/







































































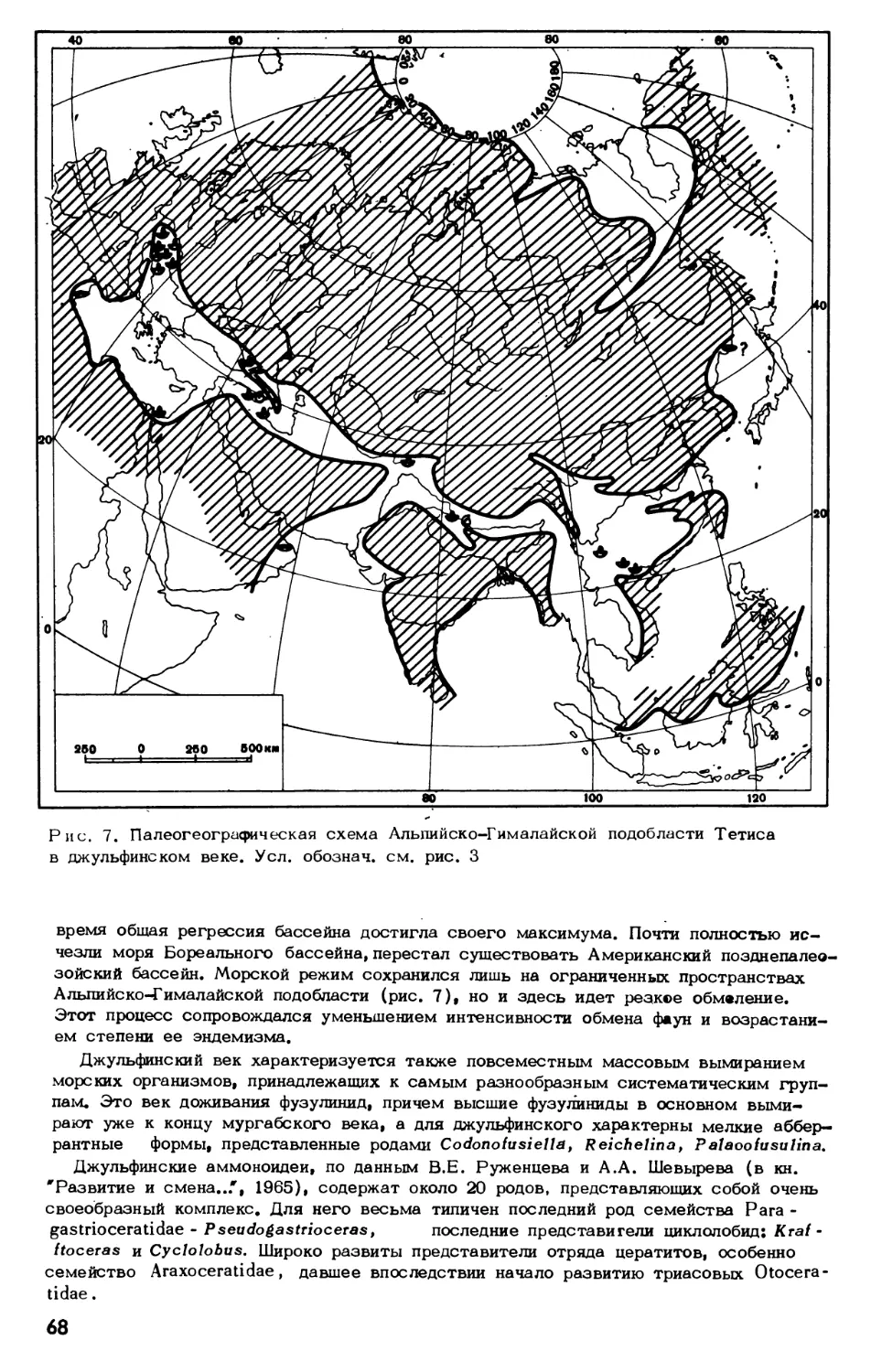



























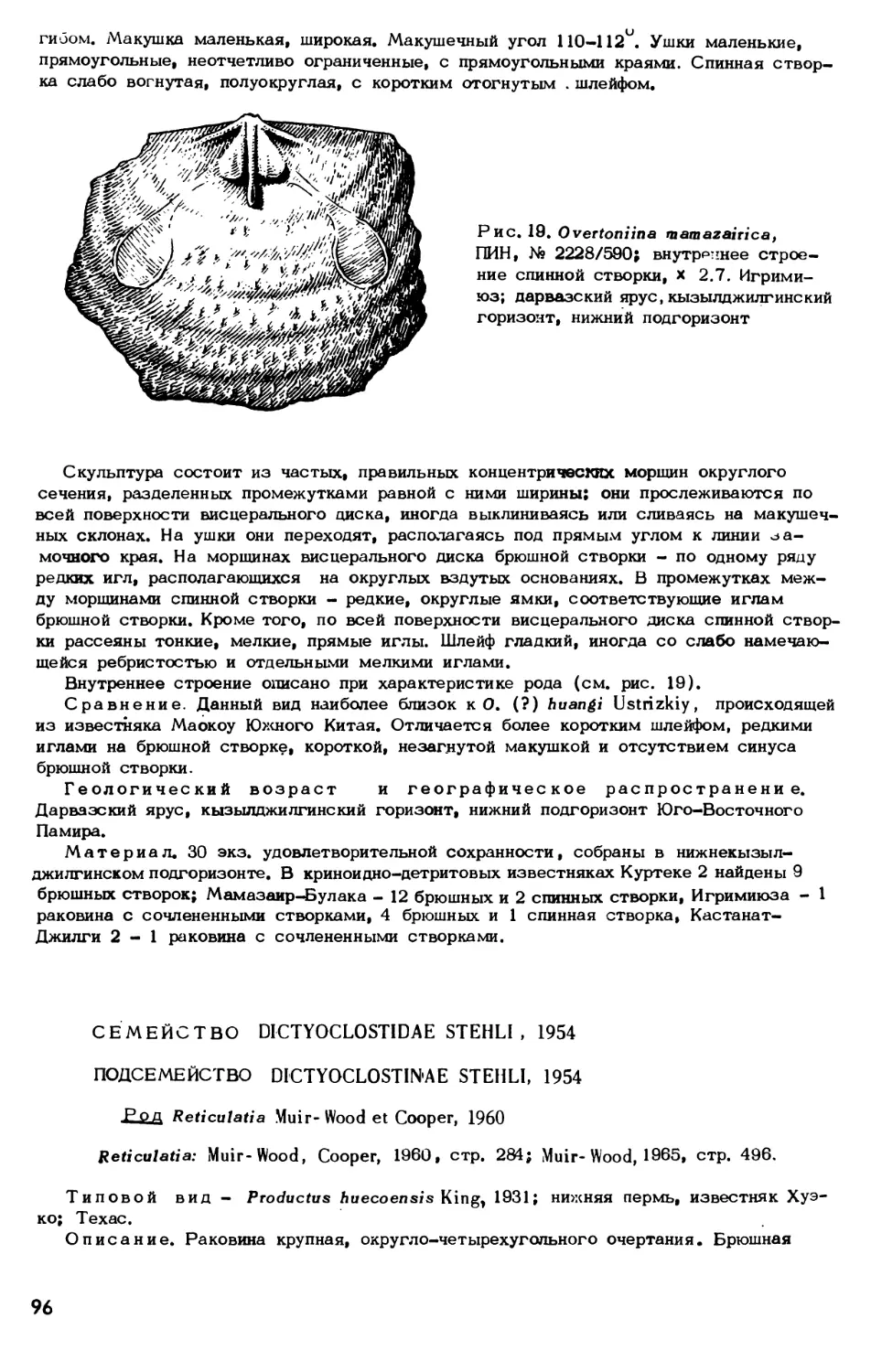
























































































































Text
Т.Л.ГГУИТ, В.Ю.ДМИТРИЕВ
ПЕРМСКИЕ
БРАХИОПОДЫ
ПАМИРА
АКАДЕМИЯ НАУК СССР
ТРУДЫ ПАЛЕОНТОЛОГИЧЕСКОГО ИНСТИТУТА
Том 136
ACADEMY OF SCIENCES OF THE USSR
TRANSACTIONS OF THE PALAEONTOLOGICAL INSTITUTE
Vol. 136
T. A. GRUNT, V. JU. DMITRIEV
PERMIAN
BRACHIOPODA
OF THE PAMIR
PUBLISHING HOUSE <NAUKA>
Moscow 1973
Т. А. ГРУНТ, В. Ю. ДМИТРИЕВ
ПЕРМСКИЕ
БРАХИОПОДЫ
ПАМИРА
ИЗДАТЕЛЬСТВО «НАУКА»
Москва 1973
УДК 564.8(113.6) (235.211)
Пермские брахиоподы Памира, Грунт Т. А., ДмитриевВ.Ю. 1973 г.
В книге впервые описаны пермские брахиоподы Юго-Восточного Па-
мира. Всего описано 53 вида, 18 из которых новые. Здесь также рас-
сматривается стратиграфия пермских отложений Юго-Восточного Пами-
ра, при этом выделяется три стратиграфических комплекса брахиопод,
дается характеристика их систематического состава и экологических
особенностей. В работе сделаны выводы о палеобиогеографическом рай-
онировании пермского Тетиса, путях миграции брахиопод в послеассель-
ское время, показана их закономерная связь с палеогеографической эво-
люцией морских бассейнов.
Книга рассчитана на широкий круг геологов-стратиграфов, палеонтоло-
гов и биологов.
Табл. 16, илл. 60, библ. 421 назв.
Ответственный редактор
Т.Г.Сарычева
_____Q21QH 818-73
(012)02 =73818'73
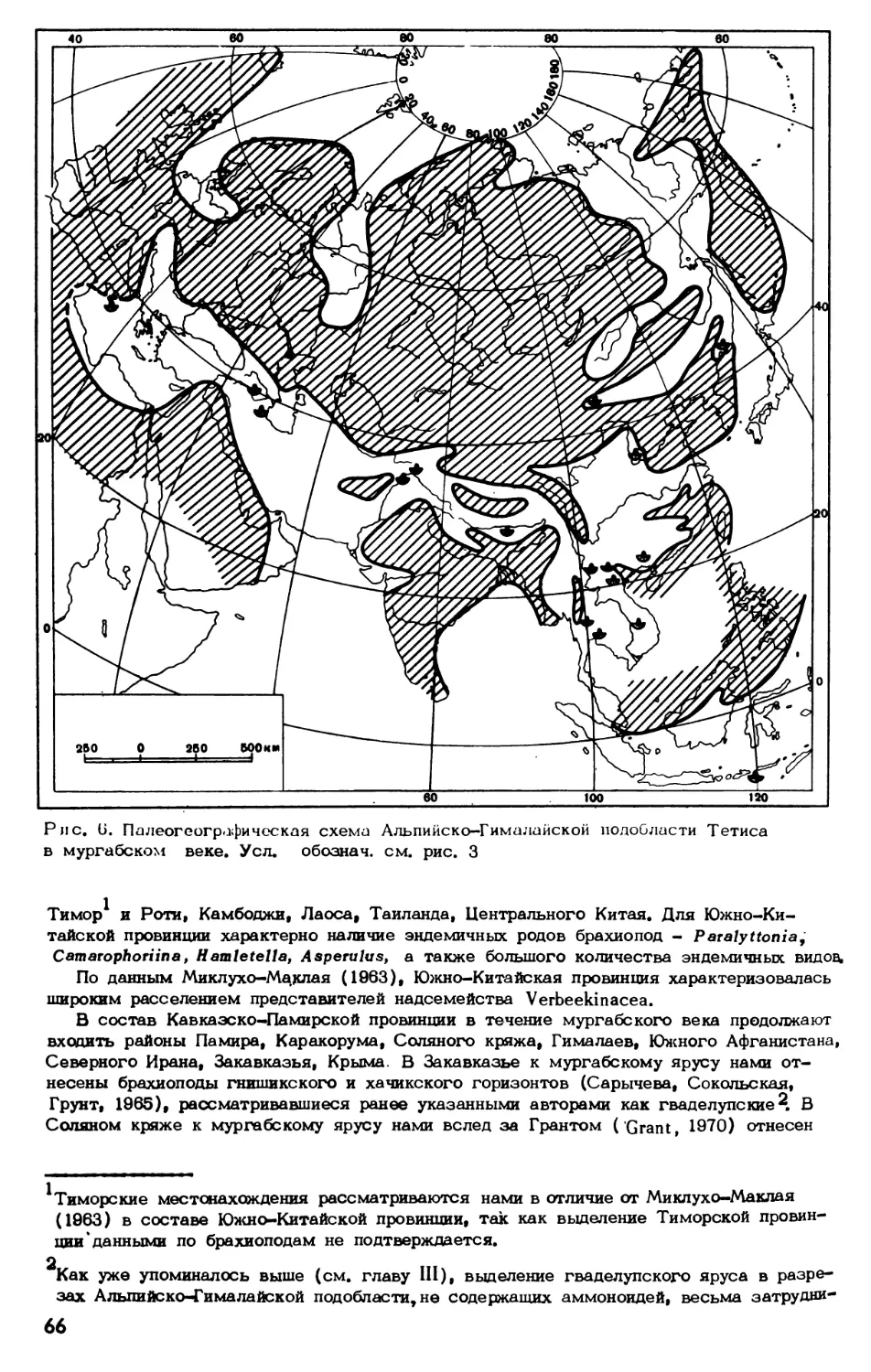
Пермские морские отложения: широко распространены на территории Советского
Союза. Многочисленная и разнообразная фауна этого возраста во многих районах
подробно изучена. Она характерна в основном для Бореальной, или Арктической,
палеобиогеографической области. Лишь сравнительно небольшие районы на юге стра-
ны были заняты в пермское время морями Тропической палеобиогеографической об-
ласти (Тетиса). В пределах СССР к ним относятся Крым, Северный Кавказ и За-
кавказье, Памир и Приморье. Из всех этих районов до последнего времени Памир
оставался наименее изученным. В то же время эта область характеризуется доста-
точно полными и хорошо обнаженными разрезами пермских морских отложений с
обильной и разнообразной фауной.
Географическое положение этого района обусловило, с одной стороны, его непос-
редственные связи с акваториями Куньлуня, Читраля, Каракорума, Соляного кряжа
и далее на восток через Тибет и Гималаи с Китаем и Индокитаем, а с другой
стороны, через Афганистан, Крым, Турцию и Кавказ - с альпийскими и сопредель-
ными акваториями.
Этим и объясняется глубокий интерес, проявляемый стратиграфами и палеонто-
логами к этому району. Но систематическое изучение богатейшей пермской фауны
Памира только начинается, число научных публикаций сравнительно невелико. Наи-
более изученной группой организмов в настоящее время являются фузулиниды, ко-
торым посвящены работы М.А. Калмыковой, Э.Я. Левена, А.Д. Миклухо-Маклая.
В ряде работ О.Г. Туманской и статьях А.М. Павлова рассмотрены пермские ам-
моноидеи некоторых районов Памира. Ругозы изучаются Т.Г. Ильиной и И.В. Пыжья-
новым, мшанки - Р.В. Горюновой.
Пермские брахиоподы Памира до настоящего времени оставались практически
неизученными. Они рассмотрены лишь в краткой монографии Ф.Н. Чернышева и от-
дельных небольших статьях В.И. Волгина. Ряд определений пермских брахиопод
Памира был сделан Б.К. Лихаревым, В.И. Волгиным, Т.В. Машковой. В настоящей
работе впервые монографически описаны пермские брахиоподы одной из структурно-
фациальных зон Памира - Мургаб-Аксуйской (Юго-Восточный Памир). На основе
изученного материала и сравнения пермских брахиопод Юго-Восточного Памира с
одновозрастными брахиоподовыми фаунами сопредельных районов сделана попытка
выявления основных этапов развития пермских брахиопод в Евро-Азиатской части
Тетиса и палеобиогеографического районирования этого бассейна.
Материалы по брахиоподам Юго-Восточного Памира собраны Т.А. Грунт в 1963,
1964, 1966 гг., Т.Г . Ильиной в 1966 г. и В Ю. Дмитриевым в 1967, 1968 гг. Всего
изучено 53 вида, принадлежащих к 43 родам, 27 семействам и 6 отрядам. Из них -
18 видов и 2 рода - новые. Подробно описаны в данной работе наиболее характерные
и редко встречающиеся роды, а также те, в морфологическую характеристику кото-
рых памирский материал вносит какие-либо дополнения. Диагнозы широко распро-
страненных и подробно описанных в литературе родов не приводятся, дается лишь
краткое описание относящихся к ним видов. Для некоторых видов, по которым в
коллекциях содержится ограниченный материал, и видов, подробно описанных в ра-
ботах последних лет, приводятся только данные о материале и распространении.
Два вида ретикуляриид: Kitakamithyris petaliformis Pavlova, sp.noy. и Permophri-
codothyris caroli (Gemm.) описаны E.E. Павловой.
Брахиоподы найдены в верхней Части базардаринской свиты, отнесенной к сак-
марс кому - нижней части артинского яруса, в нижне- и среднекызылджилгинском
5
Рис. 1. Схема расположения изученных местонахождений брахиопод
1 - северная граница Мургаб-чАксуйской структурно-фациальной зоны (Рушанско-
Пшартский разлом); 2 - Мургабский надвиг; 3 - граница Мургаб-Аксуйской и Шах-
да ринской структурно-фациальных зон; 4 - границы подзон Мургаб-чАксуйской струк-
турно-фациальной зоны (подзоны: I - Центральная; II - Промежуточная;, III -
Окраинная» объяснение см. в гл. I ); 5 - местонахождения брахиопод: 1 - Куталь1;
2 - Куталь 2; 3 - Музду -Булак; 4 - Шор-Булак; 5 - Ак-Бура; 6 - Кур-Истык 1; 7 -
Кур-Истык 2; 8 - Южный Ак-Архар; 9 - Игримиюз; 10 - Джилга-Кочусу; 11 -
Куртеке 1; 12 - Куртеке 2; 13 - Кастанат- Джилга 1; 14 - Кастанат-Джилга 2;
15 - Чакобай; 16 - Мамазаир-Булак
подгоризонтах дарвазского яруса в нижне- и среднетахтабулакской подсвитах джуль-
финского яруса. Брахиоподы происходят из следующих 16 пунктов (рис. 1)1:
1) правый борт долины Куталь, на гребне отрога хр.Ган, в 1 км к северу от
высоты 3895,3 (Куталь 1); 2) верховья левого истока долины Куталь, в 1 км к
северо-западу от высоты 4839,0 (Куталь 2); 3) истоки долины Карабелес, правый
борт (Музду-Булак); 4) левый борт долины Шор-Булак-сай, в 600 м к юго- юго-
западу от высоты 4453,5 (Шор-Булак); 5) правый борт долины Зор-Ак-Бура-сай
в 6 км от ее впадения в долину Кош-Агыл (Ак-Бура); 6) левый борт долины Кур-
Истык в 1,2 км выше озерка (Кур-Истык 1); 7) левый борт долины Кур-Истык в
0,5 км ниже озерка (Кур-Истык 2); 8) верховья р.Южный Ак-Архар, правый борт
долины, близ слияния двух составляющих реки (Южный Ак-Архар); 9) верховья
безымянного сухого сая, впадающего справа в р.Суль-Истык близ устья р. Игримиюз,
на гребне водораздела с последним (Игримиюз); 10) гребень водораздела р.Шинды
и р.Джилга-Кочусу в 400 м выше днища долины (Джилга-Кочусу); 11) верховья
второго снизу левого притока р.Куртеке близ водораздела со следующим к западу
небольшим сухим саем (Куртеке 1); 12) верховья второго снизу левого притока
р.Куртеке, левый борт (Куртеке 2); 13) левый борт первого правого безымянного
притока долины Кастанат-Джилга в 1 км от его верховьев (Кастанат-Джилга 1);
14) правый борт первого правого безымянного притока долины Кастанат-Джилга
в 1,5 км от его верховьев (Кастанат-Джилга 2); 15) седловина водораздела р.Чако-
бай и р.Салы-Ункур- сай в 1 км к западу от 459 километра Памирского тракта
(Чакобай); 16) урочище Мамазаир-Булак в 1 км к востоку от 467 километра Па-
мирского тракта (Мамазаир-Булак).
В скобках указаны сокращенные названия разрезов, употребляемые далее в тексте,
а также в объяснениях к таблицам и подписях к рисункам.
6
Изученная коллекция содержит около 2500 экземпляров удовлетворительной сохран-
ности. Многие виды представлены раковинами с двумя сочлененными створками, с
хорошо сохранившимися элементами внутреннего строения, иногда частично пере-
кристаллизованными. При изучении внутреннего строения представителей отрядов
Rhynchonellida, Spiriferida, Terebratulida применялся метод последовательных при-
шлифовок раковин с последующим снятием, лаковых пленок* Полученные пленки за-
рисовывались с помощью аппарата г Микрофот*, а также фотографировались. Детали
внутреннего строения наблюдались также на внутренних ядрах раковин.
Использованные в работе определения различных групп ископаемых организмов
Производились Э.Я. левеном (фузулиниды), Т.Г. Ильиной (ругозы)• В.Е. Руженцевым
и М.Ф. Богословской (аммоноидеи), Р.В. Горюновой (мшанки).
Ценные сведения о ряде разрезов пермских отложений Юго-Восточного Памира
были получены от геологов Горно-Бадахшанской партии Таджикского Геологического
Управления В.И. Дронова, Б.К. Кушлина, Б.М. Гущина, сотрудника МГРИ Э.Я. Ле-
вена, сотрудника Ленинградского горного института А.М. Павлова. Всем этим лицам
авторы выражают свою признательность.
Особую благодарность авторы выражают научному руководителю работы доктору
биологических наук, профессору Т.Г. Сарычевой, а также доктору гео лого-минера ло-
гических наук А.И. Осиповой и всем сотрудникам лаборатории брахиопод ПИН АН
СССР, в которой была выполнена эта работа.
Рисунки в тексте выполнены художником В.И. Дорофеевым и частично авторами;
фотографии - в фотолаборатории Палеонтологического института Н.П. Финогеновым и
Р.М. Шестаковой*
ОБЩАЯ ЧАСТЬ
Глава I
ИСТОРИЯ ИССЛЕДОВАНИЯ ПЕРМСКИХ ОТЛОЖЕНИЙ
ЮГО-гВОСТОЧНОГО ПАМИРА
Первые сведения о верхнепалеозойских отложениях Юго-Восточного Памира во-
обще и о пермских, в частности, сводятся к нескольким упоминаниям о находках
органических остатков в различных местах. Так, в коллекции индийского геолога
Фердинанда Столичко, собранной в 1874 г., К.Ридом (Reed, 1925) был обнаружен и
описан коралл Sy г in gopora sp. верхнепалеозойского облика, происходящий из извест-
няков Мамазаир-Булака.
В неопубликованных дневниках Д.Л. Иванова, проводившего работы на Юго-Вос-
точном .Памире в 1883 г., содержится указание на находку фузулинид в районе пере-
вала Ак-Бура (Ренгартен, 1935). В 1914 г. Г.Гайден (Hayden, 1916) отметил вы-
ходы пермских фузулинидовых известняков в долине Шинды. В 1915 г. Д.В. На ли в кин
(Наливкин, 1916) во время своего путешествия по Памиру обнаружил фузулинид вбли-
зи перевала Джамантал. В 1927 г. П.П. Чуенко нашел обломок гониатита Agathice-
ras sp. в сланцевой толще района Джарты-Гумбез (Наливкин и др., 1932).
С организацией Таджикско-Памирской экспедиции АН СССР начались планомер-
ные работы по изучению верхнепалеозойских разрезов Юго-Вост.очного Памира. В
1931 г. Н.В. Иониным (Ионину- 1933, 1934) проводилась геологическая съемка хр.
Базар-Дара, при которой! верхнепалеозойские отложения с фузулинид а ми были обна-
ружены в долине р.Куберганды.
В 1932 г. А.В. Хабаков описал пермский разрез близ перевала Джамантал; здесь
были собраны фузулиниды, обработанные Г.А. Дуткевичем (Хабаков, 1933; Дуткевич,
Хабаков, 1934).
В 1933-1934 гг. .на Юго-Восточном Памире производилась геологическая съемка:
В.П. Ренгартеном в районе Мургаба (Ренгартен, 1934, 1935), С.И. Клунниковым в
юго-восточной части района (Клунников и др., 1936), И.Г. Барановым в районе мес-
торождения исландского шпата Боз-Tepe (Баранов, 1935, 1936), Г.А. Дуткевичем в
массиве Мын-Хаджир и в Базардаринском хребте (Дуткевич^ 1934) 1935а; Дуткевич,
Калмыкова, 1935, 1936). Одновременно Г.А. Дуткевич посетил большинство известньрс
районов распространения пермских отложений в пределах Юго-Восточного Памира
(Дуткевич, 1934).
В процессе геологосъемочных работ В.П. Ренгартен собрал фораминифер и бра-
хиопод в органогенно-обломочных известняках Мамазаир—Булака. Брахиоподы были
определены Б.К. Лихаревым как Productus cf. aimensis Tschem., мелкая ринхонел-
лида, напоминающая Camarophoria,. Notothyris multiplicata var. minuta Broili (non Waagenj
(Ренгартен, 1935). В.П. Ренгартен отнес отложения, вмещающие эту фауну, к ниж»-
ней части верхней перми.
В результате упомянутых работ был выявлен и изучен ряд пермских разрезов
и предложены первые схемы их стратиграфического расчленения. В.П. Ренгартеном
(1935) была дана трехчленная схема деления пермских отложений, ныне представляю-
щая лишь исторический интерес, в то время как детальная схема Г.А. Дуткевича,
которому принадлежит огромная заслуга в изучении верхнепалеозойских отложений
Юго-Восточного Памира, до сих пор не утеряла своего значения. Наиболее полная
характеристика предложенных Г.А. Дуткевичем стратиграфических подразделений при-
ведена в обобщающей работе 1936 г., где выделенные пачки названы по литологи-
ческому составу (Дуткевич, 1936). Несколько позже (Дуткевич, 1937а) эти же под-»
разделения получили географические названия (см. табл. 1), и схема стратиграфичес-
кого расчленения приобрела следующий вид (снизу вверх):
8
1. Базардаринская толща черных аспидных сланцев с прослоями
песчаников.......................................................*1‘ ‘ 1500-2000 м
2. Кубергандинская известняково-сланцевая толща с фузулинидами
Pseudofusulina kraffti Schellw. et Dyhr., Misellina claudiae Depr.,Cance7-
lina primigena Hayden, Ne os ch wagerin a craticulifera Schwag. и аммоноиде-
ями Popanoceras pamiricum Toum., Sicanites mojsisovicsi Gemm., Adrianites
elegan? Gemm. и др........................................................до 60 м
3. Агалхарская известняково-сланцевая толща с Neoschwagerina ex gr.
craticulifera Schwag., N .colaniae Ozawa и Sumatrina .....................до 50 м
4. Джамантальская толща желтоватых и серых кремнистых сланцев
с прослоями известняков с N eoschwagerina craticulifera Schwag. и Sumat-
rina аппае Volz...........................................................35-40 м
5. Дейринская фиолетово—зеленая кремнистая толща без определи-
мой фауны.................................................................5-80 м
6. Карасинская толща массивных желтых и серых известняков, пере-
ходящих в известняковые конгломераты с Neoschwagerina colaniae
Ozawa, N .margaritae Deprat, Sumatrina annae Volz.........................до 95 м
7. Кутальская фиолетово-зеленая кремнистая толща с обедненным
комплексом фораминифер, известных из нижележащей пачки....................до 40 м
8. Тахтабулакская толща туфогенных и туфогенно-осадочных пород
с Colaniella sp. и Reichelina mirabilis Dutk..............................до 300 м
Перечисленные толщи были фаунистически охарактеризованы почти исключительно
фораминиферами, стратиграфическое распределение которых также было изучено Г .А. Дут-
кевичем (1937 б,см. табл. 1 в данной главе). Лишь в кубергавдинской толще долины
р.Куберганды Г.А. Дуткевичем были обнаружены аммоноидеи, изученные О.Г. Тумане—
кой (Дуткевич, Туманская, 1935), которая отнесла их к самым верхним горизонтам
нижней перми (кунгурскому ярусу). Базардаринская толща Г.А. Дуткевичем была от-
несена к верхнему карбону - нижней перми, кубергандинская - к верхней части ниж-
ней перми, а все вышележащие толщи - к верхней перми. На основании данных по
аммоноидеям и фораминиферам Г.А. Дуткевич за границу отделов пермской системы
принял основание агалхарской толщи с Sumatrina, характеризующееся, появлением
в разрезе пластов светлых известняковых песчаников.
Богатое местонахождение аМмоноидей было обнаружено И.Г. Барановым (Бара-
нов, 1935, 1936) в районе перевала Боз-Tepe. Отсюда О.Г. Туманской был определен
комплекс артинских форм, относящихся к более древним горизонтам, чем аммоноидеи
из долины р.Куберганды (монографии -ки эти аммоноидеи были описаны О.Г. Туман-
ской значительно позже - в 1949 и 1963 гг.). Г.А. Дуткевич (1936) отнес слои с
аммоноидеями, найденные И.Г. Барановым, к нижней части известняково-сланцевой
(кубергандинской) толщи.
В.П. Ренгартеном (1935) и Г.А. Дуткевичем (1936) отмечено развитие в районе
пос. Пост Памирский (ныне Мургаб) преимущественно сланцевых толщ, ранее выделяв-
шихся (Д.В. Наливкин и др., 1932) в качестве мургабской свиты. В.П. Ренгартен,
исходя из характера метаморфизма этих толщ и их предполагаемого стратиграфическо-
кого положения ниже базардарине кой свиты, отнес мургабскую свиту к каменноуголь-
ной системе, что нашло подтверждение в определениях Fusulinella ex gr. bocki Moller,
Г. ex gr. rhomboides ’Lee et Chen,Triticites sp.,сделанных Г.А. Дуткевичем (1936). Перм-
ские отложения этого района отличаются, по Г.А. Дуткевичу (1934, 1936), рядом осо-
бенностей; здесь на мощной толще глинистых сланцев залегает известняковый горизонт
с верхнепермскими фузулинидами, перекрывающийся также мощной толщей глинистых
сланцев. Верхняя толща сланцев по своему характеру близка к самой верхней части
1 ахтабулакской толщи большинства разрезов, но имеет намного большую мощность.
В то же время тахтабулакские туфогенные породы в разрезах окрестностей пос. Мур-
габ не развиты. Различия в строении разрезов Г.А. Дуткевич и другие авторы объяс-
няли фациальным переходом в районе пос. Мургаб тахтабулакских туфогенных повод
Здесь и далее родовые названия приведены в соответствии с современной класси-
фикационной номенклатурой.
9
в нижнюю часть верхней толщи глинистых сланцев, а малая мощность таких же гли-
нистых сланцев в верхней части тахтабу лаке кой толщи связывалась с наличием пред-
верхнетриасового размыва, так как известняки, перекрывающие тахтабулакскую сви-
ту, были отнесены к верхнему триасу по находкам разнообразных Halobia (Рен-
гартен, 1935). В то же время Г.А. Дуткевич не исключал возможности раннетриасо-
вого ‘возраста самой верхней части тахтабулакской толщи, так как в ее нижней
части представлен комплекс фораминифер, характеризующий самые высокие горизонты
пермской системы.
Таким образом, в результате работ Таджикско-Памирской экспедиции АН СССР
и, в первую очередь, исследований Г.А. Дуткевича было выявлено широкое распро-
странение пермских отложений на Юго-Восточном Памире, разработана детальная схема
их стратиграфического расчленения и проведена типизация разрезов, изучена фауна
фузулинид и аммоноидей. Однако фундаментальные исследования Г.А. Дуткевича не
были завершены, вследствие чего разработанная им схема не получила распростра-
нения в тех районах, где ее применение было затруднено из-кза изменчивости раз-
резов5 Состояние изученности пермских отложений района по окончании работ Тад-
жикско-Памирской экспедиции хорошо иллюстрируется обзором П.Д. Виноградова
(1959), составленным, в основном, по материалам экспедиции.
После завершения работ Таджикеко-Памирской экспедиции полевое изучение
пермских отложений^ Юго-Восточного Памира было прервано на длительный срок.
Оно возобновилось с началом работ по государственной геологической съемке, про-
водившейся Памирской экспедицией Таджикского Геологического Управления (1954-
1959 гг.). До появления первых результатов работ экспедиции материалы по стра-
тиграфии пермских отложений Юго-Восточного Памира использовались в основном
в работах А.Д. Миклухо-Маклая (1949, 1955, 195 Z) по разработке общей стратигра-
фической шкалы пермских отложений южных областей (см. табл. 1). Первые обоб-
щения материалов геологической съемки также были сделаны А.Д. Миклухо-Маклаем
(1958а, б, 1960,1961), консультировавшим съемочные работы. Одновременно геологом
Памирской экспедиции Э.Я. Левеном (1958, 1959) была предложена местная стратигра-
фическая схема, а позже (Левен, 1963, 1965, 1967) и общая стратиграфическая схема
для Тетиса. Э.Я. Левен (1958, 1959) расчленил пермские отложения Юго-Восточного
Памира на пять свит: ^базардаринс кую, шиндыйскуюЧ кубергандинскую, ганскую и ка-
рабе лесс кую По сравнению со схемой Г. А. Дуткевича новым компонентом явилась
шиндыйская свита, к которой были отнесены эффузивы основного состава, залегаю-
щие во многих разрезах в верхней части базард арине кой свиты Г. А. Дуткевича. Ган-
ская свита представляет собой объединение нескольких толщ Г.А. Дуткевича (см.
табл. 1), которые, по данным Э.Я. Левена, могут быть выделены только в наиболее
мощных разрезах Юго-Восточного Памира и не прослеживаются по всему району.
Нижняя граница ганской свиты Левеном проводится по появлению первых Neoach—
wetina, т. е. ганская свита включает слои, которые Г.А. Дуткевич откосил еще
к кубергандинской свите; последняя, таким образом, понимается Э.Я. Левеном нес-
колько уже, чем Г.А. Дуткевичем. Название *-ка рабе лесс кая свита* было предложено
вместо старого названия *тахтабулакская свита* после того, как Левеном (1959)
были изучены разрезы окрестностей пос. Мургаб и сходные с ними разрезы в
западной части района. Здесь было установлено, что маломощная па чка_ известняков,
подстилающих верхнюю толщу глинистых сланцев, объединяет аналогиганской и тах-
табулакской свит, а также кобригенской свиты нижнего. - среднего Триаса. (Ниж-
ний и средний триас были выделены в эти годы на основной части территории Юго-
Восточного Памира (Дюфур, Дронов, Кушлин, 1958) по находкам двустворок в нижней
части слоев, относимых ранее к верхнему триасу.) Поэтому толща глинистых сланцев бы-
ла отнесена к триасовой системе, а состав тахтабулакской свиты пересмот-
рен Э.Я. Левеном {1959), в связи с чем она получина новое название. Кроме
того Э.Я. Левеном (.1959) было установлено присутствие маломощного горизонта пер-
мо-триасовых известняков в средней части мургабской свиты (Наливкин и др.,1932),
после чего название *мургабская свита* для этих толщ больше не употреблялось.
Впервые свита была названа *шиндынской* (Левен, 1958); во всех более поздних
публикациях она фигурирует под названием 'шиндыйская*.
10
Сопоставление стратиграфических схем пермских отложений юго-восточного Памира
В.П.Ренгартен, 1935 Г.А. Дуткевич, 1936; Г.А. Дуткевич, М.А. Калмыкова, 1936 Г.А. Дуткевич, 1937 а Г.А. Дуткевич, 1937 6 А.Д. Миклухо- Маклай, 1949
Верхняя сланцевая свита Туфогенная порфири- то-песчано-сланце- вая свита Т ахтабулакская толща Слои с Reichelina mirabilis
Верхняя фиолетово- зеленая кремнистая свита Кутал ьская толща Слои с Parafusulina gigantea, Verbeekina verbeeki, Neoschwa— gerina margaritae, Sumatrina annae Горизонт с Sumatrina,
Свита конгломера- товидных известня- ков Карасинекая толша Polydiexodina
Известняковая свита Нижняя фиолетово- зеленая кремнистая свита Дейринская толша Слои C Jangchienia compressa pamirica, N eoschwagerina co- laniaefSumatrina annae
Кремнисто-извест- няковая свита Джамал ьс кая толша
Агалхарская толша •
Известняково- сланцевая сцита Кубергандинская толша Слои c Misellina claudiae, N eoschwa— gerina craticulifera Слои c Parafusuli— na japonica, Cancel— lina primigena Слои c Pseudofu— salina kraffti, P.ex gr. vulgaris, P.japo— nice, Misellina dy— hrenfurti, Canellina Горизонт с Misellina
Нижняя сланце- вая свита Песчано-слан- цевая свита Базардаринская толща Фаунистически охарактеризован- ные отложения неизвестны
— ... А.Д. Миклухо- Маклай, 1955 Э.Я. Левен, 1958, 1959 А.Д. Миклухо- Маклай, 1961 Ф. Р. Бенш, 1958 А .Д.Миклухо-Ме 1958а, 19586, 1961
Горизонт "мелких фораминифер" Карабелесская свита Памирская свита Горизонт мелких фораминифер и развернутых фузулинид Памирский горизонт (ярус) Lagenidae
Горизонт с Sumatrina, Polydiexodina Верхняя фио- летово-зеленая кремнистая пачка Горизонт с /V eosch wagerina, Verbeekina, Sumatrina
? Ганская свита Пачка конгло- мератовидных известняков Нижняя фиоле- тово-зеленая кремнистая пачка Известняково- кремнистая пачка Мургабская свита Мургабский гор зонт (ярус) с N eosch wagerina, Afghanella, Sumatrina, Polydiexodina
Кубергандинская свита Дарвазский гор зонт (ярус) с Parafusulina
Горизонт с Misellina Кубергандинская свита Шин- Горизонт с Misellina, А г теп in а и первыми выс1 фузулинидами
дыйская “ свита^^^ Базардаринская свита Базардаринская свита Горизонт с Parafusu line и артинскими псевдофузулинами Кар ач атырс к ий горизонт (ярус) Schwagerina
Т аблица 1
А .Д .Миклухо-Маклай 1958а, 19586, 1961 । Решения совещания по разработке уни- фицированных стра- тиграфических схем для Средней Азии, 1959 А.Д. Миклухо- Маклай, 1963 Э.Я. Левен, 1963 Э.Я. Левен, 1965, 1967 В.Е. Руженцев, Т.Г. Сарычева, А.А. Шевырев, 1965 Б.М.Гуинн 1969а, 19696 Схема, принятая в данной работе 7
Памирский горизонт (ярус) с Lagenidae Памирский горизонт - зона мелких фораминифер и развернутых фузулинид Памирский ярус - зона мелких фораминифер и Palaeofusulina, Reichelina, Codonofusiella Памирский ярус - зона Jabeina, Lepidolina Памирский ярус - зона Jabeina, Lepidolina, Codonofusiella Джульфинский ярус - слои с Codonofusiella, Palaeofusulina, Reichelina Джульфинский ярус - зона Colaniella,Palae- fusulina, Codonofusiella Памирская серия \ Тахтабу- / \ лакская / \ свита / в / ( ь / \ °/ \«0 / \ ь ( ) s \ / «о \ / ° S) 1 3 £/ V “/ \ в \ /ь \ / * Джульфинский ярус Тахтабулакская свита Верхняя под- свита
Средняя под- свита Слои с много- численными Codonofusiella, Reichelina, Palaeo fusulina, Colaniella
Нижняя под свита
Мургабский ярус Ганская свита Верхняя подсвита Слои с единичными Codonofusiella, Reichelina
Мургабский гори- зонт (ярус) с /V eosch wagerina, Afghanella, Sumatrina, Polydiexodina Мургабский гори- зонт - зона Neoschwagerina, Sumatrina, Polyd iexodina Мургабский ярус - зона Polydiexodina, Neoschwagerina, Sumatrina Мургабский ярус - зона Neoschwa — gerina Мургабский ярус - зона Neoschwagerina Подзона N.margar.itae Гваделупекий ярус Капитанский подъярус - зона Jabeina, Neoschwagerina Гваделупский ярус Зона Jabeina М уздуайрекская серия Мойнокская свита Слои c Neoschwa — gerina margaritae, Jabeina archaica
Подзона N.schuberti | Зона Neoschwagerina Подзона N.crati- culifera Дейринская свита Слои c N eoschwa- gerina schuberti
Джам анталь- ская свита Нижняя подсвита
Дарвазский гори- зонт (ярус) с Parafusulina Подзона N.simplex Подзона N.simplex Куберган- динская свита Слои c N eosch w a- gerina simplex
и первыми высшими фузулин идами Карачатырский горизонт (ярус) с Schwagerina Кубергандинс кий горизонт - зона Misellina, Armenina • Дарвазский ярус - зона Parafusulina и первых высших фузулинид Карачатырский ярус “ зона Schwagerina s.l. ргандин- ярус Зона Cancellina Кубергандинский ярус - зона Cancellina Вордекий подъярус - зона Cancellina Зона Cancellina к СО м о о. <0 X о. S Кубергандин- ская свита Слои c Cancellina
Зона Armenina
Кубе с к ИЙ Зона Misellina 1 । Артинский ярус Зона Misellina Артинский ярус ' Артинский ярус Зона Misellina ? о а <3 дыйская свита Дарвазский ярус Слои c Misellina
[ Артинский ярус Зона Brevaxina
Улукский горизонт - зона Parafusulina и артинских псевдо- фузулин Карачатырский горизонт - зона Schwagerina Зона Pseudo— fusulina и примитивных Parafusulina _ о S Карадоу- с кая свита Базардаринская свита К ызылджилгинский горизонт верхний подгори- зонт Слои c Misellina и Darvasites ?
Средний подгори- зонт
( Слои c Pseudo- fusulina и прими- тивными Parafu — sulina?)
Нижний подгори- зонт
Сакмарский ярус - зона Schwagerina Сакмарский ярус- нижнейртинскии подъярус Базардарин- ская свита (верхняя часть)
В 1958 г. в Ташкенте состоялось совещание по унификации стратиграфических схем
Средней Азии (Тезисы...., 1958; Решения ...,1959). В докладах А.Д. Миклухо-Маклая
и Ф.Р. Бенш были предложены схемы разделения пермских отложений Средней Азии
на горизонты, охарактеризованные комплексами фузулинид. А.Д. Миклухо-Маклай
предложил расчленить пермские отложения на четыре горизонта: карачатырский го-
ризонт с Schwagerina, дар вазе кий горизонт с Parafusulina и первыми высшими
фузулинидами, мургабский горизонт с N eoschwagerina, Afghanella и Sumatrina и па-
мирский горизонт с Lagenidae.
В схеме Ф.Р. Бенш не рассматриваются слои с Schwagerina, вышележащие тол-
щи она подразделяет на четыре части, из которых две верхние близко совпадают
с мургабским и памирским горизонтами, а две другие соответствуют дар вазе кому
горизонту. Нижний из горизонтов Ф.Р. Бенш, названный в принятой Совещанием ра-
бочей стратиграфической схеме улукским, характеризуется комплексом Parafusulina
и артинских псевдофузулин, верхний горизонт - кубергандинский — первыми высшими
фузулинидами Misellina,Armenina и Cancellina.
Соотношения горизонтов рабочей схемы с подразделениями пермской системы Ура-
ла и Русской платформы в работе этого совещаний не рассматривались и остались
во многом неясны.
Кубергандинский, мургабский и памирский горизонты были выделены на основе
разрезов Юго-Восточного Памира. Кубергандинский горизонт соответствует здесь
кубергандинской свите, мургабский - большей части ганской свиты, памирский -
верхней. части ганской свиты и карабелесской свитам.
Выделение на Юго-Восточном Памире кара четыре кого и у луке кого горизонтов
более сложно, так как стратотипы обоих горизонтов находятся в Фергане. Органичес-
кие остатки карачатырского горизонта на Юго-Восточном Памире не были известны.
В Решениях Совещания по разработке унифицированных стратиграфических схем Сред-
ней Азии (1959) к нему была условно отнесена большая часть базардарине кой
свиты. К улукскому горизонту отнесены шиндыйская свита и верхняя часть базар-
fl аринекой свиты, содержащие органические остатки артинского возраста. В этой час-
ти разреза были сделаны следующие находки (Дронов, Карапетов, Левен, 1959; Дро-
нов, Левен, 1961):
1. В долине р.Шинды в известняковых стяжениях среди эффузивов шиндыйской сви-
ты Э.Я. Левеном были собраны следующие верхиеартинские аммоноидеи: Parapronri-
tes timoren&is Han., Propinacoceras sp., Sicanites cf. in sulcatum (Han.), Agathiceras
sundaicum Han., A.(Gaetahoceras) martini Han., Eothinites sp., Metalegocerasadjaralen—
sis Ruzh.,Popanoceras sp. (определения B.E. Руженцева). Ниже эффузивной толщи,
в верхней части базардаринской свиты Левеном были обнаружены фузулиниды:, Tri—
iicites sp., Parafusulina ex gr. ferganica M—Mac]., Monodiexodina ex gr. wanneri (Schub.)^.
2. Верхиеартинские аммоноидеи были обнаружены при геологической съемке в
верхней части базардаринской свиты в массиве Ак-Бура. Они представлены здесь
видами, описанными О.Г. Туманской (1949) из района перевала Боз-Tepe. Более
подробно аммоноидеи из района перевала Боз-Tepe были описаны О.Г. Туманской в
1963 г. Она отнесла их к улукскому горизонту и пришла к выводу о его соответствии
артинскому ярусу. Как уже упоминалось, слои, содержащие аммоноидеи в районе
перевала Боз-Тере,были отнесены Г.А, Дуткевичем (1936) к нижней части куберган-
динской (известняково—сланцевой) толщи. Поэтому бозтеринские аммоноидеи, с одной
стороны, послужили основанием для выделения на Юго-Восточном Памире улукского
горизонта, а с другой,некоторые их виды приводятся при характеристике куберган-
динского горизонта (см. Решения ..., 1959).
При геологической съемке в 50—е годы слои с боэтеринскими аммоноидеями (
были обнаружены также по правобережью р.Суль-Истык в ее нижнем течении (водо-
раздел с долиной р.Игримиюз).
3. В долине Куртеке В.И. Дроновым, С.С. Карапетовым и Э.Я. Левеном (1959)
был изучен разрез угленосных отложений, ранее отнесенных П.Д. Виноградовым и
Н.Н. Ошурковым к перми и триасу. В.И. Дроновым и др. (1959) было установлено,
Позднее эта форма была определена Э.Я. Левеном (1967) как Monodiexodina
я hi pt on i (Dunbar).
11
что угли приурочены к базальным слоям юрских отложений, трансгрессивно перекры-
вающих пермо-триас. К пермской системе относится толща (50 м) известняков, под-
стилаемая глинистыми сланцами, с видимой мощностью 300-400 м. В своей ниж-
ней части известняки слоистые, детритовые, содержат фораминифер кубергандинского
и мургабского горизонтов;,в верхней части известняки светлые, массивные, содер-
жат фораминифер памирского горизонта. Брахиоподы Chonetes latesinuatus Schellwien,
Buxtonia aff. porrecta (Kutorga), Dictyoclostus aff. uralicus (Tschem.), Cancrinella ca-
ncriniformis (Tschem.), Linoproductus ufenais (Fredericks), L.aagardi (Toula), Leior—
hynchus ripheicus Stepanov, Neospirifer aff. nitienais (Diener), Spiriferella djigtilensis Ste-
panov, Neophricodothyris sp., Spiriferina cristata Schlotheim, Athyris royssi Waagen (non
Leveille), Dielasma el on gat urn (Schlotheim),определенные T.B. Машковой, были
собраны в известковых прослоях в верхней части толщи глинистых сланцев.
На этом основании вмещающие отложения были отнесены к артинскому ярусу нижней
перми.
В.И. Дроновым и Э.Я. Левеном (1961) на основании изучения геологических раз-
резов Юго-Восточного Памира была предложена схема фациальной зональности для
пермо-^гриасового и юрского этапов развития. Ими было показано, что зона, откуда
известны разрезы, в основном изучавшиеся Г.А. Дуткевичем, образует плавную дугу,
обращенную вершиной к северо-западу (Промежуточная подзона), в то время как
внутри этой дуги (Центральная подзона) и по ее периферии (Окраинная подзона) рас-
положены разрезы иного типа. Для внутренней части дуги свойственны разрезы, сход-
ные с разрезами долины р.Куртеке. Они были обнаружены, в нескольких выходах,
расположенных в узкой полосе, протягивающейся с юго-востока на северо-запад на
расстояние. более 100 км. По периферии дуги располагаются разрезы такого же типа,
как в окрестностях пос. ^Иургаб. В пермское время территория Юго-Восточного Па-
мира (Мургаб-чАксуйская структурно-фациальная зона) представляла собой пологий
прогиб, с удлиненным осевым поднятием, соответствующим Центральной подзоне.
Некоторая детализация этой схемы фациальной зональности была произведена
С.В. Руженцевым в связи с тем, что особенности площадного распространения раз-
резов разного типа позволяют предполагать наличие на Юго-Восточном Памире круп-
ных сдвигов (Руженцев, 1962, 1963, 1968; Руженцев, Швольман, 1963). В частности,
была выделена узкая зона разрезов, переходных по своему характеру от Промежу-
точной к Центральной подзоне (районы перевала Боз-Tepe, Игримиюза). Они харак-
теризуются наряду с другими особенностями тем, что на базардарине кой свите здесь
залегают маломощные глинистые сланцы и известняки с аммоноидеями бозтеринского
типа, а эффузивные породы шиндыйской свиты не развиты.
Вскоре после Ташкентского совещания А.Д. Миклухо-Маклай (1958а) придал ка-
рачатырскому, дарвазскому, мургабскому и памирскому горизонтам значение ярусов,
которые могут выделяться в пределах так называемой Кавказско-Синийской палео-
биогеографической области (Миклухо-Маклай, 1957). Несколько позже А.Д. Миклухо-
Маклай (1961, 1966) упоминает мургабскую и памирскую свиты, соответствующие по
своему объему мургабскому и памирскому горизонтам (ярусам) в их стратотипах,
однако обоснования этих свит он не приводит и в дальнейшем, кроме автора, никто
этих названий не использовал.
Подробное обоснование предложенного ярусного расчленения было приведено
А.Д. Миклухо-Маклаев в 1963 г. (Миклухо-Маклай, 1963).
Иная схема ярусного расчленения была предложена Э.Я. Левеном (1963, 1965,
1967). Признавая региональный характер подразделений верхнего отдела перми, он
считает возможным выделять в нижнепермских отложениях сакмарский (в широком
смысле - вместе с ассельским) и артинский ярусы.
При проведении границы отделов пермской системы Э.Я. Левен (1963) исходил
из того, что верхняя пермь Южной Евразии соответствует времени расцвета высших
фузулинид (семейство Verbeekinidae). Расчленение верхнепермских отложений на
зоны было основано на последовательности появления родов в единой филогенетической
ветви подсемейства Neoschwagerininae (семейство Verbeekinidae).
Наиболее древний представитель семейства Verbeekinidae - род Brevaxina, дал
начало филогенетическому ряду Misellina — Armenina — Cancellina — Neoschwagerina —
Jabeina - Lepidolina (подсемейство Neoschwagerininae). В разрезах Юго-Вос-
точного Памира самая нижняя часть кубергандинской свиты, содержащая редких при-
митивных представителей семейства Verbeekinidae, относящихся к единственному
12
роду Brevaxina, при доминировании швагеринид Parafusulina и Pseudofusulina
была отнесена еще к артинскому ярусу. Последовательные родовые зоны Miselli—
па, Armenina, Cancellina, охватывающие большую часть кубергандинской свиты,
были включены в состав верхней перми в качестве кубергандинского яруса, при этом
граница отделов была значительно понижена по сравнению со схемой А.Д. Миклухо-
Маклая и Решениями Ташкентского совещания.
Следует отметить, что точное стратиграфическое положение и границы зон Breva—
xina, Miisellina, Armenina и Cancellina в каком-либо из разрезов кубергандинской
свиты Э.Я. Левеном указаны не были.
Мургабскому ярусу в схеме Э.Я. Левена соответствует родовая зона Neoschwa-
gerina, памирскому - зоны Jabeina и Lepidolina. При этом Э.Я. Девен разошелся
во мнении с А.Д. Миклухо-Маклаем, который относит слои с Jabeina и Lepidolina
к мургабскому ярусу, считая, что в памирском ярусе присутствуют только мелкие
фораминиферы, преимущественно лагениды и редкие фузулиниды, причем здесь ха-
рактерны аберрантные роды Codonofusiella, Reichelina, Palaeofusulina. По мнению
Э.Я. Левена, последние в ряде мест встречаются совместно с родами Jabeina и
Lepidolina, поэтому в районах, где в силу фациальных или биогеографических причин
присутствует только комплекс лагенид и аберрантных фузулинид, его можно считать
соответствующим слоям с Jabeina и Lepidolina. Именно таким образом обстоит
дело на Юго-Восточном Памире, где выше пачки конгломератовидных известняков
с высокоразвитыми неошвагеринами залегают слои с Codonofusiella, Reichelina и др.
Редкие находки примитивных Jabeina archaica Dutk. в пачке конгломератовидных
известняков (Дуткевич, 1936, 1967; Левен, 1965, 1967) были отнесены Э.Я. Левеном
еще к зоне Neoschwagerina.
Несколько позже Э.Я. Левен (1965, 1967) изменил схему зонального расчленения
верхней перми и положение границы отделов. Зона Neoschwagerina (мургабский ярус)
была разделена на три подзоны (снизу вверх): N.siinjplex, N.schubertii и N.margari—
tae, соответствующие последовательным стадиям филогенетического усложнения пред-
ставителей этого рода. Кроме того, при ревизии родового состава и филогении се-
мейства Verbeekinidae (Левен, 1964) род Brevaxina был признан синонимом рода
Misellina, а род Armenina исключен из филогенетического ряда Neoschwa geitini—
пае. Таким образом, вместо четырех родовых зон осталось две: зона Miisellina
и зона Cancellina, из которых первая была отнесена к нижней перми, а вторая к
верхней (кубергандинский ярус). Изменение границы отделов опиралось также на
данные по аммоноидеям, так как известное в долине Куберганды местонахождение
аммоноидей (Дуткевич, Туманская, 1935) было отнесено Э.Я. Левеном к верхней
части зоны Miisellina.
Как уже упоминалось, О.Г. Туманская (1935, 1963) отнесла слои с кубергандински-
ми аммоноидеями к наиболее высоким горизонтам нижней перми. В последнее время
В.Е. Руженцев (см. книгу гРазвитие и смена.../, 1965) вновь рассмотрел состав
кубергандинских аммоноидей в связи с работой по уточнению стратиграфии верхне-
пермских отложений и также пришел к выводу об их принадлежности к наиболее вы-
соким частям нижнего отдела.
В упомянутой книге - коллективной работе сотрудников Палеонтологического ин-
ститута АН СССР на основе изучения вррхнепермских отложений Закавказья и сводки
мирового материала по стратиграфии верхней перми приводится расчленение верхней
перми на два яруса: гваделупский и джульфинский (см. табл. 1 настоящей работы).
Гваделупе кий ярус в типовых разрезах Западного Техаса объединяет зоны аммонои-
дей Waagenoceras и Tiimoidtes и залегает на слоях с Perrinites, отнесенных к
артинскому ярусу (включая - кунгурские слои), т.е. выделение его и определение
границы отделов основано преимущественно на’стратиграфическом распространении
аммоноидей. В разрезах Юго-Восточного Памира слои с Cancellina (кубергандин-
ский ярус) были отнесены к нижней части гваделупского яруса (зона Waagenoceras).
Это основывалось на их стратиграфическом положении выше слоев с кубергандин-
скими аммоноидеями, при использовании некоторых других данных о возможной синх-
роничности слоев с Cancellina и слоев с Waagenoceras. Мургабский горизонт
также был отнесен к гваделупскому ярусу, а памирский - к джульфинекому.
Некоторые данные по стратиграфии пермских отложений Юго—Восточного Памира
получены в последнее время А.М. Павловым (1965, 1967а, 19676) и Б.М. Гущиным
(1966, 1969а, 19696, 1969в). А.М. Павлов (1965, 1967а) изучил стратиграфическое
13
। распределение аммоноидей в разрезе Игримиюза. В этом разрезе выше слоев Боз-
Тере, характеризующихся присутствием родов Daraelites, Parapronorites, Propinacoce—
ras, Medlicottia, Metapeninites, Penmites, Agathiceras, Marathonites, Pamiritella^
Crimites, N eocrimites, Popanoceras, Thalassoceras, Prothalassoceras, Eothinites, Me—
talegoceras, им были выделены слои Суль-Истык с несколько иным комплексом, в
котором наряду с бозтеринскими формами появляются представители и других родов:
Sicanites, Edmedlicottia, Snndaites, Gaetanoceras, Adrianites, Epadrianites, ? Spirolego—
ceras, Pseudovidrioceras, а также родов Mikluchoceras, Pamirioceras, Murgabites,
latycbcerdafAksuites, Cardiella, впервые описанных А.М. Павловым (Павлов, 19676).
Из перечисленных родов Adrianites, Sundaites, Epadrianites и Eumedlicottia обычно
рассматриваются как верхнепермские. В то же время здесь отсутствуют такие ха-
рактерные верхнепермские роды, как Waagenoceras и Stacheoceras. Поэтому А.М.Пав-
лов (1965) в результате анализа с уль-ис тыке кого комплекса приходит к выводу, что
поскольку его*... нельзя уверенно считать верхнепермским, то эти слои можно вы-
делить как переходные между нижней и верхней пермью*. Фузулиниды из верхней час-
ти слоев Суль-Истык были отнесены Э.Я. Левеном (Павлов, 1965; Левен, 1967) к
зоне Misellina.
В другой статье А.М. Павлов (1967а) относит большую часть базардаринской сви-
ты к каменноугольной системе на основании находок в ней аммоноидей Agathiceras
и Stenopronorites?, а также нового рода каменноугольного облика. В верхней части
базардаринской свиты Промежуточной подзоны (Южный Ак-Архар) А.М. Павловым
отмечены аммоноидеи бозтеринского типа (артинский ярус).
Б.М. Гущин (1966) также относит основную часть базардаринской свиты к карбону,
в Связи с определениями в ее средней части органических остатков визе-намюрского
возрастаi •
В последнее время Б.М. Гущиным (1969а, б, в) пересмотрена местная схема стра-
тиграфии пермских отложений Юго-Восточного Памира и их ярусная принадлежность.
В частности, им выделена зона Jabeina, соответствующая подзоне Neoschwagenina
margaritae Э.Я. Левена. Верхнепермские отложения Б.М. Гущиным подразделяются
на гваделупский и джульфинский ярусы. Нижняя граница джульфинского яруса им
несколько понижена. К этому ярусу отнесена верхняя половина карасинской толщи
Г.А. Дуткевича (толщи конгломератовидных известняков), которая выделяется здесь
в самостоятельную чатскую свиту. В чате кой свите, по данным Б.М. Гущина, появ-
ляется большое количество фораминифер джульфинского яруса. Кроме того, им вы-
делены в качестве самостоятельной карадоуской свиты маломощные туфы, отмеченные
Э.Я. Левеном (1967) в верхней части базардаринской свиты в Центральной подзоне,
и предложен ряд новых названий для толщ, понимаемых Б.М. Гущиным несколько
иначе, чем другими авторами, или не подушивших ранее" собственных наименований
(например, в Окраинной подзоне).
* * *
Таким образом, в результате многолетних исследований пермских отложений Юго-
Восточного Памира разработана детальная схема их расчленения для Промежуточной
подзоны (Дуткевич, Левен), наряду с ней выделены Окраинная и Центральная под-
зоны и намечены их основные сопоставления (Левен); изучено стратиграфическое
распределение фузулинид и разработана схема родовых зон фузулинид (Дуткевич,
Левен);на основе распределения фузулинид произведены сопоставления с разрезами
других частей Южной Евразии и предложены схемы общего расчленения пермской
системы (Миклухо-Маклай, Левен) Кавказско-Синийской зоогеографической области.
Специальное изучение пермских брахиопод и других групп бентосной макрофауны
на Юго-Восточном Памире до настоящего времени не производилось, по-видимому,
вследствие сравнительной редкости находок этих организмов, и в связи с этим их
Список предварительных определений, приведенный! в статье Б.М. Гущина, содержит
большое количество форм, для которых определена только их родовая принадлеж-
ность или приблизительная видовая, причем многие формы имеют широкое стратигра-
фическое распространение. Поэтому список, на наш взгляд, не является достаточно
убедительным для установления возраста вмещающих отложений.
14
небольшого значения для решения стратиграфических вопросов. Только в последнее
время появились статьи Т.Г. Ильиной (1970) по ругозам и Р.В. Горюновой (1970,
1971) по мшанкам, посвященные описанию некоторых новых видов этих групп. Настоя-
щее исследование, посвященное монографическому описанию пермских брахиопод
Юго-Восточного Памира, частично восполняет этот пробел.
Недостаточная изученность стратиграфической и фациальной приуроченности перм-
ских брахиопод Тетиса заставляет учитывать при определении геологического воз-
раста комплексов брахиопод все данные о распределении в разрезах сопутствующих
групп ископаемых, особенно фузулинид и аммоноидей. На практике частая приурочен-
ность представителей различных групп ископаемых к разным фациям привела к
необходимости проведения детальных, по возможности послойных, сопоставлений раз-
резов различного типа. Такая работа позволила уточнить схему стратиграфии района,
которая кратко охарактеризована в следующей главе.
Глава II
СТРАТИГРАФИЧЕСКИЙ ОЧЕРК
Верхнепалеозойские отложения широко распространены в пределах Юго-Восточного
Памира (Мургаб-Аксуйская структурно-фациальная зона). Они отчетливо разде-
ляются на две части. Нижняя часть - базардаринская свита - представляет собой
мощную толщу терригенных осадков карбона и нижней перми. Верхняя часть верх-
непалеозойских отложений - относительно маломощная, представлена известняками,
глинистыми породами, эффузивами, туфами и туфогенно-осадочными породами дар-
ва зс кого, мургабс кого и джульфинского ярусов (кызылджилгинский горизонт, кубер-
гандинская, ганская и тахтабулакская свиты). На пермских отложениях залегают
тонкоплитчатые известняки и известняковые конгломераты кобригенской свиты, ох-
ватывающей нижний, средний и часть верхнего триаса (рис. 2).
Наиболее древние фаунистически охарактеризованные отложения, достоверно от-
носящиеся к пермской системе, обнаружены авторами в верхней части базардарин-
ской свиты в Промежуточной I подзоне, где они содержат ассельских аммоноидей
Boesites sp., Meta pronorites timorensis (Han.), Metapronorites sp., Synartinskia sp., Pro—
thalassoceras sp.,? Thalassoceras sp., Agathiceras (A) sp., Eoasianites sp., Proper—
rinites sp., Tabantalites sp., Marathonites (Almites) sp., ? Crimites sp., Waagenina die-
neri (Smith.) (определения B.E. Руженцева и М.Ф. Богословской). В верхней части
свиты в Центральной подзоне известны сакмарско-раннеартинские брахиоподы. В
результате сопоставления разрезов Промежуточной и Центральной подзон (Дмит-
риев, 19736) можно предполагать, что в сводном разрезе базардаринской свиты
слои с сакмарско-раннеартинскими брахиоподами должны располагаться на 200-300 м
выше стратиграфического уровня, на котором находится местонахождение ассельских
аммоноидей. Пермские брахиоподы, более древние, чем сакмарско-раннеартинские,до
настоящего времени на Юго-Восточном Памире неизвестны, поэтому ниже рассмат-
ривается разрез пермских отложений, начиная от слоев с этими брахиоподами.
Сакмарский ярус - нижнеартинский подъярус нерасчлененные
Базардаринская свита (верхняя часть).
Фаунистически охарактеризованные слои в верхней части базардаринской свиты
известны в Центральной подзоне, в разрезах долин Куртеке, Кастанат-Джилга, Ча-
ко бай и в устье долины Кур-Истык. В долине Куртеке они представлены пачкой (30-
40 м) черных битуминозных глинистых алевролитов, в различной степени известко-
вистых, с прослоями (0,5-1,5 м) буровато-серых органогенно-обломочных известня-
ков с песчано-алевритовой примесью и известковистых песчаников с базальным
пойкиллитовым цементом. Остатки ископаемых организмов приурочены к наиболее
известковистым разностям глинистых алевролитов, где встречаются преимущест-
венно ветвистые мшанки, членики криноидей, многочисленные строфомениды (Der-
by га) и более редкие крупные продуктиды ( Reticulatia), и к органогенно-об-
ломочным известнякам, содержащим брахиопод, мшанок, ругоз и многочисленные
членики криноидей. В более глинистых разностях известняков, как и в известковисть
песчаниках,те же органические остатки встречаются реже; для прослоев и лин-
зочек, обогащенных глинистым материалом, характерны в одних случаях двуст-
ворки и гастроподы, в других - мелкие хонетиды.
В разрезе Куртеке I собраны следующие остатки ископаемых организмов:
брахиоподы: Derbyia grandis Waagen, D.diversa Reed, Taeniothaerus rusticus Grunt,
16
Tb,
Центральная подзона
Промежуточная подзона
Gn2
Kb3
Kb2
Gn
Tb2
Kbj
Kz
Bz
Tbj
Вертикальный масштаб
О 20 40 60 80 100
Рис, 2. Схематический литолого-фациальный профиль пермских отложений Промежуточной и Центральной подзон Мургаб-Аксуйской струк-
турно-фациальной зоны по линии Куталь 1 (I) - Куталь 2 (II) - Кур-Истык (III) - Игримиюз (IV) - Кастанат-Джилга (V) - Кур-
теке (VI )
Буквенными индексами обозначены: Bz - базард арине кая свита; - кызылджилгинский горизонт; Kzj - нижнекызылджилгинский
подгоризонт, Kz2 - среднекызылджилгинский подгоризонт, Kz^ - верхнекызылджилгинский подгоризонт; кубергандинская сви-
та; Kbj — слои с Misellina, Kb2 — слои с Cancellina, Kb^ — слои с АГeoschwagerina simplex; Gnj_2~ ганская свита; Gni — нижне-
ганская подсвита; Gng - верхнеганская подсвита; Tb|__g- тахта б у лаке кая f свята: ТЬ| -нижнетахтабулакская подсвита, То2 - средне-
тахтабулакская подсвита, Tbg - верхнетахтабулакекая подсвита; Кг — кобригенская свита. 1 - кварцевые известковистые песчаники;
2 - известковистые алевропелиты; । 3 - глинистые алевролиты с пачками аргиллитов; 4 - аргиллиты; 5 - известняковые конгломераты
и гравелиты; 6 - песчанодетритовые известняки; 7 - криноидно—детритовые известняки; 8 - мелкодетритово-шламмовые известняки;
9 — микрозернистые известняки с незначительной глинистой и алевритовой примесью; 10 - алевритистые известняки; 11 - известняки
с туфогенной примесью; 12 - фораминиферовые известняки; 13 - гидроидно-водорослевые рифогенные известняки; 14 - тонкоплитчатые
микрозернистые известняки с тонкими прослойками и примазками глинистого материала; 15 - переслаивания аргиллитов, микрозериистых
и мелкодетритовых известняков; 16 - туфогравелиты и туфоконгломераты; 17 — туфопесчаники; 18 — туфоаргиллиты; 19 - туфы кислого
состава; 20 - эффузивы основного состава; 21 - водорослевые, табулятозые и табулято-гидроидные биогермы. Основные местонахож-
дения органических остатков: 22 - фузулинид; 23 - ругоз; 24 - аммоноидей; 25 - мшанок; 26 - брахиопод
Il
Промежуточная подзона
III
Центральная подзона
IV
VI
Gn
Kz
Kb
Bz
•«2
Bz
Рис, 2. Схематический литолого-4)ациальный профиль пермских
турно-фациальной зоны по линии Куталь 1 (I) - Куталь 2 (II)
теке (VI )
отложений Промежуточной и Центральной подзон Мургаб-Аксуйской струк-
- Кур-Истык (III) - Игримиюз (IV) - Кастанат-Джилга (V) - Кур-
Вертикальный масштаб
20 40 60 80 100 м
-1____i___I____I____I
Кг
Щтъ/
Tbj
Кг’Кг2
К21
Буквенными индексами обозначены: Bz - базард арине кая свита; - кызылджил пинский горизонт; Kzj - нижнекызылджилгинский
подгоризонт, Kz2 - среднекызылджилгинский подгоризонт, Kz^ - верхнекызылджилгинский подгоризонт; кубергандинская сви-
та: Kbj — слои с Misellina, Kk>2 — слои с Cancellina, Kb^ — слои с N eoschwagerina simplex; Gdj_2“ ганская свита; Gni — нижне-
ганская подсвита; Gr^ - верхнеганская подсвита; Tbj^g- тахта бу лаке кая ( свита: ТЬ| -нижнетахтабулакская подсвита, Т&2 - средне-
тахтабулакская подсвита, ТЬз - верхнетахтабулакская подсвита; Кг - кобригенекая свита. 1 - кварцевые известковистые песчаники;
2-известковистые алевропелиты; > 3 - глинистые алевролиты с пачками аргиллитов; 4 - аргиллиты; 5 - известняковые конгломераты
и гравелиты; 6 — песчанодетритовые известняки; 7 - криноидно—детритовые известняки; 8 — мелкодетритово-шламмовые известняки;
9 — микрозерни с тые известняки с незначительной глинистой и алевритовой примесью; 10 — алевритистые известняки; 11 — известняки
с туфогенной примесью; 12 — фораминиферовые известняки; 13 — гидроидно-водорослевые рифогенные известняки; 14 — тонкоплитчатые
микрозернистые известняки с тонкими прослойками и примазками глинистого материала; 15 — переслаивания аргиллитов, микрозернистых
и мелкодетритовых известняков; 16 — туфогравелиты и туфоконгломераты; 17 — туфопесчаники; 18 — туфоаргиллиты; 19 — туфы кислого
состава; 20 — эффузивы основного состава; 21 — водорослевые, табулятозые и та буля то-гид рои дные биогермы. Основные местонахож-
дения органических остатков: 22 - фузулинид; 23 - ругоз; 24 - аммоноидей; 25 - мшанок; 26 - брахиопод
sp.nov., Reticulatia pamiri ca 'Grunt, sp.nov., Cancnne/Ja altissima King, Geraaaimovia
bactriana Grunt, sp.nov., Punctocyrtella spin о за Plodowski, fl eoapirifer joharenaia (Die-
ner), Cleiothyridina ai la ken ai a (Reed);
мшанки: Fistulamina curtekenaia Gorjunova, Ramiporidra aaiatica Gorjunova, Dyacri-
tel la sp.nov., Streblaacopora luminosa Gorjunova, Chainodictyon lucidum Gorjunova, Fe-
neatella pamirenaia Gorjunova, F .aubquadratopora Schulga—Nesterjenko, F.biiida Eich—
wald subsp. craaaiaeptata Schulga—Nesterjenko, Polypora brevicellata Baranova, P.soahki—
пае Schulga—Nesterjenko, P.aubcrotilla Tnizna;
ругозы: Pleurophyllum auatrale Hinde, Waagenophyllum sp.
В долине р.Кастанат-Джилги доступен наблюдению более мощный разрез базар-
даринской свиты (около 250 м). Она представлена здесь однообразной толщей темных
и черных глинистых алевролитов с нечастыми прослоями бурых известковистых
песчаников и черных глинистых известняков. Известняки иногда м, содержат остатки
брахиопод, мшанок, двустворок и гастропод плохой сохранности. Многочисленны сак-
марско-раннеартинские брахиоподы: । Taeniothaerua ruaticua Grunt, sp.nov., Reticulatia
pamirica Grunt, sp.nov., Cancrinella altiaaitna King, Geraaaimova bactriana Grunt, sp. nov.,
Neospirifer joharensis (Diener), Kitalamithyria petalifonais Pavlova, sp. nov., Martinia
uralica Tschem., Cleiothyridina ailakenaia (Reed.). Они найдены в верхней части
свиты в прослое (1 м) буровато-серого сильно известковистого глинистого
алевролита. Здесь же собраны немногочисленные мшанки Feneatella sp.nov., Ж/a-
tkella orientalia(Eichw.\Ramiporidra aaiatica Gorjunova.
Близ устья сая Чакобай доступны наблюдению только фрагментарные выходы са-
мых верхних частей свиты. Здесь обнажены черные алевритистые аргиллиты с нес-
колькими прослоями (0,3-0,5 м) черных глинистых известняков и темно-серых из-
вестковистых глинистых алевролитов. Отсюда определены брахиоподы: Derbyia sp.,
Reticulatia pamirica Grunt, sp.nov., Cancrinella altiaaima King, Geraaaimovia bactriana
Grunt, sp.nov., Cleiothyridina ailakenaia (Reed).
В устье долины р.Кур-Истык непосредственно близ кровли базардаринской свиты
в черных аргиллитах располагаются тонкие прослои (10-20 см) черных битуминозных
известняков с немногочисленными двустворками, гастроподами и аммоновдеями Ме-
ta pronorites timorenaia (Haniel), Marathonitea (Almites) gracilis Smith., Waagenina aff.
dieneri (Smith.), Agathiceraa sp. Эти аммоноидей, по заключению М.Ф. Богослов-
ской, характерны преимущественно для сакмарского века, но нельзя в то же время
считать, что они не могут иметь и раннеартинский возраст.
Дарваэский ярус
Дарваэский ярус в пределах Юго-Восточного Памира рассматривается нами в
объеме кызылджилгинского горизонта (Дмитриев, 1973а) и кубергандинской свиты,
что соответствует фузулинидовым зонам Pseudofusulina и примитивных Parafusulina,
Misellina, CancelИпа и подзоне Neoschwaheitina simplex.
Кызылджилгинский горизонт залегает трансгрессивно на разновоз-
растных слоях базардаринской свиты, обычно с признаками размыва, местами на
маломощной глинистой коре выветривания, В результате последовательного сопостав-
ления разрезов кызылджилгинский горизонт может быть подразделен на три части
(подгоризонты).
В основании иижнекыз ылджилгинс кого подгоризонта залегает пласт
(1-3 м) серого глинистого микрозернистого известняка с прослоями и линзочками
криноидно-детритового известняка. В Промежуточной подзоне известняк более глинис-
тый, содержит аммоноидей, из которых наиболее характерны: iDaraelitea pamiricua
Toum., Propinacoceraa aff. ajenae (Max.), Agathiceraa mediterranean Toum., M etale goceraa
sp., Eothinitea sp., Perrinites aubcumminai (Han.)*, Marathonitea (Almites) pamiricua
Toum., Crimitea pamiricua Toum., Popanoceraa hanieli Smith.
В Центральной подзоне и переходных в ней разрезах (Северная Боз-Tepe, Зор-
Тор, Ничке-Джилга, Чакобай, Мамазаир- Булак, Куртеке, Кастанат-Джилга, Игри-
миюз, Пустан и др.) известняк преимущественно криноидно-детритовый с брахиопо-
дами, ругозами и табулятами.
Здесь встречены брахиоподы Overtoniina mamazairica Crunt, gen. et sp. nov., Probo-
lionia himalayenaia (Diener), Globiella? roaaiae Fantini Sestini, Martinia tschemyschewi
17
Grunt, sp. nov., Сашепзша murgabica Grunt, sp.nov., Labaia dmitrievi Grunt, sp.nov. ;
ругозы Polycoelia dobrolyubovae Ilyina, PleurophyHum gerthi Schdwf., P.auatrale Hinde,
Ulimia mu Ititabu latum Schdwf., U.variabile (Schdwf.), U.alternata (Huang), U .peraimmetri-
cum Schdwf., Verbeekiella gerthi Schouppe et Stac., V.auatralia (Beynich), Wannerophyllum
tubuloaum (Gerth), Timorphyllum wanneti wanneti (Gerth), Lophophyllidium spinosum
(Martin), Jatsengia aaiatica Huang.
Вышележащая часть нижнекызылджилгинского подгоризонта в Промежуточной под-
зоне образована пачкой (10-15 м) неслоистых серых известковистых алевропелитов
с характерной палочковидной отдельностью. Алевролиты содержат незначительную
примесь мелких частиц вулканического пепла-пузырчатых и игольчатых обломков
раскристаллизованного вулканического стекла кислого состава. В нижней части
пачки иногда присутствуют тонкие (10-15 см) прослои остракодовых известняков.
В разрезах, переходных к Центральной подзоне, пачка известковистых алевропели-
тов имеет меньшую мощность (3-8 м) и содержит прослои (15-20 см) серого из-
вестняка с аммоноидеями: \Daraеlitеа pamiricua Тошп., Para pronoritea timorenaia Han..
Propinacoceraa sp., Agathiceraa (A.) mediterraneum Toum., Gaetanoceraa martini (Han.),
Eothinitea sp., Perrinitea aubcumminai (Han.), Perrinitea sp., Marathonitea (Almitea)
pamiricua Toum., Crimitea sp., Cardiella graaia Pavl., Akauitea permicua Pavl.
В Центральной подзоне известковистые алевропелиты отсутствуют и непосред-
ственно на криноидно-детритовом известняке залегают среднекызылджилгинские от-
ложения.
Среднекызылджилгинский подгоризонт в Промежуточной подзоне
образован пачкой (15-35 м) темно-серых и черных аргиллитов и глинистых алевро-
литов с прослоями известковистых кварцевых песчаников, алевролитов и алевритис-
тых известняков. В прослоях наиболее крупнозернистых песчаников встречаются круп-
ные фузулиниды М onodi exodina ahi pt on i (Dunbar).
В разрезах» переходных к Центральной подзоне, феднекызылджилгинский под-
горизонт (5-25 м) слажен породами такого же типа, как в нижнем подгоризонте -
серыми известковистыми алевропелитами с прослоями известняков. Известняки сред-
него подгоризонта характеризуются значительной алевритистостью; в них содержат-
ся аммоноидеи Parapronoritea timorenaia Han., Mikluchoceraa pamiricum Pavl., Propina -
coceraa ajenae Max., Thalaaaoceraa mediterraneum Toum., Agathiceraa (A.) mediterraneum
Toum., Gaetanoceraa martini (Han.), Eothinitea sp., Perrinitea sp., Marathonitea (Almitea)
pamiricua Toum., Pseudovidrioceras dun6ari Mill. et Fum., Crimitea gemmellaroi Toum.,
Akauitea permicua Pavl., Pamiritea clinei Toum., Cardiella graaia Pavl. Приблизитель-
но к этой части разреза приурочены найденные А.М. Павловым (1965) немногочис-
ленные экземпляры Sicanitea, Eumedlicottia, Sundaitea, Adrianitea.
В Центральной подзоне среднекызылджилгинский подгоризонт представлен темно-
серыми алевритистыми известняками (2-3 м) с редкими аммоноидеями. В некоторых
разрезах Центральной подзоны (Чакобай, Кастанат-Джилга, р.Пустан) среднекызыл-
джилгинский подгоризонт сложен зелеными микроэернистыми известняками (6-12 м)
с кристаллокластами плагиоклаза, кварца и обломками игольчатого вулканического
стекла. Известняки связаны постепенными переходами с темно-зелеными туфами
(карадоуская свита Б.М. Гущина).
В известняках с кристаллокластами: встречены брахиоподы: Probolionia himalayen—
sia (Diener), Coatiferina sp., Martinia tachakobaica Grunt, sp. nov., M.tachemyschewi
Grunt, $p. nov., Cameriama murgabica Grunt, sp.nov., Spiriferella sp., Labaia dmitrievi
Grunt, sp. nov. ; ругозыzDuplophyllum achindewolii Schouppe.
К верхнекызылджилгинскому подгоризонту в Промежуточной под-
зоне относится толща (20-120 м) черных и зеленоватых глинистых сланцев» содер-
жащая мощные пласты (10-80 м) эффузивов основного состава и туфов (шиндыйская
свита Э.Я. Левена). В линзах известняка среди эффузивов были собраны верхнеар-
тинские аммоноидеи (см. список в главе I, стр. 11). Нами пачка черных и зеле-
новатых глинистых сланцев (5-15 м)» залегающих выше шиндыйских эффузивов и
относимых обычно к кубергандинской свите» отнесена к верхнему подгоризонту кы-
зылджилгинс кого горизонта» так как по своей характеристике глинистые сланцы
вполне подобны сланцам» подстилающим эффузивы и залегающим между их пластами.
В Центральной подзоне к верхнекызылджилгинскому подгоризонту относятся ма-
ломощные (4-6 м) темно—серые фузулинидовые известняки» содержащие, по данным
18
Э.Я. Левена (1967), Darva sites contractus (Schellw. et Dyhr.), Pseudofusulina curte-
kensis Leven, Pa. quasi fusuliniformis Leven, Ps. pavlovi Leven, Ps. ex gr. compacta (White),
Parafusulina dzamantalensis Leven, P.grupperaensis (Thomps. et Mill.), P.vulgariaiformis
Mor., P.globosaeformis Leven, P.poatkraffti Leven, P.annae Leven, M'isellina aliciae
(Dept.). В разрезах, переходных к Центральной подзоне, наблюдается фациальный
переход темно-серых фузулинидовых известняков в глинистую пачку (J0-15 м), со-
ответствующую толще с эффузивами Промежуточной подзоны.
Кубергандинская свита образует верхнюю часть дарваэского яруса Мур>-
-аб-Аксуйской структурно-фациальной зоны. В данной работе кубергандинская свита
рассматривается в объеме, близком к тому, который ей придавал автор свиты Г.А.Дут-
севич (1937а). Нижняя граница кубергандинской свиты проводится в Промежуточной
подзоне по изменению общего облика пород, обычно совпадающему с появлением пер-
зых известняковых прослоев с фузулинидами. Слои с N eoschwagerina simplex Ozawa, 1
юключенные Э.Я. Левеном из кубергандинской свиты и отнесенные им к ганской
гвите, нами, вслед за Г.А. Дуткевичем, рассматриваются в составе кубергандинской
;виты, так как особенности их состава и залегания свидетельствуют о более тесной
гвязи с основной частью кубергандинской свиты, чем с вышележащими слоями с
Sumatrina (подзона Neoschwagenina s ch u be it i Э.Я. Левена).
В наиболее мощных разрезах (до 110 м) Промежуточной подзоны (Куталь, Ку-
эерганды и др.) кубергандинская свита представлена чередованием серых,зеленоватых
а буроватых мелкозернистых и грубодетритово-фузулинцдовых известняков с темно-
серыми желтоватыми и зеленоватыми глинистыми сланцами и тонкими прослоями
:еро-кзеленых известковистых мелкопсаммитовых туффитов. В основании свиты нередко
обнаруживаются прослои, содержащие мелкий растительный детрит. По данным Э.Я. Ле-
вена (1967), для нижней части свиты, относящейся к зоне Misellina, характерен
следующий комплекс фузулинид: Schubertella sp., N eofusulinella cf. tan ten nisi Deprat,
Pseudofusulina pavlovi Leven, Parafusulina gruppdraenais (Thomps. et Mill.), P.dzamanta-
lensis Leven, P.schucherti Dunb. et Sk., Misellina claudiae (Depr.), M.aliciae (Depr.),
H.otakiensis Hu aim Oto, M.termieri (Depr.), M.aff. olgae Leven.
Как видно из приведенного списка, комплекс фузулинид слоев с Misellina в ниж-
пей части кубергандинской свиты отличается от комплекса верхнекызылджилгинокого
подгоризонта; особенно характерно присутствие в последнем рода Dazwaaites, не
представленного в кубергандинской свите и обилие в ней более высокоразвитых пред-
ставителей рода Мisellina.
Средняя часть свиты относится к зоне Cancellina, она характеризуется фузули-
аидами: Nan kin е Па sp., Schubertella sp., N eofusulinella lantenoi&i Depr., Jangchienia
com pres sa (Ozawa), J. haydeni longa Leven, Minojapanella (Wutuella) wutuensis Sheng, Ghu—
sen el la tieni Chen, Ch. schwageriniformis Sheng, Pseudofusulina chihsiaensis Lee, Ps.guem-
beli (Dunb. et Sk.), Ps. quasifusuliniformis Leven, Parafusulina dzamantalensis Leven,
P.densa Raich., P.skinneri Dunbar, P.kinosakii (Мог.), P. shaksgamensis Reich., Miselli —
na claudiae (Depr.), Cancellina cutalen&is Leven, C.nipponica (Ozawa), C.dutkevitchi Leven,
C.pamitica Leven, Armenina aaiatica Leven, A.cf. salgirica A.M.—Maclay, Pseudodoliolina
ozawai J a be et Hanzawa.
В верхней части кубергандинской свиты преобладают известняки серого и темно-
серого цвета, появляются мелкие кремневые конкреции, резко уменьшается роль ту-
фэге иного материала в составе пород. Отсюда Э.Я. Левеном (1967) определены Mino-
japanеПа cf. pulchra А.М.—Mad., Neofusulinella lantenoiai Depr., Jangchienia compressa
(Ozawa), Ps. cf. quasifusuliniformis Leven, Ps. ex gr. compacta (White), Misellina ovalis
(Deprat), N eoschwagerina simplex Ozawa, Prae sumatrina sp., (?) Afghan el la sp., Armenina
sp., Verbeekina verbeeki (Gein.), Pseudodoliolina sp.
Кубергацдинские аммоноидеи (Дуткевич, Туманская, 1935) были собраны, согласно
описанию Г.А. Д утке вина, все в одном слое, в 2 м выше которого была найдена
Cancellina primigena Hayden, т.е. они характеризуют самую верхнюю часть зоны
Misellina или пограничные части зон Misellina и Cancellina. Родовой состав ам-
моноидей из этого местонахождения был пересмотрен В.Е. Руженцевым (в книге
'Развитие и смена*..., 1965), который приводит следующий их список: Propinacoce-
ras, ? Artinskia, Medlicottia, Sundaites, Agathiceras, Neocrimites, ? Stacheoceras, Popano -
ceras.
В разрезах с сокращенными мощностями, располагающихся во внутренней части
Дуги Промежуточной подзоны,кубергандинская свита (15-25 м) слажена преимушест-
19
венно известняками. Обычно в нижней части свиты залегает характерный горизонт
(7-15 м) бурых и зеленоватых известняков с фузулинидами, характерными для верх-
ней части зоны Misellina и с прослоями зеленых известковистых туффитов. Места-
ми наблюдается быстрое уменьшение мощности слоев с Misellina и замещение из-
вестняков глинистыми сланцами (например, в нижнем течении р.Шор-Булак-Сай и по
правобережью р.Шинды), В долине р.Южный Ак-Архар к нижней части кубергандин-
ской свиты может быть условно отнесена пачка (9 м) темно-серых глинистых слан-
цев, в основании которой залегает прослой брахиоподового ракушника, сложенного
раковинами Тепакеа nalivkini Grunt, sp. nov.
Верхняя часть свиты слажена темно-серыми мелкодетритовыми и мелкозернистыми
известняками (5-15 м) с прослоями темно-серых глинистых сланцев и желтовато-
серых микрозернистых известняков. В известняках обычно присутствуют некрупные
кремневые конкреции, в некоторых разрезах преобладают темно-серые глинистые
сланцы. В верхней части темно-серых известняков собраны фузулиниды подзоны
Neosch wagerin a simplex.
В большинстве разрезов Промежуточной подзоны кубергандинская свита может быть
подразделена на ряд пачек, характеризующихся сходными чертами строения с зале-
ганием в основании этих пачек известняковых песчаников и гравелитов и далее
переслаиваниями мелкодетритовых известняков, глинистых сланцев и туффитов. Эти
пачки хорошо выделяются в наиболее мощных разрезах свиты; они группируются
в три более крупных ритма, прослеживающихся почти во всех разрезах Промежуточ-
ной подзоны и близко соответствующих слоям с Misellina, слоям с Cancellina и
слоям с Neoschwagerina simplex.
В разрезах, переходных к Центральной подзоне, не всегда можно с уверенностью
решить вопрос о присутствии кубергандинской свиты. Здесь наблюдается дальней-
шее сокращение мощностей свиты, изменение ее состава, иногда выпадение всей
свиты или ее части из разреза. Так, в районе перевала Боз-Tepe к кубергандинской
свите может быть отнесена пачка (до 15 м) серых глинистых сланцев с прослоями
зеленых и желтоватых пепловых туффитов и известняков, связанная постепенными ~
переходами с глинистыми породами верхнекызылджилгинского подгоризонта. В разре-
зах Игримиюза и приустьевой части долины Кур-Истык кубергандинская свита, по-
видимому, выпадает из разреза, и известняки мургабского яруса залегают не-
посредственно на верхнекызылджилгинс ких слоях.
В Центральной подзоне стратиграфические аналоги кубергандинской свиты уста-
навливаются по присутствию слоев с Cancellina и Neosch wagerin a simplex Ozawa
(Левен, 1967). В разрезах Куртеке и Кастанат-Джилги они представлены грубосло-
истыми светло-серыми детритово-фузулинидовыми известняками (20-25 м). Нижняя
часть кубергандинской свиты в Центральной подзоне неизвестна. (Не исключено ее
частичное соответствие верхней части темно-серых фузулинидовых известняков, от-
несенных нами целиком к верхнекызылджигинскому подгоризонту.) Развитие во всех раз-
резах Центральной подзоны осыпей, закрывающих часть разреза (5-10 м), от темно-
серых фузулинидовых известняков до выходов слоев с Cancellina не позволяет точ-
но решить этот вопрос.
Мургабский ярус
К мургабскому ярусу на Юго-Восточном Памире относится ганская свита, со-
ответствующая агалхарской, джамантальской, дейринской, карасинской и большей
части кутальской толщи ГА. Дуткевича. Мургабский ярус включает в себя слои с
Neoschwagerina schuberti, слои с N eoschwagerina margaritae и Jabeina archaica и слои
с первыми Codonofasiella и Reich el in а.
Ганская с вита (Левен, 1958) рассматривается в данной работе в несколько
иных границах, чем ее автором, однако общий объем свиты в типовых разрезах
хр. Ган изменен незначительно. Нижняя граница свиты проводится по кровле слоев
с N eosch wagerin a simplex Ozawa. На этой границе происходит резкое изменение общего
облика толщ, наблюдающееся по всей Промежуточной подзоне и соответствующее
началу нового этапа осадконакопления для территории Юго-Восточного Памира.
Верхняя граница свиты несколько понижена, что будет рассмотрено далее.
В Промежуточной подзоне ганская свита может быть разделена на две подсвиты.
20
Нижнеганская подсвита (агалхарская и джамантальская толщи Г.А. Дут-
кевича) в наиболее мощных разрезах Промежуточной подзоны образована чередова-
нием пластов (2,5-4 м) мелкогалечных известняковых конгломератов и песчаников
и пачек (2-5 м) темно-серых, серо-желтых и красноватых глинисто-кремнистых
сланцев с тонкими прослоями микрозернистых глинистых известняков. Снизу вверх
по разрезу подсвиты исчезают пласты известковистых конгломератов, которые за-
мещаются детритовыми известняками, и происходит уменьшение роли известняковых
песчаников. Характерны прослои с кремнистыми конкрециями среди детритовых из-
вестняков; мелкие кремневые конкреции встречаются также в связи с микрозер-
нистыми известняками в пачках глинистых сланцев. Общая мощность подсвиты до
80 м.
Известняковые песчаники и конгломераты и в меньшей мере детритовые извест-
няки содержат фузулинид: Minojapanella sp., N eofusulinella lantenoisi Dep г., Jang -
chienia sp., Kahlerina (?) sp., Neoschwagerina simplex Ozawa, N. schuberti Koch.-Dev.,
praesumatrina neoschwagerinoides (Depr.), Afghanella aff. schencki Thomps., Sumatrina an-
пае brevis Leven, Armenina salgirica A.M.-Macl., A. sphaera (Ozawa), Verbeekina verbeeki
(Gein.), V. pon tica (A.M.-Macl.), P seudodoliolina ozawai Jabe et Hanzawa.B верховьях
долины Куталь автором найден вместе с фузулинид а ми гониатит Neocrimites (So-
siocrimites) sp. В разрезах Промежуточной подзоны с сокращенными мощностями
происходит выпадение глинистых пород и уменьшение мощности известняковых сло-
ев, обычно сложенных известняковыми песчаниками, переходящими в детритовые из-
вестняки. Мощность подсвиты составляет здесь 10-12 м.
Верхнеганская подсвита (дейринская, карасинская и большая часть ку-
тальской толщи Г.А. Дуткевича) в наиболее мо^рных разрезах Промежуточной под-
зоны (на южных склонах хр. Ган) подразделяется на три части: нижняя часть под-
свиты (дейринская толща Г.А. Дуткевича) образована тонкопереслаивающимися жел-
товато-серыми, зеленоватыми и фиолетовыми микроэернистыми известняками,
глинистыми и глинисто-кремнистыми сланцами и серыми мелкозернистыми и мел-
кодетритовыми известняками (20-40 м). В известняках часто встречаются мелкие
кремневые конкреции. Комплекс фораминифер, очень редко встречающихся в мелко-
детритовых известняках, нехарактерен. Э.Я. Левеном (1967) пачка переслаиваний
предположительно отнесена к подзоне Neoschwagerina schuberti.
Средняя часть подсвиты (карасинская толща Г.А. Дуткевича) представлена мас-
сивными и грубослоистыми известняковыми конгломератами и песчаниками (15-20 м).
Обломки в конгломератах плохо окатаны, они сложены обломочно-детритовыми из-
вестняками, сходными с известняками, цементирующими их. Среди конгломератов
встречаются небольшие линзы водорослевых и гидроидных известняков, такими же
известняками образованы некоторые обломки. Снизу вверх по разрезу толщи конгло-
мераты переходят в известняковые песчаники и детритово-фузулинидовые известняки.
В верхней части толщи известняки приобретают зеленоватый оттенок и содержат при-
месь вулканогенного материала. Фузулиниды весьма многочисленны в цементе конг-
ломератов и в детритовых разностях, но, по мнению Э.Я. Левена (1967), большей
частью переотложены. Наиболее характерными формами он считает N eoschwagerina
margaritae Deprat, N. minoensis Depr., AL graticulifera (Schwager), Jabeina archaica Dutk.
Верхняя часть подсвиты (большая часть кутальской толщи Г.А. Дуткевича) сло-
жена такими же переслаиваниями, как и нижняя часть подсвиты, и имеет мощность
15-25 м. Она отличается только появлением редких тонких (5-10 см) прослоев из-
вестняковых песчаников с фиолетовым глинисто-карбонатным цементом. Форамини-
феры представлены очень редкими Colaniella parva (Colani), Palaeofusulina pamirica
Leven, Codonofusiella sp., Reichelina sp.
В пределах большей части Промежуточной подзоны, характеризующейся разрезами
с сокращенными мощностями, верхнеганская подсвита представлена монотонной тол-
щей микрозернистых плитчатых известняков (30-35 м), в то время как горизонт
известняковых конгломератов в этих разрезах полностью выклинивается. Б.М. Гу-
щиным (1969а) в некоторых более мощных разрезах этого типа (Куталь 2, ур.Ша-
карак) обнаружены пласты диабазов, которые по своему стратиграфическому уров-
ню, по-видимому, соответствуют пачке известняковых конгломератов.
В Центральной подзоне ганская свита до настоящего времени не выделялась. Как
Уже упоминалось, здесь выше черных фузулинидовых известняков верхнекызылджил-
гинского подгоризонта, отделяясь от них осыпями, залегают светлые слоистые из-
21
вестняки с фузулинидами зон Cancellina и Neosch wagerin a simpex Ozawa (Куртеке), от-
несенные нами к кубергандинской свите. Над ними располагаются массивные, свет-
лые биогенные известняки (40-50 м), образованные преимущественно гидроидными 1
и водорослями. В нижней части массивных известняков Э.Я. Левеном (1967) об-
наружены Jabeinatf) sp., в верхней части - комплекс микрофауны памирского (джуль-
финского) яруса. Все ^звестняки Центральной подзоны были объединены Левеном
в куртекинскую свиту и более дробно расчленялись только по фауне фораминифер.
Невозможность выделения в Центральной подзоне ганской свиты определялась не
только трудностью расчленения рифовых биогенных известняков, а также и предста-
влением (Левен, 1967) о том, что они непосредственно перекрываются кобригенской
свитой нижнего триаса и, таким образом, замещают тахтабулакскую свиту, охва-
тывая всю верхнюю часть верхней перми. Тем не менее ганская свита может быть
выделена в Центральной подзоне. Она включает основную часть толщ, залегающих
на слоях с Neoschwagerina simpex за исключением верхней части биогенных извест-
няков, выше которых, как нам удалось установить, залегает маломощная пачка гли-
нистых пород, соответствующая большей части тахтабулакской свиты. Параллелиза-
пия указанных слоев с ганской свитой может быть произведена в результате наблю-
дений в разрезах, переходных к Центральной подзоне, в частности, в долине
Шинды. Здесь наблюдается следующие соотношения: в разрезе левого борта долины,
напротив устья р.Кызыл- Джилга разрез ганской свиты типичен для Промежуточной
подзоны с сокращенными мощностями. Иной разрез наблюдается в устье долины
Джилга-Кочусу (левый приток р. Шинды). Он во многих отношениях сходен с раз-
резами Центральной подзоны. Здесь выше нижнеганских светлых известняков (12 м)
залегает маломощная пачка светлых микрозернистых плитчатых известняков, выше
которых располагается толща массивных и грубослоистых водорослево-гидроидных
известняков. При сопоставлении с предыдущим разрезом легко устанавливается
замещение биогенными известняками большей части верхнеганской подсвиты. Верх-
няя часть биогенных известняков в устье Джилги-Кочусу частично замещает слои,
перекрывающие верхнеганскую подсвиту в разрезе против р.Кызыл-Джилга, и отно-
сится нами так же, как и эти слои, к тахтабулакской свите джульфинского яруса.
Точно так же к джульфинскому ярусу должна быть отнесена верхняя часть рифо-
генных известняков в большинстве разрезов Центральной подзоны, где они содержат
характерный комплекс фузулинид.
Брахиоподы в ганской свите почти совершенно неизвестны. Находки единствен-
ного вида Martinia triquetra Gemm. приурочены к нижнеганской (?) подсвите в раз-
резе Игримиюза. Их стратиграфическое положение может быть установлено лишь
предположительно, вследствие особого характера этого разреза, в котором на верх-
некызылджилгинском подгоризонте с признаками размыва залегают известковые
конгломераты с линзами врдорослево-гидроидных известняков и органогенно-детри-
товые известняки мощностью 19 м. Верхняя часть известняковой толщи по комплек-
су фузулинид, определенных Э.Я. Левеном, должна быть отнесена к джульфинскому
ярусу (нижнетахтабулакской подсвите). Она отделяется от нижней части горизонтом
(5-6 м) известняковых конгломератов, который, несмотря на быструю фациальную
изменчивость слоев, прослеживается в районе Игримиюза почти повсеместно. Нижнюю
часть известняковой толщи можно сопоставить с ганской свитой, исходя из сравнения
с разрезами долины Кур-Истык (Промежуточная подзона с сокращенной мощностью)
и стратиграфического положения ниже нижнетахтабулакских известняков с фузули-
нидами. Линза микрозернистого светлого известняка, содержащая цельные рако-
вины и разрозненные створки Martinia triquetra Gemm., обнаружена среди водорос-
левых известняков в 2 м от подошвы известняковой толщи.
Наблюдения в районе Игримиюза подтверждают правильность выделения ганской
свиты в Центральной подзоне. В средней части располагающейся здесь полосы выходов
По мнению некоторых исследователей, эти организмы относятся к водорослям рода
Hikorocodium (Гущин, 1969а).
^Название 'куртекинекая свита* ранее было использовано для обозначения палеоге-
новых отложений этого же района (Наливкин и др., 1932).
22
перми происходит переход всей известняковой толщи в крупный водорослево-гидро-
идный биогерм (до 30 м)ти разрез верхней перми в целом становится очень сходным
с близко расположенным типичным разрезом Центральной подзоны в долине Кастанат-
Джилга (рис. 2).
Джульфинский ярус
Тахтабулакская свита\ В данной работе тахтабулакская свита (Дуткевич,
1937а) понимается в несколько большем объеме, чем ранее. В ее состав в качестве
нижней подсвиты включены темные известняки, подстилающие тахтабулакские туфоген—
ные породы в большинстве разрезов Промежуточной подзоны. Нижнетахтабулакская
подсвита наилучшим образом выражена в разрезах Промежуточной подзоны с сокра-
щенными мощностями в восточной части района (массив Ак-Бура, долина Кур-Ис-
тык, Сарыкольский хребет). В наиболее мощных разрезах Промежуточной подзоны
(южные склоны хр.Ган) она соответствует, по нашим данным, верхней части куталь-
ской толщи Г.А. Дуткевича. Таким образом, нижняя граница тахтабулакской свиты
в ее типовых разрезах незначительно понижена.
В основании нижнетахта булакс кой поде виты в Промежуточной подзоне с
сокращенными мощностями залегают темно-серые несортированные известняковые
конгломераты (5-10 м) с обильным карбонатным цементом и небольшими (0,3-0,8м)
биогермами черных водорослевых известняков. Цемент конгломератов содержит не-
значительную примесь туфогенного материала. В долине Кур-Истык среди водоросле-
вых известняков собраны брахиоподы Orthotichia avuschensis Sokolskaja, Parenteletes
ruzhencevi Sokolskaja,/Vote t/iyris pseudodjoulfensis Lich., sp. nov.
Над известняковыми конгломератами залегает пачка (3,5-7,5 м) серых глинистых
сланцев, переслаивающихся с темно-серыми мелкодетритовыми известняками. В верх-
ней части глинистые сланцы обычно имеют зеленоватый оттенок и содержат тонкую
пирокластическую примесь uСостав фораминифер в тёмных известняках был изучен
Э.Я. Левеном (1967) в среднем течении долины р.Шинды (напротив устья р.Кызыл-
Джилга). Из фузулинид им определены: Staffella zisongzhengensis (Scheng.), Nankinella
sp., Reichelina pulchra К. M.-Maclay, Codonofusiella sp., Palaeofusalina pamirica Leven.
Мелкие фореминиферы весьма многочисленны, особенно характерны Colaniella
ранга (Colani). Остатки макрофауны в черных известняках встречаются редко. Только
в долине Шинды Э.Я. Левеном собраны немногочисленные брахиоподы Productus sp.,
Heterelasmina schuchertiLich., Uncinunellina cf. sicula Gemm. (определения Б.К. Лихарева).
На южных склонах хр. Ган обе пачки сокращены в мощности и изменяются фациаль-
но. В разрезе Куталя известняковые конгломераты переходят в темные желто-серые
известняковые песчаники с плавающими гальками черных известняков. Мощность пес-
чаников подвержена сильным изменениям, в средней части долины Куталь она состав-
ляет 3 м, в близкорасположенном разрезе Джамантала мощность известняковых пес-
чаников меняется на коротких расстояниях от 1,2 до 0,2 м, причем в раздувах слоя
встречаются многочисленные небольшие светлые известняковые желваки (биогенные?).
Пачка глинистых сланцев и черных известняков также меняет свой характер. Она пред-
ставлена в этих разрезах тонкими переслаиваниями фиолетовых глинистых сланцев и
микрозернистых известняков, иногда с линзами известняковых конгломератов (4-6,5м).
Переименование тахтабулакской свиты в карабелесскую (Левен, 1958) было пред-
ложено вследствие того, что в состав свиты Г.А. Дуткевичем и другими были не-
верно включены верхнетриасовые толщи. Легко устанавливается, что карасинская,
кутальская и тахтабулакская толщи Г.А. Дуткевича получили свои наименования в
единой полосе выходов Джамантал-Куталь, которые должны быть приняты за страто-
типические (Гущин, 1969а). Неверное понимание стратиграфических подразделений за
пределами их стратотипов по Правилам стратиграфической номенклатуры не может
служить основанием для их переименования,в связи с чем должно быть использовано
название *тахтабулакская свита*, тем более, что оно до последнего времени приме-
няется в литературе (см. Миклухо-Маклай, 1966).
23
Разрез нижнетахтабулакской подсвиты в Промежуточной подзоне завершается пач-
кой (2,0-5,5 м) светло-серых известняковых гравелитов и песчаников» переходящих
в детритовые известняки. Известняки переслаиваются с фиолетовыми или зелеными
туфоаргиллитами. В разрезе Кур-Истыка в детритовых известняках собраны брахио-
поды Stenoaciama armenica Sok., Рermophricodothyriз caroli Gemm.
В разрезах долины р.Куберганды нижнетахтабулакская подсвита представлена в
нижней своей части массивными темно-серыми известняковыми конгломератами (Им),
в верхней части - пачкой желто-серых тонкослоистых микроэернистых известняков
с прослоями темно- и светло-серых детритовых известняков (8,5 м).
В этой долине Б.М. Гущин (1969а) выделяет темно-серые известняковые конг-
ломераты в особую чатскую свиту, содержащую Colaniella sp., Palaeofuaulina sp.,
Reichelina sp., Codonofuaiella paradoxic a Dunbar et Skinner, Staffella ziaongzhengensis
(Sheng), Neoschwagerina craticulifera (Schwager), Misellina sp., Minojapanella sp.l
В разрезах, переходных к Центральной подзоне, нижнетахтабулакская подсвита
представлена различно в разных частях района. В нижнем течении р.Шинды (устье
р.Джилга-Кочусу) верхняя часть подсвиты сложена такими же известняками,как в
упоминавшемся разрезе средней части долины, а нижняя часть подсвиты замещается
грубослоистыми и массивными светлыми водорослево-гидроидными известняками,
содержащими брахиопод Haydenella tumida Waagen, L ер to du a sp., Notothyris paeudodjoul-
fenaia Lich., sp.nov., Martinia rupicola Gemm. В разрезе Игримиюза и в районе
перевала Боз-Tepe нижнетахтабулакская подсвита представлена серыми детритовыми
известняками и известковистыми песчаниками (5-7 м). В первом из них Э.Я. Ле-
веном собраны фузулинидых Codonofuaiella sp., Рalaeofиsulina sp.f Staffella ex gr.
ziaongzhengensis (Sheng).
В Центральной подзоне к нижнетахтабулакской подсвите может быть отнесена
верхняя часть водорослево-гидроидных известняков, откуда Э.Я. Левеном (1967)
определены: Colaniella parva(Colani), Staffella ziaongzhengensis (Scheng), Nankinel—
la sp., Reichelina pulchra K.M.—Maclay, R. cf. chan gh sin gen sis Scheng et Chang,Codonofu-
aietla curtekensia Leven. В долине Куртеке в верхней части водорослево-гидро-
идных известняков найдена Еп teletella nikschitchi Licharew.
Среднетахтабулакская подсвита представлена в Промежуточной под-
зоне неяснослоистой толщей зеленых и фиолетовых туфоаргиллитов и зеленых туфо-
песчаников с прослоями мелкогалечных туфо конгломератов. Под свита повсеместно
отчетливо подразделяется на среднюю, преимущественно песчаниковую часть и ниж-
нюю и верхнюю, образованные преимущественно туфоаргиллитами. Наибольшую мощ-
ность (до 80 м) подсвита имеет на южных склонах хр. Ган, где сильно увеличена
ее средняя песчаниковая часть (50-55 м). В разрезах Промежуточной подзоны с
сокращенными мощностями среднетахтабулакская подсвита имеет мощность 25-30 м,
при мощности пачки туфопесчаников 15-20 м.
Среди туфопесчаников и туфоконгломератов среднетахтабулакской подсвиты встре-
чаются некрупные биогермы, особенно характерные для разрезов с сокращенными
мощностями. Биогермы имеют округлую или овальную форму и размеры от 0,3 до
10 м. Они сложены преимущественно водорослями, табулятами, гидроидными (стро-
матопоровдеями ?); подчиненную роль в сложении биогермов играют сфинктоэоа,
иногда также хететиды и колониальные ругозы. Наряду с рифообразующими орга-
Нижнетахтабулакская подсвита (чатская свита) долины Куберганды включает боль-
шую часть слоев, отнесенных Г.А. Дуткевичем (19356) к толще конгломератовид-
ных известняков и верхней фиолетово-зеленой кремнистой толще, которые позже
(Дуткевич, 1937а) были названы карасинской и кутальской. В связи с массовым
распространением в чатской свите джульфинских фораминифер нижняя граница джуль-
финского яруса была понижена Б.М. Гущиным (1969а) до середины карасинской тол-
щи. Однако разрезы долины Куберганды не являются типовыми для карасинской и
кутальской толщ, которые получили свои наименования по разрезам южных склонов
хр. Ган. Состав фораминифер в типовых разрезах этих толщ был хорошо изучен
Э.Я. Левеном (1967) и не дает оснований для понижения границы мургабского и
джульфинского ярусов до средней части карасинской толщи.
24
ннзмами в некоторых биогермах встречаются многочисленные брахиоподы, членики
крнноидей и редкие остатки двустворок, гастропод и наутилоидей. В каждом биогер-
ме резко преобладают представители одной-двух групп рифообразующих организмов,
вследствие чего можно выделить по основному составу рифообразователей водоросле-
вые, табулятовые и табулятово-гидроидные биогермы. В табулятовых, табулятово-
гидроидных и гораздо реже в водорослевых бигермах содержатся брахиоподы
(см. табл. 5).
В разрезах, переходных к Центральной подзоне (Шинды, Игримиюз), среднетах-
табулакская подсвита представлена пачкой (17-22 м) зеленых туфоаргиллитов с тон-
кими прослоями зеленоватых и фиолетовых известняков с туфогенной примесью. В
разрезе Игримиюза в туфоаргяллитах содержатся немногочисленные мелкие хонетиды.
В Центральной подзоне к среднетахтабулакской подсвите относятся маломощные се-
ро-желтые аргиллиты (3-5 м) с редкими тонкими (0,05-0,1 м) прослоями темно-
серых микрозернистых известняков.
Верхнетахтабулакс кая подсвита связана с среднетахтабулакской пос-
тепенными переходами. Она наиболее полно представлена в разрезах Промежуточ-
ной подзоны с сокращенными мощностями. Здесь подсвита сложена темно-серыми
и бурыми аргиллитами с редкими прослоями темно—серых микрозернистых или бурых
детритовых известняков (10-15 м).
В Промежуточной подзоне с максимальными мощностями (Джа манта л, Куталь)
верхнетахтабулакская подсвита представлена небольшой пачкой темно-серых аргил-
литов (5 м).
Подсвита с очень резкой, четкой границей перекрывается кобригенской свитой
нижнего - среднего триаса, сложенной в нижней части характерными тонкоплитча-
тыми светло-серыми известняками, которые в некоторых разрезах содержат прослои
и линзы конгломератов с гальками этих же известняков, и, кроме того, обломками
и валунами оолитовых известняков. По данным М.С. Дюфура, В.И. Дронова, Б. К. Куш-
лина (1958), нижняя часть кобригенской свиты Промежуточной подзоны содержит
нижнетриасовых двустворок Claraia cf. orientalia Richt., E итог phot is inaequicoatata
Ben., E.ex.gr. venetiana Bitt, и других и аммоноидей Flemingitea sp.
В разрезах, переходных к Центральной подзоне, наблюдаются уменьшение роли
аргиллитов в составе поде виты и увеличение количества прослоев черных микрозер-
нистых известняков. В долине р.Шинды эти слои перекрываются светлыми извест-
няковыми конгломератами нижнего триаса, содержащими характерные обломки ооли-
товых известняков. В Центральной подзоне мощность подсвиты сильно уменьшена,
из ее состава почти полностью выпадают аргиллиты и преобладают черные тонко-
слоистые известняки, обычно относимые к кобригенской свите. Таким образом, наз-
вание *кобригенская свита* применяется в Промежуточной и Центральной подзонах
к разновозрастным слоям, причем нижняя часть "кобригенской свиты* Центральной
подзоны является стратиграфическим аналогом верхнетахтабулакской подсвиты Про-
межуточной подзоны и нами отнесена к последней.
Фаунистические остатки в верхнетахтабулакской подсвите неизвестны, в связи
с чем она может быть отнесена к пермской системе лишь условно. Не исключен
нижнетриасовый возраст верхней части подсвиты.
♦ ♦ ♦
Общепринятое разделение пермских отложений Тропической палеобиогеографи-
ческой области на ярусы и отделы до настоящего времени не разработано. В гла-
ве 1 рассмотрено ярусное расчленение пермских отложений Юго-Восточного Пами-
ра, предлагавшееся различными авторами. Ниже кратко обсуждаются вопросы раз-
деления пермских отложений Юго-Восточного Памира на ярусы и отделы для по-
яснения принятой в данной работе стратиграфической схемы. Принятая номенклатура
отражает представления авторов о наиболее характерных особенностях строения
пермских отложений Юго-Восточного Памира и основана на анализе главных этапов
осадконакопления района. Это представляет некоторый материал для стратиграфичес-
Ских выводов общего характера, которые, однако, требуют привлечения фактических
Данных, далеко выходящих за рамки задач настоящего ясследпеання
25
На Юго-Восточном Памире верхне каменноугольные отложения по условиям своего
образования тесно связаны с нижнепермскими, они вместе образуют большую часть
терригенной базардаринской свиты.
Вышележащие пермские толщи Юго-Восточного Памира могут быть разделены
на две примерно равные по объему и сходно построенные крупные части (см. рис.2).
Нижняя охватывает кызылджилгинский горизонт и кубергандинскую свиту (дарвазский
ярус), верхняя - ганскую и тахтабулакскую свиты(мургабский и джульфинский ярусы).
Отложения дарвазского яруса Юго-Восточного Памира четко отграничены, по
времени своего образования они отвечают крупному естественно-историческому эта-
пу развития рассматриваемой территории.
Первая половина этого этапа (кызылджилгинское время) характеризуется раз-
витием трангрессии после непродолжительного континентального перерыва. В Про-
межуточной подзоне наблюдается переход от мелководных осадков нижнекызылд-
жилгинского под горизонта с признаками временных отклонений от нормального мор-
ского режима (остракодовые известняки) к относительно более глубоководным гли-
нистым породам, и шаровым лавам верхиекызылджилгинского подгоризонта.
Кубергандинское время характеризуется формированием мелководных обломочно-
детритовых известняков с фузулинидами и глинистых сланцев. В нижней части ку-
бергандинской свиты местами отмечен мелкий растительный детрит, свидетельствую-
щий о сравнительной близости береговой линии; верхняя часть свиты представлена,
по-видимому, более мористыми фациями. Кубергандинская свита тесно связана с под-
стилающими отложениями и в маломощных разрезах переходных к Центральной под-
зоне практически не отделяется от верхнекызылджилгинского подгоризонта, как и в
Окраинной подзоне^•
Название *дарвазский ярус* было предложено А.Д. Миклухо-Маклаем (1958а) для
толщ, объединяющих зоны Parafusulina и Misellina. В качестве стратотипа яруса
была избрана часть разреза Юго-Западного Дарваза, соответствующая основной
части дарвазской серии Г.А. Дуткевича (1937а) и объединяющая зыгарскую, челам-
чинскую, сафетдаронскую и гундаринскую свиты Н.Г. Власова (Миклухо-Маклай, 1963).
По нашим наблюдениям, на Юго-Западном Дарвазе зыгарская и вышележащие
челамчинская, сафетдаронская и гундарине кая свиты вместе образуют столь же круп-
ный самостоятельный член разреза пермской системы в ее средней части, как и на
Юго-Восточном Памире. Этим определяется большое сходство в положении толщ
дарвазского яруса внутри пермских отложений обоих районов, очень различных по
своей геологической истории и достаточно удаленных друг от друга.
Нижняя часть яруса в его стратотипических разрезах (зыгарская свита) охарак-
теризована фораминиферами: Geinitzina sp., Leella sp., Darvasites daroni M.—Macl.,
D. ex gr. contractus Schellw. et Dyhr., Pseudofusulina kraffti Schellw. et Dyhr., Ps.vu/ga—
ris Schellw. et Dyhr., Ps. globose Schellw. et Dyhr., Parafusulina ferganica M.—Macl.; ам-
моноидеями: Propinacoceras bomemani Toum., Prosicanites edelsteini Toum., Artin skia
aff. artiensis var. timorensis Han., Popanoceras lahuseni Karp., Popanoceras (?) sp., Me-
ta legoceras sp. (Миклухо-Маклай, 1963).
Перечисленные аммоноидеи характеризуют верхнеартинские отложения. Они были
описаны О.Г. Туманской (Туманская, Борнеман, 1937), которая сопоставляет их с боз-
теринскими, аммоноидеями Юго-Восточного Памира (Туманская, 1963).
В средней части дарвазского яруса (сафетдаронская свита) фузулинидыпред-
ставлены видами Nankinella orbicularis Lee, Kahlerina aff. pachytheca Kochansky—De—
vide, M inojapanella elongate Fuz. et Kan., Robustoschwagerina tumida Lich., Darvasites
contractus Schellw. et Dyhr., Pseudofusulina exigua Schellw. et Dyhr., Ps. kraffti Schellw.
et Dyhr., Ps. mengi Chen, Chusenella conicocilindrica Chen, Parafusulina aff. japonica
Gumb., Brevaxina sp., Misellina sp., Armenina sp.
В верхней части яруса (гундаринская свита) определены Nankinella aff. свис a si-
са Dutk., Darvasites contractus Schellw. et Dyhr., Pseudofusulina kraffti Schellw. et
Dyhr., Ps. vulgaris Schellw. et Dyhr., Parafusulina ex gr. japonica Gumb., Brevaxina sp.,
Misellina sp.
Разрезы Окраинной подзоны нами не рассматриваются, так как в связи с отсут-
ствием в них брахиопод и сильным уменьшением мощностей всех горизонтов (кро-
ме кызылДжилгинского) они не представляют интереса для данной работы.
26
Нижняя граница дарвазского яруса определена А.Д. Миклухо-Маклаем не совсем
точно. При выделении яруса (Миклухо-Маклай, 1958а) было принято его соответ-
ствие всей дарвазской серии Г.А. Дуткевича (Дуткевич, 1937а), нижняя часть ко-
торой в действительности отвечает значительной части карачатырского яруса. Более
точное указание на объем яруса в разрезах Юго-Западного Дарваза (Миклухо-Мак-
лай, 1963) не оставляет сомнения в том, что автор имел в виду отложения загыр-
окой и вышележащих свит.
Верхняя граница яруса не могла быть точно охарактеризована палеонтологически
в стратотипических разрезах в связи с недостаточностью фаунистической характе-
ристики гундаринской свиты, перекрывающейся конгломератовой иол лиха рекой свитой.
Поэтому положение верхней границы яруса по необходимости было установлено
А.Д. Миклухо-Маклаем (1958а) в разрезах Юго-Восточного Памира. Здесь он от-
нес к дарвазскому ярусу кубергандинскую свиту и нижележащие слои, содержащие
бозтеринских аммоноидей. Кровля дарвазского яруса определяется налеганием на
кубергандинскую свиту известняковых песчаников с Sumatrina и Neoschwagerina в
типовых разрезах выделенного А.Д. Миклухо-Маклаем (>1958а) мургабского яруса
верхней перми. Фаунистически верхняя часть кубергандинской свиты охарактеризована
комплексом фузулинид зоны Cancellina (Левен, 1965), которую А.Д. Миклухо-Маклай
не отделял от слоев с Misellina.
Самые высокие слои кубергандинской свиты относятся к подзоне Neoschwageri—
па simplex. N. simplex появляется в разрезе ниже указанной А.Д. Миклухо-Маклаем (1958а)
границы мургабского яруса (в основании агалхарской толщи), что, по-види-
мому, А.Д. Миклухо-Маклаю не было известно. Поэтому он (Миклухо-Мак-
лай, 1958а, 1963) в противоречии с особенностями распространения фузулинид в
указанном им стратотипическом разрезе рассматривал всю родовую зону Neoschwa—
germa в составе мургабского яруса.
Э.Я. Левен (1963, 1965, 1967) также относит зону Neoschwagerina полностью
к мургабскому ярусу. На Юго-Восточном Памире нижняя ’ граница мургабского
яруса проводится им по появлению первых неошвагерин в основании ганской свиты
(Левен, 1958). Нами граница дарвазского и мургабского ярусов принимается в кров-
ле кубергандинской свиты Г.А. Дуткевича, включающей подзону Neoschwagerina sim-
plex, что соответствует указаниям А.Д. Миклухо-Маклая на ее положение в стра-
тотипических разрезах.
Некоторая нечеткость в указании границ яруса, допущенная А.Д. Миклухо-Мак-
лаем, имеет большее отношение к чисто номенклатурной стороне дела и вряд ли
может быть препятствием к применению названия после соответствующих уточнений.
Названия *гваделупский*, *кубергандинский* и 'артинский* ярусы кажутся менее
удовлетворительными, так как они не отражают особенностей геологической истории
района. Кроме того, имеется ряд других трудностей для их применения, как чисто
номенклатурных, так и по существу.
Название *гваделупский ярус* было впервые введено в советскую литуратуру в
коллективной монографии 'Развитие и смена морских организмов на рубеже палео-
зоя и мезозоя* (1965) для отложений гнищикского и хачикского горизонтов Закав-
казья и их аналогов. При этом слои с Cancellina были отнесены к верхней перми
в качестве нижней части гваделупского яруса, залегающего на артинских отложениях.
Гваделупе кий ярус (Girty, 1908) был выделен в разрезах Западного Техаса, где
он охарактеризован многочисленными аммоноидеями, и в меньшей мере фузулинидами
и брахиоподами. Зональное расчленение и корреляция гваделупского яруса произво-
дится преимущественно по аммоноидеям, которые на Юго-Восточном Памире пока
неизвестны. Подробный анализ критериев, служащих для выделения этого яруса, при-
водит Э.Я. Левена (1967) к заключению о невозможности применить это название
к разрезам Евро-Азиатской части Тропической области. Действительно, положение
слоев с Cancellina в основании гваделупского яруса устанавливается чисто логичес-
ким путем в результате сложных сопоставлений и может быть принято только как
предположительное. Не исключено, что Гваделупе кий ярус соответствует в основном
более высоким слоям и не имеет стратиграфических аналогов в кубергандинской
свите.
Кубергандинский ярус выделен Э.Я. Левеном (Левен, 1963, 1965). Он относит
нижнюю часть дарвазского яруса (зону Pseudofusulina и примитивных Parafusulina,
27
а также эону Misellina) к аргунскому ярусу, а верхнюю - к верхней перми, выде-
ляя эону Cancellina в самостоятельный кубергандинский ярус. (Слои с N eoschwage-
rina simplex Э.Я. Левен относит к мургабскому ярусу.)
Кубергандинский ярус в разрезах Юго-Восточного Памира соответствует только
части кубергандинской свиты. Намечающееся при детальном изучении разрезов
кубергандинской свиты обособление слоев с Cancellina пока не дает основания для.
признания их частью разреза равноправной по своему стратиграфическому объему
ярусным подразделениям и свидетельствует скорее о соответствии другим фузули-
нидовым зонам и подзонам (см. рис. 2). В номенклатурном отношении применение
этого названия сильно затруднено вследствие того, что название 'кубергандинский
(ая)' применялось не менее чем в четырех различных значениях для одних и тех
же разрезов (см. табл. 1).
В качестве артинского яруса в разрезах Юго-Восточного Памира обычно выде-
лялись слои с бозтеринскими аммоноидеями, относимыми к верхней части базарда-
ринской свиты и нижняя часть кубергандинской свиты (зона Brevaxina схемы Э.Я. Ле-
вена 1963 г. и зона Misellina его же схемы 1965 г.). В результате уточнения
стратиграфической приуроченности бозтеринских аммоноидей они-были отнесены к
нижней части выделяемого автором горизонта, отделяющегося от базардаринской свиты
поверхностью размыва. Таким образом, артинские отложения Юго-Восточного Памира
делятся на две части, причем нижняя часть не отчленяется от сакмарских отложений,
а геологически существенная граница проходит в основании верхнеартинских отло-
жений. Верхняя граница артинского яруса в разрезах Тропической области ('Разви-
тие и смена...', 1965; Левен, 1967) определяется обычно по фауне аммоноидей как
граница эон Perninites (артинский ярус) и Waagenoceras (гвадалупский ярус).
Как уже отмечалось, положение этой границы в раэреэах Юго-Восточного Памира
остается неясным; неизвестно также, на каком стратиграфическом уровне она должна
располагаться в уральских разрезах • Поэтому в номенклатурном отношении название
'артинский ярус' Применительно к разрезам Юго-Восточного Памира также не яв-
ляется вполне удовлетворительным.
Следующий крупный этап осадконакопления, во многом сходный с дар вазе ким,
относится к мургабскому и джульфинскому векам. Начало этапа отмечено почти по-
всеместным образованием светлых известняковых песчаников и мелкогалечных конг-
ломератов с Sumatrina (подзона Neoschwagerina schuberti Э.Я.Левена) и местными
размывами подстилающих толщ. Мургабский ярус характеризуется трансгрессивными
чертами строения с переходом вверх по разрезу от мелководных обломочных осад-
ков к тонким карбонатным илам. С джульфинским временем связано резкое обмеление
бассейна (нижнетахтабулакская подсвита), затем происходит постепенное его сокра-
щение с образованием в конце перми глинистых, по-видимому, в значительной части
лагунных осадков (верхнетахтабулакская подсвита).
Распространение мелководных осадков в начале джульфинского века совпадает с
расцветом родов Codonofuaiella, Reichelina, Palaeofuaulina и других джульфинских
фораминифер, которые впервые появляются еще в мургабское время.
Джульфинские отложения (тахтабулакская свита) тесно связаны с мургабскими
(ганская свита). Граница ганской и тахтабулакской свит, по-видимому, не синхронна
по всему району, а является фациальной в пределах неширокого стратиграфического
интервала. В Окраинной подзоне тахтабулакская свита чрезвычайно маломощная (иног-
да 2-5 м) и с трудом отделяется от верхнеганской подсвиты.
Джульфинский ярус в работе коллектива авторов Палеонтологического института
('Развитие и смена...', 1965) признан синонимом памирского. По мнению А.Д. Мик-
лухо-Маклая (1958а, 1963), памирский ярус характеризуется отсутствием высших фу-
зулинид и расцветом мелких аберрантных фузулинид d лагенид, т.е. его характеристика
точно соответствует характеристике джульфинского яруса в указанной работе. В по-
нимании Э.Я. Левена (1967) памирский ярус представляет собой биозону Codonofu—
aietla и других аберрантных фузулинид, в нижней части которой еще существуют
высшие фузулиниды и швагериниды, т.е. в отличие от А.Д. Миклухо-Маклая, Э.Я.Ле-
вен не считает вымирание высших фузулинид за критерий для проведения границы
мургабского и памирского ярусов. В стратотипе памирского яруса не представлены
Neoschwagerina, Jabeina, Lepidolina, которые могут быть отнесены к нижней части
яруса (кутальская, возможно низы тахтабулакскойтолщи). Для сопоставления стра-
28
тотипов джульфинского и памирского ярусов важно, что джульфинский ярус в За-
кавказье в понимании С.Е. Розовской (см. 'Развитие* я смена...*, 1965) охва-
тывает эпиболу родов Reichelina и Codonofuaiella, а также и вышележащие слои,
где количество фузулинид постепенно уменьшается до их полного исчезновения в
конце перми. Это обстоятельство позволяет правильно сопоставлять джульфинский и
памирский ярусы, причем первый имеет несколько меньший стратиграфический объем
и соответствует на Юго-Восточном Памире тахтабулакской свите, в нижней части
которой получают широкое распространение аберрантные фузулиниды и Colaniella.
Касаясь вопроса о разделении пермских толщ Юго-Восточного Памира на отделы,
следует отметить, что в разрезах этого района не выявлены особенности строения
нижнепермских отложений, значительная часть которых, вместе с каменноугольными
толщами входит в состав монотонной и недостаточно охарактеризованной палеонто-
логически базардаринской свиты. Вследствие этого четко выделяющиеся в вышеле-
жащих толщах стратиграфические единицы не могут сравниваться со сходными под-
разделениями в нижней части пермской системы, что не позволяет составить полное
представление о строении всего разреза.
Залегающие выше базардаринской свиты толщи дарвазского, мургабского и
джульфинского ярусов в совокупности отвечают одному крупному этапу осадкона-
копления, имеющему отчетливое двучленное строение и далее подразделяющемуся
на четыре более мелких этапа (кызылджилгинское время, кубергандинское время ,
мургабский век, джульфинский век). Обращает на себя внимание соответствие дар-
вазского яруса примерно вдвое более крупному историко-геологическому подраз-
делению, чем мургабский или джульфинский ярусы. Подразделение дарвазского яруса
на две части находит подтверждение при изучении стратиграфического распростра-
нения таких важных групп, как фузулиниды и аммоноидеи, однако положение границы
между ними не определяется с достаточной ясностью. Основание кубергандинской
свиты, соответствующее изменению общего состава и облика пород, в остальном
ничем принципиально не отличается от нижней границы слоев с Cancellina или Neo—
schwagerina simplex. Ряд признаков указывает на возможность трансгрессивного
залегания каждой из этих толщ за пределами изученной площади. Действительно,
на Северном Памире подошва слоев с Cancellina фиксируется появлением песчано-
конгломератовых толщ (Левен, 1967), а в Закавказье пермская трансгрессия на-
чалась в середине времени Misellina, а наибольшего развития достигла не ранее
времени Cancellina (Левен, 1972). Поэтому для периодизации дарвазского века и
понимания взаимосвязей геологических процессов кыэылджилгинского и куберган-
динского времени необходимо выяснение строения и условий залегания соответству-
ющих слоев в других районах.
Фузулиниды, являющиеся наиболее распространенной в да рва зеком ярусе группой
ископаемых, также не определяют вполне однозначно положение границы двух его
основных частей.
Наиболее четкий рубеж в развитии фузулинид приходится на начало времени Can-
cel lina, соответствующее быстрой дивергенции семейства Verbeekinidae, появлению
родов Polydiexodina и Jangchienia и значительной вспышке видообразования у Ра-
rafastilina. Однако отложения, непосредственно подстилающие слои с Cancellina,
не получили на памирском материале достаточной характеристики, так что остается
неясным, не является ли это появление многих новых форм в слоях с Cancellina
дальнейшим развитием тенденций, проявившихся и ранее, в конце времени Misellina.
Этот вопрос пока что не может быть решен и на материале других районов.
Аммоноидеи широко распространены в нижней части дарвазского яруса на Юго-
Восточном Памире и известны также в Дар вазе ком хребте. Наиболее древние дар-
вазские аммоноидеи (нижнекыэыдджилгинские и зыгарские) сопоставляются с верх-
неартинскими (байгенджинскими) аммоноидеями Урала (Туманская, 1963; Павлов,
1965). В среднекызылджилгинское время на фоне прежнего комплекса появляются
немногочисленные представители новых родов (Adrianites, Epadrianites, Earned licottia
и др.), обычно рассматриваемых как верхнепермские, но существенного обновления
общего систематического состава комплекса не происходит. Аммоноидеи нижней
части кубергандинской свиты, собранные примерно на границе слоев Misellina и Can-
cellina (Дуткевич, Туманская, 1935), по заключению В.Е. Руженцева (см. книгу
* Развитие и смена...', 1965), связываются с тем же верхнеартинским комплексом,
29
являясь его наиболее молодыми представителями. Верхняя часть дарвазского яруса
в памирских разрезах аммоноидеями не охарактеризована, как и в большинстве дру-
гих районов, для которых применима памирская схема зонального расчленения по
фузулинвдам. Только в формации Цэюйфен Южного Китая, сопоставляющейся по
стратиграфическому положению с зоной Cancellina (нижняя часть формации Маокоу)
известны немногочисленные эндемичные аммоноидеи. Это два вида Strigogoniatites
и примитивные циклолобиды Kufengoceras и Shengoceras (ChaoKing—Koq,1965).Таким
образом, на уровне слоев с Cancellina намечается обновление комплекса аммонои-
дей, но недостаток фактического материала не позволяет полностью оценить его
стратиграфическое значение.
Стратиграфическое значение брахиопод дарвазского века пока что невелико в связи
с их недостаточной изученностью.
Таким образом, в дальнейшем, по-видимому, будет возможно подразделить дар-
вазский ярус на две части, но в настоящее время это кажется преждевременным,
так как точный стратиграфический объем его подразделений не определен.
Следует учесть также отсутствие употребительных стратиграфических названий,
по своему содержанию близко отвечающих двум частям яруса, а введение новых наз-
ваний в настоящее время вряд ли целесообразно.
Ташкентским совещанием 1958 г. (Решения..., 1959) предложены названия гулук-
скийг и гкубергандинскийг горизонты, из которых на Юго-Восточном Памире первое
понималась полностью в объеме слоев, объединенных автором в кызылджилгинский
горизонт, а второе - в объеме кубергандинской свиты Э.Я. Левена. Однако сопос-
тавление кызылджилгинского горизонта со стратотипом у луке кого горизонта, распо-
ложенным в Северной Фергане, проблематично, в связи с чем автор не считает воз-
можным использовать это название, а название 'кубергандинский* вряд ли может
употребляться в связи с разнобоем в трактовке этого подразделения.
Вопрос о разделении пермских отложений Юго-Восточного Памира на отделы
сводится к вопросу о единстве дарвазского яруса и его принадлежности к нижнему
или верхнему отделу пермской системы или самостоятельности яруса в качестве
среднего отдела. Безусловное решение этих вопросов в настоящее время вряд ли воз-
можно в связи с недостаточностью материала по синхроничным отложениям других
районов Тропической области. Поэтому укажем только, что авторы придерживаются'
точки зрения о единстве дарвазского яруса и невозможности вследствие этого про-
водить внутри него границу отделов.
Положение дарвазских толщ в пермской системе свидетельствует либо о самостоя-
тельности дарвазского яруса, либо о его близости к верхнему отделу перми, в свя-
зи с чем кажется также невозможным относить его к нижнему отделу, как это при-
нято А.Д. Миклухо-Маклаем (1958а, 1963, 1966; Лихарев, Миклухо-Маклай, 1964).
Однако синхроничные дарвазским отложениям верхняя часть артинского яруса Урала
(в широком понимании - вместе с кунгуром) и серия Лэнерд Северной Америки тра-
диционно относятся к нижней перми и для обоснования верхнепермского возраста,
дарвазского яруса необходимо рассмотреть корреляцию пермских отложений этих райо-
нов с дарвазскими толщами. Что касается корреляции дарвазского яруса с серией
Лэнерд, то нижнекызылджилгинский подгоризонт удовлетворительно сопоставляется
с ее нижней частью (Туманская, 1963; Павлов, 1965). Вопрос о положении на Па-
мире верхней части серии Лэнерд сложен, в связи с упоминавшимися трудностями
определения в памирских разрезах границы артинского и гваделупского ярусов. Гва-
делупский комплекс аммоноидей в Сицилии связывается с мургабскими отложениями
с Sumatrina и Neoschwagerina craticulifera. Росс и Насичук (Ross, Nassichuk, 1970,
Ross, 1971) считают возможным относить нижнюю чзсть гваделупского яруса (ворд-
ский подъярус - зона Waagenoceras) к слоям с Neoschwagerina margaritae, а верхнюю
часть серии Лэнерд к слоям с Neoschwagerina craticulifera (-подзона N.schuberti
Э.Я. Левена). Таким образом, вероятно, верхняя часть кубергандинской свиты отве-
чает еще лэнердским отложениям, а вордский подъярус соответствует в основном
верхней части зоны Neoschwagerina, скорее всего начиная со слоев с Neoschwage—
rina schuberti, т.е. мургабскому ярусу в нашем понимании.
Предположение о соответствии вордского подъяруса и слоев с N.schuberti подтверж-
дается находкой в нижнеганской подсвите Юго-Восточного Памира сицилийского го-
ниатита, относящегося к подроду Sosiocrimites.
30
Решающим для вопроса о возрасте дарвазского яруса является сопоставление его
со стратотипическими разрезами Урала. По аммоноидеям кызылджилгинский горизонт
должен быть отнесен к верхнеартинскому подъярусу, который в понимании В.Е. Ру-
женпева (1955) охватывает на Урале саргинский и саранинский горизонты и кунгур-
ские отложения. Поскольку последние практически не охрактеризованы аммоноидеями,
верхняя граница артинского яруса определяется на основе разрезов морских отло-
жений в Северной Америке, что возвращает нас к вопросу о сопоставлении с этими
разрезами. Что же касается положения на Урале нижней границы дарвазского яруса,
то ее точное соответствие основанию верхиеартинского (байгенджинского) подъяурса
неясно и не исключено, что она располагается выше него. Это предположение кос- *
венно подтверждается данными и по брахиоподам (см. главу IV ). Таким образом,
не удается точно определить стратиграфическое положение аналогов дарвазского
яруса в уральских разрезах; он, по-видимому, охватывает какую-то часть верхне-
артинского подъяруса, что не позволяет без ревизии стратиграфии пермских отложе-
ний Урала отнести дарваэский ярус к верхнему отделу перми. Интересно, что в Бо-
реальной палеобиогеографической области различными исследователями (Бархатова,
1964, 1970; Степанов, 1957, 1967; Устрицкий, 1960, 1961, 1963, 1971) выделяется
под разными названиями комплекс морских отложений, соответствующий эвапоритам
кунгура и уфимским континентальным отложениям. Он содержит характерную фау-
ну, позволяющую рассматривать его как самостоятельный ярус в составе верхней
перми (Степанов, 1957, 1967; Устрицкий, 1960, 1971) или даже ставить вопрос о
его соответствии самостоятельному отделу (Бархатова, 1964, 1970, 1971). Правда,
В.И. Устрицкий, настаивая на принадлежности этих отложений, названных им пайхойским
ярусом, к верхней перми, сопоставляет их с уфимской свитой, но по данным Е.А.Гу-
севой и В.П. Горского (1969) подошва пайхойского яруса в его стратотипической
местности коррелируется с основанием саранинского горизонта или даже располагает-
ся внутри саргинского горизонта. При этом существенная биостратиграфическая гра-
ница, рассматриваемая всеми указанными авторами как граница отделов в нормаль-
но морских толщах Бореальской области, проходит внутри слоев с верхнеартинским
комплексом аммоноидей. Все же пересмотр границы отделов перми в классических
разрезах Западного Урала до сих пор не произведен и упомянутые исследования лишь
поставили задачу дальнейшего изучения этого вопроса.
Возвращаясь к дарвазскому ярусу, укажем, что наилучшим решением в настоя-
щее время авторам настоящей работы представляется компромиссное, состоящее в
признании достаточной обособленности яруса, а вопрос о принадлежности его к ниж-
нему или верхнему отделу перми или равнозначности отделу требует дальнейшего
специального рассмотрения.
Глава III
ОСНОВНЫЕ КОМПЛЕКСЫ ПЕРМСКИХ БРАХИОПОД
ЮГО-ВОСТОЧНОГО ПАМИРА
Брахиоподы являются одной из наиболее распространенных групп фауны в пермских
отложениях Юго-Восточного Памира. Они характеризуют почти весь разрез пермской
системы. Inaiticulata здесь не найдены; Articulate представлены 53 видами, при-
надлежащими 43 родам, 27 семействам и 6 отрядам. Из них 18 видов и 2 рода -
новые. Такой систематический состав указывает на относительную бедность памирской
брахиоподовой фауны по сравнению, например, с закавказской, где только в верхней
перми известен 91 вид брахиопод, принадлежащих 48 родам, а тем более по сравнению
с северокавказской, где в отложениях никитинского и уруштенского горизонтов при-
сутствует 168 видов брахиопод. Памирский комплекс также значительно беднее широ-
ко известного комплекса пермских брахиопод Соляного кряжа (Пакистан), в сос-
тав которого входит более 250 видов и разновидностей.
Среди пермских брахиопод Юго-Восточного Памира преобладают продуктиды (19
видов), что вообще характерно для верхнего палеозоя. Следует отметить наличие
таких высоко специализированных и своеобразных родов, как Taeniothaerua, Stro-
phalosiina, Scacchinella, Richthofenia, Leptodua. Второе место занимает отряд Spi—
riferida (11 видов), причем особенно широко здесь распространено надсемейство
Delthytiacea. В семействе Martiimdae род Martinia представлен 6 видами. В се-
мействе Reticulariidae 2 рода: Kitakamithyria и Permophricodothyria, содержа-
щие по одному виду каждый. Представители ребристых спириферид редкие. Это роды
Punctocyrtella, Neoapirifer, Spiriferella, представленные каждый одним видом.
Особенностью комплекса брахиопод Юго-Восточного Памира является почти пол-
ное отсутствие атирндид; здесь найден лишь один вид рода Cleiothyridina, тогда
как в Закавказье известно 11 видов, принадлежащих 6 родам этого надсемейства.
Часто встречаются представители отряда Rhynchonellida (9 видов, принадлежащих 6
родам) и Terebratulida (5 видов, принадлежащих 4 родам). Своеобразны ортиды,
особенно такой редкий род, как Enteletella, до настоящего времени встреченный
лишь в верхней перми Северного Кавказа. Нужно отметить также присутствие рода
Geyerella (отряд Strophomenida). Для брахиоподового комплекса Юго-Восточного
Памира характерно многообразие родов при относительной немногочисленности ви-
дов: почти все встреченные здесь роды представлены единичными видами. Исключе-
ние составляют Martinia (6 видов), Geraaaimovia (3 вида), Enteletea (2 вида),
Derbyia (2 вида), Heterelaamina (2 вида), причем число особей, принадлежащих
большинству этих видов, очень велико.
Среди пермских брахиопод Юго-Восточного Памира можно выделить три комп-
лекса, происходящие с трех различных стратиграфических уровней (табл. 2):
I • Верхнебазардаринский, приуроченный к верхней части базардаринской свиты
сакмарского-нижней части артинского яруса (табл. 3).
П. Кызылджилгинский, приуроченный к отложениям нижнего и среднего подго-
ризонтов кызылджилгинского горизонта дарвазского яруса (табл. 4).
III. Тахтабулакский, приуроченный к нижней и средней подсвитам тахтабулакской
свиты джульфинского яруса (см. табл. 5).
Все три комплекса чрезвычайно своеобразны. Каждый характеризуется специ-
фическим набором не только видов, но и родов. Общим для всех них является лишь
широко распространенный род Martinia, а в нижнем и в верхнем присутствует род
Geraaaimovia.
Верхнебазардаринский, кызылджилгинский и тахтабулакский комплексы брахиопод
в своем вертикальном распространении в разрезах Юго-Восточного Памира характе-
32
ризуют не все стратиграфические горизонты перми: брахиоподы практически неиз-
вестны в кубергандинской| ганской свитах и в верхиекызылджилгинском подгоризонте.
При характеристике комплексов нами по мере возможности приводятся данные о
приуроченности видов к определенным типам осадков и условиям местообитания, хо-
тя обычно не удается дать полное объяснение наблюдающейся связи с фациями. Это
связано с трудностями аутоэкологической характеристики брахиопод. В большинстве
случаев состав тафоценозов достаточно разнообразен и характерен. Изучение его в
сочетании с анализом состава осадков, специфики захоронений и особенностей сох-
ранности раковин брахиопод и сопутствующих им организмов позволяет учитывать
явления посмертного переноса, раздробления и сортировки раковин и хотя бы частич-
но реконструировать состав прижизненных поселений.
* * *
ЬБрахиоподы верхнебазардаринского комплекса известны в от-
ложениях верхней части базард арине кой свиты в трех местонахождениях (Куртеке 1,
Кастанат-Джилга 1 и Чакобай), где они приурочены к органогенно-обломочным
алевритистым известнякам, глинистым известнякам и известковистым глинистым алев-
ролитам (табл. 3 ).
В этологическом отношении среди брахиопод верхнебазардаринского комплекса
может быть выделено несколько типов. Здесь присутствуют крупные плоские рако-
вины устрицеобрЛзных свободнолежащих строфоменид Derbyia grandis Waagen и D.diver-
sa Reed. Продуктиды представлены крупными, относительно тонкостворчатыми рако-
винами Taeniothaerus rusticus Grunt, sp.nov. и более массивными Reticulatia pamirica
Grunt, sp. nov., которые прикреплялись лишь на ранних стадиях онтогенетического раз-
вития. Здесь же присутствуют мелкие тонкостворчатые Cancrinella altissima King.
Многие брахиоподы, встреченные в верхнебазард арине ком комплексе, характери-
зуются якорным типом прикрепления. Они разделяются Е.А. Ивановой (1949) на три
группы. К представителям первой группы относятся: а) небольшие, одинаково двояко-
выпуклые раковины с фора мен ом, не закрывавшимся в течение всей жизни животного прй
довольно прочном замковом сочленении,что обусловливало возможность постоянного прикг-
репления раковины к субстрату при помощи ножки, выходящей через форамен (в на-
шем материале к этой группе могут быть отнесены представители надсемейства
Athyridacea—C leiothyridina ai la ken sis (Reed), характер прикрепления которых, ви-
димо, был близок к современным представителям отряда Terebratulida), и б) двоя-
ковыпуклые раковины преимущественно ринхонеллид, характеризующиеся на взрослых
стадяих отчетливо развитыми синусом и выступом, а также грубой ребристостью,
развитой на всей раковине. По данным Е.А. Ивановой (1949), на ранних стадиях они
характеризовались прикреплением к субстрату при помощи ножки и вертикальным
расположением раковины по отношению к субстрату. В дальнейшем роль ножки зна-
чительно уменьшалась или она вообще переставала функционировать, что отвечает
заметному относительному уменьшению диаметра форамена. На взрослых стадиях
развития раковины ножка, видимо, выполняла лишь роль якоря, а основную роль в
поддерживании раковины в устойчивом положении играли выступы брюшной створки
(Gerassimovia bactriana Grunt, sp.nov.). Ко второй этологической группе якорных
брахиопод относятся двояковыпуклые раковины с коротким смычным краем, неболь-
шой низкой ареей, необособленными ушками, большим дельтириальным отверстием
с узкими дельтидиальными пластинами. По данным Е.Е. Павловой (1969), наличие
у представителей этой группы перечисленных выше морфологических признаков ука-
зывает на возможность функционирования ножки на всех стадиях онтогенетического
развития раковины. К этой группе в нашем материале относятся тонкоиглистые Ki—
takamithyris petaliformis Pavlova, sp. nov., и Martinis uralica Tschern. К третьей
группе брахиопод, характеризующихся якорным типом прикрепления, Е.А. Иванова
относит крупных толстостворчатых спириферид с более массивной брюшной створкой,
отчетливо развитыми синусом и выступом и наличием длинной высокой ареи. В при-
жизненном положении они были ориентированы вертикально по отношению к суб-
страту, опираясь на макушку брюшной створки. Нависание макушки брюшной створ-
ки над ареей и сближение макушек обеих створок, видимо, препятствовали непосред-
ственному выходу ножки из дельтириального отверстия. Поэтому Е.А. Иванова де-
лает вывод о неизбежном усложнении способа прикрепления этих организмов, выра-
зившемся в образовании сильно разветвленной ножки, с чем, вероятно, оказалось
33
3 264
Таблица 2
си
Комплексы пермских брахиопод Юго-Восточного Памира
Тахтабулакский___________. Комплексы
Вид Сакмарский ярус нижне артинский подъяруС| нерасч- лененные Дарваэский ярус Мургаб- ский ярус Джульфинский ярус
базардаринская свита (верхняя часть) кызылджилгинский го- ризонт куберган- динская (?) свита нижнеган- ская (?) подсвита тахтабулакская свита
нижнетахта- булакская подсвита средне- тахтабу- лакская подсвита
нижний под- горизонт средний подгоризонт
Enteletes dzhagrensis Sokolskaja
E.meridionalis Gemmellaro
Geyerella alpina Schellwien
Scacchinella licharewi Grunt, sp.nov.
Strophalosiina multicostata Licharew
Richthofenia caucasica Licharew
Krotovia jisuensiformis Sarytcheva
Marginifera sexcostata Licharew
Anidanthus sinosus (Huang)
Compressoproductus mongolicus (Diener)
Haydenetla dutkevitchi Grunt, sp.nov.
Uncinunellina timorensis (Beyrich)
Wellerella arthaberi (Tschernyschew)
Gerassimovia pamirica Grunt, sp.nov.
G. abalakovi Grunt, sp. nov.
Camarophorinella leveni Grunt, sp.nov.
Martinia warthi Waagen
Heterelasmina lepton (Gemmellaro)
H.pygmaea Grunt, sp.nov.
Rostranteris (Notothyrina) pontica Lacharew
Верхнебазардаринский I Кызылджилгинский
Orthotichia avushensis Sokolskaja
Parenteletes ruzhencevi Sokolskaja
Enteletella nukschitchi Licharew
Scacchinella licharewi Grunt, sp.nov.
Leptodus sp.
Haydenella tumida (Waagen)
Stenoscisma armenica Sokolskaja
Notothyris pseud odj on I fen sis Licharew, sp.nov.
Scacchinella sp.
Permophricodothyris caroli (Gemmellaro)
Martinia rupicоla GemmeHaro___________________
_M^tri^e£r^(^mmelJaro_________________________
Terrakea nalivkini Grunt, sp.nov._____________
Costiferina sp.
Spiriferella sp.
Martinia tschakobaica Grunt, sp.nov.
Probolionia himalayensis (Diener)
Camerisma murgabica Grunt, sp.nov.
Martinia tschemyschewi Grunt,sp.nov.
Labaia dmitrievi Grunt, sp.nov.
Overtoniina mamazairica Grunt, gen.et sp. nov.
Globiella ? rossiae (Fantini Sestini)
Boloria garmoensis Grunt, gen.et sp.nov.______
Derbyia grandis~fyaagen
D.diverse Reed
Taeniothaerus rusticus Grunt, sp.nov.
Reticulatia pamirica Grunt, sp.nov.
Cancrinella a It is sima King
Gerassimovia bactriana Grunt, sp.nov.
Punctocyrtella spinosa Plodowski
Neospirifer joharensis (Diener)
Kitakamithyris petaliformis Pavlova,sp. nov.
Martinia uralica Tschemyschew
Cleiothyridina ailakensis_(ReedJ______________
Таблица 3
Распространение брахиопод верхнебазардаринского комплекса
в сакмарско-нккнеартинских отложениях Юго-Восточного Памира
Базар даринс кая свите (верхняя часть)
Куртеке 1 Кастанат- Джилга 1 Ч а к о б а й
органо- известко- известко- глинистые извест-
Вид генно-об- вистые вистые известняки ковистые
л омочи ые глинистые глинистые глинистые
алеврити- алевроли- алевроли- алевро-
стые из- ты ты литы
вестняки
Derbyia gran di s Waagen 50
D. di versa Reed 201
Taeniothaerus rusticus Grunt, 63 1
sp.nov. Reticulatia pamirica Grunt, 134 31 9 16
sp.nov. Cancrinella altissima King 27 9 2
Gerassimovia bactriana Grunt, 10 45 7 21
sp.nov. Punctocyrtella apinoaa Plodowski 78
Neospirifer joharensis (Diener) Kitakami thyris petaliformis 48 70
Pavlova, sp.nov. 132
Martinia uralica Tschemyschew 172
Cleiothyridina ailakensia (Reed) 33 194 14 4
1 Цифрами указано количество экземпляров, содержащихся в коллекции;
также в табл. 4 и 5.
функционально связанным развитие арен. К этой группе в нашем материале относятся
Neoapirifer joharenaia (Diener) и Punctocyrtella apinoaa Plodowski.
С этологическими особенностями брахиопод в тесной связи находятся типы их
захоронений, различающиеся по степени посмертной переработки прижизненных по-
селений. К первому типу относится захоронение в существенно известковистых гли-
нистых алевролитах местонахождения Кастанат-Джилга 1. По систематическому сос-
таву обнаруженные здесь брахиоподы мало отличаются от верхнебазардаринских
брахиопод местонахождения Куртеке 1, но общий облик комплекса, обнаруженного в
захоронении, определяется массовым количеством Kitakamithyria petaliformia
Pavlova, sp.nov., Cleiothyridina ailakenaia (Reed), Martinia uralica Tschem. Несколько
реже встречаются Neoapirifer joharenaia (Diener) и Geraaaimovia bactriana Grunt,
sp.nov. Продуктиды Taeniothaerua ruaticua Grunt, sp.nov., Reticulatia pamirica
Grunt, sp. nov. и Cancrinella altiaaima King здесь чрезвычайно редки. Раковины
захороняются в виде разрозненных створок в разнообразных положениях по отно-
шению к слоистости, причем количество более массивных брюшных створок в три-
четыре раза превышает количество более тонких и легких спинных. С сочлененными
створками захоронена только часть раковин Cleiothyridina ailakenaia Reed^ обла-
дающих мощным замковым сочленением. В то же время отсутствие заметной слоис-
тости глинистых алевролитов, характеризующихся текстурой взмучивания, хорошо
сохранившаяся тонкая скульптура раковин Kitakamithyria petaliformia Pavlova,
sp.nov., Neoapirifer joharenaia (Diener), Cleiothyridina ailakenais (Reed) и почти
полное отсутствие их обломков позволяет сделать вывод о незначительности по-
смертной переработки прижизненных поселений, ограничивавшейся в основном слабы-
ми перемещениями раковин легким волнением и выносом легких спинных створок
раковин с непрочным замковым сочленением, распадавшихся после отмирания.
36
Тафономически некоторым сходством с описанным характеризуется захоронение
в известковистых глинистых алевролитах местонахождения Куртеке 1. Для него ха-
рактерны прослои, переполненные параллельно залегающими, тесно сгруженными
брюшными и спинными створками Derbyia grand is Waagenи брюшными створками
Reticulatia pamirica Grunt, sp.nov., обращенными выпуклостью вверх. Основной
состав прижизненных поселений, по-видимому, оставался неизменным, хотя сам ха-
рактер захоронений определяется посмертным перемещением раковин под действием
гидродинамических факторов.
Иными особенностями отличается захоронение брахиопод в органогенно-обломоч-
ных алевритистых известняках (Куртеке 1). Основу комплекса брахиопод состав-
ляют относительно тонкостворчатые раковины продуктид Т aeniothaerus rusticus
Grunt,sp. nov. и Reticulatia pamirica Grunt,sp.nov., а также крупных спириферид Puncto—
cyrtella spinosa Plodowski и Neospirifer joharensis (Diener). Реже встречаются Can-
crinella altissima King и Cleiothyridina ailakensis (Reed). Раковины с сочленен-
ными створками присутствуют сравнительно редко, преимущественно в более тонко-
зернистых прослоях с существенной примесью глинистого материала. Здесь вместе
с крупными продуктидами, иногда захороняющимися в прижизненном положении,
встречаются строфомениды Derbyia diversa Reed. Для грубодетритовых разностей
известняков характерно захоронение разрозненных створок с резким преобладанием
брюшных. Створки залегают субпараллельно слоистости вместе с массой обломков
ветвистых мшанок, мелкими ругозами Pleurophyllum australe Hinde, крупными об-
ломками битых раковин. В породе присутствует также масса обломков игл продуктид.
Прижизненные поселения брахиопод характеризовались, по-видимбму, не очень плот-
ным расположением преимущественно крупных свободнолежащих форм и редкостью
брахиопод с якорным типом прикрепления, не образовывавших густых поселений,
как это наблюдалось в местонахождении Кастанат-Джилга 1. Брахиоподы, обнаружен-
ные в органогенно-обломочных алевритистых известняках Куртеке 1, обитали в раз-
личных, большей частью спокойных условиях гидродинамического режима, но иног-
да, видимо, наблюдалось усиление подвижности водной среды. Усиление волнения
приводило к гибели поселений и формированию ракушников. Дополнительная перера-
ботка ракушников волнением определила облик наблюдающегося тафоценоза.
Несколько иначе захороняются те же формы в существенно известковистых пес-
чаниках, связанных с органогенно-детритовыми известняками постепенными пере-
ходами по вертикали. Здесь раковины брахиопод крайне немногочисленны , обломки
их сравнительно редки, но обильны мшанки и ругозы. На поверхности напластования
наблюдаются прослойки глинистого материала, содержащие мелких хонетид.
Комплекс брахиопод, обнаруженный в местонахождении Чакобай, отличается от
описанных прежде всего значительной обедненностью систематического состава. Наи-
более резко она выражена в черных глинистых известняках с тонковетвистыми мшан-
ками. Здесь из брахиопод определены только Gerassimovia bactriana Grunt, sp. nov.
и Cleiothyridina ailakensis (Reed). Кроме них присутствуют немногочисленные
мелкие Chonetoidea, Spiniferinacea и Terebratulida плохой сохранности. Gerassi-
movia и Cleiothyridina захороняются большей частью в виде редких незакономер-
но ориентированных целых раковин; также встречаются разрозненные створки, преи-
мущественно брюшные, иногда обломки. Раковины и разрозненные створки, как пра-
вило, заметно потерты, с плохо сохранившейся скульптурой. Местами в породе об-
наруживается хорошо выраженная текстура взмучивания, местами послойная ориен-
тировка раковин брахиопод и их обломков совместно с редкими двустворками, об-
рывками мшанок, члениками криноидей. В этих участках слоя раковины брахиопод
подвергнуты наибольшей переработке гидродинамическими факторами. Интересно
отметить, что раковины двустворок часто несут следы сверления, в то время как
на брахиоподах они не обнаруживаются.
В известковистых глинистых алевролитах Чакобая, кроме Gerassimovia bactriana
и немногочисленных Cleiothyridina ailakensis, встречены нередкие Reticulatia
Pamirica Grunt, sp.nov., в том числе юные особи; единичные Cancrinella altissima
King и Derbyia sp.; многочисленные Chonetoidea и редкие Spiriferinacea. Захоро-
нение отличается сочетанием беспорядочно ориентирЬванных целых раковин с хорошо
с°хранной или чаще стертой скульптурой, разрозненных створок и их обломков, иног-
37
да тесно сгруженных и вложенных одна в другую, что несколько напоминает за-
хоронение в органогенно-обломочных известняках Куртеке 1.
Таким образом, захоронения в Чакобае указывают на различную степень посмерт-
ной переработки поселений брахиопод, но во всех случаях менее значительную, чем
в органогенно-обломочных известняках Куртеке 1 и более выраженную, чем в из-
вестковистых глинистых алевролитах местонахождений Куртеке 1 и Кастанат-Джилги1.
Большинство родов верхиебазардаринского комплекса: Cancrinella^ Neospirifer,
Kitakamithyris, Martinia, Cleiothyridina широко распространены географически и ох-
ватывают значительный стратиграфический интервал, начиная с нижнего карбона или
с верхнего девона (Kitakamithyris) до верхней перми. Род Derbyia известен в
верхнем карбоне - нижней перми Донбасса, Урала, Зарубежной Азии, Западной Ев-
ропы, Северной и Южной Америки^в перми Соляного кряжа, а также в верхней перми
Северного Кавказа. Род Reticulatia распространен в верхнем карбоне - нижней
перми Северной Америки. В Альпийско-Гималайской подобласти он известен только
в нижней перми: в ассельских отложениях Ферганы, Куньлуня, Каракорума и в точ-
нее не датируемых нижнепермских отложениях Читраля. Кроме того, этот род присут-
ствует в нижней перми Урала. Род Taeniothaerus известен в нижней перми Австра-
лии и Тасмании. В Альпийско-Гималайской подобласти он распространен в нижнеперм-
ских^отложениях Кашмира (аггломератовые слои) и в нижнем продуктусовом извест-
няке Соляного кряжа (Пакистан). Предположительно этот род указывается также
из карачатырских отложений (формация Джейруд, пачка Д) Северного Ирана. Род
Punctocyrtella известен из сакмарско-^артинских отложений Юго-Восточной Аравии
(формация Лузаба), а также из нижнепермских отложений Кашмира (аггломератовые
слои) и Афганистана. Род Gerassimovia имеет ограниченное распространение: кроме
Юго-Восточного Памира этот род встречен лишь в верхней перми Северного Кавказа.
Переходя к рассмотрению видового состава, нужно отметить, что из 11 видов
верхнебазардаринского комплекса четыре являются эндемичными. Из остальных се-
ми наиболее древний - Cancrinella altissima King известен из верхнего карбо-
на — нижней перми Техаса. Два вида: Derbyia grandis Waagen и Martinia uralica
Tschenu- из нижней перми Урала. Derbyia grandis, кроме того, встречается в перм-
ских отложениях Пакистана (средний и верхний лродуктусовый известняк Соляного
кряжа) и в верхней перми (местонахождение Ба зле о) Тимора. Derbyia diverse Reed —
в нижнем продуктусовом известняке Соляного кряжа (Пакистан); Neospirifer joharen-
sis Diener - в слоях Зеван Кашмира. Два вида: Punctocyrtella spinosa Plodowski
и Cleiothyridina ailakensis Reed указывается из точнее не датируемых нижнеперм-
ских отложений соответственно Афганистана и Читраля.
Проанализировав родовой и видовой состав верхнебазардаринского комплекса бра-
хиопод э можно прийти к выводу, что его состав весьма определенно указывает на
раннепермский возраст вмещающих отложений. Об этом свидетельствует наличие ро-
дов Taeniothaerus, Reticulatia, Punctocyrtella и видов: Punctocyrtella spinosa,
Martinia uralica, Cleiothyridina ailakensis.
Сравнивая верхнебазардаринский комплекс брахиопод Юго-Восточного Памира с
комплексами брахиопод, известными из слоев со Schwagerina (Pseudoschwagerina),
относящихся к ассельскому ярусу, из сопредельных районов (в частности, Южной
Ферганы, Дарвазского хребта, Западного Куньлуня и др.), можно отметить, что они
существенно различаются между собой по систематическому составу и практически
не имеют общих видов.
О более позднем, чем ассельский, возрасте верхнебазардаринского комплекса
брахиопод свидетельствует также то, что отложения, содержащие верхнебазардаринский
комплекс брахиопод,предположительно располагаются в 200-300 м выше слоев, содер-
жащих ассельских аммоноидей (Дмитриев, 19736).
Данные по мшанкам, происходящим из пермских отложений Юго-Восточного Па-
мира, полученные в последнее время Р.В. Горюновой (1971, 1970), указывают также,
что верхнебазардаринский комплекс содержит формы, характерные для нижнепермских,
преимущественно сакмарских и артинских отложений Урала, Таиланда, Индонезии и
других районов.
Нижний и средний продуктусовый известняк относится нами к дар вазе кому ярусу:
а верхний (формация Чидру) - к мургабскому ярусу (см. главу IV).
38
Верхняя возрастная граница слоев с верхнебазард арине ними брахиоп одами опре-
деляется их залеганием ниже кызылджилгинского горизонта* возраст нижней час-
ти которого определяется в основном находками в нем верхнеартинских аммоноидей,
сходных по родовому составу с байгенджинскими аммоноидеями Урала (см. главу П).
П.Брахи опод ы кызылджилгинского комплекса известны из нижнего
и среднего подгоризонтов кызылджилгинского горизонта. Брахиоподы нижнекызыл-
джилгинского подгоризонта встречены в Центральной подзоне в местонахождениях
Куртеке 2, Игримиюз, Мамазаир-Булак, Кастанат-Джилга 2 и приурочены к пласту
криновдно-детритового известняка ^залегающего в основании кызылджилгинского го-
ризонта (табл. 4). Они характеризуются преимущественно якорным этологическим
типом прикрепления. Повсеместно встречаются многочисленные Martinia tachemy—
sohewi Grunt, sp.nov., по-видимому, часто образовывавшие густые поселения. В двух
местонахождениях (Мамазаир-Булак и Игримиюз) присутствует много мелких рако-
вин теребратулид Labaia dmitrievi Grunt, sp. nov. Подчиненную роль играют мелкие
ринхонеллиды Boloria garmoenaia Grunt, sp.nov., Cameriama murgabica Grunt, sp.nov.
Последний вид принадлежит надсемейству Stenoscismatacea. По данным Е.А. Ивановой
(1949), представители этого надсемейства относятся к свободнолежащему этологичес-
кому типу. Они обладали сложным апикальным аппаратом, построенным таким об-
разом, что камарофорий и замочный отросток спинной створки входили в углубление
спондилия брюшной и при раскрывании раковины между створками не образовывалось
отверстия для выхода ножки. Этот вывод Е.А. Ивановой подтверждается тем, что
у многих представителей надсемейства Stenoscismatacea (у Cameriama margabica, в
частности) обнаружен столидий. Наличие этой морфологической структуры, видимо,
способствовало поддержанию раковин на поверхности тонкого илистого грунта. Про-
дуктиды представлены небольшими свободнолежащими раковинами Overtoniina татпа-
zairica Grunt, gen. et sp.nov., Probolionia himalayenaia (Diener), Globiella? roaaiae (Fan—
tini Sestini). Состав комплекса указывает на обитание всех его представителей в
спокойной среде, на тонких илистых грунтах. Совместно с брахиоподами существовали
многочисленные криноидеи, одиночные ругозы, принадлежащие родам Pleurophyllum,
Ufimia, V erbeekiella, Wannerophyllum, Lophophyllidium, значительно реже встреча-
ются колониальные формы, принадлежащие роду Poly сое На. Последние характеризу-
ются стелющимися колониями с кораллита ми, мало возвышающимися над поверхностью
субстрата. Нередко встречаются вытянутые или полусферические массивные ко-
лонии крупноячеистых табулят. Табуляты иногда обрастали стебли криноидей,
располагающиеся по оси колонии. Небольшие колонии хететцд, характеризующие-
ся уплощенной формой, слабо выступали над поверхностью субстрата. Свобод-
ноживущие бентосные организмы сравнительно редки, единичны находки гастро- .
под и двустворок, но довольно обычны мелкие трилобиты. В шлифах нередко
встречаются мелкие фораминиферы и остракоды. Постоянными обитателями этих
криноцдных банок были, по-видимому, аммоноидеи и наутилоцдеи.
Захоронение раннекызылджилгинских организмов происходило в условиях периоди-
ческого усиления гидродинамической активности среды. Условия залегания ископа-
емых указывают на различную степень переработки прижизненных поселений. Пре-
дельную степень переработки представляют собой грубодетритовые известняки, об-
разованные в основном в различной степени окатанными члениками криноидей и
содержащие, кроме них,лишь мелкие, часто также истертые обломки раковин брахиопод
и головоногих. Здесь сохраняются только ругозы, табуляты и хететиды, обычно за -
хороняющиеся не в прижизненном положении.
Криноидно-детритовые разности образуют многочисленные тонкие прослои и уд-
линенные линзы среди микрозернистого глинистого известняка; нередко наблюдаются
постепенные переходы между этими типами пород. В микрозерни ст ых глинистых из-
вестняках встречаются скопления мелких цельных раковин Botoria garmoenaia Grunt,
sp.nov. и Labaia dmitrievi Grunt, sp.nov. Здесь же обычны цельные раковины
головоногих, захороняюшихся в безразличных положениях по отношению к слоистости;
изредка встречаются трилобиты. В переходных типах известняков, где многочисленные
криноидеи погружены в обильный цемент микрозернистого карбоната, нередко встре-
чаются длинные части нераспавшихся стеблей. Нередки головоногие, но раковины их
обычно разбиты. Раковины брахиопод захоронялись в основном в тех разностях
криноидных известняков, которые характеризуются еще значительным участием в
39
„ Та бли ца 4 Распространение брахиопод кызылджилгинского комплекса в отложениях дарвазского яруса Юго-Восточного Памира
Вид Дарвазский ярус
Кызылджилгинский горизонт Южный А к- Архар, кубер- Ганди нс кая (?) свита*се- рые микро- зернистые известняки
нижний подгоризонт средний подгоризонт
Куртеке 2 криноид- но-детри- товые из- вестняки Мамазаир-Булак Кастанат- Джилга 2 криноид- но-детри- товые из- вестняки Игримиюз Чакобай туффитовые известняки Кастанат- Джилга 2 туффитовые известняки
криноид- но-дет- ритовые извест- няки микрозер- нистые глинистые известняки криноидно- детритовые известняки микрозер- нистые глинистые известняки
Подкомплекс Н ижнекызылджилгинский Среднекызылджилгинский ► Costiferina sp. Spiriferella sp. Martinia tschakobaica Grunt, sp.nov. Probolionia him a la у ea- sts (Diener) Camerisma murgabica Grunt, sp.nov. Martinia tscherny— schewi Grunt,sp.nov. Labaia dmitrievi Grunt, sp.nov. Overtonina mamazai— rica Grunt,gen.et sp. ПО5 Globiella?rossiae (Fantini Sestini) Boloria garmoensis Grun gen.et. sp.nov. 10 1 17 9 г. 23 t, 1 1 76 14 78 12 12 1 9 16 6 3 3 5 22 40 5 2 17 3
Terrakea nalivkini
Grunt,sp.nov.
составе пород микрозернистого карбоната. Они представлены, как правило, разроз-
ненными брюшными створками, иногда встречаются скопления битого ракушника
Martinia, но без признаков значительного истирания. Globiella? rossiae (Fantini
Sestini) образует линзу ракушника (Куртеке 2), где створки тесно сгружены и часто
вложены одна в другую, но хорошая сохранность скульптуры указывает на отсут-
ствие значительного посмертного переноса и истирания раковин.
Редкость бентосных организмов в прослоях микрозернистого известняка, содер-
жащего иногда только мелкие формы с якорным типом прикрепления, при общем
облике комплекса, указывающем на приуроченность составлявших его организмов
к неплотному илистому грунту (возможно к несколько консолидированной поверхности
грунта), заставляет.предполагать, что криноидные банки развивались в моменты
замедления или почти полной приостановки осадконакопления, предшествовавшие
моментам усиления гидродинамической активности среды, которое приводило к их
гибели и посмертной переработке. Наблюдающаяся иногда микроритмичность извест-
няка с залеганием наиболее окатанных криноидных разностей на внутренних по-
верхностях размыва с переходом вверх в микрозернистый глинистый известняк под-
тверждает этот вывод.
Описанные захоронения характерны для Центральной подзоны и близких к ней
разрезов, где формирование криноидных банок происходило в пределах обширного
мелководья с прерывистым осадконакоплением. В Промежуточной подзоне извест-
няк, залегающий в основании кызылджилгинского горизонта, отличается неясной
слоистостью и значительной глинистостью; криноидно-детритовые разности для него
не характерны. Здесь он вполне сходен с микрозернистыми разностями Центральной
подзоны и содержит также почти исключительно головоногих. С Центральной под-
зоной по составу нижнекыз ылджилгинс ко го базального известняка и его фауны сход-
ны некоторые участки Промежуточной подзоны, для которых можно предполагать
рост положительных конседиметационных структур (центральная часть хр. ।
Ак-Бура).
Среднекызылджилгинские брахиоподы известны в двух местонахождениях: Ча-
кобай и Кастанат-Джилга 2. Они представлены немногочисленными экземплярами
семи видов. Из них четыре - Probolionia himalayensis (Diener), Martinia tschemy—
schewi Grunt, sp.nov., Camerisma murgabica Grunt, sp.nov., Labaia dmitrievi Grunt, sp.
nov. являются общими с раннекызылджилгинским подкомплексом. Специфическими для
среднекызылджилгинского подкомплекса являются Martinia tschakobaica Grunt, sp.
nov. и немногочисленные Costiferina sp. и Spiriferella sp. К сожалению, точ-
ное определение видовой принадлежности двух последних родов, важных для даль-
нейших выводов, оказалось невозможным из-за недостаточности материала, имею-
щегося в коллекции. В этологическом отношении среднекызылджилгинские брахиопо-
ды не отличались какими-либо существенными чертами от раннекызылджилгинских,
Они также были обитателями илистого известкового грунта, но общий состав сопут-
ствующих организмов становится иным, так же, как и характер захоронения. Здесь
из прикрепленных организмов наряду с криноидеями существенную роль играли мшан-
ки, причем присутствуют как ветвистые, так и сетчатые формы. Ругозы представлены
немногочисленными экземплярами Duplophyllum schindewolfi Schouppe. Из микро-
организмов обнаружены немногочисленные фораминиферы, в том числе очень редкие
фузулиниды Pseudofusulina sp. и остракоды. В шлифах встречаются спикулы губок.
Головоногие представлены единичными аммоноидеями Propinacoceras sp. и прямыми
наутилоидеями. Брахиоподы, мшанки и ругозы залегают в неяснослоистых известня-
ках с пирокластической примесью. Чисто пирокластические породы не содержат ос-
татков ископаемых организмов; условия их образования,по—видимому, были небла-
гоприятны для обитания бентических групп вследствие еысокой скорости седимента-
ции. Брахиоподы нередко захоронены с сомкнутыми створками и хорошо сохра-
нившейся скульптурой. Они так же, как мшанки и ругозы,не образуют скоплений
в породе и, по—видимому,не образовывали густых поселений.
Захоронение среднекызылджилгинских организмов происходило в местах обитания
лишь при незначительном перемещении от прижизненного положения. Наблюдаемый
тафоценоз близко соответствует прижизненному сообществу скелетных форм, что
отчетливо видно также из сходства общего состава фауны в обоих местонахожде-
Ниях» которые удалены друг от друга более, чем на 60 км. Туффитовые известняки
41
формировались, по-видимому, в Центральной подзоне при замедленном, но непре-
рывном осадконакоплении, в несколько специфических условиях затишных участков.
Они связаны фациальными переходами с известковистыми кварцевыми песчаниками
Промежуточной подзоны, которые отлагались в совершенно иных условиях гидро-
динамического режима. Переходные разности пород, известные в долине р.Шинды,
содержат большое количество криноидей и редкие брюшные створки мартиний, часто
разбитые на части. Интересно, что в тех участках Центральной подзоны, где проис-
ходило формирование нормальноосадочных известняков с кварцевой алевритовой при-
месью при очень незначительном содержании пирокластического материала, прикреп-
ляющиеся и свободно лежащие скелетные организмы, по-видимому, совершенно не
обитали.
Малая представительность изученного материала не дает возможности точно ин-
терпретировать отличия нижне- и среднекызылджилгинского подкомплексов. Пока
можно указать лишь три формы, специфические для нижнекызылджилгинского под-
комплекса (Overtoniina mamazairica ^Globiella? rossiae и Boloria garmoenaia) и
три для среднекызылджилгинского (Costiferina sp., Spiriferella sp., Martinia tacha—
kobaica), Обращает на себя внимание то, что общий экологический состав и типы
поселений обоих подкомплексов близки, но ряд наблюдений заставляет с осторож-
ностью подходить к выводам о самостоятельности стратиграфического значения под-
комплексов. Вместе с нижнекызылджилгинскими брахиоподами обитали различные
кишечнополостные, иглокожие и головоногие, но совершенно ртсутствуют мшанки и
фузулиниды, которые известны только начиная со среднекызылджилгинского време-
ни. Общий групповой состав сопутствующей фауны указывает на существование в
обоих случаях нормальноморских условий, но в то же время и некоторых отличий
в типах стаций и в условиях обитания этих фаун. На возможность временных откло-
нений от нормальноморских условий в раннекызылджилгинское время косвенно ука-
зывает положение содержащего брахиопод известняка в основании трансгрессивной
серии осадков. Наличие тонких прослоев остракодовых известняков в нижнекызыл-
джилгинском подгоризонте подтверждает это предположение. Среднекызыдджилгин-
ское время отвечает более устойчивому нормально-морскому режиму, установив-
шемуся при дальнейшем развитии трансгрессии. Интересно, что именно к этому мо-
менту приурочено появление на Юго-Восточном Памире Spiriferella (см. главу IV).
Таким образом, весьма вероятно, что различия между нижне- и среднекызылджил-
гинским подкомплексами связаны с чисто местными причинами. Вследствие этого
в дальнейшем кызылджилгинские брахиоподы рассматриваются в качестве единого
комплекса.
За исключением нижне- и среднекызылджилгинского подгоризонтов, брахиоподы
в дарвазском ярусе Юго-Восточного Памира почти неизвестны. В верхнекызылджил-
гинских фузулинидовых известняках Центральной подзоны отмечено несколько не-
полных брюшных створок Martinia, В основных разрезах кубергандинской свиты бра-
хиоподы крайне редко встречаются в фузулинидовых известняках и имеют неудов-
летворительную сохранность. К кубергандинской свите предположительно отнесен
прослой ракушника, образованного цельными раковинами и разрозненными створ-
ками Terrakea nalivkini Grunt, sp. nov., известный в долине Южный Ак-^Архар.
Кызылджилгинский комплекс резко отличается от верхнебазардаринского по сис-
тематическому составу (нет ни одного общего вида и рода, за исключением Martinia),
Он содержит 10 видов (из них - 7 новых), принадлежащих 9 родам (из них - 2 но-
вых). Специфика комплекса состоит в его эндемизме.
Лишь два рода (Martinia и Cameriama) имеют широкое географическое и стра-
тиграфическое распространение. Род Martinia известен в каменноугольных и перм-
ских отложениях повсеместно в СССР и за рубежом. Род Cameriama встречается
в отложениях нижнего карбона Аляски, в нижней перми Урала и в нижней перми
Альпийско-Гималайской подобласти Тропической области. Род Spiriferella широко
распространен в верхнем карбоне, в нижней и в верхней перми Бореальной палео-
биогеографической области. В Альпийско-Гималайской подобласти он известен из
пермских отложений различных районов: Тибета,’ Юньнани, Тимора (местонахождение
Базлео), Северной Камбоджи (зона Jabeina->Lepidolina), Гималаев, Внутренней
Монголии (местонахождение Усу-Хонгор). Два вида рода Spiriferella (S.peraonata Reed
и S.kimaari Bion) указываются Ридом (Reed, 1932) из аггломератовых слоев Каш-
42
мира, но отнесение этих видов к роду Spiriferella сомнительно . Род Probolionia
известен из пермских отложений Орегона, а в Альпийско-Гималайской подобласти,
предположительно, в известняке* Малинг (ассельский ярус) Китая. В основном же
этот род встречается здесь в пермских отложениях Северной Камбоджи, Гималаев,
Каракорума. Род Overtoniina, кроме Юго-Восточного Памира, по-видимому, распро-
странен в более точно не датируемых нижнепермских отложениях Афганистана и в
верхнепермских отложениях Южного Китая (известняк Маокоу). Род Globiella, отме-
ченный в нашем материале со знаком вопроса, указывается из казанского яруса
верхней перми Русской платформы и Северо-Востока СССР, а в Альпийско-Гима-
лайской подобласти - из нижнепермских отложений Северного Ирана (пачка Д фор-
мации Джейруд). Кроме того, по данным Фантики Сестини ( Fantini Sestini, 1966),
в нижней перми Центральной Индии (слои Умариа). Род Costiferina, кроме Юго-
Восточного Памира, известен из отложений продуктусовых известняков Соляного
кряжа, где он представлен пятью видами, причем один вид (С.spiralis) присутству-
ет в нижнем продуктусовом известняке, а остальные четыре встречены в среднем
и в верхнем продуктусовом известняке. Род Labaia, кроме Юго-Восточного Па-
мира, до настоящего времени был известен лишь в верхнепермских отложениях
(уруштенский горизонт) Северного Кавказа.
Видовой состав бозтеринского комплекса также отличается чрезвычайным своеоб-
разием. Присутствует лишь два ранее известных вида: Globiella? rossiae ( Fantini
Sestini ), происходящая из нижней перми Северного Ирана (пачка Д формации Джей-
руд) и Probolionia himalayensis (Diener) - вид, широко распространенный в перм-
ских отложениях Альпийско-Гималайской подобласти Тропической области: в Север-
ной Камбодже, в Гималаях и Кашмире (слои Зеван). Род Теггакеа, указываемый
нами из кубергандинской (?) свиты, ветре чается в артинском ярусе Австралии и
в верхней перми Новой Зеландии.
Известную трудность представляет выявление возраста кызылджилгинского комп-<
лекса брахиопод. Его высокий эндемизм чрезвычайно затрудняет непосредственное
сравнение с одновозрастными комплексами Альпийско-Гималайской подобласти Тро-
пической области. Однако по присутствию родов Costiferina, Probolionia, Spiriferel-
la, Labaia я вида Probolionia himalayensis кызылджилгинский комплекс бра-
хиопод сближается с заведомо верхнепермскими некоторых районов Альпийско-
Гималайской подобласти (Тимор, Северная Камбоджа, Южный Китай, Каракорум,
Соляной кряж, Северный Кавказ), а также с обычно относимыми к верхней перми
комплексами брахиопод ряда других районов (Монголия,Тибет, Гималаи-Кашмир, Се-
верный Сикким, Непал). Таким образом, по систематическому составу брахиопод
кызылджилгинский комплекс должен быть отнесен к верхней перми. Однако он сущест-
венно отличается от мургабского комплекса, являясь более древним. С другой сто-
роны, раннекызылджилгинские брахиоподы встречены совместно с верхнеартинскими
(байгенджинскими) аммоноидеями. Находки одновозрастных аммоноидей в нижней
части зыгарской свиты Юго-Западного Дарваза (Туманекая, Борнеман, 1937) позво-
ляют отнести кызылджилгинский комплекс брахиопод к нижней части дарвазского
яруса. Кроме того, слои со среднекызылджилгинскими брахиоподами перекрываются
отложениями верхнекызылджилгинского подгоризонта, относящимися к нижней части
зоны Misellina.
Ш. Брахиоподы тахта бу ла кс кого комплекса (табл. 5) встречены в
нижне- и среднетахтабулакской подсвитах джульфинского яруса 2 е Тахтабулакский
комплекс значительно более разнообразен, чем верхнебазардаринский и кызылджил-
гинский. Он содержит 29 видов брахиопод (из них 7- новых), принадлежащих 24 ро-
дам, причем нет ни одного вид а, общего с двумя более древними комплексами и лишь
Описанные Ридом ( Heed, 1932) экземпляры Spiriferella kimsari* (Bion) скорее
всего принадлежат к роду N eospirifer, так как у них скульптура представлена
пучками ребер. Бион также указывал на непосредственное роДство описанных им
форм со Sp.fasoiger (Neospirifer по современной номенклатуре).
В мургабском ярусе (нижнеганская ? подсвита) сделана единственная находка не-
большого скопления раковин Martinia triquetra Gemmellaro.
43
Распространение брахиопод тахтабулакского комплекса в отложениях джульфинского яруса
Вид
Мургабский ярус нижнеган- ская (?) подсвита Игримиюз, водорослевые известняки Джульфинский ярус,
Н ижнетахтабулакс кая
Кур-Истык 1 темно-серые, детрито- вые известняки Кур-Истык 2 водорослевые, биогерм- ные известняки
Enteletes dzhagrensis Sokolskaja
E.meridionalis Gem me Haro
Gey ere Па alpina Schell wien
Scacchinella licharew i Grunt,sp.nov.
__Strophalosiina multicostata Licharew f
Richthofenia caucasica Licharew
Среднетахтабулакский подкомплекс
Krotovia jisuensiformis Sarytcheva
Margin if era sexcostata Licharew
Anidanthus sinosus (Huang)
Compressoproductus mongolicus (Diener)
Haydenella c/ut^evTFcTnTjrunt j^spJnov^
Uncinunellina timorensis (Beyrich)
Wellerella arthaberi (Tschemyschew)
Gerassimovia pamirica Grunt, sp.nov.
G.abalakovi Grunt, sp.nov.
Camarophorinella leveni Grunt,sp.nov.
Martinia wart hi Waagen
Heterelasmina lepton (Gemmellaro)
H.pygmaea Grunt,sp.nov.
Rostranteris {N otothyrina)pontica Licharew
Orthotichia avushensis Sokol ska pT
Parenteletes ruzhencevi Sokolskaja
Enteletella nikschitchi Licharew
Leptodus sp.
o Haydenella tumida (Waagen)
ф Stenoscisma armenica Sokolskaja
e Notothyris pseudodjoulfensis Licharew, sp.nov.
о Scacchinella sp.
§ Permophricodothyris caroli (Gemmellaro)
c Martinia rupicola 'Gemmellaro
M.triquetra Gemmellaro
19
два рода (Martinia и Gerassimovia) известны в верхнебазардаринском и кызыл-
джилгинском комплексах. В тахтабулакском комплексе можно выделить два подкомплек-
са: нижнетахтабулакский и среднетахтабулакский. В состав нижнетахтабулакского
подкомплекса входит 10 видов, из которых лишь два ( Permophricodothyris caroli
Gemm. и Martinia rupicola Gemm.) не встречаются в среднетахтабулакском подкомп-
лексе. Общими для обоих подкомплексов являются: Orthotichia avushensis Sok., En—
teletella nikschitchi Lich., Parenteletes ruzhencevi Sok., Haydenella tumida ер-
tod us sp., Stenoscisma armenica Sok., Notothyris pseudodjoulfensis Lich. Нижнетахта-
булакские брахиоподы встречены в Промежуточной подзоне с сокращенными мощ-
ностями в темносерых детритовых известняках и в располагающихся среди них
небольших водорослевых биогермах (Кур-Истык 1, Кур-Истык 2). В разрезах, пере-
44
Таблица 5
Юго-Восточного Памира
тахтабулакская свита
подсвита Среднетахтабулакская подсвита
о 3 £ Шор-Булак i Музду-Булак | , л । S J л ।
( Джилга-Кочусу водорослево-гидроидя известняки Куртеке 2 водоросле! гидроидные (?) рифо- вые известняки водорослевые био- гермные известня- ки । з . о ь 2 a g О g So ° ф Ф 2 П is I8» zhh У Оф ь ? о 2 ° 5 2 И о A 3 «2 о £ t9oa|s2>:2 я £ S 2 я S я aS Sa<p52siOs <|| Нг Й1 й!
1 s .
3 1 1
3 1 1
9
_4 г 1 1
- '
4 12 6 19
3
1
2 5 1
3 2 13
38 23 5 9 13
9 76 3 3
57 2 о
3 4 3
— 2 10 11
2 5 2
1 14 3)62
13 11 7 5
4 19 2
2
3
1 5 2 5
1 11 12.
3
4 ^53 11 30 1
5
ходных к Центральной подзоне, они приурочены к светлым водорослево—гидроидным
известнякам (Джилга-Кочусу). В Центральной подзоне, в верхней части светлых
водорослево-гидроидных известняков (Куртеке 2) встречен лишь единственный эк-
земпляр Enteletella nikschitchi Lich. Раннетахтабулакские брахиоподы характе-
ризуются преимущественно якорным типом прикрепления. Среди них выделяются две
группы:
1) небольшие раковины теребратулид Notothyris pseudojoulfensis Lich., обла-
дающие фораменом и характеризующиеся постоянным прикреплением раковины к суб-
страту при помощи ножки;
2) некрупные раковины, обладающие низкой ареей и широким дельтирием, оста-
вавшимся открытым на всех стадиях онтогенетического развития. К этой группе от-
45
носятся: из ортид Orthotichia avushensis Sok., Parenteletes ruzhencevi Sok., Entele —
tella nikschitchi Lich., из спириферид Permophricodothyris caroli Gemm. (семейство
Reticulariidae) и Martinia rupicola Gemm. (семейство Martiniidae.) Высокоарей-
ные продуктиды Scacchinella sp. относятся к этологическому типу прирастающих
брахиопод, a Stenoscisma armenica Sok., обладающие столидием, развитым по лоб-
ному и боковым краям, мелкие тонкостворчатые Haydenella turn id a Wa’ag. и сво-
еобразные Leptodus sp. - к с вободнол ежащему этологическому типу. Они прикреп-
лялись лишь на ранних стадиях своего онтогенетического развития.
Из-за малочисленности коллекции раннетахтабулакских брахиопод нельзя сделать
полноценный вывод об особенностях их фациальной приуроченности. Отметим только,
что продуктиды Haydenella tumida Waag., Scacchinella sp., Leptodus sp. отмечены
в единичных находках в светлых водорослево-гидроидных известняках местонахожде-
ния Джилга-Кочусу. К этим же светлым водорослево-гидроидным известнякам приу-
рочены находки Martinia rupicola Gemm. Теребратулиды Notothyris pseudodjoulfen -
sis Lieb. встречены как в водорослевых биогермах Кур-Истыка 2, так и в во-
дорослево-гидроидных известняках Джилги-Кочусу. Ортиды Orthotichia avuschensis
Sok. и Parenteletes ruzhencevi Sok. встречены только в водорослевых биогермных
известняках Кур-Истыка 2. Для фации серых детритовых известняков (Кур-Истык 1)
характерны Stenoscisma armenica Sok. и тонкоиглистые Permophricodothyris caroli
Gemm. Таким образом, имеющийся небольшой материал хотя и не позволяет сколь-
ко-нибудь полно охарактеризовать типы поселений раннетахтабулакских брахиопод,
все же демонстрирует тесную зависимость этологического состава поселений от осо-
бенностей местонахождений.
Среднетахтабулакский подкомплекс встречен в следующих местонахождениях: Шор-
Булак, Кур-41стык 1, Музду-Булак, Куталь 1, Куталь 2, Ак-Бура. Среднепахтабу-
лакский подкомплекс отличается от нижнетахтабулакского своим значительным
разнообразием, ь его составе насчитывается 27 видов, принадлежащих 23 родам.
Кроме видов, присутствующих в нижнетахтабулакеком подкомплексе,он содержит 20
видов брахиопод, характерных исключительно для него.
Среди брахиопод среднетахтабулакского подкомплекса можно видеть три основ«-
ные экологические группировки, приуроченные к биогермам разного типа: существен-
но табулятовым, табулятово-гидроидным и водорослевым. По систематическому сос-
таву брахиоподы, известные во всех трех типах биогермов, весьма близки. Их раз-
личия определяются значительной разницей в количественном соотношении брахиопод
разных этологических типов. В этологическом отношении брахиоподы среднетахта-
булакского подкомплекса могут быть отнесены к трем типам: якорному, прирастаю-
щему и свободно лежащему.
Брахиоподы с якорным типом прикрепления разделяются на две группы:
1) небольшие раковины, относящиеся к отрядам Terebratulida и Rhynchonel—
lid а с фораменом, функционировавшим на протяжении всей жизни организма:
Wellerella arthaberi (Tschem.), Gerassimovia pamirica Grunt, sp.nov., G.abalakovi Grunt,
sp.nov., Heterelasmina lepton (Gemm.), H.pygmaea Grunt, sp.nov., Rostranteris (N otothyrina)
pontica Lich., Notothyris pseudodjoulfensis Lich.,sp.nov;
2) раковины, характеризующиеся наличием крупного дельтирия, остававшегося
открытым на протяжении всего онтогенеза, что указывает на возможность постоян-
ного функционирования ножки. К этой группе относятся : Enteletes dzhagrensis Sok.,
E,meridionalis Gemm., Enteletella nikschitchi Lich., Martinia warthi Waag,
Брахиоподы, относящиеся к свободнолежащему этологическому типу, представлены
днумя видами надсемейства Stenosaismatacea*: Stenoscisma armenica Sok. и Саша—
rophorinella leveni Grunt, sp.nov. Можно предположить, что с вобо дно лежащий об-
раз жизни на взрослых стадиях своего онтоген ети чес кого развития вели U п cinun el—
lina timorensis (Beyrich), обладавшие столидием, располагавшимся по лобному
и боковым краям раковины. Такую морфологическую структуру могли иметь организ-
мы, обитавшие в зонах спокойного гидродинамического режима, как это было по-
казано Е.А. Ивановой для атрипид (1962). Свободнолежащий образ жизни на мягких
илистых грунтах внутренних частей биогермов вели и мелкие иглистые продуктиды
Krotovia jisuensiformis Sar., Marginifera sexcostata Lich., Haydenella dutkevitchi Grunt,
sp.nov., а также тонкоребристые Anidanthus sinosus Huang.
46
Некоторые виды продуктид характеризовались прирастанием на ранних стадиях
онтогенетического развития и лежали свободно на более поздних. К ним относятся:
Compressoproductus mongolicus (Diener), Leptodus sp., Strophalosiina multicostata
Lich. Последний вид обладал небольшой раковиной с тонкими иглами, заканчиваю-
щимися воронкообразными расширениями, располагавшимися по лобному краю рако-
вины.
К прирастающему этологическому типу брахиопод в среднетахтабулакском под-
комплексе могут быть отнесены: Geyerella alpina Schellw., Scacchinella licharewi
Grunt, sp.nov., Richthofenia caucasica Lich.
Обилие и количественные соотношения разных жизненных форм брахиопод в во-
дорослевых, табулятовых и табулятово-гидроидных биогермах различны. Водорос-
левые биогермы, встреченные в разрезах Музду-Булака, Шор-Булака и Ак-Буры,
характеризуются обедненностью систематического состава брахиопод. Они не пред-
ставляли удобного субстрата для поселений. Здесь совершенно отсутствуют свобод-
нолежащие и прирастающие формы, личинки которых, по-видимому, погибали под
наростами водорослей. Якорные брахиоподы, встречающиеся здесь, представлены
крайне немногочисленными экземплярами. Преобладают мелкие ринхонеллиды и те-
ребратулиды, реже встречаются мартинии и ортиды. Все брахиоподы захоронены с
сомкнутыми створками среди обволакивающих их водорослевых наростов (Шор-Бу-
лак, Музду-Булак, Ак-Бура).
Для табулятовых (Шор-Булак, Музду-Булак) и табулятово-гидроидных (Кур-
Истык 1, Куталь 2) биогермов характерны, наряду с якорными формами, прирас-
тающие и свободно лежащие, однако в первых они составляют около одной трети
всего комплекса, в то время как во вторых - всего лишь одну десятую. В то же
время табулятово-гидроидные биогермы характеризуются очень большим количеством
мелких теребратулид и ринхонеллид, которые не встречаются в таком изобилии в
табулятовых постройках, хотя и составляют в них около двух третей комплекса.
Таким образом, постройки, образованные ветвистыми табулятами, представляли со-
бой более удобные биотопы для обитания свободнолежащих и прирастающих брахио-
под, чем та бу л ято во- гидроидные, основное население которых составляют якорные
формы. Причиной этого, как можно предполагать, являлись биотические отношения
брахиопод с рифообразующими организмами. Для разъяснения данного предположения
рассмотрим сначала условия захоронения брахиопод.
Как правило, брахиоподы и другие рифожители захоронены в светлом микрозер-
нистом известняке, заполняющем неправильные промежутки между колониями рифо-
образователей. Якорные формы сохраняются в подавляющем большинстве случаев
с сомкнутыми створками, продуктиды со слабым замковым сочленением как в виде
цельных раковин, так и в виде разрозненных створок, причем спинные створки обыч-
но захороняются тут же. Тонкая скульптура продуктид отличается хорошей сохран-
ностью. Следовательно, щели и ниши во внутренних частях биогермов, развивавших-
ся среди подвижных грубообломочных осадков, представляли удобный субстрат для
поселения в спокойной среде свободнолежащих мелких иглистых продуктид и стено-
сцизматацей со столидием. К ним были близки продуктиды, прикреплявшиеся на ран-
них стадиях онтогенеза. В то же время на твердых поверхностях колоний рифооб-
ра зова тел ей селились различные прикрепляющиеся формы (как якорные, так и при-
растающие). Тафоценоз брахиопод в микрозернистом известняке внутренних частей
биогермов составлялся как из свободнолежащих форм, селившихся на илистом из-
вестковом грунте, так и из посмертно попадавших в него якорных брахиопод.
Возвращаясь к составу прижизненных поселений, отметим, что по характеру за-
хоронения табулятовые биогермы не отличаются какими-либо существенными черта-
ми от' табулятово-гидроидных, и, следовательно, явное преобладание продуктид в
первых вряд ли может быть связано с особенностями абиотической среды: характе-
ром субстрата или гидродинамическим режимом. Можно предположить, что поселе-
ние в окружении обволакивающих колоний гидроидных в табулятово-гидроидных био-
гермах не было вполне благоприятным для молодых особей продуктид, относящих-
ся к родам Scacchinella, Strophalosiina, Richthofenia, Mar gin if era, а также
высокоарейных строфоменид и стеноспизматацей. Мелкие теребратулиды и ринхонел-
лиды с якорным типом прикрепления, которые могли приподниматься над субстра-
том, оказывались, видимо, более приспособленными к обитанию в табулятово-гид-
47
роидных биогермах, где плотность поселений якорных брахиопод была намного вы-
ше, чем в табулятовых биогермах.
Среднетахтабулакский подкомплекс отличается от нижнетахтабулакского и зна-
чительно большим разнообразием систематического состава. Из 30 видов тахтабу-
лакских брахиопод 20 характерны исключительно для него, так что нижнетахтабу-
лакский подкомплекс можно рассматривать просто как обедненный. Однако он ха-
рактеризуется и некоторыми специфическими особенностями: присутствием немно-
гочисленных характерных для него видов, список которых, по-видимому, увеличит-
ся при дополнительном изучении (в частности, неизученными из-за недостаточности
материала оказались экземпляры ? Cryptospirifer omeishanensis Huang, встре-
ченные в водорослево-гидроидных известняках Джилги-Кочусу, и др.). Кроме того,
он в целом резко отличается иными количественными соотношениями составляющих
его систематических группировок. В частности, весьма многочисленные в средне-
тахта булакс ком комплексе и составляющие в нем более половины всего количества
брахиопод ринхонеллиды совершенно не представлены в нижнетахтабулакском. Это
не относится к Stenoscisma armenica Sokolskaja, которая известна в равной мере
из обеих подсвит. Биогермы среднетахтабулакской подсвиты по условиям сущест-
вования в них брахиопод вряд ли могли отличаться какими-либо существенными осо-
бенностями от нижнетахтабулакских, во всяком случае изучение характера захо-
ронения брахиопод в нижне- и среднетахтабулакской подсвитах не дает никаких пря-
мых указаний в этом отношении.
Различия нижне- и среднетахтабулакского подкомплексов Юго-Восточного Памира,
как можно предполагать, в значительной мере связаны с некоторыми отличиями фи-
зико-географической среды в ранне- и среднетахтабулакское время, вследствие чего
вопрос об их стратиграфическом значений за пределами этого района неясен.
Характерными для тахтабулакского комплекса признаками являются: мелкорослость
фауны, однообразие видового состава относительно редко встречающихся продуктид,
полное отсутствие атиридид (в Закавказье джульфинские атиридиды представлены де-
вятью видами рода Araxathyris), относительно большое количество представителей
отрядов Rhynchonellida, Terebratulida, Ort hid а по сравнению с другими районами
и почти полное отсутствие строфоменид.
Типичными для этого комплекса являются роды: Enteletella, Geyerella, Scacchi-
nella, Richthofenia, Strophelosiina, Compressoproductus, Heterelasmina, Haydenella ,
причем некоторые из них - Enteletella, Strophalosiina, Gerassimovia, Notothyrina ~
кроме Юго-Восточного Памира известны исключительно из верхней перми Северного
Кавказа. Из 30 видов тахтабулакского комплекса 25 видов встречено за пределами
Юго-Восточного Памира. Они приурочены преимущественно к верхнепермским отло-
жениям Евро-Азиатской части Тропической области. Из них шесть видов - Entele-
tella nikschitchi Lich., Scacchinella licharewi Grunt, sp.nov., Marginifera sexcostata
Lich., Strophalosiina multicostata Lich., Notothyris pseudodjoulfensis Lich., sp. nov.,
Rostranteris (N otothyrina) pontica Lich. встречены только в верхней перми Север-
ного Кавказа, а пять видов - в Закавказье. Orthotichia avushensis Sok. и Kroto-
via jisuenaiformis Sar. * известны из гнишикского горизонта; Parenteletes ruzhen-
cevi Sok., Stenoscisma armenica Sok. - из джульфинского яруса (слои с Araxilevis -
слои с Araxoceras — Oldhamina), a Enteletes dzhagrensis Sok. - из слоев с
Т от ро phi с eras - Bemhardites. Отдельные виды тахтабулакского комплекса (Rich-
thofenia caucasica Lich.) встречаются как в верхней перми Северного Кавказа, так
и Закавказья. Три вида, установленных Джеммелларо (Gemmellaro, 1899) — Enteletes
meridionalis, Martinia rupicola, Heterelasmina lepton - являются общими с фузули-
нидовыми известняками ( слои Созио)Сицилии. Наряду с перечисленными присутству-
ет ряд форм, распространенных в течение всего пермского периода практически на
всей территории Евро-Азиатской части Тропической области. Это Anidanthus sino-
sus Huag, Compressoproductus mongolicus (Diener), Uncinunellina timorensis (Beyrich),
Wellerella arthaberi (Tschem), Geyerella alpina Schellw.
Нужно отметить, что тахтабулакский комплекс брахиопод не отличается особым
своеобразием и эндемизмом. Проанализировав его родовой и видовой состав, можно
сделать вывод, что наличие в нем родов Enteletella, Strophalosiina, Gerassimo —
via, подрода Notothyrina, а также видов Enteletes dzhagrensis, Enteletella
nikschitchi, Parenteletes ruzhencevi, Scacchinella licharewi, Strophalosiina multicosta-
48
ta, Margini fera aexcostata, Stenosciama armenica, Notothyris paeudodjotilfenais^ Ros—
tranteris (Notothyrina) pontica позволяет с уверенностью относить его к джуль-
финскому ярусу верхней перми. Джульфинский возраст тахтабулакского комплекса
брахиопод подтверждается присутствием в отложениях тахтабулакской свиты фузу-
линид Palaeofuaulina pamirica Leven, Reichelina pulchra К.M.—Macl., Codonofusiella
lui Sheng (Левен, 1967).
* * *
Описанные комплексы брахиопод относятся к трем стратиграфическим уровням,
разделенным поверхностями размыва или толщами, не содержащими брахиопод. Поэто-
му особенности смены комплексов на протяжении всего пермского периода на на-
шем материале не могут быть выявлены полностью. Однако по систематическому
составу, положению в разрезах, сопутствующей фауне и другим признакам комплексы
брахиопод, описанные с Юго-Восточного Памира, могут быть связаны с определен-
ными комплексами брахиопод, известными из других районов Альпийско-Гималай-
ской подобласти. Сравнивая памирских брахиопод с брахиоподами из других районов
Тетиса, удается также наметить возможность выделения брахиоподовых комплексов
(ассельского и мургабского), не представленных на Юго-Восточном Памире.
К сожалению, основные сведения о комплексах брахиопод верхнего палеозоя Тро-
пической палеобиогеографической области могут быть получены преимущественно из
старых монографий, не всегда позволяющих установить родовую принадлежность опи-
санных форм, которые практически не изучены на современном уровне. Имеющиеся
сведения о типах захоронений брахиопод и их фациальной приуроченности также не-
достаточны, что сильно затрудняет стратиграфический и палеобиогеографический ана-
лиз комплексов, часто не позволяя выявить причин различия их систематического
состава.
Глава IV
ЗАКОНОМЕРНОСТИ СМЕНЫ КОМПЛЕКСОВ ПЕРМСКИХ БРАХИОПОД
АЛЬПИЙСКО-ГИМАЛАЙСКОЙ ПОДОБЛАСТИ ТЕТИСА И ИХ
БИОГЕОГРАФИЯ
Настоящая глава посвящена обзору основных комплексов пермских брахиопод Ев-
роазиатской части Тетиса, выявлению закономерностей их изменений на протяже-
нии пермского периода в связи с палеогеографическими изменениями и вотросам
палеобиогеографического районирования этого бассейна.
Пермь характеризуется крупными орогени чески ми движениями, постепенным на-
правленным сокращением эпиконтинентальных морских бассейнов возникновением
отчетливой климатической зональности, резкой аридизапией климата, начавшейся с
кунгура и постепенно нараставшей к концу перми (Руженцев, 1965). В конце перми
вымирают многие характерные для палеозоя группы морских организмов: фузулини-
ды, ругозы, трилобиты, гониатиты, некоторые отряды брахиопод, мшанок и др.
Биогеография морских бассейнов пермского периода представляет существенный
интерес.
Проблемам биогеографии позднепалеозойских морских бассейнов посвящены рабо-
ты Д.В. Наливкина (1929, 1936, 1937), А.Д. Миклухо-Маклая (1955, 1957, 1963),
Б.К. Лихарева и А.Д. Миклухо-Маклая (1964), В.И. Устрицкого (1958, 1961, 1965а,
19656, 1967а, 19676), О.Л. Эйнора (1957, 1964, 1966, 1967), О.Л. Эйнора и др. (1965),
В.М. Синицина (1962), О.В. Юферева (1967), А.Д. Григорьевой (19676), Л.С. Либ-
ровича ( 1968), Минато Macao и др. (1968), Шухерта (1957), Стели (Stehli, 1957,
1964,1971),Стели и Веллса ( Stehli, Wells, 1971), Аудлей-Чарльза (Aud ley -Charles,
1965),Мерла (Meria, 1934), Радвика (Rudwick, 1965),Радвика и Коуэна(Rudwick, Co-
wen, 1968). В работах Мерла, Радвика и Коуэна рассматривается распространение неко-
торых комплексов брахиопод и их отдельных родов в пределах пермского Тетиса. Интерес-
но отметить работу Гоббетта ( Gobbett, 1967) о биогеографическом распределении основ-
ных родов и семейств высших фузулинид. Более подробно мы остановимся на работах
А.Д.Миклухо-Маклая (1963)и Б.К. Лихарева совместное А.Д.Миклухо-Маклаем (1964),
так как они касаются интересующих нас районов Евро-Азиатской части Тетиса. На терри-
тории современной Евразии и Северной Америки для пермского периода ими выде-
ляются Европейско-Сибирская и Кавказско-Синийская биогеографические области,
которые сформировались в пределах Русско-Китайского бассейна, существовавшего
в карбоне. Это произошло в результате образования Кавказско-Тяньшанского пере-
шейка, отделившего бореальные бассейны от южных. В пределах Кавказеко-Синий-
ской биогеографической области раннепермской эпохи А.Д. Миклухо-Маклай (1963)
выделяет Карнийскую, Среднеазиатскую, Индо-Синийскую и Уссурийскую провинции;
в позднепермскую эпоху Кавказско-Синийская область подразделяется им на Крым-
ско-Памирскую подобласть, в состав которой входят Карнийская, Сицилийская, Крым-
ская, Северо-Кавказская, Закавкаэско-Дарвазская, Памирская, Северо-Индийская
провинции и Уссурийско-Малайскую подобласть; в последнюю входят Корякская, Ус-
сурийская, Индо-Синийская, Тиморская и Орегонская провинции (табл. 6).
Такое зоогеографическое районирование, по нашим представлениям, излишне дроб-
но и не всегда подтверждается анализом данных по брахиоподам1.
Биогеографическое районирование' позднепалеозойских морских бассейнов опирается
на географическое распространение эндемичных групп различных морских организмов
Сопоставление схем биогеографического районирования пермских морских бассейнов(
принятых различными авторами, и схемы, принятой в данной работе, показано в
табл. 6.
50
Сопоставление различных схем биогеогржрического районирования пермских морских бассейнов
В.И. У стриикий
1961
Ранняя пермь Поздняя пермь 1967
'2 Р2
Арктическая область Восточно- Западно- Арктич.про- А рктич.про- винция ВИНЦИЯ Тетис Монголе - Арктическая область Арктическая провинция Арктическая область Восточно- Западно- Арктич.про- Арктич.про- впнция К ВЙНЦИЯ Арктическая область Восточно- Западно- Арктич.про- Арктич.про- винция ВИНЦИЯ
Тетис Монголо-Гималайская провинция Тетис Тропическая область
А.Д.Миклухо-Маклай (1963), Б.К Лихарев, АЛ.Миклухо-Маклай (1964) Т.А.Гр у н т (схема, принятая в данной работе)
Ранняя пермь Поздняя пер мь
Евр о-С ибирская область Бореальная область
Кавказско-Синийская область Карни йс кая провинция Крымско-Памирская подобласть Карнийская провинция Тропическая область (Тетис) Альпийско-Гималайская подобласть Провинции Альпийская
Сицилийская провинция
Средне- азиат- ская провин- ция Крымская провинция Кавказско-Памирская
Северо-Кавказская провинция
Памирская провинция
Закавказско-Дарвазская провинция
Северо-Индийская провинция Юж но-К ита йс кая
И н до-Си ний - ская провин- ция У ссури йско-Л \ а л а й - ская подобласть Индо-Синийская провинция
Тиморская провинция
Уссурийская провинция Уссурийская
Корякская провинция Корякско- Орегонс- кйя подоб- ласть Корякская
Орегонская провинция Оре гонская
Северо-Американская область ? Североамериканская подобласть
Австралийская область Антибореальная область
так же, как биогеографическое районирование современной бентали. Отметим, что
биогеография пермских, так же как и других древних морских бассейнов, является,
в основном, палеобиогеографией шельфа.
Восстанавливая контуры пермских морских бассейнов (здесь мы обычно имеем
дело с мелководными осадками), мы основываемся на комплексах морских организ-
мов, большая часть которых является бентосными формами, распространенными на
континентальной отмели (брахиоподы, кораллы, многие фораминиферы, а также, по-
видимому, некоторые аммоноидеи). Районирование континентальной отмели совре-
менных морских бассейнов может быть проведено достаточно детально, с выделе-
нием провинций, округов, участков, имеющих сложные очертания, зависящие от ряда
самых разнообразных факторов.
Многообразие и качественная характеристика подобных факторов, действовавших
в. древних морях, в настоящее время в большинстве случаев не могут быть учтены
достаточно полно и точно. Поэтому задача достоверной реконструкции биогеографии
пермских морских бассейнов может быть разрешена лишь в самых общих чертах,
без выделения единиц более дробных, чем провинции.
Следует также сказать, что хотя при зоогеографическом районировании совре-
менных морских бассейнов и учитывается предшествующая история развития фаун,
в основном это районирование опирается на географическое распространение различ-
ных групп организмов в современную эпоху. Районирование древних морских бас-
сейнов производится не для одномоментного в геологическом отношении простран-
ственного распределения фаун, а для некоторого достаточно длительного периода.
Вследствие этого к вопросу об отсутствии определенных систематических групп в
тех или иных районах следует подходить крайне осторожно, так как это отсутствие
может быть связано с чисто местными фациальными причинами, неполнотой разре-
за или плохой изученностью района и типов захоронений. Недостаточное значение
перечисленных моментов может привести к неоправданному увеличению количества
палеобиогеографических подразделений. В некоторых случаях большую у бед и те ль- ,
ность эти подразделения приобретают при использовании аналогий с более ранними
и более поздними эпохами.
В последнее время большинство исследователей, занимающихся биогеографическим
районированием древних морских бассейнов, приходит к выводу, что оно должно про-
изводиться на тех же основах, что и районирование современного Мирового океана.
При районировании современных морских бассейнов С.Экман ( Ekman, 1935),
В.М. Гептнер (1936), Е.Ф. Гурьянова (1945, 195Z), Г.Латтин ( Lattin, 1967) ис-
ходят из наличия в современном мировом океане трех крупных областей или надоб-
ластей: Бореальной (области умеренных и холодных морей северного полушария),
Тропической и Анти бореальной (области умеренных и холодных морей южного полу-
шарния). Наличие их диктуется дифференциацией фауны Мирового океана по клима-
тическим поясам (закон Докучаева-Берга).
Внутри Тропической надобласти Гептнер (1936) выделяет две зоогеографические
области: Атлантическую и Индо-Тихоокеанскую (по данным распространения пелаги-
ческой фауны). Латтин, рассматривающий районирование Мирового океана на основе
бентонной фауны континентальной отмели, разделяет Тропическую область на четыре
подобласти: Индо-Тихоокеанскую, Восточно-Тихоокеанскую, Западно-Атлантическую,
Восточно-Атлантическую. В настоящее время Индо-Тихоокеанская и Восточно-Тихо-
океанская подобласти Тропической области, с одной стороны, и Западно- и Восточно-
Атлантическая - с другой, довольно слабо различаются между собой, так как их
обособление произошло, вероятно, относительно недавно- скорее всего в начале
кайнозоя. Зоогеография современных морских бассейнов объясняет это тем, что до
конца мезозоя на формирование современных фаунистических подразделений моря,
и особенно на развитие Тропической области, преобладающее влияние оказывал, ве-
роятно, субширотный океанический бассейн, известный под названием гТетисг (Гепт-
нер, 1936).
Этот термин был впервые употреблен Е.Зюссом ( Suess, 1893), который опреде-
лял Тетис, как экваториальный морской бассейн, разделявший континенты Евразии
(Ангариды) и Гондваны.
С тех пор название *Тетисг широко принято в литературе, но разные исследова-
тели придавали ему различный смысл. Палеогеографически Тетис позднего палеозоя
52
представляется зоогеографической областью, совпадающей с Тропической. В этом
смысле мы и будем употреблять это название в дальнейшем.
Тропическая область в позднем палеозое характеризовалась теплым климатом,
широким развитием рифовых построек, большим количеством разнообразных теплолю-
бивых организмов с массивным известковым скелетом.
В Трудах симпозиума, посвященного биогеографии Тетиса ("Aspects of Tethyan
Biogeography" , 1967), отмечается, что этот бассейн, существовавший с раннего
палеозоя, первоначально представлял собой мелководную морскую акваторию, тесно
связанную с Мировым океаном. Л С. Либрович (1968), рассматривая зоогеографию
каменноугольных аммоноидей, указывает, что на границе визейского и турнейского
веков обособление Тетиса (или Средиземной области) было очень четким.
Тетис позднего палеозоя представляется морским поясом, протягивавшимся суб-
щиротно и являвшимся основным путем миграции тропических морских фаун с дос-
таточно свободным сообщением внутри него. Этот бассейн охватывал области Юж-
ной Европы (Сицилия, Альпы, Венгрия, Балканский полуостров, острова Греческого
архипелага, Крым, Кавказ), северной части Африки (Тунис), Южной и Юго-Восточной
Азии (Аравийский полуостров, Турция, Закавказье, Иран, Афганистан, Памир, Фер-
гана, Кашмир, Пакистан, Гималаи, Китай, Индокитай, Индонезия), Японии, Хабаров-
ского и Приморского краев, Корякского нагорья, Орегона (западное побережье Се-
верной Америки) и, вероятно, Техаса.
Неясной остается проблема возможности морских связей между Евро-Азиатской
и Североамериканской частями Тропической области. При изучении органического
мира конца каменноугольного - начала пермского периода палеонтологи нередко при-
ходят к выводу о близости фаун Альпийского и Североамериканского (Техасского)
бассейнов. Обычно отмечается много общих родов среди фузулинид (Миклухо-Мак-
лай, 1963) и аммоноидей (Либрович, 1968). Если обратиться к ассельской брахио-
подовой фауне Северной Америки и Южной Европы, то можно увидеть, что для обеих
характерны такие своеобразные тетисные роды, как Parenteletes и Scacchinella.
Род Scacchinella широко распространен также в ассельских отложениях Урала и
Средней Азии. Эти данные указывают на возможность существования в позднем
карбоне и на протяжении ассельского века морских связей между Евро-Азиатским
и Североамериканским бассейнами. Последний Либрович (1968) условно выделил
в Американскую область, указав, что она, возможно, представляет собой лишь осо-
бую провинцию Тетиса. В Евро-Азиатской части Тетиса позднего карбона Либрович
выделил Уральскую и* Среднеазиатскую провинции, которые Миклухо-Маклай (1963)
рассматривал как единую Русско-Китайскую область.
Начиная с сакмарского века, непосредственные морские связи между Евро-Ази-
атским и Североамериканским бассейнами, вероятнее всего, оказались нарушенными.
В послеассельское время для Евро-Азиатского и Североамериканского бассейнов
характерно наличие большого количества представителей типичных южных надсемейств
Richthofeniacea и Lyttoniacea. В Евро-Азиатском бассейне из надсемейства
Richthofeniacea присутствуют роды Тeguliferina, Richthofenia, Cardinocrania, Cos—
cinarina, а в Североамериканском — Planispina и Prorichthofenia. Из над-
семейства Lyttoniacea в Евро-Азиатском бассейне присутствуют роды Keyser-
1 in gin а и О Id ham in а. В Североамериканском бассейне с ранней перми известен
род Leptodus, проникший в Азию только в начале дарвазского века. Техасский бас-
сейн пермского периода условно рассматривался Миклухо-Маклаем (1963) в качестве
самостоятельной Североамериканской области на том основании, что в нем отсут-
ствуют высшие фузулиниды. Однако, в действительности представители семейства
Neoschwagerinidae известны в этом бассейне (Skinner, Wilde, 1955; Stehli, 1964;
Bostwick, Nestell, 1967).
Учитывая вышесказанное, можно высказать предположение, что Североамерикан-
ский бассейн в течение всего пермского периода входил в состав Тропической био-
географической области, а также, что Евро-Азиатскую и Североамериканскую части
этой области можно рассматривать в качестве самостоятельных подобластей: Се-
вероамериканской и Альпийско-Гималайской. Последняя территориально совпадает с
Кавказе ко-Синийской биогеографической областью Миклухо-Маклая (1963) за исклю-
чением его Корякской и Орегонской провинций. Эти две провинции тесно связаны
между собой и характеризуются существенной обедненностью представителей тропи-
53
ческой фауны и присутствием многих бореальных родов брахиопод. Фузулиниды
многочисленны, но однообразны. Брахиоподовая фауна, сходная с корякской, описана
Мак Гуганом и Логаном ( McGugan, 1963; Logan, McGugan, 1968) из пермских отло-
жений Британской Колумбии (Канадские Скалистые горы). Авторы указывают на
наличие среди описанных брахиопод родов Jakovlevia, Spiriferella, Costiferina,
Pterospirifer и отмечают, что эта, так называемая "русская* фауна известна из
"среднепермских" отложений Орегона, Юкона, Канадской Арктики.
Можно предположить, что Корякско-Орегонский бассейн достигал высоких поляр-
ных широт и поэтому характеризовался сосуществованием значительно обедненной
тропической фауны и большого количества типичных бореальных родов, особенно сре-
ди представителей мелководного бентоса (брахиопод). Район Корякского нагорья
требует дальнейшего подробного изучения, которое еще только начинается.
В дальнейшем мы остановимся на рассмотрении лишь Альпийско-Гималайской
подобласти Тропической области . Особенностью Альпийско-Гималайской подобласти
является то, что в ней происходил пышный расцвет фауны брахиопод, которые до
конца перми остаются здесь одной из самых распространенных групп, в то время
как в Бореальной области, потерявшей с конца ассельского века связь с Те тисом
и характеризовавшейся более суровыми климатическими условиями, шло постепенное
обеднение состава брахиоподовой фауны. В сакмарско-раннеартинское время в Аль-
пийско-Гималайской подобласти насчитывается около 70 родов, тогда как в джуль-
финском веке -около 100. Это происходит, по-видимому, потому, что в благоприятных
условиях Тетиса огромное большинство родов, известных из сакмарских и артинских
отложений, продолжает существовать до конца перми. В то же время в джульфинском
и мургабском веках возникает довольно много аберрантных родов, представленных
единичными видами, которые имеют очень ограниченное время существования.
Ассельский этап развития брахиопод нами рассматривается очень кратко и обоб-
щенно на основании литературных данных, так как ассельские брахиоподы в нашем
материале отсутствуют.
Ассельский комплекс брахиопод устанавливается в результате сравнения сакмар-
ско-раннеартинского комплекса Юго-Восточного Памира и его аналогов с комплек-
сами брахиопод из слоев со Schwagerina (Pseudoschwagerina), которые по своему
систематическому составу существенно отличаются от сакмарско-раннеартинских.
Ассельский комплекс брахиопод прослеживается на обширных пространствах Аль-
пийско-Гималайской подобласти Тропической области. К нему мы относим брахиопод
карачатырского и, возможно, верхней части дастарского горизонта (Волгин, 1960)
или свит ” g" и ’’Г’ Южной Ферганы (Лихарев, 1946), брахиопод, обильно представ-
ленных в себисурхской свите Юго-Западного Дарваза (Власов, Лихарев, Миклухо-
Маклай, 1962; Волгин, 1964) и брахиопод, известных из горизонта с Pseudoschwa-
gerina в верхней части артушрикской свиты Западного Куньлуня (Устрицкий и др.,
1960). Предположительно к асеельскому комплексу нами относятся брахиоподы, опи-
санные Мерла ( Meria, 1934) и Фантини Сестини ( Fantini Sestini, 1965а) из Се-
верного Каракорума как "уральский комплекс". В Китае ассельские брахиоподы
известны в известняке Мапинг. Они описаны из провинций Южного Китая (Grabau,
1934,1936) и Юньнани {Hayasaka, 1922). Предположительно к асеельскому ярусу могут
быть отнесены брахиоподы, описанные из Центрального Таиланда Янагидой (Yanagida,
1967) и из Лаоса и Камбоджи Манси (Mansuy, 1913), Немногочисленные ассельские
брахиоподы из Афганистана (провинция Бамиан) изучены Ридом (Heed, 193la),а из севе-
ро-западной части Малой Азии (Бальи)-Эндерле (Enderle, 1901). В западной .части
Альпийско-Гималайской подобласти к ассельскому ярусу относятся брахиоподы из мест
1
Палеогеографические схемы, приводимые в настоящей работе (рис. 3-7), составлены
на основе данных В.М. Синицина (1962), Лю Хун-юна (1962), А.Д. Миклухо-Мак-
лая (1963) для Альпийско-Гималайской подобласти, Атласа литолого-палеогеографи-
ческих карт Русской платформы (1960) и В.И. Устрицкого (19676) для Бореальской
области, Шухерта (1957) для Северной Америки, а также на основе распространения
отдельных родов брахиопод в морях пермского периода. На схемах показаны пре-
делы максимального» распространения моря для каждого из рассматриваемых перио-
дов.
54
нахождения . Ауэрниг (Schellwien, 1892). Одновозрастные брахиоподы описаны К ос-
ти ч-Подгорской ( Kostic—Podgorska, 1949) и Симичем (Si.mi£Tr 1934в, 19356) из место-
нахождения Лике и Рамовшем (Ramovs, 1960) с холмов Витанье Северо-Западной Юго-
славии (рис. 3).
Брахиоподы ассельского яруса (швагеринового горизонта) Урала изучены Ф.Н.Чер-
нышевым (1902), Н.П. Герасимовым (1929), М.В. Куликовым (1938), Д.Л. Степано-
вым (1951). В монографии Ф.Н. Чернышева (1902) описаны смешанные коллекции,
собранные из отложений широкого возрастного диапазона вплоть до верхнеартинских
отложений, причем большая часть местонахождений, изученных Ф.Н. Чернышевым,
относится к более молодым, чем швагериновый горизонт, отложениям (Степанов,
1951). Тем не менее Степанов отмечает, что монография Чернышева дает наиболее
полное представление о составе брахиопод ассельского яруса.
Д.Л. Степанов (1951) приводит характеристику ассельских брахиопод и отмечает
их тесную связь с сакмарскими. Последнее наряду с недостаточной изученностью
брахиопод ассельского яруса приводит к тому, что непосредственное сопоставление
разрезов Альпийско-Гималайской подобласти с уральскими по комплексам брахиопод
затруднено, хотя в первых распространены многочисленные уральские виды.
Сопоставление осложнено еще двумя обстоятельствами. Во-первых, в Альпийско-
Гималайской подобласти практически не изучена в единых разрезах смена ассель-
ского комплекса брахиопод сакмарско-раннеартинским. Во-вторых, соответствие ас-
сельского яруса Урала кара чат ырскому ярусу, выделенному Миклухо-Маклаем (1958а)
для южных районов, является дискуссионным. По данным Миклухо-Маклая, швагери-
ны (Schwagerina s.l.) в южных провинциях существовали значительно дольше, чем
на Урале, вследствие чего содержащие их слои и были выделены в особый карача-
тырский ярус, охватывающий не только ассельский и сакмарский ярусы Урала, но
также и нижнюю часть артинского яруса. Позднее Лихарев и Миклухо-Маклай (1964)
сочли возможным сопоставить карачатырский ярус Средней Азии с ассельским и
сакмарским ярусами Урала, исключив из его состава нижнеартинские отложения.
К этой точке зрения присоединился Э.Я. Левен (1965, 1967), который справедливо
отметил, что при таком понимании стратиграфического объема карачатырского яруса
его название следует заменить названием гсакмарский ярус* в широком понимании
В.Е. Руженцева (1950). В отличие от предыдущих авторов Д.М. Раузер-Черноусова
(1965) приходит к выводу о точном соответствии ассельского яруса Урала и кара-
чатырского яруса Тропической области, что находит подтверждение в работах Р.Ф.Бенш
(1965, 1972) и Ф.Калера ( Kahler, 1962). Бенш сопоставляет карачатырские отло-
жения Ферганы с ассельским ярусом Урала, подразделяя их на три подгоризонта,
соответствующие уральским. В разрезах Карнийских Альп (Kahler, 1962; Раузер-
Черноусова, 1965) с ассельским ярусом Урала сопоставляются раттецдорфские из-
вестняки, в которых могут быть выделены те же три подгоризонта. Таким образом,
хотя в целом швагерины и близкие к ним роды, по-видимому, распространены в
Альпийско-Гималайской подобласти не только в ассельском ярусе, но все же в ряде
случаев удается по фузулинидам отделить ассельские отложения от сакмарских.
Важно, что все названные исследователи единодушны в оценке возраста трокго-
фельских известняков Карнийских Альп, сопоставляемых ими с сакмарским ярусом
Урала в его узком понимании (без ассельского). Данные о совместном нахождении
ассельских брахиопод и швагерин и намечающаяся по материалам Ферганы, К ар ни й-
ских Альп и, возможно, Юго-Западного Дарваза возможность параллелизации содер-
жащих их слоев с ассельским ярусом Урала позволяют считать ассельский комплекс
брахиопод самостоятельным, характеризующимся особым составом фауны, отличным
от сакмарско-раннеартинского. Самостоятельность и широкое географическое распро-
странение ассельского комплекса подтверждается тем, что характерный ассельский
комплекс выделяется на больших пространствах Бореальной области (Устрицкий, 1967а).
Интересно, что большинство ассельских родов брахиопод (около 70) являлось общи-
ми для Бореальной и Тропической областей.
Палеогеографическая обстановка ассельского века (рис. 3) в значительной мере
была унаследована от позднекаменноугольной эпохи, на что указывает тесная взаи-
мосвязь ассельского и позднекаменноугольного комплексов брахиопод. В ассельское
время связь Бореального и Тропического бассейнов осуществлялась через два про-
лива; .один проходил по Предуральскому прогибу, второй находился в восточной части
Аляски, причем первый играл более существенную роль. На протяжении ассельского
55
Рис. 3. Палеогеографическая схема Альпийско-Гималайской подобласти Тетиса
в ассельском веке
1 - море; 2 - суша; 3 - брахиоподы; 4 - аммоноидеи; 5 - фузулиниды; 6 - гра-
ница моря и суши
века и позднего карбона, видимо, существовали также непосредственные морские
связи между Альпийско-Гималайской подобластью и Североамериканской.
Альпийско-Гималайская подобласть в асеельское время характеризовалась наличи-
ем обширных морских связей, что обеспечивало свободную миграцию фаун. Об этом
свидетельствует широкое расселение ассельского комплекса брахиопод и швагерин.
В пределах Актюбинской области в ассельское время обособляется Южно-Ураль-
ское море (Степанов, 1951). Здесь встречаются Teguliferina и Keyserlingina, ко-
торые, как предполагает Т.Г. Сарычева (1964), являлись предковыми соответственно
для надсемейств Richthofeniacea и Lytton iacea. Впоследствии представители этих
надсемейств приобретают широкое распространение в пермских морях Альпийско-Ги-
малайской подобласти и отсутствуют в Бореальной области. В своем развитии они
пошли прямо противоположными путями: у рихтгофениид оказался редуцированным
лофофор, но гипертрофирована мускулатура, а у литтониид, наоборот, мускулатура
редуцирована, а лофофор сильно развит ( Rud wick, 1961; Сарычева, 1964). В Южно-
уральской провинции присутствуют также роды Scacchinella и Urushtenia, получив-
шие свое дальнейшее развитие в морях Альпийско-Гималайской подобласти. Разде-
ление ассельских бассейнов Альпийско-Гималайской подобласти на палеобиогеографи-
56
сакмарском веке - первой половине артинского. Усл. обознач. см. рис. 3
ческие провинции нами не производится. Можно отметить, что четкое обособление про-
винций, которое наблюдается в более позднее пермское время,в ассельском веке толь-
ко намечается.
В послеассельское время в Альпийско-Гималайской подобласти Тропической об-
ласти выделяется три основных этапа в развитии брахиоподовой фауны, отвечающие
существенным палеогеографическим особенностям этого времени:
1) сакмарско-раннеартинский; 2) дарвазский и 3) мургабско-джульфинский.
Сакмарско-раннеартинский этап
Брахиоподы сакмарско-раннеартинского этапа широко распространены в пределах
Альпийско-Гималайской палеобиогеографической подобласти. Кроме Юго-Восточного
Памира, они известны из нижнепермских отложений Читраля ( Reed, 1925), из долины
гУнц Западного Каракорума (Fantini Sestini, 1966b), из аггломератовых слоев Каш-
ира (Bion, 1928; Reed, 1932), из формации Лузаба Саудовской Аравии (Hudson,
Sudbery, 1959), из пачки ГДГ формации Джейруд Центрального Эльбурса в Северном
Иране (Fantini Sestini, 1966), из сакмарско-артинских отложений Западного Кунь-
57
луня (Устрицкий и др., I960), из серии Сакамотозава Японии ( Nakamura, 1960) и
трокгофельского известняка Карнийских Альп ( Schellwien, 1900b).
Сакмарско-раннеартинские брахиоподы представлены в Альпийско-Гималайской
подобласти приблизительно 70 родами. Из них наиболее обширную группу составляют
роды, начавшие свое существование в позднем карбоне или в ассельское время (а
некоторые даже раньше), распространенные повсеместно в Бореальной и Тропической
областях и вымирающие постепенно на протяжении пермского периода. Это Orthoti-
chia, Streptorhynchus, Meekelta, Wei level la, Terebratuloidea, Stenoscisma, Cancrinella,
Lin oprod uctus, Avonia, Krotovia, N eospirifer, Martinia, Cleiothyridina. Большин-
ство из них на рассматриваемой территории представлено эндемичными видами. Сре-
ди них значительную роль играют крупные диктиоклостиды, которые определялись
в старых работах как Productus semireticulatus и относятся теперь к родам Reti-
culatia и Chaoiella. Это Reticulatia uralica (Lich.), R.pamirica Grunt, sp.nov., Chaoi-
ella grUenewaldti (Krotov), Ch. bathycolpos (Schellw). К этой же группе примы-
кают типично тетасные роды Parenteletes и Scacchinella, расселившиеся повсе-
местно и одновременно в Альпийско-Гималайской и Североамериканской подобластях
Тропической области в течение позднего карбона- ассельского века и продолжавшие
существовать здесь до конца перми.
Вторую группу составляют эндемичные роды, впервые появляющиеся в различных
частях Альпийско-Гималайской подобласти в начале сакмарского века и часто встре-
чающиеся в рифовых фациях в течение всей перми. Это Geyerella, Overt on tin a?,
Compressoproductus, Oldhamina, Richthofenia.
К третьей группе принадлежат роды Taeniothaerus и Punctocyrtella, появившие-
ся в Альпийско-Гималайской подобласти в начале сакмарского века и существовав-
шие здесь в течение очень ограниченного времени (по-видимому, до начала дарваз-
ского века). В Антибореальном бассейне род Taeniothaerus известен до конца позд-
ней перми (Coleman, 1957).
Для сакмарско-раннеартинского комплекса брахиопод характерен более высокий
эндемизм по сравнению с ассельским, что связано с регрессией и разобщением
морских бассейнов. В эго время моря Альпийско-Гималайской подобласти отделяют-
ся от Бореальных в результате возникновения перешейка в районе Каспийского мо-
ря (рис. 4). Моря Урала и Приуралья, потеряв связь с Тропической областью, в
дальнейшем развивались в составе северных бассейнов. По мнению А.Д. Миклухо-
Маклая (1963), связь уральских морей с тропическими прекратилась уже в конце
позднего карбона. Однако, в действительности она, по-видимому, существовала и
в ассельском веке, на что указывает ряд видов фузулинид и брахиопод, общих для
Средней Азии и Урала в это время.
К началу сакмарского века Альпийско-Гималайский бассейн, видимо, замыкается
на западе, нарушаются его связи с Североамериканским бассейном.
Связь между Тропическим и Бореальным бассейнами в сакмарском веке могла
осуществляться только через пролив в районе Восточной Аляски и была крайне
затруднена. Наряду с довольно отчетливо выраженной климатической зональностью
это обусловило дальнейшее независимое развитие бореальной и тропической фаун.
Ослаблению связей Бореального и Тропического бассейнов на востоке способство-
вало возникновение китайско-корейского материка (Лю Хун-юн, 1962). По данным
В.И. Устрицкого (Устрицкий и др., 1960), в ассельское время в Куньлуне из 18
видов брахиопод с Уралом было общих 7, а в сакмарский и артинский века лишь 4
из 27. В сакмарско-нижнеартинских отложениях Юго-Восточного Памира известно
10 видов, из них только один общий с Уралом.
Однако в смене комплексов брахиопод сакмарского века и первой половины артин-
ского в Бореальном бассейне, с одной стороны, и брахиопод Альпийско-Гималайской
подобласти^ с другой, можно видеть. большое сходство. Если обратиться к стратоти-
пическим разрезам Южного Урала, то, по свидетельству Д.Л. Степанова (1951),
г...в отличие от ранее рассмотренных комплексов брахиоподовой фауны Урала, в
составе данного (сакмарского. - Т.Г.) комплекса пока нельзя с уверенностью ука-
зать руководящих форм, свойственных ему одному. Напротив, основной его особен-
ностью является совместное нахождение форм, переходящих из ассельского комплек-
са, и видов, появляющихся здесь впервые и получающих главное развитие в более
молодых отложениях* •
58
Аналогичная картина наблюдается и в Альпийско-Гималайской подобласти, как
это было показано в предыдущей главе для Юго-Восточного Памира и Устрицким
(Устрицкий и др., 1960) для Западного Куньлуня. Поэтому сакмарско-раннеартинс-
кие брахиоподы и рассматриваются в качестве единого комплекса. В начале сак-
марс кого века в Альпийско-Гималайской подобласти начинают дифференцироваться
следующие провинции: (Альпийская (объединяющая Сицилийскую и Карнийскую Миклу-
хо-Маклая) Кавказско-Памирская, Южнокитайская (объединяющая Тиморскую, Северо-
Индийскую и Индо-Синийскую Миклухо-Маклая) и Уссурийская . Эти провинции про-
должали существовать на протяжении всего пермского периода и наиболее резко обо-
собились к концу поздней перми.
Альпийская провинция, существование которой намечается уже в ассельском веке
(Карнийские Альпы), к началу сакмарского века обособляется довольно отчетливо
(трокгофельские известняки Карнийских Альп). Здесь в это время происходило фор-
мирование рифов, среди которых обитали своеобразные рифожители Scacchinella,
Teguliferina, Parenteletes. В сакмарском - начале артинского веков именно здесь
впервые появляются Oldhamina, Geyerella, Sicelia - роды, которые впоследствии
приобрели широкое развитие в Альпийско-Гималайской подобласти. Здесь же встре-
чено большое количество своеобразных видов, принадлежащих родам Enteletes, Ros-
tranterrs, Meekella. Довольно четко по составу брахиопод в сакмарско-раннеар-
тинское время обособляется Кавказско-Памирская провинция. К ней в это время
относились районы Читраля, Кашмира, Куньлуня, Памира, Саудовской Аравии, Север-
ного Ирана. Для Кавказско-Памирской провинции характерны роды Taeniothaerus и
Punctocyrtella, существовавшие здесь в течение очень ограниченного времени: сак-
марского века и первой половины артинского. Здесь же присутствуют своеобразные
виды, относящиеся к родам Reticulatia, Chaoiella, Neospirifer, Cleiothyridina .
Брахиоподы сакмарского века и первой половины артинского в Южно-Китайской и
Уссурийской провинциях известны крайне недостаточно, и названные провинции по
брахиоподам выделены быть не могут. На существование Уссурийской провинции это-
го времени указывает А.Д. Миклухо-Маклай (1963), выделивший ее по фузулинидам.
Дарвазский этап
Брахиоподы, сопоставимые с кызылджилгинским комплексом Юго-Восточного Па-
мира, известны из Пакистана (нижний и средний продуктусовый известняк Соляного
кряжа - Waagen, 1884; Reed, 1931b, 1944), Кашмира (слои Зеван - Diener, 1899,
1915), продуктусовых сланцев или сланцев Кулинг, прослеживающихся из района Спи-
та (Пенджаб) в Северные Гималаи К ума она (Diener, 1897b, 1899, 1903), Непала
(Waterhouse, 1966), Южного Тибета (Ting Pei-Chen, 1962), Северного Сиккима (се-
рия Дачи - Muir-Wood, Oakley, 1941), из области Шан Восточной Бирмы (Diener,
1911), местонахождения Та-ли Шао провинции Юньнань (Reed, 1927) и формации
Чися (J.Chao, 1927, 1928; Ching Yu—К an ,1963) Южного Китая, района Усу-Хонгор
Внутренней Монголии ( Grabau, 1931; Nonaka, 1944), чандалазской и нахбдкинско-
калузинской свит Южного Приморья (Фредерикс, 1925; Лихарев, 1960а, 1964; Кот-
ляр, 1961, 1964; Лихарев, Котляр, I960), из нижней части серии Канокура хребта
Китаками Японии (Hayasaka, 1963, 1967) и, предположительно, формации Доруд Се-
верного Ирана (Fantini Sestini, 1965 d ). Брахиоподы сафетдаронской свиты Юго-
Западного Дарваза изучались Ф.Н? Чернышевым (1914) и Б.К. Лихаревым (Власов,
Лихарев, Миклухо-Маклай, 1962) •
Роды брахиопод, наиболее характерные для провинций Альпийско-Гималайской подоб-
ласти, приведены в табл. 7.
^Комплекс сафетд а ронских брахиопод, как и брахиоподы других свит Юго-Западного
Дарваза, в настоящее время изучаются автором. Можно заметить, что сафетда-
ронские брахиоподы отличаются от других комплексов, относимых нами к дарваз-
скому ярусу значительным своеобразием.
59
Рис. 5. Палеогеографическая схема Альпийско-Гималайской подобласти Тетиса в
дарвазском веке. Усл. обознач. см. рис. 3
Особенностью рассматриваемого этапа является смешение брахиоподовых фаун
Бореальной и Тропической областей, происшедшее в результате обширной трансгрес-
сии на востоке (Синицин, 1962; Устрицкий, 19676). В результате этой трансгрессии
восстановились широкие морские связи Тропического и Бореального бассейнов. К
этому времени наряду с проливом, существовавшим в районе Восточной Аляски,
возникает новый пролив, соединивший Колымский бассейн с Южнокитайским, по-ви-
димому, через районы Приморья и Южной Монголии . Этот пролив ограничен с юга
В.А. Бобров и М.В. Куликов (1968) предполагают наличие в»пермское время также
Монголо-Забайкальского пролива, связывавшего Забайкальский бассейн с Южно-
китайским через Центральную Монголию. Этот вывод, представляющийся недоста-
точно убедительным, сделан на том основании, что в пермских отложениях Цент-
ральной Монголии встречен ряд видов, общих с Усу-Хонгором (т.е . уже с тропичес-
кой фауной). Но в перечисленном авторами списке большинство форм является вида-
ми широкого географического распространения. Кроме того, чрезвычайно интересная
и разнообразная пермская брахиоподовая фауна Монголии до сих пор полностью
монографически не обработана, что крайне затрудняет выявление палеогеографичес-
ких связей.
60
Таблица 7
Фаунистическая характеристика провинций Альпийско-Гималайской подобласти
тропической биогеографической области пермского периода
(преимущественно по брахиоподам)
Область Тропическая область - семейства Lyttoniidae, Richthofeniidae, Chonostegidae
Подобласть Альпийско-Гималайская подобласть — семейства Verbeekinidae, Tschemvschewiidae, Chonetellidae, Spirigerellinae
Провинция Альпийская Кавказско-Па- мирская Южно-Китайская Уссурийская
Джульфинский век T schemyschewia (много) Araxilevis , Araxathyris ? Comelicania, Janiceps Enteletella* Strophalosiina^ Scacchinella, Rich- thofen ia5 P ectenoproductuSy Camarophorinella, Labaia, N otothyri- na Преобладают продук тиды; редко атирид] ды> ринхонеллиды, те ре бра т у ли д ы - Состав брахиопод и- близок к Южно- китайскому
Мургабский век Gemmellarpia* Chonostegoides, Spirisosium * Septospirigerella^ Girlasia V ediproductus своеобразные ви- ды Scacchinella, Gey er ell a, Richtho- fenia Hamletella Paralyttonia Camarophoriina Asperulus , Poikilosakos Смешение тропических и бореальных родов
)вазекий век Брахиоподы не известны Enteletina, 0 vertoniina, Botoria, Costalosia , Costiferina, M arginifera, Spirigerella, Мигранты из Ан- тибореальной об- ласти: Нoskingia,i Jerrakea Whytspakia Geyerella1 Derbyella у М ongolina^ Н eterelasmina, Prorichthofenia^ Leptodus
Я Мигранты из Бореальной области: Anidanthus, Me gon si a, J akovlevia, Rhyn- chopora, Spiriferella
Сакмарский - первая половина S ф Parenteletes, Geye- Тaeniothaerus, Neo- По брахиоподам провинции не обособ- о rella Sicelia, Sea- spirifer^Chaoiella, ляются. 0 chinella, Richthofe- Punctocyrtella^Reti- Своеобразные шва- § nia, Oldhamina culatia герины, особенно g Acervoschwagerina Q. CO
| Ассель-| [с кий век Провинции обособляются слабо
древней Китайско-Корейской сущей, а с запада - Таримской. Трансгрессия захваты-
вает центральную и восточную части Альпийско-Гималайской подобласти (Японию ,
Монголию, Приморье, Гималаи, Южный Тибет, Соляной кряж, достигает Памира и Аф-
ганистана, оказывая существенное влияние на обйшй характер фаун брахиопод Уссу-
рийской, Южно-Китайской и восточной части Кавказско-Памирской провинции (рис.5).
В с пота в комплекса брахиопод в это время вошло значительное количество форм,
мигрирппАrtttwy сюда из соседних с Тетисом областей* Из бореального бассейна в
61
восточные районы Альпийско-Гималайской подобласти проникают Anidanthus, Megou -
sia, Jakovlevia, Rhynchopora, Spiriferella. Род Spiriferella в течение сакмарско-
раннеартинского этапа был представлен в Альпийско-Гималайской подобласти единст-
венным видом (Spiriferella ? personate Reed). В течение дарвазского этапа Spirife -
rella получила широкое распространение. В это время в Альпийско-Гималайской
подобласти насчитывается девять видов этого рода. Ряд родов: Whytspakia, Hoskin-
gia, Terrake а мигрирует сюда из Антибореального бассейна.
Из Американского бассейна сюда, по-видимому, переходят Leptodus и Proricht -
hofenia, которые вначале вселились в Уссурийскую провинцию, а в более позднее
время распространились по всей Альпийско-Гималайской подобласти. Таким образом,
состав фауны в значительной мере определяется брахиоподами, мигрировавшими сю-
да из других областей. Но, если американские роды продолжают развиваться в Аль-
пийско-Гималайской подобласти до конца перми, то бореальные и антибореальные
вымирают очень быстро. Одновременно появляется ряд эндемичных родов: Enteleti-
па, Derby ella, Сardinocrania, Costalosia, Costiferina, Spirigerella, Boloria, Heterelas -
лила. Продолжает развиваться группа древних родов, существующих начиная с позд-
него карбона, упоминавшихся при рассмотрении предыдущего этапа и представленных
в ряде случаев особыми видами.
Западные границы дарвазского бассейна неясны. Брахиподы этого возрастного
интервала в Альпийской провинции пока не обнаружены. В то же время представи-
тели дарвазских фузулинид распространены в западном направлении вплоть до Ту-
ниса (Миклухо-Маклай,. 1963; Левен, 1967). В.И. Устрицкий (1961) предполагает су-
ществование в рассматриваемое время перешейка, соединявшего Гондвану и Ангари-
ду в районе Памира, Каракорума и Восточного Афганистана. Однако в названных
районах мы обнаруживаем комплексы брахиопод, характеризующиеся присутствием
ряда бореальных родов, что не подтверждает указанное мнение В.И. Устрицкого.
В дар вазе ком веке отчетливо выделяется Уссурийская провинция, которая, по
данным А.Д. Миклухо-Маклая (1963),охватывает районы Советского Дальнего Вос-
тока, Южной и Юго-Восточной Монголии, Северо-Восточного Китая, Японии. Бра-
хиоподы Уссурийской провинции довольно своеобразны. Они широко представлены
в Южном Приморье и в Южной Монголии. Отмечается значительное сходство брахио-
подовых фаун этих двух районов. Присутствует около трети общих видов (Лихарев,
1960а). Основной особенностью провинции является смешение северных и южных
элементов. В Южном Приморье брахиоподы приурочены к отложениям находкинско-
калузинской свиты, представленной рифогенными известняками и чандалазской вул-
каногенно—терригенной свитой. Рифогенные отложения содержат преимущественно
южных брахиопод, представленных такими родами, как Enteletes, Streptorhynchus,
Phricodothyris, Prorichthofenia, Leptodus. Вулканогенно-терригенные отложения со-
держат Jakovlevia, Rhynchopora, Pterospirifer, Spiriferella и другие роды, типич-
ные для северных областей. В Южной и Юго-Восточной Монголии в это время фор-
мировались рифы, в которых наряду с северными формами встречается большое ко-
личество южных, а также эндемичных родов - Derbyella, Morrisina, Mongolina, He-
terelasmina. Сюда же перемещается род Geyerella- из Альпийской провинции. Бра-
хюподовая фауна нижней части формации Канокура хребта Китаками (Япония) также
обнаруживает некоторую провинциальную общность с Южным Приморьем.
Уссурийская провинция отчетливо выделяется и по мшанкам. На ее существование
указывал В.П. Нехорошев (1957), основывавший свое мнение на своеобразии родо-
вого и видового состава известных из этих районов мшанок, и назвавший ее При-
морской. И.П. Морозова (1970) также подтверждает наличие отчетливо обособленной
Уссурийской провинции в поздней перми и высказывает предположение о том, что
она, возможно, сформировалась уже в ранней, перми или даже в карбоне.
Трансгрессия, захватившая восточную часть Кавказско-Памирской и Юкно-Ки-
тайскую провинции, обусловила обширные связи их между собой и слабую степень эн-
демизма бракиоподовой фауны.
Из этих двух провинций известны следующие местонахождения брахиопод: бо-
гатый комплекс брахиопод нижнего и среднего продуктусового известняка Соляного
кряжа (Пакистан), брахиоподы слоев Зеван Кашмира, продуктусовых сланцев или
сланцев Кулинг Кумаона и Гюрхваля, Южного Тибета, Непала, Северного Сиккима,
области Шан Восточной Бирмы, местонахождения Та-^ли Шао провинции Юньнань и
формации Чися Южного Китая. К ним условно можно отнести и комплекс брахипод
62
формации Доруд Северного Ирана. Остановимся прежде всего на рассмотрении группы
местонахождений пермских брахиопод так называемых Гималаев Тетиса (Гансер,
1967). Это слои Зеван (Diener, 1809, 1915), сланцы Кулинг (Diener, 1879bf1899,
1903), пермские отложения Непала (Waterhouse, 1966), Южного Тибета (Ting Pei-
Chen, 1962) и серия Лачи Северного Сиккима (Muir-Wood, Oakley, 1941). Следует
отметить, что во всех перечисленных районах Гималаев Тетиса рассматриваемые от-
ложения залегают трансгрессивно на среднепалеозойских или более древних отложе-
ниях. Лишь в Кашмире они перекрывают слои с флорой G angamop teris, залегающие
на пермских траппах. Траппы, в свою очередь, подстилаются аггломератовыми слоя-
ми. Присутствие в продуктусовых сланцах аммоноидей, относящихся к родам Kraf-
ftoceras, Cyclolobus, Xenaspis , свидетельствует о принадлежности содержащих их
отложений к джульфинскому ярусу (Руженцев, 1965). Однако, если аммоноидеи приу-
рочены к верхней части продуктусовых сланцев, то, по указанию М.Кришнана (1954),
брахиоподы встречаются в основном в их нижней части. Все перечисленные комп-
лексы брахиопод очень близки между собой. Они характеризуются присутствием рго-
bolionia him alayen sis (Diener), Fusispirifer nitiensis (Diener), Spiriferella rajah (Salter),
Syringothyris lydekkeri (Diener).
Все эти комплексы относятся к единому стратиграфическому уровню, который со-
поставляется большинством авторов с средним продуктусовым известняком Соляного
кряжа, и, что менее вероятно, с верхним продуктусовым известняком.
Богатейшие коллекции брахиопод из продуктусовых известняков Соляного кряжа
были описаны Ваагеном (Waagen, 1884) и Ридом (Reed, 1931b, 1944). Вааген раз-
делил отложения с морской фауной на три части, названные им нижним, средним и
верхним продуктусовым известняком. Нижний продуктусовый известняк (формация
Амб по современной терминологии) залегает на преимущественно пресноводно-кон-
тинентальных отложениях группы Нилаван, относимой к нижней части пермской сис-
темы по палинологическим данным и присутствию горизонта с бедной морской фау-
ной двустворок и конулярий (Kummel, Teichert, 1970). Накопление осадков нижнего
продуктусового известняка, по-видимому, соответствует началу трансгрессии, совпа-
дающей по времени с трансгрессией в районе Юго-Восточного Памира. Это находит
подтверждение в присутствии в нижнем проду^тусовом известняке Parafusulina kat-
taensis Schwager, которая характерна для дарвазского яруса (Миклухо-Маклай, 1963).
На Юго-Восточном Памире Parafusulina ex gr. kattaensis Schwager присутствует в
верхнекызылджилгинском подгйризонте (Левен, 1967). В нижнем продуктусовом из-
вестняке впервые появляются такие роды брахиопод, как Costalosia, Edriosteres,
Costiferina, Marginifera, Notothyria^ получающие широкое распространение в выше-
лежащих отложениях среднего и верхнего продуктусового известняка, а также в от-
ложениях дарвазского и мургабского ярусов других районов Альпийско-Гималайской
подобласти. Из родов, характерных для более древних - нижнепермских отложений
здесь встречены представители Taeniothaerus и Neospirifer.
Если сопоставление формации Амб с кызылджилгинским горизонтом Юго-Восточ-
ного Памира* верно, то средний продуктусовый известняк (формация Варгаль), исклю-
чая его верхнюю часть (пачка* Калабах), должен бытьк сопоставлен с куберган-
динской свитой, т.е. брахиоподы среднего продуктусового известняка и его аналогов
могут быть отнесены ко второй половине дарвазского яруса. Отметим, что в верхней
части формации Амб встречены слои с флорой Glossopteris и Gangamopteris (Kummel,
Teichert, 1970), выше которых залегает средний продуктусовый известняк, что подт-
верждает двучленное строение отложений дарвазского яруса (см. главу II ). К со-
жалению, непосредственное сопоставление среднего продуктусового известняка и его
аналогов и кубергандинской свиты невозможно из-за отсутствия в первых - фузулинид,
а во второй — брахиопод.
Южно-Китайская и восточная часть Кавказско-Памирской провинций характери-
зуется проникновением сюда бореальных родов Spiriferella, Rhynchopora, Anidanthus,
Megousia. Наряду с этим можно отметить присутствие австралийских и новозе-
ландских родов - Whytspakia, Hoskingia, Terrakea. Эти роды представлены единич-
ными видами и существовали здесь в течение очень ограниченного времени - до
начала мургабского века. Основную же массу составляют роды, существовавшие с
начала ассельского века. Появляется большое количество эндемичных родов. Это
CQstalosia, Costiferina, Spirigerella, Enteletina.
63
Несколько обособленной в течение дарвазского века являлась памирская часть
Кавказско-Памирской провинции. Состав брахиоподового сообщества (кызылджилгин-
ского) чрезвычайно своеобразен, на что указывает присутствие родов Overtoniina,
Boloria, Labaia и ряда эндемичных видов. Кызылджилгинский компле.кс брахиопод
наиболее близко сопоставляется с брахиоподами нижней части верхней перми (см.
главу III). Однако аммоноидей,встреченные совместно с кызылджилгинскими бра-
хиоподами, всеми исследователями единодушно сопоставляются с байгенджинским
комплексом (Руженцев, 1955), характеризующим верхнюю часть артинского яруса
и, вероятно, кунгур.
Началу 'дарвазского* этапа развития брахиопод Тетиса в Бореальной области при-
мерно соответствует нижнепайхойское время. На границе пайхойского и артинского
ярусов происходит существенное обновление брахиоподовой фауны. В.И. Устрицкий
(1971) сопоставляет пайхойский ярус с уфимской свитой и относит его к верхней
перми.
Этап развития брахиоподовой фауны Арктики, соответствующий пайхойскому яру-
£?у, был впервые охарактеризован Д.Л. Степановым (1957), выделившим на основе
разрезов Шпицбергена самостоятельный свальбардский ярус, соответствующий кунгур-
скому и уфимскому ярусам. Сходное стратиграфическое подразделение предложила
В.П. Бархатова (1964, 1970, 1971), отметившая, что оно может рассматриваться в
составе верхнего отдела перми или в качестве среднего отдела. Эти авторы со-
поставляют выделенные ими ярусы с кунгурскими и уфимскими отложениями вместе
взятыми. В.П. Бархатова считает возможным также, что нижней части воркутского
яруса соответствует самая верхняя часть артинского яруса Западного Урала.
Таким образом, в Бореальной области выделяется крупный доказанский этап
развития брахиоподовых фаун.
По новым данным Е.А. Гусевой и В.П. Горского (1969), которые провели сопос-
тавление по комплексам остракод нижней части пайхойских отложений Печорского
бассейна с горизонтами перми Среднего Приуралья, основание пайхойского яруса
соответствует саранинскому или верхней части саргинского горизонта (верхиеар-
тинский подъярус). Другими словами, время появления новой фауны брахиопод в
Бореальной и Тропической областях близко совпадает. По данным Устрицкого (1971),
смена артинского комплекса брахиопод пайхойским происходит внутри слоев с бай-
генджинскими аммоноидеями. Более молодые формы аммоноидей (Spirolegocerast
Cyqlolobus, Eumedlicottia), относимые обычно к верхней перми, появляются в
верхнепайхойском подъярусе.
Рассмотренный этап развития брахиоподовых фаун принят нами в качестве 'сред-
непермского', как примерно равнозначный верхнепермскому мургабско-джульфинско-
му этапу. Как уже было сказано, дар вазе кие брахиоподы во многом близки к верх-
непермским и, возможно, что более правильно относить время их существования к
поздней перми. Недостаточная изученность 'среднепермского* этапа развития бра-
хиоподовых фаун Тропической области пока не позволяет разделять его более дроб-
но. Первая половина этапа охарактеризована здесь пока только кыз ылджилгинс ким
комплексом брахиопод Юго-Восточного Памира и, возможно, комплексом ниинего
продуктусового известняка Соляного кряжа. Все остальные, относящиеся к *сред-
непермскому* эт^ду комплексы брахиопод характеризуют его верхнюю часть. Соот-
ношения верхней границы 'среднепермского* этапа развития брахиопод с зонами
фузулинидовой шкалы не вполне ясны,(в частности вопрос о принадлежности к не-
му времени "N eoschwagerina simplex*', ) (Можно только отметить, что наиболее древ-
ние фузулиниды, встречающиеся совместно с мургабскими брахиоподами в Сици-
лии, по-видимому, относятся к подзоне N eoschwagerina schuberti (Левен, 1967).
Мургабско-джульфинский этап
Третий этап охватывает мургабский и джульфинский века. Он характеризуется об-
щей регрессией морских бассейнов, захватившей одновременно Бореальную и Тро-
пическую области. В начале мургабского века вновь закрываются проливы в районе
Аляски и ОхотскбЬо моря. Бореальные воды перестают проникать в Тетис, и роды»
мигрировавшие сюда с севера, вымирав^т в течение короткого времени. Наступает
64
общее обмеление и сокращение бассейнов, которое идет постепенно и достигает
своего максимума в конце джульфинского века. Брахиоподы мургабского и джуль-
финского веков тесно связаны между собой и представляют единый этап в общем
ходе их развития. В то же время следует обратить внимание на ряд специфичес-
ких черт, характеризующих мургабский и джульфинский века. Рассмотрим каждый
из них в отдельности.
Мургабский век. Брахиоподы мургабского яруса на Юго-Восточном Памире
практически неизвестны, однако при анализе данных по другим районам, в част-
ности, Закавказью, где мургабские брахиоподы изучались автором, представляется
возможным рассматривать комплекс брахиопод мургабского яруса как четко обо-
собленный от дарвазского и от джульфинского. Брахиоподы мургабского яруса из-
вестны из местонахождения Созио Сицилии (Gemmellaro, 1891а,Ь, 1892, 1894а,Ь,с,
1897, 1899; Meria, 1928; Greco, 1938, 1942, 1943; Di-Stefano, 1914; Parana, 1933,
Fabiani, 1933; Rudwick, Cowen ,1968), из гнишикского и хачикского горизонтов Ар-
мении и Нахичеванской АССР (Сарычева, Сокольская, Грунт, 1965), из формации
Руте Северного Ирана (Fantini, Sestini, 1965с), из Южного Ирана и Иранского Бе-
луджистана (Douglas, 1936, 1950), из формации Чидру и пачки Калабах Соляного
кряжа (Waagen, 1884; Reed, 1931b, 1944; Grant, 1970), из местонахождения Риму
Каракорума (Meria, 1934), местонахождения Читичун 1 Гималаев (Diener, 1897а,
1903), из местонахождения Базлео Тимора (Beyrich , 1965; Broili, 1916; Hamlet,
1928; Wanner, Silverts, 1935; Rothpletz, 1892). Ротплетцем описаны также брахио-
поды с о-ва Роти. Брахиоподы, относящиеся к мургабскому ярусу, происходят так-
же из Таиланда (Yanagida, 1964; Waterhouse, Piyasin, 1970) и из Северной Кам-
боджи (Tran-Thi Chi-Thuan, 1961).Брахиоподы Камбоджи, а также Лаоса известны
по работе Манси (Mansuy, 1913)., По-видимому, из описанных ста с лишним видов
брахиопод часть относится к ассельскому (?) ярусу, другая же - характеризует
верхнепермские, по-видимому, мургабские отложения. К сожалению, при обработке
Манси не разделил брахиопод на соответствующие комплексы. К мургабскому ярусу
относятся брахиоподы свиты Маокоу Центрального Китая (Ching Yu-kan, Zhang-Yan,
1961; Чжань Ли-пэй и Ли-*Ли, 1962).
Для мургабского века характерен все возрастающий эндемизм различных групп
фауны, в том числе и брахиопод. В это время шел процесс постепенного вымирания
многих родов: Reticulatia, Kochiproductus, Echinoconchus, существовавших здесь на,
протяжении длительного времени; почти полностью исчезли арктические и австра-
лийские мигранты - Whitspakia, Rhynchopora, Spiriferella, многие представители
которых вымерли еще -на протяжении дарвазского века. Одновременно происходило
сокращение количества видов и численности популяций для многих родов, продол-
жавших существовать до конца перми. В то же время ряд родов, известных ранее,
представлены в мургабеком веке весьма своеобразными видами - Entele -
tes tschemyschewi Gemm., E. obsoletus Gemm., E. subaequivalvis Gemm., E.haugi Gemm.,
E. waageni Gemm., Geyerella rex Gemm., Scacchinella depressa Gemm.
Появился ряд новых, не встречавшихся ранее родов: Hamletella, Paralyttonia,
Asperulus, Camarophoriina, Poikilosakos - в Южн о-Китайской провинции; Septospi -
rigerella, Vediproductus, Chonostegoides - в Кавказско-Памирской. А.Д. Миклухо-
Маклай (1963) для этого времени в пределах рассматриваемой территории выделил
Крымско-Памирскую и Уссурийско-Малайскую подобласти (см. табл. 6). В Уссу-
рийско-Малайскую подобласть им включены Орегонская, Корякская, Уссурийская,
Индосинийская и Тиморская провинции. Орегонская и Корякская провинции, как было
показано выше, рассматриваются нами в качестве самостоятельной подобласти. В
Уссурийской провинции мургабского века брахиоподы практически неизвестны. Для
нее, по данным А.Д Миклухо-Маклая (1963), характерны своеобразные Ozawai-
nellidae (Rauserella, Reichelina, Parareichelina ), а также малое распространение
или полное отсутствие Sumatrininae.
В начале мургабского века происходит отчетливое обособление Южно-Китайской
и Кавказско-Памирской провинций. Сообщение между ними осуществлялось в это
вРемя лишь через узкий пролив, ограниченный с одной стороны материком Гонд-
^ы, с другой - горной цепью древнего Куньлуня (рис. 6). Чжан Ли-пэй и Ли-Ли
(1962) отмечают, что Южно-Китайская провинция носит отчетливо изолированный
характер, видимо, благодаря наличию древнего Китайско-Корейского материка, и
Оделяется от Уссурийской. Южно-Китайская провинция охватывает районы о-вов
264
5
65
Рис. 6. Палеогеографическая схема Альпийско-Гималайской подобласти Тетиса
в мургабском веке. Усл. обознач. см. рис. 3
Тимор и Роти, Камбоджи, Лаоса, Таиланда, Центрального Китая. Для Южно-Ки-
тайской провинции характерно наличие эндемичных родов брахиопод - Paralyttonia,
Camarophoriina, Hamletella, Asperulus, а также большого количества эндемичных видов.
По данным Миклухо-Маклая (1963), Южно-Китайская провинция характеризовалась
широким расселением представителей надсемейства Verbeekinacea.
В состав Кавказско-Памирской провинции в течение мургабского века продолжают
входить районы Памира, Каракорума, Соляного кряжа, Гималаев, Южного Афганистана,
Северного Ирана, Закавказья, Крыма. В Закавказье к мургабскому ярусу нами от-
несены брахиоподы гнишикского и хачикского горизонтов (Сарычева, Сокольская,
Грунт, 1965), рассматривавшиеся ранее указанными авторами как Гваделупекие Я В
Соляном кряже к мургабскому ярусу нами вслед за Грантом ( Grant, 1970) отнесен
Тиморские местонахождения рассматриваются нами в отличие от Миклухо-Маклая
(1963) в составе Южно-Китайской провинции, так как выделение Тиморской провин-
ции'данными по брахиоподам не подтверждается.
2
Как уже упоминалось выше (см. главу III), выделение гваделупского яруса в разре-
зах Альпийско-Гималайской подобласти, не содержащих аммоноидей, весьма затрудии-
66
комплекс брахиопод, известный из формации Чидру и пачки Калабах. Грант отмечает^
тесную связь брахиопод пачки Калабах (верхней части среднего продуктусового из-
вестняка) и формации Чидру (нижней часта верхнего продуктусового известняка^).
Для формации Чидру и пачки Калабах общими являются: t Oldhamina, Waagenoconcha,
Stenoscisma, Kiangsiella.
В формации Чидру Грантом указываются представители следующих брахиопод:
Aulosteges sp.tCallispirina sp., Chonetella sp., Cleiothyridina cf. capillata (Waagen),
Derbyia cf. plicatella(Vtaageh)9Enteletes sp., Hemiptychina sp., Hustedia sp., Linopro-
ductus sp., Lyttonia sp.9 Martinia ? sp., N eo spirifer sp., О rthoti chi a sp., Richthofenia
sp., Spiriferella (?) sp., Spirigerella sp., Whitspakia sp.
По мнению Гранта, вышеназванные отложения относятся к Гваделупе кому яру-
су, причем не к самой его верхней части. Грант сопоставляет фауну брахиопод
формации Чидру и пачки Калабах с брахиоподами гнишикского и хачикского гори-
зонтов Закавказья, а также с брахиоподами формации Руте Ирана, устанавливая
целый ряд родов, общих для всех трех рассматриваемых комплексов: Orthotichia,
Neochonetes, Marginifera, Linoproductus, Spiriferellina, Septospirigerella, Dielasma
(Grant, 1970, стр, 129, табл. 2).
Иной точки зрения относительно возраста формации Чидру придерживается В.Е.Ру-
женцев (1965), а также Каммел и Тайхерт (Kummel, Teichert,1970)« На основании
находок в отложениях формации Чидру аммоноидей, принадлежащих к родам Eumedli-
cottia, Episageceras, Stacheoceras, Cyclolobus, Xenodiscus И Xenaspis, они относят
формацию Чидру к лжульфинскому ярусу. Однако, по мнению Гранта (Grant, 1970),
распространение рода Cyclolobus в разрезах Соляного кряжа и близких к нему не
ограничивается верхним продуктусовым известняком, а является значительно более
широким. Остальные же роды, встреченные в формации Чидру, указывают лишь на
позднепермский возраст вмещающих отложений.
Кавказско-Памирская провинция в мургабеком веке характеризуется присутстви-
ем эндемичных родов брахиопод Spirigerella, Septospirigerella, Chonostegoides, Vedip-
roductus и других. Районы Соляного кряжа и Гималаев (Читичун Обвязывают Кавказ-
ско-Памирскую и Южно-Китайскую провинции. В разрезах этих районов присутствуют
брахиоподы,характерные для обеих провинций.Для Южно-Китайской провинции характерны
такие виды брахиопод, как Orthotichia (Orthis in di с a Waag.), Derbyia regularis Waag.,W aag en-
ocon ch a (Productus abichi Waag.) ,A/exenia? (Productus gratiosus Waag.) ,M argin ifera typica
Waag., Haydenella kiangsiensis Kays, Uncinunellina timorensis Beyr., для Кавказско-
Памирской- роды, перечисленные выше.
К Альпийской провинции относится район Сицилии, где в это время происходило
формирование рифов и наблюдался пышный расцвет многих типично тетисных родов:
Geyerella, Scacchinella, Leptodus, Richthofenia, Enteletes, Rostranteris, Heterelasmi-
na9представленных своеобразными видами. Наряду с ними появились новые роды:
Gemmellaroia, Girlasia.
Джульфинский век является завершающим в развитии, позднепалеозойских
морей и населявшей их в течение длительного времени разнообразной фауны. В это
Продолжение сноски 2
гельно. Брахиоподы гнишикского и ха чине к ого горизонтов, по принятой авторами
шкале являются более молодыми, чем дарвазские, и должны быть отнесены к мур-
габскому ярусу. Это заключение подтверждается новыми данными по фузулинидам
(Левен, 1972).
3
Верхняя часть верхнего продуктусового известняка (формация Катвай) относится
к нижнему отделу триасовой системы( Kummel, Teichert, 1970).
4
Комплекс брахиопод местонахождения Читачун 1 Гималаев (Кашмир) обычно рас-
сматривается как джульфинский на основании находок на вершине Читичун 1 аммоцси-
Дей, представленных родами Stacheoceras, Crafftoceras, Xenaspis (Руженцев, Сары-
чева, Шевьхрев в кн. 'Развитие и смена..., 1965). Однако брахиоподы, известные из
этого местонахождения и находящиеся в неясных стратиграфических взаимоотношениях
с аммоноидеями, характеризуются существенной близостью к мургабским брахиопо-
дам, особенно к брахиоподам верхнего продуктусового известняка.
67
Рис. 7. Палеогеографическая схема Альпийско-Гималайской подобласти Тетиса
в джульфинском веке. Усл. обознач. см. рис. 3
время общая регрессия бассейна достигла своего максимума. Почти полностью ис-
чезли моря Бореального бассейна, перестал существовать Американский позднепалео-
зойский бассейн. Морской режим сохранился лишь на ограниченных пространствах
Альпийско-Гималайской подобласти (рис. 7), но и здесь идет резкое обмеление.
Этот процесс сопровождался уменьшением интенсивности обмена фаун и возрастани-
ем степени ее эндемизма.
Джульфинский век характеризуется также повсеместным массовым вымиранием
морских организмов, принадлежащих к самым разнообразным систематическим груп-
пам. Это век доживания фузулинид, причем высшие фузулиниды в основном выми-
рают уже к концу мургабского века, а для джульфинского характерны мелкие аббер-
рантные формы, представленные родами Codonofusiella, Reichelina, Palaoofusulina.
Джульфинские аммоноидей, по данным В.Е. Руженцева и А.А. Шевырева (в кн.
'Развитие и смена...', 1965), содержат около 20 родов, представляющих собой очень
своеобразный комплекс. Для него весьма типичен последний род семейства Рага -
gastrioceratidae - Pseudogastrioceras, последние представители циклолобид: Kraf -
ftoceras и Cyclolobus. Широко развиты представители отряда цератитов, особенно
семейство Araxoceratidae, давшее впоследствии начало развитию триасовых Otocera-
tidae.
68
В джульфинском веке заканчивается развитие гониатитов, четырехлучевых корал-
лов, сменившихся в мезозое шестилучевыми, и табулят, в процессе вымирания вер-
нувшихся к древнему аулопороидному типу строения. Полностью исчезают трилоби-
ты, а также некоторые цистоидеи, бластоидеи и криноидеи.
По данным Миклухо-Маклая (1955), Средиземноморской области этого времени
свойственно широкое распространение рифовых построек, образованных водорослями,
губками, кораллами (табулятами и ругозами), и теплолюбивая морская фауна. В не-
которых местах встречаются массовые скопления брахиопод и гастропод.
Кроме Юго-Восточного Памира, брахиоподы известны из никитинского и уруштен-
ского горизонтов Северного Кавказа (Лихарев, 1925, 1926, 1928, 1931а,б, 1932а,б,
1936, 1937, 1939, 1956; Licharew, 1930; 1935), с Юго-Западного Дарваза (Лихарев,
1939; Миклухо-Маклай, 1963), из Закавказья ( Abich,1878; Moller, 1879; Arthaber,
1900; Стоянов, 1910, 1916; Сарычева, Сокольская, Грунт, 1965), из формации Незен
Северного Ирана (Fantini Sestini, Claus, 1966; Stepanov et al., 1969), из беллерофо-
нового известняка Южного Тироля (Stache, 1878), Юлийских Альп северо-запада
Югославии ( Kossmat, Diener, 1910), из близких районов Словении ( Ramovs,1958) и
из аналогов беллерофонового известняка в Западной Сербии (Albrecht, 1924; Simic,
1933, 1934а, b, 1935а), из слоев Жажар гор Бюкк Северной Венгрии (Schretter,
1936, 1963), из формации Допинг Южного Китая (Kayser, 1883; Freeh, 1911; J. Chao,
1927, 1928; Huang, 1932, 1933), из средней части группы Майдзуру Японии ( Schimi-
zu, 1961 - 1963) и некоторых местонахождений Камбоджи (Н. Termier et G. Тег-
mier, 1970).
Однако повсюду шло резкое сокращение и обмеление бассейнов. Произошла неко-
торая перестройка палеогеографического плана. Южно-Китайская провинция в джуль-
финском веке была уже, видимо, изолирована от Кавказско-Памирской. В состав
Альпийской провинции вошли районы Северного Ирана и Закавказья, которые раньше
принадлежали Кавказско-Памирской провинции. Альпийская провинция охватывает
территорию Южного Тироля, Западной Сербии, Словении, Северной Венгрии, Юлийских
Альп, Северного Ирана и Закавказья и характеризуется широким расселением неко-
торых продуктид ( Tschemyschewia, Spinomarginifera)9 а также атиридид, которые
здесь представлены родами Araxathyris, Comelicania, Janiceps. Особенно обильны
атиридиды в закавказской части Альпийской провинции - 7 видов, принадлежащих к
роду Araxathyris. В то же время ребристые спирифериды, теребратулиды и ринхонел-
лиды крайне малочисленны.
Кавказско-Памирская провинция отличается развитием большого количества ри-
фов и, в отличие от Альпийской провинции, преимущественным распространением те-
ребратулид и ринхонеллид. Теребратулиды представлены родами Labaia, Dielasma,-
Hemiptychina, Heterelasmina, Notothyris, Notothyrina, Gefonia, a ринхонеллиды над-
семейства Rhynchonellacea - родами Wellerella, Pseudowellerella, Gersesimovia,
Denticuliphoria, Pseudopugnax, Uncinunellina, Terebratuloidea. При этом многие ро-
ды содержат большое количество видов, для них характерны широкие ареалы и об-
ширные популяции.
Кавказско-Памирская провинция в своей западной части (Кавказской) содержит
такие своеобразные роды брахиопод, как Enteletella, Tectarea, Loczyella, Pecte -
noproductus, Pseudopugnax, Camarophorinella, Notothyrina, некоторые из них распро-
странены также и в ее памирской части. На территории Юго-Восточного Памира
места обитания брахиопод характеризовались специфическими физико-географическими
условиями. Здесь наблюдается формирование небольших многочисленных биогермов,
населенных разнообразными брахиоподами. Биогермы развивались в области мелко-
водья, в Гидродинамически активной среде, среди осадков, с обильной примесью пи-
рокластического материала (туфоконгломераты и туфопесчаники).
Памирская часть Кавказско-Памирской провинции характеризуется отсутствием
крупных продуктид, количество мелких прирастающих форм также невелико. Зато ши-
роко представлены разнообразные теребратулиды (3 рода), ринхонеллиды (5 родов)
и ортиды, причем очень велико количество экземпляров, принадлежащих к отдельным
видам.
На Юго-Западном Дарвазе брахиоподы в это время населяли мелководные отмели,
сложенные раковинным песком в прибрежной зоне, характеризующейся развитием
оолитовых, онколитовых и мшанковых известняков и некрупных мшанково-водорос-
69
левых биогермов. В затишных участках обитали иглистые продуктиды рода Т scher-
nyschewia.
На территории Южно-Китайской провинции джульфинскому ярусу соответствуют
отложения формации Допинг, разделяющейся на серии Ушапин (зона Codonofusiella)
и Чансин (зона Palaeofusulina), известные также под названием 'литтониевого
известняка*. Провинции характеризуются присутствием разнообразных и многочис-
ленных представителей отряда Productida: Haydenella, Compressoproductus, Spinomar-
ginifera, а также большим количеством литтониид, о чем свидетельствует само
название содержащих их слоев. Разрезы Южного Китая, по мнению К.-К. Чао (Chao
King—Коо, 1965), являются самыми высокими фаунистически охарактеризованными
пермскими морскими отложениями. Серия Чансин содержит разнообразную фауну
аммоноидей, среди которых присутствуют роды Pseudotirolites, Pleuronodoceras,
Pachydiscoceras, Р seudogastrioceras и многие другие. К сожалению, эта богатая
аммонитовая фауна, представленная приблизительно 100 видами, относящимися к 30
родам, до настоящего времени находится в стадии обработки, результаты которой
пока не опубликованы. Также отсутствуют данные и о развитии брахиопод на гра-
нице палеозоя и мезозоя в этом районе, хотя в литературе имеются сведения о на-
личии здесь богато охарактеризованных фаунистически морских отложений, переход-
ных от палеозоя к мезозою.
Брахиоподы джульфинского яруса в Уссурийской провинции известны только из
слоев, содержащих Palaeofusulina, Codonofusiella, Jabeina в средней части группы
Майдзуру Японии. Брахиоподы здесь представлены Haydenella ? (Linoproductus kiang-
siensis Kays.), Alexenia ?[Produces (Dictyoclostus) gratiosa (Waagjj, Leptodus richtho-
feni Kays, и другими (Schimizu, 1961, 1963), Некоторые из встреченных здесь форм
близки к брахиоподам формации Допинг Южного Китая.
Судя по систематическому составу «брахиопод в разных частях Евро-чАзиатской
части Тетиса, можно высказать предположение о возможных центрах расселения ме-
зозойской брахиоподовой фауны. Вероятно, в Альпийской провинции переживали небла-
гоприятные условия конца перми - начала триаса атиридиды; теребратулиды и рин-
хонеллиды, явившиеся в мезозое и кайнозое основными представителями брахиопод,
развивались с конца палеозоя в Кавказско-Памирской провинции.
В заключение остановимся на характеристике небогатых комплексов брахиопод,
относимых нами к самым высоким горизонтам джульфинского яруса. Однако принад-
лежность этих комплексов к пермской системе дискуссионна, и не исключен их ран-
нетриасовый возраст. Сюда относятся брахиоподы из слоев с Tompophiceras - слоев
с Paratirolites Закавказья, брахиоподы нижней части формации Катвай Соляного
.кряжа (Пакистан) и верхней части слоев Зеван Кашмира. Все эти комплексы типично
пермские, но они мало характерны по систематическому составу и вызывают интерес
в основном с точки зрения принадлежности их к пермской или триасовой системе.
Граница перми и триаса в книге "Развитие и смена...* (1965) была проведена в
основании слоев с аммоноидеями Тompophiceras по сопоставлению их с отоцерасовыми
слоями нижнего триаса других районов, причем верхнепалеозойские виды брахиопод-
Spinomarginifera pygmaea Sai’., Haydenella kiangsieusis (Kays.), H.minuta Sar., Araxa-
thyris araxensis minor Grunt, Л. ogbinensis Grunt, Orthotichia parva Sok., Enteletes dzhag—
rensis Sok., Orthotetina sp., T ere bra tu lo idea sp. были признаны переходящими в ос-
нование нижнего триаса. В 1970 г. вопрос о положении границы перми и триаса
в Закавказье был вновь рассмотрен специальным совещанием, организованным в
Ереване и сопровождавшимся экскурсиями. Во время экскурсий сотрудником Па-
леонтологического института А.А. Шевыревым было обнаружено присутствие пропти-
хитид в слоях с Claraia* на основании чего им был сделан вывод о положении гра-
ницы перми и триаса в основании этих слоев и о пермском возрасте слоев с Тош-
pophiceras - слоев с Paratirolites. К этому мнению присоединились К.О. Рос-
товцев и Н.Р. Азарян (1971).
Брахиоподы основания формации Катвай Соляного кряжа описаны Грантом ( Grant,
1970), который относит их к джульфинскому ярусу* Здесь присутствуют Enteletes
sp. сходный с Enteletes dzhagrensis Sok. , Orthotetina cf. arakeljani Sok., Orthoteti •
na sp., Ombonia spM Derbyia sp., Spinomarginifera sp., Crurithyris 2 exima Grant, Martinia
sp., Whitspakia sp.
В большинстве разрезов основание слоев Катвай содержит пермских брахиопод,
а триасовые аммоноидей появляются несколько выше, в 0,3 м от подошвы слоев
70
Катвай. В двух местонахождениях джульфинские брахиоподы встречаются совмест-
но с триасовыми аммоноидеями. Грант предполагает,что в этих случаях брахиоподы
являются переотложенными. Таким образом, по его мнению, граница перми и триаса
должна проводиться по появлению триасовых аммоноидей и в большинстве случаев
основание слоев Катвай относится к джульфинскому ярусу . В противоположность ему
Б.Каммел и К.Тайхерт (Kummel, Teichert, 1970) рассматривают слои Катвай пол-
ностью в составе нижнего триаса, считая, что границу между системами необходимо
проводить по четкой смене характера осадконакопления (т.е. на границе формации
Чидру и слоев Катвай, где белые песчаники сменяются доломитами).
Граница палеозоя и мезозоя изучалась группой японских ученых (Nakazawaet all,
1970) в разрезе Гирул Равин (Guiyul Ravinе) Кашмира. Ими установлено, что в изу-
ченном разрезе слои Зеван без признаков перерыва перекрываются отложениями
нижнего триаса. Граница перми и триаса проводится ими по появлению двустворок,
отнесенных к роду Claraia , причем триасовые роды аммоноидей Otoceras и О phi се-
ras появляются несколько выше по разрезу. В самых верхних частях пермского раз-
реза присутствуют брахиоподы Dielasma sp. indet., Athyris cf. subexpansa Waag., Linop-
roductus cf. cora ( Orb.), Lissochonetes sp. indet., Waagenoconcha purdoni (Waagen), Pus-
tula 2 sp. indet., Marginifera himalayensis Diener, которые переходят также в слои
с Claraia. Можно предположить, что в данном разрезе граница между пермью и
триасом несколько занижена, и ее следует проводить по подошве слоев с триасовыми
аммоноидеями, где уже практически отсутствуют пермские брахиоподы. Этот же раз-
рез был кратко описан Тайхертом, Каммелом и Капуром (Teichert, Kummell, Kapoor,
1970).
Таким образом, в рассмотренных районах точное положение границы перми и
триаса остается спорным, и характер развития брахиопод на рубеже палеозоя и ме-
зозоя неясен.
Изучение хода эволюции брахиопод на протяжении всего пермского периода ука-
зывает на то, что процесс вымирания идет постепенно, захватывая самые разно-
образные группы. В арктических бассейнах процесс вымирания идет значительно
быстрее, чем в Тетисе, и заканчивается практически к концу казанского века (Уст-
рицкий, 19676). В Евро-Азиатской части Тетиса, где морской бассейн существовал
до конца перми, изменение фауны брахиопод шло постепенно в условиях непрерыв-
но сохраняющегося морского режима.
Учитывая данные опубликованных работ и обобщая имеющийся в нашем распоря-
жении материал, можно сделать вывод, что в течение джульфинского века и на ру-
беже палеозоя и мезозоя развитие брахиопод происходило в следующих направлениях;
1. Шло дальнейшее вымирание многих родов, существовавших в течение длитель-
ного времени и без каких-либо существенных изменений доживших до конца перми.
Их вымирание сопровождалось постепенным сокращением видового состава, ареалов
видов и численности популяций. Сюда относятся такие роды, как Enteletes, Strep -
torhynchus, Dielasma, Neospirifer, Chaoiella и др.
2. Происходила резкая вспышка видообразования среди представителей группы
'мобильных* родов - Т schernyschewia, Tyloplecta, Spinomarginifera, Heterelasmina,
Notothyris, Uncinunellina, Araxathyris. Как правило, такое интенсивное видообра-
зование сопровождалось повышением численности популяций и значительной внут-
ривидовой изменчивостью.
3. Возникало большое количество аберрантных родов, представленных, как пра-
вило, единичными видами и незначительным количеством экземпляров. Это Spin оlyt-
tonia, Gubleria, Tectarea, Loczyella, Pectenorpoductus, Enteletella, Araxilevis, Og-
binia, Kiangsiella, Janiceps и др. Некоторые роды обладают рекуррентными призна-
ками:* так род Chonopectella характеризуется сетчатой скульптурой, близкой к де-
вонскому Chonopectus; один из видов Ogbinia по форме раковины сходен с пред-
ставителями раннетурнейского рода Productina; Comelicania обладает формой рако-
вины, близкой к девонскому роду Anathyris (Сарычева, Сокольская, Грунт, 1965).
Отметим также появление столидия у некоторых пермских представителей надсе-
мейства Rhynchonellacea, например, у Памирско-Кавказских представителей рода
Uncinunellina. у него столидий, с одной стороны, сходен с опорными пластинами
атрипид (Иванова, 1962), с другой - с внутренними иглами девонского рода Unci-
nulus.
71
Интересно отметить, что рекуррентные признаки появляются, как у родов, отно-
сящихся к отрядам, вымирающим и не переходящим в триас (роды Chonopectella,
Ogbinia), так и у представителей развивающихся групп, потомки которых продол-
жают существовать в триасе; причем у последних рекуррентные признаки могут
присутствовать наряду с признаками, отмечающимися у более молодых форм. На-
пример, замочная пластина простой формы без висцерального форамена наряду с
очертаниями раковин, характерными для Anathyris, есть у Comelicania; фальциферовые
круры наряду со столидием присутствуют у Uncinunellina.
4. Некоторые формы на заключительном этапе своего развития становились угне-
тенными, мелкорослыми. Это отчетливо наблюдается на закавказском материале
(Сарычева и др., 1965),
В течение пермского периода возникали некоторые роды, являвшиеся, по-види-
мому, неотеническими формами. Это явление было особенно распространено среди
ринхонеллид, относящихся к различным надсемействам: Boloria и Paran ore Па (над-
семейство Rhyncohonellacea), Camarophoriina (надсемейство Stenosdsmatacea). Эти
формы характеризуются неребристой раковиной, а также расположением синуса на
спинной и выступа на брюшной створках. Известно, что в процессе онтогенеза у
большинства ринхонеллид происходила инверсия раковины, т.е. изменение располо-
жения синуса и выступа на обратное и развитие ребристости в разной степени. Поэто-
му появление родов, обладающих синусом на спинной и выступом на брюшной створ-
ках, мы вправе объяснить неотенией, о чем свидетельствует х^кже и отсутствие
ребристости раковины.
Наряду с отрядами Orthida и Productida, вымершими к концу палеозоя, широко
распространенными в это время являлись отряды Rhynchonellida, Terebratulida,
Spiriferida и надсемейство Athyridacea, многие представители которых известны
в мезозое, причем ряд признаков, характеризующих палеозойских брахиопод, сохранил-
ся и у мезозойских. Следует отметить, что огромное большинство прирастающих и
свободнолежащих форм оказалось неприспособленным к изменившимся условиям среды
и вымерло на рубеже палеозоя и мезозоя, в то время как брахиоподы, существовав-
шие в мезозойских морях и дожившие до современности, обладают преимущественно
якорным типом прикрепления. Непосредственно связанный с якорным типом прикреп-
ления ножной воротничок, являющийся признаком высокого уровня развития, появ-
ляется уже у пермских атиридид (род Spirigerella). Это морфологическое образо-
вание очень широко развито у некоторых триасовых атиридид (род Majkopella) и
многих теребратулид. В то же время присутствие висцерального форамена у пред-
ставителей родов Spirigerella, Araxathyris и Septospirigerella, объединенных в
подсемейство Spirigerellinae, свидетельствует о более высокой степени их специа-
лизации, чем триасовых форм, которым могла дать начало линия менее специали-
зированных родов CompQsifa - Cleiothyridina — Comelicania, характеризовавшихся
отсутствием висцерального форамена.
Из представителей отряда Terebratulida в мезозой переходят только редей е пред-
ставители семейств Heterelasminidae и Dielasmatidae (Дагис, 1963). Все мезозой-
ские теребратулиды в процессе онтогенеза проходят центронелловую стадию руч-
ного аппарата, т.е. центронелловая петля является исходной для них (Stehli, 1956).
Из отряда Spiriferidae в мезозой переходят представители надсемейств Delthyriacea
и Spiriferinacea.
Из отряда Rhynchonellida надсемейство Stenoscismatacea, характеризующееся
наличием спондилия в брюшной и септалия в спинной створках, полностью вымира-
ет к концу перми.
Из надсемейства Rhynchonellacea в мезозое продолжает развиваться семейство
Wellerellidae, представленное здесь, так же как и в верхнем палеозое (где они
достигли максимального расцвета), двумя подсемействами: Wellerellinae и Pseu—
dopugnaxinae. Они характеризуются цельной замочной пластиной, зубными пластинами
и спинной септой, развитыми в различной степени, а также фалышферовыми или
префальциферовыми крурами. Несмотря на такое сходство, Эгер ( Ager, 1965) без
достаточного основания отнес всех мезозойских веллереллид к новому подсемейству.
Другая группа мезозойских ринхонеллид - семейство Basiliolidae Cooper - об-
наруживает родство с палеозойским семейством Hypothyridiinidae Rzonsn. В.П. Мак-
ридин (1964) объединяет представителей этих двух семейств в надсемейство Basi—
72
lioloidea на основании сходного строения ручного аппарата, характеризующегося
наличием серповидных фалыщферовых крур.
Из всего вышесказанного можно сделать вывод, что для джульфинского этапа
развития брахиопод характерно интенсивное видообразование и изменчивость, обус-
ловленные высокой степенью приспособительных реакций. Это происходит на фоне
общего сокращения численности популяций ранее господствовавших групп, сужения
их адаптаций, иногда с возвратом давно утраченных признаков.
XXX
Изложенные в настоящей работе данные о распространении брахиопод в пермских
отложениях Альпийско-Гималайской подобласти позволяют сделать следующие вы-
воды.
1. Вымирание брахиопод, достигшее максимума на рубеже палеозоя и мезозоя,
происходило постепенно и являлось закономерным процессом, обусловленным всем
ходом эволюции группы, а не событием катастрофического порядка, как его рас-
сматривает Шиндевольф ( Schindewolf, 1954, 1963, 1964). Прослеживается непосред-
ственная связь между палеозойскими и мезозойскими брахиоподами, которых необ-
ходимо изучать в филогенетическом аспекте, выявляя среди палеозойских брахиопод
предковые формы для мезозойских.
2. Смена комплексов брахиопод в течение пермского периода явилась следствием
закономерного взаимодействия двух процессов: с одной стороны, эволюционного раз-
вития брахиопод, с другой - палеогеографических изменений в морях пермского
периода.
3. Выявленная этапность позволяет говорить о возможности трехчленного деления
пермской системы.
СИСТЕМАТИЧЕСКАЯ часть
КЛАСС ARTICULATA
ОТРЯД ORTHIDA
НАДСЕМЕЙСТВО ENTELETACEA
СЕМЕЙСТВО SCHIZOPHORHDAE SCHUCHERT, 1929
Род Orthotichia Hall et Clarke, 1892
Orthotichia avuahenaia Sokolskaja, 1965
Табл. I, фиг. i,2; рис. 8
Orthotichia avuahenaia: Сокольская, 1965, стр. 199, табл. XXIX, 4иг. 3, рис. 23.
Голотип - ПИН № 2072/55; раковина с сочлененными створками; Закавказье,
левый берег р.Джагры, выше селения Огбин; мургабский ярус, гнишикский горизонт;
Сокольская, 1965, табл. XXIX, фиг. 3.
Рис. 8. Orthotichia avushensis, ПИН, № 2774/73
8а - ядро брюшной створки; 86 - то же спинной створки; оба - 2.4 х. Кур-Истык 2;
джульфинский ярус, нижнетахтабулакская подсвита
Описание. Раковина небольшая, округло^ треугольного очертания, довольно силь-
но вздутая. Лобный край слабо унии ли кати ый , боковые - дуговидные. Замочный край
прямой, больше половины наибольшей ширины раковины, приуроченной к ее передней
части.
Брюшная створка слабо выпуклая, с маленькой, незагнутой макушкой. А рея низ-
кая^ отчетливо ограниченная. Дельтирий широко треугольный. Макушечный угол 100-
105 • В передней части створки слабая депрессия. Спинная створка более выпуклая,
Унипликатным называется лобный край раковины с синусом на брюшной и выступом
на спинной створках ( Thomas, 1927).
74
чем брюшная, с широкой, массивной, сильно загнутой макушкой и маленькой, неот-
четливой ареей. Ареи обеих створок анаклинные (угол между плоскостью, разделяю-
щей створки, и плоскостью ареи меньше 90 ).
Раковина покрыта тонкой радиальной струйчатостью (3-4 струйки на 1 мм на рас-
стоянии 20 мм от макушки). Концентрические линии нарастания редкие, неотчетливые.
Внутри брюшной створки - расходящиеся зубные пластины, поддерживающие мас-
сивные, овальные зубы и высокая септа, протягивающаяся до середины длины створ-
ки (см. рис. 8).
Мускульное поле удлиненно-треугольное, ограниченное зубными пластинами. По
бокам от него - отпечатки генитальных сосудов, наблюдающиеся на ядре в виде мел-
ких сосочков. Внутри спинной створки- расходящиеся- брахиофоры, охватывающие ок -
руглое мускульное поле и слабый септальный валик (см. рис. 8).
Размеры, мм
Экз., ,№ Местонахождение, возраст д Ш Т ША МУ1
2774/63 Шор-Булак, 19,5 20,5 15,5 10,0 100°
2774/64 джульфинский ярус, 18,6 18,8 15,7 9,7 103°
2774/69 среднетахтабулакская подсвита П.7 14,9 8,0 5,9 102°
1МУ - макушечный угол.
Сравнение. Рассматриваемый вид отличается от O.indica Waagen большей выпук-
лостью спинной створки, чем брюшной ( O.indicet характеризуется обратным соот-
ношением выпуклости створок); от О. dor ash a men sis Sokolskaja - более вздутой ра-
ковиной и ее изометричным очертанием; от O.minuta Sokolskaja - крупными разме-
рами и очень слабо развитым синусом.
Геологический возраст и географическое распространение.
Мургабский ярус, гнишикский горизонт Закавказья; джульфинский ярус, нижне- и
среднетахтабулакская подсвиты Юго-Восточного Памира.
Материал. 13 экз. с сочлененными створками удовлетворительной сохранности.
Из них 2 экз. собраны в водорослевых биогермных известняках нижнетахтабулакской
подсвиты Кур-Истыка 2, остальные - в отложениях среднетахтабулакской подсвиты
(из них 1 экз. - в табулятово-гидроидных биогермных известняках Кур-Истыка 1
и 9 - в табулятовых биогермных известняках . Шор-Булака).
СЕМЕЙСТВО ENTELETIDAE WAAGEN, 1884
ПОДСЕМЕЙСТВО ENTELETINAE WAAGEN,.1884
ЕРД Enteletes Fischer Waldheim, 1837
Enteletes dzhagrensis Sokolskaja, 1965
Табл. I, фиг. 6, табл. .XIV, фиг. 1; рис. 9
Enteletes dzhagrensis ‘ Сокольская, 1965, стр. 202, табл. XXIX, фиг. 7.
Г олотип - ПИН № 2072/60; спинная створка, Закавказье, левый берег р.Джаг-
Ры, ниже сел. Огбин; джульфинский ярус, слои с Tompophiceras—Paratirolites; Со-
кольская, 1965, табл. ХХЦС» фиг. 7. »
75
Рис. 9. Enteletes dzhagrensis, ПИН,
№2774/53; зарисовки последовательных по-
перечных сечений раковины в примаку-
шечной части, к 4,5
бр.с» - брюшная створка, сп.с. - спин-
ная створка, з.п. - зубные плас тин ы, с. с. -
срединная септа, б — брахиофоры. Цифра-
ми указано расстояние пришлифовок от
конца макушки брюшной створки (в мм);
Длина брюшной створки 11,5 мм. Кур-Ис-
тык 1; джульфинский ярус, среднетах-
табулакская подсвита
Описание. Раковина маленькая, шарообразная, со слабым синусом на брюшной
и выступом на спинной створку Лобный край слабо унипликальный, зигзагообразный;
боковые - дуговидные. Замочный край больше половины наибольшей ширины, распо-
ложенной в средней части раковины.
Брюшная створка умеренно выпуклая; ее макушка немного загнута; арея широкая,
низкая, слабо вогнутая, с большим дельтирием. Макушечный угол 102-107 . Спин-
ная створка немного более выпуклая, чем брюшная. Ее макушка более низкая и уз-
кая. Арея слабо развита, неясно ограничена, нототйрий маленький. Ареи обеих
створок анаклинные. С каждой стороны синуса и выступа располагается по 3-4 поло-
гих складки, развитых лишь в передней половине раковины.
Поверхность покрыта тонкими радиальными струйками (3-5 на 1 мм на расстоя-
нии 15 мм от макушки).
Внутри брюшной створки - тонкие, длинные, параллельно расположенные зубные
пластины и высокая септа, достигающая половины длины створки. Внутри спинной -
слабо расходящиеся массивные брахиофоры.* охватывающие овальное мускульное поле
(см. рис. 9). Посредине проходит низкий, широкий септальный валик, начинающийся
отступя 1-1,5 мм от макушки и не выходящий за пределы мускульного поля. На
пришлифовках септальный валик не наблюдался.
Размеры, мм
Экз., № Местонахождение, возраст д Ш Т ША МУ
2774/54 Кур-Истык 1, 8,6 9,3 8,0 4,2 105°
2774/53 джульфинский 11,5 10,5 9,2 5,0 102°
2774/55 ярус, среднетахтабулакская под- 10,1 10,1 9,9 3,2 106°
2774/58 свита 8,7 9,6 7,5 2,8 107°
Сравнение. Данный вид отличается от E.meridionalis Gemmellaro более мелкими
размерами и относительно более выпуклыми створками. От E.laevissimus Waagen -
изометричным очертанием и меньшим количеством складок, а от Е.subaequivalvis
Gemmellaro - более вздутыми створками, менее резкими радиальными складками
и большим их количеством.
Геологический возраст и географическое распространение.
Джульфинский ярус, сре дне тахта була кс кая подсвита Юго-Восточного Памира. Джуль-
финский ярус, слои с Tompophiceras — Paratirolites Закавказья.
Материал. 6 экз. с сочленными створками удовлетворительной сохранности;
часть из них представлена перекристаллизованными раковинами. Собраны в отложе-
ниях среднетахтабулакской подсвиты: в табулятово-гидроидных биогермных известня-
ках Кур-И стыка 1 - 5 экз; в табулятовых биогермных известняках Музду-Булака -1экз.
76
Enteletes meridionalis Gemmellaro, 1899
Табл. I । фиг. 3-5, табл. XIV, фиг. 2; рис. 10, И
Enteletes meridionalis: Gemmellaro, 1899, стр. 143, табл. XXVIII, фиг. 10-12;
табл. XXIX, фиг. 5.
Описание. Раковина средних размеров, умеренно выпуклая, с небольшим сину-
сом брюшной и выступом спинной створки. Лобный край слабо унипликатный, зазуб-
ренный. Боковые края дуговидные. Замочный край приблизительно равен половине на-
ибольшей ширины раковины, приуроченной к ее передней трети. Брюшная створка
умеренно выпуклая. Ее макушка слабо загнутая, арея низкая, вогнутая, нерезко очер-
ченная, с широким, треугольным, нерезко ограниченным дельтирием. Макушечный угол
Рис. 10. Enteletes meridionalis, ПИН, № 2774/75; 10а - ядро брюшной створки;
10 б - то же спинной; оба - х. 2.7 Усл. обознач. см. рис. 9. Кур-Истык 1; джульфин-
ский ярус, среднетахтабулакская подсвита
110-115°. Спинная створка немного более выпуклая, чем брюшная. Ее макушка нес-
колько шире и массивнее. Арея низкая, нототирий узкий. Ареи обеих створок анаклин-
ные. Продольные складки пологие, отчетливо развиты в передней половине створок,
по 4-5 с каждой стороны синуса и выступа. Поверхность раковины покрыта тонкими
радиальными струйками (3-5 на 1 мм на расстоянии 17 мм от макушки).
Внутри брюшной створки - длинные, субпараллельные зубные пластины и длинная,
тонкая септа (см. рис. 10,11). Внутри спинной - расходящиеся брахиофоры, охваты-
вающие округлое мускульное поле, и низкий септальный ватжк, который на пришлифов-
ках не наблюдался.
Размеры, мм
Экз., № Местонахождение, возраст д Ш Т ША МУ
2774/58 Шор-Бу ла к, джульфинский ярус, 14,7 18,4 12,0 7,2 110°
2774/72 среднетахтабулакская подсвита 14,3 17,1 11,2 7,0 112°
2774/74 Куталь 2, возраст тот же 20,7 25,8 18,4 10,7 115°
2774/75 Кур-Истык 1, возраст уот же 17,0 20,4 13,4 6,5 110°
77
Сравнение. Отличия от E.dzhagrensis Sokolskaja см. в описании последнего.
От E.tschemyschewi Diener рассматриваемый вид отличается менее вздутой ракови-
ной и менее резкими складками.
Геологический возраст и географическое распространение.
Мургабский ярус, фузулиновый известняк Сицилии; джульфинский ярус, среднетах-
табулакская подсвита Юго-Восточного Памира.
Материал. 5 экз. с сочлененными створками удовлетворительной сохранности,
причем некоторые из них перекристаллизованы. Собраны в отложениях среднетах-
табулакской подсвиты: в табулятово-гидроидных биогермных известняках Кур-Исты-
ка 1 - 1 экз.; Куталя 2-1 экз.; в табулятовых биогермных известняках Шор-Бу ла-
ка - 3 экз.
Ри с. 11. Enteletes тп erid i опа—
lis, ПИН, № 2774/59; зарисовки
последовательных поперечных
сечений раковины в примаку-
шечной части, х. 5,4 Ус л. обознач.
см. рис. 9. Длина брюшной створ-
ки 16,2 мм . Местонахождение
и возраст те же, что на рис. 10
Р °Д Enteletella Licharew, 1925
Р nteletella': Лихарев, 1925, стр. 719-720; 1939, стр. 79; Лихарев, Алихова,
1960, стр. 194; Williams, Wright, 1965, стр. 330.
Типовой вид - E.nikschitchi Licharew, 1925; джульфинский ярус, уруштенский
горизонт; Северный Кавказ.
Описание. Раковина средних размеров, шарообразная с сильно и одинаково
выпуклыми створками.Спинная макушка более массивная, чем брюшная. Брюшная арея
широкая, низкая, с широким дельтирием. Спинная арея развита слабо. Обе ареи анак-
линные. Синус и выступ не развиты, но срединная складка спинной створки и про-
межуток между складками брюшной могут быть несколько шире, чем боковые.
Скульптура состоит из резких, угловатых складок, развитых на обеих створках.
Поверхность раковины покрыта тонкими, радиальными струйками. Внутри брюшной
створки хорошо развитые зубные пластины, сливаясь с септой, образуют спондилий,
причем септа продолжается внутрь него (см. рис. 12). Спондилий такого типа назван
гспондилием с трисептой* (Лихарев, 19606).
Внутри спинной створки присутствуют брахиофоры и низкий септальный валик.
Сравнение. Рассматриваемый род внешне отличается от рода Enteletes Fischer
Waldheim, 1837 одинаково выпуклыми створками, более резкими складками, а также
отсутствием синуса и выступа. По внутреннему строению отличается наличием сеп-
талия в брюшной створке. От рода Enteletoides Stucketiberg, 1905 рассматриваемый
род отличается радиальными складками, присутствующими на обеих створках, а от
рода Enteletina Schuchert et Cooper, 1932 - отсутствием синуса и выступа (предста-
78
вители рода Enteletina имеют синус на спинной и выступ на брюшной створках). Кро-
ме того, ни у Enteletoides, ни у Enteletina нет спондилия в брюшной створке.
Видовой состав. Типовой вид.
Замечания. Автор рода Enteletella Б.К. Лихарев (1925) указывал на возмож-
ность отнесения к названному роду Enteletes microplocus Gemmellaro , близкого к Еп-
teletella nikscitchi по внешнему виду; внутреннее же строение Е .microplocus неиз-
вестно.
Распространение. Джульфинский ярус: уруштенский горизонт Северного Кав-
каза; нижне- и среднетахтабулакская подсвиты Юго-Восточного Памира.
Enteletella nikschitchi Licharew, 1925
Табл. I, фиг. 7; табл. XIV, фиг. 3; рис. 12
Enteletella nikschitchi': Лихарев, 1925, стр. 719-720, табл. V, фиг. 1-3; 1939,
стр. 79, табл. XVI,фиг. 4; Лихарев, Алихова, 1960, стр. 194, табл. XVI» фиг. 6.
Голотип - ЦНИГРМузей им. Ф.Н. Чернышева,№ 1296/1; раковина с сочлененными
створками; джульфинский ярус, уруштенский горизонт; Северный Кавказ; Лихарев,
1925, табл. V, фиг. 2.
Рис. 12, Enteletella nikschitchi,
ПИН, № 2774/79; зарисовка попереч-
ного сечения брюшной створки, х 6,3;
сп. т. - гспондилий с трисептой*. Ос-
тальные усл. обознач. см. рис. 9. Дли-
на брюшной створки 18,9 мм. Кур-Ис-
тык 1; джульфинский ярус, среднетах-
табулакская подсвита
Описание. Раковина средах размеров, шарообразная, с одинаково выпуклыми
створками, с ректимаргинатной , Зазубренной лобной комиссурой и дуговидными бо-
ковыми краями. Замочный край приблизительно равен половине наибольшей ширины
раковины, расположенной в ее средней части. Брюшная арея несколько выше спинной,
и ее макушка сильнее загнута. Макушечный угол 90 . Ареи обеих створок анаклин-
ные, нерезко ограниченные На каждой створке по 9-10 угловатых складок, начинаю-
щихся почти от самой макушки и слабо расширяющихся по направлению к лобному
краю Поверхность покрыта тонкими, однородными радиальными струйками (4 на 1 мм
на расстоянии 17-20 мм от макушки).
Внутреннее строение типичное для рода (см. рис. 12).
Сравнение. Отличается от Enteletellaf?) microplocus (Gemmellaro) наличием бо-
лее многочисленных, узких складок.
Геологический возраст и географическое распространение.
Джульфинский ярус: уруштенский горизонт Северного Кавказа; нижне- и среднетах-
табулакская подсвиты Юго-Восточного Памира.
Так как Enteletes microplocus пока невозможно с уверенностью отнести к роду
Enteletella, данные о его распространении не используются нами при установлении
стратиграфического распространения рода.
2
Ректимаргинатной передней комиссурой называется комиссура, характерная для ра-
ковин, не обладающих ни выступом, ни синусом.
79
Экз., № Местонахождение, возраст д Ш т ША МУ
2774/77 Кур-Истык 1, джульфинский ярус, 26,0 29,5 24,6 13,8 90°
2774/78 среднетахтабулакекая подсвита 17,8 20,5 17,0 10,1 90°
Материал. 4 экз. удовлетворительной сохранности, представленные двумя ра-
ковинами с сочлененными створками, одной брюшной и одной спинной створками.
Из водорослево-гидроидных (?) рифогенных известняков нижнетахтабулакской под-
свиты Куртеке 2 собран 1 экз.; из табулятово-гидроидных биогермных известняков
среднетахтабулакской подсвиты Кур-Ис тыка 1-3 экз.
Р ОД Parenteletes King, 1931
Parenteletes: King, 1931, стр. 48; Лихарев, Алихова, 1960, стр. 194; Williams,
Wright, 1965, стр. 330.
Типовой вид - P,cooperi King, 1931, верхний карбон, зона Uddenites* Техас,
Стеклянные горы.
Описание . Раковина средних размеров, треугольного очертания, спинная створ-
ка более выпуклая, чем брюшная. Брюшная макушка невысокая, умеренно загнутая>
с низкой широкой ареей и большим дельтирием. Спинная макушка массивная, высо-
кая, с низкой ареей. На спинной створке - синус; на брюшной - выступ. На обеих
створках, по бокам от синуса и выступа присутствуют широкинемногочисленные
складки. Поверхность раковины покрыта тонкими радиальными струйками. Внутри
брюшной створки - массивные зубные пластины, субпараллельные в примакушечной
части и несколько расходящиеся кпереди. Передними концами они срастаются со
срединной септой, которая начинается очень близко от макушки (см. рис. 13). Зуб-
ные пластины поддерживают удлиненные зубы. Внутри спинной створки - расходя-
щиеся брахиофоры, массивный, однолопастной замочный отросток и септальный валик.
Сравнение. Данный род отличается от Enteletes Fischer Waldheim, 1837 и Ente-
letella Licharew, 1925, наличием синуса спинной и выступа брюшной створки, а от
Enteletina Schuchert et Cooper, 1932, имеющей сходное с Parenteletes расположение
синуса и выступат- зубными пластинами, срастающимися со срединной септой свои-
ми передними концами внутри брюшной створки.
Видовой состав. Кроме типового вида, в состав рода Кингом (King, 1931)
был включен P.g lob os us (Girty, 1908) из верхней перми Техаса. К этому же роду
относится P.suessi (Schellwien, 1892) из ассельских отложений (слои Ауэрниг) и
сакмарско-^артинских отложений (трокгофельский известняк) Карнийских Альп; P.die-
neri (Schellwien, 1900b) из трокгофельского известняка Карнийских Альп; Р.sinen-
sis Huang, 1932 и P.mirabilis (Mansui, 1913) из верхней перми Южного Китая; Р.ака-
sakensis (Ozawa, 1925) из нижней перми. Японии; P.ruzhencevi Sokolskaja, 1965
из джульфинского яруса Закавказья и Юго-Восточного Памира.
Распространение. Верхний карбон Техаса, нижняя пермь Японии и Карнийских
Альп, верхняя пермь Южного Китая, Закавказья, Юго-Восточного Памира.
Parenteletes ruzhencevi Sokolskaja, 1965
Табл. I, фиг. 8; рис. 13
Parenteletes ruzhencevi: Сокольская, 1965, стр. 202-203, табл. XXIX, фиг. 9—11,
рис. 26.
Голотип - ПИН, № 2072/66; раковина с сочлененными створками; Закавказье,
селение Чанахчи; джульфинский ярус; Сокольская, 1965, табл. XXIX, фиг. 10.
80
Описание. Раковина крупная, округло-треугольная, с наибольшей шириной близ
лобного края. Лобная комиссура сулькатная!, резко зигзагообразная. Боковые -
дуговидные. Замочный край короткий, меньше половины наибольшей ширины рако-
вины, расположенной близ лобного края. Брюшная створка незначительно выпуклая,
трапецеидальной формы, с низким, но отчетливым выступом, начинающимся от ма-
кушки и имеющим расходящиеся края. Макушка широкая, низкая, слабо загнутая.
Арея нерезко ограниченная, дельтирий широкотреугольный, открытый. Макушечный
угол 120 . Спинная створка сильно выпуклая, с неглубоким, широким, угловатым
синусом. Макушка толстая, значительно выпуклая, массивная, нависающая над за-
мочным краем, с маленькой ареей. Макушечный угол 80 . Обе ареи анаклинные. По
бокам от синуса и выступа по 1-2 складки с каждой стороны. Радиальные струйки
тонкие (2-3 на 1 мм на расстоянии 17-20 мм от макушки).
Рис. 13. Parenteletes ruzhencevi, ПИН,
No 2774/52; пришлифовка макушки брюшной
створки; зубные пластины впереди за-
круглены и срастаются со. срединной
септой, х 1,8. Шор-Булак; джульфин-
ярус, среднетахтабулакская подсвита
Внутри брюшной створки - зубные пластины срастаются с септой своими перед-
ними концами (см. рис. 13). Внутри спинной - массивный однолопастной замочный
отросток и расходящиеся брахиофоры. Септальный валик не наблюдался.
Размеры, мм
Экз., № Местонахождение, возраст д Ш Т ША МУ
2774/50 Кур-Истык 2, джульфинский 40,3 46,6 31,5 20,3 80°
2774/51 ярус, нижнетахтабулакская под свита 30,2 32,2 22,2 12,0 70°
2774/52 Шор-Булак, джульфинский ярус, сред- нетахтабулакская подсвита 35,7 41,2 27,0 16,5 77°
Сравнение. Отличается от Р.sinensis Huang меньшей вздутостью раковины,боль-
шим числом складок и более короткими зубными пластинами, от Р.cooper i King —
нерезко очерченной ареей и более низкими синусом и выступом, а от P.mirabilis
(Mansui) - резко расширяющимися кпереди синусом, и выступом, а также бо-
лее грубой радиальной струйчатостью.
Геологический возраст и географическое распространение.
Джульфинский ярус Закавказья; джульфинский ярус, нижне- и среднетахтабулакская
подсвиты Юго-Восточного Памира.
Сулькатной называется лобная комиссура раковин с синусом на спинной и выступом
на брюшной створке.
6 2 64
81
Материал. 3 экз. с сочлененными створками удовлетворительной сохранности.
Одна раковина найдена в водорослевых биогермных известняках нижнетахтабулак-
ской подсвиты Кур-Истыка 2; две остальных - в табулятовых биогермных извест-
няках среднетахтабулакской подсвиты Шор-Булака.
ОТРЯД STROPHOMENIDA
НАДСЕМЕЙСТВО DAVIDSONIACEA
СЕМЕЙСТВО MEEKELLIDAE STEHLI, 1954
ПОДСЕМЕЙСТВО MEEKELLINAE STEHLI, 1954
Род Geyerella Schellwien, 1900
Geyerella: Schellwien, 1900a, стр. 12; 1900b, стр. 24; Girty, 1908, стр. 203,
Grabau, 1931, стр. 267; Лихарев, 1932a, стр. 45; Greco, 1942, стр. 150; Соколь-
ская, 1960, стр. 219; Williams, 1965, стр. 405; Rudwick, Cowen, 1968, стр.118.
Типовой вид .- G.gemmellaroi Schellwien, 1900; мургабский ярус, слои Со-
зио; Сицилия.
Описание. Раковина различных размеров, двояковыпукла^, с рожкообраэной
брюшной и умеренно выпуклой спинной створкой. На брюшной створке — высокая,
узкая, треугольная арея. Дельтирий узкий, закрыт выпуклым дельтидием, по бокам
от него - вторичная арея.
Скульптура состоит из радиальных складок, развитых в различной степени. Ра-
ковина ложнопористая, покрыта радиальной струйчатостью.
Внутри брюшной створки - тонкие зубные пластины, сходящиеся, не достигая дна
створки. Они поддерживаются высокой септой, образуя спондилий, названный двойным
(Лихарев, 19606). Внутри спинной створки - высокий двулопастной замочный отрос-
ток. Септа короткая. Круральные пластины расходящиеся (см. рис. 14).
Сравнение. Рассмотренный род отличается от родов Ombonia Caneva, 1906и Ре-
rigeyrella Wang, 1955 наличием двойного спондилия, в противоположность сидячему
спондилию этих родов. От Ombonia его отличает, кроме того, радиальная складча-
тость, отсутствующая у названного рода. Наличие спондилия отличает род Geyerella
также и от Meekella White et John, 1867.
Видовой состав. Кроме типового вида, в состав рода входят G.alpina Schellwi—
en, 1900, из сакмарско-артинских отложений (трокгофельский известняк и слои Ка-
раванкен) Карнийских Альп, мургабского яруса (слои Созио) Сицилии и джульфин-
ского яруса (среднетахтабулакская подсвита) Юго-Восточного Памира; G.mongoli-
са Grabau, 1931 из известняка Усу-Хонгор; G.rex Gregorio, 1930 из мургабского яру-
са (слои Созио) Сицилии; G.americana Girty, 1908 из Гваделупе кого яруса (форма-
ция..: Капитан) Техаса; G.tschemyschewi Licharew, 1932 из джульфинского яруса (уруш-
тенский горизонт) Северного Кавказа.
Распространение. Нижняя пермь Карнийских Альп, верхняя пермь Техаса,
Сицилии, Северного Кавказа, Юго-Восточного Памира, Монголии.
Geyrella al pin a Schellwien, 1900
Табл. I, фиг. 9,10; табл. XIV, фиг. 4; рис. 14
fieyrella alpina: Schellwien, 1900а, стр. 13, табл. I, фиг. 8.
Geyrella distorta: Schellwien, 1900b, стр. 25-26, табл.Ш, фиг. 6-13; ।
Greco ,1938, стр. 33-36, табл. II, фиг. 9-11.
Описание. Раковина небольшая, двояковыпуклая, с рожкообраэной брюшной и
округлой, умеренно выпуклой спинной створкой. Лобный край ректимаргинатный, бо-
ковые - дуговидные. Брюшная створка продольно-треугольного очертания, рожко-
82
зм.п.
кр
Рис. 14. Geyerella alpina, ПИН, № 2774/32;
зарисовки поперечных сечений раковины, Х4
сп. - спондилий, зм.п. - замочная пласти-
на; з. - зубы; кр.п. - крурапьные пластины.
Остальные усл. обознач. см. рис. 9. Музду-
’Булак; джульфинский ярус, среднетахтабулак-
ская подсвита
образная; арея высокая, слабо вогнутая, катаклинная (угол между плоскостью, раз-
деляющей створки, и плоскостью ареи равен 90 ), нерезко отделена от боковых скло-
нов створки. Дельтирий узкий, закрыт слабо выпуклым дельтидием. По бокам от него
узкая вторичная арея.
Макушка с неровной поверхностью прирастания. Спинная створка без ареи. Ма-
кушка короткая, незагнутая.
Скульптура состоит из нерезких, узких радиальных складок, незначительно рас-
ширяющихся кпереди, в количестве 10-15 на каждой створке. На боковых склонах склад-
ки отсутствуют. Раковина покрыта тонкой радиальной струйчатостью (3-4 струйки
на 1 мм по лобному краю).
Внутри брюшной створки - тонкие зубные пластины, поддерживающие треуголь-
ные гребневидные зубы. Зубы ограничивают края псевдодельтидия. Зубные пластины
срастаются на расстоянии 1/4 - 1/3 от дна створки и поддерживаются тонкой сеп-
той, образуя, таким образом, двойной спондилий. В спинной створке-тонкий высо-
кий, двулопастной замочный отросток, глубоко вдающийся в полость брюшной створ-
ки. Круральные основания тонкие, слабо расходящиеся (см. рис. 14).
Разме ры, мм
Экз., Nfe Местонахождение, возраст дбр д СП Ш т ВА ША Ш д МУ
2774/30 Шор-Бу ла к; джуль- финский ярус, 17,7 13,0 15,2 18,4 12,0 8,6 3,1 55°
2774/31 средн етахта була кс кая подсвита 16,0 14,7 17,6 15,5 Ю.О 7,6 2,5 82°
83
Сравнение. Отличается от G.gemmelteroi Schellwien меньшими размерами, менее
резкой радиальной складчатостью, незагнутостью брюшной макушки; от G.tscherny-
schewi Licharew _ пологими радиальными складками, узкой макушкой брюшной створ-
ки, меньшей выпуклостью» спинной створки, а от G.rex Gregorio - более низкой,
правильной брюшной створкой и симметрично расположенной макушкой.
Геологический возраст и географическое распространение.
Трокгофельский известняк и слои Караванкен Карнийских Альп; мургабский ярус,
слои Созио Сицилии; джульфинский ярус, среднетахтабулакская подсвита Юго-Вос-
точного Памира.
Материал. 5 экз. хорошей сохранности собраны в отложениях среднетахтабу-
лакской подсвиты. Из них найдены в табулятовых биогермных известняках Шор-
Булака - 3 раковины с сочлененными створками; Музду-Булака - 1 брюшная створ-
ка; в табулятово-гидроидных биогермных известняках Куталя 2-1 брюшная створка.
СЕМЕЙСТВО ORTHOTETIDAE WAAGEN, 1884
ПОДСЕМЕЙСТВО OERBYINAE STEHLI, 1954
Род Derbyia Waagen, 1884
Д er by ia': Waagen, 1884, стр. 576; Hall, Clarke, 1892, стр. 261; Schellwien,
1900a, стр. 3; Чернышев, 1902, стр. 202; Girty, 1908, стр. 181; Thomas, 1910,
стр. 89; Герасимов, 1929, стр. 825; Dunbar, Сondга, 1932, стр. 75; Сокольская,
1954, стр. 136-141 ( part.); 1960, стр. 219; Williams, 1965, стр. 411.
Типовой вид - D.regularis Waagen, 1884; нижний продуктусовый известняк;
Соляной кряж (Пакистан).
Описание. Раковина крупная, двояковыпуклая или выпукло-плоская, резупинат-
ная; полуокруглого или поперечно-овального очертания. Арея широко - треугольная,
низкая, с небольшим дельтидием. Скульптура состоит из неоднородных радиальных
струек.
Внутри брюшной створки - зубные пластины, поддерживающие зубы. Мускульное
поле округлое или удлиненно-овальное, обширное. Его пересекает септа, достигаю-
щая 1/3-1/4 длины створки. Внутри спинной створки - высокий массивный двуло-
пастной замочный отросток. Он поддерживается сильно расходящимися круральными
пластинами.
Сравнение. Рассматриваемый род отличается от Plicatoderbyia Thomas, 1937,
отсутствием радиальной складчатости; от Licharewiella Sokolskaja, 1960 - отсут-
ствием высокого поперечного валика, в который упирается брюшная септа. От внеш-
не сходного рода Orthotetes Fischer, 1829, относящегося к подсем. Omboniinae -
отсутствием спондилия и большим, двулопастным замочным отростком, который у
Orthotetes имеет меньшие размеры. От рода Streptorhynchus King, 1850 (подсем.
Streptorhynchinae) - наличием септы и зубных пластин в брюшной створке.
Видовой состав. Род содержит более 20 видов.
Распространение. Верхний карбон-пермь; повсеместно в СССР и за рубежом.
В сакмарско-нижнеартинских отложениях Юго-Восточного Памира - 2 вида.
Derbyia grandis Waagen, 1884
Табл. Ill, фиг. 1-4
Derbyia grandis': Waagen, 1884, стр. 597, табл. LI,J фиг. 1; табл. LII, фиг. 1,3;
табл. LII1, фиг. 3-5; Чернышев, 1902, стр. 207, табл. XXIV, фиг. 1,2; табл. XXVI,
фиг. 5; Freeh, 1911, стр. 125, табл. XVIII, фиг. 4; Broili, 1916, стр. 7; табл.
CXV (I), фиг. 9.
«
Описание. Раковина крупная, изменчивого очертания, близкого к полуокруг лому,
часто асимметричная. Лобный край ректимаргинатный; боковые дуговидные. Замоч-
ный край немного меньше наибольшей ширины, располагающейся в средней части
84
ковины. Брюшная створка слабо выпуклая иди почти плоская, полуокруглого очерта-
ния^ невысокой плоской апсаклинной ареей 1 треугольного очертания. Дельтирий
широкотреугольный, покрыт слабо выпуклым дельтидием. Макушка короткая, широкая,
иногда слабо асимметричная. Макушечный угол 130 • Спинная створка незначитель-
но и равномерно выпуклая, с краевой ареей. Радиальная скульптура представлена
тонкими, слабо приостренными струйками, одинаковыми на протяжении всей длины
раковины. Количество их увеличивается путем интеркаляции. Струйки разделены до-
вольно широкими, слабо вогнутыми промежутками с неотчетливой, тонкой поперечной
штриховкой. Количество струек 6-7 на 5 мм по лобному краю.
Наружная скульптура отражается на внутренней поверхности в виде широких, плос-
ких струек со слабой поперечной штриховкой, разделенных узкими правильными про-
межутками. Концентрические морщины редкие, неправильные, более отчетливые на
брюшной створке.
Внутри брюшной створки - массивная срединная септа, достигающая 1/4 длины
створки. Мускульные отпечатки лопастные, полуокруглые, отчетливые (табл. Ш,
фиг. 2,3). Зубные пластины расходящиеся, короткие, массивные, располагаются по
краям дельтирия. Они поддерживают небольшие удлиненные зубы, незначительно
выступающие за замочный край. Внутри спинной створки - массивный, двулопастной
замочный отросток с валиком посредине. Он поддерживается прямыми, широкорасхо-
дяшимися круральными пластинами (табл. Ill, фиг. 4). Присутствует слабый септаль-
ный валик.
Размеры, мм
Экз., № Местонахождение, возраст д Ш ША ВА Ш д
2774/1 - брюшная створка 2774/2 - Куртеке 1, сакмарский ярус - нижнеартинский подъярус нерасчлененные, базардаринска! свита (верхняя часть) 67,1 I 69,8
58,3 84,2
спинная створка 2774/3 - брюшная, створка 50,0 61,7 35,5 9,0 10,0.
2774/4 - 49,5 62,2
спинная створка
(ядро)
Сравнение. Рассматриваемый вид отличается от 0.regularis Waagen большими
размерами, неправильной формой и скульптурой. Отличия от D.diversa Reed - см. в
описании последнего.
Геологический возраст и географическое распространение.
Ассельэкий ярус Урала; сакмарский ярус - нижнеартинский подъярус нерасчлененные,
базард арине кая свита (верхняя часть) Юго-Восточного Памира. Средний, верхний
продуктусовый известняк Соляного кряжа. Местонахождение Базлео Тимора; джуль-
финский ярус (слои Допинг) Южного Китая.
Материал. 50 экз. удовлетворительной сохранности, представленных разрознён-
ными створками с сохранившимся раковинным веществом или их ядрами. Собраны
в известковистых глинистых алевролитах верхней части базардаринской свиты Кур-
теке 1 (из них 38 брюшных и 12 спинных створок).
Апсаклйнная арея - арея, плоскость которой наклонена по отношению к плоскости,
разделяющей створки, под угоном 90-180 .
85
J)erbyia diverse Reed, 1944
Табл. II, фиг. 1-4
Derbyia buriensis var. diversd: Reed, 1944, стр. 29—30, табл. IV, фиг. 2, 4, 4a.
Описание. Раковина крупная (47-57 мм длиной и 60-70 мм шириной), очер-
тание от округлого до поперечно-эллипсоидального. Лобный край ректимаргинатный;
боковые - дуговидные. Замочный край прямой, меньше наибольшей ширины раковины,
расположенной в ее средней части. Брюшная створка плоская в макушечной части и
слабо вогнутая в любной. . Макушка небольшая» широкая, прямая. Макушечный угол
140 . Арея плоская, низкая. Дельтирий небольшой, закрыт умеренно выпуклым дель-
тидием. Спинная створка равномерно и умеренно выпуклая, округлая или незначитель-
но поперечно-эллипсоидальная, с макушкой, не выступающей за замочный край.
Скульптура состоит из многочисленных, довольно грубых, правильных, приострен-
ных радиальных струек, увеличивающихся в количестве путем интеркаляции. Число
их 6-7 на 5 мм по лобному краю. Промежутки между ними приблизительно той же
ширины, что и струйки, приостренные, покрыты тонкой поперечной штриховкой. При-
сутствуют редкие, правильные концентрические линии нарастания.
Внутри брюшной створки - зубные пластины, располагающиеся по краям дельти-
рия и поддерживающие зубы. Зубы слабо выступают за замочный край. Мускульные
отпечатки очень слабо вогнутые, продольно-треугольные, с радиально расходящимися
валиками. Срединная септа достигает 1/4 длины створки, не соединяется с зубными
пластинами. Внутри спинной створки - высокий, двулопастной замочный отросток,
отогнутый вентрально. Он поддерживается массивными, слабо изогнутыми» широко
расходящимися круральными пластинами, охватывающими мульскульное поле. Мус-
кульные отпечатки вогнутые, продольно-треугольные, штриховатые. Септальный валик
слабый, короткий.
Размеры, мм
Экз., № Местонахождение, возраст д Ш Т ША ВА Ш д МУ
2774/7 Куртеке 1,’ сакмарс кий 57,4 60,6 21,4 35,1 6,2 7,0 115°
2774/8 - ярус - нижнеартинский 53,4 70,5
спинная створка подъярус нерасчлененные,
2774/10 - спинная створка базардаринская свита (верхняя часть) 47,5 65,0
Сравнение. Derbyia diverse отличается от D.buriensis Reed более высокой отог-
нутой назад ареей брюшной створки и узкими межреберными промежутками, от D.he-
misphaerica Waagen - крупными размерами и низкой, слабо отогнутой назад ареей,
а от D.grandis Waagen- меньшими размерами, изометричным очертанием, более тон-
кой ребристостью.
Геологический возраст и географичес кое распространение.
Нижний продуктусовый известняк Соляного кряжа; сакмарский ярус - нижнеартин-
ский подъярус нерачлененные, базардаринская свита (верхняя часть) Юго-Восточногс
Памира.
Материал. 20 экз. удовлетворительной сохранности; собраны в органогенно-
обломочных алевритистых известняках верхней части базардаринской свиты Куртеке1
(из них - 1 раковина с сочлененными створками» 16 брюшных и 3 спинных створки).
86
ОТРЯД PRODUCTIDA
ПОДОТРЯД PRODUCTIDIN А
НАДОЕ МЕЙ С ТВ О STROPHALOSIACEA
СЕМЕЙСТВО AULOSTEGIDAE MUIR -WOOD ET COOPER, 1960
ПОДСЕМЕЙСТВО AULOSTEGINAE MUIR-WOOD ET COOPER, 1960
Род Taeniothaerus Whitehouse, 1928
Taeniothaerus: Whitehouse, 1928, стр. 281; Prendergast, 1943, стр. 27;Hill,
1950, стр. 6;Coleman^ 1957, стр.85 (part.); Muir—Wood, Cooper, 1960, стр. 97; Muir-
Wood, 1965, стр. 454.
Типовой вид - Productus subquadratus Morris, 1845; нижняя пермь; Тасмания.
Описание. Раковина крупная, толстостворчатая, продольно-овального очертания.
Брюшная створка умеренно выпуклая, с синусом, развитым в различной степени.
Макушка короткая, массивная, загнутая. Арея низкая, вогнутая, меньше наибольшей
ширины раковины. Дельтирий узкий, закрыт элитридием и лофидием. На макушке име-
ется маленький рубец прирастания. Спинная створка коленчатая, с плоским или слабо
вогнутым висцеральным диском и коротким шлейфом, без ареи.
Скульптура брюшной створки состоит из многочисленных косых игл с продольно
вытянутыми вздутиями у основания. По переднему краю обычно присутствуют сбли-
женные концентрические пластины нарастания. Иглы здесь располагаются более час-
то, чем на висцеральном диске. Скульптура спинной створки такая же, как брюшной,
но более тонкая, а концентрические пластины нарастания выражены более отчет-
ливо.
Внутри брюшной створки - слабо вогнутые, продольно-овальные отпечатки аддук-
торов. Отпечатки дидукторов широкотреугольные. Внутри снинной створки - высокий
замочный отросток, прямой или слабо наклоненный внутрь. Он начинается кпереди
от линии смычного края, трехлопастной с наружной стороны. С внутренней стороны
его ствол разделен глубокой, продольной бороздой, так что его основание резко раз-
делено на две части, между которыми вклинивается задний конец длинной, массивной
срединной септы. Следы прикрепления аддукторов удлиненно-треугольные, сзади не-
сколько вдавленные, слабо ветвистые. Замочный отросток поддерживается валиками,
ограничивающими отпечатки аддукторов. Брахиальные валики неотчетливые, прибли-
зительно горизонтальные. Брахиальные отпечатки неясные.
Сравнение. Данный род отличается от рода Aulosteges Helmersen, 1847 крупными
размерами, массивностью раковины, правильной скульптурой, наличием пластин на-
растания у лобного края и низкой, вогнутой ареей. По внутреннему строению - бо-
лее массивным, прямым или слабо наклоненным внутрь замочным отростком, разде-
ленным с внутренней стороны на два валика, между которыми вклинивается основа-
ние септы.
Видовой состав. Кроме типового вида, к данному роду относятся Т.subquad-
ratus var. cracowensis Hill, 1950 из артинских отложений Австралии; Т.notabilis
Reed, 1944 и T.cotteri Reed, 1944 из нижнего продуктусового известняка Соляного
кряжа; T.permixtus Reed, 1932 и Т.brenenais Reed, 1932 из аггломератовых слоев
Кашмира, а также Т.rustucus Grunt,sp.nov. из нерасчлененных отложений сакмарско-
го яруса-нижнеартинского подъяруса Юго-Восточного Памира. Отнесение T.iranicus
Fantini Sestini, 1966, происходящего из карачатьфского яруса Северного Ирана к
рассматриваемому роду, вызывает некоторые сомнения. Замочный отросток T.irani-
cus, изображенный Фантии Сестини (Fantini Sestini, 1966, табл. 2, фиг. 4), несколь-
ко отличается от замочного отростка типичных Taeniothaerus характером сочле-
нения его со срединной септой: у Т.iranicus отсутствует альвеола в основании за-
мочного отростка и септа не вклинивается между его лопастями.
Замечания. Роды Aulosteges и Taeniothaerus чрезвычайно близки между собой,
поэтому среди различных исследователей имеются разногласия по поводу принад-
87
лежности ряда видов к тому или иному роду. Например, T.miniliensis Coleman и Tirwi—
nensis Coleman отнесены А.Д. Гирогорьевой (1962) к роду Aulosteges. Но другая
группа видов» к которой принадлежат Т ,subquadratus, Т.notabilis, Т. otteri, Т,permix-
tiiSf T.brenensis, относятся к роду Taeniothaerus единодушно. От W у at kina Frede-
rick^! 931» Taeniothaerus отличается отсутствием дельтириальных гребней, более ко-
роткой вогнутой ареей брюшной створки, менее отчетливыми пластинами нарастания,
несущими правильные концентрические ряды игл по переднему краю, которые у
Wyatkina имеют тенденцию располагаться радиальными радами. Некоторые авторы
(А.Д. Григорьева, 1962) считают род Wyatkina синонимом рода Aulosteges.
Г.В. Котляр (1964) выделила в составе рода Waagenoconcha семейства Echinocon—
chidae додрод Reedoconcha с типовым! видом Pr.(Taeniothaerus) permixtus Reed, 1932
из аггломератовых слоев нижней перми Кашмира. К этому подроду Г.В. Котляр от-
носит так же Т.brenensis Reed, Pr.lidorensis Diener, Pr.(Reedoconcha) sosninae Kot—
Ijar. Как было отмечено выше, названные виды Рида из аггломератовых слоев Каш-
мира большинством исследователей уверенно относятся к роду Taeniothaerus, Неяс-
ными остаются причины, по которым Г.В. Котляр рассматривает этих характерных
представителей семейства Aulostegidae в качестве подрода Waagenoconcha, сближая
их с последними.
Распространение. Нижняя пермь Австралии и Тасмании^ нижний продуктусовый
известняк Соляного кряжа (Пакистан); аггломератовые слои Кашмира; сакмарский
ярус - нижнеартинский подъярус нерасчлененные, базардаринская свита (верхняя
часть) Юго-Восточного Памира; карачатырский ярус Северного Ирана (?).
Taeniothaerus rusticus Grunt, sp.nov.
Табл. IV, фиг. 1-6; рис. 15, 16
Видовое название от rusticus (лат.) - грубый.
Голотип - ПИН, № 2228/542; брюшная створка неполной сохранности; Юго-Вос-
точный Памир» Куртеке 1; сакмарский ярус - нижнеартинский подъярус нерасчле-
ненные» базардаринская свита (верхняя часть); табл.IV» фиг 1.
Описание. Раковина крупная (размеры голотипа: 54,5 мм в длину и 52,4 мм в
ширину; размеры одного из самых крупных экземпляров, имеющихся в коллекции:
58»5 мм в длину и 77 мм в ширину), четырехугольного очертания, приблизительно
изометричная. Замочный край меньше наибольшей ширины раковины, которая приуро-
чена к ее средней части. Брюшная створка умеренно выпуклая, с узким синусом и
плоскими» треугольными» слабо обособленными ушками. Макушка массивная, с ма-
леньким рубцом прикрепления, умеренно загнутая. Макушечный угол 90 . Арея низ-
кая (3,1 мм высотой), вогнутая, треугольная, с узким треугольным дельтирием. Спин-
ная створка коленчатая» с плоским или слабо вогнутым висцеральным диском и ко-
ротким шлейфом. Арея отсутствует.
Скульптура обеих створок состоит из часто расположенных косых игл с продоль-
ными вздутиями в основании. Иглы брюшной створки более длинные» чем спинной, рас-
полагаются в относительно правильном шахматном порядке. На ушках иглы крупные,
прямые. По лобному краю наблюдаются сближенные концентрические пластины нарас-
тания с тонкими частыми иглами. На ушках присутствуют концентрические морщинки.
Внутреннее строение брюшной створки не наблюдалось. Внутри спинной - длинный»
массивный трехлопастной с внешней стороны замочный отросток, слабо наклоненный
внутрь. Он располагается немного кпереди от линии замочного края. С внутренней
стороны ствол замочного отростка разделен глубокой продольной бороздой. Замочный
отросток поддерживается расплывчатыми валиками (табл. IV »фиг. 2»3),которые ограни-
чивают с задней и с задне-боковых сторон слабо ветвистые выпуклые отпечатки ад-
дукторов. Срединная септа тонкая, достигает 2/3 длины висцерального диска и сво-
им основанием вклинивается между частями замочного отростка. Брахиальные вали-
ки короткие. Брахиальные отпечатки округло-треугольные, неотчетливые (см.рис„15).
88
Изменчивость и онтогенетические изменения. Изменчивым являет-
ся очертание раковины, которое варьирует от удлиненно-треугольного до удлиненно-
четырехугольного, высота ареи, а также степень выраженности рубца прикрепления
и синуса брюшной створки. Юные экземпляры, имеющие размеры от 13 до 20 мм в
длину и от 12 до 26 мм в ширину, характеризуются относительно тонкими створками,
отсутствием синуса брюшной створки, более отчетливой, чем у взрослых особей,пло-
щадкой прикрепления, окруженной иглами. Мускульные отпечатки у них расплывчатые,
мелко—штриховатые. Валики, ограничивающие мускульные отпечатки,более резкие
(см. рис. 16).
Рис. 15^ Т aeniothaerus rusti cus. Схема
внутреннего строения спинной створки (ре-
конструкция по номерам: ПИН, № 2228/540
и ПИН, № 2228/548). Куртеке 1; сакмарский
ярус - нижнеартинский подъярус нерасчле-
ненные, базардаринская свита (верхняя часть)
Рис. 16. Т aeniothaerus rusticus, ПИН,
№ 2228/544; внутреннее строение спинной
створки юного экземпляра, х 1.8. Местонахож-
дение и возраст те же, что и на рис. 15
Сравнение. От T,iranicus Fantini Sestini Т.rusticus внешне отличается продоль •
но вытянутой формой раковины и грубой скульптурой. Имеются также существенные
отличия во внутреннем строении этих видов, которые были рассмотрены выше и
которые позволяют высказать сомнение в отнесении последнего вида к роду Т aenio-
thaerus.От T.cotteri Reed рассматриваемый вид, кроме того, отличается наличием
синуса, а от Т .permixtus Reed и от Т.brenensis — более крупными размерами.
Геологический возраст и географическое распространение.
Сакмарский ярус - нижнеартинский подъярус нерасчленные, базардаринская свита
(верхняя часть) Юго-Восточного Памира.
Материал. 64 экз. удовлетворительной сохранности собраны в верхней части
базардаринской свиты. В органогенно-обломочных алевритистых известняках Курте-
ке 1 найдена 1 раковина с сочлененными створками, 10 брюшных и 17 спинных ство-
рок взрослых особей; 37 брюшных и 8 спинных створок юных. В известковистых гли-
нистых алевролитах Кастанат-Джилги 1 найдена 1 крупная спинная створка.
СЕМЕЙСТВО CHONOSTEGIDAE MUIR-WOOD ET COOPER, 1960
Р од Strophalosiina Licharew, 1935
Strophalosiina (Strophalosiina); Licharew, 1935, стр. 370; 1937, стр. 77.
Strophalosiina: Сарычева, Лихарев, Сокольская, I960, стр. 236; Muir—Wood, Coo-
per, 1960, стр. 115; Muir—Wood, 1965, стр. 457; Сарычева, Грунт, 1969,стр. 73.
Типовой вид - Aulosteges tibeticus Diener, 1897; верхняя пермь, известняк
Читичун , Гималаи
89
Описание. Раковина маленькая, округло-пятиугольная, коленчатая. Брюшная
створка с рубцом прикрепления и высокой треугольной ареей, несущей узкий, вы-
пуклый псевдодельтидий; имеется синус, развитый в разной степени. Спинная створ-
ка с плоским висцеральным диском и коротким шлейфом. По переднему краю брюш-
ной створки наблюдается слабая отогнутость со следами шипов на внутренней сто-
роне. По переднему краю спинной - гладкий, отогнутый назад ободок, отсутствую-
щий на боковых краях. В месте отгиба ободка - ряд наружных игл, каждая из
которых соответствует ребру, расположенному на шлейфе.
Скульптура обеих створок из округлых радиальных ребер, расположенных на шлей-
фе и в передней части висцерального диска или только на шлейфе и из правильных
концентрических морщин, занимающих весь висцеральный диск. На брюшной створке
как ребра, так и морщины выражены более отчетливо, чем на спинной. На висце-
ральном диске брюшной створки - редкие иглы с вздутыми основаниями, на спин-
ной - ямки, соответствующие этим иглам.
Внутреннее строение типового вида неизвестно. Судя по имеющемуся у нас ма-
териалу ( S.multicostata Licharew) из верхней перми Юго-Восточного Памира, внут-
ри спинной створки имеются кардинальные валики, несколько отклоняющиеся от за-
мочного края и тонкая срединная септа, достигающая края висцерального диска.
По бокам от септы небольшие, слабо выпуклые отпечатки передних и задних аддук-
торов. Между ними отходят косо расположенные брахиальные валики; они отчетли-
вые, треугольно-эллипсоидальные (см. рис. 17).
Сравнение. Род $trophalosiin а отличается от рода Chonosteges Muir—Wood et
Cooper иной формой резко коленчатой раковины, невысокой ареей и меньшим количест-
вом игл на всей раковине. Ободки обеих створок и иглы на них развиты слабее,
чем у американского рода. От рода Urush tenia Licharew род Strophalosiina отлича-
ется более высокой ареей, а также отсутствием маргинальной пластинки. От Cho—
nostegqides Sarytcheva - коленчатой формой раковины, наличием концентрических
морщин на висцеральном диске, отсутствием фестончатых пластин нарастания по
лобному краю брюшной створки, а до внутреннему строению - отсутствием марги-
нальной пластинки.
Видовой состав. Кроме типового вида, к данному роду относятся S.cauca-
sica Licharew, 1937f и S.multicostata Licharew, 1937 из уруштенского горизонта джуль-
финского яруса Северного Кавказа. Последний вид встречается так же в джульфин-
ском ярусе (среднетахтабулакская подсвита) Юго-Восточного Памира.
Замечания. В характеристике рода Б.К. Лихарев отмечает отсутствие рубца
прикрепления (Лихарев, 1937, стр. 77). Однако при описании типового вида (Ли/о-
stages tibeticus Diener) Линер указывает, что рубец прикрепления имеется или отсут-
ствует (Diener, 1^97а, стр. 35). У Мюр-Вуд и Купера (Muir— Wood,Cooper, 1960,
стр. 115) имеется указание на наличие наружного ободка по переднему краю ракови-
ны, но следы игл на нем не отмечены. Однако на приведенных фотографиях S.tibe—
tica(Diener) (там же, табл. 16, фиг. 6,8-11) отчетливо видна не только отогну-
тость, но и углубления от ряда игл по переднему краю брюшной створки в основании
отогнутого ободка, а также широкий гладкий ободок по переднему краю спинной створки.
Распространение. Известняк Читичун Гималаев; джульфинский ярус: уруш-
тенский горизонт Северного Кавказа; среднетахтабулакская подсвита Юго-Восточного
Памира.
Strophalosiina multicostata Licharew, 1937
Табл. V, фиг. 1,2; рис. 17
Strophaloaiina (Strophalosiina) multicostata: Лихарев, 1937, стр. 82, табл.
XII, фиг. 2—3.
Голотип - ЦНИГРмузей им. Ф.Н. Чернышева,№ 1617/2139, брюшная створка;
джульфинский ярус, уруштенский горизонт; Северный Кавказ; Лихарев, 1937, табл.XII
фиг. 2.
Описание. Раковина маленькая (11-14,5 мм в длину и 10,5-18 мм в ширину),
округло-пятиугольная, коленчатая, с коротким висцеральным диском. Замочный край
90
меньше наибольшей ширины, расположенной в средней части раковины. Брюшная створ-
ка с коленчатым висцеральным диском и коротким шлейфом. Макушка низкая, при-
остренная, с маленьким рубцом прикрепления. Арея невысокая (4,1 мм), треуголь-
ная, слабо асимметричная, располагается приблизительно в плоскости,разделяющей
створки (ортоклинная), с тонкой горизонтальной штриховкой, с узким дельтирием, по-
крытым псевдодельтидием. Ушки отсутствуют. На висцеральном диске и шлейфе -
слабая приплюснутость. По переднему краю - слабая отогнутость типа наружного
ободка и следы внутренних игл. Спинная створка с плоским или слабо вогнутым
висцеральным диском полуокруглого очертания и шлейфом, отогнутым под прямым
углом. По переднему краю створки - гладкий, не несущий скульптуры ободок; на
боковых краях от не развит.
Рис. 17. Strophalosiina multicostata, ПИН,
№2228/555; внутреннее строение спинной створки
(внутреннее ядро), х 3.6. Кур-Истык 1; джуль-
ский ярус, среднетахтабулакская подсвита
Скульптура состоит из отчетливых радиальных ребрышек, начинающихся на вис-
церальном диске, несколько отступя от макушки. Ребра правильные, ровные, разде-
лены промежутками равной с ними ширины. Количество их 7-8 на 5 мм поверхности
у лобного края. В области шлейфа они несут очень тонкую концентрическую штри-
ховку. В примакушечной части брюшной створки - иглы с округлыми вздутыми ос-
нованиями. Висцеральный диск спинной створки покрыт тонкими концентрическими
линиями нарастания и рассеянными ямками - следами игл брюшной створки. По краю
наружного ободка спинной створки располагаются иглы, являющиеся продолжением
ребрышек, развитых на шлейфе.
Внутреннее строение приводится при характеристике рода (см. рис. 17).
Сравнение. Рассматриваемый вид отличается от S.tibetica несколько меньшими
размерами раковины, отсутствием синуса на висцеральном диске и шлейфе брюшной
створки, а также радиальной скульптурой, располагающейся не только на шлейфе, но
частично и на висцеральном диске. От S.caucasica отличается более тонкой радиаль-
ной ребристостью на шлейфе и отсутствием удлиненных вздутых оснований игл на вис-
церальном диске, сливающихся в ребра.
Геологический возраст и географическое распространение.
Джульфинский ярус: уруштенский горизонт Северного Кавказа; среднетахтабулакская
подсвита Юго-Восточного Памира.
Материал. 7 экз. хорошей сохранности собраны в отложениях среднетахтабу-
лакской подсвиты. Из них в табулятовых биогермных известняках Шор-Булака -1 ра-
ковина с сочлененными створками, 2 брюшных и 2 спинных створки; Музду-Була-
ка - 1 раковина с сочлененными створками; в табулятово-гидроидных биогермных
известняках Кур-Истыка 1-1 раковина с сочлененными створками.
СЕМЕЙСТВО SCACCHINELLIDAE LICHAREW, 1928
Род Scacchinella Gemmellaro, 1897
Scacchinella : Gemmellaro, 1891b,стр. 22 (nom.nud.); 1892, стр.26 nom.nud.); 1897,стр. 114;
Schellwien, 1900b, стр. 33, Meria, 1928,стр. 77; Лихарев,1928, стр.271; 1960г,стр.235;
Stehli, 1954,стр. 333; Muir-Wood,Cooper,1960, стр. 129; Muir-Wood, 1965, стр. 461; Rud wick
Cowen, 1968, стр. 126. ’
Типовой вид - S.variabilis Gemmellaro, 1897; мургабский ярус, известняк
Созио; Сицилия.
91
Описание. Раковина от средних до крупных размеров, обычно асимметричная,
округло-треугольная, брюшная створка коническая, с рубцом прирастания и высокой
ареей, покрытой неправильными продольными струйками; без дельтирия.
Скульптура брюшной створки состоит из ризоидных игл, расположенных не боко-
вых склонах и в примакушечной части висцерального диска. На арее иглы отсутству-
ют. Спинная створка плоская или слабо выпуклая, с мелкими многочисленными игла-
ми, без ареи. Внутри брюшной створки - длинная срединная септа, входящая между
лопастями замочного отростка спинной створки. Замочный отросток состоит из двух
широко расходящихся лопастей, отогнутых вентрально. Боковые валики узкие. Средин-
ная септа короткая. Мускульные отпечатки дендровидные, массивные* Брахиальные
отпечатки небольшие, гладкие, округло-четырехугольные.
Рис. 18. Scacchinella licharewi, ПИН,
№ 2228/556; внутреннее строение спин-
ной створки, х1.8.Шор-Булак; джульфинский
ярус, среднетахтабулакская подсвита
Сравнение. От рода Derbyella Grabau, 1931,. род Scacchinella отличается от-
сутствием ребристости на раковине, а так же отсутствием дельтирия, покрытого дель-
тидием. Внутреннее строение Derbyella неизвестно. По внутреннему строению Scac-
chinella сходна с Тschemyschewia Stoyanow, 1910, но резко отличается конической
формой раковины.
Видовой состав. Кроме типового вида, в состав рода входят S.americana
Stehli, 1954 из нижней перми Техаса, S.depressa Gemmellaro, 1897 и S.variabilis
var. erecta Gregorio, 1930 из известняка Созио мургабского яруса Сицилии; S.gfgan-
tea Schellwien, 1900 из трокгофельского известняка Карнийских Альп й из нижней
перми Ферганы, Центральной Азии, Японии и Аляски; S.yakovlewi Licharew,1928 из
джульфинского яруса Северного Кавказа и S.Hcharewi Grunt,sp. nov. из джульфин-
ского яруса Северного Кавказа и Юго-Восточного Памира.
Распространение. Нижняя пермь Техаса, Ферганы, Центральной Азии, Японии,
Аляски; трокгофельский Известняк; мургабский ярус , известняк Созио Сицилии; джуль-
финский ярус, уруштенский горизонт Северного Кавказа; среднетахтабулакская под-
свита Юго-Восточного Памира.
Scacchinella licharewi Grunt, sp.nov.
Табл. Vt фиг. 3,4; рис. 18
Видовое название в честь Б.К. Лихарева.
Scacchinella yakovlewi var.; . Лихарев, 1928, стр. 269, табл. XXIV» фиг. 3..
Голотип - ПИН,№ 2228/556; раковина с двумя сочлененными створками; Юго-
Восточный Памир, Шор-Булак; джульфинский ярус, среднетахтабулакская подсвита ;
табл. V , фиг. 3.
О писание. Раковина средней величины (размеры голотипа: длина - 34,5 мм, ши-
рина - 34,5 мм, толщина - 22,9 мм, высота ареи - 9,5 мм, ширина ареи - 15,5 мм),
округло-треугольного очертания, с наибольшей шириной по лобному краю. Брюшная
92
створка коническая. Макушка неширокая (макушечный угол 61°), слабо загнутая, с
маленьким рубцом прирастания. Арея в виде равностороннего треугольника, незна-
чительно вогнутая, ортоклинная, без дельтидия. Посредине ареи проходит слабое про-
дольное возвышение. Наружная поверхность ее покрыта тонкой горизонтальной штри-
ховкой. Внутренний раковинный слой несет грубую вертикальную штриховку. Спинная
створка очень слабо выпуклая, поперечно-овальная, без ареи. Макушка не выражена .
На поверхности обеих створок многочисленные тонкие иглы, расположенные в бо-
лее или менее правильном шахматном порядке. На раковине имеются редкие неот-
четливые концентрические складки.
Внутри брюшной створки - длинная срединная септа, в макушечной части опираю-
щаяся на дно створки и арею и не доходящая до дна на более поздних стадиях. Ее
дистальный конец вклинивается между двумя сильно расходящимися лопастями за-
мочного отростка спинной створки. В спинной створке - короткая, тонкая, срединная
септа, достигающая половины длины висцерального диска. Лопасти замочного отрост-
ка опираются на мускульные поля. Мускульные отпечатки массивные, слабо ветвис-
тые, удлиненно-треугольные. Кардинальные валики короткие, косо отходят от замоч-
ного края сзади от основания замочного отростка. Брахиальные отпечатки округло-
квадратные, нерезкие (см. рис. 18). Внутренняя поверхность брюшной створки покры-
та многочисленными сосочками, а спинной - внутренними иглами.
Сравнение. Описываемый вид отличается от S. variabilis более крупнььми раз-
мерами, более правильной формой раковины и почти полным отсутствием концентри-
ческой скульптуры; от S. yakovlewi - отсутствием синуса и коленчатого перегиба
на брюшной створке, а так же выступа на спинной, а от S. gigantea - более сим-
метричной макушкой и меньшими размерами.
Геологический возраст и географическое распространение.
Джульфинский ярус: уруштенский горизонт Северного Кавказа; среднетахтабулакская
подсвита Юго-Восточного Памира.
Материал. 8 экз. удовлетворительной сохранности; собраны в табулятовых
биогермных известняках среднетахтабулакской подсвиты Шор-Булака. Из них 2 ра-
ковины с сочлененными створками, 3 брюшных и 3 спинных створки.
НАДСЕМЕЙСТВО RICHTHOFENLACE А
СЕМЕЙСТВО RICHTHOFENIIDAE WAAGEN, 1884
ПОДСЕМЕЙСТВО RICIITIIOFENIIN АЕ WAAGEN, 1884
Род Richthofenia Kayser, 1881
Richthofenia caucasica Licharew, 1928,
Табл. V, фиг. 5-7
Richthofenia caucasica: Лихарев, 1928, стр. 261-269, табл. XXIII, фиг. 1—4;
Simic, 1935а, стр. 147—149, 154, табл. XIV, фиг. 1-5; Сарычева, 1965, табл. XXXIV,
фиг. 4.
Лектотип - экземпляр, изображенный Лихаревым (Лихарев, 1928, табл. ХХП1,
фиг. 1), хранится в ЦНИГРмузее им. Ф.Н. Чернышева под номером 2/2553; джуль-
финский ярус, уруштенский горизонт Северного Кавказа.
Геологический возраст и географическое распространение.
Мургабский ярус, хачикский горизонт Закавказья: джульфинский ярус: зона Waage-
nophyllum indicum Западной Сербии; уруштенский горизонт Северного Кавказа; сред-
нетахтабулакская подсвита Юго-Восточного Памира.
Материал. 5 брюшных створок удовлетворительной сохранности; собраны в
отложениях среднетахтабулакской подсвиты; из них - в табулятовых биогермных из-
вестняках Шор-Булака - 3 экз.; в табулятово-гидроидных биогермных известняках
К/р-Истыка 1-1 экз.; Куталя 2-1 экз.
93
НАДСЕМЕЙСТВО PRODUCTACE'A
СЕМЕЙСТВО -AVONIIDAE SARYTCHEVA, 1960
Pод Krotovia Fredericks, 1928
Krotovia jisuensiformis Sarytcheva, 1965
Табл. V, фиг. 10
Krotovia jisuensiformis: Сарычева, 1965, стр. 216, табл. XXXIV, фиг. 5-8.
Голотип - ПИН,№ 2071/78; брюшная створка; Закавказье, правый берег р.Ве-
ди; мургабский ярус, гнишикский горизонт; Сарычева, 1965, табл. XXXIV \ фиг. 5.
Геологический возраст и географическое распространение.
Мургабский ярус, гнишикский и хачикский горизонт Закавказья; джульфинский ярус,
среднетахтабулакская подсвита Юго-Восточного Памира.
Материал. 32 экз. удовлетворительной сохранности. Собраны в среднетахта-
булакской подсвите; из них в табулятовых биогермных известняках Музду-Булака -
6 брюшных створок, Шор—Булака - 3 брюшных и 1 спинная створка; в табулятово-
гидроидных биогермных известняках Кур-Истыка 1-7 брюшных и 5 спинных ство-
рок; Куталя 1-1 брюшная створка; Куталя 2-9 брюшных створок.
СЕМЕЙСТВО OVERTONIIDAE MUIR-WOOD ET COOPER, 1960
ПОДСЕМЕЙСТВО OVERTONIINAE MUIR-WOOD ET COOPER, 1960
Р oq Overtoniina Grunt, gen. nov.
Название указывает на близость к роду Overtonia Thomas, 1914*
Типовой вид - Overtoniina mamazairica Grunt, sp.nov.; Юго-Восточный Па-
мир; дарвазский ярус, кызылджилгинский горизонт, нижний подгоризонт.
Описание. Раковина небольшая, полуокруглая. Брюшная створка с умеренно
выпуклым висцеральным диском и шлейфом, отделенным более или менее оезким
коленчатым перегибом. Спинная створка с уплощенным висцеральным диском и
коротким отогнутым шлейфом. Висцеральная полость умеренная.
Скульптура висцерального диска обеих створок состоит из правильных, невысо-
ких концентрических морщин, прослеживающихся по всему диску и переходящих на
ушки, где они сближены и более резки. Морщины разделены узкими, слабо вогну-
тыми промежутками примерно равной с морщинами ширины. Иглы брюшной створки
располагаются на морщинах в один ряд. Основания их округлые, слабо вздутые.
На висцеральном диске спинной створки иглы более тонкие, мелкие, частые, без
вздутий в основании. В промежутках между морщинами, кроме того, присутствуют
округлые ямки, отражающие места прикрепления игл брюшной створки. Шлейфы
брюшной и спинной створок обычно гладкие, иногда со слабо намечающимися реб-
рышками и тонкими линиями нарастания.
Внутри брюшной створки - округло-треугольное мускульное поле, занимающее
макушечную область и прол ягивающееся на 1/4 длины висцерального диска. Мус-
кульное поле состоит из маленьких овально-треугольных гладких отпечатков задних
аддукторов, разделенных слабым, тонким продольным ребром, передних продольно-
штриховатых треугольных отпечатков аддукторов и примыкающих к ним с боков треу-
гольных продольно-штриховатых отпечатков ди дукторов.
Внутри спинной створки - глубоко двураздельный замочный отросток. От его ос-
нования отходят массивные, изогнутые кардинальные валики, распространяющиеся
приблизительно до половины длины замочного края. Срединная септа короткая, менее
1/4 длины висцерального диска, с широким, слабо расширяющимся основанием и мас-
сивным вздутием на переднем конце. Отпечатки аддукторов гладкие, небольшие, гру-
94
шевидные, располагаются: непосредственно под макушкой, на возвышенной платформе,
особенно приподнятой спереди. Отпечатки аддукторов состоят из двух частей: перед-
них и задних отпечатков, окаймленных массивными валиками и с задне-боковых сто-
рон ограниченных глубокими ложбинками. Брахиальные валики отходят от передне-
боковых сторон мускульных отпечатков и располагаются косо под углом 40-45
к септе. Брахиальные отпечатки грушевидные, достигают 1/3 длины висцерального
диска. На внутренней поверхности спинной створки присутствуют пологие концентри-
ческие морщины, отражающие наружную скульптуру. Между ними частые следы внут-
ренних игл. Передние 2/3 поверхности усеяны удлиненными приостренными шпиками
(см. рис. 19).
Сравнение. Род Overtoniina отличается от рода Overtonia Thomas, 1914 по внеш-
нему виду присутствием более слабо развитых нерезких частых концентрических
морщин, несущих редкие иглы, отсутствием прерывистой радиальной ребристости,
характерной для Overtonia, а так же наличием коленчатого отогнутого гладкого
шлейфа на обеих створках. По внутреннему строению - короткими кардинальными ва-
ликами, не переходящими в боковые, и наличием валика, окаймляющего мускульные
отпечатки. От рода J аки topro ductus Kaschirzew, 1959. наш род внешне отличается,
кроме наличия гладкого шлейфа, скульптурой, состоящей из редких игл, распола-
гающихся на округлых основаниях (у J аки topro ductus иглы косые, располагаются
в шахматном порядке на удлиненных основаниях), а по внутреннему строению - бо-
лее короткой срединной септой и гладкими мускульными отпечатками спинной створ-
ки. От рода Avonia Thom as,1914 род Overtoniina отличается отсутствием радиаль-
ной ребристости на висцеральном диске и шлейфе, а также иным внутренним строе-
нием.
Видовой состав. Кроме типового вида, к данному роду, видимо, можно от-
нести экземпляры, описанные Хуаном (Huang, 1932) из известняка Маокоу Южного
Китая, провинция Гуйчжоу под названием Plicatiferatf) minor (Schellwien) (Huang,
1932, стр. 38, табл. Ill, фиг. 1-4). В.И. Устрицкий (Устрицкий и др., 1960) после
непосредственного изучения китайского материала предложил выделить их в само-
стоятельный вид Plicatifera huangi Ustrizkiy, I960, Этот вид характеризуется час-
тыми правильными концентрическими морщинами, несущими редкие иглы с округло
вздутыми основаниями. Внутреннее строение известно частично: замочный отросток
двухлопастной очень маленький, а срединная септа достигает почти половины длины
висцерального диска. Предположительно сюда может быть отнесена Overtonia cf.
fimbriatus Sowerby, 1824 из нижней перми Афганистана (Reed, 1931а, стр. 16, т$бл.П,
фиг. 13), характеризующаяся неотчетливыми концентрическими полосами и редкими
иглами на них.
Замечания. Развитие раннекаменноугольного рода Overtonia шло, по-видимому,
в двух направлениях. Представители Fimbriaria приобрели более резкую скульптуру,
состоящую из широких, правильных концентрических полос и частых, крупных игл.
Overtoniina, наоборот, характеризуется скульптурой из тонких, нерезко очерчен-
ных морщин с редкими, мелкими иглами на них.
Распространение. Дарваэский ярус, кызылджилгинский горизонт, нижний
подгоризонт Юго-Восточного Памира; предположительно нижняя пермь Афганис-
тана; верхняя пермь (известняк Маокоу) Южного Китая»
Overtoniina mamazairica Grunt,. sp. nov.
Табл. V, фиг. 8,9; рис. 19
Видовое название по местонахождению в Мамазаире.
Голотип - ПИЬЦЬЬ 2228/590; ядро раковины с сочлененными створками; Юго-
Восточный Памир, Игримиюз; дарваэский ярус, кызылджилгинский горизонт, нижний
подгоризонт; табл. V , фиг.8.
Описание. Раковина небольшая (размеры голотипа; 20 мм длиной и 25 мм ши-
риной), полуокруглая, с наибольшей ширийой в ее передней половине. Брюшная створ-
ка с умеренно выпуклым висцеральным диском и шлейфом, отделенным резким пере-
95
гибом. Макушка маленькая, широкая. Макушечный угол 110-112°. Ушки маленькие,
прямоугольные, неотчетливо ограниченные, с прямоугольными краями. Спинная створ-
ка слабо вогнутая, полуокруглая, с коротким отогнутым . шлейфом.
Рис. 19. Overtoniina mamazairica,
ПИН, № 2228/590; внутррпнее строе-
ние спинной створки, х 2.7. Игрими-
юз; дарвазский ярус, кызылджилгинский
горизонт, нижний подгоризонт
Скульптура состоит из частых, правильных концентрических морщин округлого
сечения, разделенных промежутками равной с ними ширины; они прослеживаются по
всей поверхности висцерального диска, иногда выклиниваясь или сливаясь на макушеч-
ных склонах. На ушки они переходят, располагаясь под прямьш углом к линии за-
мочного края. На морщинах висцерального диска брюшной створки - по одному ряду
редких игл, располагающихся на округлых вздутых основаниях. В промежутках меж-
ду морщинами спинной створки - редкие, округлые ямки, соответствующие иглам
брюшной створки. Кроме того, по всей поверхности висцерального диска спинной створ-
ки рассеяны тонкие, мелкие, прямые иглы. Шлейф гладкий, иногда со слабо намечаю-
щейся ребристостью и отдельными мелкими иглами.
Внутреннее строение описано при характеристике рода (см. рис. 19).
Сравнение. Данный вид наиболее близок к О. (?) huangi Ustrizkiy, происходящей
из известняка Маокоу Юх<ного Китая. Отличается более коротким шлейфом, редкими
иглами на брюшной створке, короткой, незагнутой макушкой и отсутствием синуса
брюшной створки.
Геологический возраст и географическое распространение.
Дарвазский ярус, кызылджилгинский горизонт, нижний подгоризонт Юго-Восточного
Памира.
Материал. 30 экз. удовлетворительной сохранности, собраны в нижнекызыл-
джилгинском подгоризонте. В криноидно-детритовых известняках Куртеке 2 найдены 9
брюшных створок; Мамазаир-Булака - 12 брюшных и 2 спинных створки, Игримиюза - 1
раковина с сочлененными створками, 4 брюшных и 1 спинная створка, Кастанат-
Джилги 2-1 раковина с сочлененными створками.
СЕМЕЙСТВО DICTYOCLOSTIDAE STEHLI, 1954
ПОДСЕМЕЙСТВО DICTYOCLOSTIN'AE STEHLI, 1954
Род Reticulatia Muir-Wood et Cooper, I960
Reticulatia: Muir-Wood, Cooper, 1960, стр, 284; Muir-Wood, 1965, стр. 496.
Типовой вид - Productus hиecoensis King, 1931; нижняя пермь, известняк Хуэ-
ко; Техас.
Описание. Раковина крупная, округло-четырехугольного очертания. Брюшная
96
створка коленчатая, с умеренно выпуклым висцеральным диском. Ушки большие, ци-
линдрически свернутые. Висцеральный диск спинной створки уплощенный.
Скульптура состоит из частых, правильных радиальных ребер и хорошо развитых
концентрических морщин. Пересечение их создает ясный сетчатый орнамент. Ни реб-
ра, ни морщины не переходят на ушки. Характерным является наличие длинного ряда
игл, располагающегося у основания ушек и отделяющего их от висцерального диска.
Кроме того, присутствует один-два ряда игл вдоль замочного края. Редкие иглы
рассеяны По всей поверхности висцерального диска и шлейфа. Иглы спинной створки
очень редки.
Внутри брюшной створки - веерообразные отпечатки мускулов-закрывателей и силь-
но выступающие овальные отпечатки мускулов-открывателей. Внутри спинной створ-
ки - небольшой трехлопастной замочный отросток. Септа с расширенным основанием
протягивается приблизительно на 3/4 длины висцерального диска. Кардинальные ва-
лики короткие, изогнутые, не достигают боковых краев створки. Мускульные отпе-
чатки массивные, выпуклые, широкотреугольные, ветвистые, состоящие из двух пар:
передних и задних отпечатков. Брахиальные валики короткие, приблизительно гори-
зонтальные, брахиальные отпечатки округло-четырехугольные.
Сравнение. Данный род отличается от рода Dictyoclostus Mui г-Wood, 1930 более
отчетливой сетчатой скульптурой, наличием косого ряда игл в основании ушек, а не
пучками игл на них. По внутреннему строению - более выпуклыми массивными мус-
кульными отпечатками спинной створки. От рода Antiquatonia Miloradovich, 1945
род Reticulata отличается более крупными размерами раковины, отсутствием резкой
складки, отделяющей ушки от висцерального диска, как правило, присутствующей у Дп-
tiquatonia. Кроме того, Reticulatia характеризуется более резкой отчетливой сетча-
той скульптурой. По внутреннему строению она отличается менее развитой горизон-
тальной частью брахиальных валиков.
Видовой состав. Кроме нижнепермских отложений (известняк Хуэко) Техаса
(типовое местонахождение) типовой вид широко распространен в верхнекаменноугольно-
нижнепермских отложениях Северной Америки. Кроме типового вида, к данному роду
относится Dictyoclostus newelli King, 1938 из формации Грахам верхнего карбона
Техаса и Reticulatia pamirica Grunt, sp. nov. из верхней части базардаринской свиты
сакмарского яруса - чижнеартинского подъяруса нерасчлененных Юго-Восточного Па-
мира. Мюр-Вуд и Купер высказывают предположение, что сюда принадлежат также
некоторые азиатские виды, в частности, Pro due tus uralicus Tschem., известный из ниж-
ней перми Урала, Средней Азии, Куньлуня и Productus transversalrs Tschem. из ниж-
ней перми Урала, Читраля и Каракорума. Возможно, к этому же роду относятся не-
которые экземпляры из группы Productus semireticulatus Martin , встречающиеся
в нижней перми Евро-Азиатской части Тетиса. Но окончательное решение этого вопро-
са требует тщательной ревизии материала.
Распространение, Верхний карбон — нижняя пермь Северной Америки; ассель—
ский ярус Ферганы, Куньлуня, Каракорума; сакмарский ярус - нижнеартинский подъ-
ярус нерасчлененные, базардаринская свита (верхняя часть) Юго-Восточного Пами-
ра; нижняя пермь Урала.
Reticulatia pamirica Grunt, sp.nov.
Табл. VI, фиг. 1-4; табл. VII, фиг. 1; рис. 20
Видовое название до местонахождению на Памире.
Голотип - ПИН, № 2228/582; раковина с сочлененными створками; Юго-Восточ-
ный Памир, Куртеке 1; сакмарский ярус - нижнеартинский подъярус нерасчлененные,
базардаринская свита (верхняя часть); табл. VI, фиг. 1.
Описание. Раковина крупная (размеры голотипа: длина 56,3 мм; ширина 64 мм),
относительно тонкостворчатая, округло-четырехугольного очертания, с наибольшей
шириной по лобному краю. Брюшная створка с умеренно выпуклым висцеральным
диском и колония то изогнутым шле;1фом. Замочный край прямой, длинный. Ушки боль-
шие, треугольные, умеренно свернутые, отделены от висцерального диска пологой,
расширяющейся к боковым краям ложбинкой. Макушка широкая, округлая, слабо заг-
97
7 264
нутая. Макушечный угол 90-05 . Синус узкий, неглубокий, начинается немного отсту-
пя от макушки и слабо расширяется по направлению к лобному краю. Спинная створ-
ка с умеренно вогнутым висцеральным диском и коротким шлейфом.
Скульптура состоит из правильных, ровных радиальных ребер, прослеживающихся
ио всей поверхности обеих створок. Количество их 2-3 на 5 мм у лобного края. Реб-
ра резкие, слабо при остренные, разделены немного более широкими, чем сами ребра
плоскими промежутками. Концентрические морщины частые, правильные, отчетливые,
выражены с той же степенью интенсивности, что и радиальные рёбра. Одинаковая
резкость радиальной и концентрической скульптуры создает отчетливый сетчатый
орнамент. Основания ушек на брюшной створке отделены от висцерального диска
рядом игл, количество которых 8-0 с каждой стороны от макушки. Кроме того, при-
сутствует ряд игл вдоль замочного края. Редкие, крупные иглы рассеяны по всей
поверхности висцерального диска и шлейфа. На спинной створке, где сетчатый ор-
намент выражен немного слабее, крупным иглам брюшной створки соответствуют ок-
руглые ямки спинной.
Рис. 20. Reticulatia pamirica.
Схема внутреннего строения спинной
створки (реконструкция по номерам:
ПИН, № 2228/580 и ПИН, N? 2228/582)
Куртеке 1; сакмарский ярус - нижне-
артинский подъярус нерасчлененные,
базардаринская свита (верхняя часть)
Внутри брюшной створки присутствуют расходящиеся веерообразные отпечатки
мускулов-замыкателей и располагающиеся кпереди от них продольно-овальные, сильно
выпуклые отпечатки мускулов-открывателей (табл. VI , фиг. 3). Они разделены неглу-
бокой узкой срединной бороздой. Внутри спинной створки - небольшой замочный отрос-
ток, наблюдавшийся только с наружной стороны, где он состоит из двух низких ло-
пастей. Замочный отросток опирается на расширенную у основания и резко суженную
по направлению к лобному краю септу. Септа достигает ЗА длины висцерального дис-
ка. Кардинальные валики короткие, изогнутые, располагаются близ замочного края,
не достигая боковых краев створки. Мускульные отпечатки выпуклые, массивные, ден-
дровидные, состоят из передних и задних отпечатков. Брахиальные валики с короткой,
горизонтальной частью, отходят от передне-боковых сторон мускульных отпечатков.
Брахиальные отпечатки округло-четырехугольные (см. рис. 20).
Изменчивость, возрастные изменения.
Вид отличается широкой индивидуальной изменчивостью, проявляющейся в следующем;
а) очертание раковины меняется от изометричного до удлиненно-четьцэехугольного;
б) варьирует степень выраженности синуса брюшной створки - иногда он резкий,
иногда почти полностью отсутствует? в) различна степень свернутости ушек.
Возрастные изменения выражаются в том, что более юные экземпляры характе-
ризуются тонкой раковиной, незначительно выпуклой брюшной створкой и плоской
спинной. Ряд игл, отделяющий основания ушек от висцерального диска, состоит из
меньшего числа игл.
Сравнение. Отличается от типового вида менее резкой ретикулятностью скульп-
туры, более широкой, округлой макушкой брюшной створки и отсутствием ламеллоз-
ной зоны по краю шлейфа. От R.uralica (Tschem.) наш вид отличается вогнутым
висцеральным диском спинной створки, узким, мелким синусом брюшной, более ши-
рокой брюшной макушкой и резкой сетчатой скульптурой. Наши экземпляры наибо-
98
лее близки к описанным Ридом (Reed, 1925) из нижней перми Читраля под назва-
нием Pr.transversalis Tschern., отличаясь более резкой, правильной скульптурой
। обеих створок и более свернутыми ушками.
Геологический возраст и географичес кое распространение.
Сакмарский ярус - нижнеартинский подъярус нерасчлененные, базардаринская свита
(верхняя часть) Юго-Восточного Памира.
Материал. Вид широко распространен в верхней части базардаринской свиты.
Имеется 183 экз. хорошей сохранности. Из них в органогенно-обломочных алеври-
тистых известняках Куртеке 1 найдено 19 раковин с двумя сочлененными створ-
ками, 108 брюшных и 7 спинных створок; в известковистых глинистых алевролитах
Куртеке 1 - 24 хбрюшных створки; в известковистых глинистых алевролитах Кас-
танат-Джилги 1 - 9 брюшных створок; в известковистых глинистых алевролитах
Чакобая - 1 раковина с двумя сочлененными створками, 13 брюшных и 2 отпечатка
спинных. створок.
Р од Costiferina Muir—Wood et Cooper, 1960
Costiferina sp.
Распространение. Нижний, средний, верхний продуктусовый известняк Со-
ляного кряжа; дарвазский ярус, кызылджилгинский горизонт, средний подгоризонт
Юго-Восточного Памира.
Материал. 3 брюшных створки плохой сохранности собраны в туффитовых из-
вестняках Чакобая.
СЕМЕЙСТВО MARGINIFERIOAE STEHLI, 1954
ПОДСЕМЕЙСТВО PLOBOLIONINAE MUIR-WOOD ET COOPER, 1960
Р од Probolionia Cooper, 1957
Probolionia: Cooper, 1957, стр. 27; Muir—Wood,Cooper, 1960, стр. 237; Mu—
ir—Wood, 1965, стр. 483.
Типовой вид: Pr.posteroreticulata Cooper, 1957; пермь Орегона.
Описание. Раковина маленькая или средних размеров, округло-четырехуголь-
ная, с наибольшей шириной по замочному краю. Брюшная створка коленчатая, с
синусом, со слабо выпуклым висцеральным диском и довольно длинным шлейфом.
Ушки большие, треугольные или обрубленные. Спинная створка вогнутая, коленча-
тая, со срединной складкой. Висцеральная полость узкая.
Скульптура брюшной створки - из простых частых радиальных ребер с узкими
меж реберными промежутками. На висцеральном диске нерезкие, частые линии на-
растания, создающие при пересечении с радиальными ребрами его ретикулятность.
На висцеральном диске присутствует 6 халтероидных, симметрично расположенных
игл и ряд игл, отделяющих ушки от висцерального диска. Кроме того, редкие иглы
рассеяны на поверхности брюшной створки. Скульптура спинной створки аналогична,
но иглы отсутствуют.
Внутри брюшной створки продольно-треугольные или продольно-овальные отпе-
чатки дидукторов. Отпечатки аддукторов маленькие, округло-треугольные или ок-
руглые, разделены тонкой срединной бороздой. Присутствует маргинальный валик.
Внутри спинной створки - маленький, двулопастной сидячий замочный отросток.
Септа тонкая, длинная, с альвеолой в основании. Отпечатки аддукторов гладкие,
состоят из передних и задних отпечатков. Брахиальные валики приблизительно гори-
зонтальные. Брахиальные отпечатки отчетливые. Маргинальный валик широкий. От
него отходят длинные дополнительные шлейфы.
Сравнение. Близкие роды отсутствуют.
Замечания. Среди представителей других подсемейств, относящихся к семейству
Marginiferidap, наиболее близкими являются роды Marginifera Waagen, Hystriculina
99
^liur—Wood et Cooper, Paramarginifera Fredericks, с которыми его объединяет наличие
маргинальных утолщений. От рода Hystriculina род Probolionia отличается наруж-
но й скульптурой: ретикулятностью висцерального диска и наличием 6 симметрично
расположенных халтероидных игл, а от Marginifera и Param ar gin ifer а • — иным
внутренним строением. Для рода Probolionia характерно так же наличие многочислен-
ных длинных шлейфов, хотя обломанные концы коротких шлейфов встречаются и у
Eomarginifera Miur—Wood и Kozlowskia Fredericks. Эти роды, так же как и Probo—
lionia^ обладают маргинальными образованиями, но отличаются от последнего скульп-
турой и иным внутренним строением. По наличию длинных повторяющихся шлейфов
Probolionia напоминает Diaphragmus Girty и Productus Sowerby но у Probolionia они
более многочисленны. Маргинальный валик у Probolionia значительно утолщен в про-
тивоположность тонкой уплощенной диафрагме каменноугольных родов.
Рис. 21. Probolionia himalay-
ensis, Схема внутреннего
строения спинной створки (ре-
конструкция по номерам: ПИН,
№ 2228/526 и ПИН, № 2228/527).
Чакобай; дарваэский ярус, кы-
зылджилгинский горизонт, сред-
ний подгоризонт
Видовой состав. Кроме типового вида в состав данного рода входят Р.е/оп-
gata Cooper, 1957, из перми Орегона, Р.himalayensis из дарвазского яруса (кызыл-
джилгинский горизонт, нижний и средний подгоризонты) Юго-Восточного Памира,
слоев Зеван Кашмира, слоев Кулинг Гималаев, верхней перми Северного Сиккима,
Каракорума, Северной Камбоджи. Предположительно к рассматриваемому роду отно-
сится Marginifera sp. rar из нижней перми (известняк Мапинг) Китая. Margini—
fera sp. ”a” , экземпляры которой изображены Хуангом (Huang, 1932, стр. 20, фир.1,2),
имеет гладкую, коленчатую раковину. Шлейфы и маргинальный валик образуют у них
широкую пластинчатую зону.
Распространение. Пермь Орегона; слои Зеван Кашмира; слои Кулинг Ги-
малаев; дарваэский ярус, кызылджилгинский горизонт, нижний и средний подгори-
зонты Юго-Восточного Памира. Верхняя пермь Северной Камбоджи, Северного Сик-
кима, Каракорума. Предположительно известняк Мапинг (нижняя пермь). Китая.
Probolionia himalayensis (Diener, 1899)
Табл. V, фиг. 13,14; рис. 21
Marginifera himalayensis : Diener, 1899, стр. 39-42, табл. П, фиг. 1-7; табл.
IV, фиг. 1,2; 1903, стр. 104, табл. V, фиг. 5,6,27; 1915, стр. 79, табл. У1П,фиг.9;
Muir— Wood, Oacley, 1941, стр. 19-21, табл. I, фиг. 1-3; Tran—Thi Chi -Thuan, 1961,
стр. 267, табл. I, фиг. 12-14.
Productus himalayensis: Meria, 1934, стр. 261, 267, табл. XXVI, фиг. 12,13.
Описание. Раковина небольшая (11,2 мм в длину и 24 мм в ширину), поперечно-
четырехугольного очертания. Наибольшая ширина по замочному краю. Брюшная
створка коленчатая. Висцеральный диск уплощенный или слабо выпуклый. Шлейф
довольно длинный. Синус отчетливый, начинается почти от макушки. Макушка ма-
ленькая, слабо выступающая за замочный край. Ушки большие, удлиненно-треуголь-
ные, выпуклые; отделены от висцерального диска довольно глубокой бороздой. Спин-
ная створка так же коленчато изогнута, повторяет изгиб брюшной. Висцеральная по-
лость умеренная.
100
Скульптура брюшной створки представлена частыми, довольно резкими радиаль-
ными ребрами, начинающимися почти от самой макушки. На висцеральном диске -
частые концентрические морщины, которые, пересекаясь с радиальными ребрами, при-
дают скульптуре ретикулятный характер. На брюшной створке имеется 6 симмет-
рично расположенных халтероидных игл. Кроме того, присутствуют 1-2 иглы с каж-
дой стороны макушки на перегибе между ушками и висцеральным диском, 1-2 иглы
с каждой стороны макушки по замочному краю и по одной игле на концах ушек.
Скульптура спинной створки отличается меньшей резкостью.
Внутри брюшной створки - продольно овальные, выпуклые, массивные отпечатки
дидукторов. Отпечатки аддукторов маленькие, округлые, располагаются в небольших
треугольных углублениях и разделены тонкой срединной бороздой.
Внутри спинной створки замочный отросток не наблюдался. Септа тонкая длин-
ная, не сочлененная с основанием замочного отростка. По бокам от септы - ма-
ленькие, гладкие, выпуклые отпечатки задних аддукторов. Кпереди от задних отпе-
чатков располагаются массивные овальные отпечатки передних аддукторов. Брахи-
альные валики отходят от места соединения отпечатков передних и задних аддук-
торов. Брахиальные валики слабо наклонные, брахиальные петли грушевидные. От
основания замочного отростка отходят гребневидные кардинальные валики, пере-
ходящие в латеральные, отделяющие ушки от висцерального диска. Маргинальный
валик широкий, слабо выпуклый (см. рис. 21). Шлейфы, отходящие от маргинального
валика, сохранились плохо, вероятно, из-за захоронения раковин в подвижной гидро-
динамической среде.
Сравнение. P.himalayensis отличается от американских видов P.posterore-
ticulata Cooper и P:elongata Cooper менее развитыми шлейфами, но возможно, это
объясняется лучшей сохранностью последних двух видов по сравнению с P.himalayen-
sis с Юго-Восточного Памира, найденной в органогенно-обломочных детритовых
известняках. От Marginifera sp. рассматриваемый вид отличается ретикулятной
скульптурой висцерального диска, а также ребристостью менее развитого шлейфа.
Замечания. К P.himalayensis мы не относим экземпляры, описанные под этим
видовым названием Д.Л. Степановым ( 1934) из нижнепермских отложений р.Колвы.
Это сделано на том основании, что уральские экземпляры характеризуются иным
расположением игл брюшной створки. Кроме того, среди экземпляров, имеющихся
у Д.Л. Степанова, нет ни одного, у которого можно было бы наблюдать маргиналь-
ные валики, характерные для представителей этого вида. Несоответствие уральских
экземпляров гималайским отмечает Н.П. Герасимов (Герасимов и др., 1953), опи-
савший первые под названием Productus cf.himalayensis Diener. Н.П. Герасимов
пишет: гМы не решаемся присоединиться к заключению Д.Л. Степанова о том, что
описанная им уральская форма действительно относится к Pr.himalayensis Diener.
В связи с этим мы фиксируем данный вид в описываемой фауне со знаком ”cf.”.
P.himalayensis из пермских отложений Юго-Восточного Памира характеризуется
тем, что септа не подходит непосредственно к основанию замочного отростка. В
диагнозе рода Probolionia Мюр-Вуд и Купер (Muir—Wood,Cooper, 1960) указывают,
что септа имеет расширенное основание, но на изображениях внутреннего строения
(там же, табл. 135, фиг. 15,17) можно наблюдать, что септа не примыкает к осно-
ванию замочного отростка.
Геологический возраст и географичес кое распространение.
Дарвазский ярус, кызылджилгинский горизонт, нижний и средний подгоризонты Юго-
Восточного Памира; слои Зеван Кашмира; слои Кулинг Гималаев; верхняя пермь
Северной Камбоджи, Северного Сиккима, Каракорума.
Материал. 19 экз. удовлетворительной сохранности; собраны в кызылджилгип-
ском горизонте. Из них в криноидно-детритовых известняках Куртеке 2 нижнекызылд-
жилгинского подгоризонта найдены 9 брюшных и 1 спинная створка; в туффитовых из-
вестняках Чакобая среднекызылджилгинского подгоризонта - 5 раковин с двумя соч-
лененными створками и 5 спинных створок.
101
ПОДСЕМЕЙСТВО MARGINIFERINAE STEHLI, 1954
Род Margin!feta Waagen, 1884
Marginifera: Waagen, 1884, стр. 713; Muir—Wood,Cooper, I960, стр. 206; Muir-
Wood, 1965, стр. 477; Grant, 1968, стр. 17.
Типовой вид - M.typi ca Waagen; Пакистан, Соляной кряж; средний, верхний
продуктусовый известняк.
Описание. Раковина небольшая, четырехугольного очертания, обычно продольно
вытянутая. Брюшная створка коленчатая. Ушки хорошо развитые, у большинства ви-
дов довольно большие. Спинная створка умеренно вогнутая.
Скульптура состоит из радиальных ребер, развитых в различной степени у раз-
ных видов, слабых концентрических морщин и многочисленных, прямых игл. Иглы
рассеяны на висцеральном диске и шлейфе. Обычно имеется так же одна или две
на макушечных склонах и у основания ушек. Скульптура спинной створки сходна с
брюшной, но менее отчетлива, а иглы отсутствуют.
Внутри брюшной створки - зазубренные латеральные валики, ограничивающие ос-
нования ушек, и маргинальный валик. Внутри спинной - сидячий замочный отросток;
двухлопастной со стороны брюшной и трехлопастной со стороны спинной створки.
От его основания отходят кардинальные валики, на боковых частях переходящие в
зазубренные латеральные, ограничивающие основания ушек, а в передней части створ-
ки - в низкий маргинальный валик. Септа тонкая, продолжается до 2/3 длины вис -
церального диска . Отпечатки аддукторов гладкие, состоят из двух пар: передних -
продольно-овальных и задних - маленьких, округло-треугольных. Брахиальные отпе-
чатки неясные.
Сравнение. Marginifera наиболее близка к Eomarginifera Muir—Wood, 1930 и
Kozkowskia Fredericks, 1933, от которых отличается отсутствием шеста крупных
симметрично расположенных игл на висцеральном диске брюшной створки. Кроме
того, от Eomarginifera ее отличает менее резко выраженная радиальная и концент-
рическая скульптура, а по внутреннему строению - отсутствие пластинчатой зоны
и гладкие отпечатки задних аддукторов. От рода Kozlowskia род Marginifera отли-
чается присутствием более частых игл на поверхности брюшной створки, зазубренных
кардинальных валиков и узким основанием срединной септы внутри спинной створки.
Видовой состав. Кроме типового вида, к данному роду достоверно можно от-
нести виды Ваагена (Waagen, 1884) из перми Соляного кряжа: М. trans versa, М.ехса—
vata, М.echinata из нижнего продуктусового известняка и M.omata из цефалопо-
довых слоев верхнего продуктусового известняка. Кроме того, к нему относится
М.magniplicata (Huang, 1932) из верхней перми Монголии и Закавказья и JM .sexcosta-
ta (Licharew, 1937) из джульфинского яруса верхней перми Северного Кавказа и
Юго-Восточного Памира.
Замечания. В вопросе понимания характеристики и объема рода Marginifera сре-
ди различных исследователей в настоящее время не существует единого мнения.Род
Marginifera был предложен Ваагеном (Waagen, 1884) для многочисленных продук-
тид, обладающих маргинальными образованиями, из пермских отложений Соляного
кряжа. В дальнейшем к этому роду относились все мелкие продуктиды, обладающие
какими бы то ни было маргинальными утолщениями ( более 100 видов). В результате
ревизии, произведенной Мюр-Вуд и Купером (Muir—Wood,Cooper, 1960) и несколько
позже Грантом ( Grant, 1968), изучившим типовой вид рода из топотипического место-
нахождения, объем рода был значительно сужен. Однако ни один из названных авто-
ров не мог произвести полной ревизии всех видов, относившихся различными иссле-
дователями к роду Marginifera, поэтому действительный его объем до настоящего
времени остается неясным.
Распространение. Джульфинский ярус: никитинский и уруштенский горизонты
Северного Кавказа, среднетахтабулакская подсвита Юго-Восточного Памира; верхняя
пермь Монголии и Закавказья; нижний, средний, верхний продуктусовый известняк
Соляного кряжа (Пакистан).
102
Marginifera sexcostata Licharew, 1937
Табл. V, фиг. 11, 12; рис. 22
Productus (Marginifera) sexcostatus; Лихарев, 1937, стр. 64-65, табл. IX,
фиг. 44-51.
Голотип - ЦНИГРмузей им. Ф.Н. Чернышева, № 1087/2139; брюшная створка;
джульфинский ярус, уруштенский горизонт; Северный Кавказ, Никитина балка; Ли-
харев, 1937, табл. IX, фиг. 48.
Рис. 22. Marginifera sexcostata
ПИН,№ 2228/587; внутреннее строе-
ние спинной створки, х6.3Шор-Булак;
джульфинский ярус, среднетахтабу-
лакская подсвита
Описание. Раковина маленькая (длина 10-12 мм, ширина 9-10 мм), прибли-
зительно изометричная, продольно-четырехугольного очертания. Наибольшая ширина ее,
располагающаяся по замочному краю, приблизительно равна ширине по лобному краю.
Брюшная створка сильно изогнута в продольном направлении, коленчатая. Шлейф
довольно длинный, нерезко отделен от уплощенного висцерального диска. Макушка
короткая, широкая, почти не выступающая за замочный край. Ушки небольшие, уп-
лощенные, нерезко отделенные от остальной поверхности висцерального диска. Спин-
ная створка слабо вогнутая, полуокруглого очертания. Ушки маленькие, нерезко от-
делены от поверхности висцерального диска.
Скульптура брюшной створки состоит из 7-8 простых резких округлых ребер, рас-
полагающихся на шлейфе. Межреберные промежутки немного шире, чем ребра, слабо
вогнутые. На висцеральном диске брюшной створки тонкие, частые иглы, располагаю-
щиеся на слабо удлиненных основаниях в более или менее правильном шахматном по-
рядке. Более крупные, редкие иглы располагаются вдоль замочного края и на ушках.
Иглы на спинной створке отсутствуют.
Внутреннее строение брюшной створки не наблюдалось. Внутри спинной створки -
маленький сидячий трехлоластной замочный отросток с более развитой срединной ло-
пастью, Кардинальные валики протягиваются от основания замочного отростка вдоль
замочного края, на боковых частях переходят в зазубренные латеральные валики,
ограничивающие ушки. У переднего края кардинальные валики протягиваются в виде
невысокого маргинального валика. Срединная септа тонкая, достигает 2/3 длины вис-
церального диска, имеет расширяющееся окончание. Отпечатки аддукторов гладкие,
разделяются на передние и задние. Отпечатки передних аддукторов продольно-оваль-
нце, располагаются почти параллельно септе. Отпечатки задних - округло-треуголь-
ные, маленькие. Брахиальные валики косо отходят от места соединения отпечатков
передних и задних аддукторов. Брахиальные отпечатки неясные, располагаются в сред-
ней части створки, (см. рис. 22). С наружной стороны ограничены рядом внутренних игл.
103
Сравнение. От видов Ваагена, относящихся к роду Marginifera, отличается
меньшими размерами , отсутствием отчетливого ряда йгл на макушечных склонах.
От М.transversa данный вид отличается, кроме того, отсутствием синуса брюшной створ-
ки и морщинистости на висцеральном диске, а от М .magniplicata (Huang1) - менее
грубыми, узкими ребрами, развитыми только на шлейфе.
Распространение. Джульфинский ярус: никитинский и уруштенский горизонты
Северного Кавказа; среднетахтабулакская подсвита Юго-Восточного Памира.
Материал. 3 экз. удовлетворительной сохранности, происходят из табулятовых
биогермных известняков среднетахтабулакской подсвиты Шср-Булака. Из них - 1
раковина с двумя сочлененными створками и 2 брюшных створки.
СЕМЕЙСТВО LINOPROOUCTIDAE STEHLI, 1954
P<lg Terrakea Booker, 1930
Terrakea Booker, 1930, стр. 66; Hill, 1950, стр. 8; Maxwell, 1956, стр. 333;
Muir—Wood,Cooper, 1960, стр. 315; Waterhouse, 1964, стр. 63; Muir—Wood, 1965,
стр. 503.
Типовой вид - Productus brachythaerus Morris, 1845; Австралия; нижняя пермь,
артинский ярус.
Описание. Раковина небольших или средних размеров, субтрапецеидальная. Брюш-
ная створка умеренно выпуклая, обычно коленчатая, со шлейфом различной длины.
Макушка короткая, незагнутая. Замочный край длинный, ушки треугольные, уплощен-
ные. Спинная створка вогнутая в различной степени, иногда коленчатая. Висцеральное
пространство узкое.
Скульптура состоит из тонких, неправильных радиальных ребер, располагающихся
по всей раковине. Иглы брюшной створки рассеяны по всему висцеральному диску
и шлейфу, длинные, тонкие, косые, со слабо вздутыми, удлиненными основаниями.
Вдоль замочного края и на ушках иглы крупные, прямые. На спинной створке иглы
мелкие, редкие. Может присутствовать ряд крупных игл на перегибе между висцераль-
ным диском и шлейфом.
Внутри брюшной створки, вдоль ее средней линии располагаются ветвистые треу-
гольно-овальные отпечатки аддукторов. По бокам от них - широкие, штриховатые
отпечатки дидукторов. Внутри спинной - двухлопастной замочный отросток. Септа
длинная, тонкая, с широким основанием. Иногда в основании септы присутствует глу-
бокая продольная бороздка. Отпечатки аддукторов округло-треугольные, разделяют-
ся на передние и задние и ограничены с боковых сторон нерезкими валиками. Бра-
хиальные отпечатки неясные.
Сравнение. Данный род отличается внешне от рода Linoproductus Chao, 1927
менее правильной, тонкой радиальной ребристостью и отсутствием складок на ушках
и кроме того, - тонкими, частыми иглами, располагающимися на каждом ребре; они
не захватывают своими основаниями нескольких ребер, как у Linoproductus. Внут-
реннее строение Тегтакеа характеризуется более высоким, не сидячим замочным
отростком. От рода Stepanoviella Zavodowsky, I960 род Terrakea отличается менее
отчетливой киленчатостью спинной створки, слабой концентрической скульптурой; но
внутреннему строению - более массивными кардинальными валиками.
Видовой состав. Кроме типового вида к данному роду относятся TJragilis
( Dana, 1847); Т./eve Booker, 1930; T pollex Hill, 1950; T,solidus ( Etheridge et Dunn,
1909) из нижнепермских отложений Австралии И Новой Зеландии, а так же T.naliv-
kini Grunt, sp.nov. из кубергандинской свиты (?) дарвазского яруса Юго-Восточного
Памира.
Замечания. Уотерхауз ( V:’aterhouse, 1964) рассматривает род Terrakea более
широко, чем автор рода Букер (Booker, 1930). В диагнозе Terrakea Уотерхауз ука-
зывает на наличие ряда игл на коленчатом перегибе спинной створки и борозды d
основании септы у некоторых видов - признаков, не отмеченных Букером в перво-
описании.
104
Распространение. Нижняя пермь Австралии и Новой Зеландии; дарвазский
ярус, кубергандинская свита (?) Юго-Восточного Памира.
Тегтакеа nalivkini Grunt, sp.nov.
Табл. VII, фиг. 2-5; рис. 23
Название в честь Д.В. Наливкина - одного из первых исследователей палео-
зоя Памира.
Голотип -ПИН № 2228/593; раковина удовлетворительной сохранности с двумя
сочлененными створками; Юго-Восточный • Памир, Южный Ак-Архар; дарвазский
ярус, кубергандинская (?) свита; табл. VII, фиг. 2.
Описание. Раковина средних размеров (размеры голотипа: длина 34,5 мм, ши-
рина 52,5 мм), округло-четырехугольного или округло-трапецеидального очертания.
Замочный край соответствует наибольшей ширине. Брюшная створка сильно выпук-
лая, коленчатая, с длинным шлейфом. Макушка широкая, короткая, не заходит за
замочный край. Ушки большие, широкотреугольные, с округлыми окончаниями, нерез-
ко отграничены от висцерального диска. Спинная створка вогнутая, слабо коленчатая
Скульптура состоит из тонких, частых, довольно правильных радиальных ребрышок
протягивающихся от макушки до лобного края. Количество их увеличивается кпереди
путем интеркаляции и составляет 5-8 на 5 мм поверхности в средней части раковины.
Межреберные промежутки плоские, немного шире самих ребер. Линии нарастания
нерезкие, неправильные, более отчетливые на спинной, чем на брюшной створке.
На ушках и боковых склонах раковины присутствуют слабые концентрические мор-
щины. Крупные, прямые иглы располагаются вдоль замочного края и на ушках брюш-
ной створки. Тонкие, косые рассеянные иглы присутствуют на всей ее поверхности.
Они расположены на ребрышках, которые обычно слабо вздуваются непосредственно
перед появлением иглы. На внутренних слоях раковины наблюдаются длинные полые
следы игл, располагающиеся вдоль соответствующих ребер. Иглы спинной створки не
наблюдались.
Рис. 23. Terrakea nalivkini, Схема внут-
реннего строения спинной створки (рекон-
струкция по номерам: ПИН, № 2228/594,
ПИН, N? 2228/595 и ПИН, N? 2228/593).
Южный Ак-Архар; дарвазский ярус, кубер-
гандинская (?) свита
Внутри брюшной створки - мускульное поле, состоящее из треугольно-овальных
отпечатков аддукторов. По бокам от аддукторов с задне-боковых их сторон распо-
лагаются продольно-штриховатые округло-треугольные отпечатки дидукторов. Внутри
спинной створки имеется замочный отросток - высокий, трехлопастной с наружной,
и двухлопастной с внутренней стороны. Он опирается на тонкую, длинную срединную
септу, достигающую более 3/4 длины висцерального диска. Септа с широким основа-
нием, несущим слабую продольную борозду. По бокам от септы располагаются ок-
ру гло-треугольные отпечатки адцукторов, разделяющиеся на передние и задние. От-
печатки задних аддукторов массивные, дендровидные, округло-треугольные; отпе-
105
чатки передних маленькие, гладкие, грушевидные. Отпечатки аддукторов ограничены
с задне-боковых сторон слабыми боковыми валиками. Брахиальные отпечатки неяс-
ные (см. рис. 23).
Сравнение. От видов, принадлежащих к роду Terra кеа, известных из пермских
отложений Австралии и Новой Зеландии, T.nalivkini отличаются более крупными
размерами, широкой макушкой, относительно более тонкой ребристостью, более тон-
кими, редкими иглами брюшной створки и отсутствием игл на спинной, а так же
слабой коленчатостью раковины.
Геологический возраст и географическое распространение.
Дарваэский ярус, кубергандинская (?) свита Юго-Восточного Памира.
Материал. В коллекции содержится более 100 экз. из кубергандинской (?) сви-
ты Южного Ак-Аркара. Собраны в прослое серых микрозернистых известняков, пере-
полненных цельными раковинами и разрозненными створками удовлетворительной сох-
ранности.
Р од Cancrinella Fredericks, 1928
Cancrinella altissima King, 1938
Табл.’VIII, фиг. 1,2
Cancrinella altissima': King, 1938, стр. 275-276, табл. 39, фиг. 27, 28; Stehli
1954, стр. 321, табл. 22, фиг. 1-5.
Описание. Раковина небольшая, цилиндрическая (размеры: 25,5 мм в длин^
19 мм в ширину). Брюшная створка довально сильно выпуклая, с крутыми субпарал-
лельными боковыми склонами,- слабо расширяющаяся кпереди. Макушка короткая, ши-
рокая, приплюснутая. Ушки небольшие, уплощенные. Спинная створка умеренно вог-
нутая, иногда с коленчатым перегибом между висцеральным диском и шлейфом.
Скульптура состоит из отчетливых радиальных ребрышек в количестве 7-8 на
3 мм поверхности у лобного края. Ребрышки тонкие, резкие, довольно отчетливые.
Присутствуют так же неправильные частые резкие концентрические морщины. Иглы
частые, располагаются на ребрах и имеют вздутые основания. В области замочного
края иглы собраны в пучки. Иглы спинной створки не наблюдались.
Внутреннее строение брюшной створки не наблюдалось. Внутри спинной - малень-
кий сидячий двухлопастной замочный отросток глинопродуктидногог типа. Замочный
отросток поддерживается короткой срединной септой, слабо расширяющейся у осно-
вания. По бокам от септы располагаются короткие боковые валики, ограничивающие
тонкие мускульные отпечатки и отходящие от основания замочного отростка под
углом 40-45 .
Сравнение. Данный вид отличается от С.boonепsis (Shumaid, 1858) несколько
более крупными размерами, более грубой ребристостью, продольно вытянутым очер-
танием раковины, от C.rugosa Cloud, 1944 и C.phosphatica (Girty, 1910) - более
тонюй ребристостью. По форме раковины С.altissima очень близка к С. к on in с к i ana
(Keyserling, 1846), но отличается сильной концентрической морщинистостью брюш-
ной створки.
Геологический возраст и географическое распространение.
Верхний карбон: формация Грахам (зона Triticites); нижняя пермь, формация Бонн
(вольфкэмл - ленэрд) Западного Техаса; сакмарский ярус - нижнеартинский подъ-
ярус нерасчлененные, базардаринская свита (верхняя часть) Юго-Восточного
Памира.
Материал. 38 экз. удовлетворительной сохранности собраны в верхней части
базардаринской свиты. Из них в органогенно-обломочных алевритистых известняках
Куртеке 1-3 раковины с двумя сочлененными створками, 4 спинных и 20 брюшных
створок; в известковистых глинистых алевролитах Кастанат-Дяшлги 1-0 брюшных
створок; в известковистых глинистых алевролитах Чакобая - 2 брюшных створки.
106
Р од Globiella Muir—Wood et Cooper, 1960
Globiella? rossiae (Fantini Sestini, 1966)
Табл. VII, фиг. 6-8
Stepanoviella rossiad: Fantini Sestini, 1966, стр. 28-33, табл. 4, фиг. la-5.
Голотип - Палеонтологический институт при Миланском университете, № 1А-
L213a; брюшная створка; Северный Иран, верховья долины Джаджеруд; нижняя
пермь, формация Джейруд; Fantini Sestini, 1966, стр. 28-33, табл. 4, фиг. 3.
Описание. Раковина средних размеров (20-25 мм в длину и 20-23 мм в ширину),
приблизительно изометричного очертания. Брюшная створка полуокруглая, равно-
мерно изогнутая в продольном и поперечном направлении. Макушка маленькая, слабо
приостренная, умеренно загнутая, немного выступающая за замочный край. Замоч-
ный край прямой, немного меньше наибольшей ширины раковины. Ушки небольшие,
уплощенные, треугольные. Спинная створка слабо вогнутая.
Скульптура состоит из довольно грубых радиальных ребер (7-8 на 5 мм поверх-
ности у лобного края). Ребра покрыты тонкой поперечной штриховкой. Концентричес-
кие морщины редкие, неправильные. Иглы довольно крупные на ушках и вдоль замоч-
ного края. На висцеральном диске и шлейфе иглы более тонкие, располагающиеся
на ребрах. Иглы спинной створки не наблюдались.
Внутри брюшной створки - узкие двальные отпечатки аддукторов, разделенные
тонкой бороздой. Дидукторы округло-треугольные, продольно штриховатые. Внутри
спинной створки - маленький сидячий двухлопастной замочный отросток, опирающий-
ся на короткую срединную септу. Кардинальные валики короткие, располагаются под
углом приблизительно 50 к септе.
Сравнение. Отличается от G.rewahensis (Reed, 1928) из слоев Умариа Централь-
ной Индии несколько более тонкой радиальной ребристостью, приуроченностью наи-
большей ширины раковины не к замочному краю, а к ее средней части, а также
менее массивной макушкой.
Замечания. У экземпляров из нижнепёрмских отложений Северного Ирана, опи-
санных Фантини Сестини (Fantini Sestini, 1966), так же как и у собранных нами на
Памире, к сожалению, не удалось полностью наблюдать внутреннего строения спинной
створки, поэтому отнесение данного вида к роду Globiella вызывает сомнение.
Геологический возраст и географичес кое распространение.
Карачатырский ярус, формация Джейруд (пачка Д) Северного Ирана; дарвазский ярус,
кызылджилгинский горизонт, нижний подгоризонт Юго-Восточного Памира.
Материал. 23 экз. удовлетворительной сохранности; собраны в криноидно—дет-
ритовых известняках нижнекызылджилгинского подгоризонта Куртеке 2 (20 брюшных
и 3 спинных створки).
Род Anidanthus Whitehouse, 1928
Anidanthus sinosus (Huang, 1932)
Табл. У1П,фиг. 3
Lin oprod и ct us sinosus : Huang,1932, стр. 43, табл. II, фиг. 15, 16.
Productus (Productus) sinosus: Лихарев, 1937, стр. 34 , 36, табл. VI, фиг. 7-15.
Anidanthus sinosud: Ching Yii—kan, Zhang Jan, 1961, стр. 409, табл. Ill, фиг. 1-5.
Геологический возраст и географичес кое распространение.
Джульфинский ярус: формация Допинг, литтониевые слои Юго—Западного Китая; уруш—
тенский и никитинский горизонты Северного Кавказа, среднетахтабулакская подсвита
Юго-Восточного Памира.
107
Материал. 1 Одна брюшная створка удовлетворительной сохранности, происходит из
табулятово-гидроидных биогермных известняков среднетахтабулакской подсвиты Кур-
Истыка 1.
Р од Compressoproductus Sarytcheva, 1960
Compressoproductus': Сарычева, 1960, стр. 231.
Striatifera (Substriatifera): Котляр, 1964, стр, 123.
Типовой вид - Product us сот pressus Waagen t1884; средний продуктусовый из-
вестняк; Соляной кряж (Пакистан).
Описание. Раковина от средних до крупных размеров, треугольного или оваль-
но-треугольного очертания, изменчивой формы. Брюшная створка сильно или умерен-
но выпуклая; спинная - слабо вогнутая или уплощенная. Поперечный изгиб ее от
слабого до достаточно резкого. Боковые склоны крутые, резко отделены ит осталь-
ной поверхности створки. Замочный край короткий. Ушки вертикальные. Брюшная ма-
кушка узкая, с рубцом прирастания. Висцеральное пространство узкое, раковина тон-
кая.
Скульптура состоит из частой тонкой правильной струйчатости и неправильных
концентрических морщин, развитых в различной степени. Jia боковых склонах скульп-
тура отсутствует или развита слабо. Здесь и вокруг рубца прирастания - редкие
крупные иглы.
Внутри брюшной створки, по данным Г.В. Котляр (1964) - две пары мускульных
отпечатков: вдоль средней линии вблизи макушки - вдавленные подковообразные денд-
ровидные отпечатки аддукторов. Отпечатки дидукторов - крупные, широкие, иногда
округлые, продольно-струйчатые. Внутренняя поверхность брюшной створки шипова-
тая. Внутри спинной створки - массивная срединная септа с широким основанием.
Замочный отросток небольшой, однолопастной 'стриатиферового' типа.
Сравнение. От Striatifera Chao, 1927.. рассматриваемый род внешне отлича-
ется более узкой свернутой макушкой брюшной створки, тонкой радиальной струй-
чатостью и наличием неправильных концентрических морщин на всей поверхности
раковины, внутреннее строение брюшной створки так же иное. Мюр-Вуд и Купер (Mu-
ir—Wood, Cooper, 1960) отмечают сходство представителей Compressoproductus (С.сотр-
ressus и C.mytiloides) с типовым видом рода Undaria, от которого названные виды
отличаются меньшими размерами, иной формой раковины и скульптурой.
Видовой состав. Кроме типового вида, встреченного в среднем продуктусо-
вом известняке Соляного кряжа, в нижнепермских отложениях Читраля, в верхне-
пермских отложениях Монголии, Южного Китая, Сицилии и Северного Кавказа, к дан-
ному роду относятся С.туtiloides (Waagen) из верхнего продуктусового известняка
Соляного кряжа - вид, йе принимаемый большинством исследователей в качестве са-
мостоятельного, так как он отличается от C.compressus лишь меньшей величиной
ушек, более грубой радиальной ребристостью и слабее развитой концентрической
скульптурой. В коллекции Ваагена присутствует лишь 2 экземпляра этого вида, поэто-
му устойчивость указанных признаков не могла быть проверена. К этому же роду
относятся С.mongolicus(Diener) из верхнепермских отложений Южного Китая, Гима-
лаев, Юго-Восточного Памира, Северного Кавказа; С.djoulfensis (Stoyanow) из джуль-
финского яруса верхней перми Закавказья и Северного Ирана; C.vladivostokensis
(Fredericks) из верхнепермских отложений Приморья; C.prinadai (Fredericks) из
верхнепермских отложений Приморья и Югославии и С.prinniformis (Girty, 1908) из
верхнепермских отложений Техаса.
3 амечания. Подрод Substriatifera, выделенный Г.В. Котляр в 1964 г., по-
видимому, является младшим синонимом рода Compressoproductus Sarytcheva, 1960
(Сарычева и др., 1960). На это указывает сам автор подрода: 'Габитус раковин,
характер скульптуры и ряд других признаков указывает на несомненное родство этой
своеобразной формы (Pr.mytiloides vladivostokensis Fred.) с Pr.(Striatifera) compres-
sus Waagen'1 (типовым видом рода Compressoproductus. - T .Г.). Отличия же Sub-
striatifera от Compressoproductus Г.В. Котляр не рассматривались.
108
Распространение. Нижняя пермь Читраля; верхняя пермь Южного Китая,
Гималаев, Кашмира, Приморья, Юго-Восточного Памира, Северного Кавказа, Закав-
казья, Северного Ирана, Сицилии, Югославии и Техаса.
Compressoproductus mongolicus (Diener, 1397)
Табл. VIII, фиг. 4,5
Productus mongolicus: Diener, 1897a, стр. 28, табл. IV, фиг. 8-10; 1899, стр. 38,
табл. VI, фиг. 1-8.
Productus compressus Waagen var. mongolica. Frech,1911, стр. 128, табл. 21,
фиг. 1.
Striatifera mongolica : J.Chao, 1927, стр, 102, табл. XI, фиг. 5-6; Huang, 1932,
стр. 51, табл. Ill, фиг. 21.
Productus (Striatifera) compressus Waagen forma mongolica : Лихарев, 1937,
стр. 50, табл. VIII, фиг. 1-3.
Описание. Раковина небольшая (16 мм в длину и 14 мм в ширину), продольно-
треугольного очертания. Брюшная створка умеренно выпуклая, слабо и равномерно
изогнутая в продольном направлении и более резко в поперечном, резко сужающаяся
по направлению к макушке. Наибольшая ширина ее g передней части раковины. За-
мочный край очень короткий, изогнут под углом 85 . Боковые склоны крутые. Ма-
кушка невысокая, округлая, с маленьким рубцом прирастания. Ушки небольшие,
треугольные, вертикальные, нерезко отделяются от остальной поверхности створки.
Спинная створка округло-треугольная, слабо вогнутая. Висцеральное пространство
узкое.
Скульптура представлена отчетливой тонкой радиальной ребристостью (4-5 ребер
на 5 мм по лобному краю), покрывающей всю створку. Между ребрами - тонкая,
густая радиальная стру^гчатость. Радиальная скульптура на ушках отсутствует. Кро-
ме ребристости на раковине присутствуют правильные концентрические морщины, пе-
реходящие на ушки. В макушечной области - редкие крупные иглы, окружающие ру-
бец прирастания.
Внутри спинной створки - тонкая срединная септа, достигающая пдловины длины
висцерального диска. Ее основание несколько расширено. По бокам от септы - не-
ясные удлиненно-овальные мускульные отпечатки.
Сравнение. Данный вид отличается от С. compressus Waagen. более резко по-
перечно свернутой брюшной створкой и более отчетливой правильной концентрической
скульптурой; от C.djoulfensis (Stoyanow) - меньшими размера ми раковины, ее бо-
лее правильной формой, узким смычным краем и отчетливой концентрической скульп-
турой, а от С.vladivostokensis (Fredericks) - меньшими размерами, более пра-
вильной формой раковины и ее тонкой радиальной скульптурой.
Геологический возраст и географическое распространение.
Известняк Читичун Гималаев; слои Зеван Кашмира; джульфинский ярус: слои До-
пинг Южного Китая; среднетахтабулакская подсвита Юго-Восточного Памира; уруш-
тенский и никитинский горизонты Северного Кавказа.
Материал. 8 экз. удовлетворительной сохранности, собраны в среднетахтабу-
лакской подсвите. В табулятово-гидроидных биогермных известняках Кур-Истыка 1
найдены: 1 раковина с сочлененными стьоркахми, 1 брюшная створка взрослого
экземпляра и 3 - юных; в табулятовых известняках Шор-Булака - 1 раковина с
сочлененныхми створками и 1 брюшная створка; Музду-Булака - 1 неполная брюш-
ная створки.
109
СЕМЕЙСТВО CHONETELLIDAE LICHAREW, I960
Род Haydenella Reed, 1944
Haydenella dutkevitchi Grunt, sp.nov.
Табл. VIII, фиг. 7,8; рис. 24
Видовое название в честь Г.А. Дуткевича - одного из первых исследовате-
лей пермских отложений Памира.
Голотип - ПИН, № 2228/505; брюшная створка; Юго-Восточный Памир; Ку-
таль 2; джульфинский ярус, среднетахтабулакская подсвита; табл. VIII, фиг. 7.
Описание. Раковина маленькая (размеры голотипа: длина 11,5 мм, ширина -
12,4 мм), округло-^греугольного или округло-четырехугольного очертания, прибли-
зительно изометричная. Брюшная створка умеренно и равномерно выпуклая, с низ-
кой ареей, иногда со слабой уплощенностью в передней части и небольшой, округ-
лой, выступающей складкой по лобному краю. Наибольшая ширина раковины - по
замочному краю. Ушки небольшие, уплощенные, с оттянутыми, остроконечными окон-
чаниями. Макушка меленькая, слабо при остренная, незначительно выступающая за
замочный край. Спинная створка умеренно вогнутая, повторяющая изгиб брюшной.
Висцеральная полость узкая. Створки очень тонкие.
Рис. 24. Haydenella dutkevitchi,
ПИН, № 2228/507; внутреннее
строение спинной створки (внут-
реннее ядро), х5.4Куталь 2;
джульфинский ярус, среднетахта-
булакская подсвита
Скульптура состоит из простых, округлых радиальных ребер, начинающихся почти
от самой макушки и несколько расширяющихся по направлению к лобному краю.
Концентрические морщины на висцеральном диске не развиты или развиты очень
слабо. На ушках они выражены отчетливо; количество их 5-7; морщинки распола-
гаются перпендикулярно линии замочного края около макушки и становятся наклон-
ными, приближаясь к боковым краям раковины. От окончания морщинок отходят длин-
ные гхонетесовидыыег иглы, образующие ряд по замочному краю. По одной длинной
игле имеется у каждого окончания ушка; 2-3 иглы образуют ряд в основании ушек,
отделяющий их от висцерального диска. Редкие иглы рассеяны по поверхности вис-
церального диска и шлейфа. Иглы на поверхности спинной створки не наблюдались.
Внутреннее строение брюшной створки не наблюдалось. Внутри спинной створки-
септа, достигающая половины длины створки. Ее сочленение с замочным отростком
не ясно. Мускульные отпечатки гладкие, овально-треугольные, длинными сторонами
обращены к септе. Брахиальные валики отходят от вершин треугольников, образо-
ванных мускульными отпечатками. Брахиальные отпечатки округло-треугольные
(см. рис. 24).
Изменчивость. Очертание раковины от округло-треугольного до округло-
четырехугольного, Изменчивой является так же и форма ушек: от прямоугольной
до оттянутой остроконечной; степень выраженности складки по л обкому краю также
непостоянна.
110
Сравнение. H.dutkevitchi отличается от остальных видов этого рода мень-
шими размерами раковины, наличием небольшой выступающей складки по лобному
краю, косо располагающимися морщинками на периферических частях ушек и нали-
чием ряда длинных хоне тес обидных игл по замочному краю.
Геологический возраст и географичес кое распространение.
Джульфинский ярус, среднетахтабулакская подсвита Юго-Восточного Памира.
Материал. 9 экз. удовлетворительной сохранности, собраны в отложениях сред-
нетахтабулакской подсвиты. В табулятово-гидроидных биогермных известняках Ку-
таля 1 найдена 1 брюшная створка, Куталя 2-3 брюшных створки; Кур-Истыка 1 -
1 раковина с сочлененными створками и 1 спинная створка; в табулятовых биогерм-
ных известняках Шор-Булака - 3 брюшных створки.
Haydenella tumida (Waagen, 1884)
Табл. УП1,фиг. 6
Productus tumidud: Waagen,1884, стр. 708-710, табл. ЬХХХ,фиг. 1, 2, 3.
Productus (Paramarginifera) kiangsiensis Kayser var. tumida': Reed, 1931 b,
стр. 14-16, табл. Ш, фиг. 5-7.
Haydenella tumidd: Сарычева, 1965, табл. XXXYIII, фиг. 9.
Геологический возраст и географическое распространение.
Джульфинский ярус Закавказья; джульфинский ярус, нижне- и среднетахтабулакская
подсвигы Юго-Восточного Памира. Средний, верхний продуктусовый известняк Со-
ляного кряжа (Пакистан).
Материал. 6 экз., представленных брюшными створками удовлетворительной
сохранности, собранных в нижне- и среднетахтабулакской подсвитах. В водорослево-
гидроидных биогермных известняках нижнетахтабулакской подсвиты Джилги-Кочусу
найден 1 экз. в табулятово-гидроидных биогермных известняках среднетахтабулак-
ской подсвиты Куталя 1-1 экз.; Куталя 2-2 экз.; Кур-Истыка 1 - 1 экз.; в
табулятовых биогермных известняках Шор-Булака - 1 экз.
ПОДОТРЯД OLDHAMINOIDEA
НАДСЕМЕЙСТВО LYTTONIACEA
СЕМЕЙСТВО LYTTONIIDAE WAAGEN, 1884
Род Leptodus Kayser, 1883
. Leptodus sp.
Табл. VIII фиг. 9,10
Распространение. Род широко распространен в Американской и Евро-Азиат-
ской частях Те тиса. Указывается из нижней и верхней перми Мексики, Техаса, Ка-
лифорнии; из верхней перми Северной Австралии, Камбоджи, Малайи, Суматры, Ти-
мора, Юго-Западного Китая, Соляного кряжа (Пакистан), Кашмира, Монголии, Япо-
нии,. Приморья, Юго-Восточного Памира, Северного Кавказа, Закавказья, Сицилии,
Югославии.
Материал. 13 экз., представленных брюшными створками с поврежденным, на-
ружным раковинным слоем плохой сохранности. В водорослево-гидроидных биогермных
известняках нижнетахтабулакской подсвиты Джилги-Кочусу найден 1 экз.; в табулято-
во-гидроидных биогермных известняках среднетахтабулакской подсвиты Кур-Истыка
1-2 экз.; Куталя 2-5 экз.; в табулятовых биогермных известняках Шорк-Бу-
лака - 5 экз.
111
ОТРЯД. RHYNCHONELLIDA
НАДСЕМЕЙСТВО RHYNCHONELLACEA
СЕМЕЙСТВО HYPOTHYRIDINIDAE RZONSNICKAJA, 1956
ПОДСЕМЕЙСТВО HYPOTHYRIDININAE RZONSNICKAJA, 1956
P од Uncinunellina Grabau, 1932
Uncinunellina': Grabau, 1932, стр. 72; Лихарев, 1939, стр. 104; 1960 д, стр.246;
Schmidt,Me Laren, 1965, стр. 566.
Типовой вид - Uncinulus theobaldi Waagen, 1884; верхняя часть среднего про-
дуктусового известняка и верхний продуктусовый известняк.(слои Чидру); Соляной
кряж (Пакистан).
Описание. Раковина средних размеров, поперечно-пятиугольная или поперечно-
треугольная с синусом на брюшной и выступом на спинной створке. Форамен очень
маленький.
Скульптура из многочисленных, хорошо развитых ребер, обычно несущих бороздки
по переднему краю. Некоторые виды обладают столидием. Наличие шиповатых вы-
ростов на концах ребер (видимо, имеется ввиду столидий) впервые описано у пред-
ставителей Uncinunellina Хуангом ( Huang, 1933, стр. 63, табл. X фиг. 356)., азатем Г.Шмидт
(Schmidt, 1937, стр. 30, рис. 14-24). Внутри брюшной створки - зубные пластины
(у различных видов развитые в различной степени), внутри спинной - разобщенная
замочная пластина. Замочный отросток и срединная септа отсутствуют. Круры длин-
ные, тонкие, фальциферовые.
Сравнение. Отличается от остальных родов подсемейства наличием столидия.
Кроме того, от рода AUorhynchus Weller, 1910 рассматриваемый род отличается
тонкой, хорошо развитой ребристостью с сильными, параллельными зубными пласти-
нами, а от родов Cerassim о via Licharew, 1956 и Tetebratuloidea Waagen, 1884 - хорошо
развитыми зубными пластинами в брюшной створке.
Видовой состав. Кроме типового вида, известного из среднего и верхнего
продуктусового известняка Соляного кряжа, а также из верхней перми Северной
Камбоджи, нижней перми Урала и Дарваза, в состав рода входит U.posterus (Wan-
gen, 1884) из верхнего продуктусового известняка Соляного кряжа; U.jabiensis
(Waagen, 1884) из верхнего продуктусового известняка (слои с Cyclolobus) Соляного
кряжа, верхней перми Тимора, нижней перми Дарваза и Урала; U.timorensis (Bey-
rich, 1865) из сакмарского и артинского ярусов Афганистана и Куньлуня и из верх-
ней перми Тимора, Юго-Западного Китая, Северного Кавказа (уруштенский горизонт)
Закавказья и Юго-Восточного Памира (джульфинский ярус); U. mongolica ^Grabau
1У31 из верхней перми Монголии; U.tschemyschewi Licharew, 1939 из шакарсевской
свиты верхней перми Дарваза; U .wangenheimi (Pander, 1861) из нижней перми Ура-
ла, Тимана, Новой Земли и сакмарско-^артинских отложений Афганистана и Кунь-
луня; U.velifer (Gemmellaro, 1899), U.siculus (Gemmellaro, 1899) и U.amor (Gemmel-
laro, 1899), из верхнепермских отложений Сицилии; U .laosensis (Mansuy, 1913) из
верхнепермских отложений Лаоса и Северной Камбоджи.
Замечания. Г.Шмидт (Schmidt, Me Laren, 1965) Uncinunellina отнесена со зна-
ком вопроса к подсемейству Uncinulinae Rzonsnickaja семейства Uncinulidae. По
внешнему виду Uncinunellina действительно напоминает девонский род Uncinulus
Bayle, 1878 и виды, в настоящее время принадлежащие к Uncinunellina, ранее мно-
гими исследователями относились к Uncinulus. В 1932 г. Гребо (Grabau, 1932), изу-
чив внутреннее строение пермских видов, относимых к Uncinulus, выделил их в
самостоятельный род Uncinunellina, характеризующийся рядом отличий; у Uncinu-
lus замочная пластина цельная, имеется двойная срединная септа в спинной створ-
ке и массивный замочный отросток, а у Uncinunellina замочная пластина разоб-
щи иная, замочный отросток и срединная септа отсутствуют. Эти признаки характер-
ны для представителей подсемейства HypothyricLininae семейства Hypothyrid ini -
dae.
112
Распространение. ? Ассельский ярус Урала, Тимана, Дарваза; сакмарский -
артинский ярусы Новой Земли, Афганистана, Куньлуня. Верхняя пермь Монголии, Си-
цилии; средний, верхний продуктусовый известняк Соляного кряжа; джульфинский
ярус Юго-Восточного Памира, Северного Кавказа, Закавказья.
Uncinunellina timorensis (Beyrich, 1865)
Табл. VIII, фиг. 16-18; табл. XIV, фиг. 5; рис. 25
Rhynchonella timorensis': Beyrich, 1865, стр. 72, 73, табл. I, фиг. 10.
Rhynchonella (Uncinulus) timorensis': Rothpleiz, 1892, стр. 84, табл. X, фиг. 6;
Freeh, 1911, стр. 171; Broili, 1916, стр. 59, табл. CXXVI, фиг. 6, 14; табл.
СХХУ11,фиг. 1-3;Hani]et, 1928, стр. 62, табл. X, фиг. 3-7; Reed, 1931а, стр. 20,
табл. III, фиг. 4-6.
Uncinulus timorensis': Diener, 1897a, стр. 69, 70, табл. X, фиг. 7-10; 1903,
стр. 90; Mansuy, 1913, стр. 85, табл. IX, фиг. 9; Tran Thi.Chi—Thuan, 1961, стр.
288, 289, табл. И, фиг. 18; табл. IV , фиг. 10; табл. VI, фиг. 5, табл. VII, фиг. 3.
Uncinunellina timorensis': Huang, 1933, стр. 61-64, табл. IX, фиг. 12, 13;
табл. X, фиг. 30-36; Ching Jii—kan, Zhang Jan., 1961, стр. 404; табл. I, фиг. 9-16;
Сокольская, 1965, табл. XL, фиг. 1.
Описание. Раковина небольшая, поперечно-пятиугольного очертания, с одина-
ково выпуклыми створками, со слабо изогнутым замочным краем, который незна-
чительно меньше наибольшей ширины, приуроченной к лобному краю. Лобный край
унипликатный, зубчатый; боковые - дуговидные, выпрямленные. Брюшная створка
поперечно-пятиугольная, уплощенная в задней части. В передней части - широкий
плоский синус, занимающий примерно половину ширины лобного^ края. Макушка
маленькая, короткая, прямостоящая. Макушечный угол 115-137 . Форамен очень ма-
ленький,круглый. Плечики макушки резко ограниченные. Спинная створка поперечно-
овальная, вздутая. В ее передней части - широкий, низкий плоский выступ, начи-
Рис. 25. Uncinunellina timorensis, ПИН, № 2228/16; зарисовки последовательных
поперечных сечений раковины в примакушечной части, х 3.6’
кр. -круры; з.я. - зубные ямки, кр.о. -круральные основания, з. - зубы; зм.п. - замоч-
ная пластина;остальные усл. обознач. см. рис. 9. Длина брюшной створки 14,4 мм. Кур-
Истык 1; джульфинский ярус, среднетахтабулакская подсвита
8 264
113
наюшийся приблизительно с половины длины створки. Макушка очень маленькая, при-
остренная, слабо обособленная.
Поверхность раковины покрыта радиальными ребрами, хорошо развитыми в передних
двух третях. Задняя треть - гладкая. Ребра резкие, треугольные в поперечном се-
чении, отделяются друг от друга глубокими межреберными промежутками. Количест-
во ребер в синусе 7-£, на выступе 8-9 , на боковых частях по 8—13 с каждой сторо-
ны синуса и выступа. Ребра, располагающиеся на боковых частях, короче и менее
отчетливы, чем на выступе и в синусе. При хорошей сохранности можно наблюдать,
что ребра несут продольные бороздки на гребнях. По лобному и боковым краям ра-
ковины развит с толи ди й. Ребристость, присутствующая на раковине, переходит и на
него.
Внутри брюшной створки - длинные, тонкие зубные пластины, располагающиеся
параллельно. Они поддерживают массивные округлые зубы, входящие в зубные ямки
спинной створки. Замочная пластина внутри спинной створки тонкая, разобщенная.
Круры тонкие, серповидно изогнутые, фальдиферовые (см. рис. 25).
Размеры, мм
Экз. № Местонахождение, возраст д Ш Т Ш с МУ Количество ребер
зинус выс- туп сбоку
бр.| СП.
2228/7 - Кур-Истык 1, джуль- 14,3 24,8 13,7 10,1 137° 8 8 20 20
фото 2228/9 финский ярус , средне- тахтабулакская под- 12,2 20,5 8,5 8,9 133° 9 8 17 25
2228/10 свита 10,0 17,2 8,2 7,9 134° 8 9 17 16
2228/12 13,7 22,4 9,8 12,1 130° 11 8 20 20
2228/14 10,1 15,7 6,7 8,7 117о 7 8 15 20
2228/19 10,6 14,4 6,9 7,0 119 7 6 10 8
Изменчивость, возрастные изменения. Очертания и форма раковины
очень характерны^ Варьирующими являются величина макушечного угла, колеблющая-
ся от 115 до 137 , степень выраженности срединного выступа и синуса, количество
ребер на выступе, в синусе и на боковых частях створок. Юные экземпляры харак-
теризуются плоской, гладкой, поперечно-треугольной раковиной со слабо развитыми
ребрами по лобному краю.
Сравнение. Uncinunellina timorensis наиболее близка к U.jabiensis (Waagen),
от которой отличается более вздутой, поперечно-пятиугольной раковиной, большим
количествОхМ ребер, а также их резкостью.
Замечания. На экземплярах U .timorensis, изображенных в работах большинства
исследователей, столидий обломан, но на некоторых из них видно, что он присутот -
вовал, так же как на экземплярах,имеющихся в нашей коллекции (Beyrich, 1865,
табл. I, фиг. 10; Diener, 1897а, табл. X, фиг. 7).
Геологический возраст и географичес кое распространение.
Сакмарский - артинский ярусы Афганистана и Куньлуня. Местонахождение Базлео
Тимора; верхняя пермь Юго—Западного Китая; джульфинский ярус Закавказья; джуль-
финский ярус: уруштенский горизонт Северного Кавказа; среднетахтабулакская под-
свита Юго-Восточного Памира.
Материал. 87 экз. с двумя сочлененными створками из отложений среднетах-
табулакской подсвиты. В табулятово-гидроидных биогермных известняках Кур-Исты- <
ка 1 собрано 22 экз.; Куталя 1 - 9 экз.; Куталя 2-13 экз.; в табулятовых био-
гермных известняках Шор-Булака - 38 экз.; в водорослевых биогермных известняках
М узду-Бу лака - 5 экз.
114
Р од Gerassimovia Licharew, 1956
Gerassimovia': Лихарев, 1956, стр. 59; 1960 д, стр. 246; Schmidt, Me Laren,
1965, стр. 592.
Типовой вид - G.gefoensis Licharew, 1956; верхняя пермь, уруштенский
горизонт; Северный Кавказ.
Описание. Раковина от небольших до средних размеров, от округло-треуголь-
ной до округло-пятиугольной формы с маленьким фораменом. Синус на брюшной
створке^ выступ - на спинной. Радиальные ребра развиты в различной степени: при-
сутствуют в синусе, на выступе и на боковых частях раковины или только в синусе
и на выступе.
Внутри брюшной створки - небольшие, косо расположенные зубы. Зубные пласти-
ны только на молодых стадиях. Внутри спинной створки - разделенная замочная
пластина. Замочный отросток и срединная септа отсутствуют. Круральные основания
слабо развиты. Круры фальциферовые.
Сравнение. Данный род отличается от Uncinunellina Grabau, 1932 и Allorhyn—
chus Weller, 1910 отсутствием зубных пластин в брюшной створке.
Видовой состав. Кроме типового вида к данному роду относятся G.bactria-
na Grunt, sp. nov. из верхней части базардаринской свиты сакмарского яруса -
ни; кн е артин с кого подъяруса Юго-Восточного Памира; G.abalakovi Grunt, sp. nov. и G.pa-
mirica Grunt, sp.nov. из среднетахтабулакской подсвиты джульфинского яруса Юго-
Восточного Памира. Предположительно сюда же относят Rh.morahensis Waagen, 1884,
из нижнего продуктусового известняка Соляного кряжа (Пакистан).
Распространение. Сакмарский - нижняя часть артинского яруса нерасчле-
ненные, базардаринская свита (верхняя часть) Юго-Восточного Памира; джуль-
финский ярус: уруштенский горизонт Северного Кавказа; среднетахтабулакская под-
свита Юго-Восточного Памира. Предположительно нижний продуктусовый известняк
Соляного кряжа.
Gerassimovia pamirica Grunt, sp.nov.
Табл. IX, фиг. 6; рис. 26
Видовое название по местонахождению на Памире.
Голотип - ПИН,М 2228/36; раковина с двумя сочлененными створками; Юго-
Восточный Памир, Кур-Истык 1; джульфинский ярус, среднетахтабулакская подсви-
та; табл. IX, фиг. 6.
Описание. Раковина небольшая поперечно-пятиугольного очертания, умеренно
двояковыпуклая, с изогнутым замочным краем, меньшим наибольшей ширины, приу-
роченной к лобному краю. Лобный край унипликатный, зазубренный, боковые - ду-
говидные. Брюшная створка немного менее выпуклая, чем спинная, пятиугольная.
Макушка короткая, утолщенная, слабо загнутая. Макушечный угол 97-126 . Форамен
удаленький. Синус широкий, неглубокий. Язычок отчетливый, трапецеидальный, боль-
шой. Спинная створка поперечно-пятиугольная. Выступ низкий.
Раковина покрыта ребрами, начинающимися немного отступя от макушки. Ребра
отчетливые, резкие, расширяющиеся по направлению к лобному краю, треугольные
в поперечном сечении. Количество ребер в синусе обычно 4-5, на выступе 4-5,
на боковых краях по 4-6 с каждой стороны синуса и выступа. Внутреннее строение
типичное для рода (см. рис. 26).
Сравнение. Наш вид отличается от G.gefoensiq Licharew несколько меньшими
размерами, поперечно вытянутой формой раковины, ребрами, начинающимися не от
самой макушки, отчетливо выраженным выступом спинной створки. От G. bactria-
na Grunt, sp.nov. он отличается значительно меньшими размерами и ребристостью,
отчетливо развитой как в синусе и выступе, так и на боковых частях раковины; от
G.abalakovi Grunt, sp.nov. - более крупными размерами, резкой ребристостью, боль-
шим количеством ребер в синусе и на выступа.
115
бр. С.
Рис. 26. Gerassimovia pamirica, ПИВ, № 2228/58; зарисовки последовательных по-
перечных сечений раковины в примакушечной части, х 3.6. Усл. обознач. см. рис. 9 и
рис. 25. Длина брюшной створки 16,4 мм. Кур-Истык 1; джульфинский ярус, сред-
нетахтабулакская подсвита
Размеры, мм
Экз., № Местонахожде- ние, возраст д Ш Т Ш с МУ Количество ребер
синус выс- туп сбоку
бр. 1 СП.
2228/36- голотип Кур-Истык 1, 15,3 18,8 10,8 10,8 126° 4 5 и 9
2228/37 джульфинский 15,4 16,6 10,1 8,7 102 4 5 11 10
2228/38 ярус, средне- 15,7 18,3 10,6 9,2 105 3 4 10 9
2228/39 тахта була кс кая 13,0 15,2 9,8 7,0 97° о 3 4 9 8
2228/41 юдсвита 14,6 18,1 9,2 11,1 119о 4 5 10 10
2228/43 13,9 17,1 9,0 9,6 114 5 6 10 9
2228/51 14,6 16,8 12,1 8,0 105° 4 5 10 10
Геологический возраст и географичес кое распространение.
Джульфинский ярус, среднетахтабулакская подсвита Юго-Восточного Памира.
Материал. 59 экз. хорошей сохранности с двумя сочлененными створками, час-
тично перекристаллизованные. Собраны в отложениях среднетахтабулакской подсвиты.
В табулятово-гидроидных биогермных известняках Кур-Истыка 1 найдено 57 экз.;
в водорослевых биогермных известняках Музду-Булака - 2 экз.
Gerassimovia abalakovi Grunt, sp.nov.
Табл. IX, фиг. 2; рис. 27
Видовое название в честь альпиниста В.Абалакова.
Голотип - ПИН, № 2228/170; раковина с двумя сочлененными створками; Юго-
Восточный Памир, Куталь 2; джульфинский ярус, среднетахтабулакская подсвита;
табл. IX, фиг. 2.
Описание. Раковина небольшая, округло-пятиугольная, приблизительно изимет-
ричная, умеренно вздутая. Замочный край резко изогнут, меньше наибольшей шири-
мы раковины, приуроченной к ее средней части. Лобный край унипликатный, зазуб-
ренный, боковые - дуговидные. Брюшная створка немного менее выпуклая, чем спин-
116
I 0.2
III 0Л
Рис. 27. Gerassimovia abalakovi, ПИН, N? 2228/172; зарисовки последовательных
поперечных сечений раковины в примакушечной части, х 3.6, Усл. обознач. см.рис. 9
и рис. 25. Длина брюшной створки 11,1 мм. Куталь 2; джульфинский ярус, средне-
тахтабулакская подсвита
ная, с широким округлым синусом, оттянутым в виде, широкого полуокруглого языч-
ка и начинающимся с половины длины створки. Макушка узкая,' прямостоящая. Ма-
кушечный угол 69-75 . Форамен небольшой, округлый. Спинная створка умеренно
выпуклая, с очень низким выступом, который хорошо наблюдается только по лобно-
му краю. Макушка короткая, широкая, округлая.
Скульптура состоит из простых, немногочисленных ребер, располагающихся на
выступе и в синусе - в передней трети раковины. Количество ребер на выступе -3;
в синусе - 2. Внутри брюшной створки зубные пластины отсутствуют. Зубы широкие,
массивные, отходят непосредственно от стенок раковины и входят в широкие ямки,
располагающиеся на замочной пластине. Внутри спинной створки - разобщенная за-
мочная пластина с субпараллельными круральными основаниями и слабым срединным
валиком (см. рис. 27).
Размеры, мм
Экз., № Местонахождени е, возраст д Ш Т МУ Количество ребер
синус выс- туп сбоку
бр. jcn.
2228/175 Кур-Истык 1, 10,8 10,2 7,4 72° о 2 3 4 4
2228/176- джульфинский 8,8 9,4 5,9 69 — 2 — —
2228/172 ярус, среднетахтабу- лакская подсвита 12,4 12,9 7,5 69° 2 3 2
2228/170 голотип Куталь 2, возраст тот же 11,5 11,6 7,0 70° 2 3 — —
2228/174 2228/170а Шор-Булак, возраст тот 9,6 10,4 9,6 10,7 5,8 6,7 75° 75° 2 3 — —
2228/171а же 9,5 9,7 5,5 71° 3 3 — —
Сравнение. Ст G.gefoensis Licharew, G.bactriana Grunt,sp.nov. и G.pamirica Grunt,
sp.nov. данный вид отличается значительно меньшими размерами; от G.gefoensis,
кроме того, - наличием хорошо выраженных синуса и выступа; а от G.pamirica и
G gtfoensis - менее отчетливой ребристостью, развитой только в передней трети
раковины.
117
Геологический возраст и географическое распространенно.
Джульфинский ярус, среднетахтабулакская подсвита Юго-Восточного Памира.
Материал. 10 экз. удовлетворительной сохранности с двумя сочлененными
створками. Собраны в среднетахтабулакской подсвите. В табулятово-гидроидных
биогермных известняках Кур-Истыка 1 найдены 4 экз.; Куталя 2-3 экз.; в таб.;-
лятовых биогермных известняках Шор-Булака - 3 экз.
Gerassimovia bactriana Grunt, sp.nov.
Табл. IX, фиг. 3-5; табл. XIV, фиг. 6, рис. 28
Видовое название от Бактрии - древней страны, располагавшейся на терри-
тории Памира Средней Азии.
Голотип - ПИН № 2228/178; раковина с двумя сочлененными створками; Юго-
Восточный Памир, Куртеке 1; сакмарский ярус - нижнеартинский подъярус нерасчле-
ненные, базардаринская свита (верхняя часть); табл. IX, фиг. 3.
Рис. 28. Gerassimovia bactriana, ПИН, № 2228/205; зарисовки последовательных по-
перечных сечений раковины в примакушечной части, х 2.7. Усл. обознач. см. рис. 9
и рис. 25. Длина брюшной створки 18,6 мм. Кастанат-Джилга 1; сакмарский ярус -
нижнеартинский подъярус нерасчлененные, базардаринская свита (верхняя часть)
Описание. Раковина крупная округло-пятиугольного очертания, с сильно изог-
нутым замочным краем, меньшим наибольшей ширины раковины, приуроченной к ее
средней части, умеренно двояковыпуклая. Лобный край унипликатный, зазубренный,
боковые края дуговидные. Брюшная створка субпятиугольная, уплощенная, незначи-
тельно менее выпуклая, чем спинная, с широким плоским синусом в передней поло-
вине створки. Язычок широкий, плоский, трапециевидный. Макушка широкая, резко
сужающаяся, с маленьким округлым фораменом. Макушечный угол 95-120 . Спинная
створка округло-треугольная, с высоким узким выступом, развитым в передней по-
ловине створки. Макушка приостренная, слабо обособленная.
Скульптура состоит из немногочисленных резких ребер, развитых лишь в перед-
ней части раковины на выступе (4) и в синусе (3). Внутри брюшной створки зубные
пластины близко примыкают к боковым стенкам, вьфажены слабо. Зубы массивные,
эллипсоидальные. Мускульное поле округлое, сильно вдавленное, небольшое. Васку-
лярные отпечатки в виде двух ветвящихся стволов (табл. IX, фиг. 4). Внутри спин-
ной створки - разобщенная замочная пластина с длинными, субпараллельными крураль-
ными основаниями. Септальный валик развит слабо или отсутствует (см. рис. 26).
118
Размеры, мм
Экз., № Местонахождение, возраст д Ш Т Ш с МУ Количество ребер
синус | выступ
2228/177 Куртеке 1, сакмарский 30,2 31,4 17,6 12,6 96о 3 2
2228/178- голотип ярус - нижнеартинский подъярус нерасчленен- 25,8 30,9 18,9 13,0 102 л 4 4
2228/179 ные, базардаринская 31,7 33,7 20,3 18,7 90о 3 3
2228/183 свита (верхняя часть) 25,4 28,2 18,2 11,6 91о 3 3
2228/185 22,6 21,7 12,5 14,8 97 з 3
2228/192 Кастанат-Джилга 1, возраст тот же 40,5 37,7 19,1 20,4 120° 3 3
Сравнение. От двух предыдущих видов данный вид отличается более крупны-
ми размерами. От G.gefoensis Licharew, к которому G. bactriana приближается по
размерам, - резко выраженными синусом и выступом и менее развитой ребристостью
обеих створок. По внешнему виду G.bactriana сходна с Leiorhynchus ripheicus Ste-
panov, 1948, но отличается от него разделенной замочной пластиной и отсутстви-
ем септалия в спинной створке.
Геологический возраст и географическое распространение.
Сакмарский ярус - нижнеартинский подъярус нерасчлененные, базардаринская свита
(верхняя часть) Юго-Восточного Памира.
Материал. 83 экз. удовлетворительной сохранности. Собраны в верхней части
базардаринской свиты. Из них.40 экз., представленных раковинами с двумя сочле-
ненными створками, найдены в органогенно-обломочных алевритистых известняках
Куртеке 1; 3 раковины с двумя сочлененными створками, 3 брюшных створки и 1
спинная створка - в глинистых известняках Чакобая. В известковистых глинистых
алевролитах Кастанат-Джилги 1 - 32 брюшных и 13 спинных створок; в известковис-
тых глинистых алевролитах Чакобая; — 10 раковин с двумя сочлененными створками;
10 брюшных и 1 спинная створка.
Рош Boloria Grunt, gen. nov.
Название рода от Болора - легендарного названия древнего Памира.
Типовой вид - В.garmoensis Grunt, sp. nov.* дарвазский ярус, кызылджилгин-
ский горизонт, нижний подгоризонт Юго-Восточного Памира, Мамазаир.
Описание. Раковина небольшая, гладкая, с синусом на спинной и выступом на
брюшной створке, с сулькатным лобным краем. Макушка брюшной створки оттяну-
тая, узкая. Форамен маленький, круглый. Внутри брюшнэй створки - длинные, тон-
кие, параллельные зубные пластины. Внутри спинной - небольшая разобщенная за-
мочная пластина. Септа отсутствует. Септальный валик слабый (см. рис. 29, 30).
Сравнение. От всех известных родов подсемейства Hypothyridininae Rzonsni—
ckaja, 1956, наш род отличается гладкой раковиной и обратным расположением сину-
са и выступа (т.е. синусом, расположенным на спинной и выступом на брюшной створ-
ке).
Видовой состав. Типовой вид.
Замечания. Раковины с обратным расположением синуса и выступа встречают-
ся в различных семействах. Так,роды Paranorella Cloud, 1944 и Sanjuania Amos, 1958
относятся к подсемейству Camarotoechiinae Schuchert et Le Vene,l929 семейства
Camarotoechiidae; JVore//a Bittner, 1890 к подсемейству Norellinae Ager, 1959 семейства
Dinierelli.iae, a Camarophoriina Licharew, 1956 к надсемейству Stenoscismatacea. C
перечисленными родами Boloria сходна по внешнему виду. Or родов Paranoretla и
Sanjuania она отличается отсутствием септалия внутри спинной створки, а также
119
длинными субпараллельными зубными пластинами внутри брюшной, а от рода Norella-
отсутствием септалия и сильных макушечных утолщений в спинной створке; от рода
Camarophoriina - отсутствием спондилия в брюшной створке и камарофория - в
спинной.
Распространение. Дарваэский ярус, кызылджилгинский горизонт, нижний под-
горизонт.
, Boloria garmoensis Grunt, sp.nov.
Табл. IX, фиг. 7,8; рис. 29; 30
Видовое название от Гармо - горного узла на Памире.
Голотип - ПИН,№ 2228/200; раковина с сочлененными створками; Юго-Вос-
точный Памир, Мамазаир; дарваэский ярус, кызылджилгинский горизонт, нижний
подгоризонт; табл. IX, фиг. 7.
Рис. 29. Boloria garmoensis, ПИН, № 2228/21 la; зарисовки последовательных по-
перечных сечений раковины в примакушечной части, х 3.6. Усл. обоэнач. см. рис. 9 и
рис. 25. Длина брюшной створки 9,0 мм. Мамазаир-Булак* дарваэский ярус, кы-
зылджилгинский горизонт, нижний подгоризонт
Рис. 30. Boloria garmoensis, ПИН, № 2228/208а; зарисовки последовательных по-
перечных сечений раковины в примакушечной части, х 3.6. Длина брюшной створки
10,9 мм. Усл. обознач., возраст и местонахождение те же, что и на рис. 29
Описание. Раковина небольшая, приблизительно изометричная, умеренно дво-
яковыпуклая, округло-пятиугольная, гладкая. Замочный край круто дуговидно изог-
нут, меньше наибольшей ширины раковины, приуроченной к ее средней части. Лобный
край сулькатный, боковые - дуговидные. Брюшная створка немного более выпуклая,
чем спинная, с низким, плоским срединным выступом, ограниченным двумя слабыми
складками. Макушка оттянутая, умеренно загнутая, резко сужающаяся. Форамен очень
маленький. Макушечный угол 91-108 . Спинная створка уплощенная с глубоким сину-
сом, который имеет округлое дно и нерезко очерченные склоны; он резко расширяет-
ся кпереди, где образует отчетливый язычок трапециевидной формы. Макушка ко-
роткая, приостренная. Внутри брюшной створки - длинные, параллельные зубные
пластины, достигающие дна створки и поддерживающие массивные, округлые зубы.
Замочная пластина в спинной створке разобщенная. Ее боковые части вогнуты; в
углубления входят зубы. Внутренние приямочные гребни отчетливые, высокие. Кру-
120
ральныо основания с маленькими ребрами. Срединная септа отсутствует. Септальный
валик развит слабо (см. рис. 29, 30).
Размеры, мм
Экз.,№ Местонахождение, возраст д Ш Т МУ
2228/200- голотип Мамазаир-Булак, дарвазский ярус, кызылджилгинский го- 12,4 13,0 7,0 108°
2228/201 ризонт, нижний подгоризонт 12,0 13,6 7,0 100°
2228/202а 10Д) 11,4 6,5 93
2228/208а 10.9 13,0 6,6 102°
2228/204а 10,5 12,0 7,1 91°
Геологический возраст и географическое распространение.7
Харвазский ярус,кызылджилгинский горизонт, нижний подгоризонт Юго-Восточного
1амира.
Материал. 16 экз., представленных раковинами с двумя сочлененными створками,
юбраны в нижне-кызылджилгинском подгоризонте. В криноидно-детритовых известня-
ках Куртеке 2 найден 1 экз.; в микрозернистых глинистых известняках Мамазаир-
Зула ка - 12 экз.; в микрозернистых глинистых известняках Игримиюза - 3 экз.
СЕМЕЙСТВО WELLERELLIDAE LICHAREW, 1956
ПОДСЕМЕЙСТВО WELLERELLINAE LICHAREW, 1956
Р од We Here Ila Dunbar et Condra, 1932
Wellerelld: Dunbar, Condra, 1932, стр. 286; Степанов, 1937, стр. 3; Лихарев,
1939, стр. 103; 1960д, стр. 247; Schmidt, McLaren, 1965, стр. 590.
Типовой вид - W.tetrahedra Dunbar et Condra, 1932; верхний карбон; Северная
Америка.
Описание. Раковина небольшая, округло-треугольная или округло-пятиугольная,
умереннодвояковылуклая, с синусом на брюшной и выступом на спинной створке.
Простые немногочисленные ребра развиты на всей поверхности обеих створок,
кроме примакушечной части, которая остается гладкой. Внутри брюшной створки -
тонкие зубные пластины; внутри спинной - цельная замочная пластина и тонкая
срединная септа, которая может быть развита у разных видов в различной степени.
Замочный отросток отсутствует. Круры длинные, фальциферовые (располагаются па-
раллельно плоскости симметрии раковины, серповидно изогнуты).
Сравнение. Рассматриваемый род отличается от Pseudowelterella Licharew, 1956
наличием зубных пластин в брюшной створке и обычно хорошо развитой срединной
септой - в спинной, О'5 Lissella Campbell, 1961 он внешне отличается более резкой
ребристостью и большим количеством ребер, а по внутреннему строению - срединной
септой, обычно соединяющейся с замочной пластиной.
Видовой состав. Кроме типового вида к данному роду относятся: W.arthabe—
ri (Tschemyschew, 1914) из нижнепермских отложений Юго-Западного Дарваза, верх-
ней перми Юго-Восточного Памира (среднетахтабулакская подсвита), Северного Кав-
каза (уруштенский горизонт), Закавказья. Сюда же входят W.wichmanni (Rothpletz,
1892) из верхнепермских отложений Тимора; W.cooperi Jochelson, 1962 из нижней
перми Канзаса; W .dekalbensis Dunbar et Condra,1932 и W.delicatula Dunbar et Condra,
1932 из верхнего карбона Небраски; W.osakensis (Swallow, 1860) из верхнего карбона
Северного Урала, Техаса, Небраски и нижней перми Канзаса; W.truncate Dunbar et
121
C end ra, |932 из верхнего карбона Башкирии и Небраски, а также нижней перми Кан-
заса.
Распространение. Верхний карбон - верхняя пермь Западного Приуралья, Ти-
мана, Северного Кавказа, Закавказья, Юго-Восточного Памира, Дар ваза, Китая, Се-
верной и Южной Америки.
Wellerella arthaberi (Tschemyschew, 1914)
Табл. IX, фиг. 1, табл. XIV, фиг. 7; рис. 31
Uncinulus wichmani: Arthaber, 1900, стр. 283, табл. XXII, фиг. 16.
Uncinulus arthaberi: Чернышев, 1914, стр. 15, табл. IV, фиг. 5-6.
Wellerella arthaberi: Лихарев, 1939, стр. 103, табл. XXV, фиг. 6; Сокольская,
1965, табл. XL, фиг. 6.
Описание. Раковина маленькая, субпятиугольного очертания, приблизительно
изометричная, довольно сильно вздутая. Замочный край изогнутый, меньше наиболь-
шей ширины раковины, расположенной по лобному краю. Лобный край унипликатный,
боковые края дуговидные. Брюшная створка менее выпуклая, чем спинная, уплощен-
ная в задней части и незначительно коленчато изогнутая в передней. Синус широкий,
плоский, начинается примерно с половины длины створки, впереди подогнут в виде
широкого плоского язычка. Макушка маленькая, узкая, прямая. Дельтирий закрыт
двумя маленькими дельтидиальными пластинами. Форамен на кончике макушки, ма-
ленький овальный. Макушечный угол 90-100 . Спинная створка округлая, с широким
низким выступом, отделенным от боковых частей раковины глубокими бороздками.
Макушка короткая, слабо приостренная.
Рис. 31. Wellerella arthaberi, ПИН, № 2228/96; зарисовки последовательных по-
перечных сечений раковины в примакушечной части,х 4.0. Усл. обознач. см.рис. 9
и рис. 25. Длина брюшной створки 6,8 см. Кур-Истык '1; джульфинский ярус, сред-
нетахтабулакская подсвита
Скульптура состоит из немногочисленных резких ребер, начинающихся прибли-
зительно с половины длины раковины. Макушечная область остается гладкой. Коли-
чество ребер в синусе 3-4 , на выступе 4-5, на боковых частях по 2-4 ребра с каж-
дой стороны синуса и выступа.
Внутри брюшной створки — тонкие, слабо расходящиеся зубные пластины, дости-
гающие дна створки и начинающиеся примерно на расстоянии 0,2 мм от макушки.
Зубные пластины поддерживают широкие, округлые зубы. Внутри спинной створки -
неглубокие округлые зубные ямки, ограниченные с внутренней стороны невысокими
122
внутренними приямочными ребрами. Замочная пластина цельная, тонкая. Присутству-
ет высокая, тонкая срединная септа,сочленение которой с замочной пластиной не-
ясно. Круры тонкие, серповидно изогнутые, фальциферовые (см. рис. 31).
Размеры, мм
Экз.,№ Местонахождение, возраст д Ш Т Шс МУ Количество ребер
си- нус выс- туп сбоку
бр. СП.
2228/98- Кур-Истык 1, джуль- 8,7 9,1 6,7 6,7 98° 6 7 7 6
фото финский ярус, сред- о
2228/95 нетахтабулакская 7,4 8,2 7,2 5,6 93о 4 5 6 6
2228/139 подсвита 8,1 8,7 6,6 5,5 97 4 5 7 8
2228/142 7,2 9,4 6,6 5,8 92 с 4 5 7 9
2228/107 6,4 7,3 5,2 5,3 101 ‘ 6 5 5 6
2228/137 6,7 7,4 6,0 4/) 98 3 3 5 6
2228/100 7,2 7,9 5,9 5,5 96° 6 7 5 7
2228/140 7,1 8,1 5,4 4,1 92° 2 3 7 7
2228/101 7,0 7,1 5,3 4,5 121С ' 4 5 4 6
2228/105 6,0 6,2 4,5 4,0 11ТС ’ 4 4 3 3
Изменчивость, возрастные изменения. Изменчивыми для данного вида
являются степень выпуклости раковины, резкость синуса и выступа, резкость ре-
бер. Более юные экземпляры характеризуются уплощенной, гладкой раковиной ок-
ругло-треугольной формы. Синус и выступ выражены слабо. Ребра менее резкие и в
меньшем количестве развиты лишь в передней части раковины или по лобному краю.
Сравнение. От близкого вида IV.osagensis Swallow данный вид отличается мень-
шими размерами и ребристостью, начинающейся ближе к макушке, а от W .wichmani
(Rothpl.) — округло-пятиугольной формой и ребристостью, развитой только в перед-
ней части раковины.
Геологический возраст игеографическое распространение.
Нижняя пермь Юго-Западного Дарваза; джульфинский ярус Закавказья; джульфин-
ский ярус; уруштенский горизонт Северного Кавказа; среднетахтабулакская подсвита
Юго-Восточного Памира.
Материал. 91 экз. хорошей сохранности с двумя сочлененными створками, соб-
ран в отложениях среднетахтабулакской подсвиты. В табулятово-гидроидных био-
гермных известняках Кур-Истыка Г найдены 76 экз.; Куталя 2-3 экз.; в табу-
лятовых биогермных известняках Шор-Булака - 9 экз.; в водорослевых биогермных
известняках Музду-Булака - 3 экз.
НАДСЕМЕЙСТВО STENOSCISMATACEA
СЕМЕЙСТВО ATRIBONIIDAE GRANT, 1965
Состав семейства. 2 подсемейства: Artiboniinae и Psilocamerinae.
Замечания. Р.Грант (Grant, 1965а) разделил надсемейство Stenoscismatacea
на 2 семейства: Atriboniidae и Stenoscismatidae. К семейству Atriboniidae принад-
лежат стеносцизматацеи без столидия. К нему относятся,в частности, роды Сата-
rophorinelta Licharew, 1936 и Camerisma Grant, 1965, у которых столидий не был отвечен.
Однако на представителях этих родов, имеющихся в нашей коллекции, это морфоло-
гическое образование было обнаружено.
123
Из отложений уруштенского горизонта джульфинского яруса Северного Кавказа
нами собраны многочисленные экземпляры C.caucasica Licharew - типового вида
рода Camarophorinetla, обладающие хорошо развитым столидием. При просмотре кол-
лекции Б.К. Лихарева из пермских отложений Северного Кавказа было обнаружено,
что некоторые экземпляры C.caucasica также обладают следами столидия, который,
по-видимому, был обломан при препаровке. Из кызылджилгинского горизонта Юго-
Восточного Памира происходят экземпляры, по всем признакам наружного' и внут-
реннего строения относящиеся к Camerisma, но имеющие следы столидия. Эти два при-
мера свидетельствуют о том, что семейство Atriboniidae содержит как рода^обла-
дающие столидием, так и лишенные его.
К семейству Stenoscismatidae Грант отнес роды, у которых столидий обычно
присутствует (но некоторые роды, например Coledium Grant, 1965, иногда и не об-
ладают им).
Таким образом, представляется недостаточно убедительным подразделение над-
семейства Stenoscismatacea на семейства по наличию или отсутствию столидия. Нуж-
но отметить также, что это образование обычно плохо сохраняется и зачастую пол-
ностью уничтожается, если не при захоронении раковин, то при их препаровке.
Поэтому установить, присутствовал ли он первоначально, бывает очень трудно.
ПОДСЕМЕЙСТВО ATRIBONIINAE GRANT, 1965
Р од Camerisma Grant, 1965
Camerisma': Grant, 1965a, стр. 63; 1965b, стр. 626.
Типовой вид - С.prava Grant, 1965; нижний карбон (верхняя часть мисси-
сипского отдела); Юго-Восточная Аляска.
Описание. Раковина средних размеров, округлая или округло-пятиугольная, с
утолщенными стенками, с синусом на брюшной и выступом на спинной створке.
Скульптура из немногочисленных, слабых ребер, располагающихся только в передней
части синуса и выступа. У некоторых видов присутствует столидий.
Внутри брюшной створки - глубокий двойной спондилий. Внутри спинной - цель-
ная замочная пластина с хорошо развитым замочным отростком. Камарофорий с
двойной межкамарофориальной пластиной. Срединная септа низкая, двойная.
Сравнение. От родов Atribonium Grant, 1965 и Sedenticellula Cooper, 1942 рас-
сматриваемый род отличается более массивной раковиной, массивными спондилием и
камарофорием, а также двойной межкамарофориальной пластиной.
Видовой состав. Кроме типового вида, к данному роду относятся; C.girtyi
Grant, 1965 из нижнего карбона Аляски; C.sagmarium Grant, 1965 из артинского яруса
нижней перми Югославии; С.sella (Kutorga, 1842) из нижней перми Юго-Западного
Дарваза, швагеринового горизонта Урала и Тимана, а также известняка Мапинг
Китая; C.murgabica Grunt, sp.nov. из нижнего и среднего под горизонтов кызылд-
жилгинского горизонта дарвазского яруса Юго-Восточного Памира.
Распространение. Нижний карбон Аляски; нижняя пермь Юго-Западного Дар-
ваза; артинский ярус Югославии; швагериновый горизонт Урала и Тимана; извест-
няк Мапинг Китая; дарвазский ярус , кызылджилгинский горизонт, нижний и сред-
ний подгоризонты Юго-Восточного Памира.
Camerisma murgabica Grunt, sp.nov.
Табл. VIII, фиг. 11, 12; табл. XV,фиг. 1; рис. 32
Видовое н а з в а н и е по местонахождению в районе г. Мургаба (Горпо-Бадах-
шанская АО).
Голотип - ПИН, № 2228/215; раковина с двумя сочлененными створками; Юго-
Восточный Памир, Чакобай; дарвазский ярус, кызылджилгинский горизонт, сродним
подгоризонт, табл. VIII, фиг. 11.
124
Рис. 32, Camerisma murgabica, ПИН, № 2228/219; зарисовки последовательных попереч-
ных сечений раковины в примакушечной части, х 2.2;
з.о. - замочный отросток, кмф - камарофорий, мк.п. - межкамарофориальная
пластина. Остальные усл. обознач. см. рис. 9 и 25. Длина брюшной створки 11,1 мм.
Чакобай; дарваэский ярус, кызылджилгинский горизонт, средний подгоризонт
Описание. Раковина средних размеров, округлая или округло-пятиугольная,
умеренно двояковыпуклая. Замочный край короткий, изогнутый, меньше наибольшей
ширины, приуроченной к лобному краю. Лобный край унипликатный, зазубренный, бо-
ковые края круто дуговидно изогнутые. Брюшная створка округло-пятиугольная,
немного менее выпуклая, чем спинная, с широким плоским синусом. Макушка широ-
кая, короткая, умеренно загнутая, с небольшим, округлым фораменом. Макушечный
угол 82-63 . Язычок небольшой, округлый, отогнутый в сторону спинной створки.
Спинная створка поперечно-треугольная, с неотчетливым выступом, развитым лишь
в передней части створки. Макушка короткая, округлая.
Скульптура состоит из простых, слабых радиальных ребер, располагающихся на
выступе и в синусе. В синусе их 4, на выступе - 3. Ребра развиты лишь в передней
половине раковины. Боковые части гладкие. По лобному краю наблюдается слабо
развитый столидий.
Внутри брюшной створки - массивные зубные пластины, соединяющиеся посредине.
Они поддерживаются низкой двойной срединной септой, образуя двойной спондилий.
Внутри спинной створки - цельная замочная пластина, низкий, но хорошо развитый
замочный отросток. Камарофорий заполнен двойной межкамарофориальной пластиной.
Срединная септа двойная, низкая (см. рис. 32).
Размеры, мм
Экз.,№ Местонахождение, возраст д Ш Т ш с МУ Количество ребер
синус 1 выступ
2228/215 - Чакобай,дарвазский ярус, 17,7 21,0 11.4 10,9 93° 4 5
голотип 2228/216а кызылджилгинский гори- зонт,средний подгоризонт 11,2 17,1 8,7 8,5 82° 2 3
2228/218а 11.1 12,9 12,0 9,5 85 2 3
Сравнен ие. Наш вид отличается от остальных ВИДОЕ > рода Camerisma наличием
слабо развитого столидия по лобному краю.
125
Геологический возраст и географическое распространение,
Дарвазский ярус, кызылджилгинский горизонт, нижний и средний подгоризонты Юго-
Восточного Памира.
Материал. 7 экз. удовлетворительной сохранности собраны в кызылджилгинском
горизонте. В криноидно-детритовых известняках нижнекызылджилгинского подгори-
зонта Куртеке 2 найдена 1 раковина с сочлененными створками; Мамазаир-Булака-
1 брюшная створка; в туффитовых известняках среднекызылджилгинского подгоризон-
та Чакобая - 5 раковин с сочлененными створками.
ПОДСЕМЕЙСТВО PSILOCAMERINAE GRANT, 1965
Р од Camarophorinella Licharew, 1936
Camarophorinella': Licharew, 1936, стр. 63; 1960 д, стр. 250; Grant, 1965а, стр.85;
1965 b,стр. 628.
Типовой вид - C.caucasica Licharew,1936;джульфинский ярус, уруштенский го-
ризонт; Северный Кавказ.
Описание. Раковина небольшая, поперечно-треугольная, умеренно двояковыпук-
лая, с синусом на брюшной и выступом на спинной створке.
Скульптура состоит из простых, резких немногочисленных ребер, занимающих всю
поверхность раковины. Столидий развит или отсутствует. Внутри брюшной створки -
двойной спондилий. Внутри спинной - раздельная замочная пластина с небольшим за-
мочным отростком. Камарофорий без межкамарофориальной пластины, поддерживается
срединной септой.
Сравнение. Рассматриваемый род отличается от рода Camarophorina Licharew,
1934 наличием ребристости, и расположением синуса на брюшной, а выступа на спин-
ной створках в противоположность Camarophoriina, характеризующейся обратным
расположением синуса и выступа. Отличие от Cyrolexis Grant, 1965 состоит в значи-
тельно более резко развитой ребристости, разделенной замочной пластине и меньших
размерах замочного отростка внутри спинной створки.
Видовой состав. Кроме типового вида к данному роду относится C.leveni
Grunt, sp.nov. из среднетахтабулавкой подсвиты джульфинского яруса Юго-Восточ-
ного Памира и, возможно, известняка Созио мургабского яруса Сицилии.
Распространен ие; Джульфинский ярус: уруштенский горизонт Северного Кав-
каза; среднетахтабулакская подсвита Юго-Восточного Памира. Предположительно,
мургабский ярус, известняк Созио Сицилии.
Camarophorinella leveni Grunt, sp.nov.
Табл.VIII,фиг. 15; табл.XIV,фиг. 9; рис. 33
Видовое название в честь Э.Я. Левена.
Голотип - ПИН, № 2228/161; раковина с двумя сочлененными створками; Юго-
Восточный Памир, Шор-Булак; < джульфинский ярус, среднетахтабулакская подсвита;
табл. VIII, фиг. 15.
Описание. Раковина небольшая, округло-треугольная или округло-четырех-
угольная, довольно сильно двояковыпуклая, с изогнутым замочным краем, меньшим
наибольшей ширины раковины, располагающейся в ее средней части. Лобный край
ушпликатный, зазубренный, боковые края круто дуговидные. Брюшная створка попе-
речно-пятиугольная, менее выпуклая, чем спинная. Синус широкий плоский, начина-
ется почти от самой макушки. Макушка короткая, узкая, слабо загнутая. Макушеч-
ный угол 90-115 , Фора мен маленький, округлый. Плечики макушки гладкие, нерезко
ограниченные. Спинная створка поперечно-треугольная, с широким, пологим выступом.
Макушка короткая, округлая.
126
Рис. 33. Camarophorinella leveni, ПИН, № 2228/159; зарисовки последовательных
поперечных сечений раковины в примакушечной части, х 5.4. Усл. обознач. см.рис.9,
25, 32. Длина брюшной створки 9,0 мм. Куталь 2; джульфинский ярус, среднетах-
табулакская подсвита
Ребра резкие, треугольного сечения, начинаются почти от самой макушки, дихо-
томирующие. Количество ребер в синусе 2-4, на выступе 3-5, .на боковых частях
по 2-4 ребра с каждой стороны синуса и выступа. Столидий не развит.
Внутри брюшной створки - тонкие зубные пластины, сходящиеся посредине и под-
держиваемые невысокой двойной срединной септой, достигающей половины длины вис-
церальной полости створки. При этом образуется низкий двойной спондилий. Зубы
большие, удлиненные. Внутри спинной створки - тонкая разобщенная замочная плас-
тина с небольшим замочным отростком. Срединная септа высокая, тонкая, двойная.
Она поддерживает эллипсоидальный камарофорий без межкамарофориальной пластины
(см. рис. 33).
Размеры, мм
Экз., № Местонахождение, возраст д Ш Т Ш с МУ Количество ребер
си- нус выс- туп сбоку бр. 1 СП.
2228/161- Куталь 2, джульфин- 7,6 8,0 5,0 3,9 90° 2 3 5 4
голотип ский ярус, средне-
2228/154 тахтабулакская 9,0 11,9 7,0 7,6 90° 4 5 8 6
2228/155 подсвита 8,2 11,2 7,2 7,0 115 2 3 7 6
2228/156 8,1 10,6 7,9 5,9 109° 4 5 7 7
2228/157 8,5 10,3 5,5 6,2 97 4 5 6 6
2228/158 8,0 9,0 6,2 4,1 90° 2 3 5 6
2228/165 Кур-Истык 1, воз- 10,0 11,6 7,5 6,6 91° 4 5 10 10
2228/106 раст тот же 9,0 10,0 6,9 5,5 90 2 3 9 10
127
Изменчивость. Изменчивой является форма раковины (она варьирует от ок-
ругло-треугольной до округло-четырехугольной), степень выраженности выстыла
спинной створки, количество ребер в синусе, выступе и на боковых склона^.
Сравнение. Рассматриваемый вид отличается от C.caucasica I icharew меньшими
размерами раковины, менее отчетливо выраженным выступом спинной створки, более
резкой ребристостью и отсутствием столидия. С. Zeven i по внешнему виду и ио внут-
реннему строению близка к экземплярам, хранящимся в коллекции Б.К. Лихарева в
ЦНИГРмузее им. Ф.Н. Чернышева (№ 36/2139 , 35/2139 , 33/2139, 34/2139) и опре-
делявшихся им как Camarophoria acuminata Gemmellaro. К сожалению, внутреннее строе-
ние этого вида из топотип и чес кого местонахождения (известняк Созио Сицилии) не-
известно.
Геологический возраст и географичес кое распространение.
Джульфинский ярус: среднетахтабулакская подсвчта Юго-Восточного Памира; пред-
положительно уруштенский горизонт Северного Кавказа; известняк Созио Сицилии
(мургабский ярус).
Материал. 23 экз. удовлетворительной сохранности, представленные раковина-
ми с двумя сочлененными створками. Собраны в отложениях среднетахтабулакской
подсвиты. В табулятово-гидроидных биогермных известняках Кур-Истыка 1 найдено
10 экз.; Куталя 2-11 экз.; в табулятовых биогермных известняках Шор-Булака -
2 экз.
СЕМЕЙСТВО STENOSCISMATIDAE GEHLERT, 1887 (1883)
ПОДСЕМЕЙСТВО STENOSCISMATINAE GEHLERT, 1887 (1883)
Род Stenoscisma Conrad, 1839
Stenoscisma armenica Sokol ska ja, 1965
Табл. VIII, фиг. 13, 14; табл. XIV, фиг. 8; рис.34
Stenoscisma armenica.: Сокольская, 1965, стр. 234-235, табл. XL, фиг. 9, рис. 39
Голотип - ПИН,№ 2072/71; раковина с сочлененными створками; Закавказье,
левый берег р.Джагры, ниже сел. Огбин’; джульфинский ярус; Сокольская, 1965,
табл. XL, фиг. 9.
О п и с а н и е. Раковина небольшая, треугольного очертания, умеренно или слабо
двояковыпуклая. Замочный край слабо изогнутый, меньше наибольшей ширины рако-
вины, приуроченной к ее средней части. Лобный край унипЛикатный, зазубренный,
боковые - дуговидные. Брюшная створка округло-треугольная, менее выпуклая, чем
спинная, с отчетливым синусом, начинающимся дочти от самой макушки и резко рас -
ширяющимся по направлению к лобному краю. Язычок трапецеидальный. Макушка
широкая, треугольная, прилегающая к спинной створке. Макушечный угол 100-120 .
Плечики макушки нерезко ограниченные. Форамен с крыт .Спинная створка округло-
треугольная, с нерезким, широким выступом, начинающимся на небольшом расстоя-
нии от макушки.
Скульптура состоит из резких, грубых, дихотомируюших ребер треугольного сече-
ния. Количество ребер . в синусе - 4, на выступе - о, на боковых частях - по
5-7 с каждой стороны от синуса и выступа» С толи ди й не наблюдался.
Внутри брюшной створки - толстые зубные пластины, сходящиеся посредине и
поддерживаемые низкой двойной срединной септой. При этом образуется двойной
спондилнй. Внутри спинной створки - цельная замочная пластина, массивный замоч-
ный отросток, низкая двойная срединная септа, поддерживающая кемарафорий с двой-
ной межкамарофориальной пластиной (см. рис. -34 ).
128
I 0.6
II 1.9
Рис. 34. Stenoscisma armenica, ПИН, лг9 2228/169; зарисовки последовательных по-
перечных сечений раковины в примакушечной части, х3.6.Усл. обознач. см. рис. 9,
25 и 32. Длина брюшной створки 13,0 мм. Шор-Булак; джульфинский ярус, средне-
тахтабулакская подсвита
Раз меры, мм
Экз., № Местонахождение, возраст д Ш Т Ш с МУ Количество ребер
си- нус выс- туп сбоку
бр. Jen.
2228/167 Кур-Истык 1, джульфлн- 10,4 15,5 9,7 7,2 120° 4 5 14 12
2228/168 ский ярус, нижнетахта- булакская подсвита 12,8 16,0 8,6 7,5 101° 4 5 11 11
2228/169 Шор-Булак, возраст 12,6 19.2 10,1 7,3 100° 4 5 10 10
тот же
Сравнение. Данный вид отличается от S.purdoni (Davidson) меньшими размерами,
меньшим количеством ребер и низким язычкОхМ, а от S.venusta (Girty) - изометрия-
ным очертанием, синусом и выступом, начинающимися не от самой хмакушки.
Геологический возраст и географическое распространение.
Джульфинский ярус Закавказья; джульфинский ярус, нижне- и среднетахтабулакская
подсвиты Юго-Восточного Памира.
Материал. 7 экз. удовлетворительной сохранности с двумя сочлененными створ-
ками. В темно-серых детритовых известняках нижнетахтабулакской подсвигы Кур-
Истыка 1 собрано 3 экз.; в габулятово-гидроицных биогермных известняках сред-
нетахтабулакской подсвиты Куталя 2-1 экз.; в водорослевых биогермных извест-
няках среднетахтабулакской подо виты Шор- Булака - 3 экз.
9 264
129
ОТРЯД SPIRIFERIDA
ЧАДСЕНЕЙСТВО SPIRIFERACEA
СЕМЕЙСТВО SYRINGOTHYRIDAE FREDERICKS, 1926
ПОДСЕМЕЙСТВО LICHAREWHNAE SLUSSAREWA, 1958
Род Punctocyrtella Plodowski, 1968
. Punctocyrtella': Plodowski, 1968, стр. 252; 1970, стр. 43
Типовой вид - P.spinosa Plodowski, 1968; нижняя пермь; Северный Афганис-
тан.
Описание. Раковина крупная, поперечно-ромбического очертания. Арея брюшной
створки высокая, широкотреугольная. Брюшной синус гладкий, широкий. Спинной выс-
туп узкий, иногда со слабой бороздой посредине.
Скульптура состоит из простых, не ветвящихся радиальных ребер, расположенных
на боковых склонах раковины. Раковина пористая. Внутри брюшной створки - вы-
сокие зубные пластины. Дельтириальная пластина и сиринкс отсутствуют. Замочная
пластина внутри спинной створки широкая, массивная.
Сравнение. Рассматриваемый род очень близок к роду Asyrinxia Campbell, 1957,
для которого характерно отсутствие сиринкса.
Видовой состав. Кроме типового вида к данному роду относится P.nagmar—
gensis (Bion, 1928), происходящая из нижнепермских отложений (аггломератовые
слои) Кашмира и Юго-Восточной Аравии (формации Лузаба)^.
Распространение . Аггломератовые слои Кашмира; сакмарский ярус - нижне-
артинский подъярус пересиленные, базардаринекая свита (верхняя часть) Юго-Вос-
точного Памира, нижняя пермь Аравии и Афганистана.
Punctocyrtella spinosa Plodowski, 1968
Табл. Х,ф?г. 1; табл. XV, фиг. 5; рис. 35
Punctocyrtella spinosd: Plodowski, 1968, стр. 253, табл. 1, фиг. 10; 1970,
стр.*43, табл. 7, фиг. 1-8.
Голотип - Палеонтологический ин-т, Бонн, № 25002; раковина с двумя сочле-
ненными створками; Афганистан; Дашт-Навар, нижняя пермь; Plodowski, 1968, табпЛ,
фиг. 10.*
Описание. Раковина крупная, поперечно-вытянутая, треугольного очертания, с
длинным, прямым смычным краем, соответствующим наибольшей ширине. Лобный край
парасулькатный (его комиссура образует один крупный изгиб в сторону спинной
створки. По бокам от него имеется по одному мелкому изгибу в сторону брюшной
створки). Брюшная створка поперечно-ромбоидальная. Брюшная макушка широкая,
незагнутая. Арея большая, широко-треугольная, покрыта продольной штриховкой.
Дельтирий большой, относительно узкий, закрыт плоским дельтидием. Края ареи
Г. Плодовский( Plodowski, 1970) указывает на распространение P.nagmargensis в верхнем
карбоне, считая в^рхнекаменноугольными аггломератовые слои Кашмира.Нами воз-
. раст аггломератовых слоев Кашмира принимается как нижнепермский (сакмарский-
артинский - см. главу IV).
^Г.Плодовский (Plodowski, 1970) указывает в географическом распространении рода
Punctocyrtella Австралию, однако при описании не приводит каких-либо видов этого
рода встреченных там. Таким образом, наличие названного рода в Австралии пока
нельзя считать доказанным.
130
з.п.
Рис. 35. Punctocyrtella spinosa, ПИН, № 2774/191; зарисовки последователь-
ных поперечных сечений брюшной створки в примакушечной частных 13. Усл. обоз—
нач. см. рис. 9. Длина брюшной створки 19,3 мм. Куртеке 1, сакмарский ярус-
- нижнеартинский подъярус нерасчлененные, базардаринская свита (верхняя
часть)
ограничены резко. Синус хорошо прослеживается от макушки, постепенно расши-
ряясь по направлению к лобному краю. Синус корытообразной формы, с пологими
боковыми склонами и округлым дном. Язычок широкий, округлый, заходит на спин-
ную створку. Спинная створка поперечно-треугольная. Вью туп на ней отчетливый,
округлый, слабо расширяющийся кпереди.
Синус и выступ гладкие. На некоторых экземплярах посредине синуса может при-
сутствовать тонкая борозда. На боковых склонах раковины с каждой стороны от
синуса и выступа - по 9-10 простых округлых ребер, разделенных межреберными
промежутками приблизительно равной с ними ширины. Внутри брюшной створки -
длинные зубные пластины, достигающие дна створки. Сильно развиты макушечные
утолщения. Сиринкс отсутствует (см. рис. 35).
Размеры, мм
Экз., № Местонахождение, возраст д Ш Т Ш а Ш д МУ
2774/150 Куртеке 1, сакмарский ярус - нижнеартинский 21,7 62,1 34,6 49 9 120°
подъярус нерасчлененные базардаринская свита л
2774/151 (верхняя часть) 14,2 — 27,0 — 9,5 122
Изменчивость. Вид является достаточно устойчивым по своим размерам и фор-
ме раковины. Изменчивыми являются: характер макушки, обычно прямостоящей, но
иногда слабо загнутой, ширина ребер и межреберных промежутков, наличие или от-
сутствие борозды в средней части выступа, а также количество ребер на боковых
склонах раковины.
Сравнение. Рассматриваемый вид отличается от Р. nagmargensis(Bion) более
продольно вытянутой формой раковины, оттянутыми ушками, большей резкостью си-
нуса брюшной и выступа спинной створок.
Геологический возраст и географичес кое распространение.
Нижняя пермь Афганистана; сакмарский ярус - нижнеартинский подъярус нерасчле-
непные, базардаринская свита (верхняя часть) Юго-Восточного Памира.
Материал. 78 экз. удовлетворительной сохранности, собранные в органогенно-
обломочных алевритистых известняках верхней части базардаринской свиты Курте-
ке 1. Из них:3 раковины с двумя сочлененными створками, 51 брюшная и 24 спинных
створки.
131
СЕМЕЙСТВО SPIRIFERIDAE KING, 1846
P од N eospirifer Fredericks, 1924
_N eospirifer joharensis (Diener, 1897)
Табл. IX, фиг. 10-13; табл. XVI, фиг. 1; рис. 36
.Spirifer joharensis': Diener, 1897 b, стр. 42-44; табл. IV, фиг. 3
Описание. Раковина от средних до крупных размеров, поперечно-треугольная,
умеренно двояковыпуклая. Лобный край - парасулькатный. Боковые края слабо ду-
говидные. Брюшная створка от поперечно-треугольной до поперечно-ромбоидальной
формы, умеренно выпуклая. Синус начинается от самой макушки и, постепенно расши-
ряясь, прослеживается до лобного края. Язычок полуокруглый, небольшой. Макушка
короткая, широкая, приостренная, умеренно загнутая. Арея длинная, совпадает с
наибольшей шириной раковины, широкая, покрыта тонкой вертикальной штриховкой.
Отчетливые ушки отсутствуют. Дельтирий небольшой, открытый. Спинная створка
поперечно-треугольная; выступ отчетливый, начинается от макушки и, незначитель-
но расширяясь, прослеживается до лобного края. Макушка слабо обособленная.
Рис. 36. Neospirifer joharensis, ПИН,№ 2774/161; зарисовка поперечного сечения
брюшной створки в примакушечной части, х 2/7.
з.п. - зубные пластины, м. - миотест. Длина брюшной створки 25,5 мм. Куртеке
сакмарский ярус - нижнеартинский подъярус нерасчлененные, базардаринская свита
(верхняя часть)
Скульптура состоит из резких радиальных ребер, развитых как в синусе и на
выступе, так и на боковых частях раковины. В синусе и на выступе присутствует
4-5 простых резких ребра. На боковых частях раковины ребра ветвятся, образуя
пучки, переходящие в слабо выраженные складки, которые состоят из 2—3 тонких
радиальных ребер, прослеживающихся от макушки до лобного края. Количество ребер
на боковых склонах раковины - по 15-17 с каждой стороны от синуса. Ребра резкие,
полуокруглые, приблизительно равной ширины с межреберными промежутками. Микро-
скульптура в виде резких черепитчатых пластин нарастания.
Внутри брюшной створки присутствуют короткие, утолщенные зубные пластины,
ограничивающие заднюю часть мускульного поля. Дельтириальная пластина не наб-
людалась. Мускульное поле узкое, вдавленное, покрыто слабой радиальной штрихов-
кой. По бокам и кпереди от него располагаются овариальные отпечатки в виде мелких
частых сосочков.
Сравнение. От N.nitiensis (Diener) рассматриваемый вид отличается меньшей
вытянутостью раковины в ширину, более низкой ареей и несколько меньшими раз-
мерами. От N.nifer (Waagen) он отличается, кроме меньшей поперечной вытянутости,
округлым язычком брюшной створки и низкой ареей, а от N .subfascifer (Licharew)
132
менее изометричным очертанием раковины и ее крупными размерами. Экземпляр
у „joharensis, изображенный Динером (Diener, 1897, табл. IV, фиг. 3), отличается
от памирских форм только несколько меньшими размерами раковины.
Размеры мм
Экз.,№ Местонахождение, возраст д Ш Т ША а Ш д МУ
2774/162 Кастанат-Джилга 1, сакмарский ярус - нижнеартинский подъя- рус нерасчлененные, 23,0 54,8 — 47,7 5,7 108° о
2774/165 базардаринская свита (верхняя часть) 26,7 53,2 17,0 • 24,3 5,2 105
Геологический возраст и географическое распространение.
Продуктусовые сланцы Кумаона; сакмарский ярус - нижнеартинский подъярус нерас-
члененные, базардаринская свита (верхняя часть) Юго-Восточного Памира.
Материал. 118 экз. удовлетворительной сохранности собраны в верхней части
базардаринской свиты. В органогенно-обломочных алевритистых известняках Курте-
ке 1 найдено 39 брюшных и 9 спинных створок; в известковистых глинистых алев-
ролитах Кастанат-Джилки 1-12 экз. с двумя сочлененными створками, 56 брюшных
и 11 спинных створок
Р од Spiriferella Tschemyschew, 1902
Spiriferella sp.
Табл. IX, фиг. 9; рис. 37
Распространение. Верхний карбон - пермь Русской платформы, Урала, Ти-
мана, Новой Земли, Таймыра, Северо-Востока и Дальнего Востока СССР, Гренландии,
Шпицбергена, Австралии, Северной Америки. В Евро-Азиатской части Тетиса пред-
ставители рода распространены преимущественно в верхней части нижней перми и в
0.1
Рис. 37.SpirifereIla sp. ПИН, № 2774/194;
зарисовка поперечного сечения брюшной створ-
ки в примакушечной части, х 4.5. Усл. обознач.
см. рис. 9. Длина брюшной стйорки 16,6 мм.
Чакобай; дарвазский ярус, кызылджилгинский
горизонт, средний подгоризонт
основании верхней, пижняя пермь: сакмарско-артинские отложения Читраля, аггло-
мератовые слои Кашмира; дарвазский ярус, кызылджилгинский горизонт Юго-Вос-
точного Памира. Верхняя пермь Непала, Северного Сиккима, Тибета, Юньнани, Ти-
мора (формация Базлео), Северной Камбоджи (зона Jabeina— Lepidolina), Гималаев
(слои Кулинг) и Усу-Хонгора.
Материал. 5 брюшных створок собраны в туффитовых известняках среднекызыл-
джилгинского подгоризонта Чакобая.
133
СЕМЕЙСТВО RETICULARIIDAE WAAGEN, 18841
ПОДСЕМЕЙСТВО ELYTHINAE FREDERICKS, 1924
P од Permophricodothyris Pavlova, 1965
Neophricodothyris: Лихарев, 1934, стр. 211 (pars).
Permophricodothyris: Павлова, 1965, стр. 134; 1969, стр. 97.
Типовой вид - P.ovata Pavlova, 1965; джульфинский ярус; Закавказье.
Описание Раковина гладкая, крупная, овальной формы, продольно вытянутая или
изометричная, со слабо развитым синусом и выступом. Ушки не обособлены.
Микроскульптура в виде тонких двуствольных игл, расположенных тангенциально
вдоль краев концентрических пластин нарастания.
Зубные пластины и септа внутри брюшной створки отсутствуют. Вершины брахи-
альных конусов направлены в сторону замочного края. Число оборотов спирали - до 28.
Сравнение. От близкого рода Phricodothyris рассматриваемый род отличается
иным строением брахиального аппарата. От остальных родов подсемейства, кроме
того, отсутствием апикальных пластин.
Видовой состав.7 видов из верхнепермских отложений Евро-Азиатской части
Тетиса: Р,оvata Pavlova, 1965, Закавказье; P.indica (Waagen), 1884, Соляной кряж,
Китай; P.waageni (Loczy, 1899), Китай; Р.caroli (Gemmellaro, 1899) , Сицилия; P.inae-
quilateralis (Gemmellaro, 1899), Сицилия; P.grandis(Chao, 1929), Китай; P.iranica
Fantini Sestini et Claus, 1966, Северный Иран.
Распространение. Верхняя пермь Закавказья, Соляного кряжа, Китая, Сици-
лии, Северного Ирана, Юго-Восточного Памира.
Permophricodothyris caroli (Gemmellaro, 1899)
Табл. XI, фиг. 1
•Reticularia caroli: Gemmellaro, 1899, стр. 198. табл.XXXIV, фиг. 11-20; табл.
XXXV, фиг. 1; табл. XLVI, фиг. 12.
Permophticodothyris caroli': Павлова, 1969, стр. 103, табл. X, фиг. 1.
Описание. Раковина крупная, округло-треугольного очертания. Наибольшая ши-
рина значительно превышает длину замочного края и располагается в средней части
раковины. Переход от замочного края к боковым частям - округлый. Лобный край
слабо унипликатный. Брюшная створка равномерно и умеренно выпуклая, с высокой,
узкой слабо загнутой макушкой. Арея высокая, неотчетливо отграниченная. Большая
ее часть занята крупным открытым дельтирием. Отчетливый синус отсутствует, но
имеется слабое углубление, постепенно расширяющееся к лобному краю. Спинная
створка субэллиптической формы, немного менее выпуклая, чем брюшная, со слабо
выступающей над смычным краем макушкой.
Скульптура раковины - в виде отчетливых концентрических пластин нарастания,
максимальная ширина которых приурочена к средней части створок и достигает 2-2,2 мм.
Внутреннее строение типично для рода.
Сравнение. Рассматриваемый вид наиболее близок к P,inaequilateralis, однако
сравнение с данным видом затруднительно, так как Джеммелларо ( Gemmellaro, 1899)
описал под этим названием асимметричные формы, считая асимметрию основным от-
* Представители семейства Reticulariidae описаны Е.Е. Павловой.
134
Размеры, мм
Экз.,№ Местонахождение, возраст д Ш Т Ва Да
2774/173 Кур-Истык 1, джуль- 44 42 23 12 21
финский ярус, нижнетах-
табулакская подсвита
личительным признаком этого вида. Возможно, что P.inaequilateralis и Р.caroli
принадлежат одному виду. От остальных видов рода наш вид отличается относительно бо-
лее высокой вентральной ареей, более высокой и узкой макушкой, а от P.indica и
P.iranica, кроме того, продольной вытянутостью раковины.
Геологический возраст и географическое распространение.
Мургабский ярус, слои Созио Сицилии; джульфинский ярус: уруштенский горизонт
Северного Кавказа; нижнетахтабулакская подсвита КУо-Вос точного Памира.
Материал. 7 экз., представленных различными фрагментами раковин удовлет-
ворительной сохранности. Собраны в темно-серых детритовых известняках нижнетах-
табулакской подсвиты Кур-Истыка 1.
Р ОД Kitakamithyris Minato, 1951
Torynifer (Kitakamithyris) : Minato, 1951, стр. 374.
Типовой вид - Т.(Kitakamithyris) tyoanjiensis Minato, 1951, нижний карбон мас-
сива Китаками, Япония.
Описание. Раковина гладкая, от мелких до крупных размеров, овальной формы.
Длина замочного края значительно меньше наибольшей ширины, которая приходится
на середину длины. Синус и выступ развиты слабо или отсутствуют.
Скульптура наружного слоя в виде двуствольных игл на концентрических пластинах
нарастания.
Внутри брюшной створки - зубные пластины и эусептоид; внутри спинной - кру-
ральные пластины, которые могут опираться на дно створки.
Сравнение. Род Kitakamithyris очень близок к роду Torynifer Hall et Clarke,
1894. Отличия заключаются лишь во внутреннем строении спинной створки, которая у
Kitakamithyris лишена срединной септы. По внутреннему строению последний близок
также к роду Elytha Fredericks, 1924, но отличается от него нескладчатой ракови-
ной и более крупными иглами. От остальных родов подсемейства четко отличается
строением апикального аппарата.
Видовой состав. Известно 14 видов из среднего и верхнего палеозоя Ев-
ропы и Азии: К. sp. - фаменский ярус Марокко; К.tyoanjiensis (Minato, 1961); K.se-
micircularis (Minato, 1951), К.hikoroitiensis (Minato, 1951) - нижний карбон Японии;
K.uniplicata (Campbell, 1955), K.triseptata (Campbell,, 1955) - турнейский ярус
Австралии; К,rouchelensis (Campbell, 1955) - визейский ярус Австралии; K.booralen—
sis (Campbell, 1961), K.immensa (Campbell,1961) - средний карбон Австралии; K.bu—
ravasi Hamada, 1960 - средний (?) карбон Таиланда; K.conquisita (Monachova, 1959) -
турнейский - визейский ярусы Казахстана; К,septata (Chronic, 1953) - средний кар-
бон Перу, Аргентины; K.complanata Pavlova, 1968 - средний карбон- нижняя пермь
Восточной Сибири, Восточного Казахстана; K.magna (Miloradovich, 1936) - нижняя
пермь Новой Земли; K.petaliformis Pavlova, sp. nov. - сакмарский ярус - нижнеартин—
ский подъярус нерасчлененные,базардаринская свита (верхняя часть) Юго-Восточно-
го Памира*
Распространение. Фаменский ярус Марокко; нижний карбон Японии, Централь-
ного Казахстана, Австралии, средний карбон Австралии, Таиланда, Перу, Аргентины;
135
верхний (?) карбон Восточной Сибири; нижняя пермь Новой Земли; средний карбон -
нижняя пермь Восточного Казахстана; сакмарский ярус - нижнеартинский подъяру<
нерасчлененные, базардаринская свита (верхняя часть) Юго-Восточного Памира.
^Kitakamithyria petaliformis Pavlova, sp.nov.
Табл. X, фиг. 2-5, рис. 38,39
Видовое название от petaliformis (лат.) - лепестковидный
Голотип - ПИН № 2774/174; Юго-Восточный Памир, Кастанат-Джилга 1, сак-
марский ярус - нижнеартинский подъярус нерасчлененные, базардаринская свита (верх-
няя часть); табл. X, фиг. 2.
Описание. Раковина крупная, незначительно поперечно вытянутая или изометрич-
ная, веерообразной формы, умереннодвояковыпуклая. Лобный край - W-образно изог-
нут, переход от замочного края к боковым - округлый. Наибольшая ширина раковины
значительно превышает длину замочного края и располагается в ее средней части^
Брюшная створка с маленькой, слабо загнутой макушкой. Макушечный угол 55-65 .
Макушечные склоны заметно обособлены от боковых частей ракови ны. Арея относи-
тельно высокая, но узкая, покрыта отчетливой вертикальной штриховкой. Дельтирий-
открытый, крупный, в виде равностороннего треугольника. Синус отчетливый, широ-
кий, но мелкий. Начинается немного отступя от кончика макушки. Вблизи лобного
края он образует язычок в форме полуэллипса. По бокам от синуса иногда развито
по одной очень пологой радиальной депрессии, значительно более мелкой, чем синус,
чаще вместо них имеются лишь уплощения. Спинная створка так же или незначи-
тельно более выпуклая, чем брюшная и близка к ней по длине благодаря довольно
высокой макушке. Седло округлое, расширяющееся и увеличивающееся по высоте по
направлению к лобному краю. По бокам от него по одной пологой широкой депрессии,
особенно заметной вблизи лобного края. \
Скульптура в виде концентрических пластин нарастания, каждая из которых несет
ряд двуствольных игл. Основания их - сердцевидной формы. Число игл - 10-13 на
5 мм в средней части раковины. Число пластин - 3-4 на 5 мм в средней части ра-
ковины и до 5-7 на 5 мм на боковых частях и вблизи макушки.
Внутри брюшной створки на протяжении 1/6 ее длины от макушки развито ма-
кушечное утолщение, на фоне которого видны зубные пластины, слабо расходящиеся
ко дну створки и клиновидный эусептоид.
Зубные пластины оконтуривают ромбовидное мускульное поле, доходя более, чем
до его половины. Мускульное поле продолжается примерно на 1 /3 длины створки.
Внутри спинной створки - вблизи макушки-круральные пластины в основании за-
мочного отростка сливаются с вторичным утолщением раковины (см. рис. 38, 39).
Васкулярные отпечатки на внутренней поверхности обеих створок в виде тонких, слег-
ка извилистых радиальных борозд.
Размеры, мм
Экз.,№ Местонахождение, возраст Дб Дс Ш Т
2774/174 - голотип Кастанат-Джилга 1, сак- марский ярус - нижнеар- тинский подъярус нерасч- лененные, базардаринская свита (верхняя часть) 29 27 38 22 22
Сравнение. Рассматриваемый вид наиболее близок к kryshtofovichi (Zavodowsky)
из верхнего карбона Колымского бассейна, отличаясь от него более глубоким сину-
136
I
II
Рис. 38. Kitakamithyris petaliformis, ПИН, № 2228/9; зарисовки последователь-
ных поперечных сечений раковины в примакушечной части, х 27 Кастанат-Джилга 1
сакмарский ярус - нижнеартинский подъярус нерасчлененные, базардаринская свита
(верхняя часть)
Рис. 39. Kitakamithyris petaliformis, ПИН, № 2228/6; зарисовки последовательных
поперечных сечений раковины в примакушечной части, х 2.7 Местонахождение и воз-
раст те же, что и на рис. 38
сом, значительно более развитыми депрессиями по бокам от синуса и выступа,
а также формой раковины.
Геологический возраст и географическое распространение.
Сакмарский ярус - нижнеартинский подъярус нерасчлененные, базардаринская свита
(верхняя часть) Юго-Восточного Памира.
Материал. 132 экз., представленных преимущественно фрагментами раковин,
разрозненными брюшными и спинными .створками, обычно деформированными. Соб-
раны в известковистых глинистых алевролитах верхней части базардаринекой свиты
Кастанат-Джилги 1.
137
СЕМЕЙСТВО MARTI NIIDAE WAAGEN, 1884
p од Martinia McCoy, 1844
Martinia': Сокольская, 1941, стр. 62; Dunbar, 1955, стр. 150; Е.А. Иванова,
1960, стр. 278; Pitrat, 1965, стр. 724; Безносова, Сокольская, 1968, стр. 190.
Типовой вид - Spirifer glaber Sowerby, 1820; нижний карбон Англии.
Описание. Раковина различных размеров, изометричного очертания, гладкая,
с коротким замочным и округлыми боковыми краями. Арея небольшая вогнутая. Си-
нус и выступ выражены в различной степени.
Микроскульптура в виде пористости на нарухшом и мелких, продолговатых уг-
лублений или струек на внутреннем слое.
Зубные пластины внутри брюшной створки отсутствуют. Дельтириальные валики
отсутствуют или развиты в различной степени. Септальные пластины внутри спинной
створки отсутствуют. Мускульные и васкулярные отпечатки различной формы. Кру-
ральные пластины присутствуют.
Сравнение. Данный род отличается от Martiniopsis Waagen,1844 отсутствием
зубных и септальных пластин соответственно брюшной и спинной створок^ от Маг—
tiniella Grabau, 1931 и от Fredericksia Paeckelmann, 1931 - отсутствием зубных
пластин, а от Elivella Fredericks, 1919, кроме того - гладкой раковиной без ребер.
От представителей рода В га chy t hyr is McCoy,1844 (сем. Brachythyrididae Fredericks,
1919), иногда конвергентно сходных c Martinia, представители последнего отлича-
ются отсутствием слабой ребристости, характерной для Brachythyris, а по внутрен-
нему строению - обычно отсутствием дельтприапьных валиков в брюшной створке.
Видовой состав. Более 50 видов, распространенных повсеместно в отложениях
карбона и перми в СССР и за рубежом. В перми Юго-Восточного Памира - 6 видов.
За меча ни я. Род Martinia, по—видимому, является сборным. Изучению его уде-
ляли внимание многие авторы. Ревизия этого рода наиболее полно произведена А.Н.Со-
кольской (1941), которая подробно осветила-историю его исследования и взгляды
различных палеонтологов на объем и систематическое положение этого рода. Дис-
куссия развернулась в основном при установлении наличия или отсутствия зубных
пластин у представителей рода Martinia и при выявлении систематического значения
этого признака для данной группы. В настоящее время всеми исследователями приз-
нается отсутствие зубных пластин у представителей рода Martinia. К сожалению, не-
достаточно большое внимание при изучении мартиниид уделялось характеру мускульных
и васкулярных отпечатков.
На материале, имеющемся в нашем распоряжении, можно выделить три группы
"мартиний", различающихся по характеру и форме мускульных^! васкулярных отпе-
чатков, а также по толщине раковины в ее макушечной части .
Группа Martinia warthi Waagen, 1884
Эта группа более всего приближается к М.glabra - типовому виду рода Marti-
nia, который характеризуется тонкими створками, наличием маленьких, продольно
расположенных, слабо выпуклых мускульных отпечатков внутри брюшной створки и
тонких правильных радиально расходящихся васкулярных отпечатков. Такое внутрен-
нее строение соответствует диагнозу рода Martinia, приведенному Холлом и Кларке
(Hall, Clarke, 1894) и рисунку ядра М. glabra из подмосковного карбона, приведенному
Е.А. Ивановой в "Основах палеонтологии", 1960.(рис. 400). Сюда же относятся V.warthi
Waagen, 1884; M.elongata Waagen, 1884; M. rupicola Gemmellaro, 1899; M.applanata Tscherhy-
schew, 1902.
К сожалению, привлечение литературных данных чрезвычайно ограничено тем, что
строение мускульных и васкулярных отпечатков бывает описано очень редко, поэто-
му произвести разделение "мартиний" на указанные группы можно только в резуль-
тате непосредственного изучения материала.
138
Группа Martinia uralica Tschemyschew,1902
Эта группа видов, как указывал Ф.Н. Чернышев (1902) и что хорошо видно на
нашем материале, характеризуется сильно утолщенной макушкой брюшной створки
и своеобразным строением ее мускульных и васкулярных отпечатков. Начиная от
самой макушки брюшной створки тянется глубокая борозда, разделяющая мускульное
поле и выходящая за его пределы. Борозда достигает 1/3 длины створки и наблю-
дается на ядрах в виде отчетливого узкого гребня. Мускульные отпечатки ромби-
ческой формы, очень глубоко вдавленные. На ядрах они видны в виде короткого
ланцетовидного выступа. Васкулярные отпечатки представлены 3-4 стволами с каж-
дой стороны от борозды, отходят от краев мускульного поля, дуговидно изогнутые
и разветвляющиеся.
К этой группе, кроме И.uralica, известной из нижней перми Урала и Тимана,
а также из верхней части базардаринской свиты Юго-Восточного Памира, относятся
М. tschakobaica Grunt, sp.nov. из кызылджилгинского горизонта дарвазского яруса
Юго-Восточного Памира; Mt tschemyschewi Grunt, sp. nov. - вид, к которому от-
несены экземпляры, описанные Ф.Н. Чернышевым (1902) под названием M.frique-
tra Gemmellaro, 1899 и экземпляры, происходящие из кызылджилгинского горизонта
дарвазского яруса Юго-Восточного Памира; М.semiglobosa Tschem., из нижней перми
Урала и Тимана. Возможно, при более детальном изучении материала эта группа
видов может быть выделена в самостоятельный род.
Группа M.triquetra Gemmellaro, 1899
К этой группе отнесен пока только один вид, встреченный в фузулиновых извест-
няках Сицилии и в отложениях мургабского яруса Юго-Восточного Памира.
Он характеризуется округло-треугольными, сильно вдавленными мускульными
отпечатками, разделенными невысоким, плоским выступом брюшной створки; на спин-
ной створке - низкий тонкий септальный валик, разделяющий большие овальные мус-
кульные отпечатки.
В заключение отметим, что род Martinia нуждается в ревизии, при которой не-
обходимо учитывать характер мускульных и васкулярных отпечатков, так как это
может оказаться полезным при разработке систематики 'мартиний*.
Распространение. Карбон-пермь; повсеместно в СССР и за рубежом. В перм-
ских отложениях Юго-Восточного Памира 6 видов.
Martinia rupicola Gemmellaro, 1899
Табл. XII, фиг. 7,8; рис. 40, 41
Martinia rupicola : Gemmellaro, 1899, стр. 168-169, табл. XXXI, фиг. 36-37,
табл. XXXII, фиг. 1-13.
Описание. Раковина средних размеров, изометричного очертания, округло-пя-
тиугольная, умеренно вздутая, с брюшной створкой, незначительно более выпуклой,
чем спинная. Замочный край очень короткий, прямой; боковые края дуговидные;
лобный - широко парасулькатныл. Брюшная створка округло-пягиугольная. Макушка
низкая, широкая, слабо загнутая, с низким дельтирием, по ширине совпадающим с
замочным краем. Ушки маленькие, тупоугольные. Макушечный угол 73-80 . Синус
отсутствует. На некоторых экземплярах развита очень слабая борозда. В передней
части створки - широкая уплощенность. Широкий, плоский, округло-трапецеидальный
язычок довольно сильно заходит на спинную створку.
Спинная створка округло-шестиугольная, с маленькой, приостренной, почти не
выступающей за замочный край макушкой . В передней части створки - очень слабое
возвышение или уплощенность. Лобный край слабо приподнят кпереди. По бокам от
него - слабые депрессии.
139
Рис. 40. Martinia rupicola, ПИН, № 2774/123; 40a - ядро брюшной створки, 406 -
ядро спинной створки, х Я2. Джилга-Кочусу; джульфинский ярус, нижнетахтабулак-
ская подсвита
Поверхность раковины гладкая. Видны только тонкие, неотчетливые линии на-
растания. Микроскульптура не наблюдалась.
Внутри брюшной створки зубные пластины и септа отсутствуют. Присутствуют
дельтириальные валики и широкие, массивные зубы. В макушечной части распола-
гаются маленькие, удлиненные, слабо выпуклые мускульные отпечатки, разделенные
короткой, тонкой,плоской бороздой. Внутри спинной створки септальные валики от-
сутствуют. Васкулярные отпечатки обеих створок тонкие, правильные, многочислен-
ные, радиально расходящиеся (см. рис. 40).
Размеры, мм
Экз.,№ Местонахождение, возраст д Ш Т Ш а МУ
2774/122 Д жилга-Кочус у, 24,7 27,2 17,5 9,8 73°
2774/123 джульфинский ярус, 193 24,3 17,5 12,3 8°о
2774/124 нижнетахтабула кс кая 24,3 23,7 17,5 10,6 72о
2774/125 подсвита 23,1 25 fi 16,6 10,0 “о
2774./126 26,8 25,6 18,5 . 13,2 81
Сравнение. М.rupicola отличается от M.elongete изометричным пятиуголь-
ным очертанием раковины (в противоположность продольно-овальному очертанию
М .elongata), более коротким прямым замочным краем и широким, плоским язычком
брюшной створки. От М.applicate Tschemyschew отли.чается более крупными
размерами, изометричным очертанием, уплощенным язычком брюшной створки и на-
личием слабо выраженных складок в передней части спинной створки.
Отличия M.rupicole от M.warthi Waagen см. при описании последнего.
Геологический возраст и географическое распространение.
Мургабский ярус, фузулиновый известняк Сицилии; джульфинский ярус, нижнетах-
табулакская подсвита Юго-Восточного Памира.
Материал. 5 раковин с сочлененными створками. Собраны в водорослево-гидро-
идных биогермных известняках нижнетахтабулакской подсвиты Джилги-Кочусу.
140
Рис. 41. Martinia rupicola, ПИН,
№ 2774/125; зарисовки последова-
тельных поперечных сечений рако-
вины, х »
з - зубы, д.в. - дельтириальные
валики, кр.п. - круральные пласти-
ны. Длина брюшной створки 23,0 мм.
Джилга-Кочусу, джульфинский ярус,
нижнетахтабулакская подсвита.
I 2.8
11 4.4
•Martinia warthi Waagen, 1884
Табл. XII, фиг. 5,6
Martinia warthi': Waagen, 1884, стр. 533-534, табл. XL III, фиг. 2,8.
ХMartinia chidruensis': Waagen, 1884, стр. 535, табл. XLIII, фиг. 3.
Описание. Раковина маленькая, поперечно-вытянутая, четырехугольного или
пятиугольного очертания, умеренно вздутая, с одинаково выпуклыми створками. За-
мочный край слабо изогнутый, немного меньше наибольшей ширины раковины, рас-
положенной в ее средней части. Боковые края круто дуговидно изогнуты; лобный -
широкопарасулькатный (его комиссура образует один крупный изгиб в сторону спин-
ной створки; по бокам от него имеется по одному мелкому изгибу в сторону брюш-
ной створки). Брюшная створка округло-пятиугольная. Ее макушка небольшая, широкая,
приостреияая, почти не загнутая. Макушечный угол 163-110 • Арея маленькая, не-
отчетливо выраженная; дельтирий низкий, треугольный, по ширине занимает прибли-
зительно 1/3 замочного края. Ушки небольшие, округлые. В передней части брюшной
створки - широкая уплощенность, иногда переходящая в слабо развитый синус. По
лобному краю располагается широкий, трапецеидальный язычок, отогнутый на спин-
ную створку. Спинная створка поперечно-овальная, с низким, плоским выступом в ее
передней части. Выступ ограничен с боков неглубокими, слабо развитыми складками.
Макушка маленькая, широкая, почти не выступает за замочный край.
Поверхность раковины гладкая. Видны лишь слабо выраженные неотчетливые ли-
нии нарастания. Микроскульптура не наблюдалась.
Зубные пластины и септа внутри брюшной створки и септальные пластины внутри
спинной - отсутствуют. На ядрах обеих створок наблюдаются правильные тонкие,
радиально расходящиеся васкулярные отпечатки.
Размеры, мм
Экз.,№ Местонахождение, возраст д Ш Т Ш а МУ
2774/140 Кур-Истык 1, джульфин- 15,4 19,2 10,8 5,5 109°
2774/139 ский ярус, среднетах- булакская подсвита 15,5 20,1 11,6 7,5 103°
2774/135 Куталь 1, возраст тот же 13,3 19,6 8,0 5,5 110о
2774/136 10 £ 18,0 8,0 5,0 ПО
141
Изменчивость выражается в том, что форма раковины варьирует от попереч-
но-четырехугольной до поперечно-пятиугольной. Иногда макушка брюшной створки
значительно выше, чем спинной; на некоторых экземплярах они почти равны по вы-
соте.
Сравнение. Рассматриваемый вид отличается от М.rupicola GemmeJJaro значи-
тельно меньшими размерами, поперечно вытянутой формой раковины и ее большей
уплощенностью. От М .elongate Waagen M.warthi отличается поперечно вытянутой формой
раковины и ее большей уплощенностьдо.
Замечания. Вааген (Waagen, 1884) отмечает, что М .chidruensis и M.warthi очень
близки между собой и, вероятно, являются родственными формами. М. chidruensis
характеризуется лишь немного более коротким, чем у M.warthi замочным краем и
несколько сильнее загнутой макушкой. Поэтому М.chidruensis, представленная в кол-
лекции Ваагена только одним экземпляром, сводится нами в синонимику M.warthi.
Геологический возраст и географическое распространение.
Нижний, средний продуктусовый известняк Соляного кряжа (Пакистан); джульфинский
ярус, среднетахтабулакская подсвита Юго-Восточного Памира.
Материал. 12 экз. удовлетворительной сохранности, собраны в среднетахта-
булакской подсвите. В водорослевых биогермных известняках Шор-Булака найдена 1
раковина с сочлененными створками; в табулятово-гидроидных известняках Кур-Ис-
тыка 1-2 раковины с сочлененными створками; Куталя 1-5 раковин с сочленен-
ными створками; Куталя 2-1 раковина с сочлененными створками, 1 спинная и
1 брюшная створки.
\Martinia uralica Tschemyschew, 1902
Табл. XL, фиг. 2-5; табл. XV, фиг. 3; рис. 42, 43
Martinia uralica: Чернышев, 1902, стр. 183, табл. XVIII, фиг. 1-4.
Лектотип - экземпляр, изображенный Чернышевым, 1902, табл. XVIII,фиг. 1;
хранится в ЦНИГРмузее им. Ф.Н. Чернышева под номером 1679/303;раковина с дву-
мя сочлененными створками, нижняя пермь; Урал, Улу-Тау.
Рис. 42. Martinia uralica, _ПИН, № 2774/118;
ядро брюшной створки, х 2.2. Кастанат-Джил-
га 1 ; сакмарский ярус - нижнеартинский подъ-
ярус нерасчлененные, базардаринская свита
(верхняя часть)
Рис. 43. Martinia uralica, ПИН, № 2774/103;
зарисовка поперечного сечения раковины, х 3.6.
Усл. обознач. см. рис. 41 . Местонахождение и
возраст те же, что и на рис. 42
Описание. Раковина средних размеров, вытянутая в длину, продольно-овальная.
Лобный край ректимаргинатный; боковые слабо дуговидно изогнуты. Замочный край
прямой, меньше наибольшей ширины раковины, расположенной в ее средней части.
Брюшная створка значительно более выпуклая, чем спинная, продольно-треугольной
142
формы, со слабой уплощены ос тью в ее передней трети. Макушка длинная, умеренно
загнутая, оттянутая, немного нависающая над замочным краем. Макушечный угол
70-85 . Арея большая, слабо вогнутая, с нерезко ограниченными краями. Дельтирия
большой, широко треугольный. Ушки небольшие, хорошо выраженные,тупоугольные.
Спинная створка округлая, уплощенная. В ее средней части проходит узкий, килевид-
ный выступ. Макушка маленькая, приостренная, плохо выраженная, не выступающая
за замочный край.
Раковина гладкая. Присутствуют лишь тонкие, неотчетливые концентрические ли-
нии нарастания. Микроскульптура не наблюдалась.
Раковина массивная; особенно утолщенная в макушечной части. Зубные пластины
и септа внутри брюшной, а также септальные пластины внутри спинной створки от-
сутствуют. Присутствуют дельтириальные валики. Внутри брюшной створки, начи-
наясь от макушки и продолжаясь до 2/3 длины створки, тянется глубокая борозда,
разделяющая мускульное поле и выходящая за его пределы. Борозда видна на ядрах
в виде отчетливого узкого гребня. Мускульные отпечатки ромбической формы, очень
глубоко вдавленные. На ядрах они видны в виде сильно выступающего, ланцетовид-
ного поля. Васкулярные отпечатки в виде 3-4 стволов, отходящих от концов мус-
кульного поля, дуговидно изогнутые, разветвляющиеся, располагающиеся под углом
20-40 к срединной борозде. Внутри спинной створки - короткая тонкая срединная
борозда, маленькие, овальные мускульные отпечатки, располагающиеся непосред-
ственно под макушкой и слабые васкулярные отпечатки.
Размеры, мм
Экз.,№ Местонахождение, возраст д ш Ш а В а Ш д МУ
2774/100- Кастанат-Джилга 1, сак- 29,0 27,2 13,0 4,0 6,8 70°
брюшная марский ярус - нижнеар-
створка тинский подъярус нерасч-
2774/118- лененные, базардаринская 25,0 21,4 13,0 85°
брюшная свита (верхняя часть)
створка г\
2774/110- 18,1 20,3 115
спинная
створка о
2774/116- 25,6 25,0 110
спинная
створка
Сравнение. М.uralica отличается от М .tschernyschewi Grunt, sp.nov. продольно-
овальной формой раковины, более вздутой брюшной створкой, оттянутой брюшной ма-
кушкой и отсутствием язычка, развитого по лобному краю у М .tschernyschewi. От
M.tschakobaica Grunt, sp.nov. отличается крупными размерами, сильно вздутой брюш-
ной створкой, продольно-овальным очертанием раковины и отсутствием язычка
по лобному краю, а от М .semiglobosa Tschemyschew- более крупными размерами, от-
сутствием синуса брюшной створки и язычка по лобному краю.
Геологический возраст и географическое распространение.
Ассельский ярус Урала (г.Улу-Тау); сакмарский ярус - нижнеартинский подъярус
нерасчлененные, базардаринская свита (верхняя часть) Юго-Восточного Памира.
Материал. 167 брюшных и 5 спинных створок удовлетворительной сохранности.
Собраны в известковистых глинистых алевролитах верхней части базардаринской сви-
ты Кастанат-Джилги 1 .
143
Martinia tschernyschewi Grunt, sp.nov.
Табл. XI, фиг. 6-£; рис. 44,45
Видовое название в честь Ф.Н. Чернышева, впервые описавшего этот вид под
названием М .triquetra Gemmellaro, 1899.
Martinia triquetra': Чернышев, 1902, стр. 178, табл. XVI, фиг. 1-6.
Голотип - ЦНИГРмузей им. Ф.Н. Чернышева, № 1666/303; раковина с сочленен-
ными створками, слабо деформированная; нижняя пермь; Урал, г.Улу-Тау; Черны-
шев, 1902, табл. ХУ1фиг. 3.
Рис. 44. Martinia tschernyschewi, ПИН, № 2774/133; ядро брюшной створки,х 1.8.
Мамазаир-Булак; дарвазский ярус, кызылджилгинский горизонт, нижний подгоризонт
Рис. 45. Martinia tschernyschewi, ПИН, № 2774/107; зарисовка поперечного сечения
раковины; х 5.4, Местонахождение и возраст те же, что и на рис. 44
Описание. Раковина от крупных до средних размеров, поперечно-треугольного
очертания, умеренно двояковыпуклая; брюшная створка незначительно более выпук-
лая, чем спинная. Замочный край короткий, слабо изогнутый, значительно меньше
наибольшей ширины раковины, расположенной в ее передней половине; боковые края
сильно дуговидно изогнуты; лобный край широкопарасулькатный. Брюшная створка
поперечно-треугольная, с широким, плоским синусом, выраженным в виде округло-
трапецеидального язычка, отогнутого на спинную створку. Макушка широкая, короткая,
слабо загнутая. Макушечный угол 88-110 .Арея низкая, нерезко ограниченная, ушки
округленные. Дельтирий по ширине занимает приблизительно половину длины замоч-
ного края. Спинная створка поперечно-овальная или овально-четырехугольная, с низ-
ким округлым возвышением, наблюдающимся только в передней части створки. Ма-
кушка маленькая, приостренная, почти не выступает за линию замочного края.
Раковина гладкая, с тонкими, неотчетливыми линиями нарастания. Микроскульп-
тура на наблюдалась.
Зубные пластины и септа в брюшной створке, так же как и септальные пластины
в спинной отсутствуют. Строение мускульных и васкулярных отпечатков такое же,
как у других представителей группы.
Сравнение. Отличия М.tschernyschewi от M.uralica см. в описании последнего.
От M.tschakobaica Grunt, sp.nov. отличается более крупными размерами, поперечно-
треугольной формой раковины, более узким дельтирием, а от М.semiglobosa Tscher-
nyschew - крупными размерами, поперечно-треугольным очертанием раковины и со
умеренной выпуклостью.
144
Размеры, мм
Экз.,№ Местонахождение, возраст д Ш Т ш а В а МУ
1666/303, голотип Урал, Улу-Тау, ассель- ский ярус 48,1 45,5 24,1 110°
2774/132 Юго-Восточный Памир, Игримиюз, дарваэский ярус, кызылджилгинский горизонт 28,6 38,3 18,8 12,4 2,1 88°
Геологический возраст и географическое распространение.
Ассельский ярус Урала (г.Улу-Tayj; дарваэский ярус, кызылджилгинский горизонт,
нижний и средний подгоризонты Юго-Восточного Памира.
Материал. 116 экз. удовлетворительной сохранности. Собраны в отложениях кы-
зылджилгинского горизонта. В криноидно-детритовых известняках нижнекызылджил-
гинс кого подгоризонта Игримиюза найдены: 1 раковина с сочлененными створками,
7 брюшных и 1 спинцая; в Кутреке 2-15 брюшных створок и 2 спинных; Мамазаир-
Булака — 1 раковина с сочлененными створками, 69 брюшных и 6 спинных; Кас-
танат-Джилга 2-2 раковины с сочлененными створками и 10 брюшных створок; в
туффитовых известняках среднекызылджилгинского подгоризонта Чакобая - 2 брюшных
створки неполной сохранности.
Martinia tschakobaica Grunt, sp.nov.
Табл. XIIjфиг. 1, 2; табл. XV,фиг. 2; рис. 46,47
Видовое название по местонахождению Чакобай, Юго-Восточный Памир.
Голотип - ПИН, № 2774/127; раковина с двумя сочлененными створками хоро-
шей сохранности; Юго-Восточный Памир, Чакобай, дарваэский ярус, кызылджилгин-
ский горизонт, средний подгоризонт; табл* XII,фиг. 1.
Описание. Раковина небольшая, продольно-четырехугольного или продольно-оваль-
ного очертания, умеренно двояковыпуклая с одинаково вздутыми створками. Замочный
край короткий, изогнутый, значительно меньше наибольшей ширины раковины, распо-
ложенной в ее передней части. Боковые края дуговидные, лобный - ректимаргинатный.
Брюшная створка овально-ромбическая, с широкой уплощенностью в ее Средней части.
Небольшой полуокруглый язычок отогнут на спинную створку. Макушка широкая, ок-
руглая, умеренно загнутая. Арея низкая, маленькая, нерезко ограниченная; ушки сла-
бо закругленные. Дельтирий широкотреугольный, по ширине занимает большую часть
длины замочного края. Спинная створка продольно-эллипсоидальная, со слабым, ок-
руглым возвышением в передней части. Макушка очень короткая, приостренная, поч-
ти не выступает за замочный край.
Раковина гладкая. Редкие, неотчетливые линии нарастания наблюдаются лишь по
ее переднему краю. Микроскульптура не наблюдалась.
Зубные пластины внутри брюшной створки, так же как и септальные
пластины внутри спинной створки - отсутствуют. Внутри брюшной створки присут-
ствуют дельтириальные валики, а внутри спинной круральные пластины (см. рис. 47).
В средней части брюшной створки располагается глубокое мускульное поле ромби-
ческой формы. На ядре оно выглядит в виде довольно высокого ланцетовидного выс-
тупа с косой штриховкой, протягивающегося до 1/3 длины раковины. Мускульное по-
ле разделено глубокой бороздой, которая на ядрах выступает в виде высокого гребня,
протягивающегося почти до лобного края створки. От краев мускульного поля от-
отходят дуговидные, слабоветвящиеся васкулярные отпечатки (по 3-4 ствола с каж-
дой стороны от срединной борозды).
W 2 64
145
Рис. 46. Martinia tschakobaica, ПИН, № 2774/129; брюшная створка с внутренней
стороны, х 3.1 .Чакобай; дарвазский ярус, кызылджилгинский подгоризонт, средний
подгоризонт
Рис. 47. Martinia tschakobaica, ПИН, № 2774/130; зарисовка поперечного сечения
раковины; х 6.3 < Усл. обознач. см. рис. 41. Местонахождение и возраст те же,
что и на рис. 46
Размеры, мм
Экз.,№ Местонахождение, возраст д Ш Т Ш а МУ
2774/1271 2G.0 17,5 12,4 7,9 77°
2774/129- брюшная створка Чакобай, дарвазский ярус, кызылджилгинский гори- зонт, средний подгоризонт; 15,0 14,0 — 8,0 76° о
2774/131 Кастанат-Джилга 2, возраст 22,4 19,8 14,6 8,8 78о
2774/128 тот же 20,0 17,5 15,3 10,2 82
2774/130 18,0 15,5 11,0 7,5 80°
1
голотип
Сравнение. См. описание М.uralica и М .tschernyschewi.
Геологический воэраст и географическое распространение.
Дарвазский ярус, кызылджилгинский горизонт, средний подгоризонт Юго-Восточного
Памира.
Материал. 39 экз. удовлетворительной сохранности, собраны в среднекызыл-
джилгинском подгоризонте. В туффитовых известняках Чакобая найдены 4 раковины
с сочлененными створками, 16 брюшных и 2 спинные створки; Кастанат-Джилги 2-
2 раковины с сочлененными створками и 15 брюшных створок.
146
'Martinia triquetra Gemmellaro, 1899
Табл. XII,фиг. 3,4; табл. XVI,фиг. 2; рис.48
Martinia triquetra: Gemmellaro, 1899, стр. 17, табл. XXXII, фиг. 23-28.
Описание. Раковина крупная, поперечно-треугольного или поперечно-пятиуголь-
ного. очертания, умеренно двояковыпуклая, с брюшной створкой немного более вы-
пуклой, чем спинная. Замочный край прямой, длинный, его длина немного меньше
наибольшей ширины раковины, расположенной в ее средней части. Боковые края ду-
говидные, лобный - парасулькатный. Брюшная створка поперечно-пятиугольного очер-
тания, с синусом, начинающимся почти от самой макушки в виде неглубокой бороз-
ды, резко расширяющейся по направлению к лобному краю. В передней части створки
синус глубокий, треугольного сечения. Макушка широкая, короткая, прямостоящая.
Арея большая, слабо вогнутая, нерезко ограниченная. Ушки округло-тупоугольные.
Дельтирий большой, широкотреугольный, занимает приблизительно половину длины
замочного края. Спинная створка уплощенная, поперечно-четырехугольная. Макушка
почти не выражена.
Рис. 48. Martinia triquetra, ПИН, №
2774/121 - зарисовка поперечного се-
чения брюшной створки, х 2.7,
Игримиюз, мургабский ярус,нижнеганская
(?) подсвита
Раковина гладкая. Линии нарастания тонкие, неотчетливые, наблюдаются лишь в
передней части раковины. Микроскульптура не наблюдалась. Зубные пластины и септа
внутри брюшной створки, так же как септальные пластины внутри спинной отсутствуют.
В макушечной области брюшной створки - небольшие, вогнутые удлиненно-треуголь-
ные мускульные отпечатки, разделенные широким плоским валиком. На ядре спинной
створки видны продольно-овальные мускульные отпечатки, разделенные тонким, низ-
ким валиком. Васкулярные отпечатки правильные, радиально расходящиеся.
Ра з меры, мм
Экз.,№ Местонахождение, возраст д Ш Т Ш 3 МУ
2774/119 Игримиюз, мургабский ярус, нижнеганская (?) подсви- 33,7 42,2 26,5 29,3 99°
2774/121 та 27,7 44,5 20,2 22,4 102°
Сравнение. От всех остальных видов, встреченных в пермских отложениях Юго-
Восточного Памира, данный вид отличается более крупными размерами, поперечно-
треугольным очертанием раковины, а по внутреннему строению - своеобразием мус-
147
кульных и васкулярных отпечатков. Среди многочисленных известных в литературе
видов мартиниид такое строение также нигде не было описано.
Геологический возраст и географическое распространение.
Мургабский ярус, фузулинидовый известняк Сицилии; нижнеганская (?) подсвита Юго-
Восточного Памира.
Материал. 19 экз. удовлетворительной сохранности собраны в водорослевых
известняках нижнеганской (?) подсвиты Игрихмиюза. Из них 7 экз. с двумя сочленен-
ными створками, 8 брюшных створок и 4 спинных.
INCEHTI ORDINIS
СЕМЕЙСТВО ATHYRIDIDAE McCOY, 1844
Род Cleiothyridina Buckman, 1906
Cleiothyris: King, 1850, p. 137
Cleothyridine': Buckman, 1906, стр. 324; Dunbar, Condra, 1932, стр. 359; Sta—
inbrook, 1947, стр. 326; Campbell, 1953, стр. 16; Dunbar, 1955, стр. 123; Easton,
1962, стр. 74; Gobbett, 1964, стр. 161 .
Cleiothyridina': Weller, 1914, стр. 472; Симорин, 1949, стр. 68; Литвинович,
1962', стр. 292; iBoucot, Johnson, Staton, 1965, стр. H 662; Григорьева, 1967а,
стр. 69; Грунт, 1968, стр. 201.
Типовой вид - Spirifer de roissyi Eveill^, 1835; нииний карбон, Бельгия.
On исание. Раковина различных размеров и формы, изометричная или слабо по-
перечно вытянутая, умеренно или слабо двояковыпуклая, иногда с синусом на брюш-
ной створке. Форамен маленький, округлый. Скульптура в виде концентрических
пластин нарастания, рассеченных на плоские иглы с бахромчатыми краями. Внутри
брюшной створки - маленькие, низкие зубные пластины, начинающиеся на значитель-
ном расстоянии от макушки, расходящиеся, иногда сливающиеся с боковыми стенка-
ми раковины. Зубные пластины поддерживают хорошо развитые, массивные, изог-
нутые зубы. Замочная пластина спинной створки - низкая, четырехугольная, неглу-
бокая, вдается в полость брюшной створки; у многих видов она прободена висце-
ральным фораменом и имеет неодинаковый угол наклона по отношению к вертикаль-
ной оси раковины у разных видов.
Давидсон ( Davidson, 1858-1863), описывая внутреннее строение "41hyris“
pectinifera, указывает, что по наружному краю спирали конусов покрыты тонкими
иглами. Данбар (Dunbar, 1955), исследуя внутреннее строение того же вида, не об-
наружил игл на спиралях ручного аппарата. В нашем материале они также не наблю-
дались.
Видовой состав. Род нуждается в ревизии. В настоящее время к нему отно-
сится более 50 видов, для многих из которых неизвестно внутреннее строение.
Сравнение. От всех других родов семейства отличается скульптурой, состоя-
щей из пластин нарастания, рассеченных на длинные плоские иглы.
Распространение. Нижний карбон-пермь; повсеместно в СССР и за рубежом.
В сакмарско-нижнеартинских отложениях Юго-Восточного Памира 1 вид.
Cleiothyridina ailakensis Reed, 1925
Табл. XIII,фиг. 1-5; табл. XV, фиг. 4; рис. 49
Athyris (Cleiothyridina) ailakensis': Reed, 1925i стр. 52, 53; табл. VI, фиг.15-
17; 'табл. VII, фиг. 1.
Описание. Раковина средних размеров, тюперечно-треугольного очертания, с
округленными боковыми краями и полого дуговидно изогнутым лобным краем. Створ-
ки умеренно или слабо вздуты. Синус и выступ отсутствуют. Брюшная створка не-
148
значительно менее выпуклая, чем спинная. Замочный край довольно круто изогнут,
меньше наибольшей ширины раковины, расположенной по ее переднему краю Брюшная
створка округло-треугольная, со слабой уплощенностью в ее передней части, не пе-
реходящей в синус. Уплощенность иногда образует небольшой, полуокруглый язычок,
который наблюдается не на всех экземплярах. Брюшная макушка короткая, приост-
ренная, прямостоящая. Макушечный угол 97-110 . Форамен маленький, округлый.
Скульптура из многочисленных, отчетливо выраженных пластин нарастания, рассе-
ченных на длинные, плоские иглы.
Рис. 49. Cleiothyridina ailakensis, ПИН, № 2774/201; зарисовки последовательных
поперечных сечений раковины в примакушечной части, х 2.7.
бр.с. - брюшная створка, сп.с. - спинная створка, з.п.-зубные пластины, з.-
зубы, зм.п. - замочная пластина, с.в. - септальный валик. Цифрами указаны рас-
стояния пришлифовок от макушки (в мм). Длина брюшной створки 20,7 мм. Кур-
теке 1; сакмарский ярус - нижнеартинский подъярус нерасчлененные, базардаринская
свита ( верхняя часть)
Внутренее строение типичное для рода (см. рис. 49). Зубные пластины внутри
брюшной створки короткие, иногда сливаются с ее боковыми стенками, иногда от-
делены от нее небольшим промежутком.
Размеры, мм
Экз., № Местонахождение, возраст д Ш Т МУ
2774/170 Куртеке 1, сакмарский ярус- 22,3 29,8 12,1 110° п
2774/167 нижнеартинский подъярус не- расчлененные, базардаринская свита (верхняя часть) 24,5 25,6 И.2 108 п
2774/172 Кастанат-Джилга 1, возраст 23,3 25,0 11,7 97
2774/168 тот же 16,8 23,9 - 110°
Сравнение. От широко распространенного вида С royssiana(Keyserling) рассмат-
риваемый вид отличается меньшими размерами раковины, а также ее изометричным
очертанием. От C.semiconcawa (Waagen)—отчетливо треугольной формой раковины,
149
менее выраженным или почти не выраженным язычком брюшной створки, от C.su-
bexpansa (Waagen) треугольной формой раковины, отсутствием синуса брюшной створ-
ки и ее высокой, узкой макушкой, а от С.capillata (Waagen) - меньшими размерами
и отсутствием синуса брюшной створки.
Геологический возраст и географическое распространение.
Нижняя пермь Читраля; сакмарский ярус - нижнеартинский подъярус нерасчлененные,
базардаринская свита (верхняя часть) Юго-Восточного Памира.
Материал. 245 экз. удовлетворительной сохранности, собраны в верхней части
базардаринской свиты. Из них 19 раковин с двумя сочлененными створками, 10 брюш-
ных и 4 спинных створки найдены в органогенно-обломочных алевритистых извест-
няках Куртеке 1; 8 раковин с двумя сочлененными створками и 6 брюшных створок-
в глинистых известняках Чакобая; 84 раковины с двумя сочлененными створками,
77 брюшных и 33 спинных створки - в известковистых глинистых алевролитах Кас-
танат-Джилги 1; 4 брюшных створки - в известковистых глинистых алевролитах
Чакобая.
ОТРЯД TEREBRATULIDA
ПОДОТРЯД TEREBRATULIDINA
НАДСЕМЕЙСТВО DIELASM АТ АСЕ А
СЕМЕЙСТВО LABAIDAE LICHAREW, 1956
Р од Labaia ’Licharew, 1956
Labaia: Лихарев, 1956, стр. 65; 1960е, стр. 293; Stehli, 1965, стр.Н 755.
Типовой вид - L.muirwoodae ‘Licharew, 1956; верхняя пермь, уруштенский
горизонт; Северный Кавказ.
Описание. Раковина небольшая, гладкая, овальная или овально-треугольная,
иногда со слабым возвышением в средней части брюшной створки. Макушка невы-
сокая, умеренно загнутая. Структура раковины тонкопористая.
Внутри брюшной створки - неотчетливый ножной воротничок. Зубные пластины
отсутствуют. Зубные ямки спинной створки, куда входят небольшие, изогнутые зубы
брюшной, ограничены с наружной стороны веществом раковины, а с внутренней -
довольно высокими внутренними приямочными гребнями. От их внутренних частей
отходят субпараллельные круральные основания. Замочная пластина отсутствует. Руч-
ной аппарат в виде простой, короткой центронелловой петли. В месте соединения
нисходящих ветвей присутствует вертикальная пластина. Иногда в спинной створке
- слабо развитый эусептоид.
Сравнение. Внешне отличается от близких родов Оligotbyrina Cooper, 1956 и
Pseudodielasma Bril], 1940 (сем. Labaidae) отсутствием складчатости на обеих
створках.
Видовой состав. Кроме типового вида в состав рода входит L.dmitrievi
Grunt, sp.nov. из кызылджилгинского горизонта дарвазского яруса Юго-Восточного
Памира.
Замечания. Б.К. Лихарев (Licharew, 1956) при характеристике рода Labaia
указывает на отсутствие вертикальной пластины, соединяющей нисходящие ветви
ручного аппарата у представителей рода Labaia, что отличает его от родов O/i-
gotbyrina и Pseudodielasma. Однако отсутствие вертикальной пластины у L.muirwoo-
dae, видимо, можно объяснить неполной сохранностью экземпляров. На экземпля-
рах, имеющихся в нашей коллекции, относящихся к L.dmitrievi Grunt, sp.nov., остат-
ки вертикальной пластины сохранились.
Распространение. Дарваэский ярус, кызылджилгинский горизонт, нижний и
средний подгоризонты Юго—Восточного Памира; джульфинский ярус, уруштенский
горизонт* Северного Кавказа.
150
Labaia dmitrievi Grunt, sp.nov.
Табл. ХП1, фиг. 13; рис. 50, 51
Видовое название в честь геолога В.Ю. Дмитриева, изучающего стратиграфию
пермских отложений Юго-Восточного Памира.
Голотип - ПИН, № 2228/727; раковина с двумя сочлененными створками; дар-
вазский ярус, кызылджилгинский горизонт, нижний подгоризонт; Юго-Восточный Па-
мир, Мамазаир-Булак; табл. ХШ, фиг. 13.
Рис. 50, 51. Labaia dmitrievi, 50 - ПИН, hfo 2228/720; зарисовки последовательных
поперечных сечений раковины в примакушечной части. Усл. обознач. см. рис. 49.
Длина брюшной створки 7,9 мм; 51 - ПИН, № 2228/721; зарисовка продольного се-
чения раковины в ее средней части; оба -х 5.4, Мамазаир-Булак; дарвазский ярус, кы-
зыоджилгинский горизонт, нижний подгоризонт
Описание. Раковина маленькая, овально-треугольная, слабо выпуклая, с брюш-
ной створкой немного более выпуклой, чем спинной; без синуса и выступа. Лобный
край ректимаргинатный, боковые края дуговидные. Наибольшая ширина - по лоб-
ному краю. Брюшная створка овально-треугольная. Макушка широкая, умеренно
загнутая, с небольшим, округлым фораменом. Макушечный угол 51-61 • Спинная
створка уплощенная. Макушка маленькая, слабо приостренная. Внутреннее строение
типичное для рода. Наблюдалась вертикальная пластина, соединяющая нисходящие
ветви ручного аппарата. Эусептоид отсутствует (см. рис. 50, 51).
Размеры, мм
Экз. ,№ Местонахождение, возраст д Ш Т МУ
2228/721 Мамазаир-Булак, дарвазский 9,2 7,2 — 61° о
2228/722 ярус, кызылджилгинский го- 9,5 6,4 4,6 51
2228/723 ризонт, нижний подгоризонт 9,4 7,4 5,7 62°
2228/727- 9,7 7,6 5,2 56°
голотип 2228/730 8,6 7,9 4,6 55°
2228/731 2228/732 12,7 10,5 9,0 8,5 6,8 6,1 5,2 5,6 4,5 4,8 60° 56° 61° 58° 54° 52°
2228/733 2228/734 2228/735 2228/736 10,1 9,1 8,4 8,9 z 6,3 8,9 7,0 7,5
151
Сравнение. L.dmitrievi отличается от L.muirwoodae Licharew отсутствием неболь^
шого возвышения, ограниченного слабыми уступами в задней части брюшной створки,
приплюснутостью спинной створки и расположением наибольшей ширины в передней
части раковины.
Геологический возраст и географическое распространение.
Дарвазский ярус, кызылджилгинский горизонт, нижний и средний подгоризонты Юго-
Восточного Памира.
Материал. 97 экз. с двумя сочлененными створками, собраны в кызылджилгин-
ском горизонте. Из них 78 экз. найдены в микрозернистых глинистых известняках
нижнекызылджилгинского подгоризонта Мамазаир—Булака; 3 экз,— в туффитовых из-
вестняках среднекызылджилгинского подгоризонта Кастанат—Джилги 2; 16 экз. —
в криноидно-детритовых известняках нижнекызылджилгинского подгоризонта Игри—
миюза.
СЕМЕЙСТВО HETERELASMINIDAE LICHAREW, 1956
Р од Heterelasmina Licharew, 1934
Heterelasmina: Лихарев, 19346, стр. 212; 1939, стр. 120; 1956, стр. 68;
1960е, стр. 293.
Типовой вид - H.schucherti Licharew, 1956; верхняя пермь, уруштенский
горизонт; Северный Кавказ.
Описание. Раковина гладкая, от мелких до средних размеров, продольно-оваль-
ная. Брюшная створка слабо выпуклая или плоская, с незначительно развитой ма-
кушкой и очень широким синусом. По переднему краю иногда проходит складка. Спин-
ная створка крышеообразная, с синусом или уплошенностью в передней части створ-
ки. Структура раковины тонкопористая.
Внутри брюшной створки зубные пластины отсутствуют. Имеются только неболь-
шие, изогнутые зубы, входящие в зубные ямки спинной створки. В спинной створке
замочная пластина отсутствует. Зубные ямки ограничены с наружной стороны стен-
кой раковины, а с внутренней - внутренними приямочными ребрами. Непосредственно
от боковых стенок спинной створки, не соединяясь с приямочными ребрами, отходят
круральные основания, направленные вентрально под острым углом к боковым стен-
кам и переходящие в круры. Петля ручного аппарата короткая, широкая, центронелло-
вая. Восходящие ветви резко изогнуты в сторону макушки и составляют острый
угол с нисходящими ветвями.
Сравнение. Данный род отличается от рода Afilasma Stehli, 1961 по внешнему
виду наличием синуса и выступа; по внутреннему строению - отсутствием зубных
пластин, от рода Beecheria Hall et Clarke, 1893 - отсутствием зубных пластин внутри
брюшной и замочной пластины внутри спинной створки. От Gilledia Stehli, 1961 Не—
terelasmina отличается по внешнему виду гладкой раковиной, а по внутреннему строе-
нию - отсутствием зубных пластин внутри брюшной створки. Heterelasmina по внеш-
нему виду очень близка к Heterelasma Girty, 1908 (сем. Cryptonellidae Thomson, 1926),
но по внутреннему строению резко отличается от нее отсутствием зубных пластин в
брюшной створке и замочной пластины, иногда поддерживаемой срединной септой
в спинной - признаков, характерных для Heterelasma.
Видовой состав. Кроме типового вида, в состав рода входят виды, описан-
ные Джеммелларо ( Gemmellaro, 1894а, 1899) из верхнепермских отложений Сицилии:
H.dieneri, Н.gentriflexum, H.nikitini, Н.lepton (последний известен также в отложе-
ниях джульфинского яруса Юго-Восточного Памира), а также H.pygmaea Grunt,sp.nov.
из джульфинского яруса Юго-Восточного Памира.
Замечания. Стели (Stehli, 1956) произвел ревизию рода Jistiina Grabau, 1931,
который Б.К. Лихарев (1956) считал близким к Heterelasmina. Типовым видом Ji-
suina является ].elegantula Grabau, 1931, установленный на единственном имеющем-
ся в коллекции юном экземпляре, внутреннее строение которого неизвестно. Гребо
гключил в состав своего рода следующие виды: Н emiptychina nikitini Gemmellaro,
152
1894; Hg^nuiflexa Gemmellaro, 1899; H. waageni Slacken berg, 1905; H,bokharica ГчНл'г—
nyschew, 1914.
Стели ((Stehli, 1956) , изучив внутреннее строение двух видов из перечисленных
выше - H.nikitim и И .genuiflexa - из верхнепермских отло ве .ий Сицилии, обнару-
жил, что оно сходно с внутренним строением представителей рода Heterelasmina I i—
charew, 1939 и на этом основании свел последний в синоним рода Jisttina, Мы, в
противоположность Стели, относим H.nikitini и Ч .genuiflexa к роду Heterelasmina
и вслед за Б.К. Лихаревым (1960) считаем, что вопрос о праве на существование рода
Heterelasmina не может быть решен до выявления внутреннего строения типового
вида рода Jisuina - J.elegantula Grabau на топотипическом материале.
Следует отметить, что речь идет о группе, в которой широко известны конвергент-
ные формы. Примером может служить род Heterelasma Girty, 1908, относящийся к се-
мейству Cry pt one 11 id ае. Его типовой вид - Н .shumardianum Girty, 1908 - по внешнему
виду очень близок к Heterelasmina pygm-iea Grunt,sp.no/., но существенно отличается
по внутреннему строению.
Распространение. Мургабский ярус, фузулиновый известняк Сицилии; джуль-
финский ярус: уруштенский горизонт Северного Кавказа; среднетахтабулакская под-
свита Юго-Восточного Памира.
Heterelasmina lepton (Gemmellaro, 1894)
Табл. XIII, фиг. И, 12; табл.XVI, фиг. 7; рис. 52, 53
Rhaetina lepton: Gemmellaro, 1894a, стр. 5; 1899, стр. 95, табл. XXV, фиг. 1-9;
табл. XXVII Фиг. 61.
Jisuina lepton': Stehli, 1956, стр. 104, табл. 20, рис. J.
Описание. Раковина средних размеров субпентагонального очертания, с брюш-
ной створкой несколько менее выпуклой, чем спинная. Лобный край эписулькатный.*"
лобная комиссура образует крупный изгиб в сторону спинной створки, осложненный
в средней части мелкими изгибами.По бокам от крупного изгиба имеется по одному
мелкому в сторону брюшной створки. Боковые края слабо дуговидно изогнуты. Брюш-
ная створка удлиненнопятиугольная с очень широким, пологим синусом, посредине
которого располагается складка, на разных экземплярах выраженная в различной сте-
пени. Макушка широкая, короткая, почти незагнутая. Макушечный угол 70-90 . Фо-
рамен маленький, округлый. Спинная створка удлиненно-пятиугольная, крышеобраз-
ная.
Внутреннее строение типичное для рода. Нужно только отметить, что зубы очень
короткие и располагаются близко к стенкам раковины, а круральные основания очень
низкие и начинаются значительно отступя от макушки (на расстоянии 1,4 мм).
Размеры, мм
Экз., № Местонахождение, возраст д Ш Т МУ
2228/364 Кур-Истык 1, джульфинский 21,6 12,6 10,1 70°
2228/368 ярус, среднетахтабулакская 16,2 11,9 8,3 74
2228/367 подсвита 14,2 10,4 6,8 81°
2228/752 Куталь 2, возраст тот же 18,2 14,6 8,3 90°
2228/750 17,4 11,5 8,3 72°
2228/756 Шор-Булак, возраст тот же 17,6 11.4 9,8 73°
2228/758 Куталь 1, возраст тот же 17,5 12.1 8,9 80°
2228/358 Ак-Бура, возраст тот же 17,0 13,2 8,0 80°
2228/359 18,2 13,5 8,3 90°
153
I 1.0
II 1.4
III 1.6
53
Рис. 52. Heterelasmina lepton, ПИН, № 2228/756; зарисовки последовательных по-
перечных сечений раковины в примакушечной части, х 3.6,
з. - зубы, з.я. - зубные ямки, вн.п.р. - внутренние приямочные ребра, кр.о. -
круральные основания. Цифрами указаны расстояния пришлифовок от макушки (в мм).
Длина брюшной створки 15,4 мм. Кур-Истык 1; джульфинский ярус; среднетахтабу-
лакская подсвита
Рис. 53. Heterelasmina lepton, ПИН, № 2228/368; зарисовка продольного сечения
раковины в ее средней части, хб.З.Музду-Булак; возраст тот же.
Изменчивость. Изменчивость выражается в различной степени развития средин-
ной складки в синусе брюшной створки. Иногда она почти не выражена, иногда вы-
ражена отчетливо, но только по переднему краю или достигает 2/3 длины створки
от лобного края. В различной степени бывает выражен выступ на спинной створке.
Сравнение. Рассматриваемый вид отличается от Н.ру&таеа Grunt,sp.nov. зна-
чительно более крупными размерами, отсутствием складки в синусе брюшной створки
и слабого углубления на выступе спинной.От Н.genuiflexa (Gemmellaro) он отличается наличие*
синуса брюшной створки и отсутствием двух складок, ограничивающих уплощенный
выступ спинной.
Геологический возраст и географическое распространение.
Мургабский ярус, фузулиновый известняк Сицилии; джульфинский ярус; среднетах-
табулакская подсрита Юго-Восточного Памира.
Материал. 27 экз , представленных раковинами с двумя сочлененными створ-
ками; некоторые из них частично перекристаллизованы. Собраны в отложениях сред-
нетахтабулакской подсвиты. Из них в табулятово-гидроидном биогермном известняке
Кур-Истыка 1-14 экз.; Куталя 1-1 экз.; Кута ля 2-6 экз.; в водорослевом
биогермном известняке Музду-Булака - 3 экз.; Ак-Буры - 2 экз.; в табулятовом
биогермном известняке Шор-Булака - 1 экз.
Heterelasmina pygmaea Grunt, sp.nov.
Табл. XIII, фиг. 10; рис. 54, 55
Видовое название от pygmaeus (лат.) - маленький, малорослый.
Голотип - ПИН, № 2228/718; раковина с двумя сочлененными створками; Юго-
Восточный Памир, Куталь 2; джульфинский ярус, среднетахтабулакская подсвита;
табл. XIII, фиг. 10.
Описание. Раковина очень маленькая, удлиненно-пятиугольного очертания, с
наибольшей шириной в задней трети раковины. Спинная створка более выпуклая, чем
154
III 1.4
54
I 0.1
55
Рис. 54, 55. Heterelasmina pygmaea, 54 - ПИН, № 2228/717; зарисовки после-
довательных поперечных сечений раковины в примакушечной части; условные
обозначения см. рис. 52. Длина брюшной створки 5,9 мм. Шор-Бу ла к; джульфинский
ярус, среднетахтабулакская подсвита. 55 - ПИН, № 2228/705; зарисовка продольного
сечения раковины в ее средней части; обах 4.5, Куталь 2; возраст тот же
брюшная. Лобный край эписулькатный, боковые - широкодугообразные. Брюшная створ-
ка удлиненно-пятиугольная, макушка широкая, короткая, очень слабо загнутая. Маку-
шечный угол 90-109 . Форамен точечный. Синус брюшной створки широкий, пологий,
составляющий 2/3 ширины раковины. Синус начинается на расстоянии примерно 2/3
длины раковины от макушки. Посредине синуса - короткая, неотчетливая складка.
Спинная створка овально-четырехугольная, крышеобразная, с небольшим выступом,
ограниченным с боков двумя вмятинами. Макушка маленькая, приостренная.
Внутреннее.строение типичное для рода. Круральные основания высокие, тонкие,
начинаются близко от макушки (см. рис. 54).
Размеры, мм
Экз.,№ Местонахождение, возраст д Ш 1 т ! 1 МУ
2228/700 Шор-Булак, джульфинский ярус, 6,2 5/) । 3,7 ' 1 102о
2228/701 среднетахтабулакская поде вита 5,7 4,5 зд 106 о
2228/703 6,7 5,1 4,6 98 о
2228/704 7Л 5,2 4,5 100
2228/336 Куталь 2; возраст тот же 6,0 5,8 3,4 9°о
2228/327 Кур-Истык 1, возраст тот же 6,8 5,8 4,3 104° . о
2228/342 7,4 5,7 4,3 109 о
2228/718- Куталь 2, возраст тот же 6,7 5,5 4,2 103
голотип
Сравнение. Отличия H.pygmaea от H.lepton см. в описании последнего. От H.die-
neri наш вид отличается отсутствием двух складок, разделенных слабо расширяющимся
промежутком в средней части брюшной створки; от Н.nikitini - наличием широкого
синуса брюшной створки, отсутствием двух складок, ограничивающих синус и средин-
ной складки в синусе брюшной створки; от H.schucherti Licharew - более мелкими
размерами и выпрямленной макушкой брюшной створки.
Геологический возраст и географическое распространение.
Джульфинский ярус, среднетахтабулакская подсвита Юго-Восточного Памира.
Материал. 36 экз., представленных раковинами с двумя сочлененными створка-
ми» некоторые из них перекристаллизованы; собраны в среднетахтабулакской подсви-
те. Из них в табулятовом биогермном известняке Шор-Булака - 13 экз.; в табуля-
тово-гйдроидном известняке Куталя 2-5 экз.; Кур-Истыка 1-11 экз., в водорос-
левом биогермном известняке Музду-Булака - 7 экз.
155
СЕМЕЙСТВО CENTRONELLIDAE WAAGEN, 1884
ПОДСЕМЕЙСТВО NOTOTHYRINAE LICHAREW, 1960
P од Notothyris Waagen, 1884
/Vofothyris;Waagen, 1884, стр. 375; Schellwien, 1900b, стр. 101 (pars.);Hall,
Clarke, 1892, стр. 275; Чернышев, 1902, стр. 44; Лихарев, 1960е; стр. 288; Steh-
li, 1965, стр. Н.758.
Типовой вид - Теге brat и la subvesicu laris Davidson, 1862^ пермь; Гималаи.
Описание. Раковина небольшая, удлиненно-овальная; брюшная створка немного
более выпуклая, чем спинная. Синус брюшной створки развит в различной степени
у разных видов. Форамен довольно большой. Выступ спинной створки неотчетливый
или отсутствует.
Скульптура состоит из многочисленных, грубых радиальных складок, развитых
обычно в передней части раковины, но иногда начинающихся и вблизи макушки. У
многих видов довольно отчетливо развиты ступени нарастания. Структура раковины
тонкопористая.
Внутри брюшной створки неотчетливый ножной воротничок. Зубные пластины от-
сутствуют. Зубы короткие, изогнутые, входят в зубные ямки спинной створки. За-
мочная пластина спинной створки плоская, невысокая, прободенная. Круральные ос-
нования начинаются у ее наружных концов. Петля ручного аппарата центронелловая,
короткая; состоит из двух нисходящих ветвей, соединенных посредине. В месте сое-
динения ветвей имеется тонкая вертикальная пластина. В верхней части петли от
обеих ветвей, вентрально отгибаясь, отходят маленькие дополнительные отростки. У
некоторых видов в спинной створке - слабо развитый эусептоид.
Сравнение. Род Notothyris отличается от рода Rostranteris Gemmellaro, 1899 по
внешнему виду более многочисленными резкими складками, а по внутреннему строе-*
нию - хорошо развитой, прободенной замочной пластиной и. отходящими от нее кру-
ральными основаниями. От Gefonia Licharew, 1936 внешне он отличается более разви-
тыми складками обеих створок, а по внутреннему - более простой петлей ручного
аппарата и отсутствием^ эусептоида в брюшной створке.
Видовой состав. Кроме типового вида, в состав рода входят N.multiplies—
ta Waagen, 1884; N.minuta Waagen, 1884; N.lenticularis Waagen, 1884; N.warthi
Waagen, 1884; N. inflate Waagen , 1884 преимущественно из среднего и частично из верхнего
продуктусового известняка Соляного кряжа. К этому же роду относятся N.djoul—
fensis Abich, 1878 из джульфинского яруса верхней перми Закавказья и среднего
продуктусового известняка Соляного кряжа и N .pseud odjoulfensis Licharew, sp.nov. из
джульфинского яруса верхней перми Юго-Восточного Памира и Северного Кавказа.
Замечания. Среди исследователей, изучавших верхнепалеозойских теребратулид,
не было единого мнения о том, являются ли Notothyris и Rostranteris самостоятель-
ными родами, или Rostranteris - младший синоним Notothyris. Так, Шелльвин (Schel-
lwien, 1900b), Холл и Кларк ( Hall, Clarke, 1897), Чернышев (1902), Лихарев (1960е)
высказывали предположение о том, что Rostranteris является синонимом Notothyris.
Как одно из отличий Rostranteris от Notothyris Джеммелларо указывал на незамк-
нутую петлю у Notothyris, но многие авторы объясняют это лишь неполной сохран-
К.Ридом ( Reed, 1944) описано 13 видов теребратулид (из них 11 новых), отнесенных
им к роду Notothyris. Однако ни для одного из этих видов не описано внутреннего
строения. Поэтому судить об их родовой принадлежности трудно. Но можно предпо-
лагать, что по крайней мере часть из них (например, N .praelecta Reed и его варие-
теты, а также N.lochardi Mans.var. oviformis Reed, происходящие из нижнего продук-
тусового известняка) характеризующиеся немногочисленными радиальными складками,
располагающимися в передней части раковины принадлежат к роду Rostranteris, а
не к Notothyris. Нами в разделе *видовой состав* указываются лишь виды, досто-
верно принадлежащие к роду Notothyris.
156
ностью ручного аппарата последнего. В действительности у Rostranteris ветви руч-
ного аппарата соединены, и в месте их соединения присутствует вертикальная плас-
тина. Однако все вышеуказанные исследователи отмечали, что по внешнему виду
представители этих родов отчетливо различаются. Авторами родов указывались и
некоторые различия в их внутреннем строении. Так, Вааген (Waagen, 1884 писал,что
Notothyris характеризуется наличием прободенной замочной пластины, от передних
краев которой отходят круральные основания. Rostranteris же имеет тонкую, руди-
ментарную замочную пластину, и круральные основания, отходящие от боковых стенок
спинной створки. Это хорошо видно на изображениях внутреннего строения спинной
створки, приведенных Джеммелларо ( Gemmellaro, 1899) для R.adrianense (там же,
табл. XXV, фиг. 39), R.pulchurum (табл. XXV, фиг. 62), Е.exile (табл. XXVII, фиг. 60;
табл. XXX, фиг. 42), R.ovale (табл. ХХ^фиг. 51) и особенно отчетливо у R.inflatum
(табл. XXV, фиг. 43).
Сходный рисунок внутреннего строения приводит Чернышев для R.nucleolus (Чер-
нышев, 1902, стр. 46, рис. 13-15). Шелльвин (Schellwien, 1900b) указывает, что ему,
так же как и Джеммелларо, не удалось найти прободенной замочной пластины у пред-
ставителей Rostranteris. Стели (Stehli, 1956 ) произвел ревизию рода Rostrante—
ris. Изучить его типовой вид - R.adrianense - ему не удалось, но был расшлифован
R.inflatum Gemma]laro, 1899, близкий к R.adrianense, из фузулинового известняка Си-
цилии. У этого вида было обнаружено наличие тонкой замочной пластины. Стели ука-
зывает, что круральные основания отходят от замочной пластины, а не от боковых
стенок спинной створки, как это описано у Джеммелларо (но этого не видно на се-
риальных пришлифовках, приведенных автором) 4
Таким образом, Notothyris и Rostranteris имеют достаточно четкие различия,
как по внешнему виду, так и во внутреннем строении и Стели ( Stehli, 1962) вполне
справедливо рассматривает их как самостоятельные роды.
Распространение. Средний и верхний продуктусовый известняк Соляного кря-
жа; джульфинский ярус Закавказья; джульфинский ярус: нижне- и среднетахтабулакская '
подсвита Юго-Восточного Памира; уруштенский горизонт Северного Кавказа.
N otothyris pseudodjoul fensis Licharew, sp.nov.
Табл. XIII, фиг. 6-8; табл. XVI,фиг. 5,6; рис. 56-58
Видовое название указывает на сходство с N .djoulfaensis Abich.
Голотип - ЦНИГРмузей им. Ф.Н. Чернышева, № 1386/2139; раковина с двумя
сочлененными створками; Северный Кавказ, гора Гефо, джульфинский ярус, уруштен-
ский горизонту табл. XIII, фиг. 6.
Диагноз . Раковина небольшой величины;удлиненно-овального или слегка пен-
тагонального очертания с наибольшей шириной, лежащей несколько впереди середины
длины Синус на обеих ствопках слабо развит. На брюшной створке 9 складок,
на спинной - 10
Описание.Раковина небольшая - субовальная или овально-пятиугольная, значи-
тельно выпуклая. Брюшная створка более выпуклая, чем спинная. Наибольшая1 шири-
на в средней части раковины или по лобному краю. Лобный край ректимаргинатный,
зазубренный; боковые края дуговидные, смычный край зигзагообразный. Брюшная
створка удлиненно-овальная, с синусом, начинающимся немного отступя от макушки
и занимающим примерно 1/3 ширины створки по лобному краю. Синус плоский, ог-
раничен резкими угловатыми складками, проходящими вдоль всей его длины. Ма-
кушка широкая, довольно толстая, умеренно загнутая, с большим овальным фора ме-
лом. Макушечный угол 71-81 . Спинная створка овальная, с короткой, слабо при-
остренной макушкой и уплощенностью в передней части, ограниченной с боков рез-
кими складками.
Диагноз вида дается по рукописи проф. Б. К. Лихарева.
157
56
I 2.2
Рис. 56. Notothyris pseudodjoulfensis, ЦНИГРмузей им. Ф.Н. Чернышева № 3189/
/2139; зарисовки последовательных поперечных сечений раковины в примакушечной
части; х 2.7. Северный Кавказ, гора Гефо; джульфинский
ярус, уруштенский горизонт
Рис. 57, 58. N otothyris pseudodjoulfensis,
57 - ПИН, № 2228/683; зарисовка продольного сечения раковины в ее средней
части , х 3.6.58 - ПИН, № 2228/273; зарисовка поперечного сечения раковины в при-
макушечной части, х 3.6,Усл. обознач. см. рис. 52. Длина брюшной створки 12,2 мм.
Кур-Истык 1; джульфинский ярус, среднетахтабулакская подсвита
Скульптура состоит из 2-3 складок, расположенных в синусе брюшной створки,
протягивающихся с самого начала синуса. По бокам от синуса 2-3 складки с каждой
стороны. На спинной створке 3-4 коротких складки, расположенных на уплощенной ’
средней части и по 1-3 менее отчетливых складки с каждой стороны от уплошен-
ности. Все складки резкие, угловатые. Внутреннее строение типичное для рода (см.
рис. 56-58).
Раз меры, мм
Экз.,№ Местонахождение, возраст д Ш Т МУ
1386/2139- Северный Кавказ, гора Гефо, джуль- 14,0 12,7 8,5 —
голотип 2228/253 финский ярус, уруштенский горизонт Юго-Восточный Памир, Кур-Истык 1, 13,9 11,6 10,4 71о
2228/636 джульфинский ярус, среднетахтабу- 10,8 9,2 6,6 80 о
2228/265 лакская подсвита 12,0 10,7 8,5 75
2228/240 12,1 10,0 10,0 72о
2228/241 Музду-Булак, возраст тот же 13,7 10,8 9,2 81
2228/242 12,0 10,3 8,2 78
Изменчивость. Вид довольно сильно изменчив. Размеры взрослых особей
колеблются от 8 до 14 мм в длину и от 6,5 до 11,6 мм в ширину. Очертание - от
овально-пятиугольного до почти изометричного. В различной степени выражен си-
нус брюшной и, особенно, срединная уплощенность спинной створки. Количество ребер
в синусе 2-3, а на уплощенности - 3-4. На боковых частях обеих створок присут-
ствует по 1-3 складки, развитых в различной степени.
Сравнение.От видов Ваагена отличается отчетливо выраженным синусом брюш-
ной створки, ограниченным резкими угловатыми складками и отсутствием резких сту-
пеней нарастания, развитых у видов из Соляного кряжа. От N.djoulfaensis Abich он
отличается более резкими складками и угловатым волнистым лобным краем.
158
Геологический возраст и географическое распространение.
Джульфинский ярус: уруштенский горизонт Северного Кавказа; нижне- и среднетах-
табулакская подсвиты Юго-Восточного Памира.
Материал. 127 экз., представленных раковинами с двумя сочлененными створ-
ками хорошей сохранности. Некоторые экземпляры с частично перекристаллизованной
петлей ручного аппарата. Собраны в нижне- и среднетахтабулакской подсвите. Из них
найдены в нижнетахтабулакской подсвите: в водорослевых биогермных известняках
Кур-Истыка 2-3 экз.; в водорослево-гидроидных биогермных известняках Джилги-
Кочусу - 4 экз.; в среднетахтабулакской подсвите: в табулятово-гцдроидных .био-
гермных известняках Кур-Истыка 1-53 экз.; Куталя 2-30 экз.; в табулятовых
биогермных известняках Шор-Булака - 2о экз.; Музду-Булака - 11 экз.; в водо-
рослевых биогермных известняках Ак-Буры - 1 экз.
Р од Rostranteris Gemmellaro, 1899
Rostranteris: Gemmellaro, 1899, стр. 104; Stehli, 1962, стр. 99; 1965, стр. Н 759.
Типовой вид - 0ielasma adrianense Gemmellaro, 1894; фузулиновый известняк,
Сицилия.
Описание. Раковина небольшая, округло-треугольная, с брюшной створкой немно-
го более выпуклой, чем спинная. Брюшная створка обычно с двумя короткими от-
четливыми складками, спинная - с тремя. Иногда по бокам от них имеются слабые
дополнительные складки. Лобный край зигзагообразный. Внутри брюшной створки -
неотчетливый ножной воротничок; зубные пластины отсутствуют. Зубы небольшие,
изогнутые. Внутри спинной створки - тонкая рудиментарная или слабо развитая за-
мочная пластина, которая у некоторых видов может полностью исчезать. Круральные
основания, видимо, отходят от боковых стенок спинной створки, как это указано у
Джеммелларо (Gemmellaro, 1899). Петля ручного аппарата короткая,*центронелловая.
В месте слияния нисходящих ветвей имеется тонкая, вертикальная пластина.
Сравнение. См. описание рода Notothyris.
С ос та в р од а .Данный род состоит из двух подродов. Первый подрод - Rostran—
teris (Rostranteris), к которому относится 10 видов, описанных Джеммеллэро ( Gem—
mellaro, 1899) из отложений Сицилии ( R.salomonense, R. gut tula, R. gibbosum,R.sinma—
turn, R.ovale, R.inflatum, R.mediterranean, R.exile, R.pulchrum, R.adrianense). R.mediter—
raneum кроме Сицилии найден в местонахождении Базлео Тимора, в Читичун I (Ги-
малаи) и в слоях с Yabeina— Lepidolina Северной Камбоджи; R.exile - в трокгофель-
ских известняках Карнийских Альп и в Читичун I; R.ovale - в трокгофельских из-
вестняках Карнийских Альп и в джульфинском ярусе Закавказья^. Кроме того, к '
нему относится R.nucleolus (Kutorga, 1842) - широко распространенный пермский
вид и R.simplex (Waagen, 1884) из среднего продуктусового известняка Соляного кря-
жа и нижней перми Урала. Второй подрод - это Rostranteris (Notothyrina), включаю-
щий только типовой вид R .(Notothyrina) pontica (Licharew, 1936) из верхней перми
Северного Кавказа и Юго-Восточного Памира.
Замечания. Б.К. Лихарев выделил Notothyrina в качестве подрода Notothyris,
сводя Rostranteris в синонимику рода Notothyris. Мы принимаем Notothyris и Rost-
ranteris в качестве самостоятельных родов (обоснование этой точки зрения приве-
дено выше, при характеристике рода Notothyris). Notothyrina в таком случае явля-
ется подродом Rostranteris и характеризуете я, так же как и номинативный подрод,
наличием небольшого количество складок на обеих створках и круральными основа-
ниями, отходящими от боковых стенок спинной створки.
Распространение. Нижняя пермь Урала и Карнийских Альп. Нижний, средний
и верхний продуктусовый известняк Соляного кряжа. Мургабский ярус Сицилии, Ти-
мора, Гималаев (Читичун I ), Северной Камбоджи; джульфинский ярус Закавказья
и Северного Кавказа, Юго-Восточного Памира. Здесь нами не учитывается распро-
странение R.nucleolus (Kutorga) - вида, распространенного повсеместно в пермских
отложениях и, по-видимому, являющегося сборным.
1
R.ovalis Gemm относился нами ранее (Грунт, 1965) к роду Notothyris.
159
Подрод N otothyrina Licharew, 1936
Notothyris (Notothyrina),: Licharew, 1936, стр. 271; 19606, стр. 189
Типовой вид - N .(N otothyrina) pontica Licharew, 1936; уруштенский гори-
зонт; Северный Кавказ.
Описание. По внешнему виду сходен с Rostranteris (Rostranteris), отличаясь
от него более грубыми внутренними структурами, в частности утолщенными стенками
спинной створки.
Видовой состав. Типовой вид.
Сравнение. Отличия от типового подрода крайне незначительны, но для обос-
нования нецелесообразности выделения подрода Notothyrina необходимо его изучение
на топотипическом материале.
Распространение. Джульфинский ярус: уруштенский горизонт Северного
Кавказа; среднетахтабулакская подсвита Юго-Восточного Памира.
Rostranteris (Notothyrina) pontica (Licharew, 1936)
Табл. XIII, фиг. 9; табл. XVI, фиг. 3,4; рис. 59 , 60.
Notothyris (Notothyrina) pontica: Licharew, 1936, стр. 271, табл. I, фиг. 4,
рис. 3; Лихарев, 1960е, стр. 289, табл. ’LXVIII, фИг. 5, рис. 441-442.
Голотип - ЦНИГР музей им. Ф.Н. Чернышева, № 1632/2139; раковина с соч-
лененными створками; верхняя пермь, джульфинский ярус, уруштенский горизонт;
Северный Кавказ; Licharew, 1936, табл. I, фиг. 4, рис. 3.
Описание. Раковина маленькая, округло-пятиугольная, приблизительно изомет-
ричная, умеренно выпуклая, с брюшной створкой немного более выпуклой, чем спин-
ная. Лобный край выпрямленный, зигзагообразный, боковые - дуговидные.
Брюшная створка овально-треугольная, с двумя довольно отчетливыми складками.
У некоторых экземпляров складки доходят почти до самой макушки, у других -
короткие. Кроме того, присутствует по 1-2 неотчетливых боковых складки. Макушка
широкая, толстая, умеренно загнутая, с довольно большим округлым фораменом. Ма-
кушечный угол 70-78 .
Спинная створка округло-пятиугольная, с очень маленькой приостренной макуш-
кой. В средней части створки - 2 складки, начинающиеся на небольшом расстоянии
от макушки. По бокам 1-2 неотчетливых складки. Внутреннее строение как у под-
рода (см. рис. 59, 60).
Размеры, мм
Экз., № Местонахождение,. возраст д Ш Т МУ
2228/298 Кур-Истык 1, джульфинский ярус, 11.1 9,9 8,8 76° Л
2228/ 691 среднетахтабулакская подсвита 10,0 8,9 6,5 70
2228/30 5 1’1.4 9,6 8,7 72°
2228/ 690 - 10,6 10,8 7,5 78°
2228/310 9,1 9,0 6,5 73°
Геологический возраст и географическое распространение.
Джульфинский ярус: уруштенский горизонт Северного Кавказа; среднетахтабулак-
ская подсвита Юго-Восточного Памира.
Материал. 25 экз. хорошей сохранности, представленные раковинами с двумя
сочлененными створками. Некоторые экземпляры с частично перекристаллизованными
внутренними структурами. Собраны в отложениях среднетахтабулакской подсвиты.
В табулятово-гидроидном биогермном известняке Кур-Истыка 1-19 экз.; Куталя 1 -
2 экз.; в табулятовом биогермном известняке Шор-Булака - 4 экз.
160
Рис. 59. Rostranteris (Notothyrina) pontica, ПИН, №2228/306;
зарисовки последовательных поперечных сечений раковины в при-
макушечной части, х 5,4. Дли на брюшной створки 9,7 мм. Кур-Истык 1;
джульфинский ярус, среднетахтабулакская подсвита
Рис. 60. Rostranteris (Notothyrina)
pontica, ПИН, № 2228/304; зари-
совка продольного сечения ракови-
ны в ее средней части, х 5,4. Место-
нахождение и возраст те же, что
и на рис. 59
161
п 264
ЛИТЕРАТУРА
Аракелян Р.А., Раузер-Чэрноусова Д.М., Рейтлингер Е.А., Щербович С.Ф., Ефи-
мова Н.А. 1964. Значение пермских фораминифер Закавказья для корреляции перми в пре-
делах Тетиса. Докл. сов. геологов на XXII сессии Межд. геол, конгресса. Проблема 16а,
стр. 63-75.
Атлас литолого-палеогеографических карт Русской платформы и ее геосинклинального обрам-
ления. 1960. М.-Л.
Баран ор И.Г. 1935. Массив Буз-Тере. В кн.: 'Таджикско-Памирская экспедиция 1934 года*
АН СССР, М.^Л. стр. 395-408.
Баранов И.Г. 1936. Месторождение исландского шпата Буз-Тере на Восточном Памире. В кн.:
И.Г. Баранов, С.И. К лунников. Материалы по геологии и геоморфологии Памира. - Труды
Тадж.-Памирск. экспедиции, вып. 54 , стр. 7-80.
Бархатова. В.П. 1964. Биостратиграфия карбона и нижней перми Северного Тимана.- Авто-
реферат докт. дисс., Ленинград, стр. 3-47.
Бархатова В.П. 1970. Биостратиграфия карбона и нижней перми Северного Тимана. - Труды
Всес. нефт. н.-и. геол.- разв. ин-та, выл. 283, стр. 3-227.
Бархатова В.П. 1971. К стратиграфии перми по материалам севера Европейской части СССР.
В кн.: Геология и полезные ископаемые северо-востока Европейской части СССР и северного
Урала, т. 1, Сыктывкар, стр. 196-200.
Бенш Ф.Р. 1958. Схема стратиграфии пермских отложений Средней Азии. Тезисы докладов к со-
вещанию по унификации стратиграфических схем Средней Азии. 'Госгеолтехиздат'.стр. 86-91.
Бенш Ф.Р. 1965. Верхнепалеозойские отложения. - В кн.: Стратиграфия Узбекской ССР, кн.1,
под ред. М.А. Ахмеджанова и др., стр. 259-э71.
Бенш Ф.Р. 1972. Стратиграфия и фузулиниды верхнего палеозоя. Южной Ферганы. АН Узб.ССР,
Ташкент, стр. 3-146.
Безносова Г.А., Сокольская А.Н. 1968. Семейство Martiniidae. — В кн.: Брахиоподы верх-
него палеозоя Восточного Казахстана, под ред. Т.Г. Сарычевой. - Труды Палеонтол. ин-та
АН СССР, т. 121, стр. 190-193.
Бобров В.А., Куликов М.В. 1968. Монголо-Забайкальский пролив пермского времени. - Докл.
АН СССР, т. 180» № 5, стр. 1179-1182.
В ил о г радов П.Д. 1959. Пермские отложения Юго-Восточного Таджикистана. Верхнепалеозой-
ские нерасчлененные отложения Юго-Восточного Таджикистана. Геология СССР, т. XXIV,
Таджикская ССР, ч. 1, Геологическое описание. М., стр. 149-163, 167-170.
Власов Н.Г., Лихарев Б.К., Миклухо-Маклай А.Д. 1962. К фаунистической характерис-
тике разреза нижнепермских отложений Юго-Западного Дарваза. - Докл. АН СССР, т. 144,
№ 5, стр. 1105-1108.
Волгин В.И. 1960. Брахиоподы верхнекаменноугольных и нижнепермских отложений Южной
Ферганы. - Изд-во Ленингр. ун-та, стр. 1-203.
Волгин В.И. 1964. Новые виды позднепалеозойских брахиопод Юго-Западного Дарваза. -
Вопр. палеонтологии, T.IV. Изд-во Ленингр. ун-та, стр. 100-105.
Гансер А. 1967. Геология Гималаев. М., изд-во 'Мир*, стр. 5-349.
Гептнер В.Г. 1936. Обшая зоогеография. М., изд-во 'Биомедгиз', стр. 3-548.
Герасимов Н.П. 1929. Брахиоподы Стерлитамакского известняка. - Уч. зап. Казанск. ун-та,
т. 89, кн. 5-6, стр. 779-872.
Герасимов Н.П., Девингталь В.В., Журавлева Ф.А., Каширцев А.С., Мирская М.Ф.,
Чудинова И.И., Шестакова М.Ф. 1953 (1952). Продуктиды кунгурских и артинских от-
ложений Пермского Приуралья. - Труды естеств. научн. ин-та при Пермск. гос. ун-те,т. X,
вып. 3-4, стр. 3-160.
Горюнова Р.В. 1970. Новые раннепермские мшанки Юго-Восточного Памира и Дарваза. В кн.:
Новые виды палеозойских мшанок и кораллов. М., 'Наука', стр. 60-68*
Горюнова Р.В. 1971. Пермские мшанки Юго-Западного Дарваза и Южного Памира. - Авто-
реф. канд. дисс. М., стр. 3-24.
Григорьева А.Д. 1962. Продуктиды казанского яруса Русской платформы и условия их
существования. - Труды Палеонтол. ин-та АН СССР, т. 92, стр. 3-05.
162
Григорьева А.Д. 1967а. О некоторых брахиоподах казанского яруса. - Палеонтол. журн.
3, стр. 62-75.
Григорьева А.Д. 19675. Направления и методы современной палеогеографии. - Итог» науки.
Стратигр., палеонтол. М. Ин-т н.-^гехн. инф. АН СССР, стр. 36-53.
Грунт Т.А. 1965. Надсемейство Athyridacea, отряд Terebratulida. В кн.: Развитие и смена
морских организмов на рубеже палеозоя и мезозоя, под ред. В.Е. Руженцева и Т.Г. Са-
рычевой. - Труды Палеонтол. ин-^га АН СССР, т. 108, стр. 237-254.
Грунт Т.А. 1968. Семейство Athyrididae. В кн.: Брахиоподы верхнего палеозоя Восточного
Казахстана, под ред. Т.Г. Сарычевой. Труды Палеонтол. ин-та АН СССР, т. 121, стр.200-202.
Грунт Т.А. 1970. К биогеографии брахиопод пермского Тетиса. - Бюлл. Моск об-ва исп.
природы, отд. геол., т. XLV (6), стр. 90-101.
Гурьянова Е.Ф 1945. Принципы биогеографического районирования Мирового океана. - Научн.
бюлл. Ленингр. гос. ун-та, л? 4, стр. 15-16.
Гурьянова Е.Ф. 1957. Закономерности распределения современной морской фауны и принципы
районирования Мирового океана. - В со.: Вопросы палеобиогеографии и биостратиграфии.
Труды I сессии Всес. Палеонтол. об-ва, стр. 15-24.
Гусева Е.А., Горский В.П. 1969. К вопросу о возрасте морских угленосных отложений Пе-
чорского угольного бассейна. - Докл. АН СССР, т. 189, № 3, стр. 595-59/.
Г у шин Б.М. 1966. О каменноугольных отложениях Юго-Восточного Памира (уточнение воз-
раста базардаринской свиты). -Докл. АН СССР, т. 168, № 2, стр. 407-408.
Г у шин Б.М. 1969а. Каменноугольно-пермские отложения Юго-Восточного Памира. - Изв.
АН Тадж.ССР* отд. физ-мат. и геол.-хим. наук, № 2 (32). Душанбе, стр. 72-90.
Г у шин Б.М. 19696. Литолого-стратиграфическое расчленение каменноугольно-пермских отло-
жений Юго-Восточного Памира. - Докл. АН Тадж.ССР, т. XII, № 3, стр. 50-54.
Гушин Б.М. 1969в. Ярусное расчленение каменноугольно-пермских отложений Юго-Восточного
Памира. - Докл. АН Тадж.ССР, т. XII, № 6, стр. 44-47.
Дагис А.С. 1963. Верхнетриасовые брахиоподы Юга СССР. Изд-во АН СССР, стр. 3-218.
Дронов В.И., Карапетов С.С., Левен Э.Я. 1959. О возрасте углей на Восточном Пами-
ре. - Докл. АН СССР, т. 127, № 3, стр. 634-636.
Дронов В.И., Левен Э.Я. 1961. К вопросу о геологии Юго-Восточного Памира. - Сов. геоло-
гия, И, стр. 21-36.
Дмитриев В.Ю. 1973а. Новые данные о строении нижнепермских отложений Юго-Восточного
Памира (в печати ).
Дмитриев В. Ю. 19736.К стратиграфии нижнепермских отложений Юго-Восточного Памира (в печати).
Дуткевич Г.А. 1934. Геологические исследования в бассейне Мургаба и Аличура. - В кн.:
Таджикско-Памирская экспедиция 1933г., стр. 333-368.
Дуткевич Г.А. 1935а. Геологические исследования в Шоркуль-Мынхаджирском районе ва Вос-
точном Памире в 1933 г.В кн.: К геологии Восточного Памира. - Труды Тадж.-Памирск.
экспедиции, вып. 36, стр. 3-Z9.
Дуткевич Г.А. 19356. Условия находки пермских аммоней на Восточном Памире и значение
этой находки для корреляции пермских разрезов Европы и Азии. - В кн.: Г.А. Дуткевич,
О.Г. Тумане кая. Фауна верхнего палеозоя Восточного Памира. - Труды Тадх^-Памирск.
экспедиции, вып. 31, стр. 3-41.
Дуткевич Г.А. 1936. Основные чарты строения верхнепалеозойских отложений Восточного
Памира. - В кн.: Научн. итоги Тадж.-Памирск. эксп., Изд-во АН СССР, стр. 87-163.
Дуткевич Г.А. 1937а. Пермские отложения Средней Азии. - Пробл. сов. геол., т. VII, № 7,
стр. 603-606.
Дуткевич Г.А. 19376. Значение микрофауны для корреляции перми южной части СССР. - Те-
зисы докл. XVII сессии Межд. геол, контр., стр. 95-96.
Дуткевич Г.А. 1967. Новый вид ябеин из верхнепермских отложений Юго-Восточного Пами-
ра. - Палеонтол. журн. № 1, стр. 18-21.
Дуткевич Г.А., Калмыкова М.А. 1935. Хребет Базар—Дара (краткий геологический очерк).—
В кн.: Таджике ко—Памирская экспедиция, 1934 г., стр. 371-994.
Дуткевич Г.А., Калмыков^ М.А. 1936. Восточная часть хребта Базар—Дара (геология и
геоморфология). - Труды Тадж.-Памирск. экспедиции, вып. 48, стр. 3-71.
Дуткевич Г.А., Туманская О.Г. 1935. Фауна верхнего палеозоя Восточного Памира. - Тру-
ды Тадж.-Памирск. экспедиции, вып. 31, стр. 3-104.
Дуткевич Г.А., Хабаков А.В. 1934. Пермские отложения Восточного Памира и палеогео-
графия верхнего палеозоя Центральной Азии. - Труды Тадж. компл. экспедиции 1932 г.,
вып. 8, стр. 3-112.
Дюфур М.С., Дронов В.И., Кушлин Б.К. 1958. К стратиграфии триаса Юго-Восточного Па-
мира. - Докл. АН СССР, 123. № 3, стр. 523-526.
Левадовский В.М. 1960. Новые виды пермских продуктид Северо-Востока СССР. - ВСЕГЕИ.
Новые виды древних растений и беспозвоночных СССР, ч. 1, стр. 334-340.
163
Иванова Е.А. 1949. Условия существования, образ жизни и история развития некоторых
брахиопод среднего и верхнего карбона Подмосковной котловины. - Труды Палеонтол. ин-та
АН СССР, т. 21, стр. 1-152.
Иванова Е.А. I960. Отряд Spiriferida. - В кн.: Основы палеонтологии. Мшанки, брахиоподы.
Изд-во АН СССР, стр. 264-279.
Иванова Е.А. 1962. Экология и развитие брахиопод силура и девона Кузнецкого, Мину-
синского и Тувинского бассейнов. - Труды Палеонтол. ин-та АН СССР, т. 88, стр. 3-149.
Ильина Т.Г. 1970. Некоторые новые пермские ругозы Юго-Восточного Памира. В кн.: Но-
вые виды палеозойских мшанок и кораллов. М., изд-во * Наука*, стр. 146-151.
Ионин Н.В. 1933. Хребет Базар-Дара (Центральный Памир). В кн.: Таджикская комплексная
экспедиция 1932 года. АН СССР, Л., стр. 139-148.
Ионин Н.В. 1934. Геология и полезные ископаемые Базардаринского хребта. - Труды Тадж.
комлл. экспедиции 1932 г., вып. V (полезные ископаемые), Л., стр. 1-39.
Каширцев А.С. 1959. Полевой атлас фауны пермских отложений Северо-Востока СССР.
Изд-во АН СССР, стр. 3-85.
К лунников С.И., Недзвецкий А.П., Виноградов П.Д. 1936. Геологическое строение
Юго-Восточного Памира. - Труды Тадж.компл. экспедиции 1934 г., вып. 64, стр. 3-57.
Котляр Г.В. 1961. Род Yakovlevia Fredericks. Докл. АН СССР, т. 140, № 2, стр. 459-460.
Котляр Г.В. 1964. Новые представители рода Productus из верхней перми Южного Приморья.-
Труды Всес. н.-и. геол, ин-^га, Палеонтол. и стратигр. Нов. серия, т. 93, стр. 123-133.
Кр ишнан М.С. 1954. Геология Индии и Бирмы. М., ИЛ, стр. 3-424.
Куликов М.ь. 1938. О находке казанского типа спириферов в нижнепермских отложениях
Урала. - Бюлл. Моск, об-ва исп. природы, отд; геол., 16, вып. 3, стр. 232-238.
Левен Э.Я. 1958. Пермские отложения Юго- Восточного Памира. - Тезисы докл. к Совеш.
по унификации стратигр. схем Средней Азии, изд-во *Госгеолтехиздат*, стр. 95-100.
Левен Э.Я. 1959. К вопросу о возрасте мургабской свиты на Восточном Памире. - Труды
АН Тадж. ССР, т. 104, Зап.Тадж. отд. Всес. минерал, об-ва, вып. 1, стр. 123-129.
Левен Э.Я. 1963. О филогении высших фузулинид и расчленении верхнепермских отложений
Тетиса. - Вопр. микропалеонтол., вып. 7, стр. 57-70.
Левен Э.Я. 1964. К систематике семейства Verbeekinidae (Fusulinida ). - Палеонтол. жур-
нал., X? 4, стр. 23-31.
Левен Э.Я. 1965. Стратиграфия и фузулиниды пермских отложений Памира в связи с вопро-
сами ярусного расчленения перми Тетиса. Автореф. канд. дисс., М., стр. 1-42.
Левен Э.Я. 1967. Стратиграф1я и фузулиниды пермских отложений Памира. - Труды Геологич.
ин-^га АН СССР, выл. 167, стр. 3-265.
Левен Э.Я. 1972. О пермских отложениях Закавказья. - Докл. АН СССР, т. 204, № 3, стр,
681-684.
Либрович Л.С. 1968. Распространение аммоноидей в морях карбона и проблема древнего
Средиземного бассейна. - Труды Всес. н.-и. геол, ин-та, нов. серия, т. 143, стр. 154-171.
ЛитвиновичН.В. 1962. Каменноугольные и пермские отложения западной части Централь-
ного Казахстана. - Материалы по геол. Центр. Казахстана , т. IV, Изд-во Моск. гос.
ун-та, стр. 3-389.
Лихарев Б.К. 1925. Новый представитель брахиопод из верхнепалеозойских отложений Се-
верного Кавказа. - Изв. Геол, ком., 43 (1924), X? 6, стр. 7-13.
Лихарев Б.К. 1926 (1922-1924). К морфологии и . биологии литтоний. - Ежегодн. Русск.
палеонтол. об-ва, т. 4, стр. 125-136.
Лихарев Б.К. 1928. О некоторых редких и новых представителях брахиопод из нижнеперм-
ских отложений Северного Кавказа. - Изв. Геол, ком., т. 47, № 3, стр. 261-296.
Лихарев Б.К. 1931а (1930). Некоторые дополнения к статье *К систематике подсемейства
Orthotetinae Waagen. -Ежегодн. Русск. Палеонтол. об-ва, т. IX, стр. 153-156.
Лихарев Б.К. 19316 (1930). Об одной проблематической брахиоподе из нижнепермских отло-
жений Северного Кавказа. - Ежегодн. Русск. палеонтол. об-ва, т. IX, стр. 157-161.
Лихарев Б.К. 1932а. Фауна пермских отложений Северного Кавказа. 1. Brachiopoda. Под-
семейство Orthotetinae Waagen. - Труды Всес. геол.-развед. объед., вып. 215, стр. 3-54.
Лихарев Б.К. 19326. Фауна пермских отложений Северного Кавказа. 2. Brachiopoda. Семей-
ство Lyttoniidae Waagen. Труды Всес. геол«-развед. объед.,вып. 215, стр. 55-54.
Лихарев Б.К. 1934. О некоторых новых родах верхнепермских брахиопод. - Докл. АН СССР,
нов. серия, 1, №4; стр. 210-213.
Лихарев Б.К. 1936. О некоторых представителях редких верхнепалеозойских брахиопод.
Бюлл. Моск. общ. исп. природы, отд. геол., 14, выл. 2, стр. 133-158.
Лихарев Б.К. 1937. Пермские Brachiopoda Сев.Кавказа. Семейства: Chonetidae Hall et Clarke
и Productidae Gray. - Моногр.по палеонтол. СССР, т. 39, вып. 1, стр. 1-151.
Лихарев Б.К. 1939. Класс Brachiopoda. Брахиоподы. Атлас руководящих форм ископаемых
фаун СССР, т. VI. Пермская система.' М.-Л., стр. 76-121.
Лиха ре в.Б.К. 1946. Материалы по фауне брахиопод среднего и верхнего карбона и нижней
перми Ферганы. - Материалы Всес. н.-и. геол, ин-та; Общ. серия, 7, стр. 91-109.
Лихарев Б.К. 1956. Семейство Strophomenidae, надсемейства Rhychonellacea, Spiriferacea,
Terebratulacea. - Материалы по палеонтол., нов. серия, вып. 12, стр. 52-53, 56-61, 64-70.
164
Дихарев Б.К. 1960а. О верхне пермских бра хи оп одах Южного Приморья и некоторых одновоз-
растных фаунах других районов Восточной Азии. - Докл. АН СССР, т. 132, № 2, стр. 428-
431.
Лихарев Ь.К. 19606. Общая часть. В кн.: Основы палеонтологии. Мшанки, брахиоподы. Изд-во
АН СССР, стр. 115-162.
Лихарев Б.К. 1960в. Семейство Chonetellidae. В кн.: Основы палеонтологии . Мшанки, бра-
хиоподы. Сарычева Т.Г., Лихарев Б.К., Сокольская А.Н. Отряд Productida. Изд-во АН СССР,
стр. 226.
Лихарев Б.К. 1960 г. Семейство Scacchinellidae. В кн.: Основы палеонтологии. Мшанки,
брахиоподы. Сарычева Т.Г., Лихарев Б.К., Сокольская А.Н. Отряд Productida. Изд-во
АН СССР, стр. 235.
Лихарев Б.К. 1960д. Верхнепалеозойские ринхонеллиды. В кн.: Основы палеонтологии. Мшан-
ки, брахиоподы. Ржонсницкая М.А., Лихарев Б.К., Макридин В.П. Отряд Rhynchonellida.
Изд-во АН СССР, стр. 239-257.
.Лихарев Б.К. 1960е. Верхнепалеозойские теребратулиды. В кн.: Основы палеонтологии. Мшан-
ки, брахиоподы. Лихарев БК., Макридин В.П., Ржонсницкая М.А. Отряд Terebratulida. Изд-во
АН СССР, стр. 286-305.
Лихарев Б.К. 1964. Рихтгофении из перми Южного Приморья. - Труды Всес. н.-и. геол, ин-та,
Палеонтол. и стратиграфия; нов. серия, т. 93, стр. 113-122.
Лихарев Б.К., Алихова.Т.Н. I960. Отряд 'Orth i da. В кн.: Основы палеонтологии. Мшанки,
брахиоподы. Изд-во АН СССР, стр. 193-195.
Лихарев Б.К., Котляр Г.В. 1960. Позднепермские брахиоподы Южного Приморья и их стра-
тиграфическое значение. - Материалы годичной сессии ученого совета Всес. н.-и. геол,
ин-та за 1959 г. стр. 19-20.
Лихарев Б.К., Миклухо-Маклай А.Д. 1964. Стратиграфия пермской системы. Докл. сов.
геологов на XXII сессии Межд. геол, конгресса. Проблема 16а, стр. 12-24.
Лю Хун-юн. 1962. Палеогеографический атлас Китая. М., ИЛ, стр. 9-117.
Макридин В.П. 1964. Брахиоподы юрских отложений Русской платформы и некоторых при-
лежащих к ней областей. - Изд-во 'Недра*, стр. 3-338.
Миклухо-Маклай А.Д. 1949. Верхнепалеозойские фузулиниды Средней Азии: Дарваз, Фер-
гана, Памир. Изд-во Ленингр. гос. ун-та, стр. 3-126.
Миклухо-Маклай А.Д. 1955. Некоторые вопросы зоогеографического районирования морс-
кой перми СССР и корреляции верхнепалеозойских отложений Средней Азии, Кавказа, За-
кавказья и Уссурийского края. - Уч. зап. Ленингр. гос. ун-та, № 189, серия геол., вып. 6,
стр. 3-20.
Миклухо-Маклай А.Д. 1957. К вопросу о зоогеографическом районировании морских бас-
сейнов каменноугольного и пермского периодов в СССР. - Вести. Ленинград, гос. ун-та,
№ 24, серия геол.-теогр.', вып. 4, стр. 176-179.
Миклухо-Маклай А.Д. 1958а. О ярусном делении морских пермских отложений южных райо-
нов СССР. - Докл. АН СССР, т. 120, № 1, стр. 175-178.
Миклухо-Маклай А.Д. 19586. Стратиграфия морских пермских отложений Средней Азии.
Тезисы докл. к совещанию по унификации стратигр. схем Средней Азии, стр. 93-94.
Миклухо-Маклай А.Д. I960.Корреляция верхнепалеозойских отложений Средней Азии, Кав-
каза и Дальнего Востока по данным изучения фораминифер. В кн.: Дочетвертичная микро-
палеонтология. Докл. сов. геологов на XXI сессии Межд. геол, конгр., стр. 69-76.
Миклухо-Маклай А.Д. 1961. Стратиграфия пермских отложений Средней Азии. - Вести.
Ленингр. гос. ун-та, № 12, серия геол.-геогр., вып. 2, стр. 18-41.
Миклухо-Маклай А.Д. 1963. Верхний палеозой Средней Азии. - Изд-во Ленингр. гос. ун-
та, стр. 1 -329.
Миклухо-Маклай А.Д. 1966. Среднеазиатские гер цине кая, киммерийская и альпийская склад-
чатые области. В кн.: Стратиграфия СССР, т. V. Пермская система. М., изд-во 'Недра*,
стр. 326-340.
Мин ат о Macao, Гораи Macao, Фунахаси Мицуо. 1968. Геологическое развитие Япон-
ских островов.'М.» Изд-во *Мир*, стр. 5-719.
Милорадович Б. В. 1936. Нижнепермская фауна острова Me жду шаре кого. - Труды Арктич.
ин-та, т. XXXVII, стр. 37-82.
Милорадович Ь.В. 1945. Некоторые данные по морфологии раковин продуктид. - Изв. АН
СССР, отд. биол. наук, № 4, стр. 485-500.
Монахова Л.П. 1959. Визейские брахиоподы нижней части угленосной толщи Центрального
Казахстана. - Труды Лаборат. геол, угля АН СССР, вып. IX,стр. 68-152.
Морозова ИЛ. 1970. Мшанки поздней перми. - Труды Палеонтол. ин-та АН СССР, т. 122,
стр. 3-332.
Наливкин Д.В. 1916. Предварительный отчет о поездке летом 1915 г. в Горную Бухару и на
Западный Памир. - Изв. Русск. геогр. об-ва, T.LII, стр. 203-237.
Н аливкин Д.В. 1929. Палеогеография Средней Азии в палеозое. - Труды 3-го Всес. съезда
геологов, вып. 2, Ташкент, стр. 345-355.
165
Наливкин Д.Е. 1936. Палеогеография Средней Азии. В кн.: Научные итоги Таджикско-Па-
мирской экспедиции АН СССР, стр. 35-61.
Наливкин Д.В. 1937. Палеогеография перми СССР. - Тезисы докл. XVII сессии межд. геол,
конгр, стр. 81.
Наливкин Д.В., Чуенко П.П., Попов В.И., Юдин Г.Л. 1932. Геологическое строение Па-
мира. - Труды Всес. геол.-разв. объед., вып. 182, стр. 1-45.
Нехорошее и.И. 1957. Значение мшанок для палеогеографии палеозоя СССР. - Труды 1-й
сессии Всес. Палеонтол. об-ва, стр. 126-145.
Павлов А.М. 1965. Новый комплекс аммоноидей в пермских отложениях Памира. - Докл.
АН СССР, т. 164, Л? 1, стр. 168-170.
Павлов А.М. 1967а. К стратиграфии верхнего палеозоя Юго-Восточного Памира. - Зап. Ле-
нпнгр. горн, ин-та, т. L11, вып. 2, стр. 133-136.
Павлов А.М. 19676. Пермские аммоноидей Юго-Восточного Памира. - Зап. Ленингр. горн,
ин-та, т. LIII, выл. 2, стр. 69-85.
Павлова Е.Е. 1965. Ревизия рода Neophricodothyris (отряд Spiriferida ). - Палеонтол. журн.,
№ 2, стр. 133-137.
Павлова Е.Е. 1968. Семейство Reticulariidae. В ки.: Брахиоподы верхнего палеозоя восточ-
ного Казахстана. Под ред. Т.Г. Сарычевой. Труды Палеонтол. ин-та АН СССР, т. 121, стр.
187-190.
Павлова Е.Е. 1969. Развитие брахиопод семейства Reticulariidae. Труды Палеонтол. ин-та
АН СССР, т. 120, стр. 3-128.
Развитие и смена морских организмов на рубеже палеозоя и мезозоя. 1965. Под ред. В.Е. Ру-
женцева и Т.Г. Сарычевой. - Труды Палеонтол. ин-та АН СССР, т. 108, стр. 3-430.
Раузер-Черноусова Д.М. 1965. Фораминиферы стратотипического разреза сакмарского
яруса. - Труды Геологич. ин-та АН СССР, вып. 135, стр. 5-81.
Ренгартен В.П. 1934. Геологическое строение района Мургаб-Истык. В кн.: Таджикско-
Памирская экспедиция 1933 года, АН СССР, Л , стр. 369-^74.
Ренгартен В.П. 1935. Геологическое строение района Мургаб-Истык на Восточном Памире.
В кн.: К геологии и палеонтологии Юго-Восточного Памира. - Труды Тадж.-Памирск. экспе-
диции 1933 г., стр. 3-117.
Решения совещания по разработке унифицированных стратиграфических схем для Средней Азии.
1959 Ташкент, стр. 3-129.
Ростовцев О.К., Азарян Н.Р. 1971. Граница палеозоя и мезозоя в Закавказье. - Докл.
АН СССР, т. 199, А9 2, стр. 418-421.
Руженцев В.Е. 1950.Верхнекаменноугольные аммониты Урала. - Труды Палеонтол. ин-та
АН СССР, т. 29, стр. 1-223.
Руженцев В.Е. 1955. Основные стратиграфические комплексы аммоноидей пермской систе-
мы. - Изв. АН СССР, серия биол., № 4, стр. 120-132.
Руженцев В.Е. 1965. Изменение органического мира на рубеже палеозоя и мезозоя. В кн.:
Развитие и смена морских организмов на рубеже палеозоя и мезозоя. - Труды Палеонтол.
ин-та АН СССР, т. 108, стр. 117-136.
Руженцев В.Е., Сарычева Т.Г., Шевырев А.А. 1965. Биостратиграфические выводы. В кн.:
Развитие и смена морских организмов на рубеже палеозоя и мезозоя. - Труды Палеонтол.
ин-та АН СССР, т. 108, стр. 93-116.
Руженцев В.Е., Шевырев А.А. 1965. Аммоноидей. В хн.: Развитие и смена морских орга-
низмов на рубеже палеозоя и мезозоя. - Труды Палеонтол. ин-та АН СССР, т. 108, стр,
47-56.
Руженцев С.В. 1962. Смещение фациальных комплексов пермотриаса вдоль разломов Юго-
Восточного Памира. - Докл. АН СССР, т 143, № 1, стр. 198-200.
Руженцев С.В. 1963. Сдвиги Юго-Восточного Памира. В кн.: Разломы и горизонтальные
движения земной коры. - Труды Геологич. ин-та АН СССР, вып. 80, стр. 113-127.
Руженцев С.В. 1968. Тектоническое развитие Восточного Памира и роль горизонтальных
движений в формировании его альпийской структуры. - Труды Геологич. ин-та АН СССР,
вып. 192, стр. 3-200.
Р уженцев С.В., Швольман В.А. 1963. Восточно-Памирская зона сдвигов. - Изв. АН СССР,
серия геол., № 7, стр. 80-83.
Сарычева Т.Г. 1960. Семейство Linoproductidae. В кн.: Основы палеонтологии. Мшанки,
брахиоподы. Сарычева Т.Г., Лихарев Б.К., Сокольская А.Н. Отряд Productida. Изд-во АН
СССР, стр. 229-231.
Сарычева Т.Г. 1964. Ольтгаминоидные брахиоподы из перми Закавказья. - Палеонтол. журн.,
№ 3, стр. 58-72.
Сарычева Т.Г. 1965. Подотряд Productoidea. В кн.: Развитие и смена морских организмов
на рубеже палеозоя и мезозоя, под ред. В.Е. Руженцева и Т.Г. Сарычевой. - Труды Па-
леонтол. ин-та АН СССР, т. 108, стр. 211-232.
Сарычева Т.Г.,Лихарев Б.К., Сокольская А.Н. 1960. Отряд Productida. В кн.: Основы
палеонтологии. Мшанки, брахиоподы. Изд-во АН СССР, стр. 221-238.
166
Сарычева Т.Г, Грунт Т.А. 1969. Морфология и классификация продуктид семейства Cho-
ri ost egid ае. - Палеонтол. жури., № 3, стр. 70-84.
Сарычева Т.Г., Сокольская А.Н., Грунт Т.А. 1965. Тип Brachiopcda. В кн.: Развитие
и смена морских организмов на рубеже палеозоя и мезозоя. - Труды Палеонтол. ин-та АН
- СССР, т. 108, стр. 62-8L
С и морин А.М. 1949. Брахиоподы Карагандинского бассейна. Выл. 1, Spiriferacea.' Изд-во
АН КазахеК.ССР, стр. 1-82.
Синицын В.М. 1962. Палеогеография Азии. Изд-во АН СССР, стр. 3-267.
Слюсарева А.Д. 1958. Роды Licharew ia Einor и Permospirifer Kulikov в казанском море Рус-
ской платформы. - Докл. АН СССР, т. 122, №1, стр. 127-130.
Сокольская А.Н. 1941. Брахиоподы основания Подмосковного карбона, часть 1. Spiriferidae.
Труды Палеонтол. ин-та АН СССР, т. ХП, вып. 2, стр. 1-139.
Соколье кая А.Н. 1954. Строфоменвды Русской платформы. - Труды Палеонтол. йн-та АН
СССР, т. 51. стр. 1-191
Сокольская А.Н. 1960. Отряд Strophomenida. - В кн.: Основы палеонтологии. Мшанки, бра-
хиоподы. Изд-во АН СССР, стр. 206-220.
Сокольская А.Н. 1965. Отряды Orthida, Rhynchonellida. - в кн.: Развитие и смена морских
организмов на рубеже палеозоя и мезозоя, под ред. В.Е. Ру;хенцева и Т.Г. Сарычевой. -
Труды Палеонтол. ин-та АН СССР, т. 108, стр. 198-203; 233-236.
Степанов Д.Л. 1934. Брахиоподы мшанковых известняков Колвинского района (Сев. Урал),
вып. I. Strophomenacea. - Труды Нефг. геол.-разе. ин-та, серия А, вып. 37, стр. 1-63.
Степанов Д.Л. 1937. Брахиоподы мшанковых известняков Колвинского района (Сев.Урал). -
ч. П. Изд-во Ленингр. гос. ун-та, стр. 1-101.
Степанов Д.Л. 1948. Верхнекаменноугольные брахиоподы Башкирии и их стратиграфическое
значение. - Труды Всес. нефг. геол.-разв. ин-та, н.с., вып. 22, стр. 1-64.
Степанов Д.Л. 1951. Верхний палеозой западного склона Урала. - Труды Всес. н.-и. геол,
развед. ин-та, нов. серия, вып. 54, стр. 3-221.
Степанов Д.Л. 1957. О новом ярусе пермской системы в Арктике. - Вести. Ленингр. гос.
ун-та, № 24, серия геол.-геогр., вып. 4, стр. 20-24.
Степанов Д.Л. 1967. Свальбардский ярус и его положение в стратиграфической шкале перм-
ской системы. - Сов. геология, № 2, стр. 3-18.
Стоянов А.А. 1910. О новом роде Brachiopoda.- Изв. АН, серия VI, № 4, стр. 853-855.
Стоянов А.А. 1916. О некоторых пермских Brachiopoda Армении - Труды Геол, ком., нов.
серия, вып. 3, стр. 1-95.*
Тезисы докладов к совещанию по унификации стратиграфических схем Средней Азии. 1958. М.,
стр. 3-192.
Туманская О.Г. 1935. Пермские аммоноидеи р. К уберганды и их стратиграфическое значение.
В кн.: Фауна верхнего палеозоя Восточного Памира. - Труды Тадж.-Памире к. экспедиции
АН СССР, вып. XXXI, стр. 43-104.
Туманская О.Г. 1949. О пермских аммонеях Средней Азии. - Бюлл. Моск, об-ва исп.
природы, отд. геол., т. 24, вып. 6, стр. 49-84.
Туманская О.Г. 1963. Пермские аммонеи Центрального Памира и их стратиграфическое
значение. Изд-во АН СССР, стр. 5-118.
Туманская О.Г., Борнеман Б.А. 1937. О пермских аммонеях Дарваза. - Бюлл. Моск,
об-ва.исп.природы, отд. геол., нов. серия, т.>45, № 2, стр. 104-118.
Устрицкий В.И. 1958. Палеозоогеографические провинции карбона и перми СССР. - Acta Ра—
laeontologica Sinica, vol.VI, №4, р. 350-358.
Устрицкий В.И. 1960. О границе нижней и верхней перми в Печорском бассейне и в Арктике.
Труды Н.-и. ин-та геол. Арктики, т. 114, вып. 14, стр. 39-49.
Устрицкий В.И. 1961. Основные этапы эволюции морских бассейнов и фауны брахиопод Азии
в течение пермского периода. - Сов. геология, № 1, стр. 49-£4
Устрицкий В.И. 1963. Распределение основных групп фауны в пограничных горизонтах ниж-
ней и верхней перми Арктической зоогеографической области и методика установления этой
границы. - Труды Всес. н.-и. ин-т геол. Арктики, т. 136, стр. 47-57.
Устрицкий В.И. 1965а. О положении Северного полюса и границах палеоарктической области
в позднем палеозое по палеонтологическим данным. В сб.: Тезисы XI сессии Всес. Па-
леонтол. об-ва, стр. 38-39.
Устрицкий В.И. 19656. О корреляции верхнепермских отложений Арктики и Тетиса. - Уч
зап. Н.-и. ин-та геол. Арктики. Палеонтол. и стратиграфия, вып. 7, стр. 10-13
Устрицкий В.И. 1967а. Районирование арктического позднепалеозойского морского бассейна.
В кн.: Вопросы палеогеографического районирования в свете данных палеонтологии. М.,
изд-во 'Недра*, стр. 56-69.
Устрицкий В.И. 19676. Верхний палеозой Арктики (Биостратиграфгя и биогеография). - Ав—
тореф. докт. дисс., М.» стр. 3-37.
Устрицкий В.И. 1971. Биостратиграфия верхнего палеозоя Арктики. - Труды Н.-и. ин-та
геологии Арктики, т. 164, Л., стр. 3-279.
Устрицкий В.И., Ху-Бин, Чан Ан- чжи. 1960 (кит.). Каменноугольные и пермские отло-
жения западной части Куньлуня и их фауна. - Ин-т геологии Мин-ва геологии КНР, спец,
вып. серия 'Стратиграфия и палеонтология*, т. 5, № 1, Пекин, стр. 6-132.
167
Фредерикс Г.Н. 1924 (1919). Палеонтологические этюды. 2. О верхнекаменноугольных спи-
риферидах Урала. - Изв. Геол. ком.,т. XXXVIII, X? 3, стр. 295-324.
Фредерикс Г.Н. 1925. Урсурийский верхний палеозой. II. Пермские брахиоподы с мыса Ка-
лузина. Материалы/ по геол, и полезн. ископаемым Дальнего Востока, X? 40, стр. 1-30.
Фредерикс Г.Н. 1926. Таблица для определения родов семейства Spiriferidae King. _ Изв.
АН СССР, серия 6, т. 20, стр. 393-423.
Фредерикс Г.Н. 1928. Материалы для классификации рода Productus Sow.- Изв. Геол, ком.,
т. 46, № 7, стр.< 773-792.
Фредерикс Г.Н. 1931. Верхнепалеозойская фауна Хараулахских гор. - Изв. АН СССР, X? 2,
отд. матем. и естеств. наук, стр. 199-228.
Фредерикс Г.Н. 1933. Палеонтологические этюды. 4. О некоторых верхнепалеозойских бра-
хиоподах Евразии. - Материалы Центр, н.-и. геол.-разв. ин-та. В об. Палеонтол. и стра-
тиграфия, X? 2, стр. 24-32.
Хаба ков А.В. 1933. Восточная часть Музкольского хребта. - В кн.'Таджикскаякомплекс-
ная экспедиция 1932 года*. АН СССР, Л., стр. 91-109.
Чжан Ли-пэй, Ли-ли. 1962. Раннепермские брахиоподы из свиты Маокоу Восточного
участка Чинь-Линя. - Acta Paleontol. Sinica, vol. 10, № 4, стр. 484-493.
Чернышев Ф.Н. 1902. Верхнекаменноугольные брахиоподы Урала и Тимана. - Труды Геол,
ком., т. 16, № 2Г стр. 1-749.
Чернышев Ф.Н. 1914. Фауна верхнепалеозойских отложений Дарваза. - Труды Геол, ком.,
вып. 104, стр. 1-66.
Штукенберг А.АД905.Фауна верхнекаменноугольной толщи Самарской Луки. - Труды Геол,
ком., нов. серия, выл. 23, стр. 1-144.
111 ухерт Ч. 1957. Палеогеографический атлас Северной Америки. М., ИЛ, Карты 1-84.
Эдельштейн Я.С. 1907. Вэрхнепалеозойские слои Дарваза. - В кн.: Материалы для геологии
России, т. 23, стр. 319-430.
Эйнор О.Л. 1946. Брахиоподы нижнего карбена и нижней перми Западного Таймыра. - Труды
горно-геол. упр. Главсевморпути, вып. 26, стр. 1-91.
Эйнор О.Л. 1957. Основные черты палеогеографии Сибири в каменноугольном и пермском
периодах. В кн.: Вопросы палеобиогеографии и бисстратиграфии, Труды I сессии Всес.
Палеонт. об-ва, Госгеолтехиздат, стр. 81-90.
Эйнор О.Л. 1964. Вопросы биогеографии в палеогеографическом атласе СССР. В сб.: Методы
палеогеографических исследований, № 1, М., изд-во 'Недра*, стр. 192-2061
Эйнор О.Л. 1966. Содержание и связи палеоэкологии и палеобиогеографии. В сб.: Организм ,
и среда в геологическом прошлом. М., изд-во 'Наука', стр. 85-97.
Эйнор О.Л 1967. Палеозоогеография карбоновых морей Казахстана . В сб.: Материалы по гео-
логии, геофизике, геохимии Украины, Казахстана, Забайкалья, № 3, Изд-во Киевск. гос.
ун-та, стр. 85-96.
Эйнор О.Л., Бельговский Г.Л. , Смирнов Г.А. 1965. Основные черты геологического
развития и палеогеография территории СССР в каменноугольном периоде. - Сов. геология,
№ 8, стр. 3244.
Юферев О.В. 1967. Важнейшие задачи палеобиогеографии и значение фораминифер для их
решения в карбоне и перми. - В сб.: Новые данные по биостратиграфии девона и верхнего
палеозоя Сибири, М., изд-во 'Недра', стр. 61-76.
A bi ch H.W. 1878. Geologische Forschungen in den Kaukasischen landern,
T.I. Eine Bergkalkfauna aus der Araxes-Enge bei Djoulfa in Armenien,
Wien, S.VII+1 126.
Ager D.V. 1959. The classification of the Mesozoic Rhynchonelloidea. -
J.Paleontol., v.33, N 2, p. 324-331.
Ager D.V. 1965. Mesozoic and Cenozoic Rhynchonellacea. - In: ’’Treatise
on invertebrate paleontology”, Part H. Brachiopoda, v.2, p. 597-624.
A I b e r c h t J. 1924Palaontologische und stratigraphische Ergebnisse der For-
schungsreise nach Westserbien 1918. - Denkschr. Akad. Wiss. Wien.,
Math. - naturwiss. KI., Bd 99, S. 289-307.
Amos A. 1958. Algunos Spiriferacea у Terebratulacea (Brach.) del carbo-
nifero superior del sistema del Tepuel (Provincia de Chubut). - Contr.
cient. Univ. Buenos Aires, ser. geol., v.II, N 3, p 95-108.
Arthaber G. 1900. Das jungere Palaozoicum aus der Araxes-Enge bei
Djulfa. - Beitr. Palaontol. und Geol. Oesterr. Ungarns und Orients, Bd
12, H.4, S.209-308.
Aspects of Tethyan biogeography. 1967. Syst. Assoc. Publ., N 7, p. 1-336.
A u dl ey-C ha r 1 e s M.G. 1965. Permian palaeogeography of the northern
Australia - Timor region. - Palaeogeogr., Palaeoclimatol., Paleoecol., v.I,
N 4, p.297-306.
Bayle C.E. 1878. Explication de la carte geologique de la France, v.4,
pt I. Paris, p. 158.
168
В о у rich Н. 1865. Uber eine Kohlenkalkfauna von Timor.- Abhandl. Konigl.
Akad.'Wiss. Berlin, S.61-96.
Bion H.S. 1928. The fauna of the agglomeratic slate series of Kashmir. -
Palaeontol. indica, new ser., v.XII, p.3-44.
Bittner A. 1890. Brachiopoden der Alpinen Trias.- Abhandl. Kaiserl.- Ko-
nigl. Geol. Reichanstalt, Bd 14, Sb 1-325.
Booker F.W. 1930. A review of some of the Permo-Carboniferous Produc-
tidae of New South Wales with a tentative reclassification.- Roy. Soc.
N.S.Wales J., v.64, p.65-77.
Bostwick D.A., Nestell M.K. 1967. Permian Tethys Fusulinid Faunas of
the Northern United States.- In ’’Aspects of Tethyan biogeography1’.
Syst. Assoc. Publ., N 7, p.93-102.
Boucot A.J., Johnson J.G., Staton R.D. 1965. Family Athyrididae.- In:
"Treatise on invertebrate paleontology", Part H, Brachiopoda, v.2, p.659-667.
Brill K.G. 1940. Brachiopods of the Whitehorse Sandstone.- In: Newell N.D.
The invertebrate fauna of the Late Permian Whitehorse Sandstone.- Bull.
Geol.Soc. America, v.51, p. 316-319.
Broili F. 1916. Die permischen Brachiopoden von Timor.- Palaontologie
von Timor, Lief. VII, T. 12, S.1-103.
Buckman S.S. 190 6. Brachiopod Homeomorphy: Pygope, Antinomia.Pygites.-
Quart. J. Geol.Soc. London, v.LXII, p.433-455.
Campbell K.S.W. 1953. The fauna of the Permo-Carboniferous Ingelara Beds
of Queensland.- Univ. Queensland Paper, v.4, N 3, p.1-43.
Campbell K.S.W. 1955. The genus Phricodothyris in the Carboniferous of
New South Wales.- Geol. Mag., v.92, p.374-384.
Campbell K.S.W. 1957. A Lower Carboniferous brachiopod-coral Fauna
from New South Wales.- J. PaleontoL, v.31, N 1, p.34-98.
Campbell K.S.W. 1961. Carboniferous fossils from the Kuttung rocks of
New South Wales.- Palaeontology, v.4, pt 3, p.428-474.
Caneva G. 190 6. Uber Bellerophonkalkfauna, zur Frage der Perm-Trias-
grenze.-Neues Jahrb . Mineral., Bd 1, S. 52-60.
Chao J.T. 1927. Productidae of China, Pt I. Product!.- Palaeontol. sinica,
ser. B, v.5, fasc. 2, p.1-244.
Chao J.T. 1928. Productidae of China, Pt 2. Chonetinae, Productinae and
Richthofeniinae.- Palaeontol. sinica, ser. B, v.5, fasc. 3, p.3-99.
Chao J.T. 1929. Carboniferous and Permian Spiriferids of China.- Palaeon-
tol. sinica, ser. B, v.ll, fasc. 1, p. 1-133.
Chao King-Koo. 1965. The Permian Ammonoid-bearing formations of
South China.- Scientia sinica, v.XIV, N 12, p.1814-1825.
Ching Ju-kan. 1963. Urushtenia from the Lower Permian of China.- Acta
palaeontol. sinica, v.ll, N 1, p.2 4-31.
Ching Jil-kan, Zhang Jan. 1961. An Upper Permian Brachiopoda fauna from
Jingxian, Anhui Province.- Acta palaeontol. sinica, v.9, N 4, p.415-417.
Chronic J. 1953. Invertebrate paleontology (excepting Fusulinids and Co-
rals).- In Newell N.D., Chronic J. and Roberts Th.G. Upper Paleozoic of
Peru.- Mem. Geol.Soc. America, v.58, p.49-107.
Cloud P.E. 1944. Permian Brachiopods.- In: King R.E., Dunbar C.O., Clo-
ud P.E., Miller A.K. Geology and paleontology of the Permian area
northwest of Las Delicias, south-western Coahuila.- Geol. Surv. Ameri-
ca Spec. Paper, v.52, p.49-66.
Coleman P.J. 1957. Permian Productacea of Western Australia.- Bull. Bur.
Mineral. Resources, Geol., Geophys. Australia, N 40, p. 1-147.
Conrad T.A. 1839. Descriptions of new species of organic remains. - N.Y.
State Geol. Survey, «3rd Annual Rept., p.57-66.
Cooper G.A. 1942. New genera of North American brachiopods.- J.Wash.
Acad.Sci., v.32.,N 8, p.228-235.
Cooper G.A. 195 6. Pennsylvanian Brachiopods.- J.Paleontol., v.30, N 3,
p.521-530.
Cooper G.A. 1957. Permian brachiopods from Central Oregon.- Smithsonian
Misc. Collect., v.134, N 12, p,3-79.
Dana J.D. 1847. Descriptions of fossil shells of the collections of the
Exploring Expedition under the command of Charles Wilkes, U.S.N., ob-
tained in Australia, from the lower layers of the coal formation in II-
lawazra.- Amer.J. Sci., ser.2, v.4, N 10, p. 151-160.
Davidson T. 1858-1863. A monograph of the British fossil Brachiopoda.
Permian and Carboniferous species.- Palaeont. Soc. London, v.2, pt 4,5,
p.1-280.
169
Davidson T. 1862. On some Carboniferous Brachiopoda collected in Indic»’
by A.Fleming, M.D, and W.Purdon, Esq. RG.S.- Quart.J. GeoLSoc. Lon-
don, v. 18, p.25-35.
Davidson T. 1866. Notes on the Carboniferous Brachipoda collected by
captain Godwin-Austen in the Valley of Kashmere- Quart. J. Geol. Soc.
London, v.22, pt 1-2, p.39-45.
Diener C. 1897a. The Permocarboniforous fauna of Chitichun.- Palaeontol.
indica, ser.XV, v.I, pt3, p. 1-105.
Diener C. 1897b. The Permian fossils of the Productus shales of Kumaon
and Gurhwal.- Palaeontol. indica, ser.XV, v.I, pt 4, p.1-54.
Diener C.1899. Anthracolithic fossils of Kashmir and Spiti.- Palaeontol. in-
dica, ser.XV, v.I, pt 2, p.1-95.
Diener C. 1903. Permian fossils on the Central Himalayas.- Palaeontol.
indica, ser.XV, v.I, pt 5, p.1-204.
Diener C. 1911. Anthracolithic fossils of the Shan States.- Palaeontol. indi-
ca, new ser., v.3, N 4, p.1-74.
Diener C. 1915. The Anthracolithic faunae of Kashmir, Kanaur and Spiti.-
- Palaeontol. indica, new ser., v.5, N 2, p.1-135.
Di-Stefano G. 1914. Le Richthofenia dei calcari con Fusulina di Palaz-
zo Adriano Nella valle del fiume Sosio.- Palaeontogr. ital., v.XX, p.1-2 6.
Douglas J.A. 1936, A Permocarboniforous fauna from South-West Persia
(Iran).- Palaeontol. indica, new ser., v.XXH, N 6, p.1-59.
Douglas J.A. 1950. The Carboniferous and Permian faunas of South
Iran and Iranian Baluchistan.- Palaeontol. indica, new ser., v. XXII, N 7,
p.l- 56.
Dunbar C.O. 1955. Permian brachiopod faunas of central East Greonland.-
Medd. Gr^nland, Bd 110, N 3, p.l-169.
Dunbar C.O., Condra G. 1932. Brachiopoda of the Pennsylvanian sys-
tem in Nebrasca.- Nebrasca Geol. Surv., ser. 2, N 5, p.l-377.
Easton W.H. 1962. Carboniferous formations and faunas of Central Monta-
na.- U.S.Geol. Surv. Profess. Paper, N 348, p.I-IV + 1-126.
Ekman S. 1935. Tiergeographie dos Meeres. Leipzig, S. 1-542.
Enderle J. 1901. Ueber eine anthracolithische Fauna von Balia Maaden in
Kleinasien.- Beitr. Palaontol. und Geol. Oestorr.- Ungarn und Orients,
Bd XIII, S. 49-109.
Etheridge R., Dunn W.S. 1909. Notes on the Permo-Carboniferous Pro-
duct! of Eastern Australia with synonymy.- Rec.Geol. Surv. N.S. Wales,
v.8, pt 4, p.293-304.
Eveille Ch. 1835. Apercu gcologiquo de quelques localites tres riches on
coquilles sur les frontieres de France de Belgique.- Mem.Soc. geol.
France, v.2, p.29-40.
Fabiani R. 1933. Sui giacimenti permiani del Sosio (Palermo) 1 sugli stroph-
omenidi in essi trevati.- Mem.Soc. geologica ital., v.I, p.2-22.
Fantini Sestini N. 1965a. Permian fossils of the Shaksgam valley. - IV -
- Paleontology-Zoology-Botany. Italian Expeditions to the Karakorum (к )
and Hindukush, p.149-392.
Fantini Sestini N. 1965b. Permian fossils of the upper Hunza valley.-
IV Paleontology-Zoology-Botany. Italian Expeditions to the Karako-
rum (K2) and Hindukush, p.135-148.
Fantini Sestini N. 1965c. • Bryozoans, brachiopods and molluscs from
Ruteh Limestone (Permian).- Riv. ital. palaontol. e stratigr., v.71, N 1,
p.13-108.
Fantini Sestini N. 1965d. The geology of the Upper Djadjerud and
Lar Valleys (North Iran). II. Palaeontology. Brachiopods from Do.rud for-
mation.- Riv. ital. paleontol. e stratigr., v.71, N 3, p.773-788.
Fantini Sestini N. 1966. Brachiopods from Geirud Formation, Member D
(Lower Permian).- Riv. ital. paleontol. e stratigr, v.72, N 1, p.9-50.
Fantini Sestini N., Claus M. 1966. Brachiopods from the Upper Per-
mian Nesen Formation (North Iran).- Riv. ital. paleontol. e stratigr., v.72,
N 4, p.887-930.
Fischer Waldheim G. 1829. Notice sur le systeme apophysaire ou sur
la Charpente osseuge des Terebratules.- Progr. invitat. seance publ.
Soc. Imp. Nat., Moscou, p.7-18.
Fischer Waldheim G. 1837. Oryctographie du Gouvernement de Mos-
cou, p.1-202.
Freeh F. 1911. Die Dyas. - In: Richthofen F. China, Bd. 5, Berlin, S.»
103-202.
170
Gemmellaro G. 1891a. Sulle Lyttonie dei calcari con Fusulina della val-
le del Fiume Sosio.- Boll. Soc. sci. natur. ed econ. Palermo, N 1,
p.3-5.
Gemmellaro G. 1891b. Sopra un nuovo genere di brachiopodi prove-
niente dei calcari con Fusulina della Provincia di Palermo.- Boll.Soc.
sci. natur. ed econ. Palermo, N 4, p.22-23.
Gemmellaro G. 1892. Sopra due famiglie" di brachiopodi (Strophomeni -
dae e Productidae) provenienti dei calcari con Fusulina della Pro-
vincia di Palermo.- Boll. Soc.sci. natur. ed econ. Palermo, N 3,
p.22-27.
Gemmellaro G. 1894a. Sopra due famiglie di Brachiopodi (Spiriferidae,
Rhynchonellidae e Terebratulidae) provenienti dei calcari con Fusulina
della valle del Fiume Sosio nella provincia di Palermo.- Boll. Soc.sci.
natur. ed econ. Palermo, N 1, p.1-6.
Gemmellaro G. 1894b. Le Richthofenia provenienti dei calcari con Pu-
sulina della valle del Fiume Sosio nella provincia di Palermo. Boll.
Soc.sci. natur. ed econ. Palermo, N .1, p.6-7.
Gemmellaro G. 1894c. Sopra un nuovo genere di Productidae prove-
niente dei calcari con Fusulina della provincia. di Palermo.- Boll.Soc.
sci. natur. ed econ. Palermo, N 1, p.7.
Gemmellaro G. 1897. Sopra due nuovi generi di brachiopodi provenienti
dei calcari con Fusulina della provincia di Palermo.- Giorn. sci.natur.
ed econ. Palermo, v.21, p.8-10.
Gemmellaro G. 1899. La fauna dei calcari con Fusulina della valle
del fiume Sosio nella provincia di Palermo. Brachiopoda.- Giorn. sci.
natur. ed econ. Palermo, v.22, p.95-214.
Girt у G.H. 1908. The Guadalupian fauna.- U.S. Geol. Surv. Profess. Pa-
per, v.58, p. 1-651.
Girty G. 1910. The fauna of the Phosphate Beds of the Park City For-
mation in Idaho, Wyoming and Utah.- U.S. Geol. Surv. Bull., N 436,
p.1-82.
Gobbett D.J. 1964. Carboniferous and Permian brachiopods of Sval-
bard.- Norsk Polarinst. skr„ N 127, p.1-201.
Gobbett D.J. 1967. Palaeozoogeography of the Verbeekinidae (Permian
foraminifera).- In: "Aspects of Tethyan biogeography". Svst. Assoc.
Publ., N 7, p.77-91.
Grabau A.W. 1931. The Permian of Mongolia. Natural History of Cent-
ral Asia, v.4, p.I-XLIV + 1-665.
Grabau A.W. 1932. Studies for students. The Brachiopoda.- Sci. Quart.
Nat. Univ. Peking, pt 3, v.III, N 2, p.47-84.
Grabau A.W. 1934. Early Permian fossils of China. I.Early Permian bra-
chiopods, pelecypods and gastropods of Kueichow.- Palaeontol. sinica,
ser.B, v.8, fasc.3, p.1-168.
Grabau A.W. 1936. Early Permian fossils of China. Pt 2. Fauna of the
Maping limestone of Kwangsi and Kweichow.- Palaoontol. sinica, ser.
B, v.8, fasc.4, p.1-168.
Grant R. 1965a. The Brachiopod superfamily Stenoscismatacea.-Smithsonian
Misc. Collect., v.148, N 2, p.1-192.
Grant R. 1965b. Superfamily Stenoscismatacea. In: "Treatise on inverteb-
rate paleontology1’. Part H. Brachiopoda, v.2, p.62 5-633.
Grant R. 1968. Structural adaptation in two Permian brachiopod genera,
Salt Range, West Pakistan.- J.Paleontol., v.42, N 1, p.1-32.
Grant R. 1970. Brachiopods from Permian-Triassic boundary beds and aoe
of Chidru Formation, West Pakistan.- In: Stratigraphic boundary prob-
lems: Permian and Triassic of West Pakistan. Univ. Kansas Dept. Geol.,
Spec. Publ. 4, p.117-151.
Greco B. 1938. Revisione degli Strophomenidi Fermiani del Sosio concer-
vati nel Museo di Geologia della R.Universita di Palermo.- Giorn. sci.
natur. ed econ. Palermo, v.39, Mem.II, p.1-46.
Greco B. 1942. La fauna permiana del Sosio-Parte terza. Brachipoda.Fasc.
1. Fam. Orchidae, Strophomenidae. - Palaeontogr. it al., v.40 (new ser.,
v.10), p.115-159.
Greco B.1943, La fauna permiana del Sosio-Parte terza. Brachiopoda.Fasc. 2.
Fam. Productidae, Richtgofeniidae. - Palaeontogr. ital., v.41 (new ser.,
v.ll), p. 1-22.
171
Gregorio A.1930. Sul Permiano di Sicilia (fossili del calcare con Fusulina
di Palazzo Adriano non deseritti del Prof. G.Gemmellaro conservati nel
mid privato Gabinetto). - Ann. Geol. paleontol. Palermo, liv. 52,p.18-32.
Hall I., Clarke I.M. 1890-1894. An introduction to the study of the gene ra
of Palaeozoic Brachiopoda. - Pt 1,2. - Natural History of New York.Pa-
leontol., v. 8, p.1-367 (1890-1892), 1-394 (1893-1894).
Hamada T. 1960. Some Permo-Carboniferous fossils from Thailand.-Sci.Pa-
pers Coll. Gen. Educ. Univ. Tokyo, v.10, N 2, p.337-361.
Hamlet B. 1928. Permische Brachiopoden, Lamellibranchiaten und Gastropo-
den von Timor. - Jaarb. mijnwezen in NederL-Indie, bd VI, Verh.2,ptl,
S. 1-115.
Hayasaka J. 1922. Palaeozoic Brachiopoda from Japan, Korea and China.
Pts 1,2. - Sci. Rept. Tohoku Univ., ser. 2, v.6, N 1, p. 1-109.
Hayasaka J. 1963. Some Permian fossils of Southern Kitakami. IV. -Proc.
Japan. Acad., v. 39, p.753-757.
Hayasaka J. 1967. Some Permian fossils from Southern Kitakami. VI.Three
Brachiopods. - Proc. Japan Acad., v.43, N 1, (Suppl. to v.42, N 10),
p.1223-1228.
Hayden H. 1916. Notes on the geology of Chitral, Gilgit and the Pamirs.-
Rec. India Geol. Surv., v.45, p.271-335.
Helmersen G. 1847. Aulosteges variabilis, ein neuer Brachiopode mit arti-
culaten Schlosse, aus dem Zechstein Russlands,- Bull. cl. phys.-math.
Acad. sci. St. Petersbourg., t. VI, N 9, S.135-144.
Hill D. 1950. The Productinae of the Artinskian Cracow fauna of Queens-
land. - Queensland Univ. Dept. Geol. Papers, v.3, N 2, p. 1-36.
Huang T.K. 1932. Late Permian Brachiopoda of Southwestern China. - Pa-
leontol. sinica, ser. B, v.IX, fasc.l, p.1-98.
Huang T.K. 1933. Late Permian Brachiopoda of Southwestern China. Part II.-
PalaeontoL sinica, Ser. B, vJX, fasc.2, p.1-131.
Hudson R.G.S., Sudbury M. 1959. Permian Brachiopoda from South-East
Arabia. - In: “Contributions a la geologie de la Peninsula Arabique".No-
tes et Mem. Moyen-Orient, v. VII, p. 1-35.
Jochelson E.L.1962. Phyllum Brachiopoda.-In: Mudge M.R.and JochelsonE.L.
Stratigraphy and paleontology of the Uppermost Pennsylvanian and Lo-
wermost Permian rocks in Kansas. Geol. Surv. Profess. Papers, N 323,
VI,p.74-85.
Kahler F.1962. Stratigraphische Vergleiche im Karbon und Perm mit Hilfe
der Fusuliniden.- Mitt. Geol.Ges. Wien, Bd 54, S.147-161.
Kayser E.1881. Mitteilungen uber die Fauna des Chinesischen Kohlenkalks
von Lo-Ping.- Z.Dtsch. Geol, Ges., Bd 33, S. 351-352.
Kayser E. 1883(1882). Ergebnisse einiger Reisen und darauf gegrundeter
Studien. - In: Richthofen F. China, Bd 4. PalaontoL TeiL, Berlin, S.
1-288.
Keyserling A. 1846. Geognostische Beobachtungen in Wissenschaftlichen
Beobachtungen auf einer Reise in das Peyschora-Land im Jahre 1843.
I. Palaeontologische Bemerkungen. II. Geognostische Reise. St. Petersb.,
S.151-406.
King R.E. 1931. The geology of the Glass Mountains, Texas. Pt 2. Faunal
summary and correlation of the Permian formations with description of
Brachiopoda. - Bull. Univ. Texas, N 3042, p. 1-245.
King R.E. 1938. New Chonetidae and Productidae from Pennsylvanian and
Permian strata of North-Central Texas. - J.Paleontol., v.12, p.257-279.
King W. 1846. Remarks on certain genera belonging to the class Pallio-
branchiata. - Ann. and Mag. Natur. Hist., v.18, p.2 6-42.
King W. 1850. A monograph of the Permian Fossils of England. - Palaeon-
togr. Soc. London, v. Ill, p. 1-258.
Kossmat F., Diener C. 1910. Die Bellerophonkalke von Oberkrain und
ihre Brachiopodenfauna. - Jahrb. Kaiserl.-Konigl. Geol. Reichsanstalt,
S.278-309.
К о sti £-Podgo r ska V. 1949. Fauna a Brachiopodes du Carbonifere de la
Lika en Croatie. - Ann. Geol. Penins. Balkan, Belgrad, t. 17, p.73-102.
Kumm el B.,Tei chert C. 1970. Stratigraphy and paleontology of the Permi-
an-Triassic boundary beds, Salt Range and Trans-Indus Ranges, West
Pakistan. - In: "Stratigraphic boundary problems: Permian and Triassic
of West Pakistan”. Univ. Kansas Dept., Geol., Spec. Publ. 4, p.1-109.
172
Kutorga S. 1842. Boitrag zur Palaoontologic? Russlands. - Verhandl. Ges,
St. Petersb., S.l-34.
Lattin G. 1967. Grundriss der Zoogeographic. Jena, S. 13-602.
Licharew B. 1930. On two new representations of the family Productidae
from the Lower Permian of North Caucasus. - Докл. АН СССР. A.
N 16, стр. 4 3 6-4 40.
Licharew B. 1935. Bemerkungen uber einige oberpalaozoische Brachiopoden.-
Zbl. Mineral., Geol. und Palaontol., Abt. B, S. 369-373.
Licharew B. 1936.Uber einige palaeozoische Gattungen der Terebratulacea aus
Eurasien. -Проблемы палеонтологии, т.1, стр. 263-273.
Loczy L. 1899. Die Karbonischo Fauna der Urngeburg von Kan-Tschu-Fu,
Uberreste aus der Karbonperiode. - Wissenschaftliche Ergebnisse der
Raise dos Grafen Bela Szechenyi in Ostasion, Bd III, Abt. 4, S.1-228.
Logan A., Me Gugan A. 1968. Biostratigraphy and faunas of the Permian
Ishbel Group, Canadian Rocky Mountains. - J. Paleontol., v.42, N 5,
p.1123-1139.
M a n s uy H. 1913. Faunes des calcaires a Productus de 1’Indochine.- Mem.
Serv. geol. Indochine, v. II, fasc. IV, p.1-10 4.
Maxwell W.G.H. 1956. Proposed use of plenary powers to designate a
type species in harmony with accustomed usage for ’’Terrakea” Booker
(RW.), 1930. Bull.Zool. Nomenci., v.ll, pt 11, p.333-336.
Me Coy F. 1844. A synopsis of the characters of the Carboniferous Li-
mestone fossils of Ireland. Dublin, p.1-207.
Me Coy F. 1847. On the fossil botany and zoology of the rocks associa-
ted with coal in Australia.- Ann. and Mag. Natur. Hist., v.20, p.145-157,
226-236, 298-331.
Me Gugan A. 1963. A Permian brachiopod and fusulinid fauna from the
Elk Valley, British Columbia, Canada.- J.Paleontol., v.37, N 3, p.621-627.
Meria G. 1928. Contribute alia conoscenza della Fauna dei calcari a
Schwagerina della valle del Sosio (Prov. di Palermo).- Atti Soc. Tosca-
na sci. natur. resid. in Pisa, v.XXXVII, p.70-80.
Meria G. 1934. Fossili antracolitici del Caracorum.- Spedizione Italiana de
Filippi nell’Himalaya, Caracorum e Turchestan Cinese, ser.II, v. V .p.99- 31 9
M i n at о M. 1951. On some Lower Carboniferous fossils of the Kitakami
massif, Northeast Hounsyu, Japan.- J.Fac.Sci. Hokkaido Univ., ser. 4,
v.VII, N 4, p.355-382.
Moller V. 1879. Uber die bathrologische Stellung des jungeren palaozoischen
Schichtensystems von Djoulfa in Armenien.- Neues Jahrb. Mineral.,Geol.
und Palaeontol., S.225-243.
Morris J. 1845. Descriptions of fossils.- In: Strezelecki P.E. Physical des-
cription of New South Wales and Van Diemen's Land. London,p.270-291.
Muir-Wood H. 1930. The classification of the British Carboniferous Bra-
chiopod - SubfaiYiily Productinae.- Ann. and Mag. Natur. Hist., v.V, N 2 5,
p.100-108.
Muir-Wood. H. 1965. Suborder Productidina. In: ’’Treatise on invertebrate
paleontology”. Part H. Brachiopoda, v.I, p.439-510.
Muir-Wood H., Cooper G.A. 1960. Morphology, classification and life
habits of the Productoidea (Brachiopoda).- Geol.Soc. America, mem. 81,
p.1-447.
Muir-Wood H., Oakley K.P. 1941. Upper Paleozoic faunas of North Sik-
kim.- Palaeontol. indica, new ser., v.XXXI, N 1, p.6-90.
Nakamura K. 1960. Dictyoclostus derived from the Middle Permian Ka-
nokura series and the Lower Permian Sakamotozawa series of the Ki-
takami mountains, Japan.- J.Fac.Sci.Hokkaido Univ., Ser„IV, Geol. and
Mineral., v.X, N 3, p.495-511.
Nakazawa K., Kapoor H.M., Ken-ichi Ichii, Bando Y., Maegoya T.,
Shimizu D., Nogami J., Tokuoka T., Nohda S. 1970. Preliminary
report on the Permo-Trias of Kashmir.- Mem.Fac.Sci.Kyoto Univ., ser.
Geol. and Mineral., v.XXXVII, N 2, p.163-172.
Nonaka J.1944. Some Permian Brachiopods from Inner Mongolia.- Japan.J.
Geol. and Geogr., v.XIX, p.83-87.
Oehlert D.P. 1887 (1883). In: Fischer P.H. Manuel de conchyliologie et
de paleontologie conchyliologique, ou Histoire naturelie des mollusques
vivants et fossiles, pt II, (Paris), p.1189-1334.
Ozawa J. 1925. Palaeontological and st rati graphic al studies on the Permo-
carboniferous limestone of Nagata. Pt 2. Palaeontology.- J.Fac.Sci.Imp.
Univ. Tokyo, v.45, pt 6, p.1-90.
173
Pit rat Ch.W. 1965. Suborder Spiriferidina.- In: ’’Tratise on invertebrate pa-
leontology", Part H, Brachiopoda, v. 2, p. 667-728.
Pa.?ckelmann V. 1931. Die Fauna des deutschen Unterkarbons.II. Die
Brachiopoderi des deutschen Unterkarbons. Teil 2. Die Productinae
und Productus-ahnlichen Chonetinae.- Preuss, geol. Landesanst. Abh„
N.F., H.136, S.1-440.
Pander C.H. 1861. In: Helmersen G. Die geologische Beschaffenheit des
unteren Narovathals und die Versandung der Narovamundung.- Bull.
Acad.Imp.Sci. St. Petersbourg, N 3, S.46-49.
Parona C.F. 1933. Le "Lyttonia” fra i Brachiopodi della fauna permiana
di Palazzo Adriano in Sicilia.- Mem.R.Acad.sci. Torino, Ser.II, v.LXVII,
N 13, pl2-18.
Plodowski *G. 1968. Neue Spiriferen aus Afghanistan.- Senckenberg. let-
haea, Bd 49, N 2/3, S.251-259.
Plodowski G. 1970. Stratigraphie und Spiriferiden (Brachiopoda) des Pa-
laozoikums der Dascht - E - Nawar (SW Afghanistan).- Palaeontographica,
Abt.A, Bd 134, Lief. 1-3, S.l-132.
Prendergast K.L. 1943. Permian Productinae and Strophalosiinae of Wes-
tern Australia.- J.Roy.Soc. W.Austral., v.28, p.l-73.
Ram о vs A. 1958. Die Entwicklung des Oberperms im Bergland von Skofja
Loka und Polhov Gradec (W.Slovenien, N.W.Jugoslawien).- Slov. akad.
znatosti in umetnosti, Razpr. diss., kn.IV, str.455-716.
Ramоvs A. 19 60. Razvoj mlajsih paleozojskih skladov v Vitanjskem nizu
(The development of upper paleozoic strata in the Vitanje Hills - Slo-
venia, NW Jugoslavia).- Geologija - Razpr. in Porocila - 6 kniga, Ljubl-
jana, str. 170-234.
Reed F.R.C. 1925. Upper carboniferous fossils from Chitral and the Pamir.-
Palaeontol. indica, new ser., v.6, N. 4, p.l-134.
Reed F.R.C. 1927. Palaeozoic and Mesozoic fossils from Junnan.- Palaeon-
tol. indica, new ser., v.X, Mem. I, p.l-140.
Reed F.R.C. 1931a. Upper Carboniferous fossils from Afghanistan.- Pala-
eontol. indica, new ser., v.XIX, p.l-39.
Reed F.R.C. 1931b. New fossils from the Productus Limestone of the Salt
Range, with notes on other species.-Palaeontol. indica, new ser., v.XVIl,
p.l-5 6.
Reed F.R.C. 1932. New Fossils from the agglomeratic slate of Kashmir.-
Palaontol. indica, new ser., v.XX, N 1, p.l-79.
Reed F.R.C. 1944. Brachiopoda and Mollusca from the Productus Limesto-
nes of the Salt Range.- Palaeontol. indica, new ser., v.23, N 2, p.1-
678.
Rothpletz A. 1892. Die Permo-Trias- und Jura-Formation auf Timor und
Rotti im indischen Archipel.- Palaeontographica, Bd XXXIX, S.57-10 6.
Ross Ch.A. 1971. New species of Schwagerina and Jabeina (Fusulinacea)
of Wordian age (Permian- from Northwestern British Columbia).- Bull.
Geol. Surv. Canada, N 197, p.95-106.
Ross Ch.A., Nassichuk W.W. 1970. Jabeina and Waagenoceras from
the Atlin horst area, Northwestern British Columbia. J .Paleontol., v. 44,
N 4, p.779-781.
Rud wick M.J.S. 1961. The feeding mechanism of the Permian brAchiopod
Prorichthofenia.- Palaeontology, v.3, pt 4, p.450-471.
Rudwick M.JLS. 1965. L’histoire de la paleobiologie marine.-Vie et milieu,
suppl. 19, p.315-320.
Rudwick M.J.S., Cowen R. 1968. The functional morphology of some ab-
berrant Slrophomenidae Brachiopods from the Permian of Sicily.- Bull.
Soc.Paleontol. ital., v.6, N 2, p. 113-176.
Rzonsnickaja M.A. 1956. Systematization of Rhynchonellida.- XX Congr.
geol. Internat.Resumenes de los trabojos Presentados. Mexico, p.125-12 6.
Schellwien E. 1892. Die Fauna des Karnischen Fusulinenkalkes.-TJ.-
P ala eonto graphic a, Bd 39, S.l-56.
Schellwien E. 1900a. Beitrage zur Systematik der Strophomeniden des
Oberen Palaeozoicum.- Neues Jahrb. Mineral., Geol. und Palaeontol.,
Bd 1, S.l-15.
Schellwein E. 1900b. Die Fauna der Trokgofelschichten in den Karschen
Alpen und den Karawanken. T.I.- Die Brachiopoden.- Abhandl. Kaiserl.-
Konigl. geol. Reichsanst. Wien, Bd XVI, N 1, S.1-122.
Schindewolf O.H. 1954. Uber die Faunenwende vom Palaozoikum zum Me-
sozoikum.- Z.Dtsch.geol. Ges., Bd 105, T.2, S.153-182.
174
Schindewolf O.H. 1963. Neokatastrophismus? -Z.Dtsch.geol.Ges.» Bd 114,
T.2, S.430-445.
Schindewolf O.H. 1964. Uber die Jungpalaozoische Veroisung dor Salt
Range (W. Pakistan).- Neues Jahrb. Geol., Palaontol. Abh., Bd 121,
N 1, S.55-66.
Schmidt H. 1937. Zur Morphogenie der Rhynchonelliden.- Senkenber giana.
Bd 19, N 1-2, S.22-60.
Schmidt H., Me Laren D.J. 1965. Paleozoic Rhynchonellacea. In: "Treatise
on invertebrate paleontology4’, Part H. Brachiopoda, v.2, p.552-597.
Schretter Z. 1936. Lyttonia aus dem Bukk- Gebirga- Foldt kozl., v.66,
p.113-121.
Schretter Z. 1963. Die Brachiopoden aus dem oberen Perm des Bukkgo-
birges in Nordungarn.- Geol. hung. Ser. palaeontol., fasc.28, p.87-158.
Shue he nt C. 1913. Brachiopoda.- In: Zittel K.A. Textbook of Paleontology
(transl. by C.K.Eastman), v.I, London, p.355-420.
Schu chert C. 1929. Family Schizophoriidae. In: Schuchert C., Le Vene C.M.
Fossilium catalogus. I.Animalia, Pt 42. Brachiopoda (Generum et Ge-
notyporum index et bibliographia). Berlin, Junk, p.15.
Schuchert C., Le Vene C.M. 1929. Fossilium catalogus. I.Animalia. Pt 42.
Brachiopoda (Generum et Genolyporum index et bibliographia).- Berlin,
Junk, p.3- 140.
Schuchert C., Cooper G.A. 1932. Brachiopod genera of the suborder
Orthoidea and Pentameroidea.- Mem. Peabody Mus. Natur. Hist., vJV,
pt I, p.1-270.
Shimizu D. 1961. Brachiopod fossils from the Permian Maizuru Group.-
Mem. College Sci. Univ. Kyoto, Ser.B, v.XXVII, N 3, p.309-350.
Shimizu D. 1962. The Permian Maizuru Group, its stratigraphy and 8yn-
tectonic faunal succession through the Latest Paleozoic Orogeny.-Mem.
College Sci. Univ. Kyoto, Ser.B, v.XXVlII, N 4, p.571-609.
Shimizu D. 1963. Permian Brachiopod fossils of the Maizuru. Group found
on the North of Oklahoma City, Japan.- Mem. College Sci. Univ. Kyo-
to, Ser. B, v.XXX, N 2, p.69-80.
Shumard B.F. 1858. Notice of new fossils from the Permian strata of
New Mexico and Texas, collected by Dr. George G.Srhumard.-Trans.
Acad.Sci. St. Louis, v.I, N 2» p.290-297.
Silvestri A. 1933. Fusulinidi dell'antracolitico della valle del Sosio (Pa-
lermo).- Mem.Inst. Geol. Univ.Padova, v.10, p.1-45.
Simic V. 1933. Gornji Perm u zapadnoj Srbiji.- Raspr.geol. Inst. Jugoslavije,
v.I, p.3-130.
Simi<? V. 1934a. Belerofonska fauna Niksicke Zupe u Grnoj gori.- Bull.
Serv. Geol. Jougoslavie, v.3, p.45-51.
Simic V. 1934b‘. Die Dorsalklappen bei Productus yangtzeensis Chao, P.
callocreneus Heritch und Proboscidella alpina Simic.- Bull. Serv.Geol.
Jougoslavie, T.III, fa sc. I, S. I0I-III.
Simic V-1934c. Beitrag zur Kenntniss der oberkarbonischon und ober-
permischen Faunen von Westserbien.- Bull.Serv. Geol. Jougoslavie, T.III,
fasc.I, S.II2-I28.
Simic V. 1935a. Einige Versteinerungen aus dem Oberperm von Westserbi-
en.- Bull. Serv. geol. Roy. Jougoslavie, ann4e,1934, t.IV, fasc. Ц p. 147-153.
Simic V. 1935b. Oberkarbonische Versteinerungen von Lika in Kroatien.-
Bull. Serv. geol.Roy. Jougoslavie, ahnee 1934, t.IV, fasc.I, p.141-144.
Skinner J.W., Wilde J.L. 1955. New fusulinids from the Permian of West
Texas.- J.Palaeon tol., v.29, N 6, p.927-940.
Sower by J. 1812-1845. The mineral conchologie of Great Britain. v.I-VII,
London, p. 1-1290.
S lac he G. 18 78. Beitrage zur Fauna der Bellerophonkalke Sudtirols.
Pelecypoda und Brachiopoda.-Geol. Bundesansl. Wien, Bd 28, N 1» S.93-168.
Stainbrook M.A. 1947. Brachiopods of the Percha Shale of New Mexico
and Arizona.- J.Paleontol., v.21, N 4, p.297-328.
Stehli F.G. 1954. Lower Leonardian Brachiopoda of the Sierra Diablo.- Bull.
Amer. Mus. Natur. Hist., v.10 5, art.3, p.2 63-358.
Stehli F.G. 1956. Dielasma and its external Homeomorph Beecheria.- J. Pa-
leontol., v.30, N 2, p.305-313.
Stehli F.G. 1957. Possible Permian climatic zonation and its implications.-
- Amer.J.Sci., v.255, p.607-618.
Stehli F.G. 1961. New Terebratuloid genera from Australia.- J. Paleontol.,
v.35, N 5, p.451-456.
175
Stehli F.G. 1962. Notes on some Upper Paleozoic terebratuloid Brachiopods.
J. Paleontol., v.36, N 1, p.97-111.
Stehli F.G. 1964. Permian zoogeography and its bearing on climate.- In
"Problems of Palaeo climatolo a v", p.537-549, 577-579.
Stehli F.G. 1965. Paleozoic Terebratulida.- In:"Treatise on invertebrate Pa-
leontology", Part H. Brachiopoda, v.2, p.730-7 62.
Stehli F.G. 1971. Tethyan and Boreal Permian faunas and their significan-
ce.- Smithsonian Contrib. Paleobiol.; N 3, p.337-345.
Stehli F.G., Wells J.W. 1971. Diversity and age patterns in Hermatypic
corals.- Syst. Zool., v.20, N 2, p. 115-126.
Stepanov D.L., Galshani F., Stooklin J. 1969. Upper Permian and Per-
mian- Triassic boundary in North Iran.-Geol.Surv.Iran, Rept N 12, p.3-72.
SuessE. 1893. Are great ocean depths permanent? -Natur.Sci., v.2, p.180- 187.
Swallow G.S. 1860. Descriptions of new fossils from the Carboniferous
and Devonian rocks of Missouri.- Trans.Acad.Sci.St.Louis, v.I (1856-
1860), p.635-660.
Teichert C., Kummel B., Kapoor H.M. 1970. Mixed Permian-Triassic fau-
na, Guryul Ravine, Kashmir. .- Science, v.167, p. 174-175.
Termi er H., Termi er G. 1970. Les Productoides du Djoulfien (Permien su-
perieur) dans la Tethys orientale: essai sur 1’agonie d'un phylum.-
। Ann.Soc.geol. Nord, t.90, N 4, p.443-461.
Thomas H.B. 1937. Plicatoderbyia, a new Permian Brachiopod subgenus.-
J. Paleontol., v.ll, N 1, p.13-18.
Thomas Iv. 1910. The British Carboniferous Orthotetinae.- Mem. Geol. Surv.
Great Britain, Palaeontol., vj, *pt 2, p.83-134.
Thomas Iv. 1914. The British Carboniferous Producti. I.Genera Pustula
and Overtonia.- Mem. Geol. Surv. Great Britain, Paleontol., v.I, pt 4,
p.199-366.
Thomson J.A. 1926. A revision of the subfamilis of the Terebratulidae
(Brachiopoda).- Ann. and Mag. Natur. Hist., ser.9, v.18, p.523-530.
Ting Pei-chen. 1962. Some Upper Permian brachiopods from Tibet.- Acta
palaeontol. sinica, v. 10, N 4, p.459-483'.
Tran-Thi Chi-Thuan, 1961. Les brachiopodes permiens du Phnom-Tup
(Sisbphon-Cambodge).- Ann. Fac. sci. Saigon, p.267-308.
Waagen W. 1882-1887. Salt Range fossils. Productus- limestone Fossils,
Part 4. Brachiopoda.- Palaeontol. indica, ser.13, v.I, pt 4, fasc.1-5, p.329-
770.
Wang Y. 1955. New genera of Brachiopods.Scientia sinica, v.4, N 2, p.327-357.
Wanner J., Silve.rts H, 1935. Zur Kenntnis der Permischen Brachiopoden
von Timor. I. Lyttoniidae und ihre biologische und Stammesgeschicht -
liche Bedeutung.- N.Jahrb. Mineral., Geol. und Palaontol. Beil.- Bd 74,
Abt. B, S.201-281.
Waterhouse J.B. 1964. Permian stratigraphy and faunas of New Zealand.-
N.Z.Geol. Surv. Bull., new ser., v.72, p.5-101.
Water ho use J.B. 1966. Lower Carboniferous and Upper Permian Brachio-
pods from Nepal.- Jahrb. Geol. B.A., Sonderband 12, S.5-99.
Waterhouse J.B., Piyasin S. 1970. Mid-Permian Brachiopods from Khao
Phrik, Thailand.- Palaeontographica, Abt.A, Bd 135, S.83-197.
Weller S. 1910. Internal characters of some Mississippian rhinchonelliform
shells.- Bull. Geol.Soc. America, v.21, p.497-516.
Weller S. 1914. The Mississippian Brachiopoda of the Mississippi Valley
Basin. -Illinois State Geol, Surv. Monogr., v.I, p.3-508.
White C.A., Orestes J, 1867. Descriptions of new Subcarboniferous coal-
measure fossils, collected upon the Geological Survey of Iowa; to-
gether with a notice of new generic characters involved in two spe-
cies of Brachiopoda.- Trans. Chicago Acad.Sci., v.I, p.115-127.
Whitehouse F. 1928. Notes on Upper Palaepzoic Marine Horizons in
Eastern and Western Australia.- Trans.Austral.Assoc. Adv.Sci. Rept.
(1926), Sect. C, v.18, p.281-233.
Williams A. 1965. Suborder Strophomenidina.-In: "Treatise on invertebrate
paleontology1', Part H,Brachiopoda,p.363-412.
Williams A., Wright A. 1965. Order Orthida. In "Treatise on invertebra-
te paleontology". Part H. Brachiopoda, v.I, p.299-3 61.
Yanagida J. 1964. Permian Brachiopods from Central Thailand.- Mem. Fac.
Sci. Kyushu Univ., ser.D, Geol., v.XV, N 1, p.1-22.
Yanagida J. 1967. Early Permian brachiopods from North-Central Thai-
land.- Geol. and Palaeontol. Southeast Asia, v.III, p.4 6-97.
176
ОБЪЯСНЕНИЕ ТАБЛИЦ
Таблица I
Фиг. 1,2. Orthotichia avushensis Sokolskaja................стр. 74
1 - ПИН, № 2774/63; la - спинная створка; 16 - брюшная створка; 1в - раковина сбоку; 1г-
лобный край (х 1); Шор-Булак; джульфинский ярус, среднетахтабулакская подсвита; 2 - ПИН,
№ 2774/69; 2а - ядро спинной створки, 26 - ядро брюшной створки (х 1); Кур-Истык 1;
возраст тот же
Фиг. 3-5. Enteletes meridionalis Gemmellaro...............стр. 77
3 - ПИН, № 2774/73; За - спинная створка; 36 - раковина сбоку (х 1); Музду-Булак; воз-
раст тот же; 4 - ПИН, № 2774/75; 4а - ядро спинной створки с частично сохранившейся ра-
ковиной; 46 - брюшная створка (х 1); Кур-Истык 1; возраст тот же; 5 - ПИН, № 2774/62-
ядро молодой раковины; 5а - спинная створка; 56 - брюшная створка (х 1); Кур-Истык 1;
джульфинский ярус, нижнетахтабулакская подсвита
Фиг. 6. Enteletes dzhagrensis Sokolskaja. . ..............стр. 75
ПИН, № 2774/54 - ядро с частично сохранившимся раковинным веществом; 6а - спинная створ-
ка; 66 - брюшная створка; 6в - раковина сбоку (х 2); Кур-Истык 1; джульфинский ярус,
среднетахтабулакская подсвита
Фиг. 7. Enteletella nikschitshi Licharew..................стр. 79
ПИН, № 2774/77; 7а - брюшная створка; 76 - спинная створка; 7в - раковина сбоку (х 1);
Кур-Истык 1; джульфинский ярус, среднетахтабулакская подсвита
Фиг. 8. Parenteletes ruzhencevi Sokolskaja................стр. 80
8 - ПИН, № 2774/51; 8а - брюшная створка; 86 - спинная створка; 8в т раковина сбоку ;
8г - замочный край (х 1); Шор-Булак; возраст тот же
Фиг. 9, 10. СеуегеЦа alpina Schellwien ...................стр. 82
9 - ПИН, № 2774/30; 9а - брюшная створка; 96 - спинная створка; 9в - брюшная арея; 9г -
раковина сбоку (х 1); 10 - ПИН, № 2774/31; 10а - спинная створка; 106 - поверхность при-
растания брюшной створки (х 1); 10в - брюшная арея (х 2); местонахождение и возраст
те же
178
Таблица I
106
179
Таблица II
Фиг. 1-4. Derbyia diversa Reed.................стр, 86
1 - ПИН, № 2774/7; la - спинная створка; 16 - брюшная створка; 1в - раковина сбоку; 1г -
замочный край (х 1); 2 - ПИН, № 2774/12; ядро спинной створки, вид со стороны хмакушки,
(х 1); 3 - ПИН, № 2774/10; ядро спинной створки (х 1); 4 - ПИН, № 2774/8; 4а - спинная
створка (х l)j 46 - скульптура спинной створки (х 5); Куртеке 1; сакмарский ярус - ниж-
неартинский подъярус, базардаринская свита (верхняя часть)
180
Таблица II
181
Таблица III
Фиг. 1-4. Derbyia grandis Waagen........................стр. 84
1 - ПИН, № 2774 /2; спинная створка (х 1); 2 - ПИН, № 2774/1; ядро брюшной створки (х.Ц;
3 - ПИН, N? 2774/3; ядро брюшной створки (х 1); 4 - ПИН, № 2774/4 ; 4а - ядро спинной
створки (х 1); 46 - замочный отросток сверху (х 3); 4в - замочный отросток с внутренней
стороны (х 2); Куртеке 1; сакмарский ярус - нижнеартинский подъярус нерасчлененные, базар-
даринская свита (верхняя часть)
182
Таблица III
183
Таблица IV
Фиг. 1-6. Taeniothaerus rusticus Grunt, sp.nov................стр. 88
1 - голотип - ПИН, Kfe 2228/542; 1а-в - брюшная створка в различных положениях; наружная
скульптура частично повреждена (х 1); 2 - ПИН, № 2228/548 - замочный отросток, под-
держиваемый септой (х 1); 3 - ПИН, № 2228/540 - внутреннее строение спинной створки
(х 1); 4 - ПИН, № 2228/545; 4а - спинная створка ( х 1); 46 - брюшная створка, (*1); 4в-
скульптура спинной створки; поверхностный слой раковины частично поврежден (х 3); 5 -
ПИН, № 2228/551; площадка прикрепления и окружающие ее иглы (юный экземпляр, фрагмент),
(хЗ); 6 - ПИН, № 2228/550; 6а - брюшная створка юного экземпляра (х 1); 66 - макушка
с площадкой прикрепления и ареей, прорезаннной дельтирием ( х5); Куртеке 1; сакмарский
ярус - нижнеартинский подъярус нерасчлененные, базардаринская свита (верхняя часть)
184
Таблица IV
1*12 264
185
Таблица V
Фиг. 1,2. Strophalosiina multicostata Licharew.................стр. 90
1 - ПИН, № 2228/555; la - висцеральный диск брюшной створки; 16 - шлейф брюшной створ-
ки; 1в - раковина сбоку (х 1); 1г - внутреннее строение спинной створки с частично раз-
рушенной септой (хЗ); Кур-Истык 1; джульфинский ярус, среднетахтабулакская подсвита;
2 - ПИН, № 2228/554; 2а - висцеральный диск брюшной створки; 26 - шлейф брюшной створ-
ки; 2в - спинная створка (х 1); Музду-Булак; возраст тот же
Фиг. 3,4, Scacchinella licharewi Grunt, sp. nov................ стр. 92
3 - гоЛОтил— ПИН, № 2228/556; За - брюшная створка, 36 - арея брюшной створки и спинная
створка с частично нарушенной поверхностью; виден отпечаток внутреннего строения; Зв -
раковина сбоку (* 1); Шор-Булак; джульфинский ярус, среднетахтабулакская подсвита; 4 -
ПИН, № 2228/557; 4а - арея брюшной створки; 46 - брюшная створка с обломанным лобным
краем (х 1); местонахождение и йозраст те же
Фиг. 5-7. Richthofenia caucasica Licharew................... стр. 93
5 - ПИН, № 2228/562 - брюшная створка с разных сторон (х 1); 6 - ПИН, № 2228/564 -
арея брюшной створки (х . 1); 7 - ПИН, № 2228/563 - брюшная створка, видна пузырчатая
ткань (х 1); Шор-Булак, джульфинский ярус, среднетахтабулакская подсвита
Фиг. 8,9. Overtoniina mamazairica Grunt, gen.et sp.nov.........стр. 95
8 - голотип - ПИН, № 2228/590, ядро раковины; 8а - брюшная створка (х 1); 86 - спинная
створка ( х 1,5); Игримиюз; дарвазский ярус, кызылджилгинский горизонт, нижний подгори-
зонт; 9 - ПИН, № 2228/591; 9а - висцеральный диск брюшной створки; 96 - замочный край
брюшной створки; 9в - брюшная створка сбоку (х 1); Мамазаир; возраст тот же
Фиг. 10. Krotovia jisuensiformis Sarytcheva.
ПИН, № 2228/520; 10а-в - брюшная створка с разных сторон (х 1); Куталь П джульфинский
ярус, среднетахтабулакская подсвита
Фиг. 11, 12. Marginifera sexcostata Licharew................ стр. 103
11 - ПИН, № 2228/588; Па - висцеральный диск брюшной створки; 116 - шлейф брюшной створ-
ки (х2); 11в - раковина сбоку (х 1); 12 - ПИН, № 2228/587; 12а -висцеральный диск брюш-
ной створки; 126 - шлейф брюшной створки; 1эв - спинная створка с частично разрушенным
раковинным веществом (х 2); Шор-Булак; джульфинский ярус, среднетахтабулакская подсвита
Фиг. 13,14. Probolioi) ia himalayensis (Diener) . ... стр. 100
13 - ПИН, № 2228/528; 13а-в - брюшная створка с разных сторон (* 1); 14 - ПИН, №
2228/526 - отпечаток спинной створки с сохранившимся маргинальным валиком (х 1); Чако-
бай; дарвазский ярус, кызылджилгинский горизонт, средний подгоризонт
186
Таблица V
187
Таблица VI
Фиг. 1-4. Reticulatia pamirica Grunt, sp.nov...............стр. 97
1 - голотип - ПИН, № 2228/582; la - брюшная створка; 16 - спинная створка; раковинное
вещество частично повреждено, видны мускульные отпечатки и септа (х 1); 1в - раковина
сбоку ( х 1); 2 - ПИН, № 2228/583; 2а - брюшная створка; 26 - длинный замочный край и
слабо свернутое ухо; 2в - спинная створка (х 1); 3 - ПИН, № 2228/585 - внутренняя по-
верхность брюшной створки (х 1); 4 - ПИН, № 2228/586 - юный экземпляр; 4а - брюшная
створка; 46 - спинная створка (х 1); Куртеке 1; сакмарский ярус - нижнеартинский подъярус
нерасчлененные, базардаринская свита (верхняя часть)
188
Таблица VI
189
Таблица VII
Фиг. 1. Reticulatia pamirica Grunt, sp. novk....................стр. 97
ПИН, № 2228/580; la - отпечаток спинной створки с частично сохранившейся раковиной; виден
замочный отросток, изогнутые кардинальные валики и обломанная септа; 16 - замочный отрос-
ток, вид сверху (х 1); Куртеке 1; сакмарский ярус - нижнеартинский подъярус нерасчленен-
ные, базардаринская свита (верхняя часть)
Фиг. 2-5. Terrакеа nalivkini Grunt, sp. nov.................. стр. 105
2 - голотип - ПИН, № 2228/593; 2а - спинная створка; 26 - брюшная створка; 2в - раковина
сбоку; 2г - замочный край (х 1); 3 - ПИН, К? 2228/597; За - брюшная створка; 36 - спин
ная створка; Зв - замочный край (X 1); 4 - ПИН, № 2228/594 - внутреннее строение спин-
ной створки (х 2); 5 - ПИН, №2228/595; 5а - брюшная створка, 56 - спинная створка
(х 1); Южный Ак-Архар; дарваэский ярус, кубергандинская свита (?)
Фиг. 6-3. Globiella ? rossiae Fantini Sestini...................стр. 107
6 -ПИН, № 2228/570 - брюшная створка (х 1); 7 - ПИН, № 2228/571; 7а-в - брюшная створ-
ка с разных сторон (х 1); 8 - ПИН, № 2228/572 - отпечаток спинной створки с частично
сохранившейся раковиной по замочному краю (х 1); Куртеке 2; дарваэский ярус, кызылджил-
гинский горизонт, нижний подгоризонт
190
Таблица VII
191
Таблица VIII
Фиг. 1,2. Cancrinella altissima King.................стр. 106
1 - ПИН, № 2228/567; la-в - брюшная створка с разных сторон (х 1); 2 - ПИН, № 2228/
568 - спинная створка (х 1); Куртеке 1, сакмарский ярус - нижнеартинский подъярус не-
расчлененные, базардаринская свита (верхняя часть)
Фиг. 3. Anidanthus sinosus (Huang)...................стр. 107
ПИН, №2228/599; За-в - брюшная створка с разных сторон ( х 1); Кур-Истык 1; джульфински
ярус, среднетахтабулакская подсвита
Фиг, 4,5. Compressoproductus mongolicus (Diener)..........стр. 109
4 - ПИН, № 2228/521; 4а,б - брюшная створка с разных сторон (х 1); 4в - скульптура
створки (х 4); Кутал^ 2; возраст тот же; 5 - ПИН, К? 2228/600 - юный экземпляр; 5а,б-
брюшная створка с разных сторон; Кур-Истык 1; возраст тот же
Фиг. 6. Haydenella tumida (Waagen).................. стр. Ill
ПИН, № 2228/513 - брюшная створка (х 1); Куталь 2; возраст тот же
Фиг. 7,8. Haydenella dutkevitchi Grunt, sp.nov.........стр. 110
7 - голотип - ПИН, № 2228/505; 7а - брюшная створка (х 1); 76 - замочный край брюш-
ной створки с иглами (х 2); 8 - ПИН, № 2228/510; 8а-б - брюшная створка с разных сто-
рон (х 1); Куталь 2; возраст тот же
Фиг. 9,10. Leptodus sp...............................стр. Ill
9 - ПИН, № 2228/603 - брюшная створка с поврежденным наружным раковинным слоем
(х 1); Куталь 2, возраст тот же; 10 - ПИН, № 2228/602 - брюшная створка без наружного
раковинного слоя (х 1); Кур-Истык 1; возраст тот же
Фиг. 11, 12. Camerisma murgabica Grunt, sp.nov.........стр. 124
11 - голотип- ПИН, № 2228/215; 11 а - брюшная створка; 116 - спинная створка; 11в - лоб-
ный край; 11г - раковина сбоку; 12 - ПИН, № 2228/216; 12а - брюшная створка; 125 -
спинная створка; 12в -раковина сбоку; 12г - лобный край (х 1); Чакобай; дарвазский ярус
кызылджилгинский горизонт, средний подгоризонт
Фиг. 13,14. Stenoscisma armenica Sokolskaja............стр. 128
13 - ПИН, № 2228/167; 13а - брюшная створка; 136 - спинная створка; 13в - лобный крал;
13г - раковина сбоку (х 1); 14 - ПИН, № 2228/168; 14а - брюшная створка; 146 - спинная
створка; 14в - раковина сбоку; 14г - лобный край (х 1); Кур-Истык 1; джульфинский яр>с,
среднетахтабулакская подсвита
Фиг. 151 Camarophorinella leveni Grunt, sp. nov........стр. 126
Голотип - ПИН, № 2228/161; 15а - брюшная створка; 156 - спинная створка; 15в - ракови-
на сбоку; 15г - лобный край (х 1); Куталь 2; возраст тот же
Фиг. 16-18. Uncinunellina timorensis (Beyrich )........ стр. 113
16 - ПИН, № 2228/7; 16а - брюшная створка; 166 - спинная створка; 16в - раковина сбок.\;
16г - лобный край (х 1); Кур-Истык 1; возраст тот же; it - ПИН, 2228/19 - юный
экземпляр; 17а - брюшная створка; 17 б - спинная створка; 17в - лобный край (х 1); М> »•
ду-Булак; возраст тот же; 18 - ПИН, № 2228/18 - брюшная створка со шлейфом (х 2);
Шор-Булак; возраст тот же
192
Таблица VIII
’Д13 264
193
Таблица IX
Фиг. 1. Wellerella arthaberi (Tschemyschew)......................стр. 120
ПИН, № 2228/98; la - брюшная створка; 16 - спинная створка; 1в - раковина сбоку; 1г - лоб-
ный край (х 1); Кур-Истык 1; джульфинский ярус; среднетахтабулакская подсвита
Фиг. 2. Gerassimovia abalakovi Grunt, sp.nov.....................стр. 116
Голотип - ПИН, № 2228/170, 2а - брюшная створка; 26 - спинная створка; 2в - раковина
сбоку; 2г - лобный край (х 1); Куталь 2; возраст тот же
Фиг. 3-5. Gerassimovia bactriana Grunt, sp. nov..................стр. 118
3 - голотип - ПИН, № 2228/178; За - спинная створка; 36 - брюшная створка; Зв - лобный
край; Зг - раковина сбоку (* 1); 4 - ПИН, № 2228/183 ; 4а - спинная створка; 46 - ядро
брюшной створки (х 1); 5 - ПИН, № 2228/185 - юный экземпляр; 5а - спинная створка;56 -
брюшная створка (х 1); Куртеке 1; сакмарский ярус - нижнеартинский подъярус нерасчленен-
ные, базардаринская свита (верхняя часть)
Фиг. 6. Gerassimovia pamirica Grunt, sp.nov......................стр. 115
Голотип - ПИН, N? 2228/36; 6а - брюшная створка; 66 - спинная створка; 6в - лобный край;
6г - раковина сбоку (х 1); Кур-Истык 1; джульфинский ярус, среднетахтабулакская подсвита
Фиг. 7,8. Botoria garmoensis Grunt, gen. et sp. nov..............стр. 120
7 - голотип - ПИН, № 2228/200; 7a - спинная створка; 76 - брюшная створка; 7в - ракови-
на сбоку; 7г - лобный край (х 1); 8 - ПИН, № 2228/201; 8а - брюшная створка; 86 - спин-
ная створка; 8в - лобный край; 8г - раковина сбоку (х 1); Мамазаир; дарвазский ярус,
кызылджилгинский горизонт, нижний подгоризонт
Фиг. 9. Spiriferella sp..........................................стр. 133
ПИН, X? 2774/192; брюшная створка с разных сторон (х 1); Чакобай; дарвазский ярус, кызыл-
джилгинский горизонт, средний подгоризонт
Фиг. 10—13. Neospiter joharensis (Diener)........................стр. 132
10 - ПИН, № 2774/162; 10 а - спинная створка; 10 б- раковина сбоку (х 1); 11 - ПИН, №
2774/165; 11а,б -брюшная створка в различных положениях; 12 — ПИН, № 2774/164 — ядро
брюшной створки;13 - ПИН, № 2774/166—скульптура (с отпечатка) (х 10); Кастанат-Джил-
га 1; сакмарский ярус - нижнеартинский подъярус нерасчлененные, базардаринская свита (верх-
няя часть)
194
Таблица IX
195
13 264
,Таблица X
Фиг. 1.Punctocyrtella spinosa Plodowski.........................стр. 130
ПИН, № 2774/150; la - брюшная створка; 16 - арея брюшной створки и макушка спинной;
1в - лобный край; 1г - спинная створка (х 1); Куртеке 1; сакмарский ярус - нижнеартинс-
кий подъярус нерасчлененные, базардаринская свита (верхняя часть)
Фиг. 2-5. Kitakamithyris petaliformis Pavlova,sp. nov...........стр. 136
2 - голотип - ПИН, № 2774/174 ; 2а-брюшная створка; 26 - спинная створка; 2в - раковина
сбоку, (к 1); 3 - ПИН, № 2774/177; За,б - ядро брюшной створки в различных положениях
(к 1); 4 - ПИН, № 2774/175; 4а,б - брюшная створка в различных положениях; 5 - ПИН,
№2774/176 - микроскульптура брюшной створки; Кастанат-Джилга 1, возраст тот же
196
Таблица X
197
Таблица XI
Фиг. 1. Permophricodothyris caroli (Gemmellaro)................стр. 134
ПИН, № 2774/173; la - брюшная створка; 16 - раковина сбоку; 1в - спинная створка (х 1);
Кур-Истык 1; джульфинский ярус, нижнетахтабулакская подсвита
Фиг. 2-5. Martinia uralica Tschemyschew....................... стр. 142
2 - ПИН, № 2774/100; 2а-в - брюшная створка с различных сторон (х 1); 3 - ПИН, N?
2774/110 - спинная створка (х 1); 4 - ПИН, № 2774/116 - спинная створка (х 1); 5 -
ПИН, К? 2774/118; 5а, б - ядро брюшной створки с различных сторон (х 1); Кастанат-Джил-
га 1; сакмарский ярус - нижнеартинский подъярус нерасчлененные, базардаринская свита
(верхняя часть)
Фиг. 6-9. Martinia tschernyschewi Grunt, sp. nov...............стр. 144
6 - голотип - ЦНИГРмузей, № 1679/303; 6а - спинная створка; 66 - раковина сбоку; 6в -
лобный край;Урал гора Улу-Тау; ассельский ярус; 7 - ПИН, К? 2774/132; 7а - брюшная створ-
ка; 76 - спинная створка; 7в - раковина сбоку; 7г - лобный край (х 1); 8 - ПИН, № 2774/
133; 8а,б - брюшная створка с разных сторон (к 1); 9 - ПИН, № 2774/134 спинная створка
(х 1); Мамазар; дарваэский ярус, кызылджилгинский горизонт, нижний подгоризонт
198
Таблица XI
199
Таблица XII
Фиг. 1,2. Martinia tschakobaica Grunt, sp. nov.................стр. 145
1 - голотип - ПИН, № 2774/127; la - брюшная створка; 16 - спинная створка; 1в - ракови-
на сбоку; 1г - лобный край (х 1);
2 - ПИН, № 2774/129 ; 2а - брюшная створка (х 1); 26 - внутренняя поверхность брюшной
створки (х 2); Чакобай; дарвазский ярус, кызылджилгинский горизонт, средний подгоризонт
Фиг. 3,4. Martinia triquetra Gemmellaro.................... . стр. 147
3 - ПИН, № 2774/119; За - спинная створка; 36 - брюшная створка; Зв - раковина сбоку;
Зг - замочный край; Зд - лобный край; 4 - ПИН, № 2774/120 - ядро спинной створки (х 1);
Игримиюз; мургабский ярус, нижнеганская (?) подсвита
Фиг. 5,6. Мartinia warthiWaagen................................ стр. 141
5 - ПИН, № 2774/140; 5а - брюшная створка; 56 - спинная створка; 5в - раковина сбоку;
5г - лобный край (х 1); Кур-Истык 1; джульфинский ярус, среднетахтабулакская подсвита;
6- ПИН, № 2774/136; 6а - спинная створка; 66 - брюшная створка; 6в - лобный край (х 1);
Куталь 1; возраст тот же
Фиг. 7,8. Martinia rupicola Gemmellaro.........................стр. 139
7 - ПИН, № 2774/122; 7а - спинная створка; 76 - брюшная створка; 7в - раковина сбоку;
7г - лобный край (х 1); 8 - ПИН, № 2774/123 ; 8а - ядро спинной створки; 86 - ядро брюш-
ной створки (х 1); Джилга-Кочусу; джульфинский ярус, нижнетахтабулакская подсвита
200
Таблица XII
201
Таблица XIII
Фиг. 1-5. Cleiothyridina ailakensis (Reed).............................стр. 148
1 - ПИН, № 2774/170; la - спинная створка; 16 - брюшная створка; 1в - раковина сбоку;
1г- лобный край (х 1); 2 - ПИН, № 2774/167 - ядро спинной створки (х 1); Куртеке 1;
сакмарский ярус _ нижнеартинский подъярус нерасчлененные, базардаринская свита (верхняя
часть) ; 3 - ПИН, № 2774/172 - ядро брюшной створки (х 1); 4 - ПИН, № 2774/169 - брюш-
ная створка с хорошо сохранившейся скульптурой; 5 - ПИН, № 2774/168; 5а - ядро брюшной
створки (х 2); 56 - скульптура (х 10); Кастанат-Джилга 1; возраст тот же
Фиг. 6-8. Notothyris pseudodjoulfensis Licharew, sp. nov...............стр, 157
6 - голотип - ЦНИГРмузей № 1386/2139; 6а - спинная створка, 66 - брюшная створка, 6в -
лобный край (х 1); Северный Кавказ; джульфинский ярус, уруштенский горизонт; 7 - ПИН,
№ 2228/253; 7а - брюшная створка; 76 - спинная створка; 7в - лобный край; 7г - раковина
сбоку (х 1); Кур-Истык 1; джульфинский ярус, среднетахтабулакская подсвита; 8 - ПИН,
№ 2228/636 - юный экземпляр; 8а - спинная створка; 86 - брюшная створка; 8в - лобный
край (х 1); местонахождение и возраст те же
Фиг. 9. Rostranteris (Notothyrina) pontica (Licharew)..................стр. 160
ПИН, № 2228/688; 9а - брюшная створка; 96 - спинная створка; 9в - раковина сбоку; 9г -
лобный край (х 1); Шор-Булак, возраст тот же
Фиг. 10. Heterelasmina pygmaea Grunt, sp. nov..........................стр. 154
Голотип - ПИН, № 2228/718; 10 а - спинная створка (х 1,5); Юв - брюшная створка (х 1,5);
106 - раковина сбоку ( xl); Куталь 2; возраст тот же
Фиг. 11,12. Heterelasmina lepton (Gemmellaro) стр. 153
11 - ПИН, № 2228/758; 11а - спинная створка; 116 - брюшная створка; Ив - раковина сбоку
(х 1); Куталь 1; возраст тот же; 12 -ПИН, № 2228/358; 12а - спинная створка; 126 - брюш-
ная створка; 12в - раковина сбоку; 12г - лобный край (х 1); Ак-Бура; возраст тот же
Фиг. 13- Labaia dmitrievi Grunt, sp. npv..............................-стр. 151
Голотип - ПИН, N? 2228/727; 13а - брюшная створка; 136 - спинная створка; 13в - лобный
край (х 1,5); 13г - раковина сбоку (х 1); Мамазаир; дарвазский ярус, кызылджилгинский
горизонт, нижний подгоризонт
202
Таблица XIII
203
Таблица XIV
Фиг. 1. Enteletes dzhagrensis Sokolskaja..............................стр. 75
ПИН, К? 2774/53, поперечное сечение раковины на расстоянии 2,5 мм от макушки (х 5); Кур-
Истык 1; джульфинский ярус, среднетахтабулакская подсвита
Фиг. 2. Enteletes meridionalis Gemmellaro.............................?7
ПИН, № 2774/59; поперечное сечение раковины на расстоянии 2,9 мм от макушки (х 5); мес-
тонахождение и возраст те же
Фиг. 3. Enteletella nikschitchi Licharew......................... стр. 79
ПИН, N? 2774/79; поперечное сечение брюшной створки на расстоянии 4,5 мм от макушки
(х 3); местонахождение и возраст те же
Фиг. 4. Geyerella alpina Schellwien.............................. стр. 82
ПИН, N? 2774/32; 4а - поперечное сечение брюшной створки; 46 - поперечное сечение спинной
створки (х 3); Музду-Булак; возраст тот же
стр. 113
Фиг. 5. Uncinunellina timorensis (Beyrich)..........................
ПИН, N? 2228/16; поперечное сечение раковины; 5а - на расстоянии 1,5 мм от макушки; 56 -
на расстоянии 2,1 мм от макушки (х 4); Кур-Истык 1; возраст тот же
Фиг. 6. Gerassimovia bactriana Grunt, sp. nov.................... стр. 118
ПИН, N? 2228/205; поперечное сечение брюшной створки на расстоянии 3,4 мм от макушки
(х 3,5); Кастанат-Джилга 1; сакмарский ярус - нижнеартинский подъярус нерасчлененные,
базардаринская свита (верхняя часть)
Фиг. 7. Wellerella arthaberi (Tschemyschew)...................
ПИН, N? 2228/96; поперечное сечение раковины на расстоянии 0,3 мм от макушки (х 5); Кур-
Истык 1, джульфинский ярус, среднетахтабулакская подсвита
Фиг. 8. Stenoscisma armenica Sokolskaja стр. 128
ПИН, N? 2228/169; поперечное сечение раковины; 8а - на расстоянии 1,9 мм от макушки; 8 6-
на расстоянии 2,7 мм от макушки (х 4,5); Шор-Булак; возраст тот же
Фиг. 9. Camarophorinella leveni Grunt, sp. nov стр. 126
ПИН, К? 2228/159; iоперенное сечение раковины; 9а - на расстоянии 0,6 мм от макушки;96 -
на расстоянии 1 мм от макушки (х 6); Куталь 2; возраст тот же
204
Таблица XIV
7ПЧ
Т а блица XV
Фиг. 1. Camerisma murgabica Grunt, sp. nov.........................стр. 124
ПИН, № 2228/219; поперечное сечение раковины; la - на расстоянии 2 мм от макушки; 16 -
на расстоянии 2,1 мм от макушки; 1в - на расстоянии 2,5 мм от макушки (х 4,5); Чакобай;
дарваэский ярус, кызылджилгинский горизонт, средний подгоризонт
Фиг. 2. Martinia tschakobaica Grunt, sp.nov........................стр. 145
ПИН, hfe 2774/130; поперечное сечение раковины на расстоянии*4,8 мм от макушки (х 3,5);
местонахождение и возраст те же
Фиг. 3. Martini a uralica Tschemyschew.............................стр. 142
ПИН, № 2774/103; поперечное сечение брюшной створки (х 3,5); Кастанат-Джилга 1; сакмар-
ский ярус - нижнеартинский подъярус нерасчлененные, базардаринская свита (верхняя часть)
Фиг. 4. Cleiothyridina ailakensis (Reed)...........................стр. 148
ПИН, N? 2774/201; поперечное сечение раковины; 4а - на расстоянии 3,5 мм от макушки, 46 -
на расстоянии 5,2 мм от макушки (х 4); местонахождение и возраст те же
Фиг. 5. Punctocyrtella spinosa Plodowski...........................стр. 130
ПИН, № 2774/191; поперечное сечение брюшной створки; 5а - на расстоянии 1,1 мм от макуш-
ки; 56 - на расстоянии 3,3 мм от макушки (х 3); Куртеке 1; возраст тот же
206
Таблица XV
207
Т а б лица XVI
Фиг. 1.Nеоspiriter joharensis (Diener)...............................стр.
ПИН, № 2774/161; поперечное сечение брюшной створки на расстоянии 6,2 мм от макушки
(х 3); Куртеке 1; сакмарский ярус - нижнеартинский подъярус нерасчлененные, базардарин-
ская свита (верхняя часть)
Фиг. 2. Martinia triquetra Gemmellaro.....'.......................... стр.
ПИН, № 2774/121; поперечное сечение раковины в примакушечной части (х 3,5); Игримиюз;
мургабский ярус, нижнеганская (?) подсвита
Фиг. 3,4. Rostranteris (Notothyrina) pontica (Licharew)...............стр. 160
3 - ПИН, № 2228/304; продольное сечение раковины в ее средней части . (х 5); Кур-ИСтык 2;
джульфинский ярус, среднетахтабулакская подсвита; 4 - ПИН, № 2228/306; поперечное сече-
ние раковины;-4а - на расстоянии 0,7 мм от макушки; 46 - на расстоянии 0,9 мм от ма-
кушки; 4в - на расстоянии 1,3 мм от макушки (х 4); местонахождение и возраст те же
Фиг. 5,6. Notothyris pseudodjoul fensis Licharew, sp. nov............стр. 157
5 - ПИН, - 2228/685; 6 - ПИН, № 2228/683; оба - продольные сечения раковины в ее сред-
ней части (к 5); местонахождение и возраст те же
Фиг. 7. Heterelasmina lepton (Gemmellero)...........................стр. 154
ПИН, № 2228/368; продольное сечение раковины в ее средней части (х 4); Музду-Булак; воз-
раст тот же
208
Таблица XVI
209
ОГЛАВЛЕНИЕ
Введение ..........................................................5
Общая часть
Глава 1. История исследования пермских, отложений Юго-Восточного Памира
(В.Ю. Дмитриев)............................................................ 3
Глава 2. Стратиграфический очерк (В.Ю. Дмитриев)..............................
Глава 3. Основные комплексы пермских брахиопод Юго-Восточного Памира
(Т.А. Грунт, В.Ю. Дмитриев)............................................... 32
Глава 4. Закономерности смены комплексов пермских брахиопод Альпийско-Ги-
малайской подобласти Тетиса и их биогеография (Т.А. Грунт).................59
Систематическая часть (Т.А. Грунт)
Класс Articulata........................................................................74
Отряд Orthida........................................................................ 74
Семейство Schizopohoriidae Schuchert............................................... 74
Род Orthotichia Hall et Clarke................................................74
Семейство Enteletidae Waagen.......................................................75
Род Enteletes Fischer Waldheim...............................................75
Род Enteletella Licharew.................................................... /8
Род Parenteletes King...................................................... 80
Отряд Strophomenida..................................................................82
Семейство Meekellidae Stehli...................................................... 82
Род Geyerella Schellwien.....................................................82
Семейство Orthotetidae Waagen .....................................................84
Род Derbyia Waagen...............................................................
Отряд Productida.................................................................... 87
Подотряд Productidina..................................................................
Семейство Aulostegidae Muir-Wood et Cooper.........................................87
Род Taeniothaerus Whitehouse.................................................87
Семейство Chonostegidae Muir-Wood et Cooper........................................89
Род Strophalosiina Licharew................................................ 89
Семейство Scacchinellidae Licharew..................................................91
Род Scacchinella Gemmellaro..................................................91
Семейство Richthofeniidae Waagen...................................................93
Род Richthofenia Kayser •................................................ 93
Семейство Avoniidae Sarytcheva.....................................................94
Род Krotovia Fredericks......................................................94
Семейство Overtoniidae Muir-Wood et Cooper....................................... 94
Род Overtoniina Grunt, gen. nov..............................................94
Семейство Dictyoclostidae Stehli...................................................96
Род Reticulatia Muir-Wood et Cooper..........................................96
Род Costiferina Muir-Wood et Cooper........................................ 99
Семейство Marginiferidae Stehli....................................................99
Род Probolionia Cooper ......................................................99
Род M arginifera Waagen......................................................102
Семейство Linoproductidae Stehli ..................................................104
Род Terrakea Booker..................................................... 104
Род Cancrinella Fredericks............................................. , . 106
Род Globiella Muir-Wood et Cooper....................................... 107
Род Anidanthus Whitehouse................................................107
Род Compressoproductus Sarytcheva........................................108
Семейство Chonete 11 idae Licharew.............................................. ... 110
Род Haydenella Heed............................................................HO
Семейство Lyttoniidae Waagen........................................................Ill
Род Leptodus Kayser............................................................HI
Отряд Rhynchonellida...................................................................112
Семейство (lypothyridinidae Kzpnsnickaja............................................112
Род Uncinunellina Grabau......................................................112
Род Cerassimovia Licharew.....................................................115
Род Boloria Grunt, gen. nov...................................................119
Семейство Wellerellidae Licharew....................................................121
Род Wellerella Bunbar et Condra...............................................121
Семейство Atriboniidae Grant........................................................123
Род Camerisma Grant...........................................................124
Род Gamarophorine/la Licharew.................................................126
Семейство Stenoscismatidae Gehlert..................................................128
Род Stemoscisma Conrad........................................................128
Отряд Spiriferida......................................................................130
Семейство Synngothyridae Fredericks.................................................130
Pon Punctocyrtella Plodowski................................................ 130
Семейство Spiriferidae King.........................................................132
Род Neosnirifer Fredericks....................................................132
Род Spirifereella Tschemyschew................................................132
Семейство Reticulariidae Waagen.....................................................134
Род Permophricodothyris Pavlova...............................................134
Род Kitakamithyris Minato..................................................... . 135
Семейство Martiniidae Waagen........................................................138
Род Martinia Me Coy...........................................................138
Incerti ordinis.....................................................................148
Семейство Athyrididae .McCoy........................................................148
Род Cleiothyridina Buchman............................................* . . • • 148
Отряд Terebratulida....................................................................150
Семейство Labaidae Licharew.........................................................150
Род Labaia Licharew..........................................................150
Семейство Heterelasminidae Licharew.................................................152
Род Heterelasmina Licharew...................................................152
Семейство Centronellidae Waagen.................................................... 156
Род Notothyris Waagen........................................................156
Род Rostranteris Gemmellaro..................................................159
Литература ............................................................................162
Объяснение таблиц
177
Татьяна Александровна Грунт; Виктор Юрьевич Дмитриев
ПЕРМСКИЕ БРАХИОПОДЫ ПАМИРА
Труды Палеонтологического института
Том 136
Утверждено к печати Палеонтологическим институтом
Академии наук СССР
Редактор Девяткин Е.В.
Редактор издательства Ванин В.С.
Художественный редактор Литвак С.А.
Технический редактор Сурикова Л.М.
Подписано в печать 14/1У-73 г. Формат 70 х 108 1/16
Усл. печ. л. 19J6 Уч.-изд. л. 20,63 ( 20,01 + 0,62 вкл.)
Т -04832 Бумага офсетная № 1 Тираж 750
Цена 2 р. 41 к. Тип, зак. 264
Книга издана офсетным способом
Издательство 'Наука*,
Москва, ГСП, 103717, К-62, Подсосенский пер», 21.
1-я типография издательства *Наука*
Ленинград, 199034, 9-я линия, 12.