





























































































































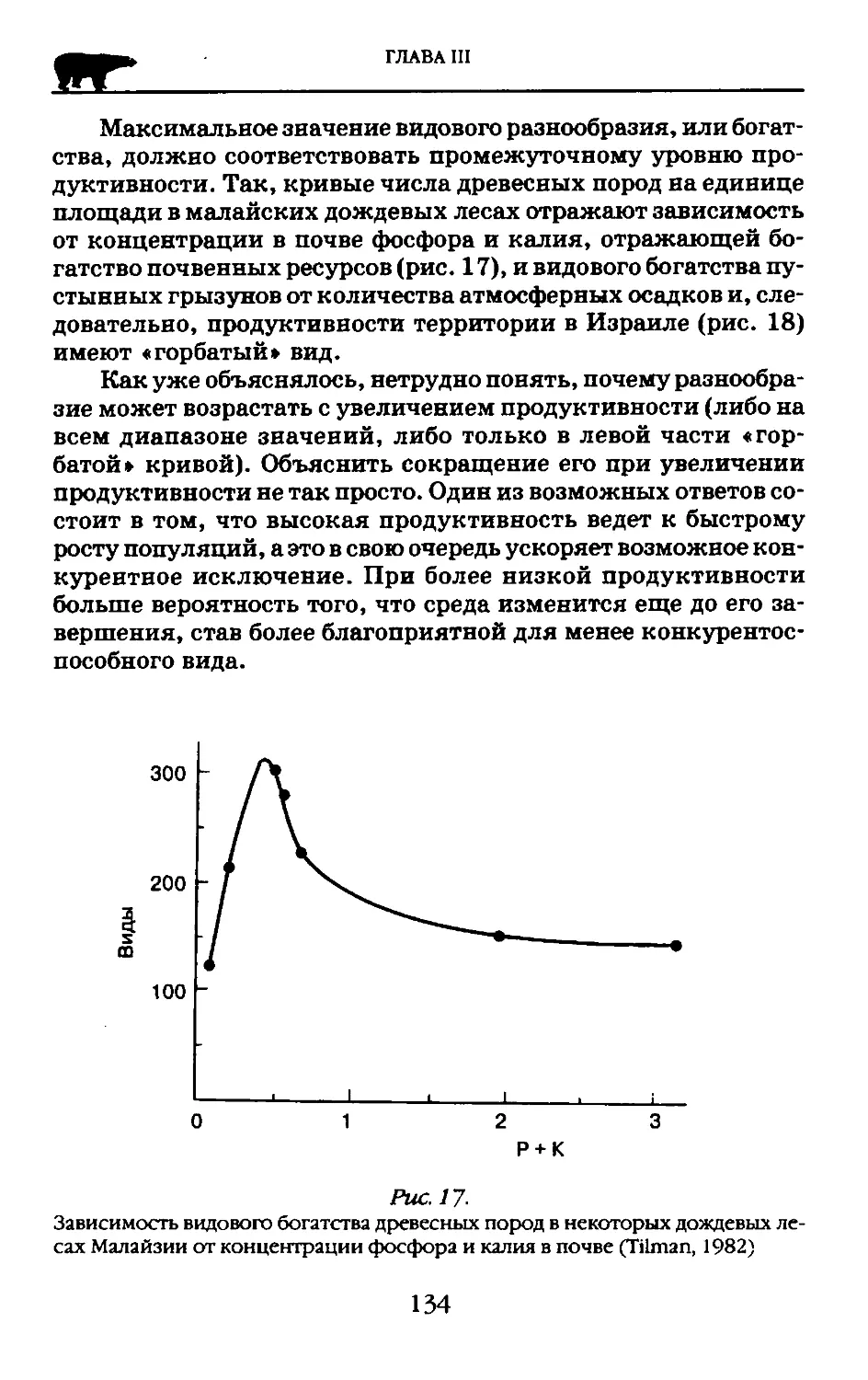









































































































































































































































































Author: Криволуцкий Д.А. Лебедева Н.В. Дроздов Н.Н.
Tags: общая экология биоценология гидробиология биогеография экология и биогеография охрана живой природы география экология учебное пособие
ISBN: 5-691-01098-0
Year: 2004




УЧЕБНОЕ ПОСОБИЕ ДЛЯ ВУЗОВ
Н.В. ЛЕБЕДЕВА, Н.Н. ДРОЗДОВ, Д.А. КРИВОЛУЦКИЙ
БИОЛОГИЧЕСКОЕ
РАЗНООБРАЗИЕ
Допущено Министерством образования Российской Федерации в качестве учебного пособия для студентов высших учебных заведений, обучающихся по направлению география и специальности география
Москва ’жммамямы! а
~\ВЛАДОС
2004
УДК 574(075.8)
ББК 28.085я73
ЛЗЗ
Лебедева Н.В., Дроздов Н.Н., Криволуцкий Д.А.
ЛЗЗ Биологическое разнообразие: Учеб, пособие для студ. высш. учеб,
заведений. — М.: Гу манит, изд. центр ВЛАДОС, 2004. — 432 с.: ил.
ISBN 5-691-01098-0.
Агентство CIP РГБ.
В учебном пособии рассмотрены основы теории биологического разнообразия и методы его оценки. Раскрыты вопросы, связанные с уровнями, географией и таксономией биоразнообразия. Большое внимание уделено взаимодействию человека н живой природы.
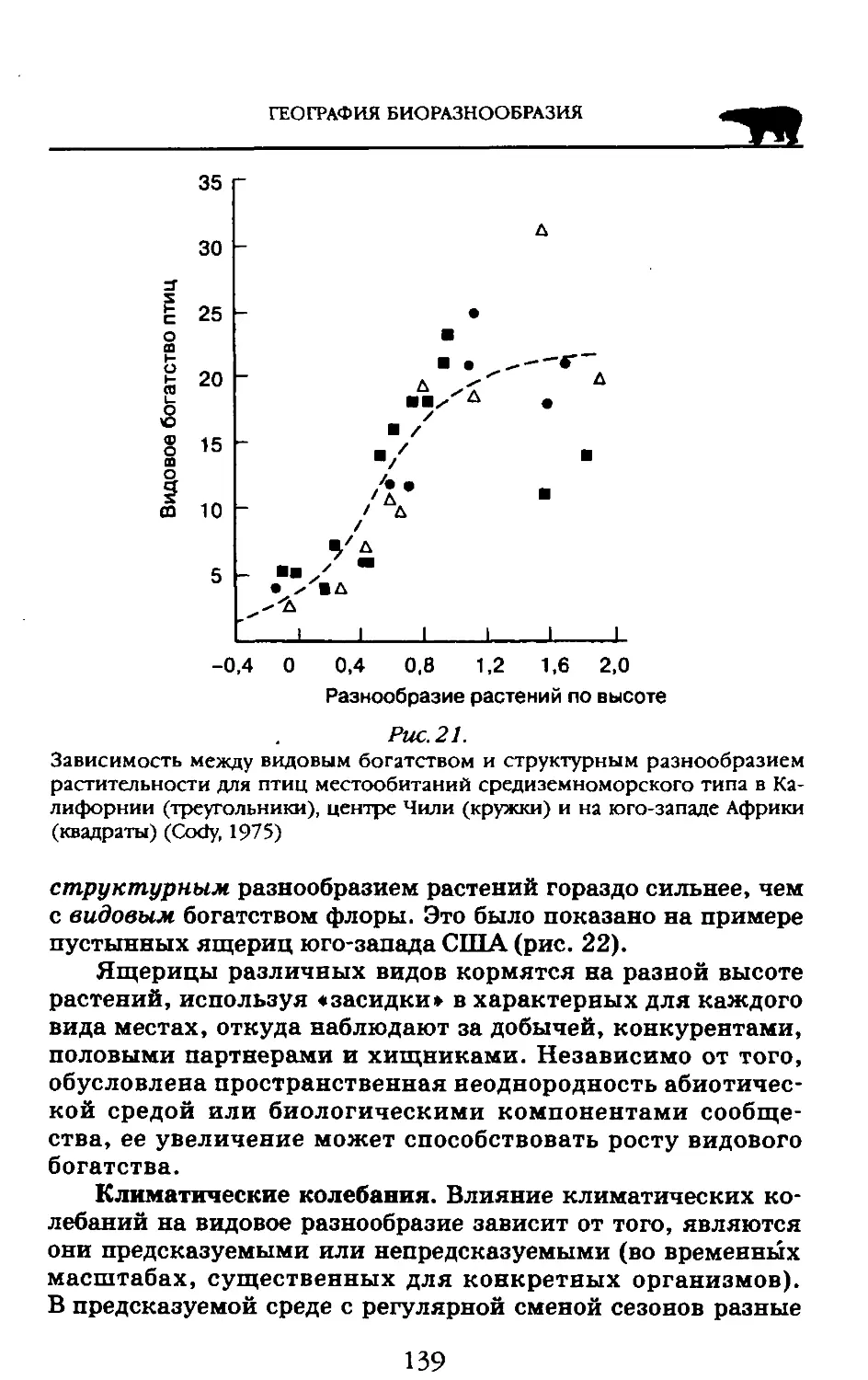
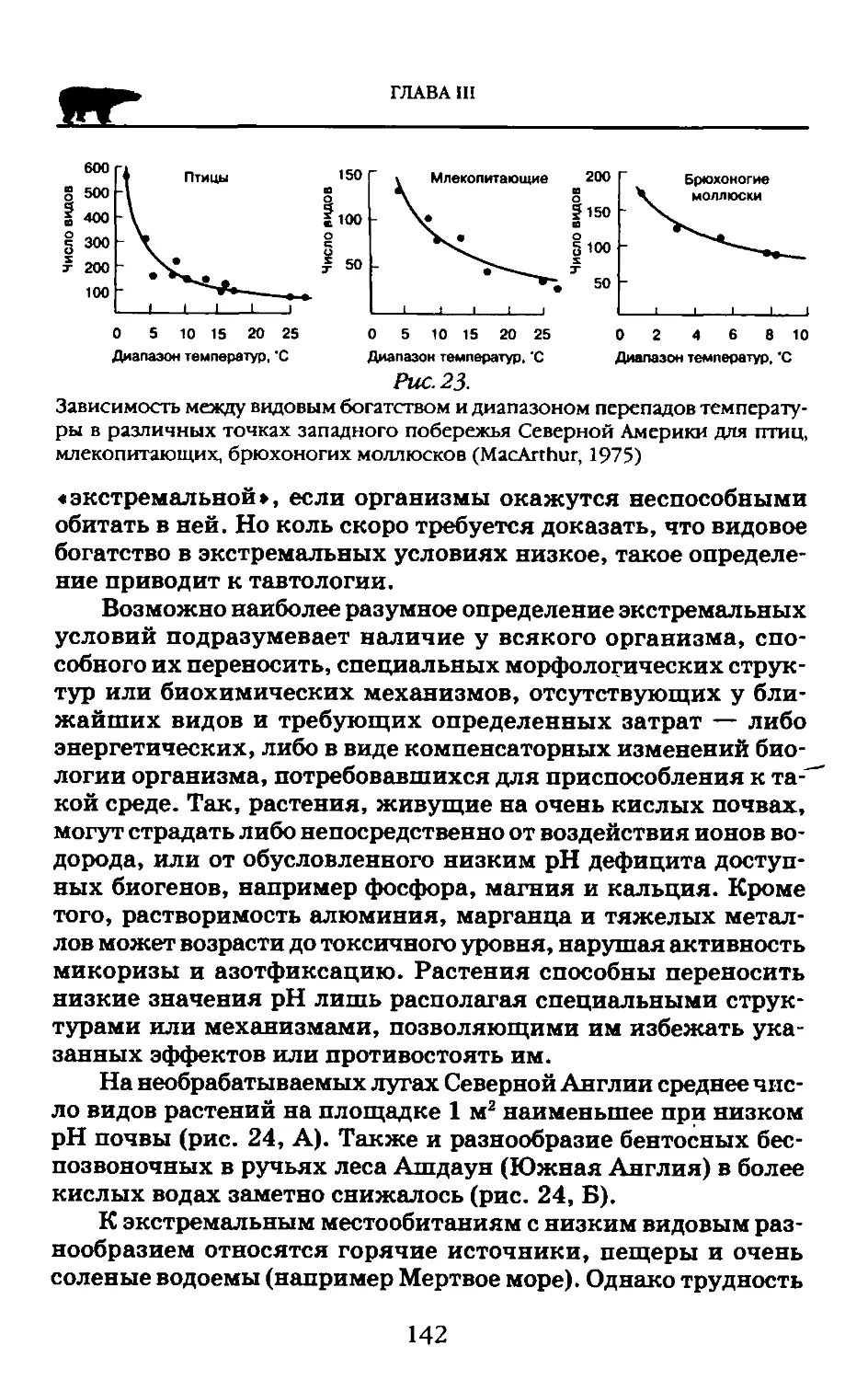
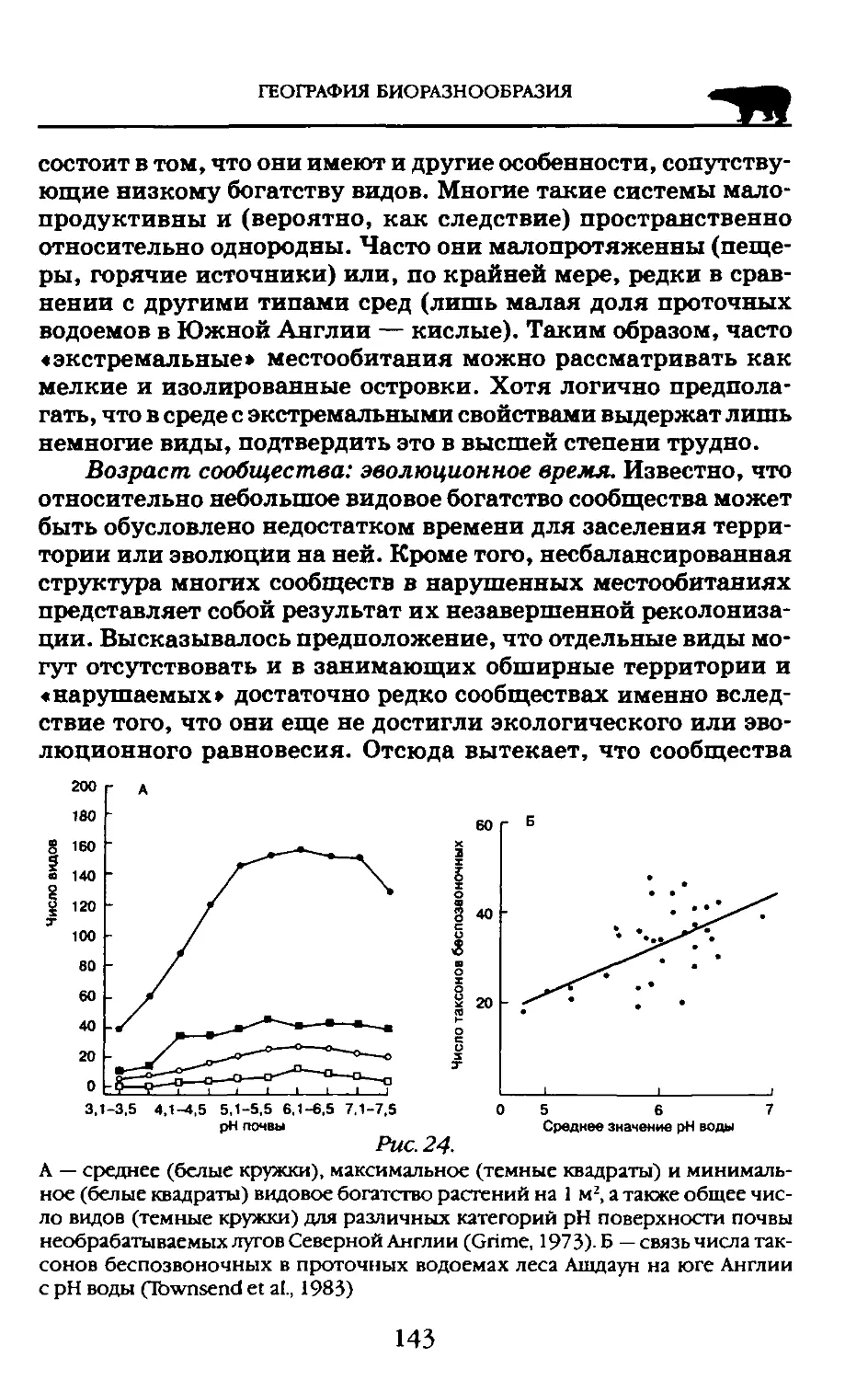
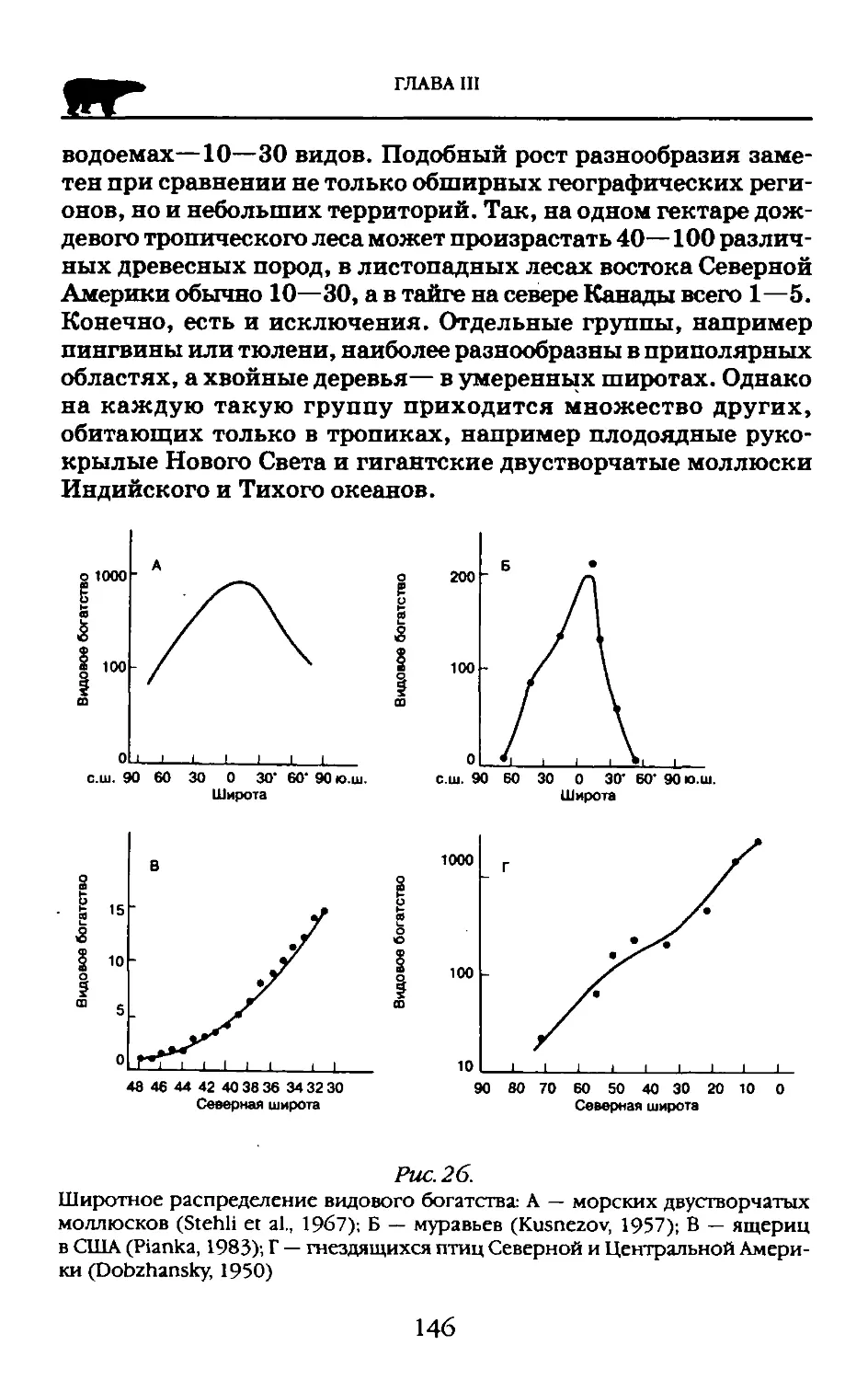
Пособие адресовано студентам высших учебных заведений обучающихся по экологическим н географическим специальностям.
УДК 574(075.8)
ББК 28.085я73
ISBN 5-691-01098-0
© Лебедева Н.В., Дроздов Н.Н., Криволуцкнй Д.А., 2004
© ООО «Гуманитарный издательский центр ВЛАДОС», 2004 © Серия «Учебное пособие для вузов» и серийное оформление.
ООО «Гуманитарный издательский центр ВЛАДОС», 2004
© Художественное оформление. ООО «Гуманитарный издательский центр ВЛАДОС», 2004
© Макет. ОСЮ^Гуманитарный издательский центрВЛАДОС», 2004
Учебное издание
Лебедева Наталья Викторовна, Дроздов Николай Николаевич, Криволуцкий Дмитрий Александрович
Биологическое разнообразие
Учебное пособие для студентов высших учебных заведений
Зав. редакцией СЛ. Платонов Редактор СЛ. Сороколетова Зав. художественной редакцией ИА Пшеничников Художники ВЛ. Филиппов, АЛ. Щетинцева, О А Филонова Художник обложкв О А Филонова. Макет АЛ. Щетинцевой Компьютерная верстка ОЛ. Емельяновой Корректор ТЯ. Кокорева
Отпечатано с диапозитивов, изготовленных ООО «Гуманитарный издательский центр ВЛАДОС».
Лицензия ИД № 03185 от 10.11.2000.
Санитарно-эпидемнологическое заключение № 77.99.02.953.Д.006153.08.03 от 18.08.2003. Сдано в набор 10.06.03. Подписано в печать 01.12.03. Формат 60x90/16. Печать офсетная. Бумага газетная. Усл. печ. л. 27. Тираж 10 000 экз. (1-й завод 1-5 000 экз.). Заказ № 1656
Гуманитарный издательский центр ВЛАДОС. 119571, Москва, просп. Вернадского, 88, Московский педагогический государственный университет. Тел. 437-11-11, 437-25-52, 437-99-98; тел./факс 735-66-25.
E-mail: vlados@dol.ru http://www.vlados.ru
ООО «Полиграфист». 160001, Россия, г. Вологда, ул. Челюскинцев, 3.
ПОНЯТИЕ БИОРАЗНООБРАЗИЯ
Разнообразие жизни издавна было предметом изучения. Первые системы живой природы, известные, например, из трудов Аристотеля (384-322 гг. до н. э.), уже относятся к анализу этого явления. Научная и методическая база для описания биоразнообразия была создана К. Линнеем в его «Системе природы» (1735). Замечательная работа Ч. Дарвина «Происхождение видов путем естественного отбора» (1859) в корне изменила представления о природе. Она завершила длительные поиски естествоиспытателей и систематиков, которые пытались найти причину для объяснения многих черт сходства и различий у наблюдаемых организмов. Тщательно и объективно подобранные им доказательства того, что виды не «неизменны», что они изменялись и изменяются, иными словами, что эволюция действительно существует, были так многочисленны, что опровергнуть их было практически невозможно. Не менее важен тот факт, что Дарвин сумел дать логическое и убедительное объяснение возникновению этих изменений, т. е. естественному отбору. Дарвин опять-таки не был первым, предложившим концепцию естественного отбора, однако именно он впервые по-настоящему
оценил связь между естественным отбором и наследственными изменениями популяции. В этом и заключался его успех; он не только сумел показать существование изменчивости, но и объяснил, каким образом она возникает.
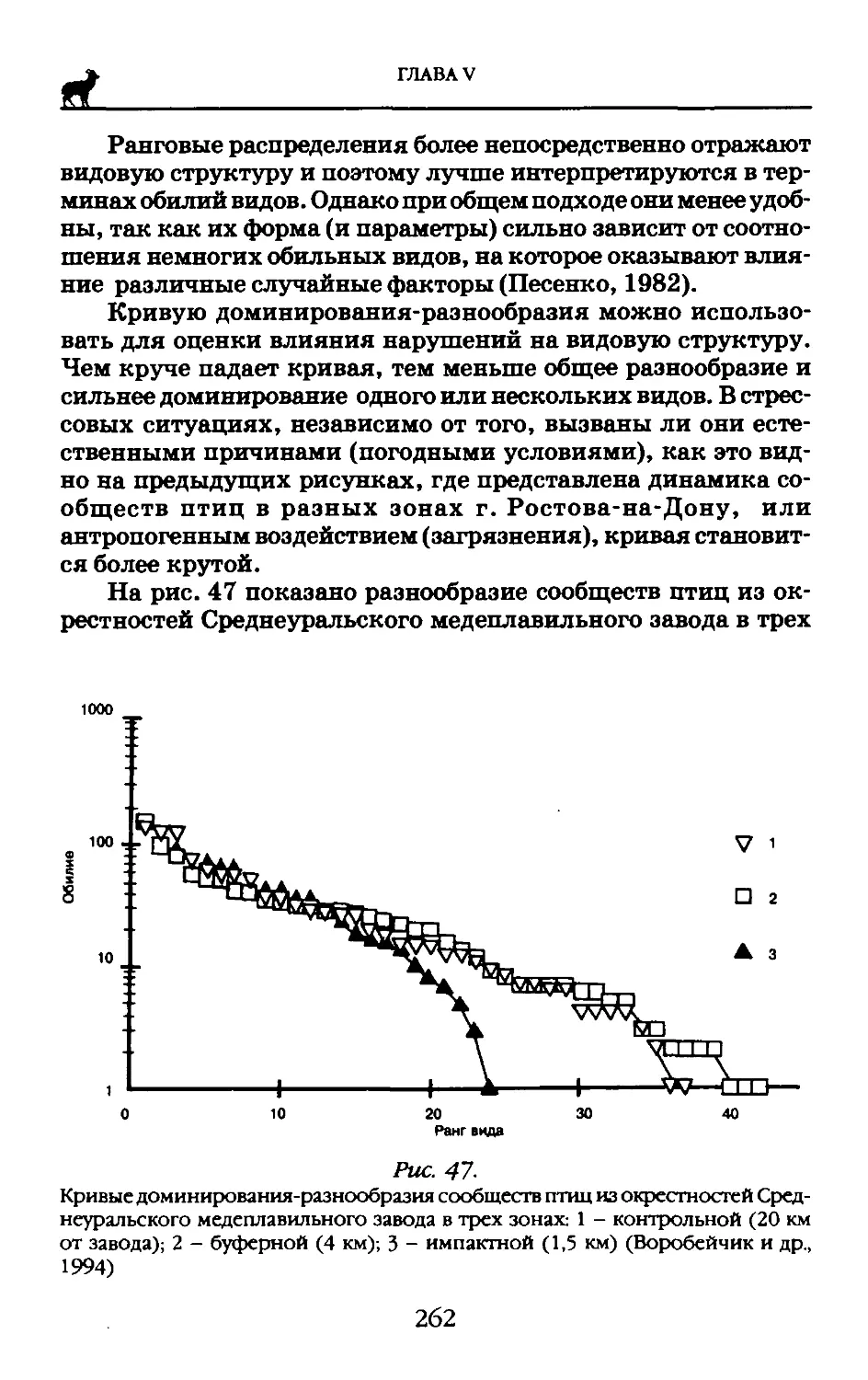
Как следствие дарвиновской теории эволюции М. Вагнером в 1868 г. была предложена концепция географического видообразования. Но еще в 1844 г. Ч. Дарвин в результате наблюдений за фауной Галапагосских островов пришел к заключению, что изоляция представляется здесь главным элементом. С тех пор ботаниками и зоологами собран огромный фактический материал, доказывающий реальность появления новых биологических форм путем пространственного разобщения.
Эволюционное учение нашего времени, которое называют «неодарвинизмом», содержит ряд положений, которых не было в первоначальной дарвиновской теории. Главной формообразующей силой по-прежнему считается естественный отбор, хотя наши понятия о корпускулярной природе генов дают нам возможность более полно представить себе возникновение изменчивости в результате мутаций, сохранение изменчивости в скрытом состоянии в диплоидных организмах, перетасовку генов в процессе генетической рекомбинации, обеспечивающую постоянный источник новых генных сочетаний, на которые мог бы действовать естественный отбор. Эволюция состоит из двух стадий: возникновения изменчивости и изменения направления этой изменчивости под действием естественного отбора.
Сейчас доказано, что изменения, возникающие под действием естественного отбора, могут иметь разные последствия в зависимости от условий существования. Один процесс наблюдается в том случае, когда условия среды, определяющие естественный отбор, весьма однородны на всем протяжении ареала вида или популяции. При этом приспособленность вида к своей среде неуклонно возрастает, а в случае изменения этой среды изменяется и вид в целом. За достаточно продолжительный период времени таким путем могут возникнуть весьма заметные изменения. Следовательно, генетическая структура отдельного ряда последовательных поколений постепенно и равномерно изменяется от поколения к поколению. Этот процесс называется филе гНичес кой эволюцией. Другой путь эволюционной дифференцировки наблюдается в том случае,
когда разные популяции одного вида тем или иным образом изолируются друг от друга и оказываются в разных условиях среды. Поскольку при этом естественный отбор действует на них по-разному, в разных популяциях возникают различные изменения. Таким образом, изолированные популяции будут все более дивергировать, пока, наконец, единый исходный вид не распадается на два или более новых вида. Описанный процесс известен под названием видообразования.
Видообразование имеет очень важное значение: оно ведет к дифференцировке, т. е. к дроблению одной генетической популяции на ряд подгрупп, каждая из которых представляет собой независимую эволюционную линию со своими возможностями для дальнейших филетических изменений. В то же время дивергенция непрерывна: она не прекращается после того, как данная группа достигла ранга вида, а продолжается дальше, приводя к возникновению более высоких таксономических категорий.
Оценки уровня биологического разнообразия Земли впервые были предприняты биогеографами, которые в XVIII-XIX вв. разработали схемы ботанико-географического и зоогеографического разделения поверхности нашей планеты по степени своеобразия флоры и фауны. В XX в. такие же схемы были составлены не только для флор и фаун, но и для сообществ растений, животных, биогеоценозов.
Само словосочетание «биологическое разнообразие» впервые применил Г. Бейтс (1892) в работе «Натуралист на Амазонке», когда описывал свои впечатления от встречи около 700 разных видов бабочек за время часовой экскурсии.
Современные представления о проблеме биологического разнообразия базируются на исследованиях популяционных генетиков 1908-1953 гг., показавших, как создается генетическое разнообразие организмов во внешне однородной популяции, и разработавших математический аппарат для его объективного описания.
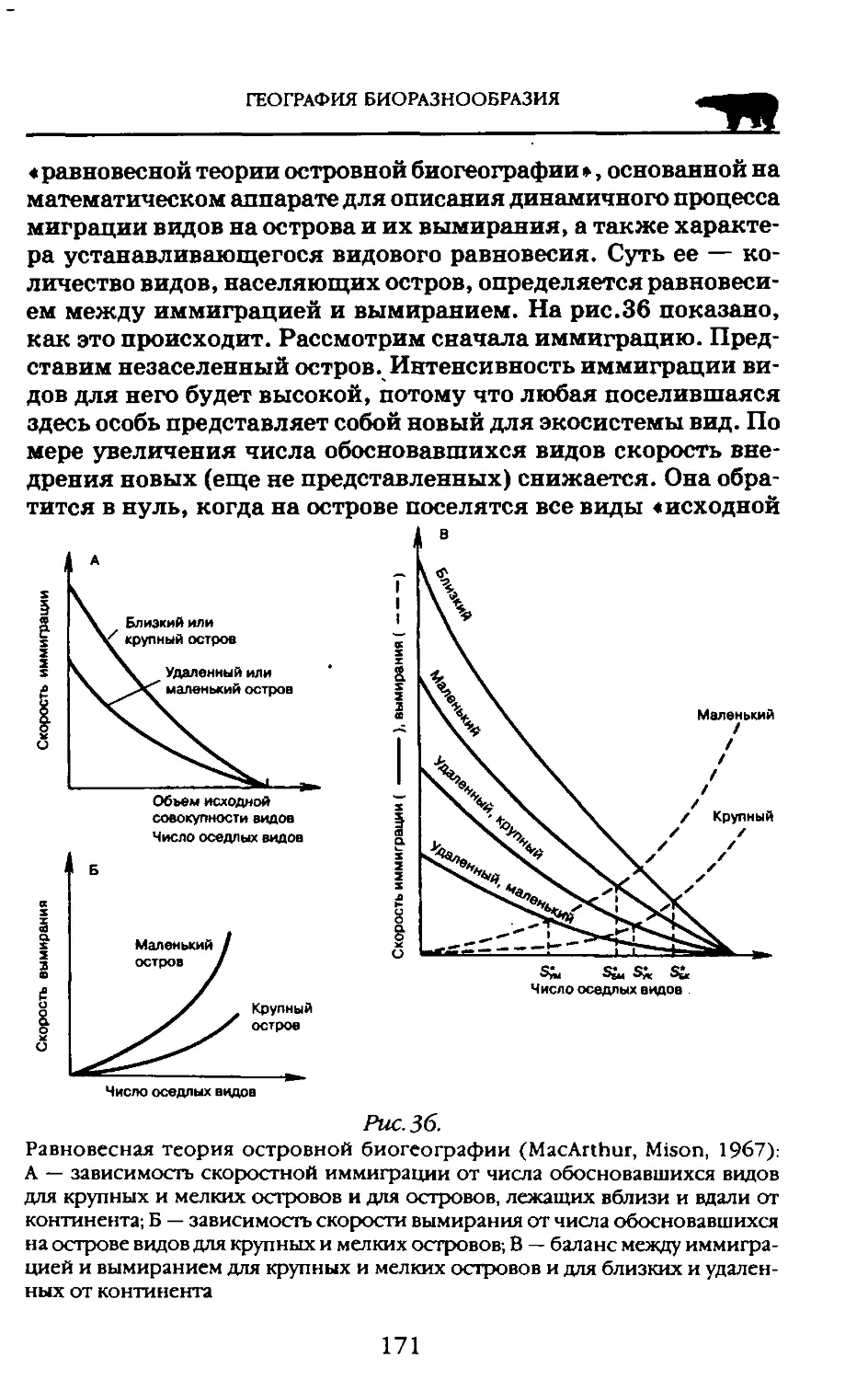
Используя математический аппарат «статистики разнообразия», Г. Хатчинсон (1962) показал эффективность его применения в полевой экологии, а затем вместе с Р. Мак-Артуром (1959) и в биогеографии. Впоследствии Мак-Артуром это представление было развито в виде «Географической экологии» (1972), а также вместе с Е. Вильсоном в «Теории островной биогеографии» (1967).
Биоразнообразие в последнее время стало одним из самых распространенных понятий в научной литературе, природоохранном движении и международных связях. Доказано, что необходимым условием нормального функционирования экосистем и биосферы в целом является достаточный уровень природного разнообразия на нашей планете. Биологическое разнообразие рассматривается как основной параметр, дающий представления о состоянии над организменных систем. В ряде стран именно характеристика биологического разнообразия выступает в качестве основы экологической политики государства, стремящегося сохранить свои биологические ресурсы, чтобы обеспечить устойчивое экономическое развитие.
Термин «биоразнообразие» является сокращением сочетания слов «биологическое разнообразие». Разнообразие — это понятие, которое имеет отношение к размаху изменчивости, или различий, между некоторыми множествами или группами объектов. Биологическое разнообразие, следовательно, имеет отношение к разнообразию живого мира. Термин «биоразнообразие» обычно используется для описания числа разновидностей и изменчивости живых организмов. В широком смысле этот термин охватывает множество различных параметров и является синонимом понятия «жизнь на Земле». В научном мире понятие разнообразия может быть отнесено к таким понятиям, как гены, виды и экосистемы, которые соответствуют трем фундаментальным, иерархически зависимым уровням организации жизни на нашей планете.
Явление разнообразия живых организмов определяется свойствами биологических макромолекул, особенно нуклеиновых кислот, — их способностью к спонтанным изменениям структуры, что приводит к трансформации геномов, к наследственной изменчивости. На этой биохимической основе разнообразие создается в результате трех независимо действующих процессов: спонтанно возникающих генетических вариаций (мутаций), действия естественного отбора в смешанных популяциях, географической и репродуктивной изоляции. Данные процессы, в свою очередь, ведут к дальнейшей таксономической и экологической дифференциации на всех последующих уровнях биологических экосистем: видовом, ценоти-ческом и экосистемном.
Термин «биологическое разнообразие» используется активно не менее полувека. За это время много сделано для по
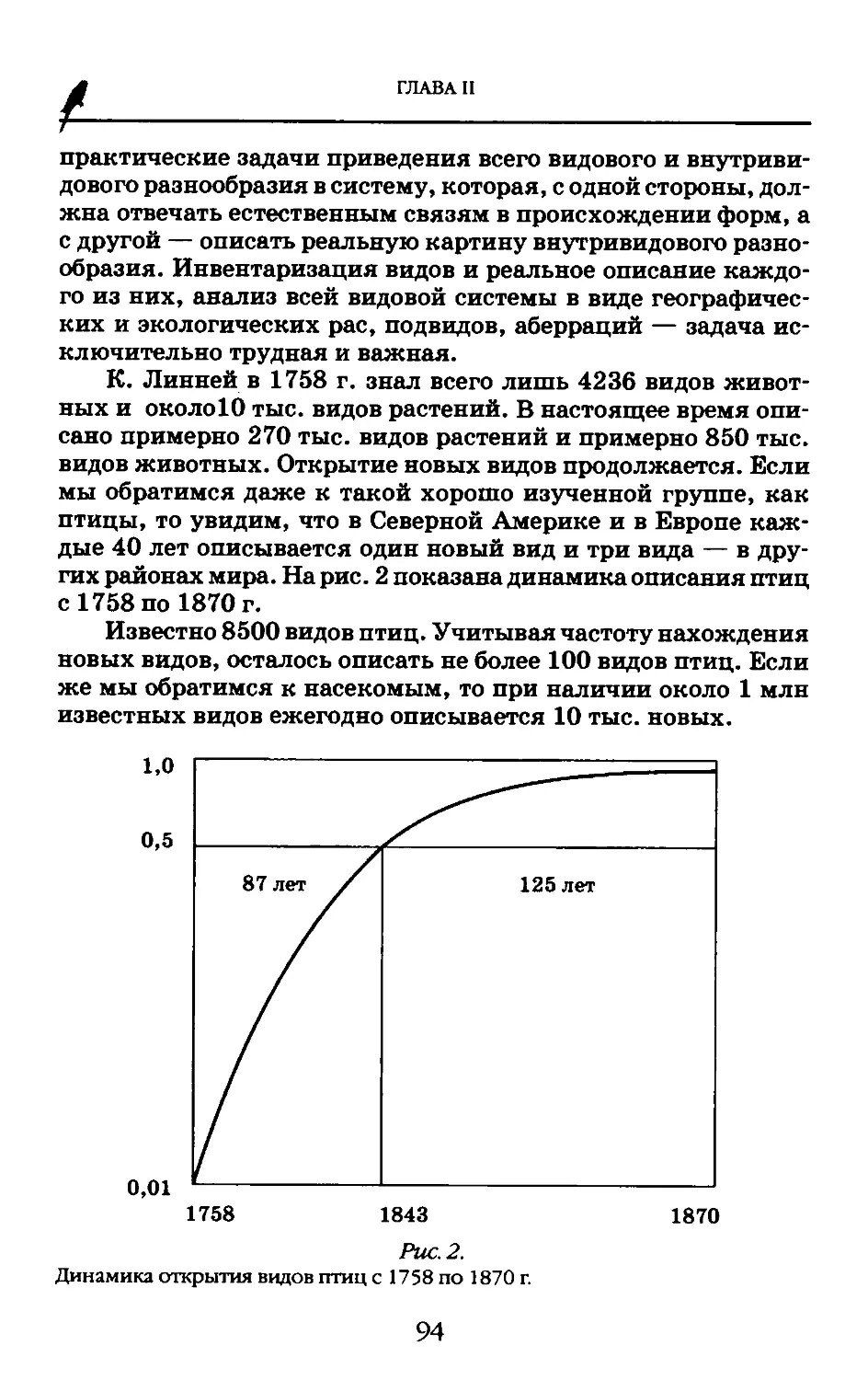
нимания самого процесса и разработки методов его измерения. Одним из существенных достижений здесь является расширение наших представлений о видовом разнообразии жизни на Земле. Если сейчас валидно описано 1,75 млн видов растений животных, микроорганизмов, то, по мнению ведущих специалистов-систематиков, их реальное число достигает не менее 10-35 млн, в том числе 1 млн видов микроорганизмов, 1 млн видов нематод, 10 млн видов насекомых и около 10 млн видов грибов. Особенно плохо изучены влажные тропики, где, как полагают, описан только 1 из 20 обитающих видов, особенно среди насекомых, грибов, а также почвенной фауны. При всей неполноте наших знаний нельзя не отметить, что в XX столетии число таксонов животных и растений увеличено по меньшей мере в 500 раз по сравнению с концом XIX в.
МЕЖДУНАРОДНАЯ ПРОГРАММА
«БИОЛОГИЧЕСКОЕ РАЗНООБРАЗИЕ»
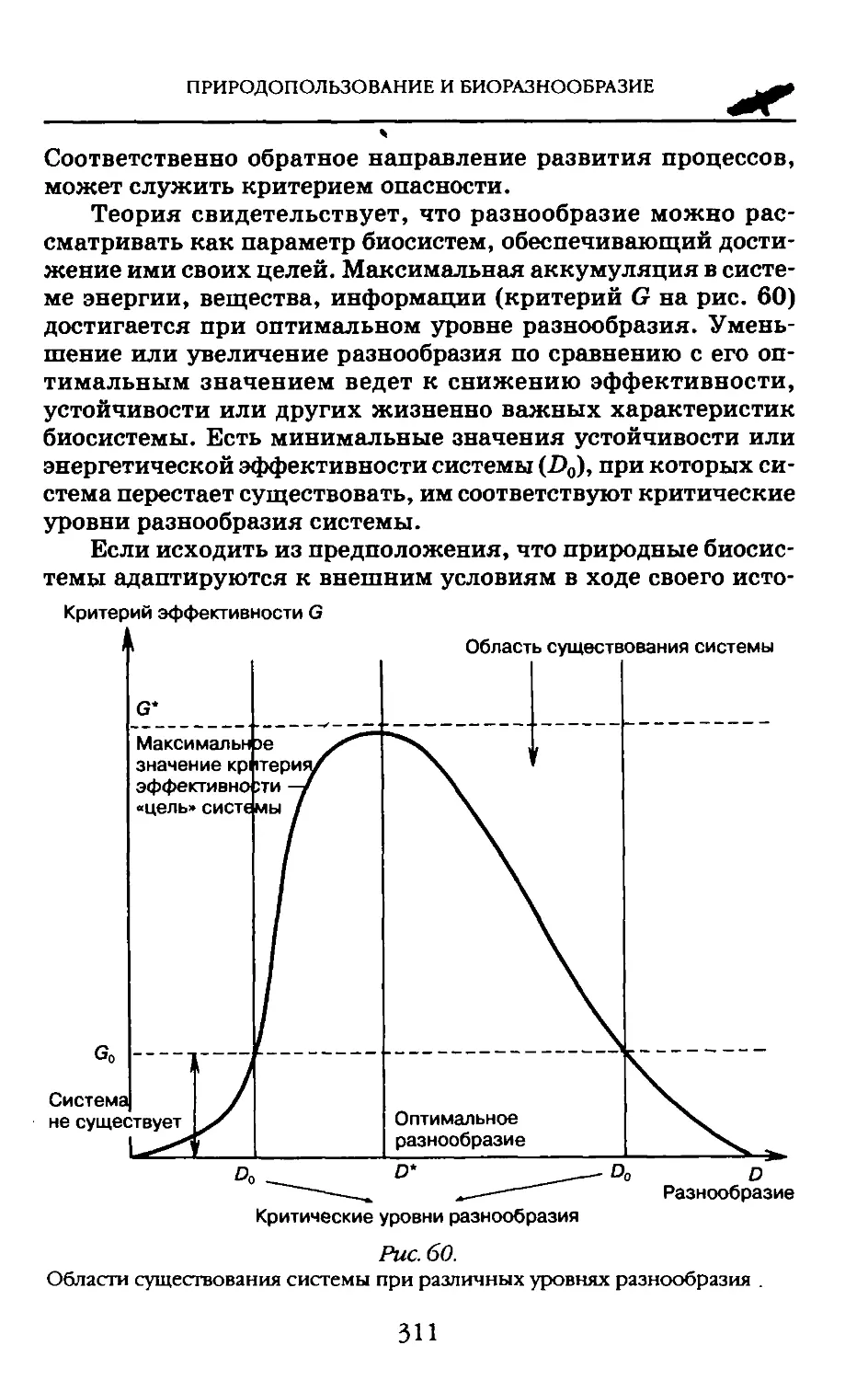
Понятие «биоразнообразия» вошло в широкий оборот только в 1972 г. на Стокгольмской конференции ООН по окружающей среде, где экологи сумели убедить политических лидеров стран мирового сообщества в том, что охрана живой природы должна стать приоритетной при любой деятельности человека на Земле.
Научную разработку программы осуществлял Международный союз биологических наук, создавший для этого в 1982 г. на Генеральной ассамблее в Канаде специальную рабочую группу. Активное участие в формировании программы исследований и первых организационных мероприятий принял академик М.С. Гиляров, ставший одним из «отцов-основателей» этого крупнейшего международного проекта. Работа по изучению биоразнообразия велась союзом с 1991 по 1997 г. в два трехлетних этапа: с 1991 по 1994 г. — первый, предварительные итоги которого проведены в Париже в 1994 г., и второй, заключительный, с 1995 по 1997 г. Итоги второго этапа, как и программы в целом, подведены в ноябре 1997 г. на 26-й сессии Генеральной ассамблеи Международного союза биологических наук в Тайбее. Исследования био-
разнообразия в других организационных формах продолжаются по более частным программам, таким как Биономенклатура, Виды-2000 — индексация известных в мире видов, Биоэтика, Систематика-2000 и др. Биоразнообразие остается одним из трех главных приоритетов исследований как в биологии наряду с биотехнологией и устойчивой агрикультурой, так и в биогеографии. Продолжаются и региональные исследования, с 1994 по 2001 г. — в рамках Государственной научно-технической программы России «Биологическое разнообразие».
Следует отметить большое значение принятия Международной конвенции о биологическом разнообразии на Конференции ООН по окружающей среде в Рио-де-Жанейро в 1992 г. Разработке и принятию Международной конвенции по биоразнообразию предшествовала активная деятельность многих организаций.
В 1975 г. вступила в силу Конвенция по международной торговле видами мировой флоры и фауны, находящимися под угрозой исчезновения. Конвенция запрещает или регулирует торговлю 20 000 видов, находящихся под угрозой исчезновения.
В1980 г. UNEP, IUCN (Международный союз охраны природы и природных ресурсов) и WWF (Всемирный фонд дикой природы) опубликовали Всемирную стратегию охраны живой природы. Более 50 стран мира использовали ее для разработки национальных стратегий охраны живых организмов.
В 1983 г. вступила в действие Конвенция по сохранению мигрирующих видов диких животных.
Создан Всемирный центр охраны и мониторинга (WCMC), целью которого является оценка распределения и обилия видов на планете, подготовка специалистов в области мониторинга биоразнообразия.
UNEP и IUCN разработали и приступили к реализации совместных планов мероприятий по сохранению африканских и индийских слонов и носорогов, приматов, кошачьих и белых медведей.
Международный Совет по генным ресурсам растений (IBPGR) в 30 странах мира организовал сеть банков генов, располагающих 40 основными мировыми коллекциями. Более 500 000 видов растений из 100 стран были собраны, оценены и размещены в хранилищах.
Международный переговорный комитет, учрежденный руководящим советом UNEP, при участии многих международных организаций подготовил Конвенцию по биологическому разнообразию. В июне 1992 г. во время Конференции ООН по окружающей среде и развитию в Рио-де-Жанейро она была подписана представителями большинства стран, включая Российскую Федерацию. Главная цель подписанного документа — сохранение биологического разнообразия и обеспечение тем самым нужд человечества.
В 1992 г. разработана Глобальная стратегия биоразнообразия, целью которой стала ликвидация условий исчезновения видов.
К настоящему времени «Конвенцию о биологическом разнообразии» подписали представители 180 стран, в том числе и России (1995 г.), взявшей на себя ответственность за сохранение живой природы 1/7 части суши нашей планеты.
Международная программа исследования биоразнообразия включает три основных уровня: генетический, таксономический и экологический (сообщества и экосистемы).
НАУЧНАЯ ПРОГРАММА «ДИВЕРСИТАС»
На 24 -й Генеральной ассамблее Международного союза биологических наук (Амстердам, сентябрь 1991 г.) при поддержке СКОПЕ и ЮНЕСКО было принято решение приступить к разработке научной программы для изучения биологического разнообразия «Диверситас». В первоначальном варианте эта программа включала четыре основных направления:
1) функционирование экосистем и поддержание биоразнообразия;
2) происхождение, сохранение и потери биоразнообразия;
3) инвентаризация и мониторинг биоразнообразия;
4) сохранение генетического разнообразия диких предков культурных растений и одомашненных животных.
Позже программа была дополнена еще тремя направлениями:
1) морское биоразнообразие;
2) разнообразие микроорганизмов;
3) роль человека в изменениях биоразнообразия.
К настоящему времени в рамках программы «Диверситас» сложились пять основных ориентаций, сфокусированных на
ключевых областях изучения биоразнообразия, и пять целевых междисциплинарных направлений.
1. Функционирование экосистем и поддержание биоразнообразия. Исследования в рамках этого направления позволят ответить на вопросы: как стабильность экосистем и их способность к восстановлению зависят от разнообразия видов; как глобальные изменения климата, землепользования, обилия чужеродных видов влияют на функционирование экосистем; какую роль играет биоразнообразие (от генетического и видового до ландшафтного уровня) в экосистемных процессах (изменения продуктивности, обеспечение преобразования и круговорота питательных веществ). Будет получена количественная оценка роли экосистем в поддержании качества и количества водных ресурсов, плодородия почв и качества атмосферного воздуха на должном уровне, а также оценка воздействия глобальных трансформаций окружающей среды на их функционирование.
2. Происхождение, сохранение и изменения биоразнообразия. Особое внимание уделяется изучению реакции видов на изменения среды обитания с учетом различных факторов, включая генетические и физиологические свойства видов, межвидовые отношения и популяционные характеристики, свойственные различным историческим эпохам. Выяснение популяционных и генетических процессов, обеспечивающих процессы видообразования и видового угасания и ведущих к увеличению и утрате мирового биотического богатства, позволит выработать эффективную стратегию по поддержанию оптимального уровня биоразнообразия.
3. Систематика: инвентаризация и классификация биоразнообразия. Цель этого направления — оценка современного биоразнообразия. Необходимо установить четкие приоритеты для проведения систематической инвентаризации видов анализа и синтеза информации, отражающей различные аспекты истории развития жизни на Земле, и создать такую систему организации работ, которая будет гарантировать качественность собранных данных и их доступность. Полученная информация послужит базой для построения прогнозов и созданию программ по улучшению системы образования.
4. Мониторинг биоразнообразия. Для принятия обоснованных решений по управлению биоразнообразием, направ
ленных на смягчение последствий его изменений, требуется обладать оперативными данными о том, где и с какой скоростью трансформируется биоразнообразие. В рамках этого направления будут разработаны эффективные и стандартные методы мониторинга и определены приоритеты, которые будут обеспечивать накопление данных, необходимых для понимания современного и будущего статуса разнообразия, в нужном объеме. Будут созданы надежные системы мониторинга и прогнозирования изменений биоразнообразия в разных частях планеты.
5. Охрана, восстановление и устойчивое использование биоразнообразия. Сохранение биологического разнообразия и устойчивое использование его компонентов определяют поддержание глобальной стабильности. Исследования, проводимые в этом направлении, позволят лучше понять, как и под воздействием каких факторов изменяется биоразнообразие. Полученные результаты будут влиять на принятие соответствующих мер по охране биоразнообразия, в том числе и на глобальном уровне. Специальные усилия будут сосредоточены на сохранении генетического разнообразия растений, окультуренных человеком, и животных, предки которых одомашнены. Будут разработаны и претворены в жизнь стратегии по восстановлению нарушенных экосистем, собран материал по демографическим и генетическим изменениям в популяциях живых организмов, которые происходят в процессе их восстановления.
6. Биоразнообразие почв и донных отложений. Биота почв и донных отложений изучена недостаточно, хотя ее важная роль в основных экосистемных процессах, включая контроль круговорота веществ, поддержание плодородия почв и влияние на состав «парниковых» газов, не вызывает сомнения. В настоящее время необходимо систематизировать накопленную информацию по отдельным видам организмов, а также определить, какие сочетания видов и структура их сообществ влияют на функционирование экосистем. Исследования будут нацелены на понимание биологических основ поддержания продуктивности почв и донных отложений, будут составлены базы данных, разработаны стандартные методы оценки и мониторинга разнообразия почв и донных отложений и проведены эксперименты для того, чтобы оценить роль различных
видов, обитающих в почвах и донных отложениях, в функционировании экосистем.
7. Морское биоразнообразие. В задачи направления входит сбор информации о том, как деятельность человека влияет на биоразнообразие океанов и прибрежных зон. Для сохранения морских экосистем требуется расширить применение моделирования процессов, контролирующих биологическое разнообразие в морях и океанах. Огромные размеры морского царства, его относительная недоступность, активные процессы по смешиванию, рассеиванию и переносу водных масс и их обитателей обусловливают сложность исследования морских сообществ. Разнообразие живых организмов в морских экосистемах огромно, но мало изучено, и поэтому процессы утраты биоразнообразия не всегда ясны и правильно оцениваются. Необходимо понять, как морское биоразнообразие меняется под воздействием рыбного промысла, эвтрофикации, физического изменения местообитаний, внедрения чужеродных видов и других форм воздействия человека. Будут созданы и развиты информационные сети для большого числа заинтересованных учреждений и организаций.
8. Биоразнообразие микроорганизмов. Микроорганизмы представляют огромное генетическое разнообразие и играют уникальную роль в экосистемах как основные компоненты пищевых цепей и биогеохимических циклов. Пока описано менее 5% существующих микроорганизмов. Будут развиваться новые методы и технологии исследования микроорганизмов, информационные базы данных по микробиологическому разнообразию.
9. Пресноводное биоразнообразие. Это направление включает исследования самых разных организмов от микроскопических бактерий до крупных птиц и млекопитающих, их структурно-функциональных связей, особенностей и процессов, определяющих потери биоразнообразия в пресноводных экосистемах. Будут проведены исследования влияния деятельности человека и глобальных изменений окружающей среды на биоразнообразие пресноводных экосистем и на их основе уточнена роль этих экосистем для человечества.
10. Роль человека в управлении биоразнообразием. Многие действия, направленные на качество жизни (производ
ство пищи и волокон, строительство и производство различных потребительских товаров, организация мест отдыха), имели негативные последствия для глобального биоразнообразия. В настоящее время человечество играет ключевую роль в изменении экосистемных процессов, и разные формы его деятельности нередко являются решающими в функционировании экосистем, сохранении биоразнообразия и его устойчивом управлении. Проводимое изучение должно способствовать четкому определению возможностей человека в управлении биоразнообразием. Это направление исследований должно активизировать получение и обобщение научной информации, требуемой: для понимания зависимостей между культурой различных групп населения и биоразнообразием; для оценки благосостояния людей во взаимосвязи с ростом населения и динамикой биоразнообразия; для анализа чрезвычайно сложных путей, которыми древнее и современное общество управляли и управляют биоразнообразием; для оценки возможностей привлечения различных слоев населения к сохранению и устойчивому управлению ресурсами биосферы.
Опираясь на рекомендации, резолюции и выводы конференций и исследовательских проектов по биоразнообразию (включая программу «Диверситас»), ведущие специалисты из ряда мировых таксономических центров, эксперты по планированию и управлению, представители различных фондов, экологи и специалисты по охране природы провели две встречи по подготовке Глобальной таксономической инициативы: в феврале 1998 г. в г. Дарвине (Австралия) и сентябре 1998 г. в Лондоне. Участники Дарвинской встречи обратились к мировой общественности с декларацией, в которой были определены четыре ключевые проблемы таксономии, от которых зависит выполнение Конвенции по биоразнообразию: установление приоритетов в таксономических проектах; развитие партнерства между таксономическими центрами для решения локальных, региональных и глобальных проблем; повышение роли таксономических центров в решении проблем биоразнообразия; обеспечение поддержки развития таксономии широким кругом организаций, озабоченных проблемой сохранения биоразнообразия.
Важной инициативой в рамках программы «Диверситас» стала подготовка к проведению в 2001 г. Международного года
наблюдений за биоразнообразием. Намечено осуществить различные проекты с региональным и глобальным охватом. Особое внимание предполагается уделить укреплению различных сетей наблюдений за компонентами биологического разнообразия и стандартизации методов и показателей, используемых разными специалистами.
ow
РЕАЛИЗАЦИЯ КОНВЕНЦИИ
О БИОРАЗНООБРАЗИИ
В РОССИИ
Россия включилась в реализацию Конвенции о биоразнообразии после ее ратификации в 1995 г. Государственной думой. Сейчас весь портфель природоохранных законов Российской Федерации напрямую увязан с выполнением обязательств нашей страны по Конвенции. По реализации Конвенции в России на федеральном уровне приняты законы «Об охраняемых природных территориях», «О животном мире», «Об экологической экспертизе», «О континентальном шельфе Российской Федерации» и др. 1 апреля 1996 г. Президент Российской Федерации подписал Указ № 440, согласно которому была утверждена «Концепция перехода Российской Федерации к устойчивому развитию » (Концепция..., 1996). В Концепции отмечена роль России в решении планетарных экологических проблем. Наша страна обладает большими по площади территориями, практически не затронутыми хозяйственной деятельностью, которые являются резервом устойчивости всей биосферы в целом. В качестве одного из важнейших направлений России в международном сотрудничестве по обеспечению устойчивого развития и охраны окружающей среды Концепция рассматривает сохранение
биоразнообразия. Впервые в нашей стране начаты работы по формированию единой системы экологического мониторинга, в которой значительное место уделено оценке и контролю состояния биологического разнообразия. Россия активно подключилась к международной деятельности по сохранению биоразнообразия, при поддержке «Глобального экологического фонда» ведутся работы по сохранению биоразнообразия, по оказанию помощи российским заповедникам, по решению проблемы охраны природы на Байкале. Проект сохранения биоразнообразия России, финансируемый Международным банком реконструкции и развития (26 млн американских долларов) стартовал в декабре 1996 г. Его цели: разработка национальной стратегии сохранения биоразнообразия России и ее региональных моделей; разработка экономических механизмов сохранения и использования ресурсов биоразнообразия, методов мониторинга его состояния; поддержка национальных парков и заповедников; реализация мероприятий по сохранению биоразнообразия и улучшение экологической обстановки Байкальского региона; поддержка общественных и образовательных программ.
Во многих зарубежных университетах курсы по биоразнообразию являются обычными. Сегодня мы наблюдаем, как результаты научных исследований в области изучения биоразнообразия определяют стратегию государственной политики, поэтому специалисты географы, биологи и экологи, выходящие из стен университетов, должны быть всесторонне подготовлены к измерению и оценке биоразнообразия. Описание разнообразия жизни и ландшафтного разнообразия среды обитания были объектами изучения экологов и биогеографов, как и объектами преподавания, задолго до 1991 г. — начала реализации программы «Биологическое разнообразие» Международного союза биологических наук. Так, разделы по разнообразию экосистем и разнообразию организмов были в учебнике П.П. Второва и Н.Н. Дроздова «Биогеография материков» (1974), раздел «Биологическое разнообразие» в учебном пособии П. Аллена «Биология — наука о жизни» (1978), изданном на русском языке в 1984 г. и т.п.
В последнее время анализ биологического разнообразия стал предметом не только научных исследований, но и преподавания в университетах. Авторы ввели преподавание анализа биоразнообразия в курсы ряда биологических дисциплин
(математические методы в биологии, экология, основы биологии, биоиндикация) в 1992 г., используя в качестве основного учебного материала программу «Биоразнообразие» Международного союза биологических наук, разрабатываемую с 1982 по 1990 г. и введенную на период с 1991 по 1997 г. в качестве приоритетной в деятельности Союза. Как учебное пособие также использовался курс Оксфордского университета «Global biodiversity» (1992). Первая учебная программа для университетов в России была разработана и опубликована Н.В. Лебедевой и А. Д. Покаржевским в 1993 г. После чего с 1994 г. отдельный курс биоразнообразия стали преподавать в РГУ и МГУ. В 1999 г. географическим факультетом МГУ опубликовано первое учебное пособие по предмету «Биоразнообразие и методы его оценки», составленное Н.В. Лебедевой, Н.Н. Дроздовым и Д.А. Криволуцким, которое используется не только для преподавания предмета, но и для проведения учебных полевых практик. Это учебное пособие стало базовым при подготовке магистров межфакультетской специализации «Эксперт в области геоэкологической безопасности» по дисциплине «Биологическое разнообразие и его оценка», осуществляемой в рамках проекта CRDF в РГУ.
Опыт преподавания курса с 1994 по 2001 г. показал, что проблема шире, чем ее можно представить. Анализируя систему оценок биоразнообразия, в учебном курсе авторы рассматривают также разнообразие на разных уровнях организации жизни: молекулярно-генетическом, клеточном, тканевом, организменном, популяционном, видовом, а также на надорганизменных уровнях (сообщества, экосистемы, биомы) и территориальных экологических подразделениях биосферы.
СИСТЕМНАЯ КОНЦЕПЦИЯ БИОРАЗНООБРАЗИЯ
По современным представлениям можно различать несколько уровней организации жизни (молекулярный, генетический, клеточный, организменный, популяционный, экосистемный, биосферный), каждый из которых обладает свойственным ему биоразнообразием. Этот подход был отражен в первой учебной программе по биоразнообразию в России (Лебедева, Покаржевский, 1993) и получил развитие в научных разработках по проблеме биологического разнообразия.
При определении сущности живого с системных позиций, живое вместе с другими его качествами нужно рассматривать как дискретные материальные системы и комплексы систем. Концепция о живом как о системах взаимодействующих частей развивалась тремя путями (Матекин, 1982):
1) формирование знаний о взаимодействии частей, слагающих организм, т. е. познания организма как целого;
2) развитие представлений о виде как взаимосвязанности индивидов;
3) развитие суждений о взаимоотношениях разных видов, обитающих совместно.
Несомненно, развитие трех этих направлений помогло становлению общей теории систем, положения которой приложимы не только к живой, но и косной материи. Основу общей теории составляет ряд частных эвристических принципов видения мира, которые позволяют открыть внутренние связи, существующие в пределах каждой формы материи и установить взаимоотношения между формами материи.
Проследим развитие понимания организма как целого.
Жорж Кювье (1769-1832), формулируя в 1817 г. принцип корреляции, или принцип конечных причин, впервые указал на целостность живого организма. Он писал: «Всякое организованное существо образует целое, единую замкнутую систему, части которой соответствуют друг другу и содействуют путем взаимного влияния одной конечной цели. Ни одна из этих частей не может измениться без того. Чтобы не изменились другие, и, следовательно, каждая из них, взятая отдельно, указывает и определяет все другие».
Столь же весомый вклад в представление о системности живого внесен физиологией. Клод Бернар (1813-1878) наблюдал изменение кровотока в ухе кролика после перерезки и раздражения симпатических нервных волокон. Так была открыта вазомоторная, т. е. сосудодвигательная, функция нервной системы и установлено значение этой функции для регуляции теплоотдачи.
Открытие вазомоторной функции нервной системы раскрывало новую сторону жизнедеятельности организма — его целостность.
Для развития идей об организме как целом чрезвычайно велико значение эмбриологических исследований. В 1924 г. Г. Мангольд, сотрудница лаборатории Г. Шпеманна, пересадила участок губы бластопора одного зародыша на спинную сторону другого зародыша. В результате у реципиента развилась вторая нервная рубка и весь осевой комплекс органов.
Дальнейшая эволюция взглядов о взаимодействии частей в развивающемся организме выразилась в установлении трех этапов морфогенеза:
1) зависимых дифференцировок;
2) само дифференцировок;
3) развития функциональных связей как заключительного периода в формировании окончательной структуры организма.
На всех этапах морфогенеза отчетливо проявляется взаимодействие возникающих структур, т.е. взаимодействие, меняющееся в своих масштабах, но, тем не менее, всегда являющееся внутренней причиной развития системы.
Существенная роль в становлении представлений о целостности организма принадлежит теории Н.И. Вавилова о гомологической наследственной изменчивости (1920). С точки зрения Вавилова, мутационный процесс, лежащий в основе наследственной изменчивости, при всей его случайности подчинен все же внутренним законам. Это выражено им в следующих словах: «Мутации в близких видах и родах идут, как правило, в одном и том же направлении». Вавилов считал, что в этом явлении находит отражение взаимодействие генов как исторически сложившихся комплексов. На основании теории Вавилова, генотип стали понимать как глубоко интегрированную систему генов, способную регулировать процессы наследственной изменчивости.
Второй путь к пониманию системности живого — анализ связи между индивидами в пределах вида.
Как известно, первое представление о виде сформулировал Джон Рей (1627-1705), который считал, что вид — это собрание особей, как дети похожие на родителей.
К. Линней, описав более 4 тыс. видов животных и растений, четко обосновал понятие о мономорфизме видов, т. е. представление о сходстве всех особей вида по всем признакам.
В 1910 г. была опубликована статья известного русского энтомолога П.П. Семенова-Тян-Шанского «Таксономические границы вида и его подразделения ». Эта статья едва ли не первая фундаментальная работа, в которой путем обобщения накопившихся данных было показано разнообразие внутривидовых категорий.
Замена взглядов о структурированности вида на представления о его системности способствовало развитию генетики. В 1931г. Н.И. Вавилов опубликовал статью «Линнеевский вид как система». В этой важнейшей для теории вида статье Вавилов рассмотрел целостность вида с позиций генетики. Он пришел к выводу, что разнообразие внутривидовых форм обусловлено неодинаковыми условиями среды с разным направлением естественного отбора. Но, вместе с тем, вид един, это единство Вавилов объясняет тем, что все структурные компоненты вида, т. е. внутривидовые формы, обмениваются генами. Таким образом, взаимосвязанность индивидов и их групп путем обмена
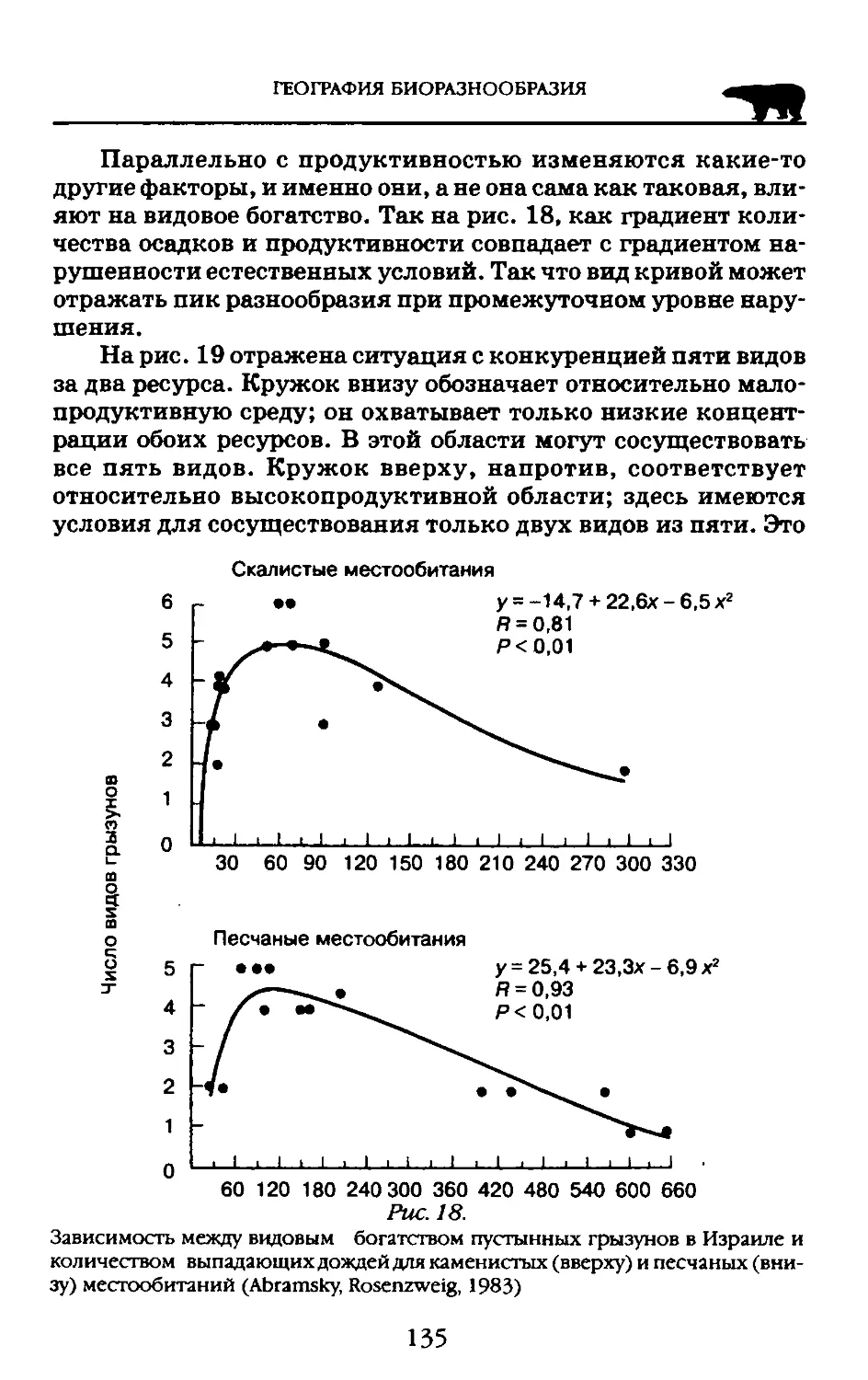
генами при скрещивании придает виду свойства системы. Но генотипические структуры аппарата наследственности ограничивают генетические связи индивидов, что и создает обособленность видовых систем.
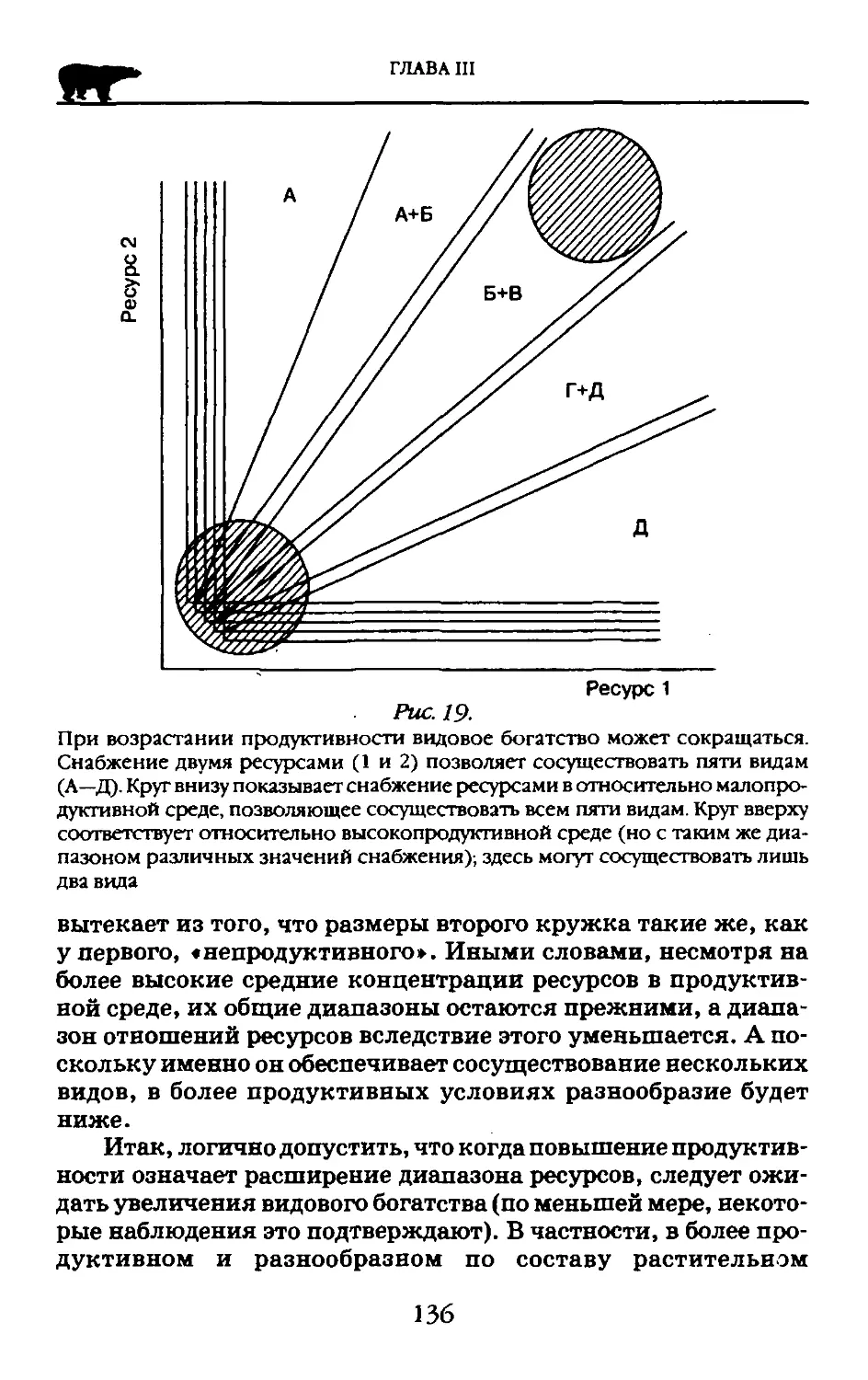
Вавилов писал: «Линнеевский вид, таким образом, в нашем понимании, — обособленная, сложная, подвижная морфофизиологическая система, связанная в своем генезисе с определенной средой и ареалом». Таким образом, вклад таксономии в развитие общих представлений о системности живого заключался в формировании взглядов о виде как ограниченной или замкнутой системе генов, стабильное существование которой возможно лишь благодаря взаимодействию частей этой обособленной системы.
Третьим направлением, которое привело к понятию системности в биологии, было развитие представлений о взаимосвязанности и взаимодействии разных видов, обитающих совместно. Этому типу взаимодействий, описываемых как альфа-, бета-, гамма-, дельта-, эпсилон- и омега-разнообразия посвящена шестая глава.
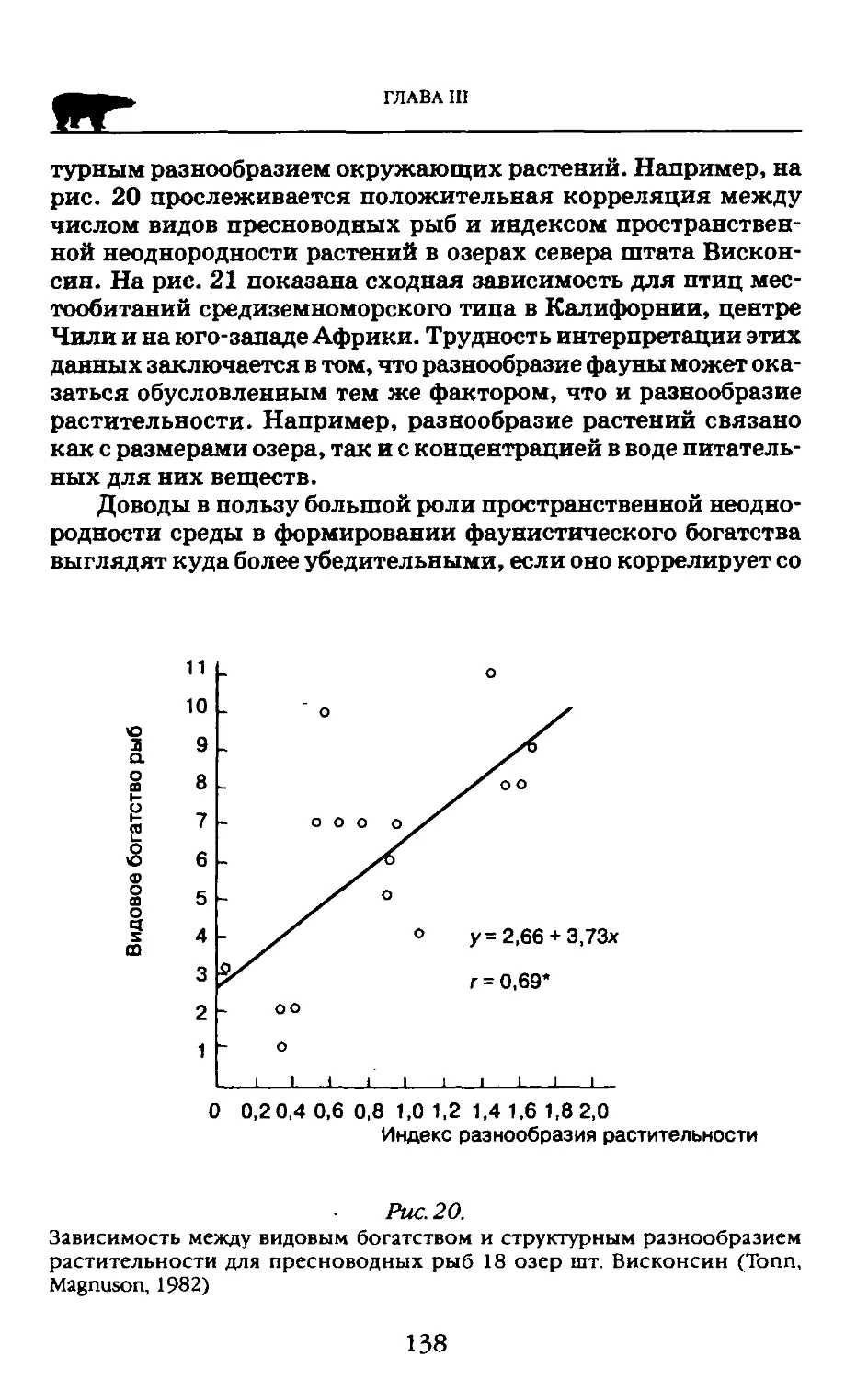
Итак, фундаментальные разделы биологии: физиология, морфология, эмбриология, генетика, экология, а также биогеография показывают не только структурированность живого, но и непременное взаимодействие между структурами. Понимание и непрерывное углубление представлений об обязательности связей между структурами живого привело к тому, что концепция системности живого, приложимая к клетке, организму, виду, биогеоценозу (экосистеме), биосфере прочно вошла в теорию биологии. Однако одним словом «система» еще не определено все то значение взаимодействий, без которых живое лишилось бы своей интегрирующей сущности. Эта теория сложилась как интеграция данных о системности самых разных форм материи.
• Философские предпосылки обоснования общей теории систем можно найти у великого английского материалиста Фрэнсиса Бэкона (1561-1626). Бэкон писал, что никто не отыщет природу вещи в самой вещи и изыскание должно быть расширено до более общего. Эта мысль может считаться едва ли не первым четким определением того, что всякое частное представляет собой элемент более общего.
В дальнейшем ученые разных стран неоднократно обращались к анализу связей между компонентами систем, при
чем и абиогенных, и биологической, и социальных. Существенное обобщение в 1950-е годы было сделано австрийским математиком Людвигом фон Берталанфи.
1. Система — это комплекс элементов, находящихся во взаимодействии, при этом степень их взаимодействия такова, что делает неправомочным аналитический подход как метод изучения системы. В то же время целое не может быть описано теми же зависимостями, какими могут быть описаны процессы в элементах системы.
Следовательно, данный тезис общей теории систем предполагает необходимость особых методов для целостного изучения системы.
2. Наличие изоморфных, т. е. одинаковых, процессов в разных категориях природных явлений требует общих законов.
3. Таким общим законом может быть известный принцип, сформулированный Ле Шателье: всякая система подвижного равновесия под действием внешнего воздействия изменяется так, что эффект внешнего воздействия сводится к минимуму.
Последний, очень важный тезис помогает представить, по крайней мере, конечные задачи исследования с помощью методов, которыми только и можно изучать целое как систему. Эти конечные задачи — определение устойчивости системы по конкретным параметрам. В биологии такими параметрами могут быть параметры гомеостазиса как организма, так и популяции, и параметры индикаторов продуктивности сообществ.
4. Свойство целого порождено свойствами элементов, в то же время свойства элементов несут свойства целого.
5. Не всегда и не только простые причинно-следственные отношения объясняют функционирование системы.
Справедливость этого тезиса подтверждается наличием и реальностью принципа обратной связи, на основании которого, как мы знаем, конечный эффект функционирования системы может изменить начальные процессы, так что новый конечный эффект будет иметь обратное значение.
6. Источник преобразования системы лежит в самой системе. В этом причина ее самоорганизованности.
7. Один и тот же материал или компонент системы может выступать в разных обличиях.
Берталанфи полагает, что развитие системного подхода заключается в переходе от вербальности (словесного описания)
к ограниченному математизированию, а далее к математическому, а не физическому рассмотрению биологических систем. Следовательно, главное — математическое выражение соотношений между переменными, описывающими поведение системы. Общая теория систем позволила, таким образом, увидеть иерархию структур в живых системах и установить наличие парциальных систем, т. е. более частных, входящих в состав общих.
Представление об иерархии систем стало той основой, на которой возникло и развилось понятие об уровнях организации живой материи. Теперь принято говорить о молекулярном, клеточном, организменном, популяционном й биоцено-тическом уровнях организации живой материи.
Далее принципы общей теории систем, как и принципы кибернетики, позволили установить, что целое воздействует на части путем определенных каналов управления. Такими каналами могут быть, прежде всего, генетическая система и системы, подобные тем, которые описываются системой регулярных синтезов, т.е. системой регуляционных метаболитов. Общая теория систем предполагает целостное понимание биологических явлений, где все биохимические процессы регулируются геномом, геном не существует вне организма, организм — вне вида, вид — вне экосистемы, а экосистема — вне географической среды.
ГЕНЕТИЧЕСКОЕ РАЗНООБРАЗИЕ
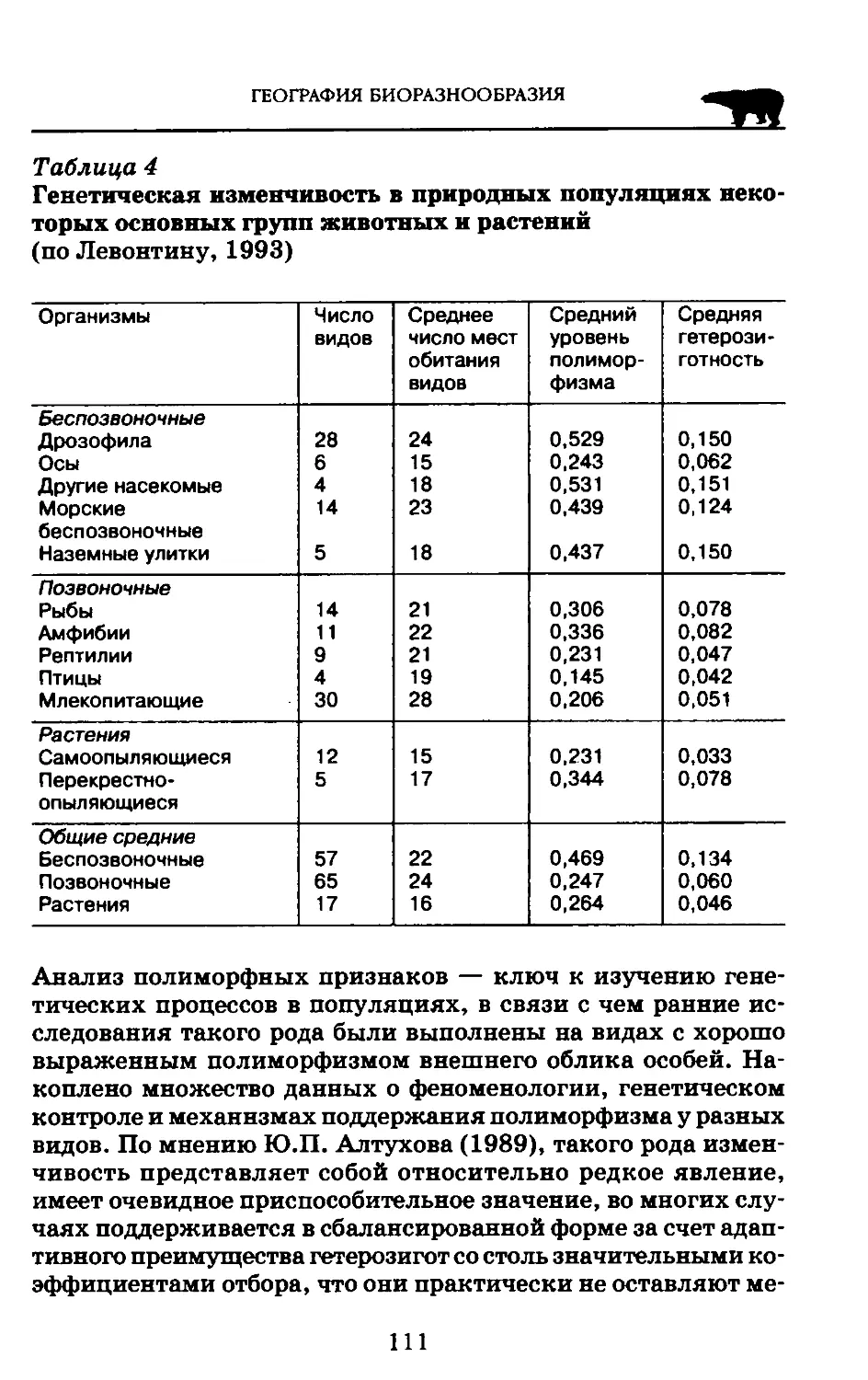
Естественное богатство нашей планеты связано с разнообразием генетических вариаций. Генетическое разнообразие, т. е. поддержание генотипических гетерозиготности, полиморфизма и другой генотипической изменчивости, которая вызвана адаптационной необходимостью в природных популяциях, представлено наследуемым разнообразием внутри и между популяциями организмов.
Как известно, генетическое разнообразие определяется варьированием последовательностей четырех комплиментарных нуклеотидов в нуклеиновых кислотах, составляющих генетический код. Каждый вид несет в себе огромное количество генетической информации: ДНК бактерии содержит около 1 000 генов, грибы — до 10 000, высшие растения — до 400 000. Огромно количество генов у многих цветковых растений и высших таксонов животных. Например, ДНК домовой мыши содержит около 100 000 генов.
Новые генетические вариации возникают у особей через генные и хромосомные мутации, а также у организмов, которым свойственно половое размножение, через рекомбинацию генов. Генетические вариации могут быть оценены у любых
организмов, от растений до человека, как число возможных комбинаций различных форм от каждой генной последовательности. Другие разновидности генетического разнообразия, например, количество ДНК на клетку, структура и число хромосом, могут быть определены на всех уровнях организации живого.
Огромное множество генетических вариаций представлено у скрещивающихся популяций и может быть осуществлено посредством селекции. Различная жизнеспособность отражается в изменениях частот генов в генофонде и является реальным отражением эволюции. Значение генетических вариаций очевидно: они дают возможность осуществления и эволюционных изменений и, если это необходимо, искусственного отбора.
Только небольшая часть (около 1%) генетического материала высших организмов изучена в достаточной мере, когда мы можем знать, какие гены отвечают за определенные проявления фенотипа организмов. Для большей части ДНК ее значение для вариации жизненных форм остается неизвестным.
Каждый из 109 различных генов, распределенных в мировой биоте, не дает идентичного вклада в формирование разнообразия. В частности, гены, контролирующие фундаментальные биохимические процессы, являются строго консервативными у различных таксонов и в основном демонстрируют слабую вариабельность, которая сильно связана с жизнеспособностью организмов.
Если судить об утере генофонда с точки зрения генной инженерии, принимая во внимание то, что каждая форма жизни уникальна, вымирание всего лишь одного дикого вида означает безвозвратную потерю от тысячи до сотен тысяч генов с неизвестными потенциальными свойствами. Генная инженерия могла бы использовать это разнообразие для развития медицины и создания новых пищевых ресурсов. Однако разрушение местообитаний и ограничение размножения многих видов приводят к опасному уменьшению генетической изменчивости, сокращая их способности адаптироваться к загрязнению, изменениям климата, болезням и другим неблагоприятным факторам. Основной резервуар генетических ресурсов — природные экосистемы — оказался значительно измененным или разрушенным. Уменьшение генотипическо-
*
го разнообразия, происходящее под воздействием человека, ставит на грань риска возможность будущих адаптаций в экосистемах.
Изучение закономерностей распределения генотипов в популяциях начато Пирсоном (1904). Он показал, что при наличии разных аллелей одного гена и действия свободного скрещивания в популяциях возникает определенное распределение генотипов, которое можно представить в виде:
р2 ДА+2рдАа + р2аа ,
где р— концентрация гена A, q — концентрация гена а.
Г.Х. Харди (1908) и В. Вайнберг (1908), специально исследовав это распределение, высказали мнение, что оно является равновесным, так как при отсутствии факторов, нарушающих его, оно может сохраняться в популяциях неограниченное время. Так стала развиваться популяционная генетика. Главная заслуга в разработке популяционной генетики, а особенно ее теоретического и математического аспектов, в этот ранний период (1920-1940 гг.) принадлежит С.С. Четверикову, С. Райту, Р. Фишеру, Дж. Холдейну, А.С. Серебровскому и Н. П. Дубинину. *
Биологическая эволюция — это процесс накопления изменений в организмах и увеличение их разнообразия во времени. Эволюционные изменения затрагивают все стороны существования живых организмов: их морфологию, физиологию, поведение и экологию. В их основе лежат генетические изменения, т. е. изменения наследственного вещества, которое, взаимодействуя со средой, определяет все признаки организмов. На генетическом уровне эволюция представляет собой накопление изменений в генетической структуре популяций.
Эволюцию на генетическом уровне можно рассматривать как двухступенчатый процесс. С одной стороны, возникают мутации и рекомбинации — процессы, обусловливающие генетическую изменчивость; с другой — наблюдается дрейф генов и естественный отбор — процессы, посредством которых генетическая изменчивость передается из поколения в поколение.
Эволюция возможна только в том случае, если существует наследственная изменчивость. Единственным поставщиком новых генетических вариантов служит мутационный процесс,
однако эти варианты могут по-новому рекомбинироваться в процессе полового размножения, т. е. при независимом расхождении хромосом и вследствие кроссинговера. Генетические варианты, возникшие в результате мутационного и рекомбинационного процессов, передаются из поколения в поколение отнюдь не с равным успехом: частота некоторых из них может увеличиваться за счет других. Помимо мутаций к процессам, изменяющим частоты аллелей в популяции, относятся естественный отбор, поток генов (т. е. их миграции) между популяциями и случайный дрейф генов.
На первый взгляд может показаться, что особи с доминантным фенотипом должны встречаться чаще, чем с рецессивным. Однако соотношение 3:1 соблюдается лишь в потомстве двух особей, гетерозиготных по одним и тем же двум аллелям. При других типах скрещивания в потомстве происходит иное расщепление признаков, и такие скрещивания также влияют на частоты генотипов в популяции. Законы Менделя ничего не говорят нам о частотах фенотипов в популяциях. Именно об этих частотах идет речь в законе Харди—Вайнберга. Основное утверждение закона Харди—Вайнберга состоит в том, что в отсутствие элементарных эволюционных процессов, а именно мутаций, отбора, миграции и дрейфа генов, частоты генов остаются неизменными из поколения в поколение. Этот закон утверждает также, если скрещивание случайно, то частоты генотипов связаны с частотами генов простыми (квадратичными) соотношениями. Из закона Харди-Вайнберга вытекает следующий вывод: если частоты аллелей у самцов и самок исходно одинаковы, то при случайном скрещивании равновесные частоты генотипов в любом локусе достигаются за одно поколение. Если частоты аллелей у двух полов исходно различны, то для аутосомных локусов они становятся одинаковыми в следующем поколении, поскольку и самцы, и самки получают половину своих генов от отца и половину — от матери. Таким образом, равновесные частоты генотипов достигаются в этом случае за два поколения. Однако в случае сцепленных с полом локусов равновесные частоты достигаются лишь постепенно.
Закон Харди—Вайнберга сформулировали в 1908 г. независимо друг от друга математик Г. X. Харди в Англии и врач В. Вайнберг в Германии. Чтобы понять смысл этого закона приведем простой пример. Предположим, что данный локус
содержит один из двух аллелей, Айа, представленных с одинаковыми для самцов и самок частотами: р для Аид для а. Представим себе, что самцы и самки скрещиваются случайным образом, или, что то же самое, гаметы самцов и самок образуют зиготы, встречаясь случайно. Тогда частота любого генотипа будет равна произведению частот соответствующих аллелей. Вероятность того, что некоторая определенная особь обладает генотипом АА, равна вероятности (р) получить аллель А от матери, умноженной на вероятность (р) получить аллель А от отца, т. е. рхр = р2 .
Закон Харди—Вайнберга гласит, что процесс наследования преемственности сам по себе не ведет к изменению частот аллелей и (при случайном скрещивании) частот генотипов по определенному локусу. Более того, при случайном скрещивании равновесные частоты генотипов по данному локусу достигаются за одно поколение, если исходные частоты аллелей одинаковы у обоих полов.
Равновесные частоты генотипов задаются произведениями частот соответствующих аллелей. Если имеются только два аллеля, А и а, с частотами р и q, то частоты всех трех возможных генотипов выражаются уравнением:
(p+g)2=p2+2pq + q2 А а АА Аа аа, где буквам во второй строке, обозначающим аллели и генотипы, соответствуют расположенные над ними частоты в первой строке.
Если имеются три аллеля, скажем, А,, А2иА3, с частотами р, q и г, то частоты генотипов определяются следующим образом:
(p + q + r)2 =р2 + q2 + г2 + 2pq+2рг + 2qr
At АгА3 А1А1АгА3А3А3А1Аг,А1А3А3А3
Аналогичный прием возведения в квадрат многочлена может быть использован для определения равновесных частот генотипов при любом числе аллелей. Заметим, что сумма всех частот аллелей так же, как и сумма всех частот генотипов, должна быть равна единице. Если имеются только два аллеля с частотами р и q, то р + q — 1, и, следовательно, р2 + 2pq + q2 = (p + q)2 =1; если же имеются три аллеля с час
тотами р, q и г, то р + q + г = 1, и, следовательно, также (p+q + rf =1 и т.д.
Организмы, обладающие удачными вариантами признаков, имеют большую вероятность по сравнению с другими организмами выжить и оставить потомство. Вследствие этого полезные вариации в ряду поколений будут накапливаться, а вредные или менее полезные вытесняться, элиминироваться. Это и называется процессом естественного отбора, который играет ведущую роль в определении направления и скорости эволюции.
Прямая взаимосвязь между степенью генетической изменчивости в популяции и скоростью эволюции под действием естественного отбора была доказана математическим путем Р. Фишером (1930) в его фундаментальной теореме естественного отбора. Фишер ввел понятие приспособленности и доказал, что скорость возрастания приспособленности популяции в любой момент времени равна генетической вариансе приспособленности в тот же момент времени. Однако прямые доказательства этого факта были получены лишь в конце 1960-х гг.
Мутационный процесс служит источником появления новых мутантных аллелей и перестроек генетического материала. Однако возрастание их частоты в популяции под действием мутационного давления происходит крайне медленно, даже в эволюционном масштабе. К тому же подавляющее большинство возникающих мутаций устраняется из популяции в течение немногих поколений уже в силу случайных причин. Неизбежность такого течения событий впервые обосновал Р. Фишер в 1930 г.
Для человека и других многоклеточных показано, что мутации обычно возникают с частотой от 1 на 100 000 (1 • 10'®) до 1 на 1 000 000 (1-Ю-8) гамет.
Новые мутанты, хотя и довольно редко, но постоянно появляются в природе, поскольку существует множество особей каждого вида и множество локусов в генотипе любого организма. Например, число особей того или иного вида насекомых обычно составляет около 100 млн (108). Если предположить, что средняя мутабильность по одному локусу равна 1 мутации на 100 000 (10-s) гамет, то среднее число вновь возникающих в каждом поколении мутантов по этому локусу для данного вида насекомых составит 2-108< 10~5=2000. (Частота возникновения мутаций умножается на число особей и еще на два, так
как любая особь представляет собой продукт слияния двух гамет.) В генотипе человека имеется около 100 000 (10s) локусов. Предположим, что у человека темп мутирования такой же, как у дрозофилы; в этом случае вероятность того, что генотип каждого человека содержит новый аллель, отсутствовавший в генотипе его родителей, равна 2- 10s* 10"s = 2. Иными словами, каждый человек в среднем несет около двух новых мутаций.
Проделанные расчеты основаны на частотах возникновения мутаций, обладающих внешним проявлением. В целом по геному темп мутирования составляет не менее 7*10“9 замен на одну нуклеотидную пару в год. У млекопитающих число нуклеотидных пар в диплоидном геноме составляет около 4*109. Следовательно, нуклеотидные замены у млекопитающих происходят с частотой не менее 4*109*7*10“9 = 28 в год на диплоидный геном. Ясно, что мутационный процесс обладает колоссальными возможностями поставлять новый наследственный материал.
Важный шаг в генетике популяций был сделан в 1926 г. С. С. Четвериковым. Исходя из закона Харди—Вайнберга, Четвериков доказал неизбежность генетической разнородности природных популяций при том, что новые мутации непрерывно появляются, но остаются обычно скрытыми (рецессивными), а в популяции идет свободное скрещивание.
Из расчетов Четверикова следовало, а впоследствии было полностью подтверждено практикой, что даже редкие и вредные для особи мутантные гены будут надежно укрыты от очищающего действия естественного отбора в гетерозиготах (организмах со смешанной наследственностью) с доминирующими безвредными генами нормального дикого типа. Мутация будет как бы поглощена популяцией, из-за чего за внешним однообразием особей одной популяции неизбежно скрывается их огромная генетическая разнородность. Четвериков это выразил так: «Вид, как губка, впитывает в себя гетерозиготные ге-новариации, сам оставаясь при этом все время внешне (фенотипически) однородным». Для жизни популяций эта особенность может иметь два разных следствия. В огромном большинстве случаев при изменении условий среды вид может реализовать свой «мобилизационный резерв» генетической изменчивости не только за счет новых наследственных изменений у каждой особи, но и благодаря «генетическому капиталу», доставшемуся от предков. Благодаря такому меха-
Г»
низму наследования популяция приобретает пластичность, без чего невозможно обеспечить устойчивость приспособлений в меняющихся условиях среды. Однако изредка возможен и другой исход: редкие скрытые вредные мутации иногда могут встретиться у потомства совершенно здоровых родителей, приводя к появлению особей с наследственными заболеваниями. И это — тоже закономерное, неистребимое биологическое явление, своего рода жестокая плата популяции за поддержание своей наследственной неоднородности.
С. С. Четверикову популяционная генетика обязана еще одним открытием, которое было изложено в маленькой, всего четыре страницы, заметке «Волны жизни», опубликованной в 1905 г. на страницах «Дневника Зоологического отделения Императорского общества любителей естествознания и этнографии» в Петербурге. Он обратил внимание, что поскольку любая природная популяция имеет конечную, ограниченную численность особей, это неизбежно приведет к чисто случайным, статистическим процессам в распространении мутаций. При этом популяции всех видов постоянно меняют численность (численность грызунов в лесу может от года к году изменяться в сотни, а многих видов насекомых — в десятки тысяч раз) из-за чего в разные годы распространение мутаций в популяциях может идти совершенно по-разному. От громадной популяции птиц, насекомых, зайцев и других животных в трудный для переживания год может остаться всего несколько особей, причем иногда совершенно нетипичных для бывшей популяции. Но именно они дадут потомство и передадут ему свой генофонд, так что новая популяция по составу генетического материала будет совершенно иной, чем прежняя. В этом проявляется генетический «эффект основателя» популяции. Постоянно изменяется и геном в популяциях человека. К. Альстрем на материале из Южной Швеции показал, что в популяции человека следующему поколению передается далеко не весь имеющийся генофонд, а лишь избранная, а то и случайно «выхваченная» часть. Так, 20% поколения здесь вовсе не оставили потомков, зато 25% родителей, которые имели трех и более детей, дали 55% численности следующего поколения.
Постоянное давление мутаций и миграции генов, а также выщепление биологически менее приспособленных генотипов по сбалансированным полиморфным локусам создает проблему так называемого генетического груза. Понятие генетичес-
*
кого груза ввел Г. Мёллер в 1950 г. в работе «Наш груз мутаций». По его расчетам, от 10 до 50% гамет у человека содержат хотя бы одну вновь возникшую мутацию. Слабо вредящие мутации, если только они проявляются в гетерозиготе, способны нанести популяции больший урон, чем полностью рецессивные летальные мутации. Каждый из нас является носителем по крайней мере восьми вредных мутаций, скрытых в гетерозиготном состоянии. Г. Мёллер в соавторстве с Н. Мортоном и Дж. Кроу (1956) произвели оценку генетического груза мутаций путем сравнения детской смертности в случайных выборках из популяций и в семьях, где имели место браки между родственниками. Они выделили собственно мутационный груз, возникающий в результате мутационного давления, и сегрегационный груз как следствие расщепления. Ими предложены расчеты летального эквивалента, соответствующего числу мутаций, дающих вместе летальный исход. Так, один летальный эквивалент может соответствовать одной летальной мутации, двум полулегальным и т.д. Было показано, что средняя величина генетического груза у человека равна ’3-5 летальным эквивалентам.
Ю. П. Алтухов с коллективом сотрудников (1989) в результате длительного изучения локальных стад рыб — больших изолированных друг от друга популяций с исторически сложившейся субпопуляционной структурой — пришел к выводу о высокой их устойчивости во времени и пространстве. Изменчивость на уровне отдельных субпопуляций не играет самостоятельной роли и отражает локальные различия действия отбора в силу гетерогенности условий обитания, а также влияние случайных факторов. К аналогичному заключению еще раньше пришел Ю. Г. Рычков с сотрудниками при исследовании изолированных групп популяций человека — коренного населения циркумполярной зоны Евразии. Американский генетик и селекционер И. М. Лернер еще в 1954 г. выдвинул представление о генетическом гомеостазе, определив его как способность популяции приводить в равновесие свою генетическую структуру и противостоять внезапным изменениям. Один из важных механизмов генетического гомеостаза — отбор в пользу гетерозигот, ведущий к сбалансированному равновесию. Вместе с тем этот же механизм служит причиной образования генетического груза, т. е. выщепляющихся гомозиготных классов особей. Такой груз был назван сбалансиро-
ванным и рассматривается как плата за поддержание гетерозигот, причисленных к генетической элите популяции.
Частоты генов в популяциях. Для описания ситуаций в популяционной генетике создано несколько математических моделей. Еще в 1928 г. Валунд установил, что если большая популяция разделена на К панмиктических групп, то в такой совокупности наблюдается эффект, подобный последствиям имбридинга в неподразделенной популяции: доля гомозигот возрастает на величину межпопуляционной вариансы частот генов за счет уменьшения доли гетерозигот.
Принципиальный вклад в описание локальной дифференциации частот генов в подразделенной популяции в терминах F-статистики был внесен С. Райтом, который обосновал несколько .^-коэффициентов как показателей меры генетической дифференциации:
1) FlT — коэффициент имбридинга особи относительно целой(Т) популяции;
2) FIS — коэффициент имбридинга особи относительно субпопуляции (S);
3) FST — коэффициент имбридинга субпопуляции относительно всей подразделенной популяции.
Соотношение между этими величинами задается равенством:
Коэффициент FST был предложен С. Райтом в 1943 г. и с тех пор неоднократно использовался при анализе распределений частот генов в природных разделенных популяциях. Коэффициент Райта представляет большой интерес, так как позволяет вычленить некоторые важные влияния популяционной подразделенности ца генетическую структуру. Для этой цели Райт предложил две оригинальные модели популяций: «островная модель» и «изоляция расстоянием».
Островная модель. Известны два варианта этой модели:
1) подразделенность вида на множество свободно скрещивающихся внутри себя субпопуляций генетически эффективного объема N, каждая из которых с равной вероятностью и с одинаковой интенсивностью т обменивается генами с любой другой;
2) большая панмиктическая популяция («материк»), окруженная множеством изолированных, генетически дифференцированных малых колоний («острова»), каждая из кото
рых получает гены с «материка» с интенсивностью т на поколение. Эффектами обратной миграции можно пренебречь.
Мерой случайной дифференциации субпопуляций в такой системе служит межгрупповая варианса генных частот:
у - ‘’('-У
4 4Nem + l и, следовательно, условие равновесия между дрейфом и миграцией генов в терминах Гет-статистики может быть записано как
F =-------.
4Ne/n-H
Более строгое решение относительно Vg задается формулой:
у - ^‘-g)______
9 4Ne-(2Ne-l)(l-m)2
Как следствие взаимодействия дрейфа и миграции мы имеем вероятностное распределение частот генов. В любой момент времени Т оно представляет функцию от Дд = -m(q -qT) как меры систематического давления миграции g(l-g) и — — выборочной вариансы частоты гена в одном
поколении за счет изоляции, т. е. случайного дрейфа:
Ф(д) = (с/У8д )exp[2j(Ag/V5q )dg].
Если мы обозначим через qt частоту гена в i-й группе (pt = = qt = 1), а через q — частоту этого же гена в подразделенной популяции как в целом, то характерные для нее средняя частота гена и ее варианса будут
к,=Е^^=(М/к)-д2.
Соответственно частоты зигот (генотипов) равны AA.£p2/K = p2=Vg ,
Aa.-£ftgf/K = 2pg-2Vg , aa.£g2/K = g2+Vg •
Сопоставляя частоты генотипов с их частотами в популяции, характеризующейся коэффициентом инбридинга F, получим соотношение
Vq = Fq(l-q) или F = Vq/q(l-q).
Поскольку величина F характеризует подразделенную популяцию в целом, то соответствующие частоты генотипов в ней равны частотам, которые были бы свойственны отдельной инбредной популяции. Иными словами, подразделен-ность популяции на отдельные скрещивающиеся группы формально эквивалентна наличию инбридинга во всей популяции.
Общая формула стационарного распределения частот генов в островной модели представляет p-функцию плотности вероятности следующего вида:
С .dq
Ф(?) = С е 9(1-9) ,
где С — нормирующий множитель, подбираемый так, чтобы
Г(4№п)
1» r(4№nq)r[4Nm(l-q)J'
В зависимости от того, каким сочетанием случайных и систематических факторов задается распределение, оно принимает вид:
а) при уравновешивании случайного дрейфа генов их миграцией
Ф(д)=Ср4Лйп₽ч(1-д)4№п(1-’Н, где р и q— частоты аллелей в субпопуляциях; pmq — средние частоты аллелей для подразделенной популяции как целого; N — эффективный размер популяции; т — коэффициент миграции;
б) при объединенном эффекте воздействия изоляции, миграции и отбора
®(q)-CW2'Vq4N'n’"1(l-g)4'Vm(1_’)-1 , где все обозначения те же, что и в предыдущем выражении, &W — средняя внутрилокусная приспособленность популя-
ft
ции, определяемая через суммирование приспособленностей генотипов с учетом их частот.
Стационарные распределения могут описывать:
1) распределения частот аллелей многих локусов в одной и той же популяции в случае нейтральности или при примерно одинаковом давлении отбора на каждый локус;
2) распределение генных частот какого-либо локуса в последовательных поколениях одной и той же стационарной популяции;
3) распределение частот аллелей одного или нескольких локусов в совокупности полностью или частично изолированных популяций.
Все эти три типа математически эквивалентны.
В островной модели величина коэффициента миграции генов не зависит от степени удаленности популяций С. Райт (1943) и Г. Малеко (1955, 1957) математически исследовали ту же популяцию, в которой интенсивность обмена между субпопуляциями зависит от расстояния. Эта модель носит название «изоляциярасстоянием» и предполагает популяцию, непрерывно распределенную на большой территории, существенно превышающей радиус индивидуальной активности особи в репродуктивный период. Особенности локальной дифференциации в такой системе зависят от репродуктивной величины или «соседства», откуда случайно происходят родители, а также от размерности ареала.
Согласно С. Райту, размер соседства приблизительно соответствует числу генетически эффективных особей внутри круга, радиус которого равен удвоенному стандартному отклонению о протяженности миграции в одном направлении в данном поколении, т. е. дистанции между местами рождения родителей и потомков. *
Дифференциация очень велика, когда Nn ~ 20, намного меньше, но все же достаточно выражена при Nn ~ 200 и почти соответствует панмиксии, когда Nn = 2000.
М. Кимура (1953) предложил другую модель популяционной структуры. Она носит название «лестничная модель» я представляет ситуацию, промежуточную между райтовской островной моделью и моделями непрерывно распределенных популяций С.Райта и Г. Малеко.
Лестничная структура миграции генов. В этой модели, как и в островной, рассматривается совокупность колоний, одяа-
*
ко обмен особями происходит только между соседними колониями, и, таким образом, непосредственно зависит от удаленности колоний друг от друга.
При равновесии межпопуляционная варианса генных частот
V ____________________________
9 2N-(2Ne -1){1-2Я1/г2/(1?! + Д2)} ’ где 1?! =^(l+a)2-(2P)2j^ ; =£(1 —a)2-(2P)2J^ , в которых
a = (l-m1 )(1-щ>,)и P = m1(l-ni00)/2; в этих уравнениях т1 — интенсивность миграций между смежными колониями, а —
давление миграции генов извне на всю совокупность колоний (соответствует коэффициенту т в островной модели С. Райта). Когда 0, то а = 1- щ>о, Р = 0,и выражение сводится к формуле Райта. Островная модель Райта, таким образом представляет собой частный случай лестничной модели в отсутствие обмена генами между соседними колониями.
Важнейшая особенность подразд елейности, также исследованная теоретически, — способность таких популяций поддерживать значительно большее генетическое разнообразие в сравнении с панмиктическими популяциями сопоставимого размера. Считается, что именно такое разнообразие и позволяет популяции более эффективно реагировать на изменения среды и вслед за ними изменять свою генотипическую структуру — тезис, играющий решающую роль в эволюционной концепции Райта, известной под названием «теория смещающегося равновесия», в которой «поверхность» Wизображается топографической картой с вершинами и долинами на едином ландшафте генных комбинаций. В этой модели важнейшее заключение состоит в том, что эволюционный процесс зависит от постоянно смещающегося баланса между факторами стабильности и изменений и что наиболее благоприятное условие для этого — наличие тонко подразделенной структуры, в которой изоляция и перекрестная коммуникация поддерживаются в соответствующем равновесии».
БИОХИМИЧЕСКАЯ СИСТЕМАТИКА
Отечественными биохимиками А.Н. Белозерским, А.С. Спириным, А.С. Антоновым и их последователями в начале 1970-х гг. XX в. были сформулированы представления об использовании генетических и биохимических показателей в оценке таксономического сходства живых организмов («Строение ДНК и положение организмов в системе», 1972), ныне получившие широкое распространение.
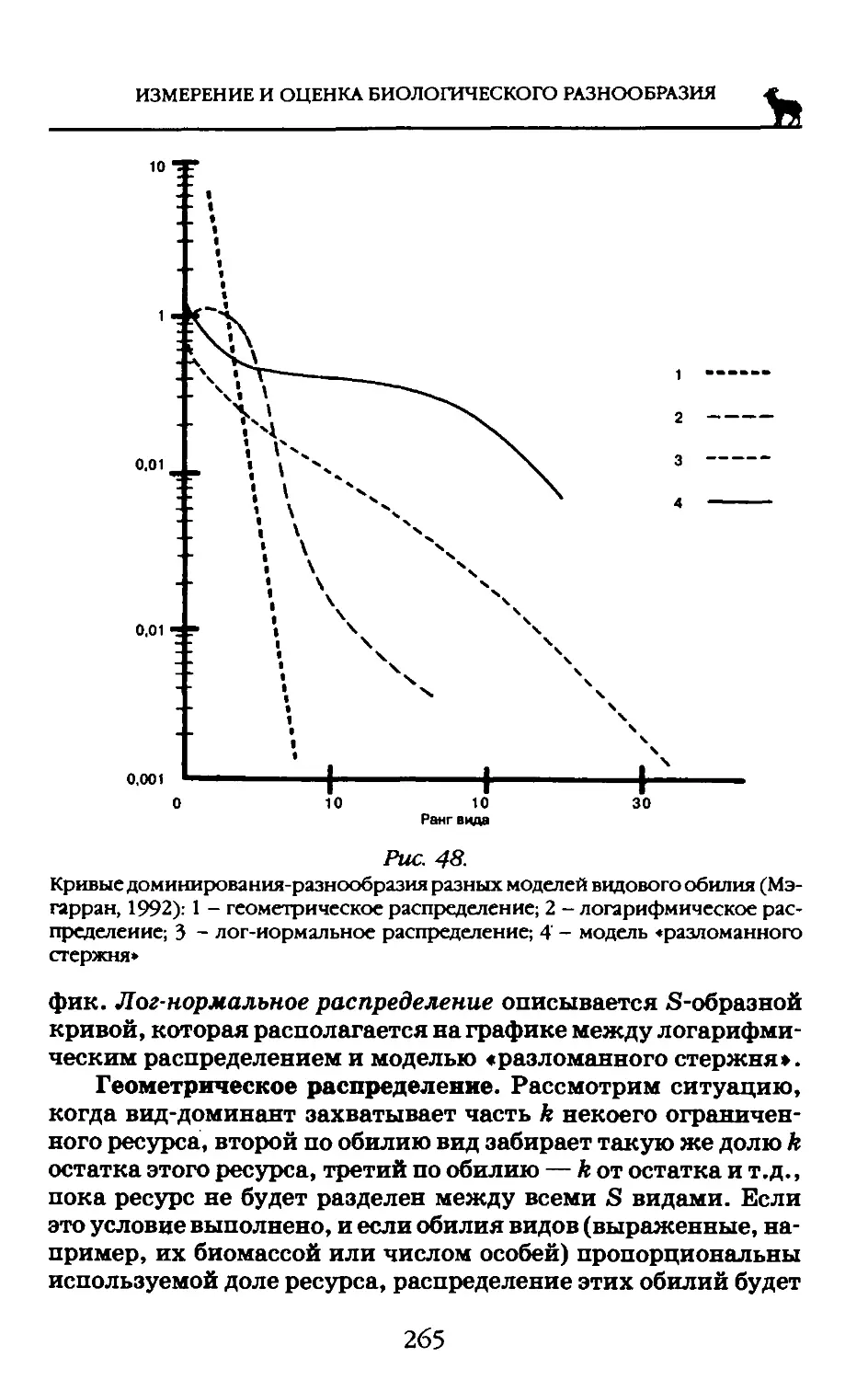
Со времен Эрнста Геккеля (1834-1919) известна триада сравнительно-филогенетических методов: морфологического, эмбриологического и палеонтологического. Исследователь, работающий классическими методами, сначала изучает морфологию и анатомию интересующих его объектов. Если черты строения органов, тканей и клеток не позволяют отнести их с достаточной уверенностью к тому или другому таксону, на помощь приходит эмбриология. В развитии организмов нередко сохраняются черты их отдаленных предков. Морфологические и эмбриологические данные там, где это возможно, дополняются палеонтологическими.
Комбинация этих методов — весьма мощный инструмент систематика-эволюциониста. Однако классические методы
пасуют именно в применении к низшим организмам. Морфологические признаки примитивных организмов за время их эволюции могли неузнаваемо измениться или, наоборот, дать многочисленные примеры конвергенции, а также параллельного развития. Кроме того, примитивные организмы именно в силу их примитивности, мало пригодны для изучения морфологии. Формы большинства бактерий (палочка, шарик, спираль) могли возникнуть самостоятельно в разных группах и многократно перейти друг в друга независимо на разных путях эволюции. То же относится к эмбриологическому методу: можем ли мы назвать «онтогенезом» деление бактериальной клетки или самосборку вирусной частицы? Палеонтологический метод также неприменим там, где речь идет об одноклеточных бесскелетных организмах: они практически не сохраняются в окаменевшем состоянии. Из этого следует, что для решения проблем систематики низших организмов необходимы принципиально новые методы, позволяющие оценить степень родства, исходя из каких-то других признаков. Такими методами могут быть биохимические.
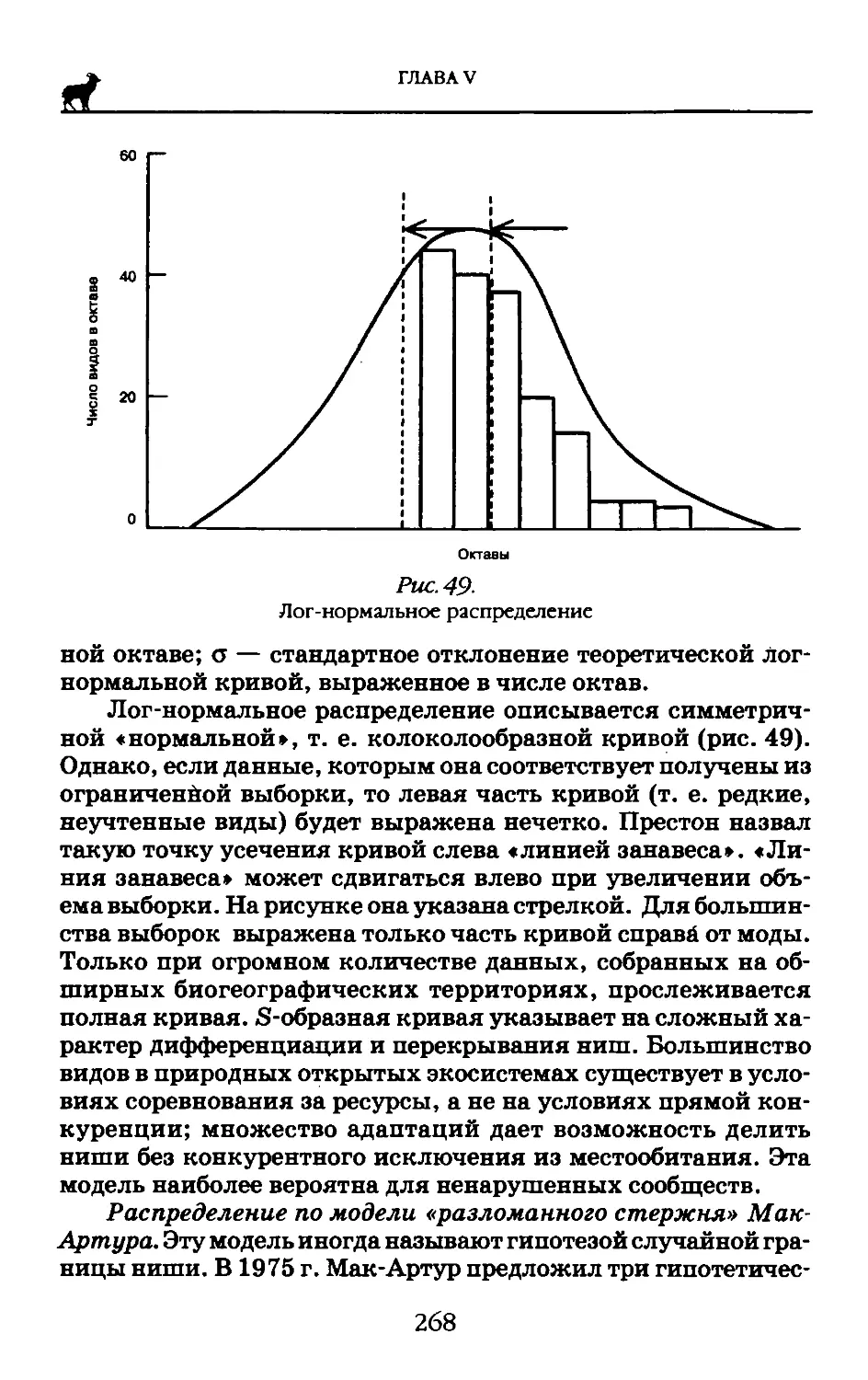
Для понимания степени филогенетического родства организмов неоднократно использовались данные о составе и свойствах их алкалоидов, жиров, белков, путях синтеза тех или иных метаболитов. Широко известны также попытки применения иммунобиологических и иммунохимических методов, а также анализа элементного состава (Лебедева, 1999).
Однако подобные исследования имели весьма ограниченный успех и не получили признания у «классических» систематиков. Дело здесь не в консерватизме систематиков. Применявшиеся биохимические показатели были так же вторичны, как и морфологические. В конечном счете нет разницы между, допустим, числом щетинок у рачка и числом изози-мов его лактатдегидрогеназы — обе величины суть признаки фенотипа, а не генотипа.
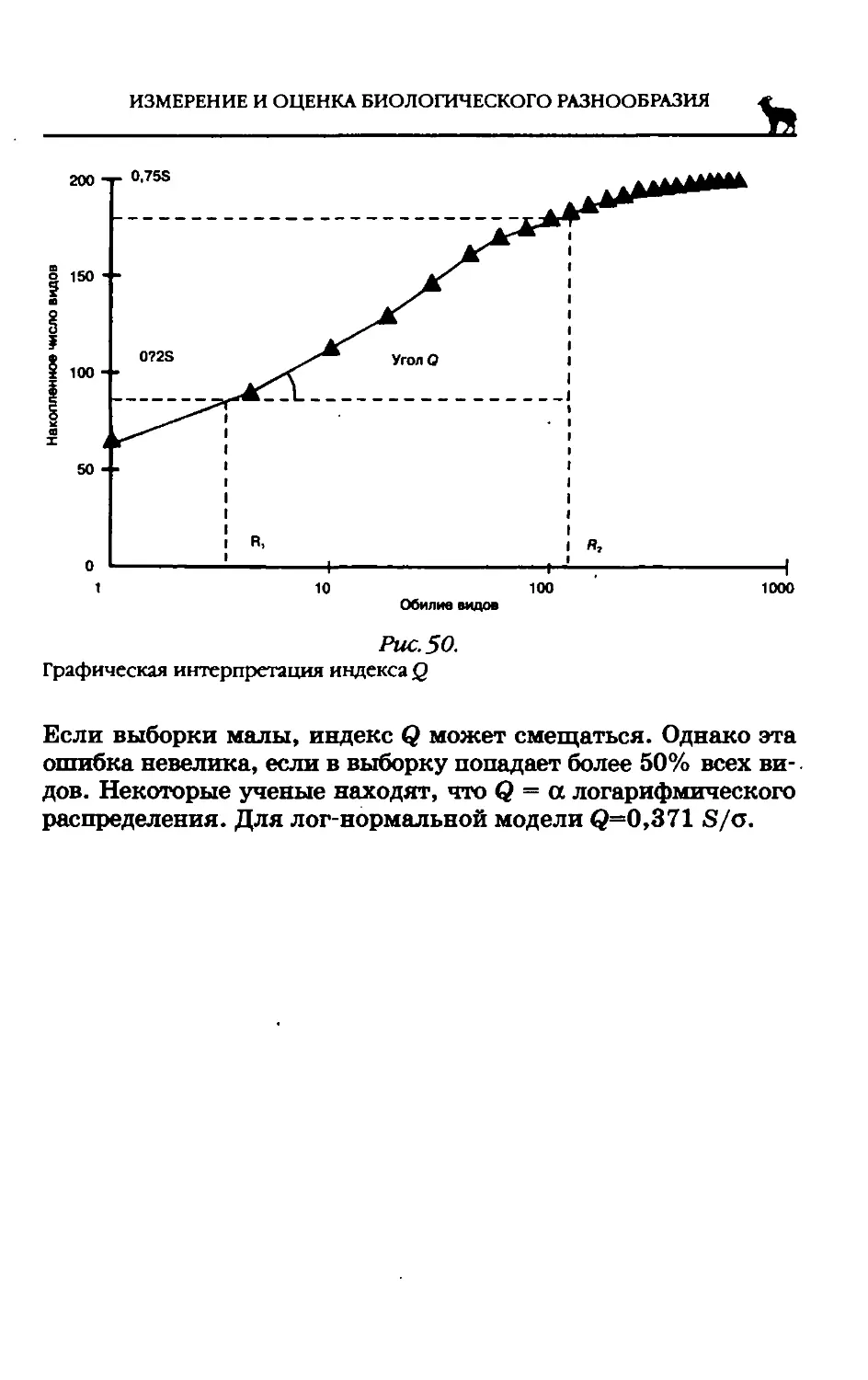
По мнению А.Н. Белозерского и его учеников, переворот в биохимической систематике произошел после того, как американский ученый Э. Чаргафф в самом начале 1950-х гг. совершил открытие кардинальной важности, установив, что нуклеиновые кислоты, так же как и белки, видоспецифичны. На протяжении последующих 20 лет были получены данные, неопровержимо доказывающие, что нуклеиновые кислоты — не только материальная форма хранения наследственной ин
формации, но и те соединения, через которые осуществляется реализация наследственности в процессах развития организма. Крайне перспективно изучение нуклеиновых кислот с точки зрения познания основ эволюционного процесса — наследственности и изменчивости.
В первую очередь речь должна идти о дезоксирибонуклеиновой кислоте — ДНК, так как именно это соединение является материальным субстратом наследственности подавляющего большинства организмов (за исключением РНК-содер-жащих вирусов). В течение более 15 лет была проведена огромная работа по изучению первичной структуры ДНК различных бактерий, актиномицетов, грибов, водорослей, высших растений, а также позвоночных и беспозвоночных животных. Полученный материал позволил сделать ряд важных выводов. Первый вывод состоит в том, что ДНК всего органического мира едина: во всех исследованных группах животных и растений она слагается из одних и тех же дезоксирибонуклеотидов, включающих два пуриновых основания (аденин и гуанин) и два пиримидиновых (цитозин и тимин). Для всего органического мира (за исключением вторичного явления — одноцепочечных ДНК некоторых вирусов) строго соблюдаются закономерности, подмеченные Чаргаффом («правила Чар-гаффа»), которые сводятся к следующему:
1. Сумма пуриновых нуклеотидов равна сумме пиримидиновых нуклеотидов:
Г+А „ пур.=пир., или ц^=1-
2. Содержание аденина равно содержанию тимина:
А=Т, или —=1.
Т
3. Содержание гуанина равно содержанию цитозина:
Г=Ц, или —=1.
Ц
4. Содержание б-аминогрупп равно содержанию 6-кето-групп:
Г+Т
А+Ц" '
5. Содержание Г+Ц и А+Т может варьировать в довольно значительных пределах.
Поэтому для ДНК наряду с единым планом строения, обусловленным ее жесткой двуспиральной структурой, присуща возможность бесконечного варьирования состава и последовательности оснований.
Несомненно, возникновение ДНК в процессе происхождения жизни на Земле было важнейшим фактором дифференциации и становления новых видов. В структуре ДНК наследственная информация не только сохраняется наилучшим образом, но и может передаваться путем репликации от поколения к поколению. В бесконечной цепи следующих друг за другом поколений естественным отбором были выбраны структуры ДНК, обеспечивавшие не только стабильность наследственной информации (консерватизм наследственности), но и возможность вариации первичной структуры ее макромолекулы.
Из пятого правила Чаргаффа вытекает второй, не менее важный вывод, сделанный одновременно и независимо друг от друга А.Н. Белозерским и А.С. Спириным и французскими исследователями Ли, Валь и Барбю, а именно: процент пар гуанин—цитозин, изменяющийся в процессе эволюции, видоспецифичен и может иметь таксономическое значение. Впер-
вые это было показано на ДНК бактерий, где мы имеем очень
большие вариации нуклеотидного состава. Показатель
А+Т у бактерий может меняться от 2,57 (сарцина) до 0,45 (возбудитель газовой гангрены). Если этот показатель равен 1 (Г+Ц=А+Т=50%), такую ДНК называют эквимолярной. В ДНК ГЦ-типа гуанина и цитозина более 50%, для ДНК АТ-типа характерно обратное соотношение оснований.
Впоследствии вариабельность нуклеотидного состава ДНК была изучена у многих групп водорослей, грибов, вирусов, а также высших растений и животных. Анализируя эти данные, А.С. Антонов пришел к выводу о том, что вариабельность соответствует эволюционному возрасту таксона. Древние группы (вирусы, бактерии, грибы, водоросли) очень варьируют по составу ДНК. То же отмечается и у животных. Так, например, у древнейшей группы простейших содержание Г+Ц может меняться почти на 39%, у губок эта величина уже 25%, у хордовых— менее 6%.
ff.
Не исключена возможность, что значительная вариабельность нуклеотидного состава ДНК микроорганизмов свидетельствует об их полифилии; в то же время она может быть лишь следствием длительности мутационного процесса. Любопытно, что ДНК практически всех высших растений и животных АТ-типа, причем нуклеотидный состав довольно стабилен. Специфичность в данном случае реализуется по линии нуклеотидной последовательности. Это обстоятельство ограничивает применение такого показателя, как нуклеотидный состав ДНК, у высших организмов. Тем не менее, использование данных о составе ДНК позволяет сделать более обоснованный выбор из числа имеющихся альтернативных схем макрофилогенеза.
А.С. Антоновым и Б.М. Медниковым (1972) методами вариационной статистики был проанализирован материал о составе ДНК около 2000 видов различных организмов. Из полученных данных сделан ряд любопытных выводов. Прежде всего можно заключить, что выделенные ранее формальные категории ДНК — ГЦ- и АТ-типа — не субъективны, а отражают реально существующую в природе закономерность: во всех наиболее изученных группах чаще всего встречаются ДНК, содержащие от 35 до 45 или от 50 до 60 мол. % ГЦ-пар нуклеотидов. Иными словами, мы имеем два модальных класса. Первый модальный класс (АТ-тип ДНК) прослеживается в группах хордовых и беспозвоночных животных, высших растений, грибов, сине-зеленых водорослей, бактерий и вирусов. ДНК второго модального класса (ГЦ-тип ДНК) отмечена у грибов, актиномицетов, водорослей, бактерий и вирусов. Самое удивительное то, что эквимолярный состав ДНК встречается относительно редко. Этот факт загадочен, так как ДНК с равным количеством всех четырех оснований должна обладать максимально возможной информационной емкостью (два бита на нуклеотид, согласно уравнению Шеннона).
Биохимики разработали методы сравнения организмов по набору аминокислот, а также по прямому сопоставлению молекул ДНК.
Второй подход сравнения последовательностей в ДНК более перспективен. Это так называемый метод молекулярной гибридизации. За последние годы он получил широкую известность среди микробиологов. Можно утверждать, что пер-
вые успешные попытки создать филогенетическую систему вирусов и бактерий основаны главным образом на применении этого метода.
Метод молекулярной гибридизации был разработан в США Хойером, Мак-Карти и Болтоном. Принцип метода сравнительно несложен. Как известно, нативная ДНК состоит из двух комплементарных цепей, соединенных водородными связями между аденином и тимином, и гуанином, и цитозином. В нормальных условиях клетки эта структура весьма устойчива. Если же повысить температуру, особенно в условиях низкой ионной силы, водородные связи между комплементарными цепями рвутся: ДНК денатурирует. При медленном охлаждении начинается процесс ренатурации, воссоединения комплементарных цепей. Ренатурация приближается к стопроцентной только у гомогенных ДНК фагов. Уже у бактерий геном настолько неоднороден, что даже за большой промежуток времени не все последовательности могут найти себе комплементарные половины.
Если денатурированную ДНК иммобилизировать осаждением на целлюлозный фильтр или включением в агаровый либо полиакриламидный гель, ренатурации не произойдет. Работая одним из основных методов, процесс гибридизации проводят в колонке с гелем, в который «вплавлена» денатурированная ДНК. Затем через колонку пропускают раствор ДНК, также денатурированной, но, кроме того, подвергнутой деградации (молекулы ДНК легко разрываются на части; этот процесс называют деградацией и обычно в качестве деградирующего агента применяют ультразвук). Короткие фрагменты деградированной ДНК легко проходят через поры геля и могут соединяться с иммобилизированной ДНК, образуя реассоциированные двойные цепи. Низкомолекулярную ДНК обычно метят радиоактивным фосфором, углеродом или тритием. В дальнейшем надо лишь вымыть из колонки неприсоединившуюся меченую ДНК. Весьма полезен прием, заключающийся в обработке препарата специфической ДНК-азой, расщепляющей одноцепочечную ДНК, но не трогающей двухцепочечную. В результате в геле остаются только гибридные цепи (в тех случаях, когда ДНК иммобилизированная и меченая происходят из разных источников). О проценте гибридизации мы можем судить по активности конечного продукта.
Болтон и Мак-Карти прекрасно отдавали себе отчет в том, насколько разработанный метод может быть цепным именно для систематики. В конечном счете это прямое определение близости геномов друг к другу. По ряду причин методу молекулярной гибридизации «нестрашны» конвергенция и параллельное развитие — проклятие всех ныне существующих методов определения сходства организмов. Например, вполне вероятно конвергентное развитие макроструктур у неродственных форм. Весьма часто и конвергентное возникновение сходных по функции белков: лизоцимы фагов и высших организмов, несомненно, не родственны. Однако конвергентное возникновение идентичных аминокислотных последовательностей уже крайне маловероятно. Еще менее вероятно подобное возникновение идентичных участков цепей ДНК, даже если они кодируют сходные по составу белки. Ведь генетический код вырожден и большинству аминокислот может соответствовать несколько триплетов. Поэтому молекулярная гибридизация — весьма многообещающий метод для выявления полифилетических таксонов.
Даже в столь хорошо изученной группировке, как млекопитающие, этим методом подтверждена полифилия отряда грызунов, многими систематиками давно разделяемого на отряды зайцеобразных и двурезцовых. Известно, что дискутируется вопрос о разделении на самостоятельные отряды зубатых и беззубых китов, а также ластоногих и насекомоядных.
Однако возникли не только технические, но и методические трудности. Уже первые работы по гибридизации ДНК показали, что у высших организмов геном неоднороден. В самом первом приближении его можно разделить на две фракции: фракция уникальных последовательностей и фракция повторяющихся последовательностей. Не менее интересны сателлитные-фракции ДНК — оказалось, что концентрации их варьируют очень сильно даже у близкородственных видов.
Комбинируя условия выделения, фракционирования и гибридизации ДНК, в принципе можно сопоставлять геномы в пределах таксонов любого ранга, вплоть до вида и в конечном счете весьма облегчить работу систематика, предоставив ему уникальные данные биохимического анализа. Методика молекулярной гибридизации, хотя и не представляет собой
*
ничего сложного, чрезмерно трудоемка. Обнадеживает то, что она поддается автоматизации, а в ряде лабораторий США и Западной Европы поставлена «на поток», стала рутинной процедурой, хотя и довольно дорогостоящей.
Впрочем, уже достаточно давно метод молекулярной гибридизации в сочетании с определением нуклеотидного состава и анализом изоплит дает результаты и не только при решении частных проблем систематики. Так, группой А.С. Антонова были проведены опыты по изучению степени гомологичности первичных структур геномов у однодольных растений и разных классов хордовых животных (главным образом птиц, рептилий). Следует остановиться на одном выводе из серии этих работ, имеющем для построения систем принципиальное значение.
Речь идет о разномасштабности таксонов в пределах различных групп растительного и животного мира. Известно, что все системы построены по иерархическому принципу: виды объединяются в роды, роды в семейства, семейства в отряды (или порядки в фитосистематике) и т. д., вплоть до царств живой природы. Возникает вопрос: соответствует ли род, например, однодольных растений роду птиц или млекопитающих? Ответить на этот вопрос трудно, так как если границы таксона систематики обычно очерчивают правильно, то ранг присваивается довольно произвольно и имеет тенденцию повышаться в связи с изученностью группы. Ясно, однако, что если таксоны равномасштабны, то и степень гомологичности первичных структур ДНК у образующих их видов должна быть одинаковой.
Однако первые опыты по гибридизации ДНК показали другое — разномасштабность таксонов в системах разных групп растительного и животного мира. Так, например, меченая ДНК домашней курицы имеет приблизительно 25—38% гомологичных последовательностей с ДНК птиц других отрядов (исследовались воробьиные, трубконосые, попугаи, голуби и дневные хищники). Приблизительно Такая же межотрядная степень гомологии ДНК установлена Мак-Карти и Болтоном для класса млекопитающих. Степень гомологичности последовательностей в ДНК представителей разных классов позвоночных, естественно, ниже (у ДНК курицы и варана, например, 10,5%). Эти участки генома, по всей вероятности, кодируют наиболее общие признаки, присущие всем хордовым.
Совсем другие цифры получены для однодольных растений. Виды, относимые к разным семействам порядка лилиецветных, имеют лишь до 10% гомологичных структур ДНК. Иными словами, семейство в группе однодольных растений по рангу должно быть приравнено к классу хордовых животных! Представители разных порядков однодольных обладают еще более гетерологичной ДНК, а между ДНК однодольных и двудольных растений иногда вообще не образуется гибридных молекул. О субъективности ранга систематических категорий говорилось уже неоднократно, но теперь мы можем количественно оценить, насколько велика эта субъективность. Во всяком случае эти данные свидетельствуют о различиях кардинального свойства в путях и темпах эволюции геномов.
ВИДОВОЕ РАЗНООБРАЗИЕ
Видовой уровень разнообразия обычно рассматривается как базовый, центральный, а вид является опорной единицей учета биоразнообразия. Рассмотрим, цитируя Б. А. Юрцева (1994), единицы биоразнообразия, на которые можно опираться, разрабатывая и реализуя систему мер по его сохранению.
Единицы учета биоразнообразия должны обладать автономным жизнеобеспечением, способностью к неограниченно длительному самоподдержанию на фоне стабильной или умеренно флуктуирующей среды, восстановлению при нарушениях и адаптивной эволюции. Особи не отвечают такой совокупности характеристик, хотя в отдельных случаях возможны исключения, когда «священные» или иные «исторические» деревья берутся под охрану как памятники природы. Перечисленным требованиям в качестве важных единиц учета и сохранения биоразнообразия удовлетворяют виды, а применительно к ограниченным территориям — представляющие вид местные популяции.
Базы разнообразной информации об организмах должны быть привязаны к конкретным видам, а виды должны иметь
четкий адрес в той или иной таксономической системе. Так, охраняя вид А, в его лице мы охраняем одного из последних представителей рода или семейства, или редкую жизненную форму. Сведения по биологии и экологии вида необходимы для выработки обязательных мер его охраны в природе и сохранении в культуре (in situ и ex situ).
Виды зачастую являются основными объектами охраны, однако природоохранная деятельность не должна строиться по таксономическому принципу. В природе виды распределены в независимости от их предполагаемого родства. Представители из разных таксонов растений, животных и микроорганизмов, взаимно дополняя друг друга, образуют биоценозы и биоты — биотические ядра экосистем; поэтому таксономические списки животного и растительного мира и специальные перечни тех их представителей, которые нуждаются в глобальной, национальной или локальной охране (Красные книги) имеют контролирующее значение. Таксономическое разнообразие любой региональной биоты слишком велико для того, чтобы могло быть охвачено Красной книгой. Чем богаче биота, тем меньше составляющих ее видов имеют шанс попасть в Красную книгу. Большая часть флоры и фауны остается без правовой защиты.
Более подробно динамика видового разнообразия и ее закономерности изложена в главе «География биоразнообразия».
БИОРАЗНООБРАЗИЕ,
СОЗДАННОЕ ЧЕЛОВЕКОМ
Человек на протяжении сотен тысяч лет своего существования активно воздействовал на окружающую его живую природу. Уже древний человек, освоив огонь, вышел победителем в соревновании с другими видами, которые заселяли природные для обитания пещеры, уничтожил многих крупных плейстоценовых млекопитающих. Но было, начиная со времени «неолитической революции», — создания производительного хозяйства, земледелия, растениеводства и животноводства — и другое глобальное воздействие: сведение естественных экосистем и замена их сельскохозяйственными угодьями, а затем и городами с их пригородными зонами. Такие экосистемы, нередко более продуктивные, чем естественные, а их биоразнообразие может быть довольно велико. Однако, говоря о созданном человеком биоразнообразии, мы имеем в виду те биологические формы, которые целенаправленно были созданы человеком путем селекции, отбора, а теперь и генной инженерии.
В нашей стране выдающиеся исследования по теории и практике создания новых сельскохозяйственных растений принадлежат плеяде замечательных генетиков и селекционе-
ров во главе с академиком Н.И. Вавиловым. Десятки тысяч сортов культурных растений были собраны в основанном им Всесоюзном институте растениеводства (ВИР) в Ленинграде, им было создано учение о центрах происхождения культурных растений.
Замечательный пример дает культура риса, которым питается около трети человечества: только в Китае известно 40 тыс. сортов риса, а на Филиппинах создан банк «Гермапласт», в котором хранится 70 тыс. культурных сортов риса и 2 тыс. его диких вариантов.
Не менее замечательно разнообразие культивируемых животных, среди которых используются сотни пород рогатого скота, пушных зверей, лошадей, рыб, птиц и не менее 2 тыс. пород собак. Инициатором изучения генетической изменчивости домашних животных был российский генетик А.С. Серебровский, создавший в 1928 г. особое научное направление — геногеографию, которая занимается картированием генетической изменчивости видов. Сам он изучал генетику кур, среди которых в начале XX в. в России были известны десятки пород. Его продолжателем стал академик Д.К. Беляев, изучавший генетическую изменчивость домашних животных, особенно в азиатской части России и организовавший на Алтае первый в мире заповедник для домашних животных.
Генетики создали немало сельскохозяйственных культур путем отдаленного скрещивания растений, не встречающихся в дикой природе: тритикале, рапс, нектарину, грейпфруты и многие другие, культивируемые в огромных масштабах.
Генетика и селекция стали основным методом отбора культивируемых видов микроорганизмов, где генная инженерия — повседневный метод создания микробов с заданными человеком свойствами и где природный генетический потенциал диких видов для селекции практически исчерпан.
Таким образом, человек не только повинен в исчезновении множества видов на нашей планете, но и создал десятки тысяч форм растений, животных, микроорганизмов, которые без его участия никогда бы не появились.
В разнообразии геномов домашних животных А.С. Серебровский еще в 20-е годы прошлого века призывал видеть та
кое же естественное богатство страны, как в запасах нефти, золота, угля и других природных ресурсов. Современное высоко производительное хозяйство без использования культурных растений и животных, без эффективных технологий их разведения уже невозможно.
ЭКОСИСТЕМНОЕ РАЗНООБРАЗИЕ
На планете мы можем наблюдать огромное разнообразие наземных и водных экосистем: от ледяных полярных пустынь до лесов и от коралловых рифов до открытого океана. Все разнообразие экосистем можно классифицировать либо по функциональным, либо по структурным признакам. Часто оно оценивается через разнообразие видового компонента. Это может быть оценка относительных обилий разных видов, общее разнообразие территории или биотопа, биомасса видов разных размерных классов на разных трофических уровнях или различных таксономических групп. Гипотетическая экосистема, состоящая только из сходных растений, будет менее разнообразна, чем экосистема, включающая такое же число особей, а также виды травоядных и хищных животных.
Миллионы видов современных организмов, насчитывающих буквально астрономическое число особей, в принципе могут группироваться в еще большее количество сочетаний. Перечислить все существующие на Земле сообщества просто немыслимо. Для того чтобы разобраться во множестве биоценозов создано несколько классификаций, группирующих их
Uo степени сходства и различия. Наиболее разработаны классификации для растительности, поскольку она выступает индикатором биоценоза.
Обычно наименьшей типологической единицей фитоценологии считают ассоциацию. Она характеризуется тем, что Объединяемые ею отдельные сообщества имеют одинаковый видовой состав организмов, причем в первую очередь отмечается сходство доминирующих видов. Кроме того, у биоценозов одной ассоциации сходное строение и по другим признакам (ярусность, синузиальность, гидротермический режим И т. п.). Например, ельник-зеленомошник-черничник или ель-иик-зеленомошник-брусничник. Сходные ассоциации образуют группы ассоциаций (ельники-зеленомошники), которые объединяются в формации. Для формации характерен общий доминирующий вид или комплекс видов, в наибольшей степени влияющий на среду обитания (вид-эдификатор). В лесных биоценозах эдификаторами являются представители древесного яруса. Обычно формации называют по наименованию видов-эдификаторов. Можно, например, говорить о формации ельников (из того или иного вида ели). Далее следуют группы формаций (например, темно-хвойные леса из формаций разных видов елей и пихт), классы формаций (хвойные леса, эди-фикаторы представлены близкими жизненными формами), типы формаций (леса).
Для каждого такого биоценоза рассматриваются виды (или группы видов, или даже биологические группы), эди-фикаторы, доминирующие (фоновые) виды и биологические группы автотрофных и гетеротрофных организмов. Функ-ционально-биоценотические группы последних — разрушители отмирающей первичной продукции (сапрофаги), потребители растительных кормов (фитофаги) и хищники (как типичные плотоядные, так и вообще гетеротрофы второго и других порядков).
Большой интерес в научном отношении представляет проблема изучения структуры и функционирования антропогенных биоценозов. Антропогенные биоценозы, формирующиеся и развивающиеся под комплексным воздействием природных и социально-экономических факторов, имеют свои особенности; специфические законы их развития еще весьма слабо изучены. Можно отметить такие свойственные антропогенным биоценозам черты, как олигодоминантность (резкое
преобладание одного или нескольких видов в растительном и животном населении), неустойчивость системы, выражающаяся в резких изменениях численности, биомассы и продукции не только по сезонам, но и по годам, повышенная уязвимость структуры ввиду относительной простоты и однозначности связей между компонентами биоценоза. Последнее объясняется исторически малым возрастом антропогенных биоценозов, строение которых обычно не достигает такой степени сложности и сбалансированности, какую мы видим в естественных биоценозах. Поэтому резкие изменения условий и воздействий на антропогенный биоценоз подчас ведут к радикальным нарушениям его структуры или к полному его разрушению. Знание закономерностей строения и жизни антропогенных биоценозов позволит регулировать и направлять развитие географической среды, все более вовлекаемой в сферу деятельности человека.
КЛАССИФИКАЦИИ
БИОРАЗНООБРАЗИЯ
Юрцев Б.А. (1992) определяет биоразнообразие как разнообразие организмов и их природных сочетаний, хотя оно прослеживается также на более низких уровнях организации живого (молекулярном, субклеточном, клеточном, тканевом и уровне органов и т.д.). При этом он рассматривает организмы, в качестве наименьших единиц биоразнообразия, обладающих автономностью, способностью к жизнеобеспечению и адаптации и являющихся носителями других форм биоразнообразия.
Разнообразие организмов можно разделить на таксономическое, или филетическое (объединение по родству), и не филе-тическое или типологическое (группировки по тем либо иным признакам, не сводимым к родству, например, структурным, функциональным, структурно-функциональным, географическим, экологическим, синэкологическим и т.д.). Круг признаков, учитываемых в анализе типологического разнообразия, может быть неограниченно широк и зависит от задач исследования. Примером могут служить жизненные формы, стратегии жизни, ценотипы, типы метаболизма, сукцессионный статус видов (т. е. место в сукцессионных рядах или системах).
ft
Таксономическое разнообразие подразделяется на иерархические уровни с серией подуровней: видовой, популяционно-генетический (подуровни — популяции разного ранга, подвиды) генотипов (фенотипы), генов и их аллелей. Можно выделять уровни надвидовых таксонов (род, семейства и т.д. вплоть до царства).
Оценки таксономического и типологического разнообразий дополняют друг друга. Описание таксономического разнообразия (филума или биоты некоторой территории), обычно представляемое длинными систематическими списками, как правило, сопровождается характеристикой каждого таксона по комплексу типологических признаков. Сопряженность таксономического и типологического разнообразий ложится в основу организации баз и банков типологической информации об организмах. Однако информация о любом типе разнообразия поступает через виды как фундаментальные единицы биоразнообразия.
Биохорологическое разнообразие подразумевает разнообразие сочетаний организмов в тех или иных территориальных выделах, частях биосферы. Разнообразие природных территориальных сочетаний организмов в свою очередь подразделяется по территориальным уровням.
Опыт геоботаники и флористики говорит о том, что с расширением площади, на которой учитывается видовое разнообразие, наступают фазы относительной стабилизации состава видов растений, когда прирост числа видов резко замедляется и даже временно прекращается. Это происходит, когда исчерпывается видовое разнообразие сообщества в рамках однородного протяженного экотопа и далее, когда состав всех экотопов на протяжении данного ландшафта выявлен достаточно полно. Очевидно, что разнообразие сообществ отражает не только разнообразие местоположений (т. е. элементов рельефа с определенным составом почвообразующей породы), но и разнообразие сукцессионных стадий на каждом местоположении. В рамках протяженного ландшафта при едином макроклимате исчерпывается разнообразие местоположений и поддерживается постоянство сукцессионных процессов, что приводит к постоянству набора и состава сообществ, а, следовательно, и к постоянству состава данной элементарной, или конкретной, флоры.
В качестве двух нижних опорных уровней оценки биохорологического разнообразия Б.А. Юрцев выделяет уровень
Г»
сообщества (приблизительные эквиваленты в терминах близких научных дисциплин: экотоп, фация в ландшафтоведении, биогеоценоз) и уровень элементарной региональной биоты. Между опорными уровнями есть ряд промежуточных ступеней. Выше — иерархия выделов биогеографического районирования (округ, провинция, область), контуры которых в разных системах биогеографического деления Земли могут существенно не совпадать, в зависимости от критериев районирования.
В зависимости от биохорологического уровня может значительно меняться и таксономический уровень единиц, по которым оценивается биоразнообразие. В качестве наиболее универсальной, обязательной единицы (для всех уровней) выступает вид. Однако виды не всегда служат лучшей единицей для оценки разнообразия, так как разные стадии жизненного цикла или различные жизненные формы одного и того же вида часто занимают разные местообитания и экологические ниши и вносят свой вклад в разнообразие (личинка и имаго чешуекрылых, стадии амфибий). При оценке биоразнообразия крупных выделов районирования, как и при оценке изменений в геологическом времени, все большее значение приобретают роды, семейства, порядки, отряды и т.д.; и только на уровне сообщества, наряду с ассоциациями видов, рассматриваются более эфемерные группировки особей (синузии, парцеллы, ценоячейки и т.д.).
Структурное разнообразие является следствием зональности, стратифицированности, периодичности, пятнистости, наличия пищевых сетей и других способов ранжирования компонентов микроместообитаний. Различные способы распределения одновременно представленных в сообществе организмов характеризуют его структурное разнообразие:
1. Стратификационные принципы (вертикальная слоистость, ярусность растительного покрова, структура почвенных профилей).
2. Зональность (горизонтальная разобщенность, вертикальная поясность в горах или литоральной зоне).
3. Характер активности (периодичность).
4. Структура пищевой сети.
5. Репродуктивные системы (ассоциации родителей и потомства, клоны растений и т.д.).
6. Социальные структуры (стада и табуны).
ft
7. Системы взаимодействия (возникают в результате конкуренции, антибиоза, мутуализма и т.д.).
8. Стохастические структуры (возникают в результате действия случайных сил).
В 1960 г. Р. Уиттекер предложил понятия а -, Р -, у-разно-образия для того, чтобы не путать разнообразие внутри одного местообитания или региона с разнообразием ландшафта либо региона, который содержит несколько местообитаний.
^-разнообразие — разнообразие внутри местообитания или одного сообщества.
^-разнообразие — разнообразие между местообитаниями.
^разнообразие — разнообразие в обширных регионах биома, континента, острова и т.д.
В 1979 г. Крюгер и Тейлор добавили к этой классификации еще Д-разнообразие.
A-разнообразие — разнообразие, определяемое изменениями климатических факторов, что выражается в смене растительных зон, провинций и т.д.
НАУЧНАЯ КЛАССИФИКАЦИЯ ОРГАНИЗМОВ
На планете Земля жизнь появилась более 3 млрд лет назад; за это время, непрерывно эволюционируя, органические формы достигли поразительного разнообразия. Благодаря трудам натуралистов и систематиков XIX и XX вв. нам уже известно более миллиона животных и более четверти миллиона растений. К этому следует добавить множество вымерших форм, обнаруженных палеонтологами. По приблизительной оценке общее число видов организмов за все время существования жизни превышает миллиард. По-видимому, не менее 40 млн из них населяют Землю в настоящее время. Хотя по некоторым группам, таким, как птицы или млекопитающие, уже имеются достаточно полные перечни видов, несомненно, остается еще много форм, которые предстоит обнаружить и формально признать; особенно это касается насекомых—группы, занимающей первое место по числу описанных форм.
Наличие сходства между особями и различий между группами особей дало возможность привести все многообразие форм органического мира в определенную систему. Были созданы и получили общеупотребительные названия категории
живых организмов. Кроме того, каждая группа, формально признанная в системе классификации, получила научное название.
Систематика, или таксономия, занимается изучением множества организмов, их отличительных признаков, их классификацией, основанной на изучении всех и каждой связей между разными организмами.
На заре систематики ее задачи ограничивались обнаружением группы, определением ее диагностических признаков и выбором названия. Считалось, что каждый вид создан независимо от других, имеет вполне конкретные морфологические признаки, неизменные во времени и пространстве, и легко отличим от иных форм. Классификация заключалась в раскладывании морфологически различающихся между собой «подлинных» сущностей, позже названных видами, в некую «естественную» систему. Предполагалось, что особи, составляющие вид, так же сходны между собой, как отливки, полученные из одной формы, а поэтому классификация форм представлялась делом несложным. Индивидуумы, отклоняющиеся от основного типа, рассматривались как исключения, и их появление объясняли ошибками развития. Черты сходства между разными видами, или родство, пытались относить за счет капризов природы или действия сверхъестественных сил в процессе независимого сотворения каждого типа.
Пионеры систематики стремились не только дать названия группам организмов, но и выяснить естественную систему, или «план творения». Наиболее удовлетворительного решения этой проблемы достиг Ч. Дарвин. Благодаря Дарвину эволюционная история популяций в конечном счете стала рациональной основой классификации, а филогенетическая интерпретация родства между различными организмами дала возможность объяснить естественную систему органического мира. Он обратил внимание на то, что принятая в таксономии иерархическая система классификации, т. е. объединение родственных популяций в один вид, родственных видов в один род, родственных родов в одно семейство, семейств в отряд, отрядов в класс и т.д., показывает существование в природе групп, подчиненных другим группам. Иерархическая классификация отражает реальную ситуацию, естественную систему, в основе которой лежат различия в уровнях филогенетической дивергенции, достигнутых в процессе эволюции.
Согласно этой гипотезе, ныне существующие виды возникли от общих предковых форм в различные периоды эволюционной истории, причем признаки отдельных форм склонны ди-вергировать путем постепенного изменения, и степень дивергенции (степень различия между группами) отражает давность их отхождения от общей предковой формы. Однако использование филогенетической концепции в классификации организмов встречает ряд серьезных трудностей. Трудности построения естественной системы связаны с необходимостью такого толкования имеющихся данных, которое подкрепляло бы филогенетическую классификацию групп. Основным критерием служит родственное происхождение. О степени родства обычно судят по сходству морфологических и анатомических признаков изучаемых групп. Прямые доказательства, т. е. данные о наличии репродуктивной связи в родословных, имеются лишь в очень редких случаях. В сущности очень многие виды организмов описываются и получают названия на основании изучения всего лишь нескольких экземпляров, хранящихся в музейных коллекциях. Лишь в немногих случаях имеются подробные сведения о популяционной структуре, биологии размножения, экологических потребностях, цитологических и физиологических признаках и распространении группы, и совсем уже редки формы, по которым собраны адекватные выборки из популяции, в полной мере отражающие изменчивость. Однако сколько бы взаимосвязей ни удалось обнаружить, филогению можно лишь постулировать. Даже построение филогенетических рядов ископаемых форм основывается на допущении существовавших между ними репродуктивных связей, которые, конечно, никто не мог наблюдать непосредственно.
Естественная система классификации в современной таксономии базируется на филогении, которая в свою очередь основывается на эволюционном принципе дивергенции форм, имеющих общее происхождение. Поэтому в любой такой классификации категории, хотя они определяются в эволюционных терминах, устанавливаются почти исключительно на основании сходства между формами, т. е. выводы о происхождении носят чисто умозрительный характер.
Утверждение о том, что биологическая классификация самая «естественная», поскольку она базируется на эволюционных принципах, не означает, однако, что не может быть
X другой системы или что хоть какая-то категория объективно отражает некую реально существующую в природе единицу. Один систематик может назвать данную группу родом, тогда как другой считает ее семейством или видит в ней два рода. Но ведущие современные систематики утверждают, что основ-ная категория классификации — вид — действительно соответствует самостоятельным независимым единицам, так же как, например, особи, клетки или хромосомы представляют собой обособленные структурные единицы.
В.Н. Беклемишев, обсуждая в 1928 г. методологию систематики, приводит относящееся к началу XX в. высказывание Г. Дриша о том, что методы построения системы принадлежат кобласти фундаментальной методологии науки. Сам В.Н. Беклемишев видел в систематике возможность решения двух задач: морфологического описания объекта и систематического описания морфопроцессов, что в полной мере требуется и при выделении жизненных форм. Корни подобных представлений содержатся в идеях Кювье о «планах строения» и Гёте об «архетипе» применительно к живым организмам. Развивая взгляды на организм как морфопроцесс, А. А. Любищев (1923) предлагал рассматривать морфологический признак (а именно такие признаки характеризуют жизненные формы) как проявление закона, управляющего органической формой, что не может не найти отражение в их систематике.
Категория вида. Перед систематиком стоит обширный ряд биологических форм, которые можно рассортировать в соответствии с разного рода системами классификации. Однако, хотя бы для одного того, чтобы стандартизировать номенклатуру, необходимо привести все это многообразие к какой-то одной официальной системе. В течение двух последних столетий иерархическая система К. Линнея (изложенная в его книге «Система природы», вышедшей в 1735 г.) принята в зоологии, а принципы этой системы — ив ботанике. Основная единица линнеевской системы — вид. По традиции каждый организм относится к определенному виду и к надвидовым категориям более высокого ранга (роду, семейству, классу и т. д.). Каждый индивидуум имеет двойное название, состоящее из названия рода и названия вида, к которому он принадлежит, — так называемая биноминальная система номенклатуры. У систематиков вошло в традицию давать видам названия и определять их в соответствии с какой-либо концепцией
категории вида. А это приводит к так называемой «проблеме» вида. Как объективно определить категорию «вид»? Иными словами, какие признаки следует считать диагностическими и как велико должно быть различие между популяциями, чтобы отнести их к разным видам? Эта проблема постепенно проясняется (Майр, 1963; Симпсон, 1961), однако объективного определения вида, которое бы содержало критерии для универсального его применения, все еще не существует. Стало понятно, что критерии распознавания (практический уровень) не могут быть установлены до тех пор, пока определение вида базируется на эволюционных концепциях и включает в себя динамические системы.
Эволюционная концепция вида. Г. Симпсон определил эволюционирующий вид как «последовательный ряд популяций родителей— потомков, эволюционирующий изолированно от других и имеющий собственную эволюционную роль и собственную тенденцию». Это определение невозможно использовать на практике ввиду отсутствия правил, на основании которых можно дать бесспорное описание «эволюционной роли и тенденции». Вместе с тем это наиболее содержательное определение — некий теоретический стандарт, с которым можно сравнивать другие определения.
При этом вид мыслится как дискретная, репродуктивно изолированная система популяций, имеющих общую эволюционную историю. Такие истории схематически изображают в виде «деревьев», что позволяет графически представить подлинную филогению, или родословную (рис.1). Концевые точки ветвей такого древа, достигающие на шкале времени современности,—это ныне существующие виды, а концевые точки, лежащие в прошлом,—вымершие популяции. О взаимосвязях родословных во времени обычно судят по родственным отношениям между ныне существующими группами; ископаемые остатки позволяют повысить вероятность правильности описания филогении.
Если бы мы располагали полной палеонтологической летописью, то могли бы проследить филетическую историю отдельного эволюционного ряда (называемого филумом) как ряда популяций родители — потомки. Но это не значит, например, что все члены ряда, прослеженного в обратном направлении от Equus через PUohippus, Merychippus, Miohippus, Mesohippus и Eohippus, принадлежат к одному виду. Согласно теории филе-
Рис. 1.
Филогенетическое древо лошади, построенное на основании палеонтологических данных Главный ствол ведет от Eohippus к современному Equus. Многие его боковые ветви вымерли. Эволюция главного ствола происходила преимущественно в Северной Америке. Hyracotherium (по-видимому, как л Eohippus) и его потомки обитали в Старом Свете, так же как Anchitherium, Hipparion и Stylohipparion. Эволюция от Pliohippus до Hippidium происходила в Южной Америке. Только те формы Equus, которые распространились в период плейстоцена по Евразии и Африке, дали современную лошадь, осла и зебру
4
тической эволюции, с течением времени могут происходить изменения, достаточные для превращения одного вида в другой в пределах одной и той же родословной. По всей вероятности, в филум от Eohippus до Equus входило большое число различных видов. На основании принципа последовательного видообразования путем филетической эволюции можно разделить любую родословную на ряд последовательных видов, или палеовидов. Но здесь можно столкнуться с одним из аспектов проблемы вида: как велико должно быть различие между популяцией родителей и популяцией потомков (т. е. между двумя ископаемыми формами), чтобы, исходя из эволюционной концепции вида, считать их разными видами?
Согласно эволюционной концепции, вид рассматривается как последовательность популяций «родители — потомки», образующая один из отрезков единой родословной. Предлагали и другие концепции вида, акцентирующие внимание на одной или нескольких связях между существующими группами организмов, но не придающие особого значения эволюционной истории видов. Е. Майр рассматривает три концепции: 1) типологическую концепцию вида; 2) концепцию безмерного вида и 3) концепцию многомерного вида.
Типологическая концепция (морфологический вид). В основе представлений о виде лежат две противоположные гипотезы: о неизменности организмов и об их превращениях. В процессе развития науки предпочтение неоднократно отдавалось то одной, то другой гипотезе. В настоящее время эволюция рассматривается как процесс, ведущий к образованию видов и обеспечивающий их постоянство. Эта последняя стадия в развитии систематики возникла под влиянием идей Ламарка, но окончательно установилась лишь после появления книги Ч. Дарвина «Происхождение видов...» (1859). В течение предшествующих 100 лет в систематике господствовали идеи К. Линнея.
В линнеевской иерархии вид был низшим рангом над уровнем особи. Предполагалось, что особи одного вида относительно однородны по анатомическому строению и четко отличаются от особей других видов. Весьма существенным для этих представлений была вера в то, что каждый вид создан независимо от других и что морфологические признаки каждого вида остаются неизменными, варьируя лишь в известных пределах, установленных Творцом.
Считалось, что поскольку вид состоит из особей, построенных по единому плану, для исследования основных свойств какой-либо группы достаточно изучить одного типичного представителя этой группы. Таким образом, типологическая концепция базируется на идее неизменности видов. Согласно этой концепции, главным критерием для описания видов служит степень морфологических различий. Описанные по такому принципу виды называются морфологическими видами, или морфовидами.
Концепция безмерного вида. Эта концепция наиболее близка к представлениям о виде натуралистов, которые изучают сосуществующие во времени и пространстве популяции различных организмов. Конспецифичные особи (члены одного вида) у перекрестнооплодотворяющихся форм объединяются в популяции. В любой момент времени каждый вид занимает в пространстве некий ограниченный ареал, состоящий из суммы всех территорий, занятых всеми популяциями этого вида. Величина ареала, а также численность и пространственное распределение в нем размножающихся колоний зависят от: 1) системы скрещивания; 2) подвижности, или способности к расселению, проявляющейся в активной (индивидуальная подвижность) или пассивной форме (расселение с помощью таких факторов, как ветер, вода или же с помощью других организмов); 3) распределения благоприятной для данного вида апологической обстановки (климатические и биотические условия, препятствующие расширению ареала) и 4) природы географических преград, мешающих расселению (таких, как океаны, горы, реки, озера, ледники и пустыни). В одних условиях виды завоевывают огромные территории, тогда как в других область их распространения сокращается, приобретая четкие Границы. Виды, приспособленные к широкому диапазону условий среды, обычно равномерно распределены по всему ареалу. Популяции, обитающие в пограничных частях ареала, Нередко представляют собой полуизолированные «группы», Которые по мере колебаний границ ареала то объединяются К остальными популяциями, то отделяются от них. Менее ши-роко распространенные формы, приспособленные к более .специфическим условиям, распределены по ареалу в виде небольших скрещивающихся колоний.
Ареалы разных видов могут перекрываться или не перекрываться. Особи всех сосуществующих в одной и той же гео-
4
графической области видов называют симпатрическими. Особи, популяции или виды, занимающие различные географические области, называют аллопатрическими. Аллопатрические популяции бывают смежными (соприкасающиеся) или разобщенными (разделенные).
Каждый вид, существующий симпатрически, представлен одной популяцией, в которой не обнаруживается ни географической изменчивости, ни дифференцировки на расы. Локальные популяции различных видов представляют собой независимые скрещивающиеся единицы; чаще всего такие популяции легко отличить от других благодаря морфологической прерывистости или « видовым разрывам ». В таких случаях «виды» характеризуются нескрещиваемостъю двух сосуществующих локальных популяций. В этом и состоит концепция безмерного вида.
Концепция многомерного вида. После того, как натуралисты получили возможность часто и много путешествовать, обнаружилось, что между многими локальными популяциями, которые в своих специфических местностях казались совершенно обособленными, существуют промежуточные группы. Следовательно, отдельную популяцию уже нельзя было рассматривать как самостоятельный вид. Стало ясно, что «вид» — групповое явление (система, состоящая из множества популяций, рассредоточенных по географическому ареалу). При описании таких систем нельзя игнорировать межпопуляционную изменчивость. Градиенты признаков, называемые кланами, служат морфологическим доказательством генетического родства сопряженных популяций, образующих надпопуляционную систему — единый вид.
Виды, обнаруживающие географическую изменчивость и состоящие из идентифицируемых групп более низкого ранга (или рас), называются политипическими. Виды, имеющие более ограниченное распространение и не обнаруживающие географической изменчивости таксономически значимых признаков, называются монотипическими.
Первые концепции многомерного вида создавались под давлением практических потребностей — необходимости давать названия различным формам жизни и определять их. Концепция вида должна была быть универсальной, т. е. приемлемой для всех встречающихся форм. Морфологическая концепция вполне соответствовала этой цели. Систематики-
практики и по сей день занимаются определением особей и именованием видов, но на иной концептуальной основе. Установление морфологических различий между индивидуумами остается необходимым и даже главным аспектом процедуры в систематике, однако сейчас вид понимают как совокупность изменчивых популяций, динамически эволюционирующих, как некую собирательную единицу, репродуктивно изолированную от остального биотического мира. Теперь, когда ясно, что единое репродуктивное сообщество может состоять из морфологически различающихся субъединиц, главным критерием определения вида служит не морфологическое различие между индивидуумами, а репродуктивная связь между популяциями. Именно на репродуктивной связи основано так называемое современное биологическое "определение вида, Это определение включает в себя четыре существенных положения.
1. Биологический вид возможен только у форм, размножающихся половым путем.
2. Вид состоит из одной или нескольких популяций родственных особей (в соответствии с многомерной концепцией).
3. Взаимоотношения между членами любой популяции вида обусловлены их зависимостью друг от друга в репродуктивном отношении (разобщенные популяции одного вида считаются потенциально способными к обмену генами).
4. Главный критерий биологического вида — репродуктивная изоляция входящих в него популяций от всех других (это как раз то, что имеется в виду при формулировании понятий «видовые разрывы» в безмерной концепции и «собственная эволюционная роль и тенденции» в эволюционной концепции вида). Короче говоря, биологический вид есть группа действительно или потенциально скрещивающихся естественных популяций, репродуктивно изолированная от других таких же групп.
Ясно, что симпатрические, морфологически отличные популяции, принадлежащие к одному виду, не могут долгое время оставаться обособленными, так как в результате скрещивания и обмена генами происходит интеграция их отличительных признаков. Отсюда следует, что различающиеся симпатрические популяции относятся к разным видам и что они различны потому, что гибридизация между ними невозможна. Человек, а также некоторые домашние животные и куль
турные растения составляют исключение. Искусственное ограничение свободного скрещивания приводит к тому, что несколько отдельных пород (рас) даже при совместном существовании сохраняют свои специфические особенности. Что касается человека, то разные расы, будучи симпатрическими, могут оставаться генетически изолированными ввиду многочисленных социально-бытовых преград, препятствующих смешанным бракам. Однако эти преграды далеко не всегда действуют, и смешанные браки довольно обычны. Межрасовые гибриды способны к воспроизводству, а, следовательно, все люди независимо от расы представляют собой (в соответствии с биологическим определением) один вид.
Наиболее существенная особенность биологического вида — репродуктивная изоляция. По той или иной причине какие-то формы оказываются неспособными скрещиваться, или производить жизнеспособное и плодовитое потомство, которое обеспечило бы передачу генетического материала последующим поколениям. Самая очевидная причина этого — пространственная изоляция видов. Однако географическая изоляция — сугубо внешняя причина репродуктивной изоляции, и при этом разобщенные формы вовсе не обязательно утрачивают потенциальную способность к скрещиванию. В то же время, даже занимая соседние территории или будучи биотически симпатрическими, популяции могут оставаться изолированными. Обмену генами между популяциями препятствует одна или несколько внутренних преград, называемых изолирующими механизмами. К изолирующим механизмам относятся такие факторы, как утрата способности к спариванию либо перекрестному опылению, резкое снижение эффективности оплодотворения, а также нежизнеспособность, или бесплодие, гибридов. Согласно био логическому определению, популяции, принадлежащие к одному виду, скрещиваются между собой и дают жизнеспособное плодовитое потомство; если такие популяции разобщены, мы подразумеваем, что они потенциально способны к эффективному скрещиванию. Популяции, принадлежащие к разным видам, репродуктивно изолированы благодаря существованию одного или нескольких изолирующих механизмов.
Ограниченность биологической концепции. Биологическое определение вида применимо к эволюционной концепции на любом отрезке времени в пределах родословной интересующих
нас форм. Филогенетические ряды остаются обособленными до тех пор, пока их популяции не смешиваются генетически.
При решении вопроса о том, принадлежат ли две формы (популяции) к разным видам, используют три главных критерия.
1. Формы настолько различны морфологически, что их трудно принять за один вид. Делается вывод, что степень репродуктивной изоляции коррелирует с величиной различий. В таких случаях решение систематика субъективно.
2. Репрезентативные представители двух форм, помещенные в экспериментальные условия, либо не скрещиваются, либо дают нежизнеспособных, или стерильных, гибридов.
3. Формы сохраняют обособленность, даже если их ареалы соприкасаются или они биотически симпатричны. Данные о скрещивании этих форм или образовании гибридов между ними отсутствуют (т. е. существует видовой разрыв).
Только третий критерий в какой-то степени приемлем. Однако даже если применять этот строгий критерий, при выделении видов часто возникают трудности, которые можно разбить на три основных типа.
1. Данные могут быть недостаточны, неубедительны или противоречивы. Степень репродуктивной изоляции между популяциями не всегда соответствует степени морфологических различий между ними. Например, существование видов-двойников. Виды-двойники обнаружены во всех группах высших животных й растений. Они очень часто встречаются среди насекомых, особенно в таких хорошо изученных родах, как Drosophila. Поскольку различия между видами-двойниками, как правило, очень невелики и весьма трудоемкие методы, необходимые для их выявления, могут быть применены только к ограниченному числу популяций, естественно предположить, что таких видов существует значительно больше, чем мы в состоянии обнаружить. Полиплоидия (кратное увеличение хромосомного набора) — весьма обычный способ видообразования у растений — привела к возникновению криптических видов во многих семействах высших растений. В некоторых местностях наряду с двумя или несколькими диплоидными видами встречаются также полиплоидные формы, возникшие в результате гибридизации от этих диплоидных форм, причем все эти формы морфологически неразли
чимы. Эта группа представляет собой полиплоидный комплекс видов-двойников, которые, тем не менее, репродуктивно изолированы. К такому комплексу морфологическая концепция вида совершенно неприменима.
Морфологические различия между группами — это внешние проявления более глубоких генетических различий, действующих на процесс развития и подверженных влиянию внешних условий. Когда таксономист определяет популяции морфологически и присваивает им ранг вида, он стремится выделить репродуктивно независимую эволюционирующую единицу. Во многих случаях степень и характер генетических различий, необходимые для возникновения репродуктивной изоляции, одновременно вызывают внешне заметные изменения в строении тела. Опытный таксономист нередко может точно предсказать генетическое различие на основании анализа коррелированной с ним изменчивости морфологических признаков. Многие из выделенных таким образом морфови-дов представляют собой настоящие биологические виды. Однако эту корреляцию можно использовать не всегда, так как степень морфологического различия между некоторыми видами так велика, что один вид можно отличить от другого даже на расстоянии. В других случаях (виды-двойники) для этого приходится прибегать к специальным тестам.
Особенно трудно решать вопрос о видовой принадлежности родственных, но дифференцированных аллопатрических популяций, поскольку в этом случае прямое определение репродуктивной изоляции невозможно.
2. Исследуемые группы находятся в состоянии эволюционной промежуточности. Процесс эволюции непрерывен, и поэтому в некоторых случаях невозможно провести строгую границу между тем, что называется расой, и тем, что называется видом. Экологические, морфологические и воспроизводительные различия между современными популяциями могут быть развиты лишь частично. Например, виды-двойники достигли репродуктивной изоляции, но почти не дифференцировались морфологически. Другие группы свободно скрещиваются и дают потомство в лабораторных условиях или на опытных участках, хотя морфологическая и экологическая дифференцировка этих групп достигла такого уровня, что их вполне можно считать отдельными мор-фовидами. Есть также формы, состоящие из ряда географи
ческих рас, между которыми возникла частичная репродуктивная изоляция.
Ситуация еще более усложняется, когда отличительные видовые признаки варьируют по степени проявления у разных популяций данного вида. Известно много примеров межвидовой гибридизации, особенно среди растений. То обстоятельство, что некоторые виды в известной мере гибридизируют друг с другом, не обязательно противоречит биологической концепции. Вряд ли можно сомневаться, что смежные популяции, свободно обменивающиеся генетическим материалом в зонах контакта, принадлежат к одному виду, а репродуктивно изолированные симпатрические формы принадлежат к разным видам. Однако критерий, основанный на репродуктивной изоляции, не всегда можно применять так объективно. Ведь репродуктивная изоляция — характеристика популяции и создается она путем постепенных эволюционных изменений. В некоторых случаях межпопуляционные преграды могут быть еще неполными, т. е. время от времени между этими популяциями может происходить скрещивание с образованием гибридов, правда, менее жизнеспособных и менее плодовитых, чем потомки от скрещивания между конспецифичными особями. Кроме того, даже хорошо развитые изолирующие механизмы могут разрушаться, допуская некоторое скрещивание. В результате то, что считалось двумя хорошими видами (полная изоляция), может оказаться пограничным случаем (частичная изоляция). Межвидовая гибридизация чаще всего дает несколько маложизнеспособных потомков Fy и потомков от некоторых возвратных скрещиваний. При этом лишь относительно небольшая часть генов одного вида соединяется с генами другого, так что «эволюционная роль» каждого вида остается относительно независимой. Этот процесс взаимного проникновения генов называется интрогрессивной гибридизацией, или интрогрессией. Чаще всего интрогрессия наблюдается между видами, изолированными экологически. Изоляция, основанная на экологической специализации, представляет собой очень слабый механизм сохранения обособленности популяций и может быстро разрушаться при самых незначительных колебаниях условий среды. Если различия между интрогрессирующими формами сохраняются, то из этого следует, что они все еще остаются различными видами.
Популяции, принадлежащие к одному виду, характеризуются фактической или потенциальной скрещиваемостью, тогда как между видами существует репродуктивная изоляция. Смежные популяции со средним уровнем гибридизационного обмена генами могут представлять собой расы с частичной репродуктивной изоляцией или виды с частично разрушенными преградами. Во всяком случае их нельзя с полным правом возвести ни в ранг расы, ни в ранг вида.
Репродуктивному критерию придается особое значение потому, что он отражает основную биологическую характеристику организмов, размножающихся половым путем (их генетическую независимость в процессе эволюции), и, таким образом, применительно к таким единицам его следует считать объективным.
3. Главный критерий биологического определения вида неприменим к бесполым формам. Два предшествующих недостатка репродуктивного критерия для классификации и номенклатуры имеют практическое значение. Существование же бесполых форм порождает теоретическую проблему, поскольку критерий «репродуктивная изоляция видов» применим только к формам, размножающимся половым путем'. Половым размножением называют любой способ размножения, обеспечивающий комбинацию и рекомбинацию генов, происходящих из различных родословных. Это чаще всего достигается за счет двух независимых процессов: 1) рекомбинации генов при мейозе и гаметогенезе; 2) сингамии, или слияния двух гаплоидных гамет для образования одной диплоидной зиготы. В результате этих процессов происходит объединение генетической информации отдельных особей, принадлежащих к одному поколению, или объединение генетической информации разных поколений. Таким образом, создается организационная единица более высокого уровня — популяция. Но среди строго бесполых организмов не существует ни генетической рекомбинации, ни генетически определенной популяционной структуры, а следовательно, не может быть и видообразования в биологическом смысле. Одна из неотложных проблем, стоящих перед систематиками, состоит в том, чтобы включить в рамки эволюционной теории многочисленные «парасексуальные» механизмы рекомбинации генов, обнаруженные у низших организмов, прежде считавшихся бесполыми.
Таким образом, термин вид используется двояко: 1) в общем смысле—как таксономическое обозначение различных организмов, которые экологически объединены, а морфологически различаются (в том числе и бесполых) и 2) в специфическом смысле — для обозначения репродуктивных изоля-тов, каждый из которых состоит из популяций фактически или потенциально скрещивающихся организмов. Использование термина в его первом значении облегчает процедуру классификации форм жизни. При этом во многих случаях происходит объединение или дробление эволюционных родословных, но, тем не менее, этот термин повсеместно применяется для описания (в первом приближении) наблюдаемой в природе упорядоченности. Второе значение термина с концептуальной точки зрения вполне приемлемо. В этом смысле он служит некой рабочей абстракцией, с помощью которой можно обобщать популяционные явления, однако в большинстве случаев применение термина затрудняется из-за отсутствия фактических данных, необходимых для принятия однозначного решения.
Внутривидовые категории (расы и подвиды). Согласно многомерной и эволюционной концепциям, виды состоят из популяций, распределенных в пространстве и во времени. Каждая локальная популяция какого-либо вида может рассматриваться как колония скрещивающихся между собой особей, приспосабливающихся к специфическому комплексу условий в их местообитании. Родственные колонии, обитающие в различных частях одной общей области обитания, по-видимому, будут занимать сравнительно сходные экологические ниши. Члены данной локальной популяции спариваются преимущественно между собой в силу территориальной близости, но в пограничных районах происходит некоторое взаимопроникновение особей и половых клеток. Поэтому для смежных популяций характерно большое число общих генов и признаков. Для более разобщенных популяций одного вида степень обмена генами ниже. Следует также ожидать, что популяции, обитающие в удаленных частях ареала, будут более своеобразны в результате приспособленности к несходным условиям среды: они обычно обладают различными генными комплексами и разными признаками. Хотя каждая популяция вида представляет собой продукт единственной в своем роде генетико-физиологической реакции на условия некой
4
локальной среды, группа родственных популяций, обитающих в одном районе, будет иметь много общих признаков, но может по некоторым признакам или частотам генетических вариантов отличаться от других групп, обитающих в других районах. Вследствие сходства и различия между популяциями иногда удобно делить их на внутривидовые группы, называемые расами, или подвидами. Человек, несомненно, принадлежит к числу политипических видов. Чаще всего политипия встречается у форм, представленных широко расселенными и разобщенными популяциями, которые приспособлены к различным условиям. Когда разные расы встречаются в природе, то наблюдается слияние их отличительных признаков в результате гибридизации. Уровень интеграции зависит, прежде всего, от степени пограничных контактов между расами.
Деление на расы — субъективная процедура, основанная на существовании двух или большего числа групп популяций, различия между которыми достигли такой величины, которая оправдывает формальное признание этого факта. Популяции являются теми реально существующими группировками особей, на которые распадаются любые виды живых организмов. Это позволяет считать популяцию элементарной единицей эволюционного процесса. Открытие популяционных явлений генетиками радикально отразилось на представлениях общей экологии. Эволюционные процессы, протекающие на разных уровнях — популяционном и видовом, принято различать.
Микроэволюция — эволюция в популяциях под влиянием ненаправленной мутационной изменчивости на подвидовом уровне, когда особи способны скрещиваться и давать плодовитое потомство. Термин «микроэволюция» ввел в науку Ю.А. Филипченко в 1927 г., чтобы разграничить два принципиально важных эволюционных явления: микро- и макроэволюцию.
Макроэволюция — эволюция на уровнях выше видового, когда нет скрещивания особей и обмена генетическим материалом между особями, но четко проявляются тенденции адаптации животных и растений к биотическим и абиотическим факторам окружающей среды. Генетики выделили удобный объект для сравнения организмов, каковым является геном — совокупность генов, характерных для гаплоидного набора хромосом любого организма. Этот термин впервые был
предложен Г. Винклером в 1920 г. Геном — функциональная единица, своего рода программа, необходимая для нормального развития и воспроизводства организмов в ряду поколений. Современная генетика располагает многочисленными средствами исследования геномов — геномным анализом.
Специальная наука — геносистематики — отрасль систематики, предметом анализа которой являются геномы организмов. Два основных ее раздела используют для изучения геномов цитологические (кариосистематика), молекулярнобиологические, биохимические и разнообразные физико-химические методы (генохемосистематика). Генотаксономия, будучи составной частью геносистематики, есть теория и практика классификации организмов, основанной на результатах изучения их генного материала.
Использование новых подходов позволяет подойти к решению проблем систематики, не решаемых при помощи классических методов. Так, например, становится возможной объективная оценка ранга таксона. Весьма перспективно применение методов геносистематики в разработке ряда проблем морфологии. Например, оценка возможности конвергентного возникновения морфологических структур разной сложности.
Методы геносистематики могут быть использованы для оценки расстояния между таксонами разного ранга. Выяснено, например, что хрящевых ганоидов, по-видимому, следует выделять в отдельный класс, связанный с костистыми рыбами весьма отдаленным родством.
Данные по молекулярной гибридизации ДНК указывают на то, что существуют дискретные степени подобия генного материала, соответствующие в классической систематике родам, семействам, отрядам и классам. Подчиненные таксоны (подкласс, надотряд и т.п.) следует, видимо, считать искусственными.
ЖИЗНЕННЫЕ ФОРМЫ
И БИОЛОГИЧЕСКОЕ РАЗНООБРАЗИЕ
Жизненная форма — это, прежде всего, биологический индикатор определенных природных условий. По набору жизненных форм, представленных на некоторой территории, можно довольно уверенно судить о степени разнообразия среды обитания и ее «узких местах» , вероятности успеха интродукции новых видов и т.п. Важным шагом вперед в разработке проблемы жизненных форм в традициях А.Н. Формозова можно считать переход к характеристике их конкретными количественными показателями — морфологическими, физиологическими и др. Эффективность подобного подхода показана в последнее время при анализе адаптации животных к радиоактивно загрязненной среде (Криволуцкий, 1996).
А.Н. Формозов считал, что вид в огромной мере несет на себе отпечаток среды, в которой он жил и живет и к которой, как правило, хорошо адаптирован. Отсюда возникновение в определенных ландшафтах специфических для них жизненных, или биологических, форм, причем в одинаковых ландшафтах разных материков могут существовать свои наборы форм, к тому же внешне и в своих повадках весьма сходных, хотя и очень
далеких в систематическом отношении. В становлении биологических форм большую роль играет конвергентная эволюция — процесс сближения морфологических, физиологических и других признаков. Этот процесс может касаться не только отдельных видов, но и в известной мере целых фаун или даже биот. В пределах одной ландшафтной зоны, например пустынь, встречается ряд специфических жизненных форм животных, по-своему решающих задачу адаптации к пустынным ландшафтам. Впервые формообразующее воздействие ландшафта на животных было показано Л.С. Бергом в 1922 г.
Г. Осборн в 1896 г. назвал конвергенцией появление сходных признаков у неродственных животных и параллелизмом появление одинаковых признаков у родственных групп животных.
Термин «жизненная форма* был введен в науку А. Гумбольдтом в 1806 г. В течение XIX в. он прочно завоевал ботанику, а затем получил и более широкое распространение. Значительной известностью по жизненным формам растений пользуются работы Раункиера, а из отечественных исследователей — В.В. Алехина, Б.А. Келлера, А.П. Щенникова, И.Г. Серебрякова и многих других.
В зоологии пропагандистами рассматриваемого понятия выступили К. Фридерикс (1930) и Д.Н. Кашкаров(1933,1945). Вначале изучение жизненных форм животных продвигалось медленно, показателем состояния проблемы в конце 1930-х гг. может служить высказывание Д.Н. Кашкарова: «У зоологов учение о “жизненной форме” совершенно не разрабатывалось. Нам не известно ни одной работы, специально этому вопросу посвященной». Успехи в изучении жизненных форм были достигнуты благодаря разработанности в зоологии за много десятилетий до рассматриваемого периода прочных основ в деле изучения процесса адаптации к конкретной среде обитания, достаточно напомнить, в частности, о ставших классическими работах К. Бэра, К.Ф. Рулье, Н.А. Северцова, М.А. Мензбира и их последователей. Адаптации животных к среде, пути и закономерности приспособительной эволюции вызывали всеобщий интерес; не случайно сложилось мнение, что «экология есть наука о приспособлениях (адаптациях) животных и растений к условиям среды*.
Натуралисты различали экологические группы животных, не называя их жизненными формами, очень давно, это
даже нашло отражение в существовании укоренившихся в обиходе названий, таких, как «нырцы», «норники», «зем-лерои» и т.п., которые соответствуют «жизненным формам» некоторых классификационных схем. Важные выводы о формообразующем влиянии среды на животных были сделаны еще в XVIII в. в отношении таких явлений, как белая окраска арктических птиц и зверей, отсутствие пигмента и глаз у пещерных и глубокопочвенных обитателей и т.п. В середине XIX в. установлены экологические правила Бергмана, Аллена, Глогера, четко сформулированные в виде общих положений Реншем в 1930-е гг. Богатейший фактический материал накоплен в ходе изучения адаптации у разных животных, образования и закономерностей изменения аналогичных и гомологичных признаков.
Все удачные попытки выделения жизненных форм были сделаны или для ограниченных групп (классов, отрядов, подсемейств), где сходные приспособления развивались на основе одинаковой схемы организации, или же в них учтены конституциональные, филогенетические признаки. Сходные формы у представителей систематически далеких групп наблюдаются исключительно редко и касаются ничтожного числа видов. Конвергентное и параллельное развитие обычно наблюдается у родственных форм. Объяснение этому дал И.И. Шмальгаузен (1946), писавший, что «для несходных организмов среда никогда не может быть одинаковой, так как разные организмы занимают в ней разное положение, т. е. сами относятся к ней по-иному», следовательно, нельзя ожидать и глубокого сходства в приспособительных реакциях у таких организмов. Подавляющее большинство жизненных форм похожи в том, что они возникают на таком этапе адаптационной эволюции группы, когда организмы, выработав важные приспособления общего порядка, переходят к более полному использованию частных особенностей среды (идиоадаптации А.Н. Северцова, алломорфозы и теломорфозы И.И. Шмаль-гаузена, адаптивная радиация Г. Осборна).
У животных жизненные формы — группы таксонов, обычно в пределах одного отряда или близких отрядов, которые обладают сходными морфоэкологическими приспособлениями для обитания в одной и той же среде. Типичным примером жизненных форм могут служить адаптивные экологические группы млекопитающих: плавающие, роющие, бегающие,
X
прыгающие, летающие и т.п, Подобные группы неоднократно описывали у птиц, насекомых, рыб, рептилий, клещей и других животных, так что можно говорить об универсальности явления адаптивного параллелизма животных, о своеобразном «четвертом правиле» адаптивной эволюции в экологии животных наряду с известными правилами Бергмана, Аллена и Глогера.
Н.П. Дубинин (1966) предлагал различать мутационные и эволюционные параллелизмы. Параллелизмы в наследственной изменчивости, выраженные законом гомологичных рядов изменчивости Н.И. Вавилова (1924), в основном описывают явления мутационного параллелизма. Что касается эволюционных параллелизмов, то здесь справедлива формулировка И.И. Шмальгаузена (1940) о том, что параллелизмы вызваны общей наследственной основой. Здесь мы встречаемся с проявлением ограниченности путей адаптивной эволюции в условиях данного типа функциональных структур. Наиболее выразительные результаты достигаются лишь при определенной перестройке системы, свойственной целой группе форм, ди-вергировавших по другим признакам, причем процесс этой перестройки может происходить на разных генетических уровнях. По-видимому, основным механизмом, обеспечивающим образование жизненных форм и адаптивную радиацию, является генетический полиморфизм, который создает предпосылки для эволюции стратегий адаптации организмов на всех стадиях жизненного цикла. В современных программах изучения биологического разнообразия рассмотрению жизненных форм уделяется немало внимания как среди основных особенностей иерархии форм жизни (Global biodiversity, 1992), так и среди механизмов, которые обеспечивают географическое разнообразие биот.
В явлении образования жизненных форм наглядно можно видеть переход от уровня стохастических, случайностных процессов, свойственных изменениям генотипа на популяционном уровне, к направленным процессам формирования адаптированного к конкретной экологической среде фенотипа. Для многих организмов, хотя далеко не для всех, фактором, направляющим отбор в генетически полиморфных популяциях, выступает географический ландшафт. Первым, кто впервые четко и определенно писал о формообразующем влиянии ландшафта и приводил множество примеров групп с аналогич-
ными признаками, которые теперь были бы названы жизненными формами, был Л.С. Берг (1922). Затем эти представления отстаивали многие зоологи, в том числе М.А. Мензбир, Г.П. Дементьев, А.К. Рустамов. Так появились представления о ландшафтных или ландшафтно-зональных, зональных жизненных формах. Подобное явление хорошо прослежено у многих наземных позвоночных, но для большинства беспозвоночных его обнаружить крайне трудно: одни из них не контактируют с ландшафтными факторами среды (многие простейшие, паразиты, обитатели толщи почвы), а другие изменяются слишком медленно, чтобы в их морфологии можно было обнаружить отклик на эфемерные в масштабах геологического времени изменения ландшафтной сферы. Например, некоторые виды панцирных клещей неизменны с миоцена, роды — с юры, а за время их жизни ландшафты, где они обитают, радикально изменялись десятки раз.
Дифференциация географических популяций в процессе адаптации к конкретной среде обитания вида нередко приводит к образованию четко различающихся по набору морфологических приспособлений экоформ у животных и растений. Тем самым выявляется связь образования жизненных форм с процессами микроэволюции. Так, на примере дождевых червей и рыб была убедительно продемонстрирована возможность образования резко различных типичных жизненных форм в пределах одного и того же биологического вида. Вероятно, и среди ранее опубликованных данных можно найти подобные факты, о чем свидетельствует, например, известная работа Л.С. Берга о яровых и озимых расах рыб, а также работа А.Н. Формозова, касающаяся цветовых форм песца*.
Ю.А. Филипченко (1929) настаивал на полном разрыве между эволюцией видов, опирающейся, по его мнению, на действие обычных факторов: изменчивости, наследственности и естественного отбора, и эволюцией родов, семейств, отрядов, классов и типов, которая совершается по другим законам. Впервые подобную идею (в 1868 г.) выдвинул Коп, хотя зачатки ее можно найти еще в высказываниях Ламарка. Ю.А. Филипченко предложил для этих двух независимых, по его мнению, потоков эволюции термины «микроэволюция» и «макроэволюция». Вполне правомочно говорить о макроэволюции как о самостоятельном этапе развития, когда мик-роэволюционные процессы, накапливаясь, получают внешнее
выражение в форме морфологически выраженных адаптаций к конкретной среде обитания. Полагают, что только при макроэволюции обнаруживаются общие тенденции, направления и закономерности (в том числе адаптивные параллелизмы), которые не поддаются наблюдению на микроэво-люционном уровне.
К макроэволюции относят и развитие основных особенностей в организации жизни, некоторые из них закладываются в процессе образования жизненных форм, когда изменяются не только эктосоматические, но и эндосоматические органы, по терминологии А.Н. Северцова. По мнению Н.П. Дубинина (1966), явления микроэволюции совершаются на протяжении ограниченного времени, они идут как бы на плоскости, внутри целой сети взаимосвязанных популяций. Изолирующие механизмы отчленяют неспециализированную популяцию, которая исходно была такой или стала ею в порядке обратимой эволюции. Через эту фазу процессы развития от уровня микроэволюции могут перейти на уровень макроэволюции, особенно в тех случаях, когда изменяются качественные особенности, появление которых было переломным моментом в морфологической эволюции определенной группы. Первопричиной того или иного типа эволюции остаются взаимоотношения организма с постоянно изменяющейся средой.
Смена экологической зоны жизни оказывает коренное влияние на формирование общих систем в строении животных. Так, переход к наземному образу жизни оказался одним из самых мощных стимулов к выработке копулятивного аппарата внутреннего оплодотворения, а вместе с тем и более сложного строения всей половой системы, вызванного выработкой приспособлений к наземному развитию в виде скорлупы яйца и т.д. Эти преобразования коснулись всех крупных групп животных, вышедших на сушу.
Действительно, обсуждение реальности различий макро-и микроэволюции с неизбежностью должно свестись к анализу различий в факторах, регулирующих эти явления. Речь идет и о факторах, которые приводят к формированию различных генных форм в пределах крупного таксона. Какие же механизмы здесь известны? Это в первую очередь те преобразования, экологические «маневры», которые возможны для организма в пределах жизненного цикла. Они позволяют выйти за пределы сложившихся трофических связей для имаги-
нальной фазы в экосистеме и занять новую экологическую нишу, зачастую свободную от врагов и конкурентов. Особенно большие возможности здесь, как говорил А.А. Любищев, дает педоморфоз, позволяющий занять новую экологическую нишу несколькими способами. Один из них — укорочение жизненного цикла и переход к размножению на все более ранних стадиях развития, к неотении. Такое изменение в далеко зашедших случаях сопровождается появлением нового тагмозиса, осей дробления в эмбриогенезе, общей архитектоники, строения нервной системы, перестройками топологии при морфогенезе, т. е. приводит именно к таким различиям, которые учитываются при делении таксонов высшего ранга.
Возможны и другие способы преобразования, например, отношения животных с «микробным звеном» трофической цепи, развитие «внутренних трофических цепей» у жвачных, моллюсков, коралловых полипов и многих других животных, что на начальном этапе трофической дивергенции связано с образованием жизненных форм. Еще один из хорошо изученных путей — изменение размеров тела, которое сразу же переводит организм в другую размерную категорию, новую систему экологических отношений, на что впервые обратил внимание М.С. Гиляров (1944). При этом измельчание, переход в категорию микрофауны, что является обычным путем образования жизненных форм почвенных обитателей, может обеспечить животным преимущества общего характера, исходя из термодинамических представлений.
Эти и другие изменения, наблюдаемые при выделении жизненных форм, не только позволяют организмам освоить новые пищевые ресурсы, избежать неблагоприятных абиотических воздействий, занять свободное от врагов и конкурентов экологическое пространство, но и приводят к усложнению, структурированности биогеоценозов и биосферы в целом, что подчеркнул Г.Н. Симкин (1969). Именно наличие подобных морфологических преобразований при дифференциации жизненных форм в самых разных группах животных позволят говорить о них как о «пусковом механизме» макроэволюции. Такие примеры можно обнаружить при рассмотрении зоологической макросистемы.
Адаптивные изменения организмов, которые находят отражение в их морфологии, так или иначе всегда учитываются при построении системы. Многие биологи в течение более чем
столетия обращали внимание на то, что таксономические группы организмов ранга отряда (порядка), семейства, а нередко и рода являются самостоятельными жизненными формами. Это стало очевидным еще при анализе «основных форм» растений А. Гумбольдта, описанных им в 1806 г., а в наше время при рассмотрении макросистем микроорганизмов, растений и животных.
Процессы образования жизненных форм очень ярко проявляются при радиации таксонов внутри классов и других высших категорий. И.И. Шмальгаузен (1940) указывает, например, что класс млекопитающих дивергентно распался на отряды вследствие того, что разные группы этого класса попали в разные условия обитания, приобрели различную трофическую специализацию (хищные, насекомоядные, копытные, китообразные, грызуны и т.д.). Отряды распались на семейства и подсемейства, которые характеризуются как морфологическими, так и экологическими особенностями (формы бегающие, скачущие, лазающие, роющие, плавающие и т.д.). Наконец, роды и виды также биологически различаются в первую очередь по образу жизни, объектам питания и т.д. И. И. Шмальгаузен (1940) всю эту группу эволюционных процессов, соответствующих идиоадаптациям А.Н. Северцова, называет алломорфозом. При алломорфозах организмы не претерпевают ни значительного усложнения, ни заметных упрощений в организации.
Насколько методологически обоснованно, корректно выделяют и описывают таксоны далекого прошлого? Очевидно, что их разнообразие и ранг могут быть объективно оценены при сравнении современных их форм, а не докембрийских с ныне живущими. Ограничимся несколькими примерами. Так, Зейлахер (1989), рассматривая разнообразие эдиакарс-ких бесскелетных беспозвоночных венда (650—570 млн лет назад), относимых палеонтологами к группе классов и даже типов, вымерших еще в докембрии и палеозое, предполагает простое и однообразное их строение. Тело этих организмов, возможно, состояло из многочисленных однотипных камер, или «пневм», с тонкой эластичной оболочкой и прочными проницаемыми перегородками, в совокупности дававшими конструкцию, напоминающую надувной матрас или стеганое одеяло, что позволяло легко трансформировать конструкцию в разнообразные жесткие объемы. Здесь типы и классы вполне
соответствуют жизненным формам, что позволило говорить об «адаптивном калейдоскопе* высших таксонов докембрия.
Еще один подобный пример просматривается в эволюции членистоногих, где недавно сделаны фундаментальные открытия в палеонтологии кембрийских и докембрийских организмов: описаны многочисленные промежуточные между червями и членистоногими червеобразные животные с мягкими, несклеротизированными конечностями, новый тип членистоногих (Schizoramnia Bergstrom, 1976) и несколько новых классов.
Очевидно, установленных филогенетически связей между разными группами животных по морфологическим, палеонтологическим, генетическим и биохимическим данным построение их макросистемы, определение ранга таксонов возможно на базе разной методологии, включая не только морфологические, генетические, фонетические системы, но и системы фенотипические, экологические (по адаптивным мор-фопроцессам), что может открыть новый путь познания разнообразия жизни.
ИНВЕНТАРИЗАЦИЯ ВИДОВ
В системе низших таксономических единиц, реально существующих в природе, центральное место занимает вид. Появление нового вида представляет собой тот узловой момент в эволюции формы, когда она приобретает характер самостоятельной единицы и, вычленившись из группы связанных рас, становится на путь самостоятельного, генетически независимого процесса эволюции.
Сколько же видов в истории Земли несли эстафету эволюции? Г. Симпсон попытался дать на этот трудный вопрос хотя бы приблизительный ответ. Для анализа, по его мнению, надо знать:
1) среднее число видов, одновременно существовавших на Земле;
2) среднюю продолжительность жизни видов;
3) время, прошедшее от начала появления видов.
Чтобы ответить на первый вопрос, надо знать число ныне существующих видов животных и растений. Описано около 1,7 млн видов растений и животных. Однако если учесть спорность многих сложных видов, то при более осторожной оценке их количество составит от 250 тыс. до 1 млн. Средняя про
должительность жизни видов, по данным палеозоологии и палеоботаники, примерно 500 тыс.— 5 млн лет. Возраст Земли равен около 5 млрд лет: она стала пригодной для жизни около 4 млрд лет назад (во всяком случае, возраст некоторых ископаемых водорослей исчисляется 2 млрд лет). Таким образом, продолжительность жизни на Земле может быть оценена в 3,8-4 млрд лет.
Исходя из этих примерных цифр, можно дать максимальную, среднюю и минимальную оценки числу видов, прошедших свою историю в эволюции (табл. 1). Для этого надо произведение среднего числа видов а на время от начала жизни b разделить на среднюю продолжительность жизни вида с, т. е. аЬ/с.
Таблица 1
Число видов в истории жизни на Земле
Оценка числа видов Число одновременно существующих видов Продолжи тельность исторической жизни вида, годы Время от начала жизни на Земле, годы Число видов и истории жизни на Земле
Максимальная 1,7 млн 5 млн 2 млрд 4 млрд
Средняя 265 тыс. 2750 тыс. 1,5 млрд 341 млн
Минимальная 250 тыс. 500 тыс. 1 млрд 50 млн
Таким образом, порядок величин, оценивающих число видов в истории жизни на Земле, равен 50 млн—4 млрд, а в среднем 400—500 млн. При этом эволюционисты и систематики всегда должны учитывать одну особенность видов, состоящую в том, что эволюция во всем объеме совершается внутри вида. Любая крупная ветвь жизни (типы, классы, семейства, отряды и роды) зарождалась в свое время в системе видов. И ныне отдельные виды таят в себе возможность эволюции, которая в будущих условиях среды приведет к грядущим преобразованиям жизни.
Поэтому к генетической системе любого вида эволюция предъявляет огромные требования. Главное из них — необходимость в носителе безграничной пластичности. Это явилось основной причиной создания корпускулярной, дискретной
наследственности для всех форм жизни. Возможность осуществления изменчивости на основе мутаций и рекомбинаций колоссальна. Количество генов у видов растений и животных исчисляется тысячами, каждый из которых через мутации может дать десятки аллелей. Рассмотрим упрощенную ситуацию кода в гаплоидном наборе, где имеется только 1000 генов, каждый из которых в состоянии путем мутаций может дать только 10 аллелей. В этом случае число генных комбинаций достигает порядка 101000, т. е. огромнейшей величины, которая больше числа электронов и протонов во Вселенной.
В реально существующих видах эта величина еще больше. Отсюда становится ясной вся гигантская, необозримая работа естественного отбора. Последний, опираясь на эту фантастическую изменчивость, в определенных условиях среды в процессе последовательной интеграции и революционных преобразований создает те сравнительно немногие генотипы, которые отвечают необходимости развития приспособленных видовых форм.
То, что именно видовые системы несут итоги эволюции и являются источником будущего в истории жизни, ярко подчеркивается фактом замкнутости генетической системы вида. Конечно, хорошо известны случаи скрещивания видов, роль гибридизации в происхождении видов, особенно растений, однако в эволюционных процессах довлеет принцип генетической замкнутости видовой системы.
Главным достижением эволюционной генетики было обнаружение тех глубоких истоков эволюции, которые в пределах популяции внутри вида зарождают процессы эволюции форм, ведут к созданию приспособительной эволюционной несовместимости генетических систем разных видов. Эволюционная генетика призвана решить еще более трудную задачу. Она должна вскрыть единство внутри вида на основе факторов эволюции и понять те силы, которые движут весь этот безграничный поток форм по путям прогресса, а также установить причины задержек в эволюции и нередкое появление регресса форм.
Вполне понятно, что эволюционист, рассматривающий историю видов; генетик, изучающий принципы генетической эволюции системы популяций внутри вида; эколог, ставящий своей задачей вскрыть прямые связи приспособительных свойств вида и действия среды, и систематик подходят к проблеме вида с разных точек зрения. Перед систематиком стоят
практические задачи приведения всего видового и внутривидового разнообразия в систему, которая, с одной стороны, должна отвечать естественным связям в происхождении форм, а с другой — описать реальную картину внутривидового разнообразия. Инвентаризация видов и реальное описание каждого из них, анализ всей видовой системы в виде географических и экологических рас, подвидов, аберраций — задача исключительно трудная и важная.
К. Линней в 1758 г. знал всего лишь 4236 видов животных и около 10 тыс. видов растений. В настоящее время описано примерно 270 тыс. видов растений и примерно 850 тыс. видов животных. Открытие новых видов продолжается. Если мы обратимся даже к такой хорошо изученной группе, как птицы, то увидим, что в Северной Америке и в Европе каждые 40 лет описывается один новый вид и три вида — в других районах мира. На рис. 2 показана динамика описания птиц с 1758 по 1870 г.
Известно 8500 видов птиц. Учитывая частоту нахождения новых видов, осталось описать не более 100 видов птиц. Если же мы обратимся к насекомым, то при наличии около 1 млн известных видов ежегодно описывается 10 тыс. новых.
Рис. 2.
Динамика открытия видов птиц с 1758 по 1870 г.
Рис.3.
Оценки количества описанных видов по разным авторам
Можно оценить динамику открытия видов животных, если сравнить списки известных видов по двум фундаментальным сводкам (Майр, 1953); (Бруска и Бруска, 1990), опубликованным с разницей всего лишь в 37 лет (рис.З).
По современным представлениям доля основных групп организмов к общему разнообразию видов на планете существенно отличается (рис.4). Однако реальное соотношение ви-
Позвоночные 3%
Др. беспозвоночные 4% Нематоды 1 %
Др. членистоногие 1%
Высшие растения 14%
Двукрылые 7%
Полужесткокрылые 8%
Чешуекрылые 9%
Жесткокрылые 24%
Рис. 4-
Доля основных групп организмов к общему разнообразию видов на планете
Др. насекомые 9%
Вирусы 1%
Бактерии 1%
Грибы 4%
Простейшие 2%/"
Водоросли 2%
Моллюски 4% ^^Ракообразные 2% Пауки 4%
Рис. 5-
Доля основных групп организмов к общему разнообразию видов на планете (прогноз)
дов растений и животных, прогнозируемое учеными, в том числе еще не известных мировой науке в мировом видовом богатстве, видимо, иное (рис.5). Многие группы организмов остаются мало изученными.
Однако анализ внутривидовых различий — дело еще более сложное. У птиц известно 26 тыс. подвидов и каждый год описываются 200 новых. Количество подвидов, рас и аберраций у насекомых поистине безгранично. Все это свидетельствует о том, что общая инвентаризация видов, и особенно инвентаризация изменчивости внутри видов, — дело исключительно сложное.
В XVIII и XIX вв. доля типовых экземпляров млекопитающих, добытых в России, составляла около 40%, а в XX в. она увеличилась до 70%> .Таким образом, большинство вновь открытых на территории бывшего СССР видов в XX в. оказались новыми и для мировой фауны млекопитающих. Хотя период активного описания новых видов, обитающих на нашей территории, заканчивается к 1950 г., позднее все же их продолжают описывать. Интересно, что новые виды описывались с территорий, где до этого проводились интенсивные ис
следования с коллектированием, что говорит о той мере условности, с которой сталкивается наблюдатель при регистрации новых видов (как новых лично для него в данном месте, так и новых на данный момент для мировой науки). Принимая во внимание непрерывный ход эволюции, можно предположить, что список видов млекопитающих, зарегистрированных на нашей территории, будет пополняться не только в результате заходов уже известных видов с сопредельных территорий, но и, возможно, в результате обнаружения новых видов для мировой фауны. При этом наибольшая вероятность таких находок может быть среди насекомоядных, грызунов и рукокрылых. Заметим, что уже в конце XIX в. были описаны два новых вида китообразных в районе о. Беринга — северный плавун (1883) и командорский ремнезуб (1885), обитающие на нашей территории.
На долю К. Линнея и П. С. Палласа выпала удача описать более 20 новых видов из общего списка. 73 зоолога описали по одному виду млекопитающих. Остальные первоописатели занимают промежуточное положение (Соколов, Шишкин, 1998).
ВИДОВОЕ
БОГАТСТВО
РОССИИ
В настоящее время на территории России зарегистрировано около 11 400 видов аборигенных и заносных сосудистых растений, принадлежащих к 1488 родам и 197 семействам. В совокупности они представляют примерно 50% состава флоры бывшего СССР. Процесс выявления состава флоры не закончен. Ежегодно на территории страны описываются десятки новых для науки видов. Обнаруживаются растения, распространенные на сопредельных территориях, и многочисленные адвентивные виды, особенно североамериканского происхождения. В Красную книгу РСФСР (1983 г.) было включено 440 видов покрытосеменных, 11 голосеменных, 10 папоротникообразных растений. Реально в той пли иной степени опасности подвергаются не менее 2 3 тыс. их видов. На охраняе мых природных территориях (государственные заповедники, национальные парки) представлено не менее 75% видов сосудистых растений флоры России. Специальных сведений о числе видов, охраняемых в заказниках, нет. Работа по инвентаризации флоры заповедников еще далека от завершения.
В России зафиксировано более 9000 видов морских, пресноводных и почвенных водорослей, что составляет примерно
1/4 мировой альгофлоры. Благодаря обширным ареалам число эндемичных видов невелико и колеблется от 2—3% во внутренних до 6—10% в морских водоемах. Наибольшей степенью эндемизма водорослей обладает оз. Байкал. Выявлено не более 1% редких, реликтовых и исчезающих видов, что связано в первую очередь с недостаточной изученностью этой группы растений.
В России насчитывается около 3000 видов лишайников и несколько тысяч видов грибов.
Фауна позвоночных животных России относительно хорошо исследована и насчитывает более 1300 видов, что составляет около 2,7% мирового разнообразия (табл.2).
Таблица 2
Разнообразие, эндемизм и состояние видов позвоночных животных РФ
Группа Известное число видов Число редких видов — на национальном уровне Число редких видов — на региональном уровне Общее число эндемичных видов В % от общего числа
Млекопитающие 320 64 90 22 7
Птицы 732 109 62 1 0,1
Рептилии 75 11 7 0 0
Амфибии 27 4 3 0 0
Рыбы* 269 9 27 57 28
Круглоротые 8 0 3 0 0
* Только пресных водоемов.
Представительна фауна птиц, млекопитающих, круглоротых (7,6; 7,0 и 40% мирового разнообразия соответственно). В России выделяется несколько регионов с высоким уровнем видового богатства: Северный Кавказ, юг Сибири и Дальний Восток. В этих регионах велик уровень эндемизма фауны, что обусловлено их исторической ролью.
Число редких и находящихся под угрозой исчезновения видов позвоночных России, согласно данным Красной книги
РСФСР (1983 г.), равно 197 (около 15%), что свидетельствует в целом о неблагоприятном состоянии фауны. В настоящее время повышается риск потери разнообразия наиболее ценной части этой группы животных.
Млекопитающие — наиболее изученная группа позвоночных животных России. Число их видов составляет около 7% мирового разнообразия этого класса. Значительное видовое богатство характерно для регионов Северного Кавказа, юга Сибири и юга Дальнего Востока. Около 23% видов млекопитающих занесено в Красную книгу России, из них 11 видов представлены только одним или несколькими своими подвидами или отдельными популяциями. Во второе издание Красной книги России планируется включить 65 видов и подвидов млекопитающих. Около 90 видов млекопитающих России (33%) находится под угрозой исчезновения на региональном (в основном в странах Центральной и Западной Европы) и мировом уровнях (39 видов, или 14%). К последним прежде всего относятся некоторые виды китов и подвиды крупных кошек.
Около 61% видового разнообразия млекопитающих (без китообразных) и около 60% видов, занесенных в Красную книгу России, встречаются на охраняемых природных территориях. В наиболее тяжелом положении находятся виды и подвиды ластоногих и копытных, для которых доля редких, охраняемых в заповедниках, животных не превышает 40%. В морях и внутренних водоемах России постоянно обитают или встречаются во время миграций 56 видов морских млекопитающих, в том числе 40 видов китообразных, 15 — ластоногих. Около 50% видов морских млекопитающих и некоторые их локальные популяции внесены в Красную книгу России и список угрожаемых видов Международного союза охраны природы (МСОП). Под угрозой полного исчезновения оказалась охотско-корейская популяция серого кита, общая численность которой не превышает 100 особей.
Фауна птиц России хорошо изучена (732 вида) и составляет 7,6% мирового разнообразия этого класса при практически полном отсутствии эндемичных видов. Подавляющее число видов (515) — гнездящиеся, а 27 видов гнездятся только в пределах России. Около 9% видов птиц занесено в Красную книгу России. Примерно 9% видов орнитофауны являются регионально редкими (в основном представители от
ряда соколообразных, а 30 видов занесены в Международную Красную книгу: кудрявый пеликан, дальневосточный аист, стерх и др.). Среди гнездящихся птиц 83% видов встречается в заповедниках (аналогичный показатель для редких видов составляет около 60%).
Фауна рептилий России немногочисленна (75 видов), что определяется достаточно суровыми климатическими условиями на большей части территории, и составляет приблизительно 1,2% мирового разнообразия этого класса позвоночных животных. Эндемичных видов нет. Наибольшее видовое богатство наблюдается на юге Дальнего Востока, на Северном и Западном Кавказе. Около 15% видов относится к редким и находящимся под угрозой исчезновения на национальном уровне, 4% видов занесено в Международную Красную книгу. Во второе издание Красной книги России планируется включить два вида рептилий.
Фауна амфибий России составляет всего 0,6% мирового разнообразия этого класса позвоночных (27 видов). Эндемичных видов нет. Около 15% видов занесено в Красную книгу России. Три вида находятся в опасдости в Европе — малоазиатский тритон, камышовая жаба, кавказская крестовка. Практически все виды амфибий (96%), в том числе включенные в Красную книгу России, встречаются на охраняемых территориях. Экономическое значение земноводных невелико. Во второе издание Красной книги России планируется включить 8 видов амфибий.
Фауна рыб России разнообразна. Многие их виды, например лососевые, карпообразные и др., в границах обширного ареала формируют многочисленные экологически и морфологически различающиеся разновидности, расы и подвиды, в том числе и эндемичные. Фауна рыб насчитывает 269 пресноводных, полупроходных и проходных видов. Не менее 400 видов встречаются в прибрежных морских водах. В целом это составляет около 2% их мирового разнообразия. Среди пресноводной фауны велик процент эндемиков. По их числу лидирует бассейн оз. Байкал. Наибольшее видовое разнообразие характерно для указанного региона и бассейна Амура.
В Красную книгу России включены 9 таксонов (около 4,5% фауны внутренних водоемов), среди которых один вид (атлантический осетр) внесен в Международную Красную книгу.
В Международную Красную книгу занесены также сахалинский осетр и белорыбица. Почти 8,5% пресноводных, полупроходных и проходных видов находится под угрозой исчезновения на региональном уровне. Во второе издание Красной книги России включены 44 таксона рыб.
Современное состояние ряда видов и подвидов вызывает серьезное опасение на национальном уровне как в связи с нарушением качества водной среды, так и из-за высокого уровня промысла, включая браконьерство. Это в полной мере относится и ко всем видам осетровых (в России сосредоточены основные мировые запасы рыб этого семейства и значительная часть ресурсов лососевых и карповых рыб).
Официальной и достоверной информации о фауне беспозвоночных животных России пока не существует. В настоящее время можно говорить только о приблизительном их количестве и оценить его в 130—150 тыс. (около 10% их мирового разнообразия) (табл.З).
Таблица 3
Разнообразие, эндемизм и состояние видов беспозвоночных животных в России
Группы животных Количество видов Количество видов, находящихся в опасности
на национальном уровне на региональном уровне
Простейшие 6500 — —
Метозои 19 — —
Губки 350 — —
Кишечнополостные 450 — —
Плоские черви 1900 — —
Круглые черви 2100 — —
Немертины 100 — —
Кольчатые черви 1 000 (13)* 5“ 1
Форониды 5 — —
Мшанки 500 (1)* —
Плеченогие 23 (1) —
Моллюски 2 000 15(42)* 15** 4
Членистоногие 12 000 34 (98)* —
в том числе:
ракообразные 2 000 (3)* 1
паукообразные 10 000 — 1
Насекомые 100 000 34 (95)* 128“ 31
в том числе:
стрекозы 150 (1)* 3
X
Окончание табл. 3
Группы животных Количество видов Количество видов, находящихся в опасности
на национальном уровне на региональном уровне
богомолы 20 1“ 1
прямокрылые 500 2(2)* 7**1 2
сетчатокрылые 400 1“ . —
ТЛИ 800 — —
полужесткокрылые 2 000 — —
жесткокрылые 22 000 13(36)* 25“ 6
чешуекрылые 12 000 12(33)* 15
двукрылые 9 000 60** —
перепончатокрылые 13 000 7 (23)* 23** 3
Иглокожие 280 —
Щетинкочелюстные 10 — 1
Погонофоры 19 — —
Полухордовые 3 — —
* Количестве видов, рекомендованных Комиссией по редким и исчезающим животным, растениям и грибам при Госкомэкологии России во второе издание Красной книги России.
* * Количество видов по Красной книге СССР (1984 г.).
Основу фауны составляют насекомые (97% всех видов). Редкие и исчезающие беспозвоночные, включенные в Красную книгу России, представлены 49 видами (0,033% их общего количества). Это свидетельствует о благополучном состоянии отечественной фауны в целом. Однако если в данный ряд показателей включить сведения из списка животных, рекомендованных во второе издание Красной книги России, то можно обнаружить четкую тенденцию роста числа таких видов (до 155).
Не меньший интерес представляет разнообразие пород домашних животных и сортов культурных растений, в том числе древних форм, которые являются объектами селекции. Так, в России имеются 30 119 сортов культурных растений (в том числе 11117 отечественных), из которых 375 сохраняются (в том числе 242 российских). Общее количество известных в России пород домашних животных — 454, а сохраняемых — 124.
ЦЕНТРЫ
ТАКСОНОМИЧЕСКОГО
РАЗНООБРАЗИЯ
Анализ распределения видов в пределах ареала рода или подвидов в ареале вида показывает, что оно чаще всего бывает неравномерным. Нередко на обширной площади род представлен всего одним или очень ограниченным числом видов, в то время как в какой-то определенной части ареала рода отмечается большое разнообразие видов. Область, в которой встречается наибольшее число видов данного рода, — это центр таксономического разнообразия (рис.6).
Повышенная концентрация видов может быть обусловлена общей благоприятностью условий для существования видов данного рода. Учитывая, что становление видов — длительный процесс, центр разнообразия рассматривается как показатель значительной давности развития рода на этой территории. Центр разнообразия, как правило, занимает значительную часть ареала, в пределах которой и происходят процессы видообразования. В связи с этим встает вопрос: не является ли центр разнообразия и центром происхождения данного таксона?
Область таксономического разнообразия иногда может совпадать с областью происхождения таксона. Возраст таксо-
Рис. 6.
Ареал рода коровяк (Verbascum) (Толмачев, 1974). Цифры обозначают количество видов
нов родового ранга относится к третичному периоду; за это время их ареалы в связи с глобальными изменениями климата претерпели многочисленные изменения (сокращение, расширение и т.д.). При этом часто возникали вторичные центры разнообразия, весьма удаленные от области происхождения. В ареале рода клен выделяются два центра таксономического разнообразия — один на востоке Северной Америки, другой — в Юго-Восточной Азии (рис.7).
Для суждения о том, какой из центров разнообразия является первичным, необходимо привлечь данные о распространении таксона в геологическом прошлом. При решении этих вопросов широко используют филогенетические методы, основанные на сравнительном изучении морфологии, кариологии и ареалов родственных форм. Особенно большое внимание уделяют хромосомной характеристике таксонов. Формы с большим числом хромосом обычно происходят от предков с низким числом хромосом (диплоидных) либо при помощи простого удвоения этого числа (автополиплоидия), либо, чаще всего, при скрещивании. Поэтому во многих родах и видах можно различать серии форм, где наряду с диплоидными
Рис. 7.
Ареал рода клен (Acer) с центрами таксономического разнообразия (наиболее темная штриховка) (Вальтер, 1982)
встречаются формы с высокой степенью полиплоидии. Это позволяет восстановить область возникновения и пути расселения данных форм. Полиплоидные формы имеют обычно большую экологическую пластичность, что позволяет им распространяться в областях, не подходящих по климату их диплоидным предкам. Случается даже, что эти формы выживают после гибели диплоидных родителей в климатически неблагоприятные периоды. Так, однолетний мятлик представлен двумя подвидами: диплоидным с числом хромосом In = 14, обитающим в Средиземноморье, и тетраплоидным с числом хромосом 2п = 28, который стал космополитным. Пырей ситниковый с прибрежных дюн представлен на Европейском побережье, от Балтики до Бискайского залива, формой с 2п = 28 хромосомами, на Средиземноморском побережье — формой с 2п = 42 хромосомами и на Черноморском — формой с 2п = 68 хромосомами. Таким образом, полиплоидия в этом случае растет с запада на восток.
ГЕНОГЕОГРАФИЯ
Популяционная генетика, зародившаяся как ветвь прикладной математики в начале XX столетия, исследует динамику наследственной информации в поколениях. Мерой такой информации служит гетерозигот ность, т. е. разнообразие генов, унаследованных от родителей потомством и определяющих различия особей внутри популяции. Например, на надкрыльях божьих коровок легко обнаружить строго индивидуальные, наследуемые пигментные узоры (пятна, точки), а популяционную принадлежность рыбок пецилий, излюбленного объекта аквариумистов, можно определять по рисунку на хвостах у самцов. Серии работ были выполнены после обнаружения методами цитологии индивидуальных различий и в строении хромосом (например, по наличию или отсутствию инверсий). Такого рода генетически детерминируемая, индивидуальная прерывистая изменчивость носит название полиморфизма и определяется как наличие в одном и том же местообитании двух или более дискретно отличающихся внутривидовых форм в таких количе ственных соотношениях, что самая редкая из них не может поддерживаться лишь давлением повторяющихся мутаций.
Однако многие виды внешне единообразны, мономорф-ны, — их гены скрыты от наблюдателя и, следовательно, генетический анализ невозможен. Как преодолеть такого рода затруднение? Развитие молекулярной биологии позволяет теперь изучать генетику любого вида, опираясь на скрытые наследственные различия, «записанные» в белковых структурах организма. Эти особенности выявляются специальными процедурами: либо иммунологическими тестами, либо при электрофорезе белков. Точно такой же полиморфизм у самых различных видов обнаружен за последние 10-15 лет при изучении первичной структуры ДНК, как ядерной, так и неядерной (например, митохондриальной), так что арсенал технических средств популяционной генетики необыкновенно расширился. Вместе с тем продолжительность и высокая стоимость ДНК-тестов пока еще ограничивают их широкое применение. Напротив, белковый полиморфизм легко выявляется при сравнительно умеренных денежных затратах на оборудование и реактивы, а общепринятая методика позволяет работать с массовым материалом (сотни образцов за день) и используется во многих лабораториях. Суть методики в следующем: кровь или экстракты различных тканей от отдельных растений или животных помещают в специальные аппараты и в нейтральных мелкопористых поддерживающих средах (гелях) сортируют молекулы в электрическом поле по заряду или другим признакам, детерминируемым наследственностью. Оказалось, что особи внутри популяции различаются составом крови, а также некоторыми особенностями строения определенных белков, и такие отличия передаются неизменными от родителей потомкам.
Достаточно посмотреть на рис. 8, где между особями тихоокеанского лосося-нерки видны различия, определяемые двухаллельным геном, кодирующим синтез фермента фосфоглюкомутазы.
Таким образом, с точки зрения генетики, популяция — смесь генотипов. Многие биологические особенности популяций, прежде всего численность и продуктивность, зависят от этой их генетической структуры. Если соотношения генотипов, а точнее, частоты генов, в популяции неизменны в поколениях, то популяция устойчива. Если соотношения меняются, то говорят об изменяющейся популяции. За время, прошедшее с конца 1960-х гг., когда метод электрофореза белков стал широко использоваться в популяционных исследовани-
Рис. 8.
Наследственный полиморфизм фермента фосфоглюкомутазы из мышечной ткани тихоокеанского лосося-нерки, Oncorhynchusnerka: 1,5 — гетерозиготы АВ; 2-4 — гомозиготы АА и ВВ, соответственно; электрофорез в крахмальном геле; стрелка указывает стартовую позицию; направление миграции молекул в электрическом поле — от катода к аноду. Расшифровка схемы такова: один ген — одна полоса на электрофореграмме; разными номерами обозначены разные рыбы из одной и той же популяции.. У рыб 2, 3 выявляется один ген, у 4 — другой, у 1 и 5 представлены оба гена, это — гетерозиготы, т. е. типы, промежуточные между двумя первыми, гомозиготными генотипами
ях, биохимическая наследственная изменчивость открыта уже у более чем 2000 различных видов — от микроорганизмов до человека.
Изучение десятков генов, разных по локализации и функции, позволило вскрыть гигантский объем биохимического полиморфизма видов и уровней гетерозиготности — показателя, жизненно важного для организма. Оказалось, что около трети генов, кодирующих синтез белков, имеют разные (аллельные) наследуемые состояния, т. е. полиморфны. А уровень индивидуальной гетерозиготности равен 10-12%. Если перенести эти оценки на геном в целом (равно 50 тыс. структурных генов), то окажется, что около 15 тыс. функционирующих генных локусов полиморфны, а «средний» индивидуум гетерозиготен по нескольким сотням своих генов. Гетерозиготность организма — не что иное, как его информационная емкость, поддающаяся строгой количественной оценке шенноновской мерой количества информации. Степень полиморфизма различна у разных групп организма (табл.4).
Обычно на практике полиморфной считают популяцию с частотой гетерозигот по некоторому локусу более 1—5%.
Таблица 4
Генетическая изменчивость в природных популяциях неко торых основных групп животных и растений
(по Левонтину, 1993)
Организмы Число видов Среднее число мест обитания видов Средний уровень полиморфизма Средняя гетерозиготность
Беспозвоночные Дрозофила 28 24 0,529 0,150
Осы 6 15 0,243 0,062
Другие насекомые 4 18 0,531 0,151
Морские беспозвоночные 14 23 0,439 0,124
Наземные улитки 5 18 0,437 0,150
Позвоночные Рыбы 14 21 0,306 0,078
Амфибии 11 22 0,336 0,082
Рептилии 9 21 0,231 0,047
Птицы 4 19 0,145 0,042
Млекопитающие 30 28 0,206 0,051
Растения Самоопыляющиеся 12 15 0,231 0,033
Перекрестно-опыляющиеся 5 17 0,344 0,078
Общие средние Беспозвоночные 57 22 0,469 0,134
Позвоночные 65 24 0,247 0,060
Растения 17 16 0,264 0,046
Анализ полиморфных признаков — ключ к изучению генетических процессов в популяциях, в связи с чем ранние исследования такого рода были выполнены на видах с хорошо выраженным полиморфизмом внешнего облика особей. Накоплено множество данных о феноменологии, генетическом контроле и механизмах поддержания полиморфизма у разных видов. По мнению Ю.П. Алтухова (1989), такого рода изменчивость представляет собой относительно редкое явление, имеет очевидное приспособительное значение, во многих случаях поддерживается в сбалансированной форме за счет адаптивного преимущества гетерозигот со столь значительными коэффициентами отбора, что они практически не оставляют ме-
ЯГ
ста эффектам случайного дрейфа генов, в ряде случаев ее следует рассматривать (из-за ограниченной фазы стабильности) как свидетельство прошедшей и продолжающейся на наших глазах дивергенции популяций в статус новых видов, она отражает запас экологической пластичности за счет постоянного выщепления и комбинации различных генотипов.
Главной единицей внутривидовой дифференцировки является популяция. Согласно С.И. Коржинскому (1892), каждая раса, или, как сказали бы сейчас, географическая популяция, характеризуется определенными морфологическими признаками и особым ареалом. Рассмотрим в качестве примера различные популяции песца. Нормальная окраска белого песца типично покровительственная: зимой снежно-белая, а летом буровато-серая, с коричневыми тонами на спине и желтоватыми на боках. В популяциях песца, обитающих в тундре на материке Евразии и на островах, соединенных зимой с материком, преобладают белые особи, однако в небольшом количестве (0,01—1,0%) встречаются и голубые. Голубой песец и зимой, и летом окрашен примерно одинаково в дымчато-серые или буроватые тона. На островах, не имеющих связи с материком, голубая окраска становится основной, а белая переходит в разряд отклонений. На Шпицбергене, в Исландии, на небольшом о. Кильдине, отделенном от материка незамерзающим проливом шириной 2—3 км, количество голубых песцов составляет 30—80%.
Сезонная белая окраска облегчает белому песцу и защиту, и, главным образом, нападение на мелких грызунов и птиц, которые являются его основной пищей в материковой тундре. Поэтому выход больших количеств голубого песца на просторы материкового ареала пресекается отбором. На островах отбор в пользу голубых форм происходит потому, что здесь белая окраска в значительной степени теряет свое приспособительное значение. На изолированных островах резко ослаблена межвидовая конкуренция, так как здесь песец — единственный наземный хищник. Однако главное состоит в изменении способа добычи пищи. Островной песец питается в основном продуктами моря (выброшенная морем рыба и млекопитающие, беспозвоночные на отливе и т.д.). В этих условиях на первый план выступают положительные особенности голубого песца, и он начинает господствовать над белыми формами, которые переходят в разряд отклонений.
Другой пример. Как известно, в популяциях большинства видов моллюсков встречаются особи с прямо противоположными по закрученности раковинками. Если у типичных особей данного вида раковинка закручена направо, по ходу часовой стрелки, то у уклоняющихся форм она закручена налево, против часовой стрелки. Частота уклоняющихся раковинок может быть различной у разных видов и в разных популяциях одного и того же вида. Так, распространение право- и левозавитковых форм у моллюсков, обитающих в окрестностях Алма-Аты, где преимущественно встречаются правозавит-ковые особи. Ограничение распространения левозавитковых форм, по-видимому, обусловлено тем, что они по сравнению с правозавитковыми обладают большей интенсивностью обмена веществ и меньшей способностью к ассимиляции пищи. Эти особенности лимитируют распространение левозавитковых форм в Семиречье, так как здесь из-за частых и длительных засух регулярно наступают периоды голодания моллюсков.
На этих примерах видно, что в процессах экологической дивергенции рас на первый план .выступает адаптация к локальным условиям среды. Как указывал В.Н. Сукачев (1928), популяции сосны представлены рядом экологических рас соответственно условиям обитания. Чем южнее происхождение сосны, тем хуже она переносит низкие температуры. Так, температуры, безвредные для скандинавского экотипа, губительны для среднеевропейских сосен. Оказалось, что основная причина этих различий — биохимические особенности тканей деревьев.
У Drosophila pseudoobscura, распространенной в западных районах Северной Америки, 27 электрофоретически различных вариантов ксантиндегидрогеназы были обнаружены в 146 независимых линиях и 26 различных вариантов фермента эстераза были найдены в 106 линиях. У людей лишь система HLA достигает такого молекулярного разнообразия. Следовательно, люди имеют несколько сниженную молекулярную изменчивость, но не настолько, чтобы их выделять среди других существ. В этом сношении они типичные млекопитающие.
ГЕНОГЕОГРАФИЯ ПОПУЛЯЦИЙ ЧЕЛОВЕКА
Человечество на ранних стадиях своей истории разделилось на множество различных по размерам популяций. Эти популяции разграничены в большей или меньшей степени географическими, политическими, религиозными и социальными барьерами. Хотя рост численности современного человека можно охарактеризовать как «популяционный взрыв» и он обитает сейчас почти во всех частях земного шара, однако все еще существуют некоторые племена и изолированные религиозные секты с очень небольшой эффективной репродуктивной величиной. Ученые считают, что даже некоторые большие популяции человека возникли сравнительно недавно от малого числа основателей.
Популяции человека полиморфны по группам крови АВО, т. е. содержат все три аллеля 1А, 1В, 1°. На основе относительных частот этих аллелей жителей различных областей земно-го шара можно характеризовать как разные «серологические» расы. Большинство рас содержат особей со всеми четырьмя группами крови О, А, В, АВ и различаются только по распространенности отдельных групп. Однако у некоторых рас те или иные группы крови вовсе отсутствуют. Например, южноаме-
риканские индейские племена были до прихода европейцев или остаются до сих пор изогенными по группе О. Аллели 1А и 1В довольно редки также среди индейцев Северной Америки. Индейцы племен блад и черноногих, обитающие на юге Канады и в штате Монтана (США), представляют исключение: они содержат аллель 1А причем частота его очень высока и достигает 80%, чего не наблюдается ни в одной другой популяции человека. Американские индейцы произошли, по-видимому, от азиатской (монголоидной) расы, небольшая группа представителей которой в свое время пересекла Берингов пролив. Хотя у азиатской расы встречаются все четыре группы крови, частота группы О относительно мала, поэтому среди основателей, пришедших из Азии, или в небольших сформировавшихся позже изолятах могли сложиться условия, благоприятствовавшие дрейфу генов, и либо просто случай, либо случай в сочетании с естественным отбором мог привести к такому необычному распределению.
Миграция и смешение людей не привели к появлению однородной массы. Одновременно со смешением происходит процесс дифференциации местных групп, поэтому всегда существуют какие-то географические различия. Некоторые из этих различий являются итогом прошлой дивергенции, которая еще не сгладилась миграцией и браками. В результате в современных частотах генов можно видеть отпечаток прошлых событий. На рис. 9 показаны клины (серии градуальных изменений) в частоте группы крови А в Японии. Тип крови А имеет высокую концентрацию на западе напротив Корейского п-ова и постепенно уменьшается к северо-востоку по направлению к Хоккайдо. Корея была традиционным источником континентальных вторжений в Японию — эта традиция воплощена в истории с камикадзе, Божественным Ветром, который рассеял флот хана Кублая в Цусимском проливе в 1281 г.
Аналогичные клины существуют в Британии и Ирландии (рис. 10). Частота группы крови А выше 50% в Восточной Англии, но к северу и западу она уменьшается до 25%. Типы В и АВ обнаруживают противоположную тенденцию: их частоты увеличиваются с 8% на востоке до 18% в Шотландии, Уэльсе и Ирландии. Эти изменения отражают то, что известно в истории как движение кельтов, пиктов, датчан и викингов через остров.
В разных географических районах существуют большие различия в цвете кожи, особенностях волос, телосложении и
Рис. 9-
Клинальиая изменчивость процента группы кровиЛ в современной Японии, расходящаяся из одного района, расположенного напротив Кореи на берегу Цусимского пролива
языке. Явные отличия между географическими группами кажутся нам гораздо большими, чем отличия между индивидами внутри групп, и касаются они различий в чертах. Это именно те характеристики, по которым мы узнаем людей и к которым мы тонко настроены, что они кажутся подходящими и для деления людей на группы. Цвет кожи жителей Ганы так очевидно отличается от цвета кожи любого датчанина, что различия между группами представляются гораздо большими, чем небольшие вариации между членами каждой группы. Типология и стереотипизация включаются в наше восприятие групп именно потому, что мы настроены на эти межгрупповые различия, оказывающиеся столь большими. Вопрос в том, являются ли эти большие различия типичными?
Чтобы ответить на этот вопрос на биологическом уровне, мы должны иметь пример изменчивости, которая может быть объективно описана, на которую не влияют особенности черт, очень заметных нам как социальным наблюдателям, и которая характерна для большой выборки географических групп, достаточной для того, чтобы дать нам общую картину. Всем этим требованиям удовлетворяет, по-видимому, простой мо-
Рис. 10.
Процент группы крови А в различных районах Британии показывает убывающую клину к северу и западу от первоначальных стоянок датчан на восточном берегу
лекулярный полиморфизм. Различия в группах крови, иммунных типах и ферментах могут быть охарактеризованы объективно; их генетика проста и вполне понятна; на протяжении последних 40 лет антропологи и генетики проверили группы крови и ферменты тысяч людей во многих местных популяциях во всех концах света. Сравнение частот генов этих полиморфизмов в разных популяциях даст нам объективную картину географического разнообразия людей.
Например, аллель 1в обычно имеет наибольшую частоту (20-25%) в Центральной и Юго-Восточной Азии и уменьшается концентрическими кругами во всех направлениях от плоскогорья Центральной Азии (рис. 11). Возможно, аллель 1В появился в Центральной Азии и затем распространился с движением степных народов из Центральной Азии в Европу, Африку и Сибирь. Полное отсутствие аллеля 1В в Новом Свете и в Австралии свидетельствует о том, что он возник после повышения уровня моря, связанного с таянием континентальных ледников 10 000 лет назад.
Популяции человека слегка отличаются друг от друга частотами генов, что подтверждается, когда исследуется более
Рис. 11.
Распределение в мире гена Is групп крови АВО. Линии соединяют места с одинаковой частотой гена. Наибольшая частота Р — в Центральной Азии. От этого района частота его уменьшается во всех направлениях
широкий набор генов. Например, исследования частоты аллелей для семи полиморфных локусов ферментов, отобранных случайным образом в выборке европейцев и африканцев, показали, что сходство между двумя группами поразительное.
К настоящему времени изучены 17 полиморфных генов человека в достаточно большом количестве наций и племен, позволяющем получить оценки сравнительного разнообразия. Небольшие изолированные группы американских индейцев, аборигенов Южной Азии, лингвистически изолированных групп, таких, как баски, или небольших островных популяций случайно теряют свое генетическое разнообразие и не могут его восстановить, потому что миграция в них из большого мира достаточно редка. Поэтому обнаруживаемые у них частоты аллелей обычно очень близки к одному или к другому концу распределения, характеризующего мир в целом. Предположим, что частота каждого аллеля в каждом известном локусе представлена отдельным измерением в многомерном
пространстве. Тогда популяцию можно представить как точку в пространстве, проекция которой на каждую из различных осей будет частотой каждого аллеля (рис. 12). Все человеческие популяции в мире образуют облако точек в таком пространстве. В центре облака будут те популяции, частота генов которых «типична» для человеческого вида, а по краям его — популяции, имеющие крайние значения в распределении по тому или иному аллелю. Маленькие изолированные острова или популяции аборигенов чаще всего будут располагаться на самых краях облака рядом с углами пространства, в то время как большие популяции Европы, Азии и Африки будут группироваться вместе в центре массива.
Чтобы дать представление о том, насколько генетически различными и насколько похожими могут быть популяции, в табл. 5 показаны три полиморфных гена, по которым больше всего различаются основные географические группы, и три, по которым они более всего похожи.
Треугольная диаграмма, показывающая состав групп крови АВО у 20 разных популяций человека из различных районов мира. Популяции 1 -3 — африканцы, 4-7 — американские индейцы, 8-13 — азиаты, 14-15 — австралийские аборигены и 16-20 — европейцы
Таблица 5
Примеры крайней дифференциации и близкого сходства в частоте аллелей групп крови в трех расах
Ген Аллель Европеоиды Негроиды Монголоиды
Duffy Fy 0,0300 0,9393 0,0985
Fy- 0,4208 0,0607 0,9015
Fy 0,5492 - -
Rhesus Яо 0,0186 0,7395 0,0409
я, 0,4036 0,0256 0,7591
я3 0,1670 0,0427 0,1951
г 0,3820 0,1184 0,0049
г' 0,0049 0,0707 0
другие 0,0239 0,0021 0
Р я, 0,5161 0,8911 0,1677
Рг 0,4839 0,1089 0,8323
Auberger Au° Аи 0,6213 0,3787 0,6419 0,3581
хд Хд" 0,67 0,55 0,54
Хд 0,33 0,45 0,46
Secretor Se Se 0,5233 0,4767 0,5727 0,4273
Самые различные — это действительно очень отличающиеся негры и азиаты: каждая из этих групп имеет разные аллели по группе крови Даффи, распространенные с очень высокой частотой, а обладание аллелем Fyb является довольно определенным указанием на европейского предка. Такая же асимметрия для негров и азиатов проявляется и для генов Auberger. Интересно, что европеоиды генетически гораздо более вариабельны по всем генам, чем любая другая «раса». Возможно, европейцы имеют больше смешанных предков, чем более «чистые» африканцы и азиаты. Каковы бы ни были генетические различия между тремя основными группами, нет таких случаев, чтобы одна группа была полностью гомозиготной по одному аллелю, а другая — по другому. Все группы имеют те же самые аллели. Не существует чистых генов расы, хотя есть аллели, которые обнаруживаются только в одной географической группе, подобно аллелю Fy* среди европеоидов или фактору группы крови Диего, который найден только
среди азиатов и американских индейцев, но он никогда не имеет большой частоты даже там.
Три полиморфных гена, которые у основных географических групп имеют наибольшее сходство, — Auberger, Xg и секретер белков — показывают, сколь вариабелен может быть ген внутри популяции при полном отсутствии различий между группами. Нужно помнить, что это три наиболее сходных полиморфных гена.
В дополнение к огромному многообразию людей, возникающему благодаря основным биохимическим полиморфизмам, существует большое разнообразие редких вариантов даже среди тех молекул, которые на самом деле не являются полиморфными. Существуют 32 «общие» и «частные» группы крови, когда различия касаются только единственной семьи. Около 25 млн различных типов HLA может быть получено из известных различных форм антигенов HLA. Для многих из тех ферментов, которые считались мономорфными на выборках небольшой величины, были обнаружены варианты при использовании электрофореза, разработанного специально для этих целей. Генетики подсчитали, что 1,75 человека на тысячу является носителем редких вариантов некоторых ферментов.
Вся популяция человеческого вида четко дифференцируется по географическим областям. Никто не спутает китайца с жителем Западной Африки или финна с австралийским аборигеном. До создания современной теории эволюции, когда считалось, что все виды были созданы сверхъестественной силой, предполагалось также и то, что эти географические группы были потомками отдельно созданных «наций». Считалось, что каждая группа представляет собой чистый законченный тип, причем, как и у других видов, эти типы оставались неизменными с момента их создания. Признание факта эволюции в конце XIX в. означало отказ от типологического подхода к видам, поскольку дарвинизм подчеркивал и наличие индивидуальной изменчивости внутри видов, и постоянное преобразование, которое претерпевает каждый вид. Однако, это изменение во взглядах биологов не чувствовалось еще много лет после выхода в свет «Происхождения видов...» Ч. Дарвина. До недавнего времени мышление антропологов было отчетливо типологическим, учебники физической антропологии содержали в основном описание и наименование че
ловеческих рас. Одни авторы («объединители») называли только дюжину человеческих рас, в то время как другие («дробители») — несметное их число.
Трудность, связанная с использованием категории расы, состоит в том, что между различными способами разделения человеческих рас существует слишком много противоречий. Являются ли турки белой расой, как свидетельствует их внешность, или они принадлежат к монголоидным племенам Центральной Азии, имеют ли они родство? Что делать с басками, которые на взгляд любого выглядят испанцами, но язык и культура которых кажутся не похожими ни на какие другие в мире? Те, кто говорит в Индии на хинди и урду, создают особую проблему. Исторически они представляют собой смешение южноазиатских дравидских аборигенов, центральноазиатских арийцев (которые явно относятся к европеоидам) и персов. Следует ли относить их к одной группе с европейцами, чьи языки ведут свое происхождение от санскрита, к которому хинди и урду очень близки, или же из-за темного цвета кожи их надо объединить в одну группу с жителями Южной Азии?
Попытка составить все более и более сложные наборы характеристик человеческих типов, которые соответствовали бы невероятному разнообразию людей, в конце концов потерпела крах, подобно эпициклической системе Птолемея, когда астрономические наблюдения стали требовать от нее все более и более изощренных объяснений. Подобно тому, как коперниковская революция в астрономии упростила астрономические объяснения, так и дарвиновская биологическая революция облегчила в антропологии наше понимание географического разнообразия людей. Антропологи уже не пытаются больше называть и определять расы и подрасы, потому что они понимают, что нет «чистых человеческих групп», которые существовали бы с момента творения как отдельные единицы. Самой поразительной чертой общей истории человечества является беспрестанная и широкая миграция и, следовательно, смешение групп из разных регионов. Массовая миграция не представляет собой нечто новое, возникшее благодаря развитию воздушного и морского транспорта, она была экономической необходимостью во все времена. Британцы являются в действительности смешением народности бикер бронзового века, индоевропейских кельтов I тысячелетия до и. э.,
англов, саксов, ютов и пиктов I тысячелетия н. э. и, наконец, викингов и их правнуков — норманнов. Смешение жителей Центральной Азии, славян, жителей Средиземноморья и Северной Европы происходило в конце существования Римской империи. Развитие ислама в VII в. н.э. имело своим следствием распространение арабского населения и его смешения с аборигенами из Кордовы в Кабул, менее чем за сто лет. Такая же ситуация была и на Дальнем Востоке. Японцы представляют собой смесь корейских завоевателей и жителей северных островов. Даже австралийские аборигены, которые рассматривались некоторыми антропологами как совершенно другой вид, имели большую долю предков папуасов и полинезийцев на восточных и северных берегах Австралии. Утверждение, что существуют стабильные чистые расы, которые только теперь подвергаются опасности смешения под влиянием современной индустриальной культуры, необосновано. Однако есть и эндогамные группы, такие, например, как пигмеи или лесные иту-ри, биологически изолированные от своих соседей географическими условиями и культурой.
Происхождение всех человеческих популяций из одного корня подтверждается и антропологическими, и генетическими данными. Исследования митохондриальной ДНК крупнейших человеческих популяций (больших антропологических рас) также поддерживают концепцию монофилитического происхождения человечества. Высказывалось мнение о том, что сначала человечество разделилось на евро-африканскую и азиатско-океанийскую расы, а в дальнейшем первая ветвь распалась на негроидов и европеоидов (кавказоидов). Однако М. Ней с соавторами (1974) показали на основании генетических расстояний по многим генетическим локусам между тремя крупнейшими человеческими расами, что отделение негроидов произошло около 110 тыс. лет назад, а разделение монголоидов и европеоидов — около 40—50 тыс. лет назад. Большую удаленность негроидов от монголоидов и европеоидов подтвердила дендрограмма расстояний морфологических пропорций тела. Вопрос о времени и месте разделения крупнейших человеческих рас пока не решен окончательно.
А.Ф. Назаровой с соавторами (2000) изучено сходство 55 человеческих популяций Европы, Азии, Америки, Африки и Океании по результатам вычисления генетических расстояний этих популяций по 28 аллелям 12 локусов белков, ферментов и
групп крови. Как видно из дендрограммы (рис. 13), раньше всех от общей азиатской прапопуляции отделились америнды — предки американских индейцев. Большой кластер, включающий 54 остальные популяции народов Европы и Азии, ранее всего разделился на ветвь индийцев и ветвь, содержащую все остальные популяции. Из истории и археологии известно, что в III тысячелетии до н.э. населяющие п-ов Индостан племена дравидов, имеющих темную кожу и ряд других негроидных признаков, начали оттесняться к югу пришедшими из более северных регионов светлокожими европеоидными племенами. В дальнейшем происходила метисация этих популяций, в результате которой образовались народности, населяющие Индию. Поэтому, видимо, индийцы и представляют на дендрограмме ветвь, ранее отделившуюся от остальных популяций Евразии. Далее от оставшихся 53 популяций обособилась ветвь лопарей (саамов). Лопари были, по-видимому, первыми древними европеоидами, заселившими север Европы. Обнаружение археологических культур верхнего палеолита в Финляндии также указывает на очень раннее заселение Северной Европы. От огромного кластера европеоидных и североазиатских популяций обособилась группа, содержащая ветвь негроидов, ранее отделившуюся от монголоидов Юго-Восточной Азии — китайцев, японцев и вьетнамцев. Из кластера европеоидов и монголоидов Северной Азии прежде всего выделилась подгруппа, содержащая популяции якутов, нганасан, американских эскимосов, чукчей и азиатских эскимосов. Дифференциация всех перечисленных популяций происходила, по-видимому, на территории Азии со среднего палеолита до неолита и сопровождалась длительной миграцией популяции от места их дифференциации до современных мест обитания. На основании обнаруженной европеоидной компоненты алтайцев по частотам полиморфных локусов белков, ферментов и групп крови, а также идентичности гаплогрупп митохондриальной ДНК у алтайцев и американских индейцев, высказано предположение, что популяции северных монголоидов, америндов и ряда европеоидных народов происходят от одной предковой популяции, обитавшей в центре Азии в среднем палеолите.
Рис.13.
Дендрограмма 55 популяций Европы, Азии, Америки, Африки и Новой Пзи-неи, построенная по матрице генетических расстояний этих популяций по 28 аллелям 12 локусов белков, ферментов и групп крови.
1 — русские, 2 — немцы; 3 — финны; 4 — алтайцы; 5 — эвенки; 6 — якуты; 7 — МОНГОЛЫ; 8 — чукчи; 9 — ЭСКИМОСЫ; 10 — ИНДИЙЦЫ; 11 — украинцы; 12 — белорусы; 13 — ПОЛЯКИ; 14 — ЭСТОНЦЫ; 15 — шведы; 16 — шотландцы; 17 — ирландцы; 18 — сербы; 19 — хорваты; 20 — болгары; 21 — чехи; 22 — венгры; 23 — марийцы; 24 — коми; 25 — манси; 26 — индейцы Америки; 27 — эскимосы Америки; 28 — японцы; 29 — китайцы; 30 — иранцы; 31 — талыши; 32 — молдаване, 33 — англичане, 34 — французы; 35 — испанцы; 36 — итальянцы; 37 — баски; 38 — грузины; 39 — вьетнамцы; 40 — ненцы; 41 — удмурты; 42 — татары; 43 — чуваши; 44 — армяне; 45 — азербайджанцы; 46 — нганасаны; 47 — осетины; 48 — чеченцы; 49 — арабы; 50 — евреи Ближнего Востока; 51 — негры Африки; 52 — греки; 53 — папуасы Новой Пзинеи; 54 — белые американцы (США); 55 — лопари
ГЕОГРАФИЧЕСКИЕ
ЗАКОНОМЕРНОСТИ
ВИДОВОГО РАЗНООБРАЗИЯ
Темпы, формы и направленность дифференциации биот определяются важнейшими факторами, среди которых выделяются географические, действующие прямо или опосредованно, через экологические процессы. Их прямое действие проявляется через изменение физических и химических параметров биосферы, на которые биота вынуждена реагировать. Таковы изменения концентрации солей в Мировом океане, региональные и локальные явления опреснения морских вод, динамика С02 и озона в атмосфере, потепления и похолодания ландшафтной сферы, оледенения, дрейф континентов и т.д. Именно эти явления определяют ♦мегауровень» дифференциации биот во времени и пространстве, включая действие факторов мегаэволюции живых организмов и их катастрофическое вымирание на рубежах геологических эпох и периодов.
Географические процессы определяют динамику биоразнообразия и опосредованно, через экологию не только сообществ, но также видов и популяций; особенно это касается регуляции структуры сообществ климатическими и почвенными факторами. Анализ картографических данных по рас-
пространению сообществ и экосистем неоспоримо свидетельствует,'что их структура и пространственное размещение находятся в теснейшей зависимости от действующих параметров абиотической среды. Но именно структура сообществ обычно сама выступает фактором, направляющим процесс эволюционной дифференциации видов через механизм конкурентного исключения и возможности натурализации мигрантов. Классические примеры здесь представляет островная география, например Мадагаскара, Новой Зеландии, Австралии, где все направления эволюции животного населения, определялись отсутствием в составе сообществ хищных плацентарных млекопитающих. Столь же эффективно, хотя и опосредованно, действуют географические факторы на эволюцию видов и популяций, определяя пространства возможностей и темпы дрейфа генов, регулируя возникновение географических популяций и пути их дальнейшей эволюции. Многие антропогенные факторы изменения биот, за исключением экоток-сикологического и прямого воздействия на виды, можно рассматривать как опосредованно-географические, поскольку механизм их действия или в устранении географических преград для расселения, или в обеднении ландшафтного разнообразия среды, что лишает многие виды их местообитаний.
Географические закономерности изменения биоразнообразия наиболее полно реализуются картографическими методами. Традиционно картографический метод используется при флористических и фаунистических работах по оценке биоразнообразия. Определение альфа-разнообразия ведется по разным показателям и индексам видового богатства. Одной из первых в этом плане следует отметить карту Е.В. Вульфа (1934) по оценке богатств флоры различных территорий земного шара. Генерализация данных о числе видов в пересчете на стандартные площади (100, 1 000, 10 000 и 100 000 км2) позволила составить серию картосхем, отражающих закономерности изменения уровней флористического богатства по широтному градиенту Северной Евразии. Информацию о бета-разнообразии традиционно содержат карты растительности разного масштаба, дающие представление о типологическом разнообразии как коренных, так и производных сообществ различных в природном отношении регионов.
Географические механизмы динамики мирового разнообразия были выявлены на примере фауны шельфовых морей
Мирового океана. Установленная связь размеров ареала таксона с его возрастом послужила основой для количественных сопоставлений разнообразия во времени на базе теории островной биогеографии. Установлена зависимость разнообразия от размеров ареала: большая площадь ареала способствует длительности жизни популяций, сокращая риск их исчезновения в критические периоды, увеличивает способы адаптаций в дифференцированной среде, в разнообразных сообществах. Ключевым моментом признана динамика площадей и числа биогеографических провинций, обладающих эндемизмом видов порядка 50%. На протяжении десятков миллионов лет происходило изменение числа провинций — главного показателя уровня глобального разнообразия. Дж. Валентин (1973) попытался объяснить явления массового вымирания видов. По его мнению, массовое вымирание обитателей моря в перми может быть объяснено не только изменениями солености Мирового океана, но и уменьшением с 14 до 8 числа провинций с их эндемичными фаунами на протяжении этого периода. На этом фоне отмечались «скачки» биоразнообразия, обусловленные присоединением новых адаптивных зон, увеличением разнообразия до локального насыщения, а также дифференциацией фауны набольших пространствах, где каждая устойчивая водная масса образует свою отдельную провинцию. Данный пример свидетельствует о том, что при рассмотрении географических аспектов биоразнообразия наряду с современным его состоянием важно учитывать изменение биоразнообразия во времени.
Рассматривая эту тему, мы попытаемся ответить на некоторые вопросы. Почему одни сообщества богаче видами, чем другие? Существуют ли закономерности или градиенты видового разнообразия? Если да, то на чем они основаны? На эти вопросы есть вполне правдоподобные и обоснованные варианты ответов, но все они имеют свои слабые стороны.
Видовое богатство сообщества можно связать с рядом факторов, относящихся к нескольким категориям. Во-первых, это — «географические» в широком смысле слова, а именно широта, высота над уровнем моря и (в водной среде) глубина. Их часто связывали с видовым богатством, как это будет рассмотрено далее, однако сами по себе они, вероятно, не могут его определять. Если оно меняется с широтой, значит, дол
жен быть еще какой-то фактор, зависящий от нее и непосредственно влияющий на сообщества.
Следующая группа факторов имеет тенденцию коррелировать с широтой и т.п., однако эта корреляция не абсолютна. В той мере, в какой она прослеживается, ими можно часто объяснить широтный и другие градиенты. Однако из-за неполноты корреляции они же затрудняют толкование зависимости от других градиентов. К таким факторам относятся продуктивность среды, климатическая изменчивость, возможно, также «возраст» местообитания и «суровость» среды.
Третья группа факторов характеризуется географической изменчивостью, не связанной с широтой, и т. п. По этой причине они, как правило, маскируют или извращают зависимость между видовым разнообразием и параметрами среды. Это относится к масштабам физических нарушений, испытываемых местообитанием, изоляции, или островному характеру, и степени его физической и химической неоднородности.
Наконец, ряд факторов представляет собой биологические свойства сообщества, но при этом оказывает существенное влияние на его структуру. Среди них особенно важны интенсивность хищничества и конкуренции, пространственная, или «архитектоническая», неоднородность, обусловленная самими организмами, а также положение сообщества в сукцессионном ряду. Эти факторы можно было бы назвать « вторичными *, поскольку сами они определяются внешними для сообщества влияниями. Тем не менее, все они способны мощно воздействовать на формирование его окончательного облика.
Продуктивность среды. Для растений продуктивность среды может зависеть от любого наиболее сильно лимитирующего рост ресурса или условия. В общем, наблюдается повышение первичной продукции от полюсов к тропикам по мере увеличения освещенности, средних температур и продолжительности вегетационного периода. В наземных сообществах снижение температуры и сокращение продолжительности вегетационного периода с высотой приводят в целом к уменьшению продукции. В водоемах продукция, как правило, падает с глубиной параллельно с температурой и освещенностью.
Часто отмечается резкое сокращение продукции в аридных условиях, где рост может лимитироваться недостатком влаги, и возрастание ее почти всегда, когда усиливается при
ток основных биогенных элементов, таких, как азот, фосфор и калий. Если говорить в самом широком смысле, то продуктивность среды для животных следует тем же самым закономерностям, поскольку зависит от количества ресурсов в основании пищевой цепи, температуры и других условий.
Если рост продукции ведет к расширению диапазона доступных ресурсов, он, вероятно, способствует и повышению видового богатства. Однако среды с разной продуктивностью могут различаться лишь количеством (интенсивностью поступления) одних и тех же ресурсов при одинаковом их ассортименте. Значит, разница между ними будет не в числе видов, а лишь в размерах популяций каждого из них. В то же время, возможно, что даже при одном и том же общем ассортименте ресурсов некоторые редко встречавшиеся их категории (или малопродуктивные участки их спектра), недостаточные для обеспечения видов в непродуктивной среде, станут настолько обильными в продуктивной, что в сообщество смогут включиться дополнительные виды. Рассуждая аналогичным образом, можно прийти к выводу, что если в сообществе преобладает конкуренция, то повышение количества ресурсов будет способствовать сужению специализации; при этом плотность популяций отдельных специализированных видов не обязательно сильно понизится (рис. 14).
Таким образом, в целом можно ожидать повышения видового богатства по мере роста продуктивности. Существует корреляция между числом видов и уровнем атмосферных осадков и у семеноядных муравьев, и у семеноядных грызунов в пустынях юго-запада США (рис. 15). В этих аридных областях среднегодовое количество осадков тесно связано с первичной продукцией и, следовательно, с количеством имеющихся запасов семян. Особенно примечательно, что в богатых видами местностях среди муравьев больше очень крупных (потребляющих крупные семена) и очень мелких (питающихся мелкими семенами) видов. Тут же встречается больше видов очень мелких грызунов. Видимо, в более продуктивных сообществах либо шире размерный диапазон семян, либо их так много, что могут прокормиться дополнительные виды консументов.
Еще какие-нибудь однозначные зависимости между видовым богатством и продуктивностью указать нелегко, потому
Непродуктивная среда
Тип ресурсов
Рис. 14.
Зависимость существования большего числа видов продуктивной среды; подгр. 1 — 5 — звенья трофической цепи
что, хотя оба этих параметра часто изменяются параллельно (например, с широтой или высотой над уровнем моря), обычно вместе с ними меняются и другие факторы, т. е. обнаруживаемая корреляция может быть обусловлена именно ими. Тем не менее, описана прямая зависимость между числом видов ящериц в пустынях юго-запада США и продолжительностью вегетационного периода — важного фактора продуктивности аридных сред (рис. 16, А), а разнообразие планктонных ветвистоусых рачков-хидорид в 14 незагрязненных озерах штата Индиана положительно коррелирует с общей продукцией этих водоемов, выраженной в граммах углерода за год (рис. 16, Б).
ЯГ
С другой стороны, рост разнообразия с увеличением продуктивности никак нельзя считать всеобщей закономерностью. Это демонстрирует, например, уникальный «газонный» эксперимент, проводимый с 1856 г. до наших дней на Ро-тамстедской опытной станции (Англия). Пастбище площадью около 2 га поделили на 20 делянок; две из них служили контролем, остальные раз в год удобрялись. С 1856 по 1949 г. менялось видовое разнообразие сообщества травянистых растений на контрольных и получавших полный набор удобрений делянках. В то время как первые оставались практически неизменными, на вторых наблюдалось постепенное сокращение видового разнообразия. Такой спад разнообразия (названный «парадоксом обогащения» среды) был выявлен и в некоторых других геоботанических исследованиях.
Аналогичным образом антропогенное эвтрофирование озер, рек, эстуариев и районов морских побережий приводит к снижению разнообразия фитопланктона (параллельно росту первичной продукции). Заметим, что два типа сообществ, относящихся в мире к наиболее богатым видам, развиваются на крайне бедных питательными веществами почвах (это южноафриканское и австралийское сообщества
Зависимость видового богатства семеноядных грызунов (треугольники) и муравьев (кружки) на песчаных почвах от выпадающих осадков и продуктивности (Brown, Davidson, 1977)
100
1000 10000 100000
Общая продукция, г С/год
Рис. 16.
А — зависимость видового богатства ящериц в 11 пунктах юго-запада США от продолжительности вегетационного периода (Pianka, 1967). Б — зависимость видового разнообразия ветвистоусых рачков-хидорид от первичной продукции озера в целом для 14 озер шт. Индиана, США. Белые кружки — три сильно загрязненные озера
зарослей жестколистных кустарников в условиях климата, близкого к средиземноморскому). Тогда как по соседству с ними, на более плодородных почвах, разнообразие растительности куда меньше.
Максимальное значение видового разнообразия, или богатства, должно соответствовать промежуточному уровню продуктивности. Так, кривые числа древесных пород на единице площади в малайских дождевых лесах отражают зависимость от концентрации в почве фосфора и калия, отражающей богатство почвенных ресурсов (рис. 17), и видового богатства пустынных грызунов от количества атмосферных осадков и, следовательно, продуктивности территории в Израиле (рис. 18) имеют «горбатый* вид.
Как уже объяснялось, нетрудно понять, почему разнообразие может возрастать с увеличением продуктивности (либо на всем диапазоне значений, либо только в левой части «горбатой* кривой). Объяснить сокращение его при увеличении продуктивности не так просто. Один из возможных ответов состоит в том, что высокая продуктивность ведет к быстрому росту популяций, а это в свою очередь ускоряет возможное конкурентное исключение. При более низкой продуктивности больше вероятность того, что среда изменится еще до его завершения, став более благоприятной для менее конкурентоспособного вида.
Рис. 17-
Зависимость видового богатства древесных пород в некоторых дождевых лесах Малайзии от концентрации фосфора и калия в почве (Tilman, 1982)
Параллельно с продуктивностью изменяются какие-то другие факторы, и именно они, а не она сама как таковая, влияют на видовое богатство. Так на рис. 18, как градиент количества осадков и продуктивности совпадает с градиентом на-рушенности естественных условий. Так что вид кривой может отражать пик разнообразия при промежуточном уровне нарушения.
На рис. 19 отражена ситуация с конкуренцией пяти видов за два ресурса. Кружок внизу обозначает относительно малопродуктивную среду; он охватывает только низкие концентрации обоих ресурсов. В этой области могут сосуществовать все пять видов. Кружок вверху, напротив, соответствует относительно высокопродуктивной области; здесь имеются условия для сосуществования только двух видов из пяти. Это
о Песчаные местообитания
О 1 1 1 I ! 1 а. 1. -L-1. J—1—1—। I । । 1 । »-J
60 120 180 240 300 360 420 480 540 600 660
Рис. 18.
Зависимость между видовым богатством пустынных грызунов в Израиле и количеством выпадающих дождей для каменистых (вверху) и песчаных (внизу) местообитаний (Abramsky, Rosenzweig, 1983)
Ресурс 1
Рис. 19.
При возрастании продуктивности видовое богатство может сокращаться. Снабжение двумя ресурсами (1 и 2) позволяет сосуществовать пяти видам (А—Д). Круг внизу показывает снабжение ресурсами в относительно малопродуктивной среде, позволяющее сосуществовать всем пяти видам. Круг вверху соответствует относительно высокопродуктивной среде (но с таким же диапазоном различных значений снабжения); здесь могут сосуществовать лишь два вида
вытекает из того, что размеры второго кружка такие же, как у первого, «непродуктивного». Иными словами, несмотря на более высокие средние концентрации ресурсов в продуктивной среде, их общие диапазоны остаются прежними, а диапазон отношений ресурсов вследствие этого уменьшается. А поскольку именно он обеспечивает сосуществование нескольких видов, в более продуктивных условиях разнообразие будет ниже.
Итак, логично допустить, что когда повышение продуктивности означает расширение диапазона ресурсов, следует ожидать увеличения видового богатства (по меньшей мере, некоторые наблюдения это подтверждают). В частности, в более продуктивном и разнообразном по составу растительном
сообществе, скорее всего, будет богаче фауна фитофагов и т. д. до конца пищевой цепи. Зато, когда повышенная продуктивность обусловлена усиленным поступлением ресурсов, а не расширением их ассортимента, теория допускает возможность, как повышения, так и сокращения видового богатства. Факты, особенно из области геоботаники, свидетельствуют, что чаще всего увеличение доступности ресурсов ведет к сокращению числа видов или же, если рассматривать весь диапазон продуктивности, к «горбатой* кривой видового богатства.
В связи со всем этим можно остановиться на свойствах света как ресурса для растений. В высокопродуктивных системах (типа тропических лесов), куда он поступает очень интенсивно, происходит его отражение и рассеяние на протяжении мощной толщи растительного покрова. Поэтому здесь существует не только высокая исходная освещенность, но и длинный плавный градиент ее снижения, а также, возможно, широкий набор частотных спектров света. Таким образом, повышение интенсивности солнечной радиации, по-видимому, связано с большим разнообразием световых режимов, за счет чего увеличивается возможность специализации и, следовательно, роста видового богатства. Еще один вывод из этого состоит в том, что наиболее высокие формы должны быть способны функционировать во всем диапазоне освещенности, поскольку они растут от уровня почвы до вершины полога.
Пространственная неоднородность. Пятнистый характер среды при агрегированном распределении организмов может обеспечить сосуществование конкурирующих видов. К тому же в средах с большей пространственной неоднородностью можно ожидать более высокого видового богатства из-за того, что в них разнообразнее микроместообитания, шире диапазон микроклиматических условий, больше типов укрытий от хищников и т. д. Словом, расширяется спектр ресурсов.
В некоторых случаях удалось показать связь между видовым богатством и пространственной неоднородностью абиотической среды. Так, растительное сообщество, занимающее ряд почв и форм рельефа, почти наверняка (при прочих равных условиях) будет богаче флористически, чем фитоценоз на ровном участке с однородной почвой.
Однако большинство работ по пространственной неоднородности связывает видовое богатство животных со струк
турным разнообразием окружающих растений. Например, на рис. 20 прослеживается положительная корреляция между числом видов пресноводных рыб и индексом пространственной неоднородности растений в озерах севера штата Висконсин. На рис. 21 показана сходная зависимость для птиц местообитаний средиземноморского типа в Калифорнии, центре Чили и на юго-западе Африки. Трудность интерпретации этих данных заключается в том, что разнообразие фауны может оказаться обусловленным тем же фактором, что и разнообразие растительности. Например, разнообразие растений связано как с размерами озера, так и с концентрацией в воде питательных для них веществ.
Доводы в пользу большой роли пространственной неоднородности среды в формировании фаунистического богатства выглядят куда более убедительными, если оно коррелирует со
0 0,20,4 0,6 0,8 1,0 1,2 1,4 1,6 1,82,0
Индекс разнообразия растительности
Рис. 20.
Зависимость между видовым богатством и структурным разнообразием растительности для пресноводных рыб 18 озер шт. Висконсин (Топп, Magnuson, 1982)
Разнообразие растений по высоте
Рис.21.
Зависимость между видовым богатством и структурным разнообразием растительности для птиц местообитаний средиземноморского типа в Калифорнии (треугольники), центре Чили (кружки) и на юго-западе Африки (квадраты) (Cody, 1975)
структурным разнообразием растений гораздо сильнее, чем с видовым богатством флоры. Это было показано на примере пустынных ящериц юго-запада США (рис. 22).
Ящерицы различных видов кормятся на разной высоте растений, используя «засидки* в характерных для каждого вида местах, откуда наблюдают за добычей, конкурентами, половыми партнерами и хищниками. Независимо от того, обусловлена пространственная неоднородность абиотической средой или биологическими компонентами сообщества, ее увеличение может способствовать росту видового богатства.
Климатические колебания. Влияние климатических колебаний на видовое разнообразие зависит от того, являются они предсказуемыми или непредсказуемыми (во временных масштабах, существенных для конкретных организмов). В предсказуемой среде с регулярной сменой сезонов разные
Рис. 22.
Зависимость между видовым богатством ящериц и тремя характеристиками растительности в пустыне юго-запада США (Pianka, 1967)
виды могут быть приспособлены к жизни в различное время года. Поэтому следует ожидать, что в сезонном климате может сосуществовать больше видов, чем в неизменных условиях среды. К примеру, разные однолетники в умеренных областях всходят, растут, цветут и дают семена в различные моменты сезонного цикла; здесь же в крупных озерах происходит сезонная сукцессия фито- и зоопланктона с поочередным доминированием то одних, то других видов — по мере того как меняющиеся условия и ресурсы становятся для них наиболее подходящими.
В то же время в несезонных местообитаниях существуют возможности специализации, отсутствующие в среде с выраженной сезонностью. Например, долгоживущему облигатно плодоядному организму было бы трудно выжить в климате, где плоды доступны лишь в определенное, весьма непродолжительное время года. А вот в несезонной тропической среде, где постоянно присутствуют плоды то одного, то другого растения, подобная специализация весьма обычна.
Непредсказуемые климатические колебания могут оказать на видовое богатство различное влияние. С одной стороны: 1) в стабильных условиях возможно существование специализированных видов, которые скорее всего не выживут там, где условия или ресурсы подвержены внезапным колебаниям; 2) в устойчивой среде более вероятно насыщение видами; 3) из теоретических соображений вытекает, что в бо
лее постоянных средах перекрывание ниш будет сильнее. Все это может увеличить видовое богатство.
С другой стороны, именно в стабильной среде выше вероятность того, что популяции достигнут своих предельных плотностей, в сообществах обострится конкуренция и, следовательно, произойдет конкурентное исключение. Поэтому логично будет считать непредсказуемые климатические колебания одной из форм нарушения, а видовое богатство, по-види-мому, будет максимально при ее «промежуточных» уровнях, т. е. оно может как возрастать, так и сокращаться при увеличении нестабильности климата.
Отдельные исследования подтверждают мнение о росте числа видов при ослаблении климатических колебаний. Например, Мак-Артур (1975), изучая птиц, млекопитающих и брюхоногих моллюсков западного побережья Северной Америки (от Панамы до Аляски), обнаружил достоверную отрицательную корреляцию между видовым богатством и диапазоном среднемесячных температур (рис. 23). Однако на этом расстоянии меняется и множество других параметров, так что такая зависимость может быть лишь косвенной. Другие исследования климатических колебаний также не привели к однозначным выводам.
Суровость среды. Среду, в которой господствует какой-то экстремальный абиотический фактор (часто ее называют суровой), распознать не так просто, как кажется на первый взгляд. С человеческой точки зрения «экстремальными» окажутся и очень холодные, и очень жаркие местообитания, и необычно соленые озера, и сильно загрязненные реки. Тем не менее, возникли виды, обитающие в таких местообитаниях, и то, что нам представляется страшно холодным и экстремальным, пингвину должно казаться подходящим и вполне обычным.
Более объективное определение можно дать, выделив для каждого фактора на непрерывной шкале крайние его значения — максимальное и минимальное. Однако будет ли относительная влажность, близкая к 100% (насыщенный водяным паром воздух), столь же «экстремальной», как нулевая? Можно ли назвать экстремальной минимальную концентрацию загрязнителя? Конечно же, нет.
Можно вовсе обойти проблему, предоставив организму «решать ее самому». Ту или иную среду мы назовем в этом случае
Диапазон температур, *С
Диапазон температур, *С
Диапазон температур, ‘С
Рис. 23.
Зависимость между видовым богатством и диапазоном перепадов температуры в различных точках западного побережья Северной Америки для птиц, млекопитающих, брюхоногих моллюсков (MacArthur, 1975)
«экстремальной», если организмы окажутся неспособными обитать в ней. Но коль скоро требуется доказать, что видовое богатство в экстремальных условиях низкое, такое определение приводит к тавтологии.
Возможно наиболее разумное определение экстремальных условий подразумевает наличие у всякого организма, способного их переносить, специальных морфологических структур или биохимических механизмов, отсутствующих у ближайших видов и требующих определенных затрат — либо энергетических, либо в виде компенсаторных изменений биологии организма, потребовавшихся для приспособления к такой среде. Так, растения, живущие на очень кислых почвах, могут страдать либо непосредственно от воздействия ионов водорода, или от обусловленного низким pH дефицита доступных биогенов, например фосфора, магния и кальция. Кроме того, растворимость алюминия, марганца и тяжелых металлов может возрасти до токсичного уровня, нарушая активность микоризы и азотфиксацию. Растения способны переносить низкие значения pH лишь располагая специальными структурами или механизмами, позволяющими им избежать указанных эффектов или противостоять им.
На необрабатываемых лугах Северной Англии среднее число видов растений на площадке 1 м2 наименьшее при низком pH почвы (рис. 24, А). Также и разнообразие бентосных беспозвоночных в ручьях леса Ашдаун (Южная Англия) в более кислых водах заметно снижалось (рис. 24, Б).
К экстремальным местообитаниям с низким видовым разнообразием относятся горячие источники, пещеры и очень соленые водоемы (например Мертвое море). Однако трудность
состоит в том, что они имеют и другие особенности, сопутствующие низкому богатству видов. Многие такие системы малопродуктивны и (вероятно, как следствие) пространственно относительно однородны. Часто они малопротяженны (пещеры, горячие источники) или, по крайней мере, редки в сравнении с другими типами сред (лишь малая доля проточных водоемов в Южной Англии — кислые). Таким образом, часто «экстремальные» местообитания можно рассматривать как мелкие и изолированные островки. Хотя логично предполагать, что в среде с экстремальными свойствами выдержат лишь немногие виды, подтвердить это в высшей степени трудно.
Возраст сообщества: эволюционное время. Известно, что относительно небольшое видовое богатство сообщества может быть обусловлено недостатком времени для заселения территории или эволюции на ней. Кроме того, несбалансированная структура многих сообществ в нарушенных местообитаниях представляет собой результат их незавершенной реколонизации. Высказывалось предположение, что отдельные виды могут отсутствовать и в занимающих обширные территории и «нарушаемых» достаточно редко сообществах именно вследствие того, что они еще не достигли экологического или эволюционного равновесия. Отсюда вытекает, что сообщества
200 г д
pH почвы
Рис. 24.
Среднее значение pH воды
А — среднее (белые кружки), максимальное (темные квадраты) и минимальное (белые квадраты) видовое богатство растений на 1 м2, а также общее число видов (темные кружки) для различных категорий pH поверхности почвы необрабатываемых лугов Северной Англии (Grime, 1973)- Б — связь числа таксонов беспозвоночных в проточных водоемах леса Ашдаун на юге Англии с pH воды (Ibwnsend et al., 1983)
могут различаться по видовому богатству из-за того, что одни ближе к состоянию равновесия, чем другие, и, следовательно, полнее насыщены видами.
Идея эта чаще всего выдвигалась в связи с восстановлением экосистем после плейстоценовых оледенений. Например, низкое разнообразие лесных пород в Европе по сравнению с Северной Америкой объясняли тем, что важнейшие горные хребты в первом случае простираются в широтном направлении (Альпы и Пиренеи), а во втором—в долготном (Аппалачи, Скалистые горы, Сьерра-Невада). Поэтому в Европе деревья оказались зажаты между ледниками и горами и, попав в своего рода западню, вымерли; а в Америке попросту отступили к югу. Прошедшего с тех пор времени в эволюционном отношении недостаточно для достижения европейскими деревьями равновесного разнообразия. По-видимому, даже в Северной Америке в межледниковые эпохи равновесие не успевало восстанавливаться; послеледниковое расселение вытесненных ледником пород идет слишком медленно.
В более широком смысле часто предполагалось, что тропики богаче, чем умеренные области, по крайней мере отчасти из-за того, что долго непрерывно эволюционируют, тогда как области ближе к полюсам до сих пор не восстановились после плейстоценовых (или даже более древних) оледенений. Однако не исключено, что экологи в прошлом сильно преувеличивали долгосрочную стабильность тропиков.
Когда климатические и природные зоны умеренных областей сдвигались во время оледенении к экватору, тропический лес, видимо, сокращался до нескольких небольших рефугий, окруженных травяными формациями (рис. 25). Поэтому упрощенно противопоставлять неизменные тропики нарушаемым и восстанавливающимся умеренным поясам нельзя. Если мы хотим хотя бы частично приписать бедность приполярной биоты состоянию, далекому от эволюционного равновесия, придется прибегнуть к сложной (и недоказанной) аргументации. (Возможно, смещение умеренных зон на совершенно иные широты приводило к вымиранию гораздо большего числа форм, нежели сокращение площади тропических систем без изменения их широтного распространения.) Проблему помогла бы решить подробная геологическая летопись, показывающая, что тропики всегда характеризовались примерно таким же видовым богатством, а
Бореальный лес
Широколиственный лес умеренного пояса Средиземноморская растительность, в основном хвойные
Средиземноморская растительность, в основном сухостеп-
Прерия (высокие травы с редкими деревьями) L——J
Рис. 25-
А — реконструкция зон растительности Европы во время максимального оледенения (18 тыс. лет назад). Основные типы растительности были сдвинуты к югу от их нынешнего положения на 10—20’, а предшественники Черного и Каспийского морей соединялись между собой (Flint, 1971). Б — плейстоценовые лесные рефугии Амазонии. В ледниковые эпохи большая часть бассейна Амазонки была занята саваннами или степями, а дождевые тропические леса встречались изолированными пятнами, как показано на карте (Prance, 1981)
в умеренных областях либо было гораздо больше видов в прошлом, либо сейчас их число здесь заметно растет. К несчастью, таких доказательств у нас нет. Итак, вероятности, одни сообщества, действительно дальше от равновесия, чем другие, но точно или хотя бы с уверенностью говорить об относительной близости к нему при современных знаниях не представляется возможным.
Градиенты видового богатства. Широта. Пожалуй, самая известная закономерность видового разнообразия — его увеличение от полюсов к тропикам. Это можно видеть на самых разных группах организмов—деревьях, морских двустворчатых моллюсках, муравьях, ящерицах и птицах (рис. 26). Кроме того, такая закономерность наблюдается и в наземных, и морских, и пресноводных местообитаниях. Обнаружено, что в небольших реках тропической Америки обитает обычно 30— 60 видов насекомых, а в умеренной зоне США в аналогичных
водоемах—10—30 видов. Подобный рост разнообразия заметен при сравнении не только обширных географических регионов, но и небольших территорий. Так, на одном гектаре дождевого тропического леса может произрастать 40—100 различных древесных пород, в листопадных лесах востока Северной Америки обычно 10—30, а в тайге на севере Канады всего 1—5. Конечно, есть и исключения. Отдельные группы, например пингвины или тюлени, наиболее разнообразны в приполярных областях, а хвойные деревья— в умеренных широтах. Однако на каждую такую группу приходится множество других, обитающих только в тропиках, например плодоядные рукокрылые Нового Света и гигантские двустворчатые моллюски Индийского и Тихого океанов.
Широта
Рис. 26.
Широтное распределение видового богатства: А — морских двустворчатых моллюсков (Stehli et al., 1967); Б — муравьев (Kusnezov, 1957); В — ящериц в США (Pianka, 1983); Г — гнездящихся птиц Северной и Центральной Америки (Dobzhansky, 1950)
Предлагался ряд объяснений этой общей закономерности, но ни одно из них нельзя принять без оговорок. Прежде всего, богатство тропических сообществ приписывали интенсивному выеданию. Высказывалось предположение, что естественные враги могут быть основным фактором поддержания высокого разнообразия древесных пород в тропических лесах: вблизи взрослых деревьев должна наблюдаться непропорционально высокая гибель подроста того же вида, поскольку родительское дерево—богатый источник видоспецифичных фитофагов. Это утверждение было подкреплено многими работами (рис.27). Если рядом со взрослым деревом вероятность возобновления этой же породы низка, возрастают шансы поселения здесь других видов, а следовательно, и повышения разнообразия сообщества. Отметим, что если специализированное на определенном типе пищи выедание и благоприятствует разнообразию тропических экосистем, оно все же не будет основной его причиной, ибо само является их свойством.
Кроме того, разнообразие связывали с возрастанием продуктивности от полюсов к тропикам. В случае гетеротрофных компонентов сообщества это, по-видимому, верно: понижение широты означает более широкий ассортимент ресурсов, т. е. больший выбор их типов, представленных в достаточном для эксплуатации количестве. Но справедливо ли такое объяснение для растений?
Если повышенная продуктивность тропических областей означает «то же самое в большем количестве» (например, свет), то здесь следовало бы ожидать сокращения, а не повышения видового богатства. В то же время большее количество света может означать и расширение диапазона световых режимов, а за счет этого возрастание разнообразия, но это всего лишь предположение. Но продукция растений определяется не только одним светом. В тропиках почвы, как правило, беднее биогенными элементами, чем в умеренном поясе, поэтому можно было бы считать тропическое богатство видов результатом низкой продуктивности среды. Тропическая почва обеднена питательными веществами, так как основная их часть заключена в огромной биомассе, а разложение и высвобождение биогенных элементов протекают здесь относительно быстро. Так что довод, связанный с «продуктивностью», следовало бы формулировать следующим образом. Освещенность, температура
Рис. 27.
Приживаемость всходов дерева дождевых тропических лесов Dipteryx panamensis возрастом от 7 месяцев до 2 лет в зависимости от расстояния до ближайшей взрослой особи (Clark D. A., Clark D. В., 1984)
и водный режим тропиков обусловливают наличие большой (но не обязательно разнообразной) биомассы растений. Это приводит к формированию бедных почв и, возможно, к широкому ассортименту световых режимов, что в свою очередь — к большому разнообразию флоры. Разумеется, это уже не просто объяснение широтных тенденций разнообразия «продуктивностью».
Некоторые экологи причиной высокого видового разнообразия в тропиках считали климат. Конечно, в экваториальных областях нет столь выраженной сезонности, как в умеренном поясе (хотя в тропиках, вообще говоря, выпадение дождей может подчиняться строгому сезонному циклу), и для многих организмов здешние условия, вероятно, более предсказуемы (это допущение чрезвычайно трудно проверить, поскольку от размеров тела и времени генерации каждого вида
в значительной степени зависит «предсказуемость» для него среды). Утверждение, что климат с меньшими сезонными колебаниями способствует более узкой специализации организмов, за последнее время неоднократно проверялось: в кустарниковых формациях и в лесах тропиков обитает намного больше размножающихся видов, чем в сравнимых экосистемах умеренного пояса, причем от 25 до 50% прироста видового богатства приходится на долю специализированных плодоядных форм, а другая его часть — на птиц, питающихся крупными насекомыми, которые только в тропиках доступны в течение всего года. Таким образом, наличие некоторых источников пищи создает дополнительные возможности для специализации тропической орнитофауны. В противоположность птицам две группы жуков, а именно короеды и древесинники (семейства Scolytidae и Platypodidae), в тропиках не столь узко специализированы по кормовым растениям, как в умеренных областях и число их видов в тропиках гораздо больше (рис. 28).
Резко различные закономерности наблюдаются среди паразитов морских рыб. У дигенетических сосальщиков специализация, если судить по проценту видов, развивающихся только на одном виде хозяина, выше поблизости от экватора (рис. 29). Зато уровень специализации моногенетических сосальщиков одинаков на всех широтах, несмотря на большее видовое разнообразие в экваториальном поясе. Итак, при любом объяснении широтного распределения разнообразия климатические различия следует анализировать, но истинная их роль остается при этом неясной.
Наконец, в качестве причины высокого видового богатства тропических сообществ выдвигали их больший эволюционный возраст. Как уже говорилось, эта теория достаточно правдоподобна, но ее справедливость еще требует доказательств.
В целом четко и однозначно объяснить наличие широтного градиента видового богатства пока не удается. Вряд ли стоит этому удивляться. Элементы возможного объяснения — тенденции, связанные с продуктивностью, стабильностью климата и т. п., сами по себе до сих пор далеко не полностью понятны нам, а ведь на разных широтах они различным образом взаимодействуют друг с другом и с другими, подчас противоположно направленными силами. Тем не менее, объяснение может оказаться очень простым и вот почему. Представим, что
семейства
Питаются Полифаги
видами
1 рода
Уровни пищевой специалимцик
>• Снижение специализация
Рис. 28.
Суммарное число видов жуков семейств Scolytidae и Platypodidae по категориям пищевой специализации. В высоких широтах специализация этих насекомых, как правило, уже (данные из Beaver, 1979)
Рис. 29.
Процент видов моногенетических и дигенетических сосальщиков, паразитирующих лишь на одном виде рыб-хозяев, для различных широт (данные из
Rohde, 1978)
существует некий внешний фактор, способствующий установлению широтного градиента видового богатства, например, среди растений. Тогда увеличение объема, разнообразия и неоднородности распределения ресурсов будет стимулировать рост видового богатства фитофагов. Следовательно, усилится их влияние на растения (вызывающее дальнейшее повышение разнообразия последних) и увеличится разнообразие ресурсов для плотоядных форм, что в свою очередь усилит давление хищничества на фитофагов и т. д. Короче, небольшая внешняя сила может породить каскадный эффект, приводящий в конечном итоге к хорошо выраженному градиенту разнообразия. Однако пока у нас нет убедительных данных о том, что могло бы запустить подобную реакцию.
Часть проблемы заключается в многочисленных исключениях из общей закономерности. Понятно, что объяснить их наличие так же важно, как и общую тенденцию. Одна из крупных категорий таких уклоняющихся сообществ — острова. Кроме того, пустыни очень бедны видами даже вблизи тропиков, возможно, из-за их крайне низкой продуктивности (связанной с недостатком влаги) и экстремальных климатических условий. Относительно бедны видами соленые марши и горячие источники, хотя продукция этих сообществ высока; судя
по всему, дело здесь в суровости абиотической среды (а в случае источников также в «островном» характере этих мелких местообитаний). Как было показано, видовое богатство соседних сообществ может различаться просто потому, что они с разной интенсивностью подвергаются физическим нарушениям.
Высота. В наземных местообитаниях сокращение видового богатства с высотой представляет собой столь же распространенный феномен, как и снижение его по мере удаления от экватора. Человек, поднимающийся в гору вблизи экватора, сначала минует тропические местообитания у подножия, затем поочередно пройдет через климатические и биотические пояса, сильно напоминающие природу Средиземноморья, умеренных и арктических областей. Если альпинист окажется к тому же экологом, он, скорее всего, заметит, как по мере подъема — сокращается количество видов. Это описано на примере птиц Новой Гвинеи и высших сосудистых растений непальских Гималаев.
Следовательно, по крайней мере, часть факторов, обусловливающих широтный градиент разнообразия, должна играть определенную роль и в формировании зависимости разнообразия от высоты (это, видимо, не относится к эволюционному возрасту и менее вероятно для стабильности климата). Разумеется, проблемы, возникающие при объяснении широтной тенденции, остаются и здесь, причем к ним прибавляется еще одно обстоятельство. Дело в том, что высокогорные сообщества практически всегда занимают меньшую площадь, чем соответствующие равнинные биомы, и, как правило, сильнее изолированы от сходных экосистем, не образуя протяженных непрерывных зон. Естественно, ограниченная поверхность и изоляция не могут не способствовать сокращению видового богатства с высотой.
На примере ландшафтов с незначительным перепадом высот установлено, что достаточно резко может различаться число видов в понижениях и на буграх сильно пересеченной местности (луг). Стоит обратить внимание на то, какие серьезные колебания в составе и разнообразии биоты могут отмечаться на очень небольшом участке, т. е., по-видимому, внутри одного сообщества.
Глубина. В водной среде изменения в разнообразии видов с глубиной происходят практически так же, как на суше с вы
сотой. Естественно, в холодных, темных и бедных кислородом глубинах крупных озер меньше видов, чем в тонком приповерхностном слое воды. Точно так же в морях растения встречаются только в эвфотической зоне (где возможен фотосинтез), редко заходящей глубже 30 м. Поэтому в открытом океане происходит быстрый спад разнообразия с глубиной, нарушаемый только некоторыми, часто причудливой формы животными, обитающими на дне. Интересно, однако, что изменение с глубиной видового богатства бентосных беспозвоночных не следует гладкому градиенту: на глубине около 2000 м наблюдается пик разнообразия, примерно соответствующий границе континентального склона. Считается, что он отражает рост предсказуемости среды от 0 до 2000 м глубины. Глубже, за пределами континентального склона, видовое богатство вновь снижается, вероятно, из-за крайне скудных кормовых ресурсов абиссальной зоны (рис.30).
Сукцессия. Каскадный эффект. В некоторых геоботанических работах указывается на постепенное возрастание видового богатства в ходе сукцессии вплоть до климакса или до определенной стадии, после которой следует обеднение флоры по мере исчезновения некоторых позднесукцессионных видов. На рис. 31 показано, как изменяется число видов в ходе залежной сукцессии с момента забрасывания поля. Немногочисленные работы, посвященные изменению фауны в процессе сукцессии, свидетельствуют о параллельном увеличении разнообразия животных.
Сукцессионный градиент видового богатства до определенной степени представляет собой закономерное следствие постепенного заселения участка видами из окружающих сообществ, находящихся на более поздних стадиях сукцессии, т. е. увеличение насыщенности видами. Однако это далеко не полное объяснение, поскольку суть сукцессии не в простом добавлении видов, а в их смене.
Как и в случае других градиентов, при сукцессии неизбежен каскадный эффект. Фактически можно представить, что она и есть этот каскадный эффект в действии. Первыми видами будут те, которые лучше других способны заселять свободные пространства и конкурировать за них. Они сразу представляют собой ранее отсутствовавшие ресурсы и обеспечивают неоднородность среды. Так, пионерные растения создают обедненные биогенными элементами участки почвы, повышая
Рис. 30.
Изменения видового богатства (в выборках из 50 особей) донных животных с увеличением глубины океана (Rex, 1981)
пространственную неоднородность концентрации питательных для растений веществ. Сами растения расширяют набор микроместообитаний и кормовой спектр для животных-фитофагов. Усиление выедания, а затем и хищничества путем обратной связи может способствовать дальнейшему росту видового богатства, обеспечивающего все больший выбор пищевых ресурсов, увеличение неоднородности среды и т.д. Вдобавок температура, влажность и скорость ветра в лесу значительно менее изменчивы, чем в раннесукцессионных сообществах, а увеличение постоянства среды может обеспечивать стабильность условий и ресурсов, что позволяет поселяться и закрепляться специализированным видам.
Как и в случае других градиентов, трудно отделить причину от следствия. И все же при формировании сукцессионного градиента разнообразия тесное переплетение причин и следствий, по всей видимости, составляет самое существо проблемы.
Видовое богатство по данным палеонтологической летописи. Неполнота палеонтологической летописи всегда была основным препятствием для изучения эволюции по ископаемым остаткам. Плохая сохранность, непропорциональное распределение находок по разным группам организмов, таксономические трудности — все это превращает раскрытие эволюционных закономерностей изменения видового разнообразия в сложнейшую задачу. И все-таки некоторые общие тенденции уже проступают.
Рис. 31.
А — изменения видового разнообразия растений (Н) по ходу залежной сукцессии на юге шт. Иллинойс, США. Б — кривые рангового распределения растений для пяти залежей разного возраста. Белые значки — травы, наполовину темные—кустарники, темные—деревья (Bazzaz, 1975)
Примерно 600 млн лет назад, всего за несколько десятков миллионов лет в палеонтологическую летопись попали почти все типы морских беспозвоночных. В течение предшествующих 2500 млн лет мир был, судя по всему, заселен только бак
териями и водорослями. Конечно, мы никогда не сможем точно сказать, отчего произошел этот кембрийский «взрыв» разнообразия. Станли (1976), полагающий, что появление более высокого трофического уровня способно повысить разнообразие на предыдущем уровне. Размышляя о причинах столь же резкого сокращения числа семейств мелководных беспозвоночных в перми, Шопф (1974) отмечал, что в конце этого периода произошло слияние материков с образованием единственного суперконтинента — Пангеи. Это заметно сократило площадь мелководных морей по окраинам материков и тем самым пространство, пригодное для беспозвоночных мелководья. Следовательно, объяснить снижение разнообразия этой фауны можно с помощью хорошо знакомого соотношения между числом видов и площадью. Аналогичным образом возрастание фаунистического богатства в послепермское время может отражать увеличение площади мелководных морей по мере нового расхождения материков.
Анализ ископаемых остатков наземных сосудистых растений показывает четыре обособленные фазы эволюции:
1) рост разнообразия древнейших сосудистых растений в силуре — среднем девоне;
2) последующее расхождение в позднем девоне и карбоне нескольких ветвей папоротниковидных;
3) появление семенных растений в позднем девоне и адаптивная радиация с формированием флоры голосеменных;
4) возникновение цветковых и завоевание ими господства в мелу и третичном периоде.
По-видимому, после выхода на сушу рост разнообразия каждой группы совпадал с его сокращением у группы, доминировавшей перед этим.
Полагают, что два подобных перехода (от древнейших форм к голосеменным и от голосеменных к цветковым) отражают конкурентное вытеснение более древних, менее специализированных таксонов более новыми и предположительно более специализированными. В то же время переход от папоротниковидных к голосеменным совпадал по времени с крупными изменениями среды обитания и мог оказаться следствием частичного вымирания первых и адаптивной радиации вторых в освободившееся экологическое пространство.
Первые насекомые, несомненно, растительноядные, известны из каменноугольного периода. После этого, постепенно
возникали современные отряды, из которых последними появились чешуекрылые, что совпало с расцветом покрытосеменных. Коэволюция растений и растительноядных насекомых почти наверняка была и продолжает оставаться важнейшим механизмом, обеспечивающим рост видового богатства обеих этих групп на протяжении всей их эволюции.
Размеры. Структура сообщества имеет еще один аспект, на который обычно не обращалось особого внимания: среди животных отмечена тенденция существования гораздо большего числа мелких видов, чем крупных. Проводилась, например, приблизительная оценка общего числа видов наземной фауны по классам физических размеров, в частности длины. Если учесть, что огромное число мелких членистоногих и других беспозвоночных до сих пор не открыто, складывается общее впечатление об удивительной редкости крупных видов животных.
В каком-то смысле это парадоксально. Согласно теории, касающейся эволюции особенностей жизненных циклов, на уровне особи естественный отбор должен способствовать возникновению крупных долгоживущих форм, и в рамках некоторых эволюционных ветвей действительно просматривается общая тенденция к увеличению размеров тела («правило Копа»), Однако этому, вероятно, противодействует обычно повышенная скорость вымирания более крупных видов с большим временем генерации. Поскольку размеры тела положительно коррелируют с последним параметром, скорость вымирания крупных видов будет, как правило, более высокой, чем у мелких форм. Из этого вытекает важное следствие: большая эволюционная пластичность присуща мелким видам с малым временем генерации. Можно ожидать, что у них не только ниже скорость вымирания, но и выше скорость видообразования.
Географические аспекты глобальной исследовательской программы «Биологическое разнообразие» составляют ее существенную часть, поскольку именно географический смысл имеют исследования альфа-, бета-, гамма-разнообразия.
РАВНОВЕСНАЯ ТЕОРИЯ ОСТРОВНОЙ БИОГЕОГРАФИИ КАК МОДЕЛЬ ДИНАМИКИ ВИДОВОГО РАЗНООБРАЗИЯ
Начало изучению островной биогеографии было положено замечательным английским натуралистом А. Уоллесом. Работая на островах Малайского архипелага, Уоллес поставил перед собой задачу найти границу между Индо-Малайской и Австралийской фаунами и в 1856 г. обнаружил ее между островами Бали и Ломбок; в 1863 г., анализируя фауны птиц на островах, он положил на карту линию разграничения между этими фаунами, получившую название «линия Уоллеса». В дальнейшем, изучая распределение в том же регионе моллюсков и млекопитающих, М. Вебер провел другую границу (так называемую «линию Вебера»), оказалось, что фауна между этими двумя линиями является смешанной (рис. 32).
Заслуга Уоллеса заключается не только в проведении разграничительной линии между двумя зоогеографическими областями, но и в объяснении причины ее образования: к западу от «линии Уоллеса» все острова расположены на мелководье континентального шельфа и в историческом прошлом они неоднократно соединялись с материком сухопутными связями. Именно Уоллесу биогеография обязана идеей рассматри-
Рис. 32.
Распространение некоторых животных в переходной зоне между Индо-Ма-лайской и Австралийской фаунистическими областями (Ibopry, 1962; Laffin, 1967): 1 — линия Уоллеса; 2 — линия Вебера; 3 — максимальная площадь суши в период последней регрессии моря
вать не только острова, но и озера, вершины гор, другие изолированные биоты в качестве «островных» экосистем среди окружающих ландшафтов, что было четко высказано им в книге «Островная жизнь» (1880).
В настоящее время изучение и анализ островных биот — активно развивающийся раздел биогеографии. Такие биоты характеризуются обилием неизвестных на материках видов, своеобразием флоры и фауны с более простой структурой экосистем. Они удобны для анализа динамики экосистем и часто являются объектами математического моделирования.
Расселение обитателей, островов. Острова Мирового океана различаются по происхождению (океанические и материковые), размерам, высоте над уровнем моря. Океанические острова никогда не соединялись с сушей, они поднялись со дна океана. К их числу относятся коралловые, вулканические и острова складчатых дуг. Материковые острова представляют собой части континентов, отделившиеся от них в ту или иную геологическую эпоху. Естественно, что условия для развития биоты на тех или других островах резко различны.
Материковые острова отделились от континента вместе со всей совокупностью живых организмов, которая этим участкам континента была свойственна. В дальнейшем часть первоначального населения вымерла, а на смену пришли виды, преодолевшие океан.
Как происходило заселение океанических островов, пытался выяснить еще Ч. Дарвин, экспериментально установивший, что плоды и семена многих видов растений сохраняют всхожесть при длительном пребывании в морской воде, и за это время могут быть доставлены течениями на многие острова. Кроме того, семена некоторых видов могут «путешествовать» в кишечном тракте птиц, не теряя всхожести. Наконец, они могут быть доставлены вместе с почвенными частицами на лапках птиц и плавающих стволах деревьев. Если рассматривать вопрос более широко, то можно сказать, что способы попадания растений и животных на острова разнообразны: морские и воздушные течения, штормы и ураганы, плавник, человек и его транспортные средства, активное передвижение по воде или по воздуху.
Установлена способность к распространению животных, относящихся к разным систематическим группам. Они указывают, что из рептилий, например, очень широко распрост
ранены сцинки и гекконы, а другие группы ящериц, как и змеи, а также пресноводные рыбы на многих океанических островах отсутствуют. Видимо, представители этих двух групп ящериц хорошо приспособлены к путешествиям на плавающих стволах деревьев или на их скоплениях — «плотах». Из других систематических групп соленую воду очень плохо переносят амфибии. Поэтому возможности их перемещения через океан крайне ограничены.
Среди беспозвоночных животных, обитающих на океанических островах, доминируют насекомые, второе по численности место занимают наземные моллюски. Насекомые распространяются всеми названными способами. Виды, распространяемые человеком, заметно отличаются от других, так как на островах они обычно являются синантропными, т. е. используют запасы пищи человека, его одежду (кожееды, моли и др.) и жилища, связаны с домашними животными и сельскохозяйственными растениями. Многие из них имеют космополитические ареалы. Перенос насекомых птицами редок. Соленая вода крайне неблагоприятно действует на них, поэтому виды, преодолевающие океанические пространства вплавь, также немногочисленны.
Взрослые растения редко остаются живыми при переносе через океан. Так могут перемещаться со стволами деревьев эпифиты. Обычно растения расселяются с помощью диаспор. Легкие семена и споры переносятся ветром. Важную роль в распространении растений играет человек, разносящий по всему миру, в том числе и по островам, сорняки.
Если два острова различной величины недавно отделились от одного и того же континента, то на большем по размерам острове материковая биота в состоянии сохраниться почти полностью, а на меньшем — может быть полностью или почти полностью исключена возможность существования целых таксономических групп, например представителей всего класса млекопитающих. Ограничивающее влияние размеров острова усиливается по мере уменьшения его территории.
Для Антильских островов выявлено следующее соотношение между размерами острова и числом видов амфибий и рептилий: при уменьшении размеров острова в 10 раз число видов этих групп уменьшается вдвое. Чем ближе остров находится к источнику миграции, тем выше степень насыщения его мигрантами. Этой закономерности подчиняются биоты материко
вых и океанических островов. Хотя расселение каждой особи носит случайный характер, однако при длительном развитии миграционных процессов они подчиняются закону больших чисел, т. е. статистической вероятности. Дарлингтон указывал, что если успешно пересекала пространство шириной в 100 миль одна особь из 1000, то при преодолении следующих 100 миль успешно выполнит эту задачу опять одна особь из 1000, миновавших первые 100 миль. Иными словами, достигнет острова, находящегося в 200 милях от источника миграции, уже только одна особь из миллиона, а острова, лежащего в 300 милях от континента, — лишь одна особь из миллиарда.
Большое влияние оказывает и положение острова по отношению к направлению ветров, которые его пересекают, так называемый «ловчий угол» для насекомых и семян растений: если остров, вытянутый в длину, расположен перпендикулярно к потоку мигрантов, то будет больше шансов, что какой-либо вид его достигнет; если же остров расположен вдоль основного направления движения, то шансов, что организмы, переносимые ветром или морскими течениями, попадут на него, будет значительно меньше.
О постепенном вымирании видов на островах свидетельствует тот факт, что небольшие по площади острова континентального происхождения имеют почти чисто океаническую фауну, в отличие от крупных островов. Так, среди островов Пирл на большом о. Рей сосредоточена примерно треть континентальных видов, а на маленьком о. Контадора — лишь 1 из 10. На островах, лежащих у побережья Южной Калифорнии, ни на одном из них не было того количества видов, которое могло бы на нем существовать, если бы этот остров являлся частью материка. Через 50 лет после первых наблюдений на этих островах число видов составляло в среднем почти половину возможного.
Распространение, дисперсия, — лишь первый этап закрепления вида на острове. Он должен пройти полный цикл развития (эцезис) от появления в данном месте до принесения жизнеспособного потомства включительно. Более толерантные (выносливые) и эвривалентные (т. е. обладающие широкими экологическими возможностями) виды легче проходят стадию эцезиса, чем менее толерантные и стенобионтные.
Эцезис осуществляется при наличии благоприятных для жизни организма условий: света, тепла, влаги и особенно
пищи. Он остается незавершенным чаще всего в результате недоступности необходимых ресурсов. Так, многие насекомые не могут обосноваться на островах из-за отсутствия кормовых растений для их личинок или взрослых особей, или в тех случаях, когда нет открытых пресных водоемов, необходимых для прохождения первых стадий развития. Многие птицы не могут выводить птенцов ввиду отсутствия на острове подходящих для гнездования мест.
Среди растений наибольшими возможностями для успешного развития и размножения обладают обитатели побережий, семена которых, попадая на остров, встречают там среду, близкую к исходной. Наоборот, обитатели высокогорий имеют небольшие возможности для переноса своих семян через океан (горы часто расположены далеко от берега) и очень мало шансов для прохождения растениями эцезиса на новой родине, где высоких гор может и не быть.
Скорость заселения островов невысока. Правда, конкретные данные о скорости этого процесса сильно расходятся. Исходя из числа видов цветковых и сосудистых споровых растений во флоре Гавайских островов и геологического возраста этого архипелага, установлено, что успешное освоение территории растениями осуществлялось здесь в среднем один раз в 20—30 тыс. лет, т. е. не чаще, чем 30—35 заселений за 1 млн лет. Однако эта скорость не может быть принята для всех островов. Так, низменные атоллы имеют чаще всего возраст 4000—6000 лет. Если принять скорость заселения, вычисленную Ф. Фосбергом для Гавайских островов, возраст низменных атоллов меньше срока заселения их одним видом. На самом деле во флоре каждого из таких атоллов насчитывается до нескольких десятков видов, из которых далеко не все были занесены человеком. На поднятый атолл Науру, возраст которого, по определениям геологов, равен примерно 100 тыс. лет, всего 4—5 видов растений могли бы проникнуть при скорости, вычисленной Фосбергом, в то время как флора атолла включает почти 100 видов. Кроме того, следует иметь в виду, что данные о флоре, несомненно, занижены, так как некоторые растения, проникшие на остров, могли впоследствии вымереть. Таким образом, для многих островов скорость их колонизации значительно выше вычисленной Фосбергом: для низменных атоллов она составляет одно заселение в 200—300 лет, для поднятого атолла Науру — примерно одно в 1000 лет.
Большинство биогеографов полагает, что проникновение видов с континента на те или иные острова могло быть облегчено существованием «каменных кладок », т. е. промежуточных островов и островков, которые на протяжении геологической истории то появлялись из вод океана, то вновь скрывались в них. Чаще всего такими временными пристанищами были острова вулканического происхождения.
Островные сообщества. Чем остров меньше, тем, как правило, однообразнее на нем жизненные условия. Этим объясняется прямая зависимость, которая наблюдается между размерами острова и числом видов, входящих в состав его биоты. Иллюстрировать это можно на примере гнездящихся птиц. Численность обитающих на острове видов зависит и от других причин, в первую очередь от возраста острова и степени его изоляции — удаленности от материка (табл. 6).
Таблица 6
Число гнездящихся птиц на островах
Острова Площадь, км2 Число видов
Новая Гвинея 758 000 495
Суматра 434 000 430
Ява 125 000 337
Шри-Ланка 85 000 251
Хайнин 34000 169
Флорес 15 000 141
Азорские 2 388 34
Бермудские 965 13
Необходимым условием видообразования на островах является изоляция. Если непрерывно осуществляется все новый и новый занос особей одного и того же вида на остров, то в результате скрещивания ранее обитавших здесь особей с особями, недавно появившимися, отмечается некоторая стабилизация особенностей вида, и процесс видообразования резко замедляется. Процесс видообразования связан также с природными особенностями островов. На высоких островах, где на небольшом пространстве наблюдается значительное разнообразие экологических условий, возможности возникновения новых подвидов и видов выше, чем на низких островах при однообразии их природных особенностей.
Отсутствие на островах в составе их биот ряда жизненных форм и систематических групп привело к тому, что некоторые виды, попадая на такие острова, проходят так называемую адаптивную радиацию: потомки одного вида, попавшего на остров или архипелаг, сильно изменяются. Так, предок птиц-цветочниц Drepanididae, американский вьюрок, проникнув на Гавайские острова, не встретил здесь конкурентов и дал начало зябликообразным, медососообразным, пищухообраз-ным, дятлообразным и дубоно-сообразным формам. Возникло несколько родов и множество видов цветочниц, которые заняли различные экологические ниши, что сделало невозможным повторение подобной адаптивной радиации в более поздние времена. Аналогичные примеры адаптивной радиации представляют пальмы на о. Куба, некоторые насекомые и моллюски на Гавайских островах.
Примером такой же адаптивной радиации, но не зашедшей столь далеко, служит дерево похутукава (Metrosideros kermadecensis) на о. Рауль (архипелаг Кермадек). В зависимости от условий местообитания оно образует формы, еще не достигшие уровня видовых различий. Это может быть приземистый, прижатый к земле кустарник в нижних частях склонов, подвергающихся действию океанического прибоя, или невысокий прямостоячий кустарник на вулканической пемзе на днище кальдеры вулкана, или прямоствольное дерево в густых насаждениях, или гигантское дерево с распростертыми горизонтальными ветвями в разреженных насаждениях, наконец, эпифит и дерево-душитель при поселении на стволах деревьев.
В других случаях в результате видообразования на островах возникают монотипные роды и даже семейства. Таковы дерево дегенерия из семейства дегенериевых Degeneriaceae на о. Фиджи и птица кагу на Новой Каледонии.
Яркая особенность островных биот — большое число эндемиков, часто высокого таксономического ранга. Количество эндемиков и уровень эндемизма зависят о? величины островов, их расстояния от континента, разнообразия экологических условий и длительности изоляции.
На островах нередко наблюдаются отклонения от обычного облика представителей тех или иных групп: гигантизм или, наоборот, карликовые размеры. Причины этого неясны. Нередко для островов характерны нелетающие птицы и насекомые. Для птиц главную роль в возникновении нелетающих видов
ЯП
играет отсутствие на островах млекопитающих, которые могли бы их истребить. Для насекомых — снос ветром и ураганами летающих видов в океан. Чтобы уцелеть, насекомые должны или обладать быстрым полетом, или, наоборот, потерять способность к полету, или затаиваться при ветре в укромных уголках. На многих, даже незначительных по площади островах имеется немало видов, характеризующихся порхающим, медленным полетом, — златоглазок, комаров, клопов, мелких дневных бабочек, молей. Их обилию способствуют особенности образа жизни, связанного с умением укрываться от порывов ветра. Таким образом, естественный отбор должен был способствовать выживанию нелетающих особей и привести в конце концов к образованию форм, утративших даже органы полета.
Наконец, острова способствуют сохранению примитивных (архаичных) форм. Примерами могут служить новозеландская гаттерия, крайне примитивный род насекомоядных — щелезуб с Антильских островов, мадагаскарская хорьковая кошка, или фосса. Объясняется это тем, что в небольших изолированных экосистемах сложившаяся структура сообществ защищена внешними географическими преградами от вторжения новых, наиболее активных групп, победивших в борьбе за существование, которые на материках, успешно расселяясь, вторгаются в уже образованные ранее экосистемы. Отмечено, что изоляция на островах способствует расхождению форм, т. е. географическому видообразованию, но в то же время эволюционный процесс здесь протекает медленнее, чем на материке.
Характерной особенностью островных биот вообще, кроме эндемизма, является ее бедность. Объясняется это вымиранием и трудностью проникновения на острова переселенцев. Численность видов подвержена по годам более или менее значительным колебаниям. Но на больших территориях эти подъемы и падения численности могут происходить лишь на какой-то части ареала вида. В то время как в одном месте численность вида падает, в другом — поднимается. Таким образом, если даже все особи известного вида в одной части ареала вымрут, то она сравнительно быстро заселится из смежных частей. На островах же вид легко может исчезнуть полностью. Ясно, что чем меньше территория острова, тем больше шансов для вида вымереть.
Изолированность биот отдельных островов — причина их легкого нарушения при изменении человеком природных условий. Вырубка лесов и замена их плантациями как древес-
ных, так и травянистых растений оказываются на островах часто необратимыми, тем более замена лесов полями. Поэтому, приводя примеры вымирания видов под воздействием человека, мы в первую очередь вспоминаем обитателей островов: стеллерову (морскую) корову, жившую у побережий Командорских островов, бескрылого чистика (о. Ньюфаундленд), моа (Новая Зеландия), дронта и др.
Наиболее катастрофическим для фауны и флоры многих островов оказывается завоз человеком (сознательный или бессознательный) новых видов на эти острова. Например, козы на многих островах истребили немало видов растений. Флора о. Св. Елены потеряла из-за коз значительное число видов ранее свойственных ей деревьев; то же отмечено на островах Кермадек и др. В настоящее время на многие острова направлены отряды охотников, цель которых резко уменьшить численность этих животных.
Растительноядное сумчатое поссум, завезенное из Австралии в Новую Зеландию, уничтожило леса во многих районах этой страны. Значительный вред фауне островов наносят попавшие туда крысы. Они истребляют яйца и птенцов птиц, гнездящихся на земле. Так, на о. Рауль (архипелаг Кермадек) они полностью ликвидировали кермадекского буревестника, ныне сохранившегося лишь на нескольких небольших по размеру островках, куда крысы не проникли. Для борьбы с крысами, причинявшими значительный ущерб сельскому хозяйству, в частности разведению сахарного тростника и риса, на Кубу и Фиджи был завезен мангуст. Однако, не ограничившись поеданием крыс, этот зверек резко сократил численность птиц, гнездящихся на земле, на Кубе почти уничтожил эндемичный вид щелезуба, а на Фиджи свел к минимуму численность фиджийской игуаны.
Огромные опустошения в составе животного населения островов производят свиньи. На Новой Зеландии они истребили представителя монотипичного эндемичного отряда — гаттерию, сохранившуюся лишь на мелких островах, лежащих у побережья Новой Зеландии; почти уничтожили нелетающих птиц — киви и совиного попугайчика и т.д.
Вселение на Новую Зеландию европейского благородного оленя привело к гибели лесов на значительной площади. Оказалось, что истребить этого зверя, завезенного человеком, очень непросто.
Таким образом, любое вселение на остров видов, ранее здесь не существовавших, должно быть тщательно про дума-
но. Необходимо всегда помнить о ранимости природы островов и трудности, а то и полной невозможности ликвидации последствий подобных акций.
Эволюция островных сообществ. Слово «остров» в данном разделе не обязательно означает участок суши, окруженный водой. Озера представляют собой «острова» воды среди суши; вершины гор — «острова» с высокогорными условиями в «океане» более низменных территорий; окна в лесном пологе, возникшие при падении деревьев, — «острова» в море древостоя. Можно говорить об «островах» с особым геологическим строением, определенным типом почв или растительности, окруженных иными горными породами, почвами или фитоценозами (рис. 33). Для всех типов «островов» также можно проследить
Рис. 33.
Центры сохранения и последующего расселения биоты (•острова во времени») на Апеннинском п-ве в период вюрмского оледенения (Мюллер, 1972). Зачерненные участки — площади, покрытые ледником; сплошные изолинии — высота снеговой линии; заштрихованные участки — прибрежные убежища средиземноморской биоты; пунктир — береговая линия
закономерное отношение между видовым богатством и площадью. Все эти вопросы очень важны для понимания структуры сообществ. Существует масса островов в океане, множество озер и горных вершин, лесов, окруженных полями, и просто отдельно стоящих деревьев (рис.34). Даже одна особь или лист представляют собой «острова» с точки зрения их паразитов. Короче говоря, не так уж много естественных сообществ, лишенных того или иного «островного» параметра.
Установлено, что число видов на острове тем меньше, чем меньше его площадь. Примеры такой зависимости для разных групп организмов приведены на рис.35. Обычно при графическом ее изображении берут логарифмическую шкалу для обоих параметров, хотя, если соотносить с логарифмом площади число видов, получаемая линия иногда ближе к прямой. Однако в простых координатах зависимость всегда имеет вид кривой, причем при увеличении площади рост числа видов замедляется.
Рис. 34.
Острова и «островные» местообитания (Мюллер, 1974): 1-2 — острова в океане; 3 — «островные» местообитания на вершинах гор; 4 — озера как «острова» на суше; 5,6 — «островные» местообитания для насекомых в кронах деревьев в лесу
Графики зависимости «число видов — площадь» для различных групп организмов на островах (MacArthur, Wilson, 1976): А — местные наземные и водоплавающие пресноводные птицы Гавайских островов; Б — сосудистые растения Азорских островов; В — земноводные и пресмыкающиеся островов Вест-Индии; Г — птицы Соломоновых островов
Применяют три подхода к изучению процессов формирования островных биот:
1) оценивают разнообразие местообитаний, пригодных на острове для заселения;
2) обращают внимание на соотношение скорости двух процессов: заселение острова новыми для него видами и вымирание уже поселившихся (теория равновесия);
3) выявляют взаимосвязь между заселением острова извне и эволюцией видов на нем.
В реальной действительности все эти процессы идут одновременно, и конкретный результат — своеобразие островной биоты — является итогом, балансом их взаимодействия.
Одним из крупных достижений биогеографии нашего времени является создание Мак-Артуром и Уилсоном (1967)
« равновесной теории островной биогеографии *, основанной на математическом аппарате для описания динамичного процесса миграции видов на острова и их вымирания, а также характера устанавливающегося видового равновесия. Суть ее — количество видов, населяющих остров, определяется равновесием между иммиграцией и вымиранием. На рис.36 показано, как это происходит. Рассмотрим сначала иммиграцию. Представим незаселенный остров. Интенсивность иммиграции видов для него будет высокой, потому что любая поселившаяся здесь особь представляет собой новый для экосистемы вид. По мере увеличения числа обосновавшихся видов скорость внедрения новых (еще не представленных) снижается. Она обратится в нуль, когда на острове поселятся все виды «исходной
Число оседлых видов
Рис. 36.
Равновесная теория островной биогеографии (MacArthur, Mison, 1967): А — зависимость скоростной иммиграции от числа обосновавшихся видов для крупных и мелких островов и для островов, лежащих вблизи и вдали от континента; Б — зависимость скорости вымирания от числа обосновавшихся на острове видов для крупных и мелких островов; В — баланс между иммиграцией и вымиранием для крупных и мелких островов и для близких и удаленных от континента
совокупности» (т. е. соседнего материка или близлежащих островов).
Конкретная кривая иммиграции зависит от степени удаленности острова от источника его потенциальных заселеннее. Нуль будет отмечаться всегда в одной и той же точке (на острове представлены все виды «исходной совокупности»), но при этом функция в целом будет иметь тем более высокие значения, чем ближе источник иммиграции, т. е. чем больше у мигрантов шансов добраться до острова. Скорость внедрения на крупные острова, как правило, выше, чем на мелкие, так как первые более доступны. Скорость вымирания равна нулю, когда на острове нет ни одного вида, и в целом низка при небольшом их количестве. Однако по мере роста числа обосновавшихся видов она, согласно теории, увеличивается.
Чтобы рассмотреть суммарный результат иммиграции и вымирания, наложим оба графика друг на друга. Число видов, соответствующее точке пересечения кривых (S*), отражает состояние динамического равновесия, т. е. потенциальное видовое богатство острова. Ниже этой точки видовое богатство растет (скорость иммиграции превышает скорость вымирания), а выше — снижается (вымирание интенсивнее иммиграции). Данная теория позволяет высказать ряд предположений о формировании островных сообществ:
1) число видов на острове в конечном итоге должно примерно стабилизироваться;
2) эта стабилизация — результат не постоянства видового состава, а непрерывной смены видов, когда одни формы вымирают, а другие вселяются;
3) на крупных островах больше видов, чем на мелких;
4) видовое богатство будет снижаться по мере удаления острова от источников заселения.
Главный вопрос островной биогеографии — существует ли «островной эффект» как таковой или же острова беднее видами просто из-за своей малой территории и небольшого количества местообитаний? В ряде исследований предпринимались попытки отделить в соотношении «число видов: площадь» для островов компоненты, полностью объясняемые неоднородностью среды обитания, от обусловленных площадью. Например, разнообразие видов рыб в озерах севера штата Висконсин (США) значимо коррелирует как с площадью озера, так и с разнообразием его растительности. Обнаружена также четкая за
висимость между видовым богатством птиц и площадью островов у побережья Западной Австралии.
Исследование зависимости между числом видов, площадью острова и разнообразием местообитаний для птиц на островах Эгейского моря показало, что разнообразие местообитаний — более важный фактор, чем площадь.
Равновесная теория применима и к насекомым-фитофагам: широко распространенные растения можно считать относительно «крупными островами», окруженными «морем» иной флоры. Кроме того, определенный вид растения можно считать «удаленным» от других, если он специфичен по своим морфологическим, биохимическим или прочим биологическим особенностям. Таким образом, из равновесной теории следует, что видовое богатство насекомых будет выше на растениях, имеющих большие ареалы, и ниже на географически изолированных или редких видах, а также на морфологически или биохимически «изолированных» растениях.
С равновесной теорией хорошо согласуются данные по островам, составлявшим в прошлом части соединявшегося с материком перешейка, который затем погрузился под воду. Если равновесное число видов в некоторой степени определяется соотношением между скоростью вымирания и площадью острова, следует предполагать, что такие недавно обособившиеся участки суши будут терять виды, пока не установится новое равновесие, соответствующее их размерам. Этот процесс принято называть «релаксацией». Так, описана зависимость между временем отделения островов Калифорнийского залива от материка и числом встречающихся на них видов ящериц (Уилкокс, 1978). Внеся поправки на площадь острова, автор продемонстрировал сокращение видового богатства с 50—75 видов примерно до 25 за 4000 лет.
Равновесная теория предполагает для «острова» не только наличие характерного видового богатства, но и постоянный обмен видов. Даже одна особь или лист представляют собой «острова» с точки зрения их паразитов. Короче говоря, не так уж много естественных сообществ, лишенных того или иного «островного» параметра.
Давно замечено, что одна из главных особенностей островных биот — «дисгармония» таксономической и трофической структуры сообщества. Под этим подразумевается иное, чем на материке, относительное участие различных таксонов в соста
ве биоты, в построении трофических сетей островных экосистем . Рассматривая отношения между числом видов и площадью, мы уже имели случай убедиться, что организмы, способные хорошо расселяться (такие, как папоротники и птицы), с большей вероятностью заселяют отдельные острова, чем виды, у которых эта способность не столь развита (большинство млекопитающих, практически все хвойные). Потенциальные возможности расселения отдельных видов, конечно, неодина-
Рис.37.
Восточные границы распространения семейств и подсемейств гнездящихся птиц, встречающихся на Новой Гвинее (Davidson, 1945): 1 — не встречаются за пределами Новой Гвинеи (пеликаны, аисты, жаворонки, коньки, райские птицы); 2 — не заходят дальше Новой Британии и архипелага Бисмарка (казуары и фазановые); 3 — не заходят дальше Соломоновых островов (совы, сизоворонки, птицы-носороги, дронго); 4 — не заходят дальше Вануату и Новой Каледонии (поганки, бакланы, скопы, вороны); 5 — не заходят дальше Тонга и Самоа (утки, дрозды, воскоклювые ткачики); 6 — не заходят дальше островов Кука и Общества (сипухи, ласточки и скворцы). 7 — к востоку от этой линии на Маркизских островах и Туамоту встречаются цапли, пастушки, голуби, попугаи, кукушки, стрижи, зимородки, славки и мухоловки
ковы и внутри таких групп. На рис. 3 7 показаны восточные границы распространения по островам Тихого океана различных наземных и пресноводных птиц, встречающихся на Новой Гвинее, фауна которых обедняется вдвое на протяжении 2600 км от области расселения. В то же время выяснилось, что большинство видов наземных улиток тихоокеанского региона, будучи очень мелкого размера, легко переносятся с острова на остров, а большинство жуков о. Святой Елены —- ксилофаги или подкоровые формы, занесенные сюда, вероятнее всего, через море вместе с плавающими стволами деревьев.
Однако разные способности к расселению — не единственная причина дисгармонии. Виды с низкой плотностью на единицу площади в естественных условиях всегда будут представлены на островах крайне малочисленными популяциями, для которых вероятность полного исчезновения в результате случайной флуктуации очень высока. На многих островах бросается в глаза отсутствие хищников, популяции которых, как правило, относительно невелики. Например, на архипелаге Тристан-да-]$унья в Южной Атлантике нет ни одного питающегося птицами хищника, не считая тех, что завезены сюда человеком.
Хищники могут отсутствовать на островах и по той причине, что их иммиграция в.состоянии привести к поселению на острове только в том случае, если там уже обосновалась их добыча (в то время, как для видов-жертв подобной зависимости от хищников не существует). Это справедливо также в отношении паразитов, мутуалистов и т.п. Иными словами, дисгармония при заселении островов возникает из-за того, что одни категории организмов более «зависимы», чем другие.
Острова и охрана живой природы. Теория островной биогеографии, возможно, наиболее важное приложение находит в области охраны природы. Большинство охраняемых территорий и заповедников окружено «океаном» местообитаний, превращенных человеком в непригодные и поэтому «враждебные» для многих видов флоры и фауны. Может ли изучение островов способствовать разработке принципов, применимых при планировании заповедных территорий? Специалисты считают, что да.
Наиболее очевидный вывод заключается в невозможности для любой территории как части континуума после превращения ее в изолированный заповедник обеспечить существо-
ванне стольких видов, сколько обитали в ней до этого. Можно также ожидать, что в крупном заповеднике сохранится больше видов (будет меньшим вымирание), чем в малом. Однако эти очевидные на первый взгляд выводы требуют некоторых оговорок. Создание изолированного заповедника означает установление границ и, следовательно, появление «пограничных» местообитаний. Значит, в заповеднике смогут обитать виды, которым подходят такие местообитания и которые прежде (т. е. до выделения заповедника из экологического континуума) могли в данной местности отсутствовать. Кроме того, некоторые виды лучше всего чувствуют себя на мелких «островах» и поэтому способны сохраниться не в больших, а в малых заповедниках. Следовательно, встает вопрос, создавать ли один крупный заповедник или несколько небольших с такой же суммарной площадью? Если в каждом из малых заповедников обитает один и тот же вид, разумеется, лучше создать один более обширный, тем самым сохраняя больше форм. Однако если регион в целом неоднороден, то в каждом из небольших заповедников будет обитать особый набор видов, и общее количество сохраняемых форм может превзойти таковое в единственном крупном заповеднике. В небольших заповедниках может сохраниться в общей сложности больше видов даже в однородной области, если учесть, что изоляцией может быть достигнута защита от эпидемий, стихийных бедствий, антропогенных воздействий, особенно в окрестностях крупных городов. Однако для крупных видов хищников, копытных, многих птиц с малой плотностью популяций один крупный заповедник явно предпочтительнее для сохранения вида.
При организации заповедников требуется учитывать многие географические особенности среды, в том числе наличие вокруг ландшафтов-аналогов, которые могли бы выполнять функции «буферных зон», «зеленых коридоров» для мигрирующих видов, возможности устойчивого самовозобновления растительного покрова, характер землепользования в округе. Однако во всех случаях теория островной биогеографии должна быть учтена.
Биом — высшая категория экологического разнообразия суши, определяемая по физиологическим признакам растительного покрова и представляющая собой совокупность биогеоценозов какой-либо климатической зоны. Подробное описание биомов проведено во многих сводках, в том числе Г. Вальтера (1968, 1982), А.Г. Воронова, Н.Н. Дроздова, Е.Г. Мяло (1989), А.Г. Воронова и др. (1999), Н.Н. Дроздова и Е.Г. Мяло (1997), Н.П. Второва и Н.Н. Дроздова (2001), откуда и взяты приводимые ниже характеристики.
Анализируя структуру растительного покрова и животного населения территорий, Г.Н. Огуреева и Н.Н. Дроздов в 1997-2000 гг. (Дроздов и др., 2002) пришли к заключению, что спектры жизненных форм и соотношение биомов на конкретном участке земной поверхности представляют собой его специфическую характеристику и потому могут рассматриваться как особый тип биоразнообразия: биомное, или омега- (завершающее) биологическое разнообразие региона (£2 — омега-разнообразие).
Элементарная единица дифференциации живого покрова суши — биогеоценоз в границах фитоценоза, который никогда не суще
ствует изолированно, а связан территориально и функционально со смежными. Результатом этого являются сочетания (комбинации) сообществ, образующие структуру живого покрова, которую можно исследовать в разных масштабах: суши в целом, материков, крупных регионов и ландшафтов. Различия в структуре живого покрова суши обусловлены многообразием изменений в пространстве климатических, орографических, почвенно-литологических условий, определяющих значительное число вариантов сочетаний разных типов сообществ друг с другом в тех или иных регионах.
Макроструктура растительного покрова суши определяется дифференциацией природной среды на планетарном уровне и находит отражение в его зональном и высотно-поясном расчленении. В основу представлений о зональной структуре живого покрова планеты легли установленные связи между растительностью крупных регионов суши и климатом.
Для выявления связей с климатом используется разработанный немецкими экологами метод климадиаграмм, позволяющий выявлять особенности того или иного климата, проводить его сравнение с климатами других географических регионов (Вальтер, 1968). При составлении климадиаграмм за основу берут чаще всего соотношение между средней месячной температурой и количеством осадков как 1:2, иными словами, 10 °C соответствует 20 мм осадков. При таком масштабе (Т : Ос = 1: 2) засушливая часть года характеризуется кривой осадков, расположенной ниже температурной кривой. Температурная кривая может служить показателем годового хода испарения. Она отражает расход влаги, а кривая осадков — ее приход: обе кривые дают представление о характере водного баланса данной территории (рис. 38). Соотношение температуры и осадков как 1 : 2 применимо для территорий, где достаточно отчетливо выражен сухой период. Там, где период засухи просматривается сдабо, используют соотношение 1 : 3, что позволяет выявить интенсивность и продолжительность полузасушливого периода.
Используя климадиаграммы, можно установить продолжительность вегетационного периода. Правда, термин «веге- <. тационный период» требует уточнения. Это может быть и безморозный период, и время года со средней суточной температурой более 5,10 или 15°. В качестве показателя лучше всего использовать период со средней суточной температурой более
Андахольо (76 м) 27,6*7089 [9-12]
Рис. 38.
Климадиаграмма влажного дождевого тропического леса (Вальтер, 1968): А - в Колумбии, Б - в Камеруне
10°, оцениваемой по кривым годового хода температуры воздуха. Важно также выявить на климадиаграммах продолжительность периода со средней суточной температурой ниже -10°, характеризующего термические условия очень холодных континентальных областей с суровыми зимами.
Таким образом, преимущество климадиаграмм состоит в том, что они помогают представить климат как единое целое, а это особенно важно при экологических исследованиях, когда требуется оценка совокупного воздействия различных элементов климата на биоту.
Тесные связи растительности с климатом еще в начале XX в. впервые были использованы немецким биоклиматологом В. Кеппеном для классификации климатов по их отношению к растительности. Подразделяя растения на экологические группы по отношению к теплу и влаге, Кеппен выделял мегатермы (обитают при постоянно высоких температурах и влажности), ксерофилы (заселяют районы с сухим климатом,, где хотя бы в отдельные сезоны отмечаются высокие температуры), мезотермы (обитатели умеренно теплых и умеренно влажных регионов с относительно теплой зимой), микротермы (произрастают в условиях низкой теплообеспеченности, обязательно с холодной зимой), гекистотермы (существуют
в термических условиях холодных полярных стран). По климатическим показателям районов распространения этих экологических групп проведено биоклиматическое деление суши на мегатермические, ксерофильные, микротермические области. Более дробную дифференциацию осуществляют по биоте. Так, среди мезотермических климатов выделяют климат оливки (средиземноморский, с сухим летом и влажной теплой зимой) и климат камелии (влажный субтропический с влажным летом и мягкой теплой зимой) и т.д.
Закономерности размещения классификационных единиц высокого ранга — типа растительности, типов биомов — в связи с изменением соотношения тепла и влаги по градиенту от низких широт к высоким выявляются при их расположении в осях влажности и температуры (рис. 39). В пределах тропических и субтропических областей, со средними температура-
Рис. 39.
Расположение основных типов биомов земного шара в осях влажности и температуры (Риклефс, 1979)
ми от 20 до 30°, типы растительности изменяются от настоящего дождевого леса, где круглый год влажность высокая, до пустыни. Между этими «крайностями* располагаются влажные листопадные леса, где в сухое время года некоторые или все деревья теряют листву, а также сухие листопадные леса и/или кустарники, часто колючие.
В умеренных широтах наблюдается смена растительности, биомов от лесов, развивающихся при достаточном увлажнении (дождевые леса умеренной зоны), к пустыням. Однако в арктических и субарктических широтах, где средние годовые температуры ниже -5°, размещается только один тип — тундра. Различия в условиях увлажнения в арктических и субарктических областях не оказывают существенного влияния. Влажная тундра перенасыщена водой, но поскольку на протяжении большей части года эта вода остается замерзшей, а на глубине 50—60 см лежит слой вечной мерзлоты, она недоступна растениям.
На «сухом» конце градиента осадков в пределах каждой температурной зоны значительную роль в господстве тех или иных типов растительности играют пожары. В африканских саваннах и прериях на среднем западе США частые пожары уничтожают проростки деревьев и препятствуют возникновению лесов, условия для. развития которых здесь вполне подходящие. Пожары благоприятствуют разрастанию многолетних травянистых растений, почки возобновления которых, размещенные ниже поверхности почвы, слабо или совсем не повреждаются огнем.
Ныне развивается представление о биомах как крупных региональных экосистемах, включающих ряд взаимосвязанных, меньших по размеру экосистем, отражающих взаимодействие климата с региональной биотой и субстратом. Биом как совокупность растительности и животного населения представляет собой сочетание конкретных экосистем разного уровня, которые наиболее эффективно используют абиотические компоненты среды (фотосинтетически активную радиацию, поступающую на участок территории, запас влаги, плодородие почв и т. д.) вследствие определенной, исторически обусловленной адаптации к этим условиям.
Итак, биом: 1) связан с зональными и высотно-поясными климатическими условиями, т. е. существует при определенном соотношении тепла и влаги; 2) включает растительные
сообщества и животное население, неразрывно связанные друг с другом; 3) как экологическая единица подразделения биосферы он, в свою очередь, может быть разделен на более мелкие подсистемы до уровня региональных биомов и далее — элементарных экосистем, или биогеоценозов, на топологическом уровне.
Биомы являются главными составляющими биосферы. Они рассматриваются как совокупности естественных экосистем со сходными структурой и физиономией сообществ, сформировавшиеся в процессе исторического развития различных в природном отношении регионов и отражающие гидротермический потенциал территории в пределах зон на равнинах и высотно-поясных спектров биокомплексов в горах. Под структурой биомов понимается закономерное сочетание и взаимное расположение экосистем, обусловленные климатическими и ландшафтными условиями. Границами биомов являются рубежи, за пределами которых экосистемы конкретных биомов не могут развиваться из-за несоответствия условиям среды, или структуре биомов, т. е. соотношение их экосистем, существенно меняется.
В целях пространственного анализа биопотенциала территории можно использовать классификацию естественных наземных экосистем (Walter, Box, 1976) с выделением единиц планетарного, зонального, регионального и локального уровня. При этом наряду с зонобиомами для характеристики высотно-поясной дифференциации биопотенциала горных территорий вводится понятие оробиома. Зональные биомы и крупные оробиомы включают субзональные биомы и оробиомы второго порядка, которые в свою очередь представлены региональными биомами.
Региональные биомы отражают биоклиматический потенциал территории на провинциальном уровне лучше, чем его биотические компоненты (растительные и животные сообщества) каждый в отдельности. В пределах биомов биота составляющих их экосистем достигает наивысшей степени адаптации к условиям внешней среды. Каждый региональный биом характеризуется специфическим составом экологических групп организмов, разнообразием, структурой растительных сообществ и животного населения. Индивидуальность регионального биома определяется пространственным расположением экосистем, обусловленным ландшафтными особен
ностями территории, включая гидрологический режим и геохимические факторы. По сути своей биом отражает биохорологическую специфику территории и соответствует в биогеографии хорологической единице типа макрокомбинации или макроценохоры.
Каждый биом имеет определенную жизненную форму доминирующих видов климаксовой растительности. Однако биомы включают не только климаксовую растительность, но и стадии сукцессий с присущими им различными видами организмов, в силу чего во многих случаях преобладают иные жизненные формы, а также эдафические климаксы. Все сообщества в данной, климатически обусловленной, совокупности биома представляют собой его естественные части. Региональные биомы как подразделения среднего уровня (между глобальным и локальным уровнями) чрезвычайно важны для изучения антропогенного воздействия на природную среду и оценки биоразнообразия.
Ключевая характеристика, позволяющая разграничивать и узнавать наземные биомы, — жизненная форма (травы, кустарники, листопадные деревья, хвойные деревья и т. д.) растительности климатического климакса. Так, климаксная растительность степного биома — злаки, хотя в разных частях биома и на различных континентах могут произрастать разные виды злаков. Следовательно, основа классификации — растительность климатического климакса, однако биом включает в себя и эдафические климаксы, и стадии развития, в которых во многих случаях доминируют иные жизненные формы. Например, степные сообщества представляют собой стадии развития лесного биома, а прибрежные леса — составная часть степного биома. Биомы мира схематически показаны на рис. 40.
Для оценки ботанического разнообразия региональных биомов хорошие результаты показало применение картографических методов. При оценке ценотического разнообразия можно использовать кадастр растительных сообществ, в котором даются описание и распространение типов подразделений растительного покрова с указанием их на аналитических картах (на конкурсной или сеточной основе). Для этого используется качественный набор фитоценотических подразделений — ценотические спектры (на уровне формаций и субформаций, классов ассоциаций) с выделением (по трехбалльной шкале)
00
IX-VIII
Рис. 40.
Основные типы биомов суши (Вальтер, 1985): I — вечнозеленые тропические дождевые леса, почти без сезонных аспектов; II — тропические листопадные леса или саванны, III — субтропическая пустынная растительность; IV — субтропические склерофильные леса и кустарники, чувствительные к морозам; V — умеренные вечнозеленые леса, чувствительные к морозам; VI — широколиственные листопадные леса, устойчивые к морозам; VII — степи и пустыни областей с холодной зимой, устойчивые к морозам; VIII — бореальные хвойные леса (тайга); IX — тундры, обычно на многолетнемерзлых почвах. Залитые контуры — высокогорная растительность
фоновых — 3 балла, сопутствующих — 2 балла и редких — 1 балл сообществ по отдельным экосистемам биома. При картографическом анализе ценотического разнообразия может быть использована также концепция разнообразия на уровне эпитаксонов, учитывающая динамическое состояние растительных ценозов с выделением коренных сообществ и их производных.
Флористическое разнообразие биомов может оцениваться по общему количеству видов с дифференциацией по спектрам отдельных формаций, по видовой насыщенности в пересчете на единицу площади для конкретных экосистем и всего биома в целом, по ценотическим комплексам или с использованием различных индексов разнообразия. В процессе работы создается база данных по каждому региональному биому, в которую входят: а) характеристики биоклиматической ситуации в регионе по отдельным экосистемам и для биома в целом; б) фитоценотические спектры; в) флористическое богатство; г) приводится филогенетическая характеристика флористических и ценотических комплексов.
ТУНДРЫ
В Евразии и Северной Америке к северу от границы древесной растительности располагаются тундры, образующие единую циркумполярную зону и занимающие наибольшие площади в Сибири и Канаде.
Сообщества тундры развиваются в условиях короткого и прохладного вегетационного периода и низкой температуры почв. Характерна высокая относительная влажность воздуха, преобладание осадков над испарением. Существенное значение для живых организмов имеют колебания температуры. Во все месяцы вегетационного периода минимальные температуры могут быть ниже нуля. Основными лимитирующими факторами служат низкие температуры и короткий сезон вегетации; осадков тоже может быть мало, но они не относятся к лимитирующим факторам, поскольку скорость испарения здесь невелика. Грунт, за исключением верхнего слоя толщиной в несколько сантиметров, на протяжении всего лета остается промерзшим. Постоянно промерзшие глубокие слои грунта называются вечной мерзлотой. Тундра — это в сущности влажная арктическая степь с растительностью, состоящей из злаков, осок, карликовых древесных растений и на более сухих
местах—лишайников («олений мох*). Для «низменной тундры* (как на прибрежных равнинах Аляски) характерны толстая рыхлая подушка из живых и очень слабо разложившихся растений, часто пропитанная водой и испещренная небольшими озерцами. «Высокая тундра», особенно если рельеф хорошо выражен, покрыта скудным покровом лишайников и злаков. Хотя вегетационный сезон краток, длинные летние фотопериоды способствуют образованию в благоприятных местах довольно большого количества первичной продукции (до 5 г сухого вещества в день). Важнейшим экологическим фактором в условиях тундры является снежный покров, защищающий зимой от низких температур и сильных ветров. Высота растений часто определяется мощностью снежного покрова. Повсеместное распространение вечной мерзлоты обусловливает формирование криогенных нано-, микро- и мезоформ рельефа, а также широкое распространение процессов заболачивания в термокарстовых понижениях.
Продуктивные водные (в том числе плодородный арктический океан) и наземные экосистемы поддерживают не только большое число перелетных птиц и насекомых, активных только в бесснежный сезон, но и постоянных обитателей, которые деятельны на протяжении всего года. Сюда относятся: овцебык, европейский северный олень, карибу, белый медведь, волки, песцы, хищные птицы, а также лемминги и другие мелкие млекопитающие, строящие ходы в толще растительности. Для крупных животных характерны обширные миграции. Особую уязвимость тундры следует иметь в виду при все возрастающей эксплуатации минеральных ресурсов и другой деятельности человека, вызывающей нарушения окружающей среды. Тонкий живой покров легко разрушается и медленно восстанавливается.
Тундровые растения — криофиты — приспособлены к короткому и прохладному вегетационному периоду и низкой температуре почв. Растения низкорослые, приземистые.
Для многих кустарничков характерны стелющиеся и подушковидные формы роста. Растениям свойственна вегетативная подвижность, что приводит к образованию куртин и латок. Благодаря низкорослости тундровые растения зимой покрываются снегом, предохраняющим их от вымерзания, а летом их надземные органы оказываются в наиболее прогретом и экологически наиболее благоприятном слое воздуха.
В тундре велико фитоценотическое значение лишайников и особенно мхов, зачастую являющихся эдификаторами сообществ. Так, в Гренландии споровых растений почти в 10 раз-больше, чем сосудистых. Сплошной моховой покров в условиях тундры существенно влияет на температурный режим почв и глубину сезонного протаивания и, следовательно, на условия обитания других растений. Лишайники оказывают на почвенные условия меньшее воздействие, но при их высоком обилии число видов трав и кустарничков снижается.
Одним из основных признаков сообществ тундры является их полидоминантность. Обычно выделяют тундры лишайниковые, моховые, кустарничковые, пушицево-осоковые и др. Как правило, все эти типы характеризуются содоминирова-нием нескольких групп растений, таких как мхи, лишайники, кустарнички, многолетние травянистые растения. Яркая черта структуры — слабовыраженная вертикальная дифференциация фитоценоза, часто и мхи, и кустарнички размещаются практически в одном высотном горизонте.
В горизонтальном строении наблюдается мозаичность, выделяются микроценозы, сложенные различными жизненными формами растений. Мозаичность тундровых сообществ обусловлена не только жизнедеятельностью растений, но и интенсивно протекающими криогенными процессами в почве, приводящими к горизонтальному расчленению ее поверхности. От типа форм нанорельефа зависит горизонтальное сложение сообществ тундры. Выделяют бугорковые, пятнистые, пятнисто-мелкобугорковые, кочкарные тундры и др. Во многих тундровых сообществах растительность далеко не полностью покрывает поверхность почвы. Характерная черта растительного покрова тундры — комплексность, обусловленная разнообразием форм микро- и мезорельефа, различиями в глубине залегания многолетнемерзлых пород и мощности снежного покрова на разных формах рельефа.
Один из важнейших признаков тундр — их безлесье. Между зоной бореальных хвойных лесов и безлесной тундрой лежит переходная полоса, в пределах которой лесные и тундровые сообщества тесно контактируют друг с другом. Это зона лесотундры. Весь комплекс экологических условий в тундре неблагоприятен для развития древесных пород. Недостаток летнего тепла, короткий вегетационный период, иссушающее действие ветров — причины, снижающие рост и замедляющие
Рис. 41.
Структура биоценоза арктической тундры о. Ляховского (Александрова, 1966). Подземные органы-. 1 — полярной ивки (SaZtxpolaris), 2 — лисохвоста альпийского (Alopecurus alpinusy 3 — крупки (Draba pseudopilosay 4 — лапчатки ги-поарктической (Potentilla hyparctica)-. 5 — неидентифицированные корневые системы; 6 — границы участков концентрации корневых систем; 7 — граница многолетнемерзлых пород в августе
развитие древесных пород, уменьшающие их конкурентную способность по сравнению с тундровыми растениями. Граница между лесом и тундрой соответствует степени континентальности климата: в наиболее континентальных районах она существенно продвигается к северу и смещается к югу в областях с господством океанических воздушных масс. На распространение леса и характер границы между лесом и тундрой весьма заметное влияние оказывает деятельность человека. В результате вырубок и пожаров граница лесов сдвигается к югу.
В связи с изменением условий от полярной границы леса по направлению к более высоким широтам тундры разделяют на субарктические, арктические и высокоарктические. Пос
ледние часто относят к полярным пустыням. В сообществах субарктических тундр доминируют или содоминируют различные полярные березки. Характерны моховые, лишайниковые, кустарничковые типы сообществ. Как правило, развит покров из мезофитных зеленых мхов. Граница между субарктическими и арктическими тундрами проходит примерно по изотерме июля +6°. Близ этого рубежа меняется облик сообществ: исчезают полярные березки, увеличивается количество пятен оголенного грунта, под которым сохраняется сомкнутость корневых систем растений (рис. 41). В арктических тундрах выделяются, как правило, два яруса: напочвенный (мохово-лишайниковый) и кустарничко-травяной. Основу растительного покрова составляют мхи, к которым на повышенных участках добавляются лишайники. Характерны осоки, пушицы, полярная ива. Ковер растительности арктических тундр нередко разорван пятнами оголенного грунта. Арктические тундры довольно сильно заболочены.
БОРЕАЛЬНЫЕ
ХВОЙНЫЕ ЛЕСА
Бореальные хвойные леса, известные под названием тайга, протянулись непрерывной полосой через Евразию и Северную Америку и образуют единую циркумбореальную зону. Ширина пояса хвойных лесов в Евразии достигает максимума в междуречье Енисея и Лены, в Северной Америке — на Тихоокеанском побережье.
Гидротермические условия характеризуются сравнительно коротким вегетационным периодом, холодной зимой, относительно прохладным летом при существенных региональных отличиях в тепло- и влагообеспеченности. Значительные площади хвойных лесов находятся в области распространения многолетнемерзлых пород. Это определяет специфику почвообразования и увлажнения почв. Как правило, количество осадков превышает расходы на испарение, с чем связано широкое распространение процессов заболачивания.
Таежные леса образованы деревьями, относящимися к ограниченному числу родов, со своеобразной (игольчатой) формой хвои. Ее наличие обеспечивает полное использование периода вегетации и предохраняет деревья от высыхания зимой.
С закрытыми устьицами многие хвойные деревья при прочих равных условиях транспирируют меньше, чем лиственные деревья в безлистном состоянии. Хвойные деревья хорошо переносят сильные зимние холода и способны к ассимиляции даже в очень прохладный сезон. Характерны узкие кроны, что снижает возможность их обламывания под тяжестью снега и позволяет растениям лучше использовать свет при низком стоянии солнца.
Таежные леса моно- или олигодоминантные, обычно с простой и четко выраженной ярусной структурой. Выделяются древесный, кустарниковый (степень выраженности которого зависит от типа леса), кустарничково-травянистый ярусы и напочвенный моховой или лишайниковый покров. Видовой состав довольно ограничен, по числу видов преобладают травянистые растения, мхи и лишайники.
В связи с изменением теплообеспеченности по широтному градиенту тайга с севера на юг подразделяется на северную, среднюю и южную. С севера на юг возрастает сомкнутость крон, увеличивается полнота насаждений. Для южной тайги характерно значительное участие в травяном покрове видов, свойственных широколиственным лесам, а также местами примесь широколиственных древесных пород,
Четыре рода хвойных — ель, пихта, сосна, лиственница — основные лесообразователи на всем протяжении тайги, представленные в Евразии и Северной Америке разными видами. В Северной Америке, кроме того, велика роль видов тсуги и дугласии (или псевдотсуги).
В зависимости от экологии лесообразователей, определяющей различия в структуре и составе сообществ, таежные леса подразделяют на темнохвойные (представленные различными видами ели, пихты, тсуги, сосны сибирской и т.д.) и светлохвойные, среди которых выделяют вечнозеленые сосновые и лиственничные леса с опадающей хвоей. Темнохвойные леса довольно просты по структуре: число ярусов обычно равно 2—3. Кроме древесного могут быть развиты ярусы травяной или травяно-кустарничковый и моховой. Иногда травяной ярус отсутствует. Кустарники единичны и выраженного яруса не образуют. Затенение значительное. В связи с этим травы и кустарнички размножаются чаще вегетативным путем, чем семенным, образуя куртины, группы. Лесная подстилка разлагается медленно, поэтому неко
торые травянистые растения не образуют хлорофилла и питаются сапрофитно (подъельник, ладьян и др.). Имеются зимнезеленые растения (брусника, грушанка). Освещение, в противоположность широколиственным лесам, одинаково в течение всего вегетационного периода, поэтому растений с цветением в ранневесенние месяцы практически нет. Венчики цветков растений нижнего яруса — белые или бледно окрашенные (бледно-розовые, бледно-голубые), именно эти цвета хорошо заметны на темно-зеленом фоне мохового покрова и в сумраке темнохвойного леса.
Светлохвойные леса характеризуются более изреженным древостоем, что связано со светолюбием лиственниц и сосен. Здесь в напочвенном покрове приобретают значительную роль лишайники, а местами сильно развит кустарниковый ярус, образованный рододендронами, ракитником, калиной, шиповником, смородиной и др. В Северной Америке в этих лесах нередко наблюдается примесь белой пихты, дугласии (псевдотсуги) и ряда других пород.
В Евразии тайга распространена в основном в России, а за ее пределами лишь в некоторых районах Северной Европы. Основные лесообразователи темнохвойных лесов — ель европейская и сибирская, пихта, сибирская кедровая сосна.
Светлохвойные леса из лиственницы сибирской и даурской обнаруживают в своем распространении четкую связь с резко континентальным климатом и замещают друг друга по мере нарастания континентальности. Облик лиственничных лесов, особенно в Сибири и на Дальнем Востоке, в значительной степени определяется пожарами, под влиянием которых в нижних ярусах получили широкое распространение багульник и брусника.
Леса из сосны обыкновенной занимают в таежной зоне большие площади, преимущественно с менее благоприятным режимом. Сосновые леса характерны для древних аллювиальных равнин, песчаных террас, развиваются на заболоченных торфянистых почвах.
Дендрофлора североамериканской области богаче евразийской, причем основное разнообразие хвойных пород сосредоточено в тихоокеанской части. Леса северной и восточной частей тайги Северной Америки близки к евразийским, что определяется сходством условий произрастания. Главные лесообразователи — три вида хвойных: ель белая,
или канадская, ель черная, лиственница американская. Местами к ним примешиваются береза бумажная и тополь осинообразный. В отличие от Евразии род сосна представлен большим числом видов, наиболее характерны леса из сосны Бэнкса, веймутовой сосны и др., приуроченные к сухим песчаным почвам. В более южных частях таежной зоны в качестве лесообразующих пород появляются тсуга канадская и туя восточная.
Притихоокеанские (западные) леса расположены между 42 и 62° с.ш. и развиваются в чрезвычайно благоприятных климатических условиях. Высокая влажность воздуха, большое количество осадков (до 1000 мм и более), часть которых выпадает в виде снега, небольшие амплитуды годовых температур обеспечивают максимальный прирост темнохвойных пород, формирование сложных высокоствольных насаждений. В этих лесах велико разнообразие хвойных пород, что обусловлено историческими причинами. Роды ель и пихта представлены здесь значительным числом видов. Наряду с ними лесообразователями являются виды родов тсуги, псевдотсуги, или дугласии, туи, кипарисовика.
Огромное воздействие на состав и структуру растительности тайги оказывают пожары и вырубки, в связи с чем на больших площадях развиваются мелколиственные леса.
Животные тайги вынуждены либо приспосабливаться к активной жизни в условиях снежной и морозной зимы, либо впадать на это время в состояние анабиоза, спячки или зимнего сна, или же откочевывать на зимний период за пределы тайги. Один из важнейших кормовых ресурсов — семена хвойных пород деревьев. Ими питаются многие птицы, грызуны и даже хищные звери. Однако урожайность ели, пихты, кедровой сосны («кедра») сильно колеблется по годам, и за годом урожайным следуют обычно несколько малоурожайных или вовсе неурожайных лет. Поэтому животные, потребляющие в основном семена хвойных, в неурожайные годы совершают дальние кочевки (белка), вылеты за пределы основного ареала (кедровка, клесты), либо значительно снижают численность (лесные полевки, оленьи хомячки, зайцы).
Общая биомасса существенно различается в разных типах леса и подзонах тайги. Больших величин она достигает в сомкнутых ельниках и пихтарниках, в богатых сообществах юж
ной и дальневосточной тайги, где составляет 350—400 т/га. Годичная продукция таких лесов достигает 8—10 т/га. Гораздо ниже показатели биомассы и продукции в разреженных лиственничках и несомкнутых лесных сообществах северной тайги. В этих районах биомасса колеблется от 50 до 200 т/га, а продукция таких лесов составляет 4—6 т/га в год.
ЛИСТОПАДНЫЕ ЛЕСА УМЕРЕННОЙ ЗОНЫ
Широколиственные, или летнезеленые, леса в умеренных широтах Северного полушария не образуют единой зоны и занимают в Европе, Азии и Северной Америке разобщенные территории с ослабленной континентальностью. Климат в ареале лесов этого типа умеренно прохладный, осадки распределены в течение года относительно равномерно, их количество варьирует в довольно широких пределах. Характерен перерыв в вегетации, обусловленный зимними холодами. В зависимости от степени континентальности климата зимы могут быть почти безморозными (приат-лантические районы Европы) или же с устойчивыми морозами при мощном снежном покрове.
Северные границы распространения широколиственных лесов зависят от продолжительности холодного периода и (или) недостатка летнего тепла. В продвижении отдельных пород на север существенную роль, по-видимому, играют поздневесенние и раннелетние заморозки, особенно губительные для подроста. На южных границах основным лимитирующим фактором является влажность, чем определяется переход к степям. Смена широколиственных лесов хвойными проис-
ходит очень постепенно, выделяется полоса смешанных хвойно-широколиственных лесов.
Леса характеризуются сомкнутым верхним древесным ярусом, нижние древесные ярусы или очень разреженные, или отсутствуют. Многие широколиственные породы дают корневую и пневую поросль, порослевый древостой более низкорослый и сомкнутый. Различают моно-, олиго- и полидо-минантные леса. Виды бука, как правило, образуют монодоми-нантный древостой. Кустарниковый ярус изменяется от хорошо развитого до крайне разреженного.
Лиан, как правило, мало, но в некоторых регионах (Восточная Азия, южная часть приатлантических лесов Северной Америки) они весьма обильны. Травянистый покров образован многолетними видами, различными в экологическом отношении. Выделяется группа мезофильных тенелюбивых трав с длительным периодом вегетации. Характерны растения, цветущие весной, когда под полог леса поступает много света, и заканчивающие свое надземное существование вскоре после распускания листьев на деревьях. Во влажном климате приморских районов в состав травянистого яруса входят некоторые вечнозеленые виды, доля участия которых понижается в континентальных районах.
Европейские широколиственные леса флористически самые бедные и относительно просты в структурном отношении. Коренных лесов практически не сохранилось. Основные лесообразователи — виды бука, дуба; роль каштана, как и граба, ясеня и липы, ограничена. В восточные, более континентальные районы и на границу со степными формациями распространяются только леса из дуба черешчатого.
Специфика лесов Азии определяется поразительным видовым богатством деревьев, кустарников и трав, причем значительная их часть относится к древним родам. По сравнению с европейскими лесами в них повышена роль лиан и папоротников-эпифитов.
В Северной Америке широколиственные леса распространены только в восточной части континента. Поразительно видовое разнообразие и богатство состава деревьев, обилие кустарников, а также лиан.
Между лесами Восточной Азии и Северной Америки обнаруживается известное сходство, усиливающееся благодаря сохранению в них большого числа представителей древней
листопадной флоры. С большим разнообразием видов деревьев связано наличие двух-трех ярусов древостоя. Доминируют виды дуба, клена, платана, вяза, ясеня, ореха, тюльпанное дерево и др. В наибольшей степени леса сохранились в предгорных районах Юго-Западных Аппалачей и в долинах рек.
В Южном полушарии широколиственные летнезеленые леса распространены крайне незначительно. В условиях очень влажного климата с ровным годовым ходом температур преобладают вечнозеленые лиственные леса, слабо отграниченные структурно и флористически от субтропических.
Хорошо выраженная ярусная структура широколиственных лесов, наличие сомкнутого древесного яруса высотой до 30 м над землей, обильная подстилка и мощный гумусовый горизонт обеспечивают полный спектр ярусов животного населения этого биома. Теплое влажное лето и холодная зима со снежным покровом обусловливают четкую сезонную динамику активности животных. На зиму пойкилотермные животные впадают в состояние анабиоза. Из гомойотермных животных (птицы и млекопитающие) одни мигрируют в более теплые регионы, другие впадают в спячку или зимний сон и лишь немногие сохраняют круглогодичную активность, переходя на специфические корма (кора и ветви деревьев, насекомые, спящие под корой, и т.п.)
Опад древесных и кустарниковых пород формирует мощный слой подстилки. Деятельность по утилизации отмершей растительной массы в широколиственных лесах осуществляет обширная и разнообразная группа животных-сапрофагов, среди которых ведущую роль играют дождевые черви семейства люмбрицид. Вместе с сапрофагами почвенный ярус населяют потребители живой корневой массы растений. К ним относятся личинки насекомых, главным образом жуков: жесткие, с плотными покровами личинки жуков-щелкунов, называемые проволочниками, белые жирные личинки пластинчатоусых жуков, живущие в почве по несколько лет до окукливания. Из них наиболее типична личинка майского жука.
В нижней и средней частях древостоя в стволах и ветвях деревьев поселяются насекомые-ксилофаги — потребители древесины. Это личинки усачей-дровосеков, пластинчатоусых (например, крупные личинки жука-оленя). Кору поедают личинки златок. В кронах лиственных пород деревьев обильны насекомые, поедающие зеленые ткани листьев. Среди них до
минируют гусеницы различных бабочек: пядениц, шелкопрядов, листоверток, личинки (ложногусеницы) пилильщиков, взрослые формы (имаго) жуков-листоедов, хрущей, в частности, упомянутые уже майские жуки. Особую группу насекомых-фитофагов образуют сокососущие виды — тли, червецы, цикады, листоблошки, цикадки, клопы.
Немало фитофагов и среди позвоночных. В наземном ярусе обитают мелкие грызуны. В евразийских лесах это лесная рыжая полевка, лесная и желтогорлая мыши, в широколиственных лесах Северной Америки — внешне похожие на мышей белоногий и золотистый хомячки.
Крупные копытные участвуют в потреблении зеленой массы листвы, травы, а зимой веточных кормов и коры деревьев. В Евразии и Северной Америке широко распространен благородный олень, известный в разных частях ареала под названием марал, изюбрь, вапити (последнее относится к американскому благородному оленю). В западноевропейском секторе к оленю присоединяется лань, а в дальневосточном — пятнистый олень. В отличие от оленей кабан питается не только надземными, но и подземными частями растений (корневищами, клубнями, луковицами), которые он выкапывает, взрыхляя носом подстилку и верхний слой почвы. Там же он находит и поедает всевозможных беспозвоночных.
Хищные животные — зоофаги населяют все ярусы широколиственных лесов. В почвенно-подстилочном живут хищные многоножки — геофилы и костянки, жуки (жужелицы и стафилины), пауки и хищные клещи. Ведущее место среди хищных беспозвоночных занимают муравьи, сооружающие наземные гнезда, но обследующие в поисках пищи все ярусы леса. Верхнюю часть почвы осваивают в поисках корма различные кроты. В европейских лесах многочислен обыкновенный крот, в лесах Восточной Азии — крот могера.
В наземном ярусе обычны амфибии и рептилии: бурые лягушки, тритоны и саламандры, особенно разнообразные в лесах Северной Америки, ящерицы и змеи.
Насекомоядные птицы собирают корм главным образом в кронах деревьев и в подстилке. В лесах Евразии обычны дрозды, мухоловки, синицы, славки и пеночки. В американских лесах дрозды и синицы также обычны, но экологическую нишу мухоловок занимают тиранны, а славок и пеночек — американские славки, или древесницы.
Хищные звери характерны как для наземного, так и для древесного ярусов. В наземном — обитают хищники, широко расселяющиеся и за пределы широколиственных лесов: лисица, волк, бурый медведь (в прошлом), горностай и ласка. На Дальнем Востоке обитают черный медведь и енотовидная собака (ныне завезенная в европейские леса), в Северной Америке — близкий к черному медведь барибал. Древесный ярус используют рысь, дикий лесной кот, там постоянно держится лесная куница, на Дальнем Востоке — харза.
Длительное и интенсивное сельскохозяйственное освоение регионов широколиственных лесов привело к резкому обеднению их животного населения, к полному исчезновению многих видов, особенно крупных позвоночных. На пахотных землях сильно размножились зерноядные грызуны. О первичном населении животных этих сообществ мы можем судить по историческим данным и по остаткам природных биоценозов на территориях заповедников и в отдаленных слабо освоенных районах.
Запасы биомассы в широколиственных лесах умеренных широт очень велики — 500—400 т/га. Продукция этих лесов также высока — от 10 до 30 и даже 50 т/га в год, что объясняется благоприятными теплыми и влажными условиями вегетационного периода. Мощность почвенного и подстилочного ярусов обеспечивает кормом громадное количество почвенных животных. В основном благодаря им зоомасса в этих лесах может достигать 1 т/га и даже более, что превышает запасы зоомассы в большинстве других биомов суши.
Сообщества листопадных лесов занимают области с большим количеством равномерно распределенных осадков (750— 1500 мм) и умеренной температурой, для которой характерны четкие сезонные колебания. Первоначально умеренные листопадные леса покрывали весь восток Северной Америки, всю Европу, часть Японии, Австралии и южную оконечность Южной Америки. Таким образом, биомы листопадных лесов в большей степени изолированы друг от друга, чем тундра и северные хвойные леса. Их видовой состав, несомненно, должен отражать эту степень изоляции. Поскольку в таких лесах деревья и кустарники в течение части года лишены листьев, контраст между зимой и летом очень велик. Травянистый и кустарниковый ярусы, а также почвенная биота развиты хорошо. Многие растения дают сочные мясистые плоды и оре
хи, например желуди или буковые орешки. Животное население первоначальных лесов Северной Америки представлено виргинским оленем, медведем, серой и черной белкой, серой лисицей, рыжей рысью и дикой индейкой. Характерные мелкие птицы зрелых стадий — красноглазый виреон, большой американский дрозд, американская хохлатая синица, желтоголовая американская славка и несколько видов дятлов.
Листопадные леса умеренной зоны представляют собой наиболее используемые человеком биотические области мира, именно в этих районах цивилизация достигла наибольшего развития. В результате этот биом сильно изменился под влиянием человеческой деятельности, его большая часть замещена культурными сообществами или сообществами лесных опушек.
САВАННЫ И СТЕПИ
Саванны — это травянистые сообщества тропического пояса, характеризующиеся наличием сомкнутого злакового покрова различной высоты с варьирующейся долей участия кустарников и деревьев. Сезонная ритмика этих сообществ связана с периодичностью выпадения осадков.
Саванны существуют в разных климатических условиях: от регионов с продолжительным сухим сезоном (как Суданские саванны в Африке и серрадос Центральной Бразилии) до регионов с достаточно влажным тропическим климатом (саванны Гвинеи). С этим связаны различия в структуре, продуктивности. Огромные площади они занимают в Африке — классической стране саванн, а также распространены в Южной Америке, Азии и Австралии.
Для злаков саванн характерны более или менее выраженный ксероморфизм, плотное укоренение, обилие семян. Высота травостоя в зависимости от увлажнения и почвенных условий варьирует весьма существенно. Особенно большой высоты достигают генеративные побеги злаков. Доминирует ограниченное число видов злаков, относящихся к нескольким
родам, широко распространенным в тропиках. Помимо злаков в травостое саванн участвуют некоторые осоковые, различные виды двудольных (многолетние и однолетние).
Деревьям и кустарникам саванн свойственна мощная корневая система, проникающая на значительную глубину даже при относительно небольших размерах надземной части. Деревья часто низкорослые с извилистыми или изогнутыми стволами, с раскидистыми кронами. Широко распространена зонтиковидная форма кроны, например у многих акаций в саваннах Африки. На стволах часто развита толстая корка. Среди деревьев и кустарников преобладают листопадные формы, но встречаются и вечнозеленые с жесткими склерофилльными листьями, например эвкалипты в саваннах Австралии.
В связи с различиями в высоте и сомкнутости травостоя, зависящими от суммы осадков и продолжительности сухого периода, различают ряд основных формаций саванн.
Влажные саванны развиваются при продолжительности сухого периода 3—5 месяцев и 800—2000 мм осадков в год. Это высокотравные (от 1,5 до 3 м и более) сообщества, зеленеющие во влажный период и высыхающие в сухой, иногда без примеси древесных видов, иногда с участием одиночных деревьев или их небольших спорадически рассеянных групп. Они располагаются на выровненных или всхолмленных пространствах междуречий; по речным долинам протягиваются галерейные леса с преобладанием вечнозеленых влаголюбивых видов. Как правило, выделяются два яруса травостоя; в сомкнутом верхнем, образованном крупными злаками с длинными и широкими листьями, число видов невелико, но они составляют основную долю фитомассы саванн. В нижнем, достаточно разреженном ярусе развиты некоторые многолетние травянистые двудольные. Вегетация растений определяется влажностью, причем начало развития приходится на период перед наступлением дождей.
Сухие саванны представлены сообществами, сформированными жестколистными и узколистными злаками высотой 1,5—2 м, растущими разреженно, не формирующими сомкнутой дернины. В областях их развития сухой период длится примерно 5—7 месяцев, сумма осадков — от 500 до 1200 мм в год. Довольно обычно участие деревьев в составе этих саванн, хотя они могут иногда и отсутствовать. Они низкорослые (5—10 м), стволы с толстой коркой и твердой древесиной. Характерны как
листопадные, так и вечнозеленые виды деревьев и деревья с сук-кулентными стволами. Сухие саванны очень бедны по видовому составу.
Колючие саванны значительно более ксероморфны, чем вышеописанные. Они развиваются при значительной продолжительности сухого периода (8—10 мес.) и варьирующем количестве годовых осадков (250—750 мм). Высота травостоя чаще всего 30—50 см и крайне редко почти 1 м. Господствуют засухоустойчивые жестколистные и узколистные злаки. Травяной покров разрежен. Обычно характерна примесь кустарников, но встречаются и отдельно растущие низкорослые деревья высотой 3—8 м, часто колючие или с суккулентными стволами, иногда безлистные стеблевые суккуленты. Велико разнообразие кустарников, преимущественно колючих, листопадных и вечнозеленых, представлены и безлистные прутьевидные формы.
Значительная часть сообществ влажных и отчасти сухих саванн в их современном распространении возникла на месте тропических лесов в результате вырубок, раскорчевки и расчистки под пашни, пожаров, сознательно используемых в целях огневой чистки облесенных участков, а также для выжигания сухой травы. Толстокорые древесные породы переносят пожары без ущерба. На многолетние злаки саванн, имеющие большое число почек возобновления в основании стебля и подземной части корневища, палы оказывают даже благоприятное влияние, способствуя разрастанию дерновины растения.
У животных четко выражены приспособления к перенесению неблагоприятного периода засухи (или двух ее периодов). Длительность анабиоза у многих насекомых, амфибий и некоторых рептилий в этих регионах заметно продолжительнее, чем в сезонно-влажных сообществах. Миграции птиц и кочевки крупных травоядных животных приобретают особый размах.
Структура животного населения значительно упрощена, фрагментарность древесного яруса не позволяет обитать в саваннах узкоспециализированным древесным животным. В то же время обилие травяных кормов обеспечивает процветание исключительно богатому и разнообразному комплексу крупных травоядных животных, в первую очередь копытным.
В саваннах, как и в других тропических типах биомов, ведущую роль в утилизации отмершей растительной массы
играют термиты. Именно здесь плотность их населения, количество и размеры надземных сооружений достигают максимальных величин. Термитники занимают до 30% поверхности почвы, а общая масса надземных сооружений термитов составляет 2400 т на 1 га. Число термитников на 1 га достигает в некоторых районах 2 тыс.
Среди птиц-фитофагов преобладают зерноядные виды семейства ткачиковых, превосходящего другие группы птиц и по численности, и по видовому составу. Некоторые виды, например красноклювая квелия и общественный ткач, образуют крупные гнездовые колонии.
Для злаковых саванн характерны крупные нелетающие птицы — страусы. Они потребляют в основном растительную пищу, а также насекомых и мелких позвоночных. В африканских саваннах обитает африканский страус — самая крупная из современных птиц (высотой до 2,7 м, массой до 90 кг). В саваннах Австралии живет страус эму ростом до 1,7 м, а в аналогичных местообитаниях Южной Америки — нанду, ростом до 1,5 м.
Среди млекопитающих саванн преобладают крупные фитофаги из отрядов парнокопытных, непарнокопытных, хоботных. Общее число видов копытных в саваннах Африки превышает 70, и большую их часть составляют антилопы. Ресурсы фитомассы распределены между всеми копытными, каждый вид использует определенный ярус растительности или даже отдельные фрагменты данного яруса, что и позволяет сосуществовать на одной площади исключительному разнообразию крупных животных.
Самые характерные группировки копытных в саванне — большие стада антилоп гну, совершающие ежегодные миграции на десятки и сотни километров в соответствии со сменами периодов вегетации травянистой растительности. Вместе с гну мигрируют и зебры. По тем же маршрутам, но в стороне от стад зебр и гну кочуют мелкие антилопы — газель Томсона, или томми, и похожая на нее, но несколько более крупная газель Гранта. Антилопы гну и томми — самые массовые виды копытных в саваннах Восточной Африки.
Древесно-кустарниковая растительность, как и травянистая, четко распределена между животными саванны. Обитающая в зарослях кустарников мелкая антилопа дик-дик обкусывает нижние веточки; на высоте примерно 1 м объедают
кусты черный носорог и антилопа импала; на высоте 2 м— антилопа геренук, или жирафовая газель. На высоте 2-4 м обламывает и обдирает ветки акаций африканский слон, который может также пастись и на злаковых пространствах, захватывая хоботом и выдирая с корнем пучки трав. На наибольшей высоте находит себе корм жираф, рост которого почти 5 м. В австралийских саваннах функцию копытных выполняют крупные кенгуру.
Хищные животные саванн также очень разнообразны. Крупные четвероногие хищники саванн кочуют за стадами антилоп, совершая вместе с ними сезонные миграции. Львы охотятся на крупных антилоп и зебр, гепарды — на мелких антилоп.
В южноазиатских саваннах помимо азиатского льва, ныне почти истребленного, распространены шакалы и полосатые гиены. В Южной Америке обитают гривистый волк и саванновая лисица. Австралийские саванны в историческое время (с появлением человека на этом материке) были заселены собакой динго.
Общие запасы сухой биомассы в различных вариантах саванн составляют от 150 до 50 т/га. Продуктивность этих сообществ довольно высокая: от 15 до 5 т/га в год, т. е. примерно 10% от общих запасов биомассы. Зоомасса обычно составляет 100—130 кг/га, из которой половина приходится на дождевых червей и четверть — на термитов. В некоторых районах скоплений крупных травоядных животных запасы зоомассы парадоксально высоки — до 50 кг/га и более.
Степи умеренной зоны расположены там, где выпадает промежуточное между пустынями и лесами количество осадков (250— 750 мм); существование степей в этих условиях зависит от температуры, сезонного распределения осадков и влагоемкости почвы. Ключевым фактором здесь является влажность почвы, в частности потому, что она лимитирует микробное разложение и возврат в круговорот биогенных элементов. Обширные степные пространства занимают внутренние части Евразийского и Северо-Американского континентов, юг Южной Америки (аргентинские пампасы) и Австралию.
В Северной Америке биом степей подразделяется (с востока на запад) на высокотравные, смешанные, низкотравные и кустисто-злаковые прерии зоны. Эта зональность опреде
ляется градиентом количества осадков, который одновременно является и градиентом уменьшения первичной продуктивности. Ниже приводится перечень некоторых наиболее важных видов многолетников, сгруппированных по высоте надземных частей.
1. Высокие злаки (от 150 до 240 см)—бородач (Andropogon gerardi), просо (Panicum virgatam), индейская трава (Sorg-hastram nutans) и в низинах Spartina pectinata.
2. Средние злаки (от 60 до 120 см) —малый бородач (Andropogon scoparius), ковыль (Stipa spartea), спороболус (Sporo-bolus heterolepis), пырей (Agropyron smithii), тонконог (Koeleria cristata), индейский рис (Oryzopsis) и многие другие.
3. Низкие злаки (от 15 до 45 см) — бизонова трава (Buchloe dactyloides), голубая грамова трава (Bouteloua gracilis), другие виды грамовой травы (Bouteloua sp.), интродуцированные мятлик и костер (Bromus sp.).
Корни большинства видов глубоко (до 2 м) проникают в почву, и масса корней климаксных многолетников в несколько раз превышает массу их надземных частей. Большое значение имеет форма роста корней. Некоторые из упомянутых видов, например бородач, бизонова трава и пырей, образуют подземные корневища и формируют дернину. Другие виды, такие, как малый бородач, тонконог, ковыль, кустятся и растут куртинами. Эти две жизненные формы можно найти во всех зонах, но кустовые злаки преобладают в более засушливых областях, где степь переходит в пустыню.
Разнотравье (сложноцветные, бобовые и т. п.) составляет обычно лишь малую часть биомассы продуцентов климаксной степи, но встречается неизменно. Некоторые виды особенно интересны как индикаторы стресса. Усиление выпаса, засуха или обе эти причины увеличивают процент разнотравья, что выражено также на ранних стадиях сукцессии. Живописные однолетние травы на обочинах дорог, такие, как солянка (Salsola) и подсолнечник (Helianthus), обязаны своим «процветанием» постоянному разрушению почвы, чему способствуют ремонтные работы на скоростных дорогах.
Обширные пространства степей, особенно высокотравных прерий, в настоящее время заняты посевами зерновых, культурными пастбищами или древесной растительностью. Сейчас трудно найти первоначальную, или девственную, высоко
травную прерию, и там, где она сохраняется для научных и учебных целей, ее нужно время от времени выжигать, чтобы сохранить специфику этого биома.
В хорошо развитом степном сообществе есть виды с различными температурными адаптациями; одна группа растет в холодные сезоны (весной и осенью), другая — в теплый (летом). Степь в целом «скомпенсирована» по температуре, что удлиняет период первичного продуцирования.
В степном сообществе развивается почва, полностью отличная от лесной, даже если оба сообщества стартовали на одной и той же материнской породе. Так как злаки по сравнению с деревьями живут недолго, почву обогащает большое количество органического вещества. Первая фаза разложения протекает быстро, в результате образуется мало подстилки, но много гумуса, иными словами, скорость гумификации велика, а минерализации мала. В итоге степная почва может содержать в 5—10 раз больше гумуса, чем лесная. Темная степная почва лучше других подходит для произрастания кукурузы, пшеницы и других зерновых, представляющих собой виды культурных злаков.
Пожары помогают степной растительности конкурировать с лесной в теплых или влажных районах. Характерная особенность степей — наличие крупных травоядных. Крупные травоядные представлены двумя «жизненными формами»: бегающей, к которой относятся только «экологические эквиваленты » бизонов, антилоп и кенгуру в степях разных биогеографических областей, и роющей — суслики и луговые собачки. Когда природные степи превращаются в пастбища, то присущие им виды травоядных животных заменяются домашними: коровами, лошадьми, овцами и козами. Поскольку степи адаптированы к мощному потоку энергии, проходящему через пастбищную пищевую цепь, такое переключение экологически безвредно. Однако люди постоянно страдают от «трагедии общин», допуская чрезмерный выпас и выпахивание. Многие степи превратились в настоящее время в созданные руками человека пустыни, которые трудно вернуть вновь к состоянию степей. Например, в аргентинских пампасах в результате интенсивного выпаса коров количество способных гореть веществ сократилось настолько, что не осталось пищи пожарам, необходимым для поддержания покрова злаков. Это привело к преобладанию колючих кустарников, рост которых прежде сдер
живался периодическими пожарами. Единственный путь восстановления продуктивности пастбищ — затрата энергии топлива для механического удаления и выжигания древесной растительности. Описанная ситуация служит примером такого антропогенного изменения растительности, которое можно сделать обратимым только дорогой ценой.
Во внутриконтинентальных регионах Северной Америки и Евразии огромные площади занимают более или менее ксеро-фильные травянистые формации, известные под названием прерии и степи. Для этих регионов характерны континентальные типы климата, часто с суровой зимой и устойчивым снежным покровом . Осадки распределены относительно равномерно в течение года, лето, особенно во второй половине, засушливое. Степная растительность связана с черноземными и каштановыми почвами, характеризующимися высоким плодородием.
В составе растительных сообществ наиболее типичны многолетние ксерофильные дерновинные злаки, относящиеся к родам ковыль (Stipa), овсяница (Festuca) и тонконог (Koeleria) и являющиеся доминантами и эдификаторами (рис. 42). В североамериканских прериях значительная роль принадлежит также видам рода бородач (Andropogon) и некоторым другим, не имеющим широкого распространения в Евразии. Характерны и корневищные злаки с единичными побегами на ползучих подземных корневищах, в целом менее засухоустойчивые и поэтому шире распространенные в более влажных частях степных областей (виды костра, пырей и др.).
Кроме злаков большую роль в сложении покрова степей играют многочисленные ксерофильные представители двудольных растений, так называемое степное разнотравье. В составе степных сообществ, особенно в более засушливых районах, представлены и коротковегетирующие растения — однолетние (эфемеры) и многолетние (эфемероиды), формирующие весенний и раннелетний аспекты и особенно обильно развивающиеся во влажные годы.
В составе степной растительности местами значительное участие принимают кустарники, иногда произрастающие группами. Это виды спиреи, степной вишни, можжевельника; в степях Монголии велика роль видов караганы, образующих своеобразные кустарниковые степи.
Для степных растений характерно наличие у многих из них глубоко проникающих и сильно разветвленных корневых
Рис. 42. Степной тип задернения (Алехин, 1957): 1 - узколистный ковыль (Stipa stenopbylla)-, 2 - обыкновенный ковыль (Stipa joannis)-, 3 - тонконог (Koeleria gracilis)-, дерновины растений сидят в более или менее глубокой чаше
систем, эффективно поглощающих влагу из почвы. В степных биоценозах запасы подземной фитомассы достигают больших величин.
Яркая особенность структуры — хорошо выраженная смена аспектов от ранней весны до глубокой осени. Так, для Стрелецкой степи под Курском насчитывается до 11 аспектов,
связанных с поочередным массовым цветением таких растений, как гиацинт, ирис, ветреница, ковыль, шалфей и др. Для степной растительности очень характерны флуктуации: в более сухие годы лучше развиваются ксерофильные виды растений, уменьшается доля эфемеров и эфемероидов; в более влажные годы преобладают виды менее засухоустойчивые.
Для нормального развития дерновинных злаков и многих видов разнотравья необходимо их очищение от отмерших побегов, сохраняющих механическую связь с живыми растениями, и отсутствие на поверхности почвы так называемого «степного войлока», покрова из отмерших частей растений. Благодаря выпасу травоядных млекопитающих происходит разрыхление скоплений ветоши на поверхности почвы, что обеспечивает развитие степных злаков. В отсутствие выпаса в течение 4—5 лет степные растения постепенно отмирают.
Не меньшую роль играет деятельность грызунов, потребляющих значительную часть травостоя и разрыхляющих почву. Устраивая глубокие норы, сурки и суслики проникают на глубину до 2—3 м; выбросы земли на поверхность образуют насыпные курганчики, часто довольно многочисленные. Возникает чередование микроповышений и микропонижений, что приводит к перераспределению осадков. В связи с этим часто развивается комплексность: к разным формам микрорельефа приурочены различные растительные сообщества.
Важный фактор в жизни степных биоценозов — пожары, уничтожающие всю надземную часть растительности. Выгорает вся ветошь и многие семена, лежащие на поверхности почвы. У дерновинных злаков, особенно крупнодерновинных, почки возобновления надежно защищены от огня основаниями листьев, благодаря чему они не погибают во время пожара. Чувствительны к воздействию огня молодые деревца, кустарники. Во время пожаров гибнут многие животные.
Условия увлажнения в пределах обширных ареалов степных биомов неоднородны, в связи с этим происходит изменение характера травостоя, его высоты, фитомассы, соотношения различных жизненных форм и т.д. По градиенту увлажнения с севера на юг степи Евразии подразделяются на подзоны или широтные полосы: луговых степей и остепненных лугов, настоящих и опустыненных степей.
Наиболее яркая особенность растительного покрова луговых степей — закономерное сочетание степных сообществ с не
большими лесными массивами, откуда, собственно, и название этой подзоны — лесостепь. На распределение растительности большое влияние оказывают перераспределение осадков по рельефу, степень промытости верхних почвенных горизонтов. В связи с этим в лесостепи европейской части России степные сообщества доминируют на междуречных равнинах, дубравы тяготеют к балкам, лощинам, изредка выходя на водоразделы. В Западной Сибири лесные биоценозы (так называемые березовые колки) приурочены к понижениям (западинам) рельефа и окружены степными сообществами.
По направлению к югу возрастает засушливость, климат степей становится теплее. На северной границе лесостепи соотношение осадков и испарения с открытой водной поверхности равняется единице, в полосе пустынных степей испарение значительно превышает количество осадков. Соответственно с севера на юг снижается видовое богатство, уменьшается число видов разнотравья, в южной части настоящих и особенно в опустыненных степях возрастает доля ксерофитов — полукустарничков, в том числе видов полыни, уменьшается число аспектов, снижаются высота травостоев и запасы биомассы.
В североамериканских прериях уменьшение количества осадков происходит с востока на запад, что и определяет субмеридиональное простирание подзон или полос: лесостепь, где чередуются фрагменты лесов (преимущественно из гикори и некоторых видов дуба); высокотравная прерия с обилием разнотравья и высоких злаков (обладают виды ковыля, бородача, овсяницы); смешанная прерия; низкотравная прерия с господством двух низкорослых видов злаков: травы грама (Bouteloua gracilis) и бизоновой травы (Buchloe dactyloides). Разнотравья здесь немного, характерно участие полыни. В связи с тем, что изменения температуры и влажности происходят в разных направлениях, в растительности каждой из субмеридиональных полос прослеживаются с севера на юг существенные различия.
В настоящее время степи и прерии в основном распаханы и заняты сельскохозяйственными культурами (особенно это относится к луговым, разнотравно-типчаково-ковыльным степям Евразии, лесостепи, высокозлаковой и смешанной прерии Северной Америки). В более засушливых полосах, где земледелие является рискованным, развито пастбищное животноводство.
В Южном полушарии пампа, а также сухие злаково-полукустарничковые формации Патагонии, находящиеся в ветровой тени Анд, чаще всего рассматриваются только как в известной степени подобные степям, .своеобразные их аналоги. Важнейшее отличие гидротермического режима областей их развития — отсутствие выраженного периода с отрицательными температурами и снегового покрова. Это оказывает существенное воздействие на состав и структуру сообществ, особенно их ритмику. Здесь характерны круглогодичная вегетация, своеобразная кустовая форма роста злаков.
Животные степей, прерий и пампы приспосабливаются к довольно жесткому гидротермическому режиму. Большинство животных вынуждены ограничивать свою активность главным образом весной, в меньшей степени — осенью. На время холодной зимы они впадают в анабиоз, а в период летних засух снижают активность, пребывают в состоянии так называемого полупокоя. Мелкие позвоночные животные (ящерицы, змеи, некоторые грызуны) впадают на зиму в спячку, крупные млекопитающие мигрируют в более южные районы с мягкой зимой, а большинство птиц совершают сезонные перелеты.
Отсутствие древесно-кустарникового яруса обусловливает простоту вертикальной структуры животного населения. Выделяется один надземный ярус, однако усиливается проникновение животных в почвенные горизонты — открытый ландшафт требует поиска убежищ, а многие грызуны способны к рытью сложных и глубоких нор.
Травянистая растительность обеспечивает обильные запасы корма животным-зеленоядам, а подземные части многих растений-геофитов (корневища, луковицы, клубни) потребляются наряду с корнями животными-ризофагами. Мощный слой подстилки и гумуса населен разнообразными сапрофагами. Таким образом, в сообществах степи, прерии и пампы значительно сильнее, чем в других, выражен подземный ярус населения животных.
Надземную зеленую массу растительности поедают разнообразные саранчовые и кузнечики. Этими же кормами питаются различные грызуны. В степях Евразии селятся большими колониями и роют сложные норы суслики. Колониальный образ жизни позволяет степным грызунам своевременно оповещать членов колонии об опасности, а норы дают им надеж
ное убежище от большинства хищников. В степной зоне сохранились еще большие поселения обыкновенного сурка, или байбака. В прериях Северной Америки обычны луговые собачки, внешне напоминающие небольших сурков. Они также роют сложные разветвленные норы на глубину до 5 м. Колонии луговых собачек достигают иногда нескольких тысяч особей. В южноамериканской пампе аналогичный образ жизни ведет крупный грызун — равнинная вискаша из семейства шиншилловых.
Грызуны-зеленояды, хотя и ведут норный образ жизни, но корм они собирают в наземном ярусе. Другая экологическая группа грызунов роет постоянные кормовые ходы, питаясь подземными частями растений — корневищами, клубнями, луковицами. Этих грызунов-ризофагов можно объединить в экологическую группу подземных землероев.
В степях Евразии обитает обыкновенная слепушонка, мелкий грызун длиной до 15 см, с маленькими глазками, вооруженный мощными резцами, которые выступают впереди губ. Этими резцами слепушонка может рыть свои кормовые ходы, не раскрывая рта, что предотвращает попадание земли в ротовую полость. Алтайские и монгольские степи населяет цокор, более крупный грызун, длиной до 25 см, также со слаборазвитыми глазами, но с мощными передними конечностями и громадными когтями. Цокор роет норы передними лапами.
Наиболее выраженные адаптации к подземному образу жизни имеют слепыши, полностью лишенные зрения (глаза скрыты под кожей), наружных ушных раковин и хвоста, с громадными резцами, постоянно торчащими изо рта, так как губы сходятся позади зубов (как и у слепушонки). Длинные и разветвленные кормовые ходы слепышей располагаются под поверхностью почвы, а гнездовая камера находится на глубине почти 3 м.
В прериях подземный образ жизни ведут грызуны семейства гоферовых. У них маленькие глаза, короткий хвост и мощные резцы, выступающие впереди губ. Они роют главную нору — галерею длиной до 140 м, от которой отходят многочисленные боковые отнорки. В южноамериканской пампе аналогичную экологическую нишу занимают грызуны туко-туко из особого неотропического семейства ктеномиид, которые роют сложные разветвленные норы с гнездовыми камерами и камерами-кладовыми. Члены колонии перекликаются меж
ду собой громкими криками «туко-туко», хорошо слышными из-под земли.
В степях Евразии несколько веков тому назад можно было увидеть пасущиеся стада диких быков туров, антилопы сайги, диких лошадей тарпанов, степных зубров. Эти копытные не только вместе с другими фитофагами потребляли зеленую массу растительности, но и активно воздействовали на структуру верхнего почвенно-подстилочного яруса.
В североамериканских прериях копытные не столь разнообразны. Фоновым ландшафтным видом здесь был лишь бизон, многотысячные стада которого паслись в прериях вплоть до появления европейцев с огнестрельным оружием. Популяция бизонов восстановлена, насчитывает тысячи особей и занимает нераспаханные участки прерии на северо-западных окраинах первичного ареала этого вида. Кроме бизона в прериях был обычным и поныне еще сохранился своеобразный вилорог, экологически замещающий отсутствующих в Новом Свете антилоп.
В пампе обитают совсем иные крупные потребители травянистой растительности. Характерный вид — безгорбый верблюд гуанако из отряда мозоленогих. Гуанако совершают сезонные миграции летом к местам водопоев и зеленым пастбищам, зимой — в районы с мягкой бесснежной погодой.
Хищные животные травянистых сообществ имеют богатый выбор кормов: от мелких насекомых и их личинок до грызунов, птиц и копытных. В наземном ярусе обычны хищные муравьи (хотя в степной зоне немало и муравьев-семеноядов), жуки-скакуны из семейства жужелиц, одиночные роющие осы, охотящиеся на разных беспозвоночных.
Мелкие хищные птицы степей (пустельга, кобчик) потребляют главным образом насекомых — саранчовых, жуков. Крупные пернатые хищники добывают грызунов соответственно своим размерам: от полевки и сусликов до сурков и луговых собачек. В степях Евразии обычны луни, канюк-курган-ник, характерен степной орел.
В прериях самая обычная птица — мелкий сокол — американская пустельга. Он питается в основном саранчовыми и другими насекомыми. И в прериях, и в пампе изредка можно увидеть ныне сильно истребленного вилохвостого коршуна.
Хищные млекопитающие питаются главным образом грызунами. Волк, лисица, горностай, ласка хотя и обычны
в степи, но не характерны для этой зоны. Наиболее свойствен светлый, или степной, хорь. С юга заходит хорек-перевязка. Представители семейства куньих легко проникают в норы грызунов и добывают их прямо в убежищах. Это лишний раз подчеркивает относительность любых защитных адаптаций: и глубокая нора не спасает ее обитателей от специализированных хищников.
В прериях группу хищных млекопитающих образуют койот, черноногий хорек, длиннохвостая ласка. В пампе в эту группу входят пампасная лисица, гривистый волк, патагонская ласка.
Таким образом, в каждом из крупных изолированных регионов травянистых сообществ складывается набор хищников от крупных наземных до мелких норных в соответствии с разнообразием фауны грызунов.
Общие запасы биомассы в ксерофильных травянистых сообществах умеренных широт варьируют в зависимости от высоты и густоты травостоя от 150 т/га сухого вещества в высокотравных прериях до 10 т/га в сухих степях и низкотравных прериях. Средние запасы в этих сообществах обычно равны примерно 50 т/га. Продукция также соответственно меняется от 30 до 5 т/га в год и составляет 20—50% от запасов годичной биомассы. Зоомасса в естественных сообществах с обилием грызунов и копытных может достигать значительных величин — 10—50 кг/га, что сравнимо с массой в тропических саваннах.
ПУСТЫНИ
Типы биоценозов, относимые к пустынным, формируются в условиях умеренного, субтропического и тропического природных поясов, что определяет существенные различия в их тепловом режиме. Однако облик, структура, динамика пустынных сообществ, уровень продукционных процессов контролируются в первую очередь увлажнением, количеством осадков, их распределением в течение года. Количество осадков в разных пустынных регионах и внутри них варьирует от 200—150 до 50—40 мм в год, вплоть до полного их отсутствия. Пустыни встречаются в тех областях, где в год выпадает меньше 250 мм осадков. Иногда к пустыням относят области и с большим количеством осадков, но эти осадки распределены очень неравномерно. Скудность осадков может быть вызвана:
1) высоким субтропическим давлением (Сахара и австралийские пустыни);
2) географическим положением в области «дождевой тени» (пустыни на западе Северной Америки);
3) большой высотой местности (пустыни Тибета и Боливии, Гоби).
В большинстве пустынь в течение года выпадает какое-то количество дождей, и имеется хотя бы редкая растительность, если эдафические условия субстрата не оказываются очень неблагоприятными (например, подвижные песчаные дюны, барханы). По-видимому, единственные места, где дождей выпадает очень мало или они не выпадают совсем, — это Центральная Сахара и пустыня на севере Чили. Чистая годовая первичная продукция составляет в пустынях менее 1000 кг сухого вещества с гектара, или около 200 ккал/м2.
В условиях орошения пустынь, когда вода перестает быть лимитирующим фактором, первостепенное значение приобретает тип почвы. Если текстура почвы и содержание элементов питания в ней благоприятны, то вследствие обилия солнечного света оводненные пустыни могут быть крайне продуктивными. Однако из-за значительных затрат на строительство и эксплуатацию ирригационных систем удельная стоимость выращенной продукции может быть очень высокой. Через такую экосистему нужно пропускать очень большие количества воды, в противном случае в почве будут накапливаться соли (вследствие высокой скорости испарения), которые станут лимитировать продуктивность. По мере «старения» орошаемой экосистемы потребности в воде растут. Это вызывает «спираль инфляции»; возникает необходимость в строительстве новых водоводов, что увеличивает стоимость продукции и ведет к более интенсивной эксплуатации подземных и горных водных ресурсов. В пустынях Старого Света легко найти развалины древних оросительных систем. Во многих случаях никто не знает, почему они пришли в упадок и почему «райские сады» опять превратились в пустыню. Эти руины должны служить нам по меньшей мере напоминанием того, что орошенная пустыня не будет процветать бесконечно, если игнорировать основные законы развития экосистемы.
Морфофизиологические адаптации растений к аридным условиям определяются дефицитом влаги, бедностью почв, часто сочетающейся с засолением, высокими температурами, действующими круглогодично или в течение сезона. Возможности развития растений в жестких условиях водного режима в сочетании с действием высоких температур обеспечиваются за счет более эффективного поглощения воды из почвы при повышении осмотического давления и развитии мощных поверхностных или глубоко проникающих корневых систем.
Многие пустынные растения характеризуются мощной корневой системой, быстрый рост в глубь почвы позволяет корням проникать в горизонты повышенной влажности, за счет чего растения могут существовать в течение довольно длительного времени. Так, семенные побеги деревьев и кустарников развивают стержневые корни, длина которых в 10 раз превышает длину надземного побега; многие злаки образуют густую корневую систему в верхних почвенных горизонтах, напоминающую войлок, а их нитевидные корни проникают в почву на метр и более. Соотношение между массой побегов и массой корней изменяется в сторону увеличения последней при возрастании засушливости.
Существуют три жизненные формы растений, адаптированные к пустыням:
1) однолетники, избегающие засухи тем, что они растут только при достаточной влажности;
2) суккуленты, такие, как кактусы, накапливающие воду;
3) пустынные кустарники, у которых многочисленные отходящие от короткого основного ствола ветви несут мелкие толстые листья.
Во время длительного засушливого сезона эти листья опадают. Пустынные кустарники всего мира весьма сходны по внешнему виду, хотя могут принадлежать к различным таксономическим группам (еще один четкий пример экологической эквивалентности).
Наиболее полно в пустынях представлены растения, относящиеся к группе ксерофитов или гиперксерофитов. Для них характерно уменьшение площади испаряющей поверхности, которое достигается за счет редукции листьев и микрофил-лии, сбрасывания ассимилирующих органов по мере нарастания сухости почвы и воздуха, замены крупных листьев первой генерации более мелкими листьями побегов последующих генераций.
Среди суккулентов, которые решают проблему водного баланса путем накопления влаги в водозапасающей паренхиме листьев и стеблей, различают стеблевые (кактусы, многие представители семейства молочайных) и листовые (например, агавы, алоэ). Особенно характерно разнообразие суккулентов и формирование специфических суккулентных формаций для пустынных регионов субтропического и тропического поясов с «двухтактным» режимом выпадения осадков (два коротких
периода дождей), что обеспечивает накопление влаги в запасающих тканях.
Особую группу представляют растения с укороченным жизненным циклом, смещенным на влажные отрезки года, что позволяет им избегать засушливых и жарких периодов. Это травянистые растения — эфемеры (терофиты) и эфемероиды (геофиты). Семена эфемеров и подземные органы эфемероидов (луковицы, клубни, корневища) сохраняют жизнеспособность в течение нескольких лет, благодаря чему легко переживают годы, когда количество осадков бывает снижено. Цикл развития может быть смещен на конец зимы — начало весны (как в пустынях Средней Азии), на весну и осень (в пустыне Сонора); связь с определенным сезоном может и отсутствовать, если осадки выпадают нерегулярно.
Для пустынных кустарников характерно очень своеобразное распределение: редкие отдельные растения разбросаны далеко друг от друга. В некоторых случаях определенную роль в поддержании дистанции между растениями играют антибиотики. Однако всегда разреженность снижает конкуренцию за скудные ресурсы, иначе интенсивная конкуренция за воду могла бы привести к гибели или задержке роста всех растений.
В формировании облика областей распространения пустынных экосистем огромное значение имеют свойства грунтов, особенности субстрата. В связи с этим выделяют пустыни песчаные, глинистые, каменистые и т.д. Так, в песчаных пустынях большую отрицательную роль играет подвижность субстрата; в каменистых и галечниковых — плотность, загипсо-ванность почв и подстилающих пород. Этим обусловлено появление у растений своеобразных приспособлений. В зависимости от них выделяются экологические группы растений: псаммофиты и гипс офиты.
Весьма жесткими экологическими условиями характеризуются глинистые пустыни, где влага атмосферных осадков по большей части испаряется с поверхности почвы, не успев в нее проникнуть. Поэтому растительный покров здесь разрежен. В песчаных пустынях, отличающихся более благоприятным водным режимом, чаще всего развиваются более сомкнутые травянистые или древесно-кустарниковые сообщества. С каменистыми субстратами связано распространение разреженно произрастающих древесно-кустарниковых форм, тяготеющих к трещинам и понижениям, где аккумулируется влага.
Своеобразный экологический режим имеют солончаковые пустыни, где развитие растений ограничено высокой концентрацией токсичных солей, в первую очередь натрия и хлора. Такие условия создаются при неглубоком залегании грунтовых вод, что особенно характерно для приморских равнин и крупных солончаковых депрессий. К этим специфическим условиям приспособилась ограниченная группа видов, относящаяся к галофитам.
В структуре растительного покрова пустынных биомов хорошо прослеживается комплексность, т. е. сочетание фрагментов различных сообществ, зависящих от тех или иных эда-фических условий. Огромную роль играет перераспределение влаги по микрорельефу, в связи с чем, как правило, проективное покрытие растительности возрастает по руслам временных водотоков, суффозионным просадкам и т.д. Это наиболее ярко проявляется в краевых частях пустынных областей (подзона полупустынь), где закономерно сочетаются фрагменты фитоценозов степей и саванн (в наиболее увлажненных местообитаниях) с пустынными сообществами.
Общими для растений и животных адаптациями к жизни в пустынях являются разреженность и мозаичность распределения по территории, наличие длительного периода покоя в неблагоприятные для активной жизни сезоны года (анабиоз у беспозвоночных, спячка у рептилий и грызунов). Сходные приспособления проявляются в освоении подземных ярусов. В отличие от растений животные ввиду своей подвижности и независимости от прямой инсоляции как источника энергии могут проводить под землей всю свою жизнь или отдельные ее циклы (термиты, мокрицы, личинки многих насекомых и других беспозвоночных).
Животные пустыни, как и растения, по-разному адаптированы к недостатку воды. Пресмыкающиеся и некоторые насекомые «преадаптированы*, так как обладают непроницаемыми покровами и выделяют сухие экскреты (мочевую кислоту и гуанин). Пустынные насекомые защищены от испарения особыми водонепроницаемыми при высокой температуре веществами. Полностью устранить испарение с поверхности дыхательных путей невозможно, однако у пустынных насекомых оно сведено к минимуму благодаря особой системе внутренних дыхалец. Заметим, что образование метаболической воды (в результате расщепления углеводов), которое часто
бывает единственным ее источником, само по себе не является адаптацией. Адаптация состоит в способности сохранять эту воду или, как наблюдается у жуков-чернотелок (характерных обитателей пустыни), в способности к усиленному образованию метаболической воды при низкой влажности. Млекопитающих как группу нельзя считать хорошо приспособленными к условиям пустыни, так как они выделяют мочевину, что связано с потерей большого количества воды. Все же некоторые виды приобрели удивительные вторичные адаптации. К таким млекопитающим в Новом Свете принадлежат грызуны из сем. Heteromyidae, особенно кенгуровая крыса (Dipodomys) и карманчиковая мышь (Perognathus), а в пустынях Старого Света — тушканчики (Dipus сем. Dipodidae). Эти животные могут неопределенно долго существовать на сухих семенах и вовсе не нуждаться в воде для питья. Днем они остаются в норах и сохраняют воду, выделяя очень концентрированную мочу и не расходуя воду на терморегуляцию. Следовательно, адаптация этих животных к жизни в пустыне носит не только физиологический, но и поведенческий характер. Другие пустынные грызуны, например древесные крысы (Neotoma), не могут существовать исключительно на сухой пище, но выживают в некоторых частях пустыни, поедая сук-кулентные кактусы или другие запасающие воду растения.
Морфофизиологические адаптации животных к экономии влаги выражаются в особенностях структуры кожных покровов, окраски и др. Отдаленной аналогией с растениями-суккулентами можно считать запасание жира в некоторых органах тела у копытных, мозоленогих и грызунов (жировые депо в хвосте у тушканчиков, песчанок, сумчатых мышей, в горбе у верблюдов). Это резерв влаги в условиях ее дефицита (путем химического разложения жировых запасов). Специфической адаптацией животных к аридным условиям среды является их миграция внутри пустынных регионов и за их пределы. Так, дальние кочевки совершают копытные и большинство видов птиц, гнездящихся в пустынях.
Для многих животных пустынь и полупустынь характерен ночной или сумеречный образ жизни, при котором активность их приходится на наиболее благоприятные часы суток. Животные, активные в дневное время, чтобы избежать прямой инсоляции и перегрева тела, перемещаются в тень кустарников. Так поступают насекомые, ящерицы, мелкие пти-
цы. Некоторые дневные ящерицы и змеи забираются в середине дня на высокие ветви кустарников, где воздух менее нагрет, чем в приземном слое. В такие часы на ветвях саксаула или акации можно увидеть сидящими агаму, шипохвоста или стрелу-змею.
Своеобразная адаптация к дефициту влаги наблюдается у типичных пустынных птиц рябков. Прилетая на водопой, расположенный нередко за многие километры от мест гнездования, эти птицы смачивают водой перья брюшка и переносят на них значительное количество воды к гнезду. Вода удерживается на перьях благодаря их особой структуре. Этой водой птицы смачивают кладку или поят птенцов.
Передвижение по сыпучему субстрату в песчаных пустынях также требует особых приспособлений. У псаммофильных ящериц и грызунов на пальцах имеются особые щеточки из роговых чешуек или удлиненных жестких волос. Некоторые ящерицы способны вибрирующими движениями тела за несколько секунд зарыться в песок. У ряда змей (рогатый гремучник, песчаная эфа, песчаная гадюка) выработался особый способ передвижения по сыпучему субстрату— так называемый боковой ход, при котором задняя часть тела приподнимается над поверхностью земли и переносится вбок, а за ней подтягиваются передняя часть тела и голова. След от движения такой змеи представляет собой не зигзаг, а ряд косых параллельных вмятин, не связанных между собой.
Животные, обитающие в глинистых пустынях и роющие там норы, обладают крепкими лапами с острыми когтями или (некоторые грызуны) мощными острыми резцами на верхней челюсти.
На основе температурного режима удобно различать два типа пустынь: жаркие и холодные, хотя такая классификация несколько произвольна. Доминирующий вид в жаркой пустыне на юго-западе Северной Америки представлен креозотовым кустарником Larrea, а в расположенных севернее холодных пустынях Большого Бассейна—полынью (Artemisia ). В южных областях широко распространен также колючий шалфей (Franseria), тогда как на высокогорьях, где влаги немного больше, характерными компонентами являются гигантский кактус Sahuaro и пало верде. Восточнее к пустынным кустарникам примешивается большое количество злаков, и пустыня переходит в полупустыню. В холодных пу
стынях, особенно на щелочных почвах внутренних водосборных бассейнов, обширные площади заняты солянками сем. Chenopodiacea, в частности, видами родов Atriplex, Grayia, Eurotia и Sarcobatus.
Солянки не менее широко распространены в аридных районах и других частей света. В мексиканской пустыне наибольшего развития достигает жизненная форма суккулентов, включая кактусы, древовидные юкки и агавы. Некоторые их виды проникают в кустарниковые пустыни Аризоны и Калифорнии, но в холодных пустынях такая жизненная форма не имеет смысла. Однолетние травы и злаки создают замечательное зрелище во всех пустынях во время короткого влажного периода. Обширные участки «голого грунта» пустынь не обязательно лишены растений. Здесь могут существовать мхи, водоросли и лишайники; на песках и других типах тонкодис-персной почвы они образуют стабилизирующую корку. Сине-зеленые водоросли (часто в составе лишайников) имеют большое значение как фиксаторы азота.
Максимальные запасы биомассы формируются в короткие периоды вегетации после выпадения дождей. В некоторых пустынях такие условия могут наступать лишь раз в два-три года. Первичная продукция в пустынных биоценозах также невелика, но может составлять до трети суммарной биомассы — 0,8—1 т/га в год. При этом, чем выше величина биомассы, тем меньшую долю ее составляет первичная продукция.
Наибольшими запасами биомассы характеризуются аридные редколесья Австралии (до 25 т/га), а также бело- и черно-саксаульники Средней Азии (до 50 т/га). Годовая продукция этих сообществ составляет примерно 20% запасов биомассы (соответственно 5 и 8 т/га), зоомасса — от 10 до 120 кг/га. Наиболее бедны по запасам биомассы и по годовой продукции пустыни Намиб, Атакама, многие районы Сахары, аравийские и центральноазиатские пустыни (Такла-Макан, Гоби).
СУБТРОПИЧЕСКИЕ
ЖЕСТКОЛИСТНЫЕ ЛЕСА
И КУСТАРНИКИ
Важнейшее влияние в субтропических широтах на структуру и функционирование биоценозов оказывают условия увлажнения, существенные и тем более продолжительные понижения температуры — явление редкое и далеко не повсеместное. Для восточных секторов материков характерно относительно равномерное распределение осадков в течение года и отсутствие ярко выраженного периода летней засухи, здесь развиты лавровые леса. Для западных секторов свойствен так называемый средиземноморский климат с зимними дождями и жарким сухим летом, с ним связано распространение жестколистных лесов и кустарников, называемых формациями средиземноморского типа. Области распространения жестколистных лесов и кустарников имеются на всех континентах, но наиболее широко они представлены в Средиземноморье и Австралии.
Наиболее яркая особенность гидротермического режима заключается в несовпадении теплого и влажного периодов во времени. Зима влажная и прохладная, лето сухое при высоких температурах воздуха. Это благоприятствует господству вечнозеленых деревьев и кустарников, относимых к своеоб-
разной группе склерофитов, для которых характерны наличие корки или пробки на стволах, начало ветвления на незначительной высоте, широкие кроны. Типичнейший их признак — жесткость листьев, сохраняющихся в течение нескольких лет. Часто листья опушены снизу и имеют матовый серо-зеленый цвет, нередко покрыты блестящим восковым налетом и содержат эфирные масла.
Для многих деревьев и кустарников характерна глубоко расположенная корневая система. Так, корни каменного дуба проникают по трещинам в породах на глубину 10—20 м. Велико разнообразие кустарников жестколистных, часто колючих.
Основная ассимиляция наблюдается весной и осенью, в середине лета и в наиболее холодный период зимы у растений этих областей отмечается относительный покой. Время цветения у многих видов — весна или осень, часто распространяется и на зиму.
Леса, как правило, довольно светлые, хорошо развиты кустарниковый и травяной ярусы. В составе последнего значительное участие принимают луковичные и клубненосные растения (геофиты), время цветения которых приходится на весну и осень.
Коренными сообществами Средиземноморья были вечнозеленые жестколистные леса с доминированием видов дуба (каменный и пробковый). До настоящего времени в составе кустарниковых сообществ встречаются отдельные деревья каменного дуба, достигающие высоты 20 м.
Исчезновение лесов Средиземноморья — многовековой процесс, результат воздействия человека. Ему сопутствовали эрозия, разрушение и смыв почвенного покрова, что привело к широкому распространению бесплодных каменистых поверхностей. Там, где нет посевов или плантаций, развиты кустарниковые типы сообществ. Эти формации, известные под названием маквис, представляют собой первую стадию деградации лесов. В состав маквиса входят различные кустарники и некоторые деревья, устойчивые к воздействию вырубок и пожаров. Особенно характерны разные виды вереска (в том числе древовидный вереск), ладанников, земляничное дерево, в Восточном Средиземноморье — дикая маслина, рожковое дерево, мирт, фисташка. Кустарники часто переплетены вьющимися растениями, нередко колючими.
Маквис также подвергается вырубкам, часто уничтожается пожарами, особенно губительными в летний период. На месте сведенного маквиса развиваются формации типа гариги — сообщества низкорослых кустарников и ксерофильных травянистых растений. В них господствует кермесовый дуб, он хорошо возобновляется после пожаров, образуя своеобразные кусты высотой до 1,5 м. Сообщества этого типа поражают обилием представителей семейств губоцветных, бобовых, розоцветных и др., выделяющих эфирные масла. На самых каменистых и скудных почвах гарига состоит из низкорослых колючих растений.
В Северной Америке распространены это жестколистные леса из вечнозеленых дубов и кустарниковые сообщества, физиономически сходные с маквисом. В Южном полушарии жестколистные леса и кустарники встречаются на всех континентах. В средней части Чили первичные жестколистные леса сохранились лишь кое-где в ущельях и по северным склонам гор.
В Африке жестколистные сообщества занимают небольшую площадь в южной и юго-западной оконечностях материка. Здесь преобладают кустарниковые сообщества, физиономически и структурно сходные с маквисом. Из деревьев отметим серебряное (леукадендрон) из семейства протейных. Кустарники достигают высоты 1—4 м. Верхний ярус, как правило, образуют различные представители протейных, в нижних ярусах преобладают бобовые и многочисленные виды рода вереск.
В Австралии леса представлены несколькими видами рода эвкалипт. Древостой разреженный, сомкнутость крон редко достигает 0,5. Нижний древесный ярус образован казуариной и видами банксии из семейства протейных. Хорошо развит кустарниковый ярус из бобовых, миртовых, протейных. Своеобразны так называемые травяные деревья из семейства лилейных, относящиеся к огнестойким породам. Жестколистные кустарники Австралии, скраб, сформированы видами эвкалиптов, казуарины с прутьевидными ветвями, здесь много бобовых и миртовых.
Сообщества субтропических лесов и кустарников граничат с тропическими сезонно-сухими лесами, саваннами и пустынями, с одной стороны, и с лесами умеренных широт —
с другой, поэтому набор видов и групп животных во многом родствен фаунам прилежащих биомов. Характерно проникновение ксерофильных видов из соседних аридных областей. Жаркое сухое лето и прохладная влажная зима обусловливают одну или две депрессии активности у большинства животных. Пики активности приходятся обычно на весенний и осенний периоды, когда сочетание условий влажности и температуры оказывается наиболее благоприятным.
По запасам биомассы различные варианты субтропических сухих лесов, редколесий и кустарников могут отличаться в зависимости от условий увлажнения. Биомасса в наиболее распространенных субтропических кустарниковых сообществах (маквис Средиземноморья, чаппараль Северной Америки) составляет примерно 50 т/га, продукция — Ют /га, что соответствует приблизительно 20% от общего запаса биомассы.
ТРОПИЧЕСКИЕ ДОЖДЕВЫЕ ЛЕСА
Разнообразие жизни достигает, по-видимому, максимума в широколиственных вечнозеленых дождевых тропических лесах, которые расположены на малых высотах полосой вдоль экватора. Эти области характеризуются ровным годовым ходом температур: средние месячные показатели составляют 1—2 . Суточная температурная амплитуда больше и может достигать 9 С. Под пологом леса, особенно на поверхности почвы, суточные амплитуды резко уменьшаются. Гидротермический режим областей распространения влажных вечнозеленых тропических лесов в течение года является оптимальным для развития живых организмов.
Влажные тропические вечнозеленые или вечнозеленые дождевые леса сконцентрированы в трех обширных регионах мира: северной части Южной (включая обширный массив в бассейне Амазонки) и прилежащей к ней части Центральной Америки; Западной экваториальной Африке; Индо-Малайском регионе. Эти области различаются по видовому составу (так как находятся в разных биогеографических областях), но сходны по структуре и экологии лесов. Различия в темге-
ратуре между зимой и летом выражены не столь сильно, как между ночью и днем. Сезонная периодичность размножения и других жизненных функций растений и животных в значительной степени связана с колебаниями количества осадков или же регулируется внутренними ритмами.
Леса этого типа относятся к самым сложным растительным формациям на Земле. Одна из ярких особенностей — их поразительное богатство видами, огромное таксономическое разнообразие. Хотя и сейчас нет уверенности в достаточной степени флористической изученности этих лесов, число видов, зафиксированных в конкретных регионах, поражает. В среднем насчитывается от 40 до 170 видов деревьев на гектар. Значительно меньше трав — 10—15 видов. Учитывая разнообразие лиан и эпифитов, число видов на относительно однородном участке леса может составлять 200—300 и более.
Преобладающей группой жизненных форм являются фанерофиты, представленные вечнозелеными гигроморфными и мегатермными кронообразующими деревьями со стройными и прямыми гладкими стволами светло-зеленого или белого цвета, не защищенными коркой, ветвящимися только в самой верхней части. Для многих характерна поверхностная корневая система, при падении стволов принимающая вертикальное положение. Яркая особенность — разновременная смена листьев на различных побегах одного и того же дерева. Смена листьев у деревьев происходит по-разному: одни растения сбрасывают их постепенно в течение года, для других характерна смена периодов листообразования и покоя. Например, некоторые деревья сем. Winteraceae растут непрерывно, тогда как для других видов этого же семейства характерен периодический рост с образованием годичных колец. Листья часто лишены почечных чешуй, иногда бывают защищены прижатыми основаниями черешков или прилистниками.
Тропические деревья могут цвести и плодоносить непрерывно или периодически, несколько раз в году, многие виды ежегодно. Среди важных эколого-морфологических особенностей следует отметить явление каулифлории — развитие цветков и соцветий на стволах и крупных ветвях деревьев, особенно расположенных в нижних ярусах леса.
Дождевой лес характеризуется очень сильно выраженной ярусностью. Деревья обычно формируют три яруса:
1) редкие очень высокие деревья, которые возвышаются над общим уровнем полога;
2) полог, образующий сплошной вечнозеленый покров на высоте 25—35 м;
3) нижний ярус, который становится густым только там, где имеются просветы в пологе.
Корни у высоких деревьев неглубокие; утолщенная нижняя часть ствола выполняет функцию «контрфорса*. Число видов растений очень велико; часто на нескольких гектарах можно встретить больше видов деревьев, чем во флоре всей Европы.
Свыше 70% видов тропического дождевого леса относятся к фанерофитам. Кроме того, здесь представлены многолетние травы и группы растений, нередко называемые внеярус-ными или межъярусными: лианы, эпифиты, полуэпифиты. Каждая из этих групп жизненных форм отличается своеобразными экологическими адаптациями.
Среди лиан, лазящих растений, преимущественно развиты деревянистые, но встречаются и травянистые формы. Многие из них обладают довольно толстыми стволами (до 20 см в диаметре), которые, поднимаясь по стволам деревьев-опор, обвивают их как канатами. Как правило, листья у лиан развиты на уровне крон деревьев, т. е. достаточно высоко над поверхностью почвы. Лианы многообразны по способу подъема на деревья-опоры. Они могут карабкаться на них, цепляясь усиками, обвиваться вокруг опоры, опираться на ствол укороченными веточками. Среди крупных лиан имеются виды, соизмеримые по высоте с наиболее высокими деревьями. Они иногда растут столь быстро и развивают такую массу ветвей и листьев в верхней части стволов, что губят поддерживающие их деревья. Зачастую лианы настолько сплетаются с ветвями крон нескольких деревьев, что отмершее дерево не падает, длительное время поддерживаемое лианами. Густые кроны лиан резко уменьшают количество света, проникающего под полог леса. Они в изобилии разрастаются по опушкам, вдоль берегов рек, по осветленным участкам.
Столь же многообразны и эпифиты, использующие в качестве субстрата для поселения стволы, ветви и даже листья деревьев, но не поглощающие воду и элементы минерального питания из живых организмов. Все они сапрофиты, получают элементы питания из мертвого органического вещества, иногда с помощью микоризы. По формам роста различают эпифиты с цистернами, гнездовые и эпифиты-бра.
Эпифиты с цистернами накапливают воду в розетках листьев и используют ее с помощью проникающих в них придаточных корней. В розетках возникают своеобразные микроценозы второго порядка, с водорослями и многочисленными водными животными. В эту группу эпифитов входят представители семейства бромелиевых, обитающие в лесах Южной и Центральной Америки.
Для эпифитов-бра и гнездовых эпифитов характерна способность накапливать почву, богатую питательными веществами, между корнями, образующими «гнездо», как, например, у папоротника «птичье гнездо», или под прилегающими к стволу дерева листьями эпифита-бра.
Эпифиты играют большую роль в жизни влажного тропического леса: они накапливают до 130 кг/га гумуса и перехватывают до 6000 л/га дождевой воды, т. е. больше, чем абсорбируют листья деревьев.
Полу эпифиты произошли от лиан, лазающих при помощи корней, путем постепенной утери связи с почвой как необходимого условия существования. Полуэпифиты остаются живыми, если все корни, связывающие их с почвой, перерезаны.
Число семейств (как и видов) травянистых растений значительно меньше, чем деревьев. Среди них хорошо представлены мареновые, постоянно присутствует незначительное число злаков, обычны селагинеллы и папоротники. Лишь на прогалинах среди леса травяной покров приобретает сомкнутый характер, обычно он изрежен.
Огромное разнообразие деревьев и межъярусных растений определяет сложную структуру леса. Высота древесного полога колеблется в разных типах леса от 30 до 50 м, над общим пологом возвышаются кроны отдельных деревьев, достигающих высоты более 60 м, это так называемые эмердженты. Деревьев, образующих основной полог от его верхних до нижних границ, много и поэтому ярусы в вертикальной структуре выражены слабо.
Кустарниковый ярус практически отсутствует. На соответствующей ему высоте представлены деревянистые растения с главным стволом, так называемые «карликовые деревья», и высокие травы. Среди последних — виды травянистых фанерофитов, т. е. трав с многолетними стеблями. Травяной покров влажного тропического леса состоит из тенелюбивых, обитающих при значительной степени затененности,
и теневыносливых, нормально развивающихся на участках с изреженным древостоем и угнетенных под сомкнутым пологом леса. Еще более усложняют структуру леса многочисленные лианы и эпифиты, вегетативные органы которых размещаются на разной высоте.
Фауна влажных тропических лесов отличается таким же богатством и разнообразием, как и флора. Здесь формируются сложные по территориальной и трофической структуре, насыщенные полидоминантные сообщества животных. Как и у растений, среди животных на всех «этажах» влажного тропического леса трудно выделить господствующие виды или группы. Во все сезоны года условия среды позволяют животным размножаться, и хотя размножение отдельных видов может быть связано с каким-либо периодом года, в целом этот процесс происходит в сообществе постоянно.
Полидоминантная структура и круглогодичное размножение способствуют плавной динамике численности животных, без резких пиков и спадов. В структуре сообщества животных можно четко выделить почвенный, подстилочный и наземный ярусы, а выше располагается серия переходящих друг в друга древесных ярусов.
Обилие животных древесных ярусов обеспечивается высокой влажностью воздуха, массой зеленых кормов, наличием «висячего» яруса почвы под эпифитами и множеством «надземных водоемов» в их розетках, пазухах листьев, дуплах и всевозможных углублениях на стволах деревьев. Поэтому в древесные ярусы широко проникают разнообразные около-водные и почвенные животные — ракообразные, многоножки, нематоды, пиявки, земноводные.
Птицы дождевого леса, по-видимому, нуждаются в периоде «покоя», поскольку размножение у них часто происходит с периодичностью, которая не связана с сезонами.
По функциональной роли в биоценозах тропического леса можно выделить ряд ведущих трофических групп, некоторые из них заметно преобладают в том или ином ярусе. Так, при общем обилии и разнообразии сапрофагов — потребителей отмершей растительной массы — доминирование этих животных наиболее выражено в почвенно-подстилочном ярусе, где обилен листовой, веточный и стволовой опад, поступающий из всех вышерасположенных ярусов. Различные группы фитофагов — потребителей живой растительной массы — гос
подствуют главным образом в средних и верхних «этажах» леса. Более равномерно, по сравнению с фитофагами и сапрофагами, распределяются по ярусам тропического леса зоофаги — разнообразные хищники в широком понимании этого слова, т.е. любые организмы, использующие в пищу животных. Это и виды, обычно называемые хищниками, и насекомоядные птицы, и хищные беспозвоночные (например, жужелицы и сколопендры), а также всевозможные экто- и эндопаразиты — клещи, пиявки, кровососущие насекомые, плоские и круглые паразитические черви.
Ведущей группой сапрофагов в тропическом лесу являются термиты. Эти общественные насекомые строят свои гнезда как на почве, так и на ветвях деревьев. Размеры наземных гнезд термитов в лесу гораздо менее внушительны, чем хорошо известные постройки термитов в саваннах. Нередко наземные термитники имеют грибовидную форму — это приспособление к защите от ежедневных тропических ливней. Гнезда термитов, расположенные на деревьях, находятся в основании толстой ветви у ствола. Такое высокое размещение гнезд у термитов служит защитой от переувлажнения.
Однако при любом поселении гнезд основной кормовой ярус термитов — почва и подстилка. Эти слои буквально пронизаны их кормовыми ходами. Поедают они растительный опад, валежник, экскременты животных и древесину растущих деревьев. Переваривание клетчатки в кишечном тракте осуществляется с помощью одноклеточных жгутиковых, разлагающих ее на более простые углеводы — сахара, которые и усваиваются термитами. Сами жгутиковые, масса тела которых составляет до трети массы тела хозяина, могут существовать только в кишечнике термита. Термиты, лишенные жгутиковых, не могут справиться с перевариванием пищи и погибают. Таким образом, между термитами и жгутиковыми складываются отношения облигатного (обязательного) симбиоза. Во влажном тропическом лесу число термитников на 1 га может достигать 800—1000, а самих термитов насчитывается от 500 до 10 тыс. особей на 1 м2.
Переработкой растительного опада заняты также разнообразные личинки насекомых (двукрылых, жуков, тлей), взрослые формы (имаго) различных мелких жуков, сеноедов и тлей, личинки растительноядных многоножек — кивсяков и сами кивсяки. Обильны в подстилке и дождевые черви. В тропических лесах Южной Африки и в Австралии обитают гигант
ские почвенные дождевые черви, достигающие нескольких метров длины, которые всюду редки и включены в Международную Красную книгу охраняемых животных.
В древесном ярусе разнообразны фитофаги: жуки, гусеницы бабочек, палочники, грызущие ткани листьев, а также цикады, сосущие соки из листьев, муравьи-листорезы. Особенностью тропического леса являются муравейники, построенные из листьев деревьев, причем не только в естественном лесу, но и на плантациях цитрусовых, гевеи, кофе.
Пыльцой и нектаром цветков наряду с листьями питаются взрослые формы жуков: хрущей, долгоносиков, усачей. Многие из них одновременно являются опылителями растений, что особенно важно в условиях сомкнутого полога леса, где опыление ветром практически исключается.
Большую группу потребителей зеленой растительной массы, а также цветков и плодов деревьев образуют живущие на деревьях обезьяны. В африканском дождевом лесу это ярко окрашенные колобусы, или гверецы, различные мартышки. В южноамериканской гилее растительные корма потребляют крупные обезьяны-ревуны, в Южной Азии — лангуры, гиббоны и орангутаны.
В отличие от лесов умеренной зоны, где жизнь приурочена к надпочвенному уровню, в дождевом лесу значительно большая доля животных обитает в верхних ярусах растительности. Например, в Британской Гвиане на деревьях живет больше 50% млекопитающих. Кроме древесных млекопитающих здесь в изобилии встречаются хамелеоны, игуаны, гекконы, древесные змеи, лягушки и птицы. Важную экологическую роль играют муравьи и прямокрылые, а также дневные и ночные бабочки. В таких лесах широко распространен симбиоз между животными и эпифитами. По отношению к фауне также справедливо утверждение, что она, как и флора, необычайно богата видами. Например, на 15 км2 в Барро-Колорад о, хорошо изученном участке дождевого леса в зоне Панамского канала, насчитывается 20 000 видов насекомых, тогда как на территории всей Франции их всего несколько сотен. Разнообразные архаичные типы животных и растений выжили в многочисленных экологических нишах неизменяющей-ся среды. Многие ученые полагают, что для тропического леса характерна высокая скорость эволюционных изменений и видообразования, поэтому такие леса стали местом возникнове
ния многих видов, вошедших позднее в состав более северных сообществ. В настоящее время ученые всего мира озабочены необходимостью сохранения больших пространств тропических лесов в качестве «ресурса генов».
Основной источник пищи животных в тропических лесах — плоды и термиты. Обилие птиц в этих лесах объясняется тем, что многие из них растительноядные; это питающиеся плодами попугаи, птица-носорог, туканы, котинги, тро-гоны и райские птицы. Поскольку «чердаки» джунглей перенаселены, многие птицы вьют себе висячие гнезда, а насекомые строят висячие коконы, что спасает их от армии муравьев и других хищников. Хотя известно несколько видов ярких птиц и насекомых, обитающих в более открытых местах, основная масса животных дождевого леса неприметна, и многие из них ведут ночной образ жизни.
В горных районах тропиков расположен горный дождевой лес, который представляет собой разновидность низинного дождевого леса с некоторыми характерными особенностями. По мере продвижения в горы лес становится все более низкорослым, и эпифиты составляют все большую долю автотрофной биомассы. Еще одна разновидность дождевого леса встречается вдоль берегов затапливаемых речных долин; это так называемый галерейный лес, или прибрежный лес.
Экосистемы влажных тропических лесов в разных регионах, несмотря на резкие различия во флористическом и фаунистическом составе, весьма сходны по структуре и представляют собой самые богатые и насыщенные сообщества в пределах биосферы Земли. При значительном разнообразии сообществ тропического леса его биомасса представлена величинами одного порядка и составляет обычно в первичных лесах 350—700 т/га (горные влажные тропические леса Бразилии), во вторичных — 140—300 т/га. Из этой биомассы, наиболее значительной по сравнению с биомассой всех сообществ суши, большая часть приходится на надземные органы растений, преимущественно деревьев, наименьшая — на корневые системы. Основная часть корневых систем деревьев располагается в почве на глубине 10—30, редко свыше 50 см. Площадь листьев колеблется от 7 до 12 га на каждый гектар поверхности почвы. Величины ежегодной продукции сильно изменяются в разных типах леса. Чистая продукция может составлять 6—50 т/га, т. е. 1—10% от биомассы.
Несмотря на кажущееся обилие животных организмов, они составляют ничтожную часть общей биомассы — примерно 1000 кг/га, т. е. 0,1% ее запасов, причем, как и в лесах умеренного климата, примерно половину зоомассы составляют дождевые черви. Влажные тропические леса, хотя и обладают мощной и равновесной структурой, легко подвергаются разрушению под антропогенным воздействием. На месте вырубленного тропического леса возникают вторичные лесные сообщества, значительно отличающиеся от коренных видовым составом и уступающие им по биомассе, продуктивности и сложности структуры. На восстановление первичного леса в благоприятных условиях требуется несколько столетий.
Замечательным свойством дождевых лесов является эффективный прямой возврат в круговороты биогенных элементов, осуществляемый мутуалистическими микроорганизмами. Именно благодаря этому на скудных почвах леса растут так же пышно, как и на более плодородных. На местах, где дождевой лес уничтожен, часто развивается вторичный лес, в состав которого входят хвойные деревья, такие как Musanga (Африка), Cecropia (Америка) и Macoranga (Малайзия).
Вторичный лес выглядит пышным, но совершенно отличается от девственного леса как экологически, так и флористически. «Климакс» возвращается обычно очень медленно, особенно на песчаных и других бедных минеральным питанием почвах, так как все элементы питания первоначального леса были утрачены в результате изъятия биомассы и нарушения микробиологической сети их возврата в круговорот.
Мангровые заросли представляют интерес прежде всего как смешение сообществ, весьма разных, если рассматривать каждое из них по отдельности. Здесь объединяются обитатели твердого и мягкого грунтов морских побережий, стволов и крон деревьев дождевого леса. Кроме того, у животных мангровых лесов, относящихся к разным эволюционным линиям, выработались многочисленные приспособления к временной или длительной жизни на суше. Значение мангровых биомов определяется также их ролью в воспроизводстве рыбных запасов — очень многие рыбы и ракообразные, имеющие большую ценность для рыболовства в примыкающих участках
моря, размножаются в манграх. Мангровые деревья с давних пор ценились человеком за высокое качество древесины, поэтому к настоящему времени они на значительных площадях уничтожены или от них сохранились небольшие фрагменты. В некоторых тропических странах, например во Вьетнаме в дельте Меконга, существуют площади мангровых лесов культурного происхождения — результат лесовосстановительных работ.
ПРЕСНОВОДНЫЕ
ЭКОСИСТЕМЫ
Пресноводные местообитания удобно подразделять на три группы: 1) стоячие воды, или лен-тические (лат. lenis — спокойный) экосистемы—озера и пруды; 2) текучие воды, или логические (лат. lotus — омывающий) экосистемы — родники, ручьи и реки; 3) заболоченные участки с колеблющимся уровнем воды часто по сезонам, но возможно и от года к году — марши и болота.
Грунтовые воды, хотя и образуют очень большой пресноводный резервуар и представляют собой важный для людей ресурс, в общем не считаются экосистемой, так как они безжизненны или населены очень слабо (иногда бактериями). Грунтовые воды связаны с тремя главными поверхностными экосистемами и, таким образом, представляют собой среду на входе и среду на выходе для этих экосистем.
По сравнению с морскими и наземными местообитаниями, пресноводные занимают сравнительно малую часть поверхности Земли, но их значение для человека поистине огромно. Это объясняется рядом причин.
1. Пресные воды — самый удобный и самый дешевый источник соды для бытовых и промышленных нужд. (Мы мо-
жем и, по-видимому, будем получать большую часть пресной воды из моря, но стоимость такой воды чрезвычайно высока, если учесть расход энергии для опреснения и возрастающее при этом засоление среды.)
2. Пресные воды — узкое место планетарного гидрологического цикла.
3. Пресноводные экосистемы наряду с лиманами и эстуариями представляют собой самую удобную и дешевую систему третичной переработки отходов. Почти все без исключения крупнейшие города мира расположены на больших реках, озерах или лиманах, которые служат бесплатными коллекторами. Человек настолько злоупотреблял использованием этого природного ресурса, что теперь уже стала очевидной необходимость прилагать значительные усилия для немедленного уменьшения возникшего стресса. В противном случае вода станет основным лимитирующим фактором для человека как биологического вида.
Вода обладает рядом уникальных термодинамических свойств, способствующих уменьшению температурных колебаний; таким образом, диапазон различных изменений в воде меньше и происходят они медленнее, чем в воздухе. Самые важные из этих свойств следующие:
1) высокая удельная теплоемкость. Это значит, что для изменения температуры воды требуется относительно большое количество тепла. Для повышения температуры 1 мл воды (1г) на 1 °C (от 15 до 16 °C) необходимо затратить 1 кал тепла. Только аммиак и немногие другие вещества обладают удельной теплоемкостью больше единицы;
2) большая скрытая теплота плавления. Для превращения 1 г льда в воду необходимо затратить 80 кал (столько же тепла выделяется при замерзании воды без изменения температуры);
3) самая высокая из известных скрытая теплота парообразования. При испарении (а оно происходит более или менее постоянно с поверхности растений, воды и льда) 1 г воды поглощается 536 кал. Значительная часть поступающей солнечной энергии в экосистемах всего мира расходуется на испарение воды. Этот поток энергии смягчает климат и делает возможным существование жизни во всем ее фантастическом разнообразии;
4) наибольшей плотностью вода обладает при 4 °C; при температурах выше или ниже 4 °C вода расширяется и,
следовательно, становится легче. Это уникальное свойство предохраняет озера от промерзания до дна.
Лектические экосистемы (озера и пруды). Большинство пресных водоемов, которые заполнены сейчас стоячей водой, в геологическом смысле молоды. Жизнь прудов продолжается от нескольких недель или месяцев (маленькие временные пруды) до нескольких сотен лет (большие водоемы). Очень немногие озера, например озеро Байкал в России, являются древними, большинство же крупных озер образовалось не раньше ледникового периода. Можно ожидать, что пресноводные экосистемы изменяются во времени со скоростью, более или менее обратно пропорциональной их размеру и глубине. Географическая разобщенность пресноводных водоемов способствует видообразованию, однако отсутствие изоляции во времени препятствует этому процессу. Вообще говоря, видовое разнообразие в пресноводных сообществах невелико, и многие таксоны (виды, роды, семейства) широко распространены в пределах континентов и даже на смежных материках.
Характерной чертой озер и крупных прудов являются четкая зональность и стратификация. В типичном случае мы можем различить литоральную зону, где встречаются прибрежные укореняющиеся растения, лимническую зону, или зону открытой воды, где доминирует планктонf и глубоководную профундальную зону, где живут только гетеротрофы. В областях с умеренным климатом озера часто становятся стратифицированными летом и зимой, что вызывается нагреванием и охлаждением воды. Более теплые верхние слои в озере, или эпилимнион (греч. limnion — озеро), временно изолируются от охлажденных глубинных вод, или гиполимниона, зоной термоклина, которая служит преградой для обмена различными веществами. Вследствие этого запасы кислорода в гиполимнионе и биогенных элементов в эпилимнионе истощаются. Весной и осенью, когда температура всей водной массы выравнивается, происходит перемешивание. «Цветение* фитопланктона часто идет именно после этих перемешиваний, поскольку в фо-тической (освещенной) зоне становятся доступными придонные биогены. В странах с теплым климатом перемешивание совершается только один раз в году (зимой), в тропиках этот процесс происходит постепенно или нерегулярно.
Первичная продукция экосистем со стоячей водой зависит от химической природы бассейна и от природы поступающих
с притоками и с суши веществ; обычно она связана с глубиной обратной зависимостью. Следовательно, урожай рыбы с единицы водной поверхности выше в мелких, чем в глубоких озерах, однако в глубоких рыба бывает крупнее. По продуктивности озера часто подразделяются на олиготрофные (малокормные) и эвтрофные (кормные). Парадокс состоит в том, что с точки зрения хозяйственного и рекреационного использования биологически бедные озера предпочтительнее высокопродуктивных. В одних частях биосферы люди увеличивают плодородие озер для нужд собственного питания, тогда как в других они его сдерживают (удаляя биогены, отравляя растения и т. д.) для сохранения красоты окружающей природы.
При строительстве искусственных прудов и озер (водохранилищ) человек изменяет ландшафт в тех регионах, где нет природных водоемов. В США почти при кажДой ферме сейчас имеется пруд, и почти на каждой реке построено водохранилище. Многие такие искусственные водоемы приносят доход, но идея строительства водохранилищ может завести слишком далеко; затопление плодородных земель, которые дают намного больше продуктов питания, чем водохранилища, не всегда лучший способ длительного землепользования. В стоячих водах загрязнения окисляются в общем менее эффективно, чем в проточных. Если водосборный бассейн не покрыт хорошей растительностью, осадки, образующиеся в результате эрозии, могут заполнить весь водоем за время жизни одного поколения. Поэтому, прежде чем строить новые водохранилища, следует проводить очень тщательный анализ всех затрат и выгод.
Тепловой режим водохранилищ может сильно отличаться 6т режима природных озер, что зависит от конструкции плотины. Если водосброс, как на плотинах гидроэлектростанций, придонный, то вниз по течению экспортируется холодная вода, богатая биогенными элементами и бедная кислородом, тогда как теплая вода задерживается в водохранилище. Водохранилище в этом случае работает как ловушка тепла и экспортер биогенных элементов в противоположность природным озерам с поверхностным водосливом, которые, таким образом, функционируют как ловушка биогенов и экспортер тепла. Следовательно, характер водосброса сильно влияет на условия вниз по течению.
На протяжении конкретного речного потока выявляются обычно две зоны:
1. Перекаты — участки с достаточно быстрым течением. Дно свободно от ила и обломочных материалов, вследствие чего поверхность его твердая. Здесь обитают специализированные организмы, которые прочно прикрепляются или прилипают к субстрату (как личинки веснянок и ручейников), или хорошие пловцы, способные плыть против течения (форель, дартер и т. д.).
2. Плёсы — глубоководные участки с медленным течением, в результате чего ил и песок оседают на дно, образуя мягкий субстрат. Такие условия благоприятны для жизни роющих и плавающих животных, укореняющихся растений и на больших плёсах — развития планктона. Сообщества плёсов крупных рек напоминают прудовые.
В верховьях реки обычно вымывают породу, врезаясь в субстрат, в результате чего дно чаще бывает твердым. В нижнем течении реки взвесь, как правило, оседает, формируются заливные долины и дельта, которые часто бывают крайне плодородными.
По химическому составу воды реки мира подразделяются на два типа: 1) жестководные, или карбонатные, содержащие 100%о или более растворенных неорганических веществ, и 2) мягководные, или хлоридные, где растворенных веществ менее 25%о. Химизм воды карбонатных рек определяется преимущественно выветриванием скальных пород, а хлоридных рек — атмосферными осадками. Еще один тип рек, характерный для теплых низменностей, — гумусовые, или «черные», реки с высокой концентрацией растворенных органических веществ.
Несмотря на малочисленность родников и источников и небольшое пространство, занимаемое ими, изучение их чрезвычайно важно.
Заболоченные пресноводные участки (марши и болота). Заболоченными участками называются любые пространства, которые хотя бы часть года покрыты пресной водой. Почвы здесь насыщены водой постоянно или на протяжении части года. Основной фактор, определяющий продуктивность и видовой состав этих сообществ, — гидропериод, т. е. периодичность колебаний уровня воды. Поэтому заболоченные местообитания можно отнести к «экосистемам, импульсная стабильность которых обеспечивается колебаниями уровня воды» (как и морские литоральные, и эстуарные экосистемы).
Болота представляют собой очень открытые системы; их удобно классифицировать по степени связи либо с более глу
бокими водоемами, либо с наземными экосистемами, либо с теми и другими вместе.
1. Речные болота, расположенные в низких депрессиях и заливных долинах, связанных с реками. Одни из самых продуктивных природных экосистем — низинные лиственные леса в долинах больших рек и пресноводные литоральные марши вдоль низовьев больших равнинных рек у берега моря.
2. Озерные болота, которые соединены с озерами, прудами или каналами запруженных рек. Они периодически заполняются водой, когда ее уровень повышается в этих глубоководных водоемах.
3. Собственно болота — это верховые и низинные болота, марши, топи, сырые луга и временные пруды, встречающиеся в депрессиях, не связанных непосредственно с озерами и реками (хотя они и могут располагаться в речных старицах и занесенных ложах прудов или озер). Таких болот много в областях бывшего оледенения. Обычно они покрыты погруженными водными макрофитами, воздушными болотными растениями, кустарниками. Часто болота служат наиболее подходящими местообитаниями для размножения водоплавающих птиц и других водных или полуводных позвоночных.
Хотя болота занимают лишь около 2% поверхности Земли, по существующим оценкам здесь содержится от 10 до 14% углерода. Некоторые болотные почвы содержат до 20% углерода по массе, а торфяники — еще больше. Дренирование таких почв и их использование для нужд сельского хозяйства способствуют высвобождению в атмосферу больших количеств СО2, что усугубляет «СО2-проблему». Несмотря на малую площадь болот, аэробно-анаэробная стратификация в болотных осадках (включая солоноводные марши) очень важна, что объясняется ролью, которую играют эти осадки в глобальных круговоротах серы, азота, фосфора и углерода.
Отношение человека к болотам коренным образом изменилось в 1970-е гг., когда исследования экологов и экономистов выявили их скрытую раньше ценность. Болота теперь уже не рассматриваются как безусловно бросовые земли, которые нужно уничтожать или переделать. В деле сохранения болот, особенно прибрежных, уже достигнуты некоторые успехи, тем не менее в этой области предстоит еще огромная работа с привлечением законодательных и политических органов.
МОРСКИЕ
ЭКОСИСТЕМЫ
Наиболее интересны, с экологической точки зрения, следующие характеристики морской среды:
1. Море занимает 70% поверхности Земли.
2. Глубина моря огромна, и жизнь обнаруживается на всей этой глубине. В океане, по-видимому, не существует безжизненных зон, однако вблизи материков и островов воды заселены значительно гуще.
3. Море непрерывно и не подразделяется на изолированные области, подобно суше и пресным водам. Все океаны соединены друг с другом. Основными барьерами для свободного передвижения морских организмов служат температура, соленость и глубина.
4. В море происходит постоянная циркуляция. Разница в температуре воздуха на полюсах и у экватора порождает сильные ветры, дующие на протяжении всего года в одном направлении (пассаты). В результате совместного действия этих ветров и вращения Земли образуются определенные течения. Заслуживают внимания экваториальные течения, направленные к востоку и западу, и прибрежные течения, направлен-
ные к северу и югу. Хорошо известны Гольфстрим и Северо-Атлантическое течение, которые несут теплую воду и смягчают климат высоких широт в Европе. Калифорнийское течение, напротив, несет на юг массы холодной воды, отчего образуется характерный для побережья Калифорнии пояс туманов (что важно для секвойи и других гигантских хвойных). Основные течения действуют подобно гигантским вертушкам (или волчкам), которые вращаются по часовой стрелке в Северном полушарии и против часовой стрелки — в Южном.
Кроме поверхностных ветровых течений имеются глубинные течения, наличие которых обусловлено различиями в плотности воды, что в свою очередь обусловлено различиями в температуре и солености. Взаимодействие ветрового давления, силы Кориолиса, термохалинных течений и физической конфигурации бассейна очень сложно и является центральной проблемой широких исследований в области физической океанографии. Перемешивание в море настолько эффективно, что часто встречающийся в пресноводных водоемах дефицит кислорода, или «стагнация», сравнительно рёдок в океанских глубинах.
5. На море постоянно господствуют разного рода волны, происходят приливы и отливы, вызванные притяжением Луны и Солнца. Роль приливов особенно велика в прибрежной зоне, где жизнь часто особенно богата и разнообразна. Именно приливы обусловливают заметную периодичность в жизни сообществ, «заводя» лунные, а не солнечные биологические «часы» литоральных животных. Эти «часы» согласовывают активность животных с периодичностью приливов. Поскольку приливы повторяются примерно через 12,5 ч, в большинстве мест они происходят дважды в сутки, сдвигаясь в последующие сутки примерно на 50 мин. Каждые две недели, когда силы солнечного и лунного притяжения складываются, амплитуда приливных колебаний достигает наибольшей величины, при этом приливы бывают особенно высоки, а отливы — особенно низки. Это так называемые сизигийные приливы. В середине этого двухнедельного периода разница между высокой и низкой водой бывает наименьшей — это, так называемые квадратурные приливы, когда силы притяжения Солнца и Луны направлены в противоположные стороны. Амплитуда приливов варьирует от менее чем 30,5 см в открытом море до 3,5 м в некоторых узких или полузамкнутых за
ливах. На приливы влияют многие факторы, поэтому картина может различаться в разных частях планеты, Самое первое, что должен сделать морской эколог, работающий в прибрежной зоне, — это получить графики приливов и отливов для данной местности.
6. Одна из характеристик морской среды — ее соленость. Средняя соленость, или содержание солей, составляет 35 частей соли на 1000 частей воды по массе, или 3,5%. Обычно это записывается так: 35%о (частей на тысячу; вспомните, что соленость пресной воды меньше 0,5%о). Около 27%о приходится на долю хлористого натрия, а остальное главным образом на соли магния, кальция и калия. Поскольку соли диссоциированы на ионы, химический состав морской воды указывают обычно в ионах. Соотношение основных компонентов в воде остается фактически постоянным, поэтому общую соленость можно рассчитать по содержанию хлорида (это легче, чем определить общую соленость). Таким образом, 19%о содержания хлора соответствует 35%о солености.
7. Морская вода имеет щелочную реакцию, сильно забу-ферена, но часто бедна жизненно важными элементами питания. Силы электролитической диссоциации катионов больше этих сил анионов (примерно на 2,4 миллиэквивалента), что и определяет щелочную природу морской воды (в норме рН=8,2). Морская вода устойчива к изменению pH. Помимо перечисленных выше ионов морская вода содержит множество других элементов (теоретически все известные). К сожалению, в морской воде возрастает содержание токсичных химических веществ промышленного происхождения. Концентрация биогенных ионов, однако, часто бывает такой низкой, что лимитирует первичную продукцию. Все эти прочие ионы составляют менее 1% солености океана.
В общем континентальный шельф простирается на некотором расстоянии от берега, на границе шельфа дно круто обрывается вниз, образуя континентальный склон; затем несколько выравнивается — это континентальное подножие и опускается к более глубокому и ровному ложу — абиссальной равнине. Мелководная зона континентального шельфа называется неритической (прибрежной), а часть берега между отметками высокой и низкой воды — зоной прилива (или литоральной зоной). Область открытого океана за пределами континентального шельфа называется океанической, область
континентального склона и подножия — батиальной зоной (может быть геологически активной, пересеченной желобами и каньонами, подверженными действию подводной эрозии и обвалов). Область океанических глубин, или абиссальная, может находиться в разных местах на глубине от 2000 до 5000 м. Абиссаль перерезают желоба, достигающие в глубину 6000 м (эти очень глубоководные области иногда называют ха-дальной зоной). Абиссаль можно рассматривать как крупнейшую в мире экологическую единицу. Это, конечно, гетеротрофная система, поскольку первичный источник энергии расположен намного выше (за исключением районов геотермальных рифтов). Считается, что срединно-океанические хребты остались после того, как разошлись континенты, прежде плотно сомкнутые.
По количеству проникающего света море подразделяют на две горизонтальные зоны: верхнюю, или эвфотическую зону первичного продуцирования, и нижнюю, простирающуюся до больших глубин, афотическую зону, где света для фотосинтеза недостаточно. Эвфотическая зона достигает больших глубин (до 100—200 м) в прозрачных водах океанической области. В более мутных (и богатых) прибрежных водах глубина эффективного проникновения света редко превосходит 30 м.
Океанические области. Из-за низкой концентрации биогенов в афотической зоне большая часть открытого океана представляет собой «пустыню» по сравнению с прибрежными водами и лиманами. Арктические и антарктические моря продуктивнее морей средних широт, на это указывает большое количество рыбы и китов, обитающих в полярных областях.
До последнего времени считалось, что первичный источник энергии в морских пищевых цепях образуют диатомеи, динофлагелляты и мелкие ракообразные, составляющие макропланктон (называемый также сетным планктоном, потому что эти организмы хорошо ловятся планктонными сетями, сделанными из шелковой ткани или нейлона). Однако позднее было обнаружено, что основу пищевых сетей в море образует микропланктон, или нанопланктон (греч. папо — карликовый, крошечный, слишком мелкий, чтобы задержаться в планктонной сети), который может быть автотрофным и гетеротрофным. В его состав входят мельчайшие зеленые и бесцветные флагеллаты и многочисленные типы бактерий. Связь между автотрофами и гетеротрофами, как и в большинстве
других типов экосистем, обеспечивается детритом — взвешенным органическим веществом (ВОВ) и растворенным органическим веществом (РОВ). Однако в море не только ВОВ может превращаться в РОВ, но и РОВ под действием волн может переходить в ВОВ. Это происходит также при образовании фекальных пеллеток и посредством других не полностью выясненных механизмов. Образующиеся агрегаты и населяющие их бактерии обеспечивают, видимо, важный источник питания для многочисленных и разнообразных фильтраторов.
Удивительно высоко разнообразие глубоководной фауны. Глубоководные рыбы во многом необычны, некоторые из них светятся (рыбы-лампы), другие способны заглатывать добычу, превосходящую по размерам их самих (рыба-гадюка, хау-лиод). Пищи мало, и она редка в темных глубинах, но рыбы наилучшим образом адаптировались к использованию счастливых возможностей.
Область континентального шельфа. Основное биоразнообразие в море сконцентрировано около берега, где благоприятны условия питания. Ни в каком другом месте, даже в дождевых тропических лесах, нет такого разнообразия жизни. Значительную часть морского прибрежного зоопланктона составляет меропланктон (временный, или сезонный, планктон), представленный пелагическими личинками донных организмов (крабов, морских червей, моллюсков и т. д.); эта особенность резко отличает его от планктона пресных вод и открытого океана, где большую часть взвешенных организмов составляет голопланктон (т. е. организмы, которые на протяжении всего жизненного цикла остаются в планктоне). Было показано, что пелагические личинки обладают способностью определять тип субстрата, пригодный для жизни прикрепленных взрослых организмов. Когда приближается время метаморфоза, личинки не оседают где попало, а только при наличии конкретных химических свойств субстрата. Бентос состоит из двух вертикальных компонентов: 1) эпифауны — организмов, которые живут на поверхности, прикрепившись к ней или свободно передвигаясь по субстрату, и 2) инфауны, представители которой закапываются в субстрат или строят трубки и норы. Агрегации бентоса распространены всюду в виде так называемых «параллельных донных сообществ», в которых доминируют экологически эквивалентные виды, часто относящиеся к одному роду.
Все крупное промысловое рыболовство мира почти полностью сосредоточено на континентальном шельфе или вблизи него, особенно в районах поднятия глубинных холодных вод либо в районах апвеллинга. Основу уловов составляют сравнительно немного видов промысловых рыб, куда входят анчоусы, сельдь, треска, макрель, сайда, сардина, камбалообразные (камбалы, палтус), лососи и тунец.
Области апвеллинга. Большую роль в жизни океана играет процесс, названный апвеллингом. Он происходит там, где ветры постоянно отгоняют поверхностную воду от крутого берегового склона. В этих местах на поверхность поднимается холодная глубинная вода, богатая накопленными биогенными элементами. Наиболее продуктивные области океанов сосредоточены в районах апвеллинга, о чем свидетельствует развитый здесь рыболовный промысел. Расположены апвеллинги вдоль западных берегов. Кроме того, они поддерживают большие популяции морских птиц, откладывающих на берегах и островах бесчисленные тонны богатого нитратом и фосфатом гуано. Приведем некоторые характерные черты биома апвеллингов:
1. Высокая концентрация биогенных элементов и организмов; преобладают пелагические, а не демерсальные (донные) рыбы.
2. Колоссальные популяции рыб (и птиц) характеризуют не только высокую продуктивность, но и короткие пищевые цепи. Некоторые виды ракообразных и рыб, которые в океанических областях хищничают, в апвеллингах становятся растительноядными. В короткой пищевой цепи доминируют диатомеи и сельдевые рыбы.
3. Отложения осадков на морском дне содержат много органических веществ и фосфата.
4. В противоположность богатству моря прилегающие районы суши часто представляют собой пустыню, так как формирующие апвеллинг ветры дуют в основном с суши, унося прочь влагу. Однако частые туманы поддерживают развитие некоторой растительности.
Иногда происходит «цветение* ядовитых динофлагеллят (красные приливы). При этом наблюдается массовая гибель рыбы, отрицательно сказывающаяся на промысловом рыболовстве. Такая катастрофа происходила несколько раз в зоне перуанского апвеллинга, который обычно оценивается как
самый продуктивный район мирового рыболовства. Человек и природа не в состоянии постоянно поддерживать очень высокий уровень продукции, циклическое поведение может быть основным свойством эвтрофицированных систем (включая индустриализованное сельское хозяйство).
Лиманы. Лиман — это полузамкнутый прибрежный водоем, свободно соединяющийся с открытым морем; таким образом, лиман сильно подвержен воздействию прилива, а морская вода в нем смешана (и обычно заметно опреснена) с пресной водой материкового стока. Примерами лиманов служат устья рек, заливы, литоральные марши и водоемы между берегом и барьерными отмелями. Лиманы характеризуются некоторыми важными свойствами, которые к тому же служат основанием для их сохранения.
1. Лиманы обычно продуктивны вследствие поступления больших энергетических субсидий в виде приливов и обилия биогенных элементов. Вертикальное перемешивание и турбулентность образуют своего рода ловушку для этих элементов, так что лиманные растения получают больше фосфора и других биогенов, чем растения из прилежащих пресноводных или морских местообитаний.
2. На протяжении всего года гетеротрофы обеспечиваются пищей за счет активности автотрофов трех типов: макрофитов (болотные и морские травы, водоросли), донных водорослей, развивающихся на поверхности осадка и в его толще (в том числе диатомей, которые живут в трубочках из ила и передвигаются в них вверх и вниз в соответствии с режимом освещенности и приливов), и фитопланктона.
3. Лиманы служат местом откорма для молоди многих морских промысловых видов. Устрицы и промысловые крабы остаются в лиманах на протяжении всего жизненного цикла, а различные креветки и многие рыбы — объекты промыслового и спортивного рыболовства, лов которых ведется вдали от берега, проводят здесь ранние стадии своего развития, где они находят изобилие пищи и защиту от хищников, благодаря чему лучше выживают и быстрее растут.
Контроль над биологическим разнообразием требует его измерения, а измерение только тогда становится возможным, когда качественные признаки могут быть описаны количественно, в величинах, которые можно сравнивать. На уровне интуиции кажется, что разнообразие сообщества тропического леса больше разнообразия сообщества тайги. Видимая простота оценки разнообразия, однако, не позволяет удовлетвориться качественными сравнениями: более разнообразное и менее разнообразное сообщество. В экологии и математике разработано множество моделей и индексов для измерения разнообразия, которые требуют различной интерпретации.
Оценивание биологического разнообразия имеет важное прикладное значение, так как:
1) позволяет контролировать сохранение генетического потенциала;
2) дает представление о состоянии экосистем на определенной территории;
3) служит основой для разработки системы менеджмента отдельных видов.
Разнообразие принято оценивать либо путем подсчета видов, измерения их относительного обилия, либо мерой, объединяющей эти два компонента. Однако оценка разнообразия только простым подсчетом видов мало информативна, так как ни одно сообщество не состоит из видов равной численности. Из общего числа видов какого-либо трофического уровня или сообщества в целом обычно лишь немногие бывают доминирующими (т. е. имеют большую биомассу, продуктивность или другие показатели), подавляющая же часть относится к редким видам (т. е. имеет низкие показатели «значимости»). Таким образом, большинство видов в сообществе малочисленны, количество других умеренно и лишь немногие обильны. В России опубликовано немало методических пособий по оценке биоразнообразия (Мэгарран, 1992; Лебедева, 1997, 1998; Лебедева и др. 1999).
АЛЬФА-РАЗНООБРАЗИЕ: ВИДОВОЕ ОБИЛИЕ
При оценке альфа-разнообразия учитываются два фактора: видовое богатство и выравненность обилий видов. Видовое богатство — число видов, для сравнения отнесенное к определенной площади. Выравненность — равномерность распределения видов по их обилию в сообществе.
Видовое разнообразие в разных местах часто зависит от шкалы измерения разнообразия (Мэгарран, 1992). Например, на 1 м2 полуестественных европейских пастбищ может быть больше видов, чем в нижнем ярусе дождевого тропического леса в бассейне Амазонки. Разнообразие видов на 1 км2 и более будет выше в тропическом лесу. Видовое разнообразие возрастает при увеличении размеров изучаемой площади.
Распределение видового богатства на Земле меняется по долготе, высоте над уровнем моря, в градиенте увлажнения, солености, содержания калия в почве и др. Уиттекер (1972) пришел к выводу, что разнообразие увеличивается от холодного к теплому климату и от морского к континентальному, а также при продвижении от высоких широт к экватору. Максимум видового разнообразия наблюдается в большинстве слу-
чаев в мезофитных сообществах. В сообществах, подвергающихся стрессовым воздействиям, видовое разнообразие уменьшается; но, кроме того, оно может снижаться в результате обострения видовой конкуренции в климаксовых сообществах, существующих в стабильной физической среде.
Высокую выравненность принято считать эквивалентной высокому разнообразию. Например, в двух выборках может быть равное число видов и особей, большая выравненность одной из выборок делает ее разнообразие более высоким. Рассмотрим теоретический пример.
На рис. 43 показаны два гипотетических сообщества, у которых равное число видов и особей (4 вида: А, В, С, В и по 12 особей в каждом). Выравненность сообщества 2 выше, чем сообщества 1. Сообщество 2, характеризующееся максимальной выравненностью и отсутствием доминирования, принято считать более разнообразным, так как все виды имеют равную численность.
Выравненность максимальна, если все виды в сообществе имеют равное обилие, и минимальна, когда один вид имеет показатели, превышающие таковые у всех остальных видов. Выравненность — это единственный серьезный показатель структуры сообщества. Выравненность, как правило, высока и постоянна среди популяций птиц (это может быть объяснено их территориальным поведением), а различия этого показателя в разных сообществах и географических зонах определяются главным образом видовым богатством. Напротив, у растений и фитопланктона выравненность в среднем низка, и оба компонента подвержены значительным вариациям.
Рис. 43.
Гипотетические сообщества, характеризующиеся разной выравненностью
МЕТОДЫ
ПОСТРОЕНИЯ ГРАФИКОВ
ВИДОВОГО ОБИЛИЯ
Наилучший способ представить оба компонента разнообразия — построить график. Перед тем как строить любой график полезно вспомнить одно неоспоримое положение, как писал Дж. Тьюки (1981): «Цели могут быть разными, но вид графика должен соответствовать цели». Можно попытаться выявить на графике либо общие тенденции, либо подробности исследуемого явления. Графический анализ биоразнообразия преследует цель выявить закономерности распределения видов в сообществе через обилие и вы-равненность.
Рассмотрим типы графиков, применяемых в анализе биоразнообразия
1. График ранг/обилие — один из способов представления данных по обилию видов. Ось абсцисс — ранг вида (порядковый номер ранжированного по обилию вида). Виды располагаются в упорядоченном ряду данных по возрастанию обилий. Ось ординат — обилие вида (число особей). Этот график используют при анализе геометрических рядов.
Линия, соединяющая точки или проходящая близко от них, называется кривой доминирования—разнообразия, или
Рис. 44.
Динамика разнообразия сообществ птиц в рекреационных зонах г. Ростова-на-Дону: на левом берегу Дона зимой (1) и весной (2), зимой в парках (3) и в Ботаническом саду (4).
кривой значимости видов. Пример такого графика представлен на рис. 44, на котором показано разнообразие сообщества птиц зимой в рекреационных зонах г. Ростова-на-Дону. Зимнее сообщество птиц наименее разнообразно.
Рис. 45-
Зависимость между количеством особей каждого вида и количеством видов
g£
2. Частотное распределение — устанавливает зависимость между числом особей каждого вида и числом видов. Ось абсцисс — число особей. Ось ординат — число видов. Пример такого распределения представлен на рис. 45.
Не у всех видов в сообществе имеется равное число особей. Обычно большинство видов представлено единичными экземплярами, в то время, как всего лишь несколько видов — очень обильны. Частотные распределения по сравнению с ранговыми в более интегральной форме отражают видовую структуру. Их форма определяется соотношением частот редких видов и видов со средним обилием, тогда как массовые виды уходят в «хвосты» распределений. Поэтому эти распределения чаще применяются при анализе экологических выборок и представляют большой интерес при описании видовых структур фаунистических коллекций (Песенко, 1982).
3. Этот же график используется при логарифмически нормальном распределении, но ось абсцисс представлена в логарифмическом масштабе.
Рис. 4б.
Распределения накопленных обилий в зимних сообществах птиц в рекреационной зоне на левом берегу Дона (1) и в Ботаническом саду (2) г. Ростова-на-Дону (Савицкий и др., 1998)
4. Типичный график, применяемый в случае модели «разломанного стержня», когда по оси ординат откладывается относительное обилие в линейном масштабе, а по оси абсцисс — порядок видов (ранг) в логарифмическом масштабе от наиболее к наименее обильным (обратное ранговое распределение).
5. Можно использовать по оси ординат накопленное обилие, выраженное в процентах, в зависимости от логарифма порядкового номера вида (ранга вида).
На рис. 46 показано видовое распределение в зимних сообществах птиц в рекреационных зонах г. Ростова-на-Дону. Сообщество птиц в Ботаническом саду менее разнообразно, чем на левом берегу Дона. Графики не пересекаются. График, отражающий менее разнообразное сообщество, располагается вверху, над графиком более разнообразного.
В табл. 7 приведены типы графического представления данных по разнообразию для различных моделей распределения видовых обилий в сообществах (см. следующий раздел).
Таблица 7
Типы графиков в анализе видового разнообразия
Тип графика Ось абсцисс Ось ординат Модель распределения
Ранг/обилие Ранг вида Обилие вида Геометрический ряд
Тоже Логарифм ранга вида Относительное обилие, % Модель «разломанного стержня»
- н - То же Накопленное обилие, % Логарифмическое распределение
Частотное распределение Число особей Количество видов Модель «разломанного стержня»
То же Логарифм числа особей То же Лог-нормальное распределение
Различные типы графических построений подчеркивают те стороны объектов, которые стремятся выделить экологи. График , наилучшим образом соответствующий модели « разломанного стержня», показывает равномерное распределение обилий; в случае геометрического ряда график должен демонстрировать присутствие немногих доминантов и многих редких видов; Гауссова кривая при логарифмически нормальном распределении привлекает внимание к видам со средним обилием.
Ранговые распределения более непосредственно отражают видовую структуру и поэтому лучше интерпретируются в терминах обилий видов. Однако при общем подходе они менее удобны, так как их форма (и параметры) сильно зависит от соотношения немногих обильных видов, на которое оказывают влияние различные случайные факторы (Песенко, 1982).
Кривую доминирования-разнообразия можно использовать для оценки влияния нарушений на видовую структуру. Чем круче падает кривая, тем меньше общее разнообразие и сильнее доминирование одного или нескольких видов. В стрессовых ситуациях, независимо от того, вызваны ли они естественными причинами (погодными условиями), как это видно на предыдущих рисунках, где представлена динамика сообществ птиц в разных зонах г. Ростова-на-Дону, или антропогенным воздействием (загрязнения), кривая становится более крутой.
На рис. 47 показано разнообразие сообществ птиц из окрестностей Среднеуральского медеплавильного завода в трех
Рис. 47.
Кривые доминирования-разнообразия сообществ птиц из окрестностей Среднеуральского медеплавильного завода в трех зонах: 1 - контрольной (20 км от завода); 2 - буферной (4 км); 3 - импактной (1,5 км) (Воробейчик и др., 1994)
зонах: контрольной (20 км от завода), буферной (4 км) и им-пактной (1,5 км). Загрязнение среды отражается на структуре сообщества птиц. Сообщества птиц в контрольной и буферной зонах сходны по разнообразию. Импактная зона характеризуется менее разнообразным сообществом за счет «выпадения» редких видов, тогда как доминирующие виды сохраняют свое обилие.
МОДЕЛИ РАСПРЕДЕЛЕНИЯ ВИДОВОГО ОБИЛИЯ
Разнообразие обычно анализируется с учетом четырех основных теоретических моделей: 1 — геометрической; 2 — логарифмической; 3 — логарифмически нормальной (лог-нормальной); 4 — распределение, описываемое моделью «разломанного стержня» Мак-Артура. Если изобразить каждую из моделей в виде графиков с осями ранг/обилие, можно увидеть переход от геометрического ряда к модели «разломанного стержня». При геометрическом распределении доминируют немногие виды при очень низкой численности большинства, при логарифмическом и лог-нормальном — виды со средним обилием становятся все более и более обычными, в распределении, описываемом моделью «разломанного стержня», обилия видов распространены с максимально возможной в природе равномерностью. Каждой из моделей соответствует характерная форма кривой на графике с осями ранг/обилие (рис. 48).
Геометрическое распределение выражается прямой линией с крутым наклоном. Логарифмическое распределение также имеет крутой наклон, но это не прямая линия, а кривая. Модель «разломанного стержня» имеет более пологий гра-
Рис. 48.
Кривые доминирования-разнообразия разных моделей видового обилия (Мэ-гарран, 1992): 1 - геометрическое распределение; 2 - логарифмическое распределение; 3 - лог-иормальное распределение, 4 - модель «разломанного стержня»
фик. Логнормальное распределение описывается 8-образной кривой, которая располагается на графике между логарифмическим распределением и моделью «разломанного стержня».
Геометрическое распределение. Рассмотрим ситуацию, когда вид-доминант захватывает часть k некоего ограниченного ресурса, второй по обилию вид забирает такую же долю k остатка этого ресурса, третий по обилию — Л от остатка и т.д., пока ресурс не будет разделен между всеми S видами. Если это условие выполнено, и если обилия видов (выраженные, например, их биомассой или числом особей) пропорциональны используемой доле ресурса, распределение этих обилий будет
описываться геометрическим рядом (или гипотезой преимущественного захвата ниши).
Пример такого ряда: наиболее обильный вид в два раза многочисленнее следующего за ним по обилию, а этот последний в свою очередь вдвое больше третьего и т.д. На графике ранг/обилие такое сообщество будет представлено прямой линией. Можно предположить, что в этом случае доминирующий вид занимает половину доступного пространства ниш, второй — половину оставшейся площади (1/4 исходной) и т.д. Таким образом, каждый вид занимает прежде всего свободную нишу, не перекрывающуюся с другими.
Модель геометрического распределения была предложена Мотомурой. Модель имеет два параметра: я, — численность самого обильного вида и k — константу геометрической прогрессии. В геометрическом ряду обилия видов от наибольшего к наименьшему выражаются формулой, разработанной Мэйем и Мотомурой:
П,:=NCkk(l-k)~\ где п, — число особей i-ro вида, N — общее число особей, Ск = [1 - (1 — k)s ]” — константа, при которой ni = N .
Распределение обилий видов по типу геометрического обнаруживается преимущественно в бедных видами место-, обитаниях или в сообществах на очень ранних стадиях сукцессии. Такое распределение характерно для некоторых растительных сообществ в суровых условиях окружающей среды (например, сообщество растений субальпийского пояса).
Логарифмическое распределение. Модель логарифмического распределения известного английского математика Фишера была первой попыткой описать отношение между числом видов и количеством особей этих видов. Особенным успехом она пользовалась в энтомологических исследованиях и была впервые применена Фишером как теоретическая модель для описания распределения видов в коллекциях.
Распределение частот видов для этой модели описывается следующей последовательностью:
ах2 ах3 ах
ах,----,----...----,
2 3 п
где ах — число видов, представленных одной особью, ах2/2 — число видов, представленных двумя особями и т.д.
Логарифмическая модель имеет два параметра а и х. Это означает, что для выборки объемом N и числом видов S существует только одно возможное распределение частот видов по их относительному обилию, так как и а, и х являются функциями N и S. Чем больше выборка, извлеченная из данного сообщества, тем больше значение х и тем меньше доля особей, относящихся к видам, представленных одной особью в выборке. Два параметра S и N связаны между собой зависимостью S=aln(l-HV/a), где a — индекс разнообразия, который можно получить из уравнения:
X где N — сумма всех особей, принадлежащих S видам:
S=Sj +S2 +•••=-aln(l-x).
Моделью логарифмического распределения, характеризующейся малым числом обильных видов и большой долей «редких», с наибольшей вероятностью можно описать такие сообщества, структура которых определяется одним или немногими экологическими факторами. Как показали исследования, проведенные Мэгарран (1992) в Ирландии, такому ряду соответствует распределение обилий видов растений наземного яруса в хвойных культурах в условиях низкой освещенности.
Логарифмически нормальное распределение. Для многих сообществ характерно лог-нормальное распределение обилий видов, но обычно эта модель указывает на большое, зрелое и разнообразное сообщество. Такое распределение характерно для систем, когда величина некоей переменной определяется значительным числом факторов.
Распределение обычно записывается в форме:
SR
тое
г/га2
где SR — теоретическое число видов в октаве, расположенной в R октавах от модальной октавы; — число видов в модаль-
Рис. 49-Лог-нормальное распределение
ной октаве; G — стандартное отклонение теоретической логнормальной кривой, выраженное в числе октав.
Лог-нормальное распределение описывается симметричной «нормальной», т. е. колоколообразной кривой (рис. 49). Однако, если данные, которым она соответствует получены из ограниченной выборки, то левая часть кривой (т. е. редкие, неучтенные виды) будет выражена нечетко. Престон назвал такую точку усечения кривой слева «линией занавеса». «Линия занавеса» может сдвигаться влево при увеличении объема выборки. На рисунке она указана стрелкой. Для большинства выборок выражена только часть кривой справй от моды. Только при огромном количестве данных, собранных на обширных биогеографических территориях, прослеживается полная кривая. S-образная кривая указывает на сложный характер дифференциации и перекрывания ниш. Большинство видов в природных открытых экосистемах существует в условиях соревнования за ресурсы, а не на условиях прямой конкуренции; множество адаптаций дает возможность делить ниши без конкурентного исключения из местообитания. Эта модель наиболее вероятна для ненарушенных сообществ.
Распределение по модели «разломанного стержня» МакАртура. Эту модель иногда называют гипотезой случайной границы ниши. В 1975 г. Мак-Артур предложил три гипотетичес
ких распределения особей по видам в сообществе, основанных на различных типах взаимоотношений ниш разных видов:
1) ниши видов в сообществе не перекрываются, но тесно прилегают друг к другу;
2) ниши видов частично перекрываются;
3) ниши видов не перекрываются и разделены промежутками.
Наиболее подробно Мак-Артур исследовал свойства первого гипотетического сообщества. Он сравнил разделение пространства ниши в пределах сообщества со случайным и одновременным разламыванием стержня на £ кусков. £ видов разделяют среду случайно между собой так, что они занимают неперекрывающиеся ниши. При этом число особей каждого вида пропорционально размеру (ширине) ниши. Эта модель рассматривает только один ресурс. Она отражает более равномерное его разделение, чем лог-нормальная, логарифмическая и геометрическая модели. Модель «разломанного стержня» характеризуется только одним параметром £ (числом видов) и сильно зависит от объема выборки.
Число особей в i-ом по порядку обилия среди £ видов (NJ получают по формуле:
Ni = NIS^\ln, n=i
где N — общее число особей, а £ — общее число видов
Эту модель можно выразить также в величинах стандартного распределения обилий видов согласно выражению, описанному Мэйом:
_ 5(5-1) fl-л?’2 л W \ W ’
Модель Мак-Артура предполагает, что пространство ниш поделено на случайные, соприкасающиеся, но неперекрывающиеся участки. Такое распределение характерно для сообществ с интенсивной межвидовой конкуренцией, территориальным поведением, например, Для лесных птиц, характер распределения которых соответствует представлению о непе-рекрывающихся случайных нишах. Лучше всего использовать модель «разломанного стержня» для доказательства большей выравненности обилий видов в определенном сообществе.
tt
Другие теоретические модели. Описанные выше модели распределения видового обилия не могут охватить всего разнообразия реальных распределений, поэтому многими исследователями предпринимались попытки подобрать к эмпирическим сообществам другие теоретические модели.
А.П.Левич, В.Д.Федоров и др. гиперболической моделью апроксимировали ранговые распределения видов в планктонных пробах. А.П.Левич предложил также смешанную дзета-модель, представляющую собой обобщение геометрического распределения и гиперболической модели. Для описания ранговых распределений видов в геоботанических выборках Ламонтом была применена экспоненциальная модель. В. Д.Федо-ров предложил модель «экспоненциально разломанного стержня», которая основана на введении в модель Мак-Артура нового параметра — плотности вероятности обилий видов, которая в исходной модели предполагается равномерным. Согласно новой модели, на степень перекрывания ниш видов, а соответственно и на соотношение их обилий, влияет плотность организмов.
ИНДЕКСЫ
ВИДОВОГО
БОГАТСТВА
Важной мерой оценки разнообразия для ограниченного в пространстве и во времени сообщества, для которого точно известно число составляющих его видов и особей, является видовое богатство. Однако в болыпи-инстве случаев исследователь имеет дело с выборкой, не располагая полным списком видов сообщества. В этом случае необходимо использовать «нумерическое видовое богатство», т. е. число видов на строго оговоренное число особей или на определенную биомассу, и видовую плотность.
Видовая плотность (например, на 1 м2) — наиболее распространенный показатель видового богатства, особенно среди ботаников и почвенных зоологов. Показатель «нумерическое видовое богатство», используется реже, хотя более популярно его применение при исследовании водных объектов. Например, при исследовании экологических воздействий на сообщества рыб, можно использовать показатель число видов на 1 000 рыб.
Не всегда можно добиться равного размера всех выборок. Но следует помнить, что при увеличении объема выборки число видов постоянно растет.
Различные сочетания S (число выявленных видов) и N (общее число особей всех S видов) лежат в основе простых показателей видового разнообразия: индекса видового богатства Маргалефа
DMg -
S-1
InW ’
индекса видового богатства Менхиника
DMn~^N‘
Например, 5 октября 1997 г. в Театральном парке г.Рос-това-на-Дону в результате экскурсии была получена выборка, которая насчитывала 17 видов птиц, представленных 149 особями. Разнообразие будет составлять: по индексу Маргалефа — DMg=3,2, по индексу Менхиника DMn=l,4. Достоинство этих индексов — легкость расчетов. Большая величина индекса соответствует большему разнообразию.
Для оценки видового богатства используется индекс Q, учитывающий распределение видовых обилий, но не требующий соответствия какой-либо модели. Этот индекс представляет собой меру межквартильного наклона кривой накопленного видового обилия и обеспечивает измерение разнообразия сообщества, не отдавая предпочтения ни очень обильным, ни очень редким видам. Несколько ранее индекс, основанный на сходной идее, был предложен Уиттекером, однако он учитывал всю кривую видовых обилий и давал ошибки на обоих концах распределения.
Индекс Q рассчитывается по эмпирическим данным:
1 1
^nR{ + 2^nr+-nR2
Q _ _____7? 1+1 f
У log(R2/Rl)
где nr — общее число видов с обилием R\S — общее число видов в выборке; Rl, R2 — вяукшяА и верхний квартили; пт— число особей в классе, соответствующем R1; — число осо-
бей в классе, соответствующем R2 (рис. 50).
По оси абсцисс откладывается обилие видов в логарифмическом масштабе (log10), а по оси ординат — накопленное число видов. Индекс Q — наклон Q между двумя квартилями.
Рис. 50.
Графическая интерпретация индекса Q
Если выборки малы, индекс Q может смещаться. Однако эта ошибка невелика, если в выборку попадает более 50% всех видов. Некоторые ученые находят, что Q = а логарифмического распределения. Для лог-нормальной модели Q=0,371 S/c.
ИНДЕКСЫ, ОСНОВАННЫЕ
НА ОТНОСИТЕЛЬНОМ
ОБР1ЛИИ ВИДОВ
Эту группу индексов называют индексами неоднородности, так как они учитывают одновременно и выравненность и видовое богатство. Индексы, основанные на относительном обилии видов, относятся к непараметрическим поскольку они не требуют никаких предположений о распределениях. Их применение углубляет оценки биоразнообразия по сравнению с индексами видового богатства. которые опираются лишь на один параметр. Выделяются два типа непараметрических индексов:
1) полученные на основе теории информации (информационно-статистические);
2) доминирования.
Индекс Шеннона. Р. Мак-Артур и Маргалеф впервые применили для исследования видовой устойчивости и разнообразия сообщества теорию информации. Теория информации основывается на изучении вероятности наступления цепи событий. Результат выражается в единицах неопределенности, или информации. Шеннон в 1949 г. вывел функцию, которая стала называться индексом разнообразия Шеннона. Расчеты этого индекса предполагают, что особи попадают в выборку слу-
чайно из «неопределенно большой» (т. е. практически бесконечной) генеральной совокупности, причем в выборке представлены все виды генеральной совокупности. Неопределенность будет максимальной, когда все события (N) будут иметь одинаковую вероятность наступления (р;= nt/N). Она уменьшается по мере того, как частота некоторых событий возрастает по сравнению с другими, вплоть до достижения минимального значения (нуля), когда остается одно событие и есть уверенность в его наступлении.
Индекс Шеннона рассчитывается по формуле:
где величина р, —доля особей i-ro вида. В выборке истинное значение р( неизвестно, но оценивается как nt/N.
Причины ошибок в оценке разнообразия с использованием этого индекса заключаются в том, что невозможно включить в выборку все виды реального сообщества. При расчете индекса Шеннона часто используется двоичный логарифм, но приемлемо также применять и другие основания логарифма (десятичный, натуральный).
Индекс Шеннона обычно варьирует от 1,5 до 3,5, очень редко превышая 4,5.
Дисперсию индекса Шеннона (Var Н") рассчитывают по формуле:
VarH, = -(^Pjlnpt)2 + S-1
N 2N2 ‘
Если значения индекса Шеннона рассчитать для нескольких выборок, то полученное распределение величин подчиняется нормальному закону. Это свойство дает возможность использовать мощнуй параметрическую статистику, включая дисперсионный анализ. Применение сравнительных параметрического и дисперсионного анализов полезно при оценке разнообразия различных местообитаний, когда есть повторности.
Для проверки достоверности различий между выборочными совокупностями значений индекса Шеннона Хатченсов предложил использовать параметрический критерий Стьюдента:
t__ н[-н'2
(VarH[-VarH'2)1/2 ‘
Число степеней свободы определяется по уравнению: (VarH[+VarH2)2
df = (VarH{)2/N1+(VarH'2)2/N2
где и jV2 — общее число видов в двух выборках.
На основе индекса Шеннона можно вычислить показатель выравненности Е (отношение наблюдаемого разнообразия к максимальному):
Е = —, IilS где Е е [0,1], причем Е=1 при равном обилии всех видов.
Индекс Шеннона оказался самым популярным в оценке данных по разнообразию и применяется чаще других.
Индекс Бриллуэна. Не всегда исследователи способны гарантировать случайный отбор объектов в выборочную совокупность или учесть все виды сообщества. Это происходит обычно из-за несовершенных методов отлова животных. Нельзя обеспечить случайность попадания объектов в выборку при отлове насекомых на свет (привлекаются виды, активные только ночью, и выпадают формы с дневной активностью). Очень разнятся списки видов паукообразных, приносимых в гнезда большой синицей и собранных в биотопе традиционными методами, рекомендуемыми при сборе беспозвоночных. Подходящей формой информационно-статистического индекса в таких случаях может быть индекс Бриллуэна, определяемый по формуле:
In АП-У In л,!
НВ =--------
Индекс Бриллуэна дает сходную с индексом Шеннона величину разнообразия, редко превышая 4,5. Однако при оценке одних и тех же данных его величина ниже индекса Шеннона. Это объясняется тем, что в нем нет неопределенности, свойственной индексу Шеннона.
Выравненность определяется по формуле:
£ = ^
НВ^ ’
max
=— bl-------7—-7----------—
N {[W/S]!} ‘ {([N/S]+l)t} •
где [2V/S] — целая часть отношения N/S, a r=N-S[N/S].
Этим индексом мало пользуются, так как он трудно вычисляется и если выборка мала, то приводит к неверным выводам. Однако этот индекс рекомендуется применять, если оценивается коллекция, а не случайная выборка, и если известен полный состав сообщества.
Меры доминирования уделяют основное внимание обилию самых обычных видов, а не видовому богатству. Лучший среди индексов доминирования считается индекс Симпсона. Его иногда называют индекс Юла, поскольку он напоминает меру, разработанную Юлом для оценки словарного запаса.
Индекс Симпсона описывает вероятность принадлежности любых двух особей, случайно отобранных из неопределенно большого сообщества, к разным видам формулой:
D=^pf , где pt — доля особей i-ro вида.
Для расчета индекса используется формула, соответствующая конечному сообществу:
у \(и,-1р| У(У-1) I где п( — число особей i-ro вида, a N — общее число особей.
По мере увеличения D разнообразие уменьшается. Поэтому индекс Симеона часто используют в форме (1—D). Эта величина носит название «вероятность межвидовых встреч» и варьирует от 0 до 1. Он очень чувствителен к присутствию в выборке наиболее обильных видов, но слабо зависит от видового богатства. Высокая или низкая величина индекса определяется типом распределения видовых обилий для случаев, когда число видов превышает 10. Многие авторы считают, что наилучшая мера — это «индекс полидоминантности»:
Sx=l/D, N(N-l)
5л (» -1)
где i=l,2,3,... S ; Sx<= [1; oo[.
Мера разнообразия Макинтоша. В 1967 г. Макинтош предложил рассматривать сообщество как точку в S-мерном гиперпространстве с координатами (п2, п2 .... я,). Тогда евклидово расстояние такого сообщества от начала координат можно использовать как меру его разнообразия:
Индекс Макинтоша U сам по себе не является индексом доминирования, однако используя его, можно рассчитать меру разнообразия D, или доминирования, которая независима от объема выборки:
в дальнейшем можно рассчитать выравненность
N-U
N-N/4S’
Индекс Бергера-Паркера — одна из мер доминирования. Его достоинство — простота вычисления. Индекс Бергера-Паркера выражает относительную значимость наиболее обильного вида:
N d = !^, N
где Nm„ — число особей самого обильного вида.
Увеличение величины индекса Бергера-Паркера, как и индекса Симпсона, означает уменьшение разнообразия и возрастание степени доминирования одного вида. Поэтому обычно используется величина, обратная индексу Бергера-Паркера 1/d. Этот индекс независим от количества видов, но на него влияет объем выборки. Некоторые ученые считают, что он служит лучшей мерой разнообразия.
СРАВНИТЕЛЬНЫЙ АНАЛИЗ
ИНДЕКСОВ РАЗНООБРАЗИЯ
Во многих работах для одних и тех же данных исследователи вычисляли равные индексы разнообразия. Было показано, что значения многих индексов сильно коррелируют друг с другом. В качестве примера можно рассмотреть сезонную динамику показателей разнообразия сообщества птиц на прудах в окрестностях г. Ростова-на-Дону (рис. 51).
Слабая разработанность общей теории мер разнообразия и методологии их практического применения затрудняет обоснованный выбор показателя для измерения разнообразия исследуемых систем, в том числе экологических сообществ и локальных фаун. Поэтому исследователи изучают любой из них, следуя личной симпатии или традиции, установившейся в работе коллег.
В идеале выбор мер разнообразия должен быть основан на продуктивности заключений, к которым приводит их применение для решения исследовательских задач.
Трудности в применении показателей разнообразия и оценке их качества возникают благодаря сложному, комплексному характеру самой измеряемой величины разнообразия, от-
Рис.51-
Сезонная динамика индексов разнообразия сообщества птиц на прудах в окрестностях г. Ростова-на-Дону. Индексы разнообразия в логарифмическом масштабе: 1 - Маргалефа; 2 - Шеннона; 3 - Бергера-Паркера; 4 - Симпсона; 5 - мера Макинтоша
сутствию какой-либо объективной шкалы отсчета, независимой от принятой концепции, а, следовательно, и от меры разнообразия. Иными словами, системы не ранжируются в какую-либо единую последовательность в соответствии с присущим им разнообразием. Например, если бы мы.хотели расположить какие-либо объекты в порядке возрастания веса, мы сделали бы это независимо от того, каким методом определяли вес или какие единицы измерения использовали.
Разнообразие системы — это ее свойство и вместе с тем то, что исследователь понимает под разнообразием.
Ю.А. Песенко (1982) провел анализ функций разнообразия, не связанных с конкретными моделями распределения видов по обилию, используя критерии, которые могут быть сформулированы в основе концепции разнообразия. По Ю.А. Песенко, функция разнообразия должна быть:
1. Не определена при S = 0 и определена для всех S > О, где S — число видов в коллекции; разнообразие не может быть установлено ни для одной выборки, не содержащей ни одной особи, но выборка, содержащая хотя бы одну особь N> 1 S> 1, имеет измерение разнообразия.
2. Непрерывна при S > 0; это подразумевает отсутствие накопленных эффектов на оценку разнообразия при измерении
обилий видов, изменения долей которых в коллекции могут быть сколь угодно малы.
3. Неотрицательна в области своего определения при S > 0.
4. Функционально не связана с объемом выборки (не имеется в виду независимость от объема выборки).
5. Возрастающей при фиксированном значении S и убывании невыравненности обилий.
6. Возрастающей при фиксированной невыравненности и увеличении S.
Трудности в применении показателей разнообразия и оценке их качества:
1. Сложный, комплексный характер самой измеряемой величины разнообразия.
2. Отсутствие какой либо объективной шкалы отсчета, независимой от принятой концепции, а, следовательно, и от меры разнообразия.
Ю.А. Песенко проверил шесть различных показателей и пришел к выводу, что среди них только индекс полидоминантности удовлетворяет всем шести критериям. Мера Макинтоша и индекс Шеннона при добавлении новых видов в выборку слабо изменяются. У вероятности межвидовых встреч и индекса Шеннона число видов является более важным фактором при небольшом числе видов, а при увеличении числа видов возрастает роль выравненности.
РЕКОМЕНДАЦИИ ДЛЯ АНАЛИЗА
ДАННЫХ ПО РАЗНООБРАЗИЮ ВИДОВ
Формирование выборок. Анализируемые выборки должны быть репрезентативны, достаточно велики и одинаковы по объему, сформированы с соблюдением правил случайного отбора.
Графический анализ данных. Необходимо построение графиков рангового распределения обилий, которые позволят получить первое представление о модели распределения.
Проверка соответствия эмпирических данных теоретической модели. В тех исследованиях, где оценка разнообразия является главной задачей, часто бывает полезно формально оценить соответствие эмпирических распределений основным моделям видового обилия. Результаты подтвердить с помощью критериев согласия, используя графики рангового распределения обилия и сравнивая их с ожидаемым распределением. Этот прием представляет наибольший интерес, когда исследуемые сообщества подвергаются действию средового стресса.
Расчет индексов разнообразия. Видовое богатство и доминирование рассчитываются по индексам Маргалефа и Бергера-Паркера. Легкость вычисления и интерпретации —
их большое преимущество. Затем определяется параметр а логарифмического распределения. Это стандартная статистическая мера разнообразия. Вместо него можно использовать индекс Q. Для сравнения с результатами исследований других авторов бывает полезным определение индекса Шеннона.
Проверка статистических гипотез. Когда выборки представлены несколькими повторностями, для проверки значимости различий между сообществами необходимо использовать дисперсионный анализ. Если сравниваются результаты двух исследований, важно использовать один и тот же индекс разнообразия. По этой причине более информативным может оказаться использование индекса Шеннона, а не поиск новых показателей, более приемлемых с теоретической и прикладной точек зрения.
АНАЛИЗ БЕТА-РАЗНООБРАЗИЯ:
СРАВНЕНИЕ, СХОДСТВО, СООТВЕТСТВИЕ СООБЩЕСТВ
Бета-разнообразие характеризует степень различий или сходства ряда местообитаний либо выборок с точки зрения их видового состава, а иногда и обилия видов. Этот термин был введен Уиттекером в I960 г. Один из общих подходов к установлению бета-разнообразия — оценка изменений видового разнообразия вдоль средового градиента. Другой путь его определения — сравнение видового состава различных сообществ. Чем меньше общих видов в сообществах или в разных точках градиента, тем выше бета-разнообразие. Этот путь используется в любых исследованиях, рассматривающих степень различий видового состава выборок, местообитаний или сообществ. Вместе с мерами оценки внутреннего разнообразия местообитаний бета-разнообразие можно использовать, чтобы получить представление об общем разнообразии условий данной территории.
Показатели сходства, основанные на мерах разнообразия. Выделено шесть мер измерения бета-разнообразия на основе данных по присутствию или отсутствию видов.
Мера Уиттекера описывается формулой:
Рж=|-Ъ где S — общее число видов, зарегистрированных в системе, а — среднее разнообразие выборок стандартного размера, измеряемое как видовое богатство.
Мера Коуди разработана для исследования изменений в сообществе птиц вдоль средового градиента:
g(H) = l(H) 2 где g(H) — число видов, прибавившихся вдоль градиента местообитаний, a 1(H) — число видов, утраченное на том же трансекте.
Меры Ратледжа. Мера /^учитывает общее видовое богатство и степень совпадения видов:
д2 рд=—-----1>
я 2r+S где S — общее число видов во всех выборках, а /— число пар видов с перекрывающимся распределением.
Мера Р1 основана на теории информации и была упрощена для качественных данных и равного размера выборок:
Р, =logfT?-(l/T)Ieilog^)-(l/T)Saylogfap, где е( — число выборок вдоль трансекта, в которых представлен i-й вид, О] — видовое богатство j -ой выборки, а Т = Zet = -Sa,. Мера РЕ — экспоненциальная форма Д:
Ре= ехр (Д) — 1.
Мера Уилсона и Шмиды. Д включает те же элементы утраты (Z) и добавления (g) видов, что и мера Коуди, но стандартизована на среднее видовое богатство выборок а, входящее в меру Уиттекера:
Д = к(Н) + ЦЯ)]/2а.
Все шесть мер были оценены Мэгарран (1992) по четырем критериям с целью определить лучший показатель:
1. Число смен сообществ (выбраны два гипотетических градиента, один из которых однороден, т. е. вдоль всей его длины присутствуют одни и те же виды, а другой состоит из неперек-рывающихся сообществ).
ft
2. Аддитивность — способность индекса давать ту же самую величину бета-разнообразия независимо от того, высчитывается ли она по данным для двух концов градиента или по сумме значений бета-разнообразия, полученных внутри градиента. Например, при трех точках сбора (а, Ь, с):
Р (а, с) = Р (а, b)+ Р (Ь, с).
3. Независимость от степени разнообразия сообществ (бедное и богатое видами сообщество).
4. Независимость от чрезмерного размера выборки.
Большинству критериев удовлетворяет мера Уиттекера Рж.
Показатели соответствия. В числовой таксономии, классификации сообществ, при сравнении фаун и флор используется большое число показателей соответствия, имеющих разнообразную математическую природу. Часть индексов основана на определенных моделях распределения видового обилия. Для того, чтобы выбрать адекватную меру сходства нужно опираться на следующие положения:
1. Теоретические представления о характере устанавливаемых связей между объектами (общность по числу совпадающих признаков, положение в многомерном пространстве признаков, корреляция, степень перекрывания, вероятность принадлежности к одной генеральной совокупности и т.д.).
2. Особенности целей сравнения (выяснение отношений близости, производности или своеобразия).
3. Кластерный анализ (свертывание информации о взаимоотношениях объектов).
4. Особенности сравниваемых объектов, в том числе тип используемых признаков (для качественных — ранги, для количественных — численность, доля видов, балл обилия, частота встречаемости и ее трансформации).
Сходство может устанавливаться по абсолютной мере с фиксированными оценкой и выражением. Степень сходства между объектами в таких случаях определяется без ссылок на какие-либо другие. Большинство показателей измеряет абсолютное сходство. Относительное сходство, степень которого определяется в связи с частным диапазоном варьирования, или с конкретным размещением объектов, или с концептуальными представлениями об их расположении.
Пример. Так, черная морфа сизого голубя может быть очень несходной с популяцией сизых голубей дикой окраски и сходной в смешанной популяции городских птиц.
Основные индексы общности для видовых списков. Самый простой способ измерения бета-разнообразия двух участков — расчет коэффициентов сходства, или индексов общности. Списки видов могут быть представлены как конечные множества (или поля), элементами которых будут составляющие их виды. Основным приемом упорядочивания данных для определения индексов общности по качественным признакам служит таблица, включающая четыре поля (табл. 8).
Таблица 8
Определение индексов общности
а (число общих видов для двух списков) b (число видов, имеющихся только во втором списке) а + b (общее число видов во втором списке)
с (число видов, имеющихся только в первом списке) d (число видов, отсутствующих в обоих списках, но имеющихся в других, в которые входит всего S видов) с + d (число отсутствующих видов во втором списке)
а + с (общее число видов в первом списке) b+d (число отсутствующих видов в первом списке) a+b+c+d=S (всего видов)
Сумма (а + d) называется числом совпадений качественных признаков; (Ь + с) — числом несовпадений; а — числом положительных nd — числом отрицательных совпадений.
Все известные индексы общности распадаются на две группы в зависимости от того, учитывают они или игнорируют число отрицательных совпадений (d).
Наибольшее значение в экологических работах имеют индексы, в формулы которых входит только число положительных совпадений. В табл. 9 приведены основные индексы общности.
Предложено огромное число индексов общности, но чаще в биоценологических, фаунистических и биогеографических работах используются индексы Жаккара и Серенсена-Чека-новского. Эти коэффициенты равны 1 в случае полного совпадения видов сообществ и равны 0, если выборки совершенно различны и не включают общих видов.
rf______
Таблица 9
Основные индексы общности, учитывающие положительные совпадения (Песенко, 1982)
ФОРМУЛА АВТОР ОТНОШЕНИЕ
/о =—— ,Ь>с В а + Ь Браун-Бланке, 1932 а к числу видов в большем списке
I&s ~ ,Ь>с а + с Шимкевич,1926, Симпсон, 1943 а к числу видов в меньшем списке
I 2а а (a + b)+(a + c) Чекановский, 1900, Серенсен, 1948 а к среднему арифметическому числу видов в двух списках
г -а( 1 1 "1 *1 2^ + 6 ' а + с ) Кульчинский, 1927 а к среднему гармоническому числу видов в двух списках
I ~ ' а °В J(a + b\a + c} Охайя,1957, Баркман,1958 а к среднему геометрическому числу видов в двух списках
a + b + c Жаккар, 1901 а к числу видов в объединенном списке
j а 53 2(a + Z> + c)— а Сокал, Снит, 1963 а к сумме числа видов в объединенном списке и числа необщих видов
j b + c Кульчинский, 1927 а к числу необщих видов
Индексы общности, учитывающие негативные совпадения, используются обычно при сравнении коллекций, когда известны полные видовые списки. Применение этой группы индексов в экологических и биогеографических исследованиях подвергалось серьезной критике. Ограниченное использование индексов, учитывающих отрицательные совпадения, связано с их большой зависимостью от редких видов, которые могут не попадать в выборки.
Объективные причины отсутствия вида были проанализированы Ю.А. Песенко (1982). Отсутствие вида в сборах может
быть результатом неподходящих условий для его существования в местах сборов, т. е. вид не может здесь жить, его ниши нет в данной местности. Вследствие некоторых исторических (географических) причин эволюция вида проходила в отдаленных от этих мест регионах, т. е. он не мог сюда попасть, хотя в данной местности и имелись подходящие для него условия. Вид может существовать в данной местности, но не попал в выборку из-за неадекватности методов сбора или из-за редкости видов. Отсутствие вида как результат первых двух причин несет ценную информацию о фауне и местообитании, но только при исключении третьей причины, что сделать, как правило, невозможно.
Наиболее распространенными индексами, учитывающими отрицательные совпадения, являются коэффициент простого совпадения, или индекс Сокала-Майченера:
_ a + d
М a+b+c+d
и индекс общности Барони-Урбани и Бюссера:
Jad + а
1вв\ = ~г= 7 ’ 0 1Вв\ 1 -
уаа +а + Ь + с
Проблема оценки достоверности этих индексов не решена. Простота вычисления, являющаяся достоинством многих индексов, оборачивается недостатком — они не включают обилие видов. Это обстоятельство привело к тому, что чаще используются модифицированные индексы, учитывающие оценку обилий.
Индекс общности для количественных данных. По мнению многих авторов (Песенко, 1982; Мэгарран, 1992), наиболее приемлемо использование в экологических исследованиях коэффициента Серенсена:
С -- 2JN N aN + bN где aN— общее число особей на участке A, bN — общее число особей на участке В, jN — сумма наименьших из двух обилий видов, встреченных на обоих участках. Так, если 12 особей вида были найдены на участке А и 29 особей того же вида на участке В, подсчитывая jN, следует взять величину 12.
ГРАФИЧЕСКИЙ АНАЛИЗ
БЕТА-РАЗНООБРАЗИЯ
Группирование и классификация выборок является следующим этапом в анализе бета-разнообразия. Эти процедуры выполняются на основе преобразования матриц, каждый элемент которой — показатель сходства между двумя выборками.
Неориентированные и ориентированные графы. Для эффективного выделения скоплений объектов существуют методы сетевого анализа. Сетевой анализ матрицы оценок сходства между объектами заключается в построении некоторых типов графов, т. е. диаграмм, где объекты изображены в виде точек (кружков) — вершин графа, которые соединяются или не соединяются линиями, называемыми ребрами графа. Степень соответствия между объектами отражается в графах или характером взаимного расположения точек, или длиной и другими особенностями линии, соединяющих точки.
При анализе матриц сходства обычно используются «неориентированные графы», в которых линии, соединяющие вершины графа, не имеют направления. Реже применяются «ориентированные графы», в которых вершины соединены стрелками. Дендрит — один из типов неориентированного
графа. Он может быть как: минимальное древо (матрица включает оценки различий между объектами) или максимальное древо (используются меры сходства). Дендрит — это ломаная линия, которая может ветвиться, но не содержит циклов. Построение дендрита заключается в нахождении для каждого объекта наиболее сходного с ним объекта и соединении их линией. В результате получается ряд отрезков, в том числе и разветвленных.
Построить дендрит (рис. 52) можно с помощью матрицы сходства выборочных совокупностей, например (А—Е), вычисленной на основе индекса сходства Жаккара (табл. 10). Для этого последовательно соединяем две наиболее сходные выборки Г и Д (0,9) отрезком. Следующая по силе величина сходства 0,85 обнаружена между выборками Г и В. Поэтому выборку В можно присоединить отрезком к уже построенной ветке Д — Г и т. д.
Рис. 52.
Последовательные этапы построения дендрита на основе матрицы сходства выборок
Таблица 10
Матрица сходства выборочных совокупностей
А Б В Г Д Е
А 0,80 0,20 0,40 0,50 0,60
Б 0,80 — 0,30 0,55 0,45 0,65
В 0,20 0,30 — 0,85 0,15 0,10
Г 0,40 0,55 0,85 — 0,90 0,75
д 0,50 0,45 0,15 0,90 — 0,25
Е 0,60 0,65 0,10 0,75 0,25 —
Основной недостаток этого графика — потеря информации, заключенной в матрице оценок сходства, в результате использования только немногих (максимальных для каждого объекта) значений показателя соответствия.
Плеяды Терентьева. Одним из видов графического анализа сходства выборок может быть построение плеяд Терентьева. Этот тип графика в отличие от дендрита учитывает всю матрицу сходства. Плеяды Терентьева (рис. 53) также можно построить с помощью матрицы фаунистического сходства, вычисленной на основе индекса сходства Жаккара (см. табл. 10).
Такой тип графика является неориентированным графом. На нем все объекты могут быть соединены линиями, отражающими связи и меру сходства объектов. Толщина или характер линий соответствуют определенному интервалу значений
Один из типов неориентированного графа - плеяд Терентьева, построенный на основе матрицы сходства выборок, где величины индекса сходства: 1 — [0,7:;0,9];
2 - [0,4; 0,7]; 3 - [0,2: 0,4]
индекса сходства. Другой графический вариант плеяд Терентьева показывает взаимосвязи между выборками на разных уровнях сходства: 0,8, 0,5 и 0,2 (рис. 54).
На уровне сходства 0,8 есть взаимосвязь между объектами Г и В, Г и Д, а также А и Б. На уровне сходства 0,5 прибавляются взаимосвязи между объектами Д и А, Г и Е, Г и Б, Аи Е, Б и Е и т. д.
Дендрограмма ( кластерный, анализ ). Если сравнивать несколько участков, хорошее представление о бета-разнообразии может дать кластерный анализ.
Кластерный анализ — один из методов многомерного анализа, сущность которого состоит в иерархической классификации объектов в разделении множества объектов на однородные группы. Графически иерархическая классификация отображается в виде дендрограммы (дерева).
Рис. 54.
Один из типов плеяд Терентьева. Взаимосвязи между объектами показаны на уровнях сходства 0,8; 0,5 и 0,2
rf.
Внутри каждой группы, получаемой в результате разбиения объектов на кластеры (группы), объекты более сходны, чем с объектами из других групп. Кластерный анализ начинается с составления матрицы сходства для каждой пары сравниваемых объектов. Затем проводится последовательное объединение объектов в группы по степени их сходства, пока все они не будут включены в одну группу. Поскольку интерпретация результатов кластерного анализа зависит от визуальной оценки дендрограммы, лучше всего использовать этот прием для небольшого количества данных.
В качестве примера рассмотрим кластеризацию выборок на основе матрицы индексов сходства (см. табл. 10). Простейшие методы кластерного анализа, применяемого в биоценологии, биогеографии и числовой таксономии разными авторами, описаны Ю.А. Песенко (1982). Эти методы могут быть с успехом использованы и в экологических исследованиях при анализе бета-разнообразия.
В наиболее простых случаях процесс группировки начинается с нахождения в матрице индексов сходства пары наиболее похожих объектов. Самыми близкими объектами в примере, рассмотренном в табл. 10, являются Г и Д (0,90). Эти объекты отображаются на графике двумя соседними точками (рис. 55).
Отходящие от точек параллельные линии соединяются отрезком на уровне величины сходства и объединяются в один
Рис. 55-Дендрограмма кластерного анализа шести объектов, построенная методом одиночного присоединения (ближайшего соседа)
кластер. Затем в матрице индексов сходства находится второй по величине индекс сходства. Если он связывает-два других, еще не объединенных в группу объекта, то их соединяют так же как и первые два, но отдельно от них на соответствующем уровне сходства. В нашем примере вторая по силе связь имеется между объектами Г и В (0,85), при этом один из этих объектов уже объединен в кластер. В этом случае присоединение связанного с ним нового объекта может быть произведено тремя разными способами: одиночным, полным и средним присоединением.
Одиночное присоединение называют также «методом ближнего соседа». Метод впервые использован Серенсеном в 1948 г. Соединение групп производится по максимальному значению сходства между объектами из каждой группы. Следуя этому методу, объект В присоединяется к объектам Д и Г, уже объединенным в кластер. Следующее по величине сходство — между объектами А и Б (0,80). Они группируются в отдельный кластер на уровне сходства. Затем идет присоединение объекта Е к группе из объединенных ранее объектов Д Г В, так как между объектами Е и Г сходство равно 0,75. На последнем этапе соединяются два сформированных ранее кластера ДГВЕиАБв один на уровне 0,60. Результаты объединения показаны на рис. 55.
Полное присоединение называют также «методом дальнего соседа». Метод был впервые предложен Свитом в 1957 г. Согласно этому правилу, после формирования кластеров ДГ
Рис. 56.
Дендрограмма кластерного анализа шести объектов, построенная методом полного присоединения (дальнего соседа)
?L_
и АБ к группе АБ присоединяется объект Е, так как минимальное сходство этого объекта с объектами этого кластера равно 0,60. Объект В присоединяется к группе ДГ только на уровне 0,15 (минимальное сходство с каждым из объектов группы). Этот объект нельзя присоединить к кластеру ЕАБ, так как минимальное сходство объекта Е с объектами этой группы всего 0,10. Результаты объединения показаны на рис. 56.
Среднее присоединение. Этот метод включает несколько вариантов. Он был предложен в 1958 г. Сокалом, Майченером и Олсоном, Миллером одновременно. Один из самых простых вариантов этого вида кластерного анализа заключается в использовании средних арифметических невзвешенных значений сходства присоединенного объекта со всеми объектами группы. Соединяются кластеры, среднее сходство между объектами которых выше. Согласно этому методу, после формирования кластеров ДГ и АБ к группе АБ присоединяется объект Е, так как среднее сходство этого объекта с объектами этого кластера (0,6 + 0,65)/2 = 0,625. Этот объект нельзя присоединить к кластеру В (сходство всего 0,10) или кластеру ДГ (среднее сходство равно 0,50). Затем к группе ДГ присоединяется объект В, так как среднее сходство его с объектами данного кластера равно 0,5, а с группой ЕАБ — лишь 0,20. Последний шаг — объединение групп ДГВ и ЕАБ в один кластер на уровне 0,49, так как эта величина показывает среднюю
Рис. 57.
Дендрограмма кластерного анализа шести объектов, построенная методом среднего присоединения
связь между объектами в двух группах. Результаты объединения показаны на рис. 57.
Более сложные и разнообразные методы кластерного анализа реализуются с помощью вычислительной техники. Разработано множество статистических пакетов программ, таких как Statgraphics, Statistics, STADIA и др., которые выполняют кластерный анализ.
ПРИМЕНЕНИЕ
ПОКАЗАТЕЛЕЙ РАЗНООБРАЗИЯ
Главные потенциальные области применения индексов разнообразия — охрана природы и мониторинг. В основе использования оценок разнообразия в этих областях лежат два положения: 1) богатые видами сообщества устойчивее бедных видами; 2) уровень загрязнения связан со снижением разнообразия и изменением характера видовых обилий. При этом в охране природы обычно используются показатели видового богатства, а в экологическом мониторинге — индексы и модели видовых обилий.
В экологических исследованиях показатели разнообразия применяются в самых различных целях. Они с успехом были использованы в работах Мак-Артура и его последователей при изучении конкуренции у птиц, насыщенности и степени перекрывания их экологических ниш. Была выяснена зависимость разнообразия птиц от разнообразия некоторых элементов местообитания и других экологических факторов.
Джейкобс в 1975 г. обобщил результаты многих исследований влияния экологических факторов на разнообразие сообществ и установил следующее.
1. Пространственная гетерогенность увеличивает разнообразие.
2. Температурная гетерогенность может уменьшать и увеличивать разнообразие в зависимости от суровости климата и других факторов.
3. Стрессовые условия среды обычно отрицательно связаны с разнообразием.
4. При повышении конкуренции в относительно небольшой период времени разнообразие может уменьшаться, но при ее наличии в течение периода, достаточного для протекания эволюционных преобразований (видообразование), разнообразие может увеличиваться.
5. Враги действуют как конкуренция, их эффект на разнообразие зависит от интенсивности их воздействия, длительности и от влияния врагов на конкуренцию среди жертв.
6. Влияние интенсивности потока энергии через сообщество и объем ресурсов питания могут быть очень важными, но степень и направление их влияния на разнообразие зависят от многих других факторов.
В период сукцессии могут протекать процессы разной направленности при изменении разнообразия.
Показатели разнообразия применяются при сравнении населения разных стаций, сезонной динамики сообществ, для экологической оценки различных видов, характера их распределения по разным местообитаниям, измерения степени пищевой специализации видов и разнокачественности пищевого рациона вида. Показатели разнообразия также успешно применяются при оценке загрязнения водоемов и территорий, в частности, при сравнений участков в градиенте загрязнения наземных экосистем.
ГА М М А- РАЗ НООБРА311Е
Г1 АЗЕМНЫХ ЭКОСИСТЕМ
Уровень гамма-разнообразия относится к более крупным пространственным единицам типа острова или ландшафта по сравнению с бета-разнообразием. Затем, если гамма-разнообразие определяется как общее разнообразие группы участков, то эпсилон-разнообразие, или ]>е-гиональное разнообразие, — общее разнообразие группы территорий гамма-разнообразия, которое относится к крупным биогеографическим областям. Наибольший интерес для изучения фитоценохор представляет именно гамма-разнообразие, относящееся к микро-, мезо- и макрокомбинациям растительного покрова, соответствующим урочищам, местностям и ландшафтам в масштабах конкретных геоботанических карт (Виноградов, 2000).
Существующие в литературе характеристики гамма-ра.з-нообразия мало способствуют его пониманию. В основополагающем документе (Global Biodiversity Strategy. 1992) отмечено только, что «экосистемное разнообразие (т. е. гамма-разнообразие) труднее измерить, чем видовое или генетическое, поскольку границы сообществ и экосистем менее дискретны». Тем не менее, как только постоянный набор
критериев используется, чтобы отличить сообщества и экосистемы, их число и распространение могут быть измерены. Б.А. Юрцев (1992) более определенно различает биохорологическое разнообразие как разнообразие сочетаний организмов тех или иных территориальных выделов, частей биосферы и выделяет биохорологические уровни в пределах экотопа, что соответствует бета-разнообразию, ландшафта — гамма-разнообразию, биогеографического района — эпсилон-разнообразию.
По определению Б.В. Виноградова, гамма-разнообразие характеризует информационную диверсификацию фитоцено-хор на надбиоценотических ландшафтных уровнях. Ввиду пространственной неоднородности и многокомпонентности сложных экосистем, наиболее показательной их характеристикой является пространственно-временное распределение биологических, геофизических, структурных и функциональных свойств, привязанных к содержательным определителям экосистем. А.А. Тишков (1998) также отмечает масштабную иерархию уровней биоразнообразия, которая может выглядеть следующим образом (рис. 58).
эпсилон-разнообразие Ей
масштаб мельче 1:1 000 000
гамма-разнообразие Г»
масштаб 1:100 000-1:1 000 000
бета-разнообразие В»
масштаб 1:10 000-1:100 000
альфа-разнообразие Ав
масштаб крупнее 1:10 000
“1 а2 Чз ап
Рис. 58.
Иерархическая (биохорологичекая) структура биоразнообразия наземных экосистем
rf_
Гамма-разнообразие может быть рассчитано так:
у = ах|3хп,
где п — общее число местообитаний или ключевых участков, где аир — среднее значение на ландшафтную единицу. Очевидно, в этом уравнении у-разнообразие может увеличиваться, когда а- и ^-разнообразия могут остаться постоянными.
Таким образом, Б.Н.Виноградов рассматривает объектом гамма-разнообразия фитоценохоры. которые, по определению академика В.Б. Сочавы, являются сложными надбиоценоти-ческими пространственными единицами растительного покрова от простых комбинаций растительности урочищ в масштабах 1:30 000 — 1:100 000 до комбинаций растительности ландшафтов в масштабах 1:1 000 000—1:3 000 000.
Основным способом исследования гамма-разнообразия фи-тоценохор является сочетание стационарных и полуста-ционарных описаний пространственных единиц растительного покрова и дешифрирование их пространственно-временных характеристик по аэрокосмическим снимкам.
Применяются различные показатели для расчета биоразнообразия растительности в пределах ландшафта (Василевич, 1992). Используются разные уравнения для оценки разнообразия, но наиболее адекватными являются нелинейные (логарифмические) функции, повышающие значение малораспространенных компонент и наоборот снижающие подавляющее значение широко распространенных (фоновых) и позволяющие вводить условные определители (Виноградов, 2000).
Привычным для расчета разнообразия является индекс Шеннона, основанный на вычислении простой энтропии.
Для изучения разнообразия фитоценохор считается эффективным использование для исследования гамма-разнообразия вычисления сложной энтропии:
N N
;=1
где Pj — априорная вероятность появления /-того класса; Pij — вероятность отнесения образца i-того класса к j-тому классу.
При многостороннем изучении сложных единиц рекомендуется вычисление условной энтропии Н„ вида:
N ял=-£р(4)Я(4), /=1
где Р(А) — условные вероятности появления i-того класса по j-той пространственной (или другой условной) характеристике:
N
Р(А,) - ’
i=i
a H(AJ— частные условные энтропии для каждого класса:
N
;=1
Применение указанного аппарата в разных направлениях, может описать различные свойства гамма-разнообразия. Нужно помнить, что все эти показатели не имеют абсолютного значения, и используется лишь их сравнение, которое дает прирост информации fAZJ в зависимости от изменения условий опыта и определяется по разности промежуточных энтропий:
Во многих работах на ландшафтном уровне биоразнообразие рассматривается как разнообразие сообществ и экосистем в терминах пространственных показателей, включая такие, как степень фрагментации, форма пятен, сложность границ, совмещенность пятен и др., связанные с измерением ландшафтной структуры. Однако такие показатели пространственной неоднородности строго не описаны в информационных терминах.
Пространственные показатели гамма-разнообразия. Наиболее корректно пространственные характеристики гамма-разнообразия выявляются по аэрокосмическим снимкам на основе измерений, произведенным по ним. Размерности частотно-пространственных характеристик (ЧПХ), а также некоторые информационные и морфометрические показатели
(энтропийная мера ландшафтного разнообразия, коэффициент ландшафтной неоднородности и др.) дают количественное выражение сложности пространственной структуры экосистем. Здесь также гамма-разнообразие сокращается с уменьшением пространственной неоднородности, укрупнением элементов пространственной структуры, выравниванием экологических условий.
Наибольший интерес представляет антропогенная дифференциация экосистем. Пространственно распределенное гамма-разнообразие антропогенных производных экосистем изменяется нелинейно: сначала ускоренно возрастает, а затем падает. На линейно возрастающем участке этой нелинейной функции рост гамма-разнообразия описывается числом возможных комбинаций п коренных составляющих и иг производных
т п!(т-п)!
Матрица ЧПХ отражает этот процесс антропизации, хотя не дает ему временной размерности. Так, например, в районе Каррыкольского стационара (Низменные Каракумы) в исходном состоянии 4 классов образуется 13 возможных комбинаций (Виноградов, 1981). При деградации на 4 стадиях дигрессии с 10 наблюдаемыми частотными разрядами возможны 1821 производные комбинации. Наконец, при полной деградации разнообразие снова падает низко, вплоть до 4 классов. По-видимому, максимум энтропии соответствует максимуму дестабилизации фитоценохоры при деградации в зоне экологического риска. Для оценки такого разнообразия предложена близкая формула, исходя из состава сегментов (S) и их частости ( F ):
Н = -log-------.
F'.(S-F)
Разномасштабные уровни гамма-разнообразия. В ряде картографических работ исследуются количественные критерии масштабной генерализации фитоценохор. При многоуровневом анализе гамма-разнообразия мерой разнообразия фитоценохор какого-либо одномерного показателя At (1= 1,2,...,1) на уровне 0П может служить размер энтропии:
0„>-Ер(4| 0n)iogP(4| 0П), i=i
где P(AJ — вероятности пространственного распределения фитоценохор At, 0П — исследуемый иерархический уровень генерализации фитоценохор. В многомерном случае, когда анализируется пространственное распределение совокупности Z значений показателей Ait BJt энтропийная мера разнообразия определяется выражением:
В ходе генерализации пространственной структуры фитоценохор происходят свертывание информации и преобразование исходной матрицы значений в матрицу меньшего размера. Отдельные группы ячеек стягиваются к сигнальным ячейкам, т. е. ядрам генерализации. Энтропия генерализованной пространственной структуры фитоценохор записывается как
Н(?\ Oi+1)=-£P^)logP(^).
Разность AZ = ZZ(Z| 0i+s)-(z| 0f) показывает, на какую величину снижается неопределенность на уровне генерализации i+S по сравнению с уровнем I. Для анализа двухмерного (и более) показателя также используется аппарат вычисления условной энтропии:
N
н(4| 0„)(j)=-£pa 10п(/)ЬвРл 10„ (j) м
ПРИ ^,|9п(/)= ,
j=l
где используется условная вероятность j- и i-ro определителей (по экологической ординации, по вероятности распознавания, относительному участию в обмене вещества и энергии, значимости информации). С увеличением уровня генерализации ус-
rf.
ловная энтропия экосистемы, как правило, повышается. По-видимому, это происходит с переходом от конкретных карт к обобщенным, что приводит к снижению информативности хорологических единиц с уменьшением масштаба. Правда, разумная тематическая генерализация препятствует здесь безграничному росту энтропии.
Информационные показатели гамма-разнообразия фитоценохор. В зависимости от разных форм обработки и анализа данных о составе и распределении фитоценохор происходит различный прирост информации при вычислении гамма-разнообразия. При этом, начиная информационный анализ, следует определить, что надо понимать под начальной энтропией системы.
Под начальной энтропией понимается сумма предварительных достоверных литературно-картографических данных о составе и структуре фитоценохор, с которыми сравнивается уменьшение неопределенности в результате проведения новых исследований. При этом нужно исходить из того, что неопределенность знаний о фитоценохорах изученной территории стремится к нулю по мере получения достаточно полной и достоверной информации.
Прирост информации (Д7,), получаемой при использовании различных форм анализа данных, определяется по разности промежуточных энтропий. Так, например, при изучении кустарниковых формаций Низменных Каракумов Б.В.Виноградовым (2000) начальная энтропия (Но) определена по пяти формациям: Haloxylonetum persici, Calligonetum dif.sp., Salsoletum richteri, Ephedretum strobilacei, Ammodendretum persici и равна 0,69. После проведения районирования территории первая промежуточная энтропия (Нг) уменьшается до 0,61. Измерение площади, занятой формациями, показывает, что вторая промежуточная энтропия (Н2) понижается до 0,59. Наибольшее снижение энтропии до 0,41 дает ландшафтное дешифрирование аэрофотоснимков (Нл), которое при вычислении условной энтропии учитывает разыную вероятность правильного распознавания растительных формаций. Это конечная энтропия нашего опыта.
По снимкам одного срока съемки для оценки гамма-разнообразия могут быть использованы частотйо-амплитудные характеристики (ЧАХ) аэрокосмического изображения.
Динамичность живой природы в пространстве и во времени осложняет оценку изменений даже в простейшей оси «хорошо—плохо». Правильный диагноз, надежный прогноз и эффективное управление живой природой требуют научных знаний, формализованных в виде теоретических моделей функционирования биологических систем. По проекту Государственного экологического фонда « Сохранение биологического разно-образия России» был проведен анализ социально-экономических факторов, влияющих на состояние биологического разнообразия (1975), который позволил определить общие принципы и направления экспертной оценки изменений биологического разнообразия в России.
При недостаточной формализации научного знания понимание изменений живой природы под влиянием человека может основываться на коллективной экспертной оценке группы ученых. Теоретическое моделирование нарушений в развитии природы, и экспертная оценка представляют собой два взаимодополняющих пути решения поставленной задачи. Экспертные оценки служат для проверки теоретических моделей, модели помогают экспертам формализовать свои знания (рис. 59).
Макроэкономические процессы
Биологическое разнообразие
Теоретическая Экспертная
модель системы оценка
Рис. 59.
Схема оценки влияния природопользования на биоразнообразие путем исследования теоретической модели системы и экспертной оценки
ВОЗДЕЙСТВИЯ ЧЕЛОВЕКА
НА БИОРАЗНООБРАЗИЕ
Какие изменения параметров живых систем следует считать опасными? Ответ на этот вопрос позволяет оптимизировать перечень наблюдаемых показателей, по которым определяется необходимость управленческого вмешательства и контролируется его эффективность. Природные системы функционируют на основе фундаментальных экстремальных принципов. Каждая биосистема (популяция, биоценоз) стремится экстремизировать (максимизировать или минимизировать) те свои характеристики, которые определяют успешность ее существования и эволюции (критерии эффективности системы). Искусственное ограничение возможностей системы в выборе значений своих параметров ведет к снижению ее жизнеспособности, различным проявлениям разбалансировки, кризисам и т.п.
В наиболее общем виде вероятные цели живых систем, движение к которым может служить критерием их благополучия, можно свести к максимальной аккумуляции в биосистеме энергии, вещества, информации, что означает одновременно максимальную степень замкнутости круговорота внутри биосистемы и минимальный поток на «выходе» из нее.
Соответственно обратное направление развития процессов, может служить критерием опасности.
Теория свидетельствует, что разнообразие можно рассматривать как параметр биосистем, обеспечивающий достижение ими своих целей. Максимальная аккумуляция в системе энергии, вещества, информации (критерий G на рис. 60) достигается при оптимальном уровне разнообразия. Уменьшение или увеличение разнообразия по сравнению с его оптимальным значением ведет к снижению эффективности, устойчивости или других жизненно важных характеристик биосистемы. Есть минимальные значения устойчивости или энергетической эффективности системы (Do), при которых система перестает существовать, им соответствуют критические уровни разнообразия системы.
Если исходить из предположения, что природные биосистемы адаптируются к внешним условиям в ходе своего исто-
Рис. 60.
Области существования системы при различных уровнях разнообразия .
рического развития, то внутрипопуляционное, видовое и пространственное разнообразие природных биосистем является оптимальным в исторически типичных условиях. Искусственное понижение или повышение исторически сложившегося уровня разнообразия неблагоприятно для будущего системы. Теоретические модели показывают, что естественный оптимум лежит ближе к минимальным допустимым значениям разнообразия. Поэтому снижение разнообразия чаще представляет большую опасность.
АНТРОПОГЕННЫЕ
ИЗМЕНЕНИЯ БИОМОВ
Человек с момента его появления на Земле оказывал и продолжает оказывать все нарастающее влияние на процесс формирования экосистем. Однако первобытный человек влиял лишь на растительный и животный мир. Затем пришли цивилизации, способные изменять педосферу, обрабатывая почву, и внедряться в литосферу, добывая из глубин Земли полезные ископаемые, изменяя при этом также и гидросферу, и атмосферу.
Многообразные проявления антропогенных воздействий на растительный покров сводятся к трем основным формам:
1) полное уничтожение растительного покрова;
2) создание культурных фитоценозов на месте естественной растительности;
3) синантропизация растительного покрова.
Синантропизация естественного растительного покрова — постепенное изменение состава и структуры растительности под давлением антропогенных факторов. Согласно П.Л. Гор-чаковскому (1979, 1998), при этом происходят:
1) обеднение видового состава флоры Земли в целом и региональных флор. Из 240 тыс. цветковых растений примерно 155 тыс. видов обитают в тропиках, где ежегодно вырубают
почти 10 млн. га тропических лесов, что ставит под угрозу исчезновения около 50 тыс. видов. Во внетропических областях из 85 тыс. цветковых растений 4,5—6 тыс. видов находятся в угрожающем состоянии и от 750 до 1500 видов — на грани исчезновения;
2) эволюционные последствия, которые включают уменьшение генетической разнородности отдельных видов, раздробление популяций и их возрастающую изоляцию, гибридизацию между ранее дифференцированными популяциями и видами из-за исчезновения экологических барьеров между ними и расселения видов-интродуцентов, появление эндемиков на техногенных субстратах (отвалы золы, горные выработки, загрязненные участки);
3) ценотические изменения, в том числе замена коренных растительных сообществ производными и синантропными, замещение эндемичных видов космополитными и стенотопных эвритопными, а автохтонных (аборигенных) видов аллохтонными (пришлыми), всеобщее обеднение и унификация региональных флор и фаун.
Трансформация экологической обстановки в результате различных антропогенных воздействий (а также их совместного влияния) приводит к изменению ценотических отношений в сообществах и внедрению видов с широкой экологической амплитудой, вытесняющих виды узкой специализации. Зачастую это сопровождается вымиранием специализированных видов, особенно имеющих небольшие ареалы.
Постоянно повторяющиеся нарушения вызывают формирование сообществ с упрощенной структурой, обедненным видовым составом, измененными соотношениями между видами (иными доминантами). Такие сообщества обладают известной устойчивостью в измененных экологических условиях, поскольку доминирующие в них виды пластичны и довольно выносливы. Все это ведет к унификации исходно различных экосистем и распространению на огромных площадях физиономически, структурно и биотически сходных сообществ, обязанных своим обликом воздействию тех или иных антропогенных факторов.
В структуре биомов в настоящее время почти везде доминируют производные сообщества, которые мы зачастую (за отсутствием эталонов) относим к коренным или условно-коренным. При этом процесс трансформации продолжается на наших глазах. Так, исследования последних десятилетий показали, что в тропических областях широкое распространение получили различные ксерофильные лесные и кустарниковые формации, а также саванны. За последние 150 лет сведено почти 40% тропических лесов. Не менее глубокие изменения охватили и пус
тынные типы биомов, причем и в тропических, и в умеренных широтах. Произошло продвижение пустынных сообществ за пределы климатически обусловленных границ. Современное состояние биомов суши определяется двумя взаимосвязанными процессами: трансформации сообществ, с одной стороны, и снижения и утраты биотического разнообразия — с другой.
Степень воздействия человека на экосистемы, уровень их изменения под этим воздействием различны. Можно различать естественные (натуральные), почти естественные (квази-натуральные), полуестественные (семинатуральные) и культивированные экосистемы.
В настоящее время антропогенные воздействия являются решающим фактором в формировании и динамике экосистем. Поэтому необходимо детально изучить закономерности синантропизации растительности и животного населения, разработать методы оценки уровня антропогенной трансформации экосистем.
Изменение окружающей среды под влиянием человека, трансформация местообитаний, разрушение квазинатурально-го растительного покрова приводят к невосполнимой утрате генетических ресурсов растительного и животного мира. Наиболее уязвимыми элементами региональных биот обычно оказываются эндемичные, реликтовые, а также используемые человеком виды. Для того чтобы своевременно принять меры по спасению редких исчезающих видов, следует знать состояние их популяций, численность, пространственную и возрастную структуру, их динамику. При оценке состояния популяций редких видов нужно принимать во внимание особенности их жизненной стратегии.
Организация заповедников, изъятие из хозяйственного использования тех или иных территорий, во многих случаях не обеспечивает осуществления задачи сохранения биоразнообразия. Знаменитые Оренбургские степи 200—250 лет назад представляли собой бескрайние открытые пространства, море волнующегося ковыля, где паслись табуны диких лошадей (тарпанов) и антилоп (сайгаков). В степях водилось множество сурков и других роющих грызунов. Для сохранения плодородия степных пастбищ местные жители, занимавшиеся скотоводством, ежегодно весной, а иногда осенью выжигали сухую траву, пуская беглые пожары (палы). Выпас копытных животных, влияние роющих грызунов, огня были необходимыми условиями поддержания состава и структуры степной растительности.
Наблюдения в Оренбургских степях и в степях Зауралья показали, что в тех местах, где воздействие выпаса и огня исключено, происходит коренное изменение степной растительности. Здесь активно расселяются кустарники (вишня степная, миндаль карликовый, таволга зверобоелистная, кизильник черноплодный и др.), оттесняющие ковыль, типчак, другие дерновинные злаки, разнотравье. Формирование зарослей кустарников на месте степей приводит к локальному вымиранию многих видов степных растений, смене видов животных.
В результате наблюдений, проведенных на пустынном стационаре в Центральных Каракумах, выяснено, что в условиях заповедного режима растительность пустынного пастбища проходит четыре периода:
1) восстановительный (7 лет);
2) наибольшей продуктивности (5 лет);
3) начального угнетения (5 лет);
4) угнетения (после 17 лет охраны).
В период угнетения значительно снижается продуктивность пастбища, поверхность почвы уплотняется, пятна мха Tortilla desertorum и некоторых лишайников занимают от 45 до 50% поверхности. Это показывает, что умеренный выпас диких или одомашненных животных — необходимое условие поддержания стабильности и продуктивности пустынной растительности; длительный заповедный режим приводит к отрицательным результатам.
Большинство скально-горностепных эндемиков Урала представлено видами, быстро расселяющимися на обнажающемся каменистом субстрате, но не выносящими конкуренции со стороны других травянистых растений, особенно дер-новинных злаков. Поэтому режим умеренного хозяйственного использования мест обитания таких эндемиков лучше обеспечивает их сохранение, чем заповедный.
Таким образом, создание заповедников не решает проблемы сохранения флоры и растительности степей, лугов и пустынь. Необходим режим охраны и использования, при котором сформировались и существовали бы соответствующие типы растительности, растительные формации и их сочетания, поэтому экологи рекомендуют обязательное включение в степные заповедные биоценозы копытных животных (лошадей), восстановление ранее существовавшего видового состава грызунов-землероев и их численности.
ОЦЕНКА ОПАСНОСТИ ИЗМЕНЕНИЙ НА УРОВНЕ ПОПУЛЯЦИЙ
Изменение естественной структуры популяций за счет выборочного изъятия одной группы (по возрасту, размеру и др.) ведет к перемене фенотипического разнообразия популяций, которое, согласно модели, более опасно для биосистемы, чем пропорциональное изъятие из всех популяционных групп. Особенно опасно изъятие редких типов (например, наиболее крупных экземпляров). В этих случаях разнообразие резко приближается к критическому порогу устойчивости биосистемы.
При росте частоты и объемов нарушений (например, промысла или разрушений среды обитания) в биосистемах чаще гибнут популяции с малыми темпами воспроизводства и длительным периодом жизни особей (К-стратегия).
На рис.61 показана зависимость типа демографической стратегии и уровня внутрипопуляционного фенотипического разнообразия (специалисты — генералисты) от интенсивности потока ресурсов всех категорий (света, энергии, минерального и органического питания, воды) — вертикальная ось (Е) и степени стабильности среды — горизонтальная ось. В левом нижнем углу схемы располагаются биосистемы, адаптированные
Интенсивность потока ресурса Е
Богатая нестабильная среда, генералнсты, г-стратеги
Одуванчик Подорожник
Крапива
Крысы Вороны
широкое разнообразие (генералнсты), г-стратегия
Богатая стабильная среда. Разнообразные варианты комбинаций популяционных характеристик, генералнсты и специалисты, К* и R-стратеги
Лось
Дуплогнездники
Широкое разнообразие (генералнсты), энергетическая неэффективность в стабильной среде, R-стратегия, устойчивость при дестабилизации
Береза Ель
Узкое разнообразие (специалисты), энергетическая эффективность в стабильной среде. К* стратегия, слабая устойчивость при дестабилизации
Болотные кустарнички
Росянка " Камнеломки
Скудная нестабильная среда. Разнообразные варианты комбинаций популяционных параметров крайне мало из-за противоречивых требований, эфемеры, «избегатели», S-стратеги
узкое
разнообразие 4X11 (специалисты), К-стратегия
Растения и
животные
олиготрофных озер
лишайники
Скудная стабильная среда Специалисты, К-стратеги
Стабильность среды Повышение трофического уровня популяции означает при прочих неизменных параметрах - рост энергозатрат особен, - рост размеров особей, - рост участков обитания, - сокращение численности, - рост толерантности особей, - сокращение внутрипопуляЦионного разнообразия, - усиление К-стратегии
Рис. 61.
Зависимость типа демографической стратегии и уровня внутрипопуляцион-ного фенотипического разнообразия от интенсивности потока ресурсов всех категорий и степени стабильности среды
к нестабильной и бедной среде. В правом верхнем углу — биосистемы стабильной и богатой среды. Соответственно в рамках этой схемы антропогенные воздействия могут интерпретироваться как сокращение/увеличение потока ресурсов или как увеличение (реже снижение) нестабильности.
Рост частоты и амплитуды изменений условий среды, а также ее гетерогенности (мозаичности), в сочетании с ростом потока ресурсов (по схеме дрейф вверх влево) опасны для специализированных видов К-стратегов и относительно мало затрагивают виды неспециализированные, с большим внутрипо-пуляционным разнообразием, короткоживущие с многочисленным потомством (R-стратегов). В условиях нестабильной (часто нарушаемой) среды роль энергетической эффективности вида (его специализации) отходит на второй план по значению для жизнеспособности популяции.
Оценка реакции биоценоза на потерю редкого вида неясна. Предположение о необходимости редких видов для адаптации к изменениям среды почти ничего не объясняет. Очевидно лишь то, что уничтожение в биоценозе ключевого вида ведет к разрушению сообщества и возникновению на его месте нового.
Последовательное сокращение размеров участков природных систем приводит к последовательному исчезновению видов (в зависимости от их требований к размерам участков обитания) и упрощению биоценотической структуры ниже оптимального уровня.
ОЦЕНКА ОПАСНОСТИ ИЗМЕНЕНИЙ
НА УРОВНЕ СООБЩЕСТВ
Структура биоценозов, относящихся к более ранним стадиям сукцессии, сходна с системами, адаптированными к менее стабильной среде, «в зрелости» их параметры массы и разнообразия соответствуют более устойчивым условиям (на рисунке это горизонтальный дрейф вправо). При определенных частоте и/или площади нарушений происходит перманентная замена зрелых стадий биосистемы на ранние стадии. На рис. 62 показано размещение биоценозов и агроценозов в тех же осях.
С использованием разработанных теоретических моделей можно производить принципиальное сравнение ра^ых типов биосистем и анализировать их реакцию на воздействия человека. Один из вариантов сопоставления эк^истем, относящихся к одному климатическому району, ноле различным типам (наземные, водные, проточные, стоячий) и разному уровню антропогенной нарушенное™ (вырубки^добрение, эвтрофикация, урбанизация), представлен на рж. 63. Рассматривая модели, можно предположить, что дрейф среды в сторону увеличения нестабильности (обобщенндевыражение антропогенных воздействий) наиболее опасен для систем прежде адаптирован-
Антропогенные возмущения среды Локальные варианты нестабильного климата Молодые системы
Протонные системы
реки (пера
Холодные зимой пустыни
море
Олиготрофные озера
реки озера
Полярные пустыни
Пещеры, глубоководные места
Стабильность среды
Возраст систем
Рис. 62.
Зависимость интенсивности потока ресурса от стабильности среды для биоценозов и агроценозов разного типа
ных к относительно стабильным и скудным условиям (правый нижний угол на схемах). Среди выделенных наземных зональных типов экосистем это степи и лесостепи, а локальных вариантов сообществ — верховые болота и олиготрофные озера. Наименее опасен этот дрейф для систем, адаптированных к нестабильным и богатым условиям (левый верхний угол на схемах).
Эфтрофнроьанныс водоемы
Эстуари
реки
азеоа
* Эстуари
оазисы
Урбакижхзанкые \
Удобряемые
болото
Орошаемые поля
орошаемые поля
Природные
Неудобряемые Вырубки < поля
реки
'Ъерховые < болота
озера
Олиготрофные озера
глубоководные
Стабильность среды
Возраст систем
Рис. 63.
Экосистемы, относящиеся к одному климатическому району, но к различным типам и разному уровню антропогенной нарушенное™
К ним относятся в первую очередь уже трансформированные системы — агроценозы и урбанизованные зоны.
Наиболее устойчивы и обеспечены ресурсами тепла и воды биомы широколиственных лесов. Эта группа сукцессионно
продвинутых экосистем формирует собственную, «ценотичес-кую» среду. В условиях, когда связи между элементами биосистем разнообразны и сильны, климатические осциляции не могут существенно влиять на структуру биома. Поэтому крупные перестройки здесь происходят лишь в случае чрезвычайно резких изменений климата (типа плейстоценовых чередований ледниковых эпох и межледниковий).
По мере продвижения как к югу, так и к северу ресурсная обеспеченность экосистем снижается и нарастает их динамичность (нестабильность). Колебания количества тепла и влаги в разные годы заметно сказываются на биомах аридного пояса и тундр (в последнем случае на животном населении), вызывая существенные изменения в соотношениях различных сообществ. Это связано с сукцессионной молодостью экосистем этих зон, определяющей слабость ценотических связей между их элементами.
Наиболее оптимальное распределение биомассы по разным структурным блокам природных экосистем характерно для биомов лесной зоны. В соответствии с теоретическими следствиями моделей разнообразия, в экосистемах с такой структурой преобладают свойства «ядра». Они отличаются повышенной устойчивостью ко всем видам воздействий. Это понятно, так как на большинство трансформаций лесные экосистемы отвечают переходом на одну из более ранних сукцессионных или демутаци-онных стадий, набор которых здесь весьма значителен. Вопреки традиционным заключениям об опасности потерь биоразнообразия, модели для некоторых степных, лесостепных и даже лесных экосистем показывают падение их устойчивости при росте разнообразия. Это связано с наличием у них не только свойств «ядра», но и значительных признаков «кризисной периферии».
Свойства кризисной периферии характеризуются падением устойчивости при увеличении разнообразия и парадоксальным ростом ее при выпадении избыточных элементов. Это сочетание свойств отражает снижение устойчивости экосистем (в первую очередь лесостепных) при появлении в них элементов, находящихся за пределами и без того очень сложных систем сукцессионных и демутационных стадий — например, массовое появление полей. Кроме того, высокая мозаичность растительного покрова этих территорий отражает неблагоприятные тенденции, когда подавляющая часть площади занята серийными местообитаниями, а климакс и предклимаксовые стадии сукцессии не достигаются.
ОСНОВНЫЕ ТИПЫ АНТРОПОГЕННЫХ НАРУШЕНИЙ И ЭКСПЕРТНАЯ ОЦЕНКА ИХ ЗНАЧИМОСТИ
Исходя из моделей были определены основные типы антропогенных нарушений, которые имеет смысл рассматривать. Первую группу составляют процессы, ведущие к сокращению, фрагментации природных сообществ или их трансформации из-за изменения эдафотопа. Наиболее сложной является вторая группа — изменения человеком среды обитания (в том числе биосистем, которые выступают факторами внешних условий для других). Третью группу составляют непосредственные трансформации биосистем (перепромысел, нарушения демографической структуры популяций, занос новых видов, изменения соотношений трофических блоков).
В силу того, что различные типы природных и антропогенных экосистем сформировались при разных уровнях нестабильности среды и потока ресурсов диагностику опасности изменения их массы и разнообразия необходимо проводить раздельно. Минимальный набор типов экосистем включает: прибрежные морские; водоемы аридной и субаридной зоны; пресноводные экосистемы; болотные, околоводные - пойменные и дельтовые комплексы; тундровые: таежные; лесные
в Европейском центре; лесные в лесостепи, долинах и горах юга России; степные и полупустынные; луговые и агроценозы; почвенные экосистемы; урбанизованные зоны. Кроме этого в отдельные типы выделены экосистемы особо охраняемых территорий и редкие виды животных и растений. Таким образом, минимальный набор изменений биосистем, которые по теоретическим и экспертным соображениям можно однозначно оценивать, представлен матрицей.
Каждый процесс в этой матрице можно понять исходя из перечисленных ранее закономерностей развития живых систем. Кроме этого были предприняты процедуры прямой экспертной оценки основных процессов по значению для сохранения или нарушения массы и разнообразия живой природы России. Результаты оценки представлены графически в виде усиления интенсивности штриховки в ячейках матрицы в соответствии с нарастанием величины экспертной оценки значимости. Наиболее важными из типов воздействий эксперты считают сокращение и фрагментацию местообитаний, нарушения экотопов (почвенно-грунтовых условий, солености и пр.) и частые возвраты к пионерной стадии сукцессионных процессов. По типам экосистем — нарушения водоемов аридной зоны, в лесах юга России и в тундре. Наименее значимыми представляются вопросы, связанные с прямым перепромыслом и подавлением хищников, а также экологические проблемы таежных регионов. Имея оценки опасности нарушений живой природы, мы можем приступать к познанию порождающих их макроэкономических и политических процессов.
ТЕХНОЛОГИЯ ЭКСПЕРТНОЙ ОЦЕНКИ
ВЛИЯНИЯ ПРИРОДОПОЛЬЗОВАНИЯ
НА БИОЛОГИЧЕСКОЕ РАЗНООБРАЗИЕ
Наибольшую трудность для анализа причин антропогенных изменений биоты представляет многосвязанность процессов, идущих в живой природе под воздействием человека. Лишь ограниченное число экспертов берется проследить цепочки событий, связывающих макрополитические шаги с конкретными последствиями для экосистем. При этом ошибки в оценках возникают на стадии политического и макроэкономического прогнозов.
Для интегральной оценки и прогноза кризисных процессов в обществе можно использовать разработанные экологами модели самоорганизации сложных систем. Полученные прогнозы очерчивают границы возможного развития, задают стенки «коридора», за пределы которого события в России не могут выходить по соображениям общих законов развития и реальной практики.
Интегральные описания имеют два основных недостатка: 1) они не дают детального анализа цепочек взаимодействий: 2) они лишены количественных оценок силы влияния разных причин и группового действия различных экономических факторов на биосистемы. Исследование детальных связей
между изменениями живой природы и макроэкономическими событиями потребовало введения промежуточного этапа, на котором эксперты давали характеристику конкретным хозяйственным процессам. Этот перечень включает семьдесят разных процессов, объединенных в десять групп: 1) сельское хозяйство; 2) лесопользование; 3) биопромыСлы; 4) добыча минеральных ресурсов и производство энергии; 5) обрабатывающая промышленность; 6) военная деятельность; 7) муниципальная сфера; 8) социально-демографические процессы; 9) природоохранная деятельность; 10) информационная структура.
Особенность перечня хозяйственных процессов в том, что с ним могут работать как эксперты-экологи, так и эксперты-экономисты. Экологи могут указать те изменения биосистем, которые связаны с разными видами хозяйственной деятельности. Экономисты же достаточно уверенно могут объединить хозяйственные процессы и их макроэкономические причины. Таким образом, появляется возможность замкнуть макроэкономику на экологию.
Для каждого включенного в матрицу экологического процесса можно получить протокол оценки его макроэкономических причин, и наоборот можно получать оцйрки экЭДргшш^ ких последствий любого макроэкономического процесса/ Такие протоколы представляют собой формализованное коллективное знанйе экспертов. При этом есть возможность оценить не только сравнительную опасность разных .процессов в прошлом, настоящем и будущем, но и увидеть^нердкознач-. ность влияния многих из них, когда экономическая йшйч'ийа, с одной стороны, способствует антропогенным ’НМф^дцеж^я^, но с другой — им же и-препятствует.
Матрицы представлены двумя основными типами. Перво* группа описывает положительные связи, когда развитию экологического нарушения способствует тот или иной макроэкономический процесс. Вторая группа характеризует отрицательные связи, когда развитию экологического нарушения препятствует тот или иной макрополитический процесс. Оценка силы положительной или отрицательной связи для всех расчетов выполняется по 100-балльной шкале. Результаты оценки могут быть представлены графически в виде усиления интенсивности штриховки в ячейках матрицы в соответствии с нарастанием величины экспертной оценки силы связи
(1 — менее 2 баллов; 2 — 2-5; 3 — 5-8; 4 — 8-13; 5 — 13-22; 6 — 22-36; 7 — 36-60; 8 — 60-100 баллов). В тех случаях, когда два процесса имеют и положительную, и отрицательную оценки связи, в матрицу включается разница этих величин.
Организация знаний экспертов по общей схеме делает их доступными для более сложных алгоритмов обработки и синтеза. Так появляется возможность анализа экологических последствий для сочетания из нескольких макроэкономических процессов, фактически экологическая оценка разных сценариев развития. Обратным ходом (от экологических процессов) можно получать интегральную оценку силы воздействий макроэкономических событий на экосистемы определенного типа. Имеется возможность определить наиболее специфичные причины любого из анализируемых экологических процессов, выделив, таким образом, макроэкономические процессы, отличающиеся наиболее направленным действием и в силу этого перспективные для использования в управлении.
Помимо исследовательского результата матрица имеет и прикладное значение. Она может служить средством экологической оценки природопользовательских проектов на стадии их планирования.
VII
ТЕХНОГЕННЫЕ
БИО-
РАЗНООБРАЗИЮ
КАТАСТРОФЫ -УГРОЗА
ж
СТАБИЛЬНОСТЬ И УСТОЙЧИВОСТЬ
БИОЛОГИЧЕСКИХ СИСТЕМ
Проблема устойчивости биотических систем является одной из актуальнейших в современной биологии. Нерациональное ведение хозяйства привело к тому, что в одних регионах наступила полная деградация естественных экосистем (например, зона европейских степей), другие — приближаются к этому состоянию. За исторический период человечество из-за эрозии потеряло 2 млрд га продуктивных земель, за относительно небольшой отрезок времени в Европе вследствие кислых дождей отмерло 7,4 млн га лесов. Прогнозы показывают, что при сохранении и в дальнейшем современных темпов развития промышленности, строительства, использования естественных ресурсов нагрузки на экосистемы возрастут в 2—3 раза. Такая ситуация вынуждает биологов искать ответы на вопрос, что такое устойчивость, чем она обусловлена, какие факторы и какой силы могут привести к ее потере.
Первым шагом к осмыслению проблемы является определение сущности понятий «устойчивость» и «стабильность». Стабильность — это общее свойство системы,раз деляющееся на два типа: резистентную устойчивость — способность оставаться в устойчивом состоянии при на-
грузках и упругую устойчивость — способность к быстрому возобновлению.
Под устойчивостью понимают внутреннюю способность системы пребывать в состоянии, близком к равновесию, и возвращаться к нему после различных нарушений, а под стабильностью — сохранять относительно неизменное состояние под влиянием некатастрофических явлений.
Р. Маргалеф (1968) рассматривает два типа стабильности: 1) стабильность, которая возникла в результате внутрен него взаимодействия; 2) устойчивость к действию внешних факторов.
Термином «устойчивость» обозначают способность системы выдерживать изменения, которые вызваны влиянием извне, и возобновляться, а «стабильность» — меру изменчивости системы. В.С. Шарашова (1989) в монографии «Устойчивость пастбищных экосистем» рассматривает отдельно устойчивость и стабильность. По ее мнению, устойчивость — это способность естественных сообществ к саморегуляции, которая обеспечивает их существование на структурно-динамической основе, она базируется не на способности противостоять различным влияниям, а на возможности адаптироваться к ним.
Ряд исследователей пришли к выводу, что устойчивость системы имеет место лишь в случае действия на нее разных факторов и проявляется в двух формах — упругой устойчивости и устойчивости сопротивления. Первая обозначает меру скорости возврата системы в исходное состояние, а вторая — способность системы избегать изменений.
Каждому биологу известны различия живых систем по их реакциям на воздействие того или иного экологического фактора, например ветра, твердых осадков засухи, вредителей, выпаса скота и др. Известно также, что живые организмы или их совокупность в различных почвенно-климатических условиях или в разных возрастных состояниях по-разному реагируют на воздействие того или иного экологического фактора. В разных условиях внешней среды также по-разному реализуется жизненная программа живой системы: взаимоотношения между ней и средой ее существования имеют и пространственную, и временную специфику.
Каждая живая система саморегулируема. В ее памяти и регуляторе заложены нормы реакции на всевозможные вне
шние возмущающие воздействия, обусловливающие отклонение от программы; т. е. каждое такое воздействие проходит оценку в регуляторных устройствах живой системы, в результате чего она избирает такую форму реакции, которая обеспечивает ей самый эффективный вариант защиты от возмущающего фактора. Следовательно, и стабильность, и устойчивость — это свойства живых систем, которые связаны с их способностью к саморегуляции. Остается лишь разграничить их сферы проявления.
Под стабильностью живой системы целесообразно понимать заложенное в ее генетической программе свойство реализовывать в изменчивых условиях внешней среды свою жизненную программу, которая, несмотря на несущественные отклонения, ведет к формированию эталонной системы (под эталонной, или гипотетической, системой в кибернетике подразумевается такое ее состояние, к которому стремится управляемая система в конкретных экологических условиях). Что касается уточнения объема упомянутых в определении ♦несущественных отклонений», то к их категории можно отнести те отклонения от программы, которые легко устраняются системой при помощи регуляции по замкнутому циклу с обратной связью.
Под устойчивостью понимается способность живой системы путем использования внутренних защитных механизмов противостоять внешним возмущающим воздействиям, адаптироваться к ним без существенного изменения структурно-функциональных особенностей или быстро возвращаться к нормальному (устойчивому, парастабильному) состоянию, если это воздействие обусловило лишь временное отклонение системы от заданной программы. Устойчивость системы может быть определена по отношению к какому-либо одному или нескольким конкретным факторам, например, к заражениям опенком, инвазиям короедов, ветровалу, засухе, загазованности, уплотнению почвы и др.
Если живая система устойчива вообще ко всем воздействиям в своей естественной среде, в таком случае она переходит в то состояние, которое характеризуется понятием ♦стабильность». Свойство стабильности присуще главным образом коренным, первичным экосистемам.
Стабильность в нашем определении представляет собой сумму разнообразных устойчивостей во времени. Иными сло
вами, стабильными могут быть только те системы, которые устойчивы во времени к разнообразным внешним возмущениям, или, как говорят экологи, к неблагоприятным воздействиям экологических факторов. Стабильные системы не могут быть не устойчивыми, но системы, устойчивые к тому или иному фактору, не обязательно стабильны. Культуры ели в поясе дубовых лесов в возрасте 10—30 лет устойчивы к ветровалам, снеголомам, короедам и грибным заболеваниям, но они нестабильны. После 40-го возраста они интенсивно разрушаются и до 60 лет погибают.
Таким образом, стабильность и устойчивость — это два различающихся понятия: стабильность — обобщенное свойство живой системы, характеризующее ее нормальное, по заданной генетической программе развитие в конкретных экологических условиях; устойчивость — частное свойство живой системы, характеризующее ее способность противостоять неблагоприятным влияниям того или иного экологического фактора. Эти понятия различаются также темпорально-пространственными показателями: стабильность связана с общей структурно-функциональной организацией системы на протяжении всего времени ее существования, устойчивость — с отдельными ее структурно-функциональными показателями и преимущественно непродолжительными отрезками времени.
В литературе преобладает мнение, что стабильность систем в значительной мере зависит от их сложности. Например, Р. Мак-Артур (1955) считал, что, чем сложнее система, тем она более стабильна. Этот вывод и много подобных ему поддавались математическому анализу. Итог этого анализа подвел Р.Мей/1987), который пришел к противоположному выводу, что увеличение числа элементов в системе ведет к ее нестабильности, т.е. к несоответствию реакции системы на незначительную смену условий. В то же время многие исследователи считают, что единой закономерности для всех систем нет и поэтому заменять одно обобщение другим не имеет смысла. Следующим условием стабильности являются способность элементов изменяться, их гетерогенность и пространственное размещение.
Стабильность систем релятивна и ограничена определенным отрезком времени, специфическим для каждой из них. В одних случаях это могут быть столетия (климаксовые ценозы), в других — годы, десятилетия (популяции эксплерен-
тов). Каждая стабильная система динамична и в зависимости от происходящих в ней процессов можно говорить об уменьшении (или увеличении) стабильности или о ее постоянном уровне. Постоянный уровень стабильности, как правило, обеспечивают флуктуационные изменения, сукцессии же ведут или к ее уменьшению, или к увеличению.
Исследования показывают, что первым, важнейшим условием стабильного существования популяции являются гетерогенность ее элементарного состава и способность элементов изменяться. Эта гетерогенность достигается за счет поливариантности онтогенеза особей. Л.А. Жукова (1988) выделяет пять типов поливариантности: размерный, морфологический, ритмологический, поливариантность по разным способам размножения и по темпам развития.
В основе поливариантности онтогенеза лежат генетический полиморфизм и модификационная изменчивость в пределах генотипа. Поливариантность онтогенеза во всех ее проявлениях — важный механизм, обеспечивающий элементарную и динамическую гетерогенность ценопопуляции, ее устойчивость к разным неблагоприятным факторам среды и стабильность.
Вторым условием стабильности существования популяции считается полночленность ее возрастной структуры.
Третье условие — пространственная гетерогенность, которая достигается за счет особенностей расселения растений и образования популяционных локусов разного возраста и жизненности, отличающихся между собой темпами развития и ролью в системе.
Четвертое условие — характер ее самоподдержания, проявляющийся в разных способах размножения (вегетативное, генеративное). Благодаря комбинациям способов размножения ценопопуляция может существовать беспредельно долго.
Одним из главных условий устойчивости живых систем надпопуляционного уровня также является гетерогенность их элементарного состава, проявляющаяся во времени их существования, пространственно-временных различиях плотности особей и массы, функций, способностей к флуктуациям, сменам и др.
Поскольку об устойчивости можно говорить лишь в случае действия определенного фактора(ов), возникает необходимость категоризации видов устойчивости. В ее основе мож
но использовать классификацию факторов, предложенную М.Ф. Реймерсом (1994). Согласно этой классификации системы могут быть эволюционно устойчивыми к действию возмущений, а также исторически и действующе устойчивы. Дальнейшую конкретизацию устойчивостей можно проводить по показателю специфики факторов (космические, абиотические, антропогенные и т.п.) и другим признакам. При этом следует иметь в виду не только прямое, но и косвенное воздействие фактора на конкретную систему путем изменения параметров системы высшего уровня организации.
Техногенные катастрофы являются одним из серьезных факторов, вызывающих изменения в структуре популяций и сообществ, нарушающих их стабильность. В оценке экологического состояния почв и водоемов основными показателями степени экологического неблагополучия являются критерии физической деградации, химического загрязнения и биологического нарушения в экосистемах. Исследуя биоразнообразие в загрязненных водных и наземных экосистемах, почвах или донных отложениях, мы имеем дело с долговременными эффектами загрязнения.
ДИНАМИКА БИОРАЗНООБРАЗИЯ
В УСЛОВИЯХ
РАДИОАКТИВНОГО ЗАГРЯЗНЕНИЯ
Одним из непредвиденных негативных последствий крупнейшего достижения науки XX в. — освоения энергии атома — стало наличие земель, загрязненных радионуклидами. В первую очередь это касается территорий полигонов по испытанию атомного оружия, земель, подвергшихся загрязнению в результате аварий на предприятиях атомной промышленности и АЭС, зон вокруг предприятий по добыче, переработке и хранению радиоактивных материалов.
Биота — наиболее динамичная составляющая любого ландшафта. При радиоактивном загрязнении она испытывает дополнительное воздействие, которое никак не затрагивает другие элементы ландшафта: подстилающие породы, воду, почву, атмосферу, поскольку выделяемая при радиоактивном распаде энергия слишком ничтожна по сравнению с потоком энергии солнечного излучения. Действие же на растения и животных при этом может быть достаточно эффективным. Однако эти же дозы даже при высоких уровнях загрязнений среды недостаточны, чтобы воздействовать на грибы и микроорганизмы. Экологу приходится учитывать несколько новых
факторов в зонах радиоактивного загрязнения, выделенных в радиационные заповедники (Криволуцкий, 1996).
В радиационных зонах, где прекращена хозяйственная деятельность человека, продолжаются естественные процессы динамики экосистем и ландшафта: сукцессионные изменения растительного покрова и животного мира, перераспределение выпавших радионуклидов по ярусам биогеоценоза, естественная динамика почвенного покрова, зарастание сначала бурьянами, а затем лесной растительностью бывших пастбищ, сенокосов и пахотных земель. Но и радиоактивное загрязнение оказывает активное воздействие на жизнь облучаемых живых организмов, их популяций, сообществ и экосистем. Такие экосистемы живут в особом режиме и отнюдь не нейтральны по отношению к окружающим территориям. Так, лесные породы здесь сильнее поражены вредителями (стволовыми, листовыми, корневой губкой), сильно поражены вредителями оставшиеся в зоне отселения насаждения фруктовых деревьев и сельскохозяйственных растений. Так, в первый год после аварии на Чернобыльской АЭС посадки картофеля в зоне отселения на 96% были уничтожены колорадским жуком. По некоторым данным, в зонах радиоактивного загрязнения из-за ослабления физиологического состояния и, возможно, иммунитета диких животных активизируются очаги природно-очаговых инфекций, таких, как клещевой энцефалит, туляремия, инфекционная желтуха. В этих зонах находят для себя убежище вредные для человека виды зверей, в частности волки. На землях радиоактивного загрязнения динамичны и создаваемые излучением «дозовые поля*, поскольку радионуклиды, выпавшие первоначально в форме аэрозолей на листву, траву и поверхность почвы, с годами оказываются почти полностью аккумулированными почвенным покровом. Это изменение дозовых полей определяющим образом воздействует на динамику растительного и животного мира. Рассмотрим это на примере изученной нами в 1986— 1999 гг. динамики животного населения почв в непосредственной близости от Чернобыльской АЭС. В обследованных биотопах численность беспозвоночных неодинакова и сильно варьирует. Минимальная (12—39 экз./м2) — на участках песчаных пляжей, крутых и обрывистых участках надпойменных террас, лишенных растительности, в молодом сосновом лесу; средняя (40—90 экз./м2) — на пастбищных лугах, в смешанном
ф
лесу, старом сосновом лесу, склонах надпойменных террас, заросших растительностью, на пастбищных лугах; максимальная (91—160, местами до 300 экз./м2) — в супралиторальной зоне, прибрежных кустарниках, сенокосных лугах, пойменных увлажненных лиственных лесах, по берегам болот, Стариц, ручьев, заросших растительностью.
Обследование почв зоны, подвергшейся радиоактивному загрязнению, проводилось с середины июня 1986 по 2001 г. сотрудниками Лаборатории биоиндикации ныне ИПЭЭ РАН им. А.Н. Северцова. Проведенные учеты показали, что почвенные животные (обитатели лесной подстилки) сильно пострадали в результате радиоактивного загрязнения среды в районе ЧАЭС на удалении 3—7 км от станции. Численность массовых групп почвенных клещей, а также ранних стадий развития животных из числа мезофауны сократилась примерно в 30 раз на середину июля 1986 г. Значительно слабее, в 2—3 раза, уменьшилась численность мезофауны на полях в толще пахотных почв, причем ни для одной группы животных, а они обитают в основном в слое почвы 5—20 см, не было отмечено катастрофического падения численности. Радиоактивное загрязнение нарушило процесс нормального воспроизводства популяций самых разных почвенных обитателей. В районе Чернобыля среди почвенного населения в сосновых лесах отсутствовали личинки и нимфы микрофауны первых возрастов. В пахотных почвах такой гибели молоди не наблюдалось, но численность молодых дождевых червей была в четыре раза ниже, чем на незагрязненных участках (Криволуцкий и др., 1996).
Поскольку мощности облучения почвенной фауны были определены, можно заключить, что дозы порядка 29 Грей вызывали катастрофические изменения в сообществе микрофауны, а около 8 Грей — регистрируемые, но незначительные изменения среди обитателей поверхности почвы. В пахотных почвах даже при суммарной экспозиционной дозе около 86 Грей на поверхности животные в толще почвы пострадали относительно слабо, вероятно, из-за того, что были хорошо защищены экранирующим слоем почвы от бета-излучения, вклад которого в общую дозу составлял около 94%. Последующие учеты показали, что уже с поздней осени 1986 г. даже на самых загрязненных участках зоны началось медленное восстановление животного населения почвы. Сначала появи-
лись отдельные личинки и нимфы, т. е. началось размножение животных. Весной 1987 г. численность микрофауны составляла еще менее половины от контроля.
Через год после аварии почвенная фауна интенсивно восстанавливалась даже в наиболее пострадавших участках. Численность дождевых червей составляла около 15% контрольной, но наличие их коконов свидетельствовало о размножении этих животных даже в наиболее загрязненных почвах.
Осенью 1987 г. общая численность почвенной фауны по многим группам сравнялась, хотя структура популяций видов еще была «ущербной»: интенсивность процессов размножения еще сильно отставала от контрольных участков. И только к осени 1988 г. (учеты были проведены в середине октября) по общей численности мезо- и микрофауна почвы практически полностью восстановилась по общему обилию, видовому разнообразию в основных чертах и по динамическим показателям структуры популяций. Можно было заключить, что остаточный уровень радиоактивности почвы уже не оказывал подавляющего влияния на восстановление животного населения почв. Это было отмечено и для тех участков прилегающего к ЧАЭС «Рыжего леса», которые еще не были сведены и засыпаны слоем песка. Таким образом, через 2,5 года после аварии почвенное население количественно и функционально полностью восстановилось, но его видовое разнообразие и через 15 лет составляло для разных групп организмов 45—75% от исходного.
Некоторые закономерности можно выделить в функционировании сообществ на этих территориях.
1. Эффекты изоляции экосистем, их развитие по принципу «островных экосистем* на местности, где живая природа функционирует в совершенно ином режиме. Специфика островных экосистем хорошо описана Р. Мак-Артуром, но общий итоговый результат происходящих здесь экологических процессов — обеднение биоты «островов» в видовом отношении, потеря биоразнообразия из-за того, что популяции тех видов, которые находятся в угнетенном состоянии постоянно или при экстремальных изменениях среды, но получают пополнения извне и имеют больше вероятности погибнуть, исчезнуть из экосистемы, чем на большом непрерывном ареале. Более того, исходный биотический потенциал заповеданной ныне территории существенно ниже, чем потенциал окружа
ющих земель, поскольку радиационные заповедники все созданы на участках, непосредственно примыкающих к крупным промышленным объектам и населенным пунктам, освоенным человеком уже несколько столетий, как это можно видеть в Чернобыле и на Южном Урале. В то же время на окружающих землях сохранялись естественные экосистемы на неудобных для освоения участках, землях лесного фонда, по болотам, поймам рек, в охотничьих хозяйствах и т.п., где разнообразие фауны и флоры все еще велико, в противоположность экосистемам на заброшенных пашнях, выпасах и поселках, где созданы радиационные заповедники.
2. Эффекты режима заповедания. Закрытые для населения, охотников, рыболовов, туристов земли, где животный мир находится в безопасности, а растительность не подвергается уничтожению через пастьбу скота, порубки и выжигание, становятся весьма привлекательными зонами для постоянного и временного концентрирования многих животных, особенно охотничье-промысловых. В закрытой зоне вокруг Чернобыльской АЭС уже в 1986 г., в первый год после аварии, появилось множество ранее здесь отсутствовавших лосей, кабанов, несколько стай волков, стадо благородных оленей, около 400 лисиц, которые все пришли в «спокойную* зону, спасаясь от охотников из Белоруссии, Брянской области России, областей Украинского Полесья. На 30 лет раньше такие же процессы отмечались в зоне загрязнения на Южном Урале, где появились оседлые популяции ранее отсутствовавших лосей, косуль, кабанов, волков, барсуков, рысей, тетеревов и т.д. Поразительный факт — только на территории Восточно-Уральского государственного заповедника, где местами уровень радиоактивного загрязнения превышает допустимый в 1000 раз, встречаются единственные во всей зауральской лесостепи гнездовья четырех видов крупных хищных птиц, внесенных в Красные книги России и СССР, здесь гнездятся и успешно размножаются такие крупные птицы, как глухарь, тетерев, тетеревятник, орлан-белохвост, большой подорлик, уральская неясыть, лебедь, серый журавль. На этой радиоактивной территории постоянно высока численность сибирской косули, лося, зайца, ондатры, барсука, горностая, степного хорька, куницы и ряда других млекопитающих, которые повсюду очень редки в освоенных под сельское хозяйство равнинных районах Челябинской и Свердловской областей.
И причина здесь» в том, что зверей и птиц не отстреливают и не беспокоят в период размножения. Такие редкие виды, как филин, скопа, большой подорлик и орлан-белохвост, чаще встречаются на территории радиационного заповедника на Южном Урале, ч.ем на соседних неохраняемых территориях. По наблюдениям: одного из авторов в июне 1991 г. на Семипалатинском полгигоне многочисленными были степные виды жаворонков, отмгечена высокая плотность серой куропатки, обычными были (балобан, болотный, степной и полевой луни, кобчик, обыкнов»енный канюк, степной орел, беркут, савка, многие из которых практически отсутствовали вне охраняемой зоны ядерно»го полигона. Последние три вида занесены в Красную книгу СССР (Криволуцкий, 1996; Лебедева, 1999; Криволуцкий и дар., 2000).
3. Эффекты радиационной динамики. Кроме упомянутых выше косвенных факторов сильнейшее влияние оказывает на биоразнообразие и прямое воздействие ионизирующей радиации, преимущественно в условиях нагрузки выше 0,5—0,7 Р/сут окспозиционной дозы. Наиболее подробно влияние динамижи радиационной обстановки на животное население изучено в зоне загрязнения Чернобыльской АЭС (Криволуцкий, 1.996). Здесь косвенные факторы определя-: ли динамику населения птиц и крупных млекопитающих, но динамика поч венной фауны в лесах вокруг ЧАЭС полно-! стью зависит от радиационного режима. При поверхностном загрязнении почга радионуклидами 1811,134Cs, 137Cs, “Sr, 239Pu и др. в апреле—ниае 1986 г. через два месяца наблюдалась ; подлинная экологическая катастрофа в зооценозах почвы, когда погибло более 90% видов беспозвоночных — обитате-' лей почвы. По меере радиоактивного распада радионуклидов i иода и лантанидов дозы облучения снизились в десятки раз ta уже с 1987 г. началось медленное восстановление популяций и сообществ.. Общая численность микроартропод вос-. становилась череез 2—3 года после аварии за счет глубоко-^почвенных мелких организмов, их биоразнообразие достигало только 50% от контроля в течение 5 лет, а с 1993 по 1995 г. началось восстановление биоразнообразия и поверхностно обитающих видов, которые стали обильны в почвах в 1995 г., хотя шх разнообразие составляло около 50% от исходного, но ко. личество видов во всех горизонтах почвы было уже 75—80 % от контроля.
Показатели биоразнообразия очень динамичны — при исчезновении повышенной радиации даже такой насыщенный видами комплекс, как почвенная фауна, восстанавливается примерно за 2 года, а при продолжении действия радиационного фактора население почвы не восстанавливается и за 100 лет, как это можно видеть в Республике Коми на почвах, загрязненных 230Th, 22eRa, 288U.
Итак, биота — наиболее динамичный компонент ландшафта в зонах аварийного или техногенного радиоактивного загрязнения. Наличие многих видов растений и животных, которых можно использовать как биоиндикаторов качества окружающей среды, позволяет успешно вести биомониторинг на загрязненных землях, оценивать риск проживания для населения. В то же время в закрытых для населения, изолированных радиационных заповедниках активно проявляются и косвенные эффекты изоляции: миграция птиц и зверей извне в «спокойные* для жизни и размножения зоны.
ВЛИЯНИЕ РАЗЛИВОВ НЕФТИ
НА РАЗНООБРАЗИЕ
МОРСКИХ СООБЩЕСТВ
Примером опасных техногенных загрязнений могут служить аварии танкеров, приводящие к разливам нефти. Такие катастрофы чрезвычайно опасны для морских сообществ.Например, приведшая к природной катастрофе авария танкера «Эксон Вальдез», произошедшая 24 марта 1989 г. близ берегов Аляски и получившая широкую известность во всем мире. Кратковременный эффект разлива нефти после этой аварии на морских птиц был тщательно исследован многими учеными и признан катастрофическим. Сразу погибло 36 115 птиц, а загрязненные нефтью особи встречались на пляжах в течение месяца после аварии. По оценкам специалистов, число морских птиц, погибших в результате разлива нефти, составило от 250 000 до 375 000 особей, из которых 74% были кайры. Более 8400 длинноклювых и короткоклювых пыжиков, самых обычных морских птиц были убиты нефтью, что составило 7% всей местной популяции. Популяции морских птиц в море и зоне разлива нефти подверглись немедленной депрессии. На 66% уменьшились популяции пыжиков в проливе Принца Уильяма по сравнению с 1970 г., так как большинство птиц, гнездящихся
здесь, не вернулись на места размножения после того, как большая часть поверхности была очищена от нефти. Вероятно, они погибли во время миграции. На 43% уменьшилась численность популяции тихоокеанского чистика.
Было установлено влияние разлива нефти в следующие несколько лет на литоральные и сублиторальные местообитания, эффективность использования морских ресурсов и социальную структуру сообществ. Нефть распространилась на сотни километров. Выброшенная на берег, она глубоко проникла на каменистые пляжи и сохранялась под камнями еще 5 лет после аварии и поверхностной очистки. Разлив нефти сразу оказал отрицательное воздействие на местообитания почти половины изученных видов. Этот эффект сохранялся в течение 2,5 лет после аварии для большинства первоначально пострадавших видов. Многие виды подверглись значительной опасности, в частности птицы, собирающие корм с поверхности моря.
На основании сравнительных данных, собранных до (1970-е гг.) и после (1989—1994 гг.) аварии, к долговременным эффектам разлива нефти на популяции были отнесены: уменьшение численности популяций, сокращение гнездового успеха и смещение сроков размножения. В районе разлива нефти были обнаружены отклонения в репродуктивном успехе белоголовых орланов. Самки черного кулика-сороки, обитающего на побережье и собирающего пищу в литоральной зоне, на загрязненных нефтью территориях в год аварии откладывали более мелкие яйца. Птенцовая смертность в 1989 г. была выше на загрязненных нефтью местах по сравнению с контрольным биотопом и положительно коррелировала с количеством нефти на кормовых участках. В 1990 г. продукция птенцов оставалась низкой на грязных участках и полностью отсутствовала на побережье, где были проведены восстановительные работы. Продуктивность обыкновенной моевки, наиболее обычного колониального вида морских птиц, гнездящегося в проливе Принца Уильяма на Аляске, упала после аварии и не улучшилась вплоть до 1994 г. Уменьшение величины кладок и успеха вылупления привело к снижению числа птенцов в загрязненных колониях. Популяции многих видов оставались в состоянии депрессии, а гнездовой успех и сроки размножения вернулись к нормальному уровню лишь в 1993 г. Исследования на уровне сообществ показали, что разлив не
фти в первый год после аварии привел к существенному снижению видового богатства и разнообразия (альфа-, лог-распределение), которые сохранялись на низком уровне спустя год.
При прогнозировании воздействия нефтяных разливов на птиц следует учитывать не только площадь загрязненных акваторий и побережья, эффективность мер очистки, роль абиотических факторов (время года, погодные условия и др.), но и видовое разнообразие сообщества, экологические ниши, численность локальных популяций отдельных видов, их охранный статус.
Возможны несколько сценариев воздействия разливов нефти на сообщества птиц. При авариях в местах скопления птиц на зимовках прогнозируемая прямая гибель составит от десятков до сотен тысяч водоплавающих в зависимости от размеров нефтяного пятна. Прогноз для локальных популяций гнездящихся редких видов с критической численностью — полное исчезновение без возможности естественного восстановления. Последствия — потеря генетического разнообразия для конкретных видов и сообщества в целом. Разливы нефти в период миграции птиц способны сказаться и на останавливающихся дальних мигрантах.
Птицы, являясь важными компонентами морских и прибрежных экосистем, замыкают трофические цепи и подвергаются прямому (загрязнение нефтью оперения, гнезд, яиц и птенцов, отравление) и косвенному воздействию загрязнения через снижение пищевых ресурсов (гибель фито-, зоопланктона, макрофитов и рыб). Косвенному воздействию могут подвергаться редкие виды хищных птиц, имеющих статус уязвимых (орлан-белохвост и скопа). При выбросах нефти в большей степени страдают птицы, которые собирают корм с поверхности моря (бакланы, чайки, крачки). Последствия — уменьшение численности за счет гибели особей, нарушения структуры популяций и репродуктивных стратегий, невозвращение на места гнездования с зимовок, депрессия численности. Долговременные эффекты — смещение сроков размножения, уменьшение величины кладки, ухудшение качества яиц и потомства, повышенная птенцовая и эмбриональная смертность, снижение репродуктивного успеха. В целом в сообществах при загрязнении происходит снижение видового богатства птиц, которое возрождается через несколько лет после принятия восстановительных мер (Лебедева, 1999).
ВЛИЯНИЕ ТЕХНОГЕННОГО ЗАГРЯЗНЕНИЯ
НА ЛЕСНЫЕ СООБЩЕСТВА
Последствия для экосистем, процесс их трансформации в результате воздействия Карабашско-го медеплавильного комбината были исследованы на Урале в Свердловской обл.
Были выявлены следующие стадии техногенной сукцессии.
1. Стадия выпадения чувствительных видов. Наблюдается элиминация наиболее чувствительных видов эпифитных лишайников. По большинству остальных параметров экосистемы не отличимы от фонового уровня.
2. Стадия структурных перестроек экосистемы. Регистрируется ухудшение санитарного состояния деревьев, но плотность древостоя и его запас не изменяются. Происходят изменения в травяно-кустарничковом ярусе (выпадают чувствительные виды лесного разнотравья). Значительно уменьшается численность дождевых червей. Замедлены процессы, осуществляемые почвенными микроорганизмами. Незначительно увеличивается толщина подстилки. Существенно уменьшаются разнообразие и обилие эпифитных лишайников. Параметры населения птиц и мелких млекопитающих остаются на уровне фона.
3. Стадия частичного разрушения экосистемы. Древесный ярус угнетен и изрежен, значительно уменьшены его запас и полнота, нарушено возобновление. В травяном ярусе почти отсутствуют лесные виды, которые заменены луговыми и видами-эксплерентами. Повышена кислотность верхних почвенных горизонтов, из них выносятся обменный кальций и магний. Активизируются эрозионные процессы. Биологическая активность почвы резко снижена. Крупные почвенные сапрофаги отсутствуют. Уменьшена скорость деструкции опада, который накапливается в виде толстого слоя подстилки. Лишайниковый покров сохраняется только у самого основания стволов, представлен одним—тремя устойчивыми видами. Происходит элиминация крупных лесных видов птиц, меньше общая плотность орнитонаселения. В то же время наблюдается вселение синантропных видов и видов, приуроченных к открытым местообитаниям (это характерно для населения птиц, мелких млекопитающих и муравьев). Беспозвоночные хортобионты увеличивают численность.
4. Стадия полного разрушения («коллапса») экосистемы. Древесный ярус полностью разрушен, сохраняются лишь отдельные сильно угнетенные экземпляры деревьев. Травяной ярус представлен одним-двумя видами злаков, в увлажненных местах встречается хвощ. В микропонижениях значительное развитие может получать одновидовой моховой покров. Лишайниковый покров отсутствует. На значительной части территории полностью смыты подстилка и верхние горизонты почвы (вплоть до горизонта В). В микропонижениях встречаются «захоронения* неразложившегосяопада. Биологическая активность почвы снижена до нуля. Типично почвенные животные отсутствуют (только под куртинами мха и в «захоронениях* опада встречаются личинки щелкунов и литобии-ды). Группировки птиц и мелких млекопитающих не поддерживают свою структуру и существуют за счет притока мигрантов с соседних участков.
Общее направление техногенных смен противоположно ходу естественных сукцессий и может быть охарактеризовано как движение вспять. Состояние экосистем в зоне техногенной пустыни аналогично стадии пионерного сообщества при демутационных сукцессиях. По мере увеличения нагрузки экосистема как бы продвигается к нему от климаксного (фонового) состояния через ряд последовательных стадий. Но
в значительной степени это сходство внешнее, поскольку механизмы, лежащие в основе техногенных и естественных смен, различны.
Техногенная сукцессия сопровождается снижением общего биологического разнообразия, падением продуктивности и упрощением структуры, замедлением и разрывом круговорота биогенов. Тормозятся как продукционные, так и деструк-ционные процессы, нарушается баланс между ними. Ряд структурных элементов полностью элиминируется (лесное разнотравье, почвенные сапрофаги, эпифитные лишайники и др.). Все это дает основание рассматривать наблюдающиеся изменения не просто как трансформацию экосистемы, а именно как ее деградацию. Две последние стадии, очевидно, патологичны. Переход к ним означает для экосистемы полную потерю устойчивости как способности возвращаться в исходное состояние.
МЕЖДУНАРОДНЫЕ ПРОГРАММЫ
МОНИТОРИНГА БИОРАЗНООБРАЗИЯ
Под мониторингом биоразнообразия понимается комплексная информационная система наблюдений за состоянием микроорганизмов, растительного и животного мира в целях выявления, анализа и прогнозирования возможных изменений на фоне естественных процессов и под влиянием антропогенных факторов. В полном объеме, т. е. с охватом всех районов и таксонов (или хотя бы основных), система мониторинга биоразнообразия сегодня не может быть реализована. Только общее исследование состояния биологического разнообразия на Земле согласно международной программе должно занять не менее 50 лет.
После подписания в 1992 г. Международной конвенции по биоразнообразию вопросы сохранения и устойчивого использования биологических ресурсов в разных странах мира приобрели еще большее значение. Решение многих из этих вопросов. весьма актуальных и для России, невозможно без укрепления международного сотрудничества. Важное место в этом сотрудничестве по праву занимает создание эффективных систем мониторинга за состоянием биоразнообразия и обмен данными, получаемыми на национальном, региональ-
ном и глобальном уровнях. В период реализации Международной программы «Биоразнообразие* в США с 1992 по 1997 г. действовала в этом направлении специальная «Биологическая информационная служба» России с 1994 по 2001 г. — департаменты Госкомитета по охране природы.
Глобальная система наземных наблюдений (GTOS). Мониторинг биоразнообразия не может проводиться в отрыве от наблюдений за другими компонентами окружающей среды и процессами, в ней происходящими, так как на биологические сообщества оказывают большое влияние и глобальные изменения климата, и уровень содержания тех или иных химических веществ в атмосфере, и особенности землепользования. Вопрос о необходимости осуществления глобальной программы мониторинга для сбора сопоставимых данных в разных типах экосистем был поставлен в 1972 г. на Стокгольмской конференции по окружающей среде, а в 1975г-в Найроби был открыт Программный центр ЮНЕП, который должен был объединить усилия разных стран и обеспечить формирование Глобальной системы мониторинга окружающей среды (GEMS). В течение последующих лет удалось добиться определенных успехов в этом направлении, и к настоящему времени эта система стала занимать центральное место в деятельности ЮНЕП по наблюдениям за нашей планетой (Earthwatch Programme). В ней в той или иной мере участвуют более 140 стран.
В 1985 г. ЮНЕП приступил к созданию Глобальной информационной базы данных о ресурсах (GRID), которая с использованием современных технологий, включая оперативную обработку спутниковой информации и создание ГИС, должна наряду с GEMS и другими системами, созданными ЮНЕП (INFOTERRA и ENRIN), обеспечивать лиц, принимающих решения, необходимыми сведениями о состоянии окружающей среды. В разных странах мира уже созданы и активно действуют 11 центров GRID, координацию деятельности которых осуществляет специальный отдел в штаб-квартире ЮНЕП. В перспективе планируется, что все страны мира через региональные центры GRID примут участие в этой глобальной сети. Важное место в программах сбора и обработки информации через центры GRID должны занять данные о состоянии биоразнообразия и ходе выполнения Конвенции по биоразнообразию. В связи с этим в ЮНЕП стали уделять больше внимания вопросам создания баз данных по биоразнообра-
зию, а для информации стран-участниц Конвенции в 1996 г. стал издаваться информационный бюллетень: «Biodiversity Data Management (BDM) Update».
При финансовой поддержке ЮНЕП и Всемирного фонда охраны природы (WWF) накопление и обработку данных по биоразнообразию осуществляет Всемирный центр мониторинга охраны природы (WCMC), созданный Международным союзом охраны природы и природных ресурсов (IUCN) в 1983 г. в Кембридже, Великобритания. При участии Комиссии МСОП по сохранению видов (IUCN/SSC) Центр регулярно издает Красные книги (Red Data books) и Красные списки (Red Lists) с данными о состоянии популяций различных видов животных и растений, которым грозит исчезновение. Важным вкладом в общую оценку состояния и степени изученности биоты на Земле стала публикация Центром сводки по глобальному биоразнообразию.
Несмотря на эти усилия и определенные успехи ряд вопросов создания и обеспечения функционирования многоцелевой глобальной системы мониторинга окружающей среды оставался нерешенным, на что было обращено внимание на Конференции ООН по окружающей среде и развитию (Рио-де-Жанейро, 1992).
По данным, представленным на Всемирном конгрессе по охране природы (Монреаль, Канада, 13—23 октября 1996 г.), в региональной сети долгосрочного экологического мониторинга (ROSELT) согласилась участвовать 21 страна Западной, Северной и Восточной Африки, которые в августе 1995 г. выделили 30 обсерваторий для наблюдений. Основная цель этих наблюдений — сбор разнообразных данных, которые должны помочь лучше понять механизмы, определяющие развитие процессов опустынивания на аридных территориях, и отобрать методы и технологии, которые благоприятны для рационального использования местных природных ресурсов и обеспечивают сохранение окружающей среды. На участках, выделенных для каждой обсерватории, предполагается обеспечить три пространственных уровня сбора данных, различающихся по своей интенсивности и объему. Каждая обсерватория может осуществлять наблюдения и исследования по отдельным или всем пяти рекомендованным направлениям: мониторинг состояния природных ресурсов, мониторинг их использования, восстановление ресурсов, определение допустимого уровня использования ресурсов и прогнозирование долговременных эко-
логических изменений. Одна из задач на будущее — по возможности согласовать методы наблюдений и обеспечить взаимодействие ROSELT с Глобальной системой наземных наблюдений (GTOS) (табл.11).
Таблица 11
Основные параметры Глобальной системы наземных наблюдений (GTOS) для мониторинга наземных и пресноводных экосистем
Категории участков и пунктов, входящих в GTOS Количество участков или пунктов Площадь обследуемой территории (кмг) Основные объекты и переменные Частота сбора данных
1. Участки для проведения крупномасштабных экспериментов и градиентных исследований 10 1000 Изменения газового состава и границ биомов на экологических мега-трансектах По специальной программе
2. Центры долговременных исследований 100 10 Изменения кругооборота энергии, воды, углерода, питательных веществ и др. переменных Стандартные измерения, регулярно в течение года с разной частотой
3. Полевые станции 1000 1 Изменения продуктивности экосистем, урожайности полей,характе ра землепользования и др. переменных Стандартные измерения, регулярно в течение года с разной частотой
4. Участки периодических наблюдений (без постоянных сотрудников) 10000 0,01-0,1 Характеристика земной поверхности и почв(содержание гумуса и др. переменные) С интервалом в 5—10 лет
5, Участки для частых дистанционных (remote sensing) наблюдений 10е 0,001-1,0 Динамика состояния крон деревьев и земной поверхности Регулярная спутниковая съемка
Согласно программе, в ближайшие годы в ключевых биомах на суше (и в ряде случаев на море) при тесном сотрудничестве с правительствами заинтересованных стран должны быть открыты долговременные (как минимум на 50 лет) исследовательские обсерватории, на базе которых будут проводиться не только наблюдения за различными параметрами окружающей среды, но и повышение квалификации местных полевых исследователей и обучение студентов и школьников. Долговременный мониторинг будет проводиться на всех обсерваториях (точное их число пока не установлено) по унифицированным программам, которые охватывают три основные категории параметров:
1) описание обследуемого участка, включая историю его освоения человеком;
2) фоновые характеристики окружающей среды: от климатических показателей до регистрации различных природных и антропогенных воздействий;
3) оценки изменений в экосистемах: влагообеспеченность, растительность, животное население, почвенный покров, отражательная способность земной поверхности и др.
Идея создания сети геосферно-биосферных обсерваторий для проведения комплексных исследований глобальных изменений окружающей среды давно привлекает внимание международных организаций. В частности, в 1987 г. в Париже было проведено специальное совещание, на котором были разработаны рекомендации по созданию таких обсерваторий разного класса и предложено размещать их для изучения пространственных и временных закономерностей на экологических мегатрансектах. Эти идеи получили дальнейшее развитие в международной программе «Диверситас* и в международной геосферно-биосферной программе.
К этому времени в 62 странах мира уже было 226 биосферных заповедников. Разработанный на Минском конгрессе и утвержденный затем ЮНЕСКО и ЮНЕП «План действий по биосферным заповедникам* (1984) должен был на многие годы определить их развитие по 9 целевым направлениям, среди которых важное внимание уделено вопросам инвентаризации таксонов растений и животных и развитию долгосрочного экологического мониторинга.
В 1996 г. общее число биосферных заповедников, утвержденных ЮНЕСКО/МАБ, достигло 337, они расположены в 85 странах. Все российские заповедники многие годы ведут на
блюдения по программе «Летопись природы* (Заповедники России, 1994) и, как видно из Справочника по биосферным заповедникам Европы и Северной Америки (ACCESS, 1996), наши биосферные заповедники наилучшим образом обеспечены постоянными участками для мониторинга различных показателей (флора, фауна, климат, почвы и др.).
Наибольшее количество биосферных резерватов (БР) создано в США (47), на втором месте находится Россия (21). Третье место занимает Болгария (17), далее следуют Китай и Испания (по 16), Германия и Великобритания (по 13), Австралия (12) и Мексика (11). В каждой из остальных стран число БР менее 10. Таким образом, пока еще наблюдается определенная диспропорция в распределении БР по отношению к площади стран и их ландшафтно-зональному разнообразию, что особенно заметно при сравнении сети БР в Болгарии и Китае. В ближайшем будущем предстоит глобальный анализ сети БР с целью их более равномерного распределения и сравнимой репрезентативности в отношении экосистемного разнообразия. В противном случае сеть БР, не сравнимых между собою по площади, принадлежности к определенной биогеографической провинции и т. д., не сможет быть основой для равновесного глобального экологического мониторинга.
Интересен опыт работы не только заповедников, но и национальных парков в деле сохранения живой природы, не изолированной от человека.
Со времени заселения Австралии европейцами (с 1788 г.) здесь вымерло 30 видов зверей и птиц и 100 видов растений. Под угрозой исчезновения (в Красной книге) находятся 60 видов зверей, птиц, амфибий и рептилий, рыб, сотни видов беспозвоночных, более 200 видов растений. Первый национальный парк (НП) в Австралии — Ройял — был создан в 1879 г., через семь лет после создания Йеллоустонского НП в США в 1872 г. Это второй НП в мире! Его посещают более 2 млн человек в год! Всего в Австралии 550 НП и 2300 других охраняемых территорий на площади 50 млн га, что составляет 6,5% территории страны! Самый большой НП страны — Какаду, его площадь — 2 млн га. Аборигены жили на его территории еще 60 тыс. лет тому назад, судя по наскальным рисункам.
Морские парки (МП) — их 160, на акватории 40 млн га. Самый большой МП в мире — Большой Барьерный Риф, площадью 35 млн га. Остальные МП — около 5 млн га.
МОНИТОРИНГ БИОРАЗНООБРАЗИЯ
В РОССИИ
В России, имеющей развитую сеть заповедников (93, включая 21 биосферных) и национальных парков (31) и значительное число экологических и агрономических станций, принадлежащих различным научным ведомствам и учреждениям, к сожалению, пока не создано собственной Программы долговременных экологических исследований (LTER), и она временно официально не присоединилась к создаваемой Глобальной системе наземных наблюдений (GTOS). Скорее всего это задача ближайшего будущего, так как уже разрабатывается Единая государственная система экологического мониторинга (ЕГСЭМ) и предлагается даже сформировать федеральную целевую программу для обеспечения ее создания и функционирования.
В последние годы активно велась разработка научных основ мониторинга природных процессов и состояния окружающей среды. Важнейшие теоретико-методологические аспекты мониторинга были разработаны в нашей стране к концу 1980-х годов. В системе бывшего Госкомгидромета были организованы базовые наблюдения по программе экологического мониторинга, а в ряде биосферных заповедников — важней-
шие элементы глобального мониторинга. В этой системе мониторинг биоразнообразия рассматривается как составная часть биологического или экологического мониторинга.
Мониторинг может проводиться с разными целями, с разной степенью полноты наблюдений; он может охватывать как отдельные популяции или виды, так и быть предметом комплексного глобального наблюдения за биосферой в целом из космоса. Такой глобальный мониторинг следит за изменениями в планетарном масштабе и требует для этого наименее измененных природных экосистем и специальных методов регистрации. На практике обычно имеют дело с региональным мониторингом, который требует иных подходов, чем глобальный мониторинг.
Известно, что биологическое разнообразие тесно связано с организацией экосистем. Стабильные и нетронутые экосистемы сохраняют высокую степень разнообразия, а в разрушенных экосистемах или находящихся под сильным влиянием антропогенных факторов обычно отмечается обеднение биоразнообразия. Особую ценность имеют длительные наблюдения за состоянием экосистем. Однако связь разнообразия и устойчивости экосистем вряд ли укладывается в простую формулу «чем больше разнообразие, тем лучше». Принято считать, что исторически сложившееся экосистемное разнообразие какого-либо региона, безусловно, способствует стабилизации круговорота веществ и энергии.
На первом этапе исследований нас интересует прежде всего таксономическое разнообразие: списки видов основных групп растений и животных, различные простые показатели разнообразия (индексы доминирования, показатели выравненности видов и т.п.). На втором этапе проводятся выделение доминантных видов и выяснение их функциональной роли в этих экосистемах, оценка биомассы и численности основных видов, анализ внутрисистемных связей и роли абиотических факторов в динамике численности популяций, определение ведущих потоков энергии. После этого возможен переход к третьему этапу работ — организации долгосрочного мониторинга.
Главная цель — слежение за состоянием биоразнообразия видов и экосистем в масштабе всей страны или крупного региона. Это подразумевает сбор данных, адекватных по репрезентативности, точности и регулярности их поступления для исследования состояния, причин изменения и принятия решений. Основное внимание здесь уделяется выбору показателей,
по которым можно судить о состоянии экосистемы. По установленным закономерностям и подмеченным тенденциям строится экологический прогноз. Стратегическая цель — предсказание возможных изменений и управление экосистемами.
Таким образом, настоящий мониторинг биоразнообразия экосистем возможен при работе определенных коллективов, и существенные успехи могут быть достигнуты при применении современных методов получения, обработки и хранения информации.
В наше время существуют предпосылки для создания единой системы мониторинга на основе объединения усилий уже работающих структур. Можно разработать и единую национальную программу по биологическому мониторингу. Объектом такой национальной программы должно быть все разнообразие животного и растительного мира. С этой точки зрения все разнообразие видов можно разбить на три группы:
1) фоновые виды, к которым можно применить стандартный подход;
2) индикаторные виды, по которым необходимо получать углубленные данные;
3) редкие виды (особо охраняемые), по которым собираются любые доступные сведения.
Мониторинг не следует рассматривать как принципиально новую систему наблюдений, он может и должен опираться на опыт и инфраструктуру уже существующих служб.
В России в прошлом был налажен мониторинг по разным группам растений и животных. И все ведомства, связанные с управлением и использованием природных ресурсов, традиционно вели мониторинг их состояния. Это, прежде всего, служба защиты растений (ВНИИЗР и др.), которая вела и ведет контроль за болезнями и вредителями растений. Фитоса-нитарный мониторинг организует такую систему наблюдений, которая гарантирует оперативное получение информации для проведения защитных мероприятий для предотвращения вреда от насекомых, грызунов и возбудителей болезней. На современном этапе все больше применяются автоматические агрометеостанции, которые стали обязательным элементом высокоразвитого растениеводства во многих странах мира.
В России имелся опыт мониторинга почв в виде сети агрохимических станций. По инициативе Д.Н. Прянишникова было организовано около 1500 таких станций для слежения
за почвой под разными группами сельскохозяйственных культур. Сейчас почвенно-экологический мониторинг проводится инфраструктурой системы Росгидромета и Роскомзема. Он служит тем экологическим фоном, на котором прослеживаются пространственно-временные изменения почвенной биоты и растительности. Его основу составляют наблюдения за гидротермическим режимом почв и химическими загрязнениями на сельскохозяйственных полях. Выбранные эталонные объекты мониторинга должны отражать состояние и динамику изменений биоразнообразия на зонально-региональном, локальном и точечном уровнях.
При активном участии Н.П. Наумова создавалась система противочумных станций, которая на большой площади степных и пустынных ландшафтов на протяжении 50 лет вела контроль за грызунами и эпидемиологической обстановкой в стране (Северо-Запад, Северный Кавказ, Прикаспий, Волго-Уральский район, Средняя Азия, Тува, Алтай, Забайкалье и Дальний Восток). Все противочумные станции в настоящее время находятся в подчинении Комитета по Госсанэпиднадзору. Большие материалы по другим природноочаговым зоозонам накоплены и эпидстанциями. Главная задача станций — выявление видового состава и оценка численности грызунов как основных носителей возбудителей чумы в одних и тех же биотопах на протяжении длительного отрезка времени с целью оценки эпизоотологической и эпидемиологической ситуации. К сожалению, сейчас эта служба находится в заброшенном состоянии.
Лесной мониторинг проводится лесотехнической службой страны давно, причем это не просто наблюдения за изменением состояния лесного покрова с разработкой кадастра типов леса, но и прогнозирование будущего состояния лесного фонда России с выходом на систему управления лесным хозяйством. Аналогичная система мониторинга водных ресурсов существует на пресноводных и морских водоемах страны (система отраслевых институтов рыбного хозяйства типа ВНИРО, ГосНИОРХ и ВНИПРХ), которая обязана не просто вести мониторинг водных ресурсов страны, но давать оценку их запасов, определять допустимые нормы вылова и предлагать рекомендации по увеличению биологической продуктивности водоемов. Мониторинг гидробионтов имеет свои традиции и особенности: здесь давно используется продукционный подход, а в оценке разнообразия применяются не только списки
видов, но и такие количественные показатели, как численность, биомасса, продукция, характеристики роста и воспроизводства, трофические связи в экосистеме.
В 1979 г. в России создана государственная служба учета охотничьих ресурсов страны, и собранные ею данные были использованы для оценки разнообразия массовых видов крупных и средних млекопитающих Центральной России и водоплавающих птиц Сибири. Эта служба довольно скоро может быть использована для мониторинга биоразнообразия наземных сообществ на большей части территории России. Уже с 1991 г. оценка численности охотничьих животных ведется в 70 субъектах Российской Федерации. Эти данные служат основой для определения норм отстрела животных и управления охотничьим хозяйством страны.
Программа ведения Красной книги курирует сбор информации по редким и исчезающим видам растений и животных. В 1978 г. опубликована «Красная книга СССР», в 1985 и 1988 гг. вышли два тома «Красной книги РСФСР», в 2001 г. — ♦Красная книга Российской Федерации».
Особо следует остановиться на системе заповедников России. Именно в заповедниках можно наиболее успешно проводить мониторинг биоразнообразия. Сейчас в России насчитывается 93 заповедника, которые занимают 1,8% территории страны. В Летописях природы находится уникальный материал, в некоторых заповедниках имеются непрерывные ряды наблюдений за 50 лет. Однако заповедники сами по себе не могут быть идеальными полигонами для мониторинга, так как они не отражают всего многообразия ландшафтов, особенно нарушенных, аихбиотавсе больше превращается в «ложноостровную». Поэтому при выборе стационаров для наблюдения необходим регионально-ландшафтный подход с учетом мозаичности разнообразия на всех уровнях, и в последние годы для этих целей все чаще стали использоваться «биосферные заповедники».
Исторически на примере заповедников выделяются следующие этапы мониторинга:
1) инвентаризация биоразнообразия;
2) выбор репрезентативных стационаров для наблюдения;
3) проведение режимных наблюдений за состоянием экосистемы;
4) создание и ведение информационной системы хранения, обработки и анализа оперативной информации об их состоянии;
5) оценка состояния биоразнообразия и прогнозирование возможных изменений.
Помимо этой системы государственных служб мониторинг ведут институты Российской Академии наук, университеты и отраслевые институты на своих биостанциях и стационарах. Отметим такие давние российские биологические подмосковные станции, как «Косино» и «Озеро Глубокое», морские станции МГУ, станции РАН «Картеж» и «Дальние Зеленцы» и многие другие.
Можно отметить следующие принципы в организации и проведении мониторинга в рамках всей страны.
1. Сохранение старой и создание новой инфраструктуры. Вопрос о сохранении старой инфраструктуры, хорошо зарекомендовавшей себя в сборе данных, стоит особенно остро в современной экономической ситуации страны. Естественно, что эти структуры должны работать по новым методическим разработкам и с новой техникой. Вместе с тем назрела необходимость создания иных структур. Одной из таких структур может быть единый всероссийский центр по мониторингу птиц: только на его основе, могут быть сделаны взвешенные выводы о состоянии вида в ареале; развито картографическое, математическое и программное обеспечение мониторинга.
2. Выбор районов зависит от целей и задач конкретного мониторинга. Различают эталонные территории (обычно заповедные), фоновые (для стандартных наблюдений) и зоны с антропогенной трансформацией. При выборе районов для мониторинга биоразнообразия целесообразно ориентироваться на «горячие точки», т. е. на участки с повышенным числом видов. Особого внимания заслуживают районы с максимальным эндемизмом, уникальные экосистемы (Байкал, ковыльные степи и др.). Несомненно, следует продолжить сбор данных по традиционным маршрутам и трансектам (например, «Кольский меридиан» в Баренцевом море и азиатско-российская ме-гатрансекта).
3. Периодика сбора данных. Выделяются два основных временных интервала для сбора и анализа данных: ежегодный и раз в 5—10 лет. Ежегодно должны собираться данные на эталонных территориях и/или по индикаторным видам. Раз в 5— 10 лет необходимо проводить полную съемку.
4. Выбор параметров. Много проблем связано с выбором показателей. Невозможно контролировать и вести наблюде-
5Г
ния за всеми видами и всеми экосистемами. Выбор модельных групп должен определяться их хорошей изученностью и возможностью слежения не только за видовым составом, но и на популяционном и внутривидовом уровнях. Из параметров прежде всего важна регистрация факта «присутствия—отсутствия» вида. Однако в большинстве случаев этого мало, поэтому для массовых видов обязательна оценка численности, размерно-возрастного состава и других популяционных параметров. При проведении массового мониторинга и на обширных территориях обычно число параметров ограничено, при стационарных исследованиях их число возрастает в зависимости от целей и задач мониторинга, а также от реальных возможностей исследовательских групп.
5. Создание баз данных и банка данных. Этим занимаются практически все научные коллективы. Основные проблемы в этом направлении — совместимость различных баз данных для создания единого банка данных и сохранение авторских прав на информацию.
6. Финансирование работ по мониторингу, особенно на первом этапе его организации, должно хотя бы в минимальном объеме выполняться из бюджетных средств как в виде грантов, так и плановых заданий. Можно ожидать, что первые же успешные шаги могут быть активно поддержаны международными природоохранными организациями, которые заинтересованы в сотрудничестве.
Новым в работах по мониторингу является выход на международный уровень. Это опыт создания станции фонового глобального экологического мониторинга на базе Центрально-лесного биосферного заповедника в рамках российско-германского сотрудничества. Здесь интересно отметить разработку методов дистанционного мониторинга состояния наземных экосистем и моделирование экологических процессов с целью оценки текущего и прогнозируемого состояния экосистем. В России созданы две сети экологического мониторинга, на которых должны выполняться международные программы: сеть биосферных заповедников (МАБ/ЮНЕСКО) и сеть станций по международной программе влияния загрязненного воздуха на экосистемы (ЮНЕП ЕЭК ООН). Несомненно, что в ближайшем будущем таких совместных международных программ будет все больше и больше, и в них видное место займет мониторинг биоразнообразия.
ГЛОБАЛЬНЫЕ
ЭКОЛОГИЧЕСКИЕ
ИЗМЕНЕНИЯ
Воздействие человека на биосферу приобрело глобальный характер, а его масштабы и темп продолжают возрастать. В результате как прямого, так и косвенного антропогенного влияния многие биологические виды исчезают или их популяции на критическом пределе численности, ставящем под угрозу возможность воспроизводства вида. Воздействие человека на сообщества живых организмов стало настолько мощным, что они уже не в состоянии противостоять процессам антропогенной трансформации и утрачивают важнейшее свойство природных сообществ — способность к самовосстановлению. В связи с этим сокращается площадь тропических лесов, идет расширение пустынных формаций за счет уменьшения площади сообществ саванн, лесов, степей, происходит так называемое антропогенное опустынивание, что отражено в ежегоднике ЮНЕП «Глобальная экологическая перспектива» (2000).
Изменение климата. В конце 1990-х гг. ежегодные выбросы двуокиси углерода почти в четыре раза превысили уровень 1950 г., а ее содержание в атмосфере достигло наивысшего за последние 160 тыс. лет уровня. По мнению Межправп-
тельственной группы по изменению климата, «имеющиеся доказательства свидетельствуют о явном влиянии человека на климат планеты». Ожидаются, в частности, смещение климатических зон, изменение видового состава и продуктивности экосистем, увеличение числа экстремальных погодных явлений и усиление влияния изменения климата на здоровье людей. В соответствии с Рамочной конвенцией ООН об изменении климата и Киотским протоколом в ближайшее время начнется реализация мер по контролю и снижению выбросов парниковых газов.
Разрушение стратосферного озонового слоя. Основные результаты по сокращению производства, потребления и выброса озоноразрушающих веществ (ОРВ) достигнуты и продолжают достигаться благодаря Монреальскому протоколу и соответствующим поправкам к нему. Максимальное содержание ОРВ в нижних слоях атмосферы наблюдалось в 1994 г., а сейчас оно постепенно снижается. Ожидается, что приблизительно к 2050 г. озоновый слой восстановится до уровня, существовавшего до 1980 г.
Проблема незаконной торговли постоянно находится в центре внимания национальных правительств. Тем не менее значительные объемы ОРВ контрабандой перевозятся через государственные границы. Многосторонний фонд и Фонд глобальной окружающей среды оказывают содействие развивающимся странам и странам с переходной экономикой в поэтапном сокращении производства ОРВ.
Избыточный азот. Мы удобряем Землю в глобальных масштабах посредством интенсивного сельского хозяйства, сжигания ископаемого топлива и широко распространенного возделывания бобовых культур. Растет число доказательств того, что громадные объемы дополнительно вносимого азота усиливают ацидификацию (закисление), приводят к изменению видового состава экосистем, непомерно увеличивают содержание нитратов в потребляемой человеком пресной воде, а также ведут к эвтрофикации многих пресноводных экосистем. Кроме того, перенасыщенные азотом воды и стоки с удобряемых сельхозугодий усиливают так называемое цветение водоемов в прибрежных зонах, что может привести к кислородному голоданию и последующему замору рыбы на больших глубинах, а также к снижению биоразнообразия морских организмов в результате межвидовой конкуренции. Выбросы азо-
та в атмосферу способствуют глобальному потеплению. Ученые все больше сходятся во мнении по вопросу о том, что современные масштабы нарушения круговорота азота могут иметь глобальные последствия, сравнимые с последствиями нарушения круговорота углерода.
Опасность химического загрязнения. В связи с глобальным распространением химических веществ и активизацией их применения возрастает воздействие пестицидов, тяжелых металлов, мелких частиц и других химических веществ, которые все больше угрожают здоровью людей и состоянию окружающей среды. Применение пестицидов служит причиной 3,5—5 млн случаев острых отравлений в год. Во всем мире ежегодно вырабатывается 400 млн т опасных отходов, причем около 75% производства пестицидов и опасных отходов приходится на развитые страны. Несмотря на существующие во многих из них ограничения на применение токсичных и стойких химикатов, таких как ДДТ, ПХД и диоксины, эти вещества до сих пор делаются на экспорт и широко используются в развивающихся странах. Прилагаются значительные усилия с целью поощрения выпуска более чистой продукции, ограничения выбросов, поэтапного отказа от применения некоторых стойких органических загрязнителей, контроля за производством и продажей отходов, а также совершенствования способов сбора и удаления отходов.
Катастрофы. Частота и последствия стихийных бедствий, таких как землетрясения, извержения вулканов, ураганы, пожары и наводнения, продолжают возрастать. Это не только воздействует на жизнь миллионов людей, приводя к их гибели, увечьям и экономическим потерям, но и создает новые проблемы окружающей среды. В качестве одного из примеров можно привести вышедшие из-под контроля лесные пожары, свирепствовавшие в 1996—1998 гг. в Бразилии, Греции, Индонезии, Италии, Канаде, в автономном районе Внутренняя Монголия на северо-востоке Китая, в Мексике, России, США, Турции и во Франции. Влияние лесных пожаров на здоровье людей может быть весьма серьезным. Эксперты считают показатель загрязнения, равный 100 мкг/м3, вредным для здоровья; в Малайзии этот индекс достигал 800 мкг/куб. м. Ущерб, нанесенный здоровью жителей Юго-Восточной Азии в результате лесных пожаров, оценивается приблизительно в 1,4 млрд долларов США. Пожары представляют также серь-
езную угрозу для сохранения биоразнообразия, особенно в случаях, когда выгорают охраняемые территории. Системы раннего оповещения и реагирования пока еще слабы, прежде всего в развивающихся странах, поэтому необходимо срочно совершенствовать информационные инфраструктуры и развивать технические возможности реагирования.
Тропическое течение Эль-Нинъо. Необычные погодные условия, наблюдавшиеся в 1997—1998 гг., объясняются приходом южного течения Эль-Ниньо (полное название: Эль-Ниньо-южная осцилляция, ЭНСО), что привело к рекордному повышению температуры в Тихом океане. До середины 1998 г. наличие этой огромной массы теплой воды влияло на климат всего мира и стало причиной существенных разрушений и ущерба во многих районах, включая и умеренный пояс. Главными факторами отрицательного воздействия стали обильные осадки и наводнения, засухи и лесные пожары.
Земельные и лесные ресурсы и биоразнообразие. Леса, редколесья и лугопастбищные угодья продолжают подвергаться деградации или уничтожению, маргинальные земли превращаются в пустыни, а площади природных экосистем сокращаются и дробятся, что повышает риск уменьшения биоразнообразия. Новые данные позволяют утверждать, что изменение климата может в ближайшие десятилетия привести к усилению почвенной эрозии во многих регионах, что грозит сокращением производства продовольствия. В развивающихся странах высокими темпами продолжается сведение лесов, вызванное главным образом потребностями в лесо- и пиломатериалах, а также необходимостью отвода земель для нужд сельского хозяйства и других целей. За период 1990—1995 гг. было вырублено около 65 млн га лесов, а до этого их общая площадь составляла 3500 млн га. Увеличение лесных земель в развитых странах на 9 млн га лишь в малой степени возмещает эту потерю. Качество остающихся лесов находится под угрозой по ряду причин, включая, в частности, вредное воздействие кислотных дождей, использование древесины на дрова, а также оскудение водоемов и лесные пожары. Сокращение или деградация мест обитания животных чреваты снижением биоразнообразия на генетическом, видовом и экосистемном уровнях, что затрудняет обеспечение основными видами товаров и услуг. Еще одна важная причина снижения биоразнообразия — широко распространенная в настоящее
время интродукция. Большинство подвергающихся опасности видов обитают на суше, причем более половины из них — в лесу. Очень уязвимы также пресноводные и морские места обитания, особенно коралловые рифы.
Морские и прибрежные зоны. Урбанизация и развитие промышленности, туризм, сельское хозяйство, сброс и слив отходов в море приводят к деградации прибрежных районов по все земному шару и вызывают разрушение таких экосистем, как водно-болотные угодья, мангровые заросли и коралловые рифы. Климатическое изменение оказывает влияние на качество морской воды и на изменение уровня моря. Низменные территории, включая множество малых островов, находятся под угрозой затопления. За период 1975—1995 гг. общемировой улов рыбы почти удвоился, и в настоящее время рыболовство находится в состоянии кризиса. Около 60% районов мирового рыболовства уже достигли или скоро достигнут такого состояния, при котором уловы начинают снижаться.
Атмосфера. Существуют значительные различия в тенденциях загрязнения воздуха в развитых и развивающихся странах. Энергичные усилия многих промышленно развитых стран привели к тому, что загрязнение атмосферы начало уменьшаться. Однако в большинстве крупных городов развивающихся стран уровень загрязнения воздуха достигает критических величин. Главные причины загрязнения воздуха — автомобильный транспорт, сжигание угля и серосодержащих видов топлива, а также лесные пожары. Население развивающихся стран также подвержено воздействию значительного загрязнения воздуха внутри помещений от открытых источников огня. В настоящее время считается, что около 50% хронических респираторных заболеваний связано с загрязнением атмосферы. Из-за кислотных дождей подвергаются деградации значительные площади лесных и сельскохозяйственных угодий.
Биоразнообразие. Все виды, так же как и все особи внутри отдельного вида, имеют конечную продолжительность жизни, и, следовательно, изменения в биоразнообразии неизбежны. Тем не менее ускорившееся и усугубившееся снижение разнообразия на генном, видовом и экосистемном уровнях не только нежелательно само по себе, но и представляет существенную угрозу материальному благополучию человечества, по-
5
скольку предполагает снижение потенциала экосистем в отношении предоставления ключевых продуктов и услуг.
Общее число видов на Земле огромно: примерно 1,7 млн было описано, но полагают, что их гораздо больше: оценки колеблются от 5 до 100 млн. В качестве разумной рабочей оценки было предложено число в 12,5 млн видов. Наиболее богатые видами экосистемы на Земле представлены влажными тропическими лесами, которые занимают более 8% площади суши и, вероятно, содержат более 90% всех видов Земли. В целом самыми богатыми по биоразнообразию являются Африка, Азиатско-Тихоокеанский регион и Латинская Америка.
Статус сохраненности большинства видов в подробностях неизвестен, но две крупные группы животных — млекопитающие и птицы — были всесторонне оценены и могут дать представление о состоянии биоразнообразия в целом. В1996 г. 25% из примерно 4630 видов, обитающих в мире млекопитающих, и 11% из 9675 видов птиц, согласно оценкам, находились под угрозой в глобальном масштабе, т. е. подвергались значительному риску полного исчезновения (IUCN, 1996). Бесчисленное количество других видов, хотя и не находятся под угрозой исчезновения в глобальном масштабе, в настоящее время существуют в меньших количествах и в виде разрозненных популяций, и многим из них угрожает исчезновение на уровне отдельных стран.
Большинство из находящихся под угрозой видов живут на суше, более половины из них обитают в лесах, но появляется все больше свидетельств уязвимости пресноводных мест обитания и таких морских мест обитания, как коралловые рифы. Например, в США пресноводные виды — почти 70% двустворчатых моллюсков, 50% речных раков и 37% рыб — подвергаются большему риску вымирания, чем сухопутные виды.
Пищевые растения служат примером основополагающих ценностей биоразнообразия. Изначально растения употреблялись в пищу непосредственно в своем природном состоянии^ и сбор природной продукции сегодня продолжается по всему миру. Только несколько из многих видов цветковых растений рассматриваются как прямые источники пищи, хотя другие служат пищей для животных, на которых, в свою очередь, охотятся или которых разводят люди.
Примерно 200 видов были окультурены как пищевые растения, и около 20 из них имеют международное экономичес-
кое значение. На относительно небольшое число семейств растений приходятся основные пищевые растения мира: Gramineae (травы, включая хлебные злаки) и Leguminosae (бобовые культуры, включая горох, бобы и чечевицу) занимают среди них ведущие места.
Благодаря эволюции пищевых культур за много веков возделывания увеличился спектр генного разнообразия, но развитие и продвижение высокоурожайных сортов для современного интенсивного земледелия в настоящее время стремительно обращает эту тенденцию вспять, что создает опасную зависимость от генетически однородных культур, часто нуждающихся для получения высоких урожаев в больших дозах удобрений и пестицидов. С широким распространением интенсивного земледелия многие местные разновидности были вытеснены, а некоторые полностью исчезли. Диким родственникам окультуренных видов часто также угрожает вымирание в результате изменений мест обитания.
Все большее сужение генетической основы, вероятно, является причиной периодических спадов производства экономически важных культур, что приводит к росту колебаний урожайности и синхронности этих колебаний на обширных территориях. Например, снижение урожайности кукурузы на 15% в 1970 г. в США приписывается широкомасштабному распространению сорта, подверженного заболеваниям.
Древесина, все еще добываемая в основном в дикой природе, является одним из наиболее важных товаров в международной торговле. Потенциально ценные ресурсы лесоматериалов во многих регионах мира деградируют из-за чрезмерных порубок, ненадлежащего управления и утраты мест обитания. Например, из более чем 600 видов крупных деревьев в Гане около 60 имеют коммерческое значение, а примерно 25 видов были признаны нуждающимися в защите из-за их чрезмерной эксплуатации или потому, что они редко встречаются. Недавно проведенный анализ примерно 10 тыс. видов деревьев (из 100 тыс. возможно имеющихся в мире) показал, что почти 6 тыс. соответствуют критериям статуса вида, находящегося под угрозой исчезновения. Из них 976 видов признаны находящимися в критическом состоянии, 1319 — находящимися под угрозой и 3609 — уязвимыми. Потеря или изменение мест обитания составляет основу риска, особенно для видов с ограниченным ареалом обитания, но из всех причин в ка-
честве угрозы чаще всего называлась вырубка леса (по 1290 видам).
В самом общем плане потери в биоразнообразии вызываются экономическими интересами, которые не придают должного значения окружающей среде и ее ресурсам, законодательными и институциональными системами, которые способствуют неустойчивой эксплуатации, а также неравномерным распределением собственности на природные ресурсы и доступа к ним, включая выгоды от их использования. В то время как некоторые виды находятся под непосредственной угрозой, например вследствие охоты, браконьерства или нелегальной торговли, основная опасность исходит от изменения землепользования, приводящего к уничтожению, трансформации или дроблению мест обитания. Например, за последние два десятилетия утрачено 80% пресноводных водно-болотных угодий Нигера. Две трети ареалов обитания диких животных и растений в Азии были уничтожены, причем наиболее существенные потери имели место на Индийском субконтиненте, в Китае, Вьетнаме и Таиланде. В Латинской Америке среднегодовые темпы обезлесения в 1990—1995 гг. составляли 2,1%, в т.ч. более 1% в Парагвае, Эквадоре, Боливии и Венесуэле.
Еще одна серьезная причина потерь биоразнообразия — широко распространенное внедрение видов растений и животных в районы, не являющиеся их естественной средой обитания, что влечет за собой изменения на уровнях сообществ и экосистем, а иногда и полное уничтожение некоторых из первоначальных видов. Например, около 18% из 119 видов млекопитающих Австралии и обеих Америк, а также 20% птиц всего мира, находящихся под угрозой исчезновения, в 1992 г. пострадали из-за внедренных в среду их обитания хищников или конкурирующих видов. Последствия внедрения чужеродных видов наиболее заметны в закрытых экосистемах, таких как озера и острова. Например, по меньшей мере 60% рыб в озере Виктория считаются вымершими в результате внедрения туда нильского окуня.
Морские и прибрежные зоны. Океаны — крупнейшие экосистемы земного шара. Они так же богаты и разнообразны, как и любая наземная экосистема, но все еще остаются в большинстве своем неисследованными. Хотя глубины океана в основном еще не загрязнены, по некоторым районам имеются све-
дения об ухудшении состояния экосистем среды и снижении численности многих морских видов. Напротив, среда морских побережий, несомненно, испытывает воздействие изменения и разрушения мест обитания, перепромысла и загрязнения. Многие из этих воздействий вызваны деятельностью, ведущейся на суше, вдали от морей. Наибольшая опасность угрожает внутренним морям. Аральское море практически мертво, а эпиконтинентальные моря, такие как Средиземное, Черное и Балтийское, сильно загрязнены. Прибрежные лагуны загрязнены повсюду в мире.
Более трети населения земного шара живет на побережьях морей — 50% в Северной Америке и 60% в Латинской Америке, где на морском побережье расположено соответственно 60 из 77 крупнейших городов, — живет в радиусе 100 км от побережья. В Азии к 2000 г. почти 500 млн человек оказались сосредоточенными в городских конгломерациях вдоль побережья. Природная среда прибрежных районов, к которой относятся водно-болотные угодья, устья рек, мангровые заросли и коралловые рифы, подвергается деградации в ходе сельскохозяйственного и городского развития, деятельности промышленных предприятий, сооружения портов и дорог, драгированная и засыпки, туризма и аквакультуры. Сооружение плотин, даже расположенных в глубине материка, может изменить режим водотока, где сосредоточены существенные запасы промысловых рыб, а также перекрыть поступление наносов, необходимых для поддержания состояния дельт и побережья.
Множество людей, живущих в прибрежных зонах и даже в глубине материка, производят большое количество отходов и прочих загрязняющих веществ, которые попадают в моря напрямую или через прибрежные водосборы, бассейны рек и выпадение осадков из загрязненного воздуха. Хотя загрязнение побережий во многих развитых странах постепенно начинает контролироваться, оно все еще увеличивается быстрыми темпами в результате прироста населения, урбанизации и промышленного роста в развивающихся регионах. Например, 38% побережья Африки и 68% принадлежащих ей морских заповедников подвергаются серьезной опасности из-за развития экономики. Более чем половине рифов земного шара потенциально угрожает деятельность человека, а в наиболее населенных районах под угрозой находятся до 80% рифов.
В состоянии прибрежной и морской среды происходят некоторые, хотя и единичные улучшения. Например, улучшилось состояние пляжей для купания во многих регионах, стали чище некоторые реки Западной Европы и снизилось содержание ДДТ в Балтийском море и у Тихоокеанского побережья Северной Америки, что повлекло за собой восстановление численности некоторых популяций животных и птиц. Тем не менее гораздо больше предстоит сделать, чтобы во всем мире масштабы восстановления превзошли масштабы разрушения, также необходимо приложить усилия для решения проблемы сброса отходов в моря, угрожающего флоре и фауне.
Растет понимание потенциального влияния изменения климата на морскую среду. Например, увеличение испарения с поверхности более теплых морей повышает атмосферную влажность и таким образом усиливает парниковый эффект. До недавнего времени внимание уделялось в основном воздействию подъема уровня моря на малые островные государства и на страны, расположенные на небольшой высоте над уровнем моря, а также увеличению частоты и интенсивности штормов вследствие изменения климата. Возможны, тем не менее, и более сложные последствия. Например, если потепление продолжится, на Норвежском и Гренландском морях может образоваться линза пресной воды от растаявших льдов Арктики, что повлечет за собой изменение структуры глубинных течений, которые могут повернуть к югу воды Гольфстрима, в настоящее время обогревающие зимой Европу.
Потепление поверхности моря и рост термической стратификации могут также сократить производительность фитопланктона, который является основой всей пищевой цепи в морях. Накопление углекислого газа в атмосфере может привести к повышенной кислотности поверхностных слоев океана, что наряду с проникновением ультрафиолетового излучения также сократит производительность фитопланктона. Может последовать и изменение содержания карбонатов в поверхностных водах, что повлияет на рост кораллов.
Общий объем улова морской рыбы в мире возрос примерно с 50 млн тв 1975 г. до более чем 97 млнтв 1995 г. Это свидетельствует о том, что планомерно эксплуатируются и истощаются все новые виды рыб и новые места лова. Производительность аквакультуры тем временем резко возросла, и теперь на ее долю приходится почти 20% всего производства рыбы и
морепродуктов. Постоянные неудачи с установлением контроля за переловом означают, что около 60% рыбных промыслов Мирового океана близки к точке, на которой уловы уменьшаются, или уже достигли ее, и многие местные рыболовные общины уже переживают катастрофическое сокращение своих ежегодных уловов.
Часто рыболовный промысел ведет к деградации морской среды обитания и видов в наиболее биологически продуктивных и коммерчески ценных морских экосистемах, таких как мангровые заросли и коралловые рифы. Интенсивные формы аквакультуры создают дополнительные проблемы в отношении окружающей среды в виде сильного загрязнения местных вод и гибели прибрежных экосистем.
Для почти 1 млрд человек рыбная продукция является основным источником белка, и прогнозируется, что спрос на рыбопродукты увеличится примерно с 75 млн т в 1994—1995 гг. до 110—120млнтв2010г.При рациональном управлении морские уловы могут устойчиво повышаться приблизительно на 10 млн т в год. Тем не менее, если в ближайшее время не будут приняты решительные меры, производительность может снизиться. По данным ФАО, большая часть прогнозируемого увеличения спроса на рыбу может быть удовлетворена только путем дальнейшего развития аквакультуры. Отмеченные процессы по-разному проявляются на отдельных континентах.
Африка обладает богатой и разнообразной флорой и фауной, в том числе основными сельскохозяйственными культурами, такими как сорго и просо. На континенте произрастает свыше 50 тыс. известных видов растений, водится 1 тыс. видов млекопитающих и 1,5 тыс. видов птиц. Традиционно многие из этих местных видов были основой существования африканских общин, причем у африканцев вошло в обычай охранять и поддерживать численность животных в целях выживания не только нынешнего, но и будущих поколений. На территории проживания отдельных племен участки с особенно богатым биоразнообразием часто выделялись в качестве священных рощ и охраняемых земель.
В Африке первые национальные парки были созданы в первой половине XX в., в том числе национальный парк Крюгера в Южной Африке (1928 г.) и природный заповедник в районе горы Тубкаль в Марокко (1944 г.). В1938 г. арабские страны провели симпозиум по проблемам охраны природы и в ре-
зультате официально признали многие из существовавших в этих странах охраняемых территорий (UNESCO, 1954).
Наибольшее число эндемичных видов млекопитающих (55%), птиц (63%), пресмыкающихся (49%) и земноводных (40%) встречается в Восточной Африке, в то время как в Северной Африке эндемиков относительно мало. Больше всего их встречается на Мадагаскаре — занимает шестое место в мире по числу эндемичных видов (более 300) среди высших позвоночных (млекопитающих, птиц и земноводных) и третье место в Африке, после Конго и Танзании, по количеству видов растений. Капская флористическая область входит в число шести областей мира, отличающихся наиболее значительным разнообразием видов растений.
Самые обширные по площади экосистемы Африки — саванны, богатейшие лугопастбищные угодья в мире. В них обитают многие местные виды растений и животных и сосредоточены самые большие в мире популяции крупных млекопитающих — слонов, буйволов, носорогов, жирафов, львов, леопардов, гепардов, зебр, бегемотов, лесных антилоп, водяных козлов и сернобыков.
Во всех районах Африки это масштабное и разнообразное биологическое наследие находится в опасности. Некоторые виды уже считаются вымершими, в том числе четыре вида антилоп в Лесото и Свазиленде, голубой гну в Малави, сасса-би в Мозамбике, эндемичный нильгау на юго-западе Капской провинции в Южной Африке и болотный козел в Танзании. В настоящее время многие другие виды находятся под угрозой исчезновения. В Мавритании опасность грозит 23% млекопитающих. В Западной и Центральной Африке среди исчезающих видов древесных растений можно отметить Guarea excelsa, Milicia excelsa, Nauclea diderric, а также лекарственные растения Voacanga africana, Zanthoxyhmm zanthoxyloides и Brucea guineensis. Под угрозой исчезновения оказались такие виды млекопитающих, как шимпанзе, сенегальская коровья антилопа (Alcelaphus bucelaphus), слоны (Loxodanta africana) и один из трех видов ламантина (Trichechus senegalensis). Сообщается, что в Эритрее исчезновение угрожает 22 видам растений. Количество находящихся под угрозой исчезновения видов может быть больше, поскольку видовое разнообразие в Африке изучено и описано частично.
ГЛАВА IX
£
Африканские водно-болотные угодья обладают богатым биоразнообразием с большим количеством эндемичных и редких видов растений, а также представителей дикой фауны, в том числе мигрирующих птиц. Водно-болотные угодья встречаются в большинстве африканских стран, самые обширные среди них — дельта р. Окаванго, дельта р. Судд в суданской провинции Верхний Нил, берега озер Виктория и Чад, а также затапливаемые земли в дельтах рек Конго, Нигер и Замбези. Несмотря на то, что водно-болотные угодья относятся к экосистемам с самой высокой в Африке биологической продуктивностью, на местном уровне их часто рассматривают как пустоши, места обитания сельскохозяйственных вредителей, представляющие угрозу для здоровья населения, или потенциальные площади для сельскохозяйственного освоения. В результате многие водно-болотные угодья исчезают. Например, за последние 20 лет XX в. в Нигере площадь незасоленных водно-болотных угодий сократилась более чем на 80%. Возрастает также угроза утраты прибрежных угодий в Египте и Тунисе и незасоленных водно-болотных угодий в Судане. Из всех экосистем наибольшая опасность угрожает, возможно, пресноводным экосистемам озер, рек и водно-болотных угодий. Они уже утратили больше характерных дЛя них видов и мест обитания, чем наземные и морские экосистемы. Неизбежны новые потери в результате строительства плотин, загрязнения, чрезмерного вылова рыбы и прочих факторов.
Снижению биоразнообразия в течение XX столетия способствовала также интродукция чужеродных видов растений — некоторые «пришельцы» вытеснили представителей местной растительности. К примеру, отдельные части «фин-бос» в Южной Африке и пастбища на восточных плоскогорьях Зимбабве оказались засорены австралийскими видами Acacia и Pinus, в результате чего местные виды Rest io, Erica и Protea оказались под угрозой исчезновения. Островные виды, например произрастающие на островах Индийского океана, особенно подвержены исчезновению, вызываемому конкуренцией или хищническим образом жизни экзотов. Однако процесс интродукции сейчас замедлился, и, если будут ужесточаться правила ввоза биоресурсов, эта положительная тенденция может сохраниться.
Гражданские конфликты и войны также наносят серьезный ущерб окружающей среде и приводят к снижению био-
разнообразия как в пределах, так и вне границ охраняемых территорий, а также создают большие трудности в работе организаций, занимающихся окружающей средой, и в реализации природоохранных программ. К 1991 г. численность диких животных в национальных парках и заповедниках Анголы жз-за гражданской войны сократилась в 10 раз по сравнению С их количеством в 1975 г.
Биоразнообразию в Африке также стало угрожать изменение климата. Уже признанным считается тот факт, что климатическое изменение, выразившееся в резком уменьшении объемов водных объектов в результате продолжительной засухи и интенсификации хозяйственной деятельности по берегам водоемов, способствовало сокращению популяций земноводных.
На Азиатско-Тихоокеанский регион приходятся части трех из восьми биогеографических областей земного шара — Палеарктической, Индо-Малайской и Океанической. В регионе находятся и высочайшая горная система мира — Гималаи, второй по площади массив влажных тропических лесов и более половины всех имеющихся в мире коралловых рифов. В Юго-Восточной Азии отмечается наибольшее разнообразие диких и культурных видов злаков и фруктовых пород.
Из 12 стран, отличающихся «мегаразнообразием», четыре расположены в данном регионе. Это Австралия, Китай, Индонезия и Малайзия. По биоразнообразию Китай занимает третье место в мире: здесь представлено более 30 тыс. видов высших растений и 6347 видов позвоночных, что составляет соответственно 10 и 14% этих видов, обитающих на земном шаре. В Австралии насчитывается примерно 1 млн видов, из которых 85% видов цветковых растений, 84% видов млекопитающих, более 45% видов птиц, 89% видов пресмыкающихся, 93% видов лягушек и 85% видов прибрежных рыб, обитающих в умеренном поясе, эндемичны.
Вследствие растущей фрагментации местообитаний в Юго-Восточной Азии сократилось разнообразие лесной продукции, служившей основным источником пищи, лекарственных средств и доходов для коренного населения. В меньшей степени эти процессы затронули бассейн Меконга, в частности труднодоступные районы вдоль государственных границ. В Южной Азии, особенно в пределах Индийского субконтинента, основное беспокойство вызывает уменьшение биоразнообра-
зия, обусловленное сочетанием таких факторов, как длительные нагрузки на луговые угодья, быстрый рост населения и поголовья скота.
Современное сельское хозяйство также ведет к уменьшению генетического разнообразия. Например, в Индонезии за 1975—1990 гг. исчезли около 1500 сортов риса, аналогичные тенденции наблюдаются в регионе повсеместно. Ожидается, что к 2005 г. 75% производимого в Индии риса будет получено всего лишь от 10 сортов, тогда как традиционно в стране выращивалось 30 тыс. сортов.
Охота, браконьерство, незаконная торговля видами, находящимися под угрозой исчезновения, оказали сильное воздействие на биоразнообразие во многих странах. В Лаоссе и во Вьетнаме, например, мясо диких животных считается деликатесом, и поэтому множество видов являются объектами охоты. Браконьерство, незаконный сбор и торговля лекарственными растениями и животными все шире распространяются. В Пакистане соколов отлавливают и контрабандой доставляют в страны Ближнего Востока, ящериц и змей убивают ради кожи, а охота на крокодилов до сих пор является там излюбленным видом спорта и отдыха.
Интродукция видов — еще одна важная причина снижения биоразнообразия, особенно в Океании. Например, бурая древесная змея, которая уничтожила многие местные популяции птиц.
Коммерческое рыболовство, в том числе с использованием отравляющих и взрывчатых веществ, приводит к истощению прибрежных биологических ресурсов. В водах Новой Зеландии недавно появились признаки разрушения экосистем отмелей рыболовными траулерами, которые занимаются ловлей глубоководных рыб, таких как австралийский ерш. Кроме того, на морскую среду отрицательно влияет загрязнение с судов, в особенности нефтью, а в некоторых районах — сбросы токсичных отходов. Прибрежные экосистемы, особенно коралловые рифы, где обитают рыбы, разрушаются под влиянием таких факторов, как сельскохозяйственные стоки, заиливание, сброс городских сточных вод, промышленное загрязнение, а в таких странах, как Мальдивы, от индустрии туризма. Уничтожение мангровых зарослей приводит к разрушению нерестилищ многих морских видов и нередко служит причиной возникновения других форм деградации прибрежных эко
систем. Два ярких примера — мангры Чакарья Сундар-бана на востоке Бангладеш, практически полностью сведенные для развития аквакультуры, и мангровые заросли Таиланда, более половины которых (около 208 220 га) было уничтожено за период 1961—1993 гг.
Количественная оценка реального масштаба изменений мест обитания и сокращения видового разнообразия в регионе до сих пор не проводилась, так как имеющиеся данные не полны или устарели. Однако установлено, что около двух третей всех мест обитания диких животных Азии уже уничтожены, а 70% основных типов растительности Индо-Малайской биогеографической области (включающей Южную Азию, бассейн Меконга и Юго-Восточную Азию) исчезли, что могло привести к потери еще до 15% наземных видов. Сухие и влажные леса утрачены соответственно на 73 и 69%, а площади переувлажненных земель, болот и мангров сократились на 55%. В целом наиболее значительные потери мест обитания имели место на Индийском субконтиненте, в Китае, Вьетнаме и Таиланде.
«Горячими точками» (где исчезновение уже находящихся под угрозой влажных тропических лесов могло бы привести к самым ощутимым потерям биоразнообразия) являются еще сохранившиеся леса на Филиппинах, в пределах полуостровной Малайзии, на северо-западе Калимантана, в Восточных Гималаях, Западных Гхатах в Индии, на юго-востоке Шри-Ланки и в Новой Каледонии.
Под угрозой вымирания находится большое число видов. Из 640 видов, включенных в список охраняемых согласно Конвенции о международной торговле видами фауны и флоры, находящимися под угрозой исчезновения (СИТЕС), 156 видов встречаются в Китае, из них 15—20% находятся под Угрозой уничтожения.
В основных экосистемах Австралии за 1788—1995 гг. произошли следующие изменения: сведено около 40% всех лесов; утрачено более 60% водно-болотных угодий прибрежной зоны на юге и востоке страны; сведено около 90% лесных массивов и зарослей кустарникового эвкалипта в умеренном поясе; утрачено более 99% низинных лугопастбищных угодий в умеренном поясе на юго-востоке страны; сведено около 75% влажных тропических лесов; погибло до 85% некоторых массивов водорослей. Поставлен печальный мировой рекорд по исчезновению видов млекопитающих. За последние 200 лет вымер-
ли 10 из 144 видов сумчатых животных и 8 из 53 местных видов грызунов; 5% высших растений и 23% млекопитающих перестали существовать; находятся под угрозой исчезновения или уязвимы 9% птиц, 7% пресмыкающихся, 16% земноводных и 9% пресноводных рыб.
Кроме того, в Австралию было завезено большое число видов, что нанесло сильный урон местным экосистемам. Это кролики (приблизительно 200 млн особей), лисы (5 млн), кошки (12 млн), козы, бизонова трава, криптостегия, пурпуровое просо, гигантская мимоза стыдливая, сиамский сорняк и грибок Phytophthora cinnamoni, являющийся патогеном, угрожающим всем местным растительным сообществам в некоторых районах на юге Австралии. Интродуцировано по меньшей мере 55 видов морских рыб и беспозвоночных, а также несколько видов морских водорослей. Некоторые из них были завезены намеренно для развития аквакультуры, а некоторые попали случайно с балластными водами морских судов или прикрепившись к их корпусу.
В Австралии около 5% покрытосеменных растений и 9% наземных позвоночных относятся к категории либо угрожаемых, либо уязвимых видов. В Новой Зеландии список видов, находящихся под угрозой исчезновения, продолжает расширяться, частично из-за того, что состояние некоторых видов и мест обитания продолжает ухудшаться. Под угрозой вымирания находятся около 800 видов и 200 подвидов животных, растений и грибов. Одной из наиболее уязвимых групп являются эндемичные наземные и пресноводные птицы Новой Зеландии, 37 видов которых (из 50) могут перестать существовать.
В настоящее время в Европе практически не осталось нетронутых природных территорий. Около трети площади занимают леса, доля которых составляет от 6% в Ирландии до 66% в Финляндии (ЕЕА, 1995).
В странах Восточной Европы почти на половине территории сохранились леса, а еще 20% общей площади приходится на естественные луга, т. е. 70% их земельных ресурсов можно считать неокультуренными. Тем не менее, на протяжении XX в. почти 10% таких земель было превращено в пашни или пастбища.
Увеличение площади лесных массивов в Западной Европе и в странах Балтии будет, вероятно, продолжаться, особенно в связи с выводом ряда территорий из интенсивного сельско-
хозяйственного использования в соответствии с новой Общей сельскохозяйственной политикой ЕС. Сокращение выбросов закисляющих газов уменьшит вредное воздействие на лесные массивы, однако для восстановления лесных почв, подвергшихся закислению, потребуется много времени.
Основную угрозу биоразнообразию в Европе и Центральной Азии представляют изменения в землепользовании, загрязнение окружающей среды, новые методы ведения сельского, лесного и водного хозяйств, интродукция чужеродных видов и пород животных и растений, чрезмерная эксплуатация ресурсов, а также развитие туризма. Важным (если не преобладающим) фактором негативного воздействия может также стать изменение климата.
Во многих европейских странах под опасностью исчезновения оказывается половина известных видов позвоночных. Особую важность для Западной Европы имеет проблема защиты мест сезонного пребывания перелетных птиц. Более трети европейских видов птиц находится на грани исчезновения главным образом из-за того, что в результате изменений в землепользовании и возрастания нагрузки со стороны сельского и лесного хозяйства наносится вред их местам обитания.
В последнее время становится все очевиднее, что природные и полуприродные места обитания уменьшаются в размерах и не могут обеспечить условий существования определенных видов растений и животных. Особенно остро эта проблема стоит в Западной Европе, где наибольшую угрозу местам обитания представляет продолжающееся развитие сети автомобильных дорог и высокоскоростных железнодорожных магистралей.
В Центральной Азии избыточное орошение в бассейне Аральского моря привело почти к полному уничтожению уникальных лесов в долинах и дельтах рек, служивших некогда средой обитания множества видов птиц, рыб и млекопитающих. Резко сократилось на этих территориях и многообразие видов.
Главнейшим препятствием для сохранения биоразнообразия является сокращение природных и полуприродных мест обитания. Одной из основных задач в масштабах Европы становится объединение различных охраняемых территорий в общеевропейскую сеть путем создания биокоридоров, или проходов, для диких животных. Обязательно надо создать более
надежную защиту ландшафтов в целом и связанных с ними специфических систем землеустройства. Например, сохранение и надлежащий уход за небольшими лесными массивами и живыми изгородями вокруг пашен обеспечивают убежища, необходимые для выживания многих видов птиц и мелких полевых животных. Модернизация методов ведения сельского хозяйства, обычно влекущая за собой увеличение размеров полей, часто приводит к уничтожению таких убежищ для диких животных.
Благодаря своему значению дДя сохранения живой природы водно-болотные угодья оказались одними из первых мест обитания, взятых под охрану. В рамках Рамсарской конвенции о водно-болотных угодьях, имеющих международное значение, в Европе и Центральной Азии охраняются около 300 таких мест обитания, помимо примерно 70 объектов мирового природного наследия и биосферных заповедников, которые также имеют важное значение для охраны диких животных. Однако водно-болотные угодья продолжают сокращаться, особенно в Южной Европе, а также и во многих сельскохозяйственных и урбанизированных районах Северо-Западной и Центральной Европы. Среди других важных мест обитания и ландшафтов чрезмерную нагрузку испытывают песчаные дюны, старые и полуприродные лесные массивы, а также районы полуприродного сельского хозяйства.
В Западной Европе почти нет признаков улучшения ситуации с исчезновением видов и потерей их мест обитания. В других регионах ситуация менее однозначна. Так, относительно мало затронутые деятельностью человека территории все еще сохранились в Центральной и Восточной Европе и в Центральной Азии. Главным препятствием к достижению природоохранных целей остается здесь невключенность проблемы биоразнообразия в другие политические сферы деятельности.
Экосистемы тропического, субтропического и умеренного поясов Латинской Америки отличаются огромным биоразнообразием. В неотропической области произрастает около 68% всех влажных тропических лесов в мире; сосредоточено 40% видов животных и растений планеты; считается, что там наблюдается наибольшее разнообразие флоры в мире.
В теплых речных долинах Амазонии, высоких и холодных Андах, приатлантических лесах Бразилии, сухих лесах Цен-
тральной Америки располагаются самые богатые экосистемы мира. Аридная и семиаридная растительность встречается в горных районах от юга Эквадора до Чили, а также на севере Колумбии, Венесуэлы, Аргентины и на северо-востоке Бразилии. В Бразилии, Парагвае и Боливии находятся обширнейшие водно-болотные угодья мира, из которых 400 тыс. км2 — болота, известные своим биоразнообразием.
Главная проблема — как предотвратить разрушение мест обитания и последующее исчезновение видов, многие из которых еще не изучены наукой. Распространение сельского хозяйства на семиаридные районы, вырубка лесов, иссушение водно-болотных угодий значительно уменьшили популяции многих видов. Потеря мест обитания является основной угрозой. Изменение условий обитания доходит до критического уровня в лесах Центральной Америки, лесах «чако», экосистемах саванн бразильских «серрадо» (где наблюдается наибольшее в мире разнообразие растительности саванн), кустарниках средиземноморского типа на Тихоокеанском побережье. В Мексике зимует 51% всех видов перелетных птиц из северных стран, и потеря жизненно важных мест перезимовки из-за обезлесения и других типов изменения землепользования может угрожать выживанию этих популяций.
Несмотря на то, что не предпринимались систематические оценки изменения среды обитания и уменьшения отдельных видов, имеющиеся данные указывают на значительное воздействие человека на животный мир. В настоящее время исчезновение грозит нескольким сотням видов позвоночных животных. Интенсификация сельскохозяйственного производства, лесозамена посредством лесопосадок, новые технологии возделывания суходолов (важного источника биоразнообразия) и изменение береговой линии свидетельствуют о том, что эти тенденции могут усугубиться в ближайшем будущем.
Под угрозой находится биота всех стран региона. Бразилия занимает второе место в мире по числу находящихся под угрозой исчезновения видов птиц (103 вида), а Перу и Колумбия — пятое место (по 64 вида в обеих странах). Трети позвоночных животных Чили (исключая морских рыб) грозит уничтожение. В Бразилии, кроме того, находится в опасности 71 вид млекопитающих (четвертое место в мире). Более чем 50% млекопитающих и птиц Аргентины также грозит исчезновение. Как правило, на территориях, где имеется большое чис-
ло находящихся под угрозой видов птиц, отмечается так же тенденция в отношении млекопитающих. Это означает, что обе названные группы животных могут быть чувствительны к сходным неблагоприятным воздействиям. Экосистемы и их растительность также находятся под угрозой. Например, по приблизительной оценке, в центральных районах Чили в период с 1978 по 1987 г. 30% естественных лесов «маулино» в Кордильера-де-ла-Коста было замещено посадками сосны. Площадь земель, охваченных различными формами охраны и защиты, продолжает расти, при этом около 6,6% земель региона относится к категории строго охраняемых.
На территории Северной Америки степень биоразнообразия увеличивается с севера на юг. Примерно 7807 видов растений, 233 вида млекопитающих и 160 видов птиц эндемичны или уникальны для Северной Америки. Особенно большой процент эндемиков приходится на Мексику. К ним относятся не менее 40—50% видов цветковых растений, более половины всех земноводных и рептилий; почти 50% видов рыб и 33% видов млекопитающих. Напротив, в Канаде эндемизм сравнительно низок. Уникальные виды в основном встречаются на островах либо в районах, избежавших оледенения. В США эндемизм наиболее распространен на островах, особенно на Гавайях, где 44% видов высших растений, 95% ракообразных и моллюсков, 43% птиц и 30% рыб внутренних водоемов являются уникальными.
В XX столетии разрушение мест обитания, нерегулируемая охота или заготовка лесопродуктов, конкуренция со стороны привнесенных видов привели к упадку и исчезновению многих североамериканских видов растений и животных. Отмечалось также сокращение генетического разнообразия сельскохозяйственных культур и животных. В последнее время опасность, приписываемая охоте и чрезмерной эксплуатации биоресурсов, уменьшилась по сравнению с вредом, проистекающим из разрушения, деградации и фрагментации мест обитания. Действительно, потеря мест обитания и их изменение стали основной угрозой сохранению разнообразия живой природы в Северной Америке.
В особо опасном положении находятся места обитания в водно-болотных угодьях, жизненно необходимые для многих форм живой природы. Начиная с XVII в. шел процесс исчезновения или деградации многих из канадских водно-болот-
ных угодий. В 85% случаев это было связано с осушением земель для нужд сельского хозяйства. С этой целью в Канаде осушено 80% болот в дельте р. Фрейзер, 70% котловин в прериях, 68% заболоченных земель на юге провинции Онтарио и 65% засоленных маршей на Атлантическом побережье. Аналогично в США было осушено, дренировано или различным образом изменено более половины всех площадей водно-болотных угодий. Большая часть этих угодий (48 млн га) утеряна в 48 граничащих между собой штатах. На Аляске потери водно-болотных угодий невелики по сравнению с их первоначальной площадью в 68 млн га. Тем не менее в целом переувлажненные земли по-прежнему занимают около 1,27 млн км2, или 13% всей территории Канады. Это четверть мировой площади водно-болотных угодий.
В Северной Америке находятся 50 массивов переувлажненных земель международного значения — так называемые «Рамсарские зоны». Страны, присоединившиеся к Конвенции о водно-болотных угодьях, имеющих международное значение главным образом в качестве мест обитания водоплавающих птиц, согласились отвести под водно-болотные заказники. Их совокупная площадь составляет 14,2 млн га, из которых 13 млн га находятся в Канаде. Сокращение площадей водно-болотных угодий, в числе прочего, способствовало в долгосрочной перспективе уменьшению численности некоторых популяций диких уток.
Закон об охране перелетных птиц, принятый в США в 1918 г., помог восстановить численность многих североамериканских видов, значительно сократившуюся в результате бесконтрольной охоты. Совместные действия в этом направлении, в том числе проведение инициативы под названием «Исследование гнездовий птиц в Северной Америке», начатое в 1966 г. США и Канадой, позволили получить дополнительную информацию о состоянии и тенденциях изменения численности мигрирующих видов североамериканских птиц. Тем не менее проблемы охраны мигрирующих видов сохраняются, и утрата мест обитания остается одной из основных опасностей, грозящих популяциям мигрирующих видов в Северной Америке. Многие птицы, летучие мыши, бабочки и морские млекопитающие проводят летний сезон в северных районах континента, а зиму — в южных, а другие мигрируют исключительно в пределах тропического пояса. В Мексику
прилетает 51 % всех видов перелетных птиц из северных стран, и потеря мест перезимовки из-за обезлесения и других изменений землепользования может угрожать выживанию этих популяций. Трансформация или деградация районов гнездования птиц на севере, наряду с сокращением в застроенной прибрежной зоне мест, необходимых для остановок на отдых в процессе миграции, способствует сокращению численности перелетных певчих птиц. С многочисленными проблемами сталкиваются и другие мигрирующие виды. Например, бабочка данаида в процессе своей исключительно своеобразной миграции подвергается угрозе гибели в связи с интенсивным хозяйственным использованием земель на побережье Калифорнии, вырубкой деревьев в зоне ее обитания во влажных тропических лесах Мексики, а также потерей мест обитания (заросли молочая) в Канаде.
В 1990—2000 гг. целенаправленная или случайная интродукция видов-«пришельцев» стала создавать растущую угрозу коренному биоразнообразию, нанося при этом не только биологический, но и экономический ущерб. Так, в США на сегодняшний день насчитывается по меньшей мере 4500 популяций видов-«пришельцев», из которых около 15% наносят тот или иной вред. К видам, оказывающим наиболее значительное воздействие, относятся непарный шелкопряд, привнесенные муравьи Рихтера (Solenopsis Invicta), дербенник иво-листный (Lythrum salicaria) и. полосатый двустворчатый моллюск.
В ноябре 1990 г. Конгресс США принял закон о некоренных видах водных растений и животных, а в мае 1993 г. — первый в мировой практике закон, регламентирующий использование балластных вод. Законом для судов, следующих в порты Великих озер, предусматривается обязательная замена балластных вод, причем производимая в открытом море. Такая замена снижает риск проникновения видов-«пришельцев», но не исключает его.
В целом в Северной Америке 430 видов животных, птиц, пресмыкающихся, земноводных и рыб находятся под угрозой исчезновения. Треть запасов пресноводных рыб относится к редко встречающимся видам либо находится под угрозой исчезновения. Водоемы США отличались наибольшим в мире разнообразием видов пресноводных моллюсков, но 55% из них уже исчезли или могут быть уничтожены. Вымирание расти-
тельных видов также вызывает обеспокоенность как на региональном, так и на глобальном уровне, особенно исходя из соображений сохранения генетического фонда. По данным подготовленного в 1997 г. отчета, посвященного оценке состояния примерно 20,5 тыс. коренных видов растений и животных США, две трети из них находятся в удовлетворительном состоянии, а треть требует принятия мер по охране. Особенно низкие оценки состояния получили организмы, распространенные в пресных водоемах, такие как моллюски, ракообразные, рыбы и земноводные. Цветочные растения также получили низкие оценки, причем треть видов, общим числом около 5144, была отнесена к проблемной группе.
В прилегающих к территории США акваториях Атлантического и Тихого океанов и Мексиканского залива насчитывается не менее 85 видов морских млекопитающих, 18 из них, в соответствии с Законом США, относятся к числу находящихся под угрозой исчезновения. Запрет китобойного промысла благоприятствовал росту популяций, но до сих пор действует ряд факторов, способствующих дальнейшему сокращению численности этих видов. К ним относятся побочный вылов животных в результате промышленного лова рыбы, браконьерство, выброс на берег, запутывание в сетях, распространение заболеваний и воздействие загрязняющих веществ.
Около 2,5 млн км2 по всей территории Северной Америки, включая наземные, пресноводные и морские угодья, были выведены из хозяйственного оборота и получили статус национальных парков и других типов охраняемых районов, чтобы сохранить редкие, находящиеся под угрозой или уязвимые экосистемы и поддерживаемые ими виды и генетические ресурсы. Эти земли составляют приблизительно 9% от всей территории Северной Америки.
Для обеспечения защиты биоразнообразия необходимо создание базы данных с целью мониторинга его состояния и установления приоритетов. Примером служит Канадская стратегия защиты биоразнообразия, разработанная правительством. Эта стратегия определяет общенациональные задачи и стратегические направления работы по охране и обеспечению устойчивого использования биоразнообразия. Таким образом, для властей федерального, провинциального и территориального уровней создается основа планирования в этой сфере.
ГЛАВА IX
£
Экосистемы Западной, Азии весьма разнообразны. Наземные экосистемы включают средиземноморские леса на севере и субтропическую горную растительность на юге и юго-западе региона. От северных до южных частей региона протянулись обширные пустыни со скудным растительным покровом, в особенности в так называемой «пустой четверти» (пустыни Руб-эль-Хали, Дехна, Нефуд и др.). Морские экосистемы включают обширные прибрежные зоны, ограничивающие полузамкнутые акватории Персидского залива. Средиземного и Красного морей, а также открытого Аравийского моря. Основные морские экосистемы представлены затопляемыми во время приливов участками прибрежной полосы, мангровыми зарослями, водорослями и коралловыми рифами. Пресноводные экосистемы сформировались вокруг малых и крупных рек Ирака, Сирии, Ливана и Иордании. На всей территории региона встречаются природные источники пресной воды.
Населяющие регион народы традиционно использовали природные места обитания и сохраняли биоразнообразие путем, например, применения системы охраны пастбищных угодий (Аль-Хема) и запретов на охоту в определенные месяцы года. Отбор по генетическим признакам с целью выведения улучшенных сортов зерновых культур и пород овец здесь начался более 10 тыс. лет назад. Однако в более поздние периоды перевыпас, обезлесение и охота привели к опустыниванию и вымиранию некоторых местных видов растений и животных. К ним относятся азиатский лев (Panthera leo persicus), который обитал на севере и исчез в 1918 г., сирийский дикий осел (Equus hemionus hemippus), уничтоженный в 1928 г., и аравийский страус (Struthio camelus syriacus), обитавший в Сирии и Аравии и истребленный в результате перепромыс-ла в 1940-е гг.
Экосистемы Западной Азии включают многочисленные виды растений и животных. Количество зарегистрированных видов растений колеблется от 301 в Катаре до более 3 тыс. в Сирии. Количество видов морских водорослей колеблется от 216 в Персидском заливе до 481 в Красном море; 21 вид млекопитающих обитает в Кувейте и 92 вида — на Западном берегу р. Иордан и в секторе Газа; видов птиц Засчитывается от 312 в Кувейте до 413 в Саудовской Аравии; видов пресмыкающихся — от 29 в Кувейте до 84 в Саудовской Аравии.
Красное и Аравийское моря отличаются богатством флоры и фауны. Здесь обитает более 330 видов кораллов, 500 видов моллюсков, 200 видов крабов, 20 видов морских млекопитающих и более 1200 видов рыб. Серьезный ущерб морскому биоразнообразию был нанесен переловом рыбы, загрязнением среды и разрушением мест обитания.
Многие виды морских животных, в том числе средиземноморский тюлень-монах, морские черепахи и морские губки, находятся под угрозой исчезновения из-за продолжающегося ухудшения качества воды в прибрежных зонах. Реальную опасность для прибрежных экосистем представляет внедрение морских вод. Массовая выемка песка для строительных целей на побережье Ливана и Сирии усугубила эту проблему и привела к разрушению мест обитания многих видов животных прибрежной зоны и прилегающей морской акватории, в том числе для морских черепах.
В регионе насчитывается более 800 эндемичных видов сосудистых растений, 7 эндемичных видов млекопитающих, 10 эндемичных видов птиц. От 20 до 23% видов кораллов являются эндемичными, а в Красном море эндемичны 17% видов рыб. Более 30% видов растений встречаются только в этом регионе, из них около 233 находятся под угрозой исчезновения. Эндемичными являются 32% видов растений на о. Сокотра, Йемен. Под угрозой исчезновения находятся такие эндемичные виды, как аравийский леопард (Panthera pardus nimr), полосатая гиена (Hyaena hyaena), аравийский тар (Hemitragus jayakari) и аравийский волк (Canis lepus arabs).
Повсеместно в регионе создаются охраняемые территории и национальные парки. В качестве примеров можно привести кедровый лес Барук, национальный заповедник Эхден и морской резерват на Пальмовом острове в Ливане, научный заповедник переувлажненных земель Азрак в Иордании, болотный заповедник Умм-Куаср в Ираке, заповедник Харрат-эль-Хар-ра, национальный парк Асир и морской заказник Эль-Джу-байль в Саудовской Аравии, заповедник аравийского сернобыка (орикса) в Джиддат-эль-Харасисе и заповедник морской черепахи в Рас-эль-Хадде в Омане, а также кедровый и пихтовый заповедник в Сирии.
Полярные регионы. В результате деятельности человека в Арктике сократились площади девственных ландшафтов,
или «дикой природы». Восемь арктических государств — членов Арктического совета в рамках Программы по охране арктической флоры и фауны (ОАФФ) обязались взять под охрану не мёнее 12% площади в каждой экологической зоне Арктики.
Эндемичные для Арктики виды исчисляются сотнями. Многие из них уникальны с генетической точки зрения, значительная часть является мигрирующими, и они часто образуют крупные скопления на ограниченных территориях, например, вдоль кромки плавучих льдов и в пределах наземных миграционных коридоров.
Занимающие крайне малые площади экосистемы Арктики вместе с населяющими их немногочисленными видами растений, животных и микроорганизмов испытывают существенные климатические колебания.
Уже доказано, по крайней мере для растений, что климатические колебания повлекли за собой значительные генетические вариации в пределах вида. Примерами могут служить сосуществование кустовой и стелющейся форм пурпурной камнеломки (Saxifraga oppositifolia) на Шпицбергене, произрастание отдельных локально адаптированных популяций дриад (Dryas octopetela) в сообществах в каменистых пустынях и на снеговых пластах, а также развитие необычайно сильной изменчивости у растений вида Draba. Подобные экотипичные вариации ценны с точки зрения выживания видов в долгосрочном плане, поскольку наделяют их свойствами, позволяющими противостоять колеблющимся климатологическим условиям.
Весьма вероятно, что гетерогенность мест обитания и популяций в Арктике наряду с длительной историей климатических колебаний в регионе высоких широт позволили современной флоре Арктики выработать определенную гибкость, необходимую для приспособления к существенным и порой быстрым изменениям внешней среды, и сохранить видовое разнообразие. Возможно, аналогичное генетическое разнообразие присуще также видам животных и микроорганизмов Севера, обеспечивая их «преадаптацию» (т. е. заранее существующую возможность приспособления) к изменению климата.
Хотя уровни антропогенного воздействия на экосистемы и биологическое разнообразие Арктики, видимо, относительно низки (по сравнению с регионами умеренного пояса и тро-
5
пиков), они могут иметь более серьезные последствия для этих относительно простых экосистем. Биоразнообразию Арктики угрожает как непосредственная деятельность человека внутри и за пределами этого региона, так и ее косвенное воздействие. Причинами фрагментации мест проживания и вреда, наносимого видам и среде их обитания, являются различные формы загрязнения, туризм, добыча полезных ископаемых, чрезмерное изъятие биологических ресурсов, внедрение чужеродных видов, а также последствия изменения климата и повышение интенсивности ультрафиолетового излучения. Следует учитывать и кумулятивный эффект этих факторов. Многие виды человеческой деятельности сами по себе не представляют опасности для биоразнообразия, но в совокупности наносят природе ощутимый вред. В частности, результатом кумулятивного воздействия лесозаготовок, туризма и разработки полезных ископаемых может стать фрагментация мест обитания.
Разнообразна растительность островов Субантарктики, которая насчитывает 30—40 видов местных сосудистых растений. На островах холодного умеренного пояса произрастает более 150 видов растений, включая кустарники и деревья. Острова Субантарктики и холодного умеренного пояса характеризуются высоким уровнем эндемизма вследствие их длительной географической и экологической изоляции друг от друга и от окружающих массивов континентальной суши. Так, генетически обособленная флора южных островов Новой Зеландии, состоящая примерно из 250 таксонов, включает 35 таксонов, эндемичных для данного региона, и несколько видов, эндемичных для отдельной группы островов.
Наземную фауну Антарктиды и соседних островов представляют почвенные клещи, коллемболы (ногохвостки), а на Антарктическом п-ве — два вида мелких бескрылых насекомых и ограниченное число других микроскопических почвенных беспозвоночных (простейших одноклеточных организмов, тихоходок, нематод и ротифер). Фауна островов Субантарктики и холодного умеренного пояса, как и их флора, более разнообразна и насчитывает множество редких видов и эндемиков, особенно среди беспозвоночных.
Некоторые острова Антарктики, на которых сохранились незатронутые антропогенным воздействием природные экосистемы, имеют международное значение для науки и охраны
окружающей среды. Многие другие острова несут на себе характерный отпечаток антропогенных воздействий, связанных в первую очередь с намеренной интродукцией хищников и паразитов животных. Особую проблему представляют завезенные сюда млекопитающие — грызуны, кошки, кролики, овцы, крупный рогатый скот, северные олени. В последние годы было проведено несколько успешных попыток очистить острова Антарктики от чужеродных млекопитающих. Эти работы продолжаются. Острова также являются жизненно важным местом отдыха и размножения морских птиц и тюленей, кормовой базой для которых служит Южный океан.
Еще одна серьезная опасность для флоры и фауны Антарктики исходит от сохраняющейся возможности непреднамеренного попадания в регион чужеродных видов либо в ходе осуществления национальных программ научных исследований, либо в связи с туризмом. В частности, случайно занесенным в Антарктику вирусом инфекционного бурсита поражены популяции пингвинов — императорского и Адели.
Морские экосистемы. В настоящее время угроза биоразнообразию Южного океана значительно ниже, чем раньше, когда китобойный промысел наносил серьезный урон популяциям китообразных. Хотя биоразнообразие Южного океана изучено далеко не полностью, по сравнению с некоторыми другими регионами, но все-таки вполне удовлетворительно. В ходе исследований выяснилось, что в Южном океане разнообразие видов в пределах многих таксономических уровней намного ниже, чем можно было предполагать, исходя из общих закономерностей распределения видов в Мировом океане. Примером могут служить брюхоногие и двустворчатые моллюски, а также бентические рыбы. Объяснения такой видовой скудности пока не найдены. В некоторых случаях ее можно разъяснить общей тенденцией к уменьшению биоразнообразия от экваториальных широт в направлении полюсов (широтное снижение разнообразия). Что касается рыб, то низкое разнообразие их видов может объясняться отсутствием в пределах Южного океана мест обитания, традиционно богатых видами рыб. В то же время некоторые группы организмов — мелкие ракообразные, бриозооны, губки — хорошо представлены в Южном океане. Наши знания о биоразнообразии Южного океана, как и большинства морей мира, в основном ограничиваются зоной континентального шельфа и материкового скло
на. Почти ничего не известно о фауне окружающих Антарктиду глубоководных морей.
В прошлом, после того как в результате перепромысла в конце XVIII — начале XIX в. морские хищники, в первую очередь южные морские котики (Arctocephalus gazella), были ликвидированы как возможный объект промысла и почти полностью истреблены физически, начался хищнический отлов гренландских китов. Единственным исключением стал строгий контроль за добычей южных морских слонов (Mirounga leonina) в районе Южной Георгии. Доказательства того, что эксплуатация ресурсов Южного океана в прошлом привела к исчезновению каких-либо биологических видов, отсутствуют. Популяции южных морских котиков, в частности, восстановились полностью.
Особое беспокойство вызывает сопровождающий рыбный промысел побочный вылов, и в первую очередь альбатросов и буревестников. Даже по самым скромным оценкам, смертность альбатросов в рыболовных сетях в Южном океане составляет 44 тыс. особей в год. Популяции этих птиц в некоторых районах сокращаются примерно на 7% в год. При таких темпах гибели самовосстановление популяций невозможно. Самой серьезной угрозой для альбатросов считается вылов рыбы крючковыми снастями.
Ведущийся другими способами рыболовный промысел также может приводить к существенным побочным последствиям. В частности, придонное траление может угрожать бентическим сообществам океана, для которых характерны очень низкие темпы естественного самовосстановления. Данные по этому вопросу в настоящее время отсутствуют. В целом можно сказать, что в будущем угроза биоразнообразию Южного океана от прямых и косвенных видов воздействия рыбного промысла сохранится. Оцениваться эти воздействия должны с учетом естественной изменчивости природной среды Южного океана, а также физического влияния на нее, например динамики льдов.
МЕЖДУНАРОДНЫЙ ОПЫТ МЕРОПРИЯТИЙ
ПО СОХРАНЕНИЮ ЖИВОЙ ПРИРОДЫ
Проблемы охраны природы, животного и растительного мира не могут быть ограничены какими-либо региональными рамками и даже границами целого материка. Для решения наиболее сложных и широких, глобальных проблем охраны природы необходимы координированные усилия ученых, администраторов, государственных руководителей как отдельных стран, так и международного сообщества в целом.
В 1948 г. был создан Международный союз охраны природы и природных ресурсов МСОП (IUCN), который сейчас объединяет представителей более 120 стран мира. Это крупная профессиональная организация, внимание которой направлено на сохранение почв, ландшафтов, вод, атмосферы и живых организмов на нашей планете, являющихся естественными ресурсами, без которых невозможно существование человечества. Создание МСОП имело свою предысторию в конце XIX в. и в первой половине XX в.
1885 г. Конвенция по ловле лососевых рыб в Рейне, Берлин.
1895 г. Конвенция по защите птиц, Париж.
1900 г. Конференция по защите африканских млекопитающих, Лондон.
1909 г. Первый Международный конгресс по защите ландшафтов. Париж, 17—20 октября.
1910 г. Пятый Международный орнитологический конгресс собрался в Берлине в июне: принято решение создать постоянную организацию с целью охраны птиц.
Восьмой Международный зоологический конгресс: образовал комитет для рассмотрения вопроса об охране природы в глобальном масштабе.
1913 г. Международная конференция по охране природы состоялась 17—19 ноября в Берне, была создана Консультативная комиссия по Международным проблемам охраны природы со штаб-квартирой в Базеле. Конвенция была подписана 19 странами.
1922 г. Международный совет по охране птиц (ICBR) образован в Лондоне.
1929 г. Образован Международный офис по охране природы со штаб-квартирой в Брюсселе. Эта организация позднее вошла в состав IUPN (первое название МСОП).
1933 г. Лондонская конвенция по сохранению флоры и фауны в естественных местообитаниях. Внесла огромный вклад в сохранение природы Африки.
1942 г. Лондонская конвенция по охране природы и сохранению дикой природы в Восточном полушарии.
1946 г. Международная конвенция по регулированию китобойного промысла.
1946—1947 гг. Серьезные усилия были предприняты Шведским союзом охраны природы, основанным в 1909 г., в создании Международной организации охраны природы; Конференции в Базеле (1946) и Брюнене (1947) были толчком к образованию временного Международного союза охраны природы.
1948 г. Во время конференции в Фонтенбло (Франция) под эгидой ЮНЕСКО и Франции был создан Международный союз охраны природы (International Union for Protection of Nature, IUPN). Это произошло 5 октября. 130 участников конференции представляли 18 правительств, 108 учреждений и организаций разных стран и 7 международных организаций.
ГЛАВА IX
€
Основные вехи деятельности МСОП
1949 г. ЮНЕСКО и МСОП была организована в США Международная техническая конференция по охране природы, которая совпала по времени проведения с Конференцией ООН по сохранению и использованию ресурсов.
1953 г. Первый выпуск Бюллетеня МСОП.
1954 г. Парижская Международная конвенция по охране птиц и Лондонская Международная конвенция по предотвращению загрязнения морей нефтью.
1954 г. На Генеральной ассамблее в Эдинбурге Международный союз охраны природы (IUPN) переименован в Международный союз охраны природы и природных ресурсов (International Union for Concervation of Nature and natural resources, IUCN).
1959 г. Резолюция 810 (XXXI) Экономического и социального совета ООН, которая утвердила список национальных парков и приравниваемых к ним резерватам по просьбе Генерального секретаря МСОП, представившего список.
1960 г. На Генеральной ассамблее МСОП в Варшаве было решено перенести штаб-квартиру из Бельгии в Швейцарию.
1961 г. Создание Всемирного фонда охраны дикой природы ВВФ (WWF).
1966 г. МСОП фокусирует внимание на проблемах окружающей среды Запада, в частности загрязнении. Генеральная ассамблея собирается в Лозанне, Швейцария.
1968 г. Подписание главами государств, входящих в состав Организации Африканского единства, Африканской конвенции по охране природы и природных ресурсов.
1969 г. Десятая Генеральная ассамблея МСОП в Нью-Дели (Индия). Глубокие изменения в направлениях деятельности МСОП, приоритетными становятся обращение к основным причинам экологических проблем, предупреждающие меры охраны.
1971 г. Совещание в Фунексе, Швейцария, привело к публикации документа «Развитие и окружающая среда: доклады и сообщения группы экспертов, созванной Генеральным Секретарем ООН, по материалам конференции “Окружающая среда человека” ».
1972 г. Конференция ООН по окружающей среде человека (Стокгольм) обратила внимание правительств на взаимоотно-
3 шения человека и окружающей среды и повлияла на возникновение UNEP — Программы ООН по окружающей среде и многочисленных Министерств по окружающей среде.
1974 г. Конвенция по международной торговле видами дикой фауны и флоры, находящимися под угрозой исчезновения (CITES), Вашингтон. Первая Всемирная конференция по народонаселению в Бухаресте, Румыния, по использованию ресурсов, окружающей среде и развитию. Стратегия принята в Мехико.
1975 г. Рамсарская конвенция по водно-болотным угодьям международного значения, в частности, как местообитаний водоплавающей дичи.
1977г. Перваямежправительственная конференция по образованию в области окружающей среды, Тбилиси, СССР.
Конференция ООН по опустыниванию, организованная МСОП.
1980 г. Мирная революция — публикация Всемирной стратегии охраны (МСОП, ВВФ, UNEP в сотрудничестве с ЮНЕСКО и ФАО).
1982 г. Принятие Генеральной Ассамблеей ООН Всемирной хартии по окружающей среде, подготовленной МСОП.
1987 г. Доклад «Наше общее будущее», опубликованный Всемирной комиссией по окружающей среде и развитию. В докладе была подчеркнута идея о глобальном экологическом кризисе и необходимости интеграции окружающей среды и развития.
В последующие годы определяющим международным документом явилась «Конвенция о сохранении биоразнообразия» (Рио-де-Жанейро, 1992).
В основе деятельности МСОП лежит Всемирная стратегия охраны природы. Ее главные принципы: сохранение важнейших экологических процессов и экосистем; сохранение генетического разнообразия; численность всех видов должна оставаться на уровне, достаточном для выживания.
Всемирная стратегия охраны природы основывается на: определении задач в сохранении живой природы; выявлении приоритетных потребностей для выполнения каждой из задач; предложении национальных стратегий охраны для выполнения приоритетных потребностей; рекомендациях по предупреждающей экологической политике, межнациональная природоохранная политика и открытая система национальных от-
четов; предложениях путей помощи сельскохозяйственным общинам в сохранении их жизненных ресурсов и защита широкого общественного участия в планировании и принятии решений в сохранении жизненных ресурсов.
Первая Конференция ООН по окружающей человека среде проходила в Стокгольме в июне 1972 г. На этой конференции был принят План мероприятий, который сыграл огромную роль в деятельности ООН, и принято решение, которое послужило основой создания Программы по окружающей среде ЮНЕП (UNEP). ЮНЕП называют «совестью ООН в вопросах окружающей среды ». Она стала первой организацией ООН, находящейся в развивающейся стране. ЮНЕП разместилась в Кении.
Деятельность ЮНЕП преследует две главные цели: удовлетворение основных потребностей человека; бережное отношение к ограниченным ресурсам биосферы.
Учитывая рекомендации Стокгольмской конференции были выделены шесть приоритетных областей деятельности ЮНЕП:
1. Развитие человеческих поселений и поддержание здоровья человека: поселения человека и среда его обитания; здравоохранение и состояние окружающей среды; борьба с вредителями.
2. Экосистемы суши: пустыни и полупустыни, тропические леса, горы, острова, побережья и т.д.; почвы; воды; биологические ресурсы; дикая природа и охраняемые территории.
3. Окружающая среда и экономическое развитие: комплексный подход к проблемам окружающей среды и экономического развития с учетом экологических факторов; использование природных ресурсов; «чистые» технологии; промышленность и окружающая среда.
4. Мировой океан и ресурсы морской флоры и фауны: загрязнение морей; биологические ресурсы морей; внутренние моря.
5. Энергетика.
6. Стихийные бедствия.
Основные направления деятельности по защите окружающей среды включают: оценку состояния окружающей среды — «надзор за Землей»; управление окружающей средой; вспомогательные мероприятия.
Функционирование системы мониторинга окружающей среды осуществляется:
1. Глобальной системой мониторинга окружающей среды ГСМОС (GEMS).
2. Международной справочной службой MCC (GRID) — использует компьютерные технологии анализа данных о состоянии окружающей среды на географической основе; используя информацию, собранную обширной сетью агентств МСС, накапливает и устанавливает взаимосвязи с другими данными, часто обобщает информацию в форме карт. Ее цель увидеть как взаимодействуют между собой различные переменные на местном, национальном, региональном и глобальном уровнях.
3. Международным регистром потенциально токсичных химических веществ ИРПТК (IRPTC) собирает и распространяет информацию о состоянии окружающей среды и влиянии химических загрязнителей на здоровье человека.
4. Международной справочной системой источников информации по окружающей среде ИНФОТЕРРА (INFOTERRA) используется специалистами многих стран.
В 1980 г. ЮНЕП, МСОП и ВВФ опубликовали Всемирную стратегию охраны живой природы.Она была использована более чем 50 странами для разработки национальных стратегий охраны живых организмов.
В июне 1992 г. во время Конференции ООН по окружающей среде и развитию была подписана Конвенция по биологическому разнообразию. Международный переговорный комитет, учрежденный руководящим советом ЮНЕП, подготовил эту конвенцию при участии ЮНЕП, ФАО, ЮНЕСКО и МСОП. Главная цель документа — сохранение биологического разнообразия и обеспечение тем самым нужд человечества.
В 1992 г. МСОП и ЮНЕП при содействии ФАО и ЮНЕСКО, разработали Глобальную стратегию сохранения биоразнообразия, цель которой ликвидировать условия исчезновения видов.
СТРАТЕГИЯ СОХРАНЕНИЯ
БИОЛОГИЧЕСКОГО PA311ООБРАЗИЯ
Одна из стратегических задач, осознанных мировым сообществом в связи с проблемой экологического кризиса биосферы, — сохранение биологического разнообразия на разных уровнях дифференциации биосферы. В настоящее время признано, что проблема охраны биологического разнообразия не сводится только к охране редких таксонов. Генеральная стратегия сохранения биоразнообразия должна быть ориентирована на сохранение систем (совокупностей) видов в их пространственном распределении — от типов экосистем глобального уровня (типов и биомов) вплоть до конкретных биогеоценозов и их сопряженных территориальных сочетаний на локальном уровне.
Стратегия сохранения биоразнообразия требует прежде всего количественной и сравнительной оценок его в природных экосистемах различного уровня.
Число видов растений пли животных в конкретном месте зависит в первую очередь от экологических условий, однако видовое богатство на обширной территории определяется в основном историческими процессами. Так. природная экосистема ранга ландшафта и выше имеет свою историю изоляции.
5
колонизации новыми видами, вымирания данных видов, и эти исторические процессы определяют альфа-разнообразие указанной экосистемы. Следовательно, чтобы понять значение этого критерия в применении к крупной экосистеме, необходимо получить информацию о гамма-разнообразии, а чтобы истолковать их отношение, нужно рассмотреть и бета-разнообразие.
Комплексная оценка всех критериев биоразнообразия позволяет ученым понять степень устойчивости изучаемой экосистемы, уровень антропогенного воздействия на ее структуру, роль и место редких видов растений и животных в данной экосистеме. Для осуществления обоснованной и взвешенной стратегии охраны природы необходимо применять методы оценки биоразнообразия по описанным критериям. Такая оценка дает возможность на количественной основе планировать перспективную сеть охраняемых территорий, намечать необходимые меры по охране и восстановлению редких видов растений и животных.
В настоящее время непосредственная угроза исчезновения нависла над 281 видом и подвидом млекопитающих, 396 видами и подвидами птиц, 105 — рептилий, 41 — амфибий, 194 видами и подвидами пресноводных рыб. Отдельными списками в каждом томе Международной Красной книги приведены виды животных, вымершие с 1600 г. до настоящего времени (так называемый «Черный список»). Информация о каждом виде приводится по стандартной системе: научное и английское название, систематическое положение, статус в Красной книге, распространение, численность, характерные местообитания, предпринятые меры охраны, предлагаемые меры охраны, дополнительные замечания, источники информации. Таким образом, в сжатом виде читатель получает нужные сведения о состоянии вида и о тех мерах, которые необходимы для его спасения и восстановления.
В Международную Красную книгу включены все виды отряда сиреновых: североамериканский, бразильский и африканский ламантины и дюгонь. Пятый вид этого семейства — стеллерова корова — был истреблен еще в середине XVHI в. На tpaHH исчезновения находится представитель монотипического семейства руконожковых — мадагаскарская руконожка, или ай-ай, вся популяция которой насчитывает несколько десятков особей. Видимо, безвозвратно исчез тасманийский
сумчатый волк. Хотя он еще сохранился на красных листах Международной Красной книги, но вот уже около полувека никому не удается обнаружить его в природе. Очевидно, придется перевести сумчатого волка в Черный список истребленных человеком животных.
В Международную Красную книгу включены все представители семейства человекообразных обезьян: орангутан, горилла, обыкновенный и карликовый шимпанзе. Редки или находятся на грани исчезновения все виды семейств лемуровых и индриевых, все пять видов носорогов мировой фауны. Под угрозой исчезновения оказались многие виды черепах и крокодилов, особенно тревожно положение всех видов семейства морских черепах, а также представителей монотипических семейств кожистой черепахи и гангского гавиала.
Среди факторов, обусловливающих исчезновение и уменьшение численности животных, основным считается разрушение естественных мест обитания человеком. Среди видов, включенных в Красную книгу, 672 оказались в угрожаемом положении именно по этой причине. Важную роль играет также чрезмерная эксплуатация ресурсов животных (37% видов из Красной книги) и воздействие интродуцированных экзотических видов (19% видов, включенных в Красную книгу). Некоторые виды, исчезнувшие в природе, удалось сохранить благодаря их содержанию в питомниках и зоопарках. Например, после того, как аравийский орикс был истреблен, провели под эгидой МСОП «Операцию Орикс». В 1981—1982 гг. из сохранившегося в неволе поголовья (более 300 экз.) были сформированы группы антилоп, которые удалось реинтродуцировать в месте их былого обитания — пустыни Омана и другие районы Аравийского п-ова. Осуществлена операция по возвращению в природу лошади Пржевальского. Для этого также использовался фонд животных, сохраненный в зоопарках. Табуны этих лошадей появились уже в Гобийском национальном парке Монголии. В результате операции по спасению маврикийской пустельги, проведенной МСОП, численность этой редчайшей птицы возросла с 6 до 15 особей и пара отловленных птиц впервые успешно вырастила птенцов в неволе. Сейчас создается резервный фонд этих птиц в питомнике для последующего выпуска их в природу.
При МСОП создана специальная комиссия по исчезающим видам растений, подготовившая Международную Красную
книгу растений (1978), в которую включено 250 таксонов. В то же время в охране, по мнению комиссии МСОП, нуждается около 20 000 видов высших растений или около 10% мировой флоры. В Международной Красной книге использованы следующие категории редкости видов: виды, исчезнувшие или, по-видимому, исчезнувшие; виды уязвимые (сокращающиеся в численности); виды редкие. Отнесение вида к определенной категории требует хорошего знания структуры его ареала, особенностей экологии и размножения.
Большую помощь при составлении Красной книги оказывают списки растений, которым грозит исчезновение, составляемые для некоторых территорий различными государствами. Флора отдельных континентов и регионов изучена еще не достаточно, а антропогенное воздействие настолько велико, что некоторые виды растений, возможно, исчезнут раньше, чем станут известны науке. Лучше изучена флора таких континентов как Европа, Северная Америка, где установлено, что около 10% от состава видов нуждается в тех или иных охранных мероприятиях.
Таксоны, включенные в Красную книгу, объединены в группы в зависимости от того, в результате каких воздействий вид получил статус угрожаемого. Ряд видов с ограниченным ареалом оказался на грани исчезновения в связи с вырубкой насаждений и раскорчевкой освободившихся участков под распашку. К этой группе отнесено в Красной книге несколько видов рода эвкалипт, подавляющая часть которого распространена в Австралии. Опасность исчезновения нависла над 120 видами, 7 из которых занесены в Красную книгу.
Загрязнение водоемов поставило под угрозу такой вид из злаковых как зизания техасская (Zizania texana), произрастающая в водоемах центральной части Техаса (США). За последние 20 лет произошло резкое сокращение численности растения, оно перешло только на вегетативный способ размножения. В результате развития туризма резко уменьшилось количество замии флоридской, ранее обычного растения п-ова Флорида. Сокращение местонахождений вида обусловлено выкапыванием его подземных клубней.
Примером растений, резко сокративших свою численность в связи с выпасом, распашкой и использованием населением, являются два вида драконова дерева, занесенные в Красную книгу. Нубийское драконово дерево (Dracaena ambit) — реликт
Джибути, Эфиопии и Судана. Возможно, что оно произрастает еще и вдоль побережья Сомали. В связи с сильным перевы-пасом коренной растительности, а также добычей сока, рубкой на дрова, использованием для плетения различных изделий сохранились только отдельные экземпляры драконова дерева на труднодоступных скалах.
Значительная часть видов оказалась под угрозой в результате различного рода коллекционирования и сборов. Так, в Красную книгу занесено алое многолистное (Aloepolythylia) — сук-кулентный многолетник из семейства лилейных. Этот эндемик Южной Африки имеет большое значение для садоводства. Запасы его сократились из-за выкапывания растений для продажи садоводам. В настоящее время осталось только около 300 экземпляров примерно в 60 пунктах. Вид строго охраняется законом и его экспорт запрещен.
Наконец, среди видов, занесенных в Красную книгу, можно выделить те, для которых характерна критически низкая численность популяции и существует угроза прекращения размножения. Так, агава аризонская (Agave arizonica), произрастающая в штате Аризона (США), известна всего в 12— 14 пунктах, в каждом из которых растет по одному или несколько экземпляров в радиусе 3—5 км.
В настоящее время выявляются целые семейства, значительная часть видов которых нуждается в охране. Например, известное семейство орхидных, одно из наиболее обширных, насчитывает около 20 тыс., а по некоторым данным до 30 тыс. видов (это около 7% всех цветковых растений), а сейчас около 17 тыс. видов этого семейства находятся на пути к исчезновению. Особую опасность для орхидей создают коллекционирование, сбор для продажи, все увеличивающиеся в объеме, а также вырубка тропических лесов, пожары, широкое распространение вторичных, более простых сообществ, которое приводит к изменению условий их существования, к сокращению ареалов, а иногда и полному исчезновению. Почти во всех странах орхидеи охраняются.
Не менее интересна судьба семейства пальмовых, включающего 2 400 видов, относящихся к 270 родам. Современное положение значительного числа пальм критическое: одни виды исчезли, другие находятся на грани исчезновения, состояние многих видов пока не определено, но часть из них близка к исчезающим видам. Высокая степень уязвимости пальм опреде-
ляется продолжающейся вырубкой тропических лесов, расширением площадей вторичных насаждений, различными мелиоративными работами, приводящими к глубокому изменению условий существования многих видов пальм. Некоторые из них, как например, африканская пальма медемия аргун (Medemia arguri), отличаются в настоящее время критически низкой численностью популяции, угрожающей прекращению размножения. Значительное число видов пальм внесено в Международную Красную книгу, еще больше в национальные Красные книги. В МСОП при Комиссии по исчезающим растениям создана специальная подкомиссия по пальмам.
Особое место в разработке охранных мероприятий уделяется эндемичным и реликтовым растениям, представляющим уникальный генетический материал. В Международную Красную книгу включено 26 монотипных родов, являющихся эндемиками. Установлено, что эндемичные растения, имеющие узкий ареал и характеризующиеся рассеянным произрастанием, оказываются наиболее чувствительными к антропогенному воздействию. Их исчезновение в отдельных частях географически ограниченного ареала быстро приводит к снижению численности вида до критической и ставит его под угрозу.
Таким образом, обзор Международной Красной книги показывает, что утрата разнообразия видов — реальная угроза, нависшая над растительным миром планет. Помимо Международной Красной книги создаются национальные и региональные Красные книги. Это научные документы первостепенной важности, которые используются деятелями охраны природы и администраторами при разработке и осуществлении мероприятий по сохранению генофонда планеты.
В связи с настоятельной необходимостью сохранения биологического разнообразия и слежения за естественными экосистемами (экологического мониторинга) возникла идея о биосферных заповедниках (резерватах).
Создание глобальной сети биосферных резерватов было одной из основных задач Программы «Человек и Биосфера», начавшей свою работу в 1970 году под эгидой ЮНЕСКО. К 2004 году в результате разносторонней деятельности многих международных и национальных организаций в рамках этой Программы создана глобальная сеть биосферных резерватов, позволяющая осуществлять экологический мониторинг в планетарном масштабе. Общее число биосферных резерва-
тов достигло к настоящему времени 380, они расположены в 91 стране мира и занимают общую площадь 268 млн га.
К началу нового тысячелетия в России сформировалась базовая сеть особо охраняемых природных территорий (ООПТ), которая может служить основой для дальнейшей работы по охране биологического разнообразия на национальном и региональном уровнях и для осуществления в пределах нашей страны программы глобального экологического мониторинга.
В первую очередь выполнению этих целей служат биосферные резерваты (ООПТ международного статуса) и природные заповедники (ООПТ национального статуса). На 1 января 2004 года в России функционирует 30 биосферный резерват под эгидой программы «Человек и Биосфера» ЮНЕСКО. Необходимо упомянуть, что почти все они были созданы на базе наиболее известных, успешно и много лет работающих государственных заповедников и наряду с новым статусом сохранили и прежние функции. Первые биосферные резерваты в России были утверждены в 1978 году. Ими стали Кавказский, Приокско-Террасный, Окский, Сихотэ-Алинский и Центрально-Черноземный государственные заповедники. Примером биосферного резервата, созданного по специальной программе на незаповедной территории, может служить Центральносибирский биосферный резерват (1986).
На 1 января 2004 года в России существует 100 природных заповедников общей площадью более 33,5 млн га, что составляет свыше 1,6% площади страны. Самый новый, 100-й заповедник создан в декабре 2000 года. Это заповедник «Эрзи», расположенный в горах Северного Кавказа на территории Ингушетии. Также в России имеется 35 национальных парков общей площадью 7 млн га, что составляет 0,4% территории России. Таким образом, общая площадь ООПТ (биосферные резерваты, природные заповедники и национальные парки) в начале XXI века превысило 40 млн га и составляет более 2% площади страны.
Для большей полноты картины к этим данным необходимо прибавить площадь наиболее крупных государственных заказников, заявленных Россией и включенных в список охраняемых территорий ООН. Это 91 государственный заказник общей площадью 14,2 млн га. Вместе с ними площадь наиболее значимых охраняемых территорий в России достигает 54,2 млн га и составляет 2,8% площади страны.
СИСТЕМА ОХРАНЯЕМЫХ
ПРИРОДНЫХ ТЕРРИТОРИЙ
В РОССИИ
К началу нового тысячелетия в России сформировалась базовая сеть особо охраняемых природных территорий (ООПТ), которая может служить основой для дальнейшей работы по охране биологического разнообразия на национальном и региональном уровнях и для осуществления в пределах нашей страны программы глобального экологического мониторинга.
В первую очередь выполнению этих целей служат биосферные резерваты (ООПТ международного статуса) и природные заповедники (ООПТ национального статуса). На 1 января 2001 г. в России функционировал 21 биосферный резерват под эгидой программы «Человек и Биосфера» ЮНЕСКО. Почти все они были созданы на базе наиболее известных, успешно и много лет работающих государственных заповедников и наряду с новым статусом сохранили и прежние функции. Первые биосферные резерваты в России были утверждены в 1978 г., ими стали Кавказский, Приок-ско-Террасный, Окский, Сихотэ-Алинский и Центральночерноземный государственные заповедники. Примером биосферного резервата, созданного по специальной программе
на незаповедной территории, может служить Центральносибирский биосферный резерват (1986).
На 1 января 2001 г. в России было 100 природных заповедников общей площадью более 30 млн га, что составляет свыше 1,5% площади страны. Самый молодой, 100-й заповедник создан в декабре 2000 г. Это заповедник «Эрзи», расположенный в горах Северного Кавказа на территории Ингушетии. На эту же дату в России было 25 национальных парков общей площадью 6,8 млн га, что составляет 0,4% территории России. Таким образом, общая площадь ООПТ (биосферные резерваты, природные заповедники и национальные парки) к началу XXI в. достигла 40 млн га и составляет 2% площади страны.
Для большей полноты картины к этим данным необходимо прибавить площадь наиболее крупных государственных заказников, заявленных Россией и включенных в список охраняемых территорий ООН. Это 91 государственный заказник общей площадью 14,2 млн га. Вместе с ними площадь наиболее значимых охраняемых территорий в России достигает 54,2 млн га и составляет 3,7% площади страны.
До последнего времени официальной Красной книгой России считалась Красная книга РСФСР (1983). В том «Животные» было включено 247 видов и подвидов животных. Они распределялись по пяти категориям: 1) находящиеся под угрозой исчезновения, 2) сокращающиеся в численности, 3) редкие, 4) неопределенные и 5) восстановленные. Из них 15 видов и подвидов моллюсков, 34 — насекомых, 9 — рыб, 4 — амфибий, 11 — рептилий, 107 — птиц и 68 видов и подвидов млекопитающих. Категория «восстановленные» была применена только к млекопитающим — это ладожская нерпа, серый кит, новоземельский северный олень и зубр.
В том «Растения» Красной книги РСФСР (1988) включено 17 видов грибов, 29 — лишайников, 4 — плауновидных, 10 — папоротникообразных, 11 — голосеменных и 440 видов покрытосеменных.
Российская Академия наук и Госкомэкологии РФ издали в электронной версии Красную книгу Российской Федерации (1999). Эта книга подготовлена с учетом материалов Красной книги РСФСР (1983), переработана и расширена. Отдельно опубликован список животных, включенных в Красную книгу Российской Федерации (Красная книга России: правовые акты, Москва, 2000). В него вошли 415 видов и около 500 подвидов
позвоночных и беспозвоночных животных. Различные типы и классы представлены следующим образом: 13 видов червей, 1 — мшанок, 1 — плеченогих, 42 — моллюсков, 3 — ракообразных, 95 — насекомых, 4 — круглоротых, 39 — рыб, 8 — амфибий, 21 — рептилий, 123 — птиц и 65 видов млекопитающих.
Согласно ст. 6 Конвенции о биологическом разнообразии все страны-участницы Конвенции должны подготовить Национальную стратегию и План действий.
По рекомендациям секретариата Конвенции о биологическом разнообразии в разных странах еще в 1995 г. была начата подготовка Национальных стратегий и Планов действий. По форме и характеру материалов их можно разделить на три группы:
1) стратегия — основные направления государственной политики в области биоразнообразия, охватывающие все стороны его использования и сохранения (принятый на государственном уровне пакет документов);
2) стратегия — система долгосрочных мероприятий по развитию сети охраняемых природных территорий и охране редких видов растений и животных (государственная программа);
3) стратегия — приоритетные направления деятельности государственных и неправительственных экологических организаций по охране живой природы (программы, рекомендации, меморандумы, планы мероприятий и пр.).
Для России имеется острая необходимость в детальной Национальной стратегии, которая на основе выделенных приоритетов определяет основные направления действий законодательной и исполнительной властей, всех групп и секторов общества, всех заинтересованных граждан и организаций. Не менее важно иметь и План действий — тактический план неотложных мероприятий по сохранению живой природы страны и отдельных регионов.
Поэтому в течение нескольких лет в России по проекту ГЭФ «Сохранение биоразнообразия» разрабатывалась национальная стратегия (Амирханов, Тишков, 1999). Актуальность этой работы заключается в том, что в условиях переходной экономики и резкого сокращения финансирования охраны живой природы есть угроза потери биоразнообразия на крупных территориях и разрушения сложившейся системы управления его сохранением. Особенностью России являются ее исключительно крупные размеры (1/7 площади суши Земли), природное
м
разнообразие (от полярных пустынь и тундр на севере до пустынь и субтропиков — на юге) и наличие крупных субъектов Федерации (всего 89), каждый из которых обычно крупнее средней европейской страны. Следовательно, не менее актуально для России разработать модели региональной стратегии сохранения биоразнообразия и внедрить их в совершенствование управления охраной природы в регионах.
Государственная политика России в сохранении биоразнообразия в настоящее время строится на правовой основе (федеральных законах и законодательных документах субъектов Российской Федерации, обеспечивающих выполнение обязательств страны по Конвенции о биологическом разнообразии — КБР) и Указах Президента Российской Федерации о переходе на модель устойчивого развития.
Объект Национальной стратегии — биоразнообразие на всех уровнях его проявления в его актуальном состоянии без регламентов в отношении процессов естественной эволюции, динамики и последствий, связанных с традиционным экстенсивным хозяйствованием в природных экосистемах, т. е. выпасом скота, оленеводством, промысловой охотой и пр. Приоритетными для сохранения объектами считаются редкие виды растений и животных, редкие и исчезающие экосистемы. Стратегия в их отношении должна быть направлена на восстановление популяций, покровительственную охрану редких видов, создание системы охраняемых природных территорий и экологическую реставрацию. Проблемы охраны живой природы, связанные с прогнозом глобальных изменений климата, решаются путем учета возможных последствий для островных экосистем, прибрежных морских экосистем и дельт, рефугиумов или «островов» природной растительности в антропогенном ландшафте, где изменения климата способны привести к утрате редких видов и их местообитаний.
Субъекты Национальной стратегии и ее ориентация. Национальная стратегия и Национальный план действий ориентированы на лиц, принимающих решения в министерствах и ведомствах, использующих природные ресурсы, осуществляющих контроль за состоянием биоразнообразия, проходящих финансирование, законодательную, научную и информационную поддержку этой деятельности, а также на всех заинтересованных в решении проблем охраны живой природы.
Традиционными партнерами в деле сохранения биоразнообразия являются министерства и ведомства природно-ресурсного сектора исполнительной власти: Министерство природных ресурсов, Минсельхозпрод России, МПР России, Миннауки России, Минздрав России, Минкультуры России, Минэкономики России, а также отраслевая, академическая и вузовская наука (РАН, РАСХН, Межведомственная ихтиологическая комиссия и др.). Координация их действий осуществляется Межведомственной комиссией по проблемам биоразнообразия. Законодательная деятельность в области сохранения биоразнообразия ведется Комитетом по природным ресурсам и Комитетом по экологии Госдумы Федерального собрания Российской Федерации. Постоянными партнерами остаются общественные природоохранные организации — российские и международные, а также средства массовой информации (хотя в последние годы, несмотря на появление массы новых периодических изданий, каналов на ТВ, интерес СМИ к проблемам охраны живой природы существенно ослаб).
Сфера партнерства в деле сохранения биоразнообразия расширяется за счет других министерств и ведомств, которые вовлекаются в процесс охраны живой природы на своих объектах или при выполнении своих функций, например, Минобороны России (охрана живой природы на военных объектах), ГТК России (контроль за экспортом и импортом объектов биоразнообразия, карантинный и ветеринарный контроль), ФПС России (охрана морских биологических ресурсов, содействие охране редких видов и функционированию пограничных ООПТ в пограничной полосе), Роскомгидромет (организация системы мониторинга), Федеральная служба почтовой связи (контроль за экспортом и импортом объектов биоразнообразия), Госкомстат России (мониторинг использования биологических ресурсов) и др.
В перспективе особое внимание должно быть обращено на относительно новых партнеров в экономический (товаропроизводители, крупные компании (РАО ЕЭС России, РАО Газпром, Лукойл и пр.), частный сектор экономики, малый бизнес, использующие биологические ресурсы и участвующие в работах по охране живой природы) и социальной сферах (общественные движения на местах, образование, СМИ, религиозные конфессии и др.).
Круг традиционных и новых партнеров в подготовке и реализации Национальной стратегии и Национального плана действий определился в процессе специально проведенного опроса и анализа отношения разных общественных групп и слоев населения к проблемам сохранения биоразнообразия, а также при подготовке проектов межотраслевых и секторальных стратегий сохранения биоразнообразия.
ЦЕЛЬ НАЦИОНАЛЬНОЙ СТРАТЕГИИ И НАЦИОНАЛЬНОГО ПЛАНА ДЕЙСТВИЙ
Целью Национальной стратегии и Национального плана действий является консолидация усилий всего общества на сохранение национальной живой природы, как составного элемента окружающей среды и ресурса, обеспечивающего условия жизни человека, его благополучие и саму возможность существования.
Они направлены на обеспечение законодательных и исполнительных органов, государственных, частных и общественных природоохранных организаций и СМИ научно и экономически обоснованными рекомендациями, долгосрочным прогнозом и планами мероприятий по управлению сохранением и устойчивым использованием биоразнообразия.
Сохранение живой природы России, осознание роли природы нашей страны в поддержании биосферных процессов на Земле, понимание масштабности роли биологических ресурсов в национальном богатстве — это может стать основой национальной идеи. Очищенная от идеологии, она рассматривается «здоровой» базой патриотизма и побудительным моментом социальной активности населения в поддержке деятельности властей по охране природы.
Именно исключительно высокая стоимостная оценка объектов живой природы и их возобновляемость делают биоразнообразие и биологические ресурсы ведущим стратегическим ресурсом для России и ставит ее в мировые лидеры по уровню национального богатства. Это позволит существенно расширить сферу внешнеполитической и внешнеэкономической активности страны.
Во всем многообразии активных и пассивных участников процесса сохранения биоразнообразия в стране (государственных органов, товаропроизводителей, разных секторов населения, научных и общественных организаций и пр.) есть реальные партнеры, имеющие свои интересы, а есть потенциально восприимчивые к действиям по формированию сознания в области охраны живой природы. Чтобы повысить эффективность реализации планируемых мероприятий их адресность должна касаться не только места, управления, характера действий, источника финансирования, но и на кого они ориентированы.
При крайне низком бюджетном обеспечении реализации государственной политики в области сохранения биоразнообразия важно ориентировать свои действия исключительно на приоритетные направления и конкретные действия, которые могут дать более широкий эффект, а также комплексно решать проблемы и согласовывать свои действия с заинтересованными ведомствами и организациями. Это вполне оправданный элемент управления, обеспечивающий соединение средств и организационных усилий участников процесса сохранения живой природы.
Важно четкое разграничение задач федерального, регионального и местного уровней. Это позволяет достичь эффективности и оперативности действий, а в ряде случаев избежать дублирования действий. Анализ многолетнего наблюдения за изменениями среды выявил среди административных методов управления предпочтение территориальных форм сохранения биоразнообразия: создание различных по форме, статусу, ведомственной принадлежности и целевому назначению охраняемых природных объектов, организация их в региональные экологические сети, введение щадящих режимов хозяйствования в ключевых регионах устойчивого развития. Внедрение комплексного подхода, обеспечивающего сохранение видов, их местообитаний, экосистем, сухопутных и водных ландшаф-
тов, одновременно позволяет решать не только природоохранные задачи, но и совершенствовать систему стабильного использования биологических ресурсов. В этом мы видим важный принцип учета интересов всех заинтересованных министерств и ведомств, участвующих в сохранении и использовании биоразнообразия страны.
Стратегия должна рассматриваться как совокупность экспертно выделенных приоритетных направлений деятельности государственных, коммерческих, научных и общественных структур по сохранению живой природы. Для реализации она представляется как пакет государственных документов долгосрочного планирования по приоритетным направлениям сохранения живой природы, учитывающим специфику природы и экономики страны, этап ее современного развития, динамику политического, экономического и социального фона.
В настоящее время намечена система приоритетов сохранения биоразнообразия, среди которых ведущими признаны: экономические (включение ценности живой природы в макроэкономические показатели страны, экономическая оценка биоразнообразия и пр.), управленческие (формирование системы партнерства, в том числе вовлечение новых партнеров в реализацию стратегии, межотраслевая координация и пр.), законодательные (обеспечение реализации стратегии законодательной и нормативной базой), научные и информационные (создание системы определения приоритетов и системы принятия решений — мониторинг, инвентаризация биоразнообразия, научное обоснование прогнозов состояния биологических ресурсов, ведение кадастров объектов живой природы и пр.), международные (внедрение категорий ценности биоразнообразия страны в практику международной деятельности страны, расширение сферы участия в конвенциях, соглашениях) и т.д.
Особенности современной России при реализации Национальной стратегии. Экспертами, привлекаемыми к подготовке Национальной стратегии и Национальному плану действий, определена специфика условий России, которые следует учитывать при реализации стратегических планов в области охраны живой природы. Это:
глубокий экономический кризис, который привел к резкому сокращению объемов природоохранных инвестиций;
относительно слабая трансформированное^ экосистем, особенно азиатской части;
снижение антропогенной нагрузки на экосистемы в последние годы в результате спада промышленного производства;
ослабление внимания населения к проблемам охраны живой природы;
отсутствие реальной оценки биоразнообразия как наиболее весомого элемента национального богатства страны;
переходная экономика, слабое развитие частного сектора, отсутствие развитого рынка купли-продажи земли;
незавершенность формирования законодательной базы в области сохранения биоразнообразия (до сих пор не приняты законы: О растительном мире. Об охоте. Об охране водных биологических ресурсов и др.);
многонациональный характер страны, наличие более 50 групп малочисленных народов, имеющих традиционные формы хозяйствования и различия в отношениях к биологическим ресурсам и биоразнообразию;
значительная протяженность сухопутных и морских границ и проблем, связанных с сохранением биоразнообразия пограничных территорий и охраной мигрирующих животных;
крупные размеры страны, разнообразие природных условий (разнообразие природных зон, наличие крупных горных систем и равнин, речных бассейнов, озер, системы арктических архипелагов);
значительная доля сохранившихся экосистем, особенно на Севере, в Азиатской части России (более 26% девственных лесов планеты находятся в России);
наличие развитой, функционирующей более 80 лет, сети ООПТ (около 2% площади страны);
глубокие научные традиции, развитая система научной поддержки мер по сохранению биоразнообразия (широкая сеть научных учреждений и учебных заведений, традиции таксономических исследований и проведения работ по инвентаризации, оценки состояния объектов живой природы и пр.);
низкая эффективность экономических и финансовых механизмов сохранения биоразнообразия.
ПРИОРИТЕТЫ
НАЦИОНАЛЬНОЙ СТРАТЕГИИ
Среди приоритетов Национальной стратегии и Плана действий по сохранению биоразнообразия можно выделить:
1. Формирование категории экономической ценности живой природы и биологическогораэнообраэия как национального богатства.
Создание политических, экономических и идеологических предпосылок для включения ценности живой природы в оценку национального богатства и макроэкономические показатели развития страны важно, прежде всего, с позиций роли живой природы России в биосфере Земли: значения в формировании климата и запасов пресных вод, балансе углерода, стабилизации состава атмосферы и пр. Основой этих стратегических действий в стране должно стать внедрение метода логии и методов оценки стоимости объектов живой приро ды, включение ее обобщенных показателей в макроэкономические параметры страны и в систему внешнеэкономических отношений. Для этого потребуются совершенствование законодательной и нормативно-методической баз в области экономики сохранения биоразнообразия, проведение масштабных
экономических оценок на основе общей стоимости применения, стоимости альтернативного применения, затрат на восстановление, оценок предотвращенного ущерба, ренты и пр. Подобные работы выполнены в отдельных регионах (Ярославская и Московская области, Республика Алтай, Ямало-Ненецкий АО и др.). Так, например, предотвращенный ущерб биологическому разнообразию, связанный с деятельностью российских заповедников (около 2% от площади страны), по разным данным (расчет через стоимость отдельных объектов живой природы и выполняемых ими функций, стоимость восстановления экосистем и пр.) оценивается некоторыми экспертами в 260—500 млрд долларов США. Стоимость восстановления популяций охотничьих животных Подмосковья оценена в 34,4 млн долларов США, затраты на экологическую реставрацию нарушенных экосистем определяются в пределах от 10,0 до 70,0 тыс. долларов США на 1 га. Даже опираясь на эти приблизительные цифры можно понять, что для поддержания действий по сохранению биоразнообразия России на современном уровне и для полноценного выполнения ею обязательств по Конвенции о биологическом разнообразии требуются средства минимум на порядок выше, чем в настоящее время (по экспертным оценкам сейчас эти затраты составляют 45—50 млн долларов США в год).
Одним из главных моментов в Национальной стратегии станет и реформирование экономики использования биологических ресурсов, включающей систему платежей, оценки ущерба, компенсацию, лицензирование и пр., а также развитие системы экономического стимулирования и поддержки инвестиций в сохранении и использовании биоразнообразия, в том числе деятельность экологических фондов, федеральные целевые программы и пр. Следует понимать, что живая природа — не нищенка с протянутой рукой, а «основа основ» жизни и практически неисчерпаемый источник благосостояния страны и населения.
2. Формирование системы партнерства в деятельности по сохранению живой природы и биологического разнообразия.
Осуществляется посредством межведомственной координации, создания и функционирования Единой государственной системы мониторинга, информационного обмена, обеспечивающего выявление приоритетов финансирования охраны живой природы.
Интеграция России в Международную систему по сохранению биоразнообразия также привлекает в нашу страну новых зарубежных партнеров, в первую очередь международные экологические организации, которые активно участвуют в деле охраны живой природы (IUCN, WWF, Wetlands International и др.) и содействуют в выполнении обязательств по различным конвенциям. В связи с этим одним из самых актуальных вопросов является и интеграция нашей страны в Пан-Европейскую стратегию в области биологического и ландшафтного разнообразия, рассматриваемую в качестве инструмента реализации Конвенции о биологическом разнообразии в Европе.
Оказание помощи в принятии Национальных стратегий и планов действий (постоянно информируются руководящие органы Пан-Европейской стратегии о ходе подготовки Национальной стратегии и плана действий в России в рамках Проекта ГЭФ, проводится обмен опытом и пр.).
3. Совершенствование системы определения приоритетов в сохранении живой природы и биологического разнообразия.
Базовыми действиями в системе определения приоритетов являются развитие научных исследований в области сохранения, восстановления и рационального использования биоразнообразия, совершенствование системы государственных учетов (земельного, лесного, охотничьего и др.), государственной статистики, ведения государственных кадастров и формирование блока контроля состояния биоразнообразия в Единой системе экологического мониторинга.
Приоритеты научной поддержки мер по охране живой природы должны складываться из: развития таксономических исследований и функционирования базовых биологических коллекций; подготовки и издания российских и региональных флористических и фаунистических сводок, в том числе «Флоры России»; проведения инвентаризации флоры и фауны отдельных регионов и ООПТ; расширения исследований по экологии редких видов для обоснования мер их охраны; создания научных основ экологической реставрации нарушенных экосистем, развития сети питомников дикой флоры и фауны, криобанков, «живых коллекций» и пр.; обоснования нормирования использования отдельных биологических ресурсов, оценок и прогнозов, квотирования и регламентирования ис
пользования по объемам, срокам, регионам и водоемам; научной и информационной поддержки мониторинга состояния биоразнообразия (на базе многолетних наблюдений) и принятия управленческих решений в области сохранения живой природы.
4. Создание системы мер и механизмов реализации Национальной стратегии.
Реализация Национальной стратегии возможна при объединение усилий федеральных и региональных органов законодательной и исполнительной власти, министерств, ведомств и организаций, НПО, СМИ, научных организаций по сохранению биоразнообразия, что достигается подготовкой и внедрением Общественного договора по охране живой природы России. Опыт в этой области имеется (например, Общественный договор в Байкальском регионе, соглашения о совместных действиях в отдельных субъектах Российской Федерации и пр.).
«Фоном» реализации стратегии должно стать завершение формирования законодательной и нормативной баз и развитие механизмов и регламентов правоприменения в области охраны живой природы.
В отраслевых стратегиях сохранения биоразнообразия приоритетным является минимизация воздействия на биоразнообразие разных форм хозяйственной деятельности (использующих биологические ресурсы или нарушающих в процессе деятельности среду обитания растений и животных), в том числе в лесном, сельском и рыбном хозяйствах, в добывающих отраслях и пр. В настоящее время собственно отраслевые программы (планы действий) имеются в Рослесхозе, Госкомры-боловстве России, в Минсельхозпроде России и его отдельных департаментах, в РАО Газпроме, РАО Норильский никель и др. В перспективе отраслевые планы действий в области сохранения живой природы, согласованные с Госкомэкологией России, будут созданы Минобороны России, МПР, МПС, в крупных добывающих отраслях и др.
Развитие территориальных форм охраны биоразнообразия рассматривается как наиболее деятельное и приоритетное направление Национальной стратегии, ориентированное на увеличение площади заповедников и национальных парков до 3% площади России. Реализация этих позиций стратегии осуществляется в соответствии с Перечнем государственных природ
ных заповедников и национальных парков, рекомендуемых для организации на территории Российской Федерации в 1994—2005 гг. (Распоряжение Правительства РФ от 23 апреля 1994 г. №572-р), Государственным кадастром ООПТ, инвентаризацией и резервированием земель для создания новых ООПТ. В перспективе предусматривается создание 72 новых заповедников и 42 новых национальных парков на площади 103,6 тыс. км. Перечнем государственных природных заказников федерального подчинения на период до 2005 г. предусмотрено создание 40 новых заказников на площади 24,0 тыс. км2. Можно прогнозировать, что в России будет расширяться система участков Всемирного природного наследия, водно-болотных угодий Международного значения, пограничных ООПТ и пр., которые наряду с действующими ООПТ разного статуса, ведомственной принадлежности и целевого назначения в перспективе составят национальную экологическую сеть, которая войдет в экологическую сеть Европы и Азии.
Наконец, в Национальной стратегии среди приоритетов поддерживаются позиции сохранения, восстановления и использования популяций редких видов растений и животных in situ и ex-situ, сохранение и разведение редких видов в зоопарках, ботанических садах и питомниках, разработку приемов и технологий воспроизводства в природе, хранения генетических материалов в криобанках и пр., совершенствование государственного контроля за использованием объектов животного и растительного мира.
Приоритетность направлений Национальной стратегии и Национального плана действий определялась экспертно и аналитически, в том числе с использованием: оценок воздействия разных отраслей и секторов общества на природу и отдельные биомы; анализа результатов социологических опросов, кон-тент-анализа публикаций в СМИ; анализа заявок на научные гранты (РФФИ, фонд Мак-Артуров и др.); оценки соответствия актуальности отдельных проблем охраны живой природы и направлений законотворчества (Госдума и Совет Федерации Федерального Собрания Российской Федерации) и подготовки Правительством подзаконных актов и федеральных целевых программ в этой области; экспертных оценок ученых-экологов и практиков охраны живой природы и др.
Разнообразие информации в глобальной компьютерной сети очень велико, почти как биологическое разнообразие в живой природе. Ниже мы приводим некоторые ссылки на ресурсы сети, касающиеся проблем биологического разнообразия.
Электронные атласы
http: / /www .sbras.nsc .ru/cgi-bin/vesta/win/elbib/bio/db / Электронный атлас: «Биоразнообразие растительного мира Сибири*. В нем есть поиск растений Сибири по критериям: семейство, род, вид, морфология, природная зона обитания, экология зоны обитания, географический район обитания, хозяйственное значение вида.
http://www.helsinki.fi/ kmus/afe/database. html
Растения Европы — Атлас (Atlas Florae Europaeae Database). База данных видового биоразнообразия. Предлагается программа для просмотра географических карт биоразнообразия.
Электронные базы данных
http://bmras.newmail.ru/bioras.html
Каталог баз данных по биоразнообразию (BIODIVERSITY DATABASES CATALOGUE) — оперативно обновляемая и постоянно пополняемая сводка по биологическим базам данных, отражающая все значимые изменения, происходящие в них. Проект впервые был анонсирован на Международном симпозиуме «Информационно-поисковые системы в зоологии и ботанике* (май 1999, Петербург). Каталог создан на основе базы сведений, отобранных в ходе электронного анкетирования держателей баз данных. Особое место в Каталоге занимает ГРНТИ — он встроен в систему и предлагается в качестве инструмента тематического поиска в соответствии с информационными потребностями пользователя. Сегодня на сайте размещен обновленный вариант «Анкеты для держателей информации по биоразнообразию*. Этот «Каталог каталогов* можно скачать в виде базы данных для Microsoft Access 97/2000.
Электронные коллекции
www.si.edu/departments/botany.html
Национальный гербарий США — Ресурсы Смитсоновского института.
Информационные сайты
www.biodiversity.uno.edu
The Biodiversity and Biological Collections Web Server содержит обширнейшую информацию по ботанике, орнитологии, энтомологии, млекопитающим и др., а также о различных биологических коллекциях, отчетах и многом другом. Имеются ссылки на соответствующие разделы.
WWW .bsponline.org/index. html
Сайт The Biodiversity Support Program позволяет получить быстрый доступ к информации по многим проектам в виде электронной библиотеки, которая содержит книги, брошюры, отчеты и другие публикации. BSP, разработан совместно с World Wildlife Fund (WWF), The National Conservansy (TNC) и World Research Institute (WRI) под руководством Consortium institution и The United States Agency for International Development (USAID).
www.nbii.gov
Национальная биолого-информационная инфраструктура США (NBII). Огромная организация, обеспечивающая ученым, представителям правительственных и частных организаций, людям, кому не безразлично состояние окружающей среды, широкий доступ к базам данных и информации о национальных биологических ресурсах. На главной странице расположены общая информация об организации и перечень ссылок на основные направления работы НБИИ: сохранение биоразнообразия; работа по исследованию вирусов; распространение примесей; сохранение биоресурсов; географические программы; образование. Работа по сохранению биоразнообразия ведется в трех направлениях: генетическое разнообразие, видовое разнообразие, экологическое или экосистемное разнообразие.
www. sci. aha.ru/biodiv/
Сайт имеет два варианта: русский и английский. Содержит несколько разделов по биоразнообразию: сводка конференций стран Европы и Евразии, общеевропейская Стратегия сохранения биоразнообразия, занимательная интернет-книга по биоразнообразию для детей и министров.
Биоразнообразие России
www. ihst. ru/org/rcmc / russian/docs /
Национальная Стратегия сохранения биологического разнообразия России: документы и процесс.
www ihst.ru/org/rcmc/russian/docs/report97/cont_r. html Национальный доклад РФ по сохранению биоразнообразия. В документе описывается: роль Конвенции о биологическом разнообразии в сохранении и устойчивом использовании биологического разнообразия России. Россия ратифицировала Конвенцию о биологическом разнообразии в феврале 1995 г. На третьем совещании Конференции сторон КБР в Аргентине в ноябре 1996 г. впервые были определены конкретные меры по реализации целей Конвенции, в частности по таким важным для России направлениям, как сохранение биоразнообразия в лесном и сельском хозяйствах. Перечислены положительные результаты национальных действий по сохранению биоразнообразия в связи с ратификацией КБР, также выделены серьезные проблемы, связанные с необходимостью выполнения Россией обязательств по КБР; современное состояние и проблемы сохранения биоразнообразия: территориальная охрана биоразнообразия, охрана редких и исчезающих видов, сохранение биоразнообразия в лесном хозяйстве, охрана и рациональное использование охотничьих животных, сохранение биоразнообразия морей и внутренних водоемов, сохранение биоразнообразия в сельском хозяйстве; проект Глобального экологического фонда «Сохранение биоразнообразия* в России.
www.sci.aha.ru/ATL/raOO.htm#Refl
Биоразнообразие и охрана природы в Web-атласе «Здоровье и окружающая среда». Содержит следующую информацию: основные особенности местоположения, природы, населения и исторического пути развития России; естественное воспроизводство населения России: численность и компоненты прироста населения, демографическая нагрузка, воспроизводство населения и его перспективы; расселение и миграции населения России: плотность населения и система расселения, жизненные миграции и тенденции урбанизации; природные условия России; биологическое разнообразие в России, особо охраняемые природные территории (ООПТ) и охрана животного и растительного мира на них; особенности взаимоотношений с окружающей средой и природопользование; леса России: состояние и динамика в 1989—1996 гг.; сельское хозяйство России: состояние и динамика в 1989—1996 гг.; природные факторы формирования здоровья населения: комфортность климата и демографические последствия дискомфортное™ климата, природно-очаговые и сельскохозяйственные зооантропонозы,
медико-экологическое районирование и медико-экологическая оценка условий жизни населения. Информация иллюстрирована картами.
Электронные журналы по биоразнообразию
http: / /books. nap. edu/books /03090522 70/html /index .html Журнал Biodiversity II. Электронная версия.
Организации
www.nysm.nysed.gov
NYS Институт исследований проблем сохранения биоразнообразия.
www.york.biosis.or
Всемирный Институт но проблемам сохранения биоразнообразия.
www. cabs. conservation, org
Сайт Центра Прикладных исследований по биоразнообразию (Center of Applied Biodiversity Science).
Международное сотрудничество
www.biodiv.org
Сайт посвящен конвенции по биоразнообразию. Конвенция — первому всестороннему договору, по всем аспектам сохранения биоразнообразия, генетических ресурсов и экосистем. На сайте можно прочитать не только о Международной конференции 1992 г. (Рио-де-Жанейро), но и о предстоящих Международных акциях вплоть до 2002 г.
www.rao.sio.rssi.ru
На этом сайте можно получить информацию о конференциях и международных акциях, которые проводились и будут проводиться международным сообществом.
http://www.biodiv.org
Biodiversity Conservation Information System. Сайт посвящен биоразнообразию и биобезопасности. Имеются различные документы по данной тематике. Рассказывается о встречах и конференциях, посвященных биоразнообразию и биобезопасности. Имеются списки прошедших, текущих и будущих встреч, конференций и многих других событий, связанных с биоразнообразием. Они представлены в виде таблиц с заголовками полей «Дата и место проведения», «Название* и «Контактная информация». Возможен поиск информации.
ЛИТЕРАТУРА
Алтухов ЮЛ. Природоохранная генетика // Экология в России на рубеже XX века (наземные экосистемы). М., 1999. С. 9—26.
Амирханов АМ„ Тишков АА. Национальная стратегия и План действий по сохранению биологического разнообразия в России //Сохранение биоразнообразия. М., 1999. С. 27—28.
Бигон М., Харнер Д., Таусенд К. Экология. Особи, популяции, сообщества. М., 1989.
Вальтер Г. Растительность земного шара. М., 1968. Т. 1.
Василевич ВЛ. Альфа-разнообразие растительных сообществ и факторы его определяющие //Биологическое разнообразие: подходы к изучению и сохранению. СПб., 1992. С. 162—170.
Виноградов Б.В. Гамма-разнообразие наземных экосистем //Биогеография. Вып. 8. География биоразнообразия. М., 2000. С. 11—20.
Воронов ГА, Дроздов Н.Н., Криволуцкий ДА, Мяло Е.Г. Биогеография с основами экологии. М., 2003. 408 с.
Воронов Г А, Дроздов НН., Мяло ЕЕ. Биогеография мира. М., 1985. 270 с.
Второв ПЛ., Дроздов Н.Н. Биогеография. М., 2001. 270 с.
Дроздов НН. Биологическое разнообразие, структура животного населения и процессы формообразования в очагах аридиости // Вести. Моск, ун-та. Сер. 5. География. 1994. № 6. С. 52—58.
Дроздов Н.Н., Мяло ЕЕ. Экосистемы мира. М., 1997. 238 с.
Дроздов Н.Н., Криволуцкий ДА, Огуреева ГН. Биомное разнообразие / / Биогеография. 2002. Вып. 10. С. 8—14.
Закон Российской Федерации № 2254 «Конвенция о биологическом разнообразии» // Собр. закоиодат. актов РФ, № 19. С. 4742—4764.
Концепция перехода Российской Федерации к устойчивому развитию. М., 1996. 12 с.
Криволуцкий ДА. Динамика биоразнообразия и экосистем в условиях радиоактивного загрязнения // Докл. РАН, 1996. Т. 347, № 4. С. 1—4.
Криволуцкий ДА. Жизненные формы и биоразнообразие животных // Бюл. МОИП. Отд. биол. 1999. Т. 347. Вып. 5.
Криволуцкий ДА, Калякин В.М., Лебедева Н.В., Рябов ИН. Земли радиоактивного загрязнения как убежища для редких видов животных // Вести. Моск, ун-та. Сер. 5. География. 2000. № 6. С. 20—23.
Лебедева Н.В. Измерение и оценка биологического разнообразия. Методическое пособие. Ч. 1. Ростов-на-Дону, 1997. 39 с.
Лебедева Н.В. Измерение и оценка биологического разнообразия. Методическое пособие. Ч. 2. Ростов-иа-Дону, 1999. 40 с.
Лебедева НЕ. Экотоксикология и биогеохимия географических популяций птиц. М., 1999. 199 с.
Лебедева НЕ., Дроздов НН., Криволуцкий ДА. Биоразнообразие и методы его оценки. М., 1999. 94 с.
Лебедева Н.В., Покаржевский АД. Биологическое разнообразие: Программа курса. Ростов-на-Дону, 1993. 9 с.
Левонтин Р. Человеческая индивидуальность, наследственность н среда. М., 1993.
Матекин П.В. История и методология биологии. М., 1982. 162 с.
ЛИТЕРАТУРА
Мэгарран Э. Экологическое разнообразие и его измерение. М., 1992.181с.
Назарова Л.Ф., Алтухов СМ., Машу ров AM. Эволюция человеческих популяций. Липецк, 2000. 16 с.
Огуреева ГЛ., Котова Т£. Классификация и картографирование биомов Сибири // Геоботаническое картографирование. СПб., 1997. С. 22—30.
Песенко ЮЛ. Принципы и методы количественного анализа в фаунистических исследованиях. М., 1982. 287 с.
Реймерс Н.Ф. Экология. М., 1994. 366 с.
Савицкий РМ„ Лебедева Н.В., Савицкая НЛ. Видовой состав и динамика разнообразия птиц в городе Ростове-на-Дону// Кавказский орнитологический вестник. 1998. № 10. С. 114—123.
Соколов В£., Шишкин В.С. Динамика первоописаний видов млекопитающих на территории Российского государства за 250 лет // Состояние териофауны в России и ближнем зарубежье/Тр. Междунар. со-вещ. М., 1996. С. 309—314.
Сохранение биологического разнообразия России. Первый национальный доклад Российской Федерации. Выполнение Россией обязательств по Конвенции о биологическом разнообразии. Под ред. А.М. Амирханова. М., 1997. 202 с.
Строение ДНК и положение организмов в системе. М., 1972. 327 с.
Терещенко В.Г., Терещенко Л.И., Сметанин ММ. Оценка различных индексов для выражения биологического разнообразия сообщества/ /Биоразнообразие: Степень таксономической изученности. М., 1994. С. 86—98.
Тишков АЛ. Устойчивость в природном разнообразии // Экология и жизнь, 1998. № 4. С. 26—33.
Тьюки Дж. Анализ результатов наблюдений. М., 1981. 693 с.
Уиттекер Р. Сообщества и экосистемы. М., 1981. 328 с.
Юрцев ВЛ. Эколого-географическая структура биологического разнообразия и стратегия его учета и охраны //Биологическое разнообразие: подходы к изучению и сохранению. СПб., 1992. С. 7—21.
Diamond JM. Assembly of species communities // Ecology and evolution of communities. Cambridge, 1975. P. 342—444.
Global biodiversity: Status of the Earth’s living resources. World Conservation Monitoring Centre. London, 1992. 594 p.
Hutchinson GJ!., MacArthure R.H. A theoretical ecological model of size distribution among species of animal // American. Natur. 1959. Vol. 93. P. 117—125.
MacArthure ЯЛ. Pattern of terrestrial bird communities // Avian biology, 1971. Vol. 1. P. 189—221.
MayrE. Evolution and the diversity of life. Cambridge, 1976.
Simpson G. G. Principles of Animal Taxonomy. New York, 1961.
Walter H., Box E. Global classification of natural terrestrial ecosystems // Vegetatio. 1976. Vol. 32, № 2. P. 75—82.
Whittaker ЯЛ. Evolution and measurement of species diversity // Taxon, 1972. Vol. 21. P. 213—251.
ОГЛАВЛЕНИЕ
ВВЕДЕНИЕ 3
Понятие биоразнообразия 4 Международная программа
♦Биологическое разнообразие* 9
Научная программа ♦Диверситас» ________________________12
Реализация Конвенции о биоразнообразии в России 18
Глава I
УРОВНИ БИОРАЗНООБРАЗИЯ" 21
Системная концепция биоразнообразия_____________________22
Генетическое разнообразие_______________________________28
Биохимическая систематика_______________________________42
Видовое разнообразие____________________________________51
Биоразнообразие, созданное человеком ___________________53
Экосистемное разнообразие_______________________________56
Классификации биоразнообразия 59
Глава II V
ТАКСОНОМИЧЕСКОЕ РАЗНООБРАЗИЕ Д 63
Научная классификация организмов_____________________64
Жизненные формы и биологическое разнообразие ________82
Инвентаризация видов_________________________________91
Видовое богатство России_____________________________98
Центры таксономического разнообразия 104
Глава III ГЕОГРАФИЯ БИОРАЗНООБРАЗИЯ 107
Геногеография 108
Геногеография популяций человека _____________________114
Географические закономерности видового разнообразия 126
Равновесная теория островной биогеографии как модель динамики видового разнообразия 158
Глава IV БИОМНОЕ РАЗНООБРАЗИЕ — ВЫСШИЙ
УРОВЕНЬ РАЗНООБРАЗИЯ ЭКОСИСТЕМ ХП 177
Тундры_____________________________________________187
Бореальные хвойные леса____________________________192
Листопадные леса умеренной зоны 197
Саванны и степи ___________________________________203
Пустыни ___________________________________________218
Субтропические жестколистные леса и кустарники_____226
Тропические дождевые леса _________________________230
Пресноводные экосистемы____________________________240
Морские экосистемы 246
Глава V ИЗМЕРЕНИЕ И ОЦЕНКА БИОЛОГИЧЕСКОГО РАЗНООБРАЗИЯ 253
Альфа-разнообразие: видовое обилие___________________256
Методы построения графиков видового обилия___________258
Модели распределения видового обилия_________________264
Индексы видового богатства 271
Индексы, основанные на относительном обилии видов____274
Сравнительный анализ индексов разнообразия 279
Рекомендации для анализа данных по разнообразию видов 282 Анализ бета-разнообразия: сравнение, сходство, соответствие сообществ_______________________________284
Графический анализ бета-разнообразия 290
Применение показателей разнообразия__________________298
Гамма-разнообразие наземных экосистем 300
Глава VI ПРИРОДОПОЛЬЗОВАНИЕ И БИОЛОГИЧЕСКОЕ РАЗНООБРАЗИЕ 307
Воздействия человека на биоразнообразие______________310
Антропогенные изменения биомов_______________________313
Оценка опасности изменений на уровне популяций 317
Оценка опасности изменений на уровне сообществ 320 Основные типы антропогенных нарушений и экспертная оценка их значимости 324
Технология экспертной оценки влияния природопользования на биологическое разнообразие 326
Глава VII
ТЕХНОГЕННЫЕ КАТАСТРОФЫ - УГРОЗА
БИОРАЗНООБРАЗИЮ 329
Стабильность и устойчивость биологических систем 330
Динамика биоразнообразия в условиях радиоактивного загрязнения 336 Влияние разливов нефти на разнообразие
морских сообществ 343
Влияние техногенного загрязнения
на лесные сообщества 346
Глава VIII
МОНИТОРИНГ БИОРАЗНООБРАЗИЯ
Международные программы мониторинга биоразнообразия ___________________________
Мониторинг биоразнообразия в России
349
350
Глава IX
ВСЕМИРНАЯ СТРАТЕГИЯ СОХРАНЕНИЯ
БИОЛОГИЧЕСКОГО РАЗНООБРАЗИЯ 363
Глобальные экологические изменения 364
Международный опыт мероприятий по сохранению живой природы____________________________________ 394
Стратегия сохранения биологического разнообразия 400
Глава X
НАЦИОНАЛЬНАЯ СТРАТЕГИЯ СОХРАНЕНИЯ
БИОЛОГИЧЕСКОГО РАЗНООБРАЗИЯ РОССИИ 407
Система охраняемых природных территорий в России 408
Цель Национальной стратегии
и Национального плана действий ___________________414
Приоритеты Национальной стратегии 418
Глава XI РЕСУРСЫ ИНТЕРНЕТ ПО СОХРАНЕНИЮ БИОРАЗНООБРАЗИЯ
428
ЛИТЕРАТУРА
Лебедева Наталья Викторовна, профессор, доктор биологических наук, академик РАЕН и Нью-Йоркской академии наук. Наталья Викторовна - инициатор разработки программ, учебных пособий и учебного курса по биоразнообразию в России. Автор фундаментальных исследований по радиоэкологии, экотоксикологии и биоразнообразию птиц, участница крупных международных экологических экспедиций в Арктике, Приазовье, Монголии, Польше, Африке.
Дроздов Николай Николаевич, профессор, доктор биологических наук, академик РАЕН и Российской экологической академии, советник Генерального секретаря ООН в области охраны природы и устойчивого развития нашей планеты.
Николай Николаевич многолетний активный деятель Международного союза охраны природы. За разработку методов сохранения биоразнообразия в России он удостоен премии ЮНЕСКО и награжден золотой медалью А. Эйнштейна.
Он неутомимый путешественник, побывавший на всех материках, выполнивший фундаментальные исследования фауны пустынь Средней Азии, Закавказья, Африки и Северной Америки.
Криволуцкий Дмитрий Александрович, член-корреспондент РАН, доктор биологических наук, профессор, лауреат Государственной премии СССР. Им обнаружено более 1100 видов ранее неизвестных в России животных микрофауны, выполнены не имеющие в мире аналогов фундаментальные исследования динамики биоразнообразия в условиях радиоактивного загрязнения.
ISSN 5-691-01098-0
О 10989
691
Пособие посвящено биоразнообразию, его уровням и таксономии, знакомит с методами его измерения. Рассматриваются механизмы воздействия человека на живую природу. Пособие адресовано студентам, обучающимся по экологическим и географическим специальностям вузов.