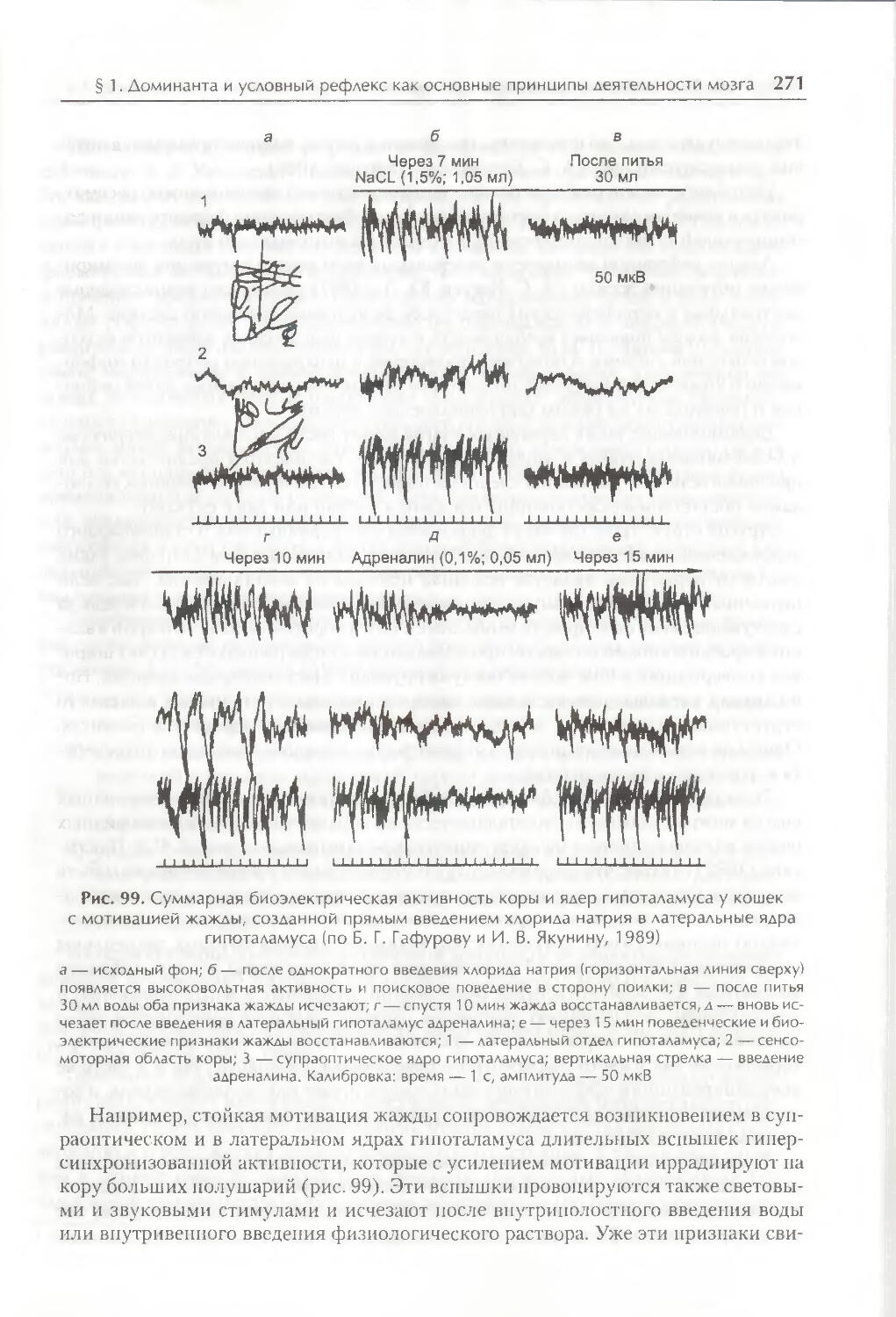
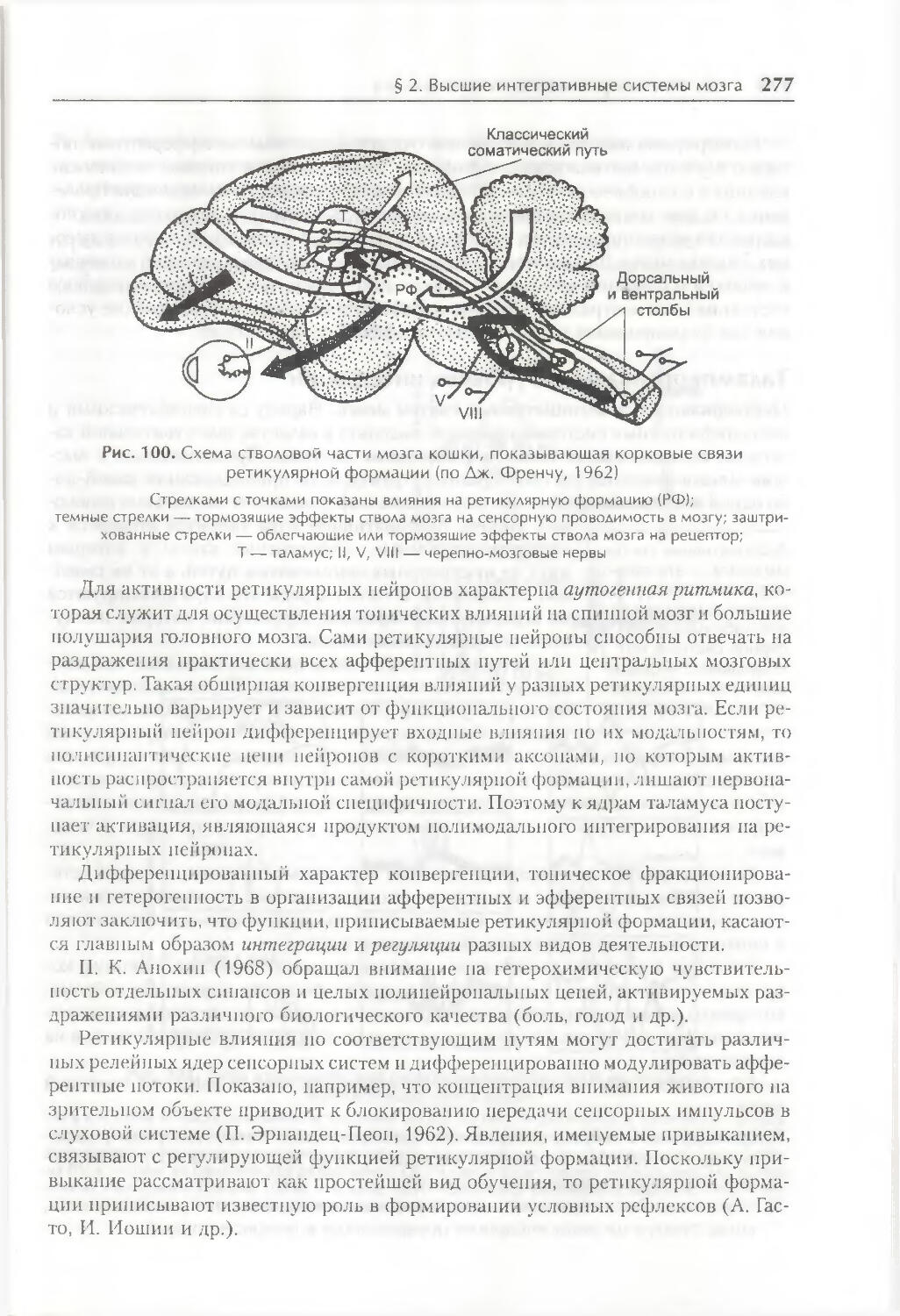
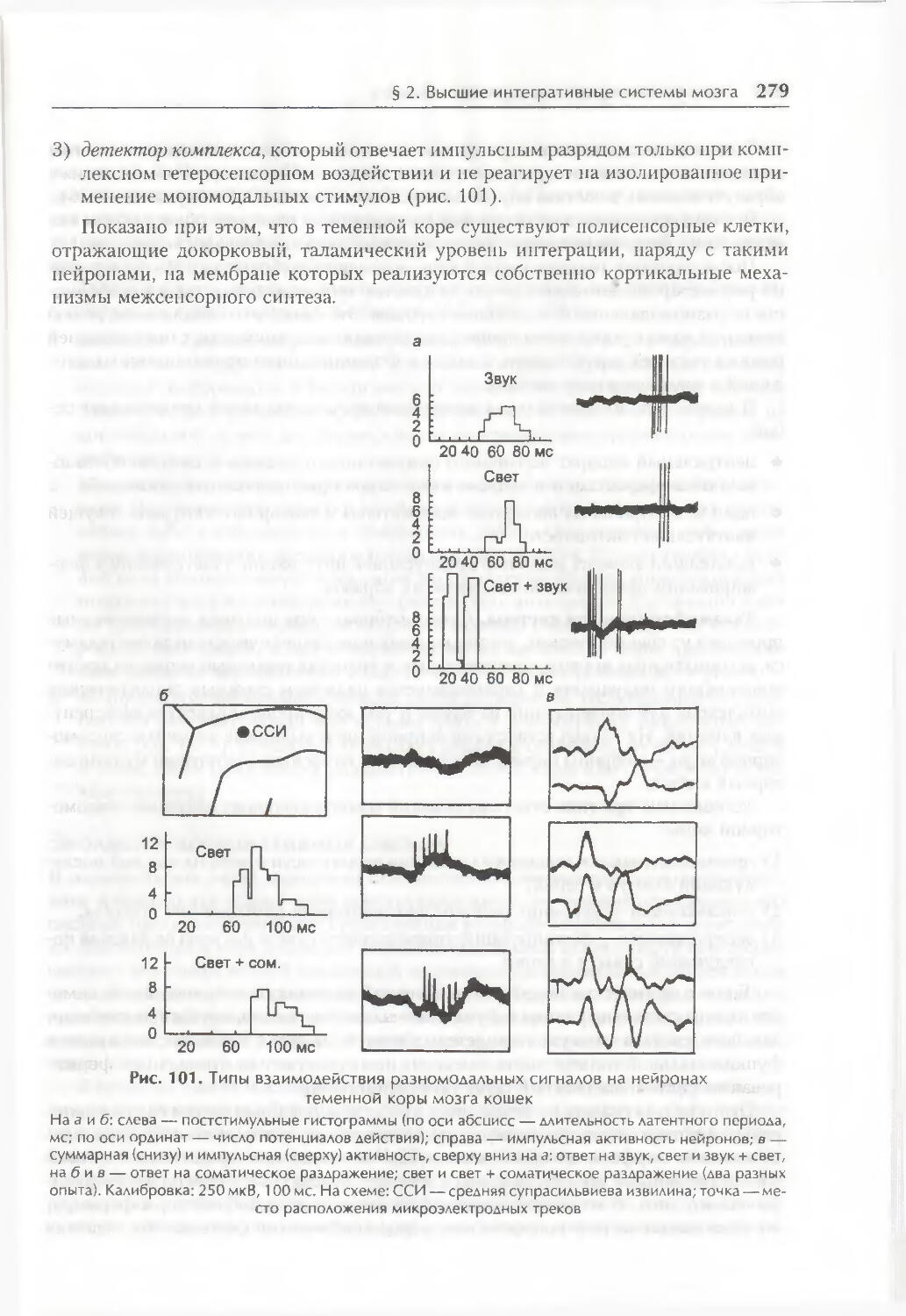
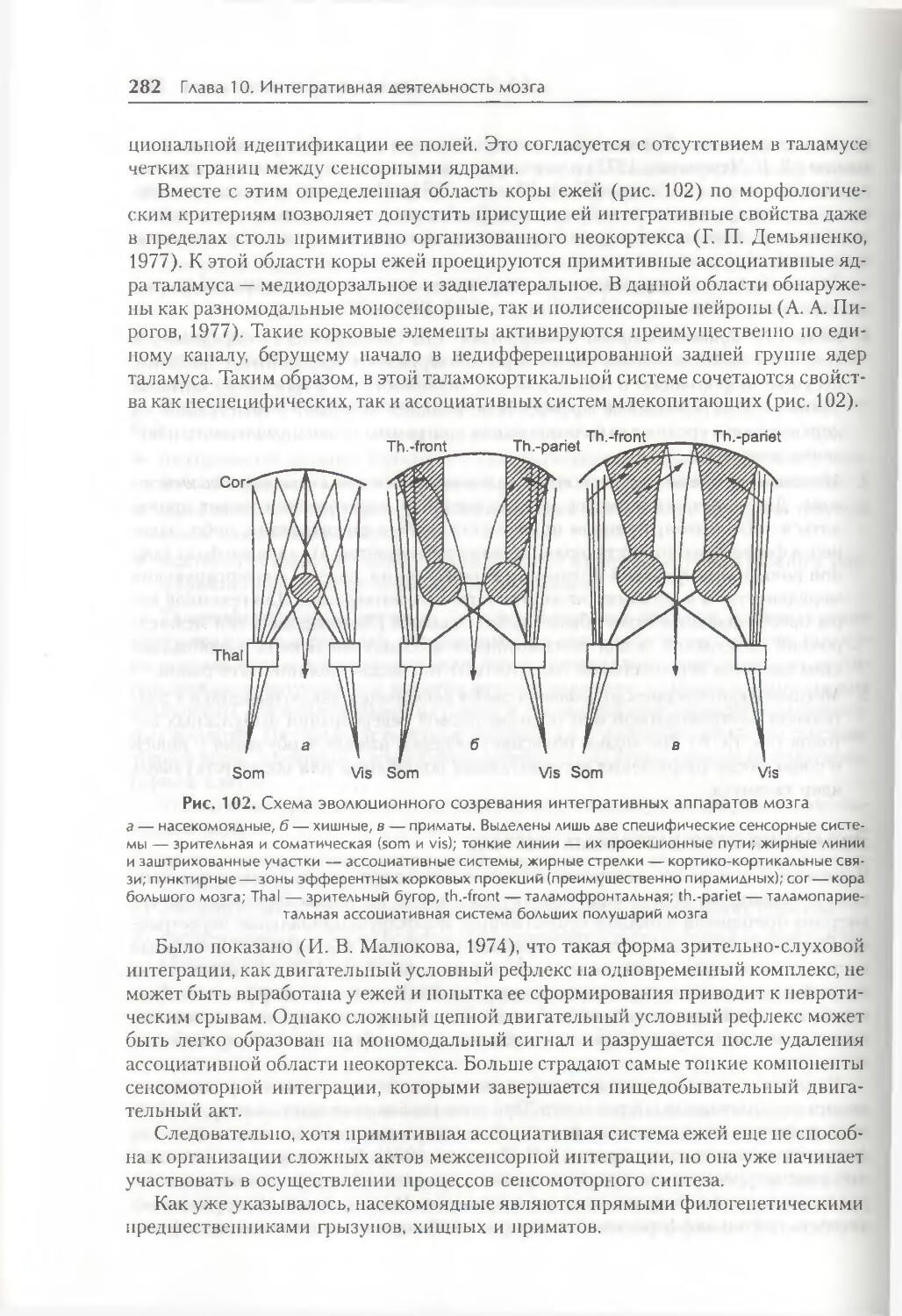
/
















































































































































































































































































































Author: Батуев А.С.
Tags: нервная система органы чувств биофизика, биохимия и физиология животных и человека физиология
ISBN: 978-5-91180-842-6
Year: 2008




Text
ЧЕБНИК С^пптсР
'ДЛЯ ВУЗОВ
А. С. Батуев
Физиология
с/ с/
высшей нервной деятельности
и сенсорных систем
3-е ИЗДЯНИе Методологические проблемы
сенсорной нейрофизиологии
Закономерности физиологии
поведения и интегративной
деятельности мозга
Соотношение сознания
и бессознательного
РЕКОМЕНДОВАНО СОВЕТОМ ПО ПСИХОЛОГИИ
УЧЕБНО-МЕТОДИЧЕСКОГО ОБЪЕДИНЕНИЯ
УЧЕБНИК
у*
ДЛЯ ВУЗОВ
А. С. Батуев
Физиология
высшей нервной деятельности
и сенсорных систем
З-е издание,
исправленное и дополненное
Рекомендовано Советом по психологии УМО
по классическому университетскому образованию
в качестве учебника для студентов высших учебных
заведений, обучающихся по направлению
и специальностям психологии
С^ППТЕР'
Москва • Санкт-Петербург ■ Нижний Новгород • Воронеж
Ростов-на-Дону ■ Екатеринбург • Самара ■ Новосибирск
Киев ■ Харьков ■ Минск
2008
ББК 28.991.782я7
УДК 612.833.8(075)
Б28
1
Федеральная целевая программа «Культура России»
(подпрограмма «Поддержка полиграфии и книгоиздания России»)
Батуев А. С.
Б28 Физиология высшей нервной деятельности и сенсорных систем:
Учебник для вузов. — 3-е изд. — СПб.: Питер, 2008. — 317 с.:
ил. — (Серия «Учебник для вузов»),
15ВЫ 978-5-91180-842-6
Издание предназначено для студентов, обучающихся по направлению
«Психология», и соответствует требованиям Государственного образова¬
тельного стандарта. В нем содержатся основные современные сведения по
анатомии сенсорных систем и физиологическим механизмам восприятия
информации, а также базовые сведения по физиологии основных поведен¬
ческих актов человека. Рассматриваются проблемы соотношения сознания
и бессознательного в психике, психологические и физиологические детер¬
минанты поведения.
Книга привлечет внимание самого широкого круга читателей — от
студентов, только что переступивших порог университета, до специали¬
стов, работающих в различных областях биологических, медицинских и
гуманитарных дисциплин.
УДК 612.833.8(075)
ББК 28.991.782я7
Все права защищены. Никакая часть данной книги не может быть воспроизведена в какой бы то
ни было форме без письменного разрешения владельцев авторских прав.
ISBN 978-5-91180-842-6
© ООО «Питер Пресс», 2008
Оглавление
Предисловие 5
От автора 6
Глава 1. Методология изучения высшей нервной деятельности 7
§ 1. Принцип рефлекса 7
§ 2. Принцип доминанты 15
§ 3. Принцип отражения 20
§ 4. Принцип системности в работе мозга 26
§ 5. Основные методы нейрофизиологии поведения. Методология, метод,
методика 33
Заключение 38
Контрольные вопросы 45
Глава 2. Сенсорная функция мозга 46
§ 1. Обшие принципы конструкции сенсорных систем 46
§ 2. Закономерности обнаружения сигналов 51
§ 3. Системная организация процессов кодирования информации 54
§ 4. Распознавание, декодирование информации 56
Заключение 58
Контрольные вопросы 59
Глава 3. Физиология сенсорных систем 61
Введение 61
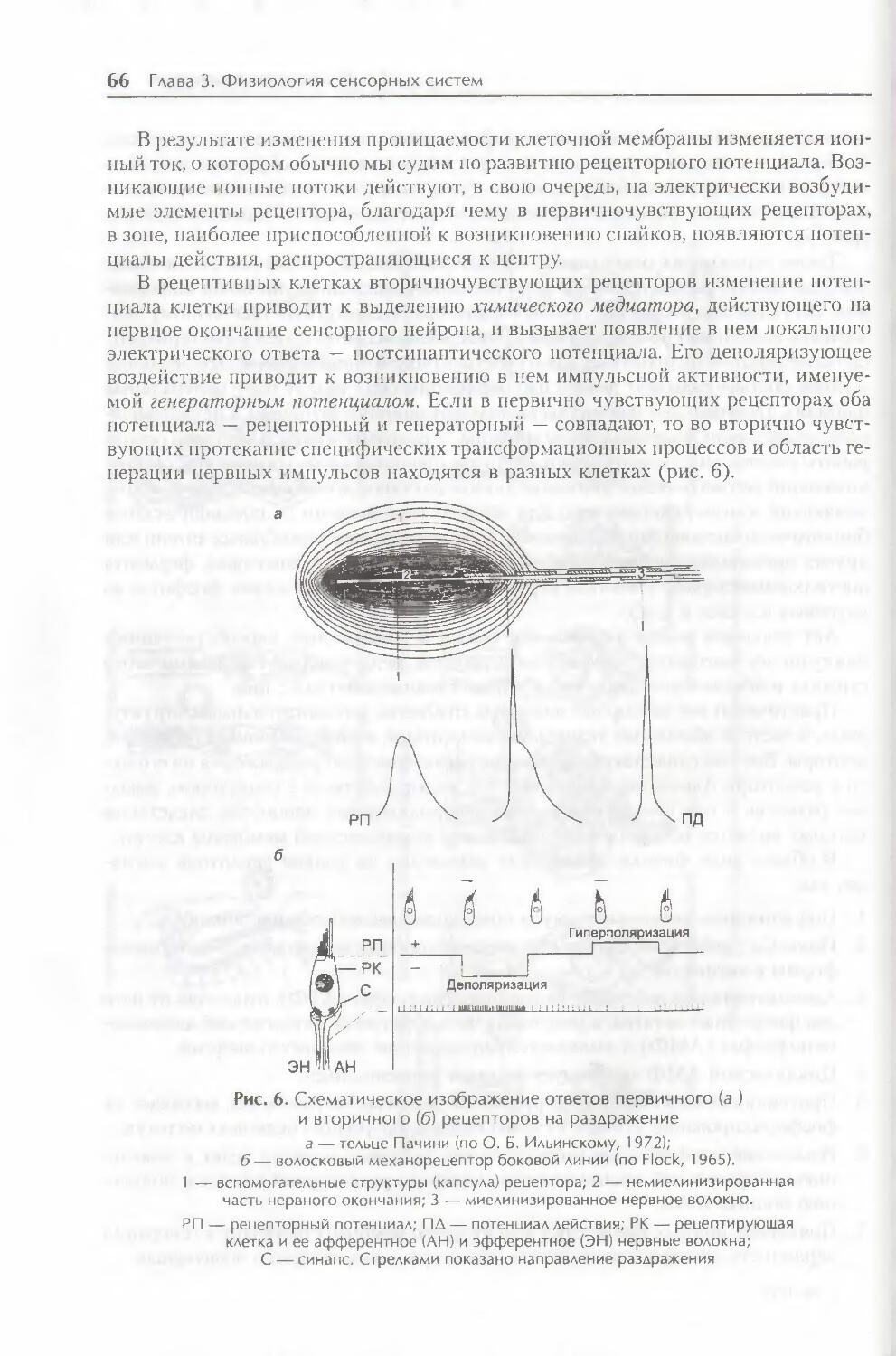
§ 1. Физиология рецепторов 62
§ 2. Кодирование видов информации 67
§ 3. Зрительная система 70
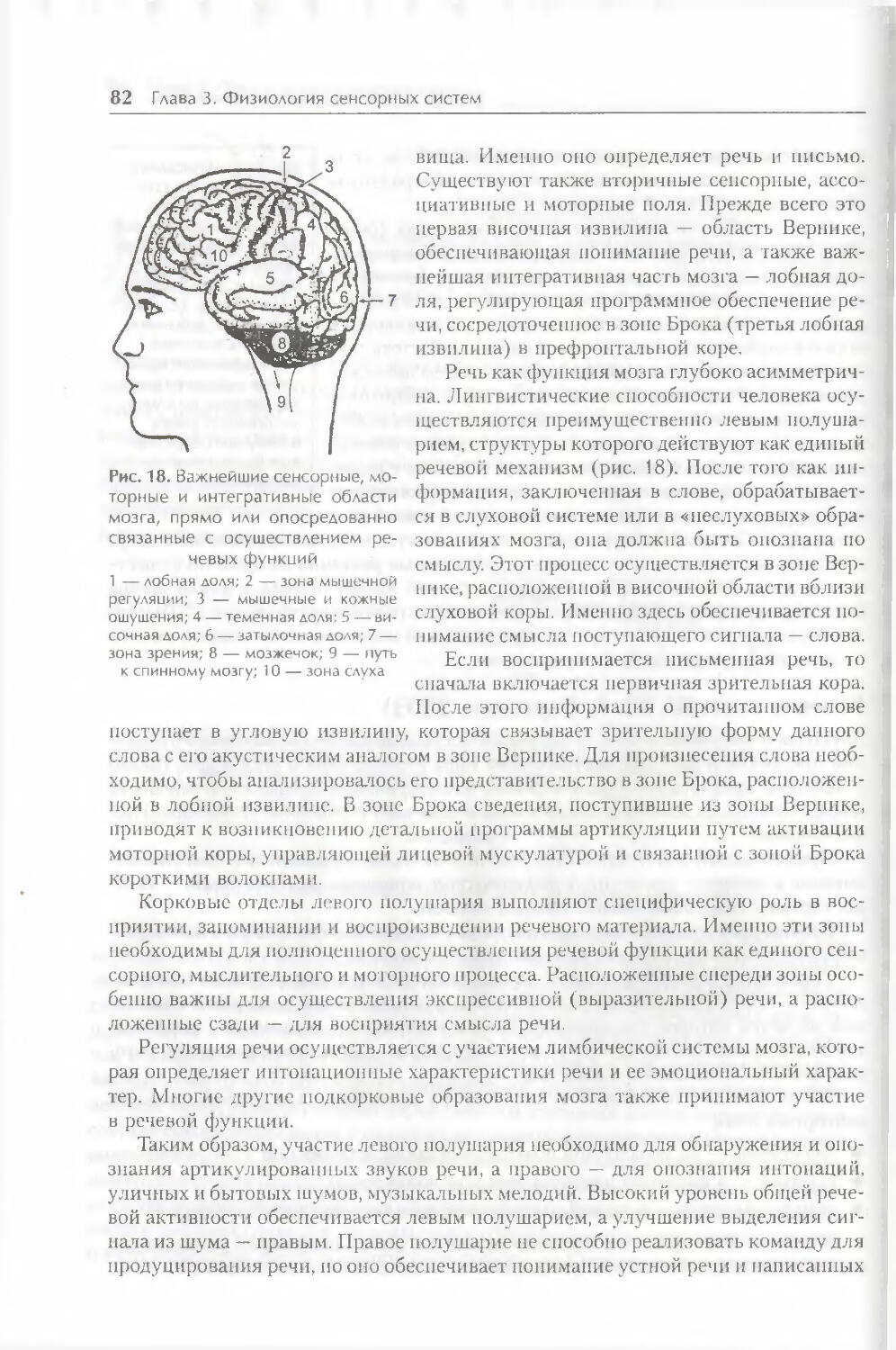
§ 4. Слуховая сенсорная система и речь 78
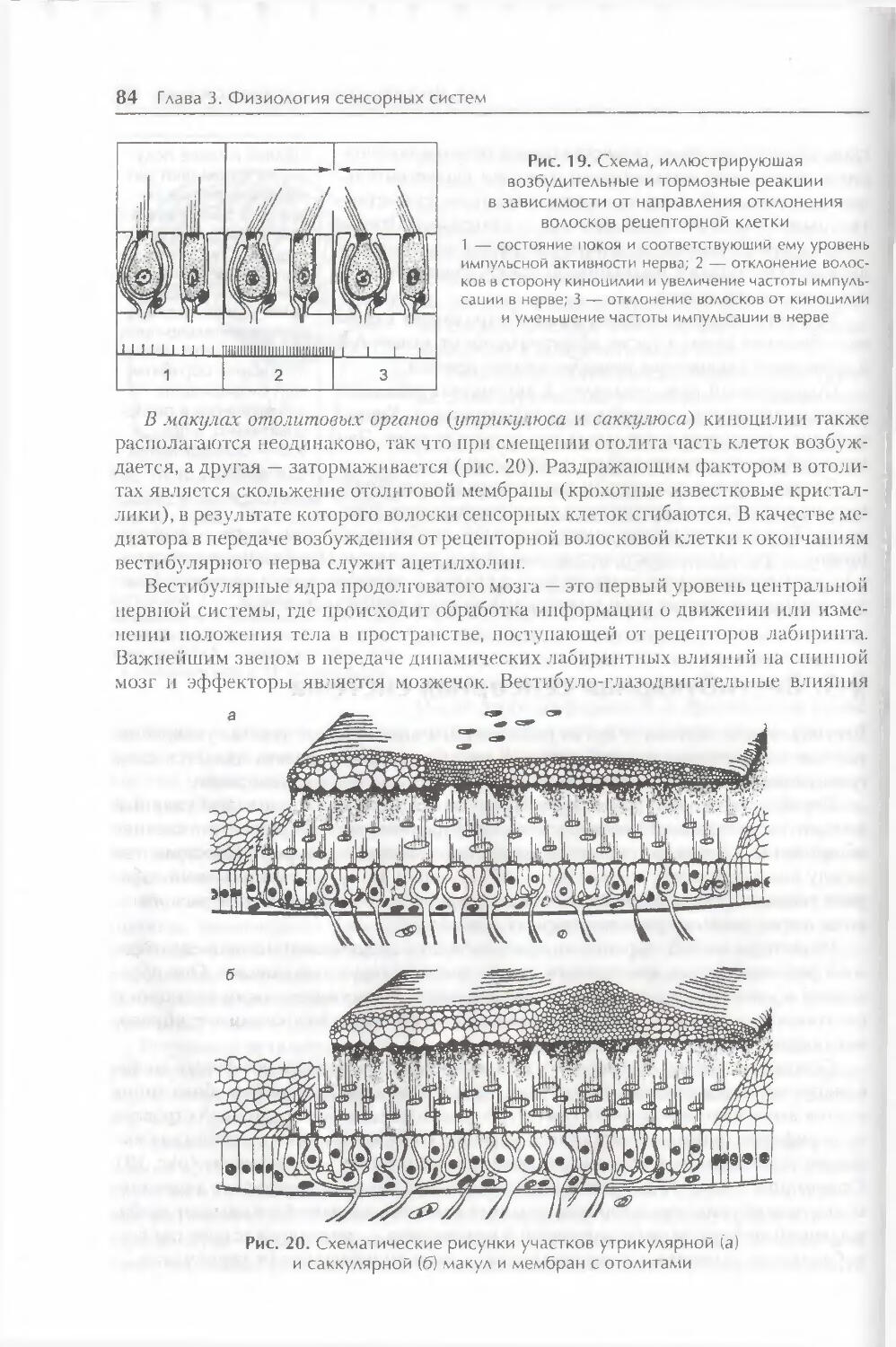
§ 5. Вестибулярная сенсорная система 83
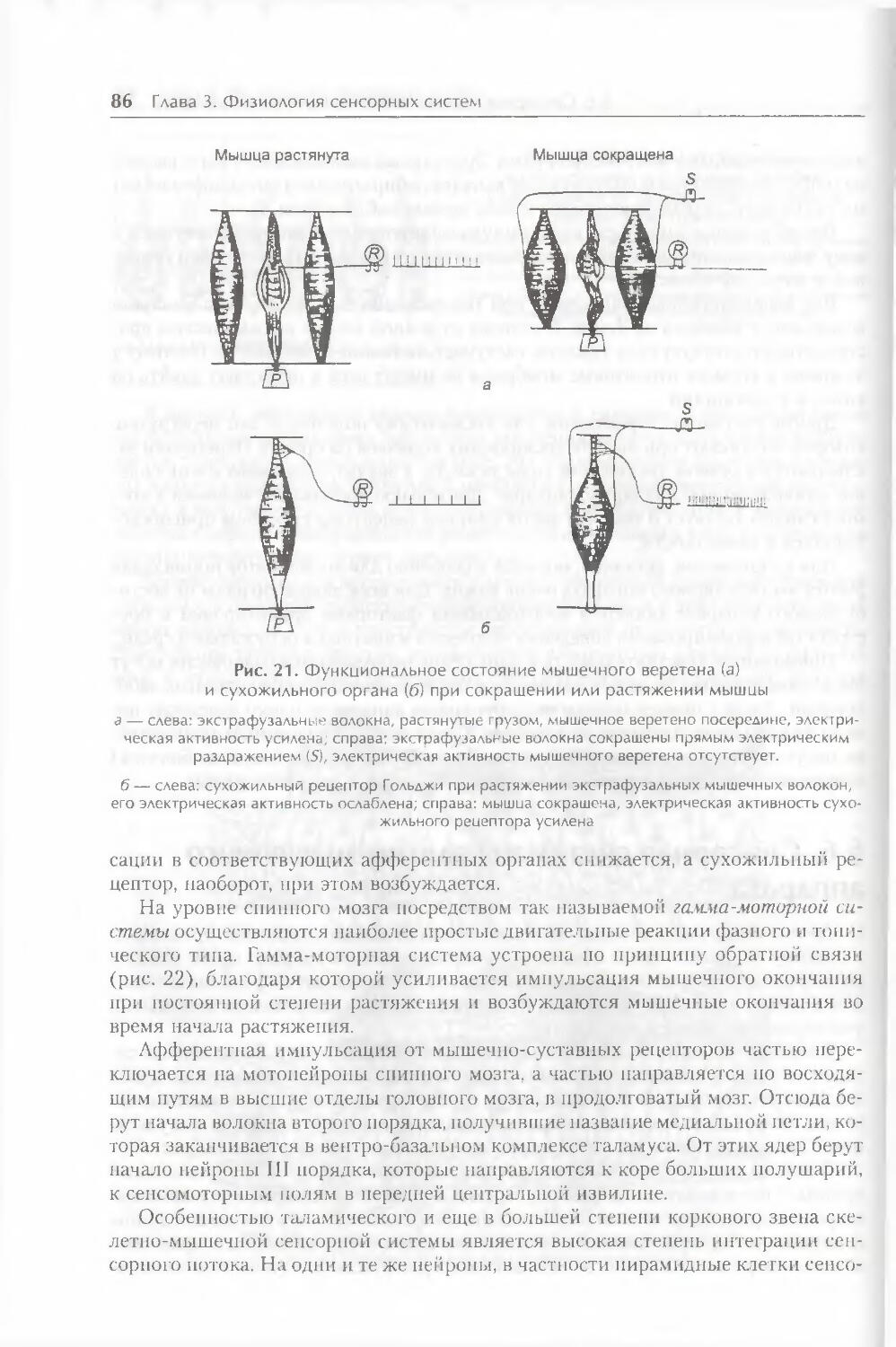
§ 6. Сенсорная система скелетно-мышечного аппарата 85
§ 7. Схема тела 88
§ 8. Кожная сенсорная система 94
§ 9. Вкусовая и обонятельная сенсорные системы 95
Заключение 98
Контрольные вопросы 98
Глава 4. Регуляция произвольных движений 100
§ 1. Обратная связь в управлении движениями 100
§ 2. Внешняя обратная связь 101
§ 3. Внутренняя обратная связь 102
§ 4. Медиальные лемниски 103
§ 5. Двигательная программа 104
Контрольные вопросы 107
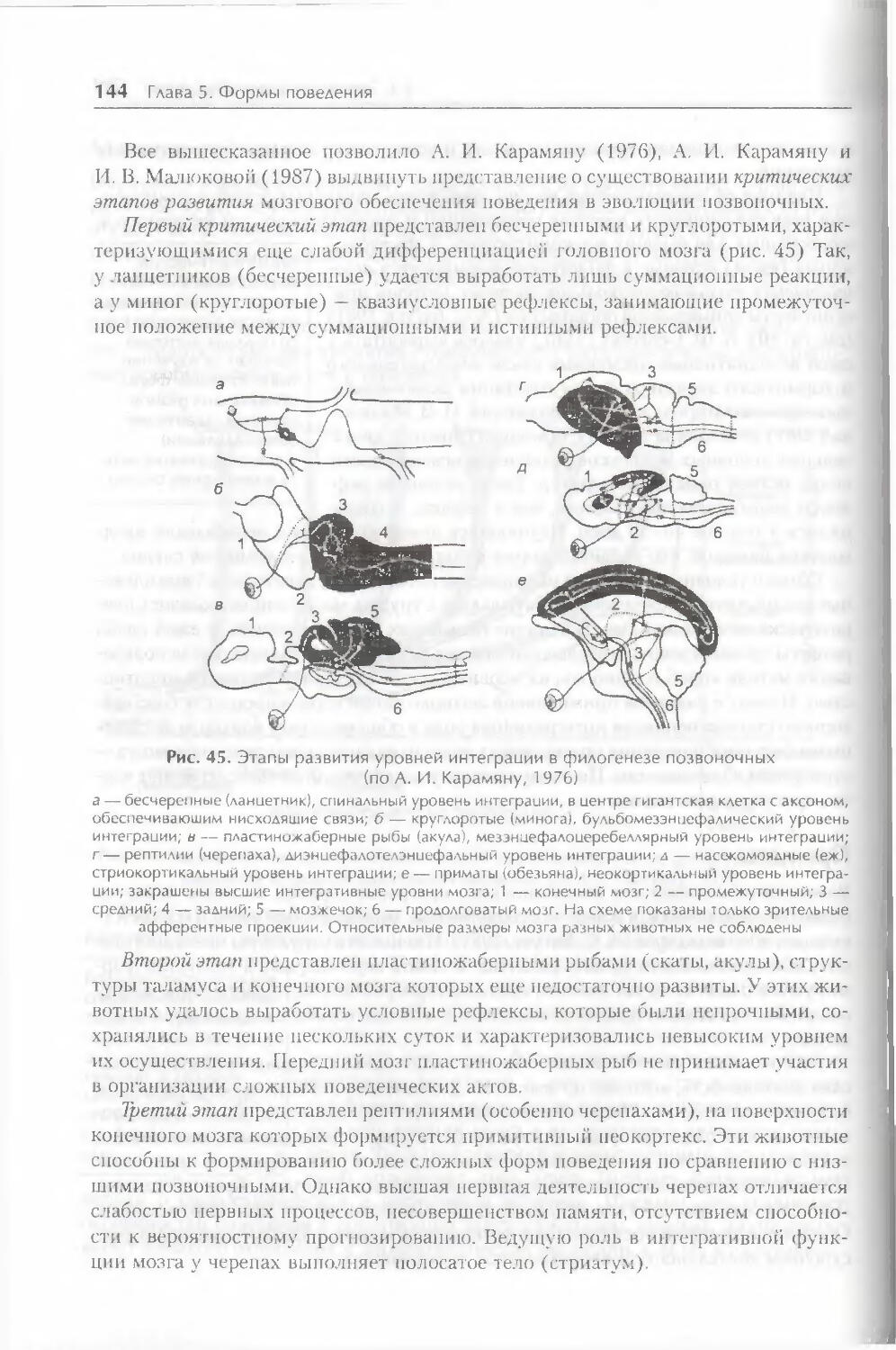
Глава 5. Формы поведения 108
§ 1. Поведение как фактор эволюции 108
§ 2. Классификация форм поведения 113
4 Оглавление
§ 3. Формы индивидуального обучения 122
§ 4. Эволюция форм поведения 135
§ 5. Формирование поведения в онтогенезе 149
Заключение 155
Контрольные вопросы 156
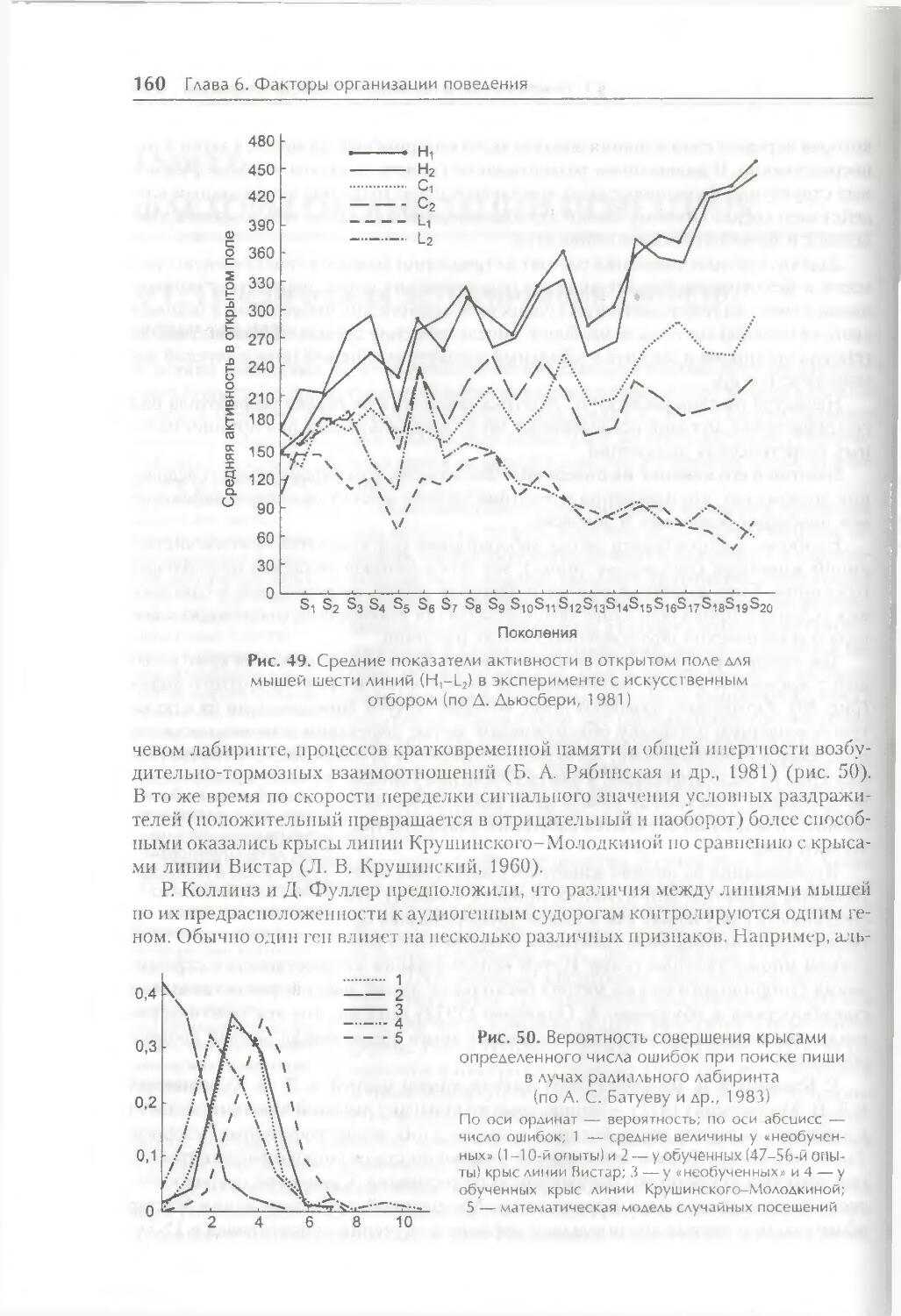
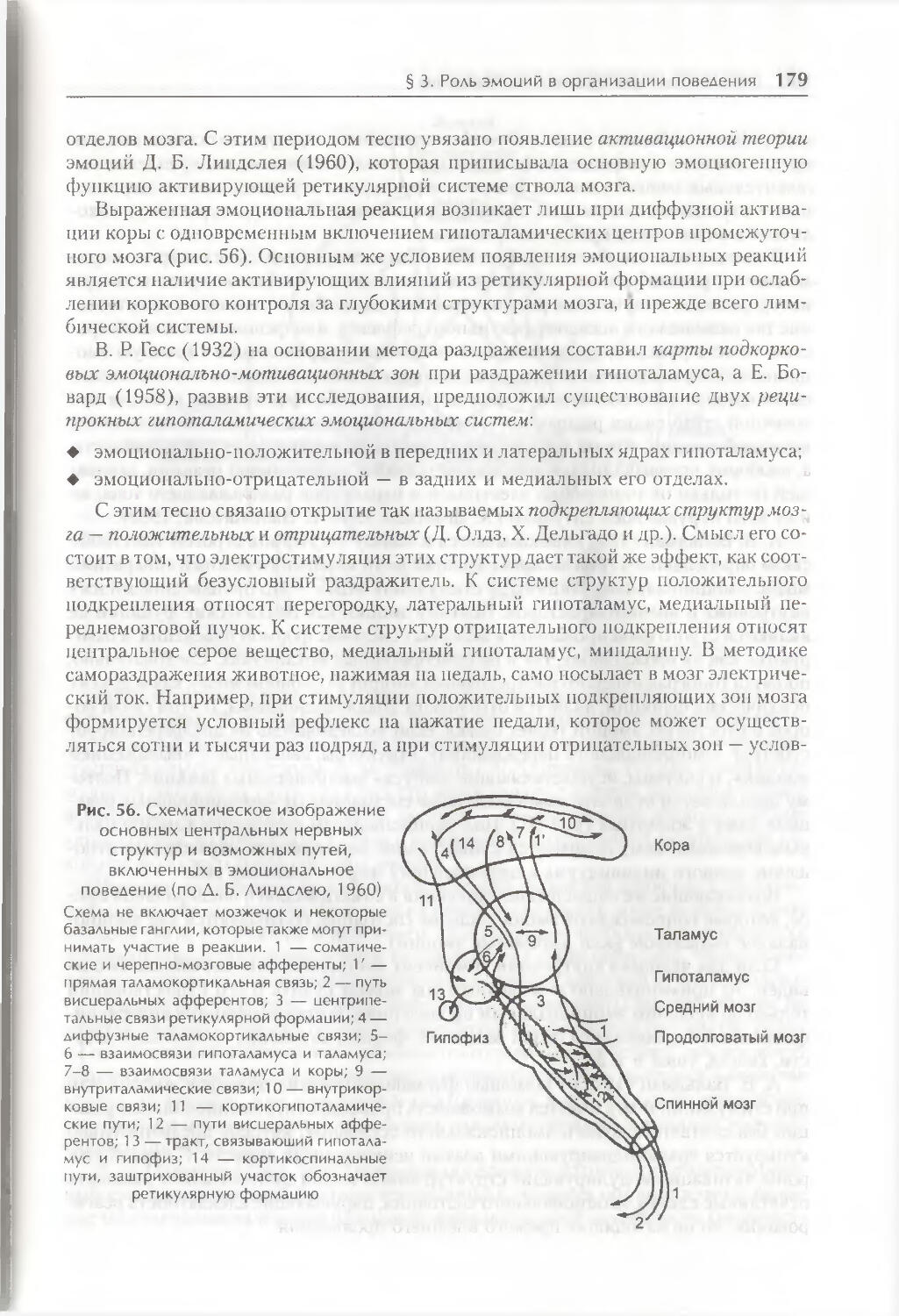
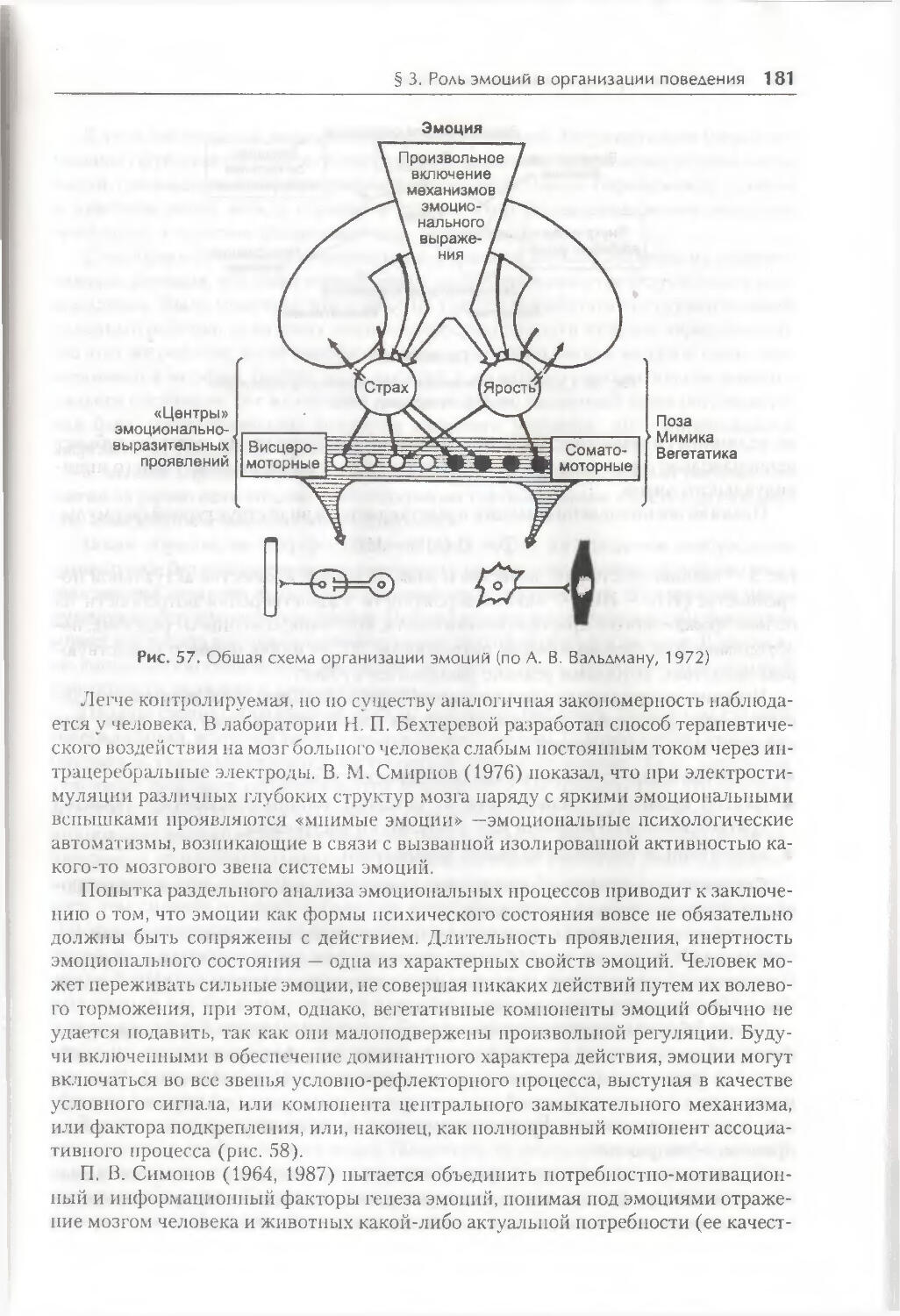
Глава 6. Факторы организации поведения 158
§ 1. Генетическая детерминация свойств поведения 158
§ 2. Биологические мотивации как внутренние детерминанты поведения ... 1 66
§ 3. Роль эмоций в организации поведения 175
§ 4. Восприятие пространства и пространственная ориентация 186
§ 5. Время как фактор организации поведения 191
Заключение 201
Контрольные вопросы 202
Глава 7. Закономерности условно-рефлективной деятельности 203
§ 1. Безусловные рефлексы 203
§ 2. Условные рефлексы 204
§ 3. Торможение условных рефлексов 212
Заключение 220
Контрольные вопросы 220
Глава 8. Механизмы формирования условных рефлексов 221
§ 1. Конвергентная теория формирования временных связей 221
§ 2. Клеточные аналоги условного рефлекса 225
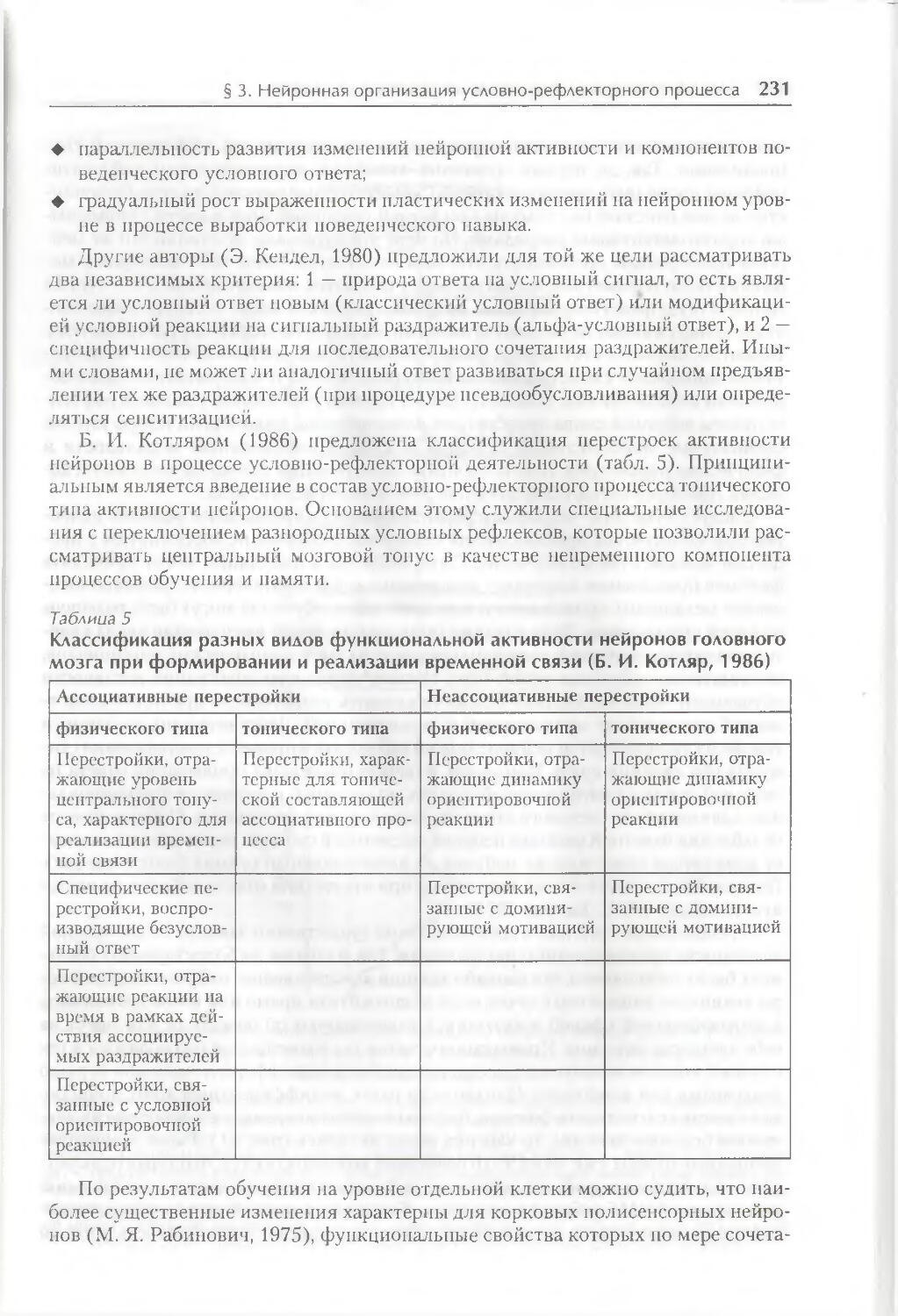
§ 3. Нейронная организация условно-рефлекторного процесса 230
§ 4. Нейронная организация условного торможения 242
Заключение 249
Контрольные вопросы 249
Глава 9. Механизмы памяти 250
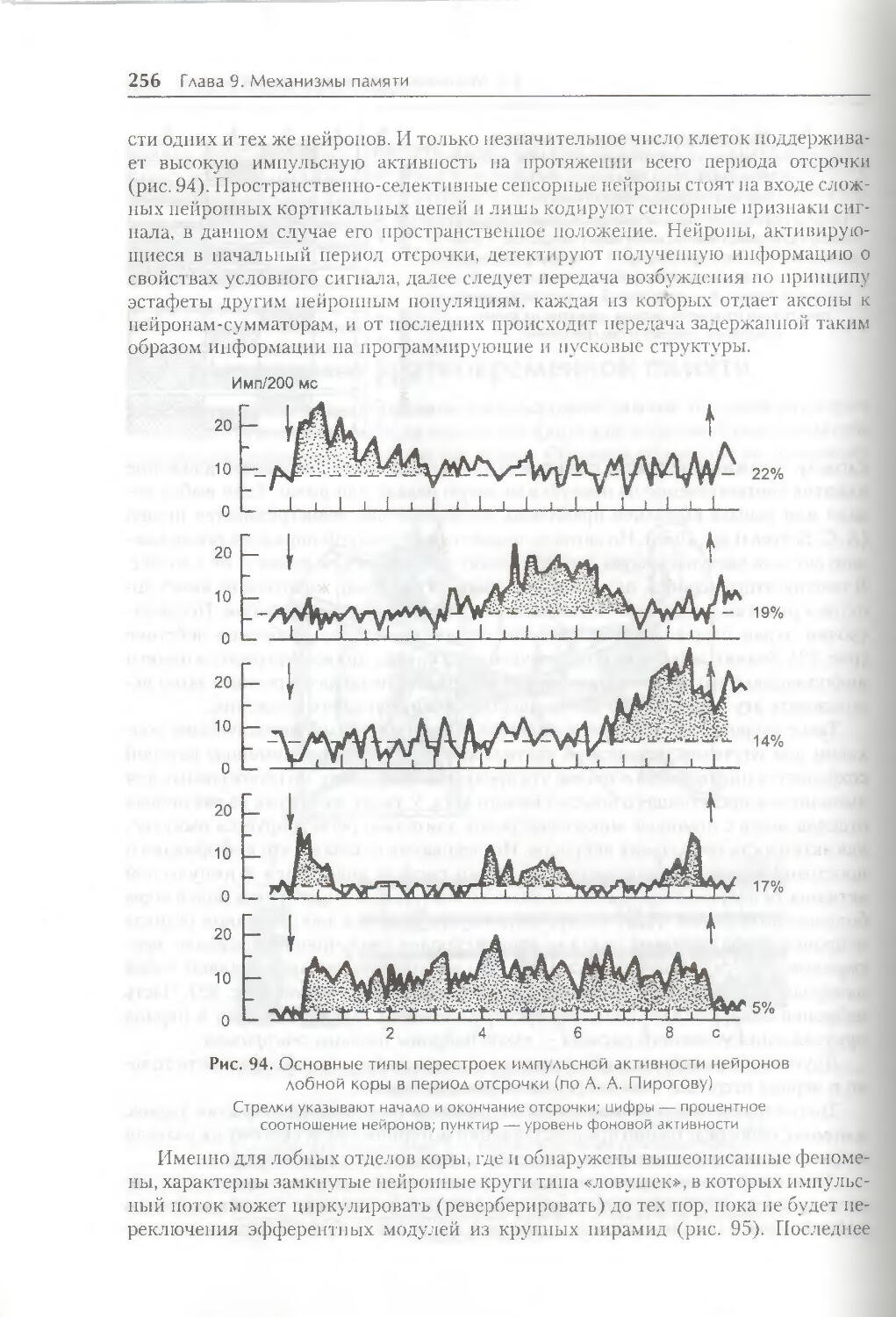
§ 1. Виды и формы памяти 250
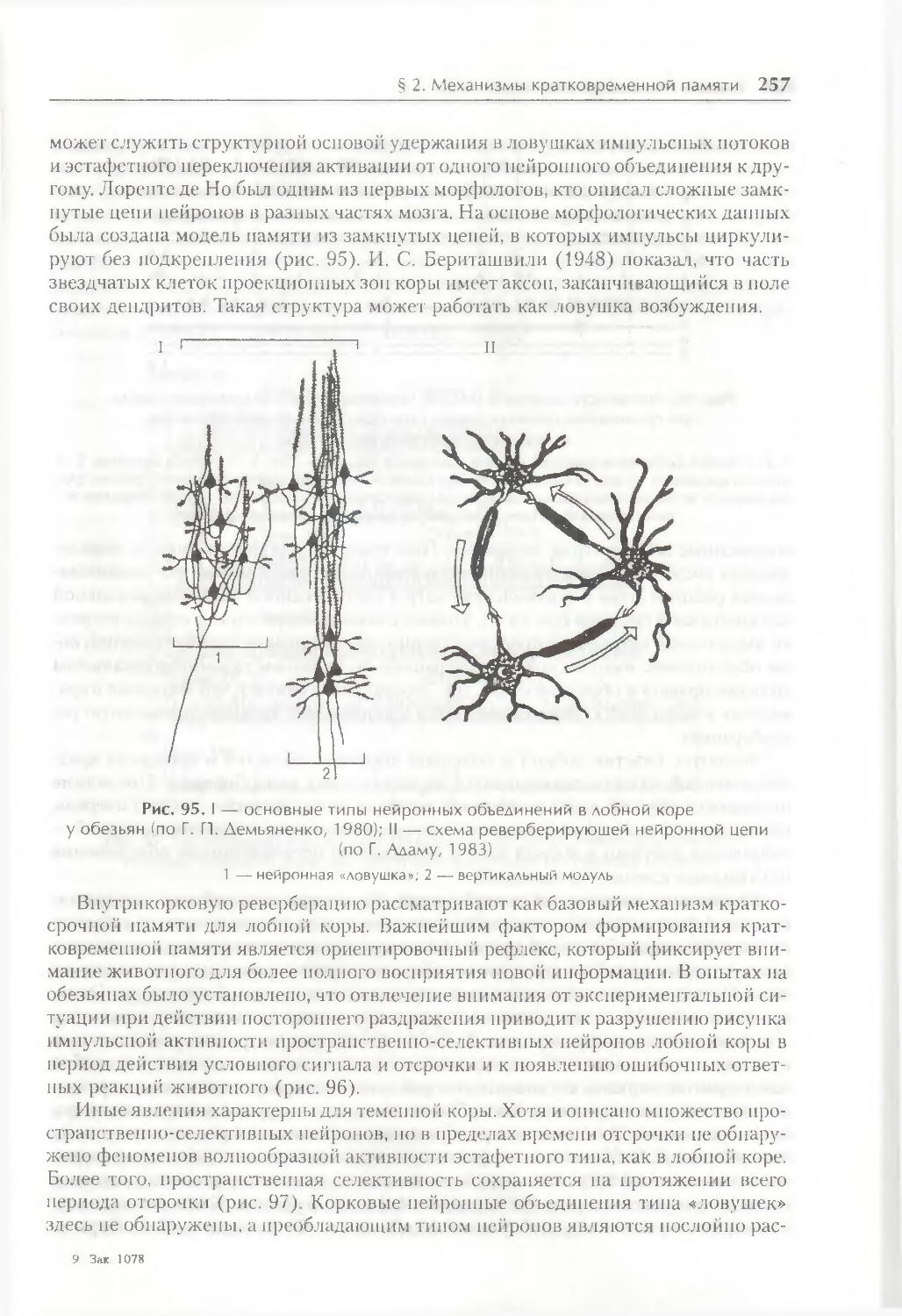
§ 2. Механизмы кратковременной памяти 254
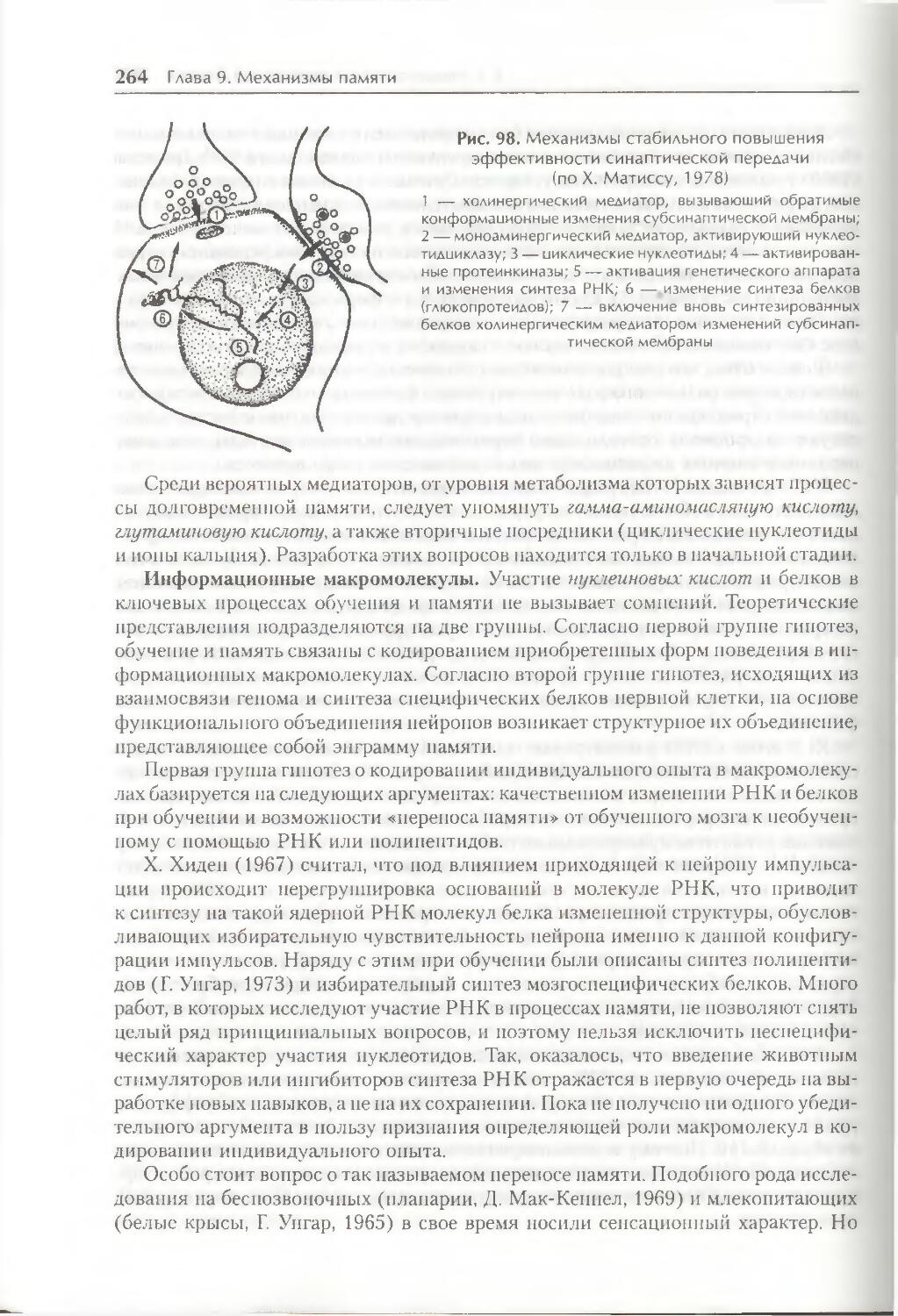
§ 3. Механизмы долговременной памяти 261
Заключение 268
Контрольные вопросы 268
Глава 10. Интегративная деятельность мозга 269
§ 1. Доминанта и условный рефлекс как основные принципы интегративной
деятельности мозга 269
§ 2. Высшие интегративные системы мозга 276
§ 3. Ассоциативные системы и сенсорная функция мозга 286
§ 4. Ассоциативные системы мозга и программирование поведения 291
Заключение 298
Контрольные вопросы 298
Глава 11. Психофизиологическая проблема 299
Контрольные вопросы 304
Заключение 305
Функциональная структура поведенческого акта 305
Литература 310
Основная 310
Дополнительная 310
Глоссарий 312
ПРЕДИСЛОВИЕ
Среди многих ныне существующих отраслей знания, посвященных изучению поведения
и психической деятельности живых организмов (этология, сравнительная и общая психо¬
логия, психофизиология и т. п.), науку о высшей нервной деятельности можно определить
как науку о мозговых механизмах поведения и психики, базирующуюся на рефлекторной
теории в ее современном виде, обогащенном достижениями целого ряда дисциплин
XX столетия. Исследованиям высшей нервной деятельности исходно присущ комплекс¬
ный междисциплинарный характер. Традиционно опираясь на различные области биоло¬
гии — генетику, биохимию, нейрофизиологию, эволюционную теорию, наука о высшей
нервной деятельности неизбежно вторгается на территорию гуманитарных дисциплин,
будь то психология, социология, философия, педагогика и т. п. Именно такому современ¬
ному статусу изучения высшей нервной деятельности максимально соответствует харак¬
тер учебника для университетов, где соседство и взаимовлияние биологического и гумани¬
тарного знания предопределено самой структурой учебного заведения.
Комплексный междисциплинарный характер науки о высшей нервной деятельности
заставил автора данного учебника проделать огромный труд по обобщению и систематиза¬
ции фактических данных и теоретических представлений, накопленных к настоящему вре¬
мени отечественными и зарубежными учеными. Два обстоятельства помогли ему спра¬
виться с этой задачей: более чем тридцатилетний опыт преподавания и собственные интен¬
сивные исследования на протяжении всех этих лет. Именно личная экспериментальная
работа, анализ ее результатов в монографиях, главах руководств и статьях позволили авто¬
ру взглянуть на излагаемый материал глазами непосредственного участника процесса по¬
знания высших и чрезвычайно сложных проявлений деятельности мозга.
Необходимо отметить, что многие вопросы, затронутые в учебнике, остаются предме¬
том острых дискуссий, не имеют общепринятых решений. Однако автор нс берет на себя
роль судьи, оценивающего объективную истинность той или иной концепции. Он посту¬
пил единственно разумным образом — поставил перед собой задачу по возможности точно
воспроизвести ныне существующие точки зрения на данную проблему, справедливо пола¬
гая, что только время и дальнейшие исследования внесут желаемую ясность в ее решение.
Вместе с тем настоящий учебник — отнюдь не бесстрастное перечисление самых разно¬
речивых мнений. В тексте отчетливо просматривается совершенно определенный идейный
стержень, который отличает труд А. С. Батуева от других попыток дать систематизирован¬
ное изложение современного состояния науки о высшей нервной деятельности. Этот стер¬
жень — творческий синтез идей двух отечественных школ: И. П. Павлова и А. А. Ухтомско¬
го, их взглядов на природу поведения и психики. Автор разделяет и разрабатывает очень
важную мысль о своеобразной дополнительности наследия Павлова и Ухтомского, по¬
скольку идеи последнего вносят принципиальнейший элемент активности как в представ¬
ления об организации поведения, так и в признание избирательности, предвзятости психи¬
ческого отражения действительности. Можно только приветствовать, что учебник, прони-
зашшй идеями А. А. Ухтомского, выходит из стен Санкт-Петербургского университета, где
трудился выдающийся биолог и мыслитель XX в.
Приближение к объективной истине, успех в познании объективных законов деятель¬
ности мозга — единственный критерий подлинного развития науки о высшей нервной дея¬
тельности.
Учебник А. С. Батуева привлечет внимание самого широкого круга читателей — от сту¬
дентов, только что переступивших порог университета, до специалистов, работающих
в различных областях биологических и гуманитарных дисциплин.
Академик П. В. Симонов
ОТ АВТОРА
Моим сотрудникам и ученикам посвящается
Наука о высшей нервной деятельности, основы которой были заложены И. М. Се¬
ченовым, И. П. Павловым, А. А. Ухтомским и другими выдающимися учеными,
традиционно является одной из основных дисциплин при подготовке современ¬
ных специалистов-психологов.
Автор настоящего учебника начиная с 1957 г. осуществляет систематическое
чтение лекций по курсу высшей нервной деятельности для студентов психологи¬
ческого факультета и факультета социологии Санкт-Петербургского университе¬
та. Основная идейная платформа построения курса базируется на теоретических
позициях двух отечественных физиологических школ — И. П. Павлова и А. А. Ух¬
томского, синтез которых позволяет формулировать ряд новых положений о моз¬
говых механизмах сложных форм поведения и психики.
Многолетний опыт чтения лекционного курса для студентов разных специаль¬
ностей убедил автора в необходимости создания специального учебного пособия
но предмету «Физиология высшей нервной деятельности и сенсорных систем».
К этому побуждает и само сущностное содержание высшей нервной деятельности,
которая может рассматриваться в цикле социально-психологических дисциплин.
Автор при подготовке учебника старался избежать неоправданного замалчи¬
вания взглядов ряда ученых, огульной критики нестандартных теоретических по¬
ложений, пытаясь использовать то рациональное зерно, которое обязательно со¬
держится в любой научной теории. При этом автор отдает себе отчет в том, что
какие-то вопросы науки о высшей нервной деятельности неизбежно оказались не¬
достаточно представленными. Оправдание этому можно видеть либо в их нераз¬
работанности к настоящему времени, либо в наличии других учебных пособий по
частным вопросам высшей нервной деятельности.
Методическая и техническая сторона исследований высшей нервной деятель¬
ности в учебнике почти не обсуждается в связи с выходом под редакцией автора
специального практикума «Методики и основные эксперименты по изучению
мозга и поведения» (Я. Буреш, О. Бурешова и Дж. Хьюстон).
Автор выражает большую благодарность за критические замечания при про¬
смотре рукописи проф. Э. А. Костапдову, доц. Ю. А. Юиатову, доц. Т. Р. Натишви-
ли, акад. Т. К Иоселиани, ст. пауч. сотр. В. Г. Каменской, ст. науч. сотр. А. Г. Смир¬
нову, ст. науч. сотр. Л. В. Соколовой.
Глава 1
МЕТОДОЛОГИЯ ИЗУЧЕНИЯ ВЫСШЕЙ
НЕРВНОЙ ДЕЯТЕЛЬНОСТИ
§ 1. Принцип рефлекса
В основе системы научного познания лежит принцип детерминизма, который по¬
зволяет выявить закономерную причинно-следственную связь явлений, опреде¬
ляемую взаимодействием материальных факторов. Современная материалисти¬
ческая наука о высшей нервной деятельности основывается па четырех принци¬
пах: принципе рефлекса, принципе доминанты, принципе отражения и принципе
системной деятельности мозга.
Механическая концепция рефлекса. Понятие о рефлексе возникло в XVI в.
в учении Р. Декарта (1596-1650) о механической картине мира. Р. Декарт жил
в эпоху расцвета механики, физики и математики. На его мировоззрение опре¬
деляющее влияние оказало открытие У. Гарвеем механизма кровообращения и но¬
ваторские идеи А. Везалия о том, что носителями психики являются «животные
духи», которые вырабатываются в желудочках мозга и передаются по нервам к со¬
ответствующим органам. Р. Декарт представлял нервные процессы по образцу
системы кровообращения, используя существовавшие в то время принципы опти¬
ки и механики.
Под рефлексом Р. Декарт понимал движение «животных духов» от мозга
к мышцам по типу отражения светового луча. Согласно его схеме, внешние пред¬
меты действуют на периферические окончания расположенных внутри «нервных
трубок» нервных «нитей», которые, натягиваясь, откры¬
вают клапаны отверстий, ведущих из мозга в нервы. По
каналам этих нервов «животные духи» перемещаются в
соответствующие мышцы, которые в результате раздува¬
ются, и, таким образом, происходит движение. Причина
двигательного акта детерминирована материальными из¬
менениями па кожной периферии тела, а нервный про¬
цесс подобен перемещению крови по сосудам. Р. Декарта
по праву можно считать родоначальником детерминисти¬
ческой психофизиологии. Важным в творчестве Р. Декарта является выработка
понятия о стимуле, необходимом для приведения в действие механизмов челове¬
ческого тела.
Исходя из рефлекторного принципа Р. Декарт пытается также объяснить и
обучаемость поведения: люди даже со слабой душой могли бы приобрести неогра¬
ниченную власть над всеми своими страстями, если бы приложили достаточно
старания, чтобы их дисциплинировать и руководить ими. Стремление Декарта
понять целостное поведение особенно четко проявилось в его учении о страстях.
Печаль и радость — это те факторы, которые формируют целесообразное отноше¬
ние организма к внешнему миру, делают реакцию координированной и сложной.
В страстях проявляется связь души с телом.
Принцип детерминиз¬
ма выявляет законо¬
мерную причинно¬
следственную связь
явлений, определяе¬
мую взаимодействием
материальных факто¬
ров.
8 Глава 1. Методология изучения высшей нервной деятельности
Основные теоретические положения Р. Декарта, ис¬
пользуемые современной физиологией, сводятся к сле¬
дующим: органом ощущений, эмоций и мыслей является
мозг; мышечный ответ порождается процессами в примы¬
кающем к мышце нерве; ощущение обусловлено изменения¬
ми в нерве, связывающем орган чувств с мозгом; движение в
сенсорных нервах отражается на моторных, и это воз¬
можно без участия воли (рефлекторный акт); вызванные
посредством сенсорного нерва движения в веществе мозга
создают готовность вновь производить такое же движе¬
ние (обучаемость).
Однако, находясь иод влиянием социально-историче¬
ских противоречий своей эпохи, Р. Декарт сделал серьез¬
ные уступки идеализму: сознание человека он рассматри¬
вал в виде субстанционного начала, способного воздейст¬
вовать посредством мозговой шишковидной железы (где,
по его мнению, располагается «объединенное чувствили¬
ще») на подчиненные рефлекторным законам телесные
процессы. Таким образом, тело и душа — самостоятель¬
ные субстанции. Дуализм Р. Декарта, его трактовка созна¬
ния препятствовали последовательному детерминизму,
ибо он допускал акты воображения, мышления, воли, про¬
исходившие от нематериальной субстанции. Поведение и сознание разводились,
превращались в два независимых ряда явлений.
В оценке общенаучного значения идей Р. Декарта, однако, важно подчеркивать
не столько механизм, сколько материалистическую сущность учения о поведении,
не столько дуализм в понимании психической деятельности, сколько первую про¬
бу ее детерминистического понимания.
Биологическая концепция рефлекса. В конце XVIII в. философия француз¬
ских материалистов завоевала широкое признание и оказывала влияние на мно¬
гих ученых Европы. Учение чешского анатома и физиолога Ииржи Прохазки
(1749-1820) — важный этап в формировании детерминистических представле¬
ний о нервно-психической деятельности.
Сущность своих взглядов па рефлекс Й. Прохазка выразил следующим обра¬
зом: внешние впечатления, возникающие в чувствительных нервах, очень быстро
распространяются но всей их длине до самого начала. Там они отражаются по
определенному закону, переходят на соответствующие им двигательные нервы и
но ним очень быстро направляются к мышцам, посредством которых производят
точные и строго ограниченные движения.
Впервые термин «рефлекс» был введен в научный язык Й. Прохазкой. Он сде¬
лал еще один шаг вперед в физиологическом утверждении стимула, ибо постули¬
ровал, что ответная рефлекторная реакция всегда проявляется в размерах, соот¬
ветствующих силе приложенного стимула.
Развивая концепцию о рефлекторной природе поведения, Й. Прохазка пытает¬
ся преодолеть вначале механистичность, а затем и дуализм картезианства. Общим
законом, по которому чувствительные раздражения переключаются на двигатель¬
ные, является присущее человеку чувство самосохранения. Й. Прохазка утверж-
Р. Декарт
Принцип рефлекса —
универсальная и свое¬
образная форма взаи¬
модействия организма
со средой, происходя¬
щая при участии нерв¬
ной системы.
§ 1. Принцип рефлекса 9
дает монистическое представление о нервной системе,
в целом относящейся к композиции «общего чувстви¬
лища», телесная часть которого локализуется в спин¬
ном мозге, а душевная — в головном. Причем для всех
нервно-психических функций характерна одна общая
закономерность: обе части «сенсориума» действуют по
закону самосохранения. Необходимые для сохранения
животного и его потомства способности — это душев¬
ные функции, а орган, служащий для этого, есть мозг,
объему и сложности которого соответствует степень
совершенства душевных функций.
Учение Й. Прохазки обогатило представление Р. Де¬
карта о рефлекторной природе поведения понятием о
биологическом (а не механическом) назначении самой й. Прохазка
структуры рефлекса, о зависимости ее усложнения от
изменения характера отношений живых существ со сре¬
дой, о пригодности ее для анализа всех уровней созна¬
тельной деятельности, о детерминирующем влиянии
чувствования.
Анатомическая концепция рефлекса. Тщательное
анатомическое изучение нервной системы явилось
сильным толчком к развитию и укреплению рефлек¬
торной концепции в XIX в. Английский анатом и врач
Чарльз Белл (1774-1842) в 1811 г. в трактате «О новой
анатомии мозга» писал, что можно произвести пере¬
резку заднего пучка нервов, исходящего из задней час¬
ти спинного мозга, без конвульсивных сокращений
мышц спины. Однако это становилось невозможным
даже при одном прикосновении кончиком ножа к пе¬
реднему корешку.
Таким образом, понятие о рефлексе как о закономерном двигательном ответе
па раздражение сенсорных нервов было превращено в естественно-научный факт.
Независимо от Ч. Белла к аналогичным выводам пришел французский физио¬
лог Ф. Мажанди (1783-1855). Переход нервного возбуждения по афферентным
нервам через спинной мозг па эфферентные нервы получил название закона Бел¬
ла- Мажанди.
Но сам Ч. Белл пошел дальше: создал теорию о «мышечной чувствительно¬
сти» и сформулировал физиологическое обоснование циклической функции нерв¬
ной системы. Между мозгом и мышцей имеется замкнутый нервный круг: один
нерв передает влияние от мозга к мышце, другой — передает в мозг чувство со¬
стояния мышцы. Если круг будет разомкнут перерезкой моторного нерва, то ис¬
чезнет движение. Если же он размыкается перерезкой чувствительного нерва,
пропадает ощущение самой мышцы, а вместе с этим исчезает и регуляция ее дея¬
тельности. Так, например, у женщины па одной руке была потеряна чувствитель¬
ность, а на другой — способность к движениям. Эта женщина могла держать ре¬
бенка на руке, потерявшей только чувствительность, до тех нор пока она на пего
смотрела. Как только она отводила взор от ребенка, сразу же возникала опасность
его падения па пол.
10 Глава 1. Методология изучения высшей нервной деятельности
Таким образом, если ранее детерминантами рефлек¬
торного акта считались лишь внешние стимулы, то
Ч. Белл показывает значение внутренней чувствительно¬
сти самих мышц, которая обеспечивает наиболее точное
и тонкое выполнение движения.
Рефлексы спинного мозга широко использовались
клиницистами, среди которых наиболее значительными
фигурами были английский врач Маршал Холл и немец¬
кий физиолог Иоганнес Мюллер. Именно М. Холлу при¬
надлежит термин «рефлекторная дуга». Рефлекторная
дуга состоит из афферентного нерва, спинного мозга и эф¬
ферентного нерва.
и. м. Сеченов М. Холл и И. Мюллер настаивали на принципиальном
отличии работы спинного мозга от головного. По их мне¬
нию, рефлекторный механизм свойствен только спинному мозгу, рефлексами мо¬
гут именоваться только такие акты, природа которых апсихична. Закономерности
протекания любого рефлекторного акта обусловливались изначально заложенны¬
ми в организме связями нервных субстратов, внешнему же стимулу отводилась
роль лишь спускового крючка. Внутренние факторы противопоставлялись внеш¬
ним. Головной мозг оказывался все дальше и дальше от сферы влияния физио¬
логии. Дистанция между физиологией и психологией становилась все более ощу¬
тимой.
Вместе с этим нельзя не видеть и прогрессивных тенденций идей Ч. Белла,
Ф. Мажанди, М. Холла, И. Мюллера. Эти ученые предприняли попытки вскрыть
впутриорганические условия протекания простейшего
рефлекторного ответа, стремились к его аналитическому
познанию как элементарной единицы нервной деятельно¬
сти, боролись против субъективно-психологических объ¬
яснений структуры рефлекса. Жесткий анатомический
характер этих теорий уже в середине XIX в. встретился с
серьезными противоречиями, которые возникали в связи
со все более широким распространением эволюционных идей, наиболее последо¬
вательно воплощенных Ч. Дарвином.
Психофизиологическая концепция рефлекса. Эволюционные идеи встрети¬
ли в России наиболее благоприятную почву, подготовленную философским уче¬
нием русских революционеров-демократов, оказавших существенное влияние на
формирование мировоззрения И. М. Сеченова (1829-1905). Само понятие о реф¬
лекторном характере нервной деятельности у И. М. Сеченова претерпело сущест¬
венные изменения.
Рассмотрим следующие основные черты сеченовской теории рефлекса (Яро-
шевский, 1961).
1. Рефлекс им понимался как универсальная и своеобразная форма взаимодейст¬
вия организма со средой, опирающаяся на эволюционную биологию. И. М. Се¬
ченов поставил вопрос о существовании двух родов рефлексов. Во-первых, по¬
стоянные, врожденные, осуществляемые низшими отделами нервной си¬
стемы. Он назвал их «чистыми» рефлексами. Во-вторых, рефлексы головного
мозга изменчивые, приобретенные в индивидуальной жизни. Эти рефлексы
Рефлекторная дуга со¬
стоит из афферентно¬
го нерва, отдела цент¬
ральной нервной
системы и эфферент¬
ного нерва.
§ 1. Принцип рефлекса 11
И. М. Сеченов представлял себе одновременно и физиологическим и психиче¬
ским явлением.
Таким образом, была впервые показана неотделимость психических про¬
цессов от мозга и одновременно обусловленность психики внешним миром.
Важнейшим для И. М. Сеченова было положение о единстве организма и усло¬
вий внешней среды. Факторы эволюции:
•Ф- определяют жизнь как.приспособление организмов к условиям существо¬
вания;
<- доказывают, что внедрение влияния способно видоизменять материальную
организацию и характер жизненных отправлений.
И. М. Сеченов был выдающимся пропагандистом дарвиновского учения в
России, он внедрил эволюционно-биологический подход в физиологию мозга и
ввел понятие об изменчивости и преобразовании рефлексов в целях успешной
адаптации, усложнения и развития. Итак, была создана материалистическая
платформа для смыкания нервных актов с психическими.
2. Физиологический субстрат рефлекторных актов характеризуется как нейро¬
динамика, отличная от динамики других систем. Открытие центрального тор¬
можения И. М. Сеченовым в 1862 г. было первым шагом к созданию им новой
физиологии головного мозга. Деятельность нервных центров мыслится теперь
как непрерывная динамика процессов возбуждения и торможения.
3. На первый план ставятся межцентральные координационные отношения. Фи¬
зиологическому анализу начинают подвергаться высшие мозговые центры. Ес¬
ли до И. М. Сеченова усиление или угнетение рефлек¬
торных реакций трактовалось не иначе как усилием
воли, сознания, разума, то И. М. Сеченов переводит
все это на строгий физиологический язык и показыва¬
ет, как центры головного мозга могут задерживать или
усиливать спинальные рефлексы.
4. Функция мозговых центров трактуется в широком
плане биологической адаптации. Центры влияют уси¬
ливающим или тормозящим образом на движения не потому, что высвобожда¬
ется заложенная в них «психическая сила», и не потому, что укорачивается или
удлиняется путь прохождения нервного импульса. И. М. Сеченов вводит поня¬
тие «физиологического состояния центра», которое непосредственно связано с
биологическими потребностями. Само же состояние центра, отражая характер
отношений со средой, представляет собой нервный субстрат потребности.
В учение о рефлексах вносится существенное дополнение. Реакция ставит¬
ся в непосредственную зависимость не только от наличных раздражений, по и
от всей суммы прежних воздействий, оставивших длительно сохраняющиеся
следы в нервных центрах.
5. Мышечная чувствительность открывает новые перспективы для детермини¬
стического анализа поведения. И. М. Сеченов считает, что мышечное чувство
при выполнении одного движения становится в порядке ассоциации рефлек¬
сов сигналом для другого движения. Принцип ассоциации рефлексов лежит в
основе обучения человека сложным формам трудовой деятельности. Устанав¬
ливается общий характер для движений и для психической деятельности — это
наличие мышечной чувствительности.
Мышечное чувство при
выполнении одного
движения становится
в порядке ассоциации
рефлексов сигналом
для другого движения
(И. М. Сеченов).
12 Глава 1. Методология изучения высшей нервной деятельности
По вопросу о соотношении физиологического и пси¬
хического И. М. Сеченов занял совершенно определен¬
ную позицию, которую выразил следующими словами:
«Для нас, как для физиологов, достаточно и того, что
мозг есть орган души, то есть такой живой механизм,
который, будучи приведен какими ни на есть причина¬
ми в движение, дает в окончательном результате тот же
ряд внешних явлений, которыми характеризуется пси¬
хическая деятельность»1.
При всей убедительности аргументов И. М. Сече¬
нова, которые он использовал для утверждения своих
взглядов на поведение и психику, ему не хватало само¬
го главного аргумента — лабораторного объективного
метода исследования.
Поднявшись до распространения рефлекторного принципа на психическую
деятельность и рассматривая рефлекс как психофизиологический феномен,
И. М. Сеченов не смог изучить конкретные механизмы поведения в связи с отсут¬
ствием надлежащего метода. Поэтому ряд его высказываний остался лишь гени¬
альными догадками, взмахом его могучей мысли.
Концепция условного рефлекса. На долю И. П. Павлова выпала чрезвычайно
ответственная миссия — блестящие догадки, предвидения и мысли И. М. Сечено¬
ва он подкрепил научной концепцией условного рефлекса. И. П. Павлов мобилизо¬
вал все свое мастерство талантливого экспериментатора, чтобы его концепция бы¬
ла введена в строгие рамки лабораторного опыта.
И. П. Павлов понимал, что он, вслед за Сеченовым, вторгается в область явле¬
ний, обычно именуемых психическими. «Вся сложная нервная деятельность, —
пишет И. П. Павлов уже в 1913 г., — которая раньше трактовалась как психическая
деятельность, представляется нам в виде двух основных механизмов: механизма
образования временной связи между агентами внешнего мира и деятельностями
организма, или механизма условного рефлекса, как мы говорим обыкновенно,
и механизма анализаторов, то есть таких приборов, которые имеют своей целью
анализировать сложность внешнего мира: разлагать его па отдельные элементы
и моменты. По крайней мере до сих пор весь добытый нами материал укладывает¬
ся в эти рамки. Но этим, конечно, не исключается возможность расширения на¬
ших теперешних представлений о деле»2.
И. П. Павлов проявил себя как последовательный материалист и детерминист.
Недаром он провозгласил, что исследование условных рефлексов опирается на
три принципа рефлекторной теории: детерминизм, анализ и синтез, структур¬
ность. И. П. Павлов целиком придерживался рефлекторной схемы Р. Декарта
и понимал значение рефлекса как одного из примеров универсального принципа
детерминирования. Уже па заре развития павловского учения становилось яс¬
ным, что условный рефлекс представляет собой закономерность более высокого
и сложного порядка, чем простые рефлексы. Условный рефлекс обеспечивает из¬
менчивость приспособительного поведения животного в отношении внешнего
1 Сеченов И. М. Избранные философские и психологические произведения. М.; Л., 1974. С. 112.
2 Павлов И. П. Собр. соч. М.; Л„ 1952.
И. П. Павлов
§ 1. Принцип рефлекса 13
мира. Условный рефлекс является важнейшим фактором биологической эво¬
люции.
Однако И. П. Павлов, захваченный полемикой с психологами и разделяя кар¬
тезианский детерминизм, стал углубленно изучать физиологические закономер¬
ности условно-рефлекторной деятельности, биологическую же сторону явления
оставил на перспективу. Отсюда и неизбежные противоречия в идее условного
рефлекса: с одной стороны, приспособительный акт целого организма, с другой —
элементарный процесс работы нервной системы. Все научное творчество И. П. Пав¬
лова было посвящено разрешению этого противоречия и созданию наименее про¬
тиворечивой идеологии в его теории высшей нервной деятельности.
Далее мы будем неоднократно рассматривать отдельные положения павлов¬
ской теории, а здесь ограничимся лишь наиболее важными ее элементами приме¬
нительно к теории рефлекса, которые были отмечены П. К. Анохиным (1979).
1. Прежде всего был создан лабораторный метод объективного изучения при¬
способительной деятельности человека и животных — метод условных рефлек¬
сов.
2. Изучая условные рефлексы на целостном организме, И. П. Павлов подчерки¬
вал их приспособительно-эволюционный смысл для животного мира.
3. И. П. Павлов предпринял попытку локализовать сам нервный процесс замыка¬
ния нервных связей в коре головного мозга у высших животных и человека. При
этом он не был категоричным и не исключал специфического участия других
отделов мозга в этом процессе. Он писал, что все наши законы всегда более или
менее условны и имеют значение только для данного времени, в условиях дан¬
ной методики, в пределах наличного материала.
4. И. II. Павлов констатировал наличие в коре головного мозга процесса тормо¬
жения, чем подкрепил сеченовские идеи о тормозящем влиянии головного мозга.
5. Было четко сформулировано учение о физиологии анализаторов, под которы¬
ми И. П. Павлов, вслед за И. М. Сеченовым, мыслил триединую конструкцию:
периферические рецепторы, проводящие пути и мозговые центры вплоть до
коры больших полушарий.
6. Были описаны феномены динамики процессов возбуждения и торможения при
условно-рефлекторной деятельности. В результате было сформировано пред¬
ставление о коре головного мозга как о мозаике из воз¬
буждений и торможений.
7. В конце своей творческой жизни И. П. Павлов выдви¬
нул принцип системности в работе коры головного
мозга, способной формировать динамический стерео¬
тип деятельности, уже в какой-то мере независимый
от качества внешних раздражений.
Идеи И. П. Павлова завоевали весь мир и продолжа¬
ют служить основой для развертывания новых научных
исследований в самых разнообразных сферах науки о по¬
ведении живых организмов.
Диалектическая концепция рефлекса. А. А. Ухтомскому (1875-1942) при¬
надлежит заслуга теоретико-физиологического плана, которая состоит в углуб¬
ленной дальнейшей разработке принципа детерминизма в рефлекторной теории.
Учение о физиологии
анализаторов — под
ним И. П. Павлов,
вслед за И. М. Сечено¬
вым, подразумевал
триединую конструк¬
цию: периферические
рецепторы, проводя¬
щие пути и мозговые
центры вплоть до коры
больших полушарий.
14 Глава 1. Методология изучения высшей нервной деятельности
Диалектичность мышления А. А. Ухтомского нашла яркое проявление в пони¬
мании им сущности рефлекса. Видя в рефлексе механизм активности, он усматри¬
вал в рефлекторном акте единство внутренних и внешних детерминант, причем
внутренние детерминанты в конечном счете также заданы и определяются внеш¬
ними условиями.
А. А. Ухтомский подчеркивал, что «...рефлекс — это такая реакция, которая мо¬
тивируется вполне отчетливо текущей обстановкой или средой. Этим, однако, не
уничтожается спонтанное действие субстрата, оно лишь
ставится в определенные границы в его противоположе¬
нии факторам среды, и от этого оно становится более
определенным по содержанию и значению. Рефлекс рису¬
ется не чисто пассивным передвижением костного шари¬
ка под влиянием полученного им удара извне; так реф¬
лекс мог изображаться, пока надо было подчеркнуть в
особенности его мотивировку из среды. Но в полноте сво¬
ей он представляется встречею во времени двух условий:
с одной стороны, деятельности заготовленной, или сложившейся в самом субстра¬
те (клетке) за предыдущую его историю, и, с другой стороны, внешних импульсов
текущего момента»1.
Следовательно, внутренние детерминанты — это накопленная история взаи¬
модействия реагирующего субстрата с фактором среды (принцип историзма).
И но происхождению, и по условиям проявления внутренние детерминанты
в конечном счете определяются факторами среды, то есть обладают лишь относи¬
тельной самостоятельностью. Внешнее выступает как комплекс условий сущест¬
вования внутреннего. Значит, среда организма — это не весь окружающий его фи¬
зический мир, а лишь та небольшая его часть, элементы которого биологически
значимы для организма. Но для организма представляет биологический интерес
лишь такое внешнее, которое может стать частью жизненного опыта, то есть ча¬
стью внутреннего, или способствовать превращению определенных внешних фак¬
торов во внутренние.
Современная теория поведения далеко ушла от простых картезианских схем.
Введение принципа историзма позволяет понять биологическую адекватность, то
есть целесообразность реакций организма па воздействие среды. Картезианское
мировоззрение базируется па жесткой однозначной причинности (жесткий детер¬
минизм Лапласа), ему чуждо признание реальных противоречий. А. А. Ухтомский
же показывает, что реальное поведение требует признания существования проти¬
воречий как непрерывного атрибута процесса развития, как движущих сил по¬
строения поведения.
Исторический подход позволил А. А. Ухтомскому выявить действительную
роль и оценить значение условного рефлекса в эволюции животного мира, а также
раскрыть одно из кардинальных свойств условного раздражителя — его превра¬
щение из индифферентного в «обязательный» компонент среды. Такой усвоенный
раздражитель начинает вызывать новую для себя реакцию. В результате такого
усвоения организм определил и зафиксировал свое отношение к данному раздра¬
жителю — определил его биологическую значимость для себя.
1 Ухтомский А. А. Собр. соч. Л., 1954. Т. 5. С. 72.
Внутренние детерми¬
нанты — это накоплен¬
ная история взаимодей¬
ствия реагирующего
субстрата с фактором
среды (приниип исто¬
ризма) (А. А. Ухтом¬
ский).
§ 2. Принцип доминанты 15
Рассматривая систему рефлексов в эволюционном
ряду, А. А. Ухтомский пишет: «...простой рефлекс клас¬
сической физиологии является не исходным и прин¬
ципиально общим типом рефлекторной активности
центров, над которой специализируется особая об¬
ласть рефлексов условных, но наоборот, — частным спе¬
циальным и поздним продуктом редукции и упроще¬
ния рефлекса условного, который становится отныне
общим типом деятельности центрального нервного ап¬
парата»'.
Индивидуальное приспособление особи с помощью
условных рефлексов служит своеобразным компасом —
ориентиром для решения проблем вида. Естественный
отбор закрепляет те мутационные приобретения, кото¬
рые соответствуют находкам особи. Таким образом, индивидуальное приспособление
идет впереди эволюционно-генетических перестроек. Принципы историзма и со¬
отношения реактивности и активности в целостном поведении как способе разре¬
шения противоречий, принесенные в физиологию А. А. Ухтомским, существенно
обогатили рефлекторную теорию, которая окончательно избавилась от дуализма
и механицизма картезианского толка, встав па твердые диалектические позиции.
§ 2. Принцип доминанты
Основные положения учения о доминанте. А. А. Ухтомский выдвинул принцип
доминанты в качестве основы складывающихся координационных отношений.
В своих теоретических построениях он активно опирался
на идеи Шеррингтона, и прежде всего на его принципы
воронки и общего пути. Шеррингтон писал, что головной
мозг, функционирующий на основе дистантной рецепции
и ведающий выполнением реакций, которые в какой-то
момент становятся реакциями предваряющими, является органом, обладающим
способностью предвосхищать ощущения, и психологическими возможностями,
вытекающими из этого преимущества. Одновременно с этим развитые Н. Е. Вве¬
денским идеи динамизма взаимоотношений в нервной системе, обусловленные
характером взаимодействия, функциональным состоянием нервных центров и ха¬
рактеристиками рабочего органа, позволили А. А. Ухтомскому сформулировать
учение о доминанте — рабочем принципе деятельности нервной системы и векто¬
ре поведения.
Во многих своих работах, подчеркивая роль доминанты как образователя
сложнейших афферентных синтезов из огромной массы текущей информации,
Ухтомский сделал вывод, что доминанта определяет вероятность возникновения
той или иной рефлекторной реакции в ответ на текущее раздражение.
Как общий принцип работы нервных центров доминанта подчиняется опреде¬
ленным закономерностям. Прежде всего в нервных сетях живого организма поч¬
ти непрерывно возникают потоки возбуждений из разнообразных центральных 1Доминанта — рабочий
принцип деятельности
нервной системы и
вектора поведения.
1 Ухтомский А. А. Указ. соч. С. 291.
16 Глава 1. Методология изучения высшей нервной деятельности
и периферических источников. Это физиологически оправдано, ибо в организме
наблюдается значительное преобладание рецепторов, улавливающих тончайшие
изменения внешней и внутренней среды, над соответствующими центрами в го¬
ловном и спинном мозге и над исполнительными органами.
Ухтомский сформулировал следующие основные свойства доминанты:
♦ повышенную возбудимость;
♦ стойкость возбуждения;
♦ инертность возбуждения;
♦ способность к суммированию возбуждения.
К этому следует добавить способность доминанты переходить в тормозное со¬
стояние и вновь растормаживаться. Ну и, конечно, доминанта только потому
и может стать таковой, что одновременно с созданием
и подкреплением самой себя она приводит к сопряженно¬
му торможению центров антагонистических рефлексов.
Что касается повышенной возбудимости, то отнюдь не
сила возбуждения в центре, а именно способность к даль¬
нейшему повышению возбуждения под влияниям нового
приходящего импульса может сделать центр доминирую¬
щим. Далее Ухтомский подчеркивает важное свойство до¬
минирующего центра достаточно интенсивно, продолжи¬
тельно и стойко накапливать и поддерживать в себе воз¬
буждение, что приобретает значение господствующего
фактора в работе прочих центров. Все это создает предпосылки для хронического
свойства доминанты, ее инертности. Наконец, относительно суммирования возбу¬
ждений Ухтомский высказывался весьма категорично, полагая, что судьба доми¬
нанты решается тем, будет ли центр способен суммировать свои возбуждения иод
влиянием доходящих до него импульсов или они застанут его неспособным к сум¬
мированию.
Определяющую роль при формировании доминантного очага играет состоя¬
ние развивающегося в нем стационарного возбуждения, в основе которого лежат
различные физико-химические процессы. Определенный уровень стационарного
возбуждения чрезвычайно важен для дальнейшего хода реакции. Если этот уро¬
вень возбуждения мал, то диффузная волна может поднять его до состояния, ха¬
рактерного для доминанты, то есть создать в нем превы¬
шенную возбудимость. Если же уровень возбуждения в
центре и без того высок, то при приходе новой волны воз¬
буждения возникает эффект торможения. Но доминант¬
ный очаг не следует рассматривать «как центр сильного
возбуждения»; определяющую роль в этом процессе иг¬
рает не количественный, а качественный фактор — повы¬
шенная возбудимость, то есть отзывчивость на приходя¬
щие волны возбуждения и способность центра суммировать эти возбуждения.
Именно такой центр — наиболее возбудимый, отзывчивый и впечатлительный
в данный момент — отвечает па приходящий стимул, анатомически даже не отно¬
сящийся к нему, именно такой центр, первым вступая в работу, предопределяет на
какой-то более или менее продолжительный промежуток времени новый ход
Сопряженное торможе¬
ние не есть подавление
всяческой деятельно¬
сти, но ее переработка
согласно с направлени¬
ем доминирующего по¬
ведения.
Приниип доминанты —
временно господст¬
вующая в нервной си¬
стеме группа нервных
иентров, определяю¬
щая характер текущей
ответной реакции ор¬
ганизма на внешние и
внутренние раздраже¬
ния и целенаправлен¬
ность его поведения.
§ 2. Принцип доминанты 17
реакции. Считая одной из основных черт доминанты ее направленность (вектор-
ность), Ухтомский вместе с тем отмечал, что векторная направленность произ¬
вольного поведения является результатом доминанты.
Не менее важную роль в процессе формирования доминанты играет процесс
сопряженного торможения. Состояние возбуждения в центре, подкрепленное воз¬
буждениями из самых отдаленных источников, будучи
достаточно стойким, то есть инертным, в свою очередь
снижает способность других центров реагировать на им¬
пульсы, имеющие к ним прямое отношение. В случае до¬
минант низшего порядка сопряженное торможение рас¬
сматривается как следствие возникновения определен¬
ного фокуса возбуждения, тогда как в случае доминант
высшего порядка (например, кортикальных) они явля¬
ются прямым фактором объединения «очагов» возбуж¬
дений в некоторый рабочий ансамбль. Но торможение в
других центрах наступает лишь тогда, когда возбуждение
в формирующем нервном центре достигает достаточной
величины. Именно сопряженное торможение играет наиболее важную роль при
формировании доминанты, и это торможение должно быть своевременным, то
есть иметь координирующее значение для работы других органов и организма в
целом. Координационное торможение — это процесс энергетически наиболее ем¬
кий, «дорогой», вырабатывающийся позднее, чем простое возбуждение. Торможе¬
ние — это наиболее тонкий, интимный процесс и вместе с тем наиболее ранимый.
Сложные рефлекторные акты осуществляются не одним анатомическим цент¬
ром, а несколькими центрами, расположенными в различных отделах спинного и
головного мозга и образующими своеобразную констелляцию центров в работе
целостного мозга. Стационарное возбуждение в констелляции центров представ¬
ляется в виде определенного цикла, когда каждый участник этой констелляции,
будучи связан с соседними и стимулируя их, в свою очередь подучает от них сти¬
муляцию. Таким образом, складывается своеобразная «циклическая машина», ко¬
торая долгое время сохраняет в себе возможность вспыхивать и усиливаться под
влиянием текущих раздражений. Состояние возбуждения в доминанте Ухтом¬
ский назвал ее симптомокомплексом.
Для образования доминантной установки необходимо нарастание в элементах
констелляции состояния возбуждения еще до того, как возникнут дискретные
волны возбуждения, способные к проведению и суммированию своих эффектов,
то есть когда слагаемые будущей констелляции заработают в единстве, когда сло¬
жится их рабочий ансамбль.
Доминантный процесс — не привилегия определенных этажей нервной систе¬
мы: он может складываться в любой центральной группе в зависимости от усло¬
вий подготовки, развития возбуждения в ней и сопряженного торможения антаго¬
нистических механизмов.
Процесс торможения не только оформляет и поддерживает доминирующую
констелляцию, но может привести и к прекращению ее функционирования, то
есть сама доминанта может тормозиться.
Часто сравнивая физиологический механизм с механизмом техническим, Ух¬
томский отмечал не только то, что он образуется по ходу самой реакции, но и то,
Торможение нейтраль¬
ное — торможение,
возникающее в нейт¬
ральной нервной систе¬
ме благодаря наличию
в ней специализирован¬
ных тормозных нейро¬
нов. Является основ¬
ным фактором коорди¬
нации деятельности
центральной нервной
системы.
18 Глава 1. Методология изучения высшей нервной деятельности
что в отличие от технического механизма он имеет не одну, а множество степеней
свободы, дающих ему возможность осуществлять множество переменных опера¬
ций. В самой природе субъекта наличие конвергенции обусловливает наделение
существующей системы исполнительных органов множеством степеней свободы,
которые обеспечивают новые функциональные разновидности, достигающиеся
новыми сочетаниями прежних анатомо-физиологических данных.
Выбор одной степени свободы при сопряженном торможении других достига¬
ется посредством доминанты, направленной на определенный рабочий вектор.
При рассмотрении вопроса о превращении доминанты из симптомокомплекса
в общий механизм действия, поведения и жизнедеятельности организма Ухтом¬
ский нередко использовал понятие функционального центра, функционального
органа. Этим подчеркивалось следующее: дело не столько в том, что в нервной
системе складываются определенные констелляции связанных между собой цент¬
ров, морфологически далеко разбросанных но мозговой массе, а в том, что они
функционально связаны единством действий, своей векторной направленностью
на определенный результат. В понятие органа может быть включено всякое соче¬
тание сил, могущее привести при прочих равных условиях всякий раз к одинако¬
вым результатам. Орган — это прежде всего механизм с определенным однознач¬
ным действием. «Громадное разнообразие морфологических черт того или иного об¬
разования нашего тела впервые приобретает значение физиологического органа
после того, как открывается значение этих морфологических деталей для рабочих
слагающих однозначно определенной физиологической равнодействующей», — пи¬
сал А. А. Ухтомский.
Единство действия достигается способностью данной совокупности приборов
взаимно влиять друг на друга с точки зрения усвоения ритма, то есть синхронизации
активности нервных центров. В этих процессах огромную роль играет усваивание
темпа и ритма возбуждений в центрах, входящих в состав
констелляции. Каждый из потенциальных компонентов
констелляции изначально обладает своим определенным
темпом и ритмом возбуждений. Именно в констелляции
более всего возможно возникновение ритмического про¬
цесса. Происходит как бы своеобразное развертывание
констелляции, когда постепенное «сыгрывание ритмов»
в отдельных ее компонентах приводит к достижению не¬
которого общего «марша» возбуждения в ней. При этом
сопряженное торможение прочих центров, не входящих в состав данной констел¬
ляции, будет развиваться тем более, чем полнее констелляция будет участвовать в
ритмической работе. Процесс синхронизации темпов и ритмов активности нерв¬
ных центров, входящих в состав определенной констелляции, делает ее домини¬
рующей.
Ее доминирование, следовательно, будет зависеть от того, насколько она будет
вовлечена в дело, насколько наладившийся ритм возбуждения будет осуществ¬
ляться без перебоев, подкрепляясь новыми текущими раздражениями, насколько
при этом будут сопряженно тормозиться другие рефлексы.
Основным аппаратом управления ритмами в тканях и органах, по Ухтомскому,
является кора головного мозга, обеспечивающая обоюдный процесс подчинения
темпов и сроков жизнедеятельности темпам и срокам сигналов из внешней среды,
равно как подчинение сроков в постепенно осваиваемой среде потребностям орга-
Выбор одной степени
свободы при сопря¬
женном торможении
других достигается
посредством доминан¬
ты, направленной на
определенный рабочий
вектор.
§ 2. Принцип доминанты 19
низма. Одновременно с поддержанием синхронизации ритмов в рабочей констел¬
ляции кора мозга принимает участие в вовлечении и других нервных центров
в этот процесс синхронизации.
Одной из черт доминирующей констелляции является способность ее отдель¬
ных компонентов или центров принимать участие в других рефлекторных актах,
входя в состав соответствующих им констелляций. Но как и чем определяется по¬
добный переход? С одной стороны, сохранение того или иного компонента — уча¬
стника рабочей констелляции — зависит от того, насколько стойко и продолжи¬
тельно он способен сохранять в себе темп и ритм, навязанные ему данной кон¬
стелляцией, то есть от его инерции. С другой стороны, переход данного нервного
центра в работу другой констелляции будет зависеть от того, насколько быстро
данный центр способен усвоить темп и ритм нового межцентрального рабочего
цикла, возникшего в нервной системе.
Постепенно формируясь, рабочая констелляция проходит несколько стадий.
Сначала доминанта в центрах вызывается ее непосредственным раздражителем
под влиянием внутренней секреции, рефлекторными влияниями и пр. На этой
стадии, названной стадией укрепления наличной доминанты по преимуществу, она
привлекает к себе в качестве поводов к возбуждению самые разнообразные внеш¬
ние раздражения. В констелляцию посредством генерализованного возбуждения
наряду с необходимыми для данного акта нервными центрами вовлекаются и по¬
сторонние клеточные группы. В этом достаточно неэкономичном процессе про¬
является диффузная отзывчивость доминирующей констелляции к различным
раздражениям. Но постепенно, в процессе повторного осуществления данного по¬
веденческого акта, диффузная отзывчивость сменяется избирательным реагиро¬
ванием только на те раздражения, которые создали данную доминанту. Подбор
биологически интересных рецепций для доминанты (стадия выработки адекват¬
ного раздражителя для данной доминанты и вместе с тем стадия предметного
выделения данного комплекса раздражителей из среды) приводит к образованию
новых адекватных поводов для той же доминанты. Теперь выполнение доминант¬
ного акта происходит более экономично, ненужные для него нервные группы за¬
тормаживаются. При повторении доминанты репродуцируется и свойственный
только ей единый ритм возбуждения.
Восстановление доминанты по кортикальному следу может быть эскизным, то
есть более экономичным ее воспроизведением через кору. При этом комплекс ор¬
ганов, участвующих в переживании восстановленной доминанты, может быть со¬
кращенным и ограничиться только кортикальным уровнем. Полное или эскизное
возобновление доминанты возможно лишь в том случае, если возобновится хотя
бы частично раздражитель, ставший для нее адекватным. Это обусловлено тем,
что между доминантой как внутренним состоянием и данным комплексом раздра¬
жителей была установлена прочная адекватная связь. На этой стадии доминанта
возобновит подбор новых, «биологически интересных поводов для себя». Поэтому
при кортикальном воспроизведении доминанты, которое является очень подвиж¬
ным сочетанием ее кортикальных компонентов, происходит иитеркортикальное
обогащение новыми рецепциями кортикальных же компонентов доминанты.
Что же представляет собой кортикальный след, по которому доминанта может
быть пережита заново? Это своего рода интегральный образ, своеобразный про-
20 Глава 1. Методология изучения высшей нервной деятельности
дукт пережитой ранее доминанты, в котором в единое целое сплетены соматиче¬
ские и эмотивные признаки доминанты с ее рецептивным содержанием, то есть
с комплексом раздражений, с которым она была связана в прошлом. При создании
интегрального образа важную роль играют как периферические, так и кортикаль¬
ные компоненты. Интегральный образ — это своеобразная памятка пережитой до¬
минанты и вместе с тем ключ к ее воспроизведению с той или иной степенью пол¬
ноты.
Если доминанта восстанавливается по своим кортикальным компонентам, то
есть более экономично, «как мимолетное воспоминание», причем с малой инерци¬
ей, то при новых условиях они всегда обходятся с помощью прежнего опыта. Био¬
логический смысл эскизно протекающих доминант состоит в том, чтобы «...по по¬
воду новых и новых данных среды очень быстро перебрать свой арсенал прежних
опытов для того, чтобы из них путем очень быстрых их сопоставлений избрать бо¬
лее или менее идущую к делу доминанту, чтобы применить ее к новому заданию.
Целесообразность или нецелесообразность выбранной доминанты прошлого ре¬
шает дело»1.
Если же доминанта восстанавливается почти с прежней полнотой, которая
предусматривает оживление работы всей соматической констелляции, то она
приживается с большей инерцией, занимая более или менее продолжительный
период жизнедеятельности организма. Подбирая при этом вновь биологически
интересные для себя раздражения из новой среды, доминанта по новым данным
переинтегрирует старый опыт.
§ 3. Принцип отражения
Отражение в биологических системах. Само понятие отражения достаточно
многогранно и сложно, а потому далеко не очевидно но своей природе. Наиболее
полным выглядит следующее определение: «Отображе¬
ние есть особый продукт действия в определенных усло¬
виях одной материальной системы на другую, воспроиз¬
ведение в преобразованном виде особенностей первой из
систем (отображаемой) в особенностях процессов второй
системы (отображающей)»2.
Итак, отражение — следствие, результат, причина же
находится вовне, она независима от своего следствия. При
этом форма отражения действительности в наших ощу¬
щениях, понятиях, теориях субъективна; следовательно, отображение зависит не
только от отображаемого объекта, но и от природы отображающей системы, ее
текущих состояний, ее истории.
Биологическая форма отражения реализуется благодаря взаимодействию
живой системы с внешней средой ее обитания. Биологическое отражение пред¬
ставляет собой сплав, получающийся от соединения отражаемого и отражающе¬
го. Организмы выработали два основных типа биологического отражения: отно¬
сительно постоянное — статическое, имеющее в качестве своих носителей раз-
1 Ухтомский А. А. Доминанта. Л., 1966. С. 256.
2 Украинцев В. С. О сущности элементарного отображения // Вопросы философии. 1960. № 2. С. 66.
Принцип отражения
представляет собой ор¬
ганическое единство
непосредственного
восприятия и следов
прошлых впечатлений,
объективного и субъек¬
тивного, формы и со¬
держания.
§ 3. Принцип отражения 21
личные морфологические образования, и динамическое, носителем которого
является психическое. Последнее также бывает двух видов: кратковременные
приспособительные реакции и реакции долговременного
приспособления.
Проблема адекватности отражения, то есть степень со¬
отнесения наших представлений с реально существующим
миром, — одна из самых древних и вместе с тем вечно но¬
вых и остро обсуждаемых проблем. От ее решения зави¬
сит сущность нашего отношения к окружающей действи¬
тельности. Прежде всего, что вкладывать в понятие адек¬
ватности образа? «Познание, — пишет В. И. Ленин, — есть отражение человеком
природы. Но это не простое, не непосредственное, не цельное отражение, а процесс
ряда абстракций, формирования, образования понятий, законов»1.
Говоря об адекватности, следует отбросить вульгарно-материалистическое по¬
нимание образа, под которым подразумевается формирование в мозге точной фо¬
тографической копии воспринимаемого объекта.
Контакт с внешним миром, воздействие его на организм возможны благодаря
высокоспециализированным аппаратам нервной системы — рецепторам. Совре¬
менная физиология располагает достаточно четкими све¬
дениями о работе органов рецепции и высших уровней
мозга, которые служат убедительной основой для постули¬
рования положения об адекватности отражения (А. С. Ба¬
туев, Г. А. Куликов, 1983).
Образный характер отражения. Под понятием «об¬
раз» мыслится такой результат познания субъектом объ¬
екта, упорядоченность элементов которого в принципе
соответствует упорядоченности свойств, связей и отно¬
шений объекта (оригинала), причем эта упорядочен¬
ность (структура) функционально отделена субъектом
от субстрата образа и непосредственно для носителя су¬
ществует в субъективно-идеальной форме.
Живой организм поддерживает состояние устойчи¬
вого неравновесия, сохраняя ряд своих параметров в огра¬
ниченных диапазонах, то есть обеспечивая гомеостаз.
Все поведенческие акты направлены в конечном сче¬
те на обеспечение относительного константного состояния организма в непрерыв¬
но меняющейся среде благодаря поступлению информации как из внешнего про¬
странства, так и из внутренней сферы самого организма.
Однако прежде чем произойдет та или иная приспособительная реакция, осу¬
ществляется процесс восприятия сигналов. Сигнал — это некая величина, от¬
ражающая определенным образом состояние физической системы. Но живой ор¬
ганизм заинтересован не в самих воздействиях как таковых, а в том, о чем они сиг¬
нализируют, и соответственно не в оценке их физических параметров, а в тех
соотношениях, которые с их помощью передаются. Вместе с тем понятно, что от¬
несение сигнала к тому или иному классу, то есть его распознавание, неизбежно
1 Ленин В. И. Поли. собр. соч. 1968. Т. 29. С. 163-164.
Образ — это такой ре¬
зультат познания субъ¬
ектом объекта, упоря¬
доченность элементов
которого в принципе
соответствует упорядо¬
ченности свойств, свя¬
зей и отношений объек¬
та (оригинала), причем
эта упорядоченность
(структура) функцио¬
нально отделена субъ¬
ектом от субстрата об¬
раза и непосредственно
для носителя существу¬
ет в субъективно иде¬
альной форме.
Отображение зависит
не только от отобра¬
жаемого объекта, но
и от природы отобра¬
жающей системы, ее
текущих состояний,
ее истории.
22 Глава 1. Методология изучения высшей нервной деятельности
Сигнал — это некая ве¬
личина, отражающая
определенным образом
состояние физической
системы.
Знак — это такой ком¬
понент психического
отражения, который не
имеет познавательно¬
образного сходства с
соответствующим ему
элементом или свойст¬
вом объекта оригинала.
основано на выделении ряда физических характеристик,
образующих область признаков данного класса сигналов.
Всякое чувственное отображение базируется па взаи¬
моотношении в нем знаковых и образных моментов. В от¬
личие от образа знак — это такой компонент психического
отражения, который не имеет познавательно-образного сходства с соответству¬
ющим ему элементом или свойством объекта-оригинала. Если образ воспроизво¬
дит структуру оригинала с определенной степенью адекватности, то знаки такой
структуры не воспроизводят. Но они воспроизводят в са¬
мом субъекте образы объектов-оригиналов, возникших
в результате предшествовавших актов отражения. Про¬
стейшим примером естественных знаков являются мо¬
дальности ощущений, из которых в результате движений
и предметных действий формируются образы объектов,
отображающие их структуру.
Основная специфика высших форм отражения, наи¬
более прогрессивно совершенствующаяся в ходе эволю¬
ции, — извлекать информацию для самих себя и в то же время абстрагироваться от
материального носителя этой информации. В отражающей системе мозга в ре¬
зультате взаимодействия с отражаемым объектом извлекается упорядоченность,
соответствующая источнику отражения, а ее материальный носитель функцио¬
нально исключается. Эта сторона отражения выражает его активный, творческий
характер.
В каждый момент времени извлекается не вся информация, а лишь та, которая
необходима для отражающей системы, ибо в процессе отражения последняя меня¬
ется, испытывает преобразования внутри себя. Эти изменения, соответствующие
какой-либо стороне воспринятого объекта, функциональ¬
но выделяются и используются отражающей системой в
качестве фактора самоуправления для сохранения ее ка¬
чественной определенности. Поэтому построение образа
определяется не субстанциональными, а функциональны¬
ми параметрами на основе извлечения информации как
неотъемлемого свойства отражаемого объекта. Сигналь¬
ный характер биологических форм отражения заключа¬
ется в том, что организм извлекает информацию в соответствии как с наследст¬
венно фиксированной программой, так и с доминирующими в данный момент по¬
требностями.
Избирательность (активность) отражения. В избирательности биологическо¬
го отражения полнее всего проявляется целесообразный характер реакций орга¬
низма, его действий, его поведения. При длительном многократном воздействии
фактора внешней среды в организме возникает качественно новое явление — оста¬
ется определенный след. Следовые процессы долговременной памяти — важнейший
компонент в системе приспособительных реакций организма.
Хотя отражение всегда вторично по отношению к своему оригиналу, П. К. Ано¬
хин (1968) выдвинул идею о существовании в животном мире «опережающего
отражения». Логика гипотезы П. К. Анохина в том, что в долговременную память
Высшие формы отра¬
жения извлекают ин¬
формацию для самих
себя и в то же время
абстрагируются от ма
териального носителя
этой информации.
§ 3. Принцип отражения 23
записываются только неоднократно повторяющиеся события. Та последователь¬
ность воздействий внешних событий, которая наблюдалась ранее, может с доста¬
точной полнотой воспроизводиться в памяти, когда вос¬
приняты еще только сигнальные признаки начальных
звеньев данной последовательности. Тогда отображение
одного из таких событий, сопоставляемое с прошлыми
отображениями других, приобретает характер отображе¬
ния не единичных объектов, а целого класса объектов.
Учитывая непременное участие прошлого жизненного
опыта в любых актах биологического отражения и при
формировании целей, планов, программ, следует признать опережающий харак¬
тер обязательным свойством отражательной деятельности мозга, которая испы¬
тывает существенные преобразования в эволюции живых существ.
Согласно распространенному представлению, образ-след записывается в дол¬
говременную память в том случае, если в прошлом вероятность встречи с этим
объектом была высокой. Использование же образов-сле¬
дов для опережающего отражения действительности, то
есть будущих событий и их последствий, с неизбежно¬
стью должно носить вероятностный характер при опери¬
ровании субъекта в непрерывно меняющейся внешней
среде. Поэтому чтобы сохранить свое приспособительное
значение, образы-следы, соответствующие элементам про¬
шлого жизненного опыта, должны характеризоваться избыточностью по отноше¬
нию к вызвавшему их объекту.
Рассматривая проблему сходства образа и отражаемого объекта, необходимо
Образ-след записыва¬
ется в долговременную
память, если в про¬
шлом вероятность
встречи с этим объек¬
том была высокой.
Избирательность био¬
логического отражения
полнее всего обеспе¬
чивает целесообраз¬
ный характер реакций
организма, его дейст¬
вий, его поведения.
учитывать два оостоятельства: во-первых, та или иная степень адекватности до¬
стигается во времени не мгновенно, а постепенно; во-вторых, окончательная оценка
адекватности производится при соотнесении нового образа с его нервной моделью,
созданной ранее на основе всего комплекса воздействий и прежнего жизненного
опыта, то есть долговременной памяти.
Иначе говоря, критерием адекватности является
опыт, практика, которые фиксируются в виде следов
и в дальнейшем служат эталонами для оценки новой
информации. «В мозгу человека отражается природа.
Проверяя и применяя в практике своей и в технике
правильность этих отражений, человек приходит к объ¬
ективной истине»1. И далее: «...практика человека и че¬
ловечества есть проверка, критерий объективности по¬
знания»1 2.
Сегодня с достоверностью установлено, что разные
внешние причины могут вызывать ощущения одной и
той же модальности, и одна и та же внешняя причина в
различных чувствительных органах производит ощу¬
щения различных модальностей.
П. К. Анохин
1 Ленин В. И. Поли. собр. соч. Т. 29. С. 183.
2 Там же. С. 193.
24 Глава 1. Методология изучения высшей нервной деятельности
Впервые на факт независимости модальности ощущений от природы раздра¬
жителя обратил внимание И. Мюллер, который сформулировал закон так назы¬
ваемой специфической энергии. Абсолютизация этого за¬
кона привела к представлениям о том, что все ощущения
зависят только от природы чувствующего аппарата, а не
от объективной реальности, которая таким образом объ¬
являлась непознаваемой. Именно на такой позиции сто¬
ял ученик И. Мюллера Герман Гельмгольц, который раз¬
работал теорию «иероглифов». Согласно последней, в ощу¬
щении и восприятии признаются лишь знаки, сигналы,
а поэтому ощущения якобы существуют параллельно
и независимо от внешнего мира.
В конце Х1Х-пачале XX в. группа исследователей из
Германии организовала научную школу гештальтпсихо¬
логов (гештальт, нем. — форма, структура), которые про¬
возгласили новую психологическую доктрину: в сознании образуются целостные
образы (гештальты), не разложимые на сенсорные первоэлементы. Гештальтам
присущи собственные характеристики и законы. Гештальтисты проповедовали
принцип изначальной упорядоченности сенсорно-интеллектуальных структур и
их динамических преобразований. Они утверждали, что образ надо изучать как
самостоятельный феномен, а не как простой эффект стимула, а потому физиоло¬
гия изолированных нервных элементов и путей должна быть заменена на физио¬
логию целостных и динамических структур-гештальтов, образование которых при
решении задач подчиняется механизмам инсайта — внезапного схватывания от¬
ношений.
Наряду с рациональным элементом этой теории, интерес к которой в связи
с изучением таких психофизиологических феноменов, как образная память и им¬
принтинг, существенно возрос, необходимо иметь в виду следующее. Образ у ор¬
тодоксальных гештальтистов выступал в виде сущности особого рода, подчинен¬
ной собственным внутренним законам и отделенной от действия. Такие методоло¬
гические предпосылки, отъединявшие сложные психофизиологические процессы
от реальной действительности, повлекли за собой критику гештальтизма.
Субъективный характер отражения. Достижения современной эволюцион¬
ной физиологии с убедительностью свидетельствуют, что наибольшей точностью
и полнотой отражения обладают высшие животные и человек с его способностью
к абстрактному мышлению. Возникает вопрос: как с физиологических позиций
оценить субъективный характер образа? Преобразование информации в какой-
либо сенсорной системе зависит не только от ее свойств и функционального со¬
стояния, но практически от всех влияний, воспринимаемых мозгом и запечатлен¬
ных в памяти. Они накладывают свой отпечаток па характер работы сенсорной
системы, меняют ее избирательность, настройку, подвижность благодаря участию
обратных связей и систем межсенсорной интеграции.
Пространственно-временная структура объекта кодируется в мозге в форме
определенной нервной модели, изоморфной внешнему воздействию. Нервная мо¬
дель есть не что иное, как физиологические основы формирующихся субъектив¬
ных образов. Последние не сводятся к нервным моделям, между субъективным
образом и нервной моделью имеется как принципиальное различие, так и опреде-
Степень адекватности
достигается во времени
не мгновенно, а посте¬
пенно; окончательная
оценка адекватности
производится при соот¬
несении нового образа
с его нервной моделью,
созданной ранее на ос¬
нове всего комплекса
воздействий и прежнего
жизненного опыта, то
есть долговременной
памяти.
§ 3. Принцип отражения 25
лепное соответствие. Образ не существует в мозге объективно в виде некоторой
уменьшенной материальной копии внешнего предмета. В то же время мозговая
нейродинамическая система существует в качестве объективной реальности. Но
она не может быть названа образом, так как не обладает предметным характером.
Она является кодом отображаемого внешнего объекта.
В нервной модели, не связанной с материальным носителем информации, про¬
изводится акт абстрагирования от конкретного объекта. Субъективный образ
воспроизводит не качество нервных процессов, состоя¬
ние рецепторов или нейронов мозга, а особенности отра¬
жаемых явлений. Иными словами, субъективный образ
возникает на базе нервных моделей при декодировании
информации и соотнесении ее с реально существующим
материальным объектом.
Этот этап отражения, получивший название декоди¬
рования, является довольно сложной и далеко не очевид¬
ной операцией в динамической структуре познания мира.
В содержании образа собственная качественная опреде¬
ленность объекта не угасает, как это имеет место в нервной модели, а демаскирует¬
ся. Субъективный образ предмета не является неизменным, статичным. Это суще¬
ствует только в абстракции, в действительном восприятии сразу же обнаружива¬
ется необходимо присущая образу динамика.
До сих пор одной из нерешенных задач остается объяснение того, как соверша¬
ется превращение материальных нейродинамических процессов в идеальное,
в субъективный образ. Справедливо замечание о том, что пытаться объяснить
идеальное анатомо-физиологическими свойствами мозга — такая же наивная за¬
тея, как объяснить денежную форму продукта труда фи¬
зико-химическими свойствами золота (Э. С. Ильенков,
1962). Только исходя из структуры акта взаимодействия,
рефлекторного по своему механизму, можно раскрыть
сложную динамическую связь между предметом, его об¬
разом и материальной основой последнего, ибо содержа¬
ние внешнего мира отражается субъектом не в формах
деятельности мозга, а в формах деятельности субъекта,
которые протекают в идеальном клаве.
Рефлекторный характер отражения. Логика приво¬
дит к необходимости анализировать различные сложные
формы рефлекторной деятельности организма. Принци¬
пиальной сущностью условно-рефлекторного акта явля¬
ется то, что он базируется па механизмах, в которых заложены не только элементы
прошлого и настоящего, но и элементы прогнозирования будущего. В этом его
главный приспособительный смысл, то есть сигнальный характер. Через сложно¬
рефлекторную деятельность организма наиболее полно проявляется субъектив¬
ный, идеальный характер отражения. При этом нельзя не учитывать, что услов¬
ный рефлекс — это лишь «частный и особый пример среди аппаратов, которыми
совершается в человеке отражение и отраженная действительность»1.
1 Ухтомский А. А. Доминанта. С. 227.
Принципиальная сущ¬
ность условно-рефлек¬
торного акта в том, что
он базируется на меха¬
низмах, в которых зало¬
жены не только элемен¬
ты прошлого и настоя¬
щего, но и элементы
прогнозирования буду¬
щего. В этом его глав¬
ный приспособитель¬
ный смысл, то есть сиг¬
нальный характер.
Пространственно-вре¬
менная структура объ¬
екта кодируется в моз¬
ге в форме определен¬
ной нервной модели,
изоморфной внешнему
воздействию. Она яв¬
ляется кодом отобра¬
жаемого внешнего объ¬
екта.
26 Глава 1. Методология изучения высшей нервной деятельности
В связи с интенсивным развитием таких дисциплин, как бионика и нейроки¬
бернетика, перед теорией отражения встают новые задачи, например изучение
отражательных свойств технических систем связи и управления, то есть искусст¬
венно создаваемых человеком устройств. Необходимо изучение соизмеримости
отражательных свойств человека и технических устройств в сложных системах
«человек—машина» для усиления интеллектуальных и практических возможно¬
стей человека.
§ 4. Принцип системности в работе мозга
Развитие проблемы «мозг и психика». Еще до нашей эры высказывались догад¬
ки о локализации психических функций в головном мозге (Гиппократ, Кротон, Га¬
лен). Не будем рассматривать средневековую историю, но лишь упомянем имя
Альберта Магнуса, предложившего концепцию о локализации психической дея¬
тельности в трех мозговых желудочках. Т. Виллис уточнил эту теорию, считая, что
общая чувствительность представлена в полосатом теле, собственные чувства —
в мозолистом теле, а память — в коре больших полушарий. Более 100 лет эта кон¬
цепция увлекала научный мир, пока оппозиция А. Галлера не отодвинула ее па
второй план. Тем не менее идеи о локализации психических свойств продолжали
существовать. Немецкий анатом М. Майер предположил, что в коре головного
мозга локализована память, в белом веществе — воображения и суждения, а в ба¬
зальных ганглиях — апперцепция и воля и все это интегрируется мозолистым те¬
лом и мозжечком.
Крупнейший австрийский анатом начала XIX в. Ф. Галль и его ученики выдви¬
нули еще более крайние френологические идеи и полагали, что умственные и мо¬
ральные качества локализуются в определенных районах поверхности мозга, при¬
чем степень развития той или иной психической способности определяется объ¬
емом ее представительства в головном мозге. Эта система Ф. Галля, хотя и
получила очень широкое распространение, была одной из самых больших ошибок
в развитии теории мозга, так как исходила из доктрины, а не основывалась на точ¬
ных наблюдениях, и стремилась жестко привязать те или иные психические явле¬
ния к определенным частям мозга. Такое направление именовалось также психо¬
морфологией или узким локализационизмом.
Фантастические взгляды френологов подвергались резкой критике со стороны
группы ученых, которые встали на позиции антилокализационных представле¬
ний (А. Галлер, Ф. Гольц, Ж.-П. Флуранс). Экспериментируя на низших живот¬
ных, они обнаружили, что та или иная форма поведения страдает не столько от то¬
го, в каком месте мозга нанесено повреждение, а от того, какой объем ткани извле¬
чен в результате операции. Поэтому эти ученые утверждали, что у всех животных
масса мозговых полушарий равноценна и однородна. Так возникла догма об экви¬
потенциальности частей мозга, а затем появилась теория универсализма, или хо¬
лизма.
Если локализационисты обращали внимание на неоднозначность мозгового
обеспечения психических функций и их прямой зависимости от работы конкрет¬
ного субстрата мозга, то эквинотенциалисты подчеркивали высокую пластич¬
ность мозга и взаимозамещаемость его частей — иными словами, динамизм —
§ 4. Принцип системности в работе мозга 27
в организации мозговых структур. Развитием этих идей явились две концепции.
Согласно первой, предложенной С. Монаковым, утрата части мозга ведет к на¬
рушению функций оставшихся мозговых структур (синдром дефекта), которые
динамически меняются во времени. Согласно второй концепции пластичности,
разработанной А. Бете (1899), любой центр, будучи поврежден, способен к функ¬
циональной реорганизации без дополнительной тренировки и обучения. Естест¬
венно, что такие взгляды затрудняли однозначное объяснение последствий ло¬
кальных поражений мозга.
Переход от умозрительных рассуждений к науке связан с именем X. Джексона
(1932) — клинициста, положившего начало соотнесению клинических симпто¬
мов с очаговыми процессами мозга. Он построил свою иерархическую систему
трех уровней функционирования мозга: нижний — уровень стабильных функций,
средний — сеисомоторный уровень и наивысший — уровень функций мышления,
присущий человеку. В обеспечении двигательного поведения эти уровни органи¬
зованы вертикально друг над другом. Джексон постулировал различные размеры
моторного и сенсорного представительства различных частей тела в зависимости
от степени их специализации и дифференциации. Скажем, область представи¬
тельства руки человека занимает в двигательной и соматосенсорной зоне коры
больших полушарий значительно большее пространство, чем все другие части
скелетно-мышечного аппарата. Джексон выдвинул правило: мозговая локализа¬
ция дефекта поведения не дает оснований для постулирования локализации пси¬
хической функции. Однако его коллеги-хирурги были менее осторожны и позво¬
лили своему воображению увести себя так далеко, что приписывали точную ло¬
кализацию в мозге умственным, эмоциональным, психическим и двигательным
функциям.
Особое внимание клиницистов было обращено на локализацию речевой функ¬
ции после открытия П. Брока «центра моторных образов слов» и находки С. Вер¬
нике аналогичного «центра сенсорных образов слов». Хотя С. Моиаков критиковал
эти попытки локализовать речь лишь па основании ее расстройств при ограничен¬
ных поражениях мозга, эти данные послужили дополнительным укреплением по¬
зиций локализационистов.
Вторая половина XIX в. ознаменовалась двумя крупнейшими событиями. Во-
первых, опытами Г. Фритча и Е. Гитцига в 1863 г. было установлено наличие «дви¬
гательных центров» в коре больших полушарий, электрическое раздражение
которых вызывает сокращение скелетной мускулатуры, а их разрушение сопровож¬
дается глубокими расстройствами движений. Во-вторых, киевский анатом В. А. Бец
в 1874 г. описал в коре передней центральной извилины мозга человека гигант¬
ские пирамидные клетки (клетки Беца), которые он связал с проведением двига¬
тельных команд в спинной мозг, а значит, с выполнением моторной функции. Эти
открытия повлекли за собой целую серию исследований: морфологических, пока¬
завших структурную неоднородность коры больших полушарий и заложивших
основы учения о цитоархитектонике мозга, и физиологических — методами уда¬
ления и раздражения.
Учение о функциональной организации мозга явилось стержневой проблемой
физиологии и психологии и развивалось в непрерывных и непримиримых проти¬
воречиях, отражающих и формирующих господствующее мировоззрение и тесно
28 Глава 1. Методология изучения высшей нервной деятельности
увязанных с общим уровнем» физиологии и смежных
наук. Одной из причин возникновения таких противо¬
речивых теорий являлось отсутствие эволюционного
подхода к изучению мозга и поведения и стремление
к широкому распространению отдельных частных за¬
ключений, выведенных на основе наблюдений в чело¬
веческой клинике (локализационисты) или на основе
экспериментов с низшими животными (эквипотен-
циалисты), на весь животный мир. Это придавало тео¬
ретическим построениям весьма однобокий характер.
И. М. Сеченов (1947) и его последователи, заложив¬
шие основы эволюционной физиологии мозга, убеди¬
в. м. Бехтерев телыю показали те преимущества, которые дает иссле¬
дователю применение исторического метода к изучению
мозга. Другой причиной непрекращающихся дискуссий, о которой подробно речь
будет идти ниже, является использование заведомо неверных методологических
предпосылок о строгой увязке отдельных частей субстрата мозга с теми или ины¬
ми целостными функциями организма.
Особенно жаркие споры с психоморфологами развернулись вокруг так назы¬
ваемых немых (а также гностических, ассоциативных, третичных) полей коры.
Еще в работах Г. Фритча и Е. Гитцига было обнаружено, что расположенные спе¬
реди от двигательной зоны мозга лобные области оказались электроневозбудимы¬
ми, то есть их стимуляция не сопровождалась сокращением скелетной мускулату¬
ры. Но в то же время разрушение лобных областей, заметно не отражаясь ни на
двигательной, ни на чувствительной сферах, приводит к глубоким расстройствам
доведения в целом. Клиницисты того времени пользовались терминологией гос¬
подствовавшей тогда ассоциативной психологии. Отсюда и появились представ¬
ления о лобных долях как об «органе абстрактного мышления», «активного вни¬
мания», «апперцепции», как о «субстрате регулирующего разума», «верховного
органа мозга» и пр.
Немецкий ученый Л. Эдингер (1904) рассматривал лобные доли как огромные
ассоциативные центры. П. Флексиг (1883) выделял в мозгу три ассоциативные
зоны: теменную, среднюю (в районе островка) и лобную, в которой возникают по¬
нятия о собственной личности, где создаются представления о «Я». И поэтому па¬
тологические процессы в области этого центра приводят к изменениям личности,
к возникновению бредовых идей величия или самоуничижения.
Близкие идеи разрабатывались итальянским ученым Л. Бианчи (1921), кото¬
рый считал, что лобные доли обеспечивают наиболее широкую координацию чув¬
ствительных и двигательных элементов, «используя продукт сенсорных зон для
психических синтезов». После разрушения лобных долей поведение распадается
на ряд фрагментов, не подчиненных единому синтезу, и перестает приспособлять¬
ся к новым условиям.
Систематический анализ богатого собственного экспериментального и клини¬
ческого материала был проведен В. М. Бехтеревым. Обсуждая результаты опытов
с лобэктомированиыми собаками, он писал: «Дело идет об утрате их высших по¬
знавательных способностей, которые устанавливают определенное отношение
между отпечатками новых внешних впечатлений и результатами прошлого опыта
§ 4. Принцип системности в работе мозга 29
и которые направляют движения и действия сообразно личной пользе»1. В совре¬
менной ему терминологии В. М. Бехтерев определяет лобные доли «как место раз¬
вития индивидуального ядра психической сферы, вследствие отложения здесь по¬
следовательных следов от внутренних раздражений». Кроме того, он отмечает, что
«вместе с этим лобные доли служат областями психорегуляторной деятельности,
обусловливающей развитие высших познавательных функций, выражающихся
правильной оценкой внешних впечатлений и выбором движений сообразно с упо¬
мянутой оценкой»1 2.
И. П. Павлов активно выступал не только против локализационизма, но и про¬
тив эквипотенциализма и сформулировал оригинальную концепцию о динамиче¬
ской локализации функций. Выступая с критикой локализационистских взглядов,
И. П. Павлов изложил свое отношение к этим идеям на примере «дыхательного
центра». Он писал: «С самого начала думали, что это — точка с булавочную голов¬
ку в продолговатом мозге. Но теперь он чрезвычайно расползся, поднялся в голов¬
ной мозг и опустился в спинной, и сейчас его границы точно никто не укажет»3.
Несмотря на критику, локализационисты не уступали своих позиций, в осо¬
бенности когда речь шла о сложных психических функциях, которые они стреми¬
лись прочно увязать с определенными морфологически¬
ми субстратами, минуя промежуточные сложные физио¬
логические процессы.
Функциональная организация мозга. Русский нейро¬
морфолог И. Н. Филимонов (1949) выдвинул принцип,
согласно которому вместо статических центров предпола¬
гается наличие одновременной и последовательной по¬
этапной локализации, и поэтому функция по самой своей
природе не может быть связана с каким-то единичным
центром. В дальнейшем была выдвинута идея о функцио¬
нальной многозначности мозговых структур, которые могут включаться в различ¬
ные функциональные системы и принимать участие в осуществлении различных
задач. В коре мозга пет узкоспециализированных зон, а каждая скорее всего пред¬
ставляет собой высший «корреляционный центр». Типичный пример таких цент¬
ров — задний (теменной) и передний (лобный) ассоциативные полюса неокор-
текса.
Специального рассмотрения заслуживают идеи А. А. Ухтомского относитель¬
но организации церебральных функций в соответствии с разработанным им уче¬
нием о нервном центре как своеобразном функциональном органе, состоящем из
обширного числа нервных аппаратов, вступающих между собой во временное це¬
ленаправленное сотрудничество и образующих физиологическую констелляцию.
«Понятие физиологической констелляции содержит в себе... не только топо¬
графический факт, что “нервный центр” может опираться на морфологически до¬
вольно широко расставленные участки нервной системы, но также физиологиче¬
ский факт, что отдельные компоненты центра могут приобретать другое функцио¬
нальное значение по связи с другими констелляциями и при участии в других
работах. А эти переходы в другие рабочие группировки и констелляции определя-
Фуншиональный ор¬
ган — это обширное
число нервных аппа¬
ратов, вступающих ме¬
жду собой во времен¬
ное целенаправленное
сотрудничество и об¬
разующих физиологи¬
ческую констелляцию.
1 Бехтерев В. М. Основы учения о функциях мозга. СПб., 1907. Т. 6-7. С. 14-64.
2 Там же. Т. 5-7. С. 14-66.
3 Павлов И. П. Собр. соч. М.; Л., 1951. Т. 3. С. 172.
30 Глава 1. Методология изучения высшей нервной деятельности
ются для центрального участка тем, насколько он способен сдвигать свою лабиль¬
ность в соответствии с темпами и ритмами возбуждения в очередной констелля¬
ции»1. Нервный центр как функциональный орган определяется следующим об¬
разом: «“Центр” определенного сложного отправления — это целое созвездие
созвучно работающих ганглиозных участков, взаимно совозбуждающих друг дру¬
га... Увязка во времени, в скоростях, в ритмах действия, а значит и в сроках выпол¬
нения отдельных компонентов реакции, впервые образует из пространственно
разных групп функционально объединенный “центр”»1 2.
Понятие о функциональном органе для современной физиологии чрезвычай¬
но ценно, ибо позволяет с новых методологических позиций рассматривать соот¬
ношение специализированных структур мозга и функций, основываясь на прин¬
ципе системной работы мозга. «Орган, — писал А. А. Ухтомский, — это прежде все¬
го механизм с определенным однозначным действием. Громадное многообразие
морфологических черт, которое мы открываем в том или ином образовании наше¬
го тела, впервые приобретает значение физиологического органа после того, как
открывается значение этих морфологических деталей как рабочих слагающих об¬
щей однозначно определенной физиологической равнодействующей. Значит,
именно физиологическая равнодействующая дает комплексу тканей значение ор¬
гана как механизма»3.
Близкие взгляды содержатся в концепции П. К. Анохина (1968) о «функцио¬
нальной системе», которая обеспечивается компонентами взаимосогласованных
актов, увязанных ради достижения запрограммированного биологического резуль¬
тата. Системное представление о высших функциях мозга позволило П. К. Анохи¬
ну по-новому рассматривать ряд неврологических синдромов. Например, так на¬
зываемый лобный синдром трактовался как распад интегративной деятельности
мозга в результате нарушения аппаратов «афферентного синтеза». Основными
звеньями последнего являются:
♦ обстановочная аффереитация;
♦ следы, записанные в памяти;
♦ пусковая аффереитация;
♦ мотивационная аффереитация.
Лобные отделы мозга связывались с геми физиологическими функциями, ко¬
торые определяют организацию двигательного поведения в обстановке, требую¬
щей выбора путем сличения «обратной афферептации» с «акцептором действия».
Так была предпринята первая попытка провести физиологический анализ уча¬
стия лобных долей мозга в организации целенаправленного поведения.
Таким образом, складывалась ситуация, в которой на смену двум крайним тео¬
ретическим представлениям (локализациопному и эквипотенциализму) выдвига¬
лись все более обоснованные концепции о системном принципе функционирования
с пересмотром старых представлений о функции и локализации.
Значительную роль в развитии теории локализации следует отвести А. Р. Лу-
рии (1973). Прежде всего А. Р. Лурия анализирует метод локальных поражений
мозга и приходит к выводу, что поражение каждой из корковых зон может привес -
1 Ухтомский А. А. Собр. соч. Л„ 1961. Т. 1. С. 97.
2 Там же. 1954. Т. 4. С. 102.
3 Ухтомский А. А. Доминанта. С. 79.
§ 4. Приниип системности в работе мозга 31
ти к распаду всей функциональной системы и «симптом» (нарушение или выпа¬
дение той или иной функции) еще ничего не говорит о ее локализации. Анализ
клинических случаев средствами нейропсихологии позволил А. Р. Лурии выдви¬
нуть концепцию о существовании трех основных функциональных блоков мозга.
Первый блок — регуляции тонуса и бодрствования, важнейшей его частью яв¬
ляются ретикулярная формация среднего мозга, неспецифическая система та¬
ламуса, гиппокамп и хвостатое ядро, работающие под мощным контролем коры
больших полушарий. Правда, многие из современных исследователей отказыва¬
ются от рассмотрения ретикулярной формации как неспецифической активирую¬
щей или тормозной системы, а скорее склонны видеть в ней структурные предпо¬
сылки для выполнения сложных интегративных процессов посредством локаль¬
ных и разнонаправленных влияний на кору и глубокие структуры мозга. Вместе с
этим критически отвергнута и другая крайняя позиция о так называемой центрэн¬
цефалической системе (У. Пенфилд, Г. Джаспер, Н. Иошии, X. Гасто), в которой
психические процессы локализуются в ретикулярной формации мозга.
Второй блок назван блоком приема, переработки и хранения информации. Сю¬
да включают все задние отделы коры, в том числе зрительную, слуховую и сома¬
тическую сенсорные области, и межпроекционную теменную область (задний ас¬
социативный полюс). Для конструкции этого блока характерны три основных за¬
кона:
♦ закон иерархического построения корковых зон, каждая из которых подразде¬
ляется на первичные, вторичные и третичные корковые поля;
♦ закон убывающей специфичности иерархически построенных зон коры, среди
которых наиболее модально специфичными являются первичные поля;
♦ закон прогрессивной асимметрии функций, который вступает в силу только на
уровне «вторичных» и «третичных» зон.
Таким образом, убывающая модальная специфичность сопровождается воз¬
растанием функциональной асимметрии. Важнейшим компонентом этого блока
являются зоны сенсорного перекрытия (теменные или теменно-височно-затылоч¬
ные), участвующие в формировании комплексных надмодальных актов высшей
нервной деятельности, и прежде всего наглядных и символических пространст¬
венных синтезов.
Третий блок назван блоком программирования, регуляции и контроля сложных
форм деятельности. В его состав входят префронтальные (лобные) отделы коры
мозга. Их нарушения сказываются в наиболее высоком уровне регуляции психи¬
ческих процессов с помощью системы речевых связей. Одним из наиболее суще¬
ственных признаков лобных поражений является нарушение сигнальной регули¬
рующей функции речи. По этому поводу А. Р. Лурия пишет: «Роль лобных отде¬
лов в синтезе предпусковых афферентаций, в формировании двигательных задач,
в сличении эффекта действия с исходными намерениями — не подлежит сомне¬
нию»1. Относя задние и передние ассоциативные корковые поля к разным функ¬
циональным блокам, А. Р. Лурия подчеркивал, что лобные доли выполняют гораз¬
до более универсальную функцию общей регуляции поведения, чем та, которую
имеет «задний ассоциативный центр» (теменная доля).
1 Лурия А. Р. Высшие корковые функции человека. М., 1962. С. 37.
32 Глава 1. Методология изучения высшей нервной деятельности
Итак, в проблеме локализации функций мозга среди прочих остается нерешен¬
ным следующее противоречие: с одной стороны — локальные поражения мозга
приводят к стабильному выпадению тех или иных физиологических функций, а с
другой — признание наличия подвижной констелляции мозговых аппаратов с по¬
лифункциональным значением каждого из них. Проще всего следует допустить
сосуществование обоих механизмов работы мозга. Потребность решать эту про¬
блему диктовалась также и следующим обстоятельством. Хорошо известно, на¬
сколько уменьшается пластичность мозговых структур с возрастом: одно и то же
по локализации и объему повреждение мозговой ткани может быть незамеченным
в раннем возрасте, но приводит к необратимым пекомненсируемым дефектам, ес¬
ли травма нанесена взрослому организму.
С одной стороны, любая даже самая простая психическая функция есть резуль¬
тат интегративной деятельности целого мозга, с другой стороны, весь опыт невро¬
логической и нейрохирургической клиники свидетельствует о том, что поврежде¬
ние целого ряда мозговых зон влечет за собой необратимый некомиенсируемый
дефект функции.
Разрешение создавшегося противоречия содержится в концепции Н. П. Бехте¬
ревой (1980) о жестких и гибких звеньях мозгового обеспечения психической дея¬
тельности. К жестким звеньям относится генетически де¬
терминированная анатомическая организация тех единиц,
которые необходимы для сохранения условий существо¬
вания вида (например, регуляция вегетативной и мышеч¬
ной систем), а также для ориентации индивида в среде
(проведение и элементарный анализ сигналов внешнего и
внутреннего мира и реализация ответов на них). Иными словами, жесткие звенья
составляют скелет системы, определяющий самое ее существование и обеспечи¬
вающий экономичность работы мозга.
Принципиально важными являются два свойства мозга:
♦ минимизация территорий мозга, необходимых и достаточных для обеспечения
сформированной и стабильно реализующейся функции;
♦ закрепление, фиксация функций в мозге в матрице долгосрочной памяти.
Имеющая место в раннем онтогенезе избыточность структурного состава моз¬
говых систем обеспечения деятельности уступает закрепленной в матрице долго¬
временной памяти минимизации функций, которая высвобождает мозг для все
более сложных видов деятельности. Например, двигательные функции обеспечи¬
ваются минимизированной структурно-функциональной матрицей, закреплен¬
ной в долгосрочной памяти. Эти матрицы зафиксированы и, с одной стороны,
поддерживают деятельность, с другой — ограничивают возможность использова¬
ния новых территорий мозга даже в случае необходимости. Поэтому поражение
таких жестких звеньев системы, закрепленных в матрице долгосрочной памяти,
может приводить к поломке функции и ее пекомпепсируемости.
Наряду с этим существуют гибкие звенья полифункциональных и полибиохи¬
мических аппаратов мозга, утрата которых может быть легко возмещена. Эти гиб¬
кие звенья мозговых систем (не участвующие в фиксированной матрице долго¬
срочной памяти) создают все то богатство возможностей, которые обеспечивают
Даже самая простая
психическая функиия
есть результат интегра¬
тивной деятельности
целостного мозга.
§ 5. Основные методы нейрофизиологии поведения. Методология, метод, методика 33
организму выполнение адаптивной деятельности в самых различных условиях су¬
ществования.
Отсутствие четких определений, что такое «функция» и «локализация», и гра¬
ниц их применения привело к неоправданно широкому их использованию — вплоть
до концепций механистического психоморфологизма. На¬
стало время отказаться от представлений о «локализации
функций». Более правильным будет использование по¬
нятия «локализация органа или системы органов». Тер¬
мин «локализация», явно имеющий анатомический смысл,
должен отражать и соответствующее содержание. Ины¬
ми словами, его следует употреблять лишь в тех случаях,
когда мы определяем строго анатомические отношения
между периферическими органами (сенсорными, сома¬
тическими, висцеральными) и теми отделами централь¬
ной нервной системы, с которыми данные органы связа¬
ны афферентно-эфферентными путями. Тогда можно сказать, что орган, то есть
его деятельность, управляется по принципу саморегуляции тем или иным мозго¬
вым аппаратом, где данный орган локализован. Этот принцип органной локализа¬
ции четко представлен в спинном мозх'е и в стволовом отделе головного мозга. Но
уже в этих структурах есть особые лабильные системы, с помощью которых уста¬
навливается межорганное взаимодействие.
Когда же речь идет о функции как о системной реакции целостного организма,
которая непременно должна быть высокоадаптивпой применительно к сиюми¬
нутной окружающей обстановке, то термин «локализация» уже неприменим, так
как любая целостная функция, поведенческая тем более, опирается на сложную
констелляцию множества пространственно разнесенных мозговых аппаратов, во¬
влекаемых симультанно и сукцессивпо для целей удовлетворения доминирующей
потребности (А. С. Батуев, 1981).
Любая поведенческая
функция организма опи¬
рается на сложную кон¬
стелляцию множества
пространственно раз¬
несенных мозговых ап¬
паратов, вовлекаемых
симультанно и сукиес-
сивно для целей удовле¬
творения доминирую¬
щей потребности.
§ 5. Основные методы нейрофизиологии
поведения. Методология, метод, методика
Обычно под методологией исследования мозга понимаются и методологические
принципы, и методические (технические приемы и средства экспериментирова¬
ния). В качестве методологических принципов служат прежде всего познанные
и конкретно примененные закономерности мозга, а также общебиологические зако¬
номерности. Таким образом, методологические принципы указывают направле¬
ние исследования, способствуют постановке проблем, влияют па истолкование
результатов эксперимента. Методики же, техника экспериментирования, по вы¬
ражению И. П. Павлова, «толкающие вперед науку», определяют конкретные
средства исследования определенного вопроса. Между методологическими прин¬
ципами исследования и методическим техническим оснащением эксперимента
существуют теснейшая связь и взаимовлияние, хотя темпы и направления разви¬
тия того и другого не одинаковы. Методологические принципы, подходы оказыва¬
ются, конечно, более стабильными, обогащаясь и видоизменяясь в деталях, тогда
как технические средства экспериментирования могут меняться очень быстрыми
2 Зак. 1078
34 Глава 1. Методология изучения высшей нервной деятельности
темпами, определяемыми успехами физики, химии, математики, кибернетики.
Эти науки оказывают на физиологию мозга столь сильное влияние, что обогаща¬
ют методологию исследования новыми идеями и подходами.
В недалеком прошлом основными методическими средствами изучения фи¬
зиологии мозга были разрушение и раздражение. С помощью этих методов полу¬
чено огромное количество фактов и в аналитическом, и в поведенческом планах.
Наблюдение за простыми или сложными поведенческими проявлениями с их де¬
тальным описанием явилось главным способом оценки целостной работы мозга.
Метод условных рефлексов, предложенный И. П. Павловым, оказал револю¬
ционизирующее влияние па физиологию, клинику, психологию и многие смеж¬
ные науки. В распоряжение ученых был дан метод объективного исследования по¬
ведения, перестраивалась вся методология научного мышления, формировалась,
говоря словами И. П. Павлова, «объективная физиология мозга и поведения».
Метод условных рефлексов был взят па вооружение во многих отечественных
и даже зарубежных лабораториях, методология же павловского научного мышле¬
ния встретила на своем пути ряд препятствий и получила в ряде случаев весьма
извращенные трактовки.
Простота и доступность методик условно-рефлекторного исследования пове¬
дения, связанная с неизбежностью, с одной стороны, искусственного вычленения
элементарных поведенческих актов, с другой — ограничения всего богатства внеш¬
ней среды нарочито избранными сигналами, создали весьма распространенное
впечатление о сводимости широкого разнообразия адаптивного поведения к эле¬
ментарным формам условно-рефлекторной деятельности. Критики павлов¬
ской методологии в запале дискуссии не хотели заметить, что главное внима¬
ние И. П. Павлова было обращено не столько па сигнальный вход и эфферентный
выход, сколько на те центральные процессы, которые имеют место при формиро¬
вании условно-рефлекторных актов. Создавалось впечатление, что павловская
методология пропагандировала редукционистский подход к пониманию механиз¬
мов мозга и поведения. Именно эта сторона условно-рефлекторной теории и была
взята па вооружение бихевиористами, доведшими до крайнего редукционизма
средства изучения поведения, регистрируя элементарные реакции животного на
искусственные стимулы и одновременно игнорируя те сложные мозговые меха¬
низмы, которые лежат в основе таких ассоциаций.
Широкое распространение бихевиористических методик, а также не всегда
верное изложение идеологии И. П. Павлова его собственными учениками послу¬
жило основой для этологов подвергнуть критике сам метод условных рефлексов.
Этологи противопоставили ему весьма ценный метод наблюдения и описания
сложных форм поведения животных в естественной среде их обитания. Следует
отметить, что как бихевиористов, так и этологов мало интересовали собственно
мозговые механизмы сложных форм поведения.
Главной же задачей самого И. П. Павлова и его школы было и есть изучение
мозговых процессов, лежащих в основе организации целенаправленного пове¬
дения. Естественно поэтому то пристальное внимание, которое всегда уделялось
вопросу о средствах и способах оперирования с живым мозгом. Наиболее распро¬
страненным приемом в школе И. П. Павлова был метод экстирпации участков ко¬
ры головного мозга (в другой крупнейшей отечественной неврологической шко¬
ле, возглавляемой В. М. Бехтеревымметод электрической стимуляции коры).
§ 5. Основные методы нейрофизиологии поведения. Методология, метод, методика 35
Техническая оснащенность физиологического эксперимента в начале XX сто¬
летия была крайне бедной. Работая с локальными разрушениями мозговых струк¬
тур при одновременной регистрации условно-рефлекторной деятельности, И. П. Пав¬
лов выдвинул представление о локализации (хотя и динамической) отдельно сен¬
сорных и моторных функций.
Как и любой крупный ученый, И. П. Павлов не избежал некоторых противоре¬
чий. Так, истово воюя против узкого локализациопизма, И. П. Павлов постулиро¬
вал локализацию корковых «пунктов» условного и безусловного раздражителей.
Это положение, переходящее из статьи в статью, из учебника в учебник, повлекло
за собой десятилетия бесплодных поисков конкретного субстрата этих «пунктов»
и самой условно-рефлекторной связи между ними. Справедливости ради следует
иметь в виду, что сам И. П. Павлов был далек от догматизации своих взглядов па
природу условно-рефлекторной связи. Во многих его последних работах такая
связь представляется им как формирование сложных динамических структур, ком¬
бинационных центров, состоящих из мозаики далеко отстоящих пунктов нервной
системы, объединенных в общей работе.
Сегодня никто не станет отрицать огромное значение методов электростиму¬
ляции мозга и экстирпации для решения общефизиологических задач. Однако
некритическое отношение к этим методам наряду с бытовавшим неверным пред¬
ставлением о мозге как конгломерате центров проникло и в неврологическую кли¬
нику, утвердив за определенными участками коры мозга или глубоких его струк¬
тур наименование «центров» памяти, сна и других сложных психических отправ¬
лений.
Внедрение в физиологию мозга методов регистрации его электрических про¬
цессов произвело революционизирующее влияние па многие аспекты учения
о мозговом обеспечении поведения. Физиологам-эксперимептаторам был дан но¬
вый метод объективного анализа собственно внутримозговой активности, сочета¬
ние которого с поведенческими актами существенно изменило представления как
о мозговой организации, так и о динамике тех процессов, которые обеспечивают
управление сложными формами поведения. Нейрофизиологическое направление
в изучении поведения и мозга базируется на регистрации электроэнцефалограм¬
мы, медленных локальных потенциалов, сверхмедленпых колебаний и, наконец,
импульсной активности одиночных нервных клеток. Возможности объективного
анализа интимных механизмов работы мозга существенно расширились.
Новый метод настойчиво формировал и новую методологию научного мышле¬
ния, путь к которой был полон противоречивых гипотез и взглядов. Казалось не
столь уж отдаленным воплощение заветной мечты — раскрытие тайн работы чело¬
веческого мозга и воспроизведение отдельных сторон его деятельности в техни¬
ческих модельных устройствах. На деле же оказалось, что объективная регистра¬
ция сверхслабых биоэлектрических процессов мозга сопряжена не только с рядом
технических трудностей, но и с проблемами общетеоретического плана. Для их
преодоления необходимо иметь методологическую платформу в виде теории си¬
стемной работы мозга, которая объединила бы в себе все накопленные знания по
изучению активного целевого поведения и в то же время позволяла проводить
корреляции с активностью отдельных мозговых субстратов с разными этапами
выполнения поведенческого теста.
36 Глава 1. Методология изучения высшей нервной деятельности
Классификация методов исследования высшей нервной деятельности. Для
удобства изложения целесообразно рассматривать отдельно методы изучения по¬
ведения и методы изучения мозга.
Методы отологического изучения поведения состоят в наблюдении за поведе¬
нием различных видов животных в естественных условиях их обитания. Главная
задача заключается в выявлении путем наблюдения основных структурных со¬
ставляющих поведения (блоков) и тех факторов, которые ответственны за их реа¬
лизацию. Эти методы были разработаны преимущественно зоологами. Особен¬
ность этих методов состоит в том, что с их помощью в основном описываются
крупные структурные единицы поведения типа инстинктов или комплексов фик¬
сированных действий и в меньшей степени те пластические компоненты сложно¬
го поведения, которые связаны с обучением. Животные в естественной среде
и при общении с соплеменниками выявляют ряд поведенческих реакций, ко¬
торые невозможно обнаружить в лабораторных условиях. Однако в связи с мно¬
жеством неучитываемых и переменных факторов зачастую невозможно выявить
определенно тот конкретный набор сигналов, которые запускают данную форму
поведения.
Методы условно-рефлекторного изучения поведения используются всецело в
лабораторных условиях при ограничении не только самого поведенческого репер¬
туара, как правило, до одной какой-либо регистрируемой реакции, по и при огра¬
ничении действующей сенсорной информации вплоть до узкого набора сигналов.
Именно эти сигналы вводятся в функциональную (временную) связь с данной по¬
веденческой реакцией. Эти методы используются в качестве основного инстру¬
мента исследования поведения в физиологических лабораториях. В отличие от
предыдущих методов при условно-рефлекторном изучении поведения выявляют¬
ся те возможные поведенческие акты, которые приобретаются в порядке обуче¬
ния. Лабораторный эксперимент позволяет моделировать различные формы по¬
ведения и подвергать их тщательному физиологическому анализу. Последнего не
удается достичь с помощью отологических методов.
Вместе с этим условно-рефлекторный эксперимент, изолирующий животное
из естественной окружающей среды, лишает исследователя возможности наблю¬
дать участие выработанных форм обучения в сложной организации целостного
поведения.
Методы когнитивного изучения поведения берут свое начало от психологии.
Главная задача этих методов, которые используются в лабораторных эксперимен¬
тах, состоит в выявлении тех сторон психики животных, которые определяют их
поведение в сложных жизненных ситуациях. Причем наряду с выявлением спосо¬
бов решения поведенческих задач путем тренировки, то есть обучения, эти мето¬
ды позволяют обнаружить в поведении животных явления так называемой эври¬
стики, то есть способности сразу находить правильный выход из новой и необыч¬
ной ситуации. Уже из самого наименования следует, что этими методами ученые
пытаются описать познавательную сторону поведения. Постепенно усложняя по¬
ставленные перед животным задачи, исследователь оценивает диапазон способно¬
стей животного к их решению, будь то лабиринт, проблемный ящик или ориента¬
ция в свободном пространстве.
§ 5. Основные методы нейрофизиологии поведения. Методология, метод, методика 37
Теперь кратко рассмотрим основные методы изучения мозга, которые в общей
форме могут подразделяться на морфологические, биохимические и физиологи¬
ческие.
Морфологические методы чрезвычайно ценны, ибо создают фундамент наших
сведений о тонком строении мозга, который используется для разработки после¬
дующих физиологических концепций. Сегодняшняя пейроморфология распола¬
гает множеством различных приемов исследования мозга с помощью световой
и электронной микроскопии. Однако успехи нейроморфологии связаны не только
с совершенствованием оптической техники, электронных микроскопов, оснащен¬
ных новейшими вычислительными устройствами, но и с успехами физико-химии,
позволяющими использовать чистые препараты для выявления тончайших эле¬
ментов нервных клеток, их отростков, синапсов и т. д. Наиболее результативно
комплексное использование приемов световой (в том числе и люминесцентной)
и электронной микроскопии вместе с приемами гистохимии, иммуноцитохимии,
флуоресцентной микроскопии и радиохимии.
Биохимические методы в последнее время начинают приобретать все большее
значение в комплексном изучении мозга. Это обстоятельство связано не только с
успехами химии и электроники, но и с изменением акцента экспериментальных
поисков, которые все больше углубляются в расшифровку метаболических про¬
цессов здорового и больного мозга в разных функциональных состояниях, фор¬
мах деятельности и пр. Несомненным шагом вперед в понимании работы мозга
явилось бурное развитие таких областей пейрохимии, как химия пептидов, медиа¬
торов и модуляторов, аминокислот и других биологически активных соединений,
приемов цитохимии, радиохимии, иммунохимии и др.
Физиологические методы включают в себя несколько экспериментальных при¬
емов. Прием разрушения отдельных участков мозга вначале был применен в лабо¬
раториях для создания моделей тех «драматических» ситуаций, в которые попадают
люди с локальными поражениями мозга. Поэтому первые сведения о функцио¬
нальной специализации отдельных участков мозга были получены благодаря их
разрушению и наблюдению за поведенческими дефектами.
Прием электрического раздражения мозга стал внедряться в эксперименталь¬
ную физиологию с середины XIX в. и долгое время наряду с разрушением был
единственно доступным для изучения мозга. Оба эти приема или подхода в наше
время не утратили своего значения, а, наоборот, стали активно применяться в свя¬
зи с внедрением в практику физиологического эксперимента стереотаксической
техники. Последняя позволяет в трехкоординатной системе вводить разрушаю¬
щий или раздражающий электрод в любой очень локальный участок мозга. Этот
же прием используется в клинической практике для терапии ряда неврологиче¬
ских и психических заболеваний.
В последнее время все активнее используются приемы хемостимуляции, термо-
и хеморазрушения (иногда обратимого блокирования) отдельных мозговых струк¬
тур, а также разрушение их ультразвуком. Эти приемы имеют ряд преимуществ
перед электростимуляцией в связи с большей локальностью и контролируемо¬
стью воздействия. Все эти методы требуют морфологического контроля и иссле¬
дования состояния мозговой ткани. Кроме локальных воздействий применяют то¬
тальное воздействие на мозг электромагнитными и звуковыми полями различной
частоты.
38 Глава 1. Методология изучения высшей нервной деятельности
Наиболее эффективным среди других является метод регистрации электриче¬
ских процессов мозга. Он стал применяться только во второй половине XX в. в свя¬
зи с бурным прогрессом в радиоэлектронике и широко распространен в физиоло¬
гических и клинических лабораториях. Самые различные формы электрических
процессов мозга — от суммарной электроэнцефалограммы до внутриклеточно ре¬
гистрируемой активности отдельной клетки бодрствующего мозга — таков широ¬
кий спектр применения этого метода. В значительной мере благодаря ему сформи¬
ровалась новая наука — нейрофизиология поведения. Нет необходимости детально
анализировать этот метод, ибо результаты его применения будут обсуждаться во
многих разделах книги. Здесь важно подчеркнуть следующее.
Современный уровень науки о высшей нервной деятельности характеризуется
высокой мультидисциплинарностыо, свидетельством чего является появление
нового объединения наук под названием нейронаука, или нейробиология. Интегра¬
ция ученых самых разных профессий, с разных позиций изучающих работу мозга
в комплексе иейроиаук, завершила процесс, начатый в физиологических и психо¬
логических лабораториях, где для решения даже самой скромной задачи, связан¬
ной с изучением мозгового обеспечения поведенческого акта, традиционно при¬
менялись многие из перечисленных методов в комплексе. Эта тенденция еще бо¬
лее проявляет себя в последнее время, когда рядом с физиологом, психологом,
нейрохимиком, пейроморфологом работают математики, инженеры, кибернети¬
ки, объединенные общим устремлением попять мозговые механизмы управления
поведением человека и животных.
В частности, наряду с регистрацией электрической активности мозга электро¬
энцефалограммы (ЭЭГ) регистрируют и вызванные потенциалы, что позволяет
выявить признаки местного или распространенного нарушения мозговой дея¬
тельности. С помощью обычного рентгеновского исследования или более слож¬
ных методов определяют размеры и форму мозга. Можно получить картину струк¬
туры мозга с расположением кровеносных сосудов — ангиограмму или с помощью
данных о потреблении мозговыми структурами глюкозы позитронную эмиссион¬
ную транскраниальную томограмму (ПЭТТ-сканирование). Таким образом, диф¬
ференцируют различные заболевания, поражающие одни и те же части мозга.
Заключение
Вся история отечественной физиологии сложилась в традициях картезианства.
Картезианское представление о рефлексе и бихевиористские идеи о поведении за¬
хватили умы многих исследователей (см. Ярошевский, 1997) прежде всего про¬
стотой схем, которые без каких бы то ни было коррективов переносились из
области теоретических представлений в сферу экспериментального анализа. Кар¬
тезианские идеи об организме как о рефлекторной машине оказали сильное влия¬
ние на мышление таких гигантов, как И. М. Сеченов и И. П. Павлов. Эта платфор¬
ма с учетом многих исторических предпосылок и послужила благодатной почвой
для появления павловской условно-рефлекторной теории, исходно предназна¬
чавшейся для объяснения высших форм поведения животных и человека, их выс¬
шей нервной деятельности (Павлов, 1952).
Поэтому принцип паирефлекторизма, активно поддерживаемый господство¬
вавшей тогда марксистско-ленинской идеологией, стал единственным в объясне¬
Заключение 39
нии всех поведенческих актов животных и человека. Причем если к непроиз¬
вольным автоматизированным актам поведения принцип рефлекса по-прежнему
более или менее применим (Бериташвили, 1975), то в случае сложных форм про¬
извольного поведения становится очевидной недостаточность рефлекторной тео¬
рии (Бериташвили, 1975; Бернштейн, 1990; Крушипский, 1986).
Рассмотрим принцип рефлекса в качестве детерминирующего для целостных
поведенческих актов. Понятно, что никакое действие не возникает без причины.
По-видимому, нет нужды оспаривать справедливость этого положения, состав¬
ляющего основу научного мировоззрения (Кругликов, 1988; Ярошевский, 1976).
Кроме того, оно оправдывается всей историей развития физиологии от Декарта
до наших дней. Действительно, соматические и вегетативные механизмы регуля¬
ции функций органов и систем базируются па рефлекторном принципе, где
в большинстве случаев с очевидностью определяется сенсорный пусковой фак¬
тор. Таковым, как правило, выступает какой-либо внешний или внутренний сиг¬
нал, который можно относительно легко подвергнуть экспериментальному анали¬
зу. Поэтому несомненно, что па основе рефлекторного принципа регулируется
деятельность отдельных органов и систем организма (Бернштейн, 1990). В схеме
рефлекторной дуги мы можем выделить афферентную, пусковую часть, центр пе¬
реключения и исполнительный орган. Все это справедливо для относительно про¬
стых и искусственно редуцированных моделей. Когда же речь идет о системном
уровне исследования организма в его активном и адаптивном взаимодействии со
средой, рефлекс (в его классическом понимании) уже не может претендовать на
роль механизма целостного поведения.
Наиболее серьезную разработку этого вопроса можно найти у А. А. Ухтомско¬
го, который, в частности, писал: «Рефлексы — это только следы прежней деятель¬
ности и прежних выработок, какие были присущи организму в его приспособле¬
нии к среде. С этой точки зрения центры, остающиеся в распоряжении обезглав¬
ленного животного и могущие осуществить еще рефлексы, являются нарочитыми
хранителями следов — аккумуляторами следов от прежних действований орга¬
низма. В нормальном организме к ним предъявляется требование осуществить
тот или иной привычный акт, тогда как высшие центры продолжают быть заняты
текущими новыми выработками по поводу задач, вновь и вновь предъявляемых
средой. С этой стороны ясно, что рефлекс как таковой, то есть остаток от прежней
целостной деятельности организма, не может содержать в себе ничего “творческо¬
го”! Он лишь инструмент в руках творческого прибора — организма в целом!..
Экспериментальному физиологу рефлекс открывается со стороны преимущест¬
венно своих мертво повторяющихся стереотипных сторон, ибо он исключил го¬
ловной мозг, то есть именно того деятеля, который варьирует, приспосабливает,
направляет рефлексы, находящиеся в его распоряжении, применяя их к потребно¬
стям текущего момента и свойствам новой среды»1.
И действительно, насколько уже усложняется картина, если мы перейдем от
аналитических моделей к естественным регуляторным процессам в организме!
Понятно, что несоизмеримому усложнению подвергнутся сигнальные и переклю¬
чательные звенья рефлекса. Это подтверждает шеррингтоновский принцип во¬
ронки и общего пути (Шеррипгтон, 1963). Увлекаясь поисками единого детерми-
1 Заметки А. А. Ухтомского на полях книги: СеллиДж. Педагогическая психология. М., 1912. Обложка.
40 Глава 1. Методология изучения высшей нервной деятельности
пирующего начала, экспериментаторы искусственно сужают афферентную часть
рефлекторной дуги. Конечно же, это не может не отразиться па существенном
ограничении и переключательных аппаратов. Получившийся в результате «абор¬
тированный» примитивный рефлекс рассматривается как основной механизм
внутрисистемной регуляции. Отнюдь не отрицая правомерности такого подхода
применительно к анализу деятельности конкретного органа, вряд ли стоит пере¬
носить его на целостный организм.
В реалиях мы сталкиваемся с тем обстоятельством, что «чем шире воронка, тем
больше потенций действий, тем более труда для того, чтобы обеспечить при этом
обилии потенций гарантию единства действий в каждый конкретный момент...»1.
Вряд ли кто-нибудь возьмется с уверенностью утверждать существование
в норме какого-то фактора, который является необходимым и достаточным усло¬
вием для запуска конкретного действия. Дело чаще всего заканчивается построе¬
нием общих схем, вынужденных считаться с полиафферентпой природой и поли-
эффекторным обеспечением любого рефлекторного акта (Гаазе-Ранпопорт, По¬
спелов, 1987). Таким образом, даже па уровне низших механизмов или «низшей
нервной деятельности» принцип рефлекса песет в себе скорее общефилософское
содержание, от которого сделать шаг в область конкретного знания о механизмах
поведения весьма непросто.
Еще А. А. Ухтомский подчеркивал, следуя принципам Ч. С. Шерриштона, что
«наиболее экономичное достижение цели в эфферентном пучке будет определять
собой вторично пределы экономизации в эфферентном пучке. Доминанта, по¬
вторно вызываясь в центрах, будет непременно подвижной в эфферентном и аф¬
ферентном пучке, пока не будет достигнута наиболее экономическая “воронка
Шерриштона”»1 2.
И далее: «Первоначально “центр” рисовался механизмом, которым определя¬
ется и объясняется функция. Впоследствии он сам стал функционально множест¬
венным прибором, единство действия которого для каждого отдельного момента
определялось условиями выполнения периферической функции. То же повторя¬
ется и для общего пути. Сначала это механизм множественного сочетания реф¬
лексов, определяющий судьбу возбуждения в каждом из компонентов. Затем это
механизм, который сам, в свою очередь, образуется из намечающегося задания
и функции... Если задание и функция приурочены лишь к целому, то и примене¬
ние общего пути по связи с целым может быть совершенно новым, никогда не осу¬
ществляющимся в нем вне связи с целым...»3
Означает ли это, что эра рефлекса уже завершена? Отнюдь пет. С помощью
экспериментально-аналитических методов современные физиологи накопили
столь огромное число фактов, что с ними невозможно не считаться (Кругликов,
1988). Однако их очень трудно, а порой и невозможно перенести па здоровый
бодрствующий организм. Есть основание полагать, что это одна из причин прямо¬
го расхождения аналитической физиологии и синтетической физиологии целост-
1 Заметки А. А. Ухтомского на полях книги: Физико-химические основы нервной деятельности. Л.;
М„ 1935. С. 5.
2 Заметки А. А. Ухтомского на полях книги: Бехтерев В. М. Общие основы рефлексологии человека.
М„ 1923. С. 228.
3 Заметки А. А. Ухтомского на полях книги: Handbuch der Normalen und Patologisehen Physiologie.
Berlin, 1927. P. 189.
Заключение 41
ноге организма (Бернштейн, 1990). Может быть, рефлекс как элементарная фор¬
ма регуляции есть просто аналитическая фикция и не более того? Ведь писал же
Ч. С. Шерриштон, что простой рефлекс — выгодная, но невероятная фикция.
А. А. Ухтомский добавлял: «Из простых рефлексов ничего образоваться не могло
по той причине, что они суть лишь абстракции, реально не существующие! Обра¬
зование могло идти из сложной доминанты дифференцированием ее сенсорных
и моторных пучков»1. И тут же А. А. Ухтомский подчеркивает значение рефлек¬
торного принципа как организующего начала: «Рефлекс — это, скорее, порядок,
в котором совершается акт, а не характер его но содержанию. Поэтому рефлектор-
ность отнюдь не исключает целесообразность. Рефлекс — целесообразный акт,
протекающий в порядке отражательной автоматичности!»1 2 А поэтому рефлекс
«не элемент, а фрагмент движения. И целое не результат складывания рефлексов,
но интеграция моментов действия»3.
Рассмотрим такой элементарный безусловный рефлекс, как чесательный, в ко¬
тором, казалось бы, легче вычленить конкретный пусковой фактор тактильной
природы. Оказывается, что такая форма поведения запускается целым набором
раздражителей, куда входят но меньшей мере тактильный, вестибулярный, про¬
приоцептивный, зрительный. Все они составляют афферентное обеспечение дан¬
ного поведенческого акта (его шеррингтоновскую воронку). Может быть, испол¬
нительная часть более локальна? Оказывается, что она непременно начинается
с торможения текущей деятельности (Ухтомский, 1966) и активной перестройки
позы, затем включается запуск ритмического движения конечности и только в за¬
вершение достигается (или пет) контакт конечности с поверхностью кожи. Не бу¬
дем забывать и о вегетативном обеспечении такого поведенческого акта. Естест¬
венно, что при этом просто невозможно думать о каком-то едином центральном
звене, обеспечивающем переключение разиомодальпой афферентации па ноли-
эффекторные аппараты данного поведенческого акта. Тогда где же располагается
(согласно классическому представлению) «центр» данного рефлекса и что высту¬
пает в качестве его рефлекторной дуги? А. А. Ухтомский так отвечает на этот во¬
прос: «Вероятно, следует говорить не о раз навсегда сорганизованных соматовеге-
тативных механизмах в коре, по о постоянно вновь и вновь организующихся со¬
четаниях во времени совершающихся событий в соматической и вегетативной
области. В зависимости от рецептивных побудителей реакции и от степени сроч¬
ности экстеро- и иитероцептивного требования будет преобладать или вегетатив¬
ный, или соматический компонент»4.
Мы привыкли эти «простые» рефлексы, пользуясь павловской терминологи¬
ей, именовать безусловными. Исследователи, как правило, довольствовались чис¬
то внешним проявлением «рефлекса» — наличным раздражителем и видимой ре¬
акцией. Именно такова идеология бихевиоризма и пеобихевиоризма (Ярошев-
ский, 1976). Представители этих широко распространенных течений стояли как
1 Заметки А. А. Ухтомского на полях книги: Бехтерев В. М. Общие основы рефлексологии человека.
С. 210.
2 Там же.
3 Заметки А. А. Ухтомского на полях журнала: Естествознание и марксизм. 1930. № 2-3. С. 86.
1 Заметки А. А. Ухтомского на полях книги: Fulton J. Physiology of the Nervous System. Oxford, 1938.
P. 485.
42 Глава 1. Методология изучения высшей нервной деятельности
раз на таких позициях, игнорируя центральные механизмы поведенческого акта
и сводя его лишь к видимым внешним проявлениям.
На деле же оказалось, что для выполнения «безусловного рефлекса» (на¬
пример, чесательпого) требуется ряд условий, и прежде всего достижение опреде¬
ленного (временного) функционального состояния системы мозговых структур
(констелляции, по Ухтомскому), которое обусловливает вовлечение в нее всех
центральных звеньев конкретного поведеическох'о акта. Возможно, в связи с труд¬
ностями идентификации функционального состояния организма этот важный
момент чаще всего ускользает от внимания исследователей.
Для определения пускового фактора того или иного рефлекса обычно исполь¬
зуют наиболее доступный и очевидный сенсорный компонент. При этом иссле¬
дователи подчас пренебрегают установленным фактом существования шеррииг-
тоновской воронки па любых уровнях центральной нервной системы. Любой
«пусковой» стимул, приходя в такую воронку, сталкивается с множеством дру¬
гих сенсорных влияний, взаимодействуя с ними весьма сложным образом.
Более того, конвергирующие сенсорные влияния создают определенное функ¬
циональное состояние нервного центра (воронки), определяя его дееспособность
и реактивность. Поэтому вариант ответной реакции организма на какое-то конк¬
ретное сенсорное воздействие определяется не только и не столько параметрами
стимула, сколько предуготовлеппостыо центрального нервного аппарата, от кото¬
рой зависит характер и степень выраженности предстоящего рефлекса. И чем вы¬
ше мы поднимаемся по «этажам» мозга или с чем более сложной филогенетиче¬
ской ступенью имеем дело, тем сильнее становится эта зависимость. А наиболее
очевидный сенсорный фактор, который мы по каким-то причинам связываем
с данной реакцией, все более приобретает не столько даже пусковое, то есть моно¬
польно запускающее, сколько «спусковое» (по терминологии Н. А. Бернштейна)
значение.
Высоко оценивая идеи Шерринггона о механизмах конвергенции и интегра¬
ции в нервной системе, А. А. Ухтомский писал: «Когда Шерриигтон высказал о
центральной иннервации мышц, что мозг мыслит не в отдельных анатомических
мышечных единицах, по в отдельных интегрированных актах, он дал зародыш
и того представления, что восприятие посредством органов чувств совершается не
в мифических “простых ощущениях”, но в интегральных образах, которые приоб¬
ретают тем более проективное значение в пространстве и времени, к чем более вы¬
сокому этажу нервной системы мы переходим»1. А поэтому «анализатор — собст¬
венно не орган чувств и не кортикальный его отдел, как... постоянный механизм,
а именно подвижное приурочение тактильное к слуховому, зрительного к слухо¬
вому, тактильного к зрительному»1 2.
Если учесть, что функциональное состояние нервного центра задается и под¬
держивается, в частности, различными гуморальными (в том числе и гормональ¬
ными) влияниями, то нетрудно себе представить, что на определенном этапе уро¬
вень готовности центра к действию будет настолько высок, что любое конверги¬
рующее к нему сенсорное воздействие может выступать в качестве «спускового»
фактора. В этом случае такое понятие, как сенсорная избирательность какого-
1 Заметки А. А. Ухтомского на полях книги: Иванов-Смоленский А. Г. Основные проблемы патофизио¬
логии высшей нервной деятельности. М„ 1933. С. 5.
2 Заметки А. А. Ухтомского на полях книги: СавичВ. В. Основы поведения человека. Л., 1924. С. 30.
Заключение 43
либо рефлекса, вообще теряет свой смысл. Тогда, естественно, возникает вопрос,
где же тот монопольный сенсорный канал, который представляет собой аффе¬
рентную составляющую данной рефлекторной дуги? Эту проблему очень хорошо
проанализировал А. А. Ухтомский, рассматривая инстинктивную деятельность
животных с позиций своего учения о доминанте (Ухтомский, 1966).
Таким образом, идея рефлекса должна оставаться лишь философским обобще¬
нием, и не следует рассматривать рефлекс как основу работы центральной нерв¬
ной системы.
С необходимостью признания сложной детерминации поведения столкнулся
еще И. М. Сеченов, пытаясь объяснить механизмы произвольных движений. В це¬
лом, оставаясь на позициях рефлекторной теории, Сеченов был вынужден ввести
понятие о «темпом мышечном чувстве», «валовом чувстве»; он первый обратил
внимание на функциональное состояние нервного центра как на важнейшее звено
в формировании ответной реакции организма (Сеченов, 1952).
Несмотря на то что биологический смысл так называемого сеченовского тор¬
можения говорил в пользу существования определенных процессов межцепт-
ралыюй интеграции, Сеченову, однако, не удалось до конца решить проблему так
называемых внутренних детерминант поведения. Он лишь поставил проблему, де¬
тальная разработка которой выдала па долю его ученика и преемника Н. Б. Вве¬
денского. Сеченовская идеология и аналитические иссле¬
дования па моделях периферического нервно-мышечного
аппарата позволили Введенскому с определенностью за¬
явить о решающей роли функционального состояния воз¬
будимой ткани (мышцы, нервного ствола, нервно-мышеч¬
ного соединения и, наконец, нервных центров) в опре¬
делении характера ее ответного реагирования. Однако
биологический смысл установленной закономерности не
был раскрыт Введенским. Эту проблему рассматривал его
преемник и ученик А. А Ухтомский. Именно Ухтомскому принадлежит идея об
определяющем значении функционального состояния мозговых центров для фор¬
мирования вектора поведения, направленного на активное решение стоящих пе¬
ред организмом задач и достижение биологически оправданного приспособитель¬
ного результата. Эти представления легли в основу учения Ухтомского о доми¬
нанте как одном из основных принципов нервной деятельности.
Доминанта базируется на трех основных факторах: преобладающей моти¬
вации, прошлом жизненном опыте и оценке вероятностной структуры среды (Ба¬
туев, 1981). Два первых фактора представляют собой внутренние детерминан¬
ты поведения, они создают мозаику функционального состояния мозговых струк¬
тур, которые в составе динамической констелляции оказываются вовлеченными
в управление предстоящей поведенческой деятельностью организма.
Любая ответная реакция организма является не только полисенсорной и осно¬
ванной на функционировании интегральных образов (психопервных образов, по
И. С. Вериташвили), но и иолиэффекторпой, то есть обеспечивается множеством
соматических и вегетативных компонентов (Черниговский, 1967). Следовательно
поиск монопольного центрального аппарата (воронки) с монопольным общим эф¬
ферентным выходом — бесперспективная задача.
Тогда какова же эфферентная часть рефлекторной дуги данного рефлекса?
В действительности дело сводится к формированию единой динамической рабо¬
Идея рефлекса долж¬
на оставаться лишь
философским обоб¬
щением, и не следует
рассматривать реф¬
лекс как механизм ра¬
боты центральной
нервной системы.
44 Глава 1. Методология изучения высшей нервной деятельности
чей констелляции. В нее сукцессивно и симультанно вовлекаются пространствен¬
но разнесенные мозговые структуры, причем каждая вносит специфическое нача¬
ло в реализацию целостного поведенческого акта. Консолидирующим фактором
формирования динамической констелляции Ухтомский считал механизм нерв¬
ной синхронизации.
Современные работы М. Н. Ливанова (1982) и его учеников подтвердили пра¬
вильность идей Ухтомского. Следует отметить, что доминанта (так же, как и реф¬
лекс) может выступать скорее в качестве методологического принципа, нежели
рассматриваться как конкретный механизм работы мозга,
ибо от формирования принципа до раскрытия реального
механизма действия — дистанция огромного размера.
Принцип доминанты Ухтомского успешно применяет¬
ся при описании активной природы целенаправленного
поведения, а принцип рефлекса остается в силе, если рас¬
сматривают конкретный поведенческий акт (короткие реф¬
лексы, по Ухтомскому) в его адаптивном значении для ор¬
ганизма. При этом надо помнить, что рефлекс (условный
или безусловный) — это не единица поведения и психи¬
ческой деятельности, а принцип реагирования. Неверно
представлять рефлекторные дуги (как было показано вы¬
ше) в виде изолированных структурных образований в центральной нервной си¬
стеме; скорее всего, это также аналитическая (по очень удобная) фикция.
А. А. Ухтомский гениально предвидел разработки продуктивных идей, осуще¬
ствленных прежде всего П. К. Анохиным и Н. А. Бернштейном. Эти ученые, каж¬
дый со своих позиций, внесли весомый вклад в развитие теории живых систем.
Нам остается отметить, что в прокрустово ложе рефлекторной теории не укла¬
дываются многие новейшие гипотезы о работе мозга, равно как и идеи когнитив¬
ной психофизиологии, которые родились в противовес классическим представле¬
ниям о рефлексе и были основаны на синтезе многих нейронаук (Прибрам, 1975).
Вместе с тем учение Ухтомского об активном характере поведения, направляемо¬
го доминантами, — это предтеча современных взглядов (Маунткасл, 1981; Эй¬
дельман, 1981) на природу когнитивных процессов мозга, а также многих законо¬
мерностей синергетики.
В основе системы научного познания лежит принцип детерминизма, который
позволяет выявить причинно-следственную связь явлений, определяемую взаи¬
модействием материальных факторов. Современная наука о высшей нервной дея¬
тельности основывается на четырех принципах: принципе рефлекса, принципе
доминанты, принципе отражения и принципе системности в работе мозга. При
прослеживании основных этапов развития принципа рефлекса видно, насколько
он отошел от простых картезианских схем. Введение принципа доминанты позво¬
лило понять целесообразный характер реакций организма на воздействия внеш¬
ней среды. Соотношение адаптивности и активности в целостном поведении как
способе разрешения реальных противоречий рассматривается как движущая сила
построения поведения. Современная теория поведения избавилась от дуализма
и механицизма, встав на диалектические позиции.
Принцип отражения основан на исследовании органического единства непо¬
средственного восприятия и следов прошлых впечатлений. Отображенный образ
Принцип доминанты
успешно применяется
при описании актив¬
ной природы целена¬
правленного поведе¬
ния, а принцип
рефлекса остается в
силе, если рассматри¬
вают конкретный пове¬
денческий акт в его
адаптивном значении
для организма.
Заключение 45
(информация) существует для его носителя в субъективно-идеальной форме. От¬
ражение носит активный, избирательный характер, оно определяется прошлым
жизненным опытом субъекта и его доминирующей потребностью. Внешний мир
отражается субъектом в формах его деятельности, носящих рефлекторный харак¬
тер и направленных па сохранение целостности организма в его активном воздей¬
ствии па среду.
Любая, даже самая простая психическая функция есть результат системной
деятельности целого мозга. Поиски локализации обучения, памяти, условных
рефлексов методологически не обоснованы. Любая целостная поведенческая
функция базируется па констелляции мозговых аппаратов, вовлекаемых в одно¬
временную и/или последовательную деятельность, направленную па удовлетво¬
рение доминирующей потребности.
Контрольные вопросы
1. В чем сущность принципа детерминизма?
2. В чем состоит дуализм учения Р. Декарта?
3. Основы биологической теории рефлекса И. Прохазки.
4. Кому принадлежит термин «рефлекторная дуга» и в чем ее сущность?
5. Основные положения сеченовской психофизиологической теории рефлекса.
6. Условно-рефлекторная концепция И. П. Павлова.
7. Принцип рефлекса в учении А. А. Ухтомского.
8. В чем сущность принципа доминанты А. А. Ухтомского?
9. Каковы основные свойства доминанты?
10. Каковы основные стадии развития доминанты?
11. Принцип отражения в биологических системах.
12. В чем суть понятия «интегральный образ»?
13. В чем суть идеи П. К. Анохина об опережающем отражении?
14. В чем сущность субъективного характера отражения?
15. Идея об узком локализациопизме и ее анализ.
16. Концепция об универсализме и ее анализ.
17. В чем суть представления о динамической локализации функций мозга?
18. В чем суть учения А. А. Ухтомского о «функциональном органе»?
19. Каковы основы концепции А. Р. Лурия о функциональных блоках мозга?
20. В чем заключается принцип органной локализации?
21. Каковы основные методы изучения мозга?
22. Каковы основные методы изучения психофизиологии поведения?
Глава 2
СЕНСОРНАЯ ФУНКЦИЯ МОЗГА
§ 1. Обшие принципы конструкции
сенсорных систем
Для понимания механизмов приема и переработки информации используют тер¬
мин «сенсорные системы», понимая под ними анатомически организованную в
структурах мозга систему ядерных образований и связей, служащую для обнару¬
жения и кодирования информации определенной модальности.
Сенсорные системы подразделяются на внешние и внутренние.
Внешние сенсорные системы снабжены экстерорецепторами, воспринимаю¬
щими раздражения из внешней среды. Экстерорецепторы делятся на дистантные
(световые, звуковые, обонятельные) и контактные (кожные, вкусовые). Внутрен¬
ние сенсорные системы имеют рецепторные аппараты — интерорецепторы — во
всей внутренней среде организма (стенки сосудов, внут¬
ренние органы, мышцы, суставы, кости скелета и пр.).
По форме энергии, которая воспринимается рецепто¬
рами, последние подразделяются на световые и звуковые
рецепторы, механорецепторы, хеморецепторы и терморе¬
цепторы. В особую группу выделяют болевые рецепторы.
В обычных условиях существования на организм пада¬
ют сложные, комплексные раздражения, которые воспри¬
нимаются одновременно различными рецепторными ап¬
паратами. Поэтому в норме сенсорные системы осуществляют свою деятельность
в тесном взаимодействии друг с другом.
Принципы многоканальности и многоуровневости. Одним из существенных
результатов эволюции сенсорных систем является постепенное формирование
многоканальное™ передачи сигнализации в высшие этажи мозга. Например,
в пределах зрительной системы можно выделить чувствительные пути, простран¬
ственно распределенные в мозговых структурах и передающие информацию о пе¬
ремещении предмета в поле зрения, о его цветовых свойствах и нр.
Поканалыюсть проведения сигнализации предполагает многоуровневый или
многоэтажный характер передачи и обработки сенсорных сообщений. Чем боль¬
шее число каналов в пределах данной сенсорной системы, тем большее число пе¬
реключений характерно для каждого капала. Многоканалыюсть, наряду с бо¬
лее детальной обработкой информации, подразумевает замедленную ее передачу
в мозговые центры (рис. 1).
Дублирование каналов связи является одним из путей обеспечения надежно¬
сти работы сенсорных систем. Поэтому существование наряду с вышеописанны¬
ми и спрямленных путей уменьшает число промежуточных звеньев переклю¬
чения импульсации и коэффициент информативности передаваемого по это¬
му каналу сигнала должен возрастать. Появление таких каналов отражает общую
тенденцию к совершенствованию конструкции мозга и повышению надежности
его сенсорных аппаратов.
Сенсорные системы •—
анатомически органи¬
зованные в структурах
мозга системы ядерных
образований и связей,
служащих для обнару¬
жения и кодирования
информации опреде¬
ленной модальности.
§ 1. Общие принципы конструкции сенсорных систем 47
Рис. 1. Схема эволюции многоканальности и многоуровневое™
в конструкции сенсорных систем (по А. А. Заварзину, 1959)
/-V — этапы усложнения конструкции сенсорных систем;
снизу — рецепторы, сверху — высшие мозговые отделы
Принцип конвергенции и дивергенции. Надежность каналов связи еще более
возрастает благодаря частичному взаимному перекрытию нейронов. Даже па ре¬
цепторном уровне, например в сетчатке глаза, концевые разветвления одного и то¬
го же нейрона контактируют с несколькими нейронами более высокого уровня
(дивергенция, мультипликация). В то же время один и тот же нейрон контактиру¬
ет, получая импульсацию, сразу с несколькими нейронами предыдущего уровня
(конвергенция). Следовательно, четкая линейность проведения информации но
независимым каналам сенсорной системы не соблюдается (рис. 2).
Поверхность, где находятся все без исключения периферические разветвления
волокна первого чувствительного (афферентного) нейрона, называется перифе¬
рическим рецептивным полем данного нейрона. Взаим¬
ное перекрытие рецептивных полей особенно отчетливо
проявляется в центральных ядрах сенсорных систем, бу¬
дучи отражением более общего принципа конструкции
центральных связей, а именно принципа «нервной ре¬
шетки». В мозговых структурах благодаря умножению
числа нейронов предпосылок для широкого пространст¬
венного распространения нервных влияний гораздо боль¬
ше, чем на периферии. Одновременно с этим действует
принцип общего пути Ч. С. Шеррингтона, то есть схожде¬
ния многих входов на одном нейроне как на «воронке».
По мере восхождения от одного этажа мозга к другому «воронка» все более рас¬
ширяется за счет присоединения все новых входов и возрастают требования к все
более точному выведению на «общий путь» биологически значимой информации.
Ч. Шеррингтон писал по этому поводу: «...если два или более афферентных ней¬
рона конвергируют па третьем, который но отношению к ним является эфферент¬
ным (исполнительным. — А. Б.) и на котором, как показано выше, сливаются или
интерферируют их влияния, то такая структура представляет собой механизм ко¬
ординации»1.
Поверхность, где нахо¬
дятся все без исключе¬
ния периферические
разветвления волокна
первого чувствитель¬
ного (афферентного)
нейрона, называется
периферическим ре¬
цептивным полем дан¬
ного нейрона.
1 Шеррингтон Ч. С. Интегративная деятельность нервной системы. Л., 1969. С. 149.
48 Глава 2. Сенсорная функция мозга
Рис. 2. Схемы обшей конструкции нейронных переключений
в сенсорных системах (по Г. И. Полякову, 1 964)
а — более простая; б — более сложная конструкция.
1 — рецепторы; 2 — периферические чувствительные нейроны; 3 — низшая
переключательная инстанция; 4 — средняя переключательная инстанция;
5 — высшая переключательная инстанция в коре
Итак, наличие в сенсорных системах ряда уровней, каждый из которых работа¬
ет но принципу дивергенции и конвергенции, — это наличие важнейших коорди¬
национных аппаратов, где происходит поэтапная обработка информации. По-ви¬
димому, оба механизма необходимы для неискаженной передачи сведений об от¬
дельных признаках и деталях наряду с объединением их при формировании
целостного образа.
Принцип обратных связей. Деятельность сенсорной системы должна рассмат¬
риваться не как процесс пассивного кодирования любого раздражения в частотно
модулированную импульсную сигнализацию, а как процесс активного восприятия и
обработки наиболее биологически существенной информации. Сенсорную систе¬
му необходимо рассматривать не просто как пассивный капал линии связи, а как
активный «сигнализатор». Сенсорная система состоит не из лестницы релейных
образований, а из аппаратов управления процессом обработки и передачи инфор¬
мации с нижележащих уровней на вышележащие. И каждый уровень системы
должен работать минимум на основе двух входов:
♦ входа информации — восходящий путь;
♦ входа управления — нисходящий путь.
Таким образом, центральная организация сенсорной системы в совокупно¬
сти прямых и обратных связей представляется в виде системы образований из
надстраивающихся нервных колец (рис. 3). Наличие нисходящих связей к раз¬
личным образованиям сенсорной системы говорит о том, что на их работу влияют
§ 1. Общие принципы конструкции сенсорных систем 49
Рис. 3. Схема построения нейронных
переключений в сенсорной системе
(по Г. И. Полякову, 1965)
Периферический чувствительный нейрон, пере¬
дающий центростремительные импульсы как на эф¬
ферентные нейроны с длинным аксоном, так и на
специальные переключательные нейроны с корот¬
ким аксоном. Наряду с восходящими проводника¬
ми (стрелки вверх) показаны нисходящие центро¬
бежные проводники (стрелки вниз).
1 — периферический чувствительный нейрон;
2 — эфферентные нейроны с длинным аксоном;
3 — переключательные нейроны с коротким
аксоном
вышерасположенные отделы многих мозговых структур. Благодаря деятельности
нисходящих связей обеспечивается регуляция пороговой чувствительности и
пропускной способности в сенсорных системах. Это естественно, ибо сенсорная
система может обработать за единицу времени объем информации во много раз
меньший, чем тот, который может поступать на вход сенсорной системы.
Направленность нисходящих влияний определяется доминирующей потреб¬
ностью как предпосылкой образования доминанты при использовании аппаратов
памяти. В связи с этим особенно важной оказывается кортикальная регуляция,
которая осуществляет три функции:
♦ пусковую;
♦ корригирующую;
♦ поддерживающую.
Значение первой из них состоит в открытии или блокировании сенсорных вхо¬
дов; второй — в трансформации нервной сигнализации или в обеспечении усло¬
вий, при которых ранее разбалансированные сенсорные системы могут быть при¬
ведены к такому режиму деятельности, который создает необходимые условия
для их наиболее полного взаимодействия и интеграции, и третьей — в поддержа¬
нии возбудимости глубоких структур дайной сенсорной системы на уровне, опти¬
мально приспособленном к восприятию и передаче сигнала в восходящем направ¬
лении.
Имеющиеся данные свидетельствуют о тормозном значении обратных связей.
Это становится попятным, если принять во внимание чрезвычайно высокую (мож¬
но сказать, критическую) чувствительность рецепторных образований сенсорных
систем. Именно на этих уровнях сенсорных систем начинается процесс активного
восприятия и обработки сигналов.
Принцип кортикализации. Процесс формирования повой коры связан с пред¬
ставительством всей совокупности сенсорных систем. Это определило принцип
функциональной многозначности коры, согласно которому все корковые облас¬
ти — это корреляционные центры, среди которых нет чисто проекционных цент¬
ров, а значит, нет места абсолютной локализации.
50 Глава 2. Сенсорная функиия мозга
Экранная структура новой коры явилась субстратом для объединения дея¬
тельности многих сенсорных систем.
Одной из общих черт корковой проекции сенсорных систем является их мно¬
жественный характер представительства в коре. В самой общей форме разграни¬
чивают первичные и вторичные проекции. Первичные корковые проекции возни¬
кают в онтогенезе человека сравнительно рано, здесь заканчиваются быстропро¬
водящие сенсорные каналы. Эти корковые зоны окружены вторичными зонами
той же сенсорной системы, импульсация к которым поступает несколько позднее,
чем к первичным зонам. Эти корковые поля принимают интегрированную инфор¬
мацию в результате взаимодействия специализированных каналов данной сен¬
сорной системы.
Наконец, выделяют зоны перекрытия разных сенсорных систем, где проис¬
ходит межсенсорпое взаимодействие (рис. 4). Эти зоны получили название тре¬
тичных, или ассоциативных, полей. Например, в зрительной системе повреждение
первичной проекционной зоны приводит к возникновению «физиологической
слепоты» — исчезает восприятие противоположной половины поля зрения (ге-
миапопсия). Повреждение же вторичных проекционных зон коры вызывает «пси¬
хическую слепоту», которая именуется зрительной агнозией (неузнавание пред¬
метов). Поэтому высшим отделом сенсорной системы (в частности, зрительной)
считают именно вторичные сенсорные поля, оставив за первичными в значитель¬
ной мере релейную, переключающую функцию.
Принцип двусторонней симметрии. Любая сенсорная система построена по
принципу билатеральной симметрии. Иными словами, рецепторные аппараты,
соответствующие им центральные проводники и центральные мозговые структу¬
ры парные, и одна половина зеркально повторяет другую. Однако, как правило,
этот принцип проявляется лишь в относительной степени, ибо даже первичный
сенсорный путь может быть связан с обоими симметричными мозговыми полуша¬
риями. Связь с контралатеральным полушарием выражена сильнее благодаря
большему числу направляющихся туда сенсорных волокон. Между симметрич¬
ными отделами сенсорных систем устанавливаются горизонтальные комиссу¬
ральные связи, обеспечивающие их взаимодействие.
4
Рис. 4. Схема, изображающая обособление в корковой
сенсорной зоне нейтрального поля, в котором концентри¬
руются прямые проекции «точка в точку» и перифериче¬
ские поля, в которые собираются преимущественно непря¬
мые, перекрестные связи (третичные, или ассоциативные,
поля) (по Г. И. Полякову, 1965)
1 — уровень рецепторов; 2 — уровень переключений в стволе
мозга; 3 — в больших полушариях; 4 — в коре мозга
§ 2. Закономерности обнаружения сигналов 51
Основным механизмом парной деятельности сенсорной системы является
механизм функциональной асимметрии при действии различным образом локали¬
зованных в пространстве объектов. Первая деятельность сенсорных систем за¬
ключается в сравнении пространственной модели ранее действовавшего стиму¬
ла с повой пространственной локализацией того же стимула. Межполушарная
асимметрия в пределах одной сенсорной системы функционирует как саморегу¬
лирующаяся система с обратной тормозной связью, осуществляя функцию свое¬
образного компаратора, выполняющего сравнение рисунков возбуждения при
раздражении симметричных рецепторов. Система каллозальных связей между ас¬
социативными полями обеспечивает тесное единство обеих половин ассоциатив¬
ной системы и создает высокую надежность ее функционирования.
Принцип структурно-функциональных корреляций. Неодинаковая степень
кортикализации сенсорных систем и большее развитие глубоких структур слухо¬
вой системы по сравнению со зрительной свидетельствует о том, что зрительная
сигнализация для некоторых животных обладает наибольшей физиологической
силой, и степень кортикализации зрительной системы настолько высока, что лю¬
бое повреждение коры прежде всего сказывается на дефектах зрительного распо¬
знавания, в меньшей степени отражаясь на анализе звуковых сигналов.
У таких животных, как дельфины, летучие мыши, слуховая система выполняет
более важную роль в пространственной ориентации, чем зрительная система.
Любая психофизиологическая функция зависит от одновременной работы не¬
скольких сенсорных систем, то есть является полисенсорной, и поэтому не может
быть локализованной в ограниченных отделах мозга. Сама же сенсорная система
выступает как определенная локализованная анатомическая система, выполняю¬
щая специализированную функцию обнаружения и преобразования информации
в нервный код, в котором заключена совокупность описания признаков восприни¬
маемого объекта или явления.
§ 2. Закономерности обнаружения сигналов
Все, что определяется понятием «живое», обладает общим свойством реактивно¬
сти, то есть способностью более или менее срочно реагировать на разнообразные
воздействия. Проявлением реактивности является раз¬
дражимость — способность организма к ответу, посредст¬
вом которого достигается приспособление к условиям
среды. Раздражителями могут быть любые физико-хими¬
ческие изменения среды, приводящие к сдвигам метабо¬
лизма.
Свойство раздражимости еще не способно обеспечить
отражение качественного своеобразия раздражителя. В отличие от раздражимости
возбудимость — это способность воспринимать существенные для организма при¬
знаки раздражений и трансформировать в специфическую нервную сигнализа¬
цию.
Специализация рецепторных элементов нервной системы связана главным об¬
разом со свойствами превращать акт внешнего раздражения в совершенную и бы¬
стро действующую нервную сигнализацию, то есть в своеобразный физиологиче¬
ский код в форме серий электрических импульсов.
Раздражимость —
способность организ¬
ма к ответу, посредст¬
вом которого достига¬
ется приспособление
к условиям среды.
52 Глава 2. Сенсорная функция мозга
Возбудимость — спо¬
собность восприни¬
мать существенные для
организма признаки
раздражений и транс¬
формировать в специ¬
фическую нервную
сигнализацию.
Следовательно, наиболее общим свойством рецепто¬
ров является генерация в них электрических процессов
при воздействии раздражителя. Они запускают нервную
сигнализацию.
На первом этане восприятия должна быть решена за¬
дача обнаружения сенсорного воздействия и определена
его модальность, что и обеспечивается рецепторами. Сущ¬
ность рецепции сводится к преобразованию энергии воз¬
действующего стимула в изменения физико-химических параметров рецептор¬
ных элементов. Иными словами, сущность первичных механизмов рецепции за¬
ключается в высвобождении потенциальной энергии рецепторных элементов.
В зависимости от силы раздражения электрические потенциалы рецепторов
преобразуются в серии импульсов различной частоты, которые подаются по чув¬
ствительным проводникам в центральную нервную систему.
Рецепторы превращают непрерывный отсчет физической величины потенциа¬
ла (являющегося аналогом внешнего стимула) в дискретный сигнал. В свою оче¬
редь, нервный проводник может быть определен как канал линии связи, работаю¬
щий на параметрах частотно-импульсной модуляции.
Приспособление рецепторов к реагированию на определенные по энергии сти¬
мулы (световые, механические и т. д.) обусловливает специфичность рецепторов
и лежит в основе их классификации по данному признаку. Степень соответствия
стимула специфичности рецепторов характеризует аде¬
кватность раздражения.
Комбинации нервных импульсов при воздействии
раздражителей на рецепторы составляют материальную
основу отражения в мозгу предметов и явлений внешне¬
го и внутреннего мира, но по своим физическим и хи¬
мическим характеристикам они совершенно несхожи со
свойствами отражаемых объектов. Комплекс внешних
раздражений и порождаемые ими биофизические и биохимические превращения
в веществе мозга служат материальной основой функционального соответствия
между сигналом и его отражением в организме. Благодаря этому обеспечивается
восприятие внешнего мира и переработка закодированной информации, посту¬
пающей в мозг через рецепторы.
Хорошо известно, что при длительном воздействии различными стимулами
ощущение постепенно уменьшается. В основе этого явления, получившего назва¬
ние адаптации, чрезвычайно характерного для различных сенсорных систем, ле¬
жат весьма сложные процессы, происходящие как в рецепторных аппаратах, так
и в центральных отделах нервной системы.
О том, что животное может менять поток сенсорной информации, поступаю¬
щей в организм, известно давно. Вначале ограничивались описанием наиболее
очевидных реакций, определяющих ориентацию рецепторных поверхностей по
отношению к источнику сигналов и изменяющих характеристики сигналов при
их прохождении через вспомогательные структуры к рецепторам. Это реакции по¬
ворота головы, глаз, ушных раковин, расширение или сужение зрачков и т. д.
Центральные управляющие влияния обнаружены почти па каждом уровне пе¬
реработки информации, что приводит к изменению порогов обнаружения тех или
иных сенсорных сигналов.
Сущность ренепиии сво¬
дится к преобразованию
энергии воздействующе¬
го стимула в изменения
физико-химических па¬
раметров рецепторных
элементов.
§ 2. Закономерности обнаружения сигналов 53
В обнаружении сенсорных сигналов важную роль выполняют так называемые
вспомогательные структуры рецепторных аппаратов, то есть всех тех дополни¬
тельных приспособлений, которые окружают собственно рецептирующий эле¬
мент (оболочки, оптическая система глаза, глазодвигательные мышцы, наружное
и среднее ухо, основная и покровная мембраны и пр.).
Нервная система влияет на работу рецепторных приборов различными спосо¬
бами:
1) изменив питание рецепторных приборов и окружающих тканей путем измене¬
ния их кровотока;
2) воздействуя па свойства вспомогательных структур;
3) непосредственно меняя функциональное состояние самих рецепторов;
4) меняя активность отходящих от рецепторов чувствительных трактов.
В общей форме задача обнаружения сигнала сводится к выделению нервной
сигнализации из собственного шума. Последний обычно связывают с фоновой
активностью нейронов, хорошо выраженной на разных
уровнях сенсорных систем. Воздействие стимула на вход
системы приводит к изменению нервной активности, а не
к ее возникновению. Так как характеристики фоновой ак¬
тивности флуктуируют и имеют определенное распреде¬
ление вероятности достижения конкретных величин, то при одних и тех же физи¬
ческих параметрах нервная активность будет меняться. При этом в зависимости
от характеристик шума распределение вероятности достижения конкретных ве¬
личии нервной активности может быть различным. Они могут и не отличаться от
величин, характеризующих активность в отсутствие раздражителя. Поэтому воз¬
можность решения задачи обнаружения сигнала основывается на введении крите¬
рия порога. При значении нервной активности больше критерия порога организм
свидетельствует о наличии сигнала. Эта теория позволяет объяснить появление
«ложных тревог» в процессе обнаружения, то есть принятия решения о воздейст¬
вии па вход сенсорной системы при фактическом отсутствии стимула.
В зависимости от положения критерия порога человек практически может не
реагировать па собственные шумы, но при этом уменьшается и вероятность обна¬
ружения сигналов. И наоборот, человек может обнаружить большинство сигна¬
лов, но при этом будет реагировать и па собственные шумы. В зависимости от не¬
обходимости обнаруживать все сигналы или, напротив, не допускать ошибок по¬
ложение критерия порога меняется. Пороги восприятия эмоционально значимых
и нейтральных сигналов также различны.
Критерий наличия или отсутствия сигналов устанавливается под влиянием
доминирующей потребности организма. Следует отличать порог возбуждения
сенсорных систем и порог ощущения. Пороги возбуждения рецепторных элемен¬
тов зависят не только от присущих им собственных свойств, но и от влияния вспо¬
могательных структур, управляющих нервно-гуморальных воздействий. Поэтому
вряд ли можно получить абсолютные величины порогов собственно рецепторных
элементов, имея в виду их непрерывную системную регуляцию.
Итак, обнаружение сенсорных сигналов зависит от различных факторов, свя¬
занных как с функционированием соответствующей сенсорной системы, так
и с интегративной деятельностью целого мозга.
Обнаружение сигна¬
ла — выделение нерв¬
ной сигнализации из
собственного шума.
54 Глава 2. Сенсорная функция мозга
§ 3. Системная организация процессов
кодирования информации
Под кодированием в нервной системе обычно понимается установление соответ¬
ствия между определенными параметрами сенсорного стимула и характеристика¬
ми импульсной активности нейрона и/или местом его расположения. Э го соответ¬
ствие может быть установлено по таким характеристикам импульсной активности,
как ее частота, величина межимпульсных интервалов, сте¬
пень их стабильности, распределение импульсов во вре¬
мени (паттерн разряда нейрона).
Для проблемы кодирования решающим является об¬
наружение зависимости частоты импульсации нейрона от
интенсивности сенсорных стимулов (Дж. Эдриан), кото¬
рая носит 5-образный характер. Следовательно, определе¬
ние амплитудных характеристик осуществляется на осно¬
ве нелинейной шкалы. Такая зависимость предполагает,
что наибольшие изменения частоты импульсации у ней¬
рона проявляются при варьировании интенсивности только в средней части кри¬
вой (динамический диапазон). Эта зависимость позволяет организму при низких
уровнях интенсивности сигналов улавливать незначительные изменения ампли¬
туды, а при высоких — обусловливает нечувствительность к тем же изменениям.
Впервые такой принцип «сжатия» информации был установлен Вебером при
изучении зависимости величины субъективного ощущения от прироста интен¬
сивности раздражения. Эта зависимость, согласно расчетам Фехнера, носит лога¬
рифмический характер (закон Вебера-Фехнера). По данным же количественных
измерений Стивенса, между величиной субъективного ощущения и амплитудой
действующего стимула существует не логарифмическая, а степенная зависи¬
мость. Функция Стивенса описывает широкий класс реакций рецепторов. Спра¬
ведливость того и/или другого закона определяется методами исследования и па¬
раметрами сенсорных стимулов.
Принято считать, что анализ интенсивности стимулов определяется деятель¬
ностью нижних уровней сенсорных систем.
Наряду с наличием определенных областей реагирования нейронов сенсорных
систем имеется возможность для их перестроек, что обеспечивает широту дина¬
мического диапазона, характерного для деятельности системы в целом.
Относительная избирательность, характерная для элементов сенсорных си¬
стем, определяет то, что точный анализ отдельных признаков сенсорных стимулов
в нервной системе не может быть основан на показании одного чувствительного
элемента. Увеличение специфичности реагирования должно осуществляться за
счет совокупной деятельности ряда элементов сенсорной системы.
Структурные особенности нервной системы создают условия для полного пе¬
рекрытия рецептивных полей нейронов. Поэтому весьма важно, что, несмотря на
потенциальную возможность такой широкой дивергенции, импульсация переда¬
ется по весьма определенным нервным каналам, то есть по принципу меченой ли¬
нии, или, как иногда говорят, кодирование сенсорной информации происходит но¬
мером капала (Б. Н. Соколов). Это, в свою очередь, соответствует топической
Кодирование — уста¬
новление соответствия
между определенными
параметрами сенсор¬
ного стимула и харак¬
теристиками импульс¬
ной активности нейро¬
на и/или местом его
расположения.
§ 3. Системная организация процессов кодирования информации 55
организации сенсорных систем, заключающейся в пространственно упорядочен¬
ном расположении нейронов на разных уровнях сенсорных систем согласно ха¬
рактеристикам их рецептивных полей; при этом расположенные рядом участки
рецепторной поверхности проецируются в соседние участки одного и того же
уровня сенсорной системы.
Наличие дивергирующих и конвергирующих нервных связей само по себе
должно было бы вызвать обширнейшую иррадиацию возбуждения, и даже слабый
сенсорный стимул запускал бы цепную реакцию нейронной сети всего мозга. При
этих условиях о тонической организации вообще не могло бы быть и речи, ибо ре¬
цептивных полей у центральных нейронов не существовало бы в принципе.
Важнейшим механизмом, ограничивающим иррадиацию возбуждения, обус¬
ловливающим наличие ограниченных рецептивных полей нейронов и, следова¬
тельно, топическую организацию сенсорных систем, является латеральное тор¬
можение. Оно основано на тормозном взаимодействии как соседних рецепторных
элементов, так и центральных нейронов.
В настоящее время известно, что в сенсорных системах и центральной нервной
системе осуществляется латеральное торможение, то есть возбужденные рецепто¬
ры или нейроны через свои возвратные коллатерали и тормозные встроенные эле¬
менты затормаживают соседние клетки. Благодаря этому возбуждение не ирра-
диирует по нервной сети и обеспечивается феномен обострения (контраста), то
есть увеличение перепада между возбужденными и певозбужденпыми нейрона¬
ми. За счет латерального торможения обеспечивается избирательный характер
реагирования нейрона при воздействии на определенные участки рецептивной по¬
верхности и ее пространственно упорядоченная поверхность в целом.
Так, на различных уровнях слуховой системы обнаружены определенные то¬
пографические соотношения между положением нейронов и частотами заполне¬
ния звуковых стимулов, реакции на которые имеют наименьшее значение порога
(тонотопия). В соматической сенсорной системе рецептивные поля кожи перед¬
них конечностей имеют значительно меньшие размеры, чем задних. Это лежит в
основе высокой разрешающей способности передних конечностей. Следствием
этого является большая протяженность кортикальной поверхности, на которую
проецируются передние конечности и голова, по сравнению с площадью проек¬
ции задних конечностей и туловища (соматотопия).
Таким образом, в сенсорных системах выявлена относительная пространст¬
венная упорядоченность расположения нейронов, соответствующая организации
рецепторной поверхности, то есть проведение возбуждения по связям «точка
в точку».
Топическая организация сенсорных систем базируется на многоканальной
передаче сигнализации в высшие этажи мозга. При этом обнаруживаются спе¬
циализированные каналы по большему числу признаков, чем подразумевает
принцип топической организации. Так, наличие быстро и медленно адаптирую¬
щихся элементов указывает па существование каналов, специализирующихся на
восприятии стационарных и изменяющихся во времени стимулов. В зрительной
и слуховой системах выделяют пространственно разнесенные каналы, связанные
с обработкой информации о локализации источника сенсорных стимулов, их пе¬
ремещении, о хроматических, частотных качествах сигналов и т. д.
56 Глава 2. Сенсорная функиия мозга
Особый интерес представляют каналы, связанные с
восприятием и проведением информации о стабильных и
строго определенных признаках сенсорного сигнала —
детекторы. Детектор — это нейрон, избирательно настро¬
енный на определенный параметр сигнала за счет фикси¬
рованной системы связей его с рецепторами или другими
нейронами более низкого уровня. На детекторе конверги¬
руют несколько каналов, но каждому из которых возбуж¬
дение поступает через синаптический контакт, эффек¬
тивность которого фиксирована. При изменении параметров стимула, действую¬
щего на детектор, его ответ уменьшается.
Из группы нейронов-детекторов выделяют инвариантные детекторы, синап¬
тические связи которых не являются постоянными, а меняются при возбуждении
вспомогательного набора детекторов. Такие детекторы обеспечивают механизм
константности восприятия.
Детекторы, свойства которых модулируются но ходу обработки информации в
сложной рецепторной системе, получили наименование пластических детекто¬
ров. Значение таких детекторов понятно, если принять во внимание, как это уже
отмечалось выше, существование наряду с жесткими сенсорными проекциями
также и пластических каналов связи, проведение и организация нейронных си¬
стем которых зависит от координирующей функции центрального торможения.
Биологическая роль такой организации может состоять в том, чтобы обеспечивать
«преднастройку» каналов сенсорной системы для более полного и адекватного
восприятия информации, возникающей при обследовании объекта (А. А. Алек¬
сандров).
Необходимо специально отметить, что разные уровни сенсорных систем полу¬
чают входы и от других «несенсорных» образований головного мозга. Следова¬
тельно, практически каждое сенсорное образование мозга, начиная с рецепторно¬
го органа, находится под непрерывным контролем как вышерасположенных отде¬
лов данной системы, так и различных других нервных образований. Этот и целый
ряд других признаков вышележащих уровней сенсорных систем опираются на
высокую эффективность процессов торможения как основного фактора, обеспе¬
чивающего как стабильный, так и пластичный характер их реагирования.
Для описания сложных стимулов рассматривается совокупная деятельность
нейронов или их структурно-функциональных объединений (модулей). Существу¬
ют убедительные свидетельства тому, что описания отдельных составляющих
сенсорных сигналов осуществляются совокупной деятельностью нейронных объ¬
единений разных уровней сенсорных систем.
Детектор — это ней¬
рон, избирательно на¬
строенный на опреде¬
ленный параметр
сигнала за счет фикси¬
рованной системы свя¬
зей его с рецепторами
или другими нейронами
более низкого уровня.
§ 4. Распознавание, декодирование информации
Прежде чем произойдет какая-либо приспособительная реакция, осуществляется
процесс восприятия сигналов. Но живой организм заинтересован не в самих воз¬
действиях как в таковых, а в том, о чем они сигнализируют, и соответственно не
в оценке их физических параметров, а в тех соотношениях, которые с их помощью
передаются. Вместе с тем понятно, что отнесение сигнала к тому или иному клас¬
§ 4. Распознавание, декодирование информации 57
Декодирование —
преобразование вход¬
ной кодовой комбина¬
ции активности нерв¬
ных элементов
сенсорных систем в
реакцию исполнитель¬
ных аппаратов.
су, то есть его распознавание, основано на выделении ряда
физических характеристик, образующих область призна¬
ков данного класса сигналов.
Из сказанного выше следует, что сенсорная функция
мозга заключается в определении сигнальной (диалогиче¬
ской) значимости сенсорных стимулов на основании ана¬
лиза их физических характеристик. Под биологической
значимостью понимается направленность реакций живо¬
го организма, которая определяется его доминирующей мотивацией, той, что воз¬
никает при отклонении параметров устойчивого неравновесия, а также информа¬
цией, извлекаемой из окружающей среды и прошлого жизненного опыта.
Для оценки биологической значимости сенсорных сигналов анализ их физиче¬
ских характеристик является необходимой, но недостаточной операцией. Био¬
логическая значимость сигнала реализуется в активации некоей совокупности
эффекторных аппаратов. Следовательно, извлечение биологически полезной
информации с целью ее использования для формирования поведенческих актов
основано на преобразовании входной кодовой комбинации активности нервных
элементов сенсорных систем в реакцию исполнительных аппаратов, что по своей
сути представляет процесс декодирования сенсорных сообщений.
Тогда оценка биологической значимости сигнала сводится к установлению со¬
ответствующей закономерности взаимодействия нервных элементов сенсорных
и двигательных систем мозга (Батуев, Куликов, 1983).
Нервные образования, входящие в систему оценки биологической значимости
сигналов, должны удовлетворять ряду требований.
Во-первых, они должны быть связаны с различными сенсорными системами.
Во-вторых, они должны быть связаны со структурами мозга, генерирующими
мотивационные состояния, ибо значимость тех или иных сигналов определяется
на основе доминирующей мотивации.
В-третьих, учитывая, что значимость одного и того же сенсорного стимула за¬
висит от всей окружающей ситуации, то есть от показаний и других сенсорных
систем, следует допустить наличие конвергенции нолимодалыюй импульсации
и способность к пластической перестройке активности нейронов.
Наконец, в-четвертых, эта система должна быть связана с регуляцией целост¬
ных двигательных актов на основе избирательного характера реагирования.
Изложенным выше требованиям удовлетворяют таламокортикальные ассо¬
циативные системы мозга.
Общим для ассоциативных систем является наличие нейронов, способных
реагировать на возбуждение различных сенсорных входов (полисенсорные нейро¬
ны). Роль ассоциативных систем в оценке биологической значимости сенсорных
сигналов отчетливее видна но отношению к зрению и слуху. Конечно, сигнальное
значение имеют все сенсорные стимулы, по способностью опережающего воздей¬
ствия обладают в первую очередь зрительные и слуховые, а также обонятельные
сигналы.
Для целей определения биологической значимости механизмы конвергенции
обеспечивают полифункциональность одного и того же сигнала, то есть зависи¬
мость его смысла от ситуации, в которой он возникает. Степень значимости лю¬
бого агента внешней среды зависит не только от определенной мотивации, но и от
58 Глава 2. Сенсорная функция мозга
сопоставления этого агента с другими факторами среды, когда он выступает в кон¬
тексте более сложного сенсорного комплекса. Именно поэтому наиболее полная
сигнальная значимость фактора окружающего пространства может реализовы¬
ваться с участием тех структур мозга, куда приходит информация о разных по сен¬
сорным качествам факторах среды, то есть по механизму одновременного гетеро¬
сенсорного сопоставления. Значит, степень совершенства интегративной деятель¬
ности мозга тесно связана со структурной дифференциацией и функциональной
специализацией ассоциативных таламокортикальных систем мозга.
Одним из проявлений динамического характера конвергенции в ассоциатив¬
ные зоны коры является свойство привыкания их электрических реакций при ис¬
пользовании однообразной ритмической стимуляции, что совпадает с закономер¬
ностями проявления ориентировочного рефлекса. Поэтому предполагается, что
«новизна» является тем информативным признаком сенсорных стимулов, с кото¬
рого начинается их оценка в ассоциативных системах
мозга. Привыкание к повторным стимулам, вероятно, свя¬
зано с процессами формирования следов кратковремен¬
ной памяти.
Система, оценивающая биологическую значимость сен¬
сорных стимулов, должна обладать избирательным харак¬
тером реагирования на их отдельные значимые признаки
и/или совокупность последних.
Закономерности избирательного реагирования могут
служить физиологическими предпосылками для трактов¬
ки механизмов сенсорного внимания. В самом общем
смысле внимание рассматривается как один из механиз¬
мов устранения избыточности сенсорных сообщений, ко¬
торый участвует как в избирательном подавлении, обеспечении сенсорных вхо¬
дов, так и в фильтрации информации, извлекаемой из систем кратковременной
и долговременной памяти.
Активный характер сенсорного восприятия, на который обращал внимание
еще И. М. Сеченов, сомнений не вызывает. И. П. Павлов также подчеркивал важ¬
ность механизма внимания.
Оно является следствием определенного состояния живого организма — доми¬
нантного, по А. А. Ухтомскому. Сам А. А. Ухтомский считал, что сформулирован¬
ный им принцип доминанты и есть физиологическая основа акта внимания, когда
из множества действующих рецепций доминанта «вылавливает» группу рецеп¬
ций, которая для нее в особенности значима.
Приведенные выше аргументы позволяют полагать, что определение вектора
поведения (по принципу доминанты) основано на взаимодействии биологически
значимых сенсорных воздействий, основным субстратом установления которого
являются высшие интегративные (в том числе и ассоциативные) системы мозга.
Заключение
Любая сенсорная функция основана на целостной деятельности сенсорных си¬
стем, и выход на исполнительные аппараты получают лишь определенные, соот¬
ветствующие доминирующей мотивации компоненты специфического импульс¬
ного воздействия.
Внимание рассматрива¬
ется как один из меха¬
низмов устранения из¬
быточности сенсорных
сообщений, который
участвует как в избира¬
тельном подавлении,
обеспечении, так и в
фильтрации информа¬
ции, извлекаемой из
систем кратковремен¬
ной и долговременной
памяти.
Контрольные вопросы 59
Классические сенсорные системы со всей сложностью их мозговых конструк¬
ций не способны обеспечить функцию опознания сигналов. Они выполняют
сложные процессы описания сигналов, осуществляя операцию кодирования ин¬
формации, развертывающуюся в многоканальной сенсорной системе — от рецеп¬
торов вплоть до высших уровней ее мозговой организации, и в значительной мере
облегчается наличием набора детекторов.
Операция опознания сигнала, то есть отнесение его к определенному классу
сигналов, записанному в аппаратах памяти, требует дополнительного участия
«внесенсорпых» структур мозга, к которым в первую очередь относятся таламо-
кортикальпые ассоциативные системы. С помощью этих структур происходит
оценка «новизны» и биологической значимости сигнала в соответствии с домини¬
рующей мотивацией, то есть сигнал оценивается как интегрированное целое, не¬
сущее значимую информацию для основной деятельности организма, — выполня¬
ется операция декодирования.
Происходит селективный отбор одних видов информации с одновременным
сопряженным торможением других сенсорных влияний. Доминирующая мотива¬
ция формируется при воздействии стимулов, ранее свя¬
занных с определенными биологическими актами. Чело¬
век способен предвидеть изменение гомеостаза с помощью
сигналов, предвосхищающих сдвиг параметров от нормы
или возвращение к ней. В связи с этим другие сенсорные
стимулы способны изменять значимость сигнала, если они
меняют доминирующую мотивацию. И тогда значимый
стимул является сигналом, вероятность реагирования на
который сопряжена с большой вероятностью достижения
желаемого результата в соответствии с доминирующей
мотивацией и прошлым жизненным опытом. Наличие со¬
пряженных признаков в комплексном сенсорном воздействии позволяет умень¬
шить неопределенность выбора поведенческих реакций, необходимых для полу¬
чения требуемого результата.
В качестве последнего выступает снижение доминирующей мотивации и как
итог — удовлетворение потребности.
Благодаря ассоциативным системам мозга осуществляется использование ши¬
рокополосного приема, подразумевающего необязательность фиксированного
выделения в сенсорных образованиях мозга определенных значимых признаков
сигналов. Потенциальная возможность широкополосного приема при наличии
специальных механизмов выделения значимых (релевантных) признаков сенсор¬
ных сигналов увеличивает адаптивность организма как системы.
Целостная сенсорная функция мозга обеспечивается содружественной дея¬
тельностью сенсорных, моторных и ассоциативных систем и направлена на орга¬
низацию адаптивных движений и действий. Последнее и лежит в основе принци¬
па «согласования движения с чувствованием», выдвинутого еще И. М. Сеченовым.
иелостная сенсорная
функция мозга обес¬
печивается содруже¬
ственной деятельно¬
стью сенсорных,
моторных и ассоциа¬
тивных систем и на¬
правлена на организа¬
цию адаптивных
движений и действий.
Контрольные вопросы
1. Что такое сенсорная система, каково ее строение и функции?
2. Принципы конвергенции и дивергенции в сенсорных системах.
60 Глава 2. Сенсорная функция мозга
3. Принцип обратной связи и его значение.
4. Двусторонняя симметрия в работе сенсорных систем.
5. Организация коркового звена сенсорных систем.
6. В чем состоит сущность рецепции?
7. От чего зависит обнаружение сенсорных сигналов?
8. В чем состоит суть кодирования информации?
9. Что такое нейрон-детектор?
10. В чем состоит основное содержание теории Е. Н. Соколова о нервной модели
стимула?
11. В чем суть декодирования информации?
12. Что такое сенсорная функция мозга?
Глава 3
ФИЗИОЛОГИЯ СЕНСОРНЫХ СИСТЕМ
Введение
Контакт с внешним миром, воздействие его па организм возможны благодаря вы¬
сокоспециализированным нервным аппаратам, получившим название сенсорных
систем. Непосредственный же субстрат, который принимает па себя воздействия
предметов и явлений окружающей среды, — это вынесенные на периферию рецеп¬
торные приборы или, как их иногда называют, органы чувств. Современная ней¬
рофизиология располагает сведениями о работе аппаратов рецепции и высших
этажей мозга, чтобы служить естественно-научной основой для постулирования
положения об адекватности отражения.
Проблема адекватности отражения — одна из самых старых и вместе с тем веч¬
но новых и остро обсуждаемых проблем. Дифференциация раздражений, их дроб¬
ный анализ происходят уже на уровне рецепторных приборов. Последние снабже¬
ны специализированными элементами — датчиками, которые активизируются
лишь в связи с изменениями определенного свойства объекта. Энергия внешнего
раздражения трансформируется благодаря физико-химическим изменениям в
частотно-модулированные и местные процессы качественно однородного харак¬
тера. Многообразие свойств объекта как бы исчезает, превращаясь в «безликий»
нервный процесс. В этом заключается важная особенность отражения для живых
организмов, в нервных клетках которых кодируется не сам материальный носи¬
тель и его энергия, а переданная информация. Частотно-модулированный им¬
пульсный процесс является важным, но не единственным фактором кодирования
пространственно-временной структуры объекта. Большую роль играют также ме¬
стные, нераспространяющиеся нервные влияния типа рецепторного потенциала.
Сенсорная система как аппарат, через который информация поступает в мозг,
функционирует посредством прямых и обратных связей, то есть как система само¬
организации и управления. Она не просто пассивно отражает воздействия, адре¬
суемые к ее рецепторному аппарату, а выступает как активный сигнализатор моз¬
га, сообщающий наиболее существенную информацию. Активная природа дея¬
тельности сенсорных систем не ограничивается центрифугальными влияниями
на рецепторы, а является общим принципом и может быть прослежена на всех си¬
наптических уровнях головного мозга, которые, испытывая на себе влияния вы¬
шележащих уровней, действуют как активные фильтры проходящего импульсно¬
го потока. Процесс фильтрации продолжается и умножается в межнейронных си¬
напсах. Как известно, на одном нейроне в центральной нервной системе могут
заканчиваться несколько тысяч окончаний аксонов других нейронов, по которым
способны одновременно поступать различные влияния. В релейных ядрах проис¬
ходит дальнейшая фильтрация импульсов, отбор из огромного их числа наиболее
важных для организма в данный момент. Обратные связи, представленные в каж¬
дой сенсорной системе, осуществляют настройку рецепторных элементов и пере¬
ключательных аппаратов к более адекватному и полному восприятию внешнего
62 Глава 3. Физиология сенсорных систем
Физиологические про¬
цессы выступают как
носители идеального
содержания лишь в том
случае, когда их ре¬
зультат соотносится
человеком с объектом
отражения. Именно от¬
несенность мозговых
процессов к объектив¬
ному миру и делает эти
процессы психически¬
ми, идеальными.
мира и в то же время обеспечивают избирательную фильтрацию биологически по¬
лезной информации из «шума», то есть комплекса разномодальных признаков.
Рассматривая проблему адекватности, то есть сходства образа и отражаемого
объекта, необходимо учитывать два обстоятельства: во-первых, та или иная сте¬
пень адекватности достигается во времени не мгновенно,
а постепенно; во-вторых, окончательная оценка адекватно¬
сти производится при соотнесении нового образа с его нерв¬
ной моделью, созданной заранее на основе всего комплек¬
са воздействий и прежнего жизненного опыта (памяти).
Естественно, что наибольшей точностью и полнотой
отражения обладает человек с его способностью к абст¬
рактному мышлению. Но тем не менее и у человека образ
предмета не является тождеством самого предмета, образ
всегда оказывается субъективной копией но отношению
к предмету как объективному началу. Образ идеален и
функционален, в нем не содержится вещественности са¬
мих предметов, а лишь их пространственно-временная
структура, упорядоченность, т. е. информация. И наконец, образ предметен, ибо
является отражением свойств определенного конкретного предмета.
Естествен вопрос: на каком этапе непрерывные физиологические преобразова¬
ния исчерпывают свое назначение и порождают психический процесс? Весь про¬
цесс абстрагирования копии от ее носителя, порождающий субъективно реальное,
идеальное, есть процесс психический. Иными словами, физиологические процес¬
сы головного мозга выступают как носители идеального содержания лишь в том
случае, когда их результат соотносится человеком с объектом отражения. Именно
отнесенность мозговых процессов к объективному миру и делает эти процессы
психическими, идеальными.
При известных допущениях правомочно говорить о локализации функций
сенсорной системы, имея в виду ее ограниченную топическую конструкцию. Иное
дело, когда речь идет о таком интегративном понятии, как «зрительная функция»,
во всей сложности и многообразии ее проявлений. Локализовать такую функцию
экспериментально невозможно и теоретически неверно, потому что всякая слож¬
ная психическая функция нолисенсорна. Локализовать мозговые структуры, не¬
обходимые и достаточные для осуществления целостной функции организма,
нельзя, и в этом нет логической необходимости. Локализованы лишь сенсорные
системы как аппараты ощущений, па базе которых возникает нервная модель, а за¬
тем — и полисенсорный субъективный предметный образ.
§ 1. Физиология рецепторов
Рецепторы — эго специализированные образования, выполняющие функцию
преобразования энергии внешнего раздражителя в специфическую активность
нервной системы, в сигналы, несущие нервным центрам информацию о раздра¬
жающем агенте. По существу, в рецепторах имеет место не энергетическое, а ин¬
формационное преобразование, с помощью которого на основе разных форм
физической энергии стимула (механической, химической и др.) формируется
§ 1. Физиология реиепторов 63
единообразный для всех сенсорных систем нервный код, представляющий собой
временную дисперсию следующих друг за другом потенциалов действия.
В зависимости от того, связаны ли рецепторные нервные окончания с внешней
средой или воспринимают раздражения из внутренней среды, различают внеш¬
ние — экстерорецепторы и внутренние рецепторы, или
интерорецепторы. Типичным примером интерорецепто-
ров являются нервные окончания, расположенные в стен¬
ках сосудов и воспринимающие химические или механи¬
ческие сигналы. Одной из разновидностей интерорецеп-
торов можно считать рецепторы опорно-двигательного
аппарата, сигнализирующие о положении или перемеще¬
нии частей тела в пространстве. Их иногда выделяют в от¬
дельную группу проприорецепторов.
В зависимости от того, возбуждаются ли рецепторы
при непосредственном контакте с источниками раздраже¬
ния или при действии раздражения па расстоянии, разли¬
чают контактные и дистантные рецепторы. К дистантным относят рецепторы
органов слуха, зрения, обоняния, к контактным — кожные и вкусовые рецепторы,
а также рецепторы внутренних органов.
Согласно наиболее удачной классификации, рецепторы подразделяются в за¬
висимости от физической энергии стимула, которая для данного рецептора явля¬
ется адекватной.
Механорецепторы. Адекватной формой энергии является механическая энер¬
гия, поэтому к мехапорецепторам относят рецепторы органов слуха, вестибуляр¬
ного аппарата, рецепторы кожи, проприорецепторы, барорецепторы сердечно-со¬
судистой системы.
Хеморецепторы. К ним относятся рецепторы вкуса и обоняния, сосудистые
и тканевые рецепторы, чувствительные к действию химических веществ, а также
хеморецепторы самой центральной нервной системы.
Фоторецепторы. К ним относятся рецепторы органа зрения, реагирующие на
электромагнитные колебания в определенном диапазоне длин воли.
Терморецепторы. Реагируют на температуру и ее изменение во времени и рас¬
полагаются в коже, внутренних органах и некоторых участках центральной нерв¬
ной системы (в частности, в гипоталамусе).
Рецепторные приборы обеспечивают каждый живой организм достаточным
количеством информации, необходимой для его нормального существования, так
как они приспособлены к восприятию именно тех сигналов, которые наиболее
важны для данного животного в данной конкретной среде обитания.
Среди рецепторных клеток сенсорных систем различают первично чувствую¬
щие и вторично чувствующие клетки. Первично чувствующие клетки отличаются
тем, что они снабжены специализированным воспринимающим раздражение пе¬
риферическим отростком, увенчанным подвижным жгутиком, или ресничкой,
и центральным отростком, по которому возникшее возбуждение передается в ви¬
де импульса в соответствующие части центральной нервной системы. Предпола¬
гают, что в процессе эволюции из первично чувствующих клеток путем их расще¬
пления развилась нервная система (Заварзин, 1941). У позвоночных зрительные
и обонятельные клетки развиваются как части самой нервной системы. Вторично
Рецепторы — это спе¬
циализированные об¬
разования, выполняю¬
щие функцию пре¬
образования энергии
внешнего раздражите¬
ля в специфическую
активность нервной
системы, в сигналы, не¬
сущие нервным цент¬
рам информацию о
раздражающем агенте.
Волосковая клетка Волосковые клетки утрикулюса
боковой линии
или утрикулюса
Рыбы
Млекопитающие Птицы
Волосковые клетки
кортиева органа
Вкусовые клетки
Млекопитающие
Птицы Насекомые Вкусовая луковица
позвоночных
Рис. 5. Схема структурной организации рецепторных клеток органов чувств
64 Глава 3. Физиология сенсорных систем
§ 1. Физиология рецепторов 65
чувствующие клетки не имеют ни периферического, ни центрального отростков.
Восприятие стимула осуществляется при помощи жгутикообразных волосков
или ресничек, а передача возбуждения — путем синаптического механизма за счет
охватывающего их конечного разветвления чувствительного нервного волокна
(рис. 5).
Таким образом, все рецепторные клетки снабжены жгутиком или ресничками,
которые можно рассматривать в качестве непосредственных датчиков информа¬
ции. Жгутики или реснички способны к автоматическому безостановочному дви¬
жению и выполняют роль подвижных универсальных антенн, они характеризуют¬
ся также сходной цитохимической и субструктурной организацией. Эти реснички
в своем составе содержат девять пар периферических и одну пару центральных
фибрилл. Центральные фибриллы выполняют опорную функцию, а перифериче¬
ские, содержащие макромолекулы миозина, — сократительную. По сути, в основе
работы рецепторных клеток имеют место два процесса: механохимический, обеспе¬
чивающий автоматическое движение антенн-датЧиков, и специфический, обуслов¬
ливающий взаимодействие того или другого вида энергии со специфическими
биологически активными веществами, локализованными в мембранах антенн или
других органоидах клетки (зрительных пигментов в фоторецепторах, фермента
ацетилхолинэстеразы в клетках кортиева органа, неспецифических фосфотаз во
вкусовых клетках и т. д.).
Акт рецепции всегда активен. Он связан с движениями, способствующими
наилучшему контакту с адекватным стимулом, непрерывными поисками этого
стимула и обеспечения наилучших условий взаимодействия с ним.
Практически все сенсорные аппараты снабжены вспомогательными структу¬
рами, а часто и обычными тканевыми элементами, в которых локализуются ре¬
цепторы. Все они существенно изменяют характеристики раздражения на его пу¬
ти к рецептору. Адекватный раздражитель взаимодействует с рецептором, вызы¬
вая развитие в нем специфических трансформационных процессов, следствием
которых является изменение проницаемости поверхностной мембраны клетки.
В общем виде физико-химические изменения па уровне рецептора выгля¬
дят так:
1. Под влиянием внешнего стимула происходит высвобождение ионов Са++.
2. Ионы Са++ действуют на фермент адепилатциклазу, переводя ее из неактивной
формы в активную.
3. Аденилатциклаза действует на аденозинтрифосфат (АТФ), отщепляя от него
два фосфорных остатка, в результате чего образуется циклический аденозип-
монофосфат (АМФ) и выделяется определенное количество энергии.
4. Циклический АМФ активирует фермент протеипкипазу.
5. Протеинкиназа, действуя па структурные макромолекулы белка, вызывает их
фосфорилирование, что ведет к изменению конформации белковых молекул.
6. Изменение конформации микротрубочек белковых молекул ведет к измене¬
нию избирательной проницаемости мембраны рецепторной клетки и появле¬
нию ионных токов.
7. Появление ионных токов через рецепторную мембрану приводит к генерации
первичного электрического ответа рецептора — рецепторного потенциала.
3 За*. 1078
66 Глава 3. Физиология сенсорных систем
В результате изменения проницаемости клеточной мембраны изменяется ион¬
ный ток, о котором обычно мы судим по развитию рецепторного потенциала. Воз¬
никающие ионные потоки действуют, в свою очередь, на электрически возбуди¬
мые элементы рецептора, благодаря чему в первичночувствующих рецепторах,
в зоне, наиболее приспособленной к возникновению спайков, появляются потен¬
циалы действия, распространяющиеся к центру.
В рецептивных клетках вторичночувствующих рецепторов изменение потен¬
циала клетки приводит к выделению химического медиатора, действующего па
нервное окончание сенсорного нейрона, и вызывает появление в нем локального
электрического ответа — постсипаптического потенциала. Его деполяризующее
воздействие приводит к возникновению в нем импульсной активности, именуе¬
мой генераторным потенциалом. Если в первично чувствующих рецепторах оба
потенциала — рецепторный и генераторный — совпадают, то во вторично чувст¬
вующих протекание специфических трансформационных процессов и область ге¬
нерации нервных импульсов находятся в разных клетках (рис. 6).
Рис. 6. Схематическое изображение ответов первичного (а )
и вторичного (б) рецепторов на раздражение
а — тельце Пачини (по О. Б. Ильинскому, 1972);
6 — волосковый механорецептор боковой линии (по Flock, 1965).
1 — вспомогательные структуры (капсула) рецептора; 2 — немиелинизированная
часть нервного окончания; 3 — миелинизированное нервное волокно.
РП — рецепторный потенциал; ПД — потенциал действия; РК — реиептируюшая
клетка и ее афферентное (АН) и эфферентное (ЭН) нервные волокна;
С — синапс. Стрелками показано направление раздражения
§ 2. Кодирование видов информации 67
Организм, получая от рецепторов сигналы о дейст¬
вующем раздражителе, в свою очередь может оказывать
на них регулирующее влияние. Оно может быть опосре¬
дованным — гуморальным и прямым — нервным. Чаще
всего сенсорные аппараты находятся под некоторым то¬
ническим тормозным влиянием. Тормозные волокна вы¬
деляют медиатор (вероятно, гамма-аминолмасляную ки¬
слоту), который приводит к изменению проницаемости
мембраны к ионам К+ и Са++. Многие рецепторы достига¬
ют максимального возбуждения при определенном на¬
правлении раздражающего воздействия, при другом же
направлении возбуждение минимально или даже тормо¬
зится. Это свойство рецепторов связано с особенностями
его строения.
Важнейшей характеристикой рецепторов являются
их рецептивные поля. Их можно рассматривать в макро-
и микроаспектах организации. Макрорецептивное поле
представляет собой чувствительную поверхность, где
располагаются окончания всех афферентных волокон первого порядка какой-то
одной сенсорной системы. Микрорецептивные ноля, или просто периферические
рецептивные поля, связаны как с одиночными нервными волокнами, так и с от¬
дельными нейронами в восходящих афферентных системах. Важную роль играет
перекрытие рецептивных нолей (одни и тот же рецептор может быть связан с дву¬
мя или более нейронами вышележащих уровней нервной системы). Это имеет
принципиальное значение для механизмов пространственной оценки сенсорных
стимулов.
В общем виде последовательность явлений, протекающих в рецепторах при
возбуждении, может быть представлена в виде следующей схемы (рис. 7).
Из повседневного опыта, подкрепленного данными точных экспериментов, хо¬
рошо известно, что при длительном воздействии различных видов стимулов (све¬
тового, температурного, звукового и др.) ощущение постепенно уменьшается. Это
явление, чрезвычайно характерное для различных сенсорных систем, носит назва¬
ние адаптация. В его основе лежат весьма сложные процессы, протекающие как
в рецепторных приборах, так и в центральных отделах нервной системы.
Рецептивные поля —
макрорецептивное по¬
ле представляет собой
чувствительную по¬
верхность, где распола¬
гаются все окончания
афферентных волокон
первого порядка какой-
либо одной сенсорной
системы. Микрорецеп¬
тивные поля, или про¬
сто периферические
рецептивные поля, свя¬
заны как с одиночны¬
ми нервными волокна¬
ми, так и с отдельными
нейронами в восходя¬
щих афферентных си¬
стемах.
§ 2. Кодирование видов информации
Количество информации зависит не только от физических характеристик дейст¬
вующего стимула, но и от состояния самого организма в данный момент времени,
от доминирующей потребности, эмоционального «фона» и т. д.
Информативными параметрами раздражителей для разных сенсорных систем
могут быть их различные свойства и качества. Наряду с этим можно выделить па¬
раметры сенсорного стимула, которые являются основными для всех сенсорных
систем. Таковыми являются:
♦ качественное своеобразие (модальность) раздражителя;
♦ амплитудные (силовые) характеристики — интенсивность;
Чувствительный
орган
Нерв
«Центр»
Вспомогательные
структуры
Рецептирующая
клетка
Синапс
Сенсорный нейрон
Дендрит Начальный сегмент Сома Аксон Терминалы
Синапс
Внешняя энергия
Рецепторный потенциал
Защищает
Проводит
Концентрирует
Анализирует
Тормозит
Повышает
чувствительность
Увеличивает
избирательность
Процесс
трансфор¬
мации
Преобразует
Процесс
генерации
Медиатор
Передает
Усиливает
Секретирует
Постсинаптический
(генераторный)
потенциал
Процесс Процесс
трансформации генерации
Преобразует
Усиливает
Генерирует
Градуальный
электрический
и химический
ответ
Г радуальный
электрический
ответ
И
Нервный импульс
Интегрирует
Проводит
Ответ«все»СекретиРУет
или «ничего»
Медиатор
Передает
*
ит. д.
Кодирует
Декодирует
Периферическое взаимодействие
Эфферентный нейрон
Рис. 7. Общая схема процессов, происходящих в сенсорном органе (модифицированная схема по Davis, 1961)
68 Глава 3. Физиология сенсорных систем
§ 2. Кодирование видов информации 69
♦ временные параметры (начало и конец действия стимула, продолжительность
действия, частота и пр.);
♦ пространственные характеристики (пространственная локализация источника
раздражения, его удаленность, размеры, форма, направление движения и пр.);
♦ пространственно-временные (скорость перемещения стимула);
♦ амплитудно-временные характеристики (изменение интенсивности действую¬
щего стимула во времени).
Модальность — поня¬
тие не физическое, а
субъективное. Под мо¬
дальностью понимают
совокупность сходных
сенсорных впечатле¬
ний, обеспечиваемых
определенным орга¬
ном. Модальность
можно определить как
качественное своеоб¬
разие сенсорного раз¬
дражителя.
Кодирование модальности. Модальность — понятие не физическое, а субъек¬
тивное. Под модальностью понимают совокупность сходных сенсорных впечатле¬
ний, обеспечиваемых определенным органом. Модаль¬
ность можно определить как качественное своеобразие
сенсорного раздражителя. Качество субъективного ощу¬
щения не столько зависит от физических характеристик
внешнего раздражителя, сколько связано со спецификой
самих органов чувств (Мюллер). За сотни миллионов лет
эволюции сформировались высокоспециализированпые
нервные образования, реагирующие оптимальным обра¬
зом на вполне определенные формы физической энергии
раздражителя.
Качественное своеобразие (модальность) раздражите¬
ля кодируется в рецепторах и связано с их специфично¬
стью и избирательностью к тому или иному виду сенсор¬
ного воздействия. При этом не следует забывать и роль центральных структур
мозга, в которых происходит окончательная переработка информации о стимуле.
Кодирование интенсивности. Каждая сенсорная система настроена на воспри¬
ятие в определенном диапазоне интенсивностей стимула. Существуют стимулы
настолько малой интенсивности, что раздражаемый ими рецептор не будет возбу¬
ждаться. Такой стимул называется подпороговъш. Стимул большей интенсивно¬
сти вызывает слабое возбуждение рецептора. Этот стимул называется пороговым.
Порог субъективного ощущения всегда выше порога возбуждения рецептора.
Интенсивность стимула, превышающая пороговый уровень, кодируется на
уровне рецептора разными способами. Как правило, с увеличением интенсивно¬
сти стимула увеличивается и частота импульсации. Сила стимула оценивается
количеством возникающих в рецепторе импульсов. Кроме того, с увеличением
интенсивности стимула в реакцию вовлекается все большее число активирован¬
ных рецепторов. При возрастании силы раздражителя увеличивается вероятность
появления возбудимых рецепторов.
Кодирование пространственных характеристик. Пространственные характе¬
ристики действующего объекта (размер, форма, удаленность, местоположение)
могут передаваться лишь упорядоченными структурами нервных окончаний. Та¬
кая упорядоченная совокупность рецепторов, связанная с одним и тем же нейро¬
ном вышележащего уровня, получила название рецептивного поля, которое может
иметь разную форму и величину, включать возбудимые и тормозные зоны и пере¬
страиваться в зависимости от стимуляции и влияний центральных структур.
Локализацию источника звука можно установить но неодинаковой силе возбу¬
ждения рецепторов правой и левой стороны. Своеобразно кодируется информа¬
70 Глава 3. Физиология сенсорных систем
ция о направлении движения действующего объекта, которая определяется на¬
правленной чувствительностью некоторых рецепторных образований.
Зрительная оценка удаленности объекта осуществля¬
ется за счет бинокулярного зрения.
Кодирование временных характеристик. Протека¬
ние процессов во времени кодируется ответами рецепто¬
ров на начало или окончание действия раздражителя,
а иногда и па то и другое. Временные интервалы боль¬
шей длительности (минуты, десятки минут) кодируются
с помощью иных механизмов. Например, слуховая сис¬
тема предназначена для восприятия сложных звуковых
сигналов, определенным образом структурированных во
времени (И. М. Сеченов). Большую роль играют ритми¬
ческие процессы, протекающие во внутренних органах.
Кодирование скорости движения. Скорость движе¬
ния, как правило, кодируется быстротой последователь¬
ного возбуждения рецепторов внутри данного рецептивного ноля. Информация
от таких рецепторов поступает к центральному нейрону, который является спе¬
циализированным детектором скорости.
Упорядоченная совокуп¬
ность рецепторов, свя¬
занная с одним и тем же
нейроном вышележаще¬
го уровня, получила на¬
звание рецептивного
поля, которое может
иметь разную форму и
величину, включать воз¬
будимые и тормозные
зоны и перестраиваться
в зависимости от стиму¬
ляции и влияний цент¬
ральных структур.
§ 3. Зрительная система
Зрительное восприятие — сложный многоступенчатый акт, который начинается
формированием изображения на сетчатке и заканчивается возникновением зри¬
тельного образа в структурах головного мозга.
Зрительная система состоит из:
♦ периферического отдела, в который входит глаз с его основными аппаратами —
оптическим, глазодвигательным и сетчаточным;
♦ подкоркового отдела, куда относят наружное коленчатое тело, верхние бугры
четверохолмия и некоторые другие образования;
♦ зрительной коры.
Все уровни зрительной системы соединены друг с другом проводящими пу¬
тями.
Глаз
Глаза снабжены большим числом вспомогательных приспособлений для их защи¬
ты. Это брови, благодаря которым стекающий со лба пот не попадает в глаза. Веки
и ресницы защищают глаза от пыли. Веки постоянно смыкаются и размыкаются
(моргание), равномерно смачивая поверхность глаза слезной жидкостью. Слезы
'образуются в слезных железах, расположенных в наружной части глазницы над
глазом. Излишки слезной жидкости стекают в носовую полость через слезный
проток. Секрет слезных желез действует не только как смазывающая, но и как дез¬
инфицирующая жидкость.
Глаз человека имеет шарообразную (или близкую к таковой) форму, что делает
возможным вращение его в определенных пределах в глазнице, а следовательно,
§ 3. Зрительная система 71
наведение в нужном направлении. По¬
следнее осуществляется с помощью 3 пар
мышц, которые иннервируются III, IV
и VI черепно-мозговыми нервами. Глаз
образован несколькими оболочками.
Склера — плотная наружная непрозрач¬
ная соединительнотканная оболочка —
в передней части глазного яблока пере¬
ходит в прозрачную роговую оболочку,
или роговицу. Под склерой лежит сосу¬
дистая оболочка, которая образована се¬
тью кровеносных сосудов. Кпереди сосу¬
дистая оболочка утолщается и переходит
сначала в ресничное тело и далее — в ра¬
дужную оболочку, которые состоят из
гладких мышечных волокон, кровеносных
сосудов и пигментных клеток. Именно эта
оболочка определяет цвет глаз. Мышеч¬
ные волокна ресничного тела прикрепле¬
ны к склере. В центре радужной оболоч¬
ки имеется отверстие — зрачок. С внут¬
ренней стороны сосудистой оболочки находится слой клеток пигментного эпите¬
лия, а к нему прилегает самая внутренняя из оболочек глаза — сетчатая оболочка,
или ретина, выполняющая основную функцию глаза — преобразование светового
раздражителя в нервное возбуждение и первичную обработку сигнала. Волокна
самой внутренней части сетчатки переходят в зрительный нерв. Между роговой и
радужной оболочками имеется полость, наполненная жидкостью, — передняя ка¬
мера глаза. За радужной оболочкой находится прозрачное тело, имеющее форму
двояковыпуклой линзы, — хрусталик, который прикреплен к ресничному телу. За
хрусталиком вся полость глазного яблока заполнена студенистым содержимым —
стекловидным телом (рис. 8).
Оптическая система глаза
Непосредственно за зрачком располагается прозрачный хрусталик, который эла¬
стичен, он может менять свою кривизну благодаря специальным мышцам. Свето¬
вые лучи от предметов проходят через зрачок, хрусталик и стекловидное тело.
У людей с нормальным зрением лучи попадают точно на сетчатку и образуют на
ней четкие изображения предметов. Но одновременно видеть с одинаковой четко¬
стью близко и далеко расположенные предметы мы не можем. В каждый момент
времени хрусталик глаза приспосабливается либо к ближнему, либо к дальнему
видению. Это достигается изменением кривизны хрусталика.
Изображение на сетчатке получается хотя и четким, но перевернутым. Почему
же тогда мы не видим все вокруг нас перевернутым вверх ногами? Один австрий¬
ский ученый изобрел специальные очки, переворачивающие изображение на сет¬
чатке. Он их носил постоянно. Первое время он видел все предметы вверх ногами,
по вскоре вновь научился видеть их нормально. В этих очках он смог даже на¬
учиться ездить на велосипеде. Но стоило ему снять очки, как первое время он сно¬
Рис. 8. Строение глазного яблока
1 — мышиа; 2 — стекловидное тело; 3 — белоч¬
ная оболочка; 4 — сосудистая оболочка; 5 —
пигментный слой; 6 — сетчатка; 7 — желтое пят¬
но; 8 — слепое пятно; 9 — зрительный нерв;
10 — радужная оболочка; 11 — хрусталик; 12 —
передняя камера; 13 — роговица; 14 -— связки
хрусталика
72 Глава 3. Физиология сенсорных систем
ва видел все окружающие предметы перевернутыми. Значит, такая особенность
нашего глаза исправляется с помощью обучения и тренировки, в которой участву¬
ют не только зрительная, по и другие сенсорные системы. Следовательно, зри¬
тельное восприятие окружающего мира основывается не только на самих зри¬
тельных ощущениях, а использует сведения и от других сенсорных систем. Среди
них главную роль выполняют органы равновесия, мышечного и кожного чувства.
В результате взаимодействия этих сенсорных систем возникают целостные обра¬
зы предметов и явлений.
У дальнозорких и близоруких людей с нарушениями оптической системы гла¬
за параллельный пучок лучей от удаленного источника сходится не на поверхности
сетчатки, а дальше или ближе ее (рис. 9). В обоих случаях изображение получает¬
ся размытым. Если же объект наблюдения приближен и лучи от него непараллель¬
ны, то вступает в действие механизм аккомодации, то есть увеличение кривизны
поверхности хрусталика, которое уменьшает его светопреломление и уменьшает
заднее фокусное расстояние, что позволяет сфокусировать объект па сетчатке.
Врожденные дефекты оптической системы глаза связаны с неправильной формой
глазного яблока, а приобретенные — с нарушением эластичности хрусталика. Эти
нарушения исправляются с помощью дополнительных линз — очков.
Рис. 9. Врожденные близорукость и дальнозоркость. Их исправление
Сетчатка глаза
Сетчатка имеет толщину 0,15-0,20 мм и состоит из нескольких слоев нервных
клеток. Первый слой сетчатки непосредственно прилегает к черным пигментным
клеткам. Этот слой образован зрительными рецепторами — палочками и колбоч¬
ками. В последних происходит трансформация световой энергии в нервное возбу¬
ждение. Это осуществляется с помощью зрительных пигментов, содержащихся в
наружных сегментах палочек и колбочек. Пигмент палочек называется родопси¬
ном, колбочек — иодопсипом. Зрительный пигмент является окрашенным бел¬
ком. Разложение пигмента после поглощения кванта света сопровождается поми¬
мо фотохимических процессов возникновением так называемого раннего рецеп¬
торного потенциала. Вслед за ним развивается поздний рецепторный потенциал
во внутреннем членике рецептора. Этот потенциал запускает всю последующую
цепь преобразований в зрительной системе. В сетчатке глаза человека палочек
в десятки раз больше, чем колбочек. Палочки возбуждаются очень быстро слабым
сумеречным светом, но не могут воспринимать цвет. Колбочки возбуждаются
медленнее и только ярким светом, они способны воспринимать цвет. Палочки
§ 3. Зрительная система 73
сравнительно равномерно распределены по сет¬
чатке. Прямо напротив зрачка в сетчатке распо¬
лагается желтое пятно, в состав которого входят
исключительно колбочки. Поэтому наиболее
отчетливо мы различаем те предметы, изобра¬
жения которых попадают прямо на желтое
пятно. С помощью глазных мышц мы можем
управлять движением глаз и изменять направ¬
ление взора. Но всегда при рассматривании но¬
вого предмета происходит перемещение взора
так, чтобы изображение частей предмета по¬
следовательно попадало па желтое пятно.
Центральная часть сетчатки представлена
биполярными клетками с двумя относительно
длинными отростками, одним из которых они
контактируют с фоторецепторами, другим —
с ганглиозными клетками сетчатки, составляю¬
щими ее внутреннюю часть (рис. 10). Таким
образом, фоторецепторы, биполяры и гангли¬
озные клетки представляют собой три последовательных звена переработки зри¬
тельной информации. Между фоторецепторами и биполярами имеются специа¬
лизированные клетки с горизонтальным расположением отростков, которые ре¬
гулируют передачу возбуждения от рецепторов к биполярам и носят название
горизонтальных. Между биполярами и ганглиозными клетками имеются так
называемые амакриновые клетки, которые управляют передачей электрических
сигналов от биноляров к ганглиозным клеткам. Согласно современным нредстав-
лениям, клетки внутреннего ядерпого слоя (биполярные, горизонтальные и амак-
риповые) являются основным источником электроретинограммы сетчатки. Нако¬
нец, аксоны ганглиозных клеток формируют зрительный нерв, который пронизы¬
вает сетчатку в противоположном направлении и входит в полость черепа. Место
его вхождения в сетчатку лишено рецепторов и называется поэтому слепым пят¬
ном. Большинство ганглиозных клеток сетчатки имеют концентрические рецеп¬
тивные поля: при освещении одной из зон ноля ганглиозная клетка возбуждается,
а при ее затемнении — тормозится. Центральную зону рецептивного поля окружа¬
ет концентрическая тормозная зона. Тормозная кайма образуется по механизму
латерального торможения — чем сильнее возбужден центр рецептивного ноля,
тем большее тормозное влияние он оказывает па периферию. Каждая ганглиозная
клетка получает возбуждающие и тормозные влияния от колбочек, имеющих раз¬
ную цветовую чувствительность. Например, если красные колбочки оказывают
возбуждающее действие па данную ганглиозную клетку, то синие ее затормажива¬
ют. Такие комбинации могут быть самыми различными. Вблизи места вхождения
в головной мозг зрительные нервы обеих глаз образуют зрительный перекрест —
хиазму, в результате чего часть волокон переходит на противоположную сторону.
После перекреста левые и правые зрительные тракты направляются к различным
ядрам среднего и промежуточного мозга. Основными центрами переработки зри¬
тельной информации являются наружное коленчатое тело, верхние бугры четве¬
рохолмия и зрительная кора.
Рис. 10. Схема синаптической
организации сетчатки
1 — рецептор; 2 — горизонтальные клет¬
ки; 3 — биполяр; 4 — амакриновые клет¬
ки; 5 — ганглиозная клетка
74 Глава 3. Физиология сенсорных систем
Наружное коленчатое тело
К наружному коленчатому телу приходят волокна только от половины сетчатки
каждого глаза. В разные слои наружного коленчатого тела приходят волокна от
разных сетчаток, эти проекции расположены одна под другой, что позволяет вы¬
делить колонкообразный участок, пересекающий все слои коленчатого тела, кото¬
рый соответствует проекции одной точки ноля зрения.
19
Рис. 11. Схема строения высших отделов зрительной системы
I—VII — слои коры; 17, 19 — корковые поля; П — подушка зрительного бугра;
НКТ — наружное коленчатое тело; ПД — переднее двухолмие; X — хиазма;
ЗТ — зрительный тракт
В структуре рецептивных полей нейронов наружного коленчатого тела отра¬
жаются свойства рецептивных полей ганглиозных клеток сетчатки, в том числе
и их цветовая чувствительность. Наружное коленчатое тело — это первый уровень
конвергенции двух сетчаток, что является необходимым условием стереоскопиче¬
ского восприятия трехмерного мира.
Верхнее двухолмие
Одной из ведущих структур, обеспечивающих ориентировочное поведение, явля¬
ется верхнее двухолмие. В верхних слоях двухолмия имеется упорядоченная про¬
екция сетчатки (ретинотопия). В нижних слоях серого вещества двухолмия име¬
ются так называемые моторные ноля, которые максимально активируются при
§ 3. Зрительная система 75
быстром движении (саккаде) глаза в определенном направлении. При рассматри¬
вании различных объектов внешнего мира глаза совершают непроизвольные бы¬
стрые саккады и медленные произвольные следящие движения. Большая часть
нейронов реагирует только на определенное направление движения, преимущест¬
венно в горизонтальной плоскости, и сила реакции зависит от скорости движения.
Нейроны промежуточных слоев имеют упорядоченные соматические проекции
передней части тела, а также упорядоченные проекции слухового пространства.
Кора больших полушарий
Зрительная кора имеет слоистую структуру и подразделяется на шесть слоев.
В состав зрительной коры входят поля 17 (первичные), 18 и 19 (ассоциативные).
Синаптические связи в зрительной коре весьма разнообразны. Особенностью ре¬
акций одиночных нейронов зрительной коры является их избирательное реагиро¬
вание на определенным образом ориентированные стимулы (лучше всего — на
светлые полосы на темном фоне или пространственные решетки, состоящие из че¬
редующихся светлых и темных полос). Большая часть нейронов отвечает лишь на
определенным образом ориентированные стимулы. Границы простых рецептив¬
ных полей имеют прямоугольную форму, состоят из центра и периферии, границы
которых параллельны друг другу. Реакция на предпочитаемое направление дви¬
жения стимула выражена сильнее всего. Нейроны со сложными рецептивными
полями лучше отвечают на полоску или решетку, оптимально ориентированную
относительно сетчатки. Нейроны сверхсложного типа могут отвечать на несколь¬
ко положений полосы, угол, кривизну контура или более сложные признаки. По-
видимому, имеет место конвергенция нейронов с простыми рецептивными поля¬
ми па сложные и далее сверхсложные нейроны. В 17-м иоле преобладают нейроны
с простыми рецептивными нолями, а в 18-м и 19-м нолях — со сложными и сверх¬
сложными полями. На основании этих фактов Хыобел и Визель выдвинули де¬
текторную гипотезу о том, что нейроны с простыми рецептивными полями, явля¬
ясь детекторами элементарных признаков зрительного изображения, конвергиру¬
ют с нейронами более высокого уровня, которые в результате этого приобретают
более сложные свойства. Сверхсложные нейроны, ответственные за опознание це¬
лостных зрительных образов (лицо, предмет и нр.), расположены за пределами
собственно зрительной коры, и в первую очередь — в пижневисочной области (ас¬
социативная кора).
Альтернативной является гипотеза Кемпбелла и В. Д. Глезера, согласно кото¬
рой нейроны зрительной коры определяют две характеристики зрительного изо¬
бражения — ориентацию стимула и его пространственную частоту. Нейроны же
ассоциативных зрительных областей формируют крупные подобразы зрительно¬
го изображения. Целостные же зрительные образы, их опознание и оценка биоло¬
гической значимости осуществляется в заднетеменпой, пижневисочной и лобной
областях коры.
Зрительное восприятие
Мы видим двумя глазами. И зрительная система состоит из двух совершенно оди¬
наковых симметричных отделов. Оба зрительных нерва вступают в соединение
с мозговыми центрами промежуточного мозга. Самые высшие зрительные центры
расположены в коре больших полушарий. В зрении участвуют оба полушария го-
76 Глава 3. Физиология сенсорных систем
ловиого мозга, каждое из которых получает информацию как от правого, так и от
левого глаза.
Важнейшим преимуществом зрения двумя глазами является способность вос¬
принимать предметы в объемном изображении и оценивать их относительную
удаленность в пространстве. Такая способность получила название стереоскопи¬
ческого зрения. Совместная работа обоих мозговых полушарий обеспечивает раз¬
личение предметов, их формы, величины, расположения, перемещения.
Один и тот же предмет внешнего мира один глаз воспринимает под одним
углом, а другой — под другим. Когда мы смотрим сначала левым, а потом правым
глазом, предмет как бы смещается в поле зрения на какое-то расстояние. Благода¬
ря работе мозга эти два изображения сливаются в одно объемное.
Эффект объемного пространства может возникнуть в тех случаях, когда мы
обоими глазами рассматриваем одну плоскую картину. В течение нескольких ми¬
нут рассматривайте одинаковые кружки (рис. 12) па расстоянии 15-20 см от глаз.
После некоторых усилий у вас появится объемное восприятие: ряд кружков ока¬
жется на переднем плане, некоторые — на среднем плане, остальные — на заднем.
По мере тренировки этот эффект начинает возникать быстрее. Вы сможете даже
перемещать взор но рисунку, а восприятие кружков на разном удалении от глаз
сохранится. А теперь попробуйте достичь этого эффекта, закрыв один глаз, — все
ваши усилия будут напрасны.
Рис. 12. Опыт, основанный на стереоскопическом зрении
Когда мы рассматриваем свои глаза в зеркале, можем обратить внимание на то,
что и крупные, и едва заметные движения оба глаза осуществляют строго одно¬
временно и в одном и том же направлении. Если попытаться зарегистрировать пу¬
ти движения глаз при рассматривании какого-либо предмета, как это сделал рус¬
ский биофизик Ярбус, то получится картина, приведенная на рис. 13. Тонкие ли¬
нии означают быстрое смещение взора, точки — это места фиксации взора во
время остановки глаз.
§ 3. Зрительная система 77
Рис. 13. Запись движения глаз при рассматривании изображения
Но всегда ли, когда мы смотрим на предметы, глаза движутся? Например, если
мы находимся в знакомой темной комнате, то даже при кратковременной вспыш¬
ке фотолампы успеваем увидеть и узнать знакомые пред¬
меты, хотя глаза в это время остаются неподвижными.
Значит, движения глаз нужны нам лишь для первоначаль¬
ного ознакомления с предметами.
С помощью движения глаз мы выделяем признаки
предметов, соотношения между ними, формируем целост¬
ные образы и все это передаем на хранение в память. Ко¬
гда же предметы хорошо знакомы, достаточно лишь от¬
дельных признаков для того, чтобы узнать, что нам пока¬
зывают. То, что мы не успеваем рассмотреть, мы можем дополнить благодаря
памяти и воображению. Следовательно, для узнавания хорошо известных предме¬
тов движение глаз необязательно.
С помощью движения
глаз мы выделяем при¬
знаки предметов, со¬
отношения между ни¬
ми, формируем
целостные образы и
все это передаем на
хранение в память.
Одной из важных характеристик зрения является острота зрения. Острота зре¬
ния определяет предельную способность глаза различать мелкие детали в поле
зрения. Острота зрения зависит от общей освещенности,
контраста деталей изображения па определенном фоне
и других причин.
Наиболее часто встречающиеся нарушения зрения —
это близорукость и дальнозоркость.
Проблема константности зрительного мира состоит в
том, что мы воспринимаем его постоянным, хотя изобра¬
жение на сетчатке постоянно смещается. Задача мозга —
вычленить инвариантные признаки объектов из непре¬
рывно меняющегося потока информации. Важная роль в
этом процессе принадлежит зрительному вниманию и на¬
личию прежнего зрительного опыта.
Для получения голов¬
ным мозгом полной
информации об окру¬
жающем мире необхо¬
димо функционирова¬
ние всей сети внутри
зрительной сенсорной
системы, включая нис¬
ходящие (обратные)
связи к первичной
зрительной коре в за¬
тылочной области.
Интеграция информации от всех корковых и подкорковых структур, связан¬
ных с обработкой зрительных сигналов, происходит лишь после тохю, как все зри¬
тельные структуры завершат свои специфические операции, то есть обработку по
соответствующим каналам. Следовательно, для получения головным мозгом пол¬
ной информации об окружающем мире необходимо функционирование всей сети
78 Глава 3. Физиология сенсорных систем
внутри зрительной сенсорной системы, включая нисходящие (обратные) связи
к первичной зрительной коре в затылочной области.
§ 4. Слуховая сенсорная система и речь
Чувство слуха — одно из главных в жизни человека. Человек слышит не все звуки
окружающего мира, а только те, которые находятся в диапазоне от 20 до 20 000 Гц.
Слух и речь вместе составляют важное средство общения между людьми, слу¬
жат основой взаимоотношений людей в обществе. Потеря слуха может привести к
нарушениям в поведении человека. Глухие дети не могут научиться полноценной
речи.
Звук, или звуковая волна, — это чередующиеся разряжение и сгущение воздуха,
распространяющиеся во все стороны от источника звука. А источником звука мо¬
жет быть любое колеблющееся тело. Звуковые колебания воспринимаются на¬
шим органом слуха.
Орган слуха построен очень сложно и состоит из наружного, среднего и внут¬
реннего уха (рис. 14).
Рис. 14. Строение органа слуха
1 — молоточек; 2 — наковальня; 3 — вестибулярный аппарат; 4 — улитка;
5 — евстахиева труба; 6 —■ стремя; 7 — барабанная перепонка;
8 — наружный слуховой проход; 9 — ушная раковина
Наружное ухо включает в себя ушную раковину и наружный слуховой проход.
Ушные раковины многих животных подвижны, это помогает животному улавли¬
вать, откуда раздается даже самый тихий звук. Ушные раковины человека также
служат для определения направления звука, хотя они и лишены подвижности.
Наружный слуховой проход соединяет наружное ухо со следующим отделом —
средним ухом.
Среднее ухо включает барабанную перепонку и три сочлененные между собой
косточки. Полость среднего уха сообщается с наружной средой через евстахиеву
трубу, которая обеспечивает поддержание в полости постоянного давления, близ¬
кого к атмосферному. При глотании происходит обмен воздухом между глоткой и
средним ухом. Изменение давления наружного воздуха, например в самолете, вы¬
зывает неприятное ощущение — «закладывает» уши. Это объясняется прогибом
барабанной перепонки из-за разницы между атмосферным давлением и давлени¬
ем в полости среднего уха. При глотании евстахиева труба открывается и давле¬
ние по обе стороны барабанной перепонки выравнивается.
§ 4. Слуховая сенсорная система и речь 79
На внутреннем конце слуховой проход перегорожен туго натянутой барабан¬
ной перепонкой. Звуковая волна, ударяя в барабанную перепонку, заставляет ее
колебаться, вибрировать. Частота вибрации барабанной перепонки тем больше,
чем выше звук. Чем сильнее звук, тем сильнее колеблется перепонка. Но если звук
совсем слабый, еле слышимый, то эти колебания очень малы. Минимальная слы¬
шимость натренированного уха находится почти па границе тех колебаний, кото¬
рые создаются беспорядочным движением молекул воздуха. Значит, человеческое
ухо — уникальный по чувствительности слуховой прибор.
Система косточек среднего уха состоит из молоточка, наковальни и стремечка.
Молоточек фиксирован своей рукояткой к барабанной перепонке, стремечко
укреплено на овальном окне, а наковальня сочленена с одной стороны с молоточ¬
ком, а с другой — со стремечком. Основная роль системы косточек состоит в пере¬
даче колебаний барабанной перепонки на структуры внутреннего уха. При этом
косточки образуют систему рычагов, которая улучшает передачу энергии колеба¬
ний из воздушной среды наружного слухового прохода. Благодаря системе косто¬
чек давление на овальное окно улитки по сравнению с давлением па барабанной
перепонке увеличивается примерно в 20 раз. Нарушение системы косточек не
приводит к полной глухоте, то есть благодаря костной проводимости слуховая
чувствительность сохраняется.
За средним ухом начинается внутреннее ухо (рис. 15). Оно расположено в глу¬
бине височной кости черепа. Внутреннее ухо представляет собой систему лаби¬
ринта и извилистых каналов, заполненных жидкостью.
Рис. 15. Внутреннее строение улитки
1 — перепончатая перегородка; 2 — чувствительные волосковые клетки;
3 — мембрана; 4 — нерв
В лабиринте находятся сразу два органа: орган слуха — улитка и орган равно¬
весия — вестибулярный аппарат. Улитка — это спирально закрученный костный
канал, имеющий у человека два с половиной оборота. Колебания перепонки оваль¬
ного окна передаются жидкости, заполняющей внутреннее ухо. И она, в свою оче¬
редь, начинает колебаться с той же частотой. Вибрируя, жидкость раздражает ре¬
цепторы, расположенные в улитке.
Базилярная и рейснерова мембраны делят канал на три части: вестибулярная
лестница, улитковый ход и барабанный ход (рис. 16). На базилярной мембране
расположен рецепторный аппарат улитки — кортиев орган, состоящий из одного
80 Глава 3. Физиология сенсорных систем
Рис. 16. Поперечный разрез завитка улитки
I — вестибулярная лестница;
II — улитковая (кохлеарная) лестница;
III — барабанная (тимпанальная) лестница.
1 — костная капсула улитки; 2 — рейснерова мембрана; 3 — базальная
(базилярная, основная! мембрана; 4 — покровная (текториальная)
мембрана; 5 — наружная волосковая клетка; 6 — внутренняя волоско-
вая клетка; 7 — биполярная клетка спирального ганглия улитки
ряда внутренних и трех-пяти рядов наружных волосковых клеток; от волосковых
клеток отходят волокна слухового нерва. Колебания жидкости, заполняющей
улитку, раздражают отдельные слуховые рецепторы. В них возникают импульсы,
которые передаются по слуховому нерву в головной мозг. На схеме (рис. 17) пока¬
заны все последовательные процессы превращения звуковой волны в нервную
сигнализацию. Контакт между нервными волокнами и волосковыми клетками
представляет собой типичный синапс с пре- и постсинаптической мембранами
и синаптической щелыо.
у
Внутреннее
ухо
Рис. 17. Схема передачи звуковых волн на слуховые рецепторы
Нейроны второго порядка входят в состав комплекса кохлеарных ядер. В них
переключаются все волокна слухового нерва. Частотная проекция кортиева орга¬
на в целом упорядоченно повторяется в пространстве каждого из подразделений
слухового комплекса ядер — тонотопия. Отростки клеток кохлеарных ядер на¬
правляются в вышележащие центры мозга, в основном противоположной сторо¬
ны, и заканчиваются на комушексе ядер верхней оливы и ядрах трапецевидного те¬
ла. Здесь уже пути от обоих ушей конвергируют на одних и тех же нейронах, обес¬
печивая бинауральный анализ звука. Аксоны слуховых клеток направляются в
составе латеральной петли к нижнему двухолмию и внутреннему (медиальному)
коленчатому телу. В нижнем двухолмии заканчивается анализ звуковых сигналов,
необходимых для обнаружения источника звука, то есть для формирования ори¬
ентировочного рефлекса на звук. Внутреннее коленчатое тело является талами¬
ческим центром слуховой системы, здесь обнаруживается тонотопия, так же как
и в слуховой коре, расположенной в височной доле. Сюда не только приходят вое-
§ 4. Слуховая сенсорная система и речь 81
ходящие волокна от внутреннего коленчатого тела, но и
берут начало нисходящие волокна ко всем центральным
образованиям слуховой системы.
Характерной особенностью коммуникативных (рече¬
вых) слуховых сигналов человека является изменение во
времени таких параметров, как амплитуда и частота, то
есть они представляют собой амплитудно- и частотно-мо-
дулированные сигналы, анализ которых осуществляется в
коре и в глубоких отделах слуховой системы. Частота сиг¬
нала оценивается с помощью тонотопии на всем протяже¬
нии слуховой системы. Интенсивность звука кодируется
не только частотой импульсации, но и количеством возбу¬
жденных нейронов. Бинауральный слух является универ¬
сальной основой локализации, человек способен с помо¬
щью движений головы определить местоположение ис¬
точника звука в горизонтальной плоскости и с помощью
Звуковая и письмен¬
ная речь — это спо¬
собность знаково¬
символического отра¬
жения предметов и яв¬
лений окружающего
мира и самих себя
в этом мире. Только
у человека признаки,
лежащие в основе
классификации пред¬
метов и внешнего ми¬
ра, связаны со слова¬
ми родного языка
и получают вербаль¬
ные (словесные) на¬
именования.
одного уха. Основой бинаурального слуха являются межушные различия стиму¬
ляции по времени и по интенсивности. Межушные различия по времени сущест¬
венны, если источник звука смещен от средней линии головы, а длина волны
больше расстояния между ушами. При частоте источника звука свыше 1,5 Гц у че¬
ловека вступает в действие другой бинауральный механизм локализации, а имен¬
но межушная разность по интенсивности.
Речевая система (И. А. Вартанян, 2003)
Звуковая и письменная речь — это способность знаково-символического отраже¬
ния предметов и явлений окружающего мира и самих себя в этом мире. Только
у человека признаки, лежащие в основе классификации предметов и внешнего
мира, связаны со словами родного языка и получают вербальные (словесные) на¬
именования.
Предпосылкой для генерации звуковой речи является развитие и совершенст¬
вование в процессе эволюции периферического аппарата звукогенерации — глот¬
ки, гортани, голосовых связок, нижней челюсти.
Важная роль принадлежит возможности координации голосового и артикуля¬
ционного аппарата, которые развиваются в онтогенезе.
Управление речевой функцией осуществляется высшими отделами мозга че¬
ловека — корой больших полушарий, значительные участки сенсорных и мотор¬
ных областей которой специализированы к восприятию, пониманию, запомина¬
нию и воспроизведению речи, а также подкорковыми образованиями мозга, кото¬
рые связаны с эмоциями и памятью.
В коре больших полушарий существует три важнейших для речевой функции
сенсорных поля:
♦ зрительное — на затылочной поверхности обоих полушарий;
♦ слуховое — в височной извилине каждого полушария;
♦ соматосенсорное — в задней центральной извилине каждого полушария.
В передней центральной извилине правого и левого полушарий расположено
первичное моторное поле, которое управляет мышцами лица, конечностей и туло¬
82 Глава 3. Физиология сенсорных систем
Рис. 18. Важнейшие сенсорные, мо¬
торные и интегративные области
мозга, прямо или опосредованно
связанные с осушествлением ре¬
чевых функций
1 — лобная доля; 2 — зона мышечной
регуляции; 3 — мышечные и кожные
ошушения; 4 — теменная доля; 5 — ви¬
сочная доля; 6 — затылочная доля; 7 —
зона зрения; 8 — мозжечок; 9 — путь
к спинному мозгу; 10 — зона слуха
вища. Именно оно определяет речь и письмо.
Существуют также вторичные сенсорные, ассо¬
циативные и моторные поля. Прежде всего это
первая височная извилина — область Вернике,
обеспечивающая понимание речи, а также важ¬
нейшая интегративная часть мозга — лобная до¬
ля, регулирующая программное обеспечение ре¬
чи, сосредоточенное в зоне Брока (третья лобная
извилина) в префронтальной коре.
Речь как функция мозга глубоко асимметрич¬
на. Лингвистические способности человека осу¬
ществляются преимущественно левым полуша¬
рием, структуры которого действуют как единый
речевой механизм (рис. 18). После того как ин¬
формация, заключенная в слове, обрабатывает¬
ся в слуховой системе или в «неслуховых» обра¬
зованиях мозга, она должна быть опознана по
смыслу. Этот процесс осуществляется в зоне Вер-
пике, расположенной в височной области вблизи
слуховой коры. Именно здесь обеспечивается по¬
нимание смысла поступающего сигнала — слова.
Если воспринимается письменная речь, то
сначала включается первичная зрительная кора.
После этого информация о прочитанном слове
поступает в угловую извилину, которая связывает зрительную форму данного
слова с его акустическим аналогом в зоне Вернике. Для произнесения слова необ¬
ходимо, чтобы анализировалось его представительство в зоне Брока, расположен¬
ной в лобной извилине. В зоне Брока сведения, поступившие из зоны Вернике,
приводят к возникновению детальной программы артикуляции путем активации
моторной коры, управляющей лицевой мускулатурой и связанной с зоной Брока
короткими волокнами.
Корковые отделы левого полушария выполняют специфическую роль в вос¬
приятии, запоминании и воспроизведении речевого материала. Именно эти зоны
необходимы для полноценного осуществления речевой функции как единого сен¬
сорного, мыслительного и моторного процесса. Расположенные спереди зоны осо¬
бенно важны для осуществления экспрессивной (выразительной) речи, а распо¬
ложенные сзади — для восприятия смысла речи.
Регуляция речи осуществляется с участием лимбической системы мозга, кото¬
рая определяет интонационные характеристики речи и ее эмоциональный харак¬
тер. Многие другие подкорковые образования мозга также принимают участие
в речевой функции.
Таким образом, участие левого полушария необходимо для обнаружения и опо¬
знания артикулированных звуков речи, а правого — для опознания интонаций,
уличных и бытовых шумов, музыкальных мелодий. Высокий уровень общей рече¬
вой активности обеспечивается левым полушарием, а улучшение выделения сиг¬
нала из шума — правым. Правое полушарие не способно реализовать команду для
продуцирования речи, но оно обеспечивает понимание устной речи и написанных
§ 5. Вестибулярная сенсорная система 83
слов. Понимание речи, осуществляемое правым полуша¬
рием, ограничено конкретными именами существитель¬
ными, в меньшей степени — отглагольными существи¬
тельными, еще в меньшей степени — глаголами. Правое
полушарие обеспечивает понимание эмоционального со¬
держания интонаций, опознание по голосу, участвует в мо¬
дуляции частот голоса.
Точность воспроизведения речи контролирует слухо¬
вая обратная связь, а также афферентация от мышечных
и суставных рецепторов речеобразующих органов.
Подкорковый путь участвует в автоматизированных
речевых процессах, не требующих обдумывания. Корко¬
вый путь участвует в сознательном контроле речи. Эти
пути действуют одновременно и параллельно.
Правое и левое полушария выполняют разные функ¬
ции, но совместно обеспечивают целенаправленное пове¬
дение. Каждое полушарие имеет отдельное особое мышление: левое — речевое,
правое — зрительно-пространственное. Левое полушарие обрабатывает информа¬
цию аналитически и последовательно, а правое — одновременно и целостно. Каж¬
дое полушарие вносит свой уникальный вклад в речь и мышление.
Правое и левое полу¬
шария выполняют раз¬
ные функции, но со¬
вместно обеспечивают
целенаправленное по¬
ведение. Каждое полу¬
шарие имеет отдель¬
ное особое мышление:
левое — речевое, пра¬
вое — зрительно-про¬
странственное. Левое
полушарие обрабаты¬
вает информацию
аналитически и после¬
довательно, а пра¬
вое — одновременно
и целостно.
§ 5. Вестибулярная сенсорная система
Вестибулярная система — орган равновесия, а иногда орган чувства ускорения,
так как адекватными раздражителями вестибулярного аппарата является сила
гравитации и силы, сообщающие телу линейное или угловое ускорение.
Периферическим органом вестибулярной системы является вестибулярный
аппарат, составляющий вместе с улиткой внутреннее ухо. Лабиринт — это костное
образование, внутри которого расположен перепончатый лабиринт. Пространство
между ними заполнено жидкостью — перилимфой. Изнутри перепончатый лаби¬
ринт также заполнен жидкостью — эндолимфой. Полукружные каналы располага¬
ются в трех взаимоперпендикулярных плоскостях.
Рецепторы вестибулярного аппарата являются вторичными механорецептора¬
ми и размещаются на кристах каждого из трех полукружных каналов. Они обра¬
зованы волосковыми клетками, которые являются чувствительными клетками и
связаны через синапсы с афферентными и эфферентными волокнами вестибуляр¬
ного нерва.
Сенсорные клетки бывают двух типов. Клетки 1-го типа имеют форму колбы
с закругленным дном, клетки 2-го типа — вид цилиндра. Из оболочек обоих типов
клеток выступают пучки волосков (стереоцилий) и один более длинный отросток
на периферии пучка — киноцилия. Смещение волосков в сторону киноцилии вы¬
зывает возбуждение, а в противоположном направлении — торможение (рис. 19).
Ориентация волосков в каждом из полукружных каналов определяет дирекцио-
нальиую возбудимость: в горизонтальном канале ток эндолимфы вызывает возбу¬
ждающий эффект, если он направлен к утрикулюсу — мешочку в основании вес¬
тибулярного аппарата, а в вертикальных — если он направлен от утрикулюса.
84 Глава 3. Физиология сенсорных систем
Рис. 19. Схема, иллюстрирующая
возбудительные и тормозные реакции
в зависимости от направления отклонения
волосков рецепторной клетки
1 — состояние покоя и соответствующий ему уровень
импульсной активности нерва; 2 — отклонение волос¬
ков в сторону киноцилии и увеличение частоты импуль-
саиии в нерве; 3 — отклонение волосков от киноиилии
и уменьшение частоты импульсации в нерве
В макулах отолитовых органов (утрикулюса и саккулюса) киноцилии также
располагаются неодинаково, так что при смещении отолита часть клеток возбуж¬
дается, а другая — затормаживается (рис. 20). Раздражающим фактором в отоли¬
тах является скольжение отолитовой мембраны (крохотные известковые кристал¬
лики), в результате которого волоски сенсорных клеток сгибаются. В качестве ме¬
диатора в передаче возбуждения от рецепторной волосковой клетки к окончаниям
вестибулярного нерва служит ацетилхолип.
Вестибулярные ядра продолговатого мозга — это первый уровень центральной
нервной системы, где происходит обработка информации о движении или изме¬
нении положения тела в пространстве, поступающей от рецепторов лабиринта.
Важнейшим звеном в передаче динамических лабиринтных влияний па спинной
мозг и эффекторы является мозжечок. Вестибуло-глазодвигательные влияния
Рис. 20. Схематические рисунки участков утрикулярной (а)
и саккулярной (б) макул и мембран с отолитами
§ 6. Сенсорная система скелетно-мышечного аппарата 85
являются сложными многонейронными. Зрительный нистагм может быть вызван
не только раздражением полукружных каналов лабиринта, но и оптокинетически¬
ми стимулами, перемещающимися в иоле зрения наблюдателя.
Вестибулярные импульсы, как и импульсы других модальностей, поступают в
кору через таламические нейроны и контактируют с корковыми клетками темен¬
ной и височной областей.
Все вышеописанные ощущения при раздражении вестибулярного аппарата
возникают у человека на Земле. В отличие от земной жизни, в космическом про¬
странстве отсутствует сила тяжести, наступает состояние невесомости. Поэтому у
человека в космосе отолитовые мембраны не имеют веса и перестают давить на
кино- и стереоцилии.
Другое состояние, характерное для космических полетов, — это перегрузки,
которые возникают при выходе космических кораблей на орбиту. Перегрузки за¬
ключаются в резком увеличении силы тяжести, а значит, оказывают очень силь¬
ное влияние па вестибулярный аппарат. Длительное пребывание человека в кос¬
мосе свидетельствует о том, что вестибулярные рецепторы способны приспосаб¬
ливаться к невесомости.
Для спортсменов, летчиков, моряков и особенно для космонавтов нормальная
работа вестибулярного аппарата очень важна. Для всех людей сигналы от вести¬
булярного аппарата являются необходимыми факторами ориентировки в про¬
странстве и формирования поведения человека и животных в окружающей среде.
Повышенная чувствительность и нарушение работы органа равновесия могут
быть врожденными, но могут возникать и после различных инфекционных забо¬
леваний. Люди с поврежденным вестибулярным аппаратом плохо переносят по¬
леты на самолетах, плавание на кораблях и даже поездки в наземном транспорте,
не могут кататься па качелях, каруселях. Явление укачивания (морская болезнь)
сопровождается головокружением, тошнотой, рвотой и даже обмороком.
§ 6. Сенсорная система скелетно-мышечного
аппарата
Скелетно-мышечный аппарат является исполнительной системой организма,
и поэтому его рецепторные элементы (проприорецепторы) играют особо важную
роль среди других чувствительных образований. Они проводят информацию о каж¬
дом моменте движения — положении суставов, длине и напряжении всех мышц,
участвующих в двигательном акте.
В составе скелетной мышцы выделяют две группы волокон. Если первые со¬
здают те усилия, которые необходимы для движений и поддержания позы (сухо¬
жильные рецепторы), то вторые входят в состав датчиков длины — мышечных ве¬
ретен, их функция сводится к формированию восходящей сенсорной импульса-
ции. Самостоятельную группу составляют рецепторы суставного угла (рис. 21).
Веретена соединены с мышечными волокнами параллельно, а сухожильные
органы — последовательно. Поэтому основные величины, измеряемые мышечны¬
ми рецепторами (веретенами и сухожильными органами), — это изменения длины
и напряжения при растяжении и сокращении мышцы. При активном сокращении
мышцы напряжение веретен ослабевает (они «разгружаются») и частота импуль-
86 Глава 3. Физиология сенсорных систем
Рис. 21. Функциональное состояние мышечного веретена (а)
и сухожильного органа (б) при сокращении или растяжении мышцы
а — слева: экстрафузальные волокна, растянутые грузом, мышечное веретено посередине, электри¬
ческая активность усилена; справа: экстрафузальные волокна сокращены прямым электрическим
раздражением (5), электрическая активность мышечного веретена отсутствует.
6 — слева: сухожильный рецептор Гольджи при растяжении экстрафузальных мышечных волокон,
его электрическая активность ослаблена; справа: мышиа сокращена, электрическая активность сухо¬
жильного рецептора усилена
сации в соответствующих афферентных органах снижается, а сухожильный ре¬
цептор, наоборот, при этом возбуждается.
На уровне спинного мозга посредством так называемой гсимма-моторной си¬
стемы осуществляются наиболее простые двигательные реакции фазного и топи¬
ческого типа. Гамма-моторная система устроена по принципу обратной связи
(рис. 22), благодаря которой усиливается импульсация мышечного окончания
при постоянной степени растяжения и возбуждаются мышечные окончания во
время начала растяжения.
Афферентная импульсация от мышечно-суставных рецепторов частью пере¬
ключается на мотонейроны спинного мозга, а частью направляется но восходя¬
щим путям в высшие отделы головного мозга, в продолговатый мозг. Отсюда бе¬
рут начала волокна второго порядка, получившие название медиальной петли, ко¬
торая заканчивается в вентро-базальном комплексе таламуса. От этих ядер берут
начало нейроны III порядка, которые направляются к коре больших полушарий,
к сенсомоторным нолям в передней центральной извилине.
Особенностью таламического и еще в большей степени коркового звена ске¬
летно-мышечной сенсорной системы является высокая степень интеграции сен¬
сорного потока. На одни и те же нейроны, в частности пирамидные клетки сенсо-
§ 6. Сенсорная система скелетно-мышечного аппарата 87
Рис. 22. Эффект фузимоторной стимуляции на
период молчания разрядов мышечных веретен
при мышечном сокращении
1 — регистрация активности афферентных волокон;
2 — механограмма.
а — фоновое напряжение (15 г) облегчает ритмическую
активность рецептора; б — прекращение разрядов при
сокращении мышцы из-за разгрузки «веретена»; в — фу-
зимоторная стимуляция снимает расслабленность вере¬
тена и поддерживает импульсный разряд в течение все¬
го периода сокращения мышцы
моторной коры, конвергируют не только афферентные входы от кожных и мы¬
шечных рецепторов, по и проекции от зрительных, слуховых, вестибулярных и дру¬
гих структур.
Большую роль в интеграции скелетно-мышечной информации и информации
от других сенсорных систем играет теменная ассоциативная область коры, где
обнаружено особенно большое количество нолисенсорпых нейронов. Здесь фор¬
мируется интегральная «схема тела» и возникает целостное представление о соот¬
несенности собственного тела с окружающим пространством. Повреждения темен¬
ной коры приводят к нарушениям скелетно-мышечной и кожной чувствительно¬
сти. При этом наблюдается значительная потеря способности к формированию
целостного образа и его локализации на площади тела и в окружающем простран¬
стве.
Деятельность мышечных веретен подвергается мощному нисходящему влия¬
нию головного мозга. В ходе двигательной реакции под влиянием нисходящих
88 Глава 3. Физиология сенсорных систем
сигналов происходит определенное перераспределение функциональной значи¬
мости восходящих систем и, следовательно, изменение приносимой ими инфор¬
мации в деятельности высших отделов мозга.
§ 7. Схема тела
Чтобы лучше объяснить, что понимается под терминами «образ тела» и «образ
пространства», обратимся к повседневной клинической практике. В клинику
Психоневрологического института поступает больная с тяжелыми расстройства¬
ми восприятия своего тела и нарушениями ориентировки в пространстве. Болезнь
длится уже три года, хотя больная продолжает работать. Во время многочасовых
приступов ей казалось, что она «ходит по воздуху», левый глаз увеличивается и
выходит из орбиты. Больная не могла найти выход из комнаты, хотя видела дверь,
но шла не в том направлении, натыкаясь на предметы. Появилось ощущение, что
левая рука и бок «чужие». Не могла надеть платье, найти правый и левый рукава,
иногда ложилась в постель, не снимая туфли с левой ноги. Ей казалось, что у нее
две левые ноги и они идут в разных направлениях. Позже появилось ощущение,
что левая рука стала большой, «ввинченной», что увеличены и подымаются вверх
ногти. Стана плохо орудовать ножом и вилкой, неправильно брала их, поворачи¬
вала не той стороной. У больной была обнаружена и затем удалена опухоль в те¬
менной области правого полушария.
У другого больного отмечен инсульт в районе артерии, снабжающей левую те¬
менно-височную область. У пего четко нарушено различение правого и левого. За¬
трудняется воспроизвести положение одних пальцев по отношению к другим, ру¬
ки или пальцев по отношению к лицу. При просьбе изобразить на рисунке челове¬
ка глаза помещает под носом, уши обращены внутрь лица. Не может правильно
нарисовать домик, при предъявлении римских цифр XI и IX их не различает. Не
может оценить углы сгибания рук в локте, точно воспроизвести пассивно задан¬
ное движение. При этом сознание у больного ясное, интеллект сохранен.
К грубым расстройствам ощущения собственного тела относят ощущение от¬
сутствия частей тела, кажущееся изменение их формы, веса, величины и, наобо¬
рот, кажущееся появление лишних конечностей, игнорирование отдельных час¬
тей тела, невнимание к ним, мнимое ощущение движений парализованных конеч¬
ностей, затруднения в словесном обозначении частей тела и их пространственных
координат и др. Если же повреждение мозга располагается в других областях, то
подобных расстройств не возникает (Трауготт и др., 1973).
Видимо, уже становится ясно, что терминами «образ тела», «схема тела»,
«трехмерная модель тела» обозначают систему обобщенных представлений и по¬
нятий человека о собственном теле в покое и при движении, о пространственных
координатах и взаимоотношении отдельных частей тела.
Теменные доли у человека сильно развиты и подразделяются на отдельные
участки. Теменные доли имеют такое богатство различных нервных связей с дру¬
гими отделами мозга, что могут выполнять свою нормальную функцию по синте¬
зу многих видов информации только во взаимодействии с другими образования¬
ми, и прежде всего с ядрами промежуточного мозга, из которых с теменными до¬
лями тесно связано заднее латеральное и в меньшей степени задпевеитралыюе
ядро. Некоторые импульсы, прежде чем достичь теменной коры, должны быть
§ 7. Схема тела 89
предварительно обработаны в этих ядрах. А поэтому и не стоит связывать такую
сложную деятельность мозга, как формирование схемы тела, с функцией исклю¬
чительно теменных долей мозга. Ярким доказательством этому служат исследова¬
ния В. М. Смирнова (1974) с применением метода интрацеребральных электро¬
дов, которые остаются в мозгу на длительное время, необходимое для постановки
диагноза и лечения. Через эти электроды врач может оказывать слабое раздра¬
жающее воздействие па глубокие структуры мозга человека и, найдя пораженный
очаг, произвести его разрушение. Так, у больного паркинсонизмом слабым элек¬
трическим током раздражалось заднее вентральное ядро таламуса правого полу¬
шария. В этот момент больной сообщал, что левая нога кажется ему «толстой»,
«расширенной». У другого больного та же стимуляция сопровождалась ощущени¬
ем холода во рту и голове, язык казался большим, толстым, «не своим». Иными
словами, раздражение, а в ряде случаев и разрушение глубоких структур мозга со¬
провождается нарушениями схемы тела.
Убедительны в этом смысле наблюдения В. М. Смирнова с фантомом ампути¬
рованных, то есть ложным восприятием (иллюзией) отсутствующей части тела,
возникшим после ее ампутации. У больного на почве ампутации левой руки воз¬
никло ощущение, что отсутствующий локоть прикреплен непосредственно к куль¬
те, то есть к верхней трети плеча, предплечье почти сливается с локтем, откуда
начинается мизинец, а остальные четыре пальца отходят прямо от культи. К суще¬
ственному лечебному эффекту привели электрические раздражения через вжив¬
ленные электроды заднего латерального ядра таламуса (фантомные боли значи¬
тельно ослабли).
Итак, ответственными за восприятие схемы тела у человека являются не толь¬
ко теменная доля, но и связанные с ней различные ядра промежуточного мозга,
которые вместе образуют единую таламо-париетальную ассоциативную систему
мозга. Хорошо известно, что детальное представительство чувствительности от¬
дельных частей противоположной половины тела, вплоть до фаланг пальцев рук,
характерно для соматосенсорной мозговой системы.
Таким образом, чувствительность всего тела, дробно топографически распре¬
деленная по поверхности коры, составляет ту основу, тот информационный мате¬
риал, из которого путем объединения формируются целостные функциональные
блоки крупных отделов тела. Эти интегративные процессы завершаются к момен¬
ту созревания человека и представляют собой закодированное описание взаимо¬
расположения частей тела, которые используются при осуществлении автома¬
тизированных стереотипных движений. А поскольку базой для этих процессов
служит автоматически закрепленная «карта» тела, логично допустить, что они со¬
ставляют основу статического образа тела. Сами процессы объединения отдель¬
ных ощущений в целостные образы взаиморасположения конечностей формиру¬
ются, скорее всего, с участием как ассоциативной соматической зоны, так и тала-
мо-париетальпой ассоциативной системы.
Для создания статического образа тела оказывается недостаточно лишь ин¬
формации от участков самого тела (кожи, суставов, мышц и т. д.). Необходимо со¬
отнести эту информацию с положением тела но отношению к силе земного притя¬
жения и взаиморасположением крупных функциональных блоков тела в системе
трех основных пространственных координат. В его создании участвует чувстви¬
тельная система вестибулярного аппарата. Перемещение всего тела вперед-назад,
90 Глава 3. Физиология сенсорных систем
вправо-влево, вверх-вниз тонко воспринимается данной сенсорной системой,
а информация о таких перемещениях поступает в те же ассоциативные районы
коры мозга, в которых происходит ее объединение с информацией от скелетно¬
мышечного аппарата и кожи. Туда же поступает импульсация от различных внут¬
ренних органов, которая также участвует в создании статического образа тела.
Причем все это происходит в здоровом организме без участия сознания. Следова¬
тельно, статический образ тела представляет собой жесткую систему связей, осно¬
ванную на системе врожденных механизмов и усовершенствованную и уточнен¬
ную в индивидуальной жизни.
Постоянно осуществляя тот или иной род деятельности, человек меняет взаи¬
морасположение частей тела, приобретает в порядке обучения новые двигатель¬
ные навыки, а значит, формирует новые трехмерные пространственные модели те¬
ла, то есть информацию о взаиморасположении частей тела в пространстве трех
координат — динамический образ тела. В отличие от первого (статического) дина¬
мический образ тела имеет значение лишь для данного конкретного момента вре¬
мени, для определенной ситуации. При изменении ситуации динамический образ
тела быстро сменяется новым. Динамический образ также имеет информативный
характер и базируется па показаниях чувствительности кожи, мышц, суставов,
вестибулярного аппарата. Динамический образ возникает при непосредственном
участии таламопариетальной системы мозга.
В мозгу происходит постоянное взаимодействие того и другого образов тела,
осуществляется сличение динамического образа с его статическим аналогом.
В результате такого сличения формируется субъективное ощущение позы, отра¬
жающее не только положение тела в данный момент, но и возможные желатель¬
ные и нежелательные его изменения в будущем. Человек может из элементов ста¬
тического образа тела запрограммировать определенную позу, то есть определен¬
ное взаиморасположение в пространстве частей своего тела. Реально же принятая
поза, оцениваемая через динамический образ тела, при сличении может совпадать
или не совпадать с запрограммированной. В случае согласования человек удовле¬
творяется принятой позой, не всегда оценивая ее субъективно. Если же такое со¬
гласование не будет достигнуто, то вступают в действие активные механизмы со¬
знательной перестройки позы, которые изменяют активность мускулатуры до тех
пор, пока не будет достигнуто желательное положение тела (рис. 23). Подробные
сведения можно найти в книге В. С. Гурфинкеля, Я. М. Коца и М. Л. Шика «Регу¬
ляция позы» (1965). Мы только укажем, что из теменной коры человека выходят
пути, с помощью которых регулируется тонус скелетной мускулатуры. Известно,
что при поражении теменной доли нарушается возможность воспроизведения за¬
данной позы.
Итак, для того чтобы субъективно оценить позу, необходимо сопоставить зако¬
дированный в памяти эталон статического образа тела с его любой конкретной ва¬
риацией (динамическим образом). Именно так трактуется возникновение фанто¬
мов, при которых внезапная утрата конечности исключает из состава динамиче¬
ского образа тела информацию об отсутствующем органе. Во время сличения со
статическим его аналогом, где эта информация привычно входила в качестве со¬
ставного элемента в схему тела, и возникает своеобразная форма рассогласования
двух информативных образов с возникновением иллюзорных ощущений, дости¬
гающих иногда патологической формы.
§ 7. Схема тела 91
Регуляция
позы
Сигнализация
с мышц, кожи,
суставов, сухожилии
Сигнализация
с вестибулярного
аппарата
Рис. 23. Схематическое изображение высших механизмов регуляции позы
Помимо уже описанных выше нарушений схемы тела у людей с поражением
теменных долей мозга отчетливо отмечаются расстройства зрительно-простран¬
ственных координаций; иными словами, больные теряют способность ориентиро¬
ваться в знакомом помещении, знакомой местности, утрачивают географические
представления (расположение частей света, соотносительное расположение горо¬
дов на карте и т. д.). Тяжелые нарушения пространственной ориентации в реаль¬
ной ситуации характерны для поражения теменной доли правого полушария.
В этих случаях расстраивается непосредственное, наглядное восприятие окру¬
жающей среды, больной не способен охватить ситуацию в целом. Вместе с этим он
не испытывает затруднений при операциях с пространственными признаками,
представленными в отвлеченной форме, обобщении объектов по этим признакам
и построении географических схем и планов. При поражении левой теменной
области больной вполне удовлетворительно ориентируется в наглядной прост¬
ранственной ситуации. Нарушения возникают преимущественно на фоне обоб¬
щенных понятий и схем, отражающих соответствующие пространственные от¬
ношения (топографические представления, работа с географическими картами,
планами и т. д.). А. Р. Лурия (1962) неоднократно подчеркивал, что нарушения
пространственных восприятий после поражения теменных долей следует связы¬
вать с расстройством механизмов одновременного синтеза различного вида ин¬
формации в единый целостный образ окружающей ситуации.
У больных с теменными поражениями (инсульт, опухоль) обнаруживали нару¬
шения в оценке различно ориентированных в пространстве отрезков прямых, уг¬
лов, дуг и пр. Требуемая для оценки этих изображений мысленная переориенти¬
ровка строится па анализе и синтезе зрительных и мышечных сигналов, возни¬
кающих при движении глаз. Расстройство этого механизма, видимо, и является
причиной нарушения зрительно-пространственной ориентировки. Помимо этого,
у больных сильно страдает пространственное восприятие звуковых сигналов.
Причем те же сигналы, но одинаково ориентированные в пространстве, распозна¬
ются больными с достаточной точностью. Итак, поражения теменных долей при¬
водят к расстройствам в оценке пространственного расположения зрительных
и звуковых сигналов.
Вместе с этим страдают и синтетические механизмы мозга. Больным предлага¬
лось, например, отличить серию последовательных сигналов (звонок, низкий тон,
92 Глава 3. Физиология сенсорных систем
высокий топ, треск) от этой же серии, в которой меняются местами второй и тре¬
тий члены. В других исследованиях больным предъявлялся ряд фигур, располо¬
женных на плоскости, с требованием отличить от другого ряда тех же фигур.
Больные либо совсем не могли справиться с этой задачей, либо решали ее с боль¬
шим числом ошибок. Кроме того, у больных наблюдались нарушения в способно¬
сти объединять различные сигналы (световые и звуковые) в единый комплекс¬
ный условный сигнал вне зависимости от их пространственного расположения
и временной последовательности.
К аналогичным доказательствам приводят и нейрофизиологические экспери¬
менты. Если источник света (или звука) располагается строго по средней линии
между глазами или ушами, то величины вызванных потенциалов в нравом и ле¬
вом полушариях будут одинаковыми. Когда раздражитель смещается влево, уве¬
личивается вызванный потенциал в правой теменной доле и уменьшается в левой,
и наоборот. Следовательно, пространственные характеристики внешнего объекта
при смещении его в горизонтальной плоскости отмечаются в электрической ак¬
тивности теменных долей мозга.
При записи электрической активности отдельных нервных клеток в теменной
коре обнаружены нейроны, которые реагируют только па комплексное действие
внешнего сигнала и не отвечают на отдельные составляющие компоненты (Таи¬
ров, 1973; Казаков и др., 1972). В исследованиях на обезьянах и кошках удалось
записать нейронную активность в ответ на строго определенные пространствен¬
ные признаки внешнего зрительного сигнала (например, клетка реагировала лишь
па перемещение черной полоски вправо, другая — лишь на смещение этой полос¬
ки па угол в 45 градусов). Такие клетки названы детекторами сверхсложных про¬
странственных признаков сигналов.
Легко представить себе, что не только образ тела является грехмерноиростран-
ствеиным, но и сам образ реальной окружающей ситуации воспринимается в том
же трехкоординатном измерении. Следовательно, если любой объект или явление
внешнего мира оценивается сознательно в терминах: нраво-лево, верх-низ, спере¬
ди-сзади, а без участия сознания — в тех же трех взаимоперпендикуляриых плос¬
костях, то таким же образом может быть оценено положение любого органа тела
но отношению к остальным.
Иными словами, внутренний образ тела является трехмерным в той же степе¬
ни, в какой трехмерным должен быть и внешний образ пространства.
Человек и любое животное оценивают окружающее пространство не абстракт¬
но, а лишь применительно к самому себе, то есть включают свое тело в качестве
непременного компонента пространства. Человек «вписывается» в окружающую
ситуацию и воспринимает ее, «привязывая», соотнося внешние объекты и собы¬
тия с положением собственного тела. Чтобы представить себе физиологические
механизмы этого сложного процесса, надо прежде всего допустить, что и внутрен¬
ний и внешний образы будут описываться одним и тем же физиологическим
«языком», в котором будет закодирован весьма ограниченный набор одних и тех
же параметров, признаков. Для этого нужно представить себе наличие, например,
какой-то вертикальной плоскости, которая делит тело на две симметричные поло¬
вины. Тогда все участки тела и предметы окружающей среды, расположенные по
одну сторону от этой плоскости, будут субъективно восприниматься как «слева»,
а по другую сторону — как «справа». Только информация о положении членов
§ 7. Схема тела 93
тела будет передаваться благодаря раздражениям с кожи, мышц, сухожилий, а ин¬
формация о положении предметов по отношению к организму — с помощью зри¬
тельного аппарата. Для правильного соотнесения внешнего объекта с положением
организма в пространстве необходимо, чтобы эти два потока информации могли
встретиться и объединиться для последующего использования в произвольной
двигательной деятельности. Как раз теменная доля и связанные с ней структуры
промежуточного мозга и представляют собой таковые образования, где сходятся
(конвергируют) на одних и тех же нейронах эти информационные потоки.
Только что рассмотренный пример относится к продольно-вертикальной плос¬
кости тела, или плоскости двусторонней симметрии. Но то же можно сказать и
про фронтальную плоскость, проходящую, но-видимому, через центры вестибу¬
лярных аппаратов. Все внешнее и внутреннее пространство по одну сторону ог
этой плоскости описывается понятием «спереди», а по другую — «сзади». Через те
же две точки проходит и горизонтальная плоскость с соответствующими поня¬
тиями — «верх» и «низ» (рис. 24). Причем последняя плоскость одновременно
проходит и через центры глазных яблок. Таким образом, наклон ли головы, содру¬
жественный ли поворот глазных яблок в ту или другую сторону от этой плоскости
способствует восприятию внешнего и внутреннего пространства в одних и тех же
параметрах. Наличие в теменной коре нейронов, реагирующих на раздражение
вестибулярного аппарата и движение зрительного объекта, говорит о том, что эти
потоки информации объединяются в одних и тех же мозговых структурах.
Итак, три воображаемые плоскости, проходящие в трех взаимно перпендику¬
лярных направлениях, пересекаются в одной воображаемой точке, которую мож¬
но условно принять за «О» отсчета. А информация, описывающая внешнее и внут¬
реннее пространство организма в параметрах трехкоордипатной системы, обраба¬
тывается в таламопариетальной системе мозга. В итоге создается целостный образ
организма в среде, который служит определенным сенсорным фоном для нача¬
ла программирования любого произвольного движения на основе сопоставления
образа внешнего пространства с образом тела.
94 Глава 3. Физиология сенсорных систем
У больных с поражением теменных долей наблюдается расстройство в форми¬
ровании новых двигательных навыков и умений, особенно пространственная ор¬
ганизация целенаправленных движений (апраксия). Движения становятся бед¬
ными, неловкими, теряется их точность. Больной не знает, как выполнить слож¬
ное движение, или не знает, что и в какой последовательности надо сделать.
Теменная доля издавна именуется дополнительным моторным нолем: при ее
раздражении возникают движения конечностей, глаз, головы; из теменной коры
берет начало целая система волокон, которые достигают двигательных аппаратов
мозга. Вся система движений, возникающих при электрораздражепии теменной
коры, свидетельствует об участии этих структур в установке тела, головы, глаз в
положение, способствующее наиболее полному восприятию окружающего про¬
странства. Иными словами, и чувствительные и двигательные механизмы темен¬
ных долей мозга нацелены на создание наиболее полного сенсорного фона для це¬
ленаправленных движений.
§ 8. Кожная сенсорная система
Кожа как орган чувств занимает особое место среди других сенсорных систем как
по первичности своего происхождения, так и по огромной сложности информа¬
ции, поступающей через обширную рецепторную поверхность, измеряемую 1,4¬
2,1 м2. Толстые миелинизированные аксоны сообщают информацию о прикосно¬
вении и от суставных рецепторов. Тонкие волокна, среди которых много немиели-
иизированных, проводят болевую и температурную чувствительность (рис. 25).
Давление
Прикосновение
Тепло
Холод
Боль
Боль
Рис. 25. Кожные рецепторы и соответствующие ощущения
Принято различать несколько видов кожной чувствительности: прикоснове¬
ния-давления, боли-зуда, тепла и холода. Рецепторы разделяют па фазные, быст¬
ро адаптирующиеся и динамические (вибрационные). Рецепторы кожи бывают
двух типов: свободные и инкапсулированные окончания нервных волокон, заверну¬
тых в капсулу или оболочку. Самым простым видом рецептора кожи является сво¬
бодный кончик тонкого афферентного волокна. Вероятнее всего, это рецепторы
повреждения и боли, зуда (щекотки), температуры. Многие свободные нервные
окончания окружают волосяные фолликулы. Они возбуждаются при смещении,
подергивании и сгибании волоса. К давлению па кожу мы привыкаем довольно
быстро. Поэтому мы очень скоро перестаем чувствовать прикосновение одежды
§ 9. Вкусовая и обонятельная сенсорные системы 95
к телу. Ощущения внутренних органов мы не можем точно определить (что и где
болит), а ощущения на коже можем определить с большой точностью. Боль — это
очень важный сигнал тревоги для организма, сигнал мобилизации на борьбу
с опасностью. К болевым ощущениям человек привыкнуть не может.
Форма кожных рецептивных полей бывает различной, обычно круглой или
овальной. Для механочувствительных волокон размеры рецептивных нолей на
тыльной поверхности руки равны 1 -2 мм2, пороговая чувствительность находит¬
ся в пределах от 0,5 до 0,6 г. Размеры рецептивных полей влияют на пороги про¬
странственной дифференциальной чувствительности, определяемые по мини¬
мальному расстоянию между двумя соседними точками, одновременное раздра¬
жение которых сопровождается отчетливым ощущением раздельности. Высокая
острота осязательной чувствительности па пальцах рук человека равна 2-3 мм.
Еще более высокая пространственная дифференциальная чувствительность име¬
ется у механореценторов языка и губ человека (0,5 мм).
Температурные рецепторы располагаются на коже неравномерно. Больше все¬
го их находится у человека па лице, шее, губах, веках, причем тепловых точек ока¬
зывается намного больше, чем холодовых. Холодовые рецепторы лежат в коже
значительно поверхностнее, чем тепловые. Температурное чувство служит в ос¬
новном одной цели — поддерживать постоянную температуру тела.
Главным органом осязания является рука. Ощупывая предметы с закрытыми
глазами, мы можем определить их форму, величину, характер поверхности, темпе¬
ратуру. Особенно тонко осязание развито у слепых. Благодаря осязанию слепые
овладевают специальным способом чтения по Брайлю. Буквы алфавита выдавли¬
ваются на плотной бумаге. Слепой человек кончиками пальцев воспринимает бу¬
квы, слова, фразы.
Сигналы от кожных рецепторов по чувствительным нервам направляются
в спинной и головной мозг. Чувствительность кожи, фасций и ощущение движе¬
ния в суставах связаны с двумя восходящими системами спинного мозга. Одна за¬
канчивается в ядрах продолговатого мозга (нежном и клиновидном), а оттуда
в составе медиальной петли возбуждение достигает веитробазалыюго комплекса
ядер таламуса. Другая направляется в составе спинноталамических трактов и за¬
канчивается в других ядрах таламуса. Третий нейрон кожной сенсорной системы
заканчивается в постцентральной области коры головного мозга. Большинство
корковых нейронов первой соматосенсорной области реагирует па раздражение
определенного ограниченного участка кожи противоположной стороны тела. Ак¬
соны нейронов других ядер таламуса проецируются преимущественно во вторую
соматосенсорную кору, которая характеризуется полным перекрытием проекций
обоих половин тела человека.
Если первая корковая зона обеспечивает различение по модальности и поло¬
жению кожной и глубокой чувствительности но принципу «точка в точку», то
вторая лишена модальной и топографической специфичности и в большей мере
связана с проведением болевой сигнализации.
§ 9. Вкусовая и обонятельная сенсорные системы
Вкус — ощущение сложное. Оно, как правило, возникает при восприятии пищи
одновременно с запахом.
96 Глава 3. Физиология сенсорных систем
Рис. 26. Строение и иннервация
вкусовой почки
1 — сенсорная клетка; 2 -— опорная
клетка; 3 — базальная клетка;
4 — эпителий; 5 — пора;
6 — афферентные волокна
По происхождению вкусовые рецепторы от¬
носятся к вторично чувствующим эпителиаль¬
ным элементам. Вкусовые почки, или лукови¬
цы, — образования круглой, яйцевидной или
колбовидной формы, расположенные на поверх¬
ности языка.
Вкусовые луковицы перпендикулярно про¬
низывают эпителий, нижним концом достигая ба¬
зальной мембраны (рис. 26), а в апикальном кон¬
це образуют вкусовой канал, соединяющийся че¬
рез вкусовую пору с ротовой полостью. Выделяют
клетки трех типов: сенсорные, опорные и базаль¬
ные. На поверхности каждой вкусовой клетки,
обращенной в сторону поры, имеются микровор¬
синки, вступающие в контакт с растворенными
веществами. Из базальных клеток в процессе он¬
тогенеза образуются сенсорные клетки.
Вкусовые луковицы тесно связаны со специ¬
альными эпителиальными структурами, образо¬
ванными складками слизистой оболочки язы¬
ка, — вкусовыми сосочками (рис. 27). Различают
четыре типа сосочков — грибовидные, листовид¬
ные, желобоватые и нитевидные. Грибовидные
сосочки рассеяны по всей поверхности; желобо¬
ватые расположены в ограниченной зоне на кор¬
не языка; листовидные образуют тесно располо¬
женные складки вдоль задних краев языка; ните¬
видные занимают остальную поверхность языка
и не содержат вкусовых луковиц, то есть не уча¬
ствуют во вкусовой рецепции.
В тесном контакте со вкусовыми рецепторами
находятся нервные окончания, образующие си¬
наптические контакты с их телами. Разные участ¬
ки языка но-разному ощущают вкус: кончик язы¬
ка более всего чувствителен к сладкому, задняя часть языка — к горькому, боковые
края — к кислому, передняя и боковые части языка — к соленому (рис. 28). На ос¬
новании этого можно предположить наличие на вкусовых рецепторах четырех ти¬
пов участков, отличающихся но ряду физико-химических параметров и отвечаю¬
щих этим модальностям. Рецептивные зоны вкусовых рецепторов сформированы
участками белковых цепей («рецептивные» белки).
В проведении сенсорной информации от вкусовых рецепторов принимают
участие волокна лицевого, языкоглоточного, блуждающего и тройничного череп¬
но-мозговых нервов. Наиболее тесно связан со вкусовой чувствительностью ниж¬
ний (латеральный) конец постцентральной извилины.
При обычном восприятии пищи работают все вкусовые рецепторы языка. Из
четырех простых вкусов мозг создает сложный вкусовой образ. Обоняние обяза-
Рис. 27. Строение вкусового
сосочка
1 — вкусовые рецепторы;
2 — вкусовой сосочек;
3 — нервные волокна
§ 9. Вкусовая и обонятельная сенсорные системы 97
телыю участвует в восприятии пищи.
В создании вкусового образа пищи обяза¬
тельно участвует и осязание, ведь мы од¬
новременно оцениваем пищу не только но
ее химическим свойствам, но и по механи¬
ческим и температурным признакам. На¬
пример, вкус мороженого возникает у нас
при одновременном появлении всех этих
ощущений. Значит, обонятельная и вку¬
совая сенсорные системы работают в тес¬
ном взаимодействии с тактильной.
Обонятельная система, как и вкусовая,
относится к хемореценторным системам.
Но в отличие от рецепторов вкусовой сис¬
темы обонятельные рецепторы являются
дистантными и способны возбуждаться
иа значительном расстоянии от источни¬
ка запаха.
Слизистая оболочка обонятельной об¬
ласти носа занимает среднюю часть верх¬
ней носовой раковины и соответствующий
ей участок слизистой оболочки носовой пе¬
регородки. Рецепторный отдел обонятель¬
ных органов представлен обонятельным эпи¬
телием. комплексом рецепторных, опорных
и базальных клеток. Поверхность обоня¬
тельного эпителия покрыта слоем слизи —
продуктом желез, расположенных в пре¬
делах эпителия и опорных клеток (рис. 29).
В типичном обонятельном рецепторе
можно выделить округлое тело с клеточ¬
ным ядром, центральный отросток и дис¬
тальный сегмент, оканчивающийся у по¬
верхности эпителия утолщенным образо¬
ванием — обонятельной булавой. Последняя
песет иа свободной поверхности различ¬
ное число респичкоподобных выростов
(обонятельных волосков), снабженных фибриллярным аппаратом. Обонятельные
волоски небольших размеров способны активно двигаться, в то время как более
крупные волоски неподвижны и выступают над слоем слизи.
Аксоны рецепторных клеток образуют немиелииизироваппый обонятельный
нерв, поступающий в обонятельные луковицы. Обонятельные луковицы лежат на
базальной поверхности лобных долей, имеют овальную форму. Иногда их включа¬
ют в состав древней коры, иногда рассматривают как особую часть обонятельного
мозга. Благодаря многочисленным конвергентным и дивергентным связям не су¬
ществует топической пространственной проекции обонятельных рецепторов на
Рис. 28. Вкусовые зоны языка
и ошушения
Рис. 29. Расположение органа
обоняния в носовой полости
1 — реснички; 2 — эпителиальные клет¬
ки; 3 — обонятельные клетки;
4 — нервные волокна
4 Зак. 1078
98 Глава 3. Физиология сенсорных систем
элементах луковицы. Существует мозаика из обонятельных рецепторов разных
типов, обладающих широкими, но различными спектрами чувствительности, ко¬
торая является вероятной основой для различения и анализа запахов.
Различают шесть основных запахов, соответствующих шести занаховым груп¬
пам (цветочный, гнилостный, фруктовый, горелый, пряный, смолистый). Эти за¬
пахи располагаются на вершинах треугольной призмы, а все остальные (промежу¬
точные) на ее ребрах и гранях (рис. 30). Однако проблема классификации и меха¬
низма восприятия запахов па сегодня является еще не решенной.
Гнилостный
Заключение
Адекватность сенсорного отражения ни в коем случае нельзя трактовать как полу¬
чение организмом точной зеркальной копии окружающего мира. Во-первых, вос¬
принимаются и оцениваются не сами по себе элементы внешнего мира, а их взаи¬
моотношения и взаимосвязи, формирующие целую структуру, которая может
иметь ту или иную биологическую значимость в данный момент и при данных об¬
стоятельствах. Во-вторых, необходимо рассматривать сенсорные системы не как
отдельные детали целостного механизма, а как тесно взаимодействующие струк¬
турно-функциональные образования, формирующие целостный неделимый образ
окружающей действительности. Наконец, в-третьих, сенсорные системы являют¬
ся не пассивными каналами линии связи, информирующими организм о событи¬
ях и изменениях в окружающем мире, а представляют собой активные преобразо¬
ватели информации, извлекающие сведения о тех событиях, которые в данный
момент являются наиболее значимыми и служат для формирования и регулиро¬
вания адекватного, целенаправленного поведения по удовлетворению домини¬
рующей мотивации.
Контрольные вопросы
1. Какова основная функция рецепторов?
2. Принципы классификации рецепторов.
3. Первичные и вторичные рецепторы.
Контрольные вопросы 99
4. Основные функции сенсорных систем.
5. Механизмы кодирования информации.
6. Вспомогательные аппараты зрительной сенсорной системы.
7. Строение глаза.
8. Оптическая система глаза.
9. Строение сетчатки.
10. Центральные механизмы зрительного восприятия.
11. Строение слуховой сенсорной системы.
12. Внутреннее ухо.
13. Центральные аппараты слухового восприятия.
14. Корковые речевые зоны.
15. Асимметрия речевой функции.
16. Слуховая обратная связь и ее функция.
17. Рецепторы скелетно-мышечной сенсорной системы.
18. Роль двигательной обратной связи.
19. Роль кожной сенсорной системы.
20. Рука как орган осязания.
21. Значение болевой рецепции.
22. Строение вестибулярного аппарата.
23. Строение вестибулярных рецепторов.
24. Почему вестибулярный аппарат называют органом равновесия?
25. К чему приводит нарушение вестибулярной функции?
26. Строение органов обоняния и вкуса.
Глава 4
РЕГУЛЯЦИЯ ПРОИЗВОЛЬНЫХ ДВИЖЕНИЙ
§ 1. Обратная связь в управлении движениями
В неврологии уже давно сложилось убеждение, что произвольные двигательные
акты связаны с функцией пирамидного и экстрапирамидного трактов, связываю¬
щих, в свою очередь, кору головного мозга и подкорковые структуры со спиналь¬
ным моторным механизмом. Было показано, что творческая аналитико-синтети¬
ческая функция принадлежит сенсорным приборам. Эфферентные же системы
являются лишь выходными воротами для соответствующих информаций, кодо¬
вый алфавит которых определяется сложным межсепсорпым синтезом.
Еще в первой четверти XIX столетия Ч. Беллом было показано, что между моз¬
гом и мышцей имеется нервный круг: один нерв несет импульсы мозга к мышце,
другой приносит ощущения состояния мышц к мозгу. Перерыв эфферентного пу¬
ти этого круга приводит к исчезновению движения; выпадение же афферентного
звена влечет за собой уничтожение ощущения состояния мышц и как следствие
этого — нарушение регулирования их функций.
В своих работах И. М. Сеченов уделял значительное место «темному мышеч¬
ному чувству в формировании элементов мысли». По его убеждениям, из этого
чувства рождаются представления о скорости движений,
об отношениях предметов в пространстве и времени.
Рецепторные приборы мышц, сухожилий и суставов
Ч. С. Шеррипгтон назвал пронриорецепторами.
П. К. Анохин применил понятие обратная связь, об¬
ратная афферентация, и эго сыграло существенную роль
во всей системе наших представлений о работе мозга. Мо¬
дальность обратной связи имеет немаловажное значение
и в большой мере определяет ее биологическую ценность
для целостного поведенческого акта, будь то, скажем, те-
лецептивная или соматическая афферентация.
Таким образом, одним из основных принципов в меха¬
низмах координации двигательных реакций является циклическая нервная связь,
в которой афферентная информация обеспечивает форму и состав эфферентного
проявления центральной интеграции.
Можно выделить ряд целевых функций, связанных с мышечной активностью:
♦ поддержание определенной позы;
♦ ориентация на источник внешнего сигнала (двигательный компонент ориен¬
тировочной реакции, глазодвигательные реакции — саккады и поддержание
равновесия);
♦ локомоция — перемещение тела в пространстве;
♦ манипулирование — куда включаются и так называемые оперантные движе¬
ния конечностей.
Одним из основных
принципов в механиз¬
мах координации дви¬
гательных реакций яв¬
ляется циклическая
нервная связь, в кото¬
рой афферентная ин¬
формация обеспечива¬
ет форму и состав
эфферентного прояв¬
ления центральной ин¬
теграции.
§ 2. Внешняя обратная связь 101
Быстрые движения
баллистического типа
относительно незави¬
симы от сигналов об¬
ратной связи, а мед¬
ленные движения в
значительной мере за¬
висят от их наличия.
Долгое время основные представления о механизмах
двигательного управления строились на положениях кон¬
цепции кольцевого управления, рассматривающей мы¬
шечную обратную связь как обязательный и решающий
фактор регулирования временных и пространственных
характеристик движения (Бернштейн, 1947). Однако в по¬
следнее время получило широкое распространение пред¬
ставление о ведущей роли в процессах управления движе¬
ниями фиксированных центральных команд (Коц, 1975; Шик, 1986). Было пока¬
зано также, что быстрые движения баллистического типа относительно независи¬
мы от сигналов обратной связи, а медленные движения в значительной мере зави¬
сят от их наличия.
Проблема трактуется так: насколько соматическая (проприоцептивная, преж¬
де всего) обратная связь является непременным фактором регуляции двигатель¬
ных актов. С одной стороны, в движениях баллистического типа классическая об¬
ратная связь не принимает участие потому, что само движение завершается рань¬
ше, чем к соответствующим мозговым центрам успеет поступить информация от
двигательных аппаратов. С другой стороны, медленные движения постоянно кор¬
ректируются благодаря показаниям с соответствующих сенсорных датчиков, рас¬
положенных в составе скелетно-мышечного аппарата и передающих информацию
в разные отделы вплоть до коры. Эти сенсорные коррекции (но терминологии
Н. А. Бернштейна) поступают к разным уровням центральной нервной системы
и способны изменить характер движения по ходу его осуществления.
Иной подход к пониманию роли обратных связей соответствует классифика¬
ции движений по характеру операционного состава. Движения, базирующиеся па
врожденных координациях, в меньшей степени требуют обратной связи от локо¬
моторного аппарата, а движения, в основе которых лежит формирование новых
координационных отношений, всецело зависят от соматической афферентации
с двигательного аппарата.
Наконец, степень участия обратных связей находится в зависимости и от уров¬
ня автоматизации двигательного акта. В первые этапы его формирования значе¬
ние информации от скелетно-мышечного аппарата значительно выше, чем на эта¬
пе его закрепления.
Таким образом, вопрос о роли соматической обратной связи в функциональной
структуре двигательного акта необходимо решать конкретно в зависимости от
временных параметров двигательного акта, его моторной дифференцированно¬
сти, сложности координационного состава, степени автоматизации и ряда других
факторов.
§ 2. Внешняя обратная связь
Движения, осуществляющиеся без помощи проприоцептивной обратной связи,
выполняются по жесткой программе. Однако формирование предварительных
программ является непременным атрибутом любого сложного движения. Поэто¬
му, особенно для медленных движений, управляемых обратными связями, необ¬
ходимо допустить сличение актуальной афферентации с тем сенсорным образом
движения, который формируется в составе программы. П. К. Анохин выделил
102 Глава 4. Регуляция произвольных движении
в общей функциональной системе двигательного акта специальный блок сличе¬
ния, который он назвал акцептором результатов действия. Результат сравнения
обратной афферентации с двигательной программой (согласование или рассогла¬
сование) является главным фактором ее перестройки. Эти влияния могут переда¬
ваться к аппаратам программирования но системе внутренней обратной связи.
Понятие о внутренней обратной связи весьма широкое и емкое, оно включает все
процессы перестроек эфферентной программы в зависимости от впутрицентраль-
ных отношений. К этому же роду явлений, несомненно, относятся и акцептор ре¬
зультатов действия, и модель предвидимого будущего (по Бернштейну), и образ
движения (по Ухтомскому).
§ 3. Внутренняя обратная связь
Если в формировании двигательных программ ведущую роль играют глубокие от¬
делы мозга и мозжечок, то кортикальный уровень связан с процессами принятия
решения и тонкого афферентного контроля сложных сеисомоторных синтезов.
Если взять для исследования три наиболее биологически адекватных движе¬
ния: прицельный прыжок, переход по перекладине и прямую побежку у кошек,
то эти движения сильно различаются по своим скоростным характеристикам, аф¬
ферентному обеспечению и координационному составу. Все эти движения имеют
полисенсорпую природу при наибольшей степени участия в прыжке зрительной
и соматической афферентации, а при переходе по перекладине — вестибулярной
и соматической афферентации. Третий вид движения — прямая побежка к кор¬
мушке — может осуществляться, будучи предварительно выработанной, без уча¬
стия зрительной, слуховой и вестибулярной афферентации. Однако непремен¬
ным условием осуществления локомоции в конкретных условиях пространства
является согласованное участие кожной и мышечной афферентации. Как бы ни
было велико значение проприорецепции от шагающих конечностей, она сама по
себе без кожной чувствительности не может обеспечить того афферентного меха¬
низма, который необходим для адаптивного характера движения. И действитель¬
но, если обученному животному, лишенному зрения, слуха, обоняния и вестибу¬
лярного аппарата, производили деафферентиацию лишь опорных частей конеч¬
ностей путем инъекции местного анестетика, направленность двигательного акта
тут же исчезала.
Другой опыт состоял в следующем. Интактная кошка обучается по сигналу
осуществлять прыжок с площадки па площадку при расстоянии между ними
100 см. Затем в подушечки конечностей вводится местный анестетик — тримека-
ин. На рис. 31 представлены результаты киноисследования динамики прицельно¬
го прыжка. Видно, что кошка, лишенная информации об опоре, не может четко
выполнять этот акт: или срывается с площадки, не допрыгнув до нее, или прома¬
хивается в результате неправильной оценки исходной нозы. Наиболее заметно на¬
рушается процесс принятия стартовой позы — и соответственно значительно уве¬
личивается латентный период. Менее страдает процесс приземления. Длитель¬
ность принятия позы приземления увеличивается незначительно, однако часто
наблюдаются промахи задних конечностей, изменение биомеханики движений
передних конечностей при приземлении, что связано с блокированием срочной
информации о контакте с поверхностью. К аналогичным последствиям приводит
§ 4. Медиальные лемниски 103
предварительное рассечение ветвей мышечных нервов конечностей при сохране¬
нии кожной чувствительности.
Рис. 31. Изменения двигательных актов у кошки после разрушения
путей соматической обратной связи (А. Г. Смирнов, 1974)
а — норма; б — после операции
Таким образом, доказана справедливость мысли П. К. Анохина о том, что про-
приорецепторы по самой своей сути не могут дать полноценную и конечную об¬
ратную афферентацию о достижении адекватного эффекта. С помощью проприо-
рецепторов мы никогда не сможем оценить результаты движения руки при взятии
какого-либо предмета, по они помогают достижению этого эффекта, создавая наи¬
более экономный тин направленного движения. Никакая саморегулирующаяся
система, если только она заканчивается полезным для организма эффектом, не
может быть построена на проприоцептивной информации.
§ 4. Медиальные леллниски
В настоящее время не вызывает сомнения определяющая роль коры в организа¬
ции произвольных движений. И в этой связи так называемая сенсомоторная кора
является объектом пристального внимания исследователей. Наряду с описанием
соматической проекции в корковую область накапливается множество данных
о ее нолисенсорпой активации по различным каналам.
Основную же роль отводят так называемому лемнисковому соматическому пу¬
ти, берущему начало от продолговатого мозга и заканчивающемуся на нейронах
вентробазального комплекса ядер таламуса (рис. 32). Создавалось представление
о лемнисковой системе как пути соматической обратной связи для кортикальных
аппаратов. В составе этой системы проходят волокна, несущие быструю инфор¬
мацию от мышц, суставов и сухожилий, которые, как принято думать, связаны
с оценкой схемы тела.
104 Глава 4. Регуляция произвольных движении
Рис. 32. Схема скелетно-мышечной
сенсорной системы
а и у — мотонейроны спинного мозга (СПМ), регулируя дея¬
тельность мыши, получают информацию от мышечных ве¬
ретен, сухожильных и суставных рецепторов по отросткам
афферентного нейрона спинального ганглия. К этим же мо¬
тонейронам по пирамидному тракту (ПТ) приходят регули¬
рующие влияния от коры больших полушарий мозга (К) и про¬
долговатого мозга (ПМ). Регуляция движений обеспечивается
содружественной деятельностью всех систем (указано стрел¬
ками), в том числе стриопаллидарной (СПС), таламуса (Т),
среднего мозга (СМ), мозжечка (М) и спинного мозга
1 — мышечные веретена, 2 — сухожильные рецепторы, 3 —
суставные рецепторы, 4 — отростки афферентного нейрона
спинального ганглия
Поэтому было проведено исследование роли лемнисковой системы в нейронной
организации сенсомоторной коры и в реализации сложных движений (А. Г. Смир¬
нов). Исследовались условно-рефлекторные акты (прицельный прыжок и переход
по перекладине) у кошек после двустороннего разрушения медиальных лемни-
сков. В результате операции латентный период условных рефлексов существен¬
но увеличился и даже спустя два месяца не достиг дооперациоиного уровня.
Стартовая поза при прыжке резко изменилась. Животные не прыгали «с ходу», а
несколько раз проделывали движения прицеливания, перебирали передними ла¬
пами, несколько раз отходили от края площадки. При переходе но перекладине
движения были замедленными, животные осторожно ощупывали перекладину,
голова опущена, ноги согнуты, тело касается перекладины. При попытке осущест¬
вить переход животные теряли равновесие и падали на бок (см. рис. 32).
У оперированных животных наблюдался повышенный мышечный тонус, лег¬
кая отвлекаемость па посторонние раздражители, иногда вегетативные и трофи¬
ческие расстройства, обедненность эмоциональных и голосовых реакций.
§ 5. Двигательная программа
Известно, что для точного осуществления движения необходима такая стартовая
позиция, которая в самом начале задала бы необходимую траекторию для переме¬
щения тела в пространстве. Помимо информации от дистантных рецепторов необ¬
ходима сигнализация об исходном положении тела в пространстве и взаиморас¬
положении суставов, которая в основном и передается по лемнисковому каналу.
Имеются указания также на существенную роль вестибулярного аппарата в фазе
инициации движения.
§ 5. Двигательная программа 105
Реализация каждого двигательного акта требу¬
ет постоянной коррекции, так как внешние реак¬
тивные силы отклоняют тело и конечности от за¬
данной траектории. Эта коррекция должна быть
очень быстрой и точно рассчитанной. В описанных
выше опытах она явно нарушена. Вероятно, поэто¬
му, чтобы не ошибиться, животные фиксируют сте¬
пени свободы и в меньшей степени освобождают
их при реализации двигательного акта, что и имеет
место при переходе но перекладине.
Таким образом, основные двигательные рас¬
стройства могут быть связаны с исчезновением бы¬
строй информации от «внутреннего пространства»
животного.
Хорошо известно, что сенсомоторные зоны ко¬
ры являются не только сомато-сенсорпыми проек¬
ционными нолями, но и содержат большое число
элементов, реагирующих на гетеромодальные воз¬
действия (Батуев, 1973). Есть основания рассмат¬
ривать сенсомоторные ноля коры в качестве «шер-
рипггоповской воронки» на кортикальном уровне,
к которой конвергирует разнообразная гетеромо-
дальпая информация и осуществляются процессы
интегративного порядка, завершающиеся форми¬
рованием эфферентного залпа. При этом основным
«входом» для данного коркового поля остается
лемнисковая импульсация. Поэтому было важно
оценить значение этого лемнискового входа в об¬
щей организации нейрональной активности сен-
сомоторпой коры, то есть представить себе, како¬
вы могут быть нейронные особенности построения
эфферентных команд в отсутствие лемписковых
влияний.
Опыты показали, что после выключения меди¬
альных лемнисков выпадают наиболее короткола¬
тентные ответы па кожную, мышечную и вести¬
булярную стимуляцию, извращается соотношение
нейронов, реактивных к телецептивпым раздраже¬
ниям (рис. 33) и в характере взаимодействия ге¬
теромодальных влияний преобладают окклюзион¬
ные отношения.
При внеклеточной регистрации мы практически
Рис. 33. Гистограмма распреде¬
ления нейронов по латентному
периоду их реакиий на разно¬
модальные стимулы до (темные
столбики) и после (белые) опе-
раиии
по горизонтали — время в миллисе¬
кундах, по вертикали — число ней¬
ронов
имеем дело с эфферентной ак¬
тивностью, и в частности с тем разрядом, который поступает но аксонным систе¬
мам к исполнительным аппаратам ствола и спинного мозга. Следовательно, по ха¬
рактеру спайковой активности нейронов можно судить о той команде, которая фор¬
мируется в коре и распространяется но путям пирамидной и экстрапирамидпой
систем. По-видимому, после разрушения медиальных лемнисков выпадают сроч-
106 Глава 4. Регуляция произвольных движении
ные корковые нисходящие влияния, увеличивается значение телецептивных ком¬
понентов в активации нейронов сепсомоторпой коры и при гетеромодалыюм тес¬
тировании вместо процессов пространственно-временной суммации преобладает
влияние какого-либо одного сенсорного входа при сопряженном торможении ос¬
тальных.
Поэтому, если перемещение конечности рассматривается как компонент обще¬
координированного акта, следует признать, что внутренняя (проприоцептивная)
и внешняя (кожная и телецептивная) афферентации выступают как содружест¬
венные компоненты системы сенсорного обеспечения движений, причем сомати¬
ческая импульсация от локомоторного аппарата для работы сегментарных спи¬
нальных механизмов может рассматриваться в качестве внешней обратной связи.
Последняя играет важную роль как для инициации сложных движений, так и для
их координированного выполнения. В первую очередь это касается медленных
компонентов двигательных актов — таких, как принятие исходной позы прицель¬
ного прыжка и перехода но перекладине, требующих непрерывных коррекций от
шагающих конечностей.
Совокупность поведенческих и нейрофизиологических исследований (Череп¬
кова, 2001) свидетельствуют о том, что на уровне сепсомоторпой коры высших
млекопитающих реализуются механизмы сенсорной и моторной координации.
В активности нейронов сепсомоторпой коры находят отражение не только про¬
цессы запуска и реализации движения, по и процессы программирования, соот¬
несения значимости и параметров сенсорных стимулов и выбора двигательной
команды о типе конкретной двигательной реакции, которая наиболее адекватно
соответствует инструкции, обусловленной этими стимулами. Эти процессы реа¬
лизуются в тех участках сепсомоторпой коры, которые принимают непосредст¬
венное участие в организации данного двигательного акта. Его проявление и на¬
ходит отражение в изменении активности на условные сигналы тех выходных пи¬
рамидных нейронов, которые имеют соматические рецептивные поля в области
исполнительных двигательных органов, а реакции во время выполнения движе¬
ния связаны либо с перемещением этих органов в целом, либо с отдельными фаза¬
ми моторного акта.
Если рассматривать корковый уровень как систему инициации адаптивного
движения сообразно пространственно-временным факторам окружающей среды,
необходимо допустить существование каналов обратной связи, по которым кора
будет информирована не об отдельных параметрах дви¬
жений, а о степени соответствия предварительно создан¬
ной двигательной программы тому наличному движению,
которое достигается в данный конкретный момент време¬
ни. Следовательно, речь идет о внутренней обратной свя¬
зи и допустимо считать медиальные лемниски одной из
важнейших систем ее осуществления. В настоящее время
описаны влияния кортикопирамидной системы на ядра
дорзальных столбов продолговатого мозга. Может быть,
последние и являются той структурой, где осуществляет¬
ся сличение какого-то двигательного цикла с копией программы, и последствия
рассогласования поступают в сеисомоторную кору для очередной коррекции кор¬
тикальной программы.
Адаптивные изменения
кортикальной двига¬
тельной программы
достигаются с помо¬
щью внутренней обрат¬
ной связи, компонен¬
том которой является
система медиальных
лемнисков.
Контрольные вопросы 107
Относительная соматотопическая организация лемписковой системы и эффе¬
рентных выходов из коры позволяет корковым аппаратам осуществлять преиму¬
щественный контроль за дистальной мускулатурой тела, причем в особенности
в моменты начала и окончания отдельного элементарного цикла движения ко¬
нечности, требующего в каждом случае коррекции с заданной интегральной про¬
граммой.
Таким образом, в конкретной организации движения внешняя обратная связь
от соматических рецепторов ограничивается спинобульбарггым уровнем, тогда
как адаптивные изменения кортикальной двигательной программы достигаются
с помощью внутренней обратной связи, компонентом которой может являться
система медиальных лемписков.
Контрольные вопросы
1. Каково значение обратной связи в управлении движениями?
2. В чем принципиальное отличие быстрых и медленных движений?
3. Какова роль «сенсорных коррекцгш» (по Н. А. Бернштейну)?
4. Суть внешней и внутренней обратной связи по П. К. Анохину.
5. Что такое двигательная программа и как огга формируется?
6. Значение коркового звегга в системе уггравлепия движениями.
7. Отличие произвольггых движеггий от непроизвольных.
8. Понятие «схема тела».
9. Статический и динамический «образ тела».
10. Феномегг фантомных болей.
И. Механизмы регуляции гюзы.
12. Что такое «сенсорггый фон движеггия»?
13. Осгговные функцигг таламоггариеталыгой сггстемы мозга в управлении движе¬
ниями.
Глава 5
ФОРМЫ ПОВЕДЕНИЯ
§ 1. Поведение как фактор эволюции
Поведение наряду со структурными признаками организма рассматривается как
важнейший фактор эволюционного процесса.
Современное развитие дарвиновской теории эволюции опирается па естест¬
венный отбор как ее краеугольный камень и рассматривает понятия макро- и микро¬
эволюции. Наследственная структура вида, его генофонд,
при развитии каждой конкретной особи выражается в на¬
боре признаков и свойств, обеспечивших ее выживание
и воспроизведение в потомстве. При изменении окру¬
жающей среды медленно меняются сам генофонд и вид
в целом. Этот процесс получил название макроэволюции.
Другой способ анализа эволюционных преобразова¬
ний организмов связан с широким использованием так
называемого популяционного подхода. Установлено, что
наиболее благоприятные условия для проявления геноти¬
пической изменчивости и соответствующего эволюцион¬
ного изменения вида возникают тогда, когда вид распада¬
ется на ряд небольших изолированных колоний — попу¬
ляций. Популяция — это совокупность особей одного
вида, давно населяющих определенную территорию, обеспечивающая свободное
скрещивание в пределах популяции, но изолированная от соседних популяций.
Микроэволюционный процесс происходит в относительно короткое время, на огра¬
ниченных территориях и в ограниченных популяциях. Таким образом, популя¬
ция — это элементарная единица процесса микроэволюции, способная реагиро¬
вать на изменение среды перестройкой своего генофонда. Комплекс представлений
о макро- и микроэволюции получил название синтетической теории эволюции.
Поведение в эволюции вида и популяции. Современная концепция вида под¬
черкивает поведенческую репродуктивную изоляцию особей разных видов и обеспе¬
чение условий для внутривидового размножения. Поведенческие механизмы, в ре¬
продуктивном отношении изолирующие один вид от другого, следует считать
наиболее важным комплексом атрибутов отдельного вида, поскольку они по опре¬
делению входят в состав видовых критериев (И. Майр,
1963).
Вид как экологическое единство взаимодействует с дру¬
гими видами как целое. Хотя вид обладает общим гено¬
фондом, между индивидуумом и видом существует про¬
межуточный уровень, который и обозначается как попу¬
ляция.
Преобладавший в течение многих десятилетий так на¬
зываемый типологический подход состоял в абстрагиро-
Репродуктивная изоля¬
ция — предотвращение
спаривания партнеров
из разных видовых по¬
пуляций на основе их
несоответствия в поло¬
вом поведении. Спо¬
собствует изоляции ви¬
довых популяций.
Поведение — совокуп¬
ность двигательных ак¬
тов организма, возни¬
кающих в ходе
взаимодействия с
внешней средой и на¬
правленных на удовле¬
творение доминирую¬
щих у организма
потребностей. Поведе¬
ние имеет адаптивное
значение и является
важнейшим фактором
эволюции вида.
§ 1. Поведение как фактор эволюции 109
ваиии от отдельности, неповторимости, изменчивости ка¬
ждого организма и рассматривании лишь черт сходства,
позволявших отнести разные организмы в одну и ту же
группу. При популяционном подходе каждый организм
рассматривается как продукт уникального в своем роде
сочетания генетических и средовых факторов.
Длительное и направленное изменение генотипа по¬
пуляции — это элементарное эволюционное явление. Ге¬
нетическая изменчивость популяции может усиливаться
благодаря мутациям, рекомбинациям и притоку новых генов, ослабляться — бла¬
годаря естественному отбору, или генетическое разнообразие популяции может
стабилизироваться благодаря чисто генетическим закономерностям, а также влия¬
нию экологических и поведенческих механизмов.
В эволюции популяции поведение играет двоякую роль: с одной стороны, с по¬
мощью поведения обеспечивается изоляция популяции, с другой — поддержива¬
ется генетическая гетерогенность популяции.
Поведение и репродуктивная изоляция. Существует много факторов, обеспе¬
чивающих изоляцию видовых популяций. К ним относятся так называемые этно¬
логические изолирующие механизмы, препятствующие межвидовому скрещива¬
нию, и прежде всего различия в коммуникативном поведении. Потенциальные
партнеры не могут встретиться в связи с разным местообитанием, несовпадением
сезонов размножения, предпочитаемым для спаривания временем суток. Предот¬
вратить спаривание на основе несоответствия в половом поведении партнеров го¬
раздо экономнее и надежнее, чем использовать другие механизмы изоляции, пре¬
пятствующие полноценному оплодотворению или появлению нежизнеспособных
гибридов.
Брачное поведение животных и птиц, представляющее собой целый комплекс
сигналов, встроенных в систему коммуникации потенциальных партнеров и жест¬
ко детерминированных генетически, получило название ритуализации. Такую ви¬
довую систему коммуникации рассматривают как динамический процесс. Каж¬
дой его фазе соответствует свой комплекс коммуникативных сигналов и правил.
Предъявление этой последовательности сигналов особи-реципиенту вызывает та¬
кое состояние, при котором ее ответная реакция будет соответствовать стереоти¬
пу коммуникаций данного вида. Приведем пример из биологии амфибий. В одной
и той же области, в одном пруду или луже могут жить несколько разных видов ля¬
гушек и одновременно возникает пение самцов нескольких видов. Самки реагиру¬
ют на крики только «своих» самцов. Видимо, видоспецифический характер их пе¬
сен возник в результате естественного отбора.
Отбор действует на уровне отдельной особи. Он сохраняет те приспособления,
которые максимизируют успех размножения данной особи на протяжении всей ее
жизни. Половой отбор — это форма естественного отбора, сведенная к неодинако¬
вым шансам на нахождение и привлечение брачного партнера, спаривания, опло¬
дотворения и выведения потомства. Поэтому отбор ведет к преобладанию геноти¬
пов с высокой приспособленностью.
Поведение и генетическое разнообразие популяции. Давно изучено явление
так называемой миграции животных из одной природной популяции в другую.
Такая миграция особей важна для формирования генофонда, и прежде всего она
Поведение особи —
это, во-первых, гене¬
тически детерминиро¬
ванная видоспецифи¬
ческая программа,
а во-вторых, лабиль¬
ная система конкрет¬
ных адаптаций к ме¬
няющимся условиям.
110 Глава 5. Формы поведения
предотвращает тесный инбридинг (близкородственное скрещивание). Например,
у павианов-анубисов половозрелые взрослые самцы и неполовозрелые самки час¬
то переходят из одной популяции в другую. То же отмечалось у горилл и шимпанзе.
Генотипическая изменчивость популяции может под держиваться существова¬
нием полиморфизма по признакам поведения, например по уровню двигательной
активности и эмоциональной возбудимости.
Наконец, важно подчеркнуть участие поведения в поддержании численности,
а также пространственной и внутренней структуры популяции в целом. В качест¬
ве одного из примеров можно упомянуть следующее. Многие птицы размножают¬
ся в колониях, причем каждая пара занимает отдельный участок определенной ве¬
личины. Если какой-либо птице не удается занять гнездовой участок в пределах
ограниченной территории для размножения, то она вовсе не размножается в дан¬
ном сезоне. Так, по достижении известного предела плотности популяции осталь¬
ные птицы «жертвуют» размножением па благо группы в целом. Значит, поведе¬
ние становится рычагом регулирования плотности популяции.
Поведение животных и современная этология. Начиная с середины 30-х гг.
одним из направлений в изучении поведения животных стала этология (от греч.
этос — характер, нравы). В задачу этой науки входит изучение «морфологии пове¬
дения животных» и в основном изучение инстинктивных
движений. Этологическая позиция характеризовалась тре¬
бованием изучать формы поведения, имеющие биологи¬
ческий смысл в естественных условиях у различных ви¬
дов домашних и диких животных. В. А. Вагнер (1914) счи¬
тал, что эволюцию поведения можно исследовать точно
так же, как изучают эволюцию органов. С его точки зре¬
ния, инстинкты подлежат тем же законам развития, ко¬
торым следуют морфологические признаки. Другие этологи (Уитмен, Юкскюль,
Хейнрот) также обращали внимание на удивительную константность инстинк¬
тивных форм поведения, которые могут, как и морфологические особенности,
служить таксономическими признаками.
В этологических исследованиях поведенческие акты рассматриваются как це¬
лостные, неделимые. Центральное место в этологическом мышлении занимают
комплексы фиксированных действий (КФД), представляющие собой сложные
стереотипные движения, образующие высокоорганизованную последователь¬
ность. Эти КФД вызываются простыми, но высокоспецифичными раздражителя¬
ми (выполняющими роль толчка) и одинаково проявля¬
ются у всех представителей данного вида. Начавшись по
таким стимулам, КФД могут продолжаться и в их отсут¬
ствие, причем эти КФД теперь уже утрачивают свой при¬
способительный смысл. Но КФД трудно вызвать повтор¬
но, поэтому их относят к самоистощающимся реакциям.
Считается, что КФД не связаны с индивидуальным опы¬
том организма и осуществляются практически с первого
же раза. К такому виду стереотипных актов относятся реакции закатывания яиц
в гнездо у серого гуся: приблизившись к яйцу, гусь вытягивает к нему шею и до¬
трагивается клювом до земли по другую сторону яйца. Совершая ряд строго сте¬
реотипных движений в вертикальной плоскости, гусь возвращает выкатившееся
Комплексы фиксиро¬
ванных действий —-
сложные стереотипные
движения, образующие
высокоорганизован¬
ную поведенческую по¬
следовательность.
Этология — наука, из¬
учающая поведение
животных, преимуще¬
ственно видоспецифи¬
ческие формы поведе¬
ния в естественных ус¬
ловиях обитания.
§ 1. Поведение как фактор эволюиии 111
яйцо в гнездо (рис. 34). Откатывание яйца в сторону, замена его на искусственную
модель сохраняет весь тот же комплекс движений.
Рис. 34. Серый гусь возвращает яйцо, выкатившееся из гнезда
(по Э. Меннингу, 1982)
а — это движение стереотипно и используется при возвращении в гнездо собственного яйца;
б — совершенно тем же способом гусь пытается подкатить и муляж яйца
Специфические стимулы, запускающие КФД, получили название «знаковых
стимулов». Самцы трехиглой колюшки, защищая свою тер]
строго специфический признак самца своего вида — крас¬
ное брюшко. Если на территорию, занятую этой рыбой,
поместить точную модель самца, но с брюшком другого
цвета, агрессивного столкновения не произойдет. Модель
же с красным брюшком будет подвергнута яростному на¬
падению. Такие стимулы-релизеры подходят к соответст¬
вующим поведенческим реакциям, как ключ к замку, вы¬
свобождая весьма сложные акты поведения, без специаль¬
ного обучения. В обычном состоянии КФД находятся иод влиянием активного
торможения. Каждый инстинктивный акт контролируется тормозным нервным
механизмом, который реализуется благодаря комбинированному действию гор¬
монов, внешних раздражений и влияний высших нервных механизмов (В. Детьер
и Э. Стеллар, 1967).
Этологи предполагают, что знаковые стимулы (релизеры, ключевые сигналы)
воздействуют на локализованный в определенном участке мозга «врожденный
разрешающий механизм», который снимает это торможение и способствует раз¬
рядке соответствующих центров. Запускаемый в результате КФД зависит не
только от стимула, по и от накопившейся «специфической энергии действия». Ес¬
ли эта энергия мала, возникают самоистощающиеся КФД; если, наоборот, велика,
КФД может возникать вообще без всякого внешнего стимула.
Инстинкты — сложнорефлекторные комплексы. Термин «инстинкт» в пере¬
воде с латинского означает внутреннее побуждение. Однако наряду с побужде¬
ниями к каким-то действиям инстинктом стали называть и сами действия.
Ч. Дарвин (1856) первым провел сравнительное описание инстинктов, свойст¬
венных как животному, так и человеку. Этим путем он доказал общность биологи¬
ческих основ поведения человека и животных и возможность изучения эволюции
поведения. Дарвин отграничивал действия, связанные с индивидуальным опытом
и способностью к его обобщению, от инстинктивных действий, переданных по на¬
следству. Инстинкты, по его мнению, — самая низкая ступень деятельности чело¬
века и животных.
Большинство последующих исследователей под инстинктами начинают пони¬
мать ту часть поведения животных, которая характерна для организмов данного
вида и закреплена за ними наследственно.
порто, реагируют па
Инстинкт — сложно¬
рефлекторный комп¬
лекс наследуемых по¬
веденческих реакций
особей одного вида,
характеризующийся
стереотипностью
и стабильностью.
112 Глава 5. Формы поведения
Немецкий зоолог Г. Э. Циглер (1964) предложил следующие критерии ин¬
стинктивного действия:
♦ побуждение и способность к действию принадлежит к числу наследственных
свойств вида;
♦ такие действия не требуют предварительного обучения;
♦ оно выполняется но существу одинаково у всех нормальных представителей
вида;
♦ оно соответствует организации животного, то есть связано с нормальным
функционированием его органов;
♦ оно приспособлено к экологическим условиям обитания вида.
Относительная стабильность инстинктивного поведения как одна из его глав¬
ных черт необходима для обязательного обеспечения жизненно важных функций,
независимо от случайных, постоянно изменчивых условий среды, в которых неиз¬
бежно оказывается любой представитель вида.
В инстинкте сосредоточен опыт поколений, это квинтэссенция видового опы¬
та. Если под инстинктом понимать лишь генетически детерминированную про¬
грамму сложных поведенческих действий, то в строгом смысле слова истинные
инстинктивные акты мы можем наблюдать у животных только при нервом их про¬
явлении. Ибо при каждой последующей реализации возникает множество новых
только что приобретенных условных рефлексов, приводящих к индивидуальной
модификации наследственно запрограммированного поведенческого акта. Так что
сложное инстинктивное поведение взрослого индивида в конечном счете — инди¬
видуальная адаптация видового опыта, то есть сплав наследственно закрепленных
стабильных и приобретенных актов поведения. Иными словами, животное рожда¬
ется с заложенной в нем наследственной поведенческой программой, но на протя¬
жении всего периода постнатального онтогенеза оно учится правильно применять
эту программу сообразно с изменчивыми факторами окружающей среды.
Стереотипность и стабильность инстинктивных действий при экстренном из¬
менении окружающей ситуации утрачивает свое приспособительное значение. Но
многие наблюдения показывают, что инстинкт не настолько уж «слепая сила»,
он способен к индивидуальной изменчивости. Принято выделять наиболее ста¬
бильные «ритуализовапные» действия и наиболее изменчивые его элементы.
К внутренним факторам инстинкта следует прежде всего отнести гумораль¬
но-гормональные отклонения в организме от обычного физиологического уровня.
Такие сдвиги во внутренней среде могут быть настолько сильными, что даже в ис¬
кусственных условиях полной изоляции животного могут привести к выполне¬
нию стереотипных инстинктивных действий. Введение лабораторным белым кры¬
сам половых гормонов вызывает у них гнездостроительную деятельность даже в
отсутствие беременности. Эндокринная система во многом определяет доминант¬
ное состояние орх'анизма: особую направленность его деятельности, когда любой
раздражитель может вызвать определенный цикл реакций (добывание пищи,
гнездостроение часто при отсутствии наличных ключевых стимулов). Однако это
характерно лишь для низших животных, так как в процессе эволюции роль гормо¬
нального компонента в регуляции полового поведения снижается, а роль нервно¬
рефлекторных факторов возрастает.
§ 2. Классификация форм поведения 113
В естественных же условиях для проявления
инстинктов чисто внутренних факторов недоста¬
точно. Кроме них необходимы ключевые или пус¬
ковые стимулы. Очень часто при отсутствии клю¬
чевых стимулов, но при наличии соответствующей
потребности животное приступает к активному
поиску этих стимулов (например, поиск партнера
в период полового возбуждения, поиск материала
для построения гнезда и пр.).
Метод использования искусственных моделей
(муляжей) при изучении инстинктивного поведе¬
ния оказался весьма эффективным. Сюда отно¬
сятся широко известные охотникам и рыболовам
приемы имитирования природных раздражителей.
Н. Тинберген экспериментальным путем устано¬
вил, что для птенцов серебристой чайки при ими¬
тации головы взрослых родителей ключевыми сиг¬
налами являются далеко не все признаки, а лишь
«красное» (имитирующее красное пятно на клю¬
ве) и «продолговатое» (имитирующее форму клю¬
ва) (рис. 35).
Таким образом, только корреляция внешних и
внутренних факторов позволяет в полной мере
реализовать инстинктивное поведение животного.
Этапы поведения. Предлагают выделять два
этапа целостного поведения: поисковое (аппетентное), или подготовительное, и
завершающее {консуматорное) поведение. К. Э. Фабри (1976) приводит убеди¬
тельный пример охотничьего поведения, начинающегося у хищника с ненаправ¬
ленного поиска жертвы. Обнаружение первого ключевого раздражителя включа¬
ет направленный поиск, а появление дополнительных ориентиров — уточнение
местоположения жертвы. Затем — подкрадывание, бросок, овладение добычей,
умерщвление, расчленение туши, поедание мяса. В этой цепи только акт поедания
относится к завершающей фазе, все предшествующее ему — это подготовка.
Поисковое поведение — наиболее пластичная изменчивая часть инстинктивно¬
го поведения, в составе которого важную роль приобретает собственный жизненный
опыт, а завершающая фаза — наиболее стабильная, генетически фиксированная.
Таким образом, число степеней свободы для реакции (А. А. Ухтомский) или
«норма» реакции (И. И. Шмальгаузеи) по мере приближения к завершающей фа¬
зе инстинктивного поведения все более снижается. Индивидуальный же опыт
(обучение) реализуется прежде всего на начальных этапах поискового поведения,
которые у высших животных становятся хотя и преобладающими, но тем не менее
предназначенными для успешного выполнения завершающей фазы инстинктив¬
ного поведения.
§ 2. Классификация форм повеления
При попытке разобраться в классификации сложнейших инстинктов И. П. Пав¬
лов положил в основу биологическую потребность, выделив среди них:
Рис. 35. Серия картонных моделей
головы серебристой чайки
(по Р. Хайнду, 1975)
Длина полоски справа показывает
число реакций, вызванных с помо¬
щью каждой модели в стандартизи¬
рованной серии тестов
114 Глава 5. Формы поведения
♦ индивидуальные — пищевой, агрессивный, активно- и пассивно-оборонитель¬
ный, рефлекс свободы, исследовательский, рефлекс игры;
♦ видовые — половой и родительский.
Ю. М. Конорски (1970) также выделил две категории безусловных рефлексов:
♦ подготовительные (драйвовые, мотивационные);
♦ исполнительные (консуматорные).
При восприятии нового стимула возникает рефлекс нацеливания и собственно
ориентировочный рефлекс па новизну. Подготовительный исследовательский
рефлекс возникает в результате «сенсорного голода», потребности в информации.
Подготовительный пищевой рефлекс (голод) выявляется в форме двигательного
беспокойства и активной настройки сенсорных систем. Страх — подготовитель¬
ный защитный безусловный рефлекс, ярость — наступательный.
А. Д. Слоним (1949) в основу своей классификации положил павловские прин¬
ципы, учение об интероцеитивной сигнализации и убеждение об отсутствии в
природе «чистых» условных или безусловных рефлексов. Эта классификация име¬
ла следующий вид;
I. Рефлексы на изменение внутренней среды организма и постоянства общества;
1) пищевые — обеспечивающие постоянство вещества;
2) гомеостатические — обеспечивающие постоянство внутренней среды.
И. Рефлексы на изменение внешней среды организма:
1) оборонительные;
2) средовые (ситуационные).
III. Рефлексы, связанные с сохранением вида:
1) половые;
2) родительский.
В настоящее время такая классификация представляется схематичной и дале¬
ко не полной, не учитывающей, например, зоосоциальные аспекты поведения.
П. В. Симонов (1986) предложил оригинальную классификацию инстинктов
как сложнейших безусловных рефлексов, составляющих потребностпо-эмоцио-
нальную основу поведения. При этом он исходит из представления о том, что спо¬
собность сохранения особи, потомства, популяции, вида представляет собой необ¬
ходимое условие развития в смысле овладения средой во все расширяющихся
временных масштабах (В. И. Вернадский, А. А. Ухтомский). С этой точки зрения
степень адаптации к среде не может быть критерием эволюционного прогресса,
сущность которого состоит в постепенном завоевании сфер; гео- и биосферы (фи¬
зическое их освоение животными и человеком) и ноосферы (интеллектуальное
освоение мира человеком).
Удовлетворение витальных потребностей. Первую самостоятельную группу
рефлексов составляют витальные рефлексы, которые обеспечивают сохранение
индивидуума и вида. Сюда относятся пищевой, питьевой, регуляции сна, обо¬
ронительные (включая рефлекс «биологической осторожности»), рефлекс эко¬
номии сил и многие другие (табл. 1). Определяющими при этом являются два
признака:
§ 2. Классификация форм поведения 115
♦ неудовлетворение соответствующей потребности ведет
к гибели особи;
♦ удовлетворение потребности не требует участия дру¬
гой особи того же вида.
Параметры тех внешних объектов, которые могут
удовлетворить соответствующую потребность уже у мо¬
лодых животных, в большей своей части генетически за¬
даны.
Таблица 1
Сложнейшие безусловные рефлексы животных (по П. В. Симонову, 1986)
Витальные
Ролевые (зоосоциальные)
Саморазвития
Пищевой
Половой
Исследовательский
Питьевой
Родительский
Имитационный
Оборонительный
Эмоциональный
Игровой
Регуляция сна — бодрство¬
вания
Резонанса, «сопережива¬
ния»
Преодоления сопротивле¬
ния, свободы
Экономии сил
Территориальный
Иерархический
Стратегии питания накладывают существенный отпечаток па характер пове¬
дения. Наиболее яркое поведение связано с хищническим образом жизни. Напри¬
мер, кошачьи — это животные засадные, которые подстерегают добычу и напада¬
ют на нее прыжком, а собачьи преследуют добычу в открытую, догоняют ее, когда
она обращается в бегство. Кошки убивают жертву, прокусывая ей затылок, а соба¬
ки, схватив жертву в зубы, размахивают ею из стороны в сторону. Насытившись,
хищник прекращает поиски пищи и охоту. Характер манипуляций, проделывае¬
мых с пищей, может быть весьма разнообразным. Например, калан ложится в воде
на спину, кладет себе на грудь плоский камень и вскрывает раковины моллюсков,
ударяя ими об этот камень (Д. Дыосбери, 1981).
Многие оборонительные рефлексы носят врожденный характер, но в своей реа¬
лизации зависят от конкретных условий. У крысы, загнанной в угол, убегание
сменяется агрессивностью. Агрессивные животные переходят к мирному сосуще¬
ствованию, когда это необходимо для выращивания потомства. Функцию защиты
от нападения выполняют и некоторые подстерегающие действия, например звуки,
издаваемые гремучей змеей, и агрессивные позы, принимаемые многими млеко¬
питающими.
У животных не существует агрессивности ради агрессивности. В основе ак¬
тивно-наступательного поведения животных лежит либо потребность в самосо¬
хранении, либо борьба за самку, за территорию, потомство и др.
Особо выделяют в группе витальных инстинктов потребность в экономии сил.
Если установить два рычага, нажатие на которые требует разных усилий, по дает
одинаковое количество пищи, крысы предпочитают легкий рычаг.
Первую самостоятель¬
ную группу рефлексов
составляют витальные
рефлексы, которые
обеспечивают сохра¬
нение индивидуума и
вида.
116 Глава 5. Формы поведения
Удовлетворение социальных потребностей. Вторую
группу поведенческих реакций образуют ролевые (зоо¬
социальные) безусловные рефлексы, которые возникают
только при взаимодействии с другими особями своего вида.
Эти рефлексы лежат в основе полового, родительско¬
го, территориального поведения, в основе феномена «со¬
переживания» (эмоционального резонанса) и формиро¬
вания групповой иерархии. При этих формах поведения
отдельная особь выступает в качестве брачного партнера, родителя или детеныша,
хозяина территории или пришельца (интрудера), лидера или ведомого.
Наиболее простые формы ролевых отношений возникают при взаимодействии
половых партнеров. Среди факторов, определяющих успешность репродуктивно¬
го поведения, выделяют в качестве основных следующие:
♦ для организации полового поведения необходимо, чтобы животное иаходи-
Вторую группу поведен¬
ческих реакций образу¬
ют ролевые (зоосоии-
альные) безусловные
рефлексы, которые воз¬
никают только при взаи¬
модействии с другими
особями своего вида.
лось в определенном гормональном состоянии;
♦ необходимо наличие соответствующих внешних стимулов;
♦ необходим индивидуальный опыт общения с особями своего вида, который
влияет на реактивность к внешним раздражениям, усовершенствование соот¬
ветствующих движений, включая готовность к спариванию.
Поведение ухаживания представляет собой сложную последовательность дви¬
гательных актов, обеспечивающую большую вероятность успешного спаривания.
К. Лоренц (1971) так описал поведение селезней при ухаживании: селезень накло¬
няет клюв к воде, выгибает тело кверху, при этом он помахивает клювом и испус¬
кает громкий свист, сопровождаемый звуком, напоминающим ворчание. Поза «го¬
лова вверх — хвост вниз» кроме свиста сопровождается резкими взмахами клю¬
вом вверх и вперед, что создает фонтан брызг (рис. 36).
Рис. 36. Десять поз, наблюдаемых у кряквы и других уток,
добывающих корм на поверхности воды (по Д. Дьюсбери, 1981)
1 — начальное покачивание клюва; 2 — вскидывание головы; 3 — помахивание хвостом; 4 — свист
и урчание; 5 — «голова вверх — хвост вниз»; 6 — поворот в сторону самки; 7 — поклоны при пла¬
вании; 8 — поворот головы затылком к самке; 9 — выпячивание груди; 10 — «вверх-вниз»
§ 2. Классификация форм повеления 117
Не меньшим разнообразием форм характеризуется поведение ухаживания
у млекопитающих. Иногда при этом ритуал ухаживания более сложен у самок,
чем у самцов. Ухаживание у дельфипов-афалин состоит в том, что самка покусы¬
вает партнера, обнюхивает его, прикасается к нему, поглаживает плавниками, то¬
пится за ним, совершает прыжки и ир.
Не менее сложен и разнообразен репертуар родительского поведения, которое
у млекопитающих начинается сразу с завершением родового акта. Забота о потом¬
стве носит видоспецифический характер. У домашних кошек, как и у большинст¬
ва животных, новорожденные котята подвергаются тщательному вылизыванию.
Инициатива кормления всегда исходит от матери: кошка ложится около котят
и изгибает тело, как бы огораживая их. Котята занимают определенное положение
при сосании, и, как правило, каждый из них сосет из одного и того же соска. На
следующей стадии взаимного приближения инициатива кормления может ис¬
ходить как от матери, так и от детенышей. А па третьей стадии уже сами голод¬
ные детеныши разыскивают самку, которая нередко стремится при этом убежать
и укрыться от них.
Забота о потомстве тесно связана с уровнем социализации. Первые же шаги со¬
циализации детеныша начинаются в общении с матерью. Особенно ярко это выра¬
жается у приматов. Например, у макак-резусов выделяют три стадии материнско¬
го поведения: 1) тесный контакт с детенышем и его защита; 2) амбивалентность;
3) отталкивание. На первой стадии контакт детеныша с матерью является посто¬
янным: новорожденный либо сосет мать, либо спит. К концу первой недели он по¬
немногу начинает самостоятельно передвигаться. К трем месяцам он покидает
самку для игры, поиска пищи, исследования окружающих предметов. К этому
времени выраженность реакции ухода за детенышем (чистка шерсти, укачивание,
перетаскивание, обнимание, удерживание возле себя) снижается, а частота угроз
и наказаний повышается. Детеныш продолжает держаться возле матери в течение
нескольких лет, но она с рождением нового детеныша уже не позволяет старшему
сосать себя.
Сколь важен для дальнейшего развития детеныша период его общения с мате¬
рью, свидетельствуют те драматические последствия, к которым приводит изо¬
ляция детеныша от матери. У изолированных от матери детенышей обезьян
(А. Г. Попугаева, 1973) характерна поза обхватывания руками своего тела и голо¬
вы, что является выражением реакции самокоптакта, компенсирующей отсутст¬
вие контакта с другой особью. Потребность в социальном контакте, в общении
и ласке имеет самостоятельное, генетически предопределенное значение.
Патогенное влияние внутривидовой изоляции на высшую нервную деятель¬
ность щенков и обезьян описал М. М. Хананашвили (1978). Значительное влия¬
ние на нормальное развитие животных оказывает их изоляция от матери, но оче¬
видна и огромная роль взаимодействия со сверстниками. Изоляция от сверстников
нарушала формирование памяти, а также полового поведения. Такие животные
оказались крайне неустойчивыми к информационным нагрузкам, у них под влия¬
нием несложных условно-рефлекторных задач возникало типичное невротиче¬
ское состояние.
Любое взаимодействие особей одного вида можно назвать общественным пове¬
дением.
118 Глава 5. Формы поведения
Но истинное сообщество — это нечто большее, чем супружеская пара или мать
с детенышем. Оно представляет собой стабильную группу, члены которой поддер¬
живают интенсивную коммуникацию и находятся в некоторых относительно по¬
стоянных отношениях друг с другом (Э. Меннинг, 1982).
Организованные сообщества животных должны отвечать определенным крите¬
риям, среди которых основными являются:
1) сложная система коммуникации между членами сообщества;
2) разделение труда, основанное на специализации;
3) стремление членов общества держаться в тесной близости друг к другу;
4) постоянство состава;
5) затруднение доступа особей того же вида, по не являющихся членами данной
группы.
Групповая жизнь животных дает несомненное преимущество: где отдельная
особь гибнет, группа выживает. Так, например, существование группами обеспе¬
чивает павианам защиту от их извечных врагов — леопардов, когда объединяются
несколько взрослых самцов, пи один леопард не может им противостоять.
Хорошо известны примеры генетически детерминированной социальной диф¬
ференциации у общественных насекомых (пчелы, термиты, муравьи). Роль, кото¬
рую играет особь в сообществах позвоночных животных, зависит от ее возраста
и пола и детерминирована значительно менее жестко, чем у насекомых.
Организация многих сообществ позвоночных базируется на двух главных фак¬
торах — доминировании и территориальности. Под доминированием большинст¬
во биологов понимают утверждение за особью в ее взаимодействии с другими чле¬
нами сообщества определенного ранга (или роли). Такое иерархическое поведе¬
ние, обеспечивающее социальную организацию, может оказаться сильнее, чем
голод, сексуальность, агрессивность и страх.
Ярко описывает поведение «господства и подчинения» у обезьян Я. Дембов-
ский (1963), который считает, что доминирование у обезьян распространяется на
все виды их поведения. Структурной основой объединения у обезьян является се¬
мья, состоящая из одного самца-вожака, нескольких половозрелых самок и боль¬
шого числа подростков. Вожак — самый сильный, наиболее агрессивный, ловкий
в драках и, вероятно, наиболее интеллектуально развитый. Доминирование вожа¬
ка признается беспрекословно остальными членами семьи, которые во всем усту¬
пают вожаку. Как правило, появление соперника в семье сопровождается его из¬
гнанием. Знаком покорности подчиненных обезьян является поза подставления,
имеющая символический характер. Чистка шерсти имеет у приматов значение
умиротворяющего жеста. Законы субординации распространяются и на других
членов стада обезьян.
Предполагают, что в основе иерархии доминирования лежат важные биологи¬
ческие тенденции. Например, наиболее жизнеспособное потомство следует ожи¬
дать от наиболее сильного и здорового самца. Или старые самки, защищающие
свое стадо, жертвуют собой (они не могут уже принести полноценного потомства)
ради сохранения жизни своих более молодых родственников.
Территориальность связана с доминированием и также является фактором,
определяющим структуру сообщества. В этом отношении территорию можно
§ 2. Классификация форм поведения 119
Третью группу пове¬
денческих актов име¬
нуют безусловными
рефлексами самораз¬
вития.
определить как область, в границах которой ее постоянный обитатель пользуется
в отношении доступа к ограниченным ресурсам правом первенства, не принадле¬
жащим ему в других областях. Таким образом, владелец
территории в ее пределах пользуется полным доминиро¬
ванием. Границы территории, например у хищных, обо¬
значаются с помощью мечения специальных мест мочой
или выделениями особых желез. Если метка, скажем, све¬
жая, то есть вероятность встречи с хозяином чужой терри¬
тории велика, группа во избежание неприятностей меняет маршрут. Таким путем
возможность встреч разных групп оказывается минимальной, а имеющееся про¬
странство эффективно и совместно используется.
Закон территориальности имеет глубокое биологическое основание: самка вы¬
бирает самца, владеющего «хорошей» территорией, что обеспечит корм их буду¬
щему потомству.
П. В. Симонов (1986) отмечает, что первобытное человеческое сообщество
вряд ли представляло собой общество равных и свободных. Изобретение орудий,
распределение совместно добытой пищи и другие черты очеловечивания наложи¬
лись на достаточно сложную и жесткую групповую организацию наших живот¬
ных предков, качественно ее трансформируя.
Удовлетворение идеальных потребностей. Третью группу поведенческих ак¬
тов именуют безусловными рефлексами саморазвития. Сюда относятся разнооб¬
разные проявления ориентировочно-исследовательского поведения, рефлексы
сопротивления (рефлекс свободы, по Павлову) и рефлексы превентивной «воору¬
женности» — имитационный и игровой. Для рефлексов саморазвития характерны
два момента:
♦ они не связаны с индивидуальной или видовой адаптацией к наличной, суще¬
ствующей в данный момент ситуации, эти рефлексы обращены к будущему;
♦ эти рефлексы самостоятельны и невыводимы из других потребностей живого
организма и несводимы к другим ранее упомянутым мотивациям.
Исследовательское поведение животных побуждается самостоятельной потреб¬
ностью в получении информации. Элементарные исследовательские реакции по¬
являются очень рано даже у незрелорождающихся животных — это, например, ма-
ятникообразпые движения головой, связанные с поиском соска матери. В даль¬
нейшем появляются более сложные ориентировочные реакции — обнюхивание,
прислушивание, присматривание и др. Наконец, еще позднее у животного возни¬
кают и подлинно исследовательские акты в связи с появлением в окружающей об¬
становке нового раздражителя: поворот глаз, ушей, головы, приближение к незна¬
комому предмету (рефлекс «что такое?», по Павлову).
В комплексе сложпорефлекторных инстинктивных поведенческих актов ори¬
ентировочно-исследовательская деятельность животного служит поиску и обна¬
ружению биологически значимых сигналов. С ориентировочного рефлекса на¬
чинается любой контакт животного с факторами среды. Эти элементарные ори¬
ентировочные реакции выполняют роль неспецифического фактора активации
мозговой деятельности, настраивающие сенсорные аппараты на более адекватное
и полное восприятие соответствующей информации (подробнее см. гл. 10).
120 Глава 5. Формы повеления
Поисково-исследовательские формы поведения как важнейший этап освоения
окружающей среды свидетельствуют о том, что нормальная жизнедеятельность
требует непрерывного поступления из внешней среды не только вещества и энер¬
гии, но и информации.
Такие формы поведения, базируясь на врожденной биологической потребно¬
сти, в значительной мере реализуются благодаря индивидуальному опыту каждо¬
го животного. Оно вынуждено самостоятельно выучивать значимые лишь для пе¬
го внешние сигналы, которые будут для разных особей того же вида весьма раз¬
личными.
Регулирование исследовательского поведения тесно связано с механизмами
эмоций. Положительные эмоции служат факторами удовлетворения исследова¬
тельской потребности, отрицательные — фактором перестройки и управления ис¬
следовательским поведением. Само по себе исследовательское поведение может
служить базой для выработки на его основе условных инструментальных рефлек¬
сов. Например, крысы обучаются выходу из лабиринта для обследования новой
территории, а собаки и обезьяны обучаются нажимать на рычаг, открывающий ок¬
но в соседнее помещение. Некоторые ученые считают, что ориентировка и обсле¬
дование нового помещения и содержащихся в нем ориентиров играют решающую
роль при формировании перцептивного обучения (см. гл. 5).
Общефизиологическое значение этой важной биологической потребности
в информации доказывается экспериментами, свидетельствующими о драмати¬
ческих последствиях «информационного голода», особенно для развивающегося
организма. Воспитание молодых животных в «обедненной» среде, изоляции и в
условиях ограничения движений не только приводит к расстройствам их поведе¬
ния, но и влияет на массу мозга, толщину коркового вещества и уровень метаболи¬
ческих процессов.
Удовлетворение самых разнообразных биологических потребностей было бы
невозможным, если бы в эволюции не получила развитие специфическая тенден¬
ция преодоления какого-либо ограничения свободы. Этот общеизвестный факт не
получил научного анализа до тех пор, пока И. П. Павлов не обратил на него специ¬
ального внимания, назвав его рефлексом свободы. В противоположность рефлексу
свободы И. П. Павлов предположил существование «прирожденного рефлекса
рабской покорности» и отмечал его биологическую значимость для сохранения
вида: пассивная поза слабейшего, естественно, ведет к падению агрессивной реак¬
ции сильнейшего.
Идея И. П. Павлова о рефлексе свободы получила свое развитие в работах
В. П. Протопопова (1950). Им был создан метод «стимульно-преградпой ситуа¬
ции», при котором на путях к овладению стимулом (например, пищевой приман¬
кой) возникала преграда в виде клетки с запертой дверцей. Животное без всякого
предварительного научения осуществляло реакцию преодоления этой преграды:
грызет зубами, рвет лапами решетку, протискивается в щели до тех пор, пока не
откроет свободный доступ к приманке.
В самостоятельную группу потребностей, а значит, и поведенческих форм для
их удовлетворения выделяют потребность в приобретении опыта, навыков, уме¬
ний, которые потребуются животным в будущей их самостоятельной взрослой
жизни. К таким формам поведения относят подражательное и игровое.
Подражательное поведение (имитационное) обеспечивает передачу опыта
от одного поколения к другому и лежит в основе «сигнальной (не генетической)
§ 2. Классификация форм поведения 121
наследственности» (М. Е. Лобашев, 1967). При этом необ¬
ходимо выделить формы имитации, которые относятся к
инстинктивному поведению, когда выполнение видоспе¬
цифичных действий одних животных является побуж¬
дающим фактором для других.
«Когда одно животное из стада, увидев и испугавшись
чего-нибудь, бросается бежать, а другие члены стада, не
видевшие сами опасности, сразу также бросаются бежать,
то мы имеем дело с инстинктивным подражанием... Так
называемая паника является примером инстинктивного
подражания. Овцы, бросившиеся в рассказе о панурговом
стаде в море, после того как бросилась одна из них, также проявили инстинктив¬
ное подражание»1. Однако такому слепому подражанию неизменно противостоит
индивидуально приобретенный опыт.
Под игровым поведением понимают совокупность двигательных актов орга¬
низма, включающим определенные мышечные группы, используемые в дальней-
Подражание (имита¬
ция) — передача ви¬
доспецифического
опыта от одного поко¬
ления к другому, от
одной особи к другой,
которая лежит в осно¬
ве «сигнальной» (не
генетической) наслед¬
ственности.
шем взрослым животным в соответствующих жизненных
ситуациях — бегстве, борьбе, добыче пищи, размножении
и т. д. (А. Д. Слоним, 1986).
Многие исследователи не без основания полагают, что
игровая деятельность является наследственно детерми¬
нированным комплексом упражнений, тестирующих и
стимулирующих развитие мышечно-опорно-двигатель¬
ной и нервной систем, а также систем дыхания и кровооб¬
ращения.
Несомненно, что это поведение имеет самостоятель¬
ные мотивационные механизмы и большое разнообразие
своих проявлений. Наряду с общими движениями —
ходьбой, бегом, прыжками — игра включает вращатель¬
ные движения, акробатику, борьбу. У приматов, и особенно у человекообразных
обезьян, игровая деятельность приобретает сложные формы манипулирования
с предметами-игрушками.
Игровая активность в целом может рассматриваться как генетически запро¬
граммированный видовой признак, характеризующийся определенным энергети¬
ческим эквивалентом выполненной работы. В процессе реализации игровой дея¬
тельности животное тренирует соответствующие поведенческие стереотипы, обо¬
гащается информацией об окружающей среде, практикуется во взаимодействии
со сверстниками, которое будет необходимым во взрослой жизни, усваивает нор¬
мы группового поведения.
Заканчивая описание основных форм поведения, направленного к удовлетво¬
рению определенной биологической потребности, мы хотим привести слова
И. П. Павлова: «Нет никакого сомнения, что систематическое изучение фонда
прирожденных реакций животного чрезвычайно будет способствовать понима¬
нию нас самих и развитию в нас способности к личному самоуправлению»1 2.
1 Кашкаров Д. Н. Современные успехи зоопсихологии. М., 1928. С. 152.
2 Физиология нервной системы (И. П. Павлов, И. М. Сеченов, Н. Б. Введенский). М.; Л., 1952.
Т. 4. С. 432.
Игровое поведение —
совокупность двига¬
тельных актов орга¬
низма, включающее
определенные мышеч¬
ные группы, исполь¬
зуемые в дальнейшем
взрослым животным в
соответствующих жиз¬
ненных ситуациях —
бегстве, борьбе, добы¬
че пиши, размноже¬
нии и т. д.
122 Глава 5. Формы поведения
Обучение — процесс,
в результате которого
организм приобретает
адаптивные изменения
индивидуального пове¬
дения, расширение
жизненного опыта,
усвоения знаний, уме¬
ний, навыков.
§ 3. Формы индивидуального обучения
Генетически детерминированные формы поведения, отражающие накопленный
в генофонде видовой опыт предшествующих поколений, оказываются недоста¬
точными, чтобы обеспечить активное существование особи в вероятностно измен¬
чивой среде. Чем меньше выражен диапазон колебаний
факторов среды, тем больше поведение животного может
опираться па опыт предшествующих поколений, записан¬
ный в его генотипе. И наоборот, чем больше изменчивость
окружающей действительности, тем меньше прагматиче¬
ская ценность видового опыта, тем в большей мере воз¬
растает необходимость в приобретении собственного, ин¬
дивидуального опыта. Последний приобретается различ¬
ными путями, в основе которых лежит общая способность
живых организмов к обучению, которое тесно увязано со
свойством фиксировать на какой-то срок элементы обучения, то есть он оснащен
памятью. Поэтому пейробиология обучения и памяти — это единая проблема для
науки.
Обучение — это процесс, состоящий в появлении адаптивных изменений ин¬
дивидуального поведения в результате приобретения опыта (В. Торп, 1963).
Следовательно, при обучении происходит отбор оптимальных реакций живот¬
ного, но сам процесс обучения, скрытый от непосредственного измерения, может
быть оценен только по результатам поведения. Иными
словами, мы оцениваем лишь то, что животные запомни¬
ли в результате обучения. Вероятно, не существует еди¬
ных общих законов обучения, а эта важнейшая способ¬
ность живых организмов должна эволюционировать как в
опто-, так и в филогенетическом отношении, чтобы обес¬
печить поведению его адаптивный характер.
Классификация форм обучения. Обучение является
предметом изучения не только физиологов, но и зоопси¬ 1 2хологов, этологов, бихевиористов. Отсюда необычайная
пестрота взглядов, противоречия отдельных школ, трудносопоставимые методи¬
Неассониативное, об¬
лигатное обучение
обусловлено набором
средовых факторов и
не требует непремен¬
ного совпадения (ассо
ииации) внешних сиг¬
налов с той или иной
целостной деятельно¬
стью организма.
ческие подходы.
Рассмотрим основные категории обучения животных (табл. 2).
1. Молодые организмы обеспечены достаточно стабильным набором внешних
стимулов, среди которых им приходится оперировать на самых ранних этапах
самостоятельного существования. Обучение в этот период носит неассоциа¬
тивный, облигатный характер, в значительной степени обусловленный набо¬
ром средовых факторов и не требующий непременного совпадения (ассоциа¬
ции) внешних сигналов с той или иной целостной деятельностью организма,
то есть является стимулзависимым.
2. На более поздних этапах онтогенеза по мере созревания организма поведение
все в большей мере приобретает активный характер. Значительно расширяется
спектр внешних факторов, могущих приобретать то или иное сигнальное зна¬
чение в зависимости от ассоциации их с целостной биологической реакцией
§ 3. Формы индивидуального обучения 123
организма (ассоциативное, факультативное обуче¬
ние). От относительно пассивного восприятия среды
организм переходит к активному процессу формирова¬
ния собственной среды путем извлечения для себя ее
функциональных составляющих, значимых для вы¬
полнения тех или иных актов поведения. Обучение в
этот период носит эффектзависимый характер, то есть
определяемый результативностью контакта организма
со средой.
3. Высшие формы обучения, свойственные в большей
степени взрослым животным с высокоразвитой нерв¬
ной системой, опираются па свойство формировать
функциональную структуру окружающей среды в кон¬
цептуальном плане, иными словами, формировать це¬
лостный образ окружающей среды. Такие формы ког¬
нитивного обучения основаны на формировании функ¬
циональной структуры среды, то есть на извлечении
законов связей между отдельными ее компонентами.
Естественно, что такие категории обучения базируют¬
ся па двух предыдущих и должны рассматриваться как продукт их прогрессив¬
ного развития в возрастном и филогенетическом плане.
Таблица 2
Классификация обучения
Категория
Основные формы
Неассоциативное, облигатное, стимул-
зависимое обучение
Суммационпая реакция
Привыкание
Запечатление (импринтинг)
Подражание
Ассоциативное, факультативное, эф-
фектзависимое обучение
Классический условный рефлекс
Инструментальный условный рефлекс
(включая обучение по типу проб и ошибок)
Когнитивное обучение
Психонервная деятельность
Рассудочная деятельность
Вероятностное прогнозирование
Иногда весьма затруднительно отнести ту или иную поведенческую адапта¬
цию к определенной категории обучения. Тем не менее богатство и разнообразие
форм индивидуального обучения живых организмов может быть ранжировано
исходя из эволюционных общебиологических закономерностей.
Рассмотрим последовательно основные формы обучения.
Ассоциативное, фа¬
культативное обуче¬
ние — активный про¬
цесс формирования
собственной среды пу¬
тем извлечения для се¬
бя ее функциональных
составляющих, значи¬
мых для выполнения
тех или иных актов по¬
ведения.
Когнитивное обучение
основано на форми¬
ровании функциональ¬
ной структуры среды,
то есть на извлечении
законов связей между
отдельными ее компо¬
нентами.
124 Глава 5. Формы поведения
Неассоциативное обучение
Суммационная реакция. Феномен суммации в центральной нервной системе из¬
вестен давно и детально изучен как на клеточном, так и на системном уровнях ор¬
ганизации. В основе этого явления лежит сенсибилизация — повышение чувстви¬
тельности нервной ткани к раздражающим агентам и фасилитация — облегчение
запуска именно данной реакции. Рядом ученых наблюдались па простейших и
низших беспозвоночных реакции, как будто бы напоминающие условные рефлек¬
сы: освоение определенного маршрута передвижения, навыки скопления в опре¬
деленных местах пространства, различение съедобных продуктов от несъедобных,
осуществление защитных двигательных реакций в ответ на прежний индиффе¬
рентный раздражитель.
Однако при углубленном анализе стало ясно, что эти навыки обладают всеми
свойствами суммационной реакции:
♦ они не могут сохраняться длительное время и обречены на исчезновение;
♦ вызывающие их раздражители не обладают специализированным сигнальным
значением;
♦ после исчезновения они самостоятельно не восстанавливаются;
♦ их исчезновение есть процесс разрушения, а не временного торможения.
Возникнув у одноклеточных организмов, суммационная реакция, основанная
па свойствах протоплазмы к сенсибилизации и фасилитации ответных реакций,
может быть рассмотрена в качестве одной из форм индивидуального приспособ¬
ления у примитивных живых существ. Однако в преобразованном, а иногда тща¬
тельно замаскированном виде суммационная реакция у высших животных высту¬
пает как важнейший элемент более сложных форм индивидуального обучения.
Клеточные механизмы суммации участвуют к процессах обучения и памяти
у высших животных.
Привыкание. В противоположность суммационной реакции привыкание пред¬
ставляет собой такую форму обучения, которая состоит в относительно устойчи¬
вом ослаблении реакции вследствие многократного предъявления раздражителя,
не сопровождающегося каким-либо биологически значи¬
мым агентом (пищевым, оборонительным, сексуальным
и пр.), именуемым обычно подкреплением. Явление при¬
выкания весьма широко распространено в животном ми¬
ре, начиная с одноклеточных организмов. Привыкание
у одноклеточных — это кратковременный процесс, не об¬
ладающий способностью к тренировке. У низших кишеч¬
нополостных с диффузно организованной нервной систе¬
мой (у гидр) выработка привыкания в каждом последую¬
щем опыте проходила быстрее, чего не наблюдалось у
инфузорий. Следовательно, у гидр, как и у инфузорий,
привыкание лежит в основе физиологических механизмов, определяющих спо¬
собность к приобретению индивидуального опыта (Н. А. Тушмалова, 1986).
Быть может, для столь низкооргапизованных живых форм, как одноклеточные
и животные с примитивной нервной системой, единственными механизмами при¬
обретения индивидуального опыта являются суммационная реакция и относи¬
тельно кратковременное привыкание. Таким образом, повышенная реактивность
Привыкание — относи¬
тельно устойчивое
ослабление реакции
вследствие многократ¬
ного предъявления раз¬
дражителя, не сопро¬
вождающегося каким-
либо биологически
значимым агентом
(подкреплением).
§ 3. Формы индивидуального обучения 125
к одним сигналам вместе со сниженной реактивностью к другим, сохраняющиеся
на протяжении относительно краткого промежутка времени, могут выступать в
качестве главных (если не единственных) механизмов адаптивных изменений ин¬
дивидуального поведения, полностью зависимого от физико-химических пара¬
метров окружающей среды. Поэтому такое поведение можно именовать стимул-
зависимым поведением. Привыкание усовершенствуется в ходе прогрессивной
эволюции и сохраняет за собой важную роль в репертуаре поведенческих адапта¬
ций. Причем привыкание возникает не в результате утомления или сенсорной
адаптации. В некотором смысле привыкание — это подавление реакций, которые
оказываются незначимыми в жизни животного.
Привыкание рассматривают как простейшую форму обучения, которая может
служить основой для развития более сложных форм обучения.
Дальнейшими исследованиями было установлено, что привыкание, присутст¬
вующее в самых разнообразных рефлекторных системах, не является утомлением
и зависит от специфичности стимула, силы раздражения и состояния организма.
Различают кратковременные и долговременные процессы привыкания, которые
используются как модель для изучения разных видов памяти (см. гл. 6).
Наиболее же распространенной формой привыкания является ориентировоч¬
ный рефлекс, который при повторении вызвавшего его раздражителя постепенно
угашается. Раздражители, которые не имеют для животного прямого биологиче¬
ского значения, постепенно утрачивают ответную реакцию организма. Такая фор¬
ма поведенческой адаптации, в основе которой лежит научение не отвечать на раз¬
дражение (негативное научение), основывается на механизмах привыкания (или
угашепия, по Павлову) ориентировочного рефлекса. Исследования показали уни¬
версальную роль ориентировочного рефлекса, с которого начинается любой вид
анализа раздражений. Биологическое значение ориентировочной реакции состо¬
ит в повышении возбудимости сенсорных систем для наилучшего восприятия
действующих на организм раздражений с целью установления их биологического
значения. Поэтому основными факторами в возникновении ориентировочного
рефлекса являются новизна, неожиданность и значимость раздражителя для ор¬
ганизма.
В составе ориентировочной реакции выделяют два процесса:
♦ начальная реакция тревоги, удивления сопровождается повышением тонуса
мышц и фиксированием позы (затаивание), генерализованным изменением
электрической активности разных отделов мозга (неспецифическая настройка);
♦ исследовательская реакция внимания, поворот головы, глаз, ориентация ре¬
цепторов по направлению к раздражителю.
Эта стадия рассматривается как период анализа раздражителя (включая услов¬
ные ориентировочные поисковые реакции) и сопровождается более локальными
изменениями биоэлектрической активности мозга.
Исследование динамики ориентировочного рефлекса у человека и животных
показало, что он возникает не на сам стимул как таковой, а в результате сличения
стимула со следом, оставленным в нервной системе предшествующими раздражи¬
телями. Если стимул и след совпадают, то ориентировочная реакция не возника¬
ет. В том случае, если информация о стимуле и хранящиеся в памяти следы от
предшествующих раздражений не совпадают, то ориентировочная реакция возни¬
126 Глава 5. Формы поведения
кает и оказывается тем интенсивнее, чем больше выражено такое рассогласование.
Следовательно, конфигурация следов, оставленных в нервной системе в результа¬
те повторения каких-то раздражителей, фиксирует все их параметры. Иными сло¬
вами, нервная система модификацией и комбинаторикой своих элементов строит
«нервную модель» стимула (Е. Н. Соколов, 1981). Динамика построения такой
нервной модели и отражается в процессах угашения ориентировочной реакции.
Отсутствие ориентировочной реакции на повторяющийся стимул свидетель¬
ствует о согласовании его нервной модели с тем реальным импульсным залпом,
который направляется в головной мозг. Недавние исследования (Э. Кеидел, 1980;
В. И. Котляр, 1986) установили тесную связь между сенситизацией и привыкани¬
ем, которые рассматриваются в качестве двух независи¬
мых процессов. В ходе многократного предъявления раз¬
дражителей отмечено первоначальное усиление ответов
(сенситизация) с последующим их подавлением (привы¬
кание). Предположили, что оба эти процесса обеспечива¬
ются разными нейронными сетями.
Импринтинг (запечатление). Среди форм индивиду¬
альных адаптаций особое место занимают процессы на
ранних этапах постнатального развития, связанные с уста¬
новлением жизненно важных контактов в гнезде, в стаде
или стае, в группе или семье, в окружении родителей.
. Комплекс поведенческих адаптаций новорожденного,
которые обеспечивают первичную связь между ним и ро¬
дителями и как бы замыкают цепь преобразований эмб¬
рионального периода, позволяя реализовать новорожденному уже сформирован¬
ные механизмы восприятия и реагирования, именуют импринтингом (запечатле-
ванием).
К. Лоренц (1937) выдвинул оригинальную теорию импринтинга. Он считал,
что молодые птицы узнают взрослых членов своего вида не инстинктивно, а с по¬
мощью имприптирования. Последнее выполняется на основе врожденной способ¬
ности следовать за движущимся объектом, который попадает в их ноле зрения
сразу после вылупления.
К. Лоренц считал, что от истинного ассоциативного обучения импринтинг от¬
личается следующими четырьмя особенностями:
♦ он приурочен к ограниченному периоду жизни, именуемому «критическим,
или чувствительным, периодом»;
♦ импринтинг необратим, то есть, возникнув в критический период, он не унич¬
тожается последующим жизненным опытом и сохраняется па всю жизнь;
♦ уникальность импринтинга определяется тем, что он происходит в тот период,
когда соответствующее (например, половое) поведение еще не развито. Ины¬
ми словами, обучение путем имприптирования не требует подкрепления.
Лоренц понимал импринтинг как форму «супериндивидуалыюго условного
рефлекса», при котором запечатлеваются не индивидуальные, а видоспецифиче¬
ские характеристики жизненно важного объекта.
Например, поведение в результате имприптирования будет направлено не
к конкретной особи, которую воспринимало животное, а к целому классу орга¬
низмов, к которому принадлежала имнринтируемая особь.
Импринтинг — комп¬
лекс поведенческих
адаптаций новорожден¬
ного, которые обеспечи¬
вают первичную связь
между ним и родителя¬
ми и как бы замыкают
цепь преобразований
эмбрионального перио¬
да, позволяя реализо¬
вать новорожденному
уже сформированные
механизмы восприятия
и реагирования.
§ 3. Формы индивидуального обучения 127
На рис. 37 показана установка для из¬
учения запечатления искусственной фи¬
гуры матери. Сохранение приобретенно¬
го опыта проверяется по реакции следо¬
вания утенка за муляжом взрослой утки.
Такая форма обучения получила название
«запечатление привязанности». Что же ка¬
сается слуховых раздражений, то пред¬
полагают, что их импринтирование может
иметь место еще раньше, то есть до рожде¬
ния или вылупления (А. Д. Слопим, 1976).
Свойством имприптирования обладают
многие животные и насекомые, а также
новорожденные дети. Причем для выра¬
ботки предпочтения оказывается несуще¬
ственным длительность экспозиции объекта. Значит, связи, возникающие при за-
печатлеваиии, шире, чем реакции следования, которую изучал К. Лоренц. Отсюда
становится понятным запоминание животными местности, положения поры, гнез¬
да и других жизненно важных ориентиров.
До сих пор спорным остается вопрос о критическом периоде имнринтирова-
ния, его продолжительности и факторах, его определяющих. Расширение спектра
воздействующих на организм раздражений, возрастание вероятностного характе¬
ра возникновения той или иной жизненной ситуации увеличивает уровень тре¬
вожности организма и побуждает его перейти от облигатных форм обучения к фа¬
культативным.
Совершенно неразработанным оказывается вопрос о возможности импринти-
рования матерью своих детенышей. Например, козы, если их лишить своих де¬
тенышей только на 15 мин после родов, принимают и допускают их к себе. При
удлинении этого времени до 3,5 ч козы отвергают детенышей. Такая же привязан¬
ность отмечена и у овец. Несомненно, что одной из основных функций имприн¬
тинга является установление контакта молодых особей с родителями и сородича¬
ми, то есть установление социальных отношений между молодыми и другими
представителями вида. Этот период первичной социализации у незрелорождаю¬
щихся животных накладывает отпечаток на всю последующую жизнедеятельность.
В самостоятельную категорию явлений запечатлевания К. Лоренц отнес «по¬
ловое импринтирование». Суть состоит в том, что самец птицы, воспитанной сре¬
ди особей другого вида, став взрослым, предпочитает в качестве половых партне¬
ров только самок этого вида, по не своего собственного. Усыновленные самцы
игнорируют самку собственного вида и ухаживают за самкой, принадлежащей ви¬
ду его приемных родителей. Тот факт, что импринтинг возникает задолго до созре¬
вания соответствующего поведения, подтверждается следующим наблюдением.
Проигрывание песни молодым птицам влияет на песню, которую они будут петь
спустя месяцы, когда достигнут половозрелости.
Это и подобного рода наблюдения — очевидное свидетельство того, что имприн¬
тинг может служить примером долговременной образной памяти (по И. С. Бери-
ташвили), возникшей без биологического подкрепления после одноразового воздей¬
ствия раздражителя. В проявлениях импринтинга используется взаимодействие
Рис. 37. Прибор, используемый для изучения
импринтинга (реакция следования) у птиц
(по А. Д. Слониму, 1976)
Движения модели утки регулируются с пульта
управления внизу. Утенок следует за моделью
128 Глава 5. Формы поведения
индивидуального опыта и врожденных свойств молодого организма к быстрой
фиксации его в механизмах памяти. Нейробиологические механизмы импринтиро-
вания как одной из форм памяти еще только начинают исследовать (Г. Хорн, 1988).
Подражание. Уже рассматривая примеры импринтинга, можно было обратить
внимание на большую роль в формировании реакции следования выводковых
птиц явлений подражания. В результате имитации (подражания) животное вы¬
полняет видотипичные действия путем непосредственного наблюдения за дейст¬
виями других животных своего вида. Эти последние являются побуждающим фак¬
тором для одновременного ухода всех членов стада или стаи от хищника, для со¬
бирания пищи, отдыха и т. д. Путем имитационного научения животные на основе
общения формируют поведение, которое вполне укладывается в рамки видового
стереотипа.
Особенно это характерно для молодых животных, которые путем имитации ро¬
дительского поведения обучаются различным проявлениям поведенческого ре¬
пертуара своего вида. Подобное имитационное научение названо облигатным.
Л. А. Орбели (1949) считал имитационное поведение главным охранителем вида,
ибо огромное преимущество заключается в том, что «зрители», присутствующие
при акте повреждения члена их же стада или сообщества, вырабатывают рефлек¬
торные защитные акты и, следовательно, могут в будущем избежать опасности.
Таким образом, облигатное, неассоциативное обучение обеспечивает жизнедея¬
тельность особи на первых этапах ее самостоятельного существования, а также за¬
кладывает основы видоспецифического характера поведения. Можно сказать, что
генетическая программа с помощью видоснецифического обучения создает осно¬
вы для сохранения целостности популяции.
Ассоциативное обучение
Классические условные рефлексы. Как уже отмечалось, формы ассоциативного
обучения характеризуются совпадением во времени (ассоциацией) какого-либо
воспринимаемого индифферентного раздражителя — внешнего или внутренне¬
го — с деятельностью самого живого организма. Биологический смысл такой ассо¬
циации — условного рефлекса — в его сигналыюсти, то есть в приобретении этим
раздражителем роли предупреждающего фактора, сигнализирующего наступле¬
ние предстоящих событий и подготавливающего организм к взаимодействию
с ними. И. П. Павлов разработал учение об условных рефлексах в основном на
примере рефлекторного слюноотделения. Индифферентный раздражитель соче¬
тался с безусловным агентом, вызывающим слюноотделительный рефлекс. В этом
случае возникала такая цепь событий: какой-то посторонний раздражитель—без¬
условное раздражение ротовой полости при попадании в нее пищи—безусловное
слюноотделение. Попадание нищи в ротовую полость именовалось подкреплени¬
ем. По мере сочетаний этих агентов первый раздражитель приобретал свойст¬
во условного сигнала, способного вызывать реакцию слюноотделения. Таким об¬
разом, условный сигнал как бы имитировал безусловно-рефлекторную реакцию,
лишь сдвинутую во времени.
Такой принцип выработки условных рефлексов был воспроизведен и в дру¬
гой экспериментальной ситуации, а именно в случае применения электрооборо¬
нительных безусловных рефлексов. Какой-либо посторонний раздражитель,
§ 3. Формы индивидуального обучения 129
например, сочетался со слабым электроболевым стиму¬
лом, приводящим к отдергиванию конечности у соба¬
ки. Спустя некоторое время отдергивание конечности
возникало уже после предъявления условного сигнала,
то есть опять же имело место сигнальное воспроизве¬
дение безусловного рефлекса. Конечно же, полного
тождества безусловной реакции по условному сигналу
не возникает. Правильнее говорить в этом случае об их
подобии, а не об идентичности. Такие условные реф¬
лексы еще называют условными рефлексами первого
типа (Ю. М. Копорски, 1970). Они получили наимено¬
вание классических. Благодаря этим условным рефлек¬
сам обеспечивается первичная ориентация животного л. А. Орбели
по признакам окружающей среды еще с самого начала
любого поведенческого акта. С помощью классических условных рефлексов жи¬
вотное адаптируется к внешней среде. Если принять во внимание модулирующее
значение физиологических потребностей, в этих случаях животное все же высту¬
пает в качестве достаточно пассивного участника событий, не имеющего возмож¬
ности кардинально изменить их последовательность. Деятельное же, активное на¬
чало сведено к достаточно простым безусловно-рефлекторным актам.
Инструментальные условные рефлексы. Некоторые ученые в самостоятель¬
ную форму ассоциативного обучения объединяют условные рефлексы, которые
строятся на основе активной целенаправленной деятельности животного. По¬
следовательность событий в данном случае зависит не
только от внешней сигнализации, но и от поведения са¬
мого животного. Например, крыса в процессе ориен¬
тировочно-исследовательского поискового поведения
в экспериментальной камере случайно нажимает лапой
на педаль, за что вознаграждается пищей. По мере та¬
ких совпадений вырабатывается прочная связь: нажа¬
тие на педаль — пища. Затем экспериментатор перед
очередным «произвольным» нажатием на педаль вклю¬
чает какой-либо посторонний раздражитель. После не¬
скольких сочетаний всей этой цепи раздражений уста¬
навливается связь: сигнал—нажатие на педаль—пища.
Активное взаимодействие животною со средой достигается благодаря включению
в эту цепь среднего звена — нажатия животным на педаль, ибо от своевременности
и качества выполнения данной реакции зависит успешность поведенческого акта
в форме достижения пищевого подкрепления. При этом следует иметь в виду, что '
в отличие от классических условных рефлексов сам по себе двигательный акт не
имеет генетической связи с получением нищи. Эта связь формируется в процессе
предварительной тренировки.
Впервые инструментальные условные рефлексы стали изучаться в павловской
лаборатории Ю. М. Конорски и Я. Миллером (1936), которые первоначально да¬
ли им название условные рефлексы второго типа. Затем их стали называть инстру¬
ментальными условными рефлексами, оперантньш поведением. По существу этот
Биологический смысл та¬
кой ассоциации — услов¬
ного рефлекса — в его
сигнальности, то есть
в приобретении этим раз¬
дражителем роли пред-
упреждаюшего факто¬
ра, сигнализирующего
наступление предстоя¬
щих событий и подготав¬
ливающего организм
к взаимодействию
с ними.
5 Зак. 1078
130 Глава 5. Формы повеления
вид обучения не отличается от обучения с помощью проб
и ошибок. Сущность инструментальной деятельности
заключается в изменении взаимоотношений организ¬
ма со средой, что происходит либо при изменении его
положения в пространстве (локомоторная деятель¬
ность), либо при воздействии организма иа окружаю¬
щие предметы (мапипуляторная деятельность).
Одним из распространенных примеров такого об¬
учения является выработка у животных навыков ори¬
ентации в лабиринте. Животное помещается в лаби¬
ринт (рис. 38), в котором находится пищевая приманка.
После неоднократного прохождения лабиринта путем
и. С. Бериташвили его тщательного исследования животное обучается
кратчайшим путем достигать местоположения приман¬
ки. Кривая обучения обычно строится па основе скорости достижения приманки
и/или количества допущенных ошибок. Лабиринтные навыки довольно быстро
становятся автоматизированными и стереотипными.
Рис. 38. Лабиринты: а — план лабиринта, применявшегося в зоопсихологических
исследованиях (лабиринт Смолла); б— лабиринт из «мостиков» (по К. Э. Фабри, 1976)
Пунктиром показана траектория побежки крысы
Было отмечено: если крысу держать в лабиринте еще до обучения, по без вся¬
кого подкрепления, то в дальнейшем она обучается быстрее и точнее. Считается,
что при обследовании лабиринта животное уже приобретает определенный опыт
(перцептивное латентное обучение), который используется для целей нахожде¬
ния кратчайшего пути в лабиринте.
Другим примером является оперантное обучение в «проблемном ящике». При
таком обучении животные начинают использовать разнообразные реакции, имею¬
щиеся в их природном репертуаре, а не навязанные экспериментатором. Напри¬
мер, кошка, помещенная в закрытую камеру, пытается убежать, безостановочно
двигается по камере и путем перебора поисковых реакций случайно поворачивает
щеколду и получает доступ к пище. Таким образом, системы классических и инст¬
рументальных условных рефлексов, участвуя в сложной конструкции адаптивно¬
го поведения, значительно расширяют приспособительные возможности живого
организма, который начинает выступать в качестве активного фактора взаимодей¬
ствия со средой.
§ 3. Формы индивидуального обучения 131
Когнитивное обучение
Образное (психонервное) поведение. И. С. Бериташвили начиная с 1926 г. из¬
учал поведение животных по разработанному им методу свободного перемещения
в экспериментальном манеже. Полученные И. С. Бериташвили (1975) факты бы¬
ло невозможно объяснить с позиций теории условно-реф¬
лекторного обучения. Суть представлений этого ученого
состоит в следующем: у высших позвоночных животных
при первом же восприятии местоположения нищи созда¬
ется образ или конкретное представление пищи и ее ме¬
стоположения в данной среде. Этот образ сохраняется,
и каждый раз, когда он репродуцируется при восприятии
данной среды или какого-либо его компонента, животное
производит такое же ориентировочное движение головы,
как и при непосредственном восприятии, ведет себя точно
так, как при восприятии, то есть идет к месту нищи, обнюхивает его и, если нахо¬
дит пищу, съедает ее. Такое психонервпое поведение, направленное образами, ста¬
ли называть произвольным в отличие от условно-рефлекторного поведения, кото¬
рое И. С. Бериташвили считал автоматизированным, непроизвольным.
И. С. Бериташвили описывает следующие характерные особенности нсихо-
нервной деятельности:
1. Психонервная активность интегрирует элементы внешней среды в одно целое
переживание, производящее целостный образ. Для этого достаточно, чтобы
животное один раз восприняло эту среду.
2. Психонервпый комплекс образа легко воспроизводится под влиянием только
одного компонента внешней среды или раздражения, напоминающего эту среду.
3. Это воспроизведение может происходить спустя длительное время после
начального восприятия жизненно важной ситуации. Иногда образ может
удерживаться всю жизнь без повторного его воспроиз¬
ведения.
4. При такой форме обучения устанавливаются времен¬
ные нервные связи между нервным субстратом образа
и двигательными центрами мозга. Индивидуально при¬
обретенные реакции, направляемые образами, легко ав¬
томатизируются при их повторении.
5. Двигательная активность животных при репродукции
образа зависит от условий его формирования, от дав¬
ности возникновения, его жизненного значения, про¬
странственных признаков ситуации.
6. Психонервпая активность высших позвоночных животных преобладает над
другими формами обучения и является определяющим фактором поведения.
7. Сознательная деятельность человека подчиняет себе двигательную активность
психопервпого процесса представления, хотя и протекает в тесном взаимодей¬
ствии со всеми другими формами поведения.
И. С. Бериташвили полагал, что нервный субстрат образной психической дея¬
тельности лежит исключительно в неокортексе.
Психонервное поведе¬
ние (образное) интег¬
рирует элементы
внешней среды в одно
нелое переживание,
производящее целост¬
ный образ, который
через ориентировоч¬
ную реакцию направ¬
ляет поведение живот¬
ного.
У высших позвоноч¬
ных животных при
первом же восприятии
местоположения пиши
создается образ или
конкретное представ¬
ление пиши и ее ме¬
стоположения в дан¬
ной среде.
132 Глава 5. Формы повеления
Система нейронов, продуцирующая образы жизненно важных объектов, мо¬
жет производить непосредственно только ориентировочную реакцию. Все осталь¬
ное поведение животного опирается па разные двигательные акты прирожденного
и условно-рефлекторного характера в соответствии с образом и окружающей си¬
туацией. Значит, образ, который лежит в основе субъективного отражения внеш¬
него мира, — это вектор предстоящего рефлекторного поведения, непосредствен¬
ная активность образа реализуется через ориентировочную реакцию, которая ре¬
гулирует направление поведения животного.
Согласно И. С. Бериташвили, образ фиксируется в памяти и извлекается отту¬
да для удовлетворения господствующей биологической потребности.
Основные отличия данной формы психонервного поведения от закономерно¬
стей условно-рефлекторной деятельности состоят в следующем (И. С. Берита¬
швили, 1984):
1. В отличие от классических условных рефлексов, которые требуют повторяе¬
мости, психопервный образ формируется сразу после одного осуществления
индивидуального поведения.
2. Условный рефлекс, не сопровождающийся безусловным подкреплением, осу¬
ществляется в течение длительного времени и гаснет постепенно. Психонерв¬
ный образ, не сопровождающийся удовлетворением соответствующей биоло¬
гической потребности, быстро перестает вызывать индивидуально направлен¬
ное поведение.
3. Если условный рефлекс может образовываться и в отсутствие коры, то целост¬
ные представления могут возникнуть только при наличии интактной коры,
и прежде всего лобных отделов коры больших полушарий.
4. Условный рефлекс подразумевает формирование временной связи между сен¬
сорными и моторными элементами. Психопервпое поведение обусловлено об¬
разованием новых действенных нервных кругов из сенсорных корковых ней¬
ронов.
5. Психонервная деятельность определяет включение и выключение условных и
безусловных рефлексов и последовательность их протекания в данном ряду.
Существенно, что индивидуальное поведение, первично направляемое психо-
иервпым образом, при повторной тренировке автоматизируется и осуществляется
по всем закономерностям условно-рефлекторного обучения.
Элементарная рассудочная деятельность животных. Поведенческие адапта¬
ции, с точки зрения Л. В. Крушинского (1977), строятся на трех основных меха¬
низмах: инстинктивном, механизме обучаемости и рассу¬
дочной деятельности (разуме). Хотя последние способно¬
сти животных противопоставляются обучению, данную
форму индивидуальной адаптации целесообразно рас¬
смотреть в общей системе форм поведения особи. На
рис. 39 приведена схема взаимосвязи основных элемен¬
тарных компонентов, участвующих в формировании по¬
веденческого акта. В результате интеграции компонен¬
тов формируется целостный поведенческий акт, который
можно определить как унитарную реакцию. Если на ранних этапах филогенетиче¬
ского развития поведение формируется под ведущим влиянием инстинктов, то по
Элементарная рассу¬
дочная деятельность —
использование предва¬
рительно воспринятой
тактики изменений в
среде для построения
стратегии своего буду¬
щего поведения.
§ 3. Формы индивидуального обучения 133
мере усложнения нервной системы большую роль начинают играть различные
формы обучения. Однако дальнейшая дифференциация конечного мозга обеспе¬
чивает элементарной рассудочной деятельности все большую роль в обучении.
Следовательно, чем выше уровень элементарной рассудочной деятельности жи¬
вотных, тем пластичнее и адаптивнее любая унитарная реакция и все поведение
в целом.
Рис. 39. Схема соотношения основных элементарных компонентов поведения
(по Л. В. Крушинскому, 1977)
Модификация поведения возможна для каждого компонента в пределах определенной нормы реакции
Наиболее характерное свойство элементарной рассудочной деятельности жи¬
вотных заключается в их способности улавливать простейшие эмпирические за¬
коны, связывающие предметы и явления окружающей среды, и возможность опе¬
рировать этими законами при построении программ пове¬
дения в новых ситуациях. Иными словами, важнейшая
функция рассудочной деятельности состоит в отборе той
информации о структурной организации среды, которая
необходима для построения программы наиболее адек¬
ватного акта поведения в данных условиях. Автор пока¬
зал, что способность к экстраполяции направления дви¬
жения раздражителя может рассматриваться как одно из
элементарных проявлений рассудочной деятельности жи¬
вотных. Основной экспериментальный подход состоит в
следующем (рис. 40). Животное должно находить кор¬
мушку А, двигающуюся прямолинейно с постоянной скоростью. Первоначальный
отрезок ее движения происходит в поле зрения животного, затем кормушка А
скрывается за преградой. Животное устремляется к месту выхода из непрозрачно¬
го коридора кормушки А с кормом. При этом животное определяет: 1) из соотно¬
шения с неподвижными точками пространства изменение положения кормушки
А, то есть направление ее движения, и 2) направление движения кормушки А по¬
сле ее исчезновения за укрытием. Таким образом, используя свою «систему отсче¬
та», животное экстраполирует направление смещения кормушки А с кормом, то
есть использует предварительно воспринятую тактику изменений в среде для по¬
строения логики своего будущего поведения. Различные тесты, предъявлявшиеся
Важнейшая функция
рассудочной деятель¬
ности состоит в отбо¬
ре той информации о
структурной организа¬
ции среды, которая
необходима для по¬
строения программы
наиболее адекватного
акта поведения в дан¬
ных условиях.
134 Глава 5. Формы поведения
3»
ai
I
-a AJ-
-А,
б Б'... „_Л!
II
±а А | -А, —
■б,;
Ill
Рис. 40. Схемы опытов
по экстраполяции с ширмой
разной степени усложнения
(по Л. В. Крушинскому, 1977)
/ — простая ширма; II — ширма с приставкой;
III — ширма со сложной приставкой; а — ис¬
ходное положение пустой кормушки; А —
кормушки с кормом; а, и Л, — положение
кормушки после их одновременного раздви-
жения; Б, и б, — ширма; Бб — шель в ширме;
пунктир — траектория движения животного.
В каждом случае животное безошибочно вы¬
полняет побежку для наиболее быстрого на¬
хождения кормушки с кормом
животным разного филогенетического уровня, выявили их способность улавли¬
вать простейшие законы пространства, времени, движения.
Л. В. Крушинский считал, что чем больше размер мозга, а значит и количество
нейронов, тем большую информацию об окружающей среде он может получить.
Наличие избыточного числа нейронов с разной функциональной активностью по¬
зволяет животному улавливать некоторые законы природы.
В процессе синтеза (эвристического уровня деятельности мозга) происходит
сличение уловленных в данный момент законов с теми, которые были познаны в
предшествующей жизни. В результате такого сравнения осуществляется выбор
наиболее адекватного пути решения задачи. Совпадение повой информации с за¬
писанной в долговременной памяти облегчает решение задачи.
Исходя из особенностей поведения животных в сообществах автор приходит
к выводу, что степень многообразия и пластичности отношений в сообществе жи¬
вотных находится в зависимости от уровня развития их
рассудочной деятельности, выполняющей особую роль
в эволюции популяций.
Вероятностное прогнозирование. Под вероятностным
прогнозированием понимается предвосхищение будуще¬
го, основанное на вероятностной структуре прошлого опы¬
та и информации о наличной ситуации. Оба эти фактора
служат основой для создания гипотез о предстоящем бу¬
дущем, причем каждой из них придается определенная
вероятность. В соответствии с таким прогнозом осуществляется подготовка к дей¬
ствиям в предстоящей ситуации, приводящим к наибольшей вероятности дости¬
жения цели (Н. А. Бернштейн, 1966).
Способность к вероятностному прогнозированию — результат биологической
эволюции в вероятностно организованной среде. Прогнозы живого организма
призваны оптимизировать результаты его действия.
В естественных условиях организм сталкивается с множеством различных
случайных раздражителей, которые нужно статистически обрабатывать, чтобы
Вероятностное прогно¬
зирование — предвос¬
хищение будущего,
основанное на вероят¬
ностной структуре про¬
шлого опыта и инфор¬
мации о наличной
ситуации.
§ 4. Эволюиия форм поведения 135
выявить повторяемость, то есть регулярность событий, на основании которой и
формируется приспособительное поведение. Теория вероятностного обучения ис¬
ходит из представлений о предсказании статистических закономерностей и выбо¬
ра оптимальных стратегий поведения при обучении субъекта независимым веро¬
ятностям стохастично возникающих раздражений.
П. В. Симонов (1986) считает, что степень информированности субъекта зави¬
сит от текущей субъективной вероятности достижения результата с учетом на¬
личных средств и оценки проблемной ситуации. В связи с этим из реальной окру¬
жающей среды выделяют и ту актуальную среду организма, от которой непосред¬
ственно зависит удовлетворение потребностей и достижение целей.
Высшие позвоночные и человек в своей деятельности непрерывно опираются
на вероятностное прогнозирование. Когда человек переходит улицу, он прогнози¬
рует ситуацию на проезжей части и интервал времени для безопасного пересече¬
ния дороги. А водитель транспорта в это время прогнозирует поведение пешехо¬
дов, смену сигналов светофора и свои собственные действия по управлению авто¬
мобилем.
Животные прогнозируют вероятность поиска пищевого объекта, одновремен¬
но прогнозируя поведение хищника, чтобы не стать жертвой собственной «бес¬
печности».
В целом вероятностное прогнозирование деятельности может иметь разные
формы (И. М. Фегепберг, Г. Е. Журавлев, 1977):
1. Прогнозирование разных форм независимых от субъекта событий.
♦ Прогнозирование своих ответных активных действий.
2. Прогнозирование целенаправленных действий не только в соответствии с их
частотой в прошлом опыте, но и с их актуальной значимостью и предполагае¬
мым результатом.
3. Поведение в общении с другими организмами предполагает использование ги¬
потез о наиболее вероятных действиях своих активных партнеров.
4. Прогнозирование действий и целей с учетом собственных энергетических за¬
трат.
Таким образом, когнитивные формы обучения, опираясь на весь широкий ре¬
пертуар более простых форм облигатного и ассоциативного обучения, выступают
в качестве одного из фундаментальных механизмов высшей нервной деятельно¬
сти животных и человека.
§ 4. Эволюция форм поведения
Проблема поведения в эволюционном плане была впервые освещена в трудах ос¬
новоположника эволюционной физиологии И. М. Сеченова, создавшего строй¬
ную систему взглядов по поводу эволюции интегративной деятельности нервной
системы. И. М. Сеченовым руководило стремление изучать исторически психиче¬
ское развитие особи с момента появления ее на свет, с тем чтобы всякую после¬
дующую фазу развития можно было бы вывести из предыдущей.
Эволюционную физиологию поведения как самостоятельную научную дисци¬
плину оформил один из представителей павловской школы — Л. А. Орбели.
136 Глава 5. Формы поведения
Отдавая себе отчет в трудностях становления эволюционной физиологии,
Л. А. Орбели выделил следующие основные подходы, на которые она должна опи¬
раться:
♦ сравнительно-физиологический;
♦ онтогенетический;
♦ клинический;
♦ экспериментально-физиологический.
Самостоятельное значение каждого из них чрезвычайно велико, но только
мультидисцинлинарные исследования способны продвинуть нас в понимании за¬
кономерностей функциональной эволюции.
Беспозвоночные
Простейшие. В этой главе уже были рассмотрены присущие донервному этапу
развития, а именно простейшим одноклеточным организмам, достаточно прими¬
тивные формы индивидуальных адаптаций. Эти виды обучения, относящиеся
к пеассоциативпым, несигнальпым, сводятся к суммации и привыканию. Анализ
различных попыток выработать у инфузорий хотя бы очень простые условные
рефлексы показал их безуспешность — все свелось к кратковременной сенсибили¬
зации. Н. А. Тушмалова и И. В. Маракуева (1986) показали, что процесс привыка¬
ния у инфузорий сопровождается существенными ультраструктурными измене¬
ниями клеток. Их данные свидетельствуют о том, что привыкание — это особое
активное состояние клетки, выражающееся в повышении активности генома, син¬
тезе матричной и рибосомалыюй РНК и возрастании цитоплазматического синтеза
белка. Эти клеточные перестройки обеспечивают индивидуальную модификацию
поведения в форме привыкания, длительность которого (до 40 мин) определяется
инерционностью систем биосинтеза и временем обратных перестроек. Значит, та¬
кие ультраструктурные перестройки относятся к обратимым пластическим реак¬
циям одноклеточного организма.
Л. Н. Серавин (1969, 1978) предостерегает против механического использова¬
ния применительно к простейшим широко распространенного термина «обуче¬
ние». Безпервные одноклеточные животные, вероятно, обладают совершенно ины¬
ми механизмами обучения, чем многоклеточные. Модификации поведения инфу¬
зорий могут носить совершенно иной характер. В качестве примера Л. Н. Серавин
предлагает разработанную им модель модификации пищевого поведения у хищ¬
ных инфузорий: путем изменения химических свойств пиши у простейших мож¬
но целенаправленно усилить пищевую реакцию за счет адекватной настройки их
хеморецепторов на определенный индуктор пищевого поведения. Соответственно
при смене корма происходит перестройка хеморецепторов на индукторные веще¬
ства, выделяемые новой жертвой. Такая форма обучения носит избирательный ха¬
рактер и может сохраняться десятки дней.
Кишечнополостные. Среди многоклеточных самое низкое положение в эво¬
люционной лестнице занимают кишечнополостные, у которых впервые появляет¬
ся хотя и слабо дифференцированная, но организованная нервная система, постро¬
енная по диффузному типу.
§ 4. Эволюция форм поведения 137
Пресноводная гидра является излюбленным объектом для изучения с позиций
сравнительной физиологии поведения. У гидры осуществление суммациопиого
рефлекса одинаково хорошо достигается при любом порядке индифферентного
и безусловного раздражений или только при одном из этих раздражений. Напри¬
мер, было показано, что у гидры при ограничении направленности движений тела
поисковые движения продолжают осуществляться преимущественно в направле¬
нии ранее разрешенного сектора еще длительное время после снятия ограничений
(Б. Ф. Сергеев, 1979) (рис. 41).
Рис. 41. Направленность поисковых движении
стебельчатой гидры (по Л. Г. Воронину, В. Ф. Сергееву, 1979)
а — в норме (в течение часа); б — после часового ограничения слева и снизу;
стрелки — направление и относительное число поисковых движений;
прерывистой линией обозначен сектор размером 90°, в направлении которого
во время тренировки могли осушествляться поисковые движения
Что касается привыкания, то новым его свойством, по сравнению с простейши¬
ми, является тренированность от опыта к опыту при интервалах между опытами,
сравнимых с временем сохранения следа (Н. А. Тушмалова, 1986).
Впервые в эволюции у червей начинается процесс централизации нервной си¬
стемы и одновременный процесс начальной цефализации. Центральная нервная
система построена по принципу лестницы с двусторонней симметрией и разрас¬
танием объема ганглиев, расположенных в головном конце тела. Однако у низших
червей, типичным представителем которых являются планарии, церебральный
ганглий имеет черты весьма примитивной организации: в нем много мышечных
клеток и элементов, выполняющих нейросекреторные функции. Примитивный
уровень «условных рефлексов» коррелирует с достаточно низкой организацией
головного ганглия нланарий.
В то же время реакция сенситизации проявляется у нланарий достаточно от¬
четливо, хотя и не обладает большой прочностью. Примитивные нестабильные
«условные рефлексы» сходны по многим параметрам с суммациопными рефлек¬
сами.
Иное дело у высших (кольчатых) червей, куда относятся и обычные дождевые
черви. Наряду с усложнением строения нервной системы, заключающемся в даль¬
нейшем увеличении окологлоточного нервного кольца — предшественника голов¬
ного мозга, расширяется разнообразие форм активного поведения этих животных.
Большую группу поведенческих реакций, которые имеют генетическую основу,
называют таксисами. Например, реакция дождевого червя на условия освещения
состоит в том, что он уползает в сторону большего затемнения (отрицательный
фототаксис).
«Условные рефлексы» характеризуются большим несовершенством. Так, дож¬
девых червей обучали поворачивать в Г-образном лабиринте в левый рукав. Без¬
138 Глава 5. Формы поведения
условным раздражением служил удар током в правом
рукаве, а условным — сам лабиринт, элементы кото¬
рого воспринимались внешней рецепцией (рис. 42).
Условный рефлекс вырабатывался медленно, потре¬
бовалось около 400 сочетаний, по мере сочетаний уве¬
личение числа правильных реакций не наступало.
Удаление надглоточного ганглия не изменяло дина¬
мику обучения, что свидетельствует о незначительной
цефализации у этих животных.
У морских кольчатых червей-полихет возникают
специальные ассоциативные центры «мозга» — грибо¬
видные тела. У полихет показана возможность выра¬
ботки стойких условных пищевых рефлексов, кото¬
рые угасали при ненодкреплении и самопроизвольно
восстанавливались (Л. Г. Воронин, 1977).
Моллюски характеризуются типом организации
нервной системы, получившим наименование разбро¬
санно-узловой, при котором проявляется как дальней¬
шая централизация нервных элементов в ограничен¬
ное число ганглиев, так и субординация с выделением
в качестве ведущего органа управления поведением
зрительных долей «мозга» у головоногих моллюсков.
Например, у осьминога (Д. Япг, 1964) зрительные до¬
ли выступают как высший интегративный нервный
аппарат, связанный со всеми остальными ганглиями.
Соответственно и поведение этих «приматов моря»
(И. Акимушкин, 1963) представляется чрезвычайно сложным, где врожденные
комплексы поведения дополняются богатым индивидуальным опытом. Так, на¬
пример, у осьминога легко и быстро вырабатывается условно-рефлекторное изме¬
нение окраски кожи при сочетании света с механическим кожным раздражением.
Наряду с инструментальными пищедобывательными условными рефлексами об¬
разуются и дифференцировочное торможение, и условный тормоз (см. гл. 6).
Более просто организованные брюхоногие моллюски являются излюбленным
объектом для исследования некоторых сторон нейронных механизмов обучения
(Э. Кепдел, 1980; Е. Н. Соколов, 1981). Сравнительно простая нервная систе¬
ма позволяет изучать внутриклеточные механизмы привыкания, сенситизации,
условных рефлексов. Например, при неожиданном стимуле у прудовика возника¬
ет рефлекс втягивания тела в раковину, которая накрывает голову, щупальца отво¬
дятся вниз и локомоция прекращается. При повторном воздействии механическо¬
го стимула латентный период уменьшается, а амплитуда рефлекса увеличивает¬
ся — возникает сенситизация, которая может быть растянута на несколько дней.
Оказалось, что облегчение может тесно взаимодействовать с привыканием это¬
го же рефлекса. Скорее всего, механизмы привыкания состоят в функциональ¬
ном изменении синаптических контактов между сенсорным и двигательным ней¬
ронами.
Условные же рефлексы у моллюсков (аплизия, виноградная улитка) выраба¬
тываются относительно легко и служат удобной моделью для изучения элемен¬
Рис. 42. Установка для
научения дождевого червя
(по К. Э. Фабри, 1976)
Движение в правый рукав лаби¬
ринта сопровождается ударом
электрического тока, что побу¬
ждает животное двигаться толь¬
ко в левый рукав лабиринта
§ 4. Эволюиия форм поведения 139
тарных клеточных механизмов обучения и памяти (Г. Г. Аракелов, 1985; П. М. Ба¬
лабан, 1985).
Другая систематическая группа животных, связанная происхождением с круг¬
лыми червями (апнелидами), представлена членистоногими, поведение которых
наиболее тщательно изучалось на представителях ракообразных и особенно насе¬
комых.
Низшим ракообразным (дафниям) свойственны лишь элементарные формы
временных связей при сочетании света с низкой температурой и затемнения —
с высокой. Избирательное реагирование животных сохранялось около одного часа.
У высших ракообразных (крабы, раки) отмечены весьма сложные формы пове¬
дения, и у них вырабатываются прочные положительные и отрицательные услов¬
ные рефлексы на одиночные и комплексные сигналы, сложные лабиринтные на¬
выки (Л. Г. Воронин, 1970). У насекомых головные ганглии образуют мощный го¬
ловной мозг в виде сложно устроенного надглоточного узла, состоящего из трех
отделов, передний из которых образует высший интегративный отдел, получив¬
ший название грибовидных тел. Продолжающаяся цефализация, выраженная
в большой концентрации нервных элементов головного ганглия, сопровождается
значительным усложнением форм поведения.
Как и у головоногих моллюсков, у насекомых можно образовать истинные
условные рефлексы спустя 15-25 сочетаний, они могут подвергаться угашению
и самовосстановлению, а также дифференцированию, переделке и пр. (М. Е. Ло-
башев, 1967). Весьма доказательными были опыты Н. Тинбергена (1969). Самки
роющих ос обучались распознавать круг из сосновых шишек, выложенных вокруг
входа в норку (рис. 43). После отлета осы кольцо из шишек вокруг норки было пе¬
ремещено в сторону. Возвратившись, оса ищет несуществующую порку внутри
кольца и находит ее лишь после повторных облетов.
Рис. 43. Поведение роюшеи осы (по К. Э. Фабри, 1976)
После отлета осы кольцо из шишек вокруг норки было перемешено в сторону.
После возвращения оса ишет норку внутри кольца и находит ее лишь после повторных облетов
(опыт Н. Тинбергена), а — кольцо из шишек вокруг норки, б — кольцо из шишек в стороне от норки
По-видимому, первые исследования научения у нчел проводились К. Фришем
(1980), который показал, что пчелы при обучении запоминают запах цветка после
первого прилета, окраску после 3-5 прилетов, а форму — примерно после 20 при¬
летов. Условно-рефлекторная же деятельность нчел в эксперименте изучалась
М. Е. Лобашевым, которому удалось описать такие интегрирующие свойства моз¬
га, которые ранее были обнаружены лишь у приматов (см. гл. 10). Хотя и мед¬
леннее, но и у пчел удалось выработать условные рефлексы па трехчленные
цепи зрительных стимулов, которые они начинали воспринимать как единый сиг¬
140 Глава 5. Формы поведения
нал после 67-158 сочетаний. У пчел обнаружены все виды внутреннего тормо¬
жения, которое обеспечивает им высокий уровень аналитико-синтетической
деятельности и достаточную пластичность поведения. У пчел допускают сущест¬
вование способности к элементарному обобщению и элементарному абстрагиро¬
ванию, что среди позвоночных присуще только высокоорганизованным млекопи¬
тающим.
Таким образом, среди беспозвоночных формируется несколько форм обуче¬
ния: привыкание, сенситизация, элементарные и истинные условные рефлексы,
возникновение которых связано со значительным усложнением центральной
нервной системы, они представляют принципиальное эволюционное приобрете¬
ние — ароморфозы в общей динамике филогенетического развития механизмов
поведения (табл. 3).
Таблица 3
Соотношение морфологических и функциональных ароморфозов на примере
эволюции повеления беспозвоночных (Н. А. Тушмалова, 1987, с дополнениями)
Группы животных
Ароморфозы
Морфологический уровень
(организация нервной систе¬
мы)
Функциональный уровень (выс¬
шая форма физиологических
механизмов поведения)
Простейшие
Допервный уровень
Привыкание, сенситизация
Кишечнополостные
Примитивная диффузная
нервная система
Привыкание + тренированность
Плоские черви
Зачатки цефализации
Нестойкие условные рефлексы
Кольчатые черви
Ассоциативный центр моз¬
га — грибовидные тела
Истинные условные рефлексы
Насекомые
Структурно-функциональная
дифференцировка головного
мозга
Положительные и отрицатель¬
ные условные рефлексы, инстру¬
ментальные условные рефлексы,
цепные и комплексные условные
рефлексы, условные рефлексы па
отношения
Позвоночные
Прежде чем перейти к анализу путей развития форм обучения у позвоночных,
вкратце остановимся на рассмотрении особенностей поведения у самых низших
хордовых — бесчерепных и круглоротых.
Бесчерепные. Б. Ф. Сергеевым (1967) показано, что у представителя бесче¬
репных — ланцетника — центральная нервная система построена по типу нервной
трубки, характеризуется эквипотенциальностью, то есть отсутствием выражен¬
ной дифференциации и цефализации, причем специализированные рецепторы
еще не развиты. Если производить сочетание слабого условного раздражителя
(света) с тем же светом, по большей интенсивности или на слабое тактильное раз¬
дражение, подкрепляемое электрическим ударом, то у этих животных удается вы¬
работать активную избегагельную реакцию. Она сохраняется в течение 10 ч, уга¬
§ 4. Эволюция форм поведения 141
сая уже на вторые сутки, ее каждый день приходится вырабатывать заново. Со¬
здается впечатление, что у бесчерепных намечаются признаки ассоциативного
условно-рефлекторного обучения, однако оно протекает по типу повышения воз¬
будимости к условному раздражителю, то есть сходно с суммационными рефлек¬
сами.
У типичного представителя круглоротых — речных миног — впервые в эволю¬
ции формируются основные рецепторные системы и в зачаточном состоянии за¬
кладываются все подразделения мозга. Головной мозг состоит из конечного, про¬
межуточного, среднего и ромбовидного вместе с мозжечком. В результате иссле¬
дований А. В. Бару, В. В. Фапарджяна и Б. Ф. Сергеева было выявлено, что у
миног вырабатываются оборонительные условные рефлексы после 3-4 сочетаний
разномодальных сигналов (свет + электрический ток), эти реакции сохраняются в
течение лишь одного дня, восстановление происходит самостоятельно или после
нескольких подкреплений. Иными словами, хотя у круглоротых ассоциативное
обучение носит характер условно-рефлекторной связи, этот тип реакций по ряду
признаков напоминает еще суммационные рефлексы.
У пластинчатожаберных рыб дифференциация структур переднего мозга
в морфофункциональном отношении еще недостаточно развита (А. И. Карамян,
И. В. Малюкова, 1987). Это отражается и па характере условно-рефлекторной
деятельности. У акул и скатов условные рефлексы вырабатываются медленно,
осуществляются с большим латентным периодом, характеризуются непрочно¬
стью, значительной вариабельностью выполнения. Функцию высшего интегра¬
тивного мозгового аппарата у этих животных выполняет сильно развитый мозже¬
чок с его богатыми межцентральными связями с остальными отделами мозга. Пе¬
редний мозг в условно-рефлекторной деятельности участия еще не принимает.
Костистые рыбы отличаются более сложной и дифференцированной организа¬
цией головного мозга и сенсорных систем. У них вырабатываются истинные пище¬
вые и оборонительные условные рефлексы на простые и сложные световые, зву¬
ковые, обонятельные сигналы, постоянное и переменное магнитное поле. Кости¬
стые рыбы характеризуются всем репертуаром форм ассоциативного научения
и сенсорного различения. Однако, как подчеркивает Б. Ф. Сергеев (1967), разоб¬
щенность центральных аппаратов сенсорных систем, а также слабость нервных
процессов препятствует образованию внутри- и межсенсорных условных рефлек¬
сов (ассоциативные временные связи), что свидетельствует о низком уровне ана¬
литико-синтетической деятельности.
В эволюции позвоночных рептилии занимают особое положение, ибо они пред¬
ставляют собой тот узловой этап эволюции, от которого формируются две незави¬
симые филогенетические линии развития мозга: стриарная, представленная пти¬
цами, и кортикальная, к которой относятся все млекопитающие вплоть до прима¬
тов и человека.
Мозг рептилий характеризуется прогрессивным усложнением структурной
организации, и впервые возникает примитивная таламокортикальная система
связей (М. Г. Белехова, 1977). Обнаружена слабая способность к фиксации следов
выработанных условных рефлексов: при обучении повой задаче стирались следы
предыдущего обучения. Стриатуму у черепах приписывается более важная роль
в интегративной деятельности мозга, чем дорсальной коре, имеющей еще доста¬
точно примитивную организацию.
142 Глава 5. Формы поведения
Млекопитающие
Обратимся к филогенетическому древу (рис. 44), из которого видно, что по мере
развития из рептилеиодобных предков основного ствола млекопитающих от него
отходит ветвь современных однопроходных, позже — сумчатых. Стволом же пла¬
центарных млекопитающих являются насекомоядные, от которых произошли
приматы, хищные и грызуны. Большая часть отрядов современных млекопитаю¬
щих — результат параллельной эволюции, а не последовательной, к которой можно
было бы отнести однопроходных, сумчатых, насекомоядных. В этом ряду и да¬
лее — вплоть до высших приматов и гоминид могут быть прослежены эволюцион¬
ные преобразования мозга и поведения.
Рис. 44. Эволюционное древо млекопитающих с выделением этапов филогенетической
эволюиии и уровней адаптивной эволюции (по А. С. Батуеву, 1979)
Филогенетический подход не исключает и другого, чрезвычайно важного
адаптационного подхода, который основан на изучении ныне живущих форм, об¬
ладающих разной степенью адаптации (специализации)
в смысле развития мозга и сенсорных систем. Такие жи¬
вотные не обязательно должны иметь общую линию эво¬
люции, и, следовательно, выводы, сделанные в результа¬
те такого сравнения, будут относиться только к общим
принципам адаптации и выживания. Эти исследования
углубляют понимание связи между развитием структур¬
ных образований мозга и их участием в целостном пове¬
дении. Филогенетический и адаптационный подходы вза¬
имно дополняют друг друга. Поэтому правомерно го¬
ворить о филогенетической и адаптивной эволюции. Понятно, что сравнение
особенностей поведения и конструкции мозга у грызунов, хищных, приматов не
имеет никакого отношения к филогенетической эволюции, а лишь к адаптивной
Большая часть отрядов
современных млекопи¬
тающих — результат
параллельной эволю¬
иии, а не последова¬
тельной, к которой
можно было бы отне¬
сти однопроходных,
сумчатых, насекомояд¬
ных.
§ 4. Эволюция форм поведения 143
в смысле усложнения организации мозга и усовершенствования биологических
адаптаций.
Начиная от насекомоядных можно проследить начальные этапы формирова¬
ния всех таламических ядерных образований и проекционных зон неокортекса,
характерных для высших млекопитающих. У насекомо¬
ядных (ежей) впервые в филогенезе формируется ассо¬
циативная таламокортикальная система, которой при¬
сущи черты примитивной организации (А. С. Батуев, 1981)
(см. гл. 10). Б. Ф. Сергееву (1967) удалось выработать у
ежей ассоциативные временные связи возбудительного
и тормозного характера путем сочетания разномодаль¬
ных звуковых и зрительных раздражений. И. В. Милюко¬
ва (1981) установила у ежей с помощью сложных двига¬
тельных условных рефлексов различение освещенности,
полос разной ориентации, фигур. Такие условные реф¬
лексы вырабатывались быстрее, чем у черепах, и сохра¬
нялись в течение 40-50 дней. Научившись новой задаче, ежи не забывали пред¬
ыдущих навыков, что свидетельствует о большой прочности храпения следов.
Однако условные рефлексы на одновременный и последовательный комплекс¬
ные раздражители, которые вырабатывались с трудом, часто сопровождались нев¬
ротическими состояниями. В отличие от высших млекопитающих, у ежей слабо
развиты ориентировочно-исследовательские реакции, не наблюдается использо¬
вания метода «проб и ошибок», их нервные процессы характеризуются инертно¬
стью. Наряду с участием примитивной ассоциативной коры в процессах сенсомо¬
торного синтеза основная интегративная роль в общемозговом контроле за слож¬
ными формами поведения принадлежит подкорковым отделам переднего мозга —
стриарным образованиям. По этому признаку насекомоядные еще сохраняют чер¬
ты, присущие рептилиям.
Приматы
Важнейшим показателем эволюционного прогресса нервной системы является
развитие неокортекса, а в нем — ассоциативных кортикальных полей и соответст¬
вующих ядер таламуса (А. С. Батуев, 1981). Именно эти структуры мозга достига¬
ют у обезьян высокого уровня развития. В самой коре впервые в эволюции фор¬
мируются длинные пучки волокон, связывающие различные области коры между
собой. Все это создает предпосылки для выполнения весьма сложных форм пове¬
дения.
Значительное место в жизни обезьян имеет ориентировочно-исследователь¬
ская деятельность, которая, но мысли Я. Дембовского (1965), является зачатком
будущего интеллекта. Убедительно показано, что обезьяны способны к формиро¬
ванию самых сложных форм условно-рефлекторной деятельности, связанной с
тонким различением зрительных объектов но самым разнообразным признакам.
Исследования следовых условных рефлексов выявили прогресс образной памяти.
Длительность хранения следов у низших обезьян исчисляется месяцами и годами.
Обезьянам присущи все формы когнитивного обучения: образное поведение, рас¬
судочная деятельность и вероятностное прогнозирование.
Филогенетически й,
подход не исключает
и другого, чрезвычайно
важного адаптаиионно-
го подхода, который
основан на изучении
ныне живущих форм,
обладающих разной
степенью адаптации
(специализации)
в смысле развития моз¬
га и сенсорных систем.
144 Глава 5. Формы поведения
Все вышесказанное позволило А. И. Карамяну (1976), А. И. Карамяну и
И. В. Милюковой (1987) выдвинуть представление о существовании критических
этапов развития мозгового обеспечения поведения в эволюции позвоночных.
Первый критический этап представлен бесчерепными и круглоротыми, харак¬
теризующимися еще слабой дифференциацией головного мозга (рис. 45) Так,
у ланцетников (бесчерепные) удается выработать лишь суммационпые реакции,
а у миног (круглоротые) — квазиусловпые рефлексы, занимающие промежуточ¬
ное положение между суммационпыми и истинными рефлексами.
Рис. 45. Этапы развития уровней интеграции в филогенезе позвоночных
(по А. И. Карамяну, 1976)
а — бесчерепные (ланцетник), спинальный уровень интеграции, в центре гигантская клетка с аксоном,
обеспечивающим нисходящие связи; б — круглоротые (минога), бульбомезэниефалический уровень
интеграции; а — пластиножаберные рыбы (акула), мезэнцефалоиеребеллярный уровень интеграции;
г— рептилии (черепаха), диэниефалотелэнцефальный уровень интеграции; л — насекомоядные (еж),
стриокортикальный уровень интеграции; е — приматы (обезьяна), неокортикальный уровень интегра¬
ции; закрашены высшие интегративные уровни мозга; 1 — конечный мозг; 2 — промежуточный; 3 —
средний; 4 — задний; 5 — мозжечок; 6 — продолговатый мозг. На схеме показаны только зрительные
афферентные проекции. Относительные размеры мозга разных животных не соблюдены
Второй этап представлен нластиножаберными рыбами (скаты, акулы), струк¬
туры таламуса и конечного мозга которых еще недостаточно развиты. У этих жи¬
вотных удалось выработать условные рефлексы, которые были непрочными, со¬
хранялись в течение нескольких суток и характеризовались невысоким уровнем
их осуществления. Передний мозг пластиножаберных рыб не принимает участия
в организации сложных поведенческих актов.
Третий этап представлен рептилиями (особенно черепахами), на поверхности
конечного мозга которых формируется примитивный неокортекс. Эти животные
способны к формированию более сложных форм поведения по сравнению с низ¬
шими позвоночными. Однако высшая нервная деятельность черепах отличается
слабостью нервных процессов, несовершенством памяти, отсутствием способно¬
сти к вероятностному прогнозированию. Ведущую роль в интегративной функ¬
ции мозга у черепах выполняет полосатое тело (стриатум).
§ 4. Эволюция форм поведения 145
Четвертый этап представлен низшими млекопитающи¬
ми — насекомоядными. Мозг ежей характеризуется увеличе¬
нием общей поверхности неокортекса и развитием дорзаль¬
ных ядер таламуса. Однако четкая дифференциация струк¬
тур таламуса и коры выражена еще слабо, хотя обнаружено
существование примитивной ассоциативной системы.
У ежей образуются сложные последовательные цепи дви¬
гательных поведенческих актов при зрительном распознава¬
нии. Впервые в эволюции позвоночных проявляется, хотя и в
слабой форме, свойство вероятностного прогнозирования.
Высшая нервная деятельность ежей характеризуется при¬
митивностью: инертностью нервных процессов, труд¬
ностью формирования межсенсорных синтезов, слабостью
ориентировочно-исследовательских реакций.
Пятый критический этап представлен приматами, где
главной особенностью организации мозга является мощное
развитие его ассоциативных систем, объединенных кортико¬
кортикальными лугами в единую интегративную систему
мозга. Таким образом, создаются все предпосылки для фор¬
мирования более сложных форм поведения, которые оказы¬
ваются специфически присущими лишь приматам.
Высшая нервная деятельность человекообразных обезь¬
ян. Некоторые исследователи поведения животных считают,
что высшие психические функции не что иное, как прими¬
тивное мышление, которым животные могут пользоваться
без предварительного научения, то есть не используя преж¬
ний жизненный опыт. Так, В. Келлер после наблюдения за
поведением шимпанзе полагал, что высшие психические
функции являются выражением изначальных свойств и спо¬
собностей мозга.
И. П. Павлов и его сотрудники, повторив опыты Келлера,
пришли к иному выводу. Шимпанзе составляет пирамиду из
ящиков, чтобы достать гроздь бананов, подвешенных к по¬
толку, на основании перенесения в новую ситуацию старого опыта, на основании
проб и ошибок (рис. 46). Сам И. Г1. Павлов писал по этому поводу: «...каждая но¬
вая ассоциация, касающаяся отношения внешних вещей, есть прибавление зна¬
ния, а пользование этим знанием есть то, что называется пониманием»1.
Еще Ч. Дарвин (1871) отмечал, что подражание сыграло значительную роль
в процессе антропогенеза, причем у родоначальников человека подражание в со¬
четании с рассудком сыграли исключительную роль в выборе оптимальных ре¬
шений.
Л. А. Фирсов (1987) выделяет подражательную деятельность в качестве само¬
стоятельной формы поведения антропоидов, которая оказывается особенно выра¬
женной у детенышей шимпанзе в возрасте 2-2,5 лет. Подражание после однократ¬
ного наблюдения решения проблемных задач или демонстрации дифференциро¬
вания предъявляемых сигналов особыо-«актером» сохраняется у «зрителя» от
Рис. 46. Решение
шимпанзе задачи
получения банана
(по В. Детьер
и Э. Стеллар, 1967)
Неопубликованные и малоизвестные материалы И. П. Павлова Л., 1975. С. 96.
146 Глава 5. Формы повеления
нескольких часов до нескольких недель. Причем кроме скорости и прочности под¬
ражания отмечается его очень широкий диапазон.
Механизм подражания следует рассматривать как особую форму поведения
или отражения действительности, где принцип «животное — окружающий мир»
заменяется принципом «животное — другое животное — окружающий мир». При¬
чем особь-«актер» воспринимается особыо-«зрителем» как обобщенная система
натуральных раздражений, которыми и являются акты поведения.
Предметная и орудийная деятельность антропоидов — это одна из базовых
проблем антропогенеза. Установлено, что манипулирование разнообразными
предметами среды — предметная деятельность — это неотъемлемая черта слож¬
ного поведения всех представителей отряда приматов. Антропоиды обладают спо¬
собностью к целенаправленному манипулированию с предметами. В процессе
этой деятельности они не только используют предмет как
таковой или при необходимости изменяют его, но способ¬
ны также сочленить несколько предметов в устойчивую
конструкцию. Использование предметов для игры моло¬
дыми шимпанзе начинается с 2,5-3-месячного возраста.
Но только в возрасте около 2,5 лет детеныши шимпанзе научались составлять но¬
вые конструкции и использовать их для приближения приманки. То же наблюда¬
ется и у других антропоидов (рис. 47).
Орудийная деятельность отчетливо проявляется у шимпанзе в опытах с огнем,
который служил препятствием для достижения приманки. В опытах И. П. Павло¬
ва Рафаэль взял бутылку с водой и вылил ее на пламя, а в опытах Л. А. Фирсова
Каролина накрыла огонь половой тряпкой и легко завладела апельсином. Из этих
опытов ясно видно свойство обобщения сигнальной ценности окружающих пред¬
метов. Наблюдения за поведением шимпанзе, проведенные Л. А. Фирсовым, дают
основание считать, что перед началом каких-либо действий у шимпанзе имеется
ближайший план, ради которого они реализуются. Вот один из примеров. Подро¬
стки шимпанзе Лада и Нева, воспользовавшись ошибкой лаборантки, достали
связку ключей, забытую ею на столе в 3 м от вольера, и оказались на свободе. Дей¬
ствия обезьян при этом были следующими:
♦ отгрызание края столешницы от стола, который уже давно находился в вольере;
♦ с помощью получившейся палки подтягивание занавеси и овладение ею;
♦ набрасывание как лассо занавеси на стол с ключами и очень осторожное подтя¬
гивание связки ключей к вольеру;
♦ открывание висячего замка и выход на свободу.
Этот пример отражает реализацию уже принятого окончательного решения,
которое сформировалось как на основе текущей деятельности, так и имеющегося
у обезьян жизненного опыта. В любом случае обезьяна способна в известной сте¬
пени предвидеть результаты своих действий.
Орудийная деятельность основана на анализе связей между предметами, яв¬
лениями и результатами манипулирования с предметами. При этом важную роль
играет уровень обобщения элементов среды с тем, чтобы можно было применить
любой предмет в любой ситуации, придерживаясь логики принятого решения.
Целенаправленность поведения с обобщенным использованием предметов внеш¬
ней среды, с частичным их изменением и составлением из них устойчивых конст¬
Предметная деятель¬
ность — манипулиро¬
вание разнообразными
предметами среды.
§ 4. Эволюция форм поведения 147
Рис. 47. Орангутан за
составлением двух трубчатых
предметов (по Л. А. Фирсову,
1987)
Орудийная деятель¬
ность основана на
анализе связей меж¬
ду предметами, явле¬
ниями и результата¬
ми манипулирования
с предметами.
рукций является наиболее поздним эволюционным
приобретением. Она обязательно проявляется в по¬
ведении ребенка, и на ней в значительной мере ос¬
новывается трудовая деятельность человека. Новая
ступень предметной деятельности — орудийная
деятельность — развивается в процессе все большей
социализации древнейшего человека.
Учитывая семейно-групповой образ жизни ан¬
тропоидов, исследователи уделяют значительное
внимание способам общения животных между со¬
бой и с экспериментатором. Основное внимание
уделяется голосовым и жестикуляторпым проявле¬
ниям. Л. А. Фирсов и В. В. Плотников (1981) отме¬
чают выраженную «нолисемаптичность» голосовых реакций обезьян и большое
значение окружающей ситуации для придания им определенной сигнальной зна¬
чимости. В силу высокоразвитых свойств подражания антропоиды легко усваи¬
вают новые звуковые проявления, обогащая их экологи¬
ческое значение. В определенных условиях эти артифи-
циальные сигналы могли оказаться более точными, чем
врожденные, и соответственно их биологическая роль воз¬
растала.
Своеобразная форма общения шимпанзе с эксперимен¬
татором была изучена А. И. Счастным (1972). Обезьян
приучали возвращать экспериментатору выданные им во
время опыта жетоны: за возвращение круглого жетона давали обезьяне конфету,
квадратного — орех, прямоугольного — компот, шестиугольного — игрушку, за
треугольник — ничего не давали. Обезьяны, используя условные «знаки», сигна¬
лизировали экспериментатору и друг другу о своей потребности в пище, питье
или игрушке.
Та же цель преследовалась опытом супругов Гарднеров, Премаков и других иссле¬
дователей па детенышах шимпанзе и горилл. В качестве символов могли исполь¬
зоваться либо специальные фишки, либо азбука глухонемых. Важно то, что вы¬
ученными жестами или фишками шимпанзе могли пользоваться между собой,
а новички по механизму подражания воспринимали их у обученных и успешно
ими пользовались.
Антропоиды обладают способностью к переносу первичного научения для
формирования обобщения с отвлечением от конкретных признаков сигнала. Ста¬
ло очевидным, что шимпанзе способны абстрагироваться
от некоторых существенных признаков предметов или яв¬
лений и обнаруживают выраженную способность к ассо¬
циативному мышлению. Обобщение и абстракция явля¬
ются результатом чувственного опыта животного, содер¬
жат информацию об истории процесса, оценке ситуации,
решении, его реализации, результатах комбинаторики
старых навыков для нового решения. Поэтому обобщение — функциональный
блок систематизированной информации, хранящейся в аппаратах памяти.
Обобщение — функ¬
циональный блок си¬
стематизированной
информации, храня¬
щийся в аппаратах па¬
мяти.
148 Глава 5. Формы поведения
Вероятно, на основе сложных обобщений у древних приматов формировался
знаковый язык. Л. А. Фирсов (1982) понимает под термином «язык» все формы
адаптивного поведения животных и человека, в том числе врожденные комму¬
никативные сигналы. Это первичный язык. Знаковая система общения у обезьян
позволяет различать во вторичных языках стадию «А» (довербальные понятия)
и стадию «Л» (вербальные понятия). Последние, как правило, реализуются с по¬
мощью речевого аппарата. Соотношение классификации языков приводится
в табл. 4.
Существенной стороной поведения антропоидов являются формы их сосуще¬
ствования в сообществах (семьях, группах, стадах). Такую кооперацию можно
рассматривать как присущий приматам тип зоосоциальиого поведения, при кото¬
ром действия нескольких особей объединяются для выполнения какой-либо опе¬
рации, направленной на достижение полезного результата. Кооперативность по¬
ведения обезьян зависит прежде всего от иерархических рангов партнеров, а затем
и от других факторов, таких как уровень мотивации, физиологическое состояние
особей и др.
Таблица 4
Классификация языков (уровни отражения) (Д. А. Фирсов, 1987)
И. П. Павлов
Л. А. Орбели
(1949)
Л. А. Фирсов (1947)
I сигнальная си¬
стема
I сигнальная си¬
стема
Дополнительный уро¬
вень
Первичный язык
Промежуточный
этап
Стадия «А»: довер¬
бальные понятия
Вторичный язык
II сигнальная си¬
стема
II сигнальная си¬
стема
Стадия «Б»: вербаль¬
ные понятия
То же
Кроме обычного наблюдения за поведением животных Л. А. Фирсовым был
использован весьма показательный объективный тест на кооперативное поведе¬
ние двух обезьян. Возле одной из кормушек находился рычаг, нажатие на который
открывало доступ к пище в противоположной кормушке, и наоборот. Кооператив¬
ное поведение устанавливается тогда, когда обезьяны по очереди подкрепляют од¬
на другую. Однако такое поведение разрушалось, как только проявилось домини¬
рование одной особи и подчиненность другой. В этом случае подчиненная особь в
несколько раз чаще нажимала на рычаг, чтобы накормить своего партнера, чем до¬
минирующая.
Наконец, хорошо изучена способность антропоидов к длительному хранению
следов. Главной отличительной особенностью шимпанзе является широкое ис¬
пользование образной памяти (рис. 48).
Вышеприведенные данные об уровне психического развития антропоидов,
включающего подражательную, орудийную, ориентировочно-исследовательскую
деятельность, способность к обобщению и абстрагированию сигналов, использо¬
ванию знаковой системы сигнализации и высокого уровня образной памяти, по¬
зволяют заключить, что обезьянообразные, ближайшие предшественники древ¬
нейшего человека, обладали не меньшим развитием перечисленных функций.
§ 5. Формирование поведения в онтогенезе 149
§ 5. Формирование поведения в онтогенезе
Типы онтогенеза. Переход от автоматизированных реакций к целенаправленным
происходит постепенно и определяется зрелостью центральной нервной системы.
Упреждающий характер развития поведения заключается в готовности ряда си¬
стем организма к определенной деятельности еще до перехода к самостоятельной
жизни.
В. Г. Кассиль (1987) отмечает, что пищевое поведение на ранних этапах жизни
регулируется иначе, чем в более поздние периоды развития. Например, у новоро¬
жденных крыс и собак отсутствует связь между активностью сосания и поступле¬
нием в организм материнского молока. При этом сигналы от рецепторов желудка
менее значимы, чем у старших особей. Потребление пищи зависит в большей сте¬
пени от сенсорных факторов, чем от метаболических. Питьевое поведение форми¬
руется позже, чем пищевое.
Даже у пезрелорождающихся животных в самые первые этапы постнатальной
жизни большое значение имеет обучение. Например, целенаправленная пищевая
реакция на обонятельный сигнал формируется по принципу импринтинга, то есть
запечатлевания. И только позже в связи с совершенствованием механизмов памя¬
ти начинают вырабатываться типичные условные рефлексы.
Существование только двух источников, определяющих видоспецифические
особенности поведения организма — генотипа и среды, не дает достаточных осно¬
ваний для разделения па две противоположные и взаимоисключающие формы
сложного адаптивного поведения. Жесткая дифференциация только двух факто¬
ров, влияющих на развитие поведения — генетических и средовых, столь же труд¬
на, как решение вопроса, от чего больше зависит площадь — от длины или от ши¬
рины (Д. О. Хебб, 1949). Еще более неплодотворна идея
деления поведения на врожденное и приобретенное.
К моменту первого своего проявления любая функ¬
циональная организация является незрелой, базирую¬
щейся на врожденных реакциях. Одни формы поведе¬
ния, например пищевое, высокооргапизовано уже к мо¬
менту рождения, другие (половое, материнское) впервые
проявляются на более поздних стадиях онтогенеза.
Л. А. Орбели (1961) следующим образом охарактери¬
зовал два тина онтогенеза животных — зрелорождающих¬
ся и пезрелорождающихся: «Одни животные уже внутри
яйца, во внутриутробной жизни достигают довольно боль¬
шой зрелости и попадают иод влияние внешней среды,
родившись или вылупившись из яйца уже вполне сфор¬
мировавшимися, и им остается только приобретать но¬
вые условные рефлексы, а другие рождаются на свет еще
не созревшими, еще не все безусловные рефлексы у них
возникли и развились, а уже на них действует внешняя
среда и устанавливает временные связи с теми реакция¬
ми, которые имеются уже налицо или которые вот-вот
только развиваются, и в зависимости от этого вся без¬
условно-рефлекторная деятельность уже дальше проте¬
Рис. 48. Состояние об¬
разной памяти (отсро¬
ченный выбор) на фоне
тренировки различения
зрительных сигналов
(по Л. А. Фирсову, 1987)
По оси ординат — отсрочка
в критическом испытании,
мин; по оси абсцисс — но¬
мера критического испыта¬
ния после образования услов¬
ного рефлекса; 1 — шимпан¬
зе; 2 — павианы; 3 -— мака¬
ки; 4 — капуцины
150 Глава 5. Формы поведения
кает иначе, чем она протекала бы, если бы животное было ограждено от влияния
внешней среды. Этот “переплет”, который создается между врожденными рефлек¬
сами и наслаивающимися на них условными рефлексами, “переплет” не просто
суммирования, накладывания друг на друга, а проникновения друг в друга и вза¬
имного влияния, представляет чрезвычайно важную сторону физиологии нерв¬
ной системы и не менее важную сторону биологического процесса приспособле¬
ния тех или иных видов к тем условиям существования, в которых они должны
жить»1.
Формирование пищевого поведения. Движения, характерные для пищевого
поведения у новорожденных млекопитающих, стереотипны и в значительной
степени не зависят от мотивационных факторов. В то же время специфическая
пищевая реакция новорожденных (сосательные движения, слюноотделение) до
первого кормления может быть вызвана тепловым воздействием. Эта реакция
усиливается при прикосновении к мордочке пучком щетинок, имитирующим
шерстный покров. В опытах X. Харлоу и Р. Циммермана (1959) детенышей макак-
резусов изолировали от матерей и кормление их производили с использованием
модели матери в форме проволочного цилиндра или такого же цилиндра, но обтя¬
нутого мягкой тканью. Независимо от того, какая модель применялась для корм¬
ления, детеныши большую часть времени проводили в контакте с «матерчатой ма¬
терью». Отсутствие стабильной температуры тела детеныша в ранний период раз¬
вития компенсируется теплом матери.
Большинство исследователей считают, что для незрелорождающихся детены¬
шей млекопитающих наиболее общим положительным раздражителем, удержи¬
вающим новорожденных около матери и обеспечивающим первый контакт с нею
путем пищевых реакций, является теплая поверхность. Иными словами, наиболее
рано проявляющейся в онтогенезе врожденной реакцией детенышей млекопитаю¬
щих является положительный термотаксис, который и обеспечивает контакт с
матерью в первые минуты жизни.
У зрелорождающихся детенышей млекопитающих (например, у копытных и
хоботных), где из-за передвижения всего стада или молодняка с самками отсутст¬
вует постоянная гнездовая территория, наряду с прикосновением к теплой по¬
верхности, которое предшествует первому кормлению, важное место среди врож¬
денных рефлексов занимают ответы па зрительные сигналы, такие как затемне¬
ние над головой, имитирующее мать, стоящую над детенышем (А. Д. Слоним,
1976), и следование за движущимся предметом. Эти рефлексы хорошо проявляют¬
ся на 1-2-й день жизни. Но если кормление происходит из рожка без затемнения
над головой или затемнение не подкрепляется нищей, то в течение первых трех
дней положительная пищевая реакция па этот сигнал исчезает.
Формирование пищевых реакций не завершается к моменту первого кормле¬
ния, а продолжается в направлении постепенного усложнения. Этот процесс об¬
лигатного обучения (Г. Темброк, 1964) зависит от критических периодов постпа-
талыюго онтогенеза. У кошек выделяют 4 таких периода: первый — от рождения
до 7-9 дней, то есть до момента открывания глаз и наружных слуховых проходов.
Второй длится около 20 дней и закапчивается к месячному возрасту. Оба эти пе¬
риода иногда объединяют в один — ранний иостнатальпый. Третий период — до
1 Орбели Л. А. Собр. соч. 1964. Т. 1. С. 382.
§ 5. Формирование поведения в онтогенезе 151
9-11 -недельного возраста и четвертый — завершается к 7 мес. постнаталыюго раз¬
вития.
1. Животные первого возрастного периода с недоразвитыми системами зритель¬
ного и слухового восприятия ориентируются главным образом с помощью
кожно-температурной и обонятельной чувствительности (К. В. Шулейкина,
1971). По термотактильным и обонятельным раздражениям котенок способен
различать признаки «своей» и «чужой» гнездовой территории. Комплекс та¬
ких реакций обозначают как хоминговую ориентацию детенышей. Успешность
достижения домашней зоны при этом крайне низка. Поэтому постоянный кон¬
такт с матерью в этих условиях имеет большое биологическое значение. Веду¬
щую роль в поддержании такого контакта играет мать, которая находит де¬
тенышей, ориентируясь по издаваемым ими крикам, возвращает их в гнездо
и кормит (Н. Н. Соколова, 1984).
2. Во второй период начинают формироваться координированные и довольно
быстрые движения глазных яблок и ушных раковин в сторону источника раз¬
дражения. Локомоторный аппарат и все сенсорные системы развиваются весь¬
ма активно. Мать, а затем и котята все чаще покидают гнездо. Именно в этом
периоде кошка впервые начинает издавать призывные материнские сигналы,
в ответ па которые котенок к ней приближается. Лактирующая самка издает
также непрерывные акустические сигналы (тихое мурлыканье), которые так¬
же служат ориентиром для котят при поиске ими гнезда.
Зрительный контроль расположения гнезда, матери и других членов семьи от¬
четливо проявляется у котят лишь к месячному возрасту. Хоминговая реакция
при этом заменяется ориентировочно-исследовательскими и игровыми формами
поведения.
Теперь уже инициатива кормления все больше начинает зависеть от детены¬
шей. Причем мать продолжает активно реагировать па призывные крики дете¬
нышей и обеспечивает непрерывный двусторонний контакт с ними. В этом перио¬
де у котят впервые начинает проявляться хищничество.
Способность к хищничеству развивается только в третьем (ювенильном, игро¬
вом) периоде жизни. Мать все чаще покидает детенышей, даже отталкивает и из¬
бегает их. Периоды кормления становятся менее продолжительными. Интенсив¬
но развивается ориентировочно-исследовательское поведение. Игра способствует
освоению окружающей территории, отрабатывает и тренирует элементы будущих
взрослых форм поведения, способствуя накоплению необходимого опыта в сен¬
сорном и моторном планах. Причем это свойство присуще только высшим млеко¬
питающим. У них в игровом периоде заметно совершенствуется способность фик¬
сации внимания, появляется реакция слежения, наблюдения за действием матери
и сопометпиков, иногда подражание им. При этом возникают первые попытки са¬
мостоятельного преследования добычи и ее умерщвления.
Четвертый период — полового созревания у кошек закапчивается в 7-9 мес.
постнаталыюго онтогенеза. Доля игровых компонентов в поведении уменьшает¬
ся. Хищническое поведение совершенствуется, стереотипизируется. Условно-реф¬
лекторная деятельность становится сложной и разнообразной. Непосредственное
преследование добычи сменяется более сложными формами поведения, вклю¬
чающими элементы вероятностного прогнозирования и экстраполяции. К концу
152 Глава 5. Формы поведения
этого периода молодые котята могут самостоятельно охотиться и защитить себя,
и семья распадается.
Взаимоотношения детенышей и родителей. Материнская забота о потомстве
может быть определена как многоактное поведение, в котором интегрированы
врожденные и индивидуально приобретенные компоненты. Формы заботы о по¬
томстве у разных отрядов и семейств млекопитающих различны. Наибольшей вы¬
раженности среди незрелорождающихся млекопитающих забота о потомстве до¬
стигает у обезьян. Детеныш сразу после появления на свет повисает на шее матери,
и она практически не расстается с ним 15-20 дней. Такому тесному контакту спо¬
собствует положительный термотаксис детенышей и свойственные им хвататель¬
ный и сосательный рефлексы. Начиная с недельного возраста хватательный реф¬
лекс ослабевает, активизируется исследовательская деятельность. Большую роль
в организации материнского поведения, особенно у человекообразных обезьян,
играют голосовые реакции детеныша, одни из которых свидетельствуют о его со¬
стоянии, другие — об угрозе нападения, что заставляет мать немедленно мобили¬
зоваться для защиты детеныша.
У большинства копытных, живущих стадами, детеныши пытаются встать и
сделать первые шаги уже через 15-20 мин после рождения и очень скоро начина¬
ют повсюду следовать за матерью. Первые 3-4 ч являются чувствительным пе¬
риодом для детеныша и матери, во время которого устанавливается тесная связь
между ними по принципу импринтинга. Детеныш запоминает на длительное вре¬
мя или на всю жизнь зрительные образы, обонятельные признаки, звуковые сиг¬
налы матери или объекта, заменившего ее. Если в течение критического периода
формирования общения детеныш не имеет возможности установить связи с осо¬
бями своего вида, впоследствии он может оказаться неспособным к общению с се¬
бе подобными и будет испытывать затруднения не только в коммуникативном, но
и в воспроизводительном поведении.
Формирование первичных коммуникативных связей устанавливается и путем
«обратного запечатлевания» детенышей их родителями. При этом существенную
роль играют акустические сигналы детенышей. Если в течение 3-4 ч после рожде¬
ния изолировать мать от детеныша, она не сможет отличить его от других. С дру¬
гой стороны, в период запечатления самки легко принимают и начинают вскарм¬
ливать чужих детенышей.
М. М. Хананашвили (1978), Е. С. Петров (1988) специально подчеркивают
значение фактора общения со сверстниками в период раннего онтогенеза для
формирования свойств высшей нервной деятельности животного. Оказалось, что
у животных, выращенных в условиях даже частичной внутривидовой изоляции,
значительно страдает способность фиксировать следы в краткосрочной памяти,
нарушается способность удержания в долгосрочной памяти нескольких систем
условных рефлексов. Отмечено, что животные, выросшие в условиях такой час¬
тичной социальной депривации, характеризуются резкими отклонениями в про¬
текании эмоциональных реакций: имеют место эмоциональная тупость, чрезмер¬
ная выраженность эмоции страха, неадекватность знака эмоции на различные раз¬
дражители.
Системогенез и теория диссолюции. Из многих теоретических представлений
о возрастной динамике формирования целостного поведения выдержали провер¬
ку временем лишь две, которые мы здесь и рассмотрим.
§ 5. Формирование поведения в онтогенезе 153
Л. А. Орбели (1964) считал, что сопоставление филогенеза с онтогенезом (за¬
кон рекапитуляции) должно быть весьма продуктивным, ибо раскрывает картину
формирования рефлекторной деятельности: как возникают, наслаиваются друг на
друга, переплетаются друг с другом, взаимодействуют, а иногда подавляют один
другого различные рефлекторные акты.
В экспериментах на эмбрионах кролика и морской свинки было показано, что
рефлекторные акты закладываются и выявляются уже в эмбриональном периоде,
но достигают полного развития в разные периоды постнатального онтогенеза. По¬
степенно включающиеся все новые и новые координации оказывают свое модули¬
рующее влияние на те рефлекторные акты, которые сформировались ранее. Путь
выявления этих влияний Л. А. Орбели видел в анализе
клинического материала. Ряд психопатологических симп¬
томов может быть истолкован «как проявление извест¬
ной рекапитуляции, как повторение пройденного ранее
этапа, как выявление того, что в норме было замаскирова¬
но иод влиянием более высоких, более молодых, более
поздних уровней развития»1. Эта система взглядов и бы¬
ла названа теорией диссолюции. Ее справедливость под¬
тверждается и экспериментальной физиологией: выклю¬
чение более молодых мозговых образований позволяет
выявить обычно маскируемые автоматизированные реф¬
лекторные акты. Способность к имитации может быть присуща как облигатным,
так и факультативным формам обучения и наиболее ярко выражена в определен¬
ный период раннего онтогенеза, а затем она не проявляется в такой степени, мас¬
кируясь множественными формами индивидуальных адаптаций. Однако удале¬
ние у взрослой собаки лобных долей коры мозга превращает ее в автоматического
имитатора, пассивно следующего за экспериментатором и повторяющего дейст¬
вия другой здоровой собаки (Н. А. Шустин, 1959).
П. К. Анохин (1968), разработав стройную систему представлений о функцио¬
нальных системах, считал, что весь процесс отражения внешнего мира живыми
организмами, закрепленный в филогенезе наследственными факторами, находит
свое выражение в развитии зародышей человека и млекопитающих. В периоде
эмбриональной жизни происходит развитие именно тех функциональных систем,
которые необходимы для осуществления жизненно важных функций новорож¬
денного, приспосабливающих его к внешней среде. Эта «подгонка» функциональ¬
ных систем к экологии данного организма достигается гетерохронньш и избира¬
тельный созреванием центральных и периферических структур, обеспечивающих
животному приспособление функции к моменту его перехода в новую внешнюю
среду. Такова наследственно закрепленная в филоэмбриогенезе особенность раз¬
вития организма. Такому разновременному созреванию различных структур заро¬
дыша предшествует также гетерохронное и избирательное развитие мозговых
структур данной функциональной системы.
Для каждого вида животного характерен свой своеобразный системогенез. Ор¬
ганизующаяся система может функционировать прежде, чем она достигнет окон¬
чательного оформления. К этому сводится суть концепции системогенеза.
Диссолюиия — теория,
основанная на законе
рекапитуляции: многие
поведенческие акты
закладываются уже в
эмбриональном перио¬
де, но достигают пол¬
ного развития в раз¬
ные периоды
постнатального онто¬
генеза.
1 Орбели Л. А. Собр. соч. 1964. Т. 1. С. 301-302.
154 Глава 5. Формы поведения
Системогенез — орга¬
низующаяся система
поведенческих актов,
которая может функ¬
ционировать прежде,
чем она достигнет
окончательного
оформления. Аля каж¬
дого вида животного
характерен свой свое¬
образный системоге¬
нез как проявление
опережающего отраже¬
ния действительности.
Например, только что вылупившиеся птенцы грача отвечают раскрытием клю¬
ва на звук «кар-р» и на движение воздуха. Оба эти фактора непременно предшест¬
вуют приему пищи в естественных условиях жизни. Оказалось, что к моменту вы-
луплепия птенцов в их слуховом аппарате созрели лишь
только те рецепторные элементы, которые способны вос¬
принимать звук «кар-р». У других птиц-дуплянок эффек¬
тивным стимулом раскрытия клюва оказалось затемне¬
ние дупла, когда мать, прилетающая с кормом, закрывает
собой единственное отверстие, через которое поступает
свет. Было установлено, что к моменту вылупления созре¬
ли те нейронные структуры мозга, которые обеспечивают
восприятие перепада освещенности и переключают воз¬
буждение на двигательные пути, управляющие раскрыти¬
ем клюва.
Как легко можно заметить, две рассмотренные кон¬
цепции онтогенеза поведения не исключают, а взаимодо¬
полняют друг друга. Причем каждая из них в большей
мере отражает одну сторону процесса развития поведения. Если концепция дис-
солюции в большей мере аргументирована клинико-физиологическими матери¬
алами, то концепция системогенеза опирается на эмбриогенетические закономер¬
ности.
Элементы адаптивного поведения в онтогенезе. Процессы, совершающиеся
в ходе онтогенеза, во многом обнаруживают те же закономерности, что и в дина¬
мике филогенеза.
Первыми реакциями, проявляющимися на начальном этапе становления
функции, являются таксисы. Таксисы считаются врожденными, генетически фик¬
сированными реакциями, проявляющимися в ответ на ключевые раздражители,
которые и определяют вектор протекания реакции. В дальнейшем они вступают
в тесные сочетания с индивидуальным опытом животного.
Другим врожденным компонентом поведения являются безусловные рефлексы,
обеспечивающие элементарный сенсомоторный уровень реагирования, способст¬
вующий подготовке висцеральных систем и скелетной мускулатуры к осуществле¬
нию определенных функций. Наличием таких врожденных компонентов поведения
обеспечивается предварительная готовность к контакту с наиболее значимыми
для выживания особи элементами внешней среды, «врожденное узнавание» этих
раздражителей (Э. К. Фабри, 1976).
На начальных этапах формирования поведенческих адаптаций проявляется
ряд стереотипных двигательных актов — комплексы фиксированных действий.
Выполнение этих действий автоматизировано, но в ходе онтогенеза происходит
уточнение, дифференцировка «узнавания» тех раздражителей, которые в даль¬
нейшем будут запускать сложные инстинктивные акты. Совершенствование взаи¬
мосвязи стимула и реакции происходит путем облигатного и факультативного
обучения. Особенностью облигатного обучения является его приуроченность к
определенным сенситивным (критическим) периодам онтогенеза. Эти периоды
приурочены к самым начальным этапам формирования функции и, как правило,
являются непродолжительными.
Заключение 155
Облигатное обучение может осуществляться путем импринтинга и играет
большую роль в становлении многих форм поведения.
Предполагается, что внутренние мозговые механизмы запечатления сходны
с механизмами доминанты (А. С. Батуев, Н. Н. Соколова, 1987). Возникновение
состояния доминанты и характеризует особенности протекания чувствительного
(критического) периода становления той или иной функции.
Формирование видовою стереотипа поведения может происходить и путем
подражания {имитации). Молодые животные путем подражания учатся выпол¬
нять некоторые жизненно необходимые действия обычного поведенческого ре¬
пертуара своего вида. Имитация может осуществляться и в процессе факульта¬
тивного обучения, идентифицирующего поведение в связи со случайными, прехо¬
дящими факторами среды.
Путем факультативного научения в онтогенезе приобретаются сложные инст¬
рументальные условные рефлексы. При этом у молодого животного-зрителя вы¬
рабатывается определенный навык в результате одного лишь созерцания дейст¬
вий другой особи, направленных па решение задачи.
Одним из средств приобретения индивидуального опыта в разные периоды он¬
тогенеза являются формы ассоциативного обучения — классические и инструмен¬
тальные условные рефлексы.
В ностнатальном онтогенезе описано явление периодизации развития услов¬
но-рефлекторной деятельности на основе созревания мозговых систем и метабо¬
лических процессов в организме.
Г. А. Образцова (1964) в исследованиях на собаках выделяет период от рожде¬
ния до 2,5-3 недель, в котором преобладают безусловные рефлексы и рефлексы
на натуральные сигналы.
В период от 3 до 6 недель появляются и упрочиваются условно-рефлекторные
связи со всех сенсорных систем.
В период от 6 до 12 педель жизни отмечена наибольшая скорость образования
условных рефлексов, их широкая генерализация в сенсорном и моторном звеньях
(см. гл. 6). В период от 6 месяцев и старше условно-рефлекторная деятельность
приобретает выраженные индивидуальные особенности. У взрослых особей от¬
четливо проявляются формы образного поведения, экстраполяции и особенно ве¬
роятностного прогнозирования, что коррелирует с высоким уровнем развития ин¬
тегративных систем мозга.
Заключение
Поведение — один из основных факторов эволюции популяций.
В основу классификации форм поведения может быть положен принцип раз¬
деления потребностей организма на биологические (витальные), социальные
(зоосоциалъные, ролевые) и идеальные (поведение саморазвития). Существуют
биологические предпосылки сложных форм потребностей у человека. Формы ин¬
дивидуального обучения подразделяются па группы:
♦ облигатное пеассоциативпое (суммация, привыкание, импринтинг, подража¬
ние);
156 Глава 5. Формы поведения
♦ факультативное ассоциативное (классические и инструментальные условные
рефлексы);
♦ когнитивное (образное психонервное поведение, элементарная рассудочная дея¬
тельность, вероятностное прогнозирование).
В динамике эволюции животного мира для допервпых форм организации
(простейшие) индивидуальное обучение проявляется в формах привыкания и сеп-
ситизации. Появление нервной системы (кишечнополостные) связано с усложне¬
нием обучения. Возникновение ЦНС (черви, моллюски) влечет за собой появ¬
ление способности к ассоциативному обучению, а четкое выделение головного
мозга в результате цефализации (насекомые) — сложные формы условно-рефлек¬
торного поведения. Приведенная динамика совершенствования форм обучения в
основном повторяется у домлеконитающих позвоночных. У млекопитающих в
связи с появлением ассоциативных систем мозга развиты все формы обучения,
особо высокого совершенства достигающие у приматов (антропоиды) с их пред¬
метной и орудийной деятельностью, подражанием, образной памятью, способно¬
стью к обобщению и абстрагированию с использованием знаковых систем сигна¬
лизации в общении.
В динамике индивидуального развития особи, подчиняющегося правилам си¬
стемогенеза и диссолюции, реализуется закон рекапитуляции: онтогенез поведе¬
ния в сжатой форме напоминает длительный процесс эволюционного созревания
поведенческих адаптации. Определяющими являются типы онтогенеза: зрелоро¬
ждающиеся и иезрелорождающиеся организмы. В последнем случае созревание
мозга и поведенческих проявлений может достигнуть дефинитивного уровня
лишь при условии постоянного общения с родителями.
Контрольные вопросы
1. Что такое репродуктивная изоляция?
2. Что такое ритуализация?
3. Популяция и полиморфизм поведения.
4. Основные постулаты этологии.
5. Что такое инстинкт?
6. Внутренние и внешние факторы инстинктивного поведения.
7. Основные этапы поведения.
8. Классификация форм поведения.
9. Витальные потребности.
10. Социальные потребности.
11. Идеальные потребности.
12. Подражательное поведение.
13. Игровое поведение.
14. Формы индивидуального обучения.
15. Неассоциативное обучение.
16. Ассоциативное обучение.
Контрольные вопросы 157
17. Когнитивное обучение.
18. Что такое импринтинг?
19. Донервпые формы поведения.
20. Особенности поведения низших беспозвоночных.
21. Особенности поведения насекомых.
22. Поведение позвоночных.
23. Мозг приматов и их поведение.
24. Типы онтогенезов.
25. Формирование пищевого поведения.
26. Взаимоотношения детенышей и родителей.
27. Особенности раннего онтогенеза человека.
Глава 6
ФАКТОРЫ ОРГАНИЗАЦИИ ПОВЕДЕНИЯ
;|, ш Iшш шт ш 111111||Ш11и |ш. ш ш — им«»»««« ш I ж /- ' адж жвжмж # л.'". 4з*.^ жив:
Генотип — вся сово¬
купность генетическо¬
го материала организ¬
ма или его часть,
то есть комплекс всех
наследственных задат¬
ков, контролирующих
развитие, строение
и жизнедеятельность
организма. Способ¬
ность к обучению де¬
терминирована, как и
любой другой признак
организма, генотипом
и обусловлена действи¬
ем множества генов.
§ 1. Генетическая детерминация свойств
поведения
Генетика поведения. Генетика поведения как самостоятельная научная дисцип¬
лина существует сравнительно недавно, ее возраст исчисляется с момента выхода
книги Д. Фуллера и В. Томпсона «Генетика поведения» в 1960 г. Главной стержне¬
вой задачей этой науки сразу же стало изучение генетиче¬
ских основ индивидуально приобретенных форм поведе¬
ния.
Всю совокупность генетического материала организма
или его часть, то есть комплекс всех наследственных за¬
датков, контролирующих развитие, строение и жизнедея¬
тельность организма, именуют генотипом. Реализация
генотипа особи происходит при ее непрерывном взаимо¬
действии с внешней средой, а продукт интеграции геноти¬
пических и средовых воздействий, заключающийся в на¬
боре морфологических, физиологических и поведенче¬
ских признаков организма, именуется фенотипом. Между
генотипом и фенотипом нет однозначного соответствия.
Генотип лишь определяет возможные пути развития орга¬
низма и его свойств при взаимодействии с внешней сре¬
дой. Последняя влияет на фенотипическую изменчивость
организма, а диапазон этой изменчивости зависит от так называемой нормы реак¬
ции, которая задается генотипом.
Напомним основные наиболее устойчивые понятия. В генах, то есть молекулах
ДНК, заключены потенции организма. Эти потенции определяют норму реакции
организма в отношении его физических и поведенческих
функций. ДНК передает закодированное генетическое со¬
общение молекулам РНК. Таким образом, РНК получа¬
ет от ДНК исходные инструкции, записанные с помощью
линейного кода, До появления надлежащих стимулов
большая часть комплексов ДНК находится в неактивной
форме. После соответствующей стимуляции ДНК акти¬
вируется и быстро синтезирует РНК так, что становится
возможным и синтез специфических белков. В большин¬
стве молекул ДНК каждый участок (локус) специализи¬
рован, поэтому его изменение приводит к фенотипическому дефекту поведения
в результате синтеза измененных РНК и белков.
Уже с момента возникновения оплодотворенной клетки в результате слияния
двух родительских клеток начинается реализация генетической информации, по¬
лученной от этих родительских клеток. На зародыш воздействует внешняя среда,
Фенотип — продукт
интеграции генотипи¬
ческих и средовых воз¬
действий, заключаю¬
щийся в наборе
морфологических, фи¬
зиологических и пове¬
денческих признаков
организма.
§ 1. Генетическая детерминация свойств поведения 159
которая передает свои влияния вначале через материнский организм, а затем и не¬
посредственно. В дальнейшем развертывание свойств генотипа в форме различ¬
ных структурно-функциональных комплексов происходит иод непрерывным воз¬
действием среды. Поэтому любой фенотипический признак — это сплав из врож¬
денных и приобретенных компонентов.
Задачи генетики поведения состоят в стремлении выделить генетическую сущ¬
ность в фенотипическом проявлении поведенческих актов, попытаться направ¬
ленно влиять на генотипическую сущность поведенческих проявлений и базовых
свойств нервной системы и, наоборот, определить силу средового воздействия па
генотип организма и оценить стабильный характер возникшей поведенческой из¬
менчивости и др.
Несмотря па свою молодость, генетика поведения не только разработала ряд
специфических методов исследования, но и сформулировала ряд принципиаль¬
ных теоретических положений.
Генотип и его влияние на поведение. Фактически все современные исследова¬
ния показывают, что изменения в генотипе глубоко влияют па внешне наблюдае¬
мое поведение животных и человека.
Наиболее распространен метод лабораторной селекции генетически чистых
линий животных (инбредные линии), все особи которой обладают одинаковым
генотипом. Если животные разных инбредпых линий, выдержанные в одинако¬
вых условиях, проявляют существенные различия в поведении, то приходят к вы¬
воду о генетической обусловленности этих различий.
Так, например, путем искусственного отбора были выведены линии крыс и мы¬
шей с высокой и низкой эмоциональной активностью в тесте «открытого поля»
(рис. 49). Активными называют крыс, которые, будучи помещенными на откры¬
тую освещенную площадку, обнаруживают частые дефекации и мочеиспускания
и, оставаясь неподвижными, часто распластываются на одном месте. У нереактив¬
ных крыс не было дефекации, и они свободно разгуливали
по площадке. Реактивные крысы из-за высокой эмоцио¬
нальности медленно обучаются избеганию удара электри¬
ческого тока.
Исследования па линиях животных с известным гене¬
тическим дефектом, или мутантах, привели к выводу, что
способность к обучению, как и любой другой признак ор¬
ганизма, детерминирована генотипом и обусловлена дей¬
ствием множественных генов. Путем использования множественного скрещи¬
вания (гибридологический метод) нескольких линий мышей, различающихся
способностями к обучению, А. Оливерио (1972) показал, что эти генетические
различия основаны на свойствах головного мозга обеспечивать разный уровень
обучения.
Р. Коллинз и Д. Фуллер (1968) вывели линию мышей, а Л. В. Крушинский
и Л. Н. Молодкина (1971) — линию крыс по признаку высокой чувствительности
к действию сильного звука, который вызывал у них эпилептиформные судороги.
Такие генуинные формы аудиогеиной эпилепсии стали широко распространен¬
ной моделью устойчивого патологического состояния. Сравнение неврологиче¬
ского статуса крыс линии Крушинского-Молодкиной с крысами линии Вистар
обнаружили у первых значительный дефицит в обучении ориентировке в 12-лу-
Способность к обуче¬
нию, как и любой дру¬
гой признак организ¬
ма, детерминирована
генотипом и обуслов¬
лена действием мно¬
жественных генов.
160 Глава 6. Факторы организации поведения
Рис. 49. Средние показатели активности в открытом поле для
мышей шести линий (Н,-12) в эксперименте с искусственным
отбором (по Д. Дьюсбери, 1981)
чевом лабиринте, процессов кратковременной памяти и общей инертности возбу¬
дительно-тормозных взаимоотношений (Б. А. Рябгшская и др„ 1981) (рис. 50).
В то же время по скорости переделки сигнального значения условных раздражи¬
телей (положительный превращается в отрицательный и наоборот) более способ¬
ными оказались крысы линии Крушинского-Молодкииой но сравнению с крыса¬
ми линии Вистар (Л. В. Крушинский, 1960).
Р. Коллинз и Д. Фуллер предположили, что различия между линиями мышей
по их предрасположенности к аудиогеиным судорогам контролируются одним ге¬
ном. Обычно один ген влияет на несколько различных признаков. Например, аль-
Рис. 50. Вероятность совершения крысами
определенного числа ошибок при поиске пиши
в лучах радиального лабиринта
(по А. С. Батуеву и др., 1983)
По оси ординат — вероятность; по оси абсцисс —
число ошибок; 1 — средние величины у «необучен¬
ных» (1-10-й опыты) и 2 — у обученных (47-56-й опы¬
ты) крыс линии Вистар; 3 — у «необученных» и 4 — у
обученных крыс линии Крушинского-Молодкиной;
5 — математическая модель случайных посещений
§ 1. Генетическая детерминация свойств поведения 161
бинизм контролируется одним геном, который влияет одновременно и на призна¬
ки поведения в открытом иоле. У человека гены, ответственные за микроцефалию
и фенилкетонурию, вызывают расстройство в строении и биохимии мозга и со¬
провождаются умственной отсталостью. Однако большая часть свойств пове¬
дения животных детерминирована действием многих генов, каждый из которых
в отдельности сравнительно мало влияет па фенотип.
Генетика инстинктов. Занимаясь генетикой поведения, ученые сталкиваются
с необходимостью и выбором для анализа «единиц поведения». Некоторые уче¬
ные считают, что наиболее удобными для этой цели являются комплексы фиксиро¬
ванных действий (КФД) благодаря их четкой выраженности и устойчивости.
В. С. Дилгер (1962) наблюдал гиездостроительпое поведение у разных видов
нопугаев-неразлучников. Особи одного вида отрывали кусочки материала для
гнезда и переносили их в клюве, а особи другого не держали их в клюве, а засовы¬
вали под боковые перья. Гибриды, полученные от скрещивания этих видов, обна¬
ружили смешанный тип поведения: птицы вначале засовывали материал для гнез¬
да в перья, а потом вынимали его и брали в клюв, доставляя к гнезду. Такая смесь
двух КФД была представлена у межвидовых гибридов. Следовательно, у гибри¬
дов в их генотипе представлены оба родительских поведенческих комплекса, од¬
нако извращена целесообразная последовательность их реализации.
Имеется много примеров, показывающих существование генов, действие кото¬
рых подобно переключателю: они определяют, появится ли данный комплекс дей¬
ствия или нет. Вероятно, естественный отбор благоприятствует группировке тех
генов, которые способствуют проявлению полезного поведенческого признака.
Эффективным путем для изучения эволюции поведения является вышеопи¬
санный прием экспериментального анализа наследования КФД. Метод получе¬
ния межвидовых гибридов оказывается в этом плане весьма убедительным.
Нервно-психические заболевания генетической природы. К настоящему
времени известно более 80 заболеваний, связанных с определенными генетиче¬
скими дефектами. И хотя очевидно, что наследственная информация влияет на
развитие психопатологических симптомов, конкретные механизмы этого влия¬
ния в большинстве случаев остаются неизвестными.
Определенные виды расстройств высшей нервной деятельности связывают
с тремя типами генетических аномалий.
1. Аномалии, связанные с рецессивньши генами, возникают в результате дефици¬
та определенного фермента в печени. Типичным примером такого рода забо¬
левания служит фепилкетонурия, которая в поведении проявляется в форме
задержки умственного развития, а иногда сопровождается эпилептическими
припадками и психозами. Данное заболевание обусловлено изменением гене¬
тического кода в одном участке хромосомы.
2. Аномалии, связанные с доминантными генами, в заболевании проявляются
при особых условиях или стрессах. Ярким примером может служить хорея Гет-
типгтона, которая начинается обычно в возрасте 30-40 лет (дегенерация коры
и базальных ганглиев) и проявляется в непроизвольных движениях, непра¬
вильной подергивающейся походке, характерных гримасах, замедленной, нев¬
нятной речи, ухудшении памяти, повышенной раздражительности, депрессии,
слабоумии.
6 Зак 1078
162 Глава 6. Факторы организации поведения
Сходны с этой болезнью и такие наследственные заболевания нервной систе¬
мы, как болезнь Альцгеймера и болезнь Паркинсона (дрожательный паралич).
Допускают, что такую же природу имеют шизофрения (грубое расстройство
интеллектуальной деятельности) и маниакально-депрессивные психозы. Вы¬
сокая наследственная предрасположенность к шизофрении не вызывает со¬
мнения. Риск заболевания шизофренией у детей, где один из родителей болен
шизофренией, 10-15 и 35-40%, если больны оба родителя. Существуют дан¬
ные о высокой наследственной предрасположенности к эпилепсии, основным
симптомом которой являются судорожные припадки.
3. Аномалии, связанные с хромосомными аберрациями, заключающимися в до¬
бавлении или утрате целой хромосомы или ее части. Типичным примером та¬
кого заболевания служит болезнь Дауна, выражающаяся в задержке умствен¬
ного и психического развития. Больной имеет 47 хромосом вместо 46, свойст¬
венных нормальному человеку. Наличие лишней хромосомы обусловливает
синтез избыточного количества фермента, необходимого для построения бел¬
ков головного мозга.
Группу заболеваний с ранним нарушением интеллекта объединяют под терми¬
ном «олигофрения». При этом хромосомные мутации могут выражаться в измене¬
нии числа хромосом, их структуры или краткого изменения гаплоидного набора.
И всегда они сопровождаются тяжелыми формами умственной отсталости в соче¬
тании с различными пороками развития.
Л. Г. Романова и Л. Г. Калмыкова (1981) выдвинули гипотезу о генетической
природе психических болезней. Они полагают, что доминантные мутации должны
приводить к поражению или дисфункции строго ограниченных структур мозга.
Приуроченность данного психического дефекта к определенным жестким мозго¬
вым системам дает возможность понять не только моз¬
говые механизмы психических расстройств, по и опреде¬
ленные аспекты нормального поведения.
Генетика свойств нервных процессов. Первым, кто
обратил внимание на необходимость изучить генетиче¬
скую природу свойств основных нервных процессов (их
силы, уравновешенности и подвижности), был И. П. Павлов в процессе разработ¬
ки им физиологической базы для определения типов нервной системы животных.
В дальнейшем отечественные ученые (Л. В. Крушинский, 1946; В. К. Красус-
ский, 1963) доказали генетическую детерминированность силы возбудительного
процесса, а В. К. Федоров (1971) показал зависимость от генотипа материнского
организма такого свойства, как подвижность нервных процессов.
М. Е. Лобашев обратил внимание на наследование такого фундаментального
свойства нервной системы, как возбудимость, которая определяет действие генов,
детерминирующих отдельные поведенческие акты.
Ученики М. Е. Лобашева создали стройную систему взглядов на нейрохимиче¬
скую и молекулярную детерминацию поведения (Н. Г. Лопатина, В. В. Понома¬
ренко, 1987).
Исследования позволили вычленить эффект отдельного гена, вносящего ос¬
новной вклад в формирование порога нервно-мышечной возбудимости, а также
установить его идентичность с определенным локусом хромосомы. Более того,
эти же гены участвуют в наследственной детерминации пороговых характеристик
Возбудимость опреде¬
ляет действие генов,
детерминирующих от¬
дельные поведенческие
акты.
§ 1. Генетическая детерминация свойств поведения 163
нервной системы в целом. У видов, пород и рас животных, имеющих высокую
нервно-мышечную возбудимость, наблюдали и более высокую пищевую возбуди¬
мость и более высокие показатели силы возбуждения. Основой такой общности
генетических механизмов могут быть функциональные или структурные особен¬
ности клеточных мембран и связанных с ними ферментов, определяющих харак¬
тер протекания внутриклеточных процессов (В. В. Пономаренко, 1970).
Генетическая детерминация особенностей обучения. Выведены линии крыс,
различающихся по обучаемости в сложном лабиринте: способные крысы ориен¬
тировались лучше по пространственным ориентирам, а неспособные — по зри¬
тельным. Большое значение при этом имеет характер мотивации: способные кры¬
сы сильнее мотивируются голодом, а неспособные — в защитно-оборонительных
ситуациях.
Вклад каждой из скрещиваемых линий в свойства обучаемости конкретного
поколения и особи неодинаков. М. П. Садовникова-Кольцова (1928) предположи¬
ла наличие у крыс трех групп генов, детерминирующих способность к обучению:
♦ гены двигательной активности;
♦ гены эмоции страха;
♦ гены ориентировочно-исследовательского инстинкта.
Однако корреляция этих свойств со способностью к обучению не всегда прояв¬
ляется, и в настоящее время рассматривают эти комбинации признаков как нося¬
щие фенотипический характер.
Наследственную основу различий в обучении М. Е. Лобашев предполагал ис¬
кать в генетически детерминированных особенностях безусловных рефлексов,
и прежде всего в уровне пищевого рефлекса как основного модулятора функцио¬
нальной активности нервной системы. Показано наличие генетических корреля¬
ций между уровнем функциональной активности нервной системы и способно¬
стью к выработке условных рефлексов (В. В. Пономаренко).
Ю. С. Дмитриевым (1981) было установлено, что порог возбудимости и ско¬
рость образования оборонительных условных рефлексов проявляют сходный ха¬
рактер наследования (рис. 51). Вероятно, в основе лежит множественный эффект
какой-то общей группы генов. В число контролируемых им признаков поведения
следует отнести ориентировочно-исследовательскую активность, общую двига¬
тельную активность и уровень эмоционального статуса. Последние могут участво¬
вать в наследственной обусловленности условно-рефлекторных форм поведения.
Ген, контролирующий вышеназванные признаки, идентичен гену, определяюще¬
му содержание биологически активных соединений в плазме крови, серотонина и
норадреналина в тканях гипоталамуса (А. Оливерио и др., 1973; Ю. С. Дмитриев,
1981).
Таким образом, множественное влияние одного и того же гена может заклю¬
чаться в контроле порога возбудимости нервной системы, содержания нейроак-
тивных соединений и способности к обучению (образованию оборонительных
условных рефлексов).
Генотип и анатомические особенности мозга. Благодаря трансплантации по¬
явилась возможность экспериментального изменения массы мозга. Пересадка аф¬
риканской рыбке-теляпии среднего мозга от донора увеличила общую массу моз¬
га реципиента и одновременно улучшила ее интеллектуальные способности: рыб-
164 Глава 6. Факторы организации поведения
Рис. 51. Корреляция между порогом возбудимости нервно-мышечного
аппарата и способностью к образованию условного рефлекса
активного избегания у мышей 16 инбредных линий
(по А. С. Дмитриеву, 1983)
а — порог возбудимости при раздражении икроножной мышцы одиночными
электрическими импульсами длительностью 2 мс; 6 — количество условных
рефлексов активного избегания в %; по горизонтали — линии мышей
ка успешно выполняла задачи по различению и переделке сигнального значения
раздражителей, что до операции делала с трудом.
К настоящему времени получено значительное число убедительных аргумен¬
тов, которые позволили приписывать особую роль структурам гиппокампа в про¬
цессах обучения и памяти. В процессе начальных этапов обучения в нейронах гип¬
покампа наблюдается более интенсивное включение меченых белков (Р. Хиден,
1972). Ухудшение способности к обучению соответственно связывают с наруше¬
нием синтеза специфических белков нервной ткани (типа 5-100). Если в наслед¬
ственной обусловленности структурных признаков и способности к обучению
участвуют общие гены, то генетическая изменчивость одного из признаков повле¬
чет за собой изменения и в уровне проявления второго. Оказалось, что изменчи¬
вость синапсов на клетках гиппокампа значима для наследственно обусловленно¬
го уровня активации этих нейронов. Причем число окончаний мшистых волокон
на дендритах пирамидных клеток находится в определенной связи со способно¬
стью мышей к образованию оборонительных условных рефлексов. Между чис¬
лом терминален на базальных дендритах и уровнем осуществления условных
рефлексов существует высокая отрицательная корреляция. Высокий уровень
условно-рефлекторной деятельности коррелирует с многочисленностью синап¬
сов на апикальных дендритах пирамид гиппокампа. Последнее связывают с тем,
что создаются лучшие условия для проявления состояния длительной активации
гиппокампа, что обеспечивает его участие в процессах обучения и запоминания.
Структурные изменения распространяются и на другие отделы мозга: у крыс
с высоким уровнем условно-рефлекторной деятельности обнаружена большая
ширина сенсомоторной области коры, большие размеры зубчатой фасции, мозо¬
листого тела с большим числом миелипизированиых волокон. Генетически детер¬
минированные структурные особенности захватывают и лимбическую систему
мозга (Н. И. Дмитриева и др., 1983, 1985):
§ 1. Генетическая детерминация свойств поведения 165
♦ у хорошо обучающихся крыс увеличивается ширина лимбической коры;
♦ увеличивается размер клеток ядер гипоталамуса и амигдалы;
♦ увеличивается число глиальных клеток свода но сравнению с животными
с низким уровнем возбудимости и скорости образования условных рефлексов.
Л. В. Крушипский (1977) считает, что избыточное количество нейронов моз¬
га — необходимое условие для полноценного восприятия окружающей среды
и формирования адекватных поведенческих реакций.
Генотип и нейрохимия мозга. Наследование свойств обучаемости по материн¬
ской линии позволяет обратить внимание па значение не только ядерного аппара¬
та нервных клеток, но и всего цитоплазматического со¬
держимого. Реализация генетической информации, зако¬
дированной в молекуле ДНК в ядре нервной клетки, осу¬
ществляется при непосредственном участии химических
факторов самой цитоплазмы клетки. Помимо широко
известных первичных химических посредников-нейроме¬
диаторов, с помощью которых информация передается к
нервной клетке и активирует ее в соответствии с прису¬
щей ей собственной генетической программой, в настоя¬
щее время в самостоятельную категорию метаболиче¬
ских факторов выделены вторичные посредники (мес¬
сенджеры). В первую очередь к ним относят циклический аденазинмонофосфат
(ЦАМФ), выполняющий функцию универсального клеточного регулятора.
Нейромедиаторы и пептидные гормоны выступают по отношению к клетке
в качестве сигналов. Эти сигналы воздействуют на мембрану клетки и способству¬
ют образованию из АТФ вторичных посредников — ЦАМФ, который стимулиру¬
ет более чем 200-кратное усиление воздействия на клетку внешнего для нее сигна¬
ла. ЦАМФ активирует соответствующие ферменты (протеинкиназы), обеспечи¬
вающие образование определенных белков. Эти рецепторные белки встраиваются
в постсинаптическую мембрану клетки и способствуют высокой эффективности
воздействия определенного сигнала на клетку.
Ионы кальция также относят к категории вторичных посредников, от которых
зависят как пре-, так и постсинаптические процессы клетки и формирование ее
электрической активности. Вслед за открытием рецептора кальция — белка каль-
модулина было установлено, что он регулирует синтез и распад ЦАМФ.
Особое внимание обращают на стероидные гормоны, которые реализуют свои
эффекты, минуя систему вторичных посредников. В отличие от пептидных гормо¬
нов стероидные гормоны уже имеют собственные возможности проникновения в
нервную клетку, где они связываются непосредственно с ее ядром. Три основных
фактора регуляции — вторичные посредники, ионы кальция и стероидные гормо¬
ны — взаимодополняют друг друга.
В. В. Пономаренко (1970) предложила гипотезу нейроэндокринной регуляции
процесса реализации генетической информации, согласно которой вторичные по¬
средники и стероидные гормоны представляют собой интеграцию функций нервной
и эндокринной систем на молекулярном уровне. Возможность регуляции активно¬
сти генов в нервной системе возрастает в эволюции за счет увеличения как нейро¬
нальных, так и гормональных звеньев этой регуляции, а также онтогенетического
влияния на эти звенья, включая весь приобретенный индивидуальный опыт.
Гипотеза нейроэндо¬
кринной регуляции
реализации генетиче¬
ской информации: вто¬
ричные посредники и
стероидные гормоны
представляют собой
интеграцию функций
нервной и эндокринной
систем на молекуляр¬
ном уровне.
166 Глава 6. Факторы организации поведения
Гетерохрония реализации генетической программы. Признаки и свойства
мозга, детерминируемые генетической программой, образуют так называемую
консервативную наследственность вида. Она может отражаться в структурных
или функциональных признаках, в константной последовательности этапов раз¬
вития поведения. Эти признаки инвариантны для целого вида и возникли под
действием стабилизирующего отбора. Динамика онтогенеза является удобной мо¬
делью для изучения закономерностей развертывания генетической программы.
Например, изучение созревания восходящих возбуждений в кору мозга от рети¬
кулярной формации, от зрительной системы и прочего позволяет представить ход
развития врожденных матриц будущих поведенческих актов, формирующихся в
процессе обучения (Ф. А. Ата-Мурадова, 1983). Особенность онтогенетического
метода заключается во вмешательстве в гетерохромную активацию генома (гл. 5).
Биогенетическая последовательность стадий развития физиологических си¬
стем отражает стадийное действие генных факторов различного эволюционного
возраста. Активность генов каждой стадии развития, формирующей соответст¬
вующий фенотипический признак, создает необходимый генетический фон и вклю¬
чает новые системы генов следующей стадии. Например, созревающие первыми
структуры восходящих корковых возбуждений прокладывают путь для других
компонентов поздно созревающих структур.
П. К. Анохин (1968) подчеркивал опережающее активирование специфиче¬
ской генетической системы, определяющей стадийность развития, и раннее вклю¬
чение нейрональных функций как системного процесса. Так, синхронизация
активности в популяциях нейронов, объединяющихся в рано созревающие ан¬
самбли, отражает системный характер активации генетической программы. Уста¬
новлена первичность созревания неспецифических восходящих в кору возбужде¬
ний. Вначале созревает более древняя холинергическая форма поведения, кото¬
рая обусловливает первичную реакцию гиперсинхронизации. Затем добавляется
адренергический механизм, заменяющий гиперсинхронизацию характерной для
взрослого десинхронизацией (Ф. А. Ата-Мурадова, 1983). При этом подчеркива¬
ется жесткость генетической запрограммированности данных феноменов, что
свидетельствует об известной инвариантности соответствующих генных инте¬
граций и об их значительном историческом возрасте.
Одной из иллюстраций такого положения могут служить эксперименты с пе¬
ресадкой (трансплантацией) мозговой ткани у амфибий. Пересадка закладки
головного мозга между зародышами разных видов бесхвостых амфибий, зна¬
чительно различающихся по поведению, показывает, что поведение в основном
определяется трансплантированным мозгом (Л. В. Полежаев, 1983). Например,
прошедшие метаморфоз остромордые лягушки с пересаженным им мозгом жабы-
чесночницы демонстрируют поведение жабы-чесночницы: ползают на брюхе, вы¬
капывают в земле ямки и зарываются в них.
§ 2. Биологические мотивации как внутренние
детерминанты повеления
Принимая в качестве необходимого фактора целенаправленного поведения нали¬
чие биологической потребности, И. П. Павлов указывал, что нелепо было бы от¬
рицать субъективный мир, как и психологию — науку, изучающую явления этого
§ 2. Биологические мотивации как внутренние детерминанты поведения 167
субъективного мира. Проблема ставилась так: каковы те внутренние состояния жи¬
вотных, которые придают их поведению целенаправленный характер? Некоторые
ученые полагали, что желания, побуждения, предшествующие действию и опреде¬
ляющие его, составляют движущие силы или мотивы действия. Термин мотивации
буквально означает то, что вызывает движение. Этот термин долгое время не ис¬
пользовался в физиологии и входил исключительно в лексикон психологов.
Мы уже отмечали (см. гл. 4), что первоначальный смысл термина инстинкт —
это побуждение к действию. Еще И. М. Сеченов полагал, что животное стремится
удержать, продлить приятное чувственное возбуждение и, наоборот, избавиться
от действия раздражений, сопровождающихся неприятными ощущениями. Таким
образом, мотивации оказывались поставленными в прямую и двустороннюю
взаимозависимость с эмоциями. И. М. Сеченов ввел категорию «желаний», куда
он относил голод, жажду, половое чувство. На этих желаниях и строятся страст¬
ные психические рефлексы», по Сеченову, которые сопровождаются томительны¬
ми отрицательными переживаниями, а сами действия, то есть удовлетворение
страстного желания, имеют положительный характер.
«Жизненные потребности, — писал И. М. Сеченов, — родят хотения, и уже эти
ведут за собой действия; хотение тогда будет мотивом или целью, а движение —
действием или средством достижения цели... Без хотения как мотива или импуль¬
са движение было бы вообще бессмысленно»1.
И. П. Павлов указывал на значение основных влечений — голода, жажды, по¬
лового чувства в возникновении «рефлекса цели». Вся жизнь, вся ее культура де¬
лается рефлексом цели, который является важнейшим
фактором жизни, начиная с капитальнейшей области —
воспитания.
Этологи назвали целенаправленное поведение аппе-
тентным, куда включались все случаи поискового пове¬
дения: поиск пищи, полового партнера, материала для по¬
стройки гнезда и т. д. В изучении эндогенных пусковых
механизмов поведения большая роль отводилась гормо¬
нальным системам. Еще в 1923 г. А. А. Ухтомский подчер¬
кивал роль гормональных механизмов в первичном возникновении некоторых до¬
минантных состояний, которые в дальнейшем могут воспроизводиться и модули¬
роваться условно-рефлекторными механизмами.
Физиологические потребности. Известно, что нормальный уровень обмена
веществ и энергии может поддерживаться только при определенном содержании
в организме ряда продуктов жизнедеятельности, при их постоянном потреблении,
накоплении и разрушении.
Увеличение или снижение содержания этих продуктов в организме существен¬
но меняет его жизнеспособность. Приобретенные в процессе эволюции механиз¬
мы саморегуляции обеспечивают организму поддержание концентрации этих ве¬
ществ во внутренней среде па относительно постоянном уровне (гомеостаз). Этот
уровень основных жизненно важных веществ в организме характеризуется как
определенные константы внутренней среды. К таковым относятся уровень содер¬
жания в крови питательных веществ, ее газового состава, осмотического давле¬
ния, температуры и пр.
Потребность — специ¬
фическая (сушностная)
сила живых организ¬
мов, обеспечиваюшая
их связь с внешней
средой для самосохра¬
нения и саморазвития
как источник активно¬
сти живых систем.
1 Сеченов И. М. Избр. произв. М„ АН СССР, 1952. Т. 1. С. 516.
168 Глава 6. Факторы организации поведения
В результате непрерывного обмена веществ внутренние константы крови име¬
ют тенденцию к колебаниям (гомеокинез), восстанавливаются до требуемого
уровня задолго до наступления существенных сдвигов в обмене веществ. Однако
такой автоматический саморегулирующийся механизм (с внутренним контуром
обратной связи) действует в конкретных пределах и с разной скоростью. Но в тех
случаях, когда отклонение от нормальных констант становится слишком значи¬
тельным и не поддается коррекции с помощью автоматической саморегуляции,
вступают в действие более высокие уровни, связанные с потреблением определен¬
ных веществ из внешней среды или выведением продуктов обмена из организма.
Эти более высокие уровни саморегуляции (с внешним контуром обратной связи)
достигаются благодаря целенаправленному поведению.
Любой живой организм может существовать лишь при периодическом потреб¬
лении из внешней среды питательных веществ, воды, минеральных солей и т. д.
Такие состояния получили название первичных физиологических потребностей.
Однако термин «потребность» может трактоваться и более широко. Исходя из
представлений В. И. Вернадского и А. А. Ухтомского о присущей живой системе
тенденции самосохранения и саморазвития П. В. Симонов (1987) предлагает иод
потребностью рассматривать специфическую (сущностную) силу живых организ¬
мов, обеспечивающую их связь с внешней средой для самосохранения и самораз¬
вития, как источник активности живых систем в окружающем мире.
П. К. Анохин (1968) и К. В. Судаков (1971,1986) в исследовании проблемы по¬
требности исходят из общебиологического принципа выживания как конечной
цели любого поведения животных.
Истинная метаболическая потребность возникает в тканях организма, однако
сигнализация о ней генерируется прежде всего в специальных высокочувстви¬
тельных рецепторах. Восприятие потребности происходит нервным и гумораль¬
ным путем, а соответствующая нервная сигнализация адресуется в специальные
подкорковые отделы мозга, и прежде всего в ядра гипоталамуса — важнейшего от¬
дела промежуточного мозга (рис. 52). Возбуждение этих и связанных с ними цент¬
ров и формирует состояние мотивации. Приведем два основных определения это¬
го термина.
Мотивация представляет собой возникающие под влиянием первичных изме¬
нений во внутренней среде эмоционально окрашенные состояния организма, харак¬
теризующиеся избирательными активирующими влияниями специальных подкор¬
ковых аппаратов на кору головного мозга и другие его отделы и направляющие по¬
ведение животного на удовлетворение исходной потребности (К. В. Судаков,
1971). Соответственно потребностно-информационный подход к пониманию дея¬
тельности мозга позволяет считать, что мотивация есть физиологический меха¬
низм активирования хранящихся в памяти следов (энграмм) тех внешних объ¬
ектов, которые способны удовлетворить имеющуюся у организма потребность,
и тех действий, которые способны привести к ее удовлетворению (П. В. Симонов,
1987).
Если первое определение мотиваций в большей мере адресуется к первичным
физиологическим потребностям, то второе объясняет целенаправленный харак¬
тер поведения во всем разнообразии его психофизиологических проявлении.
Виды мотиваций. Если исходить из определения поведения как формы жизне¬
деятельности, которая меняет вероятность и продолжительность контакта
§ 2. Биологические мотивации как внутренние детерминанты поведения 169
Результат
поведения
Рецептор]
[Мотивация'
Внутренние резервы
Гуморальная сигнализация
Целенаправленное поведение
Нервная сигнализация
Рис. 52. Принципиальная схема саморегуляторных процессов,
порождающих в центральной нервной системе мотивационное
возбуждение (по К. В. Судакову, 1972)
К — константа крови, отклонение константы от жизненно важного уровня (К, и К2)
через возбуждение рецепторов или гуморальным путем вызывает в центральной
нервной системе мотивационное возбуждение. Последнее приводит к целенаправ¬
ленному поведению и удовлетворению исходной потребности организма
с объектом удовлетворения имеющейся у организма потребности (П. В. Симонов,
1979), то вся наша жизнь представляется как непрерывная цепь формирующихся
целей и их достижений или неудач.
Превращение актуализированной потребности из побуждения в цель и состав¬
ляет содержание процесса мотивации целенаправленного поведения.
Наиболее распространенная классификация видов мотиваций предложена
Н. Миллером (1960). Он выделяет низшие, или первичные мотивации, называя их
также инстинктивными или висцеральными. Эти же со¬
стояния И. П. Павлов рассматривал как основные влече¬
ния и относил к ним голод, жажду, страх, половое чувство.
К ним же могут быть отнесены и наиболее примитивные
состояния, которые побуждают к актам мочеиспускания,
дефекации и т. д.
Вторую группу мотиваций составляют высшие, или
вторичные, мотивации, которые приобретаются в тече¬
ние индивидуальной жизни и, хотя и строятся на основе
генетически заданных потребностей, в значительной мере
опираются на накопленный индивидуальный опыт. Сюда могут быть отнесены
все виды мотиваций, которые возникают по законам условного рефлекса.
Методы изучения биологических мотиваций. Одним из основных внешних
проявлений мотиваций является целенаправленное поведение. Предложенная
П. К. Анохиным (1932) методика свободного выбора пищи животным оказалась
весьма демонстративной, ибо предпочтение животным определенного вида нищи
(мясо, молоко, солевой раствор и ир.) свидетельствовало о наличии соответствую¬
Мотиваиия — физио¬
логический механизм
активирования памят¬
ных следов тех пове¬
денческих актов, кото¬
рые способны
удовлетворить имею¬
щуюся у организма
доминирующую по¬
требность.
щей исходной мотивации.
Условно-рефлекторная методика позволяет придать различным внешним
агентам функции сигналов получения определенных видов пищи или воды и тем
170 Глава 6. Факторы организации поведения
самым определить степень соответствия внешней сигнализации естественно или
искусственно сформированной внутренней мотивации.
Например, у собаки вырабатывают условные рефлексы в форме побежки на
сигнал стука метронома 60 ударов в минуту к левой кормушке, в которой всегда
подается пресная пища, а на сигнал 120 ударов в минуту — к правой кормушке, по¬
бежка к которой подкрепляется пищей разной степени солености. Определив пре¬
дельную степень солености пищи, которую поедает животное (солевой порог), со¬
баку переводят на искусственную солевую диету с одновременным ограничением
воды. Естественно, что чем длительнее содержание животных на солевой диете, то
есть чем сильнее мотивация жажды, тем реже становятся побежки животных по
сигналу «солевого» к правой кормушке.
Наконец такие побежки прекращаются вовсе, хотя к левой «пресной» кормуш¬
ке побежки сохраняются на прежнем уровне (И. П. Никитина, 1985).
Измерить мотивацию непосредственно не представляется возможным, поэто¬
му используют косвенные приемы. Мотивация может возникнуть вследствие
длительного лишения животного воды, либо введения в желудок через фистулу
солевого раствора, либо кормления животного сухой пищей. Оценить уровень мо¬
тивации жажды можно несколькими способами (рис. 53): путем измерения обще¬
го объема выпитой животным воды, частоты нажатия на рычаг для получения
питьевого подкрепления или предельного количества хинина, добавленного в во¬
ду, при котором животное еще продолжает ее пить.
Лишение воды
(длительное)
Введение
в желудок
солевого
раствора
Кормление
сухой пищей
Мотивация
жажды
Объем
выпитой воды
Скорость
нажатия
на рычаг
Уровень хинина,
терпимый для животных
Рис. 53. Возможные способы изображения взаимосвязи
трех независимых переменных, которые позволяют
измерять потребление волы (по Э. Меннингу, 1982)
Электрическое раздражение определенных нервных центров, расположенных
в гипоталамической области, сопровождается не только возникновением первич¬
ных мотиваций, ио и соответствующим поведением, ведущим к их удовлетворе¬
нию. Так, например, у крыс предварительно вырабатывают условный рефлекс на¬
жатия на рычаг, за которым следует пищевое подкрепление. При раздражении ла¬
теральной гипоталамической области даже накормленные крысы снова начинают
нажимать на рычаг (Н. Миллер, 1960).
Особый интерес для физиологии мотиваций представляют эксперименты с са-
мораздражением (Олдс, 1958). Оказалось, что раздражение определенных струк¬
тур головного мозга (прежде всего ядер гипоталамуса) через вживленные элект¬
роды приводит к тому, что животное начинает нажатием на рычаг само включать
ток, чтобы наносить себе раздражения. Эти мотивации обладают значительной
§ 2. Биологические мотивации как внутренние детерминанты поведения 171
энергетической силой, и животные готовы преодолевать сложные препятствия
ради достижения эффекта самораздражепия.
Физиологические теории мотивации. Первая группа теорий мотивации осно¬
вана на представлении о сигнализации от периферических органов тела. В. Кеннон
(1932, 1934), Д. Хебб (1949) полагали, что мотивации определяются стремлением
организма избежать неприятных эмоциональных ощущений, сопровождающих
различные побуждения. Животное утоляет жажду, чтобы избавиться от неприят¬
ной сухости в полости рта и глотки, поедает пищу, чтобы избавиться от мышечных
сокращений пустого желудка, и т. д.
Дальнейшие поиски привели к созданию другой группы теорий, в которых
основное внимание уделялось гуморальным факторам мотиваций. Так, голод свя¬
зывался с возникновением так называемой голодной крови, то есть крови с су¬
щественным отклонением от обычной разницы в концентрации глюкозы в ар¬
териальной и венозной крови. Это и рассматривалось в качестве первопричины
«голодных» сокращений желудка. Мотивация жажды также оценивалась как
следствие изменения осмотического давления плазмы крови или снижения вне¬
клеточной воды в тканях, что может оцениваться специальными рецепторами (ос¬
мо- и волюмреценторы). Половое влечение ставилось в прямую зависимость от
уровня половых гормонов в крови.
Для оценки справедливости гуморальных теорий мотиваций существенный
интерес представляют наблюдения за двумя парами сросшихся близнецов-сакро-
пагов, выполненные Т. Т. Алексеевой (1958). У этих близнецов была общая систе¬
ма кровообращения и раздельная центральная нервная система. Верхняя часть
пищеварительного тракта у них также была раздельной. Оказалось, что кормле¬
ние одной из девочек, а значит равномерное распределение питательных веществ
в кровеносной системе обоих близнецов, никогда не приводило к насыщению вто¬
рой девочки. Наоборот, у нее длительное время могла поддерживаться потреб¬
ность в пище. Эти наблюдения, а также многочисленные наблюдения на живот¬
ных показали, что гуморальные факторы не могут быть первопричиной возникно¬
вения биологических мотиваций.
По-видимому, как возникновение мотиваций, так и их удовлетворение имеет
множественный генез, в основе которого лежит нейрогуморальпый механизм пе¬
риферического и центрального уровней.
В самих мозговых структурах обнаружены чувствительные нервные клетки,
специализированные на восприятии колебаний в содержании определенных хи¬
мических веществ, а также физических параметров питающих их жидкостей. Ос¬
новным аппаратом, содержащим такие центральные рецепторы, является гипота¬
ламус. И. Стеллар (1954), выдвинув гипоталамическую теорию мотиваций, пола¬
гал, что гипоталамус и является вместилищем «центрального мотивационного
состояния».
Рассмотрим для примера некоторые гипоталамические механизмы жажды. Во-
первых, в латеральных ядрах гипоталамуса обнаружены осмореценторы, детекти¬
рующие осмоляльность циркулирующей крови. Во-вторых, введение в латераль¬
ный гипоталамус солевого гипертонического раствора вызывает усиление потреб¬
ления воды сверх физиологических потребностей (похожую реакцию вызывает
и электрическое раздражение этих же структур). В-третьих, повреждение лате¬
рального гипоталамуса нарушает регуляцию водного баланса, а повреждение
172 Глава 6. Факторы организации поведения
переднего гипоталамуса и преоптического ядра полностью устраняет способность
к потреблению воды, ие лишая, однако, животных возможности принимать пищу.
И наконец, в-четвертых, раздражение «питьевой» зоны гипоталамуса провоциру¬
ет целый комплекс поведенческих реакций, связанных с поисками воды.
Хорошо известно, что прекращение питья наступает задолго до того, как вос¬
становятся физиологические константы крови. Показано также, что ни растяже¬
ние желудка, ни всасывание воды не имеет решающего значения в регуляции по¬
требления воды. По этому поводу высказывается наиболее вероятное предпо¬
ложение: прекращение приема воды наступает в результате обратной связи от
рецепторов ротовой полости, с помощью которой отмеряется количество погло¬
щенной воды, и через амигдалу тормозится реакция питья (Н. Мильнер, 1973).
Теперь рассмотрим некоторые теории мотивации голода, которые связывают
это состояние опять же со структурами гипоталамуса. Наиболее последователь¬
ные поиски ведутся Б. К. Анандом и Д. Р. Бробеком (1951), которые считают, что в
латеральном гипоталамусе располагается центр питания, а в медиальном гипота¬
ламусе — центр насыщения. Авторы допускают, что центры питания и насыщения
находятся в реципрокпых отношениях. Потребление пищи определяется тормоз¬
ными влияниями латеральной области па медиальную.
С. П. Гроссман (1960-1969) вводил в гипоталамус накормленных крыс биоло¬
гически активные вещества. Введение в латеральный гипоталамус адренергиче¬
ских веществ (в отличие от холинергических) вызывало усиленное потребление
пищи. Введение же холинергических веществ провоцировало питьевые реакции.
При этом возникал целый комплекс ранее заученных водо- и пищедобывательных
двигательных реакций.
Сегодня является бесспорным положение о том, что гипоталамические струк¬
туры не могут рассматриваться в качестве единственных аппаратов пищевого по¬
ведения.
Лимбический уровень (включая гипоталамические и ретикулярные механиз¬
мы) обеспечивает лишь примитивно организованные, лишенные выраженной це¬
ленаправленности типы пищевого поведения, так называемые пищевые автома¬
тизмы. Направленный же поиск нищи, ее оценка и выбор, а также формирование
избирательного аппетита требуют участия коры больших полушарий головного
мозга. Следовательно, только объединенное участие гипоталамуса, лимбических
образований и коры головного мозга формирует целостное пищевое поведение
животных.
При этом исключительная роль гипоталамуса не вызывает никаких сомнений:
этот отдел мозга как комплексный детектор физиологических дисбалансов (тем¬
пературы, питательных веществ, воды, гормонов, минеральных солей) играет до¬
минирующую роль в отношении путей использования ресурсов организма. Прак¬
тически ни одну функцию организма нельзя себе представить вне прямой или
косвенной зависимости от гипоталамуса.
Работами К. В. Судакова (1971, 1986) было показано, что каждое мотивацион¬
ное возбуждение строится па основе специфических химических механизмов, ак¬
тивирующих влияния подкорковых мотивационных центров на кору головного
мозга. Введением центральных адреполитиков (аминазина) удается избирательно
заблокировать активацию коры при болевом раздражении. Активация же коры
§ 2. Биологические мотивации как внутренние детерминанты поведения 173
головного мозга при пищевом возбуждении голодных животных избирательно
блокируется холинолитическими веществами.
Наряду с вышесказанным существует и крайняя точка зрения П. Мак-Лина
(1970) о жесткой привязке мотиваций к аппаратам лимбической системы. Он счи¬
тает, что в гипоталамусе локализованы эмоционально окрашенные поведенческие
реакции, связанные с рефлексами избегания, нападения, поиска, добывания пи¬
щи, воды и пр. В амигдале сосредоточены мотивации голода, жажды, защиты от
внешних вредных влияний. Перегородка связана с половым поведением. Проти¬
воположной позиции, которая кажется нам более справедливой, придерживается
Р. Л. Аисааксои (1978), который отмечает, что не существует одиночных анатоми¬
ческих образований для одиночных поведенческих функций, как не существует
единых поведенческих функций, которые могли бы быть приурочены к анатоми¬
ческим образованиям таким образом, чтобы эти функции оказывались полезными
в любых обстоятельствах и при всех внешних условиях.
Доминирующее мотивационное возбуждение. В каждый момент времени до¬
минирует та мотивация, в основе которой лежит наиболее важная биологическая
потребность. Сила потребности, то есть величины отклонения физиологических
констант или концентрации соответствующих гормональных агентов, получает
свое отражение в величине мотивационного возбуждения структур лимбической
системы и определяет его доминантный характер. Консервативный характер до¬
минанты заключается в ее инертности, устойчивости,
длительности. В этом ее большой биологический смысл
для организма, который стремится к удовлетворению
биологической потребности в случайной постоянно ме¬
няющейся внешней среде. В физиологическом смысле та¬
кое состояние доминанты характеризуется определенным
уровнем возбудимости центральных структур, обеспечивающей их высокую от¬
зывчивость и «впечатлительность» к разнообразным воздействиям. Но однажды
проявленная доминирующая мотивация возобновляется и живет даже в порядке
чисто нервно-рефлекторного фактора. Поэтому наряду с консервативным нача¬
лом в доминирующей мотивации как векторе поведения содержится и динамиче¬
ское начало, поскольку из множества новых подкрепляющих впечатлений проис¬
ходит непрерывный подбор «пригодного», нужного, имеющего непосредственную
связь.
Первичный очаг возбуждения при формировании доминирующих биологиче¬
ских мотиваций (голод, жажда, секс) возникает в гипоталамических образованиях
мозга (Р. А. Павлыгина, 1966). Эти первичные очаги, в свою очередь, ведут к появ¬
лению вторичных очагов в других отделах мозга, где имеются предпосылки для
продолжительного удержания следовых процессов. Кора головного мозга с ее не¬
прерывной и разномодальной активацией располагает всеми предпосылками для
создания в ней вторичных очагов стационарного возбуждения. Принципиальным
отличием таких очагов от первичных является исходная причина их возникнове¬
ния. Так, если первичный очаг в гипоталамусе возникает и поддерживается в пер¬
вую очередь за счет гуморальных факторов, то вторичные очаги в коре больших
полушарий являются проекционными по своему происхождению и поддержива¬
ются за счет широкой конвергенции к ним множества суммирующихся здесь сен¬
сорных влияний.
Консервативный ха¬
рактер доминанты за¬
ключается в ее инерт¬
ности, устойчивости,
длительности.
174 Глава 6. Факторы организации поведения
Констелляция
вторичных очагов
Рис. 54. Схема соотношения
первичных и вторичных очагов
при формировании
доминирующей констелляции
мозговых образований
(по А. С. Батуеву, 1986)
Рассмотрим несколько конкретных примеров.
Содержание животных па длительной солевой дие¬
те, формирующее у них мотивацию жажды, создает
в латеральном гипоталамусе очаг повышенной воз¬
будимости к вкусовым и висцерохимическим раз¬
дражениям (А. С. Батуев, Б. Г. Гафуров, 1988). Сни¬
мается же подобный очаг введением в организм до¬
полнительных количеств жидкости.
Итак, мотивационное возбуждение, обусловлен¬
ное преобладающей потребностью организма, фор¬
мирует в гипоталамусе первичный очаг повышен¬
ной возбудимости, обладающий свойствами доми¬
нантного очага. Благодаря этому доминантному очагу
быстро образуются условно-рефлекторные связи
с теми или иными факторами окружающей среды
(вторичные очаги), которые по своей функциональ¬
ной значимости могут быть сравнимы с эндогенны¬
ми механизмами самого мотивационного состояния
(рис. 54).
Нейрофизиология мотиваций. К. В. Судаков
(1986) сформулировал основные положения о прин¬
ципах нейрофизиологического обеспечения доми¬
нирующих мотиваций:
1. Любая биологическая мотивация обусловлена соответствующей метаболиче¬
ской потребностью и носит системный характер.
2. Потребность трансформируется нейрогуморальным путем в возбуждение ги¬
поталамических центров, которые активируют другие структуры мозга, в том
числе и кору больших полушарий.
3. Корковые и лимбические структуры мозга оказывают специфические для каж¬
дой мотивации нисходящие возбуждающие и тормозные влияния на гипотала¬
мические мотивационные центры.
4. Каждое мотивационное возбуждение представляет собой специфическую кле¬
точную и молекулярную интеграцию корково-подкорковых структур.
Избирательное возбуждение одних структурных образований сочетается с из¬
бирательным торможением других.
Мотивационные возбуждения существенно меняют конвергентные и дискри¬
минационные способности нейронов разных отделов мозга. Например, раздраже¬
ние пищевого центра латерального гипоталамуса приводило к тому, что нейроны
сенсомоторной коры, ранее не отвечавшие на световые, звуковые и гуморальные
раздражения, приобрели способность реагировать па них. С другой стороны, ней¬
роны коры, не отвечавшие ранее па введение кроликам морковного сока, начина¬
ли реагировать на эти раздражения после стимуляции центра «голода» латераль¬
ного гипоталамуса. Создается впечатление, что мотивационное возбуждение «на¬
страивает» нейроны различных областей мозга на подкрепляющие воздействия.
Мотивационное возбуждение находит свое выражение в специфическом рас¬
пределении межгшпульсных интервалов и образовании характерной пачечной
§ 3. Роль эмоиий в организации поведения 175
активности (рис. 55). Эта активность усиливается при возрастании потребности,
при возникновении препятствий к удовлетворению доминирующей мотивации.
Достижение же цели поведенческого акта сменяется в нейронограмме регулярной
активностью.
■И-НН-Н- И»1 11 И -Н1111НШЩ1 НИ'МНЩШ 11Н1Н
а 6
Рис. 55. Типичный характер нейронной активности у кролика
(по К. В. Судакову, 1986)
Под нейрограммой — гистограммы распределения межимпульсных интервалов:
а — у голодного кролика, 6 — у накормленного кролика
Возбуждения, поступающие в мозг при подкреплении, адресуются к различ¬
ным нейронам и оставляют «след» в их деятельности, формируя сложную нейро-
динамическую констелляцию. С ней и происходит срав¬
нение параметров достигнутых результатов.
В. Г. Зилов (1978) показал, что в формировании раз¬
личных биологических мотиваций участвуют одни и те
же нейромедиаторы, однако в различных комбинациях в
разных структурах мозга, что свидетельствует о специфи¬
ческой нейрохимической интеграции конкретного моти¬
вационного возбуждения. Последнее проявляется также
и в общей чувствительности отдельных нейронов к нейро¬
медиаторам и олигопептидам (К. В. Судаков, 1986). Способность ряда пептидов
(ангиотензин II, бета-липопротеип, пептогастрип и др.) при внутрижелудочковом
введении избирательно продуцировать или подавлять определенные биологиче¬
ские мотивации открывает путь к изучению молекулярной интеграции мотиваци¬
онного возбуждения.
Эмоции — субъектив¬
ные реакции живот¬
ных и человека на
внутренние и внешние
раздражители, прояв¬
ляющиеся в виде пере¬
живаний и соматове-
гетативных сдвигов.
§ 3. Роль эмоиий в организации поведения
Под эмоциями подразумевают субъективные реакции животных и человека на
внутренние и внешние раздражения, проявляющиеся в виде удовольствия или не¬
удовольствия, страха, гнева, тоски, радости, надежды, грусти и т. п.
Термин «эмоция» употребляется в разных смыслах, например:
♦ для обозначения субъективных ощущений, которые можно изучать только
путем непосредственного самонаблюдения (интроспекции);
176 Глава 6. Факторы организации поведения
♦ для обозначения экспрессивных проявлений при наблюдении за другими осо¬
бями;
♦ для описания сложного поведения типа драки, побега и т. д.
Благодаря эмоциональному возбуждению создается эмоциональная окраска
текущего поведения, происходит субъективная оценка ситуации.
Если эмоцию рассматривать как форму отражения действительности, как про¬
цесс, регулирующий отношения субъекта с внешней средой, то психическое и фи¬
зиологическое в эмоциях выступают как две стороны единой нервной деятельно¬
сти.
В эмоциях есть субъективное, но нет идеального, ибо внешний мир отражается
не в виде образов, а в виде переживаний субъективных состояний (Г. X. Шинга-
ров, 1971).
На определенном этапе эволюции возникновение ощущений обогащается
свойствами переживаемое™ и возникает то, что именуют субъективным. Отраже¬
ние психикой внутреннего состояния организма в виде субъективных пережива¬
ний служит предпосылкой возникновения субъективных образов.
Животным, так же как и человеку, присущи принципиально схожие процессы
отражения с субъективным отношением организма к окружающей его среде. Пер¬
вичные витальные эмоции (страх, ярость, боль и пр.) не являются прерогативой
человека, а унаследованы им в процессе эволюционного развития.
Эмоции и мотивации. До сих нор нет четкого мнения о том, различаются эти
два субъективных состояния организма или отражают просто разные оттенки од¬
ного и того же процесса. И. П. Павлов не проводит резкого разграничения между
эмоциями и мотивациями и часто употребляет эти термины как синонимы. И эмо¬
ции и мотивации И. П. Павлов рассматривал как проявление функционально¬
го состояния мозга в связи с действием раздражителей внешней или внутренней
среды.
Один из учеников И. П. Павлова польский физиолог Ю. М. Конорски (1970)
также не находит различий между этими явлениями и считает, что они регулиру¬
ются единой мозговой системой с помощью драйв-рефлексов (драйв — настрое¬
ние, побуждение, мотив), которые подготавливают работу исполнительной систе¬
мы мозга. Систему, контролирующую эти драйв-рефлексы, он называет эмотив-
иой или мотивационной.
И. С. Бери гашвили (1966) полагает, что эмоции теснейшим образом связаны с
удовлетворением трех основных потребностей: пищевой, защитной и половой. Он
подчеркивает, что не сам но себе голод, а то эмоциональное возбуждение, которое
возникает при представлении нищи, является побудительной причиной пищевого
поведения. Значит, в основе мотивационной деятельности лежит эмоциональное
возбуждение.
П. К. Анохиным (1968) выдвинуто, по существу, близкое представление. Он
считает, что мотивационное возбуждение сопровождается эмоциональными отри¬
цательными состояниями (жажды, голода, страха и нр.), которые и мобилизуют
организм к быстрому и оптимальному удовлетворению потребности. Удовлетво¬
ренная потребность сопровождается положительной эмоцией, которая и выступа¬
ет как конечный подкрепляющий фактор.
§ 3. Роль эмоций в организации поведения 177
В последнее время Д. Биндра (1969) развивает концепцию об общности нейро¬
физиологического содержания мотиваций и эмоций. Не найдя различий между
этими явлениями, Биндра выдвигает концепцию о «центральном мотивационном
состоянии», возникающем в результате побудительных внешних или внутренних
стимулов. Это состояние формирует избирательное внимание к определенному
классу побудительных стимулов.
Другие исследователи разграничивают мотивации и эмоции, П. Юнг (1943)
считает, что эмоция берет начало в психологической ситуации, но не является ре¬
зультатом внутриорганизменных сдвигов, побуждающих, например, к удовлетво¬
рению голода. П. В. Симонов (1970) полагает, что эмоции представляют собой са¬
мостоятельный мозговой механизм, играющий специфическую роль в организа¬
ции поведения.
А. В. Вальдман (1972) придерживается мнения, что эмоции индуцируются
преимущественно внешними, а мотивации — внутренними стимулами. Мотива¬
ционное поведение имеет организованный характер, а эмоциональное — не орга¬
низовано. Эмоции имеют психогенную природу, а мотивации — эндогенно-ме¬
таболическую. Эмоция возникает на базе сильной мотивации, если существуют
затруднения в ее удовлетворении, и может возникать как следствие взаимодейст¬
вия мотивов, конфликта в планах их осуществления. Она соответствуют такому
снижению уровня адаптивных возможностей организма, которое наступает, когда
мотивация является слишком сильной по сравнению с реальными возможностя¬
ми субъекта. Значит, существует оптимум мотивации, за пределами которого воз¬
никает эмоциональное поведение.
Теории эмоций. Ч. Дарвин (1872) создал биологическую концепцию эмоций, ос¬
нованную на сравнительном исследовании эмоциональных выразительных дви¬
жений у млекопитающих. Эти движения рассматривались как рудимент целесо¬
образных инстинктивных действий, сохраняющих в какой-то степени свой биоло¬
гический смысл и вместе с тем выступающих в качестве биологически значимых
сигналов для особей не только своего, но и других видов. Эти выразительные дви¬
жения (гнев, страх, радость и др.), и особенно мимические реакции, относились
к врожденным проявлениям эмоций.
Согласно теории II. К. Анохина (1968), эмоции возникли в эволюции как субъ¬
ективные ощущения, позволяющие животным и человеку быстро оценивать раз¬
личные внутренние потребности, действие на организм внешних факторов, ре¬
зультаты поведенческой деятельности и, наконец, удовлетворение внутренних по¬
требностей. Как правило, любая потребность сопровождается эмоциональным
переживанием отрицательного характера. Оно стимулирует животное и человека
к активной деятельности, направленной на удовлетворение данной потребности.
Удовлетворение любой потребности сопровождается эмоциональным пережи¬
ванием положительного характера, то есть вызывает чувство удовлетворения
и санкционирует успех поведенческой деятельности. Ассоциированная с успеш¬
ным завершением действия положительная эмоция закрепляется в памяти и на¬
чинает выполнять важную роль в механизмах формирования целенаправлен¬
ной деятельности. Неоднократное удовлетворение потребности, окрашенное по¬
ложительной эмоцией, способствует обучению соответствующей деятельности,
а повторные неудачи в получении запрограммированного результата вызыва¬
ют торможение неэффективной деятельности и поиски новых, более успешных
178 Глава 6. Факторы организации поведения
способов достижения цели. Эмоциональные воздействия усиливают, удлиняют
и углубляют фиксацию следов от раздражений, что помогает адекватному реаги¬
рованию па внешние сигналы.
В прошлом известной популярностью пользовалась так называемая перифери¬
ческая теория эмоций Джеймса-Ланге. Согласно этой теории, эмоция является
вторичным явлением, основанным на приходящих в мозг сигналах об изменениях
в мышцах, сосудах, внутренних органах при выполнении поведенческого акта, вы¬
званного эффективным раздражителем. Суть своей теории Джеймс выразил фор¬
мулой: «Мы чувствуем печаль, потому что плачем, мы боимся, потому что дро¬
жим». Причем каждый вид эмоционального переживания жестко детерминиро¬
ван определенным набором вегетативных реакций.
Исследования В. Кеннона, Ч. Шеррингтона, Д. Хебба и других показали, что
висцеральные проявления эмоций являются вторичными по отношению к мозго¬
вому психическому состоянию.
В качестве противопоставления «периферической» теории был предложен ряд
«центральных» теорий эмоций. В. Кеннон (1927) и В. Бард (1934) выдвинули та¬
ламическую теорию эмоций, которая помещает первичный аппарат для проявле¬
ния эмоций в таламус. Таламические эмоциогенные центры испытывают тормозя¬
щее влияние коры головного мозга и немедленно дают разряд, как только освобо¬
ждаются от кортикальных влияний. При этом условии ощущение получает
определенную эмоциональную окраску. Эти же процессы являются причиной
эмоциональных выразительных движений. Таким образом, таламус рассматрива¬
ется как резервуар эмоционального напряжения.
Под влиянием новых факторов все большую силу и убедительность приобре¬
тала лимбическая теория. Хорошо известный синдром Юновера-Быоси, выра¬
жающийся в снижении эмоциональной реактивности, наблюдался у обезьян по¬
сле разрушения некоторых глубоких структур мозга: грушевидной доли, минда¬
лины, гиппокампа. Такой же синдром «послушного поведения и бесстрашия»
наблюдался у больных с поражениями височных отделов коры и подлежащих глу¬
боких структур мозга.
В 1937 г. И. В. Пейпец, основываясь на клинических наблюдениях, предполо¬
жил, что в коре больших полушарий должен быть центр, интегрирующий пережи¬
вания и эмоциональные реакции, который он гипотетически помещал либо в по¬
ясную извилину, либо в гиппокамп. Затем он присоединил к этим образованиям
передние таламические ядра и гипоталамус. Все эти структуры и функционально
и морфологически связаны между собой и получили в дальнейшем название
«круга Пейпеца», по которому циркулируют «эмоциональные процессы».
П. Мак-Лин (1949) предложил теорию «лимбической системы», или «висце¬
рального мозга», куда он включил рад корковых, подкорковых и стволовых струк¬
тур, обладающих общими конструктивными и функциональными свойствами. По
мнению П. Мак-Лина, лимбическая система получает информацию от внутрен¬
них органов и интерпретирует ее в «терминах эмоций», то есть организует эмо¬
циональное возбуждение и экспрессии. Г1. Мак-Лин резюмировал свою идею так:
различие между новой корой и лимбической системой такое же, как различие
между «мы чувствуем» (лимбическая система) и «мы знаем» (неокортекс).
Широкое использование электроэнцефалографии в изучении мозга привело
к открытию иеспецифических функций ретикулярной формации ствола и других
§ 3. Роль эмоиии в организации поведения 179
отделов мозга. С этим периодом тесно увязано появление активационной теории
эмоций Д. Б. Линдслея (1960), которая приписывала основную эмоциогенную
функцию активирующей ретикулярной системе ствола мозга.
Выраженная эмоциональная реакция возникает лишь при диффузной актива¬
ции коры с одновременным включением гипоталамических центров промежуточ¬
ного мозга (рис. 56). Основным же условием появления эмоциональных реакций
является наличие активирующих влияний из ретикулярной формации при ослаб¬
лении коркового контроля за глубокими структурами мозга, и прежде всего лим¬
бической системы.
В. Р. Гесс (1932) на основании метода раздражения составил карты подкорко¬
вых эмоционально-мотивационных зон при раздражении гипоталамуса, а Е. Бо-
вард (1958), развив эти исследования, предположил существование двух реци-
прокных гипоталамических эмоциональных систем'.
♦ эмоционально-положительной в передних и латеральных ядрах гипоталамуса;
♦ эмоционально-отрицательной — в задних и медиальных его отделах.
С этим тесно связано открытие так называемых подкрепляющих структур моз¬
га — положительных и отрицательных (Д. Олдз, X. Дельгадо и др.). Смысл его со¬
стоит в том, что электростимуляция этих структур дает такой же эффект, как соот¬
ветствующий безусловный раздражитель. К системе структур положительного
подкрепления относят перегородку, латеральный гипоталамус, медиальный не-
реднемозговой пучок. К системе структур отрицательного подкрепления относят
центральное серое вещество, медиальный гипоталамус, миндалину. В методике
самораздражения животное, нажимая на педаль, само посылает в мозг электриче¬
ский ток. Например, при стимуляции положительных подкрепляющих зон мозга
формируется условный рефлекс на нажатие педали, которое может осуществ¬
ляться сотни и тысячи раз подряд, а при стимуляции отрицательных зон — услов-
Рис. 56. Схематическое изображение
основных центральных нервных
структур и возможных путей,
включенных в эмоциональное
поведение (по А. Б. Линдслею, 1960)
Схема не включает мозжечок и некоторые
базальные ганглии, которые также могут при¬
нимать участие в реакции. 1 — соматиче¬
ские и черепно-мозговые афференты; 1' —
прямая таламокортикальная связь; 2 — путь
висцеральных афферентов; 3 — иентрипе-
тальные связи ретикулярной формации; 4 —
диффузные таламокортикальные связи; 5-
6 — взаимосвязи гипоталамуса и таламуса;
7-8 — взаимосвязи таламуса и коры; 9 —
внутриталамические связи; 10 — внутрикор-
ковые связи; 11 — кортикогипоталамиче¬
ские пути; 12 — пути висцеральных аффе¬
рентов; 13 — тракт, связывающий гипотала¬
мус и гипофиз; 14 — кортикоспинальные
пути, заштрихованный участок обозначает
ретикулярную формацию
Кора
Таламус
Гипоталамус
Средний мозг
Продолговатый мозг
Спинной мозг
2
180 Глава 6. Факторы организации поведения
ный рефлекс избегания педали. Это обстоятельство и послужило основанием для
предположения о закреплении за структурами положительного подкрепления по¬
ложительных эмоций удовольствия, а за структурами отрицательного подкрепле¬
ния — отрицательных эмоций отвергания (аверсии). Эти структуры могут нахо¬
диться в антагонистических отношениях.
В связи с тем, что метод стимуляции и самостимуляции нашел широкое при¬
менение в разнообразных исследованиях, появляется особая осторожность при
интерпретации полученных результатов. Уже давно было отмечено существова¬
ние так называемого псевдоаффективного рефлекса, или феномена «ложной яро¬
сти», то есть реакции, напоминающей по внешнему проявлению типичную эмо¬
циональную реакцию, но не сопровождающуюся действительным возникновени¬
ем эмоционального состояния. Вероятно, сами но себе аффективные реакции при
точечной стимуляции разных зон мозга еще не являются показателями наличия
истинной эмоции. Кроме того, надо принимать во внимание не стереотипность,
а, наоборот, чрезвычайную вариабельность самой аффективной реакции, завися¬
щей не только от топографии электродов и параметров раздражающего тока, но
и от всей окружающей ситуации (X. Дельгадо, 1966; Е. Валенштейн, 1969).
А. В. Вальдман (1972) высказывается в пользу отсутствия строгой «жесткой»
связи определенного типа эмоций с конкретными морфологическими аппаратами
мозга. Эмоциональная реакция при стимуляции мозга — это производное многих
внутренних и внешних факторов. Поэтому эмоции как психические функции не
являются строго фиксированной в мозговых системах формой поведения, а бази¬
руются как на врожденных, так и на приобретенных механизмах. Следовательно,
попытка топографического распределения эмоций в головном мозгу, как и других
психических функций, является отголоском локализационизма. В этой связи во¬
прос о субстратах эмоций теряет смысл, если исследователь не дифференцирует
субстрат «эмоционального переживания», структуры, связанные с «выражением
эмоций», и системы, осуществляющие «запуск» эмоциональных реакций. Поэто¬
му предлагается отличать «эмоциональное состояние» от «эмоциональных реак¬
ций» даже у животных (рис. 57). Под эмоциональным состоянием (эмоциональ¬
ным переживанием) понимается субъективное переживание, отражающее отно¬
шение данного индивидуума к окружающему миру и к самому себе.
Возникающие же параллельно изменения в соматической и висцеральной сфе¬
ре, которые сопровождают эмоциональное состояние, обозначаются как эмоцио¬
нальное выражение (или выражение эмоции).
Если для человека внутренний компонент эмоционального переживания оче¬
виден, то применительно к животным мы можем судить о его существовании
только по внешним эмоциональным проявлениям, которые являются видоспеци¬
фичными и определяются позой, мимикой, формой движения, положением шер¬
сти, хвоста, ушей и т. д.
А. В. Вальдман (1972) с помощью фармакологических подходов доказал, что
при стимуляции мозга имеется возможность провоцировать эмоциональные реак¬
ции без соответствующего эмоционального состояния, которое предварительно
купируется транквилизирующими дозами психотропных веществ. С другой сто¬
роны, активация модулирующих структур лимбической системы может вызывать
отчетливые сдвиги эмоционального состояния, нарушающие адекватность реаги¬
рования, но не находящие прямого внешнего проявления.
§ 3. Роль эмоиий в организации поведения 181
Эмоция
«Центры»
эмоционально¬
выразительных
проявлений
Поза
Мимика
Вегетатика
Рис. 57. Обшая схема организации эмоций (по А. В. Вальдману, 1972)
Легче контролируемая, но по существу аналогичная закономерность наблюда¬
ется у человека. В лаборатории Н. П. Бехтеревой разработан способ терапевтиче¬
ского воздействия на мозг больного человека слабым постоянным током через ин¬
трацеребральные электроды. В. М. Смирнов (1976) показал, что при электрости¬
муляции различных глубоких структур мозга наряду с яркими эмоциональными
вспышками проявляются «мнимые эмоции» —эмоциональные психологические
автоматизмы, возникающие в связи с вызванной изолированной активностью ка¬
кого-то мозгового звена системы эмоций.
Попытка раздельного анализа эмоциональных процессов приводит к заключе¬
нию о том, что эмоции как формы психического состояния вовсе не обязательно
должны быть сопряжены с действием. Длительность проявления, инертность
эмоционального состояния — одна из характерных свойств эмоций. Человек мо¬
жет переживать сильные эмоции, ие совершая никаких действий путем их волево¬
го торможения, при этом, однако, вегетативные компоненты эмоций обычно не
удается подавить, так как они малоподвержены произвольной регуляции. Буду¬
чи включенными в обеспечение доминантного характера действия, эмоции могут
включаться во все звенья условно-рефлекторного процесса, выступая в качестве
условного сигнала, или компонента центрального замыкательного механизма,
или фактора подкрепления, или, наконец, как полноправный компонент ассоциа¬
тивного процесса (рис. 58).
П. В. Симонов (1964, 1987) пытается объединить потребностио-мотивацион-
ный и информационный факторы генеза эмоций, понимая под эмоциями отраже¬
ние мозгом человека и животных какой-либо актуальной потребности (ее качест-
182 Глава 6. Факторы организации поведения
Эмоциональное состояние
\
Эмоциональное выражение
Моторные \ / Вегетативные
//
Проявления
Рис. 58. Схема возникновения эмоционального состояния
(по А. В. Вальдману, 1972)
ва, величины) и вероятности (возможности) ее удовлетворения, которую субъект
непроизвольно оценивает на основе генетического и ранее приобретенного инди¬
видуального опыта.
Правило возникновения эмоции представляется в виде структурной формулы
Э - П (ИН - ИС),
где Э — эмоция, ее степень, качество и знак; П — сила и качество актуальной по¬
требности; (ИН - ИС) — оценка вероятности удовлетворения потребности на
основе врожденного и приобретенного опыта; ИН — информация о средствах, не¬
обходимых для удовлетворения потребности; ИС — информация о существую¬
щих средствах, которыми реально располагает субъект.
Перечисленные переменные являются определяющими, необходимыми и до¬
статочными факторами для возникновения какой-либо эмоции. Но кроме них
следует иметь в виду также:
♦ индивидуально-типологические особенности субъекта;
♦ фактор времени, в зависимости от которого эмоция приобретает характер
кратковременного аффекта или длительного настроения;
♦ качественные особенности самой потребности.
Соответственно эмоции, порождаемые социальными и духовными потребно¬
стями, принято называть чувствами.
Из «формулы эмоций» следует, что низкая вероятность удовлетворения по¬
требности (ИН больше, чем ИС) ведет к возникновению отрицательных эмоций.
Возрастание вероятности достижения цели в сравнении с прогнозом (ИС больше,
чем ИН) порождает положительные эмоции.
Таким образом, отражательная функция эмоций совпадает с их оценочной
функцией, поскольку цена всегда есть функция двух факторов: спроса (потреб¬
ность) и предложения (возможность удовлетворить эту потребность). Эмоции
выступают в роли своеобразной «валюты мозга», универсальной меры ценностей,
а не просто эквивалента, функционирующего по принципу: вредно — полезно,
приятно — неприятно.
С физиологической точки зрения эмоция есть активное состояние систе¬
мы специализированных мозговых структур, побуждающее изменить поведение
в направлении минимизации или максимизации этого состояния.
§ 3. Роль эмоций в организации поведения 183
В этом заключается регулирующая функция эмоций. Регулирующая (переклю¬
чающая) функция эмоций особенно ярко обнаруживается при конкуренции моти¬
ваций, при выделении доминирующей потребности. Такова борьба между страхом
и чувством долга, между страхом и стыдом и нр. Управление своими эмоциями
предстает в качестве физиологических механизмов воли.
Своеобразной и весьма существенной функцией эмоций является их подкреп¬
ляющая функция, ибо сами эмоции могут выступать в качестве безусловного под¬
крепления. Было отмечено, что у крыс не удается выработать инструментальный
условный рефлекс, если пищу вводили непосредственно в желудок через канюлю.
Но этот же рефлекс легко вырабатывается, если к вводимой в желудок пище под¬
мешивается морфин, быстро вызывающий у животного положительное эмоцио¬
нальное состояние. Тот же горький на вкус морфин, введенный через рот, переста¬
вал быть подкрепляющим фактором пищевого рефлекса, ибо сопровождался
аверсией (С. Цитава, В. Трежипьер, 1976).
У мышей хорошо вырабатывается инструментальный условный рефлекс на¬
жатия на рычаг, если это действие подкрепляется появлением в клетке другой мы¬
ши, немедленно атакуемой «исполнителем».
Таким образом, ни афферептация из полости рта, ни голодовое возбуждение
сами по себе без одновременного сочетания с механизмом генераций положитель¬
ных эмоций не может выполнить функцию подкрепления при образовании инст¬
рументального условного рефлекса. Побуждающим и подкрепляющим фактором
может выступать эмоциональное состояние другой особи того же вида. В динами¬
ке социально-исторической эволюции это могло быть биологической основой
способности человека к сопереживанию.
Первую стадию образования любого условного рефлекса, названную стадией
генерализации, именуют также эмоциональной стадией. Именно в этот период на¬
блюдается гипермобилизация вегетативной сферы организма. Если упрочение
условного рефлекса сопровождается ослаблением эмоционального напряжения
и переходом к строго избирательному реагированию па условный сигнал, то воз¬
никновение эмоций ведет ко вторичной генерализации; расширяется диапазон из¬
влекаемых из памяти эиграмм и снижаются критерии принятия решения при со¬
поставлении этих энграмм с наличными стимулами. Например, чем сильнее тре¬
вога, тем сильнее отвечает субъект на индифферентный сигнал как на опасный.
Доминантное реагирование целесообразно в условиях прагматической неопреде¬
ленности. Способствующее доминантному реагированию эмоциональное напря¬
жение зависит от размеров дефицита прагматической информации. Положитель¬
ные эмоции как бы компенсируют недостаток неудовлетворенных потребностей
и прагматической неопределенности. Компенсаторное значение эмоций заключа¬
ется в их замещающей роли. В трудной ситуаций низкой вероятностью достиже¬
ния цели даже небольшой успех (возрастание вероятности) порождает положи¬
тельную эмоцию.
В отдельных случаях переход к имитационному поведению характерен для
эмоционально возбужденного мозга. Животное, не располагающее информацией
для адекватного поведенческого акта, извлекает ее из примеров поведения других
членов сообщества. Подражательное поведение — это пример компенсаторной
функции эмоций на популяционном уровне.
184 Глава 6. Факторы организации поведения
Имея в виду системную организацию аппарата эмоций, построенного по прин¬
ципу констелляции, П. В. Симонов особую роль отводит гиппокампу, гипоталаму¬
су, миндалине и фронтальным отделам неокортекса (рис. 59). Предполагается, что
в них представлены не отдельные потребности и эмоции, а операции, необходи¬
мые для генеза самых разных функциональных состояний.
Высоковероятные > Маловероятные
события ! события
Г\ /\ ,-''1
<3
<$
Неокортекс
'ч \
\ \
ч \
\ ч
Гипоталамус
/ /ч
/ /
/ /
/ /
/ /— Гиппокамп
Миндалина
£>
£>
Доминирующая | Субдоминантные
потребность 1 потребности
Рис. 59. Принципиальная схема участия мозговых
структур в генезе эмоций и в организации
целенаправленного поведения
(по П. В. Симонову, 1987)
Все стимулы, поступающие из внешней среды, подразделяются по принципу
вероятности: с высокой вероятностью их подкрепления и с низкой вероятностью
подкрепления. Гипоталамус участвует в самых ранних стадиях организации пове¬
дения и на более поздних его этапах, когда оформляется внешне реализуемый
эмоциональный ответ. Миндалина участвует в выборе поведения путем «взвеши¬
вания» конкурирующих эмоций, порожденных конкурирующими потребностя¬
ми. Она вовлекается в процесс организации поведения на сравнительно поздних
этапах, когда актуализированные потребности уже сопоставлены с возможностью
их удовлетворения и порождены соответствующие эмоциональные состояния.
Прогнозирование вероятности удовлетворения потребности осуществляется с
участием преимущественно «информационных» мозговых структур — гиппокам¬
па и фронтальных отделов неокортекса. Однако если гиппокамп необходим для
выполнения реакций на сигналы с низкой вероятностью подкрепления, то фрон¬
тальная кора важна для организации поведения на сигналы высоковероятных со¬
бытий.
Эмоциональный стресс. Под эмоциональным стрессом в самой общей форме
понимают общую системную реакцию организма, развивающуюся на действие
стресс-факторов (стрессоров). Оценка какого-либо воздействия, ситуации как не¬
гативных (отвергаемых) при невозможности избавления от них или неподготов¬
ленности механизмов «защиты» позволяет рассматривать эти факторы в качестве
стрессоров. При этом надо иметь в виду, что не само по себе воздействие, а имен¬
но отношение к нему, негативная его оценка с сенсорной, психологической или со¬
циальной точки зрения является причиной эмоционально-стрессовой реакции
организма (Вальдман и др., 1979). Главная сущность стресс-сигнала в его инфор¬
мативном значении, поэтому он оценивается в категории субъективного, личност¬
ного к нему отношения. Следовательно, типологические свойства субъекта, осо¬
§ 3. Роль эмоции в организации поведения 185
бенности эмоционального восприятия и эмоциональных свойств личности и опре¬
деляют тип реагирования и направленность поведения.
В развитии стресса различают три стадии:
1) стадия тревоги, возникающая при внезапном действии стрессора, проходит
в форме шока как результата сильнейшего эмоционального возбуждения;
2) стадия резистентности, подготавливающая организм к повышенной устойчи¬
вости к действию вредных факторов. Адаптация в основном осуществляется
системой «гипоталамус — кора надпочечников», так называемыми адаптивны¬
ми гормонами, и прежде всего аденокортикотропным гормоном;
3) стадия истощения, возникающая при продолжающемся воздействии стрессо¬
ров. Эта стадия еще именуется состоянием психической дезадаптации.
Приведем следующий пример из экспериментов с животными (М. А. Батуева,
1985). В одном ограниченном жилом помещении находятся два однопометных
кролика, один из которых характеризуется преобладанием пассивно-оборони¬
тельных реакций, а другой — реакциями активного напа¬
дения. Иными словами, имеет место типичный случай
эмоционально-психологической несовместимости. Изо¬
ляция животных друг от друга позволила у каждого жи¬
вотного выработать двигательно-пищевые рефлексы. По
критерию стабильности рефлексов и их количествен¬
ных параметров животные не отличались друг от друга.
В один из опытных дней животные были сведены вновь
в общее жилое помещение. Спустя три дня совместного
проживания у одного из кроликов (пассивного) исчезли условные рефлексы, то¬
гда как у другого (активного) все осталось по-прежнему. Еще через три дня живот¬
ные были вновь разведены в разные жилые клетки. Ежедневное тестирование по¬
казало постепенное в течение 6 дней медленное восстановление условных реф¬
лексов у пассивного кролика. В данном случае в качестве стрессора выступает
особь того же вида при наличии антагонистических взаимоотношений.
Максимальное напряжение механизмов защиты возникает в системе при праг¬
матической неопределенности, угрожающей ситуации, при невозможности осу¬
ществления готовых программ деятельности, при необходимости срочного подбо¬
ра новых вариантов поведенческих программ.
Наиболее реактивной из структур мозга является кора больших полушарий,
а в пей — митохондрии и синапсы, особенно тормозные, а в самой нервной клет¬
ке — ферментные системы энергетического и медиаторпого обмена.
Психотропные вещества, вмешиваясь в процессы «химической интеграции»
нейронов, могут менять информационное значение сигнала для данного индиви¬
да. Длительное и эффективное психофармакологическое лечение изменяет и тот
памятный след, который является основой психотравмирующей ситуации и ведет
к полному или частичному восстановлению нормальных психических функций.
Одним из эффективных терапевтических приемов является активизация соб¬
ственных компенсаторных механизмов мозга, направленных на устранение или
ограничение патогенных процессов (М. М. Хананашвили, 1972).
Эмоциональный
стресс — обшая си¬
стемная нейрогумо-
ральная реакция цело¬
стного организма,
развивающаяся на
действие стресс-фак¬
торов (стрессоров).
186 Глава 6. Факторы организации поведения
§ 4. Восприятие пространства
и пространственная ориентация
Теории рефлекторного отражения пространства. Ориентация животного во вре¬
мени и пространстве осуществляется с помощью головного мозга с его внешними
рецепторными аппаратами.
И. М. Сеченов особую роль в пространственном восприятии отводил двига¬
тельному аппарату и создаваемому им в головном мозгу «темному мышечному
чувству». В непрерывной связи чувствования и движения И. М. Сеченов видел
суть рефлекторного отражения пространства и времени. Особая роль мышечного
аппарата обусловливается тем, что он является одновременно и рабочим органом,
и периферическим отделом «чувствующего снаряда». Другим орудием И. М. Се¬
ченов считал те раздражения, которые поступают в головной мозг из внешней сре¬
ды благодаря деятельности сенсорных органов, как сочетанную и координирован¬
ную деятельность специальных форм чувствования между собой и с двигательны¬
ми реакциями организма.
В. М. Бехтерев (1884, 1896) также рассматривал механизм отражения про¬
странственных отношений как взаимосвязанную деятельность органов равнове¬
сия с внешней рецепцией и двигательным аппаратом. Существенное значение он
придавал кожио-мышечному чувству.
А. А. Ухтомский (1954) отмечал, что конкретное восприятие совершается все¬
гда в пространстве и времени нераздельно, в хронотопе. При зрительной оценке
предмета человек руководствуется не только зрительными и проприоцептивными
(от мышц глаза) импульсами, но и одновременными рецепциями со слухового,
вестибулярного и тактильно-проприоцептивного аппаратов.
Экспериментально установлено (Э. Ш. Айрапетьяиц, А. С. Батуев, 1963), что
пространственный анализ обеспечивается комплексом динамически увязанных
между собой анализаторов и среди них в особенности — зрительного, вестибуляр¬
ного, кожного и мышечного. Интеграция комплекса и его реинтеграция в случае
повреждения обеспечиваются при непременном участии высших ассоциативных
систем мозга.
Исключительность пространственно-различительной функции какой-либо од¬
ной сенсорной системы противоречит экспериментальным данным (Б. Г. Ананьев,
1961).
Концепции И. С. Бернташвили о пространственной ориентации и А. А. Ух¬
томского об интегральном образе. И. С. Бернташвили предложил концепцию
психонервного поведения, которое регулируется целостным «представлением»
о внешней среде или образом той внешней среды, в которой находится и действу¬
ет животное. В понятие образа среды входят все объекты внешнего мира, которые
имеют связь с биологически мотивированным состоянием животного. На осно¬
ве первого знакомства со средой животное вырабатывает определенные «пред¬
ставления» («гипотезы» по Я. Кречевскому) о ее организации и использует их
в форме поведенческих тактик взаимодействия со средой с целью проверки, кор¬
ректировки и дальнейшего их усовершенствования. Это составляет тот уровень
эвристической деятельности мозга в новых ситуациях, который прогрессивно раз¬
§ 4. Восприятие пространства и пространственная ориентация 187
вивается в ходе эволюции, и на его основе формируются жесткие условно-реф¬
лекторные программы автоматизированного поведения (Т. А. Натишвили, 1887).
А. А. Ухтомский (1924) разработал концепцию об интегральном образе. Про¬
стое ощущение, отмечал А. А. Ухтомский, есть в сущности абстракция, более или
менее полезная аналитическая фикция, тогда как реальный и живой опыт имеет
дело с интегральными образами. Всякий интегральный образ является продуктом
пережитой доминанты.
Итак, имея биологическую потребность в нище, животные и человек, одно¬
кратно столкнувшись с возможностью ее получения, сразу же формируют комп¬
лексный образ местонахождения пищи (И. С. Бериташвили). По А. А. Ухтомско¬
му, это первая стадия развития доминанты, когда она «привлекает» к себе самые
разнообразные внешние рецепции. По И. П. Павлову, это стадия генерализации
условного рефлекса, которая протекает по механизмам доминанты. Генерализа¬
ция условного рефлекса создает обобщенный пространственный образ среды, ко¬
торый при наличии высокого уровня мотивации фиксируется в памяти, то есть
формирует конкретное представление.
Образное поведение — это самый первый этап индивидуального обучения.
Дальнейшее развитие поведения может происходить двумя путями. Первый —
когда организм из множества пространственных факторов среды извлекает такие,
которые с наибольшей вероятностью приведут к удовлетворению текущей биоло¬
гической потребности. В этом случае формируется прочная временная связь меж¬
ду ограниченным набором сигналов и соответствующей поведенческой реакци¬
ей — вырабатываются стабильные специализированные условные рефлексы и ор¬
ганизм переходит на режим автоматического управления.
Но допустим и второй путь развития индивидуального поведения, детерми¬
нантой в котором остается тот же образ, но прочно зафиксированный в энграмме
памяти. Что побуждает организм формировать поведение по этому, второму пути,
который представляет собой стадию закрепившейся доми¬
нанты? Во-нервых, высокий уровень мотивации, а во-вто¬
рых, низкая вероятность ее удовлетворения. При одно¬
кратном подкреплении нищей очень голодного животного
организм не в состоянии оценить вероятность повторного
получения пищи в аналогичной ситуации. Однако в связи
с тем, что естественная среда чаще всего создает именно
такую ситуацию, то есть не может гарантировать высокую
вероятность удовлетворения потребности в постоянно
меняющейся обстановке, естественный отбор жестко зафиксировал в эволюции
свойство не только воспринимать комплексное воздействие окружающей среды,
по и быстро фиксировать его в виде целостного образа. В дальнейшем будет доста¬
точно лишь одного компонента, чтобы репродуцировать целый образ и направить
поведение соответственно этому образу.
Эти два крайних механизма поведения (которые моделируются в лаборатор¬
ных условиях) подразумевают в естественной среде и наличие соответствующих
механизмов. Они строятся на сложных взаимоотношениях обоих детерминант:
как вероятностной программы (то есть интегрального образа), создающей вектор
поведения, так и автоматизированных условных рефлексов, обеспечивающих
адаптивный характер реализации этой программы в знакомой пространственной
ситуации.
Интегральный об¬
раз — продукт пере¬
житых доминант,
то есть продукт субъ¬
ективного отражения
обьективного взаимо¬
отношения организма
со средой.
188 Глава 6. Факторы организации поведения
Рис. 60. Пары изображений пространственных объектов,
предъявляемые испытуемым в опытах по мысленному
вращению объектов (по Т. А. Натишвили, 1978)
Верхняя пара — идентичные объекты, повернутые относительно
друг друга в плоскости экрана; средняя — идентичные объекты,
повернутые относительно друг друга в трех измерениях; ниж¬
няя — зеркальные конфигурации одного и того же объекта. Ис¬
пытуемые должны установить, какие пары являются идентичными
изображениями одного и того же объекта, а в каких использова¬
ны зеркальные конфигурации данного объекта
Феномен мысленного вращения объектов. В психологических исследовани¬
ях на человеке было установлено (Шепард, 1924) наличие образного представ¬
ления внешнего пространства. В основу был положен феномен мысленного вра¬
щения. Испытуемым предъявлялись пары рисунков трехмерных объектов, раз¬
личающиеся своей ориентацией (рис. 60). Испытуемые должны были решить,
являются рисунки идентичными или зеркальными копиями.
Оказалось, что время реакции возрастает с увеличением угла, на который один
из элементов пары повернут относительно другого. Испытуемый производит мыс¬
ленное вращение одного из элементов пары до геометрического его совмещения
со вторым и на основании этого заключает об их идентичности или неидентично-
сти. Причем в процессе мысленного вращения сохраняется
значительная доля образной информации об объекте. Та¬
кое образное представление реальности можно также на¬
звать аналоговым представлением (Т. А. Натишвили, 1987).
Есть основания предполагать, что аналогичные мозго¬
вые процессы могут осуществляться и высшими млекопи¬
тающими. Шепард приводит следующее наблюдение. Ов¬
чарка доставала длинную палку, переброшенную через за¬
бор, в котором отсутствовала вертикальная доска.
Собака проникла в щель, схватила в зубы палку и, повернувшись, двинулась
назад к отверстию. В последний момент она вдруг остановилась и после некото¬
рой паузы повернула голову на 90°. Держа палку в таком положении, она свободно
пролезла через щель в заборе.
Концепция когнитивных карт. Под когнитивными (познавательными) карта¬
ми понимается процесс, благодаря которому организм приобретает некое подобие
топологической карты той местности, в которой он обитает. Когнитивная карта —
это динамический образ, способный к изменениям и уточнениям на основании
информации об изменении среды либо при изменении местонахождения в ней са¬
мого субъекта (Е. Толмен, 1948).
Д. С. Олтон (1979) разработал тесты, с помощью которых было показано, что
крысы способны запоминать точное местонахождение пищи в пространстве и на¬
правление к нему независимо от исходной стартовой позиции.
Один из широко распространенных приемов для изучения когнитивного кар¬
тирования состоит в использовании двенадцатилучевого радиального лабиринта
Когнитивные карты —
процесс, благодаря ко¬
торому организм при¬
обретает некое подо¬
бие топологической
карты той местности,
в которой он обитает.
§ 4. Восприятие пространства и пространственная ориентация 189
(рис. 61). Коридоры оборудованы входными и выходными дверцами, открываю¬
щимися в центробежном направлении. В конце каждого из них помещается пища.
Каждый выбор начинается с центра платформы, затем животное идет в один из
коридоров, опускается вниз, и цикл вновь повторяется. Повторное посещение ко¬
ридора не сопровождается пищевым вознаграждением. Получение наибольшего
количества пищи достигается одноразовым посещением всех 12 коридоров, то
есть поведением, строго соответствующим когнитивной пространственной карте.
Рис. 61. Схема двенадцатилучевого радиальносимметричного
лабиринта (по Е. А. Рябинской, 1987)
а — вертикальный разрез; 6 — вид сверху, цифры — номера коридоров
Наблюдение за поведением животных в таком лабиринте позволило устано¬
вить определенные закономерности, получившие название стратегий поведения
или гипотез. Концепция поведенческих стратегий или гипотез противостоит идее
о случайном переборе животным различных реакций (метод проб и ошибок).
Я. Кречевский (1932) допускал, что животное не поступает наугад, а формирует
и проверяет серии «гипотез» для решения задачи.
Наиболее обосновано представление о существовании трех основных, страте¬
гий решения животными задач пространственной ориентации. Первая — исполь¬
зование когнитивных карт местности, вторая — поиск опорных внешних ориенти¬
ров и третья — поиск на основе последовательности собственных двигательных
реакций. Первые две стратегии являются аллоцентрическими (стратегии места),
основанными на пространственной топологии среды. Третья стратегия получила
название эгоцентрической (стратегия ответа); она основана на отношении между
элементами пространства и положением и перемещением тела животного в этом
пространстве. Если поданные стратегии и существуют, то они постоянно сменяют
друг друга и часто выступают в смешанной форме даже в процессе реализации од¬
ной поведенческой задачи.
Анализ поведения крыс в двенадцатилучевом лабиринте выявил закономерно¬
сти, именуемые поведенческими тактиками (Е. А. Рябинская, 1987). Так, напри¬
мер, крысы посещают коридор, более удаленный от коридора, который они посе¬
190 Глава 6. Факторы организации поведения
щали до этого. Кроме того, крысы чаще выбирают коридор, расположенный спра¬
ва или слева по отношению к коридору, который они только что покинули.
Э. Хониг (1978) предлагает для анализа поведения в сложных пространствен¬
ных условиях выделять рабочую память (запоминание информации, необходи¬
мой только в пределах одного опыта) и референтную память (запоминание ин¬
формации, регулярно повторяющейся во всех опытах). И тот и другой виды про¬
странственной памяти подразумевают участие обоих видов стратегий поведения,
аллоцентрической и эгоцентрической. Участие собственно двигательного ком¬
понента весьма велико, хотя животные способны прогнозировать возможные по¬
следствия самых различных движений, даже таких, которые имеются лишь
«в плане», но еще никогда не выполнялись (Т. А. Натишвили, 1987).
Нейрофизиологические корреляты пространственного восприятия. Специ¬
альными опытами на крысах было показано, что их поведение в радиальном лаби¬
ринте зависит от целостности гиппокампальной системы. Ее нарушение не приво¬
дит к дефициту аллоцентрической пространственной локализации, но приводит
к дефициту оперативной памяти, по которой отбираются коридоры лабиринта.
Заднетеменная кора представляет собой часть нервного аппарата, специализи¬
рованного для анализа и запоминания взаимного пространственного расположе¬
ния предметов.
Интересная картина выявляется при оценке активности нейронов гиппокам¬
па при ориентации крыс в коридорах радиального лабиринта (О. Кифи, 1976;
Дж. (Элтон, 1987). Отдельные нейроны гиппокампа возбуждались при нахожде¬
нии животного в определенных лучах лабиринта (рис. 62).
Рис. 62. Поля нейронов в гиппокампе у крыс (по А. И. Францевичу, 1 986)
а — участки лабиринта, в котором активируются определенные нейроны (заштрихованы) (О. Кифи,
1976); 6 — план восьмилучевого лабиринта; в-е — диаграммы активности отдельных нейронов в лу¬
чах лабиринта: окружность — уровень фоновой активности (Дж. Олтон, 1978)
Некоторые нейроны возбуждались особенно сильно, когда крыса находилась
в специфическом участке и в определенной ориентации относительно лабиринта
(рис. 63). Поворот лабиринта в системе внешних ориентиров не менял специфич¬
ности «нейронов места» в отношении идентифицируемых участков лабиринта.
На активность нейрона не влияло ни отвлечение внимания животного, ни голод¬
ное или сытое его состояние, ни тип поведения животного в этом участке лаби¬
ринта. Такие свойства клеток формируются очень быстро — не позднее чем через
10 мин после попадания животного в новую обстановку. Раз сформировавшись,
подобные клетки оказываются весьма стабильными в отношении собственного
§ 5. Время как фактор организации поведения 191
пространственного рецептивного поля. По-видимому, нейроны гиппокампа уча¬
ствуют в узнавании места и соотнесении его с координатами какой-то пространст¬
венной карты на основе аллоцентрической системы пространственной локализа¬
ции, то есть опять же в реализации оперативной памяти.
———— I <IH"m t
I 5 II 3 1 I 2 1
—,-♦■■»■4 ■■ i.. ..j|- ■ м< .»..i
| 1 1 | 4 1
— — —^ ■■ ■.■■■jı.tl" I ! i- — . ■■ .
I 5 1 ' 6 1
■4 ■..— ..»■■■■ i ı Hi Nı>и mİ \ ıMiıtt-l I
I 7 1 1 8 1 I 3 1
h M
ı 2 ' ı 4 1 I 5 1
Рис. 63. Активность нейрона дорзального гиппокампа крысы при
побежках по коридорам радиального лабиринта (по Дж. (Элтону, 1977)
Осииллографическая запись активности одиночной клетки, регистрируемой металлическим микро¬
электродом. Уровень активности клетки строго коррелирует с местонахождением животного в про¬
странстве. Импульсация клетки низка при нахождении животного в коридорах 1,4, 5, 6, 7 и 8, высока
при местонахождении животного в коридорах 2 и 3. Тем самым рецептивное поле данной клетки раз¬
деляет весь радиальный лабиринт на небольшое оп-поле в значительно большее off-поле
В исследовании на овцах с регистрацией активности нейронов нижневисочной
коры было обнаружено восприятие зрительного изображения головы представи¬
телей своего же вида, но снабженных рогами (К. Кендрик, Б. Болдвин, 1987)
(рис. 64). Из этих опытов делается вывод об участии нижневисочной коры в вос¬
приятии целостных пространственных образов, имеющих отношение к социаль¬
ному поведению животных.
Предполагают существование у млекопитающих мозговой системы, выпол¬
няющей функцию пространственной ориентации (Л. И. Францевич, 1986): ниж¬
невисочная кора — узнавание зрительных ориентиров, гиппокамп — узнавание
топологии ориентиров в пространстве, теменная кора — установление взаимного
расположения ориентиров, префронтальная кора или ее аналоги — запоминание
и расчет действий в установленной системе координат тела.
§ 5. Время как фактор организации повеления
Адаптивное значение суточных ритмов. Цикл сон—бодрствование. Вся жизне¬
деятельность организмов, обитающих на Земле, определяется ритмическими про¬
цессами, колебаниями физиологической активности систем, составляющих жи¬
вой организм.
Цикличность всех природных процессов связана с вращением Земли вокруг
собственной оси, что и определяет смену дня и ночи, то есть суточную периодич¬
ность, суточные или циркадные ритмы. Применительно к животному организму
циркадный ритм характеризуется закономерной сменой интенсивности физиоло-
192 Глава 6. Факторы организации поведения
ш
5
X
О
е
1
2
3
4
5
6
7
'^■-нит-н
Без рогов
100 мкВ [ ,
1 сек
Рис. 64. Ответы нижневисочной области коры у овцы,
отвечаюшие на предъявления изображений
(по К. М. Кендрику и Б. А. Болдвину, 1987)
Приведены ответы одиночной клетки на последовательное предъявление разных
стимулов. Момент предъявления обозначен черной горизонтальной линией.
Видно, что нейрон реагирует резким усилением импульсной активности на изо¬
бражения, в которых присутствуют рога. Величина ответа зависит от величины
рогов
§ 5. Время как фактор организации поведения 193
гических обменных процессов и поведения в целом. Таким образом, жизнедея¬
тельность организма подчиняется внешним часам, внешним датчикам времени.
Большинство млекопитающих не рождаются с готовым суточным ритмом,
а постепенно приспосабливаются к нему, то есть вырабатывают собственный
внутренний ритм. Чем быстрее и полнее животное сформирует свой внутренний
ритм, тем оно имеет больше шансов в борьбе за существование.
Из внешних факторов, которые формируют циркадную ритмику, на первое ме¬
сто ставят уровень освещенности и связанные с ним другие виды сигнализаций,
влияющие более всего на деятельность нервных центров,
регулирующие состояние гормональной сферы и интен¬
сивность обмена веществ.
Внутренние циркадные ритмы («биологические ча¬
сы») могут быть очень устойчивыми и поддерживаться
без непосредственного периодического влияния на орга¬
низм факторов внешней среды. Например, если отделить
летучую мышь (животное с четко выраженным ночным
образом жизни) от остального стада и поместить в изолированную камеру, то в ча¬
сы активности всего стада в пещере у нее можно наблюдать резкое усиление обме¬
на веществ, повышение температуры тела, учащение дыхания, общее повышение
возбудимости (А. Д. Слоним, 1970).
Таким образом, реально существует внутренний суточный ритм организма,
а смена сна и бодрствования — это одно из его проявлений.
Сон — это адаптация, проявляющаяся в подавлении активности в период наи¬
меньшей доступности нищи, угрозы резких колебаний внешних условий (темпе¬
ратуры и т. д.) и максимальной опасности со стороны хищников. Выделяют хоро¬
шо и долго спящих животных, у которых мало врагов или которые изготовляют
безопасные убежища (макаки, хищники, суслики). С дру¬
гой стороны, существуют мало спящие животные с мень¬
шей длительностью парадоксального сна. Это объекты,
которые часто служат добычей другим хищникам, ибо не
имеют безопасных убежищ (морские свинки, кролики, ко¬
пытные).
Наряду с таким объяснением ряд исследователей при¬
знает восстановительную функцию сна: сон как бы снижа¬
ет ежедневные метаболические затраты, навязывая орга¬
низму отдых, являясь выражением внутренней инстинк¬
тивной потребности (Н. И. Моисеева, 1986).
Существует ли сон у беспозвоночных, сказать трудно, хотя их активность ис¬
пытывает определенные колебания и периодически возникают состояния, похо¬
жие на сои. В подобных случаях правильно говорить о смене периодов активности
и покоя.
Сон животных бывает двух типов: монофазный — с однократным чередовани¬
ем дневного или ночного сна и периода бодрствования и полифазный — с частыми
сменами периодов сна и бодрствования в ночные и дневные часы.
У некоторых видов животных трудно выделить определенный период сна.
Ученые стали сомневаться, что обязательным внешним проявлением сна следует
считать неподвижность животных. Например, акулы всю жизнь находятся в дви-
Сон — это адаптация,
проявляющаяся в по¬
давлении активности
в период наименьшей
доступности пиши,
угрозы резких колеба¬
ний внешних условий
и максимальной опас¬
ности со стороны хищ¬
ников.
Биологические рит¬
мы —- ритмические
(периодические) коле¬
бания физиологиче¬
ских и психических
процессов (биоритмо¬
логия).
7 Заг 1078
194 Глава 6. Факторы организации поведения
жении, а дельфины вынуждены подниматься па поверхность каждые полторы ми¬
нуты для забора воздуха. Жвачные животные во время сна держат голову прямо
у них открыты глаза, и они продолжают жевать жвачку.
Эволюция сна. Весь период спа по показаниям ЭЭГ разделен на два сменяю¬
щих друг друга и резко различающихся состояния, которые именуются медленно-
волновым, медленным (ортодоксальным) сном и быстроволновым, быстрым, или
парадоксальным, сном.
В эволюции позвоночных выделяют три этапа в формировании цикла «бодрст¬
вование-сон» (И. Г. Карманова, 1977).
1этап — у рыб и амфибий обнаружены формы покоя как первичные адаптации
к дневной и ночной освещенности. Дневную и ночную формы покоя у этих живот¬
ных рассматривают как сноподобные состояния, или первичный сон. Эти формы
покоя у рыб и амфибий выполняют, вероятно, функцию защиты и пассивного от¬
дыха.
II этап связан с заменой первичной формы спа качественно новой формой по¬
коя, именуемой промежуточным сном. Этот этап характерен для рептилий и реп-
тилеподобных млекопитающих, медленноволиовой сон которых лишен еще пара¬
доксальной стадии.
III этап состоит в усложнении структуры спа у птиц и млекопитающих. Воз¬
никновение гомойотермности, значительное развитие головного мозга создали
предпосылки для очередного приобретения в виде появления двустадийного сна.
медленного и парадоксального.
Наиболее древним признаком сна являются частота сердечного ритма, тин ды¬
хания и характер тонуса соматических мышц. Биоэлектрические показатели мед-
ленноволповой и парадоксальной стадий спа возникают гораздо позднее в эво¬
люции.
Исследователи полагают, что медленноволиовой и парадоксальный сон — два
совершенно самостоятельных состояния. Парадоксальная фаза сна — это аналог
закрепившегося в эволюции состояния сторожевого бодрствования. Возбуди¬
мость мезэнцефалической ретикулярной формации в стадии парадоксального сна
оказывается повышенной по сравнению с медленным сном (Т. Н. Ониани, 1974).
Теории сна. Природа сна как наиболее загадочного состояния организма инте¬
ресовала ученых с древних времен.
Ранее других сформировалась сосудистая (циркуляторная, или гемодинами¬
ческая) теория сна. Причем одни ученые связывала наступление сна со снижени¬
ем кровотока в мозге, другие — с его усиленным кровенаполнением. Современные
исследования показали, что в течение сна действительно происходит колебание
кровенаполнения мозга.
В конце XIX в. возникла гистологическая теория сна, согласно которой чувст¬
вительные нейроны во время сна втягивают свои окончания и таким образом как
бы предотвращают воздействие внешнего мира на организм. В прямом смысле эта
гипотеза не подтвердилась.
Наиболее распространены гуморальные теории сна. Началось с поисков ве¬
ществ, накапливающихся во время деятельности организма: обеднение клеток ки¬
слородом, накопление диоксида углерода, карбоновых, молочных кислот, холе¬
стерина. Р. Лежандр и X. Пьерон (1910) считали, что сон возникает в результате
накопления токсических продуктов обмена, появляющихся вследствие утомле-
§ 5. Время как фактор организации поведения 195
иия (гипотоксины). Собакам долгое время не давали спать, а затем забивали, экст¬
рагировали вещества из мозга и вводили другим собакам. У последних развива¬
лись признаки крайнего утомления и возникал глубокий сон. То же наблюдалось
при «переносе» сыворотки крови или спинномозговой жидкости (ликвор).
В 1965 г. Ж. Монье использовал препарат с перекрестным кровообращением у
двух кроликов, то есть кровь от мозга одного кролика попадала в туловище друго¬
го и наоборот. Если у одного из кроликов раздражали участки мозга, вызывающие
сон, то второй кролик тоже засыпал. Это можно было бы объяснить переносом ка¬
ких-то «веществ спа». Действительно, во время сна в мозгу обнаружено избыточ¬
ное накопление ряда биологически активных веществ — ацетилхрлина, гамма-
аминомасляной кислоты, серотонина. Исследования в этом направлении активно
продолжаются.
Однако гуморальной теории сна противоречат наблюдения над сросшими¬
ся близнецами, обладающими общим кровообращением, но с раздельной цент¬
ральной нервной системой. Одно из таких наблюдений проведено в Москве
(Т. Т. Алексеева, 1959): сон у близнецов наступал неодновременно — одна голова
спит, а другая бодрствует.
Другое наблюдение проведено в Ленинграде (В. М. Угрюмое и др., 1964) на
близнецах-краниопагах. Авторы отмечают: если один ребенок успевал заснуть,
а другому заснуть помешали, то сон у него длительно не наступал, и он успевал
и поиграть и поплакать; другой ребенок продолжал спать. На 3-м году жизни дети
четко выражали отношение к предстоящему сну. После обеда один из малышей
охотно бы шел спать, а другой сопротивлялся и кричал: «Не хочу в комнату, не хо¬
чу спать». Однако во время сна одного из них другой вел себя тихо, как бы щадя
сон брата. Такие наблюдения сильно пошатнули гуморальную теорию сна, но от¬
вергать ее полностью все же казалось преждевременным.
Уже в середине XIX в. все большее распространение имеют нервные теории
сна.
Один путь исследований — это изучение так называемого центра сна, о сущест¬
вовании которого в ядрах гипоталамуса у животных говорил швейцарский физио¬
лог В. Гесс (1933). У больных с пораженным гипоталамусом также отмечалась по¬
вышенная сонливость: у раненого солдата осколок снаряда находился на уровне
гипоталамуса, а попытка извлечь осколок пинцетом вызывала мгновенный глубо¬
кий сон.
Наблюдения и эксперименты подобного рода пытаются объяснить и с другой
точки зрения, а именно: сои наступает в результате нарушения проведения ин¬
формации из внутренней среды, в чем решающую роль отводят именно гипотала¬
мусу.
И. П. Павлов не соглашался со взглядами Гесса о подкорковой природе сна, от¬
давая предпочтение корковым механизмам. Он рассматривал сон как следствие
иррадиации торможения, распространяющегося по коре больших полушарий,
и находил много общего между активным условно-рефлекторным сном и услов¬
ным внутренним торможением.
Сегодня с несомненностью установлена корково-подкорковая локализация
структур управления сном. П. К. Анохин (1945) считал, что лобные доли коры
мозга, заторможенные во сне, высвобождают от своего непрерывного тормозного
контроля гипоталамические структуры. Последние, активные в течение сна, бло¬
196 Глава 6. Факторы организации поведения
кируют прохождение импульсов через таламус, вызывая своеобразную функцио¬
нальную деафферентацию коры больших полушарий.
Другие ученые полагают, что блокада активирующей системы и усиление син¬
хронизирующих воздействий на кору от таламуса и ствола могут рассматриваться
как решающее условие для наступления сна.
Все большее внимание начинает привлекать так называемая информационная
теория сна (Н. Винер). Согласно этой теории, в течение дня мозг накапливает
огромную информацию, усвоение которой затруднено, а часть ее не имеет отноше¬
ния к долговременным задачам. Если кратковременная память заполняется днем,
то ночью часть содержащейся в ней информации переписывается в долговремен¬
ную память. Особенности процессов переработки информации требуют отключе¬
ния от сигналов внешнего мира. Эта теория хорошо объясняет особенности орга¬
низации психических процессов человека во время сна (см. гл. 9), хотя и остав¬
ляет без объяснения ряд противоречащих ей явлений. Исходя из этих позиций
трудно объяснить, например, почему у новорожденных детей существенно преоб¬
ладает парадоксальная стадия сна.
Структура сна высших млекопитающих. Сонное состояние мозга характери¬
зуется возникновением в ЭЭГ «сонных веретен» (12-16 колебаний в 1 с) и син¬
хронизированными крупными медленными волнами ЭЭГ в дельта-диапазоне. Та¬
кая стадия сна и получила название медленноволнового или ортодоксального спа.
Это состояние мозга периодически в течение ночи заменяется быстрой низкоам¬
плитудной десинхронизированной активностью (до 30 колебаний в 1 с), которая
напоминает ЭЭГ человека и животных во время бодрствования. Так как при этом
сои не прерывается, а по некоторым показателям становится даже более глубо¬
ким, то эта фаза спа в отличие от предыдущей получила наименование парадок¬
сального, или быстрого, сна. Смена быстрого и медленного спа происходит через
равные промежутки времени со средней длительностью 90 мин.
Парадоксальный сон ярко представлен у новорожденных, и лишь спустя не¬
сколько дней возникают симптомы медленного сна и устанавливается периодич¬
ность смены этих двух состояний мозга па протяжении ночи. Одной из характер¬
ных черт быстрого сна является возникновение быстрых движений глаз (рис. 65).
Одновременно с этим возникают изменения в скелетно-мышечной системе. Тонус
мышц особенно сильно снижается в период быстрого сна. На этом фоне возника¬
ют различные движения усов, ушей, хвоста, подергивания лап, лизательные и со¬
сательные движения и т. д.
Различия между быстрым и медленным сном отчетливо видны но анализу ве¬
гетативных функций. Так, в период медленного сна наблюдается урежение дыха¬
ния, пульса, снижение артериального давления, общие движения туловища, а при
быстром сне возникает «вегетативная буря» — учащается и становится нерегуляр¬
ным дыхание, возникает неритмичный и частый пульс, повышается артериальное
давление, усиливается гормональная активность. Весьма существенно, что при
этом активность мотонейронов спинного мозга резко заторможена.
Несомненно, что быстрый сон — это совершенно особое, но сравнению с мед¬
ленным сном, состояние мозга.
Лишение животных парадоксального сна делает их возбудимыми, раздражи¬
тельными. Это приводит впоследствии к компенсаторному увеличению продол¬
жительности парадоксального сна и «вхождению» в него, минуя обычную проме¬
жуточную стадию медлепноволиового сна.
§ 5. Время как фактор организации поведения 197
Глубина сна ^
2
Время, ч 1
Движения глаз
Движения тела
т
Г'Т" I Г”1—I—Г—I—|”Т I ! I I II!
0 12 3 4
И1«М1 шиим
II 111Н111111 1111111 I II II III
Т Г-1 I I I I I I Г~|
5 6 7
11111111111
II II II I 1111 I III
Рис. 65. Механизмы организации сна (по Г. Адаму, 1983)
Внизу — «центры» сна (гипногенные центры) в стволе мозга кошки: участки с вертикальной штри¬
ховкой — ядра, связанные со сном без сновидений; зачерненный участок соответствует располо¬
жению нейронов, контролирующих парадоксальный сон; вверху — ритмическое чередование дви¬
жений глаз и тела в течение семи часов сна, включаюших три фазы быстрых движений глаз (СВ —
сновидения)
1 — гипофиз; 2 — гипоталамус; 3 — мост; 4 — продолговатый мозг
Известная независимость быстрого и медленного сна подтверждается и разли¬
чиями в их физиологической и химической природе. Установлено, что структуры,
ответственные за медленный сон, находятся в каудальной части мозгового ствола,
главным образом — в продолговатом мозге. Наличие сходных гипногенных струк¬
тур было установлено также и в задней части моста.
В других исследованиях показано, что поведенческие и ЭЭГ-проявления пара¬
доксальной стадии сна связаны с активацией структур в области моста. Это пре¬
жде всего зона голубого пятна, а также ретикулярные и вестибулярные ядра.
В стадии быстрого сна возникают в гиппокампе периоды тега-ритма, которые свя¬
зывают с вегетативными сдвигами и движениями глаз (Т. Н. Ониаии и др., 1970).
М. Жуве (1967) развивает гипотезу о моносшинергической регуляции стадий
сна. По его представлениям, медленноволповый сои регулируется серотонинерги-
ческой системой ядер шва в продолговатом мозге (рис. 66, б ), ибо им установлена
прямая зависимость сна от функциональной активности этих мозговых образова¬
ний и общего уровня серотонина и его обмена. Катехоламипергичеекие нейроны
голубого пятна и ретикулярной формации среднего мозга выступают как антаго¬
нистическая система, ответственная за возникновение быстрого сна и состояния
бодрствования (рис. 66, а, б).
Существуют указания па особую роль глиальных элементов во время сна.
Н. Н. Демин и другие (1978) полагают, что одной из основных нейрохимических
функций медленного сна являются пластические репарационные процессы, свя¬
занные с метаболизмом белков и РНК, причем главным образом в глиальных
клетках определенных структур мозга. Активные конформационные изменения
198 Глава 6. Факторы организации поведения
в мембранах нейронов при парадоксальном сне предназначены для ликвидации
так называемой усталости синаптического проведения и являются одной из при¬
чин своеобразия ЭЭГ при данном функциональном состоянии.
Биологическое значение быстрого сна продолжает оставаться неразрешенной
проблемой. Это видно даже из краткого перечня возможных функций этого фи¬
зиологического состояния (С. Хартман, 1967):
♦ нейтрализация токсических веществ, накопившихся во время деятельности;
♦ генерация квазисенсорных импульсов, стимулирующих развитие мозга;
♦ восстановление функциональной организации рабочих механизмов мозга,
угнетенных в течение медленного сна;
♦ подготовка к переходу в состояние бодрствования;
♦ освобождение от накопившейся необработанной информации;
♦ передача сообщения из оперативной памяти в долгосрочную;
♦ организация сновидений и текущей адаптации организма к условиям среды.
Мышечный
тонус
Быстрые
корковые
волны
Мост
Ретикулярная
_ активирующая
Движение система
глаз
Мышечный
глаз
Рис. 66. Схематическое представление
о нейрофизиологии и нейрохимии сна
(по М. Жуве, 1967)
а — бодрствование; 6 — медленноволновая стадия
сна; в — парадоксальный сон
Сезонные ритмы поведения. На¬
ряду с циркадными ритмами поведе¬
ние живых организмов подчинено так¬
же сезонным колебаниям окружающей
среды, при которых изменяются усло¬
вия питания, температуры, влажности,
солнечного режима и т. д. Наиболее
известные формы сезонного поведе¬
ния проявляются в миграциях и ко¬
чевках, в явлениях зимней и летней
спячки, а также в разнообразной дея¬
тельности по постройке гнезд и убе¬
жищ. Главными факторами при этом
являются режим освещения и темпе¬
ратура, а в качестве комплекса ответ¬
ных реакций выступает изменение гор¬
мональной активности, осуществляю¬
щей запуск репродуктивного цикла.
Сезонная периодика поведения
оказывается чрезвычайно устойчи¬
вой. Так, например, было отмечено,
что австралийский страус в заповед¬
нике Аскания-Нова откладывает яй¬
ца в условиях нашей зимы, несмотря
па сильный мороз, прямо в снег в се¬
зон, соответствующий лету в Австра¬
лии (М. М. Завадовский, 1930).
Миграционная деятельность жи¬
вотных приурочена к определенным
сезонам года. Перелет птиц, миграция
§ 5. Время как фактор организации поведения 199
рыб, крупных млекопитающих связаны прежде всего с сезонными изменениями
условий питания и характеризуются всеми чертами врожденного поведения, ко¬
торому присущи некоторые общие черты, такие как изменения стадных отноше¬
ний, изменения в функции нейросекреторной системы, изменения характера тер¬
морегуляции, накопление жира в предмиграциопный период.
Понижение температуры среды рассматривают в качестве одного из внешних
сигнальных факторов для миграции птиц. Само явление разделяется на два пе¬
риода:
♦ стимуляция и поддержание собственно миграционной активности;
♦ ориентация птиц при перелете.
В. Р. Дольник предполагает, что и суточные, и сезонные ритмы имеют иод со¬
бой эндогенную основу и являются проявлением функционирования «физиоло¬
гических часов» организма. Отсчет времени в случае сезонных ритмов опирается
прежде всего на закономерные колебания активности гипоталамогипофизарной
системы, хотя детали этих механизмов остаются неясными.
Самостоятельный интерес представляет собой проблема пространственной
ориентации птиц во время перелетов. Хотя сами по себе навигационные механиз¬
мы птиц по сей день остаются загадкой, ясно, что для навигации на большие рас¬
стояния птицы используют так называемые глобальные ориентирующие поля.
Иными словами, птицы обладают способностью к сопоставлению координат ме¬
стности и координат направления (цели) перелета. Л. И. Францевич (1986) под
глобальной навигацией понимает регулирование с обратной связью в глобальных
ориентирующих полях и такую организацию своего движения, которое стремится
к уменьшению рассогласования между текущим и планируемым (эталонным)
значениями измеряемых параметров. В случаях перелетов птиц в дневное время
суток естественно предположить, что главным ориентиром является положение
солнца. Однако многие птицы, например мелкие воробьиные, перелетают ночью
и, вероятнее всего, ориентируются по картине звездного неба. В. Р. Дольник
(1981) высказал гипотезу о том, что птица запоминает картину расположения на
небе нескольких знакомых звезд в данное время суток и в новом месте стремится
повернуть голову так, чтобы в поле зрения установилась картина, сходная с эта¬
лоном. Изменение позы в сравнении с привычной для рассматривания нужных
звезд указывает направление корректирующего полета — лететь надо в ту сторону,
куда отклоняется голова. Эта же гипотеза может быть применима для понимания
ориентации по солнцу.
Помимо общебиологического значения проблемы механизмов ориентации
птиц при перелетах ее разработка принципиально важна для формулирования
теории пространственной ориентации животных.
В особую группу адаптации следует отнести сезонную цикличность периодов
спа и бодрствования, которая проявляется чаще всего в хорошо известных явле¬
ниях зимней и летней спячки. Хотя явления спячки и ночного спа животных но не¬
которым внешним признакам сходны, отмечаются и существенные различия. Ос¬
новное состоит в том, что ведущим фактором наступления зимней и летней спяч¬
ки оказывается не освещенность, а окружающая температура, которая влечет за
собой и снижение температуры тела всего организма. Это снижение может быть
настолько большим, что практически температура тела в спячке становится оди-
200 Глава 6. Факторы организации поведения
паковой с температурой среды. При этом животное принимает позу сна, выключа¬
ются высшие отделы центральной нервной системы, но усиливаются некоторые
рефлексы, регулируемые продолговатым мозгом. Резко снижается обмен веществ,
замедляются дыхание, сердечная деятельность, изменяются все циклы метаболи¬
ческих преобразований (А. Д. Слоним, 1970). Отмечено, что у летучих мышей, на¬
пример, в период зимней спячки сохраняется циркадный ритм обмена веществ.
Выработанные до спячки условные рефлексы полностью воспроизводятся и по¬
сле пробуждения, что свидетельствует о сохранении процессов долговременной
памяти в период зимней спячки.
Летняя спячка, практически совершенно неизученная, характеризуется теми
же признаками, что и зимняя, но наступает у животных — обитателей пустынь и
полупустынь в засушливое время года при наступлении неблагоприятных усло¬
вий для питания.
Развитие в последнее время биоритмологии с убедительностью показало суще¬
ствование специального механизма биологических часов, обеспечивающих про¬
явление циклических изменений в организме. Врожденным элементом этого ме¬
ханизма является способность живой системы концентрировать во времени дея¬
тельность и покой, различные уровни и формы проявления жизнедеятельности.
Однако в основе ориентации живых организмов в среде с ритмически изменяю¬
щимися параметрами лежит формирование стойких форм обучения.
Поведение в микроинтервалах времени. Одной из выраженных тенденций
адаптивного поведения является его стереотипизация, то есть регулярная повто¬
ряемость поведенческих актов, в основе которой лежит минимизация расходова¬
ния энергии при достижении определенной биологической потребности.
Выработка поведенческих стереотипов в процессе индивидуального обучения
особи базируется на системе условных рефлексов на время. В результате орга¬
низм приспосабливается к моменту возникновения определенного (ожидаемого)
события. Любой условный сигнал содержит не только информацию о том, какое
предстоит событие, но и когда оно наступит. Но посколь¬
ку в естественной среде обитания какое-либо событие не
возникает изолированно, а «вписано» в достаточно слож¬
ный континуум явлений окружающей обстановки, то
у животного формируется не одиночная рефлекторная
реакция, а их последовательная цепь — поведенческий
стереотип. И. П. Павлов, изучив это явление в лаборатор¬
ном эксперименте, назвал его динамическим стереоти¬
пом. Он установил, что в основе динамического стереоти¬
па лежит сложная система временных связей, в каждом зве¬
не которого следы от предшествующего раздражителя
могут быть использованы в качестве сигнала о вероятном
возникновении следующего раздражителя. Жесткая вре¬
менная последовательность раздражений, использованная в опытах но выработке
динамического стереотипа, отразилась и в характере следовых процессов, и в сте¬
пени их выраженности. Временные связи между элементами стереотипа были
достаточно прочными, а переделка стереотипа в целом (замена одного из сигналов
другим или исключение какого-либо компонента) требовала длительной трени¬
ровки, в итоге которой формировался новый стереотип.
Стереотип — единый
зафиксированный
комплекс поведенче¬
ских реакций, связан¬
ный под влиянием мно¬
гократных повторений
определенной последо¬
вательности раздражи¬
телей в единую цепь,
которая воспроизво¬
дится любым из раз¬
дражителей данной
системы.
Заключение 201
Тот факт, что следы нервных процессов могут выступать в качестве самостоя¬
тельного сигнального фактора, отчетливо был продемонстрирован в опытах с
условными рефлексами на «чистое время». Например, крыса обучается избегать
электрического удара через контакты в полу камер путем перехода через дверь
в аналогичную соседнюю камеру. Но в этой соседней камере спустя 2 мин через та¬
кой же электронол подается ток, заставляющий животное вернуться обратно. Та¬
кие «челночные» движения животное первое время обучается производить через
2 мин, а спустя некоторое число сочетаний крыса переходит в «безопасную» каме¬
ру, не ожидая действия тока, а за 5-10 с до его включения. Следовательно, у жи¬
вотного теперь выработан временной стереотип поведения, где в качестве услов¬
ного сигнала двигательной реакции выступает время, то есть следы возбуждения,
сформированные в ее памяти. Такие эксперименты, сопровождаемые одновремен¬
ной регистрацией электрических процессов мозга, показали существование эн¬
догенных, собственно нервных факторов организации поведения в микроинтер¬
валах времени. Они могут служить физиологическим основанием для изучения
у человека и животных «чувства времени», то есть приспособления своего поведе¬
ния к временным сдвигам окружающего пространства.
Заключение
Среди факторов, ответственных за организацию конкретного поведения, выделя¬
ют генетические, субъективные внутренние (мотивации и эмоция) и объективные
внешние (пространство и время). Генотип определяет потенциальные возможно¬
сти организма, которые реализуются во взаимодействии с внешней средой (фено¬
тип). Диапазон изменчивости организма задается генотипом и составляет норму
реакции.
Изучение генетики поведения показало возможность генетического контроля
над многими свойствами нервной системы (возбудимость, обучаемость), появле¬
ние нервно-психических заболеваний наследственной природы, возникновение
анатомических особенностей конструкции мозга, специфику метаболизма и ней¬
роэндокринной регуляции.
Мотивация рассматривается в качестве основного системообразующего фак¬
тора организации целенаправленного поведения. Биологические мотивации бази¬
руются на возникновении в гипоталамических структурах очага стационарного
возбуждения гетерохимической природы с доминантными свойствами и вторич¬
ных очагов, в том числе кортикальной и лимбической локализации, которые в со¬
вокупности образуют доминирующую констелляцию мозговых «центров».
Эмоции выступают в качестве основного фактора оценки результативности
планируемого или выполненного поведенческого акта и подразделяются на поло¬
жительные и отрицательные. Различают эмоциональные состояния (истинные
эмоции) и эмоциональные проявления, которые могут не всегда совпадать. Эмо¬
ции выполняют также отражательную, подкрепляющую и компенсаторную функ¬
ции. К мозговым аппаратам эмоций относят лимбическую систему и фронталь¬
ный неокортекс.
Организм воспринимает пространство и ориентируется в нем благодаря созда¬
нию «интегральных образов пространства» или «когнитивных карт пространст¬
ва», которым соответствуют стратегия и тактика поведения.
202 Глава 6. Факторы организации поведения
Организация поведения в макроинтервалах времени подчиняется прежде все¬
го суточным (циркадным) ритмам, которым соответствует цикл «бодрствова¬
ние — сон», а также сезонным ритмам, которые определяют зимнюю или летнюю
спячки, миграционную активность, репродуктивную деятельность. Эти формы
поведения контролируются структурами гипоталамуса. Поведение в микроинтер¬
валах времени имеет механизмы, ответственные за формирование «чувства вре¬
мени», стереотипии автоматизированного поведения и других форм адаптации.
Контрольные вопросы
1. Чем отличается генотип от фенотипа?
2. Нервно-психические заболевания генетической природы.
3. Генетическая детерминация свойств нервных процессов.
4. Генетика обучаемости.
5. Нейроэндокринная регуляция генетической информации.
6. Основные физиологические мотивации.
7. Теории физиологических мотиваций.
8. Доминирующая мотивация и поведение.
9. Что такое эмоции?
10. Чем эмоции отличаются от мотиваций?
11. Основные теории мотиваций.
12. Эмоциональное состояние и эмоциональное выражение.
13. Основные функции эмоций.
14. Эмоциональный стресс, его стадии.
15. Теории восприятия пространства.
16. Учение А. А. Ухтомского об «интегральном образе».
17. Концепция когнитивных карт.
18. Что такое поведенческие тактики?
19. Нейронные механизмы пространственного восприятия.
20. Суточные ритмы и их биологическое значение.
21. Теории и стадии спа.
22. Сезонные ритмы поведения.
23. Оценка микроинтервалов времени.
Глава 7
ЗАКОНОМЕРНОСТИ УСЛОВНО¬
РЕФЛЕКТИВНОЙ ДЕЯТЕЛЬНОСТИ
ттжтшжмттшшштшжтшштжжттжжшшшмшжшштмжтшшшштшштшшмттштжмтшшттттшшштмтмш
§ 1. Безусловные рефлексы
Безусловный рефлекс — относительно постоянная видоспецифическая, стерео¬
типная, генетически закрепленная реакция организма или отдельных его частей па
внутренние или внешние раздражители, осуществляемая при посредстве централь¬
ной нервной системы. Безусловный рефлекс формируется в ностнаталыюм онто¬
генезе под влиянием и при взаимодействии с различными формами обучения.
Термин «безусловный рефлекс» введен И. П. Павловым для обозначения реф¬
лексов, безусловно возникающих при действии адекватных раздражителей на
определенное рецептивное иоле (например, отдергивание
руки при уколе пальца). Безусловные рефлексы составля¬
ют основу низшей нервной деятельности, обеспечивающей
приспособление организма к относительно постоянным
и маломеняющимся факторам среды, и представляют со¬
бой продукт филогенетического опыта предшествующих
поколений данного вида. Поэтому безусловный рефлекс —
это видовой рефлекс. Однако непрерывно меняющаяся
внешняя среда требует от организма индивидуальных адап¬
тационных приспособлений, не предусмотренных генети¬
ческим опытом. Наследственно закрепленные безуслов¬
ные рефлексы могут возникать, тормозиться и видоиз¬
меняться в ответ на самые разнообразные раздражения,
с которыми сталкивается индивидуум. Таким путем ранее индифферентные для
определенной деятельности раздражители приобретают качества биологически
значимых сигналов. При этом образуются условные рефлексы, составляющие
основу индивидуального опыта организма, названного И. П. Павловым высшей
нервной деятельностью,
Как наследственно закрепленная форма реагирования безусловные рефлексы
выполняются на основе стабильных связей нервной системы. Любой безуслов¬
ный рефлекс осуществляется с участием многих отделов центральной нервной
системы, но в основном он связан с деятельностью низших ее этажей (спинной
мозг, мозговой ствол и др.). Важная роль в механизме безусловного рефлекса при¬
надлежит обратной афферентации — информации о результатах и степени успеш¬
ности совершенного действия.
В сравнительно-физиологическом ряду животных наблюдается тенденция
усложнения безусловных рефлексов от элементарных их проявлений у беспозво¬
ночных животных (такие реакции называются таксисами) до весьма сложных
форм генетически детерминированного поведения, к которым относят различные
проявления инстинктов или, по современной терминологии, комплексы фиксиро¬
ванных действий.
Безусловный реф¬
лекс — относительно
постоянная видоспе-
иифическая, стерео¬
типная, генетически
закрепленная реакция
организма или отдель¬
ных его частей на
внутренние или внеш¬
ние раздражители,
осушествляемая при
посредстве централь¬
ной нервной системы.
204 Глава 7. Закономерности условно-рефлективной деятельности
Безусловные рефлексы изменяются в ходе онтогенеза, зависят от влияния на
них других рефлексов, состояния центральной нервной системы, эндокринной
системы и других факторов.
Классификация безусловных рефлексов затруднена. В соответствии с характе¬
ром действующего раздражителя и биологическим смыслом ответной реакции
в школе И. П. Павлова различали пищевые, половые, оборонительные, ориенти¬
ровочные и другие безусловные рефлексы.
А. Б. Коган (1959) различает три группы безусловных рефлексов:
1) элементарные безусловные рефлексы — простые ответные реакции местного
значения, выполняемые в соответствии с жестко детерминированной програм¬
мой сегментарных центров;
2) координационные безусловные рефлексы, которые хотя и выполняются на
сегментарном уровне, но допускают коррекцию на основе положительных
и отрицательных обратных связей;
3) интегративные безусловные рефлексы, представляющие собой синтез коорди¬
нированных двигательных актов с их вегетативным обеспечением в форме
комплексных реакций определенного биологического значения; эти безуслов¬
ные рефлексы осуществляются надсегмептарными уровнями центральной
нервной системы и обеспечивают целостные ответы организма.
П. В. Симонов (1981) рассматривает в качестве сложнейших безусловных реф¬
лексов врожденную видоспецифическую форму поведения, формирование кото¬
рой завершается в постнатальном онтогенезе под влиянием условных рефлексов
и при взаимодействии с ними. По П. В. Симонову, следует различать витальные,
ролевые или социальные и безусловные рефлексы саморазвития, направленные
на удовлетворение идеальных потребностей. В основе этой классификации лежит
потребностно-информационная гипотеза организации поведения человека и жи¬
вотных.
Одним из фундаментальных безусловных рефлексов является ориентировоч¬
ный, который, по мнению Л. Г. Воронина (1979), включает три группы явлений.
Первая состоит из ряда местных реакций соматического и вегетативного характе¬
ра (расширение зрачка, снижение порогов чувствительности, поворот головы и
туловища в сторону раздражения, расширение кровеносных сосудов, изменение
частоты сердцебиений и др.), обеспечивающих наиболее полное восприятие сен¬
сорной системой нового стимула. Вторая связана со специализированными поис¬
ковыми движениями, зависящими от доминирующей в данный момент потребно¬
сти. Третья проявляется в виде исследовательской реакции, основанной на удов¬
летворении потребности организма в информации (любопытство).
В целом ориентировочно-исследовательское поведение носит черты безуслов¬
ного рефлекса и условно-рефлекторной деятельности.
§ 2. Условные рефлексы
Те рефлекторные реакции, которые могут обеспечить существование организ¬
ма лишь при относительном постоянстве окружающей (а также внутренней для
организма) среды, И. П. Павлов назвал безусловными рефлексами (см. гл. 5). А так
§ 2. Условные рефлексы 205
как условия существования весьма сложны, изменчивы
и многообразны, то приспособление организма к среде
должно обеспечиваться при помощи другого рода реак¬
ций, которые позволили бы организму адекватно реаги¬
ровать на все изменения окружающей среды. Эти реак¬
ции и были названы И. П. Павловым условными рефлек¬
сами.
Детальное изучение условных рефлексов позволило
вскрыть основные закономерности взаимодействия про¬
цессов возбуждения и торможения, определяющих при¬
способительную деятельность организма. Проявление
различных сторон высшей нервной деятельности зави¬
сит от свойств основных нервных процессов, определенная совокупность которых
характеризует те или иные индивидуальные особенности поведения.
У высших животных и человека необходимым субстратом для формирования
условных рефлексов является кора больших полушарий.
Правила образования условных рефлексов. Многолетний опыт павловских
лабораторий позволил выделить в качестве основных следующие правила образо¬
вания условных рефлексов:
1. Для образования условных рефлексов необходимо совпадение во времени (со¬
четание) какого-либо индифферентного раздражителя (условного) с раздра¬
жителем, вызывающим соответствующий безусловный рефлекс (безусловный
раздражитель).
2. Необходимо, чтобы действие условного раздражителя несколько предшество¬
вало действию безусловного.
3. Условный раздражитель должен быть физиологически более слабым по срав¬
нению с безусловным раздражителем и возможно более индифферентным,
то есть не вызывающим значительной реакции. Физиологическая сила, на¬
пример, пищевого безусловного рефлекса (слюноотделительного или иного)
определяется уровнем пищевой мотивации. Физиологическая сила защитно-
оборонительного безусловного рефлекса, избавляющего от угрозы самому су¬
ществованию организма, несомненно выше, чем пищевого безусловного реф¬
лекса. Индифферентность же условного раздражителя состоит в том, что он не
должен вызывать безусловный рефлекс, используемый в качестве подкрепле¬
ния.
4. Для образования условного рефлекса необходимо нормальное, деятельное со¬
стояние головного мозга.
5. Во время образования рефлекса должны быть исключены другие виды деятель¬
ности как ответной реакции на посторонние раздражения.
6. Эти правила составляют основу изучения условных рефлексов, хотя возмож¬
ны и их вариации для конкретных экспериментальных задач.
Общие признаки условных рефлексов. На примере хорошо известного слю¬
ноотделительного условного рефлекса в павловских лабораториях были опреде¬
лены следующие общие признаки условных рефлексов:
Условные рефлексы —
индивидуально приоб¬
ретенные системные
приспособительные ре¬
акции животных и че¬
ловека, возникающие
на основе образования
в нейтральной нервной
системе временной
связи между условным
(сигнальным) и без¬
условно-рефлекторным
актами.
206 Глава 7. Закономерности условно-рефлективной деятельности
1. Их приспособительный характер. Смысл условного рефлекса в том, что он де¬
лает поведение особенно пластичным, подогнанным к конкретным условиям
среды (времени, месту, качеству подкрепления и нр.).
2. Любые условные рефлексы образуются при участии высших отделов головно¬
го мозга (а у насекомых, например, высших головных ганглиев). Поэтому при¬
способительные реакции простейших или кишечнополостных (лишенных, как
известно, центральной нервной системы) не могут быть отнесены к реакциям
условно-рефлекторного типа.
3. Условные рефлексы приобретаются и отменяются в индивидуальной жизни ка¬
ждой конкретной особи. Этим они принципиально отличаются от безусловных
рефлексов, одинаково проявляющихся у всех особей данного вида и в опреде¬
ленной мере детерминированных генотипически. Основная масса условных
рефлексов образуется только при повторяемости сочетаний условного и без¬
условного раздражителей. Иными словами, в индивидуальную память в виде
условно-рефлекторных реакций записываются не всякие случайные совпаде¬
ния условного и безусловного раздражений, а лишь те, сочетание которых во
времени оказывается наиболее вероятным. Все остальные, случайно возник¬
шие связи затормаживаются. Однако, если вероятность получения подкрепле¬
ния (пищевого или полового, например) по данному сигналу становится близ¬
кой к нулю, такие условные рефлексы постепенно отменяются, ибо перестают
выступать в качестве фактора организаций целесообразного поведения.
4. Условный рефлекс имеет сигнальный характер, то есть всегда предшествует,
предупреждает последующее возникновение безусловного рефлекса. Смысл
условного рефлекса в том, чтобы обеспечить подготовку организма к какой-
либо биологически целенаправленной деятельности. С помощью условных
рефлексов животное может заранее избежать опасности или подготовиться
к захвату добычи, поискам полового партнера и т. д.
Таким образом, условные рефлексы — это индивидуально приобретенные си¬
стемные приспособительные реакции животных и человека, возникающие на
основе образования в центральной нервной системе временной связи между
условным (сигнальным,) раздражителем и безусловно-рефлекторным актом.
Классификация условных рефлексов. Существует несколько систем класси¬
фикации условных рефлексов, каждая из которых, естественно, не может считать¬
ся исчерпывающей. Многое в оценке этих систем зависит от тех ведущих факто¬
ров, которые положены в их основу.
1. В связи с тем, что основой условного рефлекса является безусловное подкре¬
пление, начнем рассматривать классификацию условных рефлексов с особенно¬
стей подкрепления. В зависимости от наличия или отсутствия подкрепления
условные рефлексы делят на положительные (подкрепляемые), вызывающие со¬
ответствующую реакцию организма, и отрицательные, или тормозные (непод-
крепляемые), которые не только не вызывают соответствующей реакции, но и
ослабляют ее. Эти рефлексы будут специально рассмотрены в связи с обсуждени¬
ем процессов условного торможения.
Согласно биологическому значению подкрепления различают условные реф¬
лексы в соответствии с биологическими потребностями животных и человека (см.
гл. 5, § 0). Различают витальные условные рефлексы (пищевые, оборонительные.
§ 2. Условные рефлексы 207
регуляции сна и нр.), зоосоциалъные условные рефлексы (половой, родительский,
территориальный и пр.) и, наконец, условные рефлексы саморазвития (исследо¬
вательский, имитационный, игровой и нр.).
Наиболее распространенной формой исследования условно-рефлекторных
механизмов поведения служат условные рефлексы на пищевом и оборонительном
подкреплении. Иногда в самостоятельную группу выделяют подражательные
(имитационные) условные рефлексы, при которых животное-зритель, наблюдая
за условно-рефлекторной деятельностью животного-актера, начинало приобре¬
тать аналогичные навыки поведения. Иными словами, у животпого-зрителя выра¬
батывался условный рефлекс, подкреплением которого служило лишь наблюде¬
ние за поведением особи-актера.
Подкреплением условному рефлексу может служить любая деятельность ор¬
ганизма. Поэтому достаточно грубо ее можно разделить на две большие группы:
двигательные условные рефлексы и вегетативные условные рефлексы. Такое де¬
ление, конечно, относительно, так как акцент делается на характере регистрируе¬
мого безусловного рефлекса. Например, двигательный условный рефлекс, связан¬
ный с захватом пищи, определяется обычно по виду соответствующего движения
(хватание, клевание, сосание и пр.). Однако при ехю образовании, конечно, проис¬
ходят значительные вегетативные изменения, которые обеспечивают выполнение
данного движения (изменяются просвет кровеносных сосудов, частота сердцебие¬
ний, дыхательная ритмика и т. д.). На базе безусловно-рефлекторных двигатель¬
ных реакций образуется множество инструментальных условных рефлексов, ко¬
торые служат прекрасной моделью для изучения мозговых механизмов произ¬
вольного поведения.
Образованный инструментальный условный рефлекс может в одних случа¬
ях полностью копировать безусловный рефлекс — по условному сигналу собака
поднимает одну конечность, тем самым предотвращая раздражающее действие
электрического тока на эту конечность (рис. 67). В других случаях условный дви¬
гательный рефлекс может иметь гораздо более сложную форму и не повторяет
безусловно-рефлекторное движение. Именно к такой группе рефлексов относят¬
ся, например, сложнейшие манипуляторные реакции, проявляющиеся у обезь¬
ян для добывания пищи. Инструментальные условные рефлексы в естественных
условиях существования, как правило, значительно сложнее элементарных ре¬
акций, изучаемых в лабораториях, и составляют основной фонд целенаправ¬
ленного поведения животных и человека.
Рис. 67. Графическая запись
условно-рефлекторного сгибания
ноги и условной дыхательной
реакции вслед за сочетанием звука
метронома и электрического тока
1 — движения левой передней конечно¬
сти; 2 — дыхание; 3 — отметка условного
стимула (метроном); 4 — отметка безус¬
ловного стимула (электрический ток); 5 —
отметка времени — 0,5 с
208 Глава 7. Закономерности условно-рефлективнои деятельности
И наоборот, если взять в качестве безусловного вегетативного показателя из¬
менение кровенаполнения периферических сосудов, которое может возникать на
условный сигнал и подкрепляться локальным температурным воздействием, то в
результате сочетания условного и безусловного раздражителей можно регистри¬
ровать кривую изменения тонуса сосудов (плетизмограмму), а также и другие по¬
казатели вегетативных сдвигов, например так называемую пневмограмму — изме¬
нение дыхательной ритмики (рис. 68).
Рис. 68. Образование сосудосуживающего условного рефлекса у собаки
на звонок (по А. В. Пшонику, 1952)
1 — плетизмограмма; 2 — запись дыхательных движений; 3 — отметка условного
звукового раздражителя; 4 — отметка безусловного раздражителя; 5 — отметка
времени, с. На шкале отметка кровяного давления в мм рт. ст. от изолинии
К вегетативным условным рефлексам относится классический слюноотдели¬
тельный условный рефлекс. Практически условно-рефлекторному контролю мо¬
гут подчиняться все внутренние органы, хотя и по-разному проявляться для от¬
дельных органов, с разной скоростью вырабатываться и угашаться.
Условные сердечные рефлексы вырабатываются медленно: так, учащение
сердцебиений у собаки после сочетаний звукового сигнала с введением нитрогли¬
церина возникло лишь после 30-100 сочетаний, а угашение было еще медлен¬
нее — после 296 применений гудка без подкрепления (рис. 69).
б
Рис. 69. Образование условного
сердечного рефлекса
(по М. К. Петровой)
а — электрокардиограмма собаки в нор¬
ме; б — через 30 с после применения
условного раздражителя, ставшего сиг¬
налом введения нитроглицерина; в —
электрокардиограмма через 30 с после
введения нитроглицерина, отметка вре¬
мени — 1,5 с
§ 2. Условные рефлексы 209
Целостный условный рефлекс имеет полиэффекторную природу, и его отнесе¬
ние к той или иной категории рефлексов зависит от того, какой из эффекторов из¬
бирает для регистрации экспериментатор.
В самостоятельную группу следует отнести условные рефлексы, подкреплени¬
ем которым служит прямое электрическое (или химическое) раздражение мозга.
Такие лабораторные модели могут быть полезны для изучения отдельных мозго¬
вых механизмов условно-рефлекторного процесса.
Различаются условные рефлексы и по особенностям подкрепления. Если в ка¬
честве подкрепления используется безусловный рефлекс, такой условный реф¬
лекс относят к условный рефлексам первого порядка. Если в качестве подкреп¬
ления используется ранее выработанный прочный условный рефлекс, то такой
новый условный рефлекс именуют условным рефлексом второго порядка. Соот¬
ветственно могут быть условные рефлексы третьего и последующих порядков.
Именно такого типа условные рефлексы формируются у детей и составляют осно¬
ву развития у них мыслительной деятельности (см. гл. 5).
2. Важнейшим компонентом условного рефлекса является условный сигнал,
то есть собственно рецепторная часть условно-рефлекторного процесса. По этому
признаку условные рефлексы подразделяют на две большие группы: экстероцеп-
тивные и интероцептивные.
В соответствии с тем, на какие рецепторные приборы действует раздражитель,
выделяют следующие экстероцептивные условные рефлексы: зрительные, слухо¬
вые, обонятельные, вкусовые, тактильные, температурные. Кроме того, экстеро¬
цептивные условные рефлексы делятся на дистантные (зрительные, слуховые
и др.) и контактные (тактильные, вкусовые).
Эти рефлексы играют определяющую роль в поведении животных и психиче¬
ской деятельности человека. Обучение речевому общению между людьми в осно¬
ве своей имеет специально человеческие условные рефлексы па словесные сигна¬
лы (см. гл. 4).
Интероцептивные условные рефлексы возникают в том случае, если посту¬
пивший в головной мозг афферентный залп из внутренних органов становится
условным сигналом к тем или иным изменениям в вегетативной сфере или двига¬
тельной деятельности всего организма (Э. Ш. Айрапетьянц, 1952; К. М. Быков,
1942). Эти условные рефлексы «с внутренних органов» также различаются по
следующим признакам: механические, химические, температурные, осмотические
и др. Если же в основу положить орган, с рецепторов которого формируется тот
или иной интероцептивный сигнал, то можно говорить о желудочных, кишечных,
сердечных, легочных, маточных и др. (рис. 70). Интероцептивные условные реф¬
лексы образуются медленнее экстероцептивных, а условно-рефлекторная реак¬
ция животного более диффузная, чем на экстероцептивный раздражитель.
По характеру условного раздражения условные рефлексы подразделяются на
натуральные и искусственные. К натуральным условным рефлексам относятся
такие, которые образуются на естественные признаки безусловного раздражения.
К таковым относится, например, запах пищи. Так, щенок, однажды съев сырое мя¬
со, приобретает на всю жизнь натуральный условный рефлекс на запах мяса. Лю¬
бые другие условные сигналы, не являющиеся непременным свойством пищи,
а лишь совпадающие по времени с моментом ее поедания, относятся к искусствен¬
ным условным сигналам. Эти условные рефлексы вырабатываются медленнее
и легче разрушаются.
210 Глава 7. Закономерности условно-рефлективнои деятельности
_
Рис. 70. Интероиептивный условный
рефлекс (по Э. Ш. Айрапетьяниу, 1952)
Слева: интероиептивный условный рефлекс,
проявившийся на дыхании; справа — интеро¬
иептивный условный рефлекс, проявившийся
на дыхании и на отдергивании лапы; сверху
вниз: дыхание, движение лапы, отметка вре¬
мени — 1 с, отметка безусловного раздраже¬
ния, отметка условного раздражения (разду¬
вание мочевого пузыря)
По структуре условного сигнала различают следующие четыре группы услов¬
ных рефлексов:
♦ условные рефлексы на простые раздражители (звонок, метроном, вспышки
света и пр.);
♦ условные рефлексы на одновременные комплексные раздражители, состоя¬
щие из нескольких компонентов, действующих одновременно (например,
свет + звук + кожное раздражение);
♦ условные рефлексы на последовательные комплексные раздражители, отдель¬
ные компоненты которых действуют последовательно, накладываясь друг на
друга;
♦ условные рефлексы на цели раздражителей, когда отдельные компоненты
сложного раздражителя действуют последовательно, не совпадая друг с дру¬
гом, а безусловное подкрепление присоединяется к последнему из них (рис. 71).
6 в
г I г—ц
I II II з—н
Рис. 71. Соотношение во времени между членами комплексов
сложных условных раздражителей
Одинарные линии — индифферентные раздражители; двойные — ранее выработанные сигналы,
пунктир — подкрепление; а — одновременный комплекс; 6 — суммарный раздражитель;
в — последовательный комплекс; г — иепь раздражителей
Иногда в качестве условных сигналов используют не абсолютные, а относи¬
тельные признаки раздражителей. Например, из двух одновременно предъявляе¬
мых однородных раздражителей — плоских кружков — условный рефлекс выра¬
батывается на один из них — больший по размеру. Проверка выработки такого
рефлекса производится при предъявлении животному других одинаковых гео¬
метрических фигур (скажем, треугольников), также отличающихся по размеру.
Если и в этом случае животное осуществит условно-рефлекторную реакцию на
сигнал более крупного размера, можно говорить о выработке у него условного реф¬
лекса на отношение.
3. Важнейшим признаком для классификации условных рефлексов является
соотношение во времени действия условного и безусловного раздражителей. По
одному признаку различают:
и
У
н
§ 2. Условные рефлексы 211
♦ наличные условные рефлексы, образующиеся щж совпадении во времени
условного раздражителя и подкрепления;
♦ следовые условные рефлексы, при образовании которых условный раздражи¬
тель и подкрепление отделены друг от друга «определенным временным ин¬
тервалом.
Если подкрепление почти сразу присоединяется к действию условного раздра¬
жителя, образуются так называемые совпадающие условные рефлексы. В дру¬
гих случаях подкрепление присоединяется к условному раздражителю спустя 5-
30 с от начала его действия — вырабатываются отставленные условные рефлексы
(рис. 72). Наконец, подкрепление может присоединиться к условному раздражи¬
телю после длительного изолированного действия последнего. Такие условные
рефлексы названы запаздывающими условными рефлексами.
б
1
I I 1И111[И11111111111111И1Ш1И11 I I I
4 1-И I I I I 1 I I I I I 1 I I I I I I I I I I I I I I II I I
0 12 3 4
в
1
2
3
4
||||||ц||||||||||||цц|||| I
I I I I и I I I I I I I I I I I I I I I I I I I I
0 12 3 4
Рис. 72. Схема выработки условных рефлексов
а — отставленного; 6 — запаздывающего; в — следового; 1 — слюноотделение
(в каплях слюны), 2 — условный сигнал, 3 — безусловное подкрепление,
4 — время, мин
Следовые условные рефлексы образуются в тех случаях, когда подкрепление
дается после прекращения действия условного сигнала. Условным раздражителем
образующегося рефлекса становится не сам сигнал, а след от его действия в мозго¬
вых нейронных сетях.
Особая разновидность следовых условных рефлексов — это условные рефлексы
на время. Если какой-либо безусловный рефлекс проявляется через одинаковые
интервалы времени, то в результате каждый раз по истечении данного интервала
как бы сама по себе возникает реакция, вызываемая раньше действием безуслов¬
ного раздражителя (рис. 73). Такая реакция получила название условного рефлек¬
са на «чистое» время (см. гл. 6).
Таким образом, приведенная классификация подчеркивает достаточно широ¬
кий диапазон условно-рефлекторного приспособления высших организмов и ста¬
вит условный рефлекс в качестве одного из основных феноменов высшей нервной
деятельности.
212 Глава 7. Закономерности условно-рефлективной деятельности
в
Л | / ‘
93
_1
94
1МЧЩМ1111ЦН<Ш111ШЩ1ПЧ1Ц|1Ш1П111И..1.,т....,.,
Рис. 73. Образование двигательно-оборонительного условного рефлекса
на время у собаки (по А. С. Дмитриеву, 1974)
а — первая стадия; б — вторая стадия; в — третья стадия; 1 — двигательная реакция задней лапы;
2 — отметка условного раздражителя; 3 — отметка момента электрокожного раздражителя;
4 — отметка времени (2 с), интервал между сочетаниями равен 1,5 мин, цифры —
порядковые номера сочетаний
§ 3. Торможение условных рефлексов
Объяснить поведение, признавая лишь существование возбудительного процесса,
нельзя, так как очевидно, что при возбуждении нервных аппаратов возможны
лишь судороги, а не целесообразная координированная деятельность животных и
человека. В физиологии долгое время отсутствовала теория, позволявшая объяс¬
нить координированный характер поведения. В этих условиях открытие в 1862 г.
И. М. Сеченовым центрального торможения сыграло значительную историче¬
скую роль. И. М. Сеченов рассматривал центральное торможение как деятель¬
ность (возбуждение) специальных тормозных систем по аналогии с хорошо из¬
ученными к тому времени тормозными эффектами блуждающего нерва на сердце.
Ч. Шеррингтоном (1906) развивалось представление об активном торможении
мышц-антагонистов в процессе рефлекторного акта. Согласно его представлению
в двигательном центре мышцы-антагониста при возбуждении центра другой
мышцы возникает путем индукционного наведения состояние торможения. Од¬
нако дальше констатации фактов наличия индукционных отношений между цент¬
рами мышц-антагонистов он не пошел.
А. А. Ухтомский (1923), опираясь на предшествующие взгляды, выдвинул
представление о сопряженном торможении при формировании доминанты. Цент¬
ральные механизмы наиболее биологически значимого рефлекторного акта под¬
крепляются конвергирующими к ним разнообразными возбуждениями, возни¬
кающими в центральной нервной системе, и, достигнув определенного уровня
возбудимости, эти центральные механизмы одновременно с запуском домини¬
рующей рефлекторной реакции активно тормозят биологически несовместимые
с ней любые другие формы рефлексов.
§ 3. Торможение условных рефлексов 213
Огромный экспериментальный опыт позволил выдвинуть И. П. Павлову соб¬
ственную схему классификации видов торможения при условно-рефлекторной
деятельности.
Внешнее (безусловное) торможение. Наиболее общим поведенческим при¬
знаком торможения является отсутствие ожидаемой специфической реакции при
действии порогового или даже надпорогового раздражителя или прекращение в
этих же условиях какой-либо текущей деятельности или начавшейся двигатель¬
ной или секреторной реакции.
Под внешним торможением понимается срочное подавление текущей услов¬
но-рефлекторной деятельности при действии посторонних для нее раздражителей,
вызывающих ориентировочный или другой какой-либо безусловный рефлекс.
По механизму своего возникновения этот тин торможения относят к врожден¬
ным, который осуществляется благодаря явлениям отрицательной индукции.
И. П. Павлов даже предлагал называть этот тип торможения индукционным.
А. А. Ухтомский же именовал его сопряженным торможением и видел в нем фи¬
зиологическую основу для реализации доминирующей
формы деятельности организма. Безусловное торможе¬
ние также называют внешним потому, что причина его
возникновения лежит вне структуры тормозимого реф¬
лекса.
Одним из факторов, вызывающих безусловное тормо¬
жение, является ориентировочный рефлекс. Всякий не¬
ожиданный посторонний раздражитель содержит новую
для организма информацию, и для ее более полного вос¬
приятия выполняется ориентировочный рефлекс (реф¬
лекс «что такое?», по И. Г1. Павлову). В момент возник¬
новения этого рефлекса возникает индукционное торможение конкурирующих
рефлексов. Оно может быть более или менее глубоким, кратковременным или бо¬
лее длительным, что зависит от физиологической силы ориентировочного и тор¬
мозимого рефлексов. Однако неоднократное повторное раздражение вызывает
ориентировочный рефлекс меньшей интенсивности, который затем в силу привы¬
кания к этому раздражению и вовсе пропадает. Одновременно с этим постепенно
снижается и эффект внешнего торможения. Такой вид торможения был назван
гаснущим тормозом. Механизмы гаснущего тормоза развиваются одновременно
на разных уровнях головного мозга с участием ретикулярной системы ствола.
Другой вид безусловного торможения отличается постоянством своего эффек¬
та на тот или иной тормозимый рефлекс и поэтому называется постоянным
тормозом. Стабильность внешнего торможения определяется физиологической
силой того рефлекторного акта, который становится главной причиной этого тор¬
можения. К таковым относятся оборонительные безусловные рефлексы на раз¬
ные вредящие раздражения, включая болевые. Внешнее торможение может воз¬
никнуть и при выполнении условного рефлекса, например защитно-оборонитель¬
ного.
Как и в случае гаснущего тормоза, длительность тормозного действия оборо¬
нительного рефлекса определяется его силой и характером тормозимого рефлек¬
са, и в частности степенью его упроченности. «Молодые» условные рефлексы тор¬
мозятся легче и на более длительный срок, чем более «старые», при одних и тех же
Внешнее (безусловное)
торможение — сроч¬
ное подавление теку¬
щей условно-рефлек¬
торной деятельности
при действии посто¬
ронних раздражителей,
вызывающих ориенти¬
ровочный, оборони¬
тельный или другой
безусловный рефлекс.
214 Глава 7. Закономерности условно-рефлективной деятельности
условиях. Болевые воздействия с внутренних органов обладают более длитель¬
ным тормозным влиянием на условно-рефлекторную деятельность. А иногда их
сила настолько велика, что извращает нормальное протекание даже безусловных
рефлексов. Представим себе, что собаке с выработанными пищевыми слюноотде¬
лительными условными рефлексами во время опыта наносится на одну из конеч¬
ностей электроболевое раздражение. Возникает весьма резкий оборонительный
рефлекс: животное начинает отталкиваться ногами, срывает зубами лямки, кото¬
рыми привязано в станке, скулит и пытается убежать. Условные пищевые сигналы
не сопровождаются слюноотделением, голодное животное отказывается от пред¬
ложенной ему пищи. Итак, два антагонистических рефлекса — пищевой и оборони¬
тельный — не могут сосуществовать, более слабый тормозится под влиянием бо¬
лее сильного. А чем сильнее этот рефлекс, тем ярче выражено индукционное тор¬
можение конкурирующих форм рефлекторной деятельности.
В естественных условиях обитания в определенные периоды жизни половое
поведение оказывается более сильным и тормозит защитно-оборонительные реф¬
лексы. Самцы птиц в брачный период жизни жертвуют своими перьями и други¬
ми «украшениями», становятся как бы нечувствительными к травмам.
Безусловное торможение, которое И. П. Павлов рассматривал на примере конк¬
ретных рефлексов, А. А. Ухтомский положил в основу существования целостных
поведенческих актов. Причем он подчеркивал, что такое сопряженное торможе¬
ние не есть уничтожение всяческой деятельности на местах, но ее переработка,
трансформирование согласно с направлением доминирующей деятельности.
Запредельное (охранительное) торможение. Хорошо известно, если увели¬
чивать интенсивность какого-либо раздражителя, то вызываемый им эффект уве¬
личивается. Однако это увеличение ответа будет наблюдаться лишь до определен¬
ного предела интенсивности раздражения. Дальнейшее усиление раздражения
приведет к падению или полному исчезновению эффекта. Это и есть результат не
утомления, а запредельного торможения, которое И. П. Павлов назвал охрани¬
тельным, так как оно ограждает клетки мозга от избыточного расходования энер¬
гетических ресурсов. Этот вид торможения зависит от функционального состоя¬
ния нервной системы, возраста, от типологических особенностей, состояния гор¬
мональной сферы и пр. Предел выносливости клетки в отношении раздражителей
разной интенсивности называется пределом ее работоспособности, и чем выше
этот предел, тем легче клетка переносит действие сверхсильных раздражений.
Причем речь идет не только о физической, по и об информационной силе (значи¬
мости) условных сигналов. Для корковых клеток существует предел безвредного
функционального напряжения, за которым следует вмешательство торможения.
Этот вид торможения имеет общие черты как с безусловным, так и с условным
и занимает своеобразное срединное положение между ними.
Крайним случаем запредельного торможения является оцепенение, возникаю¬
щее у животного и человека под влиянием сильного раздражения. Человек может
впасть в состояние ступора — полной неподвижности. Такие состояния возника¬
ют не только в результате действия физически сильного раздражителя (взрыва
бомбы или снаряда, например), но и вследствие тяжелых моральных потрясений
(например, при неожиданном сообщении о тяжелой болезни или смерти близкого
человека).
§ 3. Торможение условных рефлексов 215
Внутреннее (условное) торможение. К данной форме торможения текущей
условно-рефлекторной деятельности относят те случаи, когда условный раздра¬
житель перестает подкрепляться безусловным. Такое торможение возникает не
срочно, не сразу, а развивается постепенно, вырабатывается но общим законам
условного рефлекса и является столь же изменчивым и динамичным. И. П. Пав¬
лов назвал его поэтому условным торможением в отличие от безусловного. Он
считал, что такое выработанное торможение возникает внутри центральных нерв¬
ных структур самих условных рефлексов, а отсюда и его название — внутреннее
(то есть не наведенное извне, не индукционное) торможение. Правильно ли это
предположение, до сих пор неизвестно.
Выделим основные характеристики условного торможения.
1. Как уже подчеркивалось, оно развивается при неподкреплении раздражителей,
которые постепенно приобретают свойства условного тормозного раздражите¬
ля. И. П. Павлов называл подкрепляемый условный
сигнал положительным, а соответствующая реакция
именовалась положительным условным рефлексом.
Неподкрепляемый же условный сигнал снижал интен¬
сивность реакции, а иногда и вообще подавлял ее, он
именовался отрицательным сигналом. Соответствую¬
щее поведение было названо отрицательным (или тор¬
мозным) условным рефлексом.
2. Условное торможение поддается тренировке, а значит,
выработка его при повторной процедуре облегчается.
Однако заторможенный условный рефлекс может самопроизвольно восста¬
навливаться иод влиянием как внешних, так и внутренних причин. Как мы
увидим ниже, это свойство условного торможения чрезвычайно важно при
воспитании поведенческих навыков человека и животных в раннем возрасте.
3. Способность к различным проявлениям условного торможения зависит от ин¬
дивидуальных свойств нервной системы организма. У возбудимых индивиду¬
умов условное торможение вырабатывается труднее и медленнее, чем у более
спокойных.
4. Условное торможение зависит от физиологической силы безусловного реф¬
лекса, подкрепляющего положительный условный сигнал. Например, вырабо¬
тать торможение реакции, ранее подкреплявшейся электроболевым стимулом,
значительно труднее и требует гораздо больше времени, чем торможение той
же реакции на тот же сигнал, но подкреплявшийся ранее пищевым безуслов¬
ным рефлексом.
5. Условное торможение зависит от прочности ранее выработанного условного
рефлекса. Более прочные стабильные условные рефлексы значительно труд¬
нее поддаются торможению, чем только недавно сформированные условные
рефлексы.
6. Условное торможение способно взаимодействовать с безусловным, в этих слу¬
чаях возникает явлениерастормаживания, в других случаях в результате сум¬
мации условного и безусловного торможения их общий эффект может усили¬
ваться.
И. П. Павлов подразделил условное торможение на четыре вида.
Внутреннее (условное)
торможение — тормо¬
жение условно-реф¬
лекторной деятельно¬
сти, которое
развивается постепен¬
но в результате непод-
крепления условного
сигнала безусловным
рефлексом.
216 Глава 7. Закономерности условно-рефлективной деятельности
Угасательное торможение развивается при отсутствии подкрепления условно¬
го сигнала безусловным. Теперь тот же раздражитель становится сигналом отсут¬
ствия реакции. Представим себе, что у животного образовался условный рефлекс
па вид местности, который постоянно сочетался с наличием в ней необходимой
пищи. Но пищевые ресурсы местности иссякли. Не найдя пищи, то есть не полу¬
чив подкрепления, животное со временем перестает посещать знакомую мест¬
ность в результате развития угасательного торможения. Вид местности, который
раньше был положительным сигналом, теперь становится сигналом отрицатель¬
ной реакции.
Степень и скорость выработки угасательного торможения зависят от:
♦ прочности условного рефлекса (более прочно выработанные рефлексы угаша¬
ются медленнее);
♦ физиологической силы подкрепляющего рефлекса (угасить пищевой услов¬
ный рефлекс у голодной собаки значительно труднее, чем у сытой);
♦ частоты неподкрепления (при остром неподкреплении угасательное торможе¬
ние развивается в течение минут и часов, при хроническом неподкреплении —
в течение многодневных экспериментов).
Пищевые условные рефлексы угашаются значительно быстрее оборонительных.
Особенно трудно поддаются угашению двигательные оборонительные условные
рефлексы. Угасательное торможение быстрее и прочнее образуется при трениров¬
ке (повторном угашении и восстановлении), при этом угашение одного рефлекса
(первично угашенный рефлекс) ведет к ослаблению или даже исчезновению дру¬
гих рефлексов (вторично угашенные рефлексы). Угасательное торможение разви¬
вается волнообразно, и в его выработке наблюдаются индивидуальные типологи¬
ческие различия.
Дифференцировочное торможение развивается при неподкреплении раздра¬
жителей, близких к подкрепляемому сигналу. Этот вид торможения лежит в осно¬
ве различения близких по своим физическим свойствам раздражений.
Представим себе следующую экспериментальную ситуацию. У собаки выраба¬
тывается условный рефлекс на звуковой тон «ля». Все близкие к нему тоны при
первом своем применении будут вызывать аналогичную реакцию. Это явление
было названо И. П. Павловым генерализацией, то есть обобщением условных реф¬
лексов. Однако, постоянно подкрепляя топ «ля» и не сопровождая действием без¬
условного раздражителя другие тоны, можно добиться их полного различения.
Оно будет проявляться в наличии условной реакции только на тон «ля» и ее от¬
сутствии на действие прочих тонов. С помощью диффереицировочного торможе¬
ния организм из массы сходных раздражителей будет реагировать только на один
подкрепляемый, то есть биологически для него важный, а на другие сходные раз¬
дражители условная реакция будет выражена слабее или отсутствовать полно¬
стью. Таким образом достигается после первичной генерализации условных реф¬
лексов их последующая специализация.
При первом своем проявлении условный рефлекс может быть и не генерализо¬
ван. Лишь по мере своего укрепления рефлекс приобретает генерализованный ха¬
рактер. Биологическое значение генерализации видно из следующего примера.
Допустим, животному удалось избежать когтей хищника. Однако оборонитель¬
ная реакция, совпадая по времени с голосом хищника, стала носить условно-реф¬
лекторный характер на звук голоса нападающего. Генерализация, или обобщение
§ 3. Торможение условных рефлексов 217
всех вариаций голоса хищника, которые возникают в естественной среде, обеспе¬
чивает выполнение оборонительной реакции в различных ситуациях, что являет¬
ся биологически выгодным для сохранения жизни организма. Специализация в
этих условиях оказала бы пагубное влияние на животное, сильно ограничивая
возможности его самозащиты. То же справедливо и для других видов поведения
(пищевого, полового и ир.). Следовательно, свойство генерализации условных
рефлексов — неизбежный атрибут поведенческих адаптаций животных в естест¬
венной среде обитания.
Учитывая то обстоятельство, что изменчивость окружающей организм среды
происходит по вероятностному закону, колебания тех или иных биологически
значимых признаков предвидеть с высокой вероятностью невозможно, становит¬
ся биологически оправданной значительная сенсорная генерализация условных
рефлексов как стадии активного поиска жизненно важных объектов (Р. А. Павлы-
гина, 1983; П. В. Симонов, 1987). Р. А. Павлыгина считает, что в стадии генерали¬
зации условных рефлексов обнаруживается механизм доминанты, одной из ха¬
рактерных черт которой является способность доминирующей рефлекторной си¬
стемы диффузно отзываться на самый широкий репертуар внешних стимулов,
в том числе и впервые встречающихся. Однако известно, что постепенно, в про¬
цессе повторного осуществления данного рефлекторного акта, диффузная отзыв¬
чивость сменяется избирательным реагированием только на те раздражения, ко¬
торые создали данную доминанту. Наступает стадия специализации доминанты,
которая достигается благодаря механизмам дифференцировочного торможения.
Таким образом, в создании доминанты как вектора целенаправленного поведения
участвуют как процессы безусловного (сопряженного) торможения, так и процес¬
сы условного торможения.
Отметим следующие основные свойства дифференцировочного торможения:
♦ чем ближе дифференцируемые раздражители, тем труднее на один из них (не-
подкрепляемый) выработать дифференцировочное торможение;
♦ степень торможения определяется силой возбуждения, развиваемого положи¬
тельным (подкрепляемым) условным рефлексом. Как и в случае угасательного
торможения, дифференцировочное торможение легче вырабатывается при пи¬
щевом подкреплении, чем при оборонительном, легче у накормленных живот¬
ных, чем у голодных;
♦ выработка этого торможения происходит волнообразно;
♦ дифференцировочное торможение тренируемо, что лежит в основе тонкого
распознавания сенсорных факторов среды.
В самостоятельный вид условного торможения И. П. Павлов выделил услов¬
ный тормоз, который образуется при неподкреплении комбинации из положи¬
тельного условного раздражителя и индифферентного. Например, у собаки обра¬
зован пищевой условный рефлекс на звук. Если к этому сигналу присоединить
свет от лампочки и их совместное действие не подкреплять нищей, то после не¬
скольких применений этой комбинации она перестанет вызывать пищевую ре¬
акцию. Хотя изолированное применение звонка по-прежнему будет вызывать
обильное слюноотделение.
По существу, это вариант дифференцировочного торможения. Этот вид тор¬
можения часто встречается. Например, при хозяйке домашняя кошка ведет себя
218 Глава 7. Закономерности условно-рефлективнои деятельности
«благоразумно», а в отсутствие ее забирается на стол и поедает продукты. В этом
случае вид стола — условный пищевой сигнал, сочетание вида стола и хозяйки ни¬
когда не сопровождалось пищей и сделалось условным тормозом. Присутствие
хозяйки (условного тормоза) подавляет пищедобывательную реакцию кошки на
вид стола. Выработанные в течение жизни условные тормоза, как и дифференци-
ровочное торможение, уточняют условные рефлексы, делают поведение более при¬
способленным к конкретной окружающей ситуации.
Основные свойства условного тормоза следующие:
1. Он легче вырабатывается, если к слабому (световому, например) положитель¬
ному раздражителю присоединяется сильный дополнительный, например зво¬
нок.
2. Роль прибавочного раздражителя может играть след от примененного раздра¬
жения достаточно большой силы. Так, след от автомобильного гудка даже
спустя 20 с может служить в качестве условного тормоза к метрономному сиг¬
налу условного пищевого рефлекса.
3. Если прибавочный раздражитель имеет недостаточную силу, то он может пре¬
вратиться в условный раздражитель второго порядка (см. гл. 11), то есть вы¬
ступить в качестве положительного сигнала основного условного рефлекса.
4. Если прибавочный раздражитель приобрел свойства условного тормоза, то,
будучи присоединенным к любому другому положительному сигналу, он за¬
тормозит соответствующий этому сигналу условный рефлекс.
5. Прибавочный раздражитель в первый момент своего применения в комбина¬
ции с положительным сигналом вызывает ориентировочный рефлекс и индук¬
ционное торможение условной реакции, затем превращается в индифферент¬
ный раздражитель (гаснущий тормоз), и наконец на месте безусловного тормо¬
жения развивается условный тормоз.
Торможение запаздывания. При выработке такой разновидности условного
торможения подкрепление соответствующим безусловным рефлексом не отменя¬
ется (как в предыдущих видах торможения), а значительно отодвигается от нача¬
ла действия условного раздражителя. Подкрепляется лишь последний период
действия сигнала, а предшествующий ему значительный период его действия ли¬
шается подкрепления. Поэтому первый период действия условного сигнала, кото¬
рый сопровождается торможением запаздывания, называется недеятельной фа¬
зой запаздывающего условного рефлекса. По истечении его торможение прекра¬
щается и сменяется возбуждением — так называемая деятельная фаза рефлекса.
Строго говоря, в данном случае действует не один раздражитель, а два в комп¬
лексе, и второй компонент — это время. Именно время определяет момент прекра¬
щения запаздывателыюго торможения и наступление деятельной фазы рефлекса.
Допустим, у собаки выработан запаздывающий условный рефлекс на свет. Экспе¬
риментатор зажигает электрическую лампу, которая до момента подкрепления го¬
рит 3 мин. Условно-рефлекторная реакция, возникавшая по световому сигналу, не
подкрепляется и поэтому быстро гаснет, так как развивается угасателыюе тормо¬
жение. Но за некоторое время до подкрепления условная реакция проявляется
четко, она подкрепляется. Таким образом, условная реакция как бы приурочива¬
ется к моменту действия безусловного раздражителя. Начальный период дейст¬
вия раздражителя выступает в качестве сигнала отрицательной реакции, а более
§ 3. Торможение условных рефлексов 219
длительный промежуток времени, близкий ко всему периоду запаздывания, слу¬
жит положительным сигналом. В случае пищевых условных рефлексов отставле-
ние подкрепления от начала действия условного сигнала может достигать 2-
3 мин, а при электрооборонительных двигательных рефлексах этот интервал зна¬
чительно короче — 30-60 с.
Адаптивное значение торможения запаздывания состоит в тонком анализе
времени отставления раздражителя; условный рефлекс более или меиее точно
приурочивается ко времени действия сигнализируемого безусловного рефлекса.
Отметим основные свойства данного вида условного торможения:
♦ чем сильнее условный раздражитель, тем труднее выработать торможение за¬
паздывания;
♦ чем больше сила подкрепляющего рефлекса, тем труднее выработать запазды¬
вание условного рефлекса;
♦ чем медленнее удлиняется от опыта к опыту изолированное действие условно¬
го раздражителя, тем легче вырабатывается запаздывание. Если подкрепление
сразу отодвинуть от начала действия положительного сигнала на 2-3 мин, то
запаздывание выработать чрезвычайно трудно;
♦ значительное упрочение совпадающих или короткоотставлегшых условных
рефлексов препятствует выработке торможения запаздывания.
Хотя торможение запаздывания и отличается от трех других видов условного
торможения благодаря участию временного фактора, в основе его развития также
лежит пеподкрепление определенного периода действия условного сигнала.
Взаимодействие разных видов торможения. Взаимодействие условного и без¬
условного торможения может происходить чаще всего в форме феномена растор-
маживания, о котором уже упоминалось. Суть его состоит в том, что безусловное
индукционное торможение, возникающее при ориентировочном рефлексе, вре¬
менно ослабляет или полностью уничтожает эффект условного выработанного
торможения (угасателыюго, дифференцировочного, условного тормоза и тормо¬
жения запаздывания). Например, животное обучилось выполнять защитное пове¬
дение только на определенные звуки своего врага и не реагировать на все осталь¬
ные. Но выйдя на очередную охоту в привычной для себя обстановке, животное
встретилось с неожиданным новым запаховым раздражителем, который естест¬
венно вызвал ориентировочный рефлекс. Одновременно с этим настороженность
животного сильно возросла, ибо теперь уже ранее угашенные звуковые раздражи¬
тели начинали вызывать защитную реакцию (затаивание и убегание). Из такого
примера ясно, что феномен растормаживания имеет важное биологическое значе¬
ние, обеспечивая срочную перестройку привычного поведенческого стереотипа
к новой жизненной ситуации. Конечно, эффект растормаживания очень динами¬
чен, он зависит от многих факторов: силы и прочности выработанного торможе¬
ния, силы и продолжительности ориентировочного рефлекса, возраста и типа
нервной системы животного.
Другое направление взаимодействия разных видов торможения состоит в их
суммации. Например, утешение условного рефлекса может ускоряться, если кро¬
ме неподкренления применить умеренной силы посторонний раздражитель. Про¬
исходит суммация условного угасателыюго и безусловного индукционного тор¬
можения.
220 Глава 7. Закономерности условно-рефлективной деятельности
Не меньше примеров суммации разных видов условного торможения; скажем,
одновременная тренировка угасателыюго и запаздывателыюго торможения вза¬
имно усиливают друг друга. Или в другом случае один из видов условного тормо¬
жения (например, дифферепцировочное) формируется быстрее, если ему пред¬
шествовала процедура с тренировкой угасателыюго торможения. Биологическое
значение эффекта суммации торможения понятно, если иметь в виду, что трени¬
ровка торможения — одно из основных условий формирования и воспитания раз¬
личной сложности поведенческих навыков.
Сами факты взаимодействия разных видов торможения являются достаточ¬
ным основанием, чтобы предполагать их близкую физиологическую природу.
Заключение
Наряду с относительно постоянными формами реагирования на стабильные внеш¬
ние и внутренние воздействия (безусловные рефлексы) существует другой тип
реакций, позволяющих организму адекватно реагировать на все изменения окру¬
жающей среды (условные рефлексы). Условные рефлексы образуются при совпа¬
дении во времени и пространстве какого-либо индифферентного (условного) раз¬
дражителя и раздражителя, вызывающего безусловный рефлекс. Главный смысл
условного рефлекса в его сигналыюсти (предупреждающем характере) и адаптив¬
ности (приспособительном характере). Условные рефлексы разделяются по при¬
знаку особенностей безусловного подкрепления, условного сигнала или по вре¬
менному соотношению обоих сигналов.
И. II. Павлов разработал классификацию феноменов торможения условных
рефлексов. К внешнему (безусловному) торможению он отнес гаснущий и посто¬
янный тормозы, а также запредельное торможение. В первых двух случаях услов¬
ные рефлексы тормозятся при действии ориентировочного или болевого раздра¬
жений. К внутреннему (условному) торможению, которое возникает вследствие
неподкрепления условного сигнала безусловным рефлексом, относят угасатель-
ное торможение, дифферепцировочное, запаздывания, условный тормоз.
По-видимому, сходство физиологической природы разных типов и видов тор¬
можения условных рефлексов не исключает различия в их нейрофизиологиче¬
ских механизмах.
Контрольные вопросы
1. Безусловные рефлексы и их классификация.
2. Правила образования условных рефлексов.
3. Общие признаки условных рефлексов.
4. Классификация условных рефлексов.
5. Внешнее торможение и его характеристики (но Павлову).
6. Запредельное торможение.
7. Внутреннее торможение и его основные характеристики (по Павлову).
8. Взаимодействие различных видов торможения.
Глава 8
МЕХАНИЗМЫ ФОРМИРОВАНИЯ УСЛОВНЫХ
РЕФЛЕКСОВ
штжшшжшттттктжттшшттяттштштттжжттшшшттшттжштштжтттжтттюштшшшштшжшж
§ 1. Конвергентная теория формирования
временных связей
Разработке основных законов центральной нервной координации физиология
обязана исследованиям Ч. Шеррингтона, сформулировавшего фундаментальный
закон в работе нервной системы — «принцип общего пу¬
ти->. Если афферентный нейрон является индивидуаль¬
ным или частным путем, то эфферентный нейрон являет¬
ся общим путем для импульсов, возникающих в любом из
многочисленных источников рецепции. Подобная коор¬
динация складывается на основе морфологического соот¬
ношения афферентных и эфферентных путей, где первые
всегда значительно преобладают над вторыми. Так фор¬
мируется и реализуется механизм конвергенции — схождения множества аффе¬
рентных импульсов в единственный анатомически ограниченный эфферентный
канал. Частные пути, конвергируя между собой, соединяются в промежуточные
пути, и окончательно конвергенция завершается общим конечным путем в виде
воронки. Каждый этап, на котором два или более афферентных нейрона конвер¬
тируют на третьем нейроне, который по отношению к ним является эфферент¬
ным, предполагает возможность влияния или столкновения их влияний.
Идею о принципе конвергенции творчески развил А. А. Ухтомский (1954),
распространив его па всю центральную нервную систему, на все ее «этажи». Он
полагал, что в результате конвергенции наступает интерференция стимулов, ко¬
торая реализуется в определенный нервный акт. Отсюда становится понят¬
ным богатое развитие синапсов и концевых ветвлений с тенденцией увеличить
поверхность соприкосновения нервной клетки с окончаниями приходящих ак¬
сонов.
Автор принципа доминанты выдвинул новые положения об интегрирую¬
щей деятельности нервной системы. Принцип статической конвергенции пу¬
тей по отношению к исполнительному пути при дальнейшей разработке рас¬
ширяется в принцип подвижной и физиологической конвергенции нервных им¬
пульсов по отношению к области преобладающего текущего действия (принцип
доминанты).
А. А. Ухтомский находил тесную связь между учением Шеррингтона об общих
путях и теорией И. П. Павлова об условных рефлексах. В статье «Об условном от¬
раженном действии» он писал: «Если воронка Шеррингтона имела в виду посто¬
янные рефлекторные дуги, закрепившие функциональную и морфологическую
связи между собой наследственно и филогенетически, то И. П. Павлов улавливал
самое закладывание и новообразование связей в этой воронке, привлечение все
Конвергениия — схо¬
ждение множества
афферентных импуль¬
сов в единственный
анатомически ограни¬
ченный эфферентный
канал.
222 Глава 8. Механизмы формирования условных рефлексов
Рис. 74. Схема образования условного рефлекса
(по А. Гасто, 1958)
1 — ретикулярная формация нижней части ствола мозга;
II —таламическая ретикулярная формация; III — кора моз¬
га; 1 — афферентный путь безусловного раздражителя;
2 — афферентный путь условного раздражителя; 3 и 4 —
ретикулярные нейроны; 5 и 6 — коллатеральные волокна
афферентных путей, вызывающие возбуждение ретикуляр¬
ных нейронов; 7 и 8 — коллатеральные волокна афферент¬
ных путей, не вызывающие возбуждения нейронов вследст¬
вие непроходимости синапсов; 9 — восходящие пути от
ретикулярной формации, активирующие кору диффузно;
10 — восходящие пути от ретикулярной формации, активи¬
рующие кору локально; 11 — безусловно-рефлекторная ре¬
акция. Временная связь образуется между афферентными
влияниями условного и безусловного раздражителей
новых рецептивных сфер к конвергенции относительно эфферентных (исполни¬
тельных) приборов»1.
Конвергенция гетеросенсорных стимулов была положена в основу понимания
механизмов образования условных рефлексов. Так, Дж. Экклс (1953) предполо¬
жил существование нелокализованпого «центра конвергенции», где сходятся
условный и безусловный раздражители. Встреча их в центре конвергенции, взаи¬
модействие и интеграция завершаются формированием особой пространственно-
временной структуры разрядов, которая затем передается в нейронную сеть. Мно¬
гократные сочетания включают механизмы нотенциации путей условного сигнала,
который приобретает способность запускать условно-рефлекторный акт. Дли¬
тельность хранения этого эффекта обеспечивается механизмами реверберации
возбуждения в нервных сетях. А. Фессар (1962) показал, что конвергенция, то
есть сближение в пространстве мозга разномодальных сигнализаций, является
важнейшим условием формирования временной связи на уровне отдельных ней¬
ронов. В качестве двух других условий он выдвинул взаимодействие этих сиг¬
нализаций на нейронах и использование следовых процессов. Взаимодействие
квалифицирующих сигналов связано с длительным удерживанием следов актив¬
ности. В этих процессах важную роль играют механизмы тонической и посттета-
нической потенциации.
Таким образом, конвергенция и потенциация образуют некую общую цепь ди¬
намических процессов, связывающих интеграцию в пространстве (обусловлен¬
ную конвергенцией) с интеграцией во времени (обусловленную постепенным на¬
растанием облегчения на окончаниях аксонов) (А. Фессар, 1964).
Первичная конвергенция осуществляется преимущественно па клетках рети¬
кулярных образований, таламуса и ствола мозга и передается на корковые нейро¬
ны в интегративной форме («проецируемая мультивалентность»). Корковые
структуры регулируют конвергентные свойства ретикулярных структур, что обес¬
печивает специализацию условного рефлекса (рис. 74).
Эти концепции не могли объяснить свойственные условному рефлексу устой¬
чивость и приспособительную пластичность, ибо не придали должного значения
фактору биологического подкрепления. Именно это было учтено П. К. Анохиным
1 Ухтомский А. А. Доминанта. С. 175.
§ 1. Конвергентная теория формирования временных связей 223
(1968), который считал, что решающим фактором в механизмах образования ус¬
ловных рефлексов выступает биологическое значение подкрепляющего рефлекса.
Конвергирующие на нейронах сенсорные и биологические модальности опре¬
деляют организацию условного рефлекса — его сигнальные свойства и при¬
способительное значение. Стимулы различной сенсорной и биологической мо¬
дальности воздействуют при сочетаниях па синаптические входы нейронов с ге-
терохимической организацией этих входов. Свойства подкрепляющего фактора
определяются специфическими химическими реакциями, которые он вызывает,
и соответствующими молекулярными преобразованиями в пределах мембраны
и цитоплазмы нейрона. Здесь и происходит «сцепление» условной и безусловной
стимуляции с последующим выходом сложившейся интеграции на аксон в виде
клеточного разряда.
Само понятие «сцепление» следов условного и безусловного стимулов предпо¬
лагает наличие специфических ультраструктурных и молекулярных преобразований
в цитоплазме нейрона. Гипотетическая схема, предложенная М. Я. Рабиновичем
(1975), приведена на рис. 75. Сочетание стимулов двух сенсорных модальностей
(б\ и б2) приводит к образованию кратковременной связи типа сенсо-сенсорной ас¬
социации. Сочетание же каждого из сенсорных стимулов с биологически значимым
подкреплением (а) эмоционально-мотивационного характера приводит к выработ¬
ке устойчивой связи, обладающей свойствами условного рефлекса.
Рис. 75. Вероятная схема взаимодействия синоптических входов с гетерохимической
организацией для стимулов биологической и сенсорных модальностей
(по М. Я. Рабиновичу, 1975)
а — биологическая модальность; б, и 62 — сенсорные модальности.
Различные свойства входов для сенсорных и биологически значимых сигналов
условно изображены в виде размеров синапса и его активных зон
Признавая, что наиболее существенные процессы образования временных свя¬
зей реализуются па молекулярном уровне нейрона, следует иметь в виду, что
системные механизмы временных связей всецело опосредованы сложными ней¬
рофизиологическими преобразованиями. Выделяют три типа проявлений муль-
тисенсорной конвергенции (Л. Л. Воронин, 1974). Первым типом является специ¬
фический ассоциативный, при котором клетки ствола мозга, таламуса или коры
реагируют специфическим ответом на стимулы разных сенсорных модальностей.
Вторым типом является специфический проекционный, при котором клетка реаги-
224 Глава 8. Механизмы формирования условных рефлексов
рует специфической реакцией на адекватный для данной системы раздражитель и
неспецифической — на стимулы других модальностей. Наконец, третий тип име¬
нуется неспецифическим и характеризуется неспецифической ответной реакцией
нейронов на разномодальные стимулы. М. Я. Рабинович (1975) разработал гипо¬
тетическую схему, в которой синаптические входы для условного и безусловного
стимулов конвергируют на пресинаптической («входной») клетке в цепи нейро¬
нов. Сложившаяся ассоциация («интеграция») передается на постсинаптический
нейрон («выходной»), но по одному общему пути и для сигнального, и для под¬
крепляющего раздражителей (рис. 76).
УС
БС
Рис. 76. Гипотетическая схема передачи условной связи, сложившейся в процессе
сочетаний, с «входного нейрона» (а) на выходной нейрон (б) по одному обшему пути
(по М. Я. Рабиновичу, 1975)
Клетка круглой формы — вставочный нейрон возбуждающего действия; на осциллограммах — различные
формы условной активности, которые возможно регистрировать с «выходного» нейрона: 1 — обычная
реакция на условный и безусловный стимулы (УС и БС), обозначенные длинной и короткой линией под
осциллограммой; 2 — воспроизведение следовых разрядов на БС при его пропуске в сочетании; 3 — вос¬
произведение следовых разрядов на УС и БС при пропуске всего сочетания (обозначено пунктиром;
в условиях реакции на «время»
При рассмотрении разномодальной конвергенции необходимо учитывать
чрезвычайно важное свойство нейронов и нервных сетей, а именно способность
к организации пластического характера конвергенции. В этой связи следует
разграничивать конвергенцию анатомическую, или статическую, и конвергенцию
функциональную, или динамическую. Особенности организации нервных сетей го¬
ловного мозга обеспечивают связь по принципу «от каждого ко всем». Следова¬
тельно, морфологические предпосылки могут создать такие условия, когда на од¬
ной нейроне будут конвергировать самые разнообразные влияния. В процессе
эволюции и в динамике постнатального онтогенеза в головном мозге закрепляет¬
ся олигосинаптическая сеть, обеспечивающая четкий адресат проведения инфор¬
мации наряду с системой полисипаптических сетей, связи между компонентами
которой могут носить функциональный временный характер. Это обстоятельство,
то есть высокая вариативность свойств полисенсорных нейронов, и обусловливает
многообразие их реакций на разномодальную стимуляцию.
§ 2. Клеточные аналоги условного рефлекса 225
§ 2. Клеточные аналоги условного рефлекса
Выделяют две основные гипотезы обучения: синоптическую, предполагающую,
что механизмом формирования условных рефлексов является изменение эффек¬
тивности синапсов, и мембранную, утверждающую, что основным механизмом
условного рефлекса является изменение свойств возбудимой ностсипаптической
мембраны. Как мы увидим далее, такое деление весьма условно, однако представ¬
ляется удобным для первого знакомства с проблемой.
Нервные механизмы обучения обеспечиваются благодаря двум основным
свойствам нервной системы — реактивности и пластичности. Если реактив¬
ность — это способность отвечать на раздражения, то пластичность — это способ¬
ность изменять реактивность под влиянием последовательных раздражений.
В свою очередь, реактивность определяется возбудимостью ностсипаптической
мембраны и эффективностью соответствующих синапсов. Возбудимость нейрона
измеряется силой тока, необходимой для генерации потенциала действия, а синоп¬
тическая эффективность — амплитудой моносинаптического постсинаптическо¬
го потенциала.
Уже давно высказывалось предположение о том, что в основе обучения лежит
изменение эффективности межклеточных контактов, или проторение новых кон¬
тактов. На рис. 77 приведены основные схемы. Простейшим является синапс
Дж. Экклса (а), повышающий свою эффективность после собственной активации
(фасилитация). Такие синапсы могут служить для объяснения феноменов постте-
танической потенциации. Дополнением к ним являются синапсы с постепенным
снижением эффективности при повторной активации (б). Они могут обеспечи¬
вать эффекты габитуации и дегабитуации. Как мы уже отмечали, при выработке
условных рефлексов необходимо временное совпадение пресинантической акти¬
вации с активацией подкрепляющих систем, которые могут оказывать также нре-
синантическое действие (в). Д. О. Хебб (1969) главным условием синаптической
модификации считал совпадение пресинантической активности с разрядом пост¬
синаптического нейрона (г). Затем к этому была добавлена модель Альбуса, свя¬
занная не с увеличением эффективности синапса, а с ее снижением (д). Наконец,
в качестве синтетической схемы (е) рассматриваются случаи изменений синапти¬
ческой эффективности с участием трех факторов — сенсорного притока, актива¬
ции подкрепляющей системы и достижения результата.
Вопрос о характере преобразований нейрональной активности в динамике вы¬
работки условного рефлекса начал исследоваться в связи с внедрением в практи¬
ку физиологического эксперимента тонких методов микроэлектродного анализа
и последующей компьютерной обработки импульсных реакций нейронов. Попыт¬
ки использовать внутриклеточную регистрацию постсинаптических электриче¬
ских потенциалов привели к необходимости проводить исследования либо на
относительно простых нервных сетях, либо на центральных нейронах высших
млекопитающих с максимальным упрощением требуемых для анализа межней¬
ронных отношений. Такие упрощенные модели принято рассматривать в качестве
клеточных аналогов условного рефлекса.
Группой венгерских физиологов (О. Фехер, А. Барани, 1981) разработана мо¬
дель ассоциативного обучения, при котором сигнал меняет свое действие под
влиянием последующего безусловного эффекта.
8 За* 1078
226 Глава 8. Механизмы формирования условных рефлексов
а б в
+ - +
Рис. 77. Основные виды гипотетических модифицирующихся синапсов
(по Л. Л. Воронину, 1986)
а — активация синапса; б— то же; в — активация синапса и подкрепляющей системы; г— активация
синапса и постсинаптического нейрона; л — то же; е — активация синапса, постсинаптического ней¬
рона и подкрепляющей системы; квадрат — модифицирующиеся синапсы, стрелка — синапсы подкре¬
пляющей системы, кружок — постсинаптический нейрон, зачерненные знаки — элементы, активация
которых входит в условие модификации данного синапса, (+) — повышение, (-) — снижение
Рассмотрим процедуру обусловливания при «условном» раздражении вентро-
латеральпого ядра таламуса (ВЛ) и антидромной стимуляции пирамидного трак¬
та («безусловный стимул»). Сначала, как показано на рис. 78, электрическая сти¬
муляция ВЛ с частотой 1 с сопровождается только подпороговыми ПСП. После¬
дующее антидромное раздражение пирамидного тракта вызывает один потенциал
действия. По мере сочетаний раздражения таламуса с антидромной стимуляцией
амплитуда и/или длительность ТПСП увеличиваются и остаются таковыми в те¬
чение нескольких минут. Амплитуда ВГ1СП изменяется также, но к 50-му сочета¬
нию ВПСП достигает порога генерации и «условный» сигнал начинает вызывать
1-2 потенциала действия. Эффект антидромного раздражения при этом не меня¬
ется. К 100-му сочетанию амплитуда ВПСП еще больше возрастает, а число по¬
тенциалов действия достигает трех. Если проводить угашение, то есть подавать
Рис. 78. Эффекты гетеросинаптического облегчения на корковом нейроне
пирамидного тракта у кошек (по А. Барани и О. Фехеру, 1980)
УС — условный стимул; УР —- условная реакция; БС — безусловный стимул; БР — безусловная реакция
слева —до сочетания; справа — после сочетания. Приведены постсинаптические реакции нейрона на аде¬
кватную, антидромную стимуляцию пирамидного тракта и внутриклеточную поляризацию постоянным то¬
ком. ВЛ — адекватная стимуляция; ПТ — пирамидный тракт; ПОЛ — внутриклеточная поляризация
§ 2. Клеточные аналоги условного рефлекса 227
одно «условное» раздражение, не сопровождая его антидромной стимуляцией,
то амплитуда ВПСП и число генерируемых потенциалов действия уменьшается,
и спустя 350 изолированных раздражений таламического ядра ответная «услов¬
ная» реакция исчезает. Внутриклеточная поляризация исследуемого нейрона, при¬
водившая к генерации потенциалов действия, усиливала эффект раздражения
вептролатерального таламуса. Однако ни многократное применение только «услов¬
ного» стимула, ни псевдообусловливание (процедура применения условного
и безусловного раздражителей в случайном порядке) не привели к генерации по¬
тенциалов действия исследуемого нейрона. Уровень мембранного потенциала
и сопротивление мембраны при выработке временной связи менялись по-разному
в различных клетках. Это позволило думать об определяющей роли при выработке
временной связи избирательных синаптических сдвигов.
Было показано, что положительные и дифферепцировочпые связи с иссле¬
дуемым нейроном опосредуются разными синапсами независимо друг от друга.
Е. И. Соколов (1931) справедливо полагает, что элементарным локусом пластическо¬
го изменения является не нейрон, а синапс. Речь идет о синапсе Хебба, который се¬
лективно изменяется, если за синаптической посылкой следует спайковый разряд
нейрона. Такое явление получило наименование гетеросинаптического облегчения.
Этот же феномен использовался для объяснения ряда эффектов обусловлива¬
ния в другой модели (Л. Л. Воронин, 1982). Синаптические аналоги условного
рефлекса создавались в зоне коркового представительства передней конечности
бодрствующего кролика сочетаниями электрического раздражения двух точек ко¬
ры. Условным стимулом служило нодпороговое (неэффективное) раздражение
коры на некотором расстоянии от исследуемого коркового нейрона, а безуслов¬
ным — пороговое для постсинаптической реакции раздражение поверхности коры
над исследуемым корковым нейроном. При активирующих формах условной свя¬
зи в ответ па условный стимул возникали ВПСП, амплитуда которых постепенно
возрастала до критического уровня генерации потенциалов действия (рис. 79).
При отмене подкрепления, то есть при угашении, амплитуда ВПСП постепенно
уменьшалась с прекращением генерации спайковых разрядов.
Рис. 79. Усиление ВПСП и импульсных реакции на аналоге
корково-коркового условного рефлекса (по А. А. Воронину, 1986)
По оси ординат — изменение амплитуды ВПСП в ходе опыта; по оси абсцисс — номера сочетаний;
светлые кружки — амплитуда ВПСП в ответ на изолированный УС; темные кружки — то же во время
сочетаний; вертикальные черточки и стрелки отмечают генерацию частичного и полномерного потен¬
циалов действия соответственно
При тормозных формах временной связи условный стимул постепенно вызы¬
вал развивающееся угнетение механизма генерации импульсных разрядов, кото¬
рое не сопровождалось существенной гииерполяризацией мембраны клетки или
достоверным уменьшением ВПСП. При этом установлено, что дифферепцирова-
228 Глава 8. Механизмы формирования условных рефлексов
ние одним и тем же нейроном подкрепляемых и неподкренляемых стимулов реа¬
лизуется различными синаптическими входами и поддерживается благодаря ме¬
ханизмам гетеросипаптического облегчения.
Одной из моделей аналога временной связи на микропопуляции нейронов ко¬
ры больших полушарий является микроионофоретическая аппликация медиато¬
ров (глутамата и ацетилхолина), используемых в качестве аналогов ассоциируе¬
мых раздражителей (Б. И. Котляр, 1986). Установлено, что отдельные нейроны
микропопуляции в радиусе действия медиаторов обнаруживают специфические
ассоциативные перестройки своей активности. Последние могут заключаться
в следующем: изменения реактивности к сигнальному раздражителю, воспроизве¬
дение реакции на подкрепляющий раздражитель в ответ на действие условного
сигнала и реакция на время в границах действия ассоциируемых раздражителей.
В. С. Русинов (1969) пришел к выводу, что суммациопный рефлекс и доминан¬
та представляют собой начальные стадии образования любого условного рефлек¬
са и сами но себе могут служить удобной моделью для изучения простых форм
временной связи. Им была разработана модель корковой поляризационной доми¬
нанты, состоящая в том, что поляризация постоянным током участков сенсомо-
торной коры кролика при действии ранее неэффективных раздражителей — све¬
товых вспышек, приводила теперь к ритмическому сокращению передней конеч¬
ности (корковая проекция которой поляризовалась) в такт световой стимуляции.
Установившаяся под влиянием поляризации новая форма реагирования на раз¬
дражитель сохраняется до 30 мин после выключения тока.
Эта модель послужила существенным толчком для выявления пластических
характеристик корковых нейронов. Установлено, что поляризация слабым посто¬
янным током увеличивает активность нолисенсорных нейронов, что является од¬
ним из непременных механизмов возникновения движения на ранее индиффе¬
рентный раздражитель. Причем функциональные и морфологические сдвиги про¬
исходят не только в пирамидных нейронах, по и в иитернейронах и глиальных
клетках, которые также подвергаются поляризации.
Все основные свойства доминанты — повышенная возбудимость, стойкость
возбуждения, способность к суммированию возбуждений, инерции, торможению
и растормаживаншо — были обнаружены на вышеописанной модели. В доминант¬
ном очаге происходит установка на определенный уровень стационарного возбу¬
ждения, способствующий суммированию ранее нодпороговых возбуждений и пе¬
реводу на оптимальный для данных условий ритм работы, когда этот очаг стано¬
вится наиболее отзывчивым и тормозятся другие работающие очаги. Иными
словами, образуется единый функционально организованный ансамбль, или систе¬
ма. Подчеркивается особая роль нейронов нижних слоев коры, которые дольше
сохраняют ритм раздражения, чем клетки верхних слоев. Обращается серьезное
внимание па существование в системе базальных депдритов пирамидных клеток
депдродеидритических контактов, которые рассматриваются как важнейший суб¬
страт межнейронных отношений. В. С. Русинов полагает, что наиболее адекват¬
ным отражением функционального состояния системы нейронов является уро¬
вень их постоянного потенциала, а следовательно, электротонические влияния
имеют наиболее существенное значение для образования временной связи. Поля¬
ризационный очаг, созданный постоянным током, служит удачной моделью для
изучения поляризационно-электротонической гипотезы формирования доминан¬
ты и простых форм временных связей (В. С. Русинов, 1969).
§ 2. Клеточные аналоги условного рефлекса 229
Специальное внимание к нейроглиальным взаимоотношениям при формиро¬
вании клеточных аналогов условного рефлекса привлек А. И. Ройтбак (1969). Он
выдвинул глиальную гипотезу, согласно которой в основе образования и укрепле¬
ния временных связей лежит миелииизация центральных аксонов, возбуждаю¬
щихся от условного раздражения. При действии безусловного раздражения в со¬
ответствующих областях коры происходит деполяризация глиальных клеток: воз¬
буждение большого числа нервных элементов импульсами из специфических
и неспецифических путей приводит к выделению в межклеточные щели и аккуму¬
ляции ионов калия. Автор полагает, что деполяризация олигодеидроцитов слу¬
жит сигналом к образованию миелина; условием миелинизации является совпа¬
дение во времени деполяризации олигодеидроцитов и физико-химических из¬
менений в прилегающих немиелинизированиых пресииаптических терминалях
после их предшествующего возбуждения (рис. 80).
В последние годы широко развернулись исследования механизмов временных
связей на животных с относительно простой нервной системой. Наиболее удоб¬
ными для экспериментирования оказались моллюски. Простейшей моделью слу¬
жил полностью изолированный нейрон висцерального ганглия виноградной
улитки. Локальная аппликация ацетилхолина сочеталась с внутриклеточной инъ¬
екцией деполяризующего тока, в результате было показано, что изолированные
нейроны моллюсков (то есть лишенные своих сетевых свойств) не способны фор¬
мировать ассоциативный ответ в виде воспроизведения безусловной реакции сиг¬
нальным раздражителем.
Рис. 80. Схема процесса образования временной связи (по А. И. Ройтбаку,1972)
а — «потенциальный» синапс; б — образование временной связи (превращение «потенциального» си¬
напса в «актуальный»). 1 — пресинаптический участок нервного окончания; 2 — синаптическая шель;
3 — олигодендроцит; 4 — миелиновая оболочка; 5 — медиатор в синаптических пузырьках. Эффек¬
тивность проведения тока по терминальному волокну увеличивается вследствие формирования во¬
круг него оболочки из олигодендроцита
Клеточные механизмы образования условного рефлекса па морском моллюске
аплизии изучались Э. Кенделом (1980). Исследовался оборонительный рефлекс:
условный сигнал — умеренной силы раздражение сифона, вызывающее втягива¬
ние жабры в мантийную полость, безусловный раздражитель — сильная стимуля¬
ция хвоста. В результате сочетаний защитная реакция на стимуляцию сифона
усиливалась. Удобство этой модели состояло в том, что был точно определен ней¬
ронный состав рефлекторной дуги, в которую входят 24 сенсорных нейрона, обра¬
зующие прямые связи с иитернейронами и 6 моторными нейронами, иннервирую¬
щими жабру и сифон. Стимуляция хвоста возбуждает нейроны, контактирующие
с пресипантическими окончаниями сенсорных нейронов.
В данной модели обнаружились явления сенситизации, привыкания и нейро-
гуморального (субклеточно-молекулярного) воздействия безусловного раздражи¬
230 Глава 8. Механизмы формирования условных рефлексов
теля на всех этапах выработки временной связи. Суммация заключалась в неспе¬
цифическом гетеросинаптическом облегчающем эффекте — увеличении ВПСП,
связанном с увеличением выброса медиатора из пресипаптического окончания.
Суммация этих процессов приводит к возникновению длительной деполяриза¬
ции, составляющей сущность проторения путей при образовании временной свя¬
зи. Процессы привыкания связаны с ограничением потоков кальция в пресинан-
тические окончания, что приводит к гиперноляризации нейронов и снижению
их ответа па безусловный раздражитель. Нейрогуморальный эффект безусловно¬
го раздражителя достигался благодаря механизмам регуляторных нейропептидов
(см. гл. 9).
Э. Кендел (1980) считает, что в основе условной реакции втягивания жабры
у аплизии лежит механизм пресипаптического облегчения (сенситизации), кото¬
рый формируется при конвергенции раздражителей, приводящих к спайковой ак¬
тивности нейрона, и активации этого же нейрона через модулирующий вход от
безусловного раздражителя. Аналогичный механизм допускается и у позвоноч¬
ных животных, хотя прямые доказательства этого отсутствуют.
Какими бы информативными для задач общей нейрофизиологии ни были ана¬
литические подходы к исследованию наркотизированных или обездвиженных пре¬
паратов позвоночных и беспозвоночных животных с использованием различных
моделей и аналогов условных рефлексов, их конечной целью является расшифров¬
ка тех феноменов мультинейроиной активности, которые регистрируются на бодр¬
ствующем мозге при выполнении животным того или иного целостного условно-
рефлекторного акта. Сегодня мы еще не можем использовать весь арсенал сведе¬
ний аналитической нейрофизиологии для понимания клеточной активности
бодрствующего мозга. Различия двух основных подходов базируются на степени
проявления свойства пластичности, а именно — высокой изменчивости реакций
одного и того же нейрона у бодрствующего животного и значительно большей ста¬
бильности реагирования нервных клеток у наркотизированных препаратов.
§ 3. Нейронная организация условно-
рефлекторного процесса
Данные о перестройке нейрональной активности при выработке условных реф¬
лексов у бодрствующих животных представляют существенное значение для по¬
нимания функциональной организации целостной системы обучения. Такие из¬
менения реакций нейронов могут проявляться практически во всех исследован¬
ных структурах мозга. Логично допустить поэтому, что возможность образования
ассоциаций является базовым свойством нервной системы.
В связи с тем, что сигнальная сущность условного рефлекса состоит во времен¬
ном сочетании, то есть в ассоциации раздражений, представлялось необходимым
в первую очередь выделить перестройки нейрональной активности, связанные
с ассоциацией, в отличие от всех остальных сопровождающих изменений в актив¬
ности нейронов.
Р. Томпсон рекомендует для этой цели использовать три критерия:
♦ специфичность изменений активности на условный сигнал в сравнении с из¬
менениями, вызываемыми индифферентными раздражителями;
§ 3. Нейронная организация условно-рефлекторного процесса 231
♦ параллельность развития изменений нейронной активности и компонентов по¬
веденческого условного ответа;
♦ градуальный рост выраженности пластических изменений па нейронном уров¬
не в процессе выработки поведенческого навыка.
Другие авторы (Э. Кендел, 1980) предложили для той же цели рассматривать
два независимых критерия: 1 — природа ответа па условный сигнал, то есть явля¬
ется ли условный ответ новым (классический условный ответ) или модификаци¬
ей условной реакции на сигнальный раздражитель (альфа-условный ответ), и 2 —
специфичность реакции для последовательного сочетания раздражителей. Ины¬
ми словами, не может ли аналогичный ответ развиваться при случайном предъяв¬
лении тех же раздражителей (при процедуре псевдообусловливания) или опреде¬
ляться сеиситизацией.
Б. И. Котляром (1986) предложена классификация перестроек активности
нейронов в процессе условно-рефлекторной деятельности (табл. 5). Принципи¬
альным является введение в состав условно-рефлекторного процесса топического
типа активности нейронов. Основанием этому служили специальные исследова¬
ния с переключением разнородных условных рефлексов, которые позволили рас¬
сматривать центральный мозговой тонус в качестве непременного компонента
процессов обучения и памяти.
Таблица 5
Классификация разных видов функциональной активности нейронов головного
мозга при формировании и реализации временной связи (Б. И. Котляр, 1986)
Ассоциативные перестройки
Неассоциативные перестройки
физического типа
тонического типа
физического типа
тонического типа
Перестройки, отра¬
жающие уровень
центрального тону¬
са, характерного для
реализации времен¬
ной связи
Перестройки, харак¬
терные для тониче¬
ской составляющей
ассоциативного про¬
цесса
Перестройки, отра¬
жающие динамику
ориентировочной
реакции
Перестройки, отра¬
жающие динамику
ориентировочной
реакции
Специфические пе¬
рестройки, воспро¬
изводящие безуслов¬
ный ответ
Перестройки, свя¬
занные с домини¬
рующей мотивацией
Перестройки, свя¬
занные с домини¬
рующей мотивацией
Перестройки, отра¬
жающие реакции на
время в рамках дей¬
ствия ассоциируе¬
мых раздражителей
Перестройки, свя¬
занные с условной
ориентировочной
реакцией
По результатам обучения па уровне отдельной клетки можно судить, что наи¬
более существенные изменения характерны для корковых полисенсорных нейро¬
нов (М. Я. Рабинович, 1975), функциональные свойства которых по мере сочета-
232 Глава 8. Механизмы формирования условных рефлексов
иий претерпевают эволюцию, отражающую становление его афферентной спе¬
циализации. Так, до первых сочетаний стимула с подкрепляющим рефлексом
нейроны проявляют преимущественно моносенсорные реакции, то есть большин¬
ство из них отвечает на стимулы адекватной сенсорной модальности стабильны¬
ми коротколатентными разрядами. По мере последующих сочетаний эти же ней¬
роны приобретают способность отвечать на стимулы различных сенсорных мо¬
дальностей и параметров; их активность становится полисепсорной. Этот период
соответствует фазе генерализации условного рефлекса, когда положительное дей¬
ствие обнаруживают не только сигнальный, но и другие сенсорные раздражители.
Вероятно, именно в этот период данная рефлекторная система начинает приобре¬
тать доминантные свойства, выделяться среди прочих и подкрепляться разнооб¬
разными воздействиями. Наконец, при достаточно упроченном условном рефлек¬
се ответы нейронов снова приобретают моносенсорный характер, но теперь уже это
специальные моносенсорные реакции па стимул определенной модальности и
физических параметров, то есть избирательная реакция только на тот стимул, ко¬
торый приобрел сигнальное значение для поведенческого акта.
Следует отметить, что характеристики процесса привыкания реакций нейро¬
нов при монотонных предъявлениях сенсорных стимулов не коррелируют с про¬
цессом ассоциативного обучения. Одни нейроны в популяции могут проявлять
феномен привыкания, а другие — сеиситизацию, следовательно, нейрофизиологи¬
ческие механизмы привыкания и ассоциативного обучения могут быть различны
по своей организации. Если в основе привыкания лежит уменьшение числа кван¬
тов медиатора, ведущее к снижению амплитуды постсинаптических потенциалов,
ассоциативное обучение может быть связано со сдвигом эндогенной активности
обучаемого нейрона. Однако нельзя исключить возможность причинно-следст¬
венной связи между привыканием и сепситизацией. Действительно, на одном и
том же мультисенсорном нейроне можно наблюдать в процессе предъявления сти¬
мулов оба явления сразу. Например, в начале процедуры привыкания ответа па
световой стимул электрокожпый стимул, подаваемый в интервале функциональ¬
ных сдвигов после светового стимула, оказался неэффективным. Однако по мере
ослабления ответной реакции нейрона на световой стимул (привыкание) начина¬
ет появляться ответ того же нейрона на электрокожпый стимул (сенситизация).
Последний достигает своего максимума при отсутствии ответа нейрона на свето¬
вую вспышку (А. С. Батуев, 1978).
Преобладание того или иного механизма существенно зависит от сигнальной
значимости предъявляемого раздражения. Так, в опытах па бодрствующих обезь¬
янах было установлено, что сенсибилизация в реагировании нейронов лобной ко¬
ры возникает лишь в том случае, если раздражители прямо или косвенно связаны
с мотивационной сферой животного, сигнализируют об опасности или несут на
себе элементы новизны. Привыкание ответов тех же нейронов наблюдалось в тех
случаях, когда используемые раздражители были индифферентными или хорошо
знакомыми для животного. Однако если ранее индифферентный агент приобре¬
тал качества сигнального фактора, будучи сочетаем во времени с биологически зна¬
чимым болевым агентом, то картина резко менялась (рис. 81). Ранее угашенные
нейронные ответы уже через 9-10 сочетаний возобновляются, что характеризует¬
ся всеми чертами выработки условного рефлекса и подтверждается параллельной
регистрацией ЭМ Г рабочей конечности. Такой пластический характер мультисеп-
сорной конвергенции к нейронам лобной коры, диапазон и устойчивость которой
§ 3. Нейронная организация условно-рефлекторного процесса 233
i
■J.JI Ijim.i ..ДЩ. .444 ]
I
IV
10
5
о
t t
jit-jU-JKljlUU.
UU-4J
»
Рис. 81. Динамика импульсной активности коркового нейрона лобной области у обезьян
и электромиограмма (ЭМГ) мыши плеча при выработке условного рефлекса
(по А. С. Батуеву и др., 1981)
I (сверху вниз): нейронная активность, ЭМГ и график изменения средней частоты (по девяти предъяв¬
лениям) при световом раздражении; II — при кожном раздражении (уколе); III и IV — при сочетании
светового раздражения с уколом соответственно при 1 -9-м в 10-18-м их сочетаниях; V —- график из¬
менения латентного периода ЭМГ-сдвигов. На графиках I-IV'. по оси абсцисс — время в с; по оси орди¬
нат— средняя частота в имп/спри At — 100 мс; горизонтальной штриховой линией указан уровень фо¬
новой активности, на графике V — по оси ординат — порядковые номера сочетаний; по оси
абсцисс — время в с; вертикальными штриховыми линиями А и Б и стрелками указаны соответствен¬
но момент светового раздражения и момент укола. Калибровка: 500 (верхний луч) и 250 (нижний
луч) мкВ, 50 мс. Графики и осциллограммы представлены в едином временном масштабе
234 Глава 8. Механизмы формирования условных рефлексов
определяется сигнальной значимостью раздражителя, в конечном счете форми¬
руется доминирующей мотивацией и системой выработанных ранее временных
связей.
По критерию наличия или отсутствия изменений реакций нейронов нельзя
считать, что обучение характерно лишь для определенных отделов или уровней
мозга. Имея в виду неравноценность отделов мозга для процессов обучения, сле¬
дует признать, что сами по себе изменения реакций нейронов при выработке вре¬
менных связей в отсутствие дополнительных контрольных процедур еще не явля¬
ются доказательством причастности тех или иных структур мозга к процессам
обучения.
Как правило, ищут корреляцию между изменением реакций нейронов и пове¬
денческими сдвигами. Однако для заключения об организации системы, обеспе¬
чивающей условно-рефлекторный процесс, надо но крайней мере знать: любые
или вполне определенные изменения реакций нейронов соответствуют новой сиг¬
нальной значимости сенсорных стимулов и из каких образованиях мозга эти из¬
менения проявляются наиболее отчетливо. Для того чтобы установить связь ней¬
ронных перестроек именно с процессами «сцепления» условного и безусловного
стимулов, необходимо исключить возможные другие причины. К таковым могут
относиться:
♦ возможность изменения эффективной интенсивности сенсорного стимула;
♦ влияние на биоэлектрическую активность мышечных артефактов при позных
перестройках и выполнении движений;
♦ увеличение общей песпецифической возбудимости, так называемой тониче¬
ской составляющей ассоциативного процесса (по Б. И. Котляру, 1977).
При регистрации реакций нейронов разных образований мозга во время фор¬
мирования условного рефлекса возникает вопрос об их первичности. Конечные
изменения на уровне одного нейрона могут быть одинаковыми независимо от пер¬
вичности или вторичности их характера. Поиск наиболее существенных для об¬
учения структур мозга заключается в определении локализации тех нейронных
популяций, где изменения происходят в первую очередь. Критерием вовлеченно¬
сти должен являться предшествующий характер реакций нейронов относительно
поведенческой реакции, из чего логически вытекает необходимость существова¬
ния «критических синапсов», в которых прежде всего происходят изменения, об¬
условливающие распространение импульсации по новому пути. Причем наряду
с сигнально-детерминированными реакциями описан класс сдвигов импульсной
активности, которые также возникают по механизму воспроизведения следов ра¬
нее выработанных связей, но без непосредственного участия внешних сигналов —
центрально-детерминированные реакции (М. Я. Рабинович, 1975). Именно эти
реакции могут послужить удобной моделью для исследования механизмов произ¬
вольной двигательной деятельности.
Структуры мозга, в которых обнаруживаются первичные сдвиги нейрональ¬
ной активности, согласно Дж. Олдсу, представлены в разных отделах мозга. Их
можно разделить на неспецифические и специфические. К первой группе отнесе¬
ны те образования, в которых нейроны реагируют сходным образом на условные и
любые сходные стимулы той же модальности, но отличающиеся от первых по сво¬
им физическим параметрам.
§ 3. Нейронная организация условно-рефлекторного процесса 235
В специфических отделах нейроны не проявляют черт генерализации в реаги¬
ровании и не имеют сходных реакций на новые стимулы. В качестве наиболее зна¬
чимых для обучения отделов мозга отмечены фронтальная и сенсомоторная кора,
а также гиппокамп, где обнаружены изменения импульсной активности пирамид
слоя СА3 уже при первых сочетаниях условного и безусловного стимулов, и раз¬
ряд нейронов предшествовал двигательной реакции.
Группа американских ученых под руководством Ч. Вуди (1982) исследовала
внутриклеточно реакции нейронов сеисомоторпой коры кошек при выработке
мигательного условного рефлекса на звук. Безусловным стимулом служило меха¬
ническое постукивание по переносице. Реакции нейронов сеисомоторпой коры
предшествовали появлению первых мигательных движений. В результате обуче¬
ния наблюдалось, с одной стороны, избирательное снижение порогов миографи¬
ческих реакций круговой мышцы глаза, осуществляющей мигание в ответ на по¬
верхностную или внутрикорковую микростимуляцию сеисомоторпой зоны, а с
другой — избирательное усиление реакции и увеличение числа реагирующих ней¬
ронов, проецирующихся к соответствующей мышце. Было установлено, что поро¬
ги генерации спайков иа внутриклеточно приложенный импульс тока достоверно
ниже у тех нейронов, которые проецируются к мышцам, осуществляющим реак¬
цию мигания, по сравнению с проекционными нейронами других лицевых мышц.
Условные реакции нейронов на звуковой сигнал появляются с коротким латент¬
ным периодом, не превышающим латентный период реакций клеток первичной
слуховой коры, и могут сохраняться в течение длительного времени — 14 недель.
Указанные функциональные сдвиги нейронов сеисомоторпой коры были ре¬
зультатом только выработки условного рефлекса. Функциональное выключение
сеисомоторпой области аппликацией хлорида калия вызывает утрату реакций
этих нейронов и соответственно исчезновение самого акта мигания, хотя без¬
условная мигательная реакция не страдает. Результаты этих экспериментов сви¬
детельствуют о первичности пластических изменений в нейронах коркового пред¬
ставительства двигательной условной реакции.
В пользу локализации пластических изменений в сеисомоторпой коре свиде¬
тельствуют также результаты анализа нейронных механизмов условной реакции
постановки конечности на опору у кошек (Б. И. Котляр и др., 1983). Под этой ре¬
акцией понимают последовательность движений, вызванных прикосновением
предмета к дорзальной поверхности лапы, обеспечивающих постановку лапы па
поверхность этого предмета. Сигнальная условная реакция, вызванная тактиль¬
ной стимуляцией вентральной поверхности передней конечности, имела то же
двигательное проявление, что и исходная реакция, развивающаяся при раздраже¬
нии дорзальной поверхности лапы (безусловная) у необученных животных.
В результате обучения возбудительный ответ нейронов в корковом представи¬
тельстве бицепса плеча мышцы, выполняющей основную роль в сгибании локте¬
вого сустава, на условный раздражитель был в 2-2,5 раза больше, чем ответ на тот
же раздражитель у контрольных животных. Этот аффект не сопровождался изме¬
нением сенсорного притока в сенсомоторную кору.
Таким образом, реорганизация сенсомоторных связей, достигаемая путем об¬
учения, состоит в изменении эффективности связей между нейронами сенсомо-
торной коры:
♦ получающими сенсорно обусловленное эфферентное возбуждение;
♦ контролирующими сокращениями определенных групп мышц.
236 Глава 8. Механизмы формирования условных рефлексов
Внутрикорковые системные механизмы временных связей. Технические усло¬
вия ограничивают возможности одновременной регистрации активности большо¬
го числа нейронов. Чаще всего используют регистрацию активности одного, ре¬
же — двух-трех нейронов при формировании временных связей. Вместе с этим
важно при анализе характера импульсации такого нейрона исходить из предпо¬
сылки его обязательного участия в составе определенной нервной сети.
У. Г. Гасанов (1981) полагает, что методическим приемом для выявления такой
элементарной микросистемы является одновременная регистрация нескольких
клеток с последующей выборкой отдельных импульсных потоков и статистиче¬
ский анализ перекрестных интервалов между импульсами двух и трех выделен¬
ных рядов.
Он выделяет следующие особенности взаимосвязиой деятельной и нейронов
при обучении.
Деятельность микросети качественно отличается от индивидуальной активно¬
сти входящих в его состав нейронов.
Системная деятельность корковых нейронов при разномодальных длитель¬
ных раздражениях, а также на разных стадиях выработки условного рефлекса и при
угашении его основывается на разных формах взаимоотношений нейронов.
Микросистема основывается на дифференцированном участии в ней различно¬
го класса нейронов, по-разному меняющих свои сетевые свойства при обучении.
Предполагается, что в основе изменений реакций нейрона при обучении лежат
механизмы неспецифического характера. Локальные же микросети нейронов
включаются в более обширные многоуровневые системы общецеребралыюго ха¬
рактера. Для выявления собственно корковых процессов формирования времен¬
ных связей необходима либо внутриклеточная регистрация, которая но своей су¬
ти не может быть длительной, либо использование строго локальных воздействий
на исследуемый нейрон, например с помощью микроэлектрофореза различных
биологически активных соединений. Такая форма эксперимента предъявляет осо¬
бые требования к самой процедуре условно-рефлекторного обучения, состоящей
в том, что и формирование и угашение временной связи осуществляется на протя¬
жении одного и того же опытного сеанса при параллельной регистрации импульс¬
ной активности одного и того же нейрона.
Мигательный условный рефлекс отвечает таким требованиям. В. С. Сабанов
(1985) основное внимание обратил на самые ранние этапы формирования времен¬
ной связи, когда внешних проявлений условной двигательной реакции еще не на¬
блюдалось. При сочетании ритмического звукового и механического (задув воз¬
духа на роговицу глаза) раздражений наблюдалось два типа перестроек нейро¬
нальной активности: активация нейрона на условный сигнал, к которому данный
нейрон был нереактивен, и тормозной: на условный сигнал появляется тормозная
реакция, которая до сочетаний регистрировалась лишь в ответ на предъявление
безусловного сигнала (рис. 82). Эта тормозная реакция вырабатывается быстро
и угашается при изолированном действии условного сигнала. Характерно, что
клетки с активирующим типом реакции были исходно полисенсорные, то есть
реагировали на условный и безусловный сигналы, а клетки с тормозным типом
реагирования на условный сигнал были исходно моносенсорными и становились
полисенсорными лишь в процессе обучения. Исходно бисенсорные клетки с акти¬
вирующими условными реакциями отражают результат ассоциативного процес¬
са, сложившегося в результате сочетаний конвергирующих стимулов на докорко-
§ 3. Нейронная организация условно-рефлекторного процесса 237
Рис. 82. Динамика импульсной
активности коркового нейрона
с тормозным типом условной
реакции (по В. С. Сабанову, 1984)
Тонкими горизонтальными линиями отме¬
чено время нанесения звуковых щелчков,
момент нанесения безусловного стимула
(ВС) — струи воздуха на роговицу глаза;
цифры — порядковые номера сочетаний;
верхний луч — импульсная активность ней¬
рона; нижний луч — электромиограмма
мышц глаза; СБ — изолированное предъ¬
явление условного сигнала. Калибровка:
1 с и 1 мкВ
вых уровнях интеграции, — статическая проецируемая конвергенция, тогда как
тормозные условные реакции нейронов обусловлены внутрикорковыми механиз¬
мами конвергенции и интеграции — локальная динамическая конвергенция. Во¬
прос о месте формирования различного рода условных клеточных реакций имеет
принципиальное значение.
Рассмотрим схему на рис. 83, из которой видно, что таламический эфферент,
передающий безусловную стимуляцию (БС) через звездчатую клетку, вступая
в кору (2), приводит к активации тормозного иптерпейрона (3), непосредственно
контактирующего с исследуемой крупной пирамидой (1). По мере выработки вре¬
менной связи данный нейрон начинает реагировать аналогичной тормозной
условной реакцией и па условный сигнал (УС). Следовательно, ранее подпорого¬
вый вход, являющийся проекцией таламической импульсации для условного сиг¬
нала, становится пороговым для активации цепи нейронов, предпоследним зве¬
ном которой является также тормозной интернейрон (3), контактирующий с ис¬
следуемой пирамидной клеткой (1). Одновременно с этим в систему включен еще
один тормозной интернейрон (4), блокирующий проведение импульсации от без¬
условного стимула к исследуемому нейрону: ответ этого нейрона на безусловный
раздражитель исчезает. По-видимому, определяющая роль в ассоциативном об¬
учении принадлежит внутрикортикальным связям между возбуждающими и тор¬
мозными интернейронами, динамика которых может обусловливать множество
пластических перестроек, лежащих в основе системной организации условно-
рефлекторного процесса.
Рис. 83. Схема связей нейронов для объяснения
происхождения тормозных условных клеточных реакций
1 — пирамидная клетка глубоких слоев коры; 2 — звездчатая
клетка; 3 и 4 — тормозной интернейрон; 5 — ветви апикальных
дендритов в поверхностном слое коры; 6 — коллатерали аксо¬
нов; УС — путь проведения условного стимула от соседнего ней¬
ронного модуля; БС — путь проведения безусловного стимула из
таламуса
238 Глава 8. Механизмы формирования условных рефлексов
Наблюдая ряд электрофизиологических закономерностей при выработке услов¬
ного рефлекса, Г. И. Шульгина (1978) высказала гипотезу о том, что одним из
основных механизмов действия подкрепляющего стимула является ослабление
тормозных — гиперполяризационных и усиление деполяризационных процессов
в коре головного мозга. Предвозбудительное торможение в активности корковых
нейронов при ассоциативном обучении способствует последующей активации то¬
го же нейрона по механизму «отдачи», что обеспечивает синхронное срабатыва¬
ние большого числа нейронов одной популяции (автостабилизирующее превен¬
тивное торможение, по П. В. Симонову).
Рассматривая нейрон с множеством его входов в составе сложных многоней-
ронных конструкций, включающих тормозные интернейроны, наряду с механиз¬
мами локального облегчения синаптической эффективности следует признать су¬
ществование одновременно работающего механизма подавления избыточных в
данный момент синаптических входов. Оба механизма работают одновременно,
и облегчение синаптической эффективности в определенных локусах постсинап¬
тической мембраны сопровождается торможением прочих эфферентных входов,
конвергирующих на один и тот же нейрон.
Постсинаптическая мембрана представляет собой сложную динамическую мо¬
заику возбужденных и заторможенных локусов, которые, будучи сочетанными во
времени с активацией эндогенных клеточных механизмов, фиксируются, обеспечи¬
вая вовлечение нейрона в состав общецеребралыюй нейродинамической системы.
Оригинальная идея выдвинута В. М. Сторожуком (1986), который считает, что
решающая роль при обучении принадлежит песпецифическим образованиям моз¬
га, в частности центральному серому веществу. На уровне
коры основные пластические изменения происходят в си¬
напсах, воспринимающих неспецифическую афферента-
цию, тогда как специфические входы не подвергаются су¬
щественным функциональным перестройкам. Проводит¬
ся мысль о модуляции холинергических возбудительных
постсипаптических потенциалов в корковых нейронах че¬
рез систему иеспецифических облегчающих и тормозящих
влияний. При комбинации специфических и неспецифи¬
ческих входов корковый нейрон, осуществляя перестройку иеспецифического
входа, приобретает способность участвовать в точно локализованной импульсной
реакции в ответ на специфическое, но ранее индифферентное раздражение.
Динамическая констелляция центров. Разрабатывая свое учение, А. А. Ухтом¬
ский отмечал, что за каждой естественной доминантой кроется возбуждение це¬
лого созвездия (констелляции) центров, своеобразная циклическая машина,
функциональный орган, образующийся по ходу самой реакции. При этом опреде¬
ляющим фактором является усваивание ритма и темпа возбуждения в центрах,
входящих в состав констелляции.
При исследовании структур, вовлеченных в обеспечение условно-рефлекторно¬
го процесса, пришли к заключению о невозможности его привязки к узколокаль¬
ным отделам мозга. Наоборот, они имеют широкое представительство в самых
различных отделах мозга. Тем не менее подтверждается роль коры больших полу¬
шарий как высшего этажа головного мозга, ответственного за формирование но¬
вых межцентральных функциональных взаимоотношений в процессе обучения.
Одним из основных
механизмов действия
подкрепляющего сти¬
мула является ослабле¬
ние тормозных —- ги-
перполяризанионных и
усиление деполяриза¬
ционных процессов в
коре головного мозга.
§ 3. Нейронная организация условно-рефлекторного процесса 239
М. Н. Ливанов (1972) разработал концепцию о пространственной синхрониза¬
ции потенциалов различных отделов головного мозга как основе формирования
констелляции центров. По мере становления временной связи были выявлены
синхронные «усвоенные» колебания биопотенциалов.
С помощью метода корреляционно-спектрального анализа было обнаружено
значимое увеличение кросс-корреляционных коэффициентов разных структур
мозга на стадии генерализации условного рефлекса. При закреплении и специа¬
лизации условного рефлекса указанные отношения сохранялись только между те¬
ми мозговыми структурами, куда адресовались сочетаемые раздражители —
условный и безусловный. Угашение условного рефлекса сопровождалось сниже¬
нием корреляционной связи между их электрическими потенциалами.
Было установлено, что высокий уровень пространственной синхронизации со¬
провождается повышением возбудимости и облегчает процессы распространения
возбуждения в мозговых системах. Уровень пространственной синхронизации
корковых потенциалов обеспечивается регулирующими влияниями из глубоких
отделов мозга.
Р. Джон (1978) обнаружил во многих структурах мозга кошки при образовании
условного рефлекса на ритмические световые вспышки изменения формы позд¬
них компонентов вызванных потенциалов, коррелирующих с двигательным услов¬
ным ответом. Самопроизвольное возникновение выработанного движения со¬
провождалось появлением подобных же электрографических феноменов. Это
позволило думать об их связи с процессом считывания, извлечения следа памяти,
широко распределенного по многим аппаратам мозга.
В последние годы в связи с развитием представлений о модульной организа¬
ции мозга все большее число ученых соглашаются с тем, что отдельные модули
как структурно-функциональные единицы входят в состав более крупных объ¬
единений и могут вовлекаться в различные формы мозговой деятельности.
В. Маунткасл (1981) выдвинул теорию распределенных систем мозга, которая
базируется па трех основных факторах:
1. Основные структуры головного мозга построены по принципу повторения оди¬
наковых многоклеточных единиц — модулей; по своему качественному харак¬
теру функция обработки информации в коре больших полушарий одинакова
в разных ее областях.
2. Внешние связи между крупными образованиями мозга многочисленны, изби¬
рательны и специфичны.
3. Вся группа модулей разбита па подгруппы, из которых каждая соединена сво¬
ей системой связей с такими же обособленными подгруппами в других отделах
мозга. «Связанные между собой группы модулей нескольких структур мы на¬
зываем распределенными системами... Крупные структуры мозга как части
распределенных систем придают каждой из них свойство, определяемое связя¬
ми, общими для всех подгрупп модулей этой структуры, и специфическим ка¬
чеством их внутренней активности... Таким образом, модули распределенных
систем связаны между собой в “эшелонированные” параллельные и последова¬
тельные объединения. Такая распределенная система характеризуется избы¬
точностью потенциальных командных пунктов, и командная функция в разное
время может локализоваться в разных участках системы, особенно там, куда
приходит самая срочная и нужная информация. Поэтому у такой системы мно¬
240 Глава 8. Механизмы формирования условных рефлексов
го входов и выходов, а функция как свойство ее динамической активности за¬
ключена в системе как таковой»1 (выделено мной. — А. Б.).
В вышеприведенных построениях обходится молчанием вопрос о нейрофизио¬
логических механизмах «сборки», формирования и поддержания столь обширных
нейронных объединений в единой рабочей системе. На этот ключевой момент об¬
ращает внимание Дж. Экклс (1981), обсуждая механизмы корковой межмодуль¬
ной интеграции. Автор считает, что корковый модуль может участвовать в неогра¬
ниченном числе сложных систем модулей, которые он именует пространственно-
временными паттернами.
В основу своей гипотезы, в значительной мере опирающейся на морфологиче¬
ские схемы Я. Сентаготаи, Дж. Экклс положил три постулата:
1. Имеет место точная проекция из одного коркового модуля к нескольким обо¬
собленным группам модулей в том же полушарии и к зеркальному фокусу в
противоположном полушарии. Последние по обратным комиссуральным и ас¬
социативным связям проецируются на тот же начальный модуль, обеспечивая
предпосылки для колец реверберации, как внутри-, так и межполушарной.
2. Такие же кольца реципрокпых связей существуют между корой больших полуша¬
рий и таламусом, которые обеспечиваются крупными пирамидами V слоя. Эти
кольца могут служить основой для таламокортикальной реверберации, в резуль¬
тате чего активируются (с помощью таламокортикальных входов) касательные
синапсы на шипиках апикальных дендритов пирамидных клеток слоя V.
3. Апикальные дендриты крупных корковых пирамидных клеток образуют в I
и II слоях касательные синапсы с проходящими горизонтально аксонами, ко¬
торые принимают участие в механизмах формирования и считывания следов
памяти (рис. 84, а).
Эти положения позволили высказать гипотезу, согласно которой при времен¬
ном совпадении активации апикальных дендритов пирамид через касательные си¬
напсы и проведении импульсации по горизонтальным аксонам I слоя происходит
гипертрофия ограниченной популяции синапсов этих горизонтальных волокон с
апикальными дендритами тех же самых пирамид этого модуля. На рис. 84, б жир¬
ными линиями показано состояние, когда два соседних модуля активированы
кортико-кортикальными и/или таламокортикалы!ыми входами, многие из ради¬
ально расходящихся от них волокон нотенциируют синаптические контакты с
дендритами пирамидных клеток в слоях I и II в других модулях. Сама по себе ак¬
тивация синапсов в слоях I и II весьма незначительна в связи с удаленностью от
триггерной зоны, но она может быть увеличена благодаря включению пирамиды
V слоя в реверберационное кольцо. Такая продолжительная активация значитель¬
но усиливает возбуждение касательных синапсов, которые выполняют ключевую
роль в гипотезе Экклса в гипертрофии синапсов горизонтальных волокон с апи¬
кальными дендритами пирамид (возможно, это шипико-шипиковые синапсы).
В одно и то же время шипиковые звездчатые клетки переключают импульсацию
на дендриты пирамидного слоя V’ и оказывают растормаживающее действие на
нейронный модуль. Так может усиливаться слабый сигнал, поступающий к ани-
1 Эйдельман Дж., Маунткасл В. Разумный мозг. М.. 1981. С. 57—58.
§ 3. Нейронная организация условно-рефлекторного процесса 241
Рис. 84. Упрошенная схема связей в неокортексе и схема больших объединений модулей
а — схема связей в неокортексе в соответствии с теорией церебрального обучения Экклса (1981); б —
схема больших объединений модулей в горизонтальном плане. Горизонтальные волокна, возникающие
как продукт бифуркации аксонов комиссуральных (Ком) и ассоциативных (ас) волокон, а также аксо¬
нов клеток Мартинотти (Ма), образуют синапсы с апикальными дендритами звездчатых клеток в моду¬
ле В и пирамидных клеток в модулях А и Б. Внизу модуля А показана шипиковая звездчатая клетка (ЗК)
саксоном, образующим касательные синапсы (КС) со стволом апикального дендрита пирамидной клет¬
ки (ПН). Обусловленные гипертрофией (в результате совпадения импульсных влияний) ассоциативные
волокна из модуля В увеличивают синапсы на апикальных дендритах пирамидной клетки к модулю А.
Каждый модуль показан кружком. Из двух модулей образуется множество горизонтальных радиальных
волокон, которые уходят за пределы показанного участка. Каждый модуль претерпевает непременную
бифуркацию в слоях I и II так, что количество радиальных волокон удваивается в сравнении с числом
вертикальных входов — ассоциативных, каллозальных и клеток Мартинотти. В некоторых модулях
происходит гипертрофия синапсов благодаря активации проходящих горизонтальных волокон (утол¬
щенные линии). Такие горизонтальные волокна распределяются не равномерно, а преимущественно
в каком-то одном направлении
кальиым депдритам, и тем самым осуществлять избирательную активацию ней¬
ронного модуля.
Учитывая возможность связи модулей по верхнему плексиморфному слою ко¬
ры с помощью горизонтального ветвления апикальных дендритов и по глубоким
слоям коры с помощью горизонтальных коллатералей аксонов пирамидных ней¬
ронов, следует принять во внимание аксо-дендритические и деидро-дендритиче-
ские связи с апикальными и базальными дендритами пирамид, которые в основ¬
ном устанавливаются с их дендритными выростами-шиниками. Это наиболее
пластичное соединение между нейронами может быстро менять эффективность
синаптической передачи. Количество самих шипиков и синапсов на них увеличи¬
242 Глава 8. Механизмы формирования условных рефлексов
вается в онтогенезе и прямо зависит от накопления опыта, то есть от образования
энграмм памяти. Синаптическое соединение с шиииком является структурно
весьма стабильным, а функционально — подвижным. Последнее достигается с по¬
мощью изменения геометрических параметров самого шиника, что меняет сопро¬
тивление для распространяющихся по нему синаптических местных потенциалов
в направлении к стволу дендрита. Таковыми могут быть некоторые гипотетиче¬
ские механизмы формирования корковой пространственно-временной структу¬
ры — матрицы памяти.
Ч. Вуди показал, что при типичной ситуации обучения у ненаркотизирован-
ных кошек в постсинаптической клетке изменяется сопротивление мембраны, ко¬
торое сохраняется в течение недель и сопровождается устойчивым увеличением
возбудимости клеток. В. М. Смирнов, Ю. С. Бородкин (1979) также обнаружи¬
ли на целом мозге во время формирования искусственных связей при сочетан¬
ной электростимуляции мозговых структур устойчивое падение импеданса меж¬
ду ними.
Такие наблюдения позволяют допустить, что одним из механизмов создания
пространственно организованной динамической констелляции мозговых аппара¬
тов является регулирование сопротивления постсинаптических мембран нейро¬
нов, которые вовлекаются в такую констелляцию. Длительное сохранение этих
изменений свойств мембран, а следовательно, связей между отдельными нейрон¬
ными популяциями обеспечивается регулирующей ролью циклических нуклео¬
тидов (вторичных посредников) самих нейронов.
Исследованиями на бодрствующих обезьянах (А. С. Батуев и др., 1988) было
установлено, что к категории наиболее подвижных и пластичных мозговых си¬
стем следует отнести ассоциативные системы мозга, динамическая корковая кон¬
стелляция которых складывается по ходу программирования целенаправленного
действия животного и достигает наиболее высокого уровня интеграции к моменту
его выполнения. Такая динамическая констелляция и лежит в основе доминанты
вместе с ее условно-рефлекторным обеспечением.
§ 4. Нейронная организация условного
торможения
Теоретические вопросы торможения условных рефлексов. Каким же образом
неподкрепление условного сигнала приводит к ослаблению и прекращению
условно-рефлекторной реакции? Естественно, что во времена И. П. Павлова не¬
возможно было ответить на этот вопрос. Согласно его представлениям, условное
торможение возникает и первоначально локализуется в корковых нервных клет¬
ках представительства условного раздражителя. Многие данные учеников и по¬
следователей И. П. Павлова послужили свидетельством тому, что условное тормо¬
жение правильнее всего связывать с корковым «центром» безусловного рефлекса,
а Э. А. Асратян считал, что условное торможение локализуется в структурах коры,
осуществляющих временную связь представительств условного и безусловного
раздражителей.
Большинство современных исследований клеточных и системных электриче¬
ских процессов мозга при развитии условного торможения свидетельствует о весь¬
§ 4. Нейронная организация условного торможения 243
ма широком вовлечении различных мозговых структур в этот процесс. Поэтому
в настоящее время трудно говорить о приуроченности тех или иных центральных
процессов при условном торможении к работе строго ограниченных структур
мозга. И следовательно, использование термина «внутреннее торможение» как
развивающегося в пределах системы условного рефлекса вряд ли имеет сегодня
строгое логическое обоснование.
Сам И. П. Павлов говорил о сходстве физиологических механизмов безуслов¬
ного и условного торможения и о сходстве закономерностей образования поло¬
жительного и тормозного условных рефлексов.
В своей гипотезе условного торможения Э. А. Асратян (1970) исходит из пред¬
ставлений об условном рефлексе как синтезе двух безусловных рефлексов
(рис. 85). Предполагается, что возбуждение подкрепляющего безусловного реф¬
лекса производит индукционное торможение среднего звена дуги собственного
ориентировочного рефлекса условного раздражителя. После прекращения дейст¬
вия безусловного раздражителя данное торможение прекращается и сменяется
послетормознъш возбуждением. Последнее, в свою очередь, вызывает индукцион¬
ное торможение условной связи. Это торможение охранительно-восстановитель¬
ного типа предотвращает следовое перевозбуждение условной связи и утомление
соответствующих корковых клеток. При отмене подкрепления нарушается сопря¬
женная индукция и элементы условной связи испытывают длительное следовое
возбуждение. Суммация этого возбуждения при повторных неподкреплениях,
предотвращая истощение, приводит к возникновению охранительно-восстанови¬
тельного торможения в элементах временной связи. Эта гипотеза разделяется да¬
леко не всеми исследователями, ей противоречит целый ряд фактов.
Рис. 85. Схематическое изображение дуги условного рефлекса
с двусторонней связью (по Э. А. Асратяну, 1970)
А — кортикальный пункт мигательного рефлекса: а — афферентный нейрон; б — вставочный нейрон;
в — эфферентный нейрон, 1 и 2 — синаптические контакты коллатералей афферентного нейрона на
вставочный нейрон; 3 — синаптический контакт между вставочным нейроном обратной условной свя¬
зи и афферентным нейроном сигнального раздражителя; Б — кортикальный пункт пищевого рефлек¬
са; а' — афферентный нейрон; б' — вставочный нейрон; в' — эфферентный нейрон; 4 и 5 — синапти¬
ческие контакты коллатералей афферентного нейрона на вставочный нейрон; 6 — синаптические
контакты между вставочным нейроном прямой условной связи и афферентным нейроном подкреп¬
ляющего раздражителя: I — прямая, II — обратные условные связи; В — субкортикальный пункт мига¬
тельного рефлекса; Г — то же пищевого рефлекса
244 Глава 8. Механизмы формирования условных рефлексов
Согласно биологической гипотезе П. К. Анохина (1958), «в основе механизма
условного торможения лежит конкурентная борьба, столкновение различных воз¬
буждений, как это допускал И. П. Павлов в отношении безусловного торможения.
Более сильное возбуждение индукционпо тормозит текущий выработанный пи¬
щевой рефлекс. Если при безусловном торможении таким возбуждением являет¬
ся ориентировочный или оборонительный рефлекс, то при условном торможе¬
нии — биологически отрицательная реакция. Бывший прежде положительным
условный раздражитель после неподкреплеиия его едой становится сигналом
к “трудному состоянию” и потому начинает вызывать условную биологически от¬
рицательную реакцию животного. При систематическом неподкреплении данно¬
го раздражителя едой он постепенно становится тормозным, отрицательная реак¬
ция закрепляется, а пищевое возбуждение совсем тормозится»1.
Схема образования угасателыюго торможения приведена на рис. 86. Общий
механизм тормозящего эффекта определяет, по П. К. Анохину, и единую природу
коркового торможения, в основе которого может лежать пессимальиое, а иногда
и электротоническое торможение. При этом особая роль отводится поляризаци¬
онным процессам на нейронах: тормозной эффект возникает в результате того, что
гиперполяризация блокирует распространяющееся импульсное возбуждение.
100%
секреции
секреции отрицательная секреции биологически
реакция отрицательная
реакция
Рис. 86. Схема образования угасательного торможения (по П. К. Анохину, 1958)
а — нормальный условный пишевой рефлекс; б — сталия частичного торможения пишевой реакции
вследствие появления биологически отрицательной реакции; в — стадия полного торможения пише¬
вой реакции; А — анализатор; К. П. П. — корковое представительство пишевой реакции; К. П. О. —
корковое представительство биологически отрицательной реакции; М — путь для побочного тормо¬
жения
Принять гипотезу П. К. Анохина в целом нельзя, ибо возникновение «трудного
состояния» не является непременным атрибутом процедуры выработки условно¬
го торможения, а при работе на болевом подкреплении «трудное состояние» при
неподкреплении вообще отсутствует. Тем не менее положительной стороной дан¬
ной концепции является признание того, что в результате неподкреплеиия фор¬
мируется биологически новая форма деятельности, имеющая адаптивный харак¬
тер. И эта новая деятельность как биологически наиболее значимая маскирует по¬
ложительный условный рефлекс.
Ю. М. Конорски (1970) считает, что каждый безусловный рефлекс состоит из
двух антагонистических подсистем: позитивной, вызывающей данный рефлекс,
1 Анохин П. К. Биология и нейрофизиология условного рефлекса. М., 1968. С. 337.
§ 4. Нейронная организаиия условного торможения 245
и негативной, его тормозящей (рис. 87). Возбуждение одной системы по закону
отрицательной индукции тормозит другую. При неподкреплении условного сиг¬
нала образуется связь с негативной подсистемой данного рефлекса и приводит
к выработке тормозного условного рефлекса. Следовательно, и в данном случае
речь идет о формировании двух независимых условных рефлексов — положитель¬
ного и тормозного. Отдельные факты могут рассматриваться в качестве экспери¬
ментального подтверждения данной гипотезы.
I
4Ы
зм
2М
4N
ЗЫ
2Ы
1Ы
Рис. 87. Структура дуги рефлексов (по Ю. Конорски, 1970)
I — безусловный; II — условный рефлекс. Р — подсистема, ответственная за положительный рефлекс;
N — подсистема, ответственная эа отрицательный рефлекс; I, 2, 3, 4 — различные уровни нервной си¬
стемы от ствола мозга до коры; А — афферентные проводящие пути; Е — эфферентные проводящие
пути; Б — центры условных раздражителей (А — возбуждающего, С — тормозного; В — смешанного);
ЭР — положительные временные связи; БМ — тормозные связи
И. С. Бериташвили (1975) при объяснении механизма условного торможения
исходил из представления о двустороннем характере временных связей. Возник¬
новение условного торможения рассматривается как процесс усиления обратной
временной связи (от центров безусловного к центрам условного раздражителя)
вследствие отмены подкрепления, что влечет за собой активацию дендритов пира¬
мидных клеток в корковой зове двигательной реакции и одновременному блоки¬
рованию аксосоматических синапсов па теле клетки. Это последнее осуществля¬
ется путем анэлектротонического действия, заключающегося в понижении возбу¬
димости тела клетки и ее синаптических окончаний (рис. 88).
А. И. Ройтбак (1975) полагает, что при неподкреплении возбуждается восходя¬
щая таламическая ретикулярная система, волокна которой достигают верхушеч-
Рис. 88. Схема электротонического торможения корковых клеток
(по И. С. Бериташвили, 1975)
1 —тело пирамидного нейрона; 2 — верхушечный дендрит нейрона;
3 — аксон нейрона; 4 — волокно, несушее импульсы к клетке через пери-
дендритные (расположенные на верхушечных дендритах) синапсы;
5 — волокно, несушее импульсы к клетке через перикорпускулярные (за¬
канчивающиеся на теле клетки) синапсы
246 Глава 8. Механизмы формирования условных рефлексов
ных дендритов пирамидных нейронов, вызывают местное возбуждение и анэлект-
ротоническое торможение клеточных тел.
Вышеприведенные гипотезы о механизмах условного торможения построены
в значительной степени на косвенных данных, а поэтому носят умозрительный
характер, нисколько не снижающий их оригинальной ценности.
Нейронные корреляты условного торможения. Рассмотрим теоретические
положения, базирующиеся на прямых исследованиях суммарных или клеточных
процессов мозга в процессе выработки разных видов условного торможения. Сра¬
зу же отметим, что попытки выяснить механизмы условного торможения на осно¬
ве изучения фоновой ритмики ЭЭГ и отдельных компонентов вызванных потен¬
циалов дали малоубедительные результаты, ибо сама природа этих электрических
феноменов оставалась недостаточно ясной.
Е. Н. Соколов (1965) находит много общего в павловском угасателыгом тормо¬
жении и в динамике угашения ориентировочного рефлекса. Он полагает, что в ос¬
нове угашения лежит механизм усиления тормозящих влияний на ранее возбуж¬
давшихся клеточных элементах системы условного рефлекса.
Одной из первых работ по клеточному анализу условного торможения была
работа У. Г. Гасанова (1972), который пришел к выводу, что в формировании тор¬
мозной реакции участвуют все звенья положительного условного рефлекса, но
они работают в другом режиме, меняются параметры их деятельности. Отсутст¬
вие внешней реакции на отрицательный (неподкрепляемый) стимул не связано
с локальным или системным торможением структур головного мозга. В ответ на
тормозное раздражение возникала такая же многообразная активность нейронов,
как и на положительное раздражение. Таким образом, классическое представле¬
ние о нейрофизиологическом обеспечении условного торможения уже начинает
существенно видоизменяться, и это направление стало основным в последующих
исследованиях.
Многие физиологи наблюдали при угасателыюм торможении инверсию ней¬
ронных реакций: тормозящиеся на положительный сигнал нейроны усиливали
свою активность на неподкрепляемый сигнал и наоборот. Г. И. Шульгина (1978)
считает, что при угашении создаются благоприятные условия для гиперполяриза¬
ции нейронов и организации фазности в их импульсной активности. А. Б. Коган
(1979) наблюдал при образовании оборонительного условного рефлекса сниже¬
ние доли тормозящихся нейронов, а при угашении условного рефлекса возникала
обратная картина и тонкий узор ансамблевой мозаики из возбуждающихся и тор¬
мозящихся нейронов исчезал.
М. Я. Рабинович (1975) при исследовании клеточных аналогов условного реф¬
лекса описал как особую форму условные тормозные реакции клеток на положи¬
тельный сигнальный стимул, которые выступают непременным компонентом
формирования и реализации условно-рефлекторных актов. Эти тормозные услов¬
ные реакции расцениваются как одна из форм условного торможения наряду
с дифференцированием и угашением. Б. И. Котляр (1977) проявление условно¬
го торможения видит в снижении или прекращении импульсной активности клет¬
ки при продолжительном действии положительного условного сигнала (запазды-
вателыюе торможение). Л. Г. Воронин и Р. Г. Кожедуб (1970, 1971) в модельных
ситуациях обнаружили, что при утасателыюм торможении амплитуда ВПСП по¬
§ 4. Нейронная организация условного торможения 247
степенно снижалась без заметного перехода в гиперполяризацию клеточной мем¬
браны. Появилась возможность объяснить ослабление возбуждающих реакций
уменьшением выброса медиатора без активного торможения самого нейрона. По¬
добные наблюдения, также факты постепенного угнетения клеточных реакций на
угашаемый раздражитель (Н. Н. Василевский, 1979; В. М. Сторожук, 1986;
М. Я. Рабинович, 1975) могут быть связаны с понижением возбудимости корко¬
вых нейронов и дают основания предполагать, что при развитии угасателыюго
торможения имеет место «размыкание» временной связи.
При дифференцировочном торможении, как это впервые наблюдали Г. Джас¬
пер и др. (1962), часть корковых клеток снижала свою активность, а часть нейро¬
нов повышала частоту разрядов, что сопутствовало торможению условного двига¬
тельного ответа.
М. Я. Рабинович (1975) считает, что положительные и отрицательные услов¬
ные рефлексы опосредуются разными синаптическими входами активирующего
и тормозного действия. В зависимости от того, какие интернейроны включены
в полисинаптические пути условной связи, дифферепцировочное торможение
будет проявляться или как угнетение активирующих условных реакций клетки,
или как растормаживание клеточных реакций с тормозной условной активно¬
стью. Ряд клеток на неподкрепляемое раздражение усиливали импульсацию при
отсутствии их реакции на положительный условный сигнал.
При выполнении инструментальных условных рефлексов у кошек (Л. В. Че¬
репкова и др., 1989) животные обучались осуществлять пищедобывателыюе дви¬
жение на комплексный сигнал (свет + звук) и вырабатывали условное торможе¬
ние ответа при изолированном предъявлении компонентов, которые пищей не
подкреплялись (рис. 89). Авторы установили, что нейронные реакции сенсомо-
торной коры при положительном сигнале имеют паттерн ответа, существенно от¬
личающийся от такового при дифференцировочных сигналах. Предполагается,
что паттерн ответа этих нейронов на положительные или отрицательные сигналы
обусловлен той двигательной программой, которая реализуется по данному сиг¬
налу. Если угасательное торможение сопровождается распадом паттерна нейрон¬
ных ответов и/или их исчезновением, то дифферепцировочное торможение, тре-
Рис. 89. Перистимульная гистограмма активности
нейрона сенсомоторной коры кошки при выполнении
двигательного условного рефлекса захвата пиши
(по Л. В. Черенковой и др., 1988)
а — комплексный условный сигнал из одновременного предъявле¬
ния светового и звукового компонентов; 6 — отрицательный услов¬
ный рефлекс на изолированное предъявление светового компо¬
нента; в — то же, звукового компонента; по оси ординат — число
импульсов, каждый столбик соответствует времени 50 мс, по оси
абсцисс — время в секундах, под гистограммой — отклонение
вверх — условный сигнал, вниз — подача кормушки, черточки
обозначают моменты проявления двигательной реакции передней
конечности. Каждая гистограмма построена на основе накопле¬
ния 10-й реализации программы
20 -
в
20
Ш1ИЦш1Ш1)11Ы|И1)1|11|1Ш|
|11111|1|1|1||1|11]||||||[11||||||||ИМ||||||||||||| '
248 Глава 8. Механизмы формирования условных рефлексов
нируемое вместе с положительным сигналом, стабильно поддерживает новый
паттерн нейронного ответа, отражающий центральную программу поведенческой
реакции.
Г. А. Вартанян (1986) и А. А. Пирогов (1990), исследуя нейронную активность
лобной коры при разных видах условного торможения, установили три типа пере¬
строек импульсации:
1) инверсию паттерна активности нейрона, имевшего место при положительном
сигнале, в противоположный по знаку при предъявлении отрицательного сиг¬
нала;
2) рекомбинацию паттерна ответа при предъявлении неподкренляемого раздра¬
жителя в сравнении с эффектом положительного сигнала;
3) исчезновение реакций нейронов при предъявлении дифференцировочного
раздражителя.
Эти перестройки именуются как центральный нейронный эквивалент диффе¬
ренцированного торможения. В целом же условное торможение является таким
же активным системным процессом, как и условное возбуждение. По своей орга¬
низации оба процесса подобны, за исключением конечного эффекториого звена.
Поэтому невозможно искать даже отдаленное сходство в содержании понятий
«торможение» в поведенческом, павловском смысле и в его нейрофизиологиче¬
ском содержании. Вероятно, мозаика постсинаптических процессов, разыгрываю¬
щихся на корковом уровне, отражает перестройки функциональной активности
соответствующих центральных программ, эффекторпое выражение которых раз¬
ыгрывается на соответствующих мотонейронах и реализуется в форме конкрет¬
ной поведенческой реакции.
В. М. Сторожук (1986) обращает внимание на то, что при диффереицировоч-
ном торможении имеют место более сложные перестройки, более сложные отно¬
шения между структурами и популяциями нейронов, чем
при угашении. Им показана важная роль центрального се¬
рого вещества среднего мозга при условном торможении.
Установлено, что при условном торможении нейроны
центрального серого вещества резко снижают вызванную
и фоновую активность, тем самым устраняя продолжи¬
тельные тонические реакции корковых клеток и блокируя
условный рефлекс. В. М. Сторожук рассматривает услов¬
ное торможение и условное возбуждение как определенные функциональные со¬
стояния, при которых могут выполняться или блокироваться рефлекторные акты,
а также может осуществляться переход к другой индивидуально приобретенной
или врожденной деятельности. Переключение от одной формы поведения к дру¬
гой под влиянием условного торможения осуществляется на основе полной или
частичной мотивации. В этом смысле условное торможение — производное, вто¬
ричное функциональное состояние системы нервных структур, изменяющих те¬
чение приобретенных индивидуальных поведенческих реакций.
Отсутствие законченного теоретического представления о механизмах услов¬
ного торможения свидетельствует о необычайной сложности проблемы и де¬
монстрирует активный творческий поиск в плане ее решения с современных по¬
зиций.
Невозможно искать да¬
же отдаленное сходст¬
во в содержании поня¬
тий «торможение»
в поведенческом, пав¬
ловском, смысле и в
его нейрофизиологиче¬
ском содержании.
Контрольные вопросы 249
Заключение
В основе нейрофизиологических механизмов формирования условных рефлексов
лежит конвергенция к одному и тому же нейрону импульсных потоков при воз¬
действии условного и безусловного раздражений. В результате повторяющихся
сочетаний этих факторов меняется возбудимость нейрона и эффективность рас¬
положенных на нем синаптических контактов возбуждающего и тормозного зна¬
ка. Пластические свойства нейронной системы, обусловленные внутриклеточным
метаболизмом нейрона с участием вторичных посредников (мессенджеров), со¬
храняют эти взаимоотношения на длительный срок. Этому же могут способство¬
вать перестройки нейроглиальных отношений, изменения геометрии дендритных
шипиков, ответственных за эффективность аксошипиковых контактов. Выделя¬
ют на уровне неокортекса микросистему (микросеть, ансамбль, функциональный
модуль, нейронный пул) обучающихся нейронов, активность и взаимодействие
которых перестраиваются в процессе формирования условной связи. Значи¬
тельную роль в пластических перестройках отводят внутрикортикалыюму тормо¬
жению.
В результате выработки условного рефлекса формируется динамическая кон¬
стелляция мозговых структур или распределенная система гомонимпых модулей,
вовлекаемых симультанно и сукцессивно в процесс активации (синхронной или
асинхронной деятельности).
В основе феноменов условного торможения (дифференцировочного) лежит
формирование центральных альтернативных программ, связанных с реализаци¬
ей противоположных поведенческих эффектов. Взаимодействие альтернативных
центральных программ, контролируемое лобными областями коры мозга, может
обусловливать многие феномены условного торможения и растормаживания.
Контрольные вопросы
1. Что такое принцип конвергенции и общего пути Ч. С. Шеррингтона?
2. Конвергентная теория формирования временных связей.
3. Понятие о топической и посттетанической потенциации.
4. В чем суть явления гетеросипаптического облегчения?
5. Поляризационно-электротоническая гипотеза формирования доминантного
очага.
6. Глиальная теория формирования условного рефлекса.
7. Каковы основные перестройки активности нейронов в процессе условно-реф¬
лекторной деятельности?
8. Каково значение тонической составляющей условно-рефлекторного процесса?
9. Каковы внутрнкорковые механизмы временных связей?
10. Динамическая констелляция центров (по А. А. Ухтомскому) и теория распре¬
деленных систем мозга (по Маунткаслу).
11. Основные положения теории формирования временных связей Экклса.
12. В чем состоят основные теории торможения условных рефлексов?
13. Нейронные корреляты внутреннего торможения.
Глава 9
МЕХАНИЗМЫ ПАМЯТИ
ШШШШёШШШШШШШШяШШ
§ 1. Вилы и формы памяти
Формы биологической памяти. Многие из специалистов придерживаются пред¬
ставления о памяти как сложном феномене фиксации, сохранения и воспроизве¬
дения информации о взаимодействии объектов. Это определение включает широ¬
кий диапазон феноменов от элементарной памяти неживой природы до психо¬
нервной памяти человека. Принципиальное отличие живого от неживой природы
состоит в структурном воспроизведении живого, что в принципе невозможно, ес¬
ли живая система не запомнит свою стабильную организацию. Следовательно,
биологическую память можно определить как способность живых существ (или
их популяций), воспринимая воздействия извне, закреплять, сохранять и в после¬
дующем воспроизводить вызываемые этими воздействиями изменения функцио¬
нального состояния и структуры (И. П. Ашмарин, 1975). Стабилизация органиче¬
ских систем и их структурное самовоспроизведение опирается иа генетическую
память как память биологического вида. Носителем генетической памяти явля¬
ются нуклеиновые кислоты, которые позволяют обеспечить стабильность хране¬
ния информации. В основе изменчивости такой информации в филогенезе лежит
мутагенез. Воспроизведение измененных форм происходит с участием белков-
ферментов. Генетическая детерминация свойств высшей нервной деятельности
рассмотрена в гл. 5.
Второй формой биологической памяти, развивающейся позже в ходе эволю¬
ции, является иммунологическая память, которая тесно связана с генетической
памятью.
В самой общей форме иммунологическая память состоит в способности после
первой встречи с генетически чужеродными телами и веществами узнавать их при
повторной встрече, связывать и включать неспецифиче¬
ские механизмы их уничтожения. Такие чужеродные ве¬
щества именуются антигенами, а иммунные белки, обла¬
дающие способностью разрушать антигены, называются
антителами. Основными участниками иммунного ответа
являются иммунокомпетентные лимфоциты. Их поверх¬
ностная мембрана оснащена определенным набором ан¬
тител. Причем взрослый организм снабжен набором гене¬
тической информации для синтеза всего разнообразия ан¬
тител. Эти антитела на лимфоцитах служат рецепторами
для антигенов. Каждый лимфоцит имеет рецепторы к од¬
ному или к нескольким сходным антигенам. А все лимфоциты, несущие одинако¬
вый рецептор, принадлежат к одному клону, то есть являются потомками одной
материнской клетки с таким же рецептором.
Процесс селекции клонов и обусловливает иммунологическую память. Первая
встреча с антигеном вызывает увеличение числа соответствующих лимфоцитов
Биологическая па¬
мять — способность
живых существ, вос¬
принимая воздействия
извне, закреплять, со¬
хранять и в последую¬
щем воспроизводить
вызываемые этими воз¬
действиями изменения
функционального со¬
стояния и структуры.
§ 1. Виды и формы памяти 251
(формирование клопа) и их дифференцировку на эффекторпые клетки и клетки
памяти. Если первые живут несколько дней, то вторые остаются в организме на
всю жизнь и нри повторной встрече с антигеном способны вновь превращаться
в клетки обоих типов. Селективными агентами, которые обеспечивают материал
для отбора, являются антигены. Они «узнают» рецепторы антител, связываются
с ними и стимулируют их размножение. Значит, из огромного набора вариаций
антител антиген отбирает единственную и стимулирует ее количественный рост.
Эта форма памяти представляет собой эволюционное развитие генетической па¬
мяти в сторону ее большей гибкости и индивидуальной реактивности (Г. А. Варта¬
нян, М. И. Лохов, 1987).
Одним из основных свойств нервной системы является длительное хранение
информации о событиях внешнего мира и реакциях организма на эти события,
а также использование этой информации для построения текущего поведения.
Эти свойства и легли в основу формирования нервной, или нейрологической, памя¬
ти. В эволюции она возникла в связи с дифференциацией нервной системы и ока¬
залась самой сложной но проявлениям и механизмам.
В каждый момент времени животные и человек переходят из своего прошлого
к новому, ранее не испытанному состоянию, и их поведение в настоящем в значи¬
тельной мере определяется совокупностью накопленного опыта в прошлом. Вот
эту непрестанно возрастающую совокупность следов от пройденного, определяю¬
щую поведение в наступающем настоящем, и называют памятью (А. А. Ухтом¬
ский). Таким образом, память как результат обучения заключается в таких изме¬
нениях в нервной системе, которые сохраняются в течение некоторого времени и
существенно влияют на характер протекания будущих рефлекторных реакций.
Комплекс таких структурно-функциональных изменений, включающий не толь¬
ко запечатление определенной внешней ситуации, по и субъективное отношение
организма к пей, получил название процесса образования энграммы. Используя
весь ранее приобретенный опыт, а следовательно, опираясь па факторы, уже от¬
сутствующие в настоящем, энграмма оказывается избыточной. Благодаря своей
избыточности энграмма служит основой активности организма и реалистическо¬
го прогнозирования им будущих ситуаций.
Общепризнано представление о системной организации памяти, то есть ее ор¬
ганизации во времени и пространстве. И изучение элементарных механизмов па¬
мяти всегда должно ориентироваться па целостное понимание этой функции моз¬
га. Одним из трудных вопросов является отражение в памяти фактора времени.
Известно, что в памяти события размещаются по оси времени с отражением их ре¬
альной длительности. Ясно, что формирование и воспроизведение таких эиграмм
возможно лишь при допущении временного «свертывания» событий на основе су¬
ществования собственного времени мозга. На основе такой компрессии времени
создается внутренний хронотоп (по А. А. Ухтомскому), то есть внутренний про¬
странственно-временной образ внешнего единого пространственно-временного
мира. При воспроизведении такой энграммы неизбежно отражение реальной, фи¬
зической встречи с объектом. Поэтому энграмма из модели прошлого перемеща¬
ется в будущее и становится фактором, определяющим цель предстоящего пове¬
денческого акта. Значит, цель в виде реальной энграммы предшествует следствию,
то есть действию с его результатами (Р. И. Кругликов, 1987).
252 Глава 9. Механизмы памяти
Временная организация памяти. После электрического ответа рецепторной
клетки на внешнее воздействие возникают следовые процессы, продолжающиеся
некоторое время уже при отсутствии реального раздражителя. Эти первичные
следовые процессы составляют основу сенсорной памяти. Длительность хранения
следов в сенсорной памяти не превышает 500 мс, стирание следа осуществляется
за 150 мс.
Так, зрительный образ сохраняется во время мигания, при чтении, восприятии
речи и пр. На этом же виде памяти основано слитное восприятие изображений в
кино и телевидении. Предполагают, что сенсорная память человека не зависит от
его воли и не может быть подвергнута сознательному контролю. Длительность
хранения следов в сенсорной памяти продолжает оставаться дискуссионной. В ча¬
стности, требуют объяснения способности некоторых людей-эйдетиков, у кото¬
рых период сохранения зрительного образа может достигать десятков минут.
Одна из классификаций видов памяти приведена на рис. 90. Следующий за
сенсорной памятью период, связанный с хранением информации, именуется
краткосрочной памятью. И наконец, наиболее значимая
информация хранится в долгосрочной памяти. Такое де¬
ление видов памяти принимается большинством исследо¬
вателей, хотя некоторые и предлагают свою более дроб¬
ную классификацию. Кратко- и долговременная память
могут различаться по своим механизмам и оказывать друг
на друга взаимное влияние. Консолидация энграммы, то
есть ее переход из кратковременной в долговременную, — принципиально важ¬
ный момент физиологии и биохимии памяти. Некоторые полагают, что периоду
консолидации соответствует особый вид промежуточной памяти, которая как бы
встраивается между долговременной и кратковременной памятью.
Хронотоп — внутрен¬
ний пространственно-
временной образ
внешнего единого про¬
странственно-времен¬
ного мира.
Долгосрочная память
К >
Забывание путем Забывание Забывание Забывания
стирания при замене при длительном не происходит
или разрушения старой неиспользовании
информации
на новую
Рис. 90. Виды и формы памяти (по Г. М. Чайченко, 1987)
Существует мнение о том, что кратковременная и долговременная память
представляют собой последовательные этапы единого процесса. В то же время ряд
гипотез исходят из точки зрения о параллельном развитии процессов кратко-
и долговременной памяти. Предполагают, что кратковременная (нестабильная)
память образуется за счет конформационпых перестроек макромолекул и реали¬
зуется с участием синаптического и синаптосомального уровней регуляции
§ 1. Виды и формы памяти 253
(рис. 91). Перемещение ионов и/или кратковременные метаболические сдвиги во
время синаптической активности могут привести к изменению эффективности
синаптической передачи, длящейся миллисекунды и секунды. Большинство уче¬
ных усматривают в основе кратковременной памяти прежде всего электрофизио-
логические механизмы, связанные с многократным циркулированием импульса-
ции (реверберацией) по замкнутой системе нейронов.
Память
Уровни регуляции
Рис. 91. Временная организация памяти
(по X. Матиссу, 1978)
С помощью психологических тестов Эббингауза было установлено, что объем
кратковременной памяти человека измеряется 7 ± 2 единицы, то есть бессмыслен¬
ные слова после однократного их предъявления воспроизводятся испытуемым
лишь в количестве 7 ± 2.
Синаптосомальный уровень включает конформациоиные изменения струк¬
турных и ферментных белков, изменения концентрации и перемещение нейроме¬
диаторов. Это и есть длящаяся минуты и часы промежуточная память. Промежу¬
точная память, как полагают психологи, расширяет объем кратковременной па¬
мяти и увеличивает ее длительность. Долговременная память формируется на
основе синтеза макромолекул — нуклеиновых кислот и белков — и связана с акти¬
вацией генетического аппарата клетки. Следовательно, в отличие от предшест¬
вующих процессуальных видов памяти, долговременная память представляет со¬
бой новую внутримозговую функциональную структуру, базирующуюся в резуль¬
тате на изменениях в мембранах нейронов и па межнейронных связях.
Благодаря в основном исследованиям психологов в пределах краткосрочной
памяти выделена первичная память, связанная с мысленным повторением мате¬
риала с целью запоминания и его интерпретации (см. рис. 90). Длительность этой
памяти ограничивается несколькими секундами, и материал стирается при его за¬
мене на новый. В свою очередь, долговременную память подразделяют на вторич¬
ную путем создания ассоциаций, то есть взаимоотношений отдельных элементов
и явлений, которые могут храниться минуты и годы, и третичную память, куда
входят навыки, постоянно сопровождающие жизнь человека: способность к чте¬
нию, письму, профессиональные навыки и пр., которые практически не забывают¬
ся. Кроме того, различают у человека произвольное и непроизвольное запомина¬
ние; образную, моторную, логическую, эмоциональную, условно-рефлекторную
254 Глава 9. Механизмы памяти
память. Конечно, нельзя не видеть, насколько еще далеки друг от друга физиоло¬
гические и психологические схемы классификации видов памяти. Накопленные к
настоящему времени физиологические и биохимические данные позволяют с оп¬
ределенной достоверностью выделить три вида памяти:
♦ сенсорную;
♦ кратковременную;
♦ долговременную.
§ 2. Механизмы кратковременной памяти
Нейрофизиологические корреляты краткосрочной памяти. К данной категории
или стадии относится намять на только что минувшие события. Представим себе
следующую экспериментальную ситуацию. Обезьяна обучается но условному
а _Ц
Рис. 92. Методика изучения нейронной активности мозга в тестах
на краткосрочную память у обезьян
а — экспериментальная проыедура; б •— экспериментальная программа и ее
нейрональные корреляты; 1 — лампа-вспышка (предупреждающий сигнал);
2 — тубус для подачи пиши; 3 — педаль; 4 — лампа (условный сигнал); 5 —
прозрачное окно в экране (7); 6 — обшая панель; Аиб — активность нейро¬
нов лобной коры; В — двигательной коры; Г — электромиограмма дельто¬
видной мышыы плеча; Л — отметки сигналов (1 — предупреждающий; 2 и 3 —
условные; 4 — пусковой (отодвигание экрана); 5 — нажатие на клавишу). Ка¬
либровка времени — 2 с
§ 2. Механизмы кратковременной памяти 255
Рис. 93. Сравнительное распределение
пространственно-селективных нейронов лобной
и теменной коры обезьян в тестах
на кратковременную память
Цифры показывают процентное отношение к обшему
числу пространственно-селективных нейронов. Этапы
экспериментальной программы: 1 — предупреждающий
сигнал; 2 — период неспецифического ожидания; 3 —
условный сигнал; 4 — отсрочка; 5 — пусковой период
программы (открытие экрана, нажатие на рычаг,
получение пишевого подкрепления)
сигналу — зажигание лампы справа или слева на пульте — выполнять движение
нажатия соответственно на правую или левую педаль или рычаг. Если выбор пе¬
дали или рычага выполнен правильно, такое действие вознаграждается пищей
(А. С. Батуев и др., 1988). Но сигналы подаются в случайном порядке и после каж¬
дого сигнала наступает пауза, которая может длиться разное время — от 5 до 20 с.
В течение этого периода, называемого периодом отсрочки, животное не имеет до¬
ступа к рычагам и педалям: они скрыты от него непрозрачным экраном. После от¬
срочки экран открывается, и обезьяна может выполнить требуемое действие
(рис. 92). Значит, животное, получив условный сигнал, должно удержать в памяти
информацию о его местоположении в течение всего периода отсрочки, а затем ис¬
пользовать эту информацию для осуществления правильного движения.
Такое отсроченное поведение является общепризнанным методическим под¬
ходом для изучения механизмов кратковременной памяти, с помощью которой
сохраняется информация о только что прошедших событиях, но необходимых для
выполнения предстоящего поведенческого акта. У таких животных из различных
отделов мозга с помощью микроэлектродов длительно регистрируется импульс¬
ная активность нескольких нейронов. Исследования показали, что информация о
пространственном расположении условного сигнала кодируется в импульсной
активности нейронов прежде всего лобной и теменной ассоциативных полей коры
больших полушарий. Такое кодирование осуществляется либо рисунком разряда
нейронов, либо частотой импульсации нейронов, получивших название про¬
странственно-селективных. Оказалось, что эти нейроны не представляют собой
однородную популяцию, а подразделяются на несколько групп (рис. 93). Часть
нейронов обнаруживает свойство пространственной селекции только в период
предъявления условного сигнала — такие нейроны названы сенсорными.
Другая группа нейронов обнаружила различия в импульсной активности толь¬
ко в период отсрочки — их именуют нейронами памяти.
Третья группа аналогичное свойство проявила только после открытия экрана,
в момент, непосредственно предшествующий моторному акту, поэтому их назвали
нейронами моторных программ. Описаны и смешанные типы нейронов.
Детальный анализ самого периода отсрочки показал, что все ее время заполня¬
ется кратковременными вспышками импульсной активности разных нейронов
лобной коры: в начале отсрочки, в середине и в конце, либо вспышками активно¬
% 1 2
3 4 5
6,5 П
тж г I
го
о.
8 | |
щяшжтшшяшшх
о
311
| I —шшт
ГС
го
з 11
штжжжмяштх
ю
о
2 11
~шш ши
СЦ
2 I I
I шштш
зм ШШШЖШШШ
го
о.
9 11
УШ/А I I
4 ! I
I Ш I
ГО
з 11
I I жшш
I
ф
з 11
шшшшш
111
ш ~ шшт
1 И
I 1ШШШ
256 Глава 9. Механизмы памяти
сти одних и тех же нейронов. И только незначительное число клеток поддержива¬
ет высокую импульсную активность на протяжении всего периода отсрочки
(рис. 94). Пространственно-селективные сенсорные нейроны стоят на входе слож¬
ных нейронных кортикальных цепей и лишь кодируют сенсорные признаки сиг¬
нала, в данном случае его пространственное положение. Нейроны, активирую¬
щиеся в начальный период отсрочки, детектируют полученную информацию о
свойствах условного сигнала, далее следует передача возбуждения по принципу
эстафеты другим нейронным популяциям, каждая из которых отдает аксоны к
нейронам-сумматорам, и от последних происходит передача задержанной таким
образом информации на программирующие и пусковые структуры.
Имп/200 мс
Рис. 94. Основные типы перестроек импульсной активности нейронов
лобной коры в период отсрочки (по А. А. Пирогову)
Стрелки указывают начало и окончание отсрочки; цифры — процентное
соотношение нейронов; пунктир — уровень фоновой активности
Именно для лобных отделов коры, где и обнаружены вышеописанные феноме¬
ны, характерны замкнутые нейронные круги типа «ловушек», в которых импульс¬
ный поток может циркулировать (реверберировать) до тех пор, пока не будет пе¬
реключения эфферентных модулей из крупных пирамид (рис. 95). Последнее
§ 2. Механизмы кратковременной памяти 257
может служить структурной основой удержания в ловушках импульсных потоков
и эстафетного переключения активации от одного нейронного объединения к дру¬
гому. Лоренте де Но был одним из первых морфологов, кто описал сложные замк¬
нутые цепи нейронов в разных частях мозга. На основе морфологических данных
была создана модель памяти из замкнутых цепей, в которых импульсы циркули¬
руют без подкрепления (рис. 95). И. С. Бериташвили (1948) показал, что часть
звездчатых клеток проекционных зон коры имеет аксон, заканчивающийся в поле
своих дендритов. Такая структура может работать как ловушка возбуждения.
Рис. 95. I — основные типы нейронных объединений в лобной коре
у обезьян (по Г. П. Демьяненко, 1980); II — схема реверберирующей нейронной цепи
(по Г. Адаму, 1983)
1 — нейронная «ловушка»; 2 — вертикальный модуль
Виутрикорковую реверберацию рассматривают как базовый механизм кратко¬
срочной памяти для лобной коры. Важнейшим фактором формирования крат¬
ковременной памяти является ориентировочный рефлекс, который фиксирует вни¬
мание животного для более полного восприятия повой информации. В опытах на
обезьянах было установлено, что отвлечение внимания от экспериментальной си¬
туации при действии постороннего раздражения приводит к разрушению рисунка
импульсной активности пространственно-селективных нейронов лобной коры в
период действия условного сигнала и отсрочки и к появлению ошибочных ответ¬
ных реакций животного (рис. 96).
Иные явления характерны для теменной коры. Хотя и описано множество про¬
странственно-селективных нейронов, но в пределах времени отсрочки не обнару¬
жено феноменов волнообразной активности эстафетного типа, как в лобной коре.
Более того, пространственная селективность сохраняется на протяжении всего
периода отсрочки (рис. 97). Корковые нейронные объединения типа «ловушек»
здесь не обнаружены, а преобладающим типом нейронов являются послойно рас-
9 Зак. 1078
258 Глава 9. Механизмы памяти
а
1
I
2 ?
||Щ ш
4 5
ВШИН I к 1
III1 1
III1
■ I;
и ш и т щ— II 1
1 И 11
1 [
И1ЧИП1в1| 1В 1
1
1 1
1 1 1 ... 1 Г
1
2 3
4
с
11
1 1«! 1 |!»
1 1
.1 I1 1
1 1
ПК
1 1 1
МИН 1 — Ш 11111 «ИМ 1 II
Л II
1 1 1II1 1В1
В I 1
1 1
1
• А 1
1
1 ' 1 1 1
■ *■ 1 ■* 1 ■*.. 1 1 1 1 1 1 1 1 1 1 1 и.,.1 .„,.1 .
I, с
Рис. 96. Активность нейронов лобной, моторной коры и электромиограмма
при правильном решении задачи (а) и при отказе от ее выполнения
вследствие отвлечения внимания (6)
1,2 — записи активности нейронов лобной и моторной коры; 3 — ЭМГ; 4 — отметка стимулов; 5 —
отметка времени (1 с); вертикальные штриховые линии — проекции отметок на осциллограммы; сле¬
ва направо: вспышка (предупреждающий сигнал); зажигание лампы (условный сигнал); открытие эк¬
рана (пусковой сигнал); 5 — нажатие на клавишу (условная реакция)
положенные эфферентные пирамиды. Пространственная селективность, опреде¬
ляющая рисунок нейронной активности теменной коры, обусловлена взаимосвя¬
занной работой коры и таламических ядер в составе единой таламопариеталыюй
ассоциативной системы (см. гл. 9). Иными словами, сохранение в период отсроч¬
ки импульсной активности пространственно-селективных нейронов теменной ко¬
ры обусловлено импульсной реверберацией по длинным таламокортикальиым
кольцам прямой и обратной связи. Дж. Экклс (1981) считает, что корковые пира¬
мидные клетки У-У1 слоев вовлекаются в длительную таламокортикальпую ре¬
верберацию.
Вероятно, участие лобной и теменной корковых областей в процессах крат¬
ковременной памяти основывается на механизмах реверберации. Последние
отличаются друг от друга геометрией нервных колец, которые, в свою очередь,
определяются конструкцией соответствующих корковых нейронных модулей —
нейронные ловушки в лобной коре и вертикально организованные объединения
пирамидных клеток — в теменной.
Механизмы импульсной реверберации. Давно известны наблюдения клини¬
цистов о так называемой ретроградной амнезии, то есть потере памяти па события,
предшествовавшие мозговой травме. Аналогичны последствия судорог и электро¬
шока, которые приводят к стиранию лабильных кратковременных следов только
тех событий, которые непосредственно предшествовали данной процедуре. Одно¬
временно в памяти сохраняются все ранее возникшие впечатления и сведения.
Это послужило основанием для Д. Хебба (1949) рассматривать двойственную
природу следов памяти. Он предположил существование клеточного ансамбля
как закрытой системы, активность которой может реверберировать после прекра¬
щения сенсорного процесса. Ансамбли связаны друг с другом и могут активиро¬
ваться эндогенно. Они также могут образовывать связи с моторными клеточными
ансамблями. Д. Хебб считал, что фиксация следа связана со стойкими изменения¬
ми синаптической проводимости, возникающими при повторном поступлении к
синапсу импульсного разряда определенной конфигурации. К аналогичным за¬
ключениям пришли и другие ученые (Д. Джерард, Ю. Конорски, И. С. Берита-
§ 2. Механизмы кратковременной памяти 259
швили). Дж. Экклс (1981) установил, что при частом функционировании синапса
(его употреблении) возникает облегчение — длительные изменения синаптиче¬
ской проводимости, а при неупотреблении проводимость ухудшается. О двойст¬
венной природе памяти свидетельствовал и корсаковский синдром, при котором
наблюдается забывание (антероградная амнезия) при относительно сохранной
памяти о событиях и фактах, предшествующих началу заболевания (алкогольный
паралич). Операция но поводу височной эпилепсии также приводила больных к
утрате способности запоминать новую информацию при сохранении предопера¬
ционной памяти (У. Пепфильд, X. Джаспер, 1956).
Имп/200 мс
20
Прав. УС
Отсрочка (5 с)
16 18с
Рис. 97. Гистограммы импульсном активности пространственно-селективного
нейрона теменной коры при различных длительностях периода отсрочки
Явления ретроградной амнезии наблюдались в экспериментах с животными,
которым электрошок наносился сразу после обучения, по он был неэффективен,
если наносился спустя некоторое время после процедуры обучения. Многочис¬
ленные эксперименты привели к выводу о том, что все воздействия, так или иначе
отражающиеся на проводимости в синаптических межнейронных контактах, при¬
водят к нарушениям процессов кратковременной памяти. Д. Мак-Гоу полагает,
что при электрошоке разрываются функциональные реверберирующие цепи.
Несмотря на расхождения в деталях, большинство согласно с тем, что измене¬
ние проводимости в сети синапсов после многократного повторения импульсов
в результате реверберации является основой кратковременной памяти, с которой
начинается процесс обучения. Хорошо известно, что во время функционирования
нейрона происходит потеря им ионов калия и накопление ионов натрия. Внутри¬
желудочковое введение животным ингибиторов (Мг + Х)-АТФазы сказывается на
ранних этапах формирования памяти. Р. Мерк (1979) выдвинул гипотезу, согласно
которой ионные сдвиги в результате упражнения (обучения) синапсов запускают
260 Глава 9. Механизмы памяти
В основе процессов
кратковременной памя¬
ти лежит временное по¬
вышение проводимости
в синапсах, связываю¬
щих определенные ней¬
роны, и реверберация
импульсов, основанная
на ряде химических и
электрохимических ре¬
акций, не связанных с
синтезом макромолекул
длительные восстановительные процессы. Следствием их является гииерполяри-
зация мембраны, которая сопровождается поступлением аминокислот и синтезом
определенных белков. Вновь синтезированный белок используется синапсами,
стабильно меняющими свою эффективность. При этом указывается на следую¬
щие временные границы видов памяти: кратковременная — угасает через 10 мин
после обучения, лабильная (промежуточная) возникает в течение 15 мин после
обучения и угасает через 30 мин; долговременная память
возникает через 45 мин после обучения и сохраняется
неопределенно долгое время. Тогда в основе кратковре¬
менной памяти может находиться гинерполяризация
нейронов, связанная с повышением калиевой проводи¬
мости; лабильная память связана с активацией натрие¬
вого насоса; долговременная память базируется па акти¬
вации белкового синтеза.
Другие исследователи отмечают, что продолжитель¬
ность реверберации может измеряться минутами (2-
12 мин). Подчеркивается большое значение медиатора
ацетилхолина, ибо воздействие на системы, выделяю¬
щие и связывающие ацетилхолип (путем введения атропина и скополамипа, нару¬
шающих его рецепцию постсинаптической мембраной, или ядов ацетилхолипэсте-
разы), существенно влияют па кратковременную память.
Реверберация импульсов была показана в прямых исследованиях Верцеапо
при одновременной регистрации активности нескольких соседних нейронов гре¬
мя микроэлектродами. В каждом из нейронов обнаружены повторяющиеся циклы
последовательно возникающих разрядов. Видимо, эти нейроны составляют часть
какой-то цепи, где нервный импульс движется по замкнутому пути. Такая цирку¬
ляция импульсов обнаружена в нейронных цепях между клетками коры и таламу¬
са. При раздражении одной и той же частотой наблюдался один и тот же поря¬
док циркулирующих импульсов. Периодические рисунки импульсации возника¬
ют и от единичного раздражения, что доказывает наличие реверберации.
Другие авторы видели реверберацию между симметричными отделами коры
обоих полушарий, которая распространялась по волокнам мозолистого тела. Эти
межполушарные иейродинамические процессы являются одним из составных
звеньев системной организации кратковременной памяти.
С одной стороны, в цикл реверберации вовлекаются не все импульсные пото¬
ки, а лишь несущие биологически значимую информацию, с другой — ревербери¬
рующая импульсация преодолевает случайную фоновую активность и обеспечи¬
вает изменения в синаптических контактах под влиянием закономерно повторяю¬
щихся неслучайных факторов.
Итак, в основе процессов кратковременной памяти лежит временное повыше¬
ние проводимости в синапсах, связывающих определенные нейроны, и ревербера¬
ция импульсов, основанная на ряде химических и электрохимических реакций, не
связанных с синтезом макромолекул. В составе замкнутых нейронных цепей в ко¬
ре мозга участвуют тормозные интернейроны. В самоорганизации следовых про¬
цессов в пределах нервного кольца могут играть значительную роль носттормоз-
ные реакции при наличии в цепи возвратного торможения. Это создает наиболее
оптимальные условия храпения следов в течение достаточно длительного вре¬
§ 3. Механизмы долговременной памяти 261
мени. Вероятно, участие тормозных интернейронов в составе мпогоэлемеитных
внутрикорковых цепей способствует более длительному хранению следов и под¬
держанию циркулирующей активности.
§ 3. Механизмы долговременной памяти
Раньше была распространена точка зрения о том, что механизмы долговременной
памяти могут быть сведены к изучению условных рефлексов. Однако детальные
исследования показали, что феномен долговременной памяти шире и механизмы,
включающие и условно-рефлекторные процессы, разнообразнее и сложнее.
Основанием к этому служит изучение видов памяти животных и человека,
проведенное И. С. Бериташвили (1975). Он различал образную, условно-рефлек¬
торную, эмоциональную и словесно-логическую память. Кроме того, у человека
психологи и клиницисты выделяют произвольную и непроизвольную память, не¬
посредственное и опосредованное запоминание.
Под образной памятью животных и человека И. С. Бериташвили понимал со¬
хранение в памяти и репродукцию однажды воспринятого жизненно важного объ¬
екта. Под эмоциональной памятью понимают воспроизведение пережитого ранее
эмоционального состояния при повторном воздействии раздражений, обусловив¬
ших первичное возникновение этого состояния (Е. А. Громова, 1980). Эмоцио¬
нальная память обладает следующими характерными особенностями:
♦ она падмодальпа, то есть ее формирование и воспроизведение может происхо¬
дить при любых сенсорных воздействиях;
♦ она формируется очень быстро (в отличие от условно-рефлекторной) и часто
с первого раза;
♦ она характеризуется непроизвольностью запоминания и воспроизведения ин¬
формации, то есть обеспечивает пополнение подсознательной сферы человече¬
ской психики (Э. А. Костапдов, 1977).
Условно-рефлекторная память проявляется в виде воспроизведения условных
двигательных и секреторных реакций или заученных привычных движений спус¬
тя длительное время после образования. Наконец, словесно-логическая (или се¬
мантическая) — это память на словесные сигналы, обозначающие как внешние
объекты и события, так и внутренние переживания и свои собственные действия.
В любом случае имеет место фиксация предметов и явлений, включающая в свой
состав по крайней мере три этапа:
♦ формирование энграммы;
♦ сортировка и выделение новой информации;
♦ долговременное храпение значимой для организма информации.
В отличие от бытовавших в прошлом представлений о структурных основах
долговременной памяти, заключающихся в возникновении новых синаптических
контактов между нейронами, канадский ученый Хебб (1949) выдвинул другую
гипотезу. Суть ее состоит в том, что фиксация следа памяти связана со стойкими
изменениями синаптической проводимости в пределах определенного нейронно¬
го ансамбля. Многочисленные поиски были направлены па выяснение основных
факторов, обеспечивающих модуляцию эффективности синаптической передачи
262 Глава 9. Механизмы памяти
и длительное сохранение этих сдвигов в нейронных сетях. В результате этих ис¬
следований стало общепризнанным представление о том, что в основе долговре¬
менной памяти лежат весьма сложные структурно-химические преобразования
как на системном, синаптическом, так и на клеточном уровнях головного мозга.
Ниже будут последовательно рассмотрены основные участники нейрохимиче¬
ских преобразований, обеспечивающие фиксацию памятных следов.
Нейромедиаторные системы. Когда речь заходит о регуляции синаптической
эффективности, то, естественно, внимание обращается прежде всего на системы
биологически активных веществ, являющихся посредниками в синаптической пе¬
редаче. Так, было установлено увеличение содержания свободного ацетилхолина
в гиппокампе сразу после обучения оборонительному условному рефлексу. Син¬
тез ацетилхолина, зависящий от вырабатываемых навыков и индивидуальных
особенностей животных, стали связывать с процессом консолидации. Весьма чув¬
ствительным индикатором холинергической системы мозга является активность
ацетилхолинестэразы (разрушающей ацетилхолин), которая закономерно меня¬
ется в коре мозга и гиппокампе в зависимости от характера обучения и способно¬
сти животных к выработке навыков.
Дальнейшие исследования показали, что под влиянием обучения увеличивает¬
ся количество холинорецепторов, что может быть следствием либо синтеза новых
рецепторных молекул, либо демаскировки и активации уже существующих ре¬
цепторных белков. Р. И. Кругликов (1986) показал повышение чувствительности
кортикальных нейронов, вовлекаемых в формирование условного рефлекса, к мик-
роэлектрофоретически подводимому ацетилхолину. Причем чувствительность
возрастает вплоть до фиксации следа в памяти. Э. Кеидел (1980) также полагал,
что долговременная память связана со стабильным изменением чувствительности
к ацетилхолину. Активация рецепторов постсинаптической мембраны облегчает
обучение, ускоряет фиксацию, способствует извлечению следа из памяти. И на¬
оборот, антагонисты ацетилхолина нарушают обучение и воспроизведение, вызы¬
вая амнезию (Р. Ю. Илыоченок, 1977).
Системы, связанные с образованием и выделением биогенных аминов, — кате-
холаминергическая и серотонинергическая — принимают самое непосредствен¬
ное участие в механизмах долговременной памяти (Е. А. Громова, 1980; Р. И. Круг¬
ликов, 1981).
Установлено, что обучение животных в моделях с электрокожным подкрепле¬
нием условных реакций сопровождается активацией норадренергических систем
мозга, а обучение с пищевым подкреплением — снижением метаболизма и уровня
норадреналина в мозге животных.
Разработана гипотеза, в соответствии с которой норадреналин, выделяющийся
при подкреплении, пролонгирует активность нейронов, вызванную предъявлени¬
ем условного стимула, и этим облегчает формирование условного рефлекса.
Значительное снижение норадреналина его антагонистами, разрушение ней¬
ронов голубого пятна продолговатого мозга (дающего начало восходящей порад-
ренергической системе) или дорзального пучка переднего мозга замедляет обуче¬
ние, вызывает амнезию и нарушает извлечение следа из памяти (Р. И. Кругликов,
1981). Определенную роль играет и дофаминергическая система (Н. Ф. Суворов,
В. В. Суворов, 1978), ибо дофамин является предшественником образующегося из
него норадреналина.
§ 3. Механизмы долговременной памяти 263
Значительно более определенная роль в процессах, связанных с консолидаци¬
ей следов памяти, принадлежит серотонинергической системе мозга. Е. А. Громова
(1980) установила, что серотонин ускоряет обучение и удлиняет сохранение навы¬
ков, выработанных на эмоционально положительном подкреплении, нарушая вы¬
полнение и сохранение защитно-оборонительных реакций. Согласно ее концеп¬
ции, моноамины участвуют в процессах обучения и памяти опосредованно, через
нейрохимическое обеспечение положительных и отрицательных эмоциональных
состояний (см. также гл. 5). Серотонин причастен к формированию эмоциональ¬
но положительных, а норадреналин — эмоционально отрицательных состоя¬
ний. Обе моноаминергические системы находятся в рецинрокиых отношениях.
В связи с тем, что распространенным приемом исследования памяти является
использование различного рода амнезирующих (амнезия — потеря памяти) воз¬
действий стрессорного тина, было выдвинуто представление, что стрессоры дей¬
ствуют на процессы консолидации через нейроэндокринные реакции, оказывая
первичное влияние на неспецифические компоненты этого процесса.
Р. И. Кругликов (1984) разработал гипотезу, согласно которой холинергические
механизмы мозга обеспечивают информационную составляющую процесса обуче¬
ния. Роль же моноаминергнческих систем мозга сводится к обеспечению подкреп¬
ляющих и эмоционально-мотивационных составляющих процесса обучения и па¬
мяти. Причем холинергическая система находится под модулирующим влиянием
мопоаминергической системы. Стимуляция норадрепергических механизмов пе¬
рестраивает хемореактивпые свойства, которые при определенных условиях мо¬
гут закрепляться, обеспечивая создание и сохранение мпогопейропной констел¬
ляции — энграммы. Если иорадрепергические механизмы мозга в большей мере
причастны к формированию временных связей, то серотонипергическая — к их
фиксации.
X. Матисс (1979) рассматривает взаимодействие обоих типов медиаторных
систем непосредственно на мембране нейрона (рис. 98). Он полагает, что повтор¬
ная активация холинергических синапсов во время стимуляции вызывает кратко¬
временные копформациоппые перестройки постсинаптических мембран, повы¬
шающие синаптическую проводимость. Если в течение этого времени к нейрону
поступают «безусловные» влияния, опосредуемые моноамивергическими систе¬
мами, то включается цепь внутриклеточных метаболических процессов с уча¬
стием циклических нуклеотидов. В ходе этих процессов синтезируются полипеп¬
тиды или белки. Они взаимодействуют с белками постсипаитических мембран,
подвергшихся в ходе сенсорной (условной) стимуляции конформациопным пере¬
стройкам, и стабилизируют эти изменения. В результате нейрон приобретает на¬
бор синаптических входов повышенной эффективности, обеспечивающий его уча¬
стие в составе энграммы.
Данная концепция, весьма правомерная в принципе, не может быть полностью
принята, ибо показано, что 95% пресипантических моноаминергнческих термина¬
лен не образуют синапсов на нейронах, а выделяющиеся моноамины, диффунди¬
руя на значительные расстояния, оказывают модулирующие влияния на множест¬
во образований. Поэтому моноаминергические системы мозга в последнее время
стали причислять скорее к пейромодуляторным, чем к классическим медиатор-
ным системам. Объектом таких модулирующих влияний является холинергиче¬
ская система.
264 Глава 9. Механизмы памяти
Рис. 98. Механизмы стабильного повышения
эффективности синаптической передачи
(по X. Матиссу, 1978)
1 — холинергический медиатор, вызывающий обратимые
конформаиионные изменения субсинаптической мембраны;
2 — моноаминергический медиатор, активирующий нуклео-
тидциклазу; 3 — ииклические нуклеотиды; 4 — активирован¬
ные протеинкиназы; 5 — активация генетического аппарата
и изменения синтеза РНК; 6 — изменение синтеза белков
(глюкопротеидов); 7 — включение вновь синтезированных
белков холинергическим медиатором изменений субсинап¬
тической мембраны
Среди вероятных медиаторов, от уровня метаболизма которых зависят процес¬
сы долговременной памяти, следует упомянуть гамма-аминомасляную кислоту,
глутаминовую кислоту, а также вторичные посредники (циклические нуклеотиды
и ионы кальция). Разработка этих вопросов находится только в начальной стадии.
Информационные макромолекулы. Участие нуклеиновых кислот и белков в
ключевых процессах обучения и памяти не вызывает сомнений. Теоретические
представления подразделяются на две группы. Согласно первой группе гипотез,
обучение и память связаны с кодированием приобретенных форм поведения в ин¬
формационных макромолекулах. Согласно второй группе гипотез, исходящих из
взаимосвязи генома и синтеза специфических белков нервной клетки, на основе
функционального объединения нейронов возникает структурное их объединение,
представляющее собой энграмму памяти.
Первая группа гипотез о кодировании индивидуального опыта в макромолеку¬
лах базируется на следующих аргументах; качественном изменении РНК и белков
при обучении и возможности «переноса памяти» от обученного мозга к необучен¬
ному с помощью РНК или полипептидов.
X. Хидеп (1967) считал, что под влиянием приходящей к нейрону импульса-
ции происходит перегруппировка оснований в молекуле РНК, что приводит
к синтезу на такой ядерной РНК молекул белка измененной структуры, обуслов¬
ливающих избирательную чувствительность нейрона именно к данной конфигу¬
рации импульсов. Наряду с этим при обучении были описаны синтез полипепти¬
дов (Г. Унгар, 1973) и избирательный синтез мозгоспецифических белков. Много
работ, в которых исследуют участие РНК в процессах памяти, не позволяют снять
целый ряд принципиальных вопросов, и поэтому нельзя исключить неснецифи-
ческий характер участия нуклеотидов. Так, оказалось, что введение животным
стимуляторов или ингибиторов синтеза РНК отражается в первую очередь па вы¬
работке новых навыков, а не па их сохранении. Пока пе получено пи одного убеди¬
тельного аргумента в пользу признания определяющей роли макромолекул в ко¬
дировании индивидуального опыта.
Особо стоит вопрос о так называемом переносе памяти. Подобного рода иссле¬
дования на беспозвоночных (иланарии, Д. Мак-Кеннел, 1969) и млекопитающих
(белые крысы, Г. Унгар, 1965) в свое время носили сенсационный характер. Но
§ 3. Механизмы долговременной памяти 265
при их тщательном анализе оказалось, что они содержат ряд методических по¬
грешностей, существенно снижающих их доказательную силу. Вместе с этим
нельзя не считаться с достаточно убедительными наблюдениями о наличии сти¬
мулирующего влияния экстракта мозга, или ликвора, обученных допоров па спо¬
собность к обучению у реципиентов. Видимо, существует какой-то химический
фактор, обеспечивающий не прямой «перенос памяти», а облегчающий формиро¬
вание соответствующего навыка у животпых-реципиентов. Г. Адам (1983) на ос¬
новании своих исследований также приходит к выводу о неснецифическом сти¬
мулирующем эффекте экстракта мозга, отвергая за ним функцию «кода памяти».
В последнее время описаны факты прямого переноса условного сахаринового
отвращения у крыс (Г. А. Вартанян, 1986). У одной группы крыс вырабатывали от¬
вращение к сахариновому раствору при сочетаниях питья этого раствора с введе¬
нием животным хлорида лития, приводящего к интестинальному шоку. Ликвор
обученных животных вводили субокциииталыю реципиентам, у которых досто¬
верно снизилось потребление сахаринового раствора.
Другой формой «транспорта памяти» является перенос импринтинга у цып¬
лят. Получены некоторые данные, свидетельствующие об олигопептидной приро¬
де вещества-переносчика, а также о быстром включении этого вещества в форми¬
рование нового навыка у реципиента (Г. А. Вартанян, М. И. Лохов, 1987).
Для трактовки механизмов «переноса» может быть использована гипотеза об
участии иммунологических механизмов в долговременной памяти (И. П. Ашмарин,
1975). Если представить себе, что после прохождения импульсов через синапс
усиливается синтез специфических белков-антигенов, то их избыток должен вы¬
ходить в околосипантическое пространство. Эти белки взаимодействуют с рядом
расположенными клопами клеток астроцитарпой глии и индуцируют их размно¬
жение и образование антител. Последние специфически взаимодействуют с ност-
синантическими мембранами тех же нейронов и облегчают проводимость в соот¬
ветствующих синапсах. Данный клон астроцитов сохраняется в течение жизни.
В свете данной гипотезы действующим началом «переноса памяти» может быть
избыточный антиген пептидной природы, который способен автоматически най¬
ти в мозге реципиента либо соответствующую клетку глии, либо синапс.
Существует большое число данных, свидетельствующих о том, что полноцен¬
ный белковый синтез в мозге необходим для процесса консолидации и формиро¬
вания долговременной памяти. Причем при глубоком угнетении белкового синте¬
за и относительно кратковременном обучении наблюдается сохранение вырабо¬
танных рефлексов через минуты или часы обучения. Но уже через часы и сутки
после обучения выявляются глубокие нарушения в сохранении выработанных на¬
выков. Следовательно, процессы белкового синтеза не нужны в ближайшее время
после обучения, они понадобятся значительно позже па этапе консолидации эн-
граммы. Детальный анализ содержания белков в коре мозга показал, что при фор¬
мировании новой двигательной координации, связанной с предпочтением од¬
ной конечности у крыс, в сенсомоторной коре возникает выраженная асимметрия
в содержании белков в крупных пирамидных нейронах.
Убедительные данные об участии в функциях памяти получены для двух моз-
госнецифических белков 5-100 и 14-3-2 (X. Хидеи). Первый активно взаимодей¬
ствует с мембраной и сократительными белками нейрона при участии нейронов
кальция. Второй является ферментативным белком, участвующим в реакциях
266 Глава 9. Механизмы памяти
гликолиза в нейронах. Было обнаружено максимальное содержание в гиппокампе
белка 5-100 на 4-5-й дни обучения параллельно увеличению ионов кальция.
В сравнении с этим Хиден показал, что консолидация памятного следа сопровож¬
дается накоплением белка 14-3-2 прежде всего в коре мозга, а не в гиппокампе.
Ряд авторов вообще рассматривают 5-100 как преимущественно глиальный бе¬
лок, и лишь белок 14-3-2 связывают со специфическими процессами сохранения
энграммы. А уровень белка 5-100 отражает несиецифическую реакцию мозга на
усиленное функционирование церебральных структур.
Для создания устойчивости образованной энграммы должна существовать
система обновления специфических рецепторных белков, которая включает участ¬
ки генома, ответственные за синтез соответствующих белков. Либо должны воз¬
никать стабильные модификации ДНК, в результате которых в нейроне возника¬
ет и поддерживается пожизненно синтез любого нейроспецифического белка
или, наоборот, выключается необратимо синтез маскирующего белка. Г. Линч
и М. Бодри (1984) выдвинули гипотезу, сущность которой состоит в следующем.
Повторная импульсация в нейроне сопровождается увеличением концентрации
кальция в постсинаптической мембране. Это активирует фермент — кальцийзави-
симую протеипазу, которая расщепляет один из белков мембраны. Его расщепле¬
ние освобождает замаскированные ранее неактивные белковые глутаматрецепто-
ры. Число активных глутаматрецепторов возрастает, и возникает состояние повы¬
шенной проводимости синапса длительностью 3-6 суток.
Эта гипотеза имеет много прямых и косвенных доказательств в свою пользу.
Она привлекает внимание тем, что позволяет рассматривать структурную ансамб¬
левую организацию энграммы и при этом учитывает такой важный компонент
нейронной конструкции, как дендритные шипики с их белковым цитоскелетом.
Аксошиниковые контакты — наиболее пластичное соединение между нейронами,
которое может быть ответственно за эффективность синаптической передачи. Ес¬
ли представить себе, как показали многие авторы, что количество самих шипиков
и синапсов на них увеличивается в онтогенезе и прямо зависит от накопления ин¬
дивидуального опыта, то есть от образования энграмм памяти, то их использова¬
ние по поводу новых поведенческих задач и в составе новых обширных констел¬
ляций должно подразумевать наличие соответствующих механизмов. С одной
стороны, синаптическое соединение с шипиком является структурно достаточно
стабильным, ибо сохраняет свою целостность при центрифугировании сипанто-
сомальных фракций, с другой — функционально весьма недвижным. Последнее
может достигаться с помощью механизма изменения диаметра ножки шиника, ко¬
торый, в свою очередь, меняет сопротивление мембраны. Это может обеспечи¬
ваться наличием контрактильпого аппарата в ножке шипика, который представ¬
лен молекулами белка актомиозипа. Активация этих молекул может возникать
при высвобождении ионов кальция из депо, коим является содержащийся в го¬
ловке шипиковый аппарат — эндоплазматический ретикулум. Высвобождение
ионов кальция происходит при действии медиатора па постсипаптическую мем¬
брану. Сокращение молекулы актомиозипа приводит к укорочению и утолщению
ножки шипика, в результате чего меняются сопротивление и проведение электри¬
ческого тока к дендритному стволу. Эти представления носят еще весьма гипоте¬
тический характер (А. С. Батуев, В. П. Бабминдра, 1984). Значение их в том, что
они обращают внимание на построение многонейронпой энграммы и на наличие
§ 3. Механизмы долговременной памяти 267
впутрисииаптического цитоскелетного белкового комплекса регуляции синапти¬
ческой эффективности.
Нейропептиды. Г. Уигар (1977) был первым, кто обратил внимание на участие
нейропептидов в явлениях обучения и памяти, которые служат, с его точки зре¬
ния, своеобразными маркерами специфических нейронных путей.
В настоящее время основные исследования проведены на гормонах гипотала-
мо-гипофизарного происхождения или их фрагментах — коротких цепочках ами¬
нокислотных остатков.
Нейропентиды обнаружены в аксонных окончаниях нейронов вместе с медиа¬
торами. Установлено, что нейропептиды могут усилить или затормозить действие
медиатора. Такие иейропептиды-спутники вместе с медиаторами создают и под¬
держивают на постсинаптической мембране специфические рецепторные мозаич¬
ные наборы, способствующие быстрому проведению определенного вида возбуж¬
дения. Пептид-спутник повышает сродство рецептора к основному медиатору, он
более стабилен, чем основной медиатор, что обеспечивает пролонгированное об¬
легчение проведения через синапс (И. П. Ашмарин, 1987).
Так, адренокортикотропный гормон (АКТГ) и кортикостероиды и их модифи¬
кации существенно влияют па обучение и память. Их эффект зависит от интен¬
сивности обучения, он ослабевает по мере увеличения интервала между обучени¬
ем и введением пептида и является достаточно кратковременным.
Специальные исследования на животных с разрушенным гипофизом выявили
у них значительный дефицит памяти. Последнее связывали с недостатком в орга¬
низме гипофизарного гормона — вазопрессина. Дополнительные опыты на линии
крыс с генетическим дефицитом вазопрессина и нарушением памяти и улучшени¬
ем ее после инъекции дополнительных количеств вазопрессина подтвердили уча¬
стие этого пептида в формировании памятных следов. Причем у этих животных
страдал не сам процесс обучения, а именно консолидация сформировавшихся эи-
грамм. Противоположное действие оказывал другой гормон гипофиза — оксито-
цин, который рассматривают как естественный амнезирующий нейропентид. Он
нарушает сохранение выработанных навыков независимо от типа обучения у жи¬
вотных.
Среди других нейропептидов выделяют эндогенные опиоиды — эндорфипы
и энкефалины, которые оказывают выраженное влияние на обучение и память, за¬
медляют угашение условных рефлексов, улучшают их сохранение, хотя и ухудша¬
ют их формирование.
Большинство ученых склоняются к представлению о том, что нейропептиды
регулируют память через взаимодействие с медиаторами и через их влияние на
метаболизм макромолекул. Действительно, оказалось, что предварительное раз¬
рушение голубого пятна предотвращает усиливающее действие вазопрессина на
процессы консолидации. Такой же эффект возникает при предварительном разру¬
шении серотонинергических нейронов ядер шва. Но в отсутствие норадрепергиче-
ской системы чувствительность серотонинергической системы возрастает. В свою
очередь, имеются данные о том, что влияние серотонина на консолидацию эпграм-
мы опосредуется через систему опиоидпых пептидов.
Таким образом, прослеживается тесная взаимосвязь всех без исключения ней¬
рохимических механизмов обучения и памяти. По-видимому, изменение белково¬
го метаболизма является тем конечным звеном, через которое реализуются любые
268 Глава 9. Механизмы памяти
воздействия па процессы обучения и памяти. За счет изменений белкового мета¬
болизма осуществляются процессы формирования и закрепления многонейрон¬
ной эпграммы. «Хранителем всех форм нейрологической памяти служит система
межнейронных взаимодействий, и участие информационных биополимеров про¬
является лишь во включении или выключении активности различных участков
предшествующего генома, а не в синтезе новых нуклеотидных или аминокислот¬
ных последовательностей»1. Дальнейшая задача состоит в разделении специфиче¬
ских и неснецифических факторов формирования эпграммы памяти с углублен¬
ным анализом ее нейрохимических и структурных основ.
Заключение
Различают генетическую, иммунологическую и пейрологическую (нервную) фор¬
мы памяти. Последняя подразделяется на кратко- и долговременную память, по¬
мимо которых выделяют еще и промежуточную память. Перевод значимой ин¬
формации из кратко- в долговременную память называется консолидацией эн-
граммы, то есть образованием структурно-химических изменений, фиксирующих
внешнюю ситуацию и отношение к ней самого субъекта.
Кратковременная память только что минувших событий или впечатлений,
извлеченных из долговременной памяти, основывается па импульсной ревербера¬
ции в замкнутых нейронных цепях. Долговременная память формируется па
основе синтеза макромолекул — нуклеиновых кислот и белков — и связана с акти¬
вацией генетического аппарата нервной клетки, в результате чего возникают
изменения в мембранах нейронов и межнейроппых связях.
Контрольные вопросы
1. Каковы основные формы биологической памяти?
2. В чем суть временной организации памяти?
3. Учение А. А. Ухтомского о хронотопе.
4. Что такое импульсная реверберация?
5. Основные механизмы кратковременной памяти.
6. Что такое консолидация эпграмм памяти?
7. Нейромедиаторпые системы и память.
8. Что такое информационные макромолекулы?
9. Что такое пейропептиды?
1 Ашмарин И. П. Молекулярные механизмы нейрологической памяти // Механизмы памяти: Руко¬
водство по физиологии. Л., 1987. С. 61.
Глава 10
ИНТЕГРАТИВНАЯ ДЕЯТЕАЬНОСТЬ МОЗГА
§ 1. Доминанта и условный рефлекс как основные
принципы интегративной деятельности мозга
Корковый очаг стационарного возбуждения. А. А. Ухтомский экспериментально
электрическим или фармакологическим путем формировал в спинном и голов¬
ном мозге очаг стационарного возбужден! иг в результате чего он обнаружил, что
одним из его свойств является способность реагировать на посторонние для пего
возбуждения.
В лаборатории В. С. Русинова (1967) было показано, что стационарное возбу¬
ждение в коре головного мозга сопровождается сдвигом уровня постоянного по¬
тенциала. Эти же сдвиги могут быть вызваны искусственно путем поляризации
коры слабым постоянным током. Аподизация слабым постоянным током кор¬
кового представительства одной из конечностей бодрствующего кролика создает
условия, при которых ранее индифферентные световые и звуковые раздражения
начинают вызывать двигательную реакцию соответствующей конечности. Специ¬
альными опытами было показано, что такой очаг возбуждения обладает не только
способностью суммировать возбуждение, но и свойствами иптерности, торможе¬
ния при чрезмерных возбуждениях и восстановления при оптимальных условиях
раздражения.
Структурами, принимающими активное участие в образовании такого очага,
являются апикальные дендриты пирамидных клеток, интернейроны и клетки глии.
Их анодизация создает условия, при которых значительная часть нейронов коры
становится нолисенсорной.
Таким образом, в доминантном очаге происходит установка на определенный
уровень стационарного возбуждения, способствующий суммированию ранее нод-
пороговых возбуждений и переводу на оптимальный для данных условий ритм
работы, когда этот очаг становится наиболее отзывчивым и тормозятся другие ра¬
ботающие очаги.
Однако события, происходящие в поляризационном очаге, характеризуют
лишь одну из сторон процесса складывания доминанты. Оказалось, что в форми¬
ровании коркового доминантного очага участвуют различные неспецифические
образования таламуса. Дополнительная поляризация мезэнцефалических и тала¬
мических ядер ретикулярной формации облегчала формирование двигательной
доминанты, усиливала ее. Если корковый доминантный очаг тестируется свето¬
вой стимуляцией, то дополнительная оптимальная поляризация наружного ко¬
ленчатого тела подкрепляет доминанту.
На «поляризационной модели» исследовались механизмы суммации путем со¬
четания подпороговых раздражений сепсомоторной коры кошки и световых
вспышек. Замечено появление в результате суммации движения соответствую¬
щей конечности (Р. А, Павлыгипа, 1982). Этому предшествует изменение конфи¬
гурации дендритного потенциала той же зоны коры — появляется дополнитель¬
ная волна, связанная с тормозным процессом. Сразу вслед за этим наблюдается
270 Глава 10. Интегративная деятельность мозга
«отдача» в виде учащения нейронной активности, совпадающей с волнами после
разряда и движением конечности животного. Следовательно, одним из механиз¬
мов суммации является двухфазная реакция с непосредственным переходом от
торможения к возбуждению. Эти эффекты суммация исчезают сразу после пре¬
кращения сочетаний. Однако если те же сочетания проводить на фоне поляриза¬
ции коры слабым постоянным током, они сохраняются в виде следа десятки ми¬
нут после прекращения раздражений.
Сама же поляризация приводит к изменению как фоновой активности, так
и формы реагирования нейронов: учащение реакции на световое раздражение по¬
явилось у ранее нереагировавших нейронов, увеличилась интенсивность ответа
у ранее реактивных клеток коры, тормозная реакция на раздражитель сменилась
на возбудительную. Была показана возможность двух функциональных состоя¬
ний доминантного центра. Первое — длительное, скрытое, без выхода на эффек¬
тор, с повышением возбудимости не только в поляризационном участке сенсомо-
торной коры, но и в других районах, способствующее иррадиации возбуждения
из любого участка к доминантному очагу. Второе — в момент двигательной реак¬
ции угнетаются ответные реакции коры па дистантные раздражения, что является
проявлением сопряженного торможения при реализации двигательной доминанты.
Повышение возбудимости участка коры под влиянием поляризации не только
обусловлено изменениями мембранного потенциала эфферентных пирамидных
нейронов, но и само влияет на другие структуры. К таковым, в частности, относят¬
ся вставочные клетки и глия: увеличивается количество олигодендроцитов около
пирамид и интернейронов, возникают сдвиги содержания РНК в цитоплазме
и ядрах клеток. Кроме того, меняются и нейронные взаимоотношения, которые
рассматриваются как морфологический коррелят их высокой функциональной
активности.
Описывая свойства доминанты как системы центров с высокой возбудимо¬
стью, А. А. Ухтомский подчеркивал значение временного фактора ее существова¬
ния. Именно на длительности функциональных сдвигов в центрах и базируются
такие свойства доминанты, как стойкость, инертность, способность к суммации.
Иными словами, доминанта опирается в своем существовании на наличие опреде¬
ленного набора оптимальных раздражителей, но в основном на следовые процессы,
которые и отличают доминанту от частных коротких рефлексов. В противовес та¬
кому консервативному началу доминанты, которое выражается в подкреплении
ее за счет всевозможных поводов и впечатлений, А. А. Ухтомский подчеркивал ее
прогрессивную роль в процессе новообразования рефлексов на среду, когда проис¬
ходит быстрый подбор «нужных, имеющих непосредственную связь впечатлений».
Гипоталамический очаг стационарного возбуждения. С целью анализа есте¬
ственных событий в организме рассмотрим свойства отдельных хеморецентивных
зон мозга. Такие «центральные хеморецепторы» или их совокупности описаны,
обнаружена их специализация к определенному химическому соединению или
гормону. Специфические хеморецепторные нейроны, высокореактивные к коле¬
баниям «физиологических констант» организма, описаны в большом числе в яд¬
рах гипоталамуса. Поэтому первичный очаг возбуждения при формировании неко¬
торых естественных биологических доминант (голода, жажды) может возникать
именно в этих образованиях мозга. Инертность такого очага обеспечивается за
счет длительной активации соответствующих низконороговых хеморецепторных
нейронов и окружающих их глиальных клеток.
§ 1. Доминанта и условный рефлекс как основные принципы деятельности мозга 271
Через 7 мин
№С1. (1,5%; 1,05 мя)
После питья
30 мл
нм*
■ц 11 11(;‘
“I
50 мкВ
-МШ,А.Ч.к.л,И.Л.
1111111111111111 I I ’| I I I 1 I I ,1 I I I I П
г Д е
Через 10 мин Адреналин (0,1%; 0,05 мл) Через 15 мин
I 1 Ч„.и,11 111
Ш 1111 ШШ1 I ,1
Рис. 99. Суммарная биоэлектрическая активность коры и ядер гипоталамуса у кошек
с мотивацией жажды, созданной прямым введением хлорида натрия в латеральные ядра
гипоталамуса (по Б. Г. Гафурову и И. В. Якунину, 1989)
а — исходный фон; б — после однократного введевия хлорида натрия (горизонтальная линия сверху)
появляется высоковольтная активность и поисковое поведение в сторону поилки; в — после питья
30 мл воды оба признака жажды исчезают; г— спустя 10 мин жажда восстанавливается, д — вновь ис¬
чезает после введения в латеральный гипоталамус адреналина; е — через 15 мин поведенческие и био¬
электрические признаки жажды восстанавливаются; 1 —латеральный отдел гипоталамуса; 2 — сенсо-
моторная область коры; 3 — супраоптическое ядро гипоталамуса; вертикальная стрелка — введение
адреналина. Калибровка: время — 1 с, амплитуда — 50 мкВ
Например, стойкая мотивация жажды сопровождается возникновением в сун-
раоптическом и в латеральном ядрах гипоталамуса длительных вспышек гипер-
синхронизованной активности, которые с усилением мотивации иррадиируют на
кору больших полушарий (рис. 99). Эти вспышки провоцируются также световы¬
ми и звуковыми стимулами и исчезают после внутриполостного введения воды
или внутривенного введения физиологического раствора. Уже эти признаки сви¬
272 Глава 10. Интегративная деятельность мозга
детельствуют о том, что при мотивации жажды в гипоталамусе формируется стой¬
кий доминантный очаг (А. С. Батуев, Б. Г. Гафуров, 1988).
Подобного рода первичные очаги стационарного возбуждения можно рассмат¬
ривать в качестве одного из системообразующих факторов при формировании до¬
минирующей нейродинамической констелляции мозговых структур.
Анализ нейронной активности латеральных ядер гипоталамуса при формиро¬
вании мотивации жажды (А. С. Батуев, Ю. Ли, 1987) обнаружил возникновение
центральных и периферических перестроек во вкусовой сенсорной системе. Мо¬
тивация жажды повышает возбудимость входных рецепторных элементов вкусо¬
вой сенсорной системы и облегчает проведение к центральным нейронам инфор¬
мации о химических качествах пищи путем раскрытия рецептивных ролей нейро¬
нов и перехода их па режим широкополосного приема.
Возникновение таких первичных очагов может вести к появлению вторичных
и более мощных очагов в других отделах мозга, где имеются предпосылки для
продолжительного удержания следовых процессов. Например, в нейронах гиппо¬
кампа посттетаническая нотенциация длится часами или даже сутками.
Другой структурой, где могут формироваться вторичные очаги стационарного
возбуждения, является кора головного мозга. Принципиальным отличием таких
очагов от первичных является исходная причина их возникновения. Так, если
первичный очаг в гипоталамусе возникает и поддерживается в первую очередь за
счет гуморальных факторов, то вторичные очаги в коре больших полушарий явля¬
ются проекционными по своему происхождению и поддерживаются за счет широ¬
кой конвергенции к ним множества суммирующих здесь сенсорных влияний. По-
видимому, следовые процессы здесь настолько мощны, что приводят к каким-то
структурным изменениям, длительно модифицирующим передачу в синапсах.
Одним из возможных механизмов являются стационарные изменения цитоскеле¬
та в аксошиниковых синапсах.
Если применительно к биологическим мотивациям локализацию первичных
очагов можно связывать с гипоталамическими ядрами, то топография вторичных
очагов должна опираться на карту гипоталамо-кортикальных связей. Р. А. Павлы-
гина (1982) считает, что в нормально функционирующем организме должны быть
заложены механизмы, допускающие сосуществование нескольких доминант од¬
новременно. Сопряженные тормозные взаимовлияния характерны для функцио¬
нально несовместимых деятельностей, взаимоотношения которых закреплены
генетически. Образование же рецептивного поля доминанты, то есть избрание
адекватных для себя сигнальных раздражений, способных к воспроизведению и
последующему поддержанию доминанты, отражает наиболее пластическое звено
доминанты, также опирающееся на процессы центрального торможения. Значит,
торможение как в части афферентного обеспечения доминанты, так и в части ее
координированного эфферентного выполнения играет определяющую роль, и со¬
пряженное торможение первично выступает в роли организатора суммации в на¬
личном очаге возбуждения.
Таким образом, для обеспечения суммации сенсорных воздействий в пределах
рецептивного поля кортикального вторичного «очага» повышенной возбудимо¬
сти необходимо допустить торможение как второстепенных сигнализаций, так
и конкурентных рефлекторных актов.
§ 1. Доминанта и условный рефлекс как основные принципы деятельности мозга 273
Доминанта и условный рефлекс. В своей работе «Об условно-отраженном
действии» А. А. Ухтомский писал: «Шерринггоповские принципы отмечают осо¬
бо, как успело отпечатлеться уже морфологически преобладание рецептивных
приборов над исполнительными (двигательными). Но впервые И. П. Павлов со
своими учениками осветил тот механизм и принцип, которым это преобладание
формируется на ходу работы организма. Это механизм “условного рефлекса”
и принцип “временной связи”. Если воронка Шеррингтона имела в виду посто¬
янные рефлекторные дуги, закрепившие функциональную и морфологическую
связи между собой наследственно и филогенетически, то И. П. Павлов улавливал
самое закладывание и новообразование связи в этой воронке, привлечение все
новых рецептивных сфер конвергенции относительно эфферентных (исполни¬
тельных) приборов... Связь исполнительного органа с новыми рецептивными по¬
водами может закладываться, так сказать, случайно, вследствие того, что работа
данного исполнительного органа имела случаи — более или менее совпадать во
времени с данной дальней рецепцией. Кора бдительно примечает такие совпаде¬
ния, закладывает но их поводу новые связи, сначала временные, а затем могущие
закрепиться в качестве опыта и фонда, который будет использован животным для
дальнейшей жизни... перед нами новая рефлекторная дуга в процессе своего за¬
кладывания и дальнейшего закрепления»1.
Рецептивное иоле доминанты создает спектр тех сигпально значимых факто¬
ров среды, которые обеспечивают данную доминанту на разных стадиях ее суще¬
ствования. А. А. Ухтомский (1966) выделил три стадии доминанты:
1) стадия доминанты, безразлично привлекающей к себе в качестве поводов
возбуждения всевозможные рецепции;
2) стадия образования условного рефлекса, когда из множества действующих
рецепций доминанта вылавливает группу рецепций, которая для нее в осо¬
бенности биологически интересна (выработка адекватного раздражителя
для данной доминанты);
3) стадия более или менее исключительной связи данной доминанты с дан¬
ным рецептивным содержанием, когда каждый из контрагентов (внутрен¬
нее содержание и внешний образ) будут вызывать и подкреплять исключи¬
тельно друг друга.
На первой стадии достаточно устойчивой доминанты ее рецептивное поле ока¬
зывается чрезвычайно широким: разнообразные афферентации подкрепляют на¬
личную доминанту. Причем уже на этой стадии обнаруживается координирую¬
щая роль торможения, препятствующего выполнению иных форм деятельности
(Р. А. Павлыгина, 1982, 1988).
Признавая за доминантой способность подкрепляться разнообразными факто¬
рами среды, следует иметь в виду существование механизмов избирательного реа¬
гирования, то есть наиболее эффективных агентов среды, жестко связанных с под¬
держанием и реализацией данного доминантного состояния. К такого рода сигна¬
лам в первую очередь должны быть отнесены натуральные раздражители (по
павловской терминологии) или ключевые раздражители (по терминологии этоло¬
1 Ухтомский А. А. Собр. соч. Л., 1954. Т. 4. С. 225-226.
274 Глава 10. Интегративная деятельность мозга
гов). Причем совсем не обязательно, чтобы такие свойства сигналов были бы гене¬
тически детерминированы.
Например, в определенных условиях одиночные коммуникационные сигналы
животных могут приобретать характер ключевых раздражителей. Таким условием
для кошек может быть возникновение биологической доминанты, проявляющей¬
ся во время лактации (Н. Н. Соколова, 1984). В стрессорпых ситуациях котята из¬
дают сигналы, близкие по структуре к сигналам новорожденных. Такие сигналы
неизменно вызывают реакцию материнского ухаживания у самок. Если же такие
сигналы предъявляются с магнитофона, то у лактирующих самок проявляется
лишь ориентировочная реакция без активного поиска котенка. Следователь¬
но, для возникновения адекватных ответных реакций кошки-матери необходимо
комплексное воздействие разиомодальпого сигнала, звуковые же компоненты яв¬
ляются лишь частью демонстрационного поведенческого комплекса, недостаточ¬
ной для полного воспроизведения адекватных реакций.
Таким образом, рецептивное поле доминанты материнского ухаживания у лак¬
тирующих кошек представляет собой динамическую систему, в состав которой
входят жесткие компоненты (ключевые сигналы) и пластические компоненты,
биологическая значимость которых для данной доминанты оценивается в контек¬
сте всей окружающей ситуации.
Фронтальная кора у кошек-самок к моменту рождения детенышей оказывает¬
ся избирательно настроенной к тем звуковым частотам, которые являются фор¬
мантными для видоспецифических вокализаций детенышей первого месяца жиз¬
ни (Г. А. Куликов, 1989). Значит, состояние доминанты обеспечивает выделение
жестких компонентов (ключевых), связанных с восприятием наиболее значимой
в биологическом отношении сигнализации и выполнением соответствующих же¬
стко запрограммированных поведенческих актов.
Закономерности избирательного реагирования на сигнальпо значимые агенты
среды могут служить физиологическими предпосылками для трактовки механиз¬
мов сенсорного внимания. В самом общем смысле внимание рассматривается как
один из механизмов устранения избыточности сенсорных сообщений, который
участвует как в избирательном подавлении, обеспечении сенсорных входов, так и
в фильтрации информации, извлекаемой из систем кратко- и долговременной па¬
мяти.
Закрепление рецептивного поля с выделением адекватных для данной доми¬
нанты агентов внешней среды осуществляется по механизму условного рефлекса,
в котором важнейшую роль отводят фактору подкрепления. Подкрепление услов¬
ного рефлекса направлено к удовлетворению текущей доминирующей мотива¬
ции, к снижению возбудимости первичного доминантного очага. Мотивационное
поведение животных направлено прежде всего на снижение доминанты. Полное
удовлетворение биологической потребности, достигнутое в результате подкреп¬
ления систем условных рефлексов, приводит к исчезновению доминантных оча¬
гов и торможению соответствующих условных рефлексов.
Теперь обратимся к рассмотрению эндогенно вызванного доминантного состоя¬
ния (например, жажды) и его условно-рефлекторного удовлетворения (И. П. Ни¬
китина, 1983).
§ 1. Доминанта и условный рефлекс как основные принципы деятельности мозга 275
Мотивация жажды создавалась добавлением соли в пищевой рацион и ограни¬
чением воды. У животных на разные условные сигналы вырабатывались побежки
к двум разным кормушкам, в одной из которых подавалась пресная, а в другой —
соленая пища. При избыточном поступлении соли в организм возникал отказ жи¬
вотного даже от минимально соленой нищи и прекращение нобежек по сигналу
«соленого». В результате солевой перегрузки активируется система песнецифиче-
ской оценки содержания соли в предлагаемой животному пище с последующим
формированием отрицательной реакции на эту пищу. Это состояние является
стойким и гетеросенсорным по своему обеспечению, ибо не только конкретный
условный сигнал, но и вся окружающая ситуация его подкрепляла, то есть оно ха¬
рактеризовалось всеми свойствами доминантного состояния. Доминирующая мо¬
тивация жажды выступает как фактор неспецифической инициации реакций от-
вергания всего соленого и предпочтения всего пресного. Одновременно с этим мо¬
гут возникнуть и реакции неспецифического поиска предмета удовлетворения
доминирующей мотивации. Даже однократное положительное завершение такого
поведенческого акта, санкционируемое соответствующим эмоциональным сопро¬
вождением, может закрепиться в памяти.
Значение сигнальных факторов окружающей среды выступает ярко при ис¬
пользовании различных приемов противопоставления условных сигналов «пре¬
сного», которые теперь подкрепляются пищей разной степени солености. По сиг¬
налам «пресного» животное направляется к соответствующей кормушке и поеда¬
ет из нее пищу такой степени солености, которая ранее стойко отвергалась. Такое
условно-рефлекторное «опреснение» нищи (психическое опреснение) может слу¬
жить удачной моделью для исследования вкусовых шитзий.
Вероятно, пищевой условный сигнал, как и доминирующая мотивация, управ¬
ляют прежде всего вкусовыми порогами, поэтому животное и поедает неадекват¬
ную сигналу пищу. Условный сигнал приобретает свойства создавать определен¬
ный диапазон вкусовых порогов, в пределах которого данная мотивация может
быть реализована с наибольшей вероятностью. Висцеральная сигнализация, фор¬
мирующая доминирующую мотивацию, также определяет тот пороговый диапа¬
зон вкусовых сигналов, которые способны удовлетворить биологическую потреб¬
ность.
Третья стадия доминанты строится на законе «обратных отношений» А. А. Ух¬
томского: симптомокомплекс доминанты может репродуцироваться при наличии
адекватных сигналов. Такие ретенционные способности принадлежат коре боль¬
ших полушарий, которая:
♦ формирует адекватные стимулы для доминанты;
♦ запускает соответствующие доминанты по поводу этих стимулов;
♦ обеспечивает взаимодействие и переключение доминант.
На этой стадии речь должна идти не столько об элементарных биологических
доминантах (голод, жажда, половое чувство), сколько о сложном памятном следе,
носящем ситуативный характер (интегральный образ), а также о динамической
констелляции мозговых структур, обеспечивающих целенаправленное поведение
организма.
Старая доминанта возобновляется или для того, чтобы при новых данных
обойтись при помощи старого опыта, или для того, чтобы по новым данным пере-
276 Глава 10. Интегративная деятельность мозга
Комбинация механизма
доминанты с механиз¬
мами формирования
условного рефлекса
обеспечивают два фак¬
тора, необходимых и
достаточных для орга¬
низации целенаправлен¬
ного поведения: его ак¬
тивный творческий
характер (доминанта)
и точное соответствие
объективной реальности
(упроченный, тонко спе¬
циализированный
условный рефлекс).
интегрировать старый опыт (А. А. Ухтомский, 1950).
Значит, пространственно разнесенные компоненты до¬
минирующей констелляции представляют собой цент¬
ральную модель поведенческого акта, которая сформи¬
рована по закону доминанты и условного рефлекса и за¬
писана в долговременной памяти.
Справедливо заключение П. В. Симонова, который
пишет: «Комбинация (точнее — синтез) механизма до¬
минанты с механизмами формирования условного реф¬
лекса обеспечивают два фактора, необходимых и доста¬
точных для организации целенаправленного поведения:
его активный творческий характер (доминанта) и точное
соответствие объективной реальности (упроченный, тон¬
ко специализированный условный рефлекс)»1.
Таким образом, доминанта и условный рефлекс рас¬
сматриваются в качестве основных принципов организации целенаправленного
и адаптивного поведения.
§ 2. Высшие интегративные системы мозга
Ретикулярная форма¬
ция — филогенетиче¬
ски старая система
мозга, которая не пред¬
ставляет собой единого
анатомического целого
и морфологически яв¬
ляется гетерогенным
образованием.
Ретикулостволовый уровень интеграции
Ретикулярная формация — это филогенетически старая система мозга, которая
не представляет собой единого анатомического целого и морфологически являет¬
ся гетерогенным образованием. Трудно определить и ана¬
томическую протяженность ретикулярной формации, ко¬
торая занимает центральное положение на всем протяже¬
нии ствола (рис. 100).
Нейрональной основой интегрирующей функции ре¬
тикулярной формации служат длинноотростчатые
клетки, аксоны которых могут распространять влияние
как в каудальном, так и в ростральном направлении. Ре¬
тикулярные клетки соединяют разные отделы внутри ре¬
тикулярной формации, а последнюю со спинным мозгом,
большими полушариями и мозжечком.
Каждая сенсорная система направляет пути в ретикулярную формацию. Опи¬
саны мощные влияния соматических и висцеральных нервов, что позволило счи¬
тать одной из функций ретикулярной формации висцеросоматическую интегра¬
цию. Показаны пути в ретикулярную формацию от ядер тройничного нерва, вес¬
тибулярных ядер, верхней оливы.
Кора мозга оказывает регулирующие влияния на деятельность ретикулярной
формации. Пути из ретикулярной формации в нисходящем направлении — к спин¬
ному мозгу, мозжечку, ядрам специфических систем и в восходящем — к структу¬
рам больших полушарий вплоть до коры также построены по топическому прин¬
ципу, что свидетельствует о функциональной специализации ретикулярных ядер.
Симонов П. В. Взаимодействие доминанты и условного рефлекса как функциональная единица органи¬
зации поведения // Успехи физиол. наук. 1983. Т. 14, № 1. С. 18-19.
§ 2. Высшие интегративные системы мозга 277
Рис. 100. Схема стволовой части мозга кошки, показывающая корковые связи
ретикулярной формации (по Дж. Френчу, 1962)
Стрелками с точками показаны влияния на ретикулярную формацию (РФ);
темные стрелки — тормозящие эффекты ствола мозга на сенсорную проводимость в мозгу; заштри¬
хованные стрелки — облегчающие или тормозящие эффекты ствола мозга на рецептор;
Т — таламус; II, V, VIII — черепно-мозговые нервы
Для активности ретикулярных нейронов характерна аутогенная ритмика, ко¬
торая служит для осуществления топических влияний на спинной мозг и большие
полушария головного мозга. Сами ретикулярные нейроны способны отвечать на
раздражения практически всех афферентных путей или центральных мозговых
структур. Такая обширная конвергенция влияний у разных ретикулярных единиц
значительно варьирует и зависит от функционального состояния мозга. Если ре¬
тикулярный нейрон дифференцирует входные влияния по их модальностям, то
полисипантические цени нейронов с короткими аксонами, но которым актив¬
ность распространяется внутри самой ретикулярной формации, лишают первона¬
чальный сигнал его модальной специфичности. Поэтому к ядрам таламуса посту¬
пает активация, являющаяся продуктом иолимодального интегрирования на ре¬
тикулярных нейронах.
Дифференцированный характер конвергенции, топическое фракционирова¬
ние и гетерогенность в организации афферентных и эфферентных связей позво¬
ляют заключить, что функции, приписываемые ретикулярной формации, касают¬
ся главным образом интеграции и регуляции разных видов деятельности.
П. К. Анохин (1968) обращал внимание па гетерохимическую чувствитель¬
ность отдельных синапсов и целых иолипейроиальиых цепей, активируемых раз¬
дражениями различного биологического качества (боль, голод и др.).
Ретикулярные влияния по соответствующим путям могут достигать различ¬
ных релейных ядер сенсорных систем и дифференцированно модулировать аффе¬
рентные потоки. Показано, например, что концентрация внимания животного на
зрительном объекте приводит к блокированию передачи сенсорных импульсов в
слуховой системе (П. Эрпапдец-Пеон, 1962). Явления, именуемые привыканием,
связывают с регулирующей функцией ретикулярной формации. Поскольку при¬
выкание рассматривают как простейшей вид обучения, то ретикулярной форма¬
ции приписывают известную роль в формировании условных рефлексов (А. Гас-
то, И. Иошии и др.).
278 Глава 10. Интегративная деятельность мозга
Интегрированные на ретикулярном уровне размодальные афферентные по¬
сылки с учетом мотивационных факторов формируют импульсные тонические
влияния к специфическим сенсорным системам, приводящие к модуляции прове¬
дения по ним соответствующих сигнализаций. В результате создаются предпо¬
сылки для успешного синтеза различных сенсорных сигнализаций на более высо¬
ких уровнях мозга. Другая сторона восходящих ретикулярных влияний может за¬
ключаться в создании на таламокортикальном уровне такого функционального
состояния (или центрального тонуса мозга), который обеспечит адекватные усло¬
вия для формирования разиомодального афферентного синтеза.
Тадамокортикальный уровень интеграции
Нейрофизиология ассоциативных систем мозга. Наряду со специфическими и
неспецифическими системами принято выделять в качестве самостоятельной ка¬
тегории ассоциативные таламокортикалъные системы. Применительно к выс¬
шим млекопитающим это своеобразные структуры, не принадлежащие какой-ли¬
бо одной сенсорной системе, но получающие информацию от нескольких сенсор¬
ных систем. Ассоциативные ядра таламуса относятся к
«внутренним ядрам», афферентные входы к которым
идут не от сенсорных лемписковых путей, а от их релей¬
ных образований. В свою очередь, эти ядра проецируются
на ограниченные корковые территории, которые имену¬
ются ассоциативными полями.
Согласно анатомическим данным, выделяют две выс¬
шие ассоциативные системы мозга. Первая включает зад¬
нюю группу ассоциативных ядер, проецирующихся на те¬
менную область коры, и именуется таламопариетапъпой
системой. Вторая па таламическом уровне состоит из медиодорзального ядра с его
проекцией на лобную область коры и называется таламофронталъной системой.
Обе ассоциативные системы — продукт прогрессивной дифференциации неспе¬
цифического таламуса и достигают значительных размеров у приматов и чело¬
века.
Таламопариетальная система. Теменная кора является местом широкой гете-
росеисорной конвергенции по волокнам от специфических, ассоциативных и не¬
специфических ядер таламуса, а также но путям от сенсорных корковых зон
и симметричной коры противоположного полушария.
Световые воздействия оказывают наиболее сильный эффект па теменную ко¬
ру: здесь описаны наряду с мультисепсорными и собственно зрительные нейроны,
которые отвечают на перемещения в определенном направлении сложных геомет¬
рических фигур. Иногда эти же клетки отвечают и па звуковые воздействия, и на
движения глаз.
Обнаружены три типа межсенсорного взаимодействия на нейронах теменной
коры:
1) суммирующий нейрон, который при одновременном гетеросенсорном раздра¬
жении отвечает более сильным разрядом с более коротким латентным перио¬
дом, чем на мономодальные стимулы;
2) тормозящийся нейрон, латентный период ответа которого увеличивается,
когда стимул из моносеисорпого превращается в гетеросенсориый;
Ассоциативные систе¬
мы мозга — это свое¬
образные структуры,
не принадлежащие ка¬
кой-либо одной сен¬
сорной системе, но
получающие информа¬
цию от нескольких сен¬
сорных систем.
§ 2. Высшие интегративные системы мозга 279
3) детектор комплекса, который отвечает импульсным разрядом только при комп¬
лексном гетеросепсорном воздействии и не реагирует на изолированное при¬
менение мономодальных стимулов (рис. 101).
Показано при этом, что в теменной коре существуют полисенсорные клетки,
отражающие докорковый, таламический уровень интеграции, наряду с такими
нейронами, на мембране которых реализуются собственно кортикальные меха¬
низмы межсенсорпого синтеза.
г
б
20 60 100 мс
12 -
8 -
4 -
0 I-
Свет + сом.
...гч
20 60 100 мс
Звук
2^
20 40 60 80 мс
Свет
20 40 60 80 мс
4 :
П Свет + звук
—
20 40 60 80 мс
в
Рис. 101. Типы взаимодействия разномодальных сигналов на нейронах
теменной коры мозга кошек
На а и б: слева — постстимульные гистограммы (по оси абсиисс — длительность латентного периода,
мс; по оси ординат — число потенциалов действия); справа — импульсная активность нейронов; в —
суммарная (снизу) и импульсная (сверху) активность, сверху вниз на а: ответ на звук, свет и звук + свет,
на б и в — ответ на соматическое раздражение; свет и свет + соматическое раздражение (два разных
опыта). Калибровка: 250 мкВ, 100 мс. На схеме: ССИ — средняя супрасильвиева извилина; точка —ме¬
сто расположения микроэлектродных треков
280 Глава 10. Интегративная деятельность мозга
Теменная ассоциативная кора посылает мощные нисходящие связи ко многим
сенсорным, лимбическим, ретикулярным и двигательным аппаратам мозга и даже
образует волокна в составе кортикоспинального тракта (О. С. Адрианов, 1976).
Все вышесказанное дает основание рассматривать теменные области коры как
важнейший дискриминационно-интегративный аппарат больших полушарий.
После удаления теменных полей коры возникают глубокие нарушения в услов¬
но-рефлекторной деятельности как на простые мономодальные, так и в особенно¬
сти на разномодальные комплексные сигналы. Это связано со специальной ролью
теменной коры в управлении процессами селективного внимания, с оптимизацией
режима текущей деятельности, а также с формированием ориентационных дви¬
жений к опознаваемому сигналу.
В целом таламопариетальная ассоциативная система мозга представляет со¬
бой:
♦ центральный аппарат первичного симультанного анализа и синтеза обстано¬
вочной афферентации и запуска механизмов ориентационных движений;
♦ один из центральных аппаратов «схемы тела» и сенсорного контроля текущей
двигательной активности;
♦ важнейший элемент долобной предпусковой интеграции, участвующий в фор¬
мировании целостных полимодальпых образов.
Таламофронтальная система. Сенсомоторпая кора получает множественные
проекции из специфических, ассоциативных и неспецифических отделов таламу¬
са, ассоциативные кортико-кортикальные и трапскаллозальпые входы из проти¬
воположного полушария и характеризуется наличием сложных синаптических
комплексов для конвергенции па одном и том же нейроне множества афферент¬
ных влияний. Не только вставочные нейроны, по и выходные элементы сенсомо-
торной коры — нейроны пирамидного тракта — относятся к категории мультисен-
сорных клеток.
Установлено три типа ответных реакций мультисенсорпых нейронов сенсомо-
торной коры:
1) привыкание, заключающееся в снижении вероятности ответа на каждый после¬
дующий стимул в серии;
2) сенситизация — усиление реакции при повторных применениях стимула;
3) экстраполяция — формирование упреждающего ответа нейрона па каждый по¬
следующий стимул в серии.
Такие пластические перестройки ответной активности нейронов в зависимо¬
сти от характера стимуляции и функционального состояния, интенсивности и мо¬
дального состава стимулов свидетельствуют о наличии сложных механизмов
функциональной конвергенции, имеющих непосредственное отношение к форми¬
рованию системных реакций целостного организма.
Этого нельзя сказать по отношению к собственно лобным полям на дорзолате-
ралыюй поверхности прореальной извилины кошек, которые но нейрофизиоло¬
гическим и морфологическим данным еще не имеют непосредственной увязки
с интегративными процессами мозга, а скорее относятся к структурам иеспеци-
фического типа. В то же время медиобазальные отделы лобной коры формиру¬
ют нисходящие пути к таламическим ядрам лимбической системы. Последствия
§ 2. Высшие интегративные системы мозга 281
удаления нрореальной извилины у кошек состоят в дефиците зрительного опо¬
знания (Л. В. Черепкова, 1975) или в нарушениях точности зрительно контроли¬
руемых двигательных актов (Ю. А. Юнатов, 1981). Последнее связывают с тем,
что основное пространство собственно лобной коры кошек занимает глазодвига¬
тельное поле. Нарушений в других сенсорных системах или эмоционально-моти¬
вационной сферы не отмечалось.
Таким образом, можно выделить следующие основные механизмы работы ас¬
социативных систем мозга (А. С. Батуев, 1979, 1981, 1984):
1. Механизм мулътисенсорной конвергенции. Его специфичность определяется
тем, что к ассоциативным нолям коры конвергируют афферентные посылки,
несущие информацию о биологической значимости того или иного сигнала.
Такие отселектированные афферентные влияния вступают в интеграцию на
кортикальном уровне для формирования программы целенаправленного пове¬
денческого акта.
2. Механизм пластических перестроек при гетеромодалъных сенсорных воздейст¬
виях. Динамический характер мультисенсорной конвергенции может прояв¬
ляться либо в избирательном привыкании, либо в сенситизации, либо, нако¬
нец, в формировании экстраполяционного типа ответов. Установлен факт важ¬
ной роли доминирующей мотивации в определении спектра конвергирующих
модальностей и в организация впутрикорковых интеграций. Для теменной ко¬
ры преобладающим может быть горизонтальный (ламинарный) тип межней-
ронной интеграции, а для сенсомоторной ассоциативной коры преобладаю¬
щим является вертикальный (модульный) тип межнейронной интеграции.
3. Механизм краткосрочного хранения следов интеграции, заключающийся в дли¬
тельной внутрикорковой или таламокорковой реверберации импульсных по¬
токов (см. гл. 8). Последнее объясняет дефекты памяти и обучения у кошек
и собак после разрушения ассоциативных нолей коры или соответствующих
ядер таламуса.
Эволюция ассоциативных систем
В параллельных рядах, которыми шло развитие современных млекопитающих,
хотя и сохранился общий план конструкции мозга, но таламокортикальные его
системы претерпели наиболее существенные морфофункциональные перестрой¬
ки. Высокого развития достигают корковые механизмы деятельности сенсорных
систем с ясно выраженной тенденцией возрастания ассоциативных систем мозга
со свойствами полисенсорного конвергирования. В динамике морфологических
преобразований происходит обособление зон перекрытия корковых проекций,
с которыми связывают реализацию наиболее сложных форм высшей нервной дея¬
тельности.
В пределах класса млекопитающих можно выделить три основных уровня эво¬
люции ассоциативных систем мозга. При этом необходимо иметь в виду, что сте¬
пень развития ассоциативных формаций мозга рассматривается как показатель
филогенетического статуса вида (Г. И. Поляков, 1964). Причем мозг насекомояд¬
ных рассматривается как предшественник с его дальнейшим усложнением в па¬
раллельных рядах грызунов, хищных и приматов. Исследования обнаружили не¬
четкость границ дифференциации внутри новой коры и неопределенность функ¬
282 Глава 10. Интегративная деятельность мозга
циональной идентификации ее полей. Это согласуется с отсутствием в таламусе
четких границ между сенсорными ядрами.
Вместе с этим определенная область коры ежей (рис. 102) по морфологиче¬
ским критериям позволяет допустить присущие ей интегративные свойства даже
в пределах столь примитивно организованного неокортекса (Г. П. Демьяненко,
1977). К этой области коры ежей проецируются примитивные ассоциативные яд¬
ра таламуса — медиодорзальпое и заднелатеральное. В данной области обнаруже¬
ны как разномодальные моносенсорные, так и полисенсорпые нейроны (А. А. Пи¬
рогов, 1977). Такие корковые элементы активируются преимущественно по еди¬
ному каналу, берущему начало в недифференцированной задней группе ядер
таламуса. Таким образом, в этой таламокортикалыюй системе сочетаются свойст¬
ва как неспецифических, так и ассоциативных систем млекопитающих (рис. 102).
Рис. 102. Схема эволюционного созревания интегративных аппаратов мозга
а — насекомоядные, 6 — хишные, в — приматы. Выделены лишь две специфические сенсорные систе¬
мы — зрительная и соматическая (som и vis); тонкие линии — их проекционные пути; жирные линии
и заштрихованные участки — ассоциативные системы, жирные стрелки — кортико-кортикальные свя¬
зи; пунктирные — зоны эфферентных корковых проекций (преимущественно пирамидных); cor — кора
большого мозга; Thal — зрительный бугор, th.-front — таламофронтальная; th.-pariet — таламопарие-
тальная ассоциативная система больших полушарий мозга
Было показано (И. В. Малюкова, 1974), что такая форма зрительно-слуховой
интеграции, как двигательный условный рефлекс на одновременный комплекс, не
может быть выработана у ежей и попытка ее сформирования приводит к невроти¬
ческим срывам. Однако сложный цепной двигательный условный рефлекс может
быть легко образован на мопомодальный сигнал и разрушается после удаления
ассоциативной области неокортекса. Больше страдают самые тонкие компоненты
сенсомоторной интеграции, которыми завершается пищедобывательный двига¬
тельный акт.
Следовательно, хотя примитивная ассоциативная система ежей еще не способ¬
на к организации сложных актов межсепсорпой интеграции, но она уже начинает
участвовать в осуществлении процессов сепсомоторного синтеза.
Как уже указывалось, насекомоядные являются прямыми филогенетическими
предшественниками грызунов, хищных и приматов.
§ 2. Высшие интегративные системы мозга 283
Исследование конструкции ассоциативных систем мозга у крыс, кроликов, ко¬
шек, собак и низших обезьян позволили прийти к следующему заключению.
Первый уровень — грызуны, мозг которых близок к насекомоядным. У грызу¬
нов отсутствует четкая дифференциация коры и таламуса на специфические и ас¬
социативные зовы. Известная диффузиость представительства сенсорных систем
в больших полушариях головного мозга грызунов коррелирует со сравнительно
низким уровнем их аналитико-синтетической деятельности. Слабая выраженность
морфологической дифференциации и функциональной специализации полисен-
сорных структур у грызунов является фактором, определяющим несовершенство
интегративной функции мозга.
У белых крыс формирование условного рефлекса на зрительно-слуховой комп¬
лекс оказалось также безрезультатным, неоднократные тренировки этой трудной
задачи приводили к невротическим срывам. В то же время зрителыю-тактильпый
комплекс выработать удалось, хотя он характеризовался нестабильностью. При¬
мерно аналогичная картина выявилась у кроликов, у которых дифференцирова¬
ние светового и звукового компонентов от их одновременного предъявления
в комплексе не достигало выше 50-60%-ного уровня. Это отражает низкий уро¬
вень аналитико-синтетической деятельности насекомоядных и грызунов.
Второй уровень — хищные, у которых впервые появляются в коре больших по¬
лушарий развитые лобные и теменные ассоциативные поля и соответствующие
структуры таламуса. Для хищных характерны существенные структурные и функ¬
циональные различия ассоциативных таламокортикальных систем от других моз¬
говых структур. Это наличие сложных нейроносинаптических компонентов, к ко¬
торым конвергируют сенсорные потоки, несущие биологически значимую ин¬
формацию. Причем таламопариетальная система отражает усложнение актов
пространственной ориентировки и формирование механизмов, составляющих те¬
кущий сенсорный фон для выполнения целенаправленных поведенческих актов.
Таламопариетальная система — следствие усложнения конструкции и связей зри¬
тельной сенсорной системы, она обеспечивает первичный межсенсорпый синтез,
формирует сложный комплекс обстановочной афферентации и систему «схемы
тела».
Таламофроитальпая система включается в корковый отдел соматической сен¬
сорной системы с одновременной проекцией на нее лимбических аппаратов мозга.
Таламофроитальпая система участвует в организации предпусковой интеграции
и программировании сложных поведенческих актов. Здесь в большей мере отра¬
жены процессы мотивационно-эмоциональной окраски поведенческих актов бла¬
годаря прямым связям этой системы со всем лимбическим комплексом. В преде¬
лах отряда хищных лобиые отделы неокортекса усложняются, увеличиваются их
размеры и функциональная роль в организации сложных форм поведения, тре¬
бующих мобилизации механизмов краткосрочной и долгосрочной памяти.
У хищных (кошки, собаки) выработка условного рефлекса на разномодальиый
комплекс выполняется за 10-15 опытов. Уровень дифференцирования светового
и звукового компонентов от комплекса достигает 70-90%, но для этого требуется
регулярная тренировка. Иными словами, сигнальное значение комплекса поддер¬
живается при условии угашения реакций на отдельные компоненты. Следова¬
тельно, у хищных существует более высокий уровень аналитико-синтетической
284 Глава 10. Интегративная деятельность мозга
деятельности, заключающейся в способности к интеграции разпомодальных сиг¬
налов в целостный образ.
Другой важной стороной интегративной функции мозга является степень раз¬
вития процессов памяти и основанное на них свойство прогнозирования пред¬
стоящего поведения. В естественных же условиях существования животных
любая поведенческая адаптация относительна и имеет вероятностный характер.
Поэтому применительно к биологическим условиям обитания животных в веро¬
ятностно изменчивой внешней среде имеет место и вероятностное прогнозирова¬
ние, а значит, адаптивность поведенческих программ определяется степенью их
избыточности и подвижности.
Изучалось поведение в стационарных случайных средах. На звуковой сигнал
животное должно было направляться к одной из двух кормушек, причем чаще
всего к той, где наиболее вероятно получало подкрепление. Степень вероятности
подкрепления из каждой кормушки менялась (А. С. Батуев, И. В. Малюкова, 1979;
А. И. Карамян, И. В. Малюкова, 1987). Кошки способны к формированию поведе¬
ния в сторону той кормушки, вероятность пищевого подкрепления из которой
наиболее высока. Причем поведение их меняется в соответствии с изменением ве¬
роятности подкрепления. Повреждение фронтальных отделов неокортекса (в ос¬
новном сеисомоторной коры) сохраняет элементарные условные рефлексы, но
разрушает способность к вероятностному прогнозированию.
Процессы межсенсориой интеграции развиты у хищных в достаточной мере,
хотя кортикальный уровень ассоциативных систем мозга имеет перекрытие с зо¬
нами выхода эфферентных кортикальных трактов. С другой стороны, у хищных
достаточно велика роль докоркового уровня межсенсориой интеграции и кора по¬
лучает наряду с модально специфичными афферентными залпами уже перерабо¬
танную песпецифическую имнульсацию из ассоциативных ядер. Наконец, каждая
из ассоциативных систем у хищных характеризуется доминированием того или
иного сенсорного входа, что, естественно, не способствует достижению полной ге-
теросенсорной интеграции.
Третий уровень — приматы, у которых ассоциативные структуры таламуса
с их обширной и дифференцированной проекцией в лобные и теменные области
коры включаются в состав самостоятельной целостной интегративной системы
больших полушарий. Существенной особенностью приматов являются развитые
кортико-кортикальные связи, с помощью которых ассоциативные поля могут объ¬
единяться в целостную иерархически построенную систему. Благодаря компакт¬
ной системе миелинизированных ассоциативных волокон возрастает роль корти¬
кального уровня взаимодействия специфических сенсорных зон с ассоциативным
неокортексом. Ассоциативные поля неокортекса характеризуются топкой диффе¬
ренциацией с формированием из нейронных элементов целостных структурно-
функциональных ансамблей.
Спецификой ассоциативного неокортекса является конвергирование в нем
множества сенсорных сообщений о биологической значимости внешней сигнали¬
зации в соответствии с доминирующей мотивацией по независимым друг от дру¬
га афферентным каналам. При этом возрастает роль собственно кортикального
уровня межсенсориой интеграции и обеспечения процессов краткосрочной памя¬
ти. После разрушения либо лобных, либо теменных ассоциативных нолей коры
не только глубоко страдают процессы межсенсорпого синтеза, ориентировочно¬
§ 2. Высшие интегративные системы мозга 285
исследовательская деятельность, краткосрочная память, но и формирование бо¬
лее простых форм условно-рефлекторной деятельности.
Функциональная значимость отдельных ассоциативных систем расширяется
и уточняется в сравнении с хищными. Утрачивается преобладание какого-либо
одного сенсорного входа, а следовательно, расширяются возможности их интегра¬
ции. Возникает топографическая разнесенность ассоциативных полей от собст¬
венно афферентных корковых формаций, что снижает удельное значение сенсо-
моторной интеграции и расширяет роль коры в осуществлении межсенсорного
афферентного синтеза. Возникает все большая их взаимозависимость для обеспе¬
чения деятельности целостной интегративной системы полушарий.
Для приматов (низшие обезьяны) выработка условных рефлексов на одновре¬
менный комплекс представляет собой относительно легкую задачу, ибо в процессе
применения комплексного раздражителя компоненты самопроизвольно утрачива¬
ют сигнальное значение и сформированные условно-рефлекторные связи сохра¬
няются месяцами без дополнительной тренировки. У обезьян можно выработать
условный рефлекс даже на трехчленный комплекс из разпомодальных сигналов.
Это свидетельствует о более высоком уровне аналитико-синтетической деятель¬
ности мозга обезьян в сравнении с хищными, что находит отражение и в большей
структурной дифференциации и функциональной специализации ассоциативных
таламокортикальных систем.
Обезьяны легко справляются с задачами на вероятностное прогнозирование,
однако после разрушения в области ассоциативной лобной коры лишаются этой
способности, их поведение приобретает однообразный персеверативный ха¬
рактер.
Очевидно, способность использования предыдущего опыта, записанного в дол¬
госрочной памяти, для прогнозирования поведения в стационарных случайных
средах претерпевает существенные эволюционные преобразования, которые опре¬
деляются степенью развития интегративных систем мозга, уровни дифференциа¬
ции которых коррелируют со степенью совершенства аналитико-синтетической
деятельности и организации сложных форм поведения.
Онтогенез ассоииативных систем мозга
Изучение динамики формирования ассоциативных систем мозга показало гете-
рохронность этого процесса, что, по-видимому, обусловлено включением отдель¬
ных звеньев таламокортикальных систем в обеспечение разных поведенческих ак¬
тов, последовательность созревания которых определяется их необходимостью
для осуществления жизненно важных функций новорожденного животного (зре¬
ло- и незрелорождающегося).
Согласно концепции системогенеза (П. К. Анохин, 1968), неравномерность со¬
зревания нервных элементов и связей между ними объясняется их вовлечением в
структуру различных функциональных систем. Гетерохронность их созревания
определяется значимостью для выживания организма, особенно в критические
периоды жизни (см. гл. 4), когда новорожденное животное вступает в непосредст¬
венное соприкосновение с окружающей средой. Анатомически и функционально
первыми созревают те сенсорные механизмы (соматические, акустические), кото¬
рые обеспечивают выживание на начальных этапах индивидуального развития.
286 Глава 10. Интегративная деятельность мозга
В ассоциативных ядрах таламуса обнаружены проекции в кору больших полу¬
шарий уже у новорожденных котят (В. П. Бабминдра, Л. А. Васильева, 1987). В то
же время неспецифические таламические ядра формируют свои проекции на кору
несколько позже, их основная роль состоит в контроле внутриталамической ак¬
тивности.
Несмотря на раннее формирование ассоциативных систем, их окончательное
созревание происходит в течение довольно длительного периода и заканчивается
у кошек между 2-м и 3-м месяцами жизни. Именно к этому времени у котят впер¬
вые формируется полноценный условный рефлекс па одновременный комплекс,
приближающийся по своим характеристикам к аналогичному условному рефлек¬
су взрослого животного (Л. А. Васильева, Л. В. Черенкова, 1986).
Когда первый критический этап постнаталыюго развития преодолен, насту¬
пает период бурной дифференциации мозговых структур и возникновения меж¬
ду ними двусторонних связей диффузного характера. Формируется та структур¬
но-функциональная матрица, которая послужит основой для дальнейшего раз¬
вертывания процессов координации и выделения локальных функциональных
структур.
Таким образом, принцип развития — от диффузного неспецифического к ло¬
кальному специфическому (А. И. Карамяп, 1976) — общебиологическая законо¬
мерность, которой подчиняется и динамика развития ассоциативных систем мозга.
Наконец, третий этап связан с моментом формирования тормозных коорди¬
национных механизмов как в самой коре, так и в глубоких структурах. Появле¬
ние таких механизмов обеспечивает топкую специализацию как сенсорных, так
и ассоциативных систем мозга, а значит, и различных целостных поведенческих
актов.
§ 3. Ассоциативные системы и сенсорная
функция мозга
Прежде чем произойдет какая-нибудь приспособительная реакция, осуществля¬
ется процесс восприятия сигналов. Но живой орх'анизм заинтересован не в самих
воздействиях как таковых, а в том, о чем они сигнализируют, и соответственно не
в оценке их физических параметров, а в тех соотношениях, которые с их помощью
передаются. Вместе с тем понятно, что отнесение сигнала к тому или иному клас¬
су, то есть его распознавание, неизбежно основано на выделении ряда физических
характеристик, образующих область признаков данного класса сигналов.
Из сказанного выше следует, что сенсорная функция мозга заключается в опре¬
делении сигнальной (биологической) значимости сенсорных стимулов на основе
анализа их физических характеристик. Под биологической значимостью по¬
нимается направленность реакций живого организма, которая определяется его
доминирующей мотивацией, возникающей при отклонении параметров устойчи¬
вого неравновесия, а также информацией, извлекаемой из окружающей среды и
прошлого жизненного опыта. Для оценки биологической значимости сенсорных
сигналов анализ их физических характеристик является необходимой, по недо¬
статочной операцией. Биологическая значимость сигнала реализуется в актива¬
ции некоей совокупности эффекторных аппаратов. Следовательно, извлечение
§ 3. Ассоциативные системы и сенсорная функция мозга 287
биологически полезной информации с целью ее использования для формирова¬
ния двигательных актов основано на преобразовании входной кодовой комбина¬
ции активности нервных элементов сенсорных систем в реакцию исполнитель¬
ных аппаратов, что по своей сути представляет процесс декодирования сенсорных
сообщений. Тогда оценка биологической значимости сигнала сводится к установ¬
лению соответствующей закономерности взаимодействия нервных элементов
сенсорных и двигательных систем мозга.
Выход на эффекторпые элементы должен обладать свойством избирательно¬
сти, чтобы избавить систему от неадекватных реакций. Выделение экологически
значимых признаков (обычно достаточно простых) у животных низких уровней
эволюционного развития может осуществляться в самих сенсорных системах
(иногда уже на рецепторном уровне) вследствие жесткой организации нервных
связей. Однако у более высокоорганизованных животных (хищных, приматов)
эта операция уже не может ограничиться специфическими образованиями сен¬
сорных систем. Ведь по мере развития уровня организации животных рецептуар
стимулов, имеющих сигнальное значение, как правило, расширяется. Поэтому
возрастание активности организма как системы должно быть связано с возможно¬
стью перехода избирательного приема от одних сигналов к другим и в соответст¬
вии с этим — с появлением новых внутримозговых конструкций.
Нервные образования, входящие в систему оценки биологической значимо¬
сти сигналов, должны удовлетворить ряду требований. Во-первых, они должны
быть связаны с различными сенсорными системами. Во-вторых, они должны быть
связаны с мотивациогегшыми структурами лимбической системы, ибо значи¬
мость тех или иных сигналов определяется на основе доминирующей мотивации.
В-третьих, учитывая, что значимость одного и того же сенсорного стимула зави¬
сят от всей окружающей ситуации, то есть от показаний других сенсорных систем,
следует допустить наличие конвергенции нолимодальной имнульсации и способ¬
ность к пластическим перестройкам активности нейронов. Наконец, в-четвертых,
эта система должна быть связана с регуляцией целостных двигательных актов на
основе избирательного характера реагирования. Изложенным требованиям удов¬
летворяют таламокортикальиые ассоциативные системы мозга; таламопариеталь-
ная и таламофронтальпая.
Получены доказательства (А. М. Иваницкий, 1976)того, что ранние компонен¬
ты суммарного вызванного потенциала мозга человека отражают поступление
в кору информации о физических характеристиках сенсорных сигналов, а позд¬
ние — являются отражением значимости сигнала для организма. У больных с пси¬
хическими расстройствами, имеющих определенный дефицит одного из видов
информации, возникают стойкие изменения соответствующих фаз вызванных по¬
тенциалов. В особенности выделяются ассоциативные области неокортекса в свя¬
зи с выраженной конвергенцией к ним разномодальной имнульсации как основы
оценки сигнальной значимости стимула в контексте соответствующих воздейст¬
вий.
Именно этот механизм у высших позвоночных способствует селективному
отбору одних видов информации с одновременным сопряженным торможением
других сенсорных влияний. При этом у животных доминирующая мотивация воз¬
никает из биологической потребности в результате сдвигов гомеостаза. Однако
доминирующая мотивация формируется при воздействии стимулов, ранее свя¬
288 Глава 10. Интегративная деятельность мозга
занных с определенными подкреплениями. Живая система способна предвидеть
изменения гомеостаза с помощью сигналов, предвосхищающих сдвиг параметров
от нормы или возвращение к ней. В связи с этим другие сенсорные стимулы спо¬
собны изменять значимость сигнала, если они меняют доминирующую модер¬
низацию. И тогда значимый стимул является сигналом, реагирование на который
сопряжено с большей вероятностью достижения желаемого результата в соответ¬
ствии с доминирующей мотивацией и прошлым жизненным опытом. Наличие со¬
пряженных признаков в комплексном сенсорном воздействии позволяет умень¬
шить неопределенность выбора поведенческих реакций, необходимых для полу¬
чения требуемого результата.
Роль ассоциативных систем в оценке биологической значимости сенсорных
сигналов отчетливее видна но отношению к зрению и слуху. Конечно, сигнальное
значение имеют все сенсорные стимулы, но способностью опережающего воздей¬
ствия обладают в первую очередь зрительные, слуховые, а также обонятельные
сигналы. Особенностью структурной организации телецептивиых входов в ассо¬
циативные системы является множественность путей проведения к ним слуховых
и зрительных афферентаций.
Для целей определения биологической значимости механизмы конвергенции
обеспечивают полифункциональность одного и того же сигнала, то есть зависи¬
мость его смысла от ситуации, в которой он возникает. Степень значимости любо¬
го агента внешней среды зависит не только от определенной мотивации, по и от
сопоставления этого агента с другими факторами среды, когда он выступает в ка¬
честве компонента более сложного сенсорного комплекса. Именно поэтому наи¬
более полная сигнальная значимость фактора окружаю¬
щего пространства может реализовываться с участием тех
структур мозга, куда приходит информация о разных но
сенсорным качествам факторах среды, то есть по ме¬
ханизму одновременного гетеросенсорного сопоставления.
Одним из проявлений динамического характера конвер¬
генции в ассоциативные зоны коры является свойство
привыкания их электрических реакций при использо¬
вании однообразной ритмической стимуляции. Степень развития привыкания
находится в прямой зависимости от биологической значимости применяемого
сигнала. Например, стабильные ответы нейронов лобных долей у бодрствующих
обезьян возникают лишь в том случае, если раздражители прямо или косвенно
связаны с эмоционально-мотивационной сферой животного, необычны для не¬
го или сигнализируют об опасности, то есть сопровождаются четкой ориентиро¬
вочной реакцией. Привыкание ответов наблюдается в тех случаях, когда исполь¬
зуемые раздражители являются индифферентными для животного или хорошо
знакомы ему (рис. 103). Если же эти агенты сопровождаются биологически значи¬
мым подкреплением по правилам выработки условного рефлекса, то они приобре¬
тают свойства условного сигнала с одновременной стабилизацией реакций нейро¬
нов лобной коры в ответ на их применение.
Как привыкание, так и сенситизация отражают дистрибутивный характер дан¬
ных пластических реакций мозга и совпадают с закономерностями проявления
ориентировочного рефлекса. Поэтому предполагается, что «новизна» является
тем информативным признаком сенсорных стимулов, с которого начинается их
Механизмы конверген-
иии обеспечивают по¬
лифункциональность
одного и того же сигна¬
ла, то есть зависимость
его смысла от ситуа¬
ции, в которой он воз¬
никает.
§ 3. Ассоциативные системы и сенсорная функция мозга 289
I II
Рис. 103. Реакции нейрона при предъявлении раздражителей обезьяне
(по А. А. Пирогову, 1980)
/ — сохранение (эмоционально значимый раздражитель); II -— привыкание (индифферентный
раздражитель), а — изменение средней частоты импульсации на 1 -е, 3, 5 и 7-е предъявления
раздражителей; б — гистограммы ответа нейрона на соответствующие раздражения; в —
нейронограммы ответов на 1 -е и 7-е предъявления; стрелки — моменты предъявления
раздражителей
оценка в ассоциативных системах мозга. Привыкание к повторным стимулам, ве¬
роятно, связано с процессами формирования следов кратковременной памяти
(Е. Н. Соколов, 1981).
С помощью ориентировочной реакции устанавливаются соответствующие
воспринимающие поверхности для наилучшего восприятия внешнего раздраже¬
ния. При размещении источника телецептивпых стимулов но средней линии го¬
ловы наблюдается их наиболее высокое пространственное разрешение. Избира¬
тельность к этой физиологически значимой области пространства применительно
к акустическим сигналам обнаружена для нейронов мозжечка и сеисомоториой
ассоциативной коры (Я. А. Альтман, 1972; Г. А. Куликов, 1989). Участие фронталь¬
ной и теменной областей коры в обеспечении точности ориентировочных реакций
установлено и для зрительных сигналов, по отношению к которым отмечена и
пространственная, и дирекциональпая избирательная чувствительность соответ¬
ствующих нейронов. Более того, после повреждения теменной ассоциативной об¬
ласти отчетливо выступает нарушение пространственного восприятия зритель¬
ных стимулов и адекватных двигательных реакций, то есть нарушения простран¬
ственного согласования зрительно-моторной координации.
Система, оценивающая биологическую значимость сенсорных стимулов, долж¬
на обладать избирательным характером реагирования па их отдельные значимые
10 За*. 1078
290 Глава 10. Интегративная деятельность мозга
признаки и/или совокупность последних. Поэтому логичен вопрос о том, как от¬
ражаются в реакциях ассоциативных структур мозга различные физические пара¬
метры сенсорных сигналов.
Известно, что нейроны фронтальной ассоциативной коры принимают зритель¬
ную имнульсацию, не связанную с поточечным описанием контура зрительных
сигналов, но отражающую их временные и пространственные параметры. Кро¬
ме того, показано, что нейроны фронтальной коры обладают свойством избира¬
тельного реагирования на биологически значимые признаки коммуникативных
акустических сигналов. Существенно то, что для млекопитающих (в отличие от
низших позвоночных) лишь небольшое число сигналов может иметь четкое «клю¬
чевое» значение, которое в большей мере определяется доминирующим мотива¬
ционным фоном для данного животного. Поэтому становится понятным участие
в оценке биологической значимости сигнала таламофронталыюй ассоциативной
системы, тесно связанной с лимбическими структурами мозга, имеющими прямое
отношение к формированию мотивационных состояний (Г. А. Куликов, 1989).
В связи с этим закономерен вопрос, как отражается в характере электрических
реакций фронтальной коры процесс приобретения ранее индифферентным звуко¬
вым стимулом определенного сигнального значения при выработке условного
рефлекса. Специальные исследования показали, что преобразование ранее неэф¬
фективного звукового тона в мотивационно окрашенный сигнал оборонительной
реакции животного сопровождается возникновением избирательного реагирова¬
ния элементов фронтальной коры именно на данную частоту заполнения тональ¬
ного сигнала. Однако по мере стабилизации и закрепления условного рефлекса
такая избирательность выражалась слабее и наконец исчезала, что свидетельст¬
вует о динамической роли фронтального неокортекса в формировании условно-
рефлекторной сенсомоторной интеграции.
Описанные выше закономерности избирательного реагирования на сигнально
значимые агенты среды могут служить физиологическими предпосылками для
трактовки механизмов сенсорного внимания (Н. Ф. Суворов, О. П. Таиров, 1986).
В самом общем смысле внимание рассматривается как один из механизмов
устранения избыточности сенсорных сообщений, который участвует как в изби¬
рательном регулировании сенсорных сообщений, так и в фильтрации информации,
извлекаемой из систем кратковременной и долговременной памяти. Принято ду¬
мать, что процессы фильтрации осуществляются уже в специфических образо¬
ваниях сенсорных систем. Между тем в ряде работ мнение об уменьшении из¬
быточности информации основано на данных об уменьшении плотности потока
импульсации при переходе от нижележащих уровней сенсорных систем к вышеле¬
жащим и на постулировании использования в их деятельности двоичного кода.
Этот вопрос, однако, остается нерешенным с нейрофизиологических позиций, и про¬
блемой «внимания» преимущественно владеют психологи и психофизиологи.
Сам А. А. Ухтомский (1923), сформулировав принцип доминанты как физио¬
логическую основу акта внимания, писал, что из множества действующих рецеп¬
ций доминанта вылавливает группу рецепций, которая для нее в особенности био¬
логически значима.
Определение вектора поведения (по принципу доминанты) основано на взаи¬
модействии биологически значимых экстероцептивпых (окружающая среда)
§ 4. Ассоциативные системы мозга и программирование поведения 291
и интероцептивных (мотивационных) воздействий, основным субстратом уста¬
новления которых являются таламокортикальные ассоциативные системы мозга.
Следовательно, целостная сенсорная функция мозга обеспечивается содруже¬
ственной деятельностью сенсорных и ассоциативных систем и направлена на ор¬
ганизацию адаптивных движений и действий. Последнее лежит в основе согласо¬
вания движения с чувствованием (И. М. Сеченов, 1863).
§ 4. Ассоциативные системы мозга
и программирование поведения
Одним из основных атрибутов любого сложного целенаправленного движения
является формирование предварительных программ.
Роль программы в структуре двигательного акта должна рассматриваться
с учетом биологической мотивированности движения, его временных параметров,
моторной дифференцированности, степени сложности координационного соста¬
ва и уровня его автоматизированное™.
Стратегия и тактика движения. Биологическая мотивированность двигатель¬
ного акта является основным побуждающим (инициальным) фактором для его
реализации. Именно мотивации формируют целенаправленные движения, а сле¬
довательно, определяют их общую стратегию. Значит, если в основе стратегии
движения находится биологическая (или социальная) мотивация, то каждый конк¬
ретный двигательный акт будет рассматриваться как шаг к удовлетворению этой
мотивации, то есть будет решать какую-то промежуточную задачу или цель
(рис. 104). Биологические мотивации могут приводить либо к запуску «запаян¬
ных», то есть жестких, программ, устанавливать их комбинаторику, с чем мы
встречаемся у беспозвоночных и низших позвоночных и именуем инстинктами
или комплексами фиксированных действий, либо приводить к формированию
новых сложных программ, определяя одновременно степень их лабильности.
П. Тейтельбаум (1967) пишет, что в тех случаях, когда действие полностью яв¬
ляется автоматическим следствием стимула, невозможно говорить о мотивации.
При этом между стимулом и ответом существуют фиксированные взаимоотноше-
Для чего? Что? Как?
Рис. 104. Соотношение прямых и обратных связей в системе управления движением
292 Глава 10. Интегративная деятельность мозга
пия. Мотивация «ломает» эти фиксированные связи между стимулом и реакцией
с помощью процесса обучения. Например, в отличие от многих инстинктив¬
ных реакций, реакция нажатия на педаль может быть «отделена» от внутреннего
состояния животного. Операитная ситуация, сигнал, реакция, подкрепление яв¬
ляются полностью произвольными, не обладающими фиксированными связями
друг с другом.
Возьмем для примера такое биологическое адекватное движение у кошек, как
прицельный прыжок. У одной группы кошек он условно-рефлекторно закреплял¬
ся при пищевом подкреплении, у другой — при электрооборонительном (А. С. Ба¬
туев, 1960). У животных экстренно выключалась зрительная рецепция. Условные
прыжки при пищевом подкреплении исчезли полностью, а при электрооборони-
тельном — восстанавливались через две педели после операции. Следовательно,
одна и та же двигательная программа (или план движения, по Прибраму) характе¬
ризовалась разной степенью устойчивости к сенсорной депривации в зависимо¬
сти от биологического знака подкрепляющего агента. Образно говоря, если моти¬
вация определяет, для чего, с какой целью осуществляется движение, то она же,
определяя программу конкретного движения, обусловливает степень ее зависи¬
мости от сенсорных факторов. В качестве обратной афферептации здесь выступа¬
ет удовлетворение мотивации. Поэтому цель движения, его стратегия должна
быть прежде всего адаптивна но отношению к доминирующей биологической мо¬
тивации.
Программа определяет, какое конкретное движение будет осуществляться для
достижения дайной цели, и формирует его пространственно-временные парамет¬
ры в соответствии с реальной обстановкой. Следовательно, программа движения
должна быть в высокой степени адаптивной ко всем факторам окружающей среды
и строиться при их одновременном и строгом учете, то есть па основе синтеза всех
видов афферептации по закону мультисенсорной конвергенции.
В общей схеме системной организация двигательного акта существует понятие
о тактике движения, которая определяет, как будет осуществляться требуемое
движение, иными словами, каковы те эфферентные залпы к двигательным ядрам
спинного мозга, которые определяют активацию различных мышечных групп.
Последовательность включения той или иной конечности, записанная в програм¬
ме, реализуется в виде последовательности импульсных залпов надсегмеитарных
структур к различным мотопейроппым пулам.
В отличие от программы двигательные команды должны быть очень адаптив¬
ными к функциональному состоянию самого скелетно-двигательного аппара¬
та как непосредственного исполнителя этих команд. Исходя из представления
о существовании настройки перед совершением движения (Я. М. Код, 1975;
Э. Эвартс, 1975) высказано предположение о том, что срочная идентификация со¬
стояния сложной системы осуществляется путем оценки реакции системы на не¬
большое точно дозированное возмущение (тест-импульс), которое подается непо¬
средственно перед командами или с моментом их запуска. Причем тест-импульс
подается только на те мышцы, которые должны будут участвовать в следующем
движении. Реакция мышц на тест-импульс будет не настолько значительна, чтобы
существенно изменить само движение, но достаточна для возбуждения мышеч¬
ных веретен при изменении длины интрафузальных волокон на 5-10 мк. Инфор¬
§ 4. Ассоциативные системы мозга и программирование поведения 293
мация о реакции па тест-импульс конвергирует на некотором промежуточном
уровне с основным управляющим воздействием (командой), которое корректиру¬
ется в соответствии с актуальным состоянием двигательного аппарата. Учитывая
временную дисперсию проведения по пирамидному тракту, легче всего допустить
участие быстропроводящих его волокон (со скоростью 50-60 м/с) в реализации
такого тестового воздействия и коррекции последующих нисходящих импульс¬
ных залпов уже па сегментарном уровне. Таким образом, биологически мотиви¬
рованная стратегия (цель) движения, соответствующая программа конкретного
двигательного акта, иерархически построенные центральные команды и система
идентификации состояния мышечного аппарата представляют собой важнейшие
звенья организации двигательного акта, каждое из которых корректируется соот¬
ветствующими обратными связями.
Последние обеспечивают движению высокую адаптивность в конкретных усло¬
виях времени и пространства.
Участие ассоциативных систем мозга в организации движении. Роль внеш¬
них факторов, сигналов из внешней среды и соответственно роль сенсорных и ас¬
социативных систем мозга в формировании мотивированных движений весьма
значительна. Специфичность участия таламонариетальной ассоциативной систе¬
мы в организации движений определяется двумя моментами.
С одной стороны, она участвует в формировании интегральней схемы тела, все
части которого соотнесены не только друг с другом, но и с вестибулярными и зри¬
тельными сигналами.
С другой стороны, она участвует в регуляции внимания к текущим сигналам
окружающей среды с учетом ориентации всего тела относительно этих сигналов.
Таламопариетальная (как и нижневисочпая) ассоциативная система активиру¬
ется текущими сенсорными сигналами, то есть привязана в основном к настояще¬
му моменту времени, и связана с анализом главным образом пространственных
взаимоотношений разномодальных признаков.
Фронтальная ассоциативная система имеет реципрокпые отношения с двумя
функциональными системами мозга:
1) теменно-височной, которая связана с обработкой и интеграцией полимодаль-
ной сенсорной информации;
2) теленцефалической лимбической системой, включающей лимбическую кору
и связанные с ней подкорковые образования, особенно гипоталамус и районы
среднего и промежуточного мозга.
Характер связей указывает на то, что фронтальная система вовлечена и в сен¬
сорные, и в моторные механизмы мозга (рис. 105). Вентральную часть фронталь¬
ной коры рассматривают как кортикальную проекцию лимбической системы, ко¬
торая участвует в анализе и регуляции процессов внутренней среды организма.
Одно из существенных функциональных расстройств, возникших у челове¬
ка после поражения лобных долей мозга, — это распад поведенческих программ,
неспособность поддерживать нормальную временную организацию поведения.
Программы их действий, будучи однажды запущенными, повторяются, застаива¬
ются и лишь постепенно стираются, причем эта персеверация зачастую приводит
к отклонению от желаемой цели (рис. 106).
294 Глава 10. Интегративная деятельность мозга
Стратегия Связи стратегии и тактики и отбор Тактика
Мышцы
Рис. 105. Схема взаимодействия различных структур мозга, ответственных
за формирование стратегии и тактики двигательного поведения
(по X. Корнхуберу, 1974)
АМ — передние медиальные ядра таламуса; VI. — вентролатеральное ядро таламуса;
ГП — генератор плавных движений; ГС — генератор движений саккадического типа
Тот факт, что даже осознаваемое различие между целью и результатом не ведет
к смене программы, предполагает, что все эти сигналы — ошибки, обычно коррек¬
тирующие поведенческие программы, не находят выхода в эффекторную сферу.
Отсюда следует, что фронтальная ассоциативная кора опосредует мотивационные
влияния на организацию поведения в целом.
Реципрокные фронтолимбические взаимоотношения лежат в основе феноме¬
на «поведенческого предвосхищения», или экстраполяции, потеря которой яв¬
ляется одним из важных клинических симптомов фронтальных повреждений.
Обычно нормальный субъект выбирает линию поведения после сравнения боль¬
шего или меньшего числа альтернативных тактик. В конечном счете этот выбор
осуществляется при сравнении аффективных реакций на каждую из возможных
альтернатив. Выдвинутая таким образом тактика, естественно, должна пройти
«цензуру» мотивационной системы. Процессы такого предвосхищающего (экст¬
раполяционного) выбора существенно страдают при удалении лобной коры.
Утрата фронтальной коры как главного передатчика информации между корой
и лимбической системой будет сопровождаться не только нарушением выбора
программ, но также стиранием текущей или предполагаемой программы действия
самой по себе или в результате вмешательства внешних сигналов. Нарушение
процессов кратковременной памяти при лобном синдроме может быть вызвано
патологически быстрым угасанием следов сигналов, с одной стороны, и преоб¬
ладанием «теменных» механизмов переключения внимания на новые стимулы,
освобожденные от тормозных фронтальных влияний, с другой стороны (Б. Миль¬
нер, 1964, 1975).
§ 4. Ассоциативные системы мозга и программирование поведения 295
Импульсные коды моторных про¬
грамм. Программирующая деятель¬
ность мозга заключается в организации
поведенческого акта на основе доми¬
нирующей мотивации, прошлого жиз¬
ненного опыта и в соответствии с веро¬
ятностной структурой среды. Степень
адекватности программы, построенной
с учетом всех трех факторов, может
быть оценена но ее прогнозирующим
свойствам. В программе тесно взаимо¬
действуют процессы кратковременной
и долговременной памяти.
Для выявления импульсного кода
моторных программ ситуация отсрочен¬
ного пространственного выбора оказалась
весьма удобной (А. С. Батуев и др.,
1988). В опытах на обезьянах выраба¬
тывались пищедобывательные услов¬
ные рефлексы, сигналами для которых
служило вснйхивание лампы справа или слева от животного, а условной реакци¬
ей — нажатие рукой соответственно на правый или левый рычаг. Такая деятель¬
ность животного основывается на долговременной памяти. Кроме того, в задаче
фигурировали два вероятностно варьируемых фактора: чередования пространст¬
венного положения условного сигнала и продолжительность следуемой за ним
временной паузы — отсрочки. После восприятия положения сигнала животному
предстояло сохранить полученную информацию на протяжении всего периода от¬
срочки — кратковременной памяти. Степень адекватности программирования
оценивали по правильности выбора рычага, после нажатия на который следовало
пищевое подкрепление. Таким образом, у животного формировались две жесткие
моторные программы, которые извлекались из долговременной памяти и удержи¬
вались в кратковременной памяти вплоть до их реализации. Какая программа из¬
влекалась — это зависело от пространственных признаков условного сигнала и
определялось по моторному ответу. Следовательно, характер импульсной актив¬
ности нейронов от начала действия условного сигнала, на всем протяжении от¬
срочки, после открытия доступа к рычагам (пусковой сигнал) и вплоть до нажа¬
тия на рычаг может рассматриваться как импульсный код моторных программ.
Нейроны лобной и задней теменной коры характеризовались пространствен¬
ной селективностью, то есть кодированием местоположения условного сигнала
рисунком или частотой разряда (рис. 107). Такие же клетки найдены в хвостатом
ядре и вентральном переднем ядре таламуса, но они отсутствовали в бледном ша¬
ре, моторной коре, медиодорзальном и центральном латеральном ядрах таламуса.
Одна часть клеток обнаружила различия в активности только в момент действия
условных сигналов («сенсорные нейроны») или только в период отсрочки («ней¬
роны памяти»). Как отмечалось, адекватность извлеченной программы можно
оценить только по конечному результату.
„Л
> * - 8/1 ^,
б
4 р ъ ^ ^
Рис. 106. Примеры почерка больного
с локальным поражением правой лобной доли
а — видны затруднения в написании цифр и букв,
6 — навязчивые повторные движения (персевера¬
ции) при попытке написания слов
296 Глава 10. Интегративная деятельность мозга
Рис. 107. Реакции одного пространственно-селективного нейрона лобной коры
обезьян во время выполнения целенаправленного движения с альтернативным
отсроченным выбором
а — растры и соответствующие гистограммы при действии условного сигнала справа; б — то же, слева;
2-3 — период действия условного сигнала (загорание лампы после рычага); 3-4 — период отсроч¬
ки (кратковременной памяти); 4 — пусковой сигнал (открывание экрана, позволяющее осуществить
манипулирование с одним из рычагов, по вертикали — число импульсов, с, по горизонтали — время, с,
Д( = 200 мс, п = 10)
В числе пространственно-селективных клеток обнаружена популяция нейро¬
нов, которые выявляют это свойство только после открытия экрана, то есть в пус¬
ковой период, когда животное получает свободный доступ к рычагам. Их назва¬
ли нейронами «моторных программ», которые эстафетно активируются нейро¬
нами памяти и являются пусковыми для соответствующего двигательного акта
(рис. 108). Отчетливо видно, что в случаях правильного выбора рычага при одном
и том же движении, но направленном либо к правому (сверху), либо к левому
(снизу) рычагу, возникает совершенно противоположный по характеру рисунок
импульсной активности нейрона. То же наблюдается и при ошибочном выборе
рычага для нажатия.
Гистограмма правильного выбора рычага па правый сигнал отражает актива¬
цию моторной программы нажатия на правый рычаг. Но при ошибочном выборе
по условному сигналу слева животное вновь нажимает на правый рычаг. То же
справедливо и в случаях нажатия на левый рычаг. Сравнение гистограмм по диа¬
гонали убеждает в их полной идентичности. Значит, рисунок разряда нейрона при
нажатии па правый или левый рычаг отражает лишь специфику извлеченвых ра¬
нее из долговременной памяти жестких моторных программ. Рисунок разряда
нейрона отражает основное различие в этих программах — направление двига¬
тельного акта. Обнаружены также смешанные типы нейронных популяций, ко¬
торые описывают местоположение условного сигнала, сохраняя эту особенность
импульсации в период отсрочки и до реализации соответствующего двигательно¬
го акта. Такие нейроны характерны для лобной, теменной коры, хвостатого ядра
и вентрального переднего ядра.
§ 4. Ассоциативные системы мозга и программирование поведения 297
Рис. 108. Активность нейрона лобной коры обезьяны после пускового сигнала
(открывание экрана)
а — пространственный условный сигнал был дан справа; б — слева; 1 — правильный выбор рычага;
2 — ошибочный выбор; стрелка — момент открывания экрана. Видно, что рисунок гистограммы со¬
ответствует планируемому и выполняемому движению с правым (1а и 26) или с левым рычагом
(16 и 2а)
Другие исследованные структуры (моторная кора, бледный шар и др.), вероят¬
но, не имеют отношения к формированию моторных программ, по тесно связаны
с реализацией соответствующего движения.
Анализ имнульной активности нейронов позволил допустить существование
достаточно жестких форм программирования движений, при которых вероятно¬
стный сигнал сразу встраивается в уже готовую конструкцию сложного сенсомо-
торного синтеза, выработанного в процессе предварительной условно-рефлектор¬
ной тренировки. Выбор двух возможных действий выступает как извлечение из
долговременной памяти адекватной моторной программы с одновременным тор¬
можением программы альтернативной. Торможение выступает в качестве ведуще¬
го процесса ограничения степеней свободы для вывода на поведенческий уровень
единственно адекватной двигательной программы.
Таким образом, самой сложной функцией мозга является способность форми¬
ровать последовательность движений и предвидеть ее реализацию. Морфологиче¬
ски решение этой задачи связано с фронтальной ассоциативной системой. Воз¬
можность воспроизведения запрограммированной последовательности движений
связана со способностью фронтальной ассоциативной системы запоминать и хра¬
нить в памяти такие последовательности. Вероятнее всего, эти последовательно¬
сти и содержание событий кодируются некоторым сокращенным способом, па-
пример по ведущему сигнальному признаку. Высшим отражением этого кодиро¬
вания у человека является вербализация основных понятий движения. Одна из
основных характеристик лобного синдрома у человека состоит в нарушении грам¬
матической структуры речи и невозможности использования вербальных команд
для построения последовательности движений.
298 Глава 10. Интегративная деятельность мозга
Заключение
Целенаправленное поведение определяется доминирующей мотивацией, побуж¬
дающей организм к удовлетворению преобладающей потребности.
Адаптивный же характер поведения достигается с помощью множества услов¬
ных рефлексов, которые обеспечивают приспособление организма к конкретной
пространственно-временной ситуации. Неспецифическая направленность поис¬
кового поведения определяется наличием гипоталамического очага стационарно¬
го возбуждения, обладающего доминантными свойствами (инертность, высокая
возбудимость, способность к суммации); поисковая же активность в конкретной
ситуации определяется системой корковых условно-рефлекторных связей как ос¬
новы прошлого жизненного опыта, который обеспечивает направленный поиск
объекта удовлетворения потребности.
Высшие интегративные (ассоциативные) системы мозга являются основными
аппаратами управления пластичными формами поведения, которые обеспечива¬
ются механизмами:
♦ селективной конвергенции биологически значимой информации;
♦ пластических перестроек под влиянием доминирующей мотивации;
♦ краткосрочного хранения интегральных образов и программы предстоящего
поведенческого акта.
Степень развития ассоциативных систем мозга в эволюции млекопитающих
коррелирует с совершенством аналитико-синтетической деятельности и органи¬
зацией сложных форм поведения.
Способность формировать последовательность движений и предвидеть ее реа¬
лизацию как самая сложная функция мозга достигает наибольшего развития у че¬
ловека, обладающего свойствами речевого управления поведением.
Контрольные вопросы
1. Каковы свойства коркового очага стационарного возбуждения?
2. Каково происхождение гипоталамического очага стационарного возбуждения?
3. Доминанта и условный рефлекс — основные принципы интегративной дея¬
тельности мозга.
4. Особенности ретикулостволового уровня интеграции.
5. Что такое ассоциативные системы мозга?
6. Каковы основные свойства ассоциативных систем мозга?
7. Таламопариетальная система и ее функции.
8. Таламофронтальная система и ее функции.
9. В чем состоит основная линия эволюции ассоциативных систем мозга млеко¬
питающих?
10. Закономерности онтогенеза ассоциативных систем мозга.
11. Ассоциативные системы и сенсорная функция мозга.
12. Ассоциативные системы и механизмы программирования поведения.
13. Что такое импульсные коды моторных программ?
Глава 11
ПСИХОФИЗИОЛОГИЧЕСКАЯ ПРОБЛЕМА
Психофизиологическая проблема издавна привлекала внимание крупных мысли¬
телей и служила ареной ожесточенных споров, подогреваемых идеологическими
и политическими мотивами разных времен и народов. Именно эта проблема —
о соотношении физиологического и психического, биологического и социального
в природе человека — оказалась на переднем крае общественно-исторического
развития человечества, и то или иное ее решение служило идеологическим и по¬
литическим оружием в борьбе нового со старым, в решении ряда задач воспита¬
ния, обучения, организации труда и здоровья.
Кризис в науке о поведении возник в результате развития новых областей
знания: психофизики и экспериментальной психологии. В построениях неко¬
торых психологов и биологов эго направление получило название психофизиче¬
ского параллелизма. Крупнейшие представители физиологии (Ч. С. Шеррипгтон,
Дж. Экклс, Р. Гранит), изучавшие элементарные реакции и процессы мозга, попа¬
дали во власть дуалистических представлений при попытках объяснить природу
целостного поведения. Еще Э. Дюбуа-Реймон в качестве одной из семи принци¬
пиально неразрешимых мировых загадок указал на возникновение психических
процессов. Основатель экспериментальной психологии В. Вундт считал, что фи¬
зиологическими исследованиями невозможно проникнуть в тайну психики по¬
тому лишь, что психические процессы развиваются параллельно телесным и не
определяются ими. Эта идея делала бессмысленным накопление физиологиче¬
ских фактов с попыткой понять сущность и происхождение психики и механиз¬
мов поведения. Возник разрыв между физиологическими и психологическими
исследованиями — наступил кризис.
Наука настойчиво искала и нашла выход из этой ситуации -- выход из пробле¬
мы психофизического параллелизма был найден в последовательном изучении
физиологических механизмов, лежащих в основе психических феноменов.
На того, кто, не проводя серьезных исследований в качестве физиолога и пси¬
хиатра, назовет себя в будущем столетии психологом, серьезные люди будут смот¬
реть так же, как па человека, который считает себя архитектором, по не учился в
технической школе или в строительной академии, — таково было твердое убежде¬
ние В. М. Бехтерева.
А спустя еще несколько десятилетий, когда уже определились методологиче¬
ские позиции обеих наук, наш современный психолог А. Р. Лурия требовал от пси¬
хологов ясного понимания того, что знание законов работы мозга, осуществляю¬
щего психическую деятельность, так же обязательно, как и знание общественно-
исторических законов, определяющих формирование сознательной деятельности
людей.
Проблема соотношения между психическими и физиологическими процесса¬
ми (психофизиологическая проблема), и сегодня являющаяся ключевой в теории
отражения, теснейшим образом связана с основным вопросом философии — соот¬
ношение материи и сознания. Психофизиологическая проблема в сжатой форме
300 Глава 11. Психофизиологическая проблема
сводится к вопросу И. П. Павлова: «Каким образом материя мозга производит
субъективное явление?» Основная сложность при естественно-научной разработ¬
ке психофизиологической проблемы представляется в необходимости объясне¬
ния специфических особенностей психических процессов отражения на основе
механизмов деятельности нервной системы. К этим специфическим особенно¬
стям относятся предметность восприятия и его проецируемость во внешнее про¬
странство, целостность, активность и недоступность психических процессов пря¬
мому чувственному наблюдению.
В целом основная парадоксальная специфичность психического заключается
в формулируемое™ его характеристик не в терминах состояния субстрата, свой¬
ством которого оно является, а в терминах свойств его источника — объекта
(Л. М. Веккер, 1974). Сложности в объяснении свойств отражения объективного
мира, специфических черт психических процессов, как правило, и служат основой
для отрицания материалистического подхода к решению психофизиологической
проблемы. Именно абсолютизация специфичности психических процессов и не¬
состоятельность редукционизма приводила и приводит к агностицизму, идеализ¬
му, дуализму.
Широко известна гипотеза дуалистического иптеракционизма одного из наи¬
более крупных современных нейрофизиологов Джона Экклса, сформированная
под влиянием концепции «трех миров» английского философа К. Поппера. На ос¬
нове вывода, сделанного при анализе работ сторонников «научного материализ¬
ма» (сущность представления которых сводится к тому, что все психические про¬
цессы должны быть описаны в физических терминах) о невозможности создания
нейрофизиологической теории, которая объяснит, как разнообразие мозговых про¬
цессов может быть синтезировано до целостного сознания, Дж. Экклс (1979) пред¬
полагает существование разных миров, находящихся в совершенно различных
пространствах, но взаимодействующих через «связывающий мозг», представлен¬
ный фронтальной областью коры левого доминантного полушария. Несмотря на
естественную критику подобных взглядов со стороны представителей самых раз¬
ных областей научного знания (Д. И. Дубровский, 1980), сходные высказывания
появляются как в философской, так и в физиологической литературе.
Главной задачей И. П. Павлова было стремление понять физиологическую ос¬
нову психической деятельности человека и ее закономерности. Предложенный им
объективный метод условных рефлексов рассматривался как важнейший инстру¬
мент, с помощью которого можно создать фундамент психологического знания.
Вместе с этим психические процессы теснейшим образом связаны с физиологиче¬
скими явлениями, именно связаны, но несводимы к ним. Психические процессы
нетождественны тем физиологическим процессам, которые могут составлять ма¬
териальную основу первых. Л. С. Выготский (1982) писал, что следует изучать не
отдельные вырванные из единства психические и физиологические процессы, ко¬
торые при этом становятся совершенно непонятными для нас; мы должны брать
целый процесс, который характеризуется со стороны субъективной и объектив¬
ной одновременно. При этом надо иметь в виду необходимость корректного ис¬
пользования терминологии при изучении одного и того же явления средствами
разных наук.
Общепризнано, что в основе психических процессов лежат не элементар¬
ные процессы возбуждения и торможения, а системные процессы, объединяющие
отдельные физиологические проявления в интегрированное целое. Понятие
Психофизиологическая проблема 301
Системные процессы
поведенческого ак¬
та — это информаци¬
онные процессы, для
которых физиологиче¬
ские процессы высту¬
пают как материаль¬
ный носитель.
об интегральном образе как продукте пережитых доминант, то есть как продукте
субъективного отражения объективного взаимоотношения организма со средой,
явилось принципиальным в решении психофизиологиче¬
ской проблемы (А. А. Ухтомский). Важный шаг в ее раз¬
витии был сделан Г1. К. Анохиным (1968) в его концепции
функциональных систем, способствовавшей установле¬
нию концептуальной связи физиологии и психологии.
Здесь рассматривается организация физиологических
процессов в системе, которая является внутренним отра¬
жением внешней среды, что уже само но себе относится к
категории психического. Значит, системные процессы поведенческого акта —
это информационные процессы, для которых физиологические процессы высту¬
пают как материальный носитель.
Идея динамического единства физиологического и психического успешно раз¬
вивается в борьбе с дуалистическими представлениями психофизического парал¬
лелизма. Д. И. Дубровский (1980) утверждает, что психический образ и его нейро-
динамический эквивалент не только одновременные явления, по и составляют не¬
раздельную целостность.
Однако остается главная трудность, заключающаяся в невозможности тракто¬
вать результаты психологического анализа физиологическим языком, а психоло¬
гические феномены — на основе чисто физиологических закономерностей. Это
становится архисложным для конкретного психофизиологического исследова¬
ния, поскольку никакое совпадение психических феноменов и мозговых процес¬
сов проблемы не решает. Процессы восприятия, мышления, сознания нетождест¬
венны мозговым процессам и не могут быть исчерпывающе описаны понятиями
физиологии.
Это разные формы движения материи, а значит, существенно важна проблема
поиска содержательных переходов между ними.
Огромный фактический материал о поведении животных и человека не про¬
двинул проблему соотношения психики и материи, пока она решалась исходя из
представления о психике как о духовном, наджизнепном начале, реализующемся
в поведении, движении по принципу «извне — внутрь». Иные возможности воз¬
никли па основе теории отражения, представляющей психику как свойство живой
высокоорганизованной материи отражать своими состояниями независимо от
них существующую действительность. Психическое закономерно возникает па
определенной ступени развития жизни, то есть усложняющиеся условия ставят
организм перед необходимостью отражения объективной действительности в ви¬
де ощущений и восприятий, ведущих к ее осознанию. Противоположность между
субъективным и объективным не изначальна, не абсолютна, она порождается раз¬
витием определенных форм отражения, и это раздвоение мира — свойство только
сознательной деятельности человека (А. Н. Леонтьев, 1975). По мнению Л. С. Вы¬
готского (1956), ценность учения о доминанте в том, что она отвечает на основной
вопрос: как формируется единство и целостность поведения; что превращает сум¬
му отдельных реакций в целенаправленные поведенческие акты с новыми, ранее
неведомыми свойствами.
Доминирующая констелляция на уровне коры мозга способна создавать реф¬
лекторную систему, действующую как целое во всех ее двигательных и вегетатив¬
ных проявлениях. Головной мозг А. А. Ухтомский определял как удивительный
302 Глава 11. Психофизиологическая проблема
аппарат, представляющий собой множество переменных, калейдоскопически сме¬
няющих друг друга органов предупредительного восприятия, предвкушения и про¬
ектирования среды.
Таким образом, доминанта проявляется в психической деятельности, причем
не только внешней, предметно развернутой, но и внутренней. Психическое со¬
стояние и процессы «опредмечиваются» как интегральные образы, в основе кото¬
рых лежат корковые доминанты. Последние то активно действуют в поле созна¬
ния, то переходят в сферу неосознанного, где сохраняются в открытой форме как
следы прошлой деятельности. Такие доминанты могут либо поочередно «выплы¬
вать» в поле ясного внимания, либо вступать в конфликт, и тогда «душевная
жизнь будет борьбою вытесняющих друг друга несогласных доминант». Всякое
понятие и представление, всякое индивидуальное психическое содержание есть
след о пережитой некогда доминанте. Этот след, а иногда и вся доминанта в пол¬
ном ее выражении вплоть до эмоциональных состояний и вегетативных сдвигов
воспроизводится адекватным сигналом. Эти явления лежат в основе механизмов
воспоминания и мышления.
Из процессов, протекающих на любом из уровней сенсорных систем, не выво¬
дятся такие особенности сенсорных образов, как предметность, проецируемость
и т. д. Публичная наблюдаемость является существенным отличием физических
явлений от психических. Однако что считать наблюдаемостью? Если реакции от¬
дельных нейронов еще не позволяют давать однозначную интерпретацию процес¬
сов отражения, то представление о дистрибутивном характере описания сенсор¬
ных сигналов в активности многих нейронов или их совокупностей сегодня уже
позволяет предполагать принципиальную возможность такого наблюдения.
Если под предметом физиологии понимать функции и процессы, осуществ¬
ляющиеся в различных живых системах, то с полным основанием можно утверж¬
дать, что в головном мозге не существует таких нейродинамических процессов,
которые были бы принципиально недоступны физиологическим исследованиям.
Вся история физиологии XX в. являет собой убедительный пример того, как
мозг — материальный субстрат управления поведением — становился предметом
непосредственного физиологического анализа. Особенно впечатляющими явля¬
ются успехи нейрофизиологии мозга, которая не только позволила понять многие
интимные механизмы его работы, но и выработала новую систему знаний, позво¬
ливших в значительной мере преодолеть кастовую замкнутость отдельных физио¬
логических школ и направлений.
Логика наших рассуждений приводит нас к необходимости поиска на этом пу¬
ти концептуальной связи между физиологическими механизмами работы мозга
и психическими процессами, определяющими целенаправленный характер цело¬
стного поведения. П. В. Симонов (1980) считает, что такая общая концептуальная
платформа содержится в самом учении И. П. Павлова о высшей нервной деятель¬
ности: «Наука о высшей нервной деятельности не есть пи физиология, ни психо¬
логия в традиционном их понимании, ее нельзя однозначно отнести ни к биологи¬
ческим, ни к социальным наукам, ибо она включает в себя элементы всех этих от¬
раслей знания»1. Процессы отражения и деятельности можно рассматривать
в различных аспектах: со стороны нейрофизиологических, материальных меха¬
1 Симонов П. В. Наука о высшей нервной деятельности человека и психофизиологическая проблема //
Журн. высш. нерв. деят. 1980. Т. 31. С. 235-241.
Психофизиологическая проблема 303
низмов этих процессов или со стороны содержания, значения, их отношения к от¬
ражаемым объектам внешнего мира и потребностям субъекта, то есть как психи¬
ческое, субъективное.
А. М. Иваницкий (1984) полагает, что познание механизмов психики — это
изучение того, как па основе изучения мозговых механизмов, их интеграции воз¬
никает новое качество в виде психики. Под психикой по¬
нимается свойство высокоразвитого мозга воссоздавать
внутренний образ действительности, который восприни¬
мается как нечто отдельное от субъекта. Эти авторы пола¬
гают, что основным путем для решения проблемы соот¬
ношения мозга и психики является исследование, которое
строится на параллельном изучении физиологических
и психологических показателей в одном эксперименте.
А отношения между психикой и мозговыми процессами
имеют свою аналогию в соотношении информации и ее
носителя. Тогда психика — это информация, составляю¬
щая содержание определенным образом организованных
мозговых процессов (А. М. Иваницкий, 1986). При таком определении, естествен¬
но, возникает вопрос о поиске, подборе тех нервных процессов, которые имеют
определяющее значение. Исходя из системного подхода решение, возможно, сле¬
дует искать на пути сопоставления психических явлений не с отдельными нерв¬
ными процессами, а с их организацией, с их системой.
Явления, составляющие нейрофизиологический код, и информация, непо¬
средственно содержащаяся в этом коде, обладают определенным сходством внут¬
ренней организации. Поэтому задача состоит в понимании внутреннего смысла
изучаемого явления, раскрытии информационного содержания мозговых процес¬
сов, обеспечивающих психическую функцию. Примером оригинального подхода
к изучению нейрофизиологических основ психической деятельности человека,
являются исследования Н. П. Бехтеревой (1980). Разработка методических под¬
ходов к долгосрочной регистрации многих нейрофизиологических показателей,
в том числе и импульсной активности нейронов мозга бодрствующего человека
обеспечила доступ к изучению нейронных коррелятов мыслительной деятельно¬
сти. У людей при восприятии, удержании в памяти и произнесении слов структуры
импульсных реакций нейронных популяций, отражающих акустические и смы¬
словые характеристики слова, могут формироваться в мозговых образованиях,
причастных к организации движения. Н. П. Бехтерева пишет: «Обнаруженные
и исследованные нами паттерны — местные, привязанные к структуре тончайшие
нейрофизиологические корреляты психической деятельности или, точнее, нейро¬
физиологическое выражение декодирования, детерминирования в значительной
степени памятью, обучением, определяющими наряду с врожденными морфоло¬
гическими характеристиками свойство структуры в той же структуре. Это та ней¬
родинамика (или ее часть), возникновение которой находится в прямой зависи¬
мости от участия структуры в обеспечении психической деятельности».
Мышление человека как высшая форма оперирования информацией стало
предметом целенаправленного нейрофизиологического исследования. Огромная
информационная емкость человеческого мозга определяется количеством нерв¬
ных клеток, количеством связей между нервными клетками, нолифункциональ-
Психика — это свой¬
ство высокоразвитого
мозга воссоздавать
внутренний образ дей¬
ствительности, кото¬
рый воспринимается
как нечто отдельное
от субъекта. Это ин¬
формация, составляю¬
щая содержание опре¬
деленным образом
организованных моз¬
говых процессов.
304 Глава 11. Психофизиологическая проблема
ыостыо популяций и отдельных нервных клеток, обучаемостью мозга и динамич¬
ностью механизмов, лежащих в основе всех видов его деятельности (Н. П. Бехте¬
рева и др., 1985).
Основной задачей психофизиологии является объяснение специфических
особенностей психических процессов па основе механизмов деятельности моз¬
га. Ведется поиск концептуальной связи между физиологическими механизмами
работы мозга и психическими процессами, определяющими целенаправленный
характер поведения человека. Под психикой понимается свойство высокооргани¬
зованного мозга воссоздавать внутренний образ действительности, который вос¬
принимается как нечто отдельное от субъекта. Психика — это информация, со¬
ставляющая содержание организованных мозговых процессов.
Основные потребности человека (биологические, социальные и идеальные)
имеют глубокие филогенетические корни. В аналогичном плане рассматривается
феномен межполушарной функциональной асимметрии мозга человека, которая
проявляется в различии мозговых операций, выполняемых каждым полушарием.
С этим связывают также индивидуальные типологические особенности человече¬
ской психики. Структурной основой сложных форм поведения и психики высту¬
пают ассоциативные области неокортекса, имеющие отношение к развитию наи¬
более отвлеченных и обобщенных форм человеческих восприятий и действий,
а также к анализу и синтезу раздражений, связанных с речевой деятельностью.
Помимо абстрактного (словесно-логического) существует эмоциональное (на¬
глядно-действенное) мышление, понимаемое как свернутое (сокращенное) дейст¬
вие. В механизмах речепродукции и речевосприятия участвуют кортикальные
формации прежде всего левого полушария, стриопаллидарные и таламические
системы мозга.
На протяжении цикла сон—бодрствование мозг человека непрерывно работа¬
ет, периоды парадоксального (быстрого) сна заполнены сновидениями, отражаю¬
щими имеющиеся у субъекта проблемы. Материалом сновидений является нако¬
пленный жизненный опыт. Велико диагностическое значение сновидений. Гипноз
как одна из форм внушения является своеобразным состоянием бодрствования
человека с повышенной тревожностью и снижением волевых свойств личности.
Специфическая деятельность человека-оператора но управлению технически¬
ми средствами зависит в первую очередь от способности прогнозировать вероят¬
ный исход своих действий и соответственно вероятность появления тех или иных
событий. Успех такой деятельности также тесно связан с уровнем мотивации
и психофизиологической напряженности оператора.
Контрольные вопросы
1. Что такое психофизический параллелизм?
2. Соотношение понятий «психические процессы» и «информация».
3. Мышление человека.
4. Что такое наука о высшей нервной деятельности?
5. Монизм психического и физиологического в поведении человека.
ЗАКЛЮЧЕНИЕ
Функциональная структура поведенческого акта
Наиболее важным эвеном в системе адаптивных свойств организма и в динамике
эволюционных преобразований поведения является способность к формирова¬
нию планов, программ поведения. Разработка принципиальных подходов к изуче¬
нию программирующей функции мозга является одной из основных задач физио¬
логии высшей нервной деятельности.
Моделирование потребного будущего — высшее отражение активности мозга,
который сталкивается с динамически переменчивой ситуацией, ставящей его пе¬
ред необходимостью выработки свойств вероятностного
прогнозирования.
Программа поведенческого акта — это модель того,
что произойдет с организмом в будущем, ее можно рас¬
сматривать как формирование логики, алгоритма, функ¬
циональной структуры предстоящего поведенческого акта.
Такая функциональная структура опирается на про¬
шлый жизненный опыт, записанный в памяти с вероят¬
ностью, равной единице, и актуальное настоящее, куда
входит не только изменчивая окружающая среда, но и
организм с его потребностями. В результате планируется
будущий поведенческий акт, в котором организм должен
с той или иной вероятностью предвидеть возможные из¬
менения в непредсказуемо изменчивой среде. Значит, в составе вероятностного
прогнозирования находится предвосхищение будущего, основанного на структуре
прошлого опыта и информации о наличной ситуации. С расширением среды оби¬
тания и рецептивных сфер животного неопределенность окружающей среды для
организма возрастает как в динамике филогенетической эволюции, так и в инди¬
видуальном развитии. Чем больше развита способность к вероятностному про¬
гнозированию, тем больше шансы особи на выживание.
Любая поведенческая программа должна строиться но крайней мере на трех
основных детерминантах:
♦ доминирующей мотивации;
♦ прошлом жизненном опыте (долговременная память);
♦ оценке текущей ситуации и удержании ее в кратковременной памяти.
Доминирующая мотивация. В основе любого акта жизнедеятельности лежит
определенная биологическая, а для человека прежде всего социальная потреб¬
ность. Вызвав избирательную активацию определенных мозговых аппаратов, она
приводит к созданию доминирующей мотивации. Последняя является первич¬
ным системообразующим фактором, который определяет доминирующую систе¬
му (констелляцию) связанных между собой центров, временно определяющую
характер ответных реакций организма на внешние и внутренние воздействия. Эта
доминирующая центральная констелляция создает скрытую готовность организ¬
Аюбая поведенческая
программа должна
строиться по крайней
мере на трех основных
детерминантах: 1) до¬
минирующей мотива¬
ции; 2) прошлом жиз¬
ненном опыте (долго¬
временная память);
3) оценке текущей си¬
туации и удержании ее
в кратковременной па¬
мяти.
306 Заключение
ма к определенному виду деятельности при одновременном подавлении посто¬
ронних рефлекторных актов. В каждый момент времени доминирующей стано¬
вится та мотивация, в основе которой лежит наиболее важная потребность.
Доминирующая мотивация как первичный системообразующий фактор опре¬
деляет все последующие этапы мозговой деятельности. Специфика биологиче¬
ских мотиваций определяет характер и «химический статус» внутрицентралыюй
интеграции и набор вовлекаемых мозговых аппаратов. В качестве полезного ре¬
зультата определенного поведенческого акта выступает удовлетворение потреб¬
ности, приводящее к снижению уровня мотивации. При этом надо иметь в виду,
что в итоге реализации той или иной поведенческой программы организм может и
не достигнуть полезного результата. Самый смысл вероятностного прогнозирова¬
ния сводится к определенному выбору (принятие решения) в альтернативной си¬
туации наиболее вероятной поведенческой программы (гипотезы). Если нет аль¬
тернативы, то и нет необходимости принимать решение, а значит, отпадает нужда
в предварительном программировании. И живой организм опирается на уже гото¬
вые (запаянные) поведенческие решения, то есть переходит на автоматический
режим управления.
Прошлый опыт, долгосрочная память. Широкий репертуар готовых функцио¬
нальных элементов, навыков, которые либо предопределены генетически, либо
сформированы в индивидуальной жизни, но являются жесткими, стабильными,
то есть приобретшими автоматизированный характер, является фундаментом для
формирования сложных поведенческих программ. Они служат базой всякого це¬
ленаправленного поведения. На элементарном уровне поведения всякий новый
условный рефлекс — это синтез ранее сформированных жестких рефлекторных
актов, например ориентировочного и одного из безусловных рефлексов. При ана¬
лизе более сложных форм поведения его программу целесообразно рассматривать
как систему сложных (интрацентральных) условных рефлексов.
Доминирующая мотивация извлекает из долгосрочной памяти эти готовые
элементы, которые в определенных условиях могут оказаться достаточными для
достижения приспособительного эффекта. Последнее может иметь место при ста¬
бильной неальтернативной окружающей ситуации, которая в данный момент вре¬
мени обнаруживает свои жесткие связи и взаимоотношения. Вероятно, естест¬
венный отбор отселектировал целый ряд готовых поведенческих реакций на конк¬
ретные, регулярно повторяющиеся жизненные ситуации, которые при наличии
определенной биологической мотивации реализуются в поведении, не требую¬
щем предваряющего программирования. Этот тип поведенческих навыков обес¬
печивается жесткой системой связей в пределах субстрата головного мозга.
Таким образом, естественный отбор обеспечивает организму оптимальные
условия встречи с жизненно важными факторами вероятностно организованной
среды. Эта закономерность совершенствуется не только в динамике филогенеза,
но и на ранних этапах индивидуального развития. Генетически предопределенная
гетерохронность созревания основных мозговых систем обусловлена временной
потребностью их включения в целостную поведенческую реакцию. Более того,
мозг на ранних стадиях онтогенеза обладает чрезвычайно высокой запечатляемо-
стыо к биологически значимым факторам среды. Последнее обеспечивает ши¬
рокий набор жестких поведенческих навыков, которые накапливаются в памяти
Функциональная структура поведенческого акта 307
взрослого организма и являются непременным арсеналом для формирования на
их базе более сложных поведенческих актов.
Оценка текущей ситуации и краткосрочная память. Для адекватного поведе¬
ния организму необходимо следующее:
♦ оцепить собственную схему тела, вписанного в окружающую среду;
♦ извлечь биологически полезную информацию из этой среды;
♦ описать структуру среды как закон связей между ее наиболее существенными
переменными;
♦ определить ведущее кинематическое звено для реализации предстоящего по¬
веденческого акта.
Доминирующая мотивация превращает реальную среду в актуальную. Актив¬
ная природа процесса восприятия осуществляется в пределах целостной сенсор¬
ной функции мозга и состоит из двух операций:
♦ обнаружения и кодирования;
♦ распознавания и декодирования.
Высший уровень сенсорной интеграции завершается формированием образа
среды, оценкой взаимоотношений биологически значимых признаков, то есть фор¬
мированием структуры среды, логики ее построения, конструкции и закономер¬
ностей их изменений. Животные могут выполнять поведенческий акт в новой об¬
становке на основе оперирования эмпирически уловленными законами, лежащими
в основе структуры среды. Это свойство, лежащее в основе элементарной рассу¬
дочной деятельности животных, претерпевает прогрессивное развитие в эволю¬
ции и достигает наивысшего совершенства у высших приматов и человека.
Организм стремится к оптимизации вероятности встречи с биологически зна¬
чимыми предметами и явлениями окружающего мира. Организм извлекает ин¬
формацию, то есть упорядоченность для формирования логики своего поведения.
Во всех этих процессах существенная роль отводится краткосрочной памяти, ко¬
торая необходима для удержания в следах образа окружающей среды и удержа¬
ния программы предстоящего поведенческого акта вплоть до его полной реализа¬
ции.
Нейродинамическая конструкция поведенческого акта. Таким образом, в ос¬
нове организации целенаправленного поведенческого акта находится домини¬
рующая мотивация (рис. 109), которая:
1) определяет набор готовых, извлеченных из долговременной памяти поведен¬
ческих реакций;
2) формирует из реальной окружающей среды актуальное экстранерсональное
пространство с учетом системы схемы тела и сигнально значимых компонен¬
тов среды.
Последние, оцениваемые организмом в их взаимоотношениях и динамике пре¬
образований, являются базой для оценки вероятностной структуры среды. Наря¬
ду с этим из долговременной памяти (генетической и онтогенетической), состав¬
ляющей основу жизненного опыта, извлекаются уже готовые функциональные
блоки (элементы) или гипотезы. Вероятностная структура среды определяет сте¬
пень актуальности той или иной гипотезы, на основе чего и выдвигается програм¬
308 Заключение
2
=Г
О
2
т
Ф
*
О
с
о
с
Рис. 109. Функциональная структура целенаправленного поведенческого акта
ма поведенческого акта. В ней отражены взаимоотношения и динамика вовле¬
чения различных функциональных блоков. Иными словами, отдельные частные
поведенческие гипотезы (тактики) в соответствии с веро¬
ятностной структурой среды образуют новую нейродииа-
мическую систему — программу, которая должна удержи¬
ваться в краткосрочной памяти вплоть до полной реали¬
зации поведенческого акта.
Вероятностные программы формируют команды, то
есть актуализируют в строго временной зависимости ряд
гипотез, в результате чего совершается действие, которое
может привести к получению или отсутствию результата
(удовлетворения потребности). Достижение полезного
результата сопровождается стиранием данной програм¬
мы из краткосрочной памяти или переводом ее в долго-
Частные поведенческие
гипотезы (тактики) в со¬
ответствии с вероятно¬
стной структурой среды
образуют новую нейро-
динамическую систе¬
му — программу, кото¬
рая должна
удерживаться в кратко¬
срочной памяти вплоть
до полной реализации
поведенческого акта.
срочную память, а эффективность действия оценивается по степени выраженно¬
сти положительных эмоций. Последние как свидетельство успеха действия могут
привести к изменению мотивации. Система эмоций — единственная организация,
способная осуществлять оценку степени релевантности действий организма в со¬
ответствии с доминирующей мотивацией и прогнозируемой вероятностью ее удов¬
летворения.
В случае же отсутствия результата могут включаться различные мозговые
механизмы.
Функциональная структура поведенческого акта 309
При наличии высокой мотивации и при отсутствии результата программа
остается в краткосрочной памяти и действие может быть повторено.
Отрицательные эмоции, сигнализирующие о неуспехе действия, могут приве¬
сти к изменению самой мотивации (при ее недостаточной интенсивности).
Если же мотивация оказывается стойкой, перестраивается сама программа
действия, то есть выдвигаются новые гипотезы, между которыми устанавливают¬
ся новые законы связей, формируется новая функциональная структура.
Таким образом, степень адекватности поведенческой программы определяется
знаком и степенью выраженности эмоционального статуса, который сопровожда¬
ет ее реализацию.
Успех действия непременно сопровождается положительной эмоцией, которая
служит важным фактором для закрепления нового поведенческого решения в он¬
тогенетической (долговременной) памяти.
ЛИТЕРАТУРА
Основная
Анатомия. Физиология. Психология человека: Краткий иллюстрированный
словарь. СПб., 2001.
Батуев А. С. Высшая нервная деятельность. М., 1991
Батуев А. С. Куликов Г. А. Введение в физиологию сенсорных систем. М., 1984.
Блум Ф., Лайзерсон А., Хофшгпедтер Л. Мозг. Разум. Поведение М., 1988.
Данилова Н. Д., Крылова А. Л. Физиология высшей нервной деятельности. М.,
2000.
Долин А. С., Долина С. А. Патология высшей нервной деятельности. М., 1972.
Коган А. Б. Основы физиологии высшей нервной деятельности. М., 1988.
Механизмы деятельности мозга человека. Нейрофизиология человека. Л.,
1988.
Механизмы памяти. Л., 1988. Серия «Руководство по физиологии».
Физиология поведения. Нейрофизиологические закономерности Л., 1986. Се¬
рия «Руководство по физиологии».
Физиология поведения. Нейробиологические закономерности. Л., 1987. Серия
«Руководство по физиологии».
Физиология сенсорных систем. СПб., 2003.
Дополнительная
Адрианов О. С. О принципах организации интегративной деятельности мозга.
М„ 1975.
Анохин П. К. Биология и нейрофизиология условного рефлекса. М., 1968.
Асратян Э. А. Рефлекторная теория высшей нервной деятельности. М.,1983.
Батуев А. С. Высшие интегративные системы мозга. Л., 1981.
Батуев А. С. Нейрофизиология коры головного мозга. Л., 1984.
Бериташвили И. С. Избранные труды. Нейрофизиология и нейропсихология.
М„ 1976.
Бернштейн Н. А. Очерки по физиологии движений и физиологии активности.
М„ 1966.
Бехтерев В. М. Основы учения о функциях мозга. СПб., 1907. Т. 7.
Бехтерева Н. П. Здоровый и больной мозг человека. Л., 1980.
Бианки В. Л. Асимметрия мозга животных. Л., 1985.
Вартанян Г. А. Классический условный рефлекс и проблема эп граммы памяти.
Л„ 1986.
Гасанов У. Г. Системная деятельность корковых нейронов при обучении. М.,
1981.
Данилова Н. Н. Психофизиология. М., 1998.
Доминанта и условный рефлекс / Под ред. П. В. Симонова. М., 1988.
Дубровский Д. Я. Информация. Сознание. Мозг. М., 1980.
Дьюсбери Д. Поведение животных. Сравнительные аспекты. М., 1981.
Литература 311
Иваницкий А. М., Стрелец В. В., Корсаков И. А. Информационные процессы
мозга и психическая деятельность. М., 1984.
Ильин Е. П. Дифференциальная психофизиология. СПб., 2001.
Карамян А. И. Эволюция конечного мозга позвоночных. Л., 1976.
Костандов Э. А. Функциональная асимметрия полушарий мозга и неосозна¬
ваемое восприятие. М., 1983.
Котляр Б. И. Пластичность нервной системы. М., 1986.
Котляр Б. И., Майоров В. И., Тимофеева Н. О., Шулъговский В. В. Нейронная ор¬
ганизация условно-рефлекторного поведения. М., 1983.
Кругликов Р. И. Нейрохимические механизмы обучения и памяти. М., 1981.
Крушинский Л. В. Биологические основы рассудочной деятельности. М., 1977.
Леонтьев А. Н. Деятельность. Сознание. Личность. М., 1975.
Лурия А. Р. Основы нейропсихологии. М., 1973.
Мосидзе В. М. и др. Расщепленный мозг. Тбилиси, 1972.
Орбели Л. А. Избранные труды. Л., 1961. Т. 1; 1964. Т. 3.
Павлов И. П. Физиология нервной системы // И. П. Павлов, И. М. Сеченов,
Н. Е. Введенский. Л., 1952. Т. 4.
Поляков Г. И. Проблема происхождения рефлекторных механизмов мозга. М.,
1964.
Прибрсии К. Языки мозга. М., 1975.
Рабинович М. Я. Замыкательиая функция мозга. М., 1975.
Русинов В. С. Доминанта. Электрофизиологическое исследование. М„ 1969.
Сеченов И. М. Избранные произведения. М.; Л., 1952.
Симонов П. В. Мотивированный мозг. М., 1987.
Симонов П. В. Теория отражения и психофизиология эмоций. М., 1970.
Симонов II. В. Эмоциональный мозг. М., 1981.
Слоним А. Д. Среда и поведение. Л., 1976.
Соколов Е. Н. Нейронные механизмы памяти и обучения. М., 1981.
Судаков К. В. Биологические мотивации. М., 1971.
Ухтомский А. А. Доминанта. СПб., 2002.
Ухтомский А. А. Собр. соч.: В 6 т. Л., 1954.
Фабри К. Э. Основы зоопсихологии. М., 1976.
Хананашвили М. М. Экспериментальная патология высшей нервной деятель¬
ности. М., 1978.
Хорн Г. Память, запечатление и мозг. М., 1989.
Шеррингтон Ч. С. Интегративная деятельность нервной системы. Л., 1969.
ГЛОССАРИЙ
АДАПТАЦИЯ — постепенное уменьшение ощущения при длительном воздействии оди¬
наковых стимулов на рецепторную поверхность.
АССОЦИАТИВНОЕ, факультативное, обучение — активный процесс формирования соб¬
ственной среды путем извлечения для себя ее функциональных составляющих, значи¬
мых для выполнения тех или иных актов поведения.
АССОЦИАТИВНЫЕ СИСТЕМЫ — высшие интегративные структуры мозга, не принад¬
лежащие какой-либо одной сенсорной системе, но получающие информацию от не¬
скольких сенсорных систем.
БЕЗУСЛОВНЫЙ РЕФЛЕКС — относительно постоянная, видоспецифическая, стерео¬
типная, генетически закрепленная реакция организма или отдельных его частей на
внутренние или внешние раздражения, осуществляемая при посредстве центральной
нервной системы.
БИОЛОГИЧЕСКАЯ ПАМЯТЬ — способность живых существ, воспринимая воздействия
извне, сохранять и в последующем воспроизводить вызываемые этими воздействиями
изменения функционального состояния и структуры.
БИОЛОГИЧЕСКИЕ РИТМЫ — ритмические (периодические) колебания физиологиче¬
ских и психических процессов (биоритмология).
ВЕРОЯТНОСТНОЕ ПРОГНОЗИРОВАНИЕ — предвосхищение будущего, основанное
на вероятностной структуре прошлого опыта и информации о наличной ситуации.
ВИТАЛЬНЫЕ РЕФЛЕКСЫ — составляют первую самостоятельную группу рефлексов,
которые обеспечивают сохранение индивидуума и вида.
ВНЕШНЕЕ (безусловное) ТОРМОЖЕНИЕ — срочное подавление текущей условно-
рефлекторной деятельности при действии посторонних раздражителей, вызывающих
ориентировочный, оборонительный или другой безусловный рефлекс.
ВНИМАНИЕ рассматривается как один из механизмов устранения избыточности сенсор¬
ных сообщений, который участвует как в избирательном подавлении, обеспечении, так
и в фильтрации информации, извлекаемой из систем кратковременной и долговремен¬
ной памяти.
ВНУТРЕННЕЕ (условное) ТОРМОЖЕНИЕ - торможение условно-рефлекторной дея¬
тельности, которое развивается постепенно в результате неподкрспления условного
сигнала безусловным рефлексом.
ВНУТРЕННИЕ ДЕТЕРМИНАНТЫ — это накопленная история взаимодействия реаги¬
рующего субстрата с фактором среды (принцип историзма) (А. А. Ухтомский).
ВОЗБУДИМОСТЬ — способность воспринимать существенные для организма признаки
раздражений и трансформировать их в специфическую нервную сигнализацию.
ВЫСШИЕ ФОРМЫ ОТРАЖЕНИЯ — извлекают информацию для самих себя и в то же
время абстрагируются от материального носителя этой информации.
ГЕНОТИП — вся совокупность генетического материала организма или его часть, то есть
комплекс всех наследственных задатков, контролирующих развитие, строение и жизне¬
деятельность организма.
ДВИГАТЕЛЬНАЯ ПРОГРАММА кортикальная — адаптивные изменения достигаются
с помощью внутренней обратной связи, компонентом которой является система меди¬
альных лемнисков.
ДЕКОДИРОВАНИЕ — преобразование входной кодовой комбинации активности нерв¬
ных элементов сенсорных систем в реакцию исполнительных аппаратов.
ДЕТЕКТОР — это нейрон, избирательно настроенный на определенный параметр сигна¬
ла за счет фиксированной системы связей с его рецепторами или другими нейронами
более низкого уровня.
Глоссарии 313
ДЕТЕРМИНИЗМА ПРИНЦИП — выявляет закономерную причинно-следственную
связь явлений, определяемую взаимодействием материальных факторов.
диссолюция — теория, основанная па законе рекапитуляции: многие поведенческие
акты закладываются уже в эмбриональном периоде, но достигают полного развития
в разные периоды постнаталыюго онтогенеза.
ДОМИНАНТА — рабочий принцип деятельности нервной системы и вектора поведения.
ДОМИНАНТЫ ПРИНЦИП — временно господствующая в нервной системе группа нерв¬
ных центров, определяющая характер текущей ответной реакции организма на внеш¬
ние и внутренние раздражения и целенаправленность его поведения.
ЗАПЕЧАТЛЕНИЕ (импринтинг) — форма обучения, заключающаяся в том, что стимул,
предъявленный животному в особый критический период его развития, запечатлевает¬
ся животным на длительный срок и становится ключевым стимулом, запускающим опре¬
деленное поведение.
ЗВУКОВАЯ И ПИСЬМЕННАЯ РЕЧЬ — способность знаково-символического отраже¬
ния предметов и явлений окружающего мира и самих себя в этом мире.
ЗНАК — это такой компонент психического отражения, который нс имеет познавательно
образного сходства с соответствующим ему элементом или свойством объекта оригинала.
ЗРИТЕЛЬНОЕ ВОСПРИЯТИЕ — сложный многоступенчатый акт, который начинается
формированием изображения на сетчатке и заканчивается возникновением зрительно¬
го образа в структурах головного мозга.
ИГРОВОЕ ПОВЕДЕНИЕ — совокупность двигательных актов организма, включающее опре¬
деленные мышечные группы, используемые в дальнейшем взрослым животным в соот¬
ветствующих жизненных ситуациях — бегстве, борьбе, добычи пищи, размножении и т. д.
ИДЕЯ РЕФЛЕКСА - должна оставаться лишь философским обобщением, и не следует
рассматривать рефлекс как механизм работы центральной нервной системы.
ИЗБИРАТЕЛЬНОСТЬ БИОЛОГИЧЕСКОГО ОТРАЖЕНИЯ — полнее всего обеспечи¬
вает целесообразный характер реакций организма, его действий, его поведения.
ИМПРИНТИНГ — комплекс поведенческих адаптаций новорожденного, которые обеспе¬
чивают первичную связь между ним и родителями и как бы замыкают цепь преобразо¬
ваний эмбрионального периода, позволяя реализовать новорожденному уже сформи¬
рованные механизмы восприятия и реагирования.
ИНСТИНКТ — сложпорефлекториый комплекс наследуемых поведенческих реакций осо¬
бей одного вида, характеризующийся стереотипностью и стабильностью.
ИНТЕГРАЛЬНЫЙ ОБРАЗ — продукт пережитых доминант, то есть продукт субъектив¬
ного отражения объективного взаимоотношения организма со средой.
КОГНИТИВНОЕ обучение — основано на формировании функциональной структуры
среды, то есть на извлечении законов связей между отдельными ее компонентами.
КОГНИТИВНЫЕ КАРТЫ — процесс, благодаря которому организм приобретает некото¬
рое подобие топологической карты той местности, в которой он обитает.
КОДИРОВАНИЕ — установление соответствия между определенными параметрами сен¬
сорного стимула и характеристиками импульсной активности нейрона и/или местом
его расположения.
КОМПЛЕКСЫ ФИКСИРОВАННЫХ ДЕЙСТВИЙ — сложные стереотипные движе¬
ния, образующие высокоорганизованную поведенческую последовательность.
КОНВЕРГЕНЦИЯ - схождение множества афферентных импульсов в единственный
анатомически ограниченный эфферентный капал.
КРАТКОВРЕМЕННАЯ ПАМЯТЬ - в ее основе лежит временное повышение проводи¬
мости в синапсах, связывающих определенные нейроны, и реверберация импульсов,
основанная на ряде химических и электрохимических реакций, нс связанных с синте¬
зом макромолекул.
ЛЮБАЯ ПОВЕДЕНЧЕСКАЯ ФУНКЦИЯ ОРГАНИЗМА опирается на сложную кон¬
стелляцию множества пространственно разнесенных мозговых аппаратов, вовлекае¬
мых симультанно и сукцессивно для целей удовлетворения доминирующей потреб¬
ности.
314 Глоссарий
МОДАЛЬНОСТЬ — качественное своеобразие сенсорного раздражителя.
МОТИВАЦИЯ — физиологический механизм активирования памятных следов тех пове¬
денческих актов, которые способны удовлетворить имеющуюся у организма домини¬
рующую потребность.
МЫШЕЧНОЕ ЧУВСТВО — при выполнении одного движения становится в порядке ас¬
социации рефлексов сигналом к другому движению (Сеченов).
НЕАССОЦИАТИВНОЕ, облигатное, обучение — обусловлено набором средовых факто¬
ров и не требует непременного совпадения (ассоциации) внешних сигналов с той или
иной целостной деятельностью организма.
НЕЙРОЭНДОКРИННОЙ РЕГУЛЯЦИИ гипотеза реализации генетической информа¬
ции — вторичные посредники и стероидные гормоны представляют собой интеграцию
функций нервной и эндокринной систем на молекулярном уровне.
ОБНАРУЖЕНИЕ СИГНАЛА — выделение нервной сигнализации из собственного шума.
ОБОБЩЕНИЕ — функциональный блок систематизированной информации, хранящийся
в аппаратах памяти.
ОБРАЗ — это такой результат познания субъектом объекта, упорядоченность элементов
которого в принципе соответствует упорядоченности свойств, связей и отношений объ¬
екта (оригинала), причем эта упорядоченность (структура) функционально отделена
субъектом от субстрата образа и непосредственно для носителя существует в субъек¬
тивно идеальной форме.
ОБРАЗ-СЛЕД — записывается в долговременную память, если в прошлом вероятность
встречи с этим объектом была высокой.
ОБУЧЕНИЕ — процесс, в результате которого организм приобретает адаптивные измене¬
ния индивидуального поведения, расширение жизненного опыта, усвоения знаний,
умений, навыков.
ОРУДИЙНАЯ деятельность — основана на анализе связей между предметами, яв¬
лениями и результатами манипулирования с предметами.
ОТОБРАЖЕНИЕ — зависит не только от отображаемого объекта, но и от природы отобра¬
жающей системы, ее текущих состояний, ее истории.
ОТРАЖЕНИЯ ПРИНЦИП — представляет собой органическое единство непосредствен¬
ного восприятия и следов прошлых впечатлений.
ПОВЕДЕНИЕ — совокупность двигательных актов организма, возникающих в ходе взаи¬
модействия с внешней средой и направленных на удовлетворение доминирующих у ор¬
ганизма потребностей. Поведение имеет адаптивное значение и является важнейшим
фактором эволюции вида.
ПОВЕДЕНЧЕСКАЯ ПРОГРАММА должна строиться на трех основных детерминантах:
1) доминирующей мотивации, 2) прошлом жизненном опыте (долговременная память)
и 3) оценке текущей ситуации и удержании се в кратковременной памяти.
ПОДКРЕПЛЯЮЩИЙ стимул — одним из основных механизмов действия является
ослабление тормозных — гиперполяризационных и усиление деполяризационных про¬
цессов в коре головного мозга.
ПОДРАЖАНИЕ (имитация) — передача видоспецифического опыта от одного поколения
к другому, от одной особи к другой, которая лежит в основе «сигнальной» (не генетиче¬
ской) наследственности.
ПОТРЕБНОСТЬ — специфическая (сущностная) сила живых организмов, обеспечиваю¬
щая их связь с внешней средой для самосохранения и саморазвития как источник
активности живых систем.
ПРЕДМЕТНАЯ ДЕЯТЕЛЬНОСТЬ — манипулирование разнообразными предметами среды.
ПРИВЫКАНИЕ — относительно устойчивое ослабление реакции вследствие многократ¬
ного предъявления раздражителя, не сопровождающегося каким-либо биологически
значимым агентом (подкреплением).
ПРИНЦИПИАЛЬНАЯ СУЩНОСТЬ УСЛОВНО-РЕФЛЕКТОРНОГО АКТА В ТОМ, ЧТО
он базируется на механизмах, в которых заложены не только элементы прошлого и на¬
Глоссарии 315
стоящего, но и элементы прогнозирования будущего. В этом его главный приспособи¬
тельный смысл, то есть сигнальный характер.
ПРОСТРАНСТВЕННО-ВРЕМЕННАЯ структура объекта кодируется в мозге в форме
определенной нервной модели, изоморфной внешнему воздействию. Она является ко¬
дом отображаемого внешнего объекта.
ПСИХИКА — это свойство высокоразвитого мозга воссоздавать внутренний образ дейст¬
вительности, который воспринимается как нечто отдельное от субъекта. Это информа¬
ция, составляющая содержание определенным образом организованных мозговых про¬
цессов.
ПСИХИЧЕСКАЯ ФУНКЦИЯ всегда есть результат интегративной деятельности целост¬
ного мозга.
ПСИХОНЕРВНОЕ ПОВЕДЕНИЕ — интегрирует элементы внешней среды в одно целое
переживание, производящее целостный образ, который через ориентировочную реак¬
цию направляет поведение животного.
РЕПРОДУКТИВНАЯ ИЗОЛЯЦИЯ — предотвращение спаривания партнеров из разных
видовых популяций на основе их несоответствия в половом поведении. Способствует
изоляции видовых популяций.
РЕТИКУЛЯРНАЯ ФОРМАЦИЯ — филогенетически старая система мозга, которая не
представляет собой единого анатомического целого и морфологически является гетеро¬
генным интегративным образованием.
РЕФЛЕКСА ПРИНЦИП — универсальная и своеобразная форма взаимодействия орга¬
низма со средой, происходящая при участии нервной системы.
РЕФЛЕКСЫ УСЛОВНЫЕ — индивидуально приобретенные системные приспособи¬
тельные реакции животных и человека, возникающие на основе образования в цент¬
ральной нервной системе временной связи между условным (сигнальным) и безуслов¬
но-рефлекторным актами.
РЕФЛЕКТОРНАЯ ДУГА - состоит из: 1) афферентного нерва; 2) отдела центральной
нервной системы и 3) эфферентного нерва.
РЕЦЕПТИВНОЕ ПОЛЕ — упорядоченная совокупность рецепторов, связанная с одним и
тем же нейроном вышележащего уровня и имеющая разную форму и величину, вклю¬
чающая возбудимые и тормозные зоны и перестраивающаяся в зависимости от влияния
центральных структур.
РЕЦЕПТОРЫ — это специализированные образования, выполняющие функцию преобра¬
зования энергии внешнего раздражителя в специфическую активность нервной систе¬
мы, в сигналы, несущие нервным центрам информацию.
РОЛЕВЫЕ (зоосоциальные) рефлексы образуют вторую группу поведенческих реакций,
которые возникают только при взаимодействии с другими особями своего вида.
САМОРАЗВИТИЯ РЕФЛЕКСАМИ - так именуют третью группу поведенческих актов.
СЕНСОРНАЯ ФУНКЦИЯ МОЗГА обеспечивается содружественной деятельностью сен¬
сорных, моторных и ассоциативных систем и направлена на организацию адаптивных
движений и действий.
СЕНСОРНЫЕ СИСТЕМЫ — анатомически организованные в структурах мозга системы
ядериых образований и связей, служащих для обнаружения и кодирования информа¬
ции определенной модальности.
СИГНАЛ — это некая величина, отражающая определенным образом состояние физиче¬
ской системы.
СИСТЕМНЫЕ ПРОЦЕССЫ ПОВЕДЕНЧЕСКОГО АКТА - это информационные про¬
цессы, для которых физиологические процессы выступают как материальный носи¬
тель.
СИСТЕМОГЕНЕЗ — организующаяся система поведенческих актов, которая может функ¬
ционировать прежде, чем она достигнет окончательного оформления. Для каждого вида
животного характерен свой своеобразный системогенез как проявление опережающего
отражения действительности.
316 Глоссарий
СОН — адаптация, проявляющаяся в подавлении активности в период наименьшей до¬
ступности пищи, угрозы резких колебаний внешних условий и максимальной опасно¬
сти со стороны хищников.
СОПРЯЖЕННОЕ ТОРМОЖЕНИЕ не есть подавление всяческой деятельности, но ее пе¬
реработка согласно с направлением доминирующего поведения.
СПОСОБНОСТЬ К АДЕКВАТНОСТИ достигается во времени не мгновенно, а постепен¬
но; окончательная оценка адекватности производится при соотнесении нового образа с
его нервной моделью, созданной ранее на основе всего комплекса воздействий и преж¬
него жизненного опыта, то есть долговременной памяти.
СТЕПЕНИ СВОБОДЫ — выбор одной из них при сопряженном торможении других дос¬
тигается посредством доминанты, направленной па определенный рабочий вектор.
СТЕРЕОТИП — единый зафиксированный комплекс поведенческих реакций, связанный
под влиянием многократных повторений определенной последовательности раздражи¬
телей в единую цепь, которая воспроизводится любым из раздражителей данной си¬
стемы.
СХЕМА ТЕЛА (образ тела) — трехмерная модель тела, обозначающая систему обобщен¬
ных представлений и понятий человека о собственном теле в покое и при движении,
о пространственных координатах и взаимоотношении отдельных частей тела.
ТОРМОЖЕНИЕ — невозможно искать даже отдаленное сходство в содержании понятий
«торможение» в поведенческом, павловском смысле и в его нейрофизиологическом со¬
держании.
ТОРМОЖЕНИЕ ЦЕНТРАЛЬНОЕ — торможение, возникающее в центральной нервной
системе, благодаря наличию в ней специализированных тормозных нейронов. Является
основным фактором координации деятельности центральной нервной системы.
УЧЕНИЕ О ФИЗИОЛОГИИ АНАЛИЗАТОРОВ — под ним И. П. Павлов вслед за
И. М. Сеченовым подразумевал триединую конструкцию; периферические рецепторы,
проводящие пути и мозговые центры вплоть до коры больших полушарий.
ФЕНОТИП — продукт интеграции генотипических и средовых воздействий, заключаю¬
щийся в наборе морфологических, физиологических и поведенческих признаков орга¬
низма.
ФУНКЦИОНАЛЬНЫЙ ОРГАН — обширное число нервных аппаратов, вступающих ме¬
жду собой во временное целенаправленное сотрудничество и образующих физиологи¬
ческую констелляцию.
ХРОНОТОП — внутренний пространственно-временной образ внешнего единого про¬
странственно-временного мира.
ЦИКЛИЧЕСКАЯ НЕРВНАЯ СВЯЗЬ — один из основных принципов координации дви¬
гательных реакций, в которой афферентная информация обеспечивает форму и состав
эфферентного проявления центральной интеграции.
ЭЛЕМЕНТАРНАЯ РАССУДОЧНАЯ ДЕЯТЕЛЬНОСТЬ — использование предваритель¬
но воспринятой тактики изменений в среде для построения стратегии своего будущего
поведения.
ЭМОЦИИ — субъективные реакции животных и человека на внутренние и внешние раз¬
дражители, проявляющиеся в виде переживаний и соматовегетативных сдвигов.
ЭМОЦИОНАЛЬНЫЙ СТРЕСС — общая системная нейрогуморальная реакция целост¬
ного организма, развивающаяся на действие стресс-факторов (стрессоров).
ЭТОЛОГИЯ — паука, изучающач поведение животных, преимущественно видоспецифи¬
ческих форм поведения в естественных условиях обитания.
Учебное издание
Батуев Александр Сергеевич
Физиология высшей нервной деятельности
и сенсорных систем
Учебник для вузов
3-е издание
Серия «Учебник для вузов»
Заведующий редакцией
Выпускающий редактор
Литературный редактор
Художественный редактор
Корректоры
Верстка
П. Алесов
Е. Егерева
К. Ширко
К. Радзевич
Н. Баталова, Л. Комарова
Л. Егорова
Подписано к печати 24.07.07. Формат 70x100/16. Уел. п. л. 25,8. Тираж 3000. Заказ 1078
ООО «Питер Пресс», Санкт-Петербург, Петергофское шоссе, д. 73, лит. А29.
Налоговая льгота — общероссийский классификатор продукции ОК 005-93, том 2;
95 3005 — литература учебная.
Отпечатано с готовых диапозитивов в ОАО «Техническая книга»
190005, Санкт-Петербург, Измайловский пр., 29.
ЧЕБНИК
'ДЛЯ ВУЗОВ
Физиология высшей нервной деятельности и сенсорных систем
Батуев Александр Сергеевич — доктор
биологических наук, профессор, академик
РАО, заведующий кафедрой высшей нервной
деятельности и психофизиологии Санкт-
Петербургского государственного университета,
заведующий кафедрой психофизиологии
и естественных наук Санкт-Петербургского
государственного института психологии
и социальной работы, автор более 580 научных
работ, в том числе 16 монографий.
Наука о высшей нервной деятельности, основы которой были заложены
И. М. Сеченовым, И. П. Павловым, А. А. Ухтомским и другими выдающимися
учеными, традиционно является одной из основных дисциплин при подготовке
современных специалистов-психологов.
Автор данного учебника проделал огромную работу по обобщению и систематизации
фактических данных и теоретических представлений, накопленных к настоящему
времени отечественными и зарубежными учеными. Издание предназначено
для студентов, обучающихся по направлению «Психология», и соответствует
требованиям Государственного образовательного стандарта. В нем содержатся
основные современные сведения по анатомии сенсорных систем и физиологическим
механизмам восприятия информации, а также базовые сведения по физиологии
основных поведенческих актов человека. Рассматриваются проблемы соотношения
сознания и бессознательного в психике, психологические и физиологические
детерминанты поведения.
Книга привлечет внимание самого широкого круга читателей — от студентов, только
что переступивших порог университета, до специалистов, работающих в различных
областях биологических, медицинских и гуманитарных дисциплин.
Рекомендовано Советом по психологии УМО по классическому
университетскому образованию в качестве учебника для студентов высших
учебных заведений, обучающихся по направлению и специальностям психологии.
С^ППТЕР'
Заказ книг:
197198, Санкт-Петербург, а/я 619
тел.: (812) 703-73-74, postbook@piter.com
61093, Харьков-93, а/я 9130
тел.: (057) 712-27-05, piter@kharkov.piter.com
www.piter.com — вся информация о книгах и веб-магазин
Библио Глобус
Москва Мясницкая 6
http: /www .biblio-globus.ru
Тел 928-35-67
924-46-80
781-19-00