/














































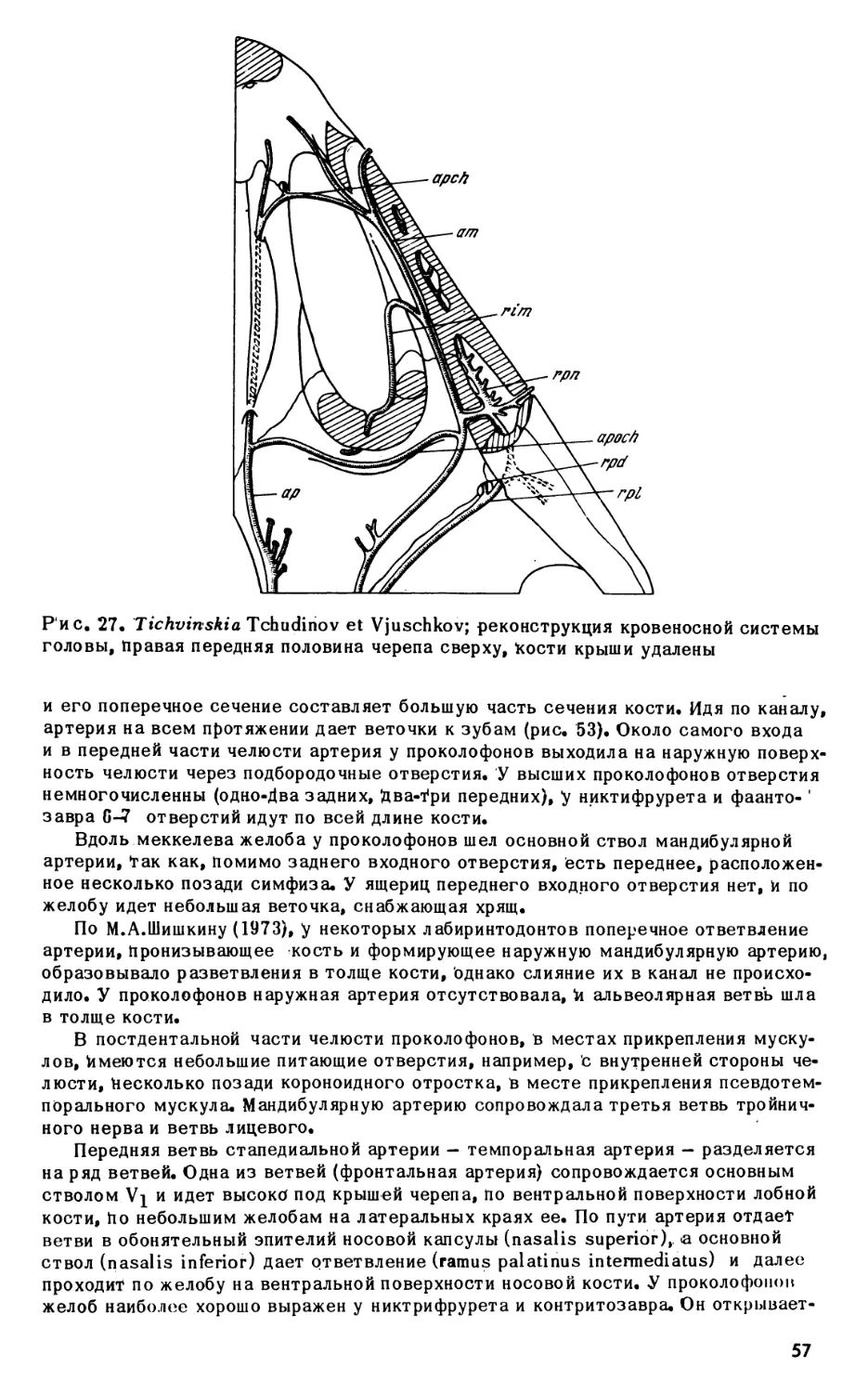




























Text
АКАДЕМИЯ НАУК СССР
М. Ф. ИВАХНЕНКО
ПЕРМСКИЕ
И ТРИАСОВЫЕ
ПРОКОЛОФОНЫ
РУССКОЙ
ПЛАТФОРМЫ
ИЗДАТЕЛЬСТВО «НАУКА»
АКАДЕМИЯ
НАУК
СССР
ТРУДЫ ПАЛЕОНТОЛОГИЧЕСКОГО ИНСТИТУТА
Том 164
М. Ф. ИВАХНЕНКО
ПЕРМСКИЕ
И ТРИАСОВЫЕ
ПРОКОЛОФОНЫ
РУССКОЙ
ПЛАТФОРМЫ
ИЗДАТЕЛЬСТВО 'НАУКА"
Москва 1 979
УДК 568.1 (47)
Ивахюнко М.Ф. Пермские и триасовые проколофоны Русской шкат-
формьь М, "Наука", 1979,80 с.
В работе приведена полная систематическая ревизия пермских и триасовых
проколофонов?уточием9&олювдояный уровень подотряда. Намечены ход эволю
ции и адаптивная радиация триасовых проколофонов. Результаты изучения
позволяют говорить о возможностях использования остатков проколофонов
для стратиграфического районирования континентальных отложений перми и
триаса.
Книга рассчитана на палеогерпетологов и биостратиграфов.
Ил. 30, библ. 140 назв.
Ответственный редактор
член-корреспондент АН СССР
Л Л. ТАТАРИНОВ
Михаил Феодосьевич Ивахненко
ПЕРМСКИЕ И ТРИАСОВЫЕ ПРОКОЛОФОНЫ РУССКОЙ ПЛАТФОРМЫ
Труды Палеонтологического института. Том 164
Утверждено к печати Палеонтологическим институтом Академии наук СССР
Редактор издательства B.C. В а н и н. Технический редактор Л.В. Русская
ИБ No 7346
Подписано к печати 4. 12. 78. Т - 17041. Формат 70 х 108 1/16
Бумага офсетная No 1. Усл.печ.л. 7,0. Уч.—издл. 7,2. Тираж 850 экз.
Тип. зак. 796 . Цена 1р. 10 к.
Книга издана офсетным способом
Издательство "Наука", 117485, Москва, В-485, Профсоюзная ул., 94а
Ордена Трудового Красного Знамени 1-я типография издательства "Наука",
199034, Ленинград, В-34, 9-я линия, 12
20801- 508
И
392-78, кн. 2
055 (02) -79
© Издательство "Наука", 1979 г.
ВВЕДЕНИЕ
Проколофоны наряду с остальными котилозаврами являются наиболее прими
тивными из известных в настоящее время рептилий. Детальное изучение их мор-
фологии, сопоставление с высшими амфибиями и остальными рептилиями дает воз
можность решить ряд вопросов, связанных с происхождением рептилий в целом и
завроморфных рептилий в частности. Однако до последнего времени проколофоны
представляли собой одну из наименее изученных групп котилозавров. В значитель
ной мере это объясняется относительно редкой встречаемостью, отсутствием, как
правило, массового материала, обычно неудовлетворительной сохранностью остат
ков проколофонов.
Из различных районов земного шара было известно 23 рода проколофонов, из
них 6 - с территории Русской платформы. Из числа пос-ледних только для одного
имелось достаточно подробное описание скелета, систематическое положение трех
родов оставалось неясным. Из экзотических форм описания лишь двух форм более
или менее удовлетворительны, остальные не отвечают требованиям, предъявляемым
в настоящее время к палеонтологическим описаниям. Подавляющее большинство
форм известно по фрагментарным остаткам.
Такая скудость сведений по морфологии и привела к тому, что родственные свя
зи проколофонов не только с другими рептилиями, но й внутри группы оставались
неясными*
Одним из факторов, затруднявших изучение морфологии проколофонов,
являются их, как правило, мелкие размеры. Лишь при широком применении оптиче
ских методов появилась возможность проводить детальное изучение остатков этих
ископаемых животных. Степень морфологической изученности определяет, в ко
нечном счете, и значение группы для стратиграфических исследований вме
щающих отложений. Разумеется, результаты морфологического аналива не
могут быть верно интерпретированы без параллельных палеоэкологических
исследований, сопоставлений фаунистических комплексов, литологического
анализа и т. д.
В этом отношении пермские и триасовые местонахождения Русской платформы
представляют особый интерес, поскольку в значительной части их проколофоны
являются преобладающим компонентом, и раскопки зачастую дают массовый мате
риал.
Изученные коллекции Палеонтологического института АН СССР содержат мате
риал по восьми родам (10 видов), из них 5 родов (6 видов) установлены автором.
Четыре рода представлены достаточно полно (скелеты, черепа, массовый фрагмен
тарный материал), остальные могут быть изучены по небольшому количеству раз
розненных костей.
В коллекцию Палеонтологического института входит Мезенская кол
лекция (сборы РЛ.Мерклина, Н J4 .Новожилова), более поздние сборы палеонто
логов Б.П.Вьюшкова, М.А.Шишкина, М.Г.Миниха, геологов Г.И.Блома, В.Р.Лозов
ского, а также автора (экспедиционные поездки 1971 и 1972 гг.). Кроме того,
В.Р.Очевым любезно предоставлена для изучения коллекция проколофонов, принад
лежащая Саратовскому университету (3 рода и 3 вида, из них 3 рода и 1 вид уста
новлены автором).
3
Работа проведена в лаборатории низших тетрапод Палеонтологического институ
та АН СССР под руководством члена-корреспондента АН СССР Л.П.Татаринова.
Автор пользуется случаем выразить руководителю сердечную признатель
ность.
Автор приносит также свою благодарность сотруднику Палеонтологического
института доктору биологических наук М.А.Шишкину, сотруднику Института эволю
ционной морфологии и экологии животных доктору биологических наук В.Б.Суха
нову и сотруднику Саратовского университета доктору геолого-минералогических
наук В.Г.Очеву за ряд ценных консультаций.
Рисунки к работе выполнены художником К.П.Мешковым.
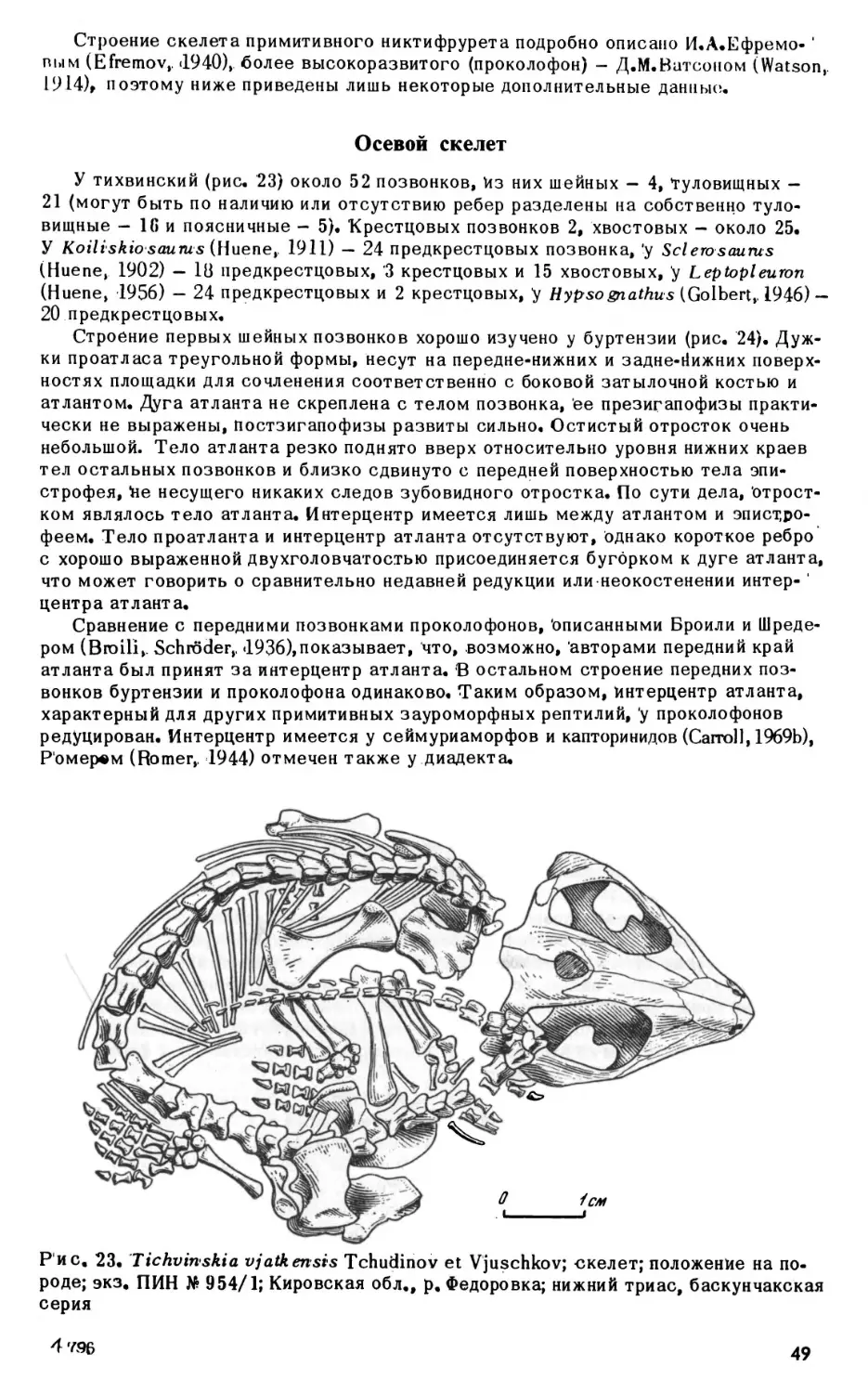
МАТЕРИАЛ
В работе были использованы коллекции Палеонтологического института АН СССР
(ПИН) и Саратовского государственного университета (СГУ), «Ниже дается распре
деление материала по отдельным видам.
Nyctiphruretus
acudens Efremov, 1938
ПИН 158; Архангельская обл., р.Мезень; верхняя пермь, нижнетатарский подь-
ярус.
Образцы: 4 - полный скелет (голотип); 5, 7, 11, 12, 13 — 24 — черепа; 16, 17, 21,
22 - неполные черепа; 14, 15, 27 - крыши черепов; 25, 37, 40, 41 - верхнечелюст
ные кости; 26, 33 - зубные кости.
ПИН 162; Архангельская обл., р. Мезень; верхняя пермь, нижнетатарский подъ-
ярус.
Образцы: 1 - череп; 7, 8, 9, 10 - зубные кости.
Tichvinskia
vjatkensis Tchudinov et Vjushkov, 1956
ПИН 954; Кировская обл.,»р. Федоровка; нижний триас, баскунчакская серия.
Образцы: 1 - полный скелет; 2, 3, 12 - части черепов; 7, 8, 9, 10-13 - верхне
челюстные кости; 4-6, 14-21
-
зубные кости.»
Tichvinskia jugensis Vjuschkov et Tchudinov, «1956
ПИН 2252; Вологодская обл., р.Шарженга, с. Вахнево; нижний триас, ветлужская
серия.
Образец: 368 - левая половина черепа (голотип).
Contritosaurus
simus Ivachnenko, «1974
ПИН 1025; Горьковская обл., р. Ветлуга, с. Спасское; нижний триас, ветлужская
серия.
Образцы: 92 - часть черепа; 50-55, 93, 98, 99-105 - верхнечелюстные кости;
56-59, 64-91, 97 - зубные кости; 60 - основная затылочная кость; 61, 95, 96 -
основные затылочные кости; 62, 63 - ушные кости; 94 - квадратная кость. »
ПИН 2482; Горьковская обл./р. Вохма, с. Спасское; нижний триас, ветлужская
серия.
Образцы: 1 - мозговая коробка; 5 - верхнечелюстная кость; 2-4
-
зубные кости.
ПИН 2890; Горьковская обл., р. Молома, с. Касьяновцы; нижний триас, ветлуж
ская серия.
Образец: 5 — часть черепа.
ПИН 3355; Горьковская обл., р. Ветлуга, с. Липово; нижний триас, ветлужская
серия.
Образцы: 1 - череп (голотип); 2,4 - верхнечелюстные кости; 3 - зубная кость.
ПИН 3356; Горьковская обл., р. Ветлуга, с. Сарафаниха; нижний триас, ветлуж
ская серия.
Образец: 1 - скелет.
5
С on tri to saurus convectorIvachnenko,
«1974
ПИН 3357; Горьковская обл., p. Ветлуга, с. Красные Баки; нижний триас, вет
лужская серия.
Образцы: 1 - постдентальная часть нижней челюсти (голотип); 2 - часть верх
нечелюстной кости.
Phaanthosaums
ignatjevi Tchudinov et Vjuschkov, «1956
ПИН 1025; Горьковская обл./ р. Ветлуга, с. Спасское; нижний триас, ветлужская
серия. •
Образцы: 1 - зубная кость (голотип); 21 - зубная кость; 2(1 - постдентальная
часть нижней челюсти,
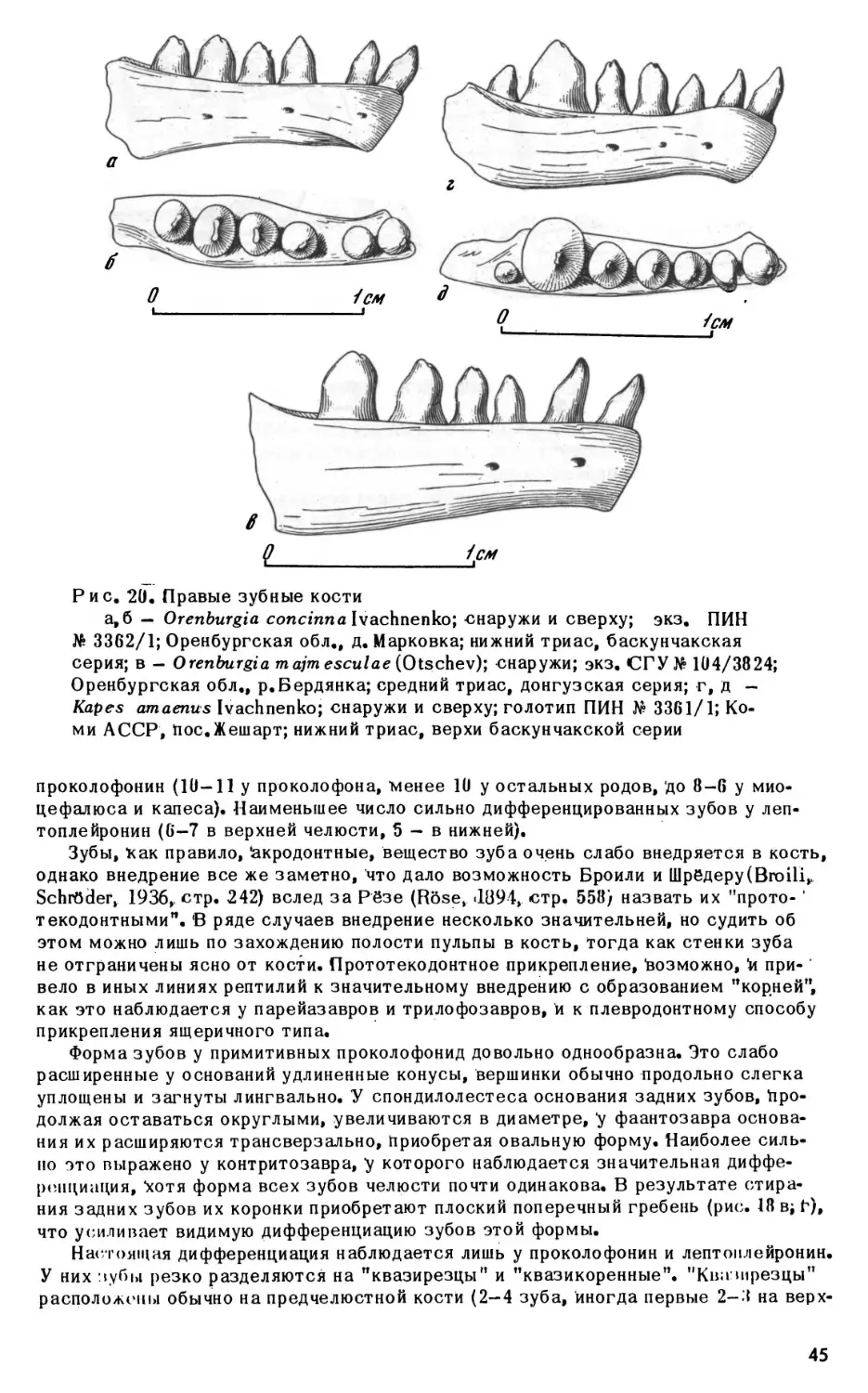
К apes am aenus Ivachnenko, «1975
ПИН 3361; Коми АССР, р.Вычегда, с.Жешарт; нижний триас, баскунчакская се
рия.
Образцы: 1 - зубная кость (голотип); 2,6-9
-
верхнечелюстные кости; 4, 5, 10,
11 - зубные кости. «
Macrophon komiensis Ivachnenko, 1975
ПИН 3361; Коми АССР, р.Вычегда, с.Жешарт; нижний триас, баскунчакская се
рия.
Образец: 3 - верхнечелюстная кость (голотип).
Orenburgia concinnaIvachnenko,
«1975
ПИН 3362; Оренбургская обл.,« с. Марковка; нижний триас, баскунчакская серия.
Образец: 1 - часть черепа (голотип).
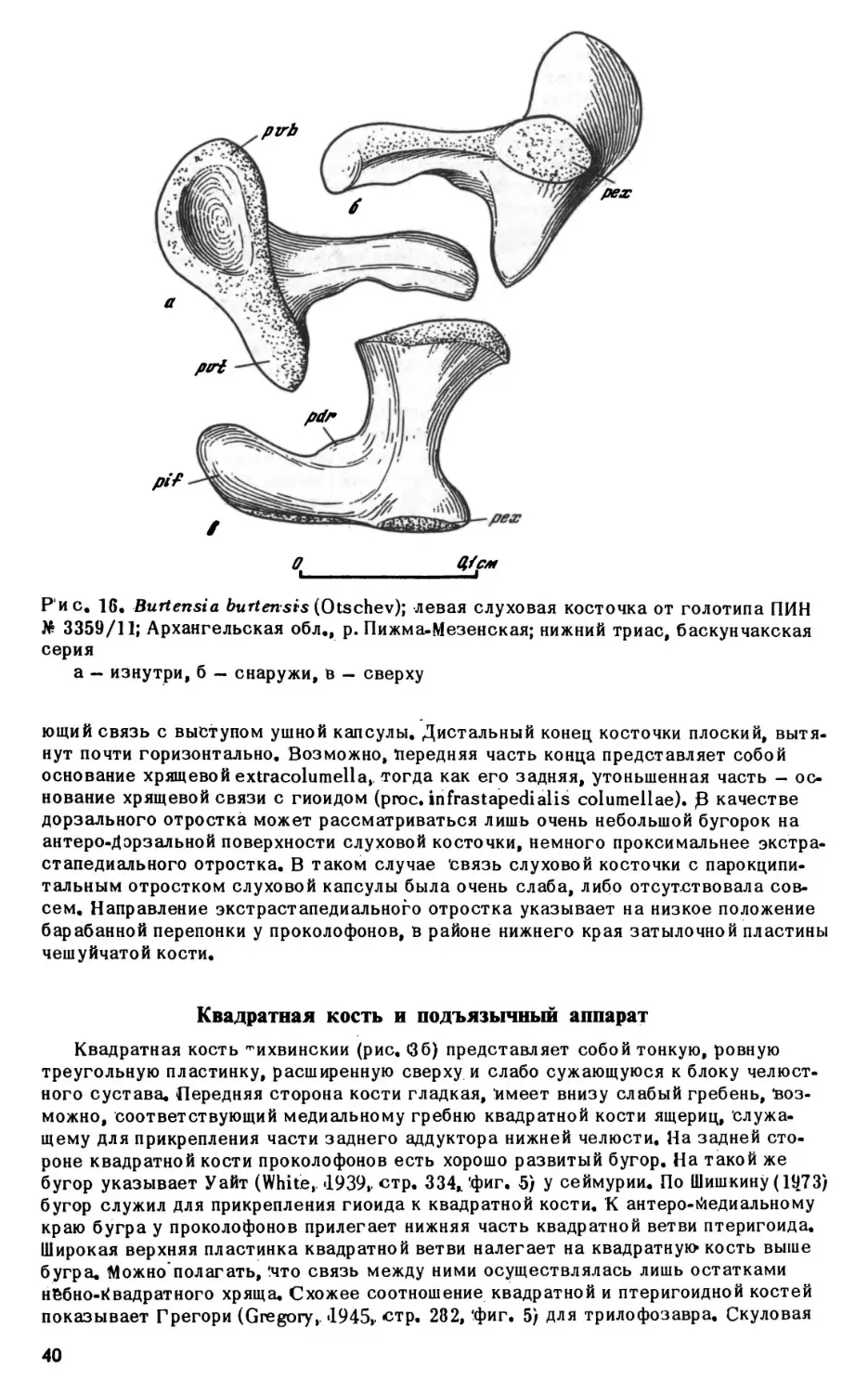
Burt ensi a burtensis (Otschev, «1958)
ПИН 3360; Архангельская обл., д. Выбор; нижний триас, баскунчакская серия.
Образцы: 1, 2, 3 — зубные кости. *
»
ПИН 3358; Архангельская обл., д.Занулье; нижний триас, баскунчакская серия.
Образец: 1 - зубная кость.
ПИН 3359; Архангельская обл./ р.Пижма-Мезенская; нижний триас, баскунчак
ская серия.
Образцы: 1—10 — челюстные кости; 11 - часть скелета с черепом.
Orenburgia majmesculae (Otschev, «1968)
СГУ 104; Оренбургская обл., р.Бердянка; средний триас, донгузская серия. <
Образец: 3824 - зубная кость.
Burtensia burtensis (Otschev, «1958)
СГУ 104; Оренбургская обл./овр. Кызыл-Сай; нижний триас, баскунчакская се
рия.
Образец: 2 - зубная кость (голотип).
Vitalia grata Ivachnenko, «1973
СГУ 104; Донская Лука, балка Липовская; нижний триас, баскунчакская серия.
Образец: 3104 - зубная кость (голотип).
В качестве сравнительного материала использовались коллекции Палеонтоло
гического института, содержащие краниальные остатки: сеймуриаморфов - Lan-
thanosuchus
(колл. «ПИН 157), Chtoniosuchus
(колл. ПИН 521, 523), Kotlassia
(крлл. ПИН 2005); котилозавров - Nycteroleter
(колл. ШН 164, 521),-
Nyctiboetus
(колл* «ПИН 161), Belebey (колл. «ПИН 164), Rhipaeosaurus
(колл. ПИН 161, 164),
Р areiasuchus (колл. ПИН 522, 2212), Scuto saurus (колл. 2005), а также Саратовско
го университета (трилофозавр Coelodontognathus,
колл. СГУ 104/3101, 3102, 3104).
ГЛАВА I
СИСТЕМАТИКА ПРОКОЛОФОНОВ
ИСТОРИЯ ИЗУЧЕНИЯ ПРОКОЛОФОНОВ
Исторически первый известный проколофон (Leptopleuron)
был описан в 1851 г.
Оуэном в "Literary Gasette". *Эту же форму переописал через год Мантелл (Mantell,
1852)». «Дав название "Telerpeton",
под которым лептоплейрон упоминается во всех
последующих работах, и лишь в 1925 г. Виллистон (Willistqn, «1925) сводит его в
синонимику. В 1857 г. Фишер де Вальдхейм (Fischer de Waldheim, 1857) опублико
вал описание склерозавра, составленное Мейером. Более подробные данные по этой
форме приведены в статье самого Мейера (Meyer,. «1859). Систематическое положе
ние склерозавра долгое время оставалось неясным, его относили и к лабиринто-
донтам ("Labyrinihodon";
Wiedersheim, «1878) и к аномодонтам
("Aristodesmus":
Seeley, «1900). Синонимичность этих родов склерозавру показал Хюне (Huene* 1932),.
однако место склерозавра среди проколофонов неясно до сих пор. В 1923 г, Нопча
(Nopcsa, 1923) выделил особое семейство склерозаврид, в состав которого вкл.кчил
склерозавра и базилеозавра. Хюне (Huene* 1956) относил склерозавра к парейазав-
ридам. Вследствие недостатка данных в настоящей работе сохраняется выделен
ное Нопчей семейство склерозаврид.
Сили, изучив описанный Оуэном (Owen, 1876) южноафриканский род Piccolo*
phon, выделил в составе отряда Cotylosauria (установленного Копом в 1880 г.} се
мейство Procolophonidae Seeley, 1888* Позже Брум (Broom, «1905а,с; 1936а, Ь; 1937)
описал ряд новых родов с территории Южной Африки — Thelegnathus,
Miсгоthel е don,
Myocephalus,
Spondylolestes.
В 1939 г. он же опубликовал описание первого поздне-
пермского рода (Owenetta).
•Продолжались находки проколофонов и на территории
Европы (Koilosk iosaurus'.WuBTiB^ «1911J Anomoiodon: Huene,. 1939).tf31928 г. был опи
сан первый проколофон из Северной Америки (Hypso&iathus:
Gilmore* 1928), «в
I960 г.
— второй (Sphodro saurus: Colbert, I960).)
В 1938-1940 гг. Ефремов (Ефремов, 1938; Efremov, «1940) описал второй род
пермских проколофонов — Nyctiphmretus.
Полагая, что ушная вырезка у никтеро-
летера развивается вторично и считая последнего проколофоном, Ефремов объеди
нил никтеролетера и никтифрурета в сем. «иктифруретид.
В работе "К познанию проколофонид" Хюне (Huene, 1943) дал краткие диагнозы
14 родов, относимых в то время к проколофонам (в том числе и род
Jrilophosaurus).
Род Myocephalus
в статье по неизвестной причине назван
"Myo&iathus".»
При переописании гипсогната Колберт (Colbert,; 1946) также дает сводку проко
лофонов. Рассматривая котилозавров.как отряд, а проколофонов как семейство,
Кольберт подразделяет их на два подсемейства — никтифруретин (никтифрурет,
никтеролетер, овенетта) и проколофонин (остальные роды).
В дальнейшем появляются описания проколофонов из различных регионов зем
ного шара - из Бразилии (Candelaria:
Price* 1947), «Мадагаскара (Barasaurus:
Pi-
veteau, «1955), «Китая (Neopiocolophon:
Young, 1957; Paoteodon: Chow et Sun, «I960).
В 1956 г. Хюне (Huene* «1956)». рассматривая проколофонов как самостоятель
ный отряд в ветви рептилиоморфов, делит его на три семейства - никтифруретид
(никтифрурет, никтеролетер, никтибет,рипеазавр),овенеттид (овенетта, спондило-
лестес), проколофонид (остальные роды). Совершенно необоснованным представля-
7
ется включение в состав проколофонов рипеозавра, отнесенного Чудиновым (1955)
к парейазаврам. Кроме того, надо отметить, что Хюне рассматривал оба рода -
Myocephalus и "Myognathus",
причем рисунок, на который он ссылался при описании "мио-
гиатуса", изображает миоцефалюса. Однако в добавлениях к работе (Huene, 1959) ошиб
ка исправлена, и оставлен лишь род Myocephalus.
Склерозавра Хюне отнес к п'арейазаври-
дам. Такой же схемы классификации придерживается и Кун (Kuhn, 1966).
Первые триасовые проколофоны с территории Русской платформы стали из
вестны в 1956 г. {Phaanthosaurus,
Tichvinskia:
Чудинов, Вьюшков, 1956). Однако
детального изучения их остатков не производилось, поэтому в дальнейшем все
формы со слабо дифференцированными зубами относились к фаантозавру ,с сильно диф
ференцированными - к тихвинский (Блом, 1968; Очев, 1958, 1968; Очев, Данилов, 1972).
Детальное изучение ранее описанного и нового материала позволило установить
ряд новых родов (Ивахненко, 1974, 1975).
В 1969 г. Дейли (Daly, 1969) опубликовала описание Acleistorhinus,
как нового
рода проколофонов. Однако наличие нижнего височного окна у этой формы дало
возможность Л.П.Татаринову (1974) объединить ее с проблематичной нижнеперм
ской формой Bolosaurus
в отряде Bolosauromorpha. рписанный В.Г.Очевым (1967)
род Соelodontognathus более близок к трилофозаврам, чем к проколофонам.
Последняя общая сводка проколофонов дана Чудиновым (1964). Приводится 21
род, среди которых роды Nycteroleter,
Nyctiboetus,
Santaisaurus.
Первые два рода
в настоящей работе исключены из состава.проколофонов (Ивахненко, 1973в). Сан-
тайзавр (Koh, 1940) при первоописании и ревизии (Young, «1948* Hoffstetter, 1953),
вероятно, вполне справедливо отнесен к лепидозаврам, во всяком случае, харак
терная для него плевродонтность зубов не наблюдается у проколофонов. Родовые
диагнозы в сводке Чудинова даны для 5 родов.
В приводимую ниже сводку входят 26 родов (из которых 7 установлены авто
ром) и вводится разделение семейства Procolophonidae на три подсемейства по
уровню развития ряда признаков (удлинение орбит и образование "ложного височ
ного окна", величина ушной вырезки, развитие выростов на костях щечного отде
ла, степень дифференциации и число зубов в челюстях).
СИСТЕМАТИКА ПОДОТРЯДА PROCOLOPHONIA
ПОДКЛАСС COTYLOSAURIA
ОТРЯД DIADECTOMORPHA
ПОДОТРЯД PROCOLOPHONIA
Диагноз. Котилозавры небольших размеров, череп скульптирован у прими
тивных форм. Орбиты более или менее сильно удлинены назад. Ушная вырезка пол
ностью смещена на чешуйчатую кость, надвисочная кость почти не участвует в ее
ограничении. Таблитчатая и заднетеменная кости сильно редуцированны, у высших
форм исчезают. В плечевом поясе два коракоида, в позвоночнике обычно сохраня
ются интерцентры.
Сост ав. Три семейства: Nyctiphruretidae Efremov, 1935; Procolophonidae Se
eley, 1888; Sclerosauridae Nopcsa, 1923.
Сравнение. От подотряда Diadectia отличаются полным смещением ушной вы
резки на чешуйчатую кость, сильной редукцией таблитчатых и заднетеменных хостей.
Распространение. Верхняя пермь, нижний, средний и верхний триас; все
материки, кроме Австралии.
СЕМЕЙСТВО NYCTIPHRURETIDAE EFREMOV, 1938
Типовой род - Nyctiphruretus
Efremov, 1938.
Диагноз. Наиболее примитивные проколофоны, с плоским с кул ьптиро ванным
черепом. Ушная вырезка маленькая. Зубы недифференцированные, многочисленные.
8
Рис. I. Nyctiphmretus
acuderts Efremov; череп; реконструкция по экз. ПИН No 158/5,
11, ПИН No 162/1; Архангельская обл., р. Мезень; верхняя пермь, нижнетатарский
подъярус
а- сверху,б - снизу,в - сбоку,г - сзади
Родовой состав. Три рода.
Сравнение. От семейства Procolophonidae и семейства Sclerosauridae отли
чаются наличием плоского скульптированного черепа, слабо удлиненными орбитами,
присутствием большого числа недифференцированных зубов.
Распространение. Верхняя пермь; Африка и Восточная Европа.
Род Nyctiphmretus
Efremov, 1938
1938. Nyctiphmretus:
Ефремов, стр. 775.
1940. Nyctiphmretus:
Efremov, S. 113.
Типовой вид - Nyctiphmretus
acuderts Efremov,. «1938; верхняя пермь; евро
пейская часть СССР.
Диагноз. Череп резко сужается кпереди. Орбиты удлиненные, пинеальное
отверстие позади уровня задних краев орбит. Небо несет многочисленные ряды мел
ких зубов, птеригоидные фланги с зубами. В челюсти свыше 30 конических зубов.
На затылочной поверхности сохраняются рудименты таблитчатой и заднетеменной .
костей.
Сравнение. От родов Owenetta и Barasaums
отличается более удлиненными
назад орбитами и присутствием зубов на птеригодных флангах.
Распространение. Верхняя пермь европейской части СССР.
9
Nyctiphmretus
acuderts Efremov,. 1938
(рис, la-г)
1938. Nyctiphmretus
acudensr. Ефремов, стр, 775. рис, 4.
1940* Nyctiphmretus
acuderts: Efremov, S. 413-440, Abb. 5-17.
Голотип — No 158/4, ПИН; скелет; Архангельская обл,, р. Мезень; верхняя
пермь.
Материал. Ряд скелетов и отдельные черепа.
Диагноз, Длина черепа не менее 5 см. Предлобные кости нависают над ор
битами. Зубы конические, заостренные, основания их слабо расширены, вершинки
загнуты лингвально.
Распространение. Верхняя пермь; европейская часть СССР, Архангель
ская обл., р.Мезень.
Род Owenetta Broom, 1939
1939, Owenetta: Broom, p. 319.
Типовой вид - Owenetta rubidgei Broom, 1939; верхняя пермь; Южная Аф
рика,
Диагноз, Череп плоский, округло-удлиненный. Орбиты слабо удлинены, не
сколько отодвинуты назад, Пинеальное отверстие позади уровня задних краев, ор
бит. Число зубов до 40 в челюсти, птеригоидные фланги без зубов.
Сравнение, Очень близок к роду Barasaums,
отличается более узкой мор
дой. От Nyctiphmretus
отличается округлыми орбитами и слабым развитием неб-
ных зубов, формой черепа.
Видовой состав. Род монотипичный.
Распространение. Верхняя пермь, зона Cistecephalus, Южная Африка,
район Нью-Бетседа.
Род Barqsmms
Piveteau, 1955
1955. Barasaums:
Piveteau, p. 1325.
Типовой вид — Barasaums
besairiei Piveteau, «1955; верхняя пермь; Мада
гаскар,
Диагноз, Череп округлый, широкий. Орбиты слабо удлинены, небольшое пи
неальное отверстие на уровне задних краев орбит. Большое число недифференци
рованных зубов.
Видовой состав. Род монотипичный.
Сравнение. От родов Owenetta
и Nyctiphmretus
отличается формой че
репа, положением теменного отверстия, положением и степенью удлине
ния орбит.
Распространение, Верхняя пермь, формация Sakamena; Мадагаскар.
СЕМЕЙСТВО PROCOLOPHONIDAE SEELEY, 1888
Типовой род - Procolophon
Owen, 1857.
Диагноз. Череп высокий, иногда слабо скульптирован, орбиты резко удлине
ны назад. Таблитчатые и заднетеменные кости отсутствуют. Зубы более или менее
дифференцированны, число их обычно невелико.
Состав. Три подсемейства,
Сравнение. От семейств Nyctiphruretidae и Sclerosauridae отличаются
наличием высокого, обычно не скульптированного черепа, удлиненными ор
битами, небольшим числом в той или иной степени
дифференцированных
зубов.
Распространение. Триас; повсеместно.
10
ПОДСЕМЕЙСТВО SPONDILOLESTINAE IVACHNENKO, SUBFAM.NOV
Типовой род - Spondylolestes
Broom, 1937*
Диагноз. Наиболее примитивные проколофониды, орбиты удлинены, как пра
вило, не сильно, зубы обычно простые, конические^относительно слабо дифферен
цированные, более 10 в челюсти.
Родовой состав. Шесть родов.
Сравнение, От остальных подсемейств отличаются слабым удлинением ор
бит и значительным числом почти не дифференцированных зубов.
Распространение. Нижний триас; Южная Африка, Южная Америка, Вос
точная Европа, Китай(?),
Род Spondylolestes
Broom, 1937
1937. Spondylolestes:
Broom, p. 299,
Типовой вид — Spondylolestes
rubidgei Broom, 1937; нижний триас, Южная Африка.
Диагноз, Череп округло-треугольный, орбиты довольно большие, округлые.
Небольшое пинеальное отверстие на уровне задних краев орбит. Зубы конические
(число их относительно велико) - не менее 13—15 в верхней челюсти.
Видовой состав. *Род монотипичный.
Сравнение, От остальных родов подсемейства отличается плоским черепом
и большим числом почти не дифференцированных зубов.
Распространение, Нижний триас, низы зоны Procolophon; Южная Африка,
район Бетесда-Роуд.
Род Phaanthosaunis
Tchudinov et Vjuschkov, 1956
1956. Phaanthosaunis:
Чудинов, Вьюшков, стр.547.
Типовой вид — Phaanthosaunis
ignatjevi Tchudinov et Vjuschkov, 1956; ниж
ний триас; европейская часть СССР.
Ди агноз. Череп широкий, округлый. Нижняя челюсть невысокая, несет 11
слабо дифференцированных зубов с загнутыми лингвально вершинками.
Видовой состав. Род монотипичный.
Сравнение. От остальных родов подсемейства отличается количеством сла
бо дифференцированных зубов. От близкого рода Spondylolestes
отличается более
округлой мордой и зубами с загнутыми лингвально вершинками.
Р ас пр остр ане ни е. Нижний триас европейской части СССР.
Phaanthosaunis
ignatjevi Tchudinov et Vjuschkov, 1956
(рис. 17б-г)
1956. Phaanthosaunis
ignatjevi: Чудинов, Вьюшков, стр. 547, рис. 2.
Голотип - No 1025/1, ПИН; зубная кость; СССР, Горьковская обл., Спасское
1, нижний триас.
Материал. Нижняя челюсть.
Диагноз. Высота передних зубов меньше, чем высота симфиза нижней челю
сти. Основания задних зубов слабо расширены. Постдентальная часть нижней челю
сти равна по длине дентальной.
Распространение» Нижний триас,низы ветлужской серии; европейская часть
СССР, Горьковская обл.
Род С on tri to saurus Ivachnenko, 1974
Типовой вид — Contritosaunissimus
Ivachnenko, 1974» нижний триас; евро
пейская часть СССР.
11
Р и с. 2. Сontritosaurus
simus Ivachnenko; череп; голотип ПИН No 3355/1; Горьков
ская обл., Д. Липово; нижний триас; низы ветлужской серии
а- сверху,б —снизу,в - сбоку
Диагноз. Череп округлый, высокий, орбиты большие, значительно удлинены
назад. На нёбе 4 ряда зубов, птеригоидные фланги без зубов, заострены вентраль-
но, в верхней челюсти 12 зубов, в нижней 9-10. Зубы дифференцированные, послед
ние 5-6 зубов с сильно расширенными основаниями. Нижняя челюсть низкая.
Видовой состав. Csimus Ivachnenko, С. convec to г Ivachnenko; нижний
триас европейской части СССР.
Сравнение, От остальных родов подсемейства отличается числом, степенью
дифференциации и формой зубов, а также формой черепа.
Распространение. Нижний триас европейской части СССР.
Contri to saurus simus Ivachnenko, 1974
(рис. 2 а-в)
1974. Contri to saurus simus: Ивахненко, стр«70^-73> рис.2, 3.
12
Голотип - No 3355/1, ПИН; череп; СССР, Горьковская обл., с.Липово; Тг:
Материал, Череп и часть скелета.
Диагноз, Длина черепа 2 см, череп высокий с тупой короткой мордой. Сош
никовый ряд небных зубов двойной. Зубы дифференцированы, но форма задних зу
бов одинакова с формой передних. Аддукторная яма нижней челюсти широкая.
Сравнение, От вида С.convector отличается более узкой нижней челюстью,
широкой аддукторной ямой, отсутствием наклона верхнечелюстных зубов.
Распространение, Нижний триас, низы ветлужской серии; европейская
часть СССР, Горьковская обл,
С ontrito saurus convector
Ivachnenko, 1974
V)7b1ontri
to saurus convector.
Ивахненко, стр.73-74, рис, 4,
Голотип — No 3357/1, ПИН; постдентальная часть нижней челюсти; СССР,
Горьковская обл,, с. Красные Баки; нижний триас.
Материал. Пост дентальная часть нижней челюсти, часть верхней челюсти.
Диагноз. Зубы верхней челюсти слабо наклонены вперед, постдентальная
часть нижней челюсти высокая, аддукторная яма узкая.
Сравнение. От вида С. simus отличается наклоном верхнечелюстных зубов,
высокой нижней челюстью, узкой аддукторной ямой.
Распространение. Нижний триас, низы ветлужской серии; европейская
часть СССР, Горьковская обл.
Род Candelaria Price, 1947
1947. Candelaria: Price* p. 17 .
Типовой вид — Сandelaria barbouri Price* 1947; средний триас Бразилии,
Диагноз. Довольно высокий округло-треугольный череп, орбиты относитель
но слабо удлинены назад, между орбитами продольная депрессия. Зубы почти не
дифференцированы, до 13 в нижней челюсти. Нижняя челюсть низкая в симфизе.
Видовой состав. Род монотипичный.
Сравнение, От остальных родов подсемейства отличается высоким черепом
со слабо удлиненными орбитами, продольной депрессией по лобным костям, боль
шим количеством почти не дифференцированных зубов.
Распространение, Средний триас, формация Santa-Maria; Бразилия, про
винция Рио-Гранде до Сул,
Род Procolophonoides
Ivachnenko, gen, no v.
1905b. Procolophon,
partim: Broom, p. 332.
1936b. Procolophon,
partim: Broom, p. 389.
Типовой вид — Pwcolophon baini Broom,
л
905; нижний триас; Южная Африка,
Диагноз*, Череп округло-треугольный, орбиты слабо удлинены, В верхней
челюсти 8 конусообразных, почти не дифференцированных зубов. Нижняя челюсть
очень высокая в симфизе.
Видовой состав. Род монотипичный.
Сравнение. От остальных родов подсемейства отличается числом зубов,
очень высокой в симфизе нижней челюстью, формой черепа.
Распространение. Нижний триас Южной Африки.
(?) Род N еор го colophon
Young, 1957
1957. Neoprocolophon: Young, p. 1.
Типовой вид — N еорю со lophon asiaticus Young, 1957; нижний триас; Китай,
Диагноз. Череп высокий, удлиненно-овальный. Орбиты сильно удлинены на
зад, между орбитами продольная депрессия, ^Крупное пинеальное отверстие вперед
13
ди уровня
задних краев орбит. На квадратно-скуловых костях небольшие
латеральные выросты. Зубы имеют слабо расширенные поперечно ко
ронки.
Видовой состав. Род монотипичный.
Сравнение. От остальных родов подсемейства отличается формой черепа,
сильно удлиненными назад орбитами, наличием выростов на квадратно-скуловых
костях..
Распространение. Нижний триас; Китай, провинция Шанси.
ПОДСЕМЕЙСТВО PROCOLOPHONINAE SEELEY,. 1888
1888* Procolophonidae: Seeley, p. 383.
1923. Procolophoninaej Nopcsa, p. 30*
Типовой род - Procolophon
Owen, 1875.
Диагноз. Черел обычно довольно высокий, орбиты сильно удлинены. Зубы
дифференцированы, часто со сложными коронками, их число, как правило, менее
10 в челюсти.
Родовой состав. 12родов.
Сравнение. От остальных подсемейств отличаются сильным удлинением ор
бит, строением и степенью дифференциации зубов.
Распространение. Нижний и средний триас; Европа и Южная Африка.
Род Procolophon
Owen, 1876
1876. Procolophon:
Owen, p. 25.
1878* Procolophon:
Seeley, p.797.
1905. Procolophon:
Seeley, p.226.
1936b. Procolophon:
Broom, p.389,
1936. procolophon:
Broili, Schroder, S.239.
1946. Procolophon:
Colbert, p. 258.
Типовой вид - Рю colophon trigoniceps Owen, 1876; «ижний триас; Южная
Африка.
Диагноз. Череп округло-треугольный, высокий, скульптирован, пинеальное
отверстие на уровне задних краев орбит. Квадратно-скуловые кости имеют широ
кие латеральные выросты. На небе 4 ряда зубов. В челюстях 10-11 зубов, 6-7 по
следних с расширенными двухвершинковыми коронками.
Видовой состав. Род монотипичный.
Сравнение. От остальных родов подсемейства отличается наличием
скульптуры, присутствием латеральных выростов на квадратно-скуловых
костях.
Распространение. Нижний триас, зона Procolophon; Южная Африка, Дон-
нибрук, Аливал-Нурт.
Род Tichvinskia Tchudinov et Vjuschkov, 1956
1956» Tichvinskia:
Чудинов, Вьюшков, стр.548.
Типовой вид - Tichvinskia
vjatkensis Tchudinov et Vjuschkov, 1956; ниж
ний триас; европейская часть СССР.
Диагноз. Череп высокий, округло-треугольный, между орбитами продольная
депрессия. Орбиты очень большие, сильно удлинены назад. Пинеальное от
верстие большое, впереди уровня задних краев орбит. На нёбе 4 ряда
зубов. Птеригоидные фланги заострены вентрально. В верхней челюсти
J0 зубов, в нижней 9. Задние зубы между латеральными буграми несут
тупой гребень.
Видовой состав. Т. ju gen sis Vjuschkov et Tchudinov, T. vjatk en sis Tchudi
nov et Vjuschkov; «ижний триас европейской части СССР.
14
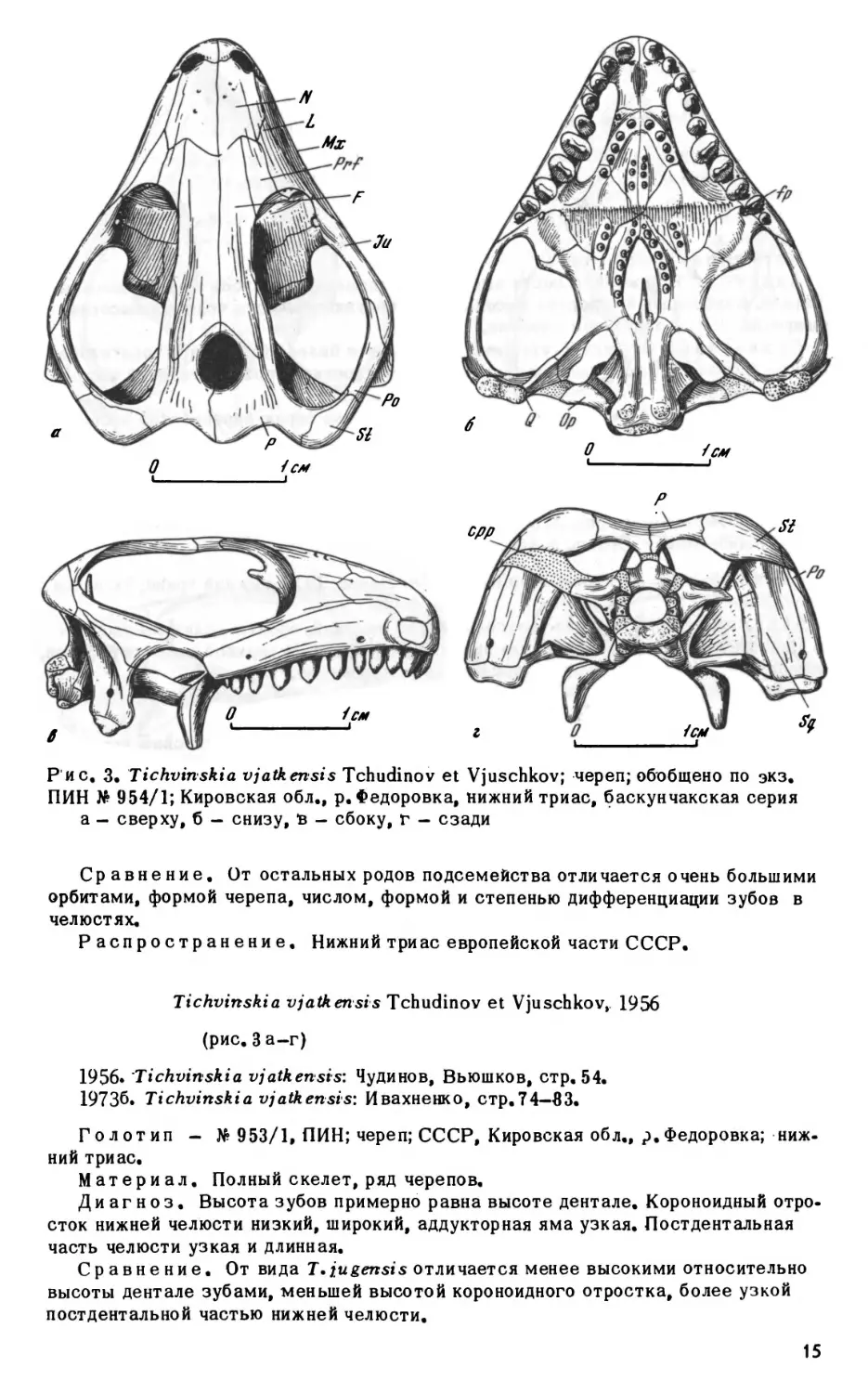
Рис. 3, Tichvinskia
vjatkensis
Tchudinov et Vjuschkov; череп; обобщено по экз.
ПИН No 954/1; Кировская обл., р.Федоровка, нижний триас, баскунчакская серия
а— сверху,б —снизу, в —сбоку, г —сзади
Сравнение. От остальных родов подсемейства отличается очень большими
орбитами, формой черепа, числом, формой и степенью дифференциации зубов в
челюстях.
Распространение. Нижний триас европейской части СССР,
Tichvinskia
vjatkensis
Tchudinov et Vjuschkov, 1956
(рис. За-г)
1956. Tichvinskia
vjatkensis:
Чудинов, Вьюшков, стр.54.
19736. Tichvinskia
vjatkensis:
Ивахненко, стр.74—33.
Голотип - No 953/1, ПИН; череп; СССР, Кировская обл., р.Федоровка; ниж
ний триас.
Материал. Полный скелет, ряд черепов.
Диагноз. Высота зубов примерно равна высоте дентале. Короноидный отро
сток нижней челюсти низкий, широкий, аддукторная яма узкая. Постдентальная
часть челюсти узкая и длинная.
Сравнение. От вида Т.jugensis отличается менее высокими относительно
высоты дентале зубами, меньшей высотой короноидного отростка, более узкой
постдентальной частью нижней челюсти.
15
Распространение. Нижний триас, верхи ветлужской — низы баскунчакской
серий; европейская часть СССР, Приуралье.
Tichvinskia jugensis Vjuschkov et Tchudinov,. 1956
1940, Microcnemus
efremovi, partim: Huene, Ss. 8— 9, t. 2, fig. 3a, b.
1956. Tichvinskia jugensis: Вьюшков, Чудинов, стр. 141, рис. 1.
Голотип - No 2252/308, ПИН; часть
черепа; СССР, р. Шарженга; ниж
ний триас.
Материал. Часть черепа.
Диагноз. В нижней челюсти высота двухвершинковых зубов меньше высоты
дентале. <Короноидный отросток высокий, постдентальная часть челюсти высокая
и широкая, аддукторная яма широкая.
Сравнение. От вида Т.vjatkensis отличается более высокими относительно
высоты дентале зубами, высоким короноидным отростком, постдентальной частью
нижней челюсти.
Распространение. Нижний триас, ветлужская серия; европейская часть
СССР, Приуралье.
Род Anomoiodon Huene, 1939
1939. Anomoiodon:
Huene,. S . 501.
1967. Anomoiodon: Ortlam, S.487.
Типовой вид - Anomoiodon lilienstemi
Huene, 1939; нижний триас; Западная
Европа.
Диагноз. Череп округло-треугольный, не высокий, орбиты удлинены умерен
но. Пинеальное отверстие на уровне задних краев орбит. В верхней челюсти 7 зубов,
в нижней — 5. Последние зубы с расширенными коронками.
Видовой состав. An. lilienstemi Huene,. средний пестрый песчаник Запад
ной Европы, An. krejcii Ortlam,. верхний песчаник.
Сравнение. От остальных родов подсемейства отличается плоским черепом,
числом и степенью дифференциации зубов.
Распространение. Нижний и средний триас Западной Европы.
Род Koiloskiosaums
Huene, 19Ц
1911. Koiloskiosaums:
Huene, S.81.
Типовой вид - Koiloskiosaums
coburgensis Huene, 1911; нижний триас; За
падная Европа.
Диагноз. Череп треугольный, Орбиты сильно удлинены назад. Пинеальное
отверстие впереди уровня задних краев орбит. Края квадратно-Скуловых костей
слегка расставлены в стороны. В верхней челюсти 9 зубов, поперечно-авальных в
сечении, в нижней - 7 зубов.
Видовой состав. Род монотипичный.
Сравнение. От остальных родов подсемейства отличается формой черепа,
числом и степенью дифференциации зубов.
Распространение. Нижний триас, средний пестрый песчаник; ФРГ, Кобург.
Род Burtensia Ivachnenko, 1975
1958. Tichvinskia:
Очев, стр. 448.
1975. Burtensia: Ивахненко, стр. 89-90.
Типовой вид - Tichvinskia
burtensis Otschev, 1958; нижний триас; европей
ская часть СССР.
Диагноз. Череп высокий, с короткой тупой мордой, Орбиты сильно удлинены.
Пинеальное отверстие несколько впереди уровня задних краев орбит. На небе 4 ко-
16
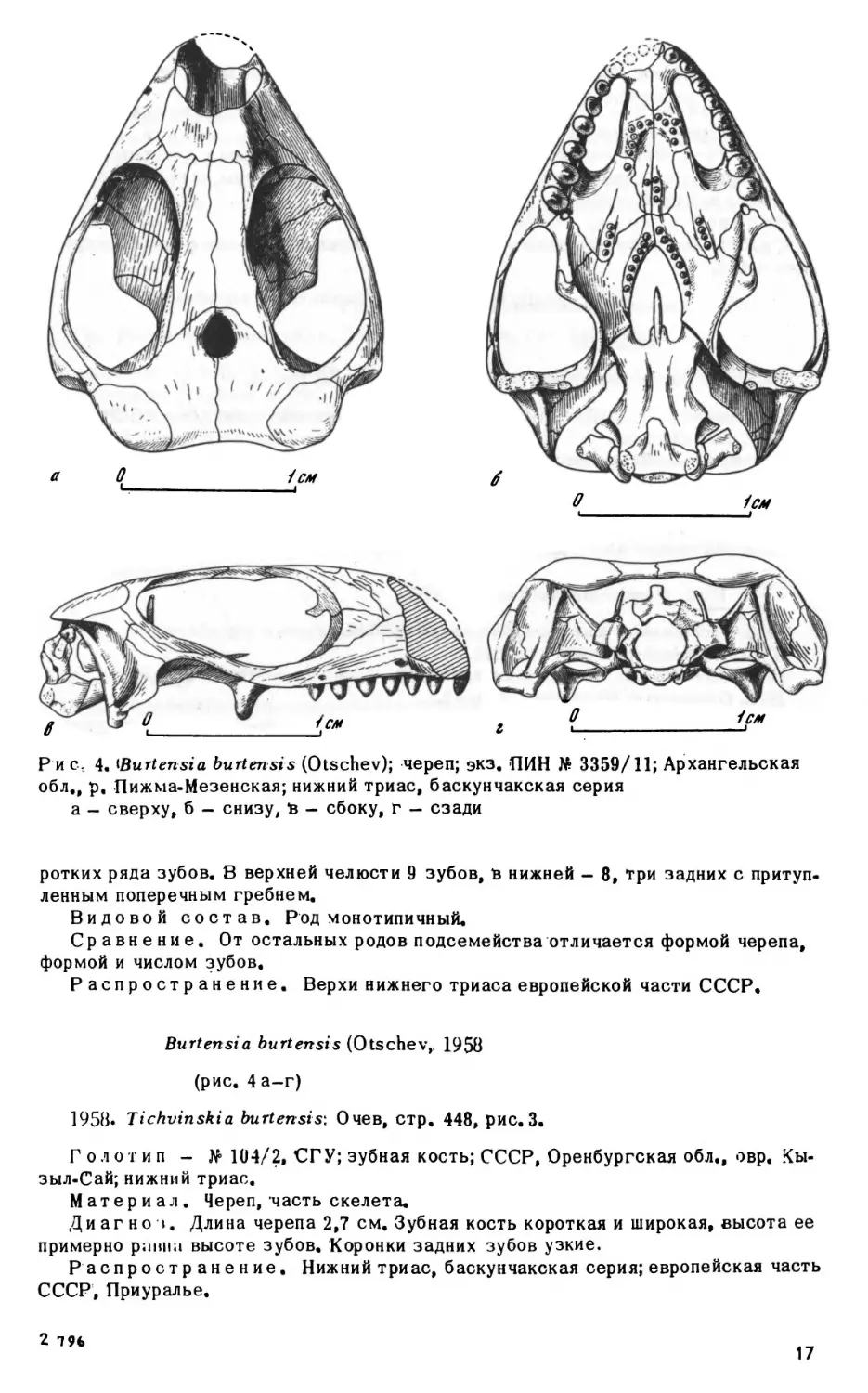
Р и с, 4, iBurtensia burtensis (Otschev); череп; экз. ПИН No 3359/11; Архангельская
обл., р, Пижма-Мезенская; нижний триас, баскунчакская серия
а- сверху,б - снизу, в - сбоку, г —сзади
ротких ряда зубов. В верхней челюсти 9 зубов, в нижней - 8, три задних с притуп
лённым поперечным гребнем.
Видовой состав. Род монотипичный.
Сравнение. От остальных родов подсемейства отличается формой черепа,
формой и числом зубов.
Распространение. Верхи нижнего триаса европейской части СССР.
Burtensia burtensis (Otschev,, 1950
(рис. 4а-г)
1958- Tichvinskia
burtensis:
Очев, стр. 448, рис.3.
Голотип - No 104/2, СГУ; зубная кость; СССР, Оренбургская обл., овр. Кы«
зыл-Сай; нижний триас.
Материал. Череп, часть скелета.
Диагно I. Длина черепа 2,7 см. Зубная кость короткая и широкая, высота ее
примерно раина высоте зубов. Коронки задних зубов узкие.
Распространение. Нижний триас, баскунчакская серия; европейская часть
СССР, Приуралье.
2 196
17
Р о д Macrophon Ivachnenko,. 1975
1975. Macrophon:
Ивахненко, стр. 91.
Тип овой вид — Macrophon komiensis Ivachnenko,. 1975; верхи нижнего три
аса; север европейской части СССР.
Диагноз. Верхнечелюстные зубы с сильно расширенными и вздутыми корон
ками, имеющими острые, широко расставленные краевые бугры, между которыми
находится режущий гребень.
Видовой состав. Род монотипичный.
Сравнение. От остальных родов подсемейства отличается формой и строе
нием зубов.
Macrophon komiensis Ivachnenko, 1975
(рис. 21а)
1975. Macrophon komiensis:
Ивахненко, Стр. 91—92, рис. 3.
Голотип — No 3361/1, ПИН; часть верхнечелюстной кости; Коми АССР, пос.
"Жешарт; нижний триас.
Материал. Часть верхнечелюстной кости.
Диагноз. Размер черепа не менее 6-7 см, на верхнечелюстной кости два
передние зуба острые, конические, задние с широкими двухбугорчатыми коронками.
Распространение, Верхи нижнего триаса, баскунчакская серия; север
европейской части СССР, Коми АССР.
Род Оrenburgia Ivachnenko, 1975
1956. Tichvinskia:
Вьюшков, Чудинов, стр. 549.
1967. Tichvinskia:
Очев, стр. 298.
1972. Tichvinskia:
Очев, Данилов, стр.82.
1975. Orenburgia: Ивахненко, стр. 92.
Типовой вид - Tichvinskia
enigmatica Vjuschkov et Tchudinov, 1956; ниж
ний триас, европейская часть СССР.
О
1с*
у
I
Рис. 5. Orenburgia concinna Ivachnenko; череп снизу; голотип ПИН # 3362/1; Орен
бургская обл., д. Марковка; нижний триас, баскунчакская серия
18
Диагноз. Череп округло-треугольный, на небе три сильно редуцированных
ряда зубов. В верхней челюсти 10 зубов, в нижней 7. Последние 4 зуба постепенно
увеличиваются кзади в размерах, несут две притуплённые вершинки на коронках.
Последний зуб обычно доминирует по размеру.
Видовой состав. Or. enigmatic a, Or. majmesculae,
Or. concinna:
нижний и
средний триас европейской части СССР.
Сравнение. От остальных родов подсемейства отличается числом ря
дов нёбных зубов, строением и способом дифференциации челюстных
зубов.
Распространение. Нижний и средний триас европейской части СССР.
Orenburgia
enigmatica(Vjuschkov,.
Tchudinov, 1956)
1956. Tichvinskia
enigmatica:
Вьюшков, Чудинов, стр. 549, рис. 4 .
Голотип - No 1043/1, ПИН; зубная кость; СССР, Донская лука; нижний триас.
Материал. Зубная кость.
Диагноз. Высота зубов примерно равна высоте зубной кости. Последний
зуб нижней челюсти не намного больше предпоследнего.
Сравнение. От Or. majmesculae
отличается относительно меньшей высотой
зубной кости, слабо доминирующим последним зубом. От Or. concinna
отли
чается большой высотой зубной кости, большей доминантностью последнего
зуба.
Распространение. Нижний триас, баскунчакская серия; европейская часть
СССР, Донская лука.
Orenbur&a
majmesculae (Otschev, 1968)
(рис. 20в)
1968. Tichvinskia
majmesculae:
Очев, Стр.298.
1972. Tichvinskia
majmesculae:
Очев, Данилов, стр. 82.
Голотип
-
No 104/3824, СГУ; зубная кость; СССР, Оренбургская обл.,
р. Бердянка; нижний триас.
Материал. Зубные кости.
Ди а г ноз. Высота зубной кости несколько больше высоты зубов, последний
зуб нижней челюсти очень велик, Доминирует.
Сравнение. От остальных видов рода отличается очень высокой зубной ко
стью, сильной доминантностью последнего зуба.
Распространение. Нижний и средний триас, баскунчакская и донгузская
серии; СССР, Приуралье.
Orenburgia concinna Ivachnenko, 1975
(рис. 20 а,б)
1975. Orenburgia concinna: Ивахненко, стр. 92-93, рис. 4.
Голотип — No 3362/1, ПИН; часть черепа; СССР, Оренбургская обл., с.Мар-
ковка; нижний триас.
Материал. Часть черепа с нижней челюстью.
Ди а г но з. Зубная кость тонкая, ее высота меньше высоты последних зубов.
Двухвершинковые зубы с широкими коронками, доминантность последнего зуба
выражена слабо.
Сравнение. От остальных видов рода отличается малой относительной вы
сотой зубной кости, слабой доминантностью последнего зуба.
Распространение. Нижний триас, баскунчакская серия; европейская часть
СССР, Приуралье.
19
Ро Д Microtheledon
Broom, 1936
1905a. Thelegnathus:
Broom, p. 275.
1936a. Micrvtheledon:
Broom, p. 850.
Типовой вид - Thelegnathus
parvus Broom, 1905; нижний триас Южной Аф
рики.
Диагноз, На верхнечелюстной кости 5 зубов, сильно расширенных попереч
но, 1с плоскими коронками.
Видовой состав. Род монотипичный.
Сравнение. От остальных родов подсемейства отличается формой зубов.
Распространение. Нижний триас, зона Cynognathus; Южная Африка, Али-
вал-Нурт.
Род Thelegnathus
Broom, 1905
1905с Thelegnathus:
Broom, p. 274.
Типовой вид - Thelegnathus
browni Broom, 1905; нижний триас Южной Аф
рики.
Диагноз. В верхнечелюстной кости б зубов. Коронки уплощенные, удлинен-
но-йвальные.
Видовой состав. Род монотипичный.
Сравнение. От остальных родов подсемейства отличается формой и числом
зубов верхней челюсти.
Распространение. Нижний триас, (зона Cynognathus; Южная Африка, Али-
вал-Нурт.
Род Myocephalus
Broom, 1936
1936b. Myocephalus:
Broom, p. 391.
Типовой вид - Myocephalus
crassidens Broom, 1936; нижний триас, Южная
Африка.
Диагноз. Череп округло-Треугольный, невысокий, орбиты удлиненные. Пине
альное отверстие овальное. В верхней челюсти 8 двухвершинковых зубов, предпо
следний зуб очень крупный, доминирует.
Видовой состав. Род монотипичный.
Сравнение. От остальных родов подсемейства отличается формой черепа,
числом и формой зубов, Наличием доминирующего зуба в верхней челюсти.
Распространение. Нижний триас, Зона Cynognathus; Южная Африка, Али-
вал-Нурт.
Род Kapes Ivachnenko, 1975
1975. Kapes: Ивахненко, стр.90.
Типовой вид — Kapes
am a enus Ivachnenko, 1974; верхи нижнего триаса;
европейская часть СССР.
Диагноз. Череп с закругленной мордой, в верхней челюсти 5-6 двухвершин
ковых зубов, в нижней — 6, последний зуб нижней челюсти доминирует по размеру.
Видовой состав. Род монотипичный.
Сравнение. От остальных родов подсемейства отличается числом, степенью
дифференциации, формой зубов, наличием доминирующего зуба в нижней челюсти.
Ра спространение. Нижний триас европейской части СССР.
Kapes
amaenus Ivachnenko, 1975
(рис. 20 г, д)
1975. Kapes
am aenus: Ивахненко, стр. 91-92, рис. 2 а, б, в.
20
Голотип - No 3361/2, ПИН; зубная кость; СССР, Коми АССР, пос. JKeiiiapT;
нижний триас.
Материал. Верхнечелюстные и зубные кости.
Диагноз. Нижняя челюсть невысокая в симфизе, зубы удлиненно-конические,
со слабо расширенными коронками, с притуплёнными латеральными буграми.
Распространение. Нижний триас, верхи баскунчакской серии, европейская
часть СССР, Коми АССР.
Род Vitali a Ivachnenko,, 1973
1973а* Vitalia: Ивахненко, стр. 133,
Типовой вид - Vitali 1 grata Ivachnenko,. 1973; нижний триас, европейская
часть СССР'.
Ди агноз. Нижняя челюсть узкая, Зубной край тонкий. Зубы со слабо уплощен
ными коронками, у передних вещество зуба значительно внедряется в кость. Зад
ние зубы акродонтные.
Видовой состав. Род монотипичный.
Сравнение. От остальных родов подсемейства отличается формой и стро
ением зубов.
Распространение. Верхи нижнего триаса европейской части СССР'.
Vitalia grata Ivachnenko,.
1973
(рис. 21б,В,Г,Д)
1973а. Vitalia grata: Ивахненко, стр. 133—144, рис. 2 а, б.
Голотип - No 104/3105, СГУ; зубная кость; СССР, Донская Лука; нижний
триас.
Материал. Часть зубной кости.
Диагноз. Нижняя челюсть невысокая в симфизе, зубы удлиненно-кони
ческие, со слабо расширенными коронками, с притуплёнными латеральными
буграми.
Распространение. Нижний триас, баскунчакская серия; европейская часть
СССР, Донская лука.
ПОДСЕМЕЙСТВО LEPTOPLEURONINАЕ IVACHNENKO, SUBFAM. NOV.
Типовой род - Leptopleuron
Owen,. 1851.
Дь агноз. Специализированные проколофоны с сильно удлиненными назад ор
битами и с шипами на костях щечного комплекса. Зубы дифференцированные, не
многочисленные.
Родовой состав. Три (четыре?) рода.
Сравнение. От остальных подсемейств отличаются наличием длинных шипов
на щечных костях, степенью дифференциации зубов.
Рас прост р ане ни е. Средний и верхний триас; Западная Европа, Северная
Америка, Китай.
Род LeptopleuronOweny
1851
1851. Leptopleuron:
Owen,, p.900 (no Huene< 1912, p. 32).
1867. Telerpeton:
Huxley,, p.77.
1904. Telerpeton: Boulenger,. p.470.
1920. Telerpetonu
Huene,. S.}89.
1925. Leptopleuron:
Williston,, p. 221.
Типовой вид - Leptopleuron
lacertinum
Owen, 1851; средний триас, Шот
ландия.
21
Диагноз. Череп невысокий, яа квадратно-скуловых, скуловых и чешуйчатых
костях развиты шипы, орбиты удлиненные, небо с тремя рядами зубов. В верхней
челюсти 5 зубов, Нижняя челюсть высокая.
Видовой состав. Род монотипичный.
Сравнение. От остальных родов подсемейства отличается относительно
слабым развитием шипов, наличием нёбных зубов.
Распространение. Средний триас, слои Stagonolepis; Шотландия, Элджин.
Род Hypso&iathus
Gilmorei, 1928
1928. Hypsognathus:
Gilmorej, p. 1.
1946. Hypsoffiathus:
Colbert, p. 237*
Типовой вид - Hypsognathus
fenneri Gilmore, 1928; верхний триас, США.
Диагноз. Череп широкий и плоский, На квадратно-скуловых костях очень силь
но развитые шипы. Орбиты сильно удлинены назад, небные зубы отсутствуют. В
верхней челюсти 7 зубов, в нижней - 5, Коронки задних сильно расширены.
Видовой состав. Род монотипичный.
Сравнение. От остальных родов подсемейства отличается сильным разви
тием шипов, сильным удлинением орбит.
Распространение. Верхний триас, серия Ньюарк, формация Брансуик;
США, Нью-Джерси.
Род Paoteodon Chow et Sun, 1960
1960. Paoteodon: Chow, Sun, p. 11.
Типовой вид — Paoteodon huangho en sis Chow et Sun, I960, верхний триас,
Китай.
Диагноз. Зубы в верхней челюсти сильно увеличены, овальные в сечении,
двухвершинковые.
Видовой состав. Род монотипичный.
Сравнение. Насколько можно судить по описанному материалу, очень бли
зок к роду Hypsognathus,
отличаясь от него размерами.
Распространение. Верхний триас; Китай, провинция Шанси.
(?) Род Sphodrosaurus
Colbert, I960
1960. Sphodrosaurus:
Colbert, p. 1.
Типовой вид — Sphodrosaurus,pennsylvanicus
Colbert,'I960;верхний триас
США.
Диагноз. Массивный скелет. Череп, возможно, широкий и плоский.
Сравнение. Единственная удовлетворительно сохранившаяся кость - бедро
мало похожа по облику на бедра проколофонов.
Распространение. Верхний триас, формация Брансуик; США, Пен
сильвания.
СЕМЕЙСТВО SCLEROSAURIDAE NOPCSA, 1923
Типовой род — Sclerosaurus
Meyer, 1857.
Диагноз. Очень крупные проколофоны, череп несет длинные шипы на квад
ратно-скуловых и скуловых костях. Орбиты маленькие, округлые. Зубы дифферен
цированы, многочисленные.
Родовой состав. Один (два?) рода.
Сравнение. От остальных семейств отличаются наличием шипов, малень
кими орбитами и многочисленными зубами.
Распространение. Средний триас Западной Европы.
22
Род Sclerosaurus
Meyer,. 1857
1857. Sclerosauius:
Meyer in Fischer de Waldheim,. p. 136.
1878. Labyrinthodon:
Wiedersheim,. p. 1.
1900. Aristodesmus:
Seeley, p.628*
1902. Sclerosauius:
Huene* p. 23.
Типовой вид - Sc/^fDsauius armatus Meyer, 1857; средний триас Западной
Европы.
Диагноз. Череп с длинными шипами на щечных костях, орбиты маленькие,
округлые. Зубы крупные, дифференцированные.
Распространение. Средний триас, верхний пестрый песчаник; Западная
Европа, Базель.
Род Basiteosaurus
Wiedersheim,. 1870
1971. Вasileosaurus:
Kuhn,. S.9.
Описание почти отсутствует, по Куну (Kuhn,. 1971) не идентичен роду Sclero*
saurus, образец потерян.
Распространение. Средний триас Западной Европы.
ГЛАВА II
СРАВНИТЕЛЬНАЯ МОРФОЛОГИЯ ПРОКОЛОФОНОВ
ЧЕРЕП
Общие данные
Проколофоны, как правило, имеют небольшие размеры. Наиболее мелкие фор
мы — это С ontrito saurus, Myocephalus,
Spondylolestes,
Owenetta.
Размер их чере
пов - 2-4 см. К наиболее крупным формам относятся Sclerosaurus,
Nеорюcolophon,
с черепами длиной 7-10 см.
Одной из наиболее примечательных черт строения черепов проколофонов явля
ются их обычно огромные, удлиненные назад орбиты. Колберт (Colbert,' 1946) и
вслед за ним Ян Чжун-чжен (Young> 1957) придавали большое значение углу
между нормалями к серединам плоскостей орбит (т.е., возможно, положению глаз
ных осей).
В работе Колберта (стр. 244, фиг. 8) приводится сопоставление таких
углов для некоторых форм и делается вывод, что в ходе эволюции шло уменьшение
угла (глаза проколофонов смещались на боковую поверхность). Сопоставление ря
да современных и ископаемых форм не свидетельствует в пользу такого предполо
жения. Чадо полагать, Дело обстояло гораздо проще. У всех форм с плоским телом
и черепом (у гипсогната, у аллигатора и других современных форм, ведущих амфи-
биотический образ жизни) угол между нормалями к плоскостям орбит невелик и
глаза животного направлены вверх. У наземных (реже водных) подвижных форм —
ящериц, высших проколофонин угол гораздо больше. Глаза направлены в стороны.
Даже у весьма примитивного никтифрурета при плоском черепе орбиты сверху
перекрываются нависающими краями предлобных костей и угол между нормалями
намного больше, чем у гипсогната.
Кольцо склеротики обнаружено лишь у никтифрурета (рис. 1в). Оно состоит из
15-18 трапециевидных пластинок, Крупных внизу и более мелких вверху. Это обра
зование интересно тем, что позволяет весьма точно определить размеры глазного
яблока (Edinger,. 1929).
Крыша черепа
Крыша черепа у примитивных форм (никтифруретиды) слабо скульптирована, у
проколофонид, Как правило, скульптура отсутствует, Юна имеется только у рода
Procolophon,
но здесь обычно очень мелка. Хюне (Huene, 1925) отмечает находку
черепа проколофона с очень крупной скульптурой, что может говорить об относи
тельной примитивности этой формы, но в то же время может быть результатом
срастания остердерм с костями. У никтифрурета (и у овенетты: Broom,, 1939) скуль
птура отсутствует на костях, образующих антеро-латеральные края черепа. Сильно
выражена скульптура (по крайней мере, у никтифрурета) на костях, образующих -
медиальный край орбиты.
Передний край черепа образуют дорзальные пластины предчелюстных костей,
никогда не срастающихся у проколофонов. Носовые кости очень широкие; у никти
фрурета (рис. 1), гипсогната (Colbert,-.
1946) и лептоплейрона (Huene,. 1920) они на
висают над ноздрями. У никтифрурета и контритозавра по внутренней поверхности
24
носовой кости от шва с лобной к ноздре идет слабо сужающийся желоб, открыва
ющийся над ноздрей (обычно под нависающей сверху частью кости) крупным опаль
ным отверстием (for. art. nasalis inferior, «рис. 26a), иногда связанным с двумя-
тремя маленькими отверстиями несколько ниже. У более высокоразвитой тихвин
ский (рис. 26а) желоб отсутствует, Имеется лишь несколько небольших отверстий
на верхней поверхности носовой кости (вероятно, для выхода ветвей латерального
этмоидного нерва).
Слезная кость обычно короткая, 'достигает ноздри лишь у примитивных форм
(никтифрурет: рис. 1)^ При этом на поверхности черепа видна узкая полоска кости,
а нижняя часть ее прикрыта снаружи дорзальной пластиной верхнечелюстной кости.
При разрастании вверх дорзальной пластины она перекрывает всю переднюю часть
слезной кости и эта часть редуцируется. Отверстие слезного протока на передней
стенке орбиты двойное. Когда кость идет до ноздри, Желоб протока сопровождает
ее по всей длине, открываясь отверстием в задней стенке ноздри, когда же кость
не достигает ноздри, двойное отверстие лишь прободает переднюю стенку орбиты,
и далее канал протока не прослеживается. Как правило, у проколофонов, даже у
проколофонид собственно, слезная кость длиннее или равна предлобной, так что
наличие короткой кости у примитивных баразавра и овенетты вызывает
удивление.
Предлобная кость ограничивает орбиту с медиального края, заходя весьма да
леко назад, иногда (контритозавр: рис. 2а) смыкаясь здесь с передним краем зад-
нелобной. У никтифрурета латеральный край предлобной кости нависает над орби
той. Задняя часть кости резко отогнута вниз, формируя переднюю часть стенки
орбиты (рис. 96). При этом (по крайней мере, у проколофонин: рис. 96) медиальный
край пластины, изгибаясь сагиттально, образует длинный плоский отросток, заост
ренный кзади. Пластинки обеих предлобных костей не смыкаются по срединной ли
нии. Вероятное назначение отростка - поддерживать хрящевые сфенэтмоидные
структуры (если они имелись) или же он входил в состав мембранной межглазнич
ной перегородки.
Лобные кости прямоугольного очертания, длчнные, вытянутые назад, Как пра
вило, образуют верхний край орбиты, хотя часто на очень небольшом участке. Лишь
у контритозавра (рис. 2 в) смыкаются передне- 'и заднелобные кости, отделяя лоб
ные от верхнего края орбиты.По лобным костям у некоторых форм (тихвинския:
рис. За; гипсогнат, канделярия) идет неглубокая продольная депрессия. На вен
тральной стороне кости гладкие, по латеральным края имеют утолщения, что де
лает их схожими с соответствующими костями ящериц, где вдоль утолщений при
крепляется расширяющаяся кзади межглазничная перегородка.
Заднелобные кости узкие. Срастание этих костей с теменными отмечается
лишь у тихвинский (где это связано, 'скорее всего, с сильно развитым кинетизмом
черепа). В этом случае шовное соединение теменных костей с лобными отмечает
ся лишь в районе проходящих вперед "заднелобных отростков" теменных костей,
а непосредственно теменные кости очень слабо соединены с лобными. Остатки шва
хорошо видны на разломах через эту область, "у буртензии (рис. 4а) срастание
отсутствует.
Пинеальное отверстие очень велико, лежит у передних краев теменных костей.
Теменные кости всегда граничат с заглазничными, так как межвисочная кость,
первично разделяющая их, исчезает. При этом наиболее вероятным представляет
ся срастание ее с окружающими костями, что иногда хорошо прослеживается у
диадекта (у молодых особей имеется свободная межвисочная кость: Чудинов, 1964).
У проколофонов, 'судя по топографии костей, она срастается либо с заглазничной,
либо с теменной. Первый случай наблюдается, например, ty никтифрурета, у которо
го заглазничная кость обходит сзади заднелобную и соединяется с почти квадрат
ной по форме теменной, второй случай — у проколофона, Когда теменная кость
имеет выступ, 'обходящий сзади заднелобную и соединяющийся с небольшой заглаз
ничной. «Характерного для лабиринтодонтов срастания межвисочной и надвисочной'
костей никогда не наблюдается, в-связи с чем вызывает удивление разделение за-
глазничных и теменных костей очень большими надвисочными у овенетты (Broom,
1939) и баразавра (Piveteau, 1955). ]
25
Заглазничная кость у примитивных форм широкая (например, у никтифрурета,
в меньшей степени у проколофона) у высших, типа тихвинский (рис, 3), очень узкая,
практически это уже не сама кость, а лишь тот край, который налегал на
скуловую.
Надвисочная кость большая, часто образует более или менее длинные "рога",
вероятно,служившие для прикрепления части депрессорного мускула. Ее участие
в ограничении ушной вырезки незначительно. Большинство авторов интерпретируют
эту кость как таблитчатую, хотя никаких данных для предположения исчезновения
или срастания межвисочной нет. (Тем более, что остатки таблитчатой кости имеют
ся у никтифрурета (рис. <12) и при сопоставлении этой формы с более высокоразви-'
тыми проколофонидами хорошо видно, что у них задний край крыши образуют имен
но теменные и надвисочные кости. Даже у примитивных ящериц Брум (Broom, 1935)
называет надвисочную кость таблитчатой. Обоснование такой точке зрения дал
Хюне (Huene, 1943)- Он считал, что у диадектоподобных форм надвисочная кость
очень мала, а у проколофонов участвует в образовании ушной вырезки, смещаясь
на задне-боковой край черепа.
Паррингтон (Parrington, 1937), на основании сопоставления окружения таблитча
тых и надвисочных костей у различных групп, делал вывод, что таблитчатые кости
на крыше черепа сохраняются лишь у амфибий и сеймуриаморфов, тогда как у кап-
торинов и терапсид они смещаются на затылок, а у ящериц исчезают. К этому на
до добавить, что у низших диадектоморфов (диадекты, парейазавры) таблитчатые
кости сохраняются, и в ходе дальнейшей эволюции редуцируются на крыше черепа,
не смещаясь на затылочную поверхность. По крайней мере, это наблюдается у наи
более типичных представителей диадектоморфных котилозавров. У других зауро-
морфов, возможно, иногда наблюдается смешение этих костей на затылочную по
верхность. Совершенно особняком стоит лантанозух, рассматриваемый в настоящее
время в составе сеймуриаморфов. Однако с сеймуриаморфами эта форма имеет
очень мало сближающих черт, и, возможно, лантанозух ближе к котилозаврам. У
лантанозуха таблитчатые и заднетеменные кости полностью смещены на затылоч
ную поверхность, почти не будучи редуцированы.
Чешуйчатая кость у примитивных проколофонов принимает небольшое участие в
построении дорзальной поверхности крыши черепа, а основная часть кости уходит
на постеролатеральную поверхность, неся пологую вогнутость — район прикрепле
ния барабанной перепонки. У прогрессивных форм (тихвинския, возможно, коило-
скиозавр: Huene, 1912) кость вообще не выходит на дорзальную поверхность черепа.
На шве между квадратной и квадратно-скуловой костями имеется небольшое пара-
квадратное отверстие.
Скуловая кость в эволюции проколофонов постепенно изгибается кверху, огра
ничивая совместно с квадратно-скуловой и верхнечелюстной костями конструкцию,
внешне сходную с нижней височной вырезкой ящериц. Квадратно-скуловая кость
сильно редуцированна. Однако это наблюдается лишь у проколофонов Русской плат
формы (и, может быть, у некоторых форм Западной Европы - аномоиодон: Huene,
1939). У рода Procolophon
на квадратно-скуловых костях появляются довольно длин
ные отростки, идущие назад и латерально. Броили и Шредер (Broili, Schroder, 1936,
стр. 241, фиг. 1) показывают на серии рисунков, как по мере роста животного уве
личиваются в размерах отростки. Хюне (Huene, 1920) описывает у лептоплейрона
шипы на надвисочных, чешуйчатых, квадратно-скуловых и скуловых костях. "Шип"
на надвисочной кости представляет ее задний, нависающий край, служивший для
прикрепления депрессора,шип на чешуйчатой принадлежит, скорее всего (судя по
его положению^квадратно-скуловой. Таким образом, шипы у лептоплейрона досто
верно имеются лишь на скуловой (небольшой выступ) и квадратно-скуловой (два
больших шипа) костях. Наибольшей степени развития шипы достигают у гипсогната.
У него имеется три направленных латерально и один вперед очень длинных шипа на
квадратно-скуловой кости. Скорее всего, расположение шипов у склерозавра почти
такое же, может быть, менее развит шип на скуловой кости и перед
ний на квадратно-скуловой, тогда как задний шип очень велик и слегка
направлен назад.
26
Орбита
Строение передней стенки орбиты изучено лишь у никтифрурета, контритозаира,
буртензии и тихвинский. У всех этих форм она построена почти идентично. Степку
слагают изгибающиеся вентрально пластины предлобной, слезной и дорзальная пла
стина небной костей. Вентральный отросток предлобной кости образует изогнутую
сагиттально узкую пластинку. Насколько можно судить, у капторина (Fox, Bowman,
1966, стр. 7, фиг. 5) передняя стенка намного уже, небная кость позади нее резко
приподнята вверх, пластинки предлобных костей отсутствуют. У проколофонов на
границе смыкания отростков предлобной и небной костей (рис. 96) имеется неболь
шое отверстие. У некоторых современных агамид {Agama) есть такое же отверстие,
пропускавшее интермедиальную артерию в носовую капсулу, тогда как у других
(Phrynocephalus)
стенка орбиты гораздо уже, поэтому отверстие отсутствует.
В нижне-боковом углу передней стенки орбиты находится крупное инфраорбиталь-
ное отверстие, в верхне-боковом - двойное отверстие слезного протока. Несколько
позади стенки орбиты, по дорзальной поверхности небной кости, к инфраорбиталь-
ному отверстию идет небольшой желобок от входного отверстия небной артерии
(возможно, здесь шла ветвь постхоанного анастомоза).
Функциональные аналоги височных окон
Аналогом верхнего окна является задняя часть орбиты. Удлинение орбиты на
чалось с простого разрастания ее назад, наблюдающегося уже у никтифруретид.
Возможно, о разрастании крыши могут свидетельствовать удлиненные лобные ко
сти. Кроме того, кости, ограничивающие орбиту сзади, у примитивных форм до
статочно большие и редуцированы крайне слабо. Лишь на уровне высших спонди-
лолестин началась интенсивная редукция костей, ограничивающих сзади орбиту.
Максимума редукция достигла у высших проколофонин (например, у тихвинский).
У них часто заглазничная кость представляет собой тонкую пластинку, почти пол
ностью лежащую на скуловой кости - скорее всего, это лишь задний край кости.
Такое удлинение орбиты привело к появлению "орбито-скулового" окна (Чудинов,
Вьюшков, 1956). По положению задняя часть этого отверстия может быть сопо
ставлена с верхним височным окном лепидозавров. Как будет показано при рас
смотрении краниальных мускулов, функционально оно, скорее всего, могло соот
ветствовать верхнему височному окну.
Можно полагать, что формирование аналогичной структуры у лептоплейронин
шло несколько иначе, по крайней мере, гипсогнат имеет очень сильно удлиненную
назад орбиту, не менее развитую, чем, например, у тихвинский, однако столь же
значительной редукции посторбитальных костей не наблюдается.
Нижнее височное окно формировалось, скорее всего, независимо в раз
личных линиях зауроморфных рептилий. У болозавроморфов имеется небольшое
окно, окруженное скуловой,, чешуйчатой и квадратно-скуловой костями. У прими
тивных диапсид (миллеретта) появляется .дель между скуловыми, чешуйчатыми и
заглазничными костями, и в дальнейшем образуется у форм типа сфенодона или
янинии в этом месте окно. На уровне прайсии и близких к ней форм (например, про-
лацерта: Parrington, 1935) происходит интенсивное разрастание окна вниз, некото
рая редукция скуловой кости и значительная — квадратно-скуловой. В результате
у высших диапсид образуется открытая снизу вырезка, ограниченная скуловой, за-
глазничной и (после окончательной редукции квадратно-скуловой) квадратной ко
стями, служащая для фиксации части аддукторной мускулатуры. Вероятно, имен
но в связи с усилением аддукторной мускулатуры у проколофонов на уровне спон-
дилолестин начинается резкое изгибание вверх и частичная редукция скуловой
кости и значительная редукция квадратно-скуловой.
Таким образом, не проходя стадии окна, у высших проколофонов образуется
структура, очень близкая по форме, и, вероятно, по функции нижней височной вы
резке ящериц. Может быть, именно такая форма связана с кинетизмом черепа, по
скольку у рода Procolophon
с почти акинетичным черепом началось образование
структуры, но далеко не зашло, и стали развиваться шипы на квадратно-скуловой
27
кости. У акинетичных лептоплейронин, вероятно, местами прикрепления мускулов
являлись мощные гребни у оснований шипов. И лишь у тихвинский с развитым ки-
нетизмом черепа образовалась структура, полностью сходная внешне со структу
рой у ящериц. У капторинид полностью отсутствует сколько-нибудь заметный из
гиб скуловых костей.
Нёбо
Нббо проколофонов представляет собой сплошную пластинку, в области перед
ней стенки орбит отогнутую вниз (по задним краям сошников и передним краям
нёбных костей). Кости небного комплекса обычно несут зубы. Костные хоаны
обычно узкие и длинные, лежат в неглубоких вдавлениях. Лишь у лептоплейрона,
•овенетты (Broom, 1939, Huene, 1912) изображают небольшие округлые хоаны.
Нёбо изучено далеко не у всех проколофонов, а описание костей небного ком
плекса с дорзальной стороны не приводилось вообще. Из всех котилозавров изо
бражение дорзальной поверхности неба приведено лишь Уотсоном (Watson, 1914b,
стр. 160, фиг. 3) для Pareiasaurus
bombidens,
однако это изображение не лишено
ряда серьезных погрешностей. Так, птеригоиды показаны продолжающимися по
медиальной линии до предчелюстных костей, не показаны желоба максиллярной
артерии и отверстия ряда других краниальных сосудов. Поэтому представляется
необходимым дать полное описание строения неба для всех изученных проколофо
нов и парейазврида Pareiasuchus
(рис. 6-9,а).
Переднюю часть неба образует вентральная пластина парной предчелюстной
кости. На вентральной поверхности пластины, на шве между костями, имеется
продольный узкий желоб, идущий от передних зубов к шву с сошниками (возможно,
для гомолога межчелюстной железы амфибий и кистеперых). В передней части
пластины имеются небольшие анастомозные отверстия медиального этмоидного
нерва.
Передние края сошников округлыми отростками налегают сверху на задние
края предчелюстных костей. Медиальные края сошников сильно утолщены, образуя
основание для межносовой перегородки. В центральной части кости это утолще
ние обычно расширяется, и в толще расширения идет канал, начинаясь у шва сош
никовой и небной костей, и кончаясь у шва сошниковой и предчелюстной костей.
Кроме того, имеется небольшое отверстие сбоку, в передней части расширения.
От этого отверстия к медиальному краю хоаны идет небольшой желобок, оканчи
вающийся иногда (тихвинския) отверстием у края кости. Надо полагать, что ка
нал в толще кости предназначался для небной артерии и нерва, а вбок отходила
ветвь предхоанного анастомоза, поскольку к противоположному краю хоаны точно
напротив подходит небольшое ответвление от желоба максиллярной артерии. У па-
рейазуха нет канала в толще кости (рис. 6), и артерия шла по поверхности. Одна
ко желоб и небольшая вырезка в районе анастомоза имеются.
От входного отверстия небной артерии назад, по краю межптеригоидной вырез
ки, идет слабо выраженный желоб, сопровождающийся частыми ответвлениями,
уходящими в толщу крыловидной кости. Возможно, сюда отходили небольшие ве
точки, питавшие небные зубы.
Межптеригоидная вырезка полностью образована птеригоидами, узкими отрост
ками смыкающимися впереди нее. На латеральных краях птеригоидов часто име
ются слабо выраженные гребни - возможно, места прикрепления птеригоидной ча
сти аддукторов. У примитивных форм (никтифрурет) птеригоидный фланг почти
полностью образован птеригоидом, развит слабо и несет зубы. У высших проколо
фонов фланг развит очень сильно, представляет собой треугольный вентральный
отросток, опускающийся вниз на расстояние, равное почти трети высоты черепа.
Фланг образован пластинами птеригоида и эктоптеригоида. Такое сильное разви
тие птеригоидного фланга характерно . шь для наиболее высокоразвитых проко
лофонов - тихвинский, гипсогната. У рода Procolophon (Seeley, 1905) фланг раз
нит слабее. У типичных ящериц фланг также развит не сильно, хотя у игуамид он
опускается довольно далеко вниз, отличаясь, однако, но строению, Лишь некото
рые агам иды {Agama, Phrynocephalus)
имеют построенный аналогично достаточно
28
теп
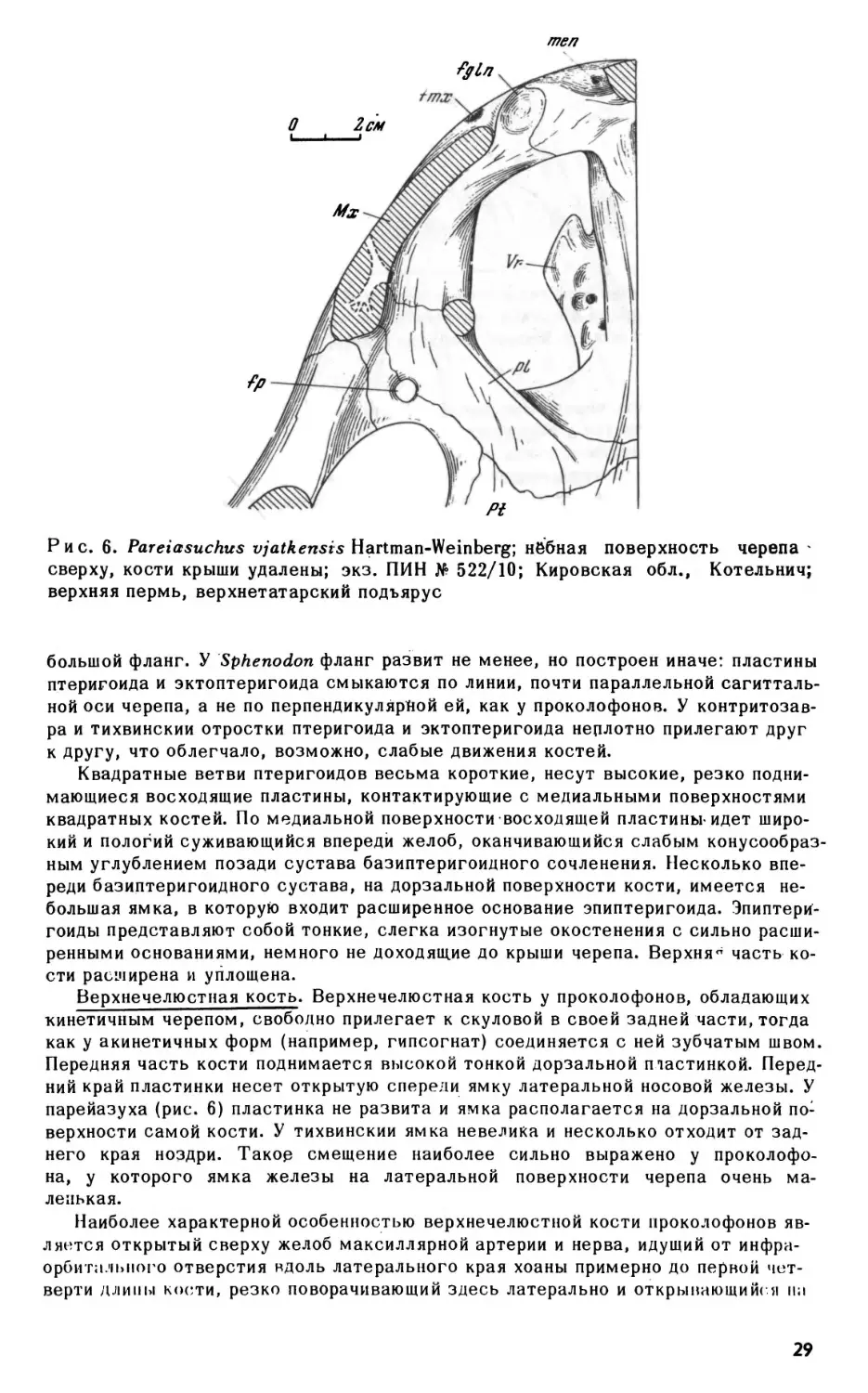
Рис. 6. Pareiasuchus
vjatkensis
Hartman-Weinberg; нёбная поверхность черепа -
сверху, кости крыши удалены; экз. ПИН No 522/10; Кировская обл.,
Котельнич;
верхняя пермь, верхнетатарский подъярус
большой фланг. У Sphenodon фланг развит не менее, но построен иначе: пластины
птеригоида и эктоптеригоида смыкаются по линии, почти параллельной сагитталь
ной оси черепа, а не по перпендикулярной ей, как у проколофонов. У контритозав-
ра и тихвинский отростки птеригоида и эктоптеригоида неплотно прилегают друг
к другу, что облегчало, возможно, слабые движения костей.
Квадратные ветви птеригоидов весьма короткие, несут высокие, резко подни
мающиеся восходящие пластины, контактирующие с медиальными поверхностями
квадратных костей. По медиальной поверхности восходящей пластины-идет широ
кий и пологий суживающийся впереди желоб, оканчивающийся слабым конусообраз
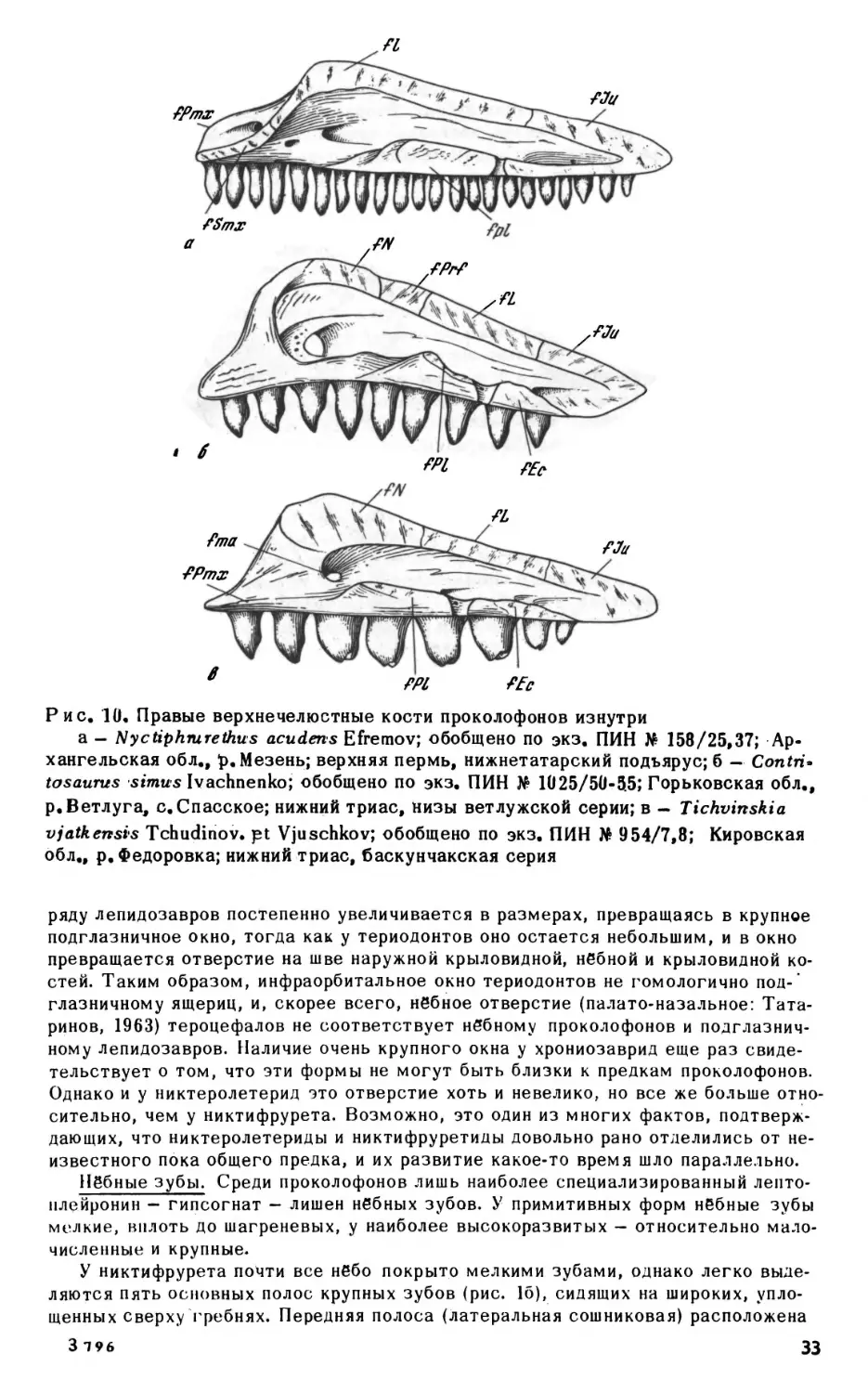
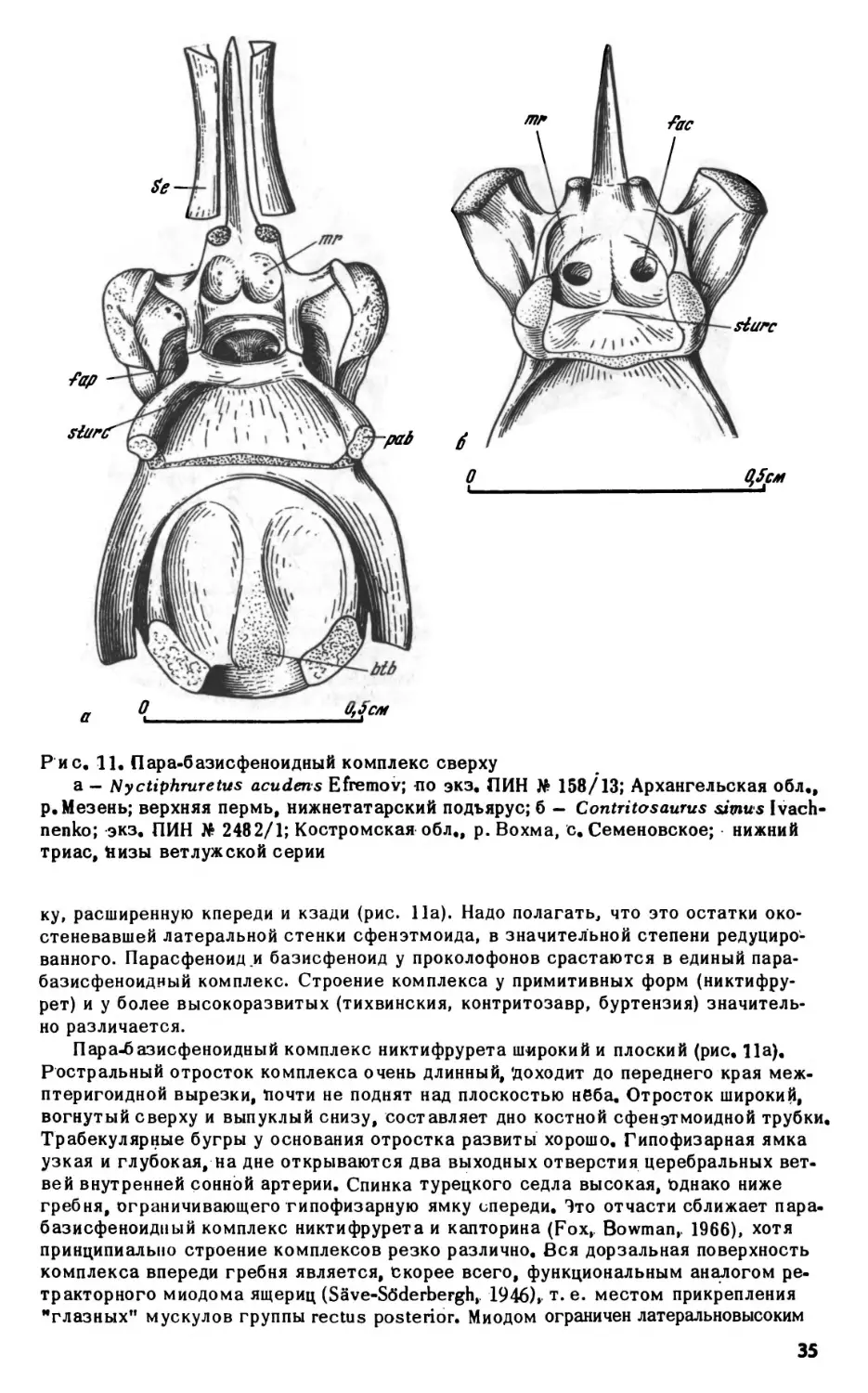
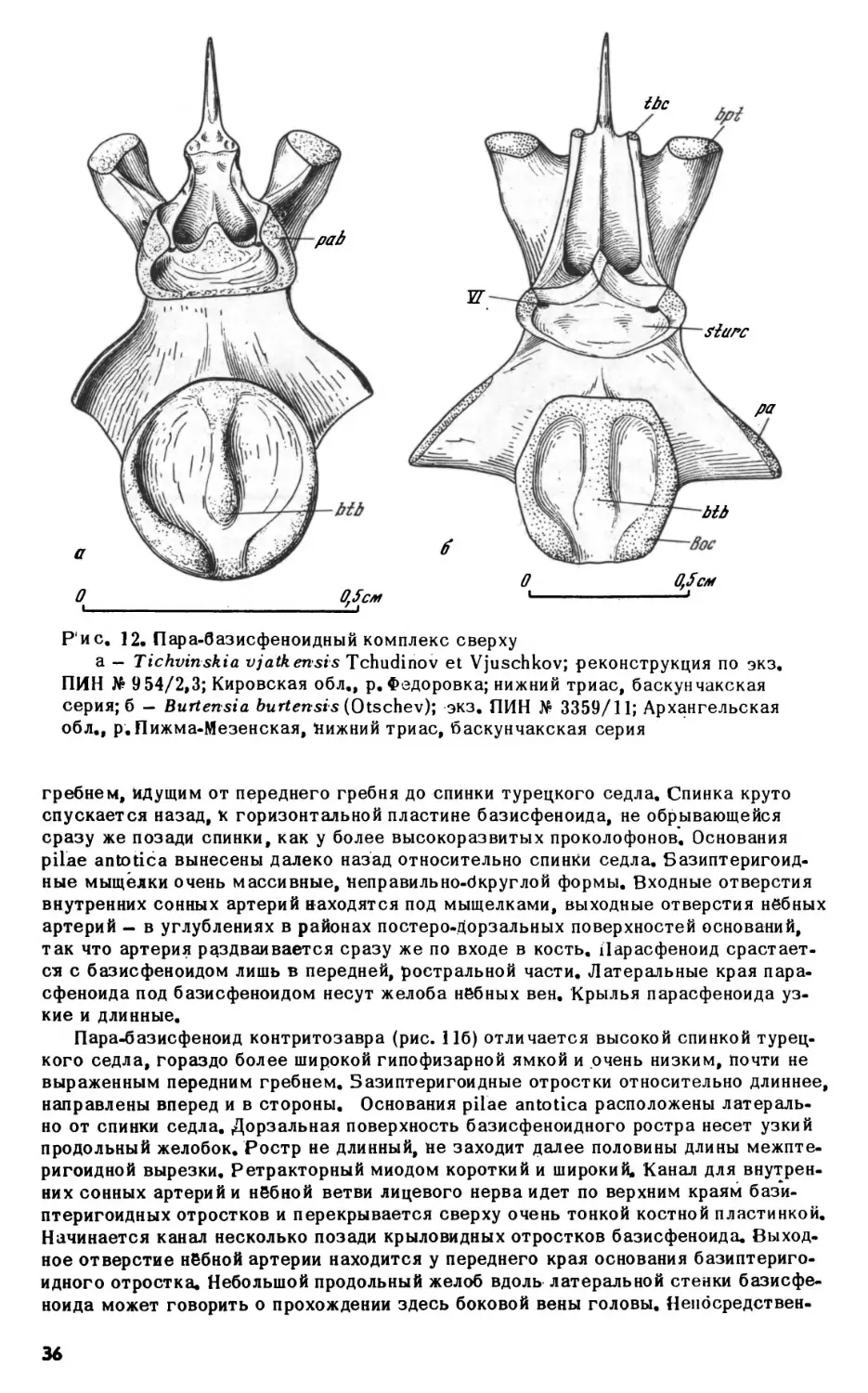
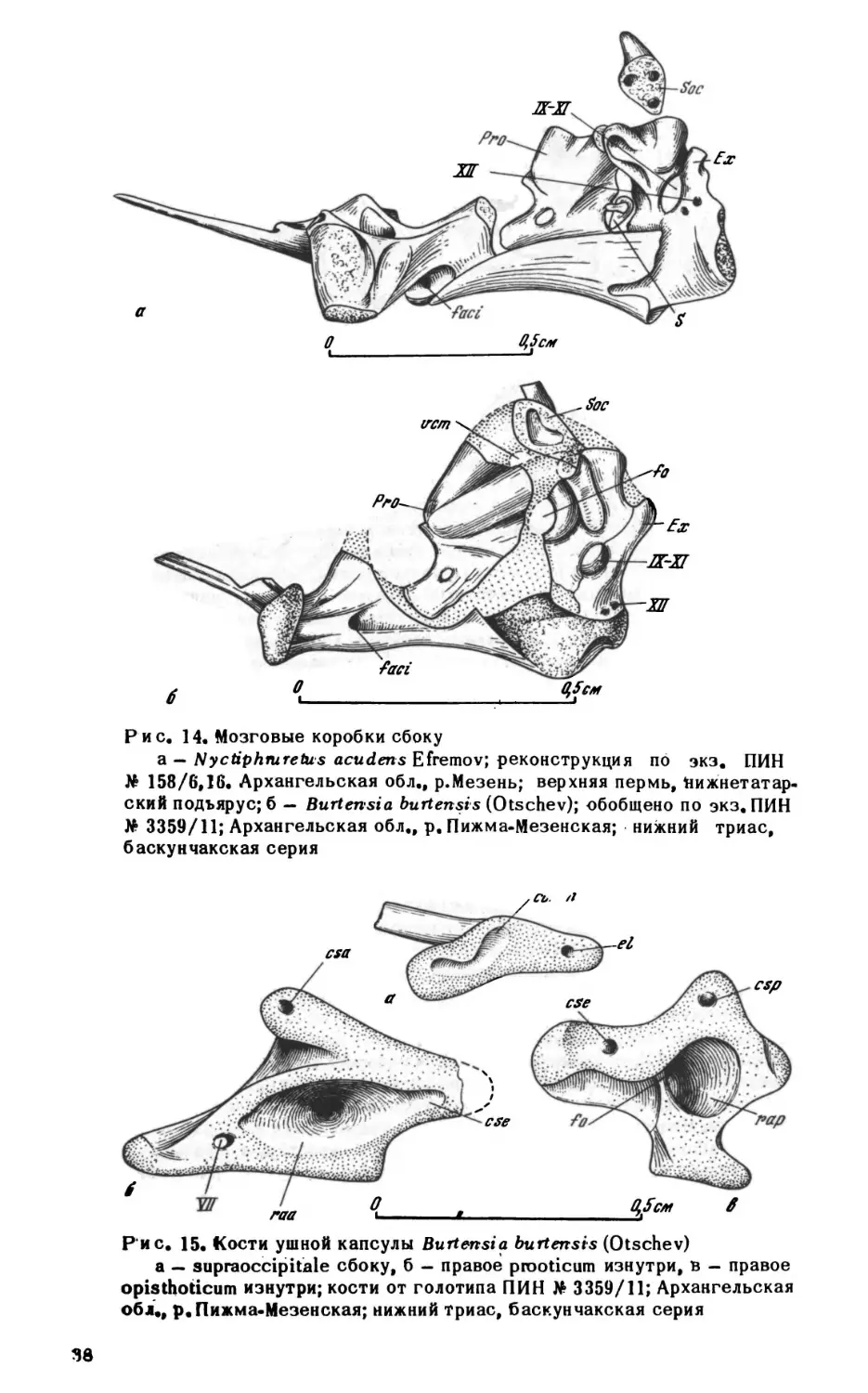
ным углублением позади сустава базиптеригоидного сочленения. Несколько впе
реди базиптеригоидного сустава, на дорзальной поверхности кости, имеется не
большая ямка, в которую входит расширенное основание эпиптеригоида. Эпиптери-
гоиды представляют собой тонкие, слегка изогнутые окостенения с сильно расши
ренными основаниями, немного не доходящие до крыши черепа. Верхня" часть ко
сти рас1чирена и уплощена.
Верхнечелюстная кость. Верхнечелюстная кость у проколофонов, обладающих
кинетичным черепом, свободно прилегает к скуловой в своей задней части, тогда
как у акинетичных форм (например, гипсогнат) соединяется с ней зубчатым швом.
Передняя часть кости поднимается высокой тонкой дорзальной пластинкой. Перед
ний край пластинки несет открытую спереди ямку латеральной носовой железы. У
парейазуха (рис. 6) пластинка не развита и ямка располагается на дорзальной по
верхности самой кости. У тихвинский ямка невелика и несколько отходит от зад
него края ноздри. Такое смещение наиболее сильно выражено у проколофо-
на, у которого ямка железы на латеральной поверхности черепа очень ма
ленькая.
Наиболее характерной особенностью верхнечелюстной кости проколофонов яв
ляется открытый сверху желоб максиллярной артерии и нерва, идущий от инфра-
орбиталмюго отверстия вдоль латерального края хоаны примерно до первой чет
верти длины кости, резко поворачивающий здесь латерально и открывающийся ил
29
О
1см
|
i
Рис. 7. Nyctiphmretus
acuderts Efremov; передняя часть нёбной поверхности чере
па сверху, кости крыши и правая септомаксилла удалены; по экз. ПИН No 158/25,
13; Архангельская обл., р.Мезень; верхняя пермь, нижнетатарский подъярус
боковую поверхность черепа (foramen maxillaris anterior, рис. 10а-б). Переднее
максиллярное отверстие у большинства форм одинарное, однако у контритозавра
оно, как правило, двойное,, очень редко это наблюдается и у тихвинский. Желоб
артерии широкий и пологий, вероятно, здесь, кроме артерии и нерва, шла вена, и
лежал край хрящевой носовой капсулы. В районе инфраорбитального отверстия, в
небольшой депрессии, образуемой желобом постхоанного анастомоза, от артерии
отходило ответвление, шедшее в толщу верхнечелюстной кости и здесь развет
влявшееся вперед и назад, образуя цепь более или менее сильно развитых пустот
в толще кости.
Обычно в этом же районе находится заднее максиллярное отвер
стие для выхода артерии на латеральную поверхность черепа, связанное каналом
с одной из пустот-лакун в толще кости. У современных ящериц в толще кости идут
два хорошо сформированных канала — сместившийся внутрь кости желоб максил
лярной артерии и соединяющийся с ним в передней части кости канал, образован
ный из слившихся лакун (рис. 28ж). У примитивного никтифрурета лакуны неболь
шие, соединены друг с другом тонкими капиллярными канальцами. У контритозав
ра и тихвинский начинается постепенное слияние лакун, наибольшего выражения
этот процесс достигает у макрофона. Однако у проколофонов не происходит сли
яния обоих каналов, как у ящериц. Именно в дне этих лакун * каналов имеются от
верстия капиллярных канальцев, ведущих в полость пульпы зубов.
Примерно на середине длины желоба максиллярной артерии от. него отходит ме
диально и несколько вперед к краю хоаны небольшой желобок, как правило, подхо
дящий к краю точно напротив желобка ветви предхоанного анастомоза нёбной ар
терии и нерва.
В передней стенке желоба, в месте поворота его латерально, у никтифрурета
имеется канал (рис. 10а), ведущий в депрессию латеральной носовой железой, а
сам желоб артерии поворачивает латерально не под прямым углом, а полого и про
ходит небольшой путь в толще кости, прежде чем открыться передним максилляр-
ным отверстием. У контритозавра желоб поворачивает круто, под прямым углом,
и в передней стенке желоба от него отходят один-два очень тонких канала, откры
вающиеся в депрессию железы. У тихвинский поворот желоба очень крутой, кана
ла в толще кости он практически не образует, ответвлений в латеральную же
лезу нет.
Таким образом, эта деталь строения схожа у никтифрурета и ящериц, у кото
рых короткий желоб полого уходит в кость, открываясь на латеральную поверх
ность несколькими отверстиями и давая крупное ответвление в депрессию железы
(foramen alveolaris inferior: Oelrich, 1956). Однако у никтифрурета канальцы, про
низывающие максиллу, отходят непосредственно от желоба, и именно они образу
ют лакуны в толще кости. Надо отметить, что у примитивного парейазуха желоб
максиллярной артерии очень пологий и широкий, также образует несколько впере
ди инфраорбитального отверстия пронизывающий макоиллу канал, от которого от-
30
о
I—
Рис. 8. С ontrito saurus simus Ivachnenko; нёбная поверхность черепа сверху, кости
крыши удалены; экз. ПИН No 2890/5. Горьковская обл., р. Молома, д. Касьяново;
нижний триас, низы ветлужской серии
ходят разветвляющиеся канальцы вперед и назад, однако-лакун в кости не образу
ется. Желоб предхоанного анастомоза у этой формы довольно широкий, находится
напротив анастомазной вырезки на переднем краю сошника. Переднее максилляр
ное отверстие большое, одинарное. Ямка железы расположена открыто впереди
желоба. У болозавра Beleb'ey желоб артерии очень пологий и широкий, под острым
углом уходит в толщу кости и проходит в ней довольно длинный путь. Однако
строение максиллы в целом схоже с таковым проколофонов, тогда как у некото
рых сеймуриаморфов такого сходства не наблюдается. К сожалению, не предста
вилась возможность изучить строение верхнечелюстной кости типичных сеймурид.
У сильно уклоняющихся хрониОзаврид (Chroniosuurus
dongusensis)
на дорзальной
поверхности максиллы нет никаких следов-желоба, поверхность ее покатая. Одна
ко в толще кости идет широкий, хорошо сформированный канал, открывающийся в
задней стенке ноздри большим отверстием и выходящий на боковую поверхность
рядом мелких отверстий. У никтеролетерид (Nyctiboetus)
желоб на дорзальной по
верхности верхнечелюстной кости выражен хорошо и имеет строение, близкое к
строению желоба у проколофонов.
Септомаксиллярная кость. У рода Procolophon
отмечают (Broili, Schroder, 1936;
Watson, 1914b) септомаксиллу, находящуюся в поверхностном положении, образу
ющую заднюю стенку ноздри. Конкретные данные о ее строении отсутствуют. Из
пермских проколофонов собственно септомаксилла обнаружена лишь у никтифру
рета, и уже у этой примитивной формы она полностью смещена в носовую полость,
однако еще не потеряла связь с передним краем верхнечелюстной кости. Верхне
челюстная кость образует горизонтально нависающий над передней частью кост
ной хоаны фланг, к которому и причленяется септомаксилла. Септомаксилла ле
жит горизонтально, медиальным краем налегая на возвышение дорзальной поверх
ности сошника. Между септомаксиллой и сошником остается обширное простран
ство, вмещавшее, безусловно, Якобсонов орган. Дорзальная поверхность септо-
максиллы продольно вогнута, и совместно с вогнутостью дорзальной поверхности
фланга верхнечелюстной кости образует широкий и пологий желоб, на котором ле
жала часть латеральной носовой железы.
У контритозавра (рис. 106) несколько впереди поворота максиллярного желоба
на латеральную поверхность черепа имеется небольшое полукруглое вдавление с
шероховатой поверхностью. Очень похожее образование имеется у современных
31
а
0
1см
i
I
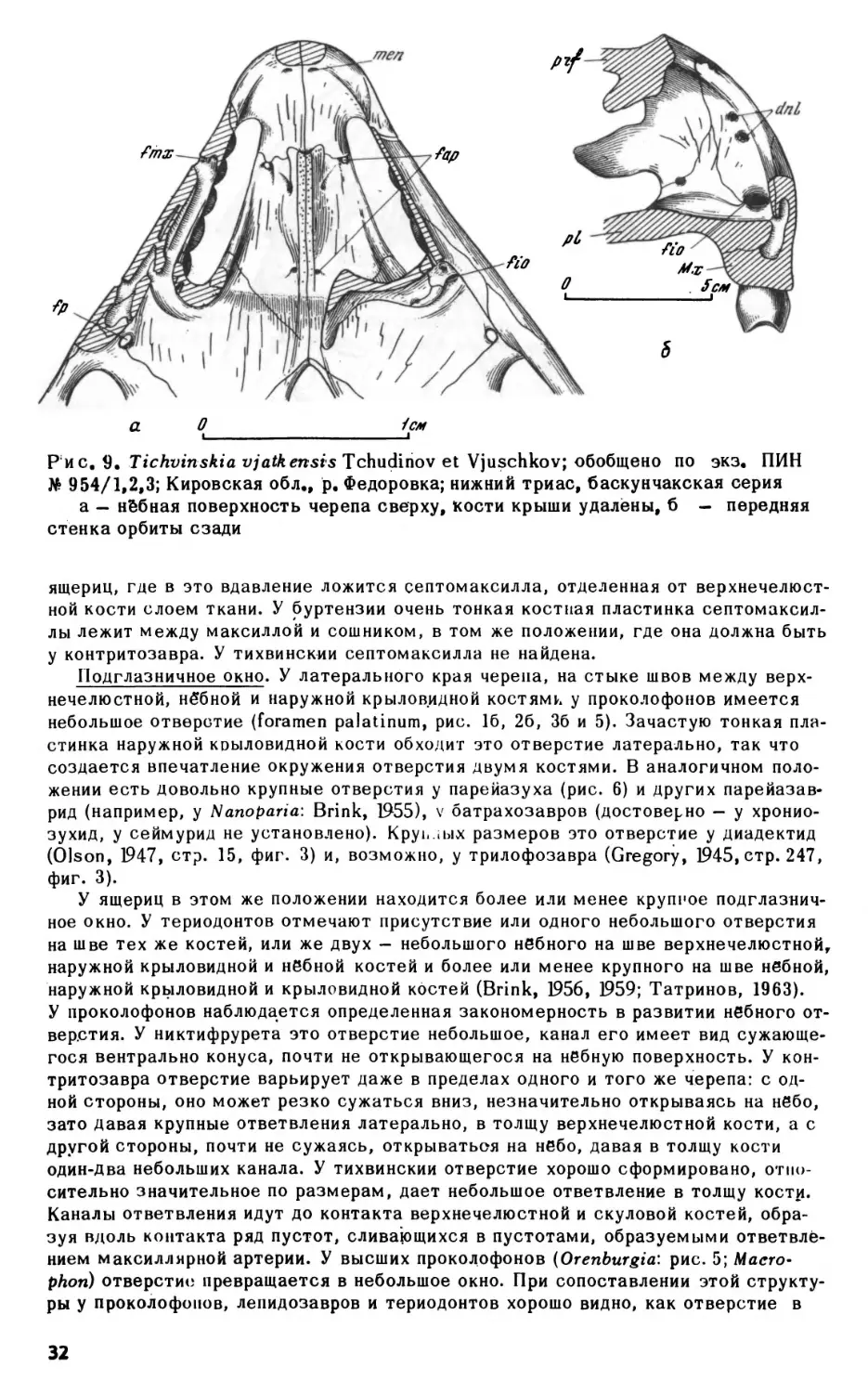
Рис. 9. Tichvinskia
vjatkensis
Tchudinov et Vjuschkov; обобщено по экз. ПИН
No 954/1,2,3; Кировская обл., р. Федоровка; нижний триас, баскунчакская серия
а — небная поверхность черепа сверху, кости крыши удалёны, б
—
передняя
стенка орбиты сзади
ящериц, где в это вдавление ложится септомаксилла, отделенная от верхнечелюст
ной кости слоем ткани. У буртензии очень тонкая костная пластинка септомаксил-
лы лежит между максиллой и сошником, в том же положении, где она должна быть
у контритозавра. У тихвинский септомаксилла не найдена.
Подглазничное окно. У латерального края черепа, на стыке швов между верх
нечелюстной, нёбной и наружной крыловидной костями у проколофонов имеется
небольшое отверстие (foramen palatinum, рис. 16, 26, 36 и 5). Зачастую тонкая пла
стинка наружной крыловидной кости обходит это отверстие латерально, так что
создается впечатление окружения отверстия двумя костями. В аналогичном поло
жении есть довольно крупные отверстия у парейазуха (рис. 6) и других парейазав-
рид (например, у Nanoparia:
Brink, 1955), v батрахозавров (достоверно - у хронио-
зухид, у сеймурид не установлено). Круьлых размеров это отверстие у диадектид
(Olson, 1947, стр. 15, фиг. 3) и, возможно, у трилофозавра (Gregory, 1945, стр. 247,
фиг. 3).
У ящериц в этом же положении находится более или менее крупное подглазнич
ное окно. У териодонтов отмечают присутствие или одного небольшого отверстия
на шве тех же костей, или же двух — небольшого небного на шве верхнечелюстной,
наружной крыловидной и нёбной костей и более или менее крупного на шве небной,
наружной крыловидной и крыловидной костей (Brink, 1956, 1959; Татринов, 1963).
У проколофонов наблюдается определенная закономерность в развитии нёбного от
верстия. У никтифрурета это отверстие небольшое, канал его имеет вид сужающе
гося вентрально конуса, почти не открывающегося на нёбную поверхность. У кон
тритозавра отверстие варьирует даже в пределах одного и того же черепа: с од
ной стороны, оно может резко сужаться вниз, незначительно открываясь на нёбо,
зато давая крупные ответвления латерально, в толщу верхнечелюстной кости, а с
другой стороны, почти не сужаясь, открываться на нёбо, давая в толщу кости
один-два небольших канала. У тихвинский отверстие хорошо сформировано, отно
сительно значительное по размерам, дает небольшое ответвление в толщу кости.
Каналы ответвления идут до контакта верхнечелюстной и скуловой костей, обра
зуя вдоль контакта ряд пустот, сливающихся в пустотами, образуемыми ответвле
нием максиллярной артерии. У высших проколофонов (Orenburgia:
рис. 5; Macro
phon) отверстие превращается в небольшое окно. При сопоставлении этой структу
ры у проколофонов, лепидозавров и териодонтов хорошо видно, как отверстие в
32
fl
Рис. 10. Правые верхнечелюстные кости проколофонов изнутри
а - Nyctiphrurethus
acudens Efremov; обобщено по экз. ПИН U 158/25,37; Ар
хангельская обл., р.Мезень; верхняя пермь, нижнетатарский подъярус; б — Contri*
tosaurus
simus Ivachnenko; обобщено по экз. ПИН No 1025/50-55; Горьковская обл.,
р.Ветлуга, с.Спасское; нижний триас, Низы ветлужской серии; в —
Tichvinskia
vjatkensis
Tchudinov. pt Vjuschkov; обобщено по экз. ПИН No 954/7,8; Кировская
обл., р.Федоровка; нижний триас, баскунчакская серия
ряду лепидозавров постепенно увеличивается в размерах, превращаясь в крупное
подглазничное окно, тогда как у териодонтов оно остается небольшим, и в окно
превращается отверстие на шве наружной крыловидной, небной и крыловидной ко
стей. Таким образом, инфраорбитальное окно териодонтов не гомологично под
глазничному ящериц, и, скорее всего, небное отверстие (палато-назальное: Тата-
ринов, 1963) тероцефалов не соответствует небному проколофонов и подглазнич
ному лепидозавров. Наличие очень крупного окна у хрониозаврид еще раз свиде
тельствует о том, что эти формы не могут быть близки к предкам проколофонов.
Однако и у никтеролетерид это отверстие хоть и невелико, но все же больше отно
сительно, чем у никтифрурета. Возможно, это один из многих фактов, подтверж
дающих, что никтеролетериды и никтифруретиды довольно рано отделились от не
известного пока общего предка, и их развитие какое-то время шло параллельно.
Нёбные зубы. Среди проколофонов лишь наиболее специализированный леито-
плейронин - гипсогнат - лишен нёбных зубов. У примитивных форм нёбные зубы
мелкие, вплоть до шагреневых, у наиболее высокоразвитых — относительно мало
численные и крупные.
У никтифрурета почти все нёбо покрыто мелкими зубами, однако легко выде
ляются пять основных полос крупных зубов (рис. 16), сидящих на широких, упло
щенных сверху гребнях. Передняя полоса (латеральная сошниковая) расположена
3196
зз
на антеро-латеральном краю сошника, она идет вдоль медиального края костной
хоаны и в передней части сошника изгибается медиально, встречаясь с аналогич
ной полосой второго сошника. Крупные зубы здесь расположены по два-три в ряд,
между ними располагаются мелкие зубики. Вторая полоса (небная) идет по меди
альному краю нёбной кости назад, несколько заходя на птеригоид. Третья полоса
(медиальная птеригоидная) идет вдоль медиального шва между сошниками, пере
ходит на крыловидную кость, продолжаясь вдоль медиальных краев межптеригоид-
ной вырезки. В передней части эту полосу составляют мелкие, очень многочислен
ные зубики, в районе межптеригоидной вырезки - более крупные, по два в ряд.
Медиальные птеригоидные ряды заходят в область базиптеригоидного сочленения.
Четвертый ряд (эктоптеригоидный) идет косо, пересекая границу птеригоида и эк-
птеригоида. Основная часть ряда на эктоптеригоиде, его составляют довольно
крупные зубы, сидящие по 1—2 в ряд. Последняя полоса находится на птеригоидном
фланге, составлена крупными зубами по одному в ряд. Вдоль границы птеригоида
и эктоптеригоида идут параллельно последней полосе 2—3 небольших ряда мелких
зубов, сидящих не на валиках, а непосредственно на поверхности кости. Аналогич
ные ряды идут также параллельно эктоптеригоидному ряду. Удивительно, что у
такой примитивной формы, как овенетта, не отмечают наличия небных зубов (кро
ме рядов вдоль межптеригоидной вырезки).
В ряду развития проколофонов Русской платформы хорошо прослеживается
судьба всех полос небных зубов. Вероятно, первыми исчезали зубы промежуточ
ных рядов, а число зубов в крупных рядах резко сократилось. Наряду с этим шло
увеличение размеров самих зубов. Так, у контритозавра лишь первый, сошниковый
ряд двойной, зубы довольно крупные, их число невелико <рис. 26). В дальнейшем
идет редукция именно переднего сошникового ряда. У тихвинский (рис. 36) этот
ряд одинарный и короткий, у оренбургии (рис. 5) от ряда остается всего два зуба,
и оба сошниковых ряда сливаются в один полукруглый ряд из четырех крупных зу
бов. Наиболее постоянными остаются ряды вдоль межптеригоидной вырезки, со
храняющиеся так же у некоторых современных ящериц.
О строении зубных рядов проколофонов иных регионов судить почти невозмож
но, так как в описаниях они не упоминаются, а на рисунках изображаются весьма
схематично. Если судить по фотографии в работе Сили (Seeley, 1905), ряды неб
ных зубов у рода Procolophon
отличаются от таковых у близких родов Русской
платформы. В частности, отсутствует медиальный сошниковый ряд, а передний
сошниковый двойной. Ряды на небных костях короткие, по 3—4 зуба, хотя у Броили
и Шредера (Broili, Schroder, 1936) изображены длинные ряды. У западноевропей
ских родов (Koiloskiosaurus,
Leptopleuron:
Huene, 1912) имеются длинные ряды,
возможно, соответствующие слившимся небным и сошниковым рядам. Надо отме
тить, что при всем разнообразии расположения зубов на небе у проколофонид в
общем наблюдается довольно схожая картина, ни в коей мере не напоминающая
расположение зубов у парейазавров. Скорее хорошо выраженные у всех парейа
завров . два медиальных ряда похожи на параллельные ряды небных зубов диа-
декта.
Мозговая коробка
Мозговая коробка изучена лишь у проколофонов Русской платформы (Nycti-
phruretus,
Contritosaurus,
Tichvinskia,
Burtensia). До сих пор в мировой литерату
ре отсутствуют данные по строению мозговой коробки у остальных проколофонов,
кроме скудных сведений о наружном виде этой структуры. Поэтому представляет
ся необходимым привести результаты изучения строения этой области у всех изу
ченных форм. Краткое описание прохождения каналов сонной артерии в парабази-
сфеноидном комплексе контритозавра дано М.А.Шишкиным (Shishkin, 1968, стр. 10,
фиг. 4d—е: "Phaanthosaurus"), строение мозговой коробки тихвинский приведено
в работе М.Ф.Ивахненко (19736).
Мозговая коробка проколофонов окостеневает гораздо слабее, чем у ящериц.
Наиболее сильные-окостенения наблюдаются у никтифрурета.
Сфенэтмоидные структуры имеются лишь у никтифрурета, где они представ
ляют собой составленную из двух несмыкающихся половинок тонкую костную труб-
34
а
0
МTM
Рис. 11. Пара-базисфеноидный комплекс сверху
а - Nyctiphruretus
acudens
Efremov; по экз. ПИН No 158/13; Архангельская обл.,
р.Мезень; верхняя пермь, нижнетатарский подъярус; б — С ontrito saurus simus Ivach
nenko; экз. ПИН Jf 2482/1; Костромская обл., р. Вохма, с. Семеновское; нижний
триас, низы ветлужской серии
ку, расширенную кпереди и кзади (рис. Па). Надо полагать, что это остатки око
стеневавшей латеральной стенки сфенэтмоида, в значительной степени редуциро
ванного. Парасфеноид и базисфеноид у проколофонов срастаются в единый пара-
базисфеноидный комплекс. Строение комплекса у примитивных форм (никтифру
рет) и у более высокоразвитых (тихвинския, контритозавр, буртензия) значитель
но различается.
Пара-базисфеноидный комплекс никтифрурета широкий и плоский (рис. 11а).
Ростральный отросток комплекса очень длинный, 'доходит до переднего края меж-
птеригоидной вырезки, Почти не поднят над плоскостью неба. Отросток широкий,
вогнутый сверху и выпуклый снизу, составляет дно костной сфенэтмоидной трубки.
Трабекулярные бугры у основания отростка развиты хорошо. Гипофизарная ямка
узкая и глубокая, на дне открываются два выходных отверстия церебральных вет
вей внутренней сонной артерии. Спинка турецкого седла высокая, однако ниже
гребня, ограничивающего гипофизарную ямку спереди. Это отчасти сближает пара-
базисфеноидный комплекс никтифрурета и капторина (Fox,. Bowman,. 1966), хотя
принципиально строение комплексов резко различно. Вся дорзальная поверхность
комплекса впереди гребня является. Скорее всего, функциональным аналогом ре-
тракторного миодома ящериц (Save-S6derbergh, 1946), т. е. местом прикрепления
"глазных" мускулов группы rectus posterior. Миодом ограничен латеральновысоким
35
Рис. 12. Пара-базисфеноидный комплекс сверху
а — Tichvinskia
vjatkensis
Tchudinov et Vjuschkov; реконструкция по экз.
ПИН No 954/2,3; Кировская обл., р. Федоровка; нижний триас, баскунчакская
серия; б - Burtensia burtensis (Otschev); экз. ПИН No 3359/11; Архангельская
обл.,
р.Пижма-Мезенская, "нижний триас, баскунчакская серия
гребнем, идущим от переднего гребня до спинки турецкого седла. Спинка круто
спускается назад, к горизонтальной пластине базисфеноида, не обрывающейся
сразу же позади спинки, как у более высокоразвитых проколофонов. Основания
pilae antotica вынесены далеко назад относительно спинки седла. Базиптеригоид-
ные мыщелки очень массивные, неправильно-йкруглой формы. Входные отверстия
внутренних сонных артерий находятся под мыщелками, выходные отверстия нёбных
артерий — в углублениях в районах постеро-дорзальных поверхностей оснований,
так что артерия раздваивается сразу же по входе в кость. Парасфеноид срастает
ся с базисфеноидом лишь в передней, ростральной части. Латеральные края пара-
сфеноида под базисфеноидом несут желоба небных вен. Крылья парасфеноида уз
кие и длинные.
Пара-базисфеноид контритозавра (рис. 116) отличается высокой спинкой турец
кого седла, гораздо более широкой гипофизарной ямкой и очень низким, почти не
выраженным передним гребнем. Базиптеригоидные отростки относительно длиннее,
направлены вперед и в стороны. Основания pilae antotica расположены латераль-
но от спинки седла. Дорзальная поверхность базисфеноидного ростра несет узкий
продольный желобок. Ростр не длинный, не заходит далее половины длины межпте
ригоидной вырезки. Ретракторный миодом короткий и широкий. Канал для внутрен
них сонных артерий и небной ветви лицевого нерва идет по верхним краям бази-
птеригоидных отростков и перекрывается сверху очень тонкой костной пластинкой.
Начинается канал несколько позади крыловидных отростков базисфеноида. Выход
ное отверстие небной артерии находится у переднего края основания базиптериго-
идного отростка. Небольшой продольный желоб вдоль латеральной стенки базисфе
ноида может говорить о прохождении здесь боковой вены головы. Ненбсредствен-
36
о
qscAt
I
.
lr
Рис. 13. Мозговая коробка Tichvinskia
vjatkensis
Tchudinov et Vjuschkov;
спереди; реконструкция по экз, ПИН No 954/1,2,3; неокостеневавшие зоны по
казаны точками; Кировская обл., р. Федоровка; нижний триас, баскунчакская
серия
но парасфеноидная часть комплекса образует идущие назад крылья, подстилавшие
антеро-медиально часть дна мозговой коробки.
Пара-базисфеноид буртензии (рис. 12а) отличается рядом существенных дета-*
лей. Ростр на дорзальной поверхности несет гребень, что свидетельствует скорее
в пользу присутствия у животного межглазничной септы, 'чем сфенэтмоида. Перед
няя часть дорзальной поверхности комплекса образует структуру, полностью срав
нимую с ретракторным миодомом ящериц. Округлые впадины для прикрепления
"глазных" мускулов разделены продольным гребнем. Гипофизарная ямка, как осо
бая структура, не выражена, отверстия церебральных ветвей внутренних сонных
артерий расположены в задней-части миодома и разделены коротким треугольным
гребнем. Спинка турецкого седла высокая, образует острый поперечный гребень,
высоко поднимающийся над плоскостью дна мозговой коробки. Относительно высо
кая спинка седла может также говорить о присутствии межглазничной септы, по
скольку у ящериц к высокой спинке прикрепляется постеро-вентральная часть меж
глазничной мембраны (membrana metoptica),. образуя расширенное краниальное дно.
Крыловидные отростки базисфеноида (основания pilae aritotica) относительно не
высокие. Базиптеригоидные отростки длинные, 'направлены вперед и слабо лате
рально.
Пар а-бази сфеноид тихвинский (рис. 126) построен в целом схоже, однако мио-
дом сильно вытянут вперед, Гипофизарная ямка выражена еще слабее, чем у бур
тензии. По краям спинки имеются небольшие вырезки, Пропускавшие отводящий
нерв. Крыловидные отростки базисфеноида довольно высокие, базиптеригоидные
отростки очень мощные, округлые в сечении, заканчиваются выпуклыми сустав
ными поверхностями. Базиптеригоидное сочленение образует настоящий сустав,
дававший" возможность для движений значительной амплитуды.
Строение ушной капсулы очень близко к таковому ящериц (например, Oelrich,.
1956; Siebenrock,. 1895)». «о степень окостенения самой капсулы меньше. Строение
ушных костей всех изученных проколофонов очень схоже. Ушную капсулу сзади,
сбоку и сверху образуют соответственно заднеушная, переднеушная и верхняя за
тылочная кости. Непосредственно друг с другом соединяются лишь ушные кости
(prooticum,. opisthoticum) в районе парокципитального отростка, в остальных местах
непосредственного контакта нет и кости отделены друг от друга хрящевыми про
слоями. Медиальная стенка капсулы не окостеневала (рис. 13, 14 а,б).
Переднеушная кость (рис. 156) представляет собой слабо окостеневшую пла
стинку, снизу лежащую на крыле парасфеноида, !а спереди прилегающую к крыло
видному отростку базисфеноида, В передней части кость несет довольно крупное
овальное выходное отверстие лицевого нерва, Над отверстием находится Г-образ-
ная система гребней для передней группы аддукторов. По латеральной поверхно-
37
Рис. 14. Мозговые коробки сбоку
а — NycUphruretus
acudens Efremov; реконструкция по экз. ПИН
No 158/6,16. Архангельская обл., р.Мезень; верхняя пермь, нижнетатар
ский подъярус; б — Burtensia burtensis (Otschev); обобщено по экз. ПИН
No 3359/11; Архангельская обл., р. Пижма-Мезенская; нижний триас,
баскунчакская серия
Рис. 15. Кости ушной капсулы Burtensia burtensis (Otschev)
а — supraoccipitale сбоку, б - правое prooticum изнутри, в - правое
opisthoticum изнутри; кости от голотипа ПИН No 3359/11; Архангельская
обл., р. Пижма-Мезенская; нижний триас, баскунчакская серия
сти кости проходит полукруглый гребень, содержащий внутри часть переднего по
лукружного канала, Горизонтальный канал идет почти перпендикулярно переднему
и выходит на задний отросток кости, смыкаясь здесь с продолжением канала в
опистотике. Кость содержит переднюю ампулу. На ее краях заметны короткие же
лобки - части латеральных стенок полукружных каналов. На антеро-медиальной
поверхности кости, Под выходом переднего полукружного канала, имеется широкий
желоб, идущий назад. У большинства ящериц именно в этом месте имеется полу
круглый отпечаток петли средней мозговой вены (Oelrich,. 1956). У агамид Phryno*
cephalus есть схожий желоб, который совместно с желобом верхней затылочной
кости образует сквозной канал. У проколофонов верхне-срединная стенка канала
была образована хрящем. Средняя мозговая вена, сделав петлю на затылочную
поверхность, по этому каналу возвращалась в мозговую коробку.
Парокципитальный отросток заднеушной кости (рис. 15в) направлен назад и
несколько вверх, т.6., находится примерно в таком же положении, как и у прола-
церты (Robinson,. 1967, 397, fig. lb). Его антеро-Латеральная поверхность -
место прикрепления хряща. Надо полагать, что широкая хрящевая пластина - про
должение отростка - шла вверх, в район контакта чешуйчатой и надвисочной ко
стей. На медиальной поверхности окостеневавшей части находится отверстие го
ризонтального полукружного канала. Заключенная в кости часть заднего полукруж
ного канала сопровождается на постеро-медиальной поверхности хорошо выражен
ным гребнем. Кость содержит заднюю ампулу и ограничивает медиально овальное
окно слуховой капсулы, край которого сужает невысокий кольцевой гребень. К
задней поверхности кости причленялась боковая затылочная, Причем между костя
ми остается широкое отверстие — часть метотикальной щели (место выхода IX—X
краниальных нервов, задней вены головы, и, возможно, перилимфатического про
тока).
Верхняя затылочная кость буртензии (рис. 15 а) и тихвинский представляет со
бой расширяющуюся к латеральным краям тонкую пластинку, слегка изогнутую
посередине. Дорзальный отросток для соединения с постеро-вентральными высту
пами теменных костей узкий и длинный, Совершенно такого же типа, как и у со
временных ящериц. Дорзальный отросток у контритозавра несколько более округ
лый в сечении, но такой же узкий. У никтифрурета дорзальный отросток отсут
ствует, верхняя часть кости по всей ширине соединяется швом с заднетеменными
костями. Верхняя затылочная кость заключает cms communis и часть канала эндо-
лимфатического протока. У буртензии канал протока отсутствует.
Основная затылочная кость плоская, округлая, у никтифрурета овально.удли-
ненная, у контритозавра трапециевидная, у тихвинский округлая. Вентральная по
верхность кости у никтифрурета несет два неглубоких конических вдавления, ко
торые у контритозавра и тихвинский почти исчезают, и непосредственно позади
них появляются сфено-окципитальные гребни для прикрепления некоторых субвер-
тебральных шейных мышц. По дорзальной поверхности кости идет невысокий про
дольный гребень (рис. 12), образующий впереди бугор (basis tuberculi basalis).
Такой же гребень, служащий для прикрепления связок медуллы, имеется у некото
рых черепах (Kesteven,. 1910, стр. 316; Gaffney,. 1972, «стр. 4). У диапсид этот гре
бень обычно находится на задней поверхности базисфеноида.
Задняя поверхность основной затылочной кости у никтифрурета несет неглубо
кое коническое вдавление для переднего конца хорды, Перекрытое сверху субме
дуллярными отростками боковых затылочных костей, срастающихся с основной за
тылочной на нижней поверхности в одно целое. Боковая затылочная кость высокая,
тонкая, у расширенного основания несет отверстие XII нерва и питающее отверстие.
Верхний край кости слабо окостеневал. Таким образом, "верхняя затылочная кость,
вероятно, представляет собой лишь продукт окостенения эмбриональной межкап-"
сулярной крыши (tectum synoticum), тогда как задняя крыша (u posterius) остава
лась у проколофонов хрящевой. Вероятно, *между обоими мостиками была фонта-
нель, так как задний край верхней затылочной кости гладкий и вогнутый.
Слуховая косточка (рис. 10) короткая, имеет массивное основание, овальное и
слегка вогнутое медиально. Передне-Нижний край основания вытянут, образуя от
росток, 'аналогичный proc. ventralis л абиринтодонтов (Шишкин, 1973), Ъеуществля-
39
О
Рис. 16. Burtensia burtensis (Otschev); левая слуховая косточка от голотипа ПИН
No 3359/11; Архангельская обл., р. Пижма-Мезенская; нижний триас, баскунчакская
серия
а — изнутри, б — снаружи, в — сверху
ющий связь с выступом ушной капсулы. Дистальный конец косточки плоский, вытя
нут почти горизонтально. Возможно, Передняя часть конца представляет собой
основание хрящевой extracolumellа, тогда как его задняя, утоньшенная часть — ос
нование хрящевой связи с гиоидом (proc. infrastapedialis columellae). Э качестве
дорзального отростка может рассматриваться лишь очень небольшой бугорок на
антеро-Дэрзальной поверхности слуховой косточки, немного проксимальнее экстра-
стапедиального отростка. В таком случае 'связь слуховой косточки с парокципи-
тальным отростком слуховой капсулы была очень слаба, либо отсутствовала сов
сем. Направление экстрастапедиального отростка указывает на низкое положение
барабанной перепонки у проколофонов, в районе нижнего края затылочной пластины
чешуйчатой кости.
Квадратная кость ^ихвинскии (рис, (36) представляет собой тонкую, ровную
треугольную пластинку, расширенную сверху и слабо сужающуюся к блоку челюст
ного сустава.
-Передняя сторона кости гладкая, 'имеет внизу слабый гребень, "воз
можно, соответствующий медиальному гребню квадратной кости ящериц, служа
щему для прикрепления части заднего аддуктора нижней челюсти. На задней сто
роне квадратной кости проколофонов есть хорошо развитый бугор. На такой же
бугор указывает Уайт (White, «1939, стр. 334,. 'фиг. 5/ у сеймурии. По Шишкину (1973)
бугор служил для прикрепления гиоида к квадратной кости. К антеро-медиальному
краю бугра у проколофонов прилегает нижняя часть квадратной ветви птеригоида.
Широкая верхняя пластинка квадратной ветви налегает на квадратную» кость выше
бугра. Можно полагать, '.что связь между ними осуществлялась лишь остатками
небно-квадратного хряща. Схожее соотношение квадратной и птеригоидной костей
показывает Грегори (Gregory, «1945,. «стр. 282, 'фиг. 5; для трилофозавра. Скуловая
Квадратная кость и подъязычный аппарат
40
и квадратно-акуловая кости у проколофонов лишь налегают снаружи на квадратную
кость. Квадратно-ушная связь, если она имелась, могла осуществляться между
верхним свободным краем квадратной кости и парокципитальным отростком ушной
капсулы. Однако этот район у проколофонов полностью лишен окостенений. Широ
кая костная полоса в этом месте есть у парейазавров, у диадекта Олсон (Olson,.
1947) также отмечает наличие квадратно-ушной связи.
Подъязычный аппарат тихвинский представлен тонкой пластинкой копулы, уз
кой посередине и расширенной на краях. Антеро-Латеральные и постеро-Латераль-
ные углы пластинки несут небольшие шероховатые площадки, Места прикрепления
дужек. У проколофона сохраняются передние дужки, прикреплявшиеся к антеро-л"а-
теральным площадкам (Broili,; SchrQder,. «1936». стр. 247, 'фиг. 7), Задние дужки не
сохранялись. У буртензии. Наоборот, Сохраняются задние дужки.
Нижняя челюсть
Нижняя челюсть у большинства проколофонов относительно узкая и тонкая,
лишь у наименее изученных форм (лептоплейрон, миоцефалюс, проколофоноидес)
отмечается очень высокая зубная кость, резко опускающаяся вниз в передней
части.
В эволюции проколофонов Русской платформы по мере усиления дифференци
ации зубов наблюдается тенденция к увеличению высоты зубов относительно den-
tale* «а также к укорочению dentale относительно пост дентальной части. Так, у
никтифрурета (рис. 17 а) зубная кость в полтора раза длиннее постдентальной ча
сти,
зуб укладывается в высоте кости (примерно в середине длины зубного ряда)
2,3 раза, У фаантозавра (рис. 176) длины дентальной и пост дентальной частей
равны, зуб укладывается в высоте два раза, у контритозавра (рис. 18 а) — 1,8; у
буртензии (рис. 18 г) - 1,4,, Однако в некоторых линиях шел противоположный про
цесс, например, у раннетриасового вида Orenburgia concinna
(рис, 20а) высота зу
ба лишь немного меньше высоты дентале, 'а у среднетриасового Or.
majmesculae
(рис. 2(i в) зуб укладывается два раза.
У проколофонов Южной Африки, вероятно, также происходило увеличение вы
соты зубной кости. Это хорошо заметно в ряду Pro со lop hono ides - Procolophon
-
Myocephalus.
Зубная кость обычно полностью образует симфиз. Возможно, "иногда (Candela°
па) пластинчатая кость доходит до симфиза,,но это нуждается в уточнении. Уорен-
бургии имеется площадка для межчелюстного мускула на нижней стороне переднего
края кости. Желоб Меккелева хряща у всех проколофонов довольно широкий (не ме
нее половины высоты кости) и идет до симфиза (например, Tichvinskia:
рис. 19 г;
Vitalia:
рис. 21 в). У трилофозавра (Gregory,. «1945; Olson,. «I960) и целодонтогната
желоб широкий сзади, резко утончается и снижается кпереди, Не доходя до симфи
за, У капторинид (например, Romer,. «1946) Меккелев канал также не доходит до сим
физа, to в построении симфиза участвует пластинчатая кость. У сеймурии (White,
1939) канал доходит до симфиза, Но симфиз образует и пластинчатая кость. Из
Меккелева желоба в толщу кости у проколофонов уходят два канала — в передней
части кости и в начале задней половины. Оба пронизывают кость и открываются
раздельно на наружной стороне в ряду нижнегубных отверстий. В толще верхнего "
края кости (рис. '19г,д) поперечные каналы соединяются хорошо сформированным
продольным, 'от которого идут веточки к зубам. Такой же продольный канал можно
видеть й у ящериц, однако он сформирован, Как правило, хуже, являясь рядом, сое
диненных между собой лакун. У лацерт и агам нет переднего входного отверстия,
только заднее. Надо отметить, что продольный канал у агамид развит гораздо луч
ше, чем у лацерт и ящурок.
Канал открывается у проколофонов на наружной стороне зубной кости рядом t
нижнегубных отверстий. У примитивных проколофонов (никтифрурет, 'фаантозавр;
число отверстий велико, Они расположены по середине высоты кости по всей ее
длине. Это схоже с расположением и числом отверстий у лацертилий - лацерт, '
тейид, тогда как у агамид, Игуанид и высших проколофонов число отверстий неве
лико.
41
д
Рис. 19.
0,SCAI
Рис, 17. Правые ветви нижней челюсти
а - Nyctiphmretus
acudens Efremov; снаружи; реконструкция по экз. ПИН No 158/5
и 162/1; Архангельская обл., р.Мезень; верхняя пермь, нижнетатарский подъярус;
б. В,г — Phaanthosaums
ignatjevi Tchudinov et Vjuschkov; снаружи, сверху и изну
три; реконструкция по экз. ПИН No 1025/1,20; Горьковская обл., р. Ветлуга, с. Спас
ское; нижний триас, низы ветлужской серии
Рис. 18. Правые ветви нижней челюсти
а, б - Contri to saums simus Ivachnenko; изнутри и сверху; обобщено по экз. ПИН
No 3355/1; Горьковская обл., д. Липово; нижний триас, низы ветлужской серии; в, г —
Burtensia burtensis (Otschev); сверху и изнутри; экз. ПИН No 3359/11; Архангельская
обл.,
р. Пижма-Мезенская; нижний триас, баскунчакская серия
Рис. 19. Tichvinskia
vjatkensis
Tchudinov et Vjuschkov; левая ветвь нижней челю
сти, обобщено по экз. ПИН No 954/1, 2,14-21; Кировская обл*, р. Федоровка; нижний
триас, баскунчакская серия
а — снаружи, б - сверху, в — изнутри, г - продольный срез зубной кости, изну
три, ft - поперечные распилы челюсти
43
Зубная кость, как правило, не участвует в построении короноидного отростка,
хотя обычно верхне-Задний край кости обходит снаружи отросток, а иногда (Contri*
tosaurus
convectoi) поднимается вверх почти до верхнего его края.
Пластинчатая кость тонкая и длинная, соединена швом с зубной в верхней ча
сти и свободно прилегает снизу, закрывая меккелев канал с внутренней стороны.
В передней части кость имеет вырезку, и далее к симфизу идет лишь нижняя часть
кости, Не доходя до симфиза. На середине длины вырезки кость несет крупное от
верстие для язычного нерва (foramen spleniale, рис. 19), канал его идет на некото
ром протяжении продольно, й лишь затем поворачивает наружу. Задний край кости
обычно образует передний край отверстия (рис. 19 в, for. Meckelianum) для комби
нированного V—VII. Форма и положение кости близки к таковым лацертилий. У ага-
мид кость резко уменьшается в размерах и даже редуцируется совсем (у кругло
головок) У капторинид (лимносцелис: Romег,. «1956) она очень узкая и почти на
всем протяжении ограничена сверху большим отверстием.
Угловая кость'занимает обычное положение, 'сходное с ее положением у всех
рептилий.
Надугловая кость (рис. 19 а) кзади резко утолщена. В этом утолщении находит
ся входное отверстие заднего ствола V3 и нижнечелюстной артерии. Нерв и арте
рия до и после входа в отверстие ветвятся, давая кожные ветви, выходящие в пе
реднее и заднее супрангулярные отверстия. Число и положение отверстий может
варьировать. Так, у тихвинский имеется крупное отверстие переднее и два задних —
над и под гребнем мускула, идущего по латеральному краю кости, у фаантозавра -
одно крупное отверстие, окруженное двумя-тремя маленькими.
Короноид небольшой, единственный, что резко отличает проколофонов от сей-
мурид (три короноида) и капторинид (два короноида). У трилофозавров короноид
сходной формы и тоже единственный, тогда как у аресцелидий {Araeoscelis:
Vaughn,
1955) два короноида. Венечный отросток у проколофонов развит слабо у примитив
ных форм (никтифрурет, тфаантозавр) и гораздо сильнее у наиболее высокоспеци
ализированных (тихвинския, лептоплейрон, гипсогнат, проколофон и др.). У капто
ринид и сеймурид короноидный отросток практически не развит. Предсочленовная
кость узкой пластиной охватывает с лабиальной стороны сочленовный отдел и зад
нюю половину верхнего края аддукторной ямы.
У фаантозавра и контритозавра по верхне-йаружному краю предсочленовной ко
сти идет небольшой гребень до половины длины аддукторной ямы. Над гребнем
предсочленовная кость резко выступает лингвально, образуя небольшой фланг.
Еще более развитый фланг предсочленовной кости имеется у большинства ящериц.
Такой же лингвальный фланг есть у склерозавра, что резко отличает его нижнюю
челюсть от таковой парейазавров.
Сочленовная кость обычно широкая, )эезко выступает медиально, Представляя
собой губчатый, не.полностью окостеневший хрящ лишь в области собственно че
люстного блока, тогда как задняя половина кости окостеневает хорошо и образует
поверхность ретроартикулярного отростка, иногда очень хорошо развитого. Верх
няя поверхность отростка несет входное отверстие chorda tympani,. «выходное зад
них стволов V3 и нижне.челюстной артерии и две продольные впадины, разделенные
невысоким гребнем. Ретроартикулярный отросток у трилофозаврид обычно также
хорошо развит, тогда как ареосцелидии, капторины и сеймурии практически его не
имеют.
Зубы
Прикус у типичных проколофонов бипрогнатический, т. е. передние зубы в
верхней и нижней челюстях наклонены вперед и верхние зубы почти на высоту ко
ронки перекрывают нижние. Такой прикус хорошо выражен у форм с сильной диф
ференциацией зубов и почти не выражен у примитивных форм.
У никтифруретид число зубов обычно весьма велико - 30—40 в челюсти, зубы
не дифференцированы, имеют одинаковую коническую форму. У спондилолестин
наблюдается слабая (спондилолестес, фаантозавр) и довольно значительная (кон-
тритозавр, возможно, проколофоноидес) дифференциация, число зубов в челюсти
резко сокращается до 12-15. Еще меньше сильно дифференцированных зубов у
44
Рис. 20. Правые зубные кости
а, б — Orenburgia concinna
Ivachnenko; «наружи и сверху; экз. ПИН
No 3362/1; Оренбургская обл., д. Марковка; нижний триас, баскунчакская
серия; в — Orenburgia
majmesculae (Otschev); снаружи; экз. СГУ No 104/3824;
Оренбургская обл., р.Бердянка; средний триас, донгузская серия; г, д —
Kapes amaenus Ivachnenko; снаружи и сверху; голотип ПИН No 3361/1; Ко
ми АССР, пос.Жешарт; нижний триас, верхи баскунчакской серии
проколофонин (10-11 у проколофона, Менее 10 у остальных родов, до 8-6 у мио-
цефалюса и капеса). Наименьшее число сильно дифференцированных зубов у леп-
топлейронин (6—7 в верхней челюсти, 5 — в нижней).
Зубы, Как правило, &кродонтные, вещество зуба очень слабо внедряется в кость,
однако внедрение все же заметно, что дало возможность Броили и Шредеру(Broili,
Schroder, 1936,, стр. 242) вслед за Рёзе (Rose, «1894, «стр. 558) назвать их "прото- '
текодонтными". 'В ряде случаев внедрение несколько значительней, но судить об
этом можно лишь по захождению полости пульпы в кость, тогда как стенки зуба
не отграничены ясно от кости. Прототекодонтное прикрепление, "возможно, "и при
вело в иных линиях рептилий к значительному внедрению с образованием "корней",
как это наблюдается у парейазавров и трилофозавров, и к плевродонтному способу
прикрепления ящеричного типа.
Форма зубов у примитивных проколофонид довольно однообразна. Это слабо
расширенные у оснований удлиненные конусы, вершинки обычно продольно слегка
уплощены и загнуты лингвально. У спондилолестеса основания задних зубов, про
должая оставаться округлыми, увеличиваются в диаметре, *у фаантозавра основа
ния их расширяются трансверзально, Приобретая овальную форму. Наиболее силь
но это выражено у контритозавра, "у которого наблюдается значительная диффе
ренциация, *хотя форма всех зубов челюсти почти одинакова. В результате отира
ния задних зубов их коронки приобретают плоский поперечный гребень (рис. 18 в^ г),
что усиливает видимую дифференциацию зубов этой формы.
Настоящая дифференциация наблюдается лишь у проколофонин и лептоилейронин.
У них :»убы резко разделяются на "квазирезцы" и "квазикоренные". "Квлшрезцы"
расположены обычно на предчелюстной кости (2-4 зуба, иногда первые 2-'.\ на верх-
45
о
Jсм
Рис. 21. Челюсти проколофонов
а — Macrophon komiensis Ivachnenko; левая верхнечелюстная кость сна
ружи; голотип ПИН No 3361/3; Коми АССР, пос. Жешарт; нижний триас, вер
хи баскунчакской серии, б, в - Vitalia grata Ivachnenko; часть левой зуб
ной кости сверху и изнутри; голотип СГУ No 104/3104, Донская Лука, овраг
Б.Липовский; нижний триас, баскунчакская серия; г, Д - предпоследний и
первый сохранившиеся зубы того же экземпляра Vitalia grata Ivachnenko
сзади
нечелюстной) и передние (1 у лептоплейронин, 2-3 у проколофонин) на нижней че
люсти. Форма "квазирезцов" аналогична форме зубов у никтифруретид.
"Квазикоренные" (рис. 19а,б;20 а-д; 22а-д) зубы имеют более или менее взду
тые коронки, расширенные поперечно основания, уплощенные вершинки, часто снаб
женные двумя латеральными бугорками. Вероятно, в связи с приспособлением к
различной пище на фоне сходного общего плана строения появляются различные
вариации. У коилоскиозавра, аномоиолона, буртензии зубы невысокие, конические,
вершинки уплощены, Двухбугорчатость почти не выражена и окончательно уничто- '
жается стиранием зуба. Наоборот, у рода Macrophon (рис. 21 а) сильно вытянутые
трансверзально коронки имеют два острых латеральных бугра и вогнутый режущий
гребень между ними. У проколофона и тихвинский строение зубов как бы промежу
точное между этими двумя типами. У рода Kapes (рис. 20 г, Д) основную роль играет
конусообразность формы, Двухбугорчатость почти не выражена, 'окончательно унич
тожается стиранием, й зуб вторично приобретает форму заостренного конуса. У
лептоплейронин зубы очень крупные, широкие, основания сильно расширены транс
верзально. Коронки имеют два острых латеральных бугра и более или менее упло
щенный гребень между ними.
Особенно интересным представляется строение зубов у рода Vitalia (рис. 21 г, Д).
Задние зубы нижней челюсти этой формы округлые, 'с куполообразными вершинка
ми, акродонтные. На передней и задней сторонах вершинок имеются небольшие бо
роздки, характерные для двухбугорчатых зубов. Вершинки притуплены вследствие
стирания. Передние зубы имеют расширенные поперечно узкие коронки, вещество
зуба сильно внедряется в кость, так что эти зубы очень схожи по строению с зуба
ми примитивного трилофозаврида — целодонтогната (особенно близки к зубам *
С. don
ensis).
Можно отметить, что у некоторых форм наблюдается небольшой наклон всех
верхнечелюстных зубов вперед (С ontrito saurus
convector).
It настоящее время не представляется возможным сказать определенно, чем
вызываются такие вариации в форме зубов проколофонов. Обычно им приписывает
ся моллюскоядность или насекомоядность. Однако современные ящерицы со сход-
46
Рис. 22. Зубы проколофонов, изображен предпоследний зуб левой стороны нижней
челюсти, *ззади
а — Tichvinskia
Tchudinov et Vjuschkov; б — Burtensia Ivachnenko; в -
Macro-
phon Ivachnenko; г - Orenburgia Ivachnenko; д- Rapes Ivachnenko
ной формой зубов (например, Tupinambus: Barbarena, Gomes,. Sanchobene, «1970) яв
ляются всеядными. В связи с этим интересно отметить, что форма зубов у диадек*
тов гораздо сложнее* Их зубы имеют оильно уплощенные основания, вздутые ко
ронки, Плоские сверху, несущие мощный латеральный бугор и слабее развитый ме
диальный. У диадекта, Диаспаракта и десматодона (Case, «1907; 1910; Vaughn,. 1972),,
кроме того, 'сильно развит средний бугор, который доминирует. Можно полагать, '
что диадекты были растительноядными формами.
Индивидуальные вариации в числе зубов у проколофонов незначительные, 'осо
бенно у примитивных видов. Например, фаантозавр имеет в нижней челюсти 11 зу
бов при изменении длины зубного ряда от 15 до 18 мм. При этом у примитивных
форм происходит смена зубов — новый зуб появляется у внутреннего основания,
впереди или сзади от замещаемого. Такая смена зубов приводит к довольно беспо-
рядочному расположению зубов на зубной кости у взрослых экземпляров (рис. 17 в).
У более высокоразвитых форм по мере роста челюсти число зубов увеличивается,
однако смена зубов наблюдается лишь изредка, и только на передних "квазирез
цах". Так, у контритозавра при удлинении зубного ряда на 0,7 мм (от G,0 до G,7)
появляется новый зуб.
Материал по Tichvinskia
vjatkensis
из местонахождения Окуневское (Кировская
обл.) дает возможность проследить для этого вида индивидуальную и возрастную
изменчивость зубной системы.
В результате изучения (данные для нижней.челюсти) выяснилось, что форма
зубов при любых вариациях размеров остается строго постоянной, а зубы могут
быть разделены по степени вариабельности в числе на две группы. Первая группа-
"квазирезцы". Число их колеблется от двух крупных до трех крупных и одного ма
ленького при изменении длины зубной кости от 9,0 до 15,0 мм. При этом число зу
бов практически не зависит от размеров челюсти. Вторая группа -
"квазикорен- '
ные".
Число этих зубов изменяется строго закономерно. При длине их ряда от 0,0
до 8,0 мм имеется 4 зуба, от 8,0 до 9,5 позади появляется еще один, Маленький,
зуб, от 9,3 до 10,0 мм - 5 крупных зубов и от 10,0 до 11,0 мм — G крупных зубов.
Таким образом, хорошо прослеживается закономерное увеличение числа зубов
(один зуб на 2 мм удлинения). Броили и Шредер(3roiII,. Schrdder,. «1936) отметили та
кое увеличение числа зубов у рода Procolophon,
однако оценили э^от признак не
правильно. Они трактовали маленькие черепа с меньшим числом зубов как черепа
самок, а большие - как самцов. Такое увеличение числа зубов при росте челюсти
характерно и для современных ящериц с акродонтными зубами, например, агам.
У целодонтогната. Первоначально отнесенного к проколофонам, смена зубов
весьма своеобразна. Замещающий зуб, формировавшийся в толще дентале, 'увели-
47
чивался в кости до такой степени, что его коронка становилась шире верхнего
края зубной кости, И при движении вверх зуб прорезал кость и края его выходили
на боковую поверхность челюсти. Судя по изображению в работе Грегори (Gregory,
1945. стр. 288, фиг. 6), абсолютно такая же смена наблюдается у трилофолавра, и
это еще один признак, который ставит целодонтогната ближе к трилофозаиру, чем
даже роды То хо lop hosaurus,
Tri cusp isaurus и Variodens,
у которых, судя л о описа
ниям, нет такой смены.
Кинетизм черепа
Одной из интересных особенностей черепов высших проколофонов (в некоторой
степени контритозавра, в гораздо большей - тихвинский и буртензии) является
хорошо выраженный кинетизм. О наличии кинетизма говорят следующие черты
строения черепа. Базиптеригоидное сочленение представляет собой сустав, допу
скавший большую амплитуду движений; верхняя затылочная кость соединялась с
теменными лишь узким и длинным восходящим отростком, >лежду отростком и те
менными костями имелась хрящевая прослойка. Челюстные кости лишь налегают
сбоку на скуловые, 'не соединяясь с ними зубчатым швом. По шву лобных и темен
ных костей вполне возможны мезокинетические движения, поскольку шовное сое
динение у тихвинский имеется лишь между "постфронтальными" отростками те
менных и лобными костями, непосредственно лобные и теменные кости лишь при
легают друг к другу. Такая конструкция могла быть достаточно гибкой. У буртен
зии срастания теменных и постфронтальных костей нет, но зато все швы' крыши
черепа резко упрощены, отсутствуют сложные зубчатые соединения.
Вполне могла изгибаться и тонкая нёбная пластинка, тем более 'что вентраль
ные отростки птеригоида и эктоптеригоида, образующие птеригоидный фланг, 'лишь
прилегают друг к другу, а не соединены швом. Подвижную зону можно предпола
гать в районе налегания передних краев сошников на предчелюстные кости, ' а
также в районе налегания предчелюстных на челюстные.
Квадратная кость прикрепляется лишь к квадратной ветви птеригоида, Причем,
возможно, 'связь осуществлялась лишь остатками небно-квадратного хряща, й квад
ратная кость могла быть достаточно подвижной.
Все перечисленные признаки говорят о том, что у проколофонов имелся раз- '
витый амфикинетизм ящеричного типа. Главным отличием является относительно
слабая подвижность квадратной кости у проколофонов. Кинетичный череп высших
проколофонов развивался из почти акинетичного черепа никтифруретид. Надо по
лагать, что развитие кинетизма у проколофонов шло параллельно с ящерицами,
при этом подвижность отдельных сегментов черепа относительно друг друга до
стигалась несколько различными путями.
ПОСТКРАНИАЛЬНЫЙ СКЕЛЕТ
Общие данные
Посткраниальный скелет проколофонид построен весьма легко* это скелет очень
подвижных, активных наземных форм. Диадектиды имеют более массивный и при
митивный скелет. Грегори (Gregory,. 1946),. сравнивая, элементы скелета парейазав-
рид и диадектид, находит очень много общего, тогда как при сходном плане стро
ения скелеты проколофонид, с одной стороны, й парейазаврид и диадектид, 'с дру
гой, Значительно различаются. В некоторой степени различия вызываются большой
разницей в размерах этих форм. Скелет наиболее крупных проколофонов (склеро-
завр) массивный и внешне схож со скелетом парейазавров, однако, как это будет
показано далее, имеет вполне определенные отличия. Возможно, "что именно сопо
ставимостью размеров объясняется сходство скелетов капторинов и проколофонов.
Наиболее схож скелет капторинида (Carroll,. <1969а). В дальнейшем скелет проколо
фонов становится более легким, исчезают срастания костей в плечевом и тазовом
поясах и идет их частичная редукция.
48
Строение скелета примитивного никтифрурета подробно описано И.А.Ефремо- '
вым (Efremov,. «1940), более высокоразвитого (проколофон) - Д.М.Ватсоном (Watson,
1914), поэтому ниже приведены лишь некоторые дополнительные данные.
Осевой скелет
У тихвинский (рис. 23) около 52 позвонков, из них шейных - 4, туловищных -
21 (могут быть по наличию или отсутствию ребер разделены на собственно туло
вищные — 1(1 и поясничные - 5). Крестцовых позвонков 2, хвостовых - около 25.
У Koiliskiosaurus
(Huene,. 1911) — 24 предкрестцовых позвонка, 'у
Sclerosaurus
(Huene, 1902) — lb* предкрестцовых, 3 крестцовых и 15 хвостовых, у
Leptopleuron
(Huene, 1956) - 24 предкрестцовых и 2 крестцовых, у Hypsognathus
(Golbert,. 1946) -
20 предкрестцовых.
Строение первых шейных позвонков хорошо изучено у буртензии (рис. 24). Дуж
ки проатласа треугольной формы, несут на передне-нижних и задне-нижних поверх
ностях площадки для сочленения соответственно с боковой затылочной костью и
атлантом. Дуга атланта не скреплена с телом позвонка, ее презигапофизы практи
чески не выражены, постзигапофизы развиты сильно. Остистый отросток очень
небольшой. Тело атланта резко поднято вверх относительно уровня нижних краев
тел остальных позвонков и близко сдвинуто с передней поверхностью тела эпи-
строфея, toe несущего никаких следов зубовидного отростка. По сути дела, отрост
ком являлось тело атланта. Интерцентр имеется лишь между атлантом и эпист,ро-
феем. Тело проатланта и интерцентр атланта отсутствуют, однако короткое ребро
с хорошо выраженной двухголовчатостью присоединяется бугорком к дуге атланта,
что может говорить о сравнительно недавней редукции или неокостенении интер- '
центра атланта.
Сравнение с передними позвонками проколофонов, 'описанными Броили и Шреде
ром (Broili,. Schroder,. «1936),показывает, что, возможно, авторами передний край
атланта был принят за интерцентр атланта. В остальном строение передних поз
вонков буртензии и проколофона одинаково. Таким образом, интерцентр атланта,
характерный для других примитивных зауроморфных рептилий, у проколофонов
редуцирован. Интерцентр имеется у сеймуриаморфов и капторинидов (Carroll, 1969b),
Р'омер^м (Romer,. 1944) отмечен также у диадекта.
Рис, 23. Tichvinskia
vjatkensis
Tchudinov et Vjuschkov; скелет; положение на по
роде; экз. ПИН No 954/1; Кировская обл., р, Федоровка; нижний триас, баскунчакская
серия
4 796
49
Ах
Рис. 24, Burtensia burtensis (Otschev); первые шейные позвонки справа; экз. ПИН
No 3359/11; Архангельская обл., р. Пижма-Мезенская; нижний триас, баскунчакская
серия
Туловищные позвонки тихвинский имеют широкие и плоские невральные дуги,
их дорзальная поверхность круто наклонена вперед, но по мере приближения к
хвостовому отделу (вследствие относительного удлинения центров) наклон умень
шается. Остистые отростки расположены в задней половине дуги, 'в сечении имеют
каплевидную, в плане заостренную вперед форму и слегка наклонены назад. Пре-
Зигапофизы тонкие, снизу поддерживаются гребнем. Постзигапофизы более ко
роткие и массивные. Поперечные отростки позвонков почти не выражены, пред
ставляют собой небольшие выступы с вертикальными фасетками прикрепления
ребер. Поперечные отростки гораздо сильнее развиты у Procolophon
(Huene* 1902).
У поясничных позвонков реберные фасетки отсутствуют, на их месте имеются
лишь треугольные бугорки. Последний предкрестцовый позвонок несет очень ши
рокий и значительно выступающий поперечный отросток с широкой фасеткой. Поз
вонки склерозавра более массивные, "между постзигапофизами имеются слабо раз
витые горизонтальные пластины.
Крестцовые позвонки тихвинский устроены также, 'лишь зигапофизы развиты
слабее. Эти позвонки имеют относительно развитые поперечные отростки с широ
кими фасетками.
Передние хвостовые позвонки устроены в общем схоже, поперечные отростки
слабо развиты, фасетки почти горизонтальные. Начиная примерно с 32-го позвон
ка, Невральные дуги утрачивают остистые отростки, 'становятся низкими, Тела по
звонков удлиняются, вплоть до середины хвоста сохраняются презигапофизы, Да
лее они представляют собой лишь фасетку сочленения на невральной дуге.
Гемальные дуги последних позвонков дорзально вогнуты, раздвоены на прокси
мальных концах, 'слегка расширены на дорзальных. Прикрепляются к фасеткам на
вентральных поверхностях тел.
Первое ребро, Находящееся у атланта, короткое двухголовчатое, расширенное
дистально. Ребро пятого позвонка Имеет характерный вид "вилочки" с раздвоен
ным дистальным концом, что может указывать на наличие лимфатического сердца.
У следующих двух ребер дистальные концы сильно расширены. Туловищные ребра
(примерно до 14—15-го позвонков) слабо изогнуты, сочленовная часть неясно под
разделяется впадинками на головку и бугорок. После сочленовной части идет не
большая перетяжка, и далее расширение, и лишь затем начинается нормальное су
жение к дистальному концу. На дорзо-медиальной плоскости расширения находится
недлинный широкий желобок.
ребер 15— 20-г*о позвонков нет разделения на голов
ку и бугорок, Нет и расширения. Последний предкрестцовый позвонок несет корот-
50
кое массивное ребро, имеющее овальную неглубокую выемку на передне-ёерхней
плоскости расширенного дистального конца. Тзкой сочленовной поверхностью это
ребро причленяется сбоку к подвздошной кости. Ребра крестцовых позвонков ко
роткие массивные, с сочленовными площадками на дистальных концах, которыми
они прилегают к внутренней поверхности подвздошной кости. Ребра передних хво
стовых позвонков сильно изогнутые, уплощенные почти горизонтально.
У тихвинский, как и у контризавра, никаких следов гастрального скелета не
обнаружено. Наличие брюшных ребер отмечено у проколофона (BroiII,t Schruder
1936), «лептоплейрона и коилоскиозавра (Ниепе* 1912). flo описанию Броили и Шре
дера, брюшные ребра проколофона начинаются за дистальным концом межключицы
и идут тесно друг к другу. Р'ебра тонкие, раздвоены на медиальном конце.
Пояса конечностей
Плечевой.пояс широкий и массивный, строение его типично для котилозавров,
однако пояс весьма облегчен за счет появления широких хрящевых прослоев между
костями и некоторой редукции самих костей. Плечевой пояс склерозавра (Seeley,
1900) построен полностью аналогично таковому проколофонов и мало похож на мас
сивный и сросшийся пояс парейазавров, к которым иногда относят склерозавра.
Лопатка плоская, расширена на нижнем и верхнем концах. Передний край кости
несет узкую площадку сочленения с клейтрумом и клавикулой. В месте их соеди
нения имеется очень слабо выраженный акромиональный выступ. По латериальной
поверхности кости в нижней части проходит гребень, отделяющий утолщенный зад-'
ний край от тонкого переднего. Задний край несет косо расположенное углубление-
BepxHe-rfepeflHFOK) часть гленоид'ной ямки.
Присутствуют оба коракоида. Прокоракоид на латеральном краю дорзальной
поверхности несет вырезку супракоракоидного отверстия. На вентральной поверх
ности кости от вырезки идет косо медиально желоб, латерально вырезку перекры- '
вает нижний край скапулы. Передне-боковой утолщенный край коракоида образует
заднюю часть гленоидной ямки. Медиальные края коракоида и прокоракоида вплот
ную прилегают к Т-с(бразной межключице. Ее вентральная сторона несет спереди
слегка изгибающийся по краям назад продольный гребень. От его середины отхо
дит поперечный гребень, Постепенно выполаживающийся и переходящий в длинный
отросток, продолжающийся каудально далеко за задние края коракоидов. Такая же харак
терная межключица и у склерозавра, резко отличная от тонкой межключицы парейазавров.
Ключицы тонкие и длинные, на внутренней стороне имеют широкий желоб и рез
ко выгнуты в латеральной трети вверх. В желоб ключиц входит передний край меж
ключицы, 'а загнутые вверх края костей прилегают к нижней половине переднего
края скапулы. Выше ключицы к этому же краю присоединяется тонкий, расширен-'
ный вверху клейтрум.
Передние конечности тихвинский относительно длинные, но массивные, 'с. корот
кой кистью (гораздо короче и шире, чем у никтифрурета). Фаланговая формула
2,3,4,4,3.
Эпифизы плечевой кости перекручены относительно друг друга на 30°. Головки
кости были в значительной степени образованы хрящем. Processus medialis раз
вит хорошо и представляет собой мощный гребень, ограничивающий с медиальной
стороны ямку для вентральных плечевых мышц. Дельтопекторальный гребень раз
вит значительно слабее, это небольшой бугорок (tuberculum majus) на наружной
стороне проксимального эпифиза. Дистальный эпифиз представляет собой тонкую
пластинку, от латерального края которой идет тонкий гребень к медиальному от- '
ростку (возможно, 'соответствует супинаторному отростку). В нижней части гребня
находится большое овальное энтэпикондилярное отверстие. Внутренний надмыщелок
для сгибателя локтя (epicondylus ulnaris) развит слабо, condyli ulnaris и radialis
представляют собой небольшие возвышения, образованные хрящем и ограниченные
сверху полукруглыми гребнями. С медиального края пластины имеется небольшой
наружный надмыщелок для разгибателя локтя.
Локтевая кость тонкая в диафизе и не сильно расширена в эпифизах. Прокси
мальный эпифиз имеет слабо развитый гребень на передней поверхности и сильнее
51
развитый на медиальном краю (возможно, соответствует олекранону). Лучевая
кость тонкая, эпифизы округлого сечения, причем сторона, обращенная к локтевой
кости, слабо уплощена.
Кисть широкая и плоская. Ossa carpalia слагает два ряда косточек, прилега- '
ющих к костям кисти и костям предплечья. Проксимальный ряд состоит из лучевой
и локтевой костей запястья и intermedium. У u In are и radi ale хорошо выражены су
ставные поверхности причленения к костям предплечья и почти не выражены суста
вы с carpalia distalia. Можно предполагать значительную гибкость кисти в запястье.
Центральная кость одна. Четыре кости дистального ряда округло прямоугольные,
суставные поверхности к метакарпусу развиты лучше, чем боковые интеркарпаль-
ные. Наибольпьая из костей находится с медиального края, наименьшая — с лате
рального. Хорошо развита округлая pisiphorme* У метакарпальных костей на про
ксимальных эпифизах - сочленовные углубления для ossa distalia,. «а дистальных —
округлые головки сочленения с фалангами. Фаланги короткие и массивные, упло
щенные. Поверхности сочленения уплощенно-бкруглые, сильно развиты флексорные
бугорки и желобки для связок на боковых поверхностях.
Таз широкий и относительно массивный. Медиальная поверхность подвздошно*
кости несет слабо выраженный гребень, разграничивающий поля присоединения
ребер крестцовых позвонков* Резко выраженные гребни — длинный передний, иду
щий наискосок от передне-н"ижнего края к середине заднего, й короткий задний,
идущий снизу вверх по заднему краю, Ограничивают спереди, 'сверху и сзади ацета-
булярную ямку с гладкой, хорошо окостневавшей поверхностью.
На расширенном латеральном краю лобковой кости имеется довольно большое
обтураторное отверстие. От этого отверстия по дорзальной поверхности кости идет
сужающийся конусообразный желоб. С переднего края желоб ограничен сильно раз
витым гребнем. Верхне-боковой край кости несет сзади гребня площадку сочлене
ния с подвздошной костью, а задне-боковой образует передне-нижнюю часть ацета-
булярной впадины. Седалищная кость широкая и длинная, узкая сзади и расширен
ная спереди, Передняя часть латерального края несет мощный бугор с двумя фа
сетками, к верхней причленяется подвздошная кость, а боковая образует задне-
нижнюю часть ацетабулярной ямки. Между костями имелись хрящевые прослои, По
этому на породе они лежат обычно на некотором расстоянии друг от друга. Насколько
можно судить, так же построены тазовые пояса всех проколофонов (в том числе и
склерозавра), тогда как у парейазаврид таз сросшийся и массивный, верхняя часть
подвздошной кости расширена и удлинена назад.
Бедро длинное и массивное, Диафиз округло-треугольного сечения. Проксималь
ный эпифиз несет слабо выраженный аддукторный гребень, на проксимальном кон
це которого в примитивном положении находится четвертый трохантер для бедрен-
но->£востовой мускулатуры. Гребень внутреннего трохантера развит, слабо. Оба
гребня разделяет широкая и глубокая межтрохантерная яма. На дистальном эпи
физе выделяются два недлинных мыщелка для причленения костей голени, большая
берцовая кость характерной формы, с массивным проксимальным эпифизом и не-'
большим дистальным. На вентро-медиальной поверхности проксимального эпифиза
имеется слабо развитый кнемиальный гребень. Малая берцовая — тонкая, сильно
изогнутая, эпифизы в сечении уплощенно-бвальной формы.
Проксимальный ряд ossa tarsalia состоит из двух костей: astragalus - резуль-
тат срастания tibiale,. intermedium и одной из centrale (имеется хорошо выраженный
шов) и calcaneus. По дорзальной поверхности таранной кости идет желоб к отвер
стию прободающей артерии между таранной и пяточной костями. Дистальный ряд
состоит из четырех костей, наибольших размеров tarsale distale IV, соединяющаяся
с двумя метатарзальными. Интертарзальный сустав выражен хорошо. Метатарзум
и фаланги ничем не отличаются от таковых кисти.
Надо отметить, что задние конечности всех проколофонид построены очень
сходно, тогда как у диадектид (Romer,. «1944) и у парейазаврид (даже у рипейазавра:
Чудинов, 1957) таранная и пяточная кости обычно срастаются.
Панцирь у проколофонов обычно отсутствует, лишь у склерозавра Хюне (Huene,.
1902) отмечает наличие панциря из трех продольных рядов крупных округлыхскуль-
птированных бляшек й нескольких мелких по сторонам.
52
МУСКУЛЫ, ЖЕЛЕЗЫ И НЕРВНО-СОСУДИСТАЯ
СИСТЕМА ГОЛОВЫ
Мускулатура головы
Реконструкция мускулатуры головы у проколофонов встречает известные труд
ности, поскольку мускулы оставляют слабые следы на костях черепа. Поэтому
часто можно лишь делать предположения о местах их прикрепления и степени раз
вития, сравнивая структуры черепов проколофонов и современных рептилий.
Первая попытка дать реконструкцию мускулов головы проколофонида была сде
лана Колбертом (Colbert,. 1946) для гипсогната. На этой реконструкции показаны
некоторые мускульные пуки аддукторов и депрессор нижней челюсти ящериц, отне
сенные к черепу гипсогната. На реконструкции показано захождение части аддук
торной мускулатуры в удлиненную орбиту,
Лингвальная мускулатура. Мускулатура дна полости рта не может быть рекон
струирована на имеющемся материале. Лишь у некоторых форм (например, Oren
burgia) можно определить положение мускула, сопоставимого с m. genioglossus
(m.ijntramandidularis: Save-Soderbergh, «1945) ящериц по наличию желоба на антеро-
вентральном краю передней части нижней челюсти.
Депрессор нижней челюсти. Депрессор (рис, 25) весьма четко фиксируется
между гребнем нависающего заднего края надвисочной кости и ретроартикуляр-
ным отростком нижней челюсти, <У тихвинский и контритозавра поле прикрепления
депрессора на ретроартикулярном отростке разделено вдоль невысоким гребнем.
Аддукторы нижней челюсти (рис, 25) у проколофонов были довольно длинные и
снабжены в большинстве случаев массивными давящими зубами. Резкое удлинение
орбиты с образованием "орбито-бкулового" отверстия говорит в пользу такого
предположения.
Орбита высших проколофонов представляет собой овально-грушевидное отвер
стие, занимающее от половины до двух третей общей длины черепа. При этом
часть, занятая глазом, Хорошо заметна, ее ограничивает сзади и сверху слабый
выступ заднелобных костей, а снизу - выступ на скуловой кости. Всю заднюю,
удлиненную часть орбиты, по-ёидимому, занимали аддукторные мускулы. Пред
ставляется почти несомненным, что у высших форм вся или большая часть верх
ней головки m.adductoris mandibularis externus входила в заднюю часть орбиты.
Обычное место прикрепления m. pseudotemporalis superficialis и т. pseudot. pro
fundus (pterigoideus posterior: Adams,. 1919) на черепе - антеро-медиальный край
верхнего височного окна, Наружная поверхность проотики (на proc. posterior) и
верхняя треть эпиптеригоида. На нижней челюсти - антеромедиальный край аддук
торной ямы, До постеро-медиального края короноида.
Нет оснований полагать, Что места прикрепления сходных мускулов у проколо
фонов значительно отличались. Верхняя часть m. pseudot. superficialis вполне могла
входить в заднюю часть удлиненной орбиты, а также прикрепляться на заднем от
ростке проотики и на верхней, расширенной и уплощенной части эпиптеригоида.
Брок (Brock,. 1938) отмечает, что прикрепление m.pseudotemporalis на внутренней
крыловидной у примитивных рептилий первично, 'а смещение на орбито-т'емпораль-
ную стенку происходит позже. Трудно судить, насколько это справедливо, однако
эпиптеригоид у примитивных проколофонов (никтифрурет) очень тонкий и округлый
в верхней части, й мало вероятно, Чтобы на нем могла прикрепляться сколько-нибудь
значительная мускульная масса.
На нижней челюсти, в районе возможного прикрепления апоневроза мускула,
имеется площадка.
Достаточно определенны места прикрепления аналогов птериго-мандибулярного
мускула. Это латеральный край квадратной ветви птеригоида и задняя часть ниж
ней челюсти. На квадратном отростке птеригоида у никтифрурета имеются ясно
выраженные гребни прикрепления мускулов. У тихвинский такие гребни развиты
гораздо слабее. Может быть, 'это связано с более сильным развитием иториго-маи-
дибуляриого мускула у примитивных форм при меньшем развитии m. add. тand. ex
ternus (в связи с относительно небольшим удлинением орбиты). Нижняя часть му-
53
Рис, 25. Tichvinskia Tchudinov et Vjuschkov; реконструкция жевательной муску
латуры головы, левая задняя половина черепа, кости щечной области удалены
скула прикреплялась к обычно хорошо выраженному гребню на наружной стороне
заднего угла нижней челюсти и обворачивалась вокруг ее нижнего края.
Мускул, соответствующий m. add. тand.posterior,мог идти от внутренней поверх
ности квадратной кости и квадратного отростка птеригоида к верхней поверхности
Меккелева хряща и внутренним краям аддукторной ямы. Внутренний край квадрат
ной кости слегка вырезан и образует небольшой гребень, аналогичный медиально
му гребню квадратной кости ящериц - месту прикрепления мускула.
Мышцы нббно-квадратного комплекса. М. levator pterygoideus (pterygo-parietalis:
Adams, 1919) у ящериц развит слабо, его обычное место прикрепления - поверх
ность птеригоида между эпиптеригоидом и базиптеригоидным суставом. У проко
лофонов (особенно у примитивных) широкое основание эпиптеригоида находится
слишком близко от сустава (у капторинид эпиптеригоид иногда участвует в обра
зовании сочленения: Warren,. 1961). Protractor pterygoideus (pterygo-sphenoidalis:
Aiams, 1919) прикреплялся на Г-образной системе гребней на латеральной поверх
ности проотики и на квадратной ветви птеригоида и был* возможно, достаточно
сильно развит.
М.Levator bulbi отходил от передней поверхности pila antotica,. «возможно, от
слабого желоба на антеро-латеральной стороне отростка и шел к глазному яблоку.
"Глазные" мускулы. Часть m. rectus superior могла прикрепляться на вентраль
ной поверхности pila antotica,. «как это наблюдается у лепидозавров (например, у
Sphenodon: Save-Soderbergh, «1946).
М. rectus posterior (m.bursalis + m.retractor bulbi) прикрепляется в районе ре-
тракторного миодома на передней части дорзальной поверхности базисфеноида,
Шейная мускулатура. Наибольший интерес представляет один из пучков шейных
мускулов, Прикреплявшихся к tuberculum spheno-occipitale.
Вой (Boy, «1971) описывает на вентральной поверхности заднего края парасфе-
ноида лабиринтодонта ("Actinodon"
latirostris)
конусообразные ямки, в которые
могли входить пучки шейных мускулов. У никтифрурета здесь имеются более глу-
54
бокие ямки, впереди которых находятся очень слабо развитые бугры. У тихвинский
бугры развиты сильнее, тогда как ямки почти исчезают. Зозможно, в связи с уве
личением подвижности шеи шло усиление мускулов. У ящериц сфено-бкципитальные
бугры служат для прикрепления вентральной группы сгибателей головы.
Железы и Якобсонов орган
Затруднения при реконструкции краниальных желез у проколофонов объясняют
ся прежде всего недостаточной изученностью соотношений костных структур че
репа и желез у современных ящериц. Поэтому может быть реконструировано рас
положение лишь некоторых желез у проколофонов.
Наиболее просто определяется положение латеральной носовой железы, которая
располагалась в более или менее глубокой ямке у заднего края ноздри. У прими
тивного парейазаврида — парейзуха (рис. 2G) ямка латеральной носовой железы
полностью располагается на дорзальной поверхности максиллы, на задней стенке
ноздри (возможно, первичное, примитивное для рептилий положение). Депрессия
железы у никтифрурета очень большая, и железа занимает значительное место на
задней стенке ноздри, несколько смещаясь при этом на боковую поверхность чере
па. Однако основная часть железы, скорее всего, лежала медиально от переднего
края максиллы, на гладкой вогнутости септомаксиллы. В ходе дальнейшей эволю
ции можно наблюдать, как ямка железы на боковой поверхности черепа проколофо
нов уменьшается в размерах и идет редукция анастомозма максиллярной и нижней
носовой артерии в районе железы (циркумнариальный анастомоз). У тихвинский
этот анастомоз полностью отсутствовал. Трудно, утверждать определенно, говорит
ли это о редукции железы или о ее смещении на постеро-медиальную поверхность
черепа. Более верным представляется все же последнее, поскольку редукция же-'
лезы плохо согласуется с рядом фактов, говорящих о возрастании наземности об-'
раза жизни проколофонов в ходе их эволюции. Кроме того, ty ящериц железа сме- '
щается на дорзо-медиальную поверхность верхнечелюстной кости (Webb, <1951),. «а
у хамелеонов заходит частично даже на внутреннюю поверхность предлобной кости
(Engelbrecht, «1951),, <не оставляя здесь более или менее ясных следов.
У некоторых проколофонов (контритозавр, тихвинскиа) на медиальном шве пред-
челюстных и сошниковых костей имеется продольное углубление с гладкими стен
ками. Для многих современных форм (Chamesaura:
du Plessis, 1944; Cordylus:
van
Pletzen,. «1946; Palmatogecko,
Oedura: Webb,. 1951) в схожем положении отмечают
сошниковую железу (glandula womeralis). Энгельбрехт (Engelbrecht, <195D для хаме
леона Micvosaura
pumila описывает продолжение железы и на нёбную кость.
О степени развития Якобсонова органа у многих проколофонов судить трудно,
поскольку прикрывающая его сверху септомаксилла обычно отсутствует, имеются
лишь отверстия веточек нббной артерии, Питавших орган. У никтифрурета сохраня
ется септомаксилла, и орган располагался в обширной полости между ней и дор
зальной поверхностью сошника (рис 7). Полость его широко открывалась в перед
ний край костной хоаны, где имеются две небольшие вырезки двойного протока
(может быть, следы слезного протока, топографически связанного с Якобсоновым
органом, как у лепидозавров и червяг).
Кровеносная система
Артериальная система. Артериальная система головы проколофонов может
быть реконструирована с известной степенью достоверности путем сравнения со
хранившихся на костях черепа отпечатков и отверстий с аналогичными следами на'
костях современных животных..При реконструировании наиболее верным представ
ляется сравнение с ящерицами. Хотя проколофоны и ящерицы, по-видимому, разви
вались весьма рано разошедшимися стволами, глубокое внутреннее морфологиче-
1
ское сходство сохранилось. Это хорошо видно по результатам реконструкции нервно-
сосудистой системы проколофонов.; Выявляется удивительное однообразие в про-
55
A
Рис. 26. Передние части черепов, сбоку
а - Nyctiphruretus
acudens
Efremov;. по экз. ПИН No 158/5,11; Архангельская
обл., р.Мезень; верхняя пермь, нижнетатарский подъярус; б — Contritosaunts
simus
Ivachnenko; по экз. ПИН No 3355/1; Горьковская обл„ д. Липово; нижний триас, ни
зы ветлужской серии; г — Tichvinskia
vjatkensis
Tchudinov et Vjuschkov; по экз.
ПИН No 954/1,2; Кировская обл., р. Федоровка; нижний триас, баскунчакская серия;
г — Agamacaucasica
Eichwald (Agamidae,. современный)
хождении не только основных артериальных и нервных стволов, но и ряда анасто
мозов и других мелких деталей. До сих пор реконструкций прохождения краниаль
ных сосудов и нервов у проколофонов не делалось.
Внутренняя сонная артерия у проколофонов до входа в базисфеноид давала от
ветвление, стапедиальную артерию, проходившую рядом со слуховой косточкой.
Однако слуховая косточка проколофонов тонкая и не несет отверстия для прохож
дения артерии,как это наблюдается у большинства примитивных форм.
У ящериц стапедиальная артерия делится на мандибулярную и темпоральную
ветви. Мандибулярная артерия у проколофонов входила в аддукторную яму нижней
челюсти и здесь давала переднюю и заднюю ветви. Задняя ветвь проходила через
два-т'ри небольших отверстия на заднем, утолщенном краю надугловой кости. Ве
точки ее выходили на наружную поверхность кости в районе гребня для птериго- '
мандибулярного мускул а'(через супрангулярные отверстия). Основная часть ветви
шла под челюстным блоком и выходила в антеро-медиальном углу ретроартикуляр-
ного отростка, снабжая депрессор нижней челюсти.
Передняя ветвь артерии давала ответвление в меккелево отверстие (рис. 19 г)
и далее входила в меккелев канал. Сразу же за короноидом она раздваивалась на
медиальную и латеральную ветви. У проколофонов (и ящериц) медиальная ветвь
выходила через сплениальное отверстие на внутреннюю поверхность челюсти. У
агам и хамелеонов, у которых пластинчатая кость сильно редуцированна, ветвь .
выходит прямо через открытый изнутри меккелев желоб. У проколофонов и всех
ящериц латеральная, альвеолярная ветвь через отверстие в зубной кости несколь
ко впереди короноида (рис. 19 д) уходит в толщу кости, проходя здесь по сквозному
продольному каналу. Этот канал у проколофонов небольшого диаметра, но сформи
рован, хорошо. У ящериц канал относительно таких же размеров, но сформирован
несколько хуже. У агамид и хамелеонов (рис. 54д) этот канал сформирован хорошо
56
Рис. 27. Tichvinskia Tchudinov et Vjuschkov; реконструкция кровеносной системы
головы, Правая передняя половина черепа сверху, кости крыши удалены
и его поперечное сечение составляет большую часть сечения кости. Идя по каналу,
артерия на всем протяжении дает веточки к зубам (рис. 53). Около самого входа
и в передней части челюсти артерия у проколофонов выходила на наружную поверх
ность челюсти через подбородочные отверстия. У высших проколофонов отверстия
немногочисленны (одно-Два задних, йва-три передних), у никтифрурета и фаанто- '
завра С-7 отверстий идут по всей длине кости.
Вдоль меккелева желоба у проколофонов шел основной ствол мандибулярной
артерии, так как, Помимо заднего входного отверстия, есть переднее, расположен
ное несколько позади симфиза. У ящериц переднего входного отверстия нет, И по
желобу идет небольшая веточка, снабжающая хрящ.
По М.А.Шишкину (1973), у некоторых лабиринтодонтов поперечное ответвление
артерии, Пронизывающее кость и формирующее наружную мандибулярную артерию,
образовывало разветвления в толще кости, однако слияние их в канал не происхо
дило. У проколофонов наружная артерия отсутствовала, и альвеолярная ветвь шла
в толще кости.
В постдентальной части челюсти проколофонов, в местах прикрепления муску
лов, имеются небольшие питающие отверстия, например, с внутренней стороны че
люсти, Несколько позади короноидного отростка, в месте прикрепления псевдотем
порального мускула. Мандибулярную артерию сопровождала третья ветвь тройнич
ного нерва и ветвь лицевого.
Передняя ветвь стапедиальной артерии — темпоральная артерия — разделяется
на ряд ветвей. Одна из ветвей (фронтальная артерия) сопровождается основным
стволом Vi и идет высоко под крышей черепа, По вентральной поверхности лобной
кости, ho небольшим желобам на латеральных краях ее. По пути артерия отдает
ветви в обонятельный эпителий носовой капсулы (nasalis superior),, а основной
ствол (nasalis inferior) дает ответвление (ramus palatinus intermediatus) и далее
проходит по желобу на вентральной поверхности носовой кости. У проколофоиои
желоб наиболее хорошо выражен у никтрифрурета и контритозавра. Он открывает-
57
ся одним-Двумя отверстиями несколько выше ноздри. У современных ящериц эта
артерия выходит через небольшую вырезку в верхне-Эаднем краю ноздри и обра
зует циркумнариальный анастомоз с выходящей из переднего альвеолярного от
верстия максиллярной артерией (Oelrich,, 1956). Лроколофоны первично (никтифру-
рет, рис. 1) имеют структуру, схожую с передним альвеолярным отверстием, по
скольку латеральная носовая железа лежит у задней стенки ноздри, однако по ме
ре смещения железы отверстие постепенно уменьшается и совсем исчезает у
тихвинский. При этом уменьшаются и отверстия глубокой носовой артерии, сме- '
щаясь дорзально. Ход редукции анастомозных структур в эволюции проколофонов
хорошо виден при сопоставлении носовых отделов (рис. 2Са-г).
Темпоральная артерия у ящериц после отхождения фронтальной артерии раз
дваивается, йавая две орбитальные ветви - art.orbitalis superior и art.orbitalis
inferior. Art.orbitalis superior у ящериц, пройдя через отверстие в орбито-т'емпо-
ральной мембране, йает ветви к глазу, глазной мускулатуре и склере.
Art.orbitalis inferior (максиллярная артерия) ящериц сопровождается V2 » У яще
риц артерия подходит к инфраорбитальному отверстию и дает здесь четыре ответ
вления. Первое, "медиальное, образует постхоанный анастомоз с ветвью нёбной
артерии. Второе идет несколько назад, прободает подглазничную мембрану в ее
переднем углу и анастомозирует на вентральной поверхности мембраны с вен-'
тральной ветвью боковой ветви небной артерии, и проходит несколько вперед по
вентральной поверхности максиллы, сопровождаясь латеральной нёбной ветвью ли
цевого нерва. Третье ответвление уходит в толщу верхнечелюстной кости и обра
зует здесь продольный канал с неровными стенками (рис. 28 ж). Из канала веточки
кровеносных капилляров идут к зубам. Четвертая ветвь — йнфраорбитальная арте
рия - проходит в инфраорбитальное отверстие, следует по короткому желобу на
поверхности максиллы и уходит в толщу кости, сливаясь с третьей ветвью,идущей
в канале. При прохождении артерии в канале она дает несколько небольших вето
чек, выходящих на боковую поверхность черепа через два-'гри передних максилляр-
ных отверстия. До входа в канал основной ствол артерии анастомозирует с интер
медиальной нёбной ветвью фронтальной артерии и несколько впереди — с медиаль
ной ветвью нёбной артерии (предхоанный анастомоз). Передняя ветвь максилляр
ной артерии выходит в переднее альвеолярное отверстие, 'образуя вокруг ноздри
анастомоз с нисходящей ветвью фронтальной артерии, и дает субнариальную ветвь,
снабжающую передние носовые структуры и анастомозирующую с нёбной артерией.
У никтифрурета (рис. 28 а) наблюдается лишь три ответвления максиллярной ар
терии перед входом в инфраорбитальное отверстие. Желобок позади передней стен
ки орбиты, идущий от инфраорбитального отверстия к желобу нёбной артерии, го
ворит о наличии постхоанного анастомоза. Вторая ветвь артерии уходила в неболь
шое отверстие в толщу верхнечелюстной кости и, образуя небольшое расширение
у шва верхнечелюстной и скуловой костей, выходила на латеральную поверхность
черепа. В толще кости, вдоль шва, имеется ряд небольших лакун, связанных друг
с другом капиллярными каналами. Основная часть артерии проходила в инфраорби
тальное отверстие, Шла по желобу и давала ответвление на предхоанный анастомоз
(хорошо выраженный желобок, идущий медиально к краю костной хоаны). Передние
ветви выходили через два крупных отверстия в задней стенке ноздри, причем ме
диальное отверстие выходит в ямку латеральной носовой железы и по положению
соответствует переднему альвеолярному отверстию ящериц (не будучи его гомо
логом), а латеральное - переднему максиллярному (переднему из ряда лабиальных
отверстий). У контритозавра (рис. 28в,г) ход артерии несколько отличается. Си
стема лакун в толще верхнечелюстной кости развита сильнее, они лучше соедине
ны между собой и уже не связаны со швом между скуловой и верхнечелюстной
костями, Медиальное отверстие в задней стенке ноздри очень небольшое, а лате
ральное смещается на боковую поверхность черепа. У тихвинский (еще лучше это
выражено у макрофона) цепь синусов в толще кости сливается в сплошной канал,
идущий вдоль кости до латерального отверстия. Однако объединения желоба ос
новного ствола и канала (как у ящериц) не происходит. Надо отметить, *что пород
нее альвеолярное отверстие ящериц не совсем соответствует таковому примитив
ных проколофонов, Поскольку у проколофонов через это отверстие проходит основ-
58
apoch
af/oc/г
Рис. 28, Схемы прохождения верхнечелюстной и части небной артерий
а, б — Nyctiphmretus
Efremov; в, г — Contritosaums
Ivachnenko; Д,.е.
—
Tichvin°
skia Tchudinov et Vjuschkov; ж, з — A gam a Daudin. а, в, д, ж - горизонтальные се
чения верхней челюсти на уровне альвеолярного канала; б, г, е, 3 — схема прохож
дения артерий
ной итвол артерии, а у ящериц - слившийся с проходящей в толще кости ветвью.
Также и переднее максиллярное отверстие проколофонов пропускает лишь часть
основного ствола артерии, а у ящериц в лабиальные отверстия выходят ветки из
канала.
Внутренняя сонная артерия у проколофонов шла в кранио-квадратном проходе,
над крыльями парасфеноида, И, пройдя базисфеноид, 'давала мозговую и нёбную
ветви. Ветвление происходит в толще базисфеноида, примерно на уровне спинки
турецкого седла.
Небная артерия у ящериц дает три ветви сразу же по выходе из видиева канала.
Небольшая задняя ветвь идет по вентральной стороне пара-базисфеноидного ком
плекса назад, восходящая ветвь поднимается по базисфеноидному ростру И про
ходит вдоль межглазничной септы в носовую капсулу, оканчиваясь в олфакторной
области. Ход основного ствола артерии у проколофонов хорошо фиксируется сла
бым желобом, сопровождаемым мелкими отверстиями, уходящими в глубь кости,
к зубам на нббе. В этом районе артерия давала крупное ответвление - латераль
ную ветвь, проходившую в небное (подглазничное) отверстие, й разветвлявшуюся
здесь на две ветви - анастомозную, уходившую в толщу кости, й нисходящую, вы-
59
ходившую па вентральную сторону нёба. Ветвь нёбной артерии, выходящую на ла
теральную поверхность черепа, 'отмечает Ярвик (Jarvik, 1942) у Polypterus.
Далее артерия у проколофонов у шва нёбных и сошниковых костей давала от
ветвление на постхоанный анастомоз с максиллярной артерией и проходила в сош
никовый канати У тихвинский в месте выхода артерии к латеральному краю хоаны
идет узкий желоб, около дистального конца которого имеется небольшое.отверстие,
пропускавшее ветку на вентральную поверхность нёба к сошниковым зубам. Желоб
оканчивается на латеральном краю хоаны, точно напротив анастомозного желобка
максиллярной артерии. Иногда (у контритозавра) выходное отверстие анастомоз-
ных ветвей нёбной артерии находится примерно на середине длины сошника, Иногда
(у тихвинский) на самом переднем краю его, у шва с предчелюстными костями, но
всегда напротив желобка максиллярной артерии.
В этом отношении интересно отметить, что у парейазуха (рис. G) нёбная арте
рия шла по поверхности сошника, давая небольшие ответвления в толщу кости, "к
нёбным зубам, а напротив желобка анастомозной ветви максиллярной артерии на
переднем краю сошника имеется вырезка, По которой и проходила эта ветвь нёб
ной артерии.
Апикальные ветви артерии, у проколофонов, Как и у всех рептилий, "могли ана-
стомозировать в передней части носовой капсулы с ветвями нижней носовой арте
рии и с су.бнариальной артерией.
Реконструкция прохождения некоторых сосудов для тихвинский дана на рис. 27.
Венозная система. Венозная система головы проколофонов может быть рекон
струирована весьма не полно. Можно с уверенностью определить положение двух-
трех венозных синусов. Это в основном объясняется тем, 'что вены почти не остав
ляли собственных отпечатков и отверстий на костях, а, кроме того, венозная си
стема современных ящериц изучена менее подробно, чем хотелось бы, особенно
плохо известно соотношение костей черепа и вен.
В желобе на дорзальной поверхности верхнечелюстной кости проколофонов
могла лежать нёбная вена; максиллярная вена, Проходящая в толще верхнечелюст
ной кости у ящериц, возможно, имелась и у проколофонов, о чем может говорить
наличие синусов. Из поперечных венозных структур может быть определено лишь
положение гипофизарной вены - по нижне-йередним поверхностям pilae antotica.
У никтифрурета, в связи с сильным развитием маргинальных гребней, ограничи
вающих трабекулярный отдел эндокрания и протягивающихся от спинки турецкого
седла до трабекулярных бугров, vena pituitaria была поднята довольно высоко над
поверхностью базисфеноида.
В шве между переднеушной и верхнезатылочной костями у проколофонов име
ется крупное отверстие, к которому по дорзо-медиальной поверхности передне-
ушной кости идет желоб. У игуан и лацерт в этом месте есть небольшой полукруг
лый желобок - след петли средней мозговой вены на внутренней поверхности моз
говой коробки. У агамид имеется здесь же отверстие, полностью схожее по поло
жению с отверстием у проколофонов. Над верхней затылочной костью средняя
мозговая вена выходит на затылочную поверхность, Делает здесь петлю и через
отверстие возвращается в мозговую коробку.
Задние мозговые вены, отходившие от медио-Дорзального синуса под крышей
черепа, Могли выходить через вагусные отверстия.
Желоба у никтифрурета, На парасфеноиде, Могут соответствовать желобам у
лабиринтодонтов, и тогда по ним проходила нёбная вена (Shishkin,. 1968). Дальней
шая редукция парасфеноида у проколофонов привела к исчезновению желобов.
Черепно-мозговые нервы
Сходство черепных структур высших проколофонов и ящериц помогает со зна
чительной степенью достоверности восстанавливать прохождение некоторых кра
ниальных нервов исследуемой группы.
N. trigeminus (V). Тройничный нерв делится на три ветви - глазную (n.ophtal-
micus),. «верхнечелюстную (п.maxillaris или infraorbitales) и нижнечелюстную
(n.mandibularis). Приблизительное место выхода ствола нерва у проколофонов на-
60
ходится несколько выше переднего края проотики (у никтифрурета и буртензии
имеется вырезка нерва).
Глазная ветвь (V^) сопровождает фронтальную артерию, Идя высоко по вен
тральной поверхности лобных костей. У ящериц, через орбито-йазальную мембрану
она входит в носовую капсулу и здесь разветвляется на две ветви - медиальный и
латеральный этмоидный нервы. У проколофонов ход латерального этмоидного нер
ва хорошо прослеживается. Он идет по желобу на вентральной поверхности носо
вых костей и выходит на латеральную поверхность черепа в области верхней части
латеральной носовой железы. Здесь, скорее всего, эта ветвь анастомозировала с
апикальной ветвью максиллярного нерва. У высших проколофонов (тихвинския) в
связи со смещением латеральной носовой железы происходит редукция анастомоза,
и нерв выходит тонкими веточками через два-три отверстия на дорзальной поверх
ности носовой кости. Медиальный этмоидный нерв, как и у всех рептилий, Вероят
но, подойдя к передней стенке носовой капсулы, Выходил на вентральную сторону
неба через небольшие анастомозные отверстия в предчелюстных костях.
Верхнечелюстной нерв (V2) проходил по дорзальной поверхности нёбного от
ростка птеригоида и подходил к инфраорбитальному отверстию, по пути давая ве
точки, уходившие под глазное дно и, вероятно, образовывавшие здесь ряд сплете
ний (инфраорбитальное и нёбное) с симпатическими волокнами лицевого нерва.
В местах этих сплетений имеются небольшие отверстия для прохождения ветвей
на вентральную сторону нёба и иннервации нёбных зубов. Далее основной ствол
верхнечелюстного нерва проходил в инфраорбитальное отверстие и сопровождал
дорзальную ветвь максиллярной артерии, Шедшую по желобу верхнечелюстной ко
сти. Апикальные ветви нерва, Вероятно, анастомозировали вокруг ноздри с ветвя
ми латерального этмоидного нерва. Перед входом в инфраорбитальное отверстие
верхнечелюстной нерв отдавал две ветви - медиальную'на постхоанный анастомоз
с нёбным нервом и латеральную, уходившую в толщу верхнечелюстной кости и раз
ветвлявшуюся здесь вперед и назад, проходя у высших проколофонов в продольном
канале максиллы и иннервируя зубы. Заднее ответвление анастомозировало с ла
теральной ветвью, Шедшей от инфраорбитального сплетения и проходившей в под
глазничное окно. Ветвление нерва в этом районе у ящериц в целом схоже, однако
латеральная ветвь, идущая от инфраорбитального сплетения на вентральную по
верхность нёба, 'не дает ответвления, "уходящего вглубь максиллы, а анастомози
рует с особой задней веточкой верхнечелюстного нерва на вентральной поверхно
сти подглазничной мембраны.
У бесхвостовых амфибий (Rana: Jarvik,. 1942) основную роль играет палато-на-
зальный ствол, Выходящий на вентральную поверхность нёба и разветвляющийся
здесь на переднюю и заднюю ветви, йннервирующие зубы. Этот стволи образует
пред- и постхоанные анастомозы с нёбным нервом. Небольшая дорзальная веточка
сопровождает слезный проток.
Сравнивая черепные структуры бесхвостых и остеолепиформов, Ярвик (Jarvik,.
1942) отмечает, что у последних инфраорбитальная ветвь также небольшая, идущая
вдоль слезного протока, а мощный ствол палато-н'азального нерва идет в толще
кости, 'сопровождаемый параллельной ветвью ("Rami Mx + Pmx"), идущей несколько
вентральнее и иннервировавшёй органы боковой линии. Пост- и предхоанные ана
стомозы образует палато-н'азальная ветвь. Однако у некоторых остеолепиформов,
возможно, палато-н'азальный ствол идет по вентральной поверхности нёба (Э.Й.Во
робьева, 1973).
У териодонтов (Татаринов, 1909) палато-назальный нерв выходит на вентраль
ную поверхность нёба через отверстие на стыке швов максиллы, трансверзума и
крыловидной кости (т.е. в положении, топографически схожем с положением под
глазничного окна ящериц и проколофонов), 'дорзальная ветвь развита слабо и па
лато-н'азальный и нёбный нервы анастомозируют на вентральной поверхности нёба.
Иная картина у лабиринтодонтов-бентозухид (Shishkin,, <1968)> «у которых анасто
мозы образует дорзальный ствол, а палато-йазальной названа ветвь, уходящая по- '
зади сублакримального отростка в толщу максиллы и разветвляющаяся вперед и
назад, тогда как у брахиопоидов (Шишкин, 1973) имеется мощная палато-н'азальная
ветвь, образующая, по крайней мере, постхоанный анастомоз, а дорзальная слезно-
61
носовом веточка слабо развита, и иногда вместо нее передние носовые структуры
иниервируют палато-максиллярная ветвь лицевого нерва.
Таким образом, Гомологизация ветвей максиллярного нерва у проколофонов и
ящериц весьма затруднительна, особенно в связи с отсутствием данных для сей-
муриаморфов. Однако представляется вполне возможным сопоставление ветви,
уходящей в толщу максиллы и иннервировавшей зубы, с палато-назальной ветвью.
Если в эволюции не происходило широкого вертикального перемещения ветвей
максиллярного нерва, То можно предполагать значительную редукцию палато-йа-
зального нерва у предков проколофонов, переход анастомозов к дорзальной ветви,
и затем вторичное усиление палато-йазального нерва без восстановления связей
с небным нервом. Такое усиление может быть хорошо прослежено в ряду никтифру-
рет - контритозавр - тихвинския. В этом случае картина ветвления максиллярно
го нерва у териодонтов по сравнению с проколофонами более близка к таковой у
бесхвостых и кистеперых.
Нижнечелюстной нерв (V3) проходит в аддукторную яму нижней челюсти и здесь
разветвляется на передний и задний стволы. Задний ствол, сопровождая ветвь ниж
нечелюстной артерии, выходит в отверстие в ретроартикулярном отростке и дает
веточки в супрангулярные отверстия. Передний ствол образует смешанные волок
на с барабанной струной, дает ответвление язычного нерва и входит в канал зуб
ной кости, йннервируя зубы и кожу (ветви, проходящие в подбородочные отверстия).
N. abducens (VI). Отводящий нерв снабжал "глазную" мускулатуру группы rec
tus posterior, отходя от вентральной поверхности продолговатого мозга. На спинке
турецкого седла этот нерв оставлял неглубокие короткие желобки, расположенные
у оснований pilae antotica. Образования закрытого сверху канала, как у ящериц,
не происходило.
N.facialis (VII). Лицевой нерв выходил из мозговой коробки через крупное
овальное отверстие в боковой стенке проотики, Под гребнем передних аддукторов.
Сразу же по выходе нерв разделялся на две ветви, Переднюю и заднюю. Задняя,
гиомандибулярная ветвь проходила несколько дорзальнее слуховой косточки. По
выходе из тимпанальной полости гиомандибулярный нерв у ящериц и других рептилий де
лится на два ствола — ramushyoideus и chorda tympani (г. mandibularis internus VII).
Гиоидная ветвь иннервирует структуры, окружающие подъязычный аппарат, и
не оставляет следов на костях.
Барабанная струна проникает в нижнюю челюсть через отверстие в передне-
наружном углу ретроартикулярного отростка и идет далее совместно с мандибу-
лярной ветвью тройничного нерва.
Небная ветвь лицевого нерва входит совместно с внутренней сонной артерией
в канал в базисфеноиде, и., пройдя его, сопровождает нёбную артерию, давая по
пути веточки на дорзальную поверхность птеригоида и нёбной кости к нёбному и
подглазничному сплетениям в области глазного дна. От нёбного сплетения отхо
дит крупное ответвление — латеральная ветвь, которая у ящериц прободает мем
брану подглазничного окна в задне-йрединном углу и выходит на вентральную по
верхность нёба, образуя здесь анастомоз с ветвью максиллярного нерва. У про
колофонов эту ветвь могло пропускать нёбное (подглазничное) окно на стыке
швов нёбной, наружной крыловидной и верхнечелюстной костей. В латеральной
стенке канала, идущего от этого отверстия, имеются небольшие канальцы в толщу
верхнечелюстной кости, Пропускавшие волокна нерва на анастомоз с задней
ветвью палато-йазального нерва.
Непосредственно позади передней стенки орбиты нёбный нерв давал анастомоз-
ную ветку (постхоанный анатомоз) с максиллярным нервом.
Далее основной ствол нёбного нерва входил в канал на дорзальной поверхности
сошника и, совместно с нёбной артерией, выходил вновь на поверхность, давая по
пути ответвления на предхоанный анастомоз и веточки на вентральную сторону к
сошниковым зубам. Апикальные ветви, как и у ящериц, Могли в районе передней
стенки носовой капсулы анастомоэировать с ветвями максиллярного нерва.
Nn.glossopharyngeus (IX)» vagus (X),. accesorius (XI). Нервы выходили из мозго
вой коробки через крупное отверстие на шве боковой затылочной и заднеушной костей.
N.byppoglossus (VII) проходил-через 1-2 отверстия в боковой затылочной кости.
62
ГЛАВА III
ПРОИСХОЖДЕНИЕ, ФИЛОГЕНИЯ И СТРАТИГРАФИЧЕСКОЕ
РАСПРОСТРАНЕНИЕ ПРОКОЛОФОНОВ
ПРОИСХОЖДЕНИЕ И ФИЛОГЕНИЯ ДИАДЕКТОМОРФНЫХ
КОТИЛОЗАВРОВ
Диадектоморфы, в состав которых входят проколофоны, помещаются некоторы
ми исследователями вслед за Ватсоном (Watson, 1917, 1957) в основание филогене
тического ствола всех зауропсидных рептилий.
Это вызывает постоянный интерес к вопросу о происхождении и дальнейшей
судьбе диадектоморфов.
В ранних работах Олсон (Olson, 1947) выводил проколофонов наряду с сейму-
риаморфами, парейазаврами и диадектами независимыми стволами из гипотетиче
ских примитивных эмболомеров. Позже (1962) Олсон предлагал иную схему, по ко-
которой диадекты имеют своими предками непосредственно сеймуриаморфов. Од^
нако пока неизвестны сеймуриаморфы, которые могли бы быть морфологически
исходной группой для диадектоморфов.
Так, хрониозухид можно исключить из числа возможных предков, что хорошо
виднопри сопоставлении строения пара-базисфеноидного комплекса и прохожде
ния ветвей внутренней сонной артерии (Shishkin, 1968), подтверждается рядом спе
цифических черт у хрониозухид (наличие предглазничной фонтанели, крупного нёб
ного отверстия, сквозного канала в верхнечелюстной кости и др.).
Сеймуриды и котлассииды высоко специализированы, но все же морфологи
чески гораздо более близки к диадектоморфам. В некотором отношении сходны с
диадектоморфами гефиростегиды. По М.А.Шишкину (1973), гефиростегиды и диа
дектоморфы представляют собой параллельные ветви, происходящие от примитив
ных сеймуриаморфов. В обеих группах в результате фетализации сохранилось де
финитивно-личиночное состояние ушной вырезки, захватывающей весь задний край
щеки, совместно с относительно вертикальной ориентировкой квадратной кости в
затылочной плоскости и горизонтальным расположением парокципитального отро
стка. В дальнейшем шло опускание щеки и дистальной части барабанной полости
и смена слуховой косточкой дорзо-латеральной ориентировки на горизонтальную.
К этому можно лишь добавить, что последовавшая затем редукция костей заднего
края крыши довершила в общих чертах формирование диадектоидного типа черепа
(рис. 29).
Таким образом, возможно, намечаются две исходные линии зауроморфных реп
тилий. Не исключено, что линий было гораздо больше — например, лантанозух име
ет ряд черт, не позволяющих сближать его ни с одной из этих линий.
Дальнейшее развитие диадектоморфов шло, вероятно, тремя независимыми
стволами - диадектоидным, парейазавроидным и никтеролетероидным. Формы, ко
торые могут быть близки к исходным для каждого из стволов, имеют одинаковую
схему строения черепа. Это стефаноспондилюс (Stappenbeck, 1905) для диадекто-.
идного ствола, рипеазавр - для парейазавроидного и никтиббт — для никтероле-
тероидного. Все три формы имеют плоские, весьма грубо скульптированные чере
па, небольшую по размерам ушную вырезку, находящуюся на чешуйчатой кости и
ограниченную сверху надвисочной. При этом на крыше черепа сохраняются таб
литчатые и заднетеменные кости, и межвисочная кость остается свободной. В
63
Рис. 29, Схема смещения барабанной перепонки в эволюции зауроморфных репти
лий. Точками показано положение квадратной кости; заштрихован район расположе
ния барабанной перепонки
а - Kotlassia
Amalitzky; б — Nycteroleter
Efremov; в - Diadectes
Соре (Olson,.
1947,. с дополнениями); г - Burtensia Ivachnenko; Д - Sphenodon Gray (Abel,. 1919,.
с дополнениями); е — Cnemidophorus
Wagler (Fisher, Tanner,. 1970)
Рис. 30. Схема редукции костей посторбитального и затылочного отделов черепа
у некоторых сеймуриаморфов и низших зауроморфов. Изображены левые половины
посторбитальных отделов крыши и затылков
а — Kotlassia prima Amalitzky; б — Diadectes
molaris Cope (Olson,. 1937, «с из- «
монениями); в —'Rhipaeosaurus talonophorus
Tchudinov; г — Scutosaurus
karpinskii
(Amalitzky); д — Nyctiboetus
liteus Tchudinov; e — Nycteroleter
bashkirikus
Efremov;
ж — Nyctiphmretus
acudens Efremov; 3 — Tichvinskia
vjatkensis Tchudinov et Vjuschkov
64
дальнейшем во всех стволах идет усиление специализации (увеличиваются разме
ры, череп становится высоким, происходит специализация и иногда слабая диффе
ренциация зубов), а также интенсивная редукция костей заднего края крыши чере
па: дорзальные п*ластины таблитчатых и заднетеменных костей уменьшаются в
размерах, от парокципитального отростка таблитчатой кости остается лишь перед
няя часть, так что весь задний край парокципитального отростка опистотики пре
вращается в свободный "фланг". Межвисочная кость часто срастается с теменной
и заглазничной (причем у молодых экземпляров диадекта отмечается свободная
межвисочная кость: Чудинов, 1964). Это хорошо видно на рис. 30.
Таким образом, намечается ряд достаточно важных черт строения, сближающих
парейазавров, диадектов и никтеролетеров, тогда как различия между этими груп
пами объясняются в основном разницей в размерах и способе питания. Это позво
ляет объединить их в один подотряд диадектий, включающий три дивергирующих
ствола.
По сравнению с диадектиями, у проколофонов ушная вырезка полностью сме
щается на чешуйчатую кость, и надвисочная кость практически не участвует в ее
ограничении сверху. Таблитчатые и заднетеменные кости сохраняются лишь у при
митивных никтифруретид в виде рудиментов - узких костных полосок под надви-
сочными и теменными костями. Следовательно, по степени развития учитываемых
признаков проколофоны представляют собой более высокую ступень.
Кроме того, очень важным признаком, отличающим диадектий от проколофоний,
представляется различие в местах прикрепления аддукторной мускулатуры ниж
ней челюсти. У диадектий пучки мускулов шли от аддукторной ямы нижней челю
сти в район посттемпорального отверстия, а у проколофоний - в заднюю часть
орбиты.
На основании всего вышеизложенного в данной работе используется следующая
схема классификации диадектоморфных котилозавров (Ивахненко, 1973в).
Отряд Diadectomorpha
Подотряд Diadectia
Надсемейство Diadectoidea
Семейство Stephanospondylidae Stappenbeck, 1905
Семейство Diadectidae Соре, 1880
Надсемейство Pareiasauroidea
Семейство Rhipaeosauridae Tchudinov, 1955
Семейство Pareiasauridae Seeley, 1888
Надсемейство Nycteroleteroidea
Семейство Nycteroleteridae Romer, 1956
Подотряд Procolophonia
Семейство Nyctiphruretidae Efremov, 1938
Семейство Procolophonidae Seeley, 1888
ПРОИСХОЖДЕНИЕ И ОБЩИЕ НАПРАВЛЕНИЯ
ЭВОЛЮЦИИ ПРОКОЛОФОНОВ
Некоторые исследователи (Parrington, 1958, 1959; Romer, 1966) выводят проко
лофонов из капториноморфов. Критика таких взглядов дана Шишкиным (1973) и в
настоящей работе нет необходимости повторять ее. Представляется верной точка
зрения, по которой капторины рассматриваются как предки тероморфного ствола
(см., например, Watson, 1954, 1957; Татаринов, 1969).
Вероятно, единственной группой, которая может претендовать на роль репти-
лийных предков проколофонов, являются никтеролетериды. Череп никтеролетерид .
весьма схож по строению с черепом примитивных проколофонов. Чтобы, базируясь
на схеме строения черепа никтеролетера, получить никтифруретидный тип чере
па, достаточно продолжить отмеченный выше процесс редукции заглазничного от
дела крыши.
A* t*oi.
65
Но никтеролетериды имеют и ряд особых черт. Главная из них - на
личие очень большой ушной вырезки. А.И.Ефремов (Efremov, 1940) полагал, что
ушная вырезка у никтеролетера развивается вторично, на фоне проколофоноидно-
го черепа. Однако череп никтеролетера отличается от черепа проколофонов рядом
признаков, ставящих его на более низкую ступень, и ушная вырезка никтеролете
ра значительно отличается от вырезки никтифрурета, и, как уже говорилось выше,
схема ее строения ближе к таковой диадектид. Но надо отметить, что вырезка у
никтеролетера продолжается далеко вперед, вследствие чего сверху ее ограничи
вает межвисочная кость, а спереди она подходит к заглазничной кости, Это под
тверждает предположение И.А.Ефремова о значительном вторичном разрастании
вырезки, но не на базе проколофоноидного черепа.
Отделение проколофонов от никтеролетерид произошло довольно рано, по край
ней мере, все известные никтеролетероиды несут черты собственной специализа
ции, так что после разделения обеих групп они прошли длительный собственный
путь развития. Так, число зубов у никтеролетерид относительно велико, но у боль
шинства форм (Nycteroleter
bashkyricus,
N.kassini)
наблюдается слабая специали
зация зубов. Нёбное (подглазничное) отверстие у никтеролетера большое и не ме
нее хорошо сформировано, чем у проколофонин. Слезная кость, если и достигает
ноздри, то очень узкой полосой (т. е. дорзальная пластина максиллы развита силь
но). Птеригоидные фланги развиты сильнее, чем у никтифрурета. Наиболее прими
тивной формой представляется никтибёт, возможно, близкий к исходной для про
колофонов форме.
До сих пор никтеролетериды из других регионов неизвестны,кроме одной про
блематичной и плохо изученной формы (Tseajaia:
Vaughn, 1964), поэтому отсут
ствуют какие-либо данные, которые могли бы говорить о происхождении проколо
фонов различными стволами, и, основываясь на сходстве группы в целом, можно
делать вывод об общности их происхождения и разделении на различные линии
лишь на уровне достаточно высокоразвитых форм (неизвестные пока примитивные
никтифруретиды).
Достаточно интересным представляется вопрос о дальнейшей судьбе диадекто-
морфных котилозавров. Нет особых сомнений в том, что диадекты и парейазавры
являются тупиковыми ветвями. Олсон (Olson, 1947, 1965) сближал с диадектами
предков черепах. Эпплби (Appleby, 1961) считает предками черепах проколофонов,
однако различие в местах прикрепления аддукторной мускулатуры говорит скорее
в пользу гипотезы Олсона.
Имеется определенное сходство в строении черепов проколофонов и трилофо-
завров, относимых в настоящее время к ареосцелидиям. Основным отличием явля
ется присутствие у трилофозавра верхнего височного окна и глубокое внедрение
зубной ткани в кость, с образованием "корней" зубов. Однако именно такое стро
ение передних зубов у проколофонида Виталии, а также у парейазавров. Черты,
сближающие трилофозавров с проколофонами,- это наличие широкой ушной вырез
ки "диадектомор'фного" типа, положение латеральной носовой железы с выходом
в неб переднего максиллярного канала, высокое поднятие ростра базинфеноида,
охват задними краями птеригоида гиоидного бугра на квадратной кости, форма и
расположение комплекса мбзговой коробки, общий план строения нижней челюсти
и другие.
Исследования показывают, что височные окна и схожие с ними структуры не
однократно возникали в ходе развития в самых разнообразных группах, поэтому
одно лишь наличие окна вряд ли может служить критерием для отнесения к раз*
личным таксонам достаточно высокого ранга (например, к подклассам). В состав
трилофозаврид в настоящее время могут быть включены следующие роды: Trilo-
phosaurus
(Gregory, 1945); Variodens,
Tricuspisaurus
(Robinson, 1957);
Toxolophosa-
urus (Olson, 1960); Anisodontosaurus
(Welles, 1947); Co elodonto gnat hits (Очев, 19 67-).
Лишь для трилофозавра имеются некоторые данные по строению черепа.
При разработке филогенетической схемы проколофонов приходится оперировать
признаками различной значимости. В качестве основного, всегда соблюдающегося
показателя уровня развития формы могут рассматриваться лишь чирло и степень
дифференциации зубов, поскольку во всех линиях имеется тенденция к увеличению
66
степени дифференциации и уменьшению числа зубов. Большое значение* имеет де
тальный морфо-структурный анализ строения черепа. К сожалению, практически
для всех экзотических проколофонов необходимые подробности строения не описа
ны в литературе, поэтому приходится ограничиваться общими признаками, как-то:
степень удлинения орбит, высота черепа, развитие или отсутствие щечных шипов.
Другими словами, здесь используются явно адаптационные признаки, весьма по
верхностно отражающие сущность эволюционных изменений. Филогенетическая
схема, построенная по таким данным, может оказаться весьма приблизительной,
так как внешне схожие адаптации могут возникнуть у весьма различных форм, а
близко родственные животные — иметь достаточно различающийся внешний
облик.
Например, большое сходство в строении зубов позволяло обычно сравнивать южно
африканского проколофона и восточноевропейскую тихвинскию. В действительно
сти эти роды принадлежат далеким, хотя и почти параллельным линиям и находят
ся на различных эволюционных уровнях.
Приводимые ниже общие закономерности морфологической эволюции проколо
фонов выводятся на основании анализа тех линий, в отношении которых имеются
достаточно полные данные (в основном линии Русской платформы и Южной Африки).
Черепа примитивных проколофонов плоские, широкие, похожие на черепа наибо
лее примитивных сеймуриаморфов. На ранних этапах эволюции происходило увели
чение высоты черепа, особенно в затылочной части. И лишь после достижения ка
кого-то порогового уровня начинается широкая адаптивная радиация, сопровожда
ющаяся выработкой различных типов черепов - высоких "ящерицеобразных" у про-
колофонин, плоских и вторично широких у лептоплейронин.
В ходе всей эволюции наблюдается прогрессивное уменьшение числа зубов в
челюстях и увеличение степени их дифференциации, связанное с дальнейшей спе
циализацией представителей разных ветвей. Такое усиление зубной системы, ве
роятно, вызванное переходом к всеядности, сопровождалось усилением челюстной
мускулатуры, что привело к значительной перестройке посторбитальной области
черепа. Известное влияние могло оказывать и развитие шейных мышц, связанное
с увеличением подвижности головы, что необходимо для активных животных на
суше. Совместное действие этих факторов и привело, вероятно, к редукции ряда
костей посторбитальной и затылочной областей черепа, к консолидации крыши —
срастанию ряда костей друг с другом, образованию "ложного височного окна". У
ящериц те же факторы привели к срастанию теменных костей, значительной редук
ции посторбитального комплекса, появлению височных окон.
В.начальный период своей адаптивной радиации проколофоны разделились на
две ветви. Одна ветвь (лептоплейронины) пошла по пути сохранения малоподвиж
ного образа жизни, их тела и черепа вторично стали широкими и плоскими. Воз
можно, начавший развиваться кинетизм исчезает. Однако процесс усиления жева
тельной мускулатуры продолжался, что вызвало интенсивное удлинение орбит.
Число зубов у представителей этой ветви также резко сократилось, зубы стали
плоскими и давящими. Интенсивного укорочения посторбитальной области черепа
не наблюдается, возможно, в связи с небольшими потребностями в движении го
ловы.
По-иному шел процесс адаптации проколофонов других линий, например, на Рус
ской платформе. Здесь наблюдается появление очень подвижных, "ящерицеподоб-
ных" форм. У них происходило значительное облегчение скелета, удлинение конеч
ностей, сильное развитие шейной мускулатуры. Это последнее вызвало активную
редукцию посторбитального отдела крыши черепа (например, полностью исчезает
дорзальная пластина чешуйчатой кости). Как и у лептоплейронин, шло усиление
зубов и дифференциация их, развитие и усиление аддукторных мускулов. Это при
вело к резкому удлинению орбит (возможно, при этом большую, чем у лептоплей
ронин, роль играла редукция костей). Скорее всего, более прогрессивным являет
ся и приобретение аналога нижней височной вырезки, а не прикрепление мус
кулатуры у оснований щечных гребней. Интересной чертой является разви
тие кинетизма черепа, вряд ли намного более примитивного, чем у современ
ных ящериц.
67
Во зсех этих отношениях проколофоны Южной Африки стоят как бы между леп-
топлейронииами Западной Европы и Америки и проколофонинами Русской платфор
мы. Здесь наблюдается смешение черт обеих линий -= появляется высокий череп
с резко укороченным затылком и несколько удлиненная орбита. Зубы резко спе
циализируются, скелет становится более облегченным. Однако при этом сохраня
ется ряд архаичных черт (например, скульптуры на черепе у проколофонов), а так
же приобретение "лептоплейронинных" черт специализации (выросты на квадратно-
скуловых костях).
Несколько особняком стоят неопроколофон и канделярия. Вероятно, их разви
тие шло особым путем. Определенный ответ на этот вопрос невозможен до более
подробного изучения форм.
Все это полностью подтверждает мнение В.Г.Очева (1967, 1972) о различных
направлениях и темпах эволюции проколофонов.
АДАПТИВНАЯ РАДИАЦИЯ ПРОКОЛОФОНОВ
Развитие проколофонов шло по более или менее параллельным линиям, ответ
вление которых произошло достаточно рано (хотя и на разных уровнях) от общего
ствола. За наличие одного исходного ствола может говорить прежде всего боль
шое сходство между представителями различных линий, возрастающее по мере
рассматривания все более древних форм. Сама глубина параллелизмов, приводящих
к формированию сходно адаптированных форм в явно различных линиях,может так
же подтверждать монофилию проколофонов. Особенно это хорошо заметно при
сравнении проколофонов Восточной Европы и Южной Африки.
Ниже рассматривается развитие проколофонов по отдельным регионам.
Проколофоны Русской платформы изучены лучше всего, и именно на их при
мере может быть прослежено, как глубокие внутренние перестройки в ходе эволю
ции сопровождались изменением внешнего вида черепа. Выяснение этой зависимо
сти интересно по той причине, что практически ничего не известно о строении, на
пример, мозговой коробки или кровеносной системы проколофонов иных регионов,
тогда как сравнительное изучение именно этих деталей может дать ключ к пост
роению систематики и филогении всей группы, а также выявить родственные свя
зи с другими группами.
Наиболее примитивной формой с территории Русской платформы (и, пожалуй,
наиболее примитивной из всех известных) является никтифрурет из низов верхней
перми. Внешние признаки его представляют смешение архаичных черт и черт до
вольно высокой специализации. К архаичным чертам могут быть отнесены такие,
как плоский и широкий, сильно скульптированный череп, недифференцированные
многочисленные (до 44 в челюсти) зубы, очень маленькая ушная вырезка, доста
точно большие размеры дорзальной пластины чешуйчатой кости, сохранение остат
ков таблитчатой и заднетеменной костей. Небные зубы развиты хорошо, образуют
многорядные полосы, птеригоидные фланги низкие и несут зубы. И в то же время
орбиты удлинены назад гораздо более, чем, например, у безусловно более высоко
организованной овенетты, пожалуй, даже более, чем у проколофона. К примитив
ным признакам в сравнении с более поздними формами должен быть отнесен и ряд
деталей внутреннего строения. Ретракторный миодом на дорзальной поверхности
пара-базисфеноида короткий, базиптеригоидные мыщелки развиты слабо и направ
лены латерально. Имеется желоб для хорды на дорзальной поверхности основной
затылочной кости. Латеральная носовая железа находится на задней стенке ноздри,
слабо смещена медиально. Дорзальная пластина верхнечелюстной кости неболь
шая, и поэтому слезная кость достигает ноздри. Небное отверстие маленькое.
Сеть кровеносных лакун в верхней челюсти развита плохо. На затылке отсутству
ют сфено-окципитальные бугры, вместо них имеются небольшие впадины, как у
лабиринтодонтов и сеймуриаморфов, что может говорить о слабом развитии шей
ных мышц.
Следующую ступень представляет фаантозавр, известный по нижней челюсти.
Длина дентале по отношению к постдентальной части уменьшается, число зубов
No
до 11 в нижней челюсти. Намечается некоторое расширение оснований задних зу
бов. Вероятно, именно на этой стадии происходит разделение ствола на ряд линий.
Одна из линий развивалась на территории "северного" триасового поля, и по
меньшей мере две — на территории "южного" триасового поля.
В северной линии происходит резкая дифференциация зубной системы. Вначале
происходит поперечное расширение оснований задних зубов, однако форма всех зу- •
бов остается одинаковой. У типичного представителя этой ступени — контитозавра-
происходит резкое удлинение назад орбиты, череп становится высоким, исчезает
скульптура, латеральная носовая железа смещается на медиальную поверхность
максиллы. Идет дальнейшая редукция покровных костей затылочного и щечного
отделов черепа, слезная кость перестает достигать ноздри в связи с сильным раз
витием дорзальной пластины максиллы. Число кровеносных лакун в верхней челю
сти увеличивается, они начинают сливаться. Пара-базисфеноидный комплекс ста
новится короче и массивнее, исчезает передний гребень турецкого седла, и моз
говая коробка приобретает "ящеричный" вид. Судя по способу причленения максил-.
лы, по строению базиптеригоидного сустава, верхнезатылочной кости, начинает
развиваться кинетизм.
Следующая морфологическая ступень представлена тихвинскией, у которой зу
бы дифференцируются на "квазирезцы" и "квазикоренные" (число последних еще
велико — 5—6 в нижней челюсти). Сильно редуцированы кости посторбитального и
щечного отделов, вследствие чего хорошо развиты "ложные височные окна". Кине
тизм черепа развит не хуже, чем, например, у современных агам. Небные зубы
нем ногочисленны.
Буртензия по ряду признаков представляет собой более высокую ступень раз
вития (дальнейшее уменьшение числа зубов, увеличение подглазничного окна и ор
биты, укорочение переднего отдела мозговой коробки). Однако внешне она ближе
к контритозавру, чем к тихвинский (небольшие размеры, короткая тупая морда и
т.д.), так что тихвинския, скорее всего, сильно аберрантная форма. У наиболее
молодой формы - капеса - последний зуб резко увеличивается в размерах и ста
новится доминирующим. Кровеносный канал в верхнечелюстной кости сформирован
лучше, чем у буртензии; небное отверстие большое.
От макрофона известна лишь часть верхнечелюстной кости, однако его крупные
размеры и строение зубов сближают его скорее с тихвинскией, чем с буртензией.
Вторая линия развития - южная - представлена лишь одним родом — оренбур-
гией. Этот род примечателен тем, что дифференциация зубов у него достаточно
сильная, хорошо отличимы одновершинковые "квазирезцы" от двухвершинковых
"квазикоренных", однако переход от одних к другим по размеру постепенный, нет
хорошо выраженного у всех северных видов промежутка с%мелкими зубами. Свое-"
образие этой линии, скорее всего, вызвано ранним отделением от общего ствола и
более быстрыми темпами эволюции. Это подтверждается наличием у наиболее при
митивного вида - Orenburgia concinna
-
сильной редукции небных зубов, хот.я дру
гие признаки (например, степень развития кинетизма, строение кровеносной си
стемы и некоторые другие) ставят ее в один ряд с тихвинскией. Если судить по из
вестным в настоящее время чертам строения, эволюция в этой линии шла в сторо
ну увеличения высоты нижней челюсти и усиления зубов, вплоть до образования
очень крупного, доминирующего последнего "квазикоренного".
Очень мало пока известно о представителях третьей линии. У единственно из
вестного рода — Виталии - отмечается сильное внедрение вещества передних зу
бов в кость, что сближает эту форму с трилофозаврами. Можно ожидать, что даль
нейшие находки позволят выяснить проблему происхождения своеобразной
группы трилофозавров, по целому ряду существенных признаков более близ-*
ких к котилозаврам, и даже к собственно проколофонам, чем к ареосце-
лидиям.
На территории Южной Африки также известны представители всех ступеней
развития проколофонов. Это, скорее всего, может говорить о распространении
всех групп в пермское и триасовое время на обоих материках.
Позднепермские роды овенетта и баразавр имеют плоские широкие черепа, ок
руглые орбиты, значительные по размерам надвисочные кости. В максилле до 30
69
недифференцированных зубов. Однако небные зубы сильно редуцированные, и на
птеригоидных флангах наличия зубов не отмечается.
Представитель следующей ступени развития — спондилолестес из верхов зоны
Lystrosaurus — низов зоны Procolophon., Он имеет более узкую морду и, возмож
но, более высокий череп4
.
Орбиты продолжаются назад за задние края лобных ко
стей, зубы более крупные, их число значительно меньше, наблюдается некоторое
расширение оснований задних зубов. По строению зубной системы эта форма близ
ка к фаантозавру.
У проколофоноидеса значительно удлиненные назад орбиты, более высокий че
реп с тупой мордой. Зубы слабо дифференцированны, верхнечелюстная кость со
держит всего около 8 зубов. К сожалению, ни точный возраст, ни место находки
этой формы не известны. Можно полагать, что по учитываемым признакам уро
вень развития проколофоноидеса соответствует уровню контритозавра. Далее про-
колофоны приобретали более удлиненную орбиту, шло уменьшение числа зубов и
разделение их на "квазирезцы" и "квазикоренные" (проколофон). Надо отметить,
что для проколофона характерно и приобретение черт "лептоплейронинной" спе
циализации (выросты на квадратно-скуловых костях).
Максимального выражения дифференциация зубов достигает у миоцефалюса.
Эта форма имеет в верхней челюсти 4— 5 квазикоренных зубов, из которых пред
последний очень крупный, доминирующий по размеру. Хорошо видно, что все рас
смотренные три линии оканчиваются формами, имеющими некоторые схожие
черты.
С территории Центральной Европы известно всего 4 рода. Склерозавр из верх
него пестрого песчаника представляет собой весьма проблематичную, скорее все
го, очень сильно уклонившуюся форму.
Два других рода - аномоиодон и коилоскиозавр - приблизительно одновозраст-
ны. Описаны они весьма поверхностно, и установление их взаимоотношений за
труднительно. По строению зубной системы более примитивным представляется
коилоскиозавр, имеющий к тому же несколько менее удлиненные орбиты, чем ано
моиодон. Кроме того, нижняя челюсть аномоиодона более высокая, а увеличение
высоты нижней челюсти наблюдается в других линиях в ходе повышения уровня
специализации. Однако коилоскиозавр имеет такой признак высокой специализации,
как наличие небольших отростков на квадратно-скуловых костях. По общему габи
тусу черепа аномоиодон стоит ближе, например, к контритозавру, тогда как коило
скиозавр - к тихвинский.
Таким образом, в настоящее время нет возможности сколько-нибудь уверенно
говорить о том, являются ли эти формы представителями одной линии развития
или различных. В случае признания одной особой линии логичнее всего видеть ее
завершение в лептоплейроне.
У лептоплейрона небольшое число сильно дифференцированных зубов и резко
удлиненные назад орбиты. Квадратнотскуловые кости несут "воротник" из мощных
шипов.
Единственный более или менее хорошо изученный североамериканский род -
гипсогнат - по целому ряду признаков близок к лептоплейрону, однако его сте
пень специализации гораздо выше. Шипы на квадратно-скуловых костях развиты
гораздо сильнее, более удлинены назад орбиты, дальше зашла редукция костей
носторбитального отдела крыши черепа. Зубы на небе полностью редуцированы.
Несмотря на большую территориальную разобщенность гипсогната и лептоплейро
на, слишком много признаков говорит в пользу их близкого родства, и вполне ве
роятно, что лептоплейрон близок к предковым для гипсогната формам.
С территории Южной Америки известна лишь проблематичная канделярия. По
степени дифференциации и числу зубов она близка к фаантозавру и другим прими-^
тивным спондилолестинам, однако общая форма черепа, некоторые черты (напри
мер, наличие продольной депрессии между орбитами) сближает ее с проколофони-
нами. Возможно, это представитель особой, неизвестной пока линии.
11с более, чем о канделярии, можно сказать и о неоироколофоне, у которого
наблюдается сочетание признаков, характерных и для спондилолестин, и для про
колофонин. Так, большое число почти не дифференцированных зубов, высокая ту-
70
пая морда сближают его со спондилолестинами, тогда как удлиненные назад ор
биты, поперечные отростки на квадратно-скуловых костях есть показатели гораз
до более высокого уровня специализации. Но надо отметить, что строение черепа
неопроколофона изучено очень плохо, и может оказаться, что он близок к формам,
исходным для лептоплейронин. Тем более что с территории Китая известен пао-
теодон, близкий к гипсогнату, как это было убедительно показано авторами рода
(Chow et Sun, I960).
Новые, еще не описанные проколофоны известны из Восточной Сахары (Lehman,
1971), Индии (Andersson, Andersson, 1970), Австралии (Bartholomai, Howie, 1970),
Канады (Carrol, Belt, Dineley, McGregor, 1972), Антарктиды (Kitching, Collinson,
Elliot, Colbert, 1972).
Можно заключить, что в настоящее время известно лишь незначительное число
из существовавшего некогда огромного разнообразия проколофонов. Например, с
территории Русской платформы известны обломки челюстей, весьма отличных от
уже описанных. Так, Tichvinskia
sp„ с р. Мутусья и из местонахождения Терюхан
на р. Вятке значительно'отличаются от остальных представителей рода и друг от
друга, но от каждой имеются лишь обломки плохо диагностирующейся верхней че
люсти. Burtensia sp. из Выбора II крупными размерами отличается от типового
вида.
Из некоторых местонахождений севера Русской платформы известны неболь
шие обломки верхних челюстей с сильно расширенными и уплощенными зубами, не
сколько похожие на челюсти телегнатуса из Южной Африки. Надо полагать, что
новые поиски принесут обширный материал по этой интересной группе, и тогда по
явится возможность разработки более детальной филогенетической и классифика
ционной схемы проколофонов.
Интересен вопрос о темпах развития и времени существования отдельных ли
ний. Представляется вероятным, что весьма быстрыми темпами шло развитие в
сторону ящерицеподобных форм типа буртензии. Эти проколофоны приобрели ряд
черт специализации, схожих с таковыми ящериц, но на базе весьма примитивной
общей организации. Надо полагать, что формирование их шло в тех же условиях,
что и ящериц, и зоны обитания перекрывались. Эти проколофоны безусловно были
экологическими предшественниками ящериц, и к началу расцвета настоящих яще
риц (средний триас) они вымерли.
Представители второго направления эволюции проколофонов (лептоплейронины)
дожили до позднего триаса, что объясняется, скорее всего, их обитанием в осо
бых условиях по отношению к ящерицам и другим рептилиям, ведущим сходный
образ жизни.
СТРАТИГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ ПРОКОЛОФОНОВ
Остатки проколофонов найдены в настоящее время.на всех материках. Наи
большее число форм описано из перми и триаса Русской платформы и из отложе
ний системы Карру Южной Африки. В остальных районах находки проколофонов
носят случайный характер. Четыре рода известны с территории Западной Европы
(ФРГ и Шотландия), два рода - из Северной Америки, один - из Южной Америки,
два - с территории Китая- Есть данные о находках неизученных пока проколофо
нов из Северной Африки, Индии, Антарктиды, Австралии, Канады.
Большинство местонахождений Русской платформы не раскапывалось, находки
представляют собой чаще всего отдельные разрозненные обломки костей. Это за
трудняет изучение фаунистического состава местонахождений. Поэтому приведен
ные ниже данные весьма неполные и нуждаются в уточнении.
На территории Русской платформы известно лишь небольшое количество косте-
носных точек позднепермского возраста, содержащих остатки никтифрурета и со
средоточенных в бассейне р.Мезень Архангельской обл. Проколофоны найдены в
красном плитчатом мергеле совместно с остатками Nycteroleter
ineptus
Efr.,
Mescnosaurus
Efr.,
не описанными пока рипейазавридами, примитивным
лантанозухидом и формой, близкой к биармозухам (по определению U.K. Чу-
динова).
71
Местонахождения низов ветлужской серии нижнего триаса расположены в основ
ном на территории Горьковской и Кировской областей. Литологически здесь пре
обладают красные и серые конгломераты из мелкой глинисто-мергелистой гальки
и песчаники, реже красные глины. Типичные местнахождения - Спасское, Красно-
Ваковское, Лииовское, Сарафаниха на реке Ветлуге, Семеновское на р. Вохме
(данные по местонахождениям по Блому, 1968). Совместно с массовыми остатка
ми контритозавра и очень редкими — фаантозавра здесь найдены остатки лабирин-
тодонта Tupilakosaurus
wetlugensis
Shishkin, Chroniosuchidae g„ ind„, разрознен
ные кости мелких псевдозухий и лепидозавров. Проколофоны в этих местонахож
дениях составляют существенный, а иногда и преобладающий элемент фауны.
В верхней половине ветлужской серии известна лишь одна находка проколофо
на Tichvinskia
jugensis
в местонахождении Вахнево на р. Шарженге (данные по
Ефремову и Вьюшкову, 1955), совместно с остатками Benthosuchus
sushkini
Efr.,
Thoosuchus
sp.,
Microcnemus
Huene,
разрозненными костями псевдо
зухий.
Отложения баскунчакской серии нижнего триаса литологически весьма разно
образны, преобладают серые конгломераты из галек мергелей и серые песчинки.
Проколофоны Tichvinskia
vjatkensis
и Burtensia burtensis
встречаются всегда раз
дельно, но с близкими сопровождающими формами (лабиринтодонтами, мелкими
псевдозухиями и лепидозаврами, остатками цератодов)„Основные местонахожде
ния — Окуневское на р. Федоровке (по Блому, 1968), ВыСор 11 (по Ефремову и
Вьюшкову, 1955), Кызыл-Сай 11 (по Гаряинову и Очеву, 1962), Липовское (по Ры
кову и Очеву, 1966). В последнем местонахождении появляются трилофозавриды -
целодонтогнаты и проколофон Vitalia grata. Местонахождения по.р. Вычегде — Же-
шарг, Гам (по Лозовскому, 1969), вероятно, более высокие по стратиграфическо
му положению, содержат остатки проколофонов Rapes amaenus и Macrophon
komi-
ensis совместно с остатками трематозавров и паротозавров. На территории "юж
ного" триасового поля в одновозрастных местонахождениях капеса и макрофона
замещает Orenburgia
concinna.
Среднетриасовое местонахождение (донгузская серия) известно в настоящее
время лишь одно, на р. Бердянке в Оренбургской обл. (Бердянка 1, по Гаряинову
и Очеву, 1962). Здесь в красных мергелях совместно с остатками Orenburgia
maj-
mesculae найдены черепа и целые скелеты дицинодонтов Rhinodicynodon
gracile
Kalandadze, остатки псевдозухий.
Из сравнения фаунистического состава местонахождений видно, что остатки
проколофонов и крупных наземных рептилий захоронялись в разных тафономиче-
ских условиях. Проколофоны захоронялись обычно в отложениях озер, болот, не
больших дельт, т. е. в местах, куда чрезвычайно редко попадали кости крупных
рептилий. Однако проколофоны, безусловно, были не настолько привязаны к воде,
как, например, лабиринтодонты, и следует ожидать появления достаточного мате
риала по ним при детальных раскопках местонахождений, содержащих остатки ди
цинодонтов и псевдозухий. Это позволит улучшить корреляцию местонахождений,
и, учитывая лучшую изученность крупных форм для различных регионов, проводить
более точные возрастные сопоставления отложений, содержащих остатки проколо
фонов.
На территории Южной Африки отложения зоны Cystecephalus представляют со
бой переслаивание пестрых красных и красно-фиолетовых глин и мергелей. Именно .
в этих отложениях в районе Нью-Бетесда был найден череп проколофона
Owenetta.
Здесь же найдены остатки дицинодонтов, горгонопсов, тероцефалов. Сопоставле
ние комплексов фаун этих местонахождений Русской платформы позволяет утвер
ждать, что отложения района р. Мезень, откуда известен никтифрурет, гораздо
более древние и, по П.К.Чудинову (1955) и В.Г.Очеву (1960),соответствуют отложе
ниям зоны Tapinocephalus, хотя в одновозрастных Мезени местонахождениях
(Очерское, Ножовка, Белебей) найдены формы, близкие к раннепермским северо
американским - Belebey (Ивахненко, 1973а), болозавр, не описанные пока диссо-
рофиды.
Barasaurus
найден на Мадагаскаре, в районе Ранохира, в отложениях серии Са-
квмена позднепермского возраста.
72
Триасовые проколофоны Южной Африки найдены в отложениях зон Lystrosau-
rus, Procolophon и Cynognathus (данные о местонахождениях по Huene, 1925).
Отложения зоны Lystrosaurus представляют собой переслаивание красных и зе
леных мергелей, песчаных банок, глин и содержат остатки крупных лабиринтодон
тов, дицииодонтов, цинодонтов, псевдозухид. Из проколофонов здесь найден череп
спондилолестеса.
Литологический состав различных местонахождений зоны Procolophon разно
образен, однако преобладают серые, зеленые, розоватые песчаники с остатками
проколофонов (Procolophon,
Procolophonoid.es),
лабиринтодонтов, псевдозухий, ле
пидозавров, редкими отдельными костями дицииодонтов. Отложения обеих рассмот
ренных выше зон представляют собой разные фации одного стратиграфического
горизонта, поскольку в последнее время появились сообщения о находках на тер
ритории Южной Африки проколофона совместно с листрозавром (Kitching, Collin-
son, Elliot, Colbert, 1972) и ряд исследователей объединяют эти зоны (например,
Kitching, 1970).
Сопоставление литологического и фаунистического состава местонахождений
зон Южной Африки и серий Русской платформы показывает, что в ряде случаев мо
жет иметь место искусственное разделение синхронных, но разнофациальных от
ложений (для Африки — зон Lystrosaurus и Procolophon, для Русской платформы,
возможно, ряда местонахождений ветлужской и баскунчакской серий "северного"
и "южного" триасовых полей).
Из местонахождений зоны Cynognathus известны остатки проколофонов (The
legnathus,
Myocephalus,
Microtheledon),
рыб, лабиринтодонтов, лепидозавров — из
песчаников, дицииодонтов и цинодонтов - из красных мергелей.
По стратиграфическому уровню отложения зоны Lystrosaurus соответствует
низам ветлужской серии (Очев, 1960; Лозовский, 1969; Пахтусова, Шишкин, 1962).
Недавно в местонахождении Асташихинское, одновозрастном ассоциации Tupila-
kosaurus — Contritosaurus, найден собственно листрозавр (Каландадзе, 1973).
Соответствие отложений зоны Cynognathus баскунчакской серии Русской пат-
формы не вызывает сомнений.
Проколофоны на территории Центральной Европы найдены в отложениях сред
него и верхнего пестрого песчаника. Средний пестрый песчаник содержит остатки
проколофонов Anomoiodon
и Koiloskiosaums
и, вероятно, должен сопоставляться
с баскунчакской серией Русской платформы.
Верхний пестрый песчаник, содержащий остатки проколофонов Sclerosaums
и
лабиринтодонта Stenoto saurus, возможно, относится уже к среднему триасу (Очев,
1966; Лозовский, 1969, ссылки на работы польских стратиграфов).
С территории Шотландии (Элджин) проколофон Leptopleuron
описан из слоев
Stagonolepis (верхний триас). Остатки лептоплейрона найдены совместно с ринхо
цефалами, ринхозаврами и псевдозухиями.
На территории Северной Америки остатки проколофонов Hypsognathus
и Spho
drosaurus найдены в отложениях низов формации Брансуик из группы Ньюарк на
территории Нью-Джерси (Пенсильвания), в пестрых красных песчаниках, пересла
ивающихся с глинами, совместно с остатками лабиринтодонта Metoposaums
и
псевдозухий Stegomus.
Эти отложения относят обычно к верхнему триасу.
Единственный известный с территории Бразилии род Candelaria описан из от
ложений формации Санта-Мария, относимых к среднему триасу. Совместно с кан-
делярией найдены остатки ринхозавров, псевдозухий, прозауропод, дицииодонтов
и цинодонтов - траверзодонтид.
С территории Китая известны два рода проколофонов - Neoptоcolophon
и Pao
teodon. Неопроколофон описан из отложений формации Ер-ма-ян, относимых к
среднему триасу (провинция Шаньси). Паотеодон найден в красном пестром песча
нике в бассейне р. Хуанхэ. Вследствие отсутствия ассоциированной фауны опре
деление возраста этих отложений затруднительно, однако горизонт лежит значи
тельно выше отложений формации Ер-ма-ян.
ЗАКЛЮЧЕНИЕ
Приведенный в настоящей работе материал по морфологии, филогении и стра
тиграфическому положению проколофонов позволяет сделать следующие выводы.
_L
Проколофоны совместно с диадектами, парейазаврами и никтеролетерами
представляют собой особую ветвь зауроморфных рептилий, происходящих от позд-
некарбоновых сеймуриаморфов (что, вероятно, сопровождалось некоторой фетализацией
структур ушной области черепа и редукцией посттемпорального ряда костей крыши).
_L
Проколофоны появляются в начале поздней перми (или в конце ранней), их
предками могли быть примитивные никтеролетероиды. В раннем триасе группа до
стигает расцвета, распространяясь на все материки. До позднего триаса дожива
ют лишь отдельные, высокоспециализированные формы.
^_По уровню эволюционного развития проколофоны распадаются на примитив
ных никтифруретид и специализированных триасовых проколофонйд. Основные
различия между этими группами заключаются в степени приспособленности к на
земному образу жизни. Переход от никтифруретид к проколофонидам сопровож
дался преобразованиями в строении носовой капсулы, звукопроводящего аппарата,
крыши черепа (возможно, побочное следствие усиления шейной мускулатуры),
посткраниального скелета.
jk Быстро, в течение поздней перми; проколофоны проходят путь морфологи
ческой эволюции от примитивных, полуназемных форм до настоящих наземных
рептилий, и к середине раннего триаса дают ряд узкоспециализированных ветвей.
По степени и направлениям специализации семейство Procolophonidae может быть
разделено на три подсемейства. Подсемейство Spondylolestinae по уровню специ
ализации (в основном по строению зубной системы) может быть исходным для
подсемейств Procolophoninae и Leptopleuroninae, хотя, возможно, такое разделе
ние произошло уже на уровне никтифруретид, и тогда семейство Procolophoninae
является гетерогенным. Однако общие направления эволюции обеих групп до от
деления лептоплейронин были одинаковы.
J3. Поздние проколофонины (по крайней мере, Русской платформы) приобрета
ют черты конвергентного сходства с лепидозаврами, выражающиеся в общем пла
не строения мозговой коробки, появлении "ложных височных окон" и амфикине-
тизма черепа, облегчении посткраниального скелета. Однако такие преобразова
ния происходили на фоне примитивной общей организации и были морфологически
менее совершенными (быстрая специализация зубной системы, своеобразное стро
ение "височных окон" и т.д.). Возможно, это делало проколофонин неконкуренто
способными по отношению к лепидозаврам и привело к вымиранию проколофонин
в начале среднего триаса, ко времени появления высокоорганизованных лепидозавров.
6. Лептоплейронины, вероятно, эволюировали в сторону вторичного приспособ
ления к амфибиотическому образу жизни, что, по-видимому, смягчило их конку
рентные взаимоотношения с ящерицами и позволило просуществовать до конца триаса.
1^ Стратиграфическое значение проколофонов достаточно велико. Это обуслов
лено ярко выраженными морфологическими отличиями, и, следовательно, легкой
диагностикой остатков, разнообразием форм, быстрыми темпами эволюции, эври-
бионтностью и, по последним данным, распространением на всех материках. Осо
бенно большую роль группа может играть при корреляции разнофациальных конти
нентальных отложений перми и триаса.
74
СПИСОК СОКРАЩЕНИЙ ЛАТИНСКИХ ТЕРМИНОВ
НА РИСУНКАХ
am
arteria maxillaris
mps — muse, pseud. ?superficialis
Ang — angulare
mpt — muse, pterygomandibularis
ар
— arteria palatina
mr
—
miodomus pro musculi retractori bulbi
apch — anastomosis praechoanalis
oculi
apoch — anast. postchoanalis
Mx
—
maxillare
Art
articulare
N
— nasale
At
-
atlas
Op — opisthoticum
Ax
—
axis
P
—
parietale
Boc
-
basioccipitale
pa
—
processus alaris parasphenoidei
btb — basis tuberculi basalis
pab — processus alaris basisphenoidei
bpt - tuberculum basipterygoideum
Part — praearticulare
с
-
coronoideum
Pat
— proatlas
Cat
-
centrum atlantis
pdr
—
processus dorsalis columellae
comm - cms communis
pex
—
proc. Bxtrastapedialis columellae
cpp
-
cartilago paroccipitalis
pif — proc. infrastapedialis columellae
csa
-
canalis semicircularis anterior
PI
—
palatinum
cse
-
can. jsemicirc. externus
Pmx — praemaxillare
csp
—
can. jsemicirc. posterior
Po
— postorbitale
cty
foramen chordae tympani
Pof
— postfrontale
D
— dentale
Pp — postparietale
dgJ — ductus glandulae Jacobsoni
РР
—
processus paroccipitalis
dm
— musculus depressor mandibulae
Prf
— praefrontale
dnl
_
ductus nasolacrimal
Pro
— prooticum
Ec
—
ectopterygoideum
Pt
— pterygoideum
el
—
foramen ductus endolymphaticum
pvb
—
processus vestibularis columellae
Ept
-
epipterygoideum
pvt
—
proc. iventralis columellae
Ex
—
exoccipitale
raa
—
recessus ampullaris anterior
F
—
frontale
rap
—
rec. lamp, posterior
faa
—
foramen alveolaris anterior
rim
— ramus intermediatus
fac
—
for. larteriae cerebralis
rpd — r. idescendens lateralis arteriae pala-
faci — for. jart. parotis interna
rpd
tinae
fani — for. iart. pasalis inferior
rpl
—
r. lateralis arteriae palatinae
fap
—
for. jart. palatina
rpn
—
r. palato-nasalis
fgln — fossa glandulae lateralis nasi.
S
— stapes
fro
_
foramen infraorbitale
Sang — suprangulare
fm
—
for. jnentale
Se
— spheirethmoideum
fmck
for. iMeckelianum
Smx — septomaxillare
fmx
_
for. jnaxillare
Soс
— supraoccipitale
fo
fenestra ovalis
Spl — spleniale
fp
_
foramen palatinum
St
— supratemporale
fpn
—
for. palato-nasale
sturc — sella turcica
fsa
—
for. jsuprangulare anterior
Sq — squamosum
fsp
—
for. jsuprang. posterior
T
—
tabulare
fspl — for. jspleniale
tbc
—
basis trabecularis cranii
Icax _ intercentrum axis
vcm — canalis venae cerebralis media
It
intertemporale
Vr
—
vomer
Ju
_
j ugale
0
—
quadra turn
L
lacrimale
Oi — quadгаtojugale
mae
musculus adductor mantibularis externus
fEc, fju, fL. fN, fPrf, fPl, fPmx, fSmx - пло-
map.
—
muse. jadd. mand. posterior
щадки причленения соответствующих ко
Mck — canalis pro cartilagne Meekeli
стей
men — foramen nervi ethmoidal is medial is
VI,. VIIV IX—XIV XII — места выхода соответ
mpp.
musculus pseudotemporalis profundus
ствующих черепномозговых нервов
75
ЛИТЕРАТУРА
Б лом Г, И. Каталог местонахождений фа-
унистических остатков в нижнетриасовых
отложениях Среднего Поволжья и При
камья, Казань: Изд-во Казанск.гос, ун-та,
1968, 375 с.
Воробьева Э.И. Остеолепиформные кис-
теперые СССР и общие проблемы морфо
логии и эволюции рипидистий. Автореф, <
докт, дисс. М., 1973.
Вьюшков Б.П., Чу ди но в П.К, О триасо
вых рептилиях — Microспemus и Tichvin*
skicu ;- Докл. АН СССР, 1956, ПО, Я 1,
с. 141-144,
ГаряиновВ,А„ ЭчевВ,Г,1962,Каталог
местонахождений позвоночных в пермских
и триасовых отложениях Оренбургского
Приуралья и юга Общего Сырта, Саратов:
Изд-во Саратовск, гос, ун-та, 1962, 61 с.
Ефремов И,А, Некоторые новые пермские
рептилии СССР, - Докл, АН СССР, 1938,
19, No 9, с, 773-781.
Ефремов И.А., Вьюшков Б.П. Каталог
местонахождений пермских и триасовых
наземных позвоночных на территории
СССР. Труды ПИН АН СССР, т, 46. М.:
Изд-во АН СССР, 1955, 185 с, •
Ивахненко М.Ф. • НовыекотилозаврыПри
уралья. -
Палеонт. журн., 1973а, 2, с. 131-134.
Ивахненко М.Ф. Строение черепа ранне-
триасового проколофона Tichvinskia vjaU
kensis Tchudinov et Vjuschkov, — Палеон-
тол. «журн., 197 36, 4^ стр. 74-83.
Ивахненко М.Ф. Систематика диадекто-
морфных котилозавров, — Бюлл. МОИП,
отд. геол., 1973в, J5, с, 15(1-151.
Ивахненко М.Ф. Новые данные по проко-
лофонидам раннего триаса СССР. — Па
леонтол. журн., 1974, 3, с, «68-74.
Ивахненко М.Ф. О родовом составе про.
колофонид раннего триаса Приуралья.
-
Палеонтол. «журн., 1975, ^ с, «88-93.
Каландадзе Н.Н. О межконтинентальных
связях фаун тетрапод в триасовом пери
оде. -
Палеонтол. журн., 1963,3, с. 75—85.
Лозовский В.Р. Триасовые отложения
осевой зоны Московской синеклизы. Авто
реферат канд. дисс, М., 1969, 53 с.
О че в В, Г, Новые данные по фауне триасо
вых прзвоночных Оренбургского При
уралья. -
Докл, АН СССР, 1958, 122.МЗ,
с. 486-488,
Очев В. Г, О континентальном перерыве
между палеозоем и мезозоем на востоке
Европейской части СССР.
-
Изв, «ысш.
уч. зав., геол. и разв., 1960,_3, с, U2-36.
Очев В. Г. К вопросу о стратиграфической
схеме континентальных триасовых отло
жений Русской платформы и Приуралья.
Докл. АН СССР, 1966, 171, No 3, с, 698-701.
Очев В. Г. Новый род проколофонов из три
аса Донской Луки. -
Изв. высш. уч. зав.,
геол. и разв,, 1967,^ с. 15-20.
Очев В. Г.» Новый представитель триасовых
проколофонов Башкирии, ^ В кн.: Ежегодн.
Всес, палеонтол. об-ва, т. 18. Л.: Недра,
1968, 298 с.
Очев В.Г., Данилов А.И. «1972. О первой
находке проколофона в среднетриасовых
отложениях СССР.
— В кн.г Вопросы гео
логии Южного Приуралья и Поволжья. *
Вып. в, ч. 1, Саратов: изд-во Саратовск.
гос, ун-та, с. 81-84.
Пахтусова Н. А., Шишкин М.А. Новые мате
риалы для установления границы перми и три
аса в бассейне Северной Двины. -
Докл. АН
СССР, 1962, 143, No 1, стр. 194-197.
Рыков СП., Очев В.Г. 1966. О местонахож- ^
дении триасовых позвоночных на Донской Л у- у
Iке.-
В кн.: Вопросы геологии Южного При
уралья и Поволжья. Вып. 2, ч. 1. Саратов:
изд-во Саратовск. гос. ун-та, с. 58-62.
Татарин о в Л, П. Новый позднепермский
тероцефал, - Палеонт. журн., 1963, 4,
стр. 76-94.
Т ат ар ино в Л.П. • Проблемы эволюции те
риодонтов. Автореферат докт. дисс. М.,
1969, «2 с.
Татаринов Л. П.' Находка болозавра в
нижней перми СССР. — Палеонтол. «журн./
1974.
с. 144-146.
Чудинов П.К. Котилозавры Шихово-Чир-
ковского местонахождения. — Докл. АН
СССР, 1955, 103, И 5, с. 913-916.
Чудинов П.К. Котилозавры из верхнеперм
ских красноцветных отложений Приуралья.
В кн.: Котилозавры и батрахозавры верх
ней перми СССР. Труды ПИН АН СССР,
т. 68. -М.: Изд-во АН СССР, 1957, с. 19-87.
Чудинов П.К. Отряд Cotylosauria.
—
В кн.?
Основы палеонтологии. Земноводные, пре
смыкающиеся, птицы. М.: Наука, 1964,
с. 213-230.
Чудинов П.К., Вьюшков Б.П. Новые дан
ные о мелких котилозаврах из перми и
76
триаса СССР.
-
Докл. АН СССР, 1956,
108. No 3, е. 347-550.
Шишкин М. А. Морфология древних земно-
водных и проблемы эволюции низших те-
траиод. Труды НИН АН СССР, т. 137, «М.:
Паука, 1973, 257 с.
Abel О. Die Stamme der Wierbeltiere. Berlin;.«
Leipzig,-1919» 354 S.
Adams L. A. A memoir on the Phylogeny of
the Jaw Muscles in recent and fossil verteb
rates. -
Ann. N.Y. Acad. Sci., 1919, 28,
p. 51-66.
Anderson H.M.,. Anderson J.A. A prelimi
nary review of the biostratigraphy of the up
permost Permian,, Triassic and lowest Juras
sic of Gondwana-land. v- Paleontol. Afr.,;
1970,. 13, p. i -22.
Appleby R.M. On the cranial morphology of
ichthyosaurs.
-
Proc. lZool. "Soc. London,.«
1961, 137, p. 330-370.
Barberena M.C., Gomes N.M.B., S a n с h o-
tene L.M.P. Osteologia craniana de Tupi-
nambus teguixin (Lacertilia,. Teiidae).
—
Publ. Espan. TESC. Geol. P. Alegre, -1970,
2b p. 1-32.
Bartholomai A., Howie A.A. Vertebrate
fauna from the lower Trias of Australia. —
Nature, 1970, 225, p. 1063.
Boulenger G. A. On the characters and af
finities of the Triassic reptile Telerpeton
elginese. - Proc. Zool. Soc. London, 1904,
p. 470-481.
Boy J.A. Ein Bemerkenswerter Schadelrest
eines unterpermischen Labyrinthodontiers
(Amphibia) aus dem Saargebiet. — Abh. Hess.
Landesamt. Bodenforsch., 1971»60»S. 31-43.
Brink A.S. On Nanoparia Broom. — Paleon
tol. Afr., 1955, J3, p. 57-63.
Brink A.S. On Aneugomphius ictidoceps
Broom et Robinson. — Paleontol. Afr., 1956,
6, p. 97-115.
Brink A.S. Notes on some whaitsiids and
moshorhinids. — Paleontol. Afr., 1959,^
p. 23-49.
Brock G.T. The cranial muscles of the
Gecko. - Proc. Zool. Soc. London, ser. B,
1938, 108, pt 4, p. 735-761.
Broil i F.; Schroder J. Beobachtungen an
Wierbeltieren der Karrooformation. XXI.
Uber Procolophon Owen. — Sitzungsber.
Bayer. Akad. Wiss. Math.-naturwiss., Abt. B,
1936, 2, S. 239-256.
Broom R. L. Preliminary notice of some new
fossil reptiles collected by Mr. Brown of
Aliwal North of South Africa. — Rec. Albany
Mus., 1905a, 1, N 4, p. 269-275.
Broom R. L. Notice of some new fossil rep
tiles from the Karroo beds of SouthAfrica. —
Rec. Albany Mus., 1905b, 1, N 5, p. 331-
337.
Broom R. L. On the affinities of the primitive
reptile Procolophon. - Proc. Zool. Soc. Lon
don, 1905c, 6, N 4-5, p. 212-217.
Broom R. L. On the structure of the temporal
region in lizard skulls. — Ann. Transvaal.
Mus., 1935, 18, pt 1, p. 13-22.
Broom R. L. On some new genera and spe
cies of Karroo fossil reptiles, with notes
on some others. — Ann. Transvaal. Mus.,
1936a, 18, pt 3, p. 349-386.
Broom R. L. The South African Procolopho-
nia.
— Ann. Transvaal. Mus., 1936b, lf^
pt 4, p. 387-391.
Broom R. L. A further contribution to our
knowledge of the fossil reptiles of the Kar
roo. — Proc. Zool. Soc. London, ser. B,
1937, 107, p. 299-318.
Broom R. L. A new type of cotylosaurian
Owenetta rubidgei. — Ann. Transvaal. Mus.,
1939, 19, p. 319-321.
Carroll R. L. A middle pennsylvanian cap-
torhinomorph, and the interrelationships of
primitive reptiles. — J. Paleontol., 1969a,
43, N 1, p. 151-170.
Carroll R. L. Problems of the origin ot Rep
tiles. -
Biol. Re.vs, 1969b, 44, p. 393-432.
Carroll R.L., Belt E.S., Dineley D. L.,
Baird D., McGregor D.C. Vortebrate
paleontology of Eastern Canada.
—
XXIV
Intern. Geol. Congr. Montreal, 1972, 113 p.
Case E.S. Restoration of Diadectes. - J.
Geol., 1907,15, p. 556-559.
Case E.S. New or little-known reptiles and amphi
bians from the Permian of Texas.—Bull- Amer.
Mus. Natur Hist., 1910,28, p. 163-181.
Chow M., Sun A.L. A new procolophonid from
North-Western Shansi. — Vertebr. Palasiati-
ca, 1960,1, N 1, p. 11-13.
Colbert E.H. Hypsognathus, a Triassic rep
tile from New Jersey.
— Bull. Amer. Mus.
Natur Hist., 1946, 86, N 5, p. 229-274.
Colbert E.H. A new Triassic Procolophonid
from Pennsylvania. — Amer,. Mus. Novit.,
1960, 2022, p. 1-19.
Daly E. A new Procolophonid reptile from the
lower Permian of Oklahoma.
— J. Palaeontol.,
1969, 43, N 3, p. 676-687.
Du Plessis S.S. Cranial Anatomy of the
South African Cordylra Chamesaura anguina. —
South Afr. J. Sci., 1944, 37> p. 1-192.
Edinger T. Uber knflcherne Scleralringe. —
Zool. Jahrb. (Anat.), 1929,51, S. 242-249.
Efremov I.A. Die Mesen-Fauna der permi-
schen Reptilien. — N. Jahrb. Mineral. Geol. und
Palaontol. Beih., 1940,84. Abu B, S. 379-466.
Engelbrecht D.Z. Contributions to the cra
nial morpholbgy of the chamaeleon Micro-
saura pumila (Daudin). — Ann. Univ. Stellen-
bosch, 1951, 27, sect. A, N 1, p. 3-31.
Fischer de Waldheim G. Uber Sclerosa
urus armatus H.v. Meyer.
— N. Jahrb. Mineral.
Geol. unci Palaontol., 1857, S. 136-140.
Fisher D.L., Tanner W.W. Osteological and
myological comparisons of the head and tho
rax regions of Cnemidophorus irgris septen
trional is Burger and Ameiva undulata parva
Barbour and Noble (Family Teiidae).— Brig-
ham Young Univ. Sci. Bull., s^r. Biol., 1970,
1L p. 1-41.
Fox R.C., Bowman M.C. Osteology and rela
tionships of Captorhinus aguti (Cope) (Repti-
lia: Captorhinomorpha). — Univ. Kansas Paleon-
77
tol. Contrib. Vertebrata, A, 1966, П, p. 1-79.
Caffney E.S." An illustrated Glossary of tur
tle skull. -
Amer. Mus. Novit., 1972,2406, p.1-33.
Gil more C. W. A new .fossil reptile from the
Triassic of New Jersey.
— Proc. UsS. Nat.
Mus., 1928, 73, art. 7, p. 1-8.
Gregory J.T. Osteology and relationships
of Trilophosaurus. - Univ. Texas Pub].,
1945, 4401, p. 273-359.
Gregory W. K. Pareiasaurus versus placo-
donts as near ancestors to the turtles. —
Bull. Amer. Mus. Natur. Hist., 1946, 86,
p. 218-323.
Hoffstetter R. Sur la position systemati-
que de Santaisaurus. - Bull. Soc. geol.
France, 1953, 6, t. 3, p.-99-103.
Huene F. von. Ubersicht (lber die Repti-
lien des Trias.
— Geol. und Pala'ontol. Abh.
N.F., 1902, 12, S. 1-83.
Huene F. von. Ueber die Procolophoniden,
mit einer neuen Form aus dem Bundsand-
stein. — Zbl. Mineral., Geol. und Palaonto].,
1911, 3, S. 78-83.
Huene F. von. Die Cotylosaurier des Trias.
-
Palaeontografica, 1912, 59, S. 80-102.
Huene F. von. Ein Telerpeton mit gut er- v
haltenem Schadel.
— Zbl. Mineral., Geol.
und Palaonto]., 1920, 11-12, S. 189-192.
Huene F. von. Die Sadafrikanische Kar
roo-Formation als geologisches und faunis-
tisches Lebensbild. — Fortschr. Geol. und
Palaeontol., 1925, 12, S. 1-124.
Huene F. von. Die fossile Reptilordnung
Saurischia, ihre Entwicklung und Geschich-
te. — Monogr. Geol. und Palaontol., 1932,
2, S. 51-92.
Huene F. von. Ein neuer Procolophonide
aus dem deutschen Bundsandstein. — N. Jahrb.
Mineral., Geol. und Palaonto]., 1939, 81,
S. 501-511.
Huene F. von. Eine Reptilfauna aus der
altesten Trias Nordrusslands. — N. Jahrb.
Mineral., Geol. und Palaonto]., Beih., 1940,
84, S. 1-23.
Huene F. von. Zur Beurteilung der Proco
lophoniden. — N. Jahrb. Mineral., Geol. und
Palaonto!., 1943, 7,.S. 192-198.
Huene F. von. Palaontologie urid Phyloge-
nie der Niederen Tetrapoden. Procolophonia.
Jena, 1956, S. 173-182.
Huene F. von. Palaontologie und Phyloge-
nie der Niederen Tetrapoden. Nachtrage und
Erganzungen. Jena, 1959, 58 S.
Huxley Т.Н. On a new specimen of Telerpe
ton elginese, - Quart. J. Geo]. Soc. London,
1867, 23, p. 77-84.
Jarvik E. On the structure of the snout of
Crossopterygians and lower Gnathostomes
in general. — Zool. bidr. Uppsala, 1942, 21,
p. 235-675.
Kesteven H. L. The anatomy of the head
of the green turtle Chelone midas, Latr.
Pi I. The skull.
-
J. Proc. Roy. Soc. N.S.W.,
1910, 44, p. 368-400.
Kitching J.W. A short review of the Bea-
fort zoning in South Africa. Symposium on
78
Gondwana. Stratigr. and Paleontol. Cape
Town; Johannesburg, Pretoria, 1970, 123 p.
Kitching J.W.,
Collinson J.W., El
liot D.H., Colbert E.H. Lystrosaurus
zone (Triassic) fauna from Antarctica.
—
Science, 1972, 175, N 4021, p. 524-527.
Koh T, P. Santaisaurus yuani gen. et sp. nov.
ein neues Reptil aus dem unteren Trias von
China. -
Bull. Geol. Soc. China, 1940, 20,
p. 73-92.
Kuhn 0. Die Reptilien. Krai]ling b. Munchen,
1966, 154 S.
Kuhn 0. Die Saurier der deutschen Trias.
Allotting, 1971, 105 S.
Lehman J. P. Nouveaux vertebres fossiles
du Trias de la serie de Zaraaitine.
—
Ann.
Paleontol., 1971, 57, p. 1-18.
Mantel] G. A. Description of the Telerpeton
elginese, a fossil reptile recently discove
red in the Old Red Sandstone of Moray. —
Quart. J. Geol. Soc. London, 1852, .8»
p. 100-105.
Meyer H. v. Sclerosaurus armatus aus dem
bunten Sandstein von Rheifelden. — Pala-
eontographica, 1859, 7_, S. 35—40.
Nopcsa F. Die Familien der Reptilien. —
Fortschr. Geol. und Palaeontol., 1923, 2!,
S. 1-210.
Оe1гiсh Т.M.Theanatomy oftheheadof
Ctenosaura pectunata (Iguanidae). — Misc.
Pubis. Mus. Zool. Univ. Michigan, 1956,
94, p. 1-122.
Olson E.C. The familly Diadectidae and
its bearung on the classification of Repti
les.
— Fieldiana: Geo!., Chicago Natur
Hist. Mus., 1947, 11, N 1, p. 1-53.
Olson E.C. A trilophosaurid reptile from
the Kootenai formation (lover Cretaceous).—
J. Paleontol., 1960, 35, p. 551-555.
Olson E.C. Les Problemes de I'origine des
Reptiles. Problemes acruels de Paleontolo-
gie (Evolution des vertebras), v. 104. Paris,
1962, p. 157-174.
Olson E.C. Relationships of Seymouria,
Diadectes and Chelonia. — Amer. Zool.,
1965, 5, N 2, p. 295-307.
Ortlam D. Fossile Boden als Leithorizonte
far die Gliederung des Hoheren Bundsand-
steins im nordlichen Schwarzwald und stld-
lichen Odenwald. — Jahresber. und Mitt.
Oberrhein. Geol. Ver., N.F.,
1967, 48,
S. 485-589.
Owen R. Descriptive and illustrated catalo-
que of the fossil reptilia of South Africa in
the collection of the British Museum. Lon
don, 1876, p. 25-26.
Parrington F.R. On Prolacerta broomi,
gen. et sp. п., and the origin of Lizards.
—
Ann. and Mag. Natur Hist., ser, 10, ~1935,
16, p. 197-205.
Parrington F.R. A note on the supratempo-
ral and tabular bones in reptiles.— Ann. and
Mag. Natur. Hist., ser. 10,1937,20, p. 69-76.
Parrington F.R. The problem of the clas
sification of reptiles. — J. Linn. Soc. (Zo
ol.), 1958, 44. P- 99 -П5.
Parringjton F.R. A note on the labyrintho-
dont middle ear. — Ann. and Mag. Natur.
Hist., ser. 13, 1959,_2, p. 24-28.
Piveteau J. Existence d'un Reptile du
groupe des Procolophonide*s a Madagascar.
Consequences stratigraphiques et paleonto-
logiques.
—
C. r. Acad, sci., 1955, 241,
p. 1325-1327.
Price L.J. Urn procolophonideo do Triassi-
co do Rio Grande do Sul. — Bol. Sev. geol.
e mineral. Brasil., 1947, 122, p. 9—26.
Robinson P.L. An unusual sauropsid den
tition. — Proc. Linn. Soc. London, 1957,
154, p. 283-293.
Robinson P.L. The evolution of the lacer-
tilia. — Problemes actuels de Paleontolo-
gie (Evolution des vertebres), v. 163. Pa
ris, 1967, p. 396-407.
Romer A.S. The permian cotylosaur Diadec
tes tenuitectes.
-
Amer. J. Sci., 1944, 242,
p. 139-144.
Romer A.S. The primitive reptile Limnosce-
lis restudied.
-
Amer. J. Sci., 1946, 244,
p. 149-188.
Romer A.S. The osteology of the Reptilia.
Ed. 3. Chicago, 1956, 772 p.
Romer A.S. Vertebrate Paleontology. Ed. 3.
Chicago, 468 p.
Rose C. Das Zahnsystem der Wierbeltiere.
Ergebnisse der Anatomie und Entwicklungs-
geschichte von Merkel und Bonnet. Abt. 2,
Berlin, 1894, S. 539-591.
Sa" ve-Sode rbe rgh G. Notes on the trigemi
nal musculature in non-mammalian tetra-
pods. — Nova Acta regiae soc. sci. upsal.,
ser. 4, 1945, 13, p. 1-48.
Sa ve-Sode rbe rgh G. On the fossa hypo-
physeos and the attachment of the retractor
bulbi group in Sphenodon, Varanus and La-
certa.
-
Arkiv zool., Bd. 38a, 1946, U,
p. 1-24.
Seeley H. G. On new species of Procolophon
from the Cape Colony preserved in Dr. Gri-
ersons Museum, Thornhill, Demfriesshire,
with some remarks on the affinites of the
genus. — Quart. J. Geol. Soc. London, 1878,
33, p. 797-807.
Seeley H. G. Researches on the structure,
organization, and classification of the fos
sil Reptilia. VI. On the Anomodont reptilia
and thier allies. — Proc. Roy. Soc. London,
1888, 44, p. 381-383.
Seeley H. G. On the anomodont reptile, Aris-
todesmus rutimeyeri (Wiedersheim) from the
Bunten Sandstone near Basel. — Quart. J.
Geol. Soc. London, 1900, 56, p. 620-645.
Seeley H. G. On the primitive reptile Proco
lophon. - Proc. Zool. Soc. London, 1905,
2x p. 218-230.
Shishkin M.A. On the cranial arterial sy
stem of the labyrinthodonts. — Acta zool.,
1968, 49, p. 1-22.
Siebenrock F. Skelet der Agamidae. —
Sitz.-Вег. Kais. Akad. Wiss. Math.-natur
wiss., Bd. 104, Bat. 1,1895,104, S. 1089-1196.
btappenbeck R. Uber Stephanospondylus
n. g. und Phanerosaurus H. v Meyer. — Z.
Dtsch. geol. Ges., 1905, 57, S. 380-437.
van Pletzen R. The cranial morphology
of Cordylus with special reference to the
cranial kineses. — Ann. Univ. Stellenbosch,
A, 1946, 24, p. 1-40.
Vaughn P.P. The permian reptile Araeos-
celis restudied. — Bull. Mus. Сотр. Zool.,
1955, 113, p. 305-467.
Vaughn P.P. Vertebrates from the Organ
Rock Shale of the Cutler Group, Permian
of Mounument Valley and Vieinty, Utah and
Arisona. -
J. Paleontol., 1964, 38, N 3,
p. 567-583.
Vaughn P.P. More vertebrates, including
a new microsaur, from the upper Pennsyl-
vanian of Central Colorado.
—
Contribs Sci.
Natur. Hist. Mus. Los Angeles, 1972, 223,
p. 1-30.
Warren J.W. The basicranial articulation
of the early Permian Cotylosaur, Captorhi-
nes. - J. Paleontol., 1961,35» N 3, p. 561-563.
Watson D. M. S. Procolophon trigoniceps, a
cotylosaurian reptile from South Africa. —
Proc. Zool. Soc. London, 1914a, p. 735—747.
Watson D. M.S. On the skull of a pareiasa-
urian reptile and on the relationship of that
type. — Proc. Zool. Soc. London, 1914b,
p. 155-180.
Watson D. M. S. A sketch classification of
the pre-Jurassic tetrapod vertebrates. —
Proc. Zool. Soc. London, 1967, p. 167—186.
Watson D. M. S. On Bolosaurus and the ori
gin and classification of reptiles. — Bull.
Mus. Compar. Zool., 1954,3, N 9, p. 299-459.
Watson D. M.S. On Millerosaurus and the
early history of the sauropsid reptiles. —
Phil. Trans. Roy. Soc. London, ser. B,
1957, 240» N 673, p. 325-400.
Webb M. The cranial anatomy of the South
African Geckoes Palmatogecko rangei (An
dersson) and Oedura carroica (Hewitt). —
Ann. Univ. Stellenbosch., A, 1951,27, N5,
p. 131-165.
Welles S. P. Vertebrates from the upper
Moenkopi Formation of Northern Arisona. —
Univ. California Pub]., 1947, 27, N 7,
p. 241-294.
White Т.Е. Osteology of Seymouria baylo-
rensis Broili. — Bull. Mus. Compar. Zool.
Cambridge, 1939, 85, N 5, p. 325-409.
Wiedersheim R. Labyrinthodon rutimeyeri.
Ein Beitrag zur Anatomie von Gesammtske-
let und Gehirn der triassischen Labyrintho-
doten. — Ber. Schweiz. Palaeontol. Ges.,
1878,^ S. 1-56.
Wil 1 is ton S.W. The osteology of the reptiles.
Cambridge: Harvard Univ. Press, 1925,300 p.
Young Chun g-c h i e n. A review of Lepido-
sauria from China. — Amer. J. Sci., 1948,
246, N 11, p. 711-719.
Young Chung-chien. Neoprocolophon
asiaticus^new cotylosaurian reptile from Chi
na. -
Vertebr. Palasiatica, 1957,j:1N 1, p. 1-7.
79
ОГЛАВЛЕНИЕ
ВВЕДЕНИЕ
:
3
МАТЕРИАЛ
5
Глава I. СИСТЕМАТИКА ПРОКОЛОФОНОВ
7
История изучения проколофонов
*
7
Систематика подотряда Procolophonia
,
8
Семейство Nyctiphruretidae
-
Семейство Procolophonidae
•
Ю
Семейство Sclerosauridae
22
Глава II. СРАВНИТЕЛЬНАЯ МОРФОЛОГИЯ ПРОКОЛОФОНОВ
24
Череп
'
-
Общие данные . . . .,
*
Крыша черепа
Орбита ..*....
27
Функциональные аналоги височных окон .
-
Нёбо
28
Мозговая коробка
34
Квадратная кость и подъязычный аппарат
40
Нижняя челюсть
41
Зубы
44
Кинетизм черепа
48
Посткраниальный скелет
Общие данные
Осевой скелет
49
Пояса конечностей
51
Мускулы, железы и нервно-сосудистая система головы
53
Мускулатура головы
Железы и Якобсонов орган
55
Кровеносная система
Черепно-мозговые нервы
•.
.
.
.
60
Глава III. ПРОИСХОЖДЕНИЕ, ФИЛОГЕНИЯ И СТРАТИГРАФИЧЕСКОЕ.РАСПРО
СТРАНЕНИЕ ПРОКОЛОФОНОВ
63
Происхождение и филогения диадектоморфных котилозавров
....
Происхождение и общие направления эволюции проколофонов ....
65
Адаптивная радиация проколофонов
68
Стратиграфическое распространение проколофонов
71
ЗАКЛЮЧЕНИЕ
*.
74
СПИСОК СОКРАЩЕНИЙ ЛАТИНСКИХ ТЕРМИНОВ НА РИСУНКАХ
75
ЛИТЕРАТУРА
76