/



























































































































































































































































Text
А.Г. Юсуфов
ЛЕКЦИИ
ПО эволюционной
ФИЗИОЛОГИИ
РАСТЕНИЙ
А.Г. Юсуфов
ЛЕКЦИИ
ПО ЭВОЛЮЦИОННОЙ
ФИЗИОЛОГИИ
РАСТЕНИЙ
Издание второе,
переработанное и дополненное
Рекомендовано Министерством общего
и профессионального образования
Российской Федерации в качестве
учебного пособия для студентов высших
учебных заведений, обучающихся
по направлению и специальности
«Биология»
Москва
«Высшая школа»
1996
ББК 28.57
Ю 91
Работа выпущена за счет частичной финансовой поддержки Республики
Дагестан.
Рецензент —д-р биол. наук,проф. В. В. Полевой (Санкт-Петер-
бургский государственный университет)
Юсуфов А. Г.
Ю 91 Лекции по эволюционной физиологии растений: Учеб,
пособие для студ. вузов по спец. «Биология».— 2-е изд.,
перераб. и доп.— М.: Высш, шк., 1996.— 255 с.: ил.
• ISBN 5-06-003335-Х
В книге освещены наиболее общие вопросы эволюционной физиологии
растений: тенденции функциональной эволюции, эволюция механизмов
автотрофного питания и аэробного дыхания, химический состав растений
и его приспособительное значение, роль периодических явлений в жизни
растений.
В настоящее издание (первое —1985 г.) включены новые разделы
«Наземная среда И приспособительная эволюция растений», «Эволюция
онтогенеза растений», дополнен фактический материал всех глав.
Пособие предназначено для студентов, специализирующихся по физи-
ологии и биохимии растений, микробиологии, ботанике и теории эволюции.
Ю
1906000000—079
001(01)—96
52—96
ББК 28.57
581.4
ISBN 5-06-003335-Х
© А. Г. Юсуфов, 1996
СПИСОК СОКРАЩЕНИЙ
АБК —абсцизовая кислота
АЛК —3-аминолевулиновая кислота
АМФ— аденозинмонофосфат
АТФ — аденозинтрифосфат
ВВМ — вещество вторичного метабо-
лизма
ВПФ—восстановительный пентозно-
фосфатный цикл, или цикл Кальвина
ГА — гиббереллины
ГАЗ — гибберелловая кислота
ГГЦ —гликолатно-глиоксилатный
цикл
ДНК —дезоксирибонуклеиновая
кислота
ИУК — индол ил-3-уксусная кислота
КоА — кофермент А
МДГ — малатдегидрогеназа
МП — мембранный потенциал
Мт митохондриальная ДНК
МтДНК — митохондрия
НАД — никотинамидадениндинуклео-
тид окисленный
НАДН — никотинамидадениндинуклео-
тид восстановленный
НАДФ — никотинамидадениндинуклео-
тидфосфат окисленный
НАДФН — никотинамидадениндину-
клеотидфосфат восстановленный
ПБК — пигмент-белковый комплекс
, ПВК — пировиноградная кислота
ПФП — пентозофосфатный путь окис-
ления глюкозы
Пх — пластохинон
Пц — пластоцианин
РБФ — рибулозобисфосфат
РБФК—О-рибулозобисфосфаткарбок-
силаза (оксигеназа)
РНК — рибонуклеиновая кислота
мРНК — матричная (информационная)
РНК
рРНК —рибосомальная РНК
тРНК—транспортная РНК
Рн — фосфор неорганический
РЦФ — реакционный центр фотосин-
теза
САМ — суккулентный (толстянковый)
тип фотосинтеза
ФГА (ЗФГА) — 3-фосфоглицериновый
альдегид
ФГК (ЗФГК) — 3-фосфоглицериновая
кислота
ФАД — флавинадениндинуклеотид
ФЕП — фосфофенолпйруват
ФЕПК — фосфофенол пироткарбокси-
лаза
ФС1 — фотосистема I
ФСП — фотосистема II
ФСЕ — фотосинтетическая единица
ФФ —фотофосфорилирование
хпДНК — хлоропластная ДНК
Цт — цитокинин
ЦТК — цикл ди- и трикарбоновых кис-
лот
ЩУК — щавелевоуксусная кислота (ок-
салоацетат)
ЭР — эндоплазматический ретикулум
яДНК — ядерная ДНК
ЭТЦ — элекгрон-транспортная цепь
ПРЕДИСЛОВИЕ
Повышение уровня подготовки специалистов в высших учебных
заведениях немыслимо без регулярного обновления учебников и
учебных пособий. Издание их особенно важно при изучении пред-
метов, находящихся на стыке наук. К ним по праву относится и
эволюционная физиология растений — комплексная дисциплина,
возникшая в пограничной области физиологии и биохимии расте-
ний с эволюционным учением, генетикой и экологией. Изложению
части ее проблем и посвящено предлагаемое пособие.
Физиология растений —одна из ведущих областей современ-
ной биологии — занимается изучением процессов жизнедеятельно-
сти растительных организмов. Проблемы и задачи физиологии
растений обширны и сложны, решение их сопряжено с примене-
нием комплекса физико-химических методов, разнообразных экс-
периментальных и теоретических подходов. В силу указанных
причин эта дисциплина постепенно входит в ранг точных наук,
усиливаются ее роль в качестве теоретической основы растениевод-
ства и связи с другими разделами биологии (агрохимия, биофизика,
биохимия, генетика развития, иммунология, микробиология, фо-
тобиология и т. д.).
Задачи повышения урожайности важнейших сельскохозяйст-
венных культур связаны с необходимостью глубокого знания фи-
зиологии растительных организмов. Закономерности же
жизнедеятельности современных растений нельзя понять без изу-
чения их прошлой истории, что определяет место и роль эволюци-
онного подхода в физиологии растений. В то же время углубление
и развитие указанного подхода имеет не только мировоззренческое
и методологическое значение, но важно в связи с растущей эколо-
гической напряженностью и задачами защиты окружающей среды
от загрязнений. Все сказанное и обусловливает существенную роль
физиологии растений и ее эволюционного направления в подготов-
ке специалистов-биологов.
Ежегодно во всем мире появляются десятки монографий и
несколько тысяч других публикаций, издаются и переиздаются
учебные пособия по физиологии растений. Отметим как положи-
тельное явление наметившийся в последние годы интерес и к
проблемам эволюционной физиологии растений. Эти вопросы на-
шли отражение в сборнике «Эволюция функций в растительном
мире» (Л., 1985) и более поздних изданиях учебной литературы по
физиологии растений (В. В. Полевой, 1989; А. Т. Мокроносов,
В. Ф. Гавриленко, 1992). Тем не менее необходимость издания
пособия по вопросам эволюционной физиологии растений все еще
велика. Это обстоятельство и побудило нас подготовить к изданию
лекции по этому предмету, читаемые в Дагестанском государствен-
ном университете с 1972 г.
В первом издании «Лекций по эволюционной физиологии рас-
тений» (М., 1985) были освещены наиболее общие вопросы —
направление и закономерности функциональной эволюции на при-
мере энергетики, химического состава и периодических явлений у
растений.' Недостатки первого издания были Связаны не только с
неполнотой охвата проблем функциональной эволюции растений,
но и с конспективностью их изложения. Сказалась и нехватка
фактического материала по генетике большей части функций рас-
тений, в результате чего изложение вопроса об эволюционном их
становлении не всегда достигало логического завершения.
Все прошедшие годы после первого издания автор продолжал
работать над рукописью. Новое издание существенно отличается от
предыдущего: 1) расширен фактический материал всех глав, кроме
гл. 6, за счет включения новых данных; 2) материал изложен с учетом
успехов изучения молекулярно-генетической природы функцио-
нальных проявлений у растений; 3) включены новые разделы
«Наземная среда и приспособительная эволюция растений» и «Эво-
люция онтогенеза растений»; 4) поставлены дискуссионные вопро-
сы; 5) усилена дидактическая ориентация текста. Несмотря на
центральное положение вопросов, касающихся эволюции энерге-
тйческих механизмов, в пособии не обойдены и другие стороны
жизнедеятельности растений (см. гл. 5, 7).
В предлагаемое пособие не включены материалы по устойчиво-
сти, росту и развитию растений. С этими вопросами студенты
знакомятся при изучении общего курса. Нам же казалось целесо-
образным обратить внимание прежде всего на наиболее трудные
разделы.
Автор приносит свои извинения за отсутствие ссылок в тексте
на целый ряд важных источников, что вызвано желанием избежать
перегрузки материала. Однако их данные во многих случаях исполь-
зованы. В список литературы вошли только обобщающие работы,
рекомендуемые для самостоятельного изучения.
По указанным причинам предлагаемое пособие не является
обзором литературы по проблемам функциональной эволюции
растений. При его написании автор стремился избежать излишней
детализации фактов, важных для понимания физико-химической
природы функции, но знакомых студентам по общему курсу. Не
вдаваясь в характеристику последовательных этапов становления
каждой из функций, автор в пособии дает оценку их роли с точки
зрения последствий для растительного обмена и жизни на Земле в
5
целом — биосферы. В этой связи общая энергетика растений рас-
сматривается как явление космического значения.
Учебное пособие «Лекции по эволюционной физиологии рас-
тений» предназначено для студентов старших курсов, специали-
зирующихся по физиологии и биохимии растений, микробиологии
и ботанике. Для его усвоения необходимо хорошее знание общих
курсов физиологии растений, биохимии, генетики и теории эво-
люции.
Автор с благодарностью примет критику, направленную на
улучшение содержания пособия и читаемого курса в целом.
ГЛАВА 1
ПРЕДМЕТ, МЕТОДЫ И ИСТОРИЯ РАЗВИТИЯ
ЭВОЛЮЦИОННОЙ ФИЗИОЛОГИИ РАСТЕНИЙ
Однозначно определить предмет и методы эволюционной фи-
зиологии растений нелегко. Усиливающиеся связи физиологии и
биохимии растений с эволюционным учением и генетикой привели
к тому, что выделился новый раздел — эволюционная фитофизио-
логия.
1.1. ПРЕДМЕТ И ЗАДАЧИ ЭВОЛЮЦИОННОЙ
ФИЗИОЛОГИИ РАСТЕНИЙ
Эволюционная физиология растений изучает развитие приспо-
собительных реакций в онто- и филогенезе, включая историческое
развитие самого онтогенеза, функции и структур, общего метабо-
лизма и отдельных его звеньев у растений. Изучение функциональ-
ных приспособлений в онто- и филогенезе — наиболее общая
задача эволюционной физиологии растений.
Конкретные задачи эволюционной физиологии сводятся к изу-
чению механизма возникновения и приспособительного значения
особенностей водообмена, фотосинтеза, дыхания, устойчивости,
роста и развития растений разных систематических и экологических
групп. Указанные задачи трудно исчерпать в рамках пособия. Поэтому
на современном этапе следует ограничиться более общими вопросами
эволюционной физиологии растений: изучением направлений и за-
кономерностей функциональной эволюции, развития в филогенезе
метаболизма и биоритмов, присущих миру растений в целом. По-ви-
Димому, здесь можно ожидать наиболее интересных обобщений,
касающихся эволюции функций у растений.
Эволюцию функций растений не всегда можно изобразить в
виде иерархического «древа», как это принято в систематике. При-
чина кроется в том, что «целевое назначение» многих функций
(фотосинтез, дыхание, механизмы роста и др.), возникших на заре
развития царства растений, не изменилось; преобразовались лишь
их интенсивность и способы реализации, не затрагивающие биохи-
мических основ. В силу этого разветвленность филогенетических
«Древ» по функциональным особенностям часто ограничена. По-
7
этому при изложении вопросов эволюционной физиологии расте-
ний чрезвычайно важно понять экологическую значимость отдель-
но взятой функции или комплекса функций.
В связи с этим следует напомнить, что растения обладают как
общими чертами, характерными для живой природы, так и специ-
фическими особенностями строения, жизнедеятельности и приспо-
собления. Отсюда при изложении проблем эволюционной
физиологии растений необходимо показать прежде всего пути
развития жизнедеятельности растений и их функциональную ди-
вергенцию в ходе эволюции. Эта общая задача далее будет конкре-
тизирована при освещении следующих вопросов: 1) классификация
функциональных приспособлений; 2) генетическая природа функ-
ций и их совершенствование под действием отбора; 3) эволюция
основных физиологических процессов в прошлом и будущем; 4)
взаимосвязь изменений функций и структур в ходе экологической
дивергенции растений; 5) связь между функциональной и морфо-
логической дивергенцией растений; 6) особенности химического
состава в связи с приспособлением их к среде обитания; 7) эволюция
онтогенеза растений и его регуляторных механизмов; 8) эволюция
ритмической деятельности и способов движения; 9) механизмы
взаимодействия растений в ценозах—физиологические основы
борьбы за существование.
Разумеется, здесь перечислена лишь небольшая часть проблем
эволюционной физиологии растений. Однако даже их изложение
означало бы существенное углубление общих представлений об
эволюции растительного мира, основанных на данных морфологии,
систематики, эмбриологии, генетики, биогеографии растений и
палеоботаники. К сожалению, уровень развития идей, методов и
направлений в физиологии растений не дает нам возможности пока
ответить на все вопросы, касающиеся эволюции функций растений.
Поэтому ниже обращено внимание прежде всего на эволюцию
энергетических механизмов растений.
Энергетика организма — основа всех процессов. На примере ста-
новления энергетических механизмов будут раскрыты и некоторые
общие черты эволюции растений. Необходимость обращения к энер-
гетическим механизмам продиктована еще и фундаментальной их
ролью в поддержании жизни на Земле. Это вытекает из ведущего
положения растений в круговороте веществ на нашей планете.
Наши знания о функциональной эволюции растений определя-
ются состоянием изученности тех или иных процессов в онтогенезе.
В зависимости от успехов в этом направлении существенно изме-
нятся и представления о функциональной эволюции растений. Но
нельзя ставить вопрос о том, что важнее изучать: механизмы
физиологических процессов у современных растений или эволю-
8
цию самих процессов. Это разные подходы к позйанию одного и
того же явления. '
Правильнее было бы рассматривать современное функциональ-
ное состояние растений с точки зрения его прошлого и изменения
жизнедеятельности в будущем. В такой постановке вопроса функ-
циональные особенности растений приобретают значение для по-
нимания их планетарной (космической) роли. Напомним, что
большая часть функций живого на Земле (энергетическая, концен-
трационная, деструктивная, средообразующая и транспортная) па-
дает на долю 'растений. Растения, хотя в целом и ведут
прикрепленный образ жизни, участвуют в переносе веществ на
дальние расстояния, как в случае перемещения водорослей, трав и
деревьев с потоком воды.
Велика роль растений в круговороте веществ и энергии на
планете. Растения своим существованием поддерживают жизнь на
Земле, в том числе и человека. Пища, энергия, запасы воды и
кислорода — продукты жизнедеятельности растений. Разве этого
одного недостаточно для серьезного изучения истории развития
«растительной жизни», уникальных и неутомимых продуцентов
веществ? При умелом подходе растения могут стать неисчерпаемым
источником жизни на планете в будущем.
Природа проявила исключительную находчивость при поиске
механизмов растительной жизни. Принципиальные новшества бы-
ли найдены путем проб и ошибок в критических ситуациях. В этих
целях производилось опробование и прежних механизмов в новых
условиях и их сочетаний между собой. Словом, имело место дости-
жение успеха в борьбе за жизнь путем преадаптации. Преадаптация
обеспечивала выживание на начальных этапах критической ситуа-
ции (создавая толерантность) до возникновения настоящей устой-
чивости на базе складывающихся совершенных механизмов.
Сказанного достаточно, чтобы понять, насколько грандиозны
задачи эволюционной физиологии растений и каковы трудности,
стоящие на этом пути. Их решение имеет значение и для управления
продуктивностью растений, и для защиты биогеоценозов. Поэтому
при описании конкретных экспериментов физиологии растений
Должны уделять внимание их эволюционной интерпретации. Однако
констатация различий в жизнедеятельности у организмов разных
видов растений и в распределении каких-то веществ у сравниваемых
объектов представляет лишь начальный этап эволюционного подхода.
По существу, эволюционный подход связан не столько с опи-
санием и выявлением различий у сравниваемых объектов (хотя и
это очень важно), сколько с объяснением их роли в борьбе за
существование. В этой связи любое отличие в жизнедеятельности
Растений нуждается в рассмотрении с точки зрения его формиро-
9
вания в процессе отбора; как оно могло быть подхвачено и усовер-
шенствовано в ходе эволюции.
При ответе на указанные вопросы мы встаем перед необходи-
мостью анализа генетических предпосылок процессов жизнедея-
тельности, оценки направлений изменчивости в процветании вида.
Разумеется, сегодня мы все еще не в состоянии судить об этом
применительно к большинству физиологических проявлений. От-
сюда и важность разработки соответствующих методов и подходов.
История становления и преобразования жизнедеятельности рас-
тений в ходе эволюции может быть воссоздана только на основе
теории Ч. Дарвина. При этом появляется возможность выяснить
пути формирования многообразия физико-химических процессов,
происходящих в растениях, и объяснить конкретные феномены их
жизни.
Анализ закономерностей функциональной эволюции растений
основан на рассмотрении конкретного экспериментального мате-
риала физиологии растений. В то же время многие детали эволюции
водного режима, питания, фотосинтеза и т. д. не могут быть объяс-
нены без применения сравнительного метода (см. ниже). Более того,
очевиден разрыв между изучением жизнедеятельности у современ-
ных растений и объяснением истории ее развития. Поэтому нельзя
сводить предмет эволюционной фитофизиологии к предмету общей
физиологии, а целесообразнее специально выделить проблемы эво-
люционной физиологии растений.
1.2. МЕТОДЫ ЭВОЛЮЦИОННОЙ ФИТОФИЗИОЛОГИИ
В современной теории эволюции для выяснения направлений
развития филогенетических групп, структуры и функций применя-
ют разнообразные методы: палеонтологический, онтогенетический
(эмбриологический) и морфологический (сравнительно-анатоми-
ческий). Заметим, что в классической филогенетике методы изуче-
ния функциональной эволюции все еще остаются слабо
разработанными. Они были предложены позже и отражены в сле-
дующей классификации методов филогенетических исследований
живой природы (Л. П. Татаринов, 1976).
I. Генетические методы:
1. Выявление различий в составе ДНК (молекулярная гибриди-
зация, определение состава и последовательности пуриновых и
пиримидиновых оснований в ДНК).
2. Выявление генного состава (гибридологический, электрофо-
ретический, иммунохимический).
II. Фенотипические методы:
ю
1. Сравнительный (сравнение структурных типов, реконструк-
ция предковых форм и т. д.).
2. Палеонтологический.
3. Эмбриологический.
III. Экстрасоматические методы:
1. Биогеографический.
2. Паразитологический.
3. Тектологический (изучение способов построения жилищ и
гнезд).
4. Этологический.
Не все из перечисленных методов одинаково «работают» при
изучении особенностей функциональной эволюции. Сложность
изучения эволюции функций связана с ограниченными возможно-
стями применения палеонтологического метода, являющегося наи-
более надежным в эволюционных построениях. В тех случаях, когда
он применяется для оценки значения формы листовой пластинки,
формы ветвления побега и т. д., чаще прибегают к экстраполяциям
фактов из жизни современных растений. Искусственность таких
экстраполяций несомненна, хотя они имеют объективные основы
и значимы для науки.
Для решения вопросов функциональной и биохимической эво-
люции растений используют комплекс методов: минералогический
(выявление возраста породы), палеонтологический (установление
состава видов и реконструкция ископаемых остатков) и сравнитель-
но-физиолого-биохимический (сравнение особенностей современ-
ных растений). В частности, биохимическими сравнениями ныне
живущих растений показано, что ранние метаболические звенья
синтеза стеринов, каротиноидов, вторичных метаболитов, реакций
гликолиза и дыхания у многих растений и животных сходны и не
нуждаются в участии кислорода (Дж. Шопф, 1981). Завершающие
же стадии многих метаболических путей, особенно у высокоорга-
низованных существ, протекают только с участием О2. Это свиде-
тельствует об их эволюционной молодости и сложении в аэробной
фазе истории Земли. Эукариотная клетка по величине больше
прокариотной. В связи с этим о времени возникновения эукариот-
ности судят косвенно по размерам ископаемых одноклеточных и ее
возраст исчисляют в 1,5 млрд. лет.
Большие надежды возлагали на возможности молекулярной
палеонтологии — изучение ископаемых органических веществ био-
генного происхождения и построение филогенетических древ бел-
ков и геномов современных растений. Н. Г. Холодный (1943)
Указывал на необходимость использования данных о химическом
составе ископаемых организмов для выявления родственных связей
Между видами растений. Так, по содержанию в органическом
веществе ископаемых организмов «правых» аминокислот представ-
11
ляется возможным датировать время их образования: чем ближе к
современности, тем больше их концентрация в биогенном веществе.
Изучение физиолого-биохимических особенностей современных
растений важно для оценки «расхождения признаков, связанных у
различных организмов общностью происхождения» (Н. Г. Холод-
ный, 1943), что ярче всего проявляется в действии естественного
отбора. Сказанное делает понятной необходимость применения
сравнительного метода для выяснения особенностей эволюции как
отдельно взятой функции, так и всей жизнедеятельности растений.
Несмотря на успехи молекулярной палеонтологии, следует при-
знать, что здесь нас ожидают еще большие разочарования, связанные
с «неполнотой геологической летописи» и отсутствием надежных
критериев для определения истинного происхождения и принадлеж-
ности найденных в пород ах биогенных веществ. Конечно, ископаемые
биогенные вещества (хорошо сохранившиеся хемофоссилии) несут
реальную информацию о былой структуре. В то же время многие из
них могли измениться и изменились после их отложения и первичного
синтеза. Направление таких изменений определялось также трофиче-
ской ценностью отложений для микроорганизмов. Так, многие белки
и углеводы, важные для метаболизма микроорганизмов, плохо сохра-
нились в ископаемом состоянии.
Чаще всего наука располагает хемофоссилиями типа отбросов
или ВВМ.
Анализ изотопного состава углерода в ископаемых отложениях
организмов, существовавших на заре жизни, показал, что до появ-
ления свободного кислорода в атмосфере мир прокариот был более
разнообразным, чем в последующем. Если успехи молекулярной
палеонтологии в какой-то мере и обозримы в будущем, то дости-
жения палеофизиологии растений пока крайне ограничены. Было
бы важно знать о величине компенсационной точки фотосинтеза
вымерших водорослей, о реакции на качество света и длину дня
растений в пермском периоде, дневном ходе фотосинтеза и транс-
пирации, ритмике развития уже вымерших растений. По-видимому,
мы никогда с точностью об этом не узнаем даже в отношении
растений, вымерших в кайнозое. На этом пути не приходится
ожидать принципиальных решений. Вряд ли удастся определить
картину жизнедеятельности ископаемых растений по их скудным
морфологическим остаткам.
Интересными и перспективными следует признать попытки
перенесения тенденций изменения современных растений на иско-
паемые формы. На примере современных деревьев показана зави-
симость камбиальной активности, размера и формы клеток
древесины, ширины годичного ее кольца и прироста от внешних
(влажность, температура, освещенность, длина дня, наличие пита-
тельных веществ в субстрате) и внутренних (наличие регуляторов
12
роста, возраст, сезонная ритмика) условий. Форма и размер клеток,
толщина кольца древесины различны у одного и того же растения
на разных сторонах кроны.
Поэтому, переходя от наблюдений на материале современных
деревьев к вымершим, можно сделать вывод, что сезонная актив-
ность камбия и роста древесины отчетливо проявляется начиная с
карбона. Она сильнее оказывается выражена в мезозое. В то же
время уже в палеозое (карбон и пермь) у древесных прослежена
зональность роста, коррелирующаяся с их географическим распро-
странением (G. Т. Greber, W. G. Chaloner, 1984). В этой связи
интересны также наблюдения над зарастанием техногенных суб-
стратов, являющихся практически стерильными. Их обычно начи-
нают обживать цианобактерии из-за своей способности
выдерживать экстремальные условия влажности и температуры,
устойчивости к химическим агентам, развивать симбиотические
отношения с другими видами и фиксировать атмосферный азот.
По-видимому, исторически они служили пионерами освоения су-
ши, как и в современных условиях.
Более или менее достоверно судят о форме и размерах листьев,
величине открытости, расположении устьиц и о внутривидовом
разнообразии у хорошо сохранившихся ископаемых видов Metase-
quoia disticanap. (В. А. Самылинаи Н. В. Лузина, 1993). Показано,
например, что в конце палеогена (в олигоцене) у метасеквой
кутикула утолщается, устьица укрупняются, ширина устьичных и
межустьичных полос уменьшается. В миоцене замечены обратные
процессы. Такие же изменения в конце палеогена отмечены у Ginkgo
orientalis (А. С. Кудышев, 1983).
Описаны изменения в анатомии древесины у многочисленных
ископаемых растений, например у ископаемых елей Восточной
Сибири. В частности, в миоцене или плиоцене наблюдаются тан-
гентальная поровость трахеид, спиральное утолщение разных типов
трахеид, утрата древесной паренхимы, изменение мест локализации
горизонтальных смоляных ходов и т. п. (Н. И. Блохина, 1983).
Подобные наблюдения, несмотря на их косвенный характер,
важны для углубления представлений о палеоэкологии и палеофизи-
ологии растений. В целом же изучение отпечатков листьев, древесины,
плодов и т. п. дает ценный материал о палеоэкологии (следовательно,
и о палеофизиологии) растений. Так, мы лучше сейчас представляем
физиологию ископаемых водорослей (К. Б. Кордэ, 1973). Если о до-
кембрийской жизни мы знаем по органогенным остаткам цианобак-
терий, то в последующем появляются многочисленные карбонатные
сооружения водорослей (А. Ю. Розанов, 1985).
В связи с ограниченными возможностями палеонтологического
метода в изучении функциональной эволюции растений особое
13
значение приобретает использование сравнительного метода —
анализ жизнедеятельности современных видов, включая и древние
организмы. Так, мембраны архебактерий состоят из изопреновых
эфиров глицерина, способных образовывать самопроизвольно
Мембраноподобные структуры. Они содержат крупные молекулы
кодирующих белков, отличающихся полифункциональностью.
Сравнивая 5S рРНК, РБФК у древних фототрофов, можно обнару-
жить наличие нескольких ветвей в их эволюции. Линия хлоропла-
стов дивергировалась рано; она берет свое начало от Clostridium
pasteurianum в сторону фотосинтезирующих прокариот. Затем па-
раллельно отходят три линии, ведущие к митохондриям: Pseudomo-
nas и Mycobacterium, архебактерий и эукариот. От последних
происходят Thermus, aquaticus, Bacillus, Mycoplasma. Примитивные
эукариоты разделяются на восемь ветвей.
Биохимические сравнения растений показали, что у древних
форм в белках доминируют аминокислоты глицин и аланин, тогда
как у филогенетически молодых — глутамин и аспарагиновая кис-
лота (Е. Г. Судьина, Г. И. Лозовая, 1982). В эволюционном про-
движении растений отмечена тенденция нарастания разнообразия
белков и их аминокислотного состава. По этой причине не все белки
могут служить маркерами для генетического анализа современных
растений. У растений выделены три группы белков по степени
консервативности молекул (В. Г. Конарев, 1983). Различают белки
древние — консервативные (гистоны ядра), молодые и функцио-
нально-активные (ферменты и запасные белки) и полиморфные
(изозимы). Первые характеризуют общность крупных таксонов,
вторые — видов, третьи — внутривидовых групп.
Использованием методов молекулярной генетики показано, что
растительные леггемоглобины (Лг) и животные глобины (Гл) ди-
вергировали 0,9—1,4 млрд, лет назад, степень их сходства составляет
30%. В то же время обнаружено сходство последовательностей
некодирующих и кодирующих участков генов Лг и Гл у животных
и растений. Основной локус Лг сформировался у растений путем
ряда дупликаций и, возможно, серии неравных кроссинговеров. Для
установления родства начали широко применять и рестрикционный
анализ хлоропластного генома (Хг). При этом показано наличие
большей идентичности Хг у Beta vulgaris и В. maritima (Mikami Т et al,
1984). Другие виды (В. trigyna, В. lomatogona, В. patellaris, В. procumkns)
отличались от В. vulgaris по набору фрагментов рестрикции.
Из приведенных выборочных данных видна перспективность
применения сравнительного метода для изучения функциональной
и биохимической эволюции растений. Однако возможности его все
еще слабо используются. Очевидно и другое, о чем в свое время
писал К. А. Тимирязев: «...ни морфология со своим блестящим и
плодотворным сравнительным методом, ни физиология со своим
14 '
еще более могущественным экспериментальным методом не покры-
вают всей области биологии, не исчерпывают ее задачи; и та, и
другая ищет дополнения в методе историческом»1. Поэтому чрез-
вычайно важно при изучении проблем эволюционной физиологии
растений использовать комплекс методов, в том числе и метода
количественного анализа, применяемые в эволюционной биохимии
и популяционной генетике при учете изменчивости (генетические
методы), а также обычные экспериментальные методы для сравне-
ния особенностей жизнедеятельности растений разных видов в
онтогенезе. Следующая задача состоит в анализе эксперименталь-
ных данных с точки зрения истории развития растений. К сожале-
нию, до сих пор сравнительный и исторический методы все еще
недостаточно применяются в физиологии растений. Этим объясня-
ется отсутствие или неполнота данных об эволюции физиологиче-
ских особенностей.
1.3. КРАТКИЙ ОЧЕРК ИСТОРИИ РАЗВИТИЯ
ЭВОЛЮЦИОННЫХ ИДЕЙ В ФИЗИОЛОГИИ РАСТЕНИЙ
Периодизация истории развития любой научной дисциплины
является важным и трудным вопросом, она достижима только после
серьезного анализа основных направлений поиска. Не вдаваясь в
подробный экскурс в эту область, ограничимся указанием на недо-
статочную ясность подхода к периодизации истории эволюционной
физиологии растений, М. X. Чайлахян (1959), сравнивая успехи
развития эволюционной морфологии и физиологии растений, пра-
вильно подметил, что эволюционная физиология растений пока
делает первые шаги и мы вынуждены искать ее зачатки «как
крупицы мысли, разбросанные в отдельных высказываниях и
статьях физиологов». Сказанное, однако, не является отрицанием
заслуг ряда отечественных и зарубежных исследователей в поста-
новке и разработке вопросов функциональной эволюции расте-
ний, что уже получило должное отражение в печати (Н. Г. Холодный,
1943; К. В. Манойленко, 1974, 1985). Далее будут приведены крат-
кие сведения об основных вехах развития эволюционной физиоло-
гии растений.
Значение эволюционного учения для объяснения сложных фено-
менов из жизни растений было показано ещё в работах Ч. Дарвина,
К. А. Тимирязева, A. G. Фаминцына. Позднее эволюционный под-
ход в фитофизиологии, к сожалению, стал занимать более скромное
место, хотя в области биохимии растений он широко применялся
(К. В. Манойленко, 1974).
1 Тимирязев К. А. Избр. соч., 1949. Т. VI. С. 61.
15
Попытаемся воссоздать историю проникновения эволюцион-
ных идей в фитофизиологию начиная с 60-х годов XIX в. В книге
«Происхождение видов» Ч. Дарвин дал эволюционное истолкова-
ние продолжительности жизни, явлению покоя, сезонности раз-
вития, опылению растений и т. д. Эти наблюдения получили
дальнейшее развитие в других его работах: «Движения и повадки
лазающих растений» (1865), «Насекомоядные растения» (1875),
«Способность к движениям у растений» (1880). В них Ч. Дарвин
пытался объяснить приспособительное значение и пути становле-
ния разнообразных движений у растений. Интерес Ч. Дарвина к
движениям растений был не случаен. В это время в физиологии
широко распространилось мнение о непознаваемости явлений раз-
дражимости не только у растений, но и у животных. В этой области
господствовал витализм. Характеризуя значение работ Ч. Дарвина,
К. А. Тимирязев писал, что «за ним остается никем неоспоримая
заслуга обобщения, подведения под одно общее начало самых
разнородных явлений движения»1.
Эволюционный анализ физиологических явлений, начатый
Ч. Дарвином, был продолжен К. А. Тимирязевым применительно
к процессам фотосинтеза и устойчивости растений к засухе. В его
работах «Значение лучей различной преломляемости в процессе
разложения углекислот растениями» (1865), «Спектральный анализ
хлорофилла» (1871), «Об усвоении света растением» (1875) и других
показан принцип действия законов естественного отбора и сохра-
нения энергии. В исследованиях по засухоустойчивости К. А. Ти-
мирязев продолжил анализ путей приспособления растений к
«физиологическому злу» — транспирации, показав ее биологиче-
ский смысл. К. А. Тимирязев указывал, что приспособления к
засухе приобретают автоматический характер, они «обращают на
пользу растения действия тех самых сил, с которыми''’оно ведет
борьбу»1 2.
Проникновение эволюционных идей в фитофизиологию про-
исходит различными путями, т. е. связано с изучением разных
явлений. Выдающуюся роль в этом сыграли работы отечественных
ученых—А. С. Фаминцына, С. П. Костычева, Н. А. Максимова,
В. Н. Любименко, Н. Г. Холодного и др. Из зарубежных исследо-
вателей, внесших вклад в разработку вопросов эволюционной фи-
тофизиологии, следует упомянуть Г. Клебса, Г. Боннье, Д. Босса,
К. Ван-Нила, Г. Фиттинга и др.
Термин эволюционная физиология впервые употреблен в работе
А. Н. Северцова «Современные задачи эволюционной теории»
1 Тимирязев К. А. Избр. соч., 1949. Т. IV. С. 229—230.
2
Там же. Т. II. С. 129.
16
(1914), где было поставлено в качестве одной из важнейших задач
изучение «эволюции физиологических и биохимических особенно-
стей организмов». Однако основателем эволюционной физиоло-
гии считается Л. А. Орбели (1961), которому принадлежат не
только определение предмета, целей и задач этой науки, но и
наиболее фундаментальные работы в этой области. Благодаря
трудам Л. А. Орбели в нашей стране и за рубежом эволюционный
подход в исследованиях физиологов животных занял ведущее место.
Эволюционные идеи в фитофизиологию вошли раньше, чем в
физиологию животных, но в дальнейшем стали разрабатываться
менее активно. Поэтому в решениях I симпозиума по эволюционной
фитофизиологии (1974) обращалось внимание на необходимость
развертывания широких исследований в этой области, регулярной
публикации их материалов и преподавания такой дисциплины в
университетах.
Тенденция развития физиологии растений в 60-х годах нынеш-
него столетия, связанная с широким использованием сложных
физико-'химических методов для изучения молекулярных основ
жизнедеятельных растений, ныне нуждается в экологизации. Воз-
вращение на новой основе к экологическому подходу, традицион-
ному для отечественной физиологии растений, позволит глубже
взглянуть на приспособленность функциональных проявлений в
мире растений. С другой стороны, он поднимет роль физиологии
растений в разработке методов защиты окружающей среды. В связи
с этим особенно велика значимость развития экологического и
эволюционного направлений в различных областях современной
физиологии растений. Это сегодня уже понимают многие исследо-
ватели и стараются подобные идеи реализовать экспериментально
(А. В. Благовещенский, М. В. Гусев, В. И. Кефели, Р. Клейтон, А.
Л. Курсанов, М. Н. Запрометов, А. Т. Мокроносов, В. В. Поле-
вой, М. X. Чайлахян, Дж. Эдвардс, Д. Уокер и др.). Значимость
подобных исследований в дальнейшем будет еще больше возрастать.
ГЛАВА 2
ОСНОВНЫЕ ТЕНДЕНЦИИ ФУНКЦИОНАЛЬНОЙ
ЭВОЛЮЦИИ РАСТЕНИЙ
2.1. ОПРЕДЕЛЕНИЕ ПОНЯТИЙ
Вместо понятия физиологический признак, предложенного
С. Л. Ивановым (1926) применительно к процессам метаболизма
растений, удобнее использовать термин функция или свойство. В то
же время употреблять термин функция в значении ассимиляции и
диссимиляции представляется нецелесообразным. Поэтому пра-
вильнее говорить об отдельных световых и темновых реакциях
фотосинтеза, роли продуктов обмена (метаболитов) в зимостойко-
сти, об отношении растений к длине дня, качеству света и т. д.
Физиологическое свойство — это элементарная и генетически
обусловленная реакция организма на условия среды, имеющая
значение для онтогенеза. Например, процесс фотосинтеза может
быть разделен на ряд этапов: поглощение энергии света, ее передача
для осуществления химических реакций и т. д. К физиологическим
свойствам растений относятся реакции на длину дня, на красный
и синий свет, сейсмонастическая активность, способы движения и
т. д. Несмотря на генетическую обусловленность физиологических
свойств, они более или менее обратимы в онтогенезе и лабильны в
темпах реализации.
Термины эволюция, приспособление и дивергенция в физиологии
растений все еще не имеют однозначного употребления. Эволюция —
эгк> возникновение каких-либо новшеств в метаболизме, морфоло-
гии и реакциях растений, популяций и видов. Функциональное
приспособление—появление в онтогенезе и филогенезе реакций,
обеспечивающих выживание и репродуктивный успех популяции.
Термин приспособление, или адаптация, физиологи чаще всего
понимают как устойчивость к стрессам. Однако устойчивость к
стрессам — лишь одно из проявлений адаптации популяций; дру-
гое, более важное проявление — их сложная жизнедеятельность в
нормальных условиях. Функциональная дивергенция — возникнове-
ние физиологических отличий между сортами, популяциями и
видами в процессе эволюции.
Функциональная эволюция — это наследственные сдвиги в жиз-
недеятельности растений разных таксономических и экологических
групп, исторически закрепленные отбором.
18
Вслед за Л. А. Орбели (1961) следует также выделить и понятие
эволюция функций, которое употребляют в более конкретном смысле,
функциональная эволюция —интегральный результат истории
развития отдельных функций. В ее основе лежит и биохимическая
продвинутость, для характеристики которой А. В. Благовещенский
(1966) предложил термин арахимоз. Существует мнение, что биохи-
мическая изменчивость растений не может быть регрессивной
(С. Л. Иванов, 1926); это в принципе спорно. Так, переход к
гетеротрофному питанию у части цветковых растений (полупарази-
ты и паразиты) можно рассматривать как регресс функции фото-
синтеза. Говоря об эволюции жизнедеятельности растений, следует
иметь в виду разнообразные случаи, приводящие к преобразованию
функций как в сторону их усиления, так и упрощения или утраты
(прогресс и регресс).
2.2. ГЕНЕТИЧЕСКАЯ ИЗМЕНЧИВОСТЬ
ФУНКЦИОНАЛЬНЫХ ПРОЯВЛЕНИЙ
Для эволюционного преобразования любого признака и свой-
ства необходима наследственная изменчивость как одна из трех
предпосылок действия естественного отбора. Поэтому анализ про-
явлений генетической изменчивости является традиционным при
рассмотрении механизмов и результатов эволюционного процесса.
Обратимся и мы к данному вопросу в рамках задач пособия.
Как известно, источники наследственной изменчивости в по-
пуляциях разнообразны. В отношении функциональных особенно-
стей необходимо иметь в виду еще большую их генетическую
вариабельность, так как они представляют собой совокупность
молекулярных (генных), метаболических (биохимических) и струк-
турных (гистологических и морфологических) изменений. Этим и
определяются сложная природа функциональных проявлений рас-
тений, их полигонная обусловленность и связь с разными локусами.
Теоретически мыслимы следующие направления генетических
изменений функций: 1) формирование принципиально нового ме-
таболического механизма как совокупности последовательных ре-
акций фотосинтеза или дыхания; 2) усиление сопряженности
разных реакций, ранее функционировавших относительно самосто-
ятельно; 3) повышение эффективности реакций в результате уси-
ления клеточной дифференциации: появление органелл и
усложнение их структуры; 4) создание дефицита или избытка в клетке
активных (ферменты, регуляторы роста) и запасных (крахмал, глюко-
зиды и т. п.) веществ; 5) усиление жизнедеятельности путем увеличе-
ния числа одноименных структур (хлоропластов, митохондрий,
19
клеток, корней, листьев и почек); 6) перенос функций с одних
морфологических структур на другие; 7) преобразование габитуса
растительного организма как физиологически целостной системы.
Надо полагать, что указанными направлениями далеко не ис-
черпываются возможности генетической изменчивости функцио-
нальных особенностей растений. В то же время из сказанного
нетрудно представить наличие неограниченных предпосылок для
функционального усовершенствования на основе генетической из-
менчивости функций. Достаточно напомнить, что генетическая
изменчивость проявляется по активности роста и морфогенеза, в
основе которой лежат сложные метаболические процессы. Так,
такой признак, как карликовость растений, обусловлен мутациями,
блокирующими синтез ГА3 и других веществ. Сам дефицит ГА3
может быть сопряжен или блокировкой ее синтеза, или повышением
АБК, проявляющей антигибберелловую активность. Выделены му-
танты типа «ксанта», отличающиеся по распределению фитогормо-
нов. У гороха описаны мутации карликовости (К-29, К-202 и т. д.),
не зависящие от содержания ауксина и отличающиеся высоким
уровнем АБК. Часть мутаций у гороха различается по типу листа
(афильные формы), которые обладают неодинаковой продуктивно-
стью и чувствительностью к регуляторам роста. Растения разных
жизненных форм обнаруживают генетически устойчивые различия
в интенсивности фотосинтеза и расходования энергетических ре-
сурсов уже на стадии прорастания семян. Все это весьма значимо
для конкурентоспособности видов.
Как подчеркивают А. Т. Мокроносов и В. Ф. Гавриленко
(1992), физиологический процесс фотосинтеза представляет чрез-
вычайно сложную систему пространственно- и временно-коорди-
нированных явлений, детерминируемых ядерными и пластидными
генами, подвергающимися мутациям. Он зависит от генетических
особенностей организации растения и листа (мезоструктуры по
А. Т. Мокроносову), а также протекания множества химических
реакций. Вклад разных генов в продуктивность фотосинтеза разли-
чен, тем не менее они значимы и наследуются в поколениях, образуя
всевозможные комбинации.
'Подтверждением сказанному отчасти служат селекционно вы-
деленные инцухт-линии у культурных растений (сахарная свекла,
кукуруза, пшеница и др.), которые отличаются разнообразным
сочетанием морфологических и функциональных признаков, опре-
деляющих продуктивность фотосинтеза. В связи с этим такие
особенности изменения, как устойчивость, рост, развитие и про-
дуктивность растений, подвергаются генетической изменчивости и
подпадают в природных условиях под действие отбора. Это видно
и на примере активности устьичных движений. Так, получены
20
мутации, характеризующиеся постоянной открытостью устьиц
(flaco), что приводит к быстрому завяданию растений в условиях
даже кратковременного дефицита воды. Описаны мутации по из-
менению числа волосков на поверхности клеток эпидермиса лис-
товой пластинки и кончика корня, по степени развитости
проводящей системы и т. д. Генотипы сорго, отличающиеся по
эффективности фотосинтеза, обнаруживали заметные различия в
активности ФЕПК, НАДФ-малик-энзима, интенсивности транспи-
рации И в содержании хлорофилла. Эти различия у генотипов
особенно резко проявлялись в условиях водного стресса.
Функциональные особенности обычно контролируются многи-
ми локусами. Так, в адаптации растений к минеральному составу
субстрата и высокому содержанию солей в почве (А. А. Жученко,
1980; Дж. Харборн, 1985) участвует большое число генов и биохи-
мических процессов, контролируемых ими. Еще большее число
генов и их комплексов принимает участие в адаптации растений к
климатическим условиям, которая выступает как совокупность взаи-
мосвязанных биохимических, физиологических и морфологических
изменений растений. На примере экотипов Agrostis tenuis и Festuca
ovina, толерантных к содержанию высоких концентраций свинца, меди
и цинка (нормально произрастающих на отвалах рудников), показано,
что данная способность обусловлена генетически, выражением чего
служит неоднородность потомства, полученного от устойчивых мате-
ринских растений (A. D. Bradshaw et al., 1965).
Чтобы дальше пояснить генетическую природу функциональ-
ных проявлений растений, остановимся на фактах мутационной
изменчивости отдельных функций. Только по синтезу ГА3 описан
ряд мутаций, связанных с блокировкой отдельных звеньев цепи,
включая и образование кауренсинтетазы. Некоторые из мутаций
блокировки синтеза ГА3 можно фенотипически имитировать обра-
боткой растений химическими ингибиторами роста. К наследствен-
ным особенностям растений относится и разная степень
содержания ГА3 в тканях, с чем коррелирует разная интенсивность
роста. Многие мутации карликовости кукурузы и гороха проявляют.
высокую чувствительность к экзогенному введению ГА3 (Г. С. Му-
ромцев, В. Н. Агнистикова, 1984). Напротив, у некоторых из му-
тантов картофеля и фасоли, а также большей части мутантов
пшеницы не обнаружена связь карликовости с низким содержанием
ГА3, а также реакция на ее экзогенное введение.
Описаны многочисленные мутации, способствующие измене-
нию устойчивости растений к патогенным агентам. Поскольку
элементами иммунной системы являются каротиноиды, антоцианы,
аминокислоты, ВВМ, фитоалексины и т. п., постольку мутацион-
ными изменениями бывает охвачен широкий круг веществ. Так,
моногенная устойчивость кукурузы к Helminthosporium turcium, обус-
21
ловленная действием гена Ht, оказывается связанной с большим
накоплением фитоалексинов—гидроксановых кислот. При этом
развивается и хлоротичность листьев. Комплексная устойчивость
растения к патогенным возбудителям связана с образованием блоков-
генов, контролирующих развитие определенных морфологических и
физиологических свойств. Такие блоки-гены ведут себя как доминан-
тные. У льна описано 25 генов, влияющих на устойчивость к ржавчине.
Отбор культурных растений на устойчивость идет по линии
усиления гомозиготности под давлением патогенов, и его результат
зависит от концентрации генов вирулентности в популяциях пато-
гена. Повышение устойчивости растения-хозяина приводит к но-
вому циклу отбора на агрессивность в популяциях патогена и т. д.
(П. М. Жуковский, 1973).
Изучение генетики функциональных особенностей значительно
продвинулось в связи с успехами в расшифровке геномов митохон-
дрий и хлоропластов (М. С. Одинцова, 1988; В. Е. Семененко,
1988; К. В. Войников и др., 1991), а также генов изоферментов части
ключевых ферментов растений (Е. В. Левитес, 1986). Описан ряд
мутаций, блокирующих разные звенья синтеза хлорофилла, функ-
цию ЭТЦ митохондрий и хлоропластов, активность ключевых фер-
ментов, что позволило составить генетические карты хромосом. В
частности, у кукурузы выявлен 91 локус, контролирующий 31
ферментную систему, у томатов —25, гороха — 12, ячменя —20
локусов, контролирующих синтез изоферментов (Е. В. Левитес,
1986). У ячменя описано более 200 ядерных генов, контролирующих
биосинтез пластидных пигментов; у гороха синтез хлорофилла и
каротиноидов регулируется примерно 170 генами, расположенными
в 15 локусах; у арабидопсиса биосинтез пластидных пигментов опре-
деляется несколькими десятками неаллельных генов (Ю. С. Насырой,
1975).
У мутаций по генам хлорофилл-недостаточности обнаружены
повышенная активность хлорофиллазы и высокое содержание по-
лифенолов (S. Sheen, Т. Hamilton, 1983). Генетические различия
описаны в строении хлоропластов (Дж. Бил, Дж. Ноузл, 1981), в
наследовании особенностей фотосинтеза (Н. Авратощукова, 1980)
и иммунитета (Я. Вандерпланк, 1981) у разных сортов и видов
культурных растений. Однако в отношении генетической природы
других функциональных проявлений растений предстоит еще мно-
гое выяснить.
Это и есть одна из важных задач сравнительной генетики
растений и генетики развития. Н. Авратощукова (1980) указывает
на генетические различия видов, сортов и растений внутри сорта
по фотосинтетической активности в целом и частным ее реакциям
(реакция Хилла —расщепление воды, активность ФЕПК, РБФК и
т. д). Так, коэффициент наследуемости фотосинтетической акгив-
22
ности для сортов табака имел среднее значение 0,85, для кар-
тофеля —0,94, для огурцов —0,53. Описаны генетические разли-
чия в фотосинтезе полиплоидов (W. Gottschalk, 1976). Установлены
внутрипопуляционные генетические различия в дыхании у плевела
и табака (J. Zelitch, 1956). Изучены мутации по синтезу хлорофилла
и некоторых веществ вторичного метаболизма у хламидомонады,
связанные с дупликацией генов. На примере водорослей и высших
растений выделены мутации по дефициту хлорофилла, элементов
ЭТЦ, каротиноидов, а также по избыточности синтеза некоторых
ферментов. У нежелтеющей мутации райграса пастбищного (Bf 986)
разобщена связь между синтезом хлорофиллазы и хлорофилла. На
примере нежелтеющей мутации (Bf 993) показано, что хлорофил-
локсидазная активность связана с ФСI. По активности указанного
фермента типичные и мутантные генотипы не различались. Однако
у Мутантов продолжительность периода хлорофиллоксидазной ак-
тивности в три раза превышала таковую в контроле. Нежелтеющая
мутация у типчака не связана с потерей активности хлорофиллок-
сидазы. У риса из желто-зеленых мутантов исследована флуорес-
ценция хлорофилла Ь. У 10 мутантов хлорофилл b полностью
отсутствовал, а у 6 — содержался в небольших количествах. У
мутантов, лишенных хлорофилла Ь, отсутствовали антенные хлоро-
филл-белковые комплексы ФС II, а также белки с молекулярной
массой 20 и 21 кД и антенные хлорофилл-белковые комплексы ФС I
(В. Malkin, 1985).
Изучены гены, контролирующие синтез части ферментов. На-
пример, у томатов эстеразный кластер генов локализован во 2-й
хромосоме в 29-м положении, а в 41-м локусе —кластеры генов
пероксидазы. Короткое плечо 7-й хромосомы у кукурузы несет 7
структурных генов зеина. У ржи и других растений длина вегетаци-
онного периода определяется наличием большого числа хиазм в
хромосомах.
Многие морфологические признаки также имеют функциональ-
ное значение и своим возникновением обязаны биохимическим
мутациям. Таковой является окраска цветков, плодов и листьев,
зависящая от состава пигментов и антоцианов. Наблюдается гео-
графическая изменчивость распространения таких мутаций. Так,
аборигенные пшеницы Грузии несут гены некроза (Ne, и Ne2) и
красного хлороза (Chr) листьев. У африканских и индийских попу-
ляций Pennisetum thyphoides распространены преимущественно оп-
ределенные типы хлорофилльных мутаций. Все это влияет на
продуктивность растений. Величина листовой поверхности и харак-
тер листорасположения как генетически детерминированные при-
знаки также оказывают влияние на продуктивность и адаптивность
культурных растений.
23
Значительна роль и явления рекомбинации наследственных фак-
торов в изменчивости физиологических особенностей. Содержание
сухих веществ, витаминов, антоцианов и т. д. зависит от сочетания
разных локусов и хромосомных перестроек. Так, только на основе
мутационной изменчивости не могла быть обеспечена быстрая
эволюция растений (G. L. Stebbins, 1968). Большая роль здесь
принадлежит гибридизации и рекомбинации. В. Грант (1984), опи-
раясь на механизм рекомбинации, объяснил возникновение супер-
гена у примулы, регулирующего тип цветка и способы опыления.
Низкая приспособленность культурных растений по сравнению
с дикорастущими и сорными видами обусловлена уменьшением их
наследственной гетерогенности в процессе селекции. Например,
дикие томаты характеризуются большей гетерозиготностью по мно-
гим локусам, чем культурные сорта (А. А. Жученко, 1989). Поэтому
необходимо повысить адаптивность культурных растений за счет
поиска соответствующих генных комплексов и создания на их
основе «синтетических» гибридов (А. А. Жученко, А. Б. Король,
1985). Накоплено много фактов генетической обусловленности
биохимических особенностей растений (см.: Генетический подход
к биохимии растений, 1991; L. Wen-Hsiung, D. Graur, 1991).
Мы еще не зИаем генетической природы многих физиологиче-
ских проявлений. Судя по функциональной их сложности и лабиль-
ности, можно предположить наличие полигенного наследования
многих функций. Например, такое важное экологическое приспособ-
ление, как засухоустойчивость, обусловлено сочетанием комплекса
генов, влияющих на биохимию, физиологию и морфологию индиви-
дуума. Причем засухоустойчивость у разных экологических типов
растений (ксерофиты, мезофиты, суккуленты и т. п.) различается по
природе (П. А. Генкель, 1983), хотя в целом она определяется спо-
собностью противостоять обезвоживанию и перегреву организма. В
отдельности же способность перенести и обезвоживание, и перегрев
имеет далеко не одинаковую наследственную основу.
На примере образцов 15 видов пшениц и 6 видов эгилопса
показано, что одни виды способны перенести обезвоживание ко-
лоса, а другие — листьев. Как правило, тетра- и гексаплоидные
пшеницы с геномами AUB и AUBD легко переносят обезвоживание
и листьев, и колосьев. В ходе эволюции пшениц повышение устой-
чивости к обезвоживанию произошло в двух случаях: при переходе
на тетраплоидный уровень с присоединением генома В от эгилопса,
а у репродуктивных органов — с переходом на гексаплоидный
уровень и после присоединения генома D (Н. М. Кожушка, 1985).
Таким образом, и внутривидовые, и внутриродовые различия в
способности переносить обезвоживание у пшениц и эгилопса оп-
ределяются спецификой генома, сложившегося в ходе эволюции.
24
С одной стороны, это свидетельствует о сложности физиологии
засухоустойчивости и ее генетической детерминации, с другой — о
несовпадении для разных видов вектора отбора по блокам генов.
Следовательно, один и тот же экологический результат (засухо-
устойчивость) достигается закреплением разного комплекса генов,
что зависит от объекта и среды обитания. Сказанное о природе
засухоустойчивости в равной мере относится и к большей части
функциональных особенностей растений. Таким образом, откры-
ваются неограниченные возможности их генетической изменчиво-
сти и эволюционного или селекционного совершенствования.
Отмечены и парадоксы в эволюции генома: морфологически
сходные организмы имеют разные величины геномов, количество
ДНК не коррелирует со сложностью организма, геномы характери-
зуются избыточностью ДНК (Ф. Айяла, 1984). Значение избыточ-
ности ДНК растений еще не ясно. (В. Грант, 1984; Э. М. Ахундова,
1986). Геномы растений имеют сложную организацию. На примере
изучения геномов пшеницы, ржи, ячменя и овса показано, что они
произошли от общего предка в результате амплификации его генома.
Так, у пшеницы более 80% оснований ДНК представляют амплифи-
кации разного типа. В зависимости от степени и типа амплификации
сравниваемые растения обнаруживают различия в повторяющихся и
неповторяющихся основаниях ДНК. Например, у пшеницы, ржи,
ячменя и овса обнаружено соответственно 16, 23, 28 и 55% повторя-
ющихся оснований (R. Travell et. al., 1979). Этим отчасти можно
объяснить и причины сходства в химизме растений (см. гл. 5).
Примеры генетической изменчивости на функциональном уров-
не будут приведены еще в соответствующих главах. Можно сделать
вывод, что нет ни одного признака, который не подвергался бы
изменчивости (Ч. Дарвин, 1939); изменчивость становится всеоб-
щим явлением в живой природе. Известные генетические механиз-
мы обеспечивают накопление резерва наследственной
изменчивости функциональных особенностей у растений, необхо-
димого для вознйкновения адаптаций под контролем отбора. В этой
связи следует остановиться на классификации физиологических
свойств и адаптаций растений.
2.3. КЛАССИФИКАЦИЯ ФИЗИОЛОГИЧЕСКИХ
СВОЙСТВ И АДАПТАЦИЙ
Физиологические свойства как наследственно дискретные осо-
бенности жизнедеятельности растений чрезвычайно разнообразны
и могут быть разделены на общие и частные. К общим относятся
способность к фотосинтезу и аэробному дыханию, зависимость
25
продуктивности и размножения растений от изменения среды,
наличие зеленого пигмента и т. д. Частные свойства проявляются
в ограничении транспирации (суккуленты) и высоком осмотиче-
ском давлении (галофиты), в предпочтительном поглощении нит-
ратного или аммонийного азота разными растениями, в специфике
фотопериодической реакции, специфике покоя и т. д.
Общие и частные (специфические) физиологические свойства
могут быть разложены на отдельные составляющие. Так, путь
метаболизма углерода у растений состоит Из множества реакций,
каждая из которых важна для синтеза органического вещества и
поддержания жизнедеятельности. Транспирация растений складыва-
ется из особенностей метаболизма устьичных клеток, состояния овод-
ненности клеток листа, выделительной деятельности клеток и т. д.
Физиологические свойства реализуются на молекулярном, кле-
точном, тканевом и органном уровнях, а также на уровне целого
растения. С переходом от молекулярного к организменному уровню,
особенно у многоклеточных, физиологические свойства, приобре-
тают более частный характер. Так, молекулярные процессы жизне-
деятельности всех растений носят более общий характер, чем
процессы на органном и организменном уровнях. Это и есть одно
из проявлений физиологической дифференциации видов. Напомним,
что общие молекулярные механизмы фотосинтеза и поглощения
веществ могут иметь специфические особенности на тканевом,
органном и организменном уровнях. Так, клетки не всех тканей и
органов ассимилируют СО2 подобно клеткам палисадной ткани
листа. Точно так же не все растения, способные к фотосинтезу,
осуществляют его в листьях.
Физиологические свойства—интегральный результат молеку-
лярных и морфологических изменений. Первичной основой функ-
циональной специфики являются молекулярные процессы,
осуществляющиеся на мембранах и в органеллах, а их конечная
Схема 1
Физиологические свойства
Общие
I
Молекулярный и клеточный
уровни
I
Структурное однообразие
(мембран, органелл)
Частные
i
Тканевой, органный
и организменный уровни
I
>• Структурное разнообразие
26
реализация зависит от сложной и разнообразной структуры (тканей,
органов, систем органов). Поэтому структурная дифференциация
оказывается предпосылкой для дальнейшей функциональной эво-
люции (см. сх. 1). Это еще одно свидетельство неправомерности
ограничения функциональной эволюции молекулярными про-
цессами.
Все это имеет отношение и к классификации функциональных
адаптаций растений. Вопрос о классификации адаптаций — один
из нерешенных в эволюционном учении. Наиболее удачной при-
знана пока классификация адаптаций по масштабу, происхожде-
нию, принадлежности к разным средам и т. д. Функциональные
адаптации растений можно разделить на генетические (устойчивые)
и фенотипические (неустойчивые).
К устойчивым, как правило, относят адаптации, сочетающиеся
с наследственными изменениями в морфологии. Например, нали-
чие обкладки пучка сочетается с изменением пути фиксации СО2
(С4-путь фотосинтеза), а устойчивость к засухе определяется ком-
плексом признаков. Так, устойчивость к обезвоживанию может
быть усилена изменением мембранной активности, осмотических
сил клеток и корня, индуцированием развития световых листьев,
часть которых является модификациями, но тем не менее они важны
для выживания растений. Нередко их обозначают термином аккли-
мация, чтобы отграничить от наследственно фиксированных адап-
таций (N. Turner, Р. Kramer, 1980).
Функциональные адаптации не всегда сопровождаются види-
мыми морфологическими перестройками, например суточная и
сезонная динамика фотосинтеза и транспирации, Изменение ин-
тенсивности дыхания при временных перепадах температуры и
т. п. Многие адаптации функционального характера морфологи-
Схема 2
1
Генотипические Фенотипические
Устойчивые Неустойчивые
4 4
Сочетающиеся с морфологией Не сочетающиеся с морфологией
Изменение метаболизма,
роста и биомассы
27
чески выражены, отражаются на росте растений и органов, со-
отношении подземной и надземной биомассы, на числе клеток и
сосудов (см. сх. 2).
По специфике и рангу функциональные адаптации могут быть
разделены на группы: 1) проявляющиеся стабильно и общие для
всех растений (реакции циклов' Кальвина и Кребса, клеточные
основы и полярность роста и т. д.); 2) развивающиеся в строго
определенных условиях и чередующиеся в онтогенезе (суточные и
сезонные ритмы, фотопериодическая реакция, покой и т. д.);
3) необходимые для переживания стрессовых условий, отражающи-
еся на структуре (последствия засухи, засоления и т. д); 4) реализу-
ющиеся в течение короткого времени для обеспечения одного из
этапов онтогенеза (функция нектарников, зародышевых корней и
семядолей, реакция на опыление и т. д); 5) ненаследственные и
обратимые свойства-модификации (временные изменения обмен-
ных процессов, морфологическая пластичность организма й т. д.),
имеющие также значение для выживания организма. Признаки
1—4-й групп наследственно закреплены и эволюционно стабили-
зированы.
Взаимоотношения между перечисленными физиологическими
свойствами изображены на сх. 3, из которой видно, что на общий
каркас организации жизнедеятельности растения (7) наслаиваются
специфические реакции, выступающие как результат экологиче-
ской дивергенции видов (2—5).
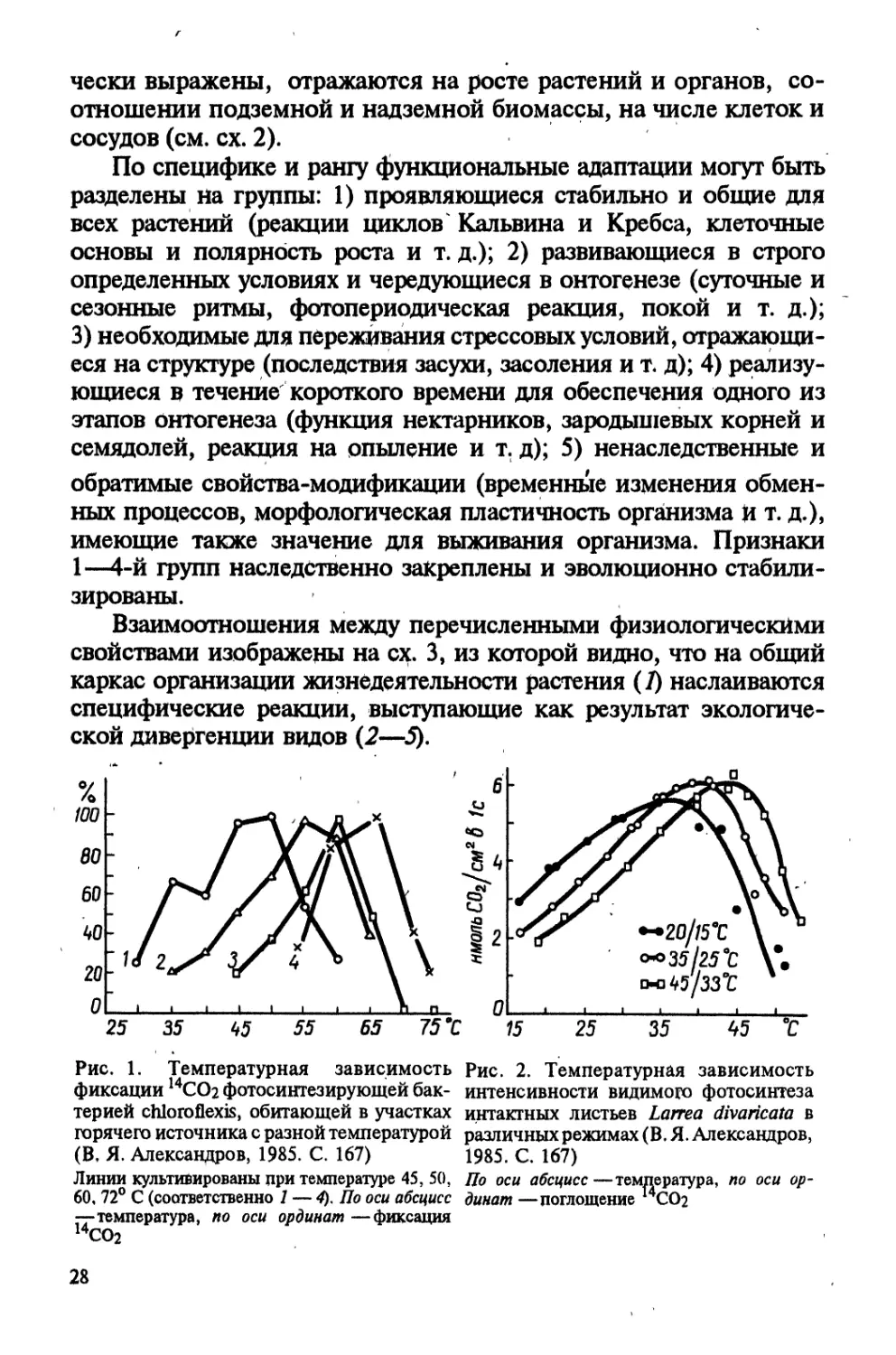
Рис. 1. Температурная зависимость
фиксации 14СО2 фотосинтезирующей бак-
терией chloroflexis, обитающей в участках
горячего источника с разной температурой
(В, Я. Александров, 1985. С. 167)
Линии культивированы при температуре 45, 50,
60, 72° С (соответственно 1 — 4), По оси абсцисс
—температура, по оси ординат —фиксация
14СО2
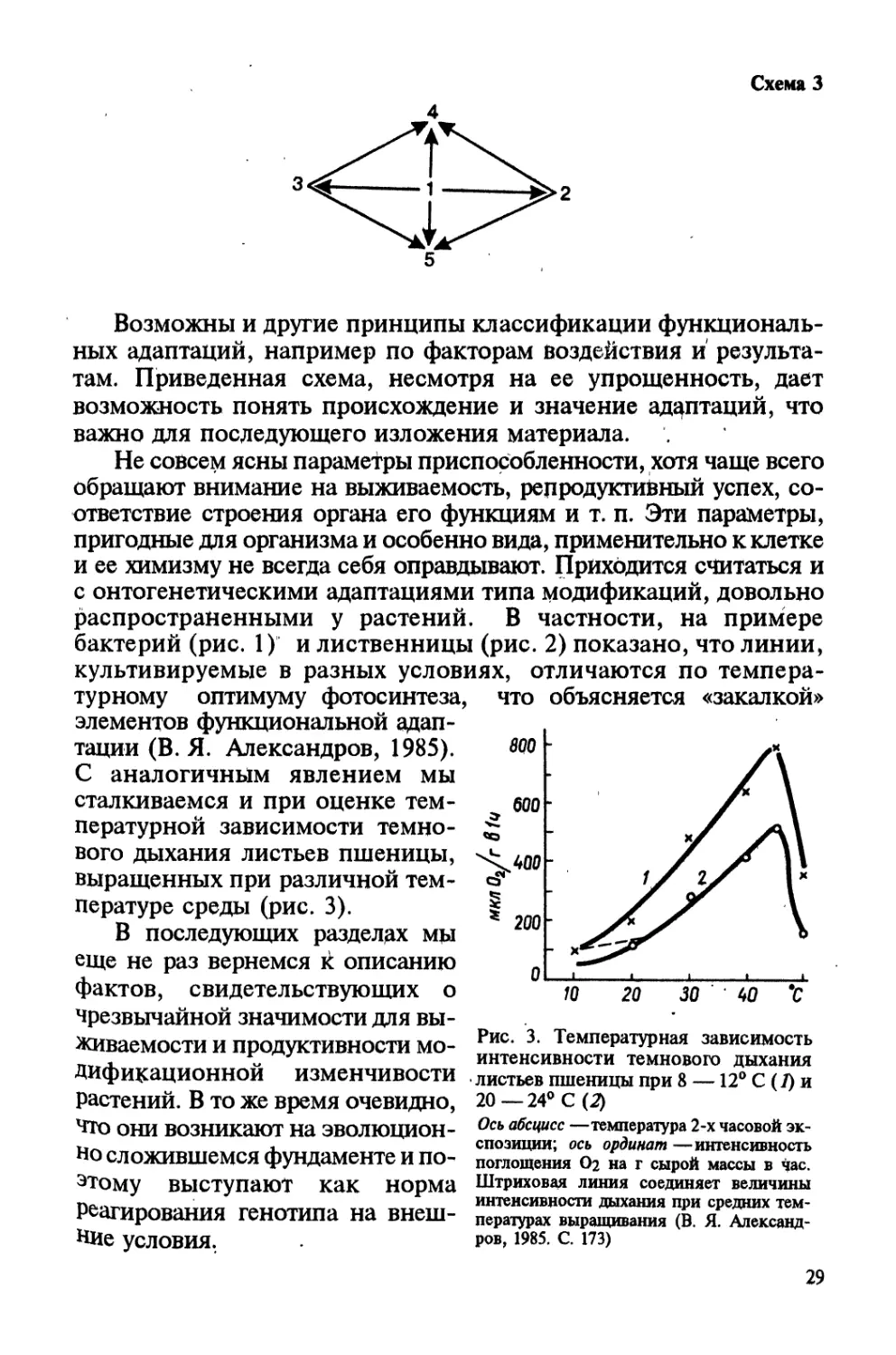
Рис. 2. Температурная зависимость
интенсивности видимого фотосинтеза
интактных листьев Larrea divaricata в
различных режимах (В, Я. Александров,
1985. С. 167)
По оси абсцисс —температура, по оси ор-
динат —поглощение 14СО2
28
Схема 3
Возможны и другие принципы классификации функциональ-
ных адаптаций, например по факторам воздействия и результа-
там. Приведенная схема, несмотря на ее упрощенность, дает
возможность понять происхождение и значение адаптаций, что
важно для последующего изложения материала.
Не совсем ясны параметры приспособленности, хотя чаще всего
обращают внимание на выживаемость, репродуктивный успех, со-
ответствие строения органа его функциям и т. п. Эти параметры,
пригодные для организма и особенно вида, применительно к клетке
и ее химизму не всегда себя оправдывают. Приходится считаться и
с онтогенетическими адаптациями типа модификаций, довольно
распространенными у растений. В частности, на примере
бактерий (рис. 1) и лиственницы (рис. 2) показано, что линии,
культивируемые в разных условиях, отличаются по темпера-
турному оптимуму фотосинтеза,
элементов функциональной адап-
тации (В. Я. Александров, 1985).
С аналогичным явлением мы
сталкиваемся и при оценке тем-
пературной зависимости темно-
вого дыхания листьев пшеницы,
выращенных при различной тем-
пературе среды (рис. 3).
В последующих разделах му
еще не раз вернемся й описанию
фактов, свидетельствующих о
чрезвычайной значимости для вы-
живаемости и продуктивности мо-
дификационной изменчивости
растений. В то же время очевидно,
что они возникают на эволюцион-
но сложившемся фундаменте и по-
этому выступают как норма
Реагирования генотипа на внеш-
ние условия.
Рис. 3. Температурная зависимость
интенсивности темнового дыхания
листьев пшеницы при 8 —12° С (Т) и
20 — 24° С (2)
Ось абсцисс —температура 2-х часовой эк-
спозиции; ось ординат —интенсивность
поглощения 02 на г сырой массы в час.
Штриховая линия соединяет величины
интенсивности дыхания при средних тем-
пературах выращивания (В. Я. Александ-
ров, 1985. С. 173)
29
2.4. МНОГООБРАЗИЕ ФОРМ И ФУНКЦИЙ РАСТЕНИЙ.
ВЗАИМОСВЯЗЬ ФОРМЫ И ФУНКЦИИ
Живые существа нашей планеты олицетворяют «уникальность
биологического вещества» (В. И. Вернадский, 1967), т. е. имеют
единую биохимическую организацию. Есть веские доказательства в
пользу того, что почти 93% всех ферментов, характерных для высших
растений, играли роль в жизни древних прокариот (Э. Брода, 1978).
Точно так же почти одинаковы механизмы фиксации СО2 и аэроб-
ного дыхания у зеленой водоросли и покрытосеменных, несмотря
на значительный морфологический разрыв между ними. Сходны и
механизмы поглощения веществ и испарения воды у наземных
растений при огромном их многообразии. В связи с этим иногда
говорят о прекращении эволюции веществ с наступлением эры
биологической эволюции.
Здесь уместно остановиться на взаимосвязи формы (структуры)
и функции. Этот вопрос является одним из древних и традиционных
в натурфилософии и биологии. Долгое время он был оплотом
идеалистических построений. В свое время Аристотель, задавая
вопрос: «Ради чего существует орган?», отвечал: «Ради выполнения
определенной цели», т. е. функции! Идеалисты исходили из тезиса,
что животные и растения с известными органами и привычками
якобы созданы для обитания в строго конкретных условиях.
Форма определяет функцию в онтогенезе, а функция — форму
в филогенезе. Значение любой функции можно оценить объективно
только в иерархии систем функций и структур. При наличии
множества функций у структуры значимость последней возрастает
при различных экологических ситуациях. Многие важные биоло-
гические функции обычно обеспечиваются несколькими структу-
рами. Например, функция фотосинтеза протекает при участии
листьев, стеблей, чашелистиков, плодов, остьев и т. п. Нередко
монофункциональность на молекулярном уровне переходит в муль-
тифункциональность на уровне организма. Так, функция молекулы
хлорофилла на уровне организма расширяется: улавливание энер-
гии света, фототаксис хлоропласта, дыхание и т. п.
Структуру (форма и орган) и функцию (физиологическое от-
правление) рассматривают в различных соотношениях, отдавая
предпочтение то одной, то другой. Обычно первенство принадлежит
изменению функции. Но это не всегда правильно. Вернее усматри-
вать взаимосвязь и взаимообусловленность их изменений в эволю-
ции. В «Происхождении видов» Ч. Дарвин писал, что «...для нас
несущественно, изменяются ли сперва привычки, а затем строение
органов...». Примеры, подтверждающие данное положение на жи-
вотных, описал сам автор и другие исследователи. Взаимосвязь
формы и функции у животных и растений представлены на сх. 4.
30
Заметим, что все эти возможности встречаются и у растений;
соответствующие примеры приведены ниже. Однако в ботаниче-
ской литературе вопросу о взаимоотношении формы и функции
уделяют мало внимания. Между тем растительное царство может
дать не менее убедительные примеры взаимообусловленности из-
менения формы и функции в ходе эволюции. Выход растений на
сушу, как известно, ознаменовался приобретением комплекса мор-
фофизиологических новшеств: защитных покровов (эпидермы й
кутикулы), проводящей системы, дифференциации тела на органы
и т. д. Подобными изменениями прежде всего было достигнуто
уменьшение потери воды от испарения и усиление ее движения по
растению. В приведенных примерах трудно сказать' что чему пред-
шествовало, морфологические или физиологические изменения,
хотя, несомненно, характер первых определял состояние организма
в новых условиях. В то же время очевидно, что «заказ» на умень-
шение отрицательных последствий недостатка воды повлек за собой
отбор растений на развитие защитных покровов и проводящей
системы в наземных условиях.
Схема 4
Цепь событий следовала одна за другой. Развитие непрерывного
слоя кутикулы должно было отрицательно сказаться на фотосинте-
зе, так как препятствовало газообмену растений со средой,, поэтому
необходимо было обеспечить газообмен и при развитии кутикулы.
Это достигается формированием устьица из двух клеток с наруж-
ными тонкими и внутренними толстыми стенками, что ведет к
изменению просвета щели между ними в зависимости от их тургор-
ного давления — открыванию и закрыванию устьиц. Хотя форма
Устьиц в ходе эволюции подвергалась изменениям (известны 10
типов устьиц по строению), принципиальный механизм их регуля-
31
ции остается неизменным у всех наземных растений (включая и
механизм поступления воды в клетки устьиц). В ходе эволюции
растений на суше менялось несколько и строение самого устьичного
аппарата (по мере аридизации климата) в сторону неравномерного
увеличения толщины стенок и возникновения побочных клеток.
При отсутствии необходимости устьичные клетки редуцируются
(риниофиты, растения-паразиты, подводные листья). Из 10 извест-
ных типов устьичного аппарата наиболее древним признан анемо-
цитный, который конвергентно может появляться и на поздних
этапах эволюции (Ал. А. Федоров, 1978).
Некоторые наиболее древним считают пароцитный тип устьиц.
Устьичный аппарат отличается полиморфизмом в пределах се-
мейств, рода и даже у отдельного растения. Так, у амарантовых
описаны почти все типы устьичного аппарата, характерные для
цветковых. Местоположение устьиц на растении (побег, верхняя и
нижняя поверхности листовой пластинки) также влияет на струк-
турный тип устьиц. Вероятно, в функциональном отношении все
типы устьиц довольно сходны, хотя их разнообразие обусловлено
особенностями замыкающих клеток и листовой пластинки. Быть
может, в этом также заключается один из парадоксов изменения
формы, на что обращают внимание С. В. Мейен, 1988; В. А. Кра-
силов, 1988.
Развитие устьиц в ходе эволюции не привело бы к заметным
успехам в борьбе за существование, если бы оно не шло параллельно
с усилением проводящей функции растений. Поэтому с повышением
роли устьиц в жизни растений происходит постепенное развитие
активности проведения воды вверх по стеблю в сторону листьев.
Как раз с выходом растений на сушу возникает необходимость
уменьшения потерь воды и усиления ее движения. В этом направ-
лении совершенствовался проводящий аппарат: выделялись сосуды
и трахеиды, возникала поровость, повышалась роль мертвых клеток
в восходящем токе и т. д. Без таких изменений развитие устьиц
сыграло бы отрицательную роль в водообмене растений. Сосуды и
их членики стали неотъемлемой частью ксилемы только у цветко-
вых, хотя первые из них были бессосудистыми. Ксилема состояла
из трахеид лестничного типа. С увеличением сухости воздуха во
флоре снижается процент растений с сосудами лестничной перфо-
рации. В результате усовершенствования проводящей системы в
нескольких направлениях скорость ксилемного транспорта дости-
гает более 10 м/ч, а флоэмного — 1 м/ч.
Такая скорость ксилемного транспорта не могла быть обеспе-
чена проведением воды и растворенных веществ по живым клеткам,
что сопряжено с большими энергетическими затратами. Поэтому в
ходе эволюции растений наблюдается постепенное удлинение пути
по мертвым клеткам. Как раз с возникновением трахеид, особенно
32
сосудов (наземные растения), с образованием сосудов лестничной
перегородки усиливается функция передвижения воды и веществ,
обеспечивается возможность переброски их на значительные рас-
стояния.
У первых цветковых описаны лишь трахеиды лестничного типа.
Несмотря на разнообразие сосудов у цветковых, они образуют
морфологический ряд перехода от члеников с многочисленными
лестничными перфорациями до члеников с простыми перфораци-
ями на поперечных стенках. Специализация сосудов и соответст-
венно усиление их проводящей функции в ходе эволюций
покрытосеменных были связаны с уменьшением длины, утолщени-
ем стенок, расширением просвета, округлением пор, возникнове-
нием поперечной стенки, уменьшением числа перекладин и
увеличением ширины сквозных отверстий сосудов. Эти и другие
структурные изменения привели к уменьшению сопротивления току
жидкости и обеспечили ее подъем на большую высоту.
В течение эволюции совершился переход ситовидных клеток
(богатых порами) в членики ситовидных трубок. В результате было
достигнуто усиление флоэмного транспорта. Элементы, аналогич-
ные флоэмным, появляются уже у некоторых водорослей. Однако
с выделением удлиненных клеток, снабженных ситовидными от-
верстиями, способность к проведению органических веществ уси-
ливается. Дальнейшая интенсификация флоэмного транспорта
оказалась возможной благодаря разрушению ядра у ситовидных
клеток. Последнее имело бы отрицательные функциональные по-
следствия, если бы ситовидные клетки были лишены клеток-
спутниц, отличающихся интенсивной жизнедеятельностью.
Отрицательные последствия потери ядра компенсируются усилени-
ем взаимодействия ситовидных клеток и их спутниц (рис. 4).
Различные виды и таксоны растений отличаются по уровню
организации проводящей системы.
Как указывает А. Л. Курсанов(1976),
различия в проводящей системе раз-
ных таксонов не всегда правомерно
рассматривать как промежуточные
этапы общей дифференциации, ско-
рее они представляют «отдельные
ветви эволюционного древа». Автор
так подытоживает представления об
эволюции системы дальнего транс-
порта в растительном мире: «Вероят-
но, исходным было передвижение
веществ в недифференцированных Рис 4 Стадии эволюции сито-
клетках с помощью конвекционных видной пластинки от сложной (А)
токов цитоплазмы, что явилось суще- к простой (Б) (по к. Эсау, 1980)
2-157
33
ственным шагом вперед по сравнению с диффузией и что и теперь
еще часто остается основным механизмом внутриклеточного пере-
распределения веществ- Однако для осуществления транспорта на
далекие расстояния этот процесс должен был быть еще более
ускорен, что и привело к возникновению экстраподвижности в виде
той, которую можно наблюдать в клетке ацетабулярии, в пыльцевой
трубке или в корневом волоске».
Сказанное можно подытожить так: у одноклеточных водорослей
транспорт веществ ограничен обменом между клеткой и средой, а
также между органеллами. У многоклеточных низших растений
появляются межклеточные потоки веществ, приводящие к трофи-
ческой интеграции тканей. У высших сосудистых формируются
механизмы быстрого и однонаправленного движения веществ меж-
ду органами. Такое эволюционное усложнение транспорта веществ
связано с возникновением специализированных структур для обес-
печения потока веществ—симпласта, апопласта, плазмодесм и
переходных клеток (д ля перекачки веществ из апопласта в симпласт
и обратно). В эволюционном ряду от мхов до цветковых растений
симпласт усложняется образованием ситовидных трубок, а апопласт
упрощается путем образования мертвых сосудов (J. МоогЬу, 1981).
В связи с этим объяснимы причины различий растений по длине
мелких жилок на листьях. Так, при площади листа дыни 1,68 дм2
. их длина составляет примерно 115 м. Изучение около 730 видов
растений показало, что переход от деревьев к травам сопровождается
изменением плазмодесменных связей между клетками мезофилла
и пучка, увеличением числа поглощающих клеток, разным сочета-
нием симпластного и апопластного путей (Ю. В. Гамалей, 1994).
Соответствующие структурные различия способствуют усилению
загрузки флоэмы и изменению состава флоэмного экссудата.
Ухудшение водообмена растений в связи с аридизацией климата
Земли в поздних геологических эпохах потребовало изменения
структуры уже самого организма как целостной системы для вос-
становления равновесия между поступлением и отдачей воды. Уже
у однолетних трав преобладает подземная (всасывающая) поверх-
ность над надземной (испаряющей). Только такой тип организации
индивидуума мог обеспечить жизнедеятельность растений в новых
условиях даже при наличии совершенной проводящей системы
(В. О. Казарян, 1990).
Связь формы и функции в ходе эволюции растений можно
продемонстрировать и на других примерах. Заслуживают упомина-
ния молекулярный и клеточный уровни. Наличие сложной внутренней
организации клетки и органелл (многочисленных крист, гран,
тилакоидов и мембран) обеспечивает непрерывность протекания
энергетических и синтетических процессов. Причем и в онтогенезе
по мере усложнения внутренней организации органелл их функци-
34
ональные возможности постепенно усиливаются. На примере хими-
ческой изомерии биологических молекул (еще со времен Л. Пастера)
показана возможность значительного изменения функциональной
активности молекул. У молекулы одного и того же фермента может
существовать несколько изомеров, отличающихся по некоторым
деталям строения («правые» и «левые» формы) и физиологической
активности.
Для дальнейшего углубления рассматриваемого вопроса пред-
ставляют интерес данные об изменении симпластических контактов
в онто- и филогенезе растений (Ю. В. Гамалей, 1994; М. В. Пахо-
мова, 1981). Специализация плазмодесм и плотность их распреде-
ления возрастают от мезофилла к ситовидным элементам. У бурых
водорослей развитие симпластических связей ограничено форми-
рованием плазмодесменного поля, у голосеменных и мхов — фор-
мированием ситовидных полей, у покрытосеменных — настоящих
ситовидных пластинок.
На примере эволюции корня также показано, что усйление его
функций шло параллельно нарастанию внутренней дифференциа-
ции и гетерогенности клеток верхушки, образованию многоклеточ-
ной паренхимы, системы межклетников, поясков Каспари
(пересекающих радиальные и поперечные стенки эндодермы) и т. д.
(М. Ф. Данилова, 1974). С усилением гетерогенности клеток вер-
хушки корня возрастала избирательность поглощения минеральных
элементов.
Особенности транспорта ионов в растениях определяются специ-
фикой структур, что прослеживается в иерархическом ряду от архи-
тектуры биологических мембран до организации ценоза (Д. Кларксон,
1978). В растениях имеются механизмы, контролирующие внутрен-
нюю ионную среду (меняющие молекулярную организацию мемб-
раны), взаимосвязь между ее элементами и метаболическим
аппаратом клетки и структурной организацией самого растения/
Соответствие структуры и функции видно и на примере расте-
ний с С3- и С4-путями фотосинтеза, подробное описание которых
дано в гл. 3. Здесь же отметим, что С4-путь имеет специфические
структурные особенности, пространственное и временное разделе-
ние мест первичной фиксации СО2 н его дальнейшее вовлечение в
Фотосинтез (рис. 5).
Не только в филогенезе, но и в онтогенезе есть соответствие
Между структурой хлоропластов и интенсивностью фиксирования
в Них СО2. Вообще адаптации растений к новому световому режиму
в онтогенезе сопровождаются перестройкой листа на тканевом и
клеточном уровнях, числа пластид и т. п. (А. Т. Мокроносов, 1981;
А. Шахов, 1993). Отдельные элементы мезоструктуры фотосин-
тетического аппарата взаимосвязаны в онтогенезе. Хотя строение
35
Рис. 5. Схема С4-пуги фотосинтеза (Дж. Эдвардс, Д. Уокер,
1986.С. 313)
ФА теснр коррелирует с его функцией (Дж. Эдвардс, Д. Уокер,
1986), не всякие структурные и функциональные изменения возни-
кают одновременно и оказываются выгодными для их носителя.
Более того, возникающие в ходе эволюции и сальтационно крупные
макромутации первично не всегда оказывались функционально
более значимыми, чем имеющиеся. Однако отбор часто «попусти-
тельствовал» их фиксации ввиду нейтральности или слабой адап-
тивной значимости. В макроэволюции высших растений
морфогенез и семифилогения (онто- и филогенез) структур часто
предшествовали функциональному совершенствованию индивиду-
умов и популяций (С. В. Мейен, 1987).
Обращалось внимание и на неполное соответствие плана орга-
низации растений принципу адаптивности. Так, К. Уордлоу (С.
Wordlaw, 1965) приводит случаи, когда развитие прилистников,
зазубренность пластинки и неравномерность ее окраски, утолщен-
ность черешка и сложное строение цветка не всегда отвечают
указанному принципу.
Удивительны примеры взаимосвязи формы и функции для
биологических молекул. Так, для гиббереллинов показано, что
изменение конфигурации и степени насыщения тетрациклической
молекулы (метильными и карбоксильными группами) в разных
положениях резко меняет их физиологическую активность. Число
гиббереллинов, описанных по этому принципу, превышает 60.
Некоторые из них характерны только для . грибов, другие—для
высших растений, третьи —для тех и других. Структура (или орга-
низация) должна быть признана характернейшей чертой живого (А-
Сент-Дьерди), как и функция. В то же время функциональные
особенности имеют некоторые опережающие возможности для
36
изменения. Так, на примере системы биосинтеза хлорофилла про-
демонстрирована возможность регуляции размеров фотосинтети-
ческой единицы в зависимости от условий —г состояния
функционирования самой молекулы.
Один из примеров влияния функциональных преобразований
на структуру органов—укорененные листья.-Их черешки после
длительного самостоятельного культивирования приобретают сход-
ство по строению со стеблем, в них усиливаются проводящая и
механическая функции (Н. И. Дубровицкая).
Структура и функция — неотъемлемые свойства живой приро-
ды, они взаимосвязаны в онто- и филогенезе. Любой орган обладает
разнообразием функций. Если из множества функций корня (про-
ведение веществ или их запасание, образование придаточных почек,
прикрепление, синтез и т. п.) одна получит преимущественное
развитие, то строение корня в филогенезе изменится сообразно этой
функции. В то же время отбор растений может происходить по
количественному выражению одной и той же функции. Такие
изменения способствуют изменению и структуры органа.
Соответствие строения органа его функциям не следует пони-
мать узко (в отношении любых морфологических особенностей),
оно относится к системе органов и организму в целом. Теория
отбора не исходит из признания пользы любых особенностей
организации.
Отбор способствует возникновению фенотипов, у которых до-
стигается использование первичных механизмов какого-либо про-
цесса в адаптивных для них структурах. С переходом на другой
структурный уровень происходит усиление процесса при сохране-
нии первичных механизмов (сх. 5).
Схема 5
ДИФФЕРЕНЦИАЦИЯ ТЕЛА У РАСТЕНИЙ
t
СПЕЦИАЛИЗАЦИЯ СТРУКТУР ( ОРГАН)
t
ВЗАИМОДЕЙСТВИЕ КЛЕТОК (ТКАНЬ)
t
ЭУКАРИОТИЧЕСКАЯ КЛЕТКА
f .
ПЕРВИЧНЫЕ МЕХАНИЗМЫ (ПРОКАРИОТЫ)
В итоге наблюдается заметная функциональная дивергенция
Растений, что также зависит от таксономического и экологического
Рангов вида. Значение подобной дивергенции для эволюции расте-
1цЙ исключительно.
37
Проблема взаимосвязи структуры и функции должна быть
разработана применительно к разным уровням живой природы.
Так, в основе фотосинтеза лежат общие физико-химические
механизмы и реакции, протекающие на мембранах. Однако в ходе
эволюции фотосинтез менялся на уровне организма, популяции
и растительности в целом (см. гл. 3). Отсюда и изменения
структуры, и функции фотосинтетического аппарата на разных
уровнях его организации возникали в разное время и с помощью
различных механизмов.
В рамках конкретного физиологического проявления необходи-
мо рассматривать его эволюцию на всех уровнях организации.
Однако физиологи растений традиционно взаимосвязь структуры
и функции ограничивают организменным и клеточным уровнями,
так как организм представляет основной предмет их интереса. В то
же время очевидно, что этим не может быть исчерпан сложный
вопрос о возникновении того или иного физиологического явления.
Следовательно, для понимания глобального значения «раститель-
ной жизни» необходимо исследовать ее основные параметры с точки
зрения их реализации на всех уровнях организации жизни.
Фотосинтез возникает в результате эволюции надорганизмен-
ных уровней для выполнения растениями космической функции.
Из этих соображений исходят при оценке роли тех или иных
организмов на Земле. Функция фотосинтеза на уровне надорганиз-
менных образований видна при оценке общей биомассы, ежегодной
и удельной продуктивности фитопланктона и фитобентоса в Ми-
ровом океане. Половина от ежегодной продукции кислорода рас-
тениями (3,2 • 10*’ т) приходится на Мировой океан. Для создания
современного фонда кислорода биосферы при этом водорослям
понадобилось бы 3700 лет (А. И. Перельман, 1979). Что прежде
всего определяет эволюционное назначение фотосинтеза в поддер-
жании жизни на планете.
Только с этих позиций проясняются некоторые факты измене-
ния структуры и функции организма и в онтогенезе. Так, мы знаем
о падении обменных (основных) процессов растений по мере
старения организма и органа, т. е. с увеличением общей массы тела,
всех его структурных компонентов.
На примере популяций Cyctoseira crinita показано, что молодые
особи в мелководьях и условиях высокой подвижности воды обес-
печивают поток 1800 г углерода/м2 дна, тогда как взрослые особи,
обитающие на большой глубине и при спокойной воде,— 200—300 г
углерода/м2 дна (К. М. Хайлов, р. П. Парчевский, 1983). Причины
таких изменений могут быть различными, в том числе и как
проявление в онтогенезе филогенетических тенденций развития
(см. гл. 8).
38
Влияние структуры на функцию не всегда однозначно. Так,
большое количество ДНК в ядре значимо для продолжительности
митоза, мейоза и жизненного цикла. Однако было бы неверно
связывать содержание ДНК в ядре только с указанными событиями.
2.5. ОСНОВНЫЕ НАПРАВЛЕНИЯ И ФОРМЫ
ФУНКЦИОНАЛЬНОЙ ЭВОЛЮЦИИ РАСТЕНИЙ
Для понимания сложной картины функциональной эволюции
растений важно выяснить ее направления. Это даст возможность
сопоставить данные об их изменении с направлением морфологи-
ческой эволюции. Направления эволюционного процесса связаны
или с повышением организации и обшей энергии жизнедеятельно-
сти организма, приводящего к расширению ареала группы с выхо-
дом в новую адаптивную зону (орогенез), или с развитием ее в
прежней зоне (аллогенез), но с резким расширением территории и
увеличением многообразия форм при сохранении общих особенно-
стей строения и функции.
' К приобретениям типа арогенеза у растений следует отнести
механизмы фотосинтеза и аэробного дыхания, формирование ядра
и многоклеточности, дифференциацию тела на органы, развитие
проводящей системы, покровов тела и устьичного аппарата в на-
земных условиях, переход, к гормональной регуляции процессов
роста и развития и т. д. Каждое из указанных приобретений спо-
собствовало выходу растений в новую адаптивную зону и процве-
танию эволюционирующей группы.
Вслед за подобными изменениями возникали и функциональ-
ные аллогенезы: возникновение разных экологических типов рас-
тений по использованию механизмов фотосинтеза и дыхания,
различие растений в транспирации, по строению и числу хлоро-
пластов и митохондрий, изменение типа листорасположения и
формы листовой пластинки, отличия видов по строению устьичного
аппарата и проводящей системы, специфические формы движения
и питания растений, изменение фотопериодической реакции и т.
п. Вследствие перечисленных приобретений растения могут полно
и разносторонне использовать адаптивную зону, сохраняя метабо-
лические механизмы. Крайнее выражение аллогенеза — специали-
зация, которая широко распространена на функциональном уровне
(С4- и CAM-типы ассимиляции углерода, насекомоядность, сукку-
ленты, галофиты, эфемеры и т. п.). После каждого функциональ-
ного арогенеза шел поиск возможностей для наиболее полного и
разнообразного его испытания в борьбе за существование (аллоге-
нез). Так что значение обоих направлений для возникновения
39
разнообразия растений нельзя умалять, хотя они и не равнозначны
в эволюционных масштабах.
Функциональная эволюция растений, как и морфологическая,
проявляется в различных формах. Так, когда говорят о филетиче-
ской эволюции, имеют в виду постепенное изменение и превраще-
ние одного вида в другой. Одним из ее примеров на
функциональном уровне следует признать возникновение аэробно-
го дыхания, а также преобразования в ряду хлорофилл —цито-
хром — ферредоксин.
Широко распространены на функциональном уровне и такие
формы эволюции, как дивергенция, конвергенция и параллелизм.
Дивергенция — это формирование многообразия в живой при-
роде под действием отбора. Дивергенция хорошо изучена на при-
мере морфологических признаков. В отношении физиологических
особенностей она также проявляется, хотя есть и кажущиеся про-
тиворечия. Например, с момента возникновения фотосинтеза не
отмечены существенные изменения в его основных реакциях. Точно
так же мало изменились механизмы аэробного дыхания и ряда
других процессов (см. гл. 4, 5). Как будет показано в гл. 3,
дивергенция растений по фототрофному питанию идет в двух
основных направлениях: усовершенствование аппарата фотосинте-
за и поиск экологических возможностей для большего накопления
ассимилятов. Отбор в указанных направлениях привел к диверген-
ции видов по эффективному сочетанию ростовых процессов и
способов фиксации С02 (А. А. Ничипорович, 1980). Такие после-
дующие изменения, как различия в числе устьиц и строении пучков,
в величине и плоидности клеток палисадной ткани и размерах
листовой пластинки (W. Gottschalk, 1976), также имели непосред-
ственное отношение к дивергенции видов по интенсивности фото-
синтеза и аэробного дыхания. Кроме того, используются и другие
возможности для стабилизации механизмов энергетики, возникших
на начальных этапах развития растений. Таким путем движущий
отбор действовал в сторону усиления экологической дивергенции
растений по эффективному использованию процессов. Поэтому на
основе одних и тех же биохимических механизмов достигаются
различия в их физиологической эффективности (М. Флоркен,
1947).
В отношении отдельных свойств можно даже вычислить темпы
дивергенции видов. Так, устойчивость подсолнечника к заразихе и
агрессивность последней заметно изменились за истекшие 80 лет.
Ныне наблюдается широкая дивергенция видов и популяций рас-
тений по устойчивости к промышленным выбросам. Вокруг про-
мышленных городов происходит смена растительности и выживают
40
преимущественно низкорослые кустарники или деревья с мелкими
листьями.
Параллелизм — независимое возникновение одинаковых при-
знаков и свойств у генетически близких видов. Н. И. Вавилов (1967)
подчеркивал, что чем блйже друг к другу виды, тем чаще прояв-
ляется параллелизм у растений; речь идет о сходстве их измен-
чивости и эволюции. Примеры параллелизма, связанные со
сходной устойчивостью к грибным заболеваниям (иммунитет),
были описаны Н. И. Вавиловым для хлебных злаков. Теперь они
широко известны и для других культур.
Напомним, что скороспелые формы встречаются в пределах
разных сортов и видов. Часто один и тот же результат — скороспе-
лость и засухоустойчивость —может быть обусловлен мутациями,
затрагивающими разные этапы онтогенеза. Представляют интерес
примеры параллелизма у разных органов одного и того же растения.
Так, антбцианы, определяющие окраску цветков (Р. Вагнер, Г.
Митчел, 1958; Б. М. Медников, 1980), образуются благодаря слож-
ным биохимическим превращениям. Оказывается, такие же пре-
вращения происходят в листьях, клубнях, плодах и побегах.
Параллелизм в изменении окраски перечисленных органов возни-
кает независимо и на разных этапах онтогенеза в силу их генети-
ческой общности.
Параллелизм в пределах вида, рода и семейства установлен по
таким признакам, как озимость и яровость, гидрофильность и
ксерофильность, устойчивость к холоду, содержание белка, продук-
тивность фотосинтеза и т. д. Н. И. Вавилов выявил параллелизм
даже между семействами и типами по «изменчивости как морфо-
логических, так и физиологических признаков».
Напомним, что древовидные формы встречаются в пределах
разных семейств покрытосеменных. Сходный ряд наследственной
изменчивости у генетически далеких форм Н. И. Вавилов назвал
аналогичным.
Конвергенция.— это явление схождения признаков у далеких
форм. Вопрос о конвергенции (схождении) функциональных осо-
бенностей растений изучен слабо. Определенно можно сказать, что
биохимия и физиология дают не меньше фактов о конвергенции
видов, чем морфология растений. На примере изучения состава
алкалоидов, эфирных масел, гормонов и т. д. выявлена конверген-
ция систематически далеких видов растений (см. § 5.5).
Функциональная конвергенция растений показана и на приме-
ре способности к движениям и насекомоядности (Ч. Дарвин, 1941;
С. П. Костычев, 1922; В. Е. Tuniper et al., 1989). Н. М. Гайдуков
(1926) описал примеры конвергенции семян и плодов по морфоло-
гическим признакам и биологическим свойствам, морфологии и
41
физиологии у молочайных и кактусовых, обитающих в одйнаковых
условиях. Точно так же в сходных условйях, например при переходе
к паразитизму у зеленых растений, стоящих на разных ступенях
филогенетического развития, отмечена постепенная утрата хлоро-
филлоносного аппарата, корней, функции фотосинтезаи, напротив,
усиление активности протеолитических ферментов. Указанные чер-
ты особенно ярко выступают у специализированных цветковых
паразитов (Cuscutaceae, Orobanchaceae).
Заслуживает внимания, что насекомоядные растения и живо-
тные обнаруживают одинаковые способности к расщеплению бел-
ков животного происхождения (Л. С. Берг, 1977). Установлены
также черты сходства в энергетическом обмене у болотных растений
и ныряющих животных (Р. М. Кроуфорд, 1981). Близок по строе-
нию и функции сократительный белок мышцы животных и подушки
листа мимозы. Концентрацией данного белка определяется способ-
ность листьев мимозы к раздражимости. В подушках листьев ми-
мозы локализованы Са2+- и Mg^-АТФазы, сходные с АТФазами
мышц и немышечных подвижных клеток животных (М. Н. Люби-
мова-Энгельгард и др., 1981). Актиноподобный белсж встречается
в проводящих пучках и во флоэме высших растений, в цитоплазме
у простейших и в плазмодии миксомицет. В немышечных клетках
актиноподобный белок принимает участие в создании сети микро-
филаментов, которая, взаимодействуя с миозинподобными белка-
ми, обеспечивает подвижность клеточных структур и цитоплазмы,
светозависимое перемещение хлоропластов.
Изучение роли белков в защитных реакциях растений против
фитовирусов показало их сходство с интерферонами животных по
вирусоспецифичности, молекулярной массе, способности сущест-
вовать в мономерных и полимерных формах. Причем индукторы
интерферонов животных в тканях растений вызывают образование
антивирусного белка, равно как фитовирусы, введенные в ткани
животных, индуцируют синтез интерферона. Здесь наглядна кон-
вергенция механизмов защитных функций на основе далекой об-
щности происхождения животных и растений.
Одним из примеров конвергенции может служить сходство защит-
ных веществ у растений и животных. Так, грибные полисахариды с
(3-связями играют индукторную роль в защитных реакциях не только
растений, но и теплокровных животных и человека (Л. В. Метлицкий,
О. Л. Озерецковская, 1985). Грибные циклические пептиды (гщкло-
спорионы) подавляют защитные реакции у высших животных.
Бесспорные факты функциональной конвергенции у растений
описаны на примере С4-пути фотосинтеза у представителей далеких
таксонов в пределах одно- и двудольных растений. Дивергенция
С4-растений проявляется по строению обкладки пучка и первичным
продуктам запасания СО2. Однако принцип пространственного
42
разделения механизма первичного запасания СО2 и его дальнейшего
вовлечения в фотосинтетический метаболизм остается общим при
конвергенции видов по С4-пути как результат отбора растений в
определенных экологических условиях (см. гл. 3).
Есть и другие примерь!. Так, Г. Балтчевским (см.: Г. Деборин и
др., 1975) показана общность элекгрон-транспортных белков у
животных и растений. Сходны строение и функция зрительного
ретинала (родопсина) и бактериородопсина с каротиноидами. В
частности, благодаря бактериородопсину Halobacterium halobium,
живущая в условиях соленой рапы, осуществляет «бесхлорофилль-
ный» фотосинтез. У растений встречаются ацетилхолин и гамма-
аминомасляная кислота —вещества, выполняющие у животных
роль нервных регуляторов.
Можно привести и другие примеры функциональной конвер-
генции. Так, устойчивость к неблагоприятным условиям (засуха,
затопление, заморозки и т. д.) характерна для представителей ни-
зших и высших, голосеменных и покрытосеменных, однодольных
и двудольных растений. В ряде случаев конвергенция наблюдается
даже в отношении одинаковых реакций, обусловливающих устой-
чивость к недостатку воды в почве: сосущей силы, осмотического
давления, интенсивности транспирации. Конвергенцию ошибочно
пытались объяснить вопреки механизму действия естественного
отбора (Л. С. Берг, 1977). Однако только направленность отбора в
близких условиях способствует конвергенции видов (Ч. Дарвин,
1939). Экологическая экспансия видов приводит к захвату сходных
ниш представителями систематически далеких форм. Это и способ-
ствует их функциональной конвергенции.
Все еще делаются попытки объяснить явления конвергенции
и параллелизмов вне механизма действия естественного отбора,
опираясь на палеонтологические, сравнительно-морфологиче-
ские (А. Б. Иваницкий, 1977; В. А, Кордюм, 1982; А. А. Любы-
шев, С. В. Мейен, 1979, 1988) и молекулярно-генетические
данные (Л. И. Корочкин, 1985, 1991). В этом же плане следует
рассматривать и гипотезу автоэволюции (А. Лима де Фариа, 1991),
где факты конвергентного и параллельного развития формы и
функции представлены как результат реализации общих закономер-
ностей неживой природы. В принципе нельзя отрицать наличие
аналогии между организацией строения стебля растений и минера-
лов (А. Лима де Фариа, 1991), проявление общих закономерностей
развития неживой природы, как в случае накопления и синтеза
веществ вторичного метаболизма (М. Н. Запрометов, 1988, 1993;
М. Е. Лоткова, 1981; М. Лукнер, 1979; В. А Пасешниченко, 1991;
К. Mothes, 1981), при схождении физиологических особенностей у
генетически неродственных форм (Т. К. Горышина, 1989; W. Lar-
43
cher, 1980; W. V. Zucher, 1983) и пигментов у растений и животных
(D. Fox, 1979). В то же время подобные факты нельзя понять вне
механизма действия естественного отбора или при ограничении его
роли «доработкой» новшеств. Явления конвергенции и параллелиз-
мов в дарвинизме получили свое объяснение (К. М. Завадский и
Э. И. Колчинский, 1977; А. С. Северцов, 1990; Т. Я. Сутг, 1977;
Л. П. Татаринов, 1988), что в равной степени относится и к функ-
циональному уровню.
2.6. ПРИНЦИПЫ ПРЕОБРАЗОВАНИЯ ФУНКЦИЙ
В ХОДЕ ЭВОЛЮЦИИ РАСТЕНИЙ
Способность приспособительного реагирования на условия не
является универсальным свойством живой системы. Отсюда в задачи
эволюционной биологии входит изучение механизмов возникновения
приспособлений, среди которых для растений наиболее типичны
функциональные проявления (Ю. Сакс, К. Гебель, Г. Клебс, К. А.
Тимирязев). Бесспорно, указанная проблема лучше разработана при-
менительно к животным, однако она давно обсуждается и в физиоло-
гии растений, хотя вопрос о способах (модусах) филогенетического
преобразования органов и функций растений еще слабо разработан.
Применительно к функциональным, явлениям выделены следу-
ющие количественные закономерности приспособительного реаги-
рования организмов на изменение среды (Г. Ф. Гаузе, 1984):
неизменность функций (сохранение важных функций при измене-
нии второстепенных), выключение функций (ингибирование фер-
мента при отсутствии субстрата), усиление функций, сочетание
усиленных функций между собой. Все это в какой-то мере прояв-
ляется и у растений (К. В. -Манойленко, 1984). Однако для них в
первую очередь можно выделить такие модусы, как множествен-
ность реализации, усиление или ослабление, компенсация, стаби-
лизация и автономизация функций, перекомпоновка функций.
Множественность реализаций функций. Это предпосылка фило-
генетического преобразования функциональных особенностей.
Здесь, как и в случае преобразования органов в филогенезе, про-
являются характерные для любой структуры мультифункциональ-
ность и способность любой функции изменяться количественно
(А. Н. Северцов, 1939). По мнению А. М. Голдовского (1975), в
пределах каждой группы веществ существует целый ряд близких по
строению и функции соединений. Подобные наблюдения легли в
основу сформулированного им закона множественности или мно-
говариантности протекания реакций.
Множественность реализации функций подтверждается: 1) воз-
можностью изменения первичных и вторичных продуктов метабо-
44
лизма; 2) участием неродственных веществ в регуляции одного и
того же явления (например, рост и развитие зависят от комплекса
энергетически и физиологически активных веществ); 3) индуциро-
ванием одной и той же реакции разными воздействиями (например,
развитию ксерофильной структуры способствуют засуха, интенсив-
ность освещения, некоторые химические агенты и т. д.).
Множественность реализации функций свидетельствует прежде
всего о сложности процессов жизнедеятельности растений, она
способствует повышению надежности онтогенеза за счет расшире-
ния нормы реагирования организма. Например, наличие разных
ферментов, катализирующих одну и ту же реакцию (Fe- и Си-про-
теины, флавопротеины и т. д.), заметно повышает интенсивность
дыхания в благоприятных условиях (Б. А. Рубин, М. Е. Ладыгина,
1974). Точно так же в качестве приспособления для улавливания
света В толще водоема возникает разнообразие пигментов у бактерий
и водорослей (А Клюйвер, К. Ван-Ниль, 1959; Е. Н. Кондратьева,
1972). Поэтому продукты метаболизма, состав пигментов и способы
дыхания у бактерий и водорослей оказываются более разнообраз-
ными, чем у цветковых. Усиление специализации метаболических
звеньев имеет значение для интенсификации процессов жизнедея-
тельности,
Блочный принцип эволюции или перекомпоновка функций. Для
блочного принципа эволюции функций (А. М. Уголев, 1985) имеет
значение мультифункциональность на уровне макромолекул, вы-
ступающая в виде их мультипотенциальности. Специализация мо-
лекул приводит к уменьшению их мультипотенциальности и
интенсификации функций. Все это проявляется на молекулярном
клеточном и тканевом уровнях.
Любая сложная функция возникает из менее простых компо-
нентов. Так, сложные процессы живых организмов складываются
из сочетания особенностей элементарных частиц, химических эле-
ментов и веществ, минералов (А. Лима де Фариа, 1991). Например,
на основе способности красящих веществ абиогенного происхож-
дения (пигментов) на свету поглощать СО2 и выделять О2 с возник-
новением живых существ складываются их фотохимические
процессы, в том числе и фотосинтез. Вся специфика живых орга-
низмов есть ничто иное, как объединение в комплексы молекул с
разным сочетанием без потери индивидуальности каждой из них
(М. Эйген, 1973; С. Э. Шноль, 1979).
Подобная «химическая предопределенность» и способствует
становлению жизни. Отбор в ходе предбиологической эволюции
Шел по принципу максимального повышения каталитической ак-
тивности не только макромолекул, но и эффективности сочетания
их в системах путем усиления пространственно-временной упоря-
45
доченности удачных сочетаний (В. А. Карасев, В. Е. Стефанов,
1991). Сочетание структур с мультифункциональными особенно-
стями —предпосылка многих физиологических явлений.
Вероятно, первично и система синтеза белка происходила на
основе простых каталитических систем (Р. Фокс, 1992). С обра-
зованием нуклеопротеинов усиливаются специфичность синтеза
белков и разнообразие их функций благодаря отбору. Так, обра-
зование молочной кислоты и в бактериях, и в мышцах человека
имеет одинаковый механизм, но роль молочной кислоты меня-
ется (Дж. Мецлер, 1980).
Усиление или ослабление функций имеет место при становлении
механизмов фотосинтеза и дыхания. Не менее убедительные данные
дает изучение эволюции процессов передвижения веществ (рис. 6).
При переходе к многоклеточной организации усиление транспорта
веществ достигается с помощью разных механизмов и развития
новых структур (К. Эсау, 1969; А. Л. Курсанов, 1976).
Тенденцию усиления функций можно проследить и на примере
механизмов водоотдачи. Первичноводные растения не обладали
сколько-нибудь развитой системой активного испарения. Излиш-
няя жидкость, накапливающаяся в организме, выводилась у них со
всей поверхности или через специальные поры —гидатоды, как у
некоторых современных наземных растений. С выходом на сушу (при
высокой влажности воздуха) у растений усиливается кутикулярная, а
в связи с аридизацией климата — устьичная транспирация. Такую же
смену способов транспирации проследил П. А. Генкель в онтоге-
незе типичного наземного растения как выражение рекапитуляции
функций.
Постепенное усиление функций можно продемонстрировать и
на примере эволюции способности к движениям у растений. В
элементарной форме она присуща всем растениям, а в особых
экологических условиях (борьба за влагу, свет, аэрацию и т. д.) эта
способность была усилена отбором в одном из следующих направ-
лений: круговые вращения стебля, лазание с помощью усиков,
листьев и т. д.
Усиление главной функции часто достигается увеличением чис-
ла одноименных компонентов, участвующих в ее осуществлении.
Например, интенсивность фотосинтеза зависит от количества хло-
ропластов и содержания в них хлорофилла, дыхание — от числа
митохондрий, рост — от числа и размеров клеток и концентрации
гормонов.
В процессе эволюции нередко наблюдается и ослабление фун-
кций. Выше было показано ослабление или полная утрата фото-
синтетической активности у цветковых полупаразитов и паразитов
(рис. 6, II). Другой пример —снижение интенсивности транспира-
ции у суккулентов и некоторых ксерофитов, ослабление способно-
46
Несосудистые
растения
Многоклеточное
водоросли
Взаимодействие
клеток
Взаимодействие
органелл
Рис. 6. Экологический ряд усиления (/) и ослабления (/7)
функций у растений
сти к движениям у цветковых даже в пределах одной и той же
систематической труппы, например у разных тыквенных.
Механизмы и причины ослабления функций не изучены. Не
исключен отбор растений по обратным мутациям (структурным и
функциональным) или выпадение какого-либо звена из сложной
цепи метаболизма. Например, ослабление транспирации у сукку-
лентов обусловлено подавлением механизма устьичных движений.
Явления усиления и ослабления функций тесно связаны с процес-
сами компенсации.
47
Компенсация функций. Известно много фактов, когда функция
редуцированного органа не утрачивается, а выполняется другими
структурами. Так, редукция листьев у части суккулентов как сред-
ство для предотвращения излишнего испарения воды могла бы
иметь отрицательные последствия для выживания вида, если бы
стебель не взял на себя функцию листьев—фотосинтез. Так, в
пустынных районах юго-западной части США встречается Fouqui-
eria splendens. Растение этого вида большую половину года из-за
засухи остается безлистным и фотосинтезирует стебель даже в очень
засушливые периоды. Поэтому его стебель имеет зеленые ткани с
хлоропластами.
Н. А. Максимов (1955) показал, что у части ксерофитов интен-
сивная потеря воды при транспирации компенсируется ее активным
поглощением из глубоких слоев почвы мощно развитой корневой
системой. Недостаток воды в окружающей среде восполняется
также образованием эндогенной воды в процессе дыхания (Б. Б.
Вартапетян, 1970).
В ходе эволюции у растений формируется ряд компенсаторных
механизмов, устраняющих нарушения функциональной целостно-
сти организма: метамерность дифференциации, способность к ре-
генерации, гипертрофия структуры и функций оставшегося
единичного органа и т. д.
В качестве примера компенсации функции укажем на «подтал-
кивание» реакций в процессах фотосинтеза. В отличие от животных
в клетках растений, в частности в хлоропластах, триозофосфатде-
гидрогеназа (ТДГ) связана с НАДФ • Н (акцептором электронов и
протонов)..Активность НАДФ • Н — зависимой ТДГ — определя-
ется наличием света и приводит к образованию НАДФ • Н. Выра-
ботка большего количества НАДФ • Н с участием ТДГ
благоприятствует «подталкиванию» реакции в сторону образования
глицеральдегид-3-фосфата (ГАФ). Без такого «подталкивания» не
удалось бы преодолеть термодинамического барьера реакции, ката-
лизируемой фосфоглицераткиназой. 1,3-дифосфоглицерат + АДФ ->
-> 3-фосфоглицерат + АТФ.
Из-за недостаточности свободной энергии (Г Дж/моль) может
возникнуть термодинамический барьер на пути осуществления фото-
синтеза. Этот барьер преодолевается накоплением большого количе-
ства АТФ (благодаря фотофосфорилированию) и расходованием
1,3-ДФГ (при участии активной НАДФ • Н —зависимой ТДГ).
Эти процессы осуществляются только при компенсации фун-
кции фосфоглицераткиназы активной НАДФ • Н —зависимой
ТДГ.
Явление компенсации функций видно на примере изменения
активности ферментов в зависимости от условий. Так, на РБФК.
48
показано, что в темноте, даже при наличии СО2 в среде, она мало
активна, тогда как после включения света резко возрастает в
считанные секунды. Постепенное уменьшение интенсивности света
приводит к соответствующему падению активности РБФК за 2—10
мин (Ю. А. Вийль, Т. П. Пярник, 1992). Этому способствует также
полное прекращение регенерации РБФ в тканях растений в темноте.
Как известно, в РЦФ использование поглощенной энергии для
переноса электронов от донора к акцептору происходит нередко
против термодинамического градиента. Благодаря этому сохраня-
ется сопряжение потока энергии и потока электронов (А. Т. Мок-
роносов, В. Ф. Гавриленко, 1992). В целом эффективность работы
РЦФ достигается непрерывной регенерацией компонентов цепи,
что сопровождается их новообразованием и восстановлением элек-
тронной структуры ранее, окисленного пигмента.
Известно, что у злаков хлорофилл содержится в остьях и коло-
сковых чешуйках. Эти структуры, находясь ближе к зерновке и в
хорошо освещенном положении, играют важную роль в наливе
зерна. Точно так же хлорофилл околоплодника многих растений
участвует в фиксации СО2, выделяющегося тканями плода при
дыхании. Показано, что хлоропласт мезокарпа плода гороха фик-
сирует СО2 воздуха, а внутреннего эпидермиса —до 50—70% СО2,
выделяемого семенем (Дж. Эдвардс, Д. Уокер, 1986).
Присутствие хлорофилла в плодах и зародышах имеет большое
значение для обеспечения их тканей продуктами фотосинтеза и
эндогенным кислородом (М. С. Яковлев, Г. Я. Жукова, 1993). На-
пример, количество хлорофилла в плодах томатов составляет всего
0,25% от содержания его в листьях; тем не менее фотосинтез плодов
оказывается в пределах 4—5% от фотосинтеза листьев (Б. Б. Вар-
тапетян, 1970). Подобные возможности расширяют норму реакции
организма, повышают надежность морфогенеза и способствуют
поддержанию гомеостаза системы в целом.
Стабилизация и автономизация функций. Пластичность растений
хорошо известна (см. § 2.8). В то же время процессы морфогенеза
и жизнедеятельности растений нельзя рассматривать как нестабиль-
ные, легко изменяющиеся. Это подтверждается данными по изу-
чению эндогенных ритмов, полярности развития, покоя,
фотопериодизма у растений и т. д. (см. гл. 6).
Эволюция ряда физиолого-биохимических процессов, как и
онтогенеза в целом, шла в направлении повышения их относитель-
ной независимости от внешних условий (L. Whyte, 1965). Поэтому
организму присуща стабильность жизнедеятельности. Так, накоп-
ление эфирных масел, имеющих значение для реализации воспро-
изводительных потенций растений, мало зависит от факторов среды
(А. А Прокофьев, 1955). Многообразие метаболических путей как
показатель дифференцировки на молекулярном уровне — предпо-
49
сылка для стабилизации жизнедеятельности организма в постоянно
меняющихся условиях среды. По этой причине лабильность отдель-
но взятой реакции в сложной цепи биохимических процессов не
приводит к преобразованию или заметному нарушению метаболиз-
ма благодаря барьерным механизмам. Так, при недостатке СО2 в
воздухе растение использует для фотосинтеза СО2, выделяемый при
дыхании; при избытке С02 в тканях ночью осуществляется карбок-
силирование.
Процессы жизнедеятельности протекают по универсальному
принципу обратной регуляции, т. е. накопление промежуточных
продуктов служит сигналом для прекращения их синтеза, тогда как
их нехватка стимулирует механизмы новообразования. Это показа-
но при изучении фотосинтеза с использованием 2-диокси-О-глю-
козы. Биосинтез хлорофилла характеризуется динамической
устойчивостью в меняющихся условиях среды, так как осуществля-
ется по принципу авторегуляции (В. Л. Калер, 1976). Эти примеры
свидетельствуют об автономности физиологических процессов.
Биосинтез хлорофилла, находящийся под контролем полифер-
ментного комплекса, регулируется сложной системой механизмов,
обеспечивающих строгую координацию последовательных реакций.
В нормальных условиях порфирины не накапливаются в клетке
из-за соответствия между скоростью их синтеза и превращения. В
этом отношении интересен пример с протохлорофиллидом, синтез
которого даже в темноте продолжается лишь до определенного
уровня, после чего его накопление идет на убыль ввиду прекращения
синтеза предшественника. Биологический смысл такой регуляции
очевиден, так как накопление порфиринов при отсутствии их
превращений было бы губительным для клетки из-за высокой их
фотосенсибилизирующей активности и бесполезной затраты исход-
ного материала (А. Т. Мокроносов, В. Ф. Гавриленко, 1992). По
этой причине синтез порфиринов находится под метаболическим,
генетическим и фитохромным контролем, что стабилизирует меха-
низм синтеза хлорофилла и фотоморфогенез растений.
Благодаря сложной регуляторной системе внутриклеточных ме-
таболических процессов, а также морфогенетических взаимодейст-
вий на уровне целого организма достигается автономизация
онтогенеза, закономерно усиливающаяся в ходе эволюции растений
(см. гл. 8). При автономизации онтогенеза внешние условия играют
роль сигналов, запускающих внутренние механизмы морфогенеза.
Внешняя среда поддерживает морфогенез энергетически, но при
сильных изменениях факторов внешней среды у растений возника-
ют тераты (уродства). Однако необходимо подчеркнуть, что тераты
не наследуются, что опять-таки показывает важность усиления
автономизации онтогенеза в ходе эволюций.
50
2.7. ДОСТИЖЕНИЯ ФУНКЦИОНАЛЬНОЙ
ЭВОЛЮЦИИ РАСТЕНИЙ
Одно из важных достижений эволюции — возникновение адап-
таций принципиального характера. Подобные изменения, по-види-
мому, возникают в метаболизме растений не часто. Так, П. Хочачко
и Дж. Семеро (1977) в становлении клеточного метаболизма вы-
деляют такие ступени: 1) изменение типа макромолекул, 2) изме-
нение количества (концентрации) макромолекул, 3) регуляция
функций макромолекул. Сообразно этому различают и пути адап-
таций организмов: 1) использование имеющихся обменных процес-
сов в онтогенезе для восстановления нарушенного состояния —
компенсаторные адаптации, 2) выработка новых механизмов в фи-
логенезе для выхода в новую зону — эксплуатативные адаптации.
Хочачко и Сомеро далее замечают, что биохимические адаптации
представляют собой «крайнее средство» и происходят лишь тогда,
когда у организма исчерпаны физиологические способы предотв-
ращения неблагоприятных воздействий среды. При выработке фун-
кциональных приспособлений чаще всего происходит опробование
ранее сложившихся физико-химических механизмов в различных
ситуациях.
В качестве примеров функциональных приспособлений расте-
ний могут служить образование световых или теневых листьев в
зависимости от условий освещения и влажности, аккомодация
роста, изменение темпов развития и потенций к регенерации при
смене условий среды, при нарушениях целостности, корреляции и
полярности организма. Так, при фотосинтезе в световых и теневых
листьях происходят одинаковые химические процессы, тем не менее
благодаря изменению структуры листа у них заметно меняется
количественное выражение основного процесса. Точно так же,
используя один и тот же механизм устьичных регуляций, растения
в течение суток активно противостоят иссушающему воздействию
среды и поддерживают фотосинтез и дыхание.
В ходе эволюции решаются важные принципиальные задачи
путем изменения количества структур и метаболистов. Так, увели-
чение числа сосудов и устьиц, густоты жилкования пластинки,
толщины слоя кутикулы и т. п. оказалось определяющим при
расселении растений в наземных условиях. Велика и роль соотно-
шения фитогормонов и ингибиторов роста в тканях (В. И. Кефели,
1974; К. 3. Гамбург, 1976;, К. Thimann, 1977). Например, изменение
их соотношения представляет собой универсальный механизм не
только для регуляции морфогенеза и роста (включая явления апи-
кального доминирования и устьичных движений), но и для осуще-
51
ствления ориентации индивидуума в пространстве. Здесь выступают
исключительно эффективные и разнообразные способы использо-
вания возникшего биохимического новшества в условиях прикреп-
ленного образа жизни.
Количественное изменение содержания веществ в онтогенезе
достигается и с помощью выработанного в ходе эволюции механиз-
ма взаимопревращения метаболитов. В качестве примера можно
указать на фитохромную систему (Рвв0 5* Р730), зарядку и разрядку
биологического аккумулятора (АТФ 52г АДФ), переход хлорофилла
из основного в возбужденное состояние и на роль аммиака как
«альфы и омеги» азотного обмена.
Одним из примеров регуляции жизнедеятельности растений
путем количественного изменения содержания вещества служит
вода. Так, влажность кукушкина льна при длительной засухе со-
ставляет 1%, сфагнума — 7% по отношению к общей сухой массе
тела. При пересыхании мхи и лишайники впадают в анабиоз.
Нередко даже высшие растения из семейства геснериевых, миро-
тамниевых и осоковых пересыхают в пустыне до такого состояния,
что их можно растереть в порошок. Тем не менее после увлажнения
их жизнедеятельность полностью восстанавливается.
Функциональная эволюция нередко связана с вычленением
основных процессов в качестве узловых в цепи превращений.
Таковым в цикле Кальвина оказалась реакция, катализируемая
РБФК:
С02 + РБФ -»(промежуточный С6-продукт) -» ЗФГА
В листьях зеленых растений на долю РБФК, как известно,
приходится до 50% всего белка, тогда как концентрация СО2 в
листьях очень низка (7 мкм). Казалось бы, парадокс. Для сравнения
укажем, что фосфофруктокиназа (главный регуляторный фермент
гликолиза) содержится в листьях в малых количествах. Несоответ- f
ствие содержания РБФК концентрации СО2 не случайно, а биоло-
гически необходимо для извлечения из атмосферы инертного
диоксида углерода, не обладающего развитой стереохимической
активностью и зарядом молекулы. Ввиду важности РБФК для
фиксации СО2 вырабатываются и специальные механизмы, регули-
рующие ее активность в онтогенезе (свет, фруктозо-6-фосфат).
Итак, структурно-функциональная дивергенция осуществляет-
ся при сохранении консерватизма основных путей метаболизма.
Для разных видов ее результаты часто не совпадают, поэтому и
способы приспособления растений к засухе и засолению оказыва-
ются весьма разнообразными. Этот определяется отбором.
52
2.8.ЕСТЕСТВЕННЫЙ ОТБОР
И ФУНКЦИОНАЛЬНАЯ ЭВОЛЮЦИЯ РАСТЕНИЙ
Выяснение способов и путей формирования того или иного
признака —очень сложная задача, так как необходимо познать,
«в чем заключается эволюционный процесс и как он сложился»
(Л. А. Орбели, 1961).
Для обсуждаемого вопроса имеют значение работы по модели-
рованию механизма действия естественного отбора в упрощенных
системах, по термодинамическим и информативным аспектам ор-
ганической эволюции (М. Эгейн, 1973; С. Э. Шноль, 1979). Они
показывают, что отбор ведет к кинетическому совершенствованию
метаболизма, т. е. к усовершенствованию аппарата матричного
воспроизведения, процессов энергетического обеспечения и био-
синтеза макромолекул.
Попытки распространить эти выводы на популяции и виды
требуют некоторого пояснения. Например, устойчивость подсол-
нечника к заразихе —функциональное проявление, зависящее от
многих сторон метаболизма растения. Она сложилась в результате
сопряженного действия естественного и искусственного отбора.
Указанное свойство можно объяснить и кинетическим совершен-
ством метаболизма. Но для понимания эволюции устойчивости
подсолнечника к заразихе этого недостаточно.
Отбор на функциональном уровне обычно действует по блоч-
ному принципу. Так, достаточно было «появиться» в клетке пор-
фиринам, чтобы автоматически сработались процессы, связанные
с поглощением, удержанием и превращением световой энергии. В
структуре и физико-химических свойствах молекулы порфиринов
предопределены почти все физические предпосылки, необходимые
для становления фототрофности. Поэтому в принципе трудно ожи-
дать наличия в клетке генов для каждого отдельно взятого фотофи-
зического процесса фотосинтеза. В то же время свойства любой
макромолекулы несколько модифицируются в зависимости от упа-
ковки. Так, в листе интенсивность флуоресценции хлорофилла
составляет всего 3% от ее уровня в органическом растворителе ввиду
использования большей части энергии света для фотосинтеза.
Различные типы взаимодействия пигмента с белками и липидами
(электростатические, координационные и гидрофобные) усили-
вают физиологическую сбалансированность процесса фотосин-
теза (А. Т. Мбкроносов, В. Ф. Гавриленко, 1992).
Отбор на функциональном уровне связан с преимущественным
размножением генотипов, обладающих комплексом биохимических
и структурных мутаций, а также усилением комплементарности
действия генов и регуляторных механизмов. Так, эволюция регуля-
торных пептидов шла не только в различных направлениях (изме-
53
нение первичной структуры самой молекулы, структуры и свойства
реципиента, коэволюция структуры регуляторных белков и рецеп-
торов и т. д.), но и привела к усилению стабильности (консерватив-
ности) участков регуляторных белков, ответственных за узнавание
рецептора (И. П. Ашмарин, 1986). В ходе эволюции регуляторных
пептидов у животных имело место вовлечение древних пептидов
для выполнения более филогенетически сложных функций, что
характерно и для растений (см. гл. 5).
Отбор привел к усилению активности многих молекул при
образовании ими комплексов с металлами. Например, карбоангид-
разы активны только при наличии цинка в молекуле (Н. И. Гулиев
и др., 1992). В то же время эволюция ферментов связана с повто-
рением функционально важных аминокислот в молекуле даже у
животных и растений. Дивергенция близких видов нередко связана
с замещением отдельных аминокислот, без существенных измене-
ний активности исходного фермента. Поэтому все карбоангидразы,
несмотря на дивергенцию их по отдельным аминокислотам, едины
в своем происхождении.
Отбор связан также с усилением функциональной специфики
молекул и выработкой механизмов регуляции их активности. На
примере эвглены показано, что тип питания организма (фототроф-
ный, фотогетеротрофный и гетеротрофный) существенно не влияет
на реакцию молекулы РБФК при внесении в среду NaCl. Оказалось,
что «собственная устойчивость» РБФК к высоким концентрациям
солей проявляется только в системе клетки и при наличии света.
При этом наблюдается резкое ингибирование активности РБФК в
условиях засоления у гетеротрофной эвглены (Л. В. Назаренко,
1992).
Функциональная дивергенция видов сопровождается возникно-
вением различий в молекуле РБФК по молекулярной активности
и физико-химическим свойствам. Так, величина температурной
полуинактивации РБФК из листьев позднеспелого клевера^ока-
зывается выше, чем из листьев раннеспелого (И. Ф. Карпилова,
А. К. Романова, 1992). Отбор растений в аридных засоленнык
условиях идет по комплексу признаков, в том числе и на поддер-
жание ионного баланса ассимилирующих органов растений, созда-
ние или усиление механизмов синтеза органических кислот,
способных нейтрализовать избыток катионов в тканях. Это позво-
ляет растениям сохранить положительный углеродный баланс в
экстремальных условиях (Т. А. Глаголева, М. В. Чулановская,
1992) и системную устойчивость к ним.
Разнообразие неблагоприятных факторов усложняет отбор на
устойчивость из-за возможности изменения его вектора. Все это
способствует возникновению системной устойчивости, с которой
коррелирует наличие сложной координации органов. При наступ-
54
лении гипоксии важной оказывается регуляция на уровне целого
организма (см. гл. 4). Такие процессы бывают выражены по-разному,
у различных видов растений. Так, у риса в отличие от пшеницы в
анаэробных условиях повышается активность МДГ, что способст-
вует восстановлению ЩУК в малат. Многие корневищные растения
в условиях затопления обходятся без О2 благодаря включению
процессов брожения. У устойчивых форм при этом меньше накап-
ливается в тканях корневища этанола (L. S. Monk et al., 1984).
Разные древесные формы, в том числе представители одного
рода, далеко не? одинаково устойчивы к гипоксии. К ней более
толерантна береза черная, нежели береза повислая. При продувании
питательного раствора азотом замечено, что в корнях у березы
черной содержание АДФ и АТФ составило 35 и 23% от контроля,
а у березы повислой, соответственно, 13 и 18%. Энергетический
баланс у первой снижался на 6—12-е сутки опыта, у второй —\
спустя 1 сут (R. R. Tripepi et al., 1984).
В последние годы возрос интерес и к возможным последствиям
ядерной катастрофы для растений, что оценивается как экологиче-
ская трагедия (А. М. Кузин, 1985). Выявлена различная чувстви-
тельность разных растений к излучениям (у-излучение). Наиболее
чувствительными к ним оказываются хвойные, а устойчивыми —
мхи и лишайники. Поэтому полагают, что хвойные леса на планете
погибнут в первые месяцы ядерного конфликта, остальные —
несколько позже, от других загрязнений. Природа разной устойчи-
вости к у-излучениям неясна. У древесных она слабо выражена по
двум причинам: из-за возможности большей концентрации радио-
активных осадков на листьях и отсутствия развитой способности к
быстрому вегетативному отрастанию (Ю. М. Свирижев, 1985).
Обеспечение устойчивости к любым.неблагоприятным услови-
ям связывают с включением механизмов защитного торможения,
компенсации и репарации на клеточном, тканевом, органном и
организменном уровнях. В устойчивости велика роль и гормональ-
ного комплекса растений (см. гл. 5). Очень часто адаптация к среде
у растений достигается за счет генетической нестабильности, т. е.
наличия множественности копий и повторов. Такие копии состав-
ляют примерно 75% всех последовательностей. За счет этого дости-
гается функционирование одних генов и подавление других у
растений, культивируемых в условиях избытка или недостатка
каких-либо элементов. При этом изменение одной из пар избыточ-
ных генов без «опаски» создает нужный для клетки генопродукт
посредством работы другого парного гена (А. С. Антонов, 1986).
Показано, что классы растений более значительно различаются
между собой по составу ДНК, чем классы животных. В связи с этим
допускают более высокие исходные темпы эволюции для первичных
55
структур ДНК у высших растений, чем у .животных. Это отчасти
объясняют повышением роли полиплоидии в эволюции растений,
причем у растений быстрые эволюционные изменения претерпева-
ют не только повторяющиеся (представленные в большом числе),
но и уникальные последовательности нуклеотидов в молекуле ДНК.
Некодирующие (повторяющиеся) последовательности нуклеотидов
в молекуле ДНК также имеют определенное значение для выжива-
ния растений ввиду их возможного участия в делении клеток и
узнавании гомологичных хромосом, регуляции жизненных процес-
сов (А. С. Антонов, 1986).
Такие последовательности неодинаково представлены у разных
высших растений, благодаря чему накопление изменений в нуклео-
тидных последовательностях в линии покрытосеменных оказалось в
10 раз выше, чем у мхов, печеночников, папоротников и голосеменных.
Приспособление к гипоксии связано и с изменением структуры
пластидного аппарата (число и величина пластид, объем пластид,
число й величина гран тилакоидной системы). Так, на примере
подводных и плавающих листьев лютиков (Batrachium eradicatum,
B.peltatum) показано наличие значительного сходства в распределе-
нии пластидного аппарата, несмотря на заметные анатомо-морфо-
логические различия самих листьев. У подводных листьев
фотосинтезирующей тканью является эпидерма, а надводных —
мезофцлл (Ю. В. Гамалей, 1985). В некоторых экстремальных ус-
ловиях (при дефиците О2) естественный отбор привел к усилению
возможностей переключения метаболических звеньев на более
древние пути. Это имеет место, в частности, у хвойных в осенне-
зимний период, когда у них преобладают гетеротрофный тип пита-
ния и анаэробное дыхание.
Во всех подобных случаях используются имеющиеся в популя-
циях предпосылки-мутации. Мутационными изменениями оказы-
ваются охвачены отдельные реакции или их комплексы,
определяющие возможности энергетики, скорость синтеза и распа-
да веществ, роста и развития индивидуума, отношение растений к
факторам среды и т. д. (см. § 2.2.). Любой, даже наиболее хорошо
отселекгированный, сорт постепенно накапливает мутации, затра-
гивающие многие признаки и свойства растений. Подобные изме-
нения даже в значительной степени встречаются в природных
популяциях, используя которые естественный отбор вырабатывает
определенный фотосинтетический аппарат. Изменчивость еще бо-
лее усиливается при сочетании результатов мутационного процесса
с генетической комбинаторикой.
В зависимости от направления естественного отбора изменяется
структура самой популяции, идет ее дифференциация по домини-
рованию тех или иных генотипов (следовательно, и фенотипов,
отличающихся по наличию соответствующих реакций и процессов),
56
отвечающих условиям среды. На основе оценки жизнеспособности
индивидуумов, их генетических различий в метаболизме и структу-
ре, происходит отбор наиболее удачных физиолого-биохимических
фенотипов, усиление или ослабление любого признака и свойства
растения. Это показано на примере дивергенции популяций и
сортов у растений по устойчивости к загрязнению среды ионами
тяжелых металлов, засухо-, соле- и морозоустойчивости и т. д.
В целом развитие функциональных приспособлений достигается
суммацией эффекта отдельных структурных и биохимических му-
таций, обеспечивающих преимущество потомству в борьбе за суще-
ствование. В случае основополагающих физиолого-биохимических
механизмов, таких, как фотосинтез и дыхание, не исключается
макроэволюционное развитие их путем симбиогенеза (см. гл. 3, 4).
Эти механизмы, однажды закрепленные в ходе эволюции у расте-
ний, в дальнейшем поддерживаются отбором как важные энерге-
тические завоевания.
Функциональная дивергенция растений основана на отборе инди-
видуумов в популяциях с наиболее полезными физиолого-биохи-
мИческими мутациями. Частота встречаемости таких уклонений в
пределах даже сорта значительна. При анализе эндемичного вида
Oenothera arganensis установлен полиморфизм в 15 ферментных
локусах, в локусе МДГ четко обнаруживались два аллеля. Система
же, обеспечивающая регулярную панмиксию, включает 45 аллелей
в локусе самонесовместимости. Р. Левонтин (1978) сообщает о
заметной генетической изменчивости аллоферментов. Так, популя-
ции Oenothera biennis обнаружили полиформность по 5 из 19 фер-
ментов. Более того, каждое растение в популяции оказалось
гетерозиготным по указанным пяти генам. Описано много му-
тантов, блокирующих синтез фитогормонов и их промежуточных
продуктов.
Эти примеры достаточны для характеристики возможности воз-
никновения генетической гетерогенности по функциональным осо-
бенностям. Попытаемся теперь понять, как действует отбор,
используя генетический полиморфизм реакций и веществ. В пре-
делах биохимических уклонений могут быть и такие, которые
способствуют или понижению (блокировка синтеза какого-либо
продукта), или повышению жизнеспособности особей. В первом
случае отбор ведет к выбраковке, во втором—к размножению
носителей соответствующих мутаций. Так, цветковые растения
оказались наиболее эволюционно продвинутыми благодаря приоб-
ретению многих новшеств, в том числе и способности к быстрому
росту и развитию. Эта способность, в свою очередь, была обуслов-
лена отбором растений по интенсивности фотосинтеза, т. е. биохи-
мическим совершенствованием. В некоторых условиях (например,
।
' 57
на Сахалине) у цветковых еще больше усиливается рост благодаря
накоплению соответствующих фитогормонов.
Другие примеры действия естественного отбора будут приведе-
ны ниже. Здесь лишь заметим, что функциональные уклонения не
возникают изолированно, а сопровождаются структурными пере-
стройками. Так, диффузия водяного пара у растений получила
биологическое завершение тогда, когда возникли устьица. В ходе
эволюции необходимость усиления потока энергии в определенном
направлении привела к выработке рациональной геометрии целых
растений, а также отдельных стволов, ветвей и листьев. Нередко
механизм, однажды закрепленный в эволюции, сохраняется без
принципиальных изменений. Однако и в этом случае эволюция
растений не прекращается, а продолжается путем совершенствова-
ния других узлов метаболизма или структуры (см. гл. 3). Развитие
функций связано с суммацией эффекта отдельных структурных и
биохимических мутаций, обеспечивающих преимущества потомст-
ву в борьбе за существование.
Интересные примеры действия естественного отбора описаны
на травянистых растениях в связи с приспособлением их к загряз-
нению среды тяжелыми металлами (A. D. Bradshaw, Т. Me. Neilly,
1981). Показано, что в ряде районов содержание металлов возросло
в 1000 раз и более по сравнению с естественным фоном. Повышение
толерантности растений к металлам связано с отбором их по
комплексу генов, определяющих особенности метаболизма и струк-
туры. Только совокупность генов, каждый из которых слабо влияет
на толерантность, определяет выживание и размножение популяций
в условиях загрязнения среды. Толерантность к тяжелым металлам
наследуется в поколениях как доминантный количественный при-
знак, подверженный фенотипической изменчивости.
2.9. ПЛАСТИЧНОСТЬ РАСТЕНИЙ
В ОНТОГЕНЕЗЕ И ЕЕ ЗНАЧЕНИЕ .
Структурная и функциональная пластичность растений в онто-
генезе имеет большое значение для перенесения ими стрессов. У
растений широко распространены модификации (ненаследствен-
ные изменения), представляющие собой норму реакции генотипа
на условия среды. Способность к ним закрепляется естественным
отбором (И. И. Шмальгаузен, 1969). В генетической теории отбора
роль модификаций в эволюции или отрицается, или рассматрива-
ется как буфер, противостоящий отбору, осуществляемому средой,
что в принципе спорно.
Пластичность растений, выражающаяся в гетерофилии (напри-
мер, стрелолист, водяной лютик, плющ и др.), поддерживается
58 - ‘
отбором как свойство, полезное для их выживания (A. Bradshaw,
1965). По этой причине у форм водяного лютика, произрастающих
в разных географических районах, обнаруживаются наследственные
сдвиги в рассеченности листьев. Вариации у разных самоопыленных
линий и простых гибридов у кукурузы в зависимости от внешних
условий также определяются генотипом.
' Многочисленные примеры неодинаковой физиологической
пластичности растений в онтогенезе при изменении условий пита-
ния и водоснабжения, освещения и температуры и т. п. представ-
ляют собой наследственную дивергенцию видов, популяций и
сортов. Различия в наследственной пластичности популяций и
сортов обусловливают и неодинаковую устойчивость к стрессам.
При резкой смене условий постепенно перестраиваются энергети-
ческие процессы, изменяются осмотические свойства клеток, соот-
ношение эндогенных регуляторов в тканях и т. д. От направления
перестройки обменных процессов зависит и устойчивость растений
к неблагоприятным условиям, их гомеостаз. В селекции очень важно
учитывать механизмы, поддерживающие гомеостаз организма: от-
бор на гомеостатичность сортов связан с повышением индивиду-
альной устойчивости растений к неблагоприятным условиям и
способностью использовать минимальные возможности для макси-
мальной продуктивности. Это достигается путем скрещивания пар
по принципу географической и генетической отдаленности, а также
сбалансированностью генрв, определяющих устойчивость к комп-
лексу неблагоприятных условий. Здесь важно изучить генетику
физиологических свойств.
Д ля рассматриваемого вопроса важно также сравнение культурных
и дикорастущих растений по содержанию эндогенных фитогормо-
нов и их реакции на введение регуляторов роста (В. И. Кёфели,
Л. Д. Прусакова, 1985; В. С. Шевелуха, 1992; В. И. Кефели, 1994),
что влияет на формирование габитуса растений. Для приспособления
растений значима способность организма поддержать благоприят-
ный баланс фитогормонов и донорно-акцепторных отношений на всех
этапах онтогенеза, особенно в экстремальных условиях (А. Т. Мок-
роносов, 1983). Изменением соотношения фитогормонов, полярности
и коррелятивных взаимодействий между органами достигаются
регуляции морфогенеза и поддержание пластичности индивидуума.
Эволюция идет в направлении формирования механизмов стабиль-
ности онтогенеза, поскольку организм выступает как целое в ин-
дивидуальном и историческом развитии (И. И. Шмальгаузен,
1968). В условиях прикрепленного образа жизни пластичность
индивидуума является важной приспособительной реакцией, выте-
кающей из нормы реагирования генотипа, без чего трудно достичь
и стабилизации продуктивности в часто меняющихся условиях.
59
* ♦ *
В главе охарактеризованы основные тенденции и направления
функциональной эволюции растений. В основе эволюции функций
лежат некоторые общие принципы: множественность проявления,
усиление или ослабление, компенсация, стабилизация, автономи-
зация и др. Важный итог функциональной эволюции — повышение
энергетической стабильности растений. Эго обусловлено включе-
нием механизма естественного отбора с использованием генетиче-
ской гетерогенности популяций.
ГЛАВА 3
ЭВОЛЮЦИЯ АВТО-
и ФОТОТРОФНОГО МЕТАБОЛИЗМА
В истории развития жизни неоднократно складывались «рево-
люционные» события, оказывавшие влияние на всю последующую
ее эволюцию. Среди них особая роль принадлежит совершенство-
ванию энергетики. Энергетические барьеры часто выступали как
факторы, ограничивающие темпы эволюции жизни на планете. Как
подчеркивает В. Грант (1991), отсутствие кислорода и медленность
насыщения им атмосферы были основной причиной длительной
задержки появления эукариот. Эукариотическая клетка с генами,
собранными в хромосомах, с распределением функций между ядром
и цитоплазмой, с включением разных органелл сыграла исключи-
тельную роль в последующем структурном и функциональном
совершенствовании живой природы. Как нехватка, так и избыток
органического вещества в экосистемах неоднократно приводили к
новым эволюционным взрывам. Поэтому эволюция сопровожда-
лась изменением самих способов питания. '
По способу питания организмы принято делить на гетеротроф-
ные (большинство бактерий, грибы, растения-паразиты, животные)
и автотрофные (фото- и хемосинтезирующие бактерии, водоросли
и зеленые растения). Однако не всегда удается четко разделить
организмы по типу питания, так как имеются переходные (миксот-
рофные) формы. Все типы питания осуществляются путем сложных
превращений веществ в клетке и организме. В этом процессе
большую роль играют мембраны.
Появление мембран у первичных организмов Земли (протоби-
онтов) усилило направленность синтетических процессов и регуля-
цию поступления веществ (А. И. Опарин, 1969). Мембраны
служили каркасом, на котором в определенной последовательности
выстраивались (фиксировались) ферменты, что придало биохими-
ческим реакциям большую организованность и повысило их эф-
фективность. Клеточный метаболизм, лежащий в основе жизни на
Земле, сходен у всех существ. Благодаря ферментам он осуществ-
ляется в 10*—1012 раз быстрее, чем обмен веществ вне клетки (без
ферментов).
61
В поддержании и дальнейшем развитии жизни на Земле фунда-
ментальную роль сыграли разнообразные автотрофные организмы,
способные синтезировать органические вещества из неорганиче-
ских. Фиксируя энергию солнечного света в продуктах фотосинтеза,
растения выполняют роль энергетического очага на Земле — кос-
мическую роль (К. А. Тимирязев). Ежегодно растения образуют до
160—200 млрд, т органических веществ и фиксируют 2 ♦ 1021 Дж
солнечной энергии. При этом растения усваивают из атмдсферы до
170 млрд, т СО2 и разлагают до 130 млрд, т Н2О, выделяя до 200—400
млрд, т свободного О2. Возникновение хлорофилла у первичных
бесцветных гетеротрофных организмов явилось йереломным эта-
пом в эволюции протобионтов и положило начало современной
биосфере на Земле. Появление фотосинтеза привело к арогенезам.
3.1. ОБЩИЕ ПРЕДСТАВЛЕНИЯ
О ВОЗНИКНОВЕНИИ ОБМЕНА ВЕЩЕСТВ
Анализ вопроса о возникновении обмена веществ в биосистемах
связан с проблемой происхождения жизни на Земле, в которой еще
много неясного. Занесена ли жизнь на Землю или она здесь
возникла? Прошел ли биологический обмен через коацерватное
состояние или первоначально возник генетический код? Почему
некоторые редкие элементы в земной коре (молибден и магний)
стали играть большую роль в биологическом обмене, чем обычные
элементы (кремний и кальций)? Подобных вопросов, ждущих сво-
его объяснения, пока еще много. Однако полагают, что общее
направление эволюционного процесса с самого начала детермини-
ровано физико-химическими свойствами атомов и молекул и самой
среды.
В принципе обмен веществ присущ не только живым существам.
На разных этапах предбиологической эволюции происходили про-
цессы, все более приближающиеся к биологическому обмену по
темпам, сложности и организованности (А. И. Опарин, 1969; Дж.
Бернал, 1969; и др.);
Возникновение сложного циклического обмена, характерного
для живых организмов, датируется возрастом более 3,2 млрд. лет.
Этот процесс можно схематически изобразить так: атомы -> моле-
кулы -> полимеризация молекул -> взаимодействие полипептидов
и полинуклеотидов -> циклический обмен (протобионты).
Синтетические процессы, протекавшие в абиогенной среде,
могли быть усилены в случае их пространственной локализации.
Так, в опытах с коацерватами показано, что при помещении их в
раствор глюкозо-1-фосфата (ПФ) они способны интенсивно син-
тезировать крахмал, если предварительно в коацерваты была вве-
дена фосфорилаза. После одновременного введения фосфорилазы
62 .
и 0-амилазы в коацерваты и помещения последних в раствор ПФ
синтезировались крахмал и мальтоза.
В связи с подобными наблюдениями полагают, что на начальных
этапах предбиологической эволюции коацерваты, в которых воз-
никла концентрация определенного набора ионов металлов и про-
стых органических веществ, стали основой для переходя к
биогенным синтезам. Эти синтезы усиливались и принимали более
направленный характер с появлением в коацерватах АТФ, синте-
зированного первоначально абиогенным путем. По современным
представлениям, даже первые живые организмы могли получать
АТФ из окружающей среды. .
С другой стороны, каталитическая активность коацерватов мог-
ла быть усилена и при возникновении комплексов между металлами
и органическими соединениями. Так, с внедрением в порфириновое
кольцо ионов железа его каталитическая активность возросла в 1000
раз по сравнению с ионами железа в растворе. В результате такого
сочетания, вероятно, появились коферменты в коацерватах, а затем
и субстратспецифические ферменты, способствовавшие биогенным
синтезам. При возникновении ферментов исходно могли быть
использованы и простые пептиды в комплексе с ионами металлов.
Химическая эволюция привела к образованию основных типов
веществ, ставших в последующем обязательными компонентами
всех организмов. Более того, при возникновении жизни имело место
частичное сужение круга органических веществ с полифункцио-
нальными свойствами. На современном этапе основу жизни состав-
ляют четыре нуклеотида, сотни органических соединений (из почти
более миллиона известных) и 20 аминокислот. Скачок от вещества
к существам был связан не только с ограничением круга первичных
соединений, но и с усилением их взаимодействия в составе сложных
комплексов. Так, взаимодополнением свойств белков и нуклеино-
вых кислот было обусловлено ускорение процесса самовоспроиз-
ведения макромолекул, что послужило толчком для возникновения
жизни. Такие системы с «комплементарным инструктированием»
оказывались выгодными в смысле воспроизведения (М. Эйген,
1973). В совокупности все эти события привели к возникновению
циклического энергообмена и биохимической универсальности жи-
вых существ.
Напомним, что в вопросе о происхождении жизни одним из
загадочных остается факт наличия абсолютной хиральной чистоты
У живых существ — содержание в молекулах белков только «левых»
аминокислот, а в нуклеиновых кислотах — «правых» сахаров. По-
добное явление Могло возникнуть только вследствие утраты пред-
биологической средой первичной зеркальной симметрии (равное
содержание «правых» и «левых» изомеров аминокислот и сахаров).
Смеси, содержащие полимеры зеркальной симметрии, называют
, 63
рацемическими. Неживой природе как раз присуща зеркальная
симметрия—рацемация.
Возникновение жизни практически исключалось до разрушения
зеркальной симметрии предбиологической среды (В. И. Гольдан-
ский, Л. Л. Морозов, 1984; В. И. Кизель, 1985; М. D. Nissinov,
1994), благодаря процессам статистической флуктуации—созда-
ния случайного избытка или недостатка одного из антиподов.
Переход к хиральной чистоте мог быть осуществлен одновре-
менно и в разных частях Вселенной более 15—20 млрд, лет назад
— «время ожидания» глобального нарушения зеркальной симмет-
рии. Современные живые существа поддерживают свою хираль-
ность исключительно внутренними процессами, главным образом
путем синтеза хирально чистых веществ в рацемической среде (А.
В. Яблоков, А. Г. Юсуфов, 1989).
По современным данным, Земля образовалась примерно 4,5—
4,6 млрд, лет назад, водоемы же 3,8 млрд, лет назад. Примерно 3,2
млрд, лет назад жизнь якобы стала полностью контролировать
земной цикл углерода. На основе таких подсчетов сделан вывод,
что для возникновения жизни в первичном «жидком бульоне»
недостаточно геологического времени для биопоэза. Поэтому Дж.
Бернал (1969) обратил особое внимание на роль коалинов в кон-
центрации органики абиогенного происхождения и межзвездного
вещества (цианистый водород, формальдегид и др.), доставляемого
на Землю кометами и метеоритами.
Земля, проходя через пылевое облако в течение 105—106 лет,
могла получить вместе с космической пылью 108—1О10 т органиче-
ского материала, что превосходит в количественном отношении
современную биомассу нашей планеты. Органическое вещество
Земли пополнялось и за счет извержения вулканов. Так, одно такое
извержение выбрасывает на Землю до 1000 т органических веществ.
Именно благодаря оседанию на поверхности ультрамикроскопиче-
ских пылинок космической органики могла возникнуть своеобраз-
ная оболочка из аминокислот, мочевины и других соединений. В
качестве энергии для этих целей использовались ультрафиолетовые
лучи.
Подобные процессы были в известной мере детерминированы
(R. Lipkin, 1994; Di. Giulio, 1994) и способствовали насыщению
первичного океана Земли органическими молекулами (предшест-
венниками живых существ), возникновению биологического обме-
на и переходу доклеточных форм к живому состоянию (В. Н.
Компаченко, 1994). Эволюция обмена в дальнейшем была связана
с постепенным усовершенствованием его механизма в критических
ситуациях, создаваемых многократными «взрывами» численности
организмов. Это прослеживается и на примере способов фиксации
СО2. Среди них самой древней являетсягетеротрофная (темновая)
64
фиксация С02. В «укороченном» Ёарианте. она возникла на заре
жизни и играла значительную роль не только у древних организмов,
но и продолжает играть у некоторых современных бактерий, живо-
тных и растений. Химизм автотрофного обмена усиливался путем
объединения катаболических и анаболических механизмов, т. е.
созданием энергетической двойственности существ. Ниже пред-
ставлены важнейшие этапы химической и биологической эволюции
(по П. Хочачко, Дж. Самеро, 1977).
Время (млн. лет назад) Этапы эволюции (что возникло в данный период)
1000 Многоклеточные организмы
Эукариотические клетки
Первые аэробные бактерии
С6Н120б + 6Н2О + 6О2 -> 6СО2 +12Н2О +30 — 40 АТФ
2000 Фотосинтез зеленых растений
6СО2 + 12Н2О С6Н12О6 + 6Н2О + 6О2
Бактериальный фотосинтез
6СО2 + 12Н2А -> СбН120б + 6Н2О + 12А
Фотофосфорилирование: трансформация световой энер-
гии в энергию АТФ
3000 Первые клетки (прокариотические)
Пентозофосфатный цикл: восстанавливающие факторы,
пентозы
6СбН120б + 6Н2О + 12 АТФ -> 12Н2 + 5C6Hi2O6 + 6СО2
Брожение: химический источник энергии
С6Н12О6 -> 2С2Н5ОН + 2СО2 + АТФ
Из приведенных данных видно, что первичные существа Земли
обладали гетеротрофным обменом. Автотрофный обмен из-за слож-
ности его механизма не мог возникнуть раньше гетеротрофного. И в
онтогенезе растений автотрофному обмену также предшествует гете-
ротрофный (А. Т. Мокроносов, 1981). Эволюционно обменные про-
цессы у растений строятся как бы на каркасе гетеротрофного
механизма, имеющего универсальное значение для живых существ. В
то же время гетеротрофная фиксация СО2 с участием ФЕП, пирувата
и ФЕП К, биосинтез пуринов и пиримидинов карбоксилирования
ацетил-КоА все еще широко используются в жизнедеятельности
растений. Могут быть и другие реакции нефотосинтетической фикса-
ции СО2, играющие роль вдыхательном метаболизме, глюконеогенезе,
поддержании pH цитоплазмы, биосинтезе липидов, метаболизме азота
и т. д. Нефотосинтетическая фиксация СО2 оказывает влияние даже
На процессы цветения растений и прорастания семян.
Первичная бескислородная среда на Земле способствовала даль-
нейшему насыщению атмосферы СО2 и привела к быстрому исчер-
панию запасов абиогенного органического вещества в среде.
3-157 65
Поэтому из-за низкой скорости абиогенного синтеза потребности
протобионтов в органическом веществе не могли быть компенси-
рованы. В этих условиях наступает конец первичной гетеротроф-
ности и отбор среди протобионтов идет в сторону выработки
автотрофного обмена.
В процессе эволюции Земля Претерпела три крупные револю-
ции: оксигенный фотосинтез, появление консумента первого по-
рядка (растительноядные многоклеточные) и консумента второго
порядка (хищники). Каждый новый этап указанной надстройки
проходил вдвое быстрее предыдущего, и с начала кембрия эволюция
земных экосистем в основных чертах завершается (Б. М. Медни-
ков, 1985).
Учитывая баланс О2 на Земле и его роль в окислении газов и
органики, nd-видимому, невелико участие первичной фототрофной
микрофлоры в окислении атмосферы Земли. Поэтому значительная
роль отводится процессам абиогенного образования О2 из
оксидного железа, находящегося в сердцевине Земли (О. Г. Соро-
хотин, 1974). Таким путем 2 млрд, лет назад концентрация О2 в
атмосфере достигла 1% от современного состояния, что, вероятно,
и способствовало появлению аэробных форм.
3.2. СУЩНОСТЬ РАСТИТЕЛЬНОЙ ЖИЗНИ. ГИПОТЕЗЫ
О СТАНОВЛЕНИИ И РАЗВИТИИ АВТОТРОФНОГО ПИТАНИЯ
Жйзнь на Земле имеет единое происхождение. Однако, говоря
об этом как об одном из крупных обобщений биологии, нельзя
забывать и о причинах ее разнообразия. Достаточно напомнить, что
число видов на Земле ныне достигает порядка 2 млн., из них
примерно 450 тыс. составляют растения. Следовательно, это разно-
образие также нуждается в объяснении, исходя из принципа исто-
рического развития живой природы.
Мы знаем, что растения и животные состоят из примерно
одинаковых химических элементов, полимерных соединений (нук-
леиновые кислоты, белки, липиды и др.) и Имеют клеточную
организацию; во многом сходно и строение их клеток. В то же время
есть и существенные различия между растениями и животными,
вытекающие из их образа жизни и организации. Достаточно запом-
нить о способности зеленых растений синтезировать органические
вещества из неорганических. Существуют и другие различия —
наличие жесткой оболочки у клеток, малая подвижность, широкое
распространение вегетативного размножения у растений и т. п.
Значение растений для поддержания жизни на Земле чрезвы-
чайно велико. Не было бы растений, вся нынешняя аэробная жизнь
вымерла бы от нехватки пищи и кислорода. Так, в среднем на 1 м2
66
площади путем фиксации СО2 за год образуют органического
вещества деревья около 1,4 кг, тропический лес —2,3 кг, травы —
0,91 кг. Общий органический продукт растений в год на земной
поверхности составляет 139 • 109 т, на долю наземных растений
приходится 110 • 109, водных — 29 • 109 т. При этом они выделяют
более 115—250 млрд, т О2.
Сами растения неоднородны по организации, продолжительно-
сти жизни, темпам размножения, продуктивности и т. д. Несмотря
на это, растения в целом выступают как средообразующий фактор
для других организмов. Поэтому в ходе эволюции живой природы
растения оказывались пионерами в освоении суши и новых терри-
торий. «Подтягивая» за собой представителей других царств на
неосвоенные территории, растения, в свою очередь, получали мощ-
ный стимул для дальнейшего эволюционного развития. При этом
складывались определенные взаимоотношения между разными ор-
ганизмами и формировались сами биогеоценозы как результат
сопряженной эволюции видов (Г. А. Заварзин, 1993; В. К. Сергеев,
1993).
Способность к синтезу органических веществ из неорганических
проявляется у разных живых существ, которые отличаются по
организации и первичным механизмам процесса. Среди них встре-
чаются одно- и многоклеточные, многоклеточные разной степени
дифференциации; железобактерии, водородные бактерии и зеленые
растения (водоросли, мхи, лишайники, папоротники, голосеменные
и цветковые). Фотосинтез — одно из проявлений автотрофного
обмена. По вопросу о возникновении автотрофного питания пока
нет единого мнения, и здесь много неясного. Заманчива гипотеза
К. С. Мережковского, А. С. Фаминцына, Б. М. Козо-Полян-
ского и других о симбиогенезе и возникновении автотрофности
путем симбиоза разных прокариот (см. § 3.7).
Показана общность молекулярных механизмов начальных ста-
дий фотобиологических процессов у живых организмов (фотосин-
тез, фотоиндукционные и фоторегуляторные процессы). Во всех
случаях имеют место поглощение света, миграция энергии, возбуж-
дение фотоактивного хромофорного комплекса и т. д. (А. Б. Рубин,
Г. Я. Фрайкин, 1987). Вообще фотоактивность заложена в природе
самой жизни, а фотосинтез является лишь более специализирован-
ным ее проявлением, результатом действия отбора «на зачаточные
стадии» полезных уклонений — фотоактивность. Последняя еще
больше усиливается при наличии в клетке порфиринов (А. Лима де
Фариа, 1991).
Одной из главных предпосылок перехода к автотрофному обме-
ну была присущая живым организмам фотохимическая активность,
67
обусловленная порфиринами. А. Л Красновский указывает, что
даже бесхлорофилльные существа синтезируют фотохимически ак-
тивные соединения — флавины и порфирины — как промежуточ-
ные продукты биосинтеза гема. Фотобиологические процессы
характерны и для грибов. У них они протекают в Юм же спектраль-
ном диапазоне и с участием тех же веществ (флавины, металлопор-
фирины, тетрапироли), что и у зеленых растений (Г. Деборин и др.,
1975).
Фотохимические реакции и возникающие свободные радикалы
(С. П. Лоч, Б. Хейлс, 1979) первично могли участвовать в создании
электрохимического потенциала ионов Н+ на поверхности мембран,
энергия которого использовалась для синтеза АТФ и выведения
избытка Ионов Na+ из клетки. В последующем указанный механизм
дополняется «протонными насосами», что привело к усилению
энергетических и транспортных возможностей Клетки (Л. Н. Во-
робьев и др., 1980). Одна из гипотез возникновения протонных
насосов исходит из возможности первичного появления пигмента
в клетке и, следовательно, использования света при транспорт?
ионов Н+ через мембрану.
С появлением пигмента в клетке начинается автотрофная эра
жизни, прошедшая через ряд ступеней совершенства. Напомним,
что среди бактерий встречаются фотоорганотрофы (несерные пур-
пурные) и фототрофы (окрашенные серные бактерии, цианеи и др.),
которые благодаря зеленому пигменту способны поглощать энер-
гию и использовать ее для ассимиляции СО2. Фототрофные бакте-
рии генетически неоднородны (ныне живущие фотосинтезирующие
бактерии — обитатели водоемов). Поэтому полагают, что фото-
трофное питание первично возникло в водоемах (куда проникал
свет) конвергентно в разных группах существ. Вероятно, по этой
причине оно встречается у разнообразных групп прокариот, далеких
в генетическом отношении. Из факта древности данного типа
биоэнергетики вовсе не следует, что любая группа примитивных
фотоавтотрофов должна быть признана филогенетически древней
(М. В. Гусев, Г. Б. Гохлернер, 1980). Наиболее примитивные из
них—пурпурные серобактерии—подвергают фотохимической
ассимиляции ацетата без поглощения СО2:
Ацетат + КоА^^ АцетилКоА -> Органическая кислота -> Ами-
нокислота (глутамин)
Другим этапом автотрофного питания можно рассматривать
способность восстанавливать СО2 при окислении органических
продуктов, в частности изопропанола, пропанола, этанола, сахаров,
органических кислот и т. п. (несерные пурпурные бактерии, серо-
бактерии). В дальнейшем, по-видимому, возникла способность
68
синтезировать органические вещества из неорганических (бактери-
альный фотосинтез), встречающаяся у,многих пурпурных и зеленых
серобактерий, которые окисляют сероводород, тиосульфат и т. д.
На стадии бактериального фотосинтеза фотоавтотрофный тип
биоэнергетики заметно стабилизируется, что видно по соответствию
величин квантового выхода бактериального фотосинтеза и фото-
синтеза зеленых растений (8—10 квантов на одну молекулу СО2).
Однако фоторедукция не связана с использованием воды в
качестве донора электронов, так как у бактерий встречаются лишь
фотосистема I (ФС1) и циклический транспорт электронов. Погло-
щенная энергия света идет у бактерий на синтез АТФ. Однако
процессы фоторедукции имеют много общего с фотосинтезом, в
том числе и восстановление СО2 при участии сильного редуциру-
ющего фактора (Н), образовавшегося на свету. У бактерий нет
ферментативных систем, способных выделять кислород в свободном
виде из окисленных продуктов, и процесс идет так:
2ОН + Н2А1 ->2Н2О + Ai
Фоторедукция отличается от фотосинтеза еще и по донору
водорода. Многие фоторедукгоры в кислородной среде переходят к
гетеротрофному типу питания, т. е. являются факультативными
автотрофами. Фоторедукция предшествовала фотосинтезу, доказа-
тельством чего отчасти может служить способность ряда Цианей,
зеленых, красных и бурых водорослей переходить к фоторедукции
в анаэробной среде. Переход от бактериального фотосинтеза к
фотосинтезу водорослевого типа следует рассматривать как важный
арахимоз в эволюции обмена. Это событие было вызвано диспро-
порцией между запасанием АТФ и восстановителя. Нехватка вос-
становителя оказалась фактором, ограничивающим бактериальный
фотосинтез, и привела к разделению функций между пигментной
и электрон-транспортной системами. Мутации, приведшие к раз-
делению функций между указанными системами, могли возникнуть
в популяциях подвижных фототрофов при перемещении их к
поверхности водоема, где давление мутагенных факторов оказалось
высоким (М. В. Гусев, Г. Б. Гохлернер, 1980).
Такой переход в биоэнергетике был сопряжен с дальнейшим
повышением энергоемкости и конкурентоспособности организмов.
Уже у синезеленых водорослей (цианей) формируются основные
элементы аппарата фотосинтеза, переносчики энергии и ферменты.
Пр некоторым данным, переход к фотосинтезу произошел еще
раньше. Так, существуют химические доказательства фотосинтеза
У организмов возраста 3,2 млрд. лет. Победе фотосинтетиков в
борьбе за существование способствовало использование ими про-
стых и легкодоступных веществ (СО2 и Н2О) для создания органи-
69
веских молекул, синтез энергетически более,богатых продуктов —
углеводов в большем количестве, выделение свободного кислорода
и т. д.
Несмотря натр что цианобактерии (Cyanophyta) из-за приобре-
тения способности к фотолизу воды достигают вершины био-
химической эволюции фототрофных прокариот (М. В. Гусев,
К. А. Никитина, 1979), они все еще сохраняют и другие метаболи-
ческие механизмы: фиксацию азота, фотосинтез по бактериальному
и растительному типу, ассимиляцию органических веществ (гете-
ротрофия) и хемосинтез. Например, у цианей (Anacystis nidulans и
др.) при высокой интенсивности света, если отношение хлорофилла
к фикоцианину составляет % СО2 фиксируется через цикл Кальви-
на, тогда как при слабом освещении или на красном свету — по
С4-пути, а при концентрациях СО2 порядка 0,03% —через цикл
карбоновых кислот. Тем не менее цианеи близки к зеленым расте-
ниям по составу пигментов, они восстанавливают НАДФ (вместо
НАД у зеленых и пурпурных бактерий), имеют каротиноиды, пла-
стоцианины, ферредоксины (близкие по составу к зеленым водо-
рослям) и ФС1. Точно так же у многих водорослей в определенных
условиях проявляются древние типы фотоавтотрофной ассимиля-
ции: они способны переходить от фотосинтеза к фоторедукции (в
атмосфере с водородом, азотом при низкой интенсивности света).
Такое переключение типов питания часто сопровождается индук-
цией синтеза дополнительных ферментов и способствует повыше-
нию экологической пластичности вида.
Таковы вероятные этапы развития автотрофного питания. В ходе
его эволюции на смену многообразным способам ассимиляции
углерода приходит более специализированный способ — фотосин-
тез (филетическая эволюция). При этом на эффективность испытаны
разные доноры электронов, источники энергии и структуры. В част-
ности, Э. Либберт(1976) указывает, что эволюция фотосинтеза прошла
три этапа: фотоорганотрофию (донор — органические вещества) ->
-> фотолитотрофию (донор—неорганические вещества) -> фотогид-
ротрофию (донор — вода). Эти ступени связаны с заметными сдвигами
в кинетическом совершенстве организмов (см. сх 6).
Таким образом, эволюцию автотрофного обмена можно
представить так: гетеротрофы -» облигатные фотогетеротрофы ->
-> факультативные фотогетеротрофы -> облигатные фстоавтогрофы ->
-> фоторедукторы -> фотосинтетики.
Наиболее резкий скачок был совершен от сбражцвателей к
фотоорганбтрофам. Между.последними и фотолитотрофами грани-
цы считаются относительными.
70
Схема б
Эта схема неполно отражает связи, существующие между раз-
ными группами фототрофов. Эти этапы характеризуют экологиче-
ское совершенство, что, вероятно, и достигалось конвергентно у
разных групп организмов. Эволюционно автотрофный обмен на-
правлен на все большую специализацию. Напомним, что пурпурные
бактерии наименее специализированы в отношении использования
субстратов, тогда как у зеленых бактерий эти возможности ограни-
чены. Как бы то ни было, в одной из групп прокариот (цианей)
путем фотосинтеза достигнута полная независимость организмов
от наличия готового органического вещества в среде.
3.3. ВОЗНИКНОВЕНИЕ И РАЗВИТИЕ ФОТОТРОФНОСТИ
Для перехода к фототрофии необходима известная подготовка.
ПожаЛуй, переход к ней не мог быть осуществлен среди форм с
явно выраженным избеганием света, т. е. с отрицательным фото-
таксисом. «Эволюционная западня» создавалась для бактерий с
положительным фототаксисом. Если в пределах одной популяции
протобионтов все особи пошли бы к свету, а у представителей
разных популяций часть особей проявляла бы фототаксис, то ни
одна из них не имела бы «преимущества перед другими» ввиду
взаимного затенения протобионтами друг друга из-за стремления
71
каждой особи занять наиболее освещенное место (Р. Клейтон, 1984).
В такой ситуации преимущества оказались бы на стороне тех особей,
которые займут место сверху огромного «протобионтного роя». Но
и такой успех был бы также сомнительным из-за губительного
влияния ультрафиолетовых лучей на доминирующие особи.
Поэтому отбор среди первичных форм жизни с положительным
фототаксисом мог быть успешным только при наличии внутриви-
довой дифференциации по способности улавливать свет разной
длины волны. В то же время допустима модификация физических
взаимодействий молекул пигментов между собой и с клеточными
компонентами, а также усиление разнообразия состава пигментов-
хлорофиллов, бактериохлорофиллов и т. д. Для решения этой
задачи необходимы были различия особей в пигментах, фотохими-
ческих реакционных центрах и связанных с ними ферментах и
коферментах (Р. Клейтон, 1984). >
На начальных этапах развития фототрофности, когда в среде еще
встречалось большое количество восстановленных органических сое-
динений абиогенного происхождения, поглощение света пигментами
как бы усиливало энергетические возможности клетки, основанные
на субстратном фосфорилировании. Энергия поглощенного света в
какой-то степени трансформировалась в химическую энергию макро-
эргических связей молекул АТФ. При этом не накапливалось большое
количество энергии, но ее было вполне достаточно при наличии в
среде активных органических соединений восстановленного типа.
Наиболее близки к этому типу создания энергии несерные пурпурные
бактерии. Запас органики в среде постепенно исчерпывался, в связи
с этим возникла необходимость их восполнения. Для этих целей
использовались разные источники углерода, в том числе и СО,. Но
инертность СО2 не могла быть преодолена только энергией АТФ и
света, которые были ранее достаточны для образования восстанови-
теля. Так сформировался нециклический транспорт электронов (у
зеленых бактерий) и осуществлен переход к использованию H,S (М.
В. Гусев, Л. А Минеева, 1984).
Одним из проявлений сложности начальных этапов эволюции
фототрофности служит наличие четыре^ типов фотосинтетических
систем у современных существ (табл. 1). Указанные типы фотоси-
стем, как показывают расчеты Р. Клейтона (1984), отличаются
энергетической эффективностью. У зеленых растений она состав-
ляет 38%, пурпурных бактерий—25%, серных и того, меньше.
Сложные преобразования процессов фототрофности у протобион-
тов выступали как результат отбора их на повышение энергетиче-
ской эффективности. Таким путем была достигнута относительная
независимость живых существ от содержания в среде готовой
органической пищи, что имело принципиальное значение для
дальнейшего расцвета жизни на Земле.
72
Таблица 1 . Сравнение фотохимических доноров и акцепторов электронов
у разных фотосинтетических систем (по Р. Клейтону, 1984)
Фотосистемы Донор электронов Акцептор электронов
Пурпурные бак- терии Зеленые бакте- рии (серные) ФС I ФС II Бхл а, специальная пара, +0,4 В Бхл о, специальная пара, +0,3 В Хл а, специальная пара, +0,4 В Хл а (мономер или специаль- ная пара), > +0,8 В Бфф а, пластохиноны (Qa, Qb) Бфф а, FeS-белки Хл а, FeS-белки Фф a, Qa и Qb
Примечание. Фф — феофитин, FeS-белки — ферредоксин, Бфф — бактериофе-
офитин, Бхл — бактериохлорофилл, Хл — хлорофилл.
Высшим итогом фотогрофности признано возникновение меха-
низма фотосинтеза. Фотосинтез как сложное физиологическое свой-
ство претерпевал эволюционные изменения на индивидуальном и
ценотическом уровнях. Нельзя даже представить эволюцию фотосин-
теза вне потребности биосферы в целом. Это видно как по специфике
физико-химических процессов, так и по характеру «вмонтирования»
их в клетке и усовершенствования всего аппарата фотосинтеза.
В связи с этим форма и число хлоропластов, величина клеток
палисадной ткани, соотношение губчатой и палисадной паренхимы
пластинки, величина и особенности строения листьев, расположе-
ние, количество и продолжительность жизни листьев на побеге —
параметры продуктивности целого растения —меняются сопря-
женно с другими признаками популяции как приспособления,
имеющие значение для успеха в борьбе за существование. Напри-
мер, в докембрии были водоросли, только стелющиеся по дну
водоема. После углубления водоемов в кембрии из-за усиления
тектонической деятельности земной коры произошло перемещение
Cyanophyta в глубокие слои водоема, где освещенность была мала.
С этим связывают возникновение комплекса пигментов, необходи-
мых для улавливания света в глубокой толще воды. С другой
стороны, ухудшение условий освещения могло привести к отбору
растений на усиление роста в направлении падания света. Это
послужило, по-видимому, причиной массового развития нитчатых
форм (К. Б. Кордэ, 1973).
Характерно, что вся последующая эволюция синезеленых водо-
рослей после кембрия не изменила существенно их морфологию
ввиду достаточной ее адаптивности для восприятия светового по-
тока и способности фиксировать положение индивидуума в водо-
еме. Возникновение вертикально растущих водорослей могло быть
также связано с постепенным засыпанием их стелющегося тела
осадочными породами в воде. В этих условиях отбор благоприят-
73
ствовал особям не только вертикально растущим, но и с выраженной
осевой активностью роста и дифференциации (К. Б. Кордэ, 1973).
На примере ископаемых красных водорослей кембрия показано,
что многие детали их организации характеризовались своей не-
устойчивостью. В раннем кембрии их эволюция шла разными
направлениями в сторону достижения прямостоячего положения.
По пути вертикального роста пошли и другие водоросли.
В развитии положения о роли отбора в формировании растений
следует указать на возникновение листостебельных форм в наземных
условиях от дихотомически ветвящихся в результате преимуществен-
ного роста одной из ветвей (перевершинивание), уплощение ветвей
при задержке роста. Такой процесс протекал параллельно во многих
линиях нижнедевонских растений. Согласно микрофильной гипотезе,
листья формируются из выростов стебля или из вторично упрощенных
ветвей, утративших проводящую функцию.
Одним из проявлений эволюции фотосинтеза как организмен-
ного явления служат пустынные растения. У видов наиболее прИт
способленных к пустынной жаре Каракумов (Haloxylon persicum,
Calligonum caput-medusae, Populus pruinosa), ассимиляционная спо-
собность изменилась путем ее удивительного сочетания с другими
особенностями индивидуума как целостной системы. Вернее, асси-
миляционная способность у указанных растений находится под
опекой тех приспособлений, которые устраняют возможность на-
ступления резкого дефицита воды в тканях. В этих целях отбор шел
также на выработку эндогенных ритмов, позволяющих пустынным
растениям накопить достаточное количество ассимилятов и био-
массы в ограниченном интервале времени года, когда наступают
благоприятные условия водоснабжения. В случае высокой интен-
сивности освещения растения пытаются «уйти» от невзгоды, сбра-
сывая листья в определенные сезоны года, или отбор индивидуумов
в популяциях пустынных растений идет в направлении редукции
листьев, усиления эфемерности, эфемероидности, низкорослости и
т. п. Таким путем пустынные растения часто избегают перегрева
тела и больших потерь воды на испарение.
О связи эволюции фотосинтеза с целостностью организма сви-
детельствуют данные об изменении фотосинтеза в онтогенезе рас-
тений (А. А. Шахов, 1993; В. И. Кефели, 1994). Так, современные
низшие фототрофы в онтогенезе проявляют древнюю способность
к хроматическим адаптациям. По этой причине у синезеленых в
онтогенезе происходят заметные сдвиги в содержании отдельных
пигментов по мере изменения освещенности. Таким путем они
настраивают себя сообразно условиям. Настройка аппарата фото-
синтеза происходит и у высших растений. Она проявляется на
уровне мембран хлоропластов, самих хлоропластов (изменением их
положения), в движении устьиц и листовой пластинки. Подобные
74
события возникают в интервале времени от нескольких секунд до
нескольких часов, они неоднократно повторяются в течение суток.
Для понимания специфики становления фотосинтеза представ-
ляют интерес организация и функционирование его ключевого
фермента РБФК, который участвует в биогеохимическом кругово-
роте СО2 на нашей планете.
3.4. ВОЗНИКНОВЕНИЕ ЭЛЕКТРОН-ТРАНСПОРТНОЙ ЦЕПИ
И МЕХАНИЗМА КАРБОКСИЛИРОВАНИЯ
Компоненты ЭТЦ фотосинтеза разнообразны и сложны, каж-
дый из них выполняет специфическую роль в восприятии световой
энергии и поддержании необходимого окислительно-восстанови-
тельного потенциала в хлоропластах (В. В. Полевой, 1992; А. Т.
Мокроносов, В. Ф. Гавриленко, 1992). Эволюция отдельных ком-
понентов ЭТЦ фотосинтеза и пути их сопряжения остаются слабо
изученными. Однако вопрос о первичности ЭТЦ фотосинтеза по
сравнению с таковой дыхания многими решается положительно. В
то же время усматривают общность происхождения ЭТЦ фотосин-
теза и дыхания, исходя из следующих соображений: 1) использова-
ние у некоторых существ одной и той же ЭТЦ в обоих процессах
(Rhodopseudomonas spheroidesy '2) сходство строения переносчика
водорода при фотосинтезе (пластохинон) и дыхании (убихинон), а
также хлорофилла и цитохромов; 3) совпадение ферментов мито-
хондрий и хлоропластов, участвующих в переносе протонов через
мембрану, и синтеза АТФ (отличие состоит в противоположной их
ориентации на поверхности мембран, что приводит к изменению
направления протонного градиента в хлоропластах и митохондриях;
4) перенос электронов в хлоропластах и митохондриях с участием
железосодержащих (цитохромы) и медьсодержащих (пластоционин
и цитохромоксидаза) белков; 5) вхождение в ЭТЦ указанных орга-
нелл флавиновых белков и хинонов.
В связи с указанными наблюдениями делается вывод о гомоло-
гичности ЭТЦ названных органелл и единстве происхождения
основных биоэнергетических процессов в ходе эволюции (А. А.
Мутускин, П. А. Колесников, 1985). На примере развития ЭТЦ
фотосинтеза и дыхания мы еще раз убеждается в том, что «природа
щедра на разнообразие, но крайне скупа на нововведение» (Ч.
Дарвин, 1939).
Для эволюции фотосинтеза чрезвычайно большое значение
имело развитие и усовершенствование механизма карбоксилирова-
ния и его ключевого фермента РБФК (А. К. Романова, 1991). Как
известно, в основе фотосинтеза лежит ВПФ-цикл, состоящий из
четырех этапов; карбоксилирование и регенерация акцептора СО,
75
(РБФ), восстановление ФГК до триозофосфата и автокатализ.
Темновое фиксирование СО2 является наиболее древним процес-
сом, присущим и гетеротрофным, и фототрофным существам. Даже
у высших фототрофных существ имеет место сочетание процессов
темнового и светового карбоксилирования. В ходе эволюции рас-
тений эффективность карбоксилирования была повышена благода-
ря возникновению механизмов постепенной регенерации акцептора
СО2 (5 из 6 молекул триозофосфатов-ФГА и ДГАФ — идут на
регенерацию РБФ, а 1 молекула —только на образование органи-
ческого продукта), автокатализа субстрата и поиска возможностей
для компенсирования низкого содержания СО2 в среде с повыше-
нием сродства фермента к ней (оно выражается величиной 10 мкМ
СО2). Заметим, что концентрация РБФ составляет около 0,6—0,75
нмоль на 1 мг хлорофилла.
В фотосинтезе есть энергетически невыгодные реакции, связан-
ные с образованием ЩУК из ПВК и СО2, а также ФЕП из ПВК,
Рн и АТФ. Поэтому эволюционно важным становится не высокое
содержание РБФ, а повышение потенциальной активности основ-
ного фермента карбоксилирования — РБФК — и его локализация.
У большинства микроводорослей, содержащих пиреноид, РБФК
локализована в пиреноиде, а у многоклеточных водорослей и
высших растений — в строме хлоропласта. Для усиления фотосин-
тетического сродства к СО2 у микроводорослей необходима полно-
стью сформированная система концентрации СО2, которая
включает не только механизмы активного транспорта и накопления
неорганического углерода, но и условия для использования внут-
реннего его пула (Н. А. Пронина, В. Е. Семененко, 1992).
У высших растений высокая активность РБФК эволюционно
фиксируется для обеспечения фотосинтетической продуктивности
в условиях падения концентрации СО2 и повышения содержания
О2 в атмосфере. Кроме того, складываются механизмы регуляции
активности РБФК. Указанный фермент использует СО2 (^„=54-15
мкМ), а ФЕПК —бикарбонат (Х,=0,34-0,4 мкМ). Сродство ука-
занных ферментов к СО2 меняется в зависимости от концентрации
О2: при концентрации О2 в среде 2% и pH 8,0 сродство РБФК и
ФЕПК к СО2 оказывается одинаковым, при наличии в атмосфере
21% О2 РБФК находится в невыгодном положении из-за возраста-
ния К„ СО2.
Отбор благоприятствовал в целом достижению ферментами
наибольшей активности при наличии концентрации СО2 и О2,
соответствующей современному уровню, а также возникновению
механизма ее регулирования для обеспечения максимального вы-
хода органического вещества. Мутантные аллели с малой активно-
стью фермента могут быть или нейтральными, или отбрасываться
76
отбором. Аминокислотные замещения в молекуле РБФК обычно
оказывают влияние на приспособленность.
Высокая интенсивность фотосинтеза эволюционно стабилизи-
руется также благодаря выработке механизма, устраняющего
перепад концентрации промежуточных продуктов фотофосфори-
лирования в хлоропластах,— прекращается утечка ФГК в цитоп-
лазму из-за поддержания высокого значения pH в строме, Mg2+ и
т. д. В ВПФ-цикле использование ФГК (ЗФГК + АТФ Mg 1,3
ДФГК + АДФ)—термодинамически невыгодная реакция из-за
необходимости АТФ, АДФ и ФГК. Поэтому возникла необходи-
мость вовлечения в качестве донора электронов НАДФН, а также
использования основной части ассимиляционной энергии для вос-
становления ФГК (что связано с повторным превращением 5 из 6
возникающих триозофосфатов). Некоторое же количество ФГК
может восстанавливаться независимо от фиксации СО2. Подобные
процессы способствуют поддержанию концентрации ФГК внутри
хлоропластов в 10 раз большей, чем в наружной среде.
Фермент РБФК рассматривают как белок номер один на Земле,
содержание его в листьях составляет примерно 50% от всего белка
в клетке, а в хлоропластах—60—70%. Фермент состоит из 8
больших и 8 малых субъединиц. Белок больших субъединиц одина-
ков у всех зеленых растений, он определяет бифункциональность,
синтезируется под контролем генома хлоропласта, структурный ген
большой субъединицы наследуется по материнской линии. Белок
малых субъединиц РБФК синтезируется под контролем ядерных
генов (на рибосомах 80). Экспрессия генов малых субъединиц
контролируется светом. Сборка же всей молекулы РБФК происходит
в хлоропласте (Ю. С. Насыров, Ю. Д. Ахмедов, 1979). Следователь-
но, каталитически активная большая субъединица РБФК невидоспе-
цифична ввиду общности ее функции у всех зеленых растений.
Для формирования функционально активного фотосинтетиче-
ского аппарата необходимо кооперативное, действие ядерных и
хлоропластных генов. Более или менее хорошо изучены ген большой
субъединицы РБФК и гены ряда полипептидов тилакоидной мем-
браны. Светом регулируется транскрипция ядерных генов, опреде-
ляющих малую субъединицу РБФК, и светособирающего
хлорофилл а, ^-связывающего белка. Среди хлоропластных генов
светозависимыми оказались и гены большой субъединицы РБФК
и ряда белков хлоропласта. Эти данные важны для понимания
сложности путей эволюции первичных механизмов фотосинтеза как
результат отбора по комплексу реакций.
На примере РБФК можно проследить некоторые общие тенден-
ции, характерные для эволюции ферментов: усиление каталитиче-
ской активной функции путем объединения разных субъединиц
77
(неравнозначных по функциям), повышение конформационной и
функциональной лабильности молекул при отборе по реакционной
активности. По этой причине РБФК у разных существ проявляет
различия. Так, у бактерий RhodospirillUm rubrum, Hydrogenomanas
eutropa, H. facillis РБФК состоит из двух субъединиц, Chromatium
thiosulfatophillum —из шести, у цианей и высших растений —из 8
больших и малых субъединиц (Е. Г. Судьина, Г. И. Лозовая, 1982),
РБФК найдена и в этиолированных листьях, где содержание малых
субъединиц в три раза превышает содержание больших.
Активность ферментов определяется многими факторами. В
частности, блокировка SH-групп и недостаток Mg2+ резко снижают
активность РБФК в тканях зеленых листьев. Все фототрофы
фиксируют СО2 с участием РБФК, и у них она равномерно распре-
делена в строме. Функция РБФК во многом зависит от концентра-
ции СО2 и О2 в среде. При этом меняется специфика «работы»
фермента:
ЗФГК.+ 02 Рибулозо-1,5-бис- С02 2 молекулы
фосфогликоле- ' фосфат ------ЗФГК
вая кислота f Т
Оксигеназная Карбоксилазная
функция функция
Говоря о функции РБФК, нельзя забывать и о том, что цикл
Кальвина складывается в геологические эпохи, когда содержание
С02 в атмосфере превышало содержание 02. Эти условия, по
мнению Р. Клейтона (1984), сказались на высоком сродстве фер-
мента к С02. У 5 видов С3- и 11 видов С4-растений была определена
активность РБФК. У С4-растений она составила 5,9 мкмоль С02 мг
белка, что вдвое превышало таковую у С3-видов (J. R. Seemann et
al., 1984).
На свету содержание РБФК в хлоропластах превышает в 100 раз
уровень ее содержания в этиопластах. При освещении этиолиро-
ванных проростков происходит быстрое и параллельное увеличение
содержания мРНК и полипептида малой субъединицы РБФК, тогда
как содержание большой субъединицы медленно меняется. В транс-
крипции мРНК малой субъединицы наблюдается три периода после
освещения: 1) быстрый, в течение 1 ч после освещения; 2) медлен-
ный, продолжительностью 36 ч, синтез мРНК возрастает постепен-
но; 3) быстрый при переносе со света в темноту и наоборот (R. J.
Ellis et al., 1984).
Интересные данные ранее были получены при сравнении тене-
выносливых растений (Aralia califomica, Disporum smithii, Viola glabella
и др.) и светолюбивых (Airiplex patula, Plantago lanceolata, Solidago
spathulata) (Bjorkman O., 1968). Так, активность РБФК (моль-IO'6
СО2/.мин) у первых в пересчете на 1 дм2 листовой пластинки
составила около 2, тогда как у вторых — свыше 10. При пересчете
78
на 1 мг хлорофилла, соответственно, 0, 38—0,76 и 1,89—5,72, на 1
мг белка 0,15—0,22 и 0,23—0,43. Низкие величины у теневыносли-
вых растений компенсировались увеличением ассимилирующей
поверхности, снижением дыхания и уменьшением доли нефотосин-
тезирующих частей.
В целом, очевидно, свойства РБФК наилучшим образом отве-
чают интересам растений, позволяют выполнить им космическую
функцию. Здесь эволюции предстояло сделать первый и принци-
пиальный выбор в усилении эффективности автотрофности, осно-
вой которой является фотосинтез. Несмотря на то что темновая
фиксация СО2 составляет первичный этап фотоассимиляции угле-
рода растениями и типы ее разнообразны (они различаются только
по характеру регенерации < акцептора СО2), все же фотосинтез
оказался основой экологического равновесия биосферных процес-
сов (А. Т. Мокроносов, В. Ф. Гавриленко, 1992). На разных уров-
нях организации фотосинтеза достигается стабилизация его
механизма ввиду значимости процесса создания органического
продукта для последующей эволюции жизни на Земле.
3.5. СТАБИЛИЗАЦИЯ МЕХАНИЗМА ФОТОСИНТЕЗА
Экспансия и увеличение разнообразия жизни в значительной
мере связаны со стабилизацией механизма фотосинтеза и его
последовательных физико-химических этапов. Без фотосинтеза
трудно представить современное состояние живой природы, обес-
печение непрерывности потока энергии и вещества на Земле.
Поэтому в ходе эволюции фототрофности наблюдается выработка
механизмов, обеспечивающих как стабильность процесса фотосин-
теза (улавливание, передача и преобразование световой энергии),
так и экологические возможности его реализации.
В сложном процессе фотосинтеза выделяют различные этапы и
происходящие при этом реакции (А Т. Мокроносов, В. ф. Гаври-
ленко, 1992). Среди них обычно различают четыре крупных карди-
нальных этапа: начальные (физические) процессы захвата энергии,
поглощение ее путем образования лабильных и богатых энергией
продуктов (фотохимические реакции), энзиматические (темновые)
реакции транспорта электронов и превращения углерода и, нако-
нец, продуктивность фотосинтеза. Энергия света поглощается со
скоростью 1015—Ю*“ с, фотохимические реакции протекают за
10*’—10’4 с, а энзиматические —за 10'4—10*2с (Б. А. Рубин, В. Ф.
Гавриленко, 1979).
Одно из важных эволюционных приобретений —стабилизация
поглощенной энергии благодаря усилению необратимости пронес-
79
сов фотосинтеза. Это достигается различными механизмами. На-
помним, что скорость передачи энергии между молекулами хлоро-
филла равна —10-’ с. За 1 с РЦФ может перенести до 50 млн.
электронов (А. Т. Мокроносов, В. Ф. Гавриленко, 1992). Все эти
события делают процесс фотосинтеза необратимым даже в условиях
короткого времени пребывания растений на свету. Эволюционные
предпосылки таких преобразований — в усилении окислительно-
восстановительного потенциала реакции фотосинтеза путем совер-
шенствования железосодержащих ферментативных систем и
вхождении марганца в состав флавопротеина, обладающего гидро-
геназными свойствами. Значение этого явления состоит в том, что
соединения марганца с галактолипидами хлоропластов на свету в
присутствии ФАД быстро окисляются с образованием пероксидов
и выделением кислорода. В процессе эволюции ферментной систе-
мы, участвующей в восстановлении СО2 при хемосинтезе, фоторе-
дукции и фотосинтезе, сохранялись восстановительные свойства
соединений негеминового железа даже в условиях, приближающих-
ся к окисленной среде. Это достигалось путем соединения восста-
новленного железа и окисленного марганца с липидами
хлоропластов. Замена флавиновых дегидрогеназ (с негеминовым
железом) дегидрогеназами с цинком, цитохромами с геминовым
железом, оксидазами с медью и т. д. имело большое значение для
повышения окислительно-восстановительного потенциала реак-
ции. При этом скорость восстановления СО2 более чем в 100 раз
превысила скорость хемосинтеза. В целом эволюция растений в
направлении повышения интенсивности фотосинтеза и дыхания
была связана с накоплением и изменением соотношения ряда
металлов в клетках, вхождением их в состав оксиредуктаз, умень-
шением содержания железа по сравнению с марганцем (табл. 2, 3).
Таблица 2. Эволюция окислительно-восстановительных
реакций фотосинтеза (по Е. А. Бойченко, 1968)
Стадии Е& при рН7 Изменение реакций в эволюции биосферы Участие в реакциях
IV +0,8 4-0,7 4-0,6 Переход к восстановлению СО2 при фо- тосинтезе с выделением О2 СО2 +>2Н2О -> (СНгО) + Н2О + 02 КоА, FeHiv пигменты, ФАД, Мп
III 4-0,5 4-0,4 4-0,3 4-0,2 4-0,1 Фоторедукция СО2 у водорослей и изо- лированных хлоропластов: СО2 4- 2Й2О 4- Ап -> (СН2О) 4- A11O2 КоА, FeHr, пигменты, ФАД
II 00 —0,1 —0,2 —0,3 —0,4 Восстановление СО2 при бактериальном фотосинтезе: СО2 4- 2Н2А1 -> (СН2О) 4- Н2О 4- 2Ai КоА, FeHr, пигменты
80
Продолжение табл. 2
Сидни при рН7 Изменение реакций в эволюции биосферы Участие в реакциях
I —0,5 Восстановление СО2 при хемосинтезе: Код
—0,6 —0,7 СО2 + 2 Н2 -> (СН2О) + Н2О FeHr
Примечание. Few — негеминовое железо, входящее в состав гидрогеназы.
Таблица 3. Изменение содержания металлов в растениях
(по Е. А. Бойченко, 1968)
Растения Число видов Среднее содер- жание Fe, % Соотношение содержания ме- таллов
Fe/Zn Fe/Cu Fe/Mn
Покрытосеменные: листья 30 1,2 • 10~2 3,8 10,0 0,2
кустарников и деревьев
целые растения 49 1,4 • 10-2 3,0 10,6 1,8
Архегониальные 13 8,2 • 10~2 9,9 58,6 3,3
Водоросли 21 1,8 • 10-1 22,8 73,6 38,3
В связи с этим интересно также отметить, что в ходе эволюции
фототрофности усиление и стабилизация функций ферредоксина в
хлоропластах у покрытосеменных шли более быстрыми темпами,
чем у синезеленых.
В то же время эти изменения сочетались с совершенствованием
(сменой) субстратов для аккумуляции энергии. Так, М. Кальвин
подчеркивает, что у пурпурных бактерий энергия аккумулируется в
пирофосфатах, у грибов — в полифосфатах, у высших растений —
в адениновых нуклеотидах и полифосфатах (Л. А. Тумерман, 1964).
Как утверждает А. А Красновский (1975), у первичных гетеротроф-
ных существ свет мог активизировать отдельные звенья цепи пере-
носа электронов еще до образования пигментной системы. Именно
необходимость использования инертных субстратов как доноров
водорода привела к включению пигментной системы в цепь пере-
носа электронов от субстрата к пиридиннуклеотиду, что имело
важное значение для стабилизации механизма фотосинтеза. Такую
же роль сыграло появление и сопряжение работы двух фотосистем,
циклического и нециклического транспорта электронов.
Компоненты ФС1 и ФСП имеют как сходство, так и различие.
При бактериальном фотосинтезе доноры электронов обладают бо-
лее восстановительными свойствами, чем вода. Поэтому бактерии
обходятся энергией ФС1. У высших растений ФС1 и ФСП взаимо-
дополняют друг друга. В эволюционном плане реакция восстанов-
ления НАДФ+ появляется на пути выделения О2 при фотосинтезе,
ее нет у фототрофных бактерий. У зеленых же растений электрон
81
от воды сначала поступает в ФСП и лишь потом в ФС1. По этой
причине в Z-схеме энергии накапливается в три раза больше, чЬм
первично высвобождается. г
Наиболее распространено мнение о равном вкладе ФС1 и ФСП
энергии в генерацию потенциала у зеленых растений. Доказана
миграция от ФСП к ФС1. Она осуществляется с максимальной
эффективностью в пространственно разобщенных тилакоидах, но
снижается при индуцировании слипания тилакоидов в граны обра-
боткой хлоропластов катионами металлов. При снижении миграции
энергии между ФСП и ФС1 усиливается ее миграция внутри ФСП
(J. М. Briantaes et al., 1982). У мутантов неактивной ФС1 редуциро-
ваны межгранные участки тилакоидов и накапливается хлорофилл
с максимумом поглощения 690, 697 и 708 нм. Неполная потеря
активности ФСП ведет к редукции числа и размеров •' гран, к
нарушению синтеза хлорофилла с максимумом поглощения 684 нм
(М. Е. Ладыгина и др., 1982). Роль указанных фотосистем зависит
от качества света. Например, при освещении красными лучами у
красных водорослей лимитирующими оказываются реакции, про-
текающие с участием ФСП, а в желтой и зеленой областях спектра,
поглощаемых фикобилинами,— ФС1.
Происхождение двух фотосистем объясняют по-разному. Н. М.
Чернявская и Д. С. Чернявский (1967) указывают, что еще до
появления системы фотолиза воды возникли ассоциаты из одина-
ковых, но последовательно работающих ФС1, видоизменение кото-
рых путем мутаций могло привести к возникновению ФСП.
Поэтому считают, что ФСП возникла позже ФС1. В онтогенезе у
этилированных листьев при позеленении вначале формируется
ФС1, а затем ФСП.
Возникновение двух фотосистем привело к усложнению реак-
ционных центров фотосинтеза. Полагают, что один тилакоид со-
держит 400 реакционных центров ФС1 и ФСП, имеет объем 1,5 х
х 1014 см3 и площадь поверхности 2 • 10“* см2. Объем внутритила-
коидного пространства и стромы в интактном хлоропласте состав-
ляет соответственно 3,3 и 2,5 мкл/мг хлорофилла (Л. Н. Воробьев
и др., 1980). Все это усложняло мембранную укладку хлоропластов
и стабилизировало фотосинтез.
Факты, накопленные при изучении основных этапов фотосин-
теза (поглощение квантов, миграция энергии возбуждения и ее
захват, роль двух фотосистем в переносе электронов и т. д.), свиде-
тельствуют о возникновении многообразных связей между его
первичными процессами в ходе эволюции. Это способствовало
стабилизации механизма фотосинтеза как универсального процесса.
Стабилизация механизма фотосинтеза привела к преобладанию
накопления органического вещества над его минерализацией,
уменьшению содержания СО2 и увеличению содержания О2 в
82
атмосфере. Таким путем уже на начальных этапах включения
Механизма фотосинтеза создается угроза для роста его эффектив-
ности. Выход был найден путем усиления потребления его продук-
тов в пределах организма, что было связано с возникновением
многоклеточности и усилением дифференциации организма (см.
гл. 4).
Для стабилизации механизма фотосинтеза эти события имели
далеко идущие последствия, о них обычно забывают, придавая
значение только его молекулярным процессам. Так, дифференци-
ация организма на вегетативные и генеративные структуры способ-
ствовала созданию фототрофами дополнительного продукта
фотосинтеза. При заселении водорослями прибрежной зоны водо-
емов (только прикрепленные формы могли там удержаться при
приливах и отливах воды, непрерывно омываясь водой, в результате
чего постоянно менялась концентрация СО2 и минеральных веществ
вокруг организма) создаются благоприятные условия для дальней-
шего действия отбора в направлении многоклеточности. Увеличе-
ние только размеров клеток ограничивало возможности усиления
фотосинтеза, хотя известно, что с увеличением размера и усложне-
нием структуры клетки более 60—70% продуктов фотосинтеза
потребляются материнской клеткой. Вероятность увеличения раз-
мера единственной клетки не беспредельна, поэтому эволюция
пошла по пути ограничения роста клеток, их измельчения, умень-
шения контактов клеток с внешней средой. Эти события (по еще
недостаточно изученным причинам) имели принципиальное значе-
ние для интенсификации процесса размножения клеток и возник-
новения многоклеточных фотоавтотрофов.
С возникновением организмов, дифференцированных на клет-
ки, ткани, органы и системы органов, появилась еще большая
потребность в продуктах фотосинтеза. Одним из доказательств этого
служит тот факт, что у многоклеточных водорослей каждая единица
массы хлорофилла способна фиксировать СО2 в большем количе-
стве, чем у одноклеточных. С выходом растений на сушу использу-
ются разные субстраты питания, в результате чего происходит
дальнейшая дифференциация тела и увеличение разнообразия ви-
дов. Как подчеркивает А. А. Ничипорович (1980), в этих условиях
возникают новые типы и механизмы взаимодействия между функ-
циями фотосинтеза и роста в целостном организме и создается
система самонастройки процессов питания, роста и размножения
на эффективное использование растениями света.
Эволюция фотосинтеза у высших растений была связана не
только с интенсификацией его механизма, но и с возникновением
ограничений в смысле возможности использования всей интен-
сивности и диапазона волн световой энергии. Дело в том, что
чрезмерное поглощение света могло оказать повреждающее дей-
83
ствие на организмы и внутриклеточные процессы. Как подчерки-
вают А. Т. Мокроносов й В. Ф. Гавриленко (1992), в эволюции
фотосинтеза высших растений одной из трудных задай было дости-
жение хорошего снабжения фотосинтетического аппарата, диокси-
дом углерода ввиду его низкого содержания в атмосфере и
необходимости преодоления ряда барьеров на пути передвижения
СО2 к фотосинтетическим центрам (устьичное сопротивление, со-
противление мезофилла и сложной системы мембран). Эти ограни-
чения компенсировались приобретением других новшеств: 1)
способность извлекать СО2 из среды при низких концентрациях и
двухступенчатой передачей диоксида углерода на карбоксилирова-
ние РБФ при участии РБФК и ФЕПК; 2) С4-пути фотосинтеза для
устранения трудности диффузии СО2 к местам карбоксилирования;
3) CAM-пути фотосинтеза, при котором ночью при открытых
устьицах много СО2 фиксируется дикарбоновыми кислотами при
участии ФЕПК; 4) фотодыхания, устранившего ингибирующее
действие О2; 5) увеличения числа устьиц на поверхности листа
(например, у С3-растений их число достигает бдлее 300, у САМ —
25—30 шт. на 1 мм2); 6) подключения к транспортировке через
мембраны СО2 карбоангидраз, роль которых особенно велика в
жизни С3-растений (поэтому у них активность карбоангидраз в
листьях в 10 раз выше, чем у растений С4-типа).
Итак, эволюция углеродного питания имеет ряд ступеней. Она
связана как с изменением первичного механизма процесса фикса-
ции СО2, так и с усилением его продуктивности при попадании
растений в разные среды. В конце концов появляются растения с
высокопродуктивным фотосинтетическим аппаратом, у которых
фотосинтез идет при низких концентрациях СО2 и высоких кон-
центрациях О2. Этому способствовало развитие определенных
структур и пигментов фотосинтеза. •
3.6. ЭВОЛЮЦИЯ ПИГМЕНТНОЙ СИСТЕМЫ ФОТОСИНТЕЗА
Эволюция фотосинтеза тесно связана со становлением его
аппарата (пигментной системы, хлоропластов, листьев и т. д.).
Данные «молекулярной палеонтологии» показывают, что порфири-
носодержащие структуры были синтезированы из пиррольных со-
единений уже в процессе химической эволюции, но только с
появлением живых организмов они нашли соответствующее при-
менение в качестве хлорофиллов, цитохромов и гемоглобинов. С
проникновением порфиринов в коацерваты создалась возможность
для обратимых фотохимических окислительно-восстановительных
реакций (А. И. Опарин, 1969). Дальнейшее усиление каталитиче-
ских функций порфиринов было достигнуто путем образования
84
комплексов с Fe, Си, Mg, Со. К тому же порфирины, обладая
антирадиационными свойствами, обеспечивали живым существам
успех в борьбе за существование.
Сложен путь эволюции молекулы фоторецептора фотосинтеза.
Первично хорошо растворимые в воде порфирины не могли быть
включены в мембраны. Поэтому отбор пошел в направлении уси-
ления растворимости их в липидах, что привело к уменьшению в
молекуле карбоксильных ipynn и увеличению гидрофобных ради-
калов — возникают молекулы с остатком фитола. Присоединение
последнего к молекуле хлорофилла и обеспечило возможность
взаимодействия пигмента с липидами мембран. В то же время в
ходе эволюции фототрофов происходит смена форм пигмента. У
зеленых бактерий (наиболее примитивных фототрофов) преоблада-
ют бактериохлорофиллы с и d, у пурпурных бактерий — бактери-
охлорофиллы а и Ъ и фитол, у цианей — хлорофилл а, у бурых и
диатомовых водорослей —хлорофилл а и с, у красных—хлоро-.
филл d. В последующем фикобилины заменяются хлорофиллом b
(эвгленовые и зеленые водоросли).
Порфирины как фоторецепторы, по-видимому, предшествовали
хлорофиллам. Они широко распространены у бактерий, растений
и животных, пути их биосинтеза очень древние. Одна из древних
форм порфиринов —уропорфирины (возникающие и абиогенно) —
обладает способностью воспринимать на свету электрон от доноров.
Такие порфирины, поглощая свет умеренной энергии, могли удов-
летворить энергетические потребности жизни на начальных этапах
ее эволюции. Но по мере увеличения суммы жизни, когда возникли
дополнительные потребности в органическом веществе, этого было
уже недостаточно. Для создания последнего требовалось усилить
фотопоглощающую способность порфиринов, что было достигнуто
отбором фототрофов на возможность включения ионов металлов в
порфириновое кольцо. Следует заметить, что порфирины имеют
более выраженное химическое сродство к ионам Fei+, Со2+ и Zn2+,
чем к Mg . Тем не менее Mg-порфирины появляются у фототрофов
предположительно несколько раньше, чем комплексы с другими
металлами. Последние стали выполнять иные функции —участие в
темновом транспорте электронов.
Порфирины могли синтезироваться на первичной Земле абио-
генно, путем конденсации четырех молекул пиррола. В восстанов-
ленной атмосфере продуктами подобной конденсации могли быть
восстановленные продукты-хлорины. Переход к окисленным фор-
мам порфиринов происходил в последующем по мере насыщения
атмосферы О2.
Возникновение хлорофилла связывают с превращением уропор-
фирина или выработкой других путей для его биосинтеза в орга-
низме. Если принять во внимание вывод о более эволюционной
8$
древности гема, чем хлорофилла (М. Кальвин), то судьба фототроф-
ности первично зависела от отбора организмов по способности
образовывать в них комплексы порфиринов с Mg2+. Именно для
них могли быть характерны наибольшие возможности к окисли-
тельно-восстановительным превращениям порфириновых соедине-
ний.
В ходе эволюции возникает сочетание функций Mg-порфиринов
и Fe-порфиронов в молекулах хлорофилла и цитохромов. Порфи-
риновое кольцо стало основой для молекул гема и хлорофилла. Эти
молекулы у животных и растений синтезируются из 5-аминолеву-
линовой кислоты, последняя у растений образуется из глутаминовой
кислоты. У животных также около 25% аминолевулиновой кислоты
образуется из глутаминовой кислоты. Итак, пути синтеза гема и
хлорофилла оказываются очень сходными, что, вероятно, связано
с общностью их происхождения. Есть указания и на то, что синтез
гема мог служить промежуточным, этапом в реакции биосинтеза
хлорофилла (Г. Гаффрон). Отбор по Mg-порфиринам среди фото-
трофов, кроме того, был обусловлен их способностью оставаться в
возбужденном состоянии и образовывать связи с другими компо-
нентами фотосинтетического аппарата. С возникновением фермен-
та метилтрансферазы (S-aдeнoзилмeтиoнин-Mg-пpoтoпopфиpин-
метилтрансфераза) окончательно складываются пути биосинтеза
хлорофилла у фототрофов.
Порфирины принадлежат к числу наиболее стабильных органи-
ческих молекул, что обусловлено их циклической структурой и
энергией резонанса (38 Дж/моль). Один из представителей Порфи-
ринов — хлорофилл — способен долго сохранять энергию погло-
щенного кванта и передавать ее идентичной молекуле. Как
указывает А. А. Красновский, благодаря этим и другим особенно-
стям порфиринов молекула хлорофилла стала основным пигментом,
участвующим в процессах фиксации энергии света.
Интересны причины, приведшие к отбору центрального металла
в порфириновом кольце. В земной коре элементы встречаются в
следующей убывающей концентрации: кремний (27,6%), алюми-
ний, железо, калий, натрий, кальций, магний и титан (0,6%).
Соединения кремния, алюминия и титана нерастворимы в воде,
поэтому у них трудно образуются порфириновые комплексы. В воде
порфириновые комплексы с натрием, калием и кальцием быстро
гидролизуются. Наиболее устойчивы и активны комплексы с маг-
нием и железом, способные к возбуждению и обратимому восста-
новлению или окислению.
Известно, что функцию «ловушек» фотонов выполняет не толь-
ко хлорофилл, но и другие пигменты. Однако в реакционный центр
86
фотосинтеза у растений входит главным образом хлорофилл, в
частности его длинноволновые формы. Роль других пигментов
(каротиноиды, фикобилины) в фотосинтезе высших растений менее
значительна, они служат для удаления из системы избыточной
энергии квантов коротковолнового света, что предохраняет хлоро-
филл от разрушения. Так, хотя каротиноиды не участвуют в пер-
вичных реакциях и фотосинтетическом транспорте электронов, они
защищают фотосинтетический аппарат от деструктивного действия
света. Роль каротиноидов в фотосинтезе усиливается с увеличением
его спектрального диапазона (Дж. Эдвардс, Д. Уокер, 1986).
В случае ингибирования процесса биосинтеза каротиноидов
специфическими ингибиторами или при возникновении соответ-
ствующих мутаций происходят необратимые нарушения фото-
синтетического аппарата (А. Т. Мокроносов, В. Ф. Гавриленко,
1992). Кроме того, каротиноиды участвуют в устранении токсич-
ности О2 (см. гл. 4), процессах оплодотворения (П. М. Жуков-
ский), защите от инфекций (см. гл. 5) и т. д. Поэтому
каротиноиды имеются у всех организмов. Разнообразен их состав
у фототрофов: только у пурпурных бактерий их обнаружено
свыше 20 (М. В. Гусев, Л. А. Минеева, 1985).
В то же время эволюция фототрофности идет в направлении
усиления роли хлорофилла. Центральное положение магния в
молекуле порфиринов предопределило их преимущественное ис-
пользование в поглощении, запасании и преобразовании световой
энергии. Как подчеркивают А. Т. Мокроносов и В. Ф. Гавриленко
(1992), циклическая структура молекулы, физико-химические свой-
ства и природа электронно-колебательных спектров, способность
к обратимым окислительно-восстановительным превращениям Mg-
порфиринов определили эффективность их участия в фотосинтезе;
они способны с 100%-ной эффективностью преобразовывать энер-
гию за счет только жесткой структуры самой молекулы. Начиная с
эвгленовых дальнейшая эволюция фототрофности была связана с
усложнением структуры молекул Mg-порфиринов и отбором в
популяциях в направлении повышения роли хлорофилла в клетке,
образование его комплексов с белками. Наконец, повышение эф-
фективности фотосинтеза достигается поддержанием гетерогенно-
сти пигментной системы (включая хлорофилл) и потоком энергии
от коротковолновых к длинноволновым формам пигментов.
Разнообразие состава пигментов у фототрофНЫх организмов
достаточно хорошо показано, что достигается за счет небольших
модификаций боковых группировок Mg-порфиринов (хлорофилл и
бактериохлорофиллы) или взаимодействием одной и той же моле-
кулы с белковыми и липидными компонентами (А' Т. Мокроносов,
В. Ф. Гавриленко, 1992). Известно более 10 нативных форм хлоро-
филла. Среди них различают хлорофилл а, обнаруженный у водо-
87
рослей и высших растений; хлорофилл b — у зеленых, харовых,
эвгленовых и высших растений; хлорофилл с — у бурых водорослей,
золотистых и диатомей; хлорофилл d— у красных водорослей.
Бактериохлорофиллы обнаружены у бактерий: а и b —у серных и
несерных пурпурных, с и d — у зеленых серобактерий. Кроме того,
у цианей и красных водорослей имеются фикоцианин и фикоэрит-
рин. У прокариот есть также каротиноиды, ксантофиллы (лютеин,
зеоксантин, виолоксантин и фукоксантин) и фикобилины (фико-
цианин и фикоэритрин), причем у разных групп в разном сочетании.
Бактериохлорофиллы поглощают лучи с большей длиной волны
(705—1100 нм), чем. хлорофиллы. У цианей фотосинтез идет с
участием хлорофилла, представленного в виде нескольких спект-
ральных форм. У них обнаружено до 30 различных пигментов,
благодаря чему цианеи способны к хроматической адаптации в
условиях пещер, куда свет проникает ограниченно. В целом разно-
образие состава пигментов фотосинтеза у бактерий и водорослей
представляет собой приспособление к условиям с ограниченным
доступом света (разнообразные водоемы с разной глубиной).
Окончательный выбор хлорофилла как фотосенсибилизатора
объясняют его способностью включаться в липопротеиновые ком-
плексы мембран (и к тому же с увеличением фотосенсибилизаци-
онной активности) и концентрироваться на мембранах (что имело
значение для упорядочения функции хлорофилла). Фотосенсиби-
лизационная активность хлорофилла в 100 раз превышает таковую
у протохлорофилла. Бактериохлорофиллы не могли стать универ-
сальными фотосенсибилизаторами для всех фототрофов ввиду низ-
кой (7,7 Дж/моль) энергии активации по сравнению с хлорофиллом
(10,5 Дж/моль). Потенциал энергии их активации оказывается
недостаточен для фотолиза воды.
Указанные обстоятельства и определили первоначальное за-
крепление хлорофилла в роли фотосенсибилизатора. Эволюцион-
ную линию, приведшую к образованию бактериохлорофиллов,
рассматривают как тупиковую в развитии фототрофности и способ-
ности выделять кислород (Р. Клейтон, 1984).
Как показал еще К. А. Тимирязев, фотосинтез осуществляется
только в лучах света, поглощаемых хлорофиллом. Указав, что
хлорофилл способствовал завоеванию растениями суши, он пишет:
«Хлорофилловая функция должна была первоначально выработать-
ся у первобытных морских водорослей, и именно в этой подводной
флоре встречаемся с наибольшим разнообразием пигментов. Из
всех этих веществ, вероятно, самое важное — хлорофилл». К важ-
ным особенностям хлорофилла, определившим его как основной
пигмент фотосинтеза, принято относить его способность к окисли^
тельно-восстановительным превращениям, поглощать квант света
и отдавать электрон акцептору. В клетках фотосинтезирующих
88
тканей этот процесс завершается за доли секунд. За 1 с молекула
хлорофилла в окислительно-восстановительном процессе оборачи-
вается в 100 раз (Р. Клейтон, 1984). Поэтому, несомненно, за
единицу времени и пространства органическое вещество создавали
наиболее продуктивно те из фототрофов, которые использовали
хлорофилл как сенсибилизатор. К другим преимуществам хлоро-
филла следует отнести его «настрой» на красный свет, способность
удерживать поглощенную энергию и донорно-акцепторную измен-
чивость. Все это определило «победу»хлорофилла в энергетической
конкуренции;
Для эволюционной стабилизации биомолекул (хлорофилл, ро-
допсин, цитохромы и др.) важное значение имеет их соответствую-
щая организация — «окружение» (Н. Т. Witt, 1983). Предполагают
такие ее эволюционные этапы: трехмерное пространство -> двух-
мерная «кювета» из жирных липидов -> анизотропия в организации
мембран -> образование замкнутых мембранных структур (вези-
кул). В итоге специфичность участия хлорофилла и реакций фото-
синтеза характеризуется: 1) наличием процессов регуляции для
утилизации энергии света — кванта; 2)кооцерацией функций двух
фоторецепторных центров, хлорофилла а и b при переносе элект-
ронов от Н2О к НАДФ+; 3) направленностью потока энергии из
возбужденных реакционных центров; 4) возникновением циркуля-
ции Н+ и накачки Н+; 5) использованием протонного потенциала
для образования АТФ; 6) использованием Н2О как источника
электронов. Говоря о необходимости организации биомолекул,
следует обратить внимание на возникновение хлоропласта для
укладки молекул хлорофилла и на усложнение структуры самой
органеллы фотосинтеза.
3.7. ХЛОРОПЛАСТЫ, ИХ ЗНАЧЕНИЕ И ЭВОЛЮЦИЯ
Фотосинтез совершается при непрерывном поступлении значи-
тельных количеств световой энергии. В связи с этим возникает
необходимость в концентрации пигмента в строго определенном
участке клетки путем соответствующей упаковки его молекул. По-
этому физико-химические свойства хлорофилла наилучшим обра-
зом реализуются в хлоропластах, имеющих довольно сложную
структуру. Формирование хлоропластов в процессе эволюции в то
же время устранило конкуренцию за электроны благодаря разделе-
нию функций синтеза АТФ в пропластидах и промитохондриях. Эти
приобретения имели значение для преобразования цикла Арнона в
Цикл Кребса, что способствовало более полному удовлетворению
потребностей клеток в АТФ.
89
Состав и структура хлоропластов сложны. Они состоят из пиг-
ментов, ДНК, ферредоксинов, рибосом, минеральных элементов и
имеют многослойную структуру, в них же находятся реакционные
центры ФС1 и ФСП. В хлоропластах ДНК содержится 2 • 10’ пар
нуклеотидов, т. е. несколько тысяч генов. Сходны ДНК хлоропла-
стов и цианей. Рибосомы состоят из 55S- и SOS-субчастиц. Генети-
ческая информация пластид обеспечивает синтез части белков ФСП
и ЭТЦ, контролирует размножение и размеры хлоропластов. В
хлоропластах около 70—80% тилакоидов концентрируются в отно-
сительно небольшом числе уложенных стопами гран, остальные
20—30% расположены в строме между отдельными гранами. Воз-
действуя на хлоропласты шпината 2—3-валентными катионами,
удается индуцировать сборку их в граны. Взаимное затенение гран
не оказывает влияния на фотосинтез ввиду высокой концентрации
ПБК в каждой отдельной гране. С увеличением продолжительности
пребывания хлоропластов на свету число единиц ФС1 и ФСП
возрастает, также происходит накопление хлорофилла и светособи-
рающего хлорофилл-белкового комплекса.
На первичной Земле абиогенным путем возникают ферредок-
сины —низкомолекулярные белки, содержащие железосерные
кластеры. Они выполняют в организме различные функции в цепях
переноса электронов: фотовосстановление НАДФ+, участие в фик-
сации азота, СО2 (при синтезе а-кетокислот у фотосинтезирующих
бактерий), декарбоксилирование пирувата (у галобактерий) и т. д.
Поэтому они встречаются у микроорганизмов, растений и живо-
тных. В одном и том же организме часто обнаруживается несколько
изоформ белка ферредоксинов. У высших растений несколько
генов, ответственных за их синтез. Полагают, что в растениях были
использованы предковые ферредоксины, которые, включаясь в
различные системы, усилили функции ферредоксинов. По этой
причине наблюдается специфика ферредоксинов хлоропластов и
бактерий (фототрофных и нефототрофных). В целом эволюция
ферредоксинов шла по пути наращивания фрагментов в одной цепи.
По этой причине ферредоксины высших растений, особенно в
пределах семейства, весьма сходны.
У прокариот реакционные центры фотосинтеза локализованы в
клеточных мембранах. У пурпурных бактерий и прохлорофитов
светособирающие пигменты в комплексе с белком входят в мемб-
раны. У других бактерий пигменты содержатся в особых структурах,
прикрепленных к поверхности мембран,—хлоросомах (зеленые
бактерии) и фикобилисомах (цианеи). Эго начальная стадия упо-
рядочивания упаковки пигментов. Организация пигмента у цианеи
более совершенна, она основана на развитой системе внутрицитоп-
лазматических мембран — тилакоидов. К последним прикрепляют-
ся и фикобилисомы.
90
Дифференциация разных сторон мембраны (в том числе и
тилакоидов хлоропластов) фиксацией пигментов привела к усиле-
нию роли мембран вообще. Так, разложение воды и выделение О2
происходят благодаря этому на внутренней стороне тилакоида, а
восстановление НАДФ — на внешней (здесь фиксируются фермен-
ты и коферменты, участвующие в восстановлении НАДФ и СО2).
Пластохиноны — посредники между ФС1 и ФСИ в переносе элек-
тронов — встречаются на обеих сторонах мембраны. Функциональ-
ная дифференциация поверхностей мембраны была нужна для
предотвращения наступления равновесия процессов (взаимное га-
шение) и обеспечения «перекачки» и перераспределения ионов и
заряда. Скорость переноса электронов через ФСП (+820 мВ) в 1,8
раза больше, чем через ФС1. Энергия последней (+250 мВ) явно
недостаточна для осуществления фотолиза воды и создания «асси-
миляционной силы» фотосинтеза — НАДФ • Н2 и АТФ:
hy ' •
АДФ + Фн + НАДФ + Н2О -> АТФ + НАДФ • Н2 + 72 О2
хлорофилл
При ассимиляции одной молекулы СО2 выделяется одна молекула
О2. Эта реакция протекает с поглощением энергии 7,8 • 10—19 Дж
(168 ккал/моль), т. е. 8—12 квантов света. Она как раз создаете^ в
результате сопряжения работы двух фотосистем.
По некоторым данным, фотохимическая реакция совершается
в РЦФ, куда входят 300—350 молекул хлорофилла. Целесообраз-
ность концентрации такой большой группы молекул хлорофилла в
1 РЦФ обусловлена тем, что каждая молекула поглощает один фотон
лишь один раз за 0,1 с (при длительности жизни возбужденного
состояния 5 • 10-9 с). Плотное расположение молекул хлорофилла
и других пигментов в хлоропластах благоприятно для переноса энер-
гии. По этой причине возникает необходимость объединения большой
группы молекул пигментов в одном фотохимическом центре. Передача
энергии между отдельными молекулами при этом весьма эффективна
и составляет примерно 80—100%. Так как в естественных условиях у
большинства видов растений концентрация хлорофилла превышает
2,5 • 10“5 г/см2, то поглощение света не лимитируется недостатком
хлорофилла (И. И. Свенитицкий, 1982).
Хлоропласты растений разных экологических групп отличаются
между собой. Усиление мощности светового потока способствует
уменьшению размеров хлоропластов, возрастанию их числа на
клетку й числа клеток хлоренхимы на единицу площади пластинки.
При низкой интенсивности света число хлоропластов на клетку
уменьшается, но содержание хлорофилла в них возрастает. Изме-
нениями в фотоактивной поверхности хлоропласта достигаются
относительное равновесие фотосинтетического аппарата и саморе-
гуляции жизнедеятельности растений при отклонении условий от
| 91
нормы (Л. В. Кахнович, 1980). Число хлоропластов на клетку в
конечном итоге генетически определено и коррелирует с содержа-
нием ДНК в ядре (А. Т. Мокроносов, 1983). Содержание хлоро-
филла на один хлоропласт по мере увеличения числа хлоропластов
на клетку падает. Хлоропласты злаков в течение 500-летней селек-
ции остались стабильными, несмотря на достигнутые успехи в
повышении продуктивности растений.
Изменением положения хлоропластов в клетке достигается
адаптивное упорядочение их деятельности при вариации освещен-
ности. При низкой и средней освещенности благодаря изменению
положения хлоропластов максимально адсорбируется световая
энергия, а при высокой интенсивности они принимают «защитную
позицию». Изменение положения хлоропластов по отношению к
свету (фототаксис хлоропластов) описано на примере многих рас-
тений и представляет адаптивную реакцию растений на направление
и интенсивность света. Фототаксисом обладают и обесцвеченные
пластиды. При относительно малых потоках энергии хлоропласты
как живая система адаптируются за короткое время к полному
использованию энергии с максимальной эффективностью для себя.
Ввиду их активного перемещения к свету содержанйе хлорофилла
растет за 30—60 мин после начала освещения. В процессе зеленения
пластид происходит становление их ламеллярной организации в он-
тогенезе и возрастание интенсивности фотосинтеза хлоропластов.
Сравнение рибосом и рибосомальных белков пластид показало,
что в строении рРНК хлоропластов и прокариот имеется много
общего (М. С. Одинцова, 1984). В то же время белки хлоропластов
оказались весьма специфичными и разнообразными (около 100
белков). Все компоненты рРНК хлоропластов синтезируются соб-
ственной ДНК, тогда как белки рибосом пластид — с участием
ядерного и хлоропластного геномов. Составлены генетические кар-
ты генома хлоропласта. По наличию интронов в генах тРНК и
спейсерной последовательности хлоропластный геном довольно
сходен с геномом эукариот.
Хлоропласты как датчик ассимилятов имеют механизмы авто-
регуляции, позволяющие поддержать баланс между первичным
синтезом и оттоком глюкозы (А. Л. Курсанов, 1988). Среди них и
контроль за деятельностью хлоропластов со стороны организма,
регуляция, активности генетического аппарата светом, метаболита-
ми и гормонами. Мы теперь в целом лучше представляем органи-
зацию и функционирование хпДНК по генетическим картам,
составленным для 20 видов растений. Геном хлоропласта имеет
полиплоидную природу, в нем содержатся две инвертированные
повторяющиеся последовательности. Мутационные изменения
обычно возникают только в уникальных ее участках, в хпДНК есть
сходные последовательности с геномами ядра и митохондрий. В то
же время хлоропластный геном оказывается более консервативным,
92
чем адерный (Ф. Ф. Литвин, 1988). Информационная емкость
хпДНК. в состоянии обеспечить синтез примерно 130 полипептидов
(В. Е. Семененко, 1988).
Генетический аппарат хлоропластов в ходе эволюции растений
также изменился, что видно из сравнения величины ДНК и после-
довательности в ней нуклеотидов низших и высших растений.
Однако подобные изменения оказались в хпДНК значительно ниже,
чем в ядерной. Обнаружены различия и в структуре мРНК хлоро-
пластов у низших и высших растений.
Все современные цианеи обладают одинаковым набором фото-
синтетических пигментов (хлорофиллы и фикобилины) и од нотипной
организацией фотосинтетических мембран (одиночные тилакоиды без
ламелл и гран). Фотосинтезирующий аппарат цианей сходен с пла-
стидами Rhodaphyta и Glaucophyta (И. М. Мирабдуллаев, 1985). Со-
гласно некоторым представлениям, в докембрии существовали восемь
типов фототрофных прокариот, использовавших в качестве доноров
электронов воду: синезеленые (считают, что они дали начало пласти-
дам красных и криптофитовых водорослей, обладали дополнительны-
ми пигментами—фикобилинами), зеленые прокариоты
(родоначальники пластид зеленых и эвгленовых водорослей, содер-
жавшие хлорофилл) и желтые прокариоты (давшие начало пластидам
динофлагеллят, золотистых, диатомовых и бурых водорослей).
Гипотеза симбиогенеза предполагает, что на ранних этапах
развития жизни имело место (в результате случайного захвата)
проникновение в прокариоты свободно живущих форм типа мито-
хондрий, хлоропластов и ядра (Л. Марголис, 1983; Л. Н. Хахина,
1979). Подобные первичные существа, найдя благоприятную эко-
логическую нишу внутри клетки, еще более активно стали выпол-
нять свойственные им функции, что оказалось обоюдно выгодным
и было поддержано отбором.
В пользу указанной гипотезы приводят доказательства: 1) нали-
чие раздельного существования прокариот, у которых все еще
разобщены процессы фотосинтеза и дыхания; 2) обнаружение зна-
чительной генетической автономности хлоропластов и митохонд-
рий в клетке, их способность к синтезам in vitro', 3) наличие
гомологичных последовательностей нуклеотидов в геномах мито-
хондрий и хлоропластов прокариот и эукариот; 4) сходство ЭТЦ
митохондрий и хлоропластов, а также наличие в их геномах генов,
кодирующих такие субъединицы, как АТФ-синтетаза, НАДН-де-
гидрогенеза и цитохромный комплекс.
Ряд авторов ставят под сомнение возможность существования
в свободном состоянии таких форм, как хлоропласты и митохонд-
рии. Поэтому их происхождение объясняют путем инвагинации
мембран клетки, а автономность—захватом генетического мате-
риала, рассеянного в полинуклеотидах прокариот. В связи с этим
ссылаются на факты неспособности геномов митохойдрий и хло-
93
ропластов обеспечить себя необходимыми для функционирования
и размножения веществами. Многие белки в них синтезируются с
участием яДНК и цитоплазмы. Более того, функционирование
мтДНК и хпДНК зависит от синтеза ферментов, кодирующих
яДНК. Кроме того, фрагменты мтДНК и хпДНК содержатся и в
составе яДНК высших растений, хотя они там малоактивны. Общим
для мтДНК, хпДНК и яДНК является наличие сходных интронов,
взаимодействие геномов органелл у высших растений при функци-
онировании.
Независимо от решения вопроса о происхождении эукариотной
и фотосинтезирующей клеток такие системы возникли в процессе
отбора и имели значение для прогресса биосферы. По-видимому,
возникновение фототрофности происходило параллельно в попу-
ляциях разных гетеротрофов, о чем отчасти свидетельствует разно-
образие форм ее проявления у современных прокариот. «Спрос» на
фототрофный метаболизм, выходя за популяционные потребности,
был связан с необходимостью обеспечения энергетическим мате-
риалом распространявшейся по Земле жизни. Это и определило
широкую дивергенцию фототрофных существ в направлении уси-
ления как физико-химических механизмов фиксации углерода, так
и поиска возможностей их эффективного использования.
Теория симбиогенеза хорошо объясняет топологию мембран
пластид зеленых и красных водорослей: тилакоиды пластид гомо-
. логичны тилакоидам фотобактерий, внутренняя мембрана оболочки
пластиды гомологична плазмолемме прокариот. Высказано пред-
положение о существовании древних цианобактерий оксифототро-
фов. Прогрессивная эволюция оксифотобактерий протекала в
направлении все большего совершенствования светособирающей ан-
тенны ФСП. Наличие ФСП обязательно для пластид и оксибактерий.
Симбиотическим происхождением хлоропластов объясняют ав-
тономность их функционирования, наличие собственной ДНК ге-
нома. Представления об эндосимбиотическом происхождении
хлоропластов и митохондрий, возможности неоднократного такого
их возникновения получили некоторые подтверждения при срав-
нительном изучении филогенетических древ ферредоксина бакте-
рий, 5S рРНК и цитохромов бактерий, водорослей, митохондрий и
хлоропластов. В специальном издании «Эндоцитобиология» (1983)
приведены данные, подтверждающие и развивающие эту гипотезу.
В частности, при сравнении последовательности нуклеотидов 16S
рРНК пластид бурой водоросли Poryphyridium обнаружена гомоло-
гия с таковыми у синезеленых Synechocystis. Гомология не отмечена
с 18S рРНК Poryphyridium. Для сравнения напомним, что гомология
в 16S рРНК у кукурузы и табака составляет 70—75%.
Хлоропласты имеют собственную белок-синтезирующую систе-
му, способную обеспечить себя ферментами и белками. Рибосомы
94
хлоропластов в 20 раз активнее и нуждаются в более высоком пуле
магния, чем рибосомы цитоплазмы. Эффективность синтеза АТФ
в хлоропластах контролируется «запросом» самого растения; при
активном ее использовании биосинтез АТФ усиливается.
В представлениях о мезоструктуре растений (А. Т. Мокроно-
сов) значительное место занимает учет числа, объема площади
проекции и поверхности хлоропластов. У большинства растений
’суммарный объем всех хлоропластов составляет 10—20%, у дре-
весных —до 35—70% от объема клетки. В одном хлоропласте
содержится (1 —2) • 10’ молекул хлорофилла, у теневыносливых
— (5 —10) • 10’ молекул (А Т. Мокроносов, В. Ф. Гавриленко,
1992). Интересно отметить, что в молодых листьях повышение
интенсивности фотосинг|еза идет параллельно с увеличением числа
хлоропластов, а в зрелых — за счет усиления функциональной
нагрузки отдельного хлоропласта.
Исследованиями последних лет (О. Ф. Монахова, Т. Е. Кисля-
кова, 1993) показано, что в ходе эволюции растений меняются число
гран хлоропластов, их диаметр, число и плотность упаковки тила-
коидов в граны, состояние стромы хлоропластов. От папоротников
к цветковым наблюдается увеличение числа клеток на единицу
поверхности листа, уменьшение размеров пластид и числа молекул
хлорофилла в хлоропластах. Однако измельчение клеток у цветко-
вых сопровождается ростом суммарной ассимиляционной поверх-
ности единицы площади листа как результат соответствующего
роста числа хлоропластов. Эволюция цветковых совпала с периодом
возрастания интенсивности освещения. Предотвращение повреж-
дения пластид действием света в ходе эволюции достигнуто умень-
шением числа молекул хлорофилла на хлоропласт.
В ходе эволюции не только возникает и совершенствуется
структура хлоропластов, но и складывается механизм для поддер-
жания их стабильности в онтогенезе — соответствие между оттоком
веществ из них и интенсивностью фотосинтеза, баланса ионов
магния и щелочного значения pH в строме хлоропласта.
Хлоропласты имеют гранальную структуру, обеспечивающую
распределение компонентов ФС1 и ФСП в разных участках мембран
и координацию миграции энергии между фотосистемами и компо-
нентами ЭТЦ фотосинтеза (компонентами ЭТЦ являются все про-
межуточные соединения на пути электронов).
Интересны различия в структуре, форме и числе хлоропластов
у разных видов зеленых растений. Так, у цианей хлоропласты
отсутствуют, а пигменты распределены на двух сближенных внут-
• риклеточных тилакоидных мембранах (имеющих двух- и трехслой-
ные ламеллы), которые составляют около 26% массы сухих клеток
и содержат до 44% белка, 17% эфирорастворимых липидов, 21%
углеводов и 8,5% хлорофилла (М. В, Гусев, К. А Никитина, 1979).
Хлоропласты (хроматофоры) водорослей отличаются большим мор-
95
фологическим разнообразием (чашевидные, лентовидные, спира-
левидные, пластинчатые, звездчатые и т. п.), большим числом,
местоположением, внутренней организацией, а также разнообрази-
ем локализации дисков. Диски могут быть одиночными (красные
водоросли) или представлены пачками по 2—4, расположенными
порознь или совместно (все другие водоросли). У зеленой водоросли
нителлы число дисков в пачке достигает 20. Увеличение числа
дисков в пачке сопровождается их укорочением.
Наиболее простая фотосинтезирующая система имеется у хло-
реллы. Она представлена всего лишь одним хлоропластом прими-
тивной (без гран) ламеллярной (слоистой) структуры. У
хламидомонады вся пластида состоит из одной грани, пигмент же
входит в состав ламелл, где осуществляются световые реакции, а
темновые — в цитоплазме. В клетке фотосинтезирующих бактерий
встречаются тилакоиды (плоские, уложенные стопками мембран-
ные структуры в виде пузырьков), образованные путем выпячивания
цитоплазматической мембраны. У высших растений чаще наблю-
даются шаровидные или дисковидные хлоропласты. У, плаунов
обнаружены ламеллярные хлоропласты (каку водорослей), граналь-
ные (как у цветковых) и переходные между ними (Т. Е. Кислякова
и др., 1976). Папоротниковидные и голосеменные по гранальной
ориентации и развитости фотоактивной поверхности хлоропластов
можно считать эволюционно продвинутыми, однако по активности
фотосинтетического аппарата, уровню энергетического обмена и
прочности связи хлорофилла с липопротеинами они уступают
цветковым растениям. В целом эволюция аппарата фотосинтеза на
клеточном уровне сопряжена с увеличением ассимиляционной
поверхности клеток мезофилла (за счет усложнения ультраструкту-
ры, величины и числа хлоропластов) и фотохимической активности
самих пластид. Все это по мере филогенетического продвижения
растений повышало эффективность механизма фотосинтеза.
Допускают полифилетическое происхождение хлоропластов
разных организмов (Э. Брода, 1978). В онтогенезе хлоропласты
формируются из протопластид — мелких образований с двумя мем-
бранами, имеющих слабо развитую внутреннюю организацию. Фор-
мирование хлоропластов в онтогенезе в какой-то мере повторяет
путь их эволюционного становления (закон рекапитуляции).
3.8. ВОЗНИКНОВЕНИЕ ЛИСТА КАК ЭТАП
БИОЛОГИЧЕСКОЙ ЭВОЛЮЦИИ ФОТОСИНТЕЗА
Экологическая дивергенция растений, представляющая резуль-
тат приспособления видов и популяций к условиям существования,
сопровождается изменением числа и формы листьев, а также топо-
графии их расположения на растениях. Несмотря на огромное число
96
видов растений и многообразие форм листьев, их функция практи-
чески мало меняется в ходе эволюции. Поэтому в процессе эво-
люции растений даже возникают запреты на изменение формы
листовой пластинки. Этим и определяется эволюционная ограни-
ченность девятью структурными типами организации листовой
пластинки (С. В Мейен, 1988).
Эволюция аппарата фотосинтеза тесно связана с появлением и
'« развитием его специального органа — листа. По вопросу о происхож-
дении листа в ходе эволюции существуют две точки зрения (А. Л.
Тахтдджян, 1964; К. Эсау, 1969; J. Richardson, 1984). Значение этого
органа для дальнейшего повышения эффективности процесса фото-
синтеза трудно переоценить. С возникновением листа происходит
значительная физиологическая и экологическая дивергенция расте-
ний по реализации механизма фотосинтеза. Особенно роль листа как
органа фотосинтеза возрастает с переходом растений к наземным
условиям и по мере аридизации среды. Тогда в пределах листа
формируются ткани и клетки, выполняющие разные функции; Как
строение листовой пластинки, так и особенности листорасположения
на растении призваны обеспечить процесс фотосинтеза.
Лист как «солнечная батарея» возникает или из выроста осей у
водорослей (микрофильная линия — плауны) или путем сращения
И утолщения теломов у псилофитов (макрофильная линия — папо-
ротники и др.). Независимо от происхождения лист имел огромное
значение для дальнейшего усиления фотосинтеза благодаря лучше-
му улавливанию световой энергии и мобилизации ее для фотосин-
теза, Развивающаяся сеть устьиц и межклетников, а также жилок
не только облегчала поступление СО2, но и создавала благоприят-
ный баланс между фотосинтезом и дыханием (из-за устранения
нагрева пластинки транспирацией). В то же время складывается
анатомо-морфологическая организация листа, позволяющая эф-
фективнее использовать энергию света: происходит разделение
функций между клетками пластинки, развивается густая сеть про-
водящих путей, разнообразятся формы пластинки, изменяется ли-
сторасположение на побеге. Однако с точки зрения физических
законов задача использования солнечной энергии листом в ходе
эволюции была решена не особенно удачно, так как его строение
рассчитано для работы при низких концентрациях СО2 и низкой
интенсивности фотосинтетической радиации. К тому же РЦФ листа
обладают низкой пропускной способностью. Но эти недостатки в
организации листа компенсируются увеличением числа и обновля-
• емости листьев на растении.
Эволюционные изменения в интенсивности фотосинтеза
обусловлены многими преобразованиями, в том числе изменени-
ем размера фотосинтетических единиц и числа реакционных
центров фотосинтеза у листьев разных растений. У филогенети-
4-157 97
чески древних видов величина фотосинтеза оказывается ниже
(2—10 мг СО2 • дм~2 • ч-1), чем у молодых (20—100 мг СО2 • дм2 • ч~
'). Единичный хлоропласт у покрытосеменных (магнолия и дримис)
по содержанию реакционных центров ФС1 и ФСП существенно не
отличается от хлоропластов папоротников и голосеменных (табл.
4). Однако у покрытосеменных из-за увеличения числа пластид и
клеток на единицу площади листа содержание реакционных центров
фотосинтеза оказывается намного больше.
Таблица 4. Содержание РЦ (на один хлоропласт), объем
и число хлоропластов в единице листовой поверхности
древних видов растений (по О. Ф. Монаховой и др., 1987)
Виды РЦ Объем хлоро- пластов, мкм3 Число хлоропла стов лис- та, 106/дм2
ФС1 ФСП
Angiopteris evecta 4,8 5,7 75,4 6,4
Encephalartos alensteinii 8,2 11,7 88,0 5,0
Ginkgo biloba 4>1 4,8 43,9 8,3
Magnolia grandiflora 2,5 3,1 20,8 17,4
Dremys winteri 3,1 3,1 90,2 7,6
Теоретическими расчетами показано, что средний хлоропласт
содержит 2,2 • 106 РЦ (А. А. Ничипорович, 1972). Повышение
интенсивности фотосинтеза у цветковых по сравнению с папорот-
никовидными выступает как результат возрастания скорости элек-
тронного транспорта и фотофосфолирования из-за увеличения
числа ЭТЦ на единицу хлорофилла и площади листа (О. Ф. Мона-
хова, Т. Е. Кислякова, 1993).
Эффективность фотосинтеза оказывается ниже 35% — величи-
ны, характерной для молекулярных процессов (А. Л. Ленивджер,
1975). Цо этой причине полагают, что фотосинтез как бы не отвечает
требованиям адаптивности и играет незначительную роль в про-
грессе жизни. При этом не учитывается комплекс изменений в
фотосинтетическом аппарате: отбор пигмента, его упаковка и кон-
центрация, образование гран в хлоропластах, увеличение размера
ФСЕ у высших растений (400 молекул против 40 у бактерий; А. Т.
Мокроносов, В. Ф. Гавриленко, 1992). В ходе эволюции формиру-
ется ЭТЦ фотосинтеза (хлоропластов) с металлопротеинами, вклю-
чаются ферредоксиноподобные соединения в транспорте
электронов, появляется ФСП цианей, возрастает число переносчи-
ков электронов и разнообразие структуры ЭТЦ хлоропластов (с
сохранением общих принципов ее работы). В итоге возникает
98 ,
современный тип ЭТЦ хлоропластов, позволяющий использовать
Н2О и СО2 в качестве исходных продуктов фотосинтеза.
Дальнейшая эволюция фотосинтеза связана с изменением листо-
вого аппарата, его структуры и функции, соотношения автотрофных
и гетеротрофных структур в пределах растения, ценотических пара-
метров растений как средообразующего фактора. На протяжении всей
эволюции жизни интенсификация фототрофности связана с сопря-
женным изменением формы и функции. В этом состоит одно из
фундаментальных требований усовершенствованию фототрофности.
Биологическая система функционирует только при наличии
структуры, что приложимо и к аппарату фотосинтеза. Все структуры
аппарата фотосинтеза (молекулы пигментов, мембраны хлоропла-
стов, хлоропласт, клетки мезофилла листа, лист, растение, расти-
тельный покров) взаимодействуют в своих функциональных
проявлениях (R. Bourdu, 1982). Поэтому естественный отбор
способствовал постепенному их совершенствованию в направлении
наибольшего выхода органического вещества путем усиления ко-
ординации деятельности фотосинтезирующих систем разной слож-
ности (фотосинтетических единиц и реакционных центров):
мембран тилакоидов, хлоропластов, зеленых клеток палисадной
ткани, листьев растений. Имел значение и отбор популяции в
фитоценозах по интенсивности фотосинтеза.
Каждую из названных систем в принципе нельзя считать иде-
альной: они только в совокупности выполняют задачу эффектив-
ного использования энергии солнечной радиации. Следовательно,
с формированием механизма фотосинтеза его эволюция как слож-
ного физиологического свойства растений и планетарного явления
не прекращалась. В ходе морфологической эволюции растений
постепенно усилилась его эффективность путем выработки слож-
ного фотосинтетического аппарата. При этом естественный отбор
был направлен на поиски возможностей для наиболее эффективной
реализации механизма фотосинтеза, возникшего уже у цианей и
одноклеточных водорослей. Задача состояла в выработке морфофи-
зиологических особенностей, необходимых для наибольшего выхо-
да органического вещества при фотосинтезе. Из этого также следует,
что фотосинтез в эволюционном плане представляет собой сложный
биологический процесс, связанный с усовершенствованием не толь-
ко физико-химического механизма, но и всей организации и жиз-
недеятельности зеленого растения как целостной системы. Поэтому
закрепление механизма фотосинтеза изменило последующий харак-
тер эволюции живой природы благодаря его конечным продуктам
— органическим (углеводы) и неорганическим (кислород). Ниже
мы вернемся к оценке эволюционной роли кислорода (см. гл. 4).
Значение органического вещества бесспорно, о чем речь шла выше.
99
3.9. ЭКОЛОГИЯ ФОТОСИНТЕЗА КАК ПОКАЗАТЕЛЬ
ЕГО БИОЛОГИЧЕСКОЙ ЭВОЛЮЦИИ
На примере растений разных экологических групп, жизненных
форм (деревьев, кустарников, трав и т. д.) и условий обитания (тро-
пики, умеренный пояс, Арктика, пустыня) накоплено достаточно
материала, характеризующего особенности фотосинтеза (О. В. Зален-
ский, 1977; К. Я. Биль, 1993). В пособиях по физиологии растений
указанной теме уделяется достаточно внимания, что делает излишним
ее широкое обсуждение. Так, разделение растений на светолюбивые
и теневыносливые основано на их способности адаптироваться к
разным световым условиям. Фотосинтетический электронный транс-
порт у светолюбивых растений интенсивнее, чем у теневыносливых.
Светолюбивые растения больше вытягиваются при недостатке
света, чем теневыносливые. В. Н. Любименко более чем на 600
видах показал, что листья у вторых по сравнению с первыми
содержат повышенное количество хлорофилла. Даже одни и те же
растения, хорошо освещенные, содержат 1,5—3 мг хлорофилла на
1 г листовой пластинки, тогда как растущие в тени —4—6 мг/г. У
теневых растений и концентрация хлорофилла на один хлоропласт
больше и сами хлоропласты крупнее. Однако интенсивность фото-
синтеза не связана с количеством хлорофилла. У освещенных
листьев хлорофилл а преобладает над хлорофиллом Ь, а у затененных
соотношение а и Ъ падает. Состав пигментов меняется и у других
растений. Так, по мере увеличения глубины водоема у водорослей
происходит увеличение содержания бурых пигментов (фукоксан-
тин), У наиболее же глубоководных (красные водоросли) —красных
пигментов (фикоэретрин). У красных водорослей отношение фи-
коэретрин/хлорофилл оказывается тем больше, чем глубже они
обитают в водоеме. Следовательно, явление «хроматической адап-
тации» — общее для зеленых растений.
Многими исследованиями показано, что растения разных эко-
логических групп содержат неодинаковое количество хлорофилла.
Очень часто содержание хлорофилла определяет энергетический
выход у листьев — накопление энергии в единицу времени (О.
Хитт, 1973). В то же время разные виды растений по числу и
размерам хлоропластов в 1 мм2 (А. Т. Мокроносов, 1981; и др.)
отличаются, что часто коррелируется с образованием сухого веще-
ства и содержанием хлоропластов. Так, у Tropaeolum major число
хлоропластов на 1 мм2 составляет 383 тыс., а сухое вещество (г/м2
листовой поверхности за 10 ч) — 4,47; у Phaseolus multiflorous —283
тыс. ц соответственно 3,22; у Ricinus communis — 495 тыс. и 5,29; у
Helianthus annuus—465 тыс. и 5,57. Число хлоропластов на 1 см2
листа варьирует от 2 до 90 млн. Много пластид в листьях ксерофитов
100 .
аридных пустынь, обитающих в условиях высокой интенсивности
света: у астрагала хивинского — 89 млн., у осоки вздутой — 72 млн.
В связи с экологией фотосинтеза растений представляют инте-
рес данные по оценке стока углерода как показателя годичной
продуктивности, выраженного величиной нетго-фотосинтеза с уче-
том затрат на дыхание (П. Ю. Вороник и др., 1995). Для этого
определяют прежде всего проективное содержание хлорофилла —
хлорофилльный индекс. Обнаружена высокая корреляция (г - 0,72)
между годичным фотосинтетическим стоком углерода и проектив-
ным содержанием хлорофилла в растительном покрове — фитоце-
нозах. Так, продуктивность растительности сухопутной России
оценивается в 1,6 Гга, тогда как планеты —50 Гга в год. Именно
наличие корреляции между проективным содержанием хлорофилла
того или иного типа фитоценоза и годичной его продуктивностью
фотосинтеза позволяет подойти к оценке приспособленности фи-
тоценозов к климатическим факторам й судить об их возможных
сукцессионных последствиях. Характерно, что биоразнообразие
мало влияет на конечный результат расчета, это облегчает задачу
оценки продуктивности фитоценоза с учетом содержания хлоро-
филла у 2—3 доминирующих видов в проективном покрытии его.
Экологические особенности видов, их, приспособленность к
среде отражаются И на продуктах фотосинтеза. В ходе эволюции
фототрофности конечные продукты становятся все более однооб-
разными («специализация» веществ). Так, у бактерий и водорослей
они более разнообразны, чем у высших растений. При сравнении
фотосинтеза лишайников и высших растений показано, что при
одинаковом наборе ассимилятов метаболизм лишайников отлича-
ется некоторыми специфическими особенностями: 1) увеличением
в продуктах фотосинтеза доли маннита и, сахарозы; 2) высокой
фотосинтетической термостабильностью; 3) восстановлением СО2
до сахароспиртов даже при температуре от —15 до —20° С.
Мир высших растений характеризуется исключительным мор-
фофизиологическим разнообразием, что отражается й на организа-
цию донорно-акцепторных отношений (А. Т. Мокроносов, 1981),
имеющих приспособительное значение. Приспособительные изме-
нения фотосинтеза можно обнаружить и при анализе положения
Координационных точек нетго-фотосинтеза при световом насыще-
нии (рис. 7), границ жизни и фотосинтеза (рис. 8) у разных
экологических групп растений; Оптимум фотосинтеза определяется
температурными границами жизни вида и популяций как результат
их приспособленности.
В пределах каждой группы растений наблюдается еще световая
адаптация. Так, водоросли в целом относятся к теневыносливым
растениям, у которых насыщение фотосинтеза происходит при
низких интенсивностях света. Например, у сифоновых водорослей,
.101
Рис. 7. Зависимость от температуры и положения кардинальных
точек нетто-фотосинтеза у растений различного типа при свето-
вом насыщении (В. Лархер, 1978. С. 120)
Рис. 8. Температурные границы жизни и нетто-фотосинтеза растений в состо-
янии активной вегетации (В. Лархер, 1978. С. 257)
Вверху —температурные границы жизни и продуктивности наиболее чувствительных к
холоду и жаре видов каждой группы растений; внизу —соответствующие величины для
наиболее устойчивых видов
обитающих в скелетах коралловых рифов, насыщение фотосинтеза
достигается при освещении 10 лк, а у хлореллы и некоторых
синезеленых — при 0,2 Лк. У одних и тех же организмов световое
насыщение фотосинтеза меняется по сезонам года: у динофлагеллят
оно происходит летом при 34,7, весной и осенью —2,5, зимой —
1 тыс. лк.
Световая адаптация фотосинтеза связана со многими события-
ми: изменением концентрации пигмента, активностью ферментных
S-4.
12 3 4 5 6 7 6 9
Содержание хлорофилла(а +д^мг/дм*
Рис. 9. Влияние содержания хлорофилла на степень исполь-
зования света (В. Лархер, 1978. С. 42)
Рис. 10. Усложнение тела у бурых водорослей: ламинария, лессония,
макроцисгис и саргассум (7 —4) (А. П. Хохряков, 1981. С. 12)
103
систем, положением и числом хлоропластов, инактивацией молекул
хлорофилла. Некоторые водоросли способны синтезировать хлоро-
филл даже в темноте, у некоторых в темноте происходит его
разрушение. Так, Scenedesmus acuta в течение 8 лет в темноте
сохранила зеленый цвет и вскоре после выставления на свет стала
фиксировать СО2. У некоторых водорослей адаптация к световым
условиям бывает растянута во времени от 2—3 сут (Cyelotella) до 2
недель (Oscillatoria rubens).
В зависимости от времени года меняется даже тип фотосинтеза
и содержание хлорофилла в листьях (рис. 9). На примере однолетних
Хлорелла Энтроморуа
Рис. 11. Морфологический ряд изменения растений, приведший к интенсификации
фотосинтеза в ходе эволюции
.104
растений из семейства марьевых, произрастающих в условиях не-
достатка влаги показано, что листья весенней генерации осущест-
вляют фотосинтез С4-типа, а летней генерации — по смешанному
С4- CAM-типу (В. Ю. Любимов, К.. Я. Биль, 1986). В фотосинте-
Лес
Рис. 11. Продолжение
105
тической адаптации растений важное значение имеет число и
распределение пластид в палисадной паренхиме. Оказалось, что
анатомический тип листа больше влияет на распределение, вели-
чину и структурные вариации пластид, чем экологические особен-
ности растений (Ю. В. Гамалей, Т. Р. Милашвилли, 1986).
В прошлом теплый климат протерозоя и палеозоя и высокая
влажность воздуха, высокое содержание СО2 и низкое содержание
О2 более благоприятствовали фотосинтезу, чем нынешние условия
(А. А. Ничипорович, 1973). Количество углерода, доступное для
фототрофов, ныне сократилось примерно в 800 раз по сравнению
с периодом возникновения жизни (В. В. Межерин, 1976).
В указанные эры масштабы фотосинтеза превосходили скорость
биологического окисления органического вещества, что способст-
вовало процветанию жизни. Одним из сдерживающих'факторов
фотосинтеза ныне стало обеднение атмосферы СО2.
Стабилизацией физико-химического механизма не завершается
эволюция фотосинтеза, а продолжается его биологический этап в
направлении поиска экологических новшеств для интенсивного
использования самого процесса. Так, с возникновением морфоло-
гического разнообразия растений в ходе эволюции параллельно
происходит экологическая их дивергенция по интенсивности фо-
тосинтеза. Переход от одноклеточных к многоклеточным и особен-
но к дифференцированным водорослям (рис. 10), а также смена
структурной организации жизненных форм и тип растительности
(рис. 11), образование многокомпонентных биогеоценозов пред-
ставляют собой ступени на пути интенсификации фотосинтеза как
планетарного явления. Подобным образом постоянно усиливается
разнообразие адаптивных возможностей фотосинтеза разных объ-
ектов и сообществ, что являетбя проявлением экологической эво-
люции растений.
3.10. ЭВОЛЮЦИЯ И ПРИСПОСОБИТЕЛЬНОЕ ЗНАЧЕНИЕ
КООПЕРАТИВНЫХ МЕХАНИЗМОВ ФОТОСИНТЕЗА
t
Процессы карбоксилирования, составляющие основы фотосин-
тетической ассимиляции СО2, широко распространены в живой
природе и представлены разнообразными реакциями. Примитив-
ные фототрофные организмы особенно отличаются многообразием
форм ассимиляции углерода. В частности, у зеленых бактерий
осуществляется восстановительный цикл карбоновых кислот (цикл
Эванса —Арнона), который в дальнейшем не имеет эволюционно-
го распространения: По соображениям, высказанным, выше, наи-
больший успех был достигнут в другом —прогрессивном —
направлении: в стабилизации цикла Кальвина как универсального
106
механизма фотосинтеза на последующих этапах развития фотосин-
теза, фототрофности.
Однако и этот цикл имеет ряд реакций, протекающих с термо-
динамическими барьерами, и также сталкивается с трудностями
снабжения фотосинтетического аппарата СО2 с учетом его низкого
содержания в атмосфере и стереохимической инертности. В то же
время поток СО2 в общем цикле Кальвина встречает ряд ограниче-
ний: устьичное сопротивление, сопротивление мезофилла и слож-
ной системы мембран (А. Т. Мокроносов, В. Ф. Гавриленко, 1992).
Достаточно напомнить, что число устьиц у CAM-типа наименьшее,
у С3-растений — наибольшее. Активность фермента карбоангидра-
зы в листьях С3-растений в 10 раз выше, чем у растений С4-типа.
По этим и другим причинам возникает эволюционная потребность
в кооперативных механизмах фотосинтезас усилением предвари-
тельного депонирования СО2. При сохранении общности первич-
ных механизмов фотосинтеза возникают различия в путях
депонирования и превращения углерода.
С4-растения не однообразны по особенностям превращения
углерода, строения обкладки пучка и организации хлоропластов.
При наличии общего механизма карбоксилирования С4-растения
отличаются спецификой транспортных метаболитов и декарбокси-
лирующих ферментов. Но для всех групп характерны наличие
кранц-анатомии, высокая интенсивность фотосинтеза и отсутствие
фотодыхания, существование высокого температурного оптимума
фотосинтеза в пределах 30—45°С, наступление углекислотно-ком-
пенсационного пункта в пределах 0—10 мкл/л СО2 (у С3-растений
—при 30—70 мкл/л), экономное использование воды (Ю. В. Га-
малей, Е. В. Вознесенская, 1986).
С4-путь обнаружен у 1500 видов, 90% из них сконцентрированы
в семействах злаковых и марьевых. Первоначальное его возникно-
вение связывают с тропиками. Число С4-видов в целом не превы-
шает 1 —2% среди высших растений, а у цветковых они обнаружены
в 16 из 320 семейств. Тем не менее значение С4-пути фотосинтеза
велико в условиях аридизации климата и ксерофилизации расти-
тельности, а также современного соотношения СО2 и О2 в атмос-
фере. Поэтому он возникает параллельно и конвергентно в разных
семействах (А. Т. Мокроносов, 1986). Он не обнаружен у водорос-
лей й водных цветковых.
Возникновение многих биохимических путей у растений связы-
вают с ответвлениями от основного пути —фотосинтетического
метаболизма. Интенсификация последнего обусловлена последую-
щей морфологической эволюцией растений. В этом отношении С4-
и CAM-типы растений являются одним из примеров эволюции в
направлении усиления подачи СО2 в цикле Бенсона—Кальвина.
Говоря о САМ-типе растений, следует напомнить, что они являются
107
8г
Рис. 12. Плотность произрастания растений различных
функционально-морфологических типов на незаселен-
ных и слабозасоленных (Л), сильнозасоленных (5) почвах
и солончаках (В) (В. Ю. Любимов и др., 1986)
СЗ —ксерофиты (/). С4 — ксерофиты (2), СЗ —суккуленты (Л,
С4 —суккуленты (4), С4 —САМ и суккуленты (J)
обитателями аридных условий, где наблюдаются резкие перепады
температур днем и ночью, большую половину/светлого периода
суток у них остаются закрытыми устьица.
К указанному типу относятся представители около 20 семейств
(Agavaceae, Bromeliaceae, Cactaceae, Crassulaceae, Liliaceae, Viticeae
и т. д.), различающиеся анатомическим строением листьев и «гиб-
костью» обмена (в зависимости от условий они функционируют как
С3-тип). Однако среди них встречаются и специфические облигат-
ные формы, у которых постоянно осуществляется CAM-тип, меня-
ясь лишь в интенсивности при сдвиге условий (Cactaceae,
Crassulaceae).
Сходство С4 и CAM-растений сострит в способе первичной
фиксации СО2 с образованием оксалоацетата (при участии ФЕПК)
и последующего декарбоксилирования последнего. Различия про-
являются не только в пространственном разобщении указанных
процессов у С4-растений, но и образовании ФЕП. Интересно
отметить, что роль ФЕП, ФЕПК и дикарбоновых кислот менее
уникальна, чем РБФ и РБФК.
По-видимому, есть основания принять адаптивность самого
явления —способности к рециклизации СО2 для повышения ус-
тойчивости фотосинтетического аппарата в условиях нарушения
газообмена при водном дефиците. Так, в аридной зоне Средней
Азии у растений семейств Роасеае и Chenopodiaceae обнаружены
разнообразные типы фиксации СО2, поэтому в районах, где наблю-
дается сильное засоление почв, преобладающими оказываются
растения С4- и САМ-типов (рис. 12).
108
15 20 25 30 35 . 40
С4-растения доминиру-
ют в условиях с высокой
годовой температурой. Они 3.
встречаются как в голаркти- |
ческом, так и палеотропи- “s 10
ческом районах (J. Vogel et £
al., 1986). При высоких тем- § .
пературах и низком освеше- I
нии С4-растения используют
энергию света эффективнее | 6
Сз-видов (рис. 13). С4- и I .
CAM-типы растения в ус- „• **
ловиях высокого гумуса в |
почве уступают по содержа- | 2
нию белка (на 1 г сухой |
массы) Сз-ксерофитным о
видам, однако на солонча-
ках они производят больше Рис. 1'
белка (В. Ю. Любимов и фотосинтеза уСз-и С4-растёний (Дж. Эдвардс *
др., 1986). Устойчивость и Д. Уокер, 1986. С. 468)
С4- и САМ-промежуточ-
ных растений в экстремальныхусловиях оказывается выше, чем С4-
и CAM-видов в отдельности. Большинство злостных сорняков
обладают С4-типом фотосинтеза. Иногда экотипы одного вида
проявляют различия в типе фотосинтеза (Alloteropis semlalata).
Сделаны попытки построить филогенетические ряды С4-типа.
метаболизма углерода (И. М. Магомедов, 1987; В. И. Пьянкой и
др., 1992).
Не вызывает сомнения, что в условиях дефицита воды, когда
устьица днем закрываются, скорость фотосинтеза у С4-растений
оказывается в 2—3 раза выше (чем у С3-видов) ввиду «челночного»
способа обеспечения ВПФ-цикла СО2. Эффективность использо-
вания воды (отношение вода/образованное сухое вещество, г/г) для
семи видов Сз-растений (Triticum aestium, Oryta sativa, Chenopodium
album и др.) колебалась в пределах 557—844, тогда как для четырех
видов С4-типа (Setaria italica, Zea mays, Amaranthus graecitens, Portu-
laca oleracea) —в пределах 260—340 (Дж. Эдвардс и Д. Уокер, 1986).
Эволюция аппарата фотосинтеза цветковых растений совпала с
периодом увеличения освещенности на планете. Это привело к
уменьшению размеров хлоропластов и количества хлорофилла в
них, но к увеличению их числа. Эти события имели значение для
предохранения хлоропластов от повреждения светом высокой ин-
тенсивности (Т. Е. Кислякова, О. Ф. Монахова, 1992). Дальней-
шая аридизация климата способствовала дифференцированию
цветковых по типам метаболизма углерода и перестройке фотосин-
109
•готического аппарата, включая строение, число и величины хло-
ропластов.
Животные предпочитают С3-растения ввиду того, что у них
крахмал более доступен. У С4-видов крахмал расположен глубоко в
клетках обкладки пучка, которые к тому же плохо прожевываются
из-за содержания большого количества лигнина. У последних мень-
ше содержится в тканях азота (Дж. Харборн, 1985). Нередко высо-
кую устойчивость С4-типа растений связывают с их способностью
синтезировать малат, который играет разнообразную роль, в том
числе и в регуляции реакций расщепления воды (в хлоропластах),
окисления по альтернативному пути (в митохондриях), углеводно-ли-
пидного метаболизма, а также устойчивости к аноксии (см. гл. 4). 1
Что касается CAM-типа метаболизма, характерного для сукку-
лентов и некоторых водных растений (Litterella uniflora), то явно
видна его способность «уходить» от дефицита воды. В нормальных
же условиях у них происходит фиксация СО2 по С3-пуги. Длитель-
ный дефицит воды ведет к переключению метаболизма на САМ-тип,
при этом накапливается и используется малат, повышается актив-
ность ФЕП. У суккулентов наблюдаются сильные перепады малата
в течение суток из-за температурных колебаний, к чему приспособ-
лены и ключевые ферменты (ФЕПК и МДГ).
У растений CAM-типа накопление СО2 в клетках разобщено во
времени, а не в пространстве. Среди них выделяют растения МДГ
и ФЕПК-типов. Первые фиксируют СО2 аналогично С3-пути, вто-
рые превращают СО2 в НСО. CAM-растения, среди которых есть
папоротники, голосеменные и покрытосеменные, в присутствии
воды открывают устьица и используют С3-путь фотосинтеза: при
засухе и засоленности почвы (устьица закрыты) идет темновая
фиксация СО2 с участием яблочной кислоты.
В течение онтогенеза один и тот же вид растений переключает
пути фиксации СО2. Например, однолетнее растение Mesembryant-
hemum crystallinum, обитатель пустынь Средиземноморья,' во влаж-
ный период сезона формирует крупные листья и использует С3-путь,
а в период засухи его рост приостанавливается и оно переходит на
CAM-путь. Это способствует не только созданию органического
вещества в условиях засухи, но и Предотвращает «тепличный»
эффект (перегрев от накопления СО2). CAM-тип характерен для
семейств толстянковых и кактусовых, он обнаружен более чем у 10
тыс. видов покрытосеменных, включая несколько тысяч орхидных.
Считают, что CAM-тип связан с засушливыми условиями, хотя он
обнаружен у водного растения Jsoetes, испытывающего в течение
дня дефицит СО2.
На примере девяти видов пиперомий показано, что степень
проявления CAM-пути коррелирует с ксероморфностью листьев и
мелкими размерами клеток. Чем меньше размер фотосинтезирую-
по -
щих клеток, тем больше накапливается в них яблочной кислоты.
Повышение температуры воздуха ночью и удлинение светового
периода способствуют снижению активности САМ-обмена. С4-путь
обмена наблюдается в сложных условиях (жара, засуха, высокая
засоленность почвы, щелочное pH). У части растений при ухудше-
нии условий происходит переключение С3-пути на С4-путь, у других
(портулак) при дефиците воды, коротком дне и низкой ночной
температуре С4-путь переключается на CAM-обмен (Т. Hultum, К.
Winter, 1982).
Несмотря на усложнения механизма фотосинтеза у С4- и САМ-
растений, преобладающее большинство Данных не оставляет сомне-
ния в приспособительном значении этих путей. Так, анализируя
тенденции изменения растительности Земли в связи с глобальным
потеплением климата, В. И. Пьянков и А. Т. Мокроносов (1993)
пришли к выводу, что в будущем произойдет перераспределение
доминант в экосистемах. При этом, как в прошлом (юрский и
меловой периоды), сухие и засоленные местообитания займут рас-
тения с признаком С4-сйцдрома и фотосинтеза, обладающие пре-
адаптацией к аридным условиям.
Ю. С. Карпилов (1970) выделяет следующие этапы в развитии
ксерофитов с С4-путем метаболизма СО2:1) появление частой сети
проводящих пучков и компактного мезофилла в листьях; 2) про-
движение хлоропластов в клетки обкладки; 3) развитие системы
транспорта ассимилятов между двумя типами хлоропластов; 4)
локализация крахмалсинтезирующей системы в пластидах клеток
обкладки; 5) нециклическое фотофосфорилирование хлоропластов
мезофилла и циклическое — хлоропластов обкладки.
Интересен вопрос о приспособлении растений к световым
условиям в онтогенезе, что также имеет значение для понимания
экологической эволюции механизма фотосинтеза. Оказалось, что в
онтогенезе существуют только три возможности адаптации расте-
ний к световым условиям (А. А. Лайск, 1982): адаптация рецепторов
СО2 (ферментативная система и диффузный аппарат листа), изме-
нение ориентации листьев и хлоропластов в соответствии с нали-
чием света и СО2. Чаще всего растения используют первые две
возможности, что повышает их пластичность и выживаемость в
онтогенезе.
Приведенные выше данные свидетельствуют о динамичности
механизмов превращения углерода и аппарата фотосинтеза в онто-
и филогенезе растений, что способствует поддержанию высокой
продуктивности фотосинтеза в разнообразных условиях и обеспе-
чению успеха в борьбе за существование — повышению энергети-
ческого потенциала и коэффициента размножаемости популяции.
Ш
3.11. К ВОПРОСУ ОБ ЭВОЛЮЦИОННОМ ПОЛОЖЕНИИ ХЕМОСИНТЕЗА
Процессы хемосинтеза — ассимиляция СО2 за счет энергии
неорганических веществ (H2S, NH3, Н2 и др.) в отсутствие света —
играли и играют значительную роль в круговороте веществ. В связи
с этим представляет интерес вопрос об эволюционном положении
хемосинтеза (литотрофии). В. Н. Любименко подчеркивал, что фо-
тосинтез никогда бы не достиг современной продуктивности, если
бы хемотрофы не ускоряли минерализацию мертвых органических
остатков, не фиксировали азот атмосферы и не выделяли продукты
окисления в среду. По этой причине он считает, что хемосинтез
должен был предшествовать фотосинтезу.
Другие авторы, исходя из сходства основного механизма усво-
ения углерода СО2 в хемо- и, фотосинтезе, полагают, что они
возникли или параллельно, или хемосинтез вторичен. У литотро-
фов, способных к фототрофии, функционирует цикл Кальвина.
Поэтому допускают существование преемственной связи между
фотоавтотрофией и хемоавтотрофией (см. сх. 6). Хемоавтотрофия
филогенетически не могла предшествовать фотоавтотрофии, так как
энергия для хемосинтеза высвобождается при дыхании. Скорее
всего хемоавтотрофы происходят от организмов, совмещающих
фотосинтез и дыхание. Неорганические субстраты при хемосинтезе
биологически окисляются тем же путем, что и органические при
дыхании. Многие из хемосинтетиков являются аэробными организ-
мами, хотя встречаются и анаэробные (хеморедукторы).
Не исключено также, что отбор на автотрофное питание в
популяциях протобионтов шел параллельно в двух направлениях:
на «проверку» эффективности сочетания хемо- и фотосинтеза. Так,
среди пурпурных бактерий встречаются виды, у которых в зависи-
мости от условий происходит или хеморедукция, или фоторедукция.
Поскольку хемосинтез связан с использованием энергии, запасен-
ной в восстановленных неорганических соединениях (сама эта
энергия в количественном отношении невелика), хемосинтез не мог
получить эволюционные преимущества перед фотосинтезом.
Для понимания эволюции и значения хемосинтеза представляют
интерес водородные бактерии и карбоксидобактерии (Г. А. Завар-
зин, 1978), способные использовать органические вещества и пере-
ходить от литотрофного к органотрофному росту за счет механизмов
превращения углеводов путем активации одних и инактивации
других ферментов. В кислородной среде они дополняют вторичные
анаэробы потребляя одни и те же субстраты, но используя другой
акцептор электронов. Продукция биомассы в год у водородных
бактерий определяется в 40—50 • 106т, а у карбоксилобактерий —
10 • 106 т. Поэтому в целом весьма отчетливо выделяется истори-
ческая роль хемосинтетиков в «очищении» среды от многих про-
112
дуктов биогенного и абиогенного происхождения, в «обживании»
нашей планеты. В эволюционном раду первичного развития жизни
механизмы хеморедукции и хемосинтеза, несомненно, следует рас-
сматривать как шаг вперед по пути усовершенствования ее органи-
зации, функционального и морфологического разнообразия.
Только теперь стало ясно, что хемосинтез представляет боковое
ответвление от магистрального развития автотрофного питания. На
начальных этапах эволюции хемосинтез выступал как важный этап
прогресса жизни. Значение хемосингетиков состояло И в том, что
они очищали среду от кислорода, который первоначально выступал
как вредный агент для живых организмов, в том числе и для
фотосинтетиков. Без хемосинтетиков вопрос «быть или не быть
фотосинтезу», вероятно, был бы решен с другими последствиями.
Появление хемосинтетиков можно отнести к первой попытке удач-
ного решения задач «охраны» живой природы. Однако с развитием
фотосинтетиков жизнь стала «захлебываться» в собственных отбро-
сах и возникла необходимость кардинальной защиты живой систе-
мы от кислорода.
* ♦ ♦
X
Процессы синтеза органического вещества из неорганического
с использованием энергии солнечного света (фотосинтез) форми-
ровались достепенно и на определенном этапе развития живой
природы. Роль и значение перехода к фотосинтезу в общей эволю-
ции жизни на Земле чрезвычайно велики. Фотосинтезирующие
растения стали самым мощным геохимическим фактором. В про-
цессе становления фотосинтеза в цепях питания взаимно приспо-
сабливались разные группы организмов.
С возникновением и стабилизацией физико-химического Меха-
низма фотосинтеза эволюция его не закончилась, а продолжалась
по пути совершенствования аппарата фотосинтеза и разнообразия
путей фиксации СО2. Благодаря количественным изменениям в
аппарате фотосинтеза усиливается, в частности, экологическое
разнообразие растений, особенно в условиях мощных антропоген-
ных факторов.
Фотосинтез создает материальную и энергетическую базу для
всех живых существ, и в этом его уникальность. Создав предпосылки
для возникновения аэробного дыхания, он оказал существенное
влияние на дальнейший прогресс жизни.
ГЛАВА 4
ЭВОЛЮЦИЯ МЕХАНИЗМОВ ДЫХАНИЯ У РАСТЕНИЙ
Энергетическая победа живых существ, несомненно, связана с
совершенствованием фотосинтетического метаболизма. Ввиду
исключительной его значимости для дальнейшего прогресса жизни
изменения в фотосинтетическом метаболизме происходили в на-
правлении постепенного повышения продуктивности и интенсив-
ности процесса фотосинтеза. В результате были усовершенствованы
системы фоторецепторов, физико-химические реакции ЭТЦ,
системы депонирования СО2, структурный аппарат и регуляторные
механизмы фотосинтеза. Возникновение фотосинтеза и его регуля-
торной системы — важнейший ароморфоз (арохимоз) в истории
развития жизни на планете. Поэтому основные этапы эволюции
биосферы Земли В. И. Вернадский так или иначе связывал с дея-
тельностью растений (первичные автотрофы, развитие животных с
кальциевым скелетом, лесные массивы и ноосфера). Несмотря на
древность механизма фотосинтеза (4—3,5 млрд, лет; Б. С. Соколов,
1971), долгое время он не мог существенно влиять на геохимические
процессы планеты из-за медленного насыщения атмосферы сво-
бодным кислородом.
' С постепенным накоплением кислорода в атмосфере прогресс
и морфологическая дифференциация живой природы сделались
необратимыми явлениями благодаря усилению энергообмена и
геохимических преобразований на Земле. В примитивной форме
механизмы энергообмена (дыхания) сложились на этапе перехода
от химической эволюции к биологической. Дыхание — важнейший
источник и составная часть биоэнергетики растений, необходимой
не только для процессов деления клеток, роста и размножения
растений, но и для образования промежуточных веществ, вовлека-
емых в дальнейшие процессы синтеза.
Как известно, кислород на Земле появился на определенном
этапе и был неравномерно распределен по ее поверхности: вначале
он сосредоточивался в верхних слоях атмосферы. Живые существа,
обитающие в воде или придонном иле, мало соприкасались с
кислородом. Только фототрофные формы (такие, как Rhodospiritlales
и др.) занимали освещенную и пограничную к кислороду толщу
воды: из их числа несерные и серные пурпурные бактерии — слои,
несколько бедные кислородом, зеленые серобактерии — еще более
114
бедные им слои. Это означает, что уже на начальных этапах
эволюции жизни существовала своеобразная дифференциация по
аэротолерантности.
В то же время организмы, выделяющие кислород, в ходе эво-
люции появляются раньше, чем организмы, потребляющие его.
Например, облигатные фототрофные синезеленые водоросли в
процессе фотосинтеза выделяют кислород, но у них нет аэробного
дыхания. Накапливающийся таким путем кислород представлял
опасность для первичных существ главным образом из-за про-
дуктов его неполного восстановления (супероксидные радикалы,
Н2О2) и окисления вообще. По этим и другим причинам возмож-
ность самозарождения жизни в кислородной среде равна нулю
(А. И. Опарин, 1969).
Однако имеются суждения и другого характера, свидетельству-
* ющие о наличии кислорода до зарождения жизни. Так полагают, что
кислород вначале образовался путем терморазложения силикатной
матрицы при ударных процессах твердых тел по поверхности планеты
(что привело к разогреву ее поверхности до температуры 30000 К). В
породах коры содержание кислорода составляет около 40%. Подсчеты
показывают, что для обогащения коры массой 2,4 • 1025 г требуется
(1—3) • 1024 г кислорода. Считают, что такое его количество не могло
быть обеспечено за счет фотосинтеза, при котором выделяется О2
только 3,4 • 1013 г/год (М. В. Герасимов, Л. И. Мухин, 1986).
Как бы то ни было, роль фотосинтетиков в последующем
обогащении атмосферы кислородом и усилении его круговорота
никем не отрицается. Несомненно, что по мере накопления О2
фототрофами в процессе эволюции у организмов вырабатывались
механизмы не только для защиты от его губительного влияния, но
и для эффективного использования реакционной активности О2. В
последнем случае благодаря увеличению выработки АТФ (38 моле-
кул. на молекулу гексозы) энергетический обмен повысился в 20
раз. Поэтому конкуренцию выдерживали существа и сообщества,
способные потреблять кислород. Они-то и дали начало новому
эволюционному взрыву гетеротрофов. Так кислород из опасного
фактора становится «творцом дифференцировок жизни на Земле»'
(В. И. Вернадский).
4.1. ОСОБЕННОСТИ ЭНЕРГЕТИКИ ОРГАНИЗМОВ РАЗНЫХ ЦАРСТВ
И ДРЕВНИЕ ФОРМЫ ЕЕ ПРОЯВЛЕНИЯ
С обогащением атмосферы свободным кислородом благодаря
все возрастающей деятельности фотосинтетиков создается озоно-
вый экран Земли (когда уровень О2 достигает примерно 1% от
современного состояния). До. возникновения озонового экрана
из-за губительного действия УФ-лучей отбор даже среди первичных
115
многоклеточных растений, в том числе и на суше, шел в сторону
безлистности.
Важным следствием насыщения атмосферы О2 было чрезвычайное
повышение энергетических процессов живых организмов путем ме-
таболического использования свободного кислорода. Это событие
имело грандиозное значение для прогрессивного развития живой
природы и сопровождалось усилением связи процессов фотосинтеза
и дыхания. Таким путем создалась возможность усиления круговорота
энергии на Земле, энергетического обеспечения хлоропластов дея-
тельностью митохондрий. И путь совершенствования живых
организмов в этом направлении был длительным и ступенчатым.
Большая часть современных живых организмов не может
обойтись без кислорода, используемого для дыхания. Напротив, на
начальных этапах развития жизни на Земле кислород выступал как
опасный для нее фактор. Это становится понятным, если принять
во внимание его способность активно поддерживать горение и
вызывать окисление разных веществ. Кислород в природе образу-
ется при разложении воды, как и в случае фотосинтеза (фотолиз
воды). Исторически сложилось так, что организмы, выделяющие
кислород, развиваются раньше, чем потребляющие его. Например,
многие даже современные цианеи в процессе фотосинтеза выделяют
О2, хотя у них нет механизмов аэробного обмена.
Существует мнение о самопроизвольном возникновении био-
энергетических механизмов обмена (R. F. Fox, 1982). В соот-
ветствии с этим энергообеспечение первых живых существ есть не
что иное, как использование механизмов, сложившихся в неживой
природе. При этом неорганический пирофосфат рассматривается
как древнейшая «конвертируемая» валюта. Возникновение протон-
ного потенциала связывают с реакцией оксиредукции между НАД
• Н и Fe2+ на мембранах. Роль трансмембранного переносчика в
последней реакции отводится убихинонам.
Как известно, ныне живущие бактерии отличаются огромным
разнообразием биоэнергетических процессов, у них значительно
варьирует ферментативный аппарат и в дыхании участвует несколь-
ко терминальных оксидаз (Э. Брода, 1978).
х На схеме 7 представлены предполагаемые этапы эволюции
генераторов энергии (М. В. Гусев, Г. Б. Гохлернер, 1980)?
Об истоках возникновения биологического обмена см. в гл. 3.
Здесь же остановимся на начальных этапах извлечения энергии
живыми существами первичной Земли. Естественно, что об этом
можно судить только по косвенным подходам —сравнению про-
цессов энергообмена у современных наиболее примитивных су-
ществ, являющихся персистированными представителями древних
форм жизни.
116
Многие данные не оставляют сомнения в том, что наиболее
древние формы жизни нашей планеты (протобионты) были гете-
ротрофами, извлекали энергию из готовых органически^ веществ в
результате субстратного фосфорилирования (гликолиза). Однако
некотррые авторы не считают процессы гликолиза самыми
примитивными проявлениями широко распространенного анаэроб-
ного энергообмена — брожения. Так, брожению могли предшест-
вовать довольно простые реакции энергетического обмена
наподобие обмена клостридий, которые для синтеза АТФ исполь-
зуют ограниченное число реакций с участием НАД, КоА, НАДФ и
ферредоксина. Наиболее примитивные организмы в ходе субстрат-
ного фосфорилирования перерабатывают органические соединения
без предварительной их активации с помощью АТФ, так как
субстраты обладают сильно выраженными восстановительными
свойствами (Э. Брода, 1978; М. В. Гусев, А. А. Минеева, 1985;
Т. В. Чиркова, Г. Ж. Валиханова, 1985).
Схема 7'
117
Дело в том, что на ранних этапах развития жизни, когда в среде
изобиловали различные органические вещества абиогенного
происхождения, было найдено простое решение для поддержания
и воспроизведения жизни — извлечение энергии из готовых
органических веществ. Смысл этого можно понять, если принять во
внимание отсутствие у протобионтов развитых собственных систем
для получения энергии. Словом, организмы не располагали воз-
можностью извлекать энергию из собственных химических реакций
ввиду их чрезвычайной упрощенности и примитивности своей
организации. Такой способ получения энергии предполагает
наличие механизма гомоферментативных реакций по типу молоч-
нокислого брожения (ключевой фермент — фосфофруктокиназа),
где гликолитический распад сахаров осуществляется (у бактерий в
цитоплазме) вне мембранных структур. Напомним, что при этом
высвобождение энергии из субстрата и фиксация ее в виде АТФ
осуществляются метаболической перестройкой органических моле-
кул, в состав которых входили не полностью окисленные (или
восстановленные) атомы углерода. Энергии при этом выделяется
мало ввиду образования в Качестве конечных продуктов сильно
восстановленных соединений. Поэтому синтезируются лишь две
молекулы АТФ на молекулу гексозы.
Ввиду опасности быстрого исчерпания субстрата для
фосфорилировании на ранних этапах развития жизни отбор
организмов среди протобионтов пошел по пути опробования разных
субстратов в энергетических целях. По принципу Гаузе эволюция
пошла в направлении использования разных субстратов как
источников энергии, что способствовало повышению емкости
первичных водных биоценозов и надежности воспроизведения
организмов. Адаптивная радиация гетеротрофных первичных
организмов сопровождалась специализацией их по переработке
разных субстратов.
Нет необходимости описывать известные реакции субстратного
фосфорилирования. Однако заметим, что наиболее важной из них
является окисление ЗФГК: здесь клетка как бы «возвращает»две
молекулы АТФ, ранее ею же затраченные на активирование субст-
рата. Эта реакция протекает вне определенных структур клетки, и
запасание Выделяемой энергии (энергетическая эффективность) в
виде АТФ составляет 40%. Другим свидетельством примитивности
субстратного фосфорилирования служит недифференцированность
донора и акцептора электронов. При этом субстрат выступает в роли
как донора, так и акцептора электронов, что ограничивает его
окислительные возможности.
Энергетические ограничения гликолиза связаны и с постепен-
ным ингибированием активности- его ключевого фермента по мере
накопление АТФ, цитрата и Н+. Торможение конечных реакций
118
обусловлено при брожении отсутствием регенерации НАД+, что
приводит к застопориванию превращения триозофосфата и обра-
зования АТФ (Т. В. Чиркова, Г. Ж. Валиханова, 1985). В то же
время субстратное фосфорилирование ввиду простоты механизма
извлечения энергии, относительно высокой энергетической эф-
фективности (даже при низком выходе энергии) и разнообразия
самих способов превращения субстратов нашло широкое исполь-
зование в последующей эволюции жизни на Земле. Оно стало
основой для развития более совершенных форм метаболизма.
Исторически наиболее древней следует признать функцию АТФ как
источника создания электрохимического потенциала на мембране
ДцН+. Позднее возникла возможность синтеза АТФ за счет ДцН'
путем изменения направления работы протонного АТФ-азного
комплекса: АТФ Дц Н+.
В эволюционном плане предшественниками АТФ служат
пирофосфаты как источники биохимических реакций. Так, на
анаэробе Desulfomaculatus nigrificans показана возможность роста в
присутствии пирофосфата:
Ацетат + пирофосфат 4- АДФ -> Ацетат + ортофосфат + АТФ
Как известно,в морской воде отсутствует пирофосфат и его
функцию там выполняет полифосфат. Многие анаэробные формы
с древнейших времен продолжают использовать пирофосфаты как
источник энергии (F. Lipmann, 1984). В этом смысле преимущест-
венное использование АТФ«=г АДФ (а не АДФ АМФ, в тер-
модинамическом отношении также выгодные) в энергетических
целях вызывает удивление. «Выбор» в ходе эволюции АТФ^АДФ
объясняют преимущественным участием в реакциях их хелатных
соединений с магнием, облегчающих взаимодействие с ферментами
(J. Е. Wilson, 1984). Возможно, что при этом сыграло роль и
химическое предрасположение — относительная легкость синтеза
адениновых нуклеотидов на стадии биопоэза.
Сказанное о роли субстратного фосфорилирования и механиз-
мов энергообмена удревних форм подтверждает мнение Ч. Дарвина
о том, что «при очень простых жизненных условиях высокая
организация не оказала бы никакой услуги...». Действительно,
Условия первичных биоценозов Земли были крайне упрощены. Им
как раз соответствовали примитивная организация протобионтов и
система их энергообеспечения. В этом проявляется целесообраз-
ность организации протобионтов, являющаяся результатом
Действия естественного отбора.
На подобной основе, вероятно, и появляется гликолиз, затем
Возникает механизм мембранного фосфорилирования, представля-
ющий собой важный этап биоэволюции. Мембранное фосфо-
119
рилирование в редокс-цепи связацо со строгой пространственной
организацией его звеньев. Впервые оно складывается у фототроф-
ных прокариот. Поэтому фототрофия эволюционно предшествовала
аэробиозу. У древних бактерий функционирует глиоксилатный путь,
представляющий собой сокращенный вариант цикла трикарбоновых
кислот (ЦТК). ЦТК также относится к древнему приобретению жизни,
который до момента появления кислорода в атмосфере не нашел
эффективного применения (М. В. Гусев, Г. Б. Гохлернер, 1980). О
Первичности анаэробного обмена свидетельствуют многробразные
метаболические превращения, даже у высших организмов соверша-
ющиеся в отсутствие О2.
На следующем этапе эволюции возникает ПФП метаболизма,
играющий важную роль в образовании нуклеиновых кислот, мета-
болически активного водорода в форме НАДФ ♦ Н и предшест-
венников ряда аминокислот. Впервые ПФП появляется также у
фототрофов. У сбраживающих бактерий ПФП не обнаружен, у
других бактерий он сформировался позже фотосинтеза.
Значение ПФП велико прежде всего в образовании активи-
рованного водорода путем расщепления воды и использовании его
для восстановления НАДФ, образовании пентоз, необходимых для
синтеза нуклеиновых кислот. В эволюционном плане эти события
способствовали усилению независимости организмов от внешней
среды, что рассматривается как один из основных результатов
прогресса живой природы (И. И. Шмальгаузен). Преимущества
ПФП обусловлены возможностью циклического его осуществ-
ления, где из шести молекул глюкозо-6-фосфата только одна рас,-
щепляется до СО2. ч
В то же время ПФП обладает рядом примитивных особенностей
(встречается у анаэробов), его ферменты (транскетолаза и трансаль-
долаза) распространены у всех живых организмов и включают некоторые
реакции брожения. Не исключено, что ПФП мог сложиться тогда, когда
организмы перешли к самостоятельному синтезу гексоз, т. е. параллель-
но с формированием фототрофии (М. В. Гусев, Г. Б. Гохлернер, 1981).
Несмотря на заметную метаболическую гибкость ПФП, он не дает
больших преимуществ в синтезе АТФ (А Ленинджер, 1974), так как у
эукариот в митохондриях НАДФ не включается в дыхательную цепь и
не используется для синтеза АТФ. Однако НАДФ • Н принимает
участие в трансгидрогеназной реакции и в генерации мембранного
потенциала:
НАДФ • н + НАД+^* НАДФ+ + НАД • Н,
что по своему направлению сходно с результатами аэробного
дыхания и гидролиза (В. П. Скулачев, 1974).
120
У цианей ЦТК выражен неполно: преобладает ПФП, а водород
вовлекается в цепь в виде НАДФ • Н. Точно так же некоторые
аэробные бактерии (уксуснокислые) осуществляют неполное
окисление органических веществ. Водород, высвобождаемый в
результате дегидрирования субстрата, вовлекаясь в процессы
окислительного фосфорилирования, превращается в воду. Полный
ЦТК впервые формируется у фототрофных анаэробных бактерий.
По указанным причинам у современных бактерий можно
встретить разнообразные сочетания процессов фотосинтеза, аэроб-
ного дыхания и гликолиза. Переход анаэробных бактерий к аэроб-
ному обмену (т. е. к дыхательной цепи) был связан не столько с
изменением структуры мембран, сколько с усилением прочности
связи ферментов с ними. Это показано на примере сравнительного
изучения локализации ряда дегидрогеназ у современных аэробных и
анаэробных бактерий. Дыхательная цепь впервые появляется у про-
кариот и исторически складывается путем замены системы бак-
териохлорофилл -> первичный акцептор цитохромами а и а3, что
привело к использованию О2 как конечного акцептора электронов.
М. В. Гусев и Г. Б. Гохлернер (1980) так представляют после-
довательность этапов энергетического обмена у бактерий: броже-
ние -> фотосинтез -> дыхание -> вторичный анаэробиоз ->
вторичный фотосинтез (микроаэрофилйя) -> нитратное «дыхание»
-> хемолитотрофия. Микроаэрофилия и нитратное «дыхание»
возникают благодаря вторичному продвижению части аэробов в бедные
кислородом экологические ниши. Перечисленные этапы обмена могли
возникать конвергентно и-неодновременно у разных прокариот путем
видоизменения реакций брожения, гликолиза и ПФП, которые и лежат
в основе дыхания представителей всех последующих царств и опреде-
ляют сходство равновесных реакций кинетического контроля их мета-
болизма.
Дыхательные ферменты растений близки к таковым животных,
У которых нет кровеносной системы и постоянной температуры
тела. У таких животных и растений на завершающем этапе дыхания
отмечена лабильность и многокомпонентность процесса. Но рас-
тения, сами вырабатывая кислород, меньше страдают от анаэро-
биоза и отличаются цианидоустойчивым дыханием (ЦУД). Это
Дыхание является следствием включения альтернативных путей
транспорта электронов с участием флавопротеинов. Кроме того,
Дыханию растений свойственна полифункциональность одних и тех
Же ферментов и связь с процессами фотосинтеза. ЦУД представляет
собой компонент общего дыхания, оно повышается у растений в
онтогенезе при переходе к репродуктивной фазе и в стрессовых
Условиях. Эволюционно возникновение ЦУД можно представить
Как результат отбора растений по активности цитохромоксидаз и
121
перехода к преимущественному использованию фруктозы и гексозы
в качестве субстратов дыхания в стрессовых условиях (Н. W. Janes,
Chin Chin-Kok, 1981).
Для понимания особенностей дыхания разных организмов следует
учитывать и результаты морфологической эволюции. Известно, что
на разных уровнях организации живых существ вопрос о снабжении
кислородом решался неоднозначно (способы обеспечения им зависят
dr толщины кутикулы и величины поверхности тела).
4.2. КИСЛОРОД КАК ФАКТОР ЭВОЛЮЦИИ ЖИЗНИ НА ЗЕМЛЕ
С появлением кислорода начал'формироваться озоновый экран
нашей планеты, что привело к отсечению УФ-лучей от солнечного
спектра. В этих условиях, естественно, отбор пошел по пути все
большего использования длинноволновой радиации в процессах
углеродного метаболизма.
Переходными между аэробами и анаэробами являются хемоав-
тотрофы И фоторедукторы, особенности которых достаточно
хорошо изучены (Э. Брода, 1978; М. В. Гусев, Г. Б. Гохлернер,
1981). Анаэробные фототрофы с сульфатным типом дыхания
появились на 1 млрд, лет раньше аэробов. Аэробные механизмы
возникают в виде дополнения к процессам брожения для
увеличения выхода АТФ (аэробное окисление у молочнокислых
бактерий) или нитратного дыхания, где нитраты (NOj) служили
акцепторами водорода (Э. Брода, 1978). В отличие от анаэробных
механизмов окислительные надстройки по существу не являются
универсальными, у различных групп современных организмов они
значительно различаются между собой. Так, в частности, у про-
кариот окислительное генерирование энергии идет на наружной
клеточной мембране и в ее впячиваниях, а у эукариот — на внут-
ренней мембране митохондрий (Т. В. Чиркова, 1988).
Разница между аэробным и анаэробным дыханием состоит
главным образом в акцепторах и продукции АТФ. Одной из важных
надстроек при возникновении аэробного дыхания является цикл
Кребса (ЦТК), протекающий в митохондриях. Возникновение его
связывают с циклом Арнона, характерным для зеленых серобак-
терий. Некоторые реакции ЦТК функционируют уже у клостридий
и метанобразующих бактерий (при синтезе глутамата из а-кетоглу-
таровой кислоты). Существует мнение о начальном самостоятель-
ном возникновении ди- и трикарбоновой части ЦТК, так как среди
цианей и зеленых бактерий встречаются формы, у которых эти
звенья еще не соединены, на уровне а-кетоглутаровой кислоты.
Электрон-транспортная дыхательная цепь, где происходит
окисление НАД * Н в аэробных условиях, складывается в резуль-
тате «инверсии» фотосинтетической ЭТЦ (Г. Б. Гохлернер, 1977).
122
Электрон в ЭТЦ дыхания передается постепенно переносчиками
на акцепторы, причем от имеющих низкий окислительный потенциал
К акцепторам с все большим окислительным потенциалом и, наконец,
к кислороду. На один атом кислорода при окислительном фосфо-
рилировании синтезируется три молекулы АТФ.
В связи с этим появление кислорода в атмосфере благодаря
деятельности фототрофов, несомненно, должно было привести к
изменению прежде всего донорно-акцепторных отношений в живой
природе. Возникла задача «отработки» системы акцептирования аг-
рессивного химического агента — кислорода. Первыми испытанию
на надежность подвергались сами организмы, выделяющие кислород;
их внутриклеточные компартменты и такие высокочувствительные к
О2 ключевые ферменты, как нитрогеназа, гидрогеназа и РБФК. Ввиду
выделения такими организмами кислорода и накопления его внутри
себя они в конце концов должны были вымереть из-за прекращения,
размножения и угнетения общего метаболизма.
Вероятность указанного процесса возрастала и по другой
причине, в частности из-за возможности перевода хлорофилла а в
сингелетное состояние, что грозило фотоокислительному
повреждению его молекулы. Поэтому необходимы были системы
внутриклеточной инактивации О2. Наиболее простой выход таился
в рамках исходной клетки. Именно тогда могли проявиться
присущие молекуле каротиноидов защитные свойства, благодаря их
способности вступать во взаимодействие с триплетным состоянием
хлорофилла и самим О2 следующим образом:
Каротиноиды + О2 -> каротиноиды (триплетное состояние) + О2 -> (основное
состояние) -> каротиноиды (триплетное состояние) -> каротиноиды (основ-
ное состояние) + теплота.
Со способностью к подобным превращениям каротиноидов^ в
том числе и у существ, не выделяющих кислород, отчасти связано
широкое распространение их в живой природе. Это обстоятельство
Делало возможным обитание таких анаэробных существ в соседстве
с кислородовыделяющими формами.
Каротиноиды выполняли функцию фотопротекторов, защищая
клетки и ткани от вредного действия видимой радиации и О2. Они
служили в качестве светособирающих пигментов в восстановленной
атмосфере в раннем периоде Земли. По мере насыщения атмосферы
02 эта функция каротиноидов постепенно утрачивалась, а фотоп-
Ротекгорная функция усиливалась (R. Maroti et al., 1984). В этом
случае они играют роль «энергетического клапана» при изменении
Интенсивности света.
В то же время, как подчеркивает Р. Клейтон (1984), первичное
Устранение токсичности О2 могло быть достигнуто его использо-
ванием для окисления Fe2+, находящегося в большом количестве в
123
водах и, следовательно, внутри самой клетки. При исчерпании
Возможностей каротиноидов и запасов Fe2+ (что наблюдалось по
мере усиления фотосинтеза самой клетки) О2 начал выходить из
клетки и поступать в окружающую среду примерно около 2 млрд, лет
назад. При этом возросла опасность для окружающей жизни. Кислород
мог выйти в атмосферу из клетки и после гибели его продуцента.
Значимость такого выхода нельзя отрицать даже если при этом
освобождалось 10“* мг О2. Агрессивность О2 могла быть значительной
в случае отмирания миллиардов существ, выделяющих кислород.
По указанным и другим причинам О2 становился все более
губительным для окружающих существ, не выделяющих его. Опас-
ность первичного О2 была связана с возможностью при его участии
образования супероксидных радикалов Oj, гидроксидного радикала
ОН, пероксида водорода Н2О2, озона О3, сингелетно-возбужденного
О2 и атомарного О состояний кислорода. В связи с этим защита
живых организмов могла быть обеспечена путем или «укрытия» их
в нишах, где нет О2, или выработки приспособлений для мета-
болической утилизации О2, Так, по имеющимся Данным, уже при
содержании в атмосфере 0,2% О2 (что соответствует 0,01% от
современного уровня его содержания) возникла необходимость
переключения процессов брожения на аэробное дыхание.
Первичная толерантность к О2, как Отмечено выше, была осно-
вана на использовании пассивных средств защиты, т. е. возможнос-
тей самой среды (внутренней и внешней). С накоплением О2 в среде
часть его под действием УФ-лучей стала превращаться в О3. Так
стал формироваться озоновый экран планеты. На значение озоно-
вого экрана в развитии жизни обращал внимание Л. С. Берг (1944),
внося коррективы в гипотезу А. И. Опарина.' В частности, Л. С.
Берг писал, что до появления фотосинтезирующих растений, когда
в стратосфере не было озонового экрана, жизнь могла возникнуть
и тем более сохраниться только в местах, защищенных от губитель-
ного действия космических лучей. В качестве таких мест автор
считал скопление воды на поверхности суши под обломками мине-
ральных пород коры выветривания. Точно так же Н. Г. Холодный
считал мелкие континентальные водоемы колыбелью земной
жизни. По мнению Л. Г. Стеббинса (1982), первые фотосин-
тезирующие бактерии появляются в руслах рек или пресных мел-
ководий. По общему признанию, озоновый экран стал играть
эффективную роль в защите многих брганизмов от губительных
компонентов солнечного света. Возможно, именно это обстоятель-
ство «вывело» жизнь из морской глубины на свет, т. е. на хорошо
освещенную поверхность воды. Такой подъем был осуществлен и
бесцветными формами жизни, что в итоге способствовало
соприкосновению разнообразных существ с О2. И тем самым повы-
шался успех отбора среди них достижением коренного решения
124
Проблемы защиты организмов от О2, метаболизаиией его в энер-
гетических целях для синтеза АТФ.
Поиск, судя по ныне живущим прокариотам, шел в разных
направлениях, в частности имел место отбор организмов по спо-
собности синтеза специальных ферментов (супероксвддисмутаза,
каталаза, пероксидаза) и клеточных метаболитов (каротиноидов),
формирования сообществ (из разных по устойчивости к О2
организмов) и структур (митохондрий и хлоропластов), а также по
использованию О2 при азотфиксации, люминесценции й т. п. По
этой причине у многих современных анаэробных прокариот найден
тот или иной путь зашиты от О2, позволяющий им жить в аэробной
среде. Сказанное можно демонстрировать и наблюдениями о пог-
лощении Ог некоторой частью современных анаэробных прокариот
при столкновении с ним (М. В. Гусев, Л. А. Минеева, 1985).
Использование О2 прокариотами в последующем пошло по пути
ферментативного и неферментативного (прямое внедрение в моле-
кулу) взаимодействия. Аэротолерантность первично возникает у
форм с субстратным фосфорилированием. Так, молочнокислые
бактерии способны, не используя О2 в, метаболизме, восстановить
его до Н2О2 благодаря наличию флавиновых ферментов. У них
пероксид накапливается в клетке из-за отсутствия каталазы. У
анаэробных пропионовокислых бактерий аэротолерантность обес-
печивается наличием супероксиддисмутазы, каталазы и перо-
ксидазы для обезвреживания супероксидного аниона и Н2О2.
Эволюция у пропионовых бактерий пошла еще дальше по пути
приспособления к аэробным условиям, у них обнаружены цитох-
ромы, реакции ПФП и ЦТК. В связи с этим у пропионовых бактерий
расширяются возможности использования разных сред и пути
вовлечения СО2 в метаболизм. С формированием ПФП становится
возможной полная деградация углеводов и начинается эра переноса
отщепленного при этом водорода на кислород.
Эволюция по пути метаболического использования О2 привела
в вовлечению его в реакции как акцептора электронов и источника
энергии для клетки. В этом как раз большую роль призваны были
сыграть складывающиеся ферментативный комплекс (для запасания
АТФ) и ЭТЦ, участвующая в переносе протонов через мембрану и
электронов на О2. Согласно представлениям П. Митчела, указанные
Два процесса могли сложиться неодновременно, причем у разных
существ. Поэтому возникает проблема их объединения в одном
существе для эффективного метаболического использования О2, что
Могло быть достигнуто или симбиозом разных существ, или
Мутационным преобразованием других звеньев метаболизма (ЭТЦ
фотосинтеза в дыхательную цепь). Мы не знаем, как обстояло здесь
Дело, хотя многие полагают наиболее вероятным второй путь. .
Как бы то ни было, возникает способность полного отщепления
Н2 от субстрата, переноса его на О2 и преобразования энергии такого
125
переноса в химическую энергию АТФ. Это стало возможным после
возникновения ЦТК. в результате «надстройки» некоторых реакций
на анаэробные энергетические механизмы клетки. Неполный, или
«разорванный» ЦТК обнаружен у ряда анаэробных фототрофных
бактерий, цианей и др. О роли и степени выраженности ЦТК у
разных прокариот достаточно сказано (М. В. Гусев, Л. А. Минеева,
1985). Для нас же важно, что указанный цикл стал функционировать
первоначально наиболее полно у фототрофов — цианей и многих
пурпурных бактерий. Именно выделение О2 последними «вдохнуло»
жизнь в «выжидающие» реакции ЦТК, т. е. подключило последние
к системе энергодобычи. Это произошло на критических этапах
эволюции жизни на Земле, когда к системе энергообеспечения
стали предъявляться повышенные требования: выжить или выме-
реть существам.
Стабилизация ЦТК в ходе эволюции связана с возможностью
получения наибольшего количества АТФ й обеспечения
значительной эффективности процесса из-за присоединения про-
межуточного метаболита к другой молекуле с низкой молекуляр-
ной массой (Н. A. Krebs, 1-981).
4.3. ПЕРВИЧНАЯ ЗАЩИТА ОРГАНИЗМОВ ОТ КИСЛОРОДА
И ВОЗНИКНОВЕНИЕ АЭРОБНОСТИ
Рассмотрим основные ступени перехода живых организмов к
использованию О2. На начальном этапе развития жизни кислород
(и то в небольшом количестве) сосредоточивался в верхних слоях
атмосферы и живые существа, обитающие в толще воды или в
придонном иле, вообще не сталкивались с ним. Только фототроф-
ные формы могли занимать освещенную или пограничную к О2
толщу воды. Из их числа несерные и пурпурные бактерии
находились в толще воды, обедненной кислородом, а зеленые
серобактерии — в бескислородных слоях. Это означает, что уже на
начальных этапах эволюции жизни возникает известная диффе-
ренциация организмов по устойчивости к О2 (аэротолерантность).
Популяционный «взрыв» фототрофов на первичной Земле
привел к накоплению кислорода в атмосфере. Этот процесс сопро-
вождался расселением фотосинтезирующих организмов и превра-
щением кислорода в «самого могущественного деятеля из всех нам
известных химических тел земной коры» (В. И. Вернадский, 1967).
Далеко не случаен весьма примечательный факт совпадения вре-
мени возникновения многоклеточных форм с появлением кисло-
рода в атмосфере, что произошло примерно в начале кембрия (600
млн. лет назад), когда уровень его составлял всего 0,6%. Отсюда О2
126
не без основания признают «творцом дифференцировок» жизни на
Земле (О. Варбург).
Как известно, О2 утилизируется как акцептор водорода при
образовании воды, при этом появляются супероксидные группы
О'г, НО2) Н2О2 и ОН. Защита аэробной клетки от кислорода достига-
ется благодаря участию ферментов супероксидцисмутазы, каталазы
и пероксидазы (Е. F. Elstner, 1982). Естественно, что переход к
аэробному дыханию не мог быть осуществлен сразу из-за отсутствия
митохондрий, ферментов, необходимых для активизации водорода
и восстановления молекулярного кислорода (оксидаз, пероксидаз,
железосодержащих порфиринов и т. д.), ступенчатого окисления
субстрата, предотвращающего излишнюю потерю энергии в виде
теплоты, и т. д. Для выживания и адаптации организмов в аэробной
среде необходима была и инактивация Н2О2. Поэтому многие
организмы первоначально занимали экологические ниши, лишен-
ные кислорода, или использовали механизмы, сочетающие анаэ-
робный и аэробный пути метаболизма.
Первичные экосистемы Земли обычно сравнивают с экосисте-,
.мами современных пещер и глубин океанов (M.dB. Гусев, Г. Б.,
Гохлернер, 1980). Уходу из зоны насыщения О2 способствовало
отчасти накопление у некоторых бактерий кристаллов магнетита,
игравших роль «стрелок компаса». Эти кристаллы обеспечивали
движение организма вдоль силовой линии магнитного поля, на-
правленного вертикально вниз (В. А. Красилов, 1986). Поэтому
бактерии опускались ниже уровня насыщения кислорода,
притягиваясь к местам скопления железа, образованного также
организмами анаэробного типа (нуждающимися в Защите от О2).
Только в ходе дальнейшей эволюции возникают анаэробные сущес-
тва, нечувствительные к О2.
। На начальных этапах защита от кислорода, по-видимому,
достигалась разными путями: 1) сочетанием в экосистемах
организмов с неодинаковым типом биоэнергетики, например
Цианей и аэротолерантных бесцветных бактерий (молочнокислых),
причем аэротолерантность бактерий могла быть даже результатом
случайного заноса в клетку гема, который, соединившись с
белком, проявлял в какой-то мере свойства каталазы разлагать
Н2О2; 2) симбиозом протоцианей и гетеротрофов (с гемом),
обеспечившим сочетание в одном существе каталазной (от гетерот-
рофов) и супероксидцисмугазНой (от автотрофов), активности; 3)
разложением Н2О2 ионами Fe2+', находящимися в первичном океане
(в составе гема каталитическая активность железа превосходит
активность его в водной среде в 103 раза, а каталазы —в 1О10 раз);
4) развитием механизмов биолюминесценции, при которой образу-
ются вещества, активно реагирующие с кислородом, в результате
чего высвобождается энергия в виде света (Л. А. Тумерман, 1964).
127
Анаэробные организмы такими путями (а возможно, и другими)
обязательно должны были приобрести определенную толерантность
к кислороду. Дальнейшее усиление толерантности к кислороду
произошло после включения в метаболизм каталазы и пероксидазы.
Основная роль в элиминации Н2О2 принадлежала каталазе, которая
безотказно действует в любых случаях и упорядочивает окисление
в клетке. Не исключается и возможность дегидрирования аро-
матических соединений пероксидазой, что могло предшествовать
этапу прямого включения кислорода в метаболизм.
Одновременно с возникновением аэротолерантности складыва-
ется фаза энергетического дыхания, где роль терминальных цито-
хромов вначале, по-видимому, выполнял какой-нибудь
самоокисляющийся двухгемовый цитохром или попарно-разные
соединения (один из цитохромов и пероксидаза). Возникновение
цитохромоксидаз связывают с преобразованием генетического
материала, ответственного за синтез хлорофилла у существ,
утративших способность к фототрофии. Вовлечение кислорода в
биоэнергетические процессы, вероятно, осуществлялось у разных
существ неодновременно и скорее всего происходило путем изме-
нения старых анаболических механизмов или замены их на более
экономные. Например, у Streptococcus facalis аэробный механизм
фиксируется в помощь сбраживающему для увеличения выхода
АТФ. В аэробных условиях эта бактерия может использовать
глицерин, предварительно дегидрируя его посредством НАД. По-
лученный при этом НАД • Н окисляется О2 через флавопротеин.
М. В. Гусев и К. А.. Никитина (1979) считают что вначале
кислород участвовал в метаболизме так же, как это происходит у
современных зеленых и пурпурных бактерий и цианей- Следующим
этапом могло быть ингибируемое светом дыхание, когда кислород
уже становился полезным для одних процессов жизнедеятельности
и вредным для других. Так, у цианобактерий защита от окисления
чувствительных структур достигается использованием кислорода
для восстановления аскорбиновой кислоты и соединений с
сульфгидральными группами.
Максимальная эффективность энергообмена в клетках растений
могла быть достигнута при условии устранения возможной конку-
ренции между фотосинтезом и дыханием путем разделения функций
коферментов НАД • Н и НАДФ • Н. При подобном разделении
первый из них специализируется на переносе электронов к кисло-
роду (дыхание), второй — на фиксации СО2 (фотосинтез). Такой
процесс был осуществлен у эукариот с появлением пропластид и
промитохондрий. Функция генерирования энергии переносится с
мембран клетки на мембраны органелл, что приводит к еще боль-
шему увеличению энергетически активной поверхности и соответ-
ственно энергетики клетки (D. Keilin, 1976).
128
В конце концов путем сложных преобразований возникает
дыхательная цепь. Для понимания ее эволюции представляют инте-
рес данные о сходстве ЭТЦ фототрофных организмов и организмов,
способных к дыханию, по составу веществ (НАД, ФАД, ферре-
доксины, цитохромы, пластохиноны). Кроме того, обнаружено
сходство строения цитохромов и хлорофилла, а также цитохромов
и ферредоксина. Однако, исходя из факта только сходства нельзя
еще делать вывод о возможности постепенного превращения ЭТЦ
фотосинтеза в дыхательную цепь.
4.4. ВОЗНИКНОВЕНИЕ МИТОХОНДРИЙ
Основной потребитель кислорода в клетке —дыхательная цепь
' митохондрий, хотя в дыхании участвуют и полиферментные системы
других органелл. Химический состав митохондрий довольно сложен:
в них нет гран, но есть кристы. Существует связь между числом крист
в митохондриях и энергетическими потребностями клетки.
Митохондрии обладают известной автономностью и способны
к саморедупликации. И митохондрии, и хлоропласты содержат
РНК, специфическую ДНК, способную к синтезу протеидов, имеют
рибосомы, фосфолипиды, сходные по строению. В митохондриях
имеются цитохромы а, Ь, а3, с, сь но отсутствуют такие компоненты
ЭТЦ, как цитохромы Ь6, присущие хлоропластам.
В препаратах МтДНК обнаружено около 95% линейных моле-
кул, хотя у части растений удается выделить до 45% кольцевой
структуры ДНК (В. И. Негрук, 1984). МтДНК состоит из попу-
ляции тех и других молекул ДНК, различающихся по размерам. В
отличие от ХпДНК каждая Из кольцевых молекул МтДНК несет
уникальные гены и содержит значительное количество некодирующей
ДНК. Содержащиеся в митохондриях пшеницы гены 26S, 18S, 5S
рРНК как в хлоропластах, так и в клетке Escherichia coli разделены
спейсерными последовательностями. Процесс вырезания — встраи-
вания нуклеотидов из ДНК гораздо чаще происходит в митохондриях,
чем в хлоропластах. Этим определяется сложность митохондриального
генома (МТГ) по сравнению с геномом хлоропласт.
Изучением генетических функций митохондрий растений пока-
зано, что они не совсем автономны в энергетическом и генетичес-
ком отношении (К. В. Войников и др., 1991). По синтезам
митохондрии зависят от ядра и других органелл. Функционирование
МтДНК также обеспечивается ферментами, кодируемыми яДНК.
МТГ растений по размерам в сотни раз превосходит (218—2500 тнм)
таковой животных, но он менее стабилен. Это, возможно, обуслов-
лено многими причинами: участием митохондрий растений не
только в дыхании, но и в устойчивости, сложностью метаболизма
растительной клетки и менее жестким ее гомеостазом.
5-157
129
Имеются данные, свидетельствующие о возможности обмена
генами между органеллами клетки в ходе онтогенеза, в том числе
и между митохондриями и ядром. Гомологию МтДНК и ХпДНК. по
содержанию 16 рРНК рассматривают как доказательство такого
обмена. Слаженным действием геномов разных органелл в онтоге-
незе растений, вероятно, обеспечивается нормальное состояние
организма, уравновешиваются процессы его жизнедеятельности
при отклонении условий от нормы. Необходимость такого уравно-
вешивания часто возникает в онтогенезе любого растения.
У прокариот нет митохондрий, транспорт электронов у них связан
с плазматической мембраной и липопротеиновой ламеллярной систе-
мой. У эукариот наблюдается большое разнообразие форм и размеров
митохондрий (обычные, нитевидные и палочковидные). Обнаружено
сходство митохондрий высших растений, дрожжей и грибов.
Гликолитический путь (эволюционно более древний) в энер-
гетическом отношении оставлял в качестве «отходов» трикарбоно-
вые кислоты, которые могли быть источником пиши для каких-то
аэробных микроорганизмов. Вероятно, такие микроорганизмы,
проникнув в прокариотическую клетку, вступали с ней в симбиоз
и могли выполнять роль митохондрий (L. Margulis, 1981). Это
предположение основано на сходстве митохондрий с микроор-
ганизмами (кольцевая ДНК, мелкие рибосомы, чувствительность к
бактериальным антибиотикам, наличие кинетопласта).
В ходе эволюции организмов митохондрии изменялись в направ-
лении увеличения размеров, сокращения генома и усиления системы
окислительного фосфорилирования. У примитивных эукариот геномы
митохондрий длиннее, чем у высших. Согласно гипотезе симбиоге-
нетического возникновения митохондрий, переход к аэробному
дыханию — результат проникновения первичной аэробной про-
кариотной формы в свободно живущее автотрофное, но анаэробное
существо. Однако теперь высказывается сомнение относительно воз-
можности самостоятельной жизни такого гипотетического аэробного
существа на первичной бескислородной Земле.
А. Ленинджер (1976), обсуждая теории биогенеза митохондрий
в онтогенезе, разделил их на три группы: синтез митохондрий de
novo из ультрамикроскопических предшественников, образование
их из других мембранных структур, рост и деление существующих
митохондрий. Каждая из этих теорий опирается на фактический
материал, имеет свои достоинства и недостатки. В . онтогенезе
митохондрии образуются, как правило, из себе подобных путем
поперечного деления, а иногда и почкования. Зачатки митохондрий,
называемые промитохондриями, передаются потомству через
яйцеклетку при половом размножении. Гетерогенность указанных
органелл рассматривается как проявление множественности путей
их дифференцировки. Эволюционно же митохондрии, вероятно,
130
возникли путем инвагинации мембран, как и хлоропласты. Это
отчасти согласуется с фактом постепенного усложнения их струк-
туры в онто- и филогенезе. '
4.5. ОСОБЕННОСТИ ДЫХАНИЯ У РАСТЕНИЙ
В дыхательную цепь митохондрий обычно входят пять
различных цитохромов, где один атом кислорода расходуется на
синтез трех молекул АТФ. Поток электронов в дыхательной цепи
напоминает таковой при фотосинтезе. Как отмечалось, дыхательная
цепь впервые появляется у прокариот и по разветвленности сходна
с митохондриальной цепью. У хемосинтетиков типичные мито-
хондрии отсутствуют, слабо сопряжены процессы окисления и
фосфорилирования, а также отмечен низкий потенциал донора
электронов. Цикл Кребса универсален для всех организмов,
дышащих кислородом.
В метаболизм растительной клетки кислород вовлекается раз-
ными путями: 1) восстановлением одно- и двухвалентными доно-
рами электронов с участием оксидаз; 2) включением продуктов его
восстановления в окисляемую молекулу с участием гидроксилаз; 3)
непосредственным включением кислорода в молекулу окисляемого •
субстрата оксигеназами.
Для растений, как и для преобладающего большинства живых
существ, характерны разнообразные превращения одного и того же
вещества. Высвобождение энергии при дыхании имеет фазовый
характер и происходит постепенно (Г. Кребс, Г. Корнберг, 1959) по
мере разукрупнения окисляемого субстрата, что предотвращает
бесполезное улетучивание энергии в виде теплоты. Этим целям как
раз служит разветвленная и многокомпонентная система фермен-
тов, участвующих в дыхании. Растительные организмы обладают и
каталитическими механизмами для Использования разнообразных
соединений в качестве субстрата для дыхания.
Механизмы аэробного дыхания и фотосинтеза у растений свя-
заны как в филогенезе, так и в онтогенезе. Они зеркально отобра-
жают и взаимодополняют друг друга (табл. 5), снабжают организм
энергией и метаболитами. Характерно, что в ассимилирующей клетке
Фотосинтез подавляет гликолиз. Но свет оказывает влияние и на
Дыхание через накопление АТФ. «Избыток» синтеза АТФ в процессе
Фотофосфорилирования оказывает ингибирующее действие на работу-'
ЭТЦ митохондрий. Образующиеся в результате цикла Кребса проме-
жуточные продукты (дикарбоновые кислоты, аминокислоты и др.)
Используются в клетке на свету, поэтому его подавление какими-либо
в°здействиями отрицательно влияет и на фотосинтез.
У растений и микроорганизмов кроме цикла Кребса как его
видоизменения встречаются и дополнительные пути окисления
131
углеводов. К ним относятся обратимое расщепление сукцината на
ацетил-КоА и НАДФ • Н2, превращение жиров в углеводы (с
образованием малата и ФЕП как промежуточных продуктов), ПФП
(его доля составляет 10—14%), ГГЦ в пероксисомах (фотодыхание
при накоплении в листьях гликолевой кислоты). В онтогенезе
растений аэробный обмен возникает на базе анаэробного. От-
сутствие узкой специализации у растений к определенному типу и
субстратам дыхания очень важно для обеспечения жизни в часто
меняющихся условиях среды. Для растений характерно цианид-ус-
тойчивое дыхание из-за наличия в митохондриях цианид-резистент-
ных оксидаз.
Таблица 5. Сравнение процессов фотосинтеза
и дыхания (по Б. А. Рубину, И. Ф. Гавриленко, 1972)
Показатели Фотосинтез Дыхание
СО2 02 н2о Энергия Органические вещества Поглощается Выделяется Разлагается Аккумулируется Синтезируются Выделяется Поглощается Синтезируется Выделяется Разлагаются
В целом механизм аэробного дыхания растений принципиально
не отличается от механизма аэробного дыхания животных и свиде-
тельствует об общности их происхождения.
4.6. ДЫХАНИЕ И ПРИСПОСОБЛЕНИЕ РАСТЕНИЙ К СРЕДЕ
Как известно, дыхательная система растений построена по
принципу множественности, т. е. на любом этапе дыхания при-
нимают участие два фермента и более, выполняющих одну и ту же
функцию. Значение этого явления становится понятным, если
принять во внимание температурный оптимум действия ферментов
и сродство их к кислороду. Так, цитохромоксидаза, ферменты
флавиновой группы и Cu-протеиды проявляют неодинаковую
активность при различном содержании кислорода в тканях. Цито-
хромоксидаза обладает наибольшим сродством к кислороду, она
активна при содержании его в среде 1—2%. Активность же фер-
ментов флавиновой группы возрастает с увеличением концентрации
кислорода.
В этой связи интересно, что наружные, хорошо аэрируемые
ткани плода мандарина имеют кислородный оптимум при 21%, а
внутренние, плохо аэрируемые,— при 10%. Ткани же мякоти, до-
ступ кислорода к которым сильно затруднен, наиболее интенсивно
дышат при содержании 5% кислорода в среде. Соответственно этому
132
в плоде распределяются ферменты: в наружных тканях преобладает
флавиновая группа (аэробные дегидрогеназы), в средних — медь-
содержащие протеиды (дифенолоксидаза, аскорбиноксидаза), во
внутренних — цитохромоксидаза. Указанные ферменты отличают-
ся И по отношению к температуре. Цитохромоксидаза функ-
ционирует при более высокой температуре, чем флавинопротеиды,
имеющие низкий температурный коэффициент активности (Q10).
Благодаря смене ферментативных систем создается возможность
•для осуществления дыхания в изменившихся условиях.
В ходе онтогенеза растений ферментативные процессы не оста-
ются постоянными. Показано, например, что при созревании пло-
дов яблонь Fe-протеиды сначала заменяются на Cu-протеиды, а
затем на более «холодостойкие» флавиновые ферменты. Интересны
наблюдения на хвойных. Известно, что хвоя лиственницы на зиму
опадает, а хвоя сосны — нет. В формирующейся хвое лиственницы
преобладает дифенолоксидаза, в период ее активного роста —
цитохромоксидаза и пероксидаза. К осени активность пероксидазы
в хвое лиственницы, постепенно падает. В хвое же сосны к осени
активность пероксидазы и дифенолоксидазы непрерывно возраста-
ет, а зимой повышается активность ряда дегидрогеназ (у лиственниц
они инактивируются). Полагают, что высокая активность дегидро-
геназ способствует повышению зимостойкости хвои сосны благо-
даря синтезу в ней масел, способных повышать эластичность
цитоплазмы и понижать температуру ее замерзания.
Обнаружены различия в ферментативных системах у водных и
полупогруженных, болотных и наземных растений. В листьях во-
дных растений преобладают гликолитические ферменты, а в листьях
наземных — ферменты аэробного цикла. Особенно поразительно
биохимическое и экологическое сходство в дыхании болотных
растений с ныряющими животными (Р. М. Кроуфорд, 1981).
Высшие растения, как правило, обеспечивают свои потребности
в кислороде не с помощью специальной системы, транспор-
тирующей кислород, а путем непосредственного газообмена со
средой, что возможно при достаточно развитой поверхности тела,
обилии устьиц и межклеточных пространств. Несмотря на это, в
Природной обстановке нередко ограничивается дыхание как у сук-
кулентов, покрытых толстой кутикулой, пробкой и восковым нале-
том, так и у водных растений, а также при затоплении и орошении,
На заболоченных и плотных почвах. Разные виды растений неодина-
ково устойчивы к анаэробиозу (D. D. Hook, Р'. М. Crawford, 1978).
Существуют приспособления к анаэробиозу, например система
воздухоносных полостей (болотные и водные растения), по которым
воздух проходит из листьев в корни и обратно. При этом листья
выполняют защитную роль как на свету, так и в темноте. В этом есть
Необходимость. Так, содержание О2 в зарослях Menyathes trifoliata на
133
глубине 25 см равно 0,47%, а на глубине 30—35 см в зарослях
Equisetum limosum — 0,34%. Воздух к корням таких растений поступает
только через надводные части (Б. Уильямс, Д. Барбер, 1964). Поэтому
растения без аэренхимы обычно не могут выжить, если их корневые
системы погружены в воду. Отсюда внутреннее строение водных
растений напоминает сложные пчелиные соты с толщиной сТенок
в одну клетку, снабженных мелкими порами для обеспечениятазом.
Стебли водных растений обычно осуществляют резервную и тран-
спортную функцию в доставке корням О2.
Как известно, у водных растений основная масса тела находится
под водой в условиях плохой аэрации. В анаэробной среде
выживание обеспечивается блокированием накопления токси-
ческих веществ в корнях (этанол, лактат и малат). Однако не только
биохимические изменения обусловливают выживание организмов
в анаэробной среде, важны и морфологические приспособления:
увеличение объема межклеточных пространств, облегчающих
диффузию газа при приливах и отливах воды между погруженными
и непогруженными частями растения. На примере осоки (гигрофит)
показано, что ее метаболизм в анаэробной среде неустойчив, при
этом наблюдается даже деструкция мембран (Б. Б. Вартапетян,
1982). Отсюда сделан вывод, что устойчивость гигрофитов к;
отсутствию кислорода обусловлена не биохимическими меха-
низмами (переключение обмена на альтернативный путь), а анатомо-
физиологическими изменениями на уровне целого организма, т. е.
развитием способности транспортировать О2 из надземной части к
подземной. В этой связи автор предлагает отнести гигрофиты к
растениям, обладающим не истинной, а кажущейся устойчивостью к
отсутствию кислорода. Данные об использовании разных механизмов
для адаптации растений к анаэробиозу свидетельствуют о взаимообус-
ловленности изменений формы и функций в эволюции.
Лишение клетки О2 резко сказывается на ее энергетике, струк-
туре и функции митохондрий. При кратковременной аноксии
происходят разбухание органелл, сокращение числа крист, дегра-
дация митохондриальных мембран. В отсутствие экзогенной глю-
козы из-за энергетического голодания аноксия приводит к
деструкции ультраструктуры клеток. Интересна отметить, что
клетки корней взрослых растений риса и осоки, живущих на
переувлажненных анаэробных почвах, не обладают большей
резистентностью к аноксии, чем корни тыквы и других растений
(обитающих на хорошо аэрируемых почвах). Адаптация к анаэ-
робиозу выступает как свойство целого растения. Из надземной
части к корням поступает лишь незначительная часть О2. Адаптация
к гипоксии и аноксии может быть обусловлена изменениями как
на молекулярном, так и на уровне целого организма. У истинно
устойчивых к аноксии растений есть молекулярные механизмы
134
резистентности. У кажущихся устойчивыми растений нет таких
механизмов, однако они способны жить в анаэробных условиях
благодаря транспорту О2 из аэрируемых органов (Б. Б. Вартапетян,
1985).
Метаболические пути приспособления растений к гипо- и
аноксии разнообразны, и на разных их уровнях осуществляется
система регуляции (Т. В. Чиркова, 1988). С усилением напряжен-
ности анаэробиоза доминирующими становятся метаболическая
регуляция, комплекс изменений для поддержания гомеостаза и
репаративных процессов. Ими затрагиваются ЭТЦ митохондрий,
осуществляются «сброс» восстановительных пиридиннуклеотидов,
трансформация дыхательных путей, возрастание доли ПФП и
синтез белков-ферментов анаэробного обмена, перестройка мемб-
ран, устраняется накопление токсических продуктов метаболизма
(лактат, этанол и др.).
Опытами установлено, что при помещении корней растений в
анаэробные условия листья их поглощают больше кислорода, чем
в аэробной среде. У гигрофитов (манник) даже в нормальных
условиях кислородного снабжения корней листья поглощают боль-
ше кислорода, чем требуется им самим. Как указывалось, роль
листьев в снабжении растений кислородом очень велика, поэтому
при лишении их листьев и света растения обнаруживают все
симптомы анаэробного обмена: Накопление молочной кислоты,
снижение дыхательного коэффициента.
В условиях, затрудняющих снабжение корней кислородом через
листья (например, под водой или ледяной коркой), растения обо-
гащаются органическими кислотами, свободными аминокисло-
тами, при этом меняется ультраструктура клеток корня,
повреждаются митохондрии, цистерны эндоплазматического
ретикулума,, уменьшаются количество рибосом в цитоплазме и
размеры ядра. Такие изменения в клетках менее выражены у
устойчивых к анаэробиозу видов (болотные растения).
В условиях анаэробиоза в результате комплекса внутриклеточных
процессов происходит угнетение роста и развития, продуктивности
растений. Как показывают исследования Г. М. Гриневой (1974), во
избежание этого осуществляются перераспределение кислорода
Между органами растений, саморегуляция энергетического обмена
путем переключения путей метаболизма в клетке, в том числе
Накопление ингибиторов роста и недоокисленных промежуточных
Продуктов (углеводы, спирты, органические кислоты), ненасыщен-
ных липидов (выполняющих роли акцепторов водорода и
заменителей кислорода), выделение токсичных веществ корнями и
Т.д. Адаптация растений к анаэробиозу связана с повышением
активности цитохромоксидаз (что поддерживает межмембранный
транспорт электронов); накоплением быстро утилизирующихся ве-
135
ществ (сукцинат и малат); усилением спиртового брожения (устра-
няющего токсическое накопление пирувата и ацетальдегида),
нитратного дыхания (при этом устраняется токсическое накопление
нитратов) и окислением глюкозы через ПФП; повышением роли
гликолиза в синтезе АТФ (проявление эволюционно древнего спо-
соба энергетики в онтогенезе). Для предотвращения отрицательного
влияния анаэробиоза имеет значение также образование дополнитель-
ных корней и газовых полостей у основания побега (сх. 8).
Схема 8
Приспособление растений выражается и в различной ус-
тойчивости к понижению или повышению температуры. Обычно
оптимум температуры для дыхания у высокогорных и полярных
растений лежит значительно ниже, чем у растений умеренной зоны,
пустынь и тропиков. Они заметно различаются и по компенсационной
точке фотосинтеза. У указанных растений меняется и качество фер-
ментов, определяемое по константам его действия (А. В. Благове-
щенский, 1966). В целом у зимостойких видов интенсивность дыхания
зимой выше, чем у незимостойких. У первых при понижении
температуры наблюдается усиление окислительно-восстановитель-
ных процессов. У растений, произрастающих в крайних условиях
существования, качество ферментов высокое, что дает им возмож-
ность сохранить активность химических реакций при резкой смене
условий. Повышение качества ферментов идет параллельно
увеличению доли низкомолекулярных белков — альбуминов и гло-
булинов. Такой параллелизм наблюдается как в онтогенезе, так и в
филогенезе (см. гл. 5).
136
В целом адаптация растений к условиям обитания сопряжена с
изменениями в метаболизме, энергетическими затратами. Как пока-
зывают многие наблюдения, в неблагоприятных условиях изме-
нения в фотосинтезе оказываются более значительными, чем в
дыхании. Это создает нехватку энергетического материала для роста
и морфогенеза, включения регенерационных процессов и
механизмов осмотических адаптаций, ведет к снижению репродук-
тивного потенциала растений. В итоге наблюдается общее истоще-
ние индивидуума и потеря устойчивости к стрессам. Поддержание
стабильности растений в неблагоприятных условиях связано с
усилением затрат на дыхание («дыхательная цена»). Сами эти
затраты можно определить количеством израсходованного углерода,
и они оказываются неравноценными у адаптированных и не
адаптированных к стрессам растений (О. А. Семихатова, 1995). Уро-
вень ингибирования растений определяется разницей между накоп-
лением (фотосинтез) и расходованием (дыхание) энергетических
ресурсов в неблагоприятных условиях. Все это делает чрезвычайно
важным учет энергетических затрат при оценке адаптации растений
и ведет к усилению давления отбора на их уменьшение при одновре-
менном обеспечении репродукционного успеха индивидуума.
Одним из примеров экологических различий в дыхании рас-
тений является фотодыхание.
4.7. РОЛЬ ФОТОДЫХАНИЯ
Эффект Варбурга — ингибирование выделения О2 и ассими-
ляции СО2 по мере увеличения концентрации О2 в среде — широко
распространен у растений с циклом Кальвина. Он обусловлен
конкуренцией СО2 и О2 за субстрат ввиду проявления РБФК
свойств, характерных для карбоксилаз и оксигеназ. Фотосинтез
даже при нормальных концентрациях СО2 и О2 в среде приводит к
образованию не только ФГК, но и гликолата.
Фотодыхание — окисление гликолата в глиоксилат на свету
через редуктивный ПФП с усиленным потреблением кислорода и
выделением СО2 —накладывает отпечаток на темновое дыхание.
При этом излишки электронов фотосинтетической системы выво-
дятся из цепи и используются для Ьинтеза АТФ. Не вызывает
сомнения наличие светозависимого потребления О2 у части рас-
тений как результат подавления темнового дыхания при
фотосинтезе. Однако фотодыхание не всегда оказывается связано с
фотосинтезом. Так, фотодыхание наблюдается у бесхлорофилло-
носных мутаций хлореллы на сильном свету, которые способны
синтезировать только каротиноиды. У таких мутантов фотодыхание
составляет 50% от уровня темнового дыхания.. В зеленых клетках
137
при отключении фотосинтеза дыхательным ядом также происходит
потребление О2 и выделение СО2.
При фотодыхании в 3—5 раз быстрее выделяется СО2, чем при
темновом дыхании (М. Зелитч, 1976). Получается так, что, с одной
стороны, в процессе фотосинтеза фиксируется СО2, с другой — она
тут же теряется на 50% при фотодыхании. Природа здесь оказалась
как бы не на уровне защиты интересов своего произведения. И
среди растений С3-типа фотосинтеза обнаружены мутанты с
блокированным фотодыханием.
В связи с подобными наблюдениями возникает вопрос, насколько
С3-растения могут обходиться без фотодыхания. При осуществлении
фотодыхания преобладает оксигеназная функция РБФК. При естес-
твенных концентрациях СО2 и О2 в атмосфере соотношение между
фотосинтезом и фотодыханием у С3-растений составляет 3; 1. Посколь-
ку соотношение указанных газов в среде (в том числе и в клетке) часто
меняется под влиянием температуры и времени суток (из-за колебания
концентрации прежде всего О2), постольку необходимым оказывается
переключениефункции РБФК на оксигеназную функцию, и наоборот.
Все зависит от того, какой газ находится в избытке.
Образование гликолата служит началом фотодыхания. Дальней-
шее его превращение происходит в хлоропластах, пероксисомах и
митохондриях. У большинства растений гликолат в хлоропластах
образуется при падении концентрации СО2 и увеличении концент-
рации О2 (сх. 9). Фотодыханию отводится некоторая положительная
роль в обеспечении клеток промежуточными веществами (глутамат,
у-глутаровая кислота, глицин, серин, глицерат и т. п.), важном для
дальнейших синтезов. Все это свидетельствует, что фотодыхание
нельзя рассматривать как бесполезный или вредный процесс.
Схема 9
Хлоропласты Пероксисомы Митохондрии Цитоплазма
Гликолат —
4
Цикл углерода
> Гликолат
Гли океплат
Глицин
Оксалоглутарат
+
Оксалоглутарат
Глутамин
г
Глутамат
[NH31
► Глутамат
138
Фотодыхание способствует поддержанию постоянства СО2
внутри листа. У растений С4-пути существует другой механизм
.поддержания концентрации СО2 (фиксация его ЩУК). Фотоды-
хание тесно связано с общим метаболизмом в зеленой клетке, оно
часто усиливается при уменьшении потребности растений в про-
дуктах фотосинтеза и, в принципе, направлено на поддержание
активности ферментативных систем и функции хлоропластов и
митохондрий (А. Т. Мокроносов, 1983). Его роль иногда связывают
с метаболизмом азотистых соединений, переносом их из одной
органеллы в другую, а также с возможностью ограничения Накоп-
ления фосфогликолата.
Явление фотодыхания открыто в 1955 г. И. Деккером. Оно в
большей мере выражено у растений С3-пути фотосинтеза и сопро-
вождается потерей углерода в цикле Кальвина из-за снижения
способности РБФК фиксировать СО2 и повышения ее активности
окислять субстрат (РБФ) в присутствии О2 (А. Лайск, 1977). В случае
окисления субстрата РБФК выступает в роли РБФ-оксигеназы. На
примере рода Panicum, в пределах которого встречаются виды с С3-,
С4- и промежуточными путями фотосинтеза, выявлены различия в
содержании органелл. У Р. riculare (С3-путь) клетки обкладки со-
держат в среднем 3 хлоропласта, 1,1 митохондрий и 0,3 пероксисом;,
у Р. milioides, Р. decipiensvi Р. schenckii (промежуточные пути) органелл
соответственно 10,6,17,7 и 3,2; у Р. р/тол1&(С4-пуп») в клетках обкладки
органелл не только больше, но они и крупнее. У указанных видов
фотодыхание снижается в направлении увеличения числа органелл в
клетках обкладки (R. Н. Brow et al., 1983).
Начальные стадии фотодыхания (образование гликолевой
кислоты) характерны для всех организмов с циклом Кальвина. Виды
же различаются механизмами метаболизма гликолевой кислоты.
Так, фотосинтезирующие бактерии и цианеи окисляют гликолат и
синтезируют глицин. Цианеи содержат фермент гликолатдегидро-
геназу и способны поглощать гликолат (в темноте и на свету), но
утилизируют его слабо. У эукариотических водорослей в процессе
фотосинтеза гликолевая кислота выделяется в окружающую среду,
хотя у них несколько лучше выражен механизм утилизации глико-
лата (при участии фермента гликолатдегидрогеназы). Подобное
выделение представляет собой способ защиты цикла Кребса от
ингибирования и на начальных этапах развития жизни было необ-
ходимо для усиления симбиотических отношений бактерий и
цианей с гетеротрофами (А. М. Степанова, А. А. Шумилова, 1980).
У высших же растений с С3-путем фиксации СО2 фотодыхание
осуществляется наиболее сложно — путем полного превращения
гликолевой кислоты, с образованием ряда важных промежуточных
продуктов (аминокислот, порфиринов и др.). Дело в том, что с
переходом растений к наземным условиям существования возмож-
.139
ности выделения гликолата в среду ограничиваются ввиду невымы-
ваемости его из организма. Поэтому защита цикла Кребса от
ингибирующего действия глиоксил ата могла быть достигнута только
путем полного превращения его в гликолат.
Несмотря на эти новшества, у высших растений потери углерода
при фотодыхании составляют 50% от его ассимилированного
количества. У них фотосинтез падает главным образом из-за кон-
куренции О2 и СО2 за молекулу РБФК и ее субстрат. Интенсивность
фотодыхания у растений С3-типа возрастает при повышении кон-
центрации О2 и температуры среды. Это ведет к ингибированию
фотосинтеза в результате снижения сродства РБФК к СО2, особенно
в аридных условиях. Поэтому отбор способствовал формированию
растений С4-типа, у которых хорошо развита система концентрации
СО2 (ФЕП, органические кислоты и др.) и выражена активность
РБФК (при одновременном подавлении оксигеназной функции
указанного фермента). Благодаря подобным и другим изменениям
у растений С4-типа потери углерода при фотосинтезе невелики и
гликрлат накапливается медленно.
Представляет интерес оценка роли фотодыхания в онтогенезе.
Как известно, в экстремальных условиях водоснабжения устьица
закрываются и резко возрастает фотодыхание. Это имеет значение
для защиты хлоропластов от разрушения (путем снабжения СО2) и
сохранения нормального соотношения НАДФ/НАДФ • Н2, в
результате чего поддерживается процесс фотосинтеза и растения
предохраняются от истощения и теплового шока. В то же время
образуемый Н2О2 при взаимодействии О2 с ЭТЦ хлоропластов может
способствовать отравлению тканей. Во избежание этого часть его
также используется в хлоропластах для не энзиматического
окисления глиоксилевой кислоты. Возникающий на промежуточ-
ных стадиях фотодыхания АТФ метаболизируется в самих мито-
хондриях, где его концентрация резко падает из-за ограничения
оттока триозофосфата из. хлоропластов и поступления СО2 в
ткани. Фотодыхание возрастает по мере старения растений и
листьев (А. Т. Мокроносов, 1981).
Фотодыхание устраняет и Избыток первичной энергии, не
используемой в фотосинтезе (J. Р. Gaudillera, 1982). Полагают, что
само по себе фотодыхание не могло служить объектом отбора, а
возникло в процессе соотбора как неизбежное следствйе эволюции
механизма фотосинтеза. Фотодыхание —адаптация к современно-
му содержанию О2 и СО2 в среде. Содержание О2 в атмосфере (21%)
превышает его потребность для темнового дыхания (2%). И фото-
дыхание предотвращает тормозящее влияние высокой концент-
рации О2 на физиологические процессы клетки и поддерживает
целостность органелл путем дополнительной его утилизации для
фиксации СО2 и окисления гликолевой кислоты. Итак, наблюдаем
140
ряд событий, ведущих к усилению фотодыхания,— выделение
гликолата и развитие механизма его использования;, появление
микротелец (пероксисомы), усиление взаимодействия клеточных
компартментов для обеспечения распределения и потока мета-
болитов. Эти события имели значение для обеспечения энер-
гетического баланса клетки, образования ряда промежуточных
продуктов метаболизма, поддержания осмотического и протонного
потенциала путем синтеза органических кислот, защиты клетки от
токсического действия О2 и хлоропластов от повреждения «выбро-
сом» СО2 из фотосинтетического цикла, устранения излишков
фитогормонов в клетке путем их окисления, окисления жирных
кислот и т. д. Значение каждого из указанных приобретений для
выживания растений не вызывает сомнения.
Ведущее место в фотодыхании принадлежит пероксисомам. Ни
один другой органоид клетки не выполняет столь разнообразные
функции, как микротельца (А. У. Игамбердиев, 1988, 1990), в том
числе и при генераций новых путей биохимических превращений.
Фотодыхание, хотя и рассматривается как ответвление фотосин-
тетического пути, играет важную роль в обеспечении нормального
функционирования цикла Кальвина, снабжая его СО2 в обход
реакций карбоксилирования. Оно также устраняет ингибирование
ЦТК метаболизацией гликолата. По этим и другим причинам
необходимо было выработать механизм утилизации гликолата, от
которого ранее (водоросли) избавлялись выделением в среду. С
появлением пероксисом у части водорослей (хлорелла) окисление
гликолата в глиоксилат с образованием Н2О2 имело значение для
рассеивания свободной энергии, поглощенной клеткой. Выделение
гликолата в среду его вымыванием имело значение для развития
симбиотических отношений автотрофов и гетеротрофов, но оно не
могло получить дальнейшего развития в наземных условиях, где
исключалась возможность этого вымывания. Отсюда и необ-
ходимость отбора растений в направлении усиления утилизации
гликолата в энергетических целях. Косвенно эти механизмы спо-
собствовали усилению транспорта воды, минеральных и органи-
ческих веществ у сосудистых растений благодаря формированию
вакуолярной системы. Интересно отметить, что в случае повторного
перехода цветковых к обитанию в воде они начинают выделять
, гликолат в среду. ,
По-видимому, роль фотодыхания наиболее велика в условиях
постоянного обезвоживания клеток растений как механизма для
предотвращения падения концентрации СО2 и истощения субстрата
дыхания (путем стабилизации фотосинтеза). Быть может, этим и
объясняется подавленность глйколатно-глиоксилатного пути у С4-
растений и при наличии в клетках обкладки соответствующих
141
ферментов—эти растения имеют механизм для поддержания
фотосинтеза даже при Закрытых устьицах. При длительном
ограничении доступа СО2 из внешней среды и у С4-растений
начинает функционировать гликолатный путь.
С энергетической точки зрения фотодыхание рассматривают как
«холостой» механизм, связанный с потерей энергии и уже
синтезированного вещества. Однако нельзя забывать его значения
для защиты окислительной системы клетки от фоторедукции в
условиях высокой интенсивности света и недостатка влаги. Найден
довольно эффективный способ для ослабления последствий подав-
ления «физиологического зла». Мы здесь еще раз сталкиваемся со
сложностью действия естественного отбора и самого процесса
эволюции.
♦ ♦ *
Выше рассмотрены пути эволюции процесса дыхания у рас-
тений. Энергетическая эволюция растений связана с увеличением
экологического разнообразия видов и форм по эффективному
использованию и сочетанию механизмов фотосинтеза и дыхания в
различных условиях. Это достигалось преимущественным
выживанием в критических ситуациях индивидуумов — носителей
благоприятных мутаций до синтезу пигментов И набору ферментов.
Поэтому энергетическая система растений развивалась постепенно,
дополняя ранее возникшие механизмы новыми реакциями обмена.
Значительны различия видов растений в экологии дыхания, в
чем проявляются характерные черты их приспособления. Показа-
телем приспособленности растений в онто- и филогенезе служит и
фотодыхание. Механизмы дыхания как энергетическая основа
жизнедеятельности оказались базисом для возникновения последу-
ющего многообразия растений, в том числе эволюции их химичес-
кого состава (см. гл. 5) и ритмики (см. гл. 6).
ГЛАВА 5
ХИМИЧЕСКИЙ СОСТАВ РАСТЕНИЙ
И ЕГО ПРИСПОСОБИТЕЛЬНОЕ ЗНАЧЕНИЕ
Учение об эволюции органического мира первоначально разра-
батывалось на основе преимущественно морфологических подхо-
дов. Уже в первых своих работах Ч. Дарвин указывал, что «многие
таинственные изменения в строении и окраске» выступают как
следствие «химических изменений в питательных жидкостях».
Однако только в 20-х годах XX в. для разработки проблем эволюции
стали использовать методы биохимии и физиологии. Значителен
вклад в изучение данного вопроса отечественных ученых (С. Л.
Иванов, 1926; А. В. Благовещенский, 1966; А. Н. Белозерский,
1976 и др.), которые, не ограничившись констатацией факта
химической дивергенции растений, начали использовать получен-
ные данные для разработки проблем таксономии и филогении
растений.
Поразительным оказался факт химического сходства микроор-
ганизмов, растений и животных. Однако эволюция веществ в
пределах единства химического состава организмов все же
происходила и имела более высокие темпы, чем на этапе абиогенеза.
Заметим, что в живой природе отсутствует автономный отбор как
веществ, так и отдельных органов. Отбор веществ осуществляется
через отбор индивидуумов в популяциях. Поэтому точнее говорить
об изменчивости химического состава видов в ходе эволюции,
условно обозначаемой термином биохимическая эволюция.
Изучение химической дивергенции имеет познавательное и
прикладное значение (Л. А. Ковалевский, 1991). Так, особенности
биоэнергетики связаны со спецификой химического состава
организмов (Э. Брода, 1978). С химической дивергенцией отчасти
связано и своеобразие функциональной деятельности растений
разных видов и отдельных их органов. Химический состав растений
выступает как итог исторического развития вида с поправкой на
условия в онтогенезе й несет приспособительный отпечаток. Боль-
шое значение имеет также изучение закономерностей химической
дивергенции ,растений для целей систематики, таксономии и
филогении. При этом такие исследования дают возможность шире
использовать растения для получения сырья и лекарств.
143 -
5.1. СРАВНИТЕЛЬНАЯ ХАРАКТЕРИСТИКА
ХИМИЧЕСКОГО СОСТАВА РАСТЕНИЙ
И ОБЩИЕ ТЕНДЕНЦИИ ЕГО ЭВОЛЮЦИИ
Как известно, теория эволюции Ч. Дарвина исходит из поло-
жения о возникновении многообразия существ на основе единства
их происхождения. Справедливость этого положения доказывается
и на примере изучения химического состава растений. Огромно
число белков (ферментов и неферментов), а также углеводов и
липидов. Разнообразны и вещества вторичного метаболизма. Одних
, только алкалоидов описано более 10 тыс. Однако для всех алка-
лоидов характерно присутствие в молекулах либо неконденсирован-
ных, либо конденсированных гетероциклов. Более'того, при их
многообразии в органическом мире они удивительно сходны в путях
биосинтеза.
В связи с указанными обстоятельствами возникают трудности
в использовании данных химической дивергенции. В этих целях
А. В. Благовещенский (1966) предложил сравнивать виды пасодер-
жанию в их семенах легкорастворимых (энергетически богатых
альбуминов) и труднорастворимых в воде (менее энергетически
емких глобулинов и глутелинов) белков. В хемосистематике рас-
тений чаще всего исследуют белковые компоненты семян ввиду их
большей видоспецифичности. Например, смена саговников
другими голосеменными была связана не только с приобретением
ими целого ряда морфологических новшеств, но и с изменением
химического состава семян. А В. Благовещенский и Е. Г. Алек-
сандрова (1974), вычислив коэффициент продвинутое™—отно-
шение содержания альбуминов к глобулинам и глутелинам — Ае
(advance of evolution) для наиболее примитивных (рода Cycas) и
продвинутых саговников (роды Macrozamia, Encephalartos), доказали
эволюционные преимущества последних. С переходом от голосе-
менных к магнолиевым содержание альбуминов в семенах повыша-
лось и соответственно возрастало Ае.
В табл. 6 представлены соответствующие данные этих авторов
для ряда семейств, среди которых розоцветные наиболее про-
двинуты морфологически и биохимически. Иногда биохимические
различия семян в пределах родов одного семейства оказываются
значительно выше, чем между представителями разных семейств.
Например, виды рода мимозовых имеют величину Ае в пределах
0,85—5,04, тогда как семейства бобовых и мимозовых — 1,20—2,20.
Биохимическая примитивность не всегда соответствует морфо-
логической примитивное™. Так, высокоспециализированная триба
Stipeae оказывается довольно примитавной биохимически. Поэтому
для суждения об эволюционной продвинутое™ рекомендуется
Использовать показатель специализации, отражающий соотно-
144
щение проламинов (определяющих устойчивость к засухе и замо-
розкам) и всех суммарных фракций белков. Эволюция злаковых как
раз шла по пути усиления биохимической специализации. И пред-
ставители рода Stipa оказались более продвинутыми биохимически,
чем виды рода Роа. Сравнения же с точки зрения морфологии не
дают столь четкой картины.
.Таблица 6. Белковые комплексы семян
у видов разных семейств (средние данные), %
Семейство и число изучен- ных видов Альбумины Глобулины Глутелины + + остаток Ае
Анноновые — 9 35.1 3,3 61,8 0,74
Лавровые — 5 15,9 0.0 84,1 ,0,18
Лютиковые —15 17,6 32,3 50,2 1,0
Розоцветные —18 31,2 43,9 24,9 3,0
В биохимическом отношении злаки специализированы на повы-
шенное содержание проламинов в эндосперме семян (В. Ф. Се-
михов, 1982). У некоторых видов злаков (Eleusine tristachya, Eragrostis
minor, Panicum miliaceum) 60% эндосперма составляют проламины,
что связано с отбором растений по легко утилизируемым белкам,
способствующим уменьшению энергетических затрат. Накопление
проламинов (проламин, зеатин, гордеин) важно для прорастания
семян ввиду их способности к быстрому гидролизу и поддержанию
осмотического давления клетки. По этой причине в небла-
гоприятных условиях семена с высоким содержанием проламинов
лучше прорастают и активнее накапливают биомассу. На примере
злаков показано отличие родов у триб между собой по аминокислот-
ному «профилю» семян. Для примитивных триб злаков характерно
как низкое содержание проламинов, так и бедность его компонент-
ного состава в семенах (В. Ф. Семихов, 1991).
Для уточнения филогенетических связей изучают последователь-
ность и соотношение нуклеотидов в молекуле ДНК (А. Н. Белозер-
ский, 1976 и др.). Установлено, что у диатомовых, зеленых и бурых
водорослей, как у бактерий, ДНК относится к АТ- и ГЦ-типам.
Если у бактерий показатель специфичности ДНК (Г+Ц/А+Т)
варьирует в пределах 0,45—2,80, у водорослей —0,58—1,76, то у
цветковых — в пределах 0,58—0,94.
Значительная изменчивость состава ДНК у бактерий объясня-
ется их большей функциональной дифференцированностью, чем
цветковых растений и высших животных. Покрытосеменные имеют
более высокое соотношение ГЦ/АУ в молекуле РНК, чем архе-
гониальные. РНК однодольных характеризуется большим отно-
шением ГЦ/АУ, чем РНК двудольных. ДНК грибов отличается от
ДНК актиномицетов. У последних она относится к ГЦ-типу с
145
показателем специфичности 2,7 и
очень мало варьирует. У низших
грибов ДНК принадлежит к сильно-
му АТ-типу, показатель специ-
фичности ее составляет 0,40. ДНК
миксомицетов и ДНК простейших
близки по составу и относятся к АТ-
типу. ДНК высших грибов сходна с
ДНК водорослей и относится к ГЦ-
типу. Эти данные позволили сделать
вывод о полифилетическом проис-
хождении грибов.
В то же время грибы по способу
обмена клетки и деления ядра
также признаны наиболее прими-
„ „ тивными эукариотами среди рас-
тений (А- Е Васильев, 1985). Как
хондрий, архебактерий и эукариот животные они способны образовы-
(<з—д) вать мочевину, гликоген и хитин
На ребрах древа указаны числа замен (Л. (ОН СОСТаВЛЯвТ 27—67% ОТ Массы
,В. Омельянчук, Н. А. Колчанов, 1987) КЛвТОЧНОЙ СТвНКИ). Среди Грибов
' только оомицеты не синтезируют
хитин. У них, как и у растений, оболочка клетки состоит из
целлюлозы. С растениями грибы сближают способы всасывания
веществ и неограниченность роста.-Систематика грибов основана
на учете особенностей жгутиковых стадий; тем самым Связывают
их происхождение со жгутиковыми.
Для выяснения скорости эволюции (Г) и филогенетических
взаимоотношений широко применяют методы молекулярной
биологии —сравнение первичной и вторичной структуры 5S, 16S,
17S РНК, секвенирование нуклеотидной последовательности
генов и т. д. Так, Идля хлоропластной линии (одно- и двудольные
растения) и эукариотической ветви оказалась почти равной и
составила-0,92 ±0,53 • 1О~10 и -0,68 ±0,43 : 10“10 замен на одну
Позицию в год..В то же время для МТГ (архебактерии, эукариоты
и эубактерии) скорость эволюции оказалась значительно выше
(« 0,8 10“10 замен на одну позицию в год; рис. 14). Подобными
сравнениями установлено наличие общего предка для цианей (Апа-
cystis nidulans) И хлоропластов (Euglena gracilis), красных и зеленых
водорослей, высших растений и даже многоклеточных животных.
Красные водоросли отнесены к самым древним эукариотам.
Геном цветковых растений по величине (67 пкг) превосходит
геномы большей части животных (6 пгк). В ДНК растений содер-
жатся минорные компоненты с высоким уровнем ГЦ-пар, она
характеризуется нестабильностью и наличием амплификаций. Вы-
сокое содержание ДНК важно для выживания и темпов развития.
146
Так, при содержании ДНК на диплоидный геном около 1—2 пкг
продолжительность мейоза при 20°С составляет 1 сут, а при более
50 пкг — около 2 недель. Возможно, по этой причине эфемеры
отличаются низким содержанием ДНК. и их почти нет среди видов
цветковых, характеризующихся высоким содержанием ДНК. В ходе
эволюции растений избыточность ДНК часто сопровождается изме-
нением ее некодирующей фракции.
Из палеонтологии известно, что около 600 млн. лет назад в
кембрии произошла «скелетизация» в мире животных и растений в
результате усиленного накопления в среде кальциевых, кальциево-
магниевых карбонатов и фосфоритов. Химический состав рас-
тений находится в зависимости от условий их жизнедеятельности
в онто- и филогенезе. Общим для живых систем является полное
использование среды и ее энергетических возможностей. В
природе происходит отбор против «нестройности и роста эн-
тропии» (Н. А. Умов) или по возрастанию «отрицательной эн-
тропии организмов» (Э. Шредингер).
Об эволюции химического состава растений чаще всего судят
по результатам изучения современных видов. Так, процент ГЦ-пар
у высших растений составляет 34—51, у бактерий :—30—70. Со-
держание 5-метилцитозина в ДНК равно 1,5—2% (мхи). ДНК
архегониальных мейее метилирована, чем цветковых. Среди цвет-
ковых растений максимум метилирования (5,4—8,0 моль %) харак-
терен для однодольных. Величина повторов в ДНК животных
составляет 10—12%, растений —до 50—70%. Структурные гены у
растений представлены 10—15% всей ДНК. Уникальные последо-
вательности у большинства эукариот составляют не более половины
гаплоидного генома и варьируют у разных животных в пределах
40—70%, тогда как у растений —всего 10—30%.
Вопрос о связи между морфофизиологическими особенностями
растений и количеством ДНК в ядре недостаточно изучен.
Увеличение кратного числа хромосом при полиплоидизации (сле-
довательно, и удвоение количества ДНК) не всегда отражается на
размерах и химическом составе растений (В. Грант, 1984). В неко-
торых случаях связь проявляется. Так, хлорофилльные мутации овса
(SM 15, St 21 и М4) имеют различия по хлорозу, амфиплоиды
пшеницы отличаются большим содержанием глиадина в эндоспер-
ме (В. Г. Конарев, 1983).
Интересные данные получены по изменению молекулярной
массы ферментов (Р. Фокс, 1992). Она возрастает в ряду микробы
-> «простые» эукариоты -> высшие эукариоты. В ходе эволюции
от нефотосинтезирующих к фотосинтезирующим прокариотам
отмечено увеличение размера молекул ферментов. Наиболее
архаичные клетки, вероятно, содержат незначительное число поли-
функциональных ферментов, обладающих низкой каталитической
147
активностью. Из них в процессе мутации и отбора складывались
клетки-потомки с высокой скоростью роста, активностью синтезов
и с множеством специализированных ферментов (В. К. Антонов,
1983). К тому же разные ферменты обладают различной эво-
люционной лабильностью. Так, наиболее консервативными у
представителей Sceale оказываются глутаминдегидрогеназа, дегид-
рогеназа, глюкозо-6-фосфат, МДГ, которые по изозимным спект-
рам совпадают у пшеницы и ржи.
Для эволюции ферментов характерно усиление их внутривидо-
вого полиморфизма (В. S. Hartley, 1979). Он выше у перекрестно-
опыляющихся видов в пределах рода, чем у самоопылителей. В ходе
эволюции меняется и активность ферментов у растений. В част-
ности, активность пероксидазы в ряду водоросли -> мхи -» папо-
ротники -> хвойные -> одно- и двудольные растения в ходе
эволюции возрастает. Морфологическое усложнение, видов у крас-
ных и бурых водорослей и у листостебельных мхов также идет
параллельно с повышением активности пероксидазы. С эво-
люционным продвижением видов замечено изменение
каталитической активности и ряда других ферментов.
Говоря об эволюции химического состава растений, следует
остановиться и на характеристике разных метаболических путей.
Так, шикиматный путь важен для синтеза ароматических
аминокислот, белков, алкалоидов, ИУК, фенольных соединений и
т. п. (М. Лукнер, 1979). Он представлен и у микроорганизмов.
Однако у высших растений шикиматный путь более сложен по
ферментам и разветвленности начальных этапов. При снижении
активности шикиматного пути остающиеся не вовлеченными в него
углеводные предшественники не отбрасываются у растений, а за-
пасаются в виде хинной кислоты, что в дальнейшем используется
для синтеза полифенолов. Происходит циклизация процесса и по
поддержанию синтеза ароматических аминокислот. Предполагают,
что метаболические пути синтеза ВВМ чаще всего возникают в
результате слияния циклов, первоначально сложившихся отно-
сительно самостоятельно (D. М. Key, 1980), с чем связана чрезвы-
чайная специфичность индивидуальных веществ.
У галофитов обнаружена связь между солеустойчивостью и
накоплением аспарагиновой кислоты, Na+ и С1— в клетках
ассимилирующих тканей. Накопление большого количества
органических кислот и аминокислот как конечных продуктов
фотосинтеза у галофитов имеет приспособительное значение для
нейтрализации избытка катионов и анионов. Подобный механизм
позволяет растениям, обитателям аридных засоленных территорий,
сохранить положительный баланс углеводов в онтогенезе (Т. А. Гла-
голева, М. В. Чулановская, 1992). Другие примеры дивергенции
растений по химическому составу, имеющие приспособительное
значение, приведены в последующих параграфах главы.
148
5.2. ИЗМЕНЕНИЕ ХИМИЧЕСКОГО СОСТАВА
РАСТЕНИЙ В ОНТОГЕНЕЗЕ
Физиология и биохимия растений располагает большим И раз-
нообразным материалом, характеризующим изменчивость химиче-
ского состава в онтогенезе в зависимости от сортовых особенностей
и условий существования. Многими авторами подмечено, что вы-
сокая влажность в сочетании с умеренной температурой ведет к
накоплению сахара, крахмала, эфирных масел, а противоположные
условия — к накоплению азотистых веществ и алкалоидов. В на-
правлении с запада на восток у возделываемых сортов пшеницы
наблюдается возрастание белковости семян, а в направлении с
севера на юг — увеличение углеводов. На химический состав рас-
тений влияют и условия репродукции семян по годам.
Химический состав растений подвержен в онтогенезе и
индивидуальной наследственной изменчивости, что показано на
примере изучения белков (D. A. Levin et al., 1979). Поэтому белки
используют как генетические маркеры для идентификации сортов
и генотипов (А. А. Созинов, 1985). Белки, какизвестно, определяют
структуру и функцию клеток и метаболические особенности орга-
низма. Отсюда и консерватизм состава части белков в течение
миллионов лет, и высокая скорость их синтеза в клетке (за считанные
секунды может быть собран полипептид из нескольких сотен
аминокислот). Особенно консервативны структурные белки и белки
мембран, они отличаются низким полиморфизмом (М. Кимура, 1985).
Сохраняя постоянство последовательности аминокислот, моле-
кулы многих белков в то же время представлены разными изофор-
мами, что обеспечивает быструю метаболическую адаптацию
организма к изменяющимся условиям среды. Гены многих
полиморфных белков обычно сформированы кластерными бло-
ками. У растений гетерозиготность каждой особи по изоферментам
составляет 17—30% (Ф, Айяла, 1984). Растения по полиморфизму
белков можно расположить в следующий убывающий ряд: космо-
политы -> тропические -> виды умеренного климата. На примере
ячменей установлено, что образцы Северной Америки и Европы
менее гетерозиготны по эстеразному локусу (0,5%), чем образцы
Среднего Востока (0,1—1,47%). Преимущественное распростра-
нение части аллелей в образцах коррелирует с устойчивостью К
болезням, климатическим невзгодам и особенностями ведения
земледелия. Под влиянием подобных факторов у растений шел
отбор по определенным кластерам генов.
Белки, будучи связанными с мембранами, играют роль в ответ-
ной реакции растений на внешние раздражители (В. Я. Александ-
ров, 1985). Специфика белков разных растений определяет
теплоустойчивость движения протоплазмы их клеток (рис. 15), и ее
149
16
Рис. 15. Теплоустойчивость движения
протоплазмы эпидермальных клеток
арктических, бореальных и субтропи-
ческих (7 —3) видов (В. Я. Александров,
1985. С. 187)
Ось абсцисс —температура 5 мин нагрева, оста-
навливающего движения протоплазмы, ось
ординат —число видов растений, у которых
движение протоплазмы останавливается при
температуре нагрева, указанной на оси абсцисс
Рис. 16. Температурная зависимость
потенциальной интенсивности фо-
тосинтеза листьев пшеницы, выра-
щенной при 8 — 12° С (7) и 20 —
24° С (2)
Ось абсцисс —температура 10 мин эк-
спозиции в присутствии 1% 14СО2; ось
ординат — интенсивность фотосинтеза.
Штриховая линия — см. рйс. 3 (В. Я. Алек-
сандров, 1985. С. 173)
показатели соответствуют исторически сложившемуся температурному
оптимуму вцца. В то же время продолжительные воздействия в онтоге-
незе не бесследны и отражаются на состоянии закалки (рис. 16).
Выделены стрессовые белки у растений (Г. И. Блехман, 1987).
Из мембран хлоропластов гороха получен белок теплового шока с
молекулярной массой 22 кД, синтез которого контролируется ядром.
Белки определяют не только способность к выживанию в не-
благоприятных условиях, но и репаративные возможности с обра-
зованием различных изоферментов (А. И. Шумская, 1987).
Изменчивы, в онтогенезе и другие органические соединения
растений (алкалоиды, фенолы, флавоноиды и др.). Растения в
пределах сортов в онтогенезе обнаруживают генетические различия
по указанным веществам. Хотя часть таких изменений нейтральна,
но некоторые из них функционально значимы.
До недавнего времени недостаточно ясна была роль диаминов
(путресцин, кадаверин) и полиаминов (спермин и спермидин).
Оказалось, что их метаболизм важен для биосинтеза и превращения
аминокислот и ряда вторичных продуктов азотного обмена. Кроме
того, полиамины влияют на нуклеиново-белковый обмен, проница-
емость мембранных структур и регуляцию гомеостаза (Н. И. Ше-
вякова, 1981). Особенно велика роль путресцина в поддержании
гомеостаза при калийном голодании растений, полиаминов —при
нарушении баланса катионов и подкислении среды. Так, при
150
исключении из среды калия содержание путресцина возрастает в
100 раз, такое же влияние наблюдается при отсутствии Mg2* и Си2*
и в условиях засоления. Защитную роль ди- и полиаминов связы-
вают с их участием в стабилизации проницаемости мембран.
Для адаптации растений к экстремальным условиям особое
значение имеет изменение в тканях соотношения регуляторов роста
(N. С. Turner, Р. J. Kramer, 1978; J. Levitt, 1980). При этом в тканях,
особенно в замыкающих клетках устьиц, увеличивается содержание
абсцизовой кислоты (АБК), снижается количество ИУК и гиббе-
реллина. Повышение содержания АБК в устьичных клетках способ-
ствует закрыванию устьиц и уменьшению транспирации. Различные
виды растений отличаются по пластичности химического состава, что
нередко кореллирует с устойчивостью к водному дефициту. Характер-
но, что накопление АБК меняется не только в ходе онтогенеза, но и
в течение суток (это также имеет приспособительное значение).
Нередко химические изменения в растении находят отражение
в его морфологии. Так, изменение содержания хлорофилла и соот-
ношения между пигментами пластид в листьях отражается на
окраске пластинки. В основе изменения окраски лепестков 'лежит
превращение флавана — вещества, имеющего два фенольных ядра.
Переход флавана во флавон обусловливает развитие желтого
пигмента у лепестков хризантемы. Иногда изменение окраски
лепестков у растений оказывается результатом не блокирования
синтеза фермента, а потери его активности—из-за колебания
температуры в окружающей среде (Б. М. Медников, 1980), как у
китайской примулы с красными цветками: при температуре 30—
35°С её цветки теряют антоциан.
Регуляцией условий в онтогенезе можно достичь значительных
изменений в химическом составе растений. Многие из них не
наследуются и выступают как норма реагирования Генотипа. На
примере изучения триглицеридов, играющих резервную и защитную
роль, показана изменчивость их состава в-онтогенезе растений.
Динамика их состава в созревающих семенах хлопчатника в онтогенезе
напоминает этапы становления триглицеридов в филогенезе. Изме-
нение химического состава в онтогенезе может быть проил-
люстрировано и на алкалоидах. Так, в семенах Catharanthus roseus
алкалоидов нет, а в период вегетации в растении обнаружено до 60 их
разновидностей (М. Е. Ловкова, 1981). Это связано с регуляцией
азотного обмена и иммунитета. У табака показана связь между изме-
нением содержания алкалоидов и чувствительностью к длине дня.
Выявление корреляции между накоплением каких-то веществ
и неблагоприятными факторами представляет интерес для познания
биологической роли отдельных групп веществ (например, изучение
связи между накоплением ряда аминокислот и фосфорных сое-
151
динений и морозоустойчивостью, растворимых углеводов и засухоус-
тойчивостью растений и т. д.). Итоги подобных исследований должны
быть оценены с применением методов эволюционнрй генетики.
5.3. ЭКОЛОГИЧЕСКАЯ РОЛЬ КАТЕХИНОВ И ПЕКТИНОВЫХ ВЕЩЕСТВ
В органическом составе растений особое место занимают
катехины и пектиновые вещества — ВВМ, широко распространен-
ные, разнообразные по составу и недостаточно изученные в функ-
циональном отношении. К кахетинам относят фенольные соединения
(полифенолы), которые найдены во всех органах высших и
большинства низших растений (М. Н. Запрометов, 1964). У фо-
тосинтезирующих бактерий Rhodospirillum гйЬгит они содержатся в
виде пара-оксибензойной, пара-оксифенилуксусной и пара-окси-
фенолпропионовой кислот. Фенольные соединения обнаружены у
водорослей, грибов, лишайников, мхов, хвощей и т. д. Эти факты
привели к выводу, что полифенолы имеют универсальное распрост-
ранение в мире растений и играют разнообразную роль в их жизни.
Например, эти соединения образуют комплексы с белками, исполь-
зуемые для заживления ран; кроме того, фенольные соединения
являются промежуточными продуктами при формировании смол и
эфирных масел, а также акцепторами водорода на конечных этапах
дыхательного процесса: хиноны -> хромогены (фенольные сое-
динения) -> хиноны. Известно также, что липидные кофакторы
фенольной природы (убихиноны, пластохиноны, витамин К и др?),
сосредоточенные в митохондриях, непосредственно принимают
участие в дыхании как связующие звенья между флавопротеинами
и цитохромами; Например, концентрация убихинона в мито-
хондриях в 20 раз превышает концентрацию цитохромов.
Не исключена и возможность того, что процесс образования
флавоноидов нужен растениям как механизм для катаболизма
фенилаланина и частично для продуцирования физиологически
активных Веществ (У. В. Маргна, 1980). Поэтому не случайно, что
в растениях обнаружено более 2000 производных флавоноидов. Их
биосинтез идет параллельно катаболическому расщеплению белков
(рис. 17), при котором и образуется большая часть фенилаланина,
являющегося предшественником и субстратом для синтеза поли-
фенолов. Образование полифенолов — фактически единственный
метаболический канал, который может принимать и пропускать
освобожденный из белков фенилаланин. Но биосинтез полифено-
лов может происходить и по шикиматному пути, хотя и в менее
значительных количествах.
Публикации последних лет (М. Н. Запрдметов, 1985, 1988, 1993)
показывают, что фенольные соединения в жизни растений выполняют
такие функции, как защита от патогенов (фитоалексины), являются
152
компонентами ЭТЦ митохондрий
и хлоропластов (убихиноны,
пластохинон и др.), участвуют в
движения^ (тургорины), разоб-
щении окислительного фосфори-
лирования (флоридзин, кварцетин
и др.) и регуляции взаимоотно-
шений между видами в ценозах
(изофлавоны, флавоны, флавоно-
лы, халконы и др.).
Полифенолы играют также
роль ингибиторов пектоли-
тических ферментов. Пектиновые
же вещества входят в состав кле-
точного сока и оболочек, играют
важную роль в газообмене и
транспирации растений. Процес-
сы, связанные с превращениями
пектиновых веществ, имеют боль-
Рис. 17. Сдвиги метаболизма в старе-
ющих листьях некоторых древесных
растений (У. В. Маргна, 1980):
А — содержание белка, Б — суммарное со-
держание флавоноидов; I — нормальные
зеленые листья, II—листья с явными
признаками старения (окрашены
антоцианом); 1—клен, 2—кизильник,
3 —свидина, 4 — черноплодная арония
шое значение в завязывании, росте и созревании плодов.
Характерной чертой фенольных соединений являются легкая
окисляемость, способность к взаимодействию с белками и комплек-
сообразованию с ионами металлов (М. Н. Запрометов, 1992). Разно-
образие строения фенольных соединений сочетается с разнообразием
их функций. Они выступают как предшественники лигнина
(оксикоричные спирты и кислоты), полисахаридных комплексов
первичной клеточной стенки (феруловая и л-кумаровая кислоты),
суберина и кутина — компонентов коры и раневой перидермы.
Среди функций фенольных соединений и такие важные свой-
ства, как защита фотосинтетического и генетического аппарата
клетки от повреждающего действия УФ-лучей (флавоноиды), сиг-
налы-аттрактанты для обеспечения взаимодействия растений и
микроорганизмов (принимают участие разные группы веществ),
роль запасных продуктов (катехины в плодах и лепестках) и субст-
рата дыхания. Из сказанного видно, что фенольные соединения
имеют важное адаптивное значение не только в онтогенезе, но и в
филогенезе — при выходе растений на сушу и защите растений от
поедания и вредителей.
К другой группе веществ вторичного обмена относят разнооб-
разные ароматические терпеновые соединения из класса углеводо-
родов (эфирные масла, смолы, каучук, гуттаперча и т. д.).
Специфический аромат цветков и плодов, необходимый для привле-
чения животных-опылителей и распространителей семян, опреде-
ляется терпеновыми соединениями. Терпеноиды, как и алкалоиды
из числа гетероциклических азотистых соединений, оказывают
153
положительное влияние на иммунитет растений и предохраняют их
от поедания животными (R. Е. Cook, 1981).
Число известных терпеноидов составляет около 4000, феноль-
ных соединений —5000, алкалоидов —10 000. Вторичный обмен
чаще всего лежит в основе клеточной специализации и не зависит
от уровня организации существ. Многие вторичные соединения не
играют определенной экологической роли, и поэтому их исчезнове-
ние в результате мутаций не сопровождается заметным ущербом
для популяций. Однако в отношении алкалоидов показано, что
полная потеря способности синтезировать их делает нежизнеспо-
собными растения алкалоидоносных родов. Алкалоиды широко
распространены среди растений пустынь и полупустынь, которые
отличаются высоким содержанием органических кислот. Возможная
роль алкалоидов в жизни растений состоит в поддержании pH клетки,
усилении иммунитета, в транспорте и резервации азота, кроме того,
алкалоиды являются субстратом для дыхания. Для филогенетически
молодых трупп растений характерно удлинение путей биосинтеза
алкалоидов, что необходимо для поддержания благоприятного баланса
азота в условиях постоянной изменчивости среды.
Биосинтез вторичных соединений имеет значение и для выве-
дения балласта. Ныне не вызывает сомнения факт использования
части вторичных соединений или для детоксикации избыточных и
ненужных соединений, или как продуктов, обеспечивающих эф-
фективное взаимодействие организмов в биогеоценозе (J. Harbome,
1978), или для биосинтеза веществ, играющих роль медиаторов. Все
это свидетельствует в пользу того мнения, что «природа не прощает
излишеств». В то же время функция некоторых продуктов
вторичного метаболизма до сих пор неизвестна.
5.4. ЭКОЛОГИЧЕСКОЕ ЗНАЧЕНИЕ
ФИТОГОРМОНОВ И ИНГИБИТОРОВ РОСТА
С момента открытия и химического выделения фитогормонов
(начиная с 30-х годов XX в.) исследования по изучению их роли и
механизма действия с каждым годом расширяются. Из фитогормо-
нбв наиболее хорошо изучены ИУК, ГА3, Цт, АБК и этилен. В
растениях встречаются и негормональные стимуляторы и инги-
биторы роста. Состав гормонов растений более ограничен, Чем
животных. Они не являются белками и такими же узкоспе-
циализированными, как гормоны животных, обладают полифунк-
циональностью (Н. Г. Холодный, 1955; В. В. Полевой, 1985). По
степени распространенности фитогормоны можно расположить в
следующий убывающий ряд: ИУК -> Цт -> ГА3 -> АБК -> этилен.
Примерно так же считают и их эволюционный возраст, о чем судят
154
по приуроченности их синтезов у видов и локализации в органах,
отличающихся своим филогенетическим возрастом.
Остается без ответа вопрос о причинах обильного синтеза части
из них микроорганизмами и низшими растениями, для которых
значение указанных веществ все еще не ясно. На сегодня же
очевидна большая значимость ИУК, ГА3, Цт, АБК и этилена для
высших, особенно цветковых, растений. Даже у архегониальных
растений спектр действия фитогормонов или ограничен или не-
четко выражен (табл. 7).
Таблица 7. Действие фитогормонов на рост и развитие мхов
(по О. Т. Демкиву, К. М. Ситнику, 1985)
Появление морфогенеза ИУК ГА, Цт АБК Этилен
Прорастание спор (+) (+) + — —
Рост протонемы + + 0 — —
Ветвление протонемы — (+) + — 0
Апикальное доминирование Дифференциация: + 0 + + 0
каулонемы + (+) 0 — —
листовидных пластинок + (+) + + +
почек гаметофитов — 0 + — 0 /
выводковых телец — 0 — — 0
архегониев (+) — + — 0
антеридиев (+) + — — 0
Образование апогамных структур 0 — + — 0
Регенерация + + + — 0
Примечание. При экзогенном введении обнаружены стимуляция [+], ингибирование [—] или слабое действие [(+)], эффект отсутствует [0].
Одним из важных результатов эволюции фитогормонов следует
признать расширение спектра действия каждого из них, возникно-
вение комплементарности и механизмов взаимодействия их для
осуществления нормального морфогенеза, а также использование
одних й тех же фитогормонов в разных соотношениях «для вклю-
чения и выключения морфогенетических и физиологических про-
грамм» (В. В. Полевой, 1985). С возникновением подобных
механизмов повышается надежность процессов дифференциации и
морфогенеза прй ограничении набора фитогормонов, что свиде-
тельствует об отборе растений в сторону усиления целостности
организма. Это Можно подтвердить фактами участия фитогормонов
в повышении устойчивости растений к экстремальным условиям
через регуляцию процессов морфогенеза и жизнедеятельности (см.
ниже), а также взаимодействия органов между собой.
В этом отношении исключение не составляет и этилен. Этилен
как один из фитогормонов играет большую роль в росте, устой-
чивости и старении растений. В частности, при подавлении синтеза
этилена слабо развиваются аэренхима и почки, но усиливается
155
эпинастия (что способствует уменьшению испарения воды расте-
ниями). В то же время накопление этилена в тканях повышает
устойчивость растений к грибной микрофлоре из-за положитель-
ного влияния его на активность пероксидазы и каталазы, синтез
лигнина, фитоалексинов и хитиназы. Известно, что высшие рас-
тения не синтезируют в норме хитин, но синтез хитиназы в них
происходит при накоплении этилена. Благодаря синтезу хитиназы
растворяется хитиновая оболочка грибов-паразитов, что предотвра-
щает распространение последних в тканях хозяина. Некоторые
используют пример синтеза хитиназы высшими растениями как
доказательство существования горизонтального переноса генов.
В плане первичного генезиса метаболических путей синтеза
фитогормонов у высших растений нельзя исключить и возможность
их перенесения из низших растений (например, пути синтеза ИУК
и ГА3). Кажется также вероятным первоначальный отбор высШих
растений^ по чувствительности к ИУК и ГА3, выделяемых парази-
тами-микроорганизмами в ткани. При этом продукты метаболизма
микроорганизмов могли влиять на изменение мембранного
потенциала клеток хозяина. В дальнейшем значение этих веществ
расширяется благодаря их участию в разнообразных процессах
Жизнедеятельности. В таком случае отбор растений должен был быть
направлен на повышение устойчивости и чувствительности к про-
дуктам своей микрофлоры.
С возрастанием физиологической роли продуктов метаболизма
микроорганизмов в жизнедеятельности растения-хозяина отбор
устранил' зависимость последних от продуцентов. Соображения
здесь таковы: микроорганизмы, как наиболее защищенные от внеш-
них невзгод и поселившиеся в ткани хозяина, могли продуцировать
соответствующие вещества и в тот период, когда растения не
нуждались в них, (например, с наступлением неблагоприятных
условий для роста растений). В этом случае обязательно должно
было произойти отмирание растения-хозяина из-за неэкономного
роста, что не могло не способствовать возникновению И стаби-
лизации их собственных метаболических путей синтеза фитогор-
монов. Только подобный исход обеспечивал уравновешенность
растений в среде. Высказанные соображения косвенно подтверж-
даются данными о сохранении симбиотйческих отношений между
некоторыми растениями и микроорганизмами-продуцентами
физиологически активных веществ, а также о морфогенетическом
значении выделений последних для части растений (возникновение
опухолей, галов).
Стабилизация собственных метаболических путей синтеза фито-
гормонов достигается у цветковых. При этом значение указанных
156
веществ в жизнедеятельности усиливается. Не исключается и парал-
лелизм в формировании путей метаболизма фитогормонов у разных
растений. Параллелизмы характерны не только для растений, но и
между растениями и животными (см. ниже).
Из сказанного видно, что эволюцию гормональной системы у
растений нельзя понять в отрыве от развития их общего метаболиз-
ма. Следовательно, эволюция растений шла по сбалансированности
синтеза физиологически активных веществ в общем метаболизме
растительного организма как целостной системы, что особенно
важно при усложнении организации индивидуума.
Данные о роли фитогормонов в морфогенезе и об изменении
их соотношения на разных этапах онтогенеза хорошо согласуются
с представлениями о механизме эксперсии генов. Многие гены
оказываются активны только на определенных этапах онтогенеза
(N. В. Broad, 1984), что и влияет на уровень содержания фитогормо-
нов в тканях. Как подчеркивает В. Грант (1991), регуляция морфо-
генеза растений достигается взаимодействием гетерогенных
комплексов генов-ингибиторов, системы оппозиционных генов,
гетерохроматиновых блоков и генов-модификаторов. Подобные
взаимодействия и обеспечивают изменение соотношения фитогор-
монов в тканях и «позиционный контроль» морфогенеза растений
(RW. Barlow, D. Е. Carr et al., 1984).
Физиология растений пока не располагает материалом, харак-
теризующим количественные соотношения фитогормонов в разных
таксонах. Но есть основания полагать, что у филогенетически
древних форм содержание ИУК выше, чем у эволюционно продви-
нутых. Минимальное содержание гиббереллинов отмечено у
представителей семейств лютиковых, кувшинковых и толстянко-
вых, а ИУК — у сложноцветных. В цветках лютиковых содержание
ИУК в 1,8 раза превышало содержание ее у сложноцветных.
- Чувствительность же к фитогормонам, наоборот, оказалось выше у
последних. Эти различия в какой-то мере отражены в табл. 8.
Предполагают, что ИУК, ГА3, Цт у низших растений (включая
микроорганизмы) выступают как функционально малозначимые
вещества. Они начинают играть заметную роль в жизнедеятельности
высших растений, что, возможно, было связано с отбором этих
растений в популяциях по чувствительности к указанным вещест-
вам, совершенствованием путей биосинтеза, передвижения и раз-
рушения регуляторов роста. В целом эволюция гормональной
системы растений шла в направлении использования все меньших
и меньших количеств веществ для достижения регуляторного эф-
фекта. Для этого в клетке вырабатываются и механизмы «узнавания»
гормонов, улавливания и усиления их эффекта. Эту роль приписы-
вают белкам.
157
Таблица 8. Различия в биосинтезе регуляторов роста у низших
и высших растений (по В. И. Кефели, Р. X. Турецкой, 1974)
Растения Содержание фитогормонов Содержание ингибиторов Система связывания фитогормонов Связь с ростом
Низшие (микроррганиз- мы, грибы) +++ , — — —
Высшие + + + 4-
Примечание. (+)—заметное, (++)—большое, (+++)—избыточное содер-
жание, (—) — отсутствие.
Биологическая стабилизация, гормонов была бы труднодо-
стижима, если бы их биосинтез и метаболизм оказались связанными
с существенной перестройкой принципиальной схемы обмена.' По-
этому в ходе эволюции было достигнуто превращение отдельных
побочных метаболитов в гормональные вещества. В принципе не
исключалась и возможность превращения веществ, вырабатывае-
мых микроорганизмами, в гормоны высших растений при их сим-
биозе. Как и в случае эволюции биогеоценозов, при таком симбиозе
отбор должен быть направлен на поиск удачных компонентов.
Гормональная система, играющая важную роль в онтогенезе
растений (М. X. Чайлахян, 1975), эволюционно возникает посте-
пенно, это обусловлено ступенчатым формированием механизмов
синтеза и рецепции, инактивации (связывания или деградации)
физиологически активных соединений, а также механизма их тран-
спорта. Все эти условия были реализованы в ходе эволюции только
у высших растений. Так, в метаболических звеньях микроорга-
низмов и низших растений нет «вилок» блокирования синтеза
индольных соединений и гиббереллинов, поэтому они на-
капливают их без видимой причины. У высших растений такие
«вилки» имеются,. что способствует синтезу из шикимовой
кислоты ИУК или и-кумаровой кислоты, а из мевалоновой
кислоты —АБК (В. И. Кефели, 1974).
С усилением давления отбора на повышение чувствительности
к отдельным метаболитам (будущим фитогормонам) необходимо
было выработать и механизмы инактивации излишков высокоак-
тивных соединений, чтобы предотвратить износ организма от не-
прерывного роста и морфогенеза. Природа решила эту задачу
синтезом их антиподов — ингибиторов роста. В отличие от рас-
тений у микроорганизмов нет метаболических ответвлений,
приводящих к синтезу ингибиторов роста.
В то же время необходимо было выработать и механизм запаса-
ния фитогормонов для периода, когда они не синтезируются.
Запасание осуществляется путем связывания гормонов с белками,
углеводами, аминокислотами, гликозидами. Выход растений из
покоя сопровождается деполимеризацией сложных органических
соединений клетки и высвобождением связанных с ними фитогор-
158
монов. Перевод в свободное состояние фитогормонов стимулирует
физиологическую активность фитогормонов, что ведет к изменению
действия генов, синтезу РНК и соответствующих белкой.
Значение образования конъюгатов фитогормонов с разными ве-
ществами состоит в облегчении транспорта фитогормонов, запасании
их для последующего использования, защите регуляторов роста от
энзиматического разрушения и поддержании гомеостаза клетки и
необходимого в ней уровня гормонов (Е. D. Cohen, R. S. Bandwsi,
1982).
Цветковые растения не только способны к синтезу регуляторов
роста, но они обладают аллостерическим центром, который, вос-
принимая уровень их содержания, ограничивает синтез фитогормо-
нов по принципу обратной связи. У микроорганизмов нет такого
центра, в результате чего наблюдается сверхпродукция фитогормо-
нов. Этому способствует также и низкая чувствительность микроор-
ганизмов к синтезируемым веществам такого типа. Для сравнения
напомним, что изменение концентрации ИУК в 10 тыс. раз не
влияет на рост гриба Taphrina epiphyНа, тогда как увеличение ее в
10 раз приводит к торможению роста отрезков колеоптилей
пшеницы (В. И. Кефели и др., 1972). Накопление фитогормонов у
микроорганизмов и грибов, возможно, имеет значение для усиления
их взаимодействия с высшими растениями. ,
Как известно, гормональная регуляция эволюционно возникает
на определенном этапе развития многоклеточных органйзмов, когда
внутриклеточные (ионные) и межклеточные (трофические и
биоэлектрические) взаимодействия оказываются недостаточными
для поддержания целостности организма. Число гормонов у живот-
ных имеет тенденцию к увеличению по мере усложнения их орга-
низации. Гормональная система у растений слабо разветвлена, нет
специальных органов для продуцирования гормонов, и гормоны
отличаются меньшей специфичностью действия, хотя в какой-то
. мере она проявляется. Так, ИУК участвует в закладке и росте корней,
рост стебля регулируется гиббереллинами, а рост листьев — преиму-
щественно цитокининами, в листопаде и переходе к состоянию
покоя большую роль играют ингибиторы. .
Таким образом, в процессе эволюции у растений появился набор
гормонов, обеспечивающий дифференциацию и регуляцию роста
на всех этапах онтогенеза. Независимо от того, идет ли речь о
фитогормонах и ингибиторах роста, механизм их инактивации
осуществляется примерно одинаково! путем окисления (разруше-
ния) соответствующими ферментами либо связывания с веществами
или структурными компонентами клетки. В некоторые, периоды
онтогенеза связывание может быть необратимым, что ведет к
превращению фитогормонов и ингибиторов роста в инертные про-
159
дукты. Это совпадает с этапом деструктивных изменений у растений
и регулируется со стороны организма как целого (К. Thimann, 1977).
Остается еще один вопрос: почему именно производные индола,
дитерпеноидов и т. д. приобретают функции гормональных факто-
ров? В. В. Полевой (1982), исходя из представлений Ч. Дарвина о
сходстве раздражимости у животных и растений, обращает внима-
ние на то, что, в частности, ИУК отбирается как фитогормон в
момент вычленения царства растений. ИУК, как и серотонин у
животных, оказывает влияние на мембранный потенциал. Серото-
нин, обнаруженный у. растений, на них не действует, так же как
ИУК, найденная у животных, на ник не влияет. Цитокинин
первично возникает как продукт распада РНК, в последующем же
складываются пути его биосинтеза (зеатин). У животных еще не
найдены аналоги цитокинина.
Гиббереллины (более 60 соединений) относятся к группе дитер-
пенов и рассматриваются как относительно молодой класс сое-
динений, ибо процесс их образования продолжается. Сходны с
гиббереллинами стероидные гормоны насекомых' и гормоны раз-
множения грибов.
АБК эволюционно могла возникнуть как путем распада ксан-
тофилов, так и путем прямого синтеза сесквитерпенов. Она обнару-
жена у высших растений. У водорослей, печеночных мхов, грибов
найдены другие вещества, обладающие абсцизовой активностью.
АБК, так же как и гиббереллины, относится к молодой группе
фитогормонов; она возникает у наземных растений как приспо-
собление к перенесению водных стрессов (участвует в регуляции
устьичных движений).
Интересны попытки осмыслить связь между изменением функ-
ций фитогормонов и их рецепторов в ходе эволюции; с расширением
функции фитогормонов (они из внутриклеточных регуляторов пос-
тепенно превращаются в межклеточные и межорганные) усилива-
ется и специализация рецепторов. Все это приводит к снижению
накопления этих гормонов и повышает регуляторную функцию.
Говоря о биологической роли фитогормонов, необходимо учи-
тывать их участие не только в эндогенной регуляции роста и
развития, но и в приспособлении растений к среде. Оказывается,
что стрессы (засуха, темнота, затопление, высокая pH и т. д.)
способствуют изменению как количества отдельных фитогормонов,
так и соотношения между ними. Так, при засухе увеличивается
содержание АБК, при затоплении — этилена, при низкой темпера-
туре снижается количество гиббереллинов и Цт (но увеличивается
содержание АБК). Подобные изменения регулируют рост и раз-
витие, снижают траты веществ, что обеспечивает переживание
организмом неблагоприятных условий. Фитогормоны оказывают
влияние также на активность ферментов ряда метаболических
звеньев < (В. В. Полевой; 1982), в том числе на деградацию
полиаминов (Н. И. Шевякова, 1981).
160
Приведенные данные представляют определенный интерес, и
нельзя считать результатом случайной конвергенции химического
состава и функций. Здесь следует усмотреть и возможность преадап-
тации растений на основе химической предопределенности свойств
самих молекул. Этим отчасти можно объяснить сходство строения
фитогормонов растений, нервных медиаторов животных и гормонов
линьки насекомых (гиббереллин сходен с экдизоном). Эволюция
фитогормонов — одно из наглядных доказательств эффективности
действия естественного отбора на функциональном уровне.
5.5. СПЕЦИАЛИЗАЦИЯ И КОНВЕРГЕНЦИЯ РАСТЕНИЙ
ПО ХИМИЧЕСКОМУ СОСТАВУ
Вопрос о специализации и конвергенции видов на функциональ-
ном уровне и по составу веществ изучен мало, но некоторые данные
в литературе все же имеются. Так, для злаков характерно содержание
проламина, для мимозовых —дженколовой кислоты, а у астрагалов
сера в аминокислотах заменена селеном. В то же время астрагалы (120
видов) могут быть разделены на две группы по наличию или отсутствию
канаванина. Для саговников характерны ядовитые вещества,
циклические спирты (инозиты) и пентол. Магнолиевые содержат
кремниевую кислоту, изохинолиновые алкалоиды, эфирные масла,
жиры, и т. д. Независимо от систематического положения у всех
древесных растений есть в составе кофейная и л-кумаровая кислоты,
а у травянистых — феруловая. У древесных имеются также лейкоан-
тоцианы, которых нет у травянистых.
Астрагалы специализированы к аккумуляции селена и использо-
ванию его для синтеза аналогов небелковых аминокислот, которые
затем депонируются в вакуолях. Селен в вакуолях безвреден для
растений, но вызывает отравление животных. У не приспособлен-
ных к селену растений он используется для синтеза белков, в
которых сера заменяется селеном (Дж. Харборн, 1985). При этом
синтезируются нефункциональные белки. Некоторые растения (Ag-
rostistenuis, Festuca ovina) произрастают на отвалах рудников, где
много свинца, меди, цинка и мышьяка. Корни таких растений
обладают высокой активностью кислой фосфатазы, содержат не-
сколько ее изозимов, белки клеточной оболочки способны связы-
вать и инактивировать ионы металлов. Иногда устойчивость связана
с ограничением их поступления в корни меди (Silene cucubalus) или
цинка (Typha latifolia). Не замечена перекрестная толерантность у
растений, т. е. одновременная устойчивость к избыточному содер-
жанию нескольких ионов в субстрате. Существуют растения-инди-
каторы на присутствие в почве металлов.
Специализация веществ и обмена связана с изменением качес-
тва ферментов. У филогенетически молодых групп качество фер-
ментов высокое, у древних —низкое. По этой причине у вторых
6-157 161
образуются продукты специализированного обмена (А. В. Благове-
щенский, 1966). Индивидуальное старение растений также связано -
с падением качества ферментов и сужением спектра накапливаемых
веществ. В связи с этим сделан вывод о том, что специализация
обмена в ходе эволюции сопровождается старением и вымиранием
видов. Специализация таксонов, как правило, сопровождается на-
коплением смол, алкалоидов, каучука, жира и т. п. Оказалось, что
чем примитивнее группа, тем больше в ней алкалоидов. У эволю-
ционно продвинутых групп преобладают гликозиды, хотя и есть
немало исключений. Нередко и таксоны со специализированным
обменом в биологическом отношении процветают.
Обнаружена определенная тенденция в распространении био-
синтетически сходных групп веществ у растений. Так, бетацианины
свойственны Centrospermae, но у них нет антоцианов. По содержа-
нию бетацианинов к этому поряжу близки представители семейства
Dictieriaceae. Состав органических кислот и красящих веществ,
содержащихся в плодах, позволяет разграничить разные виды земля-
ники, крыжовника, малины и смородины. По набору альдегидов
виды рода Prunus могут быть разделены на подроды Amygdalus,
Pruniphoro и Cerasus. Соответственно можно судить о «биохимичес-
ком профиле» семейства, рода, подрода и вида.
Обнаружена некоторая специализация растений и по содержа-
нию жира. Например, содержание жира у водорослей составляет
1,3—6,6% (в искусственных условиях — 68%), в спорах папорот-
ников и хвощей — 38, в спорах плаунов — 35, в пыльце голосемен-
ных — 46, однодольных — 9,7, двудольных — 9,4%. Масло хвощей,
плаунов и папоротников состоит из низкомолекулярных жирных
кислот, масло голосеменных и покрытосеменных — из высокомо-
лекулярных соединений. У хвощей, папоротников и плаунов нет
многих углеводородов из класса терпенов. У растений семейства
Cruciferae жир содержит глицеридные остатки эруковой кислоты, у
Umbelliferae и Araliaceae — петрозелиновую кислоту, у Palmae —
лауриновую, у Myristicaceae —миристовую, у Oleraceae —
олеиновую кислоты. Крестоцветные богаты горчичными маслами,
которые у разных видов различаются по составу аглюконов.
По химическому составу организмов иногда трудно судить об
их генетическом родстве. Так, кумарины обнаружены у 1622 видов,
относящихся к 134 семействам грибов, лишайников и цветковых./
Мало их у папоротников и голосеменных, не обнаружены они у
водорослей, мхов, хвощей и плаунов. Исключительно разнообразен
состав кумаринов у покрытосеменных. Например, из 253 видов
зонтичных выделено 171 индивидуальное вещество (из них 132
специфичны для них), из 96 видов рутовых —105 веществ (64
специфичны для них) и т. д.
. Приведенные данные в какой-то мере показывают, что «каждый
вид при неизменности внешних условий сохраняет постоянную
способность вырабатывать свойственные ему вещества, которые
162
служат его физиологическими признаками» (С. Л. Иванов, 1926).
А. М. Голдовский (1975) подчеркивает, что в организмах каждая
узкая группа органических веществ представлена набором сходных
по составу и свойствам соединений. Закон гомологической измен-
чивости (Н. И. Вавилов, 1967) имеет более общее значение, чем
обобщения упомянутых авторов. Исходя из указанного закона
удалось предсказать возможность получения из горького алкало-
идного люпина кормового негорького люпина.
Отмечена и конвергенция растений по химическому составу, в
частности по содержанию терпеноидов и алкалоидов, что имеет
значение для повышения иммунитета растений, привлечения опы-
лителей и предохраненйя от поедания животными. Так, бетаци-
анины и бетаксантины выступают как аттрактанты опыления и
предохраняют растения от вирусной инфекции. На примере
изучения жирных кислот установлено, что они регулируют несо-
вместимость видов и самоопыленных линий, влияя на рост пыль-
цевой трубки и метаболизм тканей столбика. Изопреноиды и
терпеноиды играют большую роль в иммунитете растений. Циано-
генные гликозиды обнаружены у бактерий, грибов и высших рас-
тений. У 1000 видов из 100 семейств высших растений найдено 23
цианогенных гликозида. Стероидные эстрогены, фитоэстрогены и
другие вторичные вещества у растений близки к таковым у живот-
ных и выполняют функцию антигормонов (К. Slama, 1980).
Еще более показательны примеры химической конвергенции
растений и животных (см. гл. 2). Известно, что в растущих тканях
у растений содержится много пролина (пыльца, прорастающие
глазки клубней картофеля, точки роста после яровизации и фотопе-
риодической реакции). Обычно он накапливается перед интенсив-
ным ростом, а затем используется в энергетических целях. Пролин
накапливается также в летательных мышцах насекомых и быстро
используется во время полета. Пролин и оксопролин входят в состав
белка коллагена животных, который. интенсивно образуется в
период заживления ран и в экстремальных условиях.
Напомним, что для синтеза хлорофилла а предшественником
служит о-аминолевуленовая кислота (АЛК). В то же время АЛК
встречается у животных и бактерий как предшественник тетра-
пироллов. Не менее показательны и другие случаи конвергенции
по химическому составу. Так, серотонин обнаружен в зародышах
черного ореха, где его содержание составляет 180 мкг/г сырой
биомассы. Полагают, что он важен для защиты растений от действия
рентгеновских лучей и высоких концентраций аммония. У живот-
ных же серотонин — нейромедиатор. Из проводящих пучков че-
решка борщевика Heracleum sosnowskyi выделены актино- и
миозиноподобные белки, выступающие как элементы конт-
рактильной системы и участвующие в ближнем и дальнем транс-
порте ассимилятов (М. В. Торкова и др., 1986).
163
Процессы немышечного движения энзиматически и биохи-
мически имеют много общего с механизмом мышечных сокращений
(Б. Ф. Поглазов, С. А. Бурнашева, 1989). Так, у мимозы актино- и
миозиноподобные белки, выделенные из подушек черешков (как и
соответствующие мышечные белки животных), обладают высокой
АТФазной активностью и содержание их возрастает параллельно с
повышением сейсмонастической чувствительности видов и органов
растений. Это послужило основанием для признания общности
механизмов двигательных функций у растений и животных.
Не менее интересен пример конвергенции растений и животных
по способности к синтезу пигментов (D. Fox, 1979) и гемоглобина.
Так, бобовые и другие таксоны (Betulaceae, Elaeagnaceae, Mericaceae
Ulmaceae и др.), способные к азотофиксации, имеют ген гемогло-
бина, причем степень гомологии гена гемоглобина у растений и
животных довольно велика. Корни указанной общности обнаруже-
ны в неживой природе (А. Лима де Фариа, 1991).
Так, из 15 изоформ порфиринов, известных в природе, в клетке
встречается только протопорфирин IX. На базе его комплексов с
ионами металлов возникают сходные по строению вещества —
гемоглобин, миоглобин, цитохромы и хлорофилл. Аналогичны и
представления о возникновении регуляторных пептидов, встречаю-
щихся у прокариот, но играющих специфическую роль в функциях
головного мозга и специфическом иммунитете у высших животных
(И. П. Ашмарин, 1986).
Конвергенция наблюдается и по содержанию ацетилхолина. Так,
ацетилхолин в хлоропластах участвует в регуляции проницаемости
мембран (каки в нервных клетках животных) и фотофосфорилировании.
Удивительно сходно осуществляются биогенез и функционирование
митохондрий у растений и животных при согласованном действии
ядерного и МтДНК, хотя есть и различия (Б. Б. Варгапетян, 1985).
АБК обнаружена у всех сосудистых растений и участвует в движении
устьиц, как Са2+; она регулирует сокращение гладких мышц (у крыс),
влияя на поступление в мышечные волокна Са2+. Полагают, что АБК
функционирует в качестве антагониста указанного катиона как у
прокариот, так и эукариот.
Конвергенция и специализация видов по химическому составу
часто наблюдаются в синтезе ВВМ. Указанные вещества специ-
фичны даже для отдельных видов, сортов, органов и клеток растений.
Они возникают как продукты общего метаболизма (Е. A. Bell, 1980)
и в последующем используются для разных целей. Так, в жгучих
волосках защитную роль выполняют гистамин, окситриптомин и
ацетилхолин, в иммунитете против патогенной микрофлоры пока-
зана роль фенолов, танинов, алкалоидов и фитоалексинов.
Каротиноиды участвуют в фотосинтезе, фототропизмах и процессах
старения растений. Фенольные соединения выступают не только
как ингибиторы роста, но и как регуляторы ферментов и биосинтеза
лигнина (J. Harbome, 1978).
164
Слабо изучен генный контроль синтеза специализированных
веществ. В отношении антоцианов он доказан (J. Haldane, 1956).
Присутствие того или иного гена определяет состояние окисления,
метилирования бокового фенольного кольца или введение органи-
ческой кислоты в молекулу с образованием комплексного пигмента.
Антоцианы передаются в поколениях по серии множественных
аллелей, Показана роль генного контроля в синтезе специфических
фитоалексинов у картофеля, участвующих в защите организма от
возбудителя фитофтороза. Как хозяин, так и возбудитель обладают
факторами расовой специфичности. Так, совместимые и несовме-
стимые расы возбудителя оказывают различное влияние на мембра-
ны путем накопления глюконов, взаимодействующих с рецепторами
(О. Л. Озерецковская и др., 1982).
С учетом ранее высказанных положений (см. гл. 2, 4) можно
заключить, что конвергенция по химическому составу широко
распространена в мире растений. Во многих случаях она ведет и к
преадаптации видов. Так, продукты вторичного метаболизма,
возникшие у водорослей, нашли особое применение у наземных
растений: индольные и фенольные соединения — в регуляции роста
и аллелопатии, алкалоиды, флавоноиды, фенолы — в защите от
патогенных микроорганизмов, травоядных животных, фенолы,
кроме того, имеют значение для синтеза лигнина. Этим отчасти
обусловлено постепенное усиление химической специализации рас-
тений в палеозое и мезозое. Между прочим, и в расцвете покрыто-
семенных не последнюю роль играют ВВМ.
При всем этом о роли многих других веществ в жизни растений
пока еще известно очень мало. Неизученность указанного вопроса
послужила одной из причин появления гипотез, отрицающих роль
изменения химического состава в приспособлении видов.
5.6. ИНДИВИДУАЛЬНАЯ ИЗМЕНЧИВОСТЬ ХИМИЧЕСКОГО СОСТАВА
РАСТЕНИЙ И ВОПРОС О «НЕДАРВИНОВСКОЙ» ЭВОЛЮЦИИ
А. Н. Белозерский (1976) обращал внимание на наследственные
различия в химическом составе растений в пределах сортов, на
необходимость их выявления и селекционного использования. На
примере нескольких видов Seseli и Prangos продемонстрировано
внутривидовое многообразие эфирных масел и кумаринов, изме-
нение их качественного состава и количественного содержания
отдельных компонентов в зависимости от экологических и геог-
рафических факторов. Анализ структуры цитохрома с у растений
показал, что в основе «нейтральных» аминокислотных замен лежит
отбор мутаций —трансверсий пурин — пиримидин, ведущих к
избирательному накоплению пиримидинов в смысловой цепи ДНК,
165
а пуринов —в соответствующих мРНК. Значение подобных изме-
нений связывают с увеличением помехоустойчивости трансляций.
Пожалуй, наиболее подробные наблюдения по изучению
биохимических мутаций были проведены на примере блокирования
различных этапов синтеза ГА3, ИУК (см. гл. 2) и алкалоидов. При
гибридизации кормовых сортов люпина, которые обычно не
синтезировали алкалоиды, они восстанавливали эту способность
благодаря взаимодополнению недостающих звецьев процесса
биосинтеза. И при скрещивании мутантов люпина, у которых
блокированы разные этапы единого процесса синтеза алкалоидов,
появляется потомство, способное нормально его накапливать путем
комплементарного взаимодействия генов. Разнообразие алкалоидов
настолько велико даже в пределах одного вида (Ияса rasea), что
ставит под сомнение целесообразность в адаптивном плане (К.
Mothes, Н. Schutte, 1969).
Согласно П. Зенгбушу (1982), лишь ~2% из общего количества
ДНК является носителем генетической информации, и в некоторых
ее участках последовательность нуклеотидов повторяется от 100 до
1 млн. раз. Увеличение количества ДНК обычно не сопровождается
расширением числа функций, оно характерно для примитивных и
эволюционно застывших форм.
Благодаря успехам в изучении молекулярных основ жизни,
индивидуальной изменчивости ряда веществ и ее наследственной
обусловленности широкое распространение получили идеи о нейт-
ральности эволюции на молекулярном уровне (М. Кимура, 1985).
Получены многочисленные данные, характеризующие наличие в
белках и нуклеиновых кислотах замещений, не связанных с измене-
нием функций. Подобные факты иногда противопоставляют теории
естественного отбора без достаточного основания. Скорость эволю-
ции белков часто не определяется размером популяции организмов,
и мутации в последовательности аминокислот в молекуле не всегда
затрагивают активные центры. Поэтому многие нейтральные заме-
ны в белках ограничены первичной структурой молекулы. В то же
время они не представляются безразличными в отношении к другим
компонентам аппарата синтеза и регуляции белка, что показано на
примере генов проинсулинов и глобинов (В. В. Гречко, 1985).
Сравнение вирусов, прокариот и эукариот показало, что нуклео-
тидный состав кодирующей и некодирующей частей генома харак-
теризуется наличием неодинаковых ограничений в изменчивости.
Это поддерживается естественным отбором ввиду важности для спе-
цифики структуры и функции генома. Подобное действие отбора на
молекулярном уровне необходимо для стабилизации изменчивости
популяций. Исходя из положения, что «многие особенности
организации не имеют никакого значения для их обладателей, а может
быть, не имели значения и для их предков» (Ч. Дарвин), молекулярные
нейтральные замены не вызывают удивления. Но по этой причине
166
относить молекулярные события к разряду вообще нейтральных не
правомерно. Так, лигнин первично, вероятно, бьи нейтральным вещес-
твом и использовался в наземных условиях. В последующем накопление
лигнина в почве в виде мертвой древесины создало угрозу для сущест-
вования древесных форм. Только с появлением деструктуров лигнин
вписался в наземный круговорот веществ с дальнейшим расцветом
древесных (С. М. Манская, Л. А. Кодина, 1975).
Преувеличение роли преадаптации и латериального переноса ге-
нов в эволюции видов (В. А. Кордюм) подвергнуто справедливой
критике (Д. К. Беляев, М. С. Гиляров, Л. П. Татаринов, А. В. Ябло-
ков и др.). У высших растений к результатам латерального переноса
генов склонны отнести процессы образования корончатых галов и
необычайную солеустойчивость некоторых культурных растений
(Э. С. Пирузян, М. Д. Андрианова, 1985), синтез супероксидцисму-
тазы (СОД) у представителей отдельных семейств (Р. Б. Хесин, 1984),
благодаря миграции части ДНК бактерий с помощью плазмид. Так,
из 43 исследованных семейств сосудистых растений только у пред-
ставителей трех семейств (гинкговые, нимфейные и тыквенные)
обнаружен Fe-СОД. Латеральный перенос генома допускается в
отношении гена лег-гемоглобина с помощью Ti-плазмид от Agro-
bacterium tumefaciens в клетку растений-хозяев, а также глобинового
гена от животных и бактерий к растениям (Р. Б. Хесин, 1984). Но
при этом отбор остается основным творцом эволюции как
организмов, так и перемещающихся генетических элементов. Не
исключена возможность, что в будущем число таких фактов возрас-
тет. Однако на основе латерального переноса генов в лучшем случае
можно объяснить вероятность усиления изменчивости. Ошибка
состоит в стремлении использовать примеры латерального переноса
генов для опровержения теории естественного отбора и роли борьбы
за существование в эволюции. На всех этапах химической эволюции
жизни от малых молекул до возникновения их агрегатов-макромо-
лекул проявляется принцип селективности (М. Эйген, 1973).
Эволюционные события в изменчивости химического состава
организмов могли «предварять» морфологические изменения. Так,
синтез индольных соединений низшими организмами, сыгравший
в дальнейшем важную- роль в физиологии и морфологии высших
растений, можно рассматривать как химическое предварение. Но и
для первых их биосинтез, по-видимому, имеет определенное зна-
чение, хотя бы как способ предотвращения накопления триптофана
в клетках. В отношении ключевого фермента жизни — РБФК —
показано, что в прошлом он обладал лишь карбоксилазной
активностью. С появлением О2 в атмосфере его функции расши-
рились участием в оксигеназных реакциях (Ю. С. Насыров, 1975).
Химическое предварение отмечено и в отношении молекулы гемог-
167
лрбина, которая появилась задолго до использования ее позвоноч-
ными как переносчика кислорода.
М. Кимура (1985), указывая на селективно нейтральный харак-
тер внутривидового белкового полиморфизма, подчеркнул, что
«теория нейтрализма» в принципе не отрицает возникновения
полезных и вредных изменений на молекулярном уровне. Поэтому
она не может быть рассмотрена как антагонистическая по отноше-
нию к теории отбора. Оценивая гипотезу о «недарвиновской»
эволюции в целом, следует обратить внимание еще и на то, что она
характеризует не биологическую эволюцию организмов, а специ-
фические изменения белков и особенности их молекулярной
организации (М. В. Волькенштайн, 1981, 1985). В то же время
скорость эволюции белков оказалась выше на ранних этапах разви-
тия жизни, поскольку активные центры белков находились тогда
еще на стадии своего становления. Параллельно снижению ско-
рости эволюции белков усилились их функция и взаимодействие,
что имело значение для стабилизации клеточного метаболизма.
Возможно, «нейтральные» замены у части белков являются рекапи-
туляцией самых начальных этапов их эволюции.
5.7. РОЛЬ ИЗМЕНЧИВОСТИ ХИМИЧЕСКОГО СОСТАВА РАСТЕНИЙ
В ЭВОЛЮЦИИ ЭКОСИСТЕМ
Экосистема развивается как сложная и целостная надорга-
низменная структура биосферы, где растения, животные и микроор-
ганизмы тесно связаны между собой трофическими, химическими,
средообразующими и другими взаимоотношениями. При этом ос-
новополагающими во взаимоотношениях между видами в экосисте-
мах являются химические регуляции. Для иллюстрации приведем
несколько примеров, относящихся к разным группам организмов
и отличающихся по своему характеру и назначению.
Один из таких примеров — отношения между растениями и
многими животными. Так, для привлечения насекомых-опылителей
у растений выработались разные вещества, способствующие
возникновению сложной системы запахов, оттенков окраски цвет-
ков и питательных жидкостей. У эволюционно продвинутых видов
растений в цветках преобладают красящие вещества—флавоны
(перед флавонолами). Среди высших растений циановая окраска
(фуксин) является исходной, преимущественно встречается у голо-
семенных и ветроопыляемых видов. Цианидины — наиболее при-
митивные типы пигментов — служили основой для преобразования
окраски цветков. Делеция гена, определяющего синтез цианидинов,
способствует возникновению других типов окрасок. У тропических
растений, опыляемыхt колибри, преобладает алая и оранжевая
168 7
окраска, тогда как у растений умеренных поясов, опыляемых пче-
лами,— голубая, окраска цветков (Дж. Харборн, 1985).
В этом отношении интересны данные о составе пигментов
некоторых флор. Так, при анализе флоры Австралии обнаружено,
что дельфинидиновый тип окраски характерен для 63%, циани-
диновый —для 47, пеларгонидиновый —для 2% видов растений.
Заметим, что в Австралии редко встречается опыление растений
птицами. В Индии, где растения такого способа опыления более
распространены, соотношение указанных типов окрасок соответст-
венно составляет 47, 70 и 17%. В некоторых семействах (Polemoni-
асеае), чьи представители встречаются и в тропиках и в умеренной
зоне, обнаруживают различия в окраске цветков, отвечающие
специфике опылителей. На примере Калифорнии показано, что
травянистые растения открытых прерий (опыляемые пчелами) несут
желтые цветКи, тогда как лесные (опыляемые ночными бабоч-
ками) — белые или бледно-розовые.
У растений одной и той же местности в зависимости от состава
опылителей также меняется окраска цветков (Allophyllum gilioides,
Microsteris gracilis). Узоры лепестков — чередование в них интен-
сивно и слабо окрашенных участков — также значимы для успеха
размножения растений, служат указателем нахождения нектара и
способствуют опылению растений. Подобные узоры в виде точек,
пятен или линий, не видимые человеком, воспринимаются глазом
опылителей по отражению УФ-лучей. Следовательно, внешний
колорит цветков имеет экологическое значение для борьбы за
существование растений и опылителей.
Несомненно, велико значение и запаха, и аромата для взаимо-
действия животных между собой, растений и животных. На важность
аромата цветков для опыления указывал еще Ч. Дарвин. Как теперь
установлено, запах относится к наиболее древним приобретениям и
аттрактантам растений, опыляемых насекомыми. Запах древнее
окраски (Дж. Харборн, 1985). На примере древних растений, опыля-
емых преимущественно жуками, показано, что их цветки лишены
окраски, но обладают специфическим ароматом. Часть высших цвет-
ковых растений, опыляемых ночными бабочками и летучими мышами,
выделяется своим специфическим и сильным ароматом.
Отбор растений был связан не только с усилением аромата
вообще, Но и с преимущественной его локализацией в цветках.
Усиление аромата и его специфичности определялось изменением
состава и соотношения его компонентов. Без этого трудно представить
установление узких отношений между растениями и опылителями.
Напомним, что аромат выделяют и другие части и структуры
растения—листья, лепестки, побеги, пахучие железы и т.п. Его
значение несколько иное, нежели аромата цветков. Аромат указанных
структур важен, в частности, для других целей. Так, например, муравьи
169
из рода Pseudomyrmex, питающиеся нектаром акации, защищают
последние от травоядных животных. При лишении растений акации
колоний муравьев растительноядные быстро истребляют ее заросли.
Другой пример: муравьи-листорезы {Atta cephalotes, A.octospmosus),
питающиеся грибами, снабжают последних ауксинами, извлеченными
из зеленых листьев. Это способствует быстрому росту грибов и размно-
жению самих муравьев.
Способы взаимодействия растений и животных разнообразны.
Растения вырабатывают как аттрактанты (пищевые стимуляторы),
так и репелленты (токсины). Насекомые, выделяя антитоксины
против ядовитых веществ растений, получают возможность без
конкуренции использовать данный вид растения, а последние, в
свою очередь, лучше размножаются благодаря специализации опы-
ления или зашиты от поедания. В некоторых случаях запах и
токсический нектар играют важную экологическую роль. Так, бор-
щевик обыкновенный, морозник вонючий, аронник пятнистый и
другие издают запах гниющего мяса или фекалий, что служит
своеобразной химической мимикрией. Насекомые, питающиеся
падалью, летят на запах указанных растений и, посещая цветки,
производят опыление. Химическими анализами подтверждено сход-
ство выделений названных растений и веществ, образуемых при
разлржении трупов. Как отмечает Дж. Харборн (1985), виды
аронника (Arum nigrum, A.maculatum) раскрывают свои цветки ночью
и имеют высокую активность дыхания, благодаря чему возрастает
температура тканей цветка до 30°С и усиливается улетучивание
амина. Таким путем, издавая запах фекалий, растения привлекают
опылителей — навозных жуков и мух.
Химическая специализация растений нередко связана и с кон-
вергенцией видов (см. выше). В этом отношении представляет
интерес синтез некоторыми растениями веществ, сходных с феро-
монами насекомых, что способствует привлечению к цветку насе-
комого противоположного пола в качестве опылителя. В свое время
Б. М. Козо-Полянский обратил внимание, что некоторые орхидеи
(Ophrys musciferd), как правило, опыляются самцами шмелей ввиду
их внешнего сходства с самкой. Самец, садясь на цветок орхидеи,
ведет себя, как во время копуляции с самкой. Но цветок имеет не
только внешнее сходство с'самкой, но также вырабатывает феромо-
ноподобные вещества, характерные для последней. Есть примеры и
иного характера, не менее удивительные. Так, самцы бабочек данаид
накапливают в своих андрокониях вторичные соединения (пирро-
лизидиновые алкалоиды) сорняка крестовника (Senecio) для более
активного привлечения самок. Самец, летая над самкой, разбрызгивает
эти вещества на ее усики в виде «любовной пыли» и тем самым
подготавливает ее к спариванию (Дж. Харборн, 1985). Нередко самки
пчел (Eulaema, Andrend) «пачкают» свои лапки ароматическими вещес-
твами с определенных растений. После этого поисковая активность
170
самцов повышается, что отражается на частоте их посещения
цветков соответствующих растений и величине сбора нектара.
В специализации растений по химическому составу велика роль
способности их синтезировать токсины (Р. Fenny, 1975; R. Е. Cook,
1981). Так, обследование растений клевера и лядвенца на широкой
территории Европы показало, что там, где активность улиток и
слезней (фитофаги) в течение года непрерывна и велика, способ-
ность растений к синтезу цианогенных гликозидов-токсинов доста-
точно выражена. Одновременно в популяциях фитофагов идет отбор
по устойчивости к токсину. На территориях, где отсутствует дав-
ление указанных фитофагов, токсичные формы клевера и лядвенца
оказываются вытесненными растениями того же вида, которые не
способны синтезировать токсины и проявляют большую устой-
чиврсть к неблагоприятным физическим условиям.
И насекомые нередко защищаются токсинами растений. Так,
гусеницы бабочки Danaus plexippus, питаясь листьями ваточника,
накапливают в себе его гликозиды, влияющие на повышение сер-
дечной активности и рвотной реакции их хищников (голубые
сойки). При первом поедании гусениц и бабочек у соек возникает
отрицательная реакция на жертву. По этой же причине ваточник
не поедается и крупными травоядными животными. Есть примеры
регуляции растениями численности фитофагов. Так, у тисса ягод-
ного и папоротника обыкновенного обнаружен в большом количес-
тве гормон линьки насекомых (р-экдизон). Питание указанными
растениями отражается на численности насекомых.
В ряду папоротники -> голосеменные -> древесные покрыто-
семенные -» травянистые покрытосеменные прослежена тенденция
усложнения структуры детеррентов, образование которых связано
с разными метаболическими путями синтеза ВВМ (Дж. Харборн,
1985). Велико разнообразие этих веществ (изучено более 30 тыс.
соединений), выполняющих специфическую экологическую роль —
защиту, сигналы действия, аллелопатию, устойчивость к патогенам
и т. п. Так, даже некоторые животные (многоножки, муравьи, божьи
коровки, жуки-плавунцы, лягушки) защищаются от хищников,
вырабатывая собственные алкалоиды, при этом воспроизводятся
биосинтетические пути, присущие растениям. Указанные животные
не составляют исключения в этом отношении.
Для защиты от паразитических форм растениями мобилизуется
также огромный арсенал ВВМ. Тем не менее и паразиты преуспе-
вают из-за постоянного отбора на агрессивность в их популяциях
(Г. Доувер, Р. Флейвелл, 1986).
Для выживания вида в экосистемах необходимо учитывать не
только внутривидовое разнообразие растений и животных, но и
ценотическое. Так, почвы тундры отличаются Дефицитом азота, что
Делает невозможным пышное развитие там цветковых, но они там
171
все же встречаются в ценозах со споровыми растениями, способными
фиксировать молекулярный азот и аккумулировать его в мховой
подстилке. Цветковые, в свою очередь, обеспечивают споровце
углеводами, выделяя их Через корни. Здесь фитоценоз выступает
как объект отбора на стабилизацию межвидовых отношений; в
результате сопряженной эволюции видов (Ч. Дарвин). /
Из сказанного видно, что сложные межвидовые и внутривидо-
вые взаимоотношения регулируются благодаря химическим взаимо-
действиям—синтезу ВВМ и усилению роли видов-накопителей
таких соединений (Т. С. Саламатова, О. А. Зауралов, 1991). /
Возможности приспособительной эволюций растений расширя-
ются благодаря генетической изменчивости метаболизма и дивер-
генции видов по химическому составу. Растения в «химической»
защите проявляют большую изобретательность и в случае самоот-
равления клеток пораженной ткани собственными фитоалекси-
нами, что препятствует распространению и Паразита. Конечно, мы
еще много не знаем о роли отдельных индивидуальных веществ,
звеньев метаболизма и молекулярной мимикрии в жизни растений.
Но очевидно однако, что они не могли быть подхвачены отбором
в случае ослабления успеха популяции в борьбе за существование.
Поэтому мнение об отсутствии в живой природе веществ биоло-
гически неактивных (С. А. Галактионов, 1988) правильно, хотя роль
части из них все еще неясна.
* * •
Итак, в изменчивости химического состава растений проявля-
ются некоторые общие тенденции: закон гомологичности, постоян-
ство химического состава видов, множественность представления
каждой узкой группы органических соединений, специализация и
конвергенция видов. Дивергенция видов и форм растений по хими-
ческому составу носит приспособительный характер, хотя последний
в отношении отдельного индивидуального вешества и на
внутривидовом уровне не всегда-прослеживается.
Конвергенция и специализация видов по химическому составу
чаще всего прослеживаются в отношении веществ вторичного
метаболизма. На примере этих соединений видно, что эволюция
растений сопровождается широкой адаптивной радиацией видов с
тенденцией предварения будущих требований.
Обнаружены и «нейтральные» замены в молекулах, которые,
возможно, служат рекапитуляцией ранних стадий эволюции био-
полимеров. Изменение химизма растений в онто- и филогенезе
связано с ритмическими явлениями, устойчивостью к небла-
гоприятным внешним факторам, целостностью и продуктивностью
индивидуума.
ГЛАВА 6
ПЕРИОДИЧЕСКИЕ ЯВЛЕНИЯ В ЖИЗНИ РАСТЕНИЙ
Онтогенез растения в целом, как и отдельного органа, протекает
в условиях непостоянства среды. Колебания условий могут быть
более (годовые, сезонные) или менее (суточные, дневные) значи-
тельными. С точки зрения эволюции основную роль играют посто-
янно чередующиеся (в течение суток, дня, по сезонам и годам)
условия, так как Только в этом случае возникает определенный
вектор отбора.1В конце концов складывается внутренний ритм
реагирований на подобные ситуации, что становится биологической
необходимостью. Возникновение периодичности в процессе эволю-
ции обусловлено синхронизацией жизненных явлений с геофизиче-
скими процессами, которые, в свою очередь, зависят от космических
ритмов. Изучением указанных явлений занимается новая область
науки — гелиобиология.
В связи с этим физиологи уделяют много внимания изучению
и регуляции периодических явлений в жизни растений, а также
поиску механизмов измерения биологического времени. Интерес к
периодическим явлениям обусловлен их значением для про-
дуктивности растений. Литература по этим вопросам огромна, но в
ней недостаточно рассмотрена эволюция конкретных форм прояв-
ления ритмических и периодических явлений у растений.
6.1. БИОЛОГИЧЕСКОЕ ВРЕМЯ И ЕГО ИЗМЕРЕНИЕ У РАСТЕНИЙ
Биологическое время обусловливает нормальную жизнедеятель-
ность. Разные организмы не одинаково воспринимают один и тот
же интервал физического времени, необходимый для завершения
какого-либо физиологического акта. Биологическое время харак-
теризуется спецификой геометрии, оно На разных уровнях
организации живого имеет разное выражение (A. Winfree, 1980).
Появление ядра в ходе эволюции сделало необходимым новую
временную организацию клетки. Отбор по сочетанию таких клеток
в организме в определенных условиях способствовал закреплению
эндогенных ритмов (Е. Biinning, 1978).
Д. А. Сабинин (1963) указывал, что при изучении физиологии
развития побега основой для измерения времени может быть
173
элементарный этап морфогенеза — образование узла, листа и
междоузлия (пластохрон). В частности, по изменению числа плас-;
тохронов можно объективно сравнивать скорость морфогенеза у
разных видов. 1
Для многих травянистых растений число листьев на побеге 4-
консервативный признак, генеративные органы закладываются
лишь после образования соответствующего количества листьев.
Например, у красного клевера существуют позднеспелые (однру-
косные) и скороспелые (двуукосные) формы, у которых число узлов
на побеге соответственно 7—9 и 5—7. Число листьев на главном побеге
у озимых зерновых злаков равно 7, а у яровых —4—5 (Д. А. Сабинин,
1963). Яровые и озимые, ранне- и позднеспелые формы отличаются
по числу детерминированных зачатков листьев уже в зародыше.
В физиологии растений о времени чаще всего говорят при
изучении эндогенных ритмов, фотопериодизма, скорости хими-
ческих реакций и т. д. Нередко определяют время презентации —
минимальное время для восприятия раздражителя особями разных
видов. Этот показатель позволяет оценивать степень раздражимости
растений, их реакцию на воздействие внешних условий.
В целом наши знания о биологическом времени конкретных
физиологических актов пока весьма ограничены. По замечанию
В. И. Вернадского, «во многих областях ученые изучали явление,
а не время». Ныне задача изучения специфики биологического
времени стала необходимостью, так как «бренность жизни нами
переживается как время, отличное от обычного времени физика»
(В. И. Вернадский, 1975).
Одним из проявлений измерения биологического времени у
растений служит квантированность роста и морфогенеза. Термин
квант роста был использован для обозначения единицы, или «дозы»,
прерывистого «толчкообразндго» роста растений еще в 1917 г.
А. Томпсоном (Д. А. Сабинин, 1963). Позднее выражение «кван-
тированность роста» употребляется при характеристике процесса
закладки метамеров у побегов древесных, в смысле существования
неких «ячеек» морфогенеза, включающих несколько метамеров.
Явление квантированности находит свое выражение: 1) в тенденции
соблюдения ряда Фиббоначчи или, возможно, в нескольких момен-
тах какого-либо другого ряда; 2) в соблюдении принципа кратных
отношений; 3) в многовершинных кривых варьирования размеров
органов (М. М. Магомедмирзаев, 1979).
Способы выявления и интерпретации квантов различны, но
прежде всего следует отметить два подхода. Во-первых, кванты
могут быть установлены в процессе непрерывного наблюдения за
формированием какой-либо структуры в онтогенезе, т. е. как
динамические показатели. В этом случае квантированность по сути
совпадает с понятием ритмичности физиологических процессов (см.
174
» ниже). По мнению Б. Гудвина (1966), «существует целый спектр
---------7КИХ ПрОцессов, имеющих различные периоды и образу-
рганизме сложную, но хорошо организованную временную
у». Во-вторых, кванты могут быть установлены на основе
льного анализа конечных (или промежуточных) состояний
особей в каких-либо их совокупностях (в популяциях,
г. д.), т. е. как статистические показатели.
из распространенности явления квантирования числовых
>ных признаков среди растений и животных позволил
> эволюцию его механизмов. Кванты рассматривают как
«собственного», биологического, измерения морфогенеза,
они представляют собой относительно автономные ячейки разви-
тия, между которыми лежат критические точки (фенокритические
фазы). Само возникновение этих фаз имеет свои ограничения,
связанные с необходимостью минимума вещества (массы, числа,
элементов), энергии и информации. В эволюционном плане
квантированность выступает как фактор «буферности» морфогене-
за, как предпосылка стабилизации признака. В ходе эволюции она
возникает в связи с установлением сезонности климата, а также
ритмов с продолжительным периодом смены фаз, (см. ниже).
Биологическое время в более общем смысле выражается в
темпах смены поколений (А. В. Яблоков, А Г. Юсуфов, 1981). Его
ускорение имело значение для достижения биологического и мор-
фофизиологического прогресса растений. Вообще же отсчет вре-
мени приобретает особый смысл в колеблющихся условиях и в ходе
эволюции закрепляются специальные механизмы, позволяющие
измерить его в онтогенезе («физиологические часы»).
6.2. ЦИРКАДНЫЕ РИТМЫ И ИХ ЗНАЧЕНИЕ
Для растений характерна суточная периодичность, проявляюща-
яся в движении листьев, раскрывании цветков, активности выде-
ления пасоки и нектара, изменении скорости и частоты митозов и
т. д. Такая эндогенная, суточная (циркадная) ритмика свойственна
растениям даже в темноте (рис. 18), когда они уже не подвергаются
действию света («физиологические часы»). Э. Бюннинг (1961) пола-
гает, что у растений существует временная память. Так, цветки ряда
видов начинают раскрываться незадолго до рассвета, как будто
«зная», что через несколько часов взойдет солнце. Это у них
повторяется и в темноте. В целом «растения, подобно животным,
правильно измеряют время суток, по крайней мере длительность
ночи» (Э. Бюннинг, 1961). Сейчас многие особенности жизнедея-
тельности связывают с суточной ритмикой, особенно много фактов
накоплено при изучении суточной периодичности движения
175
Рис. 18. Продолжение с незначительным ослаблением движения листьев Kalahchoe
blossfeldiana в непрерывной темноте; темные периоды заштрихованы (Э. Бюннинг,
1961. С. 20)
листьев. Для большей части изученных процессов продолжитель-
ность периода циркадных ритмов составляет 22—28 ч.
Доказаны наследственная обусловленность (Н. Mletzko, J. Mlet-
zko, 1977) суточной ритмики и ее приспособительное значение. Так,
сорта сои, возделываемые в экваториальной зоне, отличаются мень-
шей амплитудой эндогенного суточного ритма от сортов, возделы-
ваемых вне указанной зоны (рис. 19). На примере дикорастущих
злаков показана устойчивость суточной ритмики цветения, что
позволяет использовать ее как таксономический признак. Суточная
ритмика цветения выполняет важную биологическую роль, она
ускоряет дифференциацию популяций, создавая барьеры изоляции.
Рис. 19. Суточный ритм движения листьев у сортов сои в одинаковых условиях
(Э. Бюннинг, 1961. С. 35):
А —короткодневный сорт Отоотан, возделываемый в тропиках; Б —сорт Мак-РостИс
Мандарин как пример растения нейтрального дня
176
Сходная суточная ритмика цветения видов (Efytrigid) нередко
\ задерживает микроэволюционные процессы полиморфных видов
\(Е. repens), уменьшая морфологические различия благодаря естест-
венной гибридизации разных видов. Опыты по скрещиванию сортов
^неодинаковыми эндогенными ритмами показали их наследование
по промежуточному типу в Fb В то же время наследственные
эндогенные ритмы могут перестроиться в онтогенезе. Однако с
возникновением типичных условий они вновь восстанавливаются,
чтц показано в опытах по изучению влияния различной длины дня
на „движение листьев фасоли. Оказалось, что на непрерывном
освещении, как и при 3—4-часовой длине дня, листья фасоли
неактивны. Движение листьев восстанавливалось при сокращении
светового и увеличении темнового интервала времени. Так, при
16-часовом дне листья стали опускаться на свету и подниматься в
темноте на 5-й день. При 12-часовом и еще более коротком дне
листья на свету не опускались, а в темноте поднимались со зна-
чительным опережением, достигавшим нескольких часов.
В данном опыте движение листьев определялось условиями.
Наследственно закреплены не сами никтинастические движения, а
способность растений приспосабливать свой суточный метаболизм
к привычным для вида световым и другим условиям. На примере
тока пасоки в ксилеме древесных растений обнаружено, что он
меняется в зависимости от условий. Динамическую регуляцию
биоритмов рассматривают как один из универсальных адапта-
ционных механизмов, ведущих к перестройке жизнедеятельности.
Например, расход воды у растений фасоли при 24-часовом освеще-
нии оказывается в 2—3 раза меньше по сравнению с растениями,
находящимися на 14-часовом освещении. В условиях непрерывного
освещения транспирация носила постоянный характер, в то время
как в условиях 14-часового освещения она возрастала от момента
включения света до середины дня с последующим падением. При
этом в середине дня интенсивность транспирации была в 6—1 раз
выше, чем при непрерывном освещении.
У некоторых грибов и водорослей высвобождение спор из спо-
рангия представляет собой процесс с четкой суточной перио-
дичностью, регулируемый эндогенно. Свечение одноклеточной
водоросли Gonyaulaxpotyedra связано с эндогенными ритмами накоп-
ления люциферина и люциферазы ночью. У этой водоросли обнару-
жены также эндогенные ритмы деления клеток (В. М. Sweeney, 1969).
Суточный ритм физиологических функций — один из основных
и наиболее значимых для растений. Он обусловлен чередованием
дня и ночи, связан с изменением организации мембран (D. Th.
Vanden, 1979) и отражает приспособление растений к вращению
Земли и Луны вокруг оси. В ходе эволюции растений менялись
процессы, которыми управляют часы и фоторецепторы самих часов,
177
но сам принцип измерения времени на основе эндогенного ритма
природа сохранила. В естественных условиях растения «измеряют»
не длину дня, а длину ночи. Так, никтофильная перилла зацветает
только тогда, когда находится в темноте не менее 9 ч. Эволюционное
значение актиноритмизма состояло не в том, чтобы ускорить раз-
витие и спасти растения от осенних заморозков, а в том, чтобы
замедлить развитие, накопить значительную массу листьев и лишь
потом мобилизовать развитый фотосинтетический аппарат на вос-
произведение потомства. /
Суточные ритмы регулируются с участием фитохромов (Р^® и
Р7зо). У длиннодневных растений уровень Р7зо на свету низкий; а в
темноте высокий. После включения света содержание Р730 некоторое
время остается высоким. У короткодневных растений Р730 регулирует
ход биологических часов, а биологические часы — динамику
фитохрома.
Для уточнения значения циркадных ритмов представляет инте-
рес выяснение их связи с фотопериодической реакцией. Есть целый
ряд доказательств нарушения циркадных ритмов, когда ночной
период искусственно прерывался вспышкой света (в течение
минуты). При этом у длиннодневных растений даже при коротком
дне Наступает зацветание. Прерывание же световой фазы «темновым
интервалом» не дает никакого эффекта ввиду того, что решающее
значение имеет длина ночи. Отсчет фотопериодического времени
идет с участием физиологических часов, где начало освещения или
затемнения действует как датчик времени для синхронизации фаз.
Кульминационная точка фотофильной фазы у коротко- и длинно-
дневных растений разная (рис. 20). >
Циркадные ритмы у растений, несомненно, имеют приспо-
собительное значение: амплитуда их колебания расширяет норму
реакции организма в онтогенезе. Они играют важную роль и в
сезонных циклах.
Рис. 20. Схема положения фаз циркадного ритма у КД
и ДД растений (из Э. Либберта, 1976. С. 404)
Ось абсцисс —время от начала освещения
178
6.3. СЕЗОННЫЕ ЦИКЛЫ
И ИХ ПРИСПОСОБИТЕЛЬНОЕ ЗНАЧЕНИЕ
' Одно из проявлений периодичности — сезонные и годичные
циклы, связанные с чередованием периодов роста и покоя. Неза-
висимо от продолжительности жизни растения в ходе эволюции
вырабатывается определенная ритмичность, соответствующая се-
зонности климата. В формировании сезонной ритмики имеет зна-
чение обращение Земли вокруг Солнца, что ведет к смене времен
года. В то же время в жизни растений играют роль и более
продолжительные циклы. Например, в приросте многолетних рас-
тений фиксируются колебания не только климата, но и отражаются
11-летние циклы деятельности Солнца. Путем учета таких циклов
сделана запись солнечной активности по спилам секвойи за 3200
лет. Вспышки численности у ламинарии также связаны с 11 -летним
циклом Солнца. Обильное плодоношение у лиственницы в Сибири
наступает только в годы минимальной активности Солнца.
За исключением крайних типов климата (вечная мерзлота и
влажные тропики), существует чередование благоприятных и не-
благоприятных сезонов для роста и развития растений. Отрицатель-
ные воздействия среды растения вынуждены переносить путем
перестройки жизнедеятельности. Образованием органов, лишенных
активных жизненных процессов (семян, почек, укороченных побе-
гов), или сбрасыванием листьев, части побегов и плодов обеспе-
чивается выживаемость растений в неблагоприятные сезоны.
На примере водорослей из разных отделов показана сезонная
цикличность их развития (В. В. Скрипчинский, 1975). Зеленая водо-
росль Fritshiella tuberosa, обитающая в тропиках, в сезон дождей
размножается зооспорами, а в остальное время образует цисты. У
Cladophoraglomerata зимой формируются зооспоры, весной — гаметы.
У ламинарий размножение происходит с августа по март, развитие
нового таллома — весной и летом. У водорослей в разных геог-
рафических и экологических условиях определенные фазы онтогенеза
оказываются приуроченными не только к соответствующим сезонам,
но и месяцам. Нередко такая периодичность совпадает с лунными
месяцами, с приливами и отливами воды. Лунный свет в жизни
обитателей литоральной зоны приобретает сигнальное значение.
У более высоко организованных растений сезонная перио-
дичность усложняется и автономизируется по фазам онтогенеза. В
ходе эволюции прежде всего регулируются начальные этапы, т. е.
всходы развиваются с наступлением благоприятных условий, выра-
батываются механизмы, исключающие опережение или запазды-
вание в прорастании семян,—твердосемянность, послеуборочное
дозревание, глубокий покой и др. В непостоянных условиях
формируются разные типы семян, отличающиеся по срокам прора-
стания. Такое разнообразие воспроизводится в потомстве каждого
179
типа семян, что повышает устойчивость вида к неблагоприятным
условиям, так как нередко прорастание семян при этом растягивается
на несколько лет (овсюг). Интересно указать на «капризы» семян
табака, которые в летние месяцы теряют способность к прорастанию,
а в осенне-весенние месяцы восстанавливают ее. Природа регуляции
прорастания семян и ее физиологические механизмы становятся
понятными только в связи с экологией вида. Не случайно поэтому в
посевах идет отбор сорняков по темпам и срокам прорастания семян,
близким к ритму прорастания культурных растений. '
В последующем регуляция роста и развития у растений осущес-
твляется более сложным путем. Разные этапы онтогенеза отлича-
ются по чувствительности к условиям среды и выделяются как
критические по отношению к свету, влажности и температуре. С
этой точки зрения озимость и яровОсть, эфемерность развития,
однолетний, двулетний й многолетний образ жизни, тип фотопе-
риодической реакции, образование органов для переживания
периодов похолоданий и засухи выступают как разные способы
приспособления растений к неблагоприятным условиям. У тропи-
ческих же растений все почки на растении начинают и заканчивают
покой одновременно, поскольку стареющие листья вырабатывают
факторы старения и индуцируют тем самым старение других листьев
и переход почек в состояние покоя (Э. Либберг, 1976).
В средних широтах (с засушливым летом и холодными зимами)
наблюдается синхронизация активности роста растения в целом.
При этом синхронизация с климатическими условиями достигается
в пределах одного этапа развития, а все остальные берут отсчет
времени от него, причем сигнальную роль играют разные факторы.
Сезонная ритмика проявляется и в физиологических процессах,
о чем свидетельствуют материалы, характеризующие интенсивность
дыхания и фотосинтеза у растений в разные периоды года (см. гл.
3, 4). Это же подтверждают и результаты непосредственного изу-
чения эндогенного ритма темнового дыхания (рис. 21), прироста
побегов и корней у разных древесных (рис. 22).
Дыханию, как и многим другим физиологическим процессам,
свойственно > изменение суточной и сезонной активности, которое
определяется наследственностью, условиями среды и возрастным
состоянием. Эти колебания имеют эндогенную обусловленность, что
видно по сезонной активности дыхания у листопадных и вечнозеленых
растений (см. рис. 21). Так, активность дыхания у вечнозеленых
растений, обитателей более или менее постоянных условий, меняется
плавно. Эндогенный характер носят и изменения в активности роста
побегов и корней. Даже в тропиках, там, где внешние условия в целом
остаются благоприятными для роста растений в течение года, периоды
роста прерываются покоем. И для роста листопадных форм харак-
терна известная автономность (рис. 22), проявляющаяся в различиях
180
Месяцы
Рис. 21. Сезонная изменчивость интенсивности темнового
дыхания у листопадных (прерывистая линия) и вечнозеленых
(заштрихованный участок) древесных растений (В. Лархер,
1978.С. 68)
во времени начала и окончания роста побегов, в несовпадении его
пиков для разных видов. Рост корней, несмотря на большую его
зависимость от внешних условий, также является автономным
процессом, что видно при сравнении разных видов.
Цикличность проявляется и в листопаде. Он подготавливается
всем ходом развития индивидуума растений и чаше коррелирует по
времени с наступлением старения у однолетних форм или Отдельных
частей у многолетников (см. гл. 8). Деревья умеренной зоны готовятся
к зиме, а тропиков;— к летней засухе с помощью листопада. Сигналь-
ную роль при листопаде играет длина дня. Периодичность развития,
регулируемая внешними условиями, особенно хорошо изучена на
примере эфемеров, у которых все рассчитано на максимальное исполь-
зование коротких периодов благоприятных условий. Способность к
быстрому росту в течение короткого времени —отличительная осо-
бенность растений пустынь. Эфемерный тип растительности как раз
возникает в результате отбора индивидуумов, способных сокращать
время развития, что ведет к закреплению «лихорадочной поспешности
цветения». У пустынных эфемеров и эфемероидов период максималь-
ной интенсивности фотосинтеза наблюдается в марте и апреле, когда
они вступают в фазу плодоношения.
Существуют многочисленные примеры затухания роста у растений
даже в благоприятных условиях. К ним относится чередование периодов
роста с периодами покоя у побегов. Подобные циклы Д. А. Сабинин
(1963) связывал с изменением активности синтеза нуклеиновых
181
Рис. 22. Прирост в длину побега (вверх) и корней (вниз, зачернено) у различных
древесных пород в течение года (В. Лархер, 1978. С. 68). Видно совпадение
активного периода роста для разных пород
кислот. Например, если содержание нуклеопротеидов оказывается
низким, побеги (чайного куста) «заглушаются» в росте даже в летнее
время при благоприятных внешних условиях. Теперь выявлено, что
в регуляции периодичности роста растений участвуют фитогормоны
и ингибиторы (В. И. Кефели, 1974 и др.).
Заслуживает внимания также вопрос об эволюции годичных
циклов. Эти циклы исторически могли возникнуть только в те
геологические периоды, когда складывалась смена условий в
течение года, что показано на примере явлений фотопериодизма
(см. ниже). Первоначально регуляция роста, возможно, была вы-
нужденной и касалась вегетативных органов, а позднее и семян. На
основе криптобиоза и колебаний длины дня отбор формирует
механизм эндогенной регуляции роста, наиболее совершенный для
отсчета астрономического времени.
6.4. ПРОИСХОЖДЕНИЕ И ЭВОЛЮЦИЯ
ФОТОПЕРИОДИЧЕСКОЙ РЕАКЦИИ
Фотопериодизм — реакция растений на изменение соотношения
длины светового и темнового периода в течение суток — наиболее
широко распространенное и хорошо изученное ритмическое явление,
обусловливающее переход растений к размножению. Со времени
открытия фотопериодизма накоплен огромный материал по конк-
ретизации реакции разных видов (преимущественно цветковых) на
182
длину дня. Однако до сих пор явление фотопериодизма с точки
зрения происхождения, эволюции, генетики и экологии недостаточ-
но проанализировано. В. В. Скрипчинский выделяет следующие типы
фотопериодизма: физико-химический (или актиноритмизм) и
биологический (суточный, лунномесячный и сезонный).
Эти типы отражают процесс эволюции самого фотопериодизма.
Физико-химический фотопериодизм присущ предбиологическому
этапу эволюции жизни. С возникновением жизни вначале появля-
ется суточный фотопериодизм (циркадные ритмы), затем лунноме-
сячный (у обитателей литорали) и, наконец, сезонный (у наземных
высших растений).
Выяснено, что практически все низшие и высшие растения так
или иначе обладают фотопериодической реакцией, что выражается
в изменении интенсивности роста, смене фаз развития и морфоге-
неза и т. д. в зависимости от длины дня. У водорослей она прояв-
лялась в разной периодичности размножения, что совпадало с
летними и осенними месяцами, когда наступало благоприятное
чередование дня и ночи в течение суток (см. выше). У мохообразных
гаметообразование шло как при длинном (маршанцевые и юнгер-
маниевые), так и при коротком (лиственные мхи) дне. У голосе-
менных с изменением длины дня меняются прирост и сроки
наступления покоя. Изучение фотопериодизма у хвойных показало,
что северные формы и расы интенсивнее растут при длинном, тогда
как южные — при коротком дне. Среди представителей крупных
таксонов цветковых обнаружены как короткодневные, так и
длиннодневные виды, а также нейтральная группа. Поэтому можно
сказать, что у таксонов крупного масштаба отсутствует строгая связь
с определенным типом фотопериодической реакции. Это наводит
на мысль о возможности ее относительно самостоятельной эво-
люции в пределах каждого таксона, т. е. у растений шла параллель-
ная эволюция фотопериодической реакции. В пределах же . вида
разнообразие типов фотопериодической реакции значительно
меньше, чем в пределах семейств и родов.
Фотопериодическая реакция связана не столько с систематичес-
кой принадлежностью вида, сколько с зональным и широтным его
распространением. Так, давно подмечена приуроченность растений
короткого дня к низким, а длинного дня —к высоким широтам.
Однако могут быть и исключения. Например, в высокогорной
тропической зоне Анд произрастают растения, обладающие
длиннодневной фотопериодической реакцией. При понижении
температуры ночью наблюдается процветание растений длинного
Дня даже в горных условиях тропиков.
Элементарной таксономической единицей, обладающей одно-
родной фотопериодической реакцией, ныне считают биотип. На-
183
коплено много данных, свидетельствующих о строгой генетической
обусловленности фотопериодической реакции, ее полигенной
природе и передаче потомству при гибридизации. Например, при
скрещивании линий арабидопсиса, отличающихся скороспелостью,
расщепление F2 составило 3:1 при длинном дне, а при коротком —
по срокам вегетационного периода появились промежуточные
формы. Обычно гены, определяющие короткодневную реакцию,
ведут себя как рецессивные. При этом расщепление может носить
моно- и дигибридный характер. Длиннодневные формы доми-
нируют над нейтральными при скрещивании. Обнаружены мутации
по фотопериодической реакции. Нередко ее спонтанное изменение
связано с полиплоидией.
Наследственные изменения в фотопериодической реакции слу-
жат основой для отбора, так как нередко они могут иметь значение
для выживания особей в неблагоприятные сезоны года (зимы и
засухи) и особенно для процессов размножения (М. X. Чайлахян,
1981). Фотопериод служит своеобразным «астрономическим кален-
дарем», определяющим наступление благоприятных условий года.
Поэтому не случайно, что эта реакция наиболее четко выражена у
растений, обитающих в колеблющихся по сезонам условиях среды.
Если такие колебания условий наблюдаются в тропиках, то у
растения появляются соответствующие фотопериодические
реакции. Альпийские луга Эфиопии сходны по составу растений с
лугами Колумбии и Гималаев (мятлик, овсяница). В горных районах
Индо-Малайской области распространены северные растения —
рододендрон, сосна, черника. Все это свидетельствует о том, что
для миграции северных видов на юг через альпийские горные
системы фактор длины дня не мог служить непреодолимой прегра-
дой. Пониженные температуры в этих горных поясах и количест-
венная вариация фотопериодической реакции у растений
позволяют им мигрировать через горные массивы.
Учитывая распространенность фотопериодической реакции у
современных растений и ее биологическое значение, а также широт-
ные и высотные особенности распределения растений по типу
количественного и качественного варьирования данной реакции,
В. В. Скрипчинский (1975) сделал попытку представить эволюцию
фотопериодической реакции в геологическом прошлом. Автор ука-
зывает, что ввиду ограниченности материала о сезонных изменениях
климата в начале палеозойской эры (кембрий и ордовик) трудно
судить о фотопериодизме древнейших обитателей суши — лепи-
дофитов. Первые растения суши, обитавшие в постоянных условиях
(полуводные зоны теплых морей), вероятнее всего были нефото-
периодического типа.
С началом процессов горообразования, уже в силуре, появля-
ются высотные и широтные различия в климате. В этот период
формируются климатические зоны — тропическая, субтропическая
184
и умеренная. В Южной Африке и Южной Америке среди ископа-
емых девонских форм обнаружены холодолюбивые растения, а на
Новой Земле и Таймыре —теплолюбивые формы. В девоне, когда
появилась потребность в регуляции процессов роста и развития в
связи с формированием климатических зон, фотопериодическая
реакция растений усиливается. В карбоне предполагается возникно-
вецие всех ныне известных фотопериодических типов реакции, хотя
во влажных тропиках и могли еще остаться нефотопериодические
группы. Растения раннего карбона, распространенные в тропиках,
обладали преимущественно короткодневной реакцией. Это способ-
ствовало вступлению их в период покоя перед началом сухих сезонов
года. По мере проникновения растений в более высокие широты
(карбон) появляются длиннодневные формы, которые использо-
вали теплые и благоприятные сезоны года. С уменьшением зональ-
ности и сезонности климата (триас, ранняя юра) при отсутствии
необходимости строгой регуляции развития могли появиться рас-
тения с нейтральным типом реакции на длину дня.
В общих чертах эволюцию фотопериодической реакции В. В.
Скрипчинский (1975) представляет в виде следующей схемы:
Тип фотопериодизма Время возникновения реакции
Нефотопериодический
Короткодневный
Длиннодневный
Нейтральный
Кембрий, ордовик, силур
Силур (?), девон
Девон, карбон, пермь
Триас, ранняя юра (?)
В период возникновения покрытосеменных создавались условия
для сохранения коротко- и длиннодневных растений. Тип фотопе-
риодической реакции мог сыграть решающую роль при расселении
первичных покрытосеменных с их прародины. При миграции через
горные хребты преимущества были на стороне длиннодневных форм
(или при этом короткодневные превращались в длиннодневные), а
в случае миграции по низким широтам — на стороне короткоднев-
ных (или при этом длиннодневные превращались в короткоднев-
ные). При широтной миграции, в пределах тропического пояса,
преимущества оставались на стороне нефотопериодических и ко-
роткодневных растений. Широтные миграции в умеренных зонах
способствовали превращению длиннодневной реакции в нейтраль-
ную. В ледниковые эпохи растения теряли длиннодневность и
приобретали короткодневность, а в послеледниковые —наоборот.
На примере культурных растений установлено, что в центрах их
происхождения встречаются виды, обладающие различными
фотопериодическими реакциями (Н. И. Вавилов, 1967). В тропи-
ческих центрах обнаружены короткодневные (на низинах) и длин-
нодневные растения (в. горах). В своем возникновении и развитии
фотопериодическая реакция подчиняется действию отбора и имеет
приспособительное значение.
185
* * *
Таким образом, ритмические процессы связаны с экологи-
ческими факторами. По-видимому, первоначально возникновение
ритмичности было обусловлено изменением длины дня, поэтому
не случайна высокая чувствительность растений к длине дня в онто-
и филогенезе.
Сами эндогенные ритмы у разных видов выражены не одина-
ково, наиболее древние из них — циркадные (суточные). В дальней-
шем возникают ритмы с большей продолжительностью, включая
физиологический покой и фотопериодизм.
Сказанное свидетельствует о наличии приспособительных раз-
личий в проявлении физиологических процессов, что выражается
или в синхронизации периодов их активности с наступлением
благоприятных внешних условий, или в строгой автономности
регуляции. Все это, несомненно, имеет значение для достижения
репродуктивного успеха популяций. Поэтому эндогенный ритм
физиологических процессов является результатом отбора растений
в направлении автономизации онтогенеза (см. гл. 8).
ГЛАВА 7
НАЗЕМНАЯ СРЕДА И ЭВОЛЮЦИЯ РАСТЕНИЙ
Выход растений на сушу в силуре (более 400 млн. лет назад) имел
исключительное значение для дальнейшего их структурно-
функционального совершенствования. Суша для водных растений —
принципиально новая адаптивная зона, где требовалась коренная
перестройка габитуса и жизнедеятельности. Поэтому структурно-
функциональные изменения, приобретенные по комплексу признаков
в связи с выходом растений на сушу, считают одним из важных
арогенезов в эволюции растений. Значение освоения растениями суши
трудно переоценить не только для дальнейшего их прогресса, но и
всей жизни на Земле. Одной из причин возрастания дифференциации
растений, в наземных условиях следует признать большую вариабель-
ность среды и разнообразие ниш по сравнению с водной средой.
Выход растений на сушу бьи осуществлен как путем преадап-
тации, так и приобретения комплекса новшеств. К преадаптации
следует отнести возникновение многоклеточных и прикрепленных
к субстрату нитчатых водорослей, дифференцированных на органы.
Сходные изменения происходили параллельно в разных ветвях
водорослей и привели к повышению продукционного процесса (см.
гл. 3). Ряд особенностей нитчатых зеленых водорослей позволяет
рассматривать их как промежуточные между одноклеточными и
высшими растениями и как предшественники первых наземных
растений (И. М. Величко, 1989).
Для древнейших наземных растений (риниопсид) были харак-
терны дихотомическое ветвление (С. В. Мейен, 1971), отсутствие
листьев и наличие ризомоид и ризоидов (вместо корней и корневых
волосков). Фотосинтез осуществлялся в тканях стебля. Листья и
корни появляются впоследствии путем видоизменения стеблей.
Независимо от возможного предка наземные растения унаследовали
от водорослей принцип организации и функционирования клетки,
что мало меняется даже у эукариот.
Первые наземные растения (риниофиты и зостерофиты) росли на
влажных болотистых местах и по берегам водоемов и рек. Дальнейшее
повышение их устойчивости к наземной, засушливой среде было
связано с постепенной утратой функции гаметофита, возрастанием
роли спорофита (А. В. Яблоков, А. Г. Юсуфов, 1989) и вакуолярной
системы, усвоением и выделением органических веществ, развитием
187.
ЦОеткоРые
Голосеменные
Мезозойские
семенные
папоротники
Палеозойские
семенные папоротники
Рис. 23. ЭвоЛюция высших растений (Н. И. Величко, 1989. С. 92)
проводящей системы, покровных тканей и т. п. Подобные преоб-
разования происходили в разных группах растений (рис. 23).
Дивергенция и прогресс растений в наземных условиях определя-
лись успехом в обеспечении водой и светом. Поэтому морфологию
наземных растений рассматривают как геометрический компромисс
трех ограничивающих факторов: размещения относительно света,
механического стресса (рост в вертикальном положении), транспорта
воды и ассимилятов на дальнее расстояние. Отсюда в зависимости от
специфики организации экосистем формируются такие типы рас-
тений, как сфероидные, цилиндрические, стелющиеся, лианы и т. п.
Физиология наземного растения сложна, поэтому ее рассмот-
рение ограничено двумя аспектами: анализом особенностей вод ного
режима и движения растений.
188
7.1. ВОДООБМЕН И ПРИСПОСОБЛЕНИЯ РАСТЕНИЙ
В эволюционном плане наиболее изучена проблема водообмена
растений: способы добывания воды, механизмы ее передвижения,
отдачи и запасания, значение и роль воды в продуктивности и т. п.
Проблему разрабатывали применительно к растениям разных систе-
матических, филогенетических и экологических групп. Интерес к
ней связан со следующими причинами: 1) вода составляет большую
часть биомассы растений в наземных условиях (в том числе и по
мере усиления аридизации климата); 2) недостаток или избыток
воды быстро отражается на жизнедеятельности и проявляется в
морфологии (симптомы внешне легкоуловимы); 3) изучение водо-
обмена важно для решения практических задач земледелия —
регуляции урожайности возделываемых культур.
По этим и другим вопросам существует многочисленная лите-
ратура, в которой рассмотрены структурно-функциональные аспек-
ты водообмена растений. В курсе «Физиология растений» раздел
«Водный обмен» является одним из традиционных и разработанных.
Многие особенности жизнедеятельности растений, касающиеся
водообмена, уже получили эволюционную интерпретацию, соответ-
ствующие явления воспринимаются как само собой разумеющиеся
приспособления. С учетом сказанного изложение вопроса о приспо-
собительном значении особенностей водообмена растений ниже
дается конспективно.
Теперь уже не вызывает сомнения, что с выходом растений на
сушу произошла глубокая структурная и функциональная перестройка
организации и дальше она изменялась сообразно условиям обитания
конкретных видов. Нередко изменения видов носили параллельный и
конвергентный характер. Значительны и внутривидовые наследствен-
ные уклонения растений по водному обмену, что позволило выделить
экотипы по этому признаку (Г. Турресон, Б. А. Келлер и др.).
Изучение эволюции водообмена растений происходило по не-
скольким основным направлениям.
Классификация растений по экологическим типам. Пожалуй, в
исследованиях экологов впервые были выявлены приспособ-
ления растений к недостатку воды (Н. А. Базилевич и др., 1968;
К. В. Манойленко, 1974; В. В. Полевой, 1989), позволившие раз-
граничить растения по типам водообмена. В пределах наземных
выделены такие группы, как гигрофиты, мезофиты, обширная и
разнообразная по морфологии и физиологии группа ксерофи-
тов (А. Шимпер, Г. Вальтер, Е. Ф. Вотчал, В. Р. Заленский,
Н. А. Максимов, А. М. Алексеев, П. А. Генкель). В этих исследо-
ваниях получила развитие идея о приспособительном значении
различий в водообмене растений разных экологических групп
(А. Крафт и др., 1951). Характеристику особенностей водообмена
189
соответствующих групп растений проводили по комплексу призна-
ков: морфология организации корневой системы и надземных
органов, тип развития, интенсивность и продуктивность
транспирации, водоудерживающая способность тканей, способ-
ность переносить дефицит воды и т. п.
Такая комплексная характеристика не оставила сомнения в том,
что структурные и функциональные особенности, способствующие
повышению устойчивости к недостатку или избытку воды, взаимо-
дополняют друг друга (см. § 2.4) и одинаково важны для успеха в
борьбе за существование. Отбор растений был связан с оценкой
внутрипопуляционных различий в балансе водообмена. Выяс-
нилось, что один и тот же результат — устойчивость к недостатку
или избытку воды —достигался приобретением различных нов-
шеств. Так, не всегда устойчивость к дефициту воды сочетается с
ограничениями в транспирации или впадением растений в покой
при наступлении засушливого периода. Природа шла разными
путями к достижению приспособляемости. Как показал Н. А. Мак-
симов (1952), при этом растения прибегают или к уменьшению
транспирации, или к добыванию воды из глубоких слоев почвы,
или запасанию в тканях в период ее доступности. Активной или
пассивной регуляцией водообмена достигалось преодоление напря-
женной ситуации (Ф. Вент, 1972; К. Даддингтон, 1972).
Эволюция вегетативных органов и проводящей системы высших
растений. Изучение этого вопроса связано с дальнейшим углуб-
лением представлений об особенностях водообмена растений
различных экологических типов и групп. Поскольку среда обитания
водных и наземных растений принципиально отличается по эко-
логическому минимуму и оптимуму, постольку разные виды рас-
тений различаются и по степени дифференциации тела, и способам
водообмена. Именно в наземных условиях потребовалась система
тканей, предотвращающих тело растения от обезвоживания.
В наземных условиях вступает в противоречие необходимость
уменьшения потери воды с осуществлением фотосинтеза. Успеха в
завоевании суши достигли только формы, у которых удачно сочетались
фотосинтетическая деятельность (благодаря развитию листьев,
ассимилирующей ткани, клеток и органелл) с оптимальным балансом
водообмена (см. гл. 2, 3). Возникает разнообразие форм листьев,
особенно у покрытосеменных в результате нарастания их систе-
матического и экологического многообразия. Появление теломного
листа (J. В. Richardson, 1984) коренным образом изменило характер
ветвления и габитус наземных растений, что обеспечило дальнейший
их прогресс из-за увеличения ассимиляционной поверхности и
развития паренхимы. Образование крупных листьев в эволюции было
связано с такими ограничениями, как торможение закладки новых
физиологически обновленных листьев, рост боковых почек и побегов
190
и усиление потери воды (В. К. Василевская и др., 1985). Кроме
того, возникновение теломного листа способствовало диффе-
ренциации и паренхиматизации стебля — образованию прокамбия,
камбия, ксилемы, флоэмы, сосудов и т. п. Эволюция самого корня,
строение которого отвечает выполнению всасывающей и проводя-
щей функции (М. Ф. Данилова, 1974), также совпала с появлением
листьев. С паренхиматизацией стебля происходит появление все
новых и активных корней вблизи мест расположения листьев.
На примере изучения большого числа видов деревьев, кустар-
ников и кустарничков Средней Азии показано, что адаптация к
аридным условиям пустынь имеет комплексный характер и про-
исходит различными способами. Из-за возрастания засоленности
почв и сухости воздуха в среднеазиатских пустынях преобладают
виды с суккулентными листьями. Параллельная и конвергентная
эволюция разных видов растений шла в направлении сокращения
ассимилирующей и испаряющей поверхности. ПрИ этом компен-
сация ассимилирующих структур достигается развитием мелких
ксероморфных и цилиндрических листьев на брахипластах из
пазушных почек, а также быстрой дифференциацией тканей самих
листьев (А. А. Бутник, 3. А. Нигманова и др., 1991).
При далеко зашедшем процессе редукции листа (афилия) у
пустынных растений ассимилирующие функции принимает
первичная кора стебля, относящаяся к кранцевому и некранцевому
типу строения наподобие ассимилирующей ткани листьев. Аридные
растения накапливают также в тканях танниды, флавоноиды, смолы
и воски для защиты от инсоляции, развивают высокую сосущую
силу, осмотическое давление. У многих из них листья отличаются
наличием кранц-анатомйи (см. гл. 3), способны осуществлять
фотосинтез и поддерживать энергетику дыхания (О. А Семихатова,
1995) при высокой инсоляции и температуре. Фотосинтетический
аппарат пустынных и других растений (Т. К. Горышина, 1989) четко
реагирует на условия среды, претерпевает комплексные изменения
приспособительного характера к режиму освещения.
С выходом растений на сушу потребовалось создание надежной
водопроводящей системы. При сохранении общности организации
клетки (как универсальной системы поглощения веществ) в наземных
условиях происходит «макроморфологическая» эволюция в направ-
лении создания эффективных структурных и функциональных адап-
таций для поглощения, передвижения, запасания и отдачи воды (А.
А Яценко-Хмелевский, Л. А Лебеденко, 1985). Произошла не толь-
ко перестройка Покровной и проводящей систем, но и механической
ткани в целях поддержания вертикальной ориентации стебля и обес-
печения фотосинтеза и транспирации. Обеспечение нормального
водообмена растений в наземных условиях было достигнуто усовер-
шенствованием всех его этапов. Задача усложнялась еще и тем, что
191
растениям потребовалось совместить указанные процессы с потреб-
лением минеральных элементов. Поэтому возникла необходимость
формирования дополнительных структур в корн? и побеге: пропус-
кные клетки, поясок Каспари, водопроницаемые целлюлозные
оболочки, эндодерма, перицикл, трахеиды и т. п. (А. Е. Васильев,
Н. С. Воронин и др., 1978; М. Ф. Данилова, 1974).
Каждое из таких приобретений облегчало надежность водооб-
мена и минерального питания. Проводящие элементы в корне
закладываются дифференциацией прокамбия, занимающего цент-
ральное положение. В стебле из клеток прокамбия формируется
полый цилиндр. После разделения тела наземного растения на
корень и стебель наибольшие изменения отмечены в структуре
стебля. При стабильности структур, предназначенных для погло-
щения воды, в ходе эволюции возникают разнообразные типы
клеток корня и стебля, специализированные для проведения воды
по организму: кольчатые и спиральные трахеиды, ксилема
(первичная и вторичная), сосуды и их членики с разной перфо-
рацией, волокна либриформа и т. п. (см. гл. 2). В зависимости от
условий произрастания даже у представителей одной систематичес-
кой группы происходит заметная дивергенция по указанным струк-
турам, что имеет адаптивное значение. В случае же ослабления
напряженности водного режима упрощается структура водопрово-
дящей системы растений, например у трав и водных цветковых.
Значительного разнообразия достиг и устьичный аппарат (строение
замыкающих и окружающих клеток) при сохранении общности
механизма его функционирования.
Из сказанного видно, что вегетативная сфера у растений в назем-
ных условиях приобретает комплекс новшеств, благоприятных для
всех параметров водообмена. Некоторые из новшеств, однажды
появившись, сохраняются у всех растений, другие же исчезают и вновь
появляются (А А Яценко-Хмелевский, Л. А Лебеденко, 1985). В
случае перехода растений к другому образу жизни (травы, водные и
паразитические растения) отмечено упрощение вегетативной сферы
и водопроводящей системы высших растений.
Засухоустойчивость растений. Для углубления представлений о
приспособительной эволюции наземных растений значение имеют
и исследования по физиологии засухоустойчивости. Их проводили
с широким размахом на культурных и дикорастущих видах, что
позволило выявить взаимосвязь морфологических, анатомических,
биохимических и функциональных изменений в эволюции растений
(Н. А. Максимов, 1952; В. Ф. Альтергот, 1965; П. А. Генкель,
1982). В исследованиях по засухоустойчивости содержится
классификщщя способов приспособления видов к таким стрессо-
вым факторам, как перегрев, обезвоживание, почвенная и воздуш-
ная засуха на примере представителей почти всех отделов и классов,
В также характеристика границ устойчивости растений и способ-
ности переносить стрессы (К. В. Манойленко, 1983).
192
В результате разработана теория засухоустойчивости растений,
выяснены природа и механизмы устойчивости к засухе, предложены
подходы к улучшению селекционной работы по засухоустойчивости.
Засухоустойчивость — наследственное свойство, связанное с
наличием комплекса приспособлений, в том числе различий в ульт-
раструктуре и физико-химических показателях цитоплазмы. Все
подобные приобретения призваны поддержать оптимальный баланс
процессов синтеза и деструкции в условиях д лительного или кратков-
ременного дефицита воды или перегрева. При этом выявлены общие
и специфические черты приспособления растений к длительному
дефициту воды, а также связь процессов жаро- и засухоустойчивости.
Эволюция видов в условиях разной градации температуры и
влажности привела к отбору форм, обладающих способностью
переносить перегрев и обезвоживание тканей на основе разных
механизмов. Для цветковых характерна активная регуляция водо-
обмена с сохранением жизнеспособности организма, тогда как у
низших растений чаще всего складываются пассивные средства
защиты от обезвоживания — криптобиоз.
Исследования по засухоустойчивости растений внесли ясность
и в вопрос о проявлении рекапитуляции на функциональном уровне
(см. гл. 8). Подобные факты касаются устойчивости разных этапов
онтогенеза к недостатку воды («критические периоды»), последо-
вательности формирования способов транспирации и эко-
логических типов водообмена, таких, как гидрофиты, пойкило- и
гомеогидроидные (Н. И. Антипов, 1975; К. В. Манойленко, 1983).
Итак, изучение способов приспособления растений к наземным
условиям, а также направлений их дальнейшей дивергенции по
механизмам защиты от перегрева и обезвоживания дали до-
полнительный и богатый материал для познания происхождения
адаптации и общих закономерностей эволюции.
7.2. ДВИЖЕНИЯ РАСТЕНИЙ. СПОСОБЫ,
ПРЕДПОСЫЛКИ ВОЗНИКНОВЕНИЯ И ЗНАЧЕНИЕ
Способность растений к движениям давно привлекала внимание
ученых. Особенно оно усилилось после публикаций работ Ч. Дар-
вина «Движения и повадки лазающих растений» (1865), «Насеко-
моядные растения» (1875) и «Способность к движениям у растений»
(1880), в которых он рассмотрел широкий круг явлений из жизни
Растений под углом зрения их приспособительного значения. Он
также поставил проблему движений растений на эксперименталь-
ную основу, показал возможность их строгого анализа. Тем самым
Дарвинизм оказал плодотворное влияние на развитие исследований
в этой области (Н. Г. Холодный, 1941).
7-157 193
Под термином движение растений (фитодинамика) объединяют
разнородные по механизмам и значимости явления: движение
цитоплазмы и органоидов клетки, передвижение с помощью жгутиков
(монады, хлорококковые), рост растяжением, изменение положения
прорастающих семян, круговые нутации, тропизмы, ростовые и тур-
горнвде настии и т. п. (Э. Бюннинг, 1960; А. И. Меркис, 1973; Э.
Либберт, 1976; А. Гэлстон, и др., 1983; Ф. Уоринг, Ф. Филлипс,
1984). Их рассмотрение занимает большое место в учебнике
«Физиология растений» (В. В. Полевой, 1989), где они описаны не
только с точки зрения проявления и природы, но и происхождения
и эволюции.
Некоторые из названных форм движений характерны и для
животных (движение цитоплазмы и жгутиками), остальные встре-
чаются почти у всех растений. Однако в зависимости, от систе-
матического, филогенетического и ценотического положения видов
доминируют или круговые нутации, или тропизмы, или настии,
причем меняются способ и степень осуществления каждой из форм
и в онтогенезе. При доминировании одной из них другие не
исчезают, а сохраняются в виде дополнительных или в сочетаниях.
Универсальное значение для растений, особенно высших наземных,
имеют обратимые тургорные движения и настии. Сейсмонастии по
механизму близки к мышечным сокращениям (см. § 5.7).
Не останавливаясь на анализе механизмов разных форм
движений (В. В. Полевой, Т. С. Саламатова, 1985; В. В. Полевой,
1989), подчеркнем, что даже на клеточном уровне они далеко не
совпадают, не говоря уже об уровне органов и целостного растения.
В адаптивном плане разные способы движений также неравноцен-
ны. Некоторые из них (движение цитоплазмы и органоидов, кру-
говые нутации) имеют более общее значение, другие более
специфичны (лазающие, вьющиеся растения, сейсмо- и никто-
настии) в экологическом отношении.
Внимание Ч. Дарвина к движениям было вызвано и в связи с
возражением Ст. Д. Майварта о том, что принцип естественного
отбора не объясняет «начальные стадии полезных черт»1, в том числе
и у вьющихся растений. Эту способность растений Ч. Дарвин
объяснил как результат усиления под действием естественного
отбора «общей способности к движению, приобретенной ради
совершенно иных и полезных целей»1 2, которая не представляла
первоначально для них особой пользы. Далее он объясняет, что
видоизменением круговой нутации были «приобретены многие
весьма полезные и необходимые движения» растениями. Наблю-
дения над усиками Bignonia capreolata, Echinocystis lobata, Passiflora
1 Дарвин Ч. Происхождение видов. С.-Пб., 1991. С. 183.
2 Дарвин Ч. Соч. М.-Л., 1941. Т. 8. С. 202 и 503.
194
gracilis, Sicyos angulatus убедили Ч. Дарвина не только в высокой их
чувствительности, но и в специфике реакции на раздражитель.
Возможность возникновения специальных форм движений на
основе простой двигательной реакцци под действием естественного
отбора не является невероятной в случае их выгоды в борьбе за
существование. В сомкнутых фитоценозах (лес) при недостатке осве-
щения, в Поиске других ресурсов (воды, минеральных элементов и
т. п.) усилением зачаточного ростового движения в разных целях могут
сложиться вьющиеся и лазающие формы, различные способы
ориентации в пространстве (тропизмы и настии) или другие обратимые
движения у растений. В борьбе за существование путем параллелизмов
и конвергенции возникают сходные формы движений у предс-
тавителей различных семейств высших растений. Экологически
значимы нутации, настии, тропизмы, насекомояцность (В. Е. Tuniper
et al., 1989) для растений независимо от их систематического поло-
жения. Без них сложно представить стабильность растений. Несколько
труднее объяснить роль сейсмонастий у мимозы и ряда других рас-
тений, хотя рассматривают их как предотвращение повреждений от
воздействия сильных ветров и ливней.
Так или иначе способность к движениям — основа ориентации в
пространстве. Она важна и при прикрепленном к субстрату состоянии,
когда у растения нет возможности’ активно изменить положение по
отношению к односторонне действующему фактору. Поэтому почти
все формы д вижения растений представляют целесообразную реакцию
на среду (Н. Г. Холодный). Они повышают экологическую
стабильность растений, обеспечивая максимальное использование
среды путем движения органов и структур — «поисковая» активность
индивидуума. В связи с этим происходит усиление контроля целост-
ности организма над способностью к движениям растений через
механизмы гормональной регуляции и активности Н-помпы. Отбор
ведет к возникновению подобных механизмов. Заслуга Ч. Дарвина
состоит в объяснении экологической роли движений растений в связи
с действием естественного отбора.
* * *
В этой главе рассмотрены функциональные последствия выхода
растений на сушу на примере особенностей водного обмена и
способности к движениям. Несмотря на отличия сравниваемых
явлений, они имеют много общего. Благоприятный режим водооб-
мена является предпосылкой роста, а рост—предпосылкой
движений. Движения определяют активность поиска воды, мине-
ральных.элементов и света, а также ориентации в пространстве.
Особенности водного обмена и движений растений взаимодополняют
Друг друга в борьбе за существование. Поэтому в наземных условиях
У растений появляется комплекс приспособлений (см. гл. 8).
ГЛАВА 8
ЭВОЛЮЦИЯ ОНТОГЕНЕЗА РАСТЕНИЙ
Все сказанное выше о функциональной эволюции растений в
целом и о роли отдельно взятых функций имеет непосредственное
значение для онтогенеза. Адаптации нельзя представить вне конк-
ретного носителя, и таковым для функционального приспособления
является индивидуум растений с его онтогенезом. Функциональные
приспособления повышают надежность онтогенетического уровня,
а через него — и популяционно-видового.
Онтогенез (индивидуальное развитие) — срок жизни
индивидуума, комплекс его дифференцировок и реакций с момента
образования зиготы (или какого-либо другого зачатка) до полного
завершения жизненного цикла растения — основной объект изучения
ряда ботанических дисциплин. Онтогенез более или менее полно
изучен лишь у ограниченного числа цветковых растений, поэтому еще
недостаточно ясна и картина его эволюции, хотя этому вопросу
уделялось внимание (В. В. Скрипчинский, М. X. Чайлахян и др.).
Онтогенез не только результат филогенеза, но и его основа.
Онтогенетические дифференцировки имеют значение для филоге-
нетических событий — формирования новых таксонов, органов и
структур. Эволюция жизни тесно связана с преобразованием онто-
генеза, так как «развитие онтогенеза есть средство, которым была
достигнута эволюция новых, более высоких форм жизни»1.
Эволюция Онтогенеза — одна из проблем учения о макроэво-
люции. В этой связи интересно проследить роль микроэво-
люционных процессов в преобразовании онтогенеза растений, его
регуляторных механизмов. Онтогенез первично появляется на заре
жизни (А., Н. Северцов его появление связывал с колониальными
одноклеточными) и усложняется по мере ее эволюции.
8.1. ОБЩАЯ ХАРАКТЕРИСТИКА ОНТОГЕНЕЗА РАСТЕНИЙ
Число ныне живущих растений насчитывает около 500 тыс.
видов, из них 300 тыс.— цветковые. В указанное общее число
включаются и грибы, хотя в последние годы неоднократно выска-
зывалось мнение о необходимости выделения их в самостоятельное
царство.
Северцов А. Н. Собр. соч. М.-Л., 1949. Т. 5. С. 490. ।
196
Онтогенез растений, относящихся к разным таксонам, нерав-
нозначен: отличается по продолжительности, дифференцировкам,
темпам прохождения, специфике реагирования на внешние условия
и т. п. В связи с этим меняются и формы индивидуальности у
растений. По этой причине довольно условным становится срав-
нение онтогенеза у хлореллы, метасеквой и однолетнего злака,
одно- и многоклеточного растения. Но это не важно для харак-
теристики эволюции онтогенеза.
В эволюции онтогенеза растений можно проследить следующий
ряд изменений: онтогенез со сложными циклами и фазами превра-
щений (водоросли и грибы) -> преобладание диплофазы и регу-
лярное повторение вегетативных и генеративных тенденций
(Поликарпики) -> многократное повторение вегетативных и однок-
ратное повторение генеративных функций (монокарпики) -> еже-
годное обновление генеративных побегов (корневищные растения)
-> двулетники -+ однолетники -+ эфемеры. В пределах указанных
морфологических типов онтогенеза возникают различные
физиологические механизмы его реализации: автономное и
индуцированное развитие, фотопериодические и температурные
типы развития и т. д. Со сменой типа организации онтогенеза
преобразовывался и его носитель — индивидуум. В указанном ряду
развития онтогенеза целостность индивидуума усиливалась, он
вычленялся как функционально самостоятельная система — особь.
До сих пор нет единого подхода к периодизации онтогенеза
растений. Пожалуй, достичь единства взглядов в этом вопросе
трудно и периодизацию онтогенеза следует проводить сообразно
задачам и объекту исследования. У растений, как и у животных, в
онтогенезе можно выделить такие периоды, как эмбриональный,
ювенильный, активного роста, зрелости и старости. В онтогенезе
цветковых выделяют этапы: эмбриональный, молодость', зрелость,
размножение и старость (М. X. Чайлахян, Т. А. Работнов). У
низших растений в онтогенезе осуществляются сложные циклы
развития, чередование полового и бесполого поколений, т. е.
развитие по типу метаморфоза (рис. 24).
У высших растений происходит упрощение циклов развития, но
усиление степени дифференциации тела на метамерно повторя-
ющиеся органы. При этом старение разных структур не
синхронизировано. Так, у бурых водорослей осевые части и ризоиды
являются многолетними, а «листоподобные» части отпадают и
ежегодно нарастают. Среди грибов наравне с эфемерными формами
(Clavaria; Mucor, Penicillum), у которых отмирание разных структур
синхронизировано, встречаются и другие формы. Так, грибница
многих шляпочных грибов живет 15—25 лет, а в благоприятных
условиях —до 500 лет (Marasmius oreades), тогда как их плодовые
197
Рис., 24. Цикл развития улотрикса (М. В. Горленко, 1981. С. 126):
А —бесполое размножение, Б —половое воспроизведение; 1 —основная жизненная форма, 2
—образование зооспор, 3 —выходзооспор, 4 —пустая клетка, 5 —зооспоры, 6 —образование
гамет, 7—выход гамет, 8—изогамия, 9 —10—зигота, 11 —прорастание зиготы, 12 —зоос-
пора
тела — несколько месяцев. Многолетние плодовые тела характерны
для трутовиков (Forties fomentarius, Piptoporus betulinus), возраст ко-
торых достигает 25 лет. У мхов наибольшей продолжительностью
жизни обладает гаметофит, у папоротников — спорофит (кор-
невище). Независимо от систематического положения в целом
корневой системе присуща большая продолжительность жизни, чем
наземным структурам растений.
Эмбриональный период в онтогенезе растений более ограничен
дифференцировками, чем у животных. У растений эмбрион (заро-'
дыш семени) имеет упрошенное строение, особенно у
паразитических цветковых. В то же время высшие растения в
течение всей жизни сохраняют эмбриональные возможности —
образовательные ткани. Между прочим, это обстоятельство способ-
ствует повышению надежности онтогенеза растений в часто меня-
ющихся условиях, что обусловлено возможностью восстановления
целостности организма При повреждениях и повторения процессов
органообразования и развития.
198
Для онтогенеза растений (как и животных) характерна
последовательность дифференцировок. У семенных растений онто-
генез начинается с интенсивного роста корня, а затем рост корней
и надземной части идет взаимосвязанно. Закладка органов и темпы
роста определяются не только внешними условиями, но и
генетически. У цветковых растений генеративное развитие и рост
зависят от наличия благоприятных экологических условий,
специфических для разных видов.
Соответственно повторяемости дифференцировок и различиям
в формах индивидуальности не так прост вопрос о границах онто-
генеза разных растений. У многих растений со сложной системой
последовательно образующихся побегов возобновления и способ-
ных к вегетативному размножению трудно очертить границы
индивидуума и его онтогенеза. Индивидуум у таких форм состоит
из систем взаимосвязанных побегов, выступающих как части цело-
го. Лишь после отделения и перевода на самостоятельное состояние
такие побеги превращаются в новые индивидуумы. Для сложных
индивидуумов характерны малый (жизнь отдельного побега) и
большой (жизнь всего куста) жизненные циклы. При вегетативном
размножении у каждого дочернего поколения онтогенез начинается
заново, с его малыми и большими циклами.
8.2. ИНДИВИДУУМ У РАСТЕНИЙ
И ЕГО ФУНКЦИОНАЛЬНАЯ ЦЕЛОСТНОСТЬ
Понятие онтогенез тесно связано с представлениями об индиви-
дуальности. Все еще в силе остается утверждение, что одной из причин
слабой разработки проблем онтогении растений является отсутствие
ясности относительно границ их индивидуума (Б. М. Козо-По-
лянский, В. Л. Комаров). К сожалению, трудности определения
границ индивидуума у растений методически все еше не преодоле-
ны, особенно у организмов с бесполым размножением и со сложной
системой парциальных побегов. Индивид у растений — физически
обособленное и неделимое, физиологически целостное существо
(одно- или многокомпонентное), обладающее самостоятельным
онтогенезом и являющееся компонентом популяции.
При вегетативном размножении с образованием нового дочер-
него поколения начинается жизнь самостоятельного индивидуума.
Образование зарослей, кругоВин, кустов и т. д. у таких растений
является этапом становления сложного онтогенеза. Индивидуум у
Них при своем становлении проходит ряд этапов: почка -> побег ->
побег-сосунок -> побег, способный к самостоятельной жизни, ->
партикула -» самостоятельное растение (Т. А. Работнов). Поэто-
му, говоря об онтогенезе таких растений, необходимо иметь в виду
199
все эти этапы. К сожалению, фитофизиологи не приступили вплот-
ную к изучению онтогенеза таких сложных индивидуумов. Изучение
онтогенеза одно- и двулетних растений, хотя и оправдано мето-
дическими соображениями, явно недостаточно для познания зако-
номерностей эволюции онтогенеза растений.
У вегетативно размножающихся растений возрастает степень
сложности онтогенеза, хотя она не ведет к повышению уровня его
организованности (т. е. способности сохранять свою специфич-
ность и зарегулированное число признаков; В. Н. Беклемишев,
1964). Так, онтогенез, продолжающийся сотни лет и более, значи-
тельно сложнее по повторяемости циклов, чем у одно- и двулетних
растений. Но тем не менее по организации онтогенез последних не
только не уступает, но даже превосходит онтогенез многолетников.
Организация онтогенеза у растений возрастает по Пути олигоме-
ризации, с усилением целостности индивидуума и уплотнением его
дифференцировок (одно- й двулетние травы, эфемеры). Обособ-
ление индивидуума как неделимого и физиологически целостного
существа — магистральная линия эволюции растений.
Дифференциация индивида и его онтогенеза не могла бы иметь
значительного успеха, если бы она не сопровождалась процессами
интеграции. Интеграция выражена по-разному у современных рас-
тений, что связано с направлениями их соматической эволюции.
Поэтому растения, стоящие на одинаковой ступени филогенетичес-
кого развития, могут заметно отличаться по целостности организма
и его онтогенеза. Вообще экологическая эволюция накладывает
глубокий отпечаток на жизнедеятельность растений, маскируя
филогенетические отношения по биохимическим и функциональ-
ным показателям (К. М. Завадский), что следует иметь в виду при
исследованиях эволюционной физиологии растений.
Важно выяснить, что преобразуется в ходе эволюции: отдельно
взятая функция и структура или генотип в целом. Функциональная
эволюция растений прежде всего касается преобразования
индивидуума как целостной системы, его биохимической и морфо-
логической организации.
Продемонстрируем сказанное на примерах. Фотосинтез как
физиологическое явление закрепляется в интересах процветания
популяции и вида, а затем лишь жизни в целом. Это достигалось
путем преимущественного размножения особей (фенотипов), отли-
чающихся способностью накапливать избыточный продукт, необ-
ходимый для выживания и воспроизведения потомства в условиях
обеднения среда готовыми органическими веществами абиогенно-
го происхождения. Поэтому уместно говорить не только об эволю-
ции первичных механизмов фототрофности, но и о возникновении
системного контроля их реализации. Только это могло обеспечить
200
}
растениям планетарную функцию. В. Н. Любименко (1963) писал,
что «с биологической точки зрения фотосинтез есть прежде всего
физиологическая функция организма, ему подчиненная и им уп-
равляемая». С этой мыслью перекликаются современные взгляды на
эндогенную регуляцию фотосинтеза через изменение донорно-акцеп-
торных отношений в целом организме — концепция source-sink. Нам
более или менее известны механизмы этого явления и уточнены типы
донорно-акцепторных отношений (А. Т. Мокроносов, 1983).
Физико-химические реакции фотосинтеза у цианей и цветковых
Почти не отличаются между собой. Но на организменном уровне
фотосинтез имеет разные возможности для Протекания. У цианей
изменение интенсивности фотосинтеза в онтогенезе менее значитель-
но, чем у цветковых растений. На начальных этапах онтогенеза у
цветковых, когда требуется опережающий рост корневой системы,
по активности фотосинтеза доминирующими являются листья
нижнего яруса, расположенные ближе к корням. По мере роста и
развития доминирующее положение переходит к листьям вышерас-
положенных ярусов, а в фазе формирования генеративных органов
и плода таковыми являются самые верхние (молодые) листья. В то
же время и продуктивность фотосинтеза целого растения достигает
своего максимума в фазе генеративного развития. Следовательно,
наличие эволюционно сложившегося системного контроля приво-
дит к регуляции интенсивности фотосинтеза в онтогенезе как
отдельных листьев, так и растения в целом. Сказанное является
одним из следствий эволюции фотосинтеза —отбора по совокуп-
ности признаков, обеспечивающих наибольшую продуктивность
индивидуума.
Результаты селекции показывают, что более 500 лет в принципе
не менялся механизм фотосинтеза у пшеницы, хотя заметно повы-
шалась продуктивность благодаря отбору растений (выведение сор-
тов) по структуре индивидуумов, сочетанию фотосинтеза с
процессами роста, устойчивостью и т. д. Точно так же и в эволюции
имело значение, в какой системе оказывались «вмонтированными»
первичные процессы фотосинтеза. Доказательств тому много (см.
гл. 3)..Речь идет о подчинении его энергетического механизма
эволюции индивидуальности.
Отсюда следует вывод: эволюционируют не химические процес-
сы в организме, а совокупность индивидуумов, у которых эти
процессы наилучшим образом благоприятствуют воспроизведению
потомства. Здесь речь идет не о противопоставлении процесса его
носителю (организму), а о разграничении объекта (фенотипа) и
точки приложения (реакции) отбора. Это принципиально для
понимания механизма действия естественного отбора и результатов
функциональной эволюции.
201
Именно с указанных позиций должна быть оценена и диффе-
ренциация растений С3- и С4-пути, САМ и промежуточные пути
фотосинтеза. По значимости отдельных реакций фотосинтеза
подобная дифференциация видов не имеет никакого смысла. Она
важна в аспекте популяционно-видового уровня, когда необходим
наиболее устойчивый и функционально целостный индивидуум в
определенных условиях. Это же служит причиной дифференциации
растений по типам устьиц (10 типов), водорослей по структуре тела,
усиления геофилии видов в суровых условиях Арктики и т. д. Все
подобные изменения накладывают отпечаток на функциональную
дивергенцию популяций и видов.
Точно так же отбор имеет дело не с генами, а с их носителями —
фенотипами. Через выживание организмов с определенными
генами и признаками идет преобразование структуры и физиологии
индивидуума, популяций и видов. Поэтому в зависимости от
кариотипа организма изменчивость в поколениях может иметь
различный размах. Показано, что уменьшение числа хромосом
ограничивает возникновение мутаций и комбинаторику генов, тогда
как наличие большого числа хромосом в ядре усиливает эти про-
цессы в поколениях. В то же время меньшее число хромосом
предрасполагает к возникновению полиплоидных форм-рядов
(В. Грант, 1984). Генетическая система эукариот, характеризующа-
яся ростом числа генов, усиливает изменчивость за счет их
дупликаций и комбинаторики, возникновения избыточной повто-
ряемости последовательностей ДНК. Зато у высших эукариот по
сравнению с прокариотами частота кроссинговера в-10 000 раз
оказывается меньше (Н. П. Дубинин, 1985).
Общий результат эволюции —усиление дифференциации
жизни — проявляется на молекулярном, субклеточном, клеточном,
организменном и надорганизменном уровнях, Примером тому слу-
жат и различия в функции органелл, клеток, тканей и индивидуумов.
На индивидуальном уровне дивергенция может быть показана и на
примере иммунитета. Иммунитет представляет собой комплекс
новшеств, обеспечивающих устойчивость индивидуума к патоген-
ному началу. По представлениям Н. И. Вавилова, иммунитет рас-
тений следует рассматривать как «сумму из множества слагаемых».
Эта точка зрения сыграла положительную роль в развитии совре-
менной фитоиммунологии (Л. В. Метлицкий, О. Л. Озерецков-
ская, 1985). Отбор растений на устойчивость к патогенам идет не
по иммунным свойствам клеток, скорее подобные особенности
клеток меняются в связи с их значимостью для размножения и
выживания целого организма как единой функциональной системы.
В результате вырабатывается комплекс приспособлений, охра-
няющих функциональную и структурную целостность индивидуума
у растений. Среди них следует указать на наличие специальных
202
веществ для распознавания и уничтожения патогена,
конституционные и индуцированные механизмы защиты,
механические барьеры, связывание — обездвиживание паразита
(лектинами), блокирование и инактивирование токсинов и экзо-
ферментов паразита, ограничение синтеза в тканях растений ве-
ществ (используемых паразитом для своего роста и размножения),
реакция сверхчувствительности (гибель пораженных клеток вместе
с инфекцией) и т. п. Названные и другие приспособления —
результат долгой сопряженной эволюции растения и паразита. Они
проявляются в разном сочетании не только видов, но и сортов, и
штаммов.
Приведенные соображения о функциональной эволюции подт-
верждают одно из основных положений теории эволюции об отборе
генотипов через фенотипы. При этом только индивидуум как
целостная система меняется в ходе эволюции популяций и видов
(И. И. Шмальгаузен). Иммунитет прежде всего необходим для
поддержания функциональной целостности индивидуума, а через
него и для усиления конкурентоспособности популяций. Сказанное
делает понятным и изменение форм иммунитета организмов разных
таксонов в зависимости от уровня их эволюционного положения.
Растения и животные, низшие и высшие, одно- и многоклеточные
отличаются по защитным механизмам.
Структурная эволюция растений была определена необ-
ходимостью осуществления определенных функций (В. О. Каза-
рян, 1990). Так, тканевая организация имела значение для
повышения эффективности фотосинтеза (см. гл. 3), усиления пог-
лотительной функции и размножения организма (см. гл. 2). Рас-
членение тела вверх по оси способствовало возникновению
иерархии органов и взаимосвязи между ними. Вверх по оси растений
происходит как бы фильтрация ионов и других веществ. Благодаря
способности корней накапливать избыточные ионы уменьшается
их возможное отрицательное влияние на листья, цветки и заро-
дыши. Сложна структура хлоропластов, в которых одновременно
представлены и функционируют разные блоки —фотосинтез,
депонирование запасов и генетический аппарат (В. Е. Семененко,
1985). Она усложнялась в ходе эволюции для усиления
функциональной целостности растения.
Выше на примере развития фотосинтеза и иммунитета показана
приуроченность функциональной эволюции к индивидуальному
уровню, что имеет значение для размножения популяций. Инди-
видуум в ходе эволюции популяций и видов меняется как
функциональная целостная система. Совокупность таких индивиду-
умов и, позволяет популяции выполнять экологические функции,
играть роль «в экономике природы».
203
8.3. ЭВОЛЮЦИЯ ПРОДОЛЖИТЕЛЬНОСТИ ЖИЗНИ ИНДИВИДУУМА
Представления об онтогенезе связаны с необходимостью конк-
ретизировать продолжительность жизни (ПЖ) организма, т. е. с
определением индивидуальности растений. Здесь можно выделить
следующие особенности: 1) растения отличаются от животных
большей ПЖ, несмотря на ее вариации от нескольких недель до
, сотен и тысяч лет; 2) ПЖ индивидуума не является таксо-
номическим признаком, она различна даже в пределах популяций
и сортов; 3) в ходе эволюции растений наблюдается тенденция
сокращения ПЖ индивидуума, что происходит конвергентно в
разных таксонах; 4) индивидуум многолетних растений отличается
мозаичностью старения структур; 5) ПЖ индивидуума и органов
растений, хотя варьирует в широких пределах, регулируется эндо-
генно; 6) старение И отмирание индивидуума — этап онтогенеза
любого растения в экологически оптимальных условиях; 7) старение
и Отмирание органов не обязательно ведут к прерыванию индивиду-
альности растений.
Многие вопросы старения и ПЖ растений остаются
дискуссионными, несмотря на давность их постановки (Г. Молищ,
1929; Н. П. Кренке, 1950; А. Г. Юсуфов, 1992). Нередко и сейчас
дискутируется вопрос о бессмертии индивидуума у растений, что
отражает трудности в определении его границ при вегетативном
размножении. Не вдаваясь в подобные дискуссии, попытаемся
рассмотреть ПЖ и старение растений с эволюционных позиций.
Напомним, что развитие старения у всех растений связано с
ослаблением синтетических процессов тканей и клеток, падением
активности участвующих в нем ферментов, постепенной детрадацией
хлорофилла, белков и других биополимеров, накоплением инертных
веществ, ингибиторов роста и этилена, а также повышением активности
гидролитических процессов (Л. Леопольд, 1968; О. Н. Кулаева, 1973;
Т. С. Саламатова, 1983; К. Mothes, 1960; К. V. Thhnann, 1980;
Н. W. Woolhouse, 1984 и др.). Все это отражается (или протекает
параллельно) на морфологии индивидуума: замедление и остановка
роста, отмирание отдельных структур и скелетных элементов,
суховершинность и дуплистость й т. п. (В. О. Казарян, 1964;
П. И. Гупало, 1968). В принципе, развитие старческого процесса
у большей части высших растений протекает однотипно, различаясь
только в темпах. У эфемеров, одно- и двулетних растений, одно-
летних побегов многолетних трав, многолетних монокарпических
форм старческие изменения носят взрывной и необратимый харак-
тер с наступлением цветения и плодоношения (Г- Молиш, 1933). У
поликарпиков многолетних старение индивидуума растянуто во
времени и протекает постепенно. В Природе не всегда ограничение
ПЖ связано со старением индивидуума. Тем не Менее в эко-
204
логически оптимальных ситуациях старение и ПЖ индивидуума
выступают как норма реакции генотипа, т. е. генетически конт-
ролируются (Н. П. Кренке, 1950).
Поэтому важно выяснить, в чем эволюционный смысл старения
индивидуума, в связи с чем оно развивается, каковы причины
мозаичности старения органов в онтогенезе и дивергенции растений
по ПЖ индивидуума. Велись дискуссий о возможности закрепления
процесса старения естественным отбором (А. Комфорт, 1967).
Теперь очевидно, что это явление само по себе не могло быть точкой
приложения отбора. Ч. Дарвин (1939) рассматривал действие естес-
твенного отбора на долголетие опосредованно, через обеспечение
успеха в размножении.
Старение неотвратимо развивается как процесс, специфичный
для индивидуумов и органов растений по темпам и формам прояв-
ления. Даже у изолированных органов реализуется эндогенный
контроль старения, что тем более осуществляется у целого растения
и его интактных органов (А. Г. Юсуфов, 1992). Так, у изолирован-
ных листьев, семядолей, цветков и лепестков даже после развития
корней проявляются различия в ПЖ, они также специфичны для
разных видов растений. Целостное же растение состоит из комп-
лекса взаимодействующих молодых и старых структур, соотношение
которых определяется общим возрастом индивидуума (Н. П. Крен-
ке, 1950; И. И. Дубровицкая, 1961). Нарастание паренхиматизаций,
наблюдаемое у части растений и их органов в онто- и филогенезе,
сопровождается интенсификацией процессов жизнедеятельности и
сокращением ПЖ индивидуума. Между ПЖ и интенсификацией
жизнедеятельности организма обнаружена связь, реализуемая
усилением генеративных тенденций индивидуума. В этом аспекте
понятна и положительная связь между скороспелостью и
интенсивностью жизнедеятельности у культурных растений. Темпы
старения индивидуума и мозаичность старения его органов
возникают в ходе эволюции как результат отбора по репродуктивной
активности. Мозаичность старения органов — результат различий
в эволюционной специализации. По этой причине наиболее жестко
проявляется эндогенный контроль в старении цветков и лепестков.
Проявление мозаичности старения у индивидуума растений
связано с метамерностью органов и сложной пространственной и
временной расчлененностью тела. Метамерность и повторяемость
органов растения заметно снижаются в эволюционном ряду от
деревьев к однолетним травам. В этом же ряду усиливается и
синхронность старения метамерных структур, органов и
индивидуума в целом. Причиной этого является возрастание реп-
родуктивной активности благодаря необходимости «поспешного
цветения» и повышению соотношения генеративных структур.
205
В этом направлении шла эволюция растений в условиях
аридизации и похолодания климата. Сезонные и годовые резкие
колебания климата способствовали сокращению ПЖ индивидуума
и регулярному новообразованию одних и утрате других метамерных
структур. Это имело значение для повышения эффективности
использования кратковременных благоприятных условий для соз-
дания максимального органического продукта, необходимого для
полной и быстрой реализации генеративных тенденций. Один из
наиболее эффективных путей ее достижения —стабильное преобла-
дание в метамерной серии активно функционирующих молодых и
зрелых фотосинтезирующих структур (в том. числе и листьев). Такое
состояние могло быть достигнуто только при отборе особей в поп-
уляциях на частую сменяемость листьев и других структур по годам,
сезонам и этапам морфогенеза^ Коррелятивные отношения между
органами и определяют последовательность их закладки и старения.
8.4. РАЗВИТИЕ ГОМЕОСТАЗА В СВЯЗИ С ЭВОЛЮЦИЕЙ ОНТОГЕНЕЗА
Внутренняя среда любого организма—основа его жизнедея-
тельности и нормального онтогенеза, способность к ее поддер-
жанию имеет важное значение для выживания и процветания,
популяций. Она содержит воду, все запасные (углеводы, жиры,
белки, алкалоиды, дубильные вещества и т. п.) и конституционные
(постоянно входящие в состав цитоплазмы и клеточных стенок)
вещества, все химические элементы, органические кислоты, регу-
ляторы роста и т. д. Внутренняя среда разных растений различна,
меняется с возрастом и условиями, что и затрудняет ее точную
оценку. Преобладает в ней вода; которая составляет более 70%
массы растения. Особенно много воды в созревающих плодах и
жизненнЬ активных органах. Содержание большего количества
воды и сходство химического состава организмов с составом водной
оболочки планеты указывают на первичное зарождение жизни в
водной среде (В. И. Вернадский, 1967).
Как бы организмы не отличались по составу внутренней среды,
необходимым условием их существования является поддержание
его постоянства —гомеостаз, или гомеостазис (К. Бернар, 1878; У.
Кэнон, 1932). Этим мы обязаны исторически сложившимся регу-
ляторным механизмам, поддерживающим «систему в определенном
состоянии» (И. И. Шмальгаузен, 1964).
Характеристика гомеостаза занимает важное место в литературе
по физиологии животных, свидетельствуя о температурной, водно-
солевой, ионной и других формах гомеостаза и подчеркивая при
этом значение конкретных явлений для обеспечения нормальной
жизни. В то же время у растений механизмы гомеостаза слабо
изучены.
206 ,
' Несмотря на это, есть достаточные основания для признания
их’ значимости и в жизни растений. Так, накоплен достаточно
богатый материал, характеризующий наличие внутренней регу-
ляции водообмена, фотосинтеза и других процессов. В частности,
у некоторых растений (трескан, полынь, джузгун и др.) в течение 1
сут водный режим меняется 5—8 раз. В критические периоды
инсоляции для предотвращения иссушения у них увеличиваются
концентрация клеточного сока и осмотическое давление, что соп-
ровождается снижением транспирации листьев и стабилизацией
обменных процессов, усилением синтеза АБК и закрытием устьиц.
Изменение содержания солей в почве по-разному сказывается
на гомеостазе растений. На Mesembryanihemum crystallinum показано,
что при увеличении в почве концентрации NaCl содержание Na1- и
С1— в экстракте из листьев растений достигало соответственно 500
и 400 мМ. Однако в хлоропластах содержание Na+ не превышало
160—230 и СП 40—60 мМ. При содержании 20 и 400 мМ NaCl в
среде отношение Na+/K+ в хлоропластах составило соответственно
1/5. С повышением засоленности почвы содержание пролина в
экстрактах листьев (особенно в хлороПластах) возрастало, что имело
значение для осморегуляции.
Регуляция метаболизма транспортом и потреблением продук-
тов — пример гомеостаза и эндогенной регуляции процессов
(А. Л. Курсанов, 1986). Изменение в одном из звеньев механизма
транспорта, возникающее в результате разных воздействий, обычно
устраняется эндогенной регуляцией его механизма. Весьма показа-
телен в этом отношении и синтез АТФ, потребности" в котором
клетка покрывает с учетом его траты на дыхание. Немаловажна роль
и вакуоли в поддержании гомеостаза клетки.
Одним из доказательств наличия гомеостаза у растений служит
избирательное поступление катионов и анионов из почвы в корень
и растение, распределение их по органам. На примере поступления
NO3 показана его зависимость от концентрации Cl-, NHJ и
аминокислот в тканях. Все это зависит от содержания углеводов,
процесса дыхания и фотосинтеза в организме растения. Градиент
содержания веществ вдоль оси растения подвижен, происходит его
перемещение от стареющих, органов к новым путем «перекачки»
веществ из нижних к верхним, из вегетативных к генеративным
органам. Особенно велика роль указанных процессов при
ограниченном поступлении веществ из окружающей среды. Для
конкретизации этого положения нелишне еще раз напомнить и об
использовании воды, углекислоты и кислорода, образующихся в
растениях в процессах метаболизма, когда растение испытывает
недостаток их поступления извне. Если днем в тканях листа при
закрытых устьицах не хватает СО2, наблюдается отщепление его из
207
органических кислот и использование для фотосинтеза. Ночью же
при открытости устьиц и достаточном поступлении СО2 идет его
запасание в органических кислотах. Точно так же недостающие
белки у растений образуются из продуктов распада запасных белков.
Кроме того, в стрессовых условиях идет активный распад белков в
тканях растений, но отравление образующимся при этом аммиаком
предотвращается вовлечением его в новые синтезы.
Только благодаря срабатыванию механизмов поддержания го-
меостаза достигается жизнеобеспечение растений при заносе их на
ядовитые отвалы рудников. Известны растения-концентраторы
алюминия, молибдена, никеля, свинца, стронция и т. д., даже при
низком их содержании в почве (В. И. Вернадский). При этом
отравление тканей предотвращается синтезом специфических бел-
ков (определяющих устойчивость к высоким концентрациям метал-
лов), изменением количества и качества корневых выделений^
связыванием тяжелых металлов в клетках дубильными веществами
и органическими кислотами и т. д. Опыты показали, чФо до опре-
деленной концентрации в среде ионов металлов растения осущест-
вляют генетически контролируемый их транспорт из корней в
надземную часть, продолжая нормально развиваться. Если же кон-
центрация ионов оказывается очень высокой, защитные механизмы
не срабатывают и ионы тяжелых металлов, беспрепятственно пос-
тупая в растение, вызывают его отравление.
Установлено, что и обычные растения нередко оказываются спо-
собны произрастать на ядовитых отвалах рудников. В последующем
отбор среди таких растений уже по мутациям, определяющим
устойчивость к тяжелым металлам, ведет к формированию истинно
толерантных к ним популяций.
Разные галофиты, как известно, отличаются по механизму
поддержания гомеостаза. Так, способность произрастать на засо-
ленных почвах бывает обусловлена или накоплением в организме
солей в большом количестве (солянки), или выделением их листь-
ями и побегами наружу через специальные железы (кермек,
тамарикс), или ограничением поступления солей в организм
(полыни). Соответственно этому и механизм поддержания гомеостаза
по водному балансу у них не одинаков. У одних галофитов поддер-
жание гомеостаза связано с накоплением «непитательных» солей
(доленакапливающие) и повышением осмотических свойств у клеток
тканей, что создает возможность им поглощать воду из засоленного
почвенного раствора (Б. А. Келлер, 1980). У других галофитов (корни
которых непроницаемы для солей) водообеспечение достигается на-
коплением осмотически активных органических кислот и углеводов,.
У солевыводящих форм соли используются для создания осмотичес-
кого давления, затем по мере поступления их вместе с водным током
лишние ионы выводятся наружу через выделительные железки,
предотвращая отравление тканей ионами металлов.
208
Среди культурных растений неизвестны типичные галофиты, но
тем не менее они отличаются по степени солеустойчивости.
Наибольшая устойчивость из них присуща хлопчатнику, люцерне,
свекле, арбузу, доннику (они могут накапливать соли в тканях),
менее солевыносливы гречиха, кукуруза, лен, овес, пшеница и
табак. Вредное действие засоления на культурные растения прояв-
ляется не только в ухудшении их водообмена, но и в накоплении
токсических продуктов промежуточного обмена (диамина, када-
верина, путресцина и т. д.). Для синтеза указанных ядовитых ве-
ществ используются аминокислоты лизин, аргинин и глутаминовая
кислота, накопление которых связано с нарушением белкового
обмена. С увеличением концентрации солей в тканях снижается
активность диаминоксидазы, благодаря чему накапливаются
диамины ввиду блокады превращения их в алкалоиды.
У гликофитов (неустойчивых к засолению) на засоленных поч-
вах также развиваются некоторые адаптивные черты в метаболизме -
и росте, накапливаются углеводы, органические кислоты, амиды,
нуклеиновые кислоты и т. п. В конечном счете жизнеспособность
гликофитов в условиях засоления определяется сохранением
стабильности обмена путем накопления защитных веществ,
развития водоудерживающих сил и сосущей силы-клеток, эконом-
ной траты воды и образования в клетках большого числа пиноцитоз-
ных пузырьков (что ведет к выключению солей из обмена и
предохраняет ферменты от их токсического действия). В противном
случае происходит прогрессирующее обесцвечивание листьев,
отмирание участками пластинки листа (некрозы), увядание и оста-
новка роста (отмирание точек роста) растений.
Механизмы, используемые высшими растениями для поддер-
жания гомеостаза, разнообразны и не исчерпываются описанными
(А. Г. Юсуфов, 1983). Но несомненно одно: они все исторически
закреплены, направлены на повышение устойчивости организма в
онтогенезе и успеха в воспроизведении потомства. Словом, развива-
ется мощная регуляторная система онтогенеза, обеспечивающая его
устойчивость.
8.5. РЕГУЛЯТОРНАЯ СИСТЕМА ИНДИВИДУУМА,
ЕЕ РАЗВИТИЕ И ЗНАЧЕНИЕ
Проблема целостности растительного организма, с точки зрения
эволюции онтогенеза, еще не стала предметом пристального
внимания ботаников. Однако необходимость ее разработки уже
понята (М. X. Чайлахян, 1968; Н. П. Аксенова, Т. Б. Баврина,
Т. Н. Константинова, 1973; А. Г. Юсуфов, 1976; В. В. Полевой,
1982). Выделяют различные системы регуляции многоклеточных
растений, описывать которые нет необходимости. Заметим, что
209
общее направление развития регуляторной системы онтогенеза
растений шло от ионных взаимодействий на субклеточном и кле-
точном уровнях к ионным, трофическим, биоэлектрическим и
гормональным взаимодействиям на уровне тканей и органов.
Действие каждого из них проявляется постоянно внутри клетки
(ионный обмен) и дистанционно (трофические, биоэлектрические
и гормональные связи). В то же время первичный механизм
трофических, биоэлектрических и гормональных взаимодействий
также проявляется через обмен ионов на поверхности мембран,
возникновением ионных градиентов и мембранных потенциалов.
С усложнением организма и нарастанием его связей с внешней
средой регуляторная система онтогенеза растений дополняется
взаимодействием органов и метамерных структур. Здесь важную
роль играют механизмы транспорта веществ и проведения биоэ-
лектрических импульсов, а также структурная организация самого
растения (индивидуума). По этой причине у индивидуумов разных
видов и экологических групп растений система перемещения в них
даже ассимилятов оказывается неодинаково эффективной, хотя в
целом скорость самого транспорта для разных видов колеблется в
пределах 50—80 см ♦ ч—1 (А. Л. Курсанов, 1976).
Эволюция в направлении редукции размера тела, усиления диф-
ференциации проводящей системы и репродуктивной активности
индивидуума, а также сокращения жизненного цикла способство-
вала увеличению эффективности системы регуляции за счет укоро-
чения расстояния транспорта веществ, ускорения круговорота
метаболитов и повышения «запроса» к ним. Среди прочих причин,
способствовавших развитию регуляторной системы индивидуума в
ходе эволюции^ следует указать на значение стабилизации путей
синтеза, рецепции и транспорта гормонов (см. гл. 5), повышения
устойчивости физиологических процессов к неблагоприятным
условиям и т. п. Одновременно подобные изменения сыграли роль
в автономизации и усилении целостности индивидуума. Отсюда
путь к автономизации онтогенеза лежит через развитие регулятор-
ной системы индивидуума (И. И. Шмальгаузен, 1964).
Биологическое значение указанных явлений можно понять, если
принять во внимание, что онтогенез растений протекает при слож-
ных ситуациях, при циклических колебаниях условий по годам,
сезонам и в течение суток. Растениям в онтогенезе приходится
переносить недостаток воды, затенение, пониженные (включая и
отрицательные) и повышенные температуры, болезни и потравы, а
также преодолевать механические препятствия. Поэтому исто-
рически у растений возникают механизмы, предотвращающие
отрицательные эффекты подобных воздействий. Без них нельзя
представить жизнь организма, они объединяются под общим на-
званием устойчивость индивидуума.
210
Устойчивостью организма принято называть его способность
переносить внешние факторы, потенциально оказывающие вредное
действие на жизнедеятельность. Устойчивость растений имеет не-
посредственное отношение к проблемам функциональной эво-
люции, и ее изучение важно для понимания природы и механизма
приспособления видов, разработки мер для повышения про-
дуктивности культурных и природных фитоценозов.
Как известно, любой фактор среды (биотический и абиоти-
ческий) имеет свои пределы изменчивости, бывает выражен в
минимуме, оптимуме й максимуме. Указанные три точки не совпа-
дают для разных видов даже по одному фактору, т. е. виды и сорта
растений отличаются по физиологическому нулю. Отсюда и неодина-
ковая для них норма.
Понятие стресс было введено в медицину канадским ученым
Г. Селье (1972, 1979). Он обозначил агент, вызывающий стресс,
понятием стрессор, а возникающий на него ответ—понятием
общий адаптационный синдром. Последним обозначают совокуп-
ность стереотипных реакций организма на разные раздражители.
Применительно к растениям предложен термин фитостресс —
реакция организма на любые отклонения от нормы (П. А. Генкель,
1982).
Число стрессовых факторов чрезвычайно разнообразно. Услов-
но их делят на биотические (инфекции, биоповреждения, борьба с
другими существами) и физико-химические — температура (низкая
и высокая), вода (дефицит и избыток), радиация (инфракрасные,
ультрафиолетовые лучи и т. п.), химические агенты (ионы, газы,
гербициды и др.). Сообразно типам стресса различают и формы
устойчивости: иммунитет, способность к регенерации, засухоус-
тойчивость, жароустойчивость, солеустойчивость, газоустойчивость
и т.' п. Так, в отношении приспособления к низким температурам
принято делить растения на зимо-, морозо- и холодоустойчивые. В
экологическом плане одна и та же доза фактора может оказаться
стрессом для одних видов, оптимальной —для других, малозна-
чительной —для третьих. Например, пустынные условия (высокая
температура, инсоляция, недостаток воды) оказывают отрицательное
влияние на многие растения, тогда как для суккулентов они стали
нормой жизни. Некоторые растения при наступлении засухи пересы-
хают до такого состояния (мхи, лишайники, Carex, Ramondia и др.),
что их можно растереть в порошок. Однако жизнеспособность к ним
возвращается после выпадения осадков—дождей, когда их ткани
насыщаются водой. Другие растения погибают даже при меньших
потерях воды (водные лютик и кувшинки, томаты и др.). Все зависит
от истории развития вида в определенных условиях, при которых
отбор привел к выработке соответствующей нормы для вида.
211
Стрессы постоянно сопутствовали растениям в ходе их эво-
люции. Как уже указывалось, вскоре после зарождения жизни
организмы своим быстрым размножением создали для себя стрес-
совые условия, истощая запасы органической пищи (см. гл. 3). Вслед
за преодолением стресса выработкой механизма фотосинтеза
произошло ухудшение газовой ситуации на поверхности Земли
благодаря постоянному обогащению атмосферы кислородом. Мы
знаем, к каким последствиям это привело (см. гл. 4).
В сложной (многофакторной) среде обычно переплетаются раз-
нообразные стрессовые ситуации. Так, выход растений на сушу
потребовал от них выработки способов защиты от инсоляции,
низкой влажности воздуха и почвы, ветров, вредителей и т. п.
Естественно, в этой сложной обстановке в процессе Отбора растения
выработали комплекс новшеств, обеспечивающих выживаемость в
новых условиях. Гравитация Земли и направление падения света на
ее поверхность потребовали от растения выработки строгой
ориентаций в пространстве —устремление корней в грунт, а побе-
га — в сторону света. Ориентация в пространстве особенно необ-
ходимой стала в наземных условиях, где жизнь растений оказалась
' возможной только в прикрепленном состоянии. В водной среде в
ней не было столь большой потребности (за исключением донных
и прибрежных водорослей) из-за перемещения организма вместе с
движением веды. Поэтому постоянное действие силы земного
притяжения и света в определенном направлении способствовало
закреплению механизмов гео- и фототропизмов, полярности, так
как Указанные факторы стали предпосылкой морфогенеза и
выживаемости в онтогенезе.
Каковы же общие представления о функциональной ус-
тойчивости растений? Прежде всего допускают, что устойчивость у
растений может быть обусловлена избеганием неблагоприятных
факторов и толерантностью. Эволюционно наиболее прогрессивной
признана вторая форма устойчивости, так как при этом растения
сохраняют активность в борьбе за существование. В экологическом
же плане обе формы устойчивости ведут к процветанию видов (к
биологическому прогрессу).
Избегание связано с прекращением активной жизнедеятель-
ности растений в период неблагоприятных условий. Так, большая
экологическая группа растений-эфемеров, относящихся к
различным семействам и обитающих в условиях пустынь и сухих
склонов,, отличается короткой продолжительностью жизни. Они
избегают стрессовых периодов «поспешным развитием» в начале
весны или осенью, когда в почве достаточно влаги. Эфемеры не
исключения. Многие растения в состоянии покоя могут переносить
очень низкие температуры, длительные обезвоживания. Так, нос-
212
ток, пролежавший в гербарии в течение 107 лет, оказался способным
вегетировать при перенесении во влажные условия.
. Описан случай оживления тотема, хранившегося в одном из
индийских храмов. Тотем, вырезанный из толстого ствола березы
и вкопанный в землю около 100 лет назад, вдруг пустил зеленые
ростки. Специалисты квалифицировали это как редкостный фено-
мен, а индейцы —как чародейство. Споры некоторых мхов оста-
ются живыми в гербариях в воздушно-сухом состоянии в течение
50 лет и после этого способны прорастать. Семена лотоса, 2000 лет
пролежавшие в иле, сохранили всхожесть. Семена ряда мотылько-
вых, мальвовых, губоцветных и другие сохраняют всхожесть в сухом
виде более 50 лет. Семена многих растений, даже тропических,
переносят низкие отрицательные температуры в несколько десятков
градусов в сухом состоянии; так, семена кукурузы выдерживают
охлаждение до —90°С. , ,
Избегание стрессовых периодов путем впадания в физио-
логический (глубокий) или вынужденный (временный) покой
широко распространено у низших й высших растений и закреплено
как приспособительное свойство. Подобное состояние, нередко
обозначаемое как анабиоз (или криптобиоз), основано на глубокой
перестройке жизнедеятельности организма или отдельных органов
(семян, луковиц, спор, корневищ). При этом растения или отдель-
ные органы впадают в состояние «высокой пассивной устойчивости
к неблагоприятным условиям» (П. А. Генкель, 1982). Устойчивость
путем избегания возникла в ходе эволюции, при отсутствии более
активных защитных реакций организма на стресс.
Толерантность связана с сохранением жизнедеятельности при
наступлении стрессовых условий. Наиболее типичными примерами
могут служить Ксерофиты, суккуленты, галофиты и гигрофиты.
Растения, относящиеся к этим экологическим группам, отличаются
комплексом наследственно закрепленных морфофизйологических
особенностей, позволяющих переносить им недостаток воды (ксе-
рофиты и суккуленты), избыток солей (галофиты) или воды
(гигрофиты), причем они могут развиваться И в других режимах, но
выносят напряженные условия, не снижая продуктивности. .В
типичной для них естественной обстановке у указанных групп
растений в онтогенезе реализуется соответствующая норма как
результат отбора и приспособления к недостатку или избытку воды
и к повышенному содержанию солей И pH почвы.
Как ведут себя растения в онтогенезе при попадании в
нетипичные для них условия? Срабатывают механизмы,
понижающие отрицательные последствия стрессовых воздействий,
а именно; меняется активность мембранных процессов,, ряда фер-
ментов и белоксинтезирующей системы, происходит деполимери-
зация полисом, усиление связывания ядовитых веществ и активности
213
репарационных механизмов и т. д. Разумеется, направление изме-
нения в метаболизме во многом зависит от специфики воздейству-
ющего неблагоприятного фактора. Такое явление имеет место, когда
условия меняются постепенно. При резком же изменении напряжен-
ности факторов (суховеи, мгла, возрастание pH почвы или в эк-
сперименте при летальных воздействиях) реакция организма отстает
от давления мощного фактора, что приводит к необратимым повреж-
дениям тканей и органов. Исторически такие мощные повреждающие
факторы лишь временами оказывали влияние на растительные
организмы. Поэтому отбор не мог содействовать выработке соответ-
ствующих механизмов для успешной борьбы с ними.
Виды растений отличаются по степени толерантности к отдель-
ным неблагоприятным факторам или комплексу, что исторически
обусловлено. Толерантность.—сложное свойство, зависящее от
различных генов, передается по наследству в ряду поколений. Это
показано и в отношении болезнеустойчивости-растений (Я. Ван-
дерпланк, 1981). Даже при кратковременном изменении условий в
тканях растений появляются метаболические сдвиги, направленные
на нормализацию жизнедеятельности (привыкание). Такие изме-
нения могут носить известную направленность и по-разному про-
являться у наследственно устойчивых и неустойчивых форм. В
отношении неустойчивых форм рекомендуется проводить пред-
варительное закаливание семян, проростков и растений к небла-
гоприятным условиям. Эффект закаливания не передается по
наследству и постепенно исчезает в онтогенезе. Но в физиологичес-
ком отношении он важен для преодоления отрицательного воз-
действия неблагоприятного фактора в короткий период времени.
Мечтой селекционеров остается выведение сортов, комплексно
устойчивых к ряду неблагоприятных факторов среды. Вероятность
создания идеального сора мала, так как и в природе нет таких
идеальных форм. Устойчивость как проявление целесообразности
для любого вида относительна. Речь может идти лишь о защите
организма и популяций от каких-либо стрессов включением
механизмов гомеостаза на индивидуальном или популяционном
уровне. В защитных механизмах организма исключительно велика
роль гормональной системы растений, которая включает механизмы
синтеза, рецепции, инактивации и передвижения гормонов (см. гл.
5). Стрессы (засуха, неблагоприятные температуры, затопления,
засоления и т. д.) оказывают влияние на изменение количества и
соотношения гормонов и ингибиторов роста в тканях, что способ-
ствует эффективной регуляции роста и развития, траты веществ.
Все перечисленные выше механизмы повышают надежность онто-
генеза в неблагоприятных условиях, они важны для понимания
тенденций и направлений функциональной эволюции растений, а
также особенностей их изменения в будущем.
214
8.6. НАДЕЖНОСТЬ ОНТОГЕНЕЗА РАСТЕНИЙ И НЕКОТОРЫЕ
ТЕНДЕНЦИИ ИХ ФУНКЦИОНАЛЬНОЙ ЭВОЛЮЦИИ
Онтогенез — средство для достижения высших форм жизни в
ходе эволюции (А. Н. Северцов), необходимое для стабилизации
дифференцировок индивидуума. Эти события привели к повы-
шению устойчивости и надежности самого онтогенетического уровня.
Понятие надежность, заимствованное из кибернетики и техники,
нашло применение и в биологии. Введение указанного термина в
биологию не лишено значения для применения кибернетического
аппарата при объяснении явлений живой природы. В то же время
очевидна и необходимость сужения объема самого понятия, опре-
деления сферы его использования. О надежности онтогенеза следует
говорить в смысле его организованности, т. е. способности сохра-
нять свою специфику в виде зарегулированного числа признаков
(В. Н. Беклемишев, 1964). А это есть не что иное, как проявление
целостности и уплотнения дифференцировок индивидуума. Сте-
пени сложности и организованности онтогенеза не выступают как
синонимы. При высокой сложности онтогенеза его организован-
ность может оказаться недостаточной (см. выше).
Характерной чертой онтогенеза, определившей устойчивость
жизни, является наличие жесткой программы реализации его
дифференцировок и этапов развития. Структура онтогенеза
генетически детерминирована. И вопрос о соотношении наследст-
венности и условий развития (включая и внутренние условия) в
онтогенезе остается дискуссионным. Очевидно, что с упрощением
циклов развития, усилением бионтности и уплотнением диффе-
ренцировок происходит повышение роли внутренней среды в са-
морегуляции процессов морфогенезу и целостности онтогенеза
(эфемеры и эфемероиды, одно- и двулетние растения). У низших
растений, обладающих более слабой саморегуляцией морфогенеза,
зависимость хода онтогенеза от внешних условий чрезвычайно велика.
В связи с высказанными замечаниями под надежностью онто-
генеза следует подразумевать направленность и устойчивость его
основных дифференцировок и этапов развития. Все это в конечном
итоге зависит от генетической детерминации и развития регулятор-
ной системы. Генетическая детерминация индивидуального
развития очевидна у всех живых существ, даже у самых
примитивных, хотя она имеет различный временной аспект реали-
зации и зависит от степени развития регуляторной системы (гормо-
нальные и трофические взаимодействия, регенеративные процессы,
иммунный аппарат, механизмы гомеостаза, способы воспроизве-
дения и т. д.). Отсюда надежность конструкции организма выража-
ется в наличии аппарата наследственности, механизмов молекулярной
215
репарации, множественности и повторяемости процессов и струк-
тур, физиологической уравновешенности. Хотя растения отлича-
ются наличием упрощенной системы механизмов регуляции, тем
не менее регуляция у них развита (В. В. Полевой, 1982; А. Г.
Юсуфов, 1976) и выражается в высокой онтогенетической
лабильности, метамерности развития и наличии потенций к реге-
нерации.
Повышение надежности онтогенеза у растений — результат
возникновения комплекса приспособлений, органов и функций в
ходе эволюции. При однообразии условий (водная среда) расчле-
ненность организма слабо выражена, как у водорослей, или упро-
щается, как у водных высших растений. По этой же причине
оказались слабо дифференцированными первые наземные рас-
тения —риниофиты. Они не обладали четко обозначенными веге-
тативными органами, развитой проводящей системой, упорядоченным
ростом и т. п. Все это отразилось на особенностях их физиологии и
приспособленности. Усложнение среды в наземных условиях спо-
собствовало дальнейшему усилению вектора отбора на диффе-
ренциацию тела. Благодаря этому функциональная эволюция была
связана с поиском эффективных способов использования
первичных механизмов (сх. 10). ,
Схема 10
Онтогенетическая ' Экологическая
Филогенетическая
Дифференцировка
Поиск путей для эффективной
реализации механизма
t
Первичные метаболические
механизмы
Приведенная схема применима к любым функциональным
приобретениям и свидетельствует о дальнейшем углублении
физиологической дивергенции видов на основе единства мета-
болических механизмов, структурного усложнения онтогенеза и
морфологической дифференцировки индивидуума. На примере эво-
люции фотосинтеза (сх. 11) сказанное может быть представлено в
виде дифференциации как самого индивидуума на органы, так и
видов по совершенствованию некоторых звеньев механизма фотот-
рофности. . 1
216
Схема 11
С3-путь Сд-путь САМ Редукция процессов
Экологическая дифференциация
t
Дифференциация индивидуума
Эукариотическая клетка
Первичные реакции фотосинтеза
(прокариоты)
Предпосылки фототрофности
Так, уже на ранних этапах жизни отмечены стабилизация
растительного типа обмена и развитие механизмов фототрофности
и аэробности. Они оказали влияние на повышение энергетики
клеток и жизни в целом. А это, в свою очередь, способствовало
возникновению многоклеточности; следовательно, указанные
механизмы явились эрогенными приобретениями в истории живой
природы и стали предпосылкой для ускорения темпов эволюции
биосферы в целом (см. гл. 4, 5). Задача дальнейшего прогрессивного
развития жизни не могла быть решена, оставаясь на уровне про-
кариот, так как их энергетаческие ресурсы весьма ограничены.
Эволюция на усиление дифференциации привела к увеличению
разнообразия органелл и заметному повышению энергетики из-за
устранения конкуренции за электроны. В последующем возмож-
ное™ повышения энергетики в рамках одной клетки были исчер-
паны и отбор пошел в направлении развития многоклеточное™,
дифференциации организма на структуры, способные синтези-
ровать органическое вещество и структуры другого назначения. В
то же время происходила дифференциация биогеоценоза путем
поддержания определенного соотношения между продуцентами и
редуцентами. Таким путем складывалась энергетаческая основа
онтогенеза, необходимая для обеспечения его дифференцировок.
Естественно, что возникновение любых новшеств связано как
с изменением структуры (аппарата фотосинтеза), так и совершен-
ствованием функций (реакций фотосинтеза). При этом допустамы
несколько возможностей (см. § 2.3) и они реализовались при
становлении механизмов фотосинтеза, например при отборе глав-
ного пигмента и поиске способов его эффекшвного использования.
Это же видно и на примере молекулы АТФ, гормонов, мембран и
т. д. (см. гл. 3).
217
Функциональная эволюция растений связана с параллельным
преобразованием органов и функций. Предпосылкой для этого
служат также мультифункциональность и возможность количест-
венного изменения любой исходной функции (см. гл. 2). Подобные
события ведут к стабилизации процессов дифференциации в онто-
генезе (сх. 12).
Схема Г2
Стабилизация процессов
дифференциации
f
Автономизация
Стабилизация Компенсация
Усиление Ослабление
Множественность путей
реализации процесса
f
Первичные механизмы
Сходным образом происходит развитие механизмов аэробного
дыхания и гормональной регуляции; Ослабление какой-либо
функции (например, поглотительной деятельности корней или
редукция листьев) связано с возникновением компенсирующих
механизмов, что в конечном итоге также обеспечивает стабильность
процессов дифференциации и метаболизма растений (растения-
паразиты, суккуленты, ослабление опорной роли механической
ткани и т. п.). В итоге общая картина повышения надежности
онтогенеза растений носит ступенчатый характер (см. § 2.3) и
направлена на усиление структурно-функциональных особенностей
организма и развитие регуляторной системы онтогенеза. На завер-
шающей стадии эволюции регуляторной системы онтогенеза
возникают гормональные механизмы, влияющие на продук-
тивность, устойчивость (приспособленность) и воспроизведение
потомства (обновление онтогенеза). В совокупности эти процессы
определяют экологическую уравновешенность растительного орга-
низма и устойчивость его индивидуального развития. Отсюда на-
дежность онтогенеза — результат функциональной эволюции
растений. -
В то же время повышение сложности индивидуума и диффе-
ренцировок онтогенеза при переходе к наземным условиям у рас-
218
тений оказало положительное влияние на темпы функциональной
эволюции. Все это было бы невозможно без усиления меристе-
матизации индивидуума —развития первичных и вторичных
меристем, олигомеризации меристематической активности.
Меристематизация тела, создавая избыточность структур, имела
значение для завершения онтогенеза в условиях неподвижного
образа жизни и при частых повреждениях органов и тканей. Пов-
реждение части структур при их избыточности не влечет за собой
нарушения нормального хода онтогенеза (см. выше).
Дифференциация онтогенеза и индивидуума не имела бы
значительного успеха, если бы не сопровождалась интеграцией
процессов морфогенеза. Интеграция же онтогенеза выражена по-
разному у современных растений и зависит от направления их
соматической эволюции. Поэтому растения, стоящие на одинако-
вой ступени филогенетического развития, могут заметно отличаться
по целостности индивидуума и онтогенеза. Все это связано с
направлением эволюции самого онтогенеза.
8.7. ОСНОВНЫЕ НАПРАВЛЕНИЯ ЭВОЛЮЦИИ
ОНТОГЕНЕЗА РАСТЕНИЙ
Данный вопрос перекликается с предыдущим. Здесь будут поды-
тожены общие особенности эволюции онтогенеза, характерные для
высших растений. При этом выделены некоторые общие направ-
ления, которые привели к коренным изменениям самого онтогенеза
и его носителя — индивидуума. Ввиду того что пока не представ-
ляется возможным с достаточной полнотой описать пути эволюции
онтогенеза в ряду от одноклеточных до цветковых, речь может идти
только о некоторых из них.
Упрощение циклов развития и усиление дифференциации онтоге-
неза. Эволюция жизни на Земле связана с пробами и ошибками,
поисками удачных решений. И онтогенез в этом отношении не
исключение. У растений эволюционно более примитивным явля-
ется онтогенез, осуществляющийся путем сложных циклов с чере-
дованием изоморфного (ульва и диктиота) и гетероморфного
(ламинария) поколений. У грибов онтогенез также протекает раз-
нообразно, особенно с изменением условий обитания и способа
питания. У части растений онтогенез осуществляется без чередо-
вания поколений, но со сложной сменой ядерных фаз (фукус).
Чередование поколений характерно для онтогенеза мхов и папо-
ротников.
Онтогенез со сложной сменой циклов развития на ранних этапах
эволюции жизни становится одним из условий, компенсирующих
недостаток дифференциации организма. Известно, что диффе-
, 219
ренциация играет положительную роль в повышении устойчивости
организма к условиям среды. Когда организм не дифференцирован
(низшие растения), смена сложных циклов развития выступает как
приспособление для протекания онтогенеза в колеблющихся
условиях. Сложные циклы возникают, как правило, в тех условиях,
Когда ни один из них недостаточен для завершения развития, когда
частые колебания условий нарушают ход отдельных циклов. В
совокупности же эти циклы обеспечивают ход онтогенеза.
Параллельно с усилением дифференцировок в онтогенезе шло
упрощение циклов развития индивидуума. Это хорошо заметно при
переходе от низших к высшим растениям., У цветковых оно
выразилось в доминировании не только спорофита, но и однолет*
ности. Параллельно с этим происходит усиление листестебельных
тенденций и энергоемкости организма. Диплоидия не только
«защитила индивидуальное развитие от возникающих мутаций, но
и способствовала усилению соматических дифференцировок»
(Н. П. Дубинин). Поэтому не, случайно, что с преобладанием в
жизни спорофита усиливается соматическое разнообразие рас-
тений.
Дифференциация онтогенеза хорошо заметна у наземных рас-
тений. С переходом к наземным условиям отмечено постепенное
преобладание спорофита, усиление разнообразия тканей и органов,
а также повышение функциональной целостности индивидуума у
растений (см. выше).
Олигомеризация и интеграция онтогенеза. Олигомеризация го-
мологичных структур имела важное значение не только для эво-
люции животных, но и растений (В. А. Догель, 1954). Данные об
олигомеризации растений чаще всего касались генеративной сферы.
Представляет значительный интерес анализ недостаточно изучен-
ного вопроса об олигомеризации соматической сферы растений.
Известно, что олигомеризации предшествовали процессы полиме-
ризации, когда развитие обеспечивалось увеличением числа одноз-
начных, но функционально еще слабых структур. Одним из
примеров последнего является дифференциация тела на метамерно
повторяющиеся органы и образование индивидуума со сложной
иерархией побегов возобновления (рис. 25). Например, образование
колоний индивидуумов у водорослей, мхов и лишайников, с раз-
ветвленной грибницей у грибов, а также деревьев, кустарников и
многолетних трав с множеством парциальных побегов следует
рассматривать как пример все усиливающейся полимеризации рас-
тений в ходе эволюции. Здесь мы. встречаемся с образованием !
колоний путем своеобразного воссоединения «элементарных
организмов» (А. С. Фаминцын).
Последовательные процессы полимеризации привели к
возникновению древесной формы семенных растений. Процессы
полимеризации имели значение для увеличения размеров тела.
220
д
е
Рис. 25. Эволюция онтогенеза жизненных форм (А. П. Хохряков, 1975. С. 8,
12):
а —б—деревья, в—аэроксильные кустарники, г—геоксильные кустарники, д—кло-
нообразуюшие кустарники, е —клонообразующие травы
усиления приспособленности к наземным условиям, повышения
продуктивности и воспроизводительной активности растений, хотя
не всегда между этими процессами существует параллелизм.
На определенном этапе эволюции растений начинается преоб-
ладание процессов олигомеризации, т. е. редукции размеров тела с
утратой гомологичных структур, усиления функциональной
активности организма и целостности онтогенеза. Этим процессам
221
предшествует постепенное уменьшение числа, размеров и про-
должительности жизни парциальных побегов на дереве и кусте.
Словом, происходит усиление, «спрессовывание» блоков бывшей
разветвленной системы. Процессы полимеризации явились
следствием усиления эмбрионизации онтогенеза. С олигомериза-
цией происходит обрывание надставочных и виргинальных стадий
онтогенеза.
Повышение сложности индивидуума и дифференцировок в
онтогенезе у растений в наземных условиях выступало как результат
возрастания меристематизации индивидуума — развития первич-
ных и вторичных образовательных тканей. У низших растений
меристематическая активность не локализована, они растут всей
поверхностью. В ходе эволюции происходит олигомеризация
меристематической активности, что приводит к концентрации
меристем в определенных местах тела и увеличению общей их
мощности (нарастание размеров меристем и темпов диффе-
ренциации) при одновременном сохранении тотипотентности кле-
ток и других тканей. Меристематизация тела, создавая избыточность
структур, имела значение для завершения онтогенеза в условиях
неподвижного образа жизни и при частых повреждениях. Повреж-
дение части структур при их избыточности не влечет за собой
нарушения нормального хода онтогенеза.
Ускорение темпов развития в онтогенезе. Интеграция онтогенеза
имела далеко идущие последствия, одним из них является ускорение
темпов индивидуального развития — акцелерация. Акцелерацйя
онтогенеза в ходе эволюции достигалась неоднократно и
различными путями. Акцелерация онтогенеза, идущая без глубокой
дифференциации (одноклеточные) и с дифференциацией тела на
структуры (многоклеточные), имеет различные последствия.
Эволюционно более совершенной является акцелерация в онтоге-
незе многоклеточных, так как она привела к комплексной
интенсификации жизнедеятельности организма.
Акцелерация онтогенеза у растений в ходе эволюции достига-
лась путем сокращения продолжительности жизни или полного
выпадения начальных и более специализированных взрослых
стадий. Так, в эмбриональном периоде имело место сокращение
сроков развития зародыша и проростка при дивергенции цветковых
растений и в пределах жизненных форм от деревьев к травам
(L. Diels, 1906). Имеется тенденция сокращения сроков прорас-
тания семян и периода покоя у проростков (с некоторыми исклю-
чениями). Выражением этой тенденции является и живорождение,
образование «деток», в особенности с ухудшением условий, и
полное отсутствие физиологического покоя у многих растений.
Для быстроты завершения онтогенеза имела значение и тен-
денция перехода от подземного к надземному прорастанию семян
222
(А. П. Хохряков, 1975). Акцелерация у растений в ходе эволюции
связана с сокращением ряда этапов жизненного цикла и ускорением
смены скелетных образований. При этом наблюдается убыстренный
переход побегов в пределах кроны к генеративному, развитию с
последующим их отмиранием в ряду трансформации жизненных
форм по основным и боковым направлениям (И. Г. Серебряков,
1962). Ускорение онтогенеза происходит и путем выпадения конеч-
ных стадий — явление ярусной неотении при возникновении трав
из деревьев (А. Л. Тахтаджян, 1964).
Ускорение онтогенеза связано с соотбором растений на интен-
сификацию физиологических процессов. Поэтому характерной чер-
той травянистых растений по сравнению с древесными является
высокий энергетический уровень. Травянистые растения за единицу
времени и единицу поверхности накапливают больше продуктов,
чем древесные. У первых доля фотосинтезирующих структур в
пределах организма выше, чем у вторых. Это соотношение у кустар-
ников также больше, чему деревьев. Отсюда различия в их биомассе,
ежегодном приросте и продуктивности. Прирост в березняках со-
ставляет 5,4%, в тропических лесах — 6,6, саваннах — 28,4, в степях
— 35—55, эфемерных пустынях — 76% (Л. Е. Родин, Н. И. Ба-
зилевич, 1965).
Даже в пределах сортов одной культуры скороспелые харак- •
теризуются более высокой интенсивностью физиологических про-
цессов, чем позднеспелые. Поэтому возникновение скороспелости
нередко сопряжено с соотбором на высокую интенсивность жизне-
деятельности ц усиление роста в начальных этапах развития. Уско-
рение темпов развития в онтогенезе способствует повышению
относительной независимости организма от среды — авто-
номизации онтогенеза. '
Автономизация онтогенеза. Этот процесс выступает как общее
направление эволюции жизни (И. И. Шмальгаузен). Накоплен не-
который материал, характеризующий особенности автономизации
онтогенеза у растений (В. В. Скрипчинский, 1977; А. Г. Юсуфов,
1979). При автономизации онтогенеза возрастает роль внутренних
факторов развития, а внешние имеют сигнальное значение для
включения первых. Это положение может показаться несколько
парадоксальным, принимая во внимание большое значение внеш-
них факторов для развития растений. Между тем хорошо известные
факты о восприятии фотопериодического ритма растением и листом
только с достижением определенного' возраста, о подготовке к
зимнему покою растений задолго до наступления заморозков, про-
явлении эндогенных ритмов и периодичности роста растений даже
в постоянных и благоприятных условиях (в тропиках, фитотроне),
а также о возможности перехода многих растений к цветению и
морфогенезу при наличии лишь минимальной выраженности бла-
223
гоприятных экологических условий, усилении роли целостности
организма в развитии —далеко не полные доказательства авто-
номизации онтогенеза растений. Заслуживают внимания и данные
о повышении энергетического уровня ферментов в горных условиях
и стабилизации биохимически^ процессов в организме в колеб-
лющихся условиях (А. В. Благовещенский, 1966).
Автономизация онтогенеза растет в направлении от низших к
высшим растениям; наиболее отчетливо она выражена у моно-
карпических (одно- и двулетние, эфемеры). На примере эфеме-
роидных геофитов {Allium ursinum,.Anemone caucasid, Arum maculatum,
Hyacinthus orientalis, Omithogalum areuatum и др.) показано, что они
могут обходиться в онтогенезе без построения значительного
фотосинтезирующего аппарата за счет готовых резервов в кор-
невищах, клубнях и луковицах. К тому же продукты фотосинтеза,
вырабатываемые в листьях После пробуждения к росту зачатка,
используются лишь на завершающихся этапах онтогенеза, в основ-
ном для создания запасающих органов следующего поколения. У
указанных растений уже в луковицах закладываются зачатки соц-
ветий, и этот процесс происходит даже при температуре 2—6°С,
когда рост практически не идет (В. В. СкрипчиНский, 1972). Не
только снабжение вегетативных зачатков и зародышей семян запас-
ными веществами, но и развитие специальных покровов семян,
луковиц, клубней, корневищ и прчек обеспечивает защиту нового
организма от неблагоприятных условий.
Для эфемеров характерна способность к росту и развитию при
наличии благоприятной влажности и температуры в определенные
сезоны года. Несмотря на это, у них также выражена автоматизация
онтогенеза, следствием чего является очень быстрое прохождение
фаз развития при наличии минимальных условий. У них довольно
интенсивно протекают физиологические процессы. После запуска
системы (т. е. прорастания семян) максимально используются про-
дукты ассимиляции на воспроизведение потомства. Поэтому уже
отклонения условий от нормы в последующем не оказывают сколь-
ко-нибудь заметного влияния на исход онтогенеза.
Автономизация онтогенеза проявляется и у других растений,
хотя и в разной степени. Чтобы уяснить ее масштабы, укажем на
факты: глубокий покой семян и почек, способность растений
закладывать соцветия и плоды после образования определенного
числа листьев, строгая последовательность морфогенеза и всегда
равное число заложившихся структур, способность многих растений
обходиться запасами воды и минеральных веществ, содержащихся
в семенах, вегетативных зачатках и т. д.
Результаты автономизации онтогенеза закрепляются наследст-
венно как важные приспособления. При автономизации онтогенеза
внешние факторы не играют решающей роли в развитии и морфо-
224
генезё, а служат лишь как пусковые механизмы. Поясним примером.
У водяного лютика Ranunculus delphinifolius и стрелолиста Sagittaria
sagittifolia в зависимости от степени погружения растений в воду
формируются «водные» или «воздушные» листья, причем реакция
растений несколько запаздывает. Формирование разных листьев опре-
деляется не столько затоплением растений сколько их затенением.
Такая способность присуща и некоторым другим растениям (Oenanthe
aquatica, Campanularotundifolia, Limnophyllaheterophylla). Формирование
разных типов листьев с изменением условий является проявлением
нормы реагирования генотипа (И. И. Шмальгаузен).
Приведенные данные свидетельствуют о том, что даже при
наличии экологического контроля развития онтогенез растений
характеризуется жесткостью программы. Он регулируется внут-
ренними факторами и осуществляется при минимальном соот-
ветствии условий наследственной норме. Наличие внутренней
регуляции онтогенеза видно и при анализе роли явлений поляр-
ности и корреляций в морфогенезе и цветении растений. Авто-
номизацию онтогенеза следует рассматривать как важный результат
эволюции, приведший к стабилизации дифференцировок
организма и автономному контролю цветения.
8.8. РЕКАПИТУЛЯЦИЯ ПРИЗНАКОВ И ФУНКЦИЙ
В ОНТОГЕНЕЗЕ РАСТЕНИЙ
Вопрос о повторении признаков предков в онтогенезе —:
рекапитуляция — является одним из классических и дискуссионных
в биологии. Особенно, подчеркивал возможность рекапитуляции
признаков в онтогенезе Э. Геккель (1940). Возражая ему, У. Гарстанг
(1922) указывал, что онтогенез не повторяет, а творит филогенез
(Э. Н. Мирзоян, 1974). Положение о рекапитуляции признаков
применительно к растениям еще недостаточно обсуждено. Сказан-
ное справедливо в отношении как мррфологической, так и в
особенности функциональной организации растений. Ввиду отсут-
ствия зародышевых листков и выраженности метамерной
организации и повторяемости развития рекапитуляция у растений
осуществляется иначе, чем у животных (Б. М. Козо-Полянский,
1937, А. Л. Тахтаджян, 1954, 1964). В то же время палингенезы
(повторяющиеся предковые признаки) у растений почти совпадают
с ценогенезами (эмбриональными приспособлениями).
Несмотря на указанные ограничения, примеры рекапитуляции
известны и у растений. Так, все высшие растения начинают жизнь
с одноклеточной стадии и проходят нитевидную фазу при образо-
вании зародыша, что принято рассматривать как повторение пред-
ковых состояний — одноклеточных и нитчатых водорослей (рис.
Л-157 225
ff
Рис. 26. Схема развития зародыша спорофита моркови с повторением
нитевидной формы (Б. М. Козо-Полянский, 1937. С. 56):
1 — 6 —фазы развития
Рис. 27. Рекапитуляция признаков у растений (И. И. Шмальгаузен, 1969. С. 243,
370)
слева —сеянец акации с разными типами листьев; листья палеозойских (вверху) и современных
(внизу) папоротников со сходным ветвлением жилок и листовой пластинки; первые листья у
зародышей папоротников (в середине)
26). Молодые проростки и конус нарастания у высших растений до
фазы расчленения на органы напоминают слоевищных предков.
Известно много примеров, когда первичные листья отличаются от
нормальных и нижние листья на главном побеге по форме уклоня-
ются к предковым состояниям (рис. 27). Например, первые листья
у современных папоротников имеют дихотомическое ветвление, как
у их палеозойских предков и псилофитов; первые листья у зароды-
Рис. 28. Морфологическое сходство у видов солодок: голой (а), уральской (б) и
бухарской (в) как пример повторения ранних стадий развития (И. Т. Васильченко,
1965. С. 22)
227
шей папоротников формируются цельные. При развитии сеянцев
ксерофитов в затененных условиях наблюдается возврат к предко-
вым состояниям: по строению формируются листья теневого типа.
' • Поражают факты исключительного сходства проростков разных
растений й постепенное их расхождение по мере роста в онтогенезе
(рис. 28). Известны примеры увеличения числа семядолей у цвет-
ковых, что вообще характерно для голосеменных. В анатомическом
плане представляет интерес более простая организация проводяще-
го аппарата у проростков цветковых как результат уклонения к
предкам. Вообще древние .филогенетические стадии проводящей
системы сохраняются в нижней части оси побега. Юношеские
листья имеют более примитивные пучки, чем позднейшие. Парные
проводящие пучки в семядолях у Ranales, Rhoeadalespaw^r^^vyt
как примеры повторения дефинитивной (взрослой) стадии древних
голосеменных. Точно так же наличие камбия в основании
эпикотиля у проростков некоторых однодольных (чего нет у взрос-
лых форм!) служит примером рекапитуляции предковых стадий с
открытыми пучками. У многих цветковых сосуды древесины ближе к
центру (т. е. заложившиеся раньше) имеют лестничную перфорацию,
тогда как к периферии —простую. Эта смена соответствует палеон-
тологической последовательности изменения строения сосудов.
Рекапитуляция функциональных особенностей растений мало
изучена, хотя имеются суждения о довольно широком ее распрос-
транении (Ф. Д. Сказкин, 1961; П. А. Генкель, 1982). Нередко,
говоря о рекапитуляции функций растений, ограничивались указа-
нием на консервативность наследственности, последовательность
прохождения определенных фаз и стадий в онтогенезе, что в
принципе неверно. Известно, что развитие жизни шло от гетерот-
рофного К автотрофному способу. В связи с этим гетеротрофное
питание у формирующегося и прорастающего зачатка может быть
рассмотрено как рекапитуляция первичных стадий развития жизни.
Такому же объяснению подлежит тенденция к анаэробному
дыханию на начальных этапах онтогенеза у высших растений —
при прорастании семян и начальных этапах деления клеток точек
роста. В период гетеротрофного роста внутренняя газовая среда у
современных растений сходна с газовой средой Древней атмосферы
Земли, обедненной кислородом и обогащенной углекислотой, По
этой же причине в онтогенезе растений широко проявляется спо-
собность к темновой фиксации СО2.
В отношении процессов водообмена известно, что эволюционно
наиболее древней является кутикулярная транспирация, затем появ-
ляется устьичная (Н. ,И. Антипов, 1975). Соответственно этому в
онтогенезе даже цветковых наблюдается переход от кутикулярной
к устьичной транспирации. Как пример рекапитуляции указывают
на чрезвычайную чувствительность цветковых растений к неДостат-
228 .
ку воды в период гаметогенеза и оплодотворения (Ф. Д. Сказкин,
1961). Чувствительность данной фазы объясняется происхождением
цветковых от архегониальных, у которых оплодотворение обычно
осуществляется в водной среде.
Некоторые из современных растений в процессах фотосинтеза
обнаруживают родственные связи с фоторедукцией бактерий. Так,
ряд представителей синезеленых, Диатомовых и зеленых водорослей
может переходить от фотосинтеза с выделением кислорода к фото-
редукции, если их выдержать на свету в анаэробных условиях,
например в атмосфере с Н2, H2S и СО2. Способность водорослей
переключаться с одного типа усвоения СО2 на другой в зависимости
от условий легла в основу представлений об общности
фотохимических реакций зеленых растений и бактерий (At Клюйвер
и К. Ван-Ниль, 1959; X. Гаффрон,.19б2; и др.). При формировании
оптического аппарата и митохондрий в онтогенезе у растений также
проявляются черты более древних предков, происходит постепенное
усложнение их «внутренней анатомии» (см. гл. 3, 4).
Любая функция или этап жизненного цикла как бы состоит из
сочетания древних и новых приобретений (см. § 2.6): Механизм
сохранения древних особенностей связан с их значимостью в
современных условиях и наслоением на них более совершенных
реакций. По этой же причине несколько затруднено их разграни-
чение у растений. Вопросы рекапитуляции функций как у растений ,
так и животных слабо разработаны. ’Трудности сохраняются и в
применении теории филэмбриогенеза к растениям (А. Л. Тахтад-
жян, 1964).
8.9. ОНТОГЕНЕЗ — ОСНОВА ФИЛОГЕНЕЗА.
ЭВОЛЮЦИЯ РАСТЕНИЙ В БУДУЩЕМ
Выше речь шла о прошлых событиях в истории растений,
имеющих значение для современного их процветания. Очевидцем
прошлойэволюции человек, естественно, быть не мог. Он оказался
непосредственным участником только эволюции культурных рас-
тений — «управляемой эволюции» (Н. И. Вавилов, 1932). Мы до-
стоверно можем судить о скорости последней и ее результатах,
особенно когда это касается изменений в содержании каких-то
веществ (масел, белка, крахмала, алкалоидов и т. п.), иммунитета
сортов, длины волокна, сахаристости и ъ д.
В то же время человек сам стал теперь могучим фактором
эволюции, с каждым годом возрастает мощность его воздействий
на природу. Напомним несколько фактов. Подсчитано, что выбросы
только аммиака и оксидов азота в воздух ежегодно составляют около
350 млн. т, которые затем оседают на поверхность почвы и растений.
229
Загрязнение природы происходит и отбросами других отраслей
промышленности, в результате работы транспорта, применения
пестицидов, дефолиантов и т. д. (А. В. Яблоков, Л..С. Остроумов,
1983). •
Число веществ—загрязнителей природы—антропогенного
происхождения превышает уже несколько сотен и постоянно растет.
Некоторые вещества опасны не только сами по себе, но и продукты
их разрушения. Так, ДДТ и другие химикаты сохраняют в почве
свою токсичность несколько лет. В результате разрушения гексо-
хлорана образуется 25 промежуточных метаболитов, карбофоса и
хлорофоса — 14, оказывающих то или иное вредное действие и на
растения (О. Н. Прокофьев, 1983). Поэтому нас теперь интересуют
возможные эволюционные изменения растений в будущем по мере
усиления научно-технической революции и ее давления на естест-
венные ландшафты. Это как раз имел в виду В. И. Вернадский.
(1967), когда предложил выделить в биосфере область ноосферы,
где «впервые человек становится крупнейшей геологической
силой... Лик планеты — биосфера—химически резко меняется
человеком сознательно и главным образом бессознательно».
Человек своей деятельностью в течение короткого периода его
исторического существования так или иначе влиял на ход естест-
венной. эволюции. Под влиянием человека (более 50 тыс. лет)
преобразовывались биогеоценозы, возникали новые сорняки
(например, сорняк посевов льна—льняной рыжик из полевого
рыжика), появлялись более агрессивные формы заразихи и т. д. Эти
бессознательные изменения, пожалуй, меркнут перед последу-
ющими мощными воздействиями человека на природу. Речь идет
не только о последствиях изъятия части ресурсов природы для
хозяйственных целей, но и об интенсивном ее загрязнении.
Этот вопрос волновал человечество с момента развития
капиталистического общества. Так, еще в XIV—XV вв. в Германии,
Франции и Англии население обращалось с жалобами на надо-
едливую копоть в воздухе и требованиями на ограничение деятель-
ности предприятий по выбросу вредных газов в атмосферу.
Парламенты указанных стран вынуждены были обсуждать подобные
вопросы (Х.-Г. Десслер, 1981). Наш соотечественник В. И. Вер-
надский обеспокоенно предсказывал еще большие последствия
развития промышленности.
Отравление атмосферы, почвы и воды принимает во всем мире
глобальный и катастрофический характер. Источники загрязнения
природы увеличиваются, несмотря на меры, принимаемые
правительственными и межнациональными организациями. Рас-
тения первыми принимают «удар» антропогенных токсических ве-
ществ. Скопление в атмосфере газо- и пылеобразных токсических
Веществ, в почве ядовитых солей (включая тяжелые металлы)
230 • .
постепенно способствует разрушению естественных ландшафтов и
биосферы в целом (А. С. Равкин, 1981; Н. В. Гетко, 1989). В нара-
стающей критической ситуации требуется не только принятие мер
по снижению загрязнения природы, но и умение прогнозирования
его эволюционных последствий.
Вокруг крупных промышленных районов среди функциональ-
ных нарушений растений часто отмечают резкое подавление дея-
тельности ферментных, систем, интенсивности фотосинтеза,
повышение транспирации, усыхание и опадение листьев, повреж-
дение'почек и т. д. Подобные изменения в конце концов приводят
к гибели отдельных растений или исчезновению вида из загрязнен-
ного района. Наиболее очевидна при этом смена древесной расти-
тельности травянистой или возникновение форм, отличающихся
мелкими листьями, покрытыми толстой кожурой, что снижает
поступление ядовитых веществ в растения. Симптомы повреждения
ядовитыми веществами не всегда являются специфическими, что
отчасти затрудняет целенаправленный отбор на повышение ус-
тойчивости того или иного вида растений (Х.-Г. Десслер, 1981).
Испытания показали, что лиственные породы более устойчивы
к загрязнению воздуха, чем хвойные, и пылеулавливающая способ-
ность первых выше. Так, 1 га елового леса консервирует 32 т пыли,
а букового леса,— 68 т. Наиболее стойкими к пыли; даму и
загрязнениям признаны платан, гледичия, софора, дуб, бересклет,
тополь и т. д. Из хвойных —туя, кипарисовик, тисс, сосна (черная,
кедровая, горная), пихта, ель. Ель колючая не выносит загрязнения.
Айва более устойчива, чем слива. Перед селекцией растений возникла
новая задача—: обязательная оценка создаваемых сортов зерновых,
кормовых, плодовых, лесных и декоративных культур по устойчивости
к загрязнению воздуха и почвы. Будущие возможности растениевод-
ства также зависят от успеха селекционной работы на повышение
устойчивости растений к различным загрязнениям.
Результаты селекции позволяют осмыслить и возможные пос-
ледствия естественного отбора (Ч. Дарвин). Вообще растительность
обладает большими адаптационными резервами, позволяющими ей
противостоять нарастающему загрязнению. Наиболее вероятна
смена растительных формаций вокруг промышленных районов —
хвойных лесов лиственными, лиственных лесов кустарниками, кус-
тарников травянистыми растениями, особенно с преобладанием
ксероморфных видов. Следовательно, загрязнение ведет к оскуде-
нию растительности в биологическом и декоративном отношении.
Эти процессы, пожалуй, характеризуются наибольшей быстротой и
неотвратимостью. Адаптация растительности путем смены ее фор-
маций как бы является «перегруппировкой» сил и не связана с
усилением генетической толерантности видов. Скорее при этом
используются особенности разных видов, возникшие в других
231
обстоятельствах, но случайно оказавшиеся полезными в условиях
повышения загрязнения природы. Брлее активным должен быть
признан путь усиления генетической толерантности растений в
пределах вида на основе отбора мутаций, определяющих морфо-
функциональную устойчивость индивидуума к загрязнению воздуха
и почвы. Такой процесс изучен на примере заселения растениями
ядовитых отвалов рудников.
Возможности адаптации растений к загрязнению природы до-
вольно разнообразны. Но нельзя спокойно относиться к усилению
загрязнения природы, полагаясь на ее резервы. Защита раститель-
ности является частью защиты человечества, так как практически
невозможно представить цивилизацию без космической роли зеле-
ных растений. Изучение характера изменений растений под влия-
нием антропогенных воздействий — насущная задача физиологии
растений. Только это даст возможность составить эволюционно
грамотные прогнозы о масштабах нашего влияния на природу на
несколько десятков лет вперед.
Особенности Жизнедеятельности современных растений
исторически формировались постепенно, свидетельством чего
служит и постепенность их развития в онтогенезе. Любые эво-
люционные новшества первоначально возникают в онтогенезе как
мутации, и, если они' играют полезную роль, часть из них может
иметь филогенетическое значение — филэмбриогенезы (по А. Н.
Северцову). Конечно, не всякое мутационное изменение включа-
ется в морфогенетические процессы в онтогенезе и последующих
поколениях, что определяется корреляционной системой организма
(И. И. Шмальгаузен).
В принципе и функциональное усовершенствование растений
представляет собой процесс микроэволюционного преобразования
и суммирования эффекта соответствующих мутаций (см. гл. 2),
довольно частых в онтогенезе. Мутационным изменениям подвер-
гаются отдельные реакции, структурные особенности и их комплек-
сы, определяющие устойчивость, скорость роста и развития,
отношение растений к факторам среды и т. д. Любой, даже сорт
идеальной селекции постепенно накапливает мутационные изме-
нения по многим признакам и реакциям. Подобные изменения еще
чаще встречаются в природных популяциях.
На основе оценки жизнеспособности индивидуумов, их
генетических различий в метаболизме и структуре (что является
результатом сочетания мутационного процесса и генетической
комбинаторики) происходят отбор удачных физиолого-биохимиче-
ских фенотипов, усиление или ослабление любого признака и
свойства растений в поколениях. Это показано на примере дивер-
генции популяции и сортов по устойчивости к загрязнению среды
ионами тяжелых металлов, засухо-, соле- и морозоустойчивости.
232
В целом развитие функциональных приспособлений
исторически достигалось суммацией эффекта отдельных структур-
ных и биохимических мутаций в поколениях, обеспечивающих
преимущества индивидууму и популяции в борьбе за существование.
При этом нельзя исключить и значимость закрепления макрому-
таций — системных мутаций. Экспансия растений в геологическом
прошлом часто была связана с постепенным накоплением комп-
лекса новшеств, способствовавших энергетической победе или
развитию устойчивости. И эти изменения возникали в ряду онто-
генезов путем концентрации определенных генов. Подобные изме-
нения, вероятно, происходят на разных стадиях онтогенеза
растений, что показано на примере структурных преобразований
(А. Л. Тахтаджян). Однако вопрос о стадиях возникновения
физиолого-биохимических наследственных уклонений в онтогене-
зе, имеющих филогенетическое значение, нуждается в дальнейшем
изучении.
\ t
8.10. СИСТЕМНЫЙ КОНТРОЛЬ РЕГУЛЯЦИИ ЖИЗНЕДЕЯТЕЛЬНОСТИ
В ОНТОГЕНЕЗЕ РАСТЕНИЙ И ЕГО ЭВОЛЮЦИОННЫЕ ПРЕДПОСЫЛКИ
Принцип системного подхода, получивший распространение в
различных областях науки, предполагает анализ объекта во взаимо-
действии его составных элементов. Объект при этом представляется
как саморегулирующая система по принципу прямой и обратной связи
— «самонастройка» развития. В разработке теоретических основ
системного подхода применительно к индивидуальному и историче-
скому развитию живых ^организмов велика заслуга И. И. Шмаль-
гаузена, который обратил внимание не только на значение самого
явления интеграции, но и факторов и механизмов, его определя-
ющих на разных уровнях организации жизни (молекулярный, кле-
точный и т. д.). Значение указанных механизмов, по его мнению,
состоит в преодолении влияния случайных вредных воздействий и
развитии приспособлений к условиям среды для максимального
использования ее ресурсов в процессах роста и размножения. Отсюда
интеграция биологических систем выступает и как результат, и как
предпосылка их дальнейшей эволюции. В итоге И. И. Шмальгаузен
заключает, что «эволюция приобретает в определенных условиях
существования характер направленного устойчивого движения
(«филогенетический гомеорезис»1).
В плане углубления представлений о функциональной эволюции
растений важно обобщить материалы предыдущих глав с точки
зрения развития и проявления системного контроля регуляции их
Шмальгаузен И. И. Проблемы дарвинизма. Л., 1969. С. 462.
233
жизнедеятельности, т. е. онто- и филогенетических основ функци-
ональной целостности растений.
Если принять во внимание систематическое разнообразие рас-
тений и специфику жизнедеятельности в онтогенезе у предста-
вителей разных видов, то сложность решения поставленного
вопроса очевидна. Речь идет о необходимости оценки роли не только
отдельного процесса метаболической цепи и продуктов мета-
болизма в жизнедеятельности растения, но и знания того, как
устроен и работает сложный функциональный «ансамбль», каков
генезис и механизм саморегуляции на уровне клетки, тканей,
органов и целостного организма.
Ответ на вопрос связан с объективными трудностями (отсутствие
знаний о роли метаболитов и процессов в онтогенезе и о путях их
становления в филогенезе). Тем не менее функциональная целост-
ность растений априори предполагается И ее слагаемые отчасти
известны (Л. А. Курсанов, I960,' 1976; Э. Либберт, 1976; А; Т. Мок-
роносов, 1981; В. В. Полевой, 1982; О. Н. Кулаева, 1982; М. К. Чай-
лахян, 1988; Н. Е. Street, W. Cockburn, 1972; G. R. Noggle, G. J.
Fritz, 1993 и др.).
Сравнение данных о физиологии растений, относящихся к
разным систематическим и экологическим группам, показывает,
что в их онтогенезе происходит постепенная смена древних форм
метаболизма(гетеротрофный тип обмена) на фототрофный с посте-
пенным усилением (вычленением) его специфичности, характерной
для представителей определенных видов. С другой стороны, все
больше усиливается зависимость клеточного метаболизма от харак-
тера дифференциации и развития-растения в онтогенезе. В связи с
этим правомерно выделение иерархических уровней регуляции у
многоклеточных растений, где целостность организма обеспечива-
ется взаимодействием его частей и возникновением доминирующих
центров (В. В. Полевой, 1982).
В основе функциональной целостности растений в онтогенезе
лежит развитая система структурной организации и взаимосвязан-
ных Процессов. В системе регуляции доминируют донорно-акцеп-
торные отношения, основанные на сопряжении механизмов
фотосинтеза и гормонального аппарата (А. Т. Мокроносов, 1981).
Приоритетная роль сопряжения указанных систем в росте,
морфогенезе и репродукции далеко не случайна. В них замыкаются
энергетическая основа роста и морфогенеза, а также процессы,
контролирующие деление и дифференциацию клеток и экспрессию
генов (что определяет специфику метаболизма и последовательной
смены его этапов). Сопряжением механизмов фотосинтеза и гор-
монального аппарата обусловлено состояние и других процессов:
дыхания, транспорта веществ, покоя, цветения и т. д.
Переход к фототрофному метаболизму и включение гормональ-
ного контроля регуляции оказались не только предпосылкой
234
активного взаимодействия растений со средой, но и всех последующих
процессов метаболизма и морфогенеза в онтогенезе. При этом достига-
ется удовлетворение энергетических и морфогенетических потребнос-
тей самого организма и создаются запасы органического продукта,
необходимого для воспроизведения. Такие важные механизмы,
координирующие деятельность организма, как полярность корре-
ляции и процессы регенерации вытекают из донорно-акцепторных
отношений, сопряжения механизмов фотосинтеза и гормонального
баланса. Сказанное не умаляет значения других звеньев метаболизма.
Основополагающее значение процессов фотосинтеза и гормональной
регуляции определило их и как наиболее чувствительные звенья
метаболизма к колебаниям условий среды.
Естественно, что эффективность механизмов фотосинтеза и
гормонального контроля зависит от состояния дифференциации
клеток, тканей и органов, возраста, преобладания вегетативных или
генеративных тенденций индивидуума. Интенсивность и про-
дуктивность фотосинтеза регулируются не только внешними усло-
виями, но и в большей степени потребностями самого организма,
изменением в нем соотношения ассимилирующих и неасси-
милирующих структур, состоянием транспорта ассимилятов и т. п.
Поэтому на разных этапам онтогенеза меняются вклад, ярусов
листьев на побеге в создание продуктов фотосинтеза, настройка
аппарата фотосинтеза (пигментов, хлоропластов, листьев) на
восприятие света, продуктивность и интенсивность фотосинтеза
параллельно с изменением уровня потребления продуктов асси-
миляции СО2 и структурной организации индивидуума и т. д.
Очевидна и взаимосвязь между механизмами дыхания, погло-
щения и транспорта воды, органических и минеральных веществ,
процессами фотосинтеза и гормонального контроля, а также между
последними и устойчивостью растений к неблагоприятным условиям.
В онтогенезе процессы фотосинтеза и гормонального контроля
обеспечивают успех индивидуума в размножении, и популяций в
борьбе за существование. Но историческое предназначение
механизма фотосинтеза имеет глобальное значение для развития
жизни как системного явления Земли. Именно этим определяется
экологическая роль и космическая функция растений в живой
природе, в поддержании ее энергетическим материалом путем
циклического круговорота кислорода, воды и оксида углерода.
Несмотря на важность связи между механизмом фотосинтеза и
гормональным контролем у цветковых, она эволюционно и онто-
генетически возникает не сразу. Как следует из гл. 3 и 4, сами эти
явления возникают в разное время и в какой-то степени независимо.
Наиболее древним следует признать механизм фотосинтеза, кото-
рый не мог возникать из потребности самого организма по следу-
ющим причинам: 1) линия фототрофности развивалась у разных
235
групп прокариотов и этот процесс был длительным во времени {при
ограниченности ПЖ индивидуума) и постепенным из-за наслоения
новых реакций на более древние пути метаболизма; 2) фотогроф-
ность,приведшая к выделению свободного кислорода (линия
фотогидротрофии), первоначально имела летальный исход для
индивидуума из агрессивности свободного кислорода; 3) и в случае
даже развития, механизма фотосинтеза его стабилизация зависела от
потребностей жизни на Земле; 4) эффективность фотосинтеза в
геологические эпохи возрастает не в результате усовершенствования
его физико-химических реакций, а биологических потребностей
жизни.
Для удовлетворений потребностей отдельно взятого про-
кариотического организма достаточна низкая эффективность про-
цесса синтеза органического вещества из неорганических. В рамках
отдельной клетки шансы интенсификации механизма фотосинтеза
оказывались крайне ограниченными, как и в масштабах одной
фотосинтезирующей популяции. В связи с этим эволюционный
запрос на повышение эффективности механизма фотосинтеза
исходит от гетерогенности организации самой жизни в первичных
биогеоценозах с преобладанием гетеротрофных форм и все услож-
няющегося индивидуального уровня (вслед за появлением многок-
леточных). Толчком к усложнению последнего уровня опять-таки
служили потребности самой жизни как единой системы на поверх-
ности Земли, На этом этапе отбор, пожалуй, происходит на уровне
Первичных биогеоценозов со стабильной устойчивостью кругово-
рота веществ и энергии. Хотя Сегодня еще многое неясно в
механизме действия отбора на уровне экосистем, реальность самого
явления не вызывает сомнения. Именно экосистемный запрос
служил предпосылкой дальнейшей эволюции фотосинтеза как
биологического явления. В результате появляются растения с раз-
ными типами фиксации СО2, усложняется зависимость фотосинтеза
от особенностей роста, дыхания, водообмена и т. д. растений. На
этом этапе как раз складывается гормональная система растений и
усиливается ее связь с процессами фотосинтеза из-за возрастания
их биоценотической роли. Все это сопровождалось изменением
индивидуальности и геометрии, структурной организации растений.
В итоге эра включения механизма фотосинтеза становится
точкой отсчета в последующей эволюции жизни в направлении все
большей индивидуальной и филогенетической ее дифференциации.
При становлении фотосинтетического метаболизма в онтогенезе
как бы повторяются филогенетические его этапы в более быстром
и укороченном виде, подготавливается и структурная база для
реализации механизма фотосинтеза.
Сказанное свидетельствует о том, что механизмы фотосинтеза
первично возникают из запроса самой жизни на планете и из
потребностей поддержания первичных экосистем, а в последующем
236
они развиваются с усложнением и дифференциацией биоценозов.
По этой причйне на базе общности реакций фотогидротрофии
происходит эволюция фотосинтеза как сложного функцйонального
и биологического системного явления. При этом фитоценоз, явля-
ющийся основой любой экосистемы, становится объектом отбора
на устойчивость в определенных временно-пространственных
границах. Вообще появление механизма фотосинтеза и поиск путей
его эффективного использования признаны важными вехами в
процессе жизни. Фотосинтез — арахимоз принципиального харак-
тера и большого масштаба (макроэволюционное событие) в систем-
ной организации жизни на Земле.
Для понимания эволюции природы системного контроля регу-
ляции жизнедеятельности растений значение имеют и данные о
возникновении механизма взаимодействия растений между собой и с
животными в биогеоценозах, основанные на специфике химического
состава видов (гл. 5). Для их анализа, обратимся еще к примеру
коэволюции бобовых и бактерий-ризобиум, возникших независимо и
в разное время. Способность бактерий к азотофиксации является более
древним и самостоятельным приобретением, как и способность бо-
бовых образовывать «псевдоклубеньки» в виде структур для накоп-
ления и хранения крахмала (Н. А. Проворов, 1996).
В последующем в мелрвом периоде в условиях нехватки в почве
азота (столь необходимого для накопления биомассы растений,
следовательно, через них и для процветания экосистем) складыва-
ется эффективная система симбиоза бобовых растений и бактерий-
ризобиум. Эти отношения совершенствовались в ходе дальнейшей
эволюции экосистем, о чем свидетельствуют слабое их развитие у
современных наиболее древних представителей бобовых, усиление
специфичности симбиотических отношений, усложнение структу-
ры клубеньков и механизма их инфицирования у продвинутых
представителей в умеренных широтах. Бобовые постепенно стано-
вятся фактором, определяющим генетическое разнообразие,
численность и распространение популяций ризобиума, как и пос-
ледние — фактором стабильности и полиморфизма растения-
хозяина. Идет групповой и взаимный отбор на поддержание в
каждом онтогенезе эффективности азотфиксирующей системы,
состоящей из разных представителей живой природы. Результат
отбора — обеспечение оптимального уровня чувствительности рас-
тения-хозяина к инфицирующему агенту и умеренной вирулент-
ности последних, что сопровождается возрастанием концентрации
генов у бобовых, участвующих в регуляции симбиотических взаимо-
отношений на клеточном и организационном уровнях, а у бак-
терий — генов, индуцирующих развитие клубеньков бобовых.
Системный контроль регуляции в той или иной форме прояв-
ляется в онтогенезе у всех растений, что выступает как результат их
237
филогенетического развития. Отбор шел на усиление структурно-
функциональной интеграции индивидуума, что привело к выработ-
ке механизмов поддержания гомеостаза. Эти механизмы отличаются
у одно- и многоклеточных форм. У последних внутриклеточный
гомеостаз усложняется системой регуляции на уровне тканей, орга-
нов и целого растения. Интеграционные процессы индивидуума у
цветковых выражены по-разному в зависимости от степени его
расчлененности и организованности онтогенеза (§ 8.4).
Изменения в ходе эволюции онтогенеза и экосистем относятся
к макроэволюционным, в их числе и механизмы фотрсинтеза и
гормонального контроля, примеры коэволюции видов растений и
животных. Первично отбор на фототрофность выступал, по-
видимому, как межвидовой. При этом оценивался успех видов в
борьбе за существование по способности создавать органический
продукт за счет использования энергии солнечного света. Конку-
ренция видов шла за ниши в толщах водоема с хорошим осве-
щением. Однако межвидовой отбор не мог бы достичь больших
результатов, если бы не возникали генетические изменения особей
в пределах одной й той же популяции фототрофов по активности
синтеза органического вещества из простых и легко доступных
неорганических соединений среды. Именно индивидуальный отбор
особей в пределах отдельных фототрофных популяций в направ-
лении усиления тенденций фотогидротрофии сыграл решающую
роль в выработке, а затем в стабилизации физико-химического
механизма обеспечения энергией живых существ.
На примере становления механизма фотосинтеза (равно как и
других крупных звеньев метаболизма, коэволюции видов) заметна
связь микро- и макроэволюционных изменений. Исходно изме-
нения возникают в популяциях, которые подвергаются индивиду-
альному отбору —преимущественному выживанию и размножению
особей с выраженной генетической тенденцией к созданию органичес-
кого вещества из неорганических компонентов среды благодаря по-
глощению энергии света. Однако по масштабу, биологическому
назначению такие изменения выходят за пределы потребностей по-
пуляции. Они направлены на усиление взаимодействия разных видов
с образованием устойчивых экосистем, где изначально продуценты
органического вещества становятся ядром биоценоза.
Усиление же деятельности продуцентов, т. е. эффективности
использования сформированного уже механизма фотосинтеза, на
этапе развития экосистем, исключительно зависит от межвидовых
взаимоотношений — от состава и структуры экосистемы. Поэтому
экосистемы и отличаются по показателю годичной продуктивности,
выраженному величиной нетто-фотосинтеза с учетом затрат на
дыхание (§ 3.9). Устойчивость экосистем возрастает в случае пре-
обладания в них фотосинтеза над скоростью биологического
238
окисления органического вещества. Биологический этап в эволо-
юции фотосинтеза как раз связан с усилением разнообразия форм
жизни и возрастанием роли межвидовых отношений. В этой связи
вполне реален прогноз о возможности изменения самих экосистем
и перераспределения в них доминантов с потеплением климата
Земли, как результат нарушения межвидовых отношений ( § 3.10).
Такой процесс в будущем, как и в прошлом, может привести к
изменению специфики системной регуляции жизнедеятельности
растений на индивидуальном популяционно-видовом и биоце-
нотическом уровнях. В основе этого процесса окажется другое
соотношение видов с С3-, С4-, CAM-типа растений, что приведет к
изменению облика растительности и круговорота вещества в биоце-
нозах.
* * ♦
Онтогенез у растений претерпевал изменения в ходе общей их
эволюции, что выразилось в упрощении циклов развития и
усилении дифференцировок, олигомеризации процессов и струк-
тур, интеграции индивидуума, ускорении темпов развития и авто-
номизации онтогенеза. Эволюция онтогенеза растений привела к
усилению целостности, развитию механизмов гомеостаза, и ус-
тойчивости индивидуума к неблагоприятным условиям. Все это
имело непосредственное значение для адаптации и дивергенции
растений.
Проблема эволюции онтогенеза имеет много нерешенных ас-
пектов. На фоне большой литературы по морфологическому и
экологическому изучению растений разных таксонов все еще мало
сведений, касающихся эволюционных преобразований функцио-
нальных особенностей организма на разных стадиях онтогенеза. По
этой причине нет еще ясности о сфере применения некоторых
положений эволюционной теории к преобразованию функциональ-
ной деятельности растений (учений рекапитуляции, филэмбриоге-
неза, прогресса и т. п.) и использования данных сравнительной
фитофизиологии при построении филогенетических систем. '
ЗАКЛЮЧЕНИЕ
. В данном пособии известные явления и процессы из курса
физиологии растений представлены с точки зрения их историчес-
кого развития и приспособительного значения. Растительная
«жизнь» в нем рассмотрена в общем контексте эволюции живой
природы на Земле, опираясь на данные ряда смежных наук, с
которыми студенты не сталкивались при изучении курса физио-
логии растений. Поэтому пособие прежде всего призвано помочь
осмыслению ранее изученного материала. Однако пособие не является
повторением общего курса, оно содержит оригинальный материал и
выводы, касающиеся функциональной эволюции растений. Пособие —
самостоятельный курс по проблемам эволюционной физиологии рас-
тений, по которым нет учебной литературы и обобщающих научных
исследований. Его значение выходит за рамки учебных задач, так как
вопросы функциональной эволюции растений до последних лет не
служили предметом специальных обсуждений.
В пособии подведены итоги общей картины развития энерге-
тики и особенностей обмена у растений и уточнены закономерности
(постепенное усложнение, взаимосвязь формы и функции, воз-
никновение многообразия, приспособительный характер, взаимо-
дополнение механизмов), направления (дивергенция, параллелизм,
конвергенция) и принципы (множественность реализации,
усиление или ослабление, компенсация, автономизация функций)
эволюции процессов жизнедеятельности у растений. Достижения
эволюции сводились не только к возникновению совершенных
физико-химических механизмов метаболизма, но и к опробованию
их в различных условиях, к изменению соотношения тех или иных
процессов и т. п.
Энергетическое (следовательно, и адаптивное) совершенство-
вание растений в ходе эволюции происходило под контролем
естественного отбора, чему способствовали постоянно возникаю-
щие в популяциях предпосылки (генетическая изменчивость струк-
турно-функциональных особенностей, прогрессия размножения и
борьба за существование особей). Несмотря на значительную
пластичность растений в онтогенезе, в физиолого-морфологичес-
ком отношении они стабилизированы отбором. Лабильность рас-
тений — способ поддержания стабильности онтогенеза.
На примере развития автотрофного питания и аэробного дыха-
ния четко видно постеценное усложнение их механизмов в направ-
лении достижения энергетической независимости растений. Ни
одно новшество в метаболизме не было закреплено автономно как
чисто химическая реакция. Всякое химическое новшество отби-
240
ралось как составная часть общего процесса жизнедеятельности
развивающейся системы (организма), поскольку оно обеспечивало
победу популяций в борьбе за существование. Такое новшество
должно было иметь значение для успешного размножения.
С экологической дивергенцией растений связаны и различия в
химическом составе представителей разных популяций. Содержа-
ние определенных веществ и наследственная специфика популяций
и видов в их накоплении имеют приспособительное значение,
закреплены исторически и стабильно проявляются в онтогенезе.
Это показано на примере продуктов вторичного метаболизма, ка-
техинов и пектиновых веществ, фитогормонов и ингибиторов роста.
Данные о химической специализации и конвергенции популяций
и видов служат одним из подтверждений теории естественного
отбора и выводов современного дарвинизма.
Любая особенность, даже, казалось бы, самая незначительная,
подвластна отбору, если она затрагивает приспособленность попу-
ляций. С этих позиций были проанализированы и периодические
явления в жизни растений. Приспособительна сама способность
растений «измерять» время в течение суток и сезона, «следовать»
за годовыми переменами климата и т. д. В результате растения в
определенное время (специфическое для сорта и популяций) пере-
ходят к цветению и плодоношению, вступают в период покоя,
начинают листопад и стареют. Синхронизация процессов жизнеде-
ятельности растений с чередующимися условиями в онтогенезе —
результат исторического развития организмов в постоянно колеб-
лющихся условиях. Замечательно, что растения готовятся к их
наступлению, как бы предвидя будущие невзгоды. Механизм
«физиологических часов» довольно точен и имеет важное значение
для выживания популяций.
Вся жизнедеятельность растений направлена на обеспечение
процесса воспроизведения. Поэтому ни одно приобретение не
может компенсировать урон, наносимый воспроизведению, так как
«опасность не оставить потомство» представляет самую большую
угрозу для вида (Ч. Дарвин, 1939). В связи с этим обсуждение любой
проблемы эволюционной фитофизиологии и конечном итоге упирает-
ся в проблему эволюции онтогенеза и его регуляторных механизмов.
Нелишне напомнить, что на сегодня применимость к растениям
даже некоторых положений эволюционного учения об эволюции
онтогенеза остается спорной из-за слабой изученности раститель-
ных объектов в этом отношении. В связи с этим в пособии обращено
внимание на тенденции изменения онтогенеза в ходе эволюции
растений. При этом удалось выяснить, что магистральная линия
эволюции онтогенеза и у растений связана с автономизацией
процессов развития и усиления регуляторных механизмов инди-
видуумов. Однако эти особенности у растений по степени прояв-
ления отличаются от таковых у животных. И на примере растений
241
/
наблюдается взаимосвязь в изменении индивидуального и истори-
ческого развития, присущая для существ других царств.
.Несмотря на неполноту знаний о функциональной эволюции
растений, выяснилось, что становление и развитие процессор жиз-
недеятельности растений укладывается в классические постулаты
дарвинизма о постепенном усовершенствовании организации под
действием отбора. Речь идет не о формальном «согласовании»
данных физиологии растений с учением Ч. Дарвина (как с наиболее
привычной для нас теорией), а о том, что при достаточно глубоком
изучении процессов жизнедеятельности растений роль отбора в их
становлении и развитии четко вырисовывается.
Всю сложную функциональную деятельность растений можно
рассматривать с эволюционной точки зрения как цепь взаимосвя-
занных и непрерывных процессов энергетического совершенство-
вания организмов и повышения устойчивости к неблагоприятным
условиям. Наиболее архаичными в этой цепи следует считать звенья,
приведшие к развитию и стабилизации автотрофного питания. Про-
цессы аэробного дыхания, особенности химической организации и
ритмической деятельности развиваются параллельно или в непо-
средственной зависимости от специфики обмена растительной
клетки. В этом смысле процессы автотрофного обмена можно
рассматривать как предпосылки последующих эволюционных
событий в жизнедеятельности растений.
В целом в общей! и теоретической биологии вопросы
функциональной эволюции остаются наименее изученными. При
использовании методов популяционной генетики, палео-
физиологий и физико-химического анализа растений в будущем
наиболее интересных фактов можно ожидать именно в этом направ-
лений: Подобные исследования внесут изменения в привычные
представления об эволюции органического мира. При этом не надо
бояться и возможных теоретических отклонений. Так, несмотря на
большие «неудобства» представлений о недарвиновской эволюции,
они тем не менее способствовали накоплению новых фактов.
,. Материалы пособия нельзя рассматривать только как допол-
нительные доказательства.триумфа теории естественного отбора. В
ходе Изложения было обращено внимание на наличие ряда пара-
доксов в функциональной перестройке растений. Их число, несо-
мненно, увеличится по мере накопления фактов. Трудности теории
естественного отбора в объяснении функциональной эволюции
неизбежны в будущем, они о себе уже дают знать и поел ужат толчком
ее развития. Представление материала с позиции дарвинизма в
пособии не является попыткой отрицания других возможных под-
ходов. Скорее оно должно быть расценено как первое обобщение
сложного материала для акцентирования внимания на мало иссле-
дованных проблемах эволюционной физиологии растений.
242
РЕКОМЕНДУЕМАЯ ЛИТЕРАТУРА
Основная
Благовещенский А. В., Александрова Е. Г. Биохимические основы филогении
высших растений. М., 1974.
Брода Э. Эволюция биоэнергетических процессов. М., 1978.
Вернадский В. И, Биосфера. М., 1967.
Гусев М. В., Гохлернер Г. Б. Свободный кислород и эволюция клетки. М., 1980.
Дарвин Ч. Происхождение видов путем естественного отбора//Книга для
учителей. С комментариями А. В. Яблокова и Б. М. Медникова. М., 1987.
Клейтон Р. Фотосинтез. Физические механизмы и химические модели. М., 1984.
Лукнер М. Вторичный метаболизм у микроорганизмов, растений и животных.
М., 1979.
Мокроносов А. Т., Гавриленко В. Ф. Фотосинтез. Физиолого-экологические и
биохимические аспекты. М., 1992.
Полевой В. В, Физиология растений. М», 1989.
Судьина Е. Г, Лозовая Г И. Эволюционная биохимия растений. Киев, 1983.
ХарборнДж. Введение в экологическую биохимию. М., 1985.
Эволюция функций в растительном мире // Сб. статей под ред. В. В. Полевого.
Л., 1985.
Эдвардс Дж., Уокер Д. Фотосинтез Сз- и С4-растений. Механизмы и регуляция.
М., 1987.
Дополнительная
Авратощукова Н. Генетика фотосинтеза. М., 1980.
Александров В. Я. Клетки, макромолекулы и температура. Л., 1975.
Биль К. Я. Экология фотосинтеза. М., 1993.
Биогенез пигментного аппарата фотосинтеза // Под ред. Ф. Ф. Литвина. Минск,
1988.
Бутник А.А., Нигманова Р. И., Пайзиева С. А., Саидова Д. К. Экологическая
анатомия пустынных растений Средней Азии. Ташкент, 1991.
Бюннинг Э. Ритмы физиологических процессов («Физиологические часы»). М.,
1961. ‘
Вагнер Р., Митчел Г. Генетика и обмен веществ. М., 1958.
Вандерпланк Я. Генетические и молекулярные основы патогенеза у растений.
М., 1981.
Войников К. В., Константинов Ю. М., Негрук В. И. Генетические функции мито-
хондрий растений. Новосибирск, 1991.
Воробьев Л. Н., Булычев А. А., Полевой В. В., Саламатова Т. С. Ионный транспорт
в растениях // Итоги науки и техники. Физиология растений. М., 1980. Т. IV.
Гамалей Ю. В. Эндоплазматическая сеть растений. Происхождение, структура и
функции. С.-Пб., 1994.
Генкель П. А. Физиология жаро- и засухоустойчивости растений. М., 1982.
Георгиев Г. П. Гены высших организмов и их экспрессия. М., 1989.
Горизонты биохимии // Сб. статей под ред. Л. А. Тумермана. М., 1964.
Горышина Т. К Фотосинтетический аппарат растения и условия среды. Л., 1989.
Грант В. Видообразование у растений. М., 1984.
Гринева Г. М. Регуляция метаболизма у растений при недостатке кислорода. М.,
1974.
Гродзинский Д. М. Надежность растительных систем. Киев, 1983.
243
Гусев М. В., Минеева Л. А. Микробиология. М., 1985. С. 177—367.
Гэлстон А., Девис П.> Саттер Р. Жизнь зеленого растения. М.» 1983.
Деверолл Б. Дж. Защитные механизмы растений. М., 1980.
Демкиев О. Т., Сытник К. М. Морфогенез археогониат. Киев, 1985.
Жизнь растений // Под ред. Ал. А. Федорова. Т. IV. М., 1978.
Жученко А. А. Экологическая генетика культурных растений. Кишинев, 1'980.
Жученко А. А. Адаптивный потенциал культурных растений. Кишинев, 1988.
Зацрометов М. Н. Фенольные соединения. Распространение, метаболизм и
функции в растениях. М., 1993.
Земленухин А. А. и др. Глиоксалатный цикл растений. Воронеж, 1982.
Зенгбуш П. Молекулярная и клеточная биология. М.. 1982. Т. L
Инфекционные болезни растений. Физиологические и биохимические основы //
Сб. статей под ред. Ю. Т. Дьякова. М., 1985.
Казарян В. О. Физиологические аспекты эволюции от древних ктравам. Л., 1990.
Кефели В. И. Природные ингибиторы и фитогормоны. М., 1974.
Кефели В. И. Физиологический основы конструирования габитуса растений. М.,
1994.
Кефели В. И., Коф Э. М., Власов И. В., КислинЛЕ Н Природный ингибитор
роста—абсцизовая кислрта. М., 1989.
Ковалевский Л. А. Биогеохимия растений. Новосибирск, 1991.
Конарев В. Г. Белки растений как генетические маркеры. М., 1983<
Кондратьева Е. Н. Фотосинтезирующие бактерии и бактериальный фотосинтез.
М., 1972.
Курсанов А. Л. Транспорт ассимилятов в растениях. М,, 1976.
Лархер В. Экология растений. М., 1978. .
Левитес Е. В. Генетика изоферментов растений. Новосибирск, 1986,
Либберт & Физиология растений. М., 1976.
Лима де Фариа А. Эволюция без отбора. Автоэволюция формы и функций. М..
1991.
Магомедов И М. Фотосинтез и органические кислоты. Л., 1988.
Манойленко К. В. Развитие эволюционного направления в физиологии растений.
Исторические очерки. Л., 1974.
Манойленко К. В. Эволюционные аспекты проблемы засухоустойчивости рас-
тений. Л., 1983.
Маргелис Л. Роль симбиоза в эволюции клетки. М., 1983.
Метлицкий Л. В., Озерецковская О. Л. Как растения защищаются от болезней.
М.,1985.
Мокроносов А. Т. Онтогенетические аспекты фотосинтеза. М., 1981.
Мокроносов А. Т. Фотосинтетическая функция и целостность растительного
организма. М., 1981.
Муромцев Г. С., Агнистикова В, И. Гиббереллины. М., 1984,
Насыров Ю. С. Фотосинтез и генетика хлоропластов. М., 1975.
НикольсД.Дж. Биоэнергетика. Введение в хемиосмотическую теорию. М., 1985.
Орбели Л. А- Вопросы эволюционной физиологии // Избр. тр. Т. 1. М.-Л., 1961.
Палеонтология и эволюция биосферы // Сб. статей под ред. Б. С. Соколова. М.,
1983.
Пирузян Э. С., Андрианов М.Д. Плазмиды азотобактерий и генетическая инже-
нерия растений. М., 1985.
Поглазов Б. Ф., Бурнашева С. А. Немышечные двигательные системы // Итоги
науки и техники. Биологическая химия. М., 1989. Т. 29.
Полевой В. В. Фитогормоны. Л., 1982.
Полевой В. В., Саламатова Т. С. Физиология роста и развития растений. Л., 1991.
Происхождение жизни и эволюционная биохимия//Сб. статей под ред. Г.
Деборина и др. М., 1975.
Рубин Б. А., Ладыгина М. Е. Физиология и биохимия дыхания растений. М., 1974.
244
Саламатова Т. С., Зауралов О. А. Физиология выделения веществ растениями.
Л., 1991.
Семихатова О. А. Энергетика дыхания растений при повышенной температуре.
Л., 1974.
Скрипчинский В. В, фотопериодизм, его происхождение и эволюция. М., 1975.
Скрипчинский В. В. Эволюция онтогенеза растений. М.., 1977.
Созинов А. А. Полиморфизм белков и его значение в генетике и селекции. М.,
1985.
Сытник К. М., Книга Н. М., Мусатенко Л. М, Физиология корня. Киев, 1972.
Фотосинтез Ц Под ред. Говинджи. М., 1987. Т. 1 и 2.
Фокс Р. Энергия и эволюция жизни на Земле. М., 1992.
Чайлахян М. X. Регуляция цветения высших растений. М., 1988.
Чайлахян М. X,, Бутенко Р. Г.} Кулаева О. Н. и др. Терминология роста и развития
высших растений. М., 1982.
Хочачко П., Семеро Дж. Стратегия биохимической адаптации. М., 1977.
Шахов А. А. Фотоэнергетика растений и урожай. М., 1993.
Шмальгаузен И. И. Проблемы дарвинизма. Л., 1969. \
Хохряков А. П. Эволюция биоморфологии растений. М., 1981.
Чиркова Т. В. Пути адаптации растений к гипоксии и аноксии. Л., 1988,
Шноль С. Э. Физико-химические факторы эволюции. М., 1979.
Эволюция генома // Сб. статей под ред. Г. Доувера и Р. Флейвёлла. М., 1986.
Юсуфов А. Г. Функциональная эволюция растений. М., 1986.
Юсуфов А. Г. Биология старения цветковых растений. Махачкала, 1992.
Яблоков А: В., Юсуфов А. Г. Эволюционное учение. М., 1989.
Biochemical Aspects of Plant and Animal Coevolution./ Ed. J. B. Haiborrie. L.. N.-Y..
1978.
Bradshaw A. D. Me. Neilly T. Evolution and Pollution. London, 1981.
Elstner E. F Oxygen activation and oxygen toxity. Ann. Rev. Plant physiol, 33.
California, 1982.
Fox R. F. Biological energy Transduction. The Urpbarus. N.-Y., 1982.
Genome organitafiOn and expression in plant // Ed. C. Y. Leakes. L., -N.-Y., 1979.
Gottschalk W. Die Bedeutungen der Polyploidie for die Evolution der Pflanzen.
Stuttgart, 1976.
Greber G. T. Chaloner W. G. Influence of enivonnental Factors an the Wood structure
of living and Fossils Trees. The bot. rev., 1984, 50, 4, 35^—448.
Haldane J.B.S. The biochemistry of genetics. London, 1956.
Juniper В. E-, Robins R. J, Joel M. D. The comivorons plants. Acad, press. London,
1989.
Keilin D. The History of Cell Respiration an Cytochrome. Cambridge, 1976.
Levitt J. Responses of Plants to environmental Stress, v. I, II, N.-Y. London, 1980.
Malkin R. Photosysthem I. Ann. Rev. Plant Rhysiol., 33. California, 1982.
Morrby J. Transports sistems in plants. London, 1981.
Mothes K> Schutte H. Biogenese der Alkaloide. Berlin, 1969.
Plant life in Anaerobic Environments./ Eds. D. D. Hook, P. M. M. Crawford. Mi-
chigan, 1978,
Positional kontrols in "plant development // Eds. Barlow P. W., Carr D. Y. Cambridge,
1984.
Sweeney В. M. Rhytmic phenomena in plants. London, N.-Y., 1969.
X he molecular biology of plant development./ Eds. Y. Smith, D. Grierson. Oxford, 1982.
The molecular evolution^ protein polymorphism and the neutral theory./ Ed. M. Kimura.
Berlin, 1982.
Thimann К. V. Hormone action in the whole Life of Plants. Amherst, 1977.
Turner N. C, Kramer P. J. (Ed.) Adaptation of Plants to Water and hight Temperature
Stress. N.-Y.— Toronto, 1980.
245
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
Автокатализ 76
Авторегуляция 50, 92
Автотрофность
классификация 66
Автотрофная эра 68, 70
Адаптация организмов 26
— анаэробиоз 112, 116, 121, 133, 134
— антропогенные факторы 113, 229—
232
— биохимические 150
— кислороду 114
— компенсаторные 51, 190
— классификация 25—27, 218
— неустойчивые 24, 27, 214
— общие 25, 26, 104, 212
— перегруппировкой 231
— устойчивые 24, 27, 194, 211
— функциональные 7, 8, 18, 27, 144
хроматические 74, 88, 100
— частные 25, 26
— экологические 24, 100, 111
— Эксплуатационные 51
Акклимация 27
Алкалоиды 41,149, 154
—разнообразные 154
— значение 153
— мутации 166
Аминокислоты 21, 54, 149, 210
— аланин 14
— аспарагиновая кислота 14, 148
— глицин 14
— глутамин 14, 68, 84, 211
—левые 14, 35, 63
— правые 11, 35, 53
— серин 138
Анабиоз 181, 192, 213
Аноксия ПО, 134
Антоцианы 21, 41, 150, 153
Арахимоз 19, 69, 114
Архебактерии 14, 146
Ассимиляционная сила 91
Аэробность 121
Аэротолерантность 124—128
Бактериородопсин 43, 117
Бактериальный фотосинтез 69, 81 ч
Белки 149
— лабильность 150
— мультифункциональность 148
— нейтральные замены 150, 172
246
— полиморфизм 149, 168
— скорость синтеза 149
— стабильность 54, 149
— эволюция 53, 168
Бетацианины 162, 163
Биологическое вещество 30
Биохимическая примитивность 144—
146
— продвинутость 144, 169
— специализация 134, 144, 145, 165
— универсальность 63, 65
— эволюция 143
Биоэнергетика фототрофов 68, 114
Буферность морфогенеза 58, 175
Вещества биогенные 11, 12
— защитные 9
— круговорот 62
Взаимосвязь формы и функции 31—38
Взрыв популяционный 64, 115, 126
Водообмен 110, 189—193, 207
Возникновение жизни 63—66
— многоклетачности 188
Время астрономическое 178, 179
— биологическое 173,175
— ожидания 64
— геологическое 181, 185
— измерение 169, 183, 241
— презентация 174
Вторичный метаболизм 43,148,152,154,
163, 165—166, 171—172
Газообмен со средой 32
Гелиобиология 173
Гем 126
Гемоглобин 85, 164
Генераторы энергии 117
Гены блоки 22, 25, 149
— кооперативность действия 77, 94
— пластидные 91
— повторяющиеся 55, 92
— рекомбинация 24, 25
— структурные 23, 147
— экспрессия
— ядерные 22, 23, 77, 147
Геном величина 146, 147
— митохондриальный 93
— полиплоидный 56, 92, 147, 184, 202
— растений 24, 25
— хлоропластный 76, 92, 93
Генетический код 63
Гетерофилия 58
Гипоксия 55, 56
Глобины 14, 166
Гомеостаз 49, 59, 129, 135, 150, 206—208
Гормональная система 157—159, 236
Деления гена 168 • ••
Дивергенция 59
— функциональная 8, 18, 26, 37; 52,
59, 96, 97, 193
— химическая 54, 55, 143
— экологическая 25
Дифференциация тела 31, 37, 39, 67, 72,
82,203,216
— значение для фотосинтеза 82, 83,
114
— надежность 190, 191
— саморегуляция 203
Дыхание 114
— анаэробное 71, 118, 121
— аэробное 71, 115
— дыхательная цепь 121—123
— отличие от фотосинтеза 131,132 '
— примитивные способы 115,117,135
— растений 121, 131—133
— цианидоустойчивое 121
Жизнь, дифференциация 118
— коллапсирования 64
— окислительно-восстановительный
потенциал 80, 81
— растительная 9, 38, 66, 240
— фотоактивность 67, 68
Жизнедеятельность, саморегуляция 45,
52, 212—214 .
«Западня» эволюционная 71
Засухоустойчивость 24, 41, 56, 191, 211
Избыточность ДНК 25, 55
— структур 222
Изменчивость генетическая 19, 20—24
— структур 225—227
— химическая 162, 163
Изомерия химическая 35
Изоферменты 147, 149, 150
Ингибиторы роста 154, 158
Инертность COj 107
Карбоксилирование 75—77, 84, 106
Каротиноиды И, 21, 70, 87, 122—126,
137, 168
Катехины 152—154
Каурен 21
Кванты роста 174, 175
Кислород 61, 112, 122
— агрессивность 115, 116, 122
-защита 113, 115, 122, 124, 126, 127
— мёТабОЛизация 115, 116. 121,124,
126
— происхождение 38, 69, 70, 114
— тйорец дифференцировок 115, 122,
126 ?
— токсичность 113, 115, 123 1
— толерантность 114, 122; 124-428
— энергетическое использование 112.
121
Клапан энергетический 123
— метаболизм 168
Космическая роль растений 62. 66, 67.
79, 113
Коэффициент продвинутости144,
— размножаемости 111 ‘ ! :
Латеральной перенос генов'156, 167
Леггемоглобин 14; 164, 167 i ; 4
Лигнин no; 15Х 1'56. »Й5Д67
Лист
—опадение 74; 179,180, 181
—- происхождение -48, ^4; 97
— форма 97, 191* ’
— функция 97,181
Литотрофия 70, 71
Металлы в растениях* 54, 58, 68, 80, 161
— роль 83, 80, 81, 86, 119
— содержание 21, 56, 62, 80, 95, 126,
208 ?
Методы изучения эволюций 10—15
— биогеографический 10
— генетические 10,15 1
— исторический 10 :
— молекулярный 10
— палеонтологический 10—13
— сравнительный 10, 12, 14
— фенотипические 10
— экстрасоматическиеД 1
— эмбриологический 11'
— этологический И
Механизм адаптаций 21, 144—146
— аэробные 121
—защитные 55
— компенсаторные 46, 55
— стабилизации 59, 208
— фотосинтеза 57 (
Мимикрия химическая 170, 171^-172
Митохондрий 129; 130 '
— возникновение 130
— значение 129 ; 1 ’
Молекулярная генетика 16
— палеонтология И, 12, 84
Мутации 56 *
— биохимические 53, 57, 58,; 69,166
247
— дефицита ГК 20, 21
— каротиноидов 21—23
— карликовости 20
— морфологические 20, 21. 23
— нежелтеющие 23
. —устойчивости 21, 22, 208, 214
— физиологические 20—23
— хлорофильные 22, 23, 92
— хромосомные 24
Направления эволюции 39
— арогенез 39. 62, 187, 217
— аллогенез 39, 162 ?
Наследование полигонное 24
— фотосинтетической активности 23
Недарвиновская эволюция 165—168,172
Новшества морфологические 31—33
Норма реакции 45, 49, 58, 156
— специфичность 145, 146
Нуклеиновые кислоты 145, 146, 181
— значение для таксономии 145—147
— повторы кодонов 146, 147, 166,
202 у
Обмен веществ 115
— биологический 115—120
— Эволюция 146 '
Озоновый экран 115, 122, 124
Онтогенез растений 196
— автономизация 50,186, 210,218,221
— акцелерация 222
— дифференциация 197, 202, 215
— индивидуум 197—204, 210, 215
— интеграция 200, 219. 220
— микроэволюция 196, 232
— надежность 45, 192, 196, 198, 214—
219
— олигомеризация 200, 219, 220
— основа филогенеза 230—233
— организованность 200, 214 .
— периодизация 197
— полимеризация 220, 222
— регуляция 59, 208—214
— саморегуляция 206, 207
— целостность 198, 200, 202, 209, 210
— эволюция 197
— эмбрионизация 198, 222
Организация предбиологическая 61, 62
Органотрофная ветвь 94, 112
Органическое вещество 64—66, 76, 82.
85, 98, 106, 217
Отбор 73
— гомеостатичность 59
— естественный 8, 10, 12. 16, 19, 37,
43, 44. 52—58, 67, 72, 98, 194
— движущий 40
— комплекс признаков 192—194, 201.
202, 216
— механизм 119, 201
Палеофизиология 12—14
Палеоэкология 13
Пентозофосфатный путь 120, 137
— возникновение 120, 121
— значение 120
Пектиновые вещества 152—155
Пептиды регуляторные 53, 54, 164
Периодические явления 173
— значение 176—177, 181
— распространение 174, 179, 180
— эволюция 175, 179—183
Пероксисомы 131, 138, 141, 142
Пигментная система 81
— разнообразие 70, 85, 88
— эволюция 84—87
Пирбфосфаты 81, 116, 119
Пластичность формы 5Й, 59, 70, 75
Пластохиноны 73, 90, 152, 153
Позиционный контроль 157
Полифосфаты 81, 119
Порфирины 51, 53, 63—68, 84—86
Преадаптация 9, 111, 161, 165, 167, 187
Предварение химическое 45, 53, 62.
167—168, 172
Принцип Гаузе 118
Принцип блочный 45
Прокариоты фототрофные 12, 68—71,
122, 125
Протобионтный рой 61. 62. 66. 72. 112
116—119
Протонный насос 68, 71, 89, 116
Профиль биохимический 145, 162
Растения, геометрия 58, 74, 173, 188
—целостность 74,83.159,194.206.209.
219
Рацемация 64
Реакция оксиредукции 116
— подталкивания 48
— сверхчувствительности 203
Регуляторы роста 19, 20, 150, 154—156
— абсцизовая кислота 150, 154, 160
— ауксины 150, 154, 160
— гиббереллины 36, 150, 154,160
— действие 154—161
— значение 150, 154. 159, 161
— ингибиторы 158, 159, 204
— цитокинины 154, 160
— чувствительность 156, 157
— эволюция 155. 156, 160
248
— этилен 154—157
Резерв наследственности 25
Рекапитуляция 96, 151, 168, 192—193,
225—229
Рекомбинация 24
Рестрикционный анализ 14
Ритмы годовые 173
— космичЬвкие 173
— сезонные 173, 175—177, 179—180
— суточные 173, 175
— цикадные 175, 176
— эндогенные 74, 173—175
Симбиогенез 67, 68, 93, 94, 125, 130, 141
Симметрия 63, 64
Синтетические гибриды 24
Скелетизация 147 .
Скороспелость 41, 175, 183—184, 205,
221
Солеустойчивость 148, 167, 208, 211
Сопряженность эволюции 163, 164, 169
Специализация веществ 101, 162
Старение 162, 165, 180, 181, 204—206
Стерины 11, 153
Стресс 58, 59, 136, 137, 160, 192, 211
Суперген 24
Теория «нейтрализма» 168
Типы питания 54
— автотрофный 61, 64—74, 79
— гетеротрофный 54, 61—71, 81, 94
— миксотрофный 61, 66
— эволюция 66—72
Типы ДНК 145—157
— фотосинтетических систем 72, 73
— устьиц 31, 32
Толерантность, гипоксия 26, 55, 212—
214
— комплексная 134, 214
— металлы 21, 58, 162, 208
— патогены 149, 151, 154, 163, 202
— перекрестная 162
— системная 229—232
Трагедия экологическая 55
Убихиноны 75, 116, 152, 153
Углерод, земной цикл 64
— сток 101
Универсальный принцип регуляции 50,
51, 82, 106, 177, 206
Уравновешенность экологическая 212,
213,218
Уровни усиления функции 26, 34, 73
Уропорфирины 85
Ферменты
— гликолатдегидрогеназа 139
— каталаза 125, 126, 128, 158
— кауренсинтетаза 21
— малатдегвдрогеназа 55, 57, 110, 147
— малик-энзим 21
— метилтрансфераза 86
— пероксидаза 125, 126, 128, 133, 156
— супероксвдцисмутаза 115, 125, 126,
167
— РБФК-аза48, 49, 51, 54, 75—79, 84,
122, 137
-а-трансальдолаза 120
— транскетолаза 120
— ФЕПК-аза 21, 65, 76, 84, НО
— фосфофруктокиназа 52, 118
— хитиназа 156
— хлорофиллаза 22
— хлорофиллоксидаза 23
— цитохромоксидаза 121, 128, 132,
133, 135
— эволюция 54, 77, 147
Фенольные соединения 152
Ферредоксины 70, 71, 81, 90, 116
Физиологическое «зло» 16, 142
Физиологический признак 18, 163
'—свойство 18, 22, 25—27
Физиологические часы 175, 178, 240
физиологический нуль 211
Филогенетические связи ! 45, 146
Фитоалексины 21, 22, 153, 165
Фитохром 52
Флавины, флавоноиды 68, 75, 126, 133,
142
Флоэмный транспорт 33, 34
Формы эволюции 39
— дивергенция 40
— конвергенция 40—^43, 68, 71, 161
— параллелизмы 40, 41, 43, 74, 136
— филетическая эволюция 40, 70
Фосфорилирование 116
— субстратное 116—119, 126
— окислительное 119, 121, 123
Фотогидротрофия 71, 75
Фотодыхание 137
— значение 138—140
— механизмы 137—139
— специфика видов 139, 140
— эволюция 140—142
Фотолитотрофия 71, 112
Фотоорганотрофия 68, 71
Фотопериодизм 174, 182
— генетика 183—184
— значение 184—185, 186
— распространение 183, 184
249
—эдолюцияЛ83х185,: 186, , . .
Фоторедукция 69*> 7Q, 80, 112, 142
Фотосинтез 20, 67 ...
. .— биологический процесс. 96, 97
' —возникновение ЭТЦ 97, М.
— единица 97 - - ! *
— кооперативные механизмы 77,
107—110 .
— САМ-растения 84 4
— окислительнр-восстановительнЫе
реакции 77, 80, 84,112 \
— ограничения 76, 83, 84j 97, 107
— реакционные цёнтры 82, 86, 91, 97
— Сз и С4-растения 70, 78, 84,' 105—
108
—^системный контроль 74, 95
— стабилизация энергии 76, 79—84,
103
— типы 81, 82
— тропические растения 100
— уровни 99, 104—106, 121
— физиология 74, 76, 97, 101
— эволюция 69—75, 79, 97—99, 164,
201, 236
— экология 100—107
— эндогенная регуляция 174, 205, 206
— этапы 70, 71
Фотосистемы 69—73, 81, 90
— мутации 82
— эволюция 73, 82, 90, 117
Фототрофная ветвь 67—70
— эволюция 71—74
Фотохимические акцепторы 67, 68, 72
—доноры 70
— центры 89
Фосфогликолат 139
Функция живого 9
— проводящая 32, 33
Функциональные принципы 44—51
— автономизация 49, 50
— компенсация 44, 46, 47, 76
— множественность 44, 45
— ослабление 19, 44—46
— стабилизация 44, 49, 50
— усиление 19, 40—46
Хемосинтез 70, 71, 84, 112, 113
— значение 71, 112
— эволюция 112
Хемосистематика 144
Хемотрофы 112
Хемофоссилий 12
Химический состав 143
— влияние на форму 150, 168
—дивергенция > видов 143, 144
— изменение в онтогенезе 149—153
— изомерия 35, 149, 164
— индивидуальность 148, 163, 166
— конвергенция 152, 163—166, 172
— предрасположение 45,119,161,172
— Приспособленность 152, 153, 166
— скорость изменения 146, 168
— специализация 144, 145, 155, 161—
f 166, 170
—тенденция изменения 144, 172 ‘
— универсальность 143
— эволюция 143, 147
Хиральность 63
Хитин 146,156 ;
Хлоропласт 89
— адаптивность 92, 100
— ДНК 90—94
—дивергенция 91, 95, 96
— организация 35, 89—91, 95, 203
— происхождение 93, 94, 96
— функция 89, 95, 203
Хлорофилл 70
— зародышей 49
— концентрация 91
— оборачиваемость 87, 89
— плодов 49
— синтез 22, 50, 164
— функция 86—89
— эволюция 86, 89
Цикл Арнона 89, 106
— глиоксалатный 137’—141
— Кальвина 70, 78, 106, 112, 137
— Кребса 89, 119, 121, 122, 125, 139
Цитохромы 75, 89, 128, 129, 152, 166
Челночный принцип 109
Щавелевоуксусная кислота 36, 55, 75
Шикиматный путь 148, 153
Эволюция 18
— биохимическая 11, 14, 30
— механизмы 196, 201—203
— окислительно-восстановительные
реакции 79—81
— онтогенеза 8, 49, 196—208
— парадоксы 25, 32, 52, 97, 98, 242
— предбиологическая 45, 62—64,168,
183
— ситовидных пластинок 33, 34
— скорость 146, 166, 168
— сопряженная 168—170, 171, 203/
204 . '
— сосудов 32, 33, 51
250
— функциональная 7—12, 14, 18, 48,
49, 233, 239
— химическая 144—148
— экологическая 57,79,100—106,191,
196
— экосистем 98, 168—172
— этапов генераций энергии 125
— энергетическая 6, 100, 142
Эволюционный подход 9, 15,16
Эволюционная фитофизиология 4—6,
15—17
— задачи 7—9
Экспансия экологическая 43, 61, 79, 96
ЭТЦ дыхания 75, 93
— фотосинтеза 75, 79, 93, 97, 196
Энергетическая двойственность 64
— победа 57, 89, 106, 111, 114
— совершенство 217, 240
— стабильность 60, 113, 115, 132—135
— эффективность 72, 82,100,106,118,
119
Энтропия 147
Эубактерии 146
Эфемеры 74, 179, 181, 196, 212
Эффект Варбурга 137
— суммации 157
—тепличный НО
Яблочная кислота 36
Ядерная война 55
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
Agavaceae 108
Agrobacterium tumefaciens 167
Agrostis tenuius 21, 161
Allium ursinum 224
AUoteropis semialata 109
Allophyflum gilioides 169
Amaranthus graecitens 109
Amygdalis 162
Anacystis nidulans 70, 146
Andzena 171
Angiopteris evecta 98
Anemone caucasica 224
araliaceae 162 .
Aralia californica 78
Arum maculatum 170, 224
— nigrum 170
Atriplex patula 78
Atta cephalotes 170
— octospinosus 170
Bacillus 14
Batrachium
— eradicatum 56
— peltatum 56
Betulaceae 164
Beta lomatogona 14
— maritima 14
— patellaris 14
— procumlens 14
— trigyna 14
- ^vulgaris 14
Betula pendula 55, 182
Bignonia capreolata 194
Bromeliaceae 108
Cactaceae 108
Calligonum caput-medusae 74
Campanula rotundifolia 225
Carex 211
Catharanthus roseus 151
Cerasus 162
Centrospennae 162
Chloroflexis 28
Chenopodiaceae 108
Chenopodium album 109
Chromatium 78
— throsulfatophyllum 78
Chyanophyta 12, 69, 70, 73
Clavaria 197
252
Cladophora glomerata 179
Clostridium pasteurianum 14
Crassulaceae 108
Cruciferae 162
Cycas 144
Cyelotella 104
Cystoseria crinita 38
Danaus plexippus 171
Desulfomaculatus nigrificans 119
Dictieriaceae 162 '
Disporum smithii 78
Dremys winteri 98
Echinocystis lobata 194
Elaegriaceae 164
Eleusine tristachya 145
Elytriga 177
— repens 177
Encephalarto$ 98, 144 ,
— 7 alensteinii 98
Equisetum limosum 133 \‘
Eragrostis minor 145
Escherichia coli 129
Euglena gracilis 54, 146
Eulaema 170
Fagus sylvatica 182
Festuca ovina 21, 161
Fomes formentarius 198
Fouquieria splendens 46
Fritschiella tuberosa 179
Ginkgo biloba 98
— orientalis 13
Glaucophyta 93
Gonyaulax polyedra 177
Halobacterium halobium 43
Haloxylon persicum 74
Helminthosporium turcium 21
Heracleum sosriowskyi 164
Hyacinthus orientalis 224
Hydrogenomanas eutropa 77
— facillis 77
Isoetes 110
Kalahchoe blossfeldiana 176
Larix decidua 182
Larrea divaricata 28
Liliaceae 108
Limnophylla heterophylla 225
Litterella uniflora 110
Macrozamia Г44
Magnolia grandiflora 98
Marasmius oreades 197
Menyathes trifoliate 133
Mesembryanthemum crystellinum 110,
Metasequoia distica 13
Microsteris gracilis 169
Mucor 197
Mycobacterium 14 '
Mycoplasma 14
Myricaceae 162, 164
Myristicaceae 162
Oenanthe aquatica 225
Oenothera aiganensis 57
—biennis 57
Oleraceae 162
Ophrys muscifera 170
Oiyza sativa 109
Omithogalum areatum 224
Oscillatoria rubens 104
Palmae 162
Passiflora gracillis 194
Panicum 139
— decipien 139
— miliaceum 139, 145
—milioides 139
— priontis 139
— riculare 139
— schenckii 139
Penicillum 197 >
Pennisetum thyphoides 23
Phaseolus multiflorus 100
Picea abies 182
Piptoporus betulianus 198
Plantego lanceolate 78
Poa 145 1
Poaceae 108
Polemoniaceae 169
Populus pruinosa 74
— trichocappa 182
Portulaca oleracea 109
Poiyphyridium 94
Prangos 165
Pruniphora 162
Prunus 162
207 Pseudomonas 14, 170
Pseudomyrmex 170
Quercus robur 182
Ramondia 211
Ranales 228
Ranunculus delphinifolius 225
Rhoeadales 228
Rhodophyta 93
Rhodospirillales 114
Rhodospirillum rubrum 78, 117, 152
Rhodopseudomones spheroides 75
Ricinus communis 100
Sagittaria sagittifolia 225
Scataria itelica 109
Sceale 147
Scenedesmus acute 104
Senecio 170
Seseli 165
Sjcyos angulatus 195
. Silene cucubalus 162
Solidago spathulata 78
Stipa 145
Stipeae 144
Streptococcus facalis 128
Synechocystis 94
Taphrinia epiphylla 159
Thermus aquaticus 14
Thypa latifolia 162
Triticum aestium 109
Tropaellum major 100
Umbelliferae 162
Ulmaceae 164
Vinca rosea 166
Viola glabella 78
Vitaceae 108
Zea majus 109
ОГЛАВЛЕНИЕ
СПИСОК СОКРАЩЕНИЙ............................................... 3
ПРЕДИСЛОВИЕ..................................................... 4
Глава 1. ПРЕДМЕТ, МЕТОДЫ И ИСТОРИЯ РАЗВИТИЯ ЭВОЛЮЦИОН-
НОЙ ФИЗИОЛОГИИ РАСТЕНИЙ................................... 7
1.1. Предмет и задачи эволюционной физиологии растений.. 7
1.2. Методы эволюционной фитофизиологии...................... 10
1.3. Краткий очерк истории развития эволюционных идей в физиологии
растений................................................... 15
Глава 2. ОСНОВНЫЕ ТЕНДЕНЦИИ ФУНКЦИОНАЛЬНОЙ ЭВОЛЮ-
ЦИИ РАСТЕНИЙ ............................................ 18
2.1. Определение понятий . ................................. 18
2.2. Генетическая изменчивость функциональных проявлений .... 19
- 2.3. Классификация физиологических свойств и адаптаций....... 25
2.4. Многообразие форм и функций растений. Взаимосвязь формы и
функции .................................................. 30
2.5. Основные направления и формы функциональной эволюции рас-
тений ..................................................... 39
2.6. Принципы преобразования функций в ходе эволюции растений . 44
2.7. Достижения функциональной эволюции растений............. 51
2.8. Естественный отбор и функциональная эволюция растений ... 53
2.9. Пластичность растений в онтогенезе и ее значение........ 58
Глава 3. ЭВОЛЮЦИЯ АВТО- И ФОТОТРОФНОГО МЕТАБОЛИЗМА . 61
3.1. Общие представления о возникновении обмена веществ. 62
3.2. Сущность растительной жизни. Гипотезы о становлении и развитии
автотрофного питания..................... ............. . . 66
3.3. Возникновение и развитие фототрофности.................. 71
3.4. Возникновение электрон-транспортной цепи и механизма кар-
боксилирования ........................................... 75
3.5. Стабилизация механизма фотосинтеза. 79
3.6. Эволюция пигментной системы фотосинтеза................. 84
3.7. Хлоропласты. Их значение и эволюция .................... 89
3.8. Возникновение листа как этап биологической эволюции
фотосинтеза................................................ 96
3.9. Экология фотосинтеза как показатель его биологической эволюции 100
3.10. Эволюция и приспособительное значение кооперативных механиз-
мов фотосинтеза........................................... 106
3.11. К вопросу об эволюционном положении хемосинтеза... 112
Глава 4. ЭВОЛЮЦИЯ МЕХАНИЗМОВ ДЫХАНИЯ У РАСТЕНИЙ ... 114
4.1. Особенности энергетики организмов разных царств и древние формы
ее проявления............................................. 115
4.2. Кислород как фактор эволюции жизни на Земле............ 122
254
4.3. Первичная Защита организмов от кислорода и возникновение аэроб-
ности .................................................... 126
4.4. Возникновение митохондрий ............................... 129
4.5. Особенности дыхания у растений.......................... 131
4.6. Дыхание и приспособление растений к среде , s р2
4.7. Роль фотодыхания, у..............................• ? • • Щ
Глава 5. ХИМИЧЕСКИЙ СОСТАВ РАСТЕНИЙ И ЕГО ПРИСПОСОБИ-
ТЕЛЬНОЕ ЗНАЧЕНИЕ . . ... . . . . ... . . Ч ; . 143
5.1. Сравнительная характеристика химического состава растений и
общие тенденции его эволюций . . . ...... . . .'. . ... 144
5.2. Изменение химического состава растений В онтогейезе ..... 149
5.3. Экологическая роль катехинов и пектиновых веществ . . . . . " 152
5.4. Экологическое значение фигогормойов и ийгибйТороВ роста ? ч . 154
5.5. Специализация и конвергенция растений по химическому составу 161
5.6. Индивидуальная изменчивость химического состава растений и во-
прос о «недарвиновской» эволюции ............... 165
5.7. Роль изменчивости химического состава растений в эволюции
Глава 6. ПЕРИОДИЧЕСКИЕ ЯВЛЕНИЯ В ЖИЗНИ РАСТЕНИЙ. . . . 173
6.1. Биологическое время и его измерение у растений . . . . . . ?. 173
6.2. Циркадные ритмы и их значение................................\ 175
6.3. Сезонные циклы и их приспособительное значение . . 179
6.4, Происхождение и эволюция фотопериодической реакции .... 182
Глава 7. НАЗЕМНАЯ СРЕДА.И ЭВОЛЮЦИЯ РАСТЕНИЙ............., . 187
7.1. Водообмен и приспособления растений . , . . . ч ; • 189;
7.2. Движения растений. Способы, предпосылки возникновения и зна-
чение ............................................ , , 193
Глава 8. ЭВОЛЮЦИЯ ОНТОГЕНЕЗА РАСТЕНИЙ . . . . . . .-. . 196
8.1. Общая характеристика онтогенеза растений . . . ... . . 196
8.2. Индивидуум у растений и его функциональная целостность* ... 199
8.3. Эволюция продолжительности жизни индивидуума .............. . 204
8.4. Развитие гомеостаза в связи с эволюцией онтогейеза ....... 206
8.5. Регуляторная система индивидуума. Ее развитие и значение . . . 209
8.6. Надежность онтогенеза растений и некоторые тенденции их
функциональной эволюции.................................... 215 .
8.7. Основные направления эволюции онтогенеза растений.. 219
8.8. Рекапитуляция признаков и функций в онтогенезе растений ... -225
8.9. Онтогенез — основа филогенеза. Эволюция растений в будущем . 229
8.10. Системный контроль регуляции жизнедеятельности в онтогенезе
растений и его эвоюционные предпосылки.....................233
ЗАКЛЮЧЕНИЕ...................................... ... . . 240
РЕКОМЕНДУЕМАЯ ЛИТЕРАТУРА............................................. 243
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ ................................................ 246
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ ......................................... 252
Учебное издание
Юсуфов Абдул малик Гасамутдинович
ЛЕКЦИИ ПО ЭВОЛЮЦИОННОЙ ФИЗИОЛОГИИ РАСТЕНИЙ
Редактор Л. А. Савина
Художественный редактор Т. А. Коленкова
Младший редактор Г. Г. Бунина
Технический редактор В. М. Романова
Корректор Г. И. Кострикова
Компьютерная верстка Г. А. Шестакова
Оператор Г. А. Шестакова
ИБ № 10317
ЛР № 010146 от 25.J2.91. Изд. № ХЕ-154. Сдано в набор 14.02.96. Подп. в печать
30.05.96. Формат 60x88 /1$. Бум. тип. № 2. Гарнитура «Таймс». Печать офсетная.
Объем 15,68 усл. печ. л. 15,92 усЛ. кр.-отт. 16,40 уч.-изд. л. Тираж 3 000 экз.
Заказ №157.
Издательство «Высшая школа», 101430, Москва, ГСП-4, Неглинная ул., д. 29/14.
\
Набрано на персональном компьютере издательства.
Отпечатано в ОАО «Оригинал», 101898, Москва, Центр, Хохловский пер., 7