стояние на месте кобчика, пустельги или колибри. Если ввести в уравнение I/ — 0, то может быть вычислена тяга. Птица переключает тягу на подъемную силу. Чтобы птица могла лететь, не опускаясь вниз, у нее должен быть некоторый запас подъемной силы, т. е. угол атаки должен быть меньше того угла, при котором она стоит.
В рассматриваемой теории В. В. Голубева впервые установлена математическая зависимость между интенсивностью (циркуляцией) сбегающих вихрей в машущем полете и образованием тяги.
В пользу этой теории говорит ряд известных фактов получения движения в результате вихреобразования. Так, при работе кормовым веслом у рыбачьих лодок позади их образуется типичная двойная вихревая дорожка с шахматным расположением вихрей и с направлением их вращения, противоположным тому, которое имеет место в дорожках Кармана.
Наконец, чрезвычайно близко к работе кормового весла стоит механизм образования силы тяги при работе хвоста рыб. Движение хвоста рыб заставляет сходить вихри только при наибольшем отклонении хвоста.
По гидродинамической теории В. В. Голубева эти вихри, уносящие с собой некоторое количество движения жидкости, и вызывают образование пропульсивной силы.
Наличие двойной вихревой дорожки за колеблющимся крылом было подтверждено опытами инж. Я. Е. Полонского в гидроканале
ИССЛЕДОВАНИЯ Я- Е. ПОЛОНСКОГО ВИХРЕВЫХ ДОРОЖЕК ЗА МАШУЩИМ КРЫЛОМ
На рис. 105 помещена схема установки Я. Е. Полонского для эксперимента с машущим крылом-
Крыловой профиль 7, совершая поперечные колебания, двигался поступательно вместе с тележкой вдоль канала. Конструкция тележки 96
давала возможность менять величину поступательной скорости крыла, амплитуды колебания и иметь различные отношения скоростей опускания и поднятия крыла. При экспериментах наблюдалось следующее:
1. В крайних положениях крыла, т. е. в моменты перехода от опускания к поднятию или от поднятия к опусканию, от него отделялись вихри взаимно противоположного направления вращения.
Рис. 105. Установка для эксперимента с машущим крылом: 1 — газотронный выпрямитель; 2 —мотор с редуктором; 3— съемный редуктор с кулачком; 4 — подвижная рамка с роликом; 5 —пружина; 6 — направляющие рамки; 7 — державка с крылом; ведущий шкив; 9 — ведомый шкив тележки; 10 — рама тележки;
// — рельсы; 12— лоток с визирными линиями; 13 — фотоаппарат
2. После отделения вихря в одном крайнем положении крыла в процессе перехода к другому крайнему положению происходило отчетливо видимое нарастание завихренности жидкости на задней кромке крыла.
3. При равномерных колебаниях крыла отходящие вихри образуют за крылом шахматную дорожку, ось которой параллельна направлению поступательной скорости (т. е. визирным линиям лотка). При этом вихри дорожки перемещались в направлении, противоположном направлению поступательной скорости крыла. Ниже помещены фотографии таких дорожек (рис. 106).
4. При неравномерных колебаниях крыла отделяющиеся от него вихри образуют дорожку, ось которой составляет некоторый угол с направлением поступательной скорости. Дорожку отклоняется в сторону
Рис 106 Спектр вихревой зопоакч В В Голубева за машущим крылом при равномерных взмахах
Рис. 107. Спектр вихревой дорожки при неравномерных взмахах
большей скорости движения крыла. Ниже помещена фотография такой «косой» дорожки (рис. 107).
5. Вихри «косых» дорожек не имеют шахматного расположения При отклонении дорожки вниз от направления поступательной скорости вихри верхнего ряда смещены несколько влево относительно шахматного расположения; при отклонении дорожки вверх — вихри верх него ряда сдвинуты несколько вправо.
Таким образом, эксперимент полностью подтверждает гипотезы, по ложенные В. В. Голубевым в основу теории машущего крыла.
В рассматриваемом эксперименте впервые были обнаружены «ко сые» вихревые дорожки, т. е. дорожки, ось которых не совпадает < направлением общего поступательного движения. Предположение о су ществовании таких дорожек было высказано В. В. Голубевым в докла де на сессии Академии наук в январе 1946 г.
В работе Я. Е. Полонского разработана кинематика и динамика неравномерно машущего* крыла. Получена формула для угла а от 98
клонения дорожки от направления общего поступательного движения:
W су h V Z /
2 4
колебания, равная отношению
Sin ~УГ21Г] ~
1 К 1
tgO. =-----7Т-Ц
с 9лЛ
I где к—характеристика неравномерности времени поднятия крыла Тп к времени его опускания
Тп
Т„ t к — и — сред-няя скорость взмаха. _> h.
Величина -,----отноше-
ние амплитуды колебания к расстоянию между вихрями — определяется кинематикой движения крыла.
Выведенные Я. Е. Полонским формулы для подъемной силы и тяги машущего крыла приведены к обычному в аэродинамике виду:
силы в
Рис 108. Коэфициент подъемной зависимости от величины К
pV2 e 2 причем все особенности, которые вносят с собой взмахи крыла, включаются в коэ-фициенты Ср и С г . Оказывается, что1 эти величины существенным образом зависят от характеристики неравномерности колебания К (рис. 108 и 109).
Нужно иметь в виду, что в рассмотренных работах предполагается плоскопараллельное течение, т. е. не учитывается влияние конечности размаха крыла. Полученные результаты имеют первостепенное для разработки аэродинамического орнитоптера.
Г=Ст
w ?_ v
1,1
значение методов расчета
to Of Of Of- If Of Of Of Of Of Of-01- ot Of—Л____!__L—U
0 Of5 Of 0,70 to
- Of
4
2f
W V Of 0,7 Of 0,9 10
1,1
Рис. 109. Коэфициент тяги в зависимости от К
~ Of
ГЛАВА X
ЛАБОРАТОРНЫЕ ИССЛЕДОВАНИЯ КРЫЛЬЕВ ПТИЦ
ОПЫТЫ С МОДЕЛЯМИ птиц
Исследования крыла птицы, произведенные аэродинамиками совместно с орнитологами, показывают, что крыло птицы частично отбрасывает поток, создавая определенное количество движения, и частично пропускает его. Можно считать достоверно установленным: 1) отсутствие герметичности структуры крыла птицы и наличие проницания его потоком; 2) использование птицей эластичности перьев; 3) применение крыла в качестве сдвоенного парашюта; 4) управление птицей пограничным слоем своего крыла и 5) как следствие вышеуказанного, устой -чивость, управляемость и маневренность в пределах, не достижимых для самолета и планера.
Изучение птиц в полете имеет большое значение для развития аэродинамики малых скоростей. В процессе работы и опытов в этой области могут быть открыты и другие свойства, применение которых для авиации создаст новые возможности.
Для определения количественных показателей сложной физической схемы крыла необходимо проведение различных опытов в аэродинамической трубе- Во-первых, проверка всех открытых эффектов в стацио
нарном потоке в опытах с моделями птицы и, во-вторых, получение характеристик модели той же
Рис. НО. Модель чайки, испытанной в трубе
птицы при частичной или полной подвижности крыла.
До сего времени применялись два способа при проведении аэродинамических испытаний крыльев птиц: первый предложенный Марэем, относится к испытаниям на ротативной машине и описан нами в главе III1; второй способ заключается в исследовании в трубе сплошной модели птицы точно скопированной со своего оригинала в целях проверки качества внешней формы пти
1 Этот же метод применялся в 1926 г. Г. Лилиенталем для изучения крыль фрегата.
цы. Последний способ был применен также в 1944 г. при продувке модели чайки (рис. ПО). Результаты продувок показаны на диаграммах (рис. 111 и 112).
Модель чайки была сделана из гипса, армированного проволокой, задняя часть была выполнена из картона. Геометрические размеры мо-
Су \ Гб-
\5 -1,4 “
(/-1,2-
14'1
-о:в
чм
Голо fa Ns 1; Re=84700 Голова Ns Z;Re=84700 Голова NsZ;Re=120000 Голова NsZ;Re=18820
1.0 0,9 0,8 0,7 0.6 0.5 0.4
0.3 о.г o.i
Д7
п .................7-z~, , ft
О 0,04 0,08 0,12 0,16 0.20 0,24
Г Г 2° 0° 23 4° 0° 8° 10° 12? 14916'16'а
Рис. 112. Диаграмма коэфициентов Су и С5 модели чайки
Рис. 111. Поляра модели чайки при различных Рейнольдсах
1.2\0,12
1,1 0.11
1.0 -0.10^^^.-
0,3-0.09
•о—% ь
0,8-0,08
С -fife)
0,7-0,07 ______i_i____।_।___।__।__।___।_।___।__।... ।_»
0,05-10s 0,710s 0,15-Ю} Не
Рис. 114. Два варианта очертания носа чайки. Нос 2 наиболее благоприятный, с плавным переходом к туловищу
Рис. 113. Влияние числа Re на коэфициент сопротивления и подъемной силы
дели были следующие: размах L=0,9 м, площадь крыльев 8=0,017 м2, средняя хорда b =0,0916 м, длина корпуса I =0,345 м, удлинение Х=9,8.
Коэфициенты Су и Сх определялись для различных чисел Re=0, 1 • 106 до 1 - Ю6 (рис. 113).
Из продувок следует, что величина подъемной силы сохраняет свое значение постоянным от 8° до 14°. Чайка летает при числах Re, больших 100 000. Диаграмма продувок дана для двух значений Re= = 120 000 и Re = 84 700.
Рис. 115. Граница ламинарности по нижней поверхности Re =120 000
При больших значениях Re для моделей с различным очертанием носа (рис. 114) в таблице 11 приведены углы пологого планирования s и коэфициенты с у. Наряду с количественными измерениями были проведены продувки для установления границы ламинарного потока.
На рис. 115 приведены границы ламинарного потока для Re = 120 000 на верхней и нижней поверхности крыла для различных углов атаки, а на рис. 116 — для одной верхней при
Рис 116. Граница ламинарности для верхней поверхности при Re = 84 000
Таблица 11
Модель Ro ^У
№ 1 ! 118800 ! 1 :13,3 0,92
№ 1 84700 ! 1 :8,6 0,77
№ 1 84900 1 :9,8 0,73
№ 2 120000 1 :13,8 0,945
№ 2 168000 i 1:14,3 0,91
Re = 84 000. В последнем случае ламинарный поток имеет место в средней части до 75% хорды и в плечевой — минимум до 40%, причем с увеличением угла атаки область с ламинарным потоком уменьшается. На рис. 115 видно, что ламинарный поток на нижней поверхности в средней ее части занимает только 25% хорды, но зато на конце крыла, в области мануса, занимает всю поверхность.
Границы ламинарного обтекания в средней части и на фюзеляже показаны на рис. 117.
Продувка сплошного крыла дает лишь относительную оценку структуры обтекающего потока. В крыле птицы, оснащенном перьями,’ область ламинарного обтекания за счет проницания потоком может быть увеличена,
Рис. 117. Граница ламинарности по сечениям фюзеляжа
Из рассмотрения результатов продувки модели чайки видно, что они получены для жесткого крыла в случае парения. В материалах нет указания о степени закрутки крыла, профилировании его, и, кроме того, судя по эскизу (рис. НО), модель не воспроизводит концевой обратной крутки мануса.
В действительности птица при наличии перьевой структуры будет иметь иные характеристики, аэродинамика ее в общепринятом смысле окажется хуже, но возможно в большей мере будет отвечать своему назначению для машущего полета, нежели просто обтекаемое крыло.
ТРЕБОВАНИЯ К МОДЕЛЯМ ИСКУССТВЕННОЙ ПТИЦЫ
После неудачных испытаний чучел птиц в трубе задача аэродинамических исследований крыла должна быть пересмотрена на основании предложений, рассмотренных в главах II и VI.
Рис. 118. Модель птицы инж. И. Н. Виноградова с целлулоидными лопатками
Рис. 119. Модель инж. И. Н. Виноградова искусственной птицы для аэродинамических испытаний
Если на мертвой вороне проследить все явления нельзя, то, очевидно, надо проследить их на особой модели, создав такую методику испытания в трубе, при которой можно было бы проверить действие мануса, лучевого предкрылка, туннелей, парашютных отверстий, щелей и т. д.
Рис. 120. Исходная деревянная модель с металлическим предкрылком
Для более близкого подобия опытной модели крылу птицы необходимо изготовить упругие и эластичные лопатки (рис. 118). В этом случае оценка влияния пограничного слоя может быть проверена сравнительной продувкой моделей с лопатками и натурными перьями (рис. 119), причем исходная модель должна быть сплошной. На рис. 120 показан общий вид модели, параметры которой выдержаны применительно к коршуну.
Очень важным моментом испытаний является выяснение эффекта проницания потоком крыла модели при полном открытии всех щелей, туннелей, алюли и мануса- Оценка устойчивости модели в этом случае и величина коэфициента сопротивления могут служить показателем ее свойства при парашютировании.
Вторую серию испытаний в трубе должны составить опыты с моделью, имеющей машущие крылья с теми же геометрическими параметрами, в которой должны быть осуществлены колебания концевого отдела, приводимого в движение от электромотора, заключенного в фюзеляже модели. Через редуктор, трансмиссию в крыле и кривошипный механизм вращение может быть преобразовано в колебательное движение мануса с частотой 1—1,51 сек.
Основная цель испытания этой модели — получить коэфициенты тяги и подъемной силы в установившемся горизонтальном полете. Схема такой модели позволяет получить подъемную силу в средней неподвижной части крыла и тягу, создаваемую колебанием мануса.
Таким образом, модель может явиться переходной ступенью к типу летательного аппарата с машущим крылом.
ОПЫТЫ С КОЛЕБАНИЕМ НЕБОЛЬШИХ КРЫЛЬЕВ
Для изучения колебания крыла в лаборатории необходимо иметь специальную аппаратуру и измерительные устройства. Мы рассмотрим один прибор для опытов с маленькими колеблющимися поверхностями. Общий вид прибора и схемы представлены на рис. 121, 122 и 123.
Прибор имеет привод от электромотора через вал из стальной проволоки диаметром 0,2 мм, который оканчивается кривошипом 5. Передача вращения для уменьшения трения осуществлена гидравлической связью. Испытуемое крылышко из бальзы эллипсовидного очертания длиной 32 мм и шириной 8 мм устанавливается на кронштейне и монтируется таким образом, что через тягу 4 и 7 можно осуществить ударную амплитуду колебаний 6 = 68° и колебание угла атаки / в пределах от 0° до 140°.
На рис. 122 показаны шаровые шарниры 5 и 8 и тяги, создающие колебания крыла Я и шатуны 4 и 7.
Прибор снабжен масляным демпфером и стробоскопом для измерения частоты колебаний, которую можно регулировать в пределах п = = 10, 15, 20 и 25 \/сек. Вся установка собрана на переносном штативе и может быть помещена в потоке воздуха, создаваемого вентилятором.
F
Площадь, ометаемая пластинкой в амплитуде ?, равна и на-
Рис. 121. Прибор для испытаний небольших машущих крыльев
Рис. 122. Монтажная схема привода к крылу и шатунно-кривошипная схема
Рис. 124. Схема углов по отношению к потоку:
F — сметаемая площадь; о — амплитуда
Рис. 123. Схема орнитоптерного прибора и монтаж его на штативе: 1— испытуемое крылышко вращается через валик 3, колебательную тягу 4 и через кривошип 5; 10 — стальной валик диаметром 0,2 мм вращается через масляный привод; 17 — стеклянный цилиндр с маслом; 18 — цилиндр привода с желтыми и голубыми полосами для стробоскопических наблюдений; 7 — противовес; 8 — масляный демпфер; 12 — зеркало; 13 — пружина для установки в нулевое положение при отсчете сил;
15 — ролик к приводу; 20 — рычаг регулировки прибора-
правлена к потоку под углом 6 . Угол атаки а и направление подъемной силы показаны на схеме (рис. 124).
Путем поворота рычагов плоскость колебания изменялась по отношению к потоку при различных вариантах углов атаки и углов направления удара (рис. 125).
Скорость V обдува при опытах менялась в пределах от 50 до 100 см/сек.
Окружная скорость модели при опытах определялась по формуле: и = 2 s ср п, где
5 — длина крылышка (полуразмах или радиус) и п— частота Х/сек.
Особую группу испытаний представляют колебательные движения при одновременном вращении оси стержня в пределах углов / 10°, 70°, 100° и 140°.
В результате испытаний были построены диаграммы изменений тяги R's , подъемной силы к* , потребного вращательного момента /Cj и коэфициента полезного действия t]Si.
1 Соответственно в наших коэфициентах:
4as - a, 12kj — ка = С]^?
Са = Су, Cyj = Сх ,
Все кривые отнесены к выражению поступи л= так, что диаграммы напоминают винтовые характеристики, где и — окружная скорость.
Рабочие формулы будут иметь следующий вид: ASM а гт, у-. , /
q • F qF 1 qt*
где A — замеренная подъемная сила (мг), S — тяга (мг), М—момент (мг • см), ри2 q — скоростной напор ——>
к. п. д. тяги: r<s =--- • /ъ
/<1 , к* к. п. д. тяги на месте: —г
\^к\ )
Результирующий коэфициент подъемной силы:
Са = Су = ----------------
(u3+O Fj
где F[ — площадь крыла.
Для иллюстрации приводим несколько экспериментальных диаграмм, полученных на этом приборе. Диаграмма (рис. 126) составлена при отсутствии обдува (V = 0) и угле атаки (а =0°) при прямых колебаниях перпендикулярно плоскости крыла (ф ^ЭО0)1.
Рис. 125. Варианты направления плоскости колебаний и углов атаки
В этом случае возникает только тяга и притом одна и та же для всех частот колебаний, изменявшихся в пределах:
п = 10 — 25 \)сек-
При наличии обдува перпендикулярно плоскости колебания и для разных частот при V = 50 см)сек получена диаграмма тяги К s = f (/V (рис. 127).
1 На рис. 126 ф = т.
Диаграмма (рис. 128) составлена для скорости обдува V — 100 см!сек.
Из сравнения диаграмм видно, что обдув ухудшает характеристики. Перестраивая эти диаграммы по спо: собу винтовых характеристик, т. е. выражая тягу и мощность в зависимости от поступи 7 для разных углов 7, получим ряд кривых (рис. 128), откуда видно, что наименьшая тяга получается при малых углах вращения X.
Для характеристики к. п. д. при прямом ударе получены диаграммы (рис. 129). Здесь наибольший к. п. д. будет равен 0,46 для угла вращения 7 = 100°.
Изменение наклона плоскости биений или угла ф сильно влияет на характеристики колеблющегося крыла', что видно из следующих диаграмм: 1) подъемной силы ==/f/J (рис. 130); 2) тяги в зависимости от угла вращения х (рис. 131) и 3) диаграммы к. п. д. — =/(7) (рис. 132).
Последняя диаграмма показывает, что величина коэфициента подъемной силы колеблющегося крыла не превышает С а = 0,245 и что наивыгоднейшая комбинация получается при а = 20°, ф — = 70° и /= 100°.
В результате приводится полярная диаграмма (рис. 133) Ка -=f(Ks ), где нанесены кривые для различных X, пунктиром на диаграммах обозначены точки разных амплитуд вращения /.
Рис. 126. Тяга при нулевом угле атаки и прямых колебаниях крыла
Рис. 127. Тяга при обдуве, без угла атаки и прямых ударах при различной частоте п
Рис. 128. Тяга при обдуве со скоростью 100 см/сек
Следует отметить, что низкие коэфициенты Са ==С у по-лучены для малых чисел Рейнольдса: при пересчете на средние летные Re их значение увеличивается до Су = 0,55.
На основании приведенных графиков можно оценить поступательный режим, а также режим взлета с места.
Из приведенных опытов неясно, почему взяты столь большие углы поворота /
Рис. 129. Коэфициент полезного действия тяги s — f(A)
= 70— 140е. Возможно, что исследователи стремились получить от прибора все допустимые комбинации, не задаваясь вопросом практического использования результатов. Выполнение в конструкции больших углов х весьма затруднительно, хотя в природе такие случаи наблюдаются в полетах, например, колибри.
Недостатком прибора является трудность его регулировки и весьма малые габариты и вес (19 мг) испытуемой модели.
Анализируя эту работу одновременно с опытами в натуру, выполненными на орнитошерном приборе, следует отметить очень близ
кую величину, полученную для к. п. д., который для эластичного крыла равен 0,5 — 0,62.
Для проверки этой теории была построена также летающая модель орнитоптера с резиномо-тором (рис. 134), которая продержалась в воздухе 26 сек.
Данные этой модели следующие: размах -0,8 м, поверхность крыльев S = 0,07 м2, вес G = — 0,007 кг, нагрузка р = 0,1 кг/м2, мощность Лт =0,00035 л. с. Нагруз-
ка на л. с. = 20 кг*
Ударная амплитуда колебания крыла с? = 60°, средний угол взлета — 18° и максимальный — 45°, средняя частота — 2,7 1/сек; средняя скорость центра тяжести — 1,8 м/сек, угол а0 в корневом сечении + 15°, угол 7. мануса — 5°.
При другом варианте взято наоборот: в основании крыла —5° и на конце + 17°.
В результате исследований получено: средний коэфициент подъемной силы Су =0,49; средний коэфициент лобового сопротивления Сх = о,37; средний коэфициент тяги ks = 0,52; средняя часто-
Рис. 132. Коэфициент полезного действия при ф = 70° и а = 20°
Рис. 133. Зависимость Ka =^f (Ks) для разных X и углов вращения
ns та v = — = 0,6.
v
Рис. 134. Полет моделей с полным взмахом крыла
Рис. 135. Диаграмма движения центра тяжести крыла и корпуса при полете модели, полученная киносъемкой со скоростью в -эд- сек: S — полуразмах, Z —вертикальная ось, X — горизонтальная ось
На диаграмме (рис. 135) приведены колебания центра тяжести
корпуса и крыльев модели.
РАБОТЫ СОВЕТСКИХ МОДЕЛИСТОВ ДОСАВ
В 1932—1935 гг. советскими моделистами И. Трунчеиковым и М. Степченко самостоятельно были построены летающие модели с машущими крыльями.
В 1948 г. моделистом Б. Яковлевым под руководством инструктора-летчика Брылькова была построена улучшенная модель орнитоптера той же схемы, которая демонстрировалась во время доклада профессора В. В. Голубева на заседании секции машущего полета Досава (рис. 136). В 1950 г. новая модель Б. Яковлева с машущими предкрылками продержалась в воздухе 2 мин 0,4 сек.
Всесоюзное добровольное общество содействия Авиации принимает все меры к развитию нового вида летательных аппаратов.
Для успешного разрешения этой интересной проблемы необходимо проводить систематические опыты с целью изучения полетов птиц, лабораторные испытания моделей крыльев птиц в стационарном потоке колеблющихся и неподвижных.
ПОЛЕТЫ ПРИ ПОМОЩИ МАШУЩИХ КРЫЛЬЕВ
Изобретателем и конструктором Б. И. Черановским в 1921 и 1934 гг. были построены два мускуллета машущего типа. В 1935 г. им же был построен третий планер — мускуллет бипланного типа БИЧ-18
Рис. 136. Летающая модель орнитоптера В. Яковлева: вес — 10 г; площадь центре плана — 3,4 дм2; площадь машущей части — 2.4 дм2; размах — 810 мм;
длина — 540 мм; амплитуда — 48°
(рис. 137). Его испытания показали несколько лучшие качества, чем качества предыдущих планеров, однако из-за вибрации крыльев опыты были прекращены.
В 1936 г. на планерной станции Осоавиахима в с. Трикотажное были произведены испытания мускуллета П. И. Смирнова. Два крыла из бамбука, обтянутые шелковой материей, надевались на особые плечевые поручни с ограничителями взмаха. Летчик подвешивался через ролик к подвесному канату длиной 300 м и, таким образом, скользил с горы, подмахивая крыльями. Ноги летчика выполняли роль стабилизатора и руля благодаря натянутой материи между ними. Положение
Рис. 137. Планер-мускуллет конструкции Б. Н. Черановского
Рис. 138. Мускуллет П. И. Смирнова
Рис. 139. Полет на планере с машущим эластичным крылом
корпуса летчика при скольжении по тросу было горизонтальным, так что человек с крыльями напоминал гигантскую бабочку (рис. 138)-
С горы планерной станции летчик М. И. Чекалин совершил 13 скользящих спусков, во время которых он производил взмахи и при этом совершенно разгружал натянутый канат и трос своей подвески. Мускуллет показал хорошую устойчивость и управляемость.
Этот смелый опыт является своеобразной учебной тренировкой на мускуллете и, несомненно, повторение его окажется весьма полезным для изучения свойств машущих аппаратов.
На рис. 139 показан полет планера с машущими крыльями, совершившего более 30 взлетов.
Несущая поверхность планера состояла из центроплана и пары машущих крыльев длиною в 3 м (по одной трети взмаха), приводимых в движение мускульной силой. Благодаря ударам машущих крыльев удалось достигнуть увеличения расстояния до 30% и увеличения продолжительности полета до 58% по сравнению с обычным планирующим полетом.
Мускуллет-планер с машущими концами крыльев был впервые осуществлен в России в марте 1908 года летчиком А. В. Шиуковым и испытан им в гор. Тифлисе с Махатской горы. Планер имел две плоскости, на верхней плоскости по концам подвешивались машущие крылья, которые приводились в движение ногами пилота.
ОРНИТОПТЕРНЫЙ ПРИБОР
Предварительные исследования позволили создать испытательную установку для машущих крыльев в натуральную величину (рис. 140). Установка представляет собой карданно-подвешенную ферму с двухкомпонентными весами для замера подъемной силы и тяги. На корот-
Рис 140. Орнитоптерный прибор для испытания машущих крыльев в натуре
ком плече трубчатого каркаса установлен мотор трехфазного тока в 2,2 л. с. с контргрузом, а на противоположном — машущее крыло с приводом. Передаточное число между приводом машущих крыльев и мотором могло изменяться от 20,3 до 14,15. С помощью этих двух ступеней достигалось увеличение числа ударов машущих крыльев с 30 до 60 в минуту. Для того, чтобы изменять направление удара машущих
ДеистDue везВиишых сил
48 4Z ЧЧ 46 48 50 п"лшн
Рис. 141. Диаграмма испытания широких и узких крыльев
чо чг чч чб 48f п мин
крыльев, механизм крыльев присоединялся к плечу весов при помощи специального шарнира. Для замера подъемной силы были применены обычные пружинные весы, а для замера тяги и давления ветра были сконструированы специальные пружинные весы. Мощность и число ударов машущих крыльев измерялись с помощью высокочувствительных и особо защищенных от атмосферных влияний измерительных приборов.
На планерной станции было проведено до 700 отдельных испытаний при силе ветра от 4 до 12 м!сек.
Для эксперимента применялись эластичные крылья длиной около 3 м различных профилей с углами закрутки у основания до 12°. Из опытов было найдено, что машущие крылья с широкой поверхностью дают наилучший эффект, нежели длинные, узкие, как это видно из диаграммы (рис. 141).
Испытанные машущие крылья имели следующие данные: длина крыла 2,950 и, ширина — 960 мм, площадь — 2,2 м2, амплитуда — 72*, при движении вверх на 30° и вниз до 42° направления удара у = 70°. Наилучшие показатели в одном из опытов: подъемная сила У = 64 кг,
Y 64
сила тяги 7=7,5 кг при нагрузке -~-= 14,5 кг/м2, средней скоро-
2 S п „ _о .
сти на конце крыла и = = 6,78 м/сек, где S — длина дуги и п =
= 54 \/мин.
Крылья при взмахе вверх автоматически устанавливались в положение минимального лобового сопротивления.
1 I “1
У W-i la 30-zo\s/a
10
Y“30‘ t’72‘
T
«Гп kt л- 30-
л- T^80' 20-Sla^
3
'1$^^', .................
«7 W « V «7 10 ‘ <Г «f W 47 «7 «pO ' «? W W W » «7 W fam giapoB и мщ-
Y-70‘
<n. I
рис. 142. Подъемная сила и тяга в зависимости от частоты 1
п при разных углах наклона
В процессе колебания крылья выполняли управляемые ударные и поворотные движения, т. е. одновременно изменялись углы 8 и /, в крайних положениях имелись ограничители хода. В фазах подъема и опускания крыла угол атаки был положительным.
Механизм допускал перестановку машущих крыльев для измене ния направления удара (угол у). График (рис. 142) показывает влияние этого угла на увеличение силы тяги и уменьшение подъемной силы. В опыте этот угол изменялся от 70° до 105°, в последнем случае создавалась отрицательная тяга, или торможение скорости.
Рис. 143. Вид орнитоптерной установки со стороны электромотора и рычага для замера тяги
Было установлено, что при скорости обдува ветром (V = 8 м/сек) •подъемная сила при взмахе положительна. Это объясняется тем, чт« процессы обтекания и сила давления на эластичных машущих крыльях иные, чем у обычных.
Мощность, необходимая для движения машущих крыльев, определялась тормозным динамометром (рис. 143). Учитывая все потери (тепловые, электрические, на трение в подшипниках и в трансмиссии), установили, что при полной нагрузке к. п. д. машущих крыльев составляет около 60%, максимальное же его значение достигает 0,67. Данные испытаний и к. п. д. приведены в таблице 12.
Таблица 1
Испытания машущего крыла Испытание при торможении
V мсек п 1 Y Т Общая затраченная энергия Число оборотов тормозн. диска Нагрузка Q Полезная энергия крыла л. с. к. п.
мин вт I Л. с. г.
s—9 । 1 34,5 26 2,25 580 0,79 Г/0 1,0 0,3 0.3S
6 40 37,5 2,5 610 0,83 208 1,9 0,4 0,48
7—8 42 43 3,5 620 0,84 215 2,0 0,43 0,51
7—8 43,5 43 4 650 0,88 223 2,2 0,49 0,55
7 44,5 45 4 705 0,96 230 2,3 0,53 0,55
7 46 49 4,5 730 0,99 238 2,6 0,62 0,62
9-10 47,5 49,5 5 785 1,07 241 2,8 0,67 0,625
На основании отчетных материалов можно считать, что вся установка не обладала достаточной чувствительностью, так как при малых изменениях установочного угла от 2° до 10° не было обнаружено каких-либо изменений воздушных сил.
Кроме того, недостатком механизма являлось движение, противоположное крылу птицы, а именно: сверху — спереди, назад — вниз, что при встречном потоке явно невыгодно. Все же полученный материал весьма показателен с качественной стороны и может быть использован для ориентировочных расчетов орнитоптера.
ГЛАВА XI
ПРАКТИЧЕСКИЕ ЗАДАЧИ ИЗУЧЕНИЯ КРЫЛЬЕВ ПТИЦ
Совместные усилия аэродинамиков и орнитологов привели к открытиям новых свойств крыла птиц, которые заставляют нас пересмотреть как физические, так и теоретические основы машущего полета. Совершенно очевидно, что схема простого обтекания крыла птицы не объясняет ни высокой экономичности, ни совершенной управляемости и устойчивости их в полете.
Необходимо пересмотреть всю проблему с точки зрения проницаемости крыла и приспособительного нарушения обтекаемости.
Поведение подобного крыла в потоке воздуха представляет совершенно новую научную проблему. Из приведенного анализа работы крыла следует, что представляет интерес разрешение следующих задач:
1) усовершенствование крыла планера и самолета;
2) замена винта машущим крылом.
На протяжении всей истории планеризма мы встречаемся с многочисленными попытками воспроизвести внешние формы птиц.
В Советском Союзе по образцу чайки были построены планеры конструкции инж. Д. Н. Колесникова (ДК-3, ДК-4) и планер летчика А. М. Кузакова, имеющий в плане очертание орла. При сравнении характеристик этих планеров и их достижений не видно большой разницы с другими рекордными планерами обычных схем. Наоборот, можно указать, что глубоко вогнутый профиль в основании крыла планера без эластичного концевого обтекателя ухудшал полетные свойства планера, поэтому на нем рекордов не было установлено.
Не перечисляя всех возможных путей в этом направлении, можно указать на следующие радикальные преобразования, вполне достижимые при современных технических средствах.
1) Применение туннелей для внутреннего торможения потока и для создания моментов при управлении.
2) Превращение планера или самолета в парашют.
3) Применение крыльев, вращающихся относительно оси фюзеляжа.
В случае решения первого вопроса создаются новые благоприятные условия для управления бесхвостыми самолетами, разрешение второго сделает возможной посадку на любую площадку в случае остановки мотора.
Увеличение посадочного угла атаки крыла выгодно для уменьшения скорости при посадке. Можно предполагать, что осуществление птичьих методов управления пограничным слоем откроет большие перспективы для усовершенствования летательных аппаратов.
ЦЕЛЕСООБРАЗНА ЛИ ЗАМЕНА ВИНТА МАШУЩИМ КРЫЛОМ
Если произвести расчет энерговооруженности птиц и самолета по G
формуле В. С. Пышнова > то оказывается, что птицы
для своего полета затрачивают на единицу веса в несколько раз меньше энергии, чем самые совершенные самолеты.
Из приведенной сравнительной таблицы 13 видно, что нагрузка на* одну лошадиную силу у птиц больше, чем у самолета в 4—16 раз-
При переходе к моторному орнитоптеру можно принять средний, коэфициент полезного действия машущего крыла — 0,8, для редуктора— 0,85 и для трансмиссии—0,95, т. е. наибольший эффективный к. п. ,д. всего устройства будет равен 0,60.
При осуществлении орнитоптера с крылом по новой схеме мы можем допустить среднюю удельную нагрузку на единицу мощности (ог 20 до 25 кг/л. с.), т. е. повысить экономичность транспортных самолетов.
Таблица 13-
Самолеты
кг Л. с.
Птицы
кг/л. с.
Истребитель ....................
Транспортный....................
Транспортный с перегрузкой . . . .
Рекордный на дальность..........
Самолет для связи ..............
6
8,5
10
12
8,6
Ласточка.............
Грач.................
Лебедь...............
Чайка ...............
Аист черный..........
Коршун...............
30
34
37
81
42
67—95
При данном состоянии техники вполне возможна постройка легких, и небольших орнитоптеров для транспорта, связи и спортивных целей.
Конечно, здесь не ставится вопрос о прямом подражании птицам.
Обычно при переходе от моделей к натуре или от малых летательных аппаратов к большим конструктору приходится иметь дело с масштабным эффектом. В действительности не всегда оказывается возмож-
шым сохранить подобие прочности, веса конструкций, силовых и кинематических соотношений1.
ОСНОВНЫЕ ТРЕБОВАНИЯ К ОРНИТОПТЕРУ
Достижения аэродинамики за последние 30 лет сопровождались столь быстрым прогрессом самолетостроения, что- буквально на глазах одного поколения людей самолетостроение приобрело огромное значение для хозяйственной жизни и обороны страны.
Совсем другое положение мы видим в области развития маш)щего полета.
Несмотря на ряд упорных попыток многих изобретателей, полег на орнитоптере осуществить до сих пор не удалось.
К основным причинам этого обстоятельства должны быть отнесены: а) недостаточное уяснение принципа работы колеблющегося крыла; б) недостаточная разработка вопросов, связанных с управлением и устойчивостью.
В большинстве построенных аппаратов применялись жесткие и непроницаемые крылья, на движение которых затрачивалось при управлении и взмахах большое усилие, в результате коэфициент полезного действия их оказывался очень низким.
В кинематике машущих элементов допускались очень сложные виды траектории движений (восьмерки, лемнискаты и т. д.), однако не учитывались самые главные параметры, как то: изменение угла атаки и ударной амплитуды колебания.
Как теперь известно, движение концевого отдела крыла (мануса) у птиц обгоняет движение средней части, т. е. конец крыла движется с большим ускорением и имеет при спускании крыла, кроме основных угловых смещений (о и /), также смещение по горизонтали (ф).
Пренебрегая этими параметрами, невозможно добиться высокого к. п. д. тяги в колеблющейся части крыла.
Причиной, тормозящей развитие орнитоптера, является также несоответствие авиационного двигателя новым условиям работы. Много-оборотный двигатель требует применения редуктора с большим передаточным числом; кроме того, возникают конструктивные трудности уравновешивания больших колеблющихся поверхностей, движущихся возвратно-поступательно.
Мотор для орнитоптера должен быть органически связан с его конструкцией. Этому условию более удовлетворяют паровые и пневматические двигатели.
При передаче движения сжатым воздухом по гибким шлангам устраняется трансмиссия к машущим элементам и может быть достигнута большая гибкость в управлении ими.
1 Первый закон подобия ограничивает конструктивный вес, так как в больших сооружениях он растет пропорционально кубу линейных размеров, поэтому с увеличением веса надо уменьшать запас прочности и применять материалы с большой удельной прочностью.
По второму закону масштабов с увеличением размеров относительная мощность уменьшается, что вытекает из вышеприведенной формулы В. С. Пышнова для энерговооруженности летательного прибора.
ВЕРОЯТНЫЕ ПУТИ РАЗВИТИЯ ОРНИТОПТЕРА
Для создания аппаратов машущего полета нельзя ограничиваться. $<ак мы видели, одним изучением механизма колебания. Вопросы аэродинамики, управления и устойчивости должны быть разработаны комплексно и одновременно. На основе изучения полета птиц и анализа всего изложенного материала можно сформулировать основные тех-
Рис. 144. Схемы орнитоптеров
нические требования к аппарату с машущими крыльями и наметить несколько схем, удовлетворяющих этим требованиям.
От всякого летательного аппарата машущего типа необходимо требовать следующее:
1. Летательный аппарат должен обладать свойствами планера и парашюта.
2. Крыло должно быть умеренно эластичным с управляемым неграничным слоем.
3. Крыло должно иметь принудительное вращение около двух или трех пространственных осей.
4. Конец крыла должен двигаться по синусоидальной траектории.
5. Мотор должен соответствовать кинематике и динамике машущих крыльев.
6. Прибор должен развивать тягу при подъеме от 0,75 до 1,5 крат веса машины.
7. Для уменьшения пробега необходимо катапультное шасси-
Отсюда намечаются следующие три монопланные схемы орнитоптера (рис. 144 — А, Б, В) и две бипланные (рис. 144 — Г, Д).
Схема А — с раздельными функциями крыла — имеет несущий центроплан и пропеллирующий конец крыла.
Схема Б — с полным взмахом всей поверхности крыла.
Схема В — двухмаятниковая, в которой вершина крыла (манус) имеет сдвиг фаз колебания по отношению к плечевой области.
Схема Г — бипланная, типа ножниц, движущихся навстречу, «а сдвигом фаз колебаний.
Схема Д — тандемно-уступная со сдвигом фаз колебаний.
Особенностью последней схемы является разделение несущих и пропеллирующих функций, как у стрекозы.
Не следует думать, что указанные схемы исчерпывают все варианты летательных аппаратов с машущими крыльями.
Несомненно, могут быть предложены и другие рациональные схе< мы для разрешения этой трудной проблемы.
В конце концов человечество овладеет той совершенной техникой, которая заложена в гибком крыле птицы, и в результате осуществит летательный аппарат более экономичный и безопасный, нежели самолет и геликоптер.
ЛИТЕРАТУРА
1. В. Т. Горощенко и Д. В. Халезов— «К вопросу о максимальной скорости самолета». Оборонгиз. 1940 г.
2. М. К. Тихонравов — «Полет птиц и машины с машущими крыльями». ОНТИ. 1937 г.
3. Б. Н. Юрьев и П. Б. Лесникова — «Аэродинамические исследования». Изд. ЦАГИ. 1928 г. № 221.
4. Н. Е. Жуковский — «О парении птиц». Труды отделения физических наук О. Л. Е., т. IV, выпуск 2, 1892 г.
5. Д-р Арендт — «Полет птиц». Симферополь. 1898 г.
6. Е. П. Смирнов — «Новый принцип механики и динамики летания». Москва. Изд. «Новый мир». 1922 г.
7. В. Ф. Савельев — «Основы экономического полета». Журнал «Самолет» № 10 за 1936 г.
8. Н. А. Гладков — «Полеты в природе». 1948 г. Московское общество испытателен» природы.
9. Г. С. Шестакова—«Полет птиц». Научная диссертация. Труды Академии
наук СССР.
10. В. В. Голубев — «Тяга машущего крыла». Известия Академии наук № 5. Отделение технических наук. 1945 г.
11. В. В. Голубев — «Механизм образования тяги машущего крыла». Труды научной конференции 1944 г. ВВИА.
12. С. А. Чаплыгин — «Теория решетчатых крыльев». 1914 г.
13. О. Лилиенталь — «Полет птиц, как основа искусства летания». 1905 г. С.-Петербург.
14. Н. А. Гладков —«О некоторых аэродинамических свойствах птиц». Бюллетень Московского общества испытателей природы 1946 г.
15. О. Хейнрот — «Из жизни птиц». Перевод с немецкого. 1947 г. Московское общество испытателей природы.
16. С. Н. Шишкин — «О прочности крыльев птиц». Труды ЦАГИ № 258.
17. В. В. Шулейкин — «Физика моря». Академия наук СССР. 1949 г.
18. И. К. Костенко — «Бесхвостые самолеты». БИТ. ЦАГИ. 1946 г.
19. Б. Н. Юрьев — «Экспериментальная аэродинамика». ОНТК. 1936 г.
20. А. В. Столбов — «Самолетостроение последних лет». Оборонгиз. 1940 г.
21. М. К. Тихонравов — «Полет посредством машущих крыльев». Журнал «Самолет» № 1 за 1935 г. и № 5 за 1938 г.
22. Г. Шмелев — «Резонансный полет». Журнал «Самолет» № 4 за 1925 г.
23. С. П. Тимошенко — «Теория колебаний в инженерном деле». ОНТИ. 1938 г.
24. Н. Е. Жуковский — «О парении птиц». 1891 г.
25. Н. Е. Жуковский — «К теории парения». 1890 г.
26. Н. А. Закс — «Основы экспериментальной аэродинамики». ВВИА им. Жуковского. 1948 г.
_27. С. С. Туров — «Перелеты птиц». 1948 г. Московское общество испытателей природы.
28. И. А. Черемных, И. Ф. Шипилов — «Можайский — создатель первого в мире самолета». Оборонгиз. 1949 г.
29. И. Н. Виноградов — «Проектирование легких самолетов». Журнал «Самолет» №№ 10—12 за 1937 г.
30. Д. Кузьменко, Б. Висленев — «Теория авиации». Военгиз. 1936 г.
31. Журнал «Воздухоплавание». 1880 г. №№ 2—4.
32. И. Н. Виноградов — Учебник «Конструкция и расчет самолета на прочность». ОНТИ. 1935 г.
33. С. Гольдштейн — «Современное состояние гидродинамики вязкой жидкости». Т. I и II, перевод с английского под ред. проф. Н. Я. Фабриканта. 1948 г.
34. Будиг — «Применение несимметричного обдува у самолета с машущим крылом». Перевод с французского. ВВП А им. Жуковского за № 1237. 1948 г.
35. Н. А. Гладков — «Биология полета птиц». 1950 г. Московское общество испытателей природы.
36. В. В. Голубев — «Лекции по теории крыла». Гостехиздат. 1949 г.
37. А. С. Кравец—«Характеристики авиационных профилей». Оборонгиз. 1939 г.
38. С. В. Пышнов — «Оценка характеристик самолета». Журнал «Техника Воздушного Флота» № 5. 1945 г.
39. И. К. Костенко — «Летающие модели планеров». ОНТИ. 1935 г.
40. Магау Е. »Le vol des oiseaux". Paris» 1889 г.
41. G. Lilienthal. „Die Biotechnik des Fliegens“. Leipzig. 1926 г.
42. Jdrac P. „Experimentelle Untersuchungen fiber den begelflug4. Berlin. 1932 г.
ОГЛАВЛЕНИЕ
Стр
Предисловие............................................................... 3
Введение ................................................................. 5
ГЛАВА I. Как устроен летательный аппарат птиц
Основные понятия о структуре крыла.......................................9
Перьевой аппарат.......................... ...... 10
Малые маховые перья . . . ......................13
Область парашютных отверстий крыла...................................... .14
Приспособления, подобные механизации крыльев самолетов . .15
Контурные и профильные очертания крыла птицы . .... 16
Сопротивляемость крыльев нагрузкам . . . . . . .19
Углы установки и углы атаки крыльев у больших птиц........................19
Размах и удлинение крыльев птиц . . ..........................22
ГЛАВА II. Виды и приемы полетов птиц
Птицы первичной эпохи земли . . ..........................23
Виды полетов птиц ........................................................25
Приемы пилотажа у птиц . . . ................... 32
Летные характеристики птиц ...............................................32
ГЛАВА III. Результаты изучения полета птиц
Исследование живых птиц 37
Опыты по заклеиванию крыльев птиц ....................................40
О дымление крыльев .......................................................41
ГЛАВА IV. Фото-аэродинамический метод изучения крыльев
Вывод из одного поверочного расчета ...................................4
Спектр потока под крылом..................................................45
Эффект проницания крыла потоком воздуха................................46
Приспособительное нарушение обтекаемости крыла............................51
Использование вязкости воздуха............................................52
Упругий эффект больших маховых перьев.....................................56
ГЛАВА V. Как осуществляется управление пограничным слоем в крыле нтицы
Представление о пограничном слое
Методы, применяемые птицами
58
59
ГЛАВА VI. Устойчивость и управляемость птиц
Условия равновесия ...........................62
Способы сохранения устойчивости ..... 63
Путевая устойчивость птиц ........... 64
Как птица управляет полетом ........... 65
ГЛАВА VII. Силовая схема крыла птицы
Особенности движения крыла . ...................... . . 68
Синусоидальный закон движения...........................................68
Инерционные колебания в машущем полете...................................70
Маятниковое движение крыла 72
Сдала работы машущего крыла 72
Пёрвая фаза ...................... ..............................74
Вторая фаза .............................................................76
Третья фаза • • . . . ,.........................78
Четвертая фаза ..........................................................79
ГЛАВА VIII. Мускульные усилия птиц
Косвенные методы оценки усилий...........................................62
Возможен ли полет человека при помощи мускульной силы? . . . 83
ГЛАВА IX. Физическое состояние потока воздуха за крылом птицы
Резонансный полет . • . . • •.........................$6
Волнообразование за машущим крылом . ... 87
Кинематика крыла птицы...................................................90
Теория пропульсивных вихрей проф. В. В. Голубева.........................93
Исследования Я. Е. Полонского вихревых дорожек за машущим крылом . 90
. 103
. 106
. из
’13
ГЛАВА X. Лабораторные исследования крыльев птиц
Опыты с моделями птиц ............................................
Требования к моделям искусственной птицы ..... Опыты с колебанием небольших крыльев Работы советских моделистов Досава . .........................
Полеты при помощи машущих крыльев • •••••
Орнитоптерный прибор .............................................
ГЛАВА XI. Практические задачи изучения крыльев птиц
Целесообразна ли замена винта машущим крылом .... Основные требования к орнитоптеру •
Вероятные пути развития орнитоптера
Литература ... ........................................
Редактор И. Толстых________________________________Техн, редактор //. Рцьиковскш.
Г-50348. Сдано в производство 25/VII 1950 г. Подписано в печать 8/Ш 1951 г
Бумага 70X108716 = 4 бумажн. — 10,96 печати, листа. Заказ 826/124
Типография издательства ДОСАРМ, г. Тушино