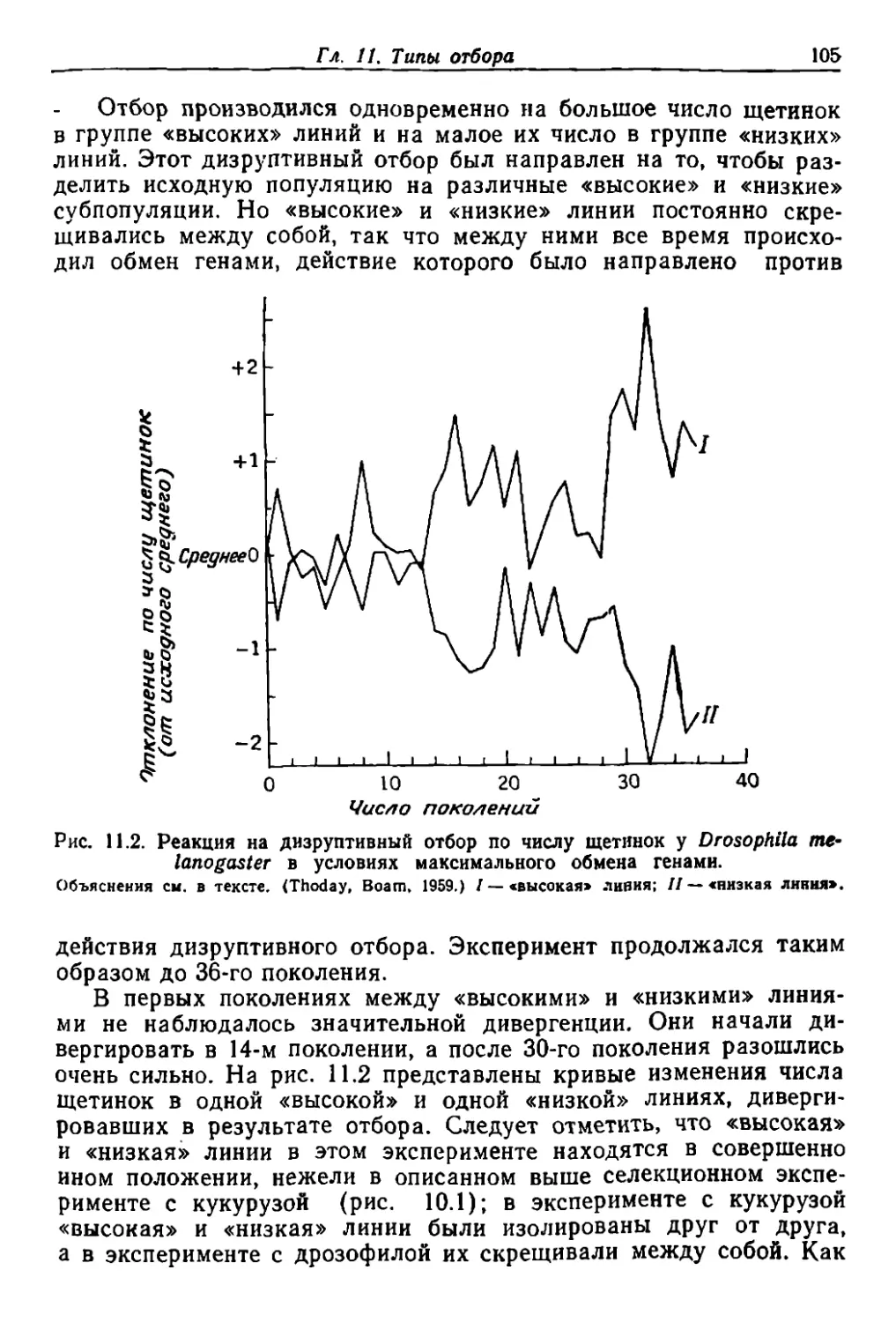
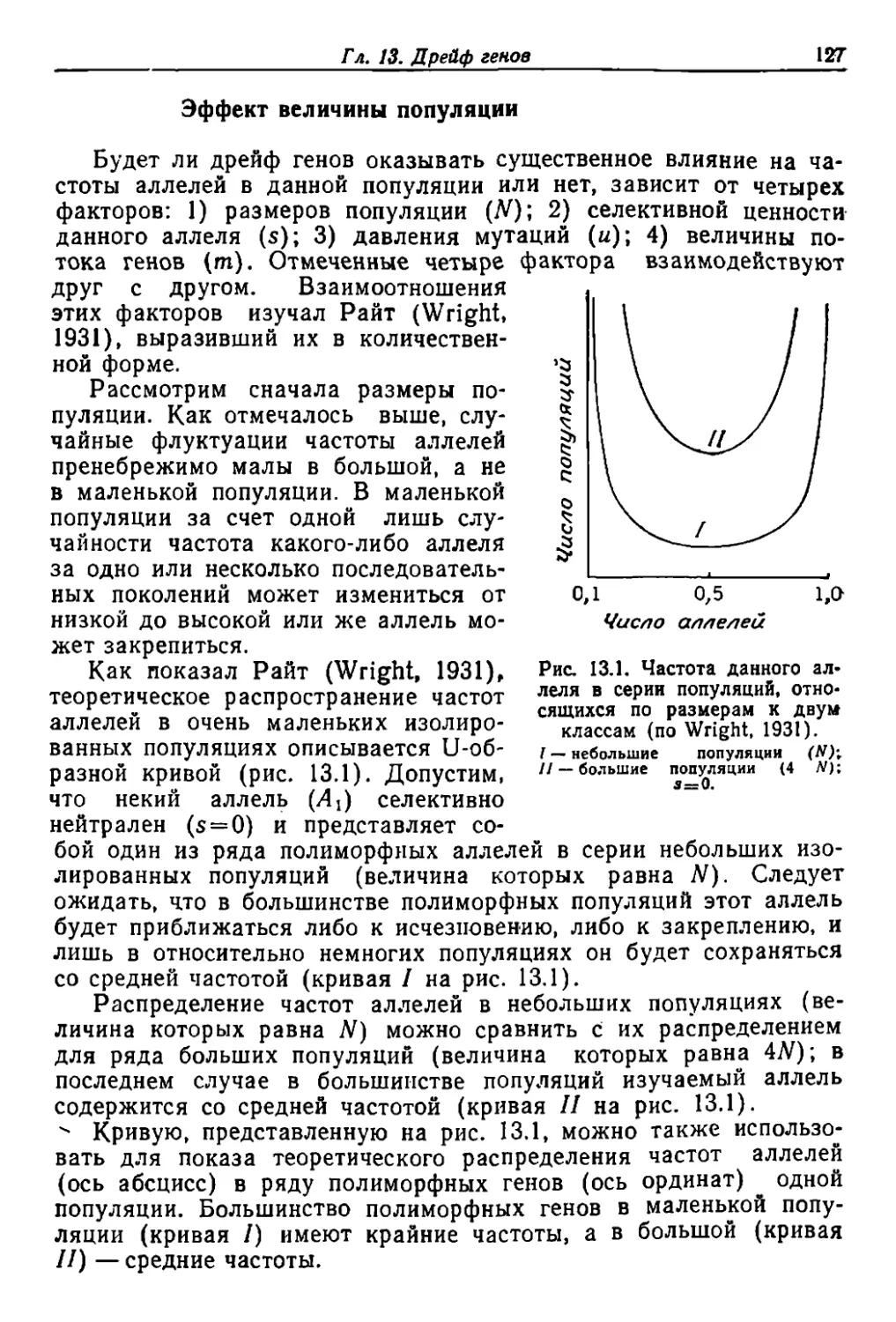
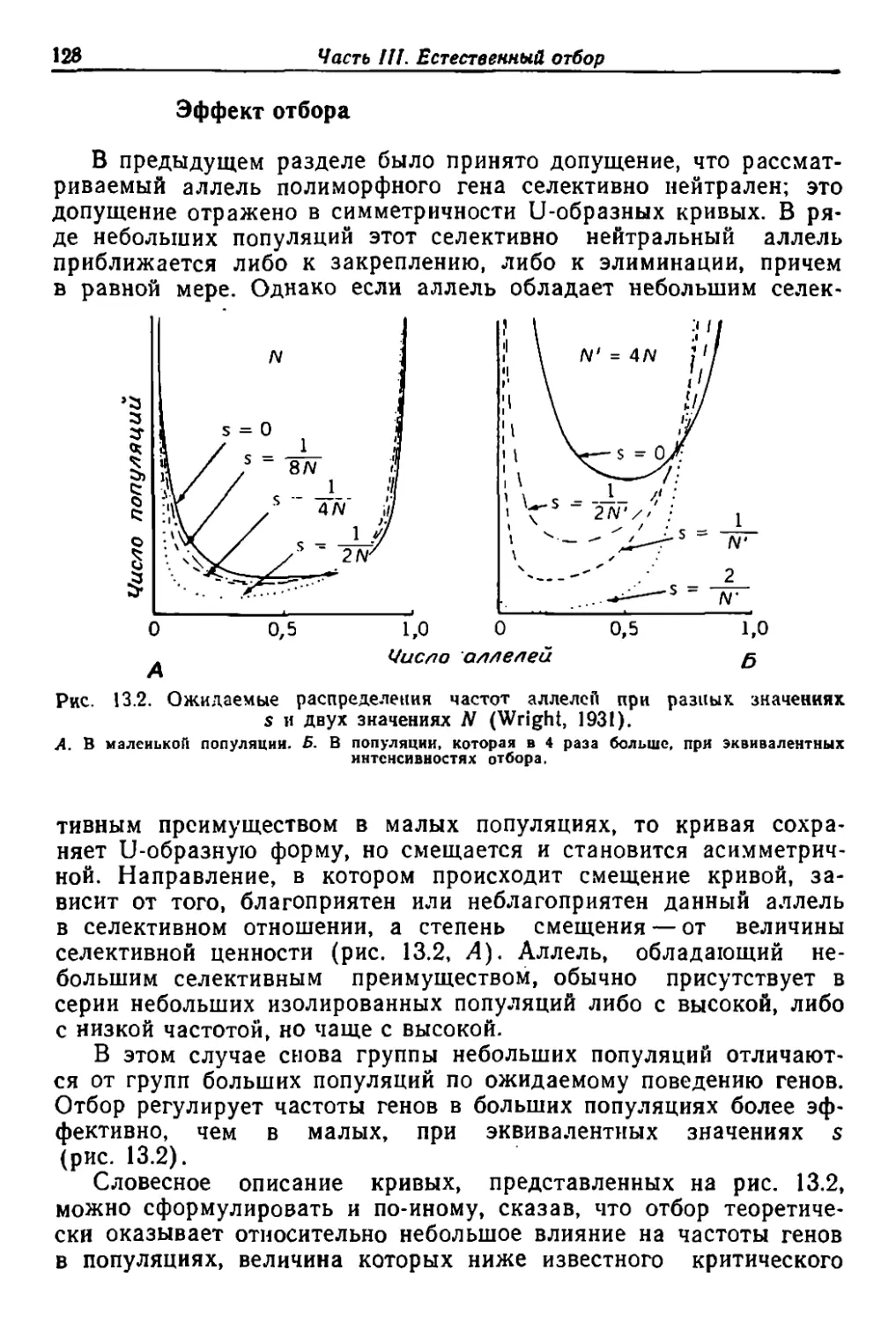
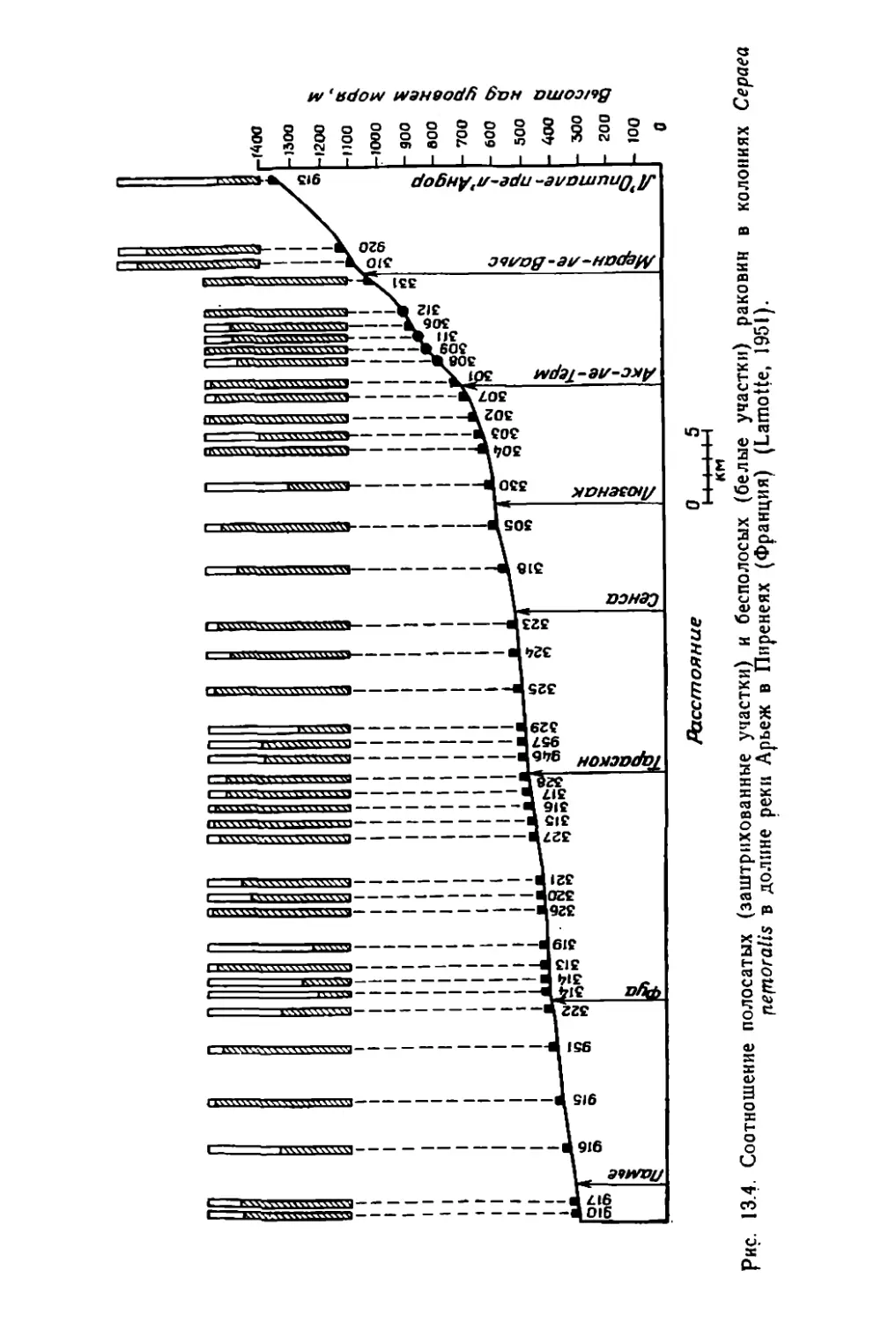
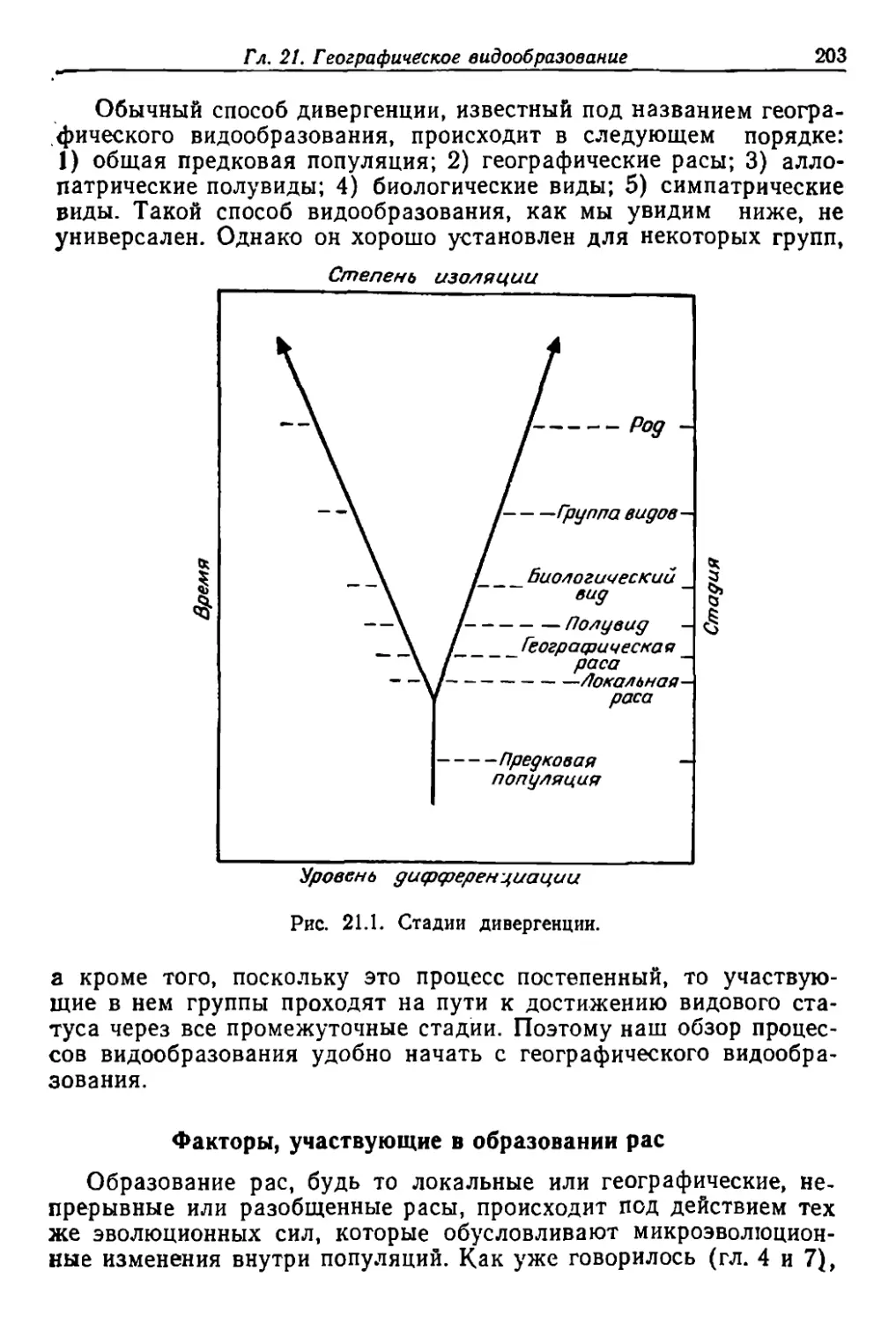
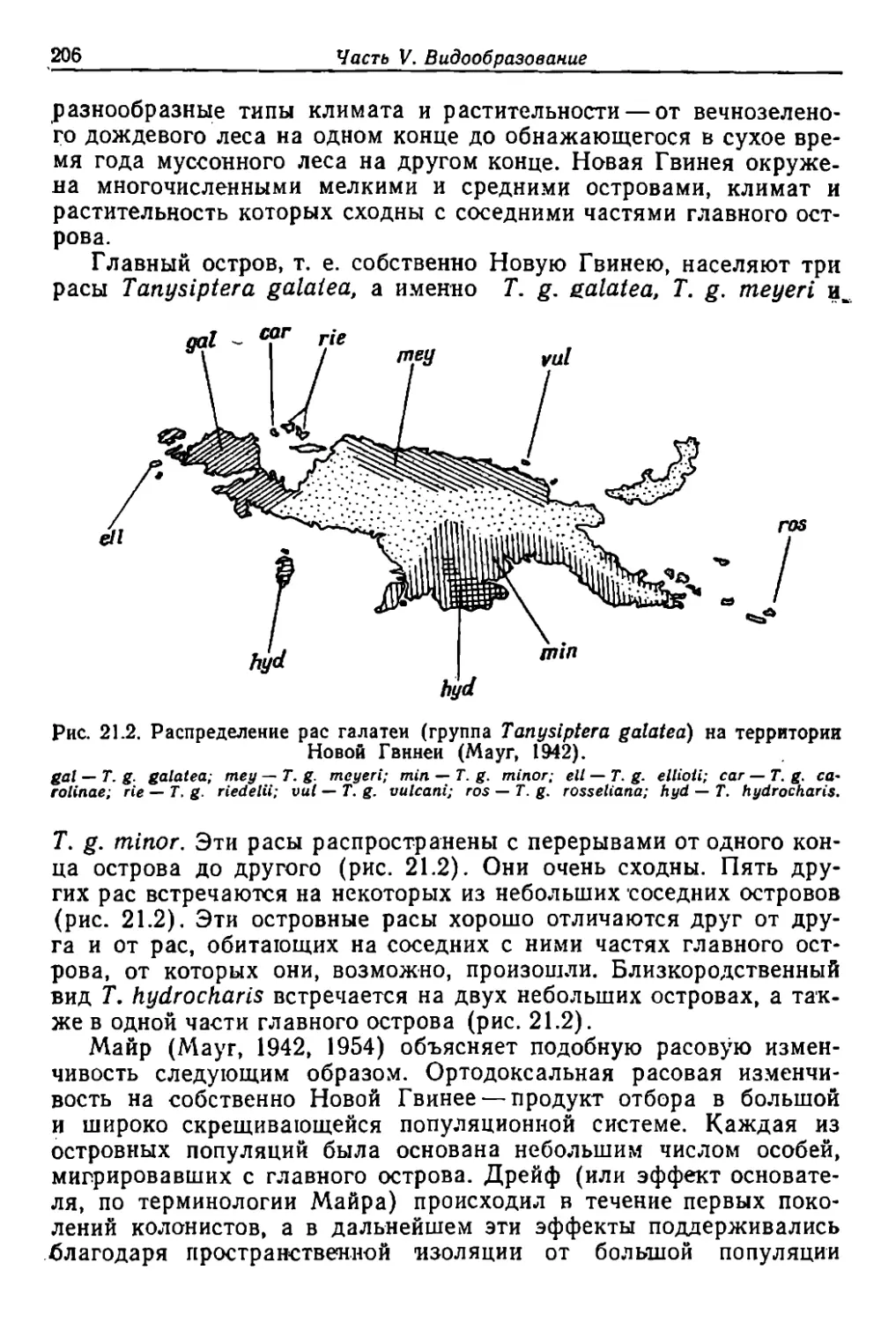
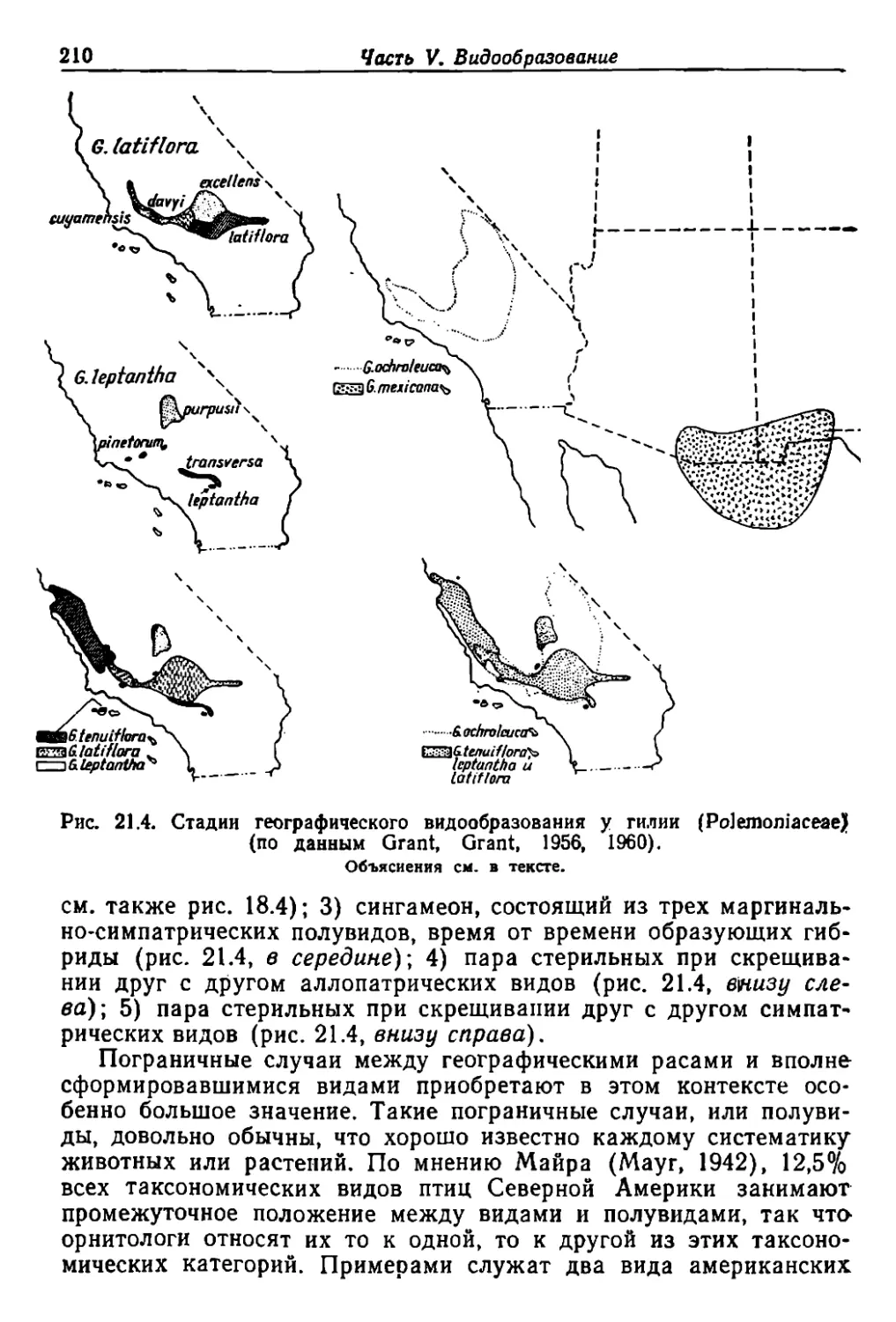
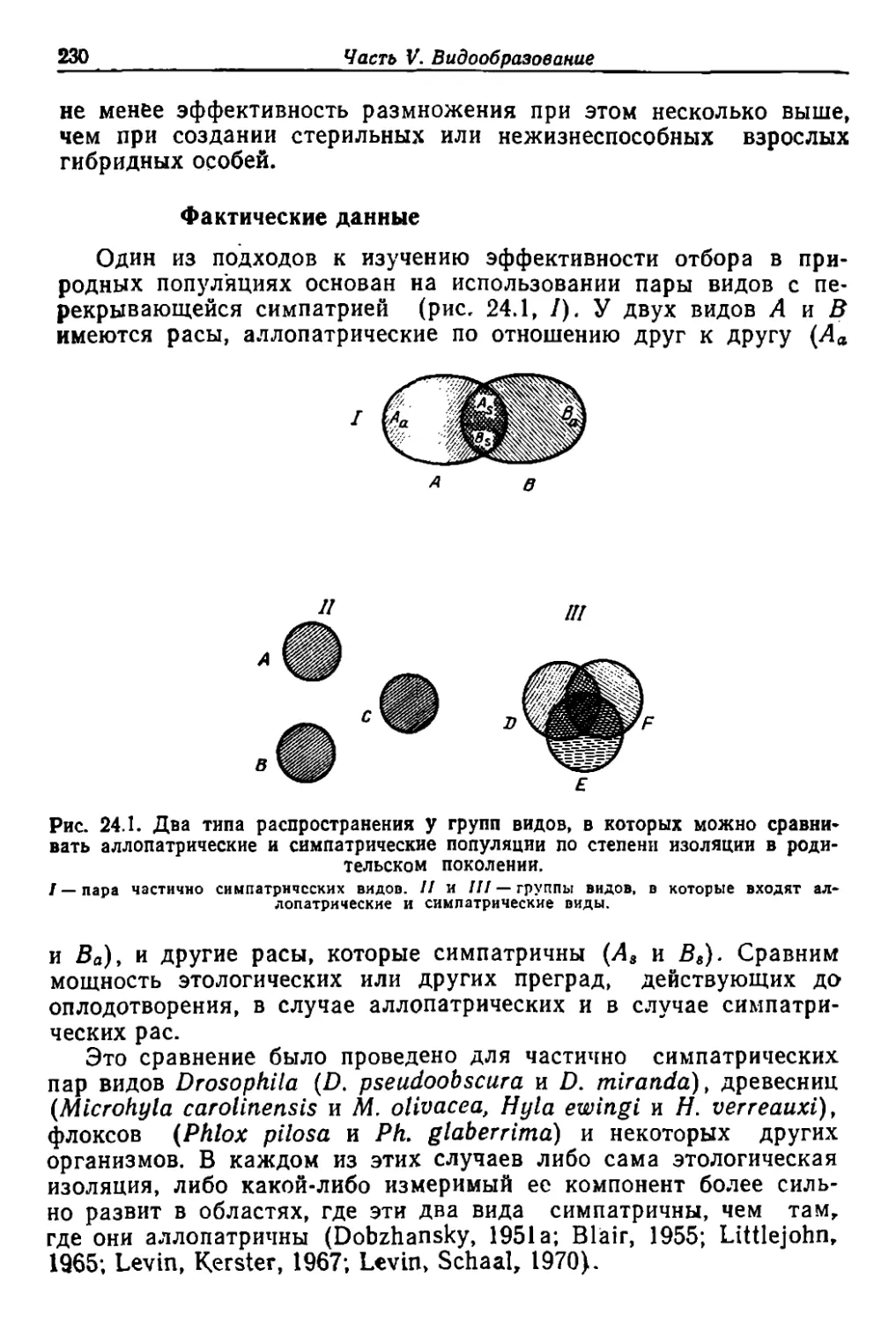
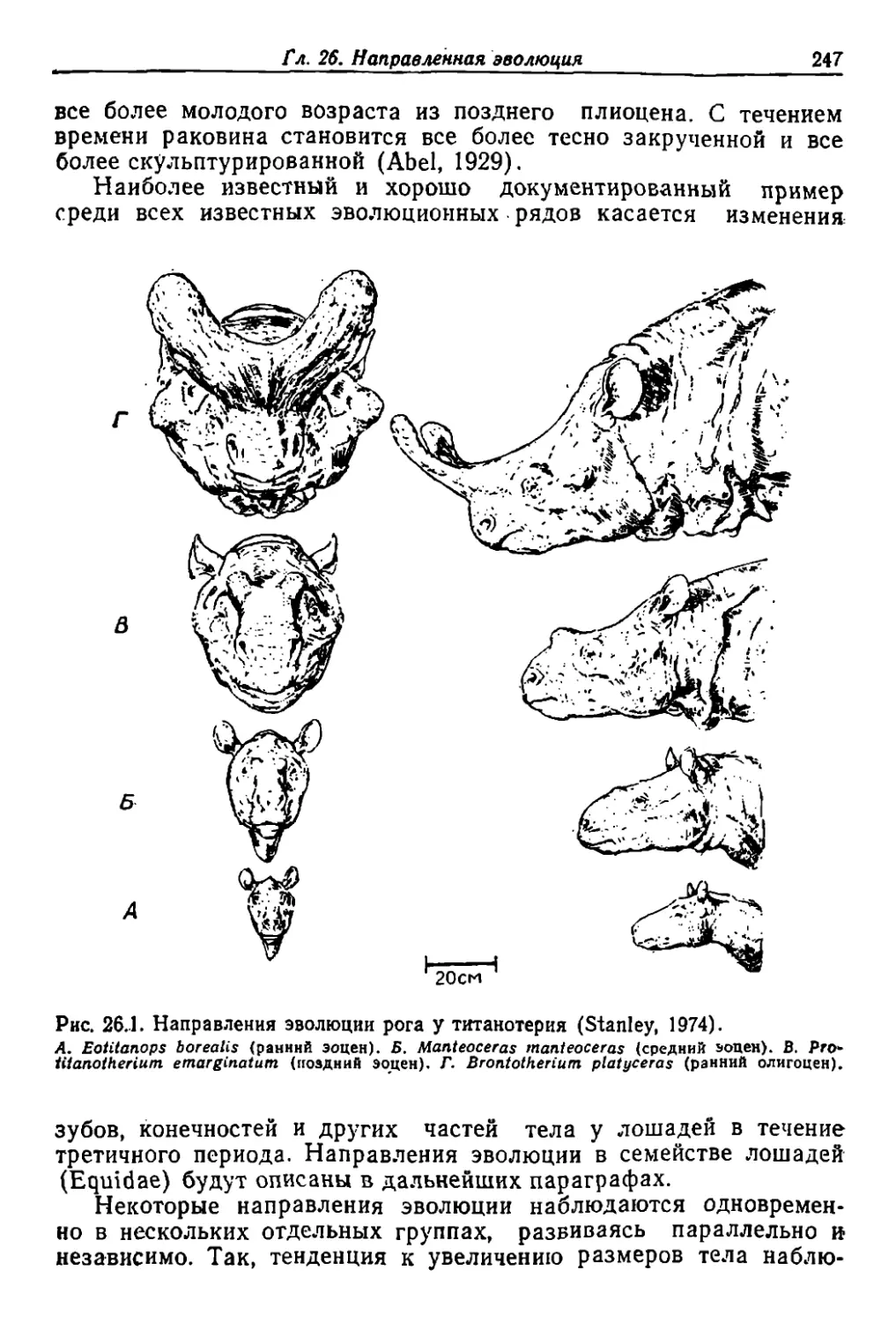
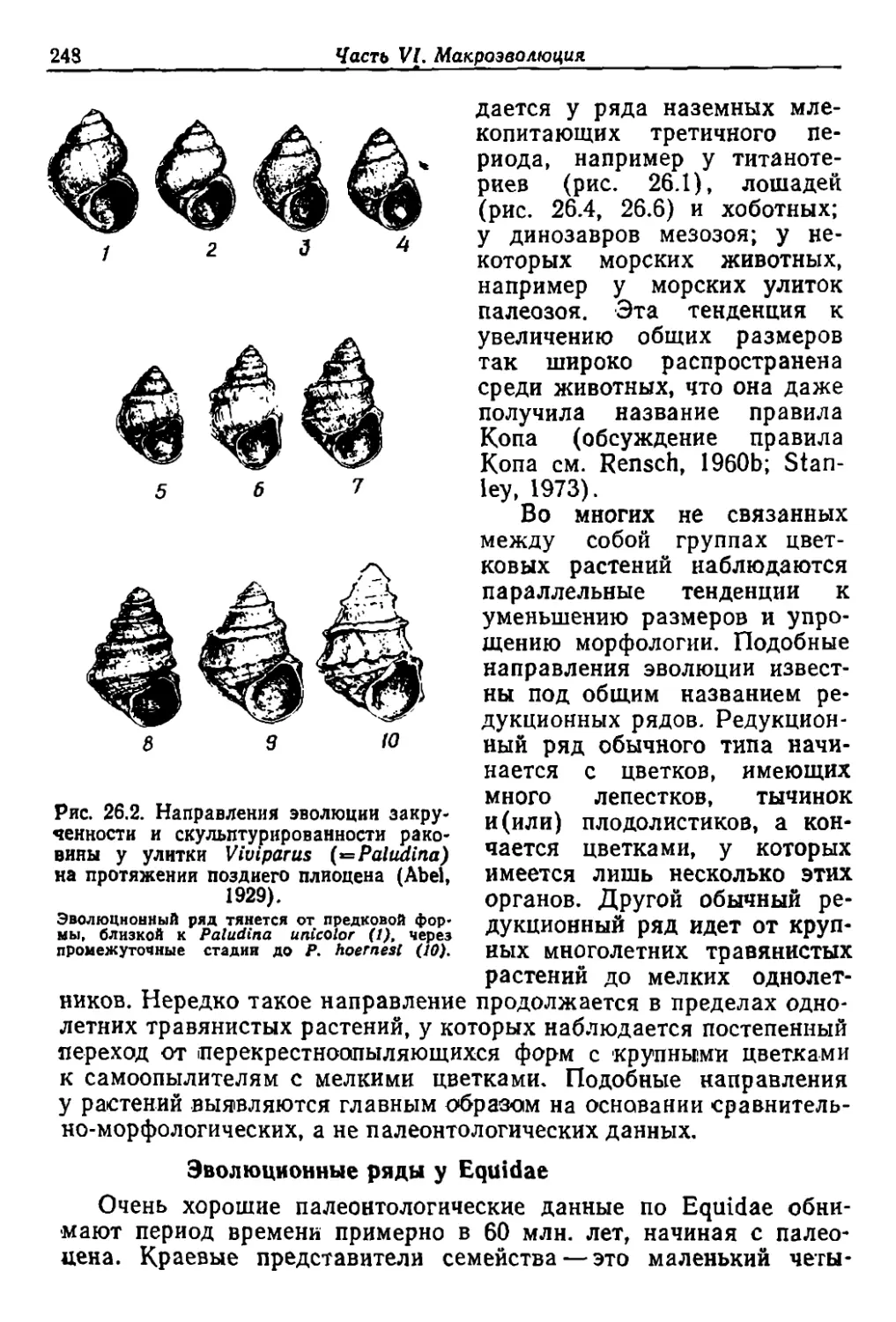
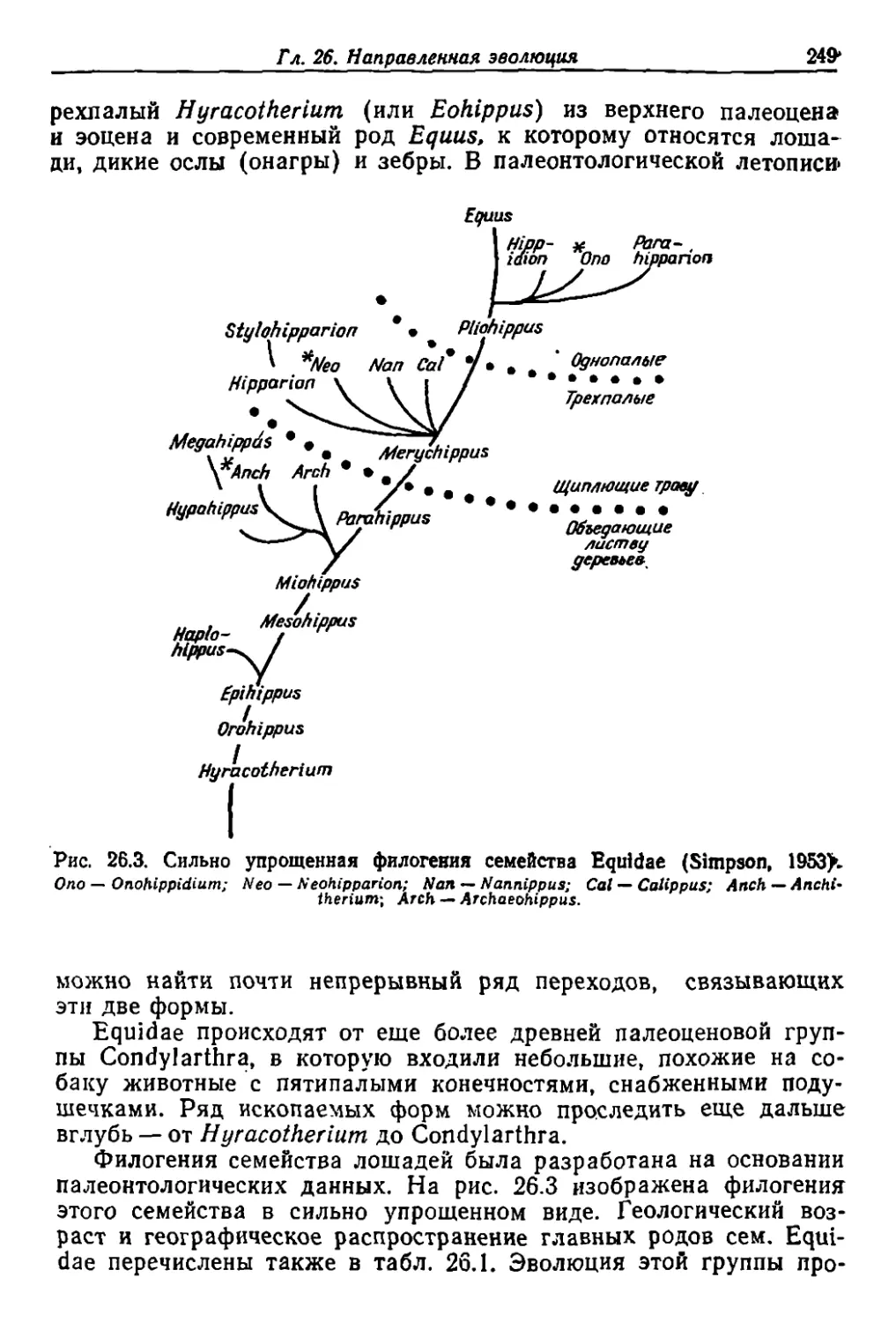
/






















































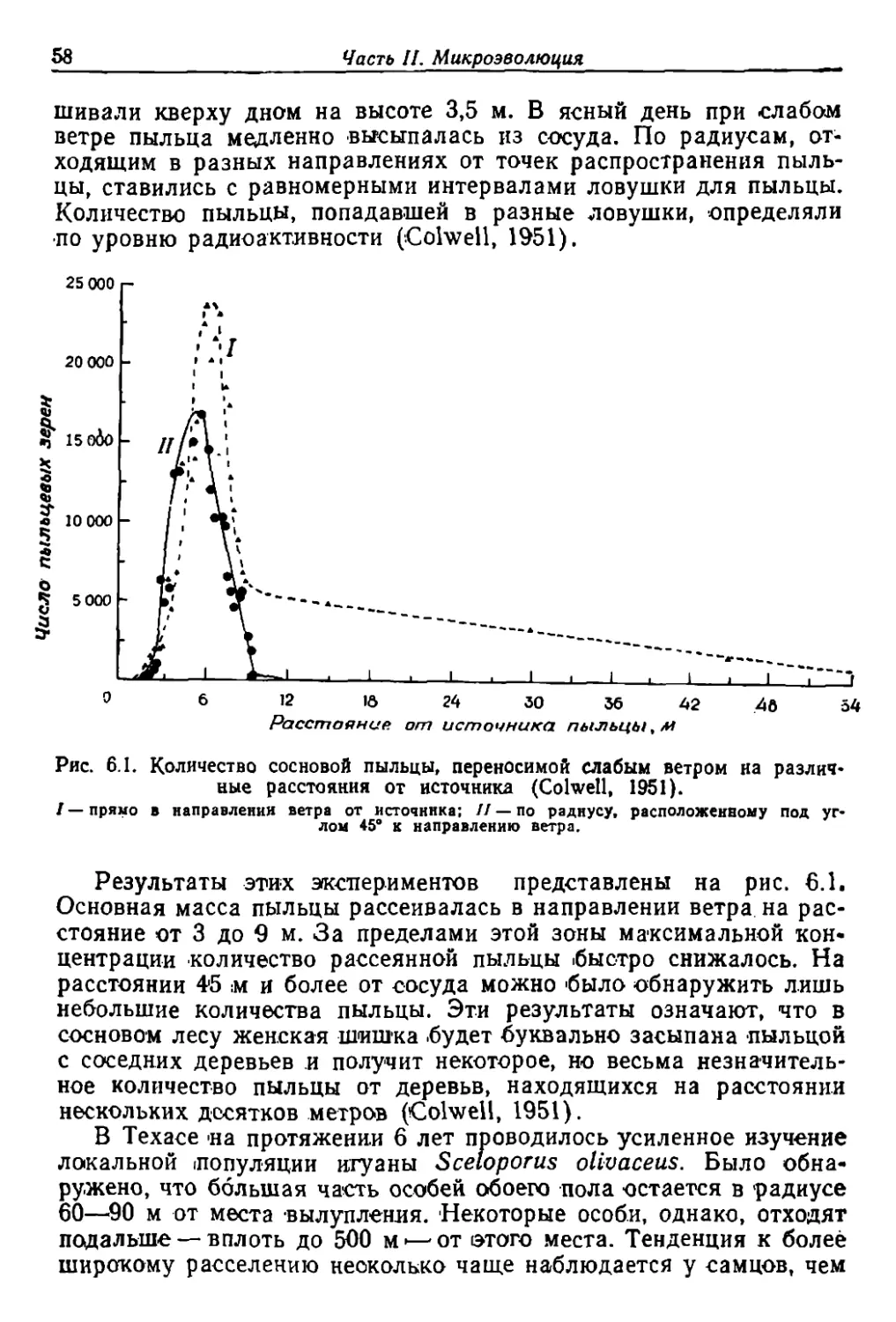
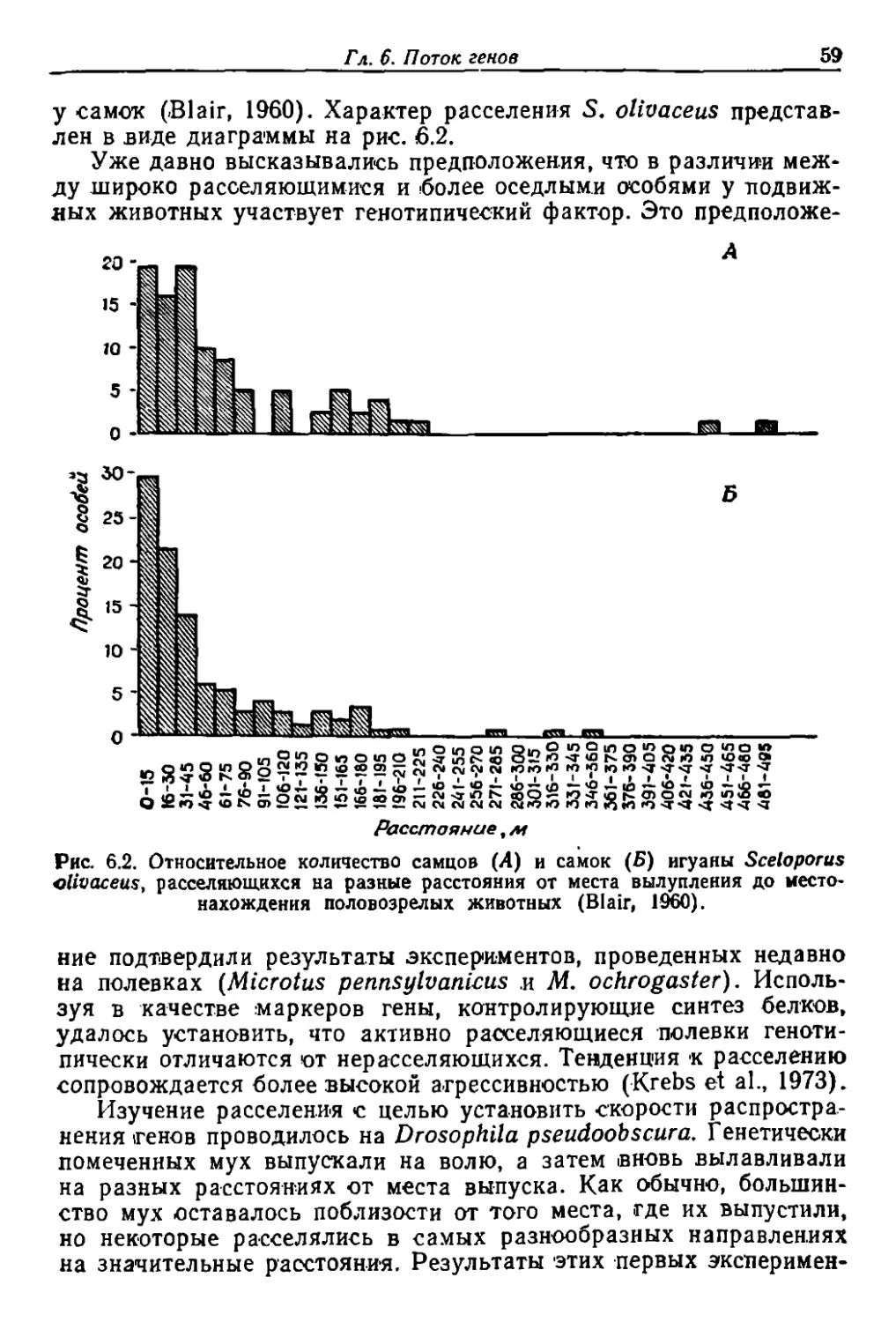


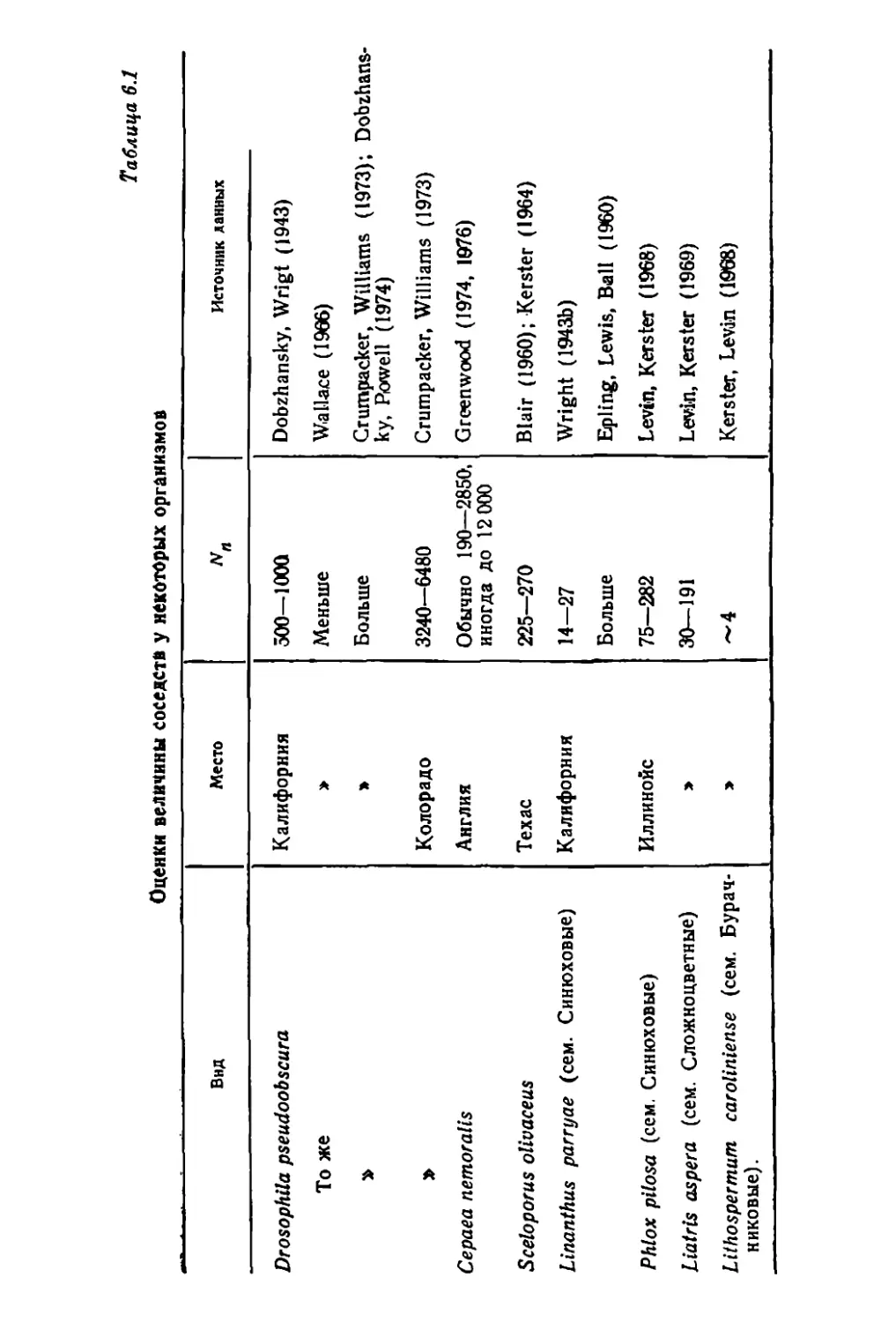


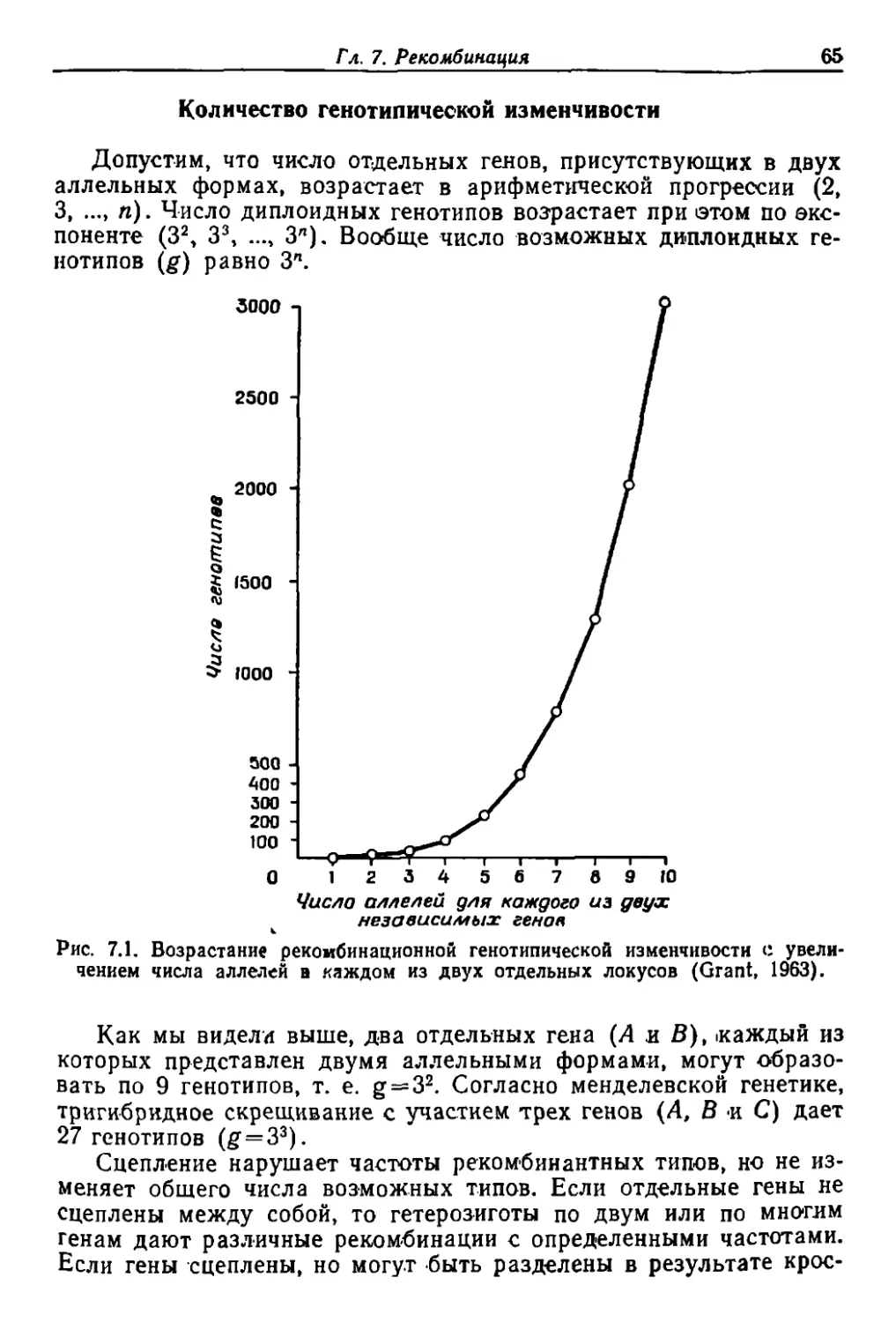



























































































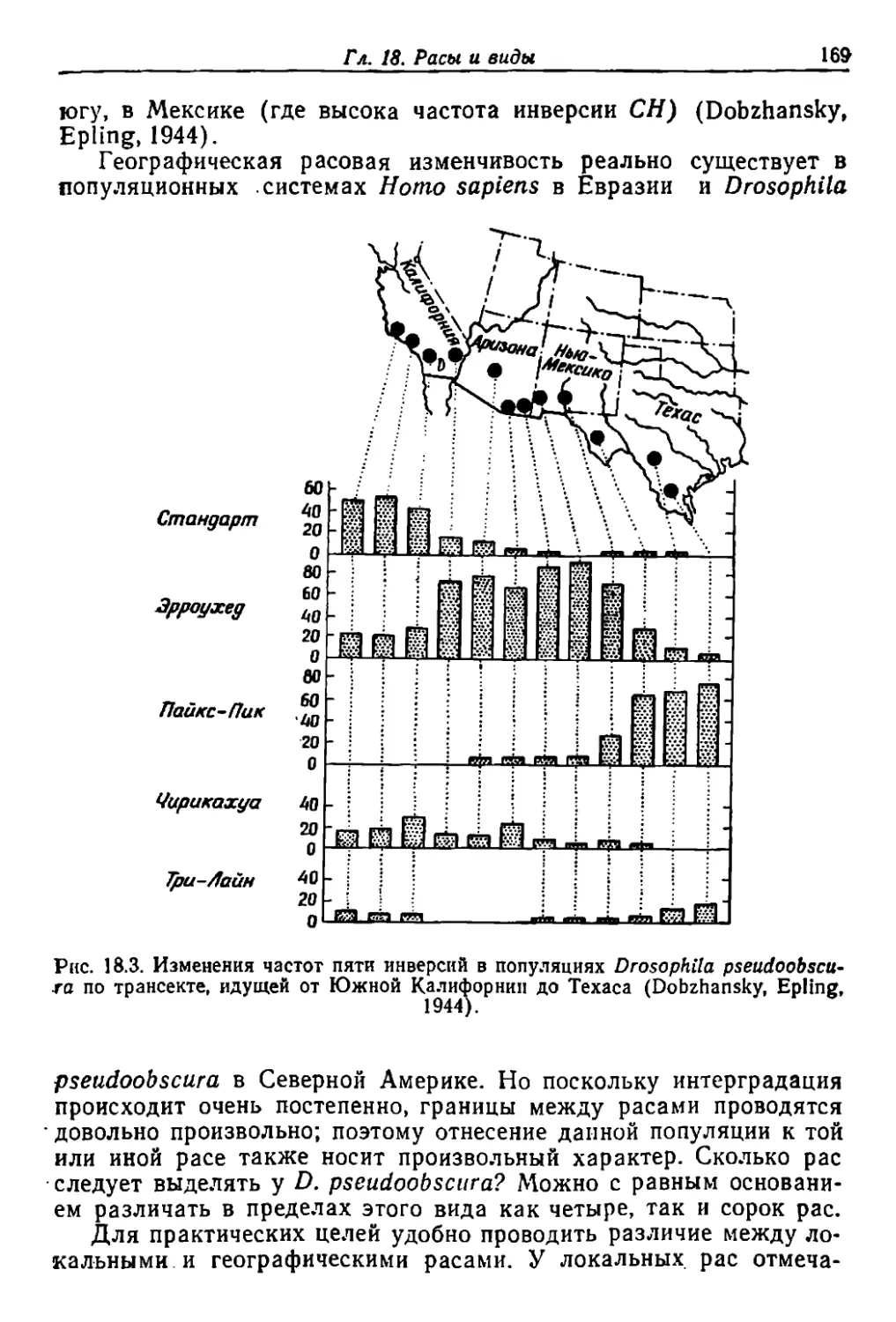

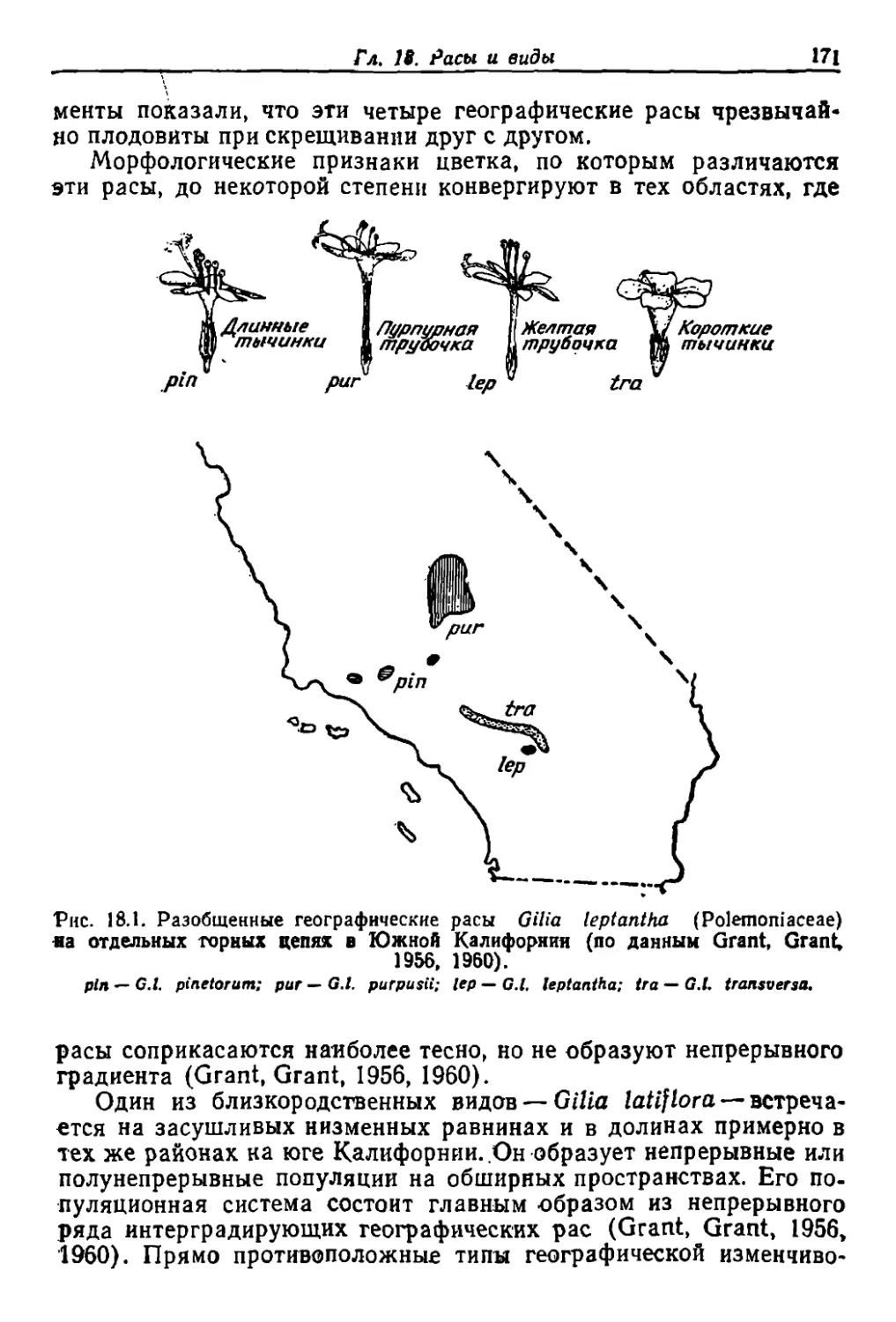
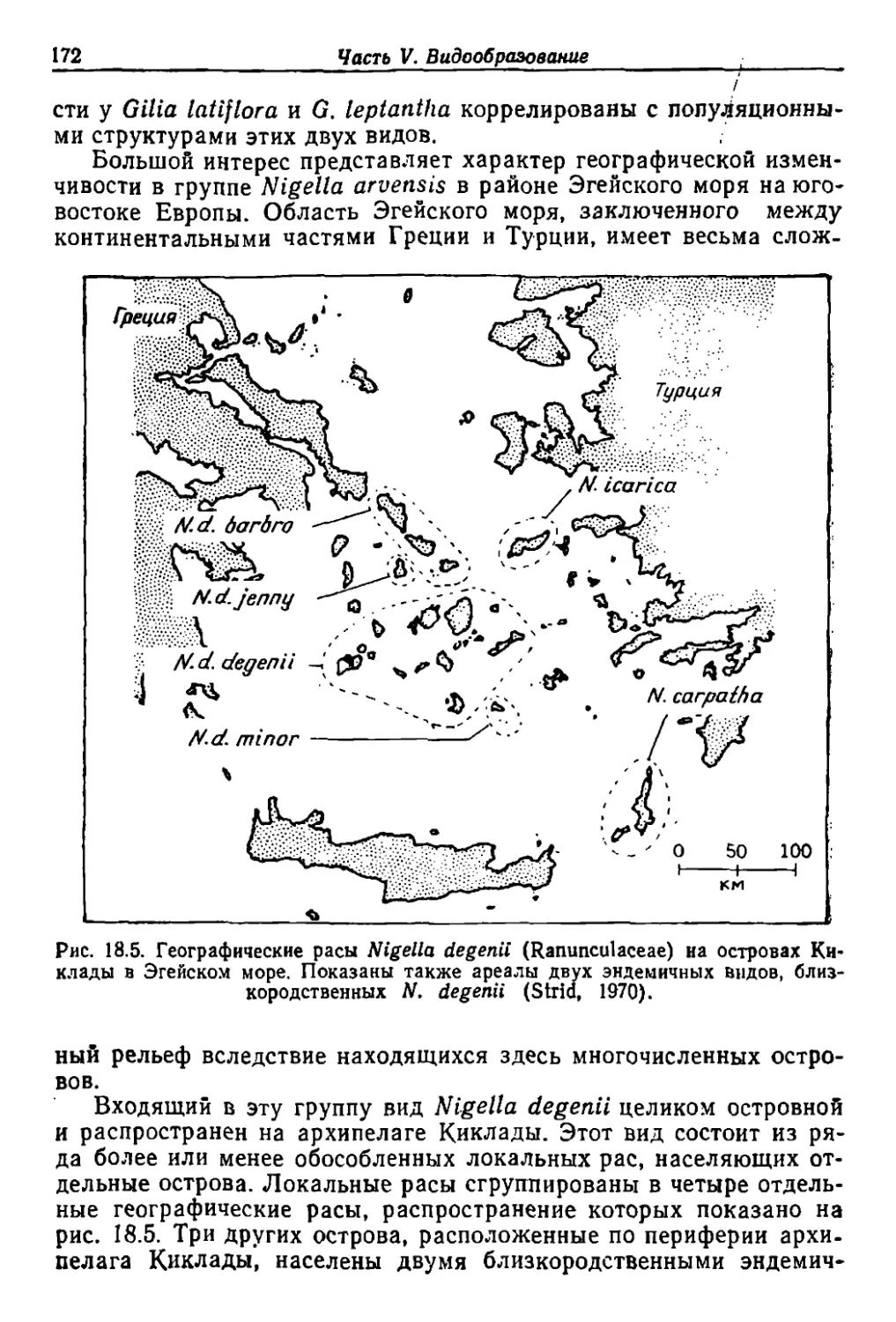



















































































































































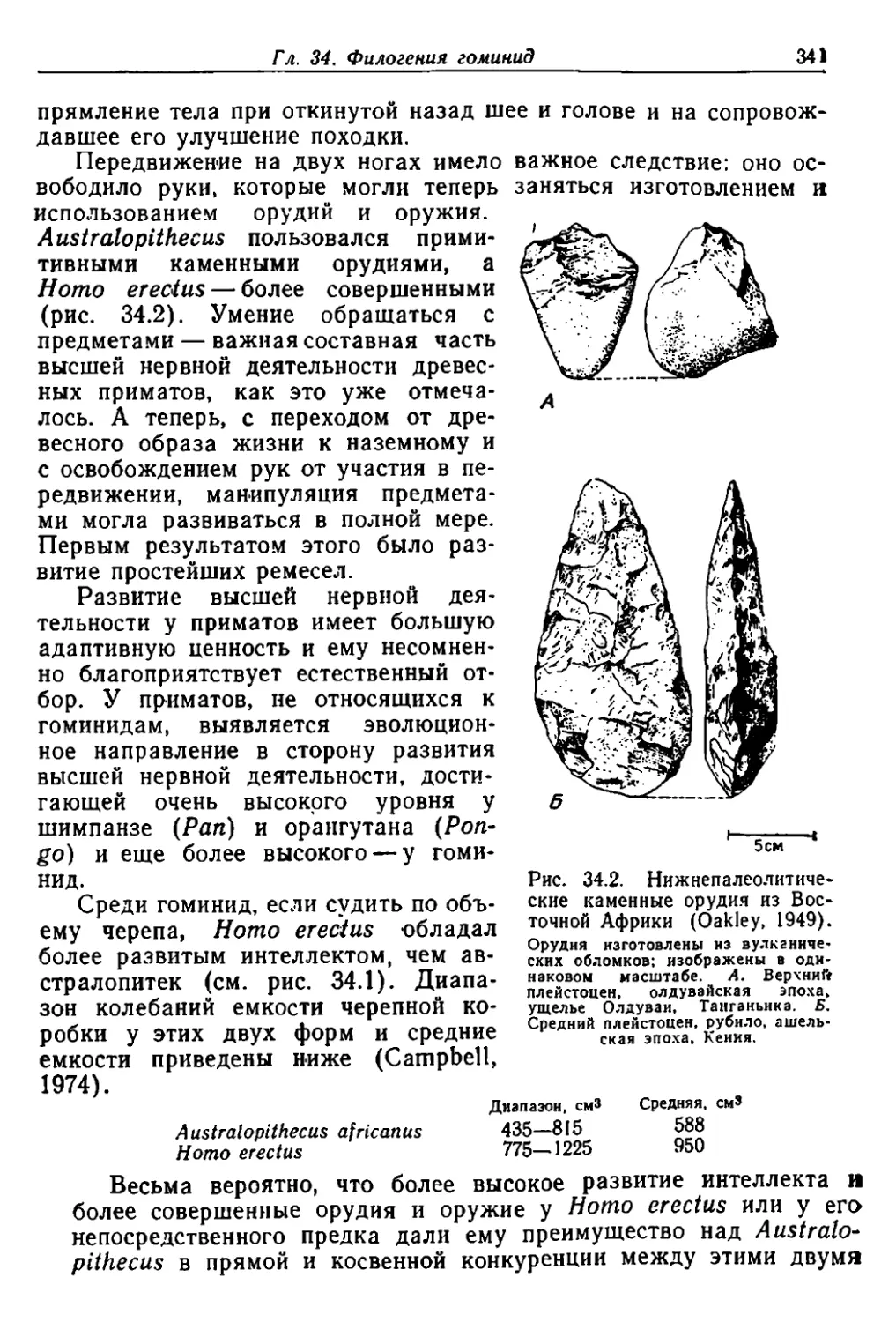



































































Author: Грант В.
Tags: биология микробиология эволюционный процесс молекулярная биология эволюция организмов издательство мир
Year: 1980




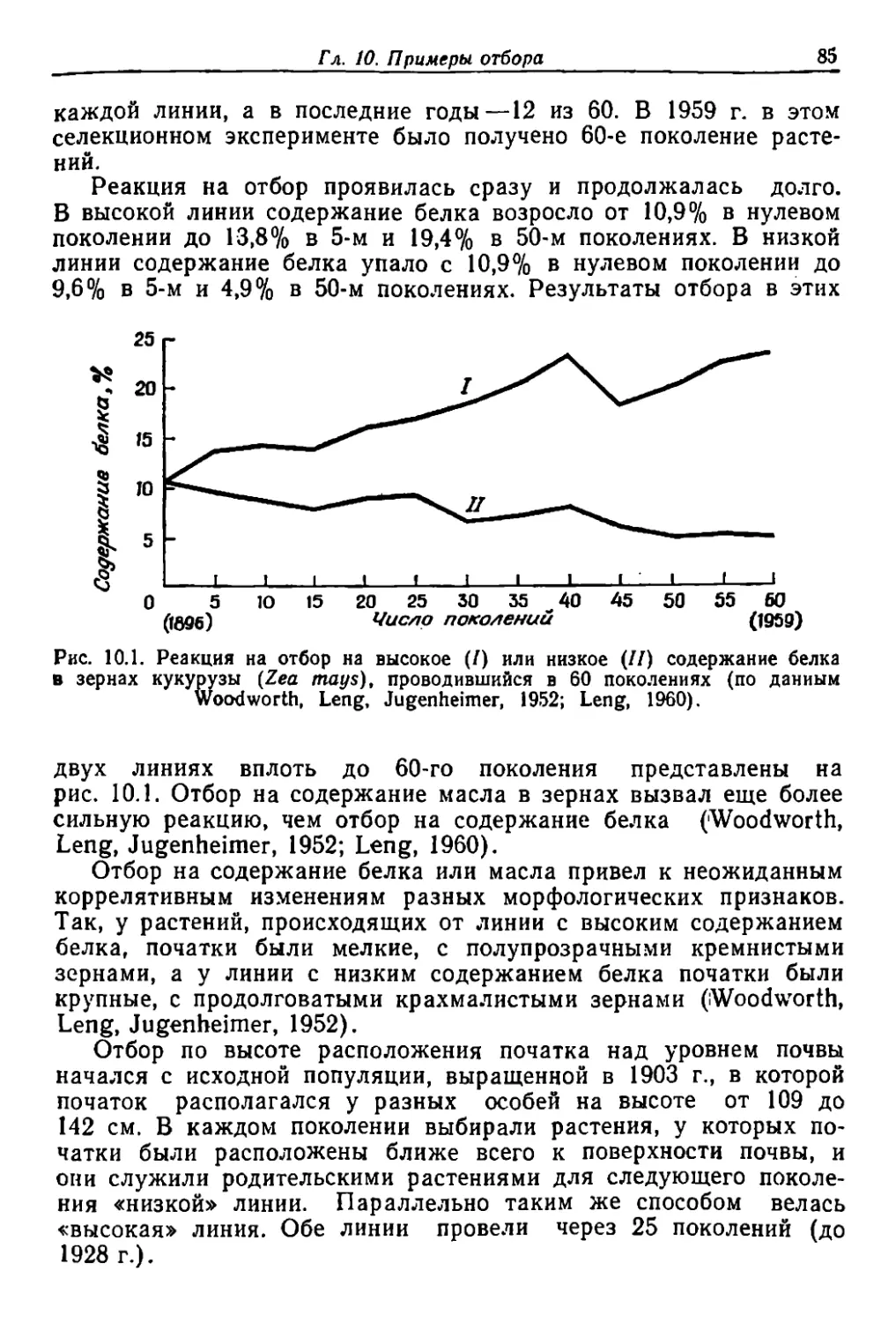
Text
B.rpaHT
ЭВОЛЮЦИЯ
орrАНИЗМОВ
1
.
.
.
I
.
.
. .
:
i
: I
, I .
·
J 1 Т }
. .
: : l'
I : ,/ t
: : : f ./
· · С' .
I f , :/
ij ., 1 '.
t s
. , ,
I l' !
:::
.
. .
I ::
t : I
,с .,
" , . :] ') , i
: '. .
r ;:"//
/
.
I : .:/
, : /"
I
J
,
i
:
t
i
.
i
.
i
I
1
.
i
OROANISMIC
EVOLUTION
VERNE GRANT.
University of Техзs
at Austin
fore.word Ьу
GEORGE GAYLORD SlMPSпJ\l;
W. Н. fREEMAN AND COMP'A-N'l
San Francisco
1977
В.rраит
эволюция
орrАНИЗМОВ
'Перевод с анrлийскоrо
Н. о. ФОМИНОй
'под ред.а1<цпей
?доктора БИО 1J. наук
;Б. .М. МЕДНИКОВА
IИЗДАТЕЛhСТВО сМИР..
.Москва J980
)'ДК 575.3/.7
Книrа известноrо ученоrо, члена Национ-альной академии наук
США представляет собой сжатый обзор современных концепций по-
важнейшим проблема м эволюционной теории на уровне орrанизма.
Блаrодаря тщатепьному отбору фактических данных и примеров,
которых, в отпичие от друrнх сводок по эвопюции, в книrе мало,
rраит сумел при отиосительио иебольшом объеме рассмотреть все
ОСНОВНЫе проблемы: микроэволюцию, естественный отбор, наслеДО-
вание приобретеНIIЫХ признаков, видообразование, макроэволlOЦИЮ
и эволюцию человека. Автор неУК.10ННО придерживается дарвинов
ской теории эволюции.
Предназначена для биопоrов всех специа.'lьностей, для студен-
тов и аспирантов биолоrических факультетов университетов и пе-
даrorических институтов, для философов и историков науки'.
Реда"цuя AurepaTyp"t по БUОАОZUUl
200 1000000
21001 121
r 041 (01) 80 J21.. 80
@ Н"1, Ьу W. Н. Freernan a.nd ComparlY
@ Перевод нз русский язык, If Мир , 1980
ПРЕДИСЛ.ОВИЕ РЕДАКТОРА ПЕРЕВОДА
в последнее .в'рем.я .э.ВQЛЮЦИЮ част.о расСМ81'рИ!вают как бы
в ,мик:роокоп, анализ'ируя ча:сти .вме.сто, ЦМООО. В мноrочиосленных
публикациях бСУЖIд'ается Э:ВОЛЮIЦия o.praBOB, 1жа!неЙ t 'клеток, а:ми.
нокисл()тных последовательностей .б J1iКОВ .и ,ну.клеотидных после.
довате.т'lыIocейй О1!делыных reHOO. Особой беды в ;подобных .nодхо--
дах HeT t OДHalКO fl1рИ этом следует помнить, 'Что ,В ст.росом CMblCJJe
слова эвюлюцио.нируют лiи.шь tПОПУ.Т'lЯ1ЦИ.И t СОС'Роящие из о.рrан'из-
МОВ. Сами по себе тены н .бе..1КИ t 'клет,ки .Н opraHbl ЭlООJIЮЦИОНИРО-
вать не MOTYT t как не MorYT .1JeTaTb .сами Ino себе 'OTдe lь'Ныe части
са:молета. Забывая '9TO t мы Пра!КТИ'ЧОСКИ не уходим дальше э.МJПе-
'ДОКiЛа (V в. до н. в.), roворивwеro. О неза'ввс.ИlМ'ОМ IВОЗНИ1кновен'ии
и .д.а.'Iьнейшем /С..1учаЙ'н'ом ,-сочетан.ии частей тела .раз.ных животных.
Л,одобные раССУЖlдения 'MorYT выlrяlд.е.тьь 'fIрюи!3мо , .однако не еле.
дует .за.бываТЬ t что тенденции -с,ведения эволюционноf.О 'процесса
к fЗамене одн'Orо аллеля ДрУ'fИiМ без учета .вза'имодейс1'ВИЯ reHOB Jf
определяемых Iими признакOIВ ,были И остают,ся Ino наостоящее в.ре-
.мя .ИJсroч.ником Iмноr.их ош.и6ач.ных теорий эволюции.
От этих 'недоста11КоОВ св060;дна .предл'а'rаемая нашему читателю
к.ни.rа В. rpaHTa. .д,.втор 'Сделал удачную Iпqпыт.ку ,pa OOMOТpeть $0-
ЛЮUИОН'RЫЙ проце.сс ВО .всем e,ro м'н.оr-ооораз.ки. И;Сn'О.!lЬЗУЯ н-овей-
шие данные rенеТИiКИ t ОН cCTpOro остается на tПозициях кл ассическ().
ro ЭЕюлюци.ониз:ма (П0-В иди-м ОМУ, эт.им объя'Сняется на'зва.ние кии-
-rи таlКИМ IНа'3JвЗ"нием автор подчер.КИJВаеТ t 'Что, .х.отя элементарными
еди'ницами .ЭВОЛЮЦИОНtНоro 'проце.сса .считаются IПОПУЛЯЦИИ t .в ко.
sечном счете они .сосroят ИЗ орrаНИiЗI:\10В). IБолее ,roro, К'о'rда совре-
менный 3Воолюционист rоОВ'ОРИТ t что .ОН Иiэучает IПОПУЛЯ1l'ИИ (!речь
идет 'О п,рирод.НЫХ t а lIе ЭКlQпер'и.меН7альных, .полностью 'контроли-
руемых ,ПОПУ.l]'Я1ЦИЯХ) t О'н :всеrда имеет ,дело лишь 1с выоор'Кам:и
орrанизм,ов. Эти лол.оженяя ,настолы<'О общеи'з:веСТ'НЫ t ЧТО t ,ка'к .пра-
ВИЛО, они 'итн()р.ирую я.
Трудно опредеЛИТЬ t IK .какой катеrории .следует .оТ'н.ести лреД-тIа-
rаемую 'Читателю книrу. Са.м автор ,считает, ЧТ.О .это учебник п()вы-
шенной т,руди'Ости, ,рассчитанный .на непЗ'с.сивное ,вос/П:риятие ИН-
формаци.и. ПРe.дlпола:rается, что. 'читате.I'JЬ станови1'СЯ BMecre с ав-
тором активным участником JП'роцеюса постижения и'Стины. Однако
В. Лра l Н1' 'не ТО.I'JЫЮ ,нообычайно lэ.рyrдирова'Н.ный :kom'Meh-татор .И ком-
пилятор, но и oJТОЛУ'ЧИIВШИЙ заслуженную известность иоследова-
тель nроцес'са эволюции. Как ни 'CTpaHoН'Ot ,в ,ПQдобном .сочетании
'6
л редисловue редактора перевода
есть IСВОИ Н6Достаl1КИ, если ,ИХ можно .назвать таковым'и. rlрзнт----
не IСТ.оронний на,блюдатель: выражаясь фиrурально, ан :не rерольд,
а ;участник турни.ра. Поэтому в :ряде случаев ОН .не избежал ()пас.
ности переаоса заlкономерностей ЭlОО...1, юци.и хорошо и-зучеlfНЫХ на
«'СБоем» материале (ЦВе11ковы-е 'ра.стен.ия), на весь ор.rанический
Мир. Но оСОО11Ношение факторов эволюции в Ipa3HblX r.руrnпах .может
быть раз.ным.
Однако Iследует IПQдчеркнуть, что r'paHT з.начителЬ'Н'о более O'c
торо жен, чем м.ноr.ие друrие авторы OOQДOK по теории эволюции
и не ;пр{)ходит ;М-ИМО точек 'зрения иссл'едователей, с которыми он
по .тем ,или .иным Iпр.и:ч'инам ,не cor ласен. Следует от.метить лоrич-
НОСть постр.оения ,I<Jниrи и UllредельнI)'Ю Чe1'lкость .И3.1J,QжеНIJIЯ. Я у;беж-
ден, .что кн'иrа В. rpaнTa ,принесет большую IПОЛЬЗУ 'советским ч.ита-
тел ЯJМ.
Хотелось Iбы оста.нов.ить'ся На \НеКQТОРЫХ вопросах, заТlрОНyrых
[рантом, ,в частности на явлении индукции. Здесь НУЖНQ соrла-
ситься с автором пре:дисл.овия -к амер.икаlНОКОМУ ,изданию СИМ'ПСО-
НОМ, кото.рый пишет: ;«...явление .индукции не останется за предела-
ми этой теории (,СИlПетиче.ской теорки эволюции ---- Б. М.), а раз-
двинет .их IJ..Hrpe». Наследст.венные Iизменения, .вознwкающие в -ре-
зультате приобретен,ия чужоrо ,rенетИ'Ческоrо 'материала, т. е.
трансдукция 'и иные формы неооловоro Iпереноса информации о/же
вошли в данные пределы, практически их не раздвинув. Ибо
тр.анс.ду.к,ция никак не а.де.кв.атна ламарк:и:ст.ской «а<:симил'Я:ции
внешних .услоаий». Это. ,не и'Н,дукция, а ,своеобразный с;поооо reHe-
тичесКОЙ ,рекомбинации. В мире \прока1РИОТИЧеских ор'rан-и.змов He
половой Iперен.ос rенетич-есж{)й инфо.рмации р'а'(ШрОСl1ранен 'Чрез-
вЬРчай.но ш'И'роко; :более To.ro, есть все .0С'Нi()вания ,полаrать, что тз(к
называемый 'полоооЙ ,процесс у баlКТерий являе'I1CЯ ero JПРОИЗБОд-
HыI.. У .ВЫОillИХ IOрrани3'МОБ ,БИ'Свк.суальный перенос rенетачес;к()й
информации практичеоки 'вытеокил !все прежние, при'митивные
способы обмена iпоследовательностя,ми днк. Роль 'редиктовых,
ЛРИМИТИlВных !Способов ТИJПа транодуК'ц.ии в эволюции еука.р'Иото.в
остается ;ДИ'ОКУiCCионной, однако ясно :О.дН.О ,непо.ТlOвОй переное
rенеТИlческой ,ИНФQР'МЗ.ци.и ,ни'Чем не м,ожет IПОМОЧЬ Iбезнадеж,но.му
делу в-озроождения ламз'ркистских теорий. Скорее он их опрове,р
raeT.
Факты, лри.водИ'мы-е rpaHTOM Б качеС'f\ве Iпр.имеров «И}{ДУ'юци.и»,
разнородны. ТЗJК, индукция r.и.бри;дн'ой стер-илыюсти у дрозофилы
.оБУСJlоОВЛИlвается оСИ'МJбиотиче'ОКИ'М ми к.роорrа'Н'И13М-ОМ , nере'даю-
щим'ся по наслеДСl1ВУ ,(нелиш'не Н3Jпомн.ить, что -еще .rербе,рт Спен-
сер тра.К70вал на 1 С.пеДС11венный ,сиф.или<: iKa,K 'наследование дри.об.ре-
TeHHoro IпрИ'знака!). Все друrnе ,п'ри.меры, та'кие, как обеcrцt8ечИlва-
иие ХЛОРофИЛЛ.ОНQС'НЫХ жryТИКOIВых, таJкже Iп.олучают 'Исчерпываю-
.щее объяс.ненне ,в pa.MKax синтетической ЭlЮлюциО'н.нюй теории, дo
полнеН'н'Ой .последними дости.жениям.и :молекулярн{)й би'оло.rии.
Предисловие редактора .nеревода
7
3.а:метим, К'стати, что структура reHOMa эукариотов такова, что
пр,и06ретение чужерОДНО'rо rен€ти'ЧеС.КQrо материала ;не дол.жно .иТ-
рать .в их ЭВОЛЮЦИИ заметн{)й ,РОJlИ. Осущеcrвлеиие а\кта.. ТР3Н{'ЛУ'К-
' эукариотических клеток .в .уело,виях лаборатории еще не может
служить тому дока'зательстоо'М.
Сложным кажется ()1'Ношение rpaHTa .к проблеме сн.м'Ц!l1.:Р..И-
:! . !! а.'30ва'ния. Вслед за Э. МаЙром он сначала прихо-
дит к выводу онеВозм ож-ности разделения одноrо JJ.ида .на Дlва
в О\дНОМ Зlреале. .однако автор отдает ,себе отчет .в том, что посту-
wТ'JaT невозмож,ности си,м'паТРИЧОС1Irorо ВИlд"ообра300вания \Вытекает
лишь из определе.ния пан.миктичес.КQЙ .менделев'ской популяции,
I(оторая, по сути дела, IВ 'реальных УСЛо/В.иях является нереализуе
мой а6страl{lцией. Недаром r,paHT везде, rде ТОЛЬКО ВОЗМ.ОЖ1НО,
И'C1Iользует терм.ин «сосеДС1UЮ» (neighborhood). Выделение С'О-
се,д.ств прои-з.'Водится .по оцеroке степени инбридинrа, а Не по сте-
пени ПЗНiМ'ИКСИИ. В конечном .счете аатор .склоняется 'к вО'зможно
сти .видообразования без rеоrраф.ичеаюй изоляции, слеrка за'вуа-
..т:Iировав ее терминами марrинально 'Си-м,пат.ричес ое .и С'меЖ'Н'о-
си;м\патр-ическое. Это и понятно: яи один э ол-оr (а rpaHT хорошо
ЗiНает IЭКОЛооrию) не оможет в \С'Воих построениях обойтись без /КОН-
цепции эколоrичесК"оr.о видообразова'ния. В последнее время и .сам
Э. Майр l'О60р'ИТ .уже не о .невозможности .сим.па'J1рИИ. а о ее недо-
статочной доказанности. ОднаlJ<О то же саМое МoQжно сказать и
() ( онцепции rеоrрафичес.коrо .видообразования. Риокуя быть обви
ненным s lКон.сервати:зме, В. rpa'HT B ЬMa с.кептически ОТНО'Сится
ко IМНlQrим /построениям автор.ов. изуча-вш-их э.волюцию на MOJIeKY-
JIЯРНОМ уровне. Gправедлива .ero криrnка так называемой «недар-
ви.новской» ЭlВол, юци-и т. е. образования новых форм Jпутем наЮ:)[I-
.т:Iения .селективно нейтра,J1Ь'НЫХ мутаций. Нель'зя не от.метИiЬ ero
TOHKOro замечания .о том, что сам Дарвин таlКУЮ ,эволюцию 'Знал Jf
писал о неи, а потому тер!м.ин «нед.арвино:вская э.волюция» попро-
сту 'неверен. Кlр'ИТИока Б. Пранто.м «моле.куляр,ной эволюции» СНИ-
детелЬ'С.'rВУ'ет не р .том, что На дroм уровне ,нельзя изучать развитие
ЖИlВой IП р.и роды , а скорее о том. что .ош.ибзлъся мож'но на любом
урооне в том числе и на tмоле,кулярно'м.
[,рант принимает лри анаJ1изе ,проб.т:Iемы Ilфоисхождения жизни
КОНЦ6111JJ:ИЮ «roлых reHoB в о.рrаничес.кюм бульоне». В 'нашей лите
ратуре более распространена теория. развитая А. И. Опариным,
с.оrлас-но .которой ,П'редб.иолоrичеока:я Эlво.дюция с caMoro качала
происходила в фазовообособленных целостных пробионтах сис
темах типа коацерватных капель. Такая теория представляется
более обоснованной. поскольку раздельное возникновение и после-
дующее сочетание компонентов белоксинтезируlOЩИХ систем на-
СТОЛЬКО маловероятны, что rраничат с чудом.
Несколько c.т:IOB ,относительно() rлав, Iпосвященных \Эволюции че-
лове.ка. .Несом нен но, эти rлавы ,ч'итаются с больш.ИJМ интересом,
"8
Предисловие редактора перевод(J
-однако .читателю вслед за ними полезно будет ознакомиться и
с ДРУl'им;и проиэведен.иям'И, i10CIВященными этому ,вопросу ----'в .пер-
вую очередь tC Iвы.шедшей 'вторым изда.нием, lIереработанной и до-
полненной новыми .да.нными СВОДКОЙ .я. я. Р,О'rинскоrо 1 .
К 'сожалению (ОИ это относи"ся не ТОЛЬ'КО 'к данной ,книre, но :и
к БОЛЬШJfНоС11ВУ aHr лоязыч.ных пу\бди каций по тео р'ИИ .эВОЛЮIЦии),
В. r.ра.ит пра<ктичеоки .не использует сове'I'ОКОЙ литератУ'РЫ' Н,О
кatКой обзор :по' теории ЭВОЛЮЦИ-И, lПо.пул,яционН'ой и 'ЭВОЛIOllJ:И'Онной
rенеТИКе может претендовать на ,ПОЛНО'"fу без yrпО'минания .и\Мен
А. Н. Северцова и С. С. Четверикова? В целом же книrа В. rpaH-
та может служить образцом сочинений подобноrо рода. Мы наде-
емся, что советский читатель найдет в ней MHoro интересноrо И
полезноrо для себя. В первую очередь это относится к студентам
и аспирантам, а также к преподавателям биолоrии и исследова-
телям, желающим пополнить свои познания в области эволюцион"
ной теории.
Б. М. Медников
J РоrинскиА Я. Я. АНТрОПОJlоrНя..... М.: Высшая школа, 1978.
ВСТУПЛЕНИЕ
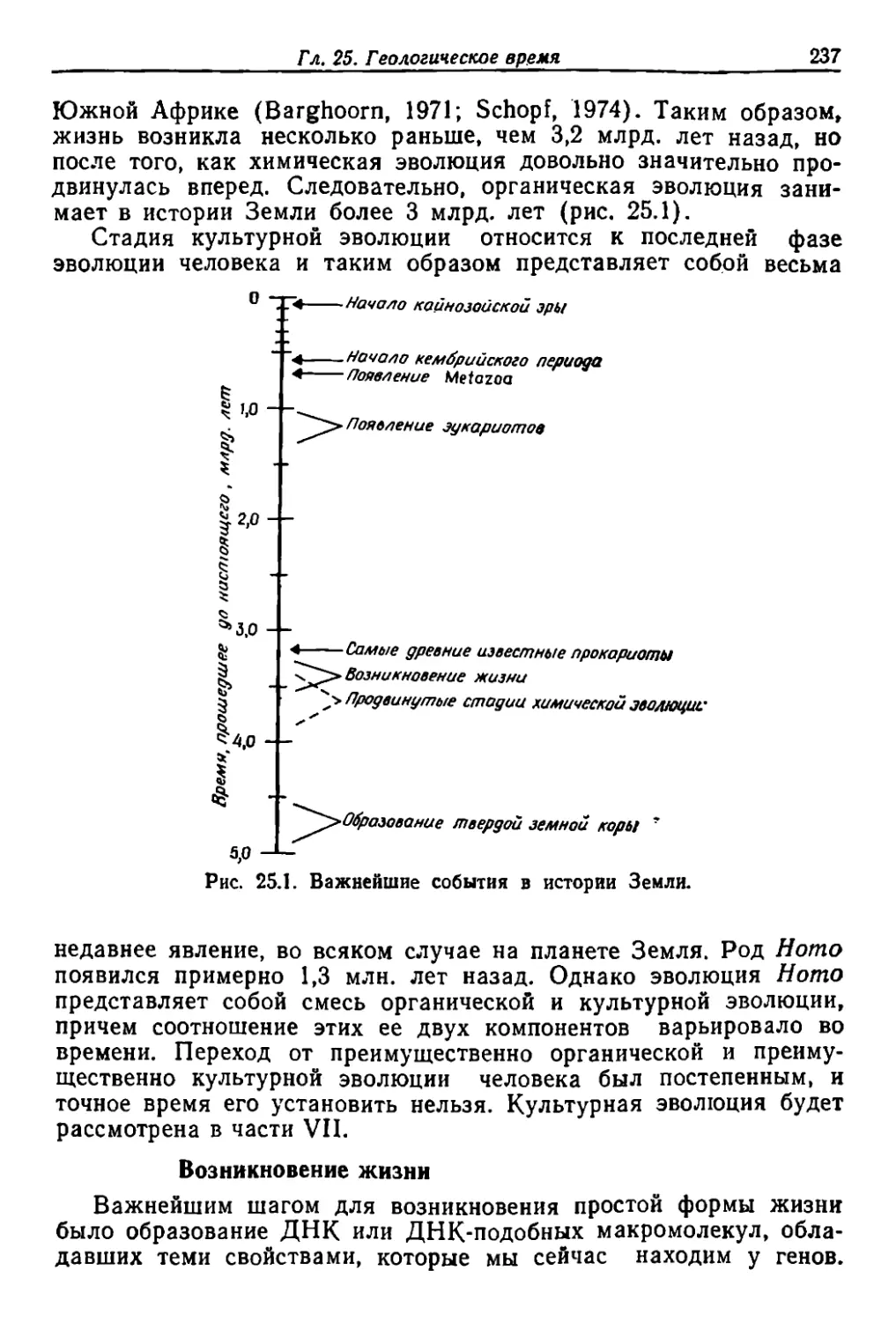
Настоящая книта 'оодер,жит общий оБЗQр ,важнейших сов.реме.н-
ных знаний об "ЭlЮJ1юции ор.rаiН.ическоrо мира. Це.пь lМ'Oero CTYn-
ления указать IMecr.o, 'зани'ма,емое ,ЭТОЙ ,кнаroй, СQпоста\вив ее
с друrи'м'И труда/М.и на ту ж тему. Для T'Oro JЧТобы ,провести такое
сопоставление 80 !Всех детаЛ1ЯХ. :понадо6илось бы на.ПИlCать еще
одну НJ1'И даже несколько 1КННJf, 'Н'О -МОЖН'О CJ].елать ЭТО .н IВ БOiЛее
сжатом в.иде. сооред'Оточи'в .вн.имание ,на некоторых И'з наи-более
зна'Ч.ительных черт IB И'СоСЛe.дJO.вании 13волюи:и:и, ха,ра.ктерных для
трех .п'etриодов: lП€lрвоrо .наЧaIВшеroся lп-рИ'Мерно сто лет назад,
BTOp:OI'Q около IПЯТИJдеСЯТJf лет ,назад .Н TpeTbero нынешнerо.
Ра'ОСМО11рение ка.ждоro из .этих Iпериодов можно еще бол е скон-
центрировать 'и однооременно 'Сократить. 'Выбрав для обсуждения
по 'Одной общ й книrе об "Эвол, юци.и ха:раlктерн'ОЙ ля С'воеro BpeMe
ни: ДЛЯ Iпервоrо ,периода .эro шедевр Дар1В.ина. lCыrrpавший в изу-
чении эволюции 'большую роль. чем любая друrая pa1OOTa; дЛЯ IВTO
poro периода кннта Л. Лалла (Lull), J<оторая была са,мым ПООУ-
лярным УЧflбн'икiOМ [10 эволюционной теории 50 лет. назад; для
TpeTbe.ro .книrа. К'ОТОрую -вы сейча."С :де'ржите ,в рука/х.
Сто лет назад Да..р!Вин еще был жиrв; он активно ,работал почти
до амой lCIМepTH. !ПостиrшеЙ ero,B 1882 r. в ,оозра.сте 73 лет. rени-
а.lIЬНЫЙ труд Дарвина. (Длинное название ](oOTOpOro обычно COКipa
щают до <<.:Проис,хожде-ния ;ВИДОВ». был' 'впервые опубликован
в 1859 r., а П()ICJIе.днее переработаннос им са,м.им .изда.н.ие (шестое)
вышло 'в 1872 ['. В 'семидесятых rодах прошл.оrо века пОЧТИ все
ком!петентные и образованные б-иолоrи .призна'ВаJ1И реа.'1ЬНОСТЬ эво
люции. Два наиболее заметных НСКЛIO'Ч.енпя 'Составляли Луи Ara.c
СИД, но ()н сконча -ся в 1873 'r.. а ero послеДQвател.и уже были ЭlВ()
Jlюцио.НJlстами. и Ричард IОуэн, который доОЖИЛ до 88 лет 'и умер
\В 1892 .r.. IHO в e-ro 'Пос.'1 дких ,работах уже СК:ВО3И..1И намеки 'на то.
что он Iвозра.жает не столько IПРОТи.в самой Iконцепции эволюци.и.
СКoQ.,ПЬКО (против KOНiКlpeT.HЫX :ПlредстаВ.IJ НИЙ Дарвина о теч.еJIIИИ это-
ТО прсщecwcа и ero причииа'х.
Таким образом, сто ileT назад каждому, '<ТО изучал биолоrию
. -под руков{)дС1'ВОМ Iквалифицированноrо Iпреподавателя. уже бы.туа
знакома ищея Э>ВОJlЮUИИ :как -QДHO 'из общих 110л,ожеиий Э'ЮI'O \Пред
мета. Насколько MIHe известно, .в то время не Iбыло rс,пециальнЫХ
курсов, 'посвященных эволюции. как са,мocr.оя.тельнаму !предмету.
Il ХОТЯ 'С'Ущес'J1В.оваJ110 немало 'Работ, iКа'Сз;в;ши.хся ,.ОТО или .ИН-o.I'о ас.-
10
Вступление
neKTa этой проблемы, НИlкакоrо друrоrо действ.ительно общеrо
ру.коВ'одС'Тва, 'кроме caMoro «ПроИ'схождени'я ,В'идов , создано не
было. Нсякий. 'кто займется 'Изучением «Происхождения .видов»,
прежде IBcero узнает, что эволюция единственное 'раЦИОllальн.ое
объяснение множества наблюдаемых фактов. Затем он узнает,
что дарвино.вская теория, cTJ}oro roворя, прИ/писывает ЭООЛЮIlИОН.
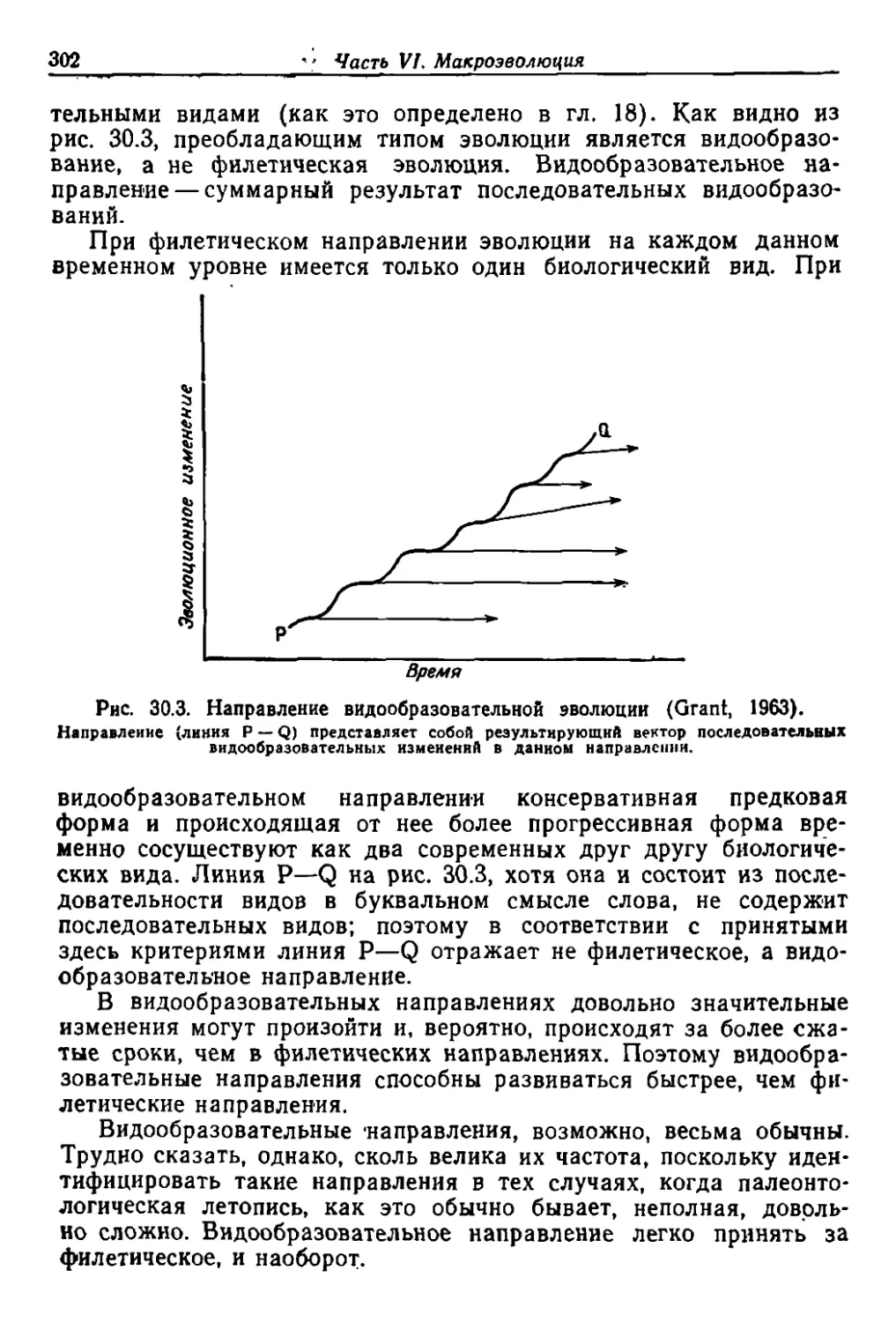
ные И3'менеwия IfлавнЫ.М образом естест.венн()му от.бору что было
H очень удачно сформули.равано ,как (<,выживание .наи.более при.
1Jсх;.обд.еIfНЫХ» .H ,в .меньшей 'степен'и на-с-ледуемым Iв-йздействиям
упраЖ'н ния .и. неу.праж'нения opraHoB, аюздейcr.виям -среды и «из-
енен'»я.м, которые 'МЫ по /сооему незнанию ,считаем ,опонтан.ныМ'И)).
рольшин'ст.ВО послсдующих авroров -объединяли .второй и. третиЙ
факторы Дарвина под общей рубр'икой .на ..1едования приобретен.
ных ,пр з.наков. Ero ;четвертый фактор 'C0011BeTc1lByeT тому, что мы
цазовали .бы .мУ.тация,ми, но 'зто В большинст:ве своем МУТ аЦIJIН ,
имеющие бол-ее обши.рные и явно выраженные эффекты, чем. обыч-
ные мута.ции. Изучающий «П'РОИlCхождение ,видов заметит также,
il1'0 Дар.вин был совершенно уозерен iВ 'рол.и ecTOCТBeHHoro отбора и
р,иве.'l ,веские данные в ,л.одтверждение эroй рол.и, .но что'.отно'Си
тель"Но Tpe друrих. факropоо он высказываЛIСЯ \менее .опреде
ленно.и выщв,Иrаемые им .доказатеЛЬС1'ва слабее.
_ На :протяжен'ии lJ1оследующих лятиде.сяти лет, т. е. вплоть до
двадцатых тодов иашеrо ,века, было от.крыто MHor,o н()вых фа кто-
ров и :сделаНО.;МНQrо 'Существенных на.блюдений. Все они указыва-
.ilИ на то, 'Что эволюция ---- единственное рационал ное и здра180е
истол ование истории ор.rаниз'мов; OT'BepraTb эволюцию -со СКОЛЬКО-
ни.будь научных пооиций стало уже неоозм'ож.ным, хотя, iКa!K ОТМС-
чае'I'CЯ в \последней части книr.И, даже .и ,по сей ,день в некоторых
неакадемических или антинаучных Kpyrax у нее находятся против-
ники. Но, .несмот,ря на та.кое единодушие ученых по ,проблеме .Б це-
лом, они 'Не достиrли -определенноrо со'rласия относительно кон-
к'ретных факroров ил.и ПрИ1fИН ,ЭВОЛЮЦИИ.
В д.вадцатые ,rOoДbl нашеrо ,столетия .в колледжах читалИ1СЬ кур-
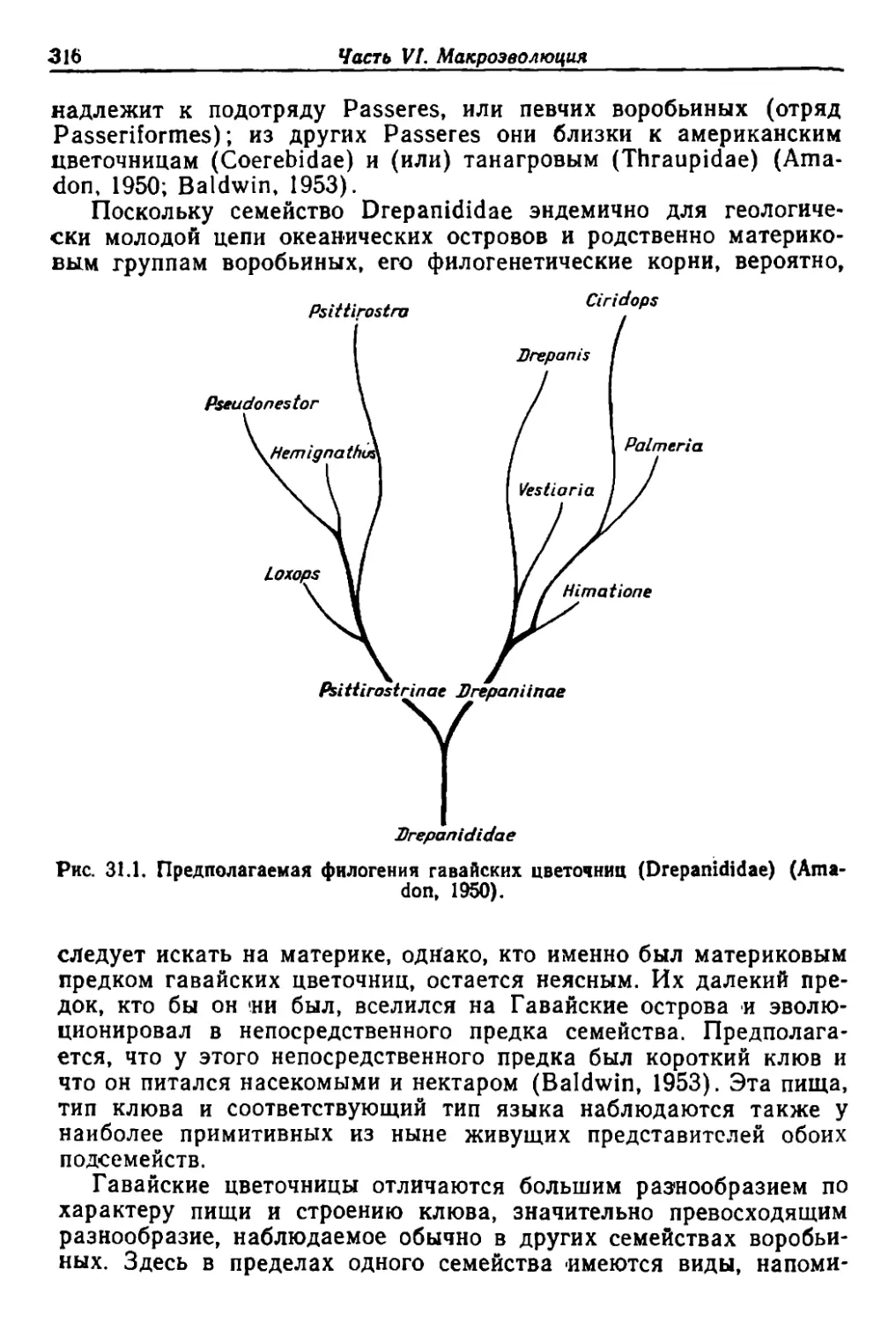
сы Jlекций по эволюции орrаничсскоrо мира. Среди них одними :иэ
наиболее ПОПУJlЯР,НЫХ были ле ции Р,ичарда Суонна ЛаЛJ1а 18 Ие:..rJЬ-
ском унwверситете. Бrо книта «.Organic .Evolution», о.п.убликован-
ная в 1917 'r., а затем 'в переработан.ном Iвиде :в 1929 r., была, ве-
роятно, наиБОJrее распространенной ИЗ неС'ко '1ЬКИХ ;имевШИХоСя в то
время книr на эту тему, и она позволяет 'судить о 'Сосroяни,и "Знаний
по !проблеме .эволюции ,и О методике ее .препода.вания пятьдесят
лет назад. Лишь небольшая ча'сть КRИ.J1И Лалла, ,менее 1-5 процен
7о.в Bcer.o Te CTa, ,была отведена 1'еориям. законам или ,(JIричина
(<<механизмам») -эволюции. Именно IB ,ЭТОЙ чаоети 1JРОЯВЛЯЮТСЯ д.ва
резких !ра.зличия ,м'ежду ИЗ.1Jожен.ием Лалла и IJIОЧТИ любым совре-
менным PYКOOOдlC1lSOM например данной книrой: мнаrочислен.н-ость
7еор-ий, которым он уделял 'Серьезное 'Внимание (некоторые из них
Вступление
11
в .насroящее !Время 'Считаются 'Заслужи.вающими лишь к.раткото
упоми,на'ния IВ ИlCТоричеоКОМ обзоре), и. 'минимальное пр.ивлечение
rенетики.
Ра'ОС'мотрение ecTecTBeHHoro от.бо:ра 'было у Лалла iкрат.ким 'и
эпизодическим и 'завершалось следующим за'ключением: «...хотя
естост.венный отбор .и можно .считать важным :фа'ктор.ом iВ .эволю",
UИИ .орrа-н-ич.ескоrо -мира, ЭТ,О, O"lевидНО, не еДИН,СТrВеН'Ный и уж, Ко-
нечно, ,не единственно важный ,фактор)). .Почти столько же внима-
ния было уделено на'следоваН'Ию приобретенных пр и'зн а ков. Лалл
пр.иходит к вы1юу,' -что ИХ наС,,1едование «не доказано», но ;добав
.'Iяет: «нельзя не 'ЧУ1ВСТВQвать», чro 'ка,ким то неизвест.ным способ.ом
приоб.ретенные Iпризнаки ;осе же 'Моryт на'Следовать'Ся. В настоящей
книre подобное на'Следо.вание .в том .смысле, каlКОЙ в Hero до СИХ
пор !Вкладывается, совершенно iCпраlведливо считается даже не
заlСЛУЖИ1вающИ'м .обсуждения.
Лалл был соrласе.н с lIроводимы.м Да:рвином рaзrра'ничением
между 'J1eC1'BeHHЫlM и 'половым отбором, .но он :пр,ида'вал несколь-
ко большее .'эП-зченйё ifбёJIё :Дне му: Ir'.Rаст.()Ящее время -половой от-
бо.р .считаетс.я одни'м из деЙ'Ст.вительно .существующих, но BTOpO
степенных случаев OCTeCTBeIlHor.o отбора. Лалл ,выделял также 'в
качестве саМQCтоятельных я!Влений ректиr,радацию п кинет.О,rенеэ.
Оба эти тер'мина, paiВHo как 'и обозначаемые .ими 'К О нцелци.и, .от-
BeprHYTbl, и сейчас о них помнят ТОЛЬКО историки науки. Ортоrенез,
I<О1'ОРЫЙ Лалл рассмзтри.вает пространно и довольно маroс.клон-
но, в на'Стоящей .КJните кратко обсуждается, однз'ко rpaHT отр.идает
эту .концепцию.
Наиболее ,важным событ.ием для эволюцион.н'ой биолоrии меж
ду первым (начавшимся сто лет назад) и вторым (пятьдесят лет
назад) периодами был-о rВозн.и.кн-овеJlие Toro, что 'часто ДОВОЛЬНО
неудачно называют Iменделевской rенетнкой. Б пе-рв-ом издании
своей КlНити (19.17 r.) Лалл дад лишь самое ,элементарное и !}{paT
кое изл.ожение «закона Менделя», сказав в за-ключение, что ОН не
имеет «осеобщеrо значения и нелрименим -ко всем случаям насле-
дования». ОБ Iперес,мотренном издании (1929 r.) изложение было
еще короче, но iВ заключение .было сказаlНО, что «-за'КQНЫ Менделя»
(на сей ,ра'з IBO .множественном числе) «и.меют универсальное зна-
чение и .приме.нимы 'ко осем 'Случаям наследования)). Никакой чет
кой с'вязи 'между «за.конз,м,и Менделя» 'Или быстро раз.вивавшейся
в то ,время rенетнкой и объяс.нением -эволюции установлено не
было. Связь, о .которой iписа,,1И в то вре.мя некоторые rенетИ'ки,
пыла антидЭ'рвин, ОВ=СКОЙ поскольку они предполаrали, что э.волю
цаей упра,вляют 'мутации, :В частности мутации со скачкообразным
феНОТИ/ПИЧеским ПРОЯIВлением, roroдa как естественному отбо.ру OT
БQДИЛ ась ОВ лучшем .случае неrативная роль.
Тем 'Самым мы !подошли к наиБО.'1ее З3'ме1'lНОМУ различию меж-
ду учебником двадцатых rодов и нынешними учебниками,. и это
12
Вступление
разЛ'ичие от.ражает CBoero 'рода 'СУ'бперевор:от 'в .э.волюциО'Н:ной био..
Jrorии, дополняющий .и расширяющий 'Тот перезороТ, ' оторый со-
вершил Да1РВИ'Н .сто с лишн'им лет назад. А.ВТОр наiCтоящей .кН.и'rи
подходит 'к основны'м .положениям эволюционн.ой теории !Прежде
Bcero енетичеСКИ !L!!Q .и..ций. Естественный отбор, хотя он и свя
заи с взаимодеИСТ8ием более обширноrо Kpyra факторов, соrласу-
ется с rенетическим подходом и ero можно обсуждать в терминах
причин и следствий в rеиетических системах, которые сейчас рас-
сматриваются rораздо шире, чем простой мендели-зм.
В своей кни'rе r'paHT .отмечает, чао новое ,на.Пlравд'ение .в 'ЭВQJIЮ-
ЦИОННОЙ теории возникло в Iконце д,вадцаты'х и .В тридцатых Irодах
в .работах ряда биолоrов, сред'И .КОТОрЫХ :внача 'Iе наИ1бол-ее .выдаю-
щи'миея были .работы аlнrЛИЙi:ко:rо статистика Р. Фише'ра (R. Fi-
sher), а'н,rлийс-коrо биолоrа Дж. ХОЛоДеЙ/на (,Наldапе), который из
nрофеос.ора физи'о ,'оrии стал I(Jрофе сором rенети.ки и биохимии, и
амеРИlкаНIQКОro те,нети'ка 'СЬЮЭЛVIа .Райта (.Sewall Wright). .в 'соро-
ковых тодах зт,от новый подх.од К ЭВОЛЮЦИОННОЙ теории дости(r рас-
цвета бла1roдаря .В'кладам, .посryпавшим от всех разделов БИО,,10rии,
среди .которых на1иболее за'метную роль .иr.рали .в то время дан'ные
популяционной rенети.ки, .си'Стематики, палеон'Т'олоrии и ботаники.
В та!ком видоиз.менсн ном и раСillи:реНII!ОМ .ВИде эволюцион,на Teo. I '
рия, .или скорее целыи .комплекс теории, Iпревратилась ,в некии СИН-
тез знаний из м.ноrих ИСТОЧНИ'КIO'.в и поэтому ее обыч,но, так же ,как
и в -этой IКIIиrе, наЗbllвают синтетической теорией.
Как и в любой друrой нау'ке, да и вообще по любому lПоводу,
существуют разноrласия .относитель.но 011ДСЛЬНЫХ ча.стей этоro
комплеюса теорий, OДjНaKO ее OQH08lHbJ i e установки и широкие обоб-
щения Iпринимаюl'СЯ в настоящее :время .orpOMHbl'M бо.тIЬШ.ИНСТВОМ
БИОЛОfОВ. По мере To..ro, ка,к совершаются новые открытия .и изу-
чают-ся новые взаим'освязи, 'Этот синтез .продолжается. rpaHT /ВЫ-
сказывает мнение (в rл. 17), ,ЧТО если будет доказана сущест.в.енная
роль rенетическаrо явления IИНДУ,кции в .процеосе эволюции, то 'эт.()
выйдет за лрсде.IJЫ .си,нтети'Ческой теории. А \м.не очеl'СЯ заметить,
что ЯВ 1ение индукции не останется за пределами 'ЭТой тео.рии, а
раздвинет их .шире.
Людя.м, и в том чtисле 'Ученым, своЙстве-нно вновь O'DKpьr,BaTb,
расширять и IПРИУ1крашивать ста.ры'е идеи, 'которые, будучи оформу
..'шро.ваны .в ,новых терминах, кажутся новыми. Большую чаi:ТЬ
ов{)ей книrи Лалл IПОСВЯТИ.Н адаптивной ра,д:иа.цни, которую. он прои-
писывал rен.ри Фэрфилду Осбо.рну, хотя упомянул мим.оходом, ЧТО
ту же идею под д.руrим назва.нием высказыва.1J Лама,рк (это невер-
но) и что ее выс'Казывал ДараИIН (это правильно). В действитель-
ности .идея об а.да:птивной радиации .была впервые со всей яrc.но-
стью высказана Дарвином, кот.орый отметил в своей автобиоrра-
фии, :в какой и-мен,но момент она 'сто .озарила. Я rоворю 06 этом
здесь именно для ToOro, чтобы ,ПОд'чер нуть, скодь искусно [,рант
Вступление
13
избеrа.ет в С'воей 'Кни'rе при.нятия новых т.ерминов за IHoBble концеп-
ции и идеи. НапрИ!Мер, в на'Ст.оящее JВремя широк-о принят н 'Исполь-
зуется термин «см.ещение п.ризнаlка)), хотя :это еще один <:лучай
примене.ния новых слов и новых пр.имеров .для описания Яiвления,
на которое ясно указал Дарвин. .Б дру.rих .случаях, как в этом
убедит-ся .сам читатель, rрЗlНТ IПрОСТО 011брасыва,ет или полностью
onpOBep'raeT тер.минолоrиче-с,кие ошибки'.
Ero кни.rа не IПрОСТО иэложени-е СИН1'ет.и-че.ской Теории. Она
н-а.писана исследователем, активно участ,вующи-м .8 'раз.витии оонов
этой теории. Автор без \кол.ебаний высказывает свои собственные
70ЧКИ зрения и вы.дв.и:rаетT новые идеи. Книrа и'меет собственное
.,1 ида.
Ha OHeц, у ,некоторых читателей может выэ.вать удивление, что
в "Этом тиро'ком и интенсивном обсуждении ряд аопектов современ-
ных эволюционных ИСС,,1едований едВа упоминае1'СЯ ;или ,даже 'BOb-се
Не затраrИlвается. ТОЛЬ'КО .для T.oro, чтобы доказать это УТlвер.жде-
ине (и ни для чеrо друrоrо), Iприведем IПрОСТОЙ пример: в кн:иrе не
рассматриваются математические модели 'изменений фаУ1НЫ на
островах и в ДрytI'их изо.тIи'рованны'x сообщеС'Рвах. Дело:в 1'ОМ, что
ни -одна книrа не ,может целик.ом объять такой 06Ш 1 И'рНЫЙ -и слож-
ный п.реДмет. как .эволюция орrанИ'змов. Дарвин считал пер,вое
издание «Происхождения IВИДОВ)). объем Koroporo дости.rал
490 сrраниц, Bcero лишь конспектом книrи об эволюции. которую
он собирался пи.сать. Для "1'oro 'Чтобы Iнаписать Од!НУ книrу .на эту
тему, нужно произвести очень тщательный отбор 'материала. Дар-
вин <:.Дела.!] это прекрасно н r:paнT Т-Qже.
Джордж fэйлорд Сuм.nсон
Таксон, Аризона
Зl д-екабря 19761'_
ПРЕДИСЛ!ОВИЕ
Эта .книrа ,посвящена процессам, которые .вывывают эволюци
онные .изменения .орrанизмо.в, и rла.в.ным факторам, 'Оказы'вающим
"Влияние IHa . 'Эти lП.роцессы. Ее название «Эволюция орrанизмов:,-
с..'1едует 'понимать ,БУ' валь:но, поскольку основ.ное в.нимание в ней
уделено эволюции орrа.нwз.мов, в ча-стности ЭВОЛЮЦИИ жи,вотных
и растений, а 'не ,молекулярной эв-олюции, 'примитивной орrаниче
екой эволюции .или .математlfЧ€СКИМ .моделям эв.олюции.
Кроме Toro, в этой книrе оановное 'мест.о аани.мают ПРИНЦИlпы,
а не детали, ,фундаментаЛJ>ные .пол-ожения, а не текущие вопросы,..
и описание .специально .выбранных примеров, а не просто переч.и-с-
ление фактов.
.«э.волюция орrанизмов» ,возн.и,к,па из ПОЛУ'rQДовоrо 'курса 'повы-
шенной сложности, .который я читаю 'начиная 'с 1952 rода. Ero.
обычно 'по'сещают студенты как мла,д.ших, так и старших курсов,.
аопиранты, а .иноrда 'на лекции п.риходят .,даже пр.офеССИоональные
биолоrи. И'менно .этот континrент читат-елей я .и-м-ел в 'виду, КО.fда
.писал «Эволюцию орrанизмов». Короче ,rоворя, книrа была 'заду-
мана и Iна:писа'на ка'к учебник ,повышенной трудности.
Поз.вольте сразу же .в'несТИ ясность. Студенты 'не особенно лю--
бят учебники, которые ЧИТc;lются и вы:rлядят как учебники, но им
нравяТоСЯ хорошие ,книrи, и я ,полностью разделяю это их nристра-
стие. Ита.к, .моей основн-ой целью было наl(JIисать хорошую 'общую'
кни.ry по эволюции, которая моrла бы предста.вить интерес Д '1я
ши-роко-rо Kpyra читате..'1ей; '.в рамках 'этой тла'вноЙ задачи я .и ст.ре..
милея 'п'ридать книrе такую 'фор.му. чтобы .она COOTBeТCTBOBa.rla CJпе..
цифическим потребностям учащих-ся.
Материал раз'бит оНа rла.вы, которые 'сr.руппи:рованы в COOTBeT
ствии 'с тремя крупными у,ровнями эволюцион,ных изменений
!микрозволюцией (iЧзсти II и HI), 'Видообразо.ванием (ча'Сть У) и
макроэв.олюцией (часть VI). 'Каждую часть IМОЖНО чита1Ъ незави
симо от ,дру.roй. Поскольку во ,'мноrих ,вышедших за по-следнее вре-
мя ниrах .по -эволюции .макроэво 'IЮЦИИ уделяется -ма.туо :внимания
Я опециально Iп-остарался дать здесь достаточно .обстоятельное /вве-
дение в эту Iпроблему. Эволюции ,челов.ека :посвящена часть УН.
Теория \эволюции дала в 'прошдом и продолжает давать .до сих пор
немало -пищи для Iра3IВИТИЯ человеч-еской мы-сли .и ,культу.ры; :эти
социальные аспекты ..эволюции вкра1"це рассматри.ваются iВ ча..
сти VIII.
п редислоsие
15
rла\Вы ко.роткие. Только. KpaТtКOCТЬ отдельных ТЛaIВ дала мне
во.змо.жность -ох.ватить очень широ.кий Kpyr вопросов В .пределах
одно.й книrи cpeдJHero. о.бъема. Из ЭIOrо. ,вытекают два следствия:
пер,вое ---- что. .каждая rлава Jп.редставляет собой ,введение. а не ис-
черпывающее рассм.от-рение то.й темы. !Которо.й ОНа посовящена. и
вто.ро.е ---- по.требность в допо.лнительно.м чтении по. то.й или иной
теме, 'затронутой 'в книrе. Я .no\пытал'Ся удовлетворить 'эту по.треб-
ность двумя /Способами: 'снабдив книrу избранно.й дополнительно.й
литературо.й .и да,в ссылки на о.риоrинаЛbJные работы в тексте. .
Специальная -термино.ло.rия в о.б.т:Iасти 'эволюционн,о.й .бао.лomи.
в ,насто.ящее время весьма обшнрона и ов зн..ачительно.й степени из
быто.чна..В дакно.й книrе я разделил .осе с.пециаль"Ные те.рмины на
две ка1'еrо.рии. К первой отнесены термины, неабх,одимые дЛЯ IПО-
нимания -осно.вных ко.нцепций. Эти термины определяются .Н иллю-
стрируютея в TeкCT . Ка второй я отношу те термины. ко.торые "я
считаю пенужными. .но. к<ото.рые друrие исследователи, очевидно.,
тако.выми не считают, поскольку эти терМИ1НЫ ,постоянно. нС!поль
ЗУЮТСЯ представителя:мн то.й И 1И иной шко.лы И "с которыми, следо.-
вательно., учащиеся неизбежно. будут стаЛ'КИ1ВЗ'!'ЬСЯ в печатных тру-
дах, на семинарах .н лекциях. Те,рми-ны, относящиеся к -это.й .вто..ро.й
катеrо.р.ии. IJ1РИВОДЯТ.СЯ в Tel<cтe в ско.бках, обычно в качестве СЯВО-
нимов или :почти СИНОНljМОВ термино.в перво.й ,катеrо.рии.
Па т:I.ный перечень спсциа lЬНЫХ терминов ---- как важней'Ш'Их,
так и вто.ростепенных ---- читатель найдет .в указателе, помещен'но.м
'в конце 'книrи.
Верне Трант.
Техасский университет
Март,.1976
Бл аrодарности
я Чlрез.вычайно мноrим обязан д-ру Джор'д'ЖУ Пэйлорду Ся'Мпсо
ну из Таксана (штат Аризона) за прочтение rлав 26, 28----32 и 37_
Высказа.н,ные д-ром Сим.псоном критичес ие замечания, ero IПредло
жения и 'Сообщенные ИМ данные lIОВЛекли за собо.й Iм,но.rочислен
'Вые ИoCJправления .и улучшения 'в атих .rлавах. Я iВOOполъзовался
также ооветз-м.и д..,ра СИ'мп.сона относительно ,разбивки материала
по. rлаl83"М.
Д-р Роберт Флей к (Fla.ke). из TeXaCCKo.r{) .университета крити-
чооки прочитал rл. 6 -и 14 и -сделал 'ряд ,полезных замечаний.
Д p Теодор Даун.с (Do.wns) из Лос-Анжелесско.rо му.зея eCTe
ствен'ной история 'предоставил мне да,иные из своих о.публ.икован-
ных и ноопу.блиtКованных работ по. иско.паемым лошадям ЭТИ ДaH
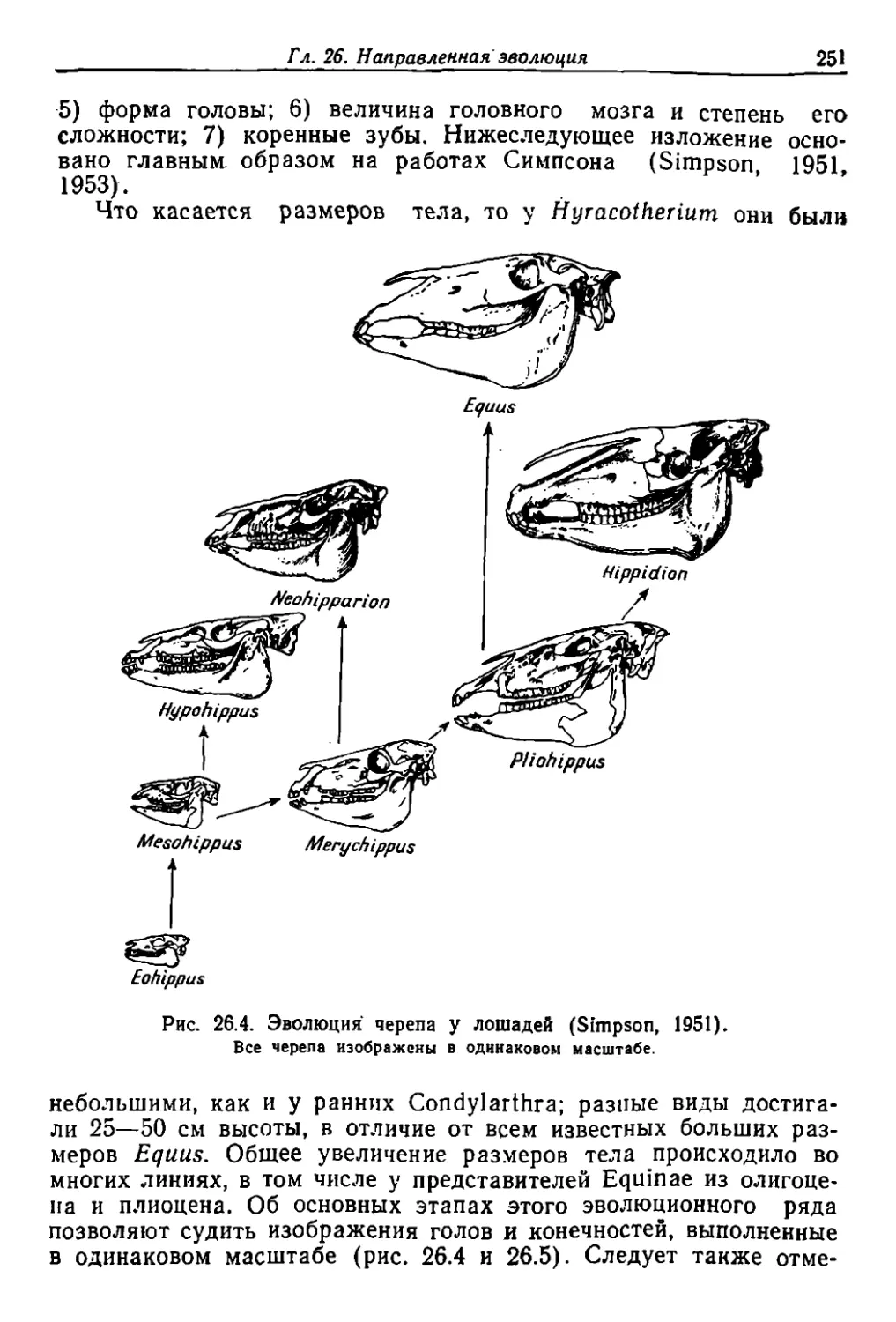
. иые 'вошли ,в .r л. 26.
r жа Карин А. rpaHT ;прочитала в ю ру.копи'Сь, iВЫИ-СКИIВая He
ясности .и устраняя 'Излиш.нее м.ноrocЛQвие. Кроме тor{), она удели
ла M'HOro ,вни'мания чтению корректур.
МОЙ сек:ретарь. r Жа Элен Барлер очень тщательно и аккуратно
перепечатала РУКQI1И-СЬ, под.обрала необходимые .материалы .в биб-
лиотеке и умело 'провела .ВСЮ <Сек'ретар.скую работу, связанную
с изданием ни и.
м..р Джан IПейнте.р из 'фирмы W. ,Н. Freeman and .сотрапу с са..
Moro начала проявил серьез-ный .и.нте'рес к 'к,ниr.е и на 'протяжени'и
Bcero ,п.роцес-са ее издания неи3'Менно оказывал всяческое содейст-
вие. r H Фред Рааб, также .из 'фирмы W. Н. Freeman and Сот'рапу
проя,вил =понимание и высокую '){Iвалификацию при редактировании
рукопи-си.
Я пользуюсь \Возможностью выразить все'м назва,нным лицам
свою ИiC.кр.ен.нюю блаroдарность.
Несколь.КО кни,rо.издателей любез.но разрешили иепольз{)вать
в 'этой книrе уже апубл'Икованные ранее иллюстрации. В подписях
к рисункам содержатся соответствующие указания.
В. Трант
Часть 1
,Введение
r л ав а 1
ПОСТАНОВКА ПРОБЛЕМЫ
Введение
Мир ЖИ'ВЫХ орrанизмо.в обладает рядом. общ.их черт, KOT.opы
в.сеrда .вызывали у чеJlOвека чувс1'В-О изумления. Пер.вая из этих
общих черт ,необычайная СЛ Ж.НOICТь crроения орrаниэмов. Вто--
ра'я очевидная целена!праlВленность ИЛИ приопособительный ха-
рактер мноrих lП"ризнако.в живых орrа,низмов. .Н наконец, т.ретья
ЯРIКО ,выражеНlная общая черта OrpOMHoe раэн.ообраэие жиз.нен..
ных форм. ТаК'им образом, проблемы би.ол.оrИоЧеской сложности и
при'Способленности объединяются самим фактом существова'ния
на 'Свете мноrих различных типов орrанизмов, обладающих этими
чертами.
Вопр.осы, порождаемые :эти,ми я,влениями. 'Совершенно очевид"
ны. Каl{lи.м оора:юм возни'Кди .СЛОЖные орrа,низмы? Под дейсmием
ка'ких -сил ()формир.ова.'IИСЬ .их .приС'пособительные ПРИЗiнаки? KaKO
во происхождение 'разнообразия орrа,ническоrо мира 'и ка'к оно под
держивается? К этому можно добавить более частные, но непо
среДС1iвенно связанные с 'предыдущими, 'Вопросы: какое Mecro за
ни.мает ,в орrаническо.м мире .человек и кто ero :предки?
Во все &еКа человек пытался :найти .на эти 'Боlпросы такие отве-.
ты, которые бы удовлеТВоря.1JИ er.o разум. В донауч,ных обществах
объя.С'нения выливались в леrекды, и некоторые из них ,перешл.и
затем в .реЛИ 1 rиозные учения. Научные объяснен.ия ,воплощены в
теории ЭВОЛЮЦИИ. Однако прежде чем .рас-сматривать эволюцион-
НУЮ теорию, очертим несколь,ко подробнее те проблемы, 'которым
эта теория ДО.'1жна дать объяснение.
Разнообразие орrаническоrо мира
.В современной .фау.не 'насчитывается п.ри:мерноI,З700Jsи.дов мле-
копитающих .Н 8600 :ВИДОВ птиц. ,Описа:но окол.о l20 ООО jБИДQВ совре-
менных рыб. Чис.1JО всех известных ныне живущих видов позвоноч-
ных достиrает примерно 1:2--000
Таблица 1.1..
11римерное число описанных современных видов в разных царствах
н основных rруппах орrанизмов
Оценки для царства животных даны по Майру (Mayr, 1969), а для
цруrих царств по fpaHTY (Grапt, 1963)
Царство и rpуrtпа
Примерное
ЧИСJlО описан.
ных видов
.иарство ЖИВОТIIЫХ
Хордовые
ЧлеНИСТОllоrие
Моллюски
Иrлокожие
Кольчатые черви
Плоские Черви
Нематоды и родственные rpYnnbl
Кишечнополостные
Мшанки и .родственные rpYnnbl
rубки
Различные мелкие rруппы
43000
838000
101 250
6000
8500
12 700
J 2 500
5:mo
3750
4800
2100
Bcero
11 043 900
:Царство растений
Цветковые растения
r олосемеНIiые
Папоротиики и родственные им rруппы
.\10хообразные
Зеленые водоросли
Бурые и красные водоросли
286 000
640
10000
23 000
5280
3400
Bcero
328 320
Царство rрибов
Настоящие rрнбы
МIfКСОМИЦСТЫ
40 000
400
Bcero
40 400
Царство протистов
Проетейшие. одноклеточные жrутиковые BOДOpOC
ли, диатомовые
30 000
Bcero
30 000
r л. 1. Постановка проблемы
19
Uapcno u rpУПП8
Примерное
число описан..
иых видов
llapcTBo монер (прокариоты)
Сине зеJ1сные водоросли
Бактерии
1400
1630
Bcero
3030
Вирусы
200
Итоrо
J 445 850
в некоторых друrих КРУlПных .rрупnах чи'Сло известных coв.pe
менных ВИДoQв rораздо 'выше описа.но 107000 в.ИДQВ 'моллюсков,
286000 видов цветковых растений и 750000 видов насекомых..
Оценки чисда ВИДОВ .В ра!зных КРУil1НЫХ трyrппах .орrанизмов приве
дены !в та1бл. 1.1. Как видно из эroй таблицы, общее число из'вест.'
ных -видов живых .существ в современном мире доститает почти.
1,.5 млн.
'СЛoQжнеЙшая задача та ксон()м.ичеокоrо изучен'ия и описания до-
вольно хорошо решена для таких rрУIПП, как птицы 'и млекопитаю-
щие, но далека от завершения .в .большинстве остальных .rрупп.
Значительное чисдо мо.рских :беспозвоночных, ЦBeT.KOIВЫx растений
н 'предстаlвителей друrих IfрУПП все еще ждет e,Boero таКоС()Iномиче-
cKoro о()писа.н'Ия. По .оценкам ихт.иодоrов, общее число 'видов COBpe .
менных 'рыб .при.ближается :к 40000, ИЗ кот()рых пока опи'Сз.НО лишь
20000. Объ-е.м ,работы, которую предстоит .проделать СИоСтематикам
насекомых, еще боль.ше. По мнению энroмолоrов, описанные дО
с.IIХ пор Iвиды насекомых (около 750 000) составляют лишь небоol'Ь
шую долю, возможно, одну ПЯ1'ую 'или одну .десятую, 'всех в.идов
насекомых, дейст,вительно существующ'их на Зе.мле в настоящее
время.
Если принять довольно скромные допущения о соотношении из-
Dt.'CTHblX и ,неизвестных 'видов .08 различных крупных r,руппах, то
можно IIIОЛУЧИТЬ tпрИ'близительные оценки общеrо .разнообразия
сущест.вующих в'идов. В своей .предыдущей книrе я указывал, что
-ЧНСЛО .ныне живущих видов составляет не менее 4,5 'мл.н. (Grant,
1963) .
Были сделаны две независwмые оценки числа 'видов, как ныне
Ж\I.В'УЩИХ, так и вымерших, существова,вших 'на Земле за .ВСЮ ее
f('ОЛoQrиче.скую исто'рию. Диа,пазоны этих оценок в значительной
МС'ре перекрываются. ,И.сходя из предварительных данных можно..
20
Часть 1. Введение
принять, что общее 'Число ,ВИдов, .населя,вших земной шар с мом-ен-
та возникновения жи'зни, составляет от QДHoro до нескольких мил-
лиардов (см. Simpson, 1952; .Grant, 1963).
п риспособлеllие
. Мноrие на-следственные !Признаки орrанизмов соот,ветствуют той
или иной особенности их ()бычной среды обитания н, таки.м обра-
зом, б.Т'Jаrоприятствуют жизни ,орrа,низ.ма в этой .среде. Эти призна-
ки называют-ся приспособитель.ными.
Известный естествоиспытатель XVIII в. Бюффон описал в С.Б-оей
«Естественной И'стории» м,ножест.во лриспособительных ,приз.наIКОВ
птиц, РЫ1б и д:руrих животных. При-ве.денное ниже тапичное утверж-
дение (Buffon, 1770, 1808) ,иллюст,рирует, 'как .был ,&веден в упо.
треблени.е :этот термин.
«Что касается .наружноrо .строе.ния Iптиц. 1'0 .ОНО уди.вительны:м
образом приспособлено ,к 'бы трому !передвижению; оно... предназ
на'Чено для 1'oro, чтобы .подни.маться в воздух... .Обширные .приопо.
собления средств к целям {наблюдаются] в общих контурах поро.
ды лернатых...»
Для Дарвина ИЗJlюбленным пр'имером приспособ.'lения с.Т']ужил
дятел. Достаточно привести один отрывок из «Происхожде,ния ви.
дов» (Dапviп, 1859, rл. 6):
«Мож.но ли ПРИiВести более разительный ПРИiмер .приопособле
ния, чем дятел, лазящий IfIО стволам деревьев и выла.вля,вающий
насекомых IB трещинах коры?»
ПредJстаlВи.м себе всю совокrупность прис<пособительных [lризна
КОВ дятла: долотообразный клюв, прочный череп. мощные мышцы
rоловы, выJвиrающийсяя язык с острыми щетинками на :конце,
лапы с за-остренными коrтями на пальцах, :причем два лальца .на-
правлены вперед и два назад. что помоrает удерживаться на Bep
'rИ 1 каJ1ЬНОЙ поверхности, и аЧ .нь Iпроч.ные рулевые ;перья, служащие
опорой д.Т']я тела 'птицы, коrда она спдит на отвесной по.верхности
СТВ{).l а.
ДруrИ:\f 'К.'1ассичоским примером приспосоf)дений служит CTpoe
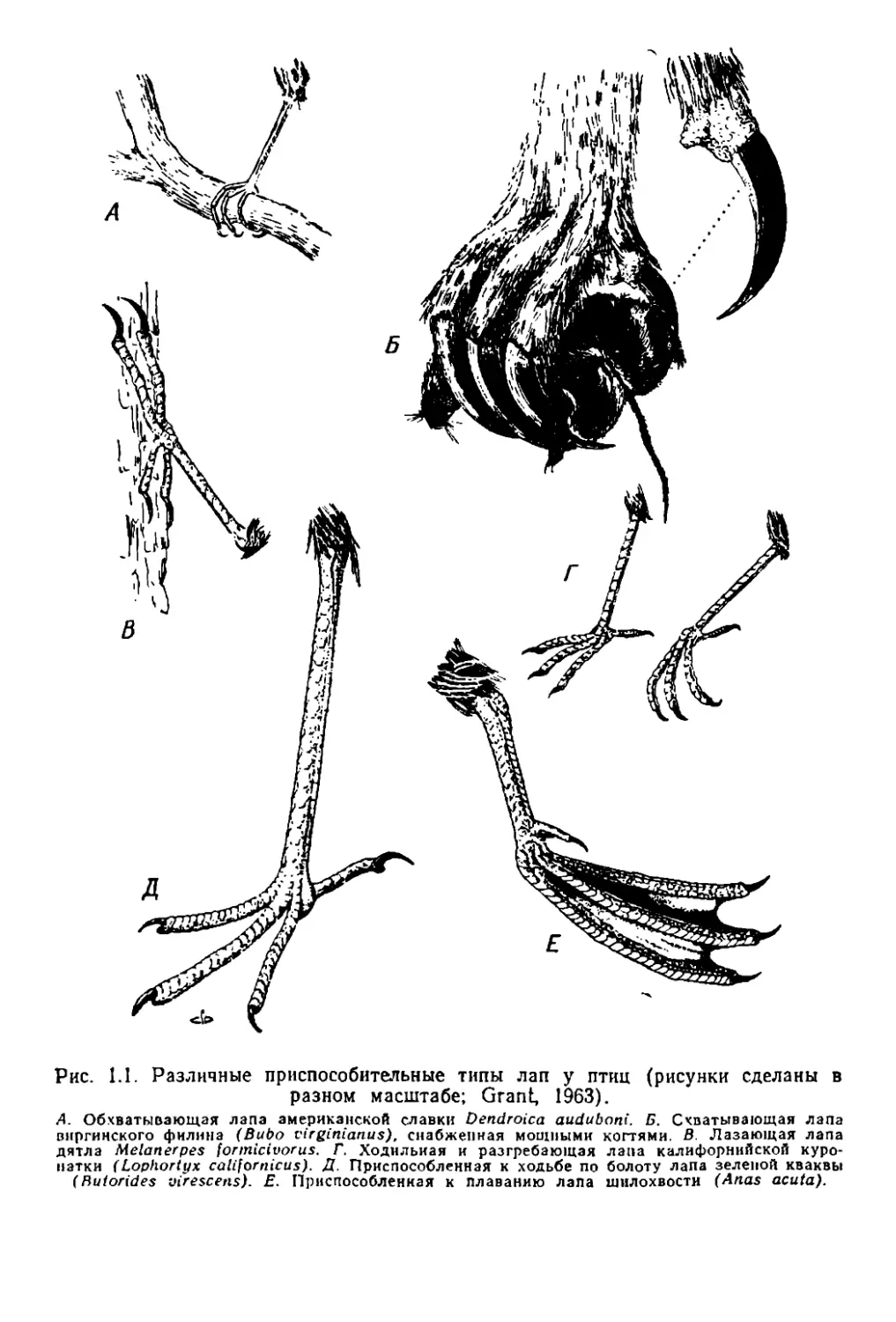
иие лапы в разных rруппах птиц (ри-с. 1.1): .приспос.обленная 'к ла
занью лапа дятла (8), оБХ1ватывающая лаlпа американс.к.ой C.тIaB
kи (А), Iсхватывающая лаlпа фи..Т'J'и-на (Б), ХОДИЛhная и разrребаю
щая лапа куропатки (Т), .при пооО'бленная к ходь:бе 'по БО 10ТУ
лапа .кваквы (Д) и перспончатая, приспособ.lJенная к П lаваниIO ла
па 'шилохвости (Е).
Следует различать общи€ и опециаЛI>ные адаптации (Sim.pson,
1953). Пер,вые оБУiCловл.ИlваlOТ триапособление ,ор'rаwизма к жизни
в :каКОЙ ЛИlбо обширной зоне 'среды, а вторые 'представляют собой
специализации к. 'l'OMY или иному ,о.пределенному образу жизни.
Так, напри.мер, .крыло птиц..... это -общая адаптация, а ДО.Т']О1'ообраз
\
,.
(/
в
Б
Рис. 1.1. Различные приспособите.льные типы лап у птиц (рисунки сделаны в
разном масштабе; Grant, 1963).
А. ОБХВ3ТI>Iвающаи лапа американской славки Dendroica aиdubпni. Б. СхпаТЫВ31Ощая J1ana
Вllрrинскоrо филина (ВиЬо t'irginianиs). Сllзбжеllная мощными коМ'ями. В. Лззающаи лапа
дятпз Melanerpes foпnicivorиs. r. ХОДИilьиая и разrребзющая J1Зllа кrМИфОРНIIПСКОП куро-
IIЗТКИ (LopllOrtyx саЩоrrziсиs). д. Лриспособленнзи к ходьбе по бопоту лапа зеJ1еной кваквы
(Rиiorides virescerts). Е. I1jJllспособленкаи к ллаванию пала ШIIJ10ХВОСТИ (Aпas acиta).
22
Часть 1. Введение
ный клюв и лазающая лапа ДЯТЛОВ адаптации специальные.
Общие адаптации входят в число 'отличительных признаков ,круп-
ных JlРУПП орrаниэ.мов.
Общие и специальные адаптации присущи всему живому. Но
приспособительны ли все признаки пли имеЮ7СЯ н епри пособи-
тельные п,ризна.ки, .встречающиеся со Iсколько нибудь значительной
частотой, друrой волрос, требующий особоrо о.бсуждения. Мне-
ние биолоrов по этому вопросу за последние сто лет колебалось
то IB .одну, то в .Д'руrую сторону. В недалеком прошлом неКО1'орые
морфолоr.и и .си.стематики сомневались в приспособительном 3Ha
чении Мlноrих предположительно .носущественных МОРфО.'Iоrичесжих
признаков, подобных тем, 'которые используются в определитель..
ных та,блицах. А ныне некоторые 'школы .биохимиков и биоматема-
тиков .считают .неприспосо6ит.ельными ;ряд из.М'енеНИЙ белка.
Вопрос о широком распространении 'нетtpис.поообительных rпр.из-
нак.ов иикоrд.а не удастся .реш.ить окоН'чатель'но, поокольку для
этоrо Iп-ришлось .бы до.казать общеотрицательное утв-ерждение, т. е.
пронзвести IПОЛНЫЙ .перебор всех признаков и доказать отсутствие
приспосО'бления IЮ всех случаях, 'однако мож.но привест.И несколько
сущеСllвен.ных соображений. Возможно 'или даже вероятно,' что.
имеются отдельные .признаки, !не .обладающие :ПРИQпособительным
значением, и что иМею1'СЯ также Iэволюционные .механизмы, закреп-
ляющие та'кие лр-изнаки. И все Же твердо .назвать т.от или иной
признак неприспо<;обительным очень трудно. Мноrие склонны счи
тать неприспособит€льным любой признак, пр.испособительные
свойства KOToporo неочевидны. Подобные антропоцентрические нс-
тол'Кования природы MorYT 'приводить к .неверным .выводам.
На первый Jвзrляд может Iпоказаться, что различие .между окра..
шенным .и белым луком не имеет n:риспособительноrо значения_
А между тем .Кlраоный .и желтый ЛУIК уст.ойчи,вы к -за'болеванию
антракнозу, вызываемому rрибом Colletotrichum cinciпaпs, тоrда
как 'белый лук чувствителен к этому -возбудителю. УСТОЙЧИВОСТЬ
к заболеван,ию создается .блаrода.ря нал.ич;ию в луковицах катехи
.на и п,ротокатехов-ой кислоты, токсичных для .в-озбудите..тrя, а эти
соединения связа.ны с крас'ным и желтым ,пиrментами, 'котО'рые со..
держатся в лу.ковицах (Jones et al., 1946; W,al.ker, Stahmann, 1955
Levin, 197.}) . .
Эволюционные объяснения
J'акие .бросающиеся .в rлаза явления. как сл.ожность живых
орrанизмов, 'их разнообразие и приспособленность, ,наука объяоС'НЯ-
ет тем, что эти я,вления возникли в проц-ессе эволюци.и. Существу
ют два 'подхода к изучению .эволюционных с.обытий: исто.рический
и прич.инный. .Пр.и :первом :прослеживают ВСЮ филоrенетическую
последовательность .событий, ,I1IРИВОДЯЩУЮ к наблюдаемому .конеч..
r л. 1. Постановка nроблеJ.fbL
. ;.
.:.
ному результату, тоrда как при втором и учают уча.ствующие в
этом причинные механизмы. Оправданы и' необходимы оба под-
хода. В данной книrе рассматриваются r.'IaBHblM обраЗО 1 причи
ны эволюции, однако при этом нередко приходится прибеrать к
-филоrенетическим данным.
Мы будем иметь дело с эволюционными явлениями, происходя
щими на трех rлавных уровнях. Это ---- эволюционные изменения
в .преде..1ах 'ПОПУ.'lяций (микроэволюция), 'эволюция подвидов и ви-
дов (видообразование) и 'ЭВОЛЮЦИЯ крупных np}'inn (м'акроэволю
дия). Каждый из этих уровней требует различных методов иссле-
дования, что в свою оч'ередь ,пр,иоодит к 'получению данных ра;зноrо
1'И'П а.
В этой книrе ,микроэволюция, видообразовани-е 'И макроэ:волю-
ция рассматриваются .по отдельности .и в том 'ПОРЯдке, iB каком они
здесь .перечис..1J6ИЫ. Порядок .изложения важен с точки зре-ния
лоr.и.ки, пос-кольку ,интерпрета.ц'Ии явлений, rПРОИ.СХОДЯЩИХ .на ypOB
не ,макроэволюции, для Toro чтобы .их можно .было считать обос-
нованными, должны ;соотв.етст,вовать данным, Iполученным !Прli изу-
чении .ми.кРОЭВOoi'IЮЦI:I'И..
Часть I1
МИКРОЗВОJlЮЦИЯ
r a:Ba 2
С РЕЩИВАЮЩАЯiCЯ rП,ОПУЛЯЦИSI
Введение
Эволюционное ,изменение 'это явление, происходящее в попу.
ляциях ,н системах ПОПУЛЯЦИЙ. iВ своей ,нзИ!БО.ТJее простой форме
эволюция имеет M TO в '-пределах локальной -с:крещивающейся по
пуляiЦИИ. Это .и есть IМИК,РОЭ.ВОЛЮЦИЯ. Таким образом, локальную
скрещивающуюся 'популяцию 'можно расс.матр.ивать как плацда1РМ
для микроэволюции. Поэтому наш обзор процессов МИКРОЭВо.пюции
необхсщим-о начать с рассмотрения общих ,признаков популяций.
Мы не можем ,п.редложить к'акое-л.ибо общее форма.,'Iьное опре-
деление локальной скрещивающейся .популяци.и, которое служило
бы эталQ.НОМ для всех -случаев. Это объясняется широ.ким lразно-
образием ,реально существующих iПОПУЛЯЦИЙ. Одни локальные по-
пуля.ции дейст,вительно имеют четкие лра.ницы, Др}"f,ие не имеют
определенных собственных r.раниц, а входят в соста:в более об-
ШИlрных .и сложных систем популяций. Удобно ,представ.'JЯТЬ себе
популяции и систе,мы ПОПУЛЯЦИЙ в .вид-е иерархии, начинающеЙся
со СЛУ'чайно скрещи.вающейся :популяции и доходящей до вида
(более .подробно эти единицы ..будут раС<:МОТрЕШЫ в дальнейшем).
Таки.М образом, локальная скрещивающаяся популяция, как под
разумевает .ее .название, 'это .популяцион.ная единица, имеющая
некую локальную протяженность в такой более или мен-ее непре:-
рывной .иерархии.
Структура ПОПУЛЯЦИИ
СкреЩИВЗlOщаяся попу.rIЯЦИЯ представляет собой репродуктив
ную единицу. У орrанизмов с половым размножением это сооб-
щество особей, связанных между собой узами скрещивания и взаи.
МООТJlошениями родители потомки (Dobzhansky, 1950). У opra-
низмов с бесполым размножением сохраняются связи родители
потомки, но перекрестные связи между особями, обусловленные
скрещиванием, сильно ослаблены; не следует, однако, ПОЛllОСТЫО
["д. 2. С"рещивающаяся популяция
25
исключать .наличие у бесполых орrанизмов таких ,е-вязей, .посколь-
ку у мноrих орrан.и.змов, которые обычно размнож ются ,бесn-олым
путем, существуют те или иные пара ексуальные .способы раЭМRО-
жения или же .время от В'ремени ,происходит .воз'врат к половому
размножению. Популяция, -ка'к правило, /Представляет сооой ,с.80-
бодн.о скрещивающуюся rруп-nу независимо от Toro, ;прои ходит Л,И
скрещивание с друrими популяциями реrУЛЯрНQ или эпизодически,
и .во ,всех ,СЛ)'Iчаях !это некая .рetnродуктивная .еди'ница.
.Популяция -представляет со.бой также эколоrичеакую еди'ницу.
Составляющие ее особи rенотипически сходны по своим эколоrи-
ческим требованиям и занимают определенную область в той или
иной ЭКОЛоrической нише или местообитании.
Pea.,1JbHble популяции весьма разно.обраоЗНЫ ,по величине и фор..
ме. Величина ,популя'ции, ПОД КQТОрОЙ .имеется .В .ВИДУ ЧИСЛО взрос-
лых размножающи.хся ос.06ей в каждом поколении, может .варьи..
ровать от одной илп неСКОJ1Ь'КИХ до мноrих миллионов особей.
Стру,ктура .fl(JПуляции слаrается из т.рех rлавных компонентов: IПрО
странственной конфиrурации, системы ,разм'ножения и -ско.рости
миrрации (последний ком!пон.е:нт .рас'сматривается в rл. 6).
Что каlCает,ся пространственноrо распrpедел'ения по.пул ЯlI.ии , то
здесь мо.жно. 'выделить три .осно.в'Ные катеroрии: 1) Болышеe ,не.ире-
рывные популяции; .2) мелкие колониальные популяции (или попу-
ляцил, .соответствующие остроВ/ному типу); 3) линейные популя..
ции. Кроме r.oro, существуют .разнообразные .состояния, промежу-
тачные между этими тремя основными т.Иinами.
Примером больших непрерывных популяций служат популяции
злаков, растущих .на ,равнинах и 'ПОКJрывающих .площади ширИ'ной
в десятки ил.и 'сотки километров. Кол()ниальные орrанизмы обра-
зуют .ряд разбросанных, разо'бщеНiНЫХ и .передка мелких популя..
ций. Пр.имерами .служат 'назем,ные животные, обитающие иа архи..
пелаrах, прес.н.оводные формы, населяющие цепь озер, обитателя
r0p'HblX ве.ршин в rорной .местности и орrани3.мы, оrран.иченные оп-
ределенным ткпом lПоч.вы ,или r,орной породы .с пятнистым lJJаспре..
делением. Линейные ПОПУ.'Iянии Iвоз.никают вдоль рек, 'на 'побережь-
ЯХ морей и в аналоrи'Чных .местооб.ита:ниях, обладающих 60ЛЬШОЙ
протяженностью ,и ,более Iили менее непрерывных 'в одном .измере-
нии, но КОРОТКИХ и {)rраниченrных в друrом.
Часro встречаются и ,раЗЛМ:'ЧiНые .промежуточные .состояния.
Большая популяция м'ожет Iбыть неп,ре'рывной ,в одних частях за-
нимаеМой ею обдасти, но ,прерыв.истой .или полунепрерывной (в 'дpy
тих. Подобным же образом колонии, населяющие систему островов,
мотут быть изолир.ованы лишь частично, а .н.е ,полностью. В сле
дующем разделе ,мы .опишем ,конк.ретный 'прим'ер ---- структуру IПОПУ-
ляци.и rиrантской секвойи, или мамонтова дерева .(Sequoiadeпdroп
gigaпteum), в .которой .сочетаются самые Iраз.нообраэные состояния,
25
Часть /l. МuкроэвОАЮЦUЯ
от . ОЛJf'Рованных , олоний на севере до прерыви-сто.rо лесноrо
пояса на юrе.
Что касается типов скрещивания, то их ,диапазон очень ши-
po .oT свО'БО'дноrо нерО'дственноrО' сжрещива'ния дО' амО'оплО'"
дО'творения. Часто встречаются такие промежутО'чные типы, как
свО'боднО'е ск.рещи;вание .м-ежду близкими соседями; .ин.бридинr,
О'существляемый иными спО'сО'бами, нежели самооплодотворение
(налр.и.мер, скрещивания :между оСкбсами у живО'тных); сочетание
неродственнО'rо оСюрещивания :с 'СамоО"пыл.ением, как у rермафрадит-
ных, НО' .самосовме тимых цветковых расте.ний.
П,ространствен.ные .канфи.rурации и системы .скрещивания .встре..
чаются 'во .всевО'зможных сачетаниях, создавая в ,реЗУ.'Iьтате чрез-
. ычайна разноО'браз:ные CTPYJKTypbl IПО'ПУЛЯЦИЙ. TaiК, БО'льшая непре-
рывная J}<JПУЛЯЦИЯ мО'жет .састоять .из своБО'днО' скрещивающихся
особей, KaJK у мнО'rих опыляемых IBeTpO'M травянистых растений ,рав-
ни.н, нО' ОНа может так,же состоять из .особей с О'rраниченной
своБО'дой скреЩ.Иiвания или .из .инбредных особей. Такая структура
популяции оказывает 'влияние на характер ее изменчивости, о чем
будет сказано:в ;дальнейшем.
Популяции мамонтова дерева
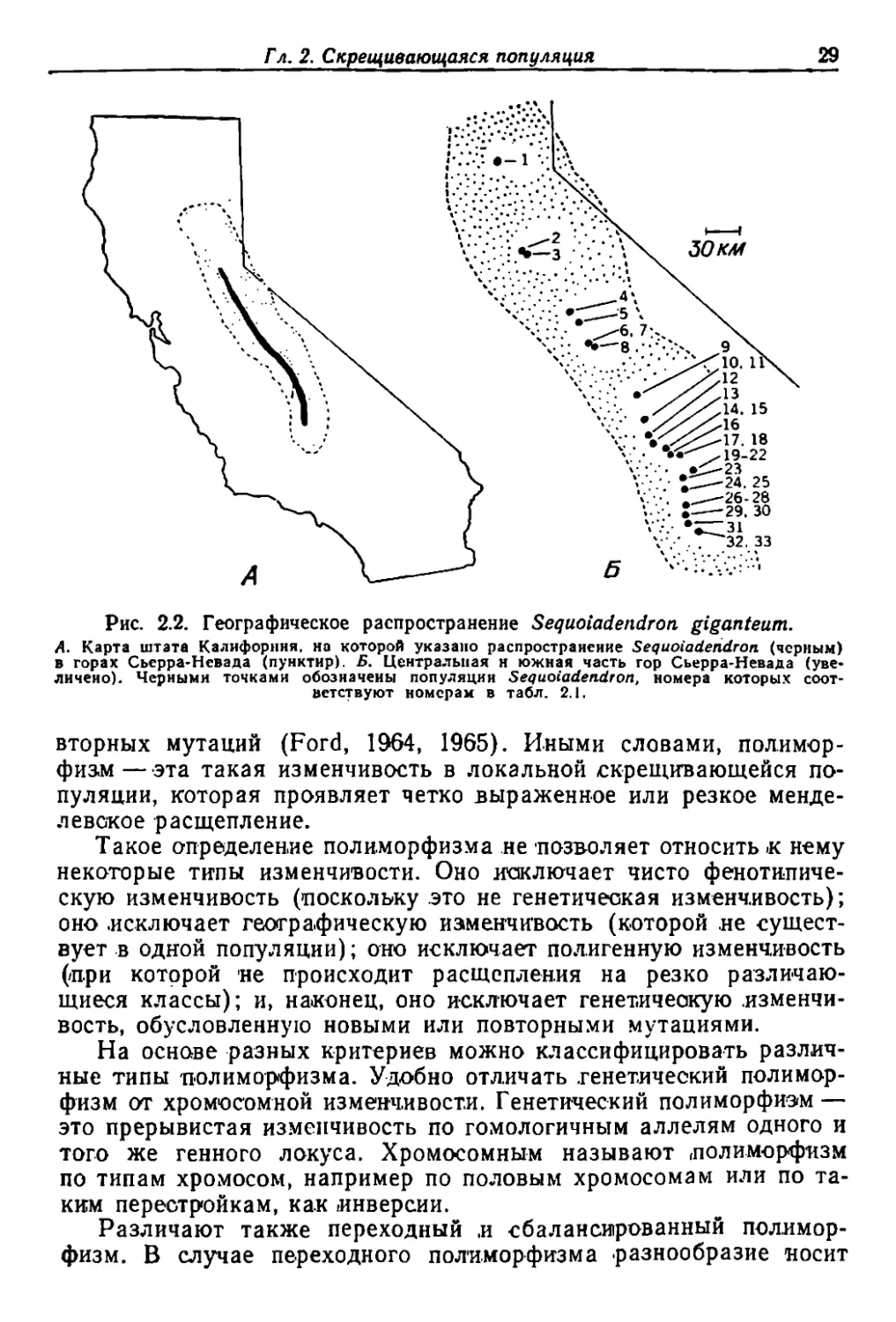
Ма.монтО'Ва дерево (Sequoiadeпdroп gigaпteum) ----о перекрестно..
О'пыляющееся IПрИ по МОЩ.И. .ветра ХВОЙНQе дерево ---- встречается
в с,оС'ново :пихтовых лесах на небольших вы.сотах (1500----.2400 м)
на западных склО'.нах rO'p Сьерра.l.Невада в КалифО'рнии .(рис_ 2.1)_
Область era .распрастранения образует узкую полосу прО'тяженнО'-
стью ОКО'ЛО' 400 км (р.ис. 2.2,А). В Iпределах \всей этО'й области ма-
мО'НТОВО' дерево ;в'стречается ,в виде 'ряда ,обособленных ,и ,БО'.,1ее ,или
менее разобщенных .популяций (рис. 2.2, Б).
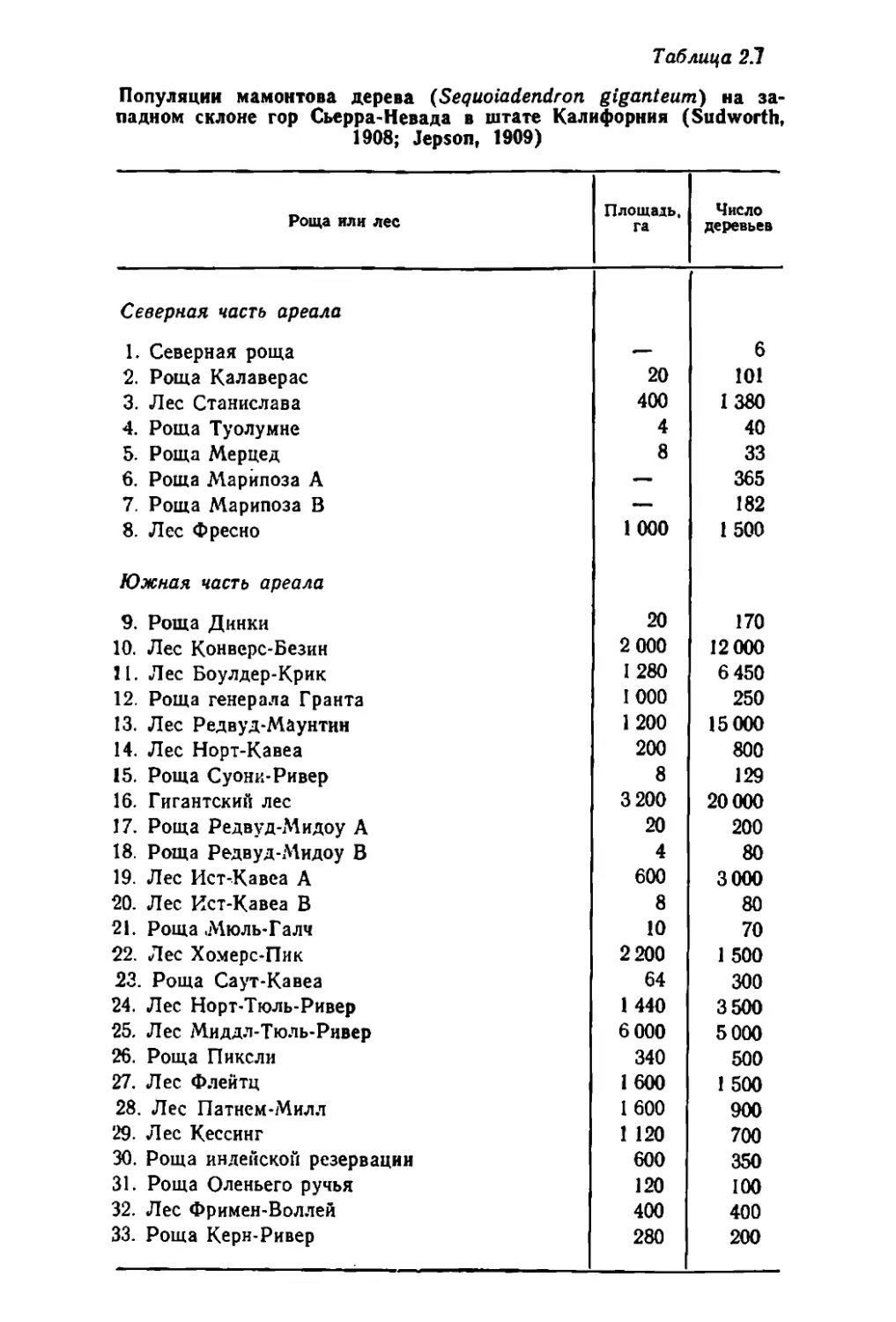
Существует ,примернО' 33 локальные популяции ма'монтО'ва дере-
Ва. В самом начале ХХ в. они ,имели размеры, yrказа'нные в
1'а'бл. 2.1. Бал'ее мелкие популяции .называют ращами, БО'лее круп-
ные лесами. В Iпериод :пр.оведения учетов, результаты каторых
представлены в табл. 2.1, эт.и леса :и.нтеасиiВ-НО вырубали.сь. Поэто
му данные табл.ицы .отнО'сятся ,к прежним, ненарушенны'М естест-
венным популяuиям. Ка,к пО'кЗ'зьrвает табл. 2.1, 'размеры пооуля..
ЦИЙ СИ.1JЬНО варьируют от ращиц, состоящих из нескО'льких де-
ревьев, да лесов, насчитывающих мнаrие тысячи особей.
Северная и южная части в'идовоrо ареала разл.ичаю'J'oCЯ по
структуре ПQПУЛЯЦИЙ. В севернО'й части ареала мамО'нтО'во дерево
абыч.н.о растет в вцде мале,ньких, СИЛЬНО' разобщенных рощиц.
Расстояния .между рощицам'и .передка соста.вляют от 15 до 80 КМ_
В южной же части БО'лее обычны, или, ве.рнее, были обычны, Iболь
шле леса, разделенные БО'лее уз.ким:и 'ПрО'странствами. В некоторых
случаях через эти разры'Вы как бы 'перекинут мост образуемый ОТ-
Таблuца 2.1
Популяции мамонтова дерева (Seqиoiadendron giganteиm) на за-
падном склоне rop Сьерра-Невада в штате Калифорния (Sudworth,
1908; Jepson, 1909)
Роща ИJlИ пес
Ппоща.IЬ, Число
ra деревьев
Северная часть ареаАа
1. Северная роща
2. Роща Капаверас
3. Лес Станислава
4. Роща Туопумне
5. Роща Мерцед
6. Роща Марипоза А
7. Роща Марипоза В
8. Лес Фресно
1000
Южная часть ареаАа
9. Роща Динки
10. Лес Конверс Безин
21. Лес Боулдер Крик
12. Роща reHepa.lfa [ранта
13. Лес Редвуд-Мзунтнн
14. Лес HopT KaBea
15. Роща Суони-Ривер
16. rиrантский лес
17. Роща Редвуд Мидоу А
18. Роща Pe-двуд Мидоу В
19. Лес Ист .Кавеа А
'20. Лес Ист-J(авеа В
21. Роща .Мюль- r алч
22. Лес XO}fepc-Пик
23. Роща Саут-Кавеа
24. Лес Hopt-Тюль Ривер
25. Лес м.идд.'J Тю.'1ь-Ривер
26. Роща Пикели
27. Лес Ф.'lейтц
28. Лес Патнем-Мил.'1
29. Лес J(ессинr
30. Роща индеЙСJ<оii резервации
31. Роща Оленьеrо ручья
32. Лес ФримеН-ВОЛ.'1ей
33. Роща Керн Ривер
20
2000
1280
1000
1200
200
8
3200
20
4
600
8
10
2200
64
1440
6000
340
1600
1600
1120
600
120
400
280
20
400
4
8
6
101
1 З8О
40
ЗЗ
365
182
1500
170
12000
6450
250
15000
800
129
20 000
200
80
3000
80
70
1500
300
3500
5000
500
1500
900
700
350
100
400
200
28
Часть 11. Микроэволюцuя
Рнс. 2.1. ]\'1aMoHTuBO дерево (SequoiacZeпtiroll gigaпtellm).
деЛЬНЫМJI деrеВЬЯМII, так что 'ПОПУЛЯЦИН ,объединены 18 по.пу.непре
рывныЙ ПОЯС.
'ОБШlIрные разрывы ,;',Н 'ЖДУ северными рОЩJ!uа t.Н соотвеТ1СТВУЮТ
ДОЛlIнам. l<ОТОРЫС во время поuюднеrо оле,денения были заняты
,'IьдаМIJ. Пред;полатают, что до наСТУlПления последнеrо оледЕ':НСНИЯ
ПОПУЛЯЦИИ aMOHTOBa дерева в центра.IJЬНОЙ части rop Cьeppa He
вада бы,rш ,бо.'1ее аБшIlрныIии и непреРЫВНЫ}"ll1, но под действием
,'IеДНIIJ\овurо КЛ,lIмата ,СОJ..:раТИЛJIСЬ 11 разБНЛIIСЬ на части и .с тех пор-
не MorYT вновь занять утраченную теРРПТОРIJЮ. В южной чаСТII
Сhе'Рра Невады олеДе'ненне не оказало столь CII.rIbHOTO 'ВЛИЯНJIЯ на
пО'пуляulJ.И мамонтова дерева (Sudworth, 1908 Axelrod, 1959).
Полиморфизм
Соrласно определению, IПОЛНМ,орфизм это сосуществование в
популяции двух ПЛII более резко различающнхся (прерывистых)
форм. при котором частота более редкоЙ формы выше частоты по
r А. 2. С"рещивающаяся nОnУАяция
29
А
!.:}< i ::[; \
\(\,, Ii, ig} \
",: . : . .............6. 7',
":: :: " 8.': ::' "" 9
,'.' ....:.......;....()Оl1
.,<.:}'.. .. l!: 15
\.: . 16
\:: ... 17. 18
\." .. /19 22
\'. :.... .!-----"" 23
,.::: ................24.25
....... ................26- 28
,"'. . 29. 30
\<. . Зl
Б \<::. .; : ::-.\з
.,...:....,
.. ,
. ..
.,
Рис. 2.2. rеоrрафическое распространение Seqиoiadeltdron giganteит.
А. Карта штата Калифорння. но которой указано распространение Sequoiaderidron (черtlЫМ)
в ropax Сьерра-Невада (пунктнр). Б. центра.,\,ыlяя н южная часть rop Сьерра-Невада (уве-
пнчено). Черными точками обозначены попу.1ЯЦИН Sequoiadendron, номера которых COOT
ветствуют номераи в табл. 2.1.
вторных мутаций (Ford, 1964, 1965). И,ными словами, пол.имо()р
физ.м 'эта такая изменчивость в .rIокальной .скрещИ'Вающейся по-
пуляции, которая проявляет четко выраженное или резкое менде-
.'1евокое 'расщепление.
Такое определение поли.морфизма .не 'позв.О.'Iяет относить ,к н-ему
некоторые типы изменчи'Вости. Оно lIOКлючает чисто феноти:пиче-
скую изменчивость (ПОСКО.'1Ьоку .это не rенетичеокая изм.енч.ивость);
оно .ис.ключает reorрафическую изменчи-вость (к,оторой .не сущест
вует .в одной популяции); оно исключ.ает по.71.иrенную изменч.И'ВОСТЬ
(/п.ри котррой 'не происходит раСЩСПJlения на резко различаю-
щиеся классы); И, на.к{)нец, оно ис.кл'ючает rенет.ичеакую .изменчи-
вость, обусловленную новыми иди повторными мутаииями.
На осно:ве .разных критериев можно классифицировать различ-
ные типы полиморфизма. Удобно отл.ичать .rенет.ический полимор
физм от ХрОМI()С'ОМ"НОЙ из Ме'НЧ,И ВОСТ.И. fенетичес.кий полиморфл'3tМ ----
это прерывистая измснчивость по rомолоrичным аллелям одноrо и
Tor.o же reHHoro локуса. ХРОМОСОМНЫМ называют rПОЛИМОрфИЗМ
ПО типам хромосом, например по половым хромосомам или по та-
ким переотprойкам, ка.к rлнвереии.
РаЗ.о1ичают также переходный .И сба.rIанс.Иlрованный пол.имор-
физм. В случае пе.рехоДноrо ПОJ1'и.морфизма ,разнообразие 'Носит
.за
Часть 11. МUКРОЭ80Аюция
'временный характер; оно на,блюдается до тех пор, пока происхо-
дит процесс замещения одной формы друrой IПрИ контролирующем
деЙствии eCTecТoBeHHoro отбора. Пр.и сбаланс.ированном полимор-
физме разные типы представляют собой более или .менее постоян-
ные ; ()Iм;поненты данной популяции бл аrодаря естественному отбо
ру, который бла,rопр.иятствует сохранению разнообразия (Ford,
1964, 1965). .
Все формы поли,морфизма reнет,ичеокий, хромосомный, пере-
ходный ,и .сбаланс,ир<>ванный весьма обычны IИ 'широко расIJoрОСТ
ранены J3 живой природе. В популяциях орrанизмов с половым раз-
множением .полиморфизм в !Сущности наблюдается всеrда. В сле
дующем разделе .мы опишем КОНlкрет.ный пр.имер пол.И'мо.рф.изм
по rруппам .крови у чеJl'ов-ека.
Явление полиморфиз.ма .подводит нас ,к концепции rенофонда,
.которая в свою очередь дает нам оозм'ожноCiЬ ,по И'ному вз,rлянуть
на локальную скрещивающуюся iПОПУЛЯЦИЮ. Рассмотрим популя
дию, ПОЛИМОРФНУЮ по reHY А 'и содержащую аллели A 1 , А 2 .и Аз.
В такой iПОПУЛЯЦИИ будут возникать дипло.идные rенот.ипы A 1 A 1 ,
А 1 А 2 , А 2 А 2 IИ т. 'л. И обусловленные ..этими rенотипами фор.мы .будут
.представлены :в любой выборке .особей, однако ясно, что одна из
.rлавных черт данной популяции '31'0 rенети'ческий !поЛ"иморфизм,
лежащий !в основе Iнаблюдаемоrо .разнообразия. ,Можно сказать,
чro rенофонд та,кой популяции содержит алле.л.и A 1 , А 2 .и Аз. Кро-
ме Toro, эти аллели nстречаю'ОСя в rенофоН!де iC олредел-енной ча
-стотой; допу-стим, что их частоты равны -соответственно 60, 30 и
1 О %. Следо'ВатеЛЬНQ, IПОПУЛЯЦИЮ .мОЖно ,описать IКО..1'ИЧGCтвен'но, 'ис
пользуя типы reHOB, содержащихся IB ее rенофонде, и .их част.оты.
Следует от.мет.ИТЬ, что концепция rенофонда !Шаре, чем концеп-
ция лодим,орфизма. rенофонд ,популящm слаrается из всех имею-
щихся в ней reHO"B. Так, rенофонд ,нашей rипотетической популя-
ции может ,быть полиморфным .по reHY А, .с<>держать ,редкий .МУ-
"антный аллель ,reHa В ,и быть МОНQМОРфНЫМ .по reHaJM С и п.
Теперь .мы .можем описать .ил.и .по крайней мере охаРЗJ теризо
ва'Ть л'ока..'lЬНУЮ ,скрещивающуюсп популядию :как rруппу особей,
имеющих ,общий теНОфОRД (Dobzhansky, '1950). Особ.и, из которых
состо'ит данная .популяция в .каждо:м данном поколе-нии, представ-
ляют .собой раЗЛИ1чные rенотипичеСЖllе ,продукты [амет, взятых из
I'енофонда этой ЛOrПУ.1ЯЦИИ .в предшествующем поколеюm.
Полиморфизм по rpynnaM .крови у человека
Давно известно, что разные люди по-разному реаrируют на пе-
реливание !кро'ви. В некоторых случаях переЛ;ИiВание крови ,пр'иво-
дил<> К аrrлютинации, или .СЛ:Иlпанию, ЭРИТрОЦИ70В, тоrда как в дру-
rих этоrо .не происходило. Такая аf1rлютинация обу.словлена ммму-
r л. 2; Скрещивающаяся популяция
3)
иолсrИ'Ческим взаимодействием ант.и-r.2НОВ,. содержащихся в эритро
цитах донора, н a-нт.итеJI. содержащиXICЯ в сыворотке крови реци....
ПНe1lта.
В зависимости от типа .с{)держащеrося в крови ант.иrена 'Раз ,и..'
чают -четыре 'rруппы крови (А, ,Б, АВ ,н О'). КажДЫЙ человек OTHO
сится к одному .из этих четырех фенотиличес.ких классов. .Инди-ви..
дуум, обладающий кровью rруппы А, может дать свою кровь длЯ!
перел.и'Вания друrому .индивидууму с той же rРУIJП{)Й крови, у ко-
Top.oro. при этом не возникнет аrrЛЮТ.инации. Подобным же обра
зам IКpOBЬ f1РYJППЫ S можно переЛ.ивать ЛЮДям, имеющим r.руппу В.
Но .пе.реЛИiвание 'Крови rрулпы А .'Iюдя'м ос rруппой В или кровк
rруппы В людя.м с rруппой А вызывает .с И'л'ЬнуЮ. аr:rлюти.на'I.J.;ИЮ..
Нет нужды описывать здесь реакции, .возникающие при осех воз
можных сочетаниях '!1'упл крови (см. Stern., 1960;. Race, Sanger,.
1962). В общем аrrлюrинация происходит в. тех случаях, КОflДЗ)
донор Н 'рецИiПИе'НТ имеют разную r.руппу крови.-
rруппы крови определяются серией трех аллелей: [.\ JB И
}О. Аллель [О рецессивен IПО отношению к а.ТIЛеЛЯМ .lA ,н [В (и-ноrда
er.o обозначают .бук'Вой i). Ал..'1е.тIИ [А и [В кодоминантны. Шесть
диллоидных rенотипов, образуемых аТIИМИ тремя а:Л.llе..1ЯМИ, фсно-
типи'Чоски проявляют-ся KalK четыре rруппы крови .в системе АВО
(табл. 2.2). На самом деле у аллеля [А имеются 'Ра'з.пич'ные, но'
hm-мунолоrИ'чееки сходные ll'зоаллели .(lAl, [А2 И Т. д.), а поэтому
число возможных rенотипОоВ БО.Т'lьше шест.и; однако в нашем обсуж
денни этими то.н.кими различиями аллеля [А .можно пренеб.речь.
Таблица 2.2
rенетические ОСНОВЫ rрупп крови системы
АВО (Stern, 1960)
rенотнп
фенотнп
(rруппа краВИ)
}Ч Л 11 }А}О
}BJ8 И }В}О
} .Ч В
}О}О
rруппа А
rруппа в
rруппа АВ
rруппа о
.Популяции человека .обычно полиморфны по :I'рулпам крови
системы АВО. Частоты разных трупп крови .и лежащих в их осно-
ве аллелей из.вестны для сотен локальных :папуляций ИЗ самых
различных 'Обла,стей зеМ"Ноrо ша1р.а. В таlбл. 2.3 Iприведены частоты
разных аЛol1елей для :rpex 'популяций. Как показывают эти .приме-
ры, разные ПОПУЛЯЦИИ человека сходны в том, что \все они по..т.и-
морфны по .rру,ппа.м крови .А!БО, однако они различаются .по часто-
, З
Часть 11. МUlсроэволюцuя
Там аллелей. У каждой локальной ПОПУЛЯЦИИ им.еется свой reHO-
фонд с хара.ктерНЫ1М для нее соотношением аллелей .reHa J.
Локальные популяции представляют собой части более крупных
реrиональных расовых rрупп. У родственных локалЬ'ных популя-
ЦИЙ, обитающих IB одН'ой и той .же ,оБЛаСТИ, rенофонды обычно не-
сколь'ко различаются. TaIK, частота аллеля fA ,среди населения
ШведС'КОО'о rор.ода Упсала iCоста,вляет, ,как указа'НО в табл. 2.3,
31,9%, а среди населения друrоrо шведскоrо rорода Фалуна ----
28,4%. И напротив, между rеоrрафическим.и ,расами на1блюдаются
устойчивые 'разЛ"ичия по частоте аллеЛей.
ТаБАица 2.3
Частоты аЛJlелей reHa 1 в трех популяциях человека
( ourant. 1954)
Частота
ЛОПУJlЯЦИЯ /А /8
/0
Упсала (Швеция) 0,319 0,019 0,602
Пенджаб (Индия) 0,181 0,259 0,560
Индейцы племени Нава-
хо (Нью Мексико ) 0,133 0,000 (\,867
П.очти во всех популяциях коренных обитателей Западной
Европы наблюд:ает.ся 'высокая частота аллеля JA и НИ3lкая (менее
10 %) частота аллеля [в. В ЦеНi'ральной Азии .на1блюдаеreя высо-
кая (20 30%) частота аллеля ]в. Среди американоких ИIFДейцев
аплель [О В"стречается с высо.кой частотой, ,тоrда IKaK аллель ]В
редок ,ил,и отсутствует вовсе (,Mourant, 1954). Равновесие меЖlДУ
разными ТИJпами несколько СДjRиrает.ся ПрИ .переходе 'из одной reo-
rрафической ,области в друrую. Мы еще .вернемся к rеоrраф.ичес.ко-
му раопределению rрупп крови .системы А;ВО в rл. 13.
Чрезвычайно интересно, что параллеЛl>н;ая полиморфная измен-
чиво.сть по rруппам крав.и .А!ВО обнаружена у чеЛ"ов.ек обрЗ'зньrх
обезьян. У lШимпа нзе найдены ,rруппы А .и О. у ора.нrутаиа и rиб-
бо'на ,имеются rруппы А, В и АВ (IMourant, 1954; Wiener, Moor-
J ankowski, 1971). Таким образом, полиморф.изм ,по rруппам ,КIРОВИ
Системы А,ВО возник в процессе эволюции раньше, чем .сам вид
Ното sapieпs и им обладают та,кже Iбл.ижайшие ,родичи челооека
в iOтряде Iпр.имаТQВ.
у .человека сущеС'Dвует еще HOCKOol'lbIKO оистем rрупп крови: си-
стема Rh, 'СИlCте.ма .MN .и друrие. Популяции человека ,ПOJlиморФны
также и .по этим системам ( R'ace, Sanger, 1962). Полиморфная из-
менчивость IПО ,системам Rh И .друrИ1М, ПО-'RИДИМОМУ, 'незаВИQима ОТ
изменчивости по сИстеме АВО.
r А. 2. Скрещивающаяся nопУАяция
зз
ПОJlИМОРФ}lЗМ по ферментам
МетОД rеЛЬ-ЭЛе<ктрофореза, 'Р.а.з.ра,батанньrй сра'Внитмьно недав"
но, дает возможность обнаруживать полиморфизм по ферментам
и 'по некоторым белкам, который не удавалось выявить обыч.ны'ми
rенет:ичеС'ки'М;и методами. МетОД элект.рофореза основан на различ-
ной ПОДВИЖ'Ност.и 'разных ферментов в .электричеоком ПОле. Экет-
ракт ткани помещают в rель и пропускают через Hero элехтриче-
Сl(ИЙ ток, чтобы создать злектр:ич-еокое поле. .При этом разлн-чные
фермеН1'Ы, содержащиеся в rеле, физически разделяются и после
окрашивания rеля выявляются в виде отдельных пятен. Таким
образом, можно .обнаруживать ТО'НКие различия между орrанизма-
Ми 'в отн.ошепли со.держащихся .в них ферментов; синтез которых
КОНl'ролирует я rенами.
ТаБАица 2.4
Полиморфные ферментные .IIOKYCbl, обнаружениые У разных ВИДОВ
(Gottlfeb, 1971; Lеwопfiп, 1973)
Вид
I чис:ло
lIЗучеН....х
. покусов
I п опиморфиые
JlОКУС:Ы. %
н ото sapieпs
Mиs тиsculиs
Peromyscиs polioпotиs
Drosophila теlалоgastеr
Drosophila pseиdoobscura
Drosophila persimilis
Drosophila willistoпi
Limиlиs polyphemus
71
40; 41
З2
19
24
24
20; 28
25
28
2()-.....ЗО
23
42
4з
25
81....86
25
Изучение выборок из природных попудяций MeroдOM !Электро-
фореза привело к неожиданным резу.тrьтатам. В -популяциях раз u
ЛИЧНЫХ видов были обнаружены удиввтельно высокие уровни по-
лиморфизма (табл. 2.4). Очевидно, полиморфизм paioopoCTpa HeH
rораздо шире он затраrивает rораздо большее число элементов re-
потапа, чем это пр-едполаталось пр.И ero .изучении классическими
методами (,НиЬЬу, Lewontin, 1966; Lеwопtiп, НиЬЬу, 1966; Lewon..
tin,1974).
Концепция ПОПУЛЯЦИИ
Концепцию rенеТIfчески изменчивых IПОПУЛЯЦИЙ ка'к репродук-
ТИвных единиц .ни .в коей мере .н.ельзя ,считать сам'оочевИ'дной. Этой
концепции не существовало .ни 'в XVIIl, ни в начале XIX В., .8 В не-
которых обл.аlСТЯХ биолоrии она отсутствует до сих пор. Соrласно
з4
Часть /1. МикроэвОАЮЦUЯ
МаЙру ('Mayr, 1972), ее ввел в биолоrию Дарвин в 18"59 [. K{),H
цепция ,популяции была одним из элементов переворота, произве
денноro Дарвином в .научном мышлении.
Концепция популя'ции прямо ПРОТИВОЛО. Т 1Ожна эссенциализму.
Сторонники .эссенциализма сч.итают, что на.блюдаемые в мире-
явления представляют собой ,выра'жения леж.ащих в их ос'Нове сущ-
ностей. Явления предстаlOТ в различных формах, сущность же их
неизменна. Следовательно, члены Toro или иноrо k.T]a-сса объе.ктов,
в том числе и Особи, составляющие ка'кую ли60 популяцию, ЭТ()
различные выражения одной и той же сущности.
Эссенциализм 'в том или и.ном варианте был традиционным фи
ЛОСОфClКим учением в Европе. Ф-ил()софия Платона, )Ср.и.стианское-
боrословие и ф.илософский идеализм представляют собой различ-
ные модиф.икации эосенциализма. Естественно, 'ЧТо эссенциал.изм
rосподст,вовал в .научном мыленииH на заре истории биолоrии..
Здесь он принял форму, которую Майр (M.ayr, 1957а, 197.2а)
назвал типолоrич-ес.ким мышлением. Соrласно 'Этим взrлядам, OT
дельные орrанизмы .представляют собой :нfocовершенны,, а поэтому
изменчивые проявлен.ия архетипа тот ВИ1да, к которому они при
'На!длежат.
Типолоrичес.кое мышление препятствует поким.аиию эволюции
для KOToporo необходимо популяционное мышление, поо ольку
эволюция это .изменение rенетическоrо состава популяций. Ве-
ликим, хотя .и очень тонким достоинством совер,шенноrо Д,арв.и.ном
переворота была, соrласно Майру (Mayr, 1972а). замена типоло-
rическоro мышления в биолоrии ПОПУЛЯЩI.OННbl.м. В.ведение популя
ционной концепции устранило даанее и сильное препятствие, Me
шавшее пониманию эволюu.ии вообще и foCTecTBeHHoro О1'бора
в частности.
rла'Ва 3
СТАТИКА (ПОПУЛЯЦИй
Закон Харди ---- Ваiiнберrа
rенофо.нд локальной популяц.и.и обычно содержит, помимо ОМО-
номорфных, такЖе и различные полиморфные reHbl. Кроме Toro.
в каждом данном .поколении алле.льные формы полиморфных re..
нов представлены .с некоторой определенной частотой. Так, напри-
мер, reH А, имеющий два алл-е.'IЯ, А и а, может быть представлен
в Irе.нофонде .одноrо ,поколения в соотношении 70 % аллелей А и
30 % аллелей а. a'КOBЫ ОБ таком .случае Iбудут 'ожидаемые чаiСТОТЫ
алл-елей в следующем поколении?
В популяции диплоидноrо орrанизма эти аллели содержатся
в ,rомозИlЛOТНЫХ и ,rетерозиrот.ных rенотипах АА, аа .и Аа, Koropble
будут встречать я также в определенных оот.ношениях в любом
данном поколении. Они служат родительскими reнотипами для
следующеr.о поколения. В связ,и с этим DоЭ'никает вопрос: ка'ковы
ожидаемые с.оотношения rенотипов во втором и в последующем
поколениях?
Ожидаемые частоты аллелей и частоты reнот.ипов можно опре
делить :по закону Харди Вайн.берrа. ЭтОТ за.кон действует при
следующих условиях: предп.олаrает-ся, что популяция достаточно
вели,ка, .с тем чтобы ,ошибки выборки не оказывали существенноrо
воздейств'ия на частоты из п.околения в поколение. и 'Что состав-
ляющие популяцию ,особи ВНОСЯТ .р.ав'Н"ое число Фун ционирующих
raMeT; иными слова.ми, разные rенот,wnы (Должны размножаться
одинаково У'Опешно. И .наlJ{,{)нец, допускае'J'lСЯ, 'что в популяции пре-
.обладает случайное крещивание.
Случайное ,скрещивание, или панмиксию, м.оЖно с ,равным у.спе-
хом определять в терм.инах особей или в терм.ина'х raMeT. Если
иметь в виду особей, то случайное скрещИ'в-ан.ие .про.и.сходит в тех
случаях, ,к()rда .особ.и, имеющие различную r-енетическую .конститу-
цию, .скрещиваются независ.имо от своих rенотипов. Напр.mмер.
сам,ка с rенотипом АА .может ,скрещroваться с самцам-и АА, Аа .или
аа, не проявляя ,ни.каlКоrо пре.д.почтения к самцам Ka oro-To адноrо
типа.
Панмиксию можно определить несколько точнее, если исходить
из наличия в raMeTHoM фонде множества raMeT. В этом смысле слу-
чайное .скреЩИlвание .означает, что каждая женС'кая ra.MeTa с оди-
на'кооой вероятностью может быть .оплодотворена МУЖСКQЙ ra.Me-
той любоrо типа и что эта ,вероятность прямо пРОПОРЦИQ-налЬ'на
частоте 'МуЖоСких raMeт дан.ноrо типа в raMeTHOM фонде. Короче
36
Часть 11. МurcрОЭ80АЮЦUЯ
.rоворя, raMeTbl, носущие разные аллели, соединяются в пары .про-
порц.иО'нально .их относительным частотам в raMeTHoM фонде. Осо-
би, состаВЛяющие популяцию в ,каЖ:ДОМ данном поколении, лред
ставляют собой в таком случае произведения разных пар raMCT
случайно извлеченных из raMeTHOTo .фонда предшествующеrо поко-
ления.
В .популяции, 'Соответ-ст.вующей указанным выше условия,м, со-
rласно закону харди----Вайнберrа, частоты аллелей будут .оставать-
ся постоянными из поколения iB поколение и в результате случай-
Horo скрещИ'Вакия в одном -поколении rенотилы достиr.нут равно..
весной частоты, которая будет сохраняться в дадьнейшем. Прнме..
ры, .иллюстри.рующие эти два обобщения, рассматриваются НИЖе
Частоты 8J1леJlеii
,
Закон .постоянства ча:стот аллелей будет проиллюстрирован K
ли'чеc-nвенны'м примером. Допустим, что .папуляция HeKoro диплонд.
Horo Бида, пол,и.морфноrо по reHY А, 'В исходном поколени.и coдep.
ЖИТ разные .rеноти.пы В следующем соот.ношении: 60% АА, 20% Аа
и 20% аа. Проследим за аллелями А шаr за шаrом на Л'ротяжеНIИИ
двух поколений.
1. Частоты аллелей 8 первом ПО1Wленuu. Поскольку ча'Стоты
rенотипов заданы .К3'К
О,60АА + О,20Аа + О,20аа,
частоты аллелей (q) в этом Же .поколения должны с.оставлять
А ---- 0,60+ 0,60 +0,20 O 7 0
q 2 ' ,
____ 0,20 + 0,20 + 0,20 ____ о 30
qa . 2 . .
2. rа.метный фонд nepBozo поколения. Предполаrается, что все
особи одинаково плодовиты; поэтому диллоидные особи будут
производить rаплоидные raMeтbl в соотношении 70% А и 30% й.
Частоты аллелей в raMeTBOM фонде такие же, как и в исходном
rенофонде.
3. Случайное скрещuвание. raMeTbl для образования зиrот ВТО-
,poro поколения извлекаются из фонда случайным образом; 'при
эт.ом :возможны такие .попарные tCочет.а'Ния:
ЖенсКJiе raмeтн
0,70 А
0,70 А
0,30 а
0,30 а
Мужские r8llеты
0,70 А
0,30 а
0,70 А
0.30 а
r л. 3. Статика популяций
4. Частоты ЗUZОТ 80 втором поколенuu. Прmеденная выше си-
стема свооодноrо скрещнвания дает следующие результаты:
О,49АА;
0,21 +0,21 ==0.42Аа;
0,0900.
СЧИ1'ается, 'что все зиrоты обладают одинаковой жизнеспособ-
ностью; следовательно, приведенные цифры дают ожидаемые рав-
новесные 'Част.оты теНIQТИПОВ во втором .поколении.
Можно заметить, что данная популяция .не находипась в рав-
нов'есии в отношении частот rенотипов в первом 'поколени.и, но
доститла pa'BHoвec.нoro состояния в результате .свободноrо скрещи-
ва'ния .B ero л.ишь в ,одном поколении.
5. Частоты аллелей во втором поколенuu. rено4юнд nToporo по-
коления, ,очевидно, будет содержать два аллеля iВ следующих
частотах:
чА
0,49 + 0,49 + о ,42 ---- о 70 '
2 "
0,42 + 0209 + 0,09 ---- 0,30.
ча
т а ки,м; образ.ом, частоты ал.пелей во втором п(ж.олении такие
Же, хаки.ми они были в пероом ;поколении.
Формула Харди ---- Вайнберrа
Формула харди----Вайнберrа дает короткий Iи ,прямой спос06
вычисления ожидаемых част-от rенотипов в 'Случайно скрещиваю-
щейся популяции ,на основании частот алле.т.ей в rенофонде.
РаlCСМОТрИМ вновь rенофонд, с-одержащий два алл-е.1Я reHa А.
Пусть робозначает чз'Стоту аллеля А, а q ---- частоту аЛ.11еля а
(приче.м p+q== 1). Случайные ,СQlЧетания raMeT А и а дадут при
.этом зиrоты в соотношении, определяемом по формуле квадрата
двучл на (p+q) 2. Иными .словами, равновесная ча.стота rеноти-пов
будет рави.а:
р2АА,
2pqAa,
q 2 aa.
Применив .эту 'формулу ,Б нашем предыдущем при-мере (rде q
используется в HeCKO.тIЬ.KO и.ном значении), можно ;путем простой
операци.и получить те же результаты: если р==О,70, то равновес-
ная частота АА == р2 == 0,49. В .тех Iслучаях, к-оrда полиморфизм свя
заи ос серией множественных аллелей, фор.мула Хард,и Вайнберrа
38
Часть /1. МU1СРОЭ80АЮЦия
lJрИRИМает :вид кв.адра1'а МНOIrО'Члена. Для трех аллелей (Al, А 2 И
Аз) е ча;СТота'ми р, q и " так что p+q+r== 1, равновесная частота
rенотипов определяется по формуле кващрата ре ЧЛена (p+q+
.+ ')2 ,
Эффекты иибридинrа
В 'большой ЛОПУ..'lяции при условии случаЙ'1Юrо скрещивания
qaCTOTbl rенoQТИПОВ быстро достwrают р'авновеспоrо состояния, II<ОТО-
рое со'Храняе'DCЯ IB дальнейшем_ Н'арушения CJlучай,ност.и .Qкрещива-
ния вызывают отклонения частот rенотипов от равновесия Xap
ди Вайнберrа.
ДОПУСТJliм, 'Ч1'о некая популяция в нулевом поколения состоит
}{ск.лючите.тIЬНО из reтер.озиr.от Аа. Ра'змнажен,ие происходит путем
самоо'пладотворения. В первом .пок.ол нии популяция будет иметь
следующий ,состав: 25% АА, 50% Аа и '25% аа. В последующих
поколениях с<жращение .класса rетерозИ\I'ОТ будет продолж.аться
с равномерной скоростью. После с,амооплодотворения в семи поко-
дениях (Т. е. в 7-м инбредно.м поколении) популяция будет содер-
жать 'почти 50% АА .и .почти 50% аа.
Доля ro м 03 Иlr от, праисходящих от преДjКОВ, rетерооиromых по
од.ному reHY (А), после т поколений са.мооплодотв-орения апрelде-
ляется по .формуле
2M l
2М
в рассматрива.емом случае поме 7 поколен,ии СЗ 1 МOQПЛ.ОД.о11ВО-
'рения ожидаемые ча1СТОТЫ rеНlQТИПОВ в 7 -М инбредиом 'покол ен;и и ,
соrлас.но этой формуле, 'составят
127/128 (АА +00);
1/128Aa.
Таким .образом, ,в результ,ате инбридинrа Ча'стотЫ rен-отипов
изменяются в сторону преобл,здания rоМ.О3Иroт. Следует, однаlКО,
отметить ,н друrой аС'пект: И'нбр.идинr ве оказывает .влияН\ия на час-
тоты а..."'Iлелей. В нашем .rнп.отетическом примере частоты аллелей
состав.'IЯЛИ -50% А и 50% а в О-м поколении; и они оставаЛJiСЬ та-
кими же как в I-M, так и в 7 -м инбредных поколениях.
ЗаКJlючение
В большой поли.морфной па,нмикт.ичес.кой популя'цИ'и, состоя-
щей из одинаково жизнеспособных и одинаково плодовитых осо-
бей, различные rомозиrотные .и rе.терозиrот.ные rеноти-пы быстро
ДОС1'иrзют 'Некоторых равновесоных частот, зависящих от .сущест-
вующих в популяци.и ча.СТОТ аллелей. Частоты rенотипов, достиr
r А. 3. Статика nопУАяций
З
вув равновесия Ха,р.ди Вайнберrа, остаются затем постоянными
на протяжеаии всех посл-едующих по.колений случаЙНОIfО "Скрещи-
вания.
Частоты аллелей в большой полиморфной популяции, состоя-
щей из одинаково жизнеспособных и одинаlКQВО плодовитых осо-
бей, обычно остаю'ОСя посroянными из по'коления ,в поколение. .эТQ
постоянство .не зависит от СJlучайноrо 'Скрещивания. Поэтому пр,и-
мен.ительн.() к чЗ!стотам аллел-ей закон ХаРДИ............jВайнбер:rа носит
более общий хар а'ктер, чем прим-ен.ительно к частот.ам rенотипов,
и в эroм своеМ аспекте он Iимеет 'оол-ее важное значение для эво-
люци.онной теории.
Полез-но изучить оообые условия, необходимые для действия
за-кoQН.э. Ха'рди Вай.Н'берrа. Эти условия ,исключают дю.бые .факто-
ры, в.оздействующие на частоту reH.OB, краме caMoro процесса БОС"
пр.оИ'зведения reHoB. До тех .пор пока уровень изменчивос'l1И iВ reHO-
фон е определяется воспроизведени-ем reHoOB как T8-KoBы1м' этот
уровень остается постоянным и неизменным в ряду ПОС.ТJедователь-
ных .ооколений.
Такова статика популяций. ,Изменение уровня изменчивости
в популяциях динамичеС;J<ая сторона проблемы т.ребует дейст-
в'ия специфических .факторов ,или сил. Это факторы МlDКрО'ЕОЛЮ--
ДИИ, которые мы раоомотрим в .следующей .rЛЗlВе.
rла а 4
ДИНАМИКА ПОПуЛЯ,ЦИй
о предеJlение МИКРО9ВОJlЮЦИИ
Как уже было отмечен.о в .предыдущей rлаве, уровень .изменчи-
вост.и в обширном rенофонде не изменяется сам по себе. Тем не
менее на.блюдения показывают. что в лока'ЛЬ'ных попупяu.иях п:ро
исходят изменения частот алделей в ряду последовательных поко
лений. С течением времени rенофо.нд .Все-та'ки изменяется.
Мы можем определить микроэволюцию как систематическое
изменение ча'Стот rомо.лоrичных аллелей. участков храмосам ил.и
целых хромосом ,в локаJIЬН<>Й популяции; иначе .п()воря. МИ1КрОЭВО-
.1Jюцией называют любое увеличение .или уменьшение частоты в re-
нофонде кЗ'кой либо вариантной IфоРМЫ, которая 'про.должает встре-
чаться в .популяци.и из по.коления ,в .поколение.
В ПрJlрОДНЫХ полуляциях можно. .найти вел.и.кое мноЖество .при-
меров М'и.КРОЭВОЛЮЦИiQННЫХ .изменений. Для ,наших целей до.статоч-
но о.писать один на.rлядный лример. ОТНQlCящийся к Drosophi1a
pseudoobscиra.
МИКРОЗВОJJюционньrе изменения
у Drosophila pseudoobscura
Для п. pseudoobscura характерна изменчивость последователь-
ности reHo.B в одном кз уч,а.с11ЮОВ ,т,ретьей хромосомы. Разные линии
мух различаются по .инверсиям tВ этом учас"ке. .Каждая ииверtCия
имеет название и бу;mенное обозначение: Стандарт (ST). Эрро.у-
хед (AR). Чирикауа (СН). Трилайн (TL). Пайкс-Пик (РР) и т. д.
У D. pseudoobscura .обнаружена 16 та:ких инверсионных форм. Они
выявляются цитолотически по xa1p,aKTepy поперечн.ой ,исчерченности
хромосом .слюнных жел-ез у личинок
Большая часть природных ПОПУ.71ЯЦИЙ п. pseudoobscura в за
падной части Северной Амер'ИlКИ полнморфна по. .третьей хромосо-
ме. ПолИ'Морфные популяции .содержат по Ba и более т.ипо'В инвер
сий в разлИiЧ'НЫХ rомозиroтных .и rетерозиrотных комбинациях (Ha
пример, ST/ST, СН/СН, SТ/СН)..Б каждой дан.НОЙ популяции раз
ные типы и.нверсий обычно вс'Лречаю'ОСя с оn-ределенньrм.и среДНIfМ.и
rодовыми ча'стот-ами.
В популяциях п. pseudoobscura, обитающих в ropax Cьeppa He-
вада (IКа.'IИфОРН.ИЯ). обы'Ч.но ,встречаю'ОСя .инверсии ST. AR .и СН.
Четыре дру.rие иН'в.ерсии (TL, OL, SC и РР) 'Встречаются в эroй
местности с низ.кой Частотой. Мы эдесь займемся rлавны1M образом
r А. 4. Динамика nопУАяций
41
одни'м из редких 11ИПОВ (РР) В ОДНОЙ нз локальных популяций
(Матер). Популяция Матера .в oKpyre й'Осемнте (Ш'I'ат Калиф.ор.
иия) на протяжении мноrих лет изучалась Добржанским и ero
сотрудниками (см. Dobzhansky, 1956, 1958, 1971).
Инверсия типа Пайкс.'П;ик встречалась, хотя и к.рай.не peДKO
в п{)пу.пяциях DrQsophila в Сьерра-'Неваде в 1945 r. и ДО этоrо, но
в популяции Матера в это .время она ,не t)ыла известна. В 1946 r..
инверсия рр стала встречаться в популяции Матера с очень низ-
кой частотой (0,3%). За ,последующие 10 лет (1947 1957 .rr.) ча-
стота .инверсии рр быстро возра тала, д<>стиrнув 10 и даже 12%
(табл. 4.1). ,Б дальнейшем (1959----1965 rr.) ее частота колооалась.
На несколь'ко более НИ3lком уровне, а затем упала довольно .с.ильно.
Повышение частоты инверсии рр в популяции Матера сопро.
вожда.ТIQСЬ понижением частоты .инверсии СН, которая раньше
была довольно обычна, до очень низких для нее уровней (2----6'%).
А сравнителыно недавно чаоеТО1'а типа СН вернулась к прежнему
уровню (табл. 4.1). .В ЭТQ же время происх-од,или аналоrичные из.
Менения в .соотношении инвер.сий ST и AR (.Dobzhansky, 1971).
ТаБАица 4.1
Изменение частоты двух типов ииверсиА (рр и СН) в популяции
Drosophila pseudoobscura в Матере (Калифорния) на протяжеиии
28 лет (Dobzhansky, 1971; Апdеrsоп et al., 1975)
rод I Частота I Частота '11П1< СП. " I Число исспедован-
типа рр. % НЫ!( XPOIlOCOM
1945 I 0,0 17 308
1946 0,3 17 336
1947 0,7 20 425
1950 3 17 812
1951 5 1l 856
1954 12 J3 1312
1957 10 4 316
1959 4 11 298
1961 6 3 350
1962 9 2 450
1963 7 6 446
1965 6 11 534
1969 2 3 312
1971 3 12 390
1972 6 17 576
РесЭIroе .повышение частоты инверсии типа Р Р наt)людалось не
только в популяции Матера. ПараЛЛeJlьн{)е увели;q ние частоты
этой инверси.и 01\мечалось в друrих обособленных популяциях
42
Часть /1. МU1СрОЭ80АЮЦия
D. pseudoobscura в .местностях, разбросанных ''по осей Калифор.
нии и Аризоне. TaIK, в ropax Сан-Джасинто .На юrе КаJLИфОрНИИ
инвер.сия рр впервые .появилаlСЬ в 1952 r., ДОСТИJfла !'ПWКОООЙ часто-
ТЫ (1 О ,И 11 %) в последующие нескольJro лет, а в шестид яты!х
rодах частота .ее вновь упала ,(Dobzhansky, 1971). Изменения ча-
СТОТЫ инверсии рр носили одинаковый хара'ктер на протяжеНИ1И
обширной rеОl1раф.ической области.
Причины lIа'блюдаемых микроэволюционных ,изменений неиз-
вестны. Имеющихся ,данных достаточно, чтобы .ИОКJIЮЧИТЬ участие
в этом НeIOО1'орых возможных фактор.ов. Полиморфные лаборатор-
ные популяции D. pseudoobscura, выращ.иваемые при блаroприят-
лых условиях, .подч.иняются .за'Кону Харди Вайнбер.rа в смысле
с()Отношения типов инверсий и сооmетствующих lДиплоидных rеио-
типов. В этом случае экС'периментальны-е данные подтверждают
теорию, исключая возм-ожность .каких-либо изменений, направ-
ляемых .изнутри. Возможность случайных эффеКТQВ исключается
большими размерами как самих популяций, так и изучавшихся
'выборок. Давление -мутаций та.кже следует ИlQКЛЮЧИТЬ, поскольку
8 сильно разобщенных локальных популяциях .наблюдал.ись парал-
лельные измеIreНИЯ. Можно :п.очт.и не сом'неваться в том, что обна-
руженные изменения обусловлены естественным от.бором. Однако
несмотря на обширные исследования, направленные На выявление
участвующеrо в ЭТОМ фактора отбора, определить ero не удалось
,.(DоЬzhапsk,у, 1956, 1958, 1971).
r Jlавные эволюционные силы
Факторы, вызывающие из'Меное.ния частоты алле..лей или часто-
ты хром-осо.мных .ин'версий, 'можно назвать rла&ными эволюцио.н.
НЫМИ с.илами. ИЗlнестН'о четы,ре таких факт{)ра: .мутационный про.
цесс, .поток rеиов, еС!fоственный отбор .и дреЙф -rенов.
ДОПУС'I'И!м, 'ЧТо rенофонд какой-либо популяции состоит rлавны'М
образом из аллелей А 1 и в меньшей стелени ........ из аЛлелей А2.
Первона'Чальные частоты reHOJВ Moryт изменяться под действием
ка.ждоrо из указанных выше четырех фаl({ТОрОВ эволюции следую-
щим образом.
АЛJlель А 1 .МОЖет М'Ут.ироватъ .в аллель А 2 однаЖДЫ .или МНОТО-
paTHO, увеличивая тем самым ча'стоту А 2 . Особи или ra.Meтbl, не-
сущие аллель А 2 , MorYT миrрировать в изучаемую популяцию из
какой-либо друrой популяции, в которой аллель А 2 встречается
чаще. Подобная миrрация ,или ,поroк reHoB та же изменит пре.дсу-
ществующие частоты reHoB в популяции-реципиеflте..
НоситеЛIJ1 алл.елей А 1 .n А 2 мотут раЗJllичатося по ,фенотипиче
ским .признакам, ,ВЛ1ИЯЮЩИМ .на ,их rCпос.обностъ к выжива.нию и ,раз-
множению, а "Тем самым .на их вклад в следующее поколение. Если
особи, .несущие аллель А2, J1ревосходят по ЭТИМ показателЯlМ осо.
r А. 4. Д ина.миlСй популяций
43
бей, Н€СУЩ1Их аллель At, 1'0 частота аЛлеля А 2 в популяции будет
постепенно ;повышаться. В данном -случае действует естественный
отбор самый ,важный из факторов, реryJlИРУЮЩlIХ частоту reHoB
в лриродных популяциях.
.На:конец, зна'чительные сдвиrи частот алл-ел-ей Al и А 2 Moryr
происхоДlПЬ в небол ших популяциях чи'Сто случайным .образом.
Этот случайный .ком:понент В изменении reHHblX частот известен
fЩД .названием дрейфа reHOВ.
И'нтенсивность дейстВ/ия каждой .из ЭТИХ четырех сил может
бы'Ть р,азличной, и ее можно охарактериэовать Iколмчест-венно.
Частота мут.ирования (и) может варыи.ровать от О до 1, rде О
ПОJшая устойчивость, а 1 полная неустоЙчивость дaHHO O reHa_
Аналоrи'Чным образом скорость .потока rеИQВ (т) изменяется
в диапазоне от т ==0, т. е. отсутствия .мШ'ра'lJJИИ, до т ==-1, т. е. по,:"I-
Horo «затопления».
Коэффициент отбора (s) служит мерой ореднеrо повышения
относительн()й ,ч.астоты одноrо а.плели ,ПО сравнению с друrими,
конкурирующими, аЛ..1е.пя'ми ,на .одно пок{)ление. Этот коэффициент
МiQжет варьиров.ать от О до 1, rде s == О .означает ОТСУТС1'в'ие отбора,
а s == 1 ---- 'полное замещение .reHa за одно поколение.
Возможность действия rенетическOfоО дреЙфа выражается за..
ВИосямостью между величиной ,паПУ 1ЯЦRИ . (N) ,и друrи'М'И перемен-
НЫМ1И. Дрейф может эффек'fJИВНО реrулировать 'ЧаСl1QТЫ lI"еноо, если
N .М ало [10 'ср.авнению с s, т rИ .и.
Взаимодействие между эволюционными силами
Первые два .из УIПОМЯНУТЫХ выше Iфакторов ---- мутационный 'про..
цесс и поток r.eHoB соз;дают ,И зм.енч;ивость. Два друтих факто-
ра ---- естесТlвенный отбор и дрейф reHOB орт,ируют эту изменч,и-
вость. Фаrкторы, создающие изменчивость, дают .начало процес.су
м Иокр.оЭВолюции, а фа,кторы, .сортирующие из-менч'ИоВОСТЬ, продолжа-
ют процесс, что пр в'Одит IK установлен.ию новых часroт .ва1риантов.
Эволюционное ,изменение :в пре:делах .ПОnYЛЯIlЩ:И м.ож-но .рассматри
вать как результат действия пр.отивоположных сил, .создающих 'и
сортирующих rенетическую .изменч:ивость.
'Одна .из старых теор.ий эволюц1flИ теория ортоrенеза, до с.их
пор имеющая ,приверженцев, ---- постулирует, что эволюционные
изменения на 1 правляю11СЯ rлавным образом мутаЦИQН'Ны.м 'процес
CO:'d. Против TaKoro взrляда существуют два веских aprYMeHTa.
Во первых, мутационные .из-менения возникают более яли менее
незавиС'и.мо от приспособитель'Ных требований орrанизма и сами
по ,себе .не ,MorYT ,создать rJIlРИСПOlCOбительные при'з.нак.и, наблюдае
мые у ор:rанизмов. .Б до.полнение к .эт.И'м изменениям необходимо-
действие ecтecтвeHHoro отбор:а, который бы перенес .процесс .< c хи
мическоr.о Yiровня ,мутации на бнолorичесК'Ий уровень адаптаЦИИ:t}
(Dar1ingt.on, 1939).
44
Часть П. Мu"роэвОАЮЦUЯ
.во-вторых, во временной последовательности М'икроэ.ВОЛЮIIlИИ
отбор .всеrДа &ступает в действие после мутаЦROююrо процесса.
Каким бы .сильным ил.и .направленным .ни был.о давлен,ие мутаций
в даllllОЙ популяции, ero бла:rоприятное .или неблаrоприятное дей-
ствие всеrда контролируется естественным отбором, за которым
и остается последнее слово.
В равной !depe справeJдЛ'ИВО, что О'J'бор не сможет деЙствовать,
если мутационный процесс не будет поставлять ему новые rенети-
чеоюие 'вар.иаЦlИ.И. Миюроэволюция .обу.словлена не действием Ка-
кой-то одной .силы, а взаимодействием двух, трех или четырех сил.
Количественная оц-енка rлавных .эоолюционных сил создает воз-
можность количе-ственноrо выражения взаимодеЙСllВИЙ между
ними. Последнее относительно просто в теоР'ИИ и и.ноrд:а достижи-
МО в создаваемых иокусственно эк-спери;менталъных популяциях,
однако в прир.одных популяциях осуществить .Э1'О райне трудно
ввиду наJ1иqия мноroчи-сл нных неконтролируемых фаIКТОрQВ. Тем
не м-енее было сделано .нескОлько .ПОПЫТО.к подойти к этой проБJIе-
ме .в реаль.ных .популяциях.
Примером взаимодействия между мутационным процес.сом и от-
бором служит rемофи.тIИЯ у чел.овека. reM-ОфИJllИЯ кровоточи-
ВОСТЬ, обусл.овл.ен.н.ая пон'Иженной свертываемостью .крови, ..... обыч-
но приводит К -смерти .в раанем возраiсте. Известно неоКОЛJ;.КО тило.в
rемофилии; мы ОI'раничимся рассмотреНИ8М классической rемофи-
лии, или .rемофил.ип А. rемофил.ией А страдают люди, rомозиrот-
ные по рецессивному аплелю (hh) reHa Hh, сцеплен'Ноrо с ПОЛОМ,
точнее, с.цепленноrо с Х-хромосо.мой. rетеРОЗ1иrотные нос.ите.льниuы
(Hh/hh) .иноrДа рОЖдают ыновей с конституцией hjO, у которых
проявляет я болезнь (Stern, 1973).
Мужчины, страдающие rемофилией, в большинстве случаев,
хотя и не всеrда, умирают до достижения половой зрелости, и По-
этому аллель hh имеет НИ3'Кое .селективное значеНие в полулеталь
нам .щиа'пазоне. rомоз'Иrоты .все ,время элиминируются ИЗ популя-
ций человека iB результате отбора. Тем не менее 'они постоянно при-
сутствуют в популяции С низкой частотой. Их сохранение объясняют
пер:иод'ическим мутир.ованием Hh в hh, причем частота мутнрова-
ния и==О,ОООО2----0,ОООО4 (Stern, 1973).
Таким образ.о.м, постоянно ,сохра'няющаяся в популяциях чело-
века низкая частота аллеля hh .представляет собой ра.в.но-весие
между периодичеоки возникающими ,мутациями и сильным .отрица-
тельным о"бором. Это ВО всяком {случае моЖно сч'итать первым
приближением к раз'р-ешению проблемы. СуlДЯ по некоторым дaH
НЫМ, rетерозн'rо'Лные женщины обладают повышеННIQЙ селектrИВliОЙ
ценностью по сраlв=нен.ию с нормальны.ми женщинами, rом()з-и.rот
ными по .одному .из д'руrих тппов rемофили'и (Rosin, Moor Jankow-
sky, S<:hneeberger, 1958). Если анало'rачным образом обстоит дело
с rеМОф:ИJI1ией типа А, то это должно сдвинуть селекци'Онн'о-'Мута.
r.1. 4. ДиНД,lщка популяций
45
ЦИО"Нiное равновесие в сторону ,бо..lее высокой равновесной частоты
в популяции ал.леля hh чем следовало бы ОЖидать в отсутствие
этоrо осложняющеrо фаlКТDpа.
Создание из.менчи:востн в результате сorчетания .мутаци:он,ноrо
.процесса и потока reH()B МОЖНО прО'иллюсrрировать Ha примере :на-
земной УЛИ1'юи Сераеа пeтorali5 в Западной Европе. В к{)лон;иях
этой ул'и,.,ки -постоянно на,блюдается ПОЛИМоОрфизм, l8ыражающийся
в .наличии .или отсутствии коричневых полос !На ра,ков.инах. Эти
фенотипичес.К!ие различия 'реrул'и.руются. rеяом, и.меющим два алле-
ЛЯ; доминаятный аллель В определяет развитие бесшолосых, а ре-
цессивный аллель Ь..... полосатых ракови'н. Мутации нз В 'в Ь и об-
ратно происходят с частотой u==O,OOOI O,0005 (Lamotte, 1951).
Структура популяnий европейской ;наземной улитки различает-
ся в разных -областях. Эти разлиЧ'ия оказывают 'влияние на ско-
рость потока тен-ов. В провинцlfИ fиень (Франция) колонИ"и распо..
ла.rаю1'СЯ дооольно близко друr от друrа и нередко связаны 'между
собой миrрирующими особями. Соrласно оценкам, поток reHoB
происх,OI.ПIит эдесь 00 скоростью m O,OO3 O,004. В пров'И'нции rиень
поток reHOB представляет собой более "сущес.твенный ИСТОЧН'1Iк И"З..
менчивост.и ПОПУЛЯЩИИ, чем мутационный процесс. А 8 провинци.и
Бретань (Франция) IКО.'IJОНИ-И улиток, на,против, располаrаются да-
леко друr от дру.rа 'и обмен м-иrpантами "между ними происхО'дит
редко. Здесь мутационный процесс и'rрает относительно б6льшую
роль, а поток .reHoB имеет меньшее значеиие, чем в ПРОВИНЦИIi
fлень (Lamotte, 19-51).
Однако() на эт.ом дело не КОНчается. Характер ПОJJосаТОСТИ рако-
вины у улиток реryл.иру.еreя, кроме Toro, отбором, а -возможно,
даже .и дреЙфом reHoB, что будет обсуждатъся в одной нз -после-
дующих rлав.
ДеЙС1\вие дрейфа reHOB целиком зависит от соотношения между
факторами N , 51 и 11 т. Напр.имер, дрейф reHOB может эффек'J1ИВ-НО
контролировать частоты reHoB при N < 1/25, если и и т настолько
малы, что .иМ'И можно прен.е6р чь. И 'наоборот, умеренные частотЫ
мутирования .и 'миrраци.и Moryт свести на нет действие дрейфа.
Чрезвычайно Iважную роль :иrрает В природе 'Сочетание дрейфа re-
нов с IОТбором. Эти взаимодей твия бол е ;подробно pa MaTp Ba.
ются в rл. 13.
ПОПУJlяционно--rенетический ПОДХОД К ЭВОЛЮЦИИ
Классические rенетикн исследовали поведение reHoB, изучая ро.
ДОСЛО.Dные семей. Однако :nОКОJIен.ия Р, Р 1 , F2 И F3 формальных re-
нети ков в-cero лишь искусственная абстракция. В 'пр.ир'оде происхо-
дит не 'просто -объединение MyraHTHoro аллеля 'с нормальным в
одной 'из особей F 1 .и -передача ero п()ловин та-мет, продуцируемых
&той .особью. В ,естественных У ЛО()ВНЯХ, вне rенетичесIroЙ ла60ра-
46
Часть 11. МUlCptJэвОАюция
то'рии ИЛИ ,опытной делянки, мута1ЦИЯ, В03НИJ{lшая у оnцельноrо.
орrанизма, Не .остаетсЯ в пределах рОДОСЛОВНОЙ одной семьи,
а .включается в rенофонд -популя.цин. И зде ь ей пред то-ит, если
ОНа f'ибельна ил.и блаrОПр'шrrна, 'Изм-енение чаcroты.
.Orсюда Iследует, что rенетнку популяций нельзя изучать долж-
.ным образом, не принимая .во .ВН'1Iмание воздействие, оказы.ваемое.
естественным rO'I'бором на на ледственные изменения. IПервоначаль-
но обособленные пути менделизма и дарвинизма ,ДОЛ1ЖНЫ сл.иты:я..
Это СJ1lИяние 'произошло ,в тридцатых r.одах и непосредcrвенно
ПрИ1вело к .современной или, 'как ее называют, си.нтеТ.иче(жой тео--
.рии эволюци.и. Начало этой теории заложил популяционно..;rенети-
ческий подХОД к эволюции. Характер происшедшеrо синтеза
01'ражают наз.вания трех ;кла'С..сичос'Ких ра.бот, О1'НQСЯЩИХ-СЯ .к -ЭТОМУ
раннему периоду: «rенетическая теория eCTeCTBeHHoro отбора
(Fisher, 1930), «Эволюция t8 менделевских популяциях» (Wright.,.
]931), «rенетика И происхождение видов» (Dobzhansky, 1937).
Популяционно.rенет.иче кий подх.од непосредственно привел к
решению (проблем, связанных с .м.икроэволюu.ией. Кроме Toro, ока-
залось, что раопространение Toro .же -популяцианноrо подхода на
друrне облаcrи БИIQлоrJm, таlJ{\ие, как .систематика яи'зших KaTero-
'рий и пал-еонтолоr.ия, дает ВQ3можность по-новому осветить про
блемы видообразован.ия и ;ма'К'роэволюци.и И тем 'самым расш"Ир'ИТь.
основы современной 'эволюционной теории.
r па,ва 5
МУТ АЦИИ
Введение
Мутацией на'Зы.вают внезапное .наследственное изменение ка.
J<оrо л,ибо ФеНQТИЛИЧес.коro призна:ка, вы.званное резким стру.ктур-
ным 'Или Iфункциональным ,изменением в ;rенетичоском материале.
rенетический матер'иаЛ .ор.rаlНизован в иерархию crру;кту;рно-функ-
циональных единиц, :нач:ииая ос О'I\д'елЬ'ных участков 'вну.три :reHa до
целых ХРOlмос{)'м ,и reH()MOB. Соотве1)ствеино этому сущесmуют раз.
яые типы мутаций ---- 'От reHHblX до 'l'eHOMHblX.
Внеза'пные .нас.ледственные изменения фенотипа .Mo.rYT быть lВы
зва.ны не 1'ольк.Q структурными 'Изменениями ,reHOB ИЛИ хромосом,
но также и друпrми тенетИ'Ч'оокими процеСоСами. Пом.им.о истинных
мутаций, существуют мутации ложные. Фенотипические изменения
.са.м'и .по .себе не 'позволяют судить о связанных с iНИМ,И rенетиче
ских процессах. На осмов-е о.дних л.ишь :пря,мых на1блюдений трудно
раЗJlичать разные ТИ1ПЫ истинных и ложных 'Му;таций.
Для 'наших целей .моЖно не вдаваться в q>азноrо IЮда сложно..
сти, которых Iнемало .в весь'ма широкой проблеме 'мутаций. Тла'вн{)е
внимание в этой rлаве сосредоточено на некоторых общих чертах
(eHHыx мутаций, имеющих Пе'plвостепен.ное значение для эволюции.
rеииые мутации
rенная мутац.ия представляет -собой .И'зменение ПОCJIедова1'ель-
:ности 'НУ'клеотидов в .пределах oдlHoro !reHa, что в .свою очередь
tИЗМеняет характер ero действия. Друrими словами, .этО молекуляр-
НОе изменение в reHe, lВызывающее .изменеННЫЙ фенот.ипический
эффект. Допустим, чro КЗIJ<{)Й-ТО .reH содержит в н€'которой с-в.оей
точке триплет ЦТТ, 'который коди,рует одну -из аминокислот в ,ПО.'1и-
лептидной 'цепи, а пмеНIНО rлутЗ"миновую 'кислоту. Триплет ЦТТ
в результате замены IBcer.o лишь ОДНОro НУ1клеотида может превра-
,-иться в триплет rTT. Такое изменение триплета пр-иводит к тому,
что в ПО. Т 1Ипептидной цепи, образующей'Ся под действие.м данноrо
тена, на .месте irлута'МиН'овой ки.слоты окажется ,rлутамнн. ИСХОД4
.ная и 'мутантная молекулы белка отлИ'Чают.ся друr от друrа, что
Б1ПОJDне может повлечь за собой друrие, вторичные фенотипкчес.кие
различия.
Стабильность r.eнoв 'на Iпротяжении Iпоследовательных \ПОКОЛе-
н'ий клеток и .особей, а следовательно, и КОКС'е'рвативность наслед-
тв'енносПI обусловлены точ'ностью процес<:а .копирования при реп.
48
Часть 1/. МUКрОЭ80ЛЮЦtJ.9
ли.кации reHa. .Однако процесс копирования не овер.шенен. Время
ОТ Bpf'MeH.)f при копировании .возникают оши,6ки. reииble мутации
можно расс'матривать ка'к та.кие ошибки копирования.
Новый 'мутантный аллель добросовестно .реплицируется до тех
пор. по.ка не !IюэниК'нет .следующее мутаЦИОННоОе изменение. Таким
образом, в реЗУJIьтате rенной мутаци.и оявляе'Лся пара или серия
rОМoQлоrwчных ллелей. И наоборот, существование аллельной .из.
меНЧ 4 И-БОСт.и по любому r-eнy в конеЧ'НIQМ счете означает, что 'Этот
reH в то ил.и н,ное в.ремя ,претерпел мутацию.
Любой тен, входящий в состав Irенотипа, по-видимому, подвер.
жен мутированию. Во веяком случае в reHax, контролирующих
.весьма разнообразные признаки, мутации наблюдаются. Напри
'мер, У Drosophila тelanogaster .известны мутанты со слеl'Ка CMOp
щенными .крыльями, с.ильно "укорочеиными рыльями 'И 1I 1И вообще
беск.рылые; мутанты с белыми или пур.пурными. rлазами; мутанты
с разн.ообразным;и и'зм-енениями щетинок и т. п. Хорош'о известны
биохимические 'мутации у 'Микроорrанизмов, 'воздействующие на те
или иные этапы их метаболиЗ'ма подобные ;мут,а.ции существуют и
у 'высших орrа.низмов, хотя в IЭТОМ случае иЗ'учать их roра'здо труд.
нее. На рис. 5.1 -п-редставлен ряд мутантных раЗНО8"Идностей с.мо-
родинноrо ;томата (Lycopersicoп piтpinellifoliuт)1 различающихся
по форме листьев.
По 'степени фенот.ипическоrо проявлснrия reHHble мутации варьи-
руют в 'широком диапазоне от мутаций со слабым.И эффектами
до мутаций, (вызывающих значительные из'мене.ния фенотипа. Эти
Два эюстремальных ТИfПа .наЗЬ1lваются -соответственно малыми мута-
циями и макромутациями. Хорошо заметные, но не об.'lадающие
сильным ей.ствием мутаци,и, nD.I<:азан.ные 'на рис. -5.1, ти.пИ'чны для
средней части диапазона. Примерами малых мутаций с.пужат му-
танты Drosophila тelaпogaster со стат.нcrrичеаки незнач.ительными
'откло.неН1ИЯМИ от 'нормальной жизнеспОСобности .или от Hop a.Т'JЬ
Horo числа щетин.ок. .Примером мак'ромута-ции слу.ЖИТ му'Тант te
traptera у D. тelaпogaster .с четырьмя крыльями вместо двух. Он
пред-ставляет собой резкое отклонение .от ДВУКРЫ.1ОСТИ, xa'PaKTep
ной для оеем. Drosophi1idae и для отряда Diptera.
у диплоидных ж.ивотных и растений Зlначительную долю новых
мутаций оостаоВЛЯЮТ рецесс.ивные мутации, а rreHbl ДИ'КQlrо типа до-
ми-нируют. .Важное .следствие рецеосивнocrи м.н<>r.их мутантных
аллел-ей за,ключается в том, ЧТО они не 'IIо.щверrаются действиJ()o
отбора нем-едленно и непосредственно, но MorYT оохранятыся IB дИ..
.плаидной ПОпуляции на протяжении МRо.rих 'ПоколеН1ИЙ.
Частота мутаций
Спонтанные частоты мутаций для разных reHOB неокольких ар..
rаниэмов пр.едставлены в табл. 5.1. Здесь следует обрат.ить внИ'Ма
нне на ряд 4моментов. Во первых, как показыв.аЮ1' данные, получен
. r/I»7
\ ?
. . "':'--.- .....
liЭJJ. . . . , /
I
: .,...' ,
'
' ff ' . (1\
! "'.;..,..;.--
А .
lOCM
б
. * , .
G! -
со- '::-'
/.: . :i'i
}f.;Д
. ......... /,....
tl:Y 4/
; ;::o. '
1
r
Рис. 5.1. Мутаитньrе формы листьев f CMOpoAHHHoro томата (Lycoperslcon plтpi.
neUifoliит) (по Stubbe, 1960).
А. HopKa.пloHaJl фор.... Б. Aecambens. В. 8Ipinn.ta. r. Diminuta. д. Carlnata. Е. DWarr.
Ж. Butlosa.
50
Часть Н. МuкроэвОАюция
ные для человека, дрозофилы и IКУ.КУРУЗЫ, /Частоты .мутирован.ия
разных reHOB у одноro и Toro же вида варь'ируют IВ довольно широ-
ких пределах. .BO-fвroРblХ, частота мутаций у 'бактерий Ifораздо
НИЖе, чем у .вы-сш.их орrанизмов. .В таблицу включены лишь tНe-
сколыкo баlктериальных тенов, представляющих собой типичные
примеры, ,выбранные из .rораздо lболее мноrочисленных данных.
Ка'к показывает 1'а:бл'Ица, 'Частота мутаЦiИЙ (и) !для .reHOB 6а,ктерий
лежит 'в 'пределах от 10 7 дО .lO 10; ,для .reHOB 'кукурузы u== lО 4----
]0----6. В этом же диапазоне находятся частоты мутаций у дрозо-
фИJlЫ, ,чело'века .Н мыши. reHbl .бактерий, по .в"ИдИ'мому, более cra-
бильны, чем rеиы lВысших 'мноrоклеroч-ных ортаниЗtМОВ.
Таблица. 5.1
. Оценка спонтанных частот мутаций дпя нескольких видов
(данные взяты из разных источников)
СРеднее
'1НСЛО НО.
Вид reH ВЫХ кутан-
тов на
I мтr.
raMeT
Мышь (Mus тusculus) { Пеrой .окраски 17
Розовоr лазости 8,5
{ Ахондроплазин 40 120
Человек (Н ото sapieпs) rемофилии А 2O 40
rемофилии в 5 IO
Хореи rентинrтона <1
{ Жептой окраски тепа 120
Плодовая мушка (Dro- Темно серой окраски тела 20
sophila тelanogaster) Различные летали 10
r Окраски растения 490
Инrибитора окраски 110
Кукуруза (Zea тays) Сахаристый эндосперм 2
1 Сморщенные семена 1
Восковидно- кр а хм алистыА
эндосперм <1
! Устойчивости к пеницилли-
ну у Staphylococcиs 0,1
Бактерин Устойчивости к фаrу у Es-
cherichia сои 0,03
Устойчивости к стрептоми-
цину у Escherichia сои 0,0004
Имеются ос.нования считать, что IЮ крайней мере некоторые
оценки частоты мутаЦIfIЙ у 'ВЫСШИХ .орrанизмов завышены. Источ-
rл. 5. Мутации
51
ии.ков ошибок может быть нес олько. Один из них ТРУ1ДНОСТЕ>
В различении истинных внутриrенных мутаций и редких 'рекомби
наций очень тесно сцепл.енных reHHblX единиц. Эти два я'вления
MOryT пр,иводить к Qдинаковому результату, а именно к 'Резкому
феНОТИ1п.и.ческому И'з-м.енению, передающемуся по наследст,ву. Лю
бой обширный !Набор мутаций какоrо-ли,бо ДИПЛQИ'дноrо ()рrаНИ3'ма
наблюдаемый на фенотипичес'ком У1р.авн.е, по всей вероятнос"и, со-
держит, 'помимо истинных rенных мутаций, некоторую ,долю 'Необ
наруженных редких рекомбинаций, что иокажает оценку частоты
мутаций 'в ,сторону повышения.
Еще ОДllН источнJfК оши.бок представляет собой ВОЗ11.еЙС,.,8Iие re
н'ов-рeryлятор.ов на проявление п'редположите..lЬНО -мута1бельных
расематриваемых reH.()'B. reH А в результате 'мутации 'может перей
ти "ИЗ домина-н.тноrо ,состояния в рецес,сивное (A a); при ЭТОМ
MOryT ,ВОЗНИ.кнуть rом,озиrоты аа, а .зате.м, по-'видимому, :происходит
обратная мутация к А. Фактически же мы наблюдаем появление
фенотипа А в инбредном :ПОТ.Qмс.тве rомозиrmноrо мутанта aa
обычно рззмножающеrося в чистоте. .одно из 1Вероятных объясне
ний возникновение обратной мутации (а ........-+А). Возможно также
что тенотип аа остае1'СЯ неизменным, но ero фенотипИ'ческое прояв
лени.е внезапнoQ подавляется rеном супрессоро'М, находящимся 8
друroм л о кУ.се, а быть может, вновь вооникшим мутантом этоro
reИЗ-С)11п.реосора. Такой процесс 'может леrко привести к завышен
ной оценке частоты мутирова'ния fенз А.
Бели даже иcrииная ча,стота мутаций у IВЫСШИХ орrанизмов на
одии или два порядка НИЖе, 'чем показывают современные оценки
ТО .все же ,эта 'частота О"кажет-ся доcrаточиой, чтобы ,поддерживаТf>
уровень мутационной изменчивости В популяциях. В популяции
средней величины, продуцирующей от 10 до 100 .млн. raMeT, на
каждый reH ,в средием будет 'возникать по крайней мере по ие
СК.Qльку КОВЫХ мутаций в ка'Ж1ДОМ ПОК-ОJIении.
rенотипический контроль
у Drosophila тelanogaster есть о.щин rеи, который вызывает
высокую 'Частоту мутаций друrих reHolB да.н:ноr.о комплемента. ЭТ,()Т"
rеи-мутзтор обозиачают символом Hi. У мух, rом.озитотных по re-
.ну Hi, частота 'мутаций в 10 раз выше обычной; у rетерозиrот П()
этому rеиу частота мутаций выше обычной в .2----7 раз. r H Hi .инду
цирует :как видимые, так и летальные мутации мноrих reHOB. Он
Вызывает также .инверсии ---- .ОДИ.н из типов "'ромосом-ных мутаций
(lves, 1950; Hinton, Ives, Еvапs, 1952).
Мы уже описывали reниые 'мутации как случайные ошибки KO
пирования, !происходящие во ;время ,воспроизведения reHa, .одна!ко,
'Как показываIО1' данные о дейс'I1ВИИ 'rеиов-мутаторов, существует
.М друrой а пект ,мутзд'Ионноrо процесса. Возникновение !Новой MY
52
Часть 11. МUlCроэвОАЮЦия
тационной изменчи.вост.и, :которая имеет 'важн.ое значение для дол-
rосрочноrо успеха данноrо ВИlДа в -эволюции, не может быть цели-
ком предоставлено .воле .случая, этому должны способствовать
reHbl-мутаторbl. Частота мутаций у да.нноrо вида частич.но может
быть одним .из контролируемых rено'J'1ИПОМ компонентов всей ero
rенетической ,с.исте:мы.
у ,близкородственных .вид.оВ Drosophila willistoni и D. prosal-
ians, обитающих в тропических областях Амер;и.ки, частота мута-
ций различна. В табл. 5.2 представ.пены частоты летальных мута-
,!.J.ий 'в хром'осомах II и 111 у ЭТИХ двух видов.
т аБАuца 5.2
Частота мутаций у двух близкородственных видов
Drosophlla willistoni и D. prosaltans
Частота мутаций
Вид
хромосома 1 I I хромосома 111
2.2'1<r 5 I 3,0 .1O 5
1 . 1 . 1 O 5 2 , 1 . 1 O 5
D. willistoni
D. prosaltans
Вид D. willistoni, у KOToporo чаСТота мутаций выше, обычен,
широко раопространен и занимает разнообразные ЭКОлоrические
ниши, тоrда .как друrой вид (D. prosaltans) встречается .редко и
.{)rраничен уЗ'кими эколоrиче.скими УСЛoQвия,ми. БЫ.'10 высказано
весьма правдоподобное мнение, что высокая частота 'мутаций
у п. willistoni, поставляя новые варианты, способствует -ПОВЫШе-
нию эколоrическоrо !раз'нообразия, а тем самым и обилию этоrо
. вида (Dobzhansky, Spassky, Spassky, 1952).
Адаптивная Ценность
Большинство .новых мутантов характеризуется .более .низкой
жизнес'По.собно'стью, чем нормальный, И",1И дикий, Т,RП. Снижение
жизнеспособности может быть выражено в разной степени от
чуть заметноrо .субвитаJlЬ"НОro состояния до полулетальности :и ле-
.тальнос1'И. П'ри оцен-ке жизнеспособности ,мутантов' Drosophila те-
laпogaster, !Возникших в результате мутаций в X-ХРОМОСOlме, 90%
-оказались менее .жизнеспособ.ным.и, чем норм.альные мухи, а 10%
были су.первитальны.ми, т. е. обладал,и повышенной жизнеспособ-
ностью. Среди 90 % мух пониженной жизнеспоообностью на.блю
да,стся весь диапазон изменений {)т слабой субвиталыности (45 %)
через :промежуточ.ные стадии понижения ж-изнеспособност,и до по.
лулетальности (6%) и летальности (14 %) (Тимофеев-Ресовский,
1940) .
Вообще адаптивная ценность новых мутантов обыqlНО бывает
:понижена. Адапти.вная цен.насть сла.rается из. ПJlОДОВИТОСТИ И Функ-
r А. 5. Мутации
53
циональной пол-езности морфол.оrичес.ких признаков, а таlкже фи.
зиолоrической жизнеспособности. Мноrие мутанты неплодовиты
незав.исимо от Toro, обладают они нормальной жизнеспособностью
иди нет. Морфолоrичос.кие макромутации обычно сопр.овождаются
наруше.ниями функциональной эффектИiВНОСТИ. Из обширной вы-
борки и'ндуциро.ванных мутантов ячменя (Hordeuт sativuт)
у 99% адапт.ивная ценность была понижена (Gustafsson, 1951).
Этим наблюдениям леrко дать 06ъя-сн-екие. Все reHbl, входящие
в состав rенотипа нормальной, или дикой, формы, на протяжени.и
мноrих lПoQколений подверrа.ТIИ-СЬ ocTecrBeHHOMY ()тбору; они пр'ошли
сквозь C TO отбора, в результате 'Чеrо ,сохра.нились те нз них, 'ко-
торые обладают маК:симальной адапти-вной ценностью. Следует
ожидать, что любые ;изменения в таких r-енах 'почти наверное ока-
жутся изменения,ми к худшему, подобио тому как .копанье науrад
в часовом механ'изме окорее нарушит, чем усовершенсrnует ero
«функциональную эффективность:..
reHHble мутации нередко называют «случайными» и'З"менениями
в reHax. В даннам контексте терми,н «случайные» требует разъяс.
Jiения. Мутационные :изменения на сам'ом деле MOryT быть неслу-
чайными на молеКу.IJЯРНОМ уровне. ,Возможно, что некоторые изме-
нен'ия последовательности ну,клеот,идов 18 цепи днк ВОзникают
чаще, 'Чем друrие. Та.к называема'я .случайность мутационноrо про-
цесса относится не к из'менениям молекулярноrо строения, а к адап-
7ИВНЫМ -свойства'м . мута'нтных reHOB. Мутации .случайны в том
.смысле, чт.о они не направлены в сторону .какоro-либо ны.нешнеrо
или будущеrо состояния приспособленнос'I'lИ данноrо орrанизма.
И все же каlКая 'Ю небольшая доля всей массы reHHblx мута-
ций у .rенетичес- и хорошо изученных орrанизм.ов в том или дру-
rOM отношении превосходит стандартный тип. В одной выборке му-
тантов ячменя примерно O,l. O,Z% -мутантов обладали 'повышен-
ной урожайностью 'в .стандартной среде !или в той среде, .из которой
происх:одил.и рсщительские формы (Gustafsson, 1951).
Мутант, обладающий пониженной адаптивной ценнос.тью:в стан-
дартных у.СЛОВИЯХ среды, может оказатьоя более риспособленнoblМ
в 'каких-л'ибо друrи.х условиях. Один .из мутантов Drosophila 'ипе-
bris мутант eversae..... при 15 ос обладает пониженной (98%)
жизнеопособностью по .сравнению ос жизнеспособностью дикоrо ти-
па, но 'при 24 ос er.o жизнес-пособность выше (1'04 %) (ПоЬzhапskу,
1951). ,Шесть мута.нтов львиноrо зева (Aпtirrhinu-m тajus) в нор-
мальных }'1СЛОВИЯХ теплицы проявляли более 'Низкие lКач-ества, чем
родительская лин'ия, но .при создании 'в 'l'епл.ице разно-rо рода ано-
мальных условий ОRИ превосходили родительскую линию .по способ-
ност.и 'к росту (Brucher. 1943; Gиstafsson, 1951).
Эти данные .об адапти,вном превосходств-е некоторых новых МУ-
7aHT B по сравнению с диким типом, будь то в стандартной или
9 новой .среде, очень важны, TaiК lКaK ОН1И .показывают, 'Что мута-
54
Часть 11. М uкроэвОАюция
ционный 'процесс может служить 011правной точкой эволюцио"Нноrо-
развития. Важное значение им-еют также данные о высокой часто
те в'редных мутаций; эти последние .дан,ные исключают rВО3МОЖ -
'ность roro, что мутационное давление иrрает роль направляющей
силы .эволюции, хотя подобное .мнени-е Ko()e rLдe еще .оохраняется
Относительное значение в 9ВОЛЮЦИИ
малых мутаций и макромутаций
БоJIыliин::ство -эволюционных !"'енетиков подчеркивают важную
роль в эвалюции малых 'Мутаций. Соrласно же м.нению .меньшин-
ства, ,к ,котораму принадлежит rольдшмидт (Golds-chmidt, 1940..
195-2, 1953, 1955) и некоторые друrие ученые, Irлавную роль в "ЭВО
людии иrрают макромутации. В прошлом между .сторонниками
двух указан.ных .противопол'ожных точ,ек зрения было MHoro с,по
ров, которые, однако, и лишни, поаколыку эти взrляды не исклю-
чают, а, .напротив, IДОПОЛНЯЮТ др}'lr друrа. .в "Эволюции иrрают 'Роль.
как малые мутации, так и макромутаци.И.
Как 'сырье для ЭВОЛЮЦИОННЫХ изменений малые мутаwии обла
дают некоторыми явными преимуществами. Каждая малая мута-
ция вызы:вает лишь небольшой фенотипичес.кий .эффект ---- к ЛУЧilIlе
му,или к худшему. Поэтому аллель, возникш'ий в результате малой
мутации 'и обладающий сл абы м !преимуществам, может в.ключить
ся в уже существующий rенот,И!II, не вызывая скалько ннбудь рез
'КОЙ дисrармон.ии. При 'помощи ряда малых мутаций, происходя
щих в разных л-аку,сах, мажно саздать тот или ,инай количествен
ный адапт.ивный эффект, не ,нарушая функцианальной эффектив
ности орrанизма во время промежуточных -стадий этаrо процесса_
Иноrда одна един'ственная мутация, затраrпвающая какую
лиоо ,ключе.вую -СТРУКТУРУ ,или функцию, может открыть .перед
своим обладателем навые возможнасти. У столь разных орrаниз-
мов, как бактерии и млекопитающие, известны примеРI?I устойчи
вости к определ'внным токси.нам, обусловленные мутацией по OДHO
МУ reHY. Устайчивая 'мутантная диния получает .возможность засе
лить тО'К!Сичную среду, закрытую для чу.вств.ите.пьноrа 'родительско..
ra ти'па. Мутантная ,бескрылая муха будет обладать преимущест
IBOM в овеваемом ветрами островном местаобитании, rде нормаль
ная крылатая.фарма не моrла ,бы выжить.
Такая единичная макромутаll.ИЯ не обязательна действует одна.
Ее действие может реrулировать'Ся -сер.ией rенов-модификаторов,
.каждый из которых сам по себе обладает незна-чительным эффек
'Том. Именно в тзlких .случаях мы имеем дела с совместным эволю-
ционным действием макромутаций и малых мутаций.
Прямые данные о типах ..reHHblx .изменений, которые :имеют ЭВО-
люционнае значение, :палучены .при rенет.ичес.ких исследовааиях
межрасовых rибри:дав. Как показывают эти данные, дифференци:'
rл. 5. Мутации
55
ровку 'каждой из близких рас обусловл'ивают .разЛ.ичные reHHble
системы. Меж'расовые разл:ичяя у растений и животных обычно
контролируются 'системами множественных reHOB. что подтверж-
дает rипотезу об эво,пюционном значении малых .мутаций. Однако
в rруппах дивер.rирующих природных популяций на6людаются
также различия -по признакам, 'контролируемым единичными reHa-
ми, хотя такие случаи довольно редки. Обыч.но rенная система,
опр-еделяющая различия !между расами у высших ор:rанизмов, со-
стоит ;из одноrО 'rла,вноrо reHa .и нескольких reHOB-МОд'ифи:каторов.
Подобноrо 'РОда if'eнная система 'поэволяет 'пред пол а тать эволюци-
онное изменение, основанное .на видимой му.тации, сопровождаю-
щейся различными малыми Iмутациями.
Вероятность сохранения новой мутации
КажlДЫЙ отдельный мутантный аллель при своем первом воз-
никновении бывает редок. Перetд ниlм встает проблема .сохранения
в родитель'с ой популяции. Со'rласно зак()ну Харди........Вайнберrа,
ча.стота HOBoro MYTaHTHor.o аллеля не может увеличиваться за счет
одноrо пяшь прouесса воспроизведения. Вероят,ность ..же чисто слу-
чайной элиминаци.и MYTaHTHoro аллеля, напротив, очень велика.
Ф.ишер (Fisher, 1930, -1958) .ВЫЧИСЛ1ИЛ вероятность сох:р анения
новпrо .MYTaHTHoro алле.ТfЯ, возникшеrо 'у одной 'особи в обширной
популяции (та , б..1. 5.3). Как показы.вают ero данные, шансы на co
хранение единичной мутации заметно ,понижают.ся :с каждым поко
лением. Е-сли Iмутантный а..'1.тIель в селективном отношении нейтра-
ден, то вероятность ero исчезновения к 3 1 му!По.колению соста'ВJIяет
94%, а к 127 My 98%. Если он обладает небольшим селектив-
ным преимуществ.ом (порядка 1 %), то .вероятность ero -исчезнове-
ния Н КOJIьк() СRижается (93 'н 97% в поколениях 31 и 127 соот..
ветст.венно), но все еще остается ,высокой.
ТаБАица 5.8
Вероятность исчезновения мутации, возникшей у одной
особи (Fisher, 1930, 1958)
Чиc.nо ПОКО.llеинА
Вероятность исчеэиовеИИJl при усповии
1 1 % HOro селектквноrо
преимущества
иеRтрапьиости
1
3
7
15
31
63
127
0,368
0,626
О ,790
0,887
0,941
0,970
0,985
0,364
0,620
0,782
0,878
0,931
0,959
0,973
rпaBa 6
.П'ОТО.К .rEHOB
Введение
Популяция мож.ет 'прнобрест.и новый аллель (например, ал..
nель А 2 тена А) в результате ли.бо мутадии, возникающей у ОДНОЙ
из особей данной 'по.пуляции, либо им.миrрации носителя "ЭТоrо. ал
леля из друrой популя ии. Этот последний процесс и есть поток
reH-DВ. .
lV\.иrрация относится к ч,ислу фа'кторов, о-казывающих влияние
на изменчивость популяции лишь в том .случае, если прише '1ЬЦЫ
01'ЛИJчаю'J'tCя от той .популяции, в которую .они всту.пают. Имм.иrрант
приносящий алл.ель А 1 'В ПОПУЛЯЦИЮ, <содержащую reH А 1 в .сБоем
rенофонде, не оказывает влияния на ее изменчивость; но имми
.rpaHT, вносящий в эту же 'самую IПОПУЛЯЦИЮ аллель А 2 , 'изменяет
ее -rенетический состав.
Носитель HOBoro аллеля А 2 MOr ,приобрести этот аллель в pe
зультате ПРОIl шедшей перед этим мутации. В .истории (Данно.rо
вида аллель А 2 , возможно, диверr.И'ровал путем мутацИ"и от предко-
вой ,формы, т. е. .от reHa А. Таким образом, для Toro чтобы п-оток
reHOB служил источником изменчи'вости, необходимо 'предвари
тельное наличие мутаций. А .поэто.му поток тенов можно расс.мат-
ривать ка;к за;лаздывающий эффект мутаци,онно.rо :процесса.
Тем не менее в каждый да.ННЫЙ момент прямым .И непосредст
венным ист.очником 'некоторо.й :измен'чиВости 'в популяции может
быть IиммИ'rрация в нее из друrих популяций носителей аллелей,
Отличных от имеющихся в ее тенофонде. Некоторые ,мутантные ал
лели, 'кикоrДа, ,nо видимо.му, не возникающие в данной популяции,
MOryT поступать .В нее из соседних популяций. Конеч.ным ист{)ч-
ником изменчивос1'И популяции-реципиента служат, слеДОвательно.,
[енные мутации, ,возникшие '8 ,каКОЙ ТО момент в ,nро.шлом п е то
в tJtpyroM месте; однако неn.оср-едствекной пр.ич.иной изменчивости,
реально действующей в данный момент в данном месте, является
пото.к lfeHoB.
Скорость иммиrрации
Скорость потока reHoB (т) варьирует 'количеСТВенНО от О До. 1,
тде т == О ОЗllачает 'полное от утств:ие .н.ммиrРИРУЮЩ1ИХ аллелей, а
т== 1 означает «затопление" иммиrрантами в течение одноrо поко-
лени,я. В общем виде
Число иммиrрантов из I ПОКOJJение
т
Общее число особей
r л. 6. ПОТОК eHoв
5
Если .на Iкаждые 1000 IralMeT в каждом п'Околении пр.иХОДИТСj)
по одному ,иммиrранту, то т==О,ООl.
Обозначим частоту данноrо аЛЛeJ1Я среди -ис,кон.ных членов по'"
пулядии Qo, а {:реди им.миrрантО'в qт. В та-ком случае 't{aCТOTa
этоrо аллеля (ч) в смешанной популяции (Falconer, 1960) соста-
вит
q==т (qrn----qo) + qo.
Изменение 'Чз-сroты тена за ОДНО IП.Qколетне (..1q) за счет !IICТOKa
тенов ,(Fakoner, 1960)
D.q;::m (qm qo).
ЕсJtи, например, ча:стота даННоrо аллеля среди исконных 'Членов
.популяция Iсоставляет qo == 0,3, среди -имм:иrранroв равна qm ==0,5,
а -скорость миrрации т==О,ООI, То ИЗМен.еКне частоты алпеля в .ре-
зультате имм-иrрации в одном поко()леl:lИИ
D.q==0,0002.
Скорость изменения частоты ..rе.иа в попуЛЯIJJiIИ, .обусловленная
lIOTOKOM rеиов, зависит, таким образом, от двух факторов: 1) от
;различия iВ частоте аллеля между :иск-онной популяцией... и им ми-
J'рзнтзмн .Н 2) -от скорости иммиrраЦ'RИ (Fakon-er, 1960).
Типы расселения
Носите.пями потока reHOB у подв.ижных высших животных яв-
дlЯЮ'f\Cя отдельные орrанизмы, у -СИДЯЧИХ морских животных сво-
бодноплавающие личинюи, у высших растений ---- пыльца и семена,
.а у ,низших растений ,и rр,ибов ---- споры. Расселение представляет
собой одну из .нормальных стадий жизненноrо ЦИК.'1З. У всех opra..
:низмов, в том 'Числе и у .сидячих, по в.идимому, имеется свободно..
жи.вущая подвижная .стадия, при помощи котороЙ происходит рас-
селение.
Но при \этом orpOMHoe большинство рас.селяющихся единиц, в
ТОМ ЧИС.'1е и 01ще.пьныe особи подвижных животных, обы'чно остают..
.ся поблизости от места -своеrо .происхождения ил.и от родной Tep
ритор.ии. Для них ха'рактериз та.кже .некоторая 'степень осе лости.
Совместное действие этих двух аспе.кто-в ---- ра'сселения и оседло-
сти определяет пространственное распред€ление особей и .их re-
нов .из поколеPrИ'Я в поколение.
ЭТИ две вза.имодополняющие -стороны 'Четко .выявил.ись при .пзу..
чении области раопространения пыльцы .с-осны. Пыльцу сосны
Культера (Pinus coulteri) метили радиоактивным фосфором, а
затем, ооздав возможность для ее ра.ссеяния ветром, выявляли при
помощи счетчика fейrера.......мЮ,,'lлера .Н методом радиоз'Втоrрафин.
Партии пыльцы объемом 1 л помещали в сосуды, которые ПОДDe-
58
Часть 11. Микроэволюция
шивади кверху дном на высоте 3,5 м. В ясный день при .слабом
ветре пыльца медленно ,вы-сыпалась 113 сосуда. По радиу-сам, от:..
ходящим В разных направлениях от точек распросrранения пыль-
цы, ставились с равномерными интервалами ЛОВУШI{И дЛЯ пыльцы.
Количеств.о пыльцЫ, попадавшей в разные ...10Вушки, 'определяли
'по уровню ради{)З'кт.ивности (Colwell, 1951).
25 000
20 000
.."
, ..
,. ..1
I ,[
I .,
, I
I IA
I
J.A
11) 15 tJUU
'tI
i 10000
5
00
r::
о
:з 5 000
..--.......
--........
......--........
........................
............... ...
... ......
о
б
12 Iб 24 30 42
PacCmORHUp. от uсmО'lНUКЙ пhlJ1ьцы. М
.4б
54
Рис. 6.1. Копичество сосновой пыпьцы, переносимоА CJIабым ветром на различ-
ные расстоянии от источника (Colwell, 1951).
1 пря о 8 иапраВ.IIеиии ветра ОТ источника; 11 по радиусу. распопожеИ80МУ под yr-
.110М 450 к направлению Ветра.
Результаты э их ЭК-СlIер.ИмеНТОВ представлены на рис. 6.1.
Основная масса пыльцы рассеивалась в направлении ветра. на рас-
стояние от 3 до 9 м. За пределами этой зоны ма'кснмальной кон-
центрации ,количество ра-ссеянной пыльцы ,быс.тро снижал ось. На
расстоянии Ф5 :м }f более от -сосуда можно ,было обнаружить л.ишь
небольшие Количества пыльцы. Эт.и результаты 0значаЮ1', 'Что в
сосново'м лесу жеН.ская .ШiИШ'ка ,будет буквально заеыпана 'пыльцой
с соседних деревьев .н полу.чит некоторое, Но весьма незна'ЧИТе.пь-
ное количест.во пыльцы от деревьв, находящихся на расстояния
носкольких ДDCЯТКОВ .метров (ColweH, 1951).
в Теха-се 'на протяжени.и 6 лет проводилось усиленное изучение
локальной IПОПУЛЯЦИИ И'I-уаны Sceloporus oUvaceus. Было обна-
ру,жено, что большая часть особей обоею 'пола 'Оста.ется в 'радиусе
60 90 м -от места 'ВЫЛУПЛ'f'ния. 'Некоторые особ.и, однако, отхо:дят
подзль.ш.е вплоть дО S{)O М.......... от rэтоrо Места. Тенденция к более
ширакому ра-сселению неоколько чаще наблюдается у -самцов, чем
r А. б. Пото" zeHOB
59
у самО'К (,Вl air, 1960). Хара.ктер расселения s. olivaceus представ
лен в .виде диаrраlММЫ На рис. 6.2.
Уже давно высказывали<сь предп.оложен.ия, что в разли'Чw.и меж
ду широко расселяющим. ася и !более оседлым.И особями у nодвиж
1IЫХ животных участвует rенотипичее:кий фактор. Это пр дположе..
2й -
А
5 -
J5
1O
о -
8
о
Е: 20
::r
15
с::::
10
Б
5
о
c o c o 8 g
c o 2 N N ..,.q
.... tt), \D....... I I . l' 1, I I I I I I I 1'" t I I I t
I I .!. .!..cb.!.8N:g;n:g = ':; t:: 5 ;; "" a; N ;;;:8;o
Q m.... .... ....N NNN qq
Расстояние, м
Рис. 6.2. Относительное копичествО самцов (А) и самок (Б) иrуаны Sceloporиs
<Jlivaceиs, рассeJtИlOЩИХСR на разные расстояния от места выпупления до место-
нахождения половозрелых животных (Blair, 1960).
иие подт!Вердили результа.ты экспери:ментов, проведенных недавно
на п{)левках (Microtus pennsylvaпi.cus .Н М. ochrogaster). Исполь
зуя в -качестве :маркеров reHbl, контролирующие синтез бел'ков,
удалось установить, что активно раttеляющиеся 'Пl()левки rеноти
пически отличаются !от нерасселяющихся. Тенденция 'к расселению
<:опр-овождается более высокой а,rрессивностью (Krebs et аl., 1973).
Изучение расселен.и,я IC целью уста.новить оС'корости распростра.
нения \reHOB проводилось на Drosophila pseudoobscura. fенетически
помеченных мух выпуС'Кали на волю, а затем IBH{)Bb вылавливали
на разных расстоян:иях ОТ места выпуска. Как обычно, большин
ство мух .оставалось лоб.низОСТИ от Toro места, rде НХ выпустили,
но нек'оторые ра.сселяли-сь в самых разнообразных направлен.ИЯХ
На знз'Чительные р'аостояни'я. Результаты 'этих .первых зксnеримен
БО
Часть 1/. Мuкроэволюция
ТОв Iпоказали, что .с,пустя 1 О мес .или через н-есколыко поколе.ниЙ1
Ilосл-е выпуска 95% ПОТОМКОВ выпущенных мух оказывается в пре
делах .окружности радиу ом 1,76 км (Dobzhansky, Wright, 1947).
Недавно эти 'эксперименты были повторены и заново лроана.'1И3И-
рованы, причем оказалось, что -скорость расселе.н.ия мух выше, чем
считали по первоначальнbl'М оценкам (Crumpacker, Williams, 1973;
Dobzhansky, Powell, 1974; см. также Johnston, Heed, 1976).
В 'природе широко ра-спространено сочетание оседлости с рассе..
лением 'некоторых особей на большие расстояния. Опи-са.нные'
выше 'Примеры достаточно типичны; аналоrичные примеры можно-
наблюдать у мышей, птиц, пчел, бабочек, растений, опыляемых на-
секомымИ или птицами, и у семян, распространяемых ветром.
Популяц.ионно-rенетиче.ские следствия, вытекающие Iиз расселе..
ния подобноro типа, совершенно очевидны. Интербридинr у сво--
бодно -скрещивающихоСЯ орrанизмов происходит rJlaBHbIM образом
между близким.и -сос-едям.и. По прошествии ряда ПОКОлений ЭТО-
приводит к значит-€-льному ,инбриди.н.rу, несмотря на систему с,во-
бодноrо -скрещивания. Тесный инбр.идинr Iдополняется, однако, от-
даленными неродствен-ными скрещиваниям.и с иммиrрантами, будь.
ТО особи или raMeTbl; это обеспечивает небольшой, но значимый
обмен rенами :между особями данной ПОПУЛЯЦИИ, находящимися.
на больших ра-сстояниях друr от друrа. Короче ,rо.воря, ВО мноrих
популяциях наблюдается сочетание значительноrо .инбридинrа
с некоторой долей .свободноrо скрещивания.
Панмиктические единицы ,н соседства
Данные о типах расселения позволяют .НаМ rлубже понять
CTpyrктypy популяций. В популяции орrанязмов с перекреС1НЫМ
оплодот.ворени.ем раз-меры rруппы, деЙствительно с рещивающейся
внутри себя, меньше, обычно .rораздо меньше. размеров caMO
популяции. ,Поэтому необходимо различать ,некоторые субш:шуля-
ционные единицы.
rруппа, в .пределах которой прои-сходит -случайное скрещ}[,ва
ние, называется паНМ1И'КТИЧООКОЙ единицей (Wright, 1943а). Эт()
реальная единица 'в популяциях с локальным св.ободным скрещи-
ванием. Число особей в такой панм.иктической единице обычно
060з.начают через N или N e , одна-ко ввиду тот,о, ч.то символ .N часто.
и-спользуют также в ,более общем .смысле, а оСимв-ол N е..... для обо-
значения друrих .в'ещей, 'мы во избежание путаницы будем обозна-
чать паllмиктиче(:кую единицу символом N п. Вел.ичина N п зави-сит
от ПЛОТНQСТИ популяции И с-корости МИI'pации. '.Следует от.метить,.
ЧТО IB паНМИlктической популяции про.исходит известнЫЙ .инбридинr,.
доля KOT.oporo составляет I/N п.
Более -общее Iприменение 'Имеет такая -субпопуляционная едини..
ца, \КаК -соседство (neighborhood; Wright, 1946). Некоторые авторы
r./1,. 6. ПОТОК zeHOB
6)
ра,ссматривают сосед.ство как синоним .панмиктической единиuы .
и при известных условиях эти две единицы действительно равно-
значны. Однако Райт (Wright, 1946) в '.с.воей первоначальной фор_.
мулиро.вке концепции сооед<:тва, которой мы здесь придерживаем-
ся. проводит тонкое различие между панмиктичеС'кими еди'ницами
и соседствами. Соседство это то же самое, что и панмиктическая
единица, там. :rде панмикоия носит локальный характер. НО ВС>.
мноrих реальных ситуациях система ..скрещивания ,приводит IK силь.
ным отклонениям от лока.l1ЬНОЙ 'панмикс.ии. ,в таких СЛУ'lаях со-
седство соответ-ствует дерива.ту чисто теоретической паН'миктиче-
ской единицы; И,,1И. выражаясь .иными словами, теоретическая пан-
микт.ичесжая единица служит неким -стандартом, из KOTopo.ro мож--
Но ,вывести величину соседства. Величи.ну соседства :МЫ здесь та.к"
же будем обозначать сим.волом N п.
В основе онцепции .соседства лежат два допущения: 1) -вероят-
ность -соединения raMeT уменьшается с увеличением расстояниЯ'
между р'Одите,,1ЬСК.ИМи особями; 2) в панмиктичес.кой популяци.и.
имеет место инбридинr, ДО.1Я KOToporo составляет, как указывалось.
выше, I/N п. В резу..lьтате подобноrо распределения по вероятности
скрещивания 'Н .большой непрерывной популяuии в каждом .поко-
.lении происходит некоторый ,,10кальный инбридинr. Ведичина пан-
миктической единицы, в которой ДО.1Я инбридин'rа из ,поколения в
поколение одинакова, соот,ветствует величине &Соседства. ИНЫМ.И'
словами, мн.ожитель ljN n используется для выведения величины
соседства в тех .случаях, 1К0rда случай.ноrо локальноrо скрещива-
Ния ,не ПРОИ-СХQДит.
Область, занимаемая соседством в непрерывной ;популяции,
это пространство, занимаемое в среднем N п особя'ми. Важный фак .
Тор, влияющий на 'Эту величину, ---- среднее квадратичное отклоне-
ние расстояний. на которые 'происходит расселение популяции (а).
Отсюда следует, ,что соседство !при распределении особей 'в двумер-
ном пространстве занимает площадь, имеющую форму окружно-
сти ,с радиусом 2<1. В ЭТОЙ ОJ\1РУЖНОСТИ сосредоточено 86,5% роди-
телей оообей, находящихся в .центре этой области.
Если .популяция имеет линейную JroнфИiryраuию, то .область со-
седства -представляет собой полосу IДЛ!ИНОЙ 3,50'. Полоса .TaKoro раз-
мера включает в себя при;мерно 92,4% родителей особей, находя-
щихся в пре.делах ,Этой полосы. [Математическое описание приве-
eHHЫX :выше рассуждений см. в работе Wright, 1946; словесные-
формулировки составлены ос п.()мощью д-ра Фл йка (Flake).]
Обширную непрерывную популяцию 'М'ожно рассматривать как
ряд пеРfIКры-вающихся соседст.в. Совершен'Но очевиДНО, что ско-
рость миrрации это факт,()р, .оказывающий влияние на величину-
tоседства. С увеличением т увеличивается .и N п. При высоких
значениях т большая непрерывная популяция приближается к'
панмиксии .в широюих масштабах {хотя не о.бязательно достиrает-
....,
tS
::r
.
о
:Е
(1)
.:с
::1:
1.
С.
О
><
2
Q,
ос)
:.с
:к:
>а
.
...
о
'v
=
::1:
-:r
.:а
е;:
111
:а::
:.:
=
о
:а
:r:
=
«1
-с
:.:
=
=
:J'
О
..
...
::s::
сп
с:
CCI
.с:
.о
о
,:)
..
м .--..
t--
с;- о)
..,.,
се
Q') CIJ .......
....... . &.. "--" ао б)
Е QJ
"6'0 t':I "i
--- ....... ti
'С ::: QJ j:Q ...... ....... ......
i i rЛ t Е;:
....... "--" . - . .
а.. s: g CI)
..:с ...... Q).... 4) t
u) ...... 4i ......
t: (.) с) ::к:;'
QJ t':I ...... i
са <..l '"" .... tIi
..с:: с.. t: ..J::: r:J r;
N Е .... tI.Q .!: . 't (/)
..1:1 са :1 .:; .;: i t
о а..;>' .... L.o i:i5 :::
с U.:.:: U О .....:1
6
8
IC"I
<=1 ....
......0
00 1:{ о
QJ QJ °Clt r-.. Q) C\J с;;
Е :3 I ; '1 t-- S
I ..Q ..Q C\J ..Q
= 8=; :210 I 1:1: I
QJ '"
о \0= . о l.t:I
J.Q C'I":) 0= ...... ta r-..
1:11: tI:
=: =
:z: о :z: u
с. 1::( с.. .:s::
о ;IIt - са О О
.g. Q, = u -е- :1::
== О с:; = =
t; t; t.. t; с;
t':I О = QJ са 1:1:
< f--o ::s;:
t':I
Q) ....... CI.
>.
QJ I.Q
:а :21
111 ::с
о 1-- :::Е
QJ
2 111 Q)
:1:: :21 :::r
= о
U IQ ::с o:u
О :Е '"
tS О
.... :i Q t; o:u
::s Q) == U :s
u =
CI) CI) ....... U "о
O.C:I :::Е
о ::s (u ....
о :Е QJ
. cj <..J
t:S
::s "i:S .2 ...
CI) ... t:S
CI) !IE .... 'о
а. о а. ... ::s .
o:u
t:S О А CI) .э Q" Q)
:::: Е- o:u ::s CI,I ....::а
(з ::s 'Q. culQ
.:::
Q" .... . Q.o
cu О )t CI)
CI) о ... 0=
о а. t: ::C
cs cj :.:l
'-3 CI) Q,. 0.4 0.4
1:
===
о
..
u
..
с:{
:r:
a:I
r л. 6. ПОТО" еен.ов
63
ее). При низ.ких же значениях .т большая непрерывная популяция
распадается .на сwрещивающиеся внутри себя rpYnnbl. т. е. на не..
большие соседства.
Теоретически в 'большой непрерывной попу..тIяции может .воз"
никнуть значительная локальная дифференциация. если размеры
случайно скрещивающейся rруппы невелики. Представим себе не..
прерывную полиморфную популяцию СО св.ободным с.крещиванием
при 'отсутствии отбора. Если N n 1000 или более, то следует ожи-
дать возникновения .небольшой :реr.иональной .расовой дифферен"
циации (пом.имо результатов действия {)тбора). .Но если N п 100.
то возм{)жно .образование реrиональных рас; если же Nn=='lO, то
.'10кальные ра,сы MorYT о'6разоваться tВ отсутствие отбора (Wright.
1943а) .
Оценки величины соседств
Различные а'вт.орЫ в разное время пытались оценить .вел.ичину
соседств у разнообразных орrаниз'Мов. Это 'нелеrкая задача. со-
пряженная с рядом методических затруднений. Несколько таких
оценок приведеНQ 'в табл. 6.1.
Для большинства перечисленных в таблице видов размеры со..
седств относительно невелики. У иrуаны Sceloporus olivaceus ве..
личина соседства Nп==225 270, пожалуй, ближе к 225, а зани-
маемая им площадь представляет собой окружность радиусом
213 м. У флокса (Phlox pilosa) величина соседства составляет
75 282 растения, а ero площадь имеет диаметр 4,4 5,2 м. Оба
эти примера довольно далеки от ширОКОЙ панмиксии.
Еще большее отступление от широкой панмиксии наблюдаетсSl
у Lithosperтит caroliniense (сем. Бурачниковые), у которых со..
седство занимает участок диаметром 4 м, а ero эффективная ве.rJИ.
чина равна примерно 4 растениям. Точно так же у некоторых
диких популяций домовой мыши (Mus тusculus) скрещивающие-
ея rруппыl представляют собой очень маленькие изолированные
tубпопуляции. состоящие примерно из 4 особей (DeFries,.
McClearn. 1972, см. также S€Iander, 1970). При таких малых раз
"ерах скрещивающихся rpynn блаrоприятные условия для ло..
'альной расовой дифференциации создаются в результате одноrо
лишь инбридинrа, без вмешательства отбора.
у наземной улитки Сераеа neтoralis н у дрозофилы соседства.
8апротив, передка достиrают значительных размеров.
rлава 7
РЕкОМБИНАЦИЯ
Введение
Мутационный процесс и поток rеИQВ мотут .создать е 'попу ляцки
язменчивость по единичным теизм. Если в результате таких пер-
вичных процессов возникает аллельная .изменчивость по ДB M или
()ольш-ему числу rеиов, то T M самым создается почва для \I!,ейст-
вия В1'оричноrо процеоса..... реКoQм'б.инации. В результате р-екомби.
нации .новые аллеЛИ t ,носителями которых первоначально, вероят-
но, были рз'зные особи, .Mo rYT .сочетаться .в одном rенотипе. .Кроме
oro, в процессе рекомбинации число различных rенотипов в попу
.JIяции может увел.ич,иться; этот процесс превращает небольшой
лервоначальный запас :изменчивости по множественным тенам в
l'ораздо более значительное ,колич.ество .rенотипической из м.еПЧИВО-
ти. '
Процесс рекомбинации
Допустим, .что в популяц.ии диплоидных орrан.измов, .размно-
жающихоСЯ половым ,путем, в двух .независимых rепах А и В ВО3-
,никд.и новые мутации. Допустим ,далее, что носителя'ми мутантиых
.аJIлелей (а и Ь) перв.оначально .были разные особи с rеноти.пами
АаВВ и ААВЬ .соответственно. При этом МОЖ'еТ ,начаться процесс
рекомбинации, слаrающийся из следующих этапов: 1) скрещива-
ние между носителями различных мутантных аллелей: АаВВ х
хААВЬ; '2) появление в Р. .rетерозиI'ОТ ПО двум reHaM АаВЬ (по
МИМQ друr.ИХ типов); 3) не:завис.имое распр-ед-еление aMe.T с обра-
:зова ни ем четырех классов raMeT ---- АВ, АЬ, аВ и аЬ; 4) образова-
ние в Р 2 девяти различных rенотипов..... ААВВ, ..., ааЬЬ.
Большую ча'сть этих девяти rеноти.пов составляют .новые reHo-
ТИЛЫ. Н .начале процесса в популяции было три rеноти.па (ААВВ,
АаВВ и ААВЬ); спустя два поколения она содержала девять J'eHo
"ИПОВ, в том числе т .кие новые 'рекомбинантные типы, .как ааВЬ
11 ааЬЬ.
Для Toro чтобы произошла рекомбинация, reHbl А и В не обя-
зательно должны быть независимы. reHbl А и В MorYT рекомбиии
1JOватьс,я, находясь в разных хромосомах или .же iВ .разных Jю'кусах
.однои хромосомы. Сцепление, если только оно не слишком тесное,
снижает rчастоту рекомбинаций. но не предотвращает их образова-
тия.
rA. 7. Рекомбинация
65
Количество rенотипическ()й изменчивости
До пуст-и м, что ЧИоСJlО от ельных reHoB, ПРИСУТСТВУЮЩИХ в ДВУХ
аллельных формах. возра-стает в арифметическoQЙ проrрессии (2.
3. ..., п). Число диплоидных rенотипов возрастает при !этом по экс-
поненте (32, 33, ..., 3 n ). Вообще число .возможных ДИ1ПJlОНДНЫХ re-
нотипов (g) равно 3".
3000
2500
... 2000
CIII
с:
::J
c:t
.500
1\)
(.)
s 1000
OO
400
300
200
100
О 1 2 3 4 5 б 7 8 9 10
Число аллелеи уля КQжgоzо из gB!I;X
незовuсиМ6/Х еено"
Рис. 7.1. Возрастание реКоибинационной rенотипическоА изменчивости (: увепн-
чением числа апле lotй в ю!ждом из двух отдепьных локусов (Grant, 1963).
Как мы видеЛ'd выше, .два отдельных rепа (А .и В). Iкаждыи из
которых пр.едставлен двумя а.,1дельными формами. MorYT образо-
вать по 9 rенотипов, т. е. g==3 2 . Соrласно менделевской rенетике,
триrи.брндное скреЩИ1зание с участием -rpex reHOB (А, В 'Н С) Дает
27 rснотипов (g == 33) .
Сцепл.ение нарушает часroты реком'бинантных ТИПоОв. н.о не из-
меняет общеrо числа возможных т.ипов. Если отдельные reHbl не
сцеплены между собой, то rетероз.иrоты по ДВУМ ИЛИ по мнО'rиМ
reHaM дают раЗ.'I,ичные рек.ом.бинации с определенными частотами.
Если reHbl 'сцеплены. но MorY.T .быть разделены в результате крое-
Часть 1/. МuкроэвОАЮЦUЯ
66
синrовера, то рекомбинанты все же образуются, но с более низки-
ми частотами, пропорциональными силе сцепления.
Полиморфные reHbl обычно пред'ста.влены в природ.ных попу-
ляциях множеств-енными аллелями. Поэтому в таких случаях для
определения числа возможных rенотипов следует ,возвести в
п ю -степень :не 3, а ка.кое нИ'будь .большее число. Общая фО'рмула
для возможноrо числа диплоидных rенот.ИПОВ (g), выражаемаJl
через п (число .отдельных reHOB) и , (числ.о аллелей каждоrо
reHa), ,имеет вид
____ [ r (, + 1) ] n
g---- 2 .
Рассмотрим применение этой формулы ДJ1Я случая Bc.ero двух
отдеol1ЬНЫХ ',reHoB с различным числ()м аллелей. Результаты пред-
ставлены rра'фически на рис. 7..t. Индив.идуальная .изменчивость,
обусловлен:ная рекомбинацией, Iбыстро возрастает п.о мере Toro,
ка/к в с{)ответствии 'с ар.ифметической Iпроrрессией увеличивается
число аллелей в 'Этих двух локусах.
ТаБАuца 7.1
Число АRПЛОИДНЫХ rенотипов, которые MorYT возникнуть в результате рекомби-
наций между раэЛИ'IНЫМ qислом отдельных reHoB, каждый из которых имеет
разное число аллелеА (Grапt, 1963)
чнс.llО Число rенотилов при рекомбинации МtЖJ(У
8J1.11епеА
кзж"оrо
rенз 2 rеиамн 3 reизми 4 rеиами 5 rtиаМR
2 9 27 81 243
3 36 216 1 296 7776..
4 100 1000 10000 100 000
5 225 3375 50 625 759 375
6 441 9261 194481 4084 101
7 784 21 952 614656 17210368
8 1296 46 656 1 679616 60466 176
9 2025 91 ] 25 4 100 625 184528 125
10 3025 166 375 9 150 625 503 284 375
r [ r(r; 1) ]2 [ r(r; 1) ]3 [ r(r:1) т [ r(r:l) 1 j
п reнами
3 n
[ r(r:1) 1"
Рассмотрим далее ,rенотипи'Ч.ескую ,изменчивость, возникающую
при 'наличии множественных аллелей более чем в ,двух докусах.
Некоторые примеры приве.дены 'В табл. 7.1. Как показы.вает таб
лица, в результате рекомбин.ации 5 reHOB, .каждый из .которых име
ет по 1 О аЛ.1Jелеи, .может возникнуть \полмиллиарда rенотипoQВ. Есл.и
выйти за преде.1ыI этой таб lИЦЫ и рассмотреть случай 6 несцеплен-
ных локу.СОВ С 10 или более аЛ 1едями в каждом, то окажется, что
число ДИiпл.ои:дных реком,бина .тов исчи.'Сляется 'в МИJ1.1и.ардах.
rA. Рекомбинация
61
Пр.и reh-ет,ичеClКОМ изучении природных популяций высших ж,и-
вот,иых И растений в больши'нстве случаев обнаруж.ивается. что
они полиморфны по разным reHaM. Допущения, .лежащие ,в основе
приведенных выше численных пр.имеров. нельзя .считать нер-еали-
стичными; скорее они СЛИШКОМ занижены.
Очевидно. что pekom-БИtllация ,представляет собой .механизм. соз-
дающий orpoMHble количества нндив.идуаЛhНОЙ reнотипичес.кой из-
менчивости. При наличии YMepeHHoro полиморфизма Bc.ero по не-
с.кольк.им нес:цеплен'ным reH'aM 'из ,эТОЙ :rен.ной .изменЧlИВОСТИ путем
рекомбинации может воэ.никнуть астрономиrческое число rенотипов.
Прн умеренной .rекной изменчивости ч.и-сло ре.комбинантов впол.не
может оказаться .выше общею чи'СЛа особей данн.оrо вида. Именно
по причине реком.оинации у орrанизмов с половым размножением
.две особи, раз-вивающиеся из раз.ных зи'rот. .Н.и'коrда Не бы.вают
совершенно одинаковыми в rеН07ИПИЧеском oQтношении.
Рекомбинация и мутация
Рекомбинация сцепленных reHo.в происходит в результате крос-
.с И. Hf. О В ер а. БСJIИ сцепленные reHN заНlимают локу.сы, находящиеся
оче.нь БЛИЗ1КО друr от друrа, то крос.синroвер наrблюдается редко.
П{)явление реком,бинан1'ОВ среди -ПОТОМКОВ в этом случае также
на.блюдается редко. Реком,бинанты .B YT себя та,к же, как и му-
танты.
Сходство .между 'редкими реком.бинантами и мутантами иллю-
-стр.иrрует следующая модель. Два тесно сцепленных reHa, А tИ В,
'реrул.ируют сходные lIроцессы и Mo'rYT замещать друr дрУ1rа. Ины-
.Ми CJIовами, нормальный .фе.нотип создается Jlи60 двумя домина,нт-
ными а.плелями (АВ), либо ОДНИМ из этих дом-инантных аллелей
.(А или В). fетеРоОзиrоты по двум reHaM :; !Имеют нормальный
.фенот,ип 'и обычно размножаются в ч.истоте. Но изредка между
...покусами А и В происходит IКросс.инrовер. в результате чеrо воз-
'Никает некоторое ЧИС..10 raMeT аЬ. Эти raMeTbl дают з'ИrотЫ ааЬЬ.
имеющие «мутаНТJIЫЙ» феНОl1ИП и 'размножающи-е.ся в чистоте по
'отклоняющемуся признаку. Есл.и wpос.оинтовер между А Jf В про-
iиСХОДИТ с частотой, сравним.оЙ .С частотой мутаций, то отличить
'Такой редкий реком'бииант ОТ мутаН1'а обычными меТОДlами не..
130ЗМОЖНО.
При помощи то.нкоr{) rенеllичеСК()IfО аиадиза у Drosophila тela-
.nogaster J Zea mays и не.которых rр.ибов было выявлено несколько
случаев, оответствующих оn.и анной выше общей модели. у дрозо-
филы кроссоверы встречались с частотой 0,00026. а возм.ожно,
даже еще реже. У кукурузы (Zea тays) имеется сложный rreH А,
'опре.деляющий окраску зерен и ДрУ'rих частей растения; этот reH
СОС1'оит .из двух лежащих 'рядом cy6reHOB, к{)торые изредка реком.
б8
Часть 11. МиКрОЭ80Аюция
бинируют, давая формы, внешне сходные с мутантами (см. обзор
Grant, 197-5, rл. 4).
Из эт.их данных 'следует, что в любой rру.ппе непроаналиэиро-
ван,ных мутантных форм, по всей ероятност.и, с{)держится, .помимо
настоящих reHHblx мутантов, некоторое число -редких рекомБИНоан-
тов. Существующие оценки ча.стоты спонтанных reHHblx мутаций,
особенно у высших растений и ЖИВО1iНЫХ, M.orYT оказаться ЛOlЭтому
слишком ,ВЫСQЮИМИ, как уже отмечалось в rл. 5. Вполне возможно,
что ис.тинные reHHble мутации у высших орrан.измов в rречаю,.ся
реже, чем это считается в настоящее время.
Значение рекомбинации
Сложные фенотипическ.ие признаки определяются Не единичны-
ми ,rе.нами, а их сочетаниями. Подобные ,сочетания reHoB состоят из
аллелей, обладающих rармоничным совместным действием. Реком-
бинация ---- '3то тот механизм, который прои'Эводит сборку reHHblX
сочетаний.
Рекомбинация иrрает -важную роль у всех орrанизмов. Во всех
царствах живоrо мира, в том Чiисле у бактерий и вируоов, сущест-
вуют те .или иные способы обмена .rенетическим М1атериалом.
у вукариотов rJIaBHblM способом, обе.спечивающим реКOIм,бинацию,
.служит пол-ов'ое размножение, а у ,прокариотов соответствующие
ФУНКЦИlи выполняют разноrо рода па'расексуальные меха,низмы,
приводящие в конечном счете к тому .же результату.
Хотя рекомбинация существует во .всех основных rРYlппах, ее
относительное значение у разных op-rа.н.ИЗМОВ весьма различно.
у Вbl'сших ЖИВО11НЫХ. rде .роль реком.б.инации .особенно .велика, она
обеспечивается облиrатным половым размножением, ,Болы1DмM чи.с.
..10М хр-омосом и друrими особенностями rенетической с.истемы.
Противоположная СИ1'уация 'имеет мест.о у бактерий .и вирусов,
которые б '1аrополучно существуют при МИНlим.альной степени ре-
комбинаци.и. Выс.шие растения занимают промеж-уточное положе
ние между 'этими двумя Iкрай.ностямя в отношении обычноrо уров-
ня Iком,бинат-и'ВНОЙ изме.нчи;воС11И.
Эти .широкие р.азличия к()ррел.ируют .со .сложкостью ортаниэ--
мов. Бактерии об ,адают относительно простой орrанизацией .и от-
нос.ительно простым rено11И'пОМ. Важные жизненные функци'и, как,
например, .способность с.интеэ.ировать некоторые необходимые ме-
таболиты, MorYT определяться у них единичными rенами; новые
простые ФУIНКЦИИ TaKoro рода нередк.о, по-.видимому, возникаЮТ
в результате rенной М'Утаuи.и в с{)четан'и.и с естественным от.бором.
В невероятно сложном J'енотипе Toro ,И 1И ИНОI"О Вbloсшеrо жи-
BOTHoro роль каждоrо единичноrо reHa в отдельности относительно
меньше. Зде.сь фенотипнческие признаки, обладающие адаптивной
ценностью, в большинстве С Т'lучаев определяются сочетаниями Ife-
r А. 7. Рекомбинация
69;
нов, причем .обычно очень сл.ожными сочетаниями. Поэтому меха-
.ниЭ!мы, при помощи которых вознИ'кают рекомбинация тенов,.
имеют существенное значение.
ЭВОЛЮЦИЯ новых .слож.ных :признаков начинается с ВО3НИl\iнове
ния Iизменчивост.и по множественным reHaM и кончается закреп-
лением HOBoro адапт.ивноrо .сочета,ния reHOB 'в да.иной популяции.
Рекомбинация Bcero лишь одна из важных точек в середине ЭТQrо
процесса.
Сначала 'ПОЯВJ1ЯЮТСЯ мутации по .одному 'или неско.Т'lЬКИМ иенам,
которые затем рассе.ивают.ся блаrодаря -потоку reHOB. Мутантные
аллели, если они рецессивны, MorYT сохраняться в популяции в ди-
пл,оидном .состоянии, це проявляя.сь на протяжении .М,ноr.иХ ПОКО.'1е
пий. Диплоидное .состояние служит Х!ранилищем для рецесс.ив.ной
,мутационной изменчивOC1lИ, а :ПОJ10IЮе размножение слу ит клю-
чом К этому хранилищу, .вЫЯВ.'1яя rомозиr.от.ных рецессивных выще
пенцев ,и разли,чные рекомб.инант.ные т.ипы. Некоторые сочетания
аллелей MorYT превысить по приспособитедьной ценности пред
.КОВЫЙ rенотип .1,ибо .в пред'ков'ой, либо в ка:к{}й-либо новой .среде.
Сборку этих сочетаний обеспечила .рекомбинация. Их закрепление
в популяци:и зависит уж:е от друrих .факторов, кот,орые нам еще
'Пред.стоит рас,смотреть.
Итак, хотя мутацио.нный процесс ,н служит исходным и.сточн.и-
ком rенетической изменчивости, rл.авным эффективным источни-
ком изменчивости у ()рrаниз,мов с половым раЗМlНожен.ием Я'вляется
рек{)м.бинация. Рекомбинация .создает большую часть т.ой инди-
ви)ДуальноЙ rенотипической изменчивости в скрещивающей.ся попу-
ляции, над которой :впоследствии предстоит работать естестве.нно-
му отбору и дрейфу тенов.
Часть IIf
Естественный отбор.
rлава 8
основ.НАЯ ТЕОРИЯ ОТБОРА
Введение
Бстественный отбор CJIа,rается -ИЗ ряда различных процессов.
С одной стороны, мы располаrаем первоначальной дарвиновской
концепцией отбора, а с дру'roй .сов.ременной rенетичес.кой тео-
рией отбора. ОБ .paMIKax последней теории мы, ,кроме Toro, должны
различать раз.ные типы -отбора. Наконец, процессы отбора дей<:т
.вуют на разных уровнях биолоrической орrанизаци.и. ПростеЙШ1ая
форма eCTeCТBeHHoro отбора отражена в однолоку,сной модели.
Однолокусная модель
До'пустим, что некая обширная по'пуляция изменчива ао reHY А.
Эта популяция содержит с высокой частотой аЛ.тIель А преДКОБоrо
или ИКОI'() ТИlпа 11 с 'н.изкой часroтой . Н"овый мутантный аллель а.
Есл,и нос.ители .аллеля а оставляют .больше потомков, т. е. -вносят
больший вклад 'в следующее .поколение, чем нос.ители аллеля А,
и если такое дифференциальное размножение этих двух аллелей
происходит систематически из поколения в поколение, то частота
аллеля а в популяции будет постепенно возрастать, а частота ал-
леля А снижаться.
Это простейшая форма естестве.нноro отбора. ОНа ,позволяет
сформулировать .определение, прим.енимое к однолоку.сной модели.
В этом смысле естественный отбор представляет собой дифферен-
циальное и н.еслучайное размножение различных альтернативных
аллелей в некой !популяции.
Естест,ве.нный отбор прои'Сходит .в том случае, коrда н-осител.и
одноrо аллеля (а) постоянно и систематически, на протяжении
ряда посдедовательных ПОJ<олений размн.ож.аются БО.'1ее успешно,
ч м носители альтернативноrо аллеля (А). Дифференциальное раз
множение альтернативных. аллелей не является случайным.
Чтобы .изм.ен.ил ась частота аллелей, .селективное преимуществ.о
алле.'1Я а по сра'внению 'с аллелем А не обязательно должно быть
r А. 8. Основная теория отбора
71
большим. Предпочитаемый аллель а может обладать лишь Iнезна
чительным преим,уществом над друr.ими кон.курирующими аллеля
М-И, а Ч1астота ero ,будет тем не менее :возра.стать.
I(оэффициент отбора
Селективное преимущества ()дноr.о аллеля иа'д альтернативным
аллеле.м (ил,и аJ1лелями) можно выразить в процентах .или в виде
коэффициента .отбора (5), .величин.а KOToporo изменяется в д.иапа
зоне от .0 )],о .1.
Коли'чествооное значение коэффициента отбора выводится из
относительных частот альтернат.ИВНЫХ аллелей. ДопустИ'м, что в
некой большой ПОПУЛЯЦИИ а ---- .предпочитаемый аллель, а А......... 'аЛ-
лель, .которому отбор не бла1rоприятствует. В этой популяции На
каждые 100 аллелей а, передаваемых .следующему поколению,
будет передаваться также некоторое число аллелей А (от 100 до О).
Коэффициент О'I1бора есть функция .этorо отношения. ВелиЧlИНУ 5
можно ()пределИ1Ъ до формуле
s == 1 Частота размножения аппепя, которому не блаrоприятствует отбор
Частота размножения алпеля, которому блаrоприятствует отбор .
Рассмотрим С.,lедующие численные примеры:
1) Orносите.'1ьиая частота размн'ожен.ия аллелей а ,и А в одном
поколении составляет 100а: 99А. Отсюда 5== 1----99/100==0'01' т. е.
аллель а обладает 1 % HЫM селективным преимуществом.
2) .вклады а.,1ьтернативных аллелей .в <:ледующее пок.ол.ение co
Сllавляют 1000 а : 999 А. Отсюда 5==0.001, а селекти.вное преимуще-
ств.о аллеля а 'равно 0.1 %.
3) Соотношение аллелей в одном ПОКО..'Iении состав.'1Яет 1000 а :
: 50 А; 5==0,5.
4) Эк.стремальиый случай ---- 100 а: О А. Здесь 5== 1; это азна-
'ч ет полное замещение reHa за ,одно поколение.
5) Противоположный экстремальный случай ----100а: 100А.
Здесь 5==0; никакоrо отбора не .про.исходит.
В.ажно отмет.ить, что селективное rrреимущество аллеля а COOT
.ветствует принципу «-в.се или Iничеrо» лишь. в четвертом случае.
Во всех друrих с,,'уч.аях оно представляет собой лишь статистиче-
ское различие в частотах размножени'я. В любом данном Iпоколе
нии скрещи,вающей'Ся популяци.и некоторые отдельные носители
аллеля а .MorYT потерпеть неудачу в разм.ножении, а некоторые
отдельные носител.и .алле.Т'IЯ А MOryT )],Ост.иnнуть .в .общем большеrо
успеха, чем носители ал..lе."Я а в .среднем. .изменения же в reHo-
фонде определяются -суммарным вкладом в .размножен.ие всех но-
сителей аллеля а.и всех носителей аJ1 еля А.
В размножени.и .пОПУ.,1ЯЦИ1И участвует случайный компонент.
Этот .случайный и;омпонент может влwять на .относительную часто-
72
Часть [/l. Естестsен.НblЙ отбор
ту ПРОДУКЦИИ аллелей А .Н а л-и60 в каких то Me Tax, либо в ОТ-
дельных поколениях. Но эти CJIучайные различия в размножении
нооителей аллелей А и а не есть отбор. Отбор происходит только
пр.и том усл.овии, что В дифферен'Циальном размя-ожении носителей
а lыернативных аллелей участ.вует также неслучайный компонент.
Именно поэтому в приведеННQМ выше .определении отбора [о-
ворится .о «дифференциальном ,и неслучаЙном» размножeйf.ИIИ; и по
этой же причине .мы специально подчеркнули, что популяция долж-
На иметь больш.ие размеры.
Скорость изменения и ero степень
Изменения в частоте 'аллелей, обусловленные отбором, MorYT
бьrrь выражены в любой степени вплоть до .полноrо замещeйlИЯ.
Новый аЛ lель а, п{)явившись за .счет мутаЦlИИ .или потока !'ен'ов,
может в.н.а,чале содержаться в популяции с низкой частотой. Но
до тех пор, пока алле.lIЬ а обладает хоть каким-то селекти.вным
преимуществом по сравнению с .существовавшим до ,Hero в 1l10пуля-
ции аллелем А, чаСТО11а аллеля а будет возраст.ать. а частота алле-
ля А снижаться. конечныJM результатом при налич.и.и доста-
точноrо времени может оказаться пол,ное замещение в популя
ции аллеля А аллелем а.
Совершенно очевидно, что эти .изменения .будут лро.исходить
быстрее пр.и BblroKOM к{).эффициенте отбора и медленнее пр.и низ-
ком коэффиц'иенте. Скорость изменеН'ия ча т{)ты аллелей прямо
пропорrци{)н.алнна величине S.
Однако для каждоrо данноrо значен.ия 5 .скорость изменения
частоты .алл.елей различна при разных частотах аЛJlелей. В слу-
чае диплоидной попул'яции и дом.инантноС11И Toro аЛJIеля, KOTOpO
МУ бла!r()lПРИЯ1'ствует отбор, 'изменение частоты аллеля происходи.т
быстро при 'н.изких ,и пром.ежуroчных ero частотах, но па.ц.ает при
выс.оких частотах. Так, частота дом'ин.антно.rо аллеля, абл'.адающе-
ro 1 %..ным селект.ИВНы.м преимущест,вом ,(5==0.01), может возрасти
с 0,01 до 98% .примерно аа 6000 поколений; но потребуется более
5000 поколений, чтобы ero частота повы<:илась с 98 до 99%, и
.примерно 1 000 000 поколен'ий. чтобы она :повысил-ась С 99 до 100%.
Е.сли же .ал ,ель, котор.ому бл а.roпр,иятствует отбор, рецессив н, то
изменение ero ча.ст.оты происходит очень медленно при ниэких ча
<:тотах, но ускоряется пр.и промежуточных Чlастотах.
В предшествующих ра.ссуждениях допускалось, что частота
алле.1JЯ, которому бла'rоприятствует отбор, будет непрерывно по--
вышаться вплоть до .полноrо er{) закрепления в популяции. Это
упрощающее .допущение. но .по ряду причин ero .нельзя считать
безусловно реалистичным.
. Коэффициент отбора не обязательно остается постоянным на
всем протяжен,ии длительноrо процесса отбора, но может .испыты-
r А. 8. Основная теория отбора
73:
вать .коле6.а'ния и резкие Iизменооия. В таких случаях тот ил.и иной
reH может обладать селективным преимуществом На одном этапе
истории данной популяции и оказаться селек11И.ВНО небла,rо.прият-
ным -на друrом ее этапе. Кр:оме TOro, с течеки€м времени в п'О.пу-
ляцию В результате мутационноrо процесса и потока renOB MorYT
включаться друrие аллели. В связи с этим допущение Bcero лишь
о двух кон'курирующих аллелях в популяции нереалистично само
по себе.
Наконец, !В диплоидной ,популяции два или большее 'числ'9 аЛ 1е-
лей включены .В ,соответствующие r.омози:rоты ил.и reтерози.rоты, на-
пример аллели А .и а в АА" .аа ,и Аа; п.ри этом отбор ока03ывает
дифференцирующ-ее действие ,не на аллели, а на rен.отнлы, в кото-
рые входят эТоИ алл.ели. Вполне возмож.но, что rенот.ип аа будет
превосходить rенот.ип АА" но Il'eтер'озиrота Аа окажеreя более цен-.
ной в селективном .отношении, чем rОМОЗИlrота аа, и в таком слу
чае популяция 6уд.ет ,перман'ентно оставаТ1.ся по..1ИМОР'Фкой по ал-
лелям А 'и а. В эroм .случае аллель а никоrда .не достиrнет полно-
то закрепления в результате от.бора. Более :подробно ,эта с:итуация
описыв.ается В rv'1. 11. Здесь же мы оrраничимся замечанием, что
папу..1яционные .rенет-ики .все ,больше ,с.клоняются .К мнению, соrлас-
но которому частота мноrих аллелей, контролируемых О'I'бо р.о м ,
очевидно, ,стабил.изи-руется на раз.ных уровнях, не достиrая
100% -:Horo уровня.
Приспособпенность
Как,ова осно.в.а этих !изменений в частоте аллелей? Почему
происходит .систематическое повышение 'частоты аллеля а .относ.и-
тельно частоты аллеля А? ,Во первых, ,в .основе селект.ивно.I1O преи-
мущества, 'п{) видимому, лежит различ.ие ,по какому-то фенотипи-
ческому пр-из.нatКу, контрол.ируемому reHoM А. В.о-вторых. такой
фен'отипический пр.изнак должен .иметь некоторое. значение для
,выжив'ания и .размножения. Носите..'tи аллеля а должны превосхо
дить носителей аллеля А по .какому-то жиз.ненно важному с.вой-
ству, влияющему на у-с-пех .размножения.
-Вопрос о сел,ективном преимуще.стве иЛи селективном дефекте
лю.боro даlН'ноrо .аллеля влечет за 'Соб.ой, таки;м .образом. постанов
ку .важных .вопросов о действия reиа, reнети'ЧeJCКИ .контролируемом
разв.итии, взаимодействии между rенами и ,взаимодейст.вии между
rенотипом .и средой. Некоторые из этих в.опрооов 06суЖJдаются
в rл. 9. Здось ,мы просто примем допущение, что нос.ител.и аллеля а
обл,адают известным преимуществом по сравнению с носителями
а.тIлеля А в отношении выживания или плодовитости.
Это подводит нас к концепции пр.wcпособлен.iюст,и (известной
также под наз.ванием адапти,вной ценности .или селекти'вной цен-
н.ости) в 'популяционной rенетике. Приспоообленность rеноТНпа.....
74
Часть 111. ЕстествеННblй отбор
это среднее чи,сло потомков, оставлен,ных .этим reнотипоы, по срав-
нен,ию со средн.им ч.ислом п,отомков У друrих конкурИ'рующих с ним
rенот.ипов. Приспособленность, таким образом, представляет собой
чисто количественную и операционную концепцию. Это количест-
веНная м.ера успеха размножения.
Пр'ИСJIlOсобленно-сть (обозначаемая .символом w) есть функция
коэффициента О'J'lбора (w == l----s). Если .отношен.ие частот размно-
жения аллелей . оС.оставляет 99 А : 100 а. тоо, как уже отмечало.сь,
5==0,01; теперь мы можем добавить IK этому, что wA==O.99. Или,
короче, WA ==99 A/IOO а ==.0,99.
Высокая ПРИ1спо'соб.пенность .в том смысле, в каком этот термин
используется в популяци-онной 'rенетике, 'не обязательно подразу
мевает «выживание наиболее приспос.обленных», а .синонимичный
ему терм.ин «.ада:пти'в.ная цен,ность» не обязательно озиач.ает, что
rен-оти.п, обладающий высокой ада'птив'ной ценностью, особенн-о
хорошо приспособлен к своей среде. Такая с.итуация возможна.
Однако возможно также, что высокая прис.пособленнО'Сть или вы-
сокая адаптивная ценность представляют -собой 'исключительно ре-
зультат высокой .п.тЮДОВИ'J'lости. Вы-со.к'ая приспоеобленность озна-
чает д'етерминированный rенетически у пех в разм'ножении; она
не указывает на какие-то определенные причины этоrо успеха, ко-
торые MorYT сильно варьировать в разных случаях.
Дарвиновская концепция отбора
популяционная rенетика и современная эволюционная теория
прираВ'ни,вают есrecтв-енный О'J'lбо.р к дифференциальному раз'мно-
жению альтернативных форм reH.oB, reHOTIJjnoB ,или дру.r,их восnpо-
изв-одимых единиц: Перв-он.ачальная дарвиновская к'О.нцепция была
несколько иной. Дарвин и ero последователи ,подч-еркивали важ-
нейшее ЗНа'ч-ение некоторых изменчивых видовых признаков, ОТ
которых зависит жизнь или смерть в «борьбе за существование».
Рассуждения Дарвина в <<lПроисхождении видов» (1.859, 1872)
Моожно суммировать .следующим 'образом. Отдельные особи данно-
ro вида .конкурируют между собой за жизненные ресурсы. Между
этими особями имеются меЛК1ие разл_ич.ия по .некоторым призна-
кам, и эти различия нередко наследуются. Некоторые варианты
лучше приспособлены к TO y, чтобы -выжить в борьбе за 'сущест-
вование, -чем д'руrие. Соответственно первые будут размножаться
преимущественно и передавать свои блаrоприятные пр.иЗ-На'I{IИ по
следующим поколениям.
Дар.вин ,иллюстр.ировал -свою точку зрен-ия ,на 'примере жирафа.
Он сделал ВПО.ТIНе 'правдоподобное допущение, что в одной из пред
ковых -популяций жираф()в отдельные особи слеrка разл;ичали'Сь по
длине шеи и передних Hor. .в периоды нед'Остатка корма в их poд
ной са:ва'Нне животные Bblc.OKoro и Rизкоrо роста должны бы..1JI
I'A. 8. Основная теория отбора
75
вступать в K-ОНlкуренцию за листву деревьев. В Тalкие периоды бо-
лее высоК'ие животные мо-rли добираться до листьев, ра.стущих на
.верхних ветвях, которые были :недоступны для ЖИВОТоных низ,коrо
роста. .пОСЛ'tЩН,ие по-этому "Поr.ибали, а ,вместе с ними и'Счезали .Н
такие их признаки, как короткая шея и короткие ноrи. И напро-
тив. длинная ш-ея .Н длин.иые Ho.rH ICOB'peMeHHor.o жирафа резуль-
тат преимущ€стве.иноro .вЫЖи:ва.ния ,и .размножения из 'пок-оления
в ,поколение более высоких -особей (.Darwin, 1872, .rл. 7).
Дарвин писал (Darwin, 1-859, .rл. 4):
< ,()жно ли... считать невероятным, чтобы друrие изменения,
'Полезны€ в каком нибу дь отношени'И для существ в ,их веJLИКОЙ и
CJlожной жизненной битве, появлял.и.сь в дли-ином -ряду последова
тельных поколений? Но если такие Iизменения п-оя,вляются, то МО-
ж-ем ли мы (помня, чт{) р.одится rораздо больше особей, чем мо-
жет выжить) сомневаться в т,ом, что особи, обладающие хотя бы
самым незначительным преимуществом перед оста."ЬНЫМИ, будут
'Иметь .больше ша'ноов на выживание и продолжение CBoero род.а?
С друrой .стороны, мы можем быть ув-ерены, что всякое .изменеН'ие,.
сколько ни,будь вредное, будет неукосните..1ЬНО подверrа.ться и.с-
треблению. Сохр'анение бл аroприятных .индивидуальных разл'И'чий
и .изменений и у.НИЧ7ожен'ие .вредных я назвал Естествеаным Отбо.
ром».
Компоненты приспособленности
Д'ифференциальное .размножение .представ..1яет собой конечный
результат мноrочи-сленных КОМ1понентов, перечисленных ниже:
Дифференциальная смертность ,в р.аннем .и молодом возрасте.
Дифференциальная смертность на стадии пол-овой зрел'ОСти.
Дифференциальная выживаемость.
Дифференци-альное стремление к с.париванию и успешность
IПОС.'Iеднеrо.
Диффере,нциальная фертильность.
Дифференциальная ,ПЛoQдов.итость.
В дарвинов.ской теорИ'и ecтecTBeHHoro отбора ПОд'черкивались
первые два или три из этих компонентов. В созданной им позднее
теор-ии полавоrо отбора (Darwin, 1871) был добавлен четвертый
КО МtlЮнеит . Что ж-е касается попу..1яционно..;rенетической теО.рJI1И от-
бора, то в ее определении есте.ствен-н.ото отбора rлавный у.пор Д -
.1Jается и'а .конечный результат ---- дифф рен иальное размнож ние;
иа практи е эта теория часто ,по.дчеркивает важную роль трех
последних компои.ентов, из которых С.1Jаrается размножение. Раз-
лиqи-е между этимtи двумя школ.ам.и отражает различия в их .про-
фе-ссиональных особе:нностях: Дарвин и ero последователи были
полевым'и био."осами, тоrда как основы /Современной теории .отбора
были за.ложены ла.бораторными исследователям'И 'и статистиками.
76
Часть 1//. Естественный отбор
РеrРОC'JIекти&но было бы желательны'м обозначить раЭ!НЫМIJI
техническими терм нами различные участвующие ,в .отборе про-
цес<сы, как это сдела,,1 сам Дар'вин в ОТн,QшеНИll половоrо отбора.
Но На самом деле мы теперь пользуемся ДНlим теРМIИНОМ, прила-
rая ero к очень ши-рокому юруry процессов. Мы ,можем юбходиться
существующей термин.олоrией, поскольку между дарвиновской \и
ПОПУJ]яционно rене'Тическ.ой концепц'Ия'Ми 01'бора нет внутреннето
пр'ОтивOtречия, но, пользуясь этой теР.Ми.нолоrией, мы должны все
время .помнить о составной природе современной концеп ии o -
.ра и быть rоroвыми, коrда lЭ1'O окажется необходимым, различать
ero .отдельные субпроцоссы.
Следует различать две ,основные формы lотбора. В первом слу-
чае селеК'J1ImНое Iпреимущества ти'па а над типом А обусловле'но
лучш'ей ,адаптирова'Нностью .к не'К'Ой Iкр'Итиltfеской ситуации JJ .среде.
В таких .сл,учаях отбор совершенствует приспо.соблен'ия орrаниз-
мов, из которых .слатается популяция, .KaIK это представлял себе
ДаlрВИН. Во втО'ром :случае типы а оставляют больше поroмк.ов,
чем типы А, вследствие более С'ильноrо стремл.ения к размноже-
нию или более выс.окой ,ПЛОДОВИ1',ост.и, незаJЗ(ИСИМО от Toro, облада-
ют они превосходстВ'Ом в отношении друrих жизненных аспектов
ил,н нет. Эта форма отоора не обязательно в'едет к по.вышению
приС'пособленоос.ти. Напротив, она может привести .к распростра-
нению призна'Ков, которые окажутся вредными для попул'Я'IJJИИ или
для Ю1да в .целом.
01'Сюда .с.".едует, что необходимо также ,различать разные Т,JfПЫ
приспос.обленности. В популяционн.ой ,rенетИ'ке tПОД высокой пр.ис.по-
собленностью понимают, KalK уже отмеч'алось, относительно боль-
шое число ,остаlвляемых поroМI{,()"В, .и само ,по себе оно не Уlказыва
ет на причины этой .вы окой приспоообленности. П.олезн{) 'Ра3JIИ
чать две ш.ир'оwие катеrори:и приспо'ообленнос'J1И: 1) адаптирован
насть, или степень приспос.о6ления особи иля опуляции к уело-
в.иям своей среды, и обусловлеНl1ryЮ Э1'им Clпособность оставлять
больше .поroМJков; 2) успех 'раЗМlЮжения ,как таковой.
r Л3Jва 9
ПРОЯ,ВЛЕНИЕ rБIiА и Ero овязь
с .ОТБОРОМ
Введение
Может возникнуть вопрос: действует ли естественный отбор
на reHbl или на фенотипы? Отбор непосредственно действует на
фенотипы и лишь косвенно на лежащие в их основе reHbl. Меж-
ДУ прямым действием отбора и возникающим в реЗУ.1Jьтате изме-
нением частоты reHa лежит, следовательно, вся сложная цепь
событий, при помощи которых действие reHa переводится в фено-
типические признаки.
Таким образом, для Toro чтобы понять деятельность eCTeCTBeH
Horo отбора, необходимо учитывать фенотипическое проявление
reHoB. Один комплекс вопросов rруппируется BOKpyr типа фено-
1ипическоrо проявления в зависимости от фона, создаваемоrо
средой; друrой комплекс касается степени изменяемости феноти
па; а третий разнообразных способов действия и взаимодейст-
вия reHOB.
Относительность селективной ценности
Рассмотрим сначала следствия, вытекающие из совершенно
очевидных взаимоотношений. Данный rенотип детерминирует про
явление одноrо и Toro же фенотипическоrо признака в ряде раз-
личных сред. В некоторых средах данный фенотип обладает Из
вестным селективным преимуществом; в друrих средах он может
иметь иное селективное преимущество, не давать никакоrо пре-
имущества или даже быть неблаrоприятным в селективном отно-
шении. Следовате..1ЬНО, в одних средах отбор будет блаrоприятст-
вовать rенотипу, детерминирующему этот фено.тип, а .в друrих
средах будет блаrоприятствовать ему меньше или даже не бла
rоприятствоват!> вовсе.
Короче rоворя, селективная ценность Toro или иноrо аллеля
или rенотипа не есть некое внутренне присущее ему свойство, а
зависит от взаимоотношений между фенотипом и средой. .
ПрОСТЫ 1 примером служит окраска раковины У европейской
наземной улитки Сераеа neтoralis. Эта улитка живет в разнооб-
разных местообитаниях от темных буковых лесов с сомкнутым
полоrом до открытых, залитых соднцем луrов. Она полиморфна
по окраске раковины, которая может быть бурой, розовой или
желтой. Эти различия в окраске раковины детерминируются од-
78
Часть 1//. ECTeCTBeHHbla отбор
ним полиморфным reHoM. Аллель бурой окраски обусловливаеr
образование бурых раковин, а аллель желтой окраски желтых
во всем диапазоне разнообразных сред, в которых обитает эта
улитка.
Улиток поедают дрозды и друrие птицы, руководствующиеся
зрением при добывании пищи. Бурые раковины обеспечивают за-
щитную окраску, затрудняющую обнаружение улиток птицами в
буковых леса , а желтые на луrах. В результате TaKoro изби
рательноrо выедания улиток птицами бурая окраска раковины
преобладает в популяциях, обитающих в лесах, а желтая в по-
пуляциях луrов. Селективные ценности аллелей коричневой и
желтой окраски связаны, таким образом, с типом местообитания
и при перемене последнеrо приобретают прямо противоположные
значения (более подробно об этом см. Cain, Sheppard, 1954;
Sheppard, 1959; Lamotte, 1959; Jones, 1973).
Drosophila pseudoobscura в западной части Северной Америки
полиморфна по инверсиям в третьей хромосоме. Каждая инверсия
имеет название и буквенное обозначение: Стандарт (ST), Чири
кауа (СН), Эрроухед (AR) и т. д. (см. rл. 4). Установлено, что
некоторые инверсии в rетерозиrотном состоянии дают своим носи-
телям селективное преимущест'во по сравнению с rомозиrотами
по этИМ инверсиям. Однако здаптивное превосходство rетсрозиrот-
ных rенотипов проявляется только в определенных условиях сре.
ды. Так, rетерозиrоты STjCH обладают более высокой се.'1ектив-
ной ценностью, чем rомозиrоты STjST и СН/СН, при темпера-
туре 21 25 ОС, но не при 16 ОС. Высокая се.пективн ая ценность
rетерозиrоты ST/CH проявляется лишь в том случае, коrда мух
выращивают в перенаселенных популяционных ящиках. При TeM
пературе 21 ос rетерозиrоты ST/CH обладают преимуществом,
если их кормят определенным видом дрожжей (Kloeckera); если
же их кормят друrими дрожжами (Zygosaccharoтyces) , TaKoro
преимущества не наблюдается (см. обзор Dobzhansky, 1970, r л. 5).
Изменяемость фенотипа
Фенотип представляет собой продукт взаимодействия rеноти-
па и среды в процеrсе развития. Каждый отдельный фенотипи-
ческий признак резу."ьтат не ТО.'1ько влияний, оказываемых во
время развития средой, но и результат действия [енов. Кроме
10ro, участие факторов среды в формировании фенотипических
признаков различается как в отношении разных признаков, так
и в отношении разных орrанизмов. У человека, например, участие
среды в формировании фенотипических признаков незначите."Ь.
но, если реЧh идет о rруппах крови, значительно в С.'1учае веса
тела и очень велико в сфере YMcTBeHHoro развития и поведения.
r л. 9. Проявление zeHa и ezo СВЯЗЬ С отбором
79
Если процесс отбора действует на какой либо фенотипический
признак, который детерминируется в основном rенотипическим
компонентом развития, тоrда как влияние среды незначительно,
то воздействие отбора на состав rенофонда будет немеД..1енным
и относите.'IЬНО прямым. Однако отбор окажется rораздо менее
эффективным ero действие будет осуществляться с большей
задержкой ---- в с.IJучае таких фенотипических признаков, которые
в значительной степени формируются средой. Способность reHo
типа реаrировать соответствующими фенотипическими МОДИфИ
кациями на широкий диапазон условий среды препятствует эф-
фективности отбора, связанноrо со средой.
Плейотропия
[ен обычно об..lадает различными и нередко не связанными
между собой фенотипическими эффектами, И.IIИ, иными словами,
каждый данный reH воздействует на бо..lее чем один фенотипиче
кий признак. Это явление известно под названием плейотропии.
Iлейотропия ус.II0жняет действие отбора.
Представьте себе, что некий П..1ейотропный а..l.ТIель оказывает
!'два раз.тшчных действия ---- одно б.IJаrоприятное, а друrое. небла
,rоприятное. Селективная ценность этоrо аЛ..1еля для всех практи
ческих целей опреде.1яется при этом чистым результатом, полу-
чаемым от сопоставления создаваемых им селективных JIреиму
ществ с ero неблаrоприятными селективными эффектами. Как
только неблаrоприятные эффекты lIачинают перевешивать пре-
имущества, отбор перестает блаrоприятствовать этому аJ1лелю и
начинает действовать против Hero.
Селекционеры И животноводы постоянно сталкиваются с этой
проблемой при искусственном отборе. Некоторые выrодные с xo
зяйственной точки зрения признаки, такие, как очень высокая ypo
жайность, просто. невозможно бывает закрепить в популяции
культурных растений или домашних животных, если они корре-
лируют с очень неблаrоприятными признаками, например с пони-
жен ной фертильностью. Несомненно, во мноrих природных попу-
ляциях П.ТIейотропные В.ТlИяния подобным же образом сдерживают
эффекты eCTecTBeHHoro отбора.
Модификаторы экспрессивности
На фенотипическое проявление данноrо reHa или сочетания
reHoB оказывают влияние друrие reHbl комплемента, известные
ПОД названием модификаторов. В этом и в следующем разделах
мы рассмотрим ро..1И rенов-модификаторов двух типов: модифи-
каторов экспрессивности и модификаторов доминантности.
80.
Часть II!. Естественный отбор
Экспрессивность это степень фенотипическоrо проявления
данноrо reHa или сочетания reHoB. У ряда особей, .имеющих оди-
наковую конституцию по данному reHY и выросших или выращи-
вавшнхся в стандартной среде, MorYT наблюдаться разные CTe
пени фенотипическоrо проявления этоrо reHa. Так, различные
особи и линии дрозофилы MorYT иметь разное число щетинок,
хотя их конституция по rлавному reHY, контролирующему ще-
тинки, одинакова и хотя они выращивались в одной и той же
среде. О таком reHe rоворят, что он обладает варьирующей экс-
прессивностыо.
Варьирующая экспрессивность может быть вызвана модифи-
каторами экспрессивности. Плюс-модификаторы усиливают фе-
нотипическое проявление rлавноrо reHa, а минус-модификаторы
подавляют ero. Отдельные орrанизмы, несмотря на их rенотипи
ческое единообразие в отношении rлавноrо reHa, rенетическJ.{ раз-
личны по своим наборам модификаторов экспрессивности. На
действие rенов-модификаrоров MorYT, кроме Toro, оказывать влия
нне условия среды, вводящие еще один осложняющий фактор.
Варьирующая экспрессивность представляет собой, таким обра-
зом, результат сложных взаимодействий между плюс- и минус-
модификаторами и факторами среды. А это взаимодействие в свою
очередь осложняется влиянием отбора.
Допустим, что некая популяция мономорфна по одному из
rлавных reHoB или сочетанию reHoB, определяющему какой либо
фенотипический признак, но содержит латентную изменчивость в
системе rеиов-модификаторов. В результате действия минус мо-
дификаторов экспрессивности данный фенотипический признак не
проявляется в одной среде (E 1 ), в которой он обладал бы адап-
тивной ценностыо, и лишь слабо проявляется в друrой среде (Е 2 ) ,
в которой он имеет определенное селективное преимущество.
Конечно, в среде El отбор был бы неэффективным. Однако можно
ожидать, что отбор создаст системы модификаторов, которые
усилят фенотипическое прояв ']ение данноrо reHa в среде Е 2 . Этот
новый набор модификаторов может, таким обр'азом, иметь эффек
ты, которые переносятся в среду El' При этом доетиrаетея про-
явление признака, и ОН может стабилизироваться под действием
отбора как в среде Е" так и в среде. Е 2 .
Уоддинrтон (Waddington, 1953, 1956) провел интересные экс
пери менты на Drosophila тelanogaster и получил данные, которые,
по видимому, подтверждают описанную выше модель. В основу
этих экспериментов БЫ 1 ноложен известный факт, а именно что
некоторые отклоняющиеся фенотипы (crossweinless, bithorax)
можно индуцировать у небольшоrо числа взрослых мух при по-
мощи резких внешних воздействий (высокая температура, эфир)
на яйца или куколки. ИндуuироваJfные таким образом фенотипи-
ческие изменения обычно не передаются по наследству, но в ЭКС-
r л. 9. П РОЯ8ление eHa и е20 связь с отбором
81
периментах Уоддинrтона они в конце концов стали наследствен-
ными.
у оддинrтон оказывал Шоковое воздействие (обработка теплом
или эфиром) на исходные популяции мух на стадиях яйца ИЛИ
КУКОw1КИ; получив соответствующие фенотипические реакции у
некоторых из взрослых мух, он проводил отбор на отклоняющие
ея фенотипы. Весь этот процесс повторялся из поколения в поко
ленке На протяжении 24 29 поколениЙ. Это был фактически ис-
кусственный отбор по ОТI(лоняющемуся фенотилическому призна
ку, проводившийся в аНО lа.,'IЬНОЙ среде, в которой этот признак
проявляется.
Отбор оказался эффективным. К концу эксперимента в проис
ходящих от этой ПОПУЛЯЦИИ поколениях мух частота особей, у
которых проявлялась феНОТИilическая реакция на шоковое B03
действие, достоверно повысил ась по сравнению с их частотой в
peДKOBЫX поко..1ениях, т. е. в нача.'Iе эксперимента. Кроме Toro
у некоторых из выведенных. таким образом мух ОТКJlОНЯlOЩИИСЯ
фенотип проявлялся не только в аномальной среде, в которой
производи.,'IСЯ отбор, но II В нормальной среде, по реакциям на
которую отбор не ПРОИЗВОДИJ1СЯ. В поздних покодениях у HeKO
торых мух наблюдались отклоняющиеся признаки, даже если их
не помещали в аномальную внешнюю среду.
Результаты, полученные Уоддинrтоном, можно интерпретиро
вать по разному. Одно из правдоподобных объяснений состоит
в том, что процесс отбора привел к созданию наборов модифика-
торов, которые УСИ.'lивают экспрессивность измененных признаков
крыла или rруди в аномальной среде. Эти новые эффективные
модификаторы влияли на фенотипическое проявление не только
в аномальной среде, но обладали, кроме Toro, побочным дейст
вием, наблюдавшимся также и в обычной среде. (Ориrинальное
описание этих экспериментов см. в работах W addington, 1953
1956; различные объяснения IW addington, 1957; Stern, 1958,
1959; Bat.eman, 1959; обзор \Grапt, 1963, стр. 206 211).
Из представленной выше модели с участием модификаторов
экспрессивности вытекают следствия, важные. для эволюционной
теории. Модель указывает, что отбор на некоторый комплекс re
нов-модификаторов, проводимый в одной среде, может создать
rенотип, который дает начало непредсказуемым феНОТИПl1ческим
лроявлениям в друrих cpeдax Новые потенциальные возможно-
сти для фенотипическоrо проявления MorYT иноrда создаваться
непреднамеренно при отборе на модификаторы эксп'рессивности.
Таким образом, ПОПУJ1ЯЦИЯ, в которой происходит отбор приме-
нительно к одной новой среде, может приобрести rенотипически
контролируемые фенотипы, которые в }<ачестве поБQчноrо резуль-
тата оказываются «преадаптированными» еще и к друrим новыМ!
средам.
'82
Часть ///. ECTeCTseHHbiu отбор
Модификаторы доминантности
Соотношение доминантности и рецессивности в паре аллелей
у диплоидноrо орrанизма может про истекать из относительной
силы действия самих этих двух аллелей, однако этим дело не or-
раничивается, поскольку доминантность и рецессивность BЫ3ЫBa
lОтся также д йствием друrих reHoB, известных как модификаторы
ДО 'lИнантности. Та или иная rетерозиrотная пара аллелей может
давать доминантный фенотип на одном rенетическом фоне и про-
.межуточный фенотип на друrом фоне. Один rенетический фон
обладает набором мощных модификаторов доминантности re-
нов модификаторов, усиливающих фенотипическое проявление
доминантноrо аллеля, тоrда как друrой rенетический фон имеет
.слабый или различающийся в каком либо друrом отношении
набор модификаторов.
Напомним, что новые мутации в большинстве случаев бывают
вредными и рецессивными. Вредное воздействие мутаций неиз
-бежное следствие случайности мутационноrо процесса в орrаниз
ме, который перед этим На протяжении мноrих поко.пений подвер-
rался отбору. Рецессивность JКe не представляет собой свойство,
внутренне присущее ,мутационному цроцоосу. Та,к почему же HO
вые мутации в бо.пьшинстве своем бывают рецессивными?
Фишер (Fisher, 1930, 1958) высказал мнение, что естествен-
ный отбор создал у диплоидных орrанизмов системы модификато-
ров, которые усиливают и стабилизуют действие нормальных ал-
делсй дикоrо типа. Эти модификаторы доминантности будут бла-
I'ОПрИЯТНЫМИ постольку, поскольку они защищают диплоидный
.орrанизм от непосредственных вредных воздействий большинства
.новых мутаций. Модификаторы доминантности дают возможность
популяции диплоидных орrанизмов держать про запас как вред-
ные, так и друrие мутации в рецессивном состоянии; при этом
такие мутации подверrаются медленному действию отбора, кото-
рый оказывает В.rIияние на относительно небольшое число rOMo-
зиrотных рецессивных выщепенцев.
Фишеровекая теория происхождения доминантности cor ласу
ется с большинством данных, относящихся к явлениям доминант-
ности и к мутационному процессу, и дает единое объяснение
различным набдюдаемым фактам, однако она не была подтверж-
дена применительно к филоrенетическим аспектам проблемы. Мы
не можем утверждать, что у Toro или Иноrо диплоидноrо вида
доминантность возникла так, как ЭТО считает Фишер. Ero теория
ПРИВ.'Iекательна, но она остается недоказанной и вызывает воз-
ражения. Теория Фишера подверrлась критике со стороны одних
авторов (Crosby, 1963; Ewens, 1965) и была взята под защиту""-
с моей точки зрения, успешно друrими (Мауо, 1966; Sheppard,
Ford, 1966).
r А. 9. П рОЯВАение zeHa и ezo свяэь с отбором
8
rенотип как единица отбора
Совершенно очевидно, что однолокусная модель отбора пред
ставляет собой сверхупрощение. Однолокусная модель биолоrи-
чески реа.ПИСТИIIна и соответствует нескольким подлинным слу-
чаям, например конкуренции между клонаl\'И бактерий, различа-
ющимися лишь по одному reHY. Но применительно к большинству
ситуаций у эукариотов эту модель следует рассматривать как
некую абстр.акцию, полезную для разъяснения более сложных
процессов, происходящих в действительности.
Фенотипические признаки, на которые действует отбор у выс-
ших орrанизмов, за редкими исключениями, детерминируются не.
единичными rенами, а сочетаниями reHoB. Отбор не имеет возмож.
ности производить выбор между аллелями одноrо reHa, а должен
действовать на альтернативные формы rенной системы, состоящей
из мноrих компонентов.
r,,'Iавная форма отбора происходит не среди самих reHoB и дa .
.JКe не среди reHHbIx систем, а среди особей данной популяции..
А отдельный орrанизм, особенно у высокоразвитых форм жизни,
uредставляет собой сложный механизм, состоящий из мноrих op
raHoB, выполняющих раЗ..тIичные функции. Эти разнообразные ор-
raHbl ДО.7JЖНЫ rармонически СОчетаться друr с друrом, а их
функции должны быть координированы. Изменение одноrо приз
нака, возможно вполне блаrоприятное в отношении специфиче-
ской функции этоrо признака, может оказать неблаrоприятные.
побочные воздействия на друrие функции орrанизма. В таких
случаях отбор сохранит новый признак или элиминирует ero в
зависимости от Toro, дает ли этот признак отде,,'1ЬНОМУ орrанизму
в це.'10М чистый выиrрыш иди чистый проиrрыш.
Конечный результат в тех С.71учаях, коrда отбор проявляет
противопо",ожные тенденции, нередко оказывается компромис .
еным. Жизнь полна таких компромиссов. Яркое оперение райских
птиц gыполняет полезную роль в брачном поведении, но вместе.
С тем дслает птицу заметной Д,,1Я хищников. Это противоречие
разрешается при помощи компромисса: ве,,1иколепное оперение
украшает самцов, которыми можно пожертвовать, а у самок OK .
раска очень неброская.
['лава 10
ПРИМЕРЫ ОТБОРА
Типы имеющихся данных
На протяжении мноrих лет эффективность eCTecTBeHHoro от-
бора ---- ero способность вызывать изменения, как это предсказы-
вает теория отбора,---- подверr.ается проверке и получает подтверж-
дение в бесчисленных исследованиях, которые проводятся на раз-
ных видах животных, растений и микроорrанизмов. В зависимости
от Toro, какой подход использовался в этих исследованиях, их
можно разбить на три rруппы: 1) селекционные эксперименты,
проводимые в лаборатории или на опытных делянках; 2) наб 1Ю-
дения внеконтролируемых природных популяциях; 3) прослежи
вание последовательных изменений. вызываемых искусственным
отбором у культурных растений и домашних животных. В этой
rлаве мы рассмотрим несколько примеров, иллюстрирующих
каждый из этих подходов.
Иллинойсский эксперимент по селекции кукурузы
На Экспериментальной сельскохозяйственной станции штата
Иллинойс начиная с 1896 r. проводится длительный эксперимент
по селекции кукурузы (Zea mays). Искусственный отбор в раз
личных линиях, полученных от изменчивой исходной популяции,
производился по нескольким признакам: по содержанию в зернах
белка и масла и по высоте расположения початка. Полученные
результаты изложены в ряде работ (Winter, 1929; Woodworth,
Leng, J.ugenheimer, 1952; Bonnett, 1954; Leng, 1960).
Отбор на содержание белка в зернах проводился параллельно
в двух линиях: в «высокой» линии (с высоким содержанием бел.
ка) и в «низкой» линии, которые велись с 1896 r. от одной и той
же исходной популяции с содержанием белка 10,9%. В каждом
следующем поколенин (кукуруза ---- однолетнее растение, дающее
по одному поколениIO в rод) собира 'IИ большое число початков и
определяли содержание бе.'Iка в некоторых их зернах. Часть
початков с наивысшим содержанием белка в зернах относили к
«высокой» линии И использовали в качестве семян для следую-
щеrо поколения. Аналоrичным образом часть початков с самым
низким содержанием белка использовали в качестве семян для
каждоrо HOBoro поколения «низкой» линии. В первые rоды про-
ведения эксперимента эта часть составляла 24 початка из 120 в
r А. 10. Примеры отбора
85
КаЖДОЙ линии, а в последние rоды 12 из 60. В 1959 r. в ЭТОМ
селекционном эксперименте было получено 60-е поколение расте-
ний.
Реакция на отбор проявил ась сразу и продолжалась до...lrо.
В высокой линии содержание белка возросло от 10,9% в нулевом
поколении до 13,8% в 5-м И 19,4% в 50-м поколениях. В низкой
линии содержание белка упало с 10,9% в HYJleBoM покодении до
9,6% в 5 M и 4,9% в 50-м ПОКО.'Iениях. Результаты отбора в этих
25
20
..
,5
"б
10
5
d
о 5 10 15 20 25 30 35 40 45 50 55 БО
(18Qб) Числ.о п.ок.оленuи (195Q)
Рис. 10.1. Реакция на отбор на высокое и) или низкое (l/) содержание белка
в зернах кукурузы (Zea тays) , проводнвшийся в 60 ПОКОJIеннях (по данным
Woodworth, Leng. Jugenheimer, 1952; Leng, 1960).
двух линиях вплоть до БО rо поколения представлены на
рис. 10.1. Отбор на содержание масла в зернах вызвал еще более
сильную реакцию, чем отбор на содержание бе.'1ка ('Woodworth,
Leng, Jugenheimer, 1952; Leng, 1960).
Отбор на содержание белка или масла привел к неожиданным
коррелятивным изменениям разных морфолоrических признаков.
Так, у растений, происходящих от ,,'1инии С высоким содержанием
белка, початки были мелкие, с полупрозрачными кремнистыми
зернами, а у линии с низким содержанием белка початки были
крупные, с продолrоватыми крахма.'1ИСТЫМИ зернами (iWoodworth,
Leng, Jugenheimer, 1952).
Отбор по высоте расположения початка над уровнем почвы
нача..1СЯ с исходной популяции, выращенной в 1903 r., в которой
початок располаrался у разных особей на высоте от 109 до
142 см. В каждом поколении выбирали растения, у которых По-
Чатки БЫJ1И расположены ближе Bcero к поверхности почвы, и
они служили родительскими растениями для следующеrо поколе
ния «низкой» линии. Параллеol1ЬНО таким же способом велась
высокая» линия. Обе линии провели через 25 поколений (до
1928 r.).
86
Часть [П. Естественный отбор
в этом случае также наб.'1юдалась хорошо выраженная peaK
ЦИЯ на отбор. В 24 ! поколении (1927 r.) средняя высота распо
ложения початка была равна 20,3 см в «НИЗКОЙ» И 306 см в «вы-
сокой» линии. Как и при отборе на содержание белка, в этих
хорошо отселектированных линиях возникали непредсказуемые
изменения, коррслирующие с признаком, по которому производил-
ся отбор. Линия с низко расподоженным почаТКО l зацветала
раньше оБЫЧRЫХ сроков, а линия с высоко расположенным почат
ком на 10 14 дней позже (Bonnett, 1954).
Zea mays ---- свободно скрещивающееся и высокоrетероэиrотное
растсние, в популяциях KOToporo содержатся большие запасы
rенстической изменчивости. Постепенные и длительные реакции
на отбор, наб.'Iюдавшиеся в ИЛЛИIlОЙССКОМ эксперименте, со ответ..
ствуют представлениям о том, что отбираемые признаки опреде-
ляются мноrими rенами, которые в исходной популяции были по
'/lИМОрфНЫМИ. Возможно, что В процессе этоrо длительноrо экспе
римента возникла новая комбинативная изменчивость, блаrодаря
которой отбор продолжал вызывать изменения в обоих направ
лениях; вподне возможно также, что за это время возникла и HO
ная мутаrщонная изменчивость. Отбор вызвал корре.1Jятивные из.
менения друrих признаков, продемонстрировав Te l самым С.1JОЖ
ность и интеrрированность rенотипа.
Жизнеспособность у дрозофилы
Добржанский и Спассхий (Dobzhansky, Spassky, 1947) поста
ви..1И на дрозофи..'1е эксперимент, спланированный таким образом,
чтобы возникающие в популяции изменения вызывались eCTeCT
венным отбором, а не сознательным в ешательством эксперимен
татора. Следовате.1JЬНО, эта работа дополняет ИЛJ1ИНОЙССКИЙ экспе
римент по селекции кукурузы, в котором ведущим фактором был
искусственный отбор.
Исходными популяциями служи.'}и линии Drosophila pseudoob
scura с пониженной жизнеспособностью. Жизнеспособность этих
линий выражалась количественно в процентах от норма.'1ЬНОЙ
жизнеспособности. Семь родственных линий были использованы
в качестве исходных популяций для закладки культуры в семи
повторностях. Каждую культуру раздели..'IИ на четыре субкульту.
ры, которые содержали в разных условиях. Вкратце эти условия
можно охарактеризовать следующим образом: 1) облучение и
скученность; 2) CKY(leHHOCTb без облучения; 3) облучение без
скученности; 4) ни скученности, ни облучения.
Линии, содержавшисся в условиях 1 и 2, переводили в rомози
rOTHoe состояние по определенным хромосомам, с тем чтобы об
.леrчить проявление новых мутаций; линии, содержавшиеся в yc
r А. 10. П pиM pЫ отбора
87
JIОВИЯХ 3 и 4, поддерживали в rетерозиrотном состоянии по этим
же хромосомам. ЛИНИИ в условиях 3 И 4 служиди КОНТРО.нем дЛЯ
ЛИНИЙ в ус.rювиях 1 и 2 соответственно. Все ЛИНИИ велись в лабо-
ратории в течение 50 покодений. Проверки на жизнеспособность
и по друrим признакам проводили через каждые несколыш поко
JIений и в. конце эксперимента.
Рассмотрим сначала изменения в одной из семи линий, содер-
жавшихся в условиях 2. Жизнеспособность этой линии в начале
эксперимента составляла 29 % (от нормальной жизнеспособности).
Самцов и самок помещали в пробирку с кормом И давали им
возможность ОТКJlадывать неоrраниченные количества яиц, что
оыстро приводило К перенаселенности культуры. Случайную BЫ
борку ИЗ этой культуры переносили в новую пробирку И давали
.ей вновь раЗ:\.lножаться. Линию вели таким образом На протяже
иии 50 поколений.
Предполаrалось, что скученность в пробирках и сильная
конкуренция за корм приведут к повышению частоты .любых му-
тантных форм, обладающих большей мощностью, чем их сибсы.
Эти ожидания оправдались. За время эксrlеримента жизнеспо-
.собность линии повысил ась с 29 до 90%.
у большей части друrих .ТlИний, содержавшихся в условиях 2,
также наб,,'lюдалось повышение жизнеспособности. В одной линии
жизнеспособность ПОВЫСИ..1ась от 60 до почти 100%, в друrой----
-от 30 до более чем 80 %. в общем жизнеспособность повысилась
у пяти из семи .ТIИний, содержавшихся в условиях 2.
Семь линий в условиях 1 содержались в общем так же, как
11 предыдущая серия, с той разницей, что самцов каждоrо HOBoro
поколения до переноса в новую пробирку подверrали peHTreHoB
-скому облучению с целью повысить число новых мутаций. В шести
из семи таких облученных линий наблюдалось повышение жизне-
.способности, иноrда довольно значите.'lьное: от 29 до 103% в од-
ной и от 65 до 115% в друrой линии.
В общей сложности в 11 из 14 линий, содержавшихся в усло-
виях интенсивной конкуренции за корм в перенаселенных культу-
рах, отмечалось заметное повышение жизнеспособности за 50
nоколений. В контрольных линиях (условия 3 и 4) аналоrичноrо
повышения жизнеспособности не происходи.'IO. Микроэволюцион-
ные изменения в первых двух сериях (условия 1 и 2) связаны с
'Частично rомозиrотной конституцией, облеrчающей проявление
новых rенетических изменений, и с дифференциа,,'IЬНОЙ смертно-
-стью, зависящей от доступности пищи. Короче rоворя, в перенасе-
ленных сериях име.1J место естественный отбор, вызвавший обна-
ружимые эффекты, тоrда как в КОНТРО"lЬНЫХ сериях изменений не
tlаблюдалось.
Часть [П. Естественный отбор
88
Мепанизм у березовой пяденицы
Березовая пяденица (Biston betularia) широко распростране
на в Анrлии. Она активна ночью, а днем отдыхает на стволах или
ветвях деревьев, rде ее поедают ма 'IИНОВКИ, дрозды и друrие на-
секомоядные птицы. Эти птицы охотятся, руководствуясь зрени-
ем. Нас интересуют в данном случае две формы этой бабочки:
так называемая типичная форма..... белая с черными пятнышка
ми ---- и меланистическая форма с черным телом и черными крыль
ями, известная под названием carbonaria. Типичная форма хорошо
сливается с фоном, образуемым корой, покрытой лишайником,
и эта защитная окраска предохраняет бабочек от уничтожения
птицами. Что касается меланистической формы, то она четко BЫ
ступает на сероватом фоне коры, поросшей ..тIишайником, но зато
сливается с корой, покрытой сажей (рис. 10.2) (Ford, 1964; r.тl. 14;
Kettlewell, 1956, 1973).
Типичная и меланистическая формы различаются по одному
reHY С. Меланистическая форма возникла в результате редкой
доминантной мутации этоrо reHa. Типичная форма имеет rенотип
ее, а меланистическая ---- Се И lИ се.
Среда, в которой обитает березовая пяденица, резко измени
лась в результате происшедшей в Анrлии промышленной револю
ции. Одним из побочных эффектов индустриа 1изации БЫ.1И дым
и сажа, изверrавшиеся на прежде не заrрязненн)'ю сельскую
местность Анrлии. Лишайники исчез.IIИ, а стволы и ветви деревьев
почернели от копоти. Эти изменения оказаJ1И rлубокое влияние на
состав популяций Biston в про:мыш."енныx районах.
Популяции березовой пяденицы R доиндустриальной Анrлии
состояли почти исключительно из типичной формы, и в незаrряз.
HeHHыx районах страны состав популяций все еще остается преж
ним.
Меланистическая форма carbonaria впервые привлекла к себе
внимание как редкий мутант в 1848 r. в Манчестере. В период
между 1848 и 1898 rr. частота этой формы в ПРОМЫШJ1енных рай-
онах быстро возрастаv1Jа; она стала обычной формой, тоrда КаК
типичная сероватая форма стада редкой. Частота а..1.'1еля C 1 обус.
ловливающеrо черную окраску, соrласно оценкам, ПОВbIсилась с ]
до 99% за 50 поколеllИЙ с 1848 по 1898 r. Ввиду TaKoro СИ.'1ьноrо
изменения частоты reHOB можно считать, что аллеv'1Ь С дает оченЬ
большое селективное преимущества (Ford. 1964; KettleweIl, 1956,
1973) .
Фактором отбора служат насекомоядные птицы, ориентирую.
щиеся при помощи зрения. Окраска типичной формы делает ее
незаметной На коре, поросшей лишайником, а меланистнческая
окраска несет защитные функции На закопченной коре (рис. 10.2).
В исследованиях с выпусканием и последующим отловом бабочек
:s:
.......
:::,
;:
с;]
;
;:::
C;]
з7:;
= :::
8=;
0'/
i--o.....
a ,
:>::: о
g c:ci
. :1'
:r: ;:
:r:
o:t:
..,
Q)
[
... :r t:
o
са е
:Q :t:
шg-=
g ;Е
'<:( \о
.:;: 52 а:
g 8=;
:i
Q) :: =
-e. u
-
:а 5
ь g-
0-&-&
-& !II:
D:: '"
u ..CI О
Q) tQ Р.
:1' Q) <1J
= о... u
..... Q)
t::t: а:
;:: ><
cj cj ;:
з
::Е tQ =
..... f-o
:: u с'о
D::.......:r
со;ЗlQ
::..........а;
== CtI
1:: О ш
= :r t:(
b::
Q)
о о о
....
u"'-&
:t
90
ЧSСТЬ 1/1. ECTuTBeHHbIiJ. отБО/l
было установлено, что в незаrрязненных районах выживаемость.
типичной формы ВДВое выше, чем выживаемость ме '1анистов;
вб '1ИЗИ промышленноrо rорода Бирминrема наблюдается обратная
картина: выживаемость меланистов вдвое выше по сравнению
с выживаемоСтыо типичной формы (Ke1tleweII, 1956). Изменение
субстрата, вызванное индустриализацией, привело к реверсии от-
носительных селективных ценностей аллелей с и с.
reH С оказывает плейотропное . воздействие на физиолоrиче-
скую мощность, что может привести к созданию вторичных се-
лективных иенностей в отношении друrих факторов. Меланистиче
ская форма оказывается более живучей, чем типичная, в CTpec
совых условиях в лаборатории, а возможно, также и в природе.
В незаrрязненных районах этот блаrоприятный плейотропный
эффект аЛ.'Iеля carbonaria сводится на нет сильным отрицательным
отбором ---- уничтожением птицами. В закопченных районах, од-
нако, популяции бабочек MorYT использовать физиолоrичеСI(ие
преимущества, предоставляемые им этим аллелем.
Параллельные изменения серой или коричневой окраски в чер-
ную наблюдались в Анrлии в тот же период и у ряда друrих ви-
дов бабочек; подобная эволюция меланизма у бабочек произошла
с некоторым СДвиrом по времени в промышленных районах Цент-
ральной Европы и на ceBepO BOCTOKe США (Ow1en, 1961; Ford,
1964; KetHewel1, 1973).
Окраска раковины ;У наземнЫх улиток
Европейская наземная улитка (Сераеа neтoralis) обычна в
разнообразных местообитаниях ---- в лесах, На луrах, в живых из
rородях ---- в Анrлии и Западной Европе. В популяциях этой улит-
ки обычно наблюдается полиморфизм по окраске раковины, KO
торая может быть бурой, розовой или желтой (рис. 10.3). Окраска
раковины детерминируется рядом множественных аллелей, среди
которых наибольшей доминантностью обладает аЛ.'lель бурой, а
наибольшей рецессивностью алде.'IЬ желтой окраски (Cain.
Sheppard, 1954; Sheppard, 1959; Lamotte, 1959; Jones, 1973).
Цвет раковины иrрает rлавную' pOJlb в защитной OKpaCI(e этих
улиток. Дрозды и друrие птицы находят улиток при помощи зре-
ния и уничтожают значительно большую долю особей, окраска
которых контрастирует с фоном, чем особей с криптической OKpac
кой. Конечно, контрастность или криптический характер окраски
цедиком зависят ОТ фона.
ОДНО из местообитапий Сераеа neтoralis"7"'""" rycTbIe буковые ле
са. Для этих лесов характерна подстилка из красно-бурых опав-
ших листьев, которая круrлый rод имеет примерно одинаI(ОВЫЙ
вид. Здесь окраска У '1иток с бурыми и розовыми раковинами ока-
зывается покровительственной, и эти формы здесь обычны. Улит
r А. 10. П рu.черы отбора
91
ки С желтой раковиной в буковых лесах встречаются редко, но
ОНИ обычны на зеленых луrах (Cain, Sheppard, 1954; Sheppard,
1959) .
Сераеа nemoralis встречается еще в одном местообитании ---- в
смешанных лиственных лесах. В этих десах характер фона изме
няется со сменой времен rода. Ранней весной подстилка бывает
бурой от опавших листьев, но к -,1ету она становится зеленой.
И селективные ценности раковин разноrо цвета здесь также из
е e . . 0 ..:...... .:.,.,.!!
. ..... {....
о' '. . . .. ..
.. . ... l'
. .
." . . .' .
. .
. "....' . , k .' :.;.
. . .
. . ,
А
6
8
Ф . е
. '. :": .
..... , . .
, . .
. . ' ..' ",:.... ,
: .
. .
. .
." о..
. .
r
А
Рис. 10.3. Полиморфизм у европейской наземной улитки Сераеа пeтoralis.
А. Желтая раКОВИllа. Б. Розовая р'зковинз. В. Бурая раКОВlIна. r. РаКОВИllа с двумя по-
.'!Осами. Д. Раковина с пятью ПО.10самн.
меняются со сменой времен rода. Ранней весной бурая и розовая
окраска лучше защищают от уничтожения птицами, а летом это
преимущество переходит к желтым улиткам. В полиморфных по-
пуляциях улиток, обитающих в ,,'Iиственных .песах, в течение rода
происходят циклические Изменения относительных частот разных
форм (Cain, Sheppard, 1954; Sheppard, 1959).
Окраска раковины несет не TOJ1bKO защитные функции. Имеют
<=я данные, свидетельствующие о различной физиолоrической вы-
носливости разных цветовых форм. Желтая форма, по видимому,
..1еrче переносит экстремальные температуры, как высокие, так и
низкие, чем розовая. Большая устойчивость желтой фор:мы к ко-
лебаниям температуры, конечно, должна давать ей преимущество
в открытых луrовых местообитаниях и в областях с жарким ле-
том. Эта форма, как уже отмечалось, деЙствительно достиrает
Высокой частоты на луrах частота ее повышается по rеоrрафиче
екой трансекте от северных районов Европы к IOry (Lamotte, 1959:
Jones, 1973).
К ЧИСollУ друrих признаков раковины относится характер по.
.,10сатости. Раковины бывают бесполосые и полосатые, а число
полос колеблется от 1 до 5 (рис. 10.3). Наличие или отсутствие
Часть "/. Естественный от60Р
92
пр.rюс детерминируется reHOM В, тесно сцепленным с reHoM, дeTep
минирующим окраску раковины. В локусе Валлель бесполосости
доминирует над аллелсм полосатоети. Число полос контролиру
ется особым локусом. В результате рекомбинации образуются Bce
возможные сочетания окраски и полосатости раковины (Cain.
Sheppard, 1954; Sheppard, 1959).
Роль eCTecTBeHHoro отбора в отношении характера полосато-
сти менее ЯСllа, чем в отношении окраски раковины. Известно,
что у друrих животных темные полосы, опоясывающие тело, участ-
вуют в образовании защитной окраски в полузатененных MeCTO
обитаниях, rде иrра света и теней создает разбитые контуры.
Возможно, что такую же функцию несут полосы на раковинах у
Сераеа nemoralis. Во всяком случае в некоторых местообитаниях
полосатая раковина лучше защищает улиток от уничтожения
птицами. Возможно также, что изменчивость по характеру поло-
сатости контролируется дрейфом reHoB; в одной из последующих
rлав мы еще вернемся к этой проблеме.
Устойчивость к токсинам
Коrда в сороковых и пятидесятых rодах в медицине впервые
стали применять пенициллин, стрептомицин и друrие антибиоти-
ки, они в небольших дозах эффективно уничтожали болезнетвор
ные микроорrанизмы. Однако вскоре после Toro, как применение
антибиот ков расширилось, их эффективность начала снижаться
и для достижения желаемых результатов приходилось использо-
вать бо..1ее высокие дозы. Существуют штаммы бактерий, устой
чивые к антибиотикам и чувствительные к ним. Появление устой-
чивых штаммов обусловлено спонтанными мутациями, возникаю-
щими с некоторой низкой частотой. Таким образом, применение
антибиотиков в низКих или умеренных дозах приводит в Действие
процесс отбора, блаrоприятствующий появлению устойчивых
штаммов.
Такие МИКРОЭВОЛIOционные изменения были обнаружены в ла
бораторных экспериментах. Примером служит селекционный экс
перимент, проведенный на одном из штаммов (NQ 209 Р) Micro
coccus pyogenes aиreus (Staphylococcus aureus) патоrенный бак
терии, вызывающей наrноение ран и пищевые отравления
(McVeigh, Hobdy, 1952).
Исходная популяция, от которой про изошел этот штамм, была
чувствительна к различным антибиотикам, и ее рост подавлялся
очень незначительными дозами этих веществ. ИЗО.тIяты, взятые из
исходной популяции, были использованы для закладки субкуль-
тур, которые выращивали на ряде культуральных сред, содер.
жавших пенициллин и друrие специфические антибиотики во все
возрастаюших концентрациях. В результате у разных штаммоВ
r л. 10. Примеры отбора
9З
возникла устойчивость к этим антибиотикам. Наибольшее по-
вышение устойчивости происходило в течение первых 1 0 20 пе-
ресевов, после чеrо устойчивость возрастала более медленно.
1-.сто"Йчивость к разным антибиотикам повысилась в разной сте..
пени:
к хлоромицетину в 193 раза
к аурсомицину в 210 раз
к N а пеницилдину в 187000 раз
к стрептомицину в 250000 раз
у возникших таким образом устойчивых штаммов наблюдают
ся и друrие изменения. Они обычно растут, медленнее, чем исход
ный чувствите.'1ЬНЫЙ штамм. УСТОЙЧИВЫЙ к ленициллину штамм в.
значительНой степени утратил свою патоrенность и СIlособность
расти в анаэробных условиях. Поэтому в обычных условиях, т. е.
при отсутствии в среде антибиотиков, чувствите..lьная форма
превосходит устойчивую. Если удалить из культуральной среды
антиuиотики, то среди бактерий начинается отбор, направленный
в противоположную сторону, т. е. на сохранение чувствите.тIЬНОЙ
формы.
Аналоrичные микроэволюционные изменения возникли у He
СКО.'Iьких видов насекомых-вредителей в результате широкоrо
применения инсектицидов. Хорошо известным примером ,служит
ПОЯВ.Т1ение штаммов комнатной мухи (Musca domestica) , устойчи-
вых к ДДТ. Друrой пример эволюция устойчивых к цианиду
штаммов щитовок (Aonidiella aurantii и друrие виды) на планта
циях цитрусовых в Калифорнии.
В живом мире встречаются разноrо рода токсины и процесс
ЭВОЛIоционноrо приспособления к ним происходит не только в yc
ловиях се.'1ьскохозяйственной и медицинской практики, н() н В
естественных условиях. Например, кактус Lophocereus schottii
растущий в l\1.ексике, содержит алкалоиды, токсичные для боль-
шинства видов дрозофилы. Однако Drosophila pachea устойчива
к этому алкалоиду и размножается в rниющих стеблях кактуса.
В этом чрезвычайно своеобразном месте размножения D. pachea
не испытывает конкуренции со стороны друrих видов Drosophila
(Kircher, Heed, 1970).
Одомашнивание
j\\ноrие виды растений и животных силыю изменяются в про-
цессе одомашнивания, при котором искусственный отбор сочета-
ется с естественным. История выведения сортов культурных расте-
Ний и пород домашних животных дает. множество хороших при-
Меров эффективности селекции. Дарвин в «Происхождении видов»
.перечисляет различия между диким скалистым rолубем и домаш-
94
Часть lJ/. Естественный отбор
ним rолубем, в том числе такими специализированными породами,
как турман и дутыш, В качестве примера rлубоких эволюционных
изменений, которые может вызвать искусственный отбор. Нетруд-
но вспомнить и друrие примеры: собака, домашний скот, томаты,
тыква и т. п. В следующем разделе мы рассмотрим один И3 та-
ких примеров ---- кукурузу_
Эволюция кукурузы
Кукуруза или маис (Zea тays) ---- один из представителей три-
бы l\1ayd,eae семейства злаков (Gramin,eae). Она резко ОТЛИЧает
ея от всех друrих членов своей трибы и семейства по ряду при
знаков, в частности по расположению opraHoB размножения и по
.строению колоса, или початка. Кукурузный початок ---- очень
сложная и высокоспециализированная часть растения; ничеrо
подобноrо нет ни у одноrо друrоrо представителя семейства зла-
ков. Но тем не менее современная кукуруза с ее своеобразным
початком и друrими признаками произошла от диких злаков,
растущих в тропических и субтропических областях Америки.
Соответствующие данные были подучены в результате сравни-
тельно-морФолоrических, таксономических, rенетических, цитоrе-
нетических, этноботанических, археолоrических и палинолоrиче-
.ских ИСС.'1едований. Ведущая роль в получении этих данных при-
надлежит Манrельсдорфу (Mangelsdorf), rалина (Galinat), Хар-
лану (Harlan) и друrим. По некоторым вопросам между разны-
ми исследователями существуют расхождения во мнениях. Более
подробно с этой проблемой можно ознакомиться в ряде интерес-
ных и важных работ (Mangelsdorf, 1958, 1959; !\'1angelsdorf,
MacNeish, Galinat, 1964; Мапgеlsdоrf, Dick, Camara-Hernandez,
1967; Anderson, Brown, 1952; .Galinat, 1970, 1971 а, 1971 Ь; De \Vet,
Harlan, Grant, 1971; De Wet, Harlan, 1972; .WИkеs, 1967, 1972).
Итоrи исследований в' этой области подведены в моноrрафии
Манrельсдорфа (Mangelsdorf, 1974).
У Zea тays тычиночные и пестичные колоски находятся в
разных соцветиях. Кукуруза, таким образом, ОДНОПШIоцветковое
,однодомное растение, что пеоБыноo для злаков. Несущая пыльцу
метелка располаrается на вершине кукурузноrо стебля, а почат
1<И, в которых находятся зерна, развиваются в пазухах листьев в
.средней части стебля (рис. 10.4). Початок }<укурузы представляет
-собой колос, состоящий из толстоrо центральноrо стержня, несу-
.1цеrо MHoro рядов пестичных колосков, а позднее ---- столько же
рядов зрелых зерен. Початок в целом покрыт модифицированны.
ми листовыми обвертками. Зерна прочно прикрепляются к стерж-
ню; зерна rолые, т. е. не покрыты пленками (шелуха ИЛИ мякина) t
:как у друrих злаков .(рис. 10.5).
r А. 10. П ри.меры отбора
(95-
у друrих злаков каждое зерно покрыто отдельной пленкой и
отделяется от соцветия в качестве самостоятельной единицы, что
сильно повышает эффективность распространения семян. Сочета"
,
Е
.
5
Рис. 10.4. Форма побеrа у двух сортов кукурузы (lea тays) зубовидной (А)
и разрывной (Б).
нне противоположных признаков у кукурузноrо початка прочно
прикрепленные rолые зерна и ..'1истовые обвертки, одевающие по..
чаток в целом, сильно снижает ВОЗ/ИОЖНОСТЬ распространен.ия.
tемян, однако зти же самые признаки очень ценны в CeJlbCKOXO
зяйственном плане, поскольку они облеrчаlOТ сбор урожая и CBO
дЯТ К минимуму потери зерна. Лучшие современные сорта куку-
рузы отличаются крупными раЗМерами ОТ'дельНЫх зерен .и Почат
96
Часть ///. Естественный отбор
ка в целом (рис. 10.5, А). У некоторых примитивных сортов Zea
тays, а также и у ее диких родичей зерна мелкие, а початки ма-
ленькие (рис. 10.5;, Б); при этом зерна у них твердые и крем-
нистые в отличие от мучнистых или сахаристых зерен лучших
сортов, }{оторые деrче размолоть или разжевать.
Высокоурожайным новейшим сортам необходим мощнЫЙ, хо-
рошо облиственный стебель, чтобы синтезировать и запасать те
А
Б
8
Рис. 10.5. Початки трех сортов кукурузы: зубовидной (А) I арrентинской разрыв
ной (Б) и пленчатой (В) (Мапgеlsdоrf, 1958).
вещества, ради которых их выращивают. Новые сорта отвечаю!
этим требованиям. Современная кукуруза rиrант по сравнению
с друrими однолетними травянистыми растениями; она не ниже
некоторых видов бамбука и rораздо выше примитивных сортов
кукурузы (рис. 10.4). Эти и друrие качества наиболее хорошо
.отселектированных сортов кукурузы полезны человеку, и он их
создавал путем искусственноrо отбора на протяжении мноrих
поколений.
Разные племена американских индейцев начали выращивать
кукурузу уже по крайней мере 7000 лет назад. Центры культу-
ры кукурузы возникли в восточной и lOro-заuадной частях Север-
r л. /0. Примеры отбора
97
ной Америки, в Мексике, Центральной Америке и в районе Анд.
у сортов кукурузы, выращиваемых в этих центрах, все еще сохра..
няются некоторые примитивные признаки; о друrих признаках
можно судить по остаткам давно вымерших сортов, найденным
при архео.поrических раскопках.
Разрывная (лопающаяся) кукуруза относительно невысокое
растение с несколькими побеrами, отходящими от основания
lсм
Рис. 10.6. Продольный разрез нижней части ископаемоrо кукурузноrо початка
из пещеры Бат-Кэйв; верхний конец початка не сохранился (Mangelsdorf, 1958).
растения, и с небольшими початками, на которых сидят мелкие
твердые зерна (рис. 10.5, Б). У пленчатой кукурузы проявляется
друrой примитивный признак зерна покрыты П.1Jенками, как у
друrих' злаков (рис. 10.5, В).
Важную находку представляет собой ископаемый кукурузный
початок, найденный в Бат-Кэйв (штат Нью-I\'\ексико), в пещере,
в которой жили первобытные люди, выращивавшие примитивный
сорт кукурузы. Этот початок маленький, меньше, чем у разрыв-
ной кукурузы. Зерна, такие же ме.'1кие, как у разрывной кукуру-
ЗЫ, сидят на длинных плодоножках, отходящих от сравнительно
TOHKoro центра..'Iьноrо стержня, и покрыты пленками, как у плен-
чатой кукурузы (рис. ]0.6). Возможно, что верхний конец колоса
нес тычиночные колоски. При помощи радиоуr.'1еродноrо и друrих
методов бы.1JО установлено, что возраст самых старых экземпля-
ров кукурузы, найденных в Бат-Кэйв, составляет 3500 4300 лет.
у БО:Iее древних ископаемых початков, найденных в Теуакан-
Воллей (l\1ексика), возраст которых достиrает почти 7000 лет,
также обнаружено сочетание признаков разрывной и пленчатой
Кукурузы; колосья бы.пи обоеполыми с тычиночными соцветиями
На конце.
98
Часть //1. Естественный отбор
Среди еще более древних ископаемых остатков из Мехико
найдены пыльцевые зерна, сходные с пыльцевыми зернами совре..
менной кукурузы. Возраст этих остатков оценивается в 80000 .'1ет.
Хотя определение на основании одних только пыльцевых зерен,
без друrих частей растения, нельзя считать вполне надежным,
весЬма вероятно, что найденная в Мехико пыльца принадлежит
какой ннбудь предковой дикой форме кукурузы.
Манrельсдорф скрестил современную разрывную кукурузу с
пленчатой, получив синтетический сорт, сочетавший в себе при-
митивные признаки каждоrо из родительских сортов. Эта ПJ1ен..
чато разрывная кукуруза имеет узкое обоеполое соцветие, типич-
ное для злаков, с женскими цветками в нижней и мужскими в
верхней части. Зерна покрыты пленчатыми чешуями и леrко от-
деляются после созревания. По видимому, синтетическая плен-
чато-разрывная кукуруза представляет собой одну из возможных
ранних стадий в эволюции кукурузы (рис. 10.7, Б).
В тропических и субтропических областях Америки есть две
rруппы диких злаков, которые близки Zea mays и считаются воз..
можными дикими предками кукурузы. К первой rруппе относит-
ся Zea mexicana (теосинте) , а ко второй небольшой род Trip..
sacuт (рис. 10.7, А и В). Из этих двух rрупп к Zea mays более
близка Z. тexicaпa.
Теосинте (Zea тexicana) однолетнее растение, широко рас..
пространенное в Мексике и Центральной Америке, rде оно часто
встречается в качестве сорняка на кукурузных полях или вблизи
них. Z. mays и Z. тexicana можно искусственно скрещивать, а в
Мексике они скрещиваются в природе, причем rибриды FI пло-
довиты. у Z. тexicana так Же, как у Z. mays, имеются отдельные
тычиночные и пестичные колоски, однако пестичные колоски
имеют у нее вид разветвленной системы тонких колосьев, окру-
женных листовой обверткой. У Z. mexicana каждый из собранных
в колос пестичных ко..l0СКОВ защищен особой отдельной обверткой
(плюской); у Z. mays таки плюски имеются, но они редуцирова-
ны и служат для повышения прочности, а не для защиты. У Z. те-
xicana стержень колоса расчленяется, так что зерна леrко отде-
ляются друr от друrа. У теосинте нет початка (рис. 10.7, В).
Разные виды Tripsacuт, в том числе Т. dactyloides, широко
распространены в теплых областях HOBoro Света. В отличие от
Zea тays и Z. тexicana Tripsacuт мноrолетники и несут обое-
полые соцветия (рис. 10.7, А). Соцветие Tripsacum мало отлича"
ется от соцветия синтетической пленчато-разрывной кукурузы,
упоминавшейся Выше. Tripsacuт успешно скрещивается с Zea
тays, но не скрещивается с Z. mexicana, и rибриды F 1 , если их
удается получить, более или менее стерильны.
Таким образом, считается, что предками Zea тays моrли быть:
1>. Tripsacuт; 2) какая то вымершая дикая форма пленчато раз"
r А. 10. Примеры отбора
А
Б
5см
8
Рис. 10.7. Колосья трех родичей современной кукурузы. обладающих примитив-
ными признаками (Mangelsdorf, 1958; Wilkes. 1967).
А. Tripsacит dactyloides верхние КОЛОСКИ мужскне. а нижние женские. Б. Синтетиче-
ская пленчато-разрывная кукуруза верхние КО.Чоски мужские, а иижние женские.
В. Zea тexicaпa ветвящкеся ряды женских колосков.
рЫБНОЙ кукурузы . (Mangelsdorf, 1958, 1974); 3) Zea mexicana
(Galinat, 1971 а; Ое Wet, Harlan, 1972). Некоторые ученые скло-
Няются к третьей rипотезе, предполаrающей происхождение ку.
курузы от теосинте (Z. mexicana) , однако эта rипотеза не под-
креП.'lена палеоботаническими данными. Что же касается второй
tипотезы, то следует отметить, что найденные в Бат Кэйв и Teya
кан Воллей ископаемые початки относятся по типу к пленчато
100
Часть 111. Естественный отбор
разрывной кукурузе. Хотя среди тех, кто изучает эволюцию ку-
курузы, все еще существуют разноrласия во мнениях, доказатель
ства в пользу второй rипотезы, по-видимому, перевешивают все
остаЛьные.
Морфолоrические различия между Zea тays и теми дикими
3J1аками, которые бы..,и ее предками, какими бы именнО они ни
оказались, очень велики. И все же эти различия можно объяснить
как результат взаимодействия между rенетической изменчивостью
с одной стороны, и естественным и искусственным отбором с
друrой. Современные сорта кукурузы обладают большими 'запаса-
ми rенетической изменчивости. Частично эта изменчивость обу-
словлена мутационным процессом, а также спонтанной rибри
дизацией между разными сортами кукурузы и между кукурузой
и ее родичами, теосинте и Tripsacuт (Mangelsdorf, 1974).
Установлено, что HeKoTopf?Ie крупные изменения по отдельным
морфолоrическим признакам Zea тays вызваны едини(!Ными re-
нами и простыми rенными системами. Так, например, пленчатые
чешуи П '1енчатой кукурузы обусловлены MYTaHTHЫ аЛ.'lелем l'eHa
Ти (tunicate), находящеrося в 4 й хромосоме. Различие между
сидячими и стебельчатыми пестичными колосками друrой при-
мер наблюдасмоrо у кукурузы различия, имеющеrо простую re
нетическую основу, но важноrо в филоrенетическом плане. Раз
личие между кремнистыми и относите.7JЬНО мяrкими зернами так-
же определяется единичным reHOM (Mangelsdorf, 1958; Galinat,
1971 а). Некоторые морфолоrические изменения, возникшие в про-
цессе эволюпии, MorYT поэтому быть реЗУ.'lьтатом искусственноrо
отбора по относительно небольшому числу rлавных reHoB и их
модификаторов. Друrие морфо.rrоrические и фИЗИО.rJоrические из-
менения, сопровождавшие эволюцию кукурузы, оБУС 10В.'Iены от-
бором по более сложным системам множественных факторов.
Заключение
1\'\ы рассмотре '1И при меры, в которых отбор мутаций, возни-
кающих в единиЧНЫХ reHax, ПрИ80ДИТ 1< относительно простым
эволюционным изменениям (меланистичсские формы березовой
пяденицы; бактерии, устойчивые к антибиотикам). В друrих рас-
смотренных случаях происходит отбор по признакам, оБУСЛОВJlен
ным мноrими rенами (содержание белка в зернах кукурузы).
Но в эволюции самой кукурузы от ее диких предков мы ста..1КИ.
ваемся с изменениями более крупноrо масштаба с изменения
ми це.rIоrо комплекса признаков, определяемоrо сочетание:\1 ряда
reHoB. Меланистическая форма березовой пяденицы остается пя-
деницей, но современная кукуруза это в сущности новый тиП
растения для семейства злаков.
r А. 10. П римеР"1 отбора
101
КРИТИI<И теории отбора давно уже высказывали мнение, что от-
бор лишен созидательноЙ силы; он может отметать неприспособ
ленных, но нельзя ожидать, чтобы в сочетании со слепым MYTa
ционным процессом он Mor создать что ..lибо новое (см., напри
мер, Koestler, Smythies, 1969).
При TaKoro рода критике упускается из виду, что комплексы
признаков детерминируются не простыми rенами, но сочетаниями
reHOB. Как мы убедились ранее, ПО.'10вое размножение механизм,
который обеспечивает сборку сочетаний а..1лелей, определяющих
новые KoMn.'IeKcы признаков. А отбор это механизм, который
может привести к закреплснию в популяциях новых сочетаний
а.Т'lлелей, если они окажутся блаrоприятными. В этом свете отбор
представляет собой механизм, способный вызывать в высшей сте.
пени маловероятные события (Fisher, 1930, 1958; Huxley, 1943,
стр. 474 475).
Кукурузный початок ма..'10I:\ероятное явление в семействе
злаков_ Каковы шансы на то. что кукурузный початок MOr бы
возникнуть без механизма отбора?
rла.ва 11
ТИПЫ ОТБОРА
Направленный отбор
Процесс отбора проявляется в различных формах и на разных
уровнях биолоrической орrанизации. В этой rлав будут рассмот-
рены основные типы отбора и уровни, на которых он происходит.
Отбор, обсуждавшийся в предшествующих rлавах, известен
под названием направленноrо (или проrрессивноrо) отбора. На-
правленный отбор вызывает проrрессивное. или направленное в
одну сторону. изменение rенетическоrо состава популяции
(рис. 11.1). Он происходит в тех случаях, коrда популяция нахо-
дится в процессе приспособления к новой среде или же коrда
происходит постепенное изменение среды, а вслед за ней и посте-
пенное изменение популяции. Хорошим примером первой ситуа-
ции служит увеличение частоты меланистической формы Biston
betularia и друrих бабочек при внезапном изменении среды, а
нменно ее заrрязнении промышленными отходами (rл. 10). Вторую
ситуацию, в которой возникает направленный отбор, ИЛЛIОСТРИРУ-
ют постепенные изменения, происходящие в среде, как, напри-
мер, изменения мноrих климатических факторов.
Стабилизирующий отбор
Совершенно иной характер носит стабилизирующий отбор.
Если популяция хорошо приспособлена к данной среде, которая
остается стабильной, то rлавное .действие отбора состоит в эли-
минации плохо пр.испособленных и периферическнх вариаций,
возникающих в результате мутационноrо процесса, потока reHoB,
расщепления и рекомбинации. Таким образом, сохраняются те
rенотипы, приспособленность которых доказана, тоrда как плохо
приспособленные формы «выпалываются». Это И есть стабилизи-
рующий отбор.
Действие стабилизирующеrо отбора rрафически представлено
на рис. 11.1. Предполаrается, что rенетическая изменчивость По-
пуляции имеет нормальное распределение, т. е. по каждому из-
менчивому и измеримому признаку основная масса особей rруппи-
руется вблизи среднеrо значения этоrо признака для данной по-
пуляции, а крайние значения наблюдаются лишь у немноrих
особей. Под действием стаБИЛИЗИРУIощеrо отбора эти особи, на-
ходящиеся по краям кривой нормальноrо распределения, элими-
нируются из поколения в поколение. Преимущественное размно'
=
:s:
::r
tJ:
Е:;
>..
с:
) о
с
ID
.о
::s ...
<.J
О
ID
Q) ::
d- :1"
;с
S CLI
:к
CI')
:5:
оа >..
:.::
<.J
Q)
Ir
=
' :, foo
CLI
:х:
CLI
'-
«1
:х:
:з .- ID
!): о
'<s с:
:1:
...
'\cs ><
!:s :)- :lI
:х:
:r
:1:
t:;
м
«1
о-
><
CLI
о-
1-0
' «1
о() о-
о
\о
....
о
Q)
==
Q) LO
Q) 1-0
<.J
o:s:
Q)
с::(
м
о
...
...
u
:1:
1"") N ..-t О.
JJпHt1VOJlOи
104
Часть 111. Естественный отбор
жение особей. у которых значения признаков близки к среднему
для данной популяции, приводит К постоянству модальных зна
чений во времени. В отличие от этоrо при направленном отборе
1IРОИСХОДИТ элиминация крайних rенетических вариантов на од-
ной стороне кривой, в результате чеrо значение средней в ряду
последовательных поколений смещается (рис. 11.1).
Праут (Prout, 1962) провел эксперимент по стаБИ '1изируюше-
му отбору у Drosophila тelanogaster. В качестве изменчивоrо
признака он использовал время развития мух. В линии, подверr
нутой стабилизирующему отбору, в каждом поколении отбирали
особей, время развития которых приближалось к среднему значе-
нию. В результате TaKoro стаБИЛИЗИРУlОщеrо отбора в ряде по-
колении дисперсия по времени развития в этой линии уменьши-
лась по сравнению с дисперсией в исходном поколении и в конт-
рольных линиях, не подверrавшихся отбору.
Стабилизирующий отбор происходит везде и всюду, хотя за
метить ero трудно; это наиболее обычный тип отбора в природе.
Мы уже ПРИВОДИ '1и в качестве яркоrо примера направленноrо от-
бора березовую пяденицу (Biston betularia) , однако при этом
ничеrо не было сказано о комплементарной роли стабилизирую
щеrо отбора в популяциях этой бабочки. До развития промыw-
ленности редкие меланистические мутанты элиминировались из
популяции стабилизирующим отбором, который, вероятно, про-
должался MHoro веков, а в измененной индустриальной среде, в
которой обитают новые ПОПУ.IJЯЦИИ, состоящие rлавным образом
из меланистав, снова действует стабилизирующий отбор, YCTpa
няя появляющиеся время от времени cBeT..lble варианты.
Дизруптивный отбор
Дизруптивный отбор (известный также как разрушающий иди
разрывающий отбор) блаrоприятствует сохранению крайних ти-
пов и элиминации промежуточных. В результате он приводит к
сохранению и усилению полиморфизма (рис. 11.1).
Рассмотрим вновь эксперимент Праута (Prout, 1962) с отбо
ром на время раЗБИТИЯ у Drosophila тelanogaster. Одна линия
дрозофилы была подверrнута дизруптивному отбору; для этоrо
в каждом ПОКОJIении отбирали мух, ВЫЛУП.IJявшихся раньше всех,
и скрещивали их с мухами, ВЫЛУП.'lявшимися позже всех. Через
ряд поколений это привело к увеличению дисперсии по вре ени
развития в сравнении с контрольными линиями, не подверrавши-
мися отбору.
Ряд экспериментов по дизруптивному отбору У D. melanogas
ter провели Тудэй и ero сотрудники. В одном из этих эксперимен
тов (Thoday, Воат, 1959), который мы здесь опишем, производи.п
ся отбор на число щетинок на теле в необычных эксперименталь-
ных условиях.
rA. 11. Типы отбора
105
Отбор производился одновременно на большое число щетинок
в rруппе «высоких» линий И на малое их число в rруппе «низких»
ЛИНИЙ. Этот ДИЗРУI1ТИВНЫЙ отбор был направлен на то, чтобы раз-
делить исходную популяцию на различные «высокие» и «НИзкие»
субпопуляции. Но «высокие» и «низкие» линии постоянно скре-
щивались между собой, так что между ними все время происхо
дил обмен rенами, действие KOToporo было направлено против
+2
t)
:t
:::s +1
O'
::,,
CpegHeeO
::s
:1'1::)
QI\.J
t:
Q,g> 1
=:I
I(t:)
......
(:"
О
30
4О
10 20
Число ПОКО/1еlfии
Рис. 11.2. Реакция на дизруптивный отбор по числу щеТJfНОК у Drosophila те-
lanogaster в условиях максимальноrо обмена rенами.
Объяснения см. в тексте. (Thoday, Boam. 1959.) I «высокая:» ивня; II «низкая ЛИRНJI .
действия дизруптивноrо отбора. Эксперимент продолжа.пся таким
образом до 36-ro поколения.
В первых поколениях между «высокими» И «низкими» линия-
ми не наблюдалось значительной диверrенции. Они начали ди-
верrировать в 14-м поколении, а после 30 ro поколения разошлись
очень сильно. На рис. 11.2 представлены кривые изменения числа
щетинок в одной «высокой» и одной «низкой» линиях, диверrи-
ровавших в результате отбора. Следует отметить, что «высокая)
И «низкая» линии В этом эксперименте находятся в совершенно
ином положении, нежели в описанном выше селекционном экспе
рименте с кукурузой (рис. 10.1); в эксперименте с кукурузой
«высокая» И «низкая» линии были ИЗОJIированы друr от друrа,
а в эксперименте с дрозофилой их скрещивали между собой. Как
106
Часть 111. Естественный отбор
показал посдедний эксперимент, дизруптивный отбор может взять
верх над потоком reHoB.
Эксперименты Стримса и Пиментела (Streams, Pimentel, 1961)
на Drosophila тelanogaster интересны количественным подходом
к взаимодействию между дизруптивным отбором и потоком reHoB.
Эти авторы использовали четыре уровня потока reHOB ----6' 20, 50
и 0% (в контрольных линиях) и дизруптивный отбор двух интен-
сивностей ---- умеренный и сильный. Сильный дизруптивный отбор
оказывал свое действие при 20 % -ном потоке reHoB, но был неэф-
фективен при 50 % -ном потоке. Что касается YMepeHHoro дизруп-
тивноrо отбора, то ero действие не моrло проявляться даже при
слабом (6%) потоке reHoB.
Более подробные сведения, касаlOщиеся экспериментов с диз-
руптивным отбором У дрозофилы, можно найти в ряде ориrиналь-
ных работ (Thoday, Воат, 1959; Millicent, Thoday, 1961; Thoday,
GiЬsол, 1970; Thoday, 1972; Streams, Pimentel, 19б 1; Soans, Pimen
tel, Soans, 1974).
Диэруптивный отбор в природе
Эффективность дизруптивноrо отбора в экспериментальных
условиях установлена достаточно хорошо. Теперь следует выяс
нить, какова ero роль в природных популяциях. Здесь мы стоим
на менее твердой почве, и нам приходится rОБОрИТЬ о некоторых
вероятностях и ВОЗМОЖНОСТЯХ, а не о бесспорных фактах.
Одна из возможных в природе ситуаций, в которых, вероятно,
вступает в действие диэруптивный отбор, возникает в тех слу
чаях, коrда хорошо дифференцированные полиморфные типы об-
ладают явным селективным преимуществом по сравнению со сла
бо дифференцированными полиморфными типами. Примером
служит половой диморфизм: самки Il самцы с хорошо дифферен-
цированными вторичными половыми признаками спариваются и
размножаются более успешно, чем различные промежуточные
типы (интерсексы и т. д.).
Вторая возможная ситуация возникает в тех случаях, коrда
полиморфная популяция занимает reTeporeHHoe местообитание.
Считается, что в таком местообитании разные типы специализи-
руются к различным нишам или субнишам и соответственно жи
вут в основном В этих специальных нишах. Так, морфа А может
обладать более высокой приспособленностью в субнише А', но
более низкой ---- в субнише В', тоrда как морфа В процветает в
субнише В', но не 8 А'. Это предполаrаемое соответствие между
разными морфами полиморфной популяции и набором субниш
в reTeporeHHoM местообитании Morпo возникнуть в результате
диэруптивноrо отбора. И наоборот, дизруптивный отбор в такоЙ
r А. 11. .Типы отбора
107
ситуации Mor бы дать возможность полиморфной популяции засе-
лить более обширный диапазон субниш в занимаемом ею место-
обитании (Levene. 1953). Э а rипотеза достаточно правдоподобна,
но твердых доказательств в ее пользу пока еще маловато.
Примером, который, по видимому, подтверждает описанную
выше модель, служит желтушка (Colias eurytheme). Самки этой
североамериканской бабочки полиморфны по окраске крыльев----
встречаются самки с оранжевыми и самки с белыми крыльями.
Этот полиморфизм обусловлен единичным reHOM.
В середине лета в нескольких местностях в Ка..1ИфОРНИИ пик
активности у белой и оранжевой форм наблюдается в разное
время дня: белая форма более активна рано утром и в позднее
послеполуденное время, а оранжевая форма ---- в полдень; по ви-
димому, эти две морфы предпочитают разную температуру и раз-
ную влажность. Кроме Toro, наблюдается и сезонная изменчивость
частоты двух цветовых морф в калифорнийских популяциях:
осенью, с понижением температуры, частота белой морфы воз-
растает (Hovanitz, 1953; Rеmiпgtоп, 1954). Как показывают фи-
зиолоrические исследования, пиrментация крыльев иrрает важ-
ную роль в реrуляции температуры у Colias eurytheтe (Watt,
1969). Таким обр.азом, создается впечатление, что полиморфизм
окраски у этоrо вида расширяет диапазон температур и продол-
жительность сезона, в пределах которых ero популяции сохра-
няют активность.
Третья ситуация, в которой проявляется действие дизруптивно-
ro отбора в природе, наблюдается у растений при некоторых осо-
бых условиях. Представим себе популяцию растений, которая, бу-
дучи прикреплена к определенному месту, свободно скрещивается
и при этом занимает участок, лежащий по обе стороны от rраницы
между двумя эколоrически различными зонами, различающимися,
например, по типу почвы или по рельефу местности. В таких
случаях у двух половин данной популяции растений нередко раз-
виваются разные комплексы приспособительных признаков.
И такая дифференциация сохраняется, несмотря на скрещивание
Между ними. Почти несомненно, что rлавным реrулирующим
фактором в подобной ситуации служит дизруптивный отбор.
Высокоrорный вид сосны Pinus albicaulis встречается в ropax
Сьерра-Невада в Калифорнии на rранице древесной раститель-
ности и непосредственно над НеЙ. Популяции, растущие на скло-
нах rop, вплоть до rраницы леса, состоят из прямостоячих де-
ревьев ---- обычная д"'Я этоrо вида форма роста. Выше rран.ицы
древесной растительности этот вид представлен низкорослым сте-
JIющимся стлаником. Древовидная и стланиковая популяции пе
реходят одна в друrую, и между ними происходит перекрестное
Опыление при помощи ветра, о чем свидетельствует наличие осо-
бей промежуточноrо типа; однако каждая из них под влиянием ди-
108
Часть 1//. Естественный отбор
BepreHTHbIx давлений отбора сохраняет соответственно в своей
субальпийской и альпийской эколоrических зонах различные
формы роста. Аналоrичные различия по обе стороны узкоrо поя-
са вдо..1Ь rраницы древесной растительности наблюдаются и у
друrих BbIcoKoropHbIX видов хвойных, а также у ив (Clausen.
1965) .
Друrая rруппа примеров. иллюстрирующих действие TaKoro
же комплекса сил, относится к пастбищным злакам. произраста-
ющим в районе свинцовых рудников в Уэльсе. В этих районах
обычная для пастбищных ,,'yroB почва. не содержащая сколько-
нибудь значительных количеств свинца. на отвалах резко пере
ходит в почву с высоким содержанием свинца. Agrostis tenuis и
друrие виды злаков образуют в этом районе непрерывные попу-
.Т]яции. область распространения которых захватывает районы с
различными типами почвы. rенетическое единство таких популя
ций поддерживается б.лаrодаря перекрестному опылению при
помощи ветра. Тем не менее между субпопу.I1ЯЦИЯМИ. растущими
на пастбищных почвах и не переносящиыи свинец, и толерантны
МИ к свинцу субпопуляциями, растущими на отвалах. возникает
с.лабая расовая диверrенция (J ain, Bradshaw. 1966; Antonovics.
1971) .
Уравновешивающий отбор
rетерозиrоты нередко превосходят соответствующие типы
rомозиrот по общей мощности и..1И по тому или иному компонен-
ту жизнеспособности, например по способности к конкуренции
или по устойчивости к заболеваниям. Таким превосходством o-
rYT об.ладать rенотипы, rетерозиrотные по единичному rCHY или
по uew'JoM)' блоку reHOB.
Если rетерозиrота Аа обладает селективным преимуществом
по сравнению с одной иди с обеими rомозиrотами, то отбор спо
собствует сохранению в популяции обоих аллелей (А и а). В re
нофонде создается равновесная частота аллелей А и а. точный
уровень которой определяется относительными селективными
ценностями альтернативных типов. Но до тех пор. пока. частоты
алле.лей контролируются отбором, ни аллель А. ни аллель а не
элиминируются. В популяции поддерживается состояние сбалан.
сированноrо полиморфизма.
Отбор. блаrоприятствующий rетерозиrотам. известен под на
званием уравновешиваroщеrо отбора (а также превосходства илli
преимущества rетерозиrот).
При уравновешивающем отборе не происходит полноrо закреп-
ления или элиминации альтернативных аллелей или блоков reHoB
как бы.ло описано выше; единицей. которой блаrоприятствует от.
бор, в данном случае оказывается rетерозиrотная пара а.л.лелеА
r А. 11. Типы отбора
109
или сочетание reHoB. Это ИМеет известное отношение к вопросу
об ОЖИДаемых результатах направленноrо отбора в сбалансиро-
ванной полиморфной системе. Полаrают, что направленный отбор
в сочетании с уравновешивающим приводит не к замене в reHo
фонде одноrо reHa или блока reHoB друrим, а к .ряду переходов от
одной rетерозиrотной комбинации к друrой, например А lA2
А2Аз АзА4 (Lerner, 1954, стр. 113 114).
Сбалансированный полиморфизм, основанный на превосходст-
не rетерозиrот, явление, которое распространено довольно ши
роко. Среди животных оно наблюдается, например, у прямокры-
лых. кур, мышей, человека и разных видов дрозофилы; среди рас-
тений оно обнаружено у кукурузы (Zea тays), ячменя (Hordeuт
sativuт), Arabidopsis и Oenothera. Наиболее подробно превосход
ство rетерозиrот изучено у Drosophila pseudoobscura, на примере
которой оно и будет описано в следующем разделе.
Превосходство rетерозиrот
у Drosophila pseudoobscura
.'
Дикий североа lериканский вид D. pseudoobscura известен
-своей изменчивостью по хромосоме 111. У Э10rо вида (как OTMe
ч.алось в rл. 4) описано 16 типов хромосомы 111, отличаlOЩИХСЯ
друr от Apyra по инверсиям. Все типы инверсий, выявляемые ци-
толоrически в хромосомах слюнных желез .пичинок, получили
названия (Стандарт, Эрроухед, Чирикауа, Тимберлайн, Пайкс
Лик и т. д.) И буквенные обозначения (ST, AR, СН, TL, рр и
Т. д.). Б6.lJьшая часть популяций полиморфна по некоторым типам
-инверсий, и в них встречаются различные возможные rомозиrоты
и rетероз иrоты.
Инвертированные участки хромосомы различаются как по
одержащимся в них reHaM, так и по своим цитолоrичсским при-
знакам. Популяция, по..1иморфная по инверсиям ST, СН и AR,
содержит Не только хромосомы трех разных типов, но и три раз-
.ных набора reHoB в инвертированных участках; поэтому rенотипы
-БТ/СН, ST/AR и CH/AR rетерозиrотны не только по инверсиям,
но и по reHaM. Но инверсии предотвращают эффективную реком-
'бинацию rCHoB в rетерозиrотах по инверсиям. Следовательно,
полиморфизм по инверсиям у D. pseudoobscura одновременно пред-
ставляет собой полиморфизм по блокам reHoB, которые остают-
-ся интактными из поколения в поколение.
Добржанский и ero шкода изучали полиморфизм по инверси
.ям в природных популяциях D. pseudoobscura по всему ареалу
этоrо вида. Наиболее обширные выборки в течение мноrих лет
. отбира.пи из некоторых популяций Сьерра-Невады и Сан-Джа
.синто (Калифорния). Кроме Toro, проводились мноrочисленные
.лабораторны-е .эксперименты в популяционных ящиках с линиями
110
Часть 1//. Естествен.н.ы.й отбор
мух, отловленных в тех же самых природиых популяциях в Ка-
лифорнии.
В этих калифорнийских ropHblX ПОПУЛЯЦИЯХ обычно встречают-
ся инверсии 8Т, СН и AR; встречаются также, но с низкой часто-
той, инверсии TL, рр и 8С. Нередко частота rетерозиrот по ин-
версиям бывает выше той, которую следовало бы ожидать на
основании равновесия Харди Вайнберrа. Действительную часто.
ту различных типов инверсий в raMeTHoM фонде определяют соот-
ветствующими методами взятия выборок из популяции; на осно-
вании полученных данных по формуле Харди Вайнберrа вы.
числяют ожидаемую частоту rетерозиrотных rенотипов. Затем эту
ожидаемую частоту сопоставляют с действительной частотой re.
терозиrот по инверсиям. При этом в определенное время rода в
природных популяциях постоянно обнаруживается статистически
значимый избыток rетерозиrот.
Так, в выборках, взятых в мае 1952 r. в ropax Сан Джасинто
из одной популяции, полиморфной по пяти инверсиям, были об-
наружены значительные различия между фактическими и ожи-
даемыми частотами rетерозиrот и rомозиrот по инверсиям
(табл. 11.1). Данные табл. 11.1 показывают, что наблюдается ус-
тойчивый избыток rетерозиrот и соответствующий недостаток ro-
мозиrот по сравнению с тем, чеrо следовало бы ожидать, исходя
из формулы Харди Вайнберrа.
1 аБАuца 11.1
Наблю.цаемые и ожидаемые частоты rетерозиrот
и rомозиrот по инверсиям в хромосоме 111
(.цанные Ерliпg, Mltchell, Mattoni, 1953)
I Наблюдаемые I Ожидаемые
частоты частоты
Твп
51/СН
5T/AR
Все rетерозиrоты
51/5Т
Все rомозиrоты
0,232
0,232
О ,830
0,134
О, 170
о , 168
0,168
0,724
0.184
0,276
.
Наблюдаемые отклонения от равновесия Харди Вайнберrа
можно объяснить на основании отбора в пользу rетерозиrот.
Один из способов проверки этой rипотезы состоит в том, чтобы
сравнить частоты rетерозиrот и rомозиrот по инверсиям в выборке
яиц, взятых из природной популяции, С ИХ частотами в выборке
взрослых мух. При этом выяснилось, что соотношение j'етерози-
rOTHblX и rомоЗиrотных rенотипов в выборках яиц соответствует
iформуле Харди ---- Вайнберrа. Недостаток rомозиrот в выборках
взрослых особей, вероятно, вызывается дифференциальной смерт-
r А. 11. Типы отбора
111
ностью В период развития от яйца до взрослых стадий, действую-
щей в пользу /rетерозиrот (Dobzhansky, Levene, 1951).
Параллельные данные о превосходстве rетеРОЗИfОТ были по
JJучены для искусственных популяций, выращенных в Полуляци
онных ящиках. Основателями этих искусственных популяций
служили линии мух, взятые из ПРИРОДНЫХ популяций Сан Джа-
синто и несущие хромосомы ST, СН и AR. Популяционные ящики
содержали в тепле и допускали возникновение в них перенасе
ленности. На стадии яйца соотношение rомозиrот и rетерозиrот
по инверсиям соответствует формуле Харди ---- Вайнберrа. Однако
на стадии взрослых особей в тех же самых искусственных попу
ляциях наблюдался достоверный избыток rетерозиrот по Инвер
еиям (Dobzhansky, 1947а).
Изучение развития и поведения мух показало, что более BЫ
сокую приспособленность rетерозиrот по инверсиям можно раз-
ложить на ряд раз.пичных компонентов: более высокая жизнеспо-
собность на стадиях, предшествующих достижению половой зре-
лости; более высокая скорость развития; большая продолжитель.
ность жизни, плодовитость И быстрота нахождения брачноrо
партнера (Moos, 1955; Dobzhansky, 1970, стр. 137----138).
Следует снова подчеркнуть, что превосходство rетерозиrот
проявляется только при определенных температуре, корме и плот-
ности популяции; в отсутствие нужных условий, будь то природ-
ные или искусственные популяции, rетерозиrоты по инверсиям
теряют свое селективное преимущество перед rомозиrотами.
Важно отметить, что природные популяции периодически дей-
ствительно сталкиваются с условиями среды, приводящими к
проявлениlO превосходства rетерозиrот. В этих популяциях если
не постоянно, то ВО всяком случае с перерывами действует урав-
новешивающий отбор. TaKoro прерывистоrо действия достаточно
для постоянноrо сохранения сбалансированноrо полиморфизма.
В прохладную поrоду в начале лета (май и июнь) в природных
популяциях возрастает частота хромосом AR и СН, а в жаркую
летнюю поrоду и осенью (с июля до сентября или октября) воз-
растает частота хромосом ST, однако эти иэменения никоrда не
доходят до полноrо закрепления или элиминации (Dobzhansky,
1943, 1947Ь, 1948). Таким образом, уравновешивающий отбор
способствует расширению периода в течение rода, на протяже-
нии KOToporo может существовать популяция, по сравнению с тем
периодом, в течение KOToporo она, вероятно, существовала бы,
будучи мономорфной.
Половой отбор
Во мноrих rруппах животных существует ясно выраженный
половой диморфизм. Этот диморфизм обычно заключается в раэ.
витии у самцов оrраниченных полом признаков признаков, ко-
112
Часть //l. Естественный отбор
торые лишь косвенно связаны с размножением. Такие вторичные
половые признаки можно разделить на два обширных кдасса:
!) признаки, относящиеся к размерам, силе и разноrо рода при
.1.аткам (например, более крупные размеры самцов у морских
львов или pora у самцов оленей); 2) украшения и демонстраци-
онное поведение (например, яркое оперение селезней, яркие lIЯТ-
на на шее у самцов колибри, особые типы песен, брачное поведе-
ние). .
В «Происхождении человека и половом отборе» Дарвин (1871)
рассматривает известные в ero время примеры вторичных поло
вых признаков у самцов животных, принадлежащих f( BeCb\fa
широкому Kpyry rрупп, и предлаrает для их объяснения теорию
ПОJlOвоrо отбора.
Дарвин представлял себе половой отбор как процесс, допол-
няющий более общий и более широко распространенный процесс
eCTecTBeHHoro отбора. Последний, по словам Дарвина, создал
адаптивные признаки данноrо вида в целом, в том числе извечные
приспособления, общие для обоих полов, а также первичные по-
ловые различия, непосредственно связанные с размножением.
Коrда все эти общие признаки получили объяснение на основе
eCTecTBeHHoro отбора в первоначальном даРВИНОВСКО 1 смысле
слова, оказалось, что за рамками этоrо объяснения остается об-
ширный класс специальных признаков, свойственных только
самцам. Такие вторичные ПО..10вые признаки, как pora оленя или
яркое оперение селезней, не представляют собой приспособлений,
блаrоприятных для вида в целом; не являются они также необ
ходимыми и для размножения. Однако можно предполаrать, что
эти признаки повышают вероятность успешноrо спаривания для
тех самцов, которые ими обладают. Теория половоrо отбора была
введена для Toro, чтобы объяснить развитие таких специальных
мужских признаков.
Дарвин описывает процесс, предложенный им в качестве объ
яснения, следующим образом (Dаrwiп, 1871): «... ПО.ТIOвой отбор...
зависит от тех преимуществ, исключительно по отношению к
воспроизведению, которыми обладают известные особи над дpy
rими особями Toro же пола и вида». Далее он объясняет (я здесь
несколько перефразировал утверждение Дарвина, сохранив ero
терминолоrию): коrда оба пода ведут совершенно одинаковый
образ жизни, но у самца opraHbl чувств и передвижения значи
тельно более развиты, чем у самки, то возможно, что ca:VluaM
нередко удавалось приобрести эти признаки не потому, что они
лучше приспособлены к выживанию в борьбе за сущест-
вование, а потому, что они обладают известными преимущсствами
над друrими самцами при размножении. «В подобных случаях
должен был вступить в действие половой отбор».
r л. 11. Типы отбора
113
История развития теории половоrо отбора со времени CBoero
возникновения в 1871 r. шла извилистым путем. Она вызывала
споры еще во времена Дарвина. Затем эта тема была предана
забвению. Коrда в период новейших эволюционных исследований
интерес к ней возродился, она предстала в новом контексте , в
котором те же самые ЯВ.'1ения рассматривались с иных альтерна
тивных точек зрения. ПО.'lОВОЙ отбор снова стал одной из проблем,
вызывающих разноrласия , пусть не очень резкие. При подобном
положении дел трудно, а по видимому, даже 11 невозможно оп.
ределить роль и значение половоrо отбора хоть с какой.то сте.
пенью точности.
Никаких особых теоретических проблем, касающихся лоловоrо
отбора, не существует. Половой отбор одна из нескольких
форм eCTecTBeHHoro отбора (а не некий альтернативный процесс,
каким ero представляли в системе концепций XIX в.). Точнее,
это некая форма отбора, опять-таки одна из нескольких, при ко-
70рОЙ селективному действию подверrаются конкурирующие чле-
ны одной из подrрупп данной популяции. В данном случае под-
rруппа это особи мужскоrо пола, а признаки, лодверrаемые OT
бору, это те признаки, которые влияют на успех спаривания.
Серьезная трудность возникает в связи с тем, что вторичные
половыс признаки образуют весьма разнородную совокупность.
Они выполняют разнdобразные функции в жизни популяции. Их
образование нельзя приписать какой либо одной форме отбора.
Рассматривая различия в размерах между особями разноrо пола,
следует иметь в виду не только половой отбор, но и отбор, спо-
собствующий эколоrической диверrенции (см. часть V). Разноrо
рода атрибуты самцов MorYT сдужить как для занятия территории
или достижения доминирующсrо положения в иерархической
структуре сообщества, так и для завоевания самок. Различные
украшения самцов, их демонстрационное поведенис, пение и за
пахи служат для стимуляции самок в процессе ухаживания, но
одновременно представляют собой видоспецифичные опознава
тельные признаки. В последнем случае они моrли быть созданы
и в результате отбора, направлснноrо на репродуктивную изоля-
цию (см. часть V), и в рсзультате ПО.rJовоrо отбора. Таким обра
зом, здесь мы имеем дело с действием целой смеси селективных
процессов, а при таких условиях трудно выявить эффекты одноrо
..1ишь половоrо отбора.
Одна rруппа случаев, в которых половой отбор определенно
иrрает важную роль, это млекопитающие и птицы с полиrамной
системой размножения. При полиrамии самые сильные самцы
собирают BOKpyr себя целый [арем из самок и охраняют ero или
ту территорию, на которой находятся их самкИ, отrоняя своих бо-
лее слабых собратьев; в результате последнИМ достается мало
самок или не достается их вовсе.
tl4
Часть l//. Естественный отбор
Мноrие млекопитающие и некоторые птицы полиrамны. Среди
млекопитающих полиrамия характерна Д.ТIЯ оленей, крупноrо po
..-атоrо скота, баранов, для большинства антилоп, для лонов, тю
леней, морских львов, моржей и павианов; среди птиц для кур,
4>азанов и павлинов. У самцов этих животных хорошо развиты
вторичные половые признаки, отсутствующие у родственных
.rрупп, которым несвойственна полиrамия.
Так, у полиtамных кур, фазанов и павлинов самцы заметно
.крупнее, более драчливы и имеют rораздо более нарядное опере
иие, чем самки, тоrда как у монотамных тетерева, кеклика и тунд-
ряной куропатки различия между особями разноrо пола относи
тельно невелики. У моржей и морских львов самцы очень круп
,ные; у мноrих копытных они украшены роrами, а у павианов они
.крупные и аrрессивные. В отличие от этоrо у MOHoraMHblx волков
и у некоторых монотамных видов обезьян, у представителей ce
мейств кошачьих, у которых детенышей выращивает в основном
мать, и у колониальных, но неполиrамных rрызунов самцы и
самки имеют примерно одинаковые размеры и силу (Dапvin,
1871, rл. 8).
rейст (Geist , 1971) тщательно изучал репродуктивное пове-
дение снежноrо барана (Ovis canadensis) в природе и в не воле.
у половозрелых самцов, как это хорошо известно, имеются БОJ1Ь-
шие тяжелые pora и кости черепа сильно утолщены. В период
rOHa между самцами происходят поединки за обладание самками.
Поединки передко бывают очень жестокими и побежденные жи-
вотные получают серьезные раны, а иноrда и rибнут. Эти наблю
дения над поведением баранов и друrих жвачных во время rOHa
не соответствуют общепринятым представлениям о том, что брач-
ное поведение самцов обычно сводится к безобидному блефу и
друrоrо рода демонстрациям. В поединках баранов, беЗУСJЮВНО,
присутствуют элементы, демонстрирующие их притязания на оп
ределенное место в иерархии, но эта демонстрация в случае He
обходимости подкрепляется дракой.
Самцы, вышедшие победителями из поединков, занимают дo
минирующее положение в иерархической структуре cBoero стада.
Они отrоняlOТ друrих самцов от самок, но сами свободно спа
риваются с самками. Система скрещивания у снежноrо барана
Не исключает иекотороrо с'паривания между самцами и самками,
занимающими подчиненное положение, но она обеспечивает преи
мущественное участие в размножении доминантных самцов. По-
этому последние, вероятно, производят больше яrнят, чем самцы,
занимающие подчиненное положение. Отсюда следует, что неко-
торые признаки poroB и черепа у самцов снежноrо барана, а так-
же их навыки нападения и защиты в битвах друr с друrом, ло
видимому, возникли в результате половоrо отбора (см. G.eist,
1971 ) .
r л. 11. Типы отбора
115
Что касается самцов неполиrамных rрупп, то объяснить про-
исхождение у них связанных с брачным поведением признаков
при помощи n0,,10BOrO отбора rораздо труднее. Это не означает,
что половой отбор, хотя бы в некоторых случ. ях, не имеет к ним
отношения. Развитие демонстрационных признаков у самцов в
некоторых неПО,,1иrамных rруппах может быть реЗУ..1ьтатом сов-
MecTHoro действия обычноrо индивидуальноrо отбора и ПОЛОВоrо
отбора. Развитию таких признаков на начальных с.тздиях Может
способствовать направленный отбор, блаrоприятствуя ухажива-
нию и спариванию; а в дальнейшем под действием половоrо от-
бора в результате преимущественноrо спаривания тех самцов, у
которых эти признаки развиты сильнее, последние MorYT достиr
нуть крайнеrо выражения (Nlayr, 1972).
Отбор на субиндивидуумных уровнях
Все обсуждавшиеся до сих пор типы отбора происходят на
уровне особи, и в сущности отбор в основном совершается именно
на этом уровне орrанизации. Однако селективные процессы MorYT
такЖе проводить различие между воспроизводимыми биолоrиче-
скими единицами, находящимися на более низкой ступени иерар-
хии биолоrической орrанизации. В этом разделе мы познакомим
читателя с несколькими типами субиндивидуумноrо отбора.
у цветковых растений мужские raMeTbI находятся в независи-
мой структуре мужском rаметофите, состоящем из пыльцевоrо
зерна и пыльцевой трубки. fетерозиrотное растение производит
rенетически различные классы пыльцы, и в некоторых случаях
среди пыльцевых зерен происходит расщепление по rенетическим
факторам, определяющим жизнеспособность пыльцы, ее способ-
ность к прорастанию или скорость роста пыльцевых трубок. Кроме
Toro, пыльца обычно продуцируется и попадает на рыльце в боль-
шем количестве, чем это необходимо Д.1JЯ оплодотворения, что BЫ
зывает конкуренцию между пыльцевыми зернами или пыльцевы-
ми трубками.
Такая конкуренция в тех случаях, коrда пыльца расщепляет-
ся по факторам роста, приводит к тому, что. некоторые классы
мужских raMeT оказываются более эффективными в смысле оп-
лодотворения, чем друrие, а следовательно, и чаще участвуют в
образовании зародышей или эндосперма. Если с фактором роста
пыльцы сцеплен какой-либо reh-маркер, определяющий видимый
морфолоrический признак, то в таком случае вмесТо ожидаемоrо
менделевскоrо соотношения по этому reHY наблюдается изменен
ное соотношение. Морфолоrический тип, детерминируемый марки-
рующим аллелем, порожденным более эффективной ПЫО'1ЬЦОЙ" 1
присутствует в поколении семян или проростков В избытке, при
недостатке противоположноrо морфолоrическоrо типа.
116
Часть J 1 J. ЕстеСТ8ен.н.ыЛ отбор
Изменение соотношения разных типов при расщеплении, обус-
.лов.пеНJlое отбором, направленным против некоторых классов
пы.fJЬЦЫ, хорошо известно у ряда видов цветковых растений. Xo
рошо проанализирован случай сцепленных reHoB: Su (определя-
ющеro тип эндосперма) и Оа (контро.fJИРУIOщеrо рост пыльцевой
.трубки) у кукурузы (Zea тays). rетерозиrоты Su/su обычно дают
зерна, расщепляющиеся по характеру эндосперма на два класса
(крахмалистый. и сахаристый эндосперм) в соотношении, соот-
ветствующем закону Менделя. Но если аЛ.,lе.пь su (сахаристый
эндосперм) сцеплен саллелем ga (медленный рост пыльцевой
.трубки) в rетерозиrоте SuGa/suga и такая rетерозиrота ИСПО.'1ь-
.зуется в качестве отцовской особи, так что сцепленный участок
.su ga передается через пыльцу, то в следующем поколении среди
.зерен наб.llюдается явная нехватка сахаристоrо типа (i\r\angels
.dorf, Jones, 1926).
Некоторые rенетически контролируемые отклонения мейоза у
Drosophila тelanogaster приводят к тому, что одна из хромосом
rомолоrичной пары вкточаетсS:l в более эффективно функциониру-
ющую сперму, чем друrая. Нарушенное расщепление хромосом
в мейозе ведет к повышению частоты хромосомы одноr.о типа R
.raMeTHoM фонде по сравнению с частотой ее rOMO.,lora. Этот про-
цесс известен под назвnнием мейотическоrо драйва.
Подобные же изменения частоты происходят у reHoB, которые
несут эти две rомолоrичные хромосомы, и у фенотипов, которые
детерминируются этими rенами. В одном случае соотношение
полов изменяется в сторону избытка самок, в друrом"""":' происхо-
дит повышеНие частоты одной рецессивной летали (Sandler, No.
vitski, 1957; Hiraizumi, Sand'loer, Crow, 1960).
Мейотический драйв это в сущности процесс дифференци.
альноrо размножения rомолоrичных XpOMOCO . Таким образом,
он, по видимому, представляет собой некую форму межхромосом-
Horo отбора.
ОДНОJ10кусная модель отбора была введена в rл. 8 как полез....
ная абстракция, как переходная ступень на пути к пониманию
процессов отбора в настоящих популяциях. Лоrически необходи-
мо, однако, предположить, что процесс отбора по одному reHY
действительно происходил на очень ранних стадиях ЭВОЛЮI{ИИ,
коrда преоб.'1адающими единицами биолоrической орrанизации
были rеноподобные частицы. Процессы, очень близкие к отбору
на уровне reHa, происходят и до сих пор при дифференциальном
размножении вирусных частиц или бактериальных клеток, раз-'
личающихся по одному вирусному или бактериальному reHY.
Междемовый отбор
Перейдем теперь к вопросу об отборе на уровне локальной
популяции или к междемовому отбору. Междемовый отбор пред-
r А. 11. Типы отбора
117
станляет собой дифференциальное размножение разных локаль-
1IЫХ попу..1ЯЦИЙ.
Здесь необходимо сделать небольшое отступление, 'касающееся
терминолоrии. «N\еждемовый отбор» общий термин. Одним из
ero синонимов является термин «популяционный отбор». Термин
<trрупповой отбор» также иноrда используют в качестве сИнонима
междемовоrо отбора, однако здесь мы cro будем применять толь
ко К особому случаю междемовоrо отбора, коrда популяции, под-
верrающиеся отбору, представляют собой сообщества. POДCTBeH
ный отбор, как указывает самый смысл слова, это отбор, осно-
ванный на родстве; он представляет собой еще более специальный
тип отбора. Родственный отбор тесно смыкается с rрупповым
отбором в тех случаях, коrда, как это часто бывает, сообщество
образовано родственными особями.
В этом разделе мы рассмотрим междеМQВЫЙ отбор в общем
виде, а затем упомянем о некоторых вероятных при мерах rруп
повоrо отбора. К rрупповому отбору мы еще раз вернемся в
rЛ.37.
Биолоrи-популяционисты далеки от единодушия в вопросе о
TO I, существует .'1и на самом деле междемовый отбор. Уильямс
(Wiiliams, 1966) и неК'оторые друrие aiBTopbl считают, что нет
никакой необходимости постулировать процесс междемовоrо от-
бора, поскольку несколько модифицированные формы отбора на
уровне особей способны вызывать эффекты, приписываемые попу
ляционному отбору. Райт (=Wright, 1931, 1956, ]960) и друrие
исследователи, в том числе и автор данной книrи, полаrают, что
междемовый отбор реа 'IЬНЫЙ процесс, происходящий в природс
при некоторых ситуациях. ПРОТИВОПО.'10жные точки зрения из..lО
жены Уильямсом (\Villiams, 1971) и Александером (Alexander,
1974) .
Райт (\Vright, 193], 1960) сравнивает попу.rтяционные системы
двух типов большую непрерывную ПОПУ..1яцию и ряд меЛI{ИХ
полуизолированных КО..10НИЙ В отношении теоретической эффек
ТИВIЮСТИ отбора. Предполаrается, что общая величина (N) обеих
популяционных систем одинакова и что орrанизмы свободно
скрещиваются между собой. .
В большой непрерывной ПОПУ..1яции отбор относительно неэф..
фективен в смысле повышения частоты блаrоприятных, 1:10 peд
ких рецессивных :мутаций. Кроме Toro, любой тенденции к по:
вышению частоты какоrо либо б",аrоприятноrо ал..1СЛЯ воднои
части данной обширной популяции противодействует скрещивание
с соседними субпопуляциями, в которых этот аллель редок. По
добным же образом б.паrоприятные новые reHHbIe сочетания, ко-
о 1'орым удалось образоваться в какой нибудь локальной доле дан-
ной популяции, разбиваются на части и элиминируются в резуль
тате с рещивания с особями соседних долей.
118
Часть /11. Естественный отбор
Все эти ТРУДНОСТИ в значительной степени устраняютс в по-
пуляционной системе, по своей структуре напоминающеи ряд
отдельных островков. Здесь отбор или отбор совместно с дрей-
фом reHOB может быстро и эффективно повысить частоту KaKoro-
либо редкоrо блаrоприятноrо аллеля в одной или нескольких
мелких колониях. Новые блаrоприятные сочетания тенов также
MorYT леrко закрепиться в одной или нескольких мелких колони-
ях. Изоляция з"ащищаст rенофонды этих колоний от «затоп..1е..
ния» В результате миrрации из друrих, не обладающих столь
блаrоприятными rенами колоний, и от скрещивания с ними. До
сих пор эта модель обходится ОДНИ 1 лишь индивидуальным от-
бором или для некоторых колоний индивидуальным отбором
в сочетании с дрейфом reHoB.
Допустим теперь, что среда, в которой находится данная по..
пуляционная система, изменилась, в результате чеrо приспособ-
ленность прежних rенотипов понизил ась. В новой среде новые
блаrоприятные reHbI или сочетания reHoB, закрепившиеся в неко-
торых колониях, обладают высокой потенциальной приспособи
тельной ценностью для популяционной системы в целом. Теперь
имеются все .условия для Toro, чтобы вступил в действие междемо
вый отбор. Менее приспособленные колонии постепенно сокраща-
ются и вымирают, а колонии, которые более приспособлены, рас-
ширяются и замещают их по всей области, занимаемой данноit
популяционной системой. Такая подразделившаяся популяцион-
ная система приобретает новый набор приспособительных при..
знаков в результате индивидуальноrо отбора в пределах HeKOTO
рых колоний, за которым следуеr дифференциальное размноже-
ние разных колоний.
Орrанический мир обладает рядом черт, которые трудно объ-
яснить на основе индивидуальноrо отбора, но которым леrко дат!.
объяснение, если рассматривать их как результат действия меж-
демовоrо отбора. Первая из них половое размножение, блаrо..
приятное для будуIЦИХ поколений данной популяции, но не для
самих родительских типов. Вторая черта..... альтруистическое по-
ведение у общественных животных, идущее на пользу сообщест-
ву, но нередко поДверrающее опасности ту особь, которая er()
проявляет. Третья черта:"" приспособительные признаки стериль-
ной касты рабочих у общественных насекомых.
Тип отбора, обусловливающий ЭВОЛlоцию пола, усиленно об-
су дался на протяжении мноrих лет и в настоящее время вновь.
привлек к себе вНимание. Некоторые эволюционисты, в том чис-
ле и автор данной книrи, склоняются к мысли, что междемовый.
или межвидовой отбор иrрает важную роль в эволюции пола_
Подобное мнение основано на том, что половое размножение дает
преимущества популяции в будущих поколениях, создавая запас
комбинативной изменчивости, но в процессе рекомбинации роди
r л. 11. Типы. отбора
119
тельские rенотипы разрушаются. Однако в проведенных недавно
модельных экспериментах было показано, что индивидуальный
отбор может блаrоприятствовать половому размножению при He
которых допускаемых условиях (WilHams, 1975; Fеlsепstеiп, у 0-
koyama, 1976). Но вопрос о том, следует ли считать развитие по-
ла в результате индивидуальноrо отбора вероятным процессом
или же это требует слишком большой натяжки, лежит в иной
плоскости и остается пока открытым.
Одна из проблем, касающаяся альтруистическоrо поведения.
.состоит 8 том, чтобы разделить селективное преимущество по
ведения TaKoro типа на ero значение для отдельной особи и для
сообщества в целом. Рассмотрим такую полезную для сообщест
ва черту поведения общественноrо животноrо, как защитные
действия против BparoB. Какая доля этой черты обусловлена
максимальным успехом в размножении отдельных обладающих
этой чертой особей, а какая доля, если таковая и меется,---- повы-
шенным успехом в размножении данноrо сообщества? У общест
венных животных трудно выделить отдельную особь и ее успех
или неудачу в размножении из Bcero сообщества, поскольку то,
что блаrоприятно д..1Я особи, часто бывает блаrоприятно для всей
rруппы и наоборот. Самому мне кажется, что альтруистическое
поведение частично возникло в результате междемовоrо отбора.
В настоящее время принято ПРИПliсывать альтруизм родствен-
ному отбору. Однако это не означает безусловноrо . иск..llОчения
междеМО80rо отбора, как иноrда думают, поскольку родственный
.отбор во мноrих реальных ситуациях очень близок к междемово.
му отбору, о чем уже rоворилось ранее.
В качестве примера общественных нзсеI{ОМЫХ рассмотрим ме-
доносную пче.'IУ (Apis тellifica). Пчелиный у'" ей или колония
представляет собой популяцию, состоящую из самок (матки),
самцов (трутни) и бесполых особей (рабочие). Матки и трутни
фертильны, но бесполезны в экономическом отношении.
Важные функции добывания пищи, защиты и поддержания
колонии и выращивания молоди выполняют рабочие особи, обла
дающие соматическими и ловсденческими адаптациями для BЫ
полнения этих функций. Носители rлавных приспособлений. бла-
rодаря которым колония существует, это рабочие особи. Между
тем рабочие особи, будучи бесполыми, не размножаются как та.
ковые, а поэтому не имеют возможности передавать reHbl, опре.
де 1]яющие их приспособите '1ьные черты. следующим поколениям.
Эту задачу выполняют матки и Tpyrпll. Следовательно, если бы
матки и трутни не несли reHoB, которые l)еспечивают развитие
умелых и прилежных рабочих пчел, то улей не Mor бы процве-
тать, а оказался бы элиминированным 8 результате конкуренции
с друrими ульями, содержащими лучше приспособленных рабочих
особей. Подобный процесс замещения одной линии пчелиных
120
Часть ///. Естественный отбор
семей друrой действитеJ1ЬНО наблюдался за последние rоды в
разных частях земноrо шара (см. Michener, 1975). Создается чет-
кое впечатление, что единицей отбора в этом случае служит ко-
лония как целое (см. Dаrwiп, 1872, а также Grant, 1963).
Общественные пчелы представляют собой один из примеров
крайней интеrрированности сообщества в мире животных. И эф-
фекты междемовоrо отбора выражены у них очень ярко. Отсюда
следует, что у животных с менее высоко интеrрированной СТРУК-
турой сообщества, как, например, у мноrих М.l1екопитающих и
птиц, эффекты междемовоrо отбора ПРОЯВ.'Iяются, вероятно, не
столь сильно, хотя некоторые их признаки все же MorYT быть ре-
зультатом отбора этоrо типа.
Отбор ба уровне вида
На видовом уровне биолоrической орrанизации действует не-
сколько типов отбора: 1) отбор, приводящий к эколоrической изо-
ляции; 2) отбор, приводящий к репродуктивной изоляции; 3) за-
мещение видов. Эти типы отбора упоминаются здесь лишь для
Toro, чтобы придать законченность нашему обзору; они обсужда-
ются в части У, посвященной явлениям видообразования.
rЛЗlВа 12
ЭФФЕКТЫ ВЗАИМОДЕйСТВИЯ ОСОБЕй
Конкуренция
На процесс отбора оказывают влияние разноrо рода взаимо
действия между особями. Наиболее обычное из этих взаимодей-
ствий ---- конкуренция за необходимое сырье, за исто ники энерrии
или жизненное пространство. Естественный отбор действует наи
более эффективно в условиях конкуренции, и, напротив, в отсут-
ствие конкуренции давление отбора ослабевает.
Это обобщение подтверждают резу.7Jьтаты мноrочисленных ce
лекционных экспериментов, проведенных на Drosophila pseudo
obscura. СедеКТИВIIЫС преимущества различных rОМОЗиrот и reTe
розиrот по инверсиям (STjST, STjCH, STjAR и т. п.) проявляют
ся ТО..1ЬКО В перенаселенных популяционных ящиках и не прояв
ляются в мадонаселенных (см. rл. 9).
Вероятно, было бы правильным сказаcrь, что отбор может про-
исходить в отсутствие конкуренции, особенно в тех случаях, коrда
селективными СИ.пами служат физические факторы среды, но что
эффективность отбора rораздо выше при наличии конкуренции.
Сукачев (1928) сравнивал жизнеспособности трех линий оду-
ванчика (Taraxacuт officinale) , размножающихся бесполым пу
тем, на экспериментальных делянках ПОД Ленинrрадом. Линии
были обозначены как биотипы А, В и С. Они выращивались в
чистых открытых насаждениях (расстояния между растениями
18 см), в чистых сомкнутых насаждениях (расстояние между
растениями 3 см) и в смешанных насаждениях (А, В и С). Естест-
венную rибель растений в различных насаждениях измеряли по
прошествии двух лет.
Нас здесь интересуют чистые насаждения двух типов. Жизне-
способность трех ЛИRИЙ, выраженная как проuент особей, вы-
живших к концу двухлетнеrо периода, представлена в табл. 12.1.
Линия С оказалась наиболее жизнеспособной в открытых насаж-
дениях. Однако в сомкнутых насаждениях, т. е. в условиях кон.
куренции, линия В превосходил а две друrие. По видимоМ'у, reHo-
типически детерминированная способность выдерживать воздей-
ствия физических факторов среды отличается от способности По-
бедить при сильной конкуренции (Сукачев. 1928).
Аналоrичные эксперименты, проведенные в дальнейшем на
растениях и на насекомых, привели к созданиЮ концепции спо-
собности к конкуренции как oc060ro свойства орrанизма, которое
контролируется rенетически. У ячменя (Hordeum sativum) и у
122
ЧlJC7Ь JJJ. ECTeCTBeHHlJlzJ отбор
Та6Аица 12.1
Относительная 1Кизнеспособность трех пиниА оду-
ванчика в чистых насаQениях (Сукачев, 1928)
Выжнвавие. "
лlllUUl открытое COMKнyroe
насаждекие насаждение
А 77 27
В 69 49
С 90 24
мучных хрущаков (Triboliuт caslaneuт и Т. confusuт) разные
rенотипы различаются по степени конкурентоспособности (Sakai,
Gotoh, 1955; Lerner, Но, 1961). Эксперименты, проведенные на
ячмене, показываlOТ, что конкурентоспособность и общая мощ-
ность разные признаки.
у высших позвоночных способность к конкуренции нередко
принимает форму аrрессивноrо поведения. Аrрессивное поведение
обычно сочетается с конкуренцией, выражается более ярко, а
иноrда и ИСКЛIочительно в условиях скученности и представляет
собой один из способов преодоления конкуренции (Wilson, 1971).
Смеси rенотипов в условиях конкуренции
Конкурентоспособность данноrо rенотипа нередко проявляется
в очень разной степени в чистом сомкнутом и в смешанном сом-
кнутом насаждении. У пшеницы (Triticuт vиlgare) и у овса
(Avena sativa) при совместном выращивании двух сортов в ус-
ловиях конкуренции число растений при сборе урожая и общая
урожайность оказались выше, чем при выращивании каждоrо
сорта по отдельности (Gustafsson, 1951). Ана.тюrичным образом
эксперимента!lьные популяции Drosophila pseudoobscиra , поли-
морфные по инверсиям, превосходят мономорфные популяции по
общей биомассе мух, развившихся на данном количестве корма
(Beardmore, Dobzhansky, Pavlovsky, 1960).
Кроме Toro, rенотип особи, успешно конкурирующей с друrи-
ми особями Toro же rенотипа. не обязательно окажется победите-
лем в полиморфной смеси rенотипов. И наоборот, конкурент, от-
носительно слабый в чИстом сомкнутом насаждении, может ока-
заться доминирующим членом смешанноrо насаждения (Gustafs-
son, 1951)_
Такое положение вещей выявилось в друrой серии экспери-
ментов, проведенных Сукачевым (1928) на Taraxacuт. Он выра-
щивал линии А, В и С на экспериментальных делянках в чистыХ
r /1.. 12. ЭффеКТIJl вэаимодеfJствия особей
123
и смешанных сомкнутых насаждениях и подсчитывал число цве-
тущих корзинок на ОДНО растение; этот признак служит хоро-
шим показателем мощности и плодоносности растений. Получен-
ные им результаты приведены в табл. 12.2. Как показывают дан-
ные, относительная конкурентоспособность rенотипа С, самая
низкая в чистом насаждении, в полиморфной смеси оказалась
самой высокой.
ТаБАица 12.2
Сравнение плодовитости трех линий одуванчика
8 сомкнутых насаждениях (Сукачев, 1928)
л ниня
Число корзинок на одно растенне
чистое насажде. I смещанное насаж"
нне дение
А
В
С
20 35
34 43
B ll
1 8
12 20
16 2З
Сходные результаты неоднократно наблюдались при односор-
товых И смешанных посевах ячменя, пшеницы, тимофеевки (Phle
ит prateпse) и друrих злаков (Gu'stafsson, 1951). Кроме Toro, и в
этом случае эксперименты с мономорфными и полиморфными
популяциями Drosophila тelanogaster и Triboliuт castaneuт (Le
wontin, 1955; Sokal, Karten, 1964) дали аналоrичные результаты.
Факторы, зависящие от плотности
Действие некоторых факторов среды, иrрающих роль селек-
тивных сил, становится более выраженным по мере увеличсния
плотности популяции. Те факторы среды, действие которых уси-
ливается с. увеличением плотности популяции, называются фак-
торами, зависящими от плотности; в отличие от них факторы,
действие которых не коррелирует с плотностью популяции, назы-
ваются факторами, не зависящими от плотност.и.
Вообще действие физических факторов среды (температуры,
влажности, наводнений, вулканических извержений и т. п.) обыч
но не зависит от плотности, тоrда как действие биотических
факторов, как правило, зависит от п..10ТНОСТИ популяции.
Хорошим примером фактора, зависящеrо от плотности, служат
инфекционные заболевания. По мере повышения плотности попу-
дяции заболеваемость и смертность увеличиваются, и на HeKO
тором уровне возро.сшей п.потности может вспыхнуть эпидемия.
Ес..1И же величина ПОПУJIЯЦИИ уменьшается, то возбудитель болез-
124
Часть /1/. EcтeCTBeNHblt'J. отбор
ни становится менее активным, а при крайней диспсрrированно
сти популяций этот фактор может совершенно прекратить свое-
действие.
Конкуренция представляет собой фактор, зависящий от П..10Т-
ности. Действие раститеJfЬНОЯДНЫХ животных на популяцию рас-
тений или действие хищников на популяцию жертвы это также
факторы, зависящие от пдотности.
Обычно зависящие от плотности факторы оказывают на чис-.
ленность популяции стабилизирующее влияние. Коrда популяция
становится слишком плотной, такой фактор вызывает сокращение
ее численности; но коrда популяция становится слишком ма.'10ЧИС"
ленной, ero действие становИтся пермиссивным, создавая возмож-
ность для восстановления численности.
Зависящие от плотности факторы обычно ПРОЯВ.1ЯЮТ свое се..
лективное .действие, коrда плотность попу.'1яции достиrает опре..
деленноrо пороrовоrо уровня или превышает ero. Под влиянием
бо",езнетворных орrанизмов в популяции, достиrшей известной
ПЛОТНОСТИ, начинается отбор на устойчивость к соответствующим
заболеваниям, и с повышением п",отности этот отбор УСИJlивается.
Иными словами, селективная ценность reHa, опреДСJlяющеrо устой
чивость к тому или иному заболеванию, не постоянна, а изменя"
ется с изменением ПЛОТНОСТИ популяции. Селективная ценность
данноrо reHa может изменяться в широком диапазоне от бу-
КRально нулевой в сильно дисперrированной популяции она до-
стиrает положите.1ьноrо значения при пороrовой плотности и
обеспечивает высокое преимущество при высоких пЛЬтностях по-
пуляции.
Отбор, зависящий от частоты
у Насекомых и у растений обнаружен рЯД случаев, в которых
селективная ценность данноrо rенз или rенотипа. варьирует в за-
висимости от ero частоты. Эта ситуация известна под названием
отбора. зависящеrо от частоты. Селективная ценность reHa обыч
но находится в обратной зависимости от ero частоты. Иными сло-
вами, данный reH И.Т'lИ rенотип обычно обладает бо.lIее высоким
селективным преимуществом, коrда частота ero невелика, чем при
ВЫсокой частоте. Известны, однако, и такие примеры, коrда низ
н:ая частота rеиа невыrодна в се..1ективном отношении.
Хорошими примерами селективной ценности, зависящей от
частоты, служит. подражательная окраска, которой обладают раз-
личные виды бабочек. Успех подражательной окраски в качестве
средства защиты от птиц, уничтожающих бабочек, связан с тем,
что подражающий вид относительно редок по сравнению с АИ
дом, служащим моделью. Насекомоядные птицы научаются избе-
fЗТЬ бабочек, служащих моделью, которых ОНИ узнают по виеш-
r л. 12. Эффекты взаимодействия особей
125'
нему виду неприятные или токсичные свойства которых они
познали на опыте. Подражающий вид извлекает пользу из CBoero
сходства с моделью косвенным образом. Если численность под
ражающеrо вида сильно повышается, то он, на собственную беду,
нарушает выработавшиеся у насекомоядных птиц реакции. По-
этому селективная IleHHoCTb подражательной окраски ве 1ика,
коrда ее обладате '1И встречаются редко, но снижается при уве.пи
чении их частоты.
В экспериментальных популяциях несколько видов Drosophila.
в том числе D. тelanogaster, п. pseudoobscura, п. paиlistoruт и
п. willistoni. сравнивался успех в спарнвании у мутантных самЦОВ
и у самцов ДИкоrо типа. Оказалось, ЧТО успешность спаривания
у самцов определенноrо типа варьирует в зависимости от их
частоты в эксперимента.'1ЬНОЙ популяции. Прсимуществом обла-
дают те самцы, которые встречаются редко, нсзависимо от Toro,
относятся ли они к мутант IOМУ или дикому типу (Ehrman, Spiess)
1969) .
Друrими примерами зависящей от частоты селективной цeHHO
сти у насекомых с.'1ужат один ИЗ ферментных локусов и одна ин-
версия у Drosophila тelanogaster. а также черный мутант у Tri..
ьоиит castaneuт (Kojima, У arbrough, 1967; Nassar, Muhs, Cook
1973; Sokal, Karten, 1964). Среди анаJl0rичных примеров у paCTe
пий можно назвать rCH, опреде '1ЯЮЩИЙ характер семенной обо
лочки У Phaseolus luпalus и раЗ 'Iичные типы венчика у Ph.lox druт..
тondii (Harding. Allard, SmeItzer, 1966; Levin, 1972).
r Ла'Ва 13
ДРЕйФ rЕИОВ
Общие соображения
Соrласно закону Харди Вайнберrа, частота аллеля, который
нейтрален в селективноМ отношении, обычно остается постоянной
из поколения в поколение. Следует, однако, помнить (см. rл. З),
что этот закон применим лишь к очень большим популяциям.
Предсказываемое им постоянство частоты аллеля представляет
собой статистическую среднюю для большоrо ЧИС,,'Iа испытаний;
размножение reHa в большой популяции удовлетворяет условию
большоrо числа испытаний. В каждом наборе из малоrо числа
испытаний, как, например, при размножении reHa в небольшой
популяции, следует ожидать отклонений от средней частоты ал-
лелей за счет одной лишь случайности.
Случайные изменения частот аллелей и rенотипов из поколе-
ния в поколение в небольшой популяции известны под названием
дрейфа reHoB. Дрейф reHoB соответствует случайному компоненту
в скорости размножения reHOB. В небольшой полиморфной попу-
ляции дрейф reHOB приводит к двум характерным явлениям: сна-
чала к флуктуациям частоты аллеля из поколения в поколение,
а в конце концов к полному закреплению или элиминации дан-
Horo аллеля.
Мы предположили, что изучаемый аллель нейтрален в селек
тивном отношении. Это упрощающее, но не обязательное допу
щение. Допустим теперь, что интересующий нас аллель обладает
небольшим преимуществом или несколько неблаrоприятен в се-
лективном отношении. Предсказанное изменение частоты аллеля
из поколения в поколение и в этих случаях представляет собой
статистическую среднюю и опять таки подвержено случайным
отклонениям.
Если, например, в популяции, полиморфной по аллелям А и
а, аллель а имеет 0,1 % Hoe селективное преимущество (5 0,00 1) ,
то, соrласно закону Харди Вайнберrа, соотношение этих двух
аЛJ1елей в rенофонде следующеrо поколения составит 1000 а: 999 А.
Это среднее соотношение действительно наблюдается в больших
ПОПУ lЯЦИЯХ; что же касается небольших популяций, то в них сле-
дует ожидать значительных отклонений от Hero, вызванных слу-
чайными факторами.
Таким образом, действие отбора само по себе не исключает
возможности действия дрейфа reHOB. В сущности есть основания
полаrать, что самая rлавная эволюционная роль дрейфа reHoB
это ero совместное действие с отбором.
r А. 13. Дреt'J.ф zeHOS
127
Будет ли дрейф reHoB оказывать существенное влияние на ча-
стоты аллелей в данной популяции или нет, зависит от четырех
факторов: 1) размеров популяции (N); 2) селективной ценности.
данноrо аллеля (5); 3) давления мутаций (и); 4) величины по-
тока reHOB (т). Отмеченные четыре фактора взаимодействуют
друr с друrом. Взаимоотношения
этих факторов изучал Райт (Wright,
193 1), выразивший их в количествен-
ной форме.
Рассмотрим сначала размеры по-
пуляции. Как отмечалось выше, слу-
чайные флуктуации частоты а.1Jлелей
пренебрежимо малы в большой, а не
в маленькой популяции. В маленькоЙ
популяции за счет одной лишь слу-
чайности частота какоrо либо аллеля
за одно или несколько последователь-
ных поколений может измениться от
низкой до высокой или же аллель мо-
жет закрепиться.
Как показал Райт (Wright, 1931),
теоретическое распространение частот
аллелей в очень маленьких изолиро-
ванных популяциях описывается U об-
разной кривой (рис. 13.1). Допустим,
что некий аллель (А 1) селективно
нейтрален (5==0) и представляет со-
бой один из ряда полиморфных аллелей в серии небольших изо
.пированных популяций (ве..1ичина которых равна N). Следует
ожидать, QTO в большинстве полиморфных популяций этот аллель
будет приб..1ижаться либо к исчезновению, либо к закреП.1Jению, и
лишь в относительно немноrих популяциях он будет сохраняться
со средней частотой (кривая / на рис. 13.1).
Распределение частот аЛ.lJелей в небольших популяциях (ве-
личина которых равна N) можно сравнить с их распределением
для ряда больших популяций (величина которых равна 4N); в
последнем С.'Iучае в большинстве попу.FJЯЦИЙ изучаемый аплель
содержится со средней частотоЙ (кривая // на рис. 13.1).
..... Кривую, представленную на рис. 13.1, можно также использо-
вать для показа теоретическоrо распределения частот аллелей
(ось абсцисс) в ряду полиморфных reHoB (ось ординат) одной
популяции. Большинство полиморфных reHoB в маленькой попу-
ляции (кривая 1) имеют крайние частоты, а в большой (кривая
/1) ---- средние частоты.
Эффект величины популяции
)::1
:::!
с:::
!;:)
с:::
!;:)
0,1 0,5
Число аллелей
1,(}
Рис. 13.1. Частота данноrо ал-
лепя в серии ПОПУЛЯl1ИЙ, отно-
сящихся по разыерам к двум
классам (по Wright, 1931).
[... небо.пьшие ПОПУ.llицни (H)
"... БО.1ьшие популяции (4 N);
з==0.
128
Часть [П. Естественн.ыа отбор
Эффект отбора
в предыдущем разделе было принято допущение, что рассмат-
риваемый аллель полиморфноrо reHa селективно нейтрален; это
допущение отражено в симметричности U образных кривых. В ря-
де неБОЛЫliИХ популяций этот селективно нейтральный аллель
приближается либо к закреплению, дибо к элиминации, причем
в равной мере. Однако если аллель обладает небольшим ce.1}eK
N
11
I
11
"
N ' = 4N
':::!
:::s
::r
Q;
t::
с)
с:::
Q
о
А
1
S 8N :.
1 ,'l
s . 4N" i"
J.
1 11.
s"'=2N
:\
: \
1\ .
11 \ 1 1.:
1 \ s 1,':
I \" 2N' /1 .: 1
\ ...... --- ./II S ='"(iiТ"
\ ,, :
, . 2
... s=
0,5 1,0 О 0,5 1,0
Цuсло ллеnей б
s O
Рис. 13.2. Ожидаемые распреде.'1е-ния частот аллелсit при разНЫх. знаqеНИRХ
s 11 двух значениях N (Wright, 1931).
А. В маленl..КОЙ популяцни. Б. В популяции. которая в 4 раза больше. при эквнвапентных
интснсивиостях отбора.
тивным прсимуществом в малых популяциях, то кривая сохра-
няет U образную форму, но смещается и становится асимметрич
НоЙ. Направление, в котором происходит смещение кривой, за
висит от Toro, блаrоприятен И.I]И неб.паrоприятен данный аллель
в селективном отношении, а степень смещения от ве..1ИЧИНЫ
селективной ценности (рис. 13.2, А). Аллель, обладающий He
большим селективным преимуществом, обычно присутствует в
серии небольших изолированных ПОПУ.,1яций либо с высокой, либо
с низкой частотой, но чаще с высокой.
В этом случае снова rруппы небольших популяциЙ отличают
ся от rрупп больших популяций ПО ожидаемому поведению reHoB.
Отбор реrулирует частоты reHoB в больших популяциях более эф
фективно, чем в малых, при эквивалентных значениях 5
(рис. 13.2). .
С..,овесное описание кривых, представленных на рис. 13.2,
можно сформулировать и по иному, сказав, что отбор теоретиче-
СКИ оказывает относительно небольшое влияние на частоты reHOB
в популяциях, величина которых ниже известноrо критическоrо
r л. 13. Дрейф еенов
129
уровня, тоrда как одна лишь случайность, по-видимому, способна
весьма эффективно реrулировать частоты reHOB при тех же ca
мых условиях ('Wright, 1931). Следует ожидать, что в маленькой
популяции дрейф reHOB возьмет верх над слабыми давлениями
отбора.
Это заставляет поставить вопрос о том, сколь мала «малень
кая» популяция и сколь велика «большая», коrда речь идет о
действии дрейфа reHOB. Критическое значение N, при котором
дрейф становится эффеКТflВНЫМ, зависит от s. Зависимость между
N и 5 представлена в табл. 13.1.
ТаБАица 13.1'
Зависимость между величиной популиции, селективной ценностыо
и дреАфом reHOB (Wrlght, 1931)
сипа. реrупирующая
часТОты аllJ1еllей
N
s
Отбор
1
:s:;;
2s
1
I
<
== 2N
1
>
== 4N
1 1"
От 4N дО 2Н
Дрейф rеиов
Отбор и дрейф reHoB
>
4s
1 I
OT ДO
4s 2s
Эти зависимости можно наrлядно изобразить, построив ли..
нейный rрафик (рис. 13.3). Как показывает rрафик, если значе
ния N низки по сравнению со значениями s, то преобладает
дрейф reHOB; при относительно высоких N преобладает отбор;
существует также область перекрывания, rде дрейф и отбор Moryr
действовать совместно.
Эти общие зависимости ..тrerKO перевести в конкретные цифры.
Допусти М, что селективная ценность данноrо аллеля s 0,0 1. Ero
частота реrулируется дрейфом reHoB при N -< 50. Но если селек..
тивная ценность аллеля s==O,OOI, то ero частота будет реrулиро .
ваться дрейфом при N -< 500. Таким образом, в общем виде при
довольно низкой селективной ценности аллеля дрейф reHoB может.
привести к erO закреплению или элиминации при совсем малой
величине популяции, но при очень низкой селективной ценности
аллеля дрейф может реrулировать ero частоту в популяции сред-
Hero размера.
Величина области перекрывания, в которой действуют дрейф.
и отбор, также варьирует в зависимости от ве.ПИЧИНЫ s. В приве
денных выше численных примерах эта област соответствует-
N 25 50 дЛЯ s O,OI и N==250 500 Дol1Я s O,OOl:
Часть 111. Естественный отбор
1130
. Возможность COBMecTHoro действия отбора и дрейфа теорети-
чески имеет очень важное значение для эволюции. Райт (Wright.
1.931, 1949. 1960) укаЗbIвает. что какой либо блаrоприятный reH
может rораздо быстрее закрепиться при помощи отбора и дрей-
фа в популяционной системе OCTpoBHoro типа. чем при помощи
ДНоrо только отбора в непрерывной большой популяции TaKoro
.же общеrо размера. Недавно была вычислен сравнительная ве-
роятность закрепления первоначально редкоrо reHa с небольшим
селективным преимуществом в популяциях ЭТИХ двух типов ----
Отбор
Дpeиtp ееНО8 Il ii;' ; ' ' ;б ;" \ """ "" ,Ш,,,,,,,,, \
", """ """"" "", "", "п
1
,
N== 100&
I I I
7 l' 1
10 4$ 2s
. I
,
q18
I
1
0,01&
I
,
0,0078
Рис. 13.3. Области действия дрейфа тенов и отбора, соответствующие различным
соотношениям N и s (Grапt, 1963).
'единой непрерывной и подразделенной на отдеЛЬНbIе колонии----
содержащих по 106 особ,ей каждая; оказалось, что в подразделен-
ной популяции эта вероятность на порядок выше (Flake, Grапt.
1974) .
Эффект потока reHoB
Райт (Wright, 1931) предложил ряд уравнений для описания
в;заимодействия между потоком и дрейфом reHoB. Частоты алле-
лей реrулируются дрейфом reHoB в том случае, коrда
1 1
N 2т или т 2N .
Частоты а.плелей реrулируются потоком reHOB в том случае,
коrда
N>
-== 4т
1
или m 4N '
а обе .силы действуют при промежуточных размерах ПОПУЛЯЦИИ:
1 1
N== по
4т ".... 2m .
1
Например. если N == 100. то критическое значение m==[V==0,005.
Таким образом. иммиrрация посторонних а.плелей со скоростями.
превышающими т == 0,005. может предотвратить дрейф reHoB в по-
fJУЛЯЦИЯХ, состоящих из 100 или менее скрещивающихся особей.
rA. 13. Дре4ф zeНOB
13f
Вообще незиачительный поток reHoB может предотвратить.
дрейф. Для Toro чтобы Mor происходить дрейф reHoB, ма.,']енькая
популяция должна быть довольно хорошо изолирована. Но при
наличии изоляции, при низких значениях как N, так и т, дрейфо
может оказывать существенное возд йствие на частоты reHoB.
Действие частоты мутаций описывается уравнениями, анало
rичными уравнениям для потока reHoB. Частота аллелей реrули-
руется давлением мутаций, KorAa N > 4 ' и дрейфом reHoB, Kor-
I
да N <:. 2и ' В небольших популяциях высокая частота мутаций
может препятствовать дрейфу reHoB.
Закрепление сочетаний reHOB
Совместное действие отбора и дрейфа reHoB в маленьких по
пуляциях, по видимому, способствует закреплению не только ОТ-.
дельных reHoB, но и их сочетаний. Последнее может иrрать очень.
важную роль в эволюции.
Допустим, что некая большая популяция содержит два ред.
ких мутантных аллеля а и Ь двух несцепленных между собой re-
иов А и В. Большинство особей в этой популяции имеет rенотип.
ААВВ; кроме Toro, в ней есть несколько особей, несущих мутант-
иые аллели АаВВ и ААВЬ. Допустим да,J1ее, что сочетание reHOB
ааЬЬ имеет высокую адаптивность в каких-то новых условиях.
среды.
В большой популяции в результате половоrо процесса изредка
будет возникать сочетание reHoB ааЬЬ, однако оно будет немед-
ленно разрушаться этим же самым процессом. Отбору трудно
«подхватить» это сочетание, и поэтому ero частота повышается
очень медленно.
Небольшой изолят этой же самой популяции может случайно-
иметь среднюю или высокую частоту в общем редких аллелей а'
и Ь. При этом в каждом поколении будет возникать и подверrать.
ся отбору пропорционально большее число зиrот ааЬЬ. После'
этоrо отбор получает возможность эффективно действовать таким
образом, что и в да."ьиейшем будет повышаться частота rенотипа
ааЬЬ. Следовательно, закрепление HOBoro сочетания reHoB может
быть достиrнуто быстрее в некоторых небольших колониях при
помощи отбора и дрейфа reHoB, чем при помощи одноrо только-
отбора в больших популяциях.
Экспериментальные данные
Керр и Райт (Ker,r. Wright, 1954) подвер.rли проверке теорию-
дрейфа reHoB в экспериментах с серией очень мелкИХ популяций пro-.
S()phila тelanogaster , полиморфных по одному из reHOB, детерми...
132
Часть 11/. Естественный отбор
ИИРУЮЩИХ щетинки, forked (1). Они создали 96 идентичных линий,
Д 1Я каждой из которых было взято по 4 самки и 4 самца. Исход
ная частота MYTaHTHoro аллеля forked в каждой линии была paB
a 0,5. Все эти. 96' линий поддержива..1И в тече.ние 16 поколений
путем случайноrо.выбора в каждом поколении родительских осо-
бей (4 самок и 4 самцов) для следующеrо поколения.
К 16 MY поколению в 41 линии произошло закрепление аллеля
.дикоrо типа; в 29 линиях закрепился аллель forked, а 26 линий
продолжали оставаться полиморфными.
.. . .Был проведен также параллельный эксперимент с использо
в.знием MYTaHTHoro аллеля Bar (В), обусловливаlOщеrо По..10СКО-
'Видные rлаза у D. тelanogaster. Здесь было заложено 108 линий,
поддерживавшихся таким же методом, как и в предыдущем экс
пери менте. Мутация Bar неблаrоприятна в селективном отноше-
нии. К 10.му поколению ллель дикоrо типа закрепился в 95 ли
ииях, аллель Bar в 3 линиях, а 10 линий оставались полиморф-
ными ('Wright, Kerr, (954).
Очевидно, что в небольшой экспериментальной популяции
значительная доля полиморфных reHOB закрепляется в результате
дрейфа. Иноrда эти reHbl закрепляются, несмотря на противодей
ствие со стороны отбора. Чаще закрепление происходит в ре-
зультате cOBMecTHoro действия дрейфа и отбора, как в случае
закрепления 95 линий дикоrо типа в эксперименте с мутацией
Bar.
Совместное действие отбора и дрейфа было продемонстриро-
вано и в друrих экспериментальных исследованиях. Одна rруппа
экспериментов была поставлена на лабораторных популяциях
Drosophila pseudoobscura, различающихся по инверсиям (Dobz-
hansky, Pavlovsky, 1957; Dobzhansky, Spassky, 1962). Друrой экс-
перимент с однолетним травянистым растением Gilia, проводив-
шийся на протяжении 17 лет, касался мощности и фертильности
в серии родственных инбредных линий (Grant, 1966а).
Дрейф в ПРИРОДНЫХ ПОПУЛЯЦИЯХ
В природе часто встречаются три ситуации, коrда величина
популяции. может быть достаточно мала, чтобы допустить эф-
фективное действие дрейфа, сопровождаемое или не сопровождае-
мое отбором: 1) популяционная система состоит из ряда изоли
рованных колоний, размеры которых постоянно остаются неболь-
шими; 2) популяция обычно имеет большие размеры, но перио-
дически сильно сокращается, а затем вновь восстанавливается
за счет нескольких выживших особей; 3) большая популяция да-
.ет начало изолированным дочерним колониям, каждая из которых
.создается одной или несколькими особями основате",ьницами.
Новые колонии проходят, таким образом, в своих первых поко-
r л. 13. Дрейф zeHOB
133
лениях сквозь «узкое rорлышко» низкой численности, хотя в
дальнейшем их размеры MorYT возрасти; это так называемый
«принцип основателя», выдвинутый Майром (Mayr, 1942, 1963).
Ес.НИ дрейф иrрает эффективную роль (опять-таки совместно
с отбором или без Hero, но скорее при ero участии), то С.'1едует
ожидать, что изменчивость колоний будет проявлять следующие
характерные черты. Во первых, небольшие колонии сестринские
колонии при ситуации 1 и дочерние колонии в первых поколениях
при ситуациях 2 и 3 должны быть rенетически довольно одно-
родными. BO BTOpЫX, между колониями должна ПРОЯВ.'Iяться до-
вольно существенная изменчивость по rснетически детерминиро-
ванным признакам. Следует ожидать, что эта локальная расовая
дифференциация должна быть выражена наиболее ясно в серии
небольших сестринских колоний (ситуация 1), но проявляется
также в некоторых сериях более крупных популяций, происходя-
щих от небольших колоний (ситуации 2 и 3). И в третьих, rpa
фик распределения межколониальной изменчивости должен но-
сить несколько незакономерный и случайный характер.
Характер изменчивости детально изучался у ряда rрупп рас-
тений с колониальной структурой популяционных систем. В неко-
1'орых из этих rpynn характер изменчивости соответствует при-
веденным выше ожиданиям, что позволяет предполаrать эффек-
тивное действие дрейфа reHOB.
Некоторые виды кипариса (Cupressus spp.) в Калифорнии
образуют ряд изолированных рощиц, причем в каждой рощице
деревья об..1адают своими отличительными морфолоrическими
признаками (,\Volf, 1948; Grant, 1958). Тот же самый тип измен-
чивости обнаружен у таких травянистых растений, образующих
популяционные системы в форме ряда колоний, как Gilia achil
Leaefolia в Калифорнии, rруппы Erysiтum candicum и Nigella
arvensis на островах Эrейскоrо моря (Grant, 1958; Snogerup,
1967; Strid, 1970). Представление о РО.lИ дрейфа во всех этих
примерах подтверждается тем, что родственные виды Juniperus,
Gilia и Nigella в друrих областях образуют большие непрерыв-
ные попу.'Iяции с иным типом изменчивости, а именно с посте..
пенной интерrрадацией по rеоrрафическим Tp HceKTaM.
Случайная локальная изменчивость набдюдается также в He
которых популяционных системах европейской наземной улитки
Сераеа neтoralis по такому признаку, как наличие или отсутствие
полос на раковине (рис. 10.3). Этот признак окраски раковины
определяется одной парой аллелей, причем аллель бесполосости
доминирует над алле..'Iем полосатости (см. rл. 10). Частота беспо-
лосоrо фенотипа и аллеля бесполосости широко варьирует в раз-
JlИЧНЫХ колониях в тех областях Франции, rде ПОПУv1яции Сераеа
neтoralis имеют колониальную структуру (рис. 13.4). Однако в
больших популяциях европейской наземной улитки частота алле..
"""
g
w 'lJdolAl Wf1Htlodli blJH DШО:J19g
oo oogOO
g g-3'
о
""
"
......., ,.....,
I'L "
....".....,.... ..................
........ ....
.",...., .....,...
. ..........., ".........".... , ,..
. n.,........".....,...,........,"',................
.............."", ,...", '''' "............"......
. ....., ,.......
.n"...."""",.... ,.....,
j9vog-аv НDtlaW
wtlaJ ..av..:J)l1;I
,"........."............ ...".......", .........
.....,....,....."....,....... ............. .............----
JiDHMOIV
....'
. ..",........, ",..................."".....
.................... ....,........................
... .........
..""....,........ ",............
. .,....................
,...'\.,.....,....,'
.................., "................
....,.... ........... ..................
D:JН8З
"..
,........,......... ................. ...... --
<:\J
cu
t)
х
11::
=
О
t:;
о
:.::
I:D
:::
=
I:D
О
:.:: .
.........
Q. ...
,,
,....Ф
=....
:.:: .
I--Q)
(..1 .
-OJ
tl'0
It) I >,
Q)...J
:а.......
t:;
:с Q),.....
\011::
1:) ...... а-
х:!:
:a
(..I '
06
t:; .......
О
СХ
(..111::
Q)Q)
\О:С
Q)
=е.
-;-а
I:D
:E
C!)
>,..:1
с...
z<
:с=
:с:.::
CI)
са Q..
040'
:c
ё5..=
1--1:";
з0
«11:1:
ID
11)
х....
:2Sё:s
1--'"
C
(..IЕ.
°cu
3
с:
Q.I
::s
:t
Q:
с)
Е:
Q)
4>
S
о
:с
1--
о
О
U
.
с.;
и.
=
Q...
t.
o
. 0 G)
00С!>
.. .80 0
ее880 80 .
...",.0 о
60\G).
.л \ 8:t:5'
с!> '... ...00000\
., 000,
'0....(;)
8 e.
o 8 G)
0
tiJ
o
@ .
0.0. .0 .
. 0.е0.
о о 0 0 ..
80.0..0. .
@ .0.
000@ 0.
8@ . .0@.8@
Е> Е> (!)@ @.
. @ ..б .. .. <:5..... .. ..
@ ,t1atJ 0 @О
O 0 О@8
..ф О е е
.. / @ @О
'@ О ... 0 О
\"..Q9........... . ...........
.@@@ ,'01 @@.
ее .Ф 'С!)О.. I
@0 0 ,'00 " "}
. @0'фО . 0
o Ф000 . оо@
0. О О
О 0 ....'0 0
..@@0 8"'... о
. . . 0 --"
o @ . 080 @
0.. о @.
.8@. .. .
@..@ . @8@
@. @ 0080Е>.Ф
@0 @80.08е.
o @@ @.0@ Ф
8@ 0 0.@0'00
..008.
Ое@ 0 ..@
@ П 0 ·
.0 ·
о . 00.0 с!)
@@ ..0.
е
о
@
.
3
:о
::Е
:
Е=
1>:
:
:Е!:;
»"'
Q,:r
:.::>->
.=
О
ltI
О
';>.
О
r-::(
>-.
IIJ)(
:а=
3
IIJ.Q
1O
О
--.о
v,1IJ
v,=
q
с:з
с')
с:з
с.
:: . :аса
u.......\;ca
g
о'> ,.,=
>-. .... о...
..:
:1) ... t
0"0 I
J::; g =>0:
>. .... :.::а
';>. oa:
о...... с,0 IIJ
:I: :<:>C
f-o UIIJ
;I: С) IIJ
:;;
o.. c,
fo-oo 6
01-
>-. ltI :.: u
::Е ::и
О CI:I ...
=0..::=0
t::( о с:
O::Et::
g CI:I 8
С) IIJ:EQ,
Е 5
с:: I!:I :е са =
::о:: О IIJ
t--a::: IQI-
О О =-е-
:с с:: ос!: 1--&
QJ с) ::::
""'.:;: 3-.оц
O 2
о.. t::{ . >->
f-oО кё:;;з
D:
:s: А:':
:r::....... о.: з о
CI) 11) IIJ 1- о:;
a:::::t ::{ о
CI) .... .......
t::{::S о::
CI) 1.,) 8
g-
Е
0.11) i I
::S о
= )(
.g. ---- "':.:
CI:I :!'
е- t
IIJ
:Еу
o:s:
:i3
а:::
CI:I
f-o
Ф
r::(
о
==
CI.I:':
:r;E
"'»
ЖQ,
8:.::
10
о -
...
:.:
:Е
»
р.
:.:-
&ci
u
:s:
I:l.
lЗn
Часть [Н. Естественный отбор
.ля беспо...10СОСТИ варьирует по rеоrрафическим трансектам еще
более постепенно (Lamotte, 1951, 1959).
Неэакономерная локальная расовая изменчивость часто встре-
чается у растений, улиток, бабочек, мышей (рис. 13.5) и в дру-
rих rруппах, в которых она коррелирована с популяционной
системой колониальноrо типа. Характер изменчивости и ero KOp
реляция со .структурой популяции представляют собой веские
доводы в пользу Toro, что в таких случаях эффективно действует
дрейф reHoB.
АллеJlИ, определяющие rpYnnbl крови
в популяциях человека
Некоторые ИЗ са мых веских данных в пользу дрейфа reHoB в
природных популяциях относятся к человеку. На всем протяже-
нии человеческой истории во мноrих частях земноrо шара су-
ществовали популяции, величина которых блаrоприятствовала
дрейфу reHOB. На стадии собирательства и охоты были обычны
небольшне изолированные или полуизолированные популяции,
СОСТОSlвшие из 200 500 взрослых особей. В разных частях света
существуют небольшие изолированные сельскохозяйственные или
рыболовецкие общины. Некоторые релиrиозные секты образуют
небольшие изолированные попу.IJ:ЯЦИИ, скрещивающиеся внутри
себя, потому что релиrиозные верования запрещают их члена :
браки с посторонними.
Б.'1аrодаря большому количеству данных, собранных по rруп-
пам крови системы АВО и друrих систем в больших и малых по
пу.I1ЯЦИЯХ человека, а также простоты reHeTH(leCKOIl основы ЭТИХ
систем, rруппы крови служат удобным показатеЛС:\1 rенетitческоrо
сходства или различия между Попу.,')яция и. Полиморфизм по
rруппам крови системы АВО был кратко описан в rл. 2.
Интересным примером служит полярное племя эскимосов, оби
тающих близ Туле на севере rренландии. Это небо.пьшое племя.
в состав KOToporo входит 271 человек, а может быть и менее, на
протяжении МНоrих поколений находилось в полной И30.,1ЯЦИИ.
ДО тех пор пока они не встретились с друrим племенем эскимо
сов, обитающим в северной части Баффиновой Земли и затратив-
шим несколько ..'1ет на поиски полярных эскимосов, последние
считали себ единственными обитателями Земли.
Оказалось, что популяция ПО.IJЯРНЫХ эскимосов заметно OT
лиqается от rлавных популяций эскимосов по чаСТоте аллелей,
олреде.,1ЯЮЩИХ rруппы крови. В более крупных популяциях rpeH
ландских ЭСКимосов частота аллеля /А состав..'Iяет 27 40%
(табл. 13.2). Сходные частоты аллеля JA обнаружены в популя
циях эскимосов Баффиновой Земли, Лабрадора и Аляски. Но пле
мя полярных эскимосов отклоняется от этой нормы, так как у
r А. 13. Дрейф еенов
137
Таблица 13.2
Частота аЛЛeJIей, опреДe.l1ЯЮЩИХ rpynnw крови системы АВО
у популяций эскимосов в rренлаидии (Lаughliп. 1950)
Частота аллепя. "
Область тА I тВ I
/"
Н норталик, oKpyr К)лианехоб (южная часть
rренландии) 27 3 70
К юrу от Нанорталика 35 5 60
Мыс Фарвель 33 3 64
Яковсхавн 29 5 66
Анrмаrссалик (восточная часть rренландии) 40 11 49
Туле (северная часть rренландии) 9 3 84
них частота аллеля /А равна 9 % (табл. 13.2). Вместе с тем у
полярных эскимосов обнаружена крайне высокая частота аллеля
/0 по сравнению с частотами 3Toro аллеля в популяциях эскимо-
СОВ в fреН..'1андии и в друrих местах (Laughlin, 1950).
Заметная локальная дифференциация по rруппам крови АВО
lIаБЛlодается также в друrих небольших изо.пированных популя-
циях че.I10века: у П.rrемен абориrенов в южной tlасти Австралии,
у релиrиозной секты данкеров в восточной части СевеРIiОЙ Аме-
рики, у еврейской общины в Риме и у некоторых ropHblx и остров-
IIЫХ деревенских общин в Японии (Birdsell, 1950; Glass, Sacks
Jahn, Hess, 1952; Dunn, Dunn, 1957; Nei, Imaizumi, 1966).
Релиrиозная секта данкеров была основана в fермании в Ha
чале XVIII века, а позднее ее члены эмиrрировали на восток
США. ЧJlены этой секты вступают в брак rлавным образом друr
с друrО:\I, в резу.'1ьтате чеrо они на протяжении мноrих поколе-
пий оставались репродуктивно изолированными от популяций
rермании и Америки, среди которых жили. Некоторые общины
данкеров очень малы; в состав одной общины на юrе Пенсильва-
"ии в период ее изучения в начале пятидесятых rодов входило
ncero 90 взрослых. Весьма примечательно, что ленсильванские
данкеры отклоняются от обычных rерманских и американских
rюпуляций по крови и по друrим признакам (Glass, Sacks, Jahn,
Hess, 1952).
В табл. 13.3 приведены частоты аллелей reHa 1 у пенсильван-
ских данкеров и у родственных им по расовой принадлежно-
<:ти ПОПУJ1ЯЦИЙ Западной fермании и восточной части США. Со-
.вершенно очевидно, что популяции Западной fермании и США
XOДHЫ по частоте разных reHoB. Что касается пенсильванских
;Данкеров, то они отличаются как от своих rермзНСКИХ предков,
138
Часть 1/1. Естественный отбор
ТаБАuца 13.3
Частота aJlJleJleA, опреДelltlЮЩИХ rруппы "рО8И системы АВО, у трех
родственных популяций, прииаАJlежащих к кавкаэскоА расе
(Glass, Sacks.. Jahn, Hess, 1952)
ЧаС'Jота аппеп". %
ПопутщlUl
JA JB
JO
Данкеры Западной rермании
Данкеры ВОСТОЧНОЙ частИ США
Данкеры В Пснсильвании
29 7
26 4
38 '2
64-
70'
50
так и от своих нынешних американских соседей: частота аллеля
JA у них существенно выше, а аллель [В близок к вымиранию
(Glass, Sacks, Jahn, Hess, 1952).
Пенсильванские данкеры отличаются от популяций своих Пред..
ков и своих теперешних соседей и по друrим признакам, напри..
мер по форме мочки уха или ПО типу волос. В этой маленькой
скрещивающейся в себе rруппе были обнаружены существенные
отлИЧия частот аллелей по пяти разным reHaM от частот, типич-
ных для окружающих популяций (Glass, Sacks, Jahn, Hess, 1952).
Аллели reHa [ не нейтральны в селективном отношении. В не-
которых популяциях человека обнаружена положительная Koppe
ляция между заболеваемостью язвой желудка и rенотипом [О/О и
между заболеваемостью раком желудка и rенотипами JA[A и /А]О.
Данные о селективном значении аллелей reHa J ИСПО,,1ЬЗУЮТ иноr..
да в качестве довода против дрейфа reHoB. Этот довод основан.
однако, на неверном представлении о том, что дрейф reHoB и OT
бор взаИмоисключающие силы, а поэтому ero нельзя считать
убедительным.
Заключение
Вывод о том, что в маленьких популяциях частоты а.п.'1елей
в значительной степени реrулируются дрейфом reHoB, вытекает
из вероятностных законов и подтверждается результатами экспе
риментальных исследований. Следующий вопрос касается воз
можной роли дрейфа reHoB в природных попу.пяциях. Иrрает ли
дрейф reHoB существенную роль в эволюции?
Этот вопрос стоя..l В центре длительной дискуссии между COB
ременными специалиста ми по теории эволюции. Участников этой
дискуссии можно rрубо разделить на две партии: «верующих» И
«неверующих». По :\lОему мнению, «не верующие» выдвинули не-
сколько слабых нозражений и неверных выводов. Например, часто
r А. 13. Лрейф ееНО8
139
утверждается. что если можно доказать действие отбора в данной
попу..1яции, то тем самым исключается дрейф reHoB. так как He
обходимость в нем отпадает (Fisher, Ford. 1947; Ford, 1955. 1964;
Mayr. 1963). Подобное рассуждение исходит из Toro, что пред-
метом спора служит дилеМ lа: дрейф reHOB или отбор, причем
отбор всеrда побеждает; к тому же оно отвлекает внимание от
более важноrо в эволюционном отношении предмета спора: отбор
и дрейф rеИОВ или же один только отбор?
Рассмотрим в качестве примера reH 1 в небольших изолиро-
ванных популяциях чеJl0века. Противники дрейфа заявляют, что
если в этих популяциях действует отбор, то дрейф уже не нужен.
Однако никто не считает дрейф единственным фактором, реrули-
рующим ЧастоТы аллелей reHa в небольших изолированных по
пуляциях утверждается лишь, что дрейф reHOB одип из фак
торов, действующих в таких ситуациях.
Еще одно возражение, часто выдвиrаемое против роли дрейфа
('еков, состоит в том, что эффективность этоrо фактора в природ-
ных популяциях не была доказана. При этоМ правила, касающие-
я получения данных в экспериментальных популяциях, неправо-
мерно распространяют на rораздо более С.'l.ожные ситуации в
природе. Мы не можем реrулировать переменные факторы в при-
родных популяциях подобно тому, как мы это делаем в экспери-
ментах, а поэтому мы не можем точно идентифицировать и оха-
рактеризовать количественно все факторы. действующие в при-
роде. Однако мы можем искать в природе ситуации, совместимые
(или несовместимые) с теоретическими или экспериментальными
данными. .
НаИ.1JУЧШИМ доказательством значения дрейфа reHoB в микро
эволюции служит характер случайной лока.пыюй дифференциации
в серии перманентно или периодически изолированных маленьких
КО..'Iоний. Дифференциация подобноrо тила MHoroKpaTHo обнару
живалась в различных rруппах животных н растений, популяuии
которых представляют собой систему колоний. Эта дифференциа
ЦИЯ, если и не доказывает, то по крайней мере сильно склоняет
к мнению о том, что дрейф reHoB иrрает важную роль вполули"
. ЦИОННЫХ си-стемах TaKoro типа.
rлава 14
ПЛАТА ЗА ОТБОР
rенетический rpуэ
БстеС1'1ВeJНIНЫЙ 01'бор .имеет как позити-аный, так и неrативиый
ас.пекты. Он .влечет за собой предпочтительное iВЫЖИfва'Ние .Н раз-
МНОЖеНlие Од'НИХ .rенот-ипов и iпреДnoQчтит-ельную эл,иминацию и не-
возможность размножения друrих. Даfиная тлава -посвящена етом.у
'второму аспекту.
Утрата 'популяцией некоторой ДОЛИ reJНQТоИПОВ '8 'результате дей-
ствия от.боjJа С()lвершенно очев.и;дна в тех случаях, 'КОI да изб'ира-
ТeJ1ьная ЭJ1-Ю.1'инация п,роя'вляется в форме смертности в дарl&ИНОО
СК'ОМ смысле, Т. е. имеется налицо ТРУ1П, который МОЖНО Уlвидеть.
Однако 016ор, .как мы .видеЛIИ ранее (см. ,rЛ. 8), 06УоСло.вливает
кроме Toro, уопех rИл.и неудачу в .одной Лишь фазе раЗlм,но.жения.
Особи" обдадающие данным rенотиП"ом. MorYT .быть фИ30и'Чеоки
крепкими, НО ввиду их низкЬй плодовитости их вклад в следующее
поколение окажется 'ПОНИЖенным. В этом случае нет 'f'JJу.па, .кото"
рый :можно было бы увидеть. Однако частота .reHOB. определяющих
ниэюую :П..1'Одо'витосrь, снижается. и они в конечном счете MorYT
соверше-НJНО lrсчез.нуть.
Н.etюз'можн,ость размножен'ия той .или иной особи. 06У'слов.:lен
ная 'От.бором в ero неrаТИ'ВНQМ а,спекте, .проявляется ли она в фор-
ме и'збиратель'ной -смертности ил.и неуда.вшеrося rраЗМiножения ка.к
TarкoBoro, извсстна ПОД .назва.нием l'енст<ичесК'ОЙ rИlбел:и (или rибе-
ли, обусловлен,ной отбором). rенеТ1ическая rJ1lбель Crни'жает репро-
ДУ,КТИiвный rпотенциал данн'Ой 'популяц.ии.
Попу.ляцня обладала .бы более высокой численностью, если бы
не потери за .счет rенетической rlfбел:и. Поэтому ,rенетическая I'и-
бель предста'вляет соБОЙ t -по крайней ме.ре п-отенциа.'IЫIO, некое
бремя д '1Я популяции. Это бремя .называют rенетичесюим I'рузом.
rенети-ческий труз определяетоСЯ (Wal1ace, 1968) ка'К l (сред.няя
ПрИСlПосооленность популяции). Ero можно 'предстЗ1В'ИТЬ себе так-
же как ,общую сумму -в<:ех случаев rенетичоской rи'бел.и на :По()КО 1е
яне. Раэл:И'Чают несКОЛЬКО ТИ'ПDВ ,тенетиче-с,}(оrо Il'руза (Brues, 1969;
Crow, 1970), в. том числе такие rлавные ero типы, как мутацион
ный, сеrреrационный и субституционный rрузы. Эти типы rруза
коррелированы с разными типами отбора.
Мутационный труз неизбежный IПобочный продукт мутацион-
НОТО Iп'роцесса. Э1'О1' 'процесс 'порождает вредные 'мутаЦИИ t которые
ДOJIЖ'ны 'быть выметены стабилиэирующим Q Op'OM.
Сеrреrаци-онный rруз сущеС'J1вует в популяциях, которые ис-
пользуют преимущества, .создаваемые ,превосходс'I'ВОМ rетероэиrот.
r А. 14. Плата за отбор
14'
При Э11ОМ В каЖДIQМ поколенИ'и ,ПРОИ:СХ,Q.дит выщеП.JIени-е менее при
способленных rомозиrот, понижающих среднюю приспосоtSлеННОСTh
rrОПУЛЯlllИИ. rомози'rоты предста'ВЛЯЮТ .собой п.пату за ур'авновеши-
вающий 01160Р.
На l J1lравленный .отбор 'ПОрОЖ'Доает .субституционный ТРУЗ. Заме-
щение .CTaporo аЛ.,l.еля новым, лревосходящим ero аллелем !Влечет'
за собой rенетнческую rибель нооителей CTapor.o алделя. суММар....
ная .rибель за 'СЧет отбора, П'РОИlCх'Одящая при полНО-М замещении
какоrо либо reHa, суммарный субституционный rруз может
быть очень .велика, .nоС' олыюу частота cTap'Oro алле.'1Я, обреченноrо
на замещение, в на'Ча.Т1е этorо ,проце,сса обычно бывает 'высокой.
Полиморфизм по белкам
Электрофорет.ическое иссл-еДlOва!н.ие ферментов в ряде популя-
пий дрозофилы, .пюзв()начных жив.отных и растен,ий показала, что
для довольно значительной ча'Сти ферментных локус.ов характерен
пол'иморфиз'м. Так, у Drosophila pseudoobscura ,из 24 ферментных
ЛОКУоС{)IВ Irюли'м'Орфизм обнаР)'1жен в 43% локусов, а у D. willistoni
до."я ПОЛИМI()РфНЫХ локусов еще 'выше (см. rл. 2). При.нято сч.и-
тать, Ч11О ,процент полиморфных л о куС.о.в , выя.вдяемыx методам.и
Э 1ектрофореза, МОЖ-НО 'принять за показатель ПОJJИМОРфности re.
потипа в цсл.ом.
Мноrочисленность полиморфных reHoB как выявлеНIIЫХ, так
и J1Iредсказываемых Iпутем 'Экстралоляции данных по выявленным
reH'aM ---- противоречит ожиданиям, основанным на раосмотреНИ 1 ]!
l'ене1'ич-ескоrо .I'pуза. Каждый 'Случай обалансированН'оrо 'пол.им,ор
физма !в данной популяции создает апределенныЙ ,се.rре'rационный
rруз, .ПОСКIQЛhКУ Т3'&ой полимор.физм влечет за собой непрерывное
выще-пление rомозиrОТ t ж'изнеспособность которых панижена.
С возраcrанием ЧИС.lJа незз,вИiС'ИМЫХ локус.ов 'со сбалансированным
полиморфизмом ;ЭТО'f .се-rреrащroнный rруз возрастает IIJО экспонен..
Те. Казалось .бы, в -какой-то м.омент популяция rOкажется не в С'о..
стоянии !неC'flИ 'этот сеrреrационный ,rруз. А между тем ВЫС'ОКDПО..
Л.llм.орфные ПОПУЛЯЦИИ дрозофилы I1Iроцв-етают.
Одна .школа исследовате.'1ей ВО rлаве с Кимур'ой -попытал ась
разрешить эту дилемму, постулировав, что 'ПО.rI.иморфизм по бел..
кам На 'Самом деле ,н-е псщдерживается О'J1бо;ром. .Соrласн'о их rипо.
тезе, аЛ.тIели reHOB, детеРМИНrИР'УЮЩИХ ферменты, нейтральны в се.
лективном 'отн,ошен:ии и дрейфуют с раЗЛIИЧНОЙ част.отой в популя.
циях (Kimura, 1968; King, Jukes, 1969). П,ри таiКОЙ Iпостановке про...
бл€ма дейст.витеЛЫfО исчезает.
НО правильно л'и rCделЗ'-нное -ими .допущение? Результаты екс)пе..
риментов на ДрОЗОфИ 1е 'пок3"зыв'ают, что IHeT. У дроз илы nОЛ ..
морфизм по беJ1lкам не проявляет случайных Iфлуктуа11l ИИ и случаи..
ной 'rеоrрафической 'изменчивости, чеr.о следовало .бы 'Ожидать, ec.rlll
:142
Часть 111. Естественный отбор
бы аллели были сел-ективно нейтральными; напротив, для этою
.полиморфизма характерна определенным образом }'Iпорядоченная
изменчивость, а это позволяет предполаrать, что в данном случае
полиморфизм поддеРЖlивается уравновешивающим .отбором (Ayala,
Anderson, 1973; Ayala, Tracey, 1974; Ayala, Gilpin, 1974).
Естественно допустить, Ч110 некоторые ,полиморфные reHbl ней-
"ральны ИЛИ почти нейтральны в отношении отбора. Однако МНО-
rие друrие ко.нТРОЛИРУЮ1'ся уравновешивающим .отбором, и, следо--
:вательно, должны ВНООИТЬ извеС1iНЫЙ вклаД .в Iсenреrацнонный rруз.
.Эту праблему неv'1ЬЗЯ отбросить, при беrну.в к та'киМ упр-ощенческим
-допущениям, :как ш.ир<жое расn:ространен'Ие селективной нейтраль-
'ности. OH1a еще ожидает cBoer<> решения.
Вероятно, следует еше раз проверить .ПР3Jвильность .предполо-
ения о том, что данные о полиморфизме ферментных локусов,
.получ-енные элеК1'Iрофоретичеоким.и методами, м'ожно считать т,и-
n:ичными для rеНО1'ипа в целом. По видимо.м'У, мы непра;вомочны
()ценивать общую частоту Iполиморфиз'Ма :путем простой экстра:по
ляции данных. о ча<:1'Оте полиморфизма 11'0 белкам. Су марный по-
.J1ИМОр.физм и сеrреrационный rруз в .популяциях м{)жет оказаться
не таким большим, как это считают М!НОl'ие авторы В настоящее
время.
Холдейновская плата за отбор
.Субституuи'Онный rpy3 тесно связан тем, .что Холдейн ('Halda
пе, 1957, 1960) .назвал ,платой за lотбор IИ что .составляет основной
предмет обсуждения этой .rлаIВЫ. По холдейновской концепции
плата за отбор это общее число смертей, ВЫЗlван,ных отбором
в пр.оцес,се .ПОЛНОfО замещения одноrо те.на друrим. След;ователь
.но, .плата за отбор это то же самое, 'ЧТО и 'КУМ'УЛЯJrИiВНЫЙ ,субсти-
туци.онный труз за ряд поколен.ий, а iCубституциоНiНЫЙ труз то же
.сам.ое, что Iприращение пдаты за {)тбор за каждое поколение.
Две первые работы Холдейна (Haldane, 1957, 1960) содержат
1.lатематичесК'ий анализ CYMMapHoro или кумуляти.вното субститу-
ционвоrо rруза в эаВИСИМ-ОСl1И ОТ веЛИЧIИНЫ поuyляци.и в :каждом
поколени.И. Он раоесматривает влияние, оказы.ваеМoQе на эту зави-
.симость И'нтеноИlВНОСТЬЮ "Отбора и .друrим:и фа.кторами, а также
Б '1,ияние платы за .отбор на скорость эволюции. Эта проблема
была вновь ИССJlедована рядом ученых, tCформулировавш'их ее по
новому (Crow, 197() Crow, Kimura, 1970 Flake, Grant, 1974).
Число особей, .элиминируемых -отбором в процесс :полноro за 4
м-ещения .одноrо аллеля друrим ( D), во MHoro ,раз больше ч,И'сла
вз.рослЬ1Х особей в любом .Q11Дельн.о взятом поколении (N.). Между
1:.D tИ N существует вп.олн-е определенная зависимость, выражаемая
коэффициентом платы (С). В общем виде если N имеет ПОСТОЯН4
ное значение, то 'f.D==C.N.
r л. 14. п Аата за отбор
14З.
Формула ДЛЯ С для QДнюr.о лоюуса ,в ,rаплоидной си теме (KOTO
рая .в алreбраическом смысле rораздо iлроще, чем для ди.плоu.ДНОЙ
системы). выводится следующим образом. Плата, приходящаяся
иа -оди.о поколение, рави'а от.иошемию
wlsq
QJ
или
WI W
W
rде WJ приспособленн'Ость ал.пеля, ,КОТОРОМУ бла'r.оприятствует
от60Р, w .средняя ,приспособленнос.ть ПОПУЛЯЦИlи, а q ----IQ,а.сТоТа
менее .ПРИICпособлеиноro аллеля. Сумм'а'рная Iплата за полН'Ое за-.
м-ещение ОДНQlrо алделя дрУ1rnм .составит при этом
C== Wl W .
W
Значение С зависит, Та'КЯМ образом, .от исходной 'ЧаСТQТЫ алле-
ля, 'которому блаrо.приятст.вует отбор, он от скорОсти замещения.
Это зна:чеНlие .велиК'О в случае н иЗ\кой ИСХОДНОЙ частоты аллеля,
которому блаrопр.иятствует отбор, и уменьшает.ся ос Iповышением
ИСХI()ДНОЙ Частоты. Кроу .и Ки.мура (Crow, Kimura, 1970) п,риводят
следующие значения С для ра'зных изменений 'Частот reHOB (q)
в ДИ'ПnОИДНОЙ ОИlCтеме при ,от.сутствии доминаНl'IНОСllИ:
q с
O,OOI O ,999 ] 3 ,81
О ,OI O,99 9,]9
О t 1 0.9 4,39
0.2....0'8 2,77
При ПОСТОЯННОЙ ИСХОДНОЙ частоте reHOB значение С изменяет-
ся ,в за'висимост.и от скорости замещения. Скорость замещения за-
виоит в свою очередь от интенсивности отбора, и поэтому 5 И.1И W
входят в формулу для вычисления вел.ичины С. .суммарное число
rибелей за счет отбор.а при замещении .одноrо ,reHa друrим о'казы-
вается практически незавнч:и мым от интеноивности отбора;. .но чlис
ЛQ по ко.лен<ий, на кoroрые ра.с.п,ределяюr.ся \rибнущие в проuесс-е
oтr60'pa особи, измеряется интенсивностью отбора (5).
ХолдеЙ1Н (Наldапе, 1957) получил численные оценки платы за.
01'бор в ти,пичаой -ситуации. Он исходил из Toro, что ,существует
некая обшщжая диплоидная .штуляц'Ия, соде'ржащая блаrопр'ият-
ный мут.знтный аллель с низкой исходной ча-стотОЙ. Предпола-
rается, Ч1'О этот новый аллель обладает YMepeНtHЫM се.'lект,ИlВНЫМ
преи:мущест,вом по сравнеиию .со -старым аллелем. При этих усл.о-.
виях сумма'рН'ое ЧИ'СЛО особей, rИ'бнущих в ,проuессе п.оЛ1НОro заме-
щения QДHoro reHa друnим, обычно в 10......J20, а иноrща даже
в 100 ,раз ,выше чи.сла размножающихся особей в -ОДНОМ поколении.
(c 10..........100). Приняв в качестве репрезенrати.вноrо KaKoe TO 'КОН-
кретное чи'сло, С.'1едует ожидать, что в среднем 'чи.сло особей, rиб-.
144
Часть 1//. Естественный отбор
иущих за счет -отбора в процеосе замещения reнa, примерно
8 30 раз больш.е числа взроCJIЫХ .особей на 'ОДНО ПОКl()ление ( D ==
==30 N; С==3().
Плата за отбор и скорость ЭВОЛЮЦИИ
БеJIlИ -скорость замещения (s) /Высока, то замещенtИе одноr:о reHa
.друrим 'будет .происходить быоСТРО, во !Всяком .случае в теории.
Одна,ко '.на -самом деле ПОПУЛЯЦИЯ мажет оказаться ,не в сocrоянии
уплатить ту цену, IВ .к,оторую обходится интен.с.ивный отбор. <: иш.
ком сильное давление от.оора .может \привести .популяцию к If1ИJбе..,'и.
Число тенетичеоких смертей, .которое мо()жет .выдержать по'пуля.
ция в каждом отдельном поколении, cTporo оrраничено. Поэтому
J'роцеее замещеНсИЯ ОДНоrр reHa дру'rим должен про.ИСХQiДИТЬ на
.протяжен'ии .мноrих поколеНlИЙ, ДЛЯ Toro чтобы !популяция моrла
постоянно с.охранять достаточную 'численность; '3то условие уста-
.навливает верхний предел скорос!'и ЭВОЛЮЦИОliJн{)'rо изменения для
тдеЛЬНQlfО reHa. В том .случае, коrда kD == 3{) N, замещение reHa
мажет .произойт.и за 300, но ника;к не за 30 .поколен,ий (Haldane,
1957) .
Плата за 011бар 'по двум ИЛIИ большему числу незаiВИС:ИМЫХ re-
:нов еще вЫШе. Эта ,плата возрастает экспонею.l.иально с увеличе.
яием чи'Сла незави.С'имых reHOB, одновременно подверrающихся
отбору (Haldane, 1957). Поэтому в популяции, непрерывно со-
храняющей Жеизнеспособность, теоретическая маКоС,има..,1ьная ск{).
рость, с которой может ПРОИСХОД:ить ЭВО"'ЮllJионное из,ме-неН1Ие по
нескольким или мноrи"М reHaM одновремен.но, должна быть rораз-
до ниже, чем 'дЛЯ любоrо отделыноrо признака, опреде..1яем.оrо €tди-
нич,ным 'reH'OM. Таким образом, если замещ н,ие еДИН-ИЧНQlfО reHa
м'Ожет прои'зойти Bcero за 100 поколен:ий, то .на замещен:ие !ДВУ'Х
lIезависимых тенов, отбо.р по ,котО'рым происходит одновременно,
может ,потребоваться МИНИМ1)'м 200 1I10колений.
Ка'к отмечалось .выше, -скорость замещения для лю60rо данноrо
-lИела незавиеимых reHOB, определяющих отдельные признаки, каж-
дый из к{)торых детеРМ'Ионrирован еДИ,НIИЧНЫМ reH'oM, 'cTpO'ro оrpани-
чена. Оrраничение скорО'Сти эволюции становится еще 'более Жест-
им, если это же -самое IЧИ-СЛО неэав.исимых reHoB ,вход,ит в качестве
составных частей в новое ада:птив.ное сочетание тенов, потому что
при этом на п-у-nи бысrроrо зз-мещения множества reHoB 'Встают
два связан,ных между обой препятствия. К IВЫСОКОЙ п.'lате за OT
бор, направлен.ный на замещение -множества reHOB, доба.вляеreя
неп'рерывное разрушение бла.roприят.ноrо сочетания IreHoB в резуль.
'Тате половоrо процееса. Действи-е ЭТ-ИХ BYX фа l КТОрОВ проЯ'вляется
с особой оилой, кorда алле..1'И, входящие 'в состав H'OBoro сочетан-ия
reHOO, все еще редки ,в даНIНОЙ по;пуля-ц'И'и.
Все эти соооражения заставляют СЧlитать, что эволюцион.ные
liЗМенения СЛ.ОЖНЫХ .признаков и сочетаний признаков обычно пр
rA. 14. ПАата эа отбор
145
ИСХОДЯТ С умеренной быстроroй иЛ,И медленно. ЭМlпи.ричеСКlи,е дан.
ные ОТlносительно окоростей эволюции в некоторых л!иниях, .эволю-
ционирующих медленно или с умеренной 'быстрот<>й, У'кладываются
в теоретические пределы, устанавливаемые платой за отбор ('Hal-
dan-e, 1957).
Извест,но, однако, неск{)лыоo пр.имеров быстро протекающей
ЭВОЛЮЦИИ, скорость которой пре.вышает те<>рети'Ческий П'Реде 'I.
Один JИз них .будет здесь описан.
Некоторые расы Miтulus guttatus (Scrophulariaceae) занима
ют в штате Юта местообитания плейстоценовоrо возраста. Счи
тается, что возраст этих местообитаний составляет 4000 лет. Сле
довательно, расовая дифференциация у Miтulus guttatus про-
исходила в штате Юта в течение 4000 лет, за которые это
травянистое растение моrло пройти через 4000 поколений (Lind
5ау, Vickery, 1967). Различия в признаках между разными расами
друrоrо травянистоrо растения Potentilla grandиlosa (Rosaceae)
обусловлено аллельными различиями по крайней мере по 100 re-
нам (Сlаusеп, Hiesey, 1958). Допустим, что rенетические различия
между расами Miтulus Toro же порядка, что и у Potentilla. В Ta
ком случае замещение 100 reHOB должно было произойти за
. 4000 поколений, т. е. в среднем за каждые 40 поколений заме-
щалось по одному reHY. эту оценку следует считать у еренной в
от.ношении как времени, так и числа reHOB.
Таким образом, ,скорость оЭВО.i1ЮЦИ:И некоторых орrа,н'из"Мов, оче
видно, .превышала поro ,()к, устанаВЛ1lfВаемый терпимой пла-rой за
отбор. .ВО.ЗНlикает естественный вопрос: 'как ЭТО им удалось?
Сцепление reHOB и взаимодействие reHOB;
ИХ влияние На оrраничение,
иалаrаемое платой за отбор
В своей первоначалыюй моде.1JИ Холдейн ПОСТУJl"Ир'овал ,боль-
шую вe.rшчину популяции, незаВИ1СИМОСТЬ IдByx или lБОJ1ьшеrо Чiисла
rеfЮВ, одновременно л.ретер.певающих замещение, .и ряд ДрУ'I1ИХ yc
повий. Эти услоВiИЯ, несом.ненно, нередко реалиэую11СЯ в прир.оде.
Столь же :верно, что \в пр.иродных попудяциях часто встречаются
отклонения .от 'этих условий. Некото.рые из таких .оТ'клонений ос-
лабляют оrраничен ие, налаrаelм.ое на скорость IЗВО 'ЮЦИИ платой з.а
отбор (ср. Grant, Flake, 1974b).
ДОII)'1сТ'им, что отдельные ,r-еНbl, входящие в новое ПРИCJrюсоби.
тельное сочетание reHoB, тесно сцеплены между .собой, О'бразуя
суперrен. [ены, входя[Цие в состав этоrо суперrена, MorYT заме
щаться .с такой же 'Ск{)ростью Iи за ту же плату, что и один reH.
Плата за отбор для одноrо суперrена .не выше, чеМ для ед.инично
ro меlfделевокоrо reHa. Если rены, составляющ,ие су.перrен, тесно
сцеплены м'ежду собой, т{) -популяция -может приобрести новое
146
Часть ///. ЕстествеюtlJlй отбор
приспосо6итеЛЫlOе сочетание reHo,B за сроки, оrранич-енные та кои
же платой за 011бор, как и при замещении oдaoro ;reHa (Grant, Fla-
ke, 1974Ь).
Некот<>рые типы взаимодействия между rенаМIИ та,кже, очев.ид
НО, .СНИЖают плату за замещение мноrих [е'НОВ. Та'К, ве]IOЯТНО, об
<';10'8Т дело В тех слу,чаях, Коrда селекти.вные преимущества или
.недостатюи отдельных reHOB, ПОдJверrающихся отбору OДHOBpeMeH
но, коррелированы между собой. Различные аллел-и, 'кот<>рым бла
rопр,иятствует отбор, MorYT ,ино,rда ветречаться вместе В ОДНОМ
И том же .rеноТ!ипе; и наоборот, разлИ'Чные аллели, которым отбор
не ,бла.rОПРИЯТС1'вует, MorYT в'стречаться также .вместе в одном ,н
том же альтернативном rенотипе. В таком случае rенетическаи.
rибель, <>6УС.'Iовленяая м,ноrочисл€НrНЫМ.Н 01'дельными тенами, .cBe
дется к rиб€л,и сравн.ительн() нооольшоrо числа ос{)бей. Это в свою
очередь делает /возможной более выоокую скорость эволюции ЛЯ
Tor<> или :иноrо Iпризнака, .контролируемоrо множественными !'ена-
ми (Mayr, 1963; Mettler, Gregg, 1969).
Влияние структуры ПОПУЛЯЦИИ на оrраничение,
налаrаемое платой за отбор
Отклонения от допущения большой Iнепрерывной пonуляЦlИИ
также MorYT снизить оrраничение, налаrаемое платой за отбор на
скорость замещения мнvжества reHOB. Рассмотрим популяционную
структуру двух типов: серию популяций-основате.пьНИЦ и подраз
деленную, или колониальную, популяционную систему (Grant.
Flake, 1974).
'Следует помнить, что плата за ,оroор ДОС'I1иrает наи-вы.сшеrо
ур.о'в'ня при низкой частоте тех аЛ.lJе.rrеи, 'которым .блаrоприятствует
отбор. Если частота этих зллелей может быть повышенз за счет
с.аучайных несе.'1е'КТИВНЫХ .фа'ктор'Ов, ТО 'это 'пО'звол'ит и беЖать наи-
более обременительной часl'И общеrо субсl1итуционн.оr<> rруза. l\1e
ханизм Д.'1я подобных изменений существует в попудяциях-основа
теv'lьницах.
у ВИда, обладающе:rо тенденцией к расселению, оБШlИрная по-
.'1им.оРфН3Я n-ре'Д'ковая популяция может .дать НЗ1ча..l0 сер-ИIИ дочер
них колоний, закладываемых QДНОЙ или неск:олькими особями_
Такие дючерние JКОЛ<>НН 1 И, или iПОПу.пяцин-основательницы, ,берут
начало от неслучайн{)й выборки из преДКО'ВQrо ,rенофон.да. Аллели,
бывшие ранее ред,КИ1МИ, MoryT досТ'ИrllУТЬ средней :Иv'lи высокой ча
стоты п<>д деЙств'ием чаСl1lIЧН-О случайных фаlКТОрОВ в период эа
крепления H K<>TOpЫX из иовых дочерmих колоний. Э И колонии
'просто из:беrаю1' значительной части субституционноrо rР'Уза.
И они мюryт Iвыдержи-вать более низкую ,плату, налаrаемую отбо
ром за IBC€ дальнейшее ПQJ3ышение частоты re-HOB, вплоть до пол-
Horo замещен'ия.
rл. 14. Плата эа отбор
147
Селекrивная .rибель ,неумолимо .собирает с популяцИ/И с.реДIНЮЮ
пошл'ину за любую данную интенсявн'ость отбора. Попу.ляц.ия, од.
иа.ко, обладает .QIIределенным репродуктивным ,ПО1'еНциалOlМ, кото-
рый ,лоз.воляет компе-нС'ировать нормальную долю случайной rибе
ли, а также известную часть селективной rибеЛtи. При некоторых
Т.Ипах .ПОПУJlЯЦИОННОЙ структуры уровень случаЙ1l'DИ смертности
может варьировать, повышаясь или ЛОllижаясь. В тех СJlучаях,
коrда смерilЮСТЬ понижается, создаЮiСЯ условия для более вы-
сокой селеКТИВНОЙ rибели.
Возьмем в качестве стандарта обширную непрерывную попу
ляцпю. Ее общая 'Сумма селеК11ИВНОЙ и случайной rибели устанав-
ливает предел сК'Орост.и эволюl.l;ИИ. Рассмотр'им теперь популяцию
Ta'J<oro те размера, 'K K crа.ндаРТlная, и подверженную тем же са-
МЫМ внешним воздействиям, н.о разделенную на части. В такой
разделенной \популяции уровень случайной смертности будет раз
JI:JfЧIНЫМ в раз,ных колониях. Колон.ии С более низк{)й случаЙной
смертностью MoryT выдер.жать ,более тяжелое, чем .в среднем, бре
мя .селекl'ИВНОЙ r.ибели, а IПОЭroму oryr по.эвоЛИ7ь себе эволlOЦИО
нИ'ровзть С более высокой ОКD!Х>СТЬЮ, чем большая стандартная
популяция.
Часть IV
Проблема приобретенных признаков
r лава 1.5
ФБНОТИПИЧЕСКИЕ М1ОДИФИКАЦИИ
Теория наследования приобретенных признаков
Вопрос о том, наследуются или не наследуются приобретенные
признаки, 'П>()чти .сто лет был основным предмеroм диС'куссий
в эволюционной биоло]'ии. Сroрон'ниюи традиu;ИоОННЫХ пред'С'Тавле-
ний "о iНа'Слetд'овании пр'Иобретенных признаков, несмотря на неодно--
кратные опровержения их теории, Не сдавали своих позиций. В не-
которых случаях эти повторные выдвижения теории были науч'-
ным анахронизмом.
.Основная 'причина такroй ДЛ1ителнн.ой дискуссии заключает-ся не
в Iнедостатке данных, поск'ольку по ряду аопектов этой "П!роб.'1емы
фак'Тическоr-о материала более чем достаточно, хотя 110 Д:РУlrИМ ее
аопектам Ta'Koro материала iCЛещовало ;бы /Иметь Irораздо боЛЬШе.
rлавным ,источником .путаницы в -прошлом :было оroут.ствие точно-
ro определения смысла термина .«приобретенный /Признак» или C!KO
рее неуме.ни.е дать Ta1<lOe определение.
При-обретен.ные (признаки мотут быть либо наследственным.и,
либо ненаследственны/Ми, о чем свидетельс11ВУЮТ та:кие .пр.и еры,
как передающиеся по 'Насле\дcrву .Иlflф кционные за,БО.1евания и
ФеНОТJlThиче(жие модификации соответственн'О. Необходимо разли-
чать два типа .приобретен'Ных ,при'знаков: фенотипические призна-
ки, опреде.'1яемыс вновь приобретенным rенетическим материа.тJOМ,
и 'ПРИЗlнаки, воз-никающ'ие в результате .феНО1'ипичеоких реа'кций
на ,внешние СТИМl)"лы, т. е. .феноти'ПичесК'ие ,модификац,и!и. Эт.и два
типа .лриобретС'нных пр.изнакюв rиrрают совершенно разную .роль
в на,слеД'Сl1венности и эволюции.
Концепция о ненаследовании фенQ ипических дификаций в
настоящее Вrремя .наСТОЛЬК-Q тв-ердо установлена и общепри.нята п
кажется 1'а I КОЙ далек'Ой от ддскуссио.нной стадии, что представ-
ЛЯе11СЯ нел'ишним lIаПQМНИТЬ .самим себе, .сколь она 'Недавнеrо про.
исх.ождения. Противоположная тачка з.рения :nредста lеНlие
о наследовании приобретенных признаков типа фенотипических
реакций разделялась всеми па протяжении большей части исто-
ри,и биолоrи'и ---- IQТ r'иппократа до Ламарка и Дар.ви'на. В частно-
r л. 15. Фенотиnичес"ие .модификации
J49
сти, у'чение Лысенко, раопростран!ившееся в СоветClКОМ Союзе
в период с 1939.по 1964 [., ,представляет .собой 80ЗрОЖ'деаие у.ста-
ревших 'ВэrлядО'в на наследственность, имевших столет.нюю дав-
ность.
,Современная точ'ка зрения, cor ласно которой фенотипичоок:ие
модификации не наследуются, имеет rораздо более короткую исто-
рию. Впервые она была Высказана Вейсманом (Weismann) в ero
кн"И'rах «=Очерки о наследственност.И (1,889 1.892) и «Зародышевая
плазма» (1893). Однако взrляды Вейсмана не были приняты сра-
зу; некоторые американские биолоrи все еще возражали против
них в тридцатых rодах, французские биолоrи продо.пжали иrнори-
ровать их в пятидесятых [одах и еще в шестидесятых [одах они
отверrаJ1ИСЬ <:ове'J1CКИМИ .биолоrам.и.
Разноrла.сия . еж..ду вейсман,измом и традициoQННЫМ-И ,взтлядам,и
на наС.тJедст:веНlIОСТЬ, которые о.бычно на.зывают ла.. арки.зМО 1.
БЫ.1И внача.1е сосре.доточены BOKpyr про.вОДИlмоrо ВеЙСI iаIЮМ раз-
rраНИочения :между СО:\-10Й И зародышевой плазмой. Вей<=ман выде-
.JIЯ.'l .катеlrори/О cOMaTo.reHHbIX вариациЙ. Под ними он имел в ВИL'1.У
реа,к,ции 'С'ОматичеС'Ких ТКаней или орrанизма в целом на внешние-
воздеЙС1\ВИЯ, в том 'Числе на ЛОIВ.режден'ия, и .реЗУЛЬ7аты у.пражне.
н:ия И неупраЖiН1ения. Он убедитель'но :И.lИ IПОЧТИ вп{мне убедитель-
но доказы,вал, ч1'О поскольку .сома отде.'Iена от русла на'СлеLl1СТВСН
ности, то и cOMaToreHHbfe вариации не MorYT попасть 'в это 'Русло.
Ра-здел.ение на зар-одышевую плазму и OMY вскоре у-ступило
место{) разделению !на ,reнО'Тип и ,фен'Отип (Johannsen, 1911). Первое
разд.еление .носит ч:исто ана1'омический .Х, ара-К1'ер rorда ка,к второе-
имеет rенетическую и морфоrенетическую основу. Концепция re..
НОТИllа феНОТИопа более .совершенна и нмеет roраздо более широ
кое приложение. ,Некоторые соматоrенные .вариации MorYT быть.
нас.Т]едственными. Однако фенотипические признаки никоrда не-
передаются -по нас.'1€ДС'I'ВУ. А \Поэтому феНОТlипические реа.кции на
внешние ,стим,vлы ненаследс,.,венны.
ПРИНЦИ1П не.наследуемости фенО"тИ'пИ'чеоких м-одификаций оста!в-
ляет, однако, за бортом те признаки, которые детерминируются
приобретенным rенетическим матер.иал-ом. Такие призяа'ки ,Mo.ryl"
стать на .Т]едственными и тем ca ЫM иrрать опре.деленную роль.
в э.в'Олюции.
В части IV наСТОЯLЦей книrи два отмеченных типа приобретен
ных пр.изнаков .РЗ1ссматриваются .по отдельности: ,в этой rлаове мы
обоудим ФеНС)1'И'шические модифика'циlИ; а эффектам Л'Риобр€те.н
нота rенетиrческоrо материала поовятим 'f.Т'f. 16 и 17.
Фенотип
fенотип это{) .вся СО'ВОК J1IНОСТЬ IreHoB данноrо орrанизма, а фе-
нотип ---- ero особый набор выражений признаков. \ежду rеноти
лом Iи фенотипом наХ'Одят-ся длите.",ьные и СЛlQжные процеС'СЫ дей-
150
Часть lУ. ПроБАема приобретенных прuзна"ов
ствия reHa и процессы развития. Эти про[ ессы протекают в опре-
делен!ной среде и находятс ЛОД ее влиянием. ФеНoQтипическое .вы-
. ражение пряэнака -представляет собой, таким 06ра:юм, результат
действия двух наборов факторов: детерминирующих rеНО1lwпи'Че-
<жих факторов си ус..100ИЙ среды, в .к.оторой пропсходит раэв'ити.е.
Каждый 'flенотиrп в разных ,средах способен дать на'ча."О опреде-
ленному С'пектру фенотипов. В этом смысле rеноти'п мож,но пред-
8'( света + /6ч темноты
2цч света
в отКР61том
ZPYHт 11
ПасаgСНl>
СМ
40
см
40
17"/I7"С
26 '117 ос
Рис. 15.1 Ростовые реакции отдепьных частей OAHOro растения Achillea borealis
из Сьюарда (Аляска) в шести разных условиях (Clauscn, Keck and Hiesey. 1948).
Первая цифра относнтся к дневноА температуре. а вторая к ночной. Пернод росТа про-
должался 3.5 мес.
.ставить .себе :как «норму реакции» (Johannsen, 1911). Иными сло-
ваМIИ, действия rенотипа ,нельзя считать -ни жестко пред-определен-
ными, ни неоrраниченными; rенотип может детерминировать ряд
фенотипических выражений в предедах, установленных самим reHO-
ти,пом (Johannsen, 1911).
Ш.ирокий спектр Iфенотипи'Ческих выражений .пеr.ко 'Продемонст-
рир.овать у растен.ий, способных к веrетатИ'&ному разм'Н'ожению.
Для этоrо достаrточно взять одно растение, разделить ero на части
и .размножать 'эти части при различных у.с.'10ВИЯХ Iсреды .в фитотро-
не или в веrетациoQННОМ домике.
Хорошим 'пример.ом служит э.КCJпер.иментадьное изучение pea'K
щий иа -внешние условия у <:.пожноцветных .rрУ'ЛПЫ Achillea millefo
Нuт (Clausen, Keck, Hiesey, 1948), мноrолетних травянистых pac
тений, которые можно размножать веrетативным путем. В Паса-
.дене (штат Калифорния) части одноrо и TOro же растения выра-
'ЩИВЗJ1И в разных камерах фитотрона, а также в открытом rруите
зимой. Все реrулируемые среды различались по продолжительно-
'сти CBeTOBoro периода, дневным и ночным температурам.
r А. 15. Фенотиnические модификации
15Ь
В ТППИЧIfО.М экеперименте шеC'l"Ь ча'Ст й о.цноrо растения Achil
lea borealis из Сьюа'рда (А.I1ЯlCка) .выlащивал-ии в течение 3,5 мес-
в пяти реrулируемых средах в фитотроне .и .в шестой Iсреде в от--
крытом трунте. Различные .ростовые реаIКЦИИ, наблюда'вшиеся'
к концу эк.сперименталь'ноrо периода, изоб.ражены на рИIC. 15.1....
r.е.ноти.п этоr.о раст ния обусловливал хороший рост в одних усло-
виях ('длинные Ten"1ble днlИ) и слаiбый рост в дlруrих (короткие'
дни). Друrие rен,отипы эТоrо же вида реа-тировал.и на эт:и условия:
по-иному (Crausen, Keck, Hiesey, 1948).
Приспособитепьные CBO CTBa фенотипических
модификаций
ФеНОТИlпичеС1кие реакции на нормальные из-менения в среде-
обычно н.осят пр.иопособительный хара.ктер. Это можно видеть на'
лриме.ре Л:ИС1'ьев растений, нахо!Дящихся на овету и .В тени. У л-и.сть--
ев, находящих я в тeJНИ (<<теневые листья»), 'площадь поверхности'
больше, что увел.ичивает их фотосwнтетическ'УЮ способность и'
компенсирует более низкую освещенность, тоrда как у листьев..
нахоtДящиXtСЯ на rCо..1J'нце (<<!солнеЧ'ные листья»), П..1:0щадь поверхно
сти меньше, что понижает транспирацию и потери воды. Один и.
тот же rенотип способен обеспечить развитие листьев этих двух
типов в ус.rIOВИЯХ сильной И слабой освещенности. Способность.
растения к подобным различным фенотипическим реакциям, об-.
условленная rенотипически, дает ему возможность приспосабли-
ваться к варьирующим условиям освещения.
Диапа:юн фенотипичеС1КОЙ модифицируемости в разных кру.п--
ных rруппах Iвысших орrанизмО'В Becь' a различен. Высшие расте-.
ния, и в частности траВЯ1нистые ра'Стения, ха1рактеризуются боль-
шой феНQтипи'Ческой пластичностью. На противопо..10Ж'НОМ KOHЦ
этоnо диапазона .находя11СЯ .на.секомые, ОТ..1ичающиеся крайне низ-
кой .фенот.ипическ.ой rибкостью. Эти различия в степени модафици-
руемости .ортан:И'зма непосреДIC'J1венно С'вя.:заны с различиями в ти-п.е.
развития указаfННЫХ двух r.рупп. Раст.ите,,1ЬНЫЙ орrанизм раэви-
l3a-е'ОСя И'3 тачек роста, котО'рые подверЖe1-lЫ сильному воздействию'
факroро.в среды, .ПJрообла'дающих в период фор ироваlНИЯ ero но-
вых частей, так что -«солнечные ..т:IИС1ЪЯ» 'появляются .на ярком све-
ту, а «теневые листья» при слабой освещенности и так далее.
В отличие от 3TOro орrанизм взрослоrо HaceKoMoro развивает-
ся внутри твердоrо наружноrо скелета, который формируется на
одной из предшествующих стадий развития. rлавные внешние при-
знаки орrанизма HaceKoMoro закладываются задолrо до Toro, как
опи приобретают функuионадьное значение. а к тому времени
коrда их можно начать использовать, они успевают принять OKOH
чательную форму, не поддающуюся изменениям.
152
Часть ТУ. Лро6леJlа приобретенных nриэнаков
Помим,о 'этих различий в 1'Иtпе развития сущеC"DВУЮТ также бо-
лее rлубокие ра,зличия в стратеrии Д-рИClIЮCобл€ния особи К ко.лоо-
JIЮЩИМСЯ усл.овиям .среды. Насекомые ПО,lJJВИЖlны, а растения
прикрепЛeJIЫ IK опреде."енном.у месТУ. Нзрослые на.секомые в из-
вестных пределах MorYT прИ\Споса:бливаться К переменным факт.о-
рам !Среды, переходя ,в более ТeJПЛoQе место в ХОЛОДНУЮ ,поrоду или
в .более 'Вла'Жное ---- 'при засухе, ,Т. е. выбирая ,себе наиболее подхо-
дящее местооби.тание. Растение же, удерживаеVIOе на ОДНО I месте
кор'нями, не имеет выбо.ра; для носо rла'Вным средством И'НДИIВJИ-
дуа.'lьноrо при<споообления к изменени'ям с.реды .служит оп-особ-
ность к фенотИ!пическим M-ОДИФИ1кациям. .
KalK ТIrПЫ феНоОТ,ипичеС1l<JИХ модификаций, так .и их ДИ3'uазон за-
висят от потребностей орrанизма. П:оэто'му весьма .вероятно, что
rенотипы подверrа:JIИСЬ отбору ,на пособность 'К ,проя;вл нию ,при-
способительных фенотипических реа ций на измеН.ение условий
среды.
Роль фенотипических модификаций в эволюции
Ф-еНОТИПlичес!кие О'ди'фи:кации охраняют 01'делыный орrаниЗlМ от
стрессовых IIOЗд.еЙ'ствий среды и IВ той мере, .В к3'К'ОЙ 'это им У дает-
я, создают буфер, .nротивостоящий отбору, 'осущеС1"Вляемому сре-
ДОЙ. Теоретически популяция, оостоящая из теноти1ЮВ, моrущих
обеспечить полный "Спектр приС'пособительных фено'Тип:и'Ческих
реаIКJJЩЙ, .вовсе не 'будет 'реа:rИ'ровать 'На даlвление eCTeC'ТBeHHoro от-
бора.
ФеНОТИlпическая "Пла.сти'ч.ность оказывает задерживающее 'В."ИЯ
иие на селеКТJflВНoQе lВоздеЙСТJ3ие peды, и этот 'задерживающий эф-
фект сказывается как на неrативных, так и на позитивных аспек-
,.ах "Отбора. Фенотнпическа-я ,пла'стичность оБУ1СЛОВJIIивает запазды-
вание Iэлиминwрующеrо действия 'отбора; она дает 1ЮЗМОЖНОСТЬ
популяции остаlваiЬ'СЯ в среде, которая изменилаlCЬ lJIеблаrоприят
ным обра'3О'М. Эта ,отс.роч'ка ' лиминации уведиtlИвает .ша-ноСЫ lПопу-
.ляции на при обретение в результате мутационноrо проuесса, пото-
ка reHoB и реКОМ1бинации новых rенетических .вариаций, которые
дадут ей rвозможность создать rеноrnпическ:ую реак.цию .на новые
неблаrОПlрияmые условия 'СредЫ.
fлава lб
ТРАНСДУКЦИЯ И ИНДУКЦИЯ
Трансформация
Пер'ВЫЙ заtCJIуживающий доверия случай наследст енноro изме-
нения, :ОЗ'НИ1кшerо IB результате прио:бретения чужоro тенетичеСIКО-
ro материала, .был .обнаружен у пневмококка. Трансфо.рмаци.я
у пневмококка была открыта rриффитом (Griffith) в 1928 r.,
а в 1944 r. Эвери, Мак-Леод и Мак-Карти (Avery, Macleod,
McCarty) ПО.'1учили ее in vitro. Все основные факты, связанные с
этими открытиями, в настоящее время общеизвестны.
Штаммы п.нев'МО'Кокка разли'чаются 'по характеру поверхности
оболоч.ки 6а'ктериальной 'к..летк:и, 'которая может 'быть rладкой 'Или
шероховатой, и степени виру..1ентности, корре 'Iированной с xapaKTe
ром поверхност.и. Эти ра'зличия ОН ас.ледственны. Если выращивать
оди'н штам'м Iба.ктерий 'на сте.РИЛЬНОМ экстракте, 'С'Одержащем днк
д-руrоro штам'ма, то кдет.очна.я поверхность некоторых дочерних
клеток пер во.rо штамма 'по своему виду .начинает на'поминать кле-
точную поверхность BToporo штамма; та'кие дочерние клетки при
обретают та,кже 'и .вирулентность, хара,ктерную для ,клеток .Бтороrо
штаММа. Эти изменения передают.ся по :на.следст.ву. Наследствен-
ные призна'ки штаМ'ма донора пе.редаются ,штамму :реЦlDпиенту
через ДН,К донора, нахоОДЯЩУЮСЯ ,в стер-ильной 'рост.авой среде
(Avery, Macleod, McCarty, 1944).
Наследственные трансформации, вызываемые стеРИ lЬНЫМИ
экстрактами, обнаружены и у бактерий друrих родов (Hemophi
lus и Streptococcus) , и притом по друrим признакам по устой
чивости К пенициллину и стрептомицину (см. Hayes, 1968, rл. 20).
ЭТИ транс-формации tiактер:ий ,на1блюдаются .в ла.борато.рии.
Возникает .вопрос: происходит lИ у бактерий а-налоrичиый Iпроцесс
в естествен'ных условиях? Природным анадоrом трансформации
ЯВiJIяется трансду.К!ция.
Трансдукция
В естественных у ловиях Iба.ктериальные ВИ'Русы, ил.я фати,
иноrда перен ят rенетический материал из одной бактериальн й
клетки :в друrую. Этот перенос Iбз-ктериаль.иоrо rенет.и'чеокоrо ма-
териала фаroм известен ПОД .названием траrн.адукции.
В -общем пр.оцесс 1'ра.нсдукци'и :протекает еледующим Qбразом:
фаr заражает каКУЮ :lибо бактериальную клетку и размножается
в ней за .счет ее дик. Затем дочерняя фа1rовая 'частица 'заражает
54
Часть /V, Проблема nрuобретенных nрuзнаКО8
.друryю бактериальную клетку и ,вносит ,при ,этом rенетический Ma
'териал пеРIВОЙ КЛе1'l}(и хозяина -во .вторую .клетку-хозяи'на. Наибо-
..пее успешную трансдукцию осуществляют умеренные фаrи, кото-
.,рые .не .вЫ'ЗЫвают полноrо разру,шения бактерnи-хозяи.на. Трансду-
.цированные бактерии лродолжают при этом свое существование
и ВОС<Пр()ИЗ1водят вои измененные 'признаки.
Установлено, что трансдукция происходит у бактерий таких
.,родов, как SalfJ1onella, Escherichia, Pseиdomonas, -Staphilococcиs,
.Н некоторых друrих. К числу признаков, передаваемых путем
.трансдукции, относятся способность к синтезу тех или иных Be
ществ, У('1'ойчивость к антибиотикам и подвижность (см. Hayes,
1968. rл. 21). в одном случае конверсии у Clostridiuт изменяется
тин токсина, вырабатываемоrо бактерией. Один из штаммов Clost
ridiuт botulinит который обычно вызывает ботулизм у человека
}f животных, при заражении определенным фаrом превращается
в друrой штамм Clostridiuт, вызывающий rазовую raHrpeHY
(Eklutld, Poysky, Meycrs, Pe[roy, 1974).
Индукция мозаицизма
у Drosophila тelanogaster
Явления, аналоrичные трансформации, обнаружены и у MHoro
клеточных ор:rанизмо.в. Для оооз.начен.ия этих я,влений .I1СПОЛЬЗУЮТ
1'ерМИll «И.llДУКЦИЯ». В этом с..lу'чае под инду,кцией 'понимают Ha
слеДС'rвенное изменение, происходящее в мноrоклеточ.ных opra
иизмах под дей'С1'вием 'чужеродной Д,НК. Имеющиеся у нас oe:Beдe
ния об индукции все еще .очень отры,вочны инеполны.
Один из неМ.ноrих у.становденных nриме.ров ОТНОСИ7СЯ .к м.озаи
ЦИЗМУ тела у D. тelaпogas.ter. Яй:ца, .содержащие зародыши
п. melanogaster дикоrо типа иа ранних ,стадиях разВ'ития, обраt6а
ты-вали 'раствором, ,содер.жащим ДНК мух со специфическими -му-
тан1'НЫМИ признака.ми. В ,качестве rеJiеТ.ических маркеров ИСПОЛЬ-
зовали -мутации, изменяющие iпиrментацию тела, форму щетин.ок
и {)краску .rлаэ (Рох, Уооп, 1966, 1970).
Из обработанных таким образом зародышей развивались взрос-
.""ые мухи, обла.дав.шие ТС"М же фенотипиче.ским признаком, что
и .мутзнтный штз,м'м, ,послуживший донорам трансформирующей
ДИК. Более Toro, транС'фоРМJrlрованные призна и аказа.'JИСЬ .нз-
следственны.ми. ОДlнз,ко транс;формированные особи были мозаич
ными, т. е. участки мутантной ткани чередова.'1ИСЬ С норма.'IЬНЫМИ
участками; полной трансформаЦИ1И Н'сеro тела ид'и какой-то ero
части не ,на1блюдалось (Fox, Уооп, 1966, 1970).
Косnен'Ные данные поз'ооляют <:читать, что траlн.с.формирующая
ДИК не ,включзется 'В хромосомы 'мух-рециrпиент.ов. Вероятно, в
некоторых клеl'ОЧНЫХ ..lИНИЯХ ИЛИ сект.орзх 1'раiноформирующая
r А. 16. Трансдукцил и индукция
155
ДНК попадает в какие-то друrие части клетки. Такое распреде-
ление позволило бы объяснить lОзаицизм фенотипицеских эффектов.
(Fox, Duggleby, Ge[bart, Уооп, 1970; Fox, Уооп, Gelbart, 1971).
Индукция rибридной стерильности
у Drosophila paulistorum
D. paulistoruт rруппа видов, состоящая из ряда зарождаю
щихся видов или полувидов, обитающих в Центральной и Южной
Америке. rибриды между некоторыми из этих полувидов стериль
НЫ. Скрещивания между друrими полувидами дают стерильных
rибридных самцов. При иных скрещиваниях все потомство оказы
вается плодовитым. Мужская стерильность в тех случаях, коrдЗ/
ОНа существует, частично оuределяется факторами, которые пере
даются через цитоплазму яйца. В конечном счете мужская CTe
рильность rибридов обуслов.llена взаимодействием между этими
цитоплазматическими факторами и хромосомами.
ПредраlС!ПОЛoQженность "к стерильности, таlКИМ образом, -пере-
дается самками мух с определенной цитоплазматической консти-
туцией их rибридны'М ПОТОМ'Ка'м ,мужС'коrо рода. А поэтому тоз-
можно такое 'положение, коrда скреЩJmание в O,!:tHOM ,на!правлении
(например, CaHTa MapTa 9 х Мезитас а) дает в FI стерильных
самцов, а реци,пракное скрещивание (Мез'итас 9 Х Сапта Ма'Рта а)
дает в F] фертильных самцов (Ehrman, Williamson. 1965; WiIliam
son, Ehrman, 1967).
Уилльямсон и Эрман (WiIriamson, Ehrman, 1967) приrотовиди
rOMoreHaT и-з цит(}плаз:мы яиц линии CaHTa- \apTa и ввели этот
rOMoreHaT самкам линии Мезитас. Обработанных таким образо,м
са'МО'К Мезитас скрестили затем .с сам'цами Cahta-]v\арта. Обрабо
танные са'миш Меэитас (в отличие от .необра'бОТaJННЫХ са'моК этой
ли.ни.и) ПРОИЗrводили стерильных мужак.их r.ибри.дов. Мужская CTe
рильность ИНДУlI;иравала,сь 'Инъекцией родителю .ЖеН'с.коrо :пола ци
топл аэмы ocoooro типа.
Какова природа цитоплазматичеСI<ИХ факторов, обусловливаю.
щих стерильность? На электронных микрофотоrрафиях репродук
тивных тканей стерильных самцов выявляется присутствие в цито-
плазме частиu, сходных с вирусоподобным орrанизмом Mycoplas
та (Ehrman, Kernaghan, 1971).
ВерОЯ'I1ная rипотеза, .п.оз.воляющая объя>сНlИТЬ ЭТИ 'ра'знородные'
rрулпы фактов, состоит (8 10М, 'Что В кле11Ках HeKoropblx полу.видов
rруппы Drosophila paulistoruт живут в качеС11Ве сим-био.нт()В Му-
coplasrпa или Ка!кие То сходные с ними ортаН!И'змы. У ,ЭТИХ [[ОЛУ'ви
дов хромосомные rепы так ХОРОШoQ 1ЛIpи.rнаны>> к -зараженной цито
. ллаз1.tе, что (нормальная rпЛf()ДО.DИТОСТЬ не Iнарушается. Однако -при
скрещи'ваниях, IB которых .эти несущие lrм,бионТоОВ ПQЛУВИДЫ ие.
156
Часть /V. Проблема nриобретеННbtХ nриэНаков
пользуются в 'каЧОС1iве родитмьокой особи жен'окоrо 'пола, а .не
содержащие сиМrБИQНl'ОВ линиИ .в Качестве особи .МУЖlCкоrо пола,
ВQЗнИ'кают ,оочетаНlИЯ цитоплазмы и хромосО'м'ных reHOB, «не пр,и
rнаШIые» друr к друrу. Фенотипическим проявлением TaKoro вза-
имодеЙСТВИ1Я между цитоплаЗМ1ОЙ и ядром является муж.ская 'Сте-
рильность rибридов (Williamson, Ehrman, 1967; Ehrman, Kernag-
Ьап, 1971).
ИНДУКЦИЯ У льна
Реа I.JjИЯ растений на K-Qличес'РВО содержащ.их,ся ОБ их .субстрате
биоrенных 'элемент,ов N, Р и К общеИЗ.вестна. Хороший рост и
БО lьшие ур.ожаи можно получать на боrатых почвах, а на бедных
почвах растения растут плохо. Хор.ошо из.вестно также, что эти
реа,кци-и предста,вляют собой феНОТИfPические модиф.ика-ции.
Исключение составляет лен (Linum usitatissiтum). у KOTOpO
ro эти .индуцирова.нные изменения, по видимому, стал'И н а'Сл ед.ст-
венными. Ра.зные растения UlbHa в исх'одных поколениях выращи-
вали при различном содержании в поч.ве N, Р 'И 1(, 'И 'они реаrиро-
ва 1И на эти разные условия обычным образом. Однако в последую-
щих поколениях потомки растений, у которых был'и индуцирова.ны
крупные размеры, сохраlНЯЛИ эти к.ру:пные раз'меры, несмотря на
то, чт,о .содержание .питательных веществ в почве было .пон'ижено;
точно так же ,пото'М,ки 'индуцированных мел'ких 'растений остава-
лись мелкими, несмотря на повышение у.ровня питательных Бе
ществ. Эти индуци'рованные .разлИ'чия в разме.рах сохра'няди,сь на
протяжении 1-0 ПоОколений. Скрещивания меж,ду полученными пу
тем индукци'и крупными и ме.'IКИМИ растениями давал.И IB FI 1f'иб
риды промежуточных размеров, что 'ПQдТtверждало заК.YJючение
о Ha.c.тIeДCT,BeHHoM характере этих изменений (.Durrant, 1962а,
1962Ь; Evans, Durrant, Rees, 1966).
Наследственные разляч'ия между индуцированным'И крупными
и меЛКИIМИ 'растениями .8 эксперименте 'Со льном, по видимому, не
связаны с цитоплазмой. Однако между этими линиями обнаруже-
ны раздичия по содержанию ДИК в ядрах: у 'индуц'Ир,ованных
круlПНЫХ растеИ1ИЙ содерж.ание днк ,Б ядрах в bJII.U е, чем у мелюих
ра.стений (Evans, Durrant, Rees, .1966).
T махорки (Nicotiaпa rustica) сходные индуцированные изме
нения размеров, корре.'Iированные с содержанием в среде пита
тельных веществ, сохранялись на протяжении трех поколений
(Hill, 1967).
Мноrие аспекты этих двух ,случаев в -настоящее ,вреbllЯ непонят-
ны и требуют дальнейших ис'('ледованнй.
FA. 16. Трансдукция II индукция
157.
Обесцвечивание у звrлены
Еще ОДНИМ при'мером нас.ледеI1венноrо и.зменения, ИНДУ'Ц:ИРО-
8aHHoro средой, .служит я.вление, .изве.стное ,как 06есцвеЧJивание и
на,блюдаемое у зелеНОrо жrути-ковоro Eugleпa. ОбесцвечИ'ваlние
оБУСЛОВ.'1ено перманентной утратой хлоропласто.в определенной
клеточной линией.
Зе..,еная окраска эвrлены создается фотосинтезируlOЩИМ пиr-
ментом хлорофиллом, который находится ,в хлоропла:стах. Хлоро-
пла.сты 'П'редставляют собой ДИК-содержащие .п ооЫ'Ч.ню самовос-
производящиеся орrанеллы цитоплазмы; это CBoer<> рода iЦитоплаз-
матические reHbl. Для нормальноrо развития и функционирования
зеленых ХJIор.опласто.в .необходимы свет и .дР)'1rие факторы.
Euglena gracilis, которая на свету сама синтезирует Необхо-
димый ей сахар, можно выращивать в темноте на искусственной
среде, содержащей сахар. В темноте хлорофилла не образуется,
а впоследствии может нарушиться и воспроизведение самих пла
стид, число которых постепенно сокращается, пока они в конце
концов вовсе не исчезают у данной клеточной линии. Эта потеря
носит перманентный характер. Культура Euglena gracilis, утра-
тившая свои хлоропласты в результате содержания в темноте, не
может восстановить их, если ее вновь перенести на свет (см. Le-
win, 1962).
Обесцвечивание у Euglena gracilis может вызываться помимо
темноты и друrими внешними факторами, в частности высокой
температурой (32----35 ОС), облучением ультрафиолетом и воздей
ствием антибиотиками, например стрептомицином и т. п. (Lewin,
1962) .
Под влиянием таких факторов среды зеленая фотосинтезирую-
щая линия Euglena может превратиться в бесцветную rетеротроф-
ную линию, способную расти ТО.'1ько на среде, содержащей сахар.
Морфолоrически такая линия Eugleпa .сходна с бесцветным эвrnе-
ноиДlOМ Astasia [anga. ,Соrлаоено одной из rи'потез, касающейся
филоrении Ж1rутиковых, лишенные <>кра.ски эВ'r.пенО'Иды возни:кл'и
от зеленых Э'вrлен в результате оОбесцвеч.и.ва.ння '(Lewin, 1962).
rлава 17
ЭВОЛЮЦИОННАЯ РОЛЬ .ИНДУКЦИИ
Введениt
Эв,олюционная pO.1Jb наследcrrвенной индукции остается на про..
тяжении МНО!1ИlC лет семейной тайной ЭВО.пюционноЙ .биолопи.и. Эта
тема peДlKO .обсуждается 'в печати, не Iподверrалась она и 'Сколько-
нибудь rлу.бокому изуче.нию. Такое \прене:6реже:ние объясняе'ТСЯ
вероятно, тем сходством, 'которое rcущест:вует 'между наследствен
ной индукцией и развенчанной ламаркистокой теорией наследова..
ния пр.ио6ретенных призна'КОБ.
А 'между тем ОТРЫ1вочные :данные, изложен.ные 'в 'предыдущей
r..lajBe, свидетельствуют о 'Существова.НИJI наследственной ин.дукции
и 'мы долж'Ны Iпопытаться 'Проанализи.ровать эти данные, куда бы
они нас ни приводил'и. ВЫ'ВОДЯТ же они нас за рам.к:и синтетнче
QКОЙ теории эволюции. Эта ,r.па,ва предстаВ.'Iяет собой Л1ИШЬ пред
варительное .обсуждение 'проб.1Jемы.
Для Ta1<OrO обсуждения важно разrраничи"Ть различные ТИIIIЫ
индуцированных 'наС.l'(едст.венных и менений. 'Суть П'роблемы co
ста!вляет не индукция наследственных и'змененrий под 'В.тIиянием
внешних фаК'Торов, как таковая. Излучение 'высок<>й энерrии и хн-
ми'ческие MYTareHbI 1'акЖе ЯБ.'1ЯЮТСЯ вие'ШНИМИ факторам'и, KOTO
рые IИНДУЦИРУЮТ reHHble мутации. Но индуцированные эти'ми фа'к-
тора'ми мутации ,ненаправленны; каЖДЫЙ мутаrенный фа'кт,ор ИН-
дуцирует 'Разнообразные ТИ1ПЫ мутац'Ий. ,На'с же интересует индук
ция направленных .наследст.венных изменений, 8ызьmаемых .внеШ
ними факторами. Направnенно'сть насл€tд'СТВ€lН'ноrо изменения
представляет собой резу..1ыат пр:и.о6ретения (или утраты) IПре-
формированноr.о rе-нетическоr{) материала определенноrо ТИПа_
Именно TalKoro рода 'и'Эме.нения мы называем здесь насл ст.веН'н'ой
индукцией.
В 'общем синтетическая теория эволюции начинается с наслед-
ственной И'3меНЧИIIЮСТИ, ОЗ'Н'И1кающей :в конечном 'Счете 'в форме
нена'праВJIенных rен-ных мутаций, и не ox.вa"ТЬDBaeT НЗiI1'р влеJННЫХ
наследcr,венных иЗ'мене.ний. Поэт.ому для Toro, чтобы эта теория
м.оrла учитывать последний класс. индуu.ированных изменений, нам,
возможно, придется .расширить наши теоретические представле-
ния и Iвыйти 'за рамки СИJlтети'Чес,]{.ой теории.
Возможные примеры
Нас интересует те.перь не то, ПРОИСХОДИТ ли вреМЯ от ,времени
наСЛед'ственная и ндую.r,и Я , а то, ,оказывает ли она оС'ко.лЫКО-НИQУДЬ
ваЖlное влия'ние на Э80nЮIl!ИЮ. Рассмотрим HeC'KO.lJbKO в'озмож-
НЫХ пркмеров.
r л. 17. Эволюционная роль инду"ции
159
Как IfОВОРИ.'ЮСЬ В .rл. 16, фотоси'нтезирующее зелеНое 'Жrутико-
вое Euglena в результате обесцвечивания превращается в .'1ишен
IIЫЙ окраски 'эвrленoQИД ТИ'Па Astasia. На этой основе Iвыд'Виrается
rИiпотеза, -что лишенные окраски '3вrленоиды произошли -в !процессе
фИ.rIоrенеза от зеленых эвrлеНiQИДОВ путем 06есI1iвечИ!Вания. 31'0
правдопадобная, но 'неlПровеРelн,ная rИIП,отеза.
Несмотря на то, что сущест.в.ова'ние трансдуюции у Iбаlктерий
твердо ус.таНQвлено, никз'ких данных, ,которые бы свидетелЬС.твова
ЛИ 06 участии ,этоrо процессз в эволюции бактерий, rПО ВИДИIМОМУ,
нет. Вместе с тем известно, что некоторые МИКРОЭIЮлюционные из-
менения у ,бактерий упраiВЛЯЮТСЯ так'ими <:та,ндартными .процес.
сами, .как мутационный ,процесх::и отбор. Развитие, .например,
УСТОЙЧ 1 ИВОС1'И к фаrу 'начинается с.о спонтанноrо появления iВ коло
ниях бактерий устойчивых 'К фаlrу мутантов. Эти мутации ВОЗИ'И-
кают с определенной ни:экой частотой, 'На КОТoQрую IHe оказывает
влия,ния нали'Ч!ие ;или отсутствие в среде фа1rа. ТаlКИМ образом
микроэволюциоиное изменение протекает по обычной схеме. К не.
rативным данным, касающимся роли трансдукции !в эволюции
ба'ктерий, не оСледу€т, однако, 011НОСИТЬСЯ .слишком .серьеЭ'но, 'по-
скольку транс.ду цию изучали мазиым образом .с Iмолекуля.р.ной,
а не с ОО.'1юциоН'ной точки 'зрения.
В цитопла'зме 'Клеток Iвсех эукариотичес'ких ор.rаниз'мов содер-
жатся митохондрии. ЦИТОП.'1азма эукариотических клеток зе..lеных
ЖlrУТIfКОВЫХ и раетений с.одержит, кроме TOro, хлоропл асты. Мито-
хондрим служат центрами дыха.ния клe-rки, а хлоропласты..... цен-т-
рами фотосинтеза. 'Ка'К те, так и Iдруrие орrа1iел.лы содержат днк
и 'пред.стЗiВЛЯЮТ собой .саМО.ВОСlllр.оизводящиеся и полуаJВТ'Он-омные
комп-о.ненты эука'риотической 'к..lеm<.И.
Соrла'сно одной да.внишней rипотезе, воэ.рожденной и усовер-
шенст-вован.и,ой 'в последние lroды, .митохондрии и хлоропласты про
исх-одят .от .uвобо'ц"НО живущих Iпрокариотов, за-селивших 'прим.иТ'ив
ные -эукариотические клетки и за:крen.И'вшихся 'в ДИТОI1JIазме IB ка1че-
Сl'ве ,постоя'н'ных :сим.бионто.в. Митохондрии ра.ссматривают ка'К
rомолоrи аэробных бактерий, от которых и проиэошли 'Эти орrа.нел-
JIbl, хлороп...lа l СТЫ 'же, ка'к Iполаrают, произошли от rCине"'зеленых
водоросл.ей. .Среди .м.иоroчислен-ных сroрОННИКQВ '9 ОЙ ,rllnOT 3bl
можно ,наз-вать ,NlарrУJIИС (Margulis, 1970) и Ра.вена (Raven, 1970);
пр.отив'оположной точ'ки зрения .ПРИдерживаются Юззел 'и ,Шполь-
скии (Uzzel1, Spolsky, 1974).
В пользу теории симбиотическоrо происхождения митохондрий
и Х.'10ропласто.в с.видетел.ьствует .структу,рное и биохимическое xoд-
СТ80 меЖду этими клеточными орrаtнеллами и с0011ВеreТ1ВУЮЩИМИ
типами современных Iпрока'р.иотов. ДНК мит.охондрий, подобно
бактериальн.ой ДIНК, состоит из двух цепочек IИ имеет ,кольцевид
ную фор.м'у. Как хлоропласты, та'к и клетки сине зеленых oдopoc-
лей, ломимо Д,НК содержат фот.оси,нтезирующий аппарат и тель.
160
Часть ТУ. П роблем.а приобретенных призн.а"оtJ
.ЦЗ, сходные с ри.босомам.и; КЗ'К те, так .и др)"rие окружены мем-
браной. К:роме 1'Oro, известно, Ч.l'О сине зеленые IВОДОРОСЛ'И имеют
тенденцию к образованию симбиотических ассоциаций с различ
ными 1Q0'временными эука'ри'Отическими орrа.ниэмами (Ris. Plaut,
1 962) .
Эти данные 'нельз.оЯ :считать }'Iбедительным'и. Сходство между
цитапла'Зiматичеакнмн орта.неллаМll кле'ftКИ 11 прокарнота'Ми может
быть .обусло'&лено общим.и предкаlМИ или конверrенц:и-ей, .лр.ичем
возможность кон,верrенции исключена \Не /была. Тем не менее си'М-
би.отич.еская 'rИ'Iютеза ,весьма :nРИlВлекаТ€JIьна. Если она Ib-ерна, 1'0
пр'иобретение ПОJIезных IВ физ'Иолоrичеоком отношен.ии про.карио-
тов примитивными эукариотами моrло быть решающим шаrом в
ЭВOJIюции жи'зни, 'сделавшим .воз.можным последующее раЗВИ1'ие
ра-стительноrо 'и Ж.ИВ:Оl'ноr.о ца.рств.
ВыС'казыва.лись 'I'аlкже Пр JIlоложения относите."ыно симбио1'И
чеекOII'О и /про'ка1риотичеекоrо !происхождения :различных друrих
орrан.ел.п .клеТ'Кll. Так, лреДЛОЛ8!таеreя, что Жlтут.ик У Ж!J'УТlIК{)ВЫХ
простейших П1рОИСХОД.ит -от спи'р.охеты (Margu1is, 1970). КаIJIпа-'Ча-
стицы ,в циroпла'зме параlмеции 'Симби.отические бактерии. Цито
плазматические фаКiОРЫ, обусловливающие .стерильность у Droso-
phila paulistoruт ----lэтО, Iбыть может, Mycoplasma 'Или сходные
с ней орrзнизмы, как уже отмечалось в 'предыдущей rлаlве.
Обсуждение
fенотип 3}'1каРИQТНЧес.ких орrанизмов мо)Кно разделить .на две
части на осно.ве степени ВНУ1'ренней 'инте.-rра1J:ИИ и подверженности
и,нваэии чужеродным rенетическим материалом. Заключенный в
хромосомах rенотип "Имеет слож.ную структуру и BЫ OKO .Иli'I'еrри-
рОБан. Чужеродной частице rенетичеС'коro материала .бы.тIO бы
очень трудно заК1репиться в хромосоме, хотя 13ТО и нельзя 'Считать
невоз.можным.
В отличие от хромосомных reHoB цитоплазматические reHbl ин-
теrрированы в меньшей степени. Кроме Toro, цитоплазма rораздо
более подвержена внешним воздействиям, чем хромосомный набор,
и, по видимому, менее устойчива к проникновению чуждых rенети-
ческих частиц. Цитоплаз а самое вероятное место для УCiпешной
наследственной индукции у эукариотов.
Для развития сложных признаков необходимо действие xopo
шо интеrрированных rеиных систем. Такие rенные системы MorYT
создаватыся и поддерживаться в хромосомном наборе, который ста-
новится штаб квартирой и информационным центром сложных
эукариотических орrанизмов. Цитоплазматич кие rеиы иrраю'I'
вторичную и 'подчиненную роль. Поэтому роль наследственной ин-
дукции в эволюции должна быть также оrраничена, и сфера ее
r л. 17. Эволюционная роль индукции
161
действия не должна ВЫХОДИТЬ за пределы тех признаков, которые
детерминируются цитоплазматическими rенами.
ПРJfобретение чужеродноrо rенетическоrо материала потенци-
ально блаrоприятно для орrанизма или для ero потомков, 11 для
осущеСТВ.'Iения rенетическоrо обмена имеются разнообразные ме-
ханизмы. К числу таких механизмов принадлежат 'Наследственная
индукция и трансдукпия. Вполне возможно, что это очень древние
процессы. Однако они неупорядоченны и действуют по принципу
«попадание промах».
Эукариотические орrанизмы обладают упорядочеННЫ 1 и точ-
ным механизмом, обеспечивающим объединение rенетическоrо Ma
териала различноrо происхождения. Этот механизм половое раз-
множение, которое действует не пзуrад, з канализировано в пре-
делах видовых популяций. Упорядоченность ПО..'Iовоrо процесса На
стадиях мейоза и ОП..'10Дотворения касается исключительно хро-
мосомных reHOB. Можно утверждать, что половое размножение
ВОЗНИКЛо вместе с эукариотическими орrанизмами и в значитель-
ной мере вытеснило неупорядоченные парасексуальные способы
обмена rенетическим материаol10М, существовавшие у прокариотов,
поскольку по достижении известной стадии орrанической сложно-
сти эти 'более примитивные и неэффективные способы оказались
иеу довлетвор ите..1ЬНЫМИ.
rенетики-эво.'IЮЦИОНИСТЫ, подобно представителям классиче-
'ской rенетИ'КИ, сосредоточивают внимание' rлавным образом на по.
ведении хромосомных reHoB, пренебреrая эффектами цитоплаЗ IЫ.
Закон Харди Вайнберrа, rенетическая теория отбора и в СУЩНО-
сти вся синтетическая теория эволюции основаны На менделирова-
нии хромосомных reHOB у орrанизмов, размножаlOЩИХСЯ половым
путем. Такой упор на хромосомный rенотип вполне оправдан. Од-
нако создающаяся в результате картина эволюционных процессов
оказывается несколько несбалансированной и однобокой. Теперь
нам необходимо побольше узнать об эволюционной рО..1и наследст-
Венной индукции.
Часть V
Видообразование
r лава 18
РАСЫ И ВИДЫ
Введение
В час-tях 11 и 111 было рассмотрено действие эволюционных си.'!
;В пределах скрещивающейся популяции. ЛокаJlьная популяция,
-.однако, представляет собой одну из составных частей более обшир-
ной популяционной системы, она принадлежит к определенной
расе и определенному подвиду. Если, как это обсуждалось в rл. 17,
.единичная скрещивающаяся популяция ЯВ.'1яется ареной, на KOTO
рой происходят эволюционные изменения, то д.,'IЯ эволюционных
сил такой ареной служит система соединяющихся между собой по-
пуляций раса и вид.
Мы обратимся теперь к изучению эволюции на уровне рас и
видов. Этот раздел эволюционных исследований известен под НаЗ-
ванием видообразования. .Мы рассмотрим видообразование на тех
.основах, которые были за..10жены в частях 11 и 111.
Прежде чем пытаться описывать эволюционные процессы, про-
:исходящие на уровне рас и видов, необходимо обсудить природу
."Этих популяционных систем. Здесь будут рассмотрены три rлав-
вые черты рас и видов: 1) типы популяционных систем и характер
'изменчивости у высших растений И животных (rл. 18); 2) преrра-
ды к скрещиванию и обмен rенами внутри популяционных систем
.и между ними (rл. 19); 3) ЭКО.'10rические взаимоотношения :\1ежду
'Видами (rл. 20). На фоне этих аспектов популяционных систем мы
'можем перейти в последующих rлавах части V к рассмотрению
:процессов, участвующих в образовании рас и видов.
Популяционные системы
у орrанизмов с половым размножением локальная популяция
представляет собой скрещивающуlOСЯ единицу, но эта единица не
находится в полно изоляции. Популяционная система..... раса и..1И
Вид..... состоит из мноrих таких .1Jокальных популяций; Все эти по-
пуляции связаны друr с друrом, так к!к блаrодаря постоянным
r А. 18. Расы и Bидьt
163
tdиrрациям и обмену rенами между ними происходит некоторое
скрещивание. Расы и виды представляют собой более обширные
скрещивающиеся rруппы, чем локальные популяции.
Члены. данной локальной популяции имеют общий rенофонд.
Реrиональная KoropTa локальных популяций ---- раса..... также име-
еТ единый rенофонд, но он обширнее. Еще более обширным и раз-
нообразным rенофондом обладает вид, представ.пяющий собой
сумму скрещивающихся между собой рас.
Видимый результат скрещивания между попу..1ЯЦИЯМИ и между
расами ---- это интерrрадация фенотипических признаков, которые
подвержены реrиональной изменчивости в преде..lах вида. Именно
эта интерrрадация фенотипических пр.изнаков дает нам первые на-
дежные указания на существование скрещивания между популя-
llИЯ 1И и на те каналы, по которым оно происходит. Прослеживая
за интерrрадациями фенотипов, мы рано или поздно ста.,'Iкиваемся
с ясно выраженной прерывистостью в характере изменчивости. Эта
прерывистость обозначает rраницу вида. Иными словами, скрещи-
вание между популяциями и расами не может выходить за извест-
ные пределы, которые соответствуют rраницам вида.
Заметим в скобках, что мы используем здесь термин «вид» В
том смысле, в каком он используется в эволюционной БИО.;lOrии.
В некоторых разделах систематики этот термин приобрел друrой
смысл. В да..lьнейшем мы еще вернемся к вопросу о различных
концепuиях вида. Но здесь мы обсуждаем биолоrический, а не так-
сономический вид.
Как уже бы.тlO сказано, rраницы БИО..10rическоrо вида COOTBeT
ствуют пределам, в которых происходит скрещивание между по-
пуляциями и интерrрадация. Установив эти внешние rраницы, мож-
но перейти к описанию некоторых типов внутривидовой изменчи
вости. Различные типы изменчивости, существующие в природе,
де.Т]ятся на три основных класса, которые будут рассмотрены по
отдельности: 1) непрерывная rеоrрафическая изменчивость,
2) прерывистая rеоrрафическая изменчивость, 2) дифференциация
ЭКО..lоrических рас. В природе разные типы изменчивости нередко
бывают перемешаны. Характер изменчивости данноrо вида может
быть раз.'1ИЧНЫМ в разных частях занимаемой им области.
Аллопатрия и симпатрия
Пространственные взаимоотношени между Jlокальными попу
ляциями и между популяционными системами представляют собой
важный фактор в определении степени происходящеrо между ни..
ми скрещивания, а следовательно, и типа развивающейся в резуль..
тате этоrо изменчивосТи. Для описания пространственных взаимо-
отношений удобно пользоваться терминами «аллопатрия» и: «сим..
патрия».
164
Часть У. Видообразование
1. Аллопатрuя. Популяции, обитающие в разных местах, назЫ-
вают аллопатрическими. Уровень скрещивания между такими по-
пу.пяциями зависит от пространственноrо расстояния между ними.
r'еоrрафические расы а.тIлопатричны. В случае смежных rеоrрафИ 4
ческих рас интерrрадация может быть более или менее непрерыв
ной; если такие расы разобщены, то степень скрещивания между
ними снижается и интерrрадация прерывается.
2. Симпатрия. Две или несколько популяций, обитающих в од-
ной И той Же" обдасти, называют симпатрическими. Нескрещиваю
щиеся между собой виды, сосуществующие в преде..1ах одной и той
же территории, симпатричны. Таким образом, термином «симпат-
рия» обозначают две различные ситуации, которые С..'Iедовало бы
различать. В первом С..'Iучае популяции сосуществуют rенетически,
но не эколоrически, тоrда как во втором они сосуществуют как re-
нетически, так и эколоrически. Первая ситуация известна под наз-
.ванием смежной, а вторая ---- биотической симпатрии.
2а. Смежная симпа7'рия. Нередко можно наблюдать случаи, Kor
.да два или несколько местообитаний примыкают друr к друrу и
между ними имеются мноrочисленные поверхности раздела на про-
'тяжении обширных rеоrрафических областей. НаПРИ! 1ер, в саванне
-пятна редколесья чередуются с участками открытых JlyroB в шах-
матном порядке; в rорной местности разные высотные зоны распо-
,лаrаются в форме концентрических колец. Популяции, которые на-
'с'еJ1ЯЮТ примыкающие друr к друrу, но различающиеся местооби-
':rания, находятся в пределах своих нормальных ареалов расселе-
ния, так что со cTporo пространственной точки зрения скрещива-
ние между ними возможно, однако они Не живут или Не растут
друr подле друrа. Это смежно симпатрические популяции. Смеж-
ная сим.патрия обычно характерна для эко..,оrических рас.
26. Биотическая сuмпатрuя. Рассмотрим теперь две или не-
'скодько популяций, живущих В одном и том же местообитании, на-
пример на пятнах редколесья в саванне. У таких популяций Наб-
людается не только перекрывание зон, в преде.ТIах которых рассе-
иваются их raMeTbI, но и возникновение контактов друr с друrом
в течение нерепродуктивных стадий цикла развития. Такие попу-
.пяции являются биотически-симпатрическими.
3. Параnатрия. Эта третья ситуация носит промежуточный ха-
рактер. Две популяции называют парапатричеСI{ИМИ, ес.'!и они за-
нимают смежные, но неперекрывающиеся области и Не скрещива
ЮТСЯ между собой.
Непрерывная rеоrрафическая изменчивость
Подобный тип изменчивости характерен для свободно скрещи
вающихся орrанизмов, образующих большие непрерывные попу
.пяции. Примерами служат человек, дрозофила и мноrие виды лес-
r А. 18. Расы и вид",
165
IIЫХ деревьев и травянистых растений равнин. Локальные популя.
ции полиморфны. Каждая локальная популяция обладает своей
характерной для нее сбалансированностью полиморфных типов,
возникающих в результате потока reHoB и различных видов от-
60ра. ИЗ 1енение частот ЭТИХ полиморфных типов обычно наблю-
дается по rеоrрафической трансекте.
Рассмотрим rруппы крови системы АВО в популяциях чслове
ка, насе..1ЯЮЩИХ Европу и Азию. Как показывает табл. 18.1, часто-
та rруппы крови В ---- фенотипическое выражение rCHa /..... изменя-
тся от почти 40% в Азии до 5 11 6% в некоторых странах Запад-
ной Европы. Частота аЛ.'lеля /В снижается аналоrичным образом
от 30% в Азии до 4.----5% в Западной Европе. rрадиенты частоты
аЛ,,'1е.ТIЯ JB изображены также на рис. 18.1. Аллель /В встречается
с высокой частотой в Центра.ТJhНОЙ Азии, но в Европе ero частота
постепенно падает, снижаясь до O 5% в разных частях Западной
Европы (рис. 18.1, А). АнаJlоrичным образом..... с востока на запад
через всю Евразию ---- наблюдается соответственное повышение ча-
стоты а.ТJ.J1елей JA и /0 Uv\ourant, 1954).
Наблюдаемый rрадиент объясняют неоднократными нашеСТВИ 4
ями монrольских П.ТJемен в западную часть Азии и в Европу. Скре-
щивания монrо.'ЮВ с народами, обитавшими на Западе, у которых,
как принято считать, алле,,1Ь fB первоначально был редок, приве-
ли к повышению частоты ЭТоrо а.'lлеля в мест.ных популяциях. ЧИС 4
'JlО монrольских завоевателей, доходивших до той И.ТJи иной обла-
сти, уменьша..'1ОСЬ по мере увеличения расстояния .от N\онrолии.
rрадиен! частоты а.ТJлеля [В, пересекающий Евразию, отражает,
таким образом, постепенное уменьшение потока reHoB нз .NloHro-
лии во все более западныс части Азии и Европы (СапdеIа, 1942).
ТаБАица 18.1
Частота rpYnnbl КРОВИ В и аллеля (В в риде популяций человека в Европе
и Азии (по данным Mourant, 1954)
ПопулЯция
Аллель /В t
О'
/0
I Чис.'10 пОЮlпь- 1 rрулпа кро",1 j
ных лопупяциА в, %
Шотпандцы
Французы
Шведы
Швейцарцы
Поляки
Юrославы
BeHrpbl
И ранцы
Пакистанцы
Индусы из ЦентрапьноА Индии
5
2
2
16
I
4
1
2
3
1
8 14
&---8
'O 1 f
5 11
17
I5 18
17
22 23
32 36
39
5 9
4----6
7 8
4 8
14-
11....14
15
16
24 26
30
IC"') Q 1() *
N N s2
1 I ""'1 1,(']
Q It") а I
О
1:
со
....
::1
О
>-
о.-
Е
>-
::ii!
О
::с
::s
41
1')
i >-
::ii!
41
I u
I IZI
I О
I t::
::
i
I Q)
t::
I о
с:..
со
(.IJ
IZI
IQ
....
с;::
t:;
41
t:;
t:;
'"
!-о
О
!-о
()
со
I D'
41
:s:
::с
41
t:;
41
1:;(
41
t::
u
'IS
о..
Q,)
о
:.::
u
41
D'
==
.g.
со
о..
'"'
о
41
"-'
...;
.о 00
....
u
aI
168
Часть У. Видообразование
Разные типы инверсий В хромосоме 111 у Drosophila pseudQ
obscura рассматривались 'Выше в качестве примеров полиморфиз-
ма, микроэволюционных изменений (rл. 4) и уравновешивающеrо
отбора (rл. 11). Эти инверсии MorYT также служить примером Не-
прерывной rеоrрафической изменчивости, наблюдаемой у них в
западной части Северной Америки.
).
f:J"
<:) 11 JO 10
5 sr 46 41 32 26 .4
АН 25 35 3б 4" 45 55 50 50
I 1
I I
I I
420 I 1: Аоиl!AlI
'
I
300 1.
r:s ... !") ..
) ' 'Oнo AeиK
150 I \&
.::
,
о
о 30 ба 90 120
Расстояние, хм
Рис. 18.2. Изменения cpeAHeroAoBblX частот инаерсий ST и AR в ПОПУJlИЦИRХ
Drosophila pseudoobscиra по трансекте через ropbl Сьерра-Невада в Калифорнии
(Dobzhansky, 1948).
На рис. 18.2 представлены среднеrодовые частоты инверсий ST
и AR через определенные интервалы по трансекте, проведснной в
центральной части rop Cьeppa HeBaдa 'в Калифорнии. .м.ожно ви
деть постепенное повышение частоты инверсии AR и понижение
частоты ST с увеличением высоты (Dobzhansky. 1948). Эта TpaH
секта представляет собой один участок rораздо более обширной
картины rеоrрафической изменчивости, предстаВ.rIенной на рис.
18.3. На трансекте, проходящей через юrо-западную часть США,
инверсия ST встречается с высокой частотой в Калифорнии, а к
востоку частота ее снижается; частота инверсии AR повышается в
направлении от Калифорнии к Аризоне и Нью Мексико, а затем
вновь снижается в Техасе. В отличие от этоrо частота инверсии
Пайкс-Пик (РР) достиrает максимальноrо уровня в Техасе, а к
западу снижается (рис. 18.3). rеоrрафическая изменчивость В еще
более широких масштабах наблюдается к северу, в районе Ска-
листых rop (rде отмечается высокая частота инверсии AR), и к
r А. 18. Расы и виды
169
юrу, в Мексике (rде высока частота инверсии СВ) (Dobzhansky,
Epling, 1944).
rеоrрафическая расовая изменчивость реально существует в
ПОПУw'lяционных .системах Ното sapiens в Евразии и Drosophila
CmaHgapm
;1ppoy;re!l
паuкс..Пик
l/UPUKQX!lQ "О
2D
О
Тpи AaЙH 40
20
О
Рис. 18.3. Изменения частот ПЯТИ инверсий в ПОПУ.'1яциях Drosophila pseudoobscи-
.ra по трансекте, идущей от Южной Ка.'1ИфОРНИIJ до Техаса (Dobzhansky, Epling,
1944) .
pseudoobscura в Северной Америке. Но поскольку интерrрадация
происходит очень постепенно, rраницы между расами проводятся
. довольно ПРОИЗВОJlЬНО; поэтому отнесение данной популяции к той
или иной расе также носит ПРОИЗВО.'lьный характер. CKO.'lЬKO рас
. следует выде.тrять у D. pseudoobsCllra? Л10ЖНО с равным основани-
ем различать в преде.'lах этоrо вида как четыре, так и сорок рас.
Для практических це.'1ей удобно проводить различие между ло-
кальными. и rеоrрафическими расами. У локальных. рас отмеча-
170
Часть у. Видообразование
ются относительно небольшие сдвиrи в частотах полимор.фных ва.
риантов (например, расы D. pseudoobscиra из Джексонвилля и
Лост-Клейма; см. рис. 18.2), тоrда как rеоrрафические расы пред.
ставляют собой реrиональные скопления локальных рас, достовер.
но раз-личающихся по своим rенофондам (наПрИ lер, популяцион.
ные системы D. pseudoobscura из Калифорнии, Аризона Нью-Мек.
сико и Техаса, см. рис. 18.3)_
Постепенность интерrрадации не всеrда бывает равномерной.
В картине изменчивости данноrо ВИДа MorYT то здесь, то там встре.
чаться крутые rрадиенты. Подобные участки резких изменений
reHHblx частот на сравнительно коротких rеоrрафических расстоя-
ниях создают естественные зоны для проведения rраниц между
расами, что несколько уменьшает ПРОИ3БОЛЬНОСТЬ при подразделе..
нии вида на rеоrрафические расы.
Разобщенные rеоrрафические расы
Популяционная система OCTpoBHoro типа раЗБиваетсн там, rде
области, приrодные для обитан"Ия данноrо вида, относительно малы.
и разделены большими промежутками. Примеры прерЫБистоrо
распределения местообитания определеННОfО типа обrцеизвестны:
острова, образующие архипелаr,..... для наземных орrаНИЗМОБ, це-
пи озер ---- для водных орrанизмов, вершины rop для альпийских
видов, выходы ropHbIx пород на травянистой равнине и т. п.
В подобных случаях создается характерная картина изменчи-
вости. В rеоrрафической изменчивости выявляются разрывы, соВ.
падающие с лрерывистостью местообитания. Колонии, если они до..
статочно хорошо изолированы, становятся специфически приспо.
собленными к условиям своей локальной среды. В результате вид
превращается в ряд разобщенных и отличающихся лруr от друrа
rеоrрафических рас. Именно такая картина изменчивости должна
возникнуть в результате уменьшения потока reHoB между колония.
ми и под действием отбора или отбора в сочетании с дрейфом
в пределах каждой колонии.
Однолетнее травянистое растение Gilia leptantha (Polemonia.
сеае) встречается на проrалинах в COCHOBbIX лесах, растущих на
ropax на юrе Калифорнии. В этих районах сосновые леса раСПОJlЗ.
rаются на средних и больших высотах ряда ИЗО,,'1ированных rop.
ных хребтов, разделенных мноrокилометровыми пространствами
лишенной лесов низменности. Соответственно популяционная си.
стема Gilia leptantha имеет разобщенное распределение, показан"
ное на рис. 18.4.
Кроме Toro, популяция, населяющие разные ropHbIe цепи, rруп-
пируются в ясно выраженные rеоrрафическпе расы. Четыре такие
расы, различающнеся ПО МОрфОJ10rичесКRМ признакам Цве1ка, си.
стематики признают отдельными подвидами (рис. 18.4). Экспери
r л. 11. i'acbl и виды
111
менты показали, что эти четыре rеоrрафические расы чрезвычай-
"о ПЛОДОВИТЫ при скрещивании друr с друrом.
Морфолоrические признаки иветка, по которым различаются
эти расы, до некоторой степени конверrируют в тех областях, тде
":..:"
ЛUНN61е
. п",чuнки
.рт pиr
ЛУРн m т K
mР9i1oч; ';;60чка тычинки
./ер tra
.
t!J рЁп
,
\.
,
,
'\
,
,
"
"
"
,
"
"
f!r
... ......... ...... -.. ...
Рис. 18.1. Разобщенные rеоrрафицеские расы ОШа leptantha (Роlemопiасеае)
"а отделЬНЫХ торных _епв 8 Южной Капифориии (по данНЫМ Grant, Grant.
) 956, 1960).
pln G.l. pinetorит; pиr... а.l. purpusii; lер... а.l. leptantha; tra... G.I. trап,vеrэа.
расы соприкасаются наиболее тесно, но не образуют непрерывноrо
rpадиента (Grant, Grant, 1956, 1960).
Один из близкородственных видов..... Gilia latiflora....... встреча.
-ется На засуш.пивых НИЗменных равнинах и в долинах примерно в
'1'ех же районах иа юrе Калифорнии. .Он .образует непрерывные и.,,"
полунепрерывные популяции на обширных пространствах. Ero по.
'пуляционная система состоит rлавныM .образом из непрерывноrо
ряда интерrрадирующих rеоrpафичес их рас (Grant, Grant, 1956,
'1960). Прямо противфположны.е ТИПD1 rеоrрафической ИЗМенчиво
172
"
I
СТИ У Gilia latiflora и а. leptant1za коррелированы с попу яционны"
ми структурами этих двух видов. .-
Большой ИIlтерес представ-пяет характер rеоrрафической измен-
чивости в rруппе Nigella arvensis в районе Эrейскоrо моря на юrо..
востоке Европы. Область Эrейскоrо моря, З3КJJюченноrо между
континентальными частями rреции и Турции, имеет весьма слож..
Часть V. Видооброзоsание
$)
.
,
rpi :!Ji:() .::.\.. . .' .
;;"if " : ;:i';:,:
......
ШiШN.; оr6:; /' "'" , ,
.:....::... :::::ж (J.. , , ' I;r' ,'i
1ЖЖ }\." g . ,., ;. 1,. , , \
:>: ',;.:. N. d jеппу . А'" -
Ш!iJР\' . ,,'0 '" '9:.:;, ."
у'" 'N. dеgепii ,"'O ,-' ,,' <.,р.
j ." ,.-,
(\.. IIJ,' '. "
N.d. тiпor '... , / ' '.,'
tJ
,.
50 100
I I
км
Рис. 18.5. rеоrрафические расы Nigella degenii (Ranunculaceae) на островах Ки-
Юlады в Эrейском море. Показаны также аре ады двух эндемичных 8I1ДОВ, 6лиз
кородственных N. degenii (Strid, 1970).
ный рельеф вследствие находящихся здесь мноrочисленных остро..
вов.
Входящий в эту rруппу вид N i.gella degenii целиком островной
и распространен на архипелаrе Киклады. Этот вид состоит из ря..
да БОJlее или менее обособленных локальных рас, населяющих от-
дельные острова. Локальные расы сrруп-пированы в четыре отдель-
ные rеоrрафические расы, распространение которых покаэано на
рис. 18.5. Три друrих острова, раСПО.,10женные по периферии архи-
пелаrа Киклады, населены двумя б lизкородственными эндемич.
\ {л. 18. Расы и виды 173
\
выми вида,и. N. icarica и -"'. carpatha (рис. 18.5). СобственнО
N. arvensis, оторый широко распространен в Европе, обладает не-
прерывной r'fоrрафической ИЗ 1СНЧИВОСТЫО на материке, Но на ост-
ровах Крит Ц Родос этот вид также представлен четко выраженной.
отдельной ра ой (Strid, 1970).
ЭКdJJоrические расы
Третий из основных типов внутривидовой изменчивости это
дифференциация эколоrических рас. fенетически и фенотипиЧ"еСКlI
различные, но П lJодовитые при скрещивании Между собой, рас-ы
приспособлены к разны:м местообитаниям в пределах одной и той
же территории. Эти ЭКО..'10rические расы симпатричны, обычно
см:ежно симпатричны. Такие расы скрещиваются между собоЙ и
интерrрадируют в мноrочисленных зонах контакта по всему ареалу
данноrо вида. Но они сохраняют свои отличитсдьные расовые при-
знаки каждый в CBoe 1 местообитании.
Gilia achilleaefolia (Polemoniaceae) встречается у подножий
Береrовых хребтов в 1\а.1JИфОРНИИ. Это очень изменчивыЙ вид.
Крайни 1И формами являются «солнеч.ные» расы и «теневые» расы,
раз..'1ичающиеся по С lJедующим признакам: у «солнечных» рас,
встречающихся на открытых, поросших травой по..'1ях, крупные си-
не фио.1Jетовые цветки собраны в малеНЬкие rоловки и перекрестно
опыляются пчелами; у «теневых» рас, растущих в лесах из вирrин-
cKoro дуба, медкие одиночные бледно окрашенные и самоопыдя-
юшиеся цветки. Тенистые леса и открытые ПО IJЯ распределены в
в районе Береrовых хребтов в Виде мозаики и такое же мозаичное
распределение характерно для различных эколоrических рас
а. achilleaefolia. «Солнечные» и «теневые» расы, дающие при скре-
щивании плодовитые rибриды, скрещиваются между собой и ин
терrрадируют в тех мноrочисленных точках, в которых соприкаса-
ются их местообитания (Grant, 1954).
Эко.поrические расы мноrообразны. К ним относятся высотные
расы ropHblx видов, расы по хозяину у насекомых и сезонные ра-
сы у орrанизмов, у которых ИМеется два или неско.'1ЬКО обособлен-
ных периодов размножения.
В ropax Cьeppa HeBaдa в Калифорнии на бо.тIЬШИХ высотах
HeCKO.тIЬKO видов деревянистых растений представлены прямо-
стоячей древовидной расой в субальпийской зоне и низкорослой
кустарниковой расой в альпийской ЗОНе. Такие соприкасающиеся
н скрещивающиеся между собой высотные расы обнаружены
у Pinus albicaulis Р. тurrayana, Tsuga тertensiana и у двух. Ви-
дов ивы (Salix). Эти виды деревьев встречаются у rраницы лесной
растительности в ropax Сьерра-Невада. Однако рост этих видов
возможен и выше верхней rраницы ..'1есной растительности, Кото-
рая является пределом лишь Д..1Я распространения субальппйских
174
Часть V. Видо06Рй3084Ние
древовидных рас, тоrда как ареал кустарниковых альпийских рас
простнрается вверх, ВЫХОДЯ за эту rраницу (Clausen,,: 1965).
Расы по хозяину обычны У насекомых. Во мноrих! rруппах на-
секомых каждый вид имеет очень узкий Kpyr хозяев, На которых
он может кормиться и размножаться. Например, в Северной Аме-
рике личинки яблонной пестрокрылки (Rhagoletis рдтоnеllа сем.
Tephritidae) заражают плоды двух деревянистых представителей
розоцветных........ боярышника (Crataegus) и культурных яблонь
(Malu ). На яблоне и боярышнике живут разные расы пестрокрыл-
ки Rhagoletis. Поскольку во мноrих местах яблони и боярышник
растут в тесной близости, расы по хозяину у Rh. ротоnеиа смежно-
симпатричны (Bush, 1969).
Пестрокрылки Rhagoletis спариваются на своих растениях-хо-
зяевах, и эта черта биолоrии упрочивает их приверженность к
определенному хозяину. Распознавание и предпочтение хозяина
у Rhagoletis и близкоrо к нему рода Procecidochares контроли-
руется одним reHOM. А следовательно, мутация, возникшая в этом
reHe, может положить начало формированию новой расы по xo
эянну (Bush, 1969; Huettel, Bush, 1972).
БИОJlоrические внды
Биолоrический вид..... общая совокупность всех рас представ-
ляет собой наиболее важную популяционную систему в природе
и в эволюционной биолоrии. Ее можно наблюдать повсеместно.....
в пределах биотических сообществ и в пределах естественных
rрупп. Так, семейство оленей (Cervidae) в Северной Америке со-
стоит из следующих хорошо выраженных видов: белохвостый
олень, чернохвостый олень, олень вапити, американский лось, ка-
рибу (Rangifer tarandиs); в Европе в это семейство входят лань,
блаrородный олень, косуля, американский лось, северный олень
(друrая раса Rangifer tarandus) , а прежде еще и Megaloceros
hibernicus.
Биолоrические виды сохраняют обособленность блаrодаря ме-
ханизмам репродуктивной изоляции, которые предотвращают или
СИЛЬНО снижают обмен rенами между ними. Разнообразные формы
репродуктивной изоляции будут рассмотрены в дальнейшем. Здесь
достаточно указать, что преrрады к размножению действительно
существуют и что они обусловливают ту явно выраженную I1pe-
рывистость в изменчивости, которая указывает rраницы между
видами.
Наличие репродуктивной изоляции между популяционными си-
стемами наиболее очевидно в тех случаях, коrда эти ПОПУЛЯllИОН-
ные системы, будучи симпатрическими, сохраняют тем не менее
различия по фенотипическим признакам. Симпатрические популя-
llионные системы орrанизмов, размножающихся половым путем,
rл. 18. Расы и виды
175
представл от собой по существу самостоятельные биолоrические
виды. Сохр нение различных сочетаний признаков в условиях сим-
латрии ---- э проверка, производимая самой природой, и наидуч-
шин критер Й, свидетельствующий о видовом статусе рассматри-
ваемых поп яционных систем.
Поскольк такое сохранение обособ..1енности при симпатрии
совершенно етко выражено в смысле видовоrо статуса рассмат-
риваемых ед ниц, удобно оформить это терминолоrически, обозна-
чив такие ед ницы, как симп трические u виды. Тем самым прово-
ди1'СЯ раЗ.ТIИчие между даннои ситуациеи и менее ясным случаем
аллопатрических видов.
Никто не п'рдверrает сомнению тот факт, что хорошие биолоrи-
ческие виды MorYT обитать на разных территориях. Желтая сосна
и Веймутова фсна обладают четкими видовыми различиями не-
зависимо от Tdro, растут .ни они вместе в одном COCHOBO 1 лесу
или же в разных лесах. Однако не всеrда обособленные аЛ.ТIOпат
рические популяционные системы представ.ТIЯЮТ собой хорошие
БИОJlоrические виды; такие системы MorYT быть также разобщен-
ными rеоrрафическими расами, обладающими сходными чертами.
Решение о TO J, к какому paHry принадлежат раЗ.ТIИчающиеся меж-
ду собой аллопатрические попу.пяции, зависит от степени различия
между ними, а это ---- понятие относите..lьное; кроме Toro, оно за-
,висит от на..1ИЧИЯ или отсутствия репродуктивной изолянии, что в
таких случаях обычно невозможно бывает установить без специ-
альных дальнейших исследований.
Степень фенотипическоrо различия между ДВУМЯ популяцион-
ными система 1И имеет меньшее значение в качестве признака,
свидетельствующеrо об их статусе био..l0rических видов, чем нади-
чие репродуктивной изоляции. В сущности пары И..1И множества
морфолоrически очень сходных, но репродуктивно изолированных
видов обнаружены во мноrих родах насекомых, цветковых расте.
ПИЙ, простейших и в друrих rруппах. Такие морфолоrически очень
сходные виды получили название видов.двойников.
Например, при более детальном анализе оказа.Т'/ось, что оби-
тающий в Европе малярийный комар, известный в старой литера-
туре под названием Anopheles maculipennis, представляет собой
комплекс из шести видов-двойников (все они были выделены в ка-
честве самостоятельных видов и получили отдельные названия:
А. тaculipennis sensu stricto, А. тelanoon, А. тesseae и друrие).
Эти виды двойники, ХОТЯ они репродуктивно изолированы, морфо-
лоrически абсолютно неразличимы. Тем не менее они различаются
по окраске ЯИЦ, МОРфО.'10rии хромосом, некоторым физиолоrичес-
ким признакам, поведению, а также по такому важному д.Т'/я Me
дицины признаку, как то, являются ли они переносчиками маля
рин или НеТ (Майр, 1963).
1.76
Часть У. Видообразование
ПОПУВНДhl
Иноrда в природе встречаются популяционные си.стемы, кото-
рые нельзя отнести НИ к хороши видам, ни к хорq'шим расам.
Примером таких систем следует считать промежутфчные стадии
дифференциации между разобщенными rеоrрафичесКjИМИ расами и
ал..l0патрическими видами или между эколоrически и расами и
симпатрическими видами. Проблематичные популяrfионные систе-
Рис. ]8.6. Перекрывающееся КОЛЬЦО рас у пчелы Hoplitis prodиcta (Megachilidae)
(Michener, 1947).
Интерrрадация от подвида Н. р. gracilis к друrик подвидак покаэана стреJlкаки. Два про-
кежуточных и TepMHHa.;tbHblx подвида этой серии (Н. р. paпarniniana и Н. р. berпardiп.a)
симпатрнчески сосуществуют С Н. Р. Kraclli$ 8 Южиой КалнфорИИU.
мы этоrо типа связаны друr с друrом разными уровнями потока
reHoB, промежуточными между УрОВНЯ IИ, типичными для рас, с
одной стороны, и типичными для видов с друrой. В результате
характер изменчивости в этих системах также оказывается про
межуточным между НН1'ерrрадацией и прерывистостью.
В некоторых случаях в одной и той же совокупности популяци-
онных систем можно найти смесь признаков, сходных с расовыми
и видовыми. Так, популяционные системы MorYT интерrрадировать
в одной части всей занимаемой ими области, но сосуществовать
симпатрически, не скрещиваясь между собой, в друrой ее чаСТII
(рис. 18.6). Или же популяционные системы MorYT быть симпат-
рическими и изолированными в большинстве мест, но скрещивать-
ся в одном ИЛИ не.скольких местах.
r А. 18. Расы u виды
177
Таблица 18.2
КпассификаЦИR попу,nяц,,"онных систем у орrанизмов, раЭмиож.аЮЩИХСR
половым путем
Уровень I Тип популяnиоииой
диверrен. системы
ЦНИ
rеоrрафические I Фенотипические I Характер
осо6ениости особеиROСТU скреlЦНВ8ННЯ
,
1 Локальные расы МlIкроrеоrрафи- Относительно Скрещиваются
ческая протя- слабая диф. между собой
женность ференциация интерrради
руют непре
рывко ил
ПОЛукепре
рывко
2 Непрерывные reo- АЮIопатрические Как на урови
rрафические ра- 1
сы
РазоБЩЕ'lIllые reo. Ал.'10патрические
rрафllческие . ра-
сы
Эколоrи((еские ра. Смежно-симпат
cы рические
3 А.'Iлопатричсские Аллопатрические Промежуточ,
полувиды ный межд
Симпатриqеские Симпатрическис уровнями 2
полувиды 4
4 _"..'1лопатриqеские Ал.'10патричеСКJlе Репродуктивно
виды Rзолированы;
Симпатрические СимпаТРllческне ! разде.,еиы
виды разрывом
5 Н здвиды rруппа аЛ.'10пат Относительно Как на уровн
рических по.'1У сн.,ьная д(-I ф 4
видов фереНlIиация
Синrамеон rруппа симпатри
ческих ПО.'lУВИ-
дов
и
е
у
и
е
Традиционных катеrорий «раса» И «вид» недостаточно для то-
ro, чтобы подвести ПОД них все встречающиеся в природе ситуа
ции. НеоБХОДИ lа какая-то ПРО lеЖУiочная катеrория Д..1Я проме-
Жуточноrо состояния. Д.'1я обозначения попу.I1ЯЦИОННЫХ систем,
промежуточных между расами и видами в 01'ношении скрещивания
друr с друrом, интерrрадации и репродуктивной ИЗО.1ЯЦИИ, исполь
эуют тер 1ИН «полувид».
Л1ножество полуви'nов представ.1Jяет собой более обширную По-
пуляционную еДИllИЦУ, чем вид. Совокупность аЛ..'10патрических по
лувидов называют наДВИДО 1; совокупность симпатрических или по
178
Часть У. ВидообраЗDвание
крайней мере марrинально симпатрических ПОЛУВИДОВ называется
синrамеоном.
Определения биолоrическоrо вида, приводившиеся ранее в этой
rлаве, применимы r..1ЗВНЫМ образом к совокупностям рас; в тех же
случаях, коrда состаВЛЯЮЩие их единицы не расы, а по.'1увиды,
они требуют некоторых уточнений. Биолоrический вид не во всех
случаях представляет собой наиболее обширную скрещивающуюся
rруппу. Это самая обширная rруппа при условии нор,иальноzо
скрещивания. Однако в иск.ry.ючительных случаях синrа меон ба.
лее всеобъемлющая популяционная система.
Разнообразие популяционных систем в природе требует более
сложной системы, чем простое деление на расы и виды. Предшест-
вующее обсуждение, проведенное в этой rлаве, попытка предста-
вить такую систему (следуя одной из более ранних работ автора;
Grant, 1963, rл. 12). На данном этапе представляется желательным
суммировать все наши рассуждения; поэтому в табл. ]8.2 приве-
ден перечень различных популяционных систем с их краткими Ха-
рактеристиками.
Вид у однополых орrанизмов
До сих пор речь шла о популяционных системах у орrанизмов
с половым размножением и перекрестным оплодотворением. Мы
рассматривали скопления скреЩU8ающUХСЯ популяций. Что каса-
ется однополых орrанизмов, как бесполых, так и парасексуальных,
то они требуют отдельноrо обсуждения. Биолоrическая концепция
вида, изложенная в этой rлаве, неприменима к однополым opra.
низмам, да н Не рассчитана на это.
Однополые орrанизмы, безусловно, образуют популяции. Их
колонии состоят из rенотипически идентичных или сходных особей,
занимающих специфическое местообитание н преДЪЯВ..1JЯЮЩИХ осо-
бые Эколоrические требования к среде. Такие КО,,10НИИ представля-
ют собой популяции, но популяции нескрещивающиеся. И этн од-
нополые популяции не объединены в дискретные биолоrические
виды.
Характер изменчивости у однополых орrаНИЗl\10В резко отлича-
ется от сочетания интерrрадации и прерывистости, наБЛIOдаемоrо
у орrанизмов с половым размножением. Вместо этоrо у них наб-
людается ряд однородных локальных (но не скрещивающихся)
популяций, возникших в результате размножения одноrо или не.
скольких адаптивных rенотипов. fенотип таких популяций нередка
слеrка изменяется в зависимости от местности. Если данная rруп-
па занимает широкий спектр местообитаний, расположенных на
обширной площади, то число rенотипически различных популяций
может достиrать нескольких сотен или тысяч, причем крайние
формы MorYT очень сильно отличаться друr от друrа. Общий диа
r А. 18. Расы u виды
179
пазон изменчивости в преде-
лах такой популяционной сис-
темы сравним с диапазоном
изменчивости в rруппе видов у
орrанизмов с половым размно
жением. Но эти диапазоны
изменчивости Не делятся eCTe
ственным образом на расы и
виды.
Биолоrическая концепция
вида в таких случаях, очевид.
но, оказывается несостоятель
ной, точно так же как и любая
друrая практическая таксоно-
мическая концепция вида, по
скольку изменчивость образу-
ет БО..'Iее И..'IИ менее непрерыв
ную сеть, не подразделенную
на четко выраженные таксо-
номические единицы.
Ботаники, сталкивающиеся
с такой ситуацией во мноrих
rруппах растений, пытались
справиться с Нею, введя спе
циальную катеrорию..... микро.
вид. МиКровид..... это rеноти
пически однородная популя
ция или популяционная систе#
ма, обладающая собственными.
отличительными фенотипиче-
скими признаками, в rруппе
однополых орrанизмов. В та-
ком сдучае rруппа родствен-
ных микровидов образует
отдельный однополый комп
лекс (см. Grant, 1971).
Таксономический вид Cre
pis occidentalis (Compositae) предстаВ.'lяет собой очень изменчи-
вую rруппу мноrолетних травянистых растений, распространенную
в западной части Север'ной Америки. Этот таксономический вид
вк..'Jючает в себя ДИШIOИдную двуполую популяционную систему----
биолоrический вид, ---- распространение которой оrы.аничено rорами
на востоке и На севере Ка..'IИфОРНИИ. По всей остальной об.тtасти,
занимае:vюй видом Crepis occidentalis sепsu lato, от Британской
Колумбии до южной части Калифорнии и от Тихоокеанскоrо побе-
режья до Великих равнин, эти популяции состоят из аrамосперм-
А
r
, ,
Б
8
, . ,
. .
. . .
. . . .
." . . .
. . .' ."
. . .
. . .
д
Е
"
ж
J
и
Рис. 18.7. Соцветии девити аrамосперм-
ных МИКРОВИДОВ Crepis occideпtalis sen.
su lato (Compositae) (по Babcock, Steb.
bins, 1938).
А. с. о. hиmilior (штат Капифоркня).
Б. С. о. пиttаlШ (штат Вашнкrтои).
В. С. о. calycиlata (штат Колорадо).
r. С. о. grayi (штат Юта). Д. с. о. elliptica
(севериая l(aCТb штата КалиФорItНR).
Е. С. о. delloidea (ПРОВННЦИR Альберта).
ж. С. о. rydbergi€ (штат J'v\oHTaKa). 3. С. о. ha.
тiltoпensis (сеаериая часть штата Капнфор'
ния). И. С. о. crassa (штат Вашин"он).
180
Часть У. Видообразование
ных типов, т. е. из растений, размножающихся семенами, которые
образуются бесполым путем (Babcock, Stebbins, 1938).
AraMocnepMHbIe и, следовательно, однополые популяции OДHO
образны. Однако разные аrа оспермные типы заметно отличаются.
друr от друrа по морфолоrическим признакам листьев, соцветий
плодов и друrих частей растения. В морфолоrической изменчиво-
сти у Crepis occidentalis sensu lato выявляются «сrустки», позво-
ляющие разбить эту rруппу на 27 различных бесполых микровидов.
каждый из которых обладает собственным rеоrрафическим apea
0I10М (Babcock, Stebbins, 1938). Соцветия нескольких из. этих мик
ровидов представлены на рис. 18.7.
Ситуация с аутоrамными растениями заслуживает особоrо упо
минания. Мноrие rруппы однолетних травянистых цветковых pac
тений и некоторые rруппы мноrолетних растений aYToraMHbI, т. е.
автоматически самоопыляются и самооплодотворяIOТСЯ И, таким об-
разом, размножаются половым путем, будучи, однако, однополы-
ми. Но аутоrамия и инбридинr обычно не бывают полными и до-
полняются некоторым перекрестным опылением. Перекрестное
опыление, несмотря на ero небольшое участие в общем размноже-
нии aYToraMHoro растения, с биолоrической точки зрения весьма
существенно и нередко оказывается достаточным Д-Т1Я объединения
особей в частично скрещивающиеся ежду собой популяции, а
этих популяций в виды.
Вид в систематике
Как и в популяционной б Ио..'lOrи и, в систематике вид также
представляет собой основную единицу. Цели систематики и попу-
ляционной биолоrии одновременно и сходны, и раЗ.1JИЧНЫ. Систе-
матика занимается формадыюй классификацией орrанизмов, а
популяционная или микросистематика ---- формальной кдассифика-
цией на уровне расы и вида. Таким образом, в систематике вид----
это прежде Bcero единица классификации. И rлавным критерием
при выделении видов в систематике с..1УЖИТ удобство и возмож-
ность практическоrо использования Д.тIЯ классификации, определе..
ния и расстановки в музейных КОol1лекциях.
Разрывы в картине изменчивости, которые оrраничивают био
лоrичесКие виды, служат также удобными поrраничными вехами
для распознавания таксономических видов. Семейство оленей в
Северной Америке удобно с таксономической точки зрения. 11 оп-
равданно биолоrически делить на следующие виды: белохвостый
олень, чернохвостый одень, олень вапити, американский лось и ка-
рибу. В таких случаях, как этот, встречающихея у эукариотических
орrанизмов очень часто и повсеместно, таксономический вид оказы-
вается синонимом биолоrическоrо вида.
r А. 18. Расы и виды,
18.
Но нередко создаются ситуации, в которых таксономическик
критерий вида не совпадает с критерием вида в популяционной
6иолоrии. В некоторых rруппах неудобно, трудно или даже невоз
можно распознавать биолоrические виды в таксономической прак .
тике. Существует три ДОВОJ1ЬНО обычных ситуации, в которых TaK
..сономический вид по необходимости расходится с био.поrичеСКИl\f
видом.
Это, во-первых, популяционные системы орrанизмов, которые-
размножаются половым nYTe 1 и которые не.IJЬЗЯ разделить На две
взаимоисключающие катеrории расы и виды, поско",ьку В них
имеются также промежуточные I{атеrории ПО..1увиды. Система
классификации, однако, содержит раз и нав-сеrда установденну»
искусственную иерархию катеrорий: рОД, .подрод, сеКllИЯ, вид, под
вид и форма (сеКllия==rруппа видов; подвид==раса). В этой
иерархии нет места д.ТУЯ rрупп, промежуточных между видом и
подвидом; в l таксономическом 'системе нет катеrории, которая COOT
ветствовала бы полувиду популяционной биолоrии. В таксономии
для Toro, чтобы были удовлетворены требования биноминальной
или триноминальной номенклатуры, ПОПУЛЯllИ"ОННУЮ систему, Ha
ходящуюся на ПО.'IУВИДОВОМ уровне диверrеНllИИ, надо рассматри-
вать либо как самостоятельный вид, .1Jибо как подвид (расу) Ka
korO-..'Iибо друrоrо вида.
BO BTOpЫX, целый ряд трудностей создают виды-двойники. TaK
сономические виды До.l'Jжны быть распознаваемы по внешним
морфолоrическим признакам, сохраняющимся у музейных экземп
JIЯрОВ. Виды-двойники не удовлетворяют этому критерию. Тем ие-
менее виды-двойники, судя по их реПрОДУКТИВНЫ 1 взаимоотноше
ниям, представ..1ЯЮТ собой настоящие биолоrические виды, неза
висимо от Toro, удается распознавать их в обычной таксономичес
кой практике или нет.
Третья об..lасть, в которой нет соответствия между двумя KOH
цепциями вида, это бесполые и парасексуальные орrанизмы.
В систематиКе принято, и этоrо требуют правила HOMeHK.lJaTypbI..
относить все орrанизмы к тем или иным видам незаВИСИ 10 от Toro..
ЯВ.'Iяются они двуполы ш и.,и нет. Но ведь у беспо..'IЫХ орrанизмоs
биолоrических видов попросту не существует.
Таl{ИМ обраЗО I, между таксономической и ПОПУ.lяционно-биоло
rической системами классификации возникают противоречия в ря-,
де важных областей. В таких случаях каждый раздед науки дол
жен сохранять верность своим собственным целям. Решение проб..
лемы заКJlючается в признании законности обеих концепций вида
таКСОНО IИческой и популяционно БИО.1]оrической. Семантической
путаНИllЫ можно избежать, называя один вид таксономическим, а
друrой БИО.тIоrическим, как это Cдe.тIaHO в предшествующем об
суждении.
182
"Часть V. Видообразование
Последовательные виды
Палеонто.поr, имеющий дело с эволюционными изменениями в
rеолоrическом времени, :вынужден различать последовательные ви.
ды. Такие виды являются узловыми точками в филетическом эво.
ЛЮЦионном ряду, достаточно сильно различающимися, чтобы им
можно было дать разные видовые названия. Например, линия ело.
IIОВ позднеrо кайнозоя состоит из пары последовательных Видов----
Elephas plaпifrons u Е. тeridionalis l которые с течением времени
постепенно интерrрадируют (см. рис. 30.2).
Последовательные виды неравноценны биолоrическим видам.
Они образуются в результате филетической эволюции, а не эволю-
ционной диверrенции. Поэтому два последовательных вида Не со-
()тветствуют двум биолоrическим видам.
rлаlва 19
ИЗОЛИРУЮЩИЕ МЕХАНИЗМЫ
Введение
Обмен rенами между разными ПОПУЛЯlI.иями или ПОП-УЛRЦИОН-
выми си темами уменьшается IlЛИ блокируется разноrо рода пре-
rрадами, известными под общим названием изолирующих меха-
низмов. Ниже приведена классификация этих весьма разнообраз-
ных изолирующих механизмов.
1. Пространственные механизмы
1. rеоrрафическая ИЗО.'lЯЦИЯ
11. Механизмы, создаваемые средой
2. Экопоrическая изоляция
111. Репродуктивиые механизмы
А. Внешние
3. Временная изоляция
4. Этолоrическая изопяция
5. еханическая изоляция
6. ИЗО.'1яция raMeT
Б. Внутренние
7. Несовместимостъ
8. Нежизнеспособность rибридов
9. Стерильиость rибридов
10. Разрушение rибридов
в этой классификации проводится rлубокое различие между
пространственной и репродуктивной. изоляцией. При пространст.
венной изоляции raMeTbl из разных популяций не встречаются друr
с друrом, потому что эти популяции живут в областях, разделен-
ных расстояниями, которые слишком велики по сравнению с по-
тенциальными возможностями орrанизмов к расселению. Короче
rоворя, пространственная изоляция это изоляция, создаваемая
rеоrрафическим расстоянием. В отличие от этоrо при репродук-
тивной изоляции преrрады к скрещиванию порождаются свойст-
вами самих орrанизмов. Блаrодаря репродуктивной изоляции по-
пуляции MorYT обитать на одной и той же территории, совсем или
почти совсем не обмениваясь между собой rен-ами.
Эти два способа изоляции входят в определения рас и видов.
Пространственная изо.ПЯЦИЯ свойственна локальным ПОnYЛЯ'ЦНЯ\f
локальным paca 1 и rеоrрафичеСКИ:\I расам. Хорошо развит.ая реп-
:tВ4
Часть V. Видообразование
родуктивная изоляция один из от личите.'1ЬНЫХ признаков биоло
rических видов. Частич'ная или неполная репродуктивная изоля
ция характерна для по.'увидов.
Изоляция. создаваемая средой, или эколоrическая изоляция.
третья и в некотором роде промежуточная катеrория. В этом слу-
чае между популяциями существуют rенетически обусловленные
различия в отношении их эколоrическнх потребностей II предпочте
ний. Способность популяций жить на одной и той же территории
-определяется наЛичием соответствующих местообитаний и ниш и
'силой межвидовой конкуренции. Если сосуществование популяций
действитеu'1ЬНО имеет место, то rибридизация между ними реrули
.руется на.'1ичием местообитаний. приrодных для их rибридноrо по
томства. Преrрады к обмену rенами имеют эколоrическую при-
роду.
Эколоrическая изоляция представляет собой универсальную
черту. характерную для всех видов. но она не ЯВ.'1яется отличи-
TeдьHыM видовым призна[(ом. Эколоrическая изоляция существу
.e-r также между эколоrическими расаМИ и между симпатрически-
ми полувидами. Весьма возможно, что между rеоrрафическими ра-
ами обычно имеется некоторая эколоrическая изоляция.
В п,риведенной выше к.'1ассификации репродуктивная изоляция
'разделена на изоляцию, создаваемую внешними и внутренними
lIреrрадами. Внешние преrрады это преrрады, препятствующие
."Встрече raMeT И.'1И (у растений) rаметофитов, тоrда как внутренние
преrрады начинают действовать после Toro, как raMeTbl или ra
:метофиты встретились друr с ApyroM.
Эколоrическая и временная изоляция
Эко.'10rическая изоляция слеДствие эколоrической дифферен-
llиации симпатрических видов. Подобная дифференциация широко
.распространена и проявляется во мноrих различных формах. Виды
дрозофилы. обитающие в одном и том же районе Калифорнии илп
БраЗИ..1I1И. И:\lеют различные пищевые предпочтения и питаются
разными видами дро.жжей. В Техасе некоторые виды дуба растут
На разных почвах: один вид (Quercus mohriana) встречается на
'известняковой почве, друrой (Q. havardi) на песчаной, а третий
(Q. grisea) на выходах маrматических пород.
Эколоrическая дифференциация на нерепродуктивных стадиях
жизни в той или иной степени снижает шансы на успешную rибри-
дизацию между. с.импатрическими видами и тем самым способст-
вует их изоляции. В r.'l. 20 эколоrическая дифференциация между
.вида 1И будет рассмотрена БО.'Iес Подробно.
у большинства ЖИВОТНЫХ н растений существуют определен-
'ВЫе сезоны размножения или цветения. Спаривание или перекре-
'oCTHO опыление происходят в определенное время rода (например,
r л. 19. ИЗОАирующие механизмы
185
летом или осенью), а нередко и в определенное вре IЯ суток (у oд
НИХ видов днем, а у друrих ночью). Б.'Iизкие виды MorYT раз.личать
ся по тому, на какое время rода или суток .приходится У них пе
риод половоro размножения. Такие межвидовые различия ведут к
временной изоляции. К временнбй изоляции ОТНОсятся сезонная.
изоляция и изоляпии, обусловленная различными суточными цик ,
лами.
Два близких вида сосны, Pinus radiata и' Р. attenuata l оказыва ,
ются в симпатрическом контакте в центральной части .I(алифор '
нийскоrо побережья. Здесь у Р. radiata пыльца осыпается рано"
в февра..'Iе, а у Р. attenuata на 6 неде.'1Ь позднее, в апреле, так
что между этими двумя видами существует сезонная изоляция
(Stebblпs, 1950). Drosophila pseudoobscura и п. persil1lilis, которые'
в западной части Северной Америки симпатричны на протяжении
обширной об,,1асти, скрещиваются в одно и то же время rода, но в.
разНое время суток: у D. pseudoobscura половая активность наб
людается по вечерам, а у D. persiтilis по утрам (Dobzhansky,.
1951Ь).
Временнjя изоляция, подобно любой друrой форме репродук
1'ИВНОЙ ИЗО.'Iяции, МОЖет быть полной иди частичной. У pOДCTBeH
ных видов растений, обитающих на одной и той же территории
пики цветения нередко приходятся на разное время, но периоды
цветения перекрываются, т. е. имеет место частичная сезонная изо
лядия.
еханическая изоляция, этолоrическая
ИЗОJIЯЦИЯ и изоляция raMeT
у высших животных и растений nмеются С.'IожНЫе репродук-
тивные системы и,,'Iи цветки, состоящие из мужских и женских op
raHoB, которые структурно коадаптированы так, чтобы об,,'Iеrчать
копуляцию, осеменение или опыление при НQрмально внутриsи
довом спаривании. Если два .вида орrанизмов различаются ПО
строению своих ПО.ТIOвых opraHoB или цветков, то это препятствует'
копуляции, осеменению или ОПЫ.'1ению между особями разных
видов. Подобные препятствия создают механическую из'оляциlO.
Механическая изоляция существует между двумя видами шаJ1
фея, Salvia тellifera ц S. apiana (Labiatae), на юrе Калифорнии.
у S. тellifera типичный двуrубый венчик, а тычинки и столбик
выступают из нижней rубы (рис. 19.1,А). Шалфей опыляют раз-
личные мелкие и среднеrо размера одиночные пче.,1Ы, которые Пе
реносят пыльцу с одноrо цветка на друrой на своих спинках (рис
19.1, Б). ДЛЯ S. apiana характерны чрезвычайно своеобразные
цветки с Д.'Iинными, СИ..'1ьно выступающими наружу тычинками и
столбиком и высокоспециализированным механиз lОМ, обеспечива
ющим опыление (рис. 19.1. В и r). Опыление у этоrо вида произ '
186
Честь У. Видообразование
ВОДЯТ пчелы-плотники и шмели, которые переносят пыльцу на
крыльях и на друrих частях тела (рис. 19,1,Д). Те насекомые, KO-
торые обычно производят опыление у S. mellife.ra l по большей час..:
ти неспособны опылять цветки S. apiana l и наоборот. В этом слу..;
чае имеет место механическая ИЗО.IJЯЦИЯ. Эта ИЗОЛяЦИЯ, однако, не,
А
А
в
r
.
Б
-
," .:8, . . . - ._ . ;
,.... ':L . I '1.
, .
. ", д
lCM
Рис. 19.1. Два вида шалфея н опыпяющие их виДЫ переПОR'Iатокрыпwх (Grant,
Grant, 1964).
А. Цветок SallJia тellifera. Б. Anthophora... обычный опылитель s. теlЩеrа. В и r. Цве-
ток S. apiana в обычно", (8) И сопрокииутом" (f) положеииях. д. nчепа пл()Тник Ху(осо-
ра brasitianorит... обычный опылитель s. apiaпa. Все рисуики 8ыпопнеиы В одинаково..
масштабе.
вполне эффективна, поскольку некоторые пчелы, имеющие проме..
жуточиые размеры, Mory" производнть межвидовое перекреС1'ное
опыление и время от времени осуществляют ero (Gr ant, Grant,
1964) .
у высших животных копуляции обычно предшествует ухажи..
вание. Ухаживание слаrается из ряда стимулов (танцы. демонст..
рации, пение, феромоны и т. п.) И реакций на эти стимулы, кото-
рые подrотавливают самок и .самцов к копуляции. У видов, при-
наД..fJежащих к той ИЛИ иной rруппе, брачное поведение варьирует
и снrналы видоспецифичны, так что самки данноrо вида не реаrи-
руют на самцов чужоrо вида. Поведенческие запреты на спари-
вание с особями чужоrо вида создают этолоrическую изоляцию.
Этолоrическая изоляция иrрает ключевую роль в предотвращении
r А. 19. ИЗОАирующие механuз.ICЫ
187
межвидовой rибридиэации во мноrих rpynnax животных, как поэ
J30НОЧНЫХ, так и беспозвоночных.
Мноrие водные орrанизмы выделяют rзметы в воду. Наружное.
оплодотворение зави-сит при этом от встречи м'ежду свободноживу
ЩИМИ яйцеклетками и сперматозоидами,. которая в свою очередь,
лровоцируется биохимическими веществами. Эти биохимические
Зiтрак1'ЗНТЫ являются видоспецифичными. В результате взаимное
привлечение и оплодотворение происходят MeJКДY яйцеклетками и
сперматозоидами одноrо и Toro же вида, но не между половыми
клетками разных видов, находящимися в одном и том же водоеме.
Это отсутствие взаимноrо привлечения у raMeT раэноrо происхож
дення н создает изоляuию ra)teT.
КлассичеСКИ 1 примерО:\-1 изоляции raMeT с.'1ужит морской еж
5trongylocentrotus 1 у KOToporo оп..тJодотворение наружное и проис
ходит в морской воде. В КОНТРО.IJируемых экспериментальных yc
oI10ВИЯХ, коrда смеси raMeT помещали в сосуды с водой, внутри
каждоrо из ДВУХ видов S. franciscanus u 5. purpuratus свобод-
но происходи.rrо оплодотворение; однако меЖВlIдовые оплодотворе
ния (5. f. Х5. р d и 5. р. 9 Х5. t о} были сильно подавлены.
Преrрады, создаваемые несовместимостью,
и нежизнеспособность rибридов
Вернемся к высшим животным п растениям, у которых имеет
место внутреннее ОП..ТIOдотворение и зародыш развивается в мате..
ринском орrанизме. Допустим, что внешние изолирующие opra..
низмы Не сработали и произошла КОПУollЯЦИЯ между особями раз
ных видов или ОПЫ..'Iение чужим видом. В таких случаях образо
взнию rибридов MorYT препятствовать разноrо рода внутренние'
преrрады, вступающие в действие либо в родительском поколении,.
либо в FI'
У покрытосеменных растений '--rежду опылением и ростом лро
ростка проходит MHoro стадий развития. Первые четыре стадии'
происходят до оплодотворения: 1) лрорастание пыльцевых зерен
на рыльцах; 2) рост пыльцевой трубки в столбике; 3) прораста
иие пыльцевой трубки до эародышевоrо мешка в семяпочке; 4) ос..
вобождение ядер спермиев; 5) их притяжение к женским raMeTaM.
Оплодотворение ---- самостоятельная стадия, которая у покрыто
семеннЫХ происходит в два этапа: 6) оплодотворение яйца; 7) оп
.Т1ОДотворение ядра эндосперма. Затем следуют первые стадии, на-
ступающие после оплодотворения: 8) первые деления зиrоты
9) развитие эндосперма; 10) развитие зародыша; 11) формирова..
ние семени; 12) прорастание семени. А сл.едуЮ1l1ие стадии относят-
ся уже к новому поколению: 13} закрепление молодоrо. пророст
ка; 14) рост проростка; 15) раЗБитие зрелоr0 растения.
188
Часть У. Видообразование
На любой из этих стадий MorYT возникать преrрады, блокиру-
ющие об11ен rенами, следующий за межвидовыми скрещиваниями.
Сходная последовательность стадий существует у млекопитаю
:lЦИХ: осеменение, миrрация сперматозоидов, зачатие, импланта
ДИЯ, развитие зародыша, рождение, рост и развитие детеныша.
у них также препятствия к успешной rибридизации MorYT возни-
'дать на любой стадии длитсльноrо процесса развития.
Все эти прспятствия известны под общими названиями: 1) пре
трады, обусловленные несовместимостью, и 2) нежизнеспособность
rибридов. rде следует проводить rраницу между этими двумя ка-
теrОрИЯ:\1И? Рассмотрим две возможности, каждая из которых име-
..ет свои преимущества и свои недостатки. r лавное, о чем следует
ПО lНить, rде бы мы ни решили провести линию раздела, это то,
'что развитие непрерывный процесс, который может быть забло-
кирован на любой из ero мноrочислснных стадий.
Мы можем произвольно приписать все препятствия, действую-
'щие до ОШlOдотворения, несовместимости, а действующие ПОС"lС
.Hero нежизнеспо.собности rибридов. Это разумно с эмБРИО..'10rи-
ческой точки зрения, но не всеrда практично. Селекционер И,,'IН
.животновод MorYT произвести определенное скрещивание, но по
'неизвестным причинам и:\t не удается ПО.IlУЧИТЬ жизнеспособных
.особей. Стадию, на КQТОрОЙ возникает препятствие, можно, конеч-
'НО, определить при помощи дальнейших ИСС.'1едований, но это не
имеет существенноrо значения для консчноrо результата искусст
"!BeHHoro (или eCTecTBeHHoro) скрещивания. Поэтому может оказать-
-ея .более удобным проводить .ТIИнию раздела между преrрадами
несовмеСТИ 10СТИ инежизнеспособностью rибридов на БО.'1ее позд-
'ней стадии.
Друrая возможность дать такое определение несовместимо-
сти, которое включало бы в себя все разнообразие преrрад, деЙст
вующих на всех стадиях, начиная с осеменения или опыления и
вплоть до рождения детеныша, откладки яиц или созревания ce
мян. В Та!{ОМ случае нежизнеспособность rибридов будет означать
заметное подаВ,,1сние мощности и пор оки развития у особей FI'
проявляющиеся после рождения, вылупления или прорастання.
Преrрады, создаваемые несовме.стимостью на ранних стадиях, c.'Ie-
дующих за осеменением или опылением, аналоrичны изоляции ra-
.мет у орrанизмов с наружнЫМ оплодотворением.
Стер.ильность rибридов
Во мноrих rруппах животных и растений при межвидовых скре-
.щиваниях образуются мощные. но стери..'1ьные rибриды Ft; хоро-
'шо известным примером таких rибридов служит мул (лошадь 9 Х
Хосел о ). Проявления стерильности, хотя они по опредеvYJению orpa-
tИичиваются 'репродуктивной стадией rибридов Fl' тем не менее
r л. 19. Н 3Dлuрующие механuзмы
189
Becь a неоднородны. Наблюдается изменчивость в отношении то-
ro, на какой именно стадии проявляется стерильность и каковы
лежащие в ее основе rенет.ические причины.
Развитие половых opraHoB и течение мейоза сложные процес
СЫ, которые MorYT быть леrко нарушены в результате неrармонич-
Horo взаимодействия reHoB у rибридов. Нарушения развития по-
ловых opra'HoB можно прои..lлюстрировать на примере некоторых
межвидовых rибридов растений, у которых образуются цветки с
абортивными пыIьниками.. У межвидовых rибридов ЖИВОТНых
процесс деления клеток зародышеnоrо пути нередко прерывается
ВСJ1едствие reHHblX нарушений: спеР)"lатоrенез может прекраТliться
до начала мейотических делений или же мейоз может протекать
аберрантно;. в люБО I с.JIучае образования сперматозоидов Не про
исходит. Нарушение сперматоrенеза на стадиях, предшествующих
мейозу, основная непосредственная причина, вызывающая сте-
рильность у самцов му..1а; нарушения мейоза причина стерильно-
сти у rибридных самцов при некоторых скрещиваниях между раз-
ными видами Drosophila (например, Р. pseudoobscuraxD. persi
milis). .
К оrраниченной полом стерильности инежизнеспособности rиб-
ридов у двудомных растений и..1и раздельнополых животных при
ЛОЖЮdО обобщсние, известное ПОД названием праВИ.rIа Холдейна.
rибриды F I ОТ межвидовых скрещиваний у раздельнополых жи-
вотных должны состоять, ВО всяком с"'учае потенциально, из reTe-
poraMeTHoro пола (несущеrо хромосомы ХУ) и rOMoraMeTHoro (ХХ)
пола. Прави..,о ХО.'1дейна rласит, что в тех случаях, коrда в ПрОSlв
лении стеРII"lЬНОСТИ ИЛИ нежизнеспособности rибридов существу
ют половые раз..1ИЧИЯ, они прояв..,яются чаще у reTeporaMeTHoro,
чем у rOMoraMeTHoro ПО.'Jа. 7 бо.пьшинства животных, в том ЧИс..lе
у ..1екопитающих и у двукры."1ЫХ, reTeporaMeTHbl самцы; мы толь-
КО что упоминали о rибридной стерильности у ЛОlпадей и у дро-
зофилы. Из правила ХО."1дейна имеются, однако, мноrочисленвые
ИСКЛIOчения (см. White, 1973).
Третья стадия -развития, на I\ОТОрОЙ может ПрОЯВ.'Iяться rибрид-
ная стеРИ.IJЬНОСТЬ, это rаметофитное поколение у растений.
у иветковых растений из ПРОДУКТОВ мейоза -непосредственно раз-
виваются rаметофиты пыльцевые зерна и зародышевые мешки,
которые содержат от двух до неско.."1ЬКИХ ядер и в которых форми-
руются raMeTbI. Нежизнеспособность rаметофитов обычная при-
чина стерильности rибридов у цветковых растений. Мейоз завер-
шается, но HopMa.1JbHOrO развития пыльцы и зародышевых мешков
не происходит.
Причины rибридной стерильности на rенетическом уровне бы-
вают троякими: rенными, хромосомными и цитоплазматическими.
Наиболее широко распространена и обычна rенная стерильность.
Не6лаrоприятные сочетания ядерных reHoB родитедьских типов,
' O
Часть. У. Видообразование
принадлежащих к разным видам, MorYT приводить и действитель-
но приводят К морфоrенетическим и цитолоrическим отклонениям
у rибридов F 1 , что лрепятствует образованию raMeT. rенетический
анализ rенной стерильности у rибридов Drosophila (D. pseudoobs
curaXD. pe,siтilis D. тelaпogasterXD. siтиlans и т. по) показы-
вает, что reHbl, обусловливающие стерильность, расположены во
всех или почти во всех хромосомах родительскоrо вида (см. OoЬ
zhansky, 1951 а, rл. 8; 1970, rл. 10). Неблаrоприятные взаимодей-
ствия между ЦИТОIlлазматическими и ядерными rенами также Be
дут к стерильности межвидовых rибридов в разных rруппах paCTe
ний и животных.
Виды растений и животных различаются также по транслока-
циям, инверсиям и друrим перестройкам, которые в rетерозиrот
ном состоянии вызывают полустерильность или стерильность. CTe
пень стерильности пропорциональна числу независимых перестро
ек: так rетерозиrотность по одной транслокации дает 50 % -НУЮ сте-
рильность, по двум независимым транслокациям 75% ную
стерильность и т. д. Стерильность растений опреде..lяется raMeTO-
фитом. У rетерозиrот по хромосомным перестройкам в результате
мейоза образуются дочерние ядра, неСУlЦие нехватки и дуплика-
ции по определенным участкам; из таких ядер не получается
функциональных пыльцевых зерен и семяпочек. Хромосомная CTe
рильность подобноrо типа очень часто встречается. у межвидовых
rибридов цветковых растений.
Течение мейоза у rибридов может быть нарушено либо rенными
факторами, либо различиями в строении хромосом. KaI< rенная, так
и хромосомная стерильность может выражаться в аберрантном Te
чении мейоза. Но типы мейотических аберраций различны. rенная
стерильность обычна у rибридов животных, а ХрО lOсомная сте-
рильность у rибридов растений. rенетический анализ некоторых
межвидовых rибридов растений показал, что ХрО:\IOсомная и reH-
ная стери.'IЬНОСТЬ часто сочетаются у одноrо и Toro же rибрида.
Разрушение rибридов
Допустим, что некий межвидовой rибрид достаточно жизне-
способен и плодовит, для Toro чтобы размножаться. В таком сду-
чае F2, BI' F з и друrие последующие поколения потомков будут
обычно содержать значительную долю нежизнеспособных, субви-
тальных, стерильных и полустерильных особей. Эти типы пред-
ставляют собой неудачные продукты рекомбинации, возникшие
при межвидовой rибридизации. Такое подавление мощности и пло-
довитости в rибридном потомстве называют разрушением rибри
дов (hybrid breakdown). Разрушение rибридов..... последнее звено
в последовательности преrрад, препятствующих обмену rенами.
Разрушение rибридов неизменно обнаруживается в потомстве
межвидовых rнбридов у растений, rде ero леrче наблюдать, чем
r А. 19. ИЗОАирующие механизмы
191
при большинстве скрещиваний у животных. fибриды F 1 от скре-
щивания Zauschneria canaxZ. septentrionalis (Onagraceae)
мощные и подуфертильные, однако большинство растений F 2 кар-
ликовые, растут медленно, восприимчивы к ржавчине или сте-
рильны. От одНоrо скрещивания между этими видами было полу-
чено 2133 растений F2' но НИ одно из них не обладало нормальной
мощностью (Clausen, Keck, Hiesey, 1940). Среди F 2 от скрещива-
ния Layia gaillardioides XL. hieracioides 80% особей были не-
жизнеспособны или субвитальны (Clausen, (951). в ПОКОJ1ениях
F 2 F б от скрещиваний Gilia тaliorX G. тodocensis (Polemo-
niaceae), cOr..lacHo оценкам, 75% зиrот были полностью или час-
тично нежизнеспособны, а среди растений, обладавших нормаль-
нои мощностью, было MHoro (70% и более) стерильных и пол усте-
рильных (Grant, 1966).
Сочетания изолирующих механизмов
Лишь в редких случаях изоляция видов обеспечивается одним
изолирующим механизмом, а возможно, таких случаев не бывает
вовсе. Обычно HecKo..'IbKO раз.тIИЧНЫХ изолирующих механизмов
действуют совместно.
Рассмотрим пару видов D. pseudoobscura и п. pe.rsiтilis. Раз-
де.7lение этих видов создается целой rруппой различных изолирую-
щих механизмов, к числу которых относятся: эколоrическая изо-
ляция, временная изоляция, этолоrическая изоляция, стерильность
rибридов и разрушение rибриДов. Ни один из этих механизмов в
отдельности Не может предотвратить rибриднзацию. Однако дей-
ствуя совместно они создают полную изоляцию этих двух сим-
патрических видов в природе (Dobzhansky, 1951Ь, 1955).
rлава 20
эколоr.ИЧЕ.СКАЯ ДИФФЕРЕНЦИАЦИЯ
Межв.идовая конкуренция
Симпатрические виды взаимодействуют не только в репродук
тивной, но и В веrетативной фазе. Эти эколоrические взаимодейст
вия проявляются в разных формах. Симпатрические виды MorYT
вступать в прямую конкуренцию за необходимые, но оrраниченные
ресурсы пищу, воду, пространство и т. д.; они MorYT избеrать
конкуренции; возможна также промежуточная ситуация косвен-
ная и неявная конкуренция. Эти раз.аичные фОр:\1Ы взаимодейст
вия приводят к различным эволюционным эффектам.
Отсутствие межвидовой конкуренции характерно для симпатри-
ческих видо.в, относящихся К разным крупным таксономическим
катеrориям, имеющих очень разные эколоrические потребности и
выполняющих разные рОJIИ в биотическом сообществе. Подобные
эколоrически хорошо дифференцированные rруппы MorYT не толь-
ко не конкурировать между собой, но даже находиться в отноше
ниях взаимной зависимости нутри CBoero биотическоrо сообщест.
ва. Биотическое сообщество эколоrическая единица, обладающая
независимой энерrетикой, а тем самым это ассоциация симпат
рических видов, иrрающих комплементарные роли в питании и
энерrетике. Мы здесь не будем вдаваться в подробности относи-
тельно взаимозависимости в биотических сообществах.
Межвидовая конкуренция превращается в действенный фактор,
Kor;x.a симпатрические виды имеют сходные эколоrические потреб
ности вообще и, в частности, нуждаются в каком-то ресурсе, кото-
рый имеется в оrраниченном количестве. Виды, участвующие в
конкуренции, часто находятся в более и..1И менее б.'IИЗКОМ родстве;
однако они MorYT также находиться в отдаленном родстве, но кон"
верrировать.
МеЖ13идовая конкуренция имеет два широких аспекта. Первый
аспект пассивное использование разными видами oДHoro и Toro
же ресурса. Так, разные виды кустарников в сообществе пустыни,
по всей вероятности, будут пассивно И"lИ неаrрессивно конкуриро..
вать за оrраниченные запасы rpYHToBbJX вод. Второй аспект, нак-
ладывающийся на первый, это прямое подаВ"lение одноrо вида
друrим, конкурирующим с ним видом.
В листьях ряда растений вырабатываются химические веще
ства, KOTo 'ыe вымываются в почву и подавляют прорастание и
рост друrих растений, находящихся поблизости (Muller, 1966,
1970). .
r А. 20. Эк,ОАоzичесlCйЯ дифференциация
193
у 1Кивотных подавление достиrается путем аrрессивноrо пове-
дения или проявления превосходства, oCHoBaHHoro на уrрозах на-
падения. В пустыне Мохаве (Калифорния и Невада) местный
сне1КНЫЙ баран (Ovis canadensis) и одичавший осел (Equus asi-
nus) конкурируют за воду и корм. При прямых столкновениях
ослы побеждают баранов; коrда ослы приближаются к источни-
кам воды, занятым баранами, последние удаляются, а иноrда
вовсе покидают данную местность (Laycock, 1974).
Замещение видов
Межвидовой отбор заключается в увеличении численности If.
эколоrическом превосходстве одноrо вида по отношению к друrо-
му, конкурирующему с ним виду. Межвидовая конкуренция ведет
к межвидовому отбору, при котором один вид обладает .каким-л'и-
бо пр исущим ему преимуществом над друrим в отношении конку-.
ренции. Так, например, после 1930 r. в Долине Смерти (Калифор.
ния) наблюдается сильное повышение численности ослов, сопро-
вождающееся заметным снижением чнсленности снежных баранов.
Процесс межвидовоrо отбора может продолжаться вплоть до-
замещения видов, при котором вид А полностью замещает вид В
в какой-либо местности, если условия среды, в которых А обла-
дает преимуществом, остаются постоянными. Таким образом дин-
ro (Canis faтiliaris dingo) на протяжении длительносо историче
.cKoro времени вытеснил су,мчатоrо ол.ка (Thylacinus) на боль
шей части территории Австралии.
Замещение видов изучалось в лабораторных экспериментах н'а
Parameciuт, Triboliuт и друrих орrанизмах. В своих классиче-
СКИХ экспериментах на парамециях rаузе (1934) выращивал
Р. aurelia и Р. caudatuт вместе в стеклянных сосудах на водной
среде, содержавшей различные соли и бактерии (Bacillus руо-
cyaneus), которые служили кормом этим простейшим; температу-
ра и состав среды поддерживались на постоянном уровне. Para-
тeciuт aurelia за несколько недель полностью заместила Р. caиda-
tuт. Все повторения этоrо эксперимента, проводившиеся в
идентичнЫХ условиях среды, всеrда давали один и тот же резуль-
тат. Но если условия изменяли, используя в качестве KopMoBoro
орrанизма друrой штамм Bacillиs, то Р. caиdatuт замещала
Р. aurelia. В целом во всех случаях, коrда два вида Parameciuт
вынуждали к конкуренции в однородной культуральной среде,
один вид в конечном счете вытеснял друrой.
Если, однако, смесь видов Paraтecium помещают в неодно-
родную среду, то полноrо замещения одноrо из видов может и не
произойти. Иллюстрацией служит эксперимент, в котором Р. aure-
иа и Р. bursaria помещали в пробирки, содержавшие вертикальи()
стратифицированные суспензии кормовых дрожжей. При этом ока-
:194
Часть У. Видообразование
залось, что Р. aurelia кормится rлавным образом в верхних слоях,
а Р. bursaria На дне. При наличии TaKoro рода разных KOpMO
.вых ниш И различных пищевых предпочтений два вида MorYT
продолжать сосуществовать долrое время (rаузе, 1935).
Замещение видов несомненно часто происходит в природе. Этот
процесс обычно трудно наблюдать и правильНО интерпретировать
в современной нам живой п.рироде, если не считать нескольких яс-
но выраженных случаев, вроде сумчатоrо волка и динrо в ABCTpa
лии. Но косвенные данные о частых замещениях видов нельзя не
заметить, рассматривая эволюционные изменения в rеО.1I0rическом
масштабе времени. Соrласно некоторым оценкам, 98% ныне жи.
вущих семеЙств позвоночных произошли от примерно 8 видов, су-
ществовавших .в раннем мезозое, а 8 эТО лишь очень небольшая
часть Toro множества видов позвоночных, которые населяли 3eM
лю в то время (Wright, 1956).
Конкурентное исключение
Эксперименты rаузе, наблюдения полевых биолоrов, в частно
стн rриннела (Grinnell) и Лэка (Lack), и математические уравне-
ния Вольтерра (Volterra) и Лотки (Lotka) привели к созданию
закона, известноrо под названием принципа KOHKypeHTHoro
исключения (а также закона rаузе, прииципа rаузе и принципа
Лотки Вольтерра; см. Hardin, 1960).
Принцип KOHKypeHTHoro исключения (как мы ero здесь будем
называть), начиная с 30 x rодов, явился той точкой, к которой
-.было приковано внимание эколоrов, и усиленно обсуждался мно-
rими авторами. Разные авторы формулируют этот принцип по-
.'р'азному. Подробное рассмотрение возникновения разных форму-
.лировок увело бы нас далеко в сторону от нашеrо изложения;
поэтому мы оrраничимся тем, что приведем цитату из Лэка
(Lack, 1947), которая отражает существо дела, а затем дадим об-
щее определение принципа.
Лэк пишет (1947) :«Теперь я совершенно изменил свою точку
зрения, оцени.в в полной мере справедливость утверждения rаузе
о том, что два вида, обладающие одинаковой эколоrией, не MorYT
сосуществовать в одной и той же области (rаузе, 1934). Это про-
стое следствие eCTecTBeHHoro отбора. Есля два вида птиц встреча-
ются совместно в одном и том же местообитании одной и той же
области, питаются одной и той же пищей и имеют одинаковые
друrие Jколоrические требования, то они будут конкурировать
друr с друrом; а поскольку вероя1'НОСТЬ Toro, что они одинаково
хорошо приспособлены, пренебрежимо мала, однн из них должен
полностью элиминировать друrой. Те.м не менее три вида зем..'IЯ-
ных вьюрков встречаются совместно в ОДНОМ и том же место-
обитании на одних и тех же rалапаrосских островах; то же самое
r А. 20. ЭКОАоzическая дифференциация
195
относится к двум видам насекомоядных древесных вьюрков. Оче
видно, существует какой.то фактор, препятствующий эффектив
ной конкуренции между этими видами.
Изложенные выше соображения заставили меня произвести
общий обзор эколоrии воробьиных птиц (Lack, 1944з). Как пока-
зал этот обзор, большинство близких видов занимают различные
местообитания И.ТIи области, однако те из них, которые встречают '
ся вместе в одном и том же местообитании, обычно отличаются
Apyr от друrа по биолоrии питания, а часто также по размер м,
в том числе по величине клюва. Установлено, что это последнее'"
различие в рЯДе с.rrучаев с;вязано С различиями в пище, и такая
корреляция, по видимому, носит общий характер, поскольку TPYД '
но представить себе, каким иным способом близкие виды моrли
бы избежать конкуренции».
Принцип KOHKypeHTHoro исключения содержит два общих по-
ложения, относящиеся к симпатрическим видам: 1) если два вида
занимают одну и ту же ЭКОJ10rическуlO нишу, то ОДИН из них прак
тически наверное п:ревосходит ApyroA в этой нише и в конце КОН-
цов вытеснит менее приспособ.'lенный вид. Или, в более краткой
форме, «сосуществование между полными конкурентами невозмож
но» (Hardin, 1960). Второе положение вытекает из первоrо: 2) ес-
ли два вида сосуществуют в состоянии устойчивоrо равновесия, то
они должны быть эколоrически дифференцированы, с тем чтобы
они моrли занимать различные ниши.
К принципу KOHKypeHTHoro искшочения можно относиться по..
разному: как к аксиоме и как к эмпирическому обобщению. Бели
рас'сматривать ero как аксиому И..1И закон природы, то он лоrичен,
ПОС.Т'Iедователен и оказывается очень эвристичным. Если paCCMaT
ривать ero как эмпирическое обобщение, то, как будет показано'
да..'1ее, он оказывается широко придожимым, но не универсальным.
Сосуществование видов
Полное замещение одноrо вида друrим Не единственный ре-
зультат межвидовой конкуренции, о чем свидетельствует Часто'
наблюдаемое в природе сосуществование близких видов со сход-
ными эколоrическими потребностями. Полноrо замещения видов
можно избежать одним ИЗ нескольких путей. Четыре таких пути.
приведены ниже (см. также Grant, 1963).
1. Замещение одноrо вида друrим длительный процесс. По
этому с..т.едует ожидать, что, производя наблюдения в любой дaH
ныИ: момент времени, мы обнаружим несколько пар конкурирую
щих видов, находящихся на стадии незавершенноrо замещения.
2. Эколоrически сходные виды MorYT сосуществовать, никоrда
не достиrая стадии прямой межвидовой конкуренции. Так будет
обстоять дело в том случае, если численность этих видов сдержи
196
Часть У. Видообразование
вается каким-то друrим фактором, а не прямой конкуренцией. На-
пример, численность популяций растительноядных животных в
природных сообществах нередко удерживается на низком уровне
хищниками И, следовательно, хищничество не допускает, чтобы
межвидовая конкуренция за пищу между растительноядными пре-
вратилась в важнЫй фактор, оказывающий влияние на сосущест-
вование этих последних.
З. Условия среды MorYT обратимо изме.няться в период дейст-
вия меж'видовоrо отбора, в результате чеrо на одной стадии селек-
тивным преимуществом обладает вид А I а на друrой вид В.
В таком случае эти два вида будут сосуществовать в условиях
циклическоrо равновесия.
4. Последний путь, имеющий важное значение в природе, воз..
можен в неоднородной среде, в разных участках которой данный
решающий фактор содержится в разных количествах. При этом
вид А может превосходить вид В в одном участке среды, тоrда
как вид В обладает преимуществом в ApyroM участке. При такой
ситуации виды А и В MorYT сосуществовать, обитая частично или
rлавным образом в соо ветствующих предпочитаемых ими участ
ках среды. Лабораторные эксперименты, проведенные с Paraтeci-
uт 1 Drosophila и друrими орrанизмами, весьма показательны в
этом отношении (rауэе, 1935; Ayala, 1969). Как уже rоворилось
выше, некоторые пары видов Paraтeciuт способны к длительному
сосуществованию, если их пища распределена по разным корма..
вым зонам, СОО'I"ветствующим различным пищевым предпочтениям
Каждоrо вида. Ес.тественные местообитания, конечно, всеrда неод-
нородны, причем часто, а может быть и как правило, такая неод-
нородность допускает некоторую эколоrическую сеrреrацию сим-
патрических видов, а следовательно, непрерывное сосуществова-
Ние.
Отбор. направленный на 9колоrическую
дифференциацию
.Мы выяе.нили, что в одних ситуациях межвидовой отбор пр'и-
водит к полному замещению видов, а в друrих ---- к эколоrическоА
диверrенции и сосуществованию. Здесь будет описана модель вто-
poro процесса, при котором результатом межвидовоrо отбора яв-
ляется эколоrическая дифференциация.
При построении этой модели были приняты следующие допу"
щения: 1) имеются два эколоrически сходных и конкурирующих
между собой вида А и В; 2) они обитают в неоднородной среде С
участками EI' Е 2 и Е з ; 3) каждый из конкурирующих видов поли-
морфен и содержит по три формы (А., А2, Аз; Вl, В2, В з ), приспо..
собленные соответственно к трем участкам среды (E 1 , Е 2 и Е з );
4) форма А 1 обладает превосходством в участке E 1 , а форма Вз
в участке Е з .
r А. 20. ЭКОАоzичеСlCая дифференциация
197
в начале процесса межвидовоrо отбора вся ситуация выrлядит
следующим образом:
Среда Et
Вид А А 1 (превосходящая
форма)
Вид В B J
Е! Е,
А, А,
82 Б з (лревосходящая
форма)
Если допустить нормальное течение межвидовоrо отбора и пре
небречь такими осложняющими факторами, как скрещивание
между полиморфными типами в пределах каждоrо вида, то полу-
чится следующий конечный результат:
Среда Е 1 Е, Е,
Вид А А 1 А 2
Вид В В, В.
Можно видеть, что между конкурирующими видами А и В воз-
никает эколоrическая диверrенция. Каждый вид становится более
узко специа.'Iизированным к тем участкам общей для этих двух ви
доn среды, в которых он обладает селективным преимуществом.
Кроме Toro в процессе специализации каждый вид теряет некото-
рую долю овоей rенетической изменчивости.
Довольно значительное количество данных, касающихся при-
родных популяций, соrласуется с этой моделью.
Лэк (Lack, 1947) указывает, что вертикальное распростране-
ние белоrл'азки (Zosterops palpebrosa) в тех районах юrо-восточ-
ной Азии, rAe она встречается одна, отличается от TaKoBoro :8
раЙонах, в которых она встречается совместно с друrими видами
Zosterops. В Бирме, rде эта rруппа видов представлена единствен
НЫМ видом Z. palpebrosa l последний размножается на всех BЫCO
1'ах ---- от уровня моря до высокоrорий. Но на Малаккском полу
:острове И на I(алимантане, rде на средних н больших высотах
обитает один из близких видов Zosterops, Z. palpebrosa оrраничеН
низменностью. На островах Яве, Бали и Флорес, rде один из близ
ких видов Zosterops занимает самые высокие места, а друrой----
прибрежные зоны, Z. palpebrosa оrраничен средними высотами.
Друrая rруппа данных связана с явлением смещения призна-
ков, которое будет описано ниже.
Смещение признаков
Смещение признаков можно наблюдать у некоторых пар ВИДОII,
-ареалы которых перекрываются, так что они оказываются сии-
патрическими в одной части своих ареалов и аллопатричеСКИМR
. друrих. Попробуем сравнить аллопатрические и симпатрические
198
Часть У. Видообразование
расы таких двух видов по их морфолоrическим, эколоrическим
или поведенческим признакам. При этом нередко оказывается,
что симпатрические расы двух таких видов различаются более
четко, чем аллопатрические. Более высокая степень дифференциа
ции симпатрических рас перекрывающихся симпатрических видов
известна под названием смещения признаков (Вrоwп, Wilson,
1956) .
Смещение признаков наблюдается у ряда rрупп животных н у
нескольких rрупп растений. Оно обнаружено у поползней (Sitta)
в Евразии в отношении общих размеров тела и размеров клюва,
у земляных Bы pKOB (Geospiza) rалапаrосских островов в отно-
шении тех же признаков, у североамериканских представителей
муравьев рода Lasius в отношении ряда признаков и у друrих
rpYnIl позвоночных и насекомых (Brown, Wi1son, 1956).
Дифференциация признаков, по которым наблюдается явление
смещения, может быть обусловлена разными типами отбора. CMe
щение признаков, относящихся к особенностям размножения, ино-
rда возникает в результате отбора, направленноrо на усиление
репродуктивной изоляции. Этот процесс нас здесь не интересует
(см. rл. 24). Но симпатрическая диверrенция признаков, оказы
вающих влияние на успех в эколоrической фазе жизненноrо цикла,
таких, как общие размеры тела и размеры клюва у птиц, скорее
представляет собой результат межвидовоrо отбора, направленно
ro на эколоrическую дифференциацию (Brown, Wilson, 1956)_
Эколоrическая ниша
«Эколоrическая ниша» термин, имеющий очень важное значе
ние при рассмотрении принципа KOHKypeHTHoro исключения. Ка}(
мы определяем этот термин? И можем ли мы дать ему такое опре-
деление, которое не зависело бы от принципа KOHKypeHTHoro иск
лючения?
Миоrо усилий было приложено к тому, чтобы найти удовлетво-
рительное формальное определение ниши, либо словесное, либо
математическое (см. Соппеl1, Orias, 1964; Hutchinson, 1965; Le-
vins, 1968; Krebs, 1973; Whittaker, Lcvin, Root, 1973; Pianka, 1974;
Rejmanek, Jenik, 1975). Это весьма скользкая тема, и поиски пока
не привели к успеху.
Хатчинсон (Hutchinson, 1965) начинает с Toro, что проводи1'
различие между так называемой фундаментальной.и реализован-
ной Н'ИШами. Фундаментальная ниша, соrласно Хатчи,нсону, ЭТО
определенный объем в MHoroMepHoM пространстве, занимаемый
данным видом, причем каждое измерение соответствует одному из
изменяющихся факторов, необходимых для жизни вида. Принима-
ется, что мноrомерный объем фундаментальной ниши не наруша"
ется присутствием конкурентных видов. А реализованная ниша........
r А. 20. Э"ОАоzическая дифференциация
199
это фундаментальная НИШа, оrраНllченная присутствием конку-
peHTHoro вида.
Однако, как нам кажется, хатчинсоновское определение фунда-
ментальной ниши относится к тому, что эколоrи растений и reo-
ботаники издавна называли «эколоrической амплитудой» или
-«rраниu.ами толерантности данноrо вида. Оно определяет поте.Н-
циальную область, которую может занимать вид, а не ero нишу.
Хатчинсоновская же «реализованная ниша» представляет собой
эколоrическую нишу. Включение эффектов межвидовой конку-
ренции в определение реализованной ниши очень важно.
Полевой натуралист знает, что такое эколоrическая ниша на
практике, независимо от Toro, может он дать ей определение или
нет. rpYHToBble воды и поверхностные воды это разные эколо-
rические ниши для корневой системы растений. Крупные семена
и мелкие семена разные кормовые ниши для зерноядных птиц.
Хорошим примером разнообразия ниш служат кормовые зоны
..американских славок в еловых лесах на ceBepO BOCTOKe США
(MacArthur, 1958). Пять симпатрических видов Dendroica ПИТа-
ются насекомыми в трех разных частях древесных крон. Они рас-
пределяются следующим образом:
.J3ерхняя часть кроны: Dendroica tigrina и п. fusca
Средняя часть кроны: черноrорлая древесная славка (D. virens)
. и каштановая древесная славка (D. castanea)
.Нижняя часть дерева: желтоrоловая древесная славка (D. coro-
nata)
Существует также более тонкая дифференциация между вида-
ми Dendroica l питающимися в одной и той же зоне. Так, в сред-
Ней зоне D. castanea ловит насекомых rлавным образом на лету,
"оrда как D. virens редко добывает себе пищу таким образом
(MacArthur, 1958).
Существует бесконечное множество подобных примеров; все
они показываlOТ, что нишу можно охарактеризовать по двум Ка..
теrория),{ факторов: 1) меетообитание или обшир.ная область сре-
ды, к которой приспособлен да.нный вид; 2) оrраничения, налаrае-
мые на использование этой среды межвидовой конкуренцией и
Межвидовым отбором. Следовательно, нишу можно рассматривать
Как тот аспект среды, к которому данный вид узкоспециализиро-
Ван.
Из этоrо лоrически вытекает представление о различиях в
ширине ниш. Эколоrические ниши MorYT быть относительно узки-
Ми или относительно широкими. И ширина ниши, по видимому,
коррелирует со степенью специализации Toro вида, который ее за-
нимает. Ряд эколоrов указывал на то. что ширина ниши умень-
шается с усилением межвидовой конкуренции. Присутствие в
200
Часть V.Видообразование
данном сообществе ряда зколоrически сходных видов способствует
созданию узкой специализации и сокращению ширины ниши
(Lack, 1944; Connell, Orias, 1964; Levins, 1968).
Влияние 9ко.nоrиЧеских требований
Межвидовая конкуренция за основные ресурсы среды приводит
в действие процесс межвидовоrо отбора, который в конеч.ном счете
может привести либо к эколоrической диверrенции, либо к полно..
му замещению одноrо вида друrим. Лоrично допустить, что меж.
видовой отбор будет протекать быстрее при наличии интенсивной
межвидовой конкуренции. Общее правило, из KOToporo имеется ряд
исключений, rласит, что крупные животные и растения предъявля-
ют к среде более высокие требования, чем мелкие орrанизмы, и
быстрее истощают ресурсы, количество которых оrраничено. Сле.
дует ожидать, что эти различия между крупными и мелкими opra..
низмами ведут к коррелятивным различиям в интенсивности меж-
видовой конкуре.нции и скорости межвидовоrо отбора.
р.осс (Ross, 1957) изучал rеоrрафическое и эколоrическое рас-
пространение шести близких видов цикадок в штате ИЛJIИНОЙС
(rруппа Erythroneura lawsoni; Jassidae, Hemiptera). Эти мелкие
насекомые питаются и размножаются на листьях платана (Plata..
nus occidentalis) и, таким образом, их эколоrическая ниша очень
оrраНИчена. Тем не менее в rруппе Е. lawsoni весьма обычна био-
тическая симпатрия и нередки случаи, коrда в одной и той же
нише сосуществует по нескольку видов. Попытки найти в такиХ
симпатрических ассоциациях признаки меж,видовой конкуренции
не увенчались успехом.
Росс (Ross, 1957) приписывает это видимое отсутствие или сла-
бость межвидовой конкуренции малым размерам цикадок, блаrо.
даря которым их популяции MorYT существовать в оrраниченной
нише, не истощая запасов пищи. Он сухо отмечает, что «было бы
трудно представить себе полдюжины видов слонов, которые все
сохранялись бы в качестве репродуктивных единиц, если бы их пи-
ща была оrраничена листьями платана».
И далее он делает общий вывод: «Число видов, которые [мо.
rYT] занимать одну и ту же нишу, обратно пропорционально пи-
щевым требованиям, а следовательно, абсолютным размерам ар.
rанизма».
Характер распространения у ряда rрупп животных соответст-
вует обобщению Росса. Знаменательно, что близкие виды крупных
млекопитающих почти всеrда ал.лопатричны, например африкан
СКИЙ и индийский слоны, виды крупных кошек и медведей в боль..
шинстве видовых комбинаций. Двуроrий и однороrий носороrи в
Южной Африке В"стречаются в одной и той же области, однако в
пределах этой области они смежно--симпатричны и заНимаЮт раз-
r Л.. 20. Э"ОАоzичес"ая дифференциация
201
ные ниши; кроме Toro, один вид редок. В отличие от эrorо у мел..
ких rРЫЗУIiОВ весьма обычны биотически-симпатрические сочета-
ния.
Противоположная крайность среди высокоорrанизованных жи-
вотных наблюдается, однако, у мелких насекомых, таких, как Ery-
throпeura l Drosophila и Anopheles, у которых обычны биотически-
симпатрические сочетания, состоящие из нескольких или мноrих
видов.
Параллельные тенденции наблюдаются у высших растений в
умеренном поясе. Близкие виды доминирующих деревьев в таких
родах, как Quercus и Pinus, и мноrолетних травянистых растений в
таких родах, как lris и Poleтoniит l в большинстве случаев ИМе-
ют аллопатрическое или смежно--симпатрическое распространение.
Но мелкие однолетние травянистые растения, подобно Gilia и
Mentzelia l образуют симпатрические сочетания из нескольких ви-
дов.
Личинки бабочек жадно поrлощают ли-ству растений, тоrда как
сами бабочки питаются на цветках в незначительной или умерен..
ной степени. Очень показательно, что данный вид бабочек часто
имеет широкую пищевую нишу на стадии имаrо, но весьма узкую
и специализированную пищевую нишу на личиночной стадии.
Взрослые бабочки питаются на цветках широкоrо спектра видов,
Но ОТК.'Iадывают яйца на специфических растениях хозяевах, rде в
дадьнейшем личинки завершают свое развитие.
у разных rрупп бабочек личинки имеют разных растений-хо-
зяев. Так, у Battlls philenor кормовым растением для личинок
служит Aristolochia, у данаиды (Danaus plexippиs) ---- ваточник
(Asclepias), а у жептуш'ки (Colias eиrytheтe) ---- люцерна, клевер
" вика (Fabaceae). Но имаrинальные стадии этих же видов бабо-
чек кормятся на широко перекрывающихся рядах видов. Парал-
JIельные различия между шириной пищевых ниш у личиночных
и имаrинальных стадий наблюдаются и во мноrих друrих rруппах
бабочек. Таким образом, у усиленно питающихся личинок мноrих
бабочек Эколоrическая дифференциация зашла rораздо дальше,
чем у взрослых бабочек, питающихся в слабой или умеренной
степени.
rлава 21
rЕоrР.АФИЧБС ОЕ .ВИДООБРАЗ-ОВАНИЕ
Эволюционная диверrенция
Дllверrенция между б..1ИЗКИМИ эволюционными линиями, если
она протекает постепенно и продолжаеreя долrо, проходит с тече-
нием времени через ряд стадий. Общая предковая популяция дает
начало двум или более локальным расам, rеоrрафическим расам,
полувидам, биолоrическим видам и rруппам видов, последователь-
но сменяющим друr друrа (рис. 21.1). Диверrенция может дасТИ-
raTb .самых высоких уровней от рода до класса и типа.
Популяционные единицы и таксономические катеrории пред..
ставляют собой отдельные стадии процесса эволюционной дивер-
rенции. Если рассматривать высшие таксономические катеrории
как продукты эволюции, то они представляют собой макроэволю-
ционные проблемы. А нас здесь интересует процесс диверrенции
на низших ступенях среднеrо уровня, т. е. на уровне рас и видов_
Диверrенция на этих уровнях относится к проблемам видообра-
зования.
Диверrенция на уровне рас и видов проявляется в трех rлав-
ных направлениях. Переход от низших к высшим уровням на вет..
вящемся филоrенетическом древе сопровождается: 1) возрастани..
ем дифференциации rенотипа; 2) возрастанием морфолоrической,
физиолоrической и поведен ческой дифференциации; 3) более СИ.ТJь-
ной изоляцией (рис. 21.1).
.Следует помнить, что все наши рассуждения носят весьма об-
щий характер. Развитие в трех перечисленных выше направлени..
ях может происходить в разных rруппах орrанизмов с разной ско-
ростью. В некоторых rруппах относительно небольшая степень re..
нотипической дифференциации выливается в резко выраже'нную
фенотипическую дифференциацию, тоrда как в друrих rруппах re..
нотипическая дифференциация выражена очень сильно, а феноти"
пическая ---- слабо. При рассмотрении обширной выборки расти-
тельных или животНЫХ rРУIПП можно Iвидеть, что репроду.ктивная
изоляция лишь .в очень слабой степени к'Оррелирооана: со степенью
фенот.и.пИ'ческой дифференциации.
Последовательность стадий видообразования также подверже-
на изменчивости. Некоторые из средних стадий видообразования
иноrда выпадают. Возможна также реверсия процесса диверrен-
ЦИИ. Подобные отклоняющиеся типы будут рассмотрены в даль..
нейше!fl. Здесь мы исключили все Э1М вариации, постулировав s
начале раздела постепенную и длительную диверrенцию.
rA. 21. rеоzрафичеС1Сое видообразование
203
. Обычный способ диверrенции, известный под названием reorpa..
.фическоrо видообразования, происходит в следующем порядке:
1) общая предковая популяция; 2) rеоrрафические расы; 3) алло-
lIатрические полувиды; 4) биолоrические виды; 5) симпатрические
виды. Такой способ видообразования, как мы увидим ниже, не
универсален. Однако он хорошо установлен для некоторых rрупп,
Степень изо/Тяции
---!руппа 8ииО8
DU0/102utleCKUU
8ug
ЛОI1У(Jug
__ fёО2раl1Juчес/(а
раса
--- --- 1I0KaA"Hal1
раса
d
преgКО{Jая
попУI1ЯЦUf{
Уровень gutpqJepeH :.{иации
Рис. 21.1. Стадии диверrенции.
а кроме Toro, поскольку это процесс постепенный, то участвую
щие в нем rруппы проходят на пути к достижению видовоrо ста-
"уса через все промежуточные стадии. Поэтому наш обзор процес
СОВ видообразования удобно начать с rеоrрафическоrо видообра-
зования.
Факторы, участвующие в образовании рас
Образование рас, будь то локальные или rеоrрафические, Не-
прерывные или разобщенные расы, происходит под действием тех
же эволюционных сил, которые обусловливают микроэволюцион-
ные изменения внутри ПОПУ.'1яций. Как уже rоворилось (rл. 4 и 7},
204
Часть V. Видообразование
к основным процессам, создающим изменчивость, относятся мута-
ционный процесс, поток reHoB и рекомбинация, а к процессам, сор'..
тирующим возникшие изменения, отбор и дрейф reHoB. Образо-
вание рас происходит в тех случаях, коrда эти процессы протека-
ют в разных направлениях (Т. е. создают различные rенофонды)
в разных rеоrрафических подразделениях области, занимаемой
видом.
Роль отбора
Естественный отбор, несомненно, служит rлавной действующей
СИ.пой, участвующей в формировании рас. О роли отбора в обра..
зованин рас свидетельствуют две rруппы явлений. Во-первых, мы
постоянно убеждаемся в том, что расовые признаки носят приспо..
собительный характер по отношению к той среде, в которой оби-
тает данная раса. BO .BTOpыx, у разных видов, на'селяющих сход-
ные диапазоны сред, нередко можно наблюдать параллельную ра..
совую изменчивость.
Одно из обобщений систематики животных, известное под наз..
ванием правила Берrмана, касается видов теплокровных позво-
ночных, которые встречаются как в теплых, так и в холодных об-
ластях. Правило Берrмана rлаСИТ t что такие виды обыq.но пред-
ставлены в областях с холодным К,,'Iиматом крупными расами, а в
областях с теплым климатом мелкими расами. Это правило под-
твердилось в отношении примерно 75 90 % видов птиц и 60 80%
видов млекопитающих в различных фаунах. Тем самым ero можно
признать справедливым обобщением, из KOToporo имеются некото-
рые исключения (см. Mayr, 1963).
Адаптивная ценность различий в размерах тела, о которых 1'0.
ворится В правиле Берrмана, зависит от термодинамических
свойств HarpeToro теnа. В холодном климате большое HarpeToe тело
обладает преимуществом, поскольку оно сохраняет тепло вслед-
ствие низкоrо отношения площади поверхности к объему. В уме-
ренно теплом климате это преимущество исчезает. А в жарком
климате небольшие размеры тела MorYT создавать определенное
преимущество в реrуляции температуры, так как высокое отно-
шение площади поверхности к объему способствует рассеянию
тепла.
Если бы подобная расовая изменчивость размеров тела была
обнаружена лишь в нескольких отдельных случаях, то основания
для утверждения о том, что эта изменчивость адаптивна, а сле..
довате.ПЬНО, контролируется отбором, были бы весьма шаткими
Однако, поскольку подобная тенденция в изменении размеров
Te,,'Ia наблюдается у мноrих различных видов в разных фаунах и
служит основанием для признания правила Берrма'на, представ-
ление о контролирующей роли отбора становится вполне оправ-
данным.
r А. 2/. rеоzрафuчес"ое видообразование
205
Друrое связанное с этим экоrеоrрафическое правило, правило
Ал..lена, относится к ДЛИне выступающих частей тела ---- ушей, но-
са, клюва, хвоста у видов теплокровных животных, обитающих
в различных климатических ЗОНаХ. Соrласно правилу Аллена, у
рас данноrо вида птиц или млекопитающих, обитающих в холод-
ных областях, выступающие части тела короче, чем у рас Toro
же вида из теп.'1ЫХ областей.
Эта общая тенденция, наБJ1юдаемая в расовой изменчивости,
подобно тенденциям в изменчивости размеров тела, связана с
проблемами термореrуляции у тепw'lОКРОВ'НЫХ животных. Си.'1ьно
выступающие части тела рассеивают тепло в окружающую среду.
Поэтому теплокровному животному, обитающему в холодном кли-
мате HeDblroAHo, чтобы эти части были длинными; в теплом же
климате, напротив, сильно выступающие части тела создают оп-
ределенную ВЫrоду.
В различных rруппах животных наблюдаются мноrие друrие
упорядоченные элементы расовой изменчивости, имеющие, возмож-
но, адаптивное значение. Более подробно об этом см. в обзорах
Ренша (Ren h, 1960а, Ь) и Майра (Mayr, 1963).
Параллельная расовая изменчивость, коррелированная с осо-
бенностями среды, обнаружена также у мноrих видов растений,
занимающих обширные ареалы. Коrда речь идет о растениях, то
rоворят обыqно не об экоrеоrрафических правилах, а об «экоти-
пах» (или адаптивных расах).
Виды 'растений, распространенные вдоль береrовой линии, мо-
rYT быть представлены стелющимися экотипами на песчаных дю-
нах и кустистыми экотипами IЩ с'калах. Виды деревянистых рас-
тений на высоких ropax MorYT быть представлены древовидными
экотипами в субальпийской зоне и кустарниковыми стланиковыми
экотипами в альпийской зоне (см. обсуждение вертикальных рас
Pinus и т. Д. В r.'I. 18). Такие экотипы или расы несомненно возни-
кают в результате отбора, производимоrо средой. Имеется ряд об-
зоров, посвященных превосходным экспериментальным исследова-
ниям ЭКОiИПОВ у растений (Turesson, ]922, 1925; Clausen, Keck,
Hiesey, 1940, 1948).
Роль дрейфа rеиов и интроrрессня
СлучаЙная расовая изменчивость наблюдается у некоторых ви
дов С колониальной популяционной системой. Под «случайной» 'мы
подразумеваем отсутствие явной корреляции между расовыми раз-
JIИЧИЯМИ и различиями в среде по трансектам, проведенным через
область. занимаемую данным видом. Иллюстрацией служит но-
воrвинейская rалатея (Tanysiptera galatea) на территории Новой
rвинеи (Маут, 1942).
Новая rвинея очень большой остров (около 2500 км в Д..1И-
ну И от 160 до 800 км В ширину). На нем можно встретить самые
206
Часть V. Видообразование
,разнообразные типы климата и растительности ---- от вечнозелено-
ro дождевоrо леса на одном конце до обнажающеrося в сухое Bpe
мя rода Myc.coHHoro леса на друrом коНце. Новая rвинея окруже-
на мноrочисленными мелкими и средними островами, климат и
растительность которых сходны с соседними частями r,,1aBHoro ост-
рова.
rлавный остров, т. е. собственно НОВУЮ rвинею, насеЛЯЮТ три
расы Tanysiptera galatea, а имен'но Т. g. I!alatea, Т. g. meyeri "'.
!Jl!l
1
ros
/
c:t
4
hgd
Рис. 21.2. Распределение рас rалатеи (rруппа Tanysiptera galatea) на территории
НОВОЙ rвннеи (Mayr, 1942).
gal Т. g. galatea; теу Т. g. meyeri; min Т. g. miпor; еи Т. g. ешои; са' Т. g. са-
,оаnае; rie Т. g. riedelii; vиt Т. g. vиtcani; ros Т. g. rosseliana; hyd Т. hydrocharis.
Т. g. minor. Эти расы распространены с перерывами от одноrо кон-
ца острова до друrоrо (рис. 21.2). Они очень сходны. Пять дpy
rих рас встречаются на некоторых из небо..1ЬШИХ 'соседних островов
(рис. 21.2). Эти островные расы хорошо отличаются друr от дру-
ra и от рас, обитающих на соседних с ними частях r..'1aBHOrO ост-
рова, от которых они, возможно, произоul.'IИ. Близкородственный
вид Т. hydrocharis встречается на двух небольших островах, а Ta'K
же в одной части r.Павноrо острова (рис. 21.2).
Майр (1'v\ayr, 1942, 1954) объясняет подобную расов);'ю измен-
чивость следующим образом. Ортодоксальная расовая изменчи-
вость на собственно Новой rвинее ---- продукт отбора в бо.тIЬШОЙ
И широко скрещиваlOщейся ПОПУ..т:Jяционной системе. Каждая из
островных популяций была основана неБО,,1ЬШИМ ЧИС,,10М особей,
миr.рировавших с r..'IaBHOrO острова. Дрейф (или эффект основате-
ля, по терминолоrии Майра) ПРОИСХОДИ,,1 в течение первых поко-
лений КО,,10НИСТОВ. а в дальнейшем эти эффекты поддерживались
.,блаrодаря пространствею.юй 'Изоляции от БО.т:JЬ1ll0Й популяции
r А. 21. rеО2раФичеС1Сое видообразование
207
rлавноrо острова. Все эти факторы привели к заметной диверrен-
ции рас. населяющих малые острова. от соседней и, возможно.
предковой популяции rлавноrо острова_ Популяция, населяющая
один из малых островов, диверrирова.ТIа так сильно, что достиrла
видовоrо paHra ---- Т. hydrocharis.
Нет необходимости допускать, что отбор не принимает участия
в образовании рас в rруппе Т. galatea. Как указывалось в rл. 13,
дрейф и отбор обычно действуют В природных популяциях COBMe
стно. Биолоrически реалистичным различием. касающимся факто
ров, ответственных 'за образование рас, следует считать различие
между .отбором в больших популяциях. с одной стороны. и взаимо
деЙJcт.вие отбор ---- дреЙф IreHoB ----.с друrой.
Известно MHOI'O дру,rих Iслучаев .расовой изменчивости. COOTBeT
сreующих тому, что наlб.1юдае1'СЯ в rР.J'1!"lJПе Tanysiptera. Одним из
них служит rруппа Nigella arvensis (Ra.nunculaceae) на островах
киклады в Эrейском море и в прилежащих материковых областях
(см. rл. 18 и рис. 18.5).
Интроrрессия пред тавляет собой особый способ естественной
rи6ридизации. известный rлавным образом у растений. Интроrрес
сия заКЛJOчается в образовании естественных rибридов F, и их
возвратном скрещивании с одним И,,1И обоими родительскими ви
дами иди ПО.'1увидами. Нередко возвратное скрещивание происхо-
дит MHoroKpaTHo в ряде последовательных поколений. В результате
возникает поток некоторых rеиов от вида или полувида, служаще-
ro донором, в популяционную систему. служащую реципиентом.
Интроrрессия ---- это доведение потока reHoB и рекомбинации до ви-
довоrо уровня.
В тех случаях, коrда два вида или полувида. А и В, марrиналь-
но сиМ'патричны и между ними происходит интроrрессия. в зоне
перекрывания возникают расы, KOHBepreHTHble по различным мор-
фолоrическим признакам и эколоrическим предпочтениям. В обла-
сти, занятой видом В, вид А в результате интроrрессии дает нача-
ло расе, которая приближается к виду В по ряду признаков. и Ha
оборот. Примеры этоrо выявлены у /ris, Tradescantia, Helianthus#
Gilia и мноrих друrих rрупп растений (см. обзор Gr ant, 1971; Hei
ser. 1973Ь).
в дaHHO 1 случае, как и в случае дрейфа, Не следует допускать
отсутствие отбора. Интроrрессия порождает изменчивость. но от-
бор сортирует возникшие вариации, элиминируя одни и сохраняя
друrие. Интроrресtивные расы ---- продукт cOBMecTHoro действия ин-
троrрессии и отбора.
3арождающаяся репродуктивная изоляция
Между разными rеоrрафическими расами одноrо и Toro же вИда
нередко cyrцecTByeT частичная репродуктивная изоляция. Расы.
обитающие в разных rеоr:рафических областяХ. скорее Bcero Имеют
208
Часть У. Видообразование
разные сезоны цветения или размножения. Столь же вероятно, что
они имеют разные эколоrические потребности.
Между rеоrрафическими раоеами MorYT также существовать ча-
стичные ВНУ1'реиние преrрады. Искусственные межрасовые rибриды
F 1 у травянистых цветковых растений обыч.но бывают полустериль-
ВЫ, а у их потомков F 2 обычно наблюдаются некоторые признаки
разрушения rибридов. Между отдельными парами rеоrрафических
рас можно наблюдать, во всяком случае в неполной или зарожда-
ющейся форме, 'все известные механизмы репродуктивной изоля-
ДИИ, описанные в rл. 19.
Репродуктивная изоляция и изоляция, создаваемая средой, мо-
жет развнвать"Ся и в самом деле развивается в качестве побочноrо
продукта эволюционной диверrенции. Рассмотрим две rеоrрафи-
'Ческие расы какоrо либо вида растений, обитающие в разных об-
.ластях, в одноЙ ИЗ которых климат прохладный и влажный, а в
Jl.руroй теплый и сухой.
Эти две расы будут подверrа1'Ь'СЯ отбору на приспособленность
'к разным климатическим, а возможно, и разным эдафическим фак-
'торам; это первая стадия эколоrической изоляции. У таких рас l1.Be-
теНне, по всей вероятности, происходит в разное время roда; в
результате возникает частичная сезонная изоляция. В прохлад-
ных влажных и теплых сухих районах, по видимому, обитают раз-
ные. rруппы пчел, а по мере Toro, как местные ра-с.ы растений при-
.спосабливаются к обычно посещающим и опыляющим их видам
пчел, в строении их цветков возникают измснения. Это приводит к
зарождению механической или этолоrической изоляции, а може}'
.быть, и обеих сразу. Некоторые изменения в строении цветка, на..
":пример изменение длины столбика, окаЗЫвают влияние на леrкость
скрещИ'вания и MorYT привести к возникновению преrрады несов-
местимости. Наконец, мноrочисленные reHHbIe различия, накапли-
.ваемые двумя такими расами за время их дифференциации, впол-
/не MorYT привести к образованию у их rибридов каких либо дие-
;rармонируlOЩИХ продуктов рекомбинации. Так возникает частич-
:ная нежизнес.пособность или полустерильность rибридов, а также.
разрушение rибридов.
Таким образом, диверrенция на ра-совом уровне неизбежно вы-
.зывает в качестве побочноrо эффекта некоторую репродуктивную
изоляциlO. Это создает основу для возмоЖноrо будущеrо видооб-
разования.
rеоrрафичес.каR теория вид,ообразования
rеоrрафическая теория видообразования была создана рядом
зоолоrов, в числе которых следует назвать М. BarHepa, работавше-
ro в конце XIX в., и К. Иордана, Д. Джордана, Рен ша, Добржзн-
CKoro, i\'\айра и друrих в первой половине ХХ в.; позднее Клау-
r л. 21. rеО2рафичесrcoе видообрDЭование
'209
зен, Стеббинс и друrие распространили эту теорию и на видооб..
разование у растений. Все теоретические рассуждения и фактичес..
кие данные по этой проблеме ясно изложены Майром в ero книrе
«Систематика И прои-схожде-
HJ{e видов» (Mayr, 1942).
Соrласно rеоrрафической
теории, биолоrические виды
обычно возникают из reorpa-
фических рас. Пространствен
ная изоляция нормальная
прелюдия к развитию репро-
дуктивной изоляции. Последо-
вательность стадий видообра
эования представлена !на
рис. 21.3. Вид, состоящий из
непрерывных rеоrрафических
рас (рис. 21.3, Б), распадается
на ряд разобщенных рас
(рис. 21.3,8), Между разоб-
щенными расами в качестве
побочноrо продукта диверrен-
ци.и развивается репродуктив-
ная изоляция. После Toro как
репродуктивная изоляция до-
ходит до известной критиче-
ской точки, д..иверrировавшие
популяции MorYT обитать на.
одной и той же территории,
не скрещиваясь между собой,
и достиrают тем самым видо-
Boro статуса (рис. 21.3, Д).
Все стадии видообразова-
ния, которых требует reorpa-
фическая теория, как проме-
жуточные, так и терминаль-
ные действительно обнаружены в разных rруппах животных и
растений. Это самый убедительный довод в пользу rеоrрафической
теории.
Ряд примеров, относящихся К одной естественной rруппе,.....
видам rилии (Polemoniaceae), представлен на рис. 21.4. Все это
однолетние диплоидные, преимущественно саМООПЫ.rIяющиеся рас..
тения пустынных и rористых районов американскоrо юrо-запада.
В этой rруппе наблюдается следующая последовательность стадий
(Grant. Grant, 1956, 1960): 1) вид, состоящий из непрерывных
rеоrрафических рас (рис. 21.4, вверху слева); 2) разобщенные и
различающиеся rеоrрафические расы (рис. 21.4, вверху справа;
А
C:::::J
cdo
!
СО О
/j
8
.r
А
СУ
Рис. 21.3. Стадии rеоrрафическоrо видо-
образовании (Mayr, 1942, с измене-
НИSIr.си) .
А. Шнроко распространениый однородный ВИ,ll..
Б. ПолитипическнА ВИД с непрерывными ра-
сами. В. По.1итипический вид с одной отде-
':швшсt\си и Оt"личаlOщеАся расой. r и Д. Миr-
рация, ведущая k восстаНОВ.1ению контакта
между О'I'де.'JИвшейся расой и основной частьЮ
вида. Т. СкреЩивание и вторИЧllая иитерrра-
дация. д. Скрещиоания не пронсходнт; по'
П}'.'lяцин, бывшие прежде разобщениыми ве-
дут себя как виовь образова шиеся внды.
210
\ G .la ,
':l i5 aQ'Iyi .).'.' lQti"o:' j '
.o
"
'\ .
"-
,
6./epfaпfпa ",
,
nllrpиs/}"
p' . ,
'neforиm. '
· · transvl!rsQ f
'?
/eptanfha
.. .......-.....
,
"
,
,
"
,
"
"
,
,
)
Часrь У. Видообразование
I
I
I
I
I I
: I
r L
I I
, I
С,... I
" I I
, I I
" , I
't I
" I
I I
r I
/ I
1 I
.. .. ...: I
..... I
,
,
,
,
!....\
'. \,
() \\'"
j
.i
\\"
,
.
" .,
'.. ,
,
,
,
,
)
Рис. 21.4. Стадии rеоrрафическоrо видообразовании у rИ.IJИИ {Po]eтoniaceae'
(по данным Grant, Grant, 1956, 1960).
Объя:снеиия СМ. в тексте.
см. также рис. 18.4); 3) синrамеон, состоящий из трех марrиналь..
но-симпатрических полувидов, время от времени образующих rиб.
риды (рис. 21.4, в середине); 4) пара стерильных при скрещива.
нии друr с друrом аллопатрических видов (рис. 21.4, 8i/tUЗУ еле.
ва); 5) пара стерильных при скрещивании друr с друrом симпат..
рических видов (рис. 21.4, внизу справа).
Поrраничные случаи между rеоrрафическими расами и вполне-
сформировавшимися видами приобретают в этом контексте осо-
бенно большое значение. Такие поrраничные случаи, или полуви-
ДЫ, довольно обычны, что хорошо известно каждому систематику
животных или растений. По мнению Майра (Mayr, 1942), 12,5%
всех таксономических видов птиц Северной Америки занимают.
промежуточное положение между видами и полувидами, так чт()
орнитолоrи относят их то к одной, то к друrой из этих таксоно.
мических каТеrорий. Примерами служат два вида американских.
r л. 21. rеО2рафичес"ое видообразование
211
ДЯТЛОВ ---- Colaptes саре, collaris и золотистый дятел (Colaptes
auratus), ---- а также два вида американских древесных славок----
Dendroica auduboni и D. coronata.
Сравнимые примеры имеются во мноrих друrих rруппах жи-
вотных и растений. В качестве одноrо TaKoro примера из царства
растений можно привести Веймутову сосну (Pinus strobus), расту-
щую в восточной части Северной Америки, и Р. тonticola, расту-
щую в западной части.
Особенно интересный тип поr.раничных с.пучаев цепочка ин-
терrрадирующих рас А, В, С, О, Е, образующих широкое кольцо
с перекрыванием :-'-Iежду формами А и Е. Расы А и Е представля-
ют собой морфолоrически и эколоrически крайние типы в данной
популяционной системе; они интерrрадируют в ряду A----B C o----
Е. Бсли исходить только из этой особенности, то рассматриваемую
популяционную систему можно считать обычным видом, состоя-
щим из rеоrрафических рас. Однако концевые и крайне различаю-
щиеся члены этоrо ряда, А и Е, сосуществуют симпатрически, а
следовательно, отношения между ними соответ.ствуют отношениям
между симпатрическими видами.
Среди классических случаев колец перекрывающихся рас сле-
дует назвать большую синицу (Parus major) в Евразии, пеночку-
весничку (Phylloscopus trochiloides) в Центра.пьной Азии, сереб-
ристых чаек (rруппа Larus argentata) на севере Евразии и Север-
ной Америки и оденью мышь (Peroтyscus тaniculatus) в западной
части Северной Америки. При новом ИСС.'1едовании некоторых из
этих давно изученных случаев (например, Parus major) обнару
живается, что изоляция между концевыми расами менее полная,
чем преДПО.ТIаrалось прежде. Сравнительно недавно было описано
MHoro друrих с.пучаев в различных родах птиц, амфибий, насе.
комых и покрытосеменных (см. обзор Майра, 1963). Кольцевое
перекрывание рас у североамериканской одиночной пчелы нори-
tis producta (Megachi1idae) предстаВ,,'Iено на рис. 18.6.
Реверсии д верrенции
Процесс rеоrрафическоrо видообразования. не.'1ЬЗЯ считать не-
поколеБИМЫ:-'-I. Эволюционная диверrенция от rеоrрафических рас
ДО биолоrических видов происходит ишь до тех пор, пока про-
должают действовать эволюционные силы, создающие диверrен-
ЦИЮ. Процесс дивсрrенции может прекратиться на любой стадии.
}-' некоторых видов rеоrрафические расы возниклИ очень давно.
Диверrенция может не только прекратиться, но и изменить свое
направление На любой стадии, вплоть до возникновения полной
етерильности между близкими видами. Естественная rибридизация,
()бычная у высших растений и хорошо известная во мноrих rpyn-
пах животных, представ.пяет собой реверсию процесса диверrенции.
rлаlВа 22
ОПОСОБЫ ВИДООБРЛJОВАНИЯ
Введение
rеоrрафич"еское видообразование. описанное в предыдущей
rлаве. представляет собой один из нескольких способов видообра-
зования. известных у высших орrанизмов. Существуют, кроме Toro,
некоторые способы видообразования. не связанные с rеоrрафичес-
кой ИЗОJ1яцией на промежуточных стадиях; эти способы теорети-
чески возможны, но не проверены и вызывают значительные разно-
rласия. Цель этой rлавы обсудить в форме KpaTKoro обзора раз-
личные способы видообразования, как установленные, так и спор-
ные.
ТеР;\fинолоrия, относящаяся к видообразовз'нию, страдает неко-
торой избыточностью. Это объясняется тем. что разные ученые не-
редко используют разные термины для обозначе,ния по существу
одноro и Toro же процесса или способа. Такое дублирование тер-
минолоrии затемняет един'ство процессов. В этой rлаве мы примем
для разных способов видообразования ряд определительных тер-
минов, которые, по нашему мнению. наиболее удачны, и в процес-
се изложения укажем также на существующие синонимы.
Квантовое видообразование
При rеоrрафическом видообразовании обычно наблюдается сле-
дующая последовательность: локальная раса----rеоrрафическая ра-
са----аллопатрический полувид----вид (как на рис. 21.1). в ОТwIJичие
от этоrо при квантовом видообразовании имеет место непосред-
ственный переход от локальной расы к новому виду. Квантовое
видообразование представляет собой, таким образом, сокращен-
ный способ формирования видов. Различиям в путях образования
соответствуют различия в контролирующих силах. В случае reor..
рафическоrо видообразования диверrенция контролируется отбо-
ром, а в случае KBaHToBoro видообразования ---- сочетанием отбора
и дрейфа reHOB.
Начальной точкой для KBaHToBoro видообразования С.1УЖИТ
обширная полиморфная свободно скрещивающаяся предковая
популяция. Допустим, что rде-то за преде.IJами области, занятой
этим предковым видом, один или несколько миrрантов основали
небольшую дочернюю колонию. Такая дочерняя КО.IJОНИЯ будет
пространственно изолирована в занимаемой ею периферической
местности.
r А. 22. Способы видообразования
213
Особи основатели приносят в эту дочернюю колонию лишь не-
большую и неслучайную выборку из rенофонда предковой попу-
ляции. Поэтому в процессе основания новой колонии имеет место
дрейф reHOB.. Далее, поскольку особям основателям вначале при-
ходится скрещиваться между собой, в течение первых поколений
в новой КО.понии происходит инбридинr и дополнительный дрейф
reHoB.
Инбридинr в такой маленькой изолированной дочерней коло-
нии ведет к образованию и закреплению новых rомозиrотных со-
четаний reHoB, которые обычно были бы разрушены в результате
свободноrо скрещивания в предковой популяции. Это само по себе
служит источником фенотипических новшеств. Однако сущест-
вует еще и дополнительный источник новшеств.
Обычно в свободно скрещивающихся популяциях наблюдается
образование rомеостатических буферных систем, обеспечивающих
развитие «нормальных» фенотипов при наJIИЧИИ высокоrетерози-
rOTHblx rенотипов. Иными с..10вами, отбор в свободно скрещиваю-
щихся популяциях блаrоприятствует а.IJлелям, которые взаимодей-
ствуют друr с друrом в rетерозиrотных сочетаниях, так, чтобы
образовались нормальные фенотипы.
Отсюда вытекает, что вынужденный инбридинr и rомозиrот"
ность снимают эти rомеостатические буферы и допускают форми-
рование новых фенотипов, часто коренным образом отличающих-
ся от нормы. Такие новые фенотипы, появляющиеся в результате
инбридинrа в популяциях, для которых нормой служит свободное
скрещивание, называют феноотклонениями. Феноотклонения хо-
рошо известны из экспериментальных исследований на дрозофиле,
курах, первоцвете, Linanthus и друrих животных и растениях.
Они постоянно возникают в небольших дочерних колониях, про-
исходящих от больших свободно скрещивающихся предковых по..
пуляций.
Следует, таким образом, раз./Iичать новые фенотипы двух уров-
ней: во-первых, простых rомозиrотных выщепенцев, которые сме..
rаются в предковых популяциях, но MorYT закрепиться в неболь-
ших изолированных дочерних колониях, и, во-вторых, фенооткло-
нения. .
Нет никаких rарантий, что новый фенотип, оказавшийся в до-
черней колонии, будь то простой rомозиrотный выщепенец или
феноотклонение, будет обладать адаптивной ценностью в той
среде, в которой находится эта колония. Если он такой ценностью
Не обладает, то колония вскоре вымрет. На самом деле большин-
ство новых фенотипов, по всей вероятности, окажутся неудачни-
ками и содержащие их колонии, вероятно, не cMorYT сохраниться.
Но в длинном ряду проб, коrда БО.'lьшая предковая популяция
основывает Мllоrочисленные дочерние колонии, в разные периоды
времени и в разных участках близ rраницы вида один или не..
214
Часть V. Видообразование
сколько раз Все же MorYT возникнуть rенотипы и фенотипы, обла-
дающие адаптивной ценностью.
Такие ценные в адаптивном отношении rенотипы, коrда они
80зникают, быстро закрепляются в дочерней колонии в резуль-
тате cOBMecTHoro действия инбридинrа, дрейфа и отбора. Эта KO
лония, новые фенотипические признаки которой. сохраняются от-
бором, может затем продолжать увеличивать свою численность и
расселяться в качестве диверrирующей расы или вида.
Такой процесс видообразования был описан в классической
работе Майра (i\'layr, 1954). Затронутые здесь идеи связаны с
более ранними концепциями rенетическоrо дрейфа (Wright, 1931)
н квантовой эволюции (Simpson, 1944), обсуждаемыми в друrих
rлавах этой книrи.
В настоящее время известно несколько примеров возможноrо
KBaHToBoro видообразования. Вспомним характер изменчивости
новоrвинейских зимородков rруппы Tanysiptera galatea. Как от-
мечалось в rл. 21, островные расы Т. galatea, происходящие, ве-
роятно, от небольшоrо числа миrрантов из популяции r.7JaBHOrO
острова, заметно ОТ 'Iичаются от нее. Степень их дифференциации
сильно отличается от обычной расовой изменчивости интерrради-
РУЮЩИХ популяций Toro же вида на rлавном острове. Одна из
островных популяций диверrировала до видовоrо уровня (Т. hy
drocharis; СМ. рис. 21.2).
Некоторые виды дрозофилы на rавайских островах, по види-
мому, возникли В результате KBaHToBoro видообразования. Воз
можную филоrению rавайских дрозофил можно ПрОС.'lедить по
сходствам и различиям инверсионных типов, наб.'lюдаемых в хро-
мосомах клеток слюнных желез. Кроме Toro, устаНОВ.'1ению отно-
сительноrо возраста разных видов способствуют данные о воз-
расте тех островов, которые они населяют.
В rруппу Drosophila planitibia входит три близкородственных
вида (п. planitibia, D. heteroneura и п. silvestris), имеющие оди-
наковую хромосомную формулу, т. е. несущие один и тот же на-
бор инверсий в rомозиrотном состоянии. Эти три вида распростра
нены на двух островах следующим образом: п. planitibia на l\1.ауи,
а D. heteroneura и п. silvestris на rавайи. Возраст острова rавайи
700000 лет, а остров Мауи возник раньше. Этот факт, в сочета
"ии с rомо.'10rичностыо хромосом, позволяет думать, что D. pla
пitibia послужила преДКОВЫ 1 видом, от KOToporo произош.'1и
п. heteroneura и п silvestris (Carson, 1970).
Последовательность событий при одном таком видовом ире-
вращении (например, п. planitibia D. Izeteroneura), вероятно,
следующая: 1) колонизация острова rавайи одним И..1И несколь-
кими миrрантами D. planitibla с острова Мауи, возможно, Bcero
ЛИШЬ одной оплодотворенной ca:\-fКОЙ; 2) закрепление reHoB, быв-
ших прежде полиморфными, в новой дочерней колонии На острове
r л.22. Способы видообразования
215
I'авайи; 3) закрепление сопровождается быстрой диверrенцией
rавайской колонии вплоть до возникновения HOBOro вида, п. he-
teroneura (Саrsоп. 1970).
Второй дочерний вид. обитающий на острове rавайи. D. sil-
f) stris, Mor возникнуть в результате независимоrо акта колониза-
ции от той же самой предковой популяции иа Мауи или же путем
диверrенции от той же самой первичной колонии основателей на
rаВ4ЙИ, от которой произошел вид п. heteroneura (Carson, 1970).
Весьма показательно, что на этих же самых двух островах
наблюдаются параллельные случаи видообразования, очевидно
KBaHToBoro, в двух друrих rруппах видов дрозофи.,1Ы (Carson,
1970) .
Среди однолетних цветковых растений uримером служит пара
видов Clarkia ьаоЬа и С. lingulata (Onagraceae) в ropax Сьерра-
Невада в Ка,,1ИфОРНИИ. Изучение хромосом и эколоrии явно сви-
детельствует о том, что широко распространенная С. biloba была
предковой формой. а узкий энде:мик С. lingulata, распространен-
ный по периферии южной части области, занимаемой С. biloba"
столь же явно представляет собой дочернюю форму. Эти два вида
различаются по двум независимым транслокациям, которые созда-
ют хромосомную стерильность, и по признакам цветка. связанным
с одной из транслокаций. По всей вероятности, полиморфная по
хромосомам популяция С. blloba закрепилась на южном краю
области, занятой этим видом. в результате дрейфа reHoB и отбора,
направленноrо на закрепление определенноrо набора транслока-
ций и reHoB, что в конечном счете приве,,10 к возникновению HOBoro
вида С. lingulata, ОТКЛОНЯlOщеrося от С. biloba (Le\vis, Roberts,
1956; Lewis. Raven, 1958; Lewis, 1962; .Gottlieb, 1974). Этот способ
видообразования. возможно, обычен у одно.петних растений.
Термин «квантовое видообразование» был предложен для
описанноrо здесь способа видообразования rpaHToM (Grant. 1963,
1971). Друrие авторы используют Д.ТIЯ этоrо друrие синонимы:
видообразование путем rенетической революции (Mayre. 1954)
видообразование путем катастрофическоrо отбора (Lewis, 1962);
видообразование в результате чередования популяционных подъе-
:\юв и спадов (Carson. 1971) И, в качестве б 1-изкоrо синонима,
rо юсеквенциальное. видообразование (Carson. 1970. 1971). Я ОТ-
даю предпочтение термину «квантовое видообразование», по
скольку он перекликается с уже известным и БО.т:Jее общим явле
нием квантовой эволюции (см. краткое обсуждение в rл. 30).
Сопоставление rеоrрафическоrо
и кваитовоrо видообразования
Как rеоrрафическое. так и квантовое :видообразование начина-
ются в больших популяциях, размножающихся половым путем.
Оба способа связаны с прохождением через период пространст-
216
Часть У. Видообразование
венной изоляции на пути к достижению видовоrо статуса. На этом
сходство кончается.
При rеоrрафическом видообразовании аллопатрическим пред-
шественником HOBoro вида служит rеоrрафическая раса; . при
квантовом видообразовании таким предшественником служит
локальная раса. В первом случае в процессе участвует отбор,
осуществляемый реrиональными факторами среды в большой
свободно скрещивающейся популяции; а во втором случае ---- от-
бор в сочетании с инбридинrом и дрейфом reHoB в маленьких
дочерних колониях. При rеоrрафическом видообразовании изме-
ения происходят постепенно. медленно и носят консервативный
характер. тоrда как квантовое видообразование происходит быст-
ро и может вызвать резкие изменения.
Различие в силах. реrулирующих эти два способа видообразо-
вания, имеет некоторые последствия. Направленный отбор на но-
вые сочетания reHoB. участвующий в rеоrрафическом видообразо-
вании. сопряжен с созданием большоrо субституционноrо rенети-
ческоrо rруза. Плата за подобный отбор. выражающаяся в числе
rенетических смертей. очень высока и ложится бременем на ре-
продуктивный потенциал популяции. Это бремя в свою очередь
оrраничивает скорость эволюции. доводя избирательную смерт-
ность до таких уровней. которые популяция может выдержать
(см. rл. 14).
Если. однако. родительская популяция перестает полаrаться
на направленный отбор. действующий в больших популяциях, а
вместо этоrо подверrает несколько маленьких популяций совмест-
ному действию отбора и дрейфа reHoB. то она может до некоторой
степени уменьшить плату за отбор. оrраничивающую скорость
этоrо процесса (см. rл. 14). Такая популяция сможет изменяться
fЭыстрее. Следовательно. при rеоrрафическом видообразовании
плата за отбор выступает в ро ']и фактора, оrраничивающеrо ско-
рость эволюции; что касается KBaHToBoro видообразования. то
оно позволяет избежать этой оrраничительной платы за отбор и,
во всяком случае теоретически. может протекать быстрее, чем
rеоrрафическое видообразование.
Видообразование путем аллополиплоидии
Образование новых видов в результате возникновения аллопо-
.nип.поидии давно известно у покрытосеменных и друrих наземныХ
растений. Более Toro, это тот способ видообразования. для кото-
poro по любой Нз крупных rрупп орrанизмов имеются хорошо до-
кументированные экспериментальные даНные. Начнем с последо-
вательноrо определения аллополиплоидии. Полиплоидными назы-
вают клетки, особи или популяции. содержащие три или более
набора хромосом (или reHOMOB); нас здесь. естественно. интере..
r А. 22. СnосоБЬL видообразования
217
сует ПОЛИПЛОИДИЯ в популяциях. Аллополиплоидия (называемая
также амфиплоидией) представляет собой rибридную полиплои..
дию или. точнее, полиплоидию, возникшую в результате межви-
довых скрещиваний.
Допустим. что хромосомные наборы или reHoMbl двух диплоид-
ных видов различаются по нескольким или по мноrим хромосом-
ным перестройкам, например транслокациям или инверсиям. Их
reHoUbl можно обозначить АА и ВВ соответственно. rде А и В
структур но дифференцированные rаплоидные наборы. У межвн..
довых rибридов АВ в первом поколении наблюдается ХРОМОСОМ-
ная стерильность, вызванная нарушениями мейоза.
Допустим далее, что у стерильноrо rибрида АВ происходит уд-
.воение 'числа хромосом. У растений оно возникает спонтанно.
Удвоение может .произойти -в сомаrrичеоких iКлет,ках, в результате
чеrо раЗВИtВаются тетра:ппоидиый побеr, а в конечном счете TeT
раплоидные цветки с reHoMoM ААВВ. Или же диплоидный rибрид
АВ ПРОИЗ80ДИТ нередуцированные raMeTbl (АВ) в качестве ко-
нечноrо результата аберрантноrо мейоза, а объединение двух та-
ких нередуцированных диплоидных raMeT дает в Ft тетраплоид-
ные зиrоты с конституцией ААВВ.
у тетраплоидноrо растения (ААВВ). имеющеrо два набора
хромосом А и два набора хромосом В, в мейозе происходит нор-
мальная конъюrация хромосом с образованием бивалентов А/А
и В/ В, после чеrо хромосомы нормально расходятся к полюсам,
так что получаются raMeTbl (АВ), сбалансированные как по хро-
мосомам, так и по reHaM. Такой аллотетраплоид плодовит. Для
Hero характерно собственное rибридное сочетание морфолоrиче-
ских, физиолоrических и эколоrических признаков, отличное от
тех сочетаний, которыми обладали один и друrой диплоидные ро-
дительские виды. И он размножается в чистоте, сохраняя сВОЮ
промежуточную .rибридную конституцию. блаrодаря внутриrе-
номной конъюrации и расхождению хромосом (А/А и В/В)
в мейозе.
Между этим аллотетраплоидом и ero диплоидными родитель-
скими видами существует преrрада, создаваемая хромосомной
стерильностью. rибриды между ними, в тех. случаях, коrда они
образуются, в FI триплоидны, а триплоиды у растений стерильны
вследствие нарушений, возникающих в мейозе. Наш новый алло-
тетраплоид ААВВ плодовит. обладает своим особым сочетанием
признаков, в отношении KOToporo он размножается в чистоте. и
стерилен при скрещивании с наиболее близкими к нему диплоид-
ными видами. Следовательно, этот аллотетраплоид представляет
собой новый биолоrический вид rибридноrо происхождения.
Аллополиплоидия весьма обычный способ видообразования
у покрытасеменных. .папоротн'И'ков и -некоторых дрyrих rруJПП рас-
тений. СоrлаlCНО недавнИiМ oцeНlKalM, 47% видов покрытосеменных
218
Часть v. Видообразование
и 95% видов папоротникообразных (папоротники и близкие к ним
rруппы) полиплоиды. Большую часть полиплоидов составляют
аллополиплоиды (см. Grant, 1971).
Среди широко распространенных культурных растений из-
вестно немало аллополиплоидов. Примерами с.."'Iужат американ-
ские сорта хлопка (Gossypium hirsutum и О. barbadense; 2п===4х==
== 52). пшеница (rriticuт aestivum; 2n == 6х==42) и табак (N icotia-
па tabacum; 2п ==4Х==48) (4х==тетраплоид; 6х==rексаплоид и т. п.).
Nicotiana tabacuт с reHOMOM SSTT аллотетраплоид. происходя-
щий от двух южноамериканских диплоидов (2п==24). близкий к
.ныне существующим видам N. toтentosiformis (ТТ) и N. silvest-
ris (SS) (Clausen, 1941).
rиБРJ:lдное видообразование у растений
Аллополиплоидия один из нескольких способов rибридноrо
видообразования у растений. Под rибридным видообразованием
подразумевается возникновение в потомстве eCTecTBeHHoro rиб-
рида новой линии, размножающейся в чистоте и изолированной
от родительских видов и ОТ своих сибсов В rибридной популяции.
Эта новая линия должна преодолеть такие препятствия, как rиб-
ридная стерильность и разрушение rибридов. Аллополиплоидия
выполняет' эту задачу. Есть и друrие механизмы, достиrающие
той же конечной цели без изменения числа хромосомных наборов.
Здесь приводится краткое описание двух таких механизмов (более
детальное изложение см. Gr ant, 1971).
Допустим, что родительские виды различаются по двум или
более независимым транслокациям (Р, Q,...). При этом один
родительский вид будет иметь набор PPQQJ а друrой ---- ppqq.
rибрид Fl от TaKoro скрещивания представляет собой rетерози-
rOTY по двум (или по мноrим) транслокациям (Р/р Q/q) и поэто-
му обладает той или иной степеныо хромосомной стерильности.
Однако в F 2 он может дать в результате расщеп..lения несколько
классов структурно rомозиrОТI1ЫХ плодовитых форм.
Два из таких классов плодовитых потомков родительские
типы, но они не являются новыми линиями. Кроме Toro, этот
rибрид может дать rомозиrотных рекомбинантов (PPqq и ppQQ),
а это уже не только плодовитые, но и новые типы. Более Toro, они
отделены от родительских видов и друr от друrа преrрадой,
создаваемой хромосомной стерильностью, хотя в данном случае
она выражена слабо.
Если число независимых транслокаций больше, то хромосом-
ная стерильность, создающая преrрады BOKpyr новых rомозиrот-
ных рекомбинантов, уси.ТIИвается, и новая линия соответственно
становится более изолированной. В основу описанной здесь моде-
ли были положены транслокации. Аналоrичное воздействие на
r А. 22. Способы видообразования
219
стерильность оказывают независимые инверсии или транспозиции.
которые можно использовать в этой модели, произведя соответст-
вующие изменения.
Описанный выше процесс был назван рекомбинационным видо
образованием. Ero можно формально определить как возникнове-
ние в потомстве межвидовоrо rибрида, обладаlOщеrо хромосом-
ной стерильностью, HOBoro структур но rомозиrотноrо рекомбина-
ционноrо типа, плодовитоrо при скрещиваниях в пределах данной
линии, но изолированноrо от друrих линий и от родительских
видов преrрадой, создаваемой хромосомной стерильностью.
Процесс рекомбинационноrо видообразования обнаружен cpe
ди потомков экспериментальных rибридов Nicotiana (Solanaceae).
Elyтus (Gramineae) и Gilia (Polcmoniaceae). Ero роль в природе
остается неясной. Вероятно, такое видообразование возникает
время от времени, но реже, чем аллополиплоидия.
Внутренние изолирующие механизмы определяются rенными
факторами rенами, обусловливающими стерильность, raMeTHbl
ми леталями, rенами несовместимости и т. п., а также cerMeHTHbl.
ми перестройками, обусловливающими хромосомную стерильность.
Теоретически, казалось бы, нет никаких причин, по которым в
основе процесса, параллельноrо рекомбинационному видообразо-
ванию, не моrли бы лежать не хромосомные перестройки, а reH-
ные факторы стерильности. Однако этот возможный способ rиб-
ридноrо видообразования еще не подверrся исчерпывающему изу-
чению.
В некоторых rруппах растений межвидовые rибриды плодови-
ты и изоляция между видами обеспечивается rлавным образом
наружными преrрадами. Эколоrическая и сезонная изоляция, а
также ИЗО 1ЯЦИЯ, обусловленная строением ЦBeTKa, rлавные
преrрады, разделяющие виды. Морфо..10rические, физиолоrические
и поведенческие различия между видами, ведущие к возникнове-
нию таких преrрад, находятся, конечно, под контролем reHoB.
у потомков естественных межвидовых rибридов, если они появ
ляются, происходит расщепление по reHHblM различиям и по со-
ответствующим признакам, определяющим внешнюю изоляцию.
Это создает возможность для возникновения продуктов межвидо-
вой рекомбинации с новыми сочетаниями признаков, заК..1адываIO
щих основу новых, внешне ИЗО '1ированных субпопуляций. Если
внешняя изоляция сохраняется и в да 1ьнейшем, то из этих суб-
популяций моrли бы возникнуть новые виды rибридноrо происхож
дения.
Вероятные примеры rибридноrо видообразования, происходя-
щеrо путем расщепления и рекомбинации по признакам, опреде
ПЯlOщим наружные преrрады, описаны в нескольких rруппах рас-
тений. Возможно, что вид живокости Delphiniит gypsophilиm,
растущий у подножий rop в Калифорнии, возник именно так, от
220
Часть У. Видообразование.
скрещивания D. recurvatuтxD. hesperium (Ranunculaceae) (Le-
wis. Epling, 1959). Аналоrичные примеры обнаружены у Amaran..
thus (Amaranthaceae). Carex (Cyperaceae) и в друrих родах.
Мы не знаем. иrрает ли rибридное видообразование существен-
ную роль в эволюции животных. Большинство эволюционистов
животных отрицает это (см.. например. Mayr, 1963; 'White, 1973).
Однако время от времени появляются сообщения о, по-видимому,
rибридном прои хождении новых видов в разных rруппах живот-
ных (например. Miller. 1955 о дятлах и Ross. 1958---- о цикад-
ках); во всяком случае о такой возможнос'rи следует помнить.
Этот вопрос требует дальнейших rлубоких исследований.
Проблема симпатрическоrо видообразования
Одной из спорных проблем эволюционной биолоrии остается
симпатрическое видообразование. Дискуссии по этому поводу
породили довольно большое число журнальных статей и книr.
Один из возникающих при этом вопросов состоит в теоретической
возможности симпатрическоrо видообразования; на этот вопрос
теперь можно ответить утвердительно. Затем встает вопрос о ре-
альности существования симпатрическоrо видообразования в
природе; мнения по этому вопросу в настоящее время резко рас-
ходятся. В высказываниях за и против немало путаницы.
Необходимо представить проблему симпатрическоrо видообра-
ЗОБан.ия 'в БО '1ее ясной .и ч-еткой форме. Сначала мы уточним yc
ловия. при которых предположительно происходит симпатрическое
видообразование, а затем можно перейти к обсуждению Toro,
осуществим ли данный процесс при этих оrраничительных усло-
виях. Для нашей темы очень важны три rруппы условий.
Во-первых. необходимо уточнить. возникает ли интересующее
нас симпатрическое видообразование в результате первичной
Диверrенции. или в результате rибридизации, или попсреме"но тем
и друrим путем. Видообразование путем аллополиплоидии явля-
ется симпатрическим и широко распространено в растительном
мире. Друrие способы rибридноrо видообразования у растений
также симпатричны и встречаются во всяком случае ИНоrда, по-
казывая тем самым. что аллополиплоидия не есть какой то особый
исключительный случай. Истинная проблема. таким образом.
сосредоточена на возможности симпатричсскоrо видообразования
в. период первичной диверrенции.
Во-вторых. надо указать систему размножения орrанизма,
участвующеrо в симпатрическом видообразовании. Если это са-
мооплодотворение. то особой проблемы не возникает. Сложная
ситуация складывается при симпатрическом видообразовании у
орrанизмов со свободным скрещиванием вследствие затопляю.
щеrо эффекта nepeKpecTHoro оплодотворения.
r А. 22. Способы видообразования
221
В-третьих, какая микроrеоrрафическзя зона имеется в виду
при употреблении термина «симпатрический»? Симпатрия может
быть либо смежной, либо биотической (см. rл. 18), и при обсуж-
дении данной проблемы следует проводить различие между этими
двумя состояниями. Симпатрическое видообразование rораздо
JIеrче осуществимо в смежно симпатрической,. чем в биотически-
симпатрической ЗОНе.
Смежно-симпатрическое видообразование
ы займемся здесь первичной диверrенцией в свободно скре-
щивающейся популяции, rде диверrИРУlощие ..'Jинии занимают
смежно симпатрические зоны. Диверrенция вызывается дизруп-
тивным отбором в соседних зонах, которые различаются по усло-
виям среды, и до некоторой степени защищена от затопления бла-
rодаря частичному пространственному разделению этих зон.
Известен ряд конкретных примеров как у растений, так и у
животных, коrда диверrенция в таких условиях доходила до уров-
ня расы. ОДНИМ из хороших примеров у раС1'ений служит, среди
прочих, свободно скрещивающийся злак Agrostis tenuis в Вели-
кобритании, образовавший различные эдафические расы на раз-
ных примыкающих друr к друrу типах почвы (Antonovics, 1971).
Интересным примером у насекомых служит пестрокрылка Rha-
goletis (Tephritidae).
Личинка яблонной пестрокрылки (Rhagoletis ротоnеиа) в Се-
верной Америке живет и кормится на плодах культурных яб-
лонь (Л1аlus) и MecTHoro боярышника (Crataegus). Популяции
HaceKOMoro на яблонях и боярышнике принадлежат к разным
расам. Пестрокрылки спариваются на растении-хозяине, после
чеrо самки откладывают яйца на плоды. Личинки развиваются в
плодах, и следующее поколение взрослых особей появляется и
скрещивается опять таки на растении-хозяине, завершая цикл
(Bush, 1969а).
В плодоводческих районах на севере США боярышник и ябло-
НИ растут в непосредственной близости друr от друrа. И расы
Rhagoletis ротоnеиа, обитающие на различных растениях-хозяе-
вах, также находятся здесь в близком соседстве. То, что каждая
раса пестрокрылки спаривается на соответствующем растении-к,?-
зяине, позволяет сохранить им расовую дифференu.иаu.ию, несмот-
ря на смежность их местообитания (Bush, 1969а).
Боярышник уроженец Северной Америки, тоrда как яблони
были ввезены на этот континент. Поэтому первоначальным расте-
нием-хозяином Rhagoletis ротоnеиа с..1УЖИЛ, вероятно, боярыш-
ник. Известно, что пестрокрылка заражала яблони в Северной
Америке начиная с 1866 f. По видимому. она переключилась с
боярышника на яблони в XIX в. (Bush, 1969а).
222
Часть V. Видообразование
Образование этой новой расы по хозяину, возможно, произо.
шло в какой либо смежно симпатрической зоне. Распознавание
и предпочтение определенноrо растения хозяина у близкоrо рода
Procecidochares# а возможно, также у Rhagoletis контролируется
единичным reHoM. Следует помнить, что спаривание происходит
на растении-хозяине. Поэтому мутация reHa, управляющеrо рас..
познаванием хозяина, может положить начало процессу смежно.
симпатрическоrо расообразования (Bush, 1969а; Huettel, Bush,
1972) .
Все эти данные служат приемлемыми косвенными доказатель.
ствами возможности диверrенции до стадии рас по хозяину. Од.
нако нас интересует проблема симпатрическоrо образования ви.
дов.
Rkagoletis ротоnеllа ---- один из четырех видов.двойников. Эти
виды симпатричны, но заражают плоды растений, принадлежащих
к разным семействам. Хозяевами представителей rруппы Rh. ро.
тоnеиа служат роды, относящиеся к следующим семействам:
Rh. pomonella
Rh. mendax
Rh. corniuora
Rh. zephyr(a
Rosaceac
Ericaceae
Cornaceae
Caprifoliaceae
Такое распределение по хозяевам моrло возникнуть в результате
перехода с представителей одноrо семейства растений на пред"
ставителей друrоrо в симпатрической ЗОНе (Bush, 1969Ь).
Существование смежно-симпатрическоrо видообразования у
пестрокрылок доказывается путем экстраполяции от образования
рас к образованию видов. Подобные экстраполяции рискованны.
Тем не менее характер распространения этих пестрокрылок по.
зволяет предполаrать наличие у них смежно симпатрической дн-
верrенции на разных таксономических уровнях. Изучение этой
проблемы продолжается.
Узкие видоспецифичные пищевые ниши обычны во мноrих
rруппах насекомых. Соблазнительно предположить, что резкие
переходы из одной пищевой ниши в друrую, подобно тому как
это было постулировано для пестрокрылок, служат обычным спо.
собом видообразования у насекомых.
Биотически.симпатрическое видообразование
Образование rенетически отличной популяции в пределах био-
тически-симпатрической зоны у какоrо-либо свободно скрещиваю.
щеrося орrанизма сопряжено с серьезными затруднениями. Но..
вая популяция подвержена затоплению и реабсорбции родитель.
r А. 22. Способы видообразования
22:1"1
екой популяцией. В обычных условиях такой процесс происходить
в природе не может.
Симпатрическая диверrенция в перекрестнооплодотворяющей-
ея популяции требует интенсивноrо дизруптивноrо отбора. Одна-
ко интенсивному отбору сопутствует тяжелый rенетический rруз,
который может оказаться непосильным для данной популяции.
В экспериментальных ПОПу.I1ЯЦИЯХ Drosophila rпelanogaster и
Musca doтestica проводили сильный дизруптивный отбор. Эти
эксперименты оказались успешными в том отношении, что между
«высокими» И «низкими» по тому или иному признаку селекцион-
ными линиями возникла сильная диверrенция. Между этими ди-
верrировавшими линиями создал ась также частичная репродук-
тивная изоляция (Thoday, Gibson, 1962, 1970; Streams, Pimentel,
1961; Pimentel , Smith, Soans, 1967; Soans, Pimentel, Soans, 1974).
Сами по себе эти результаты обнадеживают. Следует, однако,
признать, что линии, выделенные в экспериментах по дизруптив-
ному отбору, еще очень далеки от видовоrо статуса.
Роль дизруптивноrо отбора облсrчилась бы, если бы он со-
четался с ассортативным скрещиванием. Такое сочетание факто-
ров моrло бы возникнуть, если бы признак, подверrающийся диз-
руптивному отбору, вызывал позитивное ассортативное скрещи-
вание. В таких случаях дизруптивный отбор Mor бы вызвать дан-
ную степень диверrенции при пониженной плате за отбор.
Проблема биотически-симпатрическоrо видообразования Haxo
дится в настоящее время на теоретической и экспериментальной
стадиях исследования. Пока еще нет никаких данных, которые бы
свидетельствовали о том, что биотически симпатрическое первич-
Ное видообразование иrрает какую-то роль в природе.
rлава 23
ОБЩАЯ ТЕОРИЯ НИДООБРА30ВАНИЯ
Введение
В двух предшествующих rлавах мы рассмотрели несколько
способов видообразования. Попытаемся теперь найти общие
чертЫ у этих различных способов, как это было сделано в одной
ИЗ моих ранее вышедших книr (Grant, 1963).
Процесс видообразования можно разбить на четыре основных
компонента: 1) цель; 2) этапы на пути к достижению этой цели;
З) пути, состоящие из различных последовательностей этапов;
4) поля. Описав эти основные компоненты, мы сможем рассмат.
ривать разные способы видообразования как различные их со..
четания.
Цель
Симпатрические виды приспособлены к разным нишам или
местообитаниям, находящимся на общей территории. Приспособ-
ления каждоrо вида к ero нише или местообитанию основаны на
сочетаниях признаков и на сочетаниях reHoB, детерминирующих
эти признаки. Вид ---- ЭТО то поле, на котором происходит процесс
рекомбинации reHoB или, точнее, создание полезных в ПрисПО
собительном отношении reHHblx рекомбинаций.
Не все продукты рекомбинации, которые MorYT быть созданы
в результате половоrо процесса, полезны в приспособительном
отношении. Мноrие из них полностью или частично нежизнеспо-
собны и представляют собой напрасное расходование репродук-
тивноrо потенциала популяции. Функция репродуктивных изоли.
рующих механизмов состоит в том, чтобы удерживать свободное
скрещивание и создание рекомбинаций в пределах, полезных в
приспособительном отношении. Эти пределы обычно соответству.
ют rраницам биолоrических видов.
Сущность видообразования состоит, таким образом, в создании
различных репродуктивно изолированных наборов приспособи.
тельных сочетаний reHoB.
Этапы
Процесс видообразования слаrается из ряда последовательных
этапов: 1) С.Qздание изменчивости по множественным reHaM;
2) образование HOBoro сочетания аллелей; 3) закрепление этоrо
r А. 23. Общая теория видообразования
225
HOBoro сочетания аллелей в дочерней популяции; 4) защита но..
Boro сочетания алле.:тей механизмами репродуктивной изоляции.
Рассмотрим поочередно каждый из этих этапов. При этом
удобно исходить в наших рассуждениях из определенных, хотя If
сверхупрощенных изменений rенотипов. Так, предковый вид с re..
нотипом А/А В/В дает начало очередному виду а/а б/б.
1. Создание изменчивости по ,иножественным zeHaM. Предко..
вая популяция А/А В/В должна приобрести какие то аллели а и
какие то алле.пи Ь. В данном контексте нас интересуют три rлав-
ных источника такой изменчивости в природных ПОПУ..'Iяциях:
а) изменчивость, существующая обычно в полиморфных популя
циях, т. е. изменчивость, накапливающаяся за длительные перио-
ды времени; б) новая ИЗ lенчивость, возникающая в результате.
rибридизации; в) новая изменчивость, создающаяся в результате.
спорадических взрывов мутаций, reHHbIX ИJIИ хромосомных.
2. Образование новых сочетаний аллелей. В предковой попу
.I1ЯЦИИ А/А В/В в результате половоrо процесса возникает неко-
торое количество rенотипов а/а Ь/Ь. Однако эти rенотлпы вначале.
редки, а кроме Toro, они вновь разрушаются тем же ПО ТIовым
процессом.
3. Закрепление HOBOZO сочетания аллелей в дочерней популя..
ции. Закрепление HOBoro сочетания аллелей ааЬЬ в одной из до-
черних популяций представляет собой решающий шаr в процессе'
видообразования. Су.ществует два основных пути закрепления:
reHoB.
Один путь это отбор при широком свободном скрещивании
в одной из rеоrрафических рас предковоrо вида. НапраВ.'1енный
отбор в пользу rенотипа а/а Ь/Ь в большой свободно скрещиваю
щейся популяции, rенофонд которой содержит преимущественно.
аnлели А и В, постепенно повышает частоты аллелей а и Ь и в
конечном счете приводит к замещению rенотипа ААВВ rенотипом
ааЬЬ. Плата за отбор при этом процессе очень высокая, и он про-
текает медленно. Однако ПОПУ..тJяция, которая способна выдержать
rибель большоrо числа особей в процессе отбора и имеет возмож,
ность про изводить замещение в течение Д..1ите.пьноrо времени, в.
конечном счете достиrнет новой адаптивной цели. .
Друrой путь ---- инбридинr. Если редкие особи ааЬЬ MorYT CKpe .
щиваться между собой И.'1И самооплодотворяться, то число их
будет быстро уве.1ичиваться. Закрепление новых сочетаний re-
нов будет происходить быстрее и обойдется дешевле в смысле.
избирательной rибели при инбридинrе, чем при отборе и свобод..
HO}I скрещивании.
Известно два способа инбридинrа, иrрающеrо р0ЛЬ в природ-
ных популяциях, а именно скрещивание внебольших попу..'IЯЦИЯХ,.
сводящееся практически к скрещиванию между сибсами И.'IИ, са..
мое большее, двоюродными сибсами, и самооплодотворение. Tpe
226
Часть У. Видообразование
тий способ ---- ассортативное скрещивание rенетически сходных
Qсобей, возможно, таКЖе имеет важное значение, но о ero роли
в природных популяциях пока еще известно мало.
4. Защита новых сочетаний аллелей при nО.4lOщи penpoдYKTив
ной изоляции. /vlеханизмы репродуктивной ИЗО.тIяции возникают
двумя путями: в качестве побочных продуктов диверrенции reHo-
типов (см. rл. 21) и в качестве продуктов отбора, направленноrо
на репродуктивную изоляцию как таковую (см. rл. 24).
Пути
Этапы 1 и 3 бывают представлеНbl в разных формах. Эти
формы MorYT сочетаться различным образом, в результате чеrо
возникает MHoro альтернаТИВIIЫХ путей.
факторы. созgающи
изменчивость
ФактОРtJl. Jакреп.ляющuе
u3.МеНI/ШJOсть
006IЧНО существ.,/ющан v
uэмеНЧl.I(lосmь
5 Отбор (с широкuм С60боинми
скрещиванием)
Особая НlМuя иэменчuоост6
rlJБРl.IgиJOцин 11
НО86/е Щlтач"и и
\ . i
lf
'.,. i
а
Инdpиgине (и отбор)
путем сомооnлоgoт80ренип
п!lтeM оссортатионоео
!'.'ШещиОЙНUR
Рис. 23.1. Пути видообразования (Grant, 1963)..
ОБЪJrоиеКИJl СМ. в тексте.
Обозначим для удобства разные формы возникновения и эа-
крепления изменчивости буквенными символами.
Этап 1
Обычно существующая изм:еиqивость (v)
rибридизация (h)
Новые мутации (и)
Этап 3
Отбор (с широким свободным скрещиванием) (5)
Инбридинr (и отбор) (i)
В иебольших популяциях (i p )
Путем саМООШIOдотвореНlfЯ (i,)
Путем ассортативноrо скрещивания (io)
Теперь мы можем связать друr с друrом разные варианты
этапов 1 и 3 (рис. 23.1), чтобы выявить альтернативные пути.
Так, существует путь v ---- s. путь v i p и так далее. Число
r А. 23. Общая теория видообразованuя
227
возможных путей, представленных на рис. 23.1, равно 9 или 12
в зависимости оТ Toro, принимается ли в расчет ассортативное
скрещивание (ia) или нет.
Модели
Процесс видообразования может иметь место в пространст-
венных полях трех типов, а именно в аллопатрических, смежно.-
симпатрических и биотически симпатрических. Сочетая поля
этих трех типов с 9 или 12 путями, можно получить множество
возможных модеJ1ей. Конечно, большинство этих возможных мо-
делей вряд ЛИ встречаются в природе или неизвестны; однако
при этом остается достаточное число моделей, которые либо pe
альны, либо заслуживают серьезноrо внимания.
Мы перечислим здесь 8 моделей, соответствующих способам.
видообразования, обсуждавшимся в rл. 21 и 22.. Способы видооб-
разования, стоящие в начале этоrо перечня, известны, а стоя-
щие в конце представляют собой возможные способы, которые в
настоящее время изучаются.
v---s, аллопатрическая.
v ip, а.'JJJOпатрическая.
h---i, биотически симпатрическая.
h---u---i, биотичеСКИ-Сllмпатрическая.
и---; р смежно си:мпатрическая.
v---i, БИОТИЧССКИ-СИ fn:1трическая
v---s БИОТlfчеСКII-си патричсская.
v io биотичеСКИ-('IfМllатрическая.
fеоrрафичсское видообразоваиие.
Квантовое видообразование.
Рекомбинационное видообразование
rибридное видообразование с раацепл
нием в отношении наружиых преrрад.
Видообразование путем аЛ.10ПО.1ИПЛОИ-
дии.
Смежно-симпатрическое Вllдообразова
нне у свободио скрещиваlOЩИХСЯ иасе
комых и растений (вполне определенная
возможность) .
БнотичеСI<и симпатрическое видообразо.
вание у саМООПJIОДОТВОРЯЮЩkХСЯ paCTe
пиЙ и беспозвоночных (вполне опрсде
ленная возможность).
БИОТlfчески-симпатрнческое видообразо
вание у свобqдио скрещиваЮЩIIХСЯ ор-
rанизмоп при дизруптивном отборе.
(Рассматривается некоторыми учеными
как реальная ВОЗ fОЖНОСТЬ, а друrими,
к которым отношусь и я, как возмож-
ность весьма ОТД3Jlенная).
Виотически-симпаrрическое вндообраэо
вание у свободно скрещивающнхся ор-
rанизмов с ассортативным спариванием
(некая возможность).
rлава .24
ОТБОР, ,НАПРА,ВЛЕН'НЫй НА СОЗДАНИЕ
ИЗОЛЯЦИИ
Введение
Предполаrается, и к тому же с достаточным основанием, что
существует два способа возникновения репродуктивной изоля-
ции. Первый способ состоит в развитии механизмов репродуктив-
ной изоляции как побочных продуктов диверrенции от pacoBoro
до видовоrо уровня, о чем rоворилось в предыдущих rлавах. При
этом диверrировавшие виды MorYT сосуществовать симпатрически.
Однако первичные механизмы репродуктивной изоляции, раз-
вившиеся как побочные продукты диверrенции, не MorYT полно-
стью предотвратить rибридизацию. Некоторое число rибридов все
же образуется. Такие rибриды, по всей вероятности, будут не-
жизнеспособны, стерильны, слабо Ио)1И совсем плохо приспособле-
вы к условиям среды. Следовате..,ьно, образование этих rибридов
предстаВ.'1яет собой напрасную трату репродуктивноrо потенциа-
..,а родительскоrо вида.
В таких условиях родительскому виду выrодно в селективном
отношении усилить существующие первичные изолирующие меха-
низмы новыми преrрадами, которые бы эффективно предотвра-
щали rибридиэацию. В тех с.'1учаях, коrда репродуктивная изоля
ция обладает селективным преимуществом, действие отбора мо-
жет быть направлено на создание специальных механизмов ре-
продуктивной изоляции ради самой этой изоляции.
В настоящей rлаве мы кратко опишем этот второй способ Воз-
никновения репродуктивной изоляции. Отбор на создание репро
дуктивной изоляции как таковой известен также в .,1итературе
как эффект Уодлеса или процесс Уоллеса, названный так в честь
ве.;1икоrо современника Дарвина А. Р. Уоллеса; кроме Toro, ero
называют смещением признака в репродуктивной фазе.
Процесс отбора
Отбор на репродуктивную изоляцию как таковую вступает в
действие при одновременном наличии следующих условий: 1) ви-
довые популяции находятся в симпатрическом контакте; 2) между
ними происходит rибридизация; 3) rибриды и их потомки нежиз-
неспособны, стерильны или же уступают в адаптивном отноше-
иии родительским формам по каким-либо иным аспектам; 4) ени.
жение репродуктивноrо потенциала родительскими видами, вы-
r А. 24. Отбор, наnраs.л.еННtJtr1. на соэдание U30Аяцuи
229
.званное rибридизацией, неаыrодно с точки зрения отбора; иными
.словами, предотвращение rибридизации создает определенное
селективное преимущество по сравнению с существующим поло
жением вещей; 5) в одном ИЛи в обоих родительских видах су-
ществует индивидуаw'lьная изменчивость по признакам, оказываю
щим влияние на репродуктивную изоляцию,
Пятое условие затраrивает самую суть селективноrо процесса.
Мы должны допустить, что видовая популяция полиморфна в
-отношении леrкости rибридизации с чужим видом и что rибри-
дизация вызывает некоторые вредные эффекты. При таких усло-
виях те особи данной видовой популяции, которые обладают пре
rрадами, препятствующими rибридизации, привносят в .следую-
щие поколения своей популяции большее число жизнеспособных
11 плодовитых потомков, чем свободно rибридизирующие сест-
ринские особи; в результате произойдет повышение частоты reHoB
этих первых особей и их распространение по всей популяции.
Перечисленные выше условия сильно варьируют в разных па-
рах видов и в разных rруппах орrанизмов. Биотическая симпат
рия (условие 1).. например, чаще наблюдается сред,И насекомых
и ОДНО.тIетних растений, чем среди крупных наземных позвоноч-
ных и крупных деревянистых растений. А неблаrоприятные се-
.лективные последствия потери репродуктивноrо потенциала (yc
ловие 4) rораздо сильнее выражены у недолrовечных орrанизмов,
таких как однолетние растения и ме.'1кие насекомые, чем у дол-
rоживущих мноrолетних растений с высокой избыточной плодо-
витостью.
В ряде rрупп видов условия, содействующие отбору на разви-
тие изоляции, иноrда реализуются, а иноrда отсутствуют. Отбор,
.иаправ..lенный на изоляцию, не представляет собой универсаль..
Horo процесса.
В тех случаях, коrда процесс отбора на изоляцию имеет место,
-он относится rлавным образом к тем изолирующим механизмам,
которые действуют в родите..'1ЬСКОМ поколении. Конечная цель
адаптации предупреждение rибридизации. И наиболее эффек
тивным образом эта цель может быть достиrнута путем создания
.таких механизмов, как ЭТО.l'юrиче.ская изоляция или несовмести-
мость, которые действуют до оплодотворения.
Это правило можно распространить и на те орrанизмы, у КО-
70рЫХ материнская особь вынашивает и выкармливает зарОДЫШа,
как у М.'Iекопитающих и у большинства семенных растений. В та-
ких случаях теоретически возможно, чтобы отбор создавал пре-
rрады, которые дсйствуют на ранних стадиях поколения Ft. Ра-
зумное объяснение этому состоит в том, ЧТО хотя rибридный за-
родыш,- развитие KOToporo прекращается на одной из ранних ста-
ДИЙ, .в результате чеrо rибридизация блокируется, не служит
<:толь эффективной преrрадоiI, как ЭТО..10rическая изоляция, тем
230
Часть У. Видообразование
не MeH e эффективность размножения при этом несколько выше,
чем при создании стерильных или нежизнеспособных взрослых
rнбридных особей.
Фактические данные
Один из подходов к изучению эффективности отбора в при-
родных популяциях основан на использовании пары видов с пе-
рекрывающейся симпатрией (рис. 24.1, 1). У двух видов А и В
имеются расы, аJIлопатрические по отношению друr к друrу (А"
I
А
8
11
А8
/11
с
1J
8
Е
Рис. 24.1. Два типа распространения у rрупп видов, в которых можно сравни.
вать аллопатрические и снмпатрические популяции по степени изоляции в роди-
тельскоМ поколеиия.
I пара частично сиыпатрнчсских ВИДОD. 11 Н 111 rpynnъr ВИДОВ, D которые ВХОДЯТ аJl
.10паТРИlfеские и симпатрические DИДЫ.
и 8а), и друrие расы, которые симпатричны (Ав и 88)' Сравним
мощность этолоrических или друrих преrрад, действующих до
оплодотворения, в случае аллопатрических и в случае симпатри-
ческих рас.
Это сравнение было проведено для частично симпатрических
пар видов Drosophila (п. pseudoobscura и п. тiranda), древесниц
(Microhyla carolinensis и М. olivacea J Hyla ewingi и Н. verreauxi),
флоксов (Phlox pilosa и Ph. glaberriтa) и некоторых друrих
орrанизмов. В каждом из этих случаев либо сама этолоrическая
изоляция, либо какой-либо измеримый ее компонент более силь-
но развит в областях, rде эти два вида симпатричны, чем TaM
rде они а..'Iлопатричны (Dobzhansky, 1951 а; Blair, 1955; Littlejohn.
1965 Levin, I(erster, 19б7 Levin, Schaal, 1970).
r А. 24. Отбор, направленный .на со:адание изоляции
231
Так, на юrе США брачные призывы Microhyla carolinensis и
М. o.livacea, уqаствующих в создании этолоrическоЙ изоляции,
ильно различаются в зоне симпатрическоrо перекрывания, но
мало отличаются при сравнении а..1J1Опатрических рас этих двух
видов (Blair, 1955). Ана..l0rичная расовая изменчивость призыв-
ных песен обнаружена у частично симпатрической пары видов
Hyla ewingi и Н. verreauxi на юrо востоке Австралии (Littlejohn,
1965) .
Более обширные сравнения TaI<oro же рода были произведены
в надвиде Drosophila paulistoruт. Этот тропический американский
надвид СОСТОИТ из шести полувидов (Амазонский, Андский и т. д.),
TOPЫe. встречаются в различных частично симпатрических соче.
ан иях. /v\ОЩНОСТЬ этолоrической изоляции между полувидами
ожио измерить в смешанных лабораторных популяциях и вы-
азить количественно в виде коэффициента изоляции (/). Этот
оэффициент изменяется от О до 1, rде О означает свободное CKpe
ивание, а 1 полную изоляцию (Ehrman, 1965).
Коэффициент изоляции был вычислен Д..1я сочетаний аллопат-
рических рас и сочетаний симпатрических рас, представляющих
8 пар полувидов (например, АмазонскаяхАндская; АмазонскаяХ
хrвианская и т. д.) В надвиде D. paulistoruт. Ве.'IИЧИНЫ 1 для
сочетаний рас в разных парах по..1УВИДОВ показаны в табл. 24.1.
Коэффициент изоляции по всему обширному и сложному надвиду
п. paulistoruт был устойчиво выше между симпатрическими раса-
ми пары полувидов, че:м между аллопатрическими расами той же
-самой пары полувидов I (за одним исключением) (Ehrman,. 1965).
ТаБАица 24.1
разных парах
Значения
коэффициента изопяции (1) для сочетаний рас в
ПОЛУ.ИДОВ
I (пределы :олебаIlИА) I
Сочетания рас
1
(сре.!l:иее значение)
Алпопаrрические ПОЛУВИДЫ
Симпатрические полувиды
0946....0'76
0'68....0'96
0,67
0,85
Друrая ситуация, в которой МОЖНО выявить отбор, направлен-
ный на создание изоляции, представлена на рис. 24.1, 11 и 111.
На нем изображена rруппа видов, содержащая неско.'IЬКО а.,lЛО-
патрических биолоrических видов (А, В, С) и несколько симпа-
трических видов (п, Е, F). При этом сравнение силы изолиру-
ющих механизмов родите..'Iьскоrо поколения производится в меж-
. видовых сочетаниях двух классов: аллопатрический Х аллопатри-
ческиЙ и симпатрический Хсимпатрический.
ПОДХОДЯlЦей rрулпой орrанизмов для проведения TaKoro срав-
нения служит rруnпа видов rилии (Polernoniaceae). Эта естест-
232
Часть У. Видообразование
Таблица 24.2
Значение покаэателя несовместнмости (S/Fl) Апя сочетаниА разиых ВКАОВ
Coqeто... ...... I (пред,л,:I:':"б..") I (,редв, /:;:.ч...,)
)Lплопатрические
Сиипатрические
7,7 24,8
0,0 1 )2
18,1
0,2
венная rрулпа однолетних растений состоит из 9 биолоrических
видов, распространенных на ТихоокеаНСКО 1 побережье Северной
и Южной Америки. Эти виды по своему rеоrрафичеСI{ОМУ рас-
пространению делятся на два класса: пять видов, встречаlOЩИХСК
у подножий холмов и в долинах Калифорнии, в значите.тIЬНОЙ сте-
пени симпатричны; четыре друrих вида, растущие на береrовоir
линии и на б.'IИЗ.'Iежащих островах, полностью аллопатричны по-
отношению Apyr к друrу и лишь в редких случаях симпатричны
с краевыми популяциями, относящимися к первому KJIaccy. Ис-
кусственные rибриды Fl между этими девятью видами BЫCOKO
стерильны: хромосомная стерильность возникает при любых co
четаниях (Grant, 19ббЬ).
Поскольку эти растения однолетние травы и притом цве..
тущие только один раз за rод, завязывание семян имеет для них'
чрезвычайно важное значение. ПО этой же причине продукция
семян, из которых развиваются стерильные rибриды, наносит'
серьезный ущерб репродуктивному потенциа.,'lУ.
Эффективным препятствием к образованию rибридов у Gilia
служит несовместимость. Мощность преrрад между видами, соз
даваемых несов!>Лестимостью, можно определить количестве.нно в.
искусственных межвидовых скрещиваниях. Хорошей мерой для
этоrо С.'Iужит среднее число rибридных се ян, производимых на
один перекрестноопыленный цветок при бо.ТJЬШОМ числе иветков в
определенном межвидовом скрещивании (5jFI) (Grant, 19ббЬ).
Величину 51Ft опреде.'IЯЛИ для 20 межвидовых сочетаний из 9
изученных видов rи.тIИИ. Оказалось, что два rеоrрафических клас
са rибридных комбинаций резко раз. ичались по степени HeCOB 1e-
стимости (табл. 24.2). Преrрады, создаваемые несовместимостыо
очень сильно выражены между симпатрическими видами, TorAa
как между аЛ..тIопатрическими видами они незначитеЛЬНbI (Grant,.
19б6Ь) .
Этолоrическая изоляция
Видоспецифичность опознавательных признаков и особенно-'
ст-ей ухажwван.ия хорошо развита :в большин'Стве rрупп животных.
.к ним относятся брачное оперение, демонстрационное поведецие
Тл. 24. Отбор, направленны.й на соэдание иЗОАЯции
2ЗЗ
и лесни у ПТИЦ, брачные танцы и обонятельные сиrналы у насе-
комых и так далее. Разноrо рода опознавательные сиrна.тJЫ раз-
JIИЧНЫ у разных видов. Их функция состоит в том, чтобы способ
ствовать внутривидовому и препятствовать межвидовому спари-
ванию.
Происхождение видоспецифичных признаков, связанных с уха-
живанием, следует, вероятно, приписывать rлавным образом от-
бору, напраВ.'1енному на изоляцию, хотя вполне возможно, что в
этом участвуют и друrие селективные факторы. Так, признаки,
связанные с ухаживанием, действуют как факторы, стимулирую-
Iцие половое поведение, и MorYT быть, во всяком случае до не ко-
торой степени, реЗУu'lьтатами отбора, направленноrо на повышение
ПРОДУКТИВНОСТИ как таковой; но этот способ отбора не позволяет
объяснить видоспецифичность признаков, связанных с ухажива
нием. И опять-таки некоторые различия в признаках и поведении,
.связанных с ухаживанием, моrли возникну.ть как побочные про-
дукты диверrенции; однако было бы трудно объяснить более
утонченные видоспецифичные атрибуты ухаживания как побоч-
ные эффекты дифференциаu.ии, относящейся к добыванию ПИЩИ н
друrим ЭКОлоrическим аспектам жизни. Наиболее правдоподобная
'rипотеза состоит в TO I, что эти связанные с ухаживанием при
знаки, столь распространенные в животном царстве, представля-
ют собой r.,'IaBHblM образом продукты отбора, напраВJIенноrо на
этолоrическую изоляцию.
Часть УI
МаКрОЭ80JIЮЦИЯ
r ла,ва .25
rЕолоr,ИЧЕСКОЕ. ВРЕМЯ
I1рирода маКрОЭ80JlЮЦИИ
Макроэволюция подразумевает изменения rораздо большеr()
масштаба, чем те, которые происходят при М:ИКРОЭI30,пlOЦИИ и ви-
дообразовании. Изменения макроэволюционноrо уровня состоят в.
развитии признаков, по которым различаются такие крупные rруп
ПЫ, как роды, семейс.тва, отряды, класс.ы и типы. Подобные собы-
тия происходят в rео.rюrическом масштабе времени.
Методы исследования, применяемые при изучении маI{РОЭВО
люции, не MorYT не ОТ..'Iичаться от тех, которые используются при
изучении МИКРОЭI30..1IОЦИИ и видообразования, fенетика, эколоrия
и систематика на уровне низших кзтеrорий ---- r..1aBHble подходы к
изучению микроэволюции и видообразования, тоrда как все, что
нам известно. о макроэвоЛIОЦИИ,. получено в результате па..тJеонто-
лоrических и сравнительно морфолоrических исследований. Рабо-
ты по rенетике попу..1JЯЦИЙ и близким к неЙ областям позво,ляют-
получить прямые данные об эволюционных силах. Однако сведс..
иия относительно крупных событий в процессе орrанической эво
люции дают палеОНТО.'юrия и сравнительная анатомия.
Изучением эволюционной биолоrии на макроуровне и на дpy
rих уровнях обычно занимаются разные rруппы исследователей
и их представления о ро,пи эксперимента, масштабах времени и
друrих предметах нередко расходятся. .l\1ноrис палеонтолоrи и
морфолоrи старшеrо поколения недооценивали значение rене'Iиче
ских исследований для эволюционной теории; в свою очередь био-
лоrи, изучающие популяции ныне живущих орrанизмов, часто
имеют довольно смутное представление о временном элемевте
макроэволюции. Источником мноrих противоречий в эволюцион-
ной биолоrии служат характерные различия между макро- и мик
роэволюциониста и в отношении методов. исследования, образа
мышления и общей I!одrотовки.
Одна из проблем эволюционной биолоrии касается Toro, пред-
ставляет ли собой макроэволюция просто продолжение микроэво
r А. 25. ТеОАоеическ:ое время
235
люции или же между этими. двумя процессами имеется какое-то
существенное различие. Сформулируем две крайние точки зрении
по этому вопросу: 1) единственное различие М,ежду макро- и
микроэволюцией ..... продолжительность этих процессов, т. е. пер-
вый процесс представляет собой просто продолжение BToporo;
2) в макроэволюции участвуют особые факторы или процессы,
которые мы не обнаруживаем на МИКРОЭВОЛЮЦIfОННОМ уровне,
например скачкообразные изменения, системные мутации, орто-
rенетические процессы, старение рас и. Т. п. Каждая из этих ТО-,
чек зрения имеет своих приверженцев .........,...перВая мноrочисленных,
а вторая Bcero лишь ropcTKY; однако нас здесь интересуют не
.1ИЧНОСТИ, а сама проблема и пути ее решения.
Можно отметить, что первая точка зреtlИЯ находится на Bep
ном пути В cMblc.1Je экономичности rИПотез: если мы не можем вы-
явить эволюционную силу или фактор на уровне МИКРОЭВОJJЮЦИИ,
допускающем экспериментирование, мы не вправе выдумывать их
для rораздо менее ясных макроэво..'1ЮЦИОННЫХ явлений. По той И
ПО ряду друrих причин вторая точка зрения никоrда не,моrла.
ПО.'Iучить широкоrо признания среди эволюционистов.
Но укажем и на слабое :место первой точки зрения: она до-
пускае.т возможнос.ть экстрапо..1ЯЦИИ данных, ПО.Т1ученны.х .В ре..
зультате экспериментальноrо изучения 10 '50 и 100 поколений &
Jlаборатории, на события, происходившие на протяжении мцллио-
нов или десятков миллионов лет в природе. Она не учитывает
должной мере возможности, скрытые в rеолоrичеСКО I временн.
Возможна также и третья точка зрения, ПОЗВОJJЯlOщая при..
вести в соответствие то, что. нам известно о МИКРОЭВОЛIрции и ви-
дообразовании, с rеолоrическим масштабом времени. Ее BЫCKa
зал Симпсон (Simpson, 1944), и она теперь получила. : довольно
широкое распространение. Эта третья .точка зрения признает.
эволюционные Си..1Ы и факторы, разработанные популяционной.
биолоrией, но отказывается от простой экстрапо.Т"lЯЦИП. с микро-
эволюции на макроэволюцию.
Вполне возможно, что известные нам эволюционные силы и
факторы действова.'IИ в разные периоды истории данной rруппы
разными путями. Возможно, что частота мутаций в прош..'lОМ 01;'-
ЛИча..'1ась от их частоты в настоящее время. Возможно, что. в
прошлом среда И3 Iенялась быстро, а теперь она стабилизирова-
.,lacb, или наоборот. Данная rруппа моrла быть rеоrрафическя
: широко распространена на одной стадии и узко эидемиtlНа на
друrой. Или, возможно, она состояла на одной стадии из MHoro-
численных симпатрических видов, а на друrой была МОНОТИПl:fче-
екой.
rоворя о длительности времени, на протяжении KOToporo м:оrли.
происходить макроэволюционные изменения, нам следует также
.иметь в виду. возможность возникновения уникальных. событ,ий,
236
Часть V 1. Ма"роэвОАюция
имевших далеко идущие последствия. Какой-либо наземный opra
низм может колонизировать отдаленный ненаселенный остров
только однажды, после чеrо он начинает эволюционировать на
этой занятой им территории в каком-либо новом направлении.
Какой-либо примитивный микроорrанизм в результате заражения
может приобрести симбионта, дающеrо ему возможность произ
водить фотосинтез.
В пользу этой третьей точки зрения на макроэволюцию имеет..
ся довольно MHoro данных. Макроэволюция и в самом деле от ли
Чается от микроэволюции, поскольку МаКроэволюция.---- процесс
исторический.
r лавные стадии истории Земли
Историю Земли удобно разделить на четыре rлавные стадии'
в соответствии с преобладающим на каждой из. них способом
осуществления процесса космическоrо развития: 1) атомная эво
люция, в процессе которой происходили ядерные реакции, приво
дившие к образованию водорода, а затем к построению из Hero'
друrих атомов; 2) химическая эволюция, в процессе которой aTO
мы объединялись с образованием химических соединений различ
ной степени сложности, в том числе неживых орrанических MO
лекул; 3) орrаническая эволюция, к ко:rорой относятся все собы-
тия, начиная от возникновения жизни и до появления высших
животных; 4) культурная эволюция, т. е. накопление и передача.
из поколения в поколение культурноrо наследия, начавшаяся
после Toro, как человек поднялся в своем раЗБИТИИ выше живот-
Horo.
Можно установить приблизительный возраст всех этих стадий..
Образование Земли как твердоrо тела произошло 4,5----5 млрд..
лет назад. Возраст Земли установлен по количественному COOT
ношению между ураном и свинцом и между р'азличными изотопа-
ми свинца в земной коре; эти соотношения создаются в процессе
радиоактивноrо распада, скорость KOToporo известна. Самые древ..
ине известные нам вулканические породы образовались 3,0----
3,5 млрд. лет назад (см. Dott, Batten, 1971). а самые древние оса-
дочные породы ----3'4----3,8 млрд. лет назад (Cloud, 1974). Атомная
эволюция преобладала в формообразовательном периоде истории
Земли, а затем она постепенно перешла в химическую эволюцию.
Химическая эволюция в конечном счете достиrла стадии, на
которой происходило образование сложных орrанических моле
кул. Такие неживые орrанические соединения подrотовили почву
для возникновения жизни. К древнейшим известным нам ископа-
емым орrанизмам относятся бактерии и одноклеточные сине-зе"
леные водоросли, возраст которых составляет примерно 3,2 млрд.
лет; они обнаружены в допалеозой€'Кой. формации Фиr-Три в.
f А. 25. fеО.ll.оzическ.ое вр.еAl.Я
237
Южной Африке (Barghoorn, 1971; Schopf, "1974). Таким образом.
жизнь возникла несколько раньше, чем 3,2 млрд. лет назад, но
после Toro, как химическая эволюция довольно значительно про
двинул ась вперед. Следовательно, орrаническая ЭВОЛЮЦия зани
мает в истории Земли более 3 млрд. лет (рис. 25.1).
Стадия культурной эволюции относится к последней фазе
эволюции человека и таким образом представляет соб.ой весьма
о::
3.0
4,o
.
r@-
о
Начало K0/1HO.JodcKoи эр61
":: '.о
"'::
Нача/lО l<eM5puiiclrOlO пepuoga
.......... ПОЯll//енuе Mefazoa
> ПОЯ /lеНuе эgкарuотО6
.
2,0
СаМ/lIе Ilревние lJ.JBeCтH6Ie прокориоты
у BOJHUKHOBeHue жu нu
'" ",:'> ПроgSUНf/ml1lе cтogUlJ хuмuчеt:КОЦ Э60AЩ/J"
5,0
Рис. 25.1. Важнейшие событии в истории Земли.
>OOpOJOBQHUe тBepgoи .1емноu КОр6/
недавнее явление, во всяком с,,'учае На планете Земля. Род Ното
появился примерно 1,3 М..1н. лет назад. Однако эволюция Ното
представляет собой смесь орrанической и культурной эволюции,
причем соотношение этих ее двух компонентов варьировало во
времени. Переход от преимущественно орrанической и преиму-
щественно культурной эволюции человека был постепенным, и
точное время ero установить нельзя. Культурная ЭВОЛIОЦИЯ будет
рассмотрена в части VII.
Возникновение жизни
Важнейшим шаrом для возникновения простой формы жизни
было образование днк или ДИК-подобных макромолекул, обла-
давших теми свойствами, которые мы сейчас находим у reHOB.
2ЗВ
Час.ть Уl. Ма"РОЭВОАЮЦия
к этим свойствам относятся способность направлять синтез
белков и друrих веществ, дающих reHaM возможность сохранять-
ся в виде самостоятельных единиц по крайней мере в течение
kakoro-то времени, а также друrая важнейшая способность спо-
собность крепликации.
Для выпо.пнения всех этих процессов необходима пища, слу-
жащая ИСТоtlНИКОМ энерrии. Накопление боrатых энерrией соеДИ
нений уr.1ерода, происходившее в предшествующий период хими
ческой эво..тJЮЦИИ, создало запасы веществ, приrодных в качестве
потенциальной пищи.
Накоплению орrанических соединений в процессе химической
эволюции блаrоприятствова.I1И два условия, существовавшие на
этой стадии, но нарушившиеся впоследствии. Этими двумя усло-
виямИ были отсутствие живых орrанизмов и отсутствие в атмо-
сфере tсвободноrо кислорода. Именно наличием живых орrаниз-
мов И свободноrо кислорода обусловлен распад орrаническ х cq.
единений, происходящий в современном мире. Отсутствие этих
факторов в пребиотическом мире означало, что орrанические MO
лекулы, образовавшиеся в результате спонтанных химических
реакций, будут сохраняться и накап.тIИваться.
Химические реакции, в процессе которых образуются молеку-
лы орrаническ,ИХ веществ, действительно происходят спонтанно,
что было доказано экспериментально. Если создать атмосфеРУJ
состоящую из .водорода, воды, аммиака и метана, 11 подверrнуть
ее действию электрических разрядов, естественной радиоактивно.
сти, космических лучей или УJlьтрафиолета, то в ней спонтанно
образуются различные орrаничсские соединения, в том ЧИС.'1е
аминокислоты.
Поэтому лоrи но преДПОЛО!f{ИТЬ, что в тече ие стадии химиче
ской эволюции моrло пройсходить образование и накоп ение
«орrаничеСКоrо бульона», содержавшеrо запасенную энерrию.
А в какой-то момент в процессе химической эволюции, очевидно,
возникли молекулы нуклеиновых кислот, обладавшие rенетиче
екой. активностью. Эти первые частицы живоrо представляли со-
(50Й, вероятно, «fолые reHbl», жившие за счет энерrии, запасенной
в орrаническом бульоне.
,. reHbl <;>бладают способностью к репликации, размножению и
мутированниIO. Репликация создает фракцию мутантных форм.
Размножение продолжается до пределов, устанавливаемых физи-
ческими факторами среды. В результате между rенами или их но-
сителЯМИ рано или поздно возникает конкуренция за ресурсы
среды, имеющисся в оrраниченном количестве, и в этой конкурен-
ции некоторые виды будут размножаться более успешно, чем дpy
rие. Таким образом, тенденция к эвотоции один из основных
атрибутов жизни в мире оrраниченных ресурсов. Возникновение
изни положило начало процессу орrанической эволюции.
r А. 25. rеОАоzuчес"ое вреAtя
239
rJlавиьае с.тадии орrанической ЭВОЛЮЦИИ
Разделим теперь общий период орrанической эволюции, состав-
ляющий 3'2----3'5 млрд. лет, на отде.l1ьные стадии. На ОСНОВе спо-
соба питания и структурной орrаниэации наиболее ПРОдвинутых
форм, ха р актерных для каждой стадии, можно выделить 5 таких
стадий. В дальнейших разделах мы обсудим каждую стадию от.
деJ1ЬНО.
Перечислим эти стадии: 1) простые rетеротрофные частицы,
вирусоподобные или бактериоподобные; 2) прокариотические aB
тотрофные фотосинтеЗИРУlOщие или хемосинтезирующис клетки,
примерами которых служат сине зеленые ВОДОрОСJ1И и автотрофные
бактерии; 3) эукариотические аэробные клетки, примерами . кото-
рых служат ОДНОКJIеточные зеленые водоросли и жrутиковые;
ТаБАuца 25.1
Возраст разных rеОJlоrических перио,цов (KuJp, 1961; Dott,
Batten, 1971)
Эра
Период
I Возраст, м.,и.
пет до настоя-
щеrо вре)iени
о
Четверти{IНЫЙ
2,5
I(айнозой
Поэднетретичный (HeoreH)
27
Раниетретичный (палеоrеlf)
65
130
185
Мед
Мс.зозой
Юра
Триас
230
Пермь
Карбон
265
355
413
425
475
600
700
Девон
Па.'Jеозой
Силур
Ордовик
Кембрий
Докембрий I Поздиий доке"бриА
240
Часть У/. Мак.роэволюцuя
4) простые мноrоклеточные аэробные орrанизмы, примерами Ко-
торых служат мноrоклеточные зеленые водоросли, печеночники и
rубки; 5) сложно орrаНИЗ0ванные животные, например Metazoa
If наземные растения.
Стадии 1----4 зани.мают большую часть вре'1ени, в Течение КО-
Toporo проходила орrаническая эволюция,---- от 3'2----3,5 млрд. лет
назад и до начала палеозоя, т. е. 0,6 млрд. лет до настоящеrо вре-
мени (см. рис. 25.1). Древние ископаемые прокарИОТЫ ---- бактерии
и сине-зелены.е водоросли ---- обнаружены в следующих формациях
(В arghoorn, 1971):
Формация Фиr-Три, IОжная Африка
Сланцы rаНфЛИRТ, Онтарио, Канада
Формация Виттер Спринrс, Австралия
3,2 млрд. лет
1,6---1,9 млрд. лет
1 млрд. лет
в формации Биттер-Спринrс обнаружены также зеленые Во-
доросли и предполаrаемые rрибы (Barghoorn, 1971). Сообщалось
также о находках примитивных эукариотов во флоре БеК4Спринr
(Калифорния), которая имеет возраст 1,3 млрд. лет (Cloud, 1974).
Таким образом, примитивные эукариоты появились не менее
l,о l,З млрд. лет назад.
Стадия 5 начинается в позднем докембрии и развертывается
в Кембрии. Древние ископаемые Metazoa кишечнополостные и
кольчатые черви встречаются в фауне Эдиакары в Австралии,
-относящеися к позднему докембрию, абсолютный возраст KOTO
poro определяется в 0,7 млрд. лет (Simpson, 1969). Основные
rрулпы водных Metazoa стали более мноrочисленными позднее,
в кембрии (см. табл. 25.1 и рис. 25.1 и 25.2).
Начальные стадии эволюции
На протяжении примерно первых двух миллиардов лет орrЗ 4
иической эволюции было сделано несколько крупных шаrов вперед
в отношении питания и етруктурной орrанизации; эти шаrи ИМеЛИ
важное значение для последующей эволюции мноrоклеточных
орrанизмов. Такими шаrами явились: автотрофное питание, в
частности фотосинтез; аэробное дыхание; эукариотическая кле 4
точная орrанизация; половое размножение_ Точно указать на
шкале, изображенной на рис. 25.1, момент возникновения этих
свойств мы не можем; по всей вероятности, они пОявились rде-то
между 3,5 и 1,3 млрд. лет до настоящеrо времени и в той последо-
вательности, в какой они здесь перечнслены.
Непрерывный рост примитивных rетеротрофных частиц Неиз-
бежно ДОЛЖен был привести к постепенному истощению первич-
:::r
=:3
%
Cb
....
-a
'U
\1
,
"
с <:::1 <:::1 Q
N Т
806od ov.mh
ё
о
:1:
:r
=
..
со
41
са
..
4J
:r
.а
ш
::r
'"'
fВ
ф
..... о
ао;
о со
\O
c\.::s
.5
сп
...., ..
о
111
О 1:(
t'iI о
C:IiS =
о
2t
:а 1:(
:s:: !Е
I:(
о :;:
"" о:
о:;
О 1:(
с= 4J
О
CII :1:
:а t
:s:: CI.J
:s:: 111
1'1
1:(=
)(
CII 111
:= t::
4> со
g;
""" )(
о :з
&:; :1:
О t::
t=
:;:
&:;
(\s :z:'
t:
со
111
u:i
CiI
с:{
tJ
::s:: р,
p..
о
о:;
u
=
:r
о
'2:
a:I
l')
са
:;:
О
t::
2
"""
с:
х'
С::::('
u
о
242
Часть V/. МаКрОЭВОАЮЦШ!
Horo <орrаническоrо бульона». А в условиях истощения запасов
ПИLЦИ способность орrанизмов самостоятельно синтезировать себ
пищу из неорrаничеСI{оrо сырья..... воды и двуокиси уr.lIерода----
давала orpoMHoe преимущество. Развитие способности к фото-
синтезу и хемосинтезу у примитивных прокариотов было круп .
ным успехом, достиrнутым на ранних стадия)С эволю.ции. Оно.
ПрОw"'Iожило путь для BToporo важноrо шаrа развития аэробноrо
дыхания.
Первичная атмосфера Земли содержала водород, но в ней не-
было свободноrо кислорода; она обладала восстановительными,.
а не окислительными с войствю.-ш. В таких условиях примитивным
rетеротрофн м частицам и клеткам приходилось, вероятно, до-
бывать энсрrию ИЗ орrаническоrо бу.lIьона путем брожения. А бро-
жение процесс, в метаболичеСI{ОМ cMblc."'Ie не эффективный, по-
скольку при этом ббдьшая часть энерrии уr.'1еродных соединении.
остается неИСПО.'1ьзованной. В процессе аэробноrо дыхания, обес-
печивающеrо бо..lее полный распад уrо!1еродных соединений, осво.
бождается rораздо больше энерrии. Орrанизмы, по..lучающие.
необходимую им энерrиlO за счет к..lсточноrо дыхания, MorYT функ-
ционировать при rораздо более высокой скорости метабодизма,
чем орrанизмы, получающие ее за счет брожения.
Для аэробноrо дыхания необходима атмосфера, содержащая
кислород. Кислород образуется при фотосинтезе в качестве по-
бочноrо продукта. По мнению тех, IПО изучает ранние стадии-
истории Земли, переход от первоначальной бескислородной aTMO
сферы к атмосфере, содержащей кислород, произошел в резуль-
тате активности примитивных фотосинтезирующих орrанизмов.
Возникновение окислите.1ЬНОЙ аТ lОсферы, дедающей возможным
дыхание, было постепенным и очень медленным процесс ом. Имен
но медленность Этоrо процесса моrла быть причиной длительной
задержки .2 млрд. лет илп более между возникновением жиз
ни и появлением эукариотов и lv\etazoa (см. рис. 25.1). .
Эукариотическая клеточная орrанизация, при которой MHoro-
численные reHHble центры собраны в настоящие хромосомы, все
функции распределены между ядром и цитоплазмой и клетка co
держит такие орrанедлы, как хлоропласты и митохондрии, была
друrим бо.'1ЬШИМ шаrом вперед в смысле сложности структуры
и способности самостоятельно осуществлять . разнообразные жиз-
ненные процессы.
Два основных компонента половоrо размножения оплодотво-
рение и мейоз..... ста.'!и возможными б.'1аrодаря rруппировке reHoB
в настоящие хромосомы и заключению хромосом в ядро у эука-
риотов. Половое размножение как упорядоченный и симметричный
метод создания комбинативной изменчивости (см. rл. 7) возни-
кает на уровне одноклеточных эукариотов.
-.
r:::
q"
%
.
.....
OQ
g
<1
....
.
t)
в
<i
::;,-
::::J
s!
.:>.t
:ci
'\1
;:,.
с\;
:с,
:{
;:,
;:,
",,-
;;.;
"
;:,
.
":
.
с
....
с:>
. :! 9
:::;) а а С) С)
2::1 r-- 10 Ir) '1"
oo50d OV:JT/h
2
о
:с
.,.
::
f.-
...
'111
..
CI.J
Ir
.о
Q)
:7
о
1:(
,...
$ е
о) о
.... :!I:
u
.>1:
i=: ::
Q..O:;
е
..
::Е о
:ZI 111
р:; о(
1-< о
О =
aI
:s:
!Е о
...
::2 о
:!s 1::(
= iE
I::(CII
о :!I:
a:I !J:
(!) 1=;
::с 1:(
о о
t: 9
Q) j:
:i!
:ж: IQ
; :;!
)(
Q) a:I
:s: с
:.::
е-
tI'
:s: ;.о:
J-o :ZI
о :r:
1:;: t::
О ;..
f-oo
::с :.:
О
Q)
1:;:
r<j ;;;
1:: 1'3
(f) 111
ari
с'о1
!d
a:
о
t:;
и
::
:J'
t-=
2
с
:::1::
c::t
1.)
о
::
'"
....
'"
:.:
о
t:
I
...
<1
::r
::)
::)
l
о;:
i
5
1
I I
""'$1
g
::t!J
-.;.
,, c::
::)
O:::
<1
i...,
i
"
f
!
'"' .
"J
i
!II!::i '
e;s
j
.о.
;;'
l ' ,IH!
". .. ...
. : О:. }:
.1 i,'
) :.;; ;;-:
!:H]
:::: .
e
':3
i
"
';::,
o;:
::,
c::
::,
:
;j:
с)
"Q
:.
.
:::
;.'
i
CI)
'
,
.J'
С
(\s
...
с
........
J:D
О
::I!!
f')
=
:1::
(\s
J-o
а.
о
С
=
>.
а.
....
)<
::а
=
1:1
>.
а.
х
в.
(,1
с3
'<s
::s
ID
::
CIS
а.
J:D
...
u
С
u
О
""
=
са
t
CD
::t
со)
(\s
а.
:в
I!(
о
=
а.
а>
t::
'::'
i
1
.j.
u:i
ct
U
:1:
g,.
r л. 25. rеолоzuчеСlCое вреJfЯ
245
Стадия сложных. мноrоклеточных орrанизмов
Стадия сложных мноrОКJlеточных орrанизмов занимает по-
следние 700 МJlН. лет истории Земли. Такие орrанизмы редко
встречаются среди ископаемых остатков позднеrо докембрия, но
становятся весьма обильными и разнообразными в кембрии. жи-
вотные и растения на протяжении Bcero кембрия и ордовика оби-
тали в воде; в силуре и в девоне они выШЛИ на сушу, а в карбо-
не постепенно становились все лучше приспособленными к жизни
в воздушной наземной среде. Современные животные и растения
относятся к кайнозойской эре, которая началась 65 млн. лет
назад (табл. 25.2).
Таблuца 25.2
Хронолоrия каiiнозоiiской эры (Raven, Axelrod, 1974)
I Возраст, ....
Лерио.!t Эпоха .пет д.о настоя-
щеrо временн
Четвертичный Современная 0,01
Плейстоцен 2,5
Поздне'rретичный Плиоцен 10
(HeoreH) Миоцен
27
Опиrоцен 38
Раинетретичный Эоцен
(naпeoreH) 54
П апеоцен 65
Возрастающее разнообразие представителей Metazoa на про-
тяжении фанерозоя изображено rрафически на рис. 25.2 и 25.3.
Последовательное доминирование различных rрупп в биолоrиче-
ском мире показано на рис. 25.4.
r лара 26
НАПРАВЛЕННАЯ ЭВОЛЮЦИЯ
Введение
Ископаемые. остатки по мноrим rpynnaM животных и неко-
OpЫM rруппам растений ясно свидетельствуют о существовании
длительных изменений, имеющих определенную ориентацию, или
.направлений эволюции. Под направлением ЭВОЛlOции имеетея- в'ви-
ду последовательность проrрессивных изменений в какой-либо
филетическо'й л'инии, изменений, которые происходят в некото-
ром данном направлении и продолжаются на протяжении дли-
теЛьноrо периода времени. Подобные направлеЮ1Я эволюции
представляют собой факт, установленный палеонтолоrами. Вывод
об их существовании можно также сделать на основании сравни-
тельно-.морфолоrическоrо изучения мноrих современных rрупп
растений, ископаемых остатков для которых не .найдено, но среди
которых сохранились последовательности примитивных и проrрес-
сивных форм.
Фактические данные относительно напраВ.'1ений ЭВОЛЮЦИJ:l сей-
час хорошо установлены, но их объяснение долrое время вызы-
вало разноrласия, которые кое rде еще не улеrлись. Две вражду
ющие теории известны под названиеМ ортоrенеза и ортоселекции.
Преобладающее БО,1]ЬШИНСТВО данных свидетельствует в. пользу
теории ортоселекции. К вопросу об интерпретации фактов мы
вернемся в конце этой rлавы.
ПримерЬJ
Хорошим примером направления эволюции служит ув личе-
.ине «pora» у титанотерия (Brontotheriidae, Perissodactyla) в пе-
риод от эоцена до олиrоцена (Osborn, 1929; Simpson, 1967; Stan
ley, 1974). У раннеэоценовоrо титанотерия Eotitanops не было
pora (рис. 26.1, А). у позднеэоценовых титанотериев. на лицевоЙ
части черепа имелись небольшие костные выступы (рис. 26.1, Б
и В). Тупые роrообразные выступы умеренной величины были у
раннеолиrоценовоrо титанотерия Megacerops, а у друrоrо олиrо-
ueHoBoro рода Brontotherium были очень большие «pora»
(рис. 26.1, F).
Друrим классическим примером направления эво..1ЮЦИИ слу-
жит пресноводная улитка Viviparus (ранее известная под наэва..
нием Paludina, сем. Viviparidae) из плиоценовых ОтложениЙ Ев-
ропы. На рис. 26.2 изображен последовательный ряд раковин
r А. 26. Направмн",ая .ВВОАЮЦUJl
247
все более молодоrо возраста из позднеrо плиоцена. С течением
времени раковина становится все более тесно закрученной и все
более СI(у..lьптурированной (Abel, 1929).
НаиБО.1Jее извес1'НЫЙ и хорошо документированный пример
среди всех известных ЭВО.YJюционных. рядов касается изменения:
Б-
w
", '"
7, .
20см i
JV ,.
/:;.. ,:. ,).1. .
C. :; L: -,j
. .
r
8
А
Рис. 26..1. Направления эволюции pora у титанотерия (Stanley, 1974).
А. Eotitanops borealis (ранннй зоцен). Б. Manteoceras тanfeoceras (средний эоцеи). В. pro..
titanothertиm emarginatиm (rrоздниА эо.цен). r. Brontotheriиm platgceras (ранний ОJIиrоцен).
зубов, конечностей и друrих частей тела у ,,10шадеЙ в течение
третичноrо периода. Направления эволюции в семействе лошадеЙ
(Equidae) будут описаны в дальнейших параrрафах.
Некоторые направления эволюции наблюдаются одновремен-
Но в неско.пьких отде.IJЬИЫХ rрулпах, раЗБиnаясь параллельно И
независимо. Так, тенденция к увеличению размеров тела Ваблю-
248
Часть V 1. Мак.роэвОАЮЦUЯ
дается у ряда наземных мле
копитающих третичноrо пе-
риода, например у титаноте
риев (рис. 26.1) , лошадей
(рис. 26.4, 26.6) и хоботных;
у динозавров мезозоя; у не-
которых морских животных,
например у морских улиток
палеозоя. Эта тенденция к
увеличению общих размеров
так широко распространена
среди животных, что она даже
получила название правила
Копа (обсуждение правила
Копа см. Rensch, 1960Ь; Stan
ley, 1973).
Во мноrих не связанных
между собой rруппах ЦBeT
ковых растений наблюдаются
па раллельные тенденции к
уменьшению размеров и упро-
щению морфолоrии. Подобные
направления эволюции извест-
ны под общим названием ре-
ДУКЦИОННЫХ рядов. Редукцион.
иый ряд оБЫЧRоrо типа начи-
нается с цветков, имеющИХ
MHoro лепестков, тычинок
и (или) плодолистиков, а кон-
чается цветками, у которых
имеется ЛИШЬ несколько этих
opraHoB. Друrой обычный ре-
дукционный ряд идет от круп-
ных мноrолетних травянистых
растений ДО мелких однолет-
НИКОВ. Нередко такое направление продолжается в пределах одно.
летних травянистых растений, у которых наб.'Iюдается постепеНный
переход 01' Iперекрестноопыляющихся форм с 'крупными цветками
к самоопылителям с мелкими цветками. Подобные направления
у растений .выявляются rлавным -образом на ОсНоваНИИ сравнитель
но.морфолоrических, а не палеонтолоrических данных.
Эволюционные ряды у Equidae
Очень хорошие палеОНТОJIоrические данные по Equidae обни.
мают период времени примерно в 60 млн. лет, начиная с палео-
цена. Краевые представители семейства это маленький четы-
О .,...,? <j ..". . ':."' :;i'" . ':" ' 1а
. . .\' -=:\:,::01 ,'.' .
. .
. . .
2 J 4
1
t IJ :.'., t . :
. .:.' ".. .' ..
';,. . . ., ...,
.' ..
. .
. ,,'/ .
5
б
7
4 4 "' 4 'Зt
. ".';' ....i r
" -. '::" 1 . "" . ',;. ."
. . :' : ( ",..: \", _ ,'-. it
. : .. ' :" ' ,
8
9
10
Рис. 26.2. Направления эволюции закру-
чеиности и скульптурнрованности рако-
ВИНЫ у улитки Viиiparиs (а=:РаIШlinа)
на протяжении поздиеrо плиоцена (АЬе',
1929) .
ЭВОJIЮЦНОНJlЫЙ рид ТJlнеТСJI ОТ предковой фор-
МЫ. б.llиакой к Palиdiпa иnicolor (1). через
промежутоqные стадии ДО Р. hoerпesl (10).
r л. 26. Направленная эвйлючия
249'
рехпалый Н yracotheriит (или Eohippus) из BepxHero палеоцена
н эоцена и современный род Eqииs, к которому ОТНосятся лоша...
ДИ, ДИКИе ослы (OHarpbl) и зебры. В палеонтолоrической летописи.
.
.
.
.
.
MegQhippds .. .
\ * . MerychlppuS
\ Aпch Arch. .
...
. . . .
HgpalllppllS PofTihippus
Miolripp'J$
.
и.l Ne.soll/ppиs
п'Щ1,О ......... /
л/f1pиs у
Epiltippus
1.
OrohlppиS
H!lrtcotheriиm
I
. OgнопаЛ61f?
.......
ТрекпОЛfJlе
Щиплющие ТPIМ8 .
.......
lJбаеgQющие
лист8g
gереlШ!8.
Рис. 26.3. Сильно упрощенная филоreНИJl семейства Equidae (Simpson, 1953)'
Опо Oпohippidiиm; Neo Neohipparioп; Nan... Naппippиs; Cal... Calippиs; АпсЬ... АпсЫ-
theriиm; ATch... ATchaeohippиs.
МОЖНО найти поч'!и непрерывный ряд переходов. связывающих
эти две формы.
Equidae происходят от elЦe более древней палеоценовой rруп..
пы Condylarthr , в которую входили неБО.'1ьшие, похожие на со-
баку животные с пятипалыми конечностями, снабженными поду-
шечками. Ряд ископае\'\ых форм можно ПрQследить еще дальше
вrлубь ---- от Н yracotheriuт до Condylarthra.
Фипоrения семейства лошадей была разработана на основании
палеонтолоrических данных. На рис. 26.3 изображена фи.поrениЯ"
этоrо семейства в сильно упрощенном виде. fеолоrический воз-
раст И rеоrрафическое распростраНеНИе r,,1aBHblX родов сем. Equi
dae перечислены таКЖе в табл. 26.1. Эволюция этой rрулпы про-
:250
Часть У/. Макроэволюцuя.
Таблица 26.1
'Возраст раз"ых родов семейства лошаДей (по Данным Simpson, 1951 ., личноrо
сообщения Симпсона)
Эпоха
ст арий Свет
Южнаи АмеРllка
Севериа Аыерика
Современная
Плейстоцен
Ллиоnен
Мноцен
Eqиus
Е q llиS
Eqиus?
Pliohippus
Calippus
Narтippus
Neohipparion
Hipparion
Megahippus
н ypohippus
Merychippиs
Archaeohippиs
Megahippus
Hypohippus
Parahippиs
Anchitherium
Miohippus
М esohippиs
Н aplohippиs
Epihippus
Orohippиs
Н yracotherium
Поздний naJle Н yracotheriиm:::a
оцеи (== Eohipииs)
ОЛRrоцен
Эоnен
Equиs
Equиs
Stylohipparion
Eqиus
Eqииs
Н ippidion
Onohippidiит
Parahipparioп
Stylohipparion
Hipparion
Hypohippus
Anchitheriит.
н yracotheriиm
t :
исходила rлавным образом в Северной Америке, хотя в ЕвразиIt
Африке и Южной Америке в разное время в течение третичноrо
и четвертичноI'О периодов имела мес,-о эволюция боковых линиЙ.
Мы не будем пытаться дать здесь общий обзор эволюции ло-
шадей, поскольку это уже сделал Симпсон (Simpson, 1951); наше
6нимание будет сосредоточено на некоторых направлениях в эво-
люции этой rруппы. fлавными среди них были: 1) размеры .тела;
2) длина хвоста; 3) механика конечности; 4) длина конечности;
r А. 26. Н апраSАе",ная' ЭSОАюция
251
.5) форма rоловы; 6) величина rоловноrо мозrа и степень ero
сложности; 7) коренные зубы. Нижеследующее изложение осно-
вано rлавным. образом на работах Симпсона (Simpson, 1951,
1953) .
Что касается размеров тела, 'IO у f-l yracotheriuт они были
r
'PP l arioп
Hypohippиs
t
MeSjhiPPUS Merflcn/ppus
@
Eol1ippus
Нippidioп
Jf
/
Pliohippus
Рис. 26.4. Эвопюция' черепа у поwадей (Simрsоп, 1951).
Все черепа изображены в одинаковом масштабе.
небо.YJЬШИМИ, как и у ранних Condylarthra; разные виды достиrа-
ли 25 50 см высоты, в ОТЛичие от всем известных больших раз
меров Equus. Общее увеличение раЗ lеров тела происходило во
мноrих "lИНИЯХ, в том числе у представителей Equinae из 0,,1иrоце.
На и П.rIиоцена. Об основных этапах этоrо эволюционноrо ряда
позволяют судить изображения rоЛОВ и конечностей, выполненные
в одинаковом масштабе (рис. 26.4 и 26.5). Следует также отме-
252
Часть У/. Ма"роэвОАЮЦUЯ
.1'ить, что увеличение размеров, хотя оно достаточно хорошо 8Ы-
.ражено, происходило в разные периоды времени с различной
,скоростью, не распространялось на все семейство лошадей и пе-
редко обращалось вспять у разных родов, в том числе и у самих
Е q иus.
I 2 ..,
А
4..
123
Б
4
Рис. 26.5. Эволюция ноrи у пошади (Simрsоп, 1951).
1ke иоrи изображены в одинаковом масштабе. А. Задних иоrа. Б. Переднии Hora. J Hgra-
cotheriиm; 2 Mesohippиs; 3 Merychippиs; 4 Eqииs.
Хвост изменялся от длинноrо по сравнению с размерами тела
у Hyracotheriит до относительно KopoTKoro и Д.,YJинношерстноrо у
.Equus.
Наиболее ярко выраженным направлением в эволюции ноrи
лошади бы..lа редукция числа пальцев. Начальная точка этоrо
направления предковые Condylarthra, которые были пяти-
:палыми. У Н yracotheriиm на передних конечностях было по че-
.тыре, а на задних по три пальца (рис. 26.6, А). Лошади, жив-
.шие в олиrоцене, миодене и на протяжении большей части плио-
цена, были трехпалыми (рис. 26.6, Б и В). Среди трехпалых ло
шадей можно заметить разницу между преДС1'авителями более
.древних родов, у которых боковые пальцы хорошо развиты
(рис. '26,6, Б), и более поздними формами, у которых они УКОРО.
.чены (рис. 26.6, В). Завершается этот ряд однопалой конечностью
у плиоценовоrо рода Pliohippus и У ero более поздних потомков,
в том числе у Hippidion и Equus (рис. 26.6, F).
Уменьшение числа пальцев сопровождается изменением меха-
'Ники конечности. У Condylarthra и Hyracotheriит На пальцах
['л. 26. Н апраsле",нал эволюция
были подушечки, и эти животные при ходьбе опирались на эти
подушечки, как это делают собаки. Самые древние трехпалые
лошади в олиrоцене и некоторые из их прямых потомков в мио-
пене также опира..1ИСЬ на подушечки (рис. 26,6, Б). Однако у дpy
rой ветви трехпалых миоценовых лошадей и у более поздних
однопалых лошадей Hora обладала пружинящим действием. Вес
тела ложится у них не на подушечки всех пальцев, которых уже
вет, а на концевую фаланrу среднеrо па 1ьца, одетую копытом,
(рис. 26.5, 3 и 4; 26.6, В и F).
А
б
.)
в
r
Рис. :26.6. Эвопюция ноrи н пзпьцев у лошади {Simpson, 1951; Romer, 1966J.
Масштаб раэ.'1ИЧИЫЙ. А. Hyracotheriиm. Б. Mesohippиs В. Merychippиs. r. PUohippиs.
По мнению Симпсона, у трехпалых лошадей с пружинящей
ноrой два коротких боковых пальца, возможно, иноrда служили
для опоры, несколько облеrчая наrрузку на средний палец. У од-
нопа..'1ЫХ .1Jошадей эти боковые пальцы в конце концов исчезли.
I
IY них всс тела целиком ложится на копыта, одевающие средние
пальцы. Прочные связки, прикрепляющие мышцы к костям этих
пальцев, иrрают роль рессор при беrе.
Изменения в строении конечности носиди эпизодический, а не
мед..1енный и постепенный характер; эти изменения происходили
ишь в некоторых линиях и со сравнительно высокой скоростью.
Увеличение длины конечности в эволюции лошадей часТично
Qбъясняется увеличением размеров тела. Переход от конечности,
имеющей па.7JЬЦЫ, к пружинящей Hore, снабженной копытом, еще
больше увеличило эффективную длину конечности, потому что
при этом лошади ста.ТIИ ходить и беrать «на ЦЫПОЧJ<ах».
На рис. 26.4 10жн.о видеть мноrочисленные и сложные измене-
Ния как в пропорциях черепа, так и в ero размерах. Морда вытя
rивается, а кости нижней челюсти становятся шире. Эти измене.
пия в строении череп.а связаны с изменением зубов, которые бу-
дут рассмотрены в следующе 1 разделе.
Друrие изменения rоловы лошадей коррелированы с измене
Ilием объема rОЛО8ноrо моэrа. У Н yracolheriuт мозr был малень-
254
Часть V/. МакроэвОАЮЦUЯ
кий, подобно мозrу рептилий и примитивных млекопитающих
(рис. 26.7). В дальнейшей истории семейства rоловной мозr уве-
личивался в размерах и становился сложнее по мере Toro, как в
Миоцен
C08peMeHHOR и
Лllеi1сmоцен
пllиоцен
Eqиus
Pliohippus
ОАШОЦ'Н Ф Миут/рри$
3оЦ'Н М оЛiрри$
Eohippus
Рис. 26.7. ЭВОЛЮЦИЯ rO.1JOBHOrO мозrа у .10шадей (Simpson, 19=51).
rОЛОВJ10В мозr. реКОlIструнроnаllНЫЙ на OCIIOBe слепков, снятых с черепных коробок. изобра"
жен в одииаковом масштабе. Недавно получены данные (Radinsky, 1976), свидетельствую-
щнс о том. ч о у Hyracotheriит (==Eohippиs) мозr бы., БО:lее развит, чем показано на
этом рисунке.
нем развивались области, связанные с развитием высшей нервной
деятельности у современных млекопитающих. Проrрессивное раз
витие мозrа и высшей нервной деяте.1J.ЬНОСТИ у лошадей. происхо
дило постепенно на протяжении Bcero третичноrо и четвертичноrо
периодов (рис. 26.7).
Направления эволюции зубов у лошадей
Направления эволюции зубов У .ТlOшадей очень сложны и для
их подробноrо описания ПРИШ.1J.ось бы прибеrнуть к очень спе-
циальной термицолоrии. Для наших целей будет достаточно об-
щеrо очерка эволюции зубов у ..'10шадей с ИСПО..'IЬЗ0ванием мини-
мально о количества терминов.
r л. 26. НаправАе",,,,ая эвОАюци-я
255
Зубы Equus об..lадаlOТ следующими лроrрессивными ПРИЗНaI<а-
ми: 1) дифференцировка режущих передних зубов ДЛЯ откусыва-
ния и коренных зубов для пережевывания; 2) бо..lъшие размеры
коренных зубов; 3) высокая коронка у коренных зубов; 4) увели-
чение ЧIlсла функциона.IIЬПЫХ (жевательных) коренных зубов----
по шести на каждой стороне каждой чс.тIЮСТИ; 5) сильное разви-
-тие на жеватеJIЬНОЙ поверхности складок (лофодонтность), со-
стоящих из эмали; 6) развитие между СК.llадками твердоrо це.
Мента.
у l!yracotheriит этим признакам соответствуют СJlедующие
.лримитивные особенности зубов: 1) БО,,'Iее слабая дифференциров-
ка передних и боковых зубов; 2) меньшие размеры зубов; 3) низ-
.кая коронка у коренных зубов; 4) отсутствие жевательных зубов
(на каждой стороне каждой четости и:мелось по три коренных
зуба, но это не были настоящие жевательные зубы); 5) складки
эмали на жевательных поверхностях более простые и бо..Т]ее НИз-
кие; 6) отсутствие цемента (рис. 26.4 и 26.8).
Эти различия в строении зубов отражают различия в пище.
Н yracotheriuт обитал в лесах и объедаJl листву деревьев, а Eqи
us ---- тр авоядное животное, обитающее в степях.
1ноrочисленные различия в признаках зубов между Eqиus и
Hyracotheriит возникли в результате ряда направлений эволюции;
так, например, жевательные зубы ста.'1И крупнее (направление
2), коронки их ста..1И выше (направлсние 3), ск.rrадчатость ус..10Ж.
пилась (направление 5) и т. д. Уве..'1ичение чис..lа жевательных
зубов произошло в результате постепенноrо превращения у Eqиus
трех ложнокоренных зубов Н yracotheriuт в зубы, сходные с ко-
ренныМи,---- процесс, известный под названием мо..1яризации.
Эти изменения разных признаков зубов происходили не од-
новременно, а в разное время. МО.J]яризация (направление 4) яме-
JЮ leCTO rлавным образом в эоцене и Оv'Iиrоцене, а затем она
бо.ТIее И.I)И менее прекратилась. УВС.'1ичение высоты короНI\:И (на-
правление 3) произошло в основном в миоцене, причем оно кос ну.
лос(> ТО.'1ько таких вновь возникших травоядных rрупп, как Ме-
rychippus, тоrда как у миоценовых .1I0шадей, питавшихея листвой
деревьев, коронки зубов остава..'1ИСЬ низкими.
Признаки зубов изменялись постепенно, один тип переходил
в друrой. Подобную интсрrрадацию :'>tюжно наблюдать в тех слу-
чаях, коrда одна филетическая линия предста6лена ископаемыми
зубами в ряде очень близких по возрасту rеолоrических rоризон
ТОВ. Рассмотрим линию Parahippus---- 1erychippиs----Nannippus
из миоцена и плиоцена, изображенную на рис. 26.8. На рис. 26.9
показано изменение высоты коронки и жевательной поверхности
в более полной последовательности стадий в той же линии. Ясно
видно постепенное и почти непрерывное уве.'1ичение высоты ко-
ронки.
256
Часть У/. Ма/(роэволюцuя
Equиs
1" .. (
H!lpohippus llipparioп Neohipparioп Naппippиs Pliohippus
t IAr<Pa'!US 8/
. h l lMer!lChiPPUS Mer!ichif1pus
w. / prOlOI"PPUS)
Aпchifheriит Para ippиs
t
IfI
A'/esohi us
t
fI
Epil1ippus
t
") --
EoI1iPPu5
Jvtlbl 1 пpucпOCOdl/eHН6Ie.
/( OIflJegaHulO лuстtl61 I
gереБЫ!Б t
Ни8КОЯ коронка :
On:yrcr6ue цемента I
::r
\i
.
Щuплющие .зуtllJ/
В6/сокая коронка
Налuчuе цемента
Рис. 26.8. Эвопюuня зубов у пошадей (Simpson, 1951).
На рисунках (выполненных 8 одинаковоМ масштабе) показан характер складок иа жева.
тельиой повсрхности верхних коренных зубов. Друrие прнэнаки персчислены в нижней
части pIICYHKa.
Далее можно ДОПОЛНИТЬ позднемиоценовый отрезок ряда, изо-
браженноrо на рис. 26.9. Даунс (Downs, 1961) прове..l подробное
количественное изучение различий в зубах между двумя популя-
ЦИЯМИ Merychippus из формациЙ Nlаскалл и Коалинrа. Эти две
популяции разде.1Jены промежутком в 1----2 млн. лет. Черты сход-
ства и незначительные различия в складчатости зубов у этих По-
ПУЛЯЦИЙ можно видеть на рис. 26.10.
Даунс. (Downs, \961) измерил ряд признаков зубов в выбор
ках ВЗрОС.'1ых особей Merychippus из ПОПУЛЯЦИЙ формаций Мае-
CI) '"
'i,::
.
g .Е
]
CI)
.
Qj
-с:
1/)
'ё:
th
';:;
.S
а
.
s
.
.;З
'"
I .) С::
!:::I ::::S C:J
.
''''''''
-C::
с::
1:)
D
t\..
t}
JWA
lU
"'.
tD
D
't
(\,.
D
D
-.;1 '
Q'.
.,
bd;
Q
GI
Q
>:::!
DDоd :JNЭd
НО:Jби d
':::!
:::S
vvm:bWiJX а-
::,
':;:'
::::S
::t
DfjHпdO
dagпd
DdDdpoпH
D2HnVDO)l ';:]
::::S
::t
VVD>I:JDW
:::r-
с)
)fпd)( l.тт
'::,
:::J
hHOd а-
зипl/ПdJ
1:)
'IJ
()ЭVVD.J ц,
'b::t
н пlJd ::t
. ,"".
.......
t....
oqo
Q)
ё
3
.=
Ct;J
::J:
::!
е
;::
с
::
c;:s
:r:
(;J
::1
О
::
:=;z
(')
=:
-=
CIJ
r:::(
c;:s
3
е
1:;
s:
=:
с:;
а:,
I:Q.
е
с
CI)
:а
:r:
=
CI)
о..
е
:r::
:=
:с
S
о..
е
:r::
:21
Е-
е
u
CD
D:
:=
::f
.!2
t:;
о
са
(t).
а)
cj
=
Q.
258
Часть VI. Маrcроэволюция
калл и Коалинrа, и статистически обработал полученные данные_
Некоторые из ero результатов представлены в табл. 26.2. Отме-
тим, во-первых, что в пределах каждой популяции существует
значительная индивидуальная изменчивость, и, во-вторых, что
средние зиачения и диапазоны изменчивости у двух изучавшихся
популяций слеrка различаются. Различия по средним (в том числе
по некоторым средним, не включенным в табл. 26.2) по большей
Части статистически достоверны. Тем Не менее между этими дву-
Б
А
м-з М-2 M.l
КоренНI1/е
P 4 Р-З Р-2
ПреикореНlf6/е
Рис. 26.10. Складки эмапи иа верхних коренных п npeAKoptHHblX зубах двух
позднемноценовых видов Merychippus (Downs, 1961).
А. М. sefJersиs, формация Маска.llJl. Б. М. саЩоrпiсиs, формация Сев. J(озлииrа.
мя популяциями наблюдается значительное перекрывание, как
показывают пределы изменчивости и стандартные отклонения,
приведенные в табл. 26.2. Статистический характер различий
между популяциями Маскалл и Коалинrа образует мост, соеди-
няющий макроэволюционные и микроэволюционные изменения.
Известно несколько интересных примеров, Kor да тот или иной
признак зубов появлялся в какой-либо древней полиморфной
ископаемой популяции в качестве одноrо из вариантов, а затем
закреплялся в качестве постоянноrо признака в более поздней
популяции. Здесь снова можно проследить возникновение мак-
роэволюционноrо направления из микроэволюционноrо измене
ния.
Один такой случай относится к определенной складке (извест
ной под названием «кроше») на верхних коренных зубах в линии
Mesohippus ---- Equus. У Mesohippus эта складка обычно отсутство-
вала. Но в некоторых популяциях Mesohippus было по нескольку
мутантных особей с небольшими кроше. У некоторых видов-де-
r л. 26. Направленяая эволюция
259
ТаБЛllца 26.2
Изменчивость отдельных верхних коренных зубов у ВЗрОСJlЫХ лошадей
( М erychippиs) из двух позднемиоценовых формаций (Downs, 1961)
Пределы Стащ..арт- СтаТИСТЯ-
Среднее. Чнс.ло ческая
Призиак R формация нзменчнвостИ. мм ное OT зубов достовев-
ММ клоненне насть ( )
Высота коронки нестершихся
пожнокоренных и коренных
Сев. Коалинrа (МОJlоже) 30, 39,6 34,9 2,23 39 <0,01
Маска_JЛ (старше) 24,6 32,3 29,4 2,45 18'
Высота коронки стершихся ко-
ренных .
Сев. Коа.'Шlrа 23,4 33,9 27,9 2,53 57 <0,01
Маскап.'1 18, ( 27 ,5 22,9 2,37 16
Суммарная складчатость на
коренных
Сев. КоаЛliнrа 2....17 7,6 2,02 66 >0.05
Маскалп 5....11 7,3 1,52 19
риватов Miohippus из О.1Jиrоцена очень маленькое кроше пред-
ставляет собой относительно постоянный признак; у друrих, бо."ее
поздних видов-дериватов Parahippus из paHHero миоцена небо.т1Ь-
тое кроше было постоянным признаком. Позднее в линии, веду-
щей к Equus, кроше достиrло значительных размеров (Simpson,
1944; 1953). Зубной цемент впервые появляется аналоrичным об
разом в линии Parahippus ---- Merychippus. У Parahippus цемент
обычно отсутствует, но наблюдается в некоторых ero популяциях
в качестве полиморфноrо варианта, а позднее закрепляется в МО-
номорфном состоянии У Merychippus (Simpson, 1953).
Адаптивная природа направлений эволюции
в семействе лошадей
Об адаптивном характере описанных выше изменений можно
судить достаточно уверенно на основании тех условий, в которых
жили животные. Важными факторами в эволюции ..'10шадей He
сомненно были местообитание, пища и защита от BparoB. Необхо-
димо учитывать как прямые, так и косвенные воздействия этих и
друrих факторов.
В нача.'1е третичноrо периода значительная часть суши быда
покрыта .песами, однако в результате изменений климата, проис-
ходивших в миоцене, эти обширные leca частично были замещены
открытыми пространствами саванн и прерий. В плиоцене пло-
260
Часть У/. Ма1Сроэволюцuя
щадь, занимаемая луrовой растительностью, продолжала рас-
ширяться за счет лесов.
В начале третичноrо периода Equidae обитали в лесах и пи
тались древесной листвой, как уже rоворилось ВЫШе. Однако в
миоцене одна из боковых линий семейства лошадей пересели-
.лась в открытые местообитания ---- саванны и луrа ---- и перешла на
питание травой. Такое изменение местообитания и характера
питания произошло в Северной Америке; в результате эта боко-
Ba линия подверrлась, вероятно, сильным новым давлениям от-
.бора, воздействию которых можно приписать мноrие из наблюдае-
мых направлении эволюции.
Трава ---- это rрубая жесткая пища, ОТ которой зубы стира-
ются. Изменения зубов, происходившие у лошадей в миоцене и
плиоцене, представляют собой по большей. Части первоначальные
и усовершенствованные адаптации к питанию травой. Передние
режущие зубы, длинные ряды коренных зубов и складки эмали и
цемент на жевательных поверхностях этих зубов все это необ
ХОДИМО для отщипывания и пережевывания травы. Высокие ко-
ронки коренных зубов увеличивают долrовечность как самих этих
зубов, так и их обладателей.
Некоторые изменения rоловы также, по-видимому, связаны с
переходом к питанию травой. Удлинение морды способствует раз-
мещению двух дифференцированных наборов зубов ---- резцов в
передней части и коренных в задней. Широкие челюсти необхо
димы для Toro, чтобы в них моrли сидеть коренные зубы с BЫCO
кой коронкой; кроме Toro, челюсти должны быть мощны 1И, чтобы
пережевывать БО.1Jьшие количества травы.
Животные, щиплющие траву на открытых пространствах са-
ванн и равнин, rораздо БОJIее заметны для хищников, чем живот
ные, обитающие в лесу. Увеличение размеров тела и большая
сила ---- один из эффективных способов защиты от хищников у на-
земных животных. Друrой способ ---- развитие высшей нервной
деятельности, и, наконец, третий ---- способность к быстрому беrу.
Тенденции к развитию эт х признаков в семействе лошадей по
всей вероятности в значите.'1ЬНОЙ степени связаны с тем, что жи
вотному, обитающему на равнине, необходимы средства защиты.
Большие размеры тела позволяют решить некоторые проб..lе-
мы, но создают и новые. Крупной лошади, питающейся травой,
нужны, чтобы обеспечить себе питание, более крупные, бо..lее
твердые и более прочные зубы, чем маленькой лошади. также пи
тающейся травой. Поэтому. как указывает Симпсон (Simpson,
1951). направления эволюции признакоl3 зубов, вероятно, адап-
тивно коррелированы не только с из снением характера питания,
но и с изменением размеров тела.
Увеличение размеров тела оказывало также побочное влияние
на скорость беrа. Крупной лошади TPY Hee быстро беrать, чем
r л. .26. Направленная эволюция
261
-Маленькой, так как этому препятствует ее тяжелое тело; для ре-
шения этой проб.,'Jемы понадобилось усовершенствовать механику
конечности. Строение тела Н yracotheriuт было приспособлено для
45blcTporo беrа. Д.пя Toro чтобы хотя бы сохранить такую скорость
беrа, которой облада.1l предковый Н yracotheriuт, не rоворя уже
о том, чтобы превысить ее, более поздним крупным лошадям не-
.обходимо было изменить пропорции конечности и строение паль
.цев (Simpson, 1951).
Адаптивные аспекты правила Копа
Во мноrих rруппах животных наблюдается увеличение разме
ров те.'1а в течение филоrенеза. Эта довольно обычная, но не Bce
общая тенденция известна, как уже было отмечено, под названи-
ем правила Копа (Соре, 1896; Newell, 1949; R.ensch, 1960; Stanley,
1973). Среди М.'Iекопита щих такое увеличение размеров наблю
дается у сумчатых, ищных, непарнокопытных, парнокопытных,
приматов и друrих rрупп; параллельные направления эволюции
обнаружены у рептилий, члснистоноrих, моллюсков и друrих
крупных rрупп. .
Большие размеры тела создают разнообразные селективные
преимущества, во всяком случае у наземных животных. Здесь бу
дут приведены четыре таких преимущества, хотя при желании к
ним можно добавить ряд друrих.
Размеры тела иrрают важную роль во взаимоотношениях
хищник ---- жертва. Хищник должен обладать определенной вели
чиной, весом и силой, чтобы победить свою жертву; в свою оче
редь жертва Д-ОJ1жна иметь определенную величину и силу, чтобы
.спастисЬ от хищника. Мноrие сочетания хищник ---- жертва оказы
.ваются втянутыми в cBoero рода «roHKY вооружений», ведущую ко
Все возрастающей величине Te.'Ia у обоих партнеров. Возможно,
(!ТО между лошадьми и крупными хищниками сложились именно
такие взаимоотношения и с такими результатами.
Для развития высшей нервной деятельности необходим боль
шой и СJIOЖНЫЙ rоловной мозr, а для TaKoro мозrа нужна боль.
Шая rOJlOBa, что в свою очередь требует большоrо тела. Отбор на
повышение уровня развития мозrа Mor повлечь за собой отбор
На увеличение размеров тела как коррелятивноrо признака.
Сила, необходимая Д.,lЯ борьбы,---- еще один признак, Koppe
.,lированный с размерами тела. В драках между самцами за об
.lJадание самка-ми, которые происходят у мноrих видов КОПЫТНЫХ t
превосходство в размерах н силе дает седективное преимущество.
Некоторые направления эволюции, каСЗlOщиеся изменения разме-
.ров, MorYT быть отчасти результатом ПО.'IОБоrо отбора.
у крупных животных есть еще одно преимущество над мелки
.fw1и, связанное с сохранением теП.'Iа в орrанизме. Большие тела
262
Часть У/. МаКрОЭ80ЛЮЦUЯ
теряют меньше тепла, чем небольшие. вследствие более блаrопри
ятноrо соотношения между площадью поверхности и объемом.
Тенденция к увеличению размеров никоrда не была всеобщей.
В некоторых филетических линиях наблюдается такое уве.'Iиче
иие, тоrда как в друrих родственных ИМ линиях на протяжении
Toro же периода отмечается, напротив. уменьшение разм ров.
Известны и такие случаи, коrда у одной и той же филетической
линии в один период времени эволюция была направлена в сто.
рону увеличения размеров, а в друrой в сторону их уменьшения.
Оба эти отклонения от правила Копа обнаружены в семействе
лошадей, а также у слонов, оленей и в друrих rруппах.
Все это свидетельствует об относительности селективноrо пре
имущества, создаваемоrо большими размерами тела. Увеличение
этих размеров выrодно. но лишь в известных условиях. В друrих
условиях оно может не давать никаких преимуществ или даже
оказаться невыrодным. У островных животных в общем отмечает-
ся тенденция к уменьшению размеров по сравнению с их матери
ковы ми предками. ЧТО представляет собой отчасти результат от.
бора на способность к выживанию при cTporo оrраниченных пи
щевых ресурсах.
Ортоrенез или ортосеJlекция?
Соrласно старой теории ортоrенеза, направления эволюции
определяются какой-то силой. присущей самим орrанизмам.
В одном из вариантов этой теории такой внутренней направляю
Щей силой является «потребность» орrанизма. «внутреннее CTpeM
..1ение»J некий elan vital и тому подобное (Lamarck. 1815 1822;
Веrgsоп, 1911; ОsЬоrп. 1934; Teilhard, 1959 и др.). Этой таинст
венной внутренней направляющей силе дава.1JИ разные названия:
besoin (потребность; Lamarck), pouvoir de la vie (жизненная си
ла; Lamarck), еlап vital (жизненный порыв; Bergson), аристоrе
нез (Osborn), энтелехия (Driesch), телефинализм (du Nouy) и
ортоrенез (TeiIhard). Присвоение мистических наименований, KO '
нечно, никак нельзя назвать научным объяснением. Соrласно
..1.pyroMY варианту теории ортоrенеза. направление эволюции оп-
ределяется предполаrаемой тенденцией мутаций соответствовать
этому направлению (Cronquist. 1951; .Werth, 1956 и др.). Этот
второй вариант представляет собой по крайней мере научную
rипотезу. поддаlOЩУЮСЯ экспериментальному изучению, которая
.была проверена и оказалась неверной (см. rл. 5).
Короче rоворя, ни один из вариантов теории ортоrенеза не
был научно доказан. В начале XIX в. ортоrенеэ каза.'1СЯ проrрес.
сивной теорией. Однако в середине ХХ в., после создания хорошо
обоснованной синтетической теории. он превратился в анахро-
низм. в одну из неудачных попыток мыслителей прошлых пока.
ений.
r л. 26. Направле",,,,ая эволюция
263
Мы' обсуждаем здесь отжившую свой век теорию ортоrенеза,
:потому что на самом деле она еще продолжает занимать мысли
"мноrих людей. Следует помнить, что вплоть до 30 x и 40 x rодов
.нашеrо века ортоrенез был широко распространенной среди пале
-онтолоrов точкой зрения. Некоторые биолоrи старшеrо поколе
ния до сих пор все еще скл няются к ортоrенетической ИНтерпре-
,.ации макроэволюции, а друrие занимают аrностическую пози-
цию, отказываясь признать синтетическую теорию или совершен-
но отбросить теорию ортоrенеза. Кроме Toro, ортоrенез :под тем
или друrим названием (elan v-ital, энтелехия, аристоrенез, теле-
.финализм и т. д.) всеrда обладал необычайно притяrательной
силой для литераторов и боrословов, интересующихся эволюцией
(СМ., например, Teilhard, 1959; KoesHer, Smythies, 1969). Верить
R целенаправленность эволюции весьма успокоительно. Таким
образом, если rоворить о взrлядах на эволюцию среди широкой
публики и среди полупрофессионалов, то оказывается, что теория
ортоrенеза вполне здравствует.
Ортоrенез можно противопоставить одному из следствий син-
тетической теории, известному под названием ортоселекции. Со-
rласно синтетической теории направленность эволюции исходит
не от орrанизма, а от взаимодействия орrанизма со с.редой. Если
-отбор, обусловленный факторами среды, продолжает действовать
в определенном направлении в течение долrоrо времени, то такой
отбор можно назвать ортоселекцией и он приведет к возникнове-
нию HeKoro направления эволюции.
. Ортоrенез и ортоселекцию можно рассматривать в качестве
альтернативных rипотез, признав за первой право на существо
-вание, для Toro чтобы сделать возможным обсуждение. Это по
.зволит пересмотреть палеонтолоrические данные в свете обоих
возможных объяснений, а не только с точки зрения ортоrенеза.
Если внимательно исследовать палеонтолоrические данные по ка-
кой-либо rруппе, хорошо исследованной как палеонтолоrически,
так и неонтолоrически, с тем чтобы неизвестные факторы были
.-сведены до минимума, то .оказывается, что факты лучше уклады
'ваются в теорию ортоселекции, чем в теорию ортоrенеза (Simpson,
1944,1951,1953,1967).
Во-первых, мноrие направления эволюции явно адаптивны.
.К ним относятся тенденции к увеличению размеров, обобщенные
под названием правила Копа, увеличение способности к переже
'выванию и ДО.'Irовечности зубов у лошадей, мноrие редукционные
.ряды у растений и т. п.
Во-вторых, ес.ПИ .какое либо направление эволюции обуслов
.1ено ортоrенезом, то оно должно быть общим для всей филетиче-
кой rруппы и Не иметь отклонений; если же оно обусловлено
'ортоселекцией, То следует ожидать, что у этоrо направления бу
..дут проявляться признаки колебаний, что .оно будет продолжать
264
Часть V/. МаfCfJQэвпЛ,йФ/(1
ея при наличии блаrопрятных условий, но отКлоняться в сторону
при их изменении_
Данные по семейству Equidac, ИзЛоженные выше, а также дaH
ные по друrим rруппам определенно соrласуются с тем, чеrо сле
дует ожидать по теории ортоселекции. В эволюции Equidae не
было таких направлений, которые продолжалнсъ бы на всем про
тяжении истории какой либо филетической линии или имели бы
место во всех линиях одновременно. Увеличение размеров тела.
было общим направлением в ЭВО.ТIюции лошадей, но оно было
подвержено реверсиям, происходившим в некоторых ..'1иниях в
разные периоды. На самом деле филоrения .пошадей сильно раз
BeTB."'leHa, и некоторые направления, в том числе имевшие место
в линии, приведшей к Equus, развивались в боковых ветвях
(рис. 26.3) (Simpson, 1951).
В-третьих, некоторые факты индивидуальной изменчивости в
ископаемых популяциях, наблюдаемые при блаrоприятных усло-
виях массовых захоронений, трудно объяснить в свете теории
ортоrенеза, но зато они точно соответствуют тому, чеrо следовало
бы ожидать, исходя из синтетической теории. Как было показано.
в одном из предыдущих разделов, проследив за эволюцией HeKO
торых признаков зубов у лошадей в обратном напраВ.1lении, мож
Но обнаружить, что в их основе лежит полиморфизм предковых
популяций.
rлава 27
ОПЕЦИАЛИЗАЦИЯ
Введение
Несмотря на то, что все орrанизмы в той или иной мере спе
диализированы, степень их специализации очень сильно варьиру.
ет от широкой До узкой. Биолоrическая специализация это
один из аспектов адаптации, и, подобно адаптации, она может
быть относитеv'1ЬНО узкой или относительно широкой. В сущности
специализацию можно рассматривать как набор приспособ.пений
и толерантностей данноrо орrанизма применительно к той адап-
тивной зоне, которую он занимает в природе, иди, более KOHKpeT
HO, к ero нормальному местообитанию или нише. Если орrанизм
прислособлен к узкоЙ зоне, местообитаНlfЮ ИЛИ нише, то он OT
носительна высоко специа изирован; если же он приспособлен к
более ШИРОКО?vIУ диапазону условий среды, то он обладает отно-
сительно широкой специализацией. Так, всеядный енот имеет
широкую пищевую нишу по сравнению с коала, который питает-
ся одними TO bKO листьями эвкалипта; первое из этих двух мле
хопитающих имеет широкую, а второе узкую пищевую специа-
.:'Iизацию.
Я избсrаю здесь терминов «неспециализированныА» и «сверх-
специализированный», хотя они часто встречаются в работах по
эво..1ЮЦИИ, потому что оба эти термина MorYT ввести в заблужде
иие. Симпсон и друrие авторы указывают, что неспециализиро
ванный орrанизм (если таковой можно себе представить) едва
ли Mor бы существовать. Что же касается сверхспециализации,
то она требует праrматической проверки на способность выжить
в условиях непредсказуемых rрядущих изменений. Ни одну rpyn-
пу орrанизмов нельзя считать сверхспециализированной До тех
пор, пока она в состоянии выжить; любую rруппу можно назвать
сверхспеЦИЗЛIf3ИРО13ЗННОЙ после Toro, как она вымерла. Поэтому
предпочтительно rоворить о разной степени специализации.
Узкая специа..1изация создается под влиянием межвидовой
конкуренции. В биотическом сообществе, в которое входит He
сколько или MHoro видов со сходными эколоrическими потребно
стями, обычно наб..'Iюдается развитие ряда узкоспециализирован
ных форм .( см. r.1J. 20).
При изменении окружающей срсды узкоспециа..1изированному
орrанизму при прочих равных условиях вымирание уrрожает
ильнее, чем qрrанизму с широкой специа.'IИзацией. При исцезно
266
Часть V/. Ма"роэволюцuя
вении одноrо типа пищи коала вымерли бы, а еноты выжили бы..
'rроза вымирания недостаток, неразрывно связанный с узкой.
специализацией.
Саморазвитие направлений эволюции
Направления эволюции нередко ведут к повышению специали,..
зации. В предшествующей r",aBe мы пришли к выводу, что в oc
иове эволюционных тенденций нет никаких внутренних сил, KO
торые бы' их направляли. Однако в иекотором смысле та или
иная тенденция к специализации, раз возникнув, может стать.
самораззивающейся под действием каких-то внутренних факто
ров. Механизм, обусловливающий такое саморазвитие, в корне
отличен от Bcero Toro, что имеет в виду теория ортоrенеза.
Направление эволюции, связанное с каКИМ .lJибо с..l0ЖНЫМ
признаком или сочетанием признаков, однажды утвердившись
нередко становится саморазвивающимся. Эволюционисты старше
ro поколения называли такие направления необратимыми и даже-
признавали некий общий закон необратимости эволюции (из
вестный также как закон Долло); однако они допускали некото-
рое преувеличение, поскольку ииоrда эволюция бывает обрати..
мой. Тем не менее в направлениях эволюции можно усмотреть
тенденцию к саморазвитию.
Эту тенденцию можно объяснить как результат cOBMecTHoro
действия ортоселекции и специализации. Ортоселекция rлав
ная сила, под действием которой филетические линии развивают..
ся в определенном направлении. Однако по мере Toro как данная
rруппа продвиrается в этом направлении, она накапливает спе-
циализированные признаки, которые оrраничивают диапазон
функционально полезных новых мутаций. Новые мутации обла-.
дают селективной ценностью только в том случае, если они обу
словливают фенотипические изменения, rармонично вписываю
щиеся n уже имеlOщееся сочетание признаков. А с повышением
специализации диапазон ценных в селективном отношении му-
тантных форм оrраничивается. Специализация помоrает opToce
лекции поддерживать данное направление «На своем пути».
Способы, позволяющие избежать с.пециализации
Если направления, ведущие к специализации, имеют тенден--
дию становиться саморазвивающимися и при этом повышают
вероятность Toro, что в конечном счете наступит вымирание, то'
длительное существование rруппы, которая уже далеко продвину-
лась в таком направлении, может зависеть от ее способности из"
бежать специализации. Нас здесь интересует вопрос о том, каким;
образом та или иная rруппа может выйти из потенциальноrо
тупика, в который заводит специализация.
r А. 27. СпеЦUаАuэацtlJl
207
Специализированные признаки развиваются в процессе как
,онтоrенеза, так и филоrенеза. На ювенильных стадиях имеющиеся
"у взрослых особей специализации обычно отсутствуют. Изменения
.скоростей развития, которые контролируются rенами и подвер-
'жены также контролю со стороны отбора, MorYT привести к
тому, что способность к размножению достиrается на одной из
-стадий, бывших прежде ювенильными (состояние. известное под
,названием педоморфоза или неотении). Отбор, направленный на
развитие педоморфоза.. может поэтому дать какой нибудь узко
специализированной (в зрелом возрасте) rруппе новые, дополни
-тельные преимущества, способствующие выживанию. На такой
:возможный способ избежать специализации указывали мноrие
.авторы (de Beer. 1951; Hardy, 1954; Тахтаджян, 1959 и друrие).
Харди (Hardy, 1954) четко сформулировал эту мысль:
«Какой бы высокой специализации ни достиrала данная раса
животных в типичном для нее взрослом состоянии, но если у нее
.имеется ювенильная или личиночная стадия (Коtорая. eCTeCTBeH
но. уже должна быть хорошо приспособлена к своему особому
()бразу жизни). специализированная либо в меньшей степени. либо
каким-то иным образом. чем взрослая форма, и имеется rенный
комплекс. который может рано или поздно привести к развитию
неотении. то при наличии достаточноrо времени у нее естЬ шансы
избежать вымирания. У orpOMHoro большинства эволюционных
.J1иний конец, очевидно, наступает до Toro, как возникнет эта peд
кая возможность педоморфоза; но у очень небольшоrо меньшин
<.тва этот шанс возникает раньше. прежде чем станет слишком
поздно. и отбор переКЛIOчает такие линии на новые пути. откры-
вающие новые возможности для адаптивной радиации. Имеющее-
ся в распоряжении этих процессов время столь велико, что. He
<,мотря на редкость таких случаев. эти удавшиеся попытки избе
; жать специа..'Iизации, по-видимому. привели к некоторым из важ
:. нейших новшеств, возникших в проuессе эволюции».
Мне хотелось бы добавить, что нам очень мало известно о
равновесии тех факторов. от которых зависит, сумеет ли данная
rруппа избежать направления, ведущеrо к специализации, или
lieT.
rла,ва 28
СКОРОСТИ ЭВОЛЮЦИ.И
Измере,нИR скоростей
В этой rлаве излаrаются данные, свидетельствующие о раз
личиях в скорости эволюции, приводится классификация этих
скоростей, предложенная Симпсоном, а также кратко рас.смат-.
риваются факторы, определяющие разные скорости. Однако,
прежде чем обсуждать эти вопросы, следует определить единицы
измерения для скоростей ЭВО.1JЮЦИИ, а это даЛС]{Q не простая за-
дача.
Измерение скоростей эволюции сопряжено со мноrими техни..
чески ми и практическими трудностями. Скорость ЭВО..'1lОции это-
количество эво..'Iюционноrо изменения за единицу времени. Пер
вый возникающий при этом вопрос касается масштаба времени,.
KOToporo следует придерживаться. Следует ли использовать reo
лоrическое время (т. е. промежутки в миллионы лет) или биоло..
rическое время (т. е. число поколений)? Если мы хотим сравни..
вать скорости эволюции в разных rруппах, то нам следует ИСПО.l1Ь..
зовать rеолоrическое время.
Второй вопрос касается типа эволюционных изменений, ко..
10рые предполаrается изучать и измерять: будут .Т'JИ это филети .
ческие изменения, изменения типа видообразования или же CMec
тех и друrих изменений. Ответ на этот вопрос вытекает из чисто
практических возможностей. Данные, доступные нам по ОДНИ f
rруппам, относятся к фИ.'1етической эволюции, а по друrим к
видообразованию и диверrенции. Нам приходится работать с теми
данными, которые у Нас есть, а поскольку они представляют
собой смесь различных показателей, то мы до..1ЖНЫ стараться
при сравнении скоростей эволюции по разным rруппам по воз
можности не выходить за пределы какой то одной катеrории из..
менений.
Третий вопрос состоит в том, чтобы найти такую меру для'
количества морфолоrическоrо изменения, которая бы позволяла.
сравнивать скорости эволюции, например, у лошадей и опоссу
мов. Каждому, кто занимается биолоrической систематикой, иэ
вестно, как трудно сравнивать черты сходства и раз.'IИЧИЯ у rрупп,
принадлежащих к разным семействам и обладающих разными
наборами изменчивых признаков. Симпсон (Simpson, 1949, 1953..
1967) принимает в качестве заведомо несовершенной, но прак
тически полезной меры число новых родов, возникающих за мил
JlИОН лет.
r л. 28. Скорости эволюции
26
Скорости эволюции, измеренные на основе Этоrо критерия, из
вестны под названием скоростей возникновения родов. Ниже
приведены данные о средних скоростях возникновения родов. в
трех rруппах (Simpson, 1953): .
Сред,кее ЧИСЛО родов за 101 пе..
Аммониты О 105
Chaticotheriidae 0,13
Equidae (Hyracotherlит Eqиus) 0,13
Биохимические методы позволяют дать более точную оценку
количества изменений некоторых частей фенотипа и tенотипа
данной rруппы. Так можно исследовать какой нибудь rомолоrич
пый белок, например rемоr.,'lOбин, в пределах данной разнообраз-
ной естественной rруппы, с тем чтобы выявить черты сходства н
различия в пос..'Iедовательности аминокислот. При таком ИСС.1'Jедо-
вании выявляется число замещений аминокис.'IОТ, происходивших
в процесс е возникновения разнообразия в данной rруппе. И это в
сочетании с друrими Данными, касающимися фактора времени,
позволяет дать оценку скорости мо.'1еку.,'IЯРНОЙ эволюции в иссле-
дуемой rруппе (см. r..1. 29).
Можно опреде..тrить возраст ныне живущей rруппы в отсутствие
l1алеонтолоrических данных, еС.,1И эта rруппа эндемична и aBTO
хтонна для области, rеО.'lОfическая история которой известна.
Так, какая-либо раса или вид MorYT быть эндемичными для дaH
ной местности, которая стала приrодной для обитания внедавнее
время (например, изо.'lИрованная rорная цепь, которая еще He
сколько тысяч леl назад была покрыта льдами). [руппа назем-
ных животных, имеющая довольно высокий таксономический paHr,
может быть оrраничена в своем распространении каким-либо
большим изолированным островом или островом материком, на
которые она Mor.'1a попасть ТО.'1ько по сухопутным мостам, cy
lцествовавшим в какое.то известное время в прошлом, . скажем
несколько миллионов лет назад. В таких случаях представляется
l:ЮЗМОЖНЫМ сделать заключение о возрасте автuхтонной и энде-
:\-IИЧНОЙ расы, вида, рода или семейства.
Возраст данной эндемичной rруппы устанавливает пределы
для оценок скорости эволюции, но он не всеrда позволяет судить
о фактически достиrаемой максима.7JЬНОЙ скорости, потому что
рассматривае lая rруппа моrла быстро диверI'ировать от пред.
ковой линии в начальный период колонизации, а затем изменятъ
ся мало или не изменяться вовсе.
Различия в скоростях
Палеонтолоrические данные убедительно свидете.'IЬСТВУЮТ о
1 ОМ, что некоторые rруппы развивались быстро, а друrие в тече
иие Toro же caMoro периода времени оставались неизменными.
-270
Часть У/. МЙКРОЭ80ЛЮЦия
Так, в третичный период в линии лошадей последовательно воз
никло 8 родов от Hyracotheriuт до PLiohippus (в среднем по
.одному новому роду за каждые 6,25 млн. лет); аналоrичные CKO
рости эволюции наблюдаются в третичном периоде и для друrих
копытных. Между тем в линии опоссумов за rораздо более дли
тельный период времени от позднемеловоrо до cOBpeMeHHoro пе-
риода произошло очень мало изменений, и меловые опоссумы
Alphadon очень похожи на современных Didelphis (Simpson, 1967).
К классическим примерам ЭВОJlЮЦИОННО консервативных форм
или «ЖИВЫХ ископаемых» относятся опоссум (Didelphis) , fаттерия
(Sphenodon), крокодилы, устрица (Ostrea) , мечехвост (Liтulus)
и морское плеченоrое Lingula. Эти rруппы дожили от ранних reo
JIOrнческих периодов до наших дней, претерпев сравните.."'1ЬНО
.мало изменений.
Ниже перечислены консервативные rруппы, сравнительно мало
'изменившиеся за указанное для каждой из них rеолоrическое
:время. Вначале указаны линии, состоящие из двух сходных
'родов, а затем древние роды, дожившие до наших дней. Все дaH
ные заимствЬваны у Симпсона (Simpson, 1967, а также личное
-сообщение) .
А Iphadon....D idel phis
Н oтoeosaurиs S рhеnйdоn
итиШ clla Liтulus
Crocodilиs
Ostrea
Lingиla
Поздний Мел....соВреМеННый период
Н)ра современный период
Триас современный период
Поздиий мел современный период
Мел современныА: период
Силур (или ордовик?) совремеины
период
Нет нужды напоминать о том, СКОЛЬКО эволюционной воды
утекло, пока эти консервативные rруппы плелись своими вековеч
ными путями.
Палеонтолоrические данные приводят также к выводу, что
скорости эволюции не оставались постоянными в пределах каж
дой отдельной филеiической линии на всем протяжении ее из-
вестной истории. Это ЯСНО видно на rрафиках, построенных Симп
соиом для плеченоrих (рис. 28.1) и для некоторых отрядов M.1Je.
копитающих (рис. 28.2). На рис. 28.2 выяв,пяются также различия
в скоростях эволюции между разными rруппами.
Симпсон (Simpson, 1967) указывает, что такие ныне KOHcep
вативные живые ископаемые, как опоссум и мечехвост, в то вре-
мя, коrда они впервые появились в rеО..'10rической летописи, были
.проrрессивными. До cBoero nepBoro появления, т. е. в тот оrрани-
ченный период времени, коrда эти rруппы эволюционировали от
,своих предковых линий, скорости ИХ эволюции были, вероятно.
4
::"
3
::" 2
:t
11)
ос::
""
с::.
c. О
Д Мне. Пен. Р Т Ю м Трет.
Рис. 28.1. Изменение скорости ЭВОЛIОUИИ плечеlIоrих от кембрня до третичноro
времени (Sirnpson. 1967).
ПО ОСИ ординат отложено ЧИCJIО новых родов, прнХОДИЩНХСJl на мнлпион пет (CKOpOCТII
возникиовеиня); ПО ОСИ абсцисс rеопоrическне периоды.
14
,2
tt: 10
8
с;;: ь
11:>
<о
4
2
. 0"""""-'
0.../ ,/
0/ ..... o I
""O ....,(' .
" " ./....0 .. .
"" . ..
о 0/ .
" .;'0
.0.":;;- .... "/
.... /v
"
rY'
о
паnеоцен
Зоцен ОlIи20цен Muot(eH пnиоцен
РиСе 28.2. Изменен Не скоростей эвопюuии четырех отрядов млеI<опитающих в те.
чение третичноrо периода (Simpson, 1967).
По осв ордннат от.l0жено ЧНCJlО новых родоп. nРИ ОJl.ЯЩИХСИ на миМИОИ lIет (скорост.
8ОЗИlIкновеНИJl). 1 парнокопытиые: 11 хищиые: п/ непаРНОКОПЫТllые; /У иепарио.
палые (Perissodactyla).
272
Часть У/. МаКрОЭ80ЛЮЦUЯ
ДОВОЛЬНО высокими. А затем, после этоrо »ачальноrо периода, ско-
рости стабилиэировались на довольно низком или даже нулевом
уровне.
Классификация скоростей эволюции
Несколько rрупп орrанизмов, такие, как хищные, двустворча-
тые МО.1'Iлюски и диатомовые водоросли, достаточно разнообразны
н достаточно хорошо изучены палеОНТО.'Iоrически, чтобы можно
БЫoll0 построить достоверные rрафнки распределения их скоростей
ЭВО.,'IЮЦИИ. При построении таких rрафиков оказывается, что по-
.лученная кривая отклоняется от кривой нормальноrо распреде-
.ления, будучи дептокуртической, или сильно асимметричной.
у этой кривой имеется высокая вершина, J1ежащая вб.т\Изи Toro
.ее конца, который соответствует высоким скоростям. Впо.7Iне
.возможно, ЧТО лептокуртический и асимметричный характер рас-
пределения скоростей представляет соuой черту, общую Д.ня МНО-
rих, ссли не для бо..1ьшинства крупных rрупп (Simpson, 1944,
1953) .
Характер распреде..1ения скоростей С.'lужит естественной OCHO
.БОЙ дЛЯ их классификации. Различают три кэ.теrории скоростей:
ормальные скорости (rоротелия, rоротелические), низкие СКО-
.рости (брадите.IJИЯ) и высокие скорости (тахителия) (Simpson,
1944, 1953).
Высокая всршина на кривой распределения скоростей ПОЗIЮ
Jlяет сделать вывод о том, каl(УЮ скорость следует считать HOp
ма.'1ЬНоЙ для каждой данной rруппы. 'пептокуртичность означае.т,
что БО"lЬШИНСТВО членов рассматриваемой rруппы обдадаlOТ cxoд
ными характерными для нее скорос.тями ЭВОol'lюции. ТаI{ие скоро-
сти называют rоротелическими. Далее скошенность кривой рас-
преде.,1ения указывает, что эти rоротелические скорости относи-
тельно высокие.
.l\-\еньшее число особей эволюционирует с брадите.'1И 1 IССКИМИ, а
еще *lеньшее с тахителическими скоростями. Конечно, . Живые
ископае .lые находятся на нижнем конце брадителическоrо отрез-
ка. Быстрые изменения на опредеJ}енных этапах ЭБО_'IЮЦИИ лоша-
дей С..'IуЖат примером тахителичеСКIIХ скоростей.
Следует указать, что палеонтолоrические Iетоды и палеонто-
лоrlJческие данные по своему характеру наиболее приrодны ДJlЯ
изучения браДИ'1'елических и rороте..'шческих скоростей. Получение
прямых палеонтопоrических данных о тахителических скоростях
возможно ЛИШЬ при наличии очень полной rеолоrической летопи.
си, например такой, какая имеется для лошадей, и, вероятно, эти
данны(' буд.ут относиться к более медленной части тахителическо-
ro диапазона. Эволюционные ИЗ lснения, происходящие не за
миллионы, а за тысячи или сотни лет, обычно не оставляют сле.
r л. 28. CKopocru молюцuu
273
ДО В В этой летописи. Тем не менее МЫ распо.l1аrаем эксперимс.н-
та.'1ЬНЫМИ и полевыми данными относительно сдвиrов, происходя
щих в процсссе видообразования у растений на протяжении He
скольких десятков или сотен лет. Тахителические скорости MorYT
иноrда быть выше, чем можно судить по палеонтолоrическим дан-
НЫМ.
Факторы, оказывающие 8лияние
на скорости эволюции
Попробуем представить в виде некой системы те факторы,
которые влияют иди предположительно влияют на скорости эво
ЛЮЦИИ. Далее в этой rлаве мы кратко рассмотрим эти факторы,
отложив рассмотрение фактора 4, иrрающеrо очень важную роль.
!до r"l. зо.
1. Взаимоотношения между ПОПУJНщией и средой
1. Стабильность среды
JI. Характеристиюr, внутренне присущие ПОПУЛЯЦИИ
2. Запас rенеТJlqеской изменчивости
З. ПРОДОJlжительность rенерации
4. Структура популяции
5. Место, занимаемое орrанизмом в иерархии питания
.б. Место, занимаемое орrаиизмом 8 иерархии размножения.
Среда
Если среда ОТНОСИ'l'ельно стабильна, то после Toro как дaH
ная популяция становится хорошо приспособленной к данной cpe
д.е, эволюция замедляется И.'1И прекращается. Коrда популяция
д.остиrает BwcoKoro адаптивноrо уровня в стабильной среде, лю
быс новые мутации или рекомбинации, по всей вероятности, OK(:.
жутся невыrодными. Почти всякому изменению в такой популяци и
будет противодействовать стабилизирующий отбор.
Вре я от времени в каком нибудь друrом доступном местооби
тании с иными ЭКОoll0rическими условиями может возникнуть HO
вая вариаuия. обладающая ПО.ТIOжительной се.'Iективной цeHHO
стью. В таких с,1]учаях иноrда возникает боковая филетическая
.'IИИИЯ. НО центра.1ьная rруппа сохраняет при этом постоянство.
Так случилось в истории опоссума, который дал начало в Южнuй
Америке четырем крупным ветвям в paHre семейств (например.
сем. Caenolestida и т. д.), но сам при этом изменилСя мало (Sim
pson, 1953).
Брадителические, т. е. медленно эво.rllоционирующие животные
встречаются в тех местообитаниях, которые оставались BЫCOKO
стабильными на протяжении rеолоrической истории: в тропиче-
сКих лесах и в лесах умеренно теплой зоны (опоссум), в больших
274
Часть V 1. МакроэвОЛlОция
тропических реках (крокодилы), в морях (Ostrea, Lingula, Liти..
ius) и в друrих столь же длительно существующих местообита-
ниях. И напротив, во временно существующих озерах, на вы-
соких ropax, вулканических отложениях, в арктической тундре
и тому подобных местообитаниях брадителические rруппы OTCYT
ствуют.
У популяции, обитающей в изменчивой 'среде, имеется три аль
тернативы: вымирание in situ, миrрация или эволюция. В области,
среда которой подвержена изменениям, брадителия невозможна.
L(ля Toro чтобы оставаться в такой местности популяция ДОЛJКна
эволюционировать. При этом скорость ЭВО.rlюции зависит, во-пер-
вых, от скорости изменения среды, а во-вторых, от способности
данной популяции не отставать от внешних изменений.
Способность популяции эво..1юционировать с rоротелическими
или тахителическими скоростями зависит в свою очередь от раз
личных биолоrических характеристик, рассмотрениIO которых по
священ следующий раздел.
Внутренние биолоrические факторы
В качестве одноrо из возможных факторов, оrраничивающих
скорости эволюции в некоторых случаях, выдвиrалась частота
мутаций ИЛИ количество полиморфной изменчивости. Имеющиеся
данные не подтверждают такое предположение. Морфолоrические
данные свидете.:JЬСТВУЮТ о наличии в популяциях брадителическо-
ro опоссума нормальноrо или BbIcOKoro уровня rенетической из-
менчивости (Simpson, 1944). Подобным же образом э..lектрофо-
ретические данные указывают на большие запасы rенетической
изменчивости в таких брадителических rруппах, как мечехвост или
близкий к папоротникам плаун (Selander et а1., 1970; Levin,
Crepet, 1973).
Естественная межвидовая rибридизация с..1УЖИТ источником
комбинативной изменчивости 80 мноrих rруппах растений. Кроме
Toro, формы, ВознИКшие путем rибридиз ции, нередко закрепля
ются в новых местообитаниях, которые создаются в результате
нарушений среды. [руппа растений, обладающая децентрализо-
ванными запасами потенциальной изменчивости, т. е. cocpeДOTO
ченными в двух или нескольких видах, способных к rибридизации..
вероятно, обладает преимуществом по сравнению со cTporo MOHO
типической rруппой, поскольку эта первая rруппа может успешно
реаrировать на быстрые изменения среды.
Друrая присущая орrанизмам черта, которая рассматривалась
в качестве фактора, оrраничивающеrо скорость эвоJiюции, это
продолжительность rеliерации. Поско.пьку изменения среды про-
исходят в rеолоrическом или ХРОНQ.lIOrическом масштабе времени,
тоrда как естественный отбор действует ВО времени, измеряемом
r л. 28. C"opoctи МtJАЮЦUU
275
11родолжите.,1ЬНОСТЬЮ rенерации, лоrично ожидать, что у орrаниз-
мов с короткими жизненными циклами скорости эволюции MorYT
Iбыть ВЫШе, чем у орrанизмов с продолжительными циклами. Од-
.Аако па.'1еонтолоrических данных, которые подтверждали бы этот
.лоrический вывод., очень мало.
Среди млекопитающих для опоссумов с коротки жизненным
циклом характерна брадителия, тоrда как у медленно размножа
ющихся слонов эволюция протека.'1а быстро. rрызуны с KQPOT-
ким И копытные с длинным жизненным циклом, начиная с плио
цена и п.'Iейстоцена, эволюционировали в Южной Америке пример
но с одинаковой скоростью. Хищные, имеющие Дu'Iинный цикл, эво
люционировали rораздо быстрее, чем двустворчатые моллюски,
имеющие короткий цикл (Simpson, 1944).
Данные по растениям, хотя и разнородные, в общем COOTBeT
етвуют данным по животным. Например, в Калифорнии в при
6режных районах и у подножий rop однолетние травянистые pac
тения эволюционировали быстро, но в этих же самых районах
"ак же быстро эволюционировали долrоживущие деревянистые
растения (Quercus, Ceanothus и Arctostaphylos) (Stebыns, 1949.
1950). Однако деревянистые растения в калифорнийском чапар
рапе можно разделить далее на ДВе катеrории на основании спо
соба размножения и времени rенерации. У деревянистых растений
чапарраля, которые размножаются половым путем при помощи
.семян, продолжительность rенерации умеренная, тоrда как у тех.
.которые размножаются веrетативным путем, побеrами кроны,
продолжите.'IЬНОСТЬ rенерации чрезвычайно велика. У растениЙ.
относящихся к первой катеrории (например, Ceaпothus и Arcto
staphylos), скорость видообразования rораздо выше, чем у расте.
ний второй катеrории (Wells, 1969).
Существуют, вероятно, довольно высокие скорости эволюции
{измеряемые в масштабах нсторическоrо, а не rеолоrическоru
времени), которые возможны только у орrанизмов с коротким
жизненным цик..l0М. Орrанизмы с коротким и продолжительным
циклом MorYT одинаково успешно реаrировать на некоторые CKO
рости изменения среды. Но если скорость изменений среды будет
все время возрастать, то рано или поздно будет достиrнут некий
nopor, отделяющий орrаиизмы с продолжительным циклом от ор-
rанизмов с коротким циклом в отношении способности rенериро-
вать скорости эволюции, необходимые для Toro, чтобы не отста-
вать от изменений среды.
Таким образом, ПРОДО.,1житедьность rенерации представляет
собой, вероятно, фактор, оказывающий влияние на скорости эво
,1JЮЦИИ, однако этот фактор вступает в действие только на «быст-
ром» КОНЦе диапазона скоростей, rде ero нельзя выявить палеон.
ТО.10rнческими методами.
276
Часть V 1. Макроэводюция
Цепи питания и размножения
Шмальrаузен (1949) выявид корреляцию между положением..
которое занимает данный орrаиизм в пирамиде численностей. и
скоростью ero эволюции. Орrанизмы. находящиеся В основании
этой пирамиды. имеют обычно низкие скорости эволюции. а ор-
rанизмы. образующие ее вершину, высокие скорости.
Шмальrаузен разделяет пирамиду численностей. или. как он
ее называет, .цепь питания, на четыре уровня: 1) самый нижний
уровень занят орrанизмами, у которых единственным способом
защиты от хищников служит быстрота размножения (например.
планктон. бактерии. зеленые водоросли);. 2) следующий уровень
образуют орrанизмы с чисто пассивными способами защиты (Ha
пример. моллюски, животные с очень прочными покровами. BЫC
шие растения); 3) над ними стоят животные, спасающиеся от
arpeccopoB блаrодаря способности к быстрому передвижению;
4) вершина занята животными, которые самИ являются хищни.
ками.
Шмальrаузен указывает, что низкие скорости эволюции ха.
рактерны для орrаНИЗМО8, ОТН9СЯЩИХСЯ к уровням 1 и 2, а BЫCO
кие скорости часто обнаруживаются у орrанизмов. занимающих
уровни 3 и 4.
Следующая задача состоит J3 том. чтобы выявить причинные
факторы. обусловливающие набтодаемую корреляцию. По MHe
нию Шмальrаузена (1949). взаимоотношения данноrо орrанизма
с друrими орrаниз ами, отражающиеся в ero положении в цепи
питания. определяют преобладающий способ отбора, а это в свою.
очередь определяет скорость эволюции.
Орrанизмы. состав.rrяющие нача..'Iьные звенья пищевой цепи и
лишенные средств защиты, элиминируются хищниками в основ.
ном неизбирате.пьно, а поэтому их эволюция протекает медленно
и направления ее не носят проrрессивноrо характера. В отличие
от этоrо хищные животные. образующие вершину пирамиды (ypo
ВеНЬ 4). конкурируют друr с друrом за пищу. Успех ШIИ пораже
ние в этой конкуренции в значительной степени зависит от врож
денных способностей отдельных животных. Ины и словами. Э.'IИ.
минация высокоизбирательна и СОЗДает возможность для быстрой
эволюции наиболее эффективных призиаков. Высокоизбирательна
также элиминация животных, шансы которых на спасение от хищ.
ников зависят от их собственной активности (уровень 3); ДЛЯ
таких животных также характерно быстрое развитие приспособи.
тельных специализаций (Шмальrаузен. 1949).
Наблюдаемые корреляции между MecTO t, занимаемым в цепи'
питания. и скоростью эволюции можно интерпретировать и по
иному. Можно. например, допустить. что отбор действовал иа'
орrанизмы, составляющие уровни 1 и 2, но выработаJl в. качестве
r л. 28. CrcopOC7'U эволюции
27Т
защиты против уничтожения хищниками не быстроту передвиже"
ния, а высокую плодовитость, поскольку эти качества можно рас..
rматривать как альтернативные защитные стратеrии.
Кроме Toro в шмальrаузеновской корреляции между местом в
uепи питания 11 скоростью эводюции фактор величины и струк-
туры популяции представляет собой нереrулируемую переменную.
Орrанизмы, составляющие уровни 1 и 2, нередко образуют очень.
большие популяции, способные ..1ИШЬ медленно реаrировать на
отбор, тоrда как животные, принадлежащне к уровню 4, часто-
образуют популяции колониальноrо или OCTpoBHoro типа, способ
ные быс ро изменяться под действием отбора или отбора и.дрей
фа. В связи с этим возникает вопрос, лежит ли причинная связь
между уровнем, занимаемым в пирамиде, и скоростью эволюции,
и.ни между структурой популяции и скоростью эволюции, или же-
эта связь носит Сl\1ешанный характер.
Все высшие растения занимают в пирамиде ЧИС..lенности одии
и тот же низкий уровень (уровень 2), однако они заметно отли
чаются по способам размножения, ИЗ которых Стеббинс (Steb
Ыпs, 1949) построил пирамиду размножения, аналоrичную цепи
ПlIтания Шма.'1ьrаузена: 1) низший уровень, обра.зуемый растс.
ниями, у которых распространение спор, ПЫЛЬЦевых зерен ИЛИ
семян обеспечивается просто высокой численностью и пассивным.
переносом (например, ветроопыляемые хвойные, Amentiferae,.
злаки); 2) средний уровень, образуемый растениями, споры, пыль.
цсвые зерна, семена иди плоды которых защищены жесткими И.
прочными оБОol'10чками (например, некоторые виды сосны, дубы
бобовые); 3) высший уровень, образуе:\-IЫЙ растениями, у которых
.lибо опыление, либо распространение семян, ."'lибо оба эти про..
цесса обеспечиваются животными (например, некоторые бобовые,...
орхидеи) .
Скорости эволюции opraHoB, участвующих в опылении И l1И
распространении семян, в общем корреol'ШРУЮТ с ЭТИ IИ уровнями..
(St-еЬЫпs, 1949). Так, у опыляемых ветром хвойных, Amentiferae,
и злаков (уровень 1) не наб.'1юдается существенноrо ЭВОJ1ЮЦИОН.
Horo разнообразия opraHoB, продуцирующих пыльцу; однако у
иветков бобовых и орхидных, опыляемых при участии животных
(уровень -3), возникли И, по-видимому, довольно быстро, YCOBep
щснствованные специализации. Параллеollьные раз..1ИЧИЯ наблlO"
даются между семенами, шишками И..1И плодами, распространяе
мыми ветром (уровень 1) и распространяемыми животными (ypo
вень З).
rлава 29
ИЗМБНЕНИЯ НА МОЛБКУЛЯР.НОМ УРОВ.НЕ
eTOДЫ изучения
Существует 'несколько биохимических методов, позволяющих
измерить количество изменений, происшедших в rомолоrичных
макромолекулах близкородственных или далеких видов в процес-
.се их диверrенции. Обычным объектом изучения служат при этом
молекулы белка, но иноrда и молекулы ДНК. Используются rлав-
ным образом следующие методы: rибридизация молекул (для
ДНК); серолоrические тесты (для определения rрупп крови);
<lпределени-е последовательности аминокислот (для rемоrлобина,
миоrлобина, uитохрома с и т. д.) И электрофорез (д,,'IЯ разнооб-
разных ферментов).
Непосредственные результаты сравнительно биохимическоrо
,изучения макромолекул разноrо типа можно выразить колнчест
венно несколькими различными способами. Среди таких способов,.
.позволяющих измерять степень биохимической дифференцировки
между разными филоrенетическими линиями, отметим следующие:
относительную степень сродства между цепями ДНК, полученной
от различных видов (при Исследовании методом rибридизации
ДНК); число и долю замещений аминокислот в rомолоrичных по-
липептидных цепях (при изучении аМИНОКИС.."'10ТНЫХ последователь-
ностей в белках); вероятное число точковых мутаций, участвую-
щих в создании наблюдаемых различий между rомолоrичными
белками (путем экстраполяции данных об аминокислотных после-
довательностях в белках); долю ферментных локусов, оказавших-
ся электрофоретически различными (при исследовании методом
электрофореза) .
Здесь для нас представляют особый интерес косвенные резуль
таты биохимических исследований. КО.'Iичественная мера биохи
мической дифференцировки между ныне живущими видами А, В,
С и т. д. независимо от Toro, какая именно испо.пьзуется мера,
может быть наложена на филоrенетическое древо, ветви KOToporo
заканчиваются видами А, В, С, ... Известное или выведенное reo-
лоrичес"ое время, прошедшее после диверrенции соответствующих
-ветвей, служит знаменателем. Сопоставляя эти две меры, мы по
J1УЧИМ количество изменения, происходящеrо на макромолеКУJlЯР
'НОМ уровне за единицу времени, или скорость молекулярной эво
J.Iюции.
В теории все это выrлядит довольно просто, однако практиче-
.екая реализация ---- дело иноrо рода. Молекулярно биолоrические
r л. 29. Изменения на .молекуляр",о-м уровне
279
подходы к изучению эволюционных изменений и скоростей эво
люции являются новыми и пока нахоДЯТСЯ на стадии проверки.
Подходы эти еще не имели возможности принести те реЗУol1ьтаты,
которые они потенциально способны дать. Здесь мы сделаем не-
большое отступление, чтобы изложить одну связанную со всем.
этим проблему.
Критическое отступление
Эффективность любоrо научноrо метода зависит от Toro, кто:
ero применяет. Мноrие исследователи, пытающиеся в настоящее'
время применять молекулярные методы для решения ЭВD.'IЮЦИОН-
ных проблем, это специалисты в области молекулярной биоло-.
rии, а Не биолоrци орrанизмов, и, как я вынужден отметить, их.
знания в области эволюции обычно довольно поверхностны. Не-
которые ученые, компетентные в области математической биоло-.
rии, но не БИО 'Iоrии как таковой, также пытаются применять MO
деку.пярные методы для решения эволюционных проблем. Ре-.
зультаты оставляют желать луЧШеrо.
Все это постоянно приводит К возникновению серьезных проб
лем, касающихся научноrо качества хотя и н-екоторой} но доволь-
но существенной части литературы по молекулярной эволюции.
Я предпоче.1l бы вообще не rоворить об этой литературе. Однако.
количество ее стало столь значите 'IЬНЫМ, что иrнорировать ее
нельзя_ Возникла противоречивая ситуация, и мы вынуждены;
признать это. Поэтому я попытаюсь изложить все проблемы в том
виде, в каком они мне представляются. Я оrраничусь общими
рассуждениями, не указывая персонально тех, к кому относятся.
мои критические замечания, хотя это было бы нетрудно сделать.
Однако каждый, ознакомившись с литературой по молекулярной'
Э130ЛЮЦИИ, может сам убедиться в существовании этих проблем.
ОДНО неверное представление, обычно содержащееся во мно-
rих работах по молекулярной эволюции, состоит в том, что моле-
I\улярные данные более надежны или более достоверны д..1Я опре-
деления «эволюционноrо расстояния», чем морфолоrические дaH
ные. Подобное мнение можно отбросить как чистый предрассудок.
. МО.'1екулярных данных Ilмеются одни достоинства и недостат..
КИ, а у морфолоrических данных друrие.
Друrая идея, с которой связано больше шуму, чем фактических
данных, выдвинута рядом биохимиков и биоматематиков; она со-
стоит в том, что мноrие а.'lлели, обусловливающие полиморфизм.
И,7]И замещения, в се..'1еКТИ8НОМ отношении нейтральны. Что каса-
ется ПО.1иморфизма по некоторым reHaM и reHHblX замещений, то.
это может быть и верно. Однако закточение о том, что селек-
тивно нейтральные reHbl в больших ко.rlИчествах закрепляются В.
популяциях и в rенотипах, не подтверждено соответствующими
'280
. ш Часть У/. Макроэволюцuя
прямыми данны:vfИ и в настоящее время остается Bccro лишь спе-
куляцией (см. также rл. 14)..
На это накладывается еще одна идся, уже определенно HeBep
ная, идея о том, что селективную нейтральность полиморфных
reHOB можно описывать как новую, «недарвиновскую» теорию
эволюции. Термин «недарвиновская эволюция» был использован
:в этом смысле в названиях одной работы (King, Jukes, 1969) и
.одноrо симпозиума (LeCam, Neyman, Scott, 1972). Существует
яесколько недарвиновских теорий эволюции, но эта не относится
'К их числу, так как Дарвин ясно указывал на возможность су-
ществования в природе случаев нейтра.'1ьноrо в се.l1ективном OT
-ношении полиморфизма (Дарвин, 1859, r..1. 2 и 4). rоворя словами
ca oro Дарвина, «действие eCTecTBeHHoro отбора не распростра
.няется на изменения бесполезные и безвредные, они представля
.ют... колеблющийся элемент, вроде изменений, наблюдаемых нами
у некоторых полиморфных видов».
Вслед за этим идет проблема, и..1и кажущаяся проблема, аде-
'кватности методов взятия выборок, используемых при изучении
ндивидуальной из енчивости в пределах популяции. Научные
. статьи по МО.rJеку.'1ЯрНОЙ эволюции нередко бывают посвящены
.сравните.'1ЬНОЙ биохимии каКОЙ ,,'Iибо макромолекулы, имеющейся
.У нескольких видов; взять выборки необходимоrо для э-тоrо Ma
,'ериала в природе было бы трудно, однако ero леrко получить
от любой фирмы, снабжающей биолоrические учреждения, или
.даже просто в МЯСНО:\i Маrазине и на рынке. В таких статьях ни
чеrо не rоворится наличии или отсутствии индивидуальной из-
менчивости по изучаемой молекуле. И читатель такой статьи, ко-
1'oporo интересуют популяционные проблемы и который знает ав-
тора статьи как чисто лабораторноrо исследователя, не может не
задать себе вопроса о том, как производится сбор материала.
Те же самые стандартные орrанизмы фиrурируют и на фила-
тенетических древесах. На кончиках ветвей стоит: «кролик», «.'1 я-
тушка бык», «акула», «пшеница», «дрожжи» И Т. д. С чис..lОВЫМИ
индексами, указывающими на «эволюционное расстояние» между
.этими видами. Ветви соединяются внизу в опре еленных точках
'развилок, раСlJOложенных в соответствии с эволюционным рас-
стоянием. Здесь ЭВО.'IIоционисту снова приходится остановиться
и задуматься. Наши знания о фИol10rении ЛИШЬ в редких случаях
.бывают настолько ПО..1НЫМИ, чтобы дать нам право строить Tpa
фаретные филоrенетические древеса. И наоборот, филоrенетиче-
.(кие древеса, все ветви которых соединены в определенных точках
претендуют на чрезмерную точность, не. оправданную имеющими
'ся фактически и данными; именно поэтому такой способ пред-
.ставления филоrении в настоящее время мало используется D
эволюционной биолоrии, если не считать некоторых особых с.лу
' aeB.
r л. 29. Изменения на .молекулярном уровне
281
Теперь мы подош.тIИ к вопросу о скоростях. Степень молеку'"
hЯРНОЙ дифференцировки числитель для выражения CKOpO '
стей может быть установ..1ена количественно. Но знаменатель ,
ремя эволюции ---- определяется на основе филоrенетическоrо
peBa или какоrо-нибудь ана.l10rичноrо источника и точно не
liзвестен. Количественные меры скоросrей молекулярной эво.:ую
ции обычно выrлядят более точными, чем они есть на самом дe.'1e
:Кроме Toro, имеющиеся оценки скоростей используются Д..'IЯ BЫ '
'ведения общеrо заключения о скоростях молекулярной эволюции..
.к которым мы еще вернемся в дальнейшем.
l\\ежду тем друrая rруппа ученых ---- популяционные rенетики .
освоившие методы электрофореза, также С1арзlOТСЯ изучать ЭВО
люцию на биохимическом уровне и делают это, с моей пристраст-.
ной точки зрения, rораздо БОJlее успешно. r.павный недостаток В:
этом случае заключается в том, что популяционные rенетики H
об..1адают достаточной подrотовкой, чтобы ИСПОJll>зовать бо..'Iее.
С.'IОЖНые методы, предоставляемые МО.'Iекулярной биолоrией, Ha
пример методы, связанные с изучением ПОС..'1едовате..1ьностей ами
нокислот.
Само собой разумеется, что БИО.'10rи популяционисты HaCTpoe
ны чрезвычайно критически по отношению ко мноrи:м мо..lеКУ.IJЯР-
НЫМ БИО..тIоrам. Разноrласия между этими rруппами широко.
распространены, и нередко атмосфера становится beCb\-1а HaKa
ленной. Противоположные точки зрения представлены в трудах
уже упоминавшеrося симпозиума (L.eCam, Neyman, Scott, 1972) ,.
а также в текущей научной литературе.
Дискуссия ---- это один из путей к познанию истины; друrой.
путь..... сотрудничество. Один исследователь И..'IИ одна rруппа ис--
следователей с одинаКОВЫ:\НI представления ми может лишь не-
значительно продвинуться вперед в такой в значительной мере
меЖДИСЦИП.JIИнарной области, как молекулярная эволюция. rруп-
па, состоящая ИЗ молеку..1ЯРНЫХ биолоrов и популяционистов,.
непосредственно сотрудничающих друr с друrом, моrла бы избе
жать некоторых .'10вушек, в которые они попадали в недалеком:
прош.пом И рискуют попадать сейчас..
Различия в последовательностях аминокислот
Сходства и различия в послсдовательностях аминОКИСЛОТ R
полипептидных цепях rО lOлоrичных белков, приНад.....ежащих раз-
ным видам, MorYT служить опреде..'Iенной и количественной мерой.
степени молеку.'1ЯРНОЙ диффсренцировки. Сейчас уже собран()
множество данных о rомолоrичности молекул rемоrлобина, МИО
rлобина, цитохрома С, И lмуноrлобу""ина и друrих белков (см_
интересные обзоры Dayhoff, 1968, 1969, 1972). Здесь МЫ paCCMOT
рим лишь неСКО.I1ЬКО типичных примеров.
.282
Часть У/. МаКрОЭ80ЛЮЦия
Таблица 29.1
Число различий по аминокислотному составу цепей rемоrлобина Me y
человеком И разными жнвотныии (по р,анным Dayhoff, 1969)
ВIЩЫ
ЧИСЛО различиА
а.цепь -цепь
Человек шимпанэе
ЧеЛО8ек rорилла
Человек макак резус
Человек паукообразваR обезьяна
'ЧеповеК---JIоwадь
Человек---корова
.ЧеловеК....овца
Чеповек---коэа
ЧеловеК ВИНЬR
чеповек....Jl8Ма
'Человек---мышь
Человек---кролик
о
1
4
18
17
21
21
18
)6 )9
25
о
I
8
6
25
25
2 32
28---33
24
21
25
J4
rемоrлобин взрослоrо человека состоит из двух идентичных
полипептидных а-цепей, двух идентичных 13-цепей и присоединен
ных к ним rрулп reMa. Каждая а-цепь содержит по 141 амино
кислоте, а каждая цепь..... по 146. В rемоrлобине здоровоrо че
..'lOBeKa каждое' положение в цепи занято определенной аминокис-
tl10ТОЙ. ПОС.YJедовательность аминокислот известна (рис. 29.1, А,
и 29.2, А). Один reH детерминирует последовательность амино-
кислот в а-целях, а друrой, отдельный, reH ---- их последователь-
ность в -целях. Отметим попутно один интересный факт: хотя
.а- ир-цепи rемоrлобина человека различаются между собой, их
аминокислотные последовательности сходны и, вероятно, возникли
'в результате диверrенции от какой то общей предковой полипеп-
тидной цепи (Ingram, 1963).
Нас здесь больше интересует степень дифференцировки reMo-
rлобиновых цепей у разных видов. Возьмем в качестве эталона
.reMor лобин здоровоrо взрослоrо человека. Число различий по
.аминокислотам между человеком и разными друrими видами
млекопитающих представлено в табл. 29.1. Как показывает эта
таблица, у человека и у обезьяны ПОС.'1едовательности аминокис-
.лот и В "(Х-, И В -цепях идентичны. fемоrлобины че..10века и ro"
риллы различаются лишь по двум аМИНОКИС.110там. Человек и
-широконосые обезьяны довольно близки друr к друrу по строе-
,И ию rемоrлобина (рис. 29.1, Б, и 29.2, Б). Различия в строении
х-емоrлобина между человеком и представителями друrих отрядов
r л. 29. ИЗ.4Се",е",ия ",а молекуляр"'ом уровне
283'
млекопнтающих rораздо шире ---- от 14 до 33 аминокислот на каЖ
дую цепь (рис. 29.1, В, и 29.2, В; табл. 29.1).
rемоrлобины человека и карпа различаются, как и следовало
ожидать, в наибольшей степени: между их а цепями существуют
различия по 71 аминокислоте (Dayhoff, 1969).
Друrой белок дыхате,,'IЬНЫЙ фермент цитохром с ---- находится
в митохондриях эукариотических орrанизмов и очень удобен для
сравнительно-биохимических исследований представителей раз.
ных типов и разных царств. В табл. 29.2 приведены некоторые
данные относительно молекулярных различий по цитохрому
Человек служит эталоном для одной rруппы сравнений, а дрозо..
фила для друrой. Здесь снова можно отметить общую корре.
ляцию между молекулярными различиями и степенью филоrене..
тическоrо родства.
Таблица 29.2
ЧИСJlО различий по аминокислотному составу цитохрома с между
человеком и некоторыми друrими орrанизмами (по данным
Dayhoff, 1969)
ВидЫ
Число
различий
Человек --- ма как резус
Чеповек---лошадь
Человек---корова, овца
Человек---собака
ЧеловеК РО.1ИК
Человек---курица, ИНДейка
Человек---rолубь
Человек---каймановая черепаха
Человек---rремучая змея
Человек ---ляrушка бык
Чеповек---тунец
Чеповек---акупа
Человек---дрозофила
Че.'10век---мясная муха
Человек абочка TYToBoro шелкопряда
Человек---пшеница
Чсповек---нейроспора
Дрозофила мясная муха
Дрозофила бабочка тутовото шелкопряда
Дрозофила---пятиточечный бражник
Дрозофила акупа
Дрозофипа---rолубь
Дрозофипа---пшеница
1
12
10
11
9
13
12
15
14
18
21
24
29
27
31
43
48
2
15
14
26
25
47
5 10 15
УМ. "Еи SER PRO ALA А5Р .LY Т н". A A УА" L'tS At.A ALA таР CL't
1" LY:; У'!. G Y AL" НI S АlI'. GLY Ci.U Н... С"У AlA CLU ALA LEU GLU
31 ААС "'Е' P E I..е) 5ЕР. РНе PRO ТНА- ТНК L'I'S Тн" ТУ'" РНЕ PRO HIS
",. PI'IE ASP \. E ) SE !:: С\.У seR A\. r.\. t \/А" \.Y с\.., HlS GL'f \.YS
61 LVS УА\. "1." 15'- ALA Ни Tt:lR ASH ALA \/А!. Ц.I HI VAL ASP ASP
76 I"IП PRO I!S'4 H L U St AL/i Н" SER AS:' LE!J 1 S A' A II :; LYS
91 LEV .II\G YAL I P PRO \':\!. AS+4 рне L '15 H: \.EI) SER tIIS СУ5 LEU
10,. LEU YAL ТН" ни ALA ALA HIS LEU PRO AL4 CLU РНЕ ПIR PRO ALA
121 YA.L HIS ALA SER LEU ASP L'tS РНЕ lEU LA 5Е" YAL ScR 'НА. YAt.
,А 1)" LEU ТН" S.ER L'tS :ТУА. ААС *
5 10 15
YAL HU SER PRO ALA ISP L'tS 5ЕА- A5 УН LYS ALI ALI ТАР CLY
16 LYS .YAL GLY GLY'НlS 11. /i GLY GLU ТуА. GLY ALI GLV ILA "Еи (;LU
31 ЦG I"I[Т РНЕ LEU SEIt РНЕ P O 'НА 'НА LYS ТНА. ТУА. Рне PI<.O НlS
"6 F'HE 'A5 L(V :;i: 1115 GLY НiI. ALA GLN УА\. LYS GLY HIS G!\' LY5
61 LY5 Yt.l НI ASP f.LA Leu 'НА LEU ALA YAL GL'I' HIS YAL ASP ASP
Т,. МЕТ PRO ASN ALI LEU SER ALA LEU SER ASP LEV н 1 S ALA HIS "У!)
91 HU IRC YAL '.5Р PRO YAL ISN РНЕ LYS LEU LEU SEIt HIS СУ!) LEU
106 LEU YAL 'н" L U ALI ALA HIS "Еи PRQ ALA CLU РнЕ ,нА. РАО ALA
121 YAL HIS ALA 5Е" LEU ISP L У5 'НЕ LEu ALA seR YAL SER '"'" "А\.
,О В,. LEU Ун" 5Е" LYS ТУА. ARC *
S 10 .t !i
VAL LEU 5€A PRO А\.А 'ASP LYS.THR А5Н ILE L YS. 'НА. ALA TRP CLU
16 LYS. НЕ GLY SER HIS CLY GLY GЩ ПА GLY ALA GLU ILA YAL Ct.U
31 ARG.ME' РНЕ LEU CLY РнЕ РАО 'НА ТНА LYS.THR ТУ" РНЕ PRO HIS
"6 РНЕ ASP РНЕ 'НА HIS GLY SEIt GLU CLN ILE L 't'S.ALA HIS GLY lYS.
61 LYS.VIL SER GL ALA LEU HIR LYS.ALA "А" GLY HtS "Еи ASP ASP
16 LEU PRO GLY ALA LEU SER iHIt LEU SER ASP LEU HIS А\.А Н 1 S LYS.
.91',LEU ARc.YAL А5Р РАО УА!. AS" РНЕ l 'l'S.LEU LEU 5Е" HIS СУ!) LEU
'.106 LEU YAL т.'А LEU AlA ASN HIS УА" PRO SER GLU РНЕ 'н" PRO ALA
12L YAL HIS А:.А SER "Еи ASP LYS.PHE LEU ALA ASN VAL 5Е" 'НА. YAL
8 136 HU ТН" SER LYS.'YR ААС *
I>ис. 29.1. Последовательности аминокислот в а-цепях rем:оrпобина у трех видов
мпекопитающих (по Dayhoff, 1969).
А. Человек. Б. Макак.резус. В. Кролик.
5 10 15
VAL HIS LEU Ун" PRO CLU CLU LY5 5О ALA VAL Тн" ALA LEU ТА'-
16 CLY LY5 VAL ASN YAL ASP CLU YAL СО' I>i.. \. CLU ALA LEU GLY ААС
:н LEU LEU' VAL VAL ТУ" РАО ТАР ТН" CLN ААС РНЕ РнЕ .GLU SER РНЕ
46 С'" У ASP LEU SEA тнА PRO ASP AlA VAL МЕТ ЫУ ASN РRП LYS VAL
61 LYS ALA HIS CLV tYS LY ""L LEU GLY ALA РНЕ SER ASP CLV LEU
76 НА HIS ни ASP "SN Ни l VS CLV ТН'" РнЕ ALA ТНА lEU SEA GlU
91 HU HIS CVS ASP LYS LfU HIS VAL ASP РRП COLU "SN 'нЕ "АС LEU
106 LfU GLY ASN УА," LEU VAL CYS YAL LEU ALI HIS HIS РнЕ GLY tvs
.А 121 GlU РНЕ Тн" РАО РRП VAL CLN AL" ALA Т УА GLN L У$ YAL YAL ALA
136 GlV УА.. НА ASN ALA LEU AL' H(S tYS ТУА HIS '*
5 10 15
VAL HIS LEU ТН'" РRП GLU GLU t vS ASt.! ALA УАа. ТНА ТНА LEU ТА'
16 ClV ,"Y YAL -SN YAL ASP ClU УА," GLY GLY CLU ALA LEU CLV ААС
31 LEU LEU LEU УА'" ТУ" PRO TRP тн" GLN ААС PH РНЕ CLU SER РНЕ
"6 ClY А5Р LEU 5(" SEA РRП ASP AlA VAL МЕТ CLY ASN РRП LYS VAL
61 t'tS ALA HIS ctv LYS t YS УАа. ни GLY ALA РНЕ SER II.SP GLV LEU
1,. ASN HIS HU А5Р А5Н LEU LYS С'" V УН" РНЕ ALA GLN HU SEA GLU
411 LEU HIS СУ5 А5Р LYS ни HIS VAL HP РАО Ctu А5Н "НЕ LYS LEU
106 LEU CLV НН VAL \,(U VAL СУ$ VAL LEU ALA H1S HIS РНе CLV LY5
121 COLU РНЕ ТНА РАО GLN VAL COL" ALA AtA ТУ. CtN LVS VAL VAL ALA
.6 В6 GL't VAL ALA А5Н ALA LEU А\.А HIS L.YS ту. HI5 *
5 10 15
VAL HI5 LEU $ЕА НА IiLU GLU LYS SER ALA V_L ТНА ALA LEU TRP
1,. CLY LY5 VAL ASN YAt CLU CLU УАа. CLY GLY CLU AL" LEU CLt ААС
31 tE"u LEU VAL VAL ТУ" РRП ТRP 'НА CLN "RC РНЕ РНЕ CLU SER РНЕ
.6 CLY ASP ни SEA SER AlA ASN НА VAL МН ASN ASN РRП LYS VAL
61 LYS ALA HIS CLV L У5 LYS УАа. LEU ALA ALA РНЕ И" ыи GL'f LEU
16 5Н HIS LEU А5Р AS" LEU tYS GLY тн" РНЕ AlA LVS LEU SEA GLU
91 LEU HIS C'fS ASP LYS LEU HIS УА," ASD РАО GLU ASN РНЕ ARG LEU
106 LEU CLY ASt.! VAL L U VAL ILE УАа. LEU !.rA НIS t'lIS РНЕ COL'f lVS
121 CLU РНЕ 'НА PRO GLN VAL GtN Аа. А AtA ту" ClN LYS yAL УАа. ALA
/J ASt.! AtA HIS t У5 ТУ" НI S *
136 CLY Y"t ALA LEU AlA
. - ............... ..............
HC. 29.2. I10следоватenьность аминокислот в uепих rемоrпобина У трех видов
млекопитающих (по Dayhoff, 1969).
А_ Человек. Б. Макак-резус. В. кро,nик.
286
Часть V/. Макроэволюцuя
Дейхоф (Dayhoff, 1969) и ее СОТРУДНИКИ указывают, что наблю-
даемое число различий в аМИНОКИС.1JОТНОМ составе между rOMo.:IO
rЯЧНЫМИ белками не обязательно ДОw1ЖНО быть равно чисw1У
аминокислот, действительно замещенных в процессе эволюционной
диверrенции этих бе"lКОВ. В тех случаях, коrда две ПОw1ипептидные
цепи различаются по мноrим аминокислотам, эволюционное рас-
стояние становится больше, чем наблюдаемые различия. Дейхоф
и ее сотрудни){и предложили единицу эволюционноrо расстояния,
названную ими РАМ-единицей (Р AM-unit..... «accepted point ти.
tations per 100 links», т. е. число фиксированных точковых му-
таций на 100 звеньев цепи); эта единица должна служить для
Toro, чтобы можно было дать скорректированную оценку эволIO-
ционной диверrенции на молекулярном уровне.
Оiношение между наб.аюдаемым числом различий в амино-
кислотном составе на 100 звеньев цепи и ЭВОJ1ЮЦИОННЫМ расстоя-
нием, выраженным в РАМ единицах, выrлядит следующим обра-
зом (Dayhoff t 1969):
1
5
10
25
50
75
85
р АМ едниицы
1
5
11
31
8З
208
370
Чнсло аминокислотных замеН
При Э'l'ОМ эволюционное раСС'l'ояние между видами, О'1'носящи
мися К разным крупным rруплам, оказывается несколько больше,
чем можно было бы считать по данным табл. 29.1 и 29.2.
Друrие данные относительно молекулярных
изменений
Методы rибридизации ДНК СОС'l'оят в смешивании одноцепо-
чечных фраrментов ДНК двух разных видов. Доля общеrо коли
чества ДНК в смеси, которая воссоединяется с образованием
двухцепочечных спиралей, и скорость воссоединения служат ме-
рами степени rенетическоrо родства между этими видами.
Применение этоrо метода дЛЯ ДНК дрозофилы дало реЗУ"lЬ
таты, приведенные в табл. 29.3. Drosophila тelanogaster и п. si-
mulans принадлежат к одной и той же rруппе видов. Как видно
из таблицы, в опытах по rибридизации 80% их днк образует
Двойные спирали. В отличие от этоrо ДНК D. funebris, которая
r л.. 29. Из.м.енен.ия на .м.ол.екул.ярfW-М уровн.е
287
Таблица 29.3
Доля ДИ 1( разных вндов Drosophila, воссоеДННЯlOщаяся в опытах
по rибриДнэации дик (Laird, McCarthy, 1968)
I Доп. воссое-
диняющеАся
ДНК. %
Сочетание ВИДОВ
D. тelaпogasterXD. siтulaпs
О. fиnebrisXD. тelanogaster или п. siтиlans
80
25
ТIринадлежит к друrому подроду, почти не воссоединяется с днк
п. melanogaster или п. simulans (Laird, McCarthy, 1968).
Хэбби и Трокмортон (НиЬЬу, Throckmorton, 1965) использо-
вали электрофоретические методы для определения сходства или
различия белков в rруппе D. virilis. Было проведено сравнение
мноrих белков и детерминирующих их reHoB для 1 О видов, OTHO
.сящихся К этой rруппе (п. virilis, п. americana J п. texana и др.).
Среди исследованных белков 60 % оказались общими для всех
видов этой rруппы. Неско..1ЬКО меньшая доля ---- общая в пределах
близкородственных подrрупп (например, п. aтericana и п. texa
.(Ш). Остальная часть изученных белков специфична для каждоrо
отдельноrо вида эта доля равна 2,6% дЛЯ D. virilis, 5,3% ---- для
п. aтericana и состаВ.1Jяет в среднем 14 % для всех десяти видов
вместе.
В друrой серии работ, в которых использовался rель-электро-
форез, 4 вида-ДВОЙНика rруппы п. willistoni сравнивались по 14.....
28 ферментным .'IOкусам (Ayala et аl., 1970; Ayala, Tracey, 1974).
Оказалось, что эти виды различаются примерно по половине изу-
чавшихся локусов.
Тесное серолоrическое родство существует между человеком
и человекообразными обезьянами по rруппам крови системы АВО
(о системе АВО у че.повека см. rл. 2). У этих четырех видов встре-
чаются следующие rруппы крови:
Человек
rиббон
OpaHrYTaH
Шимпанзе
А. В.АВ,О
А, В,АВ
А, В, АВ
А, О
Реакции аrrлютинации крови у шимпанзе неотличимы от ана-
.Jl0rичных реакций у человека; то же самое относится к крови
тиббона. Однако rОРИ.7Jла отличается от друrих человекообразных
обезьян и от человека по rруппам крови АВО и сходна в этом
,отношении с широконосыми обезьянами (Wiener, Moor Yankowski,
1971 ) _
288
Часть У/. Ма1СРОЭ80люцu.q
Различия между человеком и шимпанзе
Степень сходства между человеI{ОМ и шимпанзе (Рап troglo-
d!ltes) на молеку.1ЯРНОМ уровне широко изучалась различными
методами, в том числе теми, о которых rоворилось в начале этой
I'Лавы. кинr и Уилсон (King, Wilson, 1975) собрали все эти об
ширные молекудярные данные. Они обнаружили, что между
человеком и шимпанзе существует значительное сходство на MaK
ромолеКУЛЯРНО:\1 уровне: мноrие белки у тех и у друrих одинако
вы, а мноrие друrие очень сходны.
Изучение последовательностей аминокислот в нескольких ro
молоrичных бе.'Iках человека и шимпанзе показало, что HeKOTO
рые из этих белков полностью или почти полностью идентичны,
различаясь лишь по одной аминокислоте или не различаясь вовсе:
rемоrпобпн о
Цитохром С О
Фибринопептид О
Лизоцим О?
oI\ цепь rемоrлобина 1
МиоrпобllН 1
Друrие белки этих двух видов отличались лишь по несколь-
КИМ аминокислотам, как, например, сывороточные альбумины,
различавшиеся по 6 аминокислотам в 580 ПО.1Jожениях (King,
Wilson, 1975).
Результаты электрофоретическоrо ана.'Iиза 44 I'омолоrичных
белков опять-таки свидетельствуют о близком СХОДстве между
человеком и шимпанзе. Примерно половина белков, изученных
этим методом, оказались идентичными у обоих видов (King, \ViI-
50П, 1975).
Степень МОJlекулярной дифференцировки между человеком И
шимпанзе соответствует той, которая существует между видами.
двойниками в друrих rруппах, таких, как комплекс Drosophila
willistoni; она rораздо меньше, чем между обычными видами в
раз.'IИЧНЫХ друrих родах (King, Wilson, 1975).
Тем не менее, как показыв.ают МОРфО.llOrические, ЭКОJ10rиче
ские и поведенческие признаки, Ното sapiens и Рап troglodytes
несомненно относятся к разным родам. Их хромосомы также от-
.:Iичаются друr от друrа по числу инверсий и транслокаций. При
надлежность этих двух видов к разным семействам признается
почти всеми. Таким образом, данные об эволюционном расстоя-
нии между человекоМ и шимпанзе, полученные на орrанизменном
уровне, отличаются от соответствующих молекулярных даНН1J1}С
(King, \Vi1sоп, 1975).
r л. 29. И A&tн.ен..uя На м.олеку!tЯРНОм'. УРОвне
289
кикr и Уилсон пытаются разрешить этот парадокс, допустив,.
что основные rенетические различия между человеком it шимпан"
зе обусловлены rлавным образом не точковыми мутациями в.
обычных или так называемых «структурных» reHax, а изменениями
Б реrуляторных системах этих двух орrаниз:'dОВ. Предполаrается,.
что изменения реrуляторных систем вызываются частично MYTa
циями в rенах-реrуляrорах, а ЧастИЧНО хромосомными перестрой
ками, изменяющими порядок reHoB и оказывающими влияние на
их выражение. «Крупные различия, существующие между чело
веком и шимпанзе на орrанизменном уровне, обусловлены, воз-
можно, относительно небольшим числом rенетических изменений
в системах, реrулирующих выражение reHa» (King, Wilson, 1975).
He хотелось бы сделать несколько замечаний по поводу Ta
КОЙ ин-rерпре1'ации данных. Она затраrивае1' ряд разных nроблем
которые следует разложить по местам.
Прежде Bcero при сравнении человека и шимпанзе сопостав"
..1яют такие черты, которые не сопоставляют при сравнениях, про
водимых между парами видов у друrlIХ высших животных. В цe
лом различия между человеком и шимпанзе обусловлены не
только биолоrической, но также и культурной эволюцией, которая
значительно усилила различия между этими двумя видами. Эти
увеличившиеся различия заставили систематиков отнести роды
Ното и Рап к разным семействам. На основании одних только
морфолоrических различий Hominidae и Pongidae нельзя было
бы считать самостоятельными семействами и соответственно нам
следовало бы, обсуждая чисто биолоrические различия между
этими rруппами, понизить их таксономический paHr. Короче ro..
БОрЯ, роды Ноmо и Рап действительно сходны на орrаниэменном
уровне, как это давно признали зооnоrн, этолоrИ и полевые
биолоrи.
Между этими двумя видами существуют также внешние фе
нотипические различия. Какова rенетическая основа этих разли
чий? ПОСКОol1ЬКУ друrие подходы к этому вопросу оказались не..
состоятельными или даже приводили к ошибкам, мы позволим
себе обратиться к обширным данным по rенетике растений в на'--
дежде, что они nOMorYT найти ключ к этой проблеме. У растений.
в отличие от высших животных нередко оказывается возможным
получать мноrочисденные потомства F 2 и F3 от межвидОвых к'
межродовых скрещиваний и подверrатЬ их факторному анализу
Работы по rибридизации во ноrих rруппах растений показыва..
ют, ЧТО виды и роды различаются по различным типам reHHblx
систем, в которых «структурные», или менделевские, reHbl иrрзют
большую роль (см. обзор Grant, 1975, rл. 7 12). Исходя из этих
данных, мы имеем основания ожидать, что между любыми двумя
видами высших прима.тов существуют мноrочисленные и слож...
ные reHHble различия.
290
Часть У/. Ма роэволюцuя
Друrая проблема, как это представляют себе Кинr и Уилсон,
касается природы реrуляторных систем, которым они уделяют
столь большое внимаllие. Эти системы reTeporeHHы и мало изу-
чены. В состав реrуляторных систем, во всяком случае у расте-
ний (см. снова Grant, 1975, rл. 8), а возможно, и у животных,
входят обычные «структурные», или менделевские reHbl. Я rOTOB
соrласиться с Кинrом и Уи.i'1СОНОМ, что реrуляторные системы
Toro или иноrо. рода, вероятно, иrрают важную роль в дифферен-
циации человека и шимпанзе. Однако из этоrо вовсе не должно
следовать, что число обычных reHHblX различий между этими
двумя видами невелико.
Несоответствие между данными по белкам и данными по внеш.
не заметным фенотипическим признакам свидетельствует, с моей
точки зрения, о том, что в опреде ении этих несоrласующихся на-
боров признаков участвуют различные reHHbIe системы. Создает-
ся впечатление, что reHbl, ВЫЯВЛЯЮЩиеся при использовании :мo
леку.i1ЯРНЫХ методов, в значите.аьной степени отличаются от тех
reHoB, которые выявляются методами классической rенетики. Во
всяком случае reHbI, определяющие белки, вопреки широко рас-
пространенному мнению, нельзя рассматривать как адекватную
выборку из rенотипа (однако см. также обсуждение во втором
разделе rл. 14). Отсюда следует, что в противоположность дpy
rOMY общепринятому допущению данные молекулярных исследо
ваний представляют собой лишь одну из нескольких обоснован-
ных -1ep rенетическоrо и эволюционноrо расстояния.
Короче rоворя, используя одни только молекулярные методы,
мы не можем приблизuться к пониманию важныx с адаптивной
ТОЧЮf зрения морфолоrических, ЭКОЛОrических И поведенческих
р.азл.ичий между человеком и шимпанзе.
Скоро ти молекулярной ЭВОJlЮЦИИ
Дейхоф (Dayhoff, 1972) вычислила средние скорости молеку-
лярных изменений в различных белках (табл. 29.4). Скорость
изменения выражена числом РАМ единиц за 100 млн. лет. Одна
РАМ-единица соответствует одной точковой мутации, вызываю-
щей замещение одной аминокислоты на 100 звеньев полипептид-
ной цепи. Использование этих единиц дает возможность сравни-
вать друr с друrом белки с разным молекулярным весом, приво-
дя их к некому общему знаменателю.
Как видно из табо!1. 29.4, разные виды белков изменяются с
различной средней скоростью. [истоны крайне консервативны.
Цитохром С, инсулин И некоторые друrие белки очень консерва-
тивны. В' отличие от этих белков фибринопептиды и ropMoHbl роста
изменяются быстро. А у rемоrлобинов наблюдается промежуточ-
ная скорость изменения.
r л. 29. Изме",е",ия на .молекулярном уровне
291
. Таблица 29.4
Средние скорости эволюционноrо изменения раэпичных белковых
макромоnекуп (Dayhoff, 1972)
ТИП белка
I Число РАМ-ед.и
ниц на 100 мпн. Ле?
Фибрииопептид
rOpMOHbl роста
Иммуноrпобуnин
Рибонукпеаза
rемоrпобин
Миоr лобик
rастрин
Энцефалитоrенные бе.'JКИ
Инсуnин
Цитохром с
r JIицеральдеrид. з фосфат деrидроrен аза
rистон
90
37
32
33
14
13
8
7
4
3
2
0,06
Некоторые специалисты по молекулярной эволюции заявляют
что скорость молекулярноrо изменения ДЛЯ белка каждоrо дан-
иоrо типа постоянна во времени (Ohta, Kimura, 1971; Kimura,
Ohta, 1971, 1972; Jukes, 1972). Предполаrается, что молекулярная
эволюция действует с paBHoMepHocTыo часовоrо механизма. По-
добное заК,,'1ючение, конечно, противоречит тому, что нам извест"
но относительно скоростей эволюции внешних фенотипических
приэнаков (см. rл. 28).
Заключение о постоянстве скоростей молекулярной эволюции
основано, однако, на элементарной ошибке, о чем писали ученые,
заннмающиеся эволюцией на уровне орrаниэмов (Stebbins, Lewon
tin, 1972; Clarke, 1973; Lеwоntiп, 1974). НеКОТ9рые исследователи
молекулярной эволюции берут общее количество молекулярных
изменений за длительный период rеолоrичеСКОrО времени (поряд-
ка 108 лет), вычисляют среднюю скорость мо..1екулярноrо измене-
НИЯ, а затем представ.'IЯЮТ эти средние скорости в качестве по-
стоянных скоростей, т. е. смешивают два совершенно разных па-
раметра. При таком подходе можно было бы сделать также не-
верный вывод о том, что внешние фенотипическне признаки эво-
люционируют с постоянными скоростями.
r J:l3lВa 30
СТРУКТУРА ,ПОПУЛЯЦИИ IB СВЯЗИ
С МАIКРОЭВQЛЮЦИЕй
Адаптивный ландшафт
Компоненты нетривиальных эводюционных изменений обсуж.
дались в предшествующих rлавах под такими названиями, как:
«адаптивные признаки и сочетания признаков»; «адаптивные со-
четания reHoB»; «широкие И узкие специа.пизации»; «направления
эволюции И скорости эволюции». Адаптивный ландшафт полез-
ная метафора, помоrающая представить взаимодействия между
перечисленными компонентами. К этой метафоре прибеrали Райт,
Добржанский, Симпсон, Стеббинс и друrие эволюционисты.
Представим себе ландшафт, состоящий 'из холмов и долин.
Рельеф местности символизирует распределение адаптивных по
лей. Вершины холмов ---- «адаптивные вершины», а долины, раз-
деляющие холмы. представляют собой для орrанизмов «ничеЙ'ную
землю» или ряд «адаптивных долин». Популяции И виды занимают
разные адаптивные вершины блаrодаря имеющимся у них соче-
таниям адаптивных признаков и лежащим в основе этих признаков
сочетаниям reHoB. Некоторые адаптивные вершины узкие, а дpy
rие широкие в соответствии с относительной широтой специализа
ции. Вершины холмов различаются также по высоте; это означает,
что на некоторые из адаП11ИВНЫХ вершин леrче взобраться и леrче
занять их по сравнению с друrими.
Эту метафору можно расширить, с тем чтобы распространить
ее на rруппы видов, роды и более крупные филетические KaTero-
рии. Вершины распределяются по адаптивному ландшафту не слу-
чайным образом, а собраны, во всяком случае до некоторой сте-
пени, в отдельные цепи. При этом цепь холмов соответствует
адаптивной зоНе Toro или иноrо рода.
До сих пор мы рассматривали адаптивный ландшафт в стати-
ческом аспекте. А как обстоит дело с ero динамикой? Каким обра
зом виды занимают свои адаптивные вершины, а роды ---- свои rop-
вые цепи? Путями, ведущими к адаптивным вершинам и цепям,
-служат направления Эволюции. А скорость восхождения соответ-
ствует скоростям эволюции. Адаптивные вершины разной высо-
ты достиrаются в результате разных сочетаний направлений эво-
.люции и ее скоростей.
Адаптивный ландшафт не всеrда остается статичным и неиз-
ыенным. Подобно настоящему ландшафту он подвержен действию
.сил, вызывающих эрозию и rорообразован'ие. Эти силы MorYT дей-
r А. 30. Структ.ура популяции в связи с JUlк.роэволюцие4
293
ствовать медленно или быстро. Изменения ландшафта 'Отражают
роль изменений среды в эволюции. Адаптивная вершина может
перемещаться с той или иной скоростью, и популяция, которая ее
занимает, должна стараться не отставать от нее.
ЭВОЛЮЦИОННЫЙ потенциал ПОПУЛЯЦИЙ раэноrо типа
Основная идея, касающаяся роли структуры популяций в вос"
хождении на адаптивные вершины, была изложена Райтом в ero
классической работе (Wright, 1931; см. также 1949, 1960). Райт
.сформулировал эволюционные характерис1'ИКИ популяций разноrо
'Типа. Для нас здесь особый интерес представляют популяции трех
типов: 1) маленькие изолированные популяции; 2) большие непре-
рывные свободно скрещивающиеся ПОПУЛЯЦИ1И; 3) особое проме-
жуточное состояние, которое Райт назвал подразделенной попу-
ляцией. Третий тип это система небоwYJЬШИХ полуизолированных
популяций; мы rоворили о ней в друrих rлавах этой книrи, назы-
вая ее популя ией колониальноrо или OCTpoBHoro типа.
Рассмотрим ожидаемое поведение популяции каждоrо типа в
условиях среды трех модальных типов: А) стабильная среда;
Б) меД.ненно изменяющаяся среда; В) быстро изменяющаяся
среда.
1. Маленькая изолированная популяция. В такой популяции
большая часть полиморфных reHOB быстро закрепляется ПОД дей-
стВ'ием дрейфа или отбора и дрейфа. В результате запас rенети-
ческой изменчивости, который обеспечивал бы возможность при.
спосабливаться к тем или иным ситуациям в будущем, оказыва-
ется очень небо.пьшим (см. rл. 13).
А. В стабильной среде такая популяция быстро достиrает
наилучшей приспособленносТ1И, возможной при имеющемся
запасе изменчивости, после чеrо наступает эволюционный
застой.
Б и В. В медленно или быстро изменяющейся среде популя-
ция оказывается неспособной к адекватной эволюционной
реакции, так как этому препятствует недостаток изменчи-
BOcrn. Она может миrрировать в какое-либо убежище
или же будет обречена на вымирание.
2. Большая неnрерЫ8ная С80бодно скреЩU8ающаяся популя-
ция. Здесь отбор относительно слабо действует на рецессивные
аллели, встречающиеся с низкой частотой, в том числе на боль.
шинство новых мутаций, а сочетание сил отбора 1}{ дрейфа совер-
шенно неэффективно. Новые сочетания reHoB не сохраняются,
а затопляются в результате широкоrо свободноrо скрещивания.
Кроме Toro, развиваются reHHbIe системы, создающие .rомеостати"
ческ е буферные свойства, и эти rомеостатические системы устой..
чивы К изменениям.
'294... _
'"Часть V/. МЙlCроэволюция
A в. стабильной ср'еде такая. _ популяция сохраняе1.' существуIO
щие адаптаЦИil но 'Не изменяется (например, ошtссум).
Б. В медленно изменяющейся среде популяция развивается Meд
ленно и непрерывно (например, лошадь).
В. В быстро изменяющейся среде п'Опуляция, однако, не поспе-
вает за изменениями среды. По ряду причин, указанных в
чредыдущем р зделе, большая непрерывная популяция не
способна -успешно реаrировать на интенсивный отбор. Она
при этом распадается на мелкие реликтовые ПОПУЛЯЦИlи (Ha
пример, секвойя) или вымирает.
Таким образом, как в очень маленьких, так и в очень больших
популяциях условия неблаrоприятны для поддержания высоких
скоростей эволюции.
Рассмотрим теперь эволюционный ПОТе'нциал популяционной
системы колониальном типа. Она лишена недостатков, присущих
мелкой изолированной и большой непрерывной популяциям, а
кроме Toro, имеет некоторые достоинства.
3. Поnуляционяая система колоН,иальноzо типа. В такой систе
ме отдельные колонии MorYT быть мономорфными ИЛИ почти MOHO
морфными, но популяция В целом обладает децентрализованным
запасом rенет ческой изменч}{вости', распределенной между раз
ными колониями, Iи эта изменчивость может распространяться от
колонии к колонии путем возникающеrо время от времени меЖ
демовоrо обмена. rенами. Каждая колония способна быстро реа.
rировать на локальные условия среды блаrодаря взаимодействию
между имеющейся в данный момент изменчивостью, с одной сто-
роны, н отбором и дрейфом reHOB..... с друrой. Блаrоприятные но-
вые сочетания reHoB, так же как и отдельные аллели, MorYT за
крепляться в той или иной колонии блаrодаря защите. от широ-
Koro свободноrо скрещивания, которую создает ПОЛj'1изоляция.
A В стабильной среде популяционная система колониальноrо
типа дифференцируется на локальные расы под действием
:отбора или отбора и дрейфа reHoB.
Б. В медленно изменяющейся среде отбор, по всей вероятно
стн, будет блаrоприятствовать некоторым локальным pacaM
которые будут расширяться, тоrда как ДРУf1ие расы будут
сокращаться. Было постулировано, что при ЭТОМ происходит
некоторый междемовый отбор (см. rл. 11).
В. Быстро изменяющаяся среда вызывает такую же реакцию.
В ней MorYT также происходить и дрyrис реакции. Некото.
рые колонии, расш"ряясь, теснее сближаются друr с дpy
rOM, что приводит к более интенсивному обмену rенами
между ними, а возможно, и к межрасовой rибридизации с
последующим выщеплением новых типов. В некоторых по-
пуляциях, которым блаrоприятствует отбор, MorYT произой
тн быстрые эволюционные изменеН!Ня в результате COBMeCT
r А. 30. СТРУICтурр популяции в связи с мшсроэволюцие4
29:
Horo действия отбора и дрейфа reHoB (см. rл. 13) и воз-.
можности избежать оrраничения, налаrаемоrо на скорость
эволюции платой за отбор (см. rл. 14).
Таким образом, популяционная система колониальноrо типа
пред<;тавляет собой на'иболее блаrоприятную основу для быстрых
и резких эволюционных изменений (Wright, 1931, ]949, 1960).
Мноrие ЭВО.тIюционисты, основываясь на теоретических соображе-
ниях, полаrают, что великие исторические эволюционные взрывы,
ПрJИводившие к образованию новых крупных rрупп, например мле-
копитающих и покрытосеменных, происходили в популяциях имен-
но TaKoro типа.
Квантовая эволюция
Следующая важная концепция, относящаяся к популяционным
аспектам макроэволlOЦИИ, ---- это квантовая эволюция (Simpson,
1944; см. также 1953). Квантовая эволюция представляет собой
особую форму филет.ической эволюции, происходящей при лереходе
данной популяции с одной адаптивной вершины на друrую (новую
или остававшуюся прежде незанятой) адаптивную вершИ'Ну. При
таком переходе она должна пересечь определенную адаптивную
ДО.'Iину.
Симпсон предполаrает, что переход на новую адаптивную вер"
шину обычно совершают небольшие популяции, эволюционирую-
щие с необычаЙно высокими скоростями. Переходный период, в
течение KOToporo популяция занимает новую вершину, сравнитель-
НО короткий. На протяжении этоrо периода популяция бывает
представлена относительно небольшим числом особей по сравне-
нию с.-их числом в более нормальные периоды истории данной
JlИНИИ. Однако количество фенотипических и rенотипичеСIQИХ изме-
нений может быть в период перехода rораздо больше, чем в тече-
ние несравненно более длительных периодов обычной. филетиче-
ской эволюции. Соотношение между скоростью эволюции и вели-
чиной популяции схематически представлено на рис. 30.1.
История каКОЙ...1ибо филетической rруппы часто состоит из
длительных периодов, в течение которых изменения происходят
с rоротелическими или брадителическими скоростями и которые
прерываются редкиМ'и короткими периодами тахителических СКО-.
ростей и кваятовых эволюционных изменений (рис. 30.1). Термин
<К8антовая эволюция» содержит в себе указание на происходя-'
щие время от времени выбросы крупномасштабной эволюцщ)нной
активности (Simpson, 1944). .
Существует несколько причин, на основании которых следует
постулировать, что квантовой эволюции подвержены небольшие
популяции. Популяция, пересекающая адаптивную долину на пути'
к но ой .вершине, по определению плохо приспособлена и под..!
296
Часть V 1. Мак.роэволюция
верrается стрессовым воздействиям а достиrнув новой адап1'ИВНОй.
вершины, она подверrается новым сильным давлениям отбора
(Simpson, 1944, 1953). Кроме roro, как отмечал ось в предыдущем
и в друrих разделах, быстрая приспособительная эволюция леrче
вызывается сочетанием отбора и дрейфа reHoB, чем одним ТОЛЬКD
отбором, а для cOBMecTHoro действия отбора и дрейфа 'необходи..
мо, чтобы п.. >пуляция Iимела небольшне размеры.
KBQHтOfJOII
.160llЮЦUII
i
Odd'"HQR qJUлетUllескйJI
.180llЮЦUR
1
/; n
1
8peMII
Рис. 30.1. Сравнение скорости эволюционных изменеиий: (1) н величины попупя-..
ции (/1) при квантовой и обычной фИJlетическоА ЭВОJlЮЦНН (.5impson, (953).
Подобноrо же рода причины позволяют предполаrать, что KBaH
товая эволюция происходит с высокой скоростью. Популяция
должна пересечь адаптивную долину и занять новую адаптивнуJOt
вершину за очень короткое время или не делать 3Toro B08ce .
В таких условиях rоротелическ"е скорости недостаточНы.
Продуктом квантовой эволюции может быть новая rруппа op
rа'низМОВ любоrо таксономическоrо paHra от вида до рода и дс>
более высоких катеrорий. Квантовая эволюция не обязатеЛЬН()I
должна участвовать в видообразовании, но в некоторых случаях:
ее участие возможно (см. rл. 22). Зато происхождение новых
крупных rрупп, таких, как семейства и отряды, вероя'rНО. связа--
но обычно с квантовой эволюцией (Simpson, 1944, 1953)\
Палеонтолоrическая история мноrих семейств и отрядов' жи-
вотных представляет собой длинную запечатленную в палеонто"
лоrической летописи последовательность, на протяжении которок
в эволюции преобладали rоротелические или брадителические CKO
рости; этой истории предшествует rораздо более короткий период
времени, о котором не сохранил ось никаких данных и во время'
KOToporo, по-видимому, возникла данная rруппа (см. табл. 31.2).
Палеонтолоrическая летопись свидетельствует о том, что в пер'ИО
ды обычной филетической эволюции популяции были большими,.
а скорости эволюции ---- rоротеличеСКИМ1И или брадителическими.
И напротив, отсутствие или редкость ископаемых данных, О'NIО-
сящихся к периоду возникновения и OCHOBHoro развития той Iили
иноi\ rpYnnbl, соответствует допущению о малых размерах попу
r л. 30. Структура популяции в связи с МйfCрОЭ80люцией
297
Jlяции при квантовой эволюции. Постулированные особенности
квантовой эволюци'и позволяют объяснить крайнюю редкость так
называемых недостающих звеньев в палеонтолоrической летописи.
По самому характеру квантовой эволюции следует ожидать.
что прямые палеонтолоrические данные относительно этоrо про-
цесса должны быть редкИМИ. Тем не менее в нескольких блаrо
приятных случаях сохранились палеонтоло ические данные, сви-
детельствующие о квантовых сдвиrах. Один из них ---- переход от
оСтопохождения к пружинящей механике конечности в семействе
Equidae, а друrой ---- переход от питания листьями деревьев к пи-
таН'ию травой у Toro же семейства (см. r.П. 26). Симпсон (Sim-
pson, 1953) приводит также примеры, относящиеся к двум друrнм
отрядам млекопитающих. Один из них касается происхождения
парнокопытных (Schaeffer, 1948), а друrой ---- происхождения
подсемейства Stylinodontinae. входящеrо в сходный с неполнозу-
быми раннетретичный отряд Taeniodonta (Patterson, 1949).
Филоrении, которые строят на основании сравН'ительноrо изу-
чения ныне живущих представителей данной rруппы, также дают
в некоторых случаях правдоподобные косвенные сведения относи-
тельно имевшей место квантовой эволюции. В своих прежних ра-
ботах (Grant, 1959, 1963) я интерпретировал происхождение двух
триб растений YMepeHHoro пояса, относящихся к семейству Po.le-
moniaceae, как примеры квантовоэволюционных сдвиrов.
rенетические революции
Следующая (в порядке публикации) важная концепция ---- это
концепция Майра (Mayr, 1954; см. также 1963, rл. 17) о rенети"
ческой перестройке, происходящей в изолированных популяциях.
Майр противопоставляет внутреннюю rенетическую среду в боль..
ших широко распространенных ПОПУ.1JЯЦИЯХ какоrо-либо вида
среде в неБОw1ЬШИХ изолятах (обычно ---- изолированных перифе..
рических популяциях) Toro же вида.
В большой непрерывной популяции в локальную субпопуляцию
все время поступает поток reHoB из соседних, а нноrда и из отда-
JleHHblX субпопуляций. Более или менее часта.я иммиrрация чужих
reHOB в данную локальную популяцию создает давления отбора,
блаrоприятствующие reHaM с хорошей комбинационнОЙ пособ.
ностью. В результате в состав ее rенофонда входят rруппы reHoB,
дающие начало нормальным жизнеспособным продуктам в весьма
разнообразных rетерозиrотных reH'HbIx сочетаниях. Фонд «хорошо
смешивающихся» reHoB, создающийся в такой rенетической среде,
обладает также, в качестве eCTecTBeHHoro следствия, rомеостати-
ческими буферными свойствами.
Это придает известный консерватизм хараК1'еру ,изценчивости
в большой непрерывной популяции. Такая популяция проявляет
298
.Чвс.ть У/. Макроэволюцuя
клинальную изменчивость и rеоrрафическую расовую изменчи-
вость (см. rл. 18). Но откло'Няющиеся или новые формы даииоrо
вида Не MorYT сохраняться 'и не сохраняются в качестве самостоя-
тельных расовых еДИНИll.
Посмотрим теперь, что происходит, коrда нескольким особям
эмиrрировавшим из большой свободно скрещивающейся популя-
ции, удается основать небольшую дочернюю колонию на перифе
рии ареала ви'да. Прежде Bcero такая популяция основателей
будет содержать лишь небольшую неслучайную выборку из reHO-
фонда родительской популяции. Ho кроме Toro, она будет изоли
рована от потока reHoB извне. Это приводит к изменению rенети-
ческой среды. Селективные ценности reHoB в этих 'изолированных
дочерних rенофондах изменены; отбор уже Не блаrоприятствует
«хорошо смешивающимся» reHaM; и, напротив, reHbl, дающие в ro
мозиrотном состоянии жизнеспособные типы, приобретают теперь
полож'ительное селективное значение (Nlayr, 1954).
Таким образом, переход от большой свободпо скрещиваю..
щейся популяции к небольшой изолированной дочерней колонии
уже сам по себе изменяет rенетическую среду и изменяет или
даж.е П9ЛН.ОСТЬЮ ревеРТlIрует селектив'ные ценности reHoB во MHO
rих локусах. Изменение внутренней rенетической среды воздейст"
вует одновременно на мноrие reHHble локусы, подобно изменению.
вызываемому rибридизацией или полиплоидией, и оно может по-
служить началом «rенетической революции» (Mayr, 1954).
Популяu я может оказаться неспособной ПрОЙiИ через узкое
rорлышко» от высокоЙ к низко и численности. Она может oQка-
заться не в состоянии выдержать rенетическую рево.l1ЮЦИЮ; в та-
КОМ случае она будет обречена на быстрое вымирание. Но при
большом числе независимых попыток основать новые колонии есть
шансы, что одной или нескольким популяциям основателей
удастся выжить. В таких случаях выживание зависит от Toro.
унаследовала ли популяция :основателей такой rенофонд, который
может выдержать rомозиrотность по мноrим локусам и который
преадаптирован к какой либо из имеюIЦИХСЯ новых эколоrических
З0Н. '
rенетическая революция в удачливой дочерней колонии при..
ВОДИТ к более или менее коренному изменению эколоrических
предпочтениЙ и морфолоrических признаков. Возникают новые
типы, приспособленные к новым эколоrическим зонам, которые
находятся З8 пределами зоны, занимаемой консервативной пред-
ХОВОЙ попу.пяцией. Таксономические даоиные подтверждают это
ожидание. В изолированных периферических популяциях некото"
рых видов и rрупп видов птиц, насекомых и однолетних растений
действительно наблюдаются новые:варианты (см. r.'1. 18,21 и 22)_
Короче rоворя, изолированные периферические популяции, воз
ннкающяе от нескольких основателей, отделившихея от большоii
r л. 30. Структура пйпуля-цuи 8 свяэu с МЙlCроэволюцuей
299
свободно скрещивающейся предковой популяции, Moryr быть те-
ми точками, rде происходит быстрая эволюция и появляются НО-
"ые формы (Mayr, 1954).
Диверrирующая ПОПуляция не обязательно навсеrда остается
аленькой, изолированной и пер'иферичеСI<ОЙ. Она может увели-
иваться в размерах, расширять занимаемую ею территорию) а в
онечном счете даже вновь вернуться на территорию предковоrо
вида. Со временем она также может стать консервативной по ха-
рактеру своей изменчивости. Но после этоrо она может дать На-
чало новому поколению золированных дочерних колоний, кото-
рые снова отрываются от предковой формы (Mayr, 1954).
Майр создал свою концепцию rенетических революций в зна-
ЧИтельной мере независимо от взrлядов Райта на подразд ен-
ную популяцию и от представлений Симлсона о квантовой эво-
люции. Тем не менее эти три концепц'ии хорошо соrласуются
между собой в отношении ожндаемых конечных результатов для
небольших популяций. Ниже рассматриваются расхождения меж-
ду ними, касающиеся реrулирующих факторов.
Майр отбросил дрейф reHoB в качестве фактора, участвующеrо
в диверrенции маленьких дочерних колоний. Вместо этоrо ОН при.
писывает начальное изменение в rенофО'нде дочерних колон'ий
тому, что он называет эффектом основателя, т. е. случайностям
выборки, возникающим в тех случаях, коrда несколь о особей,
миrрирующих из большой предковой колонии, основывают новую
дочернюю колонию (Mayr, 1954; 1963). Друrие авторы считают,
однако, что эффект основателя представляет собой, в сущности,
особый случай дрейфа reHOB (Dobzhansky, Spassky, 1962; Grant,
1963). Более Toro, в действител ьности дело здесь не в одной лишь
эффективности или неэффективности дрейфа; возможно, что про-
цесс основания новой популяции. контролируется силами, созда-
ваемыми совместным действием дрейфа и отбора.
За последние rодЫ Карсон (Carson, 1959, 1971, 1975; Carson,
Heed, 1964) ОПУ'БЛ1иковал ряд статей, в которых разрабатывается
концепция rенетических революций в периферических Ifзолятах На
основаНии изучения популяций дрозофилы. Он полаrает, что ди-
BepreHTHble изолированные периферические популяции возникают
во мноrих случаях от отдельных особей-основателей. Он вводит
также ряд новых терминов для обозначения различных аспектов
первоначальной rипотезы Майра (например, rомоселекция, циклы
расцветов и крахов популяции) (Mayr, 1954).
Квантовое видообразование
Квантовое видообразование было описано в качестве одноrо
Нз способов видообразования в rл. 22, к I<ОТОрОЙ мы И оТсылаем
читат ля. Здесь оно интересует нас в связи с ero посТулируемой
.30
Часть У/. Ма"роэвОЛЮЦUJl
ролью в макроэволюции. Высказывалось мнение, что некоторые
направления эволюции представляют собой ряд последовательных
событий KBaHToBoro видообразования (Grant, 1963). Поэтому мы
вновь коснемся здесь проблемы KBa'HToBoro видообразования в
качестве введения в рассмотрение подобных направле ий.
Напомним, что квантовое видообразование состоит в том, что
от большой ПОЛИМОРфНОЙ свободно скрещнвающейся предковой
популяции отпочковывается дочерний вид, проходящий через про
межуточиую стадию небольшой локальной расы. Локальная раса
основывается одной или несколькими особями из популяции пред
KOBoro вида. она пространственно изолирована или полуизолиро
вана от предковоrо вида; она обычно находится на периферии
области, занимаемой старым в-идом, .Н может встречаться в какой-
нибудь новой эколоrической зоНе.
В начале существования локальной расы 'на ее rенофонд OKa
зывают влияние дрейф reHoB и инбридинr, а позднее сочетание
отбора с дрейфом инередко rенетические революции. Э1'И факто-
ры MorYT ВЫзвать быстрые, а иноrда и коренные изменения в эко-
лоrических предпочтениях, морфолоrических признаках и взаимо-
оТ'ношен'иях, определяющих фертильность. Поэтому .'lOкальная
раса может быстро диверrировать до видовоrо уровня; более Toro
возникший таким образом дочерний вид может занять новую адап
тивную зону, лежащую за пределами ЭКО.'10rическоrо ареала пред
KOBoro вида.
Концепция KBaHTOBorO видообразования в Зllачительной мере
синте1'ическая концепция. Она представляет собой синтез избран
ных частей существовавших ра'нее концепций Райта (Wright
1931), Симпсона (Simpson, 1944) и Майра (Mayr, 1954), которые
были изложены выше, а также концепций, вытекающих из работ
Льюиса по эволюции растений (Lewis, Raven, 1958; Lewis, 1962)_
Однако ни одна из этих прежних концепций в отдельности не со-
ответствует в точности описанию KBaHToBoro видообразования.
Концепция отбора ---- дрейфа берется у Райта (но не у друrих aB
торов), квантовые сдвиrи (но Не видообра:ювание) ---- у Симпсона
а быстрое видообразование (но не отбор ---- дрейф) у Майра и
Льюиса; затем все эти составные части следует собрать вместе_
Синтетическая концепция KBaHToBoro видообразования была пред
ложена [рантом (1963; см. также 1971), по мнению KOToporo про
цесс этот иrрает важную роль как в видообразовании, так и в
м акроэволюци'и.
Филетнческие направления и видообраэовательные
направления
Рассмотрим теперь более подробно природу эволюционных Ha
правлений. В более ранних rлавах этой книrи мы описывали фи
летическую эволюцию в пределах отдельных линий и эволюцион"
.........
r л. 30. Структура популяции 8 связи с itакроэволюцией
зоf
.ную диверrенцию между ЛIfНИЯМИ как самостоятеЛЬНые типы эво
;ЛЮЦИИ. ЭТО есть некое упрощение Toro, что происходит на самом
Iделе, поскольку два указанных типа передко переплетаются друr'
,с друrом. Затем, рассматривая направления эволюции (rл. 26),.
мы пре ставили направления КаК явления, отgосящиеся к филе...
тическои эволюции, а это опять-таки, вероятно, ЛИШЬ частично OT .
ражает истинное положеНие вещей.
Типоло uческuе 8а96/
Е,рlапifrопs L Е, теridiопаli$
/.
CJ
с)
(fJиllетuческuе
8ug61
Е. тeridioпa/i$
Е.рlапifrОП$
4 "'У2 5 5Y:z б
l1амеЛЛЯРН61и LlHgeKc
Рис. 30.2. Филетические направления в пииии ископаемых европейских слонов
(Elephas planif,ons E. тeridionalis).
Ламел.:JЯРНЫЙ НliдeKC С.'УЖИТ мерой ко.,ичсства эмали иа кореиНЫХ зубах. фипетнческие
ВИДЫ COOTBCTCTBYIOТ пос.lедовате.1ЬНЫМ видам, а Тlfпо.,оrические виды таксономнчсскиr.l
вида м.
Для TOrO чтобы обсуждение этоrо вопроса стало яснее, следует
различать два широких типа иапраВ,,1ений эволюции: 1) филетиче.
ские направления и 2) видообразовательные направления (specia.
tional trends).
Филетическое направление представляет собой постепенное
проrрессивное эволюционное }lзменен.ие в пределах данной филети-
ческой линии. Это явление, присущее 'исключительно филетиче-
скоЙ эволюции. Отдельные стадии при этом бывают представлены
последовате.t1ЬНЫМИ видами, а Не современными Apyr друrу 6ио.
лоrическими видами (см. rл. 18). Хорошими. примерами служат
разные рОДЫ лошадей, как Merychippus в миоцене и Pliohippus
Equиs в плио плейстоцене (rл. 26). Друrим примером служит на-
правление эво.'1ЮЦИИ строения коренных зубов в ,,'Iивии ныне вы-
мерших П.,'lейстоценовых европейских слонов ---- от Elephas plaпi-
froпs дО Е. merid'ionalis (р.ис. 30.2).
Видообрззовательное направление представляет собой сту.-
пенчатое продвижение вперед, возникающее в резу.r:lьтате последо.
вателыюrо ряда изменений типа видообразования. Каждый новый
вид ряда продвиrается дальше в данном направлении эволюции.
Это направление ---- линия P----Q на рис. 30.3. Отдельные стадии
этоrо направления представлены 6иолоrическими, а не последова.
302
Часть У/. МаlCроэволюцuя
тельными видами (как это определено в rл. 18). Как видно ИЗ
рис. 30.3, преобладающим типом эволюции является видообразо-
вание, а Не филетическая эволюu.ия. Видообразовательное на-
правление суммарный результат последовательных видообразо.
Ваний.
При филетическом направлении эволюции На каждом данном
временном уровне имеется только один биолоrический вид. При
I
:t:
10)
::)
:t:
с)
::s
:::r
i
C'I)
р
Время
Рис. 30.3. Направлеиие видообразовательной эволюции (Grant, 1963).
Направление (.IIиння Р Q) представляет собоR резупьтирующнА ВЕ'ктор поспедоваТeJlЬВЫХ
вндообразоватепьных нзменений в данном напраВЛСJJIIИ.
видообразовательном направлени'и консервативная предковая
форма и происходящая от Нее более проrрессивная форма вре-
менно сосуществуют как два современных друr друrу б lIОJIоrиче-
ских вида. Линия P Q иа рис. 30.3, хотя она и состоит из после.
довательности видов в буквальном смысле слова, не содер*ит
последовательных видов; поэтому в соответствии с принятыми
здесь критериями линия P Q отражает не филетическое, а видо-
образователь'ное направление.
В видообраэовательных направлениях довольно значительные
изменения MorYT произойти и, вероятно, происходят за более ежа-
1'ые сроки, чем в филетических направлениях. Поэтому видообра-
зовате,,']ьные направления способны развиваться быстрее, чем фи-
.летичеСкие направления.
Видообразовательные 'направления, возможно, весьма обычны.
ТрУДllО сказать, однако, сколь велика их частота, поскольку идеН-
тифицировать такие направления в тех случаях, коrда палеонто-
.лоrическая летопись, как это обычно бывает, неполная, доврль-
но сложно. Видообразовательное направление леrКQ принять за
филетическое, и Hao60por.
r л. 30. Структура nОnУJ1Яции ." СВЯЗ(l С .макроэволюцией
.ЗОЗ-
Ископаемые остатки по той или иной rруппе, поскольку они
бывают собраны из различных отложений, обычно несколько раз-
бросаны по отношению к подлинному филоrенетическому древу.
В публикуемом на основании таких данных филоrене'I'ическом
древе, т. е. в филоrенетической rипотезе, имеются пробелы и неяс-
ности. В сущности на основании одних и тех же непо..1НЫХ иско-
ЭОНQ tJlJlсшеео
/// OpRg .. ,/'"
С = Аgаптuвная
зона
РИс. 30.4. Ahnenreihe (предковый ряд) и Stufenreihe (ступенчатый ряд) в про.
цессе занятия ряда адаптивных зон (Simpson, 1953).
паемых данных можно построить разные филоrении в зависимости
от Toro, как будут соединены боковые линии в тех местах, rде в
летописи имеются пробелы.
Абель (АЬе.l, 1929) и Симпсон (Simpson, 1953) ПрОВQДЯТ раз-
личие между Ahnenreihen (предковыми рядами) и Stufenreihen
(ступенчатыми рядами); такое различие помоrает решить про-
блему интерпретации данных. Эти два способа эволюции схема-
тически изображены на рис. 30.4, rде последовательность a e
соответствует Ahnenreihe, а A E Stufenreihe. Палеонтолоrиче-
ские данные по какой-нибудь rруппе передко находятся в точках
А, В, С, D и Е, TorAa как ТОЧКИ О, Ь, с, d и е неизвестны. В таких
случаях первым побуждением будет соединить точки А, В, С, D
и Е в фи..10rенетический ряд. Но это, очевидно, будет неверным,
и еСЛИ ясно представлять себе все те западни, в которые можно
попасть при интерпретации данных, то мы будем rOToBbl при-
знать А, В, С, D и Е за боковые ветви и сделать вывод о том,
что эволюция данной rруппы происходила по линии a e.
Equidae это та rруппа, ддя которой известНЫ и Stufenreihe и
Ahnenreihe. Найденный в Старом Свете палеонтолоrический ряд,
С..1аrающийся нз Hyracotheriит, Anchitherium, Hypohippus, Hip-
parion и Equиs, сначала счита..т.и филоrенетической последова-
тельностью, но позднее обнаРУЖIIЛОСЬ, что это ряд боковых вет-
4
. Часть У/. Ма5роS80ЛЮЦия
.вей, -Т. е.. Stufenreihe. ИСТИННЫЙ филоrенетический рЯД, Ahnenreihe,
найден .в Северной Америке, rде и происходила в основном эво-
.люция лошади. Североамериканский ряд Hyracotherium, Miohip-
pus, Merychippиs, Pliohippus и Equus образует Ahnenreihe этой
.rруппы (Simpson, 1953).
Совершенно очевидно, что различие между Ahnenreihen и
Stufenreihen, сходно, но вместе с тем и отличается от различия
между фИ-Т1ети.ческими и видообразовательными направлениями.
Ahnenreihe может быть синонимом как филетическоrо, так и ви-
дообраэоватеЛЫlOrо направления в зависимости от Toro, какое из
них имеет место в данном случае. Но Stufenreihe никоrда
Не может быть синонимом видообразовательноrо направления.
При видообразовательном направлении популяции образуют пря-
мой предковый ряд (подобный линии P----Q на рис. 30.3). В Stu
fenreihe (например, А, В, ..., Е на рис. 30.4) такой прямой фи-
лоrенетической связи между стадиями Не существует.
Разделение на Ahnenreihe и Stufenreihe, проведенное Абелем
(АЬе1, 1929) и Симпсоном (Simpson, 1953), предвосхитило и об-
леrчило разrраничение филетических и видообразовательных на-
правлений. Обсуждая этот вопрос, Симпсон (Simpson, 1953)
близко подходит к концепции видообразовательных направлений.
Впервые же это направление в качестве особоrо эволюциоиноrо
направления было четко выделено [рантом (Grant, 1963, 1971).
я не пользовался термином «видообразовательное направление»
в своих предшествующих рассуждениях, с тем чтобы Не вводить
лишних терминов, однако теперь этот термин, ПО ВИДИМОМУ, не-
обходим и поэтому будет здесь применяться.
Взrляды на филоrению, сходные с излаrавшимися Выше, вы-
сказывали недавно Элдредж и rоулд (Eldredge, Gould, 1972), а
также Стенли (Stanley, 1975). .
Элдредж и fоулд (1972) различают «филетический rрадуа
лизм» И «прерывистые равновесия» (punctuated equiIibria). Этими
терминами они обозначают примерно те же способы эволюции,
«оторые я соответственно называю филетическим направлением
и видоооразовательным направлением. Элдредж и rоулд в своей
CTaTLe не Сt:ылаютсs на более ранние работы, проводившиеся в
TOM же плане Симпсоном, [рантом и друrими. Элдредж и rоулд
(Eldredge, Gould, 1972) предлаrают заново интерпретировать
Все или почти все палеонтолоrические ряды как «прерывистые
равновесия» и в соответствующей степени пон'изить pO.Т'lь «фИJlети-
ческоrо rрадуализма». С моей точки зрения, это было бы слиш
ком; в моем представлении как видообра30вательные, так и филе-
iиqеские направления занимают определенное место в природе.
Я здесь коснусь двух вероятных примеров видообразовател
ных направлений у растений. В обоих случаях вывод о существо-
ВJiнии TaKoro направления делается :на основании сравнительноrо
r л. 30. Стру"тура популяции в связи с AtЙ1СрОЭ80люцией 30'
изучения биолоrии coBpeMeHHых форм. Этот ПОДХОД имеет свои
недостатки, но обладает также 'и некоторыми достоинствами, по-
скольку при эт.ом можно изучать виды в живом состоянии и опре-
делять их взаимоотношения.
Вероятное видообразовательное направление в эволюции у
Clarkia (Onagraceae) на тихоокеанском побережье Северной Аме-
рики идет от форм, приспособленных к климату с умеренной
влажностью, до ксерофитов. Предковый вид, обитавший в уме-
ренно влажных местах, был близок к современным видам
С. атоеnа или С. rubicunda. Промежуточная стадия в этом ряду
представлена ныне живущим видом С. ьиоЬа, а конечная Ксеро-
фитная стадия видом С. liпgulata. Переход от С. ьиоЬа к
С. ligulata произошел в результате KBaHToBoro видообразования
(Lewis, Lewis, 1955; Lewis, Roberts, 1956).
Вероятное направление эволюции у Polemonium (Polemonia-
сеае) в западной части Северной Америки идет от форм, присло.
сuбленных к мяrкому климату прибрежных областей YMepeHHoro
пояса, до алЬпийских форм. Отдельные стадии этоrо направле-
ния представлены следующими видами:
Polemoпiuт сатеит
Р. caeruleuт
rруппа Р. сйЩоrпiсцт Р. delicatuт
Р. pиlcherriтит
rруппа Р. ex{тiuт
Влажные при6режные низменности
Зона хвойных лесо'В
Субальпийская лесная зона
Альпийская зона
Высокие вершины в а.'JьпиfJской зоне
Этот ряд видов встречается в указанных зонах по трансектаи
от побережья до высоких rop В западной части Северной Амери-
ки. При этом названные виды Не образуют по таким трансектам
непрерывный ряд, как это Mor ло бы представляться теоретически,.
а обособлены друr от друrа, составляя ступенtlатую последова-
тельность, что позволяет предполаrать видообразовательное на-
правление (Grant, 1959, 1963).
Быстрая ЗВОJlЮЦИЯ В видообраэоватеJlЬНЫХ
направлениях
Скорость эволюции в видообразовате,,1ЬНЫХ направлениях, Воз-
можно, коррелирована с тем способом вндообразова,ния, с которым!
они связаны. Можно ож.идать, что ПОС.'1едовательности reo.-I
rрафическ'их видообразований в популяциях, постоянно сохраняю--l
щих большие размеры, будут происходить с умеренJ.lОЙ иЛИ НОР1
мальной, но не с тахителической скоростью. Успешное же кван-
товое видообразова'ние теоретически Mor .110 бы возбуждать высо-
Кие скорости у соответствующеrо эволюционноrо направления.
306
Часть У/. Ма1Сроэволюция
Видообразовательные направления этоrо последнеrо типа пред
ставляют особый интерес.
l\1ы видели прежде, что единичные случаи KBaHToBoro видооб
разования способны дать ,новые формы, быстро приспосабливаю
щиеся к новым или различным эколоrическим условиям. Кванто-
вое видообразова'ние обладает этой способностью, потому что:
1) ОНО использует сочетание сил отбора и дрейфа reHoB (см. выше
и rл. 13); 2) оно может избежать оrраничения, налаrаемоrо пла-
той за отбор при быстром замещении множества reHoB (см.
rл. 14).
Но если особые преимущества KBaHToBoro видообразования мо-
rYT быть использованы однажды, на одном временн6м уровне, ТО
их можно 'использовать снова и снова. Сдвиrи, происходящие при
квантовом видообразовании, MorYT циклически повторяться. И эти
сдвиrи MorYT иметь некое общее направление, подобно линии
P Q На рис. 30.3. Квантовое видообразователь'ное направление
подобноrо рода представляет собой способ Qбеспечения длитель-
ной быстрой эволюции в данном направлении. Быть может, это
единственный способ создания эволюционноrо направления, обла-
дающеrо тахителической скоростью.
Вернемся к метафоре адаптиВ'ноrо ландшафта. Представим се-
бе незанятую высокую адаптивную вершину и находящуюся у ее
подножия популяцию. От подножия к вершине ведет несколько
различных троп. Некоторые из них полоrие, и восхождение по этцм.
тропам происходит долrо и медленно. Они символизируют филе
тические направления и 'направления, связанные с rеоrрафическим
видообразованием. Такие тропы приrодны для восхождени,Я при
наличии достаточноrо времени.
Друrие тропы крутые, но зато короткие и потенциально обе-
спечивают быстрое восхождение. Такие тропы соответствуют на-
правлениям, связанным с квантовым видообразованием. Если по
пуляция, 'Находящаяся у подножия ropbl, должна взобраться на
вершину за КОрОТКое время, т. е. если время восхождения оrрани-
чено, то популяции придется выбрать короткую крутую тропу.
Заключение
В этой rлаве дан обзор ряда теоретических концепций, разви-
вавшихся постепенно, шаr за шаrом, на протяжении послеДН1ИХ че-
тырех или пяти десятилетий. Эти концепции позволяют объяснить
некоторые важные, но малопонятные макроэволюциО'нные явления.
Они довольно удовлетворительно соrласуются с имеющ'имися фак"
ТИЧескиМИ данными. Нельзя сказать, однако, чтобы этих данных
было слиШком MHoro; необходимо собрать rораздо больше мате..
риала относительно тех макроэволюциоН'ных процессов, которые
обсуждались в этой rлаве. ,.
rпa'Ba 31
ЭВОЛЮЦИЯ КРУПНЫХ .rРУ,ПП
Введение
Одна из важных проблем макроэволюции касается эволюции
крупных rрупп. При каких обстоятельствах в<>зникают триБЫ t се-
мейства t ОТРЯДЫ t классы? Какие эволюционные факторы участ"
вуют в происхождении и развитии rрупп среднеrо и высшеrо так-
соном'ическоrо paHra?
В этой общей проблеме содержится ряд частных проблем: Не-
обходимы ли для возникновения круп'Ных rрупп какие-то специ-
альные новые вариации? Связано ли первое появление крупных
rрупп со скачком или же с постепенной Диверrенцией от предко-
вой линии? Участвуют ли в этом какие то особые формы эволю-
ции?
Получить прямые данные по этим проблемам трудно. А отсут-
ствие убедительных фактов способствовало расцвету самых разно-
образных спекуляций. Тем не менее мы располаrаем некоторыми
косвенными данными, которые MorYT служить нитью для наших
рассуждений.
В эволюции любой крупной rруппы полезно раЗЛ1ичать две
rлавные стадии: раннюю формообразовательную стадию и после:-
дующий период расширения и распространения. Полезно такое
деление по той причине, что на этих двух стадиях, возможно, дей-
ствуют разные эволюционные процессы.
Эволюционные характеристики крупных rрупп
Прежде чем обсуждать происхождение крупных rрупп, необ-
ходимо определить основные черты, характерные для крупной.
rруппы. Крупную rруппу можно определить как филоrенетиче-
скн естественную rруппу довольно BbIcoKoro таксономическоrо
pa'Hra. Такое определение, однако, привлекает внимание Не к тому
аспекту проблемы, который нас здесь интересует. Возведение в Т0Т
или 'иной таксономический paHr производится ретроспективно,
после Toro как rруппа эволюционировала вплоть до настоящеrо
времени до эры. людей, занимающихся систематикой. Нас же
интересует данная rруппа не только на стадии достиrнутоrо ею
pacUBeTa t но и IHa самых ранних стадиях ее появл.ения.
Можно было бы также определять крупную rруппу по ее клю-
чевому пр'изнаку (или признакам). Такой подход опять-таки не.-
ЗО8
Часть VI. Макроэволюl.lUЯ
плодотворен и по той же причине. Данная крупная rруппа в мо-
мент cBoero возникновения может иметь, а может и. не иметь клю-
чевых признаков.
Более полезнbtм критерием для определения крупной rруппы
служит состояние приспособленности к той ил'и иной обширной:
ЗоНе на адаптивном ландшафте. Появление rруппы совпадает с
Занятием ею новой адаптивной зоны (Wright, 1949; Simpson,
1953). Крупной rруппой она становится позднее, в результате раз-
вития и пролиферации внутри этой зоны.
Существует два rлавных способа успешноrо пеРВОllачальноrо.
занятия адаптивной зоны.
1. Колонизируюший вид переселяется на более или менее He
обитаемую территорию. В момент колонизации эта Iновая террито-.
рия не обладает какими либо особенно новым,и условиями среды,
если не считать ее относительной незанятости. В свою очередь.
и колониз'ирующий вид не обязательно должен сильно отличаться
от своей предковой линии. Однако в новой зоне он находит эколо-
rические условия, способствующие увеличению ero численности
и возни новению разнообраз'ия.
2. Время от времени какой нибудь вид может приобрести ту
или иную черту, создающую ему возможность для HOBoro образа
жизни и преадаптирующую ero к ме.стообитанию, которое ОН
прежде не 'использовал. Так, в эволюции позвоночных развитие
J1еrких и конечностей создало возможность для заселеНИя суши,
а развитие крыльев позволило освоить воздушную среду. Преадап
тированный таким образом колонизирующий вид, используя свой
новый признак в качестве точки опоры, заселяет новое месroоби
таНие, в котором происхuдит ero дальнейшая эволюция (Huxley,.
1942; Wright, 1949).
В некоторых случаях такой новый стержневой признак возни..
кает раньше времени для тех условий среды, в которых обитает
предковая популяция, но оказывается потенциально ценным 8
местообитании HOBoro типа. Так леrк'ие, приспособленные для ды-
хания в воде, и парные плавники некоторых рыб были предшест
венниками леrких и конечностей для ползания первых наземных
позвоночных. .
Как при первом, так и при втором способе вслед за заселен'ием
новой адаптивной зоны обычно происходит адаптивная радиация
в этой зоне. Вид, первоначально заселивший !Новую зону, обычно
дает начало разнообразным линиям, занимающим различные суб-
зоны. Именно в результате адап1'ИВНОЙ радиации такая rруппа
становится распознаваемой крупной rруппой. Следует, однако,
имеТь в виду, что адаптивная радиация ---- это лишь успешное ис-
пользование на более поздней стадии той эколоrической возмож-
ности, которой воспользовался вид, первоначально заселивший HO
вую адаптивную зону.
r л. 31. Эволюция ICрчпн.rхх 2рцпп
309
Первый из описанных выше способов проще BToporo; И, напро
тив, на счету BToporo способа, вероятно, больше важных эволю
lI.ИОlfНЫХ событий, чем у первоrо. Первый способ приводит к воз
Jiикновению эндемичных rрупи среднеrо таксоном'ичеСкоrо paHra.
Хорошими примерами служат дарвиновЫ вьюрки (семейство Frin.
gillidae, подсемейство Geospizinae) на rалапаrосских островах ИI
rавайские цветочницы (семейство Drepanididae) на rавайских OCT
ровах. Вопрос о том, MorYT ли высшие катеrори'и, распространен
Jible по целому материку или по всему земному шару, возникать-
первым способом, остается открытым. Второй способ, по-видимо-
му, служит rлавяым источником настоящих крупных и широко-
распространенных rрупп, таких, как амфибии и птицы.
Скачкообразное или постепенное развитие?
Некоторые эволюционисты (например, Gol dschmidt, 1940) BЫ
сказыв ли мнение, что новые rруппы возникают в результате-
скачков, т. е. резких и внезапных или почти внезапных мутацион-
}{ых изменений у орrанизмов, существовавш'их прежде. Од'накQo
большинство эволюционистов, начиная с Дарвина, возражали про-
тив представлений о чисто скачкообразной эволюции, предпочи
тая rрадуализм в той И.'1'и иной ero форме.
Скачкообразная эволюция в своих крайних проявлениях име ..
ет примерно такие же шансы оказаться успешной, как и спонтан-
Ное возникновение жизни. Все rенетические данные склоняют К'
выводу, что rенотип состо'ит из мноrочисленных reHoB, которые
должны rармонически взаимодействовать друr с друrом, для Tor(),
чтобы орrанизм обладал нормальной жизнеспособностью и плодо-
витостью. Резкие мутации приводят к селективно неблаrоприят-
ным сдвиrам BHYTpeHHero равновесия reHoB. rенотип. который соз-
давался отбором постепенно, невозможно удачно переделать за
один раз; ero надо переделывать также постепенно.
Все эти соображения не исключают возможности перестройки-
rенотипа за относительно небольшое число поколений. Более Toro,
И 1еются экспериментальные данные о быстрой перестройке reHo
типа у растений (см. Grant, 1971, rJl. 13 и 14). Такие быстрые из-
менения некоторые авторы называют сальтаI..J:ИЯМИ. В подобных
Y fepeHHblX проявлениях сальтация может иrрать реальную роль
в эволюции.
Крайние формы rрадуализма в качестве объяснения крупных
ЭВОЛЮЦИОН'НЫХ изменений имеют друrие недостатки. rрадуализм
слишком медлоителен. rрадуализм в ctpor-ом смысле слова несов-
местим с некоторыми палеонтолоrическими данными (см. rл. 28).
Период возникновения той или иной крупной rруппы обычно co
стаВJ1яет относительно небольшую часть общей продолжительно .
сти rеолоrической IИСТОРИИ этой rруппы. К этому мы еще вернемсЯ!
8 дальнейших разделах этой rлавы.
310 .
Часть У/. Ма"роэволюция
в диапазоне от сальтации до rрадуализма ЭТИ две 1<райние
формы следует отбросить, однако между НИМИ остается обширная
промежуточная зона, и с биолоrической точки зрения вполне ре-
.ально поискать в этой зоне способы эволюции крупных rрупп.
По всей вероятности, происхождение большинства новых' rрупп
,связано с очень быстрой эволюцией. Q,П,Iако не следует считать,
что такие высокие скорости несовместимы с постепенными ступен-
чатыми изменениями. Возможно, Что в возникновении мноrих но-
вых rрупп сальтационные элементы сочетаются с элементами rpa-
Ауализма. Современные представления о квантовой эволюции и
квантововидообразовательном направлении (изложенные в
Таблица 31.1
Различия в признаках меЖАУ современными хнщными и копытными
(по Simpson, 1953)
Признак
Хищные (Fissipedia)
Копытные (Perissodaclyla
и Artiodactyla)
2
э
Наземные
Местообнтания
Наземные, древесные или
полуводные
Пальце илн стопоходящие
<:пособ передвижения
'Чиспо папьцев
Ротовые образования
4---5
Хорошо развитые коrти,
обычно втяrивающиеся
Обычно ХJIЩНИКИ, иноrда
Всеядные, редко расти-
телыlя.дныыe И.'lИ иасеко-
моядные
3 . 1 . 4 I .3. 1
3 . 1 .4---1 . 3. 1
(в исключительных случаях
4
KopeHHblX s )
Крупные
.питание
.зубная формула
.Кпыки
f1редкореиные
Обычно простые, не похо
жие на коренные
Обычно простые, без буrор-
ков искпадок
'Kope Hыe
:Хищные зубы
Р' и М. увелнчены и пре-
вращсны в хищные зубы
Брахиодонтная
Высота коронки
Обычно копытоходящие,
иноr да пальцеходящие
1 , редко 5
Хорошо развитые копыта
(кроме даманов)
Обычно раститепьнояд-
иыс, ииоrда всеядные
3---0. 1---0.4---2. 3
3.......().1---0.4 2.3
Обычно мелкие ппп от-
сутствуют; иноrда
крупные
Обычно СJIожные, часто
моляризованы
Сложные, с мноrобуrор.
чатой перетнраюшей
поверхностью
Отсутствуют
От брахиодонтной АО
rИПСОДОНТIIОЙ .
r А. 31. Эволюция крупных zpynn
311
fЛ. 30) содержат как те, так и друrие элементы и занимают тем
самым промежуточную позицию.
Образование отрядов млекопитающих
Промежуточная позиция, постулирующая постепенное измене
иие, происходящее с исключительно высокими скоростями, иллю..
стрируется историей возникновения разных отрядов млекопитаю
щих.
Современные хищные и современные копытные (непаРНОКQПЫТ-
ные и парнокопытные) представляют собой две очень хорошо диф-
ференцированные rруппы BblCUKOro paHra. В современной фауне
эти две rруппы различаются по ряду признаков. Хищники обычнс>
связываются в. нашем представлении с хищным способом пита-
ния, коrтистыми лапами, крупными клыками и резцами, а копыт-
ные ---- с раС1"ительной пищей, конечностями, снабженными копы
тами, и перетирающими коренными зубами. Эти и друrие отличи-
тельные признаки обеих rрупп приведены в табл. 31.1.
Симпсон (Simpson, 1953) показал, что предковые члены тех
же самых филетических rрупп из нижнеro палеоцена не только-
Не были хорошо дифференцированы, но и обладали большим
сходством. Предки хищников (Creodonta) и предки копытных
(Condylarthra) имели MHoro общих признаков: они одинаково-
передвиrались, имели одинаковое число пальцев (пять), одинако-
вую зубную формулу и, вероятно, питались примерно одинаково,.
будучи всеядными.
Как отмечает СИМПСОJl (Simpson, 1953), в раннем палеоцене-
хищники и копытные не были не только разными отрядами, но-
даже и разными семействами. В сущности среди всех раннепалео-
ценовых хищников и копытных, взятых вместе, разнообразие было-
меньше, чем в пределах одноrо cOBpeMeHHoro семейства хищников,
например Mustelidae или Viverridae. Один палеоценовый род----
Protogoпodon (Arctocyonidae) ---- содержал несколько видов, кото-
рых На основании всей совокупности их пр-изнаков СJlедовало бы
отнести к хищникам, и несколько друrих видов, которых следо
вало бы отнести к копытным. Друrие авторы расходятся во мне..
ниях относительно места, занимаемоrо в системе раннетретичным
семейством Arctocyonidae, оставляя ero в отряде Creodonta (на-.
прнмер, Colbert, 1955) или относя ero к Сопdуlаrthrа (например
Romer, 1966).
Таким образом, комплекс признаков, характерных д..1Я хищных,
и отряд хищных развились в процессе диверrенции от своих па-
леоценовых предков; аналоrичным образом комплекс признаков,.
характерных для копытных, и два отряда копытных (Perissodac
tyla и Artiodacty,la) развились 8 процессе диверrенции от свои,,"
паJiеоценовых предков. Крупные rруппы возникают путем дивер
rенции (Simpson, 1953).
. 12
Часть У/. Ма1СРОЭ80ЛЮЦUЯ
ПР080ДЯ зто сравнение, мы рассматриваем непарнокопытных
11 парнокопытных КаК одну монофилетичеСI<УЮ rруппу. Палеонто-
ОrИ расходятся во мнениях о()11носительно филоrенетическ,их свя-
зей между этими двумя отрядами. Отметим, однако, что если даже
ларнокопытные не относятся к той же естественной rруппе t К Ко-
торой относятся непарнокопытные, сравнение последних с хищ-
ными остается правомерным. Приэнаки, перечнсленные в правом
столбце табл. 31.1, мало изменятся, если исключить из них при
знаки парнокопытных, а общие выводы останутся прежними.
В случае хищников и копытных, очевидно, имела меСТО извест-
ная степень rрадуалнзма. Друrие данные свидетельствуют о том,
что в ра,нней истории этих .и друrих отрядов млекопитающих, Kor-
_.да происходило развитие отличительных признаков этих отрядов,
.скорости эволюции были относительно высокими. Все эти данные,
хотя они и косвенные, позволяют сделать вывод о существовании
.некоторых общих тенденций в образовании отрядов; они собраны
в табл. 31.2.
ТаБЛllца 31.2
'продолжительность существования отрядов млекопнтаlOЩНХ (по Sfmpson, 1953)1
ПродолжитеЛhНОСТь, мли" лет
Ненэвестиыlt
ОТРЯДhl известиый неизвестиыR период, " от
общая период перноц общеrо
:"Ныне жнвущие
"Marsupi.ali.a 130 80 50 38,5
1 nscctivor а J30 80 50 38,5
Edent.ata 75 60 15 20,0
Lаgооюr:р11a 75 60 15 20.0
Rode.ntia 75 60 15 20,0
Cotacea 75 45 30 40,0
Carnivora 80 75 5 6.2
Puoooscidoa 65 40 25 38,5
Sirenia 70 50 20 28,6
Perissodacty\a 65 60 5 7.7
Artiod.a-cty\ а 65 БО 5 7.7
: 8ымершне
Taeniodonta 40 з5 5 12.5
'Condy\.arthr.a 40 (55?) 35 (501) 5 21.5 (9.l?)
LiрЬоtеrnз 70 БО 10 14.3
Notoungulata 75 60 15 20,0
Pant.odonta 45 35 10 22,2
r.Средияя 73,4 55,9 17,5 23,8
I в столбцах 2 и .. приведеиы макси аJlЬИЫЕ' оценки.
[ л. 3/. ЭвОЛ1Оция "рупных ерупn
31:1.
Оценка максимальной продолжительности Toro периода в про
rIсхождении каждоrо отряда, для KOToporo не сохранилось ника..
«их ископаемЫх остатков, приведена в столбце 4 таБJl. 31.2. Как.
ожно заметить, такой период обычно составляет лишь неболь
wую часть общей оцененной продолжительности существования
a.aHHoro отряда. Эта часть составляет 6,2% для х,ищников и 7,7%
Zl.ля непарнокопытных и парнокопытных, а для всех 16 отрядов.
рместе равна в среднем 23,8%. В этих оценках, быть может, не..
Ьколько преувеличена продолжительность периода возннкиовеНИR:
Ьтрядов.
. Данные, приведенные в табл. 31.2, соответствуют rипотезе о-
быстрой эволюции и квантовой эволюции в период воз-никновения.
отрядов млекопитающих (см. также rл. 28 и 30).
Роль квантовой эволюции
Процесс ква'нтовой эволюции был описан в rл. 30, и нет нуж
ДЫ описывать ero здесь СНОВа. Следует, однако, напомнить вывод..
Симпсона (1944, 1953), основанный на совпадении разных rрупIII.
данных, что квантовая эволюция это та форма эволюции, с ко..
торой обычно связано ВО3'никновение крупных rрупп.
Это заключение можно расширить. Возможно, что изменение
nрОИСХО.llЯlllЕ'е при квантовой эволюции, обычно выражается в.
форме видообразования и тем самым превращается в случай кван..
TOBoro видообразования. Кроме тооо, ряд случаев KBaHTOBoro в'И
дообразования может соединяться во временной ряд, в результа-
Те чеrо получается видообразовательное !Направление. ВПОЛНе'воз"
.можно, что такое видообразовательное направление участвует в-
звол юции ТОЙ или ИНОЙ крупной rруппы в период ее формирова..
ния (см. rJl. 30).
Еще одии возможный вывод касается типа местности, в кота-.
.рой скорее Bcero можно ожидать возниюtовения крупной rруппы..
Для квантовой эволюции и KBaHToBoro видообразования необхо...
.ДИМЫ популяционные системы OCTpOBHoro Типа. Подобные популя
ционные сиете'МЫ обычно Нельзя встретить в районах с равнинным
рельефом. Они обычно населяют сильно пересечf'ННУЮ rОрИСТУI()
местность или rруппу островов (в случае наземных растений II
животных) .
В современном мире эволюция растений в rористых областях..
по-видимому, действительно протекает быстро и приводит К воз"
никновению БО.'Jьшоrо разнообразия. На основе этих II друrих дая
ных ряд ИССo>lедовате.l1ей эво."ЮЦИИ растении приходят к выводу,
что ropbl служат вероятным местом крупных эволюционных co
бытий В растительном царстве (например, Kerner, 1894 1895;
Axe1rod, 1970, 1974; Stebbins, 1974).
3Н
Часть У/. Aiа доэво цця
Адаптивная радиация
Коrда какому-либо виду удается обосноваться на новой тер-
рит.ории или в новом местообитании, у Hero появляется эколоrи-
ческая возможность для расселения и увеличения разнообразия.
В таких условиях первоначальный вид может дать начало ряду
дочерних видов, приспособленных к различным крупным н'ишам
в пределах данной территории или местообитания. Эти дочерние
виды становятся преДl{ами ряда боковых ветвей, коrда они в свою
очередь дают начало новым дочерним видам. При этом данная
rруппа вступает во вторую фазу cBoero развития ---- в фазу проли-
ферации.
Адаптивная радиация представляет собой тот тип эволюции,
который наблюдается в фазе пролиферации. А преобладающая
форма эволюции при адаптивной радиации ---- это видообразова-
ние. События видообразования пера.ой очереди дают начало rлав
ным ветвям растущеrо филоrенетическоrо древа. Этн rлаВНЫе
ветви соответствуют различным rлавным эколоrическим нишам.
Вторая, третья и последующие очереди видообразования в каждой
из r лавных ветвей приводят затем к подразделению имеющихся
На данной территории или в данном местооби ании эколоrических
ниш между большим числом более высоко специа.'lИэированных
.видов.
Новая территория или местообитание наполняются видами.
И пq мере Toro, как это происходит, данная rруппа превращается
Б к,рупную rруппу. А крупная rруппа приобретает таксономиче-
скую структуру, формирующуюся В соответствии с теми л,нниями,
n,о. которым происходит адаптация.
. lia одной стадии развития крупная труппа по своему TaKCO O-
ич:е.<;J'ОМУ ,paHry соответствует семейству. При этом ее подразде
лениями первоrо порядка являются трибы и роды, З3.ШЦd.ающие
COO!BeTCTBeIlHo rлавные адаптивные зоны; каждая родовая. линия
сос оит из rрупп видов, представл ющих подразделения BToporo
",орядка и занимающих соответствующие адаптивные зонР1; а
каждая rруппа в,идов состоит из видов, представляющих подраз-
д л ния TpeTbero порядка. На более продвинутой тадИИ разви-
тия такая rруппа может состоять в paHre отряда. Теперь подраз-
дел ниями первоrо порядка, опять-таки связанными с разными
адаптивными зонами, оказываютс.я семейства, а 11.0дрзздеol1ениями
BToporo, TpeTbero и четвертоrо порядков являются соответственно
роды., rруппы видов и виды.
. Концепция адаптивной радиации была сформулирована Осбор-
ном (Osborn, 1910) в связи с эволюцией млекопитающих. Осборн
различал у млеКопитающих пять rлаВRblХ линий спеЦИ8.лизаIl:ИИ в
стррении конечностей, делающих их обладателей приспособленны-
ми к различным местообитаниям Н способам передвижения:
. . r л: 31.. Э80AIOЦия ,,,:igп,,blx epynп
315
.1) быстрый беr для наземных видов; 2) роющий образ жизни для
видов, обитающих под землей; 3) плавание для видов, обитающих
'н в воде 'Н На суше, и для полнос.тью водных видов; 4) лазание
для древесных видов; 5) планирование и полет для видов, осваи-
ваЮЩfIХ воздух. Осборн различал также линии специализации :&
'строении зубов, зависящие от характера питания (всеядность, пло-
тоядность, растительноядность и т. "')' Отряды млекопнтающих
COrJ1aCHO Осборну (ОsЬоrп, 1910), обладают этими rрупnами при..
знаков в различных сочетаниях, а следовательно, относятся к раз..
ным адаптивным типам.
Адаптивная радиация у rавайских цветочниц
Прекрасным примером адаптивной радиации у современноЙ'
rруппы с.пужат птицы семейства Drepanididae fla rавайских OCT
ровах. Это семейство изучалось рядом ученых на протяжении
мноrих лет. Основные данные содержатся в моноrрафиях Амадо-
на (Amadon, 1950) и Болдуина (Baldwin, 1953); дополнительные
сведения и интерпретации приводят Бок (Bock, 1970) и Карл.
квиет (Carlquist, 1974).
Семейство Drepanididae состоит из девяти родов, объединя-е..
мых в два подсемейства (табл. 31.3). Это монофилетическое се-
мейство, эндемичное Д..,Я rавайских островов. Следует указать,
что, к сожалению, некоторое число видов сравнительно недавно
вымерло или CTa.'IO очень редким. Семейство Drepanididae при..
Таблица 31.3
Система к.пассифика.ции rавайских цветочниц
(сем. Drepanididae) (Amadon, 1950)
l'руппа
I Числ. ВИд.О
Подсемейство Psittirostrinae .(желтое
или зеленое оперение)
LDXOPS 5
Heтignathиs 4
Pseиdonesfor 1
PsittirDstra 6
Подсемейство Drcpaniinae (опереИJlе
черное с красным)
Hiтatioпe 1
I
Ciпdops I
1
Drepaпis 2
316
Часть У/. Ма"рОЭ80люция
надлежит к подотряду Passeres, или певчих воробьиных (отряд
Р asseriformes); из друrих Р asseres они 6лизки К американским
цветочницам (Coerebidae) и (или) TaHarpoBblM (Thraupidae) (Ата-
don, 1950; Baldwin, 1953).
Поскольку семейство Drepanididae эндемично для rеолоrиче
сКи молодой цепи океанических островов и родственно материко-
ВЫМ труппам воробьиных, ero филоrенетические корни, вероятно,
P5iftirostru
Ciridops
lJrepaпis
Psittirost repaпiiпae
ZJrepaпididae
Рис. 31.1. Предпопаrаемая фипоrения rаваАских цветочниц (Drepanididae) (Ата-
don, 1950).
следует искать на материке, одн.ако, кто именно был материковым
предком rавайских цветочниц, остается неясным. Их далекий пре-
док, кто бы он I.ни был, вселился на rавайские острова 'и эволю-
ционировал в непосредственноrо предка семейства. Предполаrа-
ется, что у эrоrо непосредственноrо предка был короткий клюв и
ЧТО он питался насекомыми и нектаром (BaIdwin, 1953). Эта пища,
тип клюва и соответствующий тип языка наблюдаются также у
наиболее примитивных из ныне живущих представитслей обоих
подсемейств.
rавайские цветочницы отличаются большим разнообразием по
характеру пищи и строению клюва, значительно превосходящим
разнообразие, наблюдаемое обычно в друrих семействах воробьи-
ных. Здесь в пределах одноrо семейства 'имеются виды, напоми-
r л. 31. ЭвОАюция крупных lpynn
317
lIающие вьюрков, славок, пищух, виды, питающиеся плодами или
цветками, и разные друrие формы.
Вероятная филоrения этоrо семейства вплоть до родовоrо
уровня представлена на рис. 31.1 (Amadon, 1950); Бок (Bock,
1970) предлаrает иную филоrен.ию. Два подсемейства Psittiro-
strinae и Drepaniinae образуют rлавные ветви семейственноrо
древа, а роды ---- ветви BToporo порядка. Ветви TpeTbero порядка
()бразуют виды в таких немонотипических родах, как Loxops и
Hemignathus.
Ветвление создает адаптивную радиацию, которая у rавайских
цветочн'иц связана rлавным образом с характером питания и про-
является в признаках клюва и языка, а также в друrих призна-
ках. rлавные адаптивные зоны, занятые этими птицами, соответст-
А
. .'-; ...
. ' ;; ., ' _6
-- .
р ; :.
t{.;: .
' .
.
. ."'.
. ...
'.6" .
д
.
-.: , . .
;f ;-J. '
r
_:
r
сТ'
Рис. 31.2. Типы кпювов у rавайских цветочниц (рис. Ch. Рарр по фото из Ата-
don, 1950).
А. Hiтatione saпgиiпea (питается нектаром). Б. Drepaп{s pacifica (питается нектаром).
В. Cin'dops аnпа (питается П.'!одами). r. Loxops cocciпea (питается насекомыми). Д. Heтig-
пathиs lиcidиs (питается насекомыми). Е. Psittiroslra palmeri (питается пподами и семе-
Ками).
А
319. ..
_ Часть .1[[. М а"роэвОАЮЦц,я.
вуют питанию насек..оМblМИ, нектаром, плодами и семенами. Неко-
торые из этих зон подразделяются На более мелкие зоны. Так,
среди насекомоядных цветочниц есть КОРОТКОК.rIювые формы, оби-
рающие насекомых с листьев, и длинноклювые формы, добываю-
щие их из-под коры (Baldwin, 1953).
Подсемейство Drepaniinae состоит rлавным образом из форм,
питающихся нектаром цветков. У некоторых из них клюв корот-
кий (например, Himatione, рис. 31.2, А), у друrих ---- длинный (Ha
при мер, Vestiaria и Drepanis; рис. 31.2, Б). Относящийся К этому
же подсемейству Ciridops (рис. 31.2, В) питается плодами. Неко-
торые типы клювов у представителей этоrо подсемейства, питаю-
щихся на цветках, коррелируют с цветками rавайских Lobelioi-
deae (сем. Campanulaceae).
Подсемейство Psittirostrinae состоит из видов, питающихся
насекомыми, плодами и семенами. К роду Loxops относятся 5 ви-
дов насекомоядных птиц с клювами небольших или средних раз-
меров. Виды с небольшими клювами обирают насекомых с
листьев (например, Hemignathus coccinea, рис. 31.2,r), тоrда
как виды с клювами средних размеров извлекают насекомых из
щелей в коре. У Hemignathus lucidus (рис. 31.2, Д) клюв серпо-
видной формы, и он добывает насекомых из rлубоких трещин
примерно так, как это делает пищуха. Друrие виды Hemignathus,
также имеющие серповидные клювы, соrласно сообщениям, пита-
ются на цветках.
К этому же подсемейству Psittirostrinae относится Pseиdo-
nestor, клюв KOToporo напоминает клюв попуrая и который ис-
пользует этот клюв для обламывания веточек и добывания
из под коры насекомых и их червеобразных личинок. Родствен-
ный ему Psittirostra (рис. 31.2, Е) также обладает мощным клю-
вом, НО в этом случае клюв используется для раздавливания
плодов и раскалывания семян.
Параллельным примером адаптивной радиации у друrой rруп-
пы птиц, населяющей острова, служат дарвиновы вьюрки (сем.
Fringi11idae, подсемейство Geospizinae) на rалапаrосских остро-
вах (Lack, 1947). При мер несколько иноrо рода rруппа расте-
ний, в которой адаптивная радиация связана со способом опы-
лениЯ и проявляется в разнообразии типов цветка, ---- дает семей-
ство Polemoniaceae (Grant, Grant, 1965).
Такая адаптивная радиация весьма эффективна для той rруп
пы, в которой она происход..ит, потому что эта rруппа полуqает
максимальные возможности для использования всех эколоrИ1lе.
СКих условий, предоставляемых ей новой территорией или место-
обитанием.
['лава -32
ВЫ.МИРАНИЕ
Введение
Виды, живущие в данном rеолоrическом периоде, представляют
собой прямых потомков лишь небольшой части видов, относя-
щихся К более ра'ннему периоду. OrpoMHoe большинство видов
обречено на вымирание. Вымирание yroToBaHo также большинст-
ву филетичес их rрупп низшеrо и среднеrо paHra, т. е. родам,
семействам и отрядам, хотя надвидовые rруппы обычно сохраня-
ются дольше, чем виды. Кроме Toro, в истории Земли есть перио-
ДЫ, коrда вымирание приобретает широкие ,масштабы и затраrи
nает мноrие ВИДЫ. принаД.!1ежащие к ряду разных rрупп.
Удобно различать три уровня вымирания: вымирание отдель
ных видов (напр'имер, Megaloceros gigaпteus) , вымирание надви-
довых rрупп (например, аммониты в меловом периоде) и широко
распространенное или массовое вымирание (как в позднем
мелОвом периоде).
В зависимости от характера вымирания надвидовые rруппы
МoQжно подверrнуть дальнейшему разделению. Некоторые такие
rруппы полностью вымирают, не оставляя филоrенетических по-
томков. тоrда как друrие вымирают сами, но дают начало потом-
кам, СИ.rIЬНО ОТ.пичающимся от своих предков и образующим от..
де..'JЬНУЮ надвидовую rрупnу.
В этой rлаве мы рассмотрим несколько примеров вым,ирания,
относящихся к каждому из названных выше уровней. Как мы СМО"
жем убедиться, точные причины вымирания видов и rрупп более
BblcoKoro paHra часто неизвестны или очень недосroверны. Однако
случаи, коrда вымирание происходило в очень широких масшта-
6ах, можно нередко коррелировать с rеолоrическими революция-
ми, что дает возможность объяснить вымирание какими то ,причи..
нами общеrо характера.
Megaloceros giganteus в Ирландии
Одним из знаменитых примеров вымирания в.идов служит .BЫ
мирание Megaloce.ros giganteиs rиrантскоrо оленя, жившеrо в
ппейстоцене и в начале cOBpeMeHHoro периода. Ареал этоrо вида
простирался ОТ Ирландии через всю Европу до Сибири и к. тая,
а На юrе до Северной Африки. Он процветал в Ирландии B. те-
чение периода, непосредственно следовавшеrо a отступлением
л дов в конце последнеrо 9лед не ия, и ЫMep там примерно
320
Часть У 1. Ма"роэвОЛlOция
11 000 лет назад. Возможно, что в некоторых частях материковой
Евразии он дожил до начала COBpeMeHHoro периода (GouJd, 1974).
Б6льшая часть ископаемых остатков Megaloceros giganteus
найдена в озерных отложениях и торфяных болотах Ирландии.
Это был rиrантский олень высотой около двух метров на уровне
Рис. 32.1. Pora Megaloceros giganteиs (по Mi1ais и CuvieT из Gould, 1974)'.
А. Демонстрация широких частей POroB. KOr'Aa caMeJt смотрит прямо перед собоА. Б. Де-
монстрации отростков porOB. котда rOIlOBa И8клоиеиа.
плеч. Самцы обладали оrромными роrами весом до 25 Kr и раз-
махом примерно до 3 м (рис. 32.1). Это размеры максимальные;.
подробные результаты измерений приводятся rоулдом (Gould,
1974). Ни у одноrо друrоrо представителя семейства оленей не
было таких крупных poroB. Мноrие авторы отмечали, что большие
и тяжелые pora М. giganteиs, которые олень ежеrодно сбрасывал
и котОрые отрастали вновь, вероятно, создавали тяжелую физи-
ческую наrрузку и физиолоrически истощали животных.
Среди выдвиrавш,ихся раНее ПРИЧИН вымира'Ния М. gigaпteиs
()собенно большое внимание уделялось тем неуд06ствам, которые,
r л. 32. Вымирание
32)
возможно, терпели животные из за таких больших poroB. В заВJII
С1fМОСТП от своих теоретических позиций одни авторы строили объ...
ясне ни я в терминах ортоrенеза, а друrие в терминах eCTeCT
BeHHoro отбора. Как те, так и друrие считали, что pora этих оле
ней 'не имел.и адаптивноrо значения. Вполне возможно, что pora
были фактором, способствовавшим вымиранию животноrо, но, КаК
указывал Симпсон (Simpson, 1949, 1967), нет причин считать, что
эти pora не имели адаптивноrо значения в тот период, коrда они
разв,ились и Kor да вид процветал.
Симпсон (Simpsoll, 1949, 1967) указывал далее, что величину
poroB М. gigaпteиs следует рассматривать не в качестве ИЗОJIИрО
BaHHoro признака, а как результат соотношения скоростей роста
разных частей тела в процессе филоrенеза. Среди представите.'lей
семейства оленей филоrенетическая скорость роста poroB значи--
тельно превышает скорость роста тела, так что самые крупные-
виды оленей должны иметь пропорцuонально самые большие pora.
у М. gigaпteus с ero большим телом pora были примерно TaKHe
каких и следовало бы ожидать у очень крупноrо оленя.
Следовательно, возможно, что отбор блаrоприятствовал увели
чению размеров тела М. gigaпteиs, а большие pora развились в
качестве побочноrо эффекта (Simрsоп, 1949, 1967). Увеличение
размеров тела обычная тенденция в эволюции млекопитающих,.
дающая им ряд преимуществ (см. rл. 26). Возможно также, ЧТО
половой отбор блаrоприятствовал увеличению poroB у М. gigan 06
teus, потому что это давало преимущество самцам при спарива-
нии, а побочным эффектом были большие размеры тела
(Gould, 1974).
Причины вымирания М. gigaпteus, по крайней мере в Ирлан
ДИИ и, вероятно, в друrих местах, можно искать также и в дpy
rих аспектах ero жизни. Он, по-видимому, обитал на открытых по.-
росших травой местах, перемежавшихся с участками леса. TaKoro-
рода растительность достиrла расцвета в один из периодов потеп-
ления к концу последнеrо оледенения, а именно в это время и про
цвета л М. gigaпteus. Несколько позднее в тот же период степи
смени.тIИСЬ лесами, и М. gigaпteus, который не был лесным оленем.
вероятно, не cMor выжить в rycToM лесу. Неблаrоприятные для
э оrо оленя изменения кли ата и растительности в период послед-
Hero оледенения, по видимому, и были причиной ero вымирания.
(Gou1d, 1973, 1974).
Лошадь в Северной Америке
Лошадь (Equus) вымерла в Север'ной Америке в начале со...
временной эпохи, примерно 8000 10000 лет назад. Как уже rOBO
рилось В rл. 26, эволюция Equиs происходила rлавным образом':
Часть У/. Ма"роэволюция
322
9 Северной Америке, откуда животное миrрировало на три друrих
материка. В плейстоцене в Северной Америке существовали or.
ромные табуны лошадей, принадлежавших, вероятно, к несколь.
ким видам Equиs. Затем род Бquus полностью вымер в Северной
Америке (Simpson, 1951 и личное сообщение).
Спустя MHoro тысячелетий, в 1519 r. испанцы ввезли в Север-
ную Америку европейскую домашнюю лошадь (Equus caballus).
Интродукция qказалась очень успешной, и домашняя лошадь об-
разовала дикие табуны на древней родине своих предков.
Причины вымирания Equus в Северной Америке неизвестны.
И таЙна эта усуrубляется тем, что Equus удалось вНовь успешно
закрепиться в диком состоянии на этом материке спустя несколь-
ко тысяч лет после Toro, как она там вымерла.
Высказывались различные предположения опричинах этоrо
вымирания, но все ОНИ малоубедительны. Не оказалась ли абори-
rенная североамериканская лошадь жертвой оледенения? Но ведь
Equus пережила плейстоцен, а кроме Toro, она была распростра-
нена и за пределами rраницы льдов. Была ли лошадь истреблена
первыми американскими индеЙцамИ, с которыми она ж.ила в одно
время? Такую возможность исключить нельзя. Но ведь индейцы
охотились на бизона и тем не менее не истребили ero. Возмож-
но также, что лошадь поrибла от kakoro-то эпидемическоrо забо-
левания. Однако в палеонтолоrической летописи не сохранилось
иичеrо, что свидетельствовало бы о такой эпидемии (Simp-
50П, 1951).
Те же самые события происходили параллельно в ЮЖНОЙ
Америке. Здесь род Equus также процветал в плейстоцене и вы-
мер в начале современной эпохи; затем в историческую эпоху ло-
щадь была интродуцирована из Европы, после чеrо она одичала.
Причины ее вымирания в Южной Америке также неизвестны
(Simpson, 1951).
Саблезубые кошки
Один из часто приводимых примеров вымирания надвидовой
rруппы вымирание саблезубых кошек. Саблезубые кошки бы.Т'JИ
одной из rJIaBHblX ветвей семейства Felidae, к ним относились та-
кие роды, как Hoplophoneus, Machairodus и Smilodon. Эта rруппа
отличалась от современных представителей подсемейства Felinae
большими размерами верхних клыков и друrими связанными с
этим признаками в строении челюстей и зубов (рис. 32.2).
Саблезубые кошки встречались в Северной Америке и Евра.
зии на протяжении почти Bcero третичноrо периода, 'начиная с оли-
rоцена, а в плейстоцене проникли в Южную Америку. Hoplopho-
r л. 32. Вымирание
32
пeus характерный олиrоценовый пред
ставитель этой rруппы в Северной Амери.
ке, а Smilodon более крупная плейстоце-
новая форма, жившая как в Северной, так
и в Южной Америке (рис. 32.2, А и Б).
В плейстоцене эта rруппа вымерла (iRo-
rner, 1966).
ОДНО время бы.по принятu считать, что
увеличение размеров клыков и оБLЦИХ раз.
меров тела саблезубых кошек в плейстоце-
не обусловлено ортоrенезом. Вымирание же
их считалось результатом Toro, что это op
тоrенетическое направление перешаrнуло
за пределы функциональной полезности.
Крупные клыки Smilodon, как предполаrа
ется, затрудняли кусание, что и привело к
вымиранию этой филетической rруппы.
В таком случа , отмечает Симпсон (Simp-
son, 1953), этим животным понадобилось
це..1ЫХ 40 М.,lН. лет, чтобы умереть с rолоду.
На самом деле крупные клыки не меша-
ли кусанию. Челюсти саблезубых кошек
моrли широко раскрываться и плотно сжи
маться.
Возможно, что саблезубые кошки были
специализированы к охоте На таких круп-
ных, медленно передвиrавшихся толстоко-
ЖИХ растительноядных животных, как ма-
стодонты и ленивцы. Их ДЛинные клыки
при разомкнутых челюстях моrли служить
эффективными орrанами для закалывания
и разрывания добычи; в этом им помоrали
МОLЦная шея и плечевой пояс.
В течение плейстоцена ряды толстоко-
жих растительноядных значительно поре-
дели. Снижение численности, а затем и вы.
мирание добычи этоrо типа сократило или
закрыло адаптивную зону саблезубых ко-
шек. Тем временем у представителей под-
семейства .Fe1inae в семействе кошачьих
совершенствовался альтернативный способ охоты на быстро пе.
редвиrаЮLЦиеся виды жертвы нападение из засады и преследо-
вание. Поскольку в плейстоцене адаптивная зона самих саблеэу.
бых кошек стала закрываться, а альтернативные адаптивные.
зоны уже были заняты, этим кошкам пришел конец (Simp.,
son, 1953).
А
Б
.......,.
, ...
....:0:.'..' 0_' '-( \
..:1'; .
8 .",
. .
Рис. 32.2. Черепа саБJtе
зубых кошек (по Mat
thew и Zdansky из Ro-
mer 1966, с измене.
пиями).
Черепа изображеllЫ в раз-
ном масштабе. А. Hoplo.
phoneиs, о.'Iкrоцен, д.'ина че-
репа 14 см. Б. Sтilodon.
плеАстоцен, длина черепа
30.5 см. В. Pseиdaelиrиs.
плиоцен, настовщая кошка.
изображенная ЗАесь ДЛЯ TO
ro, чтобы сопос:тавиТla раз-
..еры кпыхов.
324
Часть У/. Макрозволюцuя
Динозавры
Одним из наиболее обширных rрупповых вымираний всех вре.
мен было вымирание динозавров в позднеме.rJ.ОВОМ периоде. Ди-
нозавры, к которым относились два отряда преСМblкающихся
Saurischia н Ornitischia, rосподствоваJlИ на суше в течение Bce
ro мезозоя и поэтому мезозой называют эрой динозавров.
В течение мезозоя динозавры достиrли большоrо разнообра-
зия, о чем свидетельствует система 'их классификации, cor ласно
которой их делят на два отряда, 25 семейств и на более чем
200 родов (см. Colbert, 1961; Колберт различает 218 родов). Среди
них были такие rиrантские животные, как бронтозавры и тиран-
нозавры, а также различные формы 'небольших ,н средних разме.
ров. rруппа в целом была распространена по всему земному шару
и r.осподствовала на суше в течение более чем 100 млн. лет. Затем
в конце меловоrО периода за несколько миллионов лет вся эта
rруппа вымерла.
Причины вымирания динозавров неизвестны, и вовсе не по-
тому, что не предпри'нималось попыток объяснить это явление. Ли
ература изобилует упрощенческими и необоснованными спекуля
циями относительно причин вымирания динозавров: в позднемело
вой период климат стал слишком ХО.,'Iодным для динозавров; им
стало слишком жарко; примитивные млекопитающие поедали их
яйца; растительноядные динозавры уничтожили мезозойскую ра-
CTHTe.rtbIIOCTb; динозавры поrибли вследствие orpoMHoro усиления
излучения, сопровождавшеrо вспышку новой звезды на небе:
в результате обращения маrнитноrо поля ЗеМJ1Н смерте."ьные дозы
космическоrо излучения достиrли поверхности суши.
Два последн,их предположения, вероятно, можно исключить.
Такие астрофизические или rеофизические события, которые вы.
зывают всплески смертельных доз излучения, представляют собой
обычно факторы одноразовоrо или двухразовоrо действия, а по
этому они должны были бы немеД.,1еIlНО вызвать массовую rибель
животных; между тем вымирание динозавров продолжалось тыся-
чи и миллионы лет (Simрsоп, 1968). Постепенное изменение кли
мата в позднемеловом периоде лучше соrласуется с тем, как про-
исходило вымирание динозавров. Мноrие ученые склонны считать,
что именно неблаrоприятные изменения климата представляют
собой .наиболее вероятный фактор, как это подразумевается в двух
первых из перечисленных выше предположений, хотя они, быть
может, сформулированы в чересчур наивной форме.
Аксельрод и Бейли (Axelrod, Ваilеу, 1968) придают rлавное
значение смене pOBHoro климата неровным, которая произошла
в конце .ме.повоrо периода. На протяжении большей части мезо-
зоя преобладал ровный теплый климат, и эволюция динозавров
почти 130 млн. лет пр.отекала в таких климаТИЧесКИХ усльвиях. Во
r л. 32. Вымирание
325
второй половине меловоrо периода этот климат сменился неравно-
мерным континентальным климатом, для KOToporo характерны бо-
ee резкие колебания температуры. Друrие филетические rруппы
рептилий, жившие в меловом периоде, ящерицы, змеи и чере-
пахи Mor ли избеrать действия очень высоких или очень низких
температур, зарываясь в землю, впадая в спячку или поrружаясь
в воду, и дожили до наших дней. Но у динозавров, которые вели
Ilаземный образ жизни, не было таких способов избежать дейст-
вия крайних температур, и они вымерли (Axelrod, Ваi.lеу, 1968;
см. также Spotila et з1., 1973).
Воздействие стрессовых температур в условиях континенталь-
Horo климата позволяет объяснить MHoroe, быть может, даже
очень MHoroe, относительно вымирания динозавров. Из рептилий,
переживших ме.ТIOВОЙ период и доживших до нашеrо времени,
можно назвать крокодилов, обитающих в тропиках и субтропи
ках, и rаттерию, относящуюся к отряду хоботноrоловых и обитаю..
щую на Новой Зеландии. Почему изменение климата в конце Me
JIoвoro периода пощадило эти два отряда, повлияв только на два
отряда динозавров?
Возннкает и еще одно затруднение. Различные rипотезы, свя-
зывающие вымирание динозавров с изменениями климата, исходят
из Toro, что динозавры, поскольку они относятся к рептилиям,
БЫJlИ холоднокровными. Однако недавно были обнаружены HeKO
торые косвенные палеО'нтолоrические данные, например данные о
rистолоrии костной ткани, указывающие На то, что динозавры
были не холоднокровными, как это предполаrалось, а теплокров-
ными, 'или rомойотермными, животными (Bakker, 1975). Подтвер-
дится ли это новое заключение и если да, то удастся ли ero при-
мирить с rипотезой, связывающей вымирание динозавров со сме..
ной климата. покажет будущее. А пока эта тайна все еще
остается нераскрытой.
Массовое вымирание в позднемеловом периоде
Вымиран.ие динозавров было лишь частью rораздо более об-
ширной картины MaccoBoro вымирания в конце меловоrо периода.
В это время вымерли и друrие отряды рептилий: морские ихтио-
: aBpы. плезиозавры и мозозавры, а также летающ.ие птерозавры.
Из морских беспозвоночных в конце меловоrо периода вымерла
процветавшая прежде rруппа аммонитов.
Исчезновение нескольких rрупп морских животных в, конце ме-
.:roBoro периода трудно объяснить на основе радиационных тео-
рий, поскольку морские животные, очевядно, были защищены от
ПОСТУJ1ируемоrо в этих теориях действия излучения. Данные отно-
сительно вымирания в меловой период не противоречат климати-
ческим теориям, если эти теории прилаrаются к наземным рептн-
326
Часть V 1. Макроэволюцuя
лиям, но они вместе с тем и не подкрепляют эти теории; ведь не..
смотря на. то, что морские рептилии, казалось бы, были более или
менее ИЗОJIирова'ны от воздействия HOBoro HepoBHoro кл,им та, они
все же вымерли.
И так, в конце меловоrо периода произошло обширное обновле..
ние фауны суши, моря и воздуха. Такое обновление заставляет
предполаrать действие не каких-то отдедьных специфических эко"
лоrических факторов, а факторов общих и всепроникающих. с
мноrочисленными побочными эффектами и взаимодействиями.
Вымирание млекопитающих в плейстоцене
Друrое массовое вымирание происходило в плейстоцене. Мле..
копитающие rосподствовали на суше 1Ia протяжении Bcero третич"
Horo периода, и третичный период нередко называют эрой млеко..
питающих. Однако в позднем плейстоцене эта эра подошла к концу
или к началу конца: началось массовое вымирание млекопита.
ющих, и процесс этот продолжался в современной эпохе.
Вымирание охватило rруппы млекопитающих разноrо таксоно..
мическоrо paHra от отдельных видов (например, Megaloceros
giganteus) до надвидовых rрупп (например, саблезубые кошки).
Среди вымерших видов можно назвать М. giganteus, пещерноrо
медведя, пещерноrо льва и rиrантскоrо бобра. К вымершим Ha
двидовым rруппам относятся саблезубые кошки, меrатерий, I'и
rантский броненосец, rлиптодонты, мастодонты, мамонты, шерсти..
стый Hocopor и североамериканские дикие .."'10шади.
Вымира'ние особенно сильно затронуло виды и rруппы более
BblcoKoro paHra у крупных млекопитающих. Мелкие млекопитаю..
щие rораздо лучше пережили поздний плейстоцен и начало сов..
peMeHHoro периода. чем крупные.
Существуют две основные rипотезы относительно причин BЫ
мирания крупных млекопитающих в плейстоцене. Соrласно одной
из них, вымирание было вызвано охотой первобытноrо человека
именно на крупных млекопитающих (Martin, 1973), соrласно дру-
rой климатом 80 время оледенения (Axelrod, 1967). Обе rипо-
тезы сталкиваются с некоторыми затруднениями.
rипотеза Мартина относится rлавным образом, хотя и не ис..
ключительно, к крупным млекопитающим Северной Америки. Как
считает Мартин, 11 500 лет назад первобытный человек, находив
шийся на той стадии раЗБИТИЯ культуры, коrда он жил только за
счет охоты, проник из Сибири в Северную Америку, rде .нашел
обильные новые пищевые ресурсы в виде существовавших там в
те времена крупных млекопитающих. Эти млекопитающие, типич.
нымн представителямн которых для Северной Америки были ма..
стодонт, мамонт, лошадь, верблюд и бизон, прежде не подверrа.
.ry:ись истреблению охотившимися на них людьми. Популя.
r л. 32. Вымирание
327
ЦИЯ первобытноrо человека стала быстро разрастаться и столь же
быстро расселяться по Северной и Южной Америке блаroдаря
пище, которую обеспечивали такие крупные животные; вскоре в
результате интенсивной охоты большая часть этих животных вы-
мерла. Некоторые указанные виды и в самом деле вымерли при-
мерно в то время, коrДа, как предполаrается, произошел первый
взрыв численности популяции первобытноrо человека, т. е. пример-
но 1 О ooo 11 000 лет назад (Martin, 1973; см. также Маrtiп,
Wright, 1967).
ЭТа rипотеза опирается на широко распространенное мнение.
что человек проник в Северную Америку относительно недавно.
Однако, судя по некоторым данным, человек существует в Север-
ной Америке rораздо дольше, чем 11 ooo 12000 лет, как это по-
стулируют Мартин и друrие. С помощью одноrо из новых методов
(метод, основанный на рацемизации аспараrиновой кислоты opra
нических остатков, пропорциональной их возрасту) было установ-
лено, что возраст костей индейцев из палеолита Южной части Ка-
лифорнии равен 48000 лет (Bada, Schroeder, Carter, 1974). Таким
образом, вопрос о всех этих сроках остается открытым, поскольку
надо сначала установить, появился ли человек в Северной Амери..
ке .сравнительно недавно или давно.
Аксельрод (Axelrod, 1967) ищет причину вымирания млекопи-
тающих в изменениях климата, достнrших кульминации в плей-
стоцене. Третичный период отличался в общем ровным климатом,
и млекопитающие были приспособлены к таким К..'1иматическим
условиям. Для климата плейстоцена характерны резкие измене-
ния условий с жестоким" холодами или сильными засухами. При
очень сильных холодах отдельные особи млекопитающих rибнут
от чисто физиолоrических воздействий низких температур или от
I'олода, и эта rибель отде.ПЬНЫХ животных может в совокупности
достиrнуть уровня вымирания вида. Новый холодный (или сухой)
климат также Mor оказывать неблаrоприятные косвенные воздей..
ствия на мноrих млекопитающих, вызывая изменения раститель-
ности в их местообитаниях.
Наличие определенной зависимости между вымиранием жи-
вотных и их размерами соrласуется с климатической rипотезой.
Среди мелких животных не наблюдалось MaccoBoro вымирания.
Большинство мелких животных в холодное время MorYT укрыться
в норах или rнездах. Кроме Toro, у мелких млекопитающих репро-
дуктивный цикл обычно короткий и может быть увязан с клима-
том, подверженным резким сезонным колебаниям. У мноrих мел-
ких млеко питающих потомство достиrает зрелости в течение
KopoTKoro блаrоприятноrо сезона. Крупные млекопитающие в боль.
шинстве случаев не умеют приспосабливаться к климату, для ко..
Toporo характерны очень холодные (или очень сухие). сезоны
(Axelrod, 1967).
328
Часть V/. Макроэволюцuя
Климатическая rипотеза не может, однако, объяснить, почему
в раннем плейстоцене, коrда фактически произошел переход к
ледниковому климату, млекопитающие вымирали M3.1IO. Друrое
затруднение, с которым сталкивается климатическая rипотеза, со-
стоит в ТОМ, что мноrие крупные млекопитающие вымерли не толь-
ко на северных континентах, но и в Африке, которая не пережила
в полной мере ледниковый период (Симпсон, личное сообщение).
Эпизоды вымирания
Давно известно, что rеолоrическая история жизни отмечена
эпизодами мзссовоrо вымирания, разделенными длительными пе-
риодами nocTeneHHoro эволюционноrо изменения. Палеонтолоrиче-
ская летопись заставляет предполаrать нечто среднее между ка-
тастрофизмом Кювье и строrим униформизмом Лайеля ---- состоя..
ние эпизодической эволюции. В истории периодически возникают
эпизоды MaccoBoro вымирания одних rрупп, за которыми следует
развитие и адаптивная радиация новых rрупп (Newell, 1967).
На самом деле rраницы между rе.олоrическими эрами и перио.
дами частично Основаны на эпизодах MaccoBoro вымирания, в осо-
бенности у животных. Определяющие признаки эпизодической эво-
люции животных отражены в названиях эр (палеозойская и т. п.).
Существенная черта эпизодов вымирания ---- высокая скорость
вымирания, наблюдаемая в совершенно не связаННl1IХ между со-
бой rруппах примерно в одно и то же время. Так, у аммоноидов
и рептилий отмечается быстрое вымирание в поздней перми, позд..
нем триасе и позднем мелу (Newel.C 1967).
Ньюэлл (Ne\\'ell, 1967) сопоставил 'Н проанализировал данные
о первом и последнем появлеНИИ в палеонтоло ической летописи
2250 семейств животных, представлявших все крупные rруппы.
Полученные им результаты представлены на рИС. 32.3. На
рис. 32.3, А показана чаС,тота вымирания семейств животных на
всем протяжении rеОЛОI'ической истории по данным об их пос.пед
Нем известном появлении в nалеОНТОJIоrической "lеТОIlИСИ. Как
можно видеть, кривая вымирания очень изломана. Высuкие пики,
отражающие эп'изоды MaccoBoro вымирания, отмечаются в позд
Нем кембрии, позднем девоне, поздней перми и ПОЗД:It: М мелу;
более слабые пики приходятся на концы некоторых из промежу
точных периодов. Большинство этих эпизодов вымирания можно
коррелировать с периодами поднятия материков и установления
континентальноrо климата. Такая корреляция позволяет считать,
что возмущения. происходившие в среде, запускали сложные цеп-
ные реакции, приводившие в конечном счете к вымираниям в круп-
ных масштабах (Newell, 1967; Axelrod, 1974).
На рис. 32.3, Б видно, что первые появления в палеонтолоrиче
СКОЙ летописи новых семейств животных также распределены во
зо
20
10
А
40
.
. за
'3"
20
'а 10
6 . о
r А. 32. Вымирание
329.
к
Мис. Пен_ n т
ю
100
о
90
80
\-ot 70
..
'3БО
. 50
i
40
о
м
"100
&
90
-
80
70
БО
50
к
д Мис. Пен. П Т
ю м КОК
о
с
Рис.: 32.3. Вымирание и замещение у животиых (Newe1I, 1961)'.
..4.. Частота выu'ираНИfl семейств. Б. Частота пер-"ВЫХ появлении новых семеАств по AaRBbl"
па.nеОНТОJJоrической летописи. Объяскения сМ. в тексте.
времени очень неравномерно. Эта кривая, подобно кривой выми-
рания, также образует пики. Кроме Toro, пики кривой возникнове-
ния в общем следуют за пикам.и кривой вымирания, хотя и с не 4
которым запаздыванием; за эпизодами MaCCOBoro вымирания сле-
дует образование и развитие новых rрупп (Newell, .1967). Подоб.
ный ритм появления новых rрупп соответствует заключению, сде-
ланному е rл. 3 1. Необходимое условие для возникновения новой
rруппы наличие соответствующих эк.олоrических возможностей,
которые" создаются в результате MaCCOBoro вымирания.
Часть VII
От орrанической ЭВОJlЮЦИИ
к ЭВОJlЮЦИИ чеJlовека
rлава 33
проrРЕССИВНАЯ ЭВОЛЮЦИЯ
Концепция nporpecca в эволюции
Концепция проrресса в своей основе субъективна он обычно ис
пользуется rлавным образом применительно к человеку. Некую
тенденцию в истории человечества называют проrрессом, если одна
из более поздних стадий превосходит более раннюю по каким..
либо характеристикам, которые, по мнению наблюдате.!lЯ, имеют
особенно важное значение. Точка зрения наблюдателя и прини..
маемая им система uенностей иrрают большую роль в определе
нии проrресса. Нам нетрудно вспомнить о таких тенденциях в HO
вейшей истории, которые человек, занимающийся освоением новых
земель, называет проrрессом, а консерватор считает прискорбным
шаrом в ошибочном направлении.
Нам предстоит рассмотреть здесь следующие вопросы: 1) мож-
но ли у довлетворительиым образом переносить концепцию про
rpecca из сферы деятельности человека на эволюuионную биоло
I'ию; 2) если допустить возможность TaKoro переноса, то можно ли
превратить ero из субъективной концепции в объективную, т. е.
можно ли найти объективные критерии, позволяющие определить
понят-ие проrресса в орrанической эволюции.
В ряде r.пубоких работ, стимулирующих появление новых
идей, rексли (Huxley, 1942, 1954, 1958) высказывается положи
тельно по обоим этим вопросам. Он настаивает на объективной
реальности особоrо эволюционноrо направления, которое следует
называть проrрессивной эволюцией. Фактически rексли сначала
переносит концепцию проrресса из сферы деятельности че.'Iовека
на орrаническую эволюцию, э. затем вновь перенооит ее на чеЛове-
ка, рассматривая человечество как высшую ступень, достиrнутую
проrрессивной зволюuией.
Точку зрения rексли обсуждали мноrие эволюционисты, кото.
рые обычно не вполне соrлашаются с ним (например, Simpson
1949, 1967, 1974а; Thoday, 1958; IRensch, 1960; Stebbins, 1969;
Ayala, 1974). Сам я также заНимаю несколько двойственную по
r л. 33. П роzрессuвная эволюция
331
ЗИЦИЮ, как это станет ясно из дальнейшеrо. Тем не менее, в чем
также можно будет убедиться далее, эти положения rеКСJ1И ча-
стично выдерживают критику.
Первый из упомянутых выше вопросов не вызывает особых за-
труднений. Можно, если уrодно, а priori уравнять проrрессивную
эволюцию с эволюцией человека ил'и же, столь же произвольно,
отказаться от таких определений, как это склонен делать я. Самое
важное дать ответ на второй вопрос: представляет ли собой про-
rрессивная эволюция объективно реальный процесс, происходя-
щий в природе? Это вопрос биолоrический.
Проrресс или специализация?
rексли (Huxley, 1942) проводит различие между nporpeCCOM
п специализацией в эволюции: «Специализация... это повышение
эффективности приспособления к определенному образу жиз-
ни. . .». Но «специализация... всеrда связана с необходимостью
пожертвовать некоторыми орrанами или функциями для Toro,
чтобы друrие стали более эффективными». С ']едовательно, усовер-
шенствования, связаННые iCO 'ClпециаJтизацией ;К апре.деленной а;даn-
'ТИВНОЙ зоне, обычно оrр.аНИчи.вают возможности будущих изме-
нений, кото.рые MQrYT понадобить'Ся для продолжения суще.с1'ВО
вания в этоЙ Же аМОЙ адаПТ.ИВIIОЙ зоне. Специализации лрисуще
самооrраничение. ОРirанИ'чес'кая эволюция .своДит-ся rлавным обра
ЗОМ к раз.в.итию спецна lиза.ций. И .поэтом,у эволюция очень часто
закаич.ивае1'СЯ тупиком (Huxley, 1942).
Между тем проrресс в эволюции это усовершенствование
обll ей эффективнос.1'И механизмов жизни; это «всеобщее биолоrи-
ческое усовершенствование (Huxley, 1942). Следовательно, про
I'рессивная эволюция представляет собой направление, в котором
изменения, имевшие место в прошлом, и нынешние характеристики
не оrранич,ивают будущих возможностей. ОНа не заканчивается
тупиком (Нихlеу, 1942).
Обзор Bcero орrаническоrо мира показывает, что до сих пор
критериям проrресс.ивной эволюции удовлетворяет только вид
Ното sapieпs с ero разумом, культурой и rибким поведением.
«Лишь по одной-единственной линии проrресс продолжается и oc
тается возможным в дальнейшем по линии человека» (Hиxley.
1942) .
Смена rосподствующих ТИПОВ
К этой проблеме можно подойти с друrой стороны изучая
палеонтолоrическую историю животных. Как показывает палеон-
т<>лоrическая летопись, с течение.м времени ОДНИ I'осподствующие
ЗЗ2 ЧаСТIJ V/I. От орzанuческой эволюции к. эволюции человека
типы сменялись :друrим,и. Ниже 'перечислены ifр} n'ПЫ, rосподство
павшие в разные эры или периоды:
Морские Мо.:JЛЮСКИ, трнлобиты И друrие морскне
беспозвоночные
Рыбы
Динозавры и друrие рептипии
Млеко питающие
Человек
Ордовик
ПоздниЙ палеозой
Мезозой
Третичный период
Четвертичный период
в этой последовательности rосподствующих форм прослежи..
вается повышение общей эффективности орrаиизмов. Ее можно
рассматривать как ряд все более эффективных, и в структурном,
и в функциональном отношении, орrанизаций, предназначенных
для выполнения rлавных жизненных процессов. rлавные вехи в
этом ряду последовательное развитие KOCTHoro скелета, леrких,
теrrлокровностн и мозrа, .способноrо к мышлению (Hиxley, 1942).
Сами эти вехи rлавные стадии рядов называют rрадами
(Huxley, 1958). [рада представляет собой некий уровень YCOBep
шенствования структурной и функциональной орrанизации дaHHO
ro орrанизма. Так, в приведенном выше ряду костные рыбы соот-
ветствуют одной rраде, амфибии друrой, а наземные рептилии
третьей.
В таком ряду rрад ясно выражены черты проrрессивной ЭВО':I10
ции (Hиxley, 1942, 1958).
Настоящие последовательности rосподствующих типов, OДНlIKO,
rораздо сложнее, чем представленный выше ряд (Simpson, 1967,
1974). В одной и той же широкой адаптивной зоне одновременно
может быть два и более rосподствующих типа (например, пасе
комые и наземные позвоночные) ИJ1И же два и более rосподствуlO
щих типа MorYT существовать одновременно, но в разных адап
тивных зонах (рыбы и наземные позвоночные). Следовательно,
СУЩе<:'fIвует не ОДИН, а несколько или MHoro таких ря:д.О'В.
Критерии проrрессивной эволюции
В эволюционном ряду, идущем от Низших морских животных
и через позвоночных доходящем до человека, прослеживаются из-
менения по мноrим признакам. Если этот ряд представляет собой
проrреССоивное эволюционное направление, то такие изменяющиеся
признаки дают нам набор критериев проrрессивной эволюции, по-
лучаемых более или менее независимо от субъективных сообра-
жений.
К числу изменений в ряду ОТ низш,их животных К высшим по
звоночным относятся следующие (Hиxley, 1942; Simpson 1949
1.967; Rensch, 1960): 1) повышение энерrетическоrо уровня жиз-
r л. 33. П рО2рессuвная эвОАЮЦUЯ
333
ненных процессов, т. е. возрастание скорости метаболических про-
цессов; 2) повышение эффективности размножения, в том числе
уоиление заботы о яйцах и (или) о молоди; 3) улучшение вос.
приятия сиrналов, поступающих из внешней среды, и способности
реаrировать на внешние раздражители; 4) возрастание способно
сти управлять средой и уменьшение зависимосl'И от нее.
rексли (Нихlеу, ] 942) подчеркивает последнюю характеристи-
ку контроль над средой и, С.ТIедовательно, относительную .незави-
симость от нее как самый ва ный отличительный критерий про-
rресоивной эволюции. На основании этоrо критерия млекопитаю
щим отводится высокое место, некоторым млекопитающим, Ha
при мер бобру, очень высокое, а человеку самое высокое.
Выдвинутый rексли критерий независимости от среды и спо..
еобности управлять ею..... это почти то, что нам надо. Однако reK..
ели истолковывает «среду» В С.I1'ИШКОМ широком, а «управление»
В узком смысле. с тем чтобы сделать этот .критерий возможно бо-
лее применимым к человеку. Между тем хотелось бы иметь кри-
терий, который бы.'1 бы менее антропоцентричным и более биоло-
rичным.' ;
Можно предложить альтернативный критерий: проrресс-ивная
эволюция слаrается из приспособлений к ряду последовательных
сред, все более отдаляioщихся от первоначальной среды, в кото-
рой обитали предки. Обсуждению этоrо критерия посвящается сле-
дующий раздел.
Обсуждение
Как и у ряда друrих эволюционистов, у меня имеются некото"
рые возражения против тезиса rексли. Я считаю, что в представ
лениях rеКС.flИ о проrрессНВНОЙ ЭВО.l1юции содержится ценное зер
но истины, которое следует сохранить; однако вместе с тем я по-
лаrаю, что rексли истолковывает проrрессивную эволюцию в
слишком узком смысле, коrда подrоняет ее rлавным образом к
эволюции человека.
Вся суть орrанической эволюции в ее разнообразии. Эволюция
человека или, точнее, БИО.'10rический аспект эволюции человека,
)несомненно, представляет собой важный результат орrаническоА
эволюции; однако важные результаты достиrнуты также и в дру-
rих филетических линиях. Проrресоивное развитие наблюдается,
например, у друrих млеко питающих, у птиц, у общественных насе-
комых, а также в царстве растений, которое обычно иrнорируют
при обсуждении ЭВОЛЮlI:ионноrо проrресса.
Кроме Toro. эра человека не будет продолжаться вечно. Воз-
можно, что орrаническая эволюция достиrнет в будущем, коrда
человек сойдет со сцены, новых успех.ов. В настоящее время нель..
ая предсказать, какими будут те орrанизмы, которые займут roc
подствующее место в будущем.
.334 Часть V/I. От орzаничес/(ой эволюции к. эволюции человека
Попробуем переформулировать концепции проrресса и специа-
лизаuии, исходя из ТИПОВ эволюционных направлений. На такой
основе можно выделить три rлавных типа эволюционных направ-
лений: 1) кратковременные направления специализации, примером
которых служит одна из боковых ветвей у rавайсКИХ цветочниц
(rл. 31); 2) длительные направления специализации, примером
которых служит эволюция копыта у лошади (rл. 26) (эти два на-
правления ЯВЛS,lются каналиэированными и самооrраничиваю-
щимися); 3) проrреССИВНЫе эволюционные направления, которые
.влекут за собой общее совершенствование биолоrической орrани
зации. Ряд «наземные рептилии теплокровные млекопитаlOщие
млекопитающие, обладающие способностью к высшей вервной
деятельности, разумные rоминиды» представляет собой лишь
.один пример прQrрессивной эволюции; друrим примером служит
ряд «папоротники ---- семенные растения деревянистые покрыто-
.семенные .однолетние тр авянистые покрытасеменные».
Проrрессивные направления можно представить себе как сверх-
.длительные направления специализации, устранив тем .самым рас-
хождение между nporpeccoM и специализацией. Проrрессивные
направления это направления специализации, при которых opra
ниэмы становятся специал,изированными к адаптивным зонам,
.обычным на суше, но очень далеким от питатеJlьноrо бульона
предков.
Жизнь началась в водах, боrатых питательными веществами.
Истощение питательных веществ в морях послужило толчкоМ к
.развитию фотосинтезирующих водных форм, а также форм, п.и-
тающихся этими фотосинтезирующими формами, и хищников, пи-
тающихся последними. Последовавшее за этим переполнение мо-
.реЙ орrанизмами побудило заселеНИе несколькими филетическпми
rруппами суши, а несколькими друrими..... воздуха. Наземные ор-
rанизмы постепенно переходили из теплых влажных областей суши
13 сухие и холодные области. Человек венчает собой одно из про-
rрессивных направлений; но rрызуны, птицЫ, крылатые насеко-
мые, пустынные кустарники и однолетние растения пустыни BeH
Чают на данном этапе истории Земли друrие такие направления.
Постепенное завоевание сред, широко распространенных на
земле, но в основном малоприrодных для жизни, как, например,
бедные питательными веществами водоемы, засушливые области
и воздух, аспект, общий для истории тех длительных адаптив-
ных направлений, которые мы мазываем проrрессивными.
rлаlва 34
ФИЛОfЕНИЯ rоминид
Система классификации приматов
Отряд приматов примечателен не только своим разнообразием,
но в особе'нности диапазоном этоrо разнообразия. Расстояние
между такими формами, как тупайя или лемур, на одном конце,
и человекообразная обезьяна или человек на друrом, orpoMHO.
Разнообразие в друrих крупных отрядах млекопитающих в общем
не выходит за пределы одной rрады, однако в отряде приматов
оно соответствует ряду rрад.
Табл. 34.1 даже чисто формально дает представление об об
щей степени разнообразия среди приматов. Они делятся на 4 под..
отряда, 11 семейств и 60 родов. Но это еще не все. Попробуйте,
читая табл. 34.1, мысленно представить себе перечислеиных в ней
животных, которых вы видели в зоопарке или в книrе по естест-
венной истории М..1екопитающих. Это даст понятие как о диапазо
не, так.и о степени разнообразия в этой rруппе.
Диапазон разнообразия приматов составляет примерно пять
rрад: 1) тупайи, которых некоторые систематики выделяют из OT
ряда приматов; 2) собственно полуобезьяны (лемуры, ЛО}>И, дол..
rопяты и т. д.); 3) широконосые обезьяны; 4) узконосые обезьяны
5) человек.
Приматы, занимающие нижний конец этоrо диапазона rраду
приближаются. к несколько более примитивному отряду насекомо"
ядных. Промежуточными формами между двумя отрядами служат
тупаЙи. При маты, возможно, произошли от насекомоядных. Oc
татки насекомоядных обнаруживаются в палеонтолоrической лето-
писи вплоть до меловоrо периода, а остатки приматов вплоть до
па.ТIеоцена.
Ископаемые остатки свидетельствуют о том, что первыми при-
матами были полуобезьяны, что они были широко распространены
и что в палеоцене и эоцене у них происходила .адаптивная радиа-
ция. В зоцене и олиrоцене некоторые боковые линии вымерли, а
друrие сохранились до настоящеrо времени. ВЫЖИВШllе rлавные
ветви соответствуют различным надсемействам, перечисленным в
табл. 34.1 (Simpson, 1967).
Hominoidea
ОДНа Нз rлавных ветвей приматов, отделившаяся от остальных
в эоцене, это линия Hominoidea. Она состоит из трех. семейств:
двух ныне живущих (Pongidae и Hominid_ae. _ _ одноrо вымершеrо.
Таблuца 34.1
КJlасснфикация современных приматов (состаВJlеиа по данным
CoJbert, J969 и Walker, J964)
Крупные rpynnы
Роды It виды
rеоrpaфвчetКое
рас:простраиеНве
Подотряд Lemuroidea
НадсемеЙСТ1l0 Tupaioidea
Сем. Tupaiidae .
Тупайи
l-IадсемеАство Lemuroidea
. Сем. Lemuridae
Лемуры
Сем. Indridae
.и ндри
НадсемеЙСТ80 Daubentonioidea
Сем. Daubentoniidae
Ай-ай
1-Iадсемейство Lorisoidea
Сем. Lorisidae
Лори, raпaro .
Надсемейство Tarsioidea
. Сем. Tarsiidae
Долrопяты
Подотряд Platyrrhini
Нздсемейство Ceboidea
Сем. Cebidae
Капуцины
Сем. Callithricidae
Иrрункн
Подотряд Саtаrrhiпi
Надсемеиство Cercopithecoidea
.Сем. Cercopithecidae
Мартышки
Павианы
. Надсемейство Hominoidea
Сем. Pongidae
Человекообразные обезьяны
Сем. Hominidae
Человек
5 родов (Тираю и др.) Тропическая Азия 8
,..., 15 видов Малайзия
6 родов (Leтur и др.) MaAaracKap
,..., 16 видов 3 рода;
4 ВИда »
Монотипическое:
Daubentoпia тadagas »
carieпslS
б родов (Loris, Oa/Dgo Тропические области
и др.) ,..., 12 видов Старото Света
Единственныii род Малайзия
Tarsiиs; 3 вида
12 родов (CeЬиs, Alo- Тропические области
иatta н др.); ,..., 37 вн- HOBoro Света
Д08
4 рода (Callithrix Тропические области
и др.); ,..., 33 вида Южной Америки н
Панамы
16 родов (Масаса, Тропические и суб.
Chaeropithecиs, Papio тропические обла-
и ДР.); "-J ба виДОв сти CTaporo Света
5 родов; ,..., 10 видов Тропические обпасrи
Hylobates (rи6бон), CTaporo Света
Syтphalangus (сиз-
M3Hr), Pongo (opaH
тутан), Рап (шимпан-
зе), Gorilla (rорип-
.nа)
МОf[отипическое: Ho Весь земной шар
то sapiens
r л. 34. ФU"'ОlеНUR zо.мuнuд
331
т аблuца 34.2
становлеиныА и вероятный возраст рОАОВ и ви.l.ОВ Hominidae (СатрЬеIl, 1974)
I ПР",",ИЗNТeпЬНыА возраст, roды
РОД и вид
Эпоха
Raтapithecus
От миоцена до ПJJиоцена Вероятно. от 18 до б млн.
лет до н. э.
14 млн. пет до н. э.
9 мпн. лет до н. Э.
R. wickeri
R. puпjablcиs
Australopithecиs
А. africaпus
А. boisei
От ппиоцеиа до ппейсто- Вероитно. от 6 до 1 млн.
цена пет до н. э.
5,5 1,3 млн. пет До н. э.
3 1,2 млн. пет
Ното 1
От ппейстоцена
временной
до со- Вероятно, начиная от 1,3
мпн. пет до н. э.
1,300 000 400 000 лет до
11. 3.
250000 лет до Н. Э. И до На-
СТQящеrо времени
Н. erectиs'
Н. sapiens
1 Суд,. по некоторым IIОВЫМ предваритe.'IЬНЫМ даиным, род Ното н вид Н. r ctи,
IIOЭкнк.1Н раньше, чем здесь указано.
семейства человекообразных обезьян (сем. Oreopjthecjdae, вклю-
чающее род Oreopithecиs). Эти семейства соответствуют трем
ветвям BToporo порядка в линии Hominoidea.
Ископаемые остатки по зт,им семействам, .найденные в О.'Iиrоце-
новых и миоценовых отложениях, фраrментарны, а поэтому мно-
rие важные моменты их филоrении остаются неясными.
Jlиния Oreopithecus возникла в миоцене и вымерла в плиоцене;
вполне возможно, что она существовала еще до миоцена. Настоя
щие человекообразные обезьяны (Pongidae) представлены родом
Aegyptopithecus в олиrоцене и Dryopithecиs в миоцене и плиоцене
(МЫ здесь придерживаемся консервативных взrлядов о делении
На роды; специалисты нередко делят ЭТИ роды на несколько
друrих) .
Самым древним известным предстаsителем Hominidae является
Ramapithecus, найденный в миоцене и нижнем плиоцене. О Raтa
pithecus известно HeMHoro. По видимому, предки rоминид диверrи-
рова..1И ОТ дриопитеков. rоминиды в миоцене, очевидно, уже обо-
собились от Pongidae (см. Romer, 1966; Simpson, 1967; Colbert.
1969) .
к семейству Hominidae относятся три рода (опять таки ПРИflИ-
мая консервативную концепцию рода): Ramapithecus (древние
че",овекообезьяны), Australopithecиs (австралопитек) и Нато
ЗЗВ Часть V 11. От орzа",ичесlCOй эволюции " эволюции человеlCa
(человек). Их возрасты приведены в табл. 34.2. Эти оценки почти
непрерывно изменяются по мере обнаружения новых находок И
совершенствования методов определения их возраста.
/j
\.
Рис. 34.1. Черепа ископаемых rоминид (по Bracc, NеlS0Л. Коrл, 1971; рисунки
выполнены М. L. Brace с экземппяров, хранящихся в разных музеях).
Все рнсункн выполнены в одинаковом масштабе. А. Australopith cus a/ricanиs. иижний
ппеАстоцси. Южная Африка. Б. Ното erectus средиий ппейстоцеи. Пекии. В. Ното sapiens.
иеаидерта.1ец. верхний плеRстоцен, Ирак. r. Ното sapieпs, верхний плейстоцеи, Лекин.
Плио плейстоценовые австралопитеки известны rлавным обра
зом из Южной ,и Восточной Африки. По строению черепа, лица
и объему rоловноrо мозrа (рис. 34.1) они сходны с человекооб-
разными обезьянами, но по зубам, вертикальному положению
тела н передвижению на двух Horax близки к человеку. Сущест-
вовало по крайней мере два вида австралопитеков, живших в
одно и То же время и различавшихся по размерам (А ustralopithe
cus africanus и А. boisei), а может быть, их было и три. [Кемп
белл (Campbell, 1974) относит ископаемые формы, известные под
названиями А. robustus и Нота habilis, к А ustralopithecus boisei r
r А. 34. ФUЛО2енuЯ 20JJtинuд
339
однако некоторые друrие авторы рассматривают их как самостОЯ-
Т2льные единицы.] Австралопитеки вымерли до окончания плей-
стоцена.
Род Homo J обладающий большим rоловным мозrом, представ
J[яет друrую линию, которая, хотя и позже, перекрывалась 80
времени с Australopithecus. Различают два вида Нота: Н. erectus
(обезьяночеловек) и Н. sapiens (современный человек), к которо-
му относится и Н. neanderthaleпsis (рис. 34.1). Оба жили в Афри-
Ке и в друrих областях CTaporo Света в плейстоцене. Н. erectus
н Н. sapiens составляют пару последовательных видов, в кото-
рой первый с течением времени постепенно переходил 80 вто-
рой. На каждо отдельном временном уровне существовал только
один вид Нота, но Н. erectus или ero непосредственный предок,
по-sидимому, сосуществовал с Australopiphecus.
Нота близок к Australopithecиs и, возможно, произошел от
Hero. Точное строение филоrенетическоrо древа, однако, неизвест-
но, и разные антрополоrи придерживаются различных rипотез ОТ-
носительно филоrении этой rруппы (см. Brace et аl., 1971; РН-
Ьеаm, 1972; РilЬеаm, Gould, 1974). Одна вероятная возможность
состt)ит в том, что Нота erectus произошел от А ustralopithecиs
a/ricanus. Быть может, ископаемые находки, которые удастся об-
наружить в будущем, позволят разъяснить ero филоrению.
Наследие древесноrо образа жизни
Приматы обитают преимущественно в областях с теплым кли-
матом (см. табл. 34.1); ОНИ живут rлавным образом На деревьях,
а такие наземные формы, как па8ианы и человек, составляют ис-
ключение; приматы в основном всеядные. Древесный образ жиз-
нн привел к развитию ряда приспособлений, которые впоследствии
использовал человек.
Млекопитающее, живущее на деревьях, должны уметь точно
оценивать расстояния, фокус,ировать взrляд На определенных объ
ектах, удерживаться на ветках и схватывать предметы, rодные в
пищу. В соответствии с этими требованиями у приматов имеются
такие приспособительные особенности, как хорошо раз&итое зре-
ние, х:ватательные передние КОIIечности ос противопоставляющ.и.мся
большим пальцем и хвост, С.1'Jужащий балансиром.
Острота зрения приматов обеспечивается суммой ряда призна-
Жов. rлаза хорошо раЗБИТЫ. Они направлены вперед, что создает
возможность одновременной фокусировки обоих r лаз .на одном
и том же объекте и оценки раССТОЯНI1Я по параллаксу. Наличие
же.лтоrо пятна на сетчатке улучшает фокусировку. Наконец, при-
маты обладают цветовым зрением, KOToporo лишены мноrие rруп-
пы млек-опитающих и которое, вероятно, способствует улучшеаию
объемноrо зрения.
340 Часть Vll. От ор аничес"ой эволюции IC эволюции челове"а
Зрение служит в свою очередь одним из существенных компо-
нентов высшей нервной деяте.Т']ьности приматов, которая также
представляет собой сложную черту. Важное значение для высшей
нервной деятельности имеет прежде Bcero rоловной мозr, хорош()
развитый у всех высших приматuв. Однако то, что этот мозr мо-
жет узнать о внешнем мире, в знач,ительной степени зависит от
сочетания зрения и передних конечностей приматов. Друrие opra
ны чувств, в особенности слух, иrрают дополнительную роль.
Высшая нервная деятельность приматов основана r лавным об
разом на сочетании r лаз, способных к одновременной фокусиров-
ке, и хватательных или осязательных кистей. У друrих животных
эта деятельность имеет иную основу. У собак, например, она за-
висит прежде Bcero от сочетания способных к фокусировке r.Т'I аз,
обоняния и очень чувствительных ушей; у пчел от сочетания
СЛ()ЖНых rлаз, цветовоrо зрения и rиб их антенн, воспринимаю
щих запахи.
Сочетание rлаз рука было и остается той основой, которая
давала возможность приматам, предшествовавшим rоминидам,
концентрировать внимание на отдельных объектах. У rоминид
это же самое сочетание леrло в основу .изrотовления орудий и их
использования, экспериментирования и решения задач.
Наследие наземноrо образа жизни
Человекообразные обезьяны обитают в лесах и ведут, во вся
ком случае частично, древесный образ жизни. rиббоны проводяТ
большую часть своей жизни на деревьях и передвиrаются при по-
мощи брахиации и прыжков. Шимпанзе и rориллы полуназем
ные животные и передвиrаются по земле, опираясь на подошвы
ступней тыльную сторону пальцев передних конечностей.
В от.тшчие от этоrо Australopithecus и Ното были и остаются
животными открытых саванн и равнин. Споообом передвижения у
них служит ходьба или беr на двух Horax. Хождение на двух но-
rax и ее следствие вертикальное положение тела наиболее
яркие отличительные признаки rоминид.
В середине iи в конце третичноrо периода протяженность лес
ных местообитаний постепенно сокращалась, уступая место сте-
пям. В результате изменения климата и растительности возникали
новые местообитания. Заселение HOBoro равнинноrо местообита-
ния, возможно, определ.ило диверrенцию примитивных rоминид от
их предков человекообразных обезьян, обитавших в лесах. Пе
редвижение на двух Horax и вертикальное положение тела были
частью Toro сочетания адаптивных признаков, которое сделалО
rоминид приспособленными к ж,изни В саваннах и На равнинах.
Большая часть данных о дальнейшей эволюции rоминид в пдей-
стоцене, которыми мы располаrаем, указывает на все большее BЫ
r л. 34. Филоzе",ия 20мин.ид
341
прямление тела при откинутой назад шее и rолове и на сопровож
давшее ero улучшение походки.
Передвижен.ие на двух Horax имело важное следствие: оно ос..
вободило руки. которые моrли теперь заняться изrотовлением 11
использование орудий и оружия.
Australopithecus пользовался прими-
тивными каменными орудиями, а
Нота erec.tus более совершенными
(рис. 34.2). Умение обращаться с
предметами важная составная часть
высшей нервной деятельности древес.
ных приматов, как это уже отмеча-
лось. А теперь, с переходом от дре-
BecHoro образа жизни к наземному и
с освобождением рук от участия в пе
редвижении, манипуляция предмета-
ми Mor ла развиваться в полной мере.
Первым результатом этоrо было раз.
витие простейших peMeceJ1.
Развитие высшей нервной дея-
тельности у приматов имеет большую
адаптивную ценность и ему несомнен-
но блаrоприятствует естественный от-
бор. У пр.иматов, Не относящихся к
rоминидам, выявляется эволюцион-
ное направление в сторону развития
высшей нервной деятельности, дости-
rающей очень BblcoKpro уровня у
шимпанзе (Рап) и opaHrYTaHa (Роп-
go) и еще более BbIcOKoro..... у rоМИ-
нид.
Среди rоминид, если судить по объ-
ему черепа, Нато erectus 'Обладал
более развитым интеллектом, чем ав-
стралопитек (см. рис. 34.1). Диапа-
зон колебаний емкости черепной ко-
робки у этих двух форм и средние
емкости приведены н-иже (СаmрЬеll,
1974) .
А иstralopithecus africanus
Н ото erectиs
А
5см
Рис. 34.2. Нижнепапео.rтитиче..
ские каменные ОРУДИЯ из Вос-
точной Африки (Oakley, 1949).
Орудия изrотовлены из ву.1К2ннче.
екнх об.i10МКОВ; изображены в оди
наковом ....асшТабе. А. BepXHHff
плейетоцен, опдуваАекая эпоха,
ущелье 0.1дуваи. Таllrаньика. Б.
Средний плейетоцен. руби.10, ашель
екая эпоха. Кеиия.
Диапазон, ем3
435 815
775---1225
Средняя, см3
588
950
Весьма вероятно, что более высокое развитие интеллекта 11
более совершенные орудия и оружие у Ното erectиs или у ero
непосредственноrо предка дали ему преимущество над Australo..
pithecus в прямой и косвенной конкуренции между этими двумя
342 Часть уи. От орzа",ической эвОАюции к эволюции человека
rруппами в плейстоцене. Во всяком случае австралопитеки вы-
мерли, а Ното erectus выжил и позднее эволюционировал в
Н. sapieпs.
Расовая изменчивость
Для современной исторической эпохи характерно всесветное
распространен.ие Ното sapiens и ero диффсренцировка на ряд
rеоrрафических и .бесчисленное множество локальных рас. Джерн
(Gаrп, 1961) различает следующие rеоrрафические расы человека:
Ази aTCKO монrОЛОИДlJа я
Американские ИНДейЦЫ
Индийская (Индия, Пакистан И т. д.)
Европеоидная белая
Африканская неrроидная
Австрапийская неrроидная
Мелансзнilска SI
Микронезиiiская
Попинеэийская
Это тwпичные rеоrра.фИ'чес,кие ра.сы, подобные расам у друr.их
ВИДОВ млекопитающих. Некоторые ра,совые .признаки .разме.ров теда
и ero пропорций, ПИоr,меlJтации и т. п. коррелированы с 'Кли.матом
(ом Соол, 1955; Garn, 1961). Хорошо извоотно, например, ЧТО .ра-
совая И3менчи'вость цвета кожи КОр,релирует с КО.'Iи'чесТlВ'()М co.'1
нечноrо света в данной области.
Цвет кожи у человека реrулирует синтез витамина О.
Определенное количество этоrо витамина необходимо для нор.
мальноrо роста; недостаток ero вызывает рахит, а избыток
Qбызвествление костей. Витамин D синтезируется в коже, и ero
синтез реrулируется СО.тIнечным светом: чем больше солJ.lца, тем
больше витамина О.
Темнокожие расы живут в тех областях, rде MHoro солнца.
Темные пиrменты, содержащиеся в коже, ПJlОХО пропускают сол-
нечные лучи и предотвращают синтез избыточных количеств вита-
мина D. Напротив, белая раса, изнача.ТIЬНО обитавшая в северных
частях Европы. не моrла бы синтезировать достаточное количе.
-Сl'во витамина D, не 'имея светлой кожи, которая пропускает мак-
-симум солнечных лучей. Но в жарких странах белая кожа поrло.
щает слишком MHoro солнечноrо света, и поэтому представители
белой расы, оказавшись в таком климате, покрываются заrаром,
что представляет собой фенотипическую модификацию, препятст-
вующую избыточному поrлощению света. Что касается представ'и-
телей черных рас, то если бы им пришлось жить на севере Европы
r А. 34. Фило,е",ия zoAtиHиa
343
в естественных условиях, они вероятно, поrибли бы от рахита.
Таким образом, различия в цвете кожи представляют собой адап
тивный признак, зав'исящий от интенсивности солнечноrо излуче..
ния в данной местности (Loomis, 1967).
Подобные факты свидетельствуют о роли eCTecTBeHHoro отбора
в развитии по крайней мере некоторых расовых признаков Ното
.sapiens. Локальная расовая изменчивость иноrда носит случайный
характер, позволяя предполаrать, что в ее возникновении участ
в 'ют также дрейф reHOB 'или отбор и дрейф (см. r.11. 13). В дру"
rих случаях характер расовой изменчивости у человека указывает
на реrулнрующее влияние миrрации и потока rепов (см. rл. 18).
ВЫВОДЫ
Некоторые стороны эволюции rомИНИД, 06суждавшиеся в этой
rлаве, представляют собой продолжен-ие эволюции приматов, не
относящихся к rоминидам. Так, мы обнаружи.1И эволюционные
направления, приведшие к использованию руки, развитию высшей
нервной деятельности, изменению положения тела и т. д.; МЫ ви"
дим указания на квантовый сД"виr к новому способу передвиже
ния вместообитании HOBoro типа, на межвидовой отбор между
отдельными линиями rом,инид; образование rеоrрафических рас у
выжившеrо и широко распространившеrося вида rоминид.
Все эти аспекты эволюции rоминид предс.тавляют собой часть.
БОv1ее обширной картины ЭВО.ТIюции орrаническоrо мира. Их можно
объяснить как результат действия эволюционных факторов и
си..l, описанных в более ранних rлавах этой книrи.
В связи с этим следует указать, что за 10000 лет человек MO
JКeT пройти через 600 поколеНий. За недолrnй период отбор или
отбор в сочетании с дрейфом reHOB моrли вызвать существенные
эволюционные изменения. А за более длительные периоды, на
протяжении которых, соrласно ископаемым данным, протекала
эволюц-ия rоминид, моrли, конечно, произойти rораздо более rлу"
бокие эволюционные изменения.
rла.ва 35
эволюция ОБщвст!ввнноrо ОБРАЗА ЖИЗНИ
У .ПрИIМАТОВ
Стадность уприматов
(не относящихся к rомин дам)
Приматы ---- живОТные общественные. Отдельные их особи
<>бычно входят в состав семейных rрупп и более обширных стад.
rрупповой или стадный образ жизни у высших приматов (Т. е.
над уровнем полуобезьян) определяется двумя условиями. У ши-
роконосых и узконосых обезьян по сравнению с большинством
друrих млекопитающих начальные и ювенильные стадии развития
продолжаются сравнительно долrО t что приводит к длительной
-связи между матерью и детенышем. А отсутствие у высших прима-
70В и мноrих полуобезьян четко выраженноrо сезона размноже-
ния в свою очередь ведет к непрерывной или во всяком случае
длительн<>й связи между особями разноrо пола (более подробно
см. De Vore, 1965). Сочетание связей самка детеныш и самка
-самец приводит к образованию семенных rрупп И развитию стад-
НОСТИ.
В структуре семейных и более обширных rрупп приматов Ha
Q..'Iюдается значительная межвидовая изменчивость. Подробное
<>писание орrанизации общественной жизни у приматов выходит
за пределы темы этой rлавы (см. De Vore, 1965; Eisenberg et al.,
1972; Wilson, 1975). Айзенберr 'Н др. (Eisenberg et аl., 1972) соз-
дали полезную систему классификации разных типов rрупповой
-структуры у приматов, которую мы здесь приводим снебольшими
изменениями (таксономическая КJJасс'ификация приматов дана в
табл. 34.1).
1. Материнская се.мЬЯ 1 состоящая из матери и детеныша.
иентры активности половозрс..'Iых самок и самцов обособлены.
Наблюдается у некоторых видов лемуров и лори и у руконожек
.ай ай.
2. Семья с двумя родителями. Такая семейная rруппа состоит
из самки, самца и детенышей. Наблюдается у одноrо вида мохна-
тых индри, У некоторых мармозеток, капуцинов и rиббонов.
3. Стадо с одним самцом. Стадо СОСТОИТ из нескольких мате-
ринских семей и одноrо половозрелоrо самца, поддерживающеrо
контакт со всеми семьями. Самец Относится очень враждебно к
друrим половозрелым или созревающим сам нам. Такие стада Ha
блюдаются у некоторых капуцинов (в том числе у ревунов) и у
низших узконосых обезьян (в том числе у rамадрилов и rелад).
rл. 36. Эволюция 06щественноZО образа жизни у npu.мЙToв 345
4. Стадо со мноеими самцами раэноzо возраста. СплочеНl{ая
rруппа, состоящая из нескольких самок, нескольких самцов и дe
тенышей. Самцы проявляют дopyr к друrу некоторую терпимость.
Это делает возможным сосуществование нескольких самцов раз
Horo возраста, обычно молодых, между которыми устанавлива
ется иерархия в соответствии с их возрастом. Наблюдается у He
которых капуцинов (в том числе опять таки у ревунов), у низших
узконосых обезьян (в том числе у макак) и у rорил..1Ы.
5. Истинное мноеосаМЦО80е стадо. rруппа, подобная rруппе 4
но самцы относятся друr к друrу rораздо терпимее, что де.Ilает
возможным сосуществование нескольких половозре.ТIЫХ самцов.
Отношения между самцами носят характер кодоминантност,и и со-
трудничества, блаrодаря которым в стаде создается система rиб.
кой олиrархии. Наб.пюдается у одноrо ВИДа лемуров, одноrо вида
мохнатых индри, у некоторых низших узконосых обезьян (в том
числе у па&ианов и макак) и у шимпанзе.
Приведенные выше типы rрупповой структуры уприматов
можно прсдставить себе как некоторый эволюционный ряд. Можно
рассматривать тип 1 как наиболее примитивную структуру,
ТИП /1 как промежуточную, а типы 4 и 5 как их производные.
ПараЛ.'lе.ТIЬНО этим изменениям в структуре rруппы наблюдается
тенденция к увеличению ее размеров (Eisenberg е! а1., 1972).
Типы орrанизации стада примерно коррелируют также с ме.
стообитанием. rруппы, воэrлавляемые одним самцом (ТИП 3),
встречаются r.'IaBHbIM образом, хотя и не исключительно, у ВИДОВ,
живущих и питающихся на деревьях. В отличие от этоrо истинНО
MHorocaMUOBbIe стада (тип 5) встречаются в преобладающем БО 1Ь
шинстве случаев среди видов, ведущих ПО..1уназемный образ жизни
(Eisenberg et аl., 1972).
Адаптивное значение стадности
у высших приматов, на которых мы здесь сосредоточим свое
внимание, стадность ВЫПО.'1няет несколько важных функций. Члены
стада объединяются для удержания территории и защиты ее от
хищников. Кроме Toro, что очень важно, стадо создает ту среду,
в которой происходит передача опыта друr друrу и обучение дe
тенышей разноrо рода жизненно важным навыкам: распознаванию
съедобных плодов и листьев, распознаванию ядовитых растений,
умению обнаруживать местонахождение BparoB и т. П.
В С.Т'Iедующих двух разде.'13Х кратко рассматр.иваются HeKOTO
рые ФУНКЦИИ стадности у ПРИ)1ЭТОВ.
Язык
Потребность в передаче информации и координации деятеЛh
ности отде.Т]ЬНЫХ особей в стадах обезьян удовлетворяется при
помощи !\r1ИМИКИ И вокализации. Оба способа передачи информа
346 Часть VI1. От орzанической эволюции к эволюции человека
ции достиrли у обезьян BblcoKoro развития (см. Ое Vore, 1965).
Капуцины и мартышки особенно знамениты своей непрерывной и
шумной болтовней.
Уокер (Walker, 1964) собрал и расшифровал звуки, .издавае
мые мирикиной (Aotus trivirgatus, Cebidae). Оказалось, что эти
ночные обезьяны издают до 50 различных звуков. При помощи
таких звуков обезьяны MorYT выразить следующие наблюдения
и ощущения:
Ощущение опасности
Предупреждение об опасности
Любопытство без опасения
Любопытство с опаской
Дружепюбное приветствие
Воодуwевление
Недовольство
«Не приставай»
«Дай мне это»
«Я хочу пойти с тобой»
«Я любпю тебя»
«буук»
«вуук:,
«ююх»
«хьюх», «вью»
«чррр»
Щебетание, напоминающее птичье
«эк», «кэк»
«ук» (повторяется)
«эхе»
«ЭХ, ЭХ, эх»
«уух, УУХ»
Уошберн (Washburn, ц:ит. по De Vore, 1965) рассказывает сле-
дующий случай о павианах, живущих в Африке, в Национальном
парке Найроби. Эти павианы находятся под охраной и привыкли
к людям, разъезжающим на автомобилях. Однажды один пара
зитолоr застрелил двух павианов прямо из машины. Очевидно, Не-
сколько павианов были свидетелями этоrо события, хотя нет ни-
каких сомнений в том, что друrие ero не наблюдали. Новость
быстро распространилась по всей колонии павианов, в результате
чеrо павианы перестаЛ1И подпускать к себе людей, находившихся
в машинах, и это продолжалось по крайней мере восемь месяцев.
Уровень развития высшеii нервной деятельности
и обучение
Для сохранения и развития языка, способноrо служить для
передачи СКО.'1ько нибудь существенноrо объема информации, не-
обходимо, чтобы размеры rруппы, использующей этот язык, пре-
вышали некоторую КРИ1'ическую величину. Необходимо также,
чтобы члены такой rруппы об.'1адали определенным уровнем раз-
вития высшей нервной деятельности и способностей к обучению.
N\ежду общественными rруппами и различными аспеКТаМИ высшей
нервной деятельности существуют сложные взаимодействия rИ об
ратные связи.
r А. 35. Эволюция обществеН",ОlО образа жизни у приАЮТО8 341
Высшие приматы выделяются среди остальных Животных ypOB
нем развития высшей нервной деятельности. Размеры rоловноrо
мозrа относительно общих размеров тела у обезьян rораздо боль.
ше, чем у представителей друr.их отрядов млекопитающих. Пример.
ной мерой уровня развития высшей нервной деятельности у узко
носых обезьян может служить емкость черепа. Ниже приведены
средние данные о емкости черепа у некоторых видов (Campbell.
1974):
\aKaK (А1асаса)
Павиан (Papio)
fиббон
100 см 3
Шнмпанзе
OpaHrYTaH
rорилпа
200 »
103 »
383 »
405 »
505 »
Эта ЭВО..1юционная тенденция продолжается у rомиНИД:
Aиstralopithecиs africaпиs
н ото erectиs
Н ото sapiens
588 СМ З
950
1330
Известны мноrочисленные примеры способности узконосых
обезьян решать задачи. Большая часть таких данных относится к
шимпанзе. Шимпанзе Не только используют орудия для добывания
пищи, как это делают некоторые друrие млекопитающие и птицы,
но и в самом дe.ТIe изrотовляют. 'из палочек примитивные орудия
для выполнения каких-либо определенных целей (Goodall, цит.
по Ое Vore, 1965).
Обучение языкам и сложным навыкам требует не юлька на-
личия природных интеллектуальных способностей, но и времени.
Таблицй о З5.1
Продолжительность стадиii индивидуальноrо развития у разных при матов
(Wilson, 1975)
Продолжи- Младен- Общая продолжитель- Общая
тельность ческаfl Ю вениль- ность постнатальных продолжи-
Вид беременности. стадия, ная стадии. стад.ий ДО достижения тельность
дни roAbl roAbl половой зрелости. жизнн,
roPobl roды
Лемур 126 0,75 1,75 2,5 14
Макак-резус 168 1 ,5 6,0 7,5 27 28
rнббон 210 2,0(1) 6,5 8,5 30+
Шимпанзе 225 3,0 7,0 10,0 40
rорипла 265 3,0+ 7,0+ 10,0+ 35 ( 1)
OpaHrYTaH 275 3,5 7,0 10,5 30+
Человек 266 6,0 14,0 20,0 70---75
348 Часть 'II. От орzаНflческой , вОЛЮЦtlfl 1с эволюции человека
Друrая хорошо известная тенденция в эволюции приматов ---- YBe
JIичение ПРОДО.rIжительности стадий раЗВИТИЯ t предшествующих
наступлению половой зрелости (табл. 35.1). Отмечалось, что чело-
векообразные обезьяны, возможно, обладают способностями, Не-
Qбходимыми для Toro, чтобы выучиться основам человеческоrrо
языка, но не MorYT реал.изовать эти способности, так как слишком
быстро достиrают зрелости.
rоминиды
Все перечисленные выше адаптивные признаки, связанные со
стадностью и имеющиеся у высших обеЗЬЯН t в еще большей степе-
ни выражены у rоминид. Кроме Toro, у rомиНИД ПРОЯВ.'1яются, и
притом С еще большей аилой, некоторые друrие адаптивные пре
имущества, которые дает общественный образ жизни.
rоминиды диверrировали от предковой линии обитавших в ле-
сах ПРliматов, переселившись в саванны .и на раВНИНЫ t rде они
перешли к наземному образу жизни (СМ. r.'l. 34). Первые наземные
rОМИНИДbl, поселившиеся на О1КрЫТЫХ местах, подверrались боль-
шой опасности со стороны крупных хищников. rрупповой образ
Жизни создавал возможность успешно защищаться от таNИХ хищ-
ников, тоrда как одной или нескольким особям это было не под
си..,у. Таким образом, орrаниэоваlIпое стаДОt очеВИДНО t обладало
высокой селективной ценностью в смысле защиты от крупных
'хищников во в}{ов" заселенном открытом местообитании.
Кроме Toro, ранним rоминидам, вероятно, было леrче охотить-
ся и вообще добывать пищу rруппами, а Не поодиночке.
Поэтому есТЬ все основания считать, что австралопитеки вели
общественный образ жизни, и подобное преДПО.ТIOженне подтвер-
ждается некоторыми косвенными данными. Австралопитеки охо-
Тились на павианов, которые всеrда держатся rруппами и моrли
бы леrко оказать сопротивление отдельным охотникам, вооружен
ным камнями и дуб.инками. Отсюда явно следует вывод, что ав-
стралопитеки охотились орrанизованными rруппами (Bartholo-
mew, Birdsell, 1953).
Самая примитивная форма общественноrо объединения, из-
вестная у. Ното sapiens,..... это rруппа. занятая охотой и собира-
тельством. ЛЮД:И, живущие за счет собирательства, существовали
в разных областях земноrо шара до недавнеrо прошлоrо, а в He
которых местах существуют и до сих Пор. В качестве примеров
можно привести индейские племена ШОШонов и алrонкинов в Ce
верной Америке t бушменов и неrритосов в Африке, коренных жи-
телей Австралии и Тасманп:и, а также индийские племена t оби-
тающие в джунrлях и на Андаманских островах (BirdseIl. 1958).
В состав такой rруппы обычно входит от 20 до 100 человек,
образующих ряд семеЙ t в которых имеЮТСЯ оба родителя. fруппа
r л. 36. Эволюция общественноzо образа жизнu у npuAraToB 349
занимает определенную территорию и защищает ее. Ее члены со-
бирают в I1реде.тJах этой терр/итории ту пищу, какую им удается
найти и которая варьирует в зависиМости от времени rода. В сбо-
рах участвуют как мужчины, так и женщины. Ни запасов пищи,
ни каких.либо друrих накоплений не существует. В обществе ца-
рит равноправие; в нем нет правящеrо класса и нет половой дис-
криминаЦИ11 (Birdsel1, 1958).
Примером ныне существующеrо общества собирателей служит
племя KYHr, живущее в пустыне Калахари в Южной Африке (КО-
lata, 1974). Это кочевое племя живет здесь не менее 11 000 лет,
.занимаясь охотой и собирательством. В последнее время, однако,
ИХ жизнь начинает изменяться под действием окружающей циви-
ЛIlзации (Kolata, 1974).
в rруппах первобытных людей, занимавшихся охотой и соби-
рательством, по видимому, существовала определенная орrаниза-
ДИЯ, очень сходная с орrанизацией, наб 1юдаемой во MHorocaMUo-
вых стадах высших приматов.
Че '10веческие общества на доисторической стадии культуры
обычно имеют более сложную экономическую основу и социаль-
ную структуру и более мноrочисленны, чем rруппа собирателей.
Достаточно вспомнить племена, занимающиеся охотой На крупно-
ro зверя, пастушьи племена и ПРllмитивные земледельческие об-
щиt1ы. По мере развития циви..1ИЗ(lUИИ величина и сложность со-
обществ человека возрастает. Диапазон обществеННО ПО lитических
объединений людей крайне велик от древних сельских Посе.пе-
ний до современных промышленных национальных rосударств и
индустриальных сверхдержав.
Обсуждение
Итак, мы видим, что возникшее у приматов направление эво
пюции обществе.нноrо образа жизни продолжается у Нота sa-
pieпs. На всех ступенях от полуобезьян до COBpeMeHHoro чело-
века отмечается общая тенденция к увеличению размеров об.
щественной rруппы и сложности ее орrанизации, повышению тех-
нической мощи, усложнению языа,, развитию интеллекта н увели-
чению продолжительности пер.иода, предшествующеrо ДОСТИЖению
половой зрелости.
Все это не обособленные, а взаимосвязанные и взаимодейст-
вующие напраВJlения; кроме Toro, они носят не .тJинейный, а Bce
объемлющий характер.
При обсуждении поведения и социальных явлений обычно при-
нято проводить различие между человеком и животными. За че-
ловечеством признается способность определять собственную су дь
бу, Tor да как судьба всех друI'ИХ млекопитающих, в том числе
высших обезьян, зависит от слепых сил природы. Конкуренцию
350 Часть VI1. От ОР2анuчес"ой эволюц.ии к эволюц.ии человека
за охотничьи и rнездовые участки и ИХ защиту у позвоночных жи.
вотных называют «территориальностью»; rомолоrичные ВЗ3'имо.
действия у людей называют -борьбой между державами или в
крайних ситуациях войнами, и их изучением занимаются исто.
р-ики. У высших обезьян разноrо рода социальные и психолоrиче.
ские результаты ДОСТlIrаются за счет eCTecTBeHHoro отбора, тоrда
как rомолоrИЧllые результаты у человека приписывают «культур-
ной эволюции». .
Но в чем заключается различие между территориальностью у
позвон.очных и У человека? Чем отличается действие естественно.
[о отбора на человека и на друrих приматов? ейст&ительно ли
человек управляет своей собственной судьбой, KaI{ ему хочется
считаТЬ t и если да, то почему он делает это так плохо или, быть
можеТ t он заблуждается?
При описании эволюции общественноrо образа жизни у прима.
тов нам неоднократно приходилось пересекать rраницы, разделяю-
щие разные области науки. Мы переходили от зоопсихолоrии к
приматолоrии, палеоантрополоrии, антрополоrии, археолоrии.
истории и проблемам cOBpeMeHHoro мира, но не обнаружили при
этом Toro места, [де можно бы.по бы провести естественную rpa.
ниuу, отделяющую человека от друrих высших приматов.
Эта rлава завершается рядом вопросов об эволюции чел.овека
и ero природе, на которые очень трудно ответить. В следующей
rлаве мы попытаемся найти ответы, хотя бы частичные t на эти
вопросы.
лаlва 36
КУЛЬТУРНАЯ rЭВОЛЮЦИЯ
Культурное наСJlедие
Культурное наследие, или унаследование традиций, это вся
совокупность знаний, представлений, искусств, обычаев и тех но-
лоrических навы.ков, КОТОРЫМИ располаrае1" данное человеческое
общество в любой данный момент своей истории. Вся эта сумма
знаниЙ и традиций результат открытий и изобретений, сделан-
ных предшествующими поколениями. Она передается и будет пе-
редаваться из поколения в поколение путем обучения в широком
СМЫС.1е этоrо слова. Каждое поколение может вносить в Культур-
ное наследие что то новое, и все эт,и вклады также будут переда-
ваться последующим ПОКО.ТIеНиям при ПОМОЩИ все Toro же про-
цесса обучения.
Постепенное развитие и накопление культурноrо наследия
представляет собой культурную эволюцию. В культурной эволю-
ции наследование приобретенных признаков реальный процесс.
Различия между человеком двадцатоrо века и че.hЬвеком ка-
MeHHoro века по морфолоrическим признакам, в том числе и по
емкости черепа, относительно невелики. Однако различия в их
культуре orpOMHbl. Изменения, происшедшие с Ното sapiens
со времен палеолита до нынешней стадии ero развития, вызваны
rлавным образом культурной эволюцией.
Орrаническая эволюция моrла обусловить возникновение и пер-
вые шаrи эволюции рода Ното. Однако одна ЛИШЬ орrаническая
эволюция не моrла привести к превращению первобытноrо че.ПО-
ВеКа в cOBpeMeHHoro. Здесь вступает в силу культурная эволюция,
н она дает объяснение тем изменениям, которые нельзя объяснить
факторами орrанической эволюции.
Культурная эволюция обладает собственной движущей силой,
отличной ОТ движущих сил орrанической эволюции. И культурную
эволюц-ию можно считать совершенно самостоятельныМ процессом,
хотя на практике она взаимодействует с эволюцией орrанической.
При специальном изучении культурной эволюции ее следует изу-
чать отдельно, однако при любом изучении человечества правиль-
Нее рассматривать cOBpeMeHHoro человека как продукт совместно-
ro действия орrанической и культурной эволюции.
Бодмер и Кавалли Сфорца (Bodmer,: Cavalli-Sforza, 1976)
составили таблицу сроков основных эволЮЦflОННЫХ изменений, как
орrанических, так и культурных, ПРОИСХОДН-ВШИХ У рода Ното,
которую мы приводим на рис. 36.1.
352 Ч асть V 1 1. От орzаНU'l.цкоt2 эвОАюции " эвОАЮции чеАО8еtuZ
Обучение
Есл-и рассматривать человека как одноrо из представителей
млеко питающих и высших приматов, то ero следует считать Лро
дуктом орrанической эволюции; однако человек как предс.тавитель
человечества является, как отмечал ось выше, продуктом OДHOBpe
менно и Qрrаиичес.кой, и культурной эволюции. Кроме Toro, те
подлинные опознавательные признак-и человека, котооым" он от.
Peg!lКЦИЯ КЛЫКО
ПРlfмохожgение
Увеличение 20ЛОfJНОlО МО32а
Каменные орggия
ИСЛОЛ6:308ание OZHR, при оmО8леНtJе пищи
Я3ЬIК (?}
НаС/(QЛ6 ная живопись
ОgомаШНИ8(Jние ЖUВО;'Н6IХ и раСТении
Обработка метОАлав
....
80 HиKHbtJi!Hиe 2OpogoB
пиС6менность
r I I I
/O.s <r? /
000000 000 000
000000 tb O 000
I
/00
ООО
fOgbI go н. Э.
I I J I
/0 0$ /"
000000 000 "'00
Рис. 36.1. Примерные сроки разпнчных орrанических и культурных ИЗМеНеНИЙ
в роде Ното (Bodmer, СаvаШ-Sfоrzа).
личается от животноrо, выработались в процессе культурной эво-
Л юции.
Каждый индивидуум Нато sapiens при рождении наделен KOH
ституцией животноrо, принадлежащеrо к приматам, но лишен
какой либо культуры. Культура приобретается в процессе инди-
видуальноrо развития в результате обучения в ШИрОКОМ смысле.
В первые rоды постнатальноr{) развития каждый индивидуум из
состояния человекообразноrо животноrо (при рождении) через
стадию варвара (ребенок) переходит в человеческое состояние в
ис.тинном ИЛИ диаrностическом значении слова «человеческое».
И это умственное развитие ИНДИвидуума происходит в результате
формирующеrо влияния обучения.
r А. 36. КУЛtJтgр",ая эволюция
353
Далее индивидуум восприн"мает ту культуру. в условиях ко-
торой оН (или она) родился и был воспитан, Не считая тех редких
случаев, Коrда ИН'щивидуум оказывается способным мыслить не-
зависимо и самостоятельно и переступает rраниды своей родной
культуры; однако даже и в этих случаях он в известной степени
сохраняет эту культуру.
В таком случае наиболее существенное различие между челО-
веком и животным лежит в сфере человеческоrо разума. а не тела,
а специфически человеческие аспеКты человеческоrо разума
продукт обучения (Briffault, 1927).
Концептуальное мышление
Одна из отличительных черт умственной деятельности челове-
Ка это способность к концептуа.JlЬНОМУ мышлению, способность
мыслить отвлеченными концепциями и создавать представления.
А для концептуальноrо мышлен,ия необходим язык. И более
Toro язык, имеющий rораздо более сложную структуру, чем язык
животных, в котором каждая эмоция или приказание выражаются
своим особым сиrналом. Язык человека в отличие от языка живот-
ных содержит слова, обозначающие обобщенные классы объектов,
т. е. классы предметов и.пи абстрактных понятий. Кроме Toro, язык
человека позволяет создавать разнообразные сочетания слов
фразы и предложения; такие сочетания C.'lOB дают возможность
rоворящему выражать взаимосвязи между ИДеями и формулиро-
вать сложные концепции. Язык, обладающий такими свойствами,
МОЖет служить основой для концептуальноrо мышления.
Для сохранения подобноrо языка и передачи ero последующим
поколения м необходима общественная rруппа, размеры которой
превышали бы некоторую КРИТИ 1 1ескую величину, а для Toro чтобы
каждое новое поколение моrло овладеть таким языком, требуется
д..lите..1ЬНЫЙ период обучения. Н аличие общественной rруппы, раз-
: меры котороЙ превышают критические и которая состоит из под-
ДЗlOщихся обучению IIНДИВИДУУМОВ, ИМеет важное значение для
: развития и поддержания !{онцептуальноrо мышления. А концеп-
туальное мышление в свою очередь предстаВ,I1яет собой важный
компонент высших форм человеческой культуры.
Эволюционная ilрирода человека
На извечный вопрос «Что есть человек?:. лредлаrались разно-
образные ответы. Человек это политичное животное; это живот-
ное, изrотовляющее орудия; релиrИОЗlJое животное и тому подоб-
Ное. ЭВО."'1юционная биолоrия дает возможность подойти к этому
вопросу исторически, а не отделываться афоризмами.
з54 Часть V/I. От орzйничесtro4 эволюции к. эвОАюции чело8е/t4
в свете ЭВОЛЮЦИОННОЙ биолоrип человек ---- это млекопитающее,
точнее, примат, еще точнее человекообразное, еще точнее ...
лредставитель rоминид весьма высокоразвитоrо типа. Эти ПОСЛе-
довательные ступени иnи rраДЫ орrанической эволюции заложены
в человеческом орrанизме.
НО человек это нечто большее, чем животное; это продукт
не только орrанической, но также и культурной эволюция. Куль-
турная эволюция добавляет еще один слой, или, если уrодно, ряд
слоев к природе человека. Двойственная конституция частью
биолоrическая, а частью культурная ---- закладывается в человече-
ство процессом ero эволюционноrо развития.
Самый верхний СЛОЙ стратифицированной конституции челове-
ка, слои, наложенный культурной эволюциеЙ, и есть тот решаю
щий слой, которым человек отличается от животноrо. Мы можем
добавить к нашему перечню афоризмов еще один: «Человек ---- это
животное, надеЛ'енное культурой».
Следующий вопрос касается T-oro, rде в процессе эволюцион-
110ro развития следует проводить rраннцу между животным и че-
ловеком? В какой именно момент в процессе филоrенеза человек
стал человеком? Коrда Ното или один из. более ранних предста-
вителей rоминид стал человекоМ в том CTporoM смысле слова, оп-
ределение KOToporo было приведено выше?
Следует напомнить, что элементы, делающие человека челове-
ком, имеются также и у друrих высших приматов: общественный
образ жизни, удлинение периода развития, предшествующеrо на-
ступлению половой зрелости, высшая нервная деятельность, язык
i}I сп-особность к изrотовлению орудий (см. rл. 35). Эти элементы
развивались на протяжении животной стадии в эволюции прима-
тов; они уже имелись у ранних rоминид; и рни развивались даль-
ше, ведя человека к новым высотам эволюции.
Переход ОТ ЖИВОТНоrо к ИСТИННО человеческому СQСТОЯНИЮ про-
исходил постепенно на протяжении не особенно длительной фило-
rенетической истории. Подобный же постепенный переход проис-
ходит в онтоrенезе каЖдоrо отдельноrо человека. Поrраничную
линию в процессе онтоrенеза можно проводить лишь очень пронз-
вольно, как это делается при юридических определениях зрелости,
и весьма сомнительно, чтобы в филоrении можно было найти
какую то rраницу, которая не была бы произвольной. Эволюцион-
ное изменение состояния было постепенным и историческим про-
цессом.
rЛ8ва 37
РЕШАЮЩИЕ ФАКТОРЫ .ЭВОЛЮЦИИ ЧЕЛОВВКА
Помски еще не закончены
Современный человек завершение ряда эволюционных на-
прав..lений, которые начинаются у приматов, не принадлежащих к
rоминидам. Наиболее важные направлеJt-ия эволюции человека
обсуждались в общей форме в предшествующих rлавах (rл. 34
36). Более подробное описание этих и друrих; направлений можно
найти в работах по антрополоrии и по ЭВ-ОЛЮЦИои человека (см.,
например, Campbel1, 1974). Каковы движущие силы этих направ-
лений?
Все Е общем соrласны с тем, что эволюция человека слаrается
из двух процессов: орrанической эволюции и культурной эволю
ции. Направления орrанической эволюции приводятся в движение
длительным отбором либо ортоселекцией (rл. 26), либо MHoro
кратными воздействиями отбора в сочетании с дрейфом reHoB
(rл. 30). Культурная эволюция имеет собственные движущие
силы, позволяющие объяснить постепенное накопление культур-
Horo насле,д.ия.
Но .какова за'виси.мость между естественным от.бором и на-
ПРЭiвлениями орrанпческой .3волюци.и, с одной стороны, и КУЛЬ
турной эволюцией..... С друrой?
Прост-ейшим и .и.сториче'скИ Cal:\1blM ста,рым ответом на этот воп
рос служит положени о ТОМ, что орrанИ'ческая эволюцин довела
филетиче.ский ряд человека до известноr-о nop-or.a до Toro УРОВ-
ня, на котором стала возможНой культу.рная ЭDОЛЮЦИЯ. С это1'о
момента фи.Т'(О'rения человека ПОШ.'1а по пути культурной эв'олюции,
которая и обеспечила все последующее развитие. Орrаническая -и
культурная эволюция считаются разделенными во времени и Не-
зависимыми в своем действии. Подобная точка зрения характерна
для работ старых антропо.поrов (например, BriffauIt, 1927) .И все
еще высказывается в некоторых современных исследованиях (на-
пример, Kraus, 1973).
Значительное развитие как эволюционной биолоrии, так и ан-
трополоrии, происшедшее за последние 20 30 лет, а также воз-
росшие связи между этими двумя прежде обособленными обла-
стями науки привели к появлению более современной точки зре-
ния. Соrласно этой точке зрения, в эпоху культурной эволюции
естественный отбор продолжал действовать и орrаническая эволю-
356 Часть V 11. От Ор2ан.ичес1СОЙ эвОАюции " ввОАюции 'Iелове"а
ЦИЯ не прекращалась (см., например, Darlington, 1969; Bajema,
1971). Просто результаты культурной эволюции более заметны, чем
результаты продолжающейся орrанической эволюции.
Дальнейшее усовершенствование представляет собой концеп.
ция непрерывноrо взаимодействия между орrанической и куль-
турной эволюцией. Важным эффектом TaKoro взаимодействия MO
жет быть отбор на обучаемость и на способность воспринимать
культуру. При этом орrаническая и культурная эволюция шли бы
рука об руку. Большое внимание этой концепции уделяет Добр
жанекий (Dobzhansky, 1962).
Несмотря на значительные успехи, достиrнутые нами в пони-
мании решающих факторов эволюции человека, остается сделать
еще очень MHoroe. Существующие объяснения иноrда носят слиш-
ком общий характер, чересчур неконкретны, а в некоторых слу
чаях, возможно, и недостаточны.
Передо мной леж'ит несколько недавно опубликованных работ
по эволюции человека, написанных с антрополоrической точки
зрения. Во всех этих работах описываются эволюционные направ-
ления. В них указываются важнейшие факторы: местообитание,
использование рук, разв,итие разума и т. п. Однако, коrда дело до.
ходит до определения движущих сил, стоящих за этими направ-
лениями, авторы оrраничиваются общими ссылками на «eCTeCTBeH
НЫЙ отбор'>. Это, конечно, само по себе верно, но далеко не до-
статочно.
В друrой работе по эволюции человека, написанной специали-
стом по биолоrической эволюции (Dobzhansky, 1962), роль естест-
BeHHoro отбора обсуждается' rораздо более подробно. Но Добр-
жанекий, рассматривая разные способы индивидуальноrо отбора
применительно к популяциям человека, Не затраrивает rрупповоА
отбор. Между тем уместно задать вопрос, можно ли все rенетиче-
ски детермипированнце признаки cOBpeMeHHoro человека отнести
за счет действия индивидуальноrо отбора или же некоторые из
них представляют собой результат rрулповоrо отбора?
Некоторые признаки человека очень сложны сами по себе.
Одним из таких признаков является ,интеллект. Можно ли объяс-
нить развитие человеческоrо интеллеКта действием отбора лишь
одноrо типа? Следовало бы рассмотреть возможность Toro, что
некоторые аспекты этоrо сложноrо приэнака возникли под дейст-
вием ИНдивидуальноrо, а друrие под действием rрупловоrо от-
бора.
Обратившись к друrому компоненту двоАствеННоrо набора
движущих сил..... к культурной эволюции, MЫ обнаружим, что
наши представления о ней также носят весьма общий характер и
нуждаются в аналитическом 'исследовании.
Цель настоящей rлавы состоит в том, чтобы рассмотреть не-
которые из этих пробпем более пристально.
r л. 37. Решающие факторы эволюции "еловеl«l
357'
rрупповой отбор
у приматов, ведущих общественный образ жизни, в том числе
, человека, существуют все условия для эффективноrо действия
rРУППОБоrо отбора. В rл. 11 уже быдо дан-о представление о меж-
дем:овом и rрупповом отборе_ Напомним, что термин «rрупповой
uтбор» используется для обозначения особой формы междемо-
80ro отбора, при котором в качестве конкурирующих ПОПУJIЯЦИЙ
выступают сообщества.
Стадо или rруппа у приматов, в том числе у человека, это
более или менее связная единица орrанизации. Это единица, обес-
llсчивающая сохранение rруппы и размножение.
Разные стада или rруппы, вероятно, нередко вступают в кон-
куренцию за кормовую территорию или какой нибудь друrой не-
обходимыЙ ресурс. Успех или неудача данноrо стада или rруппы
при конкурентных стычках или борьбе за территорию с соперни-
ками определяется наряду с друrими факторами уровнем интел-
лекта членов и (или) вожаков rруппы, развитием технолоrических
навыков и способов .общения, а также численностью. В тех слу-
чаях, коrда между конкурирующими стадами или rруппами суще-
ствуют rенетичеСl{oие различия по этим признакам, может вступать
в действие rрупповой отбор.
С биолоrической точки зрения разумно предполаrать, что как у
rоминид, так и у друrих высших приматов разлиЧИЯ между rрул-
пами в отношении ;интеллекта И ero производных технолоrиче
ских навыков и способов общения частично определяются reHe-
тическими различиями. В настоящее время (в США) считается
IIдеолоrически неприемлемым серьезно рассматривать эту предпо-
сылку применительно к rруппам людей, особенно если в том же
контексте упоминается слово «раса». Это оrраничение не распро-
страняется, однако, на друrих приматов. А поэтому, дабы избе-
жать дискриминации по отношен.ию к обезьянам, мы будем исхо..
дить из биолоrич ски реалистичной предпосылки о существовании
rснетических различий в умственных .и поведенческих способно-
стях между некоторыми rруппами rоминид и друrих приматов.
Следует ожидать, что при непрерывном rрупповом отборе у
приматов, в том числе у rоминид, сообщества с наиболее высоко
развитым .интеллектом, координацией и технолоrией в среднем
будут обладать преимуществом и их rенетически детерминирован-
ные способности будут распространяться в пределах данноrо вида.
Длительный rрупповой отбор вызовет постепенное повышение
уровня развития интел.пекта, языка и технолоrическ,их навыков.
rрупповой отбор дает возможность объяснить происхождение
ряда черт общественной жизни приматов.
Одна из таких черт это язык. Следует подчерК1iуть, чт МЫ
используем здесь термин «язык» в широком смысле. включающем
з58 Часть VJJ. От ореан.ической эволюции" эволюции чеАовеа
и язык ЖИВОТНЫХ (см. rл. 35 и 36). Обмен информацией представ.
ляет собой ценный в адаптивном отношении признак стада или
rруппы, причем это rрупповой признак. Весьма вероятно, чтО язы
ковые способности приматов, включая человека, развились ПОД
действием rрупповоrо отбора.
Интеллект..... адаптивно полезный признак как для отдельноrо
животноrо. так и для сообщества. По-видимому. индивидуальный
отбор иrрает важную роль в ero развитии, однако это не исклю-
чает дополнительноrо действия rрупповоrо отбора. Стадо, состоя-
щее rлавным образом из смышленых 06езьян, вероятно, будет луч-
ше справляться с проблемами, возникающими в их жизни, чем
соседнее конкурирующее с ними стадо r лупых обезьян.
Даже численность rруппы, представляющая собой один из фак-
торов, от которых зависит ее блаrополучие, может до некоторой
степени реrулироваться rрупповым отбором. Образование более
мноrочисленных стад у полуназемных и наземных приматов кор-
релирует с уменьшением нетерпимости самцов Apyr к друrу (см.
rл. 35). Большие размеры rpYnnbl адаптивно выrодны полуназем-
ным и наземным приматам, облеrчая защиту, конкуренцию и кол-
лект»вную охоту. Если нетерпимость самцов друr к друrу препят-
ствовала расширению предсуществовавших ыелких стад и еc.n.и
в некоторых стадах самцы обладали rенетически детерминирован.
Ной более высокой терпимостью к себе подобным, чем в друrих,
то такие стада и reHbl, обусловливающие терпимость самцов, по-
лучили бы преимущество.
В семи сравнительно недавно вышедших (1962 1974) книrах
по эволюции человека, с которыми я ознакомился, rрупповой от-
бор не рассматривается. Среди современных авторов Майр (Mayr,
1963) и Уилсон (Wilson, 1975) упоминают о rрупповом отборе в
связи с эволюцией человека, но лишь мимоходом, не развивая эту
тему в сколько-нибудь значительной степени. Между тем Дарвин
в «Происх-ождении человека» (Darwin, 1883) придавал большое
значение тому, что мы сейчас назвали бы rрупповым отбором.
В своем исследовании, начатом в последарвиновскую эпоху, но
написанном и опубликованном rораздо позднее, Кейт (Keith, 1948)
развивал концепцию о том. что сообщество. в частности иеболь..
шая rруппа у прнмитивноrо человека, представляет собой основ-
ную эволюционную единицу в эволюция 'человека. По ero мнению,
ЭВОЛЮЦiИя человека..... результат конкурентных взаимодействий
Между маленьким" скрещивающимися внутри себя сообществами.
Кейт создал BOKpyr этой ИДеи эволюционную теорию, которая. ка.
залась устаревшей даже в 1948 r., не rоворя уже о сеrодняшнем
дне. Современные антрополоrи отверrают концепцию Кейта, и в
большинстве новых книr по эволюции человека ero книrа (Keith,
1948) не цитируется. Возможно, однако, что это один из тех слу-
чаев, коrда мы рискуем вместе с водой выплеснуть и ребенка.
r л. 37. Решающие фа"ТОРtJl э,о ции 'leAoselcii
SO
и нтеппект
Общеизвестно, что интеллект слаrается из нескольких раЗJIИЧ-
ifbIX умственных способностей: способности к решению задач, сло-
обности К механическому запоминанию, обучению, общению
if т. п.
Некоторые компоненты интеллекта приматов дают преимуще
:тво rлавным образом отдельной особи и ero или ее потомкам.
Это .относится к способности усваивать опыт, к памяти, к способ-
ости предсказывать результаты тех или иных действий и к уме-
f!ию пользоваться орудиями. Развитие таких слаrаемых интеЛлек-
ra в процессе ЭВОJIЮЦИИ можно в значительНой мере отнести за
счет индивидуальноrо отбора.
Преимущества, доставляемые друrими компонентами интеллек-
ra, проявляются на уровне сообщества. К таким компонентам от-
носятся умение пользоваться языком для координации деятельно
стн rруппы и обучения молодых особей. Вполне возможно, что
умственные способности TaKoro рода вырабатываются rлавным
образом за счет rрупповоrо отбора.
Следовательно, лоrическ,и оправданно предположить, что ин-
Теллект приматов создавался в результате сочетания индивиду аль-
Horo и rрупповоrо отбора.
Способность приматов к обучению зависит от продолжитель-
ности тех стадий нндивидуальноrо развития, которые предшест-
вуют достижению зрелости. Сравнительные данные о продолжи-
тельности ЭТИХ стадий пок зывают, что период, в течение I{OTOPO-
ro может происходить обучение, возрастает ОТ низших приматов к
высшим (см. табл. 35.1). Скорость индивидуальноrо развития
представляет собой rенетически детерминирова,нный признаК. Тем
самым естественный отбор можно считать движущей с,илой этоrо
эволюционноrо направления. Кроме Toro, поскольку обучение у
приматов процесс общественный, rрупповой отбор, вероятно,
иrрал большую роль в создании таких скоростей развития, кото-
рые блаrоприятствовали бы большим возможностям для обу-
чения.
rрупповой отбор, по всей вероятности, вел к повышению уров-
ня развития интеллекта во всех л.нннях приматов, ведущих обще-
ственный образ жизни. Однако у rоминид это направление, оче-
видно, продвинулось дальше, чем у друrих прнматов. Быть может,
особые биолоrические и социальные факторы, описанные в преды-
дущих rлавах (rл. 34 36),..... передвижение на двух Horax, ис-
пользование рук и т. п...... создали для более BblCOKOro развития ин-
теллекта у rоминиД iаwие потенциальные возможности, каких нет
у друrих приматов, ведущих древесный .образ жизни и передви-
rающихся на четырех Horax.
З6О Часть V/J. От ОР2аническо(J эволюции" Э80Аюции челОвека
Межвидовой отбор
В ЭВОЛЮЦИИ rоминид, очевидно, участвовал еще один способ
отбора на уровне выше индивидуальноrо, а именно .межвидовой
отбор, ведущий к замещению видов (см. rл. 20). "
Ното и Australopithecus лерекрывались во времени и были
симпатрически и. Возможно, что между Н. erectus ил.и ero непо-
средстве.иным предком и видами, принадлежавшими к роду
Australopithecus, происходил а прямая ил,и по меньшей мере KOC
венная конкуренция за пищу иди кормовые территории. Australo
pithecus вымер, тоrда как Нота erectus выжил и занял домини
рующее положение в адаптивной зоне rоминИД. Н. erectus превос
ходил австралопитеков по величине rоловноrо мозrа и по качест
ву каменных орудий. Эти два фактора, вероятно, входили в число
тех, к{)торые определили течение процесса межвидовоrо отбора.
Межвидовой отбор у rоминиД отличается от межвидовоrо от-
бора у большинства животных тем, что он сцеплен с rрупповым
отбором. Соревнование происходило между сообществами Н. етес-
tus или «pre erec.tus» и конкурирующими rруппами австралопите-
ков. Поскольку эффективными единицами размножеlIИЯ у конку-
рирующих Видов были сообщества, межвидовой отбор принял
особую форму форму межвидовоrо rрупповоrо отбора.
Механизмы культурной эволюции
Принято считать, что культурная эволюция совершается путем
постепенноrо накопления кулыурноrо наследия. Сначала происхо-
дит приобрете.ние полезных знаний, затем эти знания включаются
в общую массу знаний данноrо сообщества, после чеrо передаются
последующим поколениям пуТем обучения. Каждое следующее
поколение добавляет к этому запасу знаний то, что ему удается
открыть самому, и такие новые знания передаются дальше тем же
путем. А следовательно, культурное наследие носит кумулятивный
характер.
Несомненно, в известной степени культурная ЭВОЛЮl1.ия проте
кает именно таким образом. Но происходит ли ОНа только так,
всеrда ли она следует столь прямыми путями? Ознакомление с
историей показывает, что неТ. Наши нынешние взrляды на куль
турную эволюцию носят 'столь же общий характер и столь же ту..
манны, как с{)временные представления о роли eCTecTBeHHoro ОТ-
бора в эволюции человека, и, подобно последним, нуждаются в
критической переоценке.
Вернемся вновь к величине rруппы. rрупповой отбор, действуя
на rенетически обусловленные различия между rруппами в OTHO
шении терпимости CaMl1.oB друr к друrу, Mor привести к увс..lиче.
пию rруппы до уровня мноrосамцовоrо стада. TaKoro уровня до.
r л. 97. Решающие фак1'ОРЫ эволюции ч-еловеlCЙ
Збl
rтиrают сообщества некоторых примитивных rоминид и ДРУI'ИХ
приматов (см. r 1. 35).
Однако тендеция к уве.тшчению размеров сообщества и ero
сложности, очевидно, продолжалась в доисторическую и 'истори
ческую эпоху. rруппы сливались в племена, а племена в кланы
и rосударства. rосударства увеличивались в размерах, превра
щаясь из rосударств rородов в малые, а затем и большие нацио
нальные rосударства.
Степень терпимости самцов друr к друrу не имеет никакоrо
отношения к этим поздним стадиям эволюции, ведущим к увели-
чению размеров rруппы. Терпимость мужчин друr к друrу у не-
андертальцев и у жителей США не отличается заметным образом.
Но зато ВОЙНЫ :между племенам'и и между народами сильно зави-
сят от тенденции к увеличению rруппы. Вплоть до известноrо
уровня более МIIоrочисленные племена или народы в среднем, при
прочих равных УСЛОВИЯХ, выходят победителями в войнах с более
малочисленными племенами ;или народами. В результате ряда
последовательных войн ме..1кие обществеННО ПОv1иrические едини-
цы обычно перестаlOТ существовать в качестве незаВисимых rрупп,
сохраняясь в какой либо зависимой форме ---- в качестве колонии,
одной из составных частей федерации или сателлита, тоrда как
более крупные rруппы становЯтся независимы:ми политическими
державами. Тенденция к увеличению размеров rруппы до некото-
рой степени определяется выживанием самых крупных rрупп.
Я сказал «до некоторой степени», потому что чересчур боль-
шие пол,итические единицы, примером которых с.ТIужат империи,
не бывают долrовечными и в конце концов неизмеННО распадают-
ся на более мелкие части. Тенденции к увеличению размеров
rруппы подвержены реверсиям, особенно На своем верхнем уровне.
Увеличение размеров проявляется более определенно НИJКе этоrо
уровня.
Здесь мы имеем дело с социальной тенденцией, определяемой
неrенетическими различиями между конкурирующими rруппами.
В данной модели речь идет о неrенетических различиях в размерах
rруппы. Однако можно представить себе раз.ТIИЧИЯ между rруппа-
ми по друrим признакам культуры ---- такие различия, которые в
результате ряда последовательных конкурентных межrрупповых
столкновений моrли бы создать определенное направ.1Jение куль
турной ЭВОЛЮII'ИИ..
Например, неКОТОрЫе направления в развитии техНИКИ возник
ли, возможно, В результате длительной конкуренции между rруп-
памп с разными техническими способностями. Аналоrичным обра
зом моrли бы возникнуть и идеолоrические тенденции. Нетрудно
предска ать результаты, к которым привели бы в среднем MHoro-
числеНные столкновения между rруппами, верящими в чудеса, и
rруппами с праrматическим подходом к решению проблем.
362 Часть V/I. От ореаничес"о4 эволюции" эволюции человека
В этом разделе маятник резко отклонился от ортодоксальной
точки зрения (которую мы считаем упрощенческой) па культур-
ную э.волюu.ию, как на ЭВОЛЮЦИЮ, происходящую общим широким
фронтом, к представлению о культурной эволюции, происходящей
в результате различий в культуре между конкурирующими сооб-
ществами. Конечно, эта последняя картина также чересчур про-
ста. Ввиду недостатка места мы ВЫНуждены пренебречь мноrочис-
ленными сложностями, наблюдаемыми в реальных взаимодейст-
виях между сообществами. Здесь, вероятно, достаточно привлечь
внимание к альтернаТIIВНЫМ косвенным способам культурной эво
ЛЮЦИИ, В центре которых стоит rруппа.
Взаимодействия
Взаимодействие между отбором по морфолоrическ,им и морфО
rенетическим признакам, с одной стороны, и отбором по социаль-
ным признакам ---- с друrой, представляет собоЙ существенный ас-
пект эволюции человека.
Эволюция способности к обучен ю повлекла за собой ряд ин
тересных осложнений. Увел,ичению размеров мозrа сверх некото.
рых пределов препятствовала ширина родовых путей, через кото-
рые должна пройти rоловка ребенка при родах. Отбор на высо-
кий интеллект вступил в противоречие с отбором па выживание
женщины при родах. Для Toro чтобы выйти из этоrо тупика, попа-
добилось два компромисса.
Первым компромиссом было расширение таза у женщины. Это
повлекло за собой потерю скорости при беrе. Женщина беrает
медленнее, чем мужчина. Способность к беrу была при несена в
жертву, с тем чтобы женщина моrла рожать детей с более объеми-
стым мозrом.
Второй компром,исс состоял В снижении скорости развития
Мозrа. У человека к моменту рождения моэr еще не вполне сфор-
мирован, и новорожденный младенец совершенно беспомощен;
развитие мозrа происходит rлавным образом в период младенче-
ства и детства. Люди в этом возрасте заметно отличаются от
друrих примаrов, которы.е в момен1' рождения более самостоятель-
ны. Вторичным результатом этоrО BToporo компромисса было
увеличение продолжительности стадий индивидуальноrо развития,
предшествующих наступлению зрелости (BriffauH, 1927; Wash-
burn, 1960).
Оба компромиссных разрешения возникшей проблемы умень-
шили независимость женщины. Она оказалась обремененной за-
ботами о ребенке в течение более ДЛl1:тельноrо времени. А частич
иая утрата способност.и к беrу сократила ее возможности пол
ностью обеспечивать себе пропитание за счет OXQTbl н увеличила
ее зависимость от мужчин.
r л. 97. Решающие факторы Э80Аюции чеАовеlС4
З63
ЭТII эффекты TpeTbero порядка вызвали в сообществах пер во..
бытноrо человека эффекты четвертоrо порядка. rруппе пришлось
брать на себя известную заботу о женщинах и детях. Это в свою
очередь усилило сотру ДНоичество и общение между члена ми сооб.
щества. Тем самым причинно следственная цепь замкнул ась.
Отбор на повышение способности к обучению..... черта, полезная
для сообщества при общественном образе жизни'..... вызвал раз..
..'1ичные морфолоrические морфоrенетнческие изменения, следст-
вием которых было еще более тесно спаянное сообщество.
Заключительные замечания
Мы перечислили несколько эволюционных сил, которые, воз.
можно, участвуют в эволюции человека. К факторам орrанической
эволюции относятся: индивидуальный отбор, внутр.ивидовой rруп-
повой отбор, межвидовой rрупповой отбор и сочетание отбора с
дрейфом reHOB. К факторам культурной эволюции относятся: об-
щее накопление культурноrо наследия и тенденции в развитии куль-
туры, возникающие в результате конкуренции между сообщества-
ми, различающимися в культурном отношении (но не rенетически)'.
Некоторые из этих факторов обычно рассматриваются при об-
суждении эволюции человека, а друrие, как правило, не упоми-
наются. Мы уде.'1ИЛИ этим последним несКолько больше внима-
ния, но не потому, что считаем их более важными, а просто для
полноты картины. Мне трудно было бы оценить относительное
значение различных упомянутых выше факторов. Точнее, я в са-
мом деле считаю, что rрупповой отбор иrрает важную роль в ор-
rанической эволюuии человека и что дифференциальный успех раз-
ных сообществ имеет важное значение для ero культурной эволю-
ЦИИ, однако я Не берусь судить об относительном значении этих
rрупповых процессов по сравнению с друrими факторами.
Одна из движущих СИЛ эволюции человека, которой уделяется
довольно MHoro внимания в текущей литературе, это закрепле-
ние селективно нейтральных мутантных reHoB ПОД действием reHe-
1'ическоrо дрейфа. По этому поводу мне хочется высказать одно
соображение. Вполне ВОЗМОЖНО, что такое закрепленяе иноrда и
происходило. Однако я сомневаtOсь в том, что оно моrло nриве.
стн к 80ЗНИI{новению какой-либо существенной характеристики в
ЭВОЛЮUИИ человека. Различные эволюционные силы взаимодейст
вуют друr с друrом, что сильно осложняет анализ эволюции че-
ловека. Быть может, мы вправе УТвержда1Ь, что, несмотря на су-
ществование значительных пробелов. филоrения человека извест-
На нам сейчас rораздо лучше, чем ученым прошлоrо поколения, и
что соответствующие эволюционные фактоРЫ по большей части
определены; однако наши представления об эволюционных силах,
участвующих в эволюции человека, все еще очень неполны.
Часть VIII
Социальное значение эволюционной
теории
rлзва 38
ПОЛБМИКА ,С КРЕАЦИОНИЗМОМ
Сотворение мира
История божественноrо сотворения мира изложена в rл. 1 и 2
Книrи бытия, автором которой считается Моисей.
В rл. 1 Книrи бытия сказано, что ,на третий день боr заставил
Землю произвести растения, на пятый день животных.., а на ше
стой день человека. Боr повелел, чтобы человек владычествова,,1J
«над рыбами морскими, и над Пl'ицами небесными, и над скотом,
и над всею землею...:' (Бытие, 1 : 26).
Боr «создал человека из праха земноrо и вдунул в лицо ero
дыхание жизни...» (Бытие, 2 : 7). Затем боr поселил созданноrо ИМ
человека в саду эдемском, в котором были всяк,не деревья, даю-
щие съедобные плоды, вода и «все животные ПО.1Jевые н все птицы
небесные» (Бытие, 2: 8----20). И, наконец, пока человек спал, боr
взял одно из ero ребер п создал из Hero женщину (Бытие,
2 : 21----22) .
Этим человеком был Адам, а жеНlЦИНОЙ Ева. Адам и Ева
явил'ись прародителями человечества; «Ева... стала матерью всех
живущих» (Бытие, 3: 20). Все это произошло в 4004 r. до н. э.,
В соответствии с хронолоrией Ашера, которая была основана на
библейских данных.
Креационизм ИJIИ эволюционная теория?
На протяжении мноrих веков в западной цивилизации рассказ
оисея о сотворении мира принимался в качестве единственноrо
объяснения возникновения жизни и происхождения человека. Та.
кое исключительное положение креационизма в области мышле-
ния поддерживалось ортодоксальной церковью и релнrиозным
учением.
Этому незыблемому положению вещей наступил конец после
Toro, как в 1859 r. было опубл,иковано «Происхождение видов»
Дарвина. Традиционная ортодоксальная точка зрения СТОЛКНУ-
лась теперь с серьезным противником. Прошло время, коrда на
r А. 88. Лолемuа с "реацuониэ.мом
365
идеолоrической арене безраздельно rосподствовала одна точка
зрения; теперь на этой арене начала разыrрываться борьба между
двумя альтернативными доктринами, одной из которых была ре-
лиrиозная доrма, а друrой научная теория.
Общая -идея об орrанической эволюции существовала и до
1859 r. Ее выдвиrа.'IИ Ламарк в своей «Философии ЗООJJоrии»
(Lamarck, 1809), Чэмберс в «Следах творения» (Chambers, 1844)
и друrие. Труд Ламарка был неубедительным в части как факти-
ческих данных, так и теоретических построений; то же самое OT
носится и К трудам друrих ученых начала XIX в. Их работы,
однако, подrотовили почву для принятия «Происхождения видов'>
Дарвина.
Некоторые из rлавных событ-ий в жизни И творчестве Дарви-
на перечис.'1ены В таб.п. 38.1 (см. также de Beer, 1964).
Таблица 38.1
Ключевые события в жизни и творчестве Чарлза Дарвина
1809
1827....183 1
18З1 1836
1839
1842
1851
1859
1868
1871
1872
1875....1881
1882
Родился в Шрусбери .(Аиrлия)
Студент Ксмбриджскоrо университета
Натуралист на борту корабля «Биrль», совершающеrо Kpyro.
светнос путешествие
Женитьба на Эмме Веджвуд. Выше.') D CRCT .«Дневник изысканий]
(==«Путешсствие на корабле Биrль») (2-е иэд. в 1845 r.)
"Строение и распространение коралповых рифов:. (2-е ИЗД. 8
1874 r.)
Моноrрафия об усоноrих рачках
Вышло в свет .1 e изд. «Происхождение видов» (6 e изД. в 1872 r.)
ItИзменение животных и растений при одомашнивании:. (2-е изд.
R 1875 r.)
сПроисхождеllие че.'lОвека и ПО.'lОвой отбор» (2.е И3Д. в 1874 r.)
«Выражение ЭМОЦJIЙ у человека и животных»
Выходили В свет разные работы по ботанике. 8 том числе «Дей-
ствие перекреСТlfOl'О опыления и самоопыления в растительном
мире» ([876 r.)
Сконча.1СЯ в свосм доме в Дауне. Похоронен в ВестминстеРСI<ОМ
аббатстве
в «Происхождении видов» Дарвин систематизировал orpoMHoe
количество данных, добытых исследованиями в нескольких незави-
С-ИМЫХ направ.llениях и указывающих на существование орrани-
ческой эволюции, происходнвшей в прошлом и продолжающейся
в настоящее время. Он разработал теорию eCTecTBeHHoro отбора
чтобы объяснить силы, вызывающие эволюционные изменения.
Подход Дарвина к этой проб.'Iеме, в отличие от подходов предше-
ствовавших ему авторов, оказался убедительным для эоолоrов,
ботаников и rсолоrов. [оды и десятилетия, прошедшие после
1859 r., были отмечены постепен"ым и все расширяющимся при-
знанием ЭВО.'Iюционной теории, эахват-ившИМ весь научный мир.
Зб6 Часть V/П. СОЦШl.l1ьное значение Э80люцио",ноiJ теории
Конечно, в научной среде после 1859 r. возникали споры и
разноrласия. Однако rлавные дискуссии происходили в Kpyrax,
не имевших отношения к науке, и были связаны с попытками бо-
rословов и представителей церкви отстаивать доктрину об акте
божественноrо творения. Столкновения между ними и сторонни-
ками эволюционноrо учения в конце XIX в. составляют одну iиз
интересных, но подчас rорьких rлав в истории человеческой MЫC
ли (см. среди прочих, F. Darwin 1958; Fothergill, 1952; Eiseley,
1958; G1ick, 1974).
Б США разноrласия между сторонниками креационизма .и сто-
ронниками эволюционной теории продолжаются до сих пор.
Б 60 x и 70-х rодах нашеrо века наблюдалась даже вспышка этих
разноr ласий. Конечно, в настоящее время их можно расценивать
л,ишь как анахронизм.
В современных западных странах ряды сторонников креацио-
Низма сильно поредели по сравнению с XIX в. Одна из причин
этоrо состоит в отходе части населения от релиrии; друrая причи-
на наблюдаемая в настоящее время либерализация взrлядов во
мноrих релиr!tозных сектах. Мноrие релиrиозные rруппы и ОТ-
дельные верующие придерживаются несколько аллеrорической ин
терпретации Книrи бытия, что позволяет им признавать заключе-
ния эволюционной биолоrии. Однако в США фундаменталисты до
сих пор настаивают на cTporo буквальном толковании Книrи бы-
тия. Именно эти rруппы поддерживают анахроническую полемику.
Б центре происходящих в настоящее время в США дискуссий
находится вопрос о преподавании биолоrии в средних школах.
Сторонники креац,ионизма настаивают на том, чтобы в учебниках
биолоrии содержалось беспристрастное изложение как описания
сотворения мира, даНRоrо Моисеем, так и теории Дарвина. Кроме
Toro, они хотят преуменьшить значение эволюции, представляя ее
«как теорию, а не как установленный факт». Б ряде штатов, в том
числе в Калифорнии iИ Техасе, rруппы фундаменталистов, являю-
щиеся сторонниками креационизма, оказывают влияние на школь-
ные советы, с тем чтобы изложенные выше принципы были поло-
жены в основу школьных проrрамм. Поднятые в результате Bcero
этоrо вопросы относительно пр,инципов обучения обсуждаются на
заседаниях советов и в судах.
Критические замечания
Позиция, занимаемая сторонниками креационизма в современ-
ных условиях, ошибоч:на в ряде отношений. Прежде Bcero средняя
школа Не место для распространения релиrиозных доктр-ин тех
или иных сект.
Требование о том, чтобы в учебниках и на К.пассных занятиях
отводилось одинаковое время креационизму и эволюции, IИСХОДИТ
r А. 38. n оле.мика с ItреациОНUЭ.fЮАе
Зб7
из представления о равнозначности этих двух предметов, т. е. ИЗ
мнения о том, что они заслуживают одинаковоrо вниман,ия, а та-
кое представление ошибочно. Креационизм и эволюционное учение
НfJц:ак :нельзя ставить на одну доску. Креационизм ---- это не науч-
пая теария, К01'Орую можно было бы оцен,И'вать, сра,вн.ивая ее с
эволюционной теорией. Это ре 1иrиозная доrма. Нет НИкаких объ
ективных данных, которые бы подтверждали положение о том, что
растения, жиоотные и человек возни!кл.и .в результате акта боже
CTBeHH()rO творения. п.р,изнание эrorо ПoQлож-ения опирается на
.веру, а не на разум
Рассмотрим часто высказываемое сторонниками креационизма
утверждение о том, что эволюция ---- это теория, а не факт. П ри-
мем временно наив:ное допущение, что любой научный вопрос
можно рассматривать либо как теорию, либо как факт. И воздер-
ж,имся от утверждений, что креационизм нельзя считать ни тео-
рией, ни фактом.
ЭВО.,'1юция и в самом деле коrда-то было теорией, однако с
L859 r. утекло MHoro воды. Ббльшая часть Bcero, что относится
к эволюции перешла теперь в катеrорию провереННЫХ фактов.
Эволюционные изменения на уровнях микроэволюции и видообра
зован,ия наблюдались компетентными биолоrами, и их можно
теперь рассматривать как доказанные факты.
Изменения макроэволюционноl'O уровня занимают HeCKOJIbKO
иное ПО.110жение, поскольку это явления исторические и COOTBeTCT
nенно ни один человек не Mor наблюдать их непосредственно. От-
сутствие свидетельств. очевидцев проблема, возникающая при
любой попытке воссоздать события прошлоrо. Рассказ Моисея о
сотворении мира, который был написан спустя MHoro времени
после Toro, как возник мир, сталкивается именно с этой труд-
ностью.
Простая альтернатива теория илои факт Не распространя-
ется на макроэволюцию. Макроэволюцию нельзя наблюдать He
посредственно, о ней можно лишь судить На основании палеон-
толоrичеtких данных. Сами такие данные это факты, а макро-
эволюция ---- необходимое умозаключение, сделанное на основаниИ
этих фактов.
Данные в ПОJlЬ3У существования маКРОЭВОJlЮЦИИ
Представляется целесообразным вкратце изложить разные
rруппы доказательств существования макроэволюции. Это. жела-
тельно в связи с продолжающимися кое rде выступлениями против
теории эволюции. Ббльшая часть приведенных ниже направлений
доказательств бьта ИСПО.1Jьзована самим Дарвином в «Происхож-
дени и видов». Однако в настоящее время количество данных, со-
бранных по каждому из этих направлений, СИ."FJЬНО возросло по
сравнению с их количеством Во времена Дарвина.
368 Часть VIl/. СоциаАьное ЗНJJчение Э80Аюционнй4 теории
1_ Экономия zиnотез. Существование мпкроэволюционных из
менений и некоторых способов видообразования установлено пря-
мыми наблюдениями и (или) экспериментальными данными. Cy
ществует Iнепрерывный ряд уровней от локальной популяции и
rеоrрафической расы через вид к rруппе видов, подроду и роду.
Каждому, кто изучал живую природу и проникся чувством един
ства природы, невозможно представить себе, что орrаническое
разнообразие на. низших уровнях возникло в результате эволю-
ции, а на надвидовых уровнях ---- какими то друrими способами.
2. Данные палеонтолоzии. Для мноrих rрупп орrанизмов, xo
рошо представ..1енных в палеонтолоrической летопи-си, можно на-
блюдать последовательный ряд форм, сменяющих друr друrа в
rеолоrическом времени (см. rл. 26). В некоторых случаях сохра-
нились ископаемые остатки форм, образующих переходы между
двумя крупными rруппами (см. rл. 31 и 34).
3. Таксономический характер взаимосвязей между ныне живу
щи.чи видами. Виды естественным образом объединяются в роды,
роды ---- в семейства, семейства ---- в отряды и так далее. Это «ес-
тественное распределение орrанизмов по rруппам, подчиненным
одна друrой:., как писал Дарвин, представляет собой результат
ветвящейся филоrении. Естественный род У состоит из видов,
имеющих общих предков; родственный ему род Z слаrается из
друrоrо набора видов; роды У и Z представляют собой две ветв'и,
отходящие от более старой ветви ---- семейства, и так далее по
всей таксономической иерархии. Иерархический характер таксо-
номических взаимосвязей Не был придуман биолоrами эволIOUИО-
нистами, он был открыт систематиками доэволюционноrо перио-
да, которые признавали креационизм, а позднее был правильно
интерпретирован ЭВО f[юционистами. Вряд ли можно ожидать, что
ныне живущие виды объединялись бы в rруппы, входящие в БО.1ее
круп.ные rруппы, если бы каждый из них был продуктом незави.
симоrо акта творения. .
4. rеоzрафическое распространение. Мноrие роды, трибы, се-
мейства и друrие rруппы среднеrо таксономическоrо paHra orpa-
ничены какой-то одной rеоrрафической областью ---- определенным
архипелаrом, частью материка или материком и т. п., В которой
находится центр распространения данной rруппы. Из этоrо лоrи-
чески вытекает, что принадлежащий к такой rруппе ныне живу-
щий вид или несколько таких видов раэвивались в той области,
rде они сейчас обильны или разнообраз-ны (см. конец r.rт. 31).
Доктрина креационизма не может объяснить такой характер reo-
rрафическоrо распространения надвидовых rрупп.
5. rомоло uи. Сравнивая представителей какой-либо крупной
rруппы, можно обнаружить, что они обладают сходным общим
планом структурной ор.rа'низации, Но различаются по некоторым
rомолоrичным частям тела. В качестве примера можно привести
r /1,. 88. П О/l,еАШ1Са с "реаЦUО",UЭАСОА&
369
разные формы, которые принимают rомолоrичные передние конеч-
ности у представителей разных отрядов млекопитающих, rомоло-
rИЧllые задн.ие конечности также у представителей разных отрядов
млекопитающих, конечности в разных классах наземных позво
ночных, части венчика цветков у представителей разных семейств
иJш порядков покрытосеменных и т. д.
Приведем слова caMoro )lарвина относительно значения rOMo-
ЛОI'ИЧНЫХ opraHoB (Darwin, 1872, rл. 14):
«Что может быть любопытнее Toro, что приrодная для хвата-
ния рука человека, роющая лапа крота, 'Hora лошади, ласт дель-
фина и крыло .тIетучей мыши построены по тому же самому об-
разцу и содержат одинаковые кости с одним и тем же относи-
тельным расположением?.
Ничто не может быть более безнадежным, как попытки объ
яснить эту общность строения у членов одноrо и Toro же класса
с точки зрения полезности или учения о конечных причинах. Без-
надежность такой попытки была ясно показана Оуэном в ero в
высшей степени интересной работе «Nature of Limbs» (<<Природа
конечностей:.). Следуя обычному взrляду, признавая независимое
творение каждоrо существа, мы можем только утверждать, что
это так, что творцу yroAHo было построить всех животных и pa
стения каждоrо большоrо класса по единому плану; НО это не
научное объяснение.
Зато очень простое объяснение дается теорией отбора последо-
вательных слабых изменений, из которых каЖДое ДО известной
степени полезно изменяющейся форме, но часто обусловливается
корреляцией с друrи и частями орrанизации. При изменениях по-
добноrо рода лишь в очень слабой степени или совсем не обнару-
ЖИВается наклонность к изменению первоначальноrо плана или к
перемещенню частей. Кости конечности Moryr до известной степе-
ни стать короче и площе, одеваясь в то же время более толстым
покровом, и служить плавником; 'или же у снабженной перепон-
кой передней конечности все или некоторые кости MorYT до из.
вестной степени УДЛИНИТЬСЯ, связывающая их перепонка можеl
увеличиться, и конечность может стать крылом; но все эти ,изме.
нения не изменят OCHoBHoro П",1ама строения костей или соотноше-
ния частей. Если мы предположим, что у очень древнеrо праро-
дителя архетипа, как ero можно назвать, всех млекопитаю-
щих, пт,иц и рептилий конечности были построены по ныне сущест-
вующему общему плану, для какой бы цели они ни служили, мы
сразу поймем все значение rомолоrичноrо строения конечностей
во всех K",laccax».
б. Рудиментарные opzaHbt. У некоторых членов какой-либ{)
крупной rруппы нередко имеются атрофированные или нефункцио-
нирующие opraHbl. Такой рудиментарный орrаи rомолоrичен хо-
рошо развитому функционирующему opraHY У друrих представи-
370 Часть VIIL Социаль",ое значение эвОAJоцио",ной теории
телей той же самой rруппы. Так, у некоторых нелетающих птиц
имеются рудиментарные крылья, у некоторых китов рудимен-
тарные кости таза, а у некоторых змей, в том числе у питона,
рудиментарные задние конечност,и. Эти структуры интерпретиру
ются как редуцированные зачатки их rомолоrов, хорошо разви.
Тых у друrих членов той же самой крупной rруппы. Подrруппа,
обладающая таким рудиментарным opraHoM, перешла в новое ме-
стообита,ние или к новому образу жизни, в условиях которых
прежде функционировавший opraH оказался бесполезным и под
действием отбора сильно редуцировался, но зачатки ero сохрани.
лись в качестве филоrенетических остатков. Креационизм не мо-
жет дать убедительноrо объяснения существованию бесполезных
рудиментарных opraHoB.
7. Биохимическое сходство. Современная линия доказательств,
которой не было 80 времена Дарвина: близкое сходство биохими-
ческоrо состава и молекулярной структуры rомолоrичных белков
у членов разных, но родственных семейств иди отрядов. Хороши-
МИ примерами служат rомолоrнчные формы rемоrлобина 'и цито-
хрома с у человека и друrих приматов (см. rл. 29).
r лава 39
ПРИЛОЖВНИЯ ЭВОЛЮЦИОННаЯ Би.олоrии
Наука приносит плоды двух сортов: она дает возможность по-
нять природу и управлять пронсходящими в ней процессами. Нау-
ка питает как теорию, так и практику. Этими двумя rлавными
аспектами -науки обладает и эволюционная биолоrия.
Эволюционная биолоrия, понимаемая широко, имеет MHoro
ценных прнложений в сельском хозяйстве и медицине. Улучшение
сортов культурных растений и пород домашних животных пред
ставляет собой, по сути дела, прикладную эволюционную биоло-
rию. Знание эволюционной биолоrии необходимая составная часть
любой успешной nporpaMMbl химической или биолоrической борьбы
с патоrенными орrанизмами или насекомыми вредителями.
Однако наиболее важный вклад, по мнению мноrих эволюцио-
нистов, включая меня caMoro, эволюционная БИОJlоrия вносит в
философию и интеллектуальную сферу. Эволюционная биолоrия
дает возмо}Кность удовлетворять одну из основных потребностей
людей ---- желание узнать, как развивалась жизнь на Земле, и же-
лаНИе понять природу и I1роисхождение человека. Освобождение
человеческоrо мышления от традиционных доrм по этим вопро-
сам представляет собой заслуrу эволюционной биолоrии и продол-
жает ли'нию, Начало которой положили nроисшедшие ранее науч-
ные революции в rеолоrии и астрономии.
Эволюционная биолоrия создает перспективу, позволяющую
представить место человека в природе. Мноrие научные проблемы,
доставляющае массу затруднений, если рассматривать их вне био-
JlоrичесКоrо контекста. становятся rораздо яснее. в свете этой перс-
пективы. Приведем два примера ---- один старый, а друrой новый.
В среДНИе века широко обсуждалась в философском пла'не
проблема боли. Почему справедливый и всемоrущий боr допускает
существование в мире TaKoro неприятноrо явления, как боль? Или,
если ПОДОЙТ-II к этому вопросу с противоположноrо конца, позво-
ляет ли сушествование боли усомниться в совершенстве боrа?
Само собой разумеется, что вопрос этот не был разрешен в рам-
ках теолоrичеСКQrо подхода. Между тем с точки зрения эволюци..
онной биолоrии боль получает объяснение: это адаптявный пря-
з-нак, поскольку болевые ощущения служат сиrиалам,и, предупреж.
дающими ЖИВОТНЫЙ орrанизм о потенциально вредных воздей..
ствнях.
372 Часть V/lI. Соцuа.Аьное з",ачение МО.Аюцuон",оil теории
На сеrоднЯ вопрос об официальном разрешенИ'И абортов широ-
ко обсуждается как на юридическом, так и на политическом
уровне. Сторонники запрещения абортов rоВОрЯТ о правах плода,
сторонники разрешения ---- о правах женщин. Здесь мы снова за
ходим в тупик. И снова эволюционная биолоrия позволяет поста-
вить этот вопрос в более ш,ирокой перспективе. Одно из длитель-
ных эволюционных направлений у позвоночных, заходящее весь-
ма далеко у млекопитающих и у приматов, касается плодови-
тости. У этих животных наблюдается тенденция к уменьшению
числа детенышей и к усилению заботы о потомстве, т. е. к повы-
шению качества молодоrо поколения за счеТ сокращения ero коли-
чества. Следствия, вытекающие из этоrо направления, не поддер-
живают сторонников запрещения абортов.
В современном мире условия для жизни человека не оче'нь хо-
роши. Перенаселение, разрушение окружающей среды, нищета,
rолод, аrрессивность, франкенштейновский характер развития тех-
ники и скверное управление создают уrрозу для людей; и над
миром постоянно висит опас,ность ядерной войны. Блаrоразумные
rраждане Bcero земноrо шара, принадлежащие ко всем слоям об-
щества, озабочены в настоящее время судьбами че"ТIOвечества.
Однако биолоrи-эволюционисты старшеrо поколения Mory'" с пол-
ным правом заявить, что они были в рядах тех, кто первыми
осознали надвиrающийся кризис и забили TpeBory; если бы к тем
предостережениям, которые они высказывали в первой половине
нашеrо века, отнеСЛlIСЬ с должным вниманием, то мноrих из ны-
нешних бедствий удалось бы избежать.
Теперь, коrда пишутся эти строки, перед нами стоят пробле-
мы, возникшие к середине 70-х rодов. И эволюционная биолоr,ия
снова может прийти на помощь. Рассматривая человека как часть
природы, а точнее как rосподствующий в природе вид живот-
ных, эволюционная биолоrия понимает основную проблему
растущую дисrармонию между человеком и той средой, в кото-
рой он обитает, и может указать rлавную причину возникнове-
ния этой проблемы: безудержный рост населения. Эволюционная
биолоrия может указать также на несостоятельНость полумер
(таких, как одно лишь увеличение производства продуктов пита-
ния) .или косметических решений вопроса (конференции по вопро-
сам продовольствия) и на необходимость принятия подлинных
решений, направленных на устранение причин, а не симптомов.
Будущее человеческой uивилизации, а возможно, он caMoro
вида Ното sapieпs может зависеть от тех мер, которые будут
приняты для Toro, чтобы выправить существующее сейчас нерав-
новесие между человеком и ero земной средой. В число таких
мер должна входить стабилизация, а в конеч.ном счете и сокра-
щение населения земноrо шара путем реrуляции рождаемости.
Однако здесь Не место обсуждать конкретные меры. Достаточно
r л. 39. п рUАожения эволюционной биолоеuu
З7З
подчеркнуть, что при поисках полноценных решений необходимо
использовать все то, что может дать эволюционная биолоrия.
Мы уже указывал.и, что эволюционная биолоrия, подобно дpy
rим областям наук", имеет как практические,так н теоретические
аспекты. Различия между этими двумя аспектами достаточно
ясны, коrда речь идет о каких либо краткосрочных задачах.
Однако, коrда дело касается процессов, связанных с длительными
сроками, то практнческое значение философских перспектив иноr-
да оказывается более важным, чем праКТl!ческие приложения как
таковые. Перспективы, открываемые теоретической эволюционной
биолоrией, которые помоrл.и бы предотвратить rибель человече-
ства, окажутся практическими приложениями первостепенной
важности.
Знания, накопленные эволюционной биолоrией, -вытекающие из
них следствия 'и предостережения доступны. людям уже На протя
жении мноrих лет, 'но ими пренебреrали как те, кто стоит у вла-
сти, так и значительная часть rраждан. А тем временем старые
традиционные правительстgа не обеспечивают улучшения yc.'to
вий жизни людей и решения насущных проблем. Очевидно,. эво
люционная 6иолоrия должна вмешаться в дела общества. Ибо
предостережения эволюционной биолоrи» касаются каждоrо
из нас.
СПИСОК JIИТЕРАТУ,РЫ
АЬеl О., 1929. Palaobiologie und Stammesgeschichte, Gustav Fischer Verlag,
Jena.
Alexander R. D., 1974. The evolution of social behavior, Апп. Rev. Ecol. 5yst.,
5, З25 38З.
Aтadon D., 1950. The Hawaiian honeycreepers (Aves, Drepaniidae), Bull. Атет.
Mus. Nat. Hisl, 95 (4), 15t 252.
Anderson Е., Brown W. L., 1952. Origin of corn belt maize and its genetic sig-
пШсаl1се. 'п: Heterosis, ed. Ьу J. W. Gowen, lowa Statc College Press,
Аmез.
Anderson W., Dobzhansky Th., Pavlovsky О., Powell 1., Yardley D., 1975. аепе.
tics of natural populations. XLIII. Three decades of genetic change in Dro-
sophila pseudoobscиra, Evolution. 29, 24....36.
Andrewartha Н. а., Birch L. С., 1,954. ТЬе Distribution and Abundance of Ani-
таlз, UniveTsity of Chicago Ртезз, Chicago.
AntonotJics /., 1971. ihe еПес.ts oI а heterogeneous environment оп the genetks
of natural populations, Аmет. Sci., 59. 59З 599.
Avery О. Т., Macleod С. М., McCarty М., 1944. Studies оп the chemical nature of
the substance inducing transformation of pneumococcal types, J. Ехр. Med.,
79, 137 158.
Axelrod D. 1., 1959. Late Cenozoic evolution of the Sierran bigtree forest, Evolu-
tion, 1 3, 9 23.
Axelrod D. /., 1967. Quaternary extinctions of large таmmаlз, Univ. СаШ. РоЫ.
Geol. Sci., 74, 1 42.
Axelrod О. Т., 1970. Mesozoic paleogeography and earty angiosperm history, Bot.
Rev., 36, 277 ЗI9.
Axelrod D. 1., 1972. Ocean-floor spreading in relation (о ecosystematic probIems.
In: А symposium оп Ecosysternatics, ed. Ьу R. Т. Allen and F. С. James,
University of Arkansas Мизеиm, Fayetteville, Occasional Рарет No. 4.
Axelrod D. 1., 1974. Revolutions in the plant world, Geophytology, 4, 1 6.
Axelrod D. 1., Bailey Н. Р., 1968. Cretaceous dinosaur extinction, Evolution, 22,
595 611.
Ayala Р. 1., 1969. Experimental invalidation of the principIe of competitive excIu-
sion, Nature, 224, 107&-----1079.
Ayala F. /. 1974. The concept of biologicaI progress. Iп: Studies in the Philosophy
of Вiology, ed. Ьу Р. J. Ayala and ТЬ. Dobzhansky, МасmШап, New York
and London.
Ayala Р. 1., Anderson W. W. 1973. Evidence of natural selection in molecular
evolution, Nature, 241, 274 276.
Ayala Р. J., Gilpin М. Е., 1974. Оепе frequency СОПlрапзопз between taxa: sup
port for the natural selection of protein polymorphisms, Proc. Nat. Acad. 5ci.
USA, 71, 4847 4849.
Ayala Р. 1., Moиrao С. А., Perez 8alas 8., Richтoпd R., Dobzhaпsky Th., 1970.
Enzyme variabi1ity in the Drosophila willistoni roHp. 1. Genetic differentia'
Ноп arnong sibIing specics., Proc. Nat. Acad. SC1. USA, 67, 225 232.
Ауша Р. 1., Tracey М. L., 1974. Genetic differentiation within and between species
of the Drosophila willistoпi group, Proc. Nat. Acad. 5ci. USA, 71, 999----
1003.
Список литературы.
315'
Babcock Е. B. Slebbins О. L.. 1938. ТЬе American species оЕ Crepis: their inter.
relationships and distribution as affected Ьу polyploidy and apomixis. Car-
negie Iпstitutiоп of Wаshiпgtоп, Wаshiпgtоп. О. С., РиЫ. 504. .
Bada J. L.. Schroeder R. A. Carter О. Р.. 1974. New evidence for the antiquity
of тап in North Americ8 deduced from aspartic acid rасеmizаtiоп, Science,
184, 791 793.
Bajeтa С. J.. 1971. Natural Selection in Нитап Populations, John Wiley and
Sons, New York.
Bakker R. Т., 1975. Dinosaur renaissance, Sci. Amer., 232 (4), 58 78.
Baldwiп Р. Н.. 1953. Аппиаl сусlе, environment and evolution in the Hawaiian
hопеусrсереrs (Avcs: Drepanidae), Uпiv. СаШ. PubI. 2001., 52, 285 З98.
Barghoorn Е. S.. 1971. ТЬе oldest ЕоззНз, Sci. Amer., 224 (5), з{).........42.
Bartholoтew О. А.. Birdsell J. В.. 1953. Ecology and the protohominids, Amer.
Anthropol., 55, 481----498.
Baleтan А. 1.. 1950. Is gene disреrsiоп normal? Heredity, 4, 353.....з6З.
Bateтan 1(. а.. 1959. ТЬе genetic аssirnШаtiоп of four venation phenocopies, J.
Genet.. 56, 44З 474.
Beardтore 1. А.. Dobzhansky Th.. Pavlovsky О.. 1960. Afl attempt to compare the
fitncss of polymorphic al1d monomorphic experimental populations оС Droso
phila pseиdoobscиra. Heredity, 14, 19 33.
8eaиdry 1. R. 1900. ТЬе species concept: Нз еvоlutюп and present status, Rev.
Canad. Biol., 19, 219 240.
Bergsoп Н.. 1911. Creative Evolution, Translation, Henry Ноlt and Со., New
York, Various later reprints.
Birdsell 1. В.. 1.950. Some implications of the genetical concept of race in terms
oj spatial analysis. Cold Spring Harbor Symp. Quant. Biol., 15, 259 314.
Birdsell 1. В.. 1958. Оп population structure in generalized huпtiпg and collecting
рориlаНопз, Evolution, 12, 189 205.
Blair W. F., 1955. Mating саН and stage of speciation in the Microhyla оииасеа
М. carolinensis coтplex. Evolution, 9, 469 80.
8lair W. F.. 1960. ТЬе Rusty Lizard: А Population 5tudy, University оЕ Texas
PresS, Аustiп.
Bock W. 1., 1970. Microevolutionary sequences as а fundamental concept iп mic-
roevolutionary models. Evolution, 24, 704 722.
Bodmer W. Е" СаvаШ Sfоrzа L. L.. 1916. Genetics, Evolution and Man,
W. Н. Freeman and Со., 5ап Francisco.
Bonnett О. T. 1954. The inflorescences of maize, Science, 120, 77 87.
8race С. L.. Nelson Н.. Korn N., 1971. Atlas оУ Fossil Мап, Ноlt, Rinehart апd
Winston, New York.
Briffaиlt R.. 1927. ТЬе Mothers, 3 vols., МастШап, London.
Brower L. Р.. 1970. Plant poisons in а terrestrial food chain and imрliсаtiопs for
mimicry theory. In: Biochemical Соеvоlutiоп, Оrеgоп State University Press,
Corvallis.
Brower L. Р.. Brower 1. V. Z.. 1964.. Birds, ЬuttеrШe.s and plant poisons: а study
in ecological chemistry, Zoologica (New York), 49, lЗ7 159.
Browп W. L., Wilson Е. О,. 1956. Character disрlасemепt, Syst. ool., 5, 49:---64.
Brucher Н., 1-943. Experimentelle Uпtеrsuсhuпgеп uber den Selektlonswert kunst-
lich erzeugter Mutanten von Antirrhinиm majиs. Z. Botan., 39, 1---47.
Brиes А. М.. 1969. Genetic load and its varieties, Science, 164. 1130 1136.
Brиssard Р. Р., Ehrlich Р. R.. 1970. The population structure of Erebla epipsodea
(Lepidoptera: Satyrinae), Ecology, 51, 119....129.
Bиffon О. L. L., 1770. Histoire Naturelle des Oiseaux. Paris. .
Bиffon. О. L. L.. 1808. Natural History 01 Birds, Fish, lnsects and Rept1les. Trans-
lаНопз, 6 vols., Symопds, London. .
Bиs" а. L.. 19698. 5ympatric host race formation d speciation .in frugivorous
flies of the genus Rhagoletis (Diptera: Tephntldae}, ЕvоlutlОП, 23, 237
251.
37б
Сnисо" литератур'"
Bush О. L.. 1969Ь. Mating behavior, host specificity and the ecological sigпifiсап-
се of siЫiпg species in irugivorous flles of the gепus Rhag"oletis Dipter<J.-
ТерhrШdае), Атет. Nat., 103, 669----612.
Cain А. 1., Sheppard Р. М" 1954. Natural sеlесtiоп iп Сераеа. Gепеtiсs, 39, 89
116.
Caтpbell В., 1966, 1974. Нuтап Evolution. Eds. 1 and 2, Аldiпе-Аthеrtоп, Chicago
апd New York.
СатрЬеи В. (ed.), 1972. Sexual Selection and the Oescent о' Мап 1871 1971,
Аldiпе-Аthеrtоп, Chicago.
Candela Р. В., 194 . The introduction of bIood-grоuр В into Europe. Human 8iol.,
14, 4l3 443. "
Carlqиist S., 1914. lsland Bioiogy. Columbia Uni\fersity Press. New York.
Carson Н. L., 1957. ТЬе species аз а field (ос gene recomblnation. In: The Spe-
cies ProbIcm, ed. Ьу 1:.. Mayr. Аmеriсап Association for the Аdvапсеmепt of
Science, Washington. О. С.
Carson Н. L., 1959. ОепеНс conditions which promote or retard the (onnation of
species, Cold Spring Harbor Symp. Quапt. Biol., 24, 87....105.
Carson Н. L.. 1970. Chromosome tracers 01 the origin 01 species. Science, 168.
1414....1418.
Carson Н. L.. 1971. Speciation and the founder principle, Stadler Оепе!. Symp.,
3. 51....70.
Carson Н. L., 1975. ТЬе genetics о( speciation 8t the diploid level, Amer. Nat.,
109. вз-.--..92.
Carsoп Н. L., Heed W. В., 1964. Structural homozygosity iп mаrgiпаl populations
of nearctic and neotropical species оС Drosophila iп Florida. Proc. Nat. Acad.
Sci. USA, 52, 427---430. .
СаvаШ-Sfоrzа L. L., Bodтer W. F.. 1971. ТЬе Genetics of Ниmап Populations.
W. Н. Freeman апd Со., San Francisco.
Chaтbers R., 1844. Vestiges о' the Natural History of Сrеаtiоп, London.
Clarke В., 1973. Neutralists vs. selectionists, Science, 180. 6О()......601.
Claиsen 1., 1951. Stages iп the Evolution of Plant Species, Cornell Uпiversitу
Press. Ithaca, N. У.
Claиsen J., 1965. Рорulаtiоп studies о' аlрiпе and subalpine тасез of conifers and
wШоws in the Саlifоrпiа high Sierra Nevada, Еvоlutiоп, 19. 56----68.
Claиsen 1.. Hiesey W. М.. 1958. Experimental studies оп the n8ture of species.
lV. ОenеНс structure 01 ecological races, Carnegie lnstitution of Washington.
Washington, О. С.. РоЫ. 615.
Claиseп J., I(eck D. D., Hiesey W. М., 1940. Experimental studies оп the nature
of species. 1. Effect of varied епvirопmепts оп western North American plants,
Саrпеgiе lnstitution 01 Washington, Washington, О. С.. РоЫ. 520.
Claиsen /.. I(eck D. D., Hieseg W. М., 1948. Experimental studies оп the паturе
01 species. 111. Епvirопmепtаl rеsропsеs о' climatic races of Achillea, Came-
gie Iпstitutiоп of Wаshiпg1оп. Washington, О. С., РоЫ. 58[.
Claиsen R. Е., 1941. Polyploidy iп NiсоtiапаJ Arner. Nat., 75, 291---306.
Cloиd Р., 1974. Еvоlutiоп of ecosystems. Атес. Sci., 62. 54....66.
Colbert Е. Н., 1955. 1969. Evolution о' the Vertebrates. Eds. 1 апd 2, JOhD Wiley
and Sons, New York.
Colbert Е. Н., 1961. Dinosaurs: Their Discovery and Their World. Е. Р. Dutton
and Со., New York.
Colwell R. N.. 1951. The use of radioactive {soюрез in determining spore distri-
bution patterns. Аmет. J. Bo1., 38, 511 523.
Connell J. Н., Orias Е., 1964. 1'he есоlоgicзl regulation о! species di'lersity, Amer.
N а L, 98, 399---414.
Сnисо1С литератур",
377
Сооn С. 5., 1955. 50mе problems of human varlability and natural selection in
climate and cu1ture, Аmет. Nat., 89, 257 279.
Сооn С. 5., 1962. The Story of Мап. Ед. 2, Alfred А. Knopf, New York.
Соре Е. D. 1896. TI1C Primary Factors оС Organic Evolution, Ореп Court, Chi-
cago.
Cronquist А., 1951. Orthogenesis in evo1ution, Res. Stud. State C<l1I. Wash., 19,
3....18.
Cronqиist А., 1968. The Evo1ution апd Classification of Flowering Plants, Hough
{оп МiПНп, B05ton.
Cronqиist А., 1976. The taxonomic sigпifiсапсе of the structure of plant protefns:
а classical taxonomist's view, Brittonia, 28, 1....27.
Crosby 1. L., 1963. The еvоlutiоп and паturе of dominance, J. Theor. 8101., 5,
35-----5] .
Crow 1. F., 1970. Genetic loads апd the c05t of паturаl se[ection. In: Mathematical
Topics in Population Genetics, ed. Ьу К. Kojima, Springer Verlag, New
York.
Crow 1. F., Kiтиra М., 1970. Ап Introduction to Population Genetics Theory, Har-
per апd Row. New York.
Crumpacker D. W., Williaтs 1. 5., 1973. Density, dispersion and population struc.
ture ,п Drosophila pseиdoobscиra, Есо\. Monogr., 43, 499 538.
Darlington С.' D., 1939', 1958. ТЬе Evolution of Qenetic Systems. Eds. t and 2.
Ed. 1, Cambridge Uпivеrsitу Press, Cambridge, Ed. 2, Basic Books., New
York.
DarUngton С. D., 1969. The Evolution of Мап and Society, Simon and Schuster,
New York.
Darwin С., 1859, 1872. Оп the Origin of Sp'ecies Ьу Means of Natural Selection.
Eds. 1 and 6, John Murray, London. LИмеется перевод: Дарвин Ч. Проис.
хождение видов путем ecтecTseHHoro отбора. Соч., т. 3..... М. .... п.: Иад-во
АН СССР, 1951.]
Darwin С., 1871, 1874. ТЬе Descent о! Мап and Selection In Relation to Sex.
Eds. 1 and 2, John Murray, London. [Имеется перепод: Дарвин Ч. Проис.
хождение чеJ10века и ПО.'10ВОЙ отбор. Соч., т. 5. М. .... п.: Изд-во АН СССР,
J 953.]
Darwin С., 1875. The Variation of Animals апд Plants Under Domestication.
Ed. 2, 2 vo\s, John Мштау. London. [Имеется пере80Д: Дарвин Ч. Изменение
животных и растений при одомашнивании. Соч., т. 4..... М. .... п.: Изд-во
АН СССР, 1951.]
Darwin F. (ed.), 1958. ТЬе Autobiography of Charles Darwin and Selected Latters.
Re..erint, Dover РuЫiсаtiопs, New York.
Dayhorf М. О. (ed.), 1968. Atlas of Protein Sequence апd Structure, Уо[. З, 1967
1968. Nationa[ Biomedical Research Fоuпdаtiоп, Si1ver Sрriпg, Md.
Dayhoff М. О. (ed.), 1969. Atlas of Protein Sеquепсе and Structure, Уо]. 4, 1969.
National Biomedical Research Foundation, Silver Spring, Md.
Daghoff М. О. (ed.), 1972. Atlas о! Protein Sequence and Stтucture, Уоl. 5,
1972. National Biomedical Research Foundation, Silver Sprlng, Md.
De Вее, а. R., 1951. ЕmЬтуоз and Ancestors. Ed. 2, Oxford University Рrеsз,
Oxford.
De Вее! а. R., 1964. Chrales Darwin, Doubleday and Со., Garden City City.
N. У.
DeFrles J. С., McClearn а. Е., 1972. Behaviora1 genetics and tbe fine structure
оп mouse r)()puJ;jtion : а st1..1dv in microevolution, Еуо1. Biol., 5, 279 291.
DeVore 1. (ed.), 1965. Primate 8ehavior: Field Studies of Monkeys and Арез,
НоН, Riпеhаrt 8пd Wiпstоп, New York.
De Wet 1. М. 1., Harlan 1. R., Н172. Origin of malze: the tripartite hypothesis.
Euphytica, 21, 271....279.
De Wet J. М. 1., Harlan 1. R., Grant С. А., 1971. Origin апд evolution of teosfnte
lZea тexlcaпa (Schra .) Kuntze], ЕирhуНса, 20, 255....255. .
378
Список литературы
Dobzhansky Th., 1937, 1951а. Genetics and the Origin оЕ Species. Eds. 1 and 3,
Со]иmЫа University Press, New York.
Dobzhansky Th., ]943. Genetics of natural рориlаНопз. IX. Temporal changes iп
the composition о' рорulаtiопs of Drosophila pseиdoobscиra, Genetics, 28,
162....] 86.
Dobzhansky Th., 1947а. Genetics of natural populations. XIV. А response of cer
tain gene arrangements in the third chromosome of Drosophila pseudoobscиra
to natural selection, Genetics, 32, 142....160.
Dobzhansky Т., 1947Ь. А directional change in the genetic constitution of а natu-
ral population of Drosophila pseudoobscиra, Heredity, 8, 53 64.
Dobzhansky Th., 1948. Genetics of natural populations. XVI. AItitudinal and
seasol1a] сllэпgсs produced Ьу паturаl se]ection in ccrtain popula tiol1s о!
Drosophila pseиdoobscиra and Drosophila persiтilis, Genetics, 33, 15S-----
176. .
Dobzhansky Th., 1950. Мепdеliап populations and their evolution, Arner. Nal, 84,
401......-.418.
Dobzhansky Th., 1951а. (See Dobzhansky, 1937, 1951а, above.)
Dobzhansky Th., 1951b. Experiments оп зе.хиаl isolation in Drosophila. Х. Repro-
ductive isolation Ьеtwееп Drosophila pseudoobscиra and Drosophila persimUis
uпdеr паturаl and under laboratory conditions, Proc. Nat. Acad. Sci. USA,
37, 792....796.
Dobzha.nsky Th., 1955. ТЬе genetic basis of systematic categorles. Iп; Biological
Systematics. Bio]ogy Colloquium, Oregon State College, Corvallis.
Dobzhansky Th., 1956. Gепеtiсs -of паturаl рорulаtiопs. XXV. Genetic changes in
populations of Drosophila pseиdoobscиra апd Drosophila persimilis in some
10calities in Саlifоrпiа. Evolution, 1 О, 82....92.
Dobzhansky Th., 1958. Gепеtiсs of natural populations. ХХУII. The genetic chan-
ges iп populations of Drosophila pseиdoobscиra in the Am rican soulliwest,
Evolution, 12, З85 401.
Dobzhaп.sky Th., ] 962. Мапkiпd Evolving, Yale University Press, New Haven,
Сопп.
Dobzhansky Th., 1970. Genetics of the Evolutionary Process, Columbia University
Press, New York. .
Dobzhansky Th., 1971. Evolutionary oscillations in Drosophila pseиdoobscиra. Iп;
Ecological Gепеtiсs and Еvоlutiоп, ed. Ьу R. Creed, Blackwell Sсiепtifiс РиЬ-
lications, Oxford.
Dobzhansky Th., Ehrтan L., PavlofJsky О., Spassky В., 1964. The superspecies
Drosophila paиlistorит, Proc. Nat. Acad. Sci. USA, 51, 3 9. .
Dobz/zansky Th., Epliflg С., 1944. СопtriЬuLiОl1S to tlle gCl1etics. {ахопоmу and
ecology of Drosophila pseиdoobscиra and its relatives, Carnegie Institution
of Wаshiпgtоn, Washington, О. С., РиЫ., 554.
.Dobzhansky Th., Levene Н., 1951. Development of heterosis through natural зе-
lесtiоп in experimental рорulаtiопs of Drosophila pseиdoobscиra, Amer. Nat.,
85. 24 7 264.
.Dobzhansky Th., Pavlovsky О., 1957. Ап experimental study of interaction between
gспсtiс drift and natura\ selection, I:::volution, 18, И I ЗI9.
.Dobzhansky Th., Powell J. R., 1974. Rates оЕ dispersal of Drosophila pseиdoob-
sct.tra and its relatives, Pruc. Roy. Soc. Londun, В 887. 2 H 298.
Dobzhansky Т., Spassky В., 1947. Evolutionary сhапgеs in laboratory cultures of
Drosophlla pseиdoobscиra, Evolution, 1, 191....216.
Dobzhansky Th., Spassky В., Spassky N., 1952. А comparative study of mutation
rates in l.wo ecologically diverse spccies oi Drosophila, GeneLics. 37, б50.
664.
Dobzhansky Th., Spassky N. Р., 1962. Genetic drift and natural selection in ехре-
rimental populations 01 Drosophila pseиdoobscиra, Proc. Nat. Acad. ScL USA,
48, 148 1б6.
Сnисок литературы
379
Dobzhansky Th., Wright S., 1943. Genetics а! natural populations. х. Disреrsiоп
rates in Drosaphila pseudoobscura, Genetics, 28, 304 340.
Dobzhansky Th., Wright 5., 1947. Genetics of natural рорulаtiопs. ХУ. Rate of
dШusiоп af а mutапt gene through а population of Drosophila pseudoobscиra,
Gcnetics, 32, 303....324.
Dott R. Н., Ваиеn R. L., 1971. Evolution of the Earth, McGraw-НШ, New Yark.
Downs Т., 1961. А study af variation апd еvоlutiоп in Мiосепе Merychippиs.
СопtriЬutiопs in Science, Los Angeles соuпtу Museum. Las Апgelеs, Na. 45.
пиnn L. С., пиnn 5. Р., 1957. The Je\vish community af Rame, Sci. Arnет., 196,
(3), 118 128.
Dиrrallt А., 1962а. ТЬе епvirопmепtаl induction о! heritabIe change in иnит,
Heredity, 17, 27....61.
Dиrra:t.t А., 1962Ь. Induction, reversion and epitrophism of Пах genotrophs, Natu-
те, 196, 1302....1304.
Ehrlich Р. R., 1961. lпtriпsiс barriers to dispcrsal in checkerspat butterffy, Scien-
се, 134, 108----109.
Ehrlich Р. R., Harriтan R. L., 1971. How to Ве а Survivar, Bal1antine Books,
New York.
Ehrlich Р. R., Raven Р. Н., 1964. Butterfljes a.nd plants: а study in caevolution,
Evolution, 18, 586---608.
Ehrlich Р. R., White R. R., Singer М. С., McKechnie S. W., Gilbert L. В., 1975.
Chcckerspat buttcrflies: а historical pcrspective, Science, 188, 221 228.
Ehrтan L., 1965. Direct аЬsеrvаtiоп af sexual isolation between allopatric and
bct\v en sympaLric strains of the djffcrcnt Drosophilи paиlistoruт races. Evo.
1 u tiоп, 19. 459....464.
Ehrman L., Kernaghan R. Р., 1971. Microorganismal basis of infectiaus hybrid
male sterility in Drosophila paиlistoruт, J. Hered., 62, 66....71.
Ehrтan L., Spiess Е. В., 1969. Rare type mаtiпg аdvапtаgе in Drosophila, Аmет.
Nat., 103, 675 O.
Eltrтa1l L., Williamson О. 1.., 1965. Transmission Ьу injectiol1 оУ hybrid stcfility
to попhуЬrid males iп Drosophila paulistorит: рrеНmiпаrу repart, Proc. Nat.
Acad. Sci. USA. 54, 481....483. ,
Eiseley L., 1958. Darwin's Сепturу, Doubleday апd Со., Garden City, N. У.
Eisenberg /. F., Muckenhirn N. А., Rиdran R., 1972. The relation between ecology
and social structure in primates, Sсiепсе" 1 76, 863....874.
Eklиnd М. w., Poysky F. Т., Meyers /. А., Pelroy а. А., 1974. Iпtеrsресiеs сап-
version of Clostridiuт botиlinит type С to Clostridiит novyi type А Ьу
bacteriophage, Science, 186, 45 458.
Eldredge. N., Goиld 5. 1., 1972. Punctuated еquШЬriа: ап аltеrпаtivе to phyletic
gradualism. In: Models in Paleobiolagy, ed. Ьу Т. J. М. Schopf, Freeman.
Cooper апd Со., 5ап Francisca.
Eтlen J, М., 1973. Ecology: Ап ЕуоlиНапату Appraach, Addison- Wesley, Reading.
Mass.
Epling С., Lewis Н., Ваи F. М., 1960. The Ьrееdiпg graup апd sced starage:
а study in рорulаtiап dynamics, Evalutian, 14, 238....25:5. .
Epliп.g С., Mitchell D. F., Mattoni R. Н. Т., 1953. Оп fue role of inversions in
\\'ild populatians of Drosophila pseиdoobscиra, Evolution, 7, 342....365.
Evans а. М., Dиrrant А., Rees Н., 1966. Associated пuсlеаr changes in the induc-
tiап af flах genotraphs, Nature, 212, 697 99.
Ewens W. J., 1965. Further поtеs оп the evalutian of dominance, Heredity, 20,
44З 50.
Faegri К., vaп. der Pijl L., 1971. The Principles оС Роlliпаtiоп Ecology. Ed. 2,
Pergaman Press, Oxford.
Falconer D. 5., 1960. Iпtroduсtiоп to Quantitative Gепеtics. Ranald Press, New
York.
380
Список литературы
Farb Р., 1968. Man's Rise to Civilization as Shown Ьу the Iпdiапs of North Ате-
rica [rom Primeval Times 10 the Соmiпg of the Iпdustriаl State, Е. Р. Dut-
ton and Со., New York.
Felsenstein /., Yokoyaтa 5., 1976. ТЬе evolutionary advantage of recombination.
11. Iпdividuаl se[ection for recombination, Genetics, 83, 845---859.
Fisher R. А., 1930, 1958. ТЬе Genetical Theory of Natural Selection. Eds. 1 and 2.
Ed. 1, Clarendon Press, Oxford, Ed. 2, Dover РubJiсаtiопs, New York.
Fisher R. А., Ford Е. В., 1947. ТЬе spread of а gепе iп natural conditions iп а
соl0ПУ о! the moth Panaxia doтinиla L, Heredity. 8, [43....174.
Flake R. Н., Grcint У., 1974. Ап analysis о( the cost of selection concept, Ртос.
Nat. Acad. Sci. USA, 78, 3716 3720.
Ford Е. В., 1955. Moths, Collins, London.
Ford Е. В., [964, 197[. Ecological Genetics. Eds. 1 and 2. Ed. 1, Methuen, London,
Ed. 3, Chapman and НаН, London.
Ford 1.::. В., 19ь5. GE'nctic Роlушurрhism. Faber al1d Fabcr London.
Fothergill Р. а., 1952. Historica[ Aspects of Оrgапiс Evolution, Ноlliз and Carter,
London.
Fox А. 5., Dиggleby W. F., Gelbart W. М., Уооn S. В., 1970. DNA induced trans.
formation in Drosophila: evidence for transmission without iпtеgrаtiоп, Proc.
Nat. Acad. Sci. USA, 67, 1834....1838.
Fox А. S., Уооn 5. В., 1966. Specific genetic effects of DNA in Drosophila тela
nogaster, Genetics, 53, 897....911.
Fox А. 5., Уооn 5. В., 1970. DNA-induced transformation in Drosophila: 10cus-
sресШсitу апd the establishment of trапsfоrmеd stocks, Proc. Nat. Acad. Sci.
USA, 67, 1608....1615.
Fox А. 5., Уооn S. В., Gelbart W. М., 1971. DNA-induced transformation in Dro
sophila: genetic analysis of transformed stocks. Proc. Nat. Acad. Sci. USA,
68, 342---346.
Galinat W. С., 1970. ТЬе cupule апd its тоlе in the origin and evo[ution of maize.
Agricultural Experiment Station, Uпivеrsitу of Massachusetts, Amherst. BuH.
585.
Galinat W. С., 1971а. ТЬе origin of maize, Ann. Rev. Gепеt., 5, 447---478.
Galinat W. С., 1971 Ь. ТЬе evolution of sweet сотп. Agricultural Experiment Stati-
оп, University of Massachusetts, Amherst. BuB. 591.
Garn 5. W., 1961. Human Races, Charles С. ТЬотаз, Springfield, Ш.
rаузе т. Ф., 1934. ТЬе Struggle for Existcnce. Wil1iams and \Vilkins, Baltimo1"E'.
Fаузе [. Ф., 1935. Experimental demопstrаtiоп of Volterra's periodic оsсillаtiопs
. iп the numbers of animals. J. Ехр. Biol., 12, 44---48.
(]eist У., [971. Mountain Sheep: А Study in Behavior and Еvоlutiоп, University
of Chicago Press, Chicago.
йШ Е. D., 1975. Evolution of Australia's unique Поrа and fauna in relation to
the 'plate tectonics theory. Proc. Roy. Soc. Victoria, 87, 215....234.
Glass В., 5асЬ М. 5., /ahn Е. F., Hess С., 1952. Genetic drift in а religious iso-
1 ate: ап analysis о! the causes of variation in blood groups and other gепе
frequencies in а зmаll population, Атб. Nat.. 86, 145----159.
Glick Т. Р. (ed.) , 1974. ТЬе Comparative Reception of Dаrwiпism, University of
Техаз Prcss, Austin.
Goldschтidt R. В., 1940. ТЬе Material Basis of Evolution, Уаlе Univcrsity Press,
. New Haven, Conn.
Goldschтidt R. В., 1952. Homoeotic mutanis and c\'olution, Acta Biotheor.. 10.
87....104.
Goldschmidt R. В., 1953. Experiments with а bomoeotic mutant, bearlng оп. evo-
lution, J. Ехр. Zool., 123, 79----[ 14.
Goldschтidt R. В., 1955. Theoretical ОепеНсз, University of California Press,
Berkeley.
Gottlieb L. D., 1971. Оеl e1ectrophoresis: new approach to the study о' evolution.
Bioscience, 21, 939....943.
Список литературы.
381
Gottlieb L. D., ]974. Genetic confirmation of the origin of Clarkia lingиlata, Evo-
lution, 28, 244 250.
(}ould S. J., 1973. The mi5named, mistreated and misunderstood Irish elk, Nal
Hist.,82 (3), ]O 19.
Gould S. J., 1974. The origin and function of cbizarr9 structures: antler size
and skull size in the clrish elk , Megaloceros giganteиs, Evolution, 28, 191
220.
Graпt А., Graпt V., 1956. Genetic and taxonomic 5tudies iп Gilia. VH 1. The СоЬ-
webby gilias, AIiso, З, 2ОЗ 287.
Oraпt К. А., Grant У., 1964. Mechanical isolation of Salvia apiana and Salvia
тellifera (Labiatae). Evolution, 18, 196 212.
Oraпt К. А., Grant У., 1968. Hummingbirds and Their Flowers, СоlиmЫа Univer-
sitv Pres5, New У ork.
Grant 'У., 1954. Genetic and taxonomic studies in ОШа. IV. ОШа achilleaefolia,
Aliso, З, 1 18.
Grant У., 1958. ТЬе regulation of recornbination in рlапts, Cold Spring Harbor
Symp. Quant Biol., 23, 331 36З.
Graпt У., 1969. Natural History of the Phlox Family. Systematic Botany, М. Hij-
hоП, The Hague.
Grant У., 1960. Genetic and taxonomic studies iп ота. XI. FertiIity relationships
of the diploid Cobwebby gilias, Aliso, 4, 43 481.
Orant V., 1963. The Origin of Аdарtаtiопs, Columbia Uпivеrsitу .Press, New
У ork.
Orant У., 19б6а. Selection for vigor апd fertiIity in thc progeny of а highly sterile
species hybrid in ОШа, Genetics, 53, 757 775.
Grant V., 1966Ь. The selective origin of incompatibility barrier5 in the рlапt genus
ОШа, Amer. Nat., 100. 99 118.
Orant V., 1971. Рlапt Speciation, СоlиmЫа University Press, New York.
Grant V., 1975. Genetics of Flowering Plants, Columbia University Press. New
у ork.
Grant V., Flake R. Н., 1974а. Population structure in relation to cost of selection,
Рroс. Nat. Acad. Sci. USA, 71, 1670 lб71.
Orant V., Flake R. Н., 1974Ь. Solutions to the cost o( selection dHemrna, Proc.
Nat. Acad. Sci. USA, 71, 3863 3865.
Grant V., Grant К. А., 1965. F(ower Pollination in the РЫох Fami(y, СоlurnЫа
University Press, New York.
Gray А. Р., 1954. Mammalian Hybrids, Commonwea(th Agricultura( Bureaux,
Farnham Royal, England_ .
Gray А. Р., 1958. Bird Hybrids. Commonwealth Agricultural Витеаих. Farnham
Royal, Eng(and.
Oreenwood J. 1. D., 1974. Effectfve population. numbers fn the snail Сераеа nв-
тoralis, Еvо(utiоп, 28, 51 526. .
Greenwood J. 1. D., 1976. Effective population number In Сераеа: а modification,
Evo(ution, 30, 186.
Oиstafsson А., 1951. Mutations, environment and evolution, Cold Spring Harbor
Symp. Quanl Biol., 16, 263 281.
Haldane J. В. 8., 1932. The Causes of Evolution; Longmans, Green and Со., Lon-
don.
HaCdane J. В. 8., 1957. The <:05t of natural se(ection. J. Genet., 55, 511 524.
Haldane J. В. 8., 1960. More precise expressions for the cost of natural 5electlon,
J. Genet., 57, 351----360.
Наllат А. (ed.), 1973. Atlas of Pa1eobiogeograph}'. Аmсriсап Else\'ier. New York.
НатШоn W. D., 1964. The genetical evolution of social behavior. l and 11, J.
Theor. 8io(., 7. 1.....-.52.
Hardiп а. 1960. The competitive excluslon principle, Science, 131. 1292 1297.
Hardiп О.' 1972. Exploring New Ethics for Survival. Vilking Press, New York.
Hardiпg J:, Allard R. W., Sтeltzer D. а., 1966. Population studies {п predominant-
382
Список литературы
Iy sеlf роШпаtеd species. IX. Frequency-dependent selection in Phaseolus lи
natu"s, Ртос. Nat. Acad. Sci. USA, 56, 99 104.
Hardy А. С., 1954. Escapc from spccialization. 'п: E,,'olution as а Proc('ss. ed. Ьу
J. Huxley, А. С. Hardy and Е. В. Ford, George АНеп and Unwin, London.
Hayes W., 1968. ТЬе Genetics of Bacteria and Their Viruses. Ed. 2, John Wiley
and Sons, New York. [Имеется перевод l-ro НЗД.: Хэйс У. rенетика бак-
l'срий и бактерНОф2fОВ. М.: «Мир , 1965.1.
Heiser С. В., 1973а. Seed 10 Civilization, W. I-l. Freeman and Со., San Francisco.
Heiser С. В., 1973Ь. If!trogression re-examined, Bot. Rev., 39, 347-----366.
НШ 1., 1967. Тl1е environmental induction of heritabIe changcs in Nicotiana rиs-
tica parental and selection lines, Genetics, 55, 735 754.
Hinton Т., Ives Р. Т., Evans А. Т., 1952. Changing the gene order and number in
l1atural populations, Evolution, 6, 19-----28.
Hiraizиmi У., Sandler L., Crow 1. F.. 1960. Meiotic drive in natural populations
of Drosophila melanogaster. III. Populational implications of the segrega-
tiondistorter 10cus, Evolution, 14, 433---444.
Hovanitz W., 1953. Polymorphism and evolution, Symp. Soc. Ехр. Biol., 7, 24
253. .
НиЬЬу 1. L., Lewontin R. С., 1966. А mоlесиlат approach 10 the study of genic
heterozygosity in natural populations. 1. The number of al1eles at dШеrепt
loci in Drosophila pseudoobscura, Genetics, 54, 577 594.
НиЬЬу J. L., Throckmorton L. Н., 1965. Protcin differences in Drosophila.
11. Comparative species genetics and evolutionary probIems, Genetics, 52.
203---215.
Hиettel М. D., Bиsh О. L., 1972. The gепеtiсs of host selection and its bcaring оп
sym f. atric speciation in Procecidochares (Diptera: Tephritidae), Entomol. Ехр.
Арр., 15, 4 480.
Hutchinson а. Е., 1965. The Ecological Theater and the Evolutionary Play, Yate
University Press, New Haven, Conn.
Huxley 1. S., 1932. ProbIems of Relative Gro\vth, Methuen, London.
Huxley J. S., 1942. Evolution: The Modcrn Synthcsis, George AlIen and Unwin.
Loridon.
Huxley J. S., 1954. ТЬе evolutionary process. In: Evolution as а Process, ed. Ьу
J. Huxley, А. С. Hardy and Е. В. Ford, George Al.len and Unwin. r.ondol1.
Huxlcy J. S'J 1958. Evolutionary processes апd taxonomy with special reference
to grades. In: Systematics of Today, ed. Ьу О. Hedberg, Uppsala Universi-
tets АrsskrШ, Uppsala.
lпgrarп У. .М., 1963. The Hemoglaыns in Genetics and Evolution, Columbia Uni-
versity Press, New York.
/ves Р. Т., 1950. Thc importance of mutation rate genes [п evolution, Evolution,
4, 236---252.
lain S. К., BradS/law А. D., 1966. Evolutionary divergence among adjacent plant
populations. 1. The evidence and its thcoretical analysis, Heredity. 21, 407
441.
Jepson W. L., 1909. А Flora of СаШоrпiа. Уоl. 1, Part 1, Associated Students
Store, University of C8lifornia. Berkeley.
Johaпnsen W., 1911. The genotype conccption of heredity, Amer. Nat., 45, 129----
159.
Johnsfoп J. S., Нее([ w. В., 1976. Dispersa! of desert adapted Drosophila: the
saguaro breeding D. nigrospiracиla, Аmет. Nat., 110, 629---651.
Jones Н. А., Walker J. С., Little Т. М., iarson R. М., 1946. Relation of colo-
rinhibiting factor to smudge rcsista11ce in onion. J. Agric. Res., 72, 259
264.
lones 1. S., 1973. Ecological ge.netics and natural selection in molluscs, Science.
182. 546---552.
Jukes Т. Н., 1972. Comparison of polypeptide sequences. In: Darwinian, Neo-Dar-
winfan and Non-Darwinian Evolution, ed. Ьу L. М. LeCam, J. Neyman and
Список лuтературы
383
Е. L. Scott, Berkeley Symposia оп Mathematical Statistics and Probability,
Proceedings of the Sixth Symposium, Уоl. 5, University of California Press,
Berkeley.
J<.east А.. Erk F. С.. Glass В. (eds.). 1972. Evolution, Mammals and Southern
Continents, State University of New York Press, Albany.
J(eith А.. 1948. А Ne\v Theory of Human Evolution, Watts, London.
Кетет А., 1894 1895. ТЬе Natural History of Plants. Translation, 2 vols., Вlасюе
and Son, London.
кет, W. Е., Wright S.. 1954. Experimental studies of the distribution of gene fre-
quencies in very srnall populations of Drosophila melanogaster. 1. Forked,
Evolution, 8, 172 177. .
l(erster Н. W., 1964. Neighborhood size in the Rusty lizard, Sceloporus olitJaceus,
Evolution, 18, 445 457.
J(erster Н. W., Levin D. А., 1968. Neighborhood size in Lithospermum caroliпiense,
Genetics, 60, 577 587.
Kettlewell Н. В. D., 1956. Further selectiol1 experiments оп industrial rnelanism in
the Lepidoptera, Heredity, 10, 287 3Ql.
КеШешсll Н. В. D., 1973. ТЬе E'v"Olution of Melanism, Oxford University Prcss,
Ne\v York.
l(imura М., 1968. Genetic variabi1ity maintaincd in а finite population due to mu-
tational production of neutral and nearly neutral isoaBelcs, GCllct. Res., 11,
247 269.
l(imиra М., Ohta Т., 1971. Theorctical Aspects of Population Gcnctics, Princeton
University Press, Princeton. N. J.
l(imura М.. Ohta Т., 1972. Population genetics, molecular biometry and evolution.
ln Darwinian, New Oarwinian and Non-Oarwinian Evotution, ed. Ьу
L. М. LeCam, J. Neyman and Е. L. Scott, Berkeley Symposia оп Mathemati-
са' Statistics and J.>rоЬаыlitу. Proceedings of tllе Sixth Symposium, Vol. 5,
University of California Press, Berkeley.
I<..ing J. L., Jиkes Т. Н., 1969. Non-Darwinian evolution, Science, 164, 788 798.
J(iпg М. С., Wilson А. С., 1975. Evolution at two levels in humans and chimpan-
zees, Science, 188, 107 1I6.
Kircher Н. W., Heed W. В., 1970. Phytochemistry and host plant specificity in
Drosophila. Recent Adv. Phytochem., 3, 191---209.
J(oestler А., Smythies J. R. (eds.), 1969. Beyond Reductionism: Ne\v Perspectives
in the Life Sciences, Alpbach Symposium, 1968, Macmillan, New York.
l(ojiтa К., Yarbroиgh К. N., 1967. Frequency dependent selection at the esterase-6
10cus in Drosophila тelanogaster, Ртос. Nat. Acad. Sci. USA, 57, 645---
649.
Kolata а. В., 1974. Kung huпtеr gаthеrеrs: feminism, diet and birth control, Scien-
се, 185, 932---934.
Коорrnаn К. F., 1950. Natural selection for reproductive isolation between Droso-
phila pseиdoobscura and Drosophila persimilis, Evolution, 4, 135---148.
.l(raиs О., 1973. Homo sapiens in ОесНпе, New Diffusionist Press, Bedfordshire,
Engl and.
J(rebs С. J., 1913. Ecology: ТЬе Experimental Analysis of Distribution and АЬип-
dance, Harper and Row, New York.
Krebs С. J., Gaines М. 5., Keller В. L., Megers J. Н., Tamariп R. Н., 1973. Рори-
lation cycles in smaII rodents. Science, 179, 35 41.
Kulp J. L., 1961. Geologic time scale, Science, 133, 1l05 1114.
Lack D., 1944. Ecological aspects of species formation in passerine birds, Ibis,
86, 260---286.
Lack D., 1947. Darwin's Finches, Cambridge University Press, Cambridge. -[Имеет-
ся перевод: ЛЭК Д. Дарвиновы: вьюрки. М.: ИЛ, 1949.]
. Laird С. D.. McCarthg В. J., 1968. Magnitude of interspecific nucleotide sequence
variability in DrosophJla, Genetics, 50, ЗОЗ 322. .
384
СписоlC литературы
Lamarck J. В. Р. de., 1809. PhHosophie Zoologie, Paris. [Имеется перевод: Ла-
марк Ж.-Б. ФИ.'10СОфИЯ зоолоrии. т. I lI. ИЗД ВО АН СССР. М. Л.,
1 935.]
Lamarck J. В. Р. de., 1815-----1822. Histoire NatureIle des Animaux sans Vertebres,
Paris.
Lamotte М., 1951. Recherches sur la structure genetique des population's naturel-
les de Сераеа nemoralis (L.), BuIl. biol. France Belg. (supp.), 35, I 238.
Lamotte М., 19 9. Polymorphism of natural populations оС Сераеа пemoralis,
Cold Spring H rbor Symp. Quant. Biol., 24. 86.
Lau.ghliп W. S., 1950. Blood groups, morphology and population size оС the Eski.
mos, Cold Spring Harbor Symp. Quant. Biol., 15, 165 17З.
Laycock о., 1974. Оilетmа in the desert: bighorns or burros? Audubon Magazine.
September 1974.
LeCam L. М., Neyman J., Scott Е. L. (eds.), 1972. Darwinian, Neo Dar\\'inian.
and Non Darwinian Evolution, Berkeley Symposia оп Mathematical StaListics
and Probability. Proceedings of the Sixth Symposiurn, Val. 5, University о,
СаШоrпiа Press, Barkeley.
LeOros Clark W. Е., 1960. ТЬе Antecedents оЕ Мап, Quadrangle Books. Chicago.
Leng Е. R., 1960. Long term selection of corn for аН and protein content. Mimeo-
graphed аппиаl report, IШпоis Agricultural Ехреrirnелt Station, Urbana.
[еrnе, 1. М., 1954. Genetic Homeostasis, Oliver and Boyd, Edinburgh and Lon-
don.
Lerner 1. М., 1958. ТЬе Genetic Basis of Selection, John Wiley and Sons, New
у ork.
Lerner 1. М., Но F. к., 1961. Genotype and competitive аЫlitу of Triboliu.т specie5,
Amer. Nat., 95, З29 343.
Levene Н., 1953. Genctic' сquШЬriшn where more than опе ecological niche is
availabIe, Amer. Nat., 87, ЗЗl ЗЗ3.
Leuin D. А., 1970. Reinforcement of reproductive isolation: plants versus animals,
Amer. Nat., 104, 571----581. .
Levin D. А., 1971. Plant phenolics: ап ecological perspective, Amer. Nat., 105.
157 181. '
.Levin D. А., 1972. Low frequency disadvantage in the exploitation оС роШпаtоrs
Ьу corol1a variants in РЫох, Лmет. Nat.. 106, 453----460.
Levin D. А., Crepet W. L., 1973. Genetic variation in Lycopodiu.т lиcidulиm;
а phylogenetic relic, Evolution, 27, 622 632.
Levin D. А., Kerster Н. W., 1967. Natural selection for reproductive isolation in
PhIox. Evolution, 21, 679........б87.
Levin D. А., Kerster Н. W., 1968. Loca. gene dispersal in Phlox, Evolution, 22,
1зQ......-.139.
Levin D. А., Kerster Н. W., 1969. Density-dependent gene dispersal in Liatris,
Arner. Nat., 103, 61 74.
Levin D. А., Kerster Н. W., 1971. Neighborhood structure in plants under diverse
reproductive methods, Amer. Nat., 105. 345----354.
Levin D. А., Kerster Н. W., 1974. Gene flow in seed plants, EvoJ. Biol.. 7,
1З9 220.
Levin D. А., Schaal В. А., 1970. Corol1a color as ап inhibitor of intcrspecific hybri
dization in РЫох, Amer. Nat., 104, 273 283.
Levins R., 1968. Evolution in Changing Environments, Princeton University Press,
Princcton, N. J.
Lewin R. А. (ed.), 1962. Physiology and Biochemistry of AJgae, Academic. Press,
New York.
Lewis Н., 1962. Catastrophic selection 85 а factor in speciation, Evoltition, 16,
257 271.
Lewis Н., Epliпg С., 1969. Delphinium gypsophilum, а diplaid species 01 hybrid
origin, Evolution, 13, 511 25.
Список лuтературы
385
Lewis н., Lewis М. Е., 1955. Thc genus CJarkia. Univ. СаШ. PubI. Bot.. 20. 241
392.
Lewis Н., Raven Р. Н., 1958. Rapid evolution in Clarkia, Evolution, 12, 319----
. 336.
Lewis Н., Roberts М. R., 1956. ТЬе origin Qf Clarkia liпgиlata, Evolutian 10.
126 138. ·
Lewontiп R. С., 1955. ТЬе effects af population density and composition оп viabi-
Шу in Drosophila тelanogast r, Evalution. 9. 27 1.
Lewontin R. С., 1970. ТЬе units of selection. Ann. Rev. Есо(. Syst., 1, 1....18.
Lewoпtin R. С., 1973. Population genetics, Апп. Rev. Genet., 7. ' '7.
Lewoпtin R. С., 1974. ТЬе Genetics Basis of Evolutionary Change, Co(umbia Uni-
versity Press, New York. [Имеется перевод: Левонтин Р. rенетическне ос-
новы эвопюЦ,ни. ---- М.: Мир, 1978.]
Lewontin R. С., НиЬЬу 1. L., 1966. А molecular approach to the study of gепlс
heterozygosity in natura( populatians. Н. Amount оС variation and degree of
heterozygosity in natural populations af Drosophila pseu.doobscura, Genetics,
54. 595 609. .
Lillegraven J. А., 1974. Biogeographic considerations of the marsupial placenta(
dichatomy, Ann. Rev. Ecol. Syst., 5, 263 283.
Lindsley D. L., Grell Е. Н., (968. Gene1ic Variations af Drosophila тelaпogaster,
Carnegie Institution of Washington, Washington, О. С.. РиЫ., 627.
Lindsay D. W., Vickery R. К., 1967. Comparative evolution in Miтu.lиs guttatu.s
оС the ВоппеvШе basin, Evolution, 21. 439......0156.
Littlejohn М. J., 1965. premating iso(ation in the Hyla ewingi complex (Anura:
Hylidae), Evolution, 19. 234....243.
Looтi." W. Е., 1967. Skin-pigment regulation of vitamin D blosynthesis in тап.
Science, 157. 501 .
MacArthu.r R. Н., 1958. Populatian ecology of some warbIers of northeastern со.
niferous forests, Eco(agy. 39, 599 619.
McVeigh 1., Hobdy С. J., 1952. Deve(opment of resistance Ьу Micrococcus pyogenes
var. aиreu.s to antibiotics: morphological and physiological changes, Amer.
J. Bot., 39, 352 359.
Mangelsdorf Р. С., 1958. Reconstructing the ancestor of carn, Proc. Атет. Phil.
Soc.. 102, 454-----463.
Mangelsdorf Р. С. (ed.), 1959. The origin of corn, Bot. Mus. Leafl. Harvard
Univ., 18, 329-----440.
Mangelsdorf Р. С., 1974. Corn: Hs Origin, Evolution and Improvement, Harvard
University Press, Cambridge, Mass.
Mangelsdorf Р. С., Dick Н. W., Caтara Herпande2 J., 1967. Bat Cave revisited,
Bot. Mus. Leafl. Harvard Uni\,., 22, 1 31.
Maпgelsdorf Р. С., lones D. Р., 1926. The ехрrеssiоп of Mendelian factors in the
gametophytc of maize, Genetics, 1 " 423 455.
Maпgelsdorf Р. С., MacNeish R. S., Galiпat W. С., 1964. Domestication of соrп,
Science, 143. 538 545. .
Marllulis L., 1970. Origin af Eukaryatic Cells, Уа'е University Press. Nвw Науеп,
Conn.
MarUn Р. 5., 1973. ТЬе discovery of America. Science, 179, 969....974.
Marlin Р. 5., Wright Н. Е., Jr. (eds.), 1967. Pleistocene E,xtinctions: The Search
for а Cause, Уа(е University Press. New Haven, Conn.
Mayпard Smith 1., '938. Sexua' selection. In: А Century of Darwin, ed. Ьу
S. А. Barnett, Hcinemann, London.
Миуо О. 19б6, Оп the P.\'oJution оС dоmiпапсе. Heredity, 21. 499 51'.
Мау, Е.; 1942. Systematics and the Origin of Species, СоlитЫа University Press,
Nev.' York. [Имеется перевод: Майр Э. Систематнка и происхождение ВИ-
ДОВ. М.: ИЛ, 1947.] .
386
Список литературы
Мау, Е., 1954. Change of genetic епvirоnmеl1t апd еvоlutiоп. Iп: Evolution as
а Process, ed. Ьу J. Нихlеу, А. С. Hardy and Е. В. Ford, George Al1en and
Unwin, London.
Мау, Е., 1957з. Spccies сопссрts зпd dсfiпitiопs. Iп: Тl1С Species Probl"m. <,d. Ьу
Е. Маут, Arnerican Association (от the Аdvапсеmепt оС Science, Washington..
О. С.
Мау, Е., 1957Ь. Difficu1ties апd imроrtапсе of the biological species сопсеоt. Iп:
ТЬе Species Problem, ed. Ьу Е. Маут, American Association for the Advan.
сеmепt of Sсiепсе, Washington, О. С.
Мау, Е., 1963. Animal Species and Evolution, Harvard University Press, cambrid.
ge, Mass. f Имеется перевод: Майр Э. 300поrический вид и эволюция. --- М.:
Мир, 1968.
Мау, Е., 1969. Principles оС Systematic Zoology, McGraw-НШ, New York. [Имеет-
ся перевод: Майр э. Принципы зоолоrической систематики. --- М.: Мир.
1971. ]
Мау, Е., 1970. Populations, Species 6iпd Evolution, Harvard Uпivсrsitу Press..
Cambridge, Mass. .
Мау, Е., 1972а. ТЬе nature оС the Darwinian rcvolution, Science, 176, 981----
989.
Мау, Е., 1972Ь. Sexual selection and natural setection. In: Sexual Sеlееtiоп and
the Dеsсепt of Man 1871---1971, ed. Ьу В. СаmрЬеll, Aldine-Atherton, сы-
cago.
Mettle, L. Е., aregg Т. а., 1969. Population Genetics and Evolution, Prentic
НаН, Englewood Cliffs, N. J. [Имеется перевод: Меттпер Л., rperr т. re-
нстика попупящrй И Э80.'1ЮЦИЯ. ---М.: Мир, 1972.]
Michener С. D. j 1947. А revision of the American species of Hoplitis (Нуmепор-
tera, Megachilidac), ВиН. Amer. Mus. Nat. Hist., 89, 257---318.
Michener С. D., 1975. ТЬе Brazilian Ьее problem, Ann. Rev. Entomol., 20, 399----
416.
мте, А. Н., 1955. А hybrid woodpecker and its sigпifiсапсе in speciation iп the
gcnus Dendrocopos, Evolution, 9, 317---321.
Millicent Е., Thoday J. М., 1961. Effects оС disruptive selection. IV. Gene flow
and divеrgепсе, Heredity, 16, 199-------217.
Moos J. R., 1955. Comparative physiology of some chromosomal types in Droso-
phila pseudoobscиra, Еvоlutiоп, 9, 141---151.
Mou,ant Л. Е., 1954. Tlle DistriЬutiоп of tlle Нumап I3100d Groups, Black\\'ell
SсiепШiс Рubliсаtiопs, Oxford.
Mиller С. Н., 1966_ The role of chemical inhibition (allelopathy) in vеgеtаtiопаt
соmроsitiоп, BuH. Terrey Bot. Club, 93, 332---351.
Миие, С. Н., 1970. The role of aHelopathy iп the evolution of vegetation. In: Вio-
chemical Coevolution, Оrеgоп Statc Uni'..ersity Press, Cot'\'a1lis.
Nassa, R., Muns /1. J., Cook R. D., 1973. Fre(Jtlcl1cy dcpcndent sеl('сtiоп at the
Раупе inversion in Drosophila тelanogaster, Evolution, 27, 558---564.
Nei М., lтaizuтi У., 19б6. Gcnctic !Ф-\1сtшс of hшпап populations. 1 апd 11.
Heredity. 21, 9---35. 183---190.
Neuffer М. О., Jones L., Zuber М. S., 1968. The Mutants of Maize, Crop Sсiепсе-
Society of America, Madison, Wis.
NeflJell N. D., 1949. Phyletic size increase an important trend Шustrаtеd Ьу fos-
sil invertebrates, Еvоlutiоп, 3, 103 124.
Newell N. D., 1967. Revolutions in the history of Ше, Geol. Soc. Аmет., Spec.
Рар., 89, 63---91.
Oakley К. Р., 1949. Мао the Tool Maker, British Museum (Natural History).
Lопdоп.
Ohta Т., Klтura М., 1971. Оп the constancy of the еvоlutiопаrу rate of ,cistrons_
J. Моlес. Evol., 1, 18---25.
Опарин, А. Н., 1962, 1964. Life= Its Nature, Origin алd DeveJopment, ОНуи and
Boyd. Edinburgh (1962), Aca-demic Press, New York (1964).
Списо" литературы
387
Osborn Н. F., 1910. ТЬе Age of Mammals in Еиторе, Asia and North America,
МасmШап, New York.
Osboт Н. F., 1929. The titanothefes of ancient Wyoming. Dakota and Nebraska.
U. S. Gcological Survey, Washington, О. С., Monograph 55.
Osborп А. Р., 1934. Aristogenesis, the creative principle in the origin of species,
Атет. Nat., 68, 193 235.
Owen D. Р., 1961. Industrial me(anism in North American moths, Amer. Nat., 95,
2'Л 233.
Paterniaпi Е., 1969. Selection fOT reproductive isolation between two popu(ations
of rnaize, Zea mays L., Evo(ution, 23, 5З4 547. .
Patterson В., 1949. Rates of evolution in taeniodonts. In: Genetics, Paleontology
and Evolution, ed. Ьу G. L. Jepsen, Е. Маут and G. а. Simpson, Princeton
University Press. Princeton, N. J.
Pianka Е. R., 1974. Evolutionar r Ecology, Натрет and Row, New York.
Pilbeam D., 1972. The Asccnt о Man. М8сrnШап, New York.
раЬеат D., Goиld S. 1., 1974. Size and scaling in human evoJution, Science, 186.
892 90 1.
Pimentel D., Smith О. 1. С., Soaпs J., (967. А population model of sympatric spe-
ciation, Аrnет. Nat., 101, 493....504.
Proctor М., Уео Р., 1973. The РОШП8tiоп of Flowers, СоШпs, London.
Prout Т., 1962. ТЬе еН ects of stabilizing selection оп the Нmе of development in
Drosophila тelaпogaster, Genet. Res., 3, 364----382.
Race R. R., Sanger R., 1962. Blood Groups in Man. Ed. З, B(ackwe(J SсiепШiс
Pubtications, Oxford.
Radil1sky L., 1976. O(dest horse brains: mоте advanced than previous(y rea[ized,
Science, 194, 626 627.
Raven Р. Н., 1970. А muШрlе origin (от p(astids and mitochondria, Science, 169,
641 646.
Rauen Р. Н., Axelrod D. 1., 1974. Angiosperm biogeography and past continental
movernents, Ann. Missouri Bol Gard.. 61, 539....673.
Rejтanek М., J enik J., 19i5. Niche, habitat and related ecological concepts, Acta
Biotheor., 24, 100....107.
Reтington С. L., 1954. The genetics of Colias (Lepidoptera), Adv. Genet., 6,
403-----450.
Rensch В., 19БОа. The 18\\'s of evolution. In: Evolution After Darwin, Vol. 1,
ed. Ьу S. Тах, University of Chicago Press, Chicago.
Rensch В., 1 960Ь. Evo(ution Above the Species Level, Translation, СоlиmЫа Uni-
versity Press, New York.
Rieger R., Michaelis А., Green М. М., 1976. А Glossary оС Genetics and cytoge.
netics: Cla..sical апd Molecular. Ed. 4. SDringer Verlag. Ne\v York.
Ris Н., Plaut W., 1962. Ultrastructure оС DNA containing areas in the chloroplast
of Chlaтydomonas, J. Сеl1 Bio1., 13, 383 391.
Roтer А. S.. 1966. Vcrtcbrate Paleontolog\'. Ed. 3. Universitv of Chicago Press, Chi-
cago. [Имеется переnод J ro И .J..: Ро'мср Л. Палеонто. оrия позвоночных, roc.
научно'техн. IIЗ.I'{-ВО неdJтяноii и rорно топ.'JИВНОЙ ПрОМЫШ l. М.....л.: 1939.'
Rosin 5., Moor Jankowski J. К., Schneeberger М., 1958. Die FеrtilШit im Bluter-
stamm уоп Tenna (НаmорhШе В), Acta Genet., 8, 1....24.
Ross Н. Н., 1957. Principles of natura1 coexistence indicated Ьу leafhopper рори-
lations, Evo(ution, 11, 113 129.
Ross Н. Н., 1958. Evidence suggesting а hybrid origin for certain leafhopper
species, Е vo( ution, 12, 337....346.
Rothschild М., 1970. Toxic Lepidoptera, Toxicon, 8, 293....299.
Rothschild М., 1975. Remarks оп carotenoids in thc evolution оС signals. Iп:
Coevolution of Апimаls and Plants. ed. Ьу L. Е. GПЬеrt and Р. 1-1. Raven,
University of Texas Press, Austin.
Sakai к., Gotoh к.. 1955. Studies оп competition in plants. IV. Competitive аЬШtу
of F) hybrids in barley, J. Hcred., 46, J39 ]43.
з88
Сnисо" литературы
Saпdler L., Novitskl Е., 1957. Meiotic drive as ап evolutionary force, Аmет. Nat..
91, 105 1l0.
Schaeffer В., 1948. ТЬе origin о! а mammalian ordinal character, Evo[ution, 2.
164 175.
Ш м,альzаgзеп Н. Н., 1946. Факторы эволюции, изд-во АН СССР.
Schopf 1. W., 1974. Paleobio(ogy of {Ье Precambrian: the age of blue-green algae.
Evo(. Bio(., 7, 1.......-43.
Selander R. K. 1970. Behavior and genetic variation in natural populations. Аrnи_
Zool., 10, 5з-...-.об.
Selaпder R. К., Yang S. У., Lewontin R. С., /ohnsoп W. Е., 1970. Genetic varia-
tion in the hors'eshoe crab (Liтulиs polypheтus), а phy(ogenetic crelic , Evo-
lution, 24, 402.......-414.
Sheppard Р. М., 1959. Natural Se(ection and Heredity, Philosophical Library.
New York.
Sheppard Р. М., Ford Е. В., 1966. Natural selection and the evolution of dorninan-
се, Heredity, 21, 139....147.
Siтpson а. а., 1944. Теmро and Mode in Evolution, Columbia University Press,
New York. rИмеется перевод: Симпсои Дж. Темпы и формы э80пюции......
М.: ИЛ, 1948.]
Simpson а. а., 1949, 1967. ТЬе Meaning of Evolution. Eds. 1 and 2, Yale Uni-
versity Press, New Haven, Conn.
Simpson а. О., 1951. Horses: ТЬе Story of the Horse Family in the Modern
World and through Sixty МШiоп Years 01 Evo(ution. Oxford University Press.
New York, Paperback edition, Amcrican Museurn of Natura( History, New
у ork.
Siтpson О. О., 1952. How mапу species? Еvоlutiоп. 6, 342.
Siтpson а. О., 1953. ТЬе Major Features of Evolution, СоlиmЫа University Press.
New York.
Simpson О. а., 1961. Principles of Animal Тахопоmу, Columbia University Press.
New York.
Siтpson а. О., 1967. (See Simpson, 1949, 1967, above).
Simpson О. О., 1968. Evo(utionary effects of cosmic radiation, Science, 162.
140....141 .
Siтpson О. О., 1%9. The first three billion years of cornmunity eco(ogy, Brook-
haven Syrnp. Bio(., 22, 162....177.
Sjmpsoп а. а., 1974а. ТЬе солсер1 of prQgress in orgaпic evolиtioп, Social Res..
41, 28....51.
Siтpson а. а., 1974Ь. Recent advances in methods of phylogenetic inference.
Wenner Gren Cent. Int. Syrnp. Ser., 61, 1---35.
Siтpson а. О., Beck W. S., 1965. Lifc: Ап Introduction to Biology. Ed. 2, Нат-
court, Brace and World, New York.
Snogerup S., J 967. Studies in the Aegean Поrа. IX. Erysiтum sect, Cheiranthu$.
В. Variation and evolution in the smal( population system, Opera В01., No. 14,
1....86.
Soans А. В., Piтentel D., Soans 1. S., 1974. Evolution of reproductive isolation
in allopatric and sympatric populations, Аmет. Nat., 108, 117....124.
Sokal R. R., Кат/ел 1., 1964. Cornpetition among genotypes in Tribo/ium castaпeuт
at varying densities and gene frequencies (the bIack lQCUS), Genetics, 49.
195 21l.
Spotila J. R.. Lommen Р. W., Bakken а. S., Gates D. М., 1973. А mathematical
rnodcl for body tempcraturcs оС larger reptiles: iП1рliсЗliопs for dinosaur ccolй
gy, Arner. Nat., 107, 391---404.
Stanley S. М., 1973. An explanation for Cope's ru(e, Evolution, 27, 1....26.
Stanley S. М., 1974. Relative growth оС the titanothere horn: а new approach to
an old probIem, Evolution, 28, 447---457.
Stanley S. М., 1975. А theory of evolution above the species level, Proc. Nat.
Acad. Sci., USA, 72, 646---650.
Список лuтературbl.
389
Stebblns й: L., 1949. Rates of evolution in plants. In: Genetics, Paleontology and
Evolutlon, ed. Ьу а. L. Jepsen. Е. Mayr and а. а. Simpson Princeton Unl.
versity Press, Princeton, N. J. '
Stebblns й. L., 1950. Variation and Evolution in Plants, СоlитЫа University
Press, New York.
Stebblns G. L., 1969. The Basis of Progressive EvoIution, University of North
carolina Press, СЬареl НilI.
Stebblns а. L., 1974. Flowerfng Plants: Evolution Above the Species Level Нат-
vard University Press, Carnbridge, Mass. '
Stebblns а. L., Lewontin R. С., 1972. Comparative evolution at the I vels о! то-
lecules. organisms and populations. In: Darwinian, Neo-Darwinian and Non-
Darwinian Evolution, ed. Ьу L. М. LeCam, J. Neyman and Е. L. Scott, Bar-
keley Symposia оп Mathematical Statistics and РrоЬаЬШtу, Proceedings 01
the Sixth Symposium, Vol. 5, University оС СаШоmiа Press, Berkeley.
Stern С., 1958. Selection [or subthreshold differences and the origin о! pseudo-
exogenous adaptations, Amer. Nat., 92, 313 316.
Stern С., 1959. Variation and hereditary transmission, Proc. Amer. Phil. Soc., 103,
183 189.
Stern С., 1960. 1973. Principles of Human Genetics. Eds. 2 and 3,. W. Н. Freeman
апd Со., San Francisco.
Stirton R. А., 1947. ОЬsеrvаtiопs оп еvоlutiопаrу rates in hypsodonty, Е\'оlutiоп,
1, 32 1.
Streaтs F. А.. Piтentel D., 1961. Effects of imrnigration of the evolution of ро-
pulations, Amer. Nat., 95, 20l 210.
Strid А., 1970. Studies in the Aegean Поrа. XVI. Biosystematics o the Nigella
arvensis соrnрlех with special reference to the ртоЫет о! non-adaptive radia-
tion. Opera Bot., No. 28, рр. 1----169.
Stubbe Н., 1960. Mutanten der Wildtomate Lycopersicon pimpinellifolium (Jusl:}
МШ., I<ulturpflanze. 8. 110 137.
Sudworth а. В., 1908. Forest оС the РасiПс Slope, U. S. Departrnent of Agricul-
ture, WаsЫпgtол, О. С.
Сукачев В. Н., 1928. Einige experimentelle Untersuchungen iiber den Karnpf итэ
Dasein zwischen Biotypen derselben Art., Z. indukt. Abstamrnungs- Verer-
Ьuпgsl., 47, 54 74.
Тахтаджян А. Л., 1954. Вопросы эволюционной морфопоrии высших растений.
Л.
Teilhard de Chardin Р., 1959. ТЬе Phenomenon of Man, Translation, Harper and
Ro\v, New York.
Thoday J. М., 1958. Natural selection апd biological progress. In: А Сепturу 01
Darwin, ed. Ьу S. А. ВатпеН, George Allen and Unwin, Lопdоп.
Thoday J. М., 19i2. Disrupti\le scleclion, Proc. l oy. Soc. Lопdоп. В 182. 109
143.
Th(}day J. М., Воат Т. В., 19!19. Effects of disruptive sсlесtiоп. 11. Polymorphism
and divergencc v.'ithout isolation, Heredity, 13, 205 218.
Thoday J. М., Gibson J. В., 1962. Isolation Ьу disruptive selection, Nature, 193,
1l64 1166.
Thoda!/ J. М., Gibson J. В.. 1970. The probability оС isolation Ьу disruptive selec-
tion, Amer. Nat., 104. 219 2ЗО.
Тiтofeeff Ressovsky N. W., 1940. Мutаtiопs апd geographical vаriаtiоп. In: The
New Systematics. ed. Ьу J. Huxley. Сlаrепdоп Press, Oxford.
Turesson G., 1922. Th(' genotypical rеsропsс of the plant species to its habitat,
Hereditas, З, 211 350.
Turesson О., 1925. The plant spe<:ies in relation to habitat and climate: contribu-
tions to the knowledge of genecological units, Hereditas, 6, 1 47 236.
Uzzell Т., Spolsky С., 1974. Mitochondria апd plastids as endosymbionts: а re.
vival of special creation? Arner. Sci., 62, 334----343.
390
Список лuтературbI.
Waddington С. Н., 1953. Genetic assimilation of an acquired character, Еvоlutiоп,
7, 118 126.
Waddington С. Н., 1956. Genetic assimHation of the bithorax phenotype, Evo1uti-
оп, 10, 1 13.
Waddington С. Н., 1957. The Strategy of the Genes, George Аllеп and Unwin.
London.
Walker Е. Р., 1964. Mammals of the World. 2 vo[s. The Johns Hopkins University
Press, ВаШmоте.
Walker J. С., Stahmann М. .А., 1955. Chemica' nature o disease rеsi5tапсе iп
plants, Ann. Rev. Plant Physiol., 6, 3бl 366. .
Wallace В., 1966, Оп the dispersal of Drosophila, Amer. Nat., 100, 551 563.
Wallace В., 1968. Topies in Poptl1ation Gепеtiсs, W. W. Norton апd Со., New
York.
Washbllfn S. L., 1960. Тоо15 and human e\loluti-on, Sci. АПlеr., 203 (3), 62 75.
Washburn S. L., Moore R., 1974. Аре I"to Man, Little, Brown and Со., Boston.
Watt W. В., 1969. Adaptive sigпifiсапсе of pigment po[ymorphisms iп Colias but-
terf1ies. 11. Thermoregulation and pJ1otoperiodicaHy controlled melanin varia-
tion in Colias eиrytheme, Proc. Nat. Acad. Sci. USA. 63, 767 774.
Weismaпп А.. 1889 1892. Essays ироп Heredity апd Кindrcl Bio[ogica[ Problems.
Тrапslаtiоп, 2 vols, Oxford University Press. Oxford.
Weismann А., 1893. ТЬе Germ plasm: А Thcory of Heredity. Trans[ation, Wa1ter
Scott, London.
Wells Р. V., 1969. The re[ation between mode of reproduction and extent of specia-
tiоп iп woody gcncra oi the Саlifоrпiа chaparral. ЕvоlL1tiоп, 23, 264 267.
Werth Е., 1956. Bau uпd Leben der В[иmсп: Die bliitenbiologischen Bautypen in
Entwicklung uпd Anpassung. Епkе Ver[ag. Stuttgart.
White М. 1. D., 1973. Animal Cyto10gy and Evolution. Ed. 3, Cambridge Univer-
sity Press, Cambridge and London.
Whittaker R. Н., Levin 5. А., Root R. В., 1973. Nich habitat and ecotype, Аmес.
Nat., 107, 321 338.
Wienere А. 5., Moor Jankowski J., 1971. Blood groups of поп Ьиmап primates and
their relationship to the blood groups of man. In: Comparative Genetics' in
Мопkсуs, Apcs Зlld Мап, ed. Ьу А. В. СЫатеlli, Academic Press, Lопdоп апd
New York.
Wilkes Н. О., 1967. Teosinte: The closest Relative ос Maize. Bussey Institution,
Harvard Uпivеrsitу, Cambridge, Mass.
Wilkes Н. О., 1972. Maize and its wi1d relatives, Science, 177, 1071 1077.
Williaтs а. С., 1966. Adaptation and Natural Selection. Princeton Univcrsity
Pres5, Princeton, N. J.
Williaтs О. С. (cd.), 1971. Group Sc1ection. А[diпе-Аthеrtоп, Chicago.
Williams а. С., 1975. Sex and Evo[ution, Princeton University Press, Princeton,
N. J.
Williaтson D. L., Ehrтan L., 1967. Induction of hybrid steri1ity in nonhybrid ma-
les of Drosophila paulistorиm, Genetics, 55, 131 140.
Wills С., 1973. Iп defense оС naive pan-selectionism, Атет. Nat., 107, 23 34.
Witson Е. О., 1971. COl11petitive and aggressive behavior. In: Man and Beast:
Comparative Socia[ Behavior, ed. Ьу J. F. Eisenberg and W. S. Dillоп, Smith-
sопiап Iпstitutioп. Washington, О. С.
Wilson Е. О., 197.5. Sociobiology: ТЬе New Synthesis, Harvard University Press.
cambridge, Mass.
Wiпter Р. L., 1929. Tl1C теап апd variability as afft>cted Ьу сопtiпuоus selectiol1
for composition in corn, J. Agric. Res., 39, 451 476.
Wolf С. В., 1948. Taxonomic and distribution 5tudies 01 the New World cypresses,
Aliso, 1, 1 250.
V'oodworth С. М., Leng Е. R., 1 ugenheimer R. W., 1952. Fifty gепеrаtiопs of se.
lection (от protein апd оН iп corn. Аgrоп. J., 44. 60 65. '
Wright 5., [931. Evolution in Mendelian populations, Genetics, 16, 97 159.
Список литературы
391
Wright 5., 1943а. Isolation Ьу distance, Genetics, 28, 1I4 IЗ8.
Wright 5., 1943Ь. An ana'ysis of local variability of flowef соl0Т in Linanthиs
parryae, Genetics, 28, 13 156.
Wright 5., 1946. Iso(ation Ьу distапсе under diverse systems of mating, Genetics,
31, 39 59.
Wright 5., 1949. Adaptation and se(ection. Iп: Genetics. Paleontology and Evolu-
tion, ed. Ьу G. 1... J('pscn. Е. .\'\ayr зпd а. G. Simpsol1. Рriпсеtоп Uпivсrsitу
Press, РriпсеtQП. N. J.
Wright 5., 1956. Modes о! selection, Amer. Nat., 90, 5----24.
Wright 5., 1960. PhysiQIQgical genetics, eco[ogy of populations and natural selec-
. tiоп. In: Еvоlutiоп After Dаrwiп, vol. 1, ed. Ьу S. Тах, University of Chicago
Press, Chicago.
Wright 5., Ке" W. Е., 1954. Experimenta( studles of the distributiQn о! gепе fre-
quепсiеs iп very sma(} рорu(аtiопs Qf Drosophila melanogaster. 11. Bar. Evo
(ution, 8, 225 240.
Wynne-Edwards V. С., 1962. Anima( Dispersion in Re'ation to Social Behaviour,
Oliver апd Boyd, ЕdiпЬurgh and London.
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИй
А chillea borealis 1'50, 151
тlllefoliuт 150
Aegyptopitecu.s 337
Agrostis teпиis 108, 221
Alou.atta З36"
Alphadon 270
Amaranthaceae 220
Aтaranthu.s 220
Amentiferae 277
Anas acиta 21
Anchitheriu.т 249, 250, 256. зоз
Aпopheles 201
macuHpennis 175
melanoon 175
messeae 175
Anthophora 186
Antirrhinu.т тaju.s 53
А onidiella aurantii 93
Aotиs trivirgatиs 346
А pis тelliflca 119
Arabidopsis 109
А r.chaeohippиs 249, 250, 256
Arctocyonidae 311
Arctostaphylos 275
Aristolochia 201
Artiodactila 311 312
Asclepias 201
Astasia 159
longa 157
Aиstralopithecus 337, 339, 340, 341,
360
afrlcanиs 331, 338, 339, 341, 347
boisei 337, 338
robustu.s 338
А vena sativa 122
Bacillus pyocyaneиs 193
Baitиs philenor 201
Biston betularia 88. 89, 104
Brontotheriidae 2"46
Brontotheriит platyceras 247
ВиЬо virginianиs 21
Butorides virescens 21
Caenolestidae 273
Calippu.s 249, 250
Callithricidae 336
Callithrix 336
Саmрапи1асеае 318
Canis faтiliaris dingo 193
СартНоНасеае 222
Carex 220
Carl1ivofa 312
Catarrhini 336
Ceanothиs 275
Cebidae 336, 346
Ceboidea 336
Cebиs 336
Сераеа neтoralis 45, 62, 63, 77, 90, 9
92, 133, 134
Cercopithecidae З36
Cercopithecoidea 336
Cervidae 174
Cctacea 312
cha1icotheriidae 269
Ciridops 315. 316, 318
аnnа 317
Clarkia 305
атоеnа 305
biloba 215, 305
lingиlata 215, 305
rubicипda 305
Clostridiит botиlinuт 154
Coefebidae 316
Colaptes aиratиs 211
саре, collaris 211
Colias eurytheтe 107, 201
Colletotrichит cincinans 22
Compositae 179
Condy1arthra 249, 251, 252, 311, 312
Cornaceae 222
Crataegus 174, 221
Creodonta 311
Crepis occidentalis 179, 180
Crocodylиs 270
Cиpressus 133
Cyperaccae 220
Danaus plexippus 201
Daиbentonia тadagascariensls 336
Daubentoniidae 336
Daubentonioidea 336
Указатель латинских назsа",uа
З9З
Delphiniит gypsophilит 219
hesperiит 220
.... recиrvatum 220
Dendroica 199
.... aиduboni 21, 211
castanea 199
.... coronata 199. 211
.... fusca 199
.... tigriпa 199
.... virens 199
Dldelphis 210
Diptera 48
Drepanididae 309, 315, 316
Drepaniinae 315, 316. 318
Drepanis 315, 316, 318
.... pacifica 317
Drosophila 41, 93, 125. 196,201,230
.... americana 281
.... fиnebris 53, 286, 287
heteroneura 214, 215
.... melanogaster 33, 50, 51, 52, 67, 80.
104, 105. 106, 116, 123, 125, 131, 132,
154. 190, 223, 286, 287
.... тiranda 230
.... pachea 93
..... paulistorum 125, 155. 160, 231
---- persiтilis 33, 185, 189, 190, 191
.... рlаnШЫа 214
..... prosaltans 52
pseudoobscura 33, 40---42, 48, 59,
62, 78. 86, 109, 121, 122, 125. 132,
141, 168, 169, 170, 185, 189, 190 191,
230
silvestris 214, 215
.... simulans 190, 286, 287
(ехаnа 287
.... tJirilis 287
.... willistoni 33, 52, 125, 287, 288
Drosophilidae 48
Dryopithecus 337
Edentatao 312
Elephas тeridionalis 182, 301
.... planifrons 182. 301
Elyтus 219
Eohippиs 249. 250, 251, 254
Eotitaпops borealis 247
Epihippus 249. 250. 256
Equida,e 248, 260. 264, 269
Eqиus 249. 250. 251, 252, 254, 255.
256. 258, 259, 264, 269, 301, 303, 304.
321. 322
.... asiпиs 193
.... caballus 322
Ericaceae 222
Erysimum candicum 133
Erythroneиra 201
.... lawsoni 200
EscherCchia 154
.... соН 50
Euglena 152, .157, 159
.... gracilis 157
Fabaceae 201
Felidae 322
Felinae 323
Fissipedia 310
FriпgШidае 309, 318
O!alago 336
Geospiza 198
Geospizinae 309, 318
GШа 132, 133, 201, 207. 219
.... achilleaefolia 133, 173
--- latiflora 171, 172,210
..... leptantha 170, 171, 172,210
..... таио, 191
.... тodocensis 191
GоrШа 336
Gossypium barbadense 218
.... hirsutum 218
Gramineae 218
НарСоЫрри! 249. 250
Н aplophoneus 323
Н elianthus 207
Heтignathus 315, 316, 317,318
..... lucidus 317, 318
Hemiptera 200
Н emophilиs 153
Himatione 315. 316, 318
--- sanguinea 317
Hipparion 249. 250, 256. 303
Hippidion 249. 250. 251, 252
Hominidae 289, 335, 336. 337
Hominoidea 335, 336. 337
Ното 237, 289, 331, 338. 339, 340, 351,
352. 354. 360
.... erectиs 337, 338, 339, 341, 342, 347,
360
.... habilis 338
neanderthalensis 339
.... sapiens 33. 50. 169, 288, 336. 337.
338, 339, 342, 343, 347, 348, 349,
351,352,312
Н omoeosaurus 270
Hoplitis producta 176,211
hernardina 176
gracilis 176
.... . panamintana 176
394
Указатель латинских ",азва",ий
н oplophoneиs 323
Н ordeu.m satiuum 53 109 121
Н yla ewingi 230, 23 i '
.... uerreauxi 230 231
Hipohippus 249, '250, 251, 256, 303
Н yracotherium 249---254 261 270 303
304 ' , · ·
Indridae 336
Insectivora 312
1 ris 20 1, 207
Jassidae 200
/uniperus 133
Kloeckera 78
Labia tae 185
Lagomorpha 312
Larus argeпtata 211
Lasius 198
Layia gaillardioides 191
.... hieracioides 191
Lemu.r 336
Lemuridae 336
Lemuroidea 336
Liatris aspera 62
Limulitella 270
Liтulиs 270, 274
.... polypheтus 33
Linanthus 213
.... parryae 62
Lingula 270, 274
Ыnит usitatissimu.m 156
Liptoterna 312
Lithospermиm caroliniense 62 63
Lobclioideac 318 '
Lophocereus schottii 93
Lophortyx саЩоrniсиs 21
Loris 336
Lori!\i dac 336
Lorisoidea 336
Loxops 315, 316,317.318
--- coccinea 317, 318
Licopersicon pimpinellifolium 48, 49
Масаса 336. 347
Machairodиs 322
Malиs 174, 221
Manteoceras manteoceras 247
Marsupialia 312
Maydeae 94
Megacerops 246
Megachi1idae 176
Megahippus 249, 250
Megaloceros gigan.teus 319 320 32)
326 ' , ..
Melanerpes formicivorus 2 t
Mentzelia 201
Merychippus 249, 250, 251, 252, 253.
254, 255, 256. 257, 258 259 3 01
304 · , ·
.... californicus 258
.... seversus 258
Mesohippus 249, 250, 251. 252, 253.
254, 256. 258
Metazoa 240, 241. 242
Micrococcus pyogenes aureu.s 92
М icrohyla carolinensis 230 231
.... оиаасеа 230, 231 '
Microtus
.... ochrogaster 59
--- pennsylvanicus 59
Miтulus guttatus 145
Miohippus 248,250, 259,304
М и! muscиlus 33, 50, 63, 135
Musca domestica 93,223
Mustelidae 311
Mycoplasma 155, 150
Nannippиs 249, 250, 25 " 256 257
Neohipparion 249, 250, 256 '
N icotiana 219
.... rиstica 156
--- silvestris 218
--- tabacum 218
.... tomentosiformis 218
Nigella 133
--- arvensis 133, 172, 173, 207
--- carpatha 172, 173
--- degenii 172
--- icarica 172. 173
Notoungulata 312
Oenothera 109
Onagraccae 191. 305
Onohippidium 249, 250
Oreopithccidac 337
Oreopithecus 337
Ornithischia 324
Orohippus 249. 250
Ostrea 270, 274
Ovis canadensis 114, 193
Palmeria 315,316
Paludina 246, 248
--- hoernesi 248
u.nicolor 248
Указатель aTиHCKиx названuй
395
Рап 289, 336, 34 J
.... troglodites 288
Pantodonta 312
РаРЁО 336, 347
Parahipparion 249, 250
Parahippus 249, 250. 256, 257, 259
РататесЁит 193, 196
.... aurelia 193, 194
.... bursaria 193, 194
.... caudatum 193
Parus major 211
Passeres 316
PasserHormcs 316
Perissodactyla 246, 270, 311, 312
Peroтyscus тan;cula/us 2] 1
polionotus 33
Phaseolus lиnatus 125
РЫеит pratense 123
РЫох drummondii 125
glaberrima 230
---. pilosa 62, 63, 230 .
Phylloscopus trochiloides 211
Pinus 201, 205
.... alblcaulis 107, 173
--- attenuata 185
.... coulteri 57
--- monticola 211
.... murrayana 173
radiQ/a 185
.... strobus 211
Platanus occidentalis 200
Platyrrhini 336
Pliohippus 249, 250, 251, 252, 253,
254, 256, 270, 301, 304
- Роlсmопiасеае 171, 173, 209, 210, 219,
305
Polemonium 201, 305
caeruleum 305
.... саЩотiсum 305
саrnеит 305
.... delicatuт 305
.... eximium 305
.... pu/cherriтu.m 305
Ропgidае 289. 335, 336, 337
Pongo 336, 341
Potentilla glandulosa 145
Proboscidea 312 .
Procecidochares 174,222
Protitanotheriuт emarginatиm 247
Protogonodon 311
Protohippus 256
Pseudaelurus 323
Pseudomonas 154
.Pseudonestor 315,316,318
Psittirosfra 315,316,318
.... paCmeri 317
Psjttirostrjпae 315, 316, 318
Quercиs 201. 275
.... grisea 184
.... havardi 184
--- mohriana 184
R hamapithecus 337
.... punjabicus 337
wickeri 337
Rangifer tarandus 174
Rапuпсulасеае 207
Rhagoletis 174.221,222
--- corniuora 222
--- mendax 222
--- ротоnеllа 174. 221, 222
--- zepltyria 222
Rodentia 312
Rosaceac 145, 222
Salix 173
S almonella 154
Salvia аРЁаnа 185, 186
mellifera 185, 186
Saurischia 324
Sceloporus olivaceus 58, 59, 62, 6з
Scrophulariaceae 145
Sequoiadendron giganteum 25. 26, 27,
28, 29
Sirenia 312
Sitta 198
Sтilodon 322, 323
Solanaceae 219
Sphenodon 270
Staphylococcus 50, 154
аитеи.') 92
Streptococcus 153
. Strongylocentrotus franciscatzus 187
. pиrpuratus J 87
Stvliпоdопtiпае 297
Stylohipparion 249, 250
S yтphalangus 336
Taeniodonta 297, 312
Tanysiptera 207
galatea 205 207, 214
--- --- сатоиnае 206
.... elliot 206
.... .... galatea 206
.... --- meyeri 206
--- тinor 206
.... --- riedelii 206
.... rosseliana 206
.... --- vulcani 206
.... hydroharis 206. 207, 214
Т атахасит 122
officiпale 121
396
Указатель латинских ",азва",и4
Tarsiidae 336
Tarsioidea 336
т arsiиs 336
Tephritidae 174
Thraupidae 316
Thylaciпus 193
Tradescantia 207
Tribolium 193
.... cas/aпeиm 122, 123. 125
--- confusuт 122
Trlpsacит 98, 100
.... dac/!/loides 98. 99
rriticuт aestitJum 218
--- vulgare 122
Tsuga mertensiana 173
т upaia 336
Tupaiidae 336
Tupaioidea 3Зб
Vestiaria 315. 316. 318 '
Viverridae 311
Viviparidae 246
Viviparus 246, 248
Х ylocopa brasiliaпorum 186
Zauschпeria сапа 191
sepientrioпalis 191
Zea mays 50. 67, 84 86, 94, 95, 96.-
98, 100, 109. 116
тexicaпa 98. 99
Zosterops palpebrosa 197
Zygosaccharoтyces 78
ПРЕДМЕТНЫй УКАЗАТЕЛЬ
Аrрессивное поведение J 22, 193
Адаптации общие 20
..... специальные 20
Адаптивнаst зона 265. 308
радиация З08, 314----318
ценность 73---74
Адаптивный ландшафт 292 29Э, 306,
308
Алпопатрия 164
Аллопопиплоидия 217
Альтруистическое поведение 118 119
Ам инокиспОТllые поспедоватепъиости
в rомолоrичных белках 281---286,
288
Аристоrенез 262
Лlfаrеиез с'м. Проrрессивная эвопюuия
Атомная ЭВОЛЮI{ИЯ 236
Аутоrамия 180
Ahnenreihe 303---304
Брадитепия (брадитеJ1ические скоро-
сти) 272 274, 295
Величина популяции 25
Вид (63, (74....17 , 180....181
аЛ.10патрический 17'5
биолоrический 163. 174, 184
симпатрический 17'5
.... таксономический 179....181
ВИ.l100бразование биотически-симпат.
pHI/ecKoe 222---223, 227
--- в результате чередования популя-
ционнЫХ под'ьемов н спадов 215
rеоrрафнческая теория 303, 20в....-
211, 21'5 216, 227
--- rибридное 218....220, 227
rO foceKBeHUIlaJ'bHoe 215
квантовое 212......216' 227
общая теория 224
--- путсм аллополиплоидии 21 218,
227
.... reнетической революции 215
катастрофическоrо отбора 215
рекомбинационное 218 219, 227
симпатрическое 22 221
---- смежно.симпатрическое 221
ВидообразоватеJlьное иаправление 300
ВИДЫ Д80Аники 175, 181
..... последовательные 182
Возникновение жизни 237----238
Вторичные половые признаки 112
Вымирание 319----329
Высотные расы rop"HblX ВИДов 173
Высшая нервная деятельность у при-
матов 340, 341, 34().....з47
rибридизация 207, 220, 2 , 294
l'ибрИДнаЯ стерипьность (стериль-
ность rибридов), индукция 15s......
156, 188 190
rемофилия 44
rенетическая rибепь 140
rенетические революции 297.....299
rенетическиА rруз 140 141
.... мутационный 140
.......... сеrреrаuионныА 140
.... субституционныА 140
reHHble мутации 47 48
rеl10ТИП 149 151
rенофонд 31
rены-модификаторы доминантности
79. 82
экспрессивности 7g. 80
rеоrрафическая изменчивость 164....
173. 343
rеоrрафические расы у че.10века 342
rеопоrическое время 236, 237, 239, 245
fлавные ЭВОЛlоuионные СИJ1Ы 42....43
rомопоrИR 368----369
rоротелия (rоротепические скорости)
272 214, 295"
rрада 332, 335, 354
rраиицы толерантности 199
rрупповая структура у приматов З58
360
...... --- --- типы 344 345
rруппы крови системы АВа 31 32
Дем C-"f. Популяция покапЫiая скре-
щивающаясSl
Дрейф reHOB 43, 126----139, 205 207.
293, 363
Есrественный отбор см. Отбор
Еlап vital 262
398
п ред.метны.й указатель
Живые ископаемые 270, 272
Жизнеспособность у дрозофипы 86----
87
Закон (принцип) rаузе 194---195
До.'1ло 266
Харди Вайнберrа 35-----36
Замещение аплелеА 72
видов 193 194
Изолирующие механизмы 183---184.
См. также Изопяция
Изоляция временная 184 185
raMeT 187
м еха н ическа я 185----186
обусловленная суточными циклами
185
пространственная 183, ) 84
репродуктивная 183, 184
..... репродуктивные механизмы внеш-
ние 183. 185
внутренние 183, 184
сезонная 185
создаваемая средой 184
эколоrическая 18 185
..... этолоrическ а я 186 187, 232---233
Инбридинr 38, 2[3, 226
Инверсии у дрозофилы 40---42, 78,
1 09----111. 168----169
Индукция 154. 16 161
Инте.'1.1ект 358----'359, 362
Интроrрессня 205, 207
I(атастрофизм 328
Катастрофически А отбор СМ. Ква нто.
вое видообразование
Квантовая ЭВОЛIоция 295---297, 310,
315
Квантовое видообразование 212 216,
227
Кладоrенез см. Эвопюционная дивер-
rснuия
Ko.'JbueBoe перскрывакие рас 211
Конкурентнос ИСК.'JIочение 194---195
Конкуренция 121 124, 192 195, 200
Креационизм 370
Кроше на корснных зубах в пннии
Mesohippиs---Eqииs 258
Культурная эвопIOЦИЯ 236, 237, 351,
360 3б2
Купьтурное наС.'Jедие 351
Jlамаркиэм 149. c. ,. также ОРl'оrенеэ.
Макромутации 54
МаКРОЭВО.1JЮЦИЯ 23, 234----236, 367....
370
Массовое вы ирание 328
ейотический драйв llб
Меланизм у березовой пяденицы 88.....
90
Местообитание 199
иrраuия 56----60
Микровид 179, 180
МикроэвопlOЦИЯ 23, 24, 40
Митохондрии. теория происхождения
1'59
МолеКУЛSlрная ЭВО.'1юция .281
--- скорости 281, 290 291
Мутантные раЗllОВИДl10СТИ смородин-
Horo томата, ассumЬепs 49
--- .... Ырiппаtа 49
--- --- .... bu(]osa 49
.... --- carinata 49
--- dimintlta 49
dwarr 49
Мутации 48 55, 67---68
reHHble 47, 53
малые 48, .54
Надвид 177
Нежизнеспособность rибридов 187....
188
Необратимость эвопюции 266
Неотення 267
Обесцвеч.ивание у эвrлены \57, 159
Обучение 352
Общсственные rруппы уприматов
344----345' 357 58
Орrаническая эволюция 236
ОрrаническиА Уир. разнообразие 17
19
Ортоrснез 43, 262---264. 321, 323
ОРТОССJIСКlIИЯ 262 264
Отбор raMeT 115
--- rрупповой: 117---120, 357....359, 363
дарвиновская концепция 74 75
---- дизруптивный 1 04 108, 223
---- естественныА 042. 70
..... зависящий от частоты 124 125
.... индивидуапьный 115
...... катастрофический c,tt. Квантовое
видообразование
...... коэффициент 43, 71
.... межвидовой 193---1 . 360
междемовыЙ 11 117
напраВ.'1еIlНЫЙ 102
ПредА,етны4 уteазатеЛh
399
Отбор
--- на репродуктивную изоляцию 22в......
230
--- однолокусная модель 71, 116
основная теория 70---76
--- поповой 111---115. 261
--- популяционныА 1 17
--- при образовании рас 204---205
--- проrреесивный 102
--- родственный 117
--- сочетание с дрейфом reHOB 129---
130, 206---207, 212, 293, 294, 295,
ЗОО, 306, 363
стабипизируlОЩИЙ 102---104
уравновешивающий 108---109
Панминсня 35
Панмиктическая единица 60
Парапатрия }'.(54
Педоморфоз 267
Плата за отбор 142---147, 149. 225,
З06
--- --- --- коэффициент 142, 143
--- --- --- приращение 142
П.lJеЙотропия 70
Подражательная окраска 124---125
Полиrамия 113---114
Полиморфизм 2 30
--- rеllетический ,29---30
--- переходный 29 30
--- по бе.'1кам 141, 142
--- окраске раковины у Сераеа nе-
тoralis 45, 77---78, 9 92, 133
--- сбалансированный 29---30
хромосомныЙ 29........зо
Полип.'10ИДИЯ 217
Полувиды 176---178, 208
ПОПУJIЯЦИЯ КОJ10ниапьпая 25, 294
--- аrамоспермная 180
--- КОllпепция в БИО.lJоrип -34
--- линейная 25
Jl0ка.'1ьная СК'рещивающаяся 24 26,
ЗО
--- непрерывная 25
---- У однополых орrаниз:мов 178
Поепедовате.1ы\ые виды 182, 301
Поток reHoB 56 63. 130
Правило А.rrлена 205
--- Берrмана .204
--- Копа 261---262
ХолдсАна 189
Превосходство (преимущество) rCTe-
розиrот 108, 109
Преrрады, оБУСЛОВ.'Jснные lIесовмссти.
мостью 1 87 188
Прерывистые равновесия 304
Принцип Лотки---Во.1ьтерра 194 195
--- основатсля (эффект основате.1Я)
133, 299
Приобретенные признакн 14 149
--- --- наспедование 14 149
Приспособлсния (адаптации) 20---22
265
Лриспособпенность 73---74
.... компоненты 15
Проrрессивная эвопюция 330---334
Продолжитепьность rенерации 274
2..75
РАМ-единицы 286, 290---291
Равновесие между мутациями и от.
бором 44
--- Харди---Вайнберrа 38, 39
Разрушение rибридов 19 191
Расселение 57---60
Расы 163. 203---204
--- rеоrрафическне 164, 169
--- .'10ка.1ьные 169
--- по хозяину 173, 222
--- эколоrические 173
Реко:мбинация 64 9
Репродуктивная изоляция 18З---184
207, 208, 226. 228
Рудиментарные opraHbl 369
Са.llьтация СА«. Скачкообразная эво"
.1ЮЦНЯ
Сба.lансировашrый поли:морфизм 29,.
30
Сверхсnециа.lизация 262. См. также
Специапизация
Сезонные расы 173
Селективная нейтра.1ЬНОСТh 142, 279---
280, 363
Снмпатрия 164
--- биотическая 164
--- смежная 164
CliНraMeoH 178, 208
Синтетиt\еская теория эволюции 46..
158. 161
Скачкообразная эвопюция 309---310
Скорости возникновения родов 269
Скорость эво.'1юцни 144---145. 268-----
,277. 290----291, 305---306, 31 О
СкрещивающаJlСЯ популяция 2-5.
с.lучайное скрещивание 35, 36, 60
Смещсние прюнаков 197....198. 228
Соматоrеииые вариации 149
Соседство 60---63
--- ве.lичина 61....63
.... занимаемая оБJlасrь 61
400
п редметн.ы4 указатеАЬ
Сосуществование видов 195....196
Специапнзация 199,265, 331,334
Стери.,ьность rибридов reHHaR 189
.... ---- ХрО fосомиая 189
........ цитоппазматическая 189
Структура попупяции 24----26, 146----
147. 293
.... пространстsеннзя конфиrурации
25
.... ---- система размножения 25....26
.... .... скорость миrрации 25, 57
Субституционный rруэ 140....141
Суперrен 145
Stufenreiche З03 304
Тахителия (тахитепические скоростн)
.272.... 27 4
Телефинализм 262
Типолоrическое мышление в бнолоrии
34
Транс.з.укция 153, 154, 159, 161
Трансформация 153
Униформизм 328
Устойчивость к токсинам 92....93
Факторы, зависящие от плотности
123 124
Феноотклонения 213
Фенотип 149....1-51
Фенотипические модификации 79,
14 1049
.... приспособительиыil характер
151....152 .
.... .... роль в ЭВОЛIОЦИИ 152
Ферменты, полиморфизм 33, 141....142
Филетический rрадуалнзм 304
Формула харди....ВаАнберrа 37....38
Хим нческая эвопюция 236, 2з8
Хпоропласты. теория происхождения
159
Цепи питания 275----277
Частота аллелей 35.......{39
---- мутаций 43. 48, 5(}-",51
Частоты rенотипов 35. 37....39
Ширина ниши 199
Эволюционная диверrенция 302....303
Эволюционное направпение (направ-
ление эволюции) 246. 300
.... .... видообразоватепьное 300
.... .... длительное направпение специа-
лизации 334
.... .... кратковременное направление
специализации 334
.... ---- проrрессивной эвопюции 331....
334
---- .... фипетическое ЗОО
Эвопюционные силы 42....43
Эвопюция ато:мная fl36
.... в семействе пошадей 248....260.
321....322
.... rаваАских цветочниц 315----318
.... квантовая 295....297
.... кукурузы 94....100
.... культурная 236, 237, 351, 360....362
.... мопекупярная 281
.... на островах 31'5....318
.... начапьные стадии 240----242
---- орrаническая 236. 239....240
.... проrрессивная 330---334
---- сабпезубых кошек 322....323
.... филетическая 182, 300----305
---- химическая 236
.... эпизодическая 328----329
.... человека 335, 337....339. 348-----349
.... Hominoidea 335, 337....339
Эколоrическая амппитуда 199
Эколоrическая дифференциация меж-
ду видами 195. 196....197
ниша 198----200
........ реапиэованная 198
..... .... фундаментапьная 198
.... .... ширина 199
.... раса 1 73....174
Экотип 205
Экспрессивность 80
Энтелехия 262
ЭссеНl{иализм 34
Эффект УОЛ.lееа 228
Язык 345, 353, 357.....358
.... животных 346, 353
.... человека 353
оrЛАВЛЕНИЕ
Предисловие редактора перевода .
Вступление .
Предисловие. .
Блаrодарности
5
9
14
16
ЧАСТЬ 1. ВВЕДЕНИЕ
r лава 1. Постановка проБJlемы
Введение . . . . .
Разнообразие орrаиическоrо мира
Приспособление . . .
Эволюционные объяснения
. .
17
17
17
20
22
ЧАСТЬ 11. МИКРОЭВОЛЮЦИЯ
rJlaBa 2. Скрещивающаяся ПОПУJlЯЦИЯ
Введение . . . .
Структура популяпии. . .
Популяции мамонтова дерева .
Попиморфизм . . . . . .
Полиморфизм по rpynnaM крови у человека
Полиморфизм по ферментам .
Концепция популяции .
24
24
24
26
28
за
33
33
rлава З. Статика популяций .
Закои Харди ВаАнберrа .
Частоты аппе.лей . . .
Формула Харди Вайнберrа .
Эффекты инбридинrа
Заключение .
35
з5
36
37
38
38
rлава 4. Динамика ПОПУJlЯЦИЙ
Опреде.'Iение микроэволюnии .
МИКРОЭВО.1юционные измеиения у D,o opi1ila' тeiaпo'gastel .
rпзвные эволюционные силы . ...
Взаимодействие между эволюционными снлами .
ПОПУЛЯll.ионно-rенетический ПОДХОД к эволюnии .
40
40
40
42
43
45
fлава 5. Мутации
Введеиие
47
47
402
02АаВАен.ue
reflHble мутаци 47
Частота мутацин 48
r енотипический контродь 51
АдаnТИ8ная иеннасть . . . . 52
Относите.'lЬное зиачеиие 8 эвопюции Ma.'IblX мутаций и макромутаций 54
Вероятность сохранения новой мутации . 55
fлава 6. ПОТОК reHOB .
Введ.ение .
Скорость иммиrраuии..
Типы р"асселеиия . . . . . .
Панмнктические единицы и соседства.
Оцеики величины соседств .
56
56
5fi
57
60
63
r.,aBa 7. Рекомбинация"
Введение .
Процесс рекомбинации.
Колнчество rенотипической изменчивости
Рекомбинация и мутаllНЯ .
Зиачеflие рекомбинации
. I .
64
64
64
65
67
68
ЧАСТЬ 111. ЕСТЕСТ8ЕННЫЯ ОТБОР
fлава "8. Основная теория отбора
Введение. ..
Однолокусн"ая модель .
Коэффициент отбора . . . .
Скорость изменения и era степень.
Приспособленность. . . .
Дарвиновская концепция отбора .
Компоиенты приспособлеиности
70
70
70
71
72
73
74
75
fлава 9. Проявление reHa и ero связь с отбором .
В ведение. . .
относителыIстьь се.lJективНОЙ ценности
Изменяемость фенотипа
Плейотропия. ...
Модификаторы экспрессивиости
Модификаторы доминантности
rенотип как сдиница отбора
77
77
77
78
79
79
82
83
fлава 10. Примеры отбора
Типы имеющихся данных. .
Иплинойсский эксперимент по селекции кукуружы .
Жизнеспособиость у ДРОЗОфИЛЫ .
Меланизм у" березовой ПЯДеницы.
Окраска раковины у наземиых улиток _
Устойчивость к токсИнам .
Одомашиивание. .
Эволюция кукурузы
Заключение.
84
84
84
86
88
90
92
93
94
100
02АавАен.ие
403
fлава 11. Типы отбора.
НаПрав енн й отбор .
Стабилизирующий отбор
ДИЗРУПТИ8НЫЙ отбор . . .
Дизруптивный отбор в прнроде .
Уравновешивающий отбор. . . . . . . . .
Превосходство rетерозиrот у Drosophila pseиdoobscиra .
Половой отбор. . . . . . .'
Отбор на субиндивидуумных уровнях
Междемовый отбор .
Отбор на уровне вида .
102
102
tOZ
104
106
108
10
111
115
116
12G
fлава 12. Эффекты взаимодеАствия особей .
Конкуренция . . . . . . . .
Смеси rеНОТИПО8 в УС.110ВИЯХ конкуренции
Факторы, зависящие от ппОТfIОСТИ
Отбор, зависящий от частоты .
121
121
122
123
124
fлава 13. Дрейф rеиов .
Общие соображения . .
Эффект ВeJJИЧИИЫ попу.'1яции
Эффект отбора. .
ЭффекТ потока reHoB . .
Закрепление сочетаний reHoB
Экспериментальные данные . .
Дрейф в природных популяциях ..
АЛ.lе.:ш, опрсде.'Iяющне rруппы крови в ПОПУ_1ЯЦИЯХ человека .
ЗаК lючение
12&
12&
127
\28
lЗО
131
131
132
136
138
fлава 14. Плата за отбор . 140
rенетический rруз . . 140
Полиморфизм по белкам. . 141
ХолдеЙНОRская П_lата за отбор. . . 142
Ппата за отбор и скорость эволюции . . . . 144-
СцеП lение reHOB и flзаимодействие reHOB; их влияние на оrраничение,
Ha_laraeMoe П.lатой за отбор . 145
Впияние структуры популяиии на оrраничение, налаrаемое п.:татой
за отбор 146
ЧАСТЬ IV. ПРОБЛЕМА ПРИОБРЕТЕННЫХ ПРИ3НАКОВ .
fлава 15. Феиотипичес:кне модификации .
Теория нас едования приобретенных признаков .
Фенотип. . ., . . ......
Прнспособите.']ьные свойства фенотипических модификаций
Ропь фенотипических fOдификаи.ий в эвопюции .
148
148
149
151
152
fлава 16. Трансдукция н индукция
Трансформация .
Трансдукция . ...... . .
Индукция мозаицизма у Drosophila тelanogaste1
153
153
153
154
<404
ОиавАен.ие
ИНДУКц'ия rибридной стеРИЛЬИОС11l у Drosophila paиlisforиm
Индукция у льна. .
Обесцвечивание у эвrлены .
155
156
157
fлава 17. Эволюционная роль ИНАУКЦИИ .
Введение . . .
Возможные примеры
Обсуждение .
158
158
158
160
ЧАСТЬ V. ВИДООБРАЗОВАНИЕ
f"лава 18. Расы и виды. 162
Введение . 162
ПОПУ lяциоиные системы . 162
Аллопатрия и симпатрия . . 16З
Непрерывная rеоrрафическая измеичивость 164
Разобщенные rеоrрафические расы . . 170
Эколоrические расы 173
Виолоrические виды 174
ПО lУВИДЫ .. 176
Вид у однопо.'1ЫХ орr.аниз тв . J 78
Вид в систе fатике. . 180
Последовательные виды 182
....а.а 19. Изолирующие механизмы 183
В ве.gение . . . . . . . . 183
Эколоrическая и I;'ременная ИЗОЛЯ1U{я . 184
Механическая изоляция, этолоrическая изоляция и изопяция raMeT 185
Преrрады, создаваемые "есовместимостью; и нежизнеспособиость rиб
ридов . . . . . 187
Стерипьность rибридов . 188
Разрушение rибридов . 190
Сочетания изопирующих механизмов 191
{'лава 20. ЭКОJlоrическая дифференциация 192
Межвидовая конкуренu.ия . 192
Замещение видов . . 193
Конкуреитиое ИСК.'1ючение 194
Сосуществование видов . . . . . . . . . 195
Отбор, направпенный иа экопоrическую диффереНц'иацию . 196
Смещение признаков 197
Эколоrическая ниша . . . . . 198
Влияние эколоrических требований . 200
r..aBa 21. rеоrрафическое видообразование 202
ЭволюЦ,ионная диверreиЦ,ия . . . . 202
Факторы, участвующие в образовании рас . 203
Ропь отбора . . . . . . 204
Роль дрейфа rеиов и ннтроrрессня . . 205
3арождающаяся репродуктивная изоляция . 207
rеоrрафическая теория видообразования . 208
Реверсии диверreнции . 211
02лаВАен.U8
405
"лава 22. Способы ВИАообразования .
Введение . . . . .
Квантовое видообразование . . . . . . . . . .
Сопоставление rеоrрафическоrо и кваитовоro видообразования .
Видообразованне путем алпополиппоидии . . . . . .
rнбридное видообразование у растений. .
Проблема симпатрическоrо видообразования.
Смежно-симпатрическое видообразование. .
Биотически-симпатрllческое видообразование .
"лава 23. Общая теория видообразования
Введение
Цепь
Этапы
Пути
Моде.'!И .
rпaBa 24. Отбор, направленны" на созданИе изопяции .
Введение . .
Процесс отбора.
Фактические данные .
Этолоrическая изоляция .
ЧАСТЬ VJ. МАКРОЭВОЛЮЦИЯ
fпaBa 25. rеолоrическое время.
Природа макроэволюu.ни . .
r лаВНые стадии истории Земли
Возникновеиие жизни . . . . .
rлавные стадни орrаиической эвопюции .
Начальиые стадии эволюции . . . . .
Стадия сложных мноrоклеточных орrаНИЭblОВ
I"лава 26. Направленная 9вопюция
В ве!l.еиие
Примеры . . '"
Эволюцнонные ряды у Equidae . . .
Направлеиия эволюции зубов У лошадей.
Адаптивная природа иаправпений эволюции
Адаптивные аспекты правила Копа
Ортоrенез и.'IИ ортоселекция? .
...... .
в семействе лошадей .
rпaBa 27. Специализация
Введение . . . . . . . . . . .
Саморазвитие каправ.lJениА эвопюцин. . . .
Способы, ПОЗВО.lяющие избежать специапизацни .
('лава 28. Скорости 9ВОЛЮЦИИ
Измерения скоростей
Различия в скоростях .
212
212
212
215
216
218
220
221
222
224
224
224
224
226
227
228
228
228
. .. 230
232
234
234
236
237
239
240
245
246
246
246
248
254
259
261
262
264
264
266
266
268
268
269
406
ОzлаВАение
Классификаuия скоростей ЭВО.'lюцни ." . . . .
Факторы, оказывающие влияние на ск.орости ЭВОЛЮЦИИ
Среда . . . . . . . .
Внутренние био.'lоrические факторы .
Цепи питання и р,азмножения .
rлава 29. Изменения на молекулярном уровне
Методы изучения. . .
Критическое отступление . .
Раз.'JИЧИЯ в последовательиостях аминокислот
Друrие данные относительно мопекулярных изменений .
РЗз.'JИЧИЯ между человеком и wнмпанзе
Скорости молекудярной ЭВО.IJЮЦИИ
r лава 30. Структура популяции в связи с макроэволюциеА .
АдаПТИRНЫЙ ландшафт . . . . . . .
Эволюционный потенциал популяций разноrо типа
к.ваl11"uвая эволюция .
rенеТИ1[сские революции .
Квантовое видообразование . . . . . .
Филетические направления и видообразовательныс направлеиия
Быстрая ЭВОЛЮЦИЯ в видообразователы[ых направпениях' .
ЗаК.i1Iочение
r.,aBa 31. Эволюция крупных rpупп
Внедение . . . . . . . .
Эl:Iолюuионные характеристики крупных rрупп .
Скачкообразное или постепениое развитие? _
Образоваиие отрядов млекопитающих
Рuль квантовой ЭВО.1ЮЦИИ .
Адаптивная р,адиация .
АдаIlтивная радиация у rавайских цветочниц .
rлава 32. Вымирание
Введение . .. ...
Megaloceros gigaпteиs в Ирландии
Лошадь в Северной Америке .
Саблезvбые кошки. . . .
Диноза.вры . . .
aCCOBoe вымирание в позднемеповом периоде
Вымирание млекопитающих в плейстоцеке .
Эпизоды вымнраиия
ЧАСТЬ VJI. ОТ орrАНИЧЕСКОА ЭВОЛЮЦИ}J К ЭВОЛЮЦИИ
ЧЕЛОВЕКА
rлава 33. Проrpесснвная эволюция
КОНЦСПЦИЯ проrресса в эволюции .
Проrресс или специализация? .
Смена rосподствующих типов .
Критерии проrреССИDНОЙ эволюции .
Обсуждение .
272
273
273
274-
276
27S
278
279
281
286
288
290
292
292
293
295
297
299
зоо
305
ЗО6
307
301
307
309
311
313
314
315
319
319
319
321
322
324
325
326
" 328
ззо
330
331
. 331
332
333
ОZАавлен.ие
407
rлава 34. Филоrения rОМИНИА .
Система классификацни приматов .
Hominoidea. . . . . . .
Наследие ApeBecHoro образа жнзни
Наспедие н"аземноrо образа жизни
Расовая изменчивость
Выводы .
"лава 35. Эволюция общественноrо образа жИзни уприматов .
Стадность уприматов (Ht: i:'тносящихся к rоминндам) .
Адаптивное значение стадности . . . . . . .
Язык. . . .
YpOBellb развития высшей нервной деЯтепьности и обучение .
rоминиды .
Обсужден не .
"лава 36. Культурная эволюция
Культурное наследие
Обучение .
Концептуа.'JЬное мышление
ЭВОЛЮl1иониая природа че,'lOвека
I"лава 37. Решающие факторы эволюции человека
Поиски еще не закончены .
rрупповой отбор
Интеллект . . .
Межвидовой отбор. . . .
Механизмы ку.'1ьтурной ЭВО.I1ЮЦИИ
Взаимодействия . . .
3аКЛЮЧИТe.'Iьные замечания
ЧАСТЬ VIII. СОЦИАЛЬНОЕ ЗНАЧЕНИЕ ЭВОЛЮЦИОННОА ТЕОРИИ
("лава 38. Полемика с креационизмом
Сотворение мира . . . . . .
КреаЦИОНИЗ f ипи ЭВО.'1юционная теория?
Критические замечания. . . . . . . .
Даниые в пользу существования макроэволюции
rлава 39. Приложення эволюционной БИОJlоrни .
Список .'1итературы. . . .
Указатель латинских названий
Предметиый указатель .
ЗЗ5
з35
335
339
340
342
343
344
344
345
345
346
348
349
зы
351
352
353
353
355
355
357
359
360
360
362
363
364
364
364
366
367
371
374
392
397
УВАЖАЕМЫй ЧИТАТЕЛЫ
Ваши замечания о содержании книrИ, ее ОфОРlllпе-
нан, качестве перевода и друrие просим прнсыпать п()
адресу:
129820, ,москва, И-IIО, rсп,
1 й Рижский пер., д. 2,
издаJепьство c p
в. rpaHT
ЭВОЛЮЦИЯ орrАНИЗМОВ
Научный редактор М. Б. Николаева
Мл. научн. редактор З. В. СOJL7lертиискаJl
Художннк В. r. Ежков
Художественный редактор Б. Н. Юдинн
Техннческий редактор Н. И. МаНОХИElа
Корректор К. Л. Водяницкаи
ИВ К, 2503
Сдано в .нaCSор 23.11.79. Подписано к печатн 05.03.80.
Формат БОХ90 I /... Бумаrа типоr.pафская М 2. rарнитура латИН-
скаЯ. Печать высокая. Объем 12.75 бум. л. Уел. печ. .11. 25.50.
У..-изд. 11. 26.92. Иэд. Н. 4/0403. т.ираж 1'2500 эКа. Зак. 1991.
Цена 2 р. 10 к.
ИЗДАТЕЛЬСТВО сМИР
Москва, 1.1 РIIЖСХИЙ пер., 2.
Московская типоrpафия Nt 11 СОЮЭПОllиrpафпрока пр..
fосударствеИИОII комитете ссср по делаМ издатuьств.
попиrрафии и книжиой торrовли,
113105. Москва, Наrатинская уп., 1.