/







































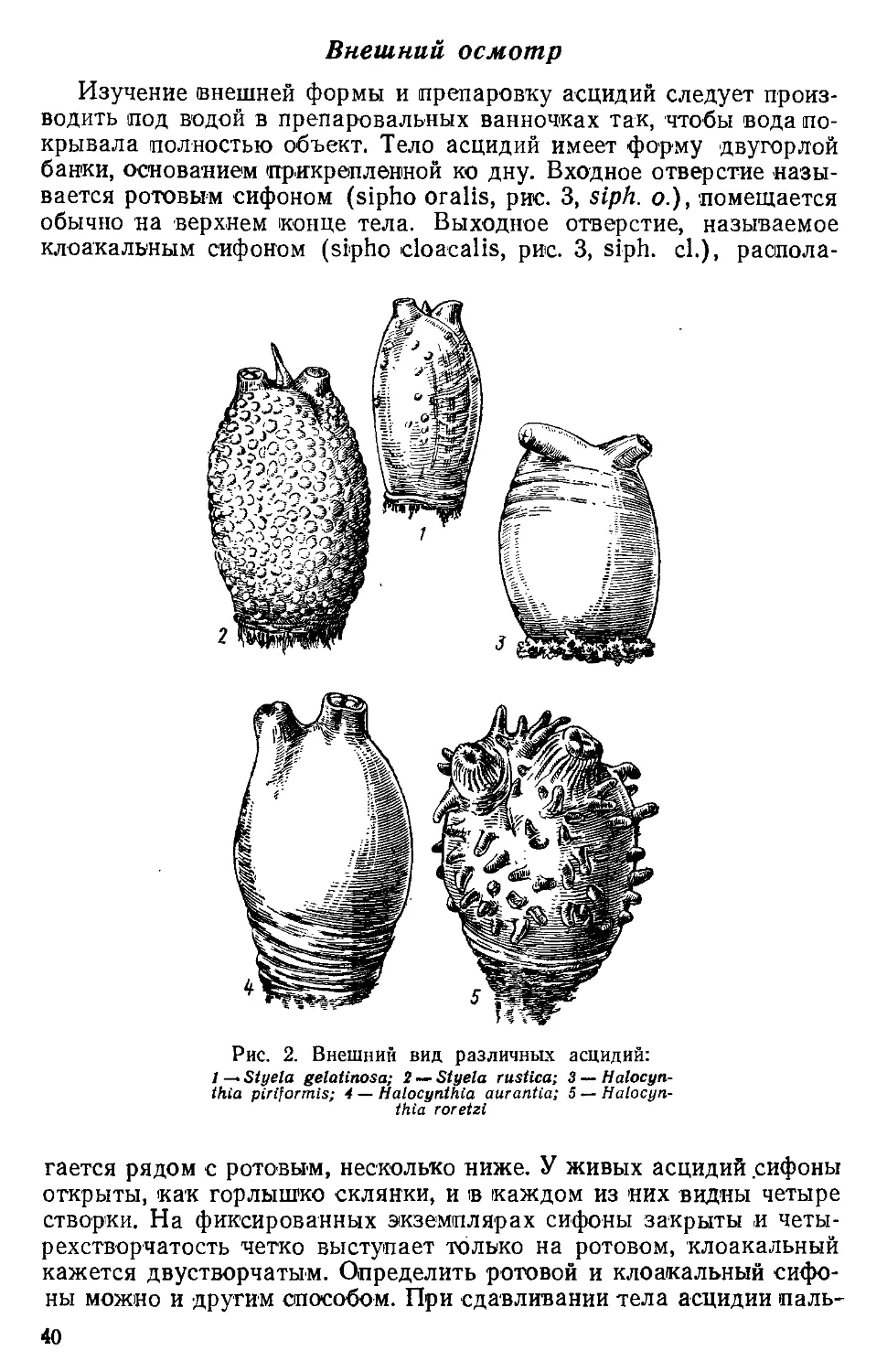








































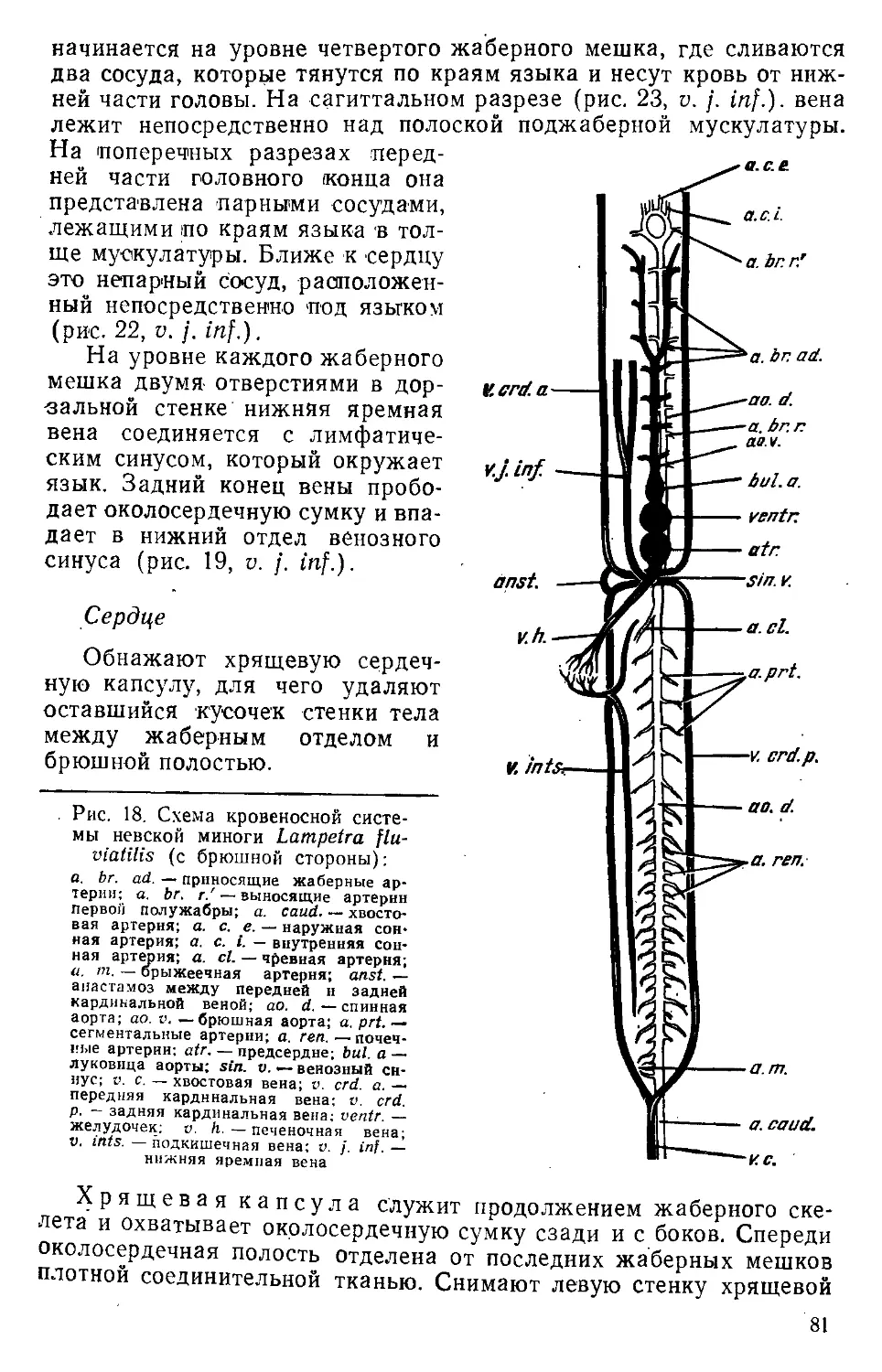


























































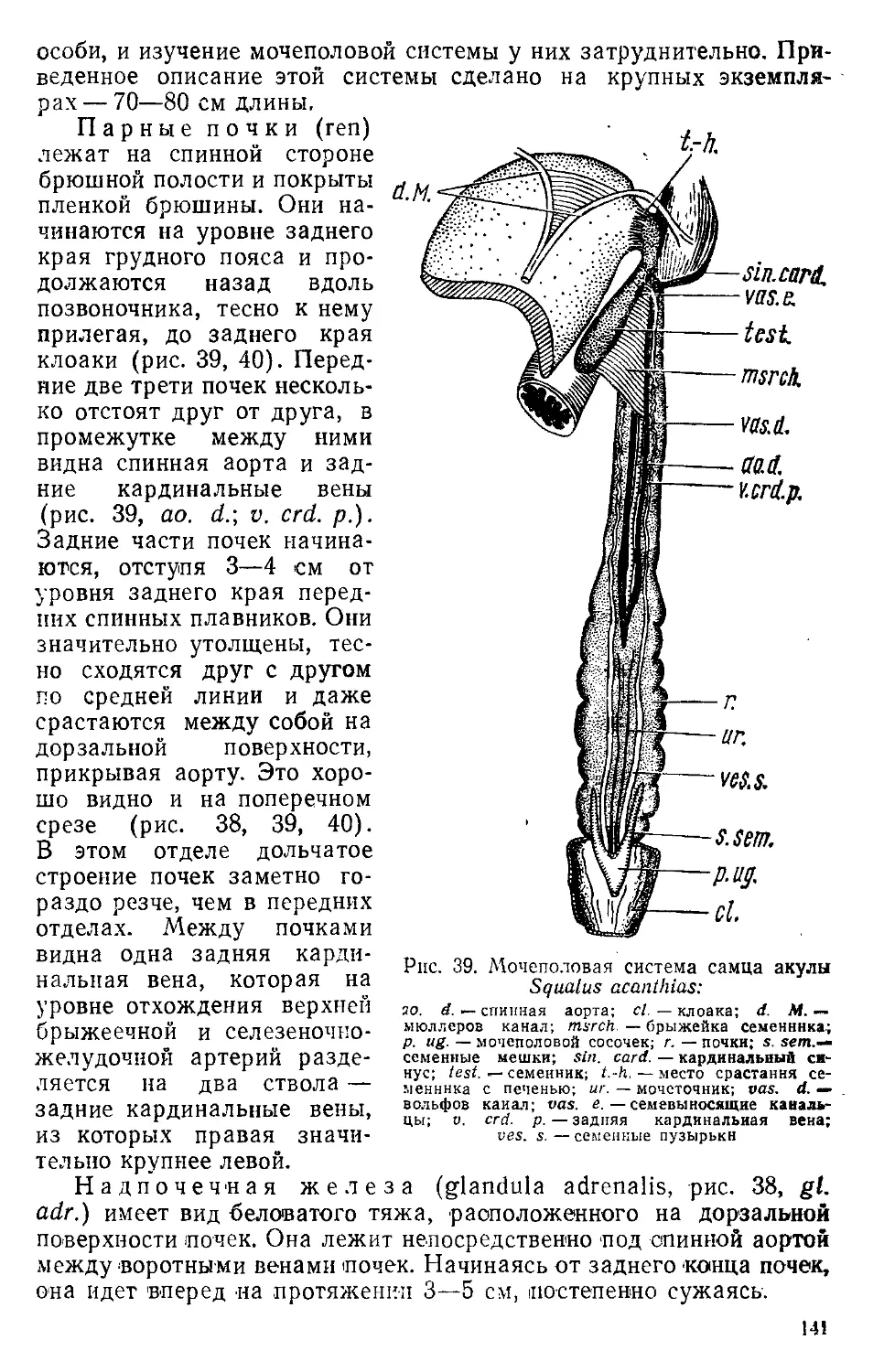



















































































































































































































Text
Н. Н. ГУРТОВОЙ, Б. С. МАТВЕЕВ, Ф. Я. ДЗЕРЖИНСКИЙ
ПРАКТИЧЕСКАЯ ЗООТОМИЯ ПОЗВОНОЧНЫХ
НИЗШИЕ ХОРДОВЫЕ, БЕСЧЕЛЮСТНЫЕ, РЫБЫ
Под редакцией
Б. С. Матвеева и Н. Н. Гуртового
Допущено
Министерством высшего и среднего специального образования СССР в качестве учебного пособия для студентов биологических специальностей университетов
59 Г95
Гуртовой Н. Н., Матвеев Б. С., Дзержинский Ф. Я.
Г95 Практическая зоотомия позвоночных (низшие хордовые, бесчелюстные, рыбы). Учеб, пособие для биологических специальностей ун-тов. М., «Высш, школа», 1976.
351 с. с ил. Библиогр. 79
Книга — практическое руководство по вскрытию, последовательности препарирования и анатомическому изучению организации всех систем органов представителей каждого класса животных. Во введении приводятся методические указания для организации самостоятельных исследований. Для правильного понимания эволюционного процесса дается анализ анатомического строения оболочников и бесчерепных. Кинга хорошо иллюстрирована.
Кроме студентов биологических факультетов университетов и педагогических институтов может быть широко использована специалистами-биологами и медиками.
21008—096
001(01)—76
82—76
59
© Издательство «Высшая школа». 1976
ПРЕДИСЛОВИЕ
Настоящее пособие представляет собой практическое руководство по анатомии хордовых животных. Предполагается издать его в трех книгах. В первой книге рассматривается строение низших хордовых, бесчелюстных и рыб. Вторая книга посвящена земноводным и пресмыкающимся. И, наконец, анатомия птиц и млекопитающих будет рассмотрена в третьей книге.
В пособии помимо описания строения всех систем органов ряда представителей позвоночных животных в последовательном порядке от низших форм к высшим даются также практические указания, как вести работу по изучению анатомии того или иного представителя.
В любой исследовательской работе каждый специалист по зоологии должен стремиться научиться читать изучаемый объект, т. е. научиться видеть, как отражаются на анатомическом строении каждого организма те особенности его биологии, условий существования и систематического положения, которые выделяют его как особый вид и род, и какие черты определяют его как представителя более крупных систематических категорий (семейства, отряда, класса). Анализ особенностей организации последовательного ряда позвоночных животных является хорошей школой исследовательского метода изучения, необходимого каждому специалисту в практической работе. Умение читать объект позволяет находить неизвестное на новых объектах исследования. Такие цели ставятся перед студентами старших курсов биологических факультетов университетов, специализирующимися по зоологии позвоночных, при выполнении ими больших практикумов по зоологии и сравнительной анатомии, а также при выполнении курсовых и дипломных работ.
Рассчитанное в первую очередь на студентов государственных университетов, это пособие может быть использовано целиком или частями на других кафедрах биологических факультетов (ихтиологии, физиологии, гистологии и др.), а также в педагогических, сельскохозяйственных, рыбохозяйственных и зоотехнических институтах. Пособие может служить справочником для студентов, аспирантов, преподавателей, научных сотрудников, сталкивающихся в своей работе с вопросами анатомии позвоночных. Практическое руководство по зоотомии позвоночных может быть использовано и при экспериментальных исследованиях по физиологии, медицине, биохимии и био
физике в случаях, когда объектами служат позвоночные животные, и анатомические данные необходимы, как материал для эксперимента.
Настоящее руководство отличается от других, широко распространенных в биологических вузах, руководств к практическим занятиям по зоологии в первые годы обучения (П. В. Терентьев; М. М. Беляев; Н. А. Аверинцев; Н. Н. Карташов, В. Е. Соколов, И. А. Шилов и др.), задачами, которые оно преследует. Там ставится задача продемонстрировать на отдельных представителях разных классов особенности строения, характеризующие каждый класс позвоночных животных. Наше руководство стремится научить студента самостоятельно разбираться в сложном организме позвоночного животного, научить понимать, как особенности образа жизни, биологии и поведения животного отражаются на его анатомическом строении. При самостоятельных занятиях необходимо научиться так пользоваться книгой, чтобы все описанное в руководстве строение той или иной системы можно было бы увидеть на препарате, изготовленном своими руками. Наше руководство не претендует занять место теоретического руководства к самостоятельному изучению курса зоологии позвоночных. Оно не затрагивает вопросов теории, а только учит студента или аспиранта самостоятельно работать над объектом с книгой. Стремясь осуществить эту задачу, авторы руководства не только использовали свой опыт работы со студентами, но при составлении текста изготовляли препараты, проделывая все те операции, которые рекомендуют студентам. С этих препаратов были сделаны рисунки, которые и помещены в данном руководстве. В редких случаях использованы рисунки из других источников. Часть рисунков была выполнена по эскизам авторов студентом В. Иваницким.
Практическое руководство * по зоотомии позвоночных составлено коллективом преподавателей лаборатории морфологии позвоночных им. А. Н. Северцова при кафедре зоологии позвоночных биологического факультета Московского государственного университета на основании многолетнего опыта проведения занятий на большом практикуме по зоологии и сравнительной анатомии под руководством проф. Б. С. Матвеева.
Первая попытка создания такого руководства была предпринята довольно давно. Описание анатомии низших хордовых, хрящевых и костных рыб и амфибий, разработанное в пределах программы большого практикума по зоологии позвоночных, проводимого в Московском университете, было издано небольшим тиражом, десятью отдельными выпусками научно-методическим кабинетом по заочному и вечернему обучению МГУ в период с 1963 по 1966 г.
Издание отдельных выпусков позволило проверить на практике работы со студентами пригодность составленных макетов.
На основе этого опыта все главы настоящего руководства подверглись не только существенным исправлениям и дополнениям, но во многих случаях и значительной переработке. Тексты глав, посвященные вскрытию хрящевых рыб и скелету костистых рыб, написанные при жизни доцентом А. Н. Дружининым и старшим преподавателем Л. В. Ганешиной, были взяты за основу и существенно переработаны. Изменения внесены также в текст по скелету и покровам осетровых рыб, соста!влен-ный доцентом В. А. Раковым.
Учитывая, что вузы располагаются в различных областях Советского Союза, и это определяет доступность тех или иных объектов, в руководстве, как правило, наряду с описанием одного представителя имеются ссылки на другие объекты. Это делает возможным не только более широкий выбор объектов для изучения, но и позволяет сравнить особенности строения представителей различных таксономических групп.
В предлагаемой книге в связи с ее основным назначением как руководства к анатомической части большого практикума даются общие методические указания к ведению большого практикума. Изложена методика работы с объектом и техника приготовления анатомических препаратов: оборудование рабочего места, техника вскрытия, подготовка материала и техника изготовления препаратов тех или иных систем и органов. Даются общие сведения о понятиях и терминах, необходимых при изучении сравнительной анатомии. Приведены общая характеристика типа хордовых, система типа хордовых, общая характеристика эволюции хордовых, а также сравнительный метод исследования и термины сопоставления органов при сравнении.
Далее изложен материал по анатомии водных позвоночных. На примере миноги рассматривается строение круглоротых, относящихся к группе бесчелюстных. Анатомия хрящевых рыб разобрана на примере акулы и ската. Дается описание анатомии костных рыб, из которых разобраны костно-хрящевые ганоиды на примере осетровых, и костистые рыбы на примере щуки, окуня, судака, трески и сазана (карпа).
В «Практикуме по зоотомии позвоночных», однако, в начале книги даны разделы, посвященные низшим хордовым животным. На примере асцидий студенты знакомятся с подтипом оболочников, или личиночнохордовых, а на примере ланцетника — с подтипом бесчерепных. Для понимания систематического и филогенетического положения позвоночных знакомство с низшими представителями хордовых обязательно.
Авторы приносят глубокую благодарность всем лицам, помогавшим при создании данного Практикума, и в первую очередь проф. Я. Я. Рогинскому и сотрудникам кафедры зоологии позвоночных МГУ Н. В. Хмелевской и М. М. Ястребцовой, принимавшим самое деятельное участие в подготовке рукописи к изданию.
Большой вклад в создание этого практикума внес Борис Степанович Матвеев. Более 50 лет Борис Степанович проводил и возглавлял большой практикум по зоологии позвоночных в Московском университете. Предлагаемая книга — результат его многолетней педагогической и научной деятельности, и большая часть книги написана им лично. Рукопись была отредактирована Борисом Степановичем, но, к сожалению, он ие дожил до выхода книги в свет.
Проф. Б. С. Матвеевым написана основная часть текста разделов: введение; оболочники; бесчерепные; скелет, мускулатура, нервная система и органы чувств хрящевых рыб, а также раздел по осетровым рыбам. Основная часть текста остальных разделов написана Н. Н. Гуртовым.
Н. Н. Гуртовой, Ф. Я- Дзержинский
ВВЕДЕНИЕ
Общие методические указания к ведению большого практикума
В системе университетского преподавания на биологических факультетах государственных университетов Советского Союза большие практикумы занимают центральное место в подготовке биолога-исследователя, ищущего новые пути в разрешении теоретических и практических вопросов. Задачей большого практикума по зоологии позвоночных является изучение организации позвоночных в последовательном порядке от низших форм к высшим, т. е. изучение особенностей строения органов и их функций у животных, составляющих тип хордовых в том порядке, как сложилась их организация в историческом развитии. Поэтому основную часть изучения на большом практикуме позвоночных животных составляют самостоятельные лабораторные занятия по анатомии отдельных представителей низших хордовых и разных классов позвоночных.
Впервые в России подобные самостоятельные работы студентов-биологов были организованы акад. П. П. Сушкиным, когда он был доцентом Института сравнительной анатомии Московского университета. После Великой Октябрьской социалистической революции эти занятия легли в основу большого практикума по зоологии позвоночных при подготовке специалистов-зоологов. В дальнейшем анатомическая часть большого практикума разрабатывалась и изменялась проф. Б. С. Матвеевым и доц. А. Н. Дружининым. В настоящее время большой практикум по зоологии позвоночных в Московском университете состоит из трех разделов: 1) изучения анатомического строения на ряде позвоночных от низших групп к высшим; 2) общего обзора организации каждой группы в теоретическом семинаре; 3) практических занятий по систематике позвоночных, по коллекциям с освоением методики определения для познания многообразия фауны нашей Родины.
Первый раздел посвящен самостоятельному изучению анатомического строения представителей всех классов позвоночных, вернее всего типа хордовых. Студенты на конкретных препаратах, изготовленных ими же, изучают все системы органов каждого представителя.
Студент должен научиться читать объект, т. е. разбираться на животном во всех деталях его строения, и понимать, какую функцию несет каждая часть организма. Важнейшим методом для этого служит метод сравнения организации последовательного ряда форм позвоночных животных от низших групп к высшим, что позволяет
при завершении большого практикума получить представление о путях и закономерностях эволюционного развития позвоночных животных в историческом прошлом.
Сравнительное изучение последовательного ряда животных имеет значительные преимущества по сравнению с всесторонним изучением отдельных объектов, избранных студентом для исследования, как это делается в некоторых университетах. Большой практикум должен дать не только нужный запас фактических данных, но и воспитать в сознании учащегося материалистическое естественнонаучное мировоззрение.
Отсюда следует второй раздел большого практикума — знакомство с многообразием животных по систематическим коллекциям путем определения. Для этого на кафедре должны быть заготовлены коллекции шкурок птиц и млекопитающих и спиртовые коллекции рептилий, амфибий, рыб и круглоротых, представителей не только фауны СССР, но и мировой фауны. Для воспитания эволюционного мировоззрения важно, чтобы студенты собственными глазами видели тех редких представителей реликтовых групп мировой фауны, которые связывают ныне живущие группы между собой.
Третьим разделом большого практикума являются теоретические обзоры — семинары, проводимые руководителем практикума еженедельно. На таком семинаре преподаватель должен стремиться на конкретном материале коллекций музея и препаратах учебной коллекции дополнить знания студента, полученные при изучении объекта, показом сравнительных данных, отображающих многообразие приспособительной эволюции изучаемых групп. Конечно, всего показать по коллекциям и препаратам нельзя, поэтому приходится пользоваться также и литературными данными и таблицами. В этом обзоре ставится задача — осмыслить отработанный фактический материал путем сопоставления анатомических данных с материалами по сравнительной анатомии, сравнительной эмбриологии и палеонтологии, используя также и данные по экологии.
Важным разделом в подготовке специалиста является также и умение пользоваться научной литературой.
К сожалению, в нашей отечественной литературе мало сводных монографий, которые могли бы служить теоретическими руководствами по большому практикуму зоологии позвоночных. Здесь можно-только назвать «Основы ихтиологии» Е. К. Суворова, «Руководство по зоологии» (т. VI, «Птицы» Г. П. Дементьева) и «Орнитологию» Л. М. Шульпина. Перед советскими биологами большой долг — заполнить этот важный пробел в подготовке специалиста — зоолога позвоночных.
Важнейшим разделом самостоятельных занятий на большом практикуме служит не только самостоятельная работа над препаратом и книгой, но и зарисовка отработанного препарата по ходу анатомирования. В исследовательской работе биолога рисование является методом последовательного анализа специфических особенностей в организации животного, оно помогает студенту восстановить по отдельным частям целое представление об изучаемой си
стеме органов. В таком рисунке в первую очередь должны быть отражены общие особенности организации исследуемого представителя той пли иной группы. При сопоставлении рисунков различных животных студент получает возможность делать заключение о преобразовании организации различных систем органов в ряду позвоночных от низших форм к высшим. Рисунки — также весьма удобная форма отчета, позволяющая установить при их проверке и беседе со студентом, насколько полно он разобрался в препарате. После выполнения каждого недельного задания преподаватель должен проверять отработанные студентами препараты и полноту их изображения на рисунках. Эти недельные зачеты весьма четко организуют труд студента.
По современным учебным планам большим практикумам по зоологии позвоночных отводится 15 ч в неделю. Из них 4 ч уходит на занятия по систематике, 9 ч — по анатомии и 2 ч — на теоретические семинары. Одновременно студенты слушают ряд спецкурсов по биологии позвоночных животных. При нормальной же работе общее количество часов достаточно для выполнения программы большого практикума, но всегда может возникнуть необходимость дополнительной работы студентов (отработка пропущенных занятий, более детальное изучение отдельных систем и т. п.). Поэтому при прохождении большого практикума студентам должны быть представлены в лаборатории постоянные рабочие места и выдано на все время оборудование, посуда, инструменты, чтобы они в любое время, свободное от других занятий, могли приходить на свое рабочее место и вести самостоятельную работу по выполнению заданий.
Методика и техника приготовления анатомических препаратов
Оборудование
Успешное проведение занятий по курсу большого практикума зависит, в частности, от наличия соответствующего оборудования. Все оборудование, необходимое для занятий, можно разделить на две группы: 1) инструменты и предметы, необходимые студенту на каждом занятии, ,и 2) оборудование, используемые студентом редко или используемое несколькими студентами поочередно в течение занятия.
Для каждого студента необходимо следующее оборудование.
1. Ванночка, на которой производится вскрытие животного. Желательно иметь ванночки двух размеров: одну большую 24X30 см для крупных объектов (акула, голубь, крыса) и одну маленькую 9X12 см для мелких (асцидия, ланцетник). Для этого вполне подходят имеющиеся в продаже эмалированные фотографические кюветы соответствующих размеров. Можно также изготовить ванночки из оцинкованного железа или толстой жести; при этом их лучше сделать более узкими, но длинными, примерно 18X35 см. Высота бортов около 4 см. Борта делаются не вертикальными, а наклонны
ми, так, чтобы верх ванночки был несколько шире ее дна. По узким сторонам, к внутренней части бортов, на расстоянии 1—1,5 см от дна, нужно припаять небольшие полоски жести, которые будут удерживать заливочный материал (воск) на дне ванночки. Ванночку следует окрасить эмалевой краской. В стандартных эмалированных ванночках такие приспособления сделать трудно, однако заливочная масса отстает от дна обычно только после длительного употребления, и это легко устранить, расплавив ее над горелкой в этой же ванночке и дав снова застыть.
Ванночка заливается расплавленной горячей заливочной смесью толщиной 2—3 см. Образовавшиеся при заливке пузырьки тщательно удаляются, пока масса еще горячая.
Рекомендуются следующие рецепты заливочных масс:
парафин — 400 г, воск — 200 г, вазелин технический — 4 столовые ложки, голландская сажа или жженая кость — 4 столовые ложки
воск — 400 г, канифоль — 200 г, вазелин технический — 4 столовые ложки, сажа голландская — 4 столовые ложки;
парафин — 400 г, воск — 200 г.
Последний рецепт дает не черную, а светло- или темно-коричневую окраску.
2. Две пары ножниц: большие с закругленными концами и маленькие — остроконечные.
3. Два анатомических (с насечкой, а не зубцами на конце) пинцета— большой и маленький (глазной).
4. Два скальпеля, лучше узких, с острыми тонкими концами — маленький (глазной) и большой или средний.
5. Иглы препаровальные (2—3 шт.). Можно пользоваться обычными препаровальными иглами с деревянной ручкой. Кончик одной иглы можно нагреть над горелкой или спичкой и загнуть. Образовавшимся небольшим крючком очень удобно производить допрепа-ровку мелких деталей. УдобнЪ! также зубоврачебные иглы с штыкообразным или изогнутым под углом концом.
6. Булавки для прикрепления вскрытого объекта ко дну ванночки. Более удобны булавки с петлей, а не с головкой на конце. Для прикрепления очень мелких объектов (асцидии, ланцетника) лучше пользоваться энтомологическими булавками № 0, 1, 2.
7. Лезвие от безопасной бритвы иногда хорошо применять вместо скальпеля. Полезно одно из лезвий разломить так, чтобы, образовалась узкая полоска с одним режущим краем и острым тонким кончиком. Широким концом кусочек лезвия вставляется в ручку от препаровальной иглы. Такой импровизированный скальпель облегчает тонкую препаровку, тем более, что затупившееся лезвие всегда легко заменить новым.
1 Последние два компонента используются для того, чтобы придать заливочному материалу темный цвет. На его качество они заметного влияния не оказывают.
8. Часовые стекла или чашки Петри, в них удобно помещать отдельные мелкие части объекта (например, вырезанный головной мозг).
9. Халат или нарукавники для работы с крупными объектами, содержащими большое количество жира.
10. Рисовальные принадлежности: альбом, лучше большого размера, и набор карандашей «Т» и «2Т».
11. Сосуды для хранения материала. Желательно, чтобы каждый студент был обеспечен отдельным сосудом для хранения материала в процессе работы.
12. Настольная лампа необходима каждому занимающемуся, так как дневного света обычно не хватает, особенно на местах, удаленных от окон.
При отработке некоторых тем необходимо специальное оборудование, которым студенты могут пользоваться поочередно.
1. Микроскопы обычно применяются при отработке задания по ланцетнику и при изучении микропрепаратов по покровам, если таковое проводится. Для занятий пригодны любые модели биологических микроскопов (М-9, М-10, МБИ-1, МБИ-4; РБИ-1 и т. д.).
2. Осветители. При работе с микроскопами, имеющими зеркало, лучше всего пользоваться специальными осветителями для микроскопирования, такими как ОИ-9, ОИ-7, ОИ-19, или осветителями, вставляющимися вместо зеркала микроскопа (ОИ-31).
3. Стереоскопические микроскопы. При работе с мелкими, а также и с крупными объектами часто возникает необходимость более детальной препаровки или изучения какого-нибудь мелкого органа или его части без вычленения данного участка из объекта. Большую помощь при этом могут оказать выпускаемые нашей промышленностью стереоскопические микроскопы МБС-1 и МБС-2.
Более удобным для работы с крупными объектами нужно считать МБС-2. Его оптическая часть крепится на горизонтальной штанге, что позволяет помещать ее над любым местом даже очень крупного объекта.
При работе с микроскопом МБС-1 нужно снять его с подставки, тогда под него удобней подставлять ванночку с исследуемым объектом.
Желательно иметь по одному микроскопу МБС-1 на одного-двух студентов. Если же невозможно обеспечить каждого студента микроскопом, необходимо иметь хотя 'бы 2—3 микроскопа на группу. Микроскопы в этом случае помещают на удобных рабочих местах. По мере необходимости каждый студент может пользоваться микроскопом, рассматривая под ним свой объект.
4. Нехватка микроскопов МБС-1 может быть восполнена или бинокулярными лупами более старых конструкций, или штативными лупами со сменным 6-, 10- и 20-кратным увеличением. Кроме того, можно использовать бинокулярную призматическую лупу (БПЛ) или лупу козырьком (БЛ-1), в крайнем случае, простую ручную лупу с 4—5-кратными увеличением, или окуляр, применяемый часовыми мастерскими.
5. Кроме выше перечисленного, необходимо иметь следующие инструменты в количестве 1—2 шт. на группу: щипцы для отламывания или откусывания небольших кусочков костей при вскрытии черепной коробки и очистке головного мозга; бритвы опасные или старые микротомные ножи, используемые при изготовлении продольных и поперечных срезов; бруски грубые и тонкие, а также ремень для правки бритв, необходимые для точки и правки инструмента во время занятий; трубочки стеклянные тонкие для поддувания легких и воздушных мешков при вскрытии наземных позвоночных и птиц; зонды хирургические с головками на конце; щетинки или моток лески диаметром 0,2, 0,3, 0,5 мм, которые применяются для введения в тонкие каналы и протоки при прослеживании их хода и мест впадения; пинцет большой или корнцанги для доставания материала из банок и сосудов; перчатки резиновые для работы с неот-мытым формалиновым материалом; набор для инъекций, если последние производятся студентами самостоятельно (о составе набора см. ниже, в разделе об инъецировании); марля, вата, тряпки, нитки.
Техника вскрытия
Подробные описания порядка вскрытия и препаровки отдельных органов и систем даются в руководстве по ходу рассмотрения объекта. Однако при любом вскрытии нужно придерживаться общих правил.
Всякое вскрытие свежего нефиксированного материала нужно начинать только после окончания трупного окоченения. Ускорить этот процесс можно осторожным разминанием мышц животного, для чего необходимо несколько раз согнуть и разогнуть конечности, шею и туловище до тех пор, пока они не станут мягкими.
Первые стадии вскрытия удобнее производить, не прикрепляя животное, так как приходится часто менять положение объекта. Наоборот, после того как вскрытие произведено, объект лучше всего укрепить на дне ванночки. Для этого его прикалывают булавками, втыкаемыми наклонно в верхнюю челюсть, в края отвернутых в стороны стенок брюшной полости и в конечности (если таковые имеются).
Рекомендуемое в литературе заполнение ванночки с объектом водой обязательно только при вскрытии мелких объектов (асцидии, ланцетник), в остальных случаях оно нежелательно, так как очень трудно бывает придать плавающим в воде органам нужное положение. Кроме того, если при вскрытии задет какой-нибудь сосуд, то сочащаяся из него кровь все время окрашивает воду, которую постоянно приходится менять. Вскрывая животное без воды, необходимо все время следить за тем, чтобы внутренние органы не подсыхали. Для этого их смачивают водой. Избыток воды (а также и кровь, выделяющаяся из порезанного сосуда) удаляются ватным тампоном. При вскрытии под водой используется дистиллированная или кипяченая вода. В водопроводной воде и в воде, взятой из во-12
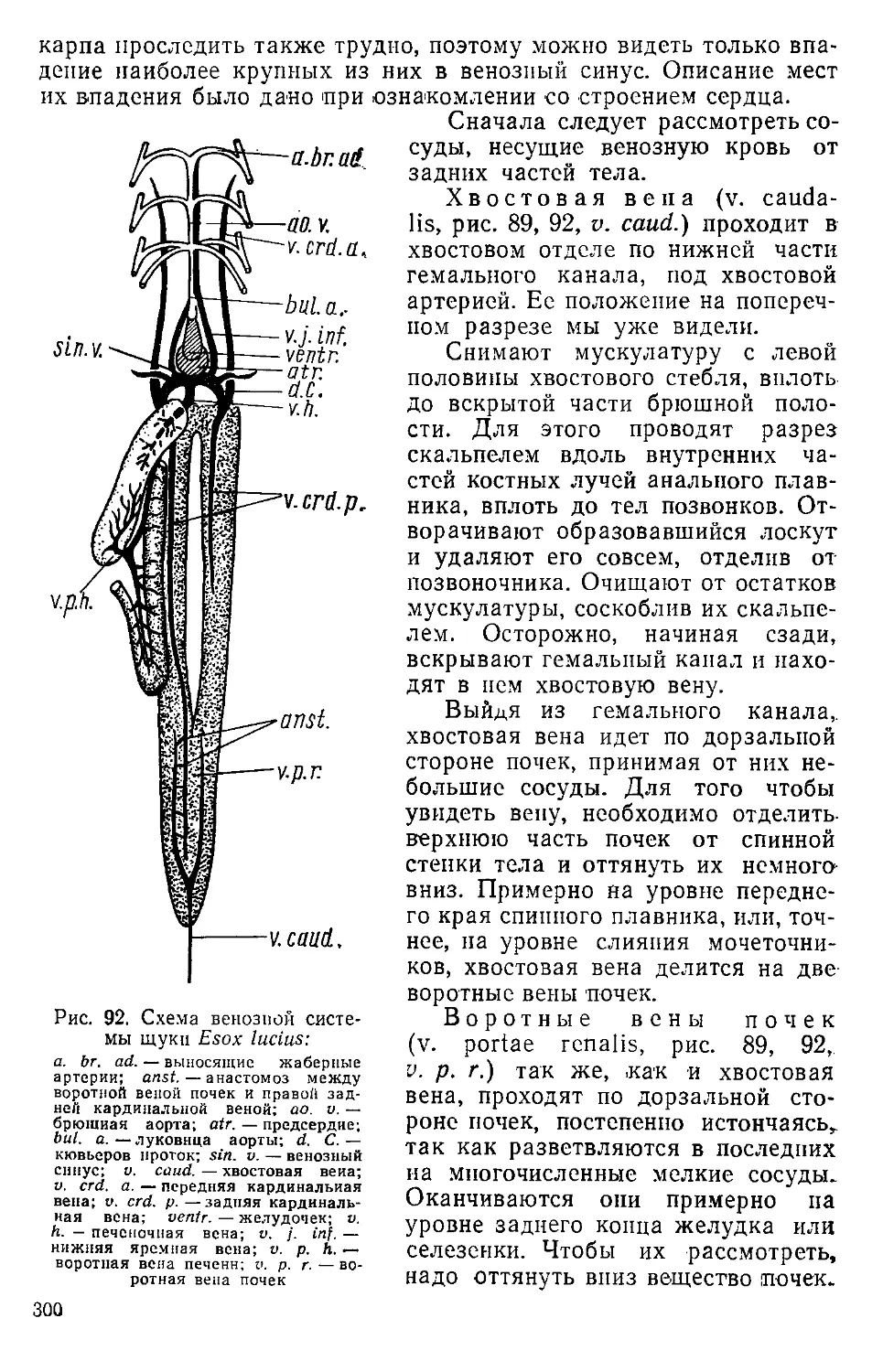
доема, выделяется большое количество пузырьков, которые оседают на объекте и сильно мешают изучению его.
Вскрывая полость тела, нужно следить за тем, чтобы стенки тела были по возможности сильнее оттянуты от внутренних органов. Это предотвращает повреждение последних. Вскрытие лучше всего производить ножницами, вводя в полость тела их тупой конец. Производя разрез, нужно держать ножницы под возможно меньшим углом к вскрываемой поверхности. Перерезку нервов, сосудов и мышц проводят только после их тщательной отпрепаровки от других органов. При этом нужно следить, чтобы подведенный конец ножниц вышел с другой стороны органа и был свободен, а не погружался в ткань, в противном случае можно повредить окружающие органы. Разрез мышцы или нерва надо производить не в местах их прикрепления или отхождения, а в средней части. Это даст возможность при необходимости легко восстановить их форму и местоположение.
Вскрывая свежее животное, необходимо избегать удаления органов или их отдельных частей. Это обычно ведет к повреждению кровеносных сосудов и усиленному кровотечению. Обычно для возможности свободного перемещения органов бывает достаточно перерезать брыжейку в местах, лишенных кровеносных сосудов.
При изучении нервной системы мелких позвоночных препаровка начинается с постепенного срезания скальпелем крышки черепной коробки, а затем ее края подрезаются ножницами или скальпелем. Для вскрытия крупных толстостенных черепов употребляются ЩИПЦЫ.
Подготовка материала
Хлороформирование животных. Для выполнения некоторых заданий довольно легко получить свежий (живой) материал. Это надо учитывать, так как вскрытие особенно удается на свежих объектах. Умерщвление животных, предназначенных для вскрытия, лучше производить накануне дня занятий или, по крайней мере, часов за 10—12 до их начала. За это время кровь в сосудах успевает свернуться и не так сильно вытекает из прорезов. Мелких животных помещают в большую банку с притертой пробкой, в которую кладут кусочек ваты, смоченный эфиром или хлороформом. Крупных млекопитающих и птиц можно просто накрыть большим сосудом или ящиком, лучше всего с фанерными стенками, под который также помещается ватка с хлороформом. Для хлороформирования крупных рыб кусочки смоченной эфиром ваты помещают под жаберные крышки. На холоднокровных животных лучше действует эфир, на теплокровных — хлороформ. Холоднокровных животных нужно выдерживать в парах наркотика значительно дольше, чем теплокровных. Особенно это касается ящериц и черепах. Последние остаются живыми даже после многочасового пребывания в парах эфира. Умерщвленных животных нужно вынуть из сосуда, лягушек обмыть и до наступления трупного окоченения свободно разложить на ван-
ночке, придав позу, удобную для последующей работы с ними. Сохранять материал до начала занятий следует при пониженной температуре, лучше всего в холодильнике, но при этом нужно следить за тем, чтобы трупы не замерзли.
Фиксация. Для изучения мускулатуры, нервной системы и др., а также при заготовлении материала впрок его необходимо фиксировать. В большинстве случаев в качестве фиксаторов применяют формалин. Обычный, имеющийся в продаже формалин, представляющий собой 40%-ный насыщенный раствор формальдегида в воде, дополнительно разводят водой из расчета 1 часть формалина на 19 частей воды.
У всех животных (кроме асцидий и ланцетника) перед фиксацией для лучшего проникновения фиксатора к внутренним органам вскрывается брюшная полость, для чего делается разрез ее брюшной стенки длиной в несколько сантиметров. Разрез нужно вести не по средней линии, а несколько отступя от нее. Можно обойтись и без разреза, но тогда необходимо инъецировать пищеварительный тракт, вводя фиксатор при помощи шприца или резиновой груши в ротовое и анальное отверстия, или вводить раствор шприцем в брюшную полость. Последнее обязательно при фиксации рептилий.
Зафиксированный материал обычно сохраняет ту форму, которая была придана ему при погружении в фиксатор, поэтому перед фиксацией животному необходимо придать форму, удобную для дальнейшей работы. Так, например, птиц, на которых будет отрабатываться мускулатура, фиксируют с расправленными крыльями и разведенными в сторону лапами. Для этого концы расправленных крыльев привязывают к деревянной планке, проходящей поперек тушки по спинной стороне. Таким же образом можно расправить и задние конечности. Описанный способ фиксации относится и к другим объектам: лягушкам, агамам, крысам, на которых будет изучаться мускулатура.
При фиксации формалином целых тушек птиц и млекопитающих их покровы плохо смачиваются, что затрудняет проникновение формалина к поверхности тела. Чтобы этого избежать, нужно предварительно окунуть тушку в 70° спирт, который хорошо смачивает перья и шерсть, вынуть из него, дать спирту стечь и только после этого опустить в формалин. При фиксации мозга крупных животных вместе с черепной коробкой необходимо вскрыть последнюю для доступа фиксатора непосредственно к головному мозгу. Отношение объема материала к объему фиксатора должно быть 1 : 10. Первую порцию фиксатора, обычно загрязненную кровью и помутневшую, через 2—3 дня следует заменить свежей, той же концентрации. В ней материал выдерживается еще одну-две недели, в зависимости от величины объекта. После окончательной фиксации материал складывают на хранение. Теперь его можно уложить поплотнее и залить более концентрированным формалином, для приготовления которого берут 1 часть формалина и 9 частей воды.
При хранении материала в металлических баках («гробах») он очень легко покрывается ржавым налетом от соприкосновения со 14
стенками бака и от частиц ржавчины, которые со временем проникают в весь раствор формалина. Для предотвращения этого стенки бака выстилаются бумагой или сам объект завязывается в несколько 'Слоев марли. Необходимо объекты сверху закрывать куском марли, препятствующей высыханию частей объекта, выступающих из раствора. Для того чтобы предотвратить появление на поверхности пленки плесени, рекомендуется добавлять в раствор формалина немного фенола.
Объекты, размеры которых невелики, нужно хранить в стеклянной посуде, лучше всего в банках с притертыми пробками. Если таковые отсутствуют, то сосуд можно закрыть куском стекла. Края сосуда при этом следует обильно смазать техническим вазелином, что значительно ослабляет испарение фиксатора. Не менее двух-трех раз в год нужно производить доливку или полную смену фиксатора. Хранить материал в 70° спирте следует только в сосудах с притертой пробкой.
Промывка материала. После любого фиксатора материал перед занятиями промывают в проточной воде в течение 1—3 суток. Для этого удобен бак с крышкой, изготовленный из оцинкованного железа. Размеры бака 50X30 см при высоте 40 см (их можно изменить как в ту, так и в другую сторону). В одну из стенок бака на расстоянии 1—2 см от дна впаиваются 2—3 тонкие металлические трубки, один конец которых запаян, в самой трубке просверлен ряд мелких отверстий. Запаянными концами трубки далеко (до противоположной стенки) вдаются в бак. Наружу торчат только небольшие кусочки, на которые можно будет надеть резиновые трубки. В ту же стенку на 5—10 см ниже верхнего края впаиваются две широкие трубки. Они могут совсем не выступать внутрь бака. Во избежание засорения внутренних отверстий толстых трубок их лучше оградить неплотной сеткой. На выступающие концы нижних трубочек надеваются резиновые трубки. Они соединяются вместе при помощи стеклянных тройников в одну трубку, свободный конец которой надевается на водопроводный кран. Две толстые резиновые трубки надеваются на наружные концы верхних металлических трубок и опускаются в раковину. Бак устанавливается рядом с раковиной на подставке. В таком баке даже большое количество материала равномерно омывается водой, текущей снизу вверх, и хорошо промывается.
Декальцинация. При препаровке нервной системы, в частности головного и спинного мозга, бывает необходимо размягчить одевающие мозг кости черепа и позвонки. Это достигается в процессе декальцинации, который сводится к превращению нерастворимых в воде солей Са, пропитывающих кости, ,в растворимые и вымыванию их из объекта. Кратковременное выдерживание препарата в кислоте полезно и перед препаровкой мускулатуры. В этом случае мышцы становятся более плотными, несколько сокращаются и хорошо отделяются друг от друга.
Чаще всего для декальцинации употребляется 5—7%-ный раствор азотной кислоты. Более концентрированные растворы дефор-
мируют ткань, менее концентрированные — декальцинируют слишком медленно, что влечет за собой мацерацию объекта еще до окончания декальцинации. Для предотвращения мацерации азотную кислоту разводят не на воде, а на растворе формалина 1 : 19. Кроме формалина можно разводить азотную кислоту и 70° спиртом. В 5—7%-ной азотной кислоте декальцинация заканчивается через 2—3 суток. О ее окончании можно судить по проникновению иголки в наиболее толстые костные части; если иголка входит легко, то можно считать декальцинацию оконченной. Если на одном и том же объекте отрабатывается мускулатура и нервная система, то нужно не доводить декальцинацию до конца, чтобы не размацери-ровались мышцы. Наоборот, если материал идет только на изготовление анатомических препаратов нервной системы, то его по возможности более полно очищают от мускулатуры и внутренних органов и декальцинируют несколько часов в 30%-ной азотной кислоте. В результате такой декальцинации все ткани, кроме нервной, размягчаются настолько, что легко удаляются при помощи пинцета, иголки и кисточки.
Наряду с препаратами, которые изготовляют сами студенты, в процессе занятий довольно часто приходится использовать готовый материал. С одной стороны, это бывает необходимо для экономии времени, с другой — специально подготовленные и смонтированные препараты помогают студентам разобраться в их собственном объекте. Такие препараты обычно изготовляются заранее и могут служить много лет. Их необходимо только профилактически ремонтировать и частично подновлять.
Подробно ознакомиться с техникой изготовления постоянных анатомических препаратов можно по руководствам или отдельным работам, список которых приведен ниже.
Здесь мы кратко остановимся только на тех методиках, которые студенты могут освоить во время большого практикума в процессе отработки той или другой темы и изготовить самостоятельно полноценные демонстрационные препараты.
На занятиях по большому практикуму требуется значительное количество микроскопических препаратов. Изготовление их связано с затратой времени, а главное, необходимы определенные навыки. Обычно студентам выдаются уже готовые препараты. Методику изготовления таких препаратов можно найти в любом руководстве ио микроскопической технике, наиболее распространенные из них приведены в списке литературы.
Приготовление скелетного материала
Существует несколько способов очистки скелета от мягких тканей, но, применяя любой из них, необходимо соблюдать некоторые общие правила. Лучшим для изготовления скелета нужно считать свежий нефиксированный материал. Сохранять собранный материал до начала его обработки можно двумя способами. При первом способе тушки животных, скелет которых содержит мало хряща, 16
можно высушить. Для этого нужно грубо очистить тушку от внутренних органов и мускулатуры, обильно посыпать солью и положить сохнуть на открытый воздух, лучше всего на солнце. Перед окончательной очисткой скелета тушка размачивается в воде, а затем обрабатывается. При втором способе хранения целые неочищенные тушки животных помещаются в 5%-ный раствор фенола; перед обработкой их нужно промывать в проточной воде.
Перед началом окончательной обработки любой скелетный материал должен быть по возможности более полно очищен, для чего с тушки снимается кожа, вынимаются внутренние органы и срезается мускулатура. По возможности скелет расчленяется по суставам на отдельные части: череп, скелет конечностей, пояса конечностей, у рыб отделяются также непарные и парные плавники. При обработке сразу нескольких объектов, чтобы избежать путаницы, отдельные части каждого объекта завязываются в марлевый мешочек, в котором и идет дальнейшая обработка. Особенно аккуратно нужно вести обработку скелетов, которые содержат много хряща. Ниже приводится несколько способов очистки скелетного материала.
Мацерация. Естественная мацерация — наиболее надежный, хотя относительно длительный способ получения скелетных препаратов, позволяющий сохранять не только отдельные кости, но и связки. Последовательность обработки следующая.
Очищенный и расчлененный скелет заливают теплой (35—40° С) водой и оставляют вымачивать на длительное время. Время выдерживания очень различно и зависит как от величины объекта, так и от культуры микроорганизмов. Так, для очистки скелета собаки или песца требуется 1—2 недели, для скелета птицы достаточно нескольких дней. Мацерировать лучше 'всего в банках с притертой пробкой во избежание неприятного запаха. Банки следует держать в теплом месте. Следующие порции материала лучше класть не в чистую воду, а уже в развившуюся культуру микробов. После мацерации следует 1—2-часовая промывка в проточной воде, окончательная очистка и высушивание.
Вываривание. Очищенный и расчлененный скелет погружают в холодную воду, которую постепенно доводят до кипения. Погружение сразу в кипяток приводит к развариванию костей, в то время как сухожилия и связки остаются еще плотными. После того как вода закипит, нужно вынуть объект и очистить с помощью пинцета и скальпеля.
Если очистка идет с трудом, то процесс кипячения повторяют опять, начав с погружения в холодную воду.
Скелеты мелких грызунов, ящериц, лягушек, рыб обычно разваливаются, если они пробыли в кипящей воде дольше 1—2 ммн. Только скелеты крупных млекопитающих выдерживают кипячение без особого для них вреда в течение 20—30 мин. При обработке скелетов в воду можно добавить едкое кали или едкий натр до крепости раствора 0,1% (но не больше!). После окончательной очистки следуют промывка и высушивание.
Антиформиновая обработка может быть рекомендована для крупных животных. При этом способе хорошо сохраняются связки, так как раствор действует .в первую очередь на мягкие ткани. Обработка идет по следующей схеме.
Приготавливают (50°) 0,5—1%-ный раствор каустической соды, в котором растворяют хлорную известь из расчета ее концентрации в растворе 0,5%. В полученную смесь погружают предварительно очищенный начерно скелетный материал. Срок действия, в зависимости от величины объекта, от нескольких часов до нескольких суток. Не следует держать до полного удаления мягких частей, так как они удаляются при следующей обработке. Промывка в течение суток, а затем сушка.
Приготовленный выше изложенными методами материал может быть использован для работы. Однако для того чтобы придать ему лучший вид, следует провести обезжиривание и отбелку. В качестве обезжиривающих веществ для мелких объектов можно применять спирт, эфир или бензин, куда сухой материал помещается на 3—4 суток. Крупный материал можно обезжиривать 5—10%-ным раствором соды.
Отбеливание мелких объектов лучше всего проводить раствором перекиси водорода. В зависимости от объекта используют 2— 3%-ный или 10%-ный растворы, куда помещают сухой скелетный материал после обезжиривания. Только для крупных костей в качестве отбеливателя можно рекомендовать раствор хлорной извести (2—3 ложки на ведро воды). В растворе материал выдерживается от 6 до 24 ч. Крупные кости можно отбелить, выдерживая их долгое время на солнце.
Окраска скелетного материала. При изучении отделов скелета, в которых костное вещество прочно связано с хрящом, выявление костных участков механическим путем невозможно. В этом случае костные элементы легко выделяются при помощи окраски ализарином, которая проводится следующим образом.
Готовится насыщенный раствор ализарина в 95° спирте. Одна часть раствора разбавляется 19 частями 70° спирта, разведенного на водопроводной воде. Полученный раствор темно-вишневого цвета титруют уксусной кислотой до приобретения им цвета крепкого чая. В этот раствор погружают промытый после спиртовой или формалиновой (последняя предпочтительней) фиксации материал. Скелет может быть очищен только начерно. Окраска около 24 ч. После этого промывка в 70° спирте, если необходимо, то и дочистка. Такие препараты следует хранить или в спирте, или в слабом формалине. Перекрашенные препараты, в которых окрасился и хрящ, можно дифференцировать в 95° спирте.
В некоторых случаях возникает необходимость в окраске хрящевых элементов скелета. Окраску в этом случае ведут в следующем растворе. В 100 см3 70° спирта растворяют 0,25 г метиленового синего или толлуидинового голубого, добавляют 1 см3 соляной кислоты, в раствор помещают объект и окрашивают в термостате при температуре 40° С. Дифференцировка в 95° спирте, подкисленном 18
соляной кислотой (до 1,0%). Промывка несколько дней в сменяемом 96° спирте. Хранение такое же, как и ализариновых препаратов.
Просветленные окрашенные препараты. При изучении деталей строения отдельных участков скелетной системы, включающих мелкие окостенения, не всегда представляется возможным тщательно очистить их от окружающих мягких тканей без нарушения порядка расположения костных элементов. В этом случае хорошие результаты дает следующая методика. Нужный участок скелета (для его получения используют фиксированный материал) очищают от мягких тканей, оставляя связки. Затем следует ализариновая окраска по вышеприведенной схеме, и далее проводка по спиртам возрастающей крепости 70,80,96,100° с .выдерживанием в каждом не менее 12 ч. Затем пропитывание смесью (1:1) 100° спирта и бензола (ксилола, толуола), чистый бензол (в каждом растворе по несколько часов до просветления) и заключение под покровное стекло в канадский бальзам. При значительной толщине препарата под покровное стекло необходимо подложить спички или узкие полоски стекла. Препараты должны храниться в горизонтальном положении. Через некоторое время необходимо добавить канадский бальзам. При такой обработке соединительная ткань, окружающая костные элементы, становится прозрачной, а окрашенные костные участки четко видны. Изучать такие препараты лучше всего под бинокулярным микроскопом.
Инъекция кровеносной системы
При изучении кровеносной системы на свежем или фиксированном материале обычно удается рассмотреть только наиболее крупные артериальные и венозные сосуды. Более детальное изучение кровеносной системы вплоть до препаровки отдельных мелких сосудов может быть осуществлено только при специальной предварительной подготовке материала, которая заключается в инъекции сосудов, т. е. наполнении их тем или иным красящим веществом.
Инъекционные массы. В зависимости от назначения препарата для заполнения кровеносных сосудов можно использовать различные инъекционные массы.
1. Раствор туши в воде 0,5—1%-ный применяется в тех случаях, когда желательно рассмотреть тончайшие, вплоть до капилляров, разветвления кровеносных сосудов. Недостатком является то, что раствор все время остается жидким и легко вытекает из разрывов сосудов, которые обычно неизбежны при дальнейшем препарировании. Кроме того, после фиксации и при высыхании тушь коагулирует и особенно в крупных сосудах скапливается только в отдельных участках, так что весь сосуд трудно проследить.
2. Растворы желатины в воде хорошо проникают в тонкие сосуды, а жидкие растворы даже в капилляры. Инъекция раствора желатины проходит по большей части успешно, поэтому ее можно в первую очередь рекомендовать для проведения инъекции во время
занятий. В зависимости от величины объекта и толщины наполняемых сосудов можно применять 2, 5 и 10%-ные растворы. Для объектов средних размеров (лягушка, ящерица, голубь, крыса) лучшим является 5%-ный раствор.
Необходимое количество сухой желатины (лучше пищевой) отвешивают на весах, заливают холодной водой и оставляют стоять для набухания на 1—2 ч. При определении необходимого количества инъекционной массы можно исходить из расчета, что на инъекцию артериальной системы крысы уходит 2—3 см3 раствора. После набухания массу помещают в термостат (37—40°) до полного растворения желатины. Затем в раствор добавляют гуашевую краску соответствующего цвета (красная для артериальной, синяя для,венозной) и гуашевые белила. Последние добавляют для придания массе меньшей прозрачности, что важно при препаровке мел-’ ких сосудов. Лучше (всего краску предварительно растворить в небольшом количестве воды. Количество краски зависит от интенсивности ее цвета, но на 100см3 раствора берут не менее чем */2 чайной ложки каждой из красок. Массу хорошо размешивают. При длительном стоянии размешивание повторяют, так как гуашь легко оседает на дно.
Необходимо учесть, что добавление значительного количества гуаши, также содержащей большое количество клеящих веществ, делает раствор более густым, что препятствует его проникновению в мелкие сосуды. Соотношение густоты желатины и количества краски в разных случаях может быть различным и подбирается опытным путем.
Раствор желатины готовят незадолго до инъецирования и хранят в закрытом сосуде во избежание подсыхания желатины в верхнем слое. При проведении инъекции сосуд с раствором желатины должен находиться в горячей воде. Шприц также должен быть предварительно хорошо пригрет. При этом выбирают шприц со свободным ходом поршня, так как тугие металлические поршни в стеклянных шприцах обычно заклинивает при нагревании. Желатина в шприце должна быть теплой во все время инъекции. При застывании желатины в тонкой игле, введенной в сосуд, ее можно попытаться растопить, капая на иглу теплую воду.
5°/о-ный раствор желатины долгое время остается жидким и при комнатной температуре, что обеспечивает его проникновение в мелкие сосуды, однако в крайних случаях при необходимости заполнения мелких сосудов, расположенных на периферии, тушку животного необходимо погрузить в теплую воду и хорошенько прогреть.
После введения инъекционной массы полностью препарат следует поместить в холодную воду для быстрого застывания желатины, а затем или начинать препаровку, или фиксировать в 2—4%-ном формалине.
При работе с густыми (10%-ный и выше) растворами, которые легко застывают, точку разжижения желатины можно понизить, прибавляя к раствору КД- Так, прибавление к 10%-ному раствору 20
на 100 г 5—6 г йодистого калия делает этот раствор жидким и при комнатной температуре. Если раствор не использован весь сразу, его можно некоторое время сохранять, добавив на 100 см3 0,25 г тимола или 0,5 г фенола.
3. Масса Тейхмана может быть рекомендована для заполнения крупных сосудов на значительных по размеру объектах, а также для изготовления демонстрационных препаратов.
Недостатком массы является ее плохое, по сравнению с желатиной, проникновение в тонкие сосуды и значительное уменьшение в объеме при застывании. Поэтому при инъекции этой массой следует добиваться значительного1 расширения сосудов, которые затем уменьшатся в объеме.
Масса готовится путем тщательного и длительного смешивания растиранием зубного порошка, масляной художественной краски и натуральной олифы или льняного масла. Вместо масляной краски можно взять краску в порошке. В этом случае примерное соотношение компонентов следующее: зубной порошок — 500 г, краска (сухая) — 100г, масло— 100—120 г. Сухую краску обычно смешивают с зубным порошком, масляную можно развести в олифе. В порошок постепенно добавляют олифу и растирают. Смесь должна приобрести консистенцию замазки и на ее поверхности выступят капли масла. Такая смесь скатывается в небольшие шарики, и они могут длительное время храниться под водой.
Для проведения инъекции масса разводится эфиром, бензином или сернистым углеродом. Чем толще инъецируемые сосуды, тем гуще должна быть масса, в противном случае при испарении растворителя масса сильно усядет, что особенно скажется на наполнении крупных сосудов.
4. Целлоидиновые смеси в первую очередь можно рекомендовать для изготовления коррозионных препаратов, когда после застывания инъекционной массы препарат помещают в 30%-ный раствор соляной кислоты. В этом случае через несколько суток все ткани мацерируются настолько, что могут быть отмыты под струей воды. При необходимости для окончательной дочистки можно перенести в 70%-ную кислоту. В результате обработки остается только дерево кровеносных сосудов.
Недостатком массы является ее очень сильная усадка при испарении растворителя, поэтому следует наполнять сосуды по возможности более густой массой. В то же время густая смесь плохо проникает в тонкие сосуды и легко закупоривает иголку из-за испарения растворителя на ее конце. Поэтому наполнение рекомендуется производить в несколько приемов. Первая порция — 3%-ный целлоидин, вторая — 5%-ный, третья—10—15%-ный. При этом через несколько часов повторяют доливку последней густой порцией целлоидина. При значительной толщине инъецируемых сосудов последняя порция может быть очень густой, вплоть до 40%. Густая смесь очень плохо проходит через иголку или канюлю для инъекции, поэтому их нужно брать предельно большого диаметра, насколько позволяет сосуд, через который проводится инъекция.
Наиболее простой способ приготовления инъекционной массы — растворение в ацетоне или смеси 100° спирта с эфиром (1:1) отмытой под горячей водой от эмульсии и высушенной рентгеновской пленки. В последнее время рентгеновская пленка стала выпускаться на негорючей подложке, которая не растворяется в приведенных выше веществах. Проверить это можно, поднеся небольшой кусочек пленки, зажатый в пинцете, к огню. Целлоидиновая пленка сразу вспыхивает и горит очень интенсивно. Для получения рабочих растворов лучше всего сначала приготовить раствор наибольшей концентрации, а затем разбавлять его растворителем. Подкраску раствора лучше производить художественными масляными красками. При обесцвечивании красителя при изготовлении коррозионных препаратов их можно подкрасить сверху масляной краской.
Порядок проведения инъекций. Несмотря на различные составы инъекционных масс и некоторые особенности их введения в зависимости от конкретных свойств, существуют общие правила и приемы инъецирования, с которыми необходимо познакомиться прежде, чем приступать к наполнению кровеносной системы.
Излагаемые в различных руководствах методы инъекции кровеносной системы обычно приводятся для крупных объектов, имеющих толстые кровеносные сосуды. В этих случаях применяются шприцы большого объема и канюли, вставляемые в разрез сосуда. Инъекция объектов мелких и средних размеров, описываемых в данном руководстве, не требует специального оборудования.
Иглы для инъекции — обычные инъекционные иглы, применяемые в медицинской практике. Лучше пользоваться иглами большего диаметра, как только позволяет диаметр сосуда, тогда масса идет через иглу легко. Игла вводится в сосуд путем прокола его стенки, поэтому она должна быть достаточно острой. Иглу .можно заточить на микрокорундовом бруске, а остроту проконтролировать под бинокулярным микроскопом.
Соединение иглы со шприцем лучше проводить не непосредственно, а через достаточно длинный отрезок ниппельной резины. Это позволяет легко манипулировать иглой отдельно от шприца; кроме того, раздуваясь, трубка несколько смягчает резкое повышение давления, которое может возникнуть. Сильное раздутие трубки указывает обычно на закупорку иглы. Трубка довольно легко надевается на конец шприца; для того чтобы так же легко надеть ее на иголку, необходимо конец на несколько минут опустить в бензол. Разбухшая и ставшая мягкой трубка легко наденется на иглу, бензол быстро испарится и трубка плотно обхватит основание иглы. При этом не требуется дополнительного закрепления ее ниткой.
При выборе шприца для инъекции не следует увлекаться шприцами большого объема. При большой поверхности поршня требуются значительные усилия при проталкивании инъекционной массы через тонкую иглу. Вполне достаточны шприцы объемом 5—10 см3. Обычно для наливки артериальной системы животного средних размеров этого объема вполне хватает. При необходимости шприц 22
легко отсоединить от резиновой трубки, снова наполнить его и продолжать инъекцию.
При заполнении шприца лучше всего набирать инъекционную массу через иглу и резиновую трубку, соединенные со шприцем. Это гарантирует от попадания в шприц крупных частиц, не проходящих через отверстие иглы, и обеспечивает заполнение иглы и трубки массой. Нужно учесть, что при заполнении шприца через тонкую иглу масса поступает в него очень медленно, и не стоит форсировать этот процесс.
Из многочисленных способов инъекций, описанных и разных руководствах, мы остановимся только на тех, которые были проверены нами на мелких объектах, не очень сложны и дают хорошие результаты.
Инъекцию лучше проводить на свежезахлороформированных животных до наступления момента трупного окоченения. Если же таковое наступило, то нужно восстановить подвижность отдельных частей тела, сгибая и разгибая их в суставах.
Рекомендуемое обычно в руководствах вымывание крови из сосудистой системы водой или физиологическим раствором нами никогда не проводилось и не является необходимым. При инъекции в первую очередь заполняется артериальная система, сосуды которой, особенно крупные, после смерти животного обычно лишены крови и легко заполняются инъекционной массой. Оставшаяся в артериальной системе кровь выдавливается при инъекции в венозную систему, делает ее контрастной и вполне пригодной для изучения даже без дополнительной инъекции. По этой же причине не рекомендуется вскрывать какой-либо крупный венозный сосуд для оттока крови. Вообще инъекция получается тем удачнее, чем меньше повреждено сосудов при вскрытии.
Инъекция через желудочек сердца является наиболее простым и легче всего выполнимым способом. У забитого животного строго по средней линии с вентральной стороны вскрывается грудная область. Стенки тела разводятся в стороны, так чтобы обнажилось сердце. В желудочек сердца или в его артериальную половину вводится довольно толстая инъекционная игла. При ее введении желудочек нужно немножко сжать ножками анатомического пинцета и удерживать в определенном положении. Иглу вводить в вершину желудочка, прокалывая стенку между ножками пинцета. После введения иглы сразу можно начинать инъекцию. Толстые мускульные стенки желудочка плотно охватывают иглу и не дают массе вытекать через прокол, дополнительной фиксации иглы не требуется. Метод при всей своей простоте дает хорошие результаты, но имеет и существенные недостатки. Отсутствие фиксации иглы не позволяет создать значительное давление, а тем самым и гарантию заполнения мелких сосудов (крупные артериальные стволы заполняются^ хорошо). Этот недостаток можно исправить, затягивая лигатурой вершину желудочка вместе с иглой, однако в этом случае инъекционная масса обычно попадает в предсердия и частично заполняет крупные венозные стволы, что нежелательно, особенно при
изготовлении демонстрационных препаратов. Этот способ инъекции хорошо удается на большинстве объектов, но особенно его можно рекомендовать для амфибий, ящериц и млекопитающих.
Хорошее наполнение артериальной системы достигается при введении массы через основания крупных сосудов, отходящих от сердца. Вскрытие производят так же, как указано выше, но после этого отпрепаровывают основания крупного сосуда, через который будет производиться наливка, таким образом, чтобы под него можно было подвести маленький пинцет. Концами пинцета захватывают предварительно смоченную нитку, подводят ее под сосуд и завязывают узлом, не затягивая его. Инъекционная игла вводится в желудочек, как указано выше, и через него в просвет сосуда. Конец иглы должен выходить за уровень наложения лигатуры. После этого затягивают узел, и можно приступать к инъекции. После наполнения сосудов иголку вынимают, и одновременно туже затягивают узел на сосуде, препятствуя вытеканию инъекционной массы. Этот способ можно рекомендовать также для всех объектов, но, как и предыдущий, он отличается одним существенным недостатком. При вскрытии вентральной стенки тела в области сердца повреждается значительное количество кровеносных сосудов. Особенно это относится к птицам, у которых приходится снимать грудину и повреждать грудные артерии. Через поврежденные даже мелкие сосуды вытекает большое количество инъекционной массы, что мешает созданию повышенного давления и полному наполнению артериальной системы. Особенно это сказывается при использовании жидких, хорошо проникающих, масс: тушь, желатина. Масса Тейхмана или густая целлоидиновая масса легче закупоривает тонкие сосуды и вытекает значительно меньше.
Вышеприведенных недостатков можно избежать, проводя инъекцию через какой-либо поверхностно расположенный сосуд на не вскрытом животном. Лучше всего инъецировать таким способом через сонную артерию или сосуды, проходящие в области бедра и голени. В этом случае делается продольный разрез на коже в области прохождения сосуда, кожу отворачивают в стороны и, раздвигая мышцы, находят нужный сосуд. Сосуд отпрепаровывают так, чтобы на некотором расстоянии он был свободным. Под него подводят нитку и путем прокола стенки вводят иглу по направлению от периферии к центру. Затем на ней затягивают узел и проводят инъекцию. По окончании иглу вынимают с одновременным затягиванием узла. Введение иглы в тонкостенный сосуд требует большой осторожности и некоторого навыка. При тупой игле можно легко оборвать сосуд или проколоть его насквозь, т. е. обе стенки. Для большей неподвижности сосуда, что очень важно при введении в него иглы, можно рекомендовать подведение под него спички: располагаясь поперек хода сосуда, она, кроме того, приподнимает его над мускулатурой, что значительно облегчает манипулирование с ним. Инъекцию через сонную артерию можно рекомендовать для млекопитающих и черепах, так как снять у
последних брюшной панцирь практически невозможно без повреждения сосудов.
Недостатком метода является его проведение «вслепую»: степень наполнения сосудов не видна, и контроль можно проводить только по наполнению мелких сосудов на концах конечностей. Кроме того, при введении большего, чем нужно, количества массы может произойти разрыв сосудов и вытекание массы в полость тела. Поэтому желательно первоначально на двух-трех экземплярах установить точное количество массы, необходимое для заполнения всей системы, и в дальнейшем заканчивать инъекцию после введения этого количества.
Для рыб, у которых при введении массы в брюшную аорту она доходит только до жаберных капилляров, можно рекомендовать наполнение кровеносной системы через хвостовую артерию. Для этого скальпелем отрезают хвост недалеко от его конца, вскрывают гемальный канал и освобождают небольшой кусочек хвостовой артерии, в которую и вводят иглу, закрепленную лигатурой.
Инъекция венозной системы целиком практически невозможна из одного места, этому препятствуют воротные системы почек и печени. Кроме того, ее инъекция от сердца, т. е. против тока крови, затруднена из-за наличия в венах клапанов, препятствующих продвижению массы в этом направлении. Как указывалось выше, в инъекции венозной системы обычно нет необходимости. Если все же такая необходимость возникает, то проводить инъекцию венозной системы нужно так же, как и инъекцию артериальной системы, через сосуды. Наиболее удобными в этом случае являются у рыб хвостовая вена, у других позвоночных — бедренные, яремные и брюшная вены.
Литература по технике приготовления препаратов
Богуславская Т. Б. Изготовление топографо-анатомических препаратов и методика некоторых анатомических исследований. М., Изд-во МОЛМИ им. Сеченова, 1959.
Давыдов И. С. Опыт применения советского синтетического латекса для инъекции кровеносных сосудов, полых органов и лимфатической системы. Жури. «Архив анатомии, гистологии и эмбриологии», т. 32, № 1, стр. 64—66, 1955.
Ковешникова А. К., Клебанова Е. А. Способы изготовления анатомических препаратов. М., Учпедгиз, 1954.
Р о м е й с. Микроскопическая техника, 1959.
Роек ин Г. И., Левинсон Л. Б. Микроскопическая техника. М., «Советская наука», 1957.
Современные методы и техника морфологических исследований. Под ред. Д. А. Жданова, М, —Л., 1955.
Субботин М. Я., Л агу ч ев С. С. и др. Гистологическая техника. М., Медгиз, 1954.
Ярославцев Б. М. Анатомическая техника. Руководство по изготовлению анатомических и биологических препаратов. Фрунзе, 1961.
Adam Н„ Czihak G. Arbeitsmethoden der makroskopischen und mikros-kopischen Anatomie. Stgt., G. Fischer, 1964.
Kiikenthal W., Mattes E. Leitfaden fur das Zoologische Praktikum. 13 Aufl. Stgt. Jena, G. Fischer, 1953.
Piechocki R. Macroscopische Praparationstechnik. Teil I. DDR, Akade-mische Verlagsgeselschaft, Leipzig, 1961.
ТИП ХОРДОВЫЕ —CHORDATA
Общая характеристика хордовых
Позвоночные вместе с человеком принадлежат к типу хордовых (Chordata). По строению и функции нервной системы тип хордовых занимает высшее место в мире животных и является наиболее молодым в геологической летописи Земли. Все позвоночные объединяются в высший подтип типа хордовых — подтип позвоночных — Vertebrata или черепных — Craniota. Бесчерепные — Acrania и оболочники — Tunicata выделяются в самостоятельные подтипы низших хордовых; они являются древнейшими и наиболее примитивными, но сильно измененными представителями хордовых. Самый древний и низший подтип хордовых — полухордовые — Hemichorda вместе с погонофорами в настоящее время выделяют в самостоятельный тип погонофора — Pogonophora, занимающий промежуточное положение между беспозвоночными и позвоночными животными.
Место хордовых в системе мира животных определяется следующими семью признаками.
1. Д в у с т о р о н н е - с и м м ет р и ч но е строение (Bilateralia) с разделением тела на голову, туловище и хвост. Это указывает на то, что по типу своей организации представители типа хордовых активно подвижные' животные, двигающиеся передним концом вперед. В связи с этим на переднем конце тела обособляется головной отдел, где прогрессивно развиваются органы чувств, с помощью которых животные ориентируются в окружающей среде. Концентрация на переднем отделе тела органов чувств вызывает прогрессивную дифференцировку переднего отдела центральной нервной системы — головного мозга — и защищающего его черепа. Этот процесс обособления головного отдела, где сосредоточены высшие органы чувств, головной мозг и защищающий их мозговой череп, носит название цефализации. При цефализации у первичных водных позвоночных на переднем конце тела образовались ротовое* отверстие и жаберные отверстия, выполняющие вегетативные функции — питание и дыхание. Хвостовой же отдел обособился как часть тела, выполняющая двигательную функцию при плавании.
2. Вторичноротость — по этому признаку позвоночных животных вместе с полухордовыми, погонофорами и иглокожими объединяют в раздел вторичноротых — Deuterostomia. Другие типы 2ft
двусторонне-симметричных животных, куда относятся разные типы червей и членистых животных, составляют группу первичноротых животных — Protostomia, их ротовое отверстие соответствует первичному рту — бластопору. Моллюски занимают особое положение.
3. Наличие вторичной полости тела — целома, т. е. полости, где помещаются органы питания, дыхания, кровообращения и выделения. Существование целома объединяет всех хордовых с высшими беспозвоночными, имеющими вторичную полость тела, в группу целомных животных — Coelomata.
4. Метамерность — расчленение тела на ряд повторяющих друг друга по продольной оси тела сегментов. Этот признак, как и первые, также относит хордовых к группе высших типов мира животных, обладающих наиболее совершенным способом передвижения тела в пространстве, — двусторонне-симметричных животных. Наиболее постоянное метамерное строение тела позвоночных выражено в скелете и нервной системе, а у низших классов водных позвоночных— также в мускулатуре и кровеносной системе.
5. Наличие первичного осевого скелета — спинной струны — хорды (chorda dorsalis), замещаемой хрящевым и костным позвоночником, что резко выделяет всех хордовых в особый тип мира животных.
6. Наличие трубчатой нервной системы с нервным каналом — невроцелем внутри, располагающейся над хордой на спинной стороне тела. Этот признак также выделяет хордовых в высший тип мира животных. У всех остальных животных нервная система представлена нервными тяжами с утолщениями — ганглиями.
7. Существование жаберных щелей, пронизывающих с обеих сторон ротоглоточную полость, у низших водных позвоночных во взрослом состоянии, а у высших наземных классов — в эмбриональном состоянии. У низших водных хордовых в жаберной области развиваются органы дыхания в виде жабр, у наземных позвоночных жаберные щели — провизорные органы зародыша.
Указанные выше признаки определяют свойственный позвоночным план строения — расчленение тела на голову, туловище и хвостовой отдел — и характерное расположение органов: нервная система в виде мозговой трубки на спинной стороне тела, под ней осевой скелет в виде хорды или позвоночника, под позвоночником основные артериальные и венозные сосуды тела и на брюшной стороне целом и лежащие в нем внутренние органы. Вентральнее пищеварительной трубки лежит сердце.
Таким образом, мы видим, что все хордовые животные являются особой ветвью двусторонне-симметричных животных, обладающих целомом и сегментацией тела. Основной ствол хордовых — позвоночные животные — в своем эволюционном развитии пошли по пути приспособления к активному подвижному образу жизни и приспособления к жизни в самых разнообразных условиях существования.
Изучая организацию разных представителей хордовых в последовательном ряду от низших групп к высшим, можно видеть постепенное повышение активности их жизнедеятельности, давшее им возможность заселить не только все водоемы земного шара, ио и выйти на сушу и постепенно заселить материки.
При сравнительном изучении организации позвоночных животных и сопоставлении их с низшими хордовыми и беспозвоночными выработана общая терминология, которая помогает сопоставлять разных животных друг с другом.
Передний конец двусторонне-симметричного животного носит название рострального отдела — от слова рыло (rostrum), задний отдел — каудального от слова хвост (cauda). Спинная поверхность тела называется дорзальной стороной от слова спина (dorsum), брюшная — вентральной от слова брюхо (venter).
Продольная плоскость, делящая тело животного на две неодинаковые по строению половины (спинную — д о р з а л ь ну ю и брюшную — вентральную), носит название фронтальной плоскости. На дорзальной стороне тела у позвоночных располагаются центральная нервная система и хорда или позвоночник с мускулатурой; на вентральной помещаются ротоглоточная и брюшная полости с внутренними органами. Продольная плоскость, делящая тело двусторонне-симметричного животного на две равные половины, правую и левую, из которых каждая является зеркальным отражением другой, носит название медиальной или сагиттальной.
По схеме строения непарные органы у позвоночных помещаются по средней линии, но могут смещаться вправо или влево; парные обычно располагаются по отношению к этой срединной линии справа и слева.
Плоскость сечения, делящая тело животного на передний и задний концы, носит название'трансверзальной. Органы, рас полагающиеся наиболее близко к средней, медиальной плоскости, носят название проксимальных, а органы, наиболее удаленные от этой плоскости в стороны, — дистальных. Так, в конечностях наземных позвоночных плечо и бедро будут занимать проксимальное положение, а кисть и стопа — дистальное положение по отношению к телу животного.
Латинские или латинизированные названия различных органов являются международной анатомической номенклатурой. Без знания этих названий трудно пользоваться анатомической литературой, как русской так и иностранной. Многие латинские термины не имеют перевода или являются основой для образования анатомического термина, как это имеет место, например, в английском языке. Поэтому рекомендуется с первых же занятий для обозначения того или иного органа пользоваться только латинскими названиями, которые приводятся в Практикуме. Для облегчения запоминания терминов они приводятся для всех органов в именительном падеже единственного числа.
Система типа хордовых
Систематика, или классификация, дает возможность привести все многообразие животного мира в определенный порядок, систему, отражающую не только черты сходства и различия, но и направление исторического развития в процессе эволюции. Начиная с Ч. Дарвина, классификация в биологии стремится быть генеалогической — естественной в отличие от искусственной классификации до-эволюционного периода в истории биологии.
Для биолога, стремящегося познать многообразие мира животных и растений Земного шара не только в современном его состоянии, но и в историческом развитии, во взаимной связи разных групп животных друг с другом недостаточно сопоставления современных групп животных. Необходимо знание и древних форм. Последние мы находим не только в виде ископаемых остатков, но и в современной фауне. Поэтому принятые в классификации современных животных систематические подразделения на виды, роды, семейства, отряды и классы часто объединяют весьма неравномерные по количеству представителей группы животных.
Так, один только надотряд рыб Teleostei из класса костных рыб Osteichthyes объединяет около 19,5 тыс. видов современных рыб из 20 тыс. видов всех рыб, в то время как к надотряду кистеперых рыб Crossopterygii относится лишь один род современных рыб — лати-мерия (Latimeria). Однако в девонский период кистеперых рыб было много, а костных еще не было. Вместе с кистеперыми рыбами в девонский период жили многие двоякодышащие рыбы (Dipnoi), а в настоящей ихтиофауне они представлены лишь тремя разрозненными родами: австралийский, цератод, дли рогозуб. (Neoceratodus), африканский протоптер (Protopterus) и американский чешуйчат-ник, лепидосирен (Lepidosiren).
Точно так же класс млекопитающих в современной фауне содержит 3200 видов, а вместе с ископаемыми известно около 6,5 тыс. видов. Из современных видов громадное большинство относится к подклассу плацентарных млекопитающих Eutheria, а в юрском и меловом периодах мезозойской эры большинство млекопитающих было представлено подклассами первозверей (Prototheria), многобугорчатыми Multituberculata (Allotheria), а также сумчатыми (Marsupialia), составляющими подкласс Metatheria, в то время как в современной фауне от этих древних подклассов млекопитающих сохранились только утконос (Ornithorhynchus) и ехидна (Tachyg-lossus) и около 200 видов сумчатых.
Для эколога-фауниста эти единичные представители древних фаун не представляют большого интереса, но для эволюционного морфолога они являются редчайшими документами, позволяющими восстанавливать условия жизни и биологию животных в прошлые этапы жизни Земли. Однако нужно помнить, что ныне сохранившиеся древние группы животных — не живые предки, остановившиеся в своем развитии. Наравне с древними малоизмененными признаками в их организации развилось много новых приспособле
ний к современным условиям жизни, которых у родоначальных предков не было. Для того чтобы правильно понимать место современных животных в истории развития мира животных и их происхождение в процессе эволюции, необходимо научиться различать, какие признаки организации у современных животных древние, т. е. сохранились от родоначальных предков, а какие новые, т. е. развились самостоятельно как приспособление к современным условиям существования.
Возникновение более современных групп животных в эволюционном развитии происходило не постепенно, а ступенчато, скачками, и эта ступенчатость исторического развития находит отражение в распределении типа хордовых на крупные систематические подразделения. Первой ступенью является деление их на два подтипа — бесчерепных — Acrania и черепных, или позвоночных — Craniota или Vertebrata. Позвоночные в свою очередь делятся на бесчелюстных— Agnatha, из которых в настоящей фауне сохранился класс круглоротых — Cyclostomata, и чёлюстноротых — Gnathostomata, которые объединяют все остальные классы позвоночных.
Обычно в учебной литературе позвоночных подразделяют на шесть классов: круглоротые, рыбы, амфибии, рептилии, птицы и млекопитающие, из которых первые два сохраняют первично водный образ жизни, а представители четырех последующих являются наземными животными и носят название четвероногих — Quadru-peda (от лат. quatuor — четыре), или Tetrapoda (от греч. tetra — четыре). В настоящее время среди палеонтологов и зоологов, учитывающих многообразие рыб в предшествующие геологические эпохи, получило широкое распространение представление, что термин Рыбы — Pisces является таким же сборным понятием, как тетраподы для наземных позвоночных. Рыбы столь разнородны по своей организации, что они должны быть подразделены на ряд самостоятельных классов, равноценных классам наземных позвоночных (земноводным, пресмыкающимся, птицам и млекопитающим). Учитывая ископаемых рыб, группу Pisces обычно подразделяют на три-четыре класса: панцирные рыбы — Placodermi, хрящевые рыбы — Chondrichthyes, акантоды — Acanthodei и костные рыбы — Osteichthyes. Действительно, глядя на распределение позвоночных и низших хордовых на геологической летописи (рис. 1), такая классификация рыб оказывается обоснованной.
Существует еще одно важное разграничение позвоночных на группы. Рыбы и земноводные, как правило, связанные по биологии размножения с водной средой, откладывающие икру и имеющие наружное оплодотворение, объединяются в группу анамний — Апагп-nia, т. е. не имеющих амниона. Пресмыкающиеся, птицы и млекопитающие, размножающиеся на суше или путем откладки яиц в скорлуповых оболочках при внутреннем оплодотворении в выводных путях самки или же как млекопитающие, имеющие внутриутробное развитие, объединяются в группу амниот — Amniota, т. е. имеющих амнион и другие зародышевые органы (так называемые зародышевые оболочки).
Наконец, рептилии и птицы, как яйцекладущие, нередко противопоставляются живородящим млекопитающим, как группа ящерообразных — Sauropsida.
Таким образом, учитывая современные и ископаемые группы животных, система типа хордовые, доведенная до расчленения на классы и подклассы, может быть представлена в виде следующей таблицы.
Тип Сhо гdatа — хордовые
Подтип Urochorda (Tunicata) — Личиночнохордовые (Оболочники)
Класс Ascidia — Асцидии
Подкласс — Enterogona
Подкласс — Pleurogona
Класс Salpae — Сальпы
Класс Appendicularia — Аппендикулярии
Подтип Acrania — Бесчерепные
Класс Cephalochorda — Головохордовые
Подтип Vertebrata (Craniota) — Позвоночные (Черепные)
Раздел Agnatha — Бесчелюстные
Класс Diplorhini (Pteraspidomorphi) —Двуноздревые Подкласс Thelodontia (Coelolepida) — Телодонты Подкласс Heterostraci — Разнощитковые
Класс Monorhini (Cephalaspidomorphi) — Одноноздревые Подкласс Osteostraci — Костнощитковые Подкласс Anaspida — Бесщитковые
Класс Cyclostomata — Круглоротые
Подкласс Petromyzones — Миноги
Подкласс Mixini — Миксины
Раздел Gnathostomata — Челюстноротые
Надкласс Pisces — Рыбы
Класс Placodermi — Панцирные рыбы
Подкласс Antiarchi (Pterichthyes) — Антиархи
Подкласс Arthrodira (Coccostei) — Артродиры
Класс Chondrichthyes — Хрящевые рыбы
Подкласс Holocephala — Цельноголовые (химеры)
Подкласс Elasmobranchii — Пластиножаберные (акулы, скаты)
Класс Acanthodei — Акантоды
Класс Osteichthyes — Костные рыбы
Подкласс Sarcopterygii — Мясистолопастные (кистеперые, двоякодышащие)
Подкласс Brachiopterygii (Polypteri) —Многоперые
Подкласс Actinopterygii — Лучеперые (палеонисциды, хрящевые и костные ганоиды, костистые рыбы)
Надкласс Tetrapoda — Четвероногие (наземные позвоночные) Класс Amphibia — Земноводные
Подкласс Apsidospondyli — Дугопозвонковые (лабиринтодонты и бесхвостые)
Подкласс Lepospondyli — Листопозвонковые (хвостатые и безногие)
Подкласс Batrachosauria (Seymouriamorpha — Сеймурио-образные
Класс Reptilia — Пресмыкающиеся
Подкласс Cotylosauria — Котилозавры
Подкласс Progonosauria — Прогонозавры (мезозавры) Подкласс Synaptosauria (Plesiosauria) — Плезиозавры Подкласс Ichthyopterygia — Ихтиоптеригии (ихтиозавры) Подкласс Testudinata — Черепахи
Подкласс Archosauria — Архозавры (текодонты, динозавры, летающие ящеры, крокодилы)
Подкласс Lepidosauria (Squamata) — Чешуйчатые
Подкласс Synapsida (Therapsida) — Зверообразные
Класс Aves — Птицы
Подкласс Aschaeornithes — Древние птицы (археоптерикс) Подкласс Neornithes — Новые птицы
Класс Mammalia — Млекопитающие
Подкласс Allotheria — Многобугорчатые
Подкласс Prototheria — Первозвери (клоачные)
Подкласс TricOnodonta — Триконодонты
Подкласс Panthotheria — Пантотерии (докодонты, симетро-донты)
Подкласс Metatheria — Низшие или сумчатые звери
Подкласс Eutheria (Placentalia) — Высшие или плацентарные
Сравнительный метод исследования
и типы сопоставления органов при сравнении
Сравнительный метод исследования дает возможность объяснить причины возникновения сходства и различий между животными. При сравнении друг с другом животных, произошедших в прошлом от общего родоначального предка, черты сходства между ними подтверждают их родство, а черты различия свидетельствуют о приспособлении к различным условиям существования (различном образе жизни, питании и т. д.).
Органы разных животных, имеющие общий план строения, развивающиеся из сходных зачатков, служат свидетельством родства животных и происхождения их в процессе эволюции от общего родоначального предка. Они могут выполнять одну и ту же функцию (как конечности четвероногих животных при хождении по земле), но могут иметь и различные функции. Так, например, парные конечности наземных позвоночных у одних служат для передвижения по суше, у других —для полета в воздухе, у третьих являются органами плавания в воде — ластами. Такое морфологическое сходство органов с одинаковой или различной функцией носит название г о-32
мологии и является одним из основных доказательств родства между животными.
Если сопоставляются друг с другом сходные органы у одного и того же животного, например передние и задние конечности позвоночного животного, или различные сегменты тела метамерного животного, то такие сериально гомологичные органы одного и того же животного носят название органов гомодинамиых.
Сопоставление друг с другом гомологичных органов родственных групп животных дает возможность судить о направлениях их эволюционного развития. В одних случаях при возникновении новых видов мы сталкиваемся с процессом, названным Ч. Дарвином расхождением признаков от одной точки в двух направлениях, шт дивергенцией, когда на фоне сходных признаков строения, унаследованных от общего родоначального предка, возникают ново-приобретенные различия; в других случаях вновь приобретенные приспособления возникают одновременно у целого ряда новых видов, и они расходятся в различных направлениях — как бы по радиусам от одного родоначального центра. Такое направление названо В. О. Ковалевским и р р а д и а ц и е й, а Г. О. Осборном — адаптивной радиацией.
Черты сходства у разных животных не всегда обусловлены одинаковым происхождением органов. При жизни в сходных условиях существования могут вырабатываться сходные приспособления у животных, по своему происхождению далеких друг от друга. Такое соответствие органов, выполняющих одинаковую функцию в сходных условиях существования, — функциональное сходство, возникающее в органах разного происхождения, носит название аналогии. Аналогичные органы являются свидетельством эволюционного развития в сходных условиях вследствие выполнения этими органами сходных функций. Так, самые различные животные для полета в воздухе имеют.крылья, которые выполняют одинаковую функцию, но развились они независимо друг от друга из различных зачатков и при функциональном сходстве имеют различное строение. Такое функциональное сходство свидетельствует о совершенно другом процессе, чем морфологическое сходство гомологических органов. Здесь наблюдается схождение признаков у современных форм в отличие от их родоначальных предков, живших в различных условиях существования. Процесс схождения признаков в эволюционном развитии носит название конвергенции.
В некоторых случаях одинаковые изменения строения органов возникают под влиянием приспособления к сходным условиям существования у родственных животных независимо друг от друга как аналогичные признаки. Такие явления параллелизма носят название гомойологии или гомоплазии.
Пользуясь сравнительным методом исследования при изучении многообразия мира животных, зоолог должен уметь различать разнородность черт сходства и различия у разных животных. Только тогда он сможет понять законы развития мира животных на Земле и научиться управлять их развитием.
$7 3?
Рис. 1. Родословное древо хордовых, наложенное на геологическую летопись. Отдельные филогенетические ветви обозначены в виде листьев, ширина которых означает относительное количество представителей в данной эпохе. Кружками обозначены гипотетические группы. Черные места означают отсутствие ископаемых остатков. Ст—кембрий, 70 млн. лет; 5i — нижний силур, 90 млн. лет; S2— верхний силур, 40 млн. лет; D — девон, 50 мли. лет; С] — нижний карбон, 50 млн. лет; С2— верхний карбон, 35 мли. лет; Р — пермь, 25 млн. лет; Т — триас, 35 млн. лет; J — юра, 35 млн. лет; Сг—мел, 65 млн. лет; К — кайнозойская эра (третичный, четвертичный и современный периоды), 55 млн. лет:
Родословное древо хордовых
Взаимоотношение современных и ископаемых групп животных в историческом развитии особенно наглядно выражено в виде родословного древа, наложенного на геологическую летопись Земли.
На рис. 1 представлено такое родословное древо исторического развития хордовых животных, восстановленное на основании сопоставления данных палеонтологии для ископаемых групп, данных сравнительной анатомии и систематики для современных групп и данных сравнительной эмбриологии при восстановлении низших хордовых животных, не сохранившихся в ископаемом состоянии. Горизонтальные линии отделяют периоды геологической летописи, выраженные в миллионах лет в одинаковом масштабе, отчего толщина периодов на таблице разная.
Каждая ветвь родословного древа в виде языка выражает начало, расцвет и вымирание группы животных по палеонтологическим данным. Верхний горизонт соответствует систематическим подразделениям современной фауны, толщина каждой ветви в схематической, условной форме отражает количество видов, составляющих каждую группу. Отдельные ветви соответствуют надотряду по шкале классификации, иногда с некоторым отклонением. При такой форме выражения естественной классификации животных, современных и вымерших, на рисунке видно, что в истории развития органического мира на Земле все время происходит замена одних фаун животных другими, более совершенными.
На рис. 1 слева представлены более древние низшие подтипы, классы, подклассы хордовых животных, а по направлению направо ответвляются более поздние группы позвоночных животных. Крайний правый верхний угол занимает высший класс позвоночных — млекопитающие с их подразделением на вымершие и современные подклассы и отряды. В тех случаях, когда между современными и ископаемыми группами не сохранилось палеонтологических данных,
взаимосвязь их выражена пунктиром, а древние гипотетические группы низших хордовых, не имеющих твердого скелета и не сохранившихся в ископаемом состоянии, оставлены незачерченными.
При прохождении большого практикума студентам рекомендуется, изучая организацию низших хордовых и позвоночных в последовательном систематическом ряду от низших к высшим, возвращаться к рассмотрению данного родословного древа при изучении каждого нового представителя животных и анализировать взаимоотношения, начало, расцвет и конец ветвей, составляющих генеалогическую систему мира животных. Выражение исторического развития органического мира на Земле в виде родословного древа от единого корня с постепенным ветвлением ствола на ветви в разные стороны, установленное Ч. Дарвином, носит название монофилии (единостволие). При изучении многообразия приспособлений животных к различным условиям существования зоологи часто встречаются с появлением сходных приспособлений к одинаковым условиям, возникающих у разных групп, родственных между собой, которые возникли у них независимо друг от друга — параллельно. Параллельное, независимое появление сходных приспособлений к одинаковым условиям жизни у разных групп носит название п а-р афилии.
Материалистическому учению о монофилии противопоставляется представление о многостволии — п о л и ф и л и и, т. е. развитии отдельных групп животных независимо друг от друга параллельными рядами в виде отдельных кустов, как поле хлебных злаков, где каждый куст развился из отдельного зерна. Согласно учению о полифилии общие черты организации у животных возникают как аналогичные органы, независимо друг от друга, руководимые общими закономерностями развития.
Литература по зоологии и сравнительной анатомии позвоночные
Беляев М. М. Зоотомия позвоночных. Учпедгиз, 1947.
Бобринский Н. А., Матвеев Б. С., Банников А. Г. Курс зоологии, т. II, «Зоология позвоночных». М., «Высшая школа», 1966.
Карташов Н. Н., Соколов В. Е., Шилов И. А. Практикум по зоологии позвоночных. М., «Высшая школа», 1969.
Ливанов Н. А. Пути эволюции животного мира (Анализ организации главнейших типов многоклеточных животных). М., «Советская наука», 1955.
Основы палеонтологии. Справочник для палеонтологов и геологов СССР в 15 томах (под ред. Ю. А. Орлова). Том Бесчелюстные, рыбы; том Земноводные, пресмыкающиеся и птицы. М„ «Наука», 1964.
Паркер Т. Курс зоотомии (позвоночные). М., 1904.
Паркер Т. и Гасвелл В. Руководство к зоологии, т. 2. «Позвоночные», 1908.
Ромер А. Палеонтология позвоночных. М., ГИЗ, 1939.
Северцов А. Н. Морфологические закономерности эволюции. Изд-во АН СССР, 1949; то же, собр. соч., т. 5. М„ 1955.
Северцов А. Н. Главные направления эволюционного процесса (морфологическая теория эволюции). Изд. 3, Изд-во МГУ, 1967.
Терентьев П. В. Практикум по зоологии позвоночных. М., «Советская наука», 1956.
Шимкевич В. М. Курс сравнительной анатомии позвоночных животных. Л., 1922.
Шмальгаузен И. И. Основы сравнительной анатомии. Изд. 4. М., «Советская наука», 1947.
Шмальгаузен И. И. Происхождение наземных позвоночных. М., «Наука», 1963.
Bolk L., Goppert Е., Kalius Е., Lubosch W. Handbuch der verg-leichenden Anatomie der Wirbeltiere. 5 Bde. Urban und Wien, 1931—1938.
Giersberg H., Rietschel P. Vergleichende Anatomie der Wirbeltiere. 2 Bde. Verl. G. Fischer, Jena, 1967, 1968.
Goodrich E. E. Studies an the structure and development of Vertebrates, London, 1930.
Grasse E. C. Traite de Zoologie. t. XI—XVII, 1948—1955.
I hie J. E. van Katnpen P. N., Nierstrasz H. F., Versluys J. Vergleichende Anatomie der Wirbeltiere. Ubers, von G. Ch. Hirsch. Berlin, Verl. Springer, 1927.
Kukenthal W., Krumbach Th. Handbuch der Zoologie, 1929—1955.
Marinelli W., Strenger A. Vergleichenden Anatomie und Morphologie der Wirbeltiere, 1953 u ff.
Plate L. Allgemeine Zoologie und Abstammunslehre. 2 Bde, Jena, G. Fischer Verl., 1922, 1924.
Portmann A. Einfuhrung in die vergleichende Morphologie der Wirbeltiere. 3 Aufl. Basel Schwabe, 1965.
Romer A. S. Vertebrate Paleontology. Chicago, 1947.
Romer A. S. The vertebrate body. Philadelphia, 1957.
Romer A. S. Vergleichende Anatomie der Wirbeltiere (Ubersetzt H. Frick), Hamburg, 1959.
ПОДТИП ЛИЧИНОЧНО-ХОРДОВЫЕ, ИЛИ ОБОЛОЧНИКИ, — UROCHORDA, S.TUNICATA
Общая характеристика подтипа
Оболочники принадлежат к низшим хордовым, вторично утерявшим типичное строение хордовых животных. Тело оболочников покрыто особой оболочкой, туникой, откуда они и получили свое название. В личиночном состоянии эти животные имеют спинную струну — хорду и трубчатую нервную систему, но у асцидий во взрослом состоянии хорда и нервная система подвергаются дегенерации. Все оболочники — морские животные.
Оболочники Tunicata разделяются на три класса: 1. Асцидии — Ascidiae. 2. Сальпы — Salpae, s. Thaliacea. 3. Аппендикулярии— Appendicularia. Типичными и наиболее распространенными представителями являются асцидии.
Оболочники представляют большой интерес. Несомненно, эта группа отделилась от первичных хордовых чрезвычайно давно, так как они сохранили самый примитивный тип питания и дыхания путем фильтрации воды через жаберные отверстия при помощи мерцательных ресничек клеток эпителия, выстилающих ротоглоточную полость. Оболочники — единственная группа среди хордовых, имеющая бесполое размножение почкованием, вследствие чего у них распространена колониальность, как это имеет место у беспозвоночных с бесполым размножением.
КЛА С С А СЦИДИИ — ASCIDIAE
Общая характеристика класса
К классу асцидий принадлежат одиночные и колониальные оболочники, ведущие сидячий образ жизни на дне морей и широко распространенные по всем морям земного шара (около 1500 видов). Асцидии — единственная группа среди хордовых, ведущая неподвижный образ жизни, отсюда в их организации отмечается ряд дегенеративных черт по сравнению с ланцетником и позвоночными животными. Тело асцидий, как и других оболочников, снаружи по-38
крыто оболочкой — туникой. Туника состоит из особого вещества, близкого к растительной клетчатке, — туницина, которое образуется как продукт выделения стенок тела. Стенки тела состоят из кожных покровов, сросшихся с мускулатурой тела в общий слой, называемый мантией.
Долгое время асцидий причисляли к моллюскам, и только в 1866 г. А. О. Ковалевский, изучивший строение личинок асцидий, обосновал принадлежность их к хордовым животным.
Изучать строение оболочников следует на одиночных асцидиях. В морях Советского Союза встречается около 150 видов асцидий и среди них имеются крупные одиночные формы.
Класс асцидий разделяется на два подкласса: Enterogonia и Pleurogonia. Большинство асцидий принадлежит к отряду Stoli-dobranchiata подкласса Enterogonia. Два других отряда (Aplousob-ranchiata и Phlebobranchiata) этого подкласса встречаются гораздо реже. В состав отряда Stolidobranchiata входят четыре семейства: Botrylidae, Styelidae, Pyuridae, Molgulidae. Все они довольно часто встречаются, но для изучения анатомии асцидий рекомендуется добывать представителей рода Styela (Goniocarpa) из семейства Styelidae, которые будут основным объектом при описании аспидий, или представителей рода Halocynthia (Tethyum) из семейства Pyuridae. На особенности строения этих представителей в тексте будут даны ссылки.
Представители класса асцидий. Род Styela (Flemming 1822), s. Goniocarpa (Hunstman) (рис. 2)
Виды рода Styela (Goniocarpa) широко распространены в северных морях. Это довольно крупные асцидии с толстой кожистой туникой, цилиндрической формы, с характерным крупным шипом между входным и выходным сифонами. Для рода характерны парные яичники, по одной гонаде на каждой стороне тела. В жаберном мешке с каждой стороны по 4 складки. В Белом море широко распространены два близких вида рода Styela: Styela rustica L. и St. gelatinosa L. Оба вида очень близки между собой. Вскрытие удобнее вести на St. gelatinosa, так как туника ее легко отслаивается от стенок тела, но St. rustica — более распространенный вид, и ее легко достать.
Для изучения особенностей строения оболочников необходимо заготовить фиксированный материал. При фиксации асцидии конвульсивно сокращают свою мускулатуру и сморщиваются. Поэтому рекомендуется перед фиксацией в сосуд с асцидиями постепенно приливать пресную воду до тех пор, пока асцидии перестанут сокращаться. Фиксировать животных следует в 5—10%-ном формалине (от продажного). Через 1—2 суток после фиксации асцидий фиксатор рекомендуется сменить и залить их 5%-ным формалином. За сутки до начала работы с асцидиями их необходимо вымочить в воде, чтобы отмыть формалин. Только после этого материал можно .выдавать студентам для работы.
Внешний осмотр
Изучение внешней формы и препаровку асцидий следует производить под водой в препаровальных ванночках так, чтобы вода покрывала полностью объект. Тело асцидий имеет форму двугорлой банки, основанием прикрепленной ко дну. Входное отверстие называется ротовым сифоном (sipho oralis, рис. 3, siph. о.), помещается обычно на верхнем конце тела. Выходное отверстие, называемое клоакальным сифоном (sipho cloacalis, рис. 3, siph. cl.), распола-
Рис. 2. Внешний вид различных асцидий: l~Styela getatinosa; 2 — Styela rustica; 3 — Halocyn-thia piriformis; 4— Halocynthia aurantia; 5 — Hatocyn-thia roretzl
гается рядом с ротовым, несколько ниже. У живых асцидий сифоны открыты, как горлышко склянки, и в каждом из них видны четыре створки. На фиксированных экземплярах сифоны закрыты и четы-рехстворчатость четко выступает только на ротовом, клоакальный кажется двустворчатым. Определить ротовой и клоакальный сифоны можно и другим способом. При сдавливании тела асцидии паль-
цами жидкость выделяется обычно через клоакальный сифон. Пространство между сифонами можно считать спинной стороной, остальную часть — брюшной. Нижним концом брюшной поверхности, или подошвой, асцидия прикрепляется ко дну или к какому-
сог.
Рис. 3. Общая топография внутренних органов асцидий Styela gela-tinosa. Полость глотки вскрыта:
м. о. к — мерцательное окологлоточное кольцо; п. с. — продольные складки глоткн; р. щ. — ротовые щупальца; с. п. — спинная пластинка; сог. — сердце; end.—эндостиль; gast. — желудок; gl. sbn. — подмозговая железа; g. nr.— мозговой ганглнй; ints — кишечник; mnt. — мантия; oes. — пищевод; ои. — яичник; oud. — яйцевод; г. — выделительные органы; siph. cl. — клоакальный сифон;
siph. о. —- ротовой сифон; test. — семенник; tnc. — туника
либо твердому предмету (камню, скале, раковине). Подошва уплощена для прикрепления к субстрату, и туника на подошве более утолщена, чем на теле.
Асцидия Styela rustica (Goniocarpa rustica, рис. 2. 2). Представители этого вида — одиночные асцидии цилиндрической формы с
сифонами, но иногда они сидят группами по нескольку штук. В живом состоянии животные окрашены в оранжевый цвет, края входного и выходного отверстий ярко-красные. Они имеют мощную мускулатуру оранжево-красного цвета, которая и обусловливает окраску тела. Фиксированный материал довольно быстро обесцвечивается.
Наружная поверхность у асцидии St. rustica неровная, бугристая, в поперечных складках и покрыта шипиками различной формы. Кроме того, она обычно покрыта различными животными и водорослями, для которых туника является субстратом— ветвистыми колониями коралловых полипов, колониями мшанок, мелкими ракушками мидий и других моллюсков, которые довольно легко удаляются с тела при соскабливании.
При раздражении асцидий мускулатура резко сокращается и фонтаном выпрыскивает воду из выходного отверстия. При сдавливании тела фиксированной асцидии вода также выпрыскивается из выходного отверстия.
Асцидия Styela gelatinosa (Goniocarpa gelatinosa, рис. 2, /). Очень сходна с St. rustica как по внешним признакам, так и по внутреннему строению, но не имеет такой яркой окраски. Стенки тела более прозрачные, откуда и название вида St. gelatinosa. Мускулатура стенок тела развита значительно слабее. Наружная поверхность более гладкая, имеются лишь небольшие бугорки. Через стенку тела просвечивают органы, что у St. rustica не наблюдается.
Асцидия Halocynthia piriformis (рис. 2, 3) — представитель семейства Pyuridae (Tethyidae Hunstmann, 1911; Редикорцев, 1916), широко распространена в Белом море, имеет боченковидную форму и окрашена в розово-оранжевый цвет. Клоакальный сифон коленообразно изогнут. Туника тонкая, но очень плотная, резинообразной консистенции, с мелкими бисеровидными бугорками. Она плотно сращена со стенкой тела и совершенно непрозрачная.
Асцидия Halocynthia aurantia (рис. 2, 4) по внешнему виду очень напоминает вышеописанную, но значительно более крупных размеров, достигает 150 мм длины. Туника очень плотная, сохраняющая и после фиксации своп ярко-оранжевый цвет. Подошва сильно утолщена. Ротовой сифон обычно имеет четыре створки, клоакальный — две. Оба сифона расположены на верхней части тела и часто одинаковы по величине.
Асцидия Halocynthia roretzi (рис. 2, 5) по размерам и форме близка к вышеописанной, но плотная туника имеет многочисленные крупные заостренные выросты.
Вскрытие асцидий
Вскрытие асцидий проводят двумя способами, в зависимости от времени, которое выделено на занятие. Кроме того, дополнительно к вскрытию рекомендуется на другом объекте сделать бритвой поперечный разрез через середину тела асцидии так, чтобы на одной половине остались сифоны, а на другой — подошва. На поперечных 42
разрезах очень удобно разбираться в строении стенок тела и в расположении органов в атриальной полости, и в полости глотки.
При первом упрощенном способе тело асцидии разрезают бритвой на две половины по сагиттальной плоскости. Для этого кладут животное в ванночку с водой на вату. Далее вместе с ватой берут ее в левую руку так, чтобы брюшная сторона была направлена вправо, ротовой сифон вверх, а клоакальный налево. Бритву следует держать в правой руке и начинать продольный разрез таким образом, чтобы ротовой и клоакальный сифоны были разрезаны пополам по средней линии. Разрез через сифоны требует некоторого усилия, затем бритва идет очень легко. Разрез необходимо проводить до самой подошвы, разрезав тело на две равные части. При удачно прошедшем разрезе жаберная полость делится пополам, и при сопоставлении обеих половинок без всякой препаровки легко разобраться в строении внутренних органов асцидии.
Однако продольный разрез может пройти косо, и тогда разбираться в препарате становится трудно, поэтому рекомендуется пользоваться более трудоемким, но более надежным методом последовательного вскрытия.
Чтобы вскрыть тунику, животное берут в руку и делают ножницами небольшой разрез туники несколько ниже клоакального сифона. У Styela gelatinosa прозрачная туника легко отстает от стенки тела, и ее можно разрезать одну. У других видов она плотно срастается со стенкой тела, которая также может быть разрезана. Если разрезать одну тунику не удается, то следует поступать, как описано ниже (при вскрытии стенки тела — мантии). Разрез туники продолжают вниз до подошвы, а потом параллельно краю подошвы до противоположной стороны. Теперь асцидию помешают в ванночку с водой и, придерживая пинцетом тунику около клоакального сифона, осторожно проводят ножницами разрез туники от клоакального сифона к^ ротовому, стараясь не повредить органов, лежащих между клоакальным и ротовым сифонами. Доведя разрез около ротового сифона до брюшной стороны и отвернув тунику направо, тело и отвернутую в сторону тунику прикрепляют булавками к восковому дну ванночки так, чтобы ротовой сифон расположился кверху, а клоакальный — налево.
Для вскрытия стенки тела (мантии асцидии) ножницами проводят продольный разрез от клоакального сифона вниз до подошвы, а затем у основания подошвы и под сифонами до брюшной стороны асцидии. Приподняв край разреза, можно увидеть околожа-берную а тр и а л ь ну ю полость (cavum atrii, см. рис. 4 га-’, а.), куда открываются наружные жаберные отверстия глотки и задн -й отдел кишки. Глотка прикрепляется к мантии тонкими мезентериальными спайками, в чем легко убедиться, рассматривая погруженные в воду поперечные разрезы асцидий (см. рис. 4).
Чтобы отпрепарировать глотку, имеющую вид тонкой сетки, пронизанной многими рядами жаберных отверстий — стигм (foramen branchiale, см. рис. 6, f. br.), надо, придерживая пинцетом стенку тела, осторожно подрезать мезентериальные спайки, подвеши
вающие глотку к стенкам околожаберной полости, стараясь при этом не повредить как глотку, так и органы, прикрепленные к внутренней поверхности мантии. После этого рекомендуется развернуть вскрытую асцидию и приколоть ее булавками к восковому дну ванночки.
Теперь можно рассмотреть общий план расположения органов. Левая часть препарата почти полностью занята глоткой. Под ней внизу около подошвы темнеет в виде непрозрачного тела желудок. На отвернутой стенке тела видны одна или несколько, в зависимости от того, какой вид вскрывается, желтых или бурых трубочек яичников. Кроме того, здесь располагаются многочисленные темные почечные пузырьки. По стенке тела, а также по другим органам могут быть разбросаны отдельные беловатые дольки мужских половых желез — семенников.
Строение туники, мантии и мускулатуры можно рассмотреть на отвернутой стенке тела, а затем уже приступить к рассмотрению строения внутренних органов (рис. 3).
Туника (tunica, рис. 3, tno.) у St. gelatinosa кожистая, полупрозрачная, незначительной толщины, на наружной стороне ее видны редко расположенные шипики. На подошвенной стороне туника утолщена и образует подошву, которой асцидия прикреплена к субстрату. Около ротового и клоакального сифонов туника сращена со стенкой тела. У разных родов и видов асцидий туника имеет различную толщину и консистенцию.
Мантией (mantia, рис. 3, mnt.) называется стенка тела, которая состоит из наружного кожного покрова, плотно сращенного с мускулатурой тела в общий кожно-мускульный мешок, одевающий все тело.
Мускулатура (рис. 3) у асцидии St. gelatinosa по сравнению со St. rustica развита слабо. От ротового сифона продольными пучками веерообразно идут мышечные волокна, замыкающие ротовой сифон. Они кончаются у основания ротового сифона на кольцевидном валике, являющемся границей жаберной области. Такая же мускулатура имеется и у клоачного сифона. Общий мускульный мешок состоит обычно из двух перекрещивающихся слоев мускулатуры: снаружи располагаются волокна продольных мышечных пучков (у St. gelatinosa очень тонкие), а внутри — кольцевая поперечная мускулатура.
У асцидий семейства Styelidae на внутренней поверхности кольцевой мускулатуры при рассмотрении в лупу поперечного разреза через асцидию можно видеть еще тонкие продольные волокна третьего, внутреннего, слоя мускулатуры.
На поперечном разрезе через тело асцидии St. rustica (рис. 4) хорошо видны все четыре слоя, составляющие стенку тела: наружный слой — мощная туника, плотно сращенная с эктодермой;, второй слой — мощно развитые отдельные пучки продольной мускулатуры; третий слой — кольцевидные волокна поперечной мускулатуры; четвертый — внутренние продольные волокна, у St. rustica они значительно крупнее, чем у St. gelatinosa.
У асцидии Halocynthis piriformis, имеющей очень плотную кожистую тунику розового цвета и неспособной сильно сокращаться, мускулатура развита очень слабо и отличается как бы обратным расположением мускульных слоев: снаружи под туникой располагается тонкий слой кольцевидной мускулатуры, плотно сращенной с туникой, а ко внутри од1 нее идет ряд пучков продольной мускулатуры. Такое впечатление создается потому, что у этого вида асцидий наружный продольный слой редуцирован, внутренний развит более сильно. То же самое относится и к двум другим представите
лям этого рода.
Рис. 4. Поперечный разрез тела асцидии Styela gelatinosa:
в. м. — внутренние продольные мышечные пучки; к. м. — пучки кольцевой мускулатуры; п. м. — наружные продольные мышечные пучки; п. с. — продольные складки глотки; с. — мезентериальные спайки; с. п. — спинная пластинка; cav. а. — атриальная полость; end.—эндостнль; ints.— кишечник; ои. — яичник; ph.— глотка; г.—выделительные органы; s. Ьг. — жаберный мешок; tnc. — туинка
Строение покровов и мантии асцидий более подробно можно изучить на микропрепаратах поперечных разрезов стенки тела и туники, изготовленных по целлоидиновой методике и окрашенных гематоксилинэозином. Лучше всего такие препараты делать толщиной 10—15 мк из мелких асцидий.с тонкой туникой, например Styela gelatinosa.
На таких препаратах даже при небольшом увеличении микроскопа видно, что наружный слой составляет туника, наружные слои ее гомогенны, а в основании наблюдается четкое слоистое строение. Под туникой располагается эпидермальный слой мантии, клетки которого лежат в один ряд и несколько отличаются от
обычных эпидермальных
клеток в связи с присутствием в нем мерокриновых бокаловидных клеток, которые секретируют кислые мукополисахариды, образуя вещество туники. Ниже располагается рыхлая мезенхиматозная ткань с разбросанными в ней отдельными клетками. В мезенхима-тозную ткань включены волокна двух слоев продольной и лежащего между ними одного слоя кольцевой мускулатуры. Как указывалось выше, у разных видов мощность этих слоев может быть различной. Внутренняя, обращенная к атриальной полости поверхность стенки тела покрыта однослойным эпителием, который, поскольку атриальная полость образуется путем впячивания, имеет эктодермальное происхождение. Иногда на срезах попадаются прилежащие
к стенке тела разрезы семенников и «почек». Расположение внут-
ренних органов можно рассмотреть на вскрытой асцидии.
Атриальная полость (cavum atrii, рис. 4, cav. а.) заменяет собой сохранившуюся у взрослых асцидий только в области сердца вторичную полость тела. Она открывается наружу через клоакальный сифон. Большая часть атриальной полости занята глоткой. К стенкам тела — мантии — она прикреплена отдельными мезентериальными спайками, которые проходят по атриальной полости и особенно густы на брюшной и спинной сторонах тела. Положение глотки в атриальной полости хорошо видно на поперечном разрезе асцидии. К мантии прикрепляются также лежащие в атриальной полости гонады и выделительные органы — почечные пузырьки.
Женские гонады — яичники (ovarium, рис. 3, ov.) у асцидий р. Styela парные и имеют форму длинных извитых железистых мешков ярко-желтого или коричневого цвета, заполненных яйцами. Располагаются они на правой и левой сторонах тела и прикреплены к стенкам мантии. У St. rustika они прямые, а у St. gela-tinosa нижние концы их загнуты. На спинной стороне тела около клоакального сифона каждый из яичников открывается в атриальную полость коротким яйцеводом (рис. 3, ovd.). У асцидий р. Halocynthia яичники также парные, но количество трубочек в каждом из них обычно равно 4, хотя может варьировать от 3 до 5.
Мужские гонады — семенники (testis, рис. 3, test) представлены в виде многочисленных беловатых долек, расположенных в нижнем отделе атриальной полости. Они или прикреплены к стенке мантии, или лежат на других внутренних органах: желудке, кишечнике, яичниках. Семяпроводов у представителей двух описываемых родов нет.
Таким образом, у асцидий существует гермафродитная половая система, т. е. у каждой особи имеются и мужские и женские половые железы. Однако самооплодотворения не происходит, так как мужские и женские половые жёлезы созревают не одновременно.
Половая система у асцидий, как и вообще у туникат, отличается большим разнообразием строения у разных видов, родов и семейств. Их отличительные особенности служат систематическими признаками при классификации асцидий.
Выделительные органы (рис. 3, г.) у асцидий представлены многочисленными почечными пузырьками, наполненными жидким секретом. Они рассеяны по стенкам мантии и свисают в атриальную полость. У мольгулид имеется одна крупная почка в виде крупного овального пузыря на правой стороне тела.
Пищеварительная система начинается пищеводом ^oesophagus, рис. 3, oes), соединяющимся с жаберным мешком на его спинной стороне на высоте нижней трети. Пищевод имеет вид трубки, располагается с левой стороны жаберного мешка и входит в желудок, лежащий под жаберным мешком в углублении, образованном подошвой.
Желудок асцидий (gaster, рис. 3, gast.) характеризуется продольной складчатостью слизистой оболочки. Он имеет веретеновид-46
ную изогнутую форму, загибается кверху и переходит в кишку (intestinum, рис. 3, ints.). Кишка поднимается прямо кверху по направлению к клоакальному сифону и, не доходя до него, открывается в артериальную полость на высоте верхнего конца яичника. Наружное анальное отверстие имеет характерные пальцевидные выросты в виде воронки. Чтобы рассмотреть кишечник, надо приподнять жаберный мешок.
Печень. У Styelidae на стенках желудка имеются складки двух сортов. Одни—продольные, другие, более подчеркнутые,— ко
сые, сходящиеся к горлу желудка. Эти более выраженные складки называют печеночными складками. У асцидий из семейств Molgulidae и Pyuridae печеночные складки образуют разветвленные дивертикулы и группируются в массу долек, которые лежат на верхней стороне желудка и получают название печени.
Пилорические железы присутствуют у всех асцидий в виде системы тонких трубочек,
Рис. 5. Мозговой ганглий и подмозговая железа асцидии Styela gelatinosa:
м. о. к. — мерцательное окологлоточное кольцо; п. с. — продольные складкн глотки; р. щ. — ротовые щупальца; с. п. — спинная пластинка; f. or. — жаберные отверстия; gl. sbn. — подмозговая железа; g. nr. — мозговой ганглий
разветвленных в стенках желудка.
После рассмотрения пищеварительной системы вскрывают жаберный мешок и ротовой сифон. Для
этого вставляют ножницы
в ротовой сифон, разрезают его ближайшую к препарирующему стенку по средней линии. Продолжая разрез, вскрывают также и жаберный мешок по средней линии вплоть до его нижнего конца. Затем делают разрез и клоакального сифона. Осторожно отворачивают края жаберного мешка и сифонов, прикалывают их булавками и рассматривают их строение. Рассматривают также строение жаберного мешка на поперечном разрезе (см. рис. 4).
Ротовой сифон (sipho oralis, рис. 3, siph. о.) имеет форму конусовидного кратера, у основания которого есть утолщенный кольцевой валик, окружающий вход к глотку. При вскрытии ротового сифона видно, что на кольцевом валике располагаются длинные щупальца разной величины (рис. 3,5, р. щ.) в количестве до 30 шт. у р. Styela. У представителей сем. Styelidae щупальца простые, у сем. Molgulidae и Pyuridae — разветвленные. Щупальца ог-
раждают вход в глотку.
Жаберный мешок, или глотка (pharynx), представляет тончайшую сетку многих рядов жаберных щелей — стигм с вертикальными ячеями, ограниченными прямыми продольными и поперечными перекладинами, в которых проходят кровеносные сосуды, обеспечивающие газообмен в жаберном аппарате. Такое расположение жаберных щелей — стигм — прямыми рядами в виде лестниц, сцепленных более крупными продольными сосудами,— очень характерный тип строения, свойственный асцидиям сем. Stye-lidae.
Вторым важным признаком строения жаберного аппарата асцидий этого семейства является наличие 8 продольных складок внутри жаберной полости, по четыре с каждой стороны. Рассмотрите их положение на вскрытой асцидии, а также на ее поперечном разрезе (продольные складки, рис. 3, 4, 5, п. с.).
На спинной стороне тела асцидии над отверстием пищевода на внутренней поверхности жаберного мешка в виде продольной складки помещается спинная пластинка (рис. 3, 4, 5, с. п.), тянущаяся кверху до околоротового валика с ротовыми щупальцами. Эта спинная пластинка распределяет складки мешка на две стороны, по четыре с каждой.
Эндостиль (endostil, рис. 3, 4, end.). На внутренней поверхности жаберного мешка, на его вентральной стороне против спинной пластинки, располагается железистая борозда в виде беловатого валика с бороздкой. Она тянется от ротового валика (около ротового сифона) по вентральной стороне мешка до самого желудка, где оканчивается на дне глотки около отверстия пищевода. Эндостиль играет важную роль в процессе питания асцидии. Мельчайшие организмы, проникающие в жаберный мешок из ротового сифона, при фильтрации воды через жаберные стигмы попадают в желобок эндостиля, склеиваются слизью, выделяемой железистыми клетками, и мерцательными волосками эндостиля передвигаются вверх к окологлоточному кольцу, образованному мерцательными клетками. По окологлоточному кольцу пищевые частицы попадают на спинную сторону глотки, где они спускаются по спинной пластинке до отверстия пищевода. Таким образом, жаберный мешок выполняет не только дыхательную функцию, но и функцию питания.
Строение дыхательного аппарата у асцидий чрезвычайно разнообразно и служит одним из основных систематических признаков при их классификации. Так, у большинства асцидий отр. Stolido-branchiata жаберный мешок со складками, у представителей отрядов Phlebobranchiata и Aplousobranchiata жаберные мешки лишены складок, а у последнего нет и продольных сосудов. Как было указано выше, у асцидий из сем. Styelidae жаберные щели прямые, а из сем. Molgulidae — изогнутые в виде прерывистой или непрерывистой спирали. Количество складок в жаберном мешке также различно у разных видов и родов: у разных видов р. Molgula число складок может быть 10, 12 или 14; у асцидий Halocynthia pirijor-48
mis из сем. Pyuridae количество жаберных складок достигает 16—18.
Важным систематическим признаком разных асцидий является также строение ротовых щупалец и их количество. Они могут быть прямыми, как у Styela, или разветвленными, как у Molgulidae и Pyuridae. Наконец, спинная пластинка может быть цельной (у Styela) или бахромчатой (у Halocynthia). Подсчет количества складок, изучение формы щупалец и спинной пластинки лучше всего проводить на верхней половине поперечного разреза асцидии.
Взаимоотношения различных органов, лежащих в области основания ротового сифона, хорошо можно разобрать на поперечном разрезе. Для этого верхнюю половину асцидии еще раз разрезают поперек ближе к ротовому сифону и рассматривают ее под бинокуляром, погрузив в сосуд с водой.
Как указывалось выше, около ротового валика с щупальцами эндостиль переходит в мерцательное окологлоточное кольцо (рис. 3, 5, м. о. к.), которое переходит на дорзальную сторону между ротовым и клоакальным сифонами. Там находятся важные дорзальные органы — мерцательная ямка и подмозговая железа, связанные с дорзальным нервным ганглием и нервной системой. Рассмотрите эти органы при большом увеличении лупы.
Мерцательная ямка, или теменной орган, соединена с подмозговой железой (рис. 3, 5 gl. sbn.), имеющей у Styela вид округлого продолговатого мешочка, лежащего на спинной стороне между ротовым и клоакальным сифонами. У р. Halocynthia подмозговая железа имеет вид двух соприкасающихся конусов, вершины которых направлены в глотку. Эти органы гомо-логизируются с гипофизом позвоночных животных и с инфундибулярным органом ланцетника.
Нервный узел, или мозговой ганглий (ganglion neurale, рис. 3, 6, g. nr.),— остаток мозговой трубки личинки асцидии, редуцированный после перехода к сидячему образу жизни,— имеет вид белого узелка, который лежит под мозговой железой. От мозгового ганглия белыми нитями в разные стороны идут разветвления нервов, иннервирующих различные органы. Два боковых из них идут по окологлоточному кольцу и дают веточки к ротовым щупальцам.
Сердце (сот, рис. 3, сог.) и кровеносную систему на фиксированном материале разобрать трудно. Однако при тщательной препаровке удается увидеть прозрачную трубку сердца, располагающуюся на вентральной стороне желудка около пищевода. От сердца один сосуд идет по спинной пластинке, давая к жаберному мешку целую серию продольных сосудов, проходящих вдоль жаберных складок; а другой — по вентральной стороне тела асцидии в основании эндостиля. Он сообщается с дорзальным сосудом многими поперечными жаберными сосудами, идущими по перегородкам жаберного мешка. У живой асцидии видно биение сердца последовательными волнами от одного конца к другому. При этом порядок биения через короткие промежутки времени меняется: сна
чала ток крови идет в одном направлении, а затем — обратно. Таким образом, одни и те же сосуды функционируют поочередно: то как артерии, то как вены.
Строение личинки асцидии
Все признаки, характеризующие асцидий как хордовых животных, полностью выражены только у личинки, поэтому желательно ее рассмотреть. Однако добывание свободноплавающих личинок — дело совсем не простое. Если этим не заниматься специально, то их приходится отбирать из гидробиологических проб. При возможно-
Рис. 6. Схема строения личинки асцидии («Жизнь животных», т. II): саи. at. — атриальная полость; ch. — хорда; епс. — мозговой пузырь; end. — эндостиль; med. sp.—спинной мозг; ос. — глазок; ph.— глотка; st. — статоцист; stom. — рот
сти достать личинок асцидий из них нужно сделать постоянные препараты. Фиксированные личинки тотально окрашиваются гема-токсилинэозином или борным кармином и после проведения по спиртам заключаются в канадский бальзам под покровное стекло. На основании нашего опыта можно сказать, что в большинстве случаев такие препараты бывают не очень удачными. Как правило, на них четко видны только расположенная в хвостовом отделе толстая хорда (chorda dorsalis, рис. 6, ch.) и лежащая над ней нервная трубка (medulla spinalis, рис. 6, med. sp.). В туловищном отделе два пигментных пятна, одно из которых соответствует глазку (oculus, рис. 6. ос.), а второе — статоцисту (рис. 6, st.). Поэтому мы здесь не будем давать подробное описание строения личинки, а ограничимся приведением взятого из литературы рисунка личинки, по которому легко разобраться и в остальных деталях, если они будут видны на препарате.
ПОДТИП БЕСЧЕРЕПНЫЕ —ACRANIA
КЛАСС ГОЛОВОХОРДОВЫЕ — CEPHALOCHORD А
Общая характеристика класса
Ланцетник, или амфиокс,— представитель подтипа бесчерепных— Acrania, названных так потому, что они в отличие от всех позвоночных — Vertebrata, или черепных — Craniata (или Сга-niota), не имеют ни головных органов чувств (обоняния, зрения и слуха), ни обособленного головного мозга, ни черепа. Род ланцетников Branchiostoma, или Amphioxus, объединяет около 20 видов, характеризующихся наличием парных гонад. Второй род бесчерепных Asymmetron отличается непарной гонадой с правой стороны тела и распадается на 6 разных видов. Оба рода объединяются в единственное семейство — Branchiostomidae — жаберноротые, которые составляют единственный класс современных бесчерепных головохордовых Cephalochorda, названных так потому, что хорда у них простирается вперед дальше переднего конца нервной трубки.
Все бесчерепные — морские, в подавляющем большинстве донные животные, обитают на песчаном грунте, а ланцетники зарываются в песок так, что только передний конец тела с ротовым отверстием высовывается наружу.
Ланцетники, как и оболочники, характеризуются пассивным питанием и дыханием, т. е. вода, снабжающая органы дыхания кислородом и содержащая пищевые объекты, продвигается в ротоглоточной области лишь движением ресничек мерцательного эпителия, которым выстланы ее стенки.
Второй особенностью бесчерепных, объединяющих их с оболочниками, является то, что наружные жаберные отверстия открываются у них не наружу, а в особую околожаберную, или атриальную, полость.
Бесчерепные встречаются на морском дне в прибрежных песках западного побережья южной Европы, у северных берегов Средиземного моря, в морях на восточных берегах Аравийского полуострова, Азии, Африки и Австралии, а также у восточных берегов Атлантического океана Центральной и Южной Америки. В СССР ланцетники обитают в прибрежной зоне Черного, моря, и их можно получить на Севастопольской биологической станции.
Представитель класса — ланцетник (Branchiostoma lanceolatum Pallas, s. Amphioxus lanceolatus)
Внешний осмотр
Рассмотрите внешний вид фиксированного ланцетника (рис. 7) под бинокулярным микроскопом или штативной лупой, поместив его в чашечку Петри или часовое стекло с дистиллированной или кипяченой водой. В сырую воду класть не рекомендуется, так как осмотру будут мешать пузырьки воздуха.
Рис. 7. Просветленный тотальный препарат ланцетника Branchiostoma lanceolatum:
г. Г. — глазки Гессе; м. о. — мерцательный орган; я. Г. — ямка Гатчека; ап. — анальное отверстие; at. — атриопор; ch.—хорда; err.—околоротовые щупальца; col.—толстая кишка; end.—эндостиль; gast.— желудок; h. — печеночный вырост; med. sp. — спинной мозг; тт, — миомер; ms. — миосепта; mtp.— метаплевральная складка; рпп. ab.—брюшной плавник; рпп. с. — хвостовой плавник; рпп. d. — спинной плавник; rm. Ьг. — жаберная щель; vel. — парус
Ланцетник имеет вытянутое в длину и сжатое с боков плоское тело, хвост в виде лезвия ланцета, откуда и его название. В живом состоянии тело ланцетника прозрачное, так что на просвет видны все органы. В фиксированном состоянии оно менее прозрачное, желтовато-белое. Передний и задний концы тела заострены.
Вдоль спинной стороны тянется кожная складка — спинной плавник (pinna dorsalis, рис. 7, рпп. d.). Он огибает передний конец тела и доходит до верхнего края ротового отверстия. На заднем конце тела плавник дает стреловидное расширение — хвостовой плав ни и (pinna caudalis, рис. 7, рпп. с.) с равными лопастями на спинной и брюшной сторонах. На брюшной стороне непарный плавник продолжается вперед в виде брюшного плавника (pinna abdominalis, рис. 7, рпп. ab.) до отверстия атриальной полости, которое лежит на расстоянии одной трети длины тела от заднего конца. Впереди от атриопора (atrioporus, рис. 7, at.) по брюшной поверхности тела тянутся вплоть до ротового отверстия парные кожные складки, носящие название металле-вральных складок (metapleura, рис. 7, mtp.). Вся плавниковая оторочка расчленена как бы на отдельные членики, выполняющие опорную функцию. Как это будет видно на микроскопических препаратах, это отдельные камеры, заполненные студенистым содержимым.
Через прозрачные кожные покровы хорошо видна мускулатура; ланцетника, разделенная белыми косыми линиями на ряд мышечных сегментов — миомеров (рис, 7, тт.), покрывающих все тело животного от самого переднего конца головы до кончика хвоста. Белые линии образованы межсегментными перегородками — миосептами (рис. 7, ms.), а между ними натянуты волокна, составляющие отдельные миомеры. Каждый миомер образует угол, направленный вперед, причем дорзальный конец его более короткий, а вентральный — более длинный. Метамеры правой и левой сторон несколько сдвинуты относительно друг друга, так что онц чередуются. Нижний конец миомеров не доходит до брюшной поверхности тела приблизительно на одну треть.
На брюшной стороне тела видны три отверстия: 1) на переднем-конце тела помещается удлиненно-овальное отверстие пред ротовой воронки (cavum praeorale), окруженное венчиком около-ротовых щупалец (cirri, рис. 7, err.). По бокам предротовой воронки начинаются парные метаплевральные складки, тянущиеся назад на протяжении двух третей тела. На заднем конце они сливаются и переходят в брюшной плавник; 2) перед брюшным плавником около заднего конца метаплевральных складок расположено округлое отверстие околожаберной атриальной полости, носящее название атриопора (atrioporus, рис. 7, at.); 3) наконец,, в начале стреловидной лопасти хвостового плавника помещается небольшое заднепроходное, или анальное, отверстие (anus,, рис. 7, ап.), сдвинутое на левую сторону тела. У половозрелых форм на брюшной стороне вдоль нижних концов миомеров просвечивают через стенки тела сегментально расположенные гонады в виде округлых тел, прижатых друг к другу на всем протяжении околожаберной атриальной полости.
Для изучения внутреннего строения ланцетника желательно иметь несколько препаратов и проводить его тремя способами. Во-первых, желательно провести анатомическое изучение ланцетника, для чего проводится его вскрытие. Во-вторых, изучаются тотальные препараты ланцетников, окрашенные борным кармином, просветленные и заключенные в канадский бальзам. Такие просветленные препараты изготовляет в Москве фабрика «Медучпособие», но их легко сделать и своими силами. В-третьих, изучаются микроскопические препараты поперечных разрезов ланцетника в жаберной области и позади атриального отверстия.
Вскрытие
Препараты вскрытого ланцетника готовят обычно из довольно крупных экземпляров. Для того чтобы провести вскрытие ланцетника, нужны некоторые навыки в анатомировании. Поэтому рекомендуется вскрытие одного-двух экземпляров ланцетника провести преподавателю. Такие препараты помещают под бинокулярный микроскоп и по ним студент при необходимости может уточ-
нить расположение и взаимосвязь отдельных органов, которые не всегда четко видны на других препаратах.
Для вскрытия ланцетника животное кладут в ванночку с во-
дой спинной стороной на дно,
Рис. 8. Общая топография внутренних органов ланцетника Branchiostoma lanceolatum (из
Беляева):
ап. — анальное отверстие; саи. or.-ph. — ротоглоточная полость; ch. — хорда; coel. — целом; err. — предротовые щупальца; end. — эндостиль; gns. — гонады; h. — печеночный вырост; inis. — кишечная труб-
ка
а брюшной — кверху и, придерживая в таком положении пальцами левой руки, делают тонкими ножницами продольный разрез через брюшную стенку тела, начиная от предротового отверстия, между метаплевральными складками назад до анального отверстия. Затем отворачивают тонким пинцетом края разреза вправо и влево, прикрепляют их тонкими булавками, лучше всего энтомологическими, ко дну ванночки и рассматривают внутреннее строение под лупой.
По средней линии вскрытой атриальной полости (са-vum peribranchiale) тянется пронизанная многочисленными косо расположенными жаберными щелями (rima branchialis) рото-глоточная полость (cavum oro-pharyngeum, рис. 8, cav. or-ph.). Впереди от нее лежит предротовая воронка (cavum praeorale), прикрытая от наблюдателя лежащими по ее краям предротовыми щупальцами (cirri, рис. 8, err.). При вскрытии часто передний отдел рото-глоточной полости повреждается и тогда на ее дорзальной стороне виден просвечивающий белый тяж — хорда (chorda dorsalis, рис. 8, ch.). По средней линии вентральной стороны глотки идет тонкая полоска эндостиля (endostyl, рис. 8, end.). Сзади глотка переходит в тонкую кишечную трубку (intesti-num, рис. 8, ints.). Почти в месте этого перехода от кишечника отходит слепой печеночный вырост (hepar, рис. 8, Л.), лежащий в атриальной полости с правой стороны животного. К наруж
ным стенкам атриальной полости прикреплены, в виде тесно прилегающих друг к другу пакетов, половые железы гонады (gonas,.. рис. 8, gns.). На уровне заднего конца гонад располагается и задний конец атральной полости, которая открывается здесь наружу атриопором. Лежащая позади нее полость, в 'которой расположена большая часть кишечника, является вторичной полостью тела— целомом (coelom, рис. 8, coel.), которая тянется вплоть до-уровня анального отверстия (anus, рис. 8, ап.).
Вследствие малого размера ланцетника строение его внутренних органов гораздо лучше можно разобрать на готовом просветленном препарате под лупой.
Изучение просветленных тотальных препаратов
Приготовление препаратов. Для приготовления просветленного' препарата из ланцетника фиксированный материал обрабатывают следующим образом. Если фиксатором служил формалин, то перед обработкой препарат промывают в проточной воде в течение 1—2 суток. Данный материал окрашивают тотально в течение одних суток в растворе борного кармина. После окраски его раскрашивают в 70%-ном спирте с соляной кислотой (одна капля крепкой соляной кислоты на 25 куб. см спирта) в течение 3—5 суток до бледно-розового цвета; подкисленный спирт меняют несколько-раз. Далее материал уплотняют крепким спиртом 90°, 96° и 100° в продолжение 24 ч в каждом из них и просветляют в ксилоле также в течение 24 ч. После этого на предметном стекле делают рамочку из кусочков предметного стекла, в которую ланцетника переносят из ксилола, заливают канадским бальзамом и покрывают большим покровным стеклом или тонким предметным. Для приготовления просветленных препаратов лучше использовать мелких, молодых, неполовозрелых ланцетников, не имеющих зрелых гонад. У взрослых половозрелых ланцетников плотные малопрозрачные гонады настолько застилают все внутреннее строение;. что разобрать их строение трудно.
Изучение просветленного тотального препарата. Рассматривают просветленный препарат под штативной лупой или бинокулярным микроскопом при проходящем свете и разбирают на нем внешние признаки так, как они были описаны при внешнем осмотре в начале занятия: плавниковую складку, ротовое отверстие, атриопор, заднепроходное отверстие, сегмантацию мускулатуры наряд миотомов. На хорошо приготовленном препарате тело ланцетника прозрачное, желтоватого цвета, в красный цвет окрашены только 'пищеварительная трубка и гонады. В дополнение к рассмотрению внешних признаков рассматривают общее расположение внутренних органов.
Скелетная система. Спинная струна — хорда (chorda dorsalis, рис. 9, ch.) —прозрачный продольный тяж на дорзальной стороне тела — выполняет функцию осевого скелета. Хорда у лан
цетника начинается на самом переднем конце тела и кончается на хвостовом отделе, разделяя стреловидный хвостовой плавник на равные лопасти: дорзальную и вентральную. Такой равнолопастный плавник носит название протоцеркального (см. рис. 7, рпп. с.).
Кроме хорды, выполняющей роль осевого скелета, у ланцетника на просветленном тотальном препарате сквозь стенки атриаль-
Рис. 9. Головной конец ланцетника Branchiostoma lanceolatum. Просветленный препарат:
в. щ. —• велярные щупальца; г. Г. — глазки Гессе; м. о. — мерцательный орган; п. к.— плавниковые камеры; п. п. — пигментное пятно; я. Г.— ямка Гатчека; я. К.—ямка Келликера; arc. Ъг. — главная жабериая дужка; arc. Ьг' — вторичная жаберная дужка (слева); cav. prr. — предротовая воронка; ch. — хорда; err. — предротовые щупальца; med. sp. — спинной мозг; тт. — мномер; ms. — мносепта; пгс. — иевроцель (его расширенная часть, соответствующая желудочкам головного мозга); mr. Ьг. — жаберная щель (справа); vel.— парус
яой полости видна скелетная решетка жаберного висцерального скелета в виде наклоненных, темноокрашенных эластиновых стержней, расположенных между жаберными щелями.
В плавниковой складке выше были отмечены сегментальные плавниковые камеры со студенистым веществом, которые играют роль плавниковых лучей, поддерживающих плавники (рис. 9, п. к.)
Центральная нервная система. Спинной мозг (medulla spinalis, рис. 9, med. sp.) располагается в виде тонкой трубки непосредственно над хордой. На просветленном препарате под лупой ее положение легко определить по ряду черных точек пигмента, так называемых глазков Гессе (рис. 9, г. Г.). На переднем конце она окрашена в красный цвет и дает небольшое расширение внутренней полости — невроцеля. Передний конец нерв
ной трубки далеко не доходит до переднего края хорды, что является важным систематическим признаком всех головохордовых. Дальнейшее изучение нервной системы следует отложить до рассмотрения препарата под микроскопом.
Пищеварительная и дыхательная системы. Для общего обзора рекомендуется рассмотреть пищеварительную и дыхательную системы сначала при слабом увеличении штативной лупы, а затем отдельными частями под микроскопом.
Пищеварительная система начинается предротовой воронкой, окруженной предротовыми щупальцами (cirri, рис. 9, err.) в виде нитевидных выростов, функционирующих как органы чувств, так как они покрыты чувствующими почками. На задней стенке предротовой полости располагается ротовое отверстие, впереди которого по стенке предротовой воронки тянутся полоски утолщенных клеток, снабженных ресничками. На просветленном препарате они имеют вид пальцевидных выростов розоватого цвета. Это так называемый мерцательный орган (рис. 9, м. о.). При рассмотрении под микроскопом на спинной стенке можно видеть особый орган, носящий название ямки Гатчека (рис. 9, я. Г.). На фоне прозрачных розово-желтых стенок выделяется полый вырост, окрашенный в более красный цвет с просветом посередине. Ямка Гатчека открывается в предротовую полость. Рядом с ямкой можно видеть второй характерный для ланцетника орган, имеющий вид тонкостенного выроста, головной нефридий — нефридий Гатчека.
Парус (velum, рис. 9, vel.) располагается на задней части предротовой чаши и окаймляет ротовое отверстие со стороны ротоглоточной полости. На нем сидят пальцевидные выросты, направленные от ротового отверстия назад (рис. 9, в. щ.).
Жаберная область на просветленном ланцетнике хорошо просвечивает через стенки атральной полости. Она занимает две трети переднего отдела пищеварительной трубки и представляет ее расширенный отдел, пронизанный справа и слева большим количеством (около 100—125) косо расположенных просветов жаберных щелей (rima branchialis, рис. 9, rm. br.), разделенных темными жаберными дужками.
Жаберные дуги (arcus branchialis) при внимательном рассмотрении под большим увеличением микроскопа имеют неодинаковое строение. Одни дужки, получившие название главных, отличаются тем, что на нижних вентральных концах их скелетные элементы вильчато раздвоены (рис. 9, arc. br.). Вторичные дужки не имеют таких разветвлений (рис. 9, arc. br'.). Во всей жаберной решетке эти дужки чередуются.
На микроскопическом препарате поперечного разреза ниже будут описаны другие отличия главных и вторичных дужек.
На брюшной стенке жаберной области проходит в виде темной полосы поджаберная борозда — эндостиль (endostyle, рис. 7, end.), имеющая большое значение в улавливании и передвижении пищевых частиц. Более подробно эндостиль будет рассмотрен при
•изучении микроскопических препаратов поперечных разрезов ланцетника. На заднем конце глотки жаберные щели несколько укорачиваются и освобождают верхнюю часть глотки, от которой отходит кишечная трубка.
Акт дыхания. Акт дыхания у ланцетника сходен с актом дыхания асцидий, так как наружные жаберные щели открываются в • околожаберную полость, а не наружу, как у водных позвоночных.
Атриальная полость имеет брюшную мускулатуру, сокращение которой и выгоняет воду из атриальной полости наружу через атриопор. Движение воды в ротоглоточную полость обеспечивается выстилающим ее реснитчатым эпителием. Развитие атриальной полости обусловливается приспособлением ланцетника к зарыванию в песок, у личинки ланцетника первично жаберные щели открываются наружу.
Печеночный вырост (hepar, рис. 7, h.) отходит от пищеварительной трубки позади последней жаберной щели и в виде толстой слепой кишки направляется вперед по атриальной полости с правой стороны тела животного.
Пищеварительная трубка позади печеночного выроста дает расширение, называемое по аналогии с позвоночными желудочным расширением (рис. 7, gast.), а далее назад на кишечной трубке выделяется темная полоса с менее прозрачными стенками, так как они покрыты густыми ресничками. Это темное кольцо кишечной трубки получает название толстой кишки (ileo-co--lon, рис. 7, col.). После этого кишечника трубка идет назад до заднепроходного отверстия, помещающегося у Br. lanceolatum на 50-м сегменте, у китайских видов Br. belcheri — на 54—55-м, а у южноамериканского Br. elongatum — на 68-м сегменте тела.
Акт питания. Пищевые объекты движением мерцательных ресничек эпителия, выстилающего ротоглоточную полость, попадают •в нее вместе с водой. Обволакиваясь слизью, они оседают на эндо-•стиле и движением его ресничек гонятся вперед. Оттуда по около-ротовому кольцевому каналу они поднимаются кверху и по надглоточной борозде мерцательными же ресничками продвигаются .назад в кишечную трубку. Расположение жаберных дужек, атриальной полости и наджаберной борозды более подробно рассматривают дальше на препарате поперечного разреза под микроскопом.
Нервная система и органы чувств. Более детально изучают передний конец просветленного ланцетника под микроскопом (рис. 9). Нервная трубка, как указано выше, видна в туловище лишь как скопление точек черного пигмента над хордой, которые являются светочувствительными органами, называемыми глазками Гессе (рис. 9, г. Г.). Головной конец их не имеет, но на переднем конце видно черное пигментное пятно (рис. 9, п. п.)—рецептор, функция которого не выяснена. Позади него виден просвет головного расширения внутримозгового канала — невроцеля, соответствующего желудочкам головного мозга. На дне этого головного расширения мозговой трубки помещается так на-58
зываемый инфундибулярный орган, соответствующий нижнемозговой железе — гипофизу позвоночных животных. Он состоит из высоких цилиндрических чувствительных клеток с мерцательными волосками. На левой стороне головного конца ланцетника над передним концом нервной трубки под микроскопом можно видеть углубление, выстланное эпителиальными клетками,— это обонятельная ямка Кёлликера (рис. 9, я. К-), которая соответствует переднему концу нервной борозды — нейропору.
Для более четкого представления о форме переднего отдела нервной системы и органов чувств рекомендуется, в дополнение к изучаемым, изготовить серию гистологических поперечных срезов переднего конца ланцетника, окрасив препараты по Малло-рии. Просматривая всю серию, легко отобрать препараты, на которых строение вышеописанных органов видно наиболее полно.
Кровеносная система, выделительная система, строение жабр, гонады и целом на просветленном тотальном препарате ланцетника не видны. Их изучение требует рассмотрения полных серий разрезов на микроскопических препаратах. Поэтому переходим к изучению отдельных поперечных разрезов, чтобы познакомиться с отдельными частями строения этих органов.'
Изучение поперечных разрезов ланцетника
Поперечный разрез ланцетника в жаберной области
Наружные покровы ланцетника образованы очень тонким слоем кожи, который надо рассмотреть под большим увеличением микроскопа на отдельном маленьком участке на спине или боках тела.
Кожа (рис. 10, cut.) состоит из тонкого эпидермиса эктодермального происхождения и бесклеточного волокнистого слоя мезодермального происхождения.
Эпидермис у ланцетника однослойный, цилиндрический, среди его клеток разбросаны отдельные чувствительные и железистые клетки. Снаружи эпидермис покрыт тонкой гомогенной пленкой— кутикулой. Это единственный пример присутствия кутикулы у хордовых. Под эпидермисом расположен соединительнотканный слой — кориум. Он имеет волокнистую структуру, но состоит из бесклеточной межуточной ткани. Между, эпидермисом и кориумом имеется базальная мембрана.
По медиальной линии микроскопического препарата поперечного разреза помещаются осевые органы, дорзальный непарный плавник, спинномозговой канал с разрезом центральной нервной системы, хорда и разрез ротоглоточной полости.
Хорда — спинная струна (chorda dorsalis, рис. 10, ch.) занимает центральное положение разреза. На разрезе — это округлое тело, заполненное плотным студенистым веществом, обусловливающим внутренний тургор хорды как осевого скелета. Сна-
ружи она одета плотной эластической оболочкой (membrana elastica, см. рис. 13).
Спинной мозг, а также хорда окружены бесклеточной студенистой тканью, образующей спинномозговой канал. Эта ткань лроникает также во все промежутки между миомерами и играет
Рис. 10. Схема поперечного разреза ланцетника Branchiostoma lanceolatum в жаберной области (из Беляева):
б. м. — брюшная мускулатура; п. к, — плавниковые камеры; arc. br. — жаберная дужка; ао. и. — брюшная аорта; cav. or.-ph. — ротоглоточная полость; cav. prb. — атриальная полость; ch.— хорда; сое/.— целом; cut.—кожа; end. — эндостиль; h. — печеночный вырост; gns. — гонада; med. sp.— спинной мозг; тт. —миомер; ms. — мносепта; пгс. — невроцель; rd. а. — корин спинной аорты; s. epb. — наджаберная борозда
роль межуточной ткани. Она образует соединительнотканную опорную систему ланцетника.
Центральная нервная система (рис. 10, med. sp.) в виде мозговой трубки с неполностью сросшимися краями нервной бороздки с тонким узким каналом — невроцелем в середине — помещается в. спинномозговом канале непосредственно над хордой. На микроскопическом поперечном разрезе видна не-60
однородность состава клеток нервной системы. Клетки, окружающие невроцель, сохраняют эпителиальную структуру. На некоторых разрезах среди нервных клеток видны крупные пигментные клетки черного цвета, представляющие так называемые глазки Гессе — специфические органы восприятия световых раздражений. Каждый глазок Гессе состоит из сочетания двух клеток: крупной чувствующей клетки и примыкающей к ней второй пигментной клетки. Среди нервных клеток встречаются особые гигантские нервные клетки, исполняющие роль коллекторов.
Плавниковые камеры или плавниковые лучи (рис. 10, п. к.) — пузыревидные образования над спинномозговым каналом, лежащие против каждого миомера и заполненные студенистым веществом, как это было видно при рассмотрении тотального препарата. Они выполняют функцию поддержания непарных плавников.
Миомеры (рис. 10, пгт.)—мышечные сегменты — на поперечном разрезе образуют боковые части и составляют главную массу туловищной мускулатуры. Вследствие зигзагообразной формы миомеров, как это видно на целом ланцетнике, на каждый разрез попадают пять-шесть миомеров. Мускулатура миомеров располагается только на дорзальной половине поперечного разреза. Вентральные отделы стенок околожаберной полости имеют тонкий слой брюшной мускулатуры (рис. 10,6. м.), не расчлененной на отдельные миомеры, и соответствуют соматическому листку боковой пластинки.
Жаберный отдел кишечной трубки, или ротоглоточная полость (cavumoro-pharyngeum, рис. 10, cav. or.-ph.), занимает центральное положение в полости поперечного разреза. На поперечном разрезе ротоглоточная полость состоит из отдельных отрезков многочисленных жаберных перегородок (рис. 10, arc. br.), разрезанных поперечно в косом направлении. Это происходит потому, что, как видно на просветленном препарате ланцетника, жаберные щели и дуги расположены близко друг от друга в сильно наклонном положении.
В каждой перегородке при большом увеличении микроскопа видна скелетная дужка треугольной формы. На некоторых препаратах удается отметить, что жаберные дужки не одинаковы по своему составу. В главных дужках кнаружи от скелета располагается полость целомического канала, а на второстепенных — целомических каналов нет. В каждой дужке с наружной и внутренней •стороны располагаются приносящий и выносящий кровеносные сосуды.
Эндостиль (endostyle, рис. 10, end..). На внутренней стороне вентральной части разреза ротоглоточной полости помещается подглоточная бороздка, носящая название эндостиля. Эта бороздка образована высокими эпителиальными клетками двоякого рода: мерцательными и железистыми (рис. 11). Середина бороздки заполнена слизистым веществом с пищевыми частичками.
Наджаберная бороздка (sulcus epibranchi alis, рис. 10, •s. epb.) помещается под хордой и занимает дорзальное положение
Рис. 11. Эндостиль ланцетника (по Кю-кеиталю):
ж. к. — железистые клетки; м. к. — мерцательные клетки; ао. v. — брюшная аорта; coel. — целом
относительно глотки. Надглоточная борозда также имеет эпителиальное строение из высоких мерцательных клеток, передвигающих своими ресничками пищу назад в кишечную трубку.
Околожаберная, или атриальная, полость (cavum peribranchiale, рис. 10, cav. prb.) — внутренняя полость, где располагаются ротоглоточный отдел пищеварительной трубки и другие внутренние органы. Околожаберная полость окружена стенками тела, подстилается с вентральной стороны вышеописанной брюшной мускулатурой.
Печеночный вырост (hepar, рис. 10, h.) имеется не на всех поперечных разрезах. В передних разрезах ротоглоточной полости его нет. Он располагается на правой стороне тела рядом с разрезом жаберной области кишечника. Печеночный вырост имеет толстые эпителиальные стенки железистого характера и густо оплетен сетью кровеносных сосудов.
Гонады (gonas, рис. 10' gns.). Половые железы занимают боковые стороны тела животного. Справа и слева они примыкают к боковым стенкам атриальной полости. У самки в половой железе хорошо видны со
ставляющие яичник яйцевые клетки; у самца гонада имеет компактное строение с нитевидной структурой половых клеток. Никаких выводных протоков ни у самки, ни у самца нет. Половые клетки, выделяются в атриальную полость прорывом стенки гонад.
Вторичная полость тела — целом (coelom). В жаберной области единой брюшной полости у ланцетника нет, так как она вытеснена околожаберной полостью, в которой мы и рассматривали все органы. Однако на поперечном разрезе можно видеть сохранившиеся отдельные участки вторичной полости тела — целома: 1) под хордой по бокам надглоточной борозды справа и слева располагаются парные дорзальные участки целома — субхордальные целомы (рис. 10, coel.), куда открываются сегментальные выделительные канальцы — нефридии. Рассмотреть на отдельных поперечных разрезах строение нефридий не удается. Для этого надо изучить всю последовательную серию поперечных разрезов; 2) как было упомянуто выше, в основных жаберных дугах около скелетных дужек имеются целомические каналы жаберных целомов; 3) печеночный вырост окружен печеночным целомом. Стенки целома в виде тонкой пленки брюшины окружают 62
эпителиальные стенки печеночного выроста; 4) целом половых желез виден в виде узкой щели, ограничивающей гонады только со стороны атриальной полости тонкой пленкой; 5) су бэндостильный целом — неболь-
шой участок в виде непарного остатка — располагается под эндостилем; 6) остатки целома сохраняются также по бокам тела в метаплевральных складках.
Кровеносная система. На поперечном разрезе можно видеть просветы основных кровеносных сосудов ланцетника: 1) под хордой справа и слева над субхордальными целомами видны разрезы парных корней спинной аорты (radix aortae рис. 10, 12, rd. а.)\ 2) под эндостилем в субэндостильном целоме виден сосуд, приносящий венозную кровь к жаберным дужкам,— брюшная аорта (aorta ventralis, рис. 10, 12, ао. ».).
Кроме описанных выше участков, другие отделы кровеносной системы на препаратах не видны, поэтому при ее изучении приходится ограничиться разбором схемы, приведенной на рис. 12.
Артериальная система представлена проходящей по вентральной стенке глотки брюшной аортой, которая отдает приносящие жаберные артерии к каждой жаберной перегородке. Сердце у ланцетника отсутствует, а движение крови осуществляется за счет пульсации брюшной аорты и расширенных участков приносящих жаберных сосудов, лежащих в их основаниях. Выносящие жаберные артерии несут уже артериальную кровь от жаберных перегородок в парные продольные сосуды— корни спинной аорты (radix aortae, rd. а.), которые сзади объединяются в непарный, идущий под хордой сосуд — спинную аорту (aorta dorsalis, ао. d.).
Рис. 12. Схема кровеносной системы ланцетника (по Францу с изменениями):
a. br. ad. — приносящие жаберные артерии; a. br. г. — выносящие жаберные артерии; а. с. с. — сонные артерии; ао. d. — спинная аорта; ао. v. — брюшная аорта; d. с. — кювьеровы протоки; rd. а. — корни спинной аорты; V. с. — хвостовая вена; о. crd. а.— передние кардинальные вены; v. crd. р. — задние кардинальные вены; v. ints. — под* кишечная вена; v. р. h. — воротная вена печени; а. с. — хвостовая артерия
Отходящие посегментно от спинной аорты сосуды распадаются на капилляры, которые затем объединяются в мелкие венозные сосу-
ды, впадающие в хвостовую или подкишечную вену. Хвостовая вена (vena caudalis, v. с.) начинается в хвостовом отделе ланцет-
ника и продолжается вперед в виде подкишечной вены (vena
intestinalis, v. inis.), лежащей, как указывалось, под кишечником.
Рис. 13. Схема поперечного разреза ланцетника Branchiostoma lanceolatum позади атриопора: г. Г. — глазки Гессе; п. к. — плавниковая камера; ch. — хорда; coel. — целом; cut. — кожа; Inst. — кишечная трубка; mb* е. — эластическая оболочка хорды; med. sp. — спинной мозг; mes. d. — дорзальный мезентерий; mes. v. — вентральный мезентерий;
тт. — миомер; ms. — миосёпта; пгс. — невроцель
Продолжаясь вперед, она входит в печеночный вырост уже под названием воротной вены печени (venae portae hepa-tis, v. p. h.), которая распадается в ткани печеночного выроста на сеть капилляров, образуя воротную систему. Из выносящих капилляров образуется печеночная вена (vena hepatica), впадающая сзади в брюшную аорту.
Кроме этих основных сосудов, по бокам спинной аорты посегментно располагаются венозные лакуны, соединяющиеся в парные кардинальные вены (vena cardi-nalis anterior et posterior). Передние кардинальные вены (n. crd. а.) несут кровь от переднего конца тела, на уровне отхождения печеночного выроста они соединяются с задними кардинальными венами (у. ord. р.). Передняя
и задняя кардинальные вены каждой стороны образуют проток Кювье (ductus Cuvieri, d. С.). Правый и левый протоки Кювье
впадают в брюшную аорту.
Поперечный разрез ланцетника позади атриопора (рис. 13)
На поперечном разрезе тела ланцетника позади атриопора расположение органов наиболее типично для хордовых животных. Центр разреза занимает хорда, одетая эластичной оболочкой (mb. е.). Над ней располагается нервная трубка, причем в ней хорошо виден щелевидный невроцель. Боковые части разреза заняты мускулатурой, имеющей здесь вид отдельных пучков, разделенных
миосептами из бесклеточной межуточной ткани. Поскольку каждый миомер имеет, как это мы видели на тотальном препарате, изогнутую форму, на поперечный разрез попадает несколько их участков, которые располагаются друг над другом. Картина расположения миомеров на правой и левой сторонах не совпадает, так как правые и левые миомеры сдвинуты относительно друг друга; Под хордой помещается разрез кишечника, наполненный пищей. По бокам от кишечной трубки располагается брюшная полость — целом. Правая и левая его половины отделены друг от друга лежащим сверху от кишечника дорзальным мезентерием (tries. d.) и снизу от него вентральным мезентерием (тез. и.). На этих мезентериях кишечник подвешен к дорзальной и вентральной стенкам полости тела.
Над спинномозговым каналом в спинном плавнике видна плавниковая камера, заполненная студенистым веществом. Ниже вентрального мезентерия в брюшном плавнике лежит точно такая же камера.
Литература по низшим хордовым
Беклемишев В. Н. Основы сравнительной анатомии беспозвоночных, т. 1, 2, АН СССР. М., «Наука», 1964.
Жизнь животных в 6 томах (под ред. Л. А. Зенкевича), т. 2. Беспозвоночные, тип Полухордовые, тип Оболочники. «Просвещение», 1968.
Редикорцев В. В. Оболочники. В кн. «Фауна России и сопредельных стран», т. 1, вып. 1, 1916.
Северцов А. Н. Современные задачи эволюционной теории. М., «Наука», 1914. То же, собр. соч., т. 3, М.— Л., Изд-во АН СССР, 1945.
Северцов А. Н. Исследования об эволюции низших позвоночных, ч. II. Журнал «Архив анатомии, гистологии и эмбриологии», т. 1, 1917. То же. Организация ближайших предков современных позвоночных, Собр. соч., т. 4. Изд-во АН СССР, 1948.
Delage I., Herouard Е. Les Procordes Traite de zoologie concrete les Procordes. 8, Paris, 1898.
Grasse P. Traite de zoologie. T. XI. Echinoderms, Stomocordes, Procordes. Paris, 1948.
3—3467
ПОДТИП ПОЗВОНОЧНЫЕ —VERTEBRATA
РАЗДЕЛ БЕСЧЕЛЮСТНЫЕ —AGNATHA
КЛАСС КРУГЛОРОТЫЕ — CYCLOSTOTATA
Общая характеристика класса
Класс круглоротые — Cyclostomata — единственный современный класс некогда большой группы бесчелюстных позвоночных — Agnatha.
Согласно работам А. Н. Северцова qt первичных бесчерепных, наряду с настоящими бесчерепными и оболочниками берет начало и группа первичночерепных животных (Protocraniata, см. рис.1), у которых наряду, с прогрессивным развитием висцерального скелета, появляются элементы черепной коробки. Они отчасти защищают развившиеся в связи с активным образом жизни органы чувств (зрения, обоняния, статоакустики), а также головной мозг, развитие определенных отделов которого связано с развитием того или иного анализатора.
Первичные черепные делятся затем на две группы: у одной из них жаберные лепестки в связи с усложнением органов дыхания, развиваются ко внутри от висцерального скелета, у другой —снаружи. Отсюда каждая из этих групп соответственно называется внутренне- и наружножаберными (Efito- et Ectobranchiata, см. рис. 1). Первая группа фактически представляет собой группу бесчелюстных позвоночных, отличающуюся пассивным способом питания. Вторая группа перешла к активному способу питания, в связи с чем у них вырабатывается челюстной аппарат. Эти первичночелюстные животные дают начало всем остальным группам позвоночных (см. рис. 1).
Представители группы, Agnatha отличаются от вышестоящих-челюстных Gnathostomata рядом примитивных признаков, из которых главнейшими являются отсутствие челюстей и парных плавников, наличие жаберных мешков с жаберными лепестками эктодермального происхождения, непарный теменной орган зрения позади ноздри (кроме двух парных по бокам головы). Вместе с тем у бесчелюстных мы находим и ряд черт, прогрессивных по сравнению с рассмотренным нами представителем бесчерепных Acrania — 66
ланцетником. Сравнительно с последним это формы, хорошо плавающие, активно разыскивающие, пищу, в связи с чем у них прогрессивно развиты органы чувств: парные органы зрения, непарная ноздря, внутреннее ухо (правда, только с двумя полукружными каналами), сейсмо-сензорные органы. Передний отдел спинномозговой трубки расширяется в головной мозг с типичными для позвоночных отделами. Появляется настоящий хрящевой скелет, охватывающий головной мозг снизу и с боков, отчасти спинной мозг, а также поддерживающий жаберные, мешки и ротовую воронку.
Расцвет группы бесчелюстных приходится на силур и девон (см. рис. 1). В большинстве своем Agnatha — это придонные формы, покрытые более или менее развитым костным панцирем. Они распадаются на три класса. Первый более древний, известный с ордовика, содержит два подкласса Thelodonti et Heterostraci. Одним из отличительных признаков этого класса является наличие парных ноздрей, отсюда и его название Diplorhini.
Другой класс Monorhini — одноноздревые, более молодой, известен с верхнего силура (см. рис. 1). Из двух его подклассов Osteostraci et Anapsida в последний входили свободноплавающие в толще, воды формы. Отсюда предполагается, что современный класс круглоротых произошел или от этого подкласса, или от общих с ним предков.
Представители современного класса круглоротых Cyclostomata относятся к двум подклассам: миноги Petromyzones и миксины Mixini. Подкласс миноги Petromyzones содержит только один отряд Petromyzoniformes с единственным семейством Pet'romy-zonidae, которое объединяет семь родов. Четыре из них (Petromy-son, Caspiomyzon, Entosphenus, Lampetra) представлены в СССР девятью видами, три из которых являются объектами промысла, отличаются довольно крупными размерами и поэтому могут быть рекомендованы для практических занятий.
Распространение и места добычи этих видов следующие:
1. Каспийская, или волжская, минога Caspiomyzon wagneri Kessler распространена по всему бассейну Каспийского моря. Промысел ведется в низовьях Волги и на Куре.
2. Тихоокеанская, или ледовитоморская, минога Lampetra japonica Mart обитает в морях Ледовитого и Тихого океанов. Промышляется-на реках Онеге, Северной Двине, Мезени, а также по западному берегу Белого моря.
3. Балтийская, или невская, минога Lampetra fluviatilis L. встречается в бассейне Балтийского моря и служит объектом про-, мысла на реке Неве, а также в Эстонии, Латвии, Литве.
Добывают миног во время их'хода в реки на икрометание, т. е. в осенние и весенние месяцы.
Кроме указанных трех видов миног можно использовать и другие, встречающиеся у нас. Все виды миног довольно мало отличаются по своему строению, поэтому дальнейшее описание делается по невской миноге. Некоторые отличия, имеющиеся у других видов, будут оговорены в тексте.
Представитель класса круглоротых — невская минога — Lampetra fluviatilis L.
Как представитель класса круглоротых Cyclostomata невская минога — Lampetra fluviatilis L. характеризуется присутствием округлой присасывательной воронки, голой кожей, висцеральным скелетом в виде решетки, с не разделенными на отдельные членики дугами.
Образ жизни миног еще плохо изучен. Постоянным местом их обитания являются моря, где они, вероятно, ведут малоподвижный образ жизни, возможно, зарываясь в ил, чему способствует общая угревидная форма тела. Питаются соками и кровью рыб, к которым присасываются. Нападению в первую очередь подвергаются малоподвижные или снулые рыбы. Возможно, что некоторую роль в питании играет и растительная пища.
Одна группа миног входит в реки для нереста осенью, другая — идет в реки в мае, нерест происходит в начале июля. С момента входа в реки до нереста миноги не питаются. В связи с этим наблюдается дегенерация кишечника и желчного пузыря, начинающаяся еще в море. После нереста производители гибнут. Из икринки вылупляется личинка, которая живет 4 года в реке, зарывшись в грунт (отсюда — название пескоройка), после чего превращается в маленькую взрослую миногу и скатывается в море.
Миног лучше всего фиксировать раствором формалина 1 :9 (одна часть формалина и девять частей воды) в течение 2—3 суток, затем промыть в проточной воде (сутки) и перенести для хранения в 70° спирт. Можно хранить и в формалине, но более' слабой концентрации— 1 : 19. Непосредственно перед занятиями материал промывается проточной водой в течение 1—2 суток.
Для отработки заданий по изучению миноги желательно иметь на каждого студента по две-три целые миноги и по одному сагиттальному разрезу. Для сагиттальных разрезов нужно отобрать ровные экземпляры, без изгибов в горизонтальной плоскости. Особенно нежелательны изгибы в области головы. Разрез делается острой опасной бритвой строго по средней линии тела. Лучше всего разрезать со спинной стороны тела, начиная с переднего конца головы. Бритву нужно стараться вести под острым углом к телу миноги. Разрез можно делать, держа миногу в левой руке спиной кверху или положив ее на бок на стол и прижимая к нему левой рукой. Последний способ наиболее приемлем для искривленного объекта. Кроме того, на -всю группу готовится серия поперечных разрезов из одного-двух экземпляров (в зависимости от количества студентов).
Поперечные разрезы делаются также опасной бритвой. Для этого, положив миногу на стол или лучше на дощечку, нарезают ее, начиная с головы, на ломтики толщиной 1—1,5 см. В жаберной области разрезы нужно проводить как по жаберным отверстиям, так и в промежутках между ними. Срезы лучше разделить на четыре группы, поместив каждую в отдельный сосуд с 70° спиртом: 68
1) срезы жаберной области; 2) срезы передней половины туловища; 3) срезы задней половины туловища; 4) срезы хвостовой области.
Вскрытие
Внешние признаки
Тело миноги (см. рис. 15) продолговатое, червеобразное, округлое в передней части и постепенно сжимающееся с боков к заднему концу. Головной, туловищный и хвостовой отделы незаметно переходят друг в друга. Граница между головным и туловищным отделами проходит по последнему жаберному отверстию; между туловищным и хвостовым — сзади мочеполового сосочка.
Кожа (cutis) миноги совершенно гладкая, лишенная каких бы то ни было чешуй. При внешнем осмотре можно отметить только наличие богатого железами эпидермиса, который на фиксированных экземплярах имеет вид мутной пленки, покрывающей тело миноги, и легко соскабливается скальпелем.
Более детально строение кожи можно изучить на поперечных срезах кожи, но для этого необходимо иметь микроскопические препараты.
При увеличении микроскопа 200-х можно видеть, что кожа состоит из двух слоев: наружного — эпидермиса и внутреннего — соединительнотканного, который является собственно кожей. Над эпидермисом имеется еще тонкий бесклеточный поверхностный слой — кутикула (cuticula), который является продуктом выделения верхнего слоя клеток эпидермиса.
Эпидермис (epidermis) в отличие от такового ланцетника многослойный. Непосредственно под Кутикулой лежит один ряд уплощенных клеток с редуцирующимися ядрами. Самый нижний — мальпигиев слой — состоит из одного ряда вытянутых цилиндрических клеток, способных к Делению в течение всей жизни животного. Он дает начало вышележащим слоям клеток. Между этими двумя слоями располагается железистый слой, составляющий основную массу эпидермиса. В нем можно различить клетки трех типов.
1. Главные клетки, наиболее многочисленные, которые в верхнем слое сменяются плоскими клетками. Эти клетки выделяют слизь, которая накапливается в надъядерной зоне, поэтому ядра смещаются в базальном направлении.
2. Зернистые железы — крупные клетки округлой или несколько вытянутой формы с зернистым содержимым. Секрет белкового характера выделяется в межклеточные пространства.
3. Колбовидные железы — хорошо заметные, крупные, вытянутые клетки, тянущиеся почти через весь эпидермальный слой от нижней границы эпидермиса до поверхности. В них хорошо различимы одно или два ядра и сгустки плазмы, занимающие только центральную часть. Основная масса клеток заполнена белковым
Рис. 14. Предротовая вороика невской миноги Lampetra fluviatilis:
в. б. з. — внутренние боковые губные зубы; в. г. з. — верхние губные зубы; в. з. п. — верхнечелюстная зубная пластинка; в. с. — внутренняя кожная складка; з. я. п. — задняя язычная пластинка; к. з. — краевые зубы; ’к. с. — наружная кожная складка; м. з. — отдельные мелкие зубчики; и. з. п. —.нижняя зубная пластинка; п. я. п. — передняя язычная пластинка; с. с. — сосудистые сосочки
секретом, который выделяется в направлении базальной мембраны. Тонкая основная перепонка (membrana basalis) — продукт совместного выделения эпидермиса и собственно кожи — лежит на границе между этими двумя слоями.
Собственно кожа, кориум или дерма (corium, derma) состоит из плотных соединительнотканных волокон, расположенных в двух взаимноперпендикулярных направлениях.
Пигмент у миноги в отличие от других позвоночных расположен между кориумом и подкожной клетчаткой. Это обнаруживается и макроскопически, если снять небольшой кусок кожи с препарата. Пигмент при этом остается на теле миноги. Рассмотрев кожу на просвет, убеждаемся, что она совершенно бесцветная и прозрачная.-Предротовая воронка ..{cavum praeorale, см. рис. 15, cav. prr.) располагается на переднем конце головы. Ее наружное отвер-' стие косо наклонено вниз. У фиксированных экземпляров она может иметь форму продольной щели, у живых присосавшихся миног — округлая. Для того чтобы расправить воронку, нужно плотно прижать ее к предметному стеклу и рассматривать через это стекло. Сзади и с боков отверстие
воронки окружено кожной складкой (рис. 14, к. с.), которая может расправляться и тогда воронка вытягивается 'вперед. Края воронки окружены бахромой из тонких сосочков. Внутрь от сосочков, непосредственно у их оснований, лежит тонкая кожная складка (рис. 14, в. с.), окружающая всю ротовую воронку.
Внутренняя поверхность воронки имеет вид конуса, на дне которого расположено ротовое отверстие. Она усажена желтыми роговыми зубами, которые вместе с таким же вооружением языка помогают миноге разрушать ткани жертвы во время питания. Зубы миноги не гомологичны зубам остальных позвоночных. Каждый зуб миноги представляет собой соединительнотканный сосочек,, одетый ороговевшим эпидермисом, в чем можно легко убедиться,, сняв роговую часть зуба иголкой.
У невской миноги непосредственно над ротовым отверстием лежит широкая верхнечелюстная зубная пластинка, с 70
двумя острыми вершинами по краям (рис. 14, в. з. п.). Лежащие над ней в один ряд пять одновершинных зубов носят название верхних губных (рис. 14, в. г. з.). По бокам от ротового отверстия с каждой стороны по три внутренних боковых губных зуба (рис. 14, в. б. з.). Верхние и нижние зубы в каждом ряду двураздельны, средние — с тремя вершинами.
-Нижняя зубная п л а с ти н к а (рис. 14, н. з. п.) находится непосредственно под ротовым отверстием и несет по верхнему краю семь зубов (количество их может варьировать от 5 до 9). По внешнему краю воронки у основания кожной складки тянется один ряд краевых зубов (рис. 14, к. з.). Кроме того, местами могут встречаться отдельные мелкие зубчики (рис. 14, м. з.).
Передний край языка, видимый в глубине ротовой воронки, также несет роговые пластинки: непарную переднюю язычную пластинку (рис. 14, п. я. п.) с мелкими зубчиками по верхнему краю и парные, лежащие более глубоко задние язычные пластинки (рис. 14, з. я. п.). При сильно втянутом языке они могут быть не видны.
Количество и форма роговых зубов варьируют у разных видов и служат систематическими признаками. У тихоокеанской миноги увеличено количество верхнегубных зубов и имеется ряд нижегуб-ных. • У каспийской вся воронка покрыта рядами округлых зубов.
Перед тем как приступить к рассмотрению органов, располагающихся на голове, необходимо удалить непрозрачный слой слизи, соскабливая его скальпелем.
Глаза (oculus, рис. 15, ос.) —парные органы зрения, лежащие по бокам головы и прикрытые тонкой прозрачной кожей. Веки у миноги отсутствуют.
Более детально со строением глаза можно будет ознакомиться при изучении органов чувств. Поэтому при дальнейшей работе осторожно вычленяют глазное яблоко у миног, которые предназначены для изучения мускулатуры и скелета, и сохраняют их в пробирке с формалином.
На дорзальной части головы, между глазами, у миноги имеется светлое, лишенное пигмента пятно. Под ним располагаются п и-неальный и париетальный органы (corpus pineale, corpus parietale, рис. 15, c. prt.), называемые также теменными глазами.
Непарная ноздря (naris, рис. 15, паг.)—наружное непарное обонятельное отверстие, назогипофизарного мешка, которое открывается на конце короткой трубочки, высовывающейся над поверхностью тела впереди теменного органа. Задний край трубочки выше переднего, поэтому отверстие направлено не только вверх, но и вперед. Между отверстием ноздри и теменным глазом видно небольшое вздутие. Оно образовано верхней стенкой обонятельной капсулы.
Органы боковой линии, или сейсмосензорные органы (linea lateralis, рис. 15, tin., I.),— мелкие светлые бугор
ки, особенно хорошо заметные на головном конце, где они концентрируются в несколько рядов. На одних экземплярах они хорошо видны, на других для их рассмотрения следует счистить скальпелем эпидермис. Верхний ряд начинается несколько позади от теменного органа и редкими точками тянется вдоль дорзальной поверхности тела, проходя у основания спинных плавников до конца хвостового отдела. Следующий ряд берет начало у верхнего края ротовой воронки, доходит до глаза, опускается вниз по переднему краю глазницы, образует за глазом небольшое скопление близко расположенных бугорков и продолжается назад по латеральной поверхности тела редкими точками, расположенными не по одной прямой линии, а то выше, то ниже. Этот ряд не доходит до конца
Рис. 15. Общая топография внутренних органов невской миноги Lampetra fluvia-tilis:
cav. prr.—предротовая воронка; c. prt.—тененной орган; f. br. — жаберное отверстие; gast. — желудок; gon. — половая железа; К. — печень; inis. — кишка; Un. I. — органы боковой линии; mes. — брыжейка; паг. — ноздря; ос. — глаз; рпп. а. — анальный плавник; рпп. с. — хвостовой плавник; рпп. d. — спиниые плавники; рп. — предпочка; р. urg. — мочеполовой сосочек; г. — почка; tph. — спиральный клапан; иг. — мочеточник
тела и заканчивается иа уровне переднего края второго спинного плавника. Короткий ряд образован бугорками, сидящими по одному или по два в промежутках -между отверстиями жаберных мешков. Самый нижний ряд окаймляет ротовую воронку сбоку и снизу тянется по вентральной поверхности головы. В самом конце он поворачивает кверху и подходит к последнему жаберному отверстию.
Жаберные отверстия (foramen branchiale, рис. 15,/. br.). Семь пар наружных отверстий жаберных мешков помещаются по бокам задней части головного отдела. Они имеют форму треугольника с округленными углами и вершиной, направленной назад. По заднему краю отверстия сидят небольшие ворсинки, играющие роль фильтра при засасывании воды в жаберные мешки. На передней стенке помещается широкий наружный клапан, играющий роль в акте дыхания при вбирании воды в жаберные мешки и при ее выбрасывании, так как оба эти процесса у присосавшейся миноги совершаются через наружные жаберные отверстия.
Мочеполовой сосоче'К (papilla urogenitalis, рис. 15, р. urg.) и з а д непр о хо д но е о т в ер ст и е (anus) помещаются на вентральной стороне тела, на границе туловищного и хвостового отделов, в неглубокой продольной щели, образованной складкой кожи. Раздвинув складки кожи, можно видеть конусовидный вырост— мочеполовой сосочек — с отверстием на конце. Анальное 72
отверстие открывается на передней стенке щели и на фиксированном материале плохо заметно.
Плавники (pinna, рис. 15)—непарные кожные складки, окаймляющие задний отдел тела миноги. Рассматривая их на просвет, можно видеть тонкие хрящевые лучи (см. стр. 105), которые поддерживают плавник. На спинной стороне тела два спинных плавника (pinna dorsalis, pun. d.) —передний и задний. Задний спинной плавник переходит непосредственно в хвостовой, который разделен осью тела на две равные лопасти: верхнюю й нижнюю. Такой тип хвоста называется протоцеркальным, т. е. первично равнолопастным. На вентральной стороне тела нижняя лопасть хвостового плавника сливается впереди с анальным плавником (pinna analis, рпп.а), который в виде низкой складки тянется вперед почти до мочеполового сосочка. Анальный плавник не поддерживается хрящевыми элементами и сильно развивается только у самок в период нереста. В остальное время он имеет вид невысокой, треугольной в поперечном сечении складки.
Общая топография внутренних органов (рис. 15)
Дальнейшее изучение внутренних органов миноги можно вести двумя путями. Метод последовательного препарирования, который лежит в основе дальнейшего описания, позволяет наиболее полно познакомиться со всеми системами, особенно если в дополнение к нему рассматривать сагиттальный и поперечный срезы. Этот метод трудоемкий, и при недостатке времени можно ограничиться только рассмотрением сагиттального и поперечных разрезов, описание которых будет дано ниже, используя данные по строению отдельных органов, приведенные при описании вскрытия.
Ножницами разрезают брюшную стенку тела по средней линии. Разрез следует начать, отступая на 1 см от последнего жаберного отверстия, и продолжать назад почти до анального отверстия. При этом нельзя вводить ножницы очень глубоко, чтобы не повредить внутренних органов. От конца разреза проводят небольшие вертикальные разрезы по левой стороне тела. Отворачивают левую стенку тела и удаляют ее совсем, отрезав ножницами или скальпелем на уровне верхней части брюшной полости. Особенно осторожно следует подрезать в хвостовой части, чтобы не повредить почки и мочеполовой синус.
Необходимо отметить, что внутренняя сторона срезанной брюшной стенки выстлана тонким пигментированным листком — брюшиной (peritoneum) так же, как и вся брюшная полость. Эта часть брюшины, выстилающая стенки полости тела, носит название париетального листка (somatopleura).
Брюшная полость, или вторичная полость тела — целом (coelom), простирается от заднего конца жаберной области до анального отверстия. У половозрелых миног она почти целиком занята половой железой (gon.), которая оттесняет к краям все остальные органы. В передней части брюшной полости
лежит треугольная печень (А.). Вдоль вентральной стенки брюшной полости тянется кишка (ints.), которая на большем своем протяжении окружена лопастями половой железы. Лентовидные почки (г.), занимают дорзальную часть задней половины брюшной полости. По их вентральному краю тянется тонкий белый канал — мочеточник (иг). Почки подвешены к дорзальной стенке на тонком листке брюшины.
Дыхательная система
Бритвой или острым скальпелем срезают левую стенку тела в области наружных жаберных отверстий примерно на одну четвертую часть толщины тела миноги. Срез начинают от первого жаберного отверстия и заканчивают у последнего. Рассматривают внутреннее строение жаберных мешков, разрезанных примерно пополам по вертикальной плоскости.
Жаберные мешки (saccus branchialis, рис. 16). Каждый из семи пар жаберных мешков имеет по два отверстия: наружное (см. рис. 15, f.br.), соединяющее мешок с'внешней средой, и внутреннее (рис. 16, f. br. in.); ведущее в дыхательную трубку. Причем внутреннее смещено вперед, относительно наружного, поэтому мешки расположены под углом к продольной оси тела. Оттянув вперед переднюю стенку жаберного мешка, можно рассмотреть внутреннее жаберное отверстие с мускульным сфинктером по краю. Внутренняя поверхность жаберного мешка покрыта многочисленными выростами, или складками, — жаберными лепестками (filum branchiale, рис. 16, fit. br.), которые тянутся параллельно друг другу от внутреннего к наружному жаберному отверстию. Каждая из таких складок покрыта мелкими лепестками второго порядка, которые тянутся от основания к свободному краю лепестка первого порядка. Жаберные лепестки первого порядка соприкасаются друг с другом почти на' всем протяжении и только у основания, где вторичные лепестки невысоки, образуются своеобразные каналы (рис. 16,ж. к.), которые расширяются по направлению к латеральной стенке жаберного мешка.'
В отличие от жабр других водных позвоночных жаберные мешки миноги энтодермального происхождения. Снаружи стенки жаберного мешка покрыты тонким мускулом — сжимателем жаберного мешка, который не имеет самостоятельного значения в акте дыхания, а только поддерживает стенки мешка и жаберные лепестки в определенном тонусе.
Околожаберные синусы (sinus peribranchialis, рис. 16, sin. prb.) —полости, которые находятся между стенкой жаберного мешка и межжаберными перегородками. В зависимости от того, сжаты или растянуты жаберные мешкр у фиксированного экземпляра, они могут быть или очень широкими, или имеют вид узкой щели. Синусы связаны с венозной системой и обычно заполнены жидкостью из смеси лимфы и крови, которая на фиксированном материале имеет вид беловатой студнеобразной массы. Околожа-74
~ верные синусы играют роль в акте дыхания, так как заполняющая их* жидкость, благодаря своей несжимаемости, передает воздейст-
вие наружной мускульной стенки при ее сокращении или сжатии на стенку жаберных мешков.
Межжаберная перегородка (septum interbranchiale, рис. 16, spt. ibr.) развивается во время метаморфоза из жаберной перего
родки пескоройки в результате обособления от нее внешнего слоя вместе с жаберными лепестками. Внутренний ее край растет в медиальном и ростральном направлении до слияния с впереди лежащей перегородкой. Таким образом, у взрослой миноги межжаберные перегородки составляют переднюю, заднюю и медиальную стенки камеры, в которой лежит, жаберный мешок. Наружный край межжаберной перегородки связан с хрящами висцеральной решетки, так что наружная стенка камеры образована латеральной стенкой жаберного аппарата.
Как видно на препарате, каждый жаберный мешок состоит из двух полужабр — передней и задней, разделенных узкой полоской стенки жаберного мешка без жаберных лепестков. Изучение эмбрионального развития показывает, что целая
Рис. 16. Продольный разрез жаберной области невской миноги Lampetra fluviatilis:
ж. к. —' каналы жаберных мешков; а. Ьг. г. — выносящая жаберная артерия; с. — хрящи жаберной решетки; f. br. in. — внутреннее отверстие жаберного мешка; fit. br. — жаберные лепестки; т. с. — констриктор жаберного мешка; т. с. d. — констриктор жаберного аппарата; sin. prb. — околожаберный синус; sph. т. — мускульный сфинктер; spt. ibr. — межжаберная перегородка
жабра состоит из двух по-
лужабр с заключенной между ними жаберной перегородкой. Таким образом, целая жабра у миноги складывается нз задней полужабры переднего мешка, межжаберной перегородки и передней полужабры заднего жаберного мешка. Первый и последний жаберные мешки^несут по одной самостоятельной полужабре.
В нижней и верхней частях промежутков между жаберными мешками видны перерезанные поперек хрящи скелета жаберной решетки (рис. 16,с.). В верхней части к их каудальной стороне примыкают небольшие отверстия, выносящие жаберные
артерии (a. branchialis revehens, рис. 16, a. br.r.). Сверху и снизу каждого жаберного мешка непосредственно к жаберной перегородке прилегают довольно мощные мышечные волокна к о н-стриктора жаберного аппарата (m. constrictor branchialis, рис. 16, т. с.. Ь.).
Осторожно, при помощи пинцета можно удалить жаберные мешки вместе с жаберными перегородками, очистить лежащие глубже органы от сгустков крови и рассмотреть их положение в головном конце животного, одновременно рассматривая их и на сагиттальном разрезе.
Дыхательная трубка (tuba respiratoria, см. рис. 23, tub. г.)—орган, характерный только для миног. Дыхательная трубка начинается от задней части ротовой полости, тянется назад над языком и слепо оканчивается за последним жаберным мешком. В нее открывается семь пар внутренних жаберных отверстий (f. br. in.). В самом начале дыхательная трубка отделена от ротовой полости двухстворчатым клапаном, или парусом (velum, vel.). Передний край каждой створки прикреплен к краю входного отверстия, а задний свободен и направлен назад так, что вода может легко проходить из ротовой полости в дыхательную трубку. При ее обратном движении створки клапана сжимаются и закрывают тем самым вход в ротовую полость.
Положение створок паруса хорошо видно на поперечном разрезе. Для этого следует провести поперечный разрез, отступая примерно на 1 см от заднего края глаза, а затем срезать тонкие слои с передней части головы по направлению вперед, пока в просвете дыхательной трубки не покажутся задние края клапанов. На сагиттальных срезах видны также сидящие по переднему краю дыхательной трубки пять тонких выростов слизистой оболочки, поддерживаемых хрящевыми стерженьками и направленных в ротовую полость. Они играют роль фильтра, препятствующего попаданию крупных частиц в дыхательную трубку.
Акт дыхания. У плывущей миноги обмен воды в жаберных мешках может проходить благодаря ее току через ротовую воронку, дыхательную трубку, жаберный мешок и наружные жаберные от-’ верстия. У присосавшейся миноги он осуществляется более сложно, так как набор воды в жаберный мешок и ее выталкивание оттуда происходит только через наружное жаберное отверстие. Внутренние жаберные отверстия могут перекрываться или через них может осуществляться связь между отдельными жаберными мешками, тогда как доступ воды в ротовую полость прегражден парусом. Действия описанного выше наружного и еще пары внутренних клапанов, лежащих в наружном жаберном отверстии, сводится к тому, что вода, попавшая внутрь жаберного мешка, при его сокращении не может выйти обратно по тому же пути, по которому она поступила, поскольку он перекрывается клапаном. Поэтому она просачивается мимо лепестков в каналы жаберных мешков, из которых выход наружу свободен. Этим обеспечивается принудительное омывание жаберных лепестков во время акта дыхания.
Органы пищеварения
Переднюю часть пищеварительного тракта удобнее рассматривать на сагиттальном разрезе (см. рис. 23). Начинается пищеварительная система предротовой воронкой, которая была описана выше (см. рис. 15 и 23, cav. prr.).
Ротовая полость (cavum orale, см. рис. 23, cav. or.) соединяется в передней части узким ротовым отверстием (stoma, рис. 23, st.) с предротовой воронкой. На дорзальной стороне в средней части ротовой полости имеется расширение — гидросинус (г.), отграниченное от остальной полости выростом — ростродорзальным клапаном (к.). Сзади ротовая полость переходит в пищевод, отверстие которого лежит непосредственно над отверстием дыхательной трубки (oes.).
Язык (lingua, см. рис. 10) в виде плотного белого мускульного тяжа подстилает дно ротовой полости и продолжается назад под дыхательной трубкой вплоть до сердечной хрящевой капсулы, к передней, части которой и прикрепляется. На сагиттальном разрезе видно, что основу языка составляет подъязычный хрящ (cartilago glossae, с. gl.), который поддерживает переднюю часть языка. Задняя часть языка образована тонким, длинным мускулом— ретрактором языка (m. cardio-apicalis, т.с.-ар.), который задним концом крепится к нижней комиссуре скелета жаберной решетки и к хрящевой околосердечной капсуле, а передней сухожильной частью — к небольшому апикальному хрящику в переднем конце языка. Этот мускул втягивает язык в глубь воронки. В средней части видны разрезанные поперек мышечные волокна m. mandibularis posterior (т.т.р.), которые, как муфта, охватывают язык. При сокращении этот мускул, вероятно, проталкивает язык вперед. Всего в системе языка насчитывается 10 мускулов, из них 4 — парные и 2 — непарные.
Пищевод (oesophagus, см. рис. 23, oes.) на нашем препарате имеет вид тонкостенной трубки, лежащей непосредственно над дыхательной трубкой. Его следует найти на сагиттальном разрезе и еще раз рассмотреть место отхождения от ротовой полости. Внутренняя поверхность пищевода, как и кишечника, покрыта продольными складками. В области сердечной капсулы пищевод незаметно переходит в кишечную трубку.
Кишечник (intestinum, рис. 15, ints.), чтобы рассмотреть его начало, на препарате следует слегка оттянуть вниз печень и приподнять половую железу. Передний отдел кишечника лежит в углублении на дорзальной поверхности печени. Вентральная стенка его соединена с печенью. По печени кишечник спускается с дорзальной стороны полости тела на вентральную и тянется назад вдоль брюшной стенки тела. Оттянув кишку книзу, можно убедиться, что почти на всем протяжении она лежит свободно и только в задней части прикреплена мезентериальными тяжами к дорзальной стенке брюшной полости (рис. 15, mes.). По этим тяжам подходят к кишечнику артериальные кровеносные сосуды. Сзади кишечная
трубка заканчивается а н а л ь н ы м отверстием (anus, рис. 17, ап.), которое располагается в складке перед мочеполовым сосочком. Его можно рассмотреть, отпрепарировав окончание кишечника, для чего срезают стенку тела впереди от мочеполового сосочка; положение анального отверстия хорошо видно также на сагиттальном разрезе. На всем протяжении кишечной трубки можно отметить три слабо выраженных расширения: первое — в том месте, где она соприкасается с печенью, принято считать желудком (gaster, рис. 15, gast.)-, второе — в средней части; третье расположено в самой задней части и носит название прямой кишки (rectum, рис. 17, ret.).
Сделав продольный разрез на одной из стенок кишечной трубки, можно видеть ее внутреннюю поверхность, покрытую многочисленными невысокими складками. Кроме того, одна крупная складка— спиральный клапан (tiphlosolis, рис. 15, tph.) тянется вдоль всей кишечной трубки, огибая ее по пологой спирали, и служит для увеличения всасывающей поверхности.
Из пищеварительных желез у миноги можно видеть только п е-чень, так как поджелудочная железа (pancreas) рассеяна по стенкам кишечнйка.
Печень (hepar, рис. 15, h.) имеет форму конуса, основание которого прилегает к задней стенке хрящевой капсулы, окружающей сердце, а вершина направлена назад.
Желчный пузырь (vesica fellea) и желчный проток (ductus choledochus) развиты только у миног, живущих в море. У идущих на нерест, а следовательно, непитающихся миног они редуцируются. По связке, соединяющей печень с кишечником, проходит воротная вена печени (v. portae hepatis). Она образована слиянием двух вен, несущих кровь от переднего и заднего отделов кишечной трубки. Воротная вена печени, распадаясь в печени на капилляры, образует воротную систему печени. Из нее кровь направляется в сердце по печеночной вене.
Печеночная вена (v. hepatica) — темный венозный сосуд, просвечивающий на вентральной поверхности печени.
Мочеполовая система
Полов_ая железа (рис. 15, gon.) — семенник у самцов и яичник у самок — непарная многодольчатая железа, которая у половозрелых особей занимает почти всю брюшную полость. Каждая долька одета листком-брыжейки, на которой вся железа подвешена к дорзальной стенке брюшной полости. Специальных половых протоков нет. Половые продукты у самок и самцов через разрывы стенок железы выпадают в полость тела, из которой через половые поры (рис. 17, por.g.) попадают в мочеполовой Синус, а оттуда через отверстие мочеполового сосочка — наружу.
Семенник (testis) беловатый, по виду напоминает жир.
Яичник (ovarium) имеет зернистую структуру от огромного количества видимых простым глазом икринок — яиц.
Почки (ren, рис. 15 и 17, г.)—парные лентовидные органы, подвешенные на брыжейке к дорзальной стенке тела. Функционирующие у взрослой миноги туловищные (мезонефрические) почки (mesonephros) начинаются примерно на уровне середины туловища и тянутся назад вдоль дорзальной стенки брюшной полости, прикрывая на препарате верхнюю часть половой железы. По нижнему краю каждой почки тянется беловатый мочеточник (ureter), узкий в передней части почек и постепенно расширяющийся кзади (рис. 15 и 17, иг.). Вперед от головного конца почек тянется более или менее заметная у разных экземпляров складка брыжейки, доходящая почти до уровня сердечной капсулы. У экземпляров со слабо развитыми половыми железами в ней можно видеть отдельные клубочки почечных канальцев. Это остатки головной почки — пронефроса. У особей с вполне развитой половой железой видна только складка брыжейки с жировыми включениями (рис. 15, р.п.).
Осторожно срезая левую стенку тела в районе мочеполового сосочка, следует отпрепарировать общий мускульный сфинктер (рис. 17, sph.m.), охватывающий кольцом задний отдел кишечника и мочеполовой синус. Перерезая мускульные волокна, сфинктора, удаляют левую его стенку и обнажают задний конец прямой кишки и мочеполовой синус.
Мочеполовой синус (sinus urogenitalis, рис. 17)—небольшой яйцевидный мешок с.узким внутренним просветом, образованный в результате слияния расширенных задних отделов мочеточников. Его задний конец выступает над брюшной поверхностью тела в виде м о че п о л о в ого сосочка (papilla urogenitalis), на вершине которого имеется открывающееся наружу мочеполовое отверстие.
Перерезав один из мочеточников недалеко от его впадения в синус, вводят в его просвет щетинку так, чтобы она вышла через мочеполовое отверстие. Следуя направлению щетинки, вскрывают стенку синуса и подрезают ее края. В передней части мочеполового синуса видны два широких отверстия мочеточников (рис. 17, иг.').
Половые поры (porus genitalis, рис. 17, por.g.). На препарате остается только одна правая пора (левая удаляется вместе со стенкой синуса). Она имеет вид узкой щели в боковой стенке средней части синуса. Пропуская через нее щетинку, убеждаются в связи мочеполового синуса с брюшной полостью.
Иногда строение мочеполового синуса хорошо видно на сагиттальных разрезах.
Кровеносная система
Кровеносная система миноги наиболее трудная для изучения. Полностью проследить местоположение и ход сосудов можно только после их инъекции, которую выполняют лишь на свежих миногах. На фиксированных экземплярах сосуды видны довольно плохо. К тому же они тонкостенны и непрочны, особенно вены, поэтому
во многих случаях целесообразнее не пытаться выделить сосуд, а лучше вскрыть его и очистить от сгустков крови. Такая операция позволяет довольно точно проследить ход крупных сосудов. Чтобы полностью и наиболее точно представить ход и расположение тех или иных сосудов, необходимо постоянное сопоставление препарата с сагиттальными и поперечными разрезами. При дальнейшей работе следует постараться разобрать сначала строение сердца, потом артериальную систему, а затем венозную. Однако не всегда это
Рис. 17. Мочеполовой сииус невской миноги Lampetra fluviatilis:
an. — анальное отверстие; рог. g. — подовые поры; г. — почка; rd. — прямая кишка; sph. т. — мускульный сфинктер; иг. — мочеточник; иг/ — отверстия мочеточников в мочеполовой сннус
удается, так как часть сосудов венозной системы приходится уда-лять, чтобы рассмотреть артерии (например, нижнюю яремную вену). Отсюда неизбежны отступления от общего порядка и в описании.
Осторожно снимают мускулатуру с жаберной области, для чего отделяют ее от скелета, начав с нижней части среза жаберной области, а затем переходят на правую сторону. Мускулатуру можно отделить вплоть до жаберных отверстий, а затем отрезать. Пинцетом снимают по частям скелет жаберной решетки. Прямо под ним лежит очень тонкостенный сосуд — нижняя яремная вена (v. jugularis inferior, см. рис. 18, 22, 23, v. j. inf.) которая собирает венозную кровь из нижней части головного отдела, а также связана с венозными синусами, окружающими язык. Отпрепарировать ее очень трудно. Ход ее легче всего проследить, вскрыв вентральную стенку и вычистив из полости сосуда сгустки крови. Яремная вена
начинается на уровне четвертого жаберного мешка, где сливаются два сосуда, которые тянутся по краям языка и несут кровь от нижней части головы. На сагиттальном разрезе (рис. 23, v. j. inf.), вена лежит непосредственно над полоской поджаберной мускулатуры.
На поперечных разрезах передней части головного конца она представлена парными сосудами, лежащими по краям языка в толще мускулатуры. Ближе к сердцу это непарный сосуд, расположенный непосредственно под языком (рис. 22, V. j. inf.).
На уровне каждого жаберного мешка двумя отверстиями в дорзальной стенке нижняя яремная вена соединяется с лимфатическим синусом, который окружает язык. Задний конец вены прободает околосердечную сумку и впадает в нижний отдел венозного синуса (рис. 19, v. j. inf.).
Сердце
Обнажают хрящевую сердечную капсулу, для чего удаляют оставшийся кусочек стенки тела между жаберным отделом и брюшной полостью.
Рис. 18. Схема кровеносной системы невской миноги Lampetra flu-viatilis (с брюшной стороны): a. br. ad. — приносящие жаберные артерии; a. br. г/— выносящие артерии первой полужабры; a. caud. — хвостовая артерия; а. с. е. — наружная сонная артерия; а. с. i. — внутренняя сонная артерия; a. cl. — чревная артерия; а. т. — брыжеечная артерия; anst. — анастамоз между передней и задней кардинальной веной; ао. d.—спинная аорта; ао. v. — брюшная аорта; a. prt. — сегментальные артерии; а. геп. — почечные артерии; air. — предсердие; bul. а — луковица аорты; sin. v. — венозный синус; V. с. — хвостовая вена; v. crd. а. — передняя кардинальная вена: v. crd. р. — задняя кардинальная вена; ventr. — желудочек; v. h. — печеночная вена; V. infs. — подкишечная вена; о. /. inf. — нижняя яремная вена
а. с. а
а.сл.
a. br. г!
a. br ad.
Herd, а
а о. d.
ae.v.
V.J. inf.
bul. a.
ventr.
atr.
Sin. v.
ansi.
a. cl.
V.h.
a. prt.
К inf
an. d.
a. ren.
v. crd.p.
v.c.
a. caud.
Хрящевая капсула служит продолжением жаберного скелета и охватывает околосердечную сумку сзади и с боков. Спереди околосердечная полость отделена от последних жаберных мешков плотной соединительной тканью. Снимают левую стенку хрящевой
капсулы, стараясь не повредить при этом печеночной и нижней яремной вен, и рассматривают отделы сердца.
Предсердие (atrium, рис. 18, 19, atr.) —большой мешок с -тонкими мускульными стенками. Если вскрытие ведется с левой стороны, то этот отдел сердца прежде всего ’бросается в глаза, так
Рис. 19. Сердце невской миноги Lampetra fluviatilis (большая часть предсердия удалена):
ао. и.— брюшная аорта; atr.—предсердие; but. а.—луковица аорты; о. s.-a. — отверстие между венозным синусом н предсердием; sin. и. — венозный сннус; v. crd. а. — передняя кардинальная вена; v. crd. р.—задняя кардинальная вена; ventr. — желудочек; v. h, — печеночная вена; v. j. inf. — нижняя яремная веиа; vl. a.-v. — атриовентрикулярные клапаны
как занимает всю левую половину сердца и закрывает все остальные отделы.
Для того чтобы лучше рассмотреть другие отделы сердца, срезают большую часть левой стороны предсердия и очищают его полость от сгустков крови. Оставшейся медиальной стенкой предсердие соприкасается с венозным синусом и желудочком. В ней можно видеть два отверстия. Заднее имеет вид вертикальной щели с двумя тонкими синусо-атриальными клапанами, которые располагаются по ее передней и задней стенкам. Через это отверстие (рис. 19, f. s.-a.) предсердие соединяется с венозным синусом. Впереди от него — небольшое круглое отверстие с двумя перепончатыми а т р и о - в е н т р и к уг лярными клапанами (valvula atrio-ventricularis, рис. 19, vl.
a.-v.); отверстием предсердие соединяется с желудочком.
Венозный с и н у с, или ве ноз н а я пазуха (sinus veno-sus, рис.-18, 19, sin.v.),— тонкостенная трубка, идущая-в вертикальном направлении посередине между предсердием и желудочком. Нижний отдел синуса образован нижней яремной веной (рис. 18, 19, 22, 23, v.j.inf.) и печеночной веной (рис. 18, 19, 23, v.h.). Последняя несет кровь из печени и проходит через заднюю стенку хрящевой капсулы.
В верхний отдел синуса впадают передние и задние кардинальные вены, которые будут рассмотрены ниже.
Желудочек (ventriculus, рис. 18, 19, ventr.) округлой формы с толстыми мускулистыми стенками занимает переднюю правую часть сердца. Вперед от него отходит брюшная аорта.
Артериальная система
Перерезав нижнюю яремную вену около сердечной капсулы, отделяют задний конец языка в месте его прикрепления к скелету
жаберной решетки. Оттягивая его на себя и подрезая скальпелем соединительную ткань, удаляют весь язык, перерезав его в самой передней части на уровне глаза. Положив объект на спинную сторону, раздвигают образовавшуюся щель, отвернув жаберные мешки и закрепив их в таком положении булавками. Осторожно очистив полость от сгустков крови, рассматривают лежащую на дне ее брюшную аорту. Следует также уточнить ее положение на сагиттальном разрезе.
Брюшная аорта (aorta Ventralis, рис. 18, 19, ao.v.) отходит от переднего конца желудочка и еще в околосердечной капсуле имеет расширение — луковицу аорты (bulbus aortae, рис. 18, 19, bul. а.); аорта тянется вперед к голове и располагается между дыхательной трубкой и мускулатурой языка. На уровне четвертого мешка делится на два сосуда. На сагиттальном разрезе брюшная аорта также имеет вид трубки, доходящей до четвертого жаберного мешка, с отверстиями перерезанных приносящих сосудов (рис. 23, ao.v.). На поперечном разрезе головы брюшная аорта лежит между языком и дыхательной трубкой (рис. 22, ao.v.).
Приносящие жаберные артерии (a. branchialis advehens, рис. 18, a. br.ad.) отходят от брюшной аорты справа и слева в числе восьми пар на уровне межжаберных перегородок. Четыре пары отходят от непарного сосуда и четыре — от парных. Каждая из приносящих жаберных артерий, кроме первой и последней, делится на два сосуда, которые отходят к двум полужабрам соседних жаберных мешков. Около внешней стенки жаберного мешка каждый из сосудов распадается на большое количество мелких сосудиков, которые идут по внешней поверхности жаберного мешка и дают ответвления к жаберным лепесткам. Первый и последний приносящие жаберные сосуды обслуживают только по одной полужабре.
Выносящие жаберные артерии, (a. branchialis ге-vehens, рис. 18, br. г.) хорошо видны на препаратах с инъецированной кровеносной системой. На обычных препаратах обнаружить их довольно трудно. Для того чтобы их рассмотреть, перерезают приносящие жаберные артерии у их основания, отделяют жаберные мешки от дыхательной трубки, перерезают дыхательную трубку вместе с пищеводом в месте их отхождения от ротовой полости. Отпрепарировав, отворачивают эти органы назад. Вскрывают ротовую полость по вентральной стенке, разворачивают в стороны края образовавшейся щели. По медиальным стенкам жаберных мешков тянутся тонкие кровеносные сосуды. Это — выносящие жаберные артерии. Каждый из выносящих жаберных сосудов образован двумя сосудами, несущими кровь от двух полужабр соседних жаберных мешков. Каждый из этих мелких сосудов в свою очередь сливается из двух сосудов: один из них несет кровь от верхней половины полужабры, другой — от нижней. Только самый последний. и самый первый выносящие жаберные сосуды несут кровь от одной полужабры. От третьей пары выносящих жаберных артерий отходит в голову сосуд, несущий кровь к ее нижним частям и мус-
кулатуре языка. Выносящие жаберные артерии каждой стороны впадают в спинную аорту, причем сосуды левой стороны- впадают несколько позади от соответствующих правых сосудов.
Спинная аорта (aorta dorsalis, рис. 18, ao.d.)-—непарный сосуд, тянущийся по медиальной линии спинной части тела, непосредственно под хордой. На препарате она лежит между верхними частями жаберных мешков правой и левой стороны. На сагиттальном разрезе в области головы это тонкая трубка, лежащая непосредственно под хордой (рис. 23, ao.d.), на латеральной стенке которой можно видеть остатки выносящих жаберных артерий. На уровне передней стенки первого жаберного мешка спинная аорта делится на два сосуда, в каждый из которых впадают выносящие сосуды первой полужабры (рис. 18, a. br.r.). Пройдя немного вперед, эти сосуды опять сливаются, образуя круг, от передней части которого самостоятельно отходят четыре сонные артерии: две внутренние сонные (a.carotis interna, рис. 18, а. с. i.) и две наружные (a. carotis externa, рис. 18, а.с.е.). В туловищном отделе спинная аорта тянется по дорзальной стенке брюшной полости между двумя кардинальными венами и дает целый ряд ответвлений к внутренним органам. Она четко видна на поперечных срезах в виде относительно толстостенного сосуда, лежащего под хордой мёжду двумя задними кардинальными венами (рис. 20, ao.d.).
Чревная артерия (a. coeliaca, рис. 18, a. cl.) отходит от аорты на уровне переднего края печени. Оттянув вниз печень и переднюю часть кишечника, следует обнаружить этот крупный сосуд, несущий кровь к стенкам кишечника.
Перерезав печеночную вену у стенки сердечной капсулы, отделяют печень от сердечной капсулы, перерезают кишечную трубку и, перерезав брыжейку, отворачивают назад внутренние органы вплоть до почек. На всем протяжении опинной аорты от нее вправо и влево соответственно мышечным сегментам отходят тонкие кровеносные сосуды (a. parietalis, рис. 18, a. prt.). Кроме того, большое количество мелких почечных артерий (a. renalis, рис. 18, а. геп.) тянутся по брыжейкам, на которых подвешены почки, снабжая кровью последние. В заднем конце от спинной аорты отходят три-четыре сосуда (a. mesenterica) к заднему отделу кишечника (рис. 18, а. т.). Они проходят по мезентериям, соединяющим кишечник с дорзальной стенкой брюшной полости.
'Хвостовая артерия (a. caudalis, рис. 18, a. caud.). Задний конец спинной аорты, идущий по хвостовому отделу, носит название хвостовой артерии. Ее лучше рассмотреть на поперечном разрезе хвостовой области. Сосуд лежит в гемальном канале, располагаясь между хордой и хвостовой веной (рис. 21, a. caud.).
Венозная система
Сначала следует отпрепарировать дорзальный отдел венозного синуса и рассмотреть впадающие в него сосуды. Тонкостенные 84
венозные сосуды очень плохо отделяются от окружающей их ткани, поэтому ход их лучше всего проследить, вскрывая одну из стенок и очищая просвет сосуда от сгустков крови. Параллельно с препаровкой полезно рассмотреть положение венозных сосудов на поперечных разрезах через туловищную, хвостовую и жаберную области.
Задние кардинальные вены (v. cardinalis posterior, рис. 18, 20, v. crd. р.) —парные широкие тонкостенные сосуды, впадающие сзади в венозный синус (кювьеровы протоки у миноги отсутствуют), вернее, образующие вместе с передними кардинальными венами, несущими кровь из головы, верхнюю часть синуса. На препарате они имеют вид двух темных полос, тянущихся по дорзальной стенке брюшной полости непосредственно под хордой. На сагиттальном разрезе чаще всего просвет одной из вен бывает вскрыт. На поперечном разрезе туловищной области это два крупных сосуда, лежащие по обеим сторонам под хордой (рис. 20, v.crd.p.).
Хвое тов а я в ей a (v. caudalis, рис. 18,21, v.caud.)—непарный широкий тонкостенный сосуд. Она начинается капиллярами в хвостовой области и тянется вперед по средней линии тела, под хвостовой артерией. Передний конец вены раздваивается, давая начало задним кардинальным венам. Хвостовую вену и место ее распадения на две кардинальные вены удобнее рассмотреть на серии поперечных срезов через хвостовую область. Место распадения хвостовой вены на кардинальные находится на уровне переднего края второго спинного плавника.
Передние кардинальные, или верхние яремные, вены (v. cardinalis anterior, s. v. jugularis superior, рис. 18, 19, v. crd. a.) — парные сосуды, впадающие спереди® верхний отдел венозного синуса. Они несут кровь из головы и тянутся над жаберными мешками по бокам хорды. Правая яремная вена коротким сосудом (рис. 18, anst.) связана с правой задней кардинальной веной. Эти сосуды трудно отпрепарировать и лучше рассмотреть на поперечном разрезе головы, где их просветы лежат под мускулатурой на уровне середины хорды (рис. 22, v. crd. а.).
Нижняя яремная вена (v. jugularis inferior, рис. 18, u. j. inf.), печеночная вена (v. hepatis, рис. 18, v.h.) и кишечная вена (v. intestinalis, рис. 18, v. ints.) были рассмотрены при вскрытии.
Поперечные разрезы
Разрез туловищной области (рис. 20.)
Основную массу поперечного среза составляют мышечные сегменты— мио меры, которые занимают все боковые части среза’ (тт.).
В центре среза лежит голубоватая спинная струна, или хорда (chorda dorsalis, ch.). Снаружи она одета двумя оболочками: верхняя очень тонкая, темного цвета носит название наруж-
ной оболочки (elastica externa, el.e.); под ней лежит толстая волокнистая оболочка (el.e'.).
Над хордой расположен спинномозговой канал, на дне которого лежит плоский спинной мозг (medulla spinalis, med.). При
Рис. 20. Поперечный разрез миноги Lampetra flu-viatilis в туловищной области:
меняя лупу, можно увидеть в его центре небольшое отверстие — невроцель (neurocoel, nr с.).
Спинномозговой канал, а . также отчасти хорда окружены скоплением жировой фиброзной ткани (ж. т.), которая продолжается вверх по средней линии до основания плавников. В этой ткани по бокам от спинномозгового канала на некоторых срезах можно видеть розоватые хрящи — верхние дуги (arcus neuralis, arc. п.). Под хордой, непосредственно по медиальной линии, лежит спинная аорта (ао. d.), а по бокам от нее — два просвета задних кардинальных вен (и. crd. р.). Вент-ральнее от сосудов по-
- мещается полость тела— целом (coel.). В ее верхней части видны парные почки (г.). Они прилегают к латеральным стенкам брюшной полости и
ж. т. — соединительнотканная оболочка хорды н спинномозгового канала; п. л. — плавниковые лучн; a. ints.— подкншечная артерия; ао. d. — спннная аорта; аге. п. — верхние дуги позвонков; ch. — хорда; coel.— целом; еД е. — наружная оболочка хорды; el. е.' — волокнистая оболочка; int. — кишка; med. sp. — спинной мозг; т. т. — миомеры; пгс. — невроцель; ov. — яичннк; г. — почка; мг. — мочеточник; о. crd. р. — задняя кардинальная вена;
V. ints. — подкншечная вена
подвешены на брыжейках к ее дорзальной стороне. Длинные, изогнутые вверху щели в вентральной части почек являются ничем иным, как просветом мочеточников (иг.). Основную массу брюшной полости занимает половая железа (у самок — крупнозернистый яичник, у самцов — семенник). Оттянув ее книзу, можно убедиться, что она так же, как и почки, подвешена на брыжейке к дорзальной стенке брюшной полости. Нижние части половой железы окружают кишечную трубку (ints.). Так как последняя не подвешена на брыжейке, то из тонких срезов легко может выпасть.
S6
Внутренняя стенка кишечной трубки покрыта многочисленными выростами. Кроме того, в ее просвет в виде крупного выроста вдается спиральный клапан. В его плотном основном веществе видны (под лупой) отверстия двух сосудов: более тонкостенный— кишечная ве^а (v. ints.) и толстостенный — подкишечная артерия (a. ints).
По медиальной линии плавника видны перерезанные поперек, кожные плавниковые лучи (п.л.). С помощью лупы можно рассмотреть два слоя кожи: более светлый верхний — эпидермис и нижний — кориум.
Поперечный разрез в передней части туловища отличается тем, что здесь отсутствуют почки. На их месте может лежать более или менее развитое жировое тело.
Разрез хвостовой области (рис. 21)
В центре среза, так же как и в области туловища, видна хорда (ch.). Над ней — спинномозговой канал со спинным мозгом (med.sp.). Под хордой просветы двух сосудов, расположенных один над другим: верхний — хвостовая артерия (a. caud.), нижний, более широкий — хвостовая вена (v. caud.). Фиброзная соединительная ткань окружает не только' спинной мозг и хорду, но также и кровеносные сосуды, напоминая по форме настоящий позвонок. Канал, в котором проходят кровеносные сосуды; носит название гемального канала (canalis haemalis). Лучи спинного плавника (п. л.) глубоко вдаются в тело и доходят до верхнего отдела соединительнотканного «позвоночника». В основании анального плавника (рпп. а.) нет никаких лучей, а имеется только плотная соединительная ткань. Большая часть среза занята мускулатурой (тт.).
Разрез жаберной области (рис. 22)
Разрез делится на дорзальную, двигательную, и вентральную, дыхательную, части. Вся дорзальная часть занята сконцентрированными здесь миомерами мускулатуры (тт.). На вентральную сторону тела переходит только узкая полоска поджаберной мускулатуры, лежащая непосредственно под кожей (тт.).
Хорда (ch.), спинной мозг (med. sp.) и спинномозговой канал выглядят так же, как и на срезах туловищной и ' хвостовой областей.
По бокам хорды видны довольно крупные просветы передних кардинальных вен (v. crd. а.), непосредственно под хордой— толстостенный непарный сосуд — с п и н н а я аорта (ao.d.). Еще ниже, также на- средней линии, видна тонкостенная трубка — пищевод (oes.). Он хорошо отличим, благодаря наличию направленных внутрь выростов стенок. Под пищеводом хорошо' заметно треугольное отверстие дыхательной трубки
(tub. г.). На некоторых срезах можно видеть ее соединение с жаберными мешками. Под дыхательной трубкой расположен толстостенный сосуд — брюшная аорта (ao.v.). На срезах, сделанных ближе к переднему концу, брюшная аорта может быть парной.
Еще ниже видны, в виде плотного белого образования, перерезанные поперек мышцы языка (ling.*), причем на срезах через
Phc. 21. Поперечный развез миноги Lampetra flu-viatilis в хвостовой обла-
сти:
п. л. — плавниковые лучи; a. cattd. — хвостовая артерия; ch. — хорда; med. sp. — спинной мозг; тт. — миомеры; ргт. а. — анальный плавник;
v. caud. — хвостовая вена
vj.lnf. rr>'
сап. п. med. sp. ch.
V. crd. CL. sin. v. ao.d.
-sin. v. oes.
c. a. bead.
tub. r.
ao.v.
ling.
Рис. 22. Поперечный разрез миноги Lampetra flu-viatilis в жаберной области:
a. br, ad. — приносящие жаберные артерии; ао. d. — спинная аорта; ао. v. — брюшная аорта; с. — хрящи •жаберной решетки; сап, п. — спинномозговой канал; ch. хорда; ling. — язык; med. sp, — спинной мозг; тт, — мно-меры; тт.' — поджаберная мускулатура; oes. — пищевод; sin. v. — венозный сииус; tub. г. — дыхательная трубка; v. crd, а. — передняя кардинальная вена; v. j.
inf. — нижняя яремная вена
переднюю часть жаберного отдела они занимают довольно большую площадь, а кзади постепенно уменьшаются. Нижняя яремная вена (v. jug. inf.) занимает самое вентральное положение и лежит на границе с поджаберной мускулатурой. Боковые области нижней части разреза заняты большими жаберными мешками. Пространство между отдельными органами и стенками жаберных мешков занято обширными полостями лимфатических •синусов (sinv.). Большинство синусов парные и располагаются вокруг аорты, пищевода и мускулатуры языка. Верхние синусы <88
соединяются с передними кардинальными ведами. Нижние так же, как и околожаберные, с нижней яремной веной.
Жаберные мешки, как уже говорилось, расположены косо по отношению к продольной оси тела, поэтому на поперечном разрезе с каждой стороны обычно видна передняя часть мешка и задняя часть другого. Очистив полости околожаберных синусов от сгустков лимфы, на некоторых срезах можно видеть сеть приносящих жаберных сосудов (a. br. ad.), которые тянутся от брюшной аорты к медиальной стенке жаберного мешка. Рассмотрите отверстия, ведущие из жаберных мешков наружу.
По периметру жаберной области видны перерезанные поперек розоватые хр ящ и жаберной решетки (с.).
Продольный разрез (рис. 23)
На продольном разрезе, примерно по средней линии, почти вдоль всего тела тянется (чуть фиолетового цвета) довольно толстый тяж — хорда (ch). Суженный передний конец хорды доходит до половины хрящевой черепной коробки и составляет часть ее
Рис. 23. Сагиттальный разрез головного конца миноги Lampetra fluviatilis (по Маринелли и Стренгеру):
г. — гндросинус; ж. т. — жировая ткань; к. — ростро-.дорзальный клапан; ао. d. — спинная аорта; ао» v. — брюшная аорта; can. n.-h.—полость назо-гипофизарного мешка; cap. о.— обонятельная капсула; cav. or. — ротовая полость; cav. рг. г. — предротовая воронка; с. gl. — подъязычный хрящ; ch. — хорда; сог. — сердце; епс. — головной мозг; f. br. in — внутренние жаберные отверстия; т. с.~ар. — m. cardio-apicalis: med. sp. — спинной мозг; т. т. — мускулатура; т. т. р. — mandibularis posterior; nar. — ноздря; oes. — пищевод; oec.' — его отверстие в ротовую полость; st. — ротовое отверстие; tub. л — дыхательная трубка; и. с. р. ~ задняя кардинальная вена; —парус; v. j. inf. — нижняя яремная вена; и. h. — печеноч-
ная вена
дна. Сзади хорда продолжается до конца тела. Сверху и снизу от хорды видны ее оболочки—наружная, в виде тонкой темно-коричневой наружной полосы, и волокнистая, лежащая под ней и имеющая более светлую окраску, чем основное вещество хорды.
Над хордой тянется спинномозговой канал, на дне которого лежит тонкая светлая полоска спинного мозга (med. sp.). В передней части спинномозговой канал расширяется, образуя полость черепной коробки, в которой лежит головной мозг (ens.). Над спинномозговым каналом тянется соединительно
тканная прослойка (ж. т.). Она тонкая в _самом начале у -черепной коробки и наибольшего развития достигает в средней части тела. В хвостовой области такая же прослойка лежит и под хордой. Над соединительнотканной прослойкой расположены миомеры (пгт) и еще выше — тонкая линия кожи.
В тулЪвищной области под хордой, если разрез прошел строго по медиальной линии, виден разрезанный вдоль беловатый сосуд — спинная аорта (ао. d.). Если разрез несколько отклонился в сторону, то видна широкая задняя кардинальная вена (левая или правая) с пигментированными тонкими стенками. В хвостовой области артериальный сосуд—хвостовая артерия и венозный — хвостовая »е н а лежат под хордой один под другим внутри соединительнотканного футляра.
В области туловища вентральнее от кровеносных сосудов расположена б р ю ш н а я полость, сплошь заполненная п о л-о в о й железой. Удалив часть железы в заднем отделе брюшной полости, рассматривают медиальную стенку почки с широким белым мочеточником, тянущимся по нижнему краю, который особенно хорошо виден именно с внутренней стороны почки.
По дну брюшной полости тянется местами перерезанная к и-шечн-а я трубка. В передней части брюшной полости виден треугольник разрезанной вдоль печени. По ее нижнему краю тянется небольшая полость — это просвет печеночной вены (о. hepatis, v. h.). Впереди от печени лежит сердце (сот.), причем при левой половине тела остается предсердие, а при правой — желудочек, в стенке которого.можно видеть отверстие, соединяющее предсердие с желудочком. Обычно на правой стороне остается и тонкостенная трубка венозного синуса, в верхней части которого видны два отверстия — места владения передней и задней кардинальных вен. В нижней части видно впадение в венозный синус печеночной и нижней яремной вен. Перед передней стенкой желудоч.ка, внутри околосердечной капсулы, брюшная аорта имеет, небольшое утолщение —луковицу аорты. Сама брюшная аорта (ао. и.) редко бывает разрезана вдоль и на препарате обычно имеет вид трубки, тянущейся вперед от сердца, стенка которой пронизана отверстиями на местах' отхождения приносящих жаберных артерий.
Для того чтобы лучше рассмотреть органы, располагающиеся в • головной части, нужно очистить полости окружающих их синусов от сгустков лимфы. Непосредственно над брюшной аортой видна разрезанная вдоль относительно широкая дых а те л ьн а я тру б-ка (tub. г.). Стенка ее пронизана семью отверстиями, ведущими в жаберные мешки (f.br.in.). Сзади дыхательная трубка оканчивается слепо, а в передней части, около места соединения ее с ротовой полостью в ней располагаются направленные назад лепестки паруса (ие/.). От верхнего и боковых краев дыхательной трубки вперед в ротовую полость направлены шупальца — выросты слизистой оболочки. Над дыхательной трубкой тянется пищевод (oes.). Следует найти отверстие, которым пищевод открывается в
ротовую полость (oes'.). Между пищеводом и лежащей дорзальнее хордой лежит спинная аорта (ao.d.). Отодвигают пищевод вниз и рассматривают впадающие в спинную аорту выносящие жаберныеартерии.
Непосредственно под брюшной аортой видна продольная и кольцевая мускулатура я з ы к а (т.с-ар., т. т. р.), а под ней — нижняя яремная вена. В переднем отделе языка внутри мускулатуры лежит длинный голубоватый хрящ, поддерживающий язык (с. gl.).
Передний конец языка, на котором хорошо видны роговые п е-редняя и задняя языковые пластинки, разграничивает две полости: лежащую впереди и вниз от него — полость ‘предротовой воронки (cav. prr.) и сзади и вверху—ротовую полость (cav. or.). Соединяющая их щель является ротовым отверстием (stoma, st.). В передней части ротовая полость образует слепой мешок — гидросинус (г.). Над ротовой полостью видна узкая полость назогипофизарного мешка (сап. п.-h.). Его передний конец открывается наружу ноз д р ей, к задней стенке которой прилегает обонятельная капсула с органом обоняния (capsula offactoria, cap. о.). Задний конец оканчивается слепо под передним концом хорды и носит название питуитарного выроста. В верхней стенке назогипофизарного мешка имеется отверстие, затянутое только тонкой пленкой, через которое внутрь мешка вдается мозговая железа — гипофиз (hypophysis). Большое количество хрящей, имеющихся в головном отделе, будут разобраны при описании скелета.
Мускулатура (рис. 24)
Изучение мускулатуры, нервной системы и скелета при недостатке материала можно провести на одном экземпляре миноги, тогда последовательность должна быть следующей: мускулатура, скелет, нервная система. Однако очистку скелета лучше проводить на отдельном экземпляре и придерживаться того порядка работы, который описан в руководстве, тем более, что хорошо очищенные скелеты легко сохранить в формалине, и они могут быть использованы на следующий год.
Всю мускулатуру позвоночных животных' можно разделить на соматическую (париетальную), которая обусловливает, движения тела, и висцеральную, производящую движения внутренних органов.
Соматическая и висцеральная мускулатура отличается друг от друга также по своему происхождению и иннервации.
Соматическая мускулатура позвоночных развивается из мускульного листка — миотома — и иннервируется нервными волокнами, входящими в состав брюшных корешков спинномозговых нервов. Вся-эта мускулатура поперечно-полосатая.
Висцеральная мускулатура развивается из боковой пластинки мезодермы и иннервируется нервами, входящими у низших позво-
ночных в состав спинных корешков. Она может быть как гладкой, таки поперечно-полосатой.
Снимают кожу с одной стороны передней части туловища миноги. Для этого ножницами или скальпелем делают два разреза — по медиальной линии дорзальной и нейтральной сторон. Разрезы следует начинать несколько сзади от последнего жаберного отверстия и продолжать вперед до края ротовой воронки. Соединяют задние края разрезов по одной из сторон (лучше левой) и снимают кожу, захватив задний край получившегося лоскута пинцетом или пальцами. Удаляют подкожную клетчатку, осторожно соскоблив ее скальпелем. Соскабливание лучше всего начинать с переднего конца.
Рис. 24. Поверхностная мускулатура миноги Lampetra fluviatilis:
tn. с. — m. cornealis; tn. con. — m. constrictor; m. m. — миомеры; ms. — миосепты; m. sbo. — m, subocularis; m. spo. — m. supraocttlaris
Мускулатура туловища (соматическая мускулатура) составляет основную массу тела миноги. Снаружи она имеет вид ряда лент—мномеров (myomer, тт.), разделенных соединительнотканными прослойками — миосептами (myoseptum, /ns.).
Фактически каждый миомер представляет собой вертикально--стоящую пластинку, наружный край которой выходит на латеральную поверхность тела, а внутренний достигает соединительнотканного «позвоночника» и стенки полости тела. Пластинка миомера имеет небольшие изгибы и, кроме того, каждый впереди лежащий миомер накладывается каудальным краем на лежащий сзади, благодаря чему на поперечных срезах мы видим их лежащими в несколько слоев (рис. 21, 22, 23, тт.). Мускульные волокна миомера лежат параллельно продольной оси тела.
Мускулатура туловищной области в отличие от вышестоящих позвоночных еще не разделена горизонтальной перегородкой на дорзальную и вентральную. Миомеры правой и левой сторон соединяются друг с другом по средней линии дорзальной и вентральной поверхности. Только в области жаберных отверстий соматическая мускулатура разделяется на дорзальную и. вентральную, причем 92
здесь в головном конце происходит- и ее дальнейшая дифференцировка. В районе последнего жаберного отверстия дорзальные и вентральные миомеры соответствуют друг.другу, но по мере продвижения вперед вентральные миомеры делаются шире, так что на каждый из них приходится два дорзальных.
Несколько передних миомеров дорзальной части мускулатуры правой и левой сторон не срастаются друг с другом. Их разделение начинается примерно за обонятельной капсулой. Передними концами эти миомеры прирастают к заднему верхнему хрящу, их называют m. supraocularis (tn. spo). При одновременном действии мускулов обеих сторон происходит приподнимание заднего верхнего хряща, при одностороннем действии вместе с другими мускулами — поворачивание головы вбок.
Ростральная часть вентральной мускулатуры образует m. subo-cularis (tn. sbo.). Он занимает всю вентральную часть головы, начиная от заднего края ротовой воронки, к которому он прикрепляется, и тянется назад до уровня заднего края глаза, где ясно отделен от остальной мускулатуры септой. Сам мускул состоит из большого числа мускульных сегментов, разделенных миосептами. При своем сокращении m. subocularis оттягивает кольцевой хрящ назад.
Между первым жаберным отверстием и задним краем глаза лежит особый мускул m. cornealis (tn. с.). Каудальным концом он прикрепляется к* жаберной решетке, а ростральным — соединен с дисцеметовой оболочкой роговицы (см. стр. 102). При сокращении мускула роговица уплощается, давит на хрусталик, и тот перемещается ближе к сетчатке. Таким образом мускул участвует в аккомодации глаза.
Производными миотомов головы являются также лежащие под глазным яблоком шесть глазных мышц, которые обеспечивают подвижность глазного яблока.
Висцеральная мускулатура представлена у миног мускулами жаберного аппарата. На препарате видны только небольшие участки этой мускулатуры в промежутках между жаберными отверстиями и дорзальной и вентральной частями париетальной мускулатуры. Это участки общего жаберного сжимателя (m. constrictor, tn. соп.), который окружает жаберную область.
Сложная мускулатура присасывательного аппарата, связанная со стенками ротовой воронки и языком, специфична для круглоротых. Частично она была описана выше, а более подробный разбор не входит в задачи данного руководства.
Нервная система
Нервная система круглоротых, как и всех бесчелюстных, имеет наиболее примитивное строение среди позвоночных животных. Туловищные и головные нервы сохраняют, как у ланцетника, отдельные брюшные (двигательные) и спинные (смешанные) корешки, не
образуя спинно-мозговых нервов. Головной мозг расчленен на четыре отдела, так как мозжечок не развит. Головных изгибов нет. Передний мозг имеет общую полость, он состоит главным образом из обонятельных долей и является центром обонятельных функций. В промежуточном мозге мощно развиты вторичные обонятельные центры (габенулярные ганглии) и парные теменные* органы (париетальный и пинеальный), а в дне мозга расположен сложно устроенных гипофизарный орган. Орган статоакустики развит слабо, глаза также. Главные мозговые центры, координирующие функции мозга, расположены в столовой части мозга (сетевидная формация, под-бугровая область). Имеется десять пар головных нервов, среди которых сложно разветвленные нервы, снабжающие висцеральный аппарат.
Удаляют мускулатуру с верхей части головы, вплоть до крыши черепной коробки и соединительнотканной трубки, в которой заключен спинной мозг. Для этого остро отточенным скальпелем или лезвием безопасной бритвы срезают ее тонкими ломтиками до тех пор, пока не обнажится крышка черепной коробки. Срезы нужно начинать, несколько отступя назад от ноздри, и продолжать до уровня 4—5-го жаберного отверстия. Осторожно срезают дорзальную стенку футляра спинного мозга на уровне 4-го жаберного отверстия. Вставляют конец ножниц в образовавшееся отверстие и, ведя разрез вперед, вскрывают спинномозговой канал и крышку черепной коробки по средней линии вплоть до обонятельной капсулы. Не следует вводить ножницы слишком глубоко, чтобы не повредить нервную систему! Образовавшуюся щель расширяют, постепенно срезая крышку черепной коробки, но так, чтобы осталась целой одна из слуховых капсул. Особенно осторожно нужно подрезать в передней части, где головной мозг вплотную прилегает к дорзальной стенке черепа. Снимают одевающие мозг мозговые оболочки, одновременно удаляют одну из боковых стенок черепной коробки, срезав ее скальпелем или ножницами примерно до половины высоты. Затем промывают препарат под слабой струей воды и знакомятся более подробно с отделами центральной нервной системы, которые хорошо видны при рассматривании мозга сверху..
Спинной мозг
Спинной мозг (medulla spinalis, рис. 25, 26, med. sp.) имеет вид ленты и лежит в спинномозговом канале. Срезают верхнюю стенку канала на небольшом протяжении. Пользуясь лупой, можно рассмотреть в боковых стенках канала спинного мозга перерезанные поперек отдельные хрящики. Это зачатки передних и задних верхних дуг позвонков. Между ними выходят вентральные корешки спинномозговых нервов. В головном отделе передние и задние дужки срастаются так, что вентральные корешки проходят через отверстие в центре единой хрящевой пластинки. Ориентируясь по этим отверстиям, находят вентральные корешки спинномозговых нервов и рассматривают их отхождение от спинного мозга (рис. 25, 94
rd. v.). Дорзальные корешки спинномозговых нервов очень тонкие
и при препаровке обычно обрываются.
Следует рассмотреть также строение спинного мозга на попе
речном разрезе. Для этого можно воспользоваться препаратами
разрезов через туловищную область минОги, на которых при 20-кратном увеличении лупы хорошо видна уплощенная форма спинного мозга. В центре имеется небольшое отверстие — невроцель (neurocoel), окруженное более темным веществом, чем все-остальное вещество мозга.
Вентрально от центрального канала расположены группы относительно крупных прозрачных полостей. Это перерезанные поперек мюллеровские волокна — отростки крупных нервных клеток, которые осуществляют связь, двигательного аппарата головного и хвостового отделов тела миноги.
Головной мозг сверху (рис. 25)
Прежде чем познакомиться с отделами головного мозга, следует обратить внимание на то, что мозг занимает не всю полость черепной коробки, а лежит в ней довольно свободно. В отличие от других позвоночных головной мозг миноги имеет только четыре хорошо развитых отдела и не имеет изгибов в вертикальной плоскости.
Продолговатый мозг (medulla oblongata, med. о.) располагается в черепной коробке на уровне слуховых капсул. Дно и стенки продолговатого мозга образованы нервной тканью, крыша состоит из тонкой эпи-
med. sp.
Рис. 25. Головной мозг мн-ногн Lampetra fluviatilis сверху:
aq. s. — сильвиев водопровод; с. prt. — ножка теменного органа; crb. — мозжечок; dnc. — промежуточный мозг; fos. rh. — ромбовидная ямка; g. hab. — га-бенулярные ганглии; hem. — полушария переднего мозга; I- olf. — обонятельные доля; I. opt. — зрительные доли; med. о. — продолговатый мозг; med. sp. —спянной мозг; pl. ch. р. — место расположения заднего мозгового сплетения кровеносных сосудов; rd. v. — брюшные
корешки спинномозговых нервов; s. с. I. — центральная борозда, tine. — передний мозг; v. trt. — полость третьего желудочка; / — обонятельный нерв; II— зрительный нерв; VII — лицевой нерв; ViII — слуховой нерв; IX — языкоглоточный нерв; X — блуждающий иерв
телиальиой пленки, которая плотно срастается с мозговыми оболочками и обычно удаляется вместе с ними при препаровке, открывая отверстие, через которое видна широкая полость четвертого мозгового желудочка, или ром-
бовидная ямка (fossa rhomboidea,
fos. rh.). Вместе с эпителиальной крышей удаляется и расположенное в ней крупное сплетение кровеносных сосудов (plexus choriodeus posterior, pl. ch. p.). Место соединения этого
сплетения с головным мозгом имеет вид двух утолщений на заднем крае ромбовидной ямки.
По медиальной части дна ромбовидной ямки проходит центральная борозда (sulcus centralis longitudinalis, s. с. I.).
Мозжечок (cerebellum; crb.) у миноги развит слабо и не выделяется как самостоятельный отдел. Ему соответствует верхняя часть передней стенки четвертого желудочка, которая имеет вид поперечного валика на дорзальной стороне передней части продолговатого мозга.
Средний мозг (mesencephalon) лежит впереди от продолговатого мозга на уровне переднего края слуховой капсулы. На его дорзальной поверхности резко выделяются два яйцевидных вздутия— зрительные доли (lobi optici, I. opt). Между зрительными долями имеется отверстие, окаймленное небольшим валиком. Нормально оно закрыто тонкой эпителиальной пленкой с развитым в ней средним мозговым сплетением (plexus cho-rioideus medius). Однако, так же жак и крыша продолговатого мозга, она легко удаляется при препаровке вместе с мозговыми оболочками. Через это отверстие видна внутренняя полость среднего мозга, носящая название сильвиева водопровода (aquaeductus Sylvii, aq. S.). Переднюю стенку отверстия образует задняя спайка (commissura posterior, сот. р.), связывающая правый и лев'ый зрительные бугры.
Промежуточный мозг (diencephalon, dnc.).. Впереди средний мозг без резкой границы переходит в промежуточный мозг. Крыша последнего сильно утолщена, однако также не полностью образована нервным веществом, поэтому в медиальной части видно отверстие треугольной формы, ведущее в полость третьего желудочка (ventriculus tertius, v. trt.). На каудальной стенке отверстия можно различить два _ небольших бугорка— г а б е н у-ларные ганЬлии (ganglion habenulae, g. hab.). С правым, более крупным ганглием, связан при помощи длинной и тонкой ножки пинеальный орган (corpus pineale), располагающийся под кожей и имеющий вид небольшого диска. У рептилий он превращается в эпифиз, отсюда его другое название epiphysis cerebri. Этот орган так же, как и лежащий под ним пузырек п ар ие-тального (corpus parietale) или пар апи не а ль ног о орга-н а, называемого также теменным глазом, можно сохранить только при тщательной препаровке под бинокулярным микроскопом. Обычно они удаляются при снятии оболочек мозга, и на препарате остается только небольшой стебелек (рис. 25, с. prt.)—остаток связи париетального органа с левым ганглием.
Передний мозг (telencephalon, tine.) своей передней стенкой примыкает непосредственно к обонятельной капсуле, а сзади отграничен от промежуточного мозга довольно четко выраженной бороздой. Он почти полностью разделен на два полушария спереди задней стенкой обонятельной капсулы, а сзади — глубоко вдающимся в него передним отделом промежуточного мозга. Правая и левая половины в свою очередь делятся на две неравные доли: не-96
большие задние доли, лежащие по бокам промежуточного мозга и являющиеся собственно большими полушариями переднего мозга (hemisphaera, hem.), и располагающиеся впереди от них крупные обонятельные доли (lobus olfactorius, I. olf.), передние отделы которых, как показывает внутреннее строение и развитие, правильнее называть обонятельными луковицами (bulbus olfactorius). К оро-медиальному углу каждой из них подходит обонятельный нерв (рис. 25,1).
Головные нервы
Следующим этапом изучения нервной системы является знакомство с черепномозговыми нервами. Для этого отодвигают головной мозг от срезанной наполовину стороны черепной коробки. Отходящие от мозга нервы становятся видимыми. Рассматривать их лучше всего под простой или бинокулярной лупой с небольшим увеличением. Не оттягивайте мозг слишком сильно, так как тонкие корешки головных нервов легко обрываются. Даже при очень тщательной препаровке трудно сохранить все нервы, поэтому удается рассмотреть только часть. Однако в дальнейшем описании будут даны все черепно-мозговые нервы и соответственно оговорены те, которые обычно отсутствуют на препарате. Все без исключения головные нервы — парные. Обычно черепно-мозговые нервы обозначаются определенными римскими цифрами. Эти обозначения взяты из анатомии человека, где порядок нумерации соответствует порядку отхождения нервов от головного мозга. Однако у позвоночных животных места отхождения нервов несколько изменены, поэтому часто нерв, имеющий больший порядковый номер, отходит впереди нерва с меньшим порядковым номером.
Обонятельный нерв, I (nervus olfactorius, рис. 25, I). Этот нерв представлен отростками рецепторных клеток, лежащих в полости обонятельной капсулы. Собираясь вместе, они выходят из капсулы через два отверстия в задней ее стенке (foramen pro nervi olfactorii) и входят в обонятельные луковицы переднего мозга. На препарате видны только небольшие отрезки нерва, лежащие между стенкой капсулы и передним мозгом.
Зрительный нерв, или зрительный тракт, II (nervus opticus, s. tractus opticus, рис. 25, II) отходит от дна промежуточного мозга, прободает стенку черепной коробки примерно на уровне заднего края переднего мозга, выходит в так называемую «орбиту», а затем входит в глазное яблоко через его медиальную стенку.
Глазодвигательный нерв, III (nervus oculomoto-rius, рис. 26, III) отходит от вентральной части латеральной поверхности среднего мозга и проходит через стенку черепной коробки на уровне границы промежуточного и среднего мозга. Он очень тонок, и обычно его часть, отходящая от мозга, обрывается при препаровке, зато довольно легко проследить его ход вне черепа. По выходе из черепной коробки он сейчас же делится на две ветви.
Одна ветвь более толстая (ramus inferior), тянется вперед и дает две ветви — одну к перед ней косой мышце глаза (m. ob-liquus anterior) и вторую, идущую вниз, к передней прямой мышце (m. rectus internus). Косые мышцы глаза у миноги, в отличие от других позвоночных, прикрепляются к черепу не одна над другой, а на довольно значительном расстоянии. Причем, одна — лежит впереди глазного яблока, а другая — позади него, поэтому их удобней называть передней и задней, а не верхней и нижней.
Вторая ветвь (г. superior) иннервирует верхнюю и нижнюю прямые мышцы (m. rectus superior, т. rectus inferior) .
Блоковый нерв, IV (nervus trochlearis) отходит от вентральной поверхности зрительных долей, проходит между средним и продолговатым мозгом и через отверстие, расположенное перед слуховой капсулой, выходит наружу. Он иннервирует заднюю косую мышцу глаза (m. obliquus posterior). При препаровке легко обрывается (рис. 26, IV). К нервам, иннервирующим глазные мышцы, относится также и отводящий нерв.
Отводящий нерв, VI (nervus abducens, рис. 26, VI) берет начало от переднего бокового угла продолговатого мозга, рядом с корешками n. trigeminus, вместе с последними выходит из черепа через отверстие перед слуховой капсулой и делится на две ветви. Одна из них подходит к m. rectus posterior, а другая — кт. rectus inferior. Корешки его обычно трудно отделить от корешков V нерва.
Тройничный нерв, V (nervus trigeminus, рис. 26, V) начинается на боковой поверхности переднего конца продолговатого мозга несколькими корешками, выходит из черепа через названное выше отверстие перед слуховой капсулой и по выходе из черепа сразу делится на две ветви: 1) г. ophthalmicus, тянется вперед в направлении нижнего края обонятельной капсулы и иннервирует кожу верхних и боковых частей головы и головные органы боковой линии; 2) г. maxillo-mandibularis иннервирует кожу по бокам головы и по краям ротовой воронки, а также мускулатуру боковых частей головы и языка.
Лицевой нерв, VII (nervus facialis, рис. 25, VII). Его корешки отходят несколько впереди от корешков VIII нерва, от латеральной поверхности средней части продолговатого мозга и вместе с n. acusticus входят в слуховую капсулу, из которой выходят через отверстие в медиально-оральном углу. По выходе лицевой нерв образует ганглий, прилегающий к V нерву, что при беглом рассмотрении создает впечатление выхода VII нерва вместе с корешками IV, V и VI нервов через отверстие впереди слуховой капсулы. От его основной ветви (г. hyomandibularis), идущей вниз и внутрь жаберного скелета, отделяется г. lateralis, которая идет назад вдоль наружной поверхности слуховой капсулы, сливается позади с ветвью блуждающего нерва, образуя латеральную ветвь, иннервирующую туловищные органы боковой линии.
Слуховой нерв, VIII (nervus acusticus, рис. 25, VIII) отходит от боковой поверхности продолговатого мозга и входит в слуховую капсулу через отверстие в середине ее медиальной стенки. Как показывает само название, этот нерв иннервирует орган слуха.
Языкоглоточный, IX (nervus glossopharyngeus) и блуждающий, X (nervus vagus) нервы (рис. 25, IX, X). Большая группа корешков, которыми отходят эти нервы, хорошо видна на препарате даже при небольшом увеличении. Начинаясь на латеральной поверхности продолговатого мозга, оба нерва выходят из спинномозгового канала уже за пределами черепа через отверстие между задним краем слуховой капсулы и передними верхними дужками. Эти нервы иннервируют жаберный аппарат, а п. vagus, кроме того, внутренние органы и вместе с г. lateralis VII нерва участвуют в иннервации органов боковой линии на туловище.
Головной мозг снизу
Перерезав скальпелем корешки головных и передних спинномозговых нервов с обеих сторон, несколько отступя от мозга, осто-
рожно, стараясь не повредить передний мозг и обрезав обонятельные нервы, отворачивают мозг назад. Для этого аккуратно, захватив передний край пинцетом, поднимают мозг вверх, отпрепа-ровывая его от дна черепной коробки. Можно также перерезать спинной мозг и тогда совсем вынуть головной мозг из черепной коробки и поместить на часовое стекло с водой. Очищают нижнюю поверхность мозга от мозговых оболочек и находят снизу границы четырех отделов мозга. На нижней поверхности мозга бросается в глаза вырост дна промежуточного мозга.
Мозговая воронка (infundibulum, рис. 26, in.). Боковые части ее носят название д о-лей воронки (lobus infundi-buli). С внутренней стороны к мозговой воронке прилегает нижняя мозговая железа —гипофиз (hypophysis), обычно отры-
Рис. 26. Головной мозг миноги Lam-pelra fluvialilis сбоку и снизу:
с. prt. — теменной орган; g. hab. — габену-лярный ганглий (правый); hem. — большие полушария; infd. — мозговая воронка; lob. о/. — обонятельные доли; med. о. — продолговатый мозг; med. sp. — спинной мозг; I — обонятельный нерв; II — зрительный нерв: /// — глазодвигательный нерв; IV — блоковый нерв; V — тройничный нерв; VI — отводящий нерв; VII— лицевой нерв;
VIII — слуховой нерв
вающаяся от нее при препаровке
и остающаяся в затянутом соединительнотканной пленкой отвер-стии дна черепной коробки, где ее можно рассмотреть, пользуясь лупой.
Полость мозговой воронки имеет вид продолговатой щели. Утолщенные края ее носят название saccus infundibuli. Дорзальная часть воронки связана отверстием с полостью промежуточного мозга.
, С помощью лупы рассматривают корешки головных нервов, отходящих от вентральной поверхности головного мозга. Впереди от infundibulum видны относительно крупные парные зрительные нервы. В отличие от таковых у других позвоночных они не образуют зрительного перекреста и направлены не вперед, а назад (рис. 26, II). Несколько позади заднего края воронки можно рассмотреть корешки глазодвигательного нерва. На вентральной поверхности зрительных долей в их задней части иногда остаются небольшие, очень тонкие корешки IV пары головных нервов (рис. 26, IV). Места отхождения V и VI, а также VII и VIII нервов легко найти, ориентируясь по их корешкам, которые были хорошо видны и при рассмотрении мозга сверху (рис. 25).
Органы чувств
Сейсмосензорные органы, или органы боковой линии; их расположение было подробно описано выше (см. рис. 15). Более детально их можно рассмотреть только на микроскопических препаратах.
При удачно прошедшем
через середину срезе можно видеть, что каждый бугорок состоит как бы из двух половинок, разделенных желобком, на дне которого и помещается чувствующая почка, состоящая из рецепторных и опорных клеток.
Орган слуха у мино-
Рис. 27. Перепончатый лабнрнит миноги Lampetra fluviatilis с латеральной стороны:
а. с. sem. а. — ампула переднего полукружного канала; а. с. sem. р.—ампула заднего полукружного канала; can. sem. ant. — передний полукружный канал; can. sem. post. — задний полукружный канал; с. сот. —« общая ножка полукружных каналов; сот. — комиссура; utr. — преддверие
ги представлен внутренним ухом, расположенным внутри слухо-вых капсул. На нетронутой при изучении головного мозга стороне
черепа очищают слуховую капсулу от окружающей мускулатуры. Маленькими ножницами вскрывают ее латеральную стенку и, оттягивая хрящи капсулы, расширяют разрез. Внутри капсулы находится перепончатый лабиринт. Отодвигая его иголкой, срезают всю верхнюю часть капсулы, а затем при помощи иголки вычленяют лабиринт. Это удается довольно легко, поскольку приходится подрезать только слуховой нерв, входящий в медиальную стенку перепончатого лабиринта. Вычленив лабиринт, помещают его в сосуд с водой и рассматрива
ют с латеральной, противоположной месту вхождения слухового нерва поверхности под бинокулярным микроскопом. В сложно устроенном внутреннем ухе можно различить следующие части. Наиболее латерально располагаются два вертикальных полукружных канала (canalis semicircularis anterior et posterior, рис. 27, can. sem. ant., can. sent. post.). Горизонтальный канал у миноги отсутствует. Дорзальные концы вертикальных каналов сливаются в общую ножку (crus communis, рис. 27, с. сотт.), которая переходит в так называемую комиссуру (рис. 27, сот.). Нижние концы полукружных каналов оканчиваются расширениями-ампулами (ampulla canalis semicircularis anterioris et posterioris, рис. 27, a. c. sem. a., a. c. sem. p.). Между каналами располагаются два вздутия преддверия (utriculus, рис. 27, utr.).
Другие образования, такие, как лежащие на основании утрику- ' люса выпячивания (переднее более мелкое соответствует саккулю-су, а заднее лагене) и отходящий от верхней части утрикулюса эндолимфатический проток, на приготовленных препаратах рассмотреть обычно не удается. Из таких временных препаратов можно изготовить постоянные, окрасив их борным кармином и заключив, после обезвоживания, в канадский бальзам.
Парные органы зрения — глаза, как уже отмечалось, располагаются по бокам передней части головы миноги (см. рис. 15, ос.). Разрезают специально оставленное для этой цели глазное яблоко на две части. Разрез следует провести так, чтобы он прошел через середину зрачка и через место вхождения в глазное яблоко зрительного нерва. Хрусталик, который становится очень плотным под влиянием фиксирующей жидкости, обычно разрезать не удается. Одну из половин глазного яблока (ту, при которой остался хрусталик) кладут на дно небольшой ванночки, залитой водой, и рассматривают внутреннее строение глаза при 15—20-кратном увеличении лупы. Более четко строение глаза видно на специально приготовленных микроскопических срезах, прошедших через середину глазного яблока. Самая наружная оболочка глазного яблока — склера (рис. 28, sc.). Ее легко отличить по слегка голубоватому цвету. Она толще на медиальной поверхности глаза и постепенно истончается к его наружному краю. В области глазного нерва склера переходит в окружающую его твердую оболочку. В передней части склера переходит вдисцеметовую оболочку (рис. 28, д. о.) —тонкую пленку, одевающую глазное яблоко спереди. У высших позвоночных дисцементова оболочка является одним из слоев роговицы, но у миноги роговица еще не вполне отдифференцирова-лась от кожных покровов и представлена участком кожи над глазным яблоком, который отличается от остальных покровов толщиной, а также гистологическим строением (рис. 28, cut.). Соответственно с этим она и удаляется при удалении кожи.
Под склерой лежит сосудистая оболочка (chorioidea, рис. 28, chor.), имеющая черный цвет от присутствия большого количества пигментных клеток, которые покрывают стенки многочисленных
кровеносных сосудов. Следует отметить также глазную артерию (a. ophthalmica), ход которой внутри твердой оболочки глазного нерва хорошо заметен благодаря покрывающим ее пигментным клеткам.
Впереди сосудистая оболочка продолжается в радужную оболочку (iris, рис. 28, ir.). Отверстие в ее центре носит название зрачка (pupilia).
Изнутри полость глазного яблока выстлана сетчатой оболочкой, или сетчаткой (retina, рис. 28, ret.). При достаточ
Рис. 28. Схема строения глаза миноги Lampetra fluviatilis (по Гирсбергу и Рит-челу):
д. о. — дисцеметова оболочка; ц. с. — циннова связка; chor.—сосудистая оболочка; cut. — кожа; fr. — радужная оболочка; lens. — хрусталик; т. с. — m. cornealis; ret. — сетчатка; sc. — склера; II — зрительный нерв
ном увеличении лупы можно видеть ее слоистое строение. У живого животного она гладкая, а складки на ее поверхности появляются в результате-фиксации. В передней части полости глазного яблока лежит хрусталик (lens crystallina, рис. 28, lens.). Он имеет типичную почти для всех первичноводных животных шарообразную форму. Полость глазного яблока заполнена стекловидным телом, которое обычно вытекает при разрезе. Тонкая волокнистая пленочка, отходящая от основания радужной оболочки и сливающаяся за-
тем с плотным слоем хрусталика, носит название цинновой связки (рис. 28, ц. с.).
Зрительный нерв прободает с медиальной стороны стенку глазного яблока. При удачном разрезе можно видеть, что он проходит через наружные (по отношению к глазному яблоку) слои сетчатки и распадается на волокна только в самом внутреннем ее слое (рис. 28, II).
Орган обоняния. На сагиттальном разрезе необходимо более детально рассмотреть три отдела, из которых состоит орган обоняния.
Носовая трубка — короткий носовой проход, открывающийся наружу отверстием ноздри (см. рис. 23, паг.). Стенки его состоят из хрящевой ткани и довольно сильно пигментированы. Как уже указывалось, передний отдел несколько выступает над уровнем дорзальной поверхности головы.
В задней стенке носового хода имеется отверстие, ведущее во второй отдел органа обоняния — обонятельную капсулу (см. рис. 23, cap. о.), перед входом в которую носовая трубка образует две складки, способные отгибаться вперед и назад в зависимости от тока воды.
Внутри обонятельной капсулы помещается обонятельный мешок. На внутренней поверхности мешка имеется 15 радиально ориенти
рованных складок из соединительной ткани, которая богата пигментными клетками и на препарате выглядит черной. Последнее наиболее четко видно на .поперечном разрезе обонятельной капсулы, который можно приготовить заранее при изготовлении поперечных разрезов головного конца. На вершинах и боковых стенках складок располагается беловатый обонятельный эпителий. Непарная медиальная складка делит обонятельную капсулу на две симметричные части. Ее нижний конец выступает в полость носового хода в виде отростка.
Носовая трубка продолжается назад в длинный питуитарный вырост (рис. 23, can. n.-h.). Этот вырост тянется вниз и назад, прободает на своем пути задний верхний хрящ у его заднего конца и, проходя под основанием черепа и передним концом хорды, заканчивается слепо. Приблизительно в средней части питуитарный вырост соединяется с полостью черепной коробки отверстием, которое, правда остается затянутым тонкой соединительнотканной перепонкой. Через него в полость выроста вдается нижняя мозговая железа — гипофиз, отчего весь канал называют также назо-гипофизарным каналом. Носовой канал вместе с питуитарным выростом является своеобразной пипеткой, насасывающей и выталкивающей воду из органа обоняния.
Скелет
Скелетная система круглоротых отличается от скелета других позвоночных рядом особенностей, указывающих на их особое положение как представителей бесчелюстных — Agnatha и особого класса круглоротых — Cyclostomata. Осевой скелет представлен спинной струной — хордой, одетой плотным соединительнотканным футляром, образующим в туловище соединительнотканный — перепончатый позвоночник. В его верхней части лежат зачаточные верхние дуги. Скелет головы состоит из трех отделов: 1) осевого черепа— хрящевой мозговой коробки; 2) системы хрящей, образующих присасывательный аппарат ротовой воронки; 3) жаберного скелета, состоящего из тонкой хрящевой решетки, одевающей жаберные мешки с наружной стороны. Кроме того, непарные плавники поддерживаются тонкими хрящевыми нитями, также относящимися к осевому скелету туловища.
Основу скелета миноги составляет хорда, одетая соединительнотканным футляром, с включенными в него отдельными хрящевыми элементами. Скелет головного отдела почти полностью состоит нз хрящевых элементов. Хрящи скелета очень тонкие и плотно сросшиеся с соединительной тканью, поэтому препаровка скелета затруднительна. При препаровке лучше всего сохранять и соединительную ткань, удаляя ее лишь там, где она мешает рассмотрению хрящевых элементов скелета. Для облегчения препаровки полезно выдержать миногу в течение 2—3 дней в 10 %-ной азотной кислоте, а затем хорошо промыть в проточной воде. Вместо азотной кислоты частичной мацерации можно добиться помещением объекта на 5—
10 дней в тепловатую воду, ежедневно сменяемую. Хранить очищенный скелет нужно в мокром виде, лучше всего в 70° спирте. Обычно скелет сначала очищают начерно, а затем дочищают по мере изучения отдельных отделов. Препаровку скелета можно проводить на тех же объектах, на которых изучали строение мускулатуры.
Удаляют кожу со всего тела миноги, а также с одной стороны плавников. Вторую сторону оставляют нетронутой. Снимают всю массу мускулатуры с обеих сторон тела, для чего пальцами раздвигают мускулатуру на спинной стороне, несколько отступя назад от последнего жаберного отверстия. Отделяют мускулатуру с каждой стороны от соединительнотканного футляра и затем, захватив пальцами или пинцетом, снимают целыми пластами в направлении назад. Особенно осторожно следует снимать мышечные пласты у основания плавников. Необходимо следить за тем, чтобы не отделить вместе с мышцами и тонких хрящевых лучей плавника. Вместе с мускулатурой удаляют и внутренние органы, вплоть до жаберной решетки. С верхней части головы мускулатуру также легко снять пинцетом. Удаляют мускулатуру с жаберной решетки так, чтобы не повредить лежащих под ней тонких хрящей жаберного скелета.
На таком грубо очищенном препарате рассматривают отделы, на которые делится скелет миноги.
1. Осевой скелет туловища — хорда и плотный соединительнотканный футляр, одевающий ее и спинной мозг.
2. Скелет плавников, непарных и хвостового, — тонкие хрящевые нити, поддерживающие это образование.
3. Скелет ротовой воронки — хрящевые образования, поддерживающие ротовую воронку, называемые губными хрящами.
4. Скелет языка — хрящи, поддерживающие язык.
5. Скелет осевого черепа, нейрокраниум, — хрящевой скелет мозговой коробки вместе со слуховыми и обонятельной капсулами.
6. Висцеральный скелет, скелет жаберного аппарата, — тонкие-хрящи жаберной решетки.
Затем знакомятся более подробно с каждым отделом, одновременно производя допрепаровку.
Осевой скелет
Осевой скелет миноги можно разделить только на два отдела: туловищный и хвостовой. Необходимо тщательно очистить небольшие участки осевого скелета в каждом из этих отделов, соскабливая остатки мускулатуры с одной и другой стороны скальпелем, рассмотреть эти участки на просвет, разобрать строение скелета отдельного сегмента тела. Границы сегмента можно определить по поперечным полоскам на соединительнотканном футляре, представляющим места прикрепления миосепт — соединительнотканных прослоек между миомерами.
Спинная струна, хорда (chorda dorsalis, рис. 29,. ch.) —цилиндрический упругий стержень, состоящий из особой пу-104
дырчатой ткани и имеющий две оболочки — наружную (elastica externa) и волокнистую. Эти оболочки хорошо видны на поперечных разрезах (см. рис. 20, el. е., el. е'.). Со всех сторон хорда одета плотным волокнистым футляром, дорзальная часть которого образует невральный канал, заключающий в себе спинной мозг. Вскрывают на небольшом участке и рассматривают хорду. Хорда тянется вдоль всего тела миноги. Впереди она оканчивается в основании осевого черепа, а назад тянется до конца хвоста, постепенно утончаясь.
Верхние невральные дуги (arcus neuralis, рис. 29, •arc. п.). Единственные элементы позвонка, развивающиеся у миноги— маленькие хрящевые палочки, лежащие в толще соединительной ткани справа и слева над хордой и принимающие участие в образовании спинно-мозгового канала. Их довольно легко обнаружить, проведя вдоль верхнего края хорды концом иголки, которая при этом издает характерный трещащий звук, цепляясь за основания верхних дуг. Чтобы рассмотреть верхние дуги, соскабливают скальпелем соединительную ткань над хордой. Соскабливание следует вести спереди назад. Лучше всего хрящи верхних дуг видны в области спинных плавников, где они наиболее длинные.
Пользуясь лупой, рассматривают положение верхних дуг. На каждый мышечный сегмент одной стороны приходится по две верхние дужки: одна лежит против места прикрепления к соединительнотканному футляру миосепты, другая — против середины миомеры. Все дужки несколько наклонены назад. В головном конце дужки более крупные и могут сливаться друг с другом. В хвостовой области и в области плавников они тонкие и длинные.
Гемальный канал (canalis haemalis). В хвостовом отделе соединительная ткань образует канал также и под хордой, в котором помещаются хвостовая артерия и хвостовая вена. Он хорошо виден на поперечном разрезе (см. рис. 21). Этот канал носит название гемального канала.
Настоящие нижние дуги позвонков у миноги отсутствуют, однако скопления скелетогенной ткани, образующие гемальный канал, соответствуют нижним дугам позвоночника. Такие же, но менее ясно выраженные скопления имеются и в туловищной области.
Скелет плавников
Скелет спинных и хвостового плавников состоит из длинных тонких сплющенных хрящевых плавниковых лучей, верхние части которых поддерживают кожную складку плавника, а основания проходят через слой мускулатуры и доходят до дорзальной стороны соединительнотканного футляра хорды. На спинной стороне они соответствуют дорзо-спинальным элементам (dorsospi-nalia), на брюшной — вентроспинальным (ventrospina-Иа) хрящевых рыб. Хвостовой плавник имеет равные лопасти и до самого конца разделен хордой. Такой хвост называется первичноравнолопастным — прото цер ка ль ны м,
Скелет ротовой воронки
Скелет ротовой воронки — своеобразные хрящи, которые поддерживают мягкие части ротовой воронки, передней части головы и языка. Вместе с мускулатурой они выполняют тройную функцию: 1) присасывание воронкой к коже жертвы; 2) разрушение языком и зубами ротовой воронки кожи и мягких частей добычи; 3) сосательные движения ротовой полости.
Рис. 29. Скелет головного отдела миноги Lampetra fluviatilis:
в. — вентральная комиссура; г. — гипотрематическая комиссура; э. —эпитрематическая комиссура; acr. Ьг.—жаберная дужка; arc. br., 1 — первая жаберная дужка; асг Ьг., 8 — восьмая жаберная дужка; аге. п. — верхняя дуга позвонка; с. ап. — кольцевой хрящ; cap. а. — слуховая капсула; cap. о. — обонятельная капсула; с. сор.—средний нижний хрящ; с. crd. — околосердечный хрящ. с. gl.—подъязычный хрящ; ch. — хорда; с. //. — передний боковой хрящ; с. тп.—стилевидиый хрящ; с. гЛ. — задний боковой хрящ; с. so.—подглазничная дуга; с. sp. — палочковидный хрящ; с. tec. — передний верхний хрящ; с. tr.—задний верхний хрящ; f. so. —подглазничное отверстие; f- II — отверстие для II нерва; f. IV, V, VI — отверстие для IV, V, VI нервов; пгег. —черепная коробка
При очистке скелета ротовой воронки нужна особая осторожность. Мускулатура, которую приходится удалять, очень плотная, кроме того, отдельные хрящи и мускулатура связаны плотной соединительной тканью, поэтому мягкие хрящи легко могут быть удалены вместе с мускулатурой и соединительной тканью. Чтобы не повредить хрящей, следует ориентироваться по рисунку. Одни и те же хрящи миноги у разных авторов имеют разные названия. Здесь приведены наиболее употребимые сейчас русские названия хрящей, а в латинских названиях использована терминология Д. К. Третьякова. В его работе, посвященной скелету и мускулатуре головы миноги, имеется и таблица синонимов названий хрящей.
Кольцевой хрящ (cartilago annularis, рис. 29, с. ап.) — массивное хрящевое кольцо, которое поддерживает ротовую воронку. К нему прикрепляются верхнечелюстные и нижнечелюстные зубы.
Передний верхний хрящ (cartilago tectoria, рис. 29 с. tec.) —широкая непарная хрящевая пластинка. Передний конец ее располагается над дорзальной частью кольцевого хряща. Задний край прикрыт передним концом заднего верхнего хряща. С каждой стороны от него отходят направленные вниз отростки.
Задний верхний хрящ (cartilage trabecularis, рис. 29, с. tr.) лежит над передним верхним хрящом вперед от обонятельной капсулы. Задним концом он срастается с передней частью черепной коробки. Имеет форму хрящевой пластинки с широким и раздвоенным передним краем и довольно узким, но значительно более толстым задним концом. Этот хрящ в виде щита покрывает дорзальную поверхность предчерепной области головы.
Передние боковые хрящи (cartilago lateralis, рис. 29, с. et.)—парные палочкообразные хрящики, лежащие под передним верхним хрящом впереди от его нисходящих отростков в толще соединительной ткани. При препаровке хрящей следует соблюдать осторожность, так как их легко можно удалять вместе с мускулатурой. К этим хрящам прикрепляется мускулатура ротовой воронки.
Задние боковые хрящи (cartilago rhomboidea, рис. 29, с. rh.) —парные относительно широкие и плоские хрящевые пластинки, лежащие под задним верхним хрящом между нисходящими отростками переднего верхнего хряща и передним боковым отростком подглазничной дуги.
К скелету ротовой воронки относятся также хрящи, поддерживающие ее вентральную часть.
Палочковидные хрящи (cartilago spinosa, рис. 29, с. sp.) —.парные, узкие, заостренные на заднем конце хрящи. Передними утолщенными концами они прикрепляются к латеральным частям заднего края кольцевидного хряща.
Средний нижний хрящ (cartilago copularis, рис. 29, с. сор.) Т-образной формы, непарный хрящ с длинным заостренным на конце средним отростком, направленным назад. Он лежит под подъязычным хрящом между выше названными палочковидными хрящами.
Скелет языка (рис. 29)
Подъязычный хрящ (cartilago glossae. с. gl.)—длинный, сплющенный с боков хрящ, поддерживающий мускулатуру языка. Передний конец его лежит в области кольцевидного хряща, далее он, постепенно утончаясь, идет назад и проникает внутрь жаберного аппарата. Средняя и боковые лопасти языка поддерживаются рядом мелких хрящиков, располагающихся около переднего конца подъязычного хряща. Правая и левая лопасти языка поддерживаются желобковидным апикальным хрящом (cartilago apica-lis), который небольшим эластическим хрящом (cartilago interglossa) связан с подъязычным хрящом. Кроме апикального хряща передний конец языка поддерживается небольшими супра-апикальными хрящами (cartilago supraapicalis) и непарным преапикальным хрящом (cartilago praeapica-lis). Первые лежат на дорзальной, а второй — на вентральной стороне апикального конца.
Осевой череп
Очищают мозговую коробку от остатков мускулатуры и соединительной ткани со всех сторон. При этом особенно осторожно следует очищать подглазничную дужку и лежащий позади нее стилевидный отросток. Плотные соединительнотканные перепонки полностью удалить обычно не удается, так как при этом можно повредить хрящи. Точные границы хрящей легко установить, прощупывая их концом препаровальной иголки.
Осевой череп, нейрокраниум, мозговая короб-к a (neurocranium, рис. 29, пгсг.) является непосредственным продолжением спинномозгового канала и продолжается вперед до задней стенки обонятельного мешка. Хрящевыми являются только основания и боковые стенки осевого черепа. Крыша мозговой коробки остается перепончатой и только в ее задней части на уровне слуховых капсул имеется тонкая хрящевая перемычка. Вторая неполная перемычка лежит несколько впереди от уровня переднего края слуховых капсул. К боковым стенкам мозговой коробки в задней части прирастают яйцевидные хрящевые слуховые капсулы (capsula auditiva, рис. 29, cap. а.). Передняя стенка нейрокра-ниума образована обонятельной капсулой (capsula olfac-toria, s. nasalis, рис. 29, cap. о.).
От переднего и заднего концов вентральной части боковых стенок отходит косо вниз и в сторону хрящевая подглазничная дуга (cartilage subocularis, рис. 29, с. so.), которая служит ложем для глазного яблока и ограничивает снизу подглазничное отверстие (foramen suboculare, рис. 29, f. so.). Подглазничная дуга по своему происхождению относится к висцеральнойу скелету.
От внутренней части каудального конца подглазничной дуги отходит направленный вниз стилевидный хрящ (cartilago mandibularis, рис. 29, с. тт.). Его нижняя часть несет вытянутый вперед тонкий хрящевой отросток (cartilago cornualis). На стилевидный отросток накладывается один из передних хрящей скелета жаберной решетки. Сами хрящевые боковые стенки прободены рядом отверстий.
Срезают ножницами крышу черепной коробки, а также верхнюю стенку спинномозгового канала, удаляют головной мозг и очищают черепную коробку с внутренней стороны.
Примерно в средней части дна мозговой коробки имеется крупное отверстие f. palato-nasalis, которое делит осевой череп на два отдела: задний — хордальный и передний — прехордальный. Само отверстие соединяет полость черепной коробки с полостью питуитарного выроста. В нем, как уже указывалось, лежит нижняя мозговая железа — гипофиз.
Рассматривают более детально строение основания черепа.
Основание заднего, хордального, отдела разделено на две половины, вдающимся в него вплоть до небно-носового отверстия передним концом хорды (рис. 30, ch.). Расположенные по бокам от нее хрящевые пластинки носят название парахордаль-108
них пластинок, или задних парах ордалий (рис. 30, pch. р.). На вентральной поверхности черепа они срастаются под хордой, а на спинной стороне дна черепа только примыкают к хорде по бокам.
Передний, п р е х о р д а л ь-ный, отдел осевого черепа характеризуется положением впереди небно-носового отверстия и носит название передних парахордалий (рис. 30, pch. а.). По аналогии с черепом других позвоночных этот отдел назывался раньше трабекулами. Он образует переднюю часть дна мозговой коробки и в основании черепа продолжается вперед в задний верхний хрящ. Передняя стенка мозговой коробки образована обонятельной капсулой (рис. 29, cap. о.).
На уровне отверстия в на-зо-гипофизарный канал в боковых стенках черепной коробки имеются крупные отверстия, которые служат для выхода зрительного (рис. 30, f. II) и глазодвигательного (рис. 30, f. Ill) нервов. Это отверстие как бы подразделяется на две неравные части спускающимся сверху выростом: через переднюю часть проходит зрительный нерв, через заднюю — глазодвигательный. Следующее отверстие перед слуховой капсулой значительно меньших размеров, но через него выходит несколько нервов: IV, V, VI (см. рис. 29, f. IV, V, VI). Отверстие в средней части медиальной стенки слуховой кап
сулы служит для прохождения внутрь капсулы VII и VIII нервов (рис. 30, f. VII, VIII). Че-
рез пространство между кау-
дальной стенкой слуховой
капсулы и первыми верхними дугами —
зачатками верхних дуг позвонков, уже вне черепа выходят IX и X пары нервов (рис. 30, IX, X).
c. ап.
с.tec.
c.tr.
c.sp.
c.rhm.
pc fi. a.
c.s o.
ШЕ
f. ВГ, HU
c. and.
pchp {
c.ii.
Рис. 30. Скелет головы миноги Lampetra fluviatilis сверху:
c. an. — кольцевой хрящ; с. aud. — слуховая капсула; ch..— хорда; с. sp. — передний боковой хрящ; с. rhm. — задний боковой хрящ; с. so. — подглазничная дуга; с. tec. — передний верхний хрящ; с. tr. — задний верхний хрящ;
f. 11, 111 — отверстие для выхода II и Ш нервов; f. IV, V, VI — отверстие для выхода IV, V и VI нервов; [. VII, VIII — отверстие дли выхода VII и VIII нервов; IX, X — место выхода IX и X нервов; pch.a. — передние пара-хордалии; pch. р. — задние парахордалии
Слуховые капсулы (capsula auditiva) полностью хрящевые и целиком входят в состав стенок черепной коробки, так что отделить их без повреждения последней невозможно.
Обонятельная капсула (capsula olfactoria, см. рис. 29, cap, о.), наоборот, соединена с черепной коробкой только при помощи связок. После их удаления капсула легко отделяется от черепа. Она почти целиком соединительнотканная и только задняя стенка хрящевая. В хрящевой части имеются два больших отверстия, затянутые по краям перепонкой, через которые проходят обонятельные нервы. Очистить отверстия очень трудно, поэтому лучше всего их местоположение обнаружить, прощупывая концом препаровальной иглы.
Висцеральный скелет
Висцеральный скелет миноги состоит из жаберной решетки и вышеописанной подглазничной дуги. Жаберный скелет имеет вид сложной хрящевой сетки, лежащей с наружной стороны от жаберных мешков. Он состоит из восьми поперечных хрящевых жаберных дужек, лежащих в жаберных перегородках на их наружной стороне, в отличие от рыб, где скелет дужек помещается на внутренней стороне. Они соединены друг с другом несколькими продольными перемычками. Спереди жаберная коробка соединяется со стилевидным отростком подглазничной дуги, а позади оканчивается полусферой хрящевой капсулы, окружающей перикардиальную сумку.
Хорошо очистить скелет жаберной решетки можно только, применяя сложные способы химической обработки, поэтому разбирают строение жаберного скелета, лишь слегка иголкой очищая красноватые хрящевые дужки и перемычки между ними и ориентируясь в их расположении по рисунку..
Жаберные дужки (arcus branchialis, см. рис. 29, arc. Ьг.) имеют вид изогнутых поперечных нитей между жаберными отверстиями. Каждая жаберная дужка дает от себя несколько отростков. В дорзальном отделе каждой дужки можно видеть три отростка. Два из них направлены вперед и вниз, а один — назад и вверх. Ниже поджаберной связи жаберная дужка также дает три выроста — два вперед и один назад.
Передняя жаберная дужка имеет особое строение. Она короче всех остальных, не имеет выростов, и от ее верхней части отходит отросток, направляющийся сначала вверх, а затем загибающийся книзу, который налегает на стилевидный отросток подглазничной дуги и плотно с ней связан. Отростком, направленным назад, она соединена с двумя продольными комиссурами, которые здесь сливаются вместе (см. рис. 29, arc. br., 1).
Восьмая жаберная дужка также имеет особое строение. У нее нет боковых выростов, и позади она связана с околосердечным хрящом (см. рис. 29, аге. Ьг., 8).
Продольные комиссуры — продольные хрящевые тяжи, связывающие жаберные дуги между собой с каждой стороны. Их четыре: две проходят дорзальнее жаберных щелей, две — нейтральнее.
1. Околохордовая — субхордальная, связывает дорзальные концы дужек под хордой. Она лежит под хордой по ее медиальной линии и поэтому на наших препаратах не видна.
2. Наджаберная — эпитрематическая, связывает дуги над жаберными отверстиями (см. рис. 29, э.). Она имеет несколько выростов, направленных вверх.
3. Поджаберная — гипотрематическая, связывает дужки под жаберными отверстиями (см. рис. 29, г.).
4. Вентральная комиссура (см. рис. 29, в.) образована слиянием вентральных концов жаберных дужек каждой стороны и частичным срастанием дужек разных сторон по медиальной линии.
Околосердечный хрящ (cartilago cardiobranchialis, см. рис. 29, с. crd.) имеет форму конической чаши, дном направленной назад. Он окружает снаружи перикардиальную сумку и спереди соединяется с восьмой жаберной дужкой тремя хрящевыми мостиками с каждой стороны.
Литература по классу круглоротых и бесчелюстных
Балабай П. П. Морфология и филогенетическое развитие группы бесчелюстных. Киев, Изд-во АН УССР, 1956.
Кольцов Н. П. История развития головы миноги. «Бюллетень императорского общества естествоиспытателей», т. 15, 1905.
Северцов А. Н. Исследования об эволюции позвоночных (морфология скелета и мускулатуры головы циклостом). Журн. «Русский архив анатомии, гистологии и эмбриологии», вып. 1, 1916, то же, соб. соч., т. 4, 1948.
Третьяков Д. К. Органы чувств речной миноги. Одесса, 1915.
Marinelli М., Strenger A. Vergleichende Anatomie und Morphologie der Wirbeltiere. Ill, Wien, 1953.
Mezojko A. Cyclostomen. Bronns Klassen und Ordnungen, Bd. VI, Abt. I, 1901—1924.
РАЗДЕЛ ЧЕЛЮСТНОРОТЫЕ — GNATHOSTOMATA
НАДКЛАСС РЫБЫ PISCES
КЛАСС ХРЯЩЕВЫЕ РЫБЫ — CHONDRICHTHYES
Общая характеристика класса
Хрящевые рыбы являются древнейшими челюстными позвоночными — Gnathostomata. От бесчелюстных Agnatha они отличаются парными плавниками, позволяющими активно плавать в воде, и активным питанием, так как схватывают добычу челюстями.
Современные хрящевые рыбы, как представители челюстноротых животных, обладают целым рядом особенностей, приобретенных этой группой позвоночных в процессе эволюции, но одновременно с этим они отличаются от древних хрящевых рыб, известных с девона (см. рис. 1) целым рядом новоприобретенных признаков в строении и образе жизни, позволивших им сохраниться до настоящего времени, тогда как все остальные хрящевые рыбы вымерли и были заменены более совершенными костными рыбами.
До настоящего времени среди зоологов и ихтиологов нет единого мнения о таксономическом ранге группы рыбы — Pisces. Некоторые авторы считают рыб единым классом, другие возводят их в ранг надкласса, разбивая на ряд классов, или вообще ликвидируют понятие рыбы, выделяя отдельные группы как самостоятельные классы надкласса челюстноротых. Соответственно с этим меняются и таксономические ранги более мелких систематических единиц, а также их количество.
В настоящем издании мы будем придерживаться принятого среди зоологов и палеонтологов деления группы первичноводных челюстноротых животных, объединяемых в надкласс рыбы (Pisces), на четыре самостоятельных класса: панцирные рыбы (Placodermi), хрящевые рыбы (Chondrichthyes), акантодии (Acanthodii) и костные рыбы (Osteichthyes). Взаимоотношения между ними можно представить следующим образом (см. рис. 1). Первичночелюстноротые Protognathostomata дали начало по крайней мере двум ветвям позвоночных животных. Представителями одной из них являются панцирные рыбы Placodermi, ископаемые остатки которых встречаются с конца силура до конца девона (см. рис. 1). Имея хрящевой скелет, эта группа отличалась и целым рядом черт, свойственных костным рыбам. Так, в позвоночнике имелись поверхностные окостенения верхних и нижних дуг, также по всей поверхности окостеневал эндокраниум. Имелся наружный костный скелет, образующий панцирь. Правда, кости покровов слились из первоначальных мелких окостенений, трудно гомологизируются с костями костных рыб и поэтому получили особые названия. Имелся челюстной аппарат, 112
но он был образован покровными костями. В эволюции этой группы наблюдался переход от придонных форм к свободноплавающим.
Вторая ветвь дает начало трем остальным классам рыб Chondr-ichthyes, Acanthodii, Osteichthyes (см. рис. 1). Из них наиболее примитивными нужно считать хрящевых рыб.
Современные хрящевые рыбы Chondrichthyes представлены двумя подклассами: пластиножаберными, или эласмобранхиями — Elasmobranchii, и цельноголовыми, или химеровыми, рыбами —• Holocephali.
Все эласмобранхии — морские, широко распространенные рыбы, отличаются от других рыб наличием поперечного рта, плакоидной чешуей, головой, вытянутой в рыло, или рострум, наличием наружных жаберных щелей, не прикрытых жаберной крышкой, и хрящевым скелетом. Характерной особенностью их является внутреннее осеменение, поэтому самцы имеют копулятивные органы, а самки или откладывают крупные яйца, покрытые рогоподобной оболочкой, или рождают живых детенышей.
Подкласс Elasmobranchii объединяет около 12 отрядов. Разбираемые в качестве представителей хрящевых рыб виды принадлежат к одному из них — Squatinida. Анатомию хрящевых рыб можно изучать на различных объектах. Наиболее удобны и доступны катран: 'Squalus acanthias L., встречающийся почти во всех морях Советского Союза, и кошачья акула — Scyliorhinus caniculus L., встречающаяся в Черном море. Оба вида относятся к подотряду настоящих акул Selachoidei (Squaloidei) и являются хорошо плавающими нектонными формами.
Можно использовать также относящихся к подотряду скатов Batoidea (Rajoidei) шиповатого ската — Raja clavata L. и морского кота — Trygon pasiinaca L. Оба вида живут у нас в Черном море. Благодаря придонному образу жизни, они имеют сплющенное в дорзо-вентральном направлении тело и сильно развитые грудные плавники, которые придают им ромбовидную форму.
В дальнейшем при изучении анатомии хрящевых рыб мы будем базироваться на описании наиболее доступного объекта — катрана, давая ссылки на особенности строения отдельных систем у других указанных видов. Некоторые особенности строения хрящевых рыб при возможности нужно рассмотреть на голубой акуле — Carchari-ntis glaucus L. и химере — Chimaera monstrosa L. На них будут даны ссылки в тексте.
Представители класса — колючая акула кокотница, или катран (Squalus acanthias L.), и шиповатый скат (Raja clavata L.)
Внешний осмотр акулы
Половозрелые экземпляры черноморской акулы катрана достигают длины метра и даже больше. На занятиях обычно используют неполовозрелые особи 50—60 см длины.
Сначала знакомятся с внешним видом катрана. Форма тела удлиненно-веретенообразная; голова, заостренная спереди, без резких границ переходит в туловище, которое, постепенно сужаясь, образует хвостовой отдел, заканчивающийся неравнолопастным (гетероцеркальным) хвостовым плавником. Тело акулы в поперечном разрезе представляет собой треугольник, вершина которого несколько закруглена. В хвостовом отделе оно значительно суживается к заднему концу и имеет в поперечном разрезе округло-овальную форму.
Непарные плавники у катрана представлены двумя спинными и хвостовым, огибающим задний конец тела. Передний спинной плавник (pinna dorsalis anterior) располагается посередине туловища, з а дни й (pinna, dorsalis posterior) — на хвостовом отделе. На переднем крае каждого спинного плавника имеется свободный колючий шип. Хвостовой плавник (pinna cauda-lis) — гетероцеркальный, верхняя лопасть большая, окаймляет задний отдел тела, нижняя — значительно короче.
Парные плавники — передние и задние парные конечности — представлены грудными и брюшными плавниками.
Поворачивают рыбу брюхом кверху и рассматривают парные плавники. Передняя граница грудных плавников одновременно является задней границей головного отдела. Как в грудном, так и в брюшном плавниках следует отметить две части: мясистую лопасть при основании плавника, которая содержит внутренний хрящевой скелет и мускулатуру, и тонкую оторочку, представляющую собой дубликатуру кожи, в которой расположены роговые лучи — эласто-трихии (elastotrichii).
Грудные плавники (pinna thoracalis). Для того чтобы придать грудным плавникам нормальное положение, следует оттянуть их в стороны и вперед. Грудные плавники имеют приблизительно треугольную форму с широким основанием. Основание переходит в задний, или постаксиальный, край кожной лопасти. Передний, или преаксиальный, край плавника обращен вперед и несколько кнаружи.
Основания грудных плавников занимают почти горизонтальное положение и являются продолжением плоской брюшной поверхности. Передний, краниальный, край плавника поднят вверх по сравнению с задним, каудальным.
Брюшные плавники (pinna abdominalis) лежат на брюшной поверхности туловища, соприкасаясь между собой своими внутренними, медиальными краями. У самцов на заднем конце постаксиального края каждого брюшного плавника развит особый мясистый вырост — копулятивный орган — птеригоподия (pterygo-podia).
Поступательное движецие акулы в воде обеспечивается сильными изгибами хвостового отдела вправо и влево. Грудные и брюшные плавники, по существу, не являются органами поступательного движения, а лишь выполняют функции несущих плоскостей и управления движением вверх, вниз и в сторону.
Клоака (cloaca, рис. 31, cl.). На брюшной стороне тела между задними краями оснований брюшных плавников у хрящевых рыб располагается продольная широкая щель — это отверстие клоаки. Чтобы ее рассмотреть, нужно раздвинуть плавники в стороны.
Брюшные поры (porus abdominalis, рис. 31, рог. а.) два щелевидных косых отверстия, лежащие по бокам заднего края клоаки и ведущие в брюшную полость.
Следует более детально рассмотреть голову катрана. Передний заостренный отдел — рыло, р о с т р у м (rostrum), лежит впереди двух расположенных на вентральной стороне носовых ямок, образующих входное и выходное отверстия обонятельной капсулы. Верхнее, меньшее, имеет округлую форму; нижнее, большее, — форму длинной носовой щели, обрамленной кожными складками. По бокам головы располагаются глаза (oculus) удлиненноовальной формы, сверху они частично прикрыты кожными складками — веками pal-pebra). По переднему и заднему краям глаза выступает белая
Рис. 31. Общая топография внутренних органов акулы Squalus acanthias:
ао. d. — спинная аорта; a. tn. i. — нижняя брыжеечная артерия; cl. —клоака; col. — толстая кишка; d. chol. — желчный проток; dens. —зубы; duod. —двенадцатиперстная кншка; gast. — желудок; gl. ret. — ректальная железа; h. — печень; I. — селезенка; Л1 — Мюллеров канал; msg. — передний дорзальный мезентерий; т. s. г. — мезентерий прямой кишки; паг. — ноздря; oes. — пищевод; о. та/. — большой сальник; о. min. — малый сальник; pner. d. — дорзальная доля поджелудочной железы; pner. v. — ее вентральная доля; рог. а. — брюшные поры; г.—почка; ret.—прямая кишка; st. — ротовое отверстие; tph. — спиральный клапан; ves. f. — желчный пузырь; и. tat. — боковая вена; at. — маточная часть яйцевода
белковая оболочка — склера (sclera), остальная часть его снаружи покрыта прозрачной оболочкой — роговицей (cornea), из-под которой просвечивает радужная оболочка (iris) с блестящим пигментом. Последняя прорезана в середине зрачком, окаймленным черной пигментной линией. За глазом лежит косоовальная щель—брызгальце (spiraculum), соединяющее полость, глотки с внешней средой. Кзади от брызгальца на границе боковой и брюшной поверхности тела расположены пять поперечных жаберных щелей. Каждая жаберная щель прикрыта спереди широкой кожной складкой. Последняя жаберная складка прикрывает собой не только жаберную щель, но и частично среднюю ча.сть пояса грудных конечностей.
Поворачивают акулу брюхом кверху. Теперь хорошо видны передние и задние ноздри (naris, рис. 31, паг.) и поперечно расположенное р о т о в о е отверстие (stoma, рис. 31, si.) полулунной формы. Обращают на себя внимание заостренные зубы верхней челюсти, расположенные в несколько рядов. Зубы нижней челюсти снаружи не видны. Верхняя челюсть окружена свободной кожной складкой, которая, дойдя до углов рта, отделяется снаружи от остальной кожи особой щелью и охватывает снаружи и изнутри скрытые в ней особые образования — губные хрящи.
Следует обратить внимание на кожные органы чувств, представленные небольшими отверстиями на поверхности кожи. Особенно отчетливо они видны в виде темных точек на вентральной поверхности рыла, на предротовой области и между ротовой и первой жаберной щелями. Хорошо видны они также на дорзальной стороне головы, на рыльной части и на надглазничной области. На туловище их легче рассмотреть чуть ниже горизонтальной септы — продольной линии, разделяющей мускулатуру туловища на спинную и брюшную части.
По внешнему виду кошачья акула — Scyliorhinus caniculus отличается от катрана несколько большей тупорылостью, отсутствием шипов в спинных плавниках и наличием непарного анального плавника, расположенного на вентральной стороне хвостового отдела.
Внешний осмотр ската
У скатов, ведущих придонный образ жизни, форма тела, форма н расположение парных и непарных плавников сильно изменены по сравнению с акулами. Тело ската имеет форму ромба, боковые острые углы которого образованы громадными разросшимися грудными плавниками, передние концы которых доходят до конца рыла, прирастают к боковым поверхностям головы и затрудняют проведение четкой границы между ней и туловищем. Пять жаберных отверстий расположены на брюшной поверхности. Наоборот тонкий, сплющенный в дорзо-вентральном направлении хвостовой отдел хорошо отличается от туловищного. У морского кота примерно от середины хвостового отдела тела отходит направленный вверх шип.
Парные плавники. Грудные плавники лежат в горизон
тальной плоскости и имеют сильно разросшуюся мясистую основную часть, окаймленную по краю широкой кожной складкой, поддерживаемой кожными лучами. Брюшные плавники значительно меньших размеров и располагаются между задними концами грудных на границе туловищного и хвостового отделов. Каждый ив них состоит из двух лопастей: передней узкой, направленной кнаружи, и задней, более широкой, вытянутой назад. У самцов на внутренней стороне брюшных плавников также развиты копулятивные органы.
Следует обратить внимание на расположенную между брюшными плавниками продольную щель клоаки и лежащие за ней два отверстия брюшных пор.
Непарные плавники у скатов редуцированы. У шиповатого ската имеются два небольших спинных плавника, отнесенные на конец хвоста, и небольшая лопасть хвостового плавника, анальный плавник отсутствует. У морского кота хвостовой отдел заканчивается тонкой нитью. В отличие от акул движение скатов осуществляется за счет волнообразных изгибов грудных плавников, так что хвостовой отдел выполняет лишь функцию руля и органа защиты.
У скатов в связи с придонным образом жизни и уплощенной формой тела глаза сдвинуты на дорсальную поверхность головы. Лежащие позади глаз отверстия брызгалец относительно велики-У лежащего на дне ската через них в ротоглоточную полость поступает необходимая для дыхания вода. Передняя стенка брызгал ь-ц а покрыта мелкими складочками, которые представляют собой ложную жабру (pseudobranchia).Жаберные отверстия в количестве пяти пар расположены полукругом на вентральной стороне тела. Чтобы их рассмотреть, нужно положить животное на спину. Несколько впереди от жаберных щелей лежит ротовое отверстие, имеющее вид поперечной, слегка изогнутой щели. Зубы у скатов также сидят в несколько рядов, но в отличие от акулы они уплощены и служат для раздваивания раковин моллюсков и панцирей ракообразных. Между отверстиями ноздрей и углами рта тянется кожная складка. Приподнимают ее и рассматривают лежащую под ней носо-ротовую борозду (sulcus oro-nasalis). Эта борозда способствует обмену воды в носовых капсулах при забирании скатом воды через рот во время плавания.
Кожные органы чувств достигают наибольшего развития на вентральной поверхности головы. Пользуясь лупой, можно обнаружить разветвленную сеть каналов органов боковой линии. Особенно' сильно каналы ветвятся в области рыла, но некоторые из них переходят и на вентральную поверхность плавников. Такое сильное развитие органов боковой линии на брюшной стороне тела связано с придонным образом жизни.
По окончании'внешнего осмотра можно более подробно ознакомиться со строением покровов. Микроскопическое строение кожи и развитие плакоидной чешуи приводится в любом учебнике по сравнительной анатомии, поэтому мы здесь на нем останавливаться не будем. Проведя рукой сзади наперед по средней линии спины, лег-
ко убедиться в том, что кожа акулы имеет шероховатое (шагреневое) строение, что зависит от присутствия мелких кожных зубчиков, или плакоидных чешуи. Вычленив одну из чешуй, рассматривают ее под лупой. Каждая чешуя состоит из основной пластинки и загнутого назад зубца. Последний покрыт снаружи твердым Слоем эмали.
У ската спинная поверхность покрыта плакоидными чешуями разной величины. Более крупные чешуи лежат вдоль средней линии тела и хвоста. Кроме того, отдельные крупные чешуи разбросаны между мелкой чешуей по всей спинной поверхности тела. По строению чешуя ската такая же, как и у акулы, в чем можно убедиться, вычленив ее и рассмотрев под лупой. Поворачивают ската брюшной стороной вверх, обращают внимание на то, что на этой стороне чешуя мелкая и расположена преимущественно по переднему краю рыла и грудных плавников.
Вскрытие
Общая топография внутренних органов (рис. 31)
Общая топография внутренних органов у описываемых акул и скатов сходная, поэтому при работе со скатами можно пользоваться тем порядком вскрытия и описания, которые будут даны для катрана. Однако следует учитывать, что брюшная полость скатов значительно короче, чем у акулы; это приводит к некоторым отличиям в расположении и форме внутренних органов. В том случае, когда эти отличия значительны, в тексте будут сделаны соответствующие оговорки.
Вскрытие хрящевых рыб обычно ведут на фиксированных экземплярах. По ним же сделано описание, поэтому не имеет значения повреждение даже крупных кровеносных сосудов, но таких повреждений следует избегать при работе на свежем объекте.
Акулу помещают на ванночку спинной стороной книзу, прощупывают на брюшной стороне, чуть кпереди от преаксиальных краев грудного и брюшного плавников, грудной и брюшной пояс.
Делают продольный разрез по средней линии брюха, начиная от заднего края грудного пояса до переднего края заднего или тазового, пояса. Затем в обе стороны от переднего и заднего концов продольного разреза делают два поперечных разреза. Передний разрез ведут вдоль заднего края грудного пояса до основания плавника, задний — вдоль переднего края тазового пояса. Отворачивают обе стенки тела и отмечают хорошо заметную сегментацию мускулатуры. Примерно по середине отвернутой стенки тела проходит темная полоса — это заполненная кровью боковая вена.
Для скатов такого вскрытия недостаточно, поскольку передняя и задняя части полости остаются прикрыты поясами передних и задних конечностей. Чтобы рассмотреть все внутренние органы, необходимо удалить вентральные участки поясов конечностей. Для этого приподнимают пинцетом плечевой пояс, расположенный меж-118
ду последними жаберными щелями, перерезают его по бокам на уровне жаберных щелей и удаляют вентральную часть. При этом следует учитывать, что непосредственно перед поясом лежит сердце; поэтому стараются его не повредить при вычленении хрящевых элементов пояса. Точно так же, сделав два разреза по краям, удаляют вентральную часть тазового пояса и только после этого приступают к рассмотрению общего расположения внутренних органов ската.
Обращают внимание на то, что вся брюшная полость акулы выстлана блестящей пленкой — брюшиной (peritoneum), у скатов она сильно пигментирована на спинной стороне. Большую часть брюшной полости занимает трехлопастная печень (Л). Отвернув в сторону ее левую и среднюю доли, можно увидеть лежащий под ними широкий пищевод (oes.) и нерезко отграниченный от него У-образно изогнутый желудок (gast.). У акул на месте изгиба к его дну тесно прилегает фиолетово-бурая заостренная на заднем конце селезенка (/.). У скатов последняя располагается между ветвями петли желудка.
Заметным перехватом желудок отделяется от начальной части кишечника — тонкой кишки, носящей название двенадцатиперстной (duod.). Чтобы ее рассмотреть, нужно отвернуть правую долю печени, которой она прикрыта. К вентральной стенке двенадцатиперстной кишки плотно прилегает брюшная доля поджелудочной железы (рпсг. о.). Далее идет расширенный отдел кишечника — толстая кишка (col.). На ее стенке хорошо заметны поперечные полосы — места прикрепления складки спирального клапана. Еще дальше следует более узкий отдел — прямая кишка (ret.), от дорзальной стенки которой отходит пальцевидная, или ректальная, железа (gl. ret.). Прямая кишка впадает в клоаку (cl.), открывающуюся наружу клоакальным отверстием. Отвернув печень и желудок вверх и влево, следует обратить внимание на то, что задний отдел пищевода, желудок, селезенка, двенадцатиперстная кишка и передняя половина толстой кишки подвешены к спинной стенке тела при помощи широкого мезентерия.
Под местом прикрепления мезентерия к спинной стенке тела, вдоль вентральной поверхности позвоночника, проходит обычно за-пустевающий сосуд — спинная аорта (ао. d.). По бокам от нее расположены два темных широких сосуда — задние кардинальные вены.
Кнаружи от задних кардинальных вен располагаются почки (г.), прикрытые брюшиной. У акул они тянутся вдоль всей брюшной полости, расширяясь в задней части, у скатов имеют дольчатое строение и занимают заднюю половину брюшной полости. На уровне переднего края печени, по бокам от нее и частично соединяясь с ней при помощи брыжеек, помещаются половые органы самца — семенники или самки — яичники. У кошачьей акулы семенники и яичники лежат в средней части брюшной полости на уровне каудального конца желудка.
Следует отметить, что на уровне переднего конца печени задние кардинальные вены расширяются, образуя обширные синусы, наполненные кровью.
Пищеварительная система
Пищеварительная система хрящевых рыб как хищников, питающихся сложной белковой пищей, имеет ярко выраженное расчленение на отделы и хорошо обособленные пищеварительные железы.
Как уже указывалось, у акул и скатов пищеварительная система имеет сходное строение и отличается только незначительными де-
Рис. 32. Пищеварительная система акулы Squalus acanthias: и ее кровоснабжение: a. cl. — чревная артерия; a. ints. — кишечная артерия; a. h. — печеночная артерия; a. g. st.—желудочная артерия; а. т. i — нижняя брыжеечная артерия; а. т. s. — верхняя брыжеечная артерия; col. — толстая кишка;
crd. — кардиальная часть желудка: d. chol. — желчный проток: fund. — дно желудка; gast. — желудок; gl. ret. — ректальная железа; К. — печень; I. — селезенка; msr. — мезентерий прямой кишки; oes. — пищевод; pner. d.— дорзальная часть поджелудочной железы; рпег. V.—ее вентральная часть; pyl. — пилорическая часть желудка; ret. — прямая кишка; ves. f. — желчный пузырь; v. р. h. — воротная вена печени
талями, поэтому, несмотря на то, что описание дается по катрану, им вполне можно пользоваться и при изучении пищеварительной системы на скатах.
Для того чтобы рассмотреть начальный отдел пищеварительной системы — ротовое отверстие, зубы и ротоглоточную полость — необходимо перерезать жаберные дуги. Это повредит рассмотрению строения дыхательной и кровеносной системы, поэтому изучение начала пищеварительной системы следует отложить и рассмотреть ее после изучения органов дыхания. Рассмотрение пищеварительной системы начинают прямо с пищевода. Для этого отворачивают левую долю печени, под которой он расположен.
Пищевод (oesophagus, рис. 31, 32, oes.) — довольно короткая трубка с толстыми мышечными стенками. Внешне без резких границ пищевод переходит в длинный желудок. Наружная перетяжка не является истинной границей между ними, а объясняется различием во внутреннем строении проксимальной и дистальной частей желудка (см. ниже описание внутренних стенок желудка).
Желудок (gaster, рис. 31, 32, gast.) имеет V-образную форму. Нисходящая часть его, называемая кардиальной (cardium, рис. 32, crd.), тянется приблизительно до половины брюшной полости (у скатов почти до ее конца), где образует значительно меньшее, краниально направленное колено, называемое пилорус, привратник (pylorus, рис. 32, pyl.). Место изгиба называется дном желудка (fundus, рис. 32, fund.). Пилорическая часть заканчивается сильным утолщением мышечной оболочки, образующей сфинктер.
Кишечник (intestinum). У хрящевых рыб отделы кишечника трудно гомологизировать с таковыми наземных позвоночных. Поэтому названия отделов кишечника, соответствующие названиям отделов высших позвоночных, условны.
Передний отдел тонкой кишки — это так называемая двенадцатиперстная кишка (duodenum, см. рис. 31, duod.). Она очень коротка. В нее открываются желчный проток и проток поджелудочной железы.
Двенадцатиперстная кишка переходит в расширенный отдел кишечника, получивший название «толстой» кишки (colon, рис. 31, 32, col.). На ее стенках хорошо заметны поперечные полосы— просвечивающие складки содержащегося в ней спирального клапана. В каудальном направлении толстая кишка постепенно сужается и близ переднего края брюшного пояса переходит в значительно более узкую и лишенную перетяжек спирального клапана прямую кишку (rectum, рис. 31, 32, ret.).
Перерезав у акулы пояс задних конечностей по средней линии, продлить разрез до передней стенки клоаки и, разведя в стороны половинки тазового пояса, слегка оттягивают в любую сторону кишку и рассматривают отходящую от нее ректальную железу.
Ректальная железа (glandula rectalis, рис. 31, 32, gl. ret.) имеет овально-удлиненную форму. Железа подвешена на складке брюшины — м е з е н т е р и и (mesorectum, рис. 31, 32, msr.), поддерживающей также и прямую кишку. Каудальным концом ректальная железа открывается на спинной поверхности в средней части прямой кишки. Она выделяет жидкость,-представляющую собой раствор NaCl. Ректальная железа способна поглощать из крови сравнительно большие количества NaCl, что и является ее функцией. Прямая кишка оканчивается несколько расширенной частью — клоакой (cloaka, см. рис. 31, cl.). Последняя открывается наружу вышеописанным клоакальным отверстием.
Печень (hepar, рис. 31, 32, h.) желтовато-бурая, занимает почти всю переднюю половину брюшной полости. Цвет ее зависит от степени наполнения жиром. В печени различают три лопасти,
расходящиеся в каудальном направлении и сливающиеся в краниальном. Правая лопасть занимает почти всю правую половину брюшной полости, левая — в значительной мере прижата к стенке тела и на вентральную сторону выходит узким краем. Центральная лопасть меньше предыдущих. В каудальном направлении она вытянута в резко суженный отросток.
Желчный пузырь (vesica fellea, рис. 31, 32, ves. f.)—зеленоватого цвета, тесно прилежит к правой стороне центральной доли печени. Его проток (ductus cysticus), приняв протоки от печени (ductus hepaticus) и именуясь теперь желчным протоком (ductus choledochus, рис. 31, 32, d. chol.), проходит по краю большого сальника. Дойдя до двенадцатиперстной кишки, он на значительном расстоянии проходит внутри ее стенки и открывается на дорзальной поверхности двенадцатиперстной кишки.
Поджелудочная железа (pancreas) имеет две лопасти: дорзальную и вентральную. Вентральная лопасть поджелудочной железы лежит на вентральной поверхности двенадцатиперстной кишки (рис. 31, 32, рпсг. v.). Дорзальная лопасть имеет форму плоского, вытянутого тела (рис. 31, 32, рпсг. d.). Ее передняя половина лежит в петле желудка, простираясь в краниальном направлении почти до самого краниального края желудка. Задняя ее половина лежит в петле, образованной восходящим коленом желудка с одной стороны, и кишечником — с другой. Ее каудальный конец располагается на уровне конца передней трети толстой кишки. Большая часть заднего отдела поджелудочной железы у катрана прикрыта селезенкой. Чтобы его увидеть, отворачивают селезенку направо. На месте разделения поджелудочной железы на переднюю и заднюю части от нее отходит тонкая перемычка к брюшной лопасти той же железы. Она охватывает дугообразно внутренний край левой половины двенадцатиперстной кишки непосредственно под пилорусом.
Проток поджелудочной железы проходит по внутренней стороне вентральной лопасти железы и открывается на вентральной стороне двенадцатиперстной кишки. Перед впадением в нее он, как и желчный проток, на значительном расстоянии проходит внутри ее ст.енки.
Мезентерии, или брыжейки. Стенки брюшной полости выстланы тонкой серозной оболочкой — париетальным листком брюшины (peritoneum), который переходит на многие внутренние органы и одевает их в виде висцерального листка. Место перехода одного листка в другой и называется брыжейкой, или мезентерием. У хрящевых рыб имеются два дорзальных и два вентральных мезентерия, связанных с пищеварительным трактом.
Передний дорзальный мезентерий (mesogaster, рис. 31, msg.) отходит от спинной стенки тела и прикрепляется к заднему отделу пищевода, желудку, тонкой кишке и началу толстой.
Задний дорзальный (mesorectum, рис. 31, 32, msr.) идет от спинной стенки тела к прямой кишке.
Два вентральных мезентерия связывают различные участки внутренних органов между собой и называются большим и малым 122
сальниками, так как в них могут располагаться отложения жира.
Малый сальник (omentum- minus s. omentum gastro-hepa-ticum, рис. 31,o. min.) — складка брюшины, которая начинается от дорзальной стороны печени, идет в каудальном направлении и прикрепляется к дорзальной стороне желудка в области его петли. Для того чтобы, лучше рассмотреть сальник, следует оттянуть желудок направо. Вы увидите, что он прикрывает полностью лежащую в дорзальном мезентерии дорзальную лопасть поджелудочной железы.
Большой сальник (omentum majus, s. omentum gastro-duodenum, рис. 31, о. maj.) начинается одновременно с малым сальником в виде складки брюшины, идущей от печени к двенадцатиперстной кишке, и далее в области толстой кишки, сливающейся со спинным мезентерием. Между листками сальников проходят основные артерии и вены внутренних органов.
Следует рассмотреть еще особую брыжейку, которая у акул прикрепляет селезенку ко дну желудочка, и брыжейку, натянутую между вентральной долей поджелудочной железы и пилорической частью желудка.
Строение внутренней стенки пищеварительной трубки. Чтобы познакомиться со строением внутренних стенок пищеварительного канала, разрезают его по средней линии брюшной поверхности и промывают водой. В толстой же кишке срезают часть ее брюшной стенки, чтобы рассмотреть строение спирального клапана.
Пищевод. На внутренней стенке пищевода отмечают обилие белых роговых сосочков, сидящих продольными рядами на складчатой поверхности пищевода. От дорзальной поверхности пищевода, внутрь него, вдается мощная ороговевшая складка, на которой сидят в несколько рядов тоже ороговевшие сосочки. Обращают внимание на значительную толщину стенки пищевода.
Желудок. Внешне без всяких границ пищевод переходит в У-образный желудок. Проксимальный отдел желудка с помощью утолщенной кольцевой складки делится на краниальную и каудальную половины. Стенка краниальной половины значительно толще каудальной. Внутренняя стенка краниальной половины выстлана продольными складками. Количество крупных продольных складок в каудальной части несколько меньше, но зато здесь появляются более мелкие складки, лежащие преимущественно на вентральной стороне желудка. Все складки заканчиваются в области дна желудка, откуда возникают продольные складки дистального колена желудка. Это — высокие и широкие складки с характерными поперечными перетяжками. Слизистая оболочка между ними, особенно ближе к пилорусу, также собрана многочисленными мелкими поперечными складками. Привратник представлен значительным утолщением мышечной оболочки, образующим сфинктер. На внутренней поверхности сфинктера имеются 4-—5 гладких складочек. Поперечные складки отсутствуют.
Кишечник. Недалеко за пилорусом в кишке начинаются обороты спирального клапана (tiphlosolis, рис. 31, tph.). Они пред-
ставляют собой как бы вставленные друг в друга конусы, обращенные суженной, вершинной частью в краниальном направлении. По периферии стенки конусов приращены к внутренней стенке кишки. Участок кишки от пилоруса до места прирастания стенки переднего конуса к стенке кишки условно может быть назван двенадцатиперстной кишкой, так как здесь открываются протоки пищеварительных желез. Проток поджелудочной железы открывается на внутренней стенке вентральной поверхности кишки, кпереди на несколько миллиметров от места прирастания переднего конуса к стенке кишки. Печеночный проток открывается на дорзальной поверхности на расстоянии 1 —1,5 см впереди от места прирастания конуса к стенке кишки. Следует заметить, что оба описанных притока прежде, чем открыться в кишечник, на значительном расстоянии проходят внутри стенки двенадцатиперстной кишки. Последний оборот спирального клапана не образует полного конуса, а, начавшись от предыдущего, вдается, петлеобразно изгибаясь, в полость кишечника, срастаясь с его стенкой.
Прямая кишка. Суженный отдел кишечника, начинающийся непосредственно за окончанием спирального клапана, называется прямой кишкой. Это относительно тонкостенная часть кишечника с высокими продольными складками. Примерно на середине дорзальной поверхности прямой кишки можно видеть отверстие протока ректальной железы, который вначале идет в стенке кишки сзади наперед. Задний отдел прямой кишки, открывается в клоаку. Клоака открывается наружу вышеописанным клоакальным отверстием.
Жаберная область
Теперь рассмотрим жаберную область, где расположены органы дыхания — жабры, центральный орган кровообращения — сердце, брюшная аорта с приносящими жаберными артериями и щитовидная железа.
Для этого делают неглубокий разрез по средней линии от грудного пояса до заднего края нижней челюсти и от него проводят разрезы в стороны. Отпрепарируют кожу, отвернув ее в стороны до нижнего края наружных жаберных щелей.
Далее перерезают по середине тонкий слой поперечной мускулатуры, вентральную часть сжимателя гиоидной дуги (см. описание мускулатуры стр. 168) и отворачивают обе половины 'мышцы в стороны. Под ними видны мощные продольные пучки мускулатуры, расположенные по центру, а по бокам от них — жаберные мешки, прикрытые мускульными волокнами вентральной части поверхностного жаберного сжимателя (см. описание жаберной мускулатуры, стр. 168). Обращают внимание на то, что волокна мускула срастаются с наружными краями жаберных перегородок.
Подрезают подъязычную мускулатуру у плечевого пояса, а также отдельные пучки у челюстной и подъязычной дуг и удаляют их. Под ними находится околосердечная сумка, одевающая сердце, а 124
впереди лежит брюшная аорта (рис. 34, ао. и.) с отходящими от нее приносящими жаберными артериями. Впереди от места раздвоения брюшной аорты на первые две приносящие жаберные артерии на нижней стороне гладкой слизистой оболочки рта находится щитовидная железа (glandula thyreoidea, рис. 34, gl. thyr.) в виде скопления мелких пузырьков.
Органы дыхания
spt.br.
a.br.r.----
a. brad.—; hembr.
ar.br
hol.br.
Рис. 33. Поперечный разрез жабр акулы Squalus acanthias (по Маринелли и Стреи-> геру):
аг. Ьг. — жаберная дуга; а. Ьг. ad. — приносящая - а. Ьг. г.—выносящая жабер-—полужабра; hoi. br.-~n.e-жабра; spt. br. — жаберная перегородка
жаберная артерия; с ная артерия; hembr. лая ; ' - ' '
Органы дыхания хрящевых рыб представлены жаберными мешками, открывающимися внутренними жаберными отверстиями в ротоглоточную полость, а наружными — по бокам головы. Чтобы детально ознакомиться со строением органов дыхания, нужно вскрыть жаберные мешки. Для этого вставляют одну половину ножниц в наружное жаберное отверстие, делают разрез в медиальном направлении, как бы продолжая вентральную часть жаберной щели. Разрезанный при вскрытии всех жаберных мешков на отдельные части поверхностный жаберный сжиматель можно удалить весь или по, крайней мере с одной жаберной дуги. При этом следует обратить внимание на лежащие по периферии
каждой жаберной перегородки тонкие хрящевые элементы — extrabranchiale. Рассматривают отделяющие жаберные мешки друг от друга — жаберные перегородки (septum branchiale, рис. 33, spt. br.). Каждая жаберная перегородка начинается на жаберной дуге, тянется до наружного жаберного отверстия и кончается конечной складкой, покрывающей жаберную щель. На жаберных перегородках сидят складки слизистой оболочки — жаберные лепестки. Необходимо отметить, что жаберные лепестки не доходят до конца перегородок, особенно в передних полужабрах, поэтому образуется свободный от них край, в котором ясно заметны мышечные волокна (m. interbran-chialis), кончающиеся на внутренней поверхности (extrabranchiale). Совокупность жаберных лепестков, приращенных к одной стороне перегородки, образует полужабру (hemibranchia,
рис. 33, hembr.). Две полужабры, сидящие на одной перегородке, образуют целую жабру (holobranchia, рис. 33, holbr.), а две полужабры, ограничивающие спереди и сзади жаберную щель, образуют жаберный мешок.
Рассматривают и пересчитывают жабры, жаберные перегородки с жаберными лепестками и жаберные щели, идущие в косом направлении изнутри кнаружи и несколько спереди назад. По переднему краю первой жаберной щели расположена первая полужабра, сидящая на задней стороне подъязычной дуги (гиоидная полужабра). Следующие четыре жаберные дуги (1—4) несут каждая по две полужабры, сидящие по переднему и заднему краям жаберных перегородок; 5-я жаберная дуга жаберных лепестков не несет.
Таким образом, если говорить о жабрах, то имеется четыре с половиной жабры, причем передней полужаброй является гиоидная. Если же считать жаберные мешки, то их окажется тоже четыре с половиной, но только половина мешка будет помещаться позади 4-й жаберной дуги. Раздвигают жаберные дуги и отмечают, что их внутренние вогнутые поверхности, обращенные в ротоглоточную полость, вооружены заметными выростами — жаберными тычинками, расположенными на всех дужках в один ряд, за исключением 4-й дуги, на которой тычинки сидят в два ряда.
Проводя пальцем по обеим поверхностям жаберной дуги, можно почувствовать шероховатость как на самой дужке, так и на тычинках, что обусловлено наличием дентиновых зубов, хорошо видимых под лупой. Тычинки, сидящие на внутренних поверхностях жаберных дуг, служат для задерживания пищевых частиц в ротоглоточной полости.
В основании жаберных лепестков через слизистую оболочку просвечивают хрящевые лучи, поддерживающие жаберную перегородку. Считают жаберные лепестки на одной из жаберных дуг и обращают внимание на то, что лучи отходят от жаберной дуги.
Теперь изучают строение жабры на поперечном срезе и ротоглоточную полость, куда открываются внутренние отверстия жаберных мешков. Для этого разрезают сочленение между верхней и нижней челюстями, перерезают гиоидную дугу и первые четыре жаберные дуги на уровне середины жаберных щелей, но только с одной стороны (удобнее перерезать левую сторону). С одной из перерезанных жабр делают поперечный срез и рассматривают его в воде под лупой. В основании жабры находится хрящевая жаберная дуга, в которой прикрепляется перегородка с сидящими на ней жаберными лепестками. Под жаберной дугой в основании межжаберной перегородки можно видеть три отверстия жаберных сосудов: непарный широкий сосуд —это приносящая жаберная артерия, по которой венозная кровь идет к жабрам, два маленьких парных сосуда — выносящие жаберные артерии, по которым артериальная кровь оттекает из жаберных лепестков.
Отворачивают влево вентральную часть висцерального аппарата и рассматривают передний отдел пищеварительной трубки — ротоглоточную полость.
Ротоглоточная полость
Ротоглоточная полость (cavum oro-pharyngeum)— расширенный передний отдел пищеварительной системы — начинается ротовым отверстием и продолжается назад до пищевода. Боковые стенки собственно глоточной полости пронизаны внутренними жаберными отверстиями, почему этот передний отдел и получил название ротоглоточной полости.
Ротовое отверстие (stoma, рис. 31, si.) полулунной формы в виде поперечной щели расположено на брюшной поверхности головы. Оно ограничено верхней и нижней челюстями, усаженными в несколько рядов зубами.
Зубы (dens, рис. 31, dens.) имеют форму острых дентиновых конусов. Они сидят на хрящах верхней и нижней челюстей; по мере снашивания и выпадения наружные зубы сменяются постоянно подрастающими зубами с внутренней стороны. По своему строению и происхождению они являются видоизмененными плакоидными че-шуями. У скатов зубы имеют такое же строение, но они уплощены.
Язык (lingua) находится на дне ротовой полости. Это складка слизистой оболочки, поддерживаемая непарным элементом — копулой подъязычной дуги (basihyale). Такой язык не имеет собственной мускулатуры, и поэтому его движения связаны с движениями подъязычного и жаберного аппаратов.
Рассматривают пять внутренних жаберных отверстий, отмечают, что наружные жаберные отверстия значительно меньше, чем внутренние отверстия.
Брызгальце (spiraculum). Впереди от первой жаберной щели, между челюстной и подъязычной дугами, находятся парные отверстия — брызгальца, которые открываются в ротоглоточную полость. Особенно сложно устроено брызгальце у скатов. Задняя его стенка ограничена гиоидной дугой, передняя расположена непосредственно позади глаза. Поворачивают ската спинной стороной к себе и на передней стенке брызгальца рассматривают складку — это клапан, способный закрывать все наружное отверстие брызгальца. По свободному краю складки расположены мелкие валики, которые образуют ложную жабру (pseudobranchia). Перерезают брызгальце со спинной стороны и рассматривают в основании ложной жабры особый хрящ, который называется спиракулярным. Такое сложное строение брызгальца, его значительные размеры, наличие клапана, препятствующего обратному току воды, связаны с тем, что у ската в связи с придонным образом жизни вода для дыхания поступает только через брызгальце. У акул — пелагических форм — вода поступает через ротовое отверстие.
Кровеносная система
Для описания кровеносной системы хрящевых рыб нами использованы инъецированные экземпляры. Инъекция производилась на свежих акулах и скатах на месте лова, поэтому следует учесть, что
на обычных фиксированных экземплярах кровеносных сосудов будет видно значительно меньше, чем это дано в описании.
Положение центрального органа кровообращения — сердца —; впереди от грудного пояса вы уже видели при вскрытии жаберной области.
Сердце (сот) хрящевых рыб, как и других рыб, называется двухкамерным, но состоит из четырех отделов. Задний отдел, куда впадают все вены, называется венозным синусом. Он широким отверстием соединяется с предсердием, имеющим вид тонкостенного мешка, наполненного кровью. Затем идет желудочек, центральный отдел сердца, с толстыми мышечными стенками, сокращением которого кровь выталкивается из сердца. Впереди желудочка в виде толстой трубки лежит передний отдел сердца — артериальный конус, переходящий в брюшную аорту. Сердце расположено в особой околосердечной полости.
Околосердечная, или перикардиальная, полость тела (cavum pericardiale) имеет форму треугольника, широкое основание его прилегает к грудному поясу. Эта полость представляет собой переднюю часть целома, отделенную от брюшной полости поперечной перегородкой (рис. 34, рте.). Она выстлана изнутри тонким париетальным листком брюшины, называемым перикардием (pericardium). Висцеральный листок плотно облегает сердце снаружи, являясь внешним слоем его стенок, и называется эпикардием (epicardium). Под ним расположен толстый слой мускулатуры, составляющий основную массу сердечной стенки — миокардий (myocardium). Внутренние же полости сердца выстланы тонким листком, называемым эндокардием (endocardium).
Вскрывают околосердечную сумку, начиная от основания луковицы аорты, примерно до переднего края перегородки. Удаляют ее брюшную стенку и рассматривают отделы сердца.
Желудочек (ventriculus cordis, рис. 34, ventr.) расположен непосредственно на вентральной стороне перикардия, занимая его большую часть. Он имеет форму закругленного треугольника с очень плотными мышечными стенками.
Артериальный конус (conus arteriosus, рис. 34, con. а.) — удлиненный, суженный отдел, находящийся впереди желудочка и сходный с ним по цвету. Его стенки, как и стенки желудочка, образованы поперечно-полосатыми мышцами, так как он является отделом сердца, обособившимся от желудочка.
Предсердие (atrium, рис. 34, atr.) — более тонкостенный отдел сердца, темного цвета от просвечивающей крови — выступает с боков от желудочка.
Венозный синус (sinus venosus, рис. 34, sin. v.) расположен в заднем отделе перикардия, служит резервуаром для венозной крови, собирающейся со всего тела. Чтобы его обнаружить, оттягивают желудочек вперед. Из венозного синуса вся венозная кровь переходит сперва в предсердие, а затем в желудочек и в артериальный конус. Таким образом, через сердце рыб проходит только 128
венозная кровь. Чтобы рассмотреть венозный синус, следует удалить вентральную часть грудного пояса. Синус имеет вид тонкостенной трубки, переходящей по краям в более узкие сосуды — кювьеровы протоки (ductus Cuvieri, рис. 34, d. С.). Разрезав вдоль брюшную стенку венозного синуса и кювьерова протока
а. fiyp.br.
s.br.
gl. thyr.
a. in. a.
a.brad.
~Ьу.
air.
ao.v. but. a. con. a.
-pro.
- cer. br. 4
- v.Jug.i
~ ventr. sin. v.
Рис. 34. Вскрытая жаберная область акулы Squalus acanthias (по Маринелли и Стренгеру):
а. br. ad. — приносящие жаберные артерии; a. hyp. br. — поджаберная артерия; a. in. а. — безымянные артерии; ао. V. — брюшная аорта; atr. — предсердие; bid. а. — луковица аорты; соп. а. — артериальный конус; d. С. — кювьеров проток; gl. /hyr. — щитовидная железа; hy. — гноид; сег. Ъг. 4 — ceratobranchiale 4; pre.— перикардий; s. br. — жаберный мешок; sin. о.—венозный синус; ventr. — желудочек; о. jug. I. — нижняя яремная вена
с одной левой стороны и промыв полость водой, рассматривают отверстия вен, впадающих в венозный синус и кювьеров проток. Прежде всего на середине задней стенки венозного синуса находят два отверстия печеночных вен, которые несут кровь из общего печеночного синуса (рис. 35, v. h.). В заднем углу начала кювьерова 5—3467 129
протока лежит крупное отверстие задней кардинальной вены (рис. 35, V. crd. р.), которая перед впадением в кювьеров проток образует расширение — кардинальный синус (sinus cardinalis, рис. 35, sin. с.). На передней стенке кювьерова протока, против отверстия задней кардинальной вены, находят место впадения передней кардинальной вены (рис. 35, v. crd. а.)-, в сущности слияни-
Рис. 35. Вскрытые венозный синус и сердце акулы Squalus acanthias:
etr. — предсердие; con. а. — артериальный конус; d. С. — кювьеров проток; f. a.-ven. — предсердно-желудочковое отверстие; sin. с. «— кардинальный синус; sin. h. — печеночный сииус; sin. v. — венозный синус; и. соп. а. — клапаны артериального конуса; о. crd. а. — передняя кардинальная вена; и. crd. р. — задняя кардинальная вена; р. h. — печеночная вена; ventr. — желудочек; и. j. inf. — нижняя яремная вена;
о. lai. — боковая вена; v. scl. — подключичная вена
ем передних и задних кардинальных вен и образован кювьеров проток. Медиальнее от места впадения передней кардинальной вены в переднюю стенку кювьерова протока впадает нижняя яремная вена (v. jugularis inferior, рис. 35, v. j. inf.). На нашем рисунке, на котором правый кювьеров проток не вскрыт, видно, что подключичная вена (v. subclavia, рис. 35, v. scl.) впадает в него с вентрально.й стороны. На препарате место впадения подключичной вены не всегда удается рассмотреть, так как оно повреждается при вскрытии венозного синуса и кювьерова протока. Наконец, находят на передней стенке венозного синуса большое синусо-предсердное
отверстие (ostium sinatriale), которое ведет из венозного синуса в предсердие. Оно окружено двумя тонкими клапанами.
Продольным разрезом вскрывают вентральную стенку желудочка, продолжив разрез впереди и на артериальный конус вплоть до луковицы аорты. Обращают внимание на толстые мышечные стенки желудочка. Раздвигают в стороны края разреза, освобождают полость желудочка и артериального конуса от сгустков крови. На дорзальной стенке желудочка видно предсердно-желудочковое отверстие (ostium atrio-ventriculare, рис. 35, о. a-о.), снабженное двумя клапанами. На внутренней стенке артериального конуса под лупой можно видеть два ряда клапанов, по три в каждом ряду. Они имеют вид тонкостенных карманчиков, позволяющих крови течь из конуса только вперед в брюшную аорту.
Вскрытие сердца удобнее произвести, выделив его из препарата и поместив в ванночку с водой, но это можно проделать только после изучения брюшной аорты с приносящими жаберными артериями или даже всей кровеносной системы.
Артериальная система
Артериальная система слагается из артерий, по которым венозная кровь идет из сердца по брюшной аорте и приносящим жаберным артериям к жабрам, где и окисляется. Затем уже окисленная кровь по выносящим жаберным артериям собирается в спинную аорту. Отходящие от спинной аорты артерии снабжают артериальной кровью все внутренние органы, кожу, мышцы и т. д. Изучение артериальной системы удобнее начать с брюшной аорты.
Брюшная аорта (aorta ventralis, см. рис. 34, ао. v.). Чтобы рассмотреть брюшную аорту, необходимо разрезать соединительнотканную пленку, покрывающую ее с брюшной стороны до околосердечной сумки. Брюшная аорта отходит от переднего конца артериального конуса и идет вперед к голове. Она белого цвета и значительно уже в диаметре, чем артериальный конус. Такое резкое различие в цвете обусловлено тем, что стенки брюшной аорты не
содержат поперечнополосатой мускулатуры.
Приносящие жаберные артерии (a. branchialis advehens, рис. 34, a. br. ad.) — ряд сосудов, отходящих от брюшной аорты вправо и влево. Они идут по жаберным перегородкам и при-
носят венозную кровь к жабрам.
Следует отпрепарировать пять приносящих жаберных артерий, отходящих от брюшной аорты, для чего приподнимают корако-бранхиальные мышцы. Первые две приносящие жаберные артерии представляют собой ветви безымянной артерии (a. innominata, s. anonyma anterior, рис. 34, a. in. а.). Передняя идет к передней полужабре гиоидной дуги, задняя снабжает две полужабры 1-й жаберной дуги, 3-я приносящая жаберная артерия направляется ко 2-й жаберной дуге. Она отходит от брюшной аорты на 1—1,5 см каудальнее места отхождения безымянной артерии. Чтобы ее рассмотреть, необходимо приподнять первую и вторую коракобранхи-
альные мышцы. Примерно на 1,5 см кзади лежит 4-я приносящая жаберная артерия, снабжающая полужабры 3-й жаберной дуги. Чтобы ее лучше рассмотреть, отгибают вперед 3-ю коракобранхи-альную мышцу. Последняя 5-я приносящая жаберная артерия 4-й жаберной дуги отходит с дорзальной поверхности брюшной аорты непосредственно над предыдущей. На препарате она лежит глубже. Она направляется в сторону и назад, прикрытая 4-й коракобран-хиальной мышцей. Последнюю удаляют. В области отхождения 4-й и 5-й приносящих жаберных артерий аорта образует заметное расширение, иногда называемое луковицей аорты (bulbus aortae, рис. 34, bul. а.). У ската три последних приносящих сосуда отходят от брюшной аорты общим стволом; получившим название задней безымянной артерии (a. innominata posterior, s. a. anonyma posterior).
Выносящие жаберные артерии (a. branchialis reve-hens, рис. 36, a. br. г.) собирают окисленную в жабрах кровь и об-р азуют систему артериальных дуг корней аорты (radix aortae), составляющих центральный артериальный ствол — спинную аорту. Они располагаются в дорзальной стенке ротоглоточной полости. Для того чтобы изучить выносящие жаберные артерии, перерезают правую половину висцерального аппарата (левая сторона была перерезана раньше, при изучении дыхательной системы) и отворачивают отрезанную часть назад. На основании черепа через слизистую оболочку крыши ротоглоточной полости слегка просве
чивают идущие косо спереди и снаружи ко внутри четыре пары выносящих жаберных артерий. Для того чтобы их рассмотреть как следует, необходимо удалить слизистую оболочку крыши глотки. На фиксированных экземплярах выносящие сосуды обычно не наполнены кровью и видны довольно плохо, поскольку не отличаются ио цвету от окружающих тканей. Контрастность их можно повы
Рис. 36. Выносящие жаберные артерии и начало спинной аорты акулы Squalus acanthias:
в. br. г, — выносящие жаберные арте-рви; а. с. с. >—общая сонная артерия; а. е. <. — наружная сонная артерия; а. а. 7. — внутренняя сонная артерия; е. d. — чревная артерия; a. hy. d. — дорзальная гиоидная артерия; а. т. — мускульная артерия; ао. d. — спинная аорта; a. opht. — глазная артерия; а. зЛ — подключичная артерия; f. о.— основное отверстие; hm. — гномандибу-лярный хрящ
сить, если ввести в них подкрашенный раствор желатины или тушь. В некоторых случаях такая инъекция на фиксированном материале бывает достаточно удачной.
Первая жаберная артерия обнаруживается вдоль переднего края pharyngobranchiale 1-й жаберной дуги (рис. 36, a. br. г., 1). Она сливается с сосудом своей пары на уровне дорзального конца pharyngobranchiale 2-й жаберной дуги, образуя начало спинной аорты (aorta dorsalis). Сходным образом расположены выносящие сосуды 2-й, 3-й, 4-й жаберных дуг. Все они идут по передней поверхности глоточных элементов жаберных дуг и последовательно впадают в спинную аорту (рис. 36, ао. d.). Каждый выносящий сосуд собирает кровь от двух полужабр жаберного мешка так: 1-й выносящий з виде основного ствола проходит по передней полужабре 1-й жаберной дуги и получает в области нижнего конца эпиэлемента ветку от гиоидной полужабры; аналогичным образом 2-й выносящий сосуд несет кровь из 2-го жаберного мешка, 3-й — из 3-го, 4-й — из 4-го жаберного мешка. Причем к 4-му выносящему сосуду присоединяется ветка из задней полужабры 4-й жаберной дуги.
При снятии слизистой оболочки с основания черепа впереди от первого выносящего жаберного сосуда следует соблюдать осторожность, так как в него на уровне сочленения pharyngo — epibranchiale первой жаберной дуги впадает нижняя гиоидная артерия (a. hyoidea ventralis, рис. 36, a. hy. щ), несущая кровь от гиоидной полуж»збры. Примерно от места впадения гиоидной артерии или прямо от последней (могут быть варианты) отходит общая сонная артерия (a. carotis communis, рис. 36, а. с. с.), которая тянется вперед сначала вдоль латерального края основания черепа, а затем поворачивает к его середине. В месте поворота она распадается на два сосуда: наружную и внутреннюю сонные артерии.
Внутренняя сонная артерия (a. carotis interna, рис. 36, а. с. г.) идет к медиальной, линии черепа, сливается здесь с одноименной артерией противоположной стороны, и общий сосуд входит внутрь черепа через непарное основное отверстие (foramen basila-ге, рис. 36, f. b.), снабжая кровью головной мозг.
Наружная сонная артерия (a. carotis externa, рис. 36, а. с. е.), наоборот, поворачивает в латеральном направлении, выходит в область глазницы, где распадается на ряд мелких ветвей, снабжающих органы, расположенные в глазнице. Одна из них тянется далеко вперед, снабжая кровью вентральные части рострума (рис. 36).
Иногда на препаратах удается рассмотреть верхнюю гиоидную артерию (a. hyoidea dorsalis, рис. 36, a. hy. d.), которая собирает кровь преимущественно от дорзальной поверхности гиоидной полужабры. Она поворачивает вперед, ложится по наружному краю сустава между подвеском и гиоидом и направляется в ложную жабру. Покинув ее, артерия идет вниз и несколько вперед и Делится на две ветви. Верхняя проникает внутрь глаза, распола
гаясь между прямыми мышцами глаза. Это a. ophtalmica (рис. 36, a. opht.). От нее отходит ветка к косым мышцам глаза. Нижняя a. hyoidea проходит по боковой поверхности черепа и проникает внутрь через foramen a. hyoideae, располагающееся несколько впереди межглазничного венного канала. Внутри черепа она сливается с разветвлениями внутренней сонной артерии.
От медиальных участков первой выносящей жаберной артерии, непосредственно перед их слиянием в спинную аорту, отходят позвоночные артерии (a. vertebralis, рис. 36, a. vrt.), которые тянутся по краям позвоночного столба и по задней части основания черепа, постепенно истончаясь. На своем пути они отдают многочисленные тонкие ветви в область спинномозгового канала и внутрь черепа.
У скатов общее расположение и взаимосвязь выносящих жаберных сосудов соответствует таковым акул, однако имеются и некоторые особенности. Так, за счет слияния некоторых выносящих жаберных артерий недалеко от жаберных отверстий спинная аорта образована не четырьмя, как у акулы, а тремя или даже двумя стволами. Позвоночные артерии значительно короче, чем у акул. Имеются и некоторые другие, более мелкие отличия в длине и расположении отдельных сосудов.
Спинная аорта (aorta dorsalis) располагается на дорзальной стороне полости тела непосредственно под позвоночником. Чтобы рассмотреть отходящие от нее артерии, отворачивают в сторону внутренние органы. Последние выносящие жаберные артерии впадают в спинную аорту примерно на уровне переднего края грудного пояса. На линии его задней границы от боковой или даже дорзальной поверхности спинной аорты отходят парные подключичные артерии (a. subclavia, рис. 36,*а. scl.), несущие кровь к грудным плавникам.
Несколько кзади от подключичных артерий от вентральной поверхности спинной аорты отходит крупный ствол чревной артерии (рис. 36, a. cl.), снабжающей кровью желудок, печень, поджелудочную железу и двенадцатиперстную кишку. В самом начале от чревной артерии отходит ряд мелких сосудов к пищеводу. Несколько отступя назад от места отхождения чревной артерии, также от вентральной поверхности спинной аорты, отходит крупный сосуд— передняя брыжеечная артерия (рис. 32, а. т. s.). У скатов бывают достаточно хорошо видны отходящие от нее в самом начале передние артерии половых желез (a. genitalis anterior). У акул рядом с верхней брыжеечной артерией отходит также селезеночно-желудочная. Далее назад от спинной аорты отходит задняя брыжеечная артерия (а. те-senterica inferior), которая идет по переднему краю мезоректума и ветвится в ректальной железе. На препаратах кровеносной системы скатов бывают видны отходящие от нее парные задние половые артерии (a. genitalis posterior). На всем протяжении от спинной аорты отходят сегмен т а р н ые артерии (a. parie-talis), которые в области почек дают небольшие почечные артерии.
У ската эти почечные артерии (a. renalis) особенно сильно развиты в области задних концов почек. Близ заднего края полости тела от спинной аорты отходят две подвздошные артерии (a. iliaca), они идут кнаружи к брюшным плавникам.
Отрезают левую сторону клоаки от стенки тела и отворачивают клоаку влево. На уровне задней половины клоаки видны идущие от спинной аорты подвздошные артерии. В самом начале от них отходят основные п о ч еч н ы е артерии (a. renalis), которые направляются вперед, проходят по внутреннему краю брюшной поверхности почек и снабжают их артериальной кровью. После отхождения подвздошных артерий спинная аорта продолжается назад как хвостовая артерия (a. caudalis), которая проходит в гемальном канале хвостовых позвонков.
Теперь следует рассмотреть ветвление наиболее крупных сосудов, отходящих от спинной аорты.
Чревная артерия (a. coeliaca, рис. 36, a. cl.), отойдя от спинной аорты, выходит на вентральную сторону брюшной полости, проходя между печенью и пищеводом. Чтобы ее рассмотреть, необходимо отвернуть печень вправо или вперед. Дойдя до переднего края сальника, она на небольшом промежутке тянется параллельно желчному протоку, а затем распадается на два сосуда: правый— кишечную артерию и левый — желудочно-печеночную.
Кишечная артерия (a. intestinalis, рис. 36, a. ints.) у акул и скатов ветвится несколько по-разному.
У акул кишечная артерия тянется параллельно желчному протоку и в самом начале от нее отходит тонкий сосуд, который идет вниз параллельно кишечной артерии. Он снабжает кровью краниальный конец спинной доли поджелудочной железы, а также частично пилорический отдел желудка, его дорзальную поверхность.
Главный ствол кишечной артерии направляется к пилорической части желудка, дугообразно охватывает внутреннюю поверхность двенадцатиперстной кишки и в этом месте от него отходят следующие сосуды: 1) к наружной поверхности пилорического отдела желудка; 2) к самому пилорусу; 3) совместная веточка к двенадцатиперстной кишке и брюшной лопасти поджелудочной железы и 4) самостоятельная веточка к двенадцатиперстной кишке. После отхождения упомянутых ответвлений кишечная артерия снабжает кровью дорзальную поверхность толстой кишки, отдавая парные сосуды, идущие по перетяжкам, образованным спиральным клапаном.
У скатов кишечная артерия, несколько не доходя до двенадцатиперстной кишки, делится на два ствола. Один меньший — артерия двенадцатиперстной кишки (a. duodenalis) снабжает кровью часть двенадцатиперстной кишки, пилорус с дорзальной стороны и брюшную долю поджелудочной железы. Второй, очень крупный сосуд — нижняя желудочная артерия (a. gast-rica inferior) проходит по каудальному краю пилорической и донной частям желудка, отдавая им ветки.
Желудочно-печеночная артерия (a. gastro-hepatica) делится на печеночную и желудочную артерии.
Печеночная артерия (a. hepatica, см. рис. 32, a. h.) идет вперед, пересекает кишечную артерию с вентральной стороны и направляется к средней лопасти печени.
Желудочная артерия (a. gastrica, см. рис. 32, a. gst.) тянется в каудальном направлении и на уровне кардиальной части желудка делится на три ствола: передний снабжает задний отдел пищевода и кардиальную часть желудка; оба задних ствола направляются назад и снабжают кровью желудок по его малой кривизне. От правого заднего ствола вначале отходит ветка к краниальной половине дорзальной лопасти поджелудочной железы. Для того чтобы проследить разветвления чревной артерии, следует снять верхнюю пленку большого сальника.
Верхняя брыжеечная артерия (a. mesenterica superior, см. рис. 32, а. т. s.) у акул и скатов тянется в вентральную часть брюшной полости к стенке толстой кишки, которую и снабжает кровью с помощью парных мелких сосудов, проходящих по бороздам кишки, образованным складками спирального клапана. Сосуд оканчивается на стенке прямой кишки. Недалеко от места отхождения верхней брыжеечной артерии от нее же отходят ветви к половым железам.
У скатов от верхней брыжеечной артерии отходят также сосуды к дорзальной лопасти поджелудочной железы и к селезенке.
С е л е з е н оч н о-ж е л у д оч н а я артерия (a lieno-gastrica) имеется только у акул и отходит от спинной аорты практически общим стволом с верхнебрыжеечной. Она проходит близ заднего края мезентерия и подходит приблизительно к середине селезенки. Несколько не доходя до селезенки, она делится на два ствола — верхнюю и нижнюю селезеночные артерии. От верхней селезеночной артерии отходит, направляясь вперед, ветка к каудальной половине дорзальной лопасти поджелудочной железы. Обе селезеночные артерии входят в селезенку, где и ветвятся. Каждая из артерий помимо селезеночных веток отделяет артерии, снабжающие донную часть желудка.
Венозная система
Венозная система состоит из сосудов, по которым венозная кровь возвращается в сердце. На препарате вены легко отличить от артерий, так как это широкие тонкостенные сосуды с просвечивающей кровью. Можно отметить два пути, по которым венозная кровь идет к сердцу: 1) система кардинальных вен, собирающих венозную кровь со всего тела, причем задние кардинальные вены проходят через воротную систему почек, и 2) воротная система печени, через которую проходит венозная кровь от всего пищеварительного канала, пищеварительных желез и селезенки. Вся венозная кровь собирается в вышеописанный венозный синус.
Основными стволами венозной системы являются передние и задние кардинальные вены.
. Передние кардинальные вены (v. cardinalis anterior, рис. 37, v. crd. а.), или верхние яремные (v. jugularis superior), они же внутренние яремные вены (v. jugularis interna), возвращают кровь от головного конца тела, в основном от
Рис. 37. Схема венозной системы акулы (4) и ската (5):
С. — кювьеров проток; sin. с. — кардинальный синус; sin. h. — печеночный синус; V. с. — хвостовая вена; v. cl. — клоакальная вена; v. crd. а.—передняя кардинальная вена; v. crd. р. задняя кардинальная вена; v. fem. — бедренная вена; v. h. — печеночная вена; и. inf. — ннжняя яремная вена; и. lat. — боковая вена; и. р. Л.—воротная вена печени; и. р. г. — воротная веиа почек; v. s. cl. — подключичная вена
головного мозга, черепной коробки и органов чувств. Они выходят из черепной коробки через отверстие для X нерва, идут назад над верхними сегментами жаберных дуг и впадают в кювьеровы протоки. Просмотреть весь ход этих вен трудно, обычно виден только небольшой отрезок сосуда перед впадением его в кювьеров проток (рис. 35, V. crd. а).
Нижние яремные вены (v. jugularis inferior, рис. 35, 37, v. j. inf.) собирают кровь с нижней стороны головы, в основном от висцерального аппарата. Они проходят по его вентральной поверхности, по внешней стенке околосердечной полости и впадают в проток Кювье.
Для того чтобы рассмотреть хвостовую вену и сосуды, образующие воротную систему почек, следует сделать ряд поперечных срезов через хвостовой отдел и заднюю часть тела рыбы. Срезы можно сделать либо на другом экземпляре, либо на данном, но после изучения мочеполовой системы.
Хвостовая вена (v. caudalis, рис. 37, v. caud.) лежит в ге-мальных дугах позвонков под хвостовой артерией и собирает венозную кровь из хвостового отдела. На уровне задних концов почек
Рис. 38. Поперечный разрез тела акулы Squalus acanthias: ао. d. — спинная аорта; col.—толстая кишка; gl. adr.— надпочечник; gl. ret. — ректальная железа; msr, — мезентерий ректальной железы; periton. — брюшина; г. — почка; иг. — мочеточник; tut.— матка; v. crd. р. d.—задняя кардиальная вена, правая; v. lot. -— боковая вена; v. р. г. — воротная вена почек
хвостовая вена разделяется на правую и левую воротные вены йочек (v. portae renalis, рис. 37, 38 v. р. г.). Эти вены идут вперед по дорзальному краю почек, образуя их воротную систему, т. е. систему разветвлений венозных сосудов .в почках, которая служит для выделения из крови жидких продуктов распада (мочи).
Задние кардинальные вены (v. cardinalis posterior, рис. 37, 38, v. crd. р.) — выносящие сосуды воротной системы почек. Сначала они появляются в виде непарного сосуда (рис. 38, V. crd. р.), который на уровне задней трети почек делится на правую и левую задние кардинальные вены. Они идут вперед по медиальным краям вентральной поверхности почек, причем правая вена значительно крупнее левой. Между ними имеется ряд тонких ана-138
стомозов. Воротные вены почек постепенно делаются все тоньше н исчезают.
На уровне заднего края грудных плавников задние кардинальные вены образуют заметные расширения — кардинальные синусы (рис. 37, sin. с.). С каждой стороны кардинальный синус, сужаясь, сливается с передней кардинальной веной, образуя поперечно идущий сосуд — кювьеров проток (ductus Cuvieri, см. рис. 35, 37, d. С.).
У скатов непарный кардинальный синус достигает необычайно мощного развития, кроме того, располагаясь на уровне половых желез, он получает венозную кровь от последних по широким п о-ловым синусам (s. genitalis), впадающим в боковые отделы кардинального синуса. На каудальном конце кардинальный синус резко сужается, образуя довольно удлиненный отдел, который прикрепляется к ректальной железе. Вся венозная кровь от железы оттекает в кардинальный синус.
Далее следует рассмотреть вены, собирающие венозную кровь из парных плавников и боковой стенки тела.
Б о к о в ы е в е н ы (v. lateralis, рис. 31, 35, 37, v. lai.) — парные сосуды, каждый из которых начинается от бедренной вены (v. femoralis, рис. 37, v. fem.), несущей кровь от брюшных плавников, и проходит по боковым стенкам брюшной полости, принимая по ходу от них вены. Они доходят до коракоидной области плечевого пояса, где и сливаются с подключичной веной.
У акул боковые вены в своем каудальном конце соединены друг с другом анастомозом (рис. 37).
У скатов с их каудальными концами связан венозный сосуд, собирающий кровь от боковых частей прямой кишки и клоаки. Таким образом у скатов кровь от задних отделов пищеварительной системы минует воротную систему печени.
Подключичная вена (v. subclavia, рис. 35, 37, v. scl.) собирает кровь от грудного плавника и плечевого пояса, проходя по его каудальному краю. После соединения с боковой веной она впадает в кювьеров проток с его вентральной стороны.
Воротная система печени образована венами, приносящими кровь от пищеварительного тракта, пищеварительных желез и селезенки. В печени вены образуют разветвления, где кровь фильтруется и снова собирается в крупные сосуды — печеночные вены, которые впадают в венозный синус.
Теперь необходимо познакомиться с венами, собирающими венозную кровь от пищеварительной системы и образующие в печени воротную систему. В большинстве случаев эти вены идут параллельно артериям.
У акул дистальная часть воротной системы представлена двумя венами, идущими вдоль дорзальной и вентральной поверхности толстой кишки. Вентральная вена, следуя более или менее параллельно верхней брыжеечной артерии, направляется ко дну желудка, где сливается с крупной селезеночной веной (см. рис. 32); далее, принимая кровь от дорзальной лопасти поджелудочной железы, она
идет краниально вдоль ее дорзальной поверхности. Вена дорзальной поверхности толстой кишки следует параллельно кишечной артерии, принимает вену из брюшной лопасти поджелудочной железы, проходит под пилорической частью желудка и идет краниально между кишечной артерией и ее пилорической частью. От желудка кровь оттекает по нескольким венам, которые идут параллельно желудочным артериям и сливаются в общий ствол.
У ската в отличие от акулы от кишечника отходит только один крупный венозный сосуд, в который на уровне пилорической части желудка впадают две вены: одна несущая кровь от пилорической части и дна желудка; другая — от двенадцатиперстной кишки и поджелудочной железы. Образующийся таким способом крупный сосуд на уровне пищевода сливается с тремя венами, несущими кровь от желудка: верхняя от кардинального отдела, вторая от малой кривизны и третья от селезенки. Слиянием всех этих вен и образуется крупный сосуд — воротная вена печени.
Общая желудочная вена (v. gastrica) у акулы сливается с вентральной веной толстой кишки, и их общий ствол непосредственно впереди от дорзальной лопасти поджелудочной железы соединяется с дорзальной кишечной веной, образуя воротную вену печени.
Воротная вена печени (v. portae hepatis, рис. 32, 37, v. р. h.), прикрытая с вентральной стороны печеночной артерией, идет к центральной доле печени. В печени эта вена распадается на капилляры, образуя воротную систему.
Печеночные вены (v. hepatica, рис. 35, 37, v. h.) выносят венозную кровь из печени, перед выходом образуют расширение — печеночный синус (sinus hepaticus, рис. 35, 37, sin. h.). У акулы, чтобы его обнаружить, надо срезать брюшную часть печени в переднем отделе. Открываются печеночные вены в задний отдел венозного синуса, что мы видели раньше при его вскрытии.
У ската печеночный синус лежит вне печени, впереди ее переднего края, поперек брюшной поверхности пищевода. Его суженные боковые части впадают в кювьеровы протоки.
Мочеполовая система самцов хрящевых рыб
Мочеполовая система всех описываемых видов построена сходным образом. У скатов она практически отличается только меньшей протяженностью выводных путей, в связи с относительно более короткой брюшной полостью. Эта компактность выделительной и половой системы ската несколько затрудняет ее изучение, поэтому желательно изучение мочеполовой системы проводить на акулах. Исходя из этого описание мочеполовой системы дается только по катрану. Это описание легко перенести и на другие виды, при этом необходимо учесть, что у кошачьей акулы половые железы расположены не в передней части тела, как у катрана, а в середине.
Необходимо отметить, что используемые на занятиях экземпляры акулы не превышают 40—50 см длины. Это неполовозрелые 140
особи, и изучение мочеполовой системы у них затруднительно. Приведенное описание этой системы сделано на крупных экземпля-
рах— 70—80 см длины.
Парные почки (геп) лежат на спинной стороне брюшной полости и покрыты пленкой брюшины. Они начинаются на уровне заднего края грудного пояса и продолжаются назад вдоль позвоночника, тесно к нему прилегая, до заднего края клоаки (рис. 39, 40). Передние две трети почек несколько отстоят друг от друга, в промежутке между ними видна спинная аорта и задние кардинальные вены (рис. 39, ао. d.-, V. crd. р.). Задние части почек начинаются, отступя 3—4 см от уровня заднего края передних спинных плавников. Они значительно утолщены, тесно сходятся друг с другом по средней линии и даже срастаются между собой на дорзальной поверхности, прикрывая аорту. Это хорошо видно и на поперечном срезе (рис. 38, 39, 40). В этом отделе дольчатое строение почек заметно гораздо резче, чем в передних отделах. Между почками
видна одна задняя кардинальная вена, которая на уровне отхождения верхней брыжеечной и селезеночножелудочной артерий разделяется на два ствола — задние кардинальные вены, из которых правая значительно крупнее левой.
Надпочечная желе
Рис. 39. Мочеполовая система самца акулы Squalus acanthias:
20. d. — спинная аорта; cl. — клоака; d. М. — мюллеров канал; march. — брыжейка семенника; р. ug. —мочеполовой сосочек; г. —почки; s. sem.— семенные мешки; sin. card. — кардинальный синус; test. — семенник; t.~h.— место срастания семенника с печенью; иг. — мочеточник; vas. d. — вольфов канал; vas. е.—семевыносящие канальцы; о. crd. р. — задняя кардинальная вена;
ves. s.—семенные пузырьки
a (glandula adrenalis, рис. 38, gl.
adr.) имеет вид беловатого тяжа, расположенного на дорзальной
поверхности почек. Она лежит непосредственно под спинной аортой между воротными венами почек. Начинаясь от заднего конца почек, она идет вперед на протяжении 3—5 см, постепенно сужаясь.
Семенники (testis, рис. 39, test.). Приподнимают и отводят в сторону печень. Под ней видны парные семенники неправильно удлиненной формы, закругленные на концах. Они лежат с боков от пищевода, передней половиной налегая на вентральную поверхность кардинальных синусов, и прикрыты снизу и с боков печенью. При помощи брыжейки —mesorchium (рис. 39, msrch.) семен-поверхности полости тела. Внутренней поверхностью передней части семенники плотно срастаются с дорзальной поверхностью печени (рис.
иики закреплены на спиннои
Рве 40. Вскрытый мочеполовой синус самца акулы Squalus асап-thias:
с1»кдоака; г.—почка; s. setn. — се-хеявыв мешки; sin. ug. — мочеполовой синус; иг. — мочеточник; иг/ — отверстие мочеточника; vas. d/— отверстие вольфова канала; ves. s. — семенные пузырьки
39, t.-h.).
Вольфов канал (vas deferens, рис. 39, vas. d.) — выводной проток семени. Примерно от середины наружного края семенника можно проследить начало вольфова канала, который возникает из семявыносящих канальцев (vas. efferens, рис. 39, vas. е.), совокупность которых образует будущий придаток семенника, или epididymis.
Вольфов канал служит в основном для выделения семени и тянется по внутреннему краю вентральной поверхности почек. Вначале проток образует многочисленные, частые, неглубокие извилины, которые по направлению к заднему концу становятся более малочисленными и более пологими. Начиная с последней трети почек, вольфовы протоки становятся заметно шире, идут пря
мо назад и несколько ковнутри, сходясь между собой на спинной поверхности клоаки, где они и открываются в парные выросты м о ч еп о л о в о г о синуса (sinus arogenitalis, рнс. 40, sin. ug.). Перед впадением в синус каудальная часть вольфовых каналов у взрослых самцов образует замет-
ные расширения, так называемые семенные пузырьки, или ампулы (vesicula seminalis, s. ampula seminalis, рис. 39, 40, ves. s.). В мочеполовой синус несколько каудальнее отверстий вольфовых протоков открываются мочеточники (ureter, рис. 39, 40, иг.). Они начинаются от задней трети почек и лежат дорзальнее вольфова канала, несколько прикрытые веществом почки. Отпрепаровывая вольфовы каналы и мочеточники, можно легко убедиться в том, что в мочеточники открываются многочисленные моче выс юа н а лЪщ ы (рис. 40).
К вентральным поверхностям каудальных концов вольфовых каналов прилегают утончаюшиеся вперед семенные мешки,
или мужские матки (sacculus seminalis, s. uterus masculinus, рис, 39, 40, s. sem.). Они являются остатками задних частей мюллеро-вых каналов. Их задние концы, расширяясь, непосредственно пе-реходят в парные отделы мочеполового синуса, которые, сливаясь вместе, образуют непарную часть мочеполового синуса, оканчивающуюся мочеполовым отверстием на вершине мочеполового сосочка. Мочеполовой сосочек (papilla urogenitalis, рис. 39, р. ug.) расположен на дорзальной стороне клоаки. Чтобы его рассмотреть, вскрывают вентральную стенку клоаки, срезают вентральную стенку сосочка и обращают внимание на то, что из раздвоенной краниальной части мочеполового синуса легко проникнуть в семенные мешки. Дорзальнее этих отверстий находят места впадения вольфовых каналов, расположенных непосредственно сбоку от места соединения медиальных стенок парных частей (рогов) мочеполового синуса. Находят овально-щелевидные отверстия мочеточников — метанефрических протоков (рис. 40, иг'.).
Помимо остатков каудальных отделов мюллеровых каналов, образующих семенные мешки, у самцов сохраняются и передние их отделы. Они начинаются общим отверстием на вентральной поверхности проксимальной половины центральной доли печени. От общего отверстия, тесно прилегая друг к другу, на протяжении 2— 3 см эти каналы идут вперед (рис. 39, d. Л4.), расходясь далее в стороны по переднему краю печени, отделяя правую и левую ее доли от центральной. Далее они поворачивают в дорзальном направлении, прилегая с боков от пищевода к брюшной поверхности кардинальных синусов, где и теряются в соединительной ткани. От места расхождения мюллеровых каналов между ними натянута тонкая треугольная брыжейка, кончающаяся на перикардии. От него же отходит вторая складочка, которая стоит в плоскости, перпендикулярной к первой, и кончается у места сближения мюллеровых каналов.
Мочеполовая система самок хрящевых рыб
Положение и строение почек у самки сходно со строением почек самца. Следует, однако, отметить, что последние две трети почек у самки несколько тоньше и уже. Местонахождение же сосудов и надпочечника точно такое же, поэтому можно воспользоваться описанием почек, сделанным для самца. Положение половых желез также сходно — они лежат под печенью, поэтому надо поднять и отвернуть печень в сторону, под ней можно видеть парные яичники.
Яичники (ovarium, рис. 41, ov.) имеют неправильно удлиненную форму, несколько сужены на каудальных концах. Они лежат с боков от пищевода, налегая передней частью на вентральную поверхность кардинальных синусов, и прикрыты снизу и с боков печенью. При помощи брыжейки (mesovarium) яичники закреплены на спинной поверхности брюшной полости тела, а также и на пищеводе. Внутренней поверхностью передней части яичники плотно' срастаются с дорзальной поверхностью печени.
Яйцеводы (oviduclus, рис. 41, ovd.). Оба яйцевода, или мюл-
леровы каналы, начинаются общим отверстием в полость тела на вентральной поверхности центральной доли печени, в ее прокси-
мальной половине. От общего
Oh
y.crd.p.
Рис. 41. Мочеполовая система самки акулы Squalus acanthias:
cl, и» клоака; tnesev. — брыжейка яичника; og. —• скорлуповая железа; oh. — место срастания яичника с печенью; оу. — яичник; ovd. — яйцевод; р. иг. —• мочевой сосочек; г. — почка; sin. card. — кардинальный синус; иг. — метанефрический проток; ut.— матка; vas. d.--мочеточник (вольфов канал); о. crd.
р. — задняя кардинальная вена
отверстия, тесно прилегая друг к другу, на протяжении 2— 3 см, эти каналы идут вперед, расходятся далее в стороны по переднему краю печени, отделяя правую и левую ее доли от центральной, и далее поворачивают в дорзальном направлении, прилегая с боков от пищевода к брюшной поверхности кардинальных синусов. От места расхождения мюллеровых каналов между ними натянута тонкая, треугольной формы брыжейка, кончающаяся на перикарде. От него же отходит вторая складочка, которая стоит в плоскости, перпендикулярной к первой, и кончается у места сближения мюллеровых каналов. Пройдя по вентральной поверхности кардинальных синусов в каудальном направлении, на уровне их задних отделов, яйцеводы образуют небольшие расширения— скор лу новые железы (рис. 41, og.). У половозрелых самок железы достигают большой величины, и их железистые стенки выделяют «роговую» оболочку яйца. Далее назад яйце-
. воды проходят по вентральной поверхности почек и, не доходя на 1—2 см до клоаки, расширяются и образуют матки (uterus, рис. 31, 41, ut.).
Для того чтобы рассмотреть отверстия маток, разрезают вентральную стенку клоаки, раздвигают ее в стороны и -на дорзальной стенке находят мочевой сосочек (рис. 41, р. иг.), а по бокам от него щелевидные отверстия маток. Делают продольный разрез матки в краниальномца-правлении, обращая вниманиенк— складчатое строение ее стенки.
Срезают вентральную стенку мочевого сосочка и рассматривают удлиненную полость — мочевой синус, в верхний отдел которого отдельными отверстиями открываются два мочеточника — вольфовы каналы (vas. deferens, рис. 41, vas. d.). Они идут в самой ткани почек, вдоль их внутреннего края, под брюшиной. Яйцеводы же и матки проходят по вентральной поверхности почек, не прикрытые брюшиной. Чтобы рассмотреть мочеточники, отделяют матки и яйцеводы от почек, отводят их в сторону, снимают брюшину с почек и осторожно удаляют верхний слой почечной ткани. Они хорошо заметны в заднем отделе почек. Несколько не доходя до клоаки, каждый проток заметно расширяется. Перед самым впадением в верхний отдел мочевого синуса в него впадает тонкий, так называемый метанефрический проток (ureter, рис. 41, иг.). При этом обращают внимание, что в метанефрический проток впадает несколько тонких мочевых протоков.
Скелет хрящевых рыб
Изучение скелета и мускулатуры хрящевых рыб лучше проводить на акулах, а не на скатах, у которых, как известно, в связи с приспособлением к донному образу жизни тело имеет уплощенную форму, что менее типично для хрящевых рыб.
В то же время желательно ознакомиться с особенностями скелета ската, что можно сделать по постоянным, приготовленным заранее препаратам. Некоторые элементы скелета катрана вторично усложнены, поэтому необходимо иметь также препараты по скелету кошачьей или голубой акулы. Желательно иметь и постоянный препарат целого скелета химеры для ознакомления с примитивным строением осевого скелета, скелета плавников и особенностями скелета головы.
Для приготовления скелета акулы формалиновый материал следует промыть в воде, а затем выдержать в следующей смеси: 60%-ного спирта — 70 объемов, глицерина — 15 объемов, крепкой азотной кислоты —10 объемов. В этом растворе материал следует держать 1—2 суток и после снова промыть в проточной воде.
Снимают с акулы, выдержанной в вышеописанной смеси, кожу со всего тела. При снятии кожи с плавников, как непарных, так и парных, следует снимать ее только с одной стороны плавника, оставляя ее на другой стороне для сохранения тонких роговых лучей, поддерживающих кожную складку плавника. При снятии кожи около жаберных отверстий необходимо соблюдать осторожность. По краям жаберных отверстий кожу можно не снимать, а лишь подрезать ее вокруг.
На занятиях большого практикума обычно не хватает времени на то, чтобы каждый студент имел возможность очистить весь скелет акулы. Поэтому приходится изучать скелет отдельными частями по препаратам: череп, парные.и непарные плавники, отдельные части позвоночника. Желательно, чтобы препараторы и лаборанты заранее приготовили материал по скелету акул для раздачи сту
дентам уже очищенных препаратов. Такие препараты хорошо сохраняются в спирту или слабом растворе формалина и могут быть использованы в течение ряда лет. Однако при самостоятельном приготовлении скелетов студентами гораздо лучше усваиваются функциональные соотношения скелета с мускулатурой и другими органами.
Чтобы составить представление о хрящевом скелете в целом, необходимо до разбора строения отдельных частей предварительно провести общий осмотр целого скелета по препаратору, смонтированному в банке. Это даст возможность представить себе функциональное распределение частей скелета хрящевых рыб соответственно сложному комплексу двигательных функций этих примитивных и типичных представителей челюстноротых позвоночных.
Общий обзор скелета акулы катрана
Скелет акулы как представителя древнейших хрящевых рыб Chondrichthyes состоит только из хряща и не содержит костных элементов. В то же время в осевом скелете еще сохраняется хорда. Будучи способны к быстрому плаванию, акулы обладают и сравнительно прочным скелетом, дифференцированным на типичные для позвоночного отделы, каждый из которых несет определенную функцию, связанную с обеспечением активного движения или схватывания преследуемой добычи. Эти отделы следующие: 1) осевой скелет — позвоночник; 2) скелет конечностей (парных и непарных плавников); 3) скелет головы.
Осевой скелет — позвоночник (рис. 42, vrlb.) служит основной опорной осью и местом прикрепления двигательной туловищной мускулатуры. Он тянется вдоль всего тела от затылочного отдела черепа до хвостового плавника, заходя в его верхнюю лопасть, и состоит из отдельных хрящевых позвонков, внутри которых сохраняется хорда.
Скелет плавников обычно состоит из элементов двоякого рода. Внутренний состоит из хрящевых элементов, наружный представлен эластиновыми лучами кожного происхождения. Внутренний скелет непарных плавников (двух спинных и подхвостового), как правило, представлен расчлененными на членики хрящевыми лучами — радиалиями (рис. 42, rad.). Скелет хвостового плавника в связи с укреплением его за счет позвоночника имеет более сложное строение. Парные плавники, играющие основную роль рулей при плавании, отличающиеся большей подвижностью и получающие большую нагрузку, имеют и более сложно устроенный скелет, В нем различают расположенную в толще мускулатуры часть — пояс конечности (рис. 42, с. е. а. и с. е. р.) и скелет^евободнойко* ценности, имеющей также внутреннюю хрящевую часть и наруйи ный скелет из эластиновых нитей. В связи с большей функцией нальной нагрузкой все части скелета передних конечностей разви-5 ты значительно лучше, чем задних.
Скелет головы подразделяется на осевой череп — нейрокрани-ум и висцеральный скелет — спланхнокраниум. Осевой череп (рис. 42, neur.), или черепная коробка, возникает как защита головного мозга. С ее боковыми стенками сливаются обонятельные (рис. 42, cap. nas.) и слуховые (рис. 42, cap. aud.) капсулы, а между ними выемка — глазница. Висцеральный скелет, возникающий как опора переднего отдела пищеварительной трубки, представлен рядом дуг. Передняя — челюстная дуга (arcus mandibularis, рис. 42, arc.
nw.
Рис. 42. Общий вид скелета акулы Squalus acanthias:
arc. br. — жаберные дуги; arc, h. — подъязычная дуга; arc. tn. — челюстная дуга; bas. — основные элементы плавника; cap. aud. — слуховая капсула; cap. nas.—обонятельная капсула; с. е. а. — плечевой пояс; с. е. р.—тазовый пояс; с. t. ~ ребра; near. ~ черепная коробка; rad. —радиалии плавников
mnd.) выполняет функцию захватывания пищи. Следующая — подъязычная дуга (arcus hyoideus, рис. 42, ars. h.) играет роль подвеска, сочленяющего челюстную дугу с черепом. Остальные пять дуг, называемые жаберными (агсиэ branchialis, рис. 42, arc. Ьг.), выполняют дыхательную функцию, так как поддерживают 'сидящие на них жабры.
Осевой скелет — позвоночник акулы
Осевой скелет состоит из ряда сегментально расположенных позвонков; они охватывают лежащую внутри них спинную струну (chorda dorsalis, рис. 43, ch.), которая четкообразно перешнурована телами позвонков. Между позвонками находятся соединительнотканные прослойки.
Позвоночник разделяется на два отдела: 1) туловищный отдел — характеризуется наличием ребер, начинается от черепа и тянется до уровня клоаки (у некоторых скатов ребер нет); 2) хвостовой отдел —отличается отсутствием ребер и смыканием нижних дуг позвоночника в гемальный канал (рис. 43, Б).
Для того чтобы рассмотреть строение позвонков, нужно очень тщательно очистить отдельные участки позвоночника: один — в туловищной, другой — в хвостовой области. Полезно также взять в туловище часть позвоночника и разрезать его вдоль по сагиттальной плоскости.
Рассматривают строение отдельных позвонков туловищного и хвостового отделов, сравнивают их друг с другом, пользуясь при этом препаратами отдельных позвонков, очищенными участками позвоночника и разрезанными продольно позвонками.
Рис. 43. Дорзальный плавник акулы Squalus acanthias и часть позвоночника с сагиттальным разрезом четырех передних позвонков (Л); поперечный разрез позвонка из хвостовой области (5); участок позвоночника из туловищной области (В-) (из Гудрича):
а. — канал для хвостовой артерии; ас.—передние хрящи (видоизмененные радиалии); Ьс. ~~ основной -хрящ (слившиеся радиалии); b. d.— основной элемент верхней дуги; bv. — основной элемент нижней дуги; са.— кальцифицированное кольцо; сА. —хорда; ct. — ребро; с. vrt.— тело позвонка; h. а. — гемальиая дуга; id. — вставочный элемент верхней дуги; in. о, — межпозвоночная связка; iv—вставочный элемент ннжней дуги; пс. — невральный канал; sp. — спинной шип; v.канал для хвостовой вены
Каждый позвонок состоит из тела позвонка, охватывающего кольцом хорду, верхних дуг, образующих спинномозговой канал, и нижних дуг с поперечными отростками для причленения ребер-в туловищном отделе. В хвостовом отделе нижние дуги позвонков смыкаются в гемальный канал.
Тела позвонков (corpus vertebrae, рис. 43, с. угА) ^короткие хрящевые цилиндры, соединенные друг с другом прочными связками, что создает возможность сгибания позвоночника в целом. Тела позвонков охватывают кругом хорду, причем посредине позвонка хорда почти нацело перешнуровывается хрящевым кольцом тела. Благодаря этому тела позвонков оказываются двояковогнутыми — амфицельными. У некоторых акуловых рыб в телах позвонков встречаются отложения твердых известковых солей, напоминающих кость. В позвонках разных видов акул эти известковые скопления имеют форму колец или звездчатых фигур. Их можно хорошо рассмотреть, если окрасить поперечные разрезы позвонков ализарином (см. раздел «Материал и методика», стр. 18).
Верхние дуги (arcus neuralis s. neurapophysis) образуют боковые стенки спинномозгового канала и состоят в каждом позвонке из двух элементов: 1) основная дужка (basidorsale, рис. 44, bd.)—задний, каудальный элемент, прирастает основанием к. телу позвонка и имеет треугольную форму. Она характеризуется тем, что посредине пронизана брюшным корешком спинномозгового нерва; 2) вставочная пластинка (interdorsale или inter-calare, рис. 44, id.)—передний, краниальный элемент; она имеет треугольную форму, но вершиной направлена вниз и лежит против границы двух позвонков, вклиниваясь между двумя соединениями основными элементами. Вставочные дужки пронизаны отверстиями спинных корешков спинномозговых нервов.
Нижние дуги (pleurapophysis) также образованы о с н о в-ными элементами (basiventrale, рис. 43, bv.) и имеют вид боковых отростков позвонков под хордой. В туловищных позвонках они образуют идущие в стороны выступы — боковые отростки (parapophysis), к которым причленяются тонкие хрящевые пластинки— ребра, легко отрывающиеся при препаровке. Между боковыми отростками, у их основания, против миосепт, у катрана помещаются, каки в верхних дугах, нижние вставочные пластинки (interventrale, рис. 43, iv.), имеющие вид маленьких треугольных хрящиков, расположенных между основными нижними дугами. Таким образом, на каждый метамер тела приходится с каждой стороны двойная закладка скелетных элементов верхних и нижних дуг (диплоспондилия), как это имеет место и у круглоротых.
Ребра (costa, рис. 42, 43, ct.) расположены в туловищной области, но они лежат не около брюшной полости, как у костистых рыб, а в горизонтальной септе, которая делит мускулатуру на спинную и боковую.
В хвостовой области поперечных отростков нет, а нижние дуги образуют идущие вниз гемальные отростки, замыкающиеся в канал, называемый гемальным каналом (рис. 43, h. а.), где про-
ходят кровеносные сосуды (хвостовые вены и артерия). Гемальные отростки, как самостоятельные брюшные отростки нижних дуг, имеются и в задних туловищных позвонках.
В отличие от акуловых рыб у цельноголовых (химер) отсутствуют тела позвонков, и осевой скелет представлен хордой с сидящими на ней верхними и нижними дугами. Причем в верхних дугах, особенно в переднем отделе позвоночника, вставочные дужки развиты значительно сильнее, чем основные. При возможности желательно изготовить постоянный препарат осевого скелета химеры.
Скелет плавников
Для приготовления препарата скелета плавников вычленяют непарные плавники, проведя разрезы справа и слева от них. Так же выделяют и брюшные плавники вместе с тазовым поясом. Тазовый пояс легко прощупывается в виде поперечной перемычки впереди клоаки и лежит свободно в брюшной мускулатуре тела. Несколько сложнее вычленить грудной плавник. Прощупывают на вентральной стороне тела между грудными плавниками плечевой пояс. Далее следуетпроследить расположение скелета плечевого пояса на боках тела позади жаберного аппарата. Осторожно отпрепарируют справа и слева хрящевую дугу плечевого пояса вместе с грудными плавниками, начиная от спины.
Непарные плавники. У акулы Squalus acanthias имеются два непарных дорзальных плавника: передний (Д 1) и задний (Д 2). Эти плавники выполняют ответственную функцию килей при плавании. Соответственно этой функции скелет непарных плавников у акулы катрана вторично изменен по сравнению с первичным строением его у хрящевых рыб. В простейшем виде скелет непарных плавников состоит из одного ряда хрящевых лучей, носящих название радиальных лучей (radialia), или птеригиофор (ptery-giophora), каждый из которых расчленен на 2—3 хрящика. К концу каждого хрящевого луча по бокам примыкают основания тонких эластиновых нитей кожного происхождения — эластотрихий (elastotrichium), поддерживающих кожную складку плавника. Такое типичное строение скелета плавников можно видеть на препаратах плавников голубой акулы, кошачей акулы и химеры. У черноморской акулы катрана хрящевые лучи сращены друг с другом в несколько хрящевых пластинок, служащих опорой для поддерживания острого шипа (рис. 43).
Хвостовой плавник (pinna caudalis, рис. 44). Очищают скелет хвостового отдела акулы. Для этого целиком отслаивают по позвоночнику всю мускулатуру с одной стороны и постепенно снимают ее до самого заднего конца тела. Хвостовой плавник акул имеет типичное строение разнолопастного гетероцеркального плавника. Верхняя дорзальная лопасть его, по существу, является продолжением позвоночника. Нижняя вентральная более широкая лопасть поддерживается у основания удлиненными основны-150
ми хрящами нижних дуг хвостовых позвонков (basiventrale, bv.). Сама же кожная складка плавника поддерживается только кожными лучами, эластотрихиями. По средней линии скелета хвостового плавника продолжается ряд хвостовых позвонков. Они отчленены друг от друга посегментно соединительнотканными прослойками и над каждым телом позвонка хорошо различимы основные дужки (basidorsale, bd.), пронизанные брюшными корешками спинномозговых нервов. Между ними вклинены хрящевые треугольники, интеркалярии (interdorsale, id.) —вставочные промежуточные
Рис. 44. Скелет хвостового плавника акулы Squalus acanthias:
bd. — основной элемент верхней дуги; bv.—основной элемент ннжней дуги; dsp—дорзоспииальные хрящн; elast.— эластиновые лучн; id. — верхняя вставочная пластинка;
vsp. — вентроспииальные хрящн
элементы, пронизанные спинными корешками спинномозговых нер-вов. На каждой из верхних дужек позвонков располагается ряд длинных не расчлененных на отдельные членики (как это было в непарных плавниках) хрящевых плавниковых лучей. Они не соответствуют радиальным лучам непарных плавников и имеют особое название дорзоспинальиых хрящей (dorsospinale, dsp.). На вентральных концах удлиненных нижних дуг, поддерживающих нижнюю лопасть хвостового плавника, также лежит ряд дополнительных маленьких хрящиков, соответствующих dorsospinalia. Их. называют ventrospinalia. Эти дополнительные элементы в позвоночнике имеются только в области хвостового плавника, в туловище их нет. При сравнении с круглоротыми они могут быть сопоставлены с остатками скелета непарных плавников древних предков позвоночных. В туловище у акул в плавниках развился другой скелет из расчлененных на членики лучей (radiale).
Брюшные плавники (pinna abdominalis, рис. 45, Б) акулы имеют скелет, состоящий из тазового пояса и скелета свободного плавника.
Тазовый пояс, или таз (pelvis, рис. 45, Б, Р.), имеет форму почти прямой хрящевой пластинки, лежащей поперек тела не-
посредственно перед клоакой, От переднего конца этой пластинки отходят два тупых отростка (processus praepubis). По бокам и снизу располагаются сочленовные площадки для причленения скелета брюшных плавников, а сама пластинка пояса пронизана отверстиями для нервов.
Скелет свободного плавника в основании имеет большой длинный базальный луч (basipterygium, bpt.), который причле-
Рис. 45. Скелет парных конечностей акулы Squalus acatithias, эластотрихин удалены. Д — скелет левой половины плечевого пояса и грудного плавника; Б— скелет левой половины тазового пояса и брюшного плавника (по Маринелли и Стренгеру): bpt. — основной хрящ; mspt. — средний основной хрящ; mtpt. — задний основной хрящ; р. с. — коракоидный отдел; р.—тазовый пояс; ppt. — передний основной хрящ; р. s. — лопаточный отдел; rad. — радиальные хрящи
няется к поясу и лежит на внутренней стороне плавника. На наружной стороне базиптеригиума причленяется ряд хрящевых плавниковых лучей (radiate, rad.), сходных с плавниковыми лучами непарных плавников. Они также расчленены на отдельные членики. Первый из них (ppt.), самый крупный, причленяется непосредственно к тазовому поясу.
Кожная лопасть брюшного плавника, как и в непарных плавниках, поддерживается большим количеством кожных эластиновых лучей (elastotrichium), основания которых сверху и снизу охватывают концы хрящей.
У самцов акул и скатов дистальные концы базиптеригиума и задних радиальных лучей превращены в скелет копулятивного ап-.152
парата. Они приобрели ложковидную форму и сильно разрослись в длину.
Грудные плавники (pinna thoracalis, s. pinna thoracica, рис. 45, Д) также имеют пояс передних конечностей и скелет свободного плавника.
Пояс грудных плавников по аналогии с наземными позвоночными называется плечевым поясом и лежит непосредственно позади последней жаберной дуги. У акул он имеет форму хрящевой дуги, охватывающей тело с боков и замкнутой на брюшной стороне. Сочленовная поверхность для причленения скелета грудного плавника находится на боковой поверхности на утолщенном крае и делит хрящевую дугу пояса на дорзальный отдел, носящий название лопаточного (pars scapularis, р. s.), и вентральный отдел, называемый коракоидпым (pars coracoidea, р.с.).
На месте срастания правого и левого поясов на внутренней стороне образуется плоская площадка, служащая дном околосердечной сумки.
Грудной плавник акулы имеет три основных хряща: проптеригиум (propterygium, ppt.) маленький; средний мезоптеригиум (mesopterygium, mspt.) — сильный, крупный хрящ, промежуточный между передним и задним хрящами; задний — метан-теригиум (metapterygium, mtpt.)— самый длинный, тянется назад по внутреннему краю плавника и соответствует основному лучу брюшного плавника (basipterygium).
Плавниковые лучи — птеригиофоры грудного плавника (radiale, rad.) —сидят по наружному краю основных трех хрящей и каждый расчленен па 2—3 отдельных хряща различных размеров. Против проптеригиума лежит один большой луч, мезоптеригиум поддерживает 8—-9 хрящевых лучей, метаптеригиум— 12-13. Дистальная часть плавника без хрящевого скелета и поддерживается эластотрихиями.
Плечевой пояс ската сильно изменен в связи с громадным разрастанием грудных плавников вперед и назад. Передняя часть плавника поддерживается отходящим от черепа отростком — processus antorbitalis. В отличие от акул плечевой пояс на спинной стороне причленяется к позвоночнику. На его наружной стороне имеются три отдельные сочленовные фасетки для причленения трех основных отделов плавника. В плечевом поясе существуют три больших отверстия, или фоптапели: переднее из них носит название foramen scapulo-coracoideum, заднее верхнее — foramen scapu-lare, заднее нижнее — foramen coracoideum.
У грудного плавника ската передние и задние основные хрящи разрастаются в крупные массивные стержни, направленные вперед и назад. Средний хрящ (mesopterygium) плоский, сравнительно небольшой хрящик, располагается между их основаниями и причленяется к плечевому поясу особой фасеткой. Поддерживающие плавник радиали' очень многочисленны и сильно удлинены. Их дистальные концы практически доходят до наружного края плавника. Каждая радиалия состоит из большого количества члеников.
Функции парных плавников. Парные плавники у акул не являются органами движения, плавания в воде. Поступательное движение совершается боковыми изгибами хвостового отдела вправо и влево. Грудным и брюшным плавникам у акул принадлежит роль несущих плоскостей, стабилизаторов и рулей глубины.
У скатов, наоборот, хвостовой отдел утерял функцию органа движения тела вперед и превратился в придаток, выполняющий функцию руля или органа защиты. Движение скатов .вперед осуществляется за счет волнообразных колебаний расширенных грудных плавников.
Скелет головы
Череп акуловых рыб хрящевой, только у некоторых акул он пропитан известковыми солями и становится твердым как >кость. Весь скелет головы разделяется па два отдела (рис. 42): 1) осевой, или мозговой череп — пейрокраниум (рис. 46 и 47), собственно черепная коробка, является непосредственным продолжением позвоночника и представляет собой хрящевую капсулу, окружающую головной мозг и органы чувств; 2) висцеральны й скелет (спланхнокраниум— splanchnocranium) состоит из ряда расчлененных хрящевых дуг (рис. 48), отделенных друг от друга жаберными щелями и поддерживающих ротоглоточную полость, т. е. ротовую полость и жаберную часть пищеварительной трубки. Передние висцеральные дуги образуют челюстной аппарат и служат для захватывания пищи, задние, в количестве 5—7 дуг, служат жаберным скелетом, к ним прикреплены жабры, и они выполняют дыхательные движения жаберного аппарата.
При препаровке черепа не следует расчленять скелет головы на отдельные части, лучше сохранить их в естественном положении, соединенными друг с другом связками. В осевом черепе нужна аккуратность при очистке переднего отдела — рыла, нежных стенок, обонятельной капсулы и хрящевых пластинок, окаймляющих отверстия ноздрей. При препаровке висцерального скелета следует обратить внимание на соединительнотканные связки, соединяющие отдельные дужки между собой. Для удобства рассмотрения рекомендуется с одной стороны тела удалить легко отпадающие хрящевые жаберные лучи, сидящие на жаберных дужках и поддерживающие жаберные перегородки, а также внешние жаберные дуги (extrabranchiale), тонкие хрящевые стержни, лежащие параллельно основным жаберным дугам по внешнему краю жаберных перегородок.
Осевой череп — нейрокраниум
Осевой череп (neurocranium, рис. 46 и 47) представляет собой хрящевую коробку, открытую спереди, которая состоит из сплошной массы хряща и не подразделена на отдельные элементы, соответствующие отдельным костям в костном черепе. По своей функ
ции и по происхождению осевой череп может быть подразделен па
следующие отделы: а) собственно мозговая коробка, окружающая головной мозг; б) капсулы органов чувств (слуховые и обонятельные, рис. 46, cap. aud., cap. nas.) и выемка между ними — глазница,, занятая глазными яблоками; в) рыло, или рострум (rostrum,
рис. 46, rst.),— отдел, лежащий впереди обонятельных капсул и поддерживающий удлиненную переднюю часть головы.
Черепная коробка в переднем и заднем отделах имеет значительные расширения (рис. 46). Переднее из них образовано выступающими обонятельными капсулами {cap. nas.) и предорбитальными отростками. Задние расширения являются слуховыми капсулами (can. aud.), вошедшими у хрящевых рыб уже полностью в состав черепной коробки. Между обонятельными и слуховыми капсулами черепная коробка несколько сужена — это область глазниц, но между глазницами имеется значительное пространство, где располагается головной мозг. Такой тип черепа называется п л а-т и б а з а л ьп ы м. Сверху по краям глазниц, расположено 5 —6 отверстий, переднее из которых наиболее крупное. Через эти отверстия проходят ветки V и VII нервов (см. ниже стр. 179), (рис. 46, V + VII).
Обонятельная область черепа (pars ethmoidea, pars olfac-ioria) передний расширенный
Рис. 46. Осевой череп акулы Squalus acanthias сверху (по Маринелли и Стренгеру):
cap. aud.—слуховая капсула; cap. nas.— обонятельная капсула; f. d. е. — отверстие эндолимфатического канала; /. ер. — отверстие эпифиза; fn. — фоитанель; /. о. т.— затылочное отверстие; f. vst. — вестибулярное отверстие; рг. осс. — затылочный отросток; рг. ро. — заглазнячный отросток; rst. — рострум; V+Vlll, IX, X — отверстия для выхода соответствующих нервов
отдел пейрокраниума, расположенный по сторонам от переднего конца мозговой коробки и образованный двумя округлыми обонятельными капсулами (рис. 46, cap. nas.) с очень тонкими хрящевыми стенками. На нижней стороне обонятельных капсул помещаются широкие обонятельные отверстия. Медиальные стенки
обонятельных капсул пронизаны отверстиями обонятельных нервов. Между обонятельными капсулами на верхней части рострума находится крупное отверстие, затянутое лишь соединительнотканной Перепонкой, — это передняя фонта цель (рис. 46, fn.).
Рыло, или рост рум (рис. 46, rst.), поддерживает передний конец заостренной головы акулы. У Squalus acanthias рострум имеет форму тонкой хрящевой ладьевидной пластинки. У разных акул рострум имеет весьма различное строение. У голубой акулы он представлен тремя тонкими палочковидными хрящами. У некоторых скатов (например, у электрического ската) его совсем нет, и передний конец головы образован сходящимися впереди черепа передними концами грудных плавников.
Глазничный отдел черепа (рис. 47) занимает среднюю часть нейрокраниума и образован глазничными впадинами. Глазница представляет собой глубокую впадину по бокам черепа между обонятельной и слуховой капсулами. Она ограничена впереди и сзади хрящевыми отростками: передними — предглазничными (processus praeorbitalis, pr. pro.) и задними — заглазничными (processus pos-iorbitalis, pr. po.). В этой области дно черепной коробки также образует два выступа — передний, расположенный под предглаз-ничным отростком, называется processus antorbitalis (pr. auto.) и задний — основной отросток (processus basilaris, pr. bas.). Верхний, а у других акул и нижний край глазницы окаймляется выдающимися хрящевыми гребнями, называемыми надглазничным и подглазничным. Внутренняя боковая стенка глазницы пронизана рядом отверстий для выхода из черепа нервов и сосудов. Они будут описаны в дальнейшем. На крыше черепа имеются отверстия для нервов.
Слуховой отдел черепа — широкий задний участок черепа, возникший как результат прирастания к бокам мозговой коробки крупных слуховых капсул (рис. 46, cap. aud.), окружающих сложно устроенный перепончатый лабиринт внутреннего уха. Передней границей слуховой области являются заглазничные отростки, далеко простирающиеся в бока, а задней — затылочные отростки (processus occipitalis, рис. 46, pr. осс.). Между ними имеется плоская выемка. Под затылочным отростком помещается сочленовная поверхность для причленения гиомандибулярного хряща — подвеска челюстного аппарата. Позади него — широкое отверстие головного нерва X пары — блуждающего (n. vagus, рис. 46, 47, X), а кнаружи of него и несколько впереди — отверстие головного нерва IX пары—< языкоглоточного (и. glossopharyngeus, рис. 46, 47, IX).
Между слуховыми капсулами лежит большое, овальной формц углубление, называемое теменным. На дне этого углубления! находят две пары отверстий. Два крупных, расположенных в заднему отделе, вестибулярных отверстия (foramen vestibuli| рис. 46, f. vst.) служат для регуляции давления, увеличения и уменьшения объема перилимфы. Впереди от них находятся два го| раздо меньших отверстия, через которые проходят эндолимфати! ческие каналы (for. ducti endolymphatici, рис. 46, f. d. e.). По бокам от теменного углубления на крыше черепа через прозрачный хрящ просвечивают в виде двух беловатых полос, расходящихся под пря мым углом в стороны, передние и задние вертикальные полукруяя ные каналы перепончатого лабиринта.
Затылочный отдел черепа можно изучить если отчленить череп от позвоночника. В этом отделе следует рассмотреть: а) большое затылочное отверстие (foramen occipitale magnum) для прохождения спинного мозга (рис. 46, f.o.m.); б) место вхождения хорды в череп под затылочным отверстием; в) отверстия X головного нерва (n. vagus), расположенные на дне парной ямки по бокам затылочного отверстия позади слуховой капсулы; г) отверстия XII пары головных нервов, подъязычного нерва (n. hypoglossus) — два очень маленьких отверстия под отверстием X нерва.
Рис. 47. Осевой череп акулы Squalus acanthias сбоку:
a. hy. — отверстие для гиоидной артерии; pr. anto.—противоглазничный отросток; pr. bas.—основной отросток; рг. ро.—заглазничный отросток; pr. pro. — предглаз-ннчпый отросток; о. io. — отверстие для межглазяичиой вены; I, II, III, IV, V+VU, г, pont. V+VII, VI, VII, IX, X, г. hm. VII. г. р. VII — отверстия для соответствующих нервов и их ветвей
Отверстия в черепе для нервов и сосудов (рис. 46 и 47).
1. В крыше черепа, как было уже сказано, в желобках около глазницы, где проходит надглазничный канал органов чувств боковой линии, а также внутри глазницы по верхнему краю располагается целый ряд отверстий для глазничной ветви V и VII 'нервов (г. ophthalmicus superficial n. V+n. VII, рис. 46 и 47, V+VII).
2. На дне обонятельной капсулы при рассмотрении ее спереди видны отверстия I головного нерва, обонятельного (tr. olfactorius, I, рис. 47, /).
3. Внутри глазницы под предглазничным отростком видно отверстие глубокой глазничной ветви тройничного нерва (г. ophthalmicus profundus, V, рис. 47).
4. В передней части глазницы, около ее нижнего края, располагается крупное отверстие зрительного нерва (n. opticus, II, рис. 47, II).
5. Выше отверстия зрительного нерва, несколько отступя назад, в глазнице лежит отверстие IV головного нерва, (n. trochlearis, IV, рис. 47, IV).
6. Вниз и назад от IV нерва, но выше уровня зрительного нерва, в глазнице помещается отверстие для выхода III головного нерва — глазодвигательного (п. oculomotorius, III, рис. 47, ///).
7. В заднем углу глазницы под заглазиичным отростком, расположено крупное отверстие тройничного нерва V пары (n. trigeminus, V+VII).
8. Позади тройничного нерва или вместе с ним выходит лицевой нерв (n. facialis, VII, рис. 47, V+WZ).
9. Под ними выходит также и VI — отводящий нерв (n. abdu-cens, VI, рис. 47, VI).
10. Под отверстиями этих нервов, над базальным углом дна черепа (angulus basalis), располагаются два отверстия: для гиоидной артерии (a. hyoidea, рис. 47, a. hy.) и межглазничной вены (v. interorbitalis, рис. 47, v. io.).
11. На боковой стенке слуховой капсулы внизу помещаются два отверстия для ветвей лицевого нерва: небной ветви (г. palatinus, VII, рис. 47, г. р. VII) и гиомандибулярной (г. hyo-mandibularis, VIL рис. 47, г. hm. VII).
12. Позади слуховой капсулы, как указано выше, располагаются два крупных отверстия: переднее —для языкоглоточного нерва, IX (n. glossopharyngeus), а позади него — для блуждающего нерва X пары (n. vagus).
Висцеральный скелет
Висцеральный скелет — с п л а н х н о к р а н и у м (splanchnocra-nium, рис. 48 и 49) —система скелетных элементов, поддерживающих передний отдел пищеварительной трубки, носящей название ротоглоточной полости, пронизанной по бокам жаберными отверстиями. Хрящевые дуги, составляющие висцеральный скелет акуловых рыб, обычно в количестве семи парных полудуг, соединенных па брюшной стороне, охватывают по бокам ротоглоточную полость. К ним прикрепляется мускулатура, обслуживающая движения висцерального аппарата.
Чтобы рассмотреть висцеральный скелет у акулы, рекомендуется, не расчленяя скелета головы на отдельные части, вставить бумажный сверток или тампон ваты через ротовое отверстие внутрь, в ротоглоточную полость, и продвинуть его в жаберную область.
Хрящевые дуги, составляющие висцеральный скелет, неодинаковы по строению и функции. Две передние, более мощные дуги образуют челюстной аппарат. Они носят название челюстной и подъязычной дуги. Остальные пять дужек поддерживают органы дыхания и называются жаберными. Правая и левая половина каждой жаберной дуги расчленена на четыре отдела, причем два располагаются на спинной стороне глоточной трубки, а два — на брюшной. Первые две дуги имеют в каждой половине только по два элемента. Между жаберными дугами находятся жаберные щели, а между челюстной и подъязычной — брызгальце (spira-culum).
Передняя дуга — челюстная (arcus mandibularis), состоит из двух массивных хрящей, покрытых острыми зубами. Дорзальный отдел челюстной дуги служит верхней челюстью и носит название н е б в о - к в а д р а т н о г о хряща (palato-quadra-tum, рис. 48, 49, p.-q.). Правый и левый небно-квадратные хрящи срастаются друг с другом передними концами но средней линии.
Рис. 48. Висцеральный скелет акулы Squalus acanthias сбоку (по Маринелли и Стренгеру):
car. bas. — непарный базибрапхиальвый элемент (копула); ce.-br. / — цератобранхиальный хрящ; с. hy. — гиоидный хрящ; с. hytn. — гкомандибулярный хрящ; с. lab. — губные хрящи; с. М. — меккелев хрящ; сор. — копулы жаберных дуг; с. p.-q.— иебно-квадратный хрящ; с. spir. — спиракулярный хрящ; e.-br. 1 — эпибранхиальные хрящи; ecto. br. do. /— 4 — эктобранхиальные дорзальные жаберные дуги; ecto. br. ve. 1—4 — эктобранхиальные вентральные жаберные дуги; ecto. hy. се.—эктобранхиальный хрящ гиоидной дуги, вентральный; h.-br. 2 — гипобранхиальный элемент второй жаберной дуги; ph.-br. 4+5 — фарингобранхиальные хрящи 4—5; pr. orb. p.-g. — глазничный отросток небно-квадратного хряща
У большинства акул и скатов небно-квадратный хрящ контактирует с глазничной областью черепной коробки при помощи направленного вверх глазничного отростка (processus orbitalis palato-quadrati, рис. 48 и 49, pr. orb. p-q.). Вентральный отдел челюстной Дуги служит нижней челюстью и носит название меккелева хряща (cartilago МескеИ, рис. 48, 49, с. М.). Задние концы меккелевых хрящей сочленяются с задними концами небно-квадратных хрящей, а передние — друг с другом на вентральной стороне. Когда рот закрыт, нижняя челюсть лежит параллельно верхней, непосредственно под ней окаймляя снизу ротовое отверстие, имеющее вид поперечной щели.
Зубы (рис. 49, dens). На верхней и нижней челюстях в несколько рядов сидят зубы. У акул, ведущих хищный образ жизни, зубы острые, конические или с боковыми зубчиками по краям. Они сходны с кожными плакоидными чешуями. На челюстях зубы сидят
косыми рядами, на наружной стороне они более крупные, а по направлению внутрь становятся мельче и менее развитыми. У скатов, питающихся раковинными моллюсками, зубы уплощены, они приспособлены для раздавливания раковин и имеют форму мостовидных плиток.
Вторая, подъязычная дуга (arcus hyoideus), так же как и челюстная дуга, расчленена на два отдела, связанных друг с другом на вентральной стороне непарным элементом (рис. 48 и 49). Верхний отдел — короткий, крепкий хрящевой стержень—носит название подъязычно-челюстного, или гиомандибу-л я р н о го, хряща (hycmandibulare, рис. 48, 49, с. hym.). Он играет роль подвеска между челюстной дугой и черепом. Своим верхним концом этот хрящ причленяется к слуховой области черепа, а нижним — к месту сочленения верхней и нижней челюстей. Таким образом, весь челюстной аппарат имеет опору в черепе лишь при помощи гиомапдибулярного хряща. Такой тип причлене-ния челюстного аппарата называется г и о ст и л и ч е с к и м. У химер небно-квадратный хрящ прирастает к черепной коробке. Такой тип причленения висцерального скелета называется аутостили-чески м. Между небно-квадратным и гиомандибулярным хрящами располагается брызгальце. Нижний отдел подъязычной дуги в виде гораздо более слабого хрящевого стержня помещается позади и ковнутри от нижней челюсти. Он носит название подъязычного, или гиоидного, хряща (hyoideum, s. с. hyа 1 is, рис. 48, 49, с. hy.). Верхний конец подъязычного хряща сочленяется с гиомандибулярным хрящом у места его причленения к челюстям, а нижний конец— с непарным вентральным элементом. Непарный вентральный элемент подъязычной дуги имеет вид широкой хрящевой пластинки. Он соединяет вентральные концы подъязычных хрящей обеих сторон и носит название основного гиоидного хряща (basi—hyale, рис. 49, с. bhy).
У скатов имеется только, верхний отдел подъязычной дуги — hyo-mandibulare. Вентральный отдел — hyoideum — редуцируется, и вместо него впереди первой жаберной дуги находится маленькая хрящевая дужка, не сочленяющаяся с гиомандибулярным хрящом и носящая название ложной гиоидной дужки (pseudohyale). Пред-брызгальцевая, или преспиракулярная, связка в виде крепкого волокнистого тяжа с небольшими хрящами внутри идет от переднего конца слуховой капсулы впереди брызгальца к месту сочленения гиомапдибулярного хряща и верхней челюсти.
У химеры в связи с прирастанием верхней челюсти к осевому черепу подъязычная дуга не играет роли подвеска, поэтому не причленяется к осевому черепу, но состоит из трех типичных для висцеральной дуги парных элементов — фаринго, эпи- и цератоэле-мента.
Жаберные дуги (arcus branchialis) в количестве пяти (рис. 48 и 49) лежат позади подъязычной дуги и отделены друг от друга жаберными щелями так, что первая жаберная щель лежит впереди 1-й жаберной дуги, а пятая — перед 5-й жаберной дугой. 160
Каждая половинка жаберной дуги расчленена на четыре элемента, направленных друг к другу под углом: два на дорзальной стороне, два — на вентральной; с брюшной стороны между ними располагается общая непарная хрящевая пластинка — копула.
Дорзальный, или глоточно-жаберный (pharyngo-branchiale, рис. 48, ph.-br.), элемент жаберной дуги лежит в крыше глоточной области и имеет вид плоской хрящевой пластинки. Свободный верхний конец его направлен внутрь и назад и прикреплен
Рис. 49. Висцеральный скелет акулы Squalus acanthias снизу (по Маринелли и Стренгеру):
с. bhy — основной хрящ гиоидной дуги; се.-Ьг. — цератобраихиаль-ный элемент; с. hy. — гиоидный хрящ; с. hym. — гиомаидибуляр-ный хрящ; с. lab. — губные хрящн; с. М. — меккелев хрящ; сор. — базнбранхиальный элемент жаберных дуг; сор. 2 — базибранхиаль-ный элемент второй жаберной дуги; с. p.-qu. — небно-квадратный хрящ; dens. — зубы; h.-br. — гипобранхиальиые элементы; pr. orb. p.-q. — глазничный отросток небно-квадратного хряща; рг. г. — жаберные тычинки; га. Ьг. — жаберные лучи
связками к позвоночнику, а нижний — вперед и кнаружи, где сочленяется со следующим элементом. Дорзальные элементы двух последних дуг сливаются между собой и с эпи-элементом пятой дуги.
Второй дорзальный элемент носит название верхнежаберного, эпибранхиальиого (epi-branchiale, рис. 48, e.-br.). Он направлен своим нижним концом вниз и назад и под углом к следующему отделу жаберной дужки. На epi-branchiale всех дужек, кроме пятой, сидят жаберные лучи (radius branchialis, рис. 49, га. Ьг.), поддерживающие жаберные перегородки, на которых сидят лепестки.
Третий отдел жаберной дуги — основной элемент на вентральной стороне глотки. Он носит название рожковидно-жаберного, ц е р а т о - б р а н х и а л ь н о г о, элемента (cerato-branchiale, рис. 49 ce.-br.). От места сочленения с верхнежаберным элементом он направлен вперед и внутрь. На заднем крае cerato-branchiale также сидят жаберные лучи. Рожковидно-жаберный отдел пятой дуги не несет жаберных лучей, и на его заднем конце имеется вырезка для прохождения кювьерова протока.
Самые нижние элементы имеются у акул на второй, третьей и четвертой жаберных дугах и носят название поджаберных, гипо-бранхиальных, элементов (hypo-branchiale, рис. 49, h.-br.). Это маленькие хрящики, направленные своими нижними концами назад, где они сочленяются и часто срастаются с непарным вентральным элементом. Пятая дуга не имеет самостоятельного гипо-бранхиального элемента. Иногда встречается hypo-branchiale первой дуги, очень маленький хрящик в связке, направленный вперед, где он соединяется с basi-hyale.
Непарный вентральный базибранхиальный элемент, нлн копула (basi-branchiale, s. copula, рис. 49, сор.), имеет у акулы вид продолговатой плоской хрящевой пластинки, заостренной сзади. Передним концом эта пластинка сочленена с hypo-branchiale, 3-й и 4-й дуги и cerato-branchiale 4-й и 5-й дуг. Базибрапхиале поддерживает верхнюю стенку околосердечной сумки. Вторая жаберная дуга имеет самостоятельный базибранхиальный элемент (рис. 49, сор. 2).
У скатов происходит срастание basi-branchiale с hypo-branchiale, в результате чего образуется широкая неправильной формы хрящевая пластинка, покрывающая сердце, так называемая сердечно-жаберная, кардиобранхиальная пластинка (cardiobranchiale).
Жаберные лучи (radius branchialis, рис. 49, га. br.) — тонкие хрящевые стержни, часто ветвящиеся на конце, сидят на наружном заднем крае основных элементов (epi-branchiale и cerato-branchiale) жаберных дуг, кроме пятой, последней, а также на гиомандибулярпом и гиоидном хрящах подъязычной дуги. Они лежат под жаберными лепестками на жаберных перегородках и поддерживают их.
Жаберные тычинки (рис. 49, рг. г.)—хрящевые конические отростки на внутреннем переднем крае всех жаберных дуг, образуют цедиальный аппарат, удерживающий пищу в ротоглоточной полости при прохождении через нее воды при дыхании.
Внешние жаберные дуги — экстра-бранхиальные хрящи (extra-branchiale), дополнительные скелетные элементы, поддерживающие наружные края межжаберных перегородок (рис. 48, ect. br. I—4). Они имеются только у хрящевых рыб. У акулы катрана они иаходятся на подъязычной и первых четырех жаберных дугах в виде двух хрящевых стержней на каждой дужке, одна на дорзальной стороне, другая на вентральной с перерывом посредине, па месте жаберного отверстия.
Губные хрящи (cartilago labialis, рис. 48, 49, с. lab.)—три тонких палочковидных элемента в складке угла рта с каждой стороны. Их считают рудиментами двух висцеральных дуг, лежавших впереди челюстной. Единственный нижний губной хрящ прилегает вплотную к меккелеву хрящу и плотно с ним связан, а два остальных заключены в кожную складку верхней губы и могут быть легко удалены при препаровке.
Жаберный аппарат при выполнении акта дыхания действует как единый двигательный механизм. Все жаберные дуги связаны друг с другом системой связок в общую жаберную коробку. Один ряд связок связывает основные дорзальные элементы — epi-branchi-ale— у их переднего конца, так как у глоточных элементов (pharyn-go-branchiale) на нижнем конце имеются два отростка, к которым прикрепляются плотные связки. Брюшные концы основных вентральных элементов (cerato-branchiale) также соединены друг с другом связками у места причленения к ним поджаберных элементов (hypo-branchiale). В углах между основными жаберными элементами (epi- и cerato-branchiale) на внутренней стороне прикреплены пучки междужных мышц (m. m. interarcuales), сокращение которых вместе с наружными сжимателями (m. т. constricto-res) обеспечивает наполнение и опорожнение водой системы жаберных мешков при дыхании, жаберные мешки при этом сокращаются последовательно спереди назад.
Мускулатура хрящевых рыб
Мускулатура акуловых рыб сохраняет в себе много примитивных черт и может служить общей схемой для представления о происхождении сложной мускулатуры высших позвоночных животных. Она одевает внутренний хрящевой скелет толстым мускульным слоем, расчлененным на зигзагообразные мышечные сегменты — миомеры, создавая типичную веретенообразную форму тела рыбы. В голове сегментация миомеров нарушена, и мускулатура расчленена на отдельные группы: 1) затылочная группа наджаберных миомеров; 2) поджаберная группа; 3) мускулатура жаберного и челюстного аппаратов и 4) глазные мышцы.
По своему функциональному значению мускулатура может быть расчленена на: 1) мускулатуру плавников, регулирующих движение тела при плавании в воде; 2) мускулатуру головы, жаберного и челюстного аппаратов; 3) мускулатуру пищеварительной системы и внутренних органов.
По происхождению мускулатура акулы, как и всех позвоночных, делится на соматическую и висцеральную (см. стр. 166).
Мускулатура туловища
Мускулатура туловища хрящевых рыб от головы до конца хвоста сохраняет правильную сегментацию в виде изогнутых поперечных лент—миомеров (рис. 50). Однако уже у хрящевых рыб на-6* 163
мечается некоторое функциональное расчленение мускулатуры туловища по продольной оси соответственно положению миомеров в теле животного. На боковой поверхности тела намечается продольная борозда, которая делит весь ряд туловищных миомеров на две группы: дорзальную (эпаксиальную) по бокам позвоночника и вентральную (гипаксиальную), образующую в туловище боковые стенки брюшной полости тела. Каждый метамер мускулатуры носит название мио мер а (рис. 50, тт.) и с наружной стороны тела имеет вид поперечной полосы, изогнутой в виде буквы W под острыми углами. Миомеры отделены друг от друга соединительнотканными прослойками—миосептами (рис. 50, ms.). Следует
Рис. 50. Общий вид мускулатуры акулы Squalus acanthias (по Маринелли и Стренгеру):
тт. — миомер; tn. obi. а. — наружная косая мышца живота; tn. г. а.—прямая мышца живота; ms. — миосепта; s. —трапсверзальная септа
проследить направление ТГ-образного изгиба миомера, начиная от спинной стороны вниз: сначала изгиб идет назад, потом поворачивает вперед до продольной соединительнотканной прослойки, называемой поперечной или трансверзальной септой (рис. 50, s. t.), в которой залегает боковая ветка X головного нерва (ramus lateralis, п. X). От дрансверзалыюй септы изгиб снова поворачивает назад и затем на брюшной поверхности снова вперед. В каждом миомере мышечные волокна идут в продольном направлении по отношению к продольной оси тела, и их последовательное сокращение приводит к изгибам всего тела.
В дополнение к рассмотрению расположения зигзагообразных изгибов миомеров на теле акулы рассматривают расположение мускулатуры на поперечном разрезе в туловищной и хвостовой областях. На разрезе видно, что в поле зрения попадает одновременно несколько мышечных сегментов благодаря их зигзагообразным изгибам (см. рис. 38), причем они имеют вид концентрических окружностей. Это происходит потому, что каждый миомер имеет очень сложное строение, в каждом изгибе он образует конус, направленный то вперед, то назад, причем конусы соседних миомеров вложены друг в друга так, что на поперечном разрезе образуют серию колец.
Мускулатура туловища делится трансверзальной септой (septum transversale, рис. 50, s. t.) на дорзальную, или н а д-
осевую (э п а к с и а л ь н у ю), мускулатуру и боковую, или п' о д-осевую (г и и а к с и а л ь и у ю). Кроме того, между брюшными и грудными плавниками на нижней боковой поверхности тела обособляется косой бороздой, проходящей через ряд миомеров, отдельная мышечная группа, представляющая собой зачаток косой мышцы живота (m. obliquus abdominis externus, рис. 50, m.obl.a.).
На брюшной стороне тела, начиная от просвечивающего между грудными плавниками хрящевого плечевого пояса, вентральные концы миомеров обособляются в продольную мышечную полосу, которая в дальнейшем у наземных позвоночных дает прямую мышцу живота (m. rectus abdominis, рис. 50, 51, т. г. а.).
Впереди от плечевого пояса туловищная мускулатура расчленяется жаберным аппаратом на дорзальную и вентральную части.
Дорзальная часть, как
Рис. 51. Мускулатура головы акулы Squalus acanihias снизу (по Маринелли и Стренгеру):
продолжение эпаксиаль-ной мускулатуры или .мускулатуры туловищных
миомеров, доходит до черепа, где и кончается, образуя затылочную мускулатуру (рис. 50, 52). Вентральная часть образует поджабе р-
т. а. т. — m. adductor mandibulae; т. с.-а. — т. coraco-arcualis; т. c.-br. — т. coraco-bran-chlalis; tn. с. hy. — т. coraco-hyoideus; tn. c. hy. v. — m. constrictor hyo-mandibularls ventralis: m. c.-m.—m. coraco-mandibularis; tn. c. s. v. — m. constrictor superficialis ventralis; tn. r. a. — прямая мышца живота
и у ю мускулатуру, идущую вперед от плечевого пояса и дифференцированную на отдельные мускульные пучки. Спереди она прикрыта межчелюстными мышцами (см. ниже). Поджаберная
мускулатура иннервируется корешками ХП головного нерва (n. hypoglossus). Перережьте по средней линии брюшной сжиматель гиоидной дуги (рис. 51, т. с. hy. v.) и отверните его в стороны. Рассмотрите лежащие под ним крупные мышцы, связывающие плечевой пояс с отдельными висцеральными дугами. Основание этих мышц составляет m. coraco-arcualis, имеющий форму треугольника и основанием прикрепленный к коракоидной части плечевого пояса (рис. 51, т. с.-а.). От его вершины отходят мышцы к различным дугам висцерального скелета. Посредине отходит непарная мышца, прикрепляющаяся к месту соединения двух половин нижней челю-
сти. Эта мышца называется корако-мандибулярной (m. coraco-mandibularis, рис. 51, т. c.-т.). Латеральнее лежит парная ко р а ко - гио иди а я мышца (m. coraco-hyoideus, рис. 51, т. c.-hy.), прикрепляющаяся к подъязычной дуге.
Раздвинув нижнюю челюсть и подъязычную дугу, находят места прикрепления этих мышц. Далее рассматривают более короткие мышечные пучки, связывающие каждую жаберную дугу с корако-идным отделом плечевого пояса. Эти мышцы называются кора-ко-жаберными (m. coraco-branchialis, рис. 51, т.с.-Ьг.).
Мускулатура непарных плавников представлена особыми мускульными пучками, отщепившимися от миотомов с обеих сторон скелета плавников. В непарных плавниках из этих пучков образуются правая и левая плавниковые группы мышц.
Мускулатура парных плавников также образована пучками мышц, отчленившимися от нескольких миотомов (рис. 50, 52). В грудных п л а в н и к а х таким путем обособлена дорзальная мускулатура над скелетными лучами (рис. 52, tn.d.)r поднимающая плавник, и вентральная мускулатура иод скелетными лучами, опускающая плавник. Дорзальная и вентральная мускулатуры плавников не дифференцированы на отдельные мускулы, а функционируют как единое целое. У акуловых рыб в построении мускулатуры грудного плавника принимают участие 6—7 миомеров, а у скатов, имеющих громадные грудные плавники и мощную мускулатуру, количество миомеров, образующих мускулатуру грудного плавника, доходит до 13—44.
Мускулатура брюшных плавников (см. рис. 50) также состоит из отдельных пучков, отщепившихся от ряда миотомов. Проксимальным концом они прикрепляются к тазовому поясу, а дистальным— к дистальным концам хрящевых лучей. Как и у грудных плавников, мускулатура брюшных плавников располагается по обеим сторонам скелета с дорзальной и вентральной сторон, образуя дорзальную и вентральную группы мышц.
Мускулатура головы
В голове сегментальное расположение мускулатуры нарушено. Расчленение на отдельные миотомы сохраняется лишь в наджаберной группе затылочных миотомов.
Соматическая мускулатура представлена в голове только тремя группами мышц: наджаберная группа затылочных миомеров; поджаберная мускулатура, описанная выше, как продолжение брюшной мускулатуры туловища, впереди от плечевого пояса; и, наконец, шесть глазных мышц, двигающих глаз (их описание будет дано вместе с описанием глаза, см. рис. 57).
Висцеральная мускулатура головы в отличие от туловищной представлена сложной системой мышц челюстного и жаберного аппаратов. В отличие от гладкой кишечной мускулатуры мускулатура челюстного и жаберного аппаратов поперечнополосата и расчленена на отдельные мышцы, совершающие активные хвататель-166
пые и дыхательные движения. Соответственно делению висцерального аппарата па ряд сегментов (челюстная дуга, подъязычная и ряд жаберных) висцеральная мускулатура расчленена по этим дугам.
Жаберная мускулатура у акул представлена топким слоем сжимателей жаберных мешков в виде поперечных лент между жаберными щелями. В глубине под ними имеются отдельные пучки между отдельными члепиками, каждой жаберной дуги, носящие название междуговых мышц (m. interarcualis). Рассмотрите только слой поверхностных мышц, которые носят название поверхностных сжимателей (m. т. constrictores superficiales).
m.a.
Рис. 52. Мускулатура головы акулы Squalus acanthias сбоку (по Маринелли и Стренгеру):
т. а. т. — m. adductor mandlbulae; т. с. hy. d. — т. constrictor hyoi-deus dorsalis; tn. c. s. d. — m. constrictor superficialis dorsalis; d. hm. — m. depresso hyomandibularis; m. d. — дорзальная мускулатура плавника; m. I. p. qu. —;m. levator palato-quadrati; tn. tr.—трапециевидная мышца
Группа жаберных мышц над жаберными отверстиями носит название дорзальных поверхностных сжимателей (m. m. constrictores superficiales dorsales, рис. 52, т. c.s.d.). Мышцы под жаберными отверстиями называются вентральными поверхностными сжимателями (m. m. constrictores superficiales ventrales, рис. 51, т. с. s. и.). У скатов, вследствие прирастания к голове громадных разросшихся грудных плавников, группа дорзальных сжимателей располагается совершенно отдельно от вентральной группы на дорзальной стороне тела, а группа вентральных сжимателей — на вентральной стороне в виде ряда лент, идущих от жаберных дуг кнаружи, к плавникам.
Мускулатура подъязычной дуги имеет сходное строение с мускулатурой жаберных дуг и точно так же представлена дорзальным и вентральным сжимателями. Д о р з ал ьп ы й гиоидный сжиматель (m. constrictor hyoideus dorsalis (рис. 52, in. с. hy. d.) располагается между первой жаберной щелью и брызгальцем и го
мологичен опускателю нижней челюсти тетрапол. Вентральный сжиматель помещается на брюшной стороне челюстного аппарата; сжиматели правой и левой сторон срастаются между собой по средней линии и с вентральным сжимателем челюстной дуги. Образуется общий вентральный гиомандибулярный сжиматель (m. constrictor hyomandibularis ventralis s. ш. intermandi-bularis, см. рис. 51, m. c. hy. u.).
Мускулатура челюстной дуги в связи с превращением челюстной дуги в аппарат захватывания пищи расчленена на отдельные мышцы. Первая порция мышц, соответствующая дорзальному сжимателю, преобразуется в маленькую мышцу — подниматель верхней челюсти (m. levator palato-quadrati, рис. 52, т. I. p.-qu.). Эта мышца лежит впереди брызгальца между черепом и небно-квадратпым хрящом в заднем углу глазницы. Вторая группа мышц обособляется в очень сильную мышцу, приводящую нижнюю челюсть (m. adductor mandibulae, рис. 51, 52, т. а. т.). Она дорзальным концом прикрепляется к верхней челюсти, а вентральным— к нижней (меккелеву хрящу) и является главной мышцей, закрывающей рот при схватывании добычи. Третья, вентральная, порция мускулатуры челюстной дуги, соответствующая вентральным сжимателям жаберных дуг, как упомянуто выше, вместе с одноименной мышцей подъязычной дуги образует тонкую широкую мышцу, натянутую между правой и левой нижними челюстями (ш. intermandibularis).
У скатов в связи с приспособлением к питанию твердыми моллюсками мышцы челюстной дуги еще более дифференцированы. Жевательная мускулатура ската состоит из большого числа мышц, так как каждая из описанных выше мышц распадается в свою очередь на ряд мускулов.
Трапециевидная мышца (m. trapezius, рис. 52, т. tr.). Позади последней жаберной дуги расположена задняя порция висцеральной мускулатуры, которая задними пучками прикрепляется к дорзальному отделу плечевого пояса и носит название трапециевидной мышцы, как у наземных позвоночных животных, а передний конец трапециевидной мышцы сливается с жаберной мускулатурой.
Иннервация головной мускулатуры. Висцеральная мускулатура головы соответственно своему происхождению от висцеральной мускулатуры, т. е. мускулатуры пищеварительной трубки, имеет особую иннервацию висцерально-двигательными ветвями головных нервов.
Мускулатура челюстной дуги, т. е. верхней и нижней челюстей, иннервируется двигательными и чувствующими ветвями тройничного головного нерва V пары (n. trigeminus). Мускулатура подъязычной дуги иннервируется висцерально-двигательными ветвями нерва VII пары (n. facialis). Первая жаберная дуга имеет собственную мускулатуру и приводится в движение соответствующими ветвями нерва IX дары (n. glossopharyngeus). Все остальные жабер
ные дуги и их мускулатура иннервируются висцерально-двигательными ветвями блуждающего нерва (n. vagus).
Соматическая мускулатура головы, как указано выше, представлена группой наджаберных затылочных миомеров, которые иннервируются передними корешками спинномозговых нервов, и группой поджаберной мускулатуры, которая иннервируется двигательными корешками головного нерва XII пары — подъязычного нерва (п. hypogiossus).
Кроме этих мышц, являющихся продолжением на голову мускулатуры туловищных миомеров, в голове к соматической мускулатуре относятся шесть глазных мышц. Изучение их следует отложить до рассмотрения разветвлений головных нервов при изучении нервной системы и органов чувств.
Нервная система хрящевых рыб
Нервная система современных хрящевых рыб, как хищников, имеет усложненное строение по сравнению с древними костными рыбами. В туловищной области спинномозговые нервы, образованные слиянием в общие сегментальные нервы брюшных и спинных корешков, образуют сложные сплетения, снабжающие парные плавники. Органы чувств дифференцированы на сложную систему кожных органов боковой линии, глаза имеют типичное для ориентации в воде строение, стато-акустические органы имеют три полукружных канала. Головной мозг имеет прогрессивно организованный передний мозг с крышей из нервного вещества, с обособлением двураздельного желудочка. Промежуточный мозг ясно расчленен на отделы и мозговую воронку. Средний мозг становится главным центром зрения. Продолговатый мозг служит центром стато-акусти-ки и органов боковой линии, а также всего висцерального аппарата (челюстного и жаберного). Мозжечок, ведающий моторикой всего тела, прогрессивно развит. Головные нервы в количестве десяти пар имеют усложненное ветвление с включением в черепные нервы 12-й пары (подъязычного нерва).
Препаровка нервной системы ската или акулы не требует никаких предварительных обработок материала. Можно воспользоваться экземплярами, на которых изучались внутренние органы или мускулатура.
Материал должен быть фиксирован в 5%-ном формалине (очень желательно предварительно вскрыть черепную полость), затем лучше перенести объект в 70° спирт. При анатомировании необходимо следить, чтобы препарат не подсыхал, удобно даже всю работу проводить в ванночке с водой.
При описании изучения первой системы и органов чувств последовательность хода препаровки приходится давать не в систематическом порядке строения, а по ходу работы. Порядок изложения принимается нами следующий: 1) кожные органы чувств; 2) строение головного мозга ската; 3) строение головного мозга акулы;
4) отхождение от мозга головных нервов; 5) разветвление головных нервов; 6) спинной мозг и спинномозговые нервы; 7) строение органов чувств — глаз, орган слуха, орган обоняния.
Кожные органы чувств боковой линии (рис. 53)
Рассматривают расположение каналов боковой линии и лорен-циниевых ампул на голове акулы.
Органы боковой линии, сейсмосенсорные органы, представлены у данного вида акул кожными каналами (сап.) и лоренциниевыми ампулами (amp. L.). Кожные каналы идут под кожей и дают ответвления к ее поверхности.
Л о р е и ц и и и ев ы е ампулы (ampulla Lorenzini) представляют собой трубочки разной длины, лежащие под кожей и окруженные плотной соединительной тканью. Наружный конец их открывается на поверхности кожи небольшими, более или менее округлыми отверстиями. Чувствующие элементы располагаются у внутренних концов трубочек.
Рассматривают расположение лоренципиевых ампул на вентральной и дорзальной поверхностях головы акулы (amp. L.).
Лоренциниевы ампулы на голове акулы концентрируются в несколько групп. Отверстиями ампул покрыта вся вентральная часть рострума, начиная от его переднего конца, вплоть до переднего края верхней челюсти. Небольшое количество их переходит также на бока головы. Две небольшие группы располагаются на вентральной стороне тела за нижней челюстью: одна — непосредственно у ее заднего края, другая — несколько отступя назад.
На дорзальной стороне головы ампулы концентрируются по бокам передней части рострума, а затем переходят на верхнюю часть головы, образуя две группировки на уровне переднего края глаза. Небольшие группы лоренципиевых ампул лежат за задним краем глазницы и за отверстием брызгальца.
Значительно хуже видны лежащие под кожей каналы боковой линии, открывающиеся па поверхность кожи рядом отверстий. Последние обычно заполнены слизью, слабо заметны и имеют вид расплывчатых белых точек. Там, где имеются лоренциниевы ампулы, легко спутать те и другие и проследить ход канала в этих участках довольно затруднительно. Ампулы отличаются тем, что при надавливании на кожу из них выделяются капли фиксирующей жидкости.
При тщательном исследовании поверхности головы с помощью лупы можно отметить ход некоторых каналов.
Латеральный капал (can. 1.)—орган чувств боковой линии— начинается на уровне заднего конца брызгальца и тянется назад сначала по верхнему участку головы, затем переходит на латеральную поверхность тела и доходит до конца хвостового стебля.
Вперед от латерального канала тянется надглазничный, супраорбитальный, кожный канал (canalis supraorbi-170
talis, can. spo.). Он хорошо виден на дорзальной поверхности головы, где в виде ряда мелких точек проходит несколько отступя от верхнего края глазницы, как бы огибая глазное отверстие. Переходя на ростральную часть, фолликулы этого канала теряются среди отверстий лоренциниевых ампул.
Небольшой участок подглазничного, инфраорбитального, канала (canalis infraorbitalis, can. ino.) можно проследить по ряду отверстий, тянущихся сбоку от угла ротового отверстия и дальше вперед в направлении отверстия ноздри.
Рис. 53. Схема распределения кожных органов чувств боковой линии на голове акулы, сбоку:
amp. L. — лоренцнииевы ампулы; can. ino.—подглазничный канал; can. I.—латеральный канал; can. or. — околоротовой канал; can. sp. о. — надглазничный канал; sp. — брызгальце
На вентральной стороне чуть позади угла рта проходит небольшой ряд отверстий околоротового, орального, кожного канала (can. or.). Несколько отступая от него назад вдоль заднего края нижней челюсти лежит ряд гуляр ных фолликул, которыми заканчивается серия веерообразно расходящихся подкожных каналов.
Снимают кожу с одной половины головы, осторожно отделяя ее от мускулатуры и хрящей. На внутренней поверхности ее можно довольно хорошо видеть некоторые участки каналов боковой линии, которые идут непосредственно под кожей и остаются на ней при препаровке. Большая часть каналов обычно остается при мускулатуре, и их трудно обнаружить. Пользуясь рисунками и сопоставляя отдельные видимые участки кожи с каналами с поверхности и с внутренней стороны кожи, можно довольно полно проследить их ход.
У ската, вероятно, в связи с придонным образом жизни и мощным развитием грудных плавников конфигурация каналов очень сложна. Для того чтобы их рассмотреть, необходимо снять кожу с
вентральной и дорзальной поверхности тела. При этом надо стараться отделить самый верхний кожный слой, оставляя нетронутой соединительную ткань, одевающую мускулатуру, так как каналы лежат непосредственно под ней и могут быть легко повреждены. В большинстве случаев каналы имеют вид двойного беловатого тяжа.
На вентральной стороне канал, начинающийся на уровне угла рта, тянется назад вдоль наружного края жаберных отверстий, уходит до переднего конца брюшной полости, некоторое время идет вдоль него, потом круто поворачивает вперед и поднимается до ростральной области. На всем протяжении он дает многочисленные мелкие веточки, идущие по вентральной поверхности плавников. В ростральной области каналы правой и левой сторон сливаются на уровне отверстий обонятельных капсул. От места слияния каналы каждой стороны поднимаются вперед вдоль рострума до переднего конца тела. Здесь они поворачивают и опускаются назад почти параллельно восходящей ветви, снова идут вперед и опять назад, образуя фигуру, несколько напоминающую букву W. На дорзальной стороне латеральный канал начинается около медиального края отверстия брызгальца и идет назад сначала вдоль основания грудных плавников, а затем по спинной стороне хвостового стебля вплоть до его конца. На своем пути он дает несколько крупных ответвлений к спинной поверхности грудных плавников. Надглазничный канал является продолжением вперед латерального канала. Он проходит вдоль медиального края глазницы и теряется в ростральной области, распадаясь на ряд мелких веточек. Подглазничный канал проходит по узкой перемычке между глазом и брызгальцем и идет вперед в ростральную часть. Особенно четко каналы боковой линии видны на голове химеры. Они здесь еще не замкнуты в трубку, а имеют вид глубоких желобков с небольшими расширениями на всем их протяжении, поэтому картина их расположения представлена очень четко.
Головной мозг хрящевых рыб
Особенности формы тела ската облегчают препаровку нервной системы. Поэтому его надо признать более выгодным объектом для проведения этой работы, и основное описание нервной системы дается именно для него.
Головной мозг ската сверху (рис. 54)
Вскрывают осторожно черепную полость, начиная разрез между глазницами и срезая постепенно отдельными кусочками хрящевые стенки, стараясь не повредить головных нервов, прободающих боковые стенки черепа. Особенно следует быть осторожными, снимая крышку черепа в области среднего мозга.
Головной мозг почти полностью заполняет черепную полость. Он суживается между глазницами, но значительно расширяется впере-172
ди них. При рассмотрении сверху будут видны следующие его части.
Передний мозг (telencephalon, tele.)—это самый широкий отдел головного мозга у ската, имеет форму трапеции, широким основанием направленной вперед, а узким — назад, где примыкает к позади лежащему промежуточному мозгу. Легкой бороздкой сверху и снизу передний мозг поделен на две половины, называемые полушариями переднего мозга, где сосредоточены первичные и
Рис. 54. Головной мозг и периферическая нервная система ската Raja clavata сверху:
аиг. сгЬ — ушки мозжечка; bul. о. — обонятельная луковица; crb. — мозжечок; d. п. с. — промежуточный мозг; f. rh. — ромбовидная ямка; lob. о. — зрительные доли; med. obi. — продолговатый мозг; pl. br. — плечевое сплетение; г. b. I. VII — щечная боковая ветвь VII нерва; г. b. т. V// —щечная внутренняя ветвь VII нерва; г. hm. VII — подъязычно-челюстная ветвь VII нерва; г. hy. VII — подъязычная ветвь VII нерва; г. lat. X — боковая ветвь X нерва; г. md. V — нижнечелюстная ветвь V нерва; г. md. е. VII — наружная нижнечелюстная ветвь VII нерва; г. md. I. VII — внутренняя нижнечелюстная ветвь VII нерва; г. тх. V —верхнечелюстная ветвь V нерва; г. opht. р. V—глубокая глазничная ветвь V нерва;
г. opht. s. V+VIII— поверхностная глазничная ветвь V и VII нервов;
г. ph. X — глоточная ветвь X нерва; г. рг. IX — преджаберная ветвь IX нерва; г. рг. X — преджаберная ветвь X нерва; г. pr.+p. V//— небио-преспнракулярная ветвь VII нерва; г. ps. IX — зажаберная ветвь IX нерва; г. ps. X — зажаберная ветвь X нерва; г. spl. X — виутреииостная ветвь X иерва; tele. — передний мозг; tr. о. — обонятельный тракт; / — обонятельные нервы; // — зрительный тракт; /// — глазодвигательный нерв; /V — блоковый нерв; V+VII — место отхождения тройничного и лицевого нервов; VIII — слуховой нерв; IX — языкоглоточный нерв;
X — блуждающий нерв; XII — подъязычный нерв
вторичные обонятельные центры. Собственно полушария (hemisphaerium) занимают среднюю часть переднего мозга справа и слева от центральной бороздки. Боковые части переднего мозга образуют обонятельные доли (lobus olfactorius), которые продолжаются в обонятельные ножки (pedunculus olfactorius), или обонятельные тракты (tractus olfactorius, ir. о.). Впереди они дают расширение, прилегающее к обонятельным мешкам и носящее название обонятельных луковиц (bulbus olfactorius, bul. о.). От обонятельных луковиц в виде тонких нитей щеточкой отходят обонятельные нервы (n. olfactorius, I). Внутри переднего мозга имеется широкая полость полушарий головного мозга, соответствующая первому и второму желудочкам мозга наземных позвоночных. От этой полости отходят полые выросты вправо и влево в обонятельные доли. Вскрытие переднего мозга разрезом по фронтальной плоскости можно сделать после рассмотрения головного мозга сверху и снизу.
Промежуточный мозг (diencephalon)—узкий отдел мозга, начинающийся сейчас же позади переднего мозга; его крыша чрезвычайно тонка, так что через нее можно видеть полость этого отдела мозга — третий желудочек. На задней части крыши промежуточного мозга помещается в виде маленького пузыревидного выроста верхняя мозговая железка, или эпифиз (epiphysis). Боковые стенки промежуточного мозга утолщены и в виде двух столбов мозговой ткани продолжаются назад в основание среднего мозга.
Верхний отдел промежуточного мозга получает название над-бугровой области — эпиталамус — и служит центром пинеального органа (эпифаза). Средний отдел называется бугровой областью— таламусом. Нижний отдел, образующий мозговую воронку (infundibulum), называется подбугровой областью — гипоталамусом, где сосредоточены очень важные центры вегетативных функций. С нижней стороны промежуточного мозга видно отхождение очень крупных зрительных нервов, образующих зрительную хиазму. Остальные части дна промежуточного мозга будут описаны при рассмотрении головного мозга с нижней стороны.
Средний мозг (mesencephalon) с верхней стороны представлен двумя овальными телами, полыми внутри и разделенными посредине бороздой. Они носят название зрительных долей (lobus opticus, lob. о.), или двухолмия, и служат первичными центрами органов зрения. В основании среднего мозга справа и слева лежит стволовая часть головного мозга, как продолжение стволовой части продолговатого мозга. В среднем мозгу стволовая часть называется ножками мозга (crura cerebri). Полость среднего мозга получает название сильвиева водопровода (aqueduc-tus cerebri). Крыша этой полости образует зрительные доли и называется крышей (tectum opticum), а дно — покрышкой (tegmentum), где сосредоточены важные центры так 'называемой сетевидной формации.
Мозжечок (cerebellum, s. metencephalon, crb.) у хрящевых, рыб развит очень сильно. Он имеет вид полого выроста крыши заднего мозга, нависающего впереди над зрительными долями, а также назад над продолговатым мозгом, закрывая собой перепончатую крышу продолговатого мозга. На его поверхности видны борозды, продольные и поперечные, в виде креста, которые делят его на переднюю и заднюю части и на правую и левую половины.
Продолговатый мозг (medulla oblongata, s. myelencephalon, med. obi.) — задний отдел мозга, непосредственно продолжающийся в спинной мозг. В значительной части он покрыт сверху мозжечком. Крыша его чрезвычайно тонка, лишена нервных клеток и обыкновенно плотно срастается с мягкой оболочкой мозга (pia mater) и кровеносными сосудами, образуя сосудистое сплетение (plexus chorioideus posterior), питающее мозг, которое очень легко удаляется при препаровке. Под ним открывается большая полость — четвертый желудочек (fossa rhombeidea, f. rh.). Боковые части продолговатого мозга образуют особые утолщения неправильной формы, которые относятся к мозжечку и носят название ушек мозжечка (auricula cerebelli, рис. 54, aur. crb.). От боковой поверхности продолговатого мозга отходит большинство головных нервов (V, VII, VIII, IX, X и XII). В продолговатом мозгу сосредоточены центры, ведающие чувствующими и двигательными функциями челюстного п жаберного аппаратов, а также иннервацией внутренних органов (желудка, сердца и т. д.). Кроме того, там же помещаются центры, ведающие системой кожных органов чувств боковой линии (сейсмосензорных органов), а также статоакустического органа.
Головной мозг ската снизу (рис. 55)
Нижнюю поверхность головного мозга можно рассмотреть, не портя общего препарата нервной системы. Очищают слизистую оболочку с крыши ротовой полости и вскрывают осторожно брюшную стенку черепа под мозгом до тех пор, пока не будет ясно видно брюшной поверхности мозга. Рассматривают отделы мозга.
Передний мозг (telencephalon) характеризуется присутствием небольшого непарного углубления, в остальном же видны те же части, что и со спинной поверхности.
Промежуточный мозг (diencephalon) с брюшной стороны развит чрезвычайно сильно, он вытянут в воронку, которая прикрывает дно среднего мозга, и дифференцирован на несколько отделов. Рассматривают отделы промежуточного мозга:
а) зрительная хиазма (chiasma nervorum opticorum) образована пересечением зрительных нервов в передней части дна промежуточного мозга. После перекреста нервы расходятся кнаружи;
б) нижние доли (lobus inferior, s. corpus lateralis, cr.) — парные яйцевидные тела, лежащие на нижней поверхности позади перекреста зрительных нервов. Нижние доли представляют собой бо
ковые части мозговой воронки (infundibulum) — воронковидного расширения полости промежуточного мозга (третьего желудочка), загибающегося назад;
в) железистый мешочек (saccus vasculosus, s. vase.) — тонкостенное полое тело, вырост дна промежуточного мозга, состоящий из трех лопастей: непарной (в желобке между нижними долями) и двух парных выростов (на свежем мозгу красноватых, на фиксированном — черного цвета) под нижними долями;
Рис. 55. Головной мозг акулы Squalus acanthias сверху, сбоку и снизу (по Маринелли и Стренгеру):
а иг. erb, — ушки мозжечка; bul. о. — обонятельная луковица; crb. — мозжечок; cr. I. — нижние доли; апс. — промежуточный мозг; ер. — эпнфиз; hyp. — гнпофнз; lob. о. — зрительные доли; med. о. — продолговатый мозг; med. sp. — спинной мозг; msc. — средний мозг; л. term. — конечный нерв; pl. ch. р. — заднее сосудистое сплетение; s. vase. — сосудистый мешок; Нс. — передний мозг; tr. о. — обонятельный тракт; II, III, IV, V, VI, VII, VIII, IX, X X// —головные нервы
г) гипофиз, или питуитарное тело (hypophysis, hyp.), — нижняя мозговая железа, представляет собой тонкостенный, непарный мешок позади железистого мешочка, прирастающий ко дну мозговой воронки.
Средний мозг (mesencephalon) с нижней стороны почти не виден, он закрыт налегающей на него мозговой воронкой промежуточного мозга. Снизу от нижней поверхности среднего мозга против места соединения воронки с гипофизом отходят глазодвигательные нервы (n. oculomotorius, Ш). От дна продолговатого мозга вперед продолжаются, несколько расходясь в сто-176
роны, стволы проводящих путей, называемых ножками среднего мозга (crura cerebri).
Продолговатый мозг (medulla oblongata, s. myelencephalon) с нижней стороны имеет форму конуса, расширенным основанием направленного вперед и отделенного впереди перешейком (isthmus) от передних отделов головного мозга. Сбоку видны отхождения головных нервов V, VII, VIII, IX и X пары. Снизу на вентральной стороне видны тонкие корешки нерва шестой пары (n. ab-bucens), снабжающие наружную прямую мышцу глаза (m. rectus externus). Позади десятого нерва (n. vagus) можно видеть корешки подъязычного нерва (n. hypoglossus, XII).
Головной мозг акулы
Головной мозг акулы катрана Squalus acanthias при сравнении с головным мозгом ската Raja clavata отражает особенности биологии и образа жизни акул, активных хищников, схватывающих свою добычу и преследующих ее при плавании.
Передний мозг (telencephalon, рис. 55, 56, tic.) имеет значительно больше развитые средние отделы, соответствующие полушариям переднего мозга (hemisphaera). Они в виде двух мощных скоплений мозговой ткани занимают дно и боковые стенки переднего мозга, их мозговые стенки загибаются вперед до смыкания по средней линии. Особенно мощной толщины скопление мозговой ткани достигает на брюшной поверхности, что видно при рассмотрении мозга с нижней стороны. В середине переднего мозга верхняя стенка перепончатая и через нее просвечивает общая полость первого и второго мозговых желудочков.
Боковые отделы переднего мозга — обонятельные доли (lobus olfactorius), являющиеся первичными обонятельными центрами, у акул развиты значительно слабее, чем у скатов, и имеют вид тонких боковых тяжей, лежащих по бокам полушарий.
Промежуточный мозг (diencephalon, рис. 55, 56, dnc.) не имеет особых отличий. Крыша его остается перепончатой и к ней прилегает сидящая на длинной ножке верхняя мозговая железа— эпифиз (epiphysis, рис. 55, ер.). С нижней стороны дно промежуточного мозга образует типичный перекрест зрительных нервов (хиазма), позади него находится воронка (infundibulum) с прилегающим к ней гипофизом (hypophysis, рис. 55, hyp.), нижними долями (lobus inferior, s. corpus lateralis) и сосудистым мешочком (saccus vasculosus, рис. 55, s. vase.).
Средний мозг (mesencephalon) представлен, как и у скатов, крупными яйцевидными зрительными долями (lobus opticus, рис. 55, 56, lob. о.). У акул они значительно сильнее развиты по сравнению со скатами, так как зрительным рецепторам акул принадлежит более важная биологическая роль, чем обонятельным.
Наконец, мозжечок (cerebellum, рис. 55, crb.) у акул также Достигает более мощного развития, так как акулы — хорошие пловцы, тогда как скаты ведут придонный образ жизни. Наоборот, бо-
ковые отделы мозжечка, его ушки (рис. 54, 55, ayr. crb.), являющиеся центрами сейсмо-сензорных кожных органов чувств боковой линии, значительно крупнее у скатов, так как эти органы там образуют очень сложные разветвления на поверхности громадных грудных плавников.
Головные нервы хрящевых рыб
Отхождение головных нервов
Необходимо проследить места отхождения от мозга и порядок отхождения головных нервов, не разбирая и не препарируя их ответвлений.
tr.aLf.
lOb.O. ---;
Ш
он 1.0.
tel. с.
х
ш
r.lat.X nspl.X
Г- —
тт
И aur. crb. med. obi.
Г. opht.s.V+ТШ ropbt.p.v гЬЛ+т.тх.И
IS
Г.Ь.Ш r. tml r. mills r.pl. ИГ г.ргЛ , m --------r.hm. ИГ rmd.e.Vl rtuj.VL
r.pr.lX r.ps.lX
r.pr.X
r.ps.X rph.X
Рис. 56. Головной мозг и головные нервы акулы Squalus acanthias:
aur. crb. — ушкн мозжечка bul. о. — обонятельная луковица; crb. — мозжечок; dnc. — промежуточный мозг; lob. о. — зрительные Доли; med. obi. — продолговатый мозг; г. b. VII — щечная ветвь VII нерва; rb. VII+г. тх. V — щечная ветвь VII и верхнечелюстная V нервов; г. hm. V//— подъязычно-челюстная ветвь VII нерва; г. hy. V//— подъязычная ветвь VII нерва; г. lat. X — боковая ветвь X нер-' ва; г. md. V - нижнечелюстная ветвь V нерва; г. md. е. VII — наружная нижнечелюстная ветвь VII нерва; г. md. i. — внутренняя нижнечелюстная ветвь VII нерва; г. тх. V — верхнечелюстная ветвь V нерва; г. opht. р. V — глубокая глазничная ветвь V нерва; г. opht. s. V+VI1— поверхностная глазничная ветвь V и VII нервов; г. pl. V// —небная ветвь VII нерва; г. ph. X—глоточная ветвь X нерва; г. рг. VII — преспираку-лярная ветвь VII нерва; г. рг. IX — преджаберная ветвь IX нерва; г. рг. X — преджаберная ветвь X нерва; г. ps. /л —зажаберная ветвь IX нерва; г. ps. X —зажаберная ветвь X нерва; г. spl. X — внутренностная ветвь X нерва; tel. с. — нередкий мозг; tr. olf. — обонятельный тракт; II, III, IV, V, VII, VIII, IX,
X, XII — головные нервы
а
Обонятельные нервы (п. olfactorius) — I пара головных нервов (рис. 54, /), отходит от обонятельных луковиц переднего мозга. Зрительные нервы (n. opticus, рис. 54, 55, 56, II) — II пара, отходит от дна промежуточного мозга. Глазодвигательный нерв (n. oculomotorius, рис. 54, 55, 56, ///)—III пара, отходит с брюш-178
ной поверхности среднего мозга (mesencephalon) и проходит через стенки глазницы к глазным мышцам. Блоковый нерв (n. troch-learis, рис. 54, 55, 57, IV) —IV пара, очень тонкий, отходит от крыши мозга на границе между мозжечком (cerebellum) и средним мозгом. Остальные головные нервы (V, VI, VII, VIII, IX, X и XII) отходят от продолговатого мозга. Чтобы рассмотреть места их отхождения, следует осторожно отодвинуть продолговатый мозг от стенки черепа, пока не откроются корешки нервов, а затем аккуратно срезать черепную стенку и слуховую капсулу с одной стороны (оставив другую для препаровки слухового лабиринта), пока корешки нервов не будут видны совершенно ясно.
Нерв V пары — тройничный (n. trigeminus), отходит вместе с первом VII пары — лицевым нервом (n. facialis) и нервом VIII пары (n. acusticus, рис. 54, 55, 56, V, VII, VIII). Все эти нервы отходят несколькими (5—7) корешками от боковой поверхности продолговатого мозга па самом широком месте, против заднего конца мозжечка. Примерно на том же уровне, но от вентральной поверхности продолговатого мозга отходит нерв VI пары — отводящий (n. abducens, рис. 56, VI).
Языкоглоточный нерв (n. glossopharyngeus) — IX пара, отходит, несколько отступя назад от слухового нерва, ближе к брюшной поверхности мозга и непосредственно примыкает к X нерву. Блуждающий нерв (n. vagus, рис. 54, 55, 56, X) —X пара, отходит несколькими корешками от боков задней части продолговатого мозга и идет мощным пучком назад.
Разветвление головных нервов (см. рис. 14)
Изучение ветвей головных нервов можно проводить, как на скате так и на акуле. Однако в связи с различиями в образе жизни ветвление некоторых стволов головных нервов у пих различно. Поэтому описание ветвей ведется по скату (рис. 54) с указаниями на отличия, имеющиеся у акулы (рис. 56, 57).
Осторожно отпрепарируют ответвления головных нервов на правой стороне черепа, оставив левую для анатомирования слухового лабиринта и для контроля.
Сначала необходимо проследить прохождение нервных стволов по глазнице и впереди нее, в области обонятельного мешка. Рассматривают последовательно головные нервы, начиная от переднего конца мозга.
Конечный нерв (п. О, п. terminalis, рис. 55, п. term.) — очень тонкий соматически-чувствующий нерв, четко видимый у акулы. Он отходит от ростральной поверхности переднего мозга и тянется вдоль медиальной поверхности обонятельного тракта к стенке обонятельного мешка.
О б о н я т е л ь н ы е н е р в ы (п. I, п. olfactorius) — многочисленные тонкие нервные волокна, выходящие из обонятельной капсулы н впадающие в расширенные передние концы обонятельных трактов— обонятельные луковицы (bulbus olfactorius, рис. 54,
55, 56, bulb. о.). Последние у скатов имеют вид белых лепт, лежащих на обонятельных капсулах, а у акул — пузыревидных вздутий. Зрительный нерв (n. II, п. opticus) входит в глазницу через отверстие в черепе и близ его нижней поверхности. В глазни
це нерв одет в очень плот-
r.cpht.s. 7+ И
1У -
т
г.рг. хи
r.hm. УЛ
Л m.r.e.
г. тх. У г. ггЛ.У
-in.r.lr.t.
m.r.s.
Рис. 57. Разветвление головных нервов в глазнице и глазные мышцы акулы Squalus . acanthias, вид сверху:
т. о. inf. — пижияя косая мышца глаза; т. о. s.— верхняя косая мышца глаза; т. г. е. — наружная прямая мышца глаз; т. г. inf. — нижняя прямая мышца глаза; т. г. s. — верхняя прямая мышца глаза; т. г. int.—внутренняя прямая мышца глаза; г. hm. VII— подъязычно-челюстная ветвь VII нерва; г. md. V — нижнечелюстная ветвь V нерва; г. тх. V — верхнечелюстная ветвь V нерва; г. ophi. р. V — глубокая глазничная ветвь V нерва; г. opht. s. V+ VII — поверхностная глазничная ветвь V и VII нервов; г. pl. V//— небная ветвь VII нерва; г. pr. VII — преспнракулярная ветвь VII нерва; II, III, IV, VI, VIII, IX— головные нервы
ную соединительнотканную оболочку и идет прямо кнаружи в глазное яблоко (рис. 57, //).
Глазодвигательный нерв (n. Ill, п. oculo-motorius) — очень тонкий двигательный головной нерв, отходит от дна среднего мозга впереди и сбоку от гипофиза, идет кнаружи и, прободая стенку черепа, выходит в глазницу. Войдя в глазницу, он распадается сразу же на три ветви, идущие к глазным мышцам: передняя ветка идет ко внутренней прямой мышце, средняя — к верхней прямой, а задняя — к двум мышцам, нижней прямой и нижней косой (рис. 57, ///).
Блоковый нерв (п. IV, п. trochlearis) — довольно тонкий двигательный головной нерв, отходит от спинной поверхности мозга на границе между средним мозгом и мозжечком. Чтобы увидеть начало нерва, следует немножко приподнять передний конец мозжечка. Он тянется вперед и кнаружи и пронизывает стенку глазницы немножко впереди ее середины над отверстием зрительного нерва. В глазнице нерв направляется к верхней косой мышце глаза/ которую иннервирует (рис! 54, 57, IV).
Тройничный нерв (п. V, n. trigeminus) — сложный голову ной нерв, выходит из черепа в глазницу и здесь обычно распадает! ся на три ветви: глазничную, верхнечелюстную и нижнечелюстную!
почему и носит название тройничного. У ската взаиморасположение ветвей несколько иное, и поэтому в глазнице нерв первоначально дает только две ветви. Как указано выше, корешки его тесно связаны с корешками VII, лицевого нерва (n. facialis, n. VII). Осторожно отодвигают мозг от стенки черепа, чтобы видеть корешки нерва внутри черепа. Корешки отходят целым пучком и образуют большой нервный узел. Чтобы проследить отхождения в черепе, проводят вскрытие черепной коробки сбоку, срезая ломтиками прозрачный хрящ. Отпрепарируют проходящие по дну глазницы нервные стволы глазничной ветви тройничного нерва и прослеживают их направление к периферии.
Глазничная ветвь (ramus ophthalmicus V+VII) отходит от переднего края упомянутого общего узла и проникает в глазницу чуть впереди от места прикрепления прямых мышц глаза. В глазнице она распадается на две ветви: а) поверхностная глазничная ветвь (г. ophthalmicus superficialis, рис. 54, 56, 57, г. opht. s. V+VII), чувствующий нерв, составляемый волокнами V и VII нервов. Он идет по верхнему краю глазницы, отдавая тонкие веточки вверх к надглазничному каналу боковой линии, и направляется вперед, где иннервирует своими ветвями кожные органы чувств верхней поверхности рыла; б) глубокая глазничная ветвь (г. ophthalmicus profundus, рис. 54, 56, 57, г. opht. р. V) — особый, также чувствующий корешок тройничного нерва, он снабжен своим отдельным ганглием. Эта ветвь отщепляется от общего глазничного нерва в глазнице и проходит между глазными мышцами. Над обонятельными луковицами глубокая глазничная ветвь сливается с поверхностной глазничной веткой и также идет к кожным органам чувств рыла.
Отрезают глаз, осторожно удаляют глазные мышцы, на дне глазницы виден толстый тяж из нервов, 'который посредине глазницы расходится на несколько ветвей тройничного и лицевого нервов. Поэтому целесообразно эти ветви рассмотреть совместно.
Самая медиальная и латеральная ветви являются щечными ветвями лицевого нерва, иннервирующими каналы боковой линии. Медиальная ветвь называется внутренней щечной ветвью (г. buccalis medialis n. VII, рис. 54, г. b. т. VII), латеральная — боковой щечной (г. buccalis lateralis n. VII, рис. 54, г. b. I. VII).
Нижнечелюстная ветвь тройничного нерва (г. mandi-bularis V, рис. 54, г. md. V) располагается латеральнее боковой щечной. Она направляется вниз и назад, подходит к углам рта и, повернув здесь в медиальном направлении, переходит на нижнюю челюсть. Эта ветвь составлена соматически-чувствующими и висцерально-двигательными волокнами. Она иннервирует кожу и зубы нижней челюсти, а также мускулатуру челюстной дуги.
Верхнечелюстная ветвь тройничного нерва (г. maxil-laris V, рис. 54, V) у ската выражена не четко и представлена отдельными тонкими веточками, начинающимися от внутренней щеч
ной и нижнечелюстной ветвей на уровне переднего края нижней челюсти. Верхнечелюстная ветвь содержит соматически-чувствую-щие волокна и иннервируют кожу и зубы верхней челюсти.
У акулы расположение четырех перечисленных ветвей лицевого и тройничного нервов несколько иное. Ветвь, проходящая по дну глазницы, распадается у ее переднего края только на три ствола. Первый проходит наиболее латерально и может считаться боковой щечной ветвью лицевого нерва. Он иннервирует нижнюю часть головы перед верхней челюстью. От ее переднего конца отходит небольшая ветвь, которая идет назад и иннервирует верхнюю челюсть в области губных хрящей, т. е. является одной из частей верхнечелюстной ветви тройничного нерва. Общий ствол двух вышеназванных ветвей следует считать совместным стволом г. buccalis lateralis VII + r. maxillaris V (рис. 56, r. b. VII+r. mx. V). Лежащая несколько медиальней вторая ветвь является чистой боковой щечной ветвью лицевого нерва (г. buccalis lateralis VII). Самая медиальная крупная ветвь является внутренней щечной ветвью (г. buccalis medialis VII, рис. 54, г. Ь. т. VII) лицевого нерва. Она распадается на две веточки. Одна иннервирует нижнюю поверхность рыла на уровне переднего края глазницы, другая доходит до конца рыла.
Нижнечелюстная ветвь тройничного нерва (г. mandibu-laris V, рис. 54, 57, г. md. V) у акулы идет вдоль заднего края глазницы, переходит на латеральную поверхность головы, затем поворачивает на ее вентральную поверхность и иннервирует нижнюю челюсть. В месте прохождения по латеральной поверхности она отдает назад крупную ветвь к мускулу, приводящему нижнюю челюсть. Почти на том же уровне вперед отходит небольшая ветвь к верхней челюсти (часть г. maxillaris V, 56, 57, г. nix. V), которая иннервирует задний отдел верхней челюсти.
Отводящий нерв (n. VI, n. abducens, рис. 57, VI) — двигательный, очень тонкий нерв, видимый только с нижней стороны мозга. Отходит внизу под корешками тройничного и лицевого пер-' вов и направляется в виде очень тонкой нити в глазницу через отверстие вместе с V и VII нервами к наружной прямой мышце глаза, которую он иннервирует. Он хорошо виден па ее вентральной поверхности.
Лицевой нерв (п. VII, н. facialis) тесно связан корешками и ветвями с тройничным нервом. Это сложный смешанный головной нерв, распадающийся на много ветвей: а) глазничная поверхностная ветвь VII нерва идет вместе с одноименной ветвью V нерва; б) щечная ветвь (г. buccalis, рис. 56, г. b. VII) VII нерва идет по дну глазницы совместно с верхнечелюстной ветвью тройничного нерва. Эти ветви описаны выше вместе с тройничным первом; в) подъязычно-челюстной нерв, или г и о м а н д и б у л я р и ы й (г. hyomandibularis VII, рис. 54, 56, 57, г. hm. VII),— главный ствол VII нерва, идущий прямо кнаружи около передней стенки слуховой капсулы и далее под кожей по задней стенке брызгальца. В конце он делится на две ветви: одна
из них, подъязычная ветвь (г. hyoideus, рис. 54, 56, г. hy. VII), иннервирует висцеральную мускулатуру, другая, наружная нижнечелюстная (г. mandibularis externus, рис. 54, 56, г. md. е. VII), идет к органам боковой линии. Вскоре по выходе подъязычного нерва из черепа от него отходит тонкая ветвь, которая проходит впереди брызгальца и распадается на отдельные веточки. Одни из них идут к ложной жабре, другие —распределяются по слизистой ткани неба. Этот нерв включает два компонента— небную ветвь и предспиракулярную (г. palatinus+r. praetre-mancus, рис. 54, 56, г. pr.+p.VII). На уровне заднего края брызгальца от гиомандибулярного нерва отходит еще одна тоненькая веточка, которая иннервирует внутреннюю поверхность нижней челюсти,—это нижнечелюстная внутренняя ветвь (г. mandibularis internus, s. chorda tympani, рис. 54, 56, r. md. i. VII).
Чтобы увидеть сзади лежащие нервы, удаляют слуховую капсулу и перепончатый слуховой лабиринт. Далее, найдя места отхождения от мозга IX и X нервов, удаляют боковую и заднюю стенки черепа. Для того чтобы проследить прохождение нервов к отдельным жаберным дугам, удаляют на спинной поверхности жаберную мускулатуру и вскрывают верхнюю яремную вену, под которой проходят ветви десятого нерва.
Слуховой нерв (n. VIII, n. acusticus, рис. 54, 56, 57, VIII) по отхождении от мозга загибается назад и разветвляется сейчас же после вхождения в слуховую капсулу на ряд мелких веточек, идущих к различным частям внутреннего уха.
Языкоглоточный нерв (п. IX, n. glossopharyngeus) идет кнаружи и назад в хрящевом канале по дну слуховой капсулы, выходит из черепа у заднего и внешнего угла капсулы против верхнего конца первой жаберной щели. Здесь он делится на две ветви, идущие впереди и позади первой жаберной щели: а) передняя, или преджаберная, ветвь (г. praetrematicus, рис. 54, 56, г. рг. IX) идет по заднему краю подъязычной дуги; б) задняя, зажаберная, ветвь (г. posttrematicus, рис. 54, 56, г. ps. IX), идет по переднему краю первой жаберной дуги. Будучи типичным жаберным нервом, языкоглоточный нерв обладает еще и глоточной ветвью, которая здесь иннервирует слизистую оболочку неба и потому названа небной ветвью (г. palatinus).
Блуждающий нерв (n. vagus X) по отхождении от мозга толстым тяжем выходит из черепа на заднем его конце и направляется назад вдоль спинного мозга по внутреннему краю жаберного аппарата, давая от себя ряд жаберных ветвей, внутренностную ветвь и боковой нерв.
Жаберные ветви (рис. 54, 56, г. branchialis) в числе четырех идут к верхним концам четырех задних (со 2-й по 5-ю) жаберных щелей. Каждая ветвь, как и у IX нерва, разветвляется на три ветви: а) тонкую переднюю, преджаберную (г. praetrematicus, г. рг. X), идущую но задней стенке передней дужки; б) большую заднюю, зажаберную (г. posttrematicus, г. ps. X), идущую по пе
редней стенке задней жаберной дужки; в) глоточную (г. pharynge-us, г. ph. X), идущую к стенке глотки.
Внутренностная ветвь (г. splanchnicus, г. spl. X) идет назад и внутрь как продолжение общего жаберно-кишечного ствола (n. branchio-intestinalis), от которого ответвляются жаберные ветви. Она уходит в глубину, иннервируя сердце, желудок и кишечник.
Рис. 58. Слуховой лабиринт и затылочные нервы акулы Squalus acanthias (из Маринелли и Стренгера):
can. sem. — полукружные каналы, г. d. d. — спинные корешки спинномозговых нервов; г. d. V. — брюшные корешки спинномозговых нервов; X — блуждающий нерв; XII — подъязычный нерв
Боковой нерв (г. lateralis, г. lat.) отходит от общего ствола около места выхождения X нерва из черепа и идет прямо назад по телу более поверхностно, чем жабернокишечный ствол, снабжая кожные органы чувств боковой линии туловища.
Подъязычный нерв (n. XII, n. hypoglossus, рис. 54, 55, 56, 58, XII) будет виден при препаровке спинного мозга и спинномозговых нервов. Это 1—3 маленьких брюшных двигательных корешка позади X нерва, перед первым полным спинномозговым нервом.
Спинной мозг и нервы
Чтобы увидеть спинной мозг и спинномозговые нервы, следует удалить горизонтальными срезами крышу спинномозгового канала, пока не откроется спинной мозг, а затем и боковые стенки так, чтобы видеть отхождение корешков спинномозговых нервов.
Спинной мозг лежит в спинномозговом канале позвоночника. Он сплющен сверху вниз. На дорзальной и вентральной стороне посредине проходит непарная продольная борозда. Внутри мозга проходит узкий центральный канал (невроцель), который впереди непосредственно переходит в полость продолговатого мозга (четвертый желудочек).
Спинномозговые нервы отходят от мозга посегментно справа и слева. Каждый нерв отходит двумя корешками: спинным— чувствующим, с утолщением — ганглием (рис. 58, г. d. d.) и б р юш н ы м — двигательным (рис. 58, г. d. v.), более коротким, лежащим в глубине, на брюшной поверхности мозга. Каждый корешок выходит из позвоночника через особые отверстия в верхних дужках позвонков, а снаружи корешки сливаются друг с другом и образуют спинномозговой нерв.
Подъязычный нерв (hypoglossus, XII) является задним головным, так называемым затылочным нервом. Образован подъязычный нерв 1—3 самыми передними брюшными корешками спинномозговых нервов, не имеющих спинных корешков с ганглиями. Эти корешки сливаются и образуют сплетение подъязычного нерва, идущего к поджаберной мускулатуре.
Плечевоесплетение (plexus brachialis, рис. 54, pl.br.) образовано передними спинномозговыми нервами, которые справа и слева веерообразно соединяются в общий ствол, идущий к грудным плавникам, или соединены между собой анастомозами (у акул). У акул плечевое сплетение меньше (5—6 нервов), у скатов больше (13—14 нервов), что находится в зависимости от величины грудного плавника.
Пояснично-крестцовое сплетение (plexus lumbo-sacralis) иннервирует брюшной плавник и расположено в задней части полости тела. Его удобнее рассматривать со стороны брюшной полости тела у вскрытого животного.
Органы чувств хрящевых рыб
Органы зрения
Глазница и глаза (см. рис. 57). Удаляют веки правого глаза, чтобы открыть глазницу, и промывают струей воды сгустки крови из глазничного синуса. Рассматривают глаз и глазные мышцы, стараясь не повредить проходящие по глазнице нервы.
Глазное яблоко почти шаровидно, но его наружная сторона, покрытая роговицей, заметно уплощена.
Глазные мышцы в количестве шести тонких и узких мускулов приводят глаз в движение: они расположены двумя группами — прямые н косые мышцы.
Прямые мышцы глаза в числе четырех отходят почти вместе в нижнем заднем углу глазницы и расходятся крестообразно вокруг глазного яблока, прикрепляясь к нему сверху, снизу, изнутри и снаружи: 1) прямая верхняя (m. rectus superior, tn. г. s.)
поднимает глаз кверху; 2) прямая наружная (m. rectus externus, т. г. е.) отводит глаз назад; 3) прямая внутренняя (т. rectus internus, т. г. int.) отводит глаз вперед; 4) прямая нижняя (т. rectus inferior, т. г. inf.) лежит глубоко под глазом напротив верхней прямой мышцы и опускает глаз вниз.
Между четырьмя вышеназванными мышцами лежит небольшой хрящевой стержень с дискообразным расширением на конце - ре-
Рис. 59. Схема вертикального разреза глаза акулы (по Franz):
chor. — сосудистая оболочка; corn. — роговица; gl. chor. — хориоидальная железа; ir. — радужина; lens. — хрусталик; т. — глазные мышцы; р. Ь. — хрящевой стержень, поддерживающий глазное яблоко: ret. — сетчатка; sc. — склера; t. opt.— зрительный тракт
dunculus bulbi, поддерживающий глазное яблоко.
Косые мышцы глаза в числе двух начинаются в переднем углу глазницы, а оканчиваются на верхней и нижней поверхностях глазного яблока: 1) косая верхняя (m. obliquus superior, т. о. s.) лежит над глазным яблоком и отводит его вверх и вперед; 2) косая нижняя (m. obliquus inferior, т. о. inf.) лежит под глазным яблоком и отводит его вниз и вперед.
Строение глаза (рис. 59). Отрезают осторожно ножницами глазные мышцы в местах их прикрепления к глазному яблоку, перерезают зрительный нерв и вынимают глаз из глазницы. Разрезают его ножницами во фронтальной плос
кости на две половины и рассматривают его строение в ванночке с водой.
Стенки глаза состоят из трех прилегающих друг к другу слоев. Внутри глаза находится полость; близ внешней стороны расположена округлая линза — хрусталик — приблизительно с горошину величиной. Рассматривают стенки глаза.
Склера (sc.)—белковая оболочка, самый наружный скелетный слой глаза, весьма плотный, хрящевой. Она одевает глаз со всех сторон, за исключением наружной, которая прикрыта роговицей.
Роговица (cornea, corn.)—плоская пленка, которая одевает глаз с наружной стороны; через нее проходит свет. Она совершенно прозрачная и на краях соединяется со склерой.
Сосудистая оболочка (chorioidea, chor.) имеет вид черной непрозрачной перепонки, подстилающей склеру с внутренней поверхности. На месте перехода склеры в роговицу сосудистая оболочка продолжается в радужную (iris, ir.), которая лежит свободно позади роговицы и пронизана в центре продолговатой щелью — зрачком (pupilia), через который свет проходит в глаз.
Внутренняя поверхность сосудистой оболочки покрыта блестящей серебристой перепонкой (tapetum lucidum), которая обусловливает зеленоватое свечение глаз живых акул.
Сетчатка (retina, ret.) — нежная оболочка нервной ткани, выстилающая большую часть внутренней поверхности глазного яблока. Это световоспринимающий слой глаза, непосредственно соединенный с глазным нервом.
Хрусталик (lens, lens.)—шаровидное тело, лежащее непосредственно позади зрачка. У живых акул хрусталик — совершенно прозрачная линза, преломляющая свет.
Стато-акустический орган (рис. 60)
Стато-акустический орган состоит из очень нежного
перепончатого лабиринта, заключенного в хрящевую капсулу в бо-
ковых расширениях хрящевого черепа позади глазницы. Перепон-
чатый лабиринт состоит из перепончатого мешка, или преддверия, лежащего в основании капсулы и трех перепончатых полукружных каналов — трех изогнутых трубочек, поднимающихся дугой над преддверием в трех перпендикулярных друг другу плоскостях. Каждый канал
amp. I utr.
amp. р.
своими концами откры-
вается в преддверие. Перепончатый лабиринт находится в полости слуховой капсулы, заполненной жидкостью — перилимфой.
Очищают задний отдел черепа с левой сто-
Рис. 60. Левый перепончатый лабиринт акулы Squalus acanthias, вид с медиальной стороны (из Маринелли н Стренгера):
amp. I. — ампула бокового полукружного канала; amp. а. — ампула переднего полукружного канала; amp. р. — ампула заднего полукружного капала; can. sem. а. — передний полукружный канал; can. sem. I. — горизонтальный (боковой) полукружный канал; can. sem. р. — задний полукружный канал; lad. — ла-гена; sac.—круглый мешочек; utr. — овальный мешочек
роны, чтобы хорошо была видна слуховая капсула. Рассматривают
просвечивающие сквозь прозрачные хрящевые стенки слуховой капсулы два полукружпых канала. Они проходят в двух пересекающихся вертикальных плоскостях. Чуть медиальнее места их соединения в общую ножку (crus commune) лежит отверстие эндолимфатического протока (ductus endolymphaticus), соединяющего внутренний перепончатый лабиринт с наружной средой. Осторожно срезают верхнюю стенку слуховой капсулы, пока не вскроют полость, выполненную перилимфатической жидкостью, в которую погружен слуховой лабиринт, и осторожно отпрепарируют полукружные
каналы и далее, в глубине, мешковидное преддверие. Рассматривают отделы перепончатого лабиринта, поместив его в сосуд с водой.
Полукружные каналы (canalis semicircularis, рис. 58, can. sem.) —три нежные, прозрачные изогнутые трубки перепончатого лабиринта, выполняющие функцию органов равновесия:
а) передний вертикальный канал идет от общей ножки вперед, затем вниз вдоль заднего края глазницы, и его передний
конец перед впадением в преддверие расширяется в ампулу оваль
ной формы;
б) задний вертикаль
Рис. 61. Орган обоняния акулы Squa-lus acanthias
ный канал — самый длинный, образует почти полное кольцо, направляется назад и имеет ампулу на заднем нижнем конце;
в) горизонтальный канал — располагается приблизительно по наружному краю слуховой капсулы, его ампула находится рядом с ампулой переднего вертикального канала около глазницы.
Преддверие (vestibulum) представляет собой сплющенный с боков мешок, лежащий внизу под полукружными каналами в глубине слуховой капсулы. Он не вполне разделен на две части,
верхнюю — овальный мешочек (utriculus) и более крупную нижнюю — круглый мешочек (sacculus), и содержит в себе известковые включения — отолиты — молочного цвета. Внут-
ри преддверие наполнено жидкостью — эндолимфой. На внутренней стороне преддверия впадает эндолимфатический проток. Через этот проток м’ожно попробовать пропустить зонд с наружной
поверхности в преддверие.
Слуховой нерв (n. acusticus, головной нерв VIII пары) входит в слуховую капсулу на ее внутренней стороне и сейчас же оазделяется на ветки, которые, как это можно проследить, проходят в преддверие и в ампулы.
Орган обоняния (рис. 61)
Рассматривают с брюшной стороны головы ноздри и органы обоняния.
Орган обоняния представляет собой два обонятельных мешка, открывающихся впереди ротовой щели на брюшной поверхности рыла отверстиями ноздрей. Форма ноздри и характерная для многих хрящевых рыб оро-назальная борозда были описаны при внешнем осмотре.
Вынимают один из обонятельных органов из его капсулы; проводят через него посредине поперечный разрез бритвой, промывают водой и рассматривают.
Орган обоняния представляет собой мешок, стенки которого собраны в виде многочисленных поперечных складок, прикрепленных попарно к непарной пластинке. Каждая складка покрыта нежной слизистой оболочкой.
Литература по классу хрящевых рыб
Северцов А. II. Исследования об эволюции низших позвоночных. Первичное строение висцерального аппарата эласмобранхий. Соор, соч., т. 4. Изд-во АН СССР, 1917—1925.
Суворов Е. К. Основы ихтиологии. Л., 1948.
Daniel I. Е. The Elasmobranch Fisches Berkeley, 1928.
Gegenb aur C. Untersuchungen zur Vergleichenden Anatomie dec Wirbel-tiere. Heft II und III. Das Kopfskelett der Selachier. Leipzig, 1872.
Grodzinski Z. Anatomia i Embriologia Ryb. Panst Wydawn Polnicze i Leske. Warszawa, 1961.
Marinelli W., Strenger A. Vergleichende Anatomie und Morphologic der Wirbeltiere, III. Squalus acanthias, Wien, 1953.
St ens io E. Upper Devonian Vertebrates from East Greenland. Kobenhavn, 1931.
St ens io E. Anatomical Studies on the Arthrodiran Head. Part. I. Aemovis and Winsell, Stocholm, 1963.
Woskoboinikoff M. Der Apparat der Kiemenatmung bei den Fischen. Zool. Jahrb. Abt. Anat. Bd. 55, 1932.
НАДКЛАСС РЫБЫ—PISCES
КЛАСС КОСТНЫЕ РЫБЫ — OSTEICHTHYES
Общая характеристика класса
Класс костные рыбы — Osteichthyes — объединяет большинство современных рыб, обладающих костным скелетом. Он включает разнообразные группы рыб, чрезвычайно неравноценные по количеству составляющих их видов. У всех костных рыб наблюдается большая или меньшая замена внутреннего хрящевого скелета настоящей костью. Кроме того, под кожей за счет костных чешуи развиваются кожные кости на голове и плечевом поясе, образуя так называемые покровные окостенения. Помимо этого, все костные рыбы имеют жаберную крышку (operculum), которая закрывает в виде заслонки наружные жаберные отверстия и участвует в дыхательных движениях жаберного аппарата (оперкулярный тип дыхания). Тело большинства рыб покрыто костными чешуями различного строения. Плакоидная чешуя сохраняется только у некоторых видов на отдельных участках тела.
В основание ствола костных рыб, ископаемые остатки которых известны с нижнего девона, вероятно, можно поставить гипотетическую группу Osteichthyes primitivi (см. рис. 1), объединяющую в себе целый ряд черт, свойственных костным рыбам. К ним нужно отнести наличие в коже определенным образом расположенных покровных костей, преобразующихся в районе ротового отверстия во вторичные челюсти; постепенную замену роговых лучей в плавниках костными; появление кожной жаберной крышки с развивающимися в ней покровными костями.
Как 'переходную группу между хрящевыми и примитивными костными рыбами можно рассматривать акаптодий (Acanthoi-dei). Представители этой группы, выделяемой в настоящее время в, самостоятельный класс, встречаются в ископаемом состоянии с конца силура до верхнего карбона (см. рис. 1). Наряду с чертами, свойственными хрящевым рыбам (хрящевой эндокраниум, челюстная дуга из небно-квадратного и меккелева хрящей, наличие хорды в осевом скелете), имеются черты, присущие костным рыбам (кожные кости на черепе и плечевом поясе, эпдокраний, так же как и элементы челюстной дуги, могут окостеневать несколькими окостенениями, окостеневающие верхние дуги в осевом скелете, плотные ряды ганоидных чешуи на теле). Есть также признаки, сближающие эту группу с Placodermi, что дает основание рассматривать акантодий как ветвь Placodermi, которые являются тогда и непосредственными предками хрящевых и костных рыб.
Примитивные костные рыбы дали начало двум ветвям (см. рис. 1), которые в настоящее время рассматриваются как два самостоятельных подкласса: мясистолопастные (Sarcopterygii) и лу-190
чеперые (Actinopterygii). Первые — примитивная группа, объединяющая два надотряда: кистеперых (Crossopterygii) и двоякодышащих (Dipnoi). Первый надотряд в ископаемом состоянии известен с девона, а в настоящее время представлен только одним глубоководным видом — Latimeria chalumnae. Второй — также известен с девона, а в современной фауне представлен несколькими специализированными тропическими видами.
К подклассу лучеперых практически относится все огромное количество современных костных рыб. В основание этого ствола можно поставить известный только в ископаемом состоянии с девона по нижний мел (см. рис. 1) надотряд Palaeoniscoidei, представители которого отличаются рядом черт, свойственных и другим надотрядам лучеперых (Holostei, Chondrostei, Teleostei). От него берет начало надотряд костных ганоидов (Holostei), который, в свою очередь, является родоначальным (см. рис. 1) для прогрессивного, наиболее богатого видами надотряда костистых рыб (Teleostei).
Родственные связи двух других надотрядов подкласса лучеперых: хрящевых ганоидов (Chondrostei) и многоперов (Brachiopte-rygii) — не совсем ясны. Учитывая черты их сходства с некоторыми ископаемыми группами Palaeoniscoidei, а также другие морфологические особенности, их можно включить в подкласс лучеперых. Это, вероятно, вполне справедливо в отношении хрящевых ганоидов, хотя непосредственные связи этой группы с предковыми формами неизвестны.
Brachiopterygii, включающие один отряд — многоперых (Ро-lypteriformes), представлены в настоящее время 10 видами, а в ископаемом состоянии известны только с третичного периода, что затрудняет установление их родственных связей. Наряду с признаками, сближающими их с ганоидами (ганоидная чешуя, спиральный клапан в кишечнике и т. д.), имеются также признаки, свойственные мясистолопастным (грудные плавники имеют в основании мясистую лопасть, поддерживаемую, правда, разросшимися радиа-лиями, имеется «легкое», кровь в которое поступает из спинной аорты, имеется задняя полая вена). Все последние признаки можно объяснить близостью древних палеонисцид к древним мясистолопастным и, предполагая происхождение многоперых от примитивных палеонисцид, отнести их к подклассу лучеперых. В то же время совокупность своеобразных признаков позволяет выделить многоперов в самостоятельный подкласс, равноценный подклассам лучеперых и мясистолопастных.
ПОДКЛАСС ЛУЧЕПЕРЫЕ —ACTINOPTERYGII
Надотряд хрящевые гаиоиды — Chondrostei
Хрящевые ганоиды, хотя и известны в ископаемом состоянии только с триаса (рис. 1), по-видимому, являются наиболее примитивной группой из всех известных костных рыб. В некоторых отно-
шениях они обнаруживают значительное сходство с древними па-леонисцидами. Для ископаемых хрящевых ганоидов характерно наличие наружного костного панциря из настоящих ганоидных че-шуй. У современных форм он редуцируется и заменяется крупными (жучки) и мелкими костными чешуями. Имеется только небольшой участок по бокам верхней лопасти хвоста, покрытый ромбовидными чешуями, похожими на ганоидные.
Современные хрящевые ганоиды составляют отряд осетрооб-разных — Acipenseriformes, известный с юры и включающий два семейства: осетровые — Acipenseridae и веслоносы — Polyodonti-dae. В последнее семейство входят роды Polyodon (в Америке) и Pseuphurus (в Китае).
Семейство осетровые встречается только в северном полушарии и наиболее многообразно (тремя родами) представлено в Советском Союзе:
1) род осетры Acipenser Linne включает 15 видов, из которых в СССР встречаются следующие: русский осетр—Ac. guldenstadti, севрюга — Ac. stellatus, стерлядь — Ac. ruthenus, сибирский осетр — Ac. baeri, шип — Ac. nudiventris;
2) род белуги Huso Brandt с двумя видами: белуга — Н. huso и калуга — Н. daurica-,
3) род лжелопатоносы Pseudoscaphirhynchus A. Nikolski.
Большинство осетровых рыб — проходные морские формы, входящие в реки для икрометания. Из пресноводных можно указать виды: стерлядь, сибирский осетр, амурский осетр, калуга.
Характеристика представителя осетровых рыб — стерляди (Acipenser ruthenus L.)
Для изучения организации осетровых рыб взята стерлядь как наиболее распространенный вид осетровых рыб в СССР. Стерлядь населяет реки бассейнов Черного, Каспийского и Балтийского морей, а также встречается в реках Обь и Енисей. Стерляди — пресноводные рыбы, редко заходящие в море. Питаются главным образом личинками насекомых, собирая их своим выдвижным ртом на дне водоемов. Стерлядь — самый мелкий представитель осетровых рыб, максимальный размер их до 80 см. Половозрелыми становятся в 4—5 лет. Как и другие осетровые рыбы, стерляди имеют в СССР промысловое значение, однако значительно меньшее, чем осетры и севрюги.
Вскрытие
Внешний осмотр
Тело стерляди, как и других осетровых рыб, имеет типичную веретенообразную форму и делится на голову, туловище и хвост. Голова покрыта костным панцирем покровных окостенений, ее задней границей служит задний край жаберной крышки. Туловище про-192
должается до заднепроходного отверстия. Осетровые рыбы отличаются от других костных рыб наличием пяти продольных рядов костных жучек, играющих защитную функцию, и особенно крупных у мальков. На спинной стороне располагается непарный ряд жучек ромбической формы с продольным шипом в середине. На уровне боковой линии тела с обеих сторон проходит верхний ряд боковых жучек, 'предохраняющих лежащие под ними кожные органы чувств боковой линии. Между грудными и брюшными плавниками расположен брюшной ряд жучек, который впереди заканчивается крупными чешуями, покрывающими вентральные концы плечевого пояса и соответствующих ключицам. Разные виды осетровых рыб имеют различное количество жучек, число их является важным си--стематическим признаком..
Кроме пяти рядов жучек, туловище осетровых рыб покрыто мелкими костными чешуйками, щитками, имеющими различную форму у разных родов и видов. У стерляди эти щитки имеют форму острых конических чешуек, часто сросшихся группами по три зубчика. У севрюги такие же зубчики срастаются в группы рядами в виде гребешков с центральным зубцом 'посередине. У осетра щитки образованы зубчиками, сросшимися в розетки с радиальным расположением. У белуги — это отдельные костные зубчики.
На хвостовом стебле вместо этих костных щитков хорошо видны правильно расположенные косые ряды ромбических чешуй, соприкасающихся друг с другом. По своему расположению они соответствуют ганоидным чешуям настоящих ганоидных рыб. Наличие таких ромбических чешуй на хвостовом стебле определяет принадлежность осетровых рыб к надотряду хрящевых ганоидов.
Голова стерляди имеет форму конуса, направленного заостренным концом вперед и покрыта на спинной стороне костным панцирем из кожных костей; передний ее конец образует вытянутое рыло, как у акуловых рыб.
Щупальца (cirri, рис. 62, err.). На нижней стороне рыла, приблизительно на половине расстояния до его переднего конца, располагаются четыре нитевидных щупальца, играющие роль осязательных органов при поиске пищи.
Рыло (rostrum, рис. 62, rst.) у стерляди имеет форму острого конуса, уплощенного с брюшной стороны. На верхней и нижней сторонах рыла видны отверстия каналов боковой линии. У осетра рыло тоже коническое, но тупое; сходной формы рыло у шипа и белуги; форму широкой лопаты имеет рыло лжелопатоносов.
Рот (stoma, рис. 62, st.) расположен на брюшной стороне головы и имеет вид поперечной щели в закрытом состоянии или округлого отверстия овальной формы — в открытом. Он окружен мясистыми губами в виде парных валиков на верхней и нижней че-"юстях; на нижней челюсти мясистые лопасти губ раздвинуты и не скопятся друг с другом. Впереди верхней челюсти видна глубокая Щель, образованная складкой кожи выдвижного челюстного аппарата. Если потянуть за верхнюю челюсть, то выдвигается ротона я воронка вместе с челюстным аппаратом; она служит для
схватывания добычи со дна: стерлядь — типичная бентосоядная рыба.
Ноздри (naris, рис. 62, п.) располагаются па боковой поверхности головы. С каждой стороны имеется по два отверстия: переднее округлое, помещается на верхнем копне кожной складки, заднее щелевидное — позади, перед глазом. Вода входит в переднее отверстие и выходит через заднее, омывая складки обонятельного мешка.
Глаза (oculus, рис. 62, ос.) помещаются по бокам головы позади обонятельных мешков. Они овальной формы и окружены кожной складкой, допускающей повороты глаз в глазнице; зрачок круг-
Рис. 62. Общин вид стерляди Acipenser ruthenus:
б. ж. — боковой ряд «жучек»; бр. ж. — брюшной ряд «жучек»; сп. ж. — спинные «жучки»; err. — усики; f. br. е. — наружное жаберное отверстие; «. — носовые отверстия; псп,— первая увеличенная чешуя спинного ряда; ос. —глаз; ор.— жаберная крышка; рпп. а.— анальный плавник; рпп. ab. — брюшной плавник; рпп. с. — хвостовой плавник; рпп. d.—спинной плавник; рпп. th. — грудной плавник; rst — рострум;
sp. — брызгальце; st. — рот
лый. Величина глаз у различных представителей осетровых рыб различная; у стерляди диаметр глаза составляет 2,2% к длине тела; у осетра и шипа — 1,9; у севрюги и белуги—1,5; у скафирии-ха —0,8%.
Жаберная крышка (operculum, рис. 62, ор.) образует боковые поверхности головы и простирается назад до переднего края костного плечевого пояса грудных плавников как сбоку, так и снизу. Это мягкая, кожистая складка, которая поддерживается костными чешуями и покровными костями кожного происхождения.
Наружное жаберное отверстие (рис. 62, f. br. е.) помещается в углублении, образованном позади жаберной крышки поясом грудных плавников. На брюшной стороне правое и левое жаберные отверстия сходятся и отделяются друг от друга узким перешейком (isthmus), в который простирается вперед поджаберная мускулатура.
Брызгальце (spiraculum, рис. 62, sp.)—рудиментарное дыхальце— у осетровых рыб помещается по верхнему краю кожистой жаберной крышки в виде маленького отверстия, сообщающего жаберную полость с наружной средой.
Грудные плавники (pinna thoracalis, рис. 62, рпп. th.) прикреплены к мощному плечевому поясу, лежащему непосредственно позади жаберного отверстия. С наружной стороны хрящевой пояс грудных плавников одет кожными покровными костями, являющимися передними членами боковых рядов костных жучек (б. ж.); верхние кости принадлежат боковому ряду жучек, нижние — брюшному. Сами грудные плавники имеют форму треугольных пластинок, прикрепленных к туловищу почти в горизонтальной плоскости, передний край их несколько приподнят. На переднем крае плавников лежит мощный костный шип, поддерживающий плавник при опоре на дно. Кожная лопасть плавников укрепляется кожными костными лучами — л е п и д о т р и х и я м и, состоящими из мелких костных чешуек, слившихся в ряды плавниковых лучей.
Брюшные плавники (pinna abdominalis, рис. 62, рпп. v.) также лежат горизонтально по бокам от анального отверстия. Они имеют ромбическую форму и поддерживаются, как и грудные, рядами кожных лучей — лепидотрихий.
Основания грудных и брюшных плавников покрыты кожей и в них располагаются мускулатура и хрящевые лучи.
Спинной плавник (pinna dorsalis, рис. 62, рпп. d.) расположен непосредственно позади дорзального ряда жучек. Он имеет килеобразную форму и вместе с анальным плавником поддерживает тело в вертикальном положении. В основании плавника находятся хрящевые радиальные лучи, по бокам их располагаются мускульные пучки.
Фулькры. Позади спинного плавника начинается ряд сводообразных костных щитков, называемых фулькрами (fulcra, рис. 76, /.), которые образуют дорзальный киль хвостового плавника и постепенно переходят в костные лучи — лепидотрихии, поддерживающие кожную лопасть хвостового плавника.
Анальный плавник (pinna analis, рис. 62, рпп. а.) помещается почти на одной вертикали с дорзальным, позади анального отверстия. Он тоже начинается отдельными костными щитками, переходящими в ряд кожных лучей — лепидотрихий, передние из которых образуют также сводообразные пластинки — фулькры.
Хвостовой плавник (pinna caudalis, рис, 62, рпп. с.) осетровых рыб имеет типичную форму неравнолопастного гетероцеркального плавника: ось тела с хордой и хвостовой мускулатурой продолжается только в верхнюю лопасть хвостового стебля, а нижняя лопасть образована лишь удлиненными кожными костными лучами — лепидотрихиями.
Анальное и половое отверстия. Между брюшными плавниками располагаются одно за другим анальное (anus, рис. 63, ап.) и половое (foramen genitale, рис. 63, f. g.) отверстия, оба они лежат в общей щели. Раздвинув щель, можно видеть также еще два маленьких отверстия по бокам — это абдоминальные поры (porus abdominalis, рис. 63, рог. а.), сообщающие полость тела с наружной средой.
Общая топография внутренних органов (рис. 63)
Стерлядь кладут в ванночку брюхом кверху, делают скальпелем небольшой поперечный разрез брюшной стенки тела непосредственно за задним краем пояса передних 'конечностей. Затем вставляют
Рис. 63. Общая топография внутренних органов стерляди Acipenser ruthenus: an. —• анальное отверстие; сап. g. — половой проток: coel. — брюшная полость; сог. — сердце; Л. сНЫ.—желчный проток; duod. — двенадцатиперстная кишка; f. g.—половое отверстие; g. card. — кардиальный отдел желудка; g. pyl. — пилорический отдел желудка; gl. pyl.— пилорическая железа; h. — печень; I.— селезенка; pre. — перикардий; рпег. — поджелудочная железа; рог. а. — абдоминальные поры; rd. — прямая кишка; test. — семенники;
/рй.— спиральный клапан; ves. f.—желчный пузырь; oes. рп. — плавательный пузырь
Рис. 64. Общин вид органов пищеварения стерляди Acipenser ruthenus:
еЫ.—спиральная кишка: d. chol. — желчный проток; duod. — двенадцатиперстная кишка; g. card. — кардиальный отдел желудка; g. pyl. - пилорический отдел желудка; gl. pyl. — пилорическая железа; f. рп. — отверстие плавательного пузыря; h. — печень; /.—селезенка; pfer. — поджелудочная железа; rd. — прямая кишка; oes. — пищевод; ves. /.—желчный пузырь; ves. рп. — плавательный пузырь
острие ножниц в отверстие и разрезают брюшную стейку по средней линии вплоть до анального отверстия. Перед анальным отверстием, между брюшными плавниками на брюхе расположены хря-?&5
щевые пластинки тазового пояса. Проводят разрез по средней линии между правым и левым элементом тазового пояса. Далее делают разрез стенки тела, чтобы вскрыть околосердечную полость. Для этого следует провести разрез через треугольник, занятый поджаберной мускулатурой, основанием которого служит плечевой иояс, а вершиной — узкий перешеек между жабрами правой и левой сторон. Вскрывать околосердечную полость необходимо осторожно, чтобы не повредить сердца, разрез следует проводить между правым и левым поясом грудных плавников, между двумя костными ключицами.
Отворачивают в стороны боковые стенки полости тела и рассматривают общее расположение внутренних органов.
Околосердечная сумка (pre.)—перикардиальная полость с лежащим в ней сердцем (сог.) — размещается ближе к голове, в треугольнике между ключицами. Перикардиальная полость отделяется от лежащей сзади брюшной полости тела поперечной перегородкой брюшины.
Брюшная 'полость тела — целом (coel.) простирается от перикардиальной полости назад до анального отверстия, в пей помещаются все внутренние органы. Стенки тела выстланы с внутренней стороны слоем брюшины.
Печень (h.), или, вернее, сложная пищеварительная железа, занимает переднюю область брюшной полости и представляет собой многодольчатую железу, охватывающую желудок спереди и по бокам.
Желудок (g. card., g. pyl.) прикрыт с боков печенью так, что видна только его задняя часть — толстостенный пилорический отдел. На месте перехода желудка в кишечную трубку помещается дольчатая пилорическая железа (gl. pyl.), имеющая бобовидную форму.
Кишечная трубка спускается вниз до середины брюшной полости тела и образует нисходящую петлю кишечной трубки. Загибаясь кверху до нижнего края правой доли печени, опа снова образует петлю и поворачивает назад, где переходит вениральную кишку (рис. 64, col.), через стенки которой просвечивают косо расположенные складки спирального клапана (tph.). На заднем конце кишечная трубка открывается самостоятельным анальным отверстием.
Селезенка (/.). На вскрытой рыбе имеет V-образную форму и лежит справа и слева от нисходящей петли кишечника.
Яичники (ov.) —парные половые железы самки — заполняют боковые отделы брюшной полости, закрывая собой все остальные органы. Около яичника видны скопления жировой ткани (рис. 65).
Семенники (lest.) — парные дольчатые органы самца — занимают боковые отделы целома справа и слева от пищеварительной системы. Семенники начинаются впереди, немного отступая от передней границы печени, а простираются назад почти до заднего конца полости тела.
Плавательный пузырь занимает центральную часть туловища на спинной стороне брюшной полости тела. При рассмотрении общей топографии внутренних органов стерляди он не виден; только приподняв переднюю петлю кишечника, можно увидеть плавательный пузырь.
П очки и их выводные протоки располагаются в задней части
А 5
Рис. 65. Половые органы самца и самки стерляди Acipenser ruthenus (по Машковцеву):
can. ur.-g.—мочеполовой капал; test. — семенник; tub. obd. — воронка яйцевода;
ov. — яичник; oud. — яйцевод
брюшной полости на спинной стороне целома. Задние концы их видны при оттягивании заднего отдела спиральной кишки. Рассмотрение их следует отложить до изучения мочеполовой системы.
Пищеварительная система стерляди
Пищеварительная система осетровых рыб по своему строению занимает промежуточное положение между хрящевыми и костистыми рыбами. Прежде всего для всех осетровых рыб характерен выдвижной челюстной аппарат в виде ротовой воронки, способной далеко выдвигаться наружу при собирании пищи со дна. Он имеет своеобразное строение и разбирается при описании висцерального скелета. Для взрослых рыб характерно отсутствие зубов, имеющихся лишь у личинок и замещающихся у взрослых форм роговыми валиками.
Пищеварительный тракт характеризуется присутствием двух приспособлений, увеличивающих площадь всасывающей
поверхности пищеварительного эпителия: с одной стороны, кишечник образует систему петель, как у костных рыб, с другой — область средней кишки сохраняет типичный спиральный клапан, свойственный хрящевым рыбам. Клоака отсутствует и имеются отдель
ные заднепроходное и мочеполовое отверстия.
Как и у всех позвоночных, пищеварительная система разделяется на переднюю кишку, к которой относятся ротоглоточная полость, пищевод и желудок, среднюю кишку, образующую петлю двенадцатиперстной кишки и нисходящий отдел спиральной
кишки, и заднюю кишку, открывающуюся анальным отверстием.
Рассматривают сначала на вскрытой стерляди расположение органов пищеварения в брюшной полости тела, отложив анатомирование ротоглоточной полости до изучения органов дыхания. Прежде всего разбирают расположение органов пищеварения без их препаровки инструментами, не разрывая соединяющей их брыжейки, а лишь отодвигая вправо и влево органы и стараясь рассмотреть со всех сторон их форму и отдельные части (см. рис. 64).
Печень (hepar, см. рис. 63, 64, А.) занимает всю переднюю область полости тела позади соединительнотканной перегородки, отделяющей целом от околосердечной сумки. Эта железа имеет неправильную форму. Заполняя все свободные места полости, она охватывает пищеварительные органы спереди, по бокам и со спинной стороны. Соответственно этому наружная поверхность печени выпуклая, а внутренняя — вогнутая. Она может быть расчленена на три доли. Правая доля печени (со стороны животного), выпуклая с наружной стороны и вогнутая с внутренней, охватывает округлый пилорический отдел желудка и спускается вниз до петли двенадцатиперстной кишки. На ее переднем крыле помещается овальный желчный пузырь (vesica fellea, см. рис. 63, 64, ves. f.). Желчный пузырь с брюшной стороны и опереди прикрыт маленькой центральной долей печени. От желчного пузыря идет желчный проток (ductus choledochus, см. рис. 63, 64, d. chol.), который огибает пилорический отдел желудка слева и впадает в двенадцатиперстную кишку у основания пилорической железы. Левая доля печени еще более длинная, чем правая. Она начинается от места отхождения центральной доли и раополагастся на дорзальной стороне тела. Чтобы ее рассмотреть, надо отвернуть желудок налево и тогда будет видно, что левая доля печени подстилает желудок до нижнего края его петли и даже проникает через петлю желудка па брюшную поверхность перед пилорическим выростом.
Отворачивают желудок палево и рассматривают его начало и соединение с пищеводом.
Пищевод (oesophagus, см. рис. 64, oes) является непосредственным продолжением ротоглоточной полости, которая суживается з трубку, лежащую на дорзальной стороне околосердечной полости, и продолжается далее назад через перегородку околосердечной сумки в брюшную полость. Начало пищевода можно увидеть, отвернув желудок и печень. Он располагается около позвоночника, простираясь назад, постепенно расширяется и переходит в желудок. Чтобы рассмотреть границу между желудком и пищеводом и структуру внутренней поверхности пищевода, рекомендуется вскрыть продольным разрезом пищевод и начало желудка до места впадения плавательного пузыря. Внутренняя эпителиальная выстилка пищевода отличается присутствием эпителиальных ворсинок в виде листовидных сосочков, расположенных продольными рядами и разветвляющихся па вершинах на два-три дополнитель-
ных сосочка. При переходе в желудок эти сосочки постепенно изменяются в продольные волнообразные складки, идущие по всему кардиальному отделу желудка 6—7 рядами.
Плавательный пузырь (vesica pneumatica, s. natatoria, см. рис. 63, 64, ves. рп.) представляет собой полый мешок с плотными волокнистыми стенками. Он занимает всю дорзальную область брюшной полости и плотно срастается верхней стенкой с брюшиной. У всех осетровых рыб плавательный пузырь сообщается с пищеварительной трубкой широким каналом. Развернув края продольного разреза пищевода и желудка, можно видеть, что у стерляди, как и у всех осетровых, место отхождения плавательного пузыря с окружающим его мускульным сфииктором лежит не в области листовидных сосочков пищевода, а в желудке между его продольными складками.
Желудок (gaster, рис. 63, 64) у осетровых рыб состоит из двух отделов. Кардиальный отдел желудка (g. card.) начинается в области отхождения плавательного пузыря, тянется вниз до половины брюшной полости, далее загибается петлей к наружной стороне полости тела, поднимается кверху до передней стенки целома и переходит в пилорический отдел желудка. Таким образом, весь желудок имеет форму рукописной буквы «р», где началом желудка будет правый изгиб, а концом (пилорический отдел)— левый изгиб. Пилорический отдел (g. pyl.) — привратник (pylorus)—имеет форму лимона сочень плотными утолщенными стенками. Он располагается в поперечном направлении в выемке между правой и левой долями печени, загибается налево (на правую сторону рыбы), где снова суживается и переходит в среднюю кишку.
Вскройте дальше продольным разрезом стенку желудка и рассмотрите внутреннюю структуру ее. Стенки желудка покрыты продольными складками в 6—7 рядов, которые из кардиального отдела переходят непосредственно в пилорический. В пилорическом отделе, имеющем толстые мясистые стенки, складки эти выражены сильнее. На месте перехода желудка в кишечник образуется мощный мускульный сфинктор, которым он соединяется с передним отделом средней кишки, называемым двенадцатиперстной кишкой.
Пилорическая железа (glandula pylorica, см. рис. 63 и 64, gl. pyl.)—бобовидный орган, лежащий на границе пилорического отдела желудка и начала средней кишки. Она имеет структуру мпогодольчатой железы с дольками, располагающимися веерообразно. Можно думать, что пилорическая железа осетровых рыб является результатом слияния в один орган многочисленных отдельных пилорических выростов, часто встречаемых у других лучепе-рых рыб.
Двенадцатиперстная кишка (duodenum, см. рис. 63 и 64, gl. pyl.)—передний отдел средней кишки. Она начинается около пилорической железы, идет вниз до середины полости тела и, загибаясь вверх, дает восходящую петлю, которая поднимается до уровня нижнего края правой доли печени или даже до пилориче-200
ской железы, снова поворачивает налево и переходит в расширенную часть — спиральную кишку.
Вскрывают продольным разрезом стенку кишечника, рассматривают строение внутренних стенок. Они покрыты слизистой оболочкой, имеющей своеобразную структуру, из-за наличия углубленных ячеек. Отверстие пилорической железы лежит рядом со сфинктором пилорического желудка. Тут же впадает желчный проток (ductus choledochus, d. chol.) желчного пузыря.
Селезенка (lien, рис. 63 и 64, Z.) у осетровых рыб — большой железистый орган V-образной формы, огибающий справа и слева петлю двенадцатиперстной кишки и подстилающий ее, что можно видеть, приподняв кишку. Начинается селезенка отдельной долей, примыкающей к пилорической железе, и далее переходит в удлиненную долю, которая образует внизу выступ в форме клина. Затем она поднимается по восходящей петле до края правой доли печени, где уже с правой стороны, под нижней долей печени, образует отдельную треугольную долю, называемую добавочной долей селезенки. Эта добавочная доля прилегает непосредственно к печени и поджелудочной железе, так что границу между ними можно определить лишь по цвету желез. У осетра добавочная доля достигает значительных размеров. Если отвернуть петлю двенадцатиперстной кишки, то можно увидеть, что селезенка, располагающаяся под ней, имеет Z-образную форму и подстилает кишку справа и слева.
Поджелудочная железа (pancreas, рис. 63, 64, рпсг.) у осетровых рыб вместе с жировыми скоплениями располагается вдоль сосудов, питающих двенадцатиперстную кишку, и находится в тесном контакте с лопастями правой доли печени и дополнительной долей селезенки. Поэтому нередко печень осетровых рыб называют hepatopancreas. Однако (особенно у крупных осетров) можно различить отдельные лопасти поджелудочной железы. Опи располагаются на месте перехода пилорического отдела желудка в двенадцатиперстную кишку и тянутся двумя продольными лопастями: одна лежит вдоль наружной стенки вместе с подкишечной веной (v. intestinalis), другая — по внутреннему краю вдоль кишечной артерии (a. mesenterica). Обе лопасти подходят к центральной доле печени.
Спиральная кишка (colon, рис. 64, col.)—задний отдел средней кишки, нисходящий отдел кишечника — является непосредственным продолжением вниз двенадцатиперстной кишки. Этот отдел кишечника имеет толстые мускулистые стенки и даже с наружной стороны его видны внутрилежащие складки спирального клапана. Спиральный клапан образуется скрученностью кишечной трубки и созданием внутри нее закрученной высокой складки слизистой оболочки, свисающей в полость, что можно видеть па поперечном разрезе, Всего имеется 7—8 таких витков. На вскрытой кишечной трубке создается впечатление, что она разделена на 7—8 отдельных камер, сообщающихся друг с другом. Помимо спирального клапана, на внутренних стенках средней кишки находят-201
ся мелкие складки слизистой оболочки, которые в конце переходят в гладкие стенки задней кишки.
Задняя кишка (rectum, рис. 63, 64, ret.)—прямой задний отдел кишечника — оканчивается анальным отверстием. У осетровых рыб нет слепых отростков па границе средней и задней кишки, свойственных другим позвоночным.
Анальное отверстие (anus, см. рис. 63, ап.)—переднее из четырех отверстий анальной зоны, расположенное непосредственно позади пояса брюшных плавников.
Брыжейки. Кишечный тракт в брюшной полости и окружающие его органы соединены со стенками тела и друг с другом мезентериями. Пищевод до места отхождения плавательного пузыря соединен с позвоночником дорзальным мезентерием. Кардиальный и пилорический отделы желудка спаяны брыжейками с долями печени. Селезенка прикреплена брыжейками к кардиальному отделу желудка и далее к двенадцатиперстной кишке. От добавочной доли селезенки и двенадцатиперстной кишки идут мезентериальные спайки к печени, вдоль которых располагаются дольки поджелудочной железы. Спиральная кишка и прямая кишка имеют дорзальный мезентерий, связывающий их сначала с плавательным пузырем, а позади него — с позвоночником. Наконец, гонады и выводные каналы мочеполовой системы своими брыжейками (meso-varium. mesotestis) прикреплены к дорзальной стенке полости тела.
Мочеполовая система
Строение мочеполовой системы осетровых рыб занимает промежуточное положение между хрящевыми и лучеперыми рыбами. Для всех осетровых рыб характерно наружное оплодотворение и откладка большого количества икры, а также отсутствие клоаки. Оба эти признака связывают осетровых с костистыми и другими лучеперыми рыбами. С другой стороны, у них имеются яйцеводы, открывающиеся воронками в полость тела, что объединяет осетровых с хрящевыми, двоякодышащими рыбами и наземными позвоночными.
Яичники (ovarium, рис. 65, ov.) — громадные парные гонады самки — занимают боковые части полости тела почти от самого перикардия до заднего конца целома. Они прикреплены к дорзальной стенке тела брыжейками на всем протяжении справа и слева от плавательного пузыря. Яичник имеет зернистое строение, он состоит из яйцеклеток одинаковой величины и одинаковой степени развития, так как икрометание происходит одновременно.
Яйцеводы (oviductus, рис. 65, ovd.)—особые мезонефрические выводные каналы — лежат на наружной стороне гонад в виде широких трубок, открывающихся как у самок, так и у самцов в полость тела широкими воронками на уровне нижней половины гонады. Они соответствуют яйцеводам — мюллеровым каналам — хрящевых и двоякодышащих рыб, но имеют .особое происхождение от первичного почечного выводного канала, к которому присоеди-202
няется мезонефрическая воронка. Правый и левый яйцеводы на заднем конце сливаются друг с другом и открываются общим отверстием позади анального отверстия.
Семенники (testis, рис. 65 и 66, test.) — парные половые же
лезы самцов — занимают, как
и яичники, боковые отделы полости тела. Они имеют дольчатое строение и сверху, при рассмотрении вскрытой рыбы, прикрыты большими скоплениями жировых тел. Как и у самок, на наружной стороне железы располагаются воронки яйцеводов, соединяющиеся с широким тонкостенным половым протоком с правой и левой сторон. Внизу парные половые протоки сливаются вместе и открываются половым' отверстием, лежащим непосредственно позади анального отверстия.
Мочеточниками (ureter) и семяпроводами (vas deferens) служат первичные почечные протоки. Они начинаются у переднего края почки с наружной стороны отдельными почечными канальцами и образуют общий проток, к которому на уровне заднего конца плавательного пузыря присоединяется воронка яйцевода, образованная у осетровых рыб не мюллсровыми каналами, а мезонефрическим каналом. Через эту воронку и выводной капал выводится наружу целомпая жидкость.
При вскрытии общего мочеполового канала видно, как в
ирг.
v.sp.s.
and
test.
и. crd.
— tub.oad.
can. ur.-g.
-an.
can g u.sp.lnf.
Pnc. 66. Почки и выводные протоки стерляди Acipenser ruthenus и их кровоснабжение:
пч. —анальное отверстие; ао. d. —спинная аорта; can. g. — половой проток; сап. ur-g. мочеполовой канал; h g. половое огвсрстие; г. — почки; test. — семенник; tub. ovd. — воронка яйцевода; v. crd. — кардинальная вена: v. sp. inf- — нижняя вена семенника; и. sp. s. — верхняя вена семен-
ника
нем открываются отверстия многочисленных нефридиальных мочевых канальцев.
Семявыносящие канальцы (vas efferens). От семенника, лежащего кнаружи от полового протока, в брыжейках, на которых подвешен семенник (рис. 66, test.), при наблюдении под бинокулярной лупой видны тоненькие, прозрачные выносящие не-фридиальные трубочки, идущие вглубь, в почку. Они служат семявыносящими канальцами. Впереди воронки по направлению к
верхней доле семенника можно видеть целый букет прозрачных семевыносящих канальцев. Они приносят семя из семенника и общим пучком впадают на уровне воронки в общий семепровод. Почечные нефридиальиые канальцы тонкими трубочками пронизывают почку и впадают в общий проток выводного канала.
Почки располагаются в заднем отделе туловища на дорзальной стенке брюшной полости тела и прикрыты выстилкой брюшины. Для того чтобы их отпрепарировать, необходимо удалить яичники или семенники, подрезав спереди назад брыжейку, прикрепляющую их к стенкам тела. Затем промывают струей воды полость тела и постепенно отчленяют яйцеводы и перитонеальную выстилку полости тела. Отпрепарируют также плавательный пузырь и отодвигают его в сторону. Далее вскрывают ножницами задний отдел брюшной полости рядом с анальным и половым отверстиями и отпрепарируют задние отделы целома, отодвигая в стороны мускулатуру стенок тела.
Почки (геп, рис. 66, г.) имеют форму парных лентовидных железистых органов, слившихся друг с другом позади плавательного пузыря по средней липин тела. Они пронизаны кровеносными сосудами, образующими воротную систему почек. Дойдя до заднего конца 'плавательного пузыря, почки раздваиваются и простираются вперед, оплетаемые разветвлениями задних кардиальных вен, лежащих ковнутри от почек. Передний конец лентовидных почек доходит до уровня петли желудка на месте перехода кардиального отдела в пилорический.
Окончив таким образом рассмотрение внутренних органов, расположенных в брюшной полости, переходят к вскрытию ротоглоточной полости для изучения ротовой полости и жаберного аппарата.
Ротоглоточная полость (рис. 67)
Для того чтобы вскрыть ротоглоточную полость и рассмотреть передний отдел пищеварительной системы и жаберный аппарат, необходимо отчленить жаберную крышку в месте ее прикрепления к голове, по краю костного панциря, покрывающего голову сверху, предварительно ознакомившись с ее строением.
Жаберная крышка (operculum) называется оперкулярной крышкой, так как она закрывает все жабры. Изучают ее строение. Это — производная подъязычной жаберной перегородки, лежащей между спиракулярной и первой жаберной щелями. Если сравнить ее с другими жаберными дугами, то становится ясным, что жаберная крышка не что иное, как разросшаяся жаберная перегородка подъязычной дуги. На внутренней стенке жаберной крышки прикреплены жаберные лепестки оперкулярной жабры. Жаберная крышка поддерживается покровными костями и кожными костными чешуями, задний ее край имеет кожистую перепонку, которая служит клапаном всасывающего насоса в дыхательном акте. Жаберные перепонки на вентральной стороне г^овы с двух сторон ent
прирастают к межжаберному промежутку — истмусу, у белуг вин срастаются друг с другом и образуют общую складку. Позади жаберной крышки помещаются наружные жаберные отверстия, окаймленные сзади передним краем пояса конечностей грудных
плавников.
Вставляют лезвие ножниц в жаберное отверстие около костного панциря и проводят разрез сзади наперед. На середине рас
стояния до глаза можно увидеть брызгальце (spiraculum, sp.). Проводят разрез через брызгальце дальше вперед и вниз по краю окостенений глазного кольца и вниз на брюшную сторону головы. Затем удаляют с обеих сторон костный и хрящевой пояса грудных плавников. Взяв грудной плавник левой рукой и оттянув его, отчленяют от головы, отрезают мускулатуру грудного пояса, отделяют его от стенки брюшной полости и затем твердыми ножницами перерезают окостенения плечевого пояса посредине. После этого осторожно отделяют вентральные отделы плечевого пояса вместе с плавниками. Да-
Рис. 67. Жаберная область стерляди Aclpen-ser ruthenus. А — разрез ротоглоточной области, вид на небную сторону; Б — жаберная дуга с жаберными лепестками и тычинками:
лее, вставляют лезвие ножниц в ротовое отверстие и проводят разрез справа и слева так, чтобы он проходил между дор-
ж. л. — жаберные лепестки; ж. т. — жаберные тычинки; arc. br.x — arc. Ьг.4 — разрезы жаберных дуг с жаберными лепестками от первой до четвертой; Ьг. ор.—оперкулярная жабра; err.— усики; hmd. — гиомапдибуляре; «I. — мускулатура; md. — челюстной хрящ; pal. v. — небные валика;
s. — брызгальце; st. — ротовое отверстие
зальным и вентральным
отделами жаберных дуг. Оттягивая отрезанные дуги, постепенно отчленяют вентральную стенку жаберного аппарата от его дорзальной стенки по линии разреза. После этого рассматривают сначала ротоглоточную полость, а затем и строение жаберного ап-
парата.
Ротовое отверстие (stoma, st.) помещается на вентральной стороне головы и имеет форму овального отверстия, расположенного в поперечном направлении; оно окаймлено сверху и снизу мясистыми губами, поддерживаемыми покровными окостенениями.
Ротоглоточная полость (cavum oropharyngeus) резкой границей разделена в поперечном направлении на две части: пе
реднюю— ротовую полость — и заднюю — жаберную полость. Первая образует выдвижной челюстной аппарат. Взявшись рукой за верхнюю челюсть, можно вытянуть ротовую воронку из кожного мешка, причем кожная складка вытягивается па небной пластинке.
Выдвижной челюстной аппарат — своеобразное приспособление для схватывания добычи на дне. Он может перемещаться как салазки на большой покровной кости парасфеноида (parasphenoideum), подстилающего крышку ротоглоточной полости. Вытягивающаяся часть челюстного аппарата состоит из небноквадратного хряща (см. «Скелет», стр. 243) и продолжается далее назад в виде небной пластинки (см. описание скелета висцерального аппарата, стр. 243). При оттягивании жаберного аппарата хорошо видна структура небных валиков, располагающихся в поперечном направлении в количестве 7—8 полосок на небной пластинке (pal. ц.)и 2—3 ряда — на небно-квадратном хряще.
Жаберная полость и ж а б р ы. Отворачивают за нижнюю челюсть вентральную стенку ротоглоточной полости и рассматривают строение жаберной полости и жабр. Жаберная крышка костных рыб образована жаберной перегородкой подъязычной дуги, дно глоточной полости — непарными хрящевыми элементами жаберного аппарата, боковые стенки пронизаны справа и слева четырьмя жаберными щелями, отделенными друг от друга жаберными дугами. Пользуясь лупой, рассматривают более подробно строение отдельной жаберной дуги.
Жаберная дуга. Вырезают первую жаберную дугу и рассматривают ее строение более подробно. Верхние концы жаберных дуг прикреплены к небу глоточно-жаберными элементами, а нижние срастаются с подъязычным аппаратом. На внутреннем крае каждой жаберной дуги в два ряда сидят жаберные тычинки (ж. т.), входящие друг в друга; как зубцы шестерни. Один ряд сидит на переднем крае, другой — на заднем. Каждая жаберная тычинка имеет форму конического треугольника. Тычинки выполняют функцию фильтра для удержания пищи. На наружном крае жаберной дуги прикреплены жаберные лепестки (ж. л.), расположенные также в два ряда и отделенные друг от друга ж а-берными перегородками. Жаберные перегородки не достигают края жаберных лепестков (в отличие от акуловых рыб, где жаберные перегородки простираются до краев жаберных отверстий). Каждый жаберный лепесток имеет форму листовидной перегородки, в которой помещается петля жаберных капилляров, составленная приносящим и выносящим жаберными сосудами. Жаберная дуга с двумя рядами жаберных лепестков на наружной и внутренней сторонах образует одну целую жабру рыб (holobranchia).
Передний ряд жаберных лепестков располагается на внутренней поверхности жаберной крышки, является полужаброй подъязычной дуги и получает название оперкулярной жабры
(branchia opercularis, op. br.)\ она ограничивает переднюю стенку первой жаберной щели. На задней стенке передней жаберной щели лежит второй ряд жаберных лепестков, который спаян жаберной перегородкой первой жаберной дуги со следующим рядом жаберных лепестков, являющихся передней стенкой второй жаберной щели. Второй ряд жаберных лепестков вместе с третьим рядом составляет первую жабру. Вторая, третья и четвертая жабры также состоят из двух рядов лепестков каждая. Последняя, пятая, жаберная дуга не песет жаберных лепестков, вплотную прилегает к плечевому поясу и скрыта под кожей, поэтому задняя стенка пятой жаберной щели ограничена передним краем плечевого пояса.
Акт дыхания у осетровых рыб, как и у других костных рыб, носит название оперкулярного дыхания. При оттягивании жаберной крышки в стороны в околожабернон полости между жаберной крышкой и жабрами образуется пространство с пониженным давлением, куда всасывается вода из ротоглоточной полости через жаберные отверстия. В ротовую же полость вода всасывается через рот при растяжении всего висцерального аппарата. При прижимании жаберной крышки вода выталкивается из околожабер-ной полости через наружные жаберные отверстия.
Кровеносная система
Кровеносная система осетровых рыб обладает типичными чертами кровообращения рыб. Сердце подразделено на четыре отдела (артериальный конус, желудочек, предсердие и венозный синус) п содержит венозную кровь, которая по брюшной аорте и приносящим жаберным сосудам приносит кровь к жабрам, где происходит ее окисление. Артериальная кровь собирается в выносящие жаберные сосуды и вливается в спинную аорту, снабжающую своими ветвями все органы. Основными венозными стволами, приносящими кровь к сердцу, являются передние и задние кардинальные вены. Пищеварительная система имеет обособленную венозную систему— воротную вену печени, приносящую кровь к печени, где происходит фильтрация крови через систему капилляров.
Изучение кровеносной системы надо начинать с сердца, для чего следует отпрепарировать околосердечную сумку, располагающуюся впереди брюшной полости тела, между грудными плавниками.
Перикардиальная полость, или околосердечная сумка (см. рис. 63, pre.),— обособившаяся передняя часть брюшной полости тела. Она отделена от нее соединительнотканной перегородкой, лежащей в поперечном направлении впереди печени. Перикардиальная полость имеет форму треугольника, направленного вершиной вперед, что можно было видеть при общем вскрытии. В этой треугольной полости располагается сердце рыб. Полость перикардия выстлана брюшиной (перитонеумом), состоящей из двух оболочек: наружная называется перикардием, а внут
ренняя, прилегающая к сердцу и прикрепляющая его к стенке тела,— эпикардием.
Сердце (сот, рис. 68), как указано выше, состоит из четырех отделов, обладающих способностью пульсации. Сначала рассматривают отделы сердца, а потом разбираются в его внутреннем строении на разрезе. Передним отделом является артериальный конус (conus arteriosus, con. а.) в виде короткой трубки,
Рис. 68. Сердце осетра Acipenser gilldenstadti. А — в разрезе; Б — вид сбоку:
«/Л — предсердие; соп. а. — артериальный конус; sin. v. — венозный синус; ventr. — желудочек; и. si. — полулунные клапаны
лежащей впереди мускулистого желудочка. Желудочек (ventriculus, ventr.) имеет трапециевидную форму с плоским и даже вогнутым основанием, направленным вниз, а вершиной — вперед. Так же, как и артериальный конус, он имеет толстые мускулистые стенки. Наружная поверхность желудочка и артериального конуса у всех осетровых рыб покрыта почти сплошь пузыревидными расширениями железистого характера, это типичная для осетровых лимфоидная железа, представленная отдельными дольками. Это можно видеть на фронтальном разрезе сердца (см. ниже). В глубине, под желудочком, располагается задний отдел сердца рыб — предсер дие (atrium, atr.), имеющий вид сморщенного мешка с мускулистыми стенками из густопереплетенпых волокон. Совсем в глубине околосердечной сумки предсердие общим широким отверстием сообщается с самым задним отделом сердца — в е н о з-
ним синусом (sinus venosus, sin. v.), принимающим венозную кровь со всего тела рыбы. Если сделать фронтальный разрез желудочка сердца, начиная от вершины, направленной вниз, и далее вперед через артериальный конус, станут видны толстые мускулистые стенки желудочка с радиальным расположением переплетающихся мышечных волокон. Внутренняя полость желудочка узкая, с лакунарными карманами, открывающимися в желудочек. На задней стенке этой полости имеется широкое атриовентрикулярное отверстие (ostium atriovcntricularis), открывающееся в предсердие. На границе между полостью желудочка и артериального конуса стоят 3—4 листовидные лопасти, перегражда-ющие обратное возвращение крови в желудочек. В самом артериальном конусе расположены три ряда полулунных клапанов (valvula semilunaris, v. si.): два ряда внизу, а третий ряд выше — у основания брюшной аорты.
Артериальная система (рис. 69)
Артериальная система осетровых рыб слагается, как и у других рыб, из брюшной аорты с приносящими жаберными артериями, несущими из сердца венозную кровь, и системы выносящих артерий, соединяющихся на спинной стороне тела в спинную аорту, которая снабжает артериальной кровью все органы туловища и хвоста.
Описание кровеносной системы дается по музейным препаратам, где кровеносные сосуды — как артерии, так и вены — наинъециро-ваны желатиновой массой. Инъекция артериальной системы производится через желудочек и артериальный конус, а венозной системы— через хвостовую вену. Без инъекции можно видеть только главные сосуды.
Для рассмотрения брюшной аорты надо отпрепарировать ее вперед от артериального конуса, раздвигая вдоль сосуда жаберную мускулатуру. Открыв брюшную аорту, следует осторожно очистить места отхождения корней приносящих жаберных артерий, которые проходят по жаберным дугам под слизистой оболочкой жаберной области. Выносящие жаберные сосуды лежат в крыше ротоглоточной полости под слизистой оболочкой.
Брюшная аорта (aorta ventralis, ао. v.)—короткий толстый сосуд, который идет по брюшной стенке жаберного аппарата одним общим стволом и впереди дает парные разветвления жаберных артерий первой жаберной дуги (a. branchialis advehens, ci. br. ad[). Каждая приносящая жаберная артерия первой жаберной дуги загибается вперед и дает от себя веточку, несущую кровь к оперкулярной жабре (a. branchialis hyoidea). Гиоидная артерия идет к оперкулярной жабре, где распадается на сеть капилляров жаберных лепестков, соединенных по краю этой сети продольными спайками. Остальные приносящие жаберные артерии 2, 3 и 4-й жаберных дуг отходят от продолжения брюшной аорты, вгибающейся внутрь и назад петлей.
ao.ir.
asci.
a.sp.(ou)s
cor
ao.d.
a.m inf.
a.il-l
a br. ad, abrade . a. brad, a.bn.ady con. a.
a.cl-m.
f br\ f.br, fbrL
a.c.at'.d —
[l 2.orb. nas
a.hy.
—~a.bnrf
a br. r? -----d.br.Fj
----abr.Fv
U—-ao.d.
7
К
Рис. 69. Артериальная система стерляди Acipenser ruthenus. A — сердце, брюшная аорта с приносящими жаберными артериями, спинная аорта п ее разветвления; Б — выносящие жаберные артерии и начало спинной аорты:
a. br. ad.] — a, br. ad.4 — приносящие артерии от первой до четвертой дуг; a. br. r.} — a. br. г.< —выносящие артерии; a. caud. — хвостовая артерия; a. hy — гиоидная артерия; а. с/.-ш. — желудочно-кишечная артерия; a. il-l.— артерия брюшных плавников; а. т. inf. — нижняя брыжеечная артерия; ао. d. — спинная аорта; a. orb. nas.— глазнично-носовая артерия; ао. v. — брюшная аорта; a. scl. — подключичная артерия; a. sp. (ov.) inf.—нижняя половая артерия самца (самки); a. sp. (ov.) s. — верхняя половая артерия самца (самки); соп. а. — артериальный конус; сог. — сердце; f. br.^—f. br.4— жаберные отверстия пер-
вое — четвертое
Приносящие жаберные артерии (a. branchialis ad-vehens, 1, 2, 3, 4, a. br. ad\—a. br. ad4) идут по наружному краю жаберных дуг. В каждый жаберный лепесток, наподобие зубцов гребенки, отходят капиллярные сосуды, которые, дойдя почти до вершины лепестка, поворачивают к его основанию и вливаются в выносящие жаберные артерии. Окисление крови происходит в капиллярах жаберных лепестков.
Выносящие жаберные артерии (a. branchialis ге-vehens, а. Ьг. г.) начинаются парными сосудами от каждой полужабры и соединяются друг с другом в четыре ствола, слияние которых в непарный продольный сосуд образует спинную аорту. Выносящие жаберные сосуды лежат на дне черепа и прикрыты слизистой оболочкой ротоглоточной полости. Чтобы их рассмотреть, необходимо осторожно снять слизистую оболочку с крыши ротоглоточной полости. Первая выносящая артерия (а. Ьг. и), выйдя из первой жаберной дуги, дает две ветви вперед для снабжения артериальной кровью головы. Внутренняя ветка — гиоидная артерия (a. hyoidea, a. hy.) —загибается вниз и снабжает ложножабру, а вторая ветвь — глазнично-носовая (a. orbitona-salis) — идет вперед к глазнице, отдавая ответвления к небу, к мускулатуре челюстной дуги и в глазницу. Вторая выносящая артерия идет от второй жаберной дуги и также вливается справа и слева в общий ствол спинной аорты. Третья и четвертая выносящие артерии перед впадением в спинную аорту сливаются вместе и уже общим стволом вливаются в спинную аорту.
Спинная аорта (aorta dorsalis, ао. d.)— крупный сосуд, лежащий под спинной струной по средней линии тела. После образования из выносящих жаберных артерий спинная аорта идет назад и на уровне грудных плавников разветвляется на правую и левую подключичные артерии (a. subclavia, a. scl.), снабжающие кровью грудные плавники. Далее назад, подходя к плавательному пузырю, аорта расширяется и уплощается, превращаясь в широкую сплющенную лакуну, тянущуюся между плавательным пузырем и позвоночником до заднего края плавательного пузыря, давая в обе стороны мелкие посегмептные ответвления к туловищной мускулатуре. От спинной аорты также отходят артериальные сосуды, снабжающие внутренние органы.
Желудочно-кишечная артерия (a. coeliaca-mesenle-rica, a. cl.-m.)—крупный сосуд, отходит от спинной аорты перед плавательным пузырем к органам пищеварения (печени, желудку и кишечнику).
Верхние семенные артерии (a. spermatica superior) у самцов или яичниковые артерии (a. ovarica superior) у самок также отходят от спинной аорты впереди плавательного пузыря, рядом с желудочно-кишечной артерией справа и слева от нее и разветвляются в гонадах (a. sp. s.; a. ov. $.). Задняя часть гонад снабжается кровью самостоятельным кровеносным сосудом, отходящим от спинной аорты на месте сужения аортного расширения. Эта нижняя половая артерия у самца (a. spermatica
inferior) и у самки (a. ovarica inferior) отходит к задней части полости тела и идет вперед (a. sp. inf.; a. ov. inf.).
Нижнебрыжеечная артерия (a. mesenterica inferior, а. т. inf.) также отходит от спинной аорты, но уже позади плавательного пузыря, и дает ответвления вперед и назад по задней кишке. Позади плавательного пузыря спинная аорта уже пе видна на вскрытой рыбе, так как опа прикрыта почками, которые в заднем отделе полости тела объединяются в непарный орган на спинной стороне брюшной полости тела. При анатомировании внутренних органов стерляди также нс видно, как спинная аорта переходит в хвостовой отдел тела, превращаясь в хвостовую артерию.
Подвздошно-поясничная артерия (a. ilio-lumbalis) отходит от спинной аорты в самом заднем конце брюшной полости, па месте разветвления хвостовой вены на приносящие вены почек. Она снабжает артериальной кровью брюшные плавники.
Хвостовая артерия (a. caudalis) и хвостовая вена (v. caudalis) —два непарных сосуда, проходящих под хордой. Их хорошо можно видеть на поперечном разрезе через хвостовой отдел стерляди позади полости тела. Сверху под хордой располагается хвостовая артерия, а под нею ниже — хвостовая вена.
Венозная система (рис. 70)
Для того чтобы рассмотреть венозную систему, еще до вскрытия рыбы рекомендуется заполнить кровеносные сосуды окрашенной инъекционной массой (см. методические указания об инъекции кровеносной системы, стр. 22). Можно также окрасить сосуды, наполнив их тушью. Для этого отрезают хвостовой отдел тела позади брюшных плавников и позади анального отверстия и вводят в поперечный разрез туловища в хвостовую вену иглу шприца, наполненного тушью или красящим веществом для инъекции. Введя иглу, завязывают весь хвостовой отдел ниткой и очень осторожно вводят краску из шприца. Затем рассматривают венозную систему, начиная с заднего конца полости тела.
Хвостовая вена (v. caudalis, v. caud.). Венозная система начинается в хвостовом отделе капиллярами хвостовой артерии, которые, сливаясь, составляют непарный сосуд — хвостовую вену, лежащую под хордой в гемальном канале позвоночника вместе с хвостовой артерией непосредственно под нею, как это можно видеть на поперечном разрезе через хвостовой отдел. .
Воротная вена почек (v. portae renalis, v. p. г.). Хвостовая вена, войдя в полость тела, разделяется па парные приносящие вены почек (v. renalis advehens), которые входят в задние концы почек, простирающиеся назад до самого заднего конца целома. В почках сосуды разветвляются на капилляры для фильтрации жидких продуктов распада, давая густую сеть сосудов воротной вены почек.
Задние кардинальные вены (v. cardinalis posterior, v. crd. p.) начинаются в заднем конце полости тела при вхождении 212
хвостовой вены в почки, где эта вена распадается на сеть капилляров воротной вены почек. С левой стороны рыбы из почечных капилляров постепенно составляется л е-вая кардинальная в е-н a (v. cardinalis posterior sinistra, v. crd. p. $.), которая идет по внутреннему краю почек до самого переднего конца полости тела, лишь отдавая в почку ветвления. Иначе говоря, в левой стороне у осетровых рыб,- как и костистых рыб, воротная вена не полная. У заднего конца плавательного пузыря там, где хвостовая артерия расширяется .в широкую лакуну спинной аорты, от кардинальной вены отходит крупное ответвление в правую сторону рыбы — задний отдел правой кардинальной вены (у. cardinalis posterior dext-ra, v. crd. p. d.), которая в правой почке полностью составляется из сети капилляров правой воротной системы почек. Лишь в передней половине полости тела в правой почке начинает восстанавливаться на наружной стороне почки венозный сосуд переднего отдела правой кардинальной вены. На уровне переднего конца гонады правая кардинальная вена принимает в себя верхнюю половую вену и вместе с левой кардинальной веной простирается вперед. Задние кардинальные вены простираются вперед до уровня сердца, пронизывают перегородку околосердечной сумки и проникают в перикар-
u.caud
Рис, 70. Венозная система стерляди Acipenser ruthenus:
d. с. — протоки Кювье; v. caud. — хвостовая вена; v. crd. р. d.—правая задняя кардинальная вена; v. crd. р. s.—левая задняя кардинальная вена; v. j. inf. — нижняя яремная вена; v. j. s. — верхняя яремная вена; v. h. —печеночная вена; v. ИЛ. — вена брюшных плавников; v. р. h. — воротная веиа печени; о. р. г.—воротная вена почек; v. г. ad.—приносящие вены почек; v. scl. — подключичная веиа; v. s. trtf. — нижняя половая вена; v. sp. s. ~ правая верхняя половая вена
дпальпую полость. Там они сливаются с передними кардинальными, или яремными, венами в общие протоки Кювье.
Рассматривают сначала вены, впадающие в задние кардинальные вены.
Нижняя половая вена семенника у самца или я и ч-н и к а у самки (v. spermatica inferior, s. v. ovarica inferior, v. sp. inf.) впадает в непарный отдел кардинальных вен в самой задней части полости тела и приносит кровь от заднего отдела гонад правой и левой частей тела.
Верхняя половая вена семенника у самца или я и ч-н и к а у самки (v. spermatica superior, s. v. ovarica superior, v. sp. s.) впадает в передний конец кардинальных вен около мест •ответвления кишечно-желудочной артерии от спинной аорты. Она «обирает кровь от передних отделов семенников и идет вместе с одноименной артерией.
Сегментальные артерии отходят от спинной аорты, а соответствующие им сегментальные вены впадают в кардинальные вены справа и слева в виде ряда поперечных сосудов, идущих вместе друг с другом по стенкам тела.
Вены брюшных плавников, подвздошно-поясничные вены (v. ilioluinbalis, v. il.-l.) впадают с обеих сторон несколькими веточками в кардинальные вены в заднем конце полости тела.
Теперь рассматривают вены, приносящие кровь в протоки Кювье.
Протоки Кювье (ductus Cuvieri, d. С.)—поперечные трубки, лежащие в глубине околосердечной полости, образуются слиянием передних и задних кардинальных вен. В них впадают с брюшной стороны от печени печеночные вены (v. hepatis, v. h.), составленные в ткани печени из разветвленных капилляров воротной вены печени. Справа и слева в кювьеровы протоки, в месте слияния кардинальных и яремных вен, впадают подключичные вены грудных плавников (v. subclavia, v. scl.).
Верхние яремные вены (v. jugularis superior, v. j. s.), или передние кардинальные вены (v. cardinalis anterior), приносят кровь от головы, головного мозга, черепа и органов чувств и впадают со спинной стороны в протоки Кювье, сливаясь вместе с задними кардинальными венами. Впадение яремных вен в кювьеровы протоки можно видеть на той стороне рыбы, где были удалены трудные плавники при рассмотрении ротоглоточной полости. Они .лежат глубоко под сердцем, непосредственно впереди впадения кардинальных вен в протоки Кювье, и идут кверху — к черепу и головному мозгу.
Нижняя яремная вена (v. jugularis inferior, v. j. inf.) приносит кровь от вентральной части головы. У осетровых рыб это непарный сосуд, проходящий по боковой стенке околосердечной сумки с правой стороны рыбы, который впадает в верхнюю яремную вену у ее основания. С левой стороны одноименной вены обнаружить у стерляди не удалось.
Совершенно отдельную систему венозных сосудов составляют вены, собирающие кровь от пищеварительной системы. Они начинаются сетью капилляров в стенках кишечника и образуют разветвления вен в брыжейках, приносящих кровь к печени.
Воротная вена печени (v. portae hepatis, v. p. h.) — система венозных сосудов, собирающих кровь от пищеварительной системы, независимо от системы венозных сосудов, составляющих кардинальные и яремные вены туловища. Она начинается капиллярами в задней кишке и, постепенно укрупняясь, идет вдоль наружных стенок кишечной трубки, принимая веточки от стенок кишечника по всем его петлям, а также от желудка и сосудов, разветвляющихся в брыжейках. Рассматривают также разветвления венозных сосудов в стенках плавательного пузыря, а также веточки отдельных лопастей печени. В печени вся эта система сосудов распадается на систему капилляров, где происходит фильтрация крови. Внутри печени капилляры снова собираются в выносящие вены, называемые печеночными венами.
Печеночные вены (v. hepatica, v. h.)—короткие широкие сосуды, несколькими трубочками соединяющие переднюю поверхность печени с задней стенкой околосердечной сумки. Пронизывая ее, они впадают в протоки Кювье.
Нервная система (рис. 71)
Нервная система осетровых рыб обладает общими чертами строения с другими костными рыбами из надотряда лучеперых рыб (Actinopterygii), но по сравнению с костистыми рыбами она отличается особенностями, показывающими более примитивную организацию осетровых рыб. Основными органами чувств, при помощи которых осетровые ориентируются в окружающей среде, являются органы чувств боковой линии, органы обоняния и статоакустики; органы зрения развиты значительно слабее. Соответственно этому в головном мозге прогрессивно развиты центры сепсмосеизорпых органов, статоакустики и обоняния.
На изучение осетровых рыб отводится мало времени, поэтому отделы нервной системы и органов чувств изложены здесь кратко.
Для приготовления анатомического препарата нервной системы осетровых рыб требуется соответствующая подготовка материала.
Зафиксированные обычным путем в 10%-ном формалине головы стерляди следует тщательно промыть в течение двух суток в проточной воде, а затем декальцинировать костный скелет в следующей смеси: 85 частей 70%-пого спирта, 10 частей крепкой азотной кислоты и 5 частей глицерина. Держать в этой смеси надо 2—3 суток, пока покровные окостенения, покрывающие хрящевой череп, не станут мягкими. После этого головы следует промыть в проточной воде и течение суток и переложить в 70%-пый спирт.
Перед вскрытие.м черепной полости необходимо удалить декальцинированный костный панцирь черепа, срезая его острым ножом и стараясь не повредить лежащий под ним хрящевой череп. Затем
осторожно срезать хрящ плоскостными срезами, пока не вскроется черепная полость с лежащим в ней мозгом.
При вскрытии мозговой полости одновременно вскрываются и слуховые капсулы, в которых находится слуховой лабиринт с полукружными каналами.
Рис. 71. Головной мозг стерляди Acipenser ruthenus. А — сбоку; Б — свеох ху; В — снизу:
Ь. — мозжечок; с. str. — базальные ганглии, полосатые тела переднего мозга; dnc. — промежуточный мозг; ер. — эпифиз; hyp. — гипофиз; infd. — воронка промежуточного мозга; I. I. I.—доли органов чувств боковой линии; lob. inf. — нижние доли промежуточного мозга; lob. olf. — обонятельные доли переднего мозга; I. vise. t. — доли блуждающего нерва продолговатого мозга; med. об/, — продолговатый мозг; п. sp. — спинномозговые нервы; par. — парафиз крыши промежуточного мозга; sac. vase. — железистый мешочек; tgtn. — покрышка; tr. olf. — обонятельный тракт; t. opt. — зрительная крышка; tine. — передний мозг; t. ас. — слуховой бугорок; vel. tr. — поперечный парус; / — обонятельные иервы; // — зрительный тракт; /// — глазодвигательный нерв; /V — блоковый нерв; V— тройничный нерв; VI— отводящий нерв; VII— лицевой нерв; VIII — слуховой нерв; /X — языкоглоточный нерв; X. — блуждающий нерв
На переднем конце головы удаление костного панциря кожных окостенений следует производить до переднего конца рыла так, чтобы можно было отпрепарировать обонятельные капсулы, лежащие по бокам головы впереди глаз.
Общая топография нервной системы в черепной полости
Осторожно препарируют черепную полость, чтобы рассмотреть головной мозг и отхождение головных нервов. Удаляя хрящевые стенки черепа, следят за тем, чтобы не повредить головные нервы, отходящие от головного мозга вправо и влево. После этого рассматривают сверху общую топографию головного мозга и головных нервов (рис. 71).
Впереди черепной полости расположены парные органы обоняния. От каждого обонятельного мешка тянутся пучки обонятельных нервов (I), переходящие в парные обонятельные доли (lobus olfactorius, lob. olf.). Они сходятся по средней линини широким основанием друг с другом и переходят в непарный передний мозг (telencephalon, tine.). Передняя стенка переднего мозга резко повышается кверху и переходит в перепончатую крышку переднего мозга, имеющего вид пузыря с общей полостью внутри. Задний отдел крыши переднего мозга сверху прикрыт перепончатой кры-шей п р о м еж у точ н о го мозга (diencephalon, dnc.), от которого вперед вытягивается на ножке пинеальный орган (эпифиз— epiphysis, ер.). На фиксированном материале перепончатые крыши мозга находятся в спавшем состоянии и опущены в полость головного мозга. При рассмотрении сверху граница между передним и промежуточным мозгом выражена очень нечетко.
Позади промежуточного мозга резко возвышается средний мозг (mesencephalon, t. opt.), образуя мощное скопление мозговой ткани — крыши среднего мозга (tectum opticum), не расчлененной у стерляди па парные зрительные доли. К зрительным долям сзади непосредственно примыкает мозжечок (cerebellum, crb.), несколько свисающий назад парными лопастями. Он представляет собой переднюю стенку ромбовидной ямки, состоящей из мозговой ткани, дальше назад тянется перепончатая крыша четвертого желудочка продолговатого мозга (medulla oblongata, med. о.), покрывающая широкую ромбовидную ямку. Крыша продолговатого мозга прикрыта сверху мощным скоплением жироподобной ткани лимфоидного органа, заполняющего затылочную область черепа.
Срезая боковые стенки затылочного расширения мозговой полости, следует отпрепарировать заполняющий его лимфоидный орган.
Лимфоидный орган черепной полости имеет грушевидную форму и располагается над продолговатым мозгом и передним концом спинного мозга в виде большого скопления жироподобной ткани, пронизанной густой сетью кровеносных сосудов. Передний заостренный конец сливается с перепончатой крышей ромбовидной ямки, а расширенный задний простирается до конца хрящевого че
репа. По-видимому, этот лимфоидный орган выполняет функцию кроветворного органа.
Вычленяют лимфоидный орган из мозговой полости и удаляют его, чтобы увидеть продолговатый мозг и отходящие от него головные нервы.
Прежде чем разбираться в строении головного мозга, рассматривают места отхождения от мозга головных нервов и их расположение в черепной полости.
Отхождение от мозга головных нервов (рис. 71)
Осторожно срезают скальпелем хрящевые стенки черепной коробки так, чтобы не повредить головные нервы, отходящие от головного мозга справа и слева и пронизывающие ее стенки. Для того чтобы увидеть отхождение головных нервов, осторожно отодвигают ручкой пинцета головной мозг в сторону и рассматривают прохождение головных нервов внутри черепной полости.
Обонятельные нервы (n. olfactorius, n. I, /), как было видно выше, отходят от обонятельных долей к обонятельным мешкам двумя пучками; каждый из них подходит к обонятельному мешку, распадаясь па пучки нервов, иннервирующих орган обоняния.
Зрительные нервы (n. opticus, п. II. //) отходят от лна промежуточного мозга в, самой глубине мозговой полости. Их можно увидеть под передним мозгом, отодвигая в стороны нижние доли промежуточного мозга, лежащего в глубине. Рассмотрение их откладывают до изучения головного мозга с нижней стороны.
Глазодвигательные нервы (n. oculomotorius, n. III,///) отходят от дна среднего мозга и направляются в стороны и вперед вместе с впереди лежащим кровеносным сосудом к глазным мышцам.
Блоковый нерв (n. trochlearis, п. IV, /V) тянется в виде очень тоненькой белой нити, пронизывающей сзади наперед боковую стенку глазницы. Он отходит на границе между мозжечком и средним мозгом и, пронизав стенку черепа, выходит в глазницу, где иннервирует верхнюю косую мышцу глаза.
Тройничный нерв (n. trigeminus, n. V, V) отходит от продолговатого мозга в глубине черепной полости мощным корешком, направляющимся вперед, и пронизывает стенку черепа внутри глазницы.
Отводящий нерв (n. abducens, п. VI, V/) здесь не виден, он находится под тройничным первом с нижней стороны мозга.
Лицевой нерв (п. facialis, п. VII, VII) начинается мощным корешком позади тройничного нерва, несколько выше его, также направляется вперед и выходит из черепа в глазнице, позади тройничного нерва.
Слуховой нерв (n. acusticus, n. VIII, VIII) отходит от продолговатого мозга непосредственно позади лицевого нерва несколькими корешками, направляющимися веерообразно в стороны в перепончатый слуховой лабиринт.
Языкоглоточный нерв (п. glossopharyngeus, n. IX, IX) лежит непосредственно позади корешка VIII нерва и направляется назад через черепную полость позади слухового лабиринта.
Блуждающий нерв (п. vagus, п. X, А')—мощный пучок, нервов, отходящих многочисленными корешками в общее сплетение позади отхождения VIII и IX нервов. Этот сложный «пучок нервов простирается по черепной 'полости назад, прободает стенку затылочной области черепа и направляется к жаберному аппарату.
Передние спинномозговые нервы (п. sp.), также отходят в затылочной области черепа, они начинаются от спиппого мозга позади ромбовидной ямки. Внутри вскрытой мозговой полости видны два спинных корешка с ганглиями, выходящих через затылочную часть черепа; следующий спинномозговой нерв отходит уже позади черепа.
Головной мозг
Чтобы рассмотреть строение головного мозга стерляди, рекомендуется вынуть его из черепной коробки, для чего острыми, топкими ножницами перерезают корешки головных и спинномозговых нервов справа и слева. Сначала перерезают около обонятельных капсул обонятельные нервы, отпрепарируют их от дна черепной полости так, чтобы можно было приподнять мозг. Далее осторожно отодвигают мозг в стороны и последовательно, спереди назад, перерезают все корешки головных нервов, сначала блоковый, потом глазодвигательный и, наконец, в глубине впереди промежуточного мозга — зрительный нерв. Затем последовательно один за другим перерезают отдельные корешки тройничного, лицевого и слухового нервов, которые своими ветвями направлены вперед и в стороны. Позади слуховой капсулы отходят корешки языкоглоточного и блуждающего нервов, а далее назад и первые спинномозговые нервы. После этого, приподнимая пинцетом, отпрепарируют головной мозг от мелких кровеносных сосудов, подходящих к нему со всех сторон, и осторожно вынимают его из мозговой полости. Кладут мозг в ванночку или в чашку Петри, заливают водой и рассматривают его строение, начиная с переднего конца, сверху, с боков и снизу.
Передний мозг состоит из резко обособленного обонятельного мозга и переднего конечного мозга, не дифференцированного на парные полушария. Внутри переднего мозга имеется большая мозговая полость.
Обонятельный мозг представлен парными обонятельными долями. Обонятельные доли (lobus ollactorius, lob. olf.) — парные образования из мозговой ткани с просвечивающей полостью внутри. Задней границей они примыкают к возвышающемуся позади них мозговому пузырю переднего и промежуточного мозга, не расчлененного на отдельные полушария и имеющего общую полость с промежуточным мозгом.
Передний, иди конечный, мозг (telencephalon, tenc.) имеет перепончатую пузыревидную крышу, состоящую из эпендимы. В глубине переднего мозга имеется большая полость, не разделенная на боковые желудочки полушарий головного мозга, носящих у наземных позвоночных название первого и второго мозговых желудочков. Нижние боковые стенки переднего мозга, являющиеся вторичными обонятельными центрами, заняты мощным скоплением мозговой ткани; это б а з а л ь н ы е г а н г л и и (ganglion basale), или полосатые тела (corpus striatum, с. str.). Передняя стенка переднего мозга также состоит из нервной ткани, а далее она переходит в перепончатую крышу переднего и промежуточного мозга.
Поперечный парус (velum transversum, vet. tr) образует заднюю границу переднего мозга. Он начинается пальцевидным выростом, идущим внутрь от передней стенки, обозначая место отхождения вперед парафиза (paraphysis, par.) — непарного переднего выроста перепончатой крыши промежуточного мозга. Поперечный парус простирается в мозговой полости па границе переднего и промежуточного мозга по диагонали от переднего верхнего угла вниз к заднему концу полосатых тел. Поперечный парус оплетен сетью кровеносных сосудов, питающих мозг, образуя переднее сосудистое сплетение (plexus chorioideus anterior). Оно хорошо видно на просвет через перепончатую крышу в виде плотного шнура и служит границей, отделяющей переднюю полость конечного мозга от полости верхнего отдела промежуточного мозга — эпиталамуса. От верхней стенки поперечного паруса вперед вытягивается ножка пинеального органа — эпифиза (epiphysis, ер).
Промежуточный мозг (diencephalon, s. thalamenceph-alon, dnc.) состоит из трех частей: эпиталамуса, таламуса и гипоталамуса.
Эпиталамус (epithalamus, s. parencephalon)—верхний отдел промежуточного мозга — описан выше, он характеризуется перепончатой крышей, от которой отходят два непарных выроста: передний — париетальный орган и задний — пинеальный орган — эпифиз (epiphysis, ер).
Таламус (thalamus)—средний отдел промежуточного мозга— является непосредственным продолжением назад мозговой ткани полосатых тел переднего мозга. Внутренняя полость промежуточного мозга носит название третьего мозгового желудочка.
Гипоталамус (hypothalamus)—нижний отдел промежуточного мозга — образует опускающуюся глубоко вниз мозговую воронку промежуточного мозга (infundibulum, infd.). Боковые стенки мозговой воронки имеют парные вздутия мозговой ткани справа и слева, называемые нижними долями (lobus inferior, lob. inf.). Это хорошо видно при рассмотрении дна промежуточного мозга снизу. На дне воронки прикреплена нижняя мозговая железа, или гипофиз (hypophysis, hyp.), резко выделяющаяся
на воронке мозга. Мозговая воронка резко выделяется по сравнению с вентральной стенкой переднего мозга. Границей с передним мозгом служит место отхождения зрительных нервов (n. opticus, II), образующих впереди воронки перекрест зрительных нервов, или зрительную хиазму (hiasma nervorum opticorum). Соответственно придонному образу жизни осетровых рыб и второстепенной роли зрения в их биологии зрительные нервы осетровых по сравнению с другими костными рыбами развиты слабо.
Железистый мешочек (saccus vasculosus, sac, vase.) — полый вырост воронки, располагающийся между нижними долями дна промежуточного мозга. Внутри он выстлан рецепторными клетками, выполняющими у рыб не вполне разгаданную функцию. У наземных позвоночных он отсутствует.
Средний мозг (mesencephalon, msc.) у осетровых рыб дифференцирован очень слабо. На дорзальной стороне головного мозга возвышается позади промежуточного мозга скопление зрительных мозговых центров — зрительная крышка (tectum opticum, t. opt.), образующая зрительные доли (lobus opticus), расчленение которых на два яйцевидных тела, характерное для других рыб, здесь еле намечается.
Стволовая часть среднего мозга — непосредственное продолжение стволовой части продолговатого и спинного мозга —образует скопление мозговой ткани и проводящих путей, носящих название покрышки (tegmentum, tgm.), или ножек мозга (crura cerebri). От дна среднего мозга отходят вниз глазодвигатель-н ы е головные нервы (n. oculomotorius, III).
Мозжечок (cerebellum, s. metencephalon, crb.) непосредственно примыкает сзади к зрительным долям среднего мозга. При рассмотрении головного мозга сверху он резко возвышается над средним мозгом в виде скопления мозговой ткани, располагающегося в поперечном направлении, и представляет собой утолщенную переднюю стенку продолговатого мозга и его ромбовидной ямки. Передняя часть мозжечка вдается в полость среднего мозга в виде мощного впячпвания мозговой ткани, которое носит название мозжечковой складки (valvula cerebelli), являющегося мозговым центром органа слуха. Верхняя стенка мозжечка переходит в перепончатую крышу продолговатого мозга.
Продолговатый мозг (medulla oblongata, s. myelencephalon, med. о.) —самый задний отдел головного мозга, непосредственно переходящий в спинной мозг. На целом мозге с дорзальной стороны видна мощная перепончатая пленка, оплетенная кровеносными сосудами заднего сосудистого сплетения, покрывающего в виде свода широкую ромбовидную ямку (fossa rhomboidea), образованную мозговой полостью продолговатого мозга и носящую название четвертого желудочка мозга. Чтобы рассмотреть ее строение, надо удалить перепончатую крышу с кровеносным сплетением, носящим название заднего хориоидального сплетения (plexus chorioideus posterior). На боковой поверхности
продолговатого мозга видны корешки головных нервов: V, VII, VIII,. IX, X и XII.
Отпрепарируют осторожно головной мозг от покрывающих его мозговых оболочек, пронизанных разветвлениями кровеносных сосудов. Одновременно с мозговыми оболочками удаляют при препаровке и перепончатые стенки крыши желудочков головного мозга. При этом оказывается снятой перепончатая крыша переднего и промежуточного мозга и вскрывается внутренняя полость. По бокам этой полости выступают утолщения базальных ганглиев, или полосатых тел (corpus striatum, с. str.), а впереди — два отверстия в обонятельные доли. В задней части этой полости видно углубление мозговой воронки промежуточного мозга. Позади мозжечка открывается полость ромбовидной ямки в развернутом виде. В дне ромбовидной ямки видны отдельные мозговые центры продолговатого мозга. Наружные валики ромбовидной ямки дают утолщения, которые служат центрами кожных органов чувств боковой линии (lobus lineae lateralis, I. I. I.). Позади центров сейсмосен-зорных органов чувств выделяются самостоятельные центры, носящие название слуховых бугорков (tuberculum acusticum, t. ас.). Внутрь от них обособляется продольная полоска центров блуждающего нерва.
Сравнительная характеристика головного мозга разных видов осетровых рыб
Сравнивая разных представителей осетровых рыб, можно отметить следующие отличия в развитии разных отделов головного мозга, отражающие различия в образе жизни и в деятельности отдельных органов чувств у разных видов и родов осетровых рыб.
Мозг стерляди—Ac. ruthenus отличается значительным развитием обонятельных мешков и мощными, но короткими обонятельными нервами. Соответственно значительного развития достигает и передний мозг, где сосредоточены обонятельные центры. Средний мозг у стерляди также развит наиболее сильно по сравнению с другими осетровыми, хотя в его крыше и не имеется ясного расчленения на парные зрительные доли. Также велика у стерляди и степень развития мозжечка, который имеет вид значительных парных скоплений мозгового вещества.
Мозг севрюги — Ac. stellatus тоже имеет крупные обонятельные мешки и очень длинные и мощные обонятельные нервы. Передний и промежуточный мозг, как и у стерляди, развит значительно. Глаза у севрюги менее крупные и зрительные доли в среднем мозге при сопоставлении со стерлядью развиты значительно слабее. В продолговатом мозге имеется четкая дифференцировка отдельных центров, как и у стерляди.
Мозг лжелопатопосов — Pseudoscaphirhynchus kaufmanni по сравнению с другими осетровыми развит значительно слабее. Особенно это бросается в глаза при сопоставлении размеров глаз и зрительных центров среднего мозга. Но и передний мозг и обонятель-222
ный мозг у этих малоподвижных донных рыб по сравнению с мозгом других осетровых рыб имеет меньшие размеры. Зато продолговатый мозг с центрами кожных органов чувств и иннервации челюстного и жаберного аппаратов, а также ромбовидная ямка развиты у лопатоноса так же хорошо, как и у других осетровых рыб.
Органы чувств
Осетровые рыбы ведут придонный образ жизни, питаясь бентосными организмами. Соответственно этому ведущее значение в добывании пищи и биологии осетровых имеют не органы зрения, как у костистых рыб, а тактильные и хеморецепторные (обонятельные и вкусовые) органы.
Тактильные органы чувств представлены у всех осетровых характерными для них четырьмя усиками, расположенными на вентральной стороне рыла, впереди от ротового отверстия. Кроме тактильных органов, на них располагаются также вкусовые чувствующие клетки.
Орган обоняния в виде типичных для рыб обонятельных мешков расположен на рыле непосредственно впереди глаз. Обонятельный мешок в полтора раза больше глаза. С наружной стороны каждый обонятельный мешок прикрыт кожистой передней стенкой с двумя отверстиями — ноздрями (паг, рис. 62, и.). При плавании вода входит в переднее отверстие и выходит через заднее. Кожистая передняя стенка обонятельного мешка легко может быть срезана по окружности тонкими ножницами и тогда становится видным внутреннее строение органа обоняния. Дно обонятельного мешка представляет розетку, образованную расходящимися от центра по радиусам складками чувствующего эпителя. Складки увеличиваются в высоту по направлению от центра к периферии. Количество складок у стерляди доходит до 30.
Органы зрения — глаза (oculus, рис. 62, ос.) у осетровых рыб невелики по размеру (см. стр. 194). Они имеют типичное строение глаз рыб. Их аккомодация совершается путем передвижения хрусталика вперед и назад.
Статоакустический орган заключен в мощно развитом затылочно-позвоночном отделе хрящевого черепа. Перепончатый лабиринт имеет типичные для всех рыб три полукружных канала, расположенных в трех взаимно перпендикулярных плоскостях соответственно трехмерной ориентации рыбы в пространстве. Отпрепарировать их можно, сняв покровные окостенения в затылочной области и вычленяя их из хрящевого черепа при помощи тех же методов, которые применялись при препаровке слухового лабиринта акулы.
Органы системы боковой линии (рис. 72). Кожные органы чувств системы боковой линии у стерляди на туловище представлены системой различных каналов и ямок-фолликулов.
Боковой канал (canalis lateralis, с. lat.) скрыт в чешуях-жучках бокового ряда. Он простирается вдоль всего тела от уров
ня верхнего края жаберной крышки до конца нижней хвостовой лопасти, пронизывая весь ряд боковых жучек. В области хвостового плавника боковой канал закрыт нижним рядом ромбических че-шуй и проходит назад и вверх до конца хвоста. Боковой канал в промежутках между соседними жучками открывается на поверхность наружными отверстиями.
Боковой канал на туловище стерляди между спинным и боковым рядом жучек сопровождается спинным рядом чувствующих фолликулов, идущим на протяжении от вертикали плечевого пояса до
Рис. 72. Схема распределения кожных органов чувств боковой линий на голове стерляди Acipenser ruthenus:
с. ino. — подглазничный капал; с. р. ор. — предкрышечный канал; с. sorb.— надглазничный канал; с. sip. — надвисочный канал; с. tp.— височный канал; f. iorb. — подглазничные чувствующие ямки; fj. — предкрышечные (щечные) чувствующие ямки; f. ор. — крышечные чувствующие ямки; f. рг. orb. — пред-глазничные чувствующие ямки; f pt. orb. — эаглазничные чувствующие ямки; f. г. — ростральные фолликулы; f. sorb. — надглазничные чувствующие ямки; f. sr. — надростральные чувствующие ямки; f. stp. — иадвисочные чувствующие ямки; f. ip. — височные чувствующие ямкн; sp. — брызгальце
уровня брюшных плавников, и дополнительным рядом чувствующих фолликулов туловища, который проходит по вентральным поверхностям жучек бокового ряда.
На голове кожные органы чувств по строению и расположению образуют сложную систему: из чувствующих каналов с погруженными в них невромастами, из групп простых чувствующих бугорков и сложных чувствующих ямок.
При рассмотрении этой сложной системы кожных органов чувств головы следует разобраться в чувствующих каналах и сложных ямах, обязательно пользуясь оптикой.
Боковой канал выше плечевого пояса в переднем направлении
прободает надвисочную кость, несколько выше ее бокового края и далее, вступив в чешуйчатую кость, распадается на два канала — височный и надвисочный.
Височный капал (canalis temporalis, с. ip) начинается в переднем отделе надвисочной кости в месте отхождения надвисоч-пого канала (с. stp.) и проходит через нее до переднего конца. Здесь височный канал дает две ветви: одну, идущую над глазом, — надглазничный канал (с sorb.) и вторую, идущую за глазом,— подглазничный канал (с. ino).
Надвисочный канал (canalis supratemporalis, с. sip.) па уровне верхнего края наружного жаберного отверстия начинается от бокового канала в переднем отделе надвисочной кости и направляется на дорзальную сторону головы; он пронизывает дополнительные надвисочные кости и кончается между заднезатылочной, или нухале, и кожной верхнезатылочиой костями.
Надглазничный канал (canalis supraorbitalis, с. sorb.) после отхождения от переднего конца височного канала (с. ip.) проходит над глазницей в лобных костях, далее идет вниз, вперед и переходит в кости межноздревой перемычки. Ниже передней ноздри он прерывается в окостенениях предносовой области.
Подглазничный канал (canalis infraorbitalis, с. ino.) проходит в заднелобной кости и заглазничных окостенениях позади глазницы, отсюда на уровне рта он переходит в ряд мелких окостенений вентральной поверхности головы и продолжается вперед к концу рыла, предварительно огибая снаружи (латерально) усики. На переднем конце рыла канал переходит на дорзальную сторону и в медиальном направлении погружается в хрящ переднего конца рыла.
На всем протяжении надглазничный и подглазничный каналы проходят в костях дермокраниума, а на тотальных препаратах прослеживаются по отверстиям их наружу к чувствующим ямкам — фолликулам смежных частей головы.
Преоперкулярный, предкрышечпый канал (canalis praeopercularis, с. pop.) лежит в предкрышечной или щечной области на уровне и ниже брызгальца, несколько кзади от рта в предкрышечных или гулярных косточках, погруженных в кожу. По происхождению он — '.преобразованные ямковые органы или фолликулы.
Разобравшись в топографии чувствующих каналов головы стерляди, внимательно рассматривают распределение и концентрацию крупных сложных чувствующих ямок (fossa) па отдельных участках головы. (Пользуйтесь оптикой!) Они резко выделяются по сравнению с большой складчатостью кожи. Для этой цели в значительной мере может служить просветленный препарат дермокраниума.
Наибольшее количество чувствующих ямок сосредоточивается на вентральной стороне головы (рострум, впереди и вокруг рта), много их на боковых участках головы (щечная область, на жаберной крышке), меньше на дорзальной стороне.
Вентральная поверхность головы впереди рта на всем протяжении между внутренними усиками, между боковыми усиками и проходящим здесь подглазничным каналом и латеральным краем рострума несет богатую систему простых и сложных чувствующих ямок. Предротовая группа простых чувствующих ямок располагается впереди рта. Система чувствующих ямок распространяется и па вентральную поверхность рострума, впереди усикового так называемая нижперостральпая группа чувствующих ямок.
При рассмотрении головы сбоку находят и прослеживают следующие группы сложных чувствующих ямок: 1) надглазничную (f. sorb.), расположенную между глазом и надглазничным каналом; 2) п р ед г л а з и и ч н у ю (f. pr. orb.), лежащую впереди надглазничной и над передним обонятельным отверстием; 3) за глазничную группу (f. pt. orb.), расположенную за глазом впереди заглазничных окостенений (заглазничная группа ямок распространяется каудально на предкрышечную область); 4) подглазничную группу (f. iorb.)-, 5) оперкулярную, или крышечную, группу (f. op.) из сложных чувствующих ямок, располагающихся на жаберной крышке, и 6) надростральную группу (f. sr.), лежащую впереди обонятельных отверстий по Латеральному краю рострума, заходя несколько на дорзальную сторону в виде многочисленных сложных чувствующих ямок.
Па дорзальной поверхности головы легко обнаруживается надвисочная группа чувствующих ямок (f. sip.), лежащая выше височного канала по ходу и впереди пего.
Скелет
Изучение скелета осетровых рыб в ряду позвоночных животных представляет особый интерес, потому что скелет этих рыб объединяет в себе признаки древних хрящевых рыб Chondrichthyes, сохраняя в осевом скелете хорду и хрящевой внутренний скелет, в то же время имеет костный скелет, свойственный всем высшим рыбам и наземным позвоночным. Костный скелет осетровых рыб состоит из настоящей костной ткани в виде кожных покровных окостенений черепа и костей пояса грудных плавников, одевающих хрящевой скелет с наружной стороны и из костных плавниковых лучей — лепидотрихий (lepidotrichium) и чешуй различной формы.
Изучение скелета осетровых рыб обычно приходится проводить на готовых препаратах, поскольку достать свежий материал не всегда удается. Для приготовления препаратов можно пользоваться морожеными тушками стерлядей.
Для ознакомления с покровным скелетом туловища лучше всего снять кожу с одной стороны туловища рыбы, начиная от заднего края плечевого пояса и до конца хвостового отдела. Кожу следует снимать только с осевого стержня, оставив лепидотрихии лопастей плавника и фулькры при тушке. Точно так же следует ос
тавить при тушке спинной и анальный плавники. Такая кожа хорошо сохраняется в сухом виде. Д.чя более детального изучения мелких костных чешуек рекомендуется вырезать небольшой кусочек кожи и поместить его в глицерин, где он просветляется и хорошо сохраняется. Можно спять кожу и целиком, но при этом необходимо сохранить целостность непарных и парных плавников.
Затем, проведя продольный разрез брюшной стенки между грудными и брюшными плавниками, тушку освобождают от внутренних органов п аккуратно вычленяют пояса передних и задних конечностей.
Пояс задних конечностей вычленяется довольно легко. При вычленении пояса передних конечностей особенно осторожно следует отчленять его от черепа, к которому он причленен вдоль нижнего края височной кости, так как в этой области дорзальная часть пояса плотно соединена с боковыми затылочными отростками. Затем отделяют от окружающей ткани задний и передний края пояса. Для того чтобы снять покровные кости с головы, ее необходимо спустить в теплую воду, которую постепенно нагревают до 70— 80° С, по не кипятят. Предварительно па коже головы делают несколько разрезов. Первый поперечный, соединяющий вентральные края жаберных отверстий. Второй перпендикулярный ему до конца рыла. При проведении этого разреза следует обойти справа и слева верхнюю и нижнюю челюсти. Выдержав голову в горячей воде (время выдерживания указать трудно, но следует не передержать, чтобы пе развалились кости), ручкой скальпеля отчленяют весь кожный скелет крыши и боков головы вместе с кожей и жаберной крышкой от лежащего под ним хрящевого черепа и висцерального скелета. Такой снятый покровный череп высушивают, предварительно плотно набив его ватой и придав естественную форму. Сухие препараты покровного черепа могут храниться очень долго.
Остальную тушку вместе с поясами конечностей мацерируют в теплой воде и дочищают начисто внутренний скелет.
Осевой и висцеральный скелет черепа можно покрасить ализарином, так же как и пояс передних конечностей, что значительно облегчает изучение вторичных окостенений в этих отделах. Для этого очищенные скелеты головы фиксируют в формалине и далее поступают по приведенной выше схеме ализариновой окраски.
Общий обзор скелета
У стерляди хорошо различим скелет наружный (cxosceletum) и внутренний (endoscelctum).
Наружный костный скелет у осетровых рыб на голове представлен костным панцирем из покровных костей, покрывающим хрящевой череп сверху и с боков до наружных жаберных отверстий.
В туловищном отделе наружный костный скелет представлен характерными для всех осетровых рыб пятью рядами костных жучек с острыми шипами, играющими защитную функцию (один дор-
зальный, два боковых и два брюшных ряда). Передние жучки боковых и брюшных рядов, разрастаясь, покрывают снаружи хрящевой пояс грудных плавников и превращаются в покровные кости пояса передних конечностей. На хвостовом стебле боковые поверхности сплошь покрыты ромбическими пластинками, сходными с ганоидными чешуями, а верхний край — вильчатыми чешуйками, фулькрами.
Внутренний скелет у осетровых рыб сохраняет во взрослом состоянии очень древний признак — полно выраженную спинную струну — хорду, одетую плотным соединительнотканным футляром, позвоночник же представлен только хрящевыми дугами позвонков.
Скелет плавников, как непарных, так и парных (грудных и брюшных), состоит из хрящевых лучей, составляющих основания плавников, и костных лучей кожного происхождения, свойственных всем лучеперым костным рыбам. Парные плавники имеют хрящевые пояса, грудной хрящевой пояс снаружи прикрыт кожными покровными костями.
Хвостовой отдел сохраняет свойственную хрящевым рыбам форму неравнолопастного хвостового плавника, где хорда продолжается в верхнюю лопасть, а нижняя лопасть поддерживается удлиненными хрящевыми лучами.
Скелет головы состоит из двух резко обособленных друг от друга отделов: осевого черепа, или мозговой коробки (нейрокраниум), и висцерального скелета (челюстной аппарат и жаберный скелет).
Осевой череп представлен в виде замкнутой хрящевой мозговой коробки, снаружи закрытой костным панцирем.
Передний отдел висцерального скелета преобразован в сложный выдвижной челюстной аппарат, который скользит взад и вперед под основанием мозгового черепа. Он состоит из сильно видоизмененных челюстной и подъязычной дуг. Жаберный скелет обособлен от челюстного аппарата и состоит из пяти расчлененных хрящевых, частично окостеневающих жаберных дуг, прикрепленных к слуховому и затылочно-позвоночному отделам черепа. Жаберные дуги соединены друг с другом на вентральной стороне непарными хрящами — копулами.
Наружный, или костный, скелет
Наружный скелет туловища (рис. 73)
Жучки — крупные костные чешуи, составляющие основу наружного скелета туловища. Их расположение на теле осетровых в 5 рядов было рассмотрено выше при внешнем осмотре рыбы. Следует познакомиться теперь более подробно с их формой, которая несколько отличается в каждом из рядов.
Дорзальный ряд жучек (В) идет по средней линии спины и поэтому отдельные чешуйки имеют вид двускатной крыши. Вершина ее (конек) к заднему концу приподнимается, утолщается и перехо-228
дит в 'направленный назад тупой шип. Обращают внимание на то, что между жучками кожа лишена скелетных образований и возможны изгибы тела.
Жучки бокового ряда (Л) меньших размеров, чем спинного, н имеют не совсем правильную ромбическую форму. Спереди назад, по наружной поверхности чешуи, проходит невысокий тупой валик— гребешок. На просветвленном препарате видно, что в про-
Рис. 73. Типы чешуй кожных покровов стерляди Acipenser ruthenus. А — жучки из бокового ряда; Б — жучки из брюшиого ряда; В — жучки из спинного ряда; Г—межрядные чешуйки
дольном направлении под продольным гребнем, чешую пронизывает канал боковой линии. Вдоль заднего края чешуи он дает вниз ответвление, открывающееся на нижнем заднем краю жучки. Передние жучки бокового ряда дают верхние элементы покровного плечевого пояса.
В брюшном ряду (Б) количество жучек меньше и сами они мельче, чем в боковом ряду. Жучки имеют форму пластинок с неправильными очертаниями, вытянутых в передне-заднем направлении. В передней части жучка несет продольный сглаженный гребеиь.
Обращают внимание на то, что жучки каждого ряда погружены в кожу.
Количество жучек в рядах различно у разных видов осетровых и служит важным систематическим признаком.
Рассмотрев жучки из всех рядов, можно перейти к изучению мелких чешуи между рядами жучек, при этом следует найти различные типы их (Г): одно-, двух-, трех- и многозубчатые, гребенчатые чешуи и переходы в более сложные чешуи-жучки, рассмотренные выше (пользуйтесь оптикой!).
Наружный скелет головы — дермокраниум (рис. 74)
Мозговая часть хрящевого черепа—(chondrocranium) снаружи закрыта костным кожным черепом—(dermocranium), состоящим из большого ряда покровных костей. Кости, составляющие дермо-
Рис. 74. Костный панцирь головы стерляди Acipenser ruthenus:
cl. — ключица; clt. — клейтрум; d. soc. — кожная верхнезатылочная кость; fr.—лобные кости; j. — предкрышечный ряд костей (скуловые костн); med. — медиальная кость; п. ~ носовая кость; nch. — нухальная кость; op. v. inf. — нижний вентральный элемент жаберной крышки; ор. и. з. — верхний вентральный элемент жаберной крышки; р. — теменные кости; р. clt. — лостклейтру.м; р. fr. — заднелобная кость; рг. orb.— предглазничная кость; pt. orb. — заднеглазничиая кость; s. clt. — надклейтрум; sb. ор. — подкрышечная кость; sb. orb. — подглазничная кость; squ.— чешуйчатая кость; sp. — брызгальце; s. orb. — надглазничная кость; sr. I. — надростральная боковая кость; sr. m. — надростральная средняя кость; s. tp. — падвисочные кости
краниум .стерляди, по величине, количеству, форме и положению подвержены большим возрастным и индивидуальным изменениям. Здесь имеются как кости типа простых чешуи, покрывающих туловище, так и крупные кости черепа сложной топографии и формы, соответствующие костям черепа других позвоночных.
Во всех случаях для костей характерно присутствие на них шипов-гребней, сходных с таковыми же на простых чсшуях и чешуях-жучках туловища. В дермокраниуме стерляди хорошо прослеживаются кости, более постоянные для всех Chondrostei.
При изучении дермокраниума прежде всего следует разобраться в покровных костях, составляющих крышу наружного костного панциря, покрывающего хрящевсй череп, далее — в окостенениях глазничной области, потом — в щечной, предкрышечпой области и, наконец, — в костях жаберной крышки.
Впереди увеличенной первой жучки спинного ряда, которую многие авторы рассматривают в составе черепа и называют затылочной (nucliale, nch.)t а А. Н. Северцов трактует как задне-затылочную кость (postoccipitale), медиально лежит небольшая треугольная кость, направленная вершиной вперед, — кожная верхнезатылочпая кость (dermosupraoccipitale, d. soc.).
Латерально от верхнезатылочной кости на уровне наружных жаберных отверстий лежат парные падвисочные кости (supratemporale, s. Zp.), имеющие форму неправильного ромба. В нижнюю часть этой кости вступает канал боковой липни. На заднебоковой границе верхпезатылочной и заднезатылочпой костей часто имеются дополнительные парные или непарные падвисочные кости, по которым проходит надвисочный канал (см. рис. 72, с. stp.).
Парные теменные кости (parietale, р.) лежат впереди затылочной и надвисочных костей, между ними, и простираются вперед и латерально. Они имеют форму плоских пластинок с радиальной исчерчепностью, как жучки, впереди граничат с лобными костями.
Чешуйчатые (squamosum, squ.) или крылоушные (pteroti-cum) кости лежат по бокам теменных костей па всем протяжении последних. Сбоку к ним примыкают верхние края жаберных крышек. Продольно через чешуйчатую кость проходит большая часть височного канала (см. рис. 72, с. tp.) системы кожных органов чувств боковой линии.
Лобные кости (frontale, fr.) парные, лежат над глазницами впереди теменных костей. Медиальный продольный гребень иа лобной кости несколько сглажен и составляет как бы продолжение гребня теменной кости. Внутри лобной кости над глазом идет надглазничный капал (см. рис. 72, с. sorb.) боковой линии.
Заднелобная кость (postfrontal, р. fr.) в виде неправильного ромба лежит кнаружи от лобной кости над глазницей и впереди чешуйчатой кости (squ.). Ее латеральный край граничит с верхнезаглазпичпой костью (pt. orb.) и надглазничными окостенениями (s. orb.). Заднелобная кость прободается подглазничным каналом (см. рис. 72, с. ino.).
Медиальные кости (mediate, med.) располагаются на уровне ноздрей между передними концами лобных костей. Медиаль
ные кости (по А. Н. Северцову) некоторые авторы рассматривают как серию центральных лобных окостенений.
Средние надростральные окостенения, по А. Н. Се-верцову (suprarostrale mediate, sr. т.), лежат впереди медиальных окостенений и переходят в ряд мелких косточек, которые простираются до конца рострума.
После рассмотрения костного панциря крыши черепа следует разобраться в покровных окостенениях, окружающих глазницу, расположение которых связано с прохождением внутри них подглазничного канала органов чувств боковой линии.
Глазница сзади ограничена заглазничным окостенением (postorbitale, pt. orb.), состоящим из двух костей — верхней и нижней заглазничной.
Кожная клиноушная (dermosphenoticum), или верхняя заглазничная (postorbitale superior), кость лежит ниже или лате-рально от заднелобной кости и впереди чешуйчатой. Эта значительных размеров Т-образная кость сзади граничит с передпеверхним краем преджаберной, или щечной, области, а спереди — с задневерхним сектором глазницы. Верхняя часть ее расширена и дает переднюю и заднюю ветви, подпирающие чешуйчатую и заднелобную кости. Вентрально она продолжается одним отростком и несколько ниже уровня вентрального края глазного отверстия сочленяется с нижней заглазничной костью.
Нижняя заглазничная (postorbitaleventrale) кость вентрально расширена и органичивает нижнезадний сектор глазного отверстия.
Надглазничные окостенения (supraorbitale, s. orb.) — ряды мелких чешуевидных костей, которые ограничивают глазное отверстие сверху.
Подглазничные окостенения (suborbitale, sb. orb.) лежат под глазным отверстием в коже и, как бы продолжая нижнюю заглазничную кость, за вертикалью ноздрей переходят в мелкие окостенения бокового ряда надростральных окостенений.
Медиально нижняя заглазничная кость переходит в ряд удлиненных трубковидных косточек, расположенных в коже на вентральной стороне головы. Они идут параллельно подглазничным и боковым надростральным окостенениям до наружного усика, постепенно уменьшаясь кпереди. Здесь этот ряд окостенений огибает спереди усик и идет далее к концу рыла параллельно базиростраль-ному и латеральному рядам надростральных окостенений. Ряд мелких трубковидных косточек окружает продолжение подглазничного канала боковой линии.
Предглазничное окостенение (praeorbitale, pr. orb.) отделяет передний край глазного отверстия от заднего обонятельного отверстия и плотно прилежит к наружному краю предглазнич-ного отростка (pr. praeorbitalis) хрящевого черепа. Оно верхним расширенным концом граничит с латеральным краем надглазничной кости, а нижний конец постепенно сужается и не доходит до переднего подглазничного окостенения.
Носовые окостенения (nasale, п.). У стерляди выше ноздрей имеется несколько увеличенных костных пластинок, которые по ‘положению, видимо, можно сравнить с носовыми окостенениями.
В толще кожистой перемычки между передним и задним носовыми отверстиями залегает ряд трубочковидиых косточек. Они окружают надглазничный канал боковой линии (см. рис. 72, с. sorb.), продолжающийся в предобонятельную область.
Дно хрящевой черепной коробки подостлано громадной пластинчатой покровной костью — парасфеноидом — и ростральными костями, которые будут описаны при рассмотрении хрящевого ней-рокраниума (стр. 239).
Скелет кожи щечной области представлен, главным образом, чешуями и костными пластинками разной формы, располагающимися в верхних слоях ее, и более крупными окостенениями, погруженными в толщу кожи. <
Пред'крышечные кости (praeoperculare, s. jugale, j) располагаются на середине латеро-вентральной половины кожи щечной области в виде поперечного ряда косточек. Этот ряд окостенений известен как гулярный, или предкрышечный. Он вытянут в дор-зовентральном направлении, его косточки имеют очень сложные контуры и рельеф наружной поверхности, форма и количество их варьируют. На окрашенных и просветленных препаратах с применением оптики можно видеть, что каждый костный элемент этого ряда пронизан сложной сетью каналов кожных органов чувств с выходами на наружную поверхность.
Жаберная крышка — задневерхняя часть жаберной крышки ниже чешуйчатой кости вплоть до верхнего края наружного жаберного отверстия, узкая, кожистая с разбросанными в ней чешуйками и костными пластинками. Основная часть жаберной крышки поддерживается тремя костными элементами. Из них дорзальный, самый массивный, есть собственно подкрышечная кость (suboperculare, sb. ор.). Ниже подкрышечной кости располагаются два других вентральных окостенения жаберной крышки меньших размеров и неправильной формы: верхний костный элемент (op. v. s.) несколько больше нижнего (op. v. inf.). Из их положения видно, что они с гиоидной дугой не связаны, а по своему происхождению, видимо, являются ее кожными лучами.
Элементы наружного костного скелета имеются и в поясах передних конечностей, но их лучше рассматривать при общем изучении поясов.
Внутренний скелет
При изучении внутреннего скелета стерляди рассматривают сначала общее строение осевого скелета туловища—хорду и позвоночник, далее детально разбирают строение отдельного скелетного сегмента в туловище и хвостовом отделе, строение хвостового плавника, непарных плавников и парных плавников и их поясов.
После этого переходят к изучению хрящевого черепа — нейрокра-ииума и, наконец, висцерального скелета — челюстно-подъязычного и жаберного аппаратов.
Осевой скелет туловища (рис. 75)
составляет основу осевого скелета сплошного круглого тяжа на всем
Рис. 75. Туловищный отдел позвоночника осетра Acipenser sturio (из Гудрича):
bd. — базидорзалиа; bv.— базивентралиа; еап. А. — гемальный канал; сап. п. — пев-Г льны й канал; ch. — хорда; ci.— ребра;
rd. rv. — отверстие для выхода дорзального н вентрального корешков спинномозгового нерва; id. — ннтердорзалиа; iv. — интервеитралиа; lg. v. — продольная позвоночная связка; meh.—оболочка хорды; pr. sp. d. — дорзальный остистый отросток;
pr. tr. — поперечный отросток
Хорда, или спинная струпа (chorda dorsalis, рис. 75 ch.), туловища. Она тянется в виде протяжении осевого скелета от гипофизарного отдела черепа до конца хвоста, заходя в верхнюю лопасть хвостового плавника. Она полупрозрачна и снаружи одета соединительнотканной и эластической оболочками (рис. 75, meh.). Последние вокруг хорды образуют своеобразный, плотно прилегающий к ней футляр, и вместе со структурными особенностями хорды определяют ее гибкость и упругость.
Позвоночник (colum-na vertebralis) представлен только верхними и нижними дугами позвонков,тела позвонков отсутствуют.
Чтобы более подробно познакомиться с деталями строения осевого скелета осетровых
рыб, следует взять участок туловища стерляди впереди уровня брюшных плавников.
При рассмотрении сбоку между верхними и нижними хрящевыми элементами позвонков хорошо видна хорда (ch.), над хордой на ее оболочке сидят массивные хрящики — в е р х н и е задние (каудальные) дуги (arcus superior caudalis), или базидорзалии (basidorsale, bd.).
Верхние каудальные дуги правой и левой сторон, сливаясь, окружают спинной мозг и над ним дают сужающийся вырост, который окостеневает и служит местом прикрепления непарного дорзального или спинного о ст и сто гб отростка (processus spinosus dorsalis, pr. sp. d.); последний в средней части окостеневает, а на концах сохраняет хрящевое состояние.
Между основаниями соседних верхних задних дуг располагаются небольшие, примерно треугольной формы хрящики — верхние передние (краниальные) дуги (arcus superior cranialis), или интердорзалии (interdorsale).
С вентральной стороны хорды напротив верхней задней дуги
располагается нижняя задняя дуга, или базивептра-л и я (arcus ventralis posterior, s. basiventrale, bv.).
По бокам между соседними нижними задними дугами залегают небольшие хрящики — нижние передние дуги (arcus ventralis anterior), или вставочные пластинки — и н т е р в е и т р а л и и (interventrale, iv.).
В туловищном отделе нижние дуги позвонков образуют поперечные отростки (processus transversus, pr., tr.), к которым прикрепляются ребра (costa, ct.). На вентральной стороне в туловище они дают г е м а л ьн ы е отростки, между которыми проходит спинная аорта. Гемальпые отростки хвостовой части вентрально смыкаются и образуют гемальный канал (canalis haema-lis, can. h.), в котором залегают хвостовые артерия и вена.
При рассмотрении положения верхних и нижних дуг позвонков относительно друг друга, мускульных сегментов--миомер, соединительнотканных перегородок — миосепт и прикрепления последних к дугам можно видеть, что у стерляди хорошо выражена дипло-спондилия, т. е. на каждый сегмент мускулатуры приходится две пары верхних и две пары нижних дуг. При этом следует особо обратить внимание на то, что через верхнюю переднюю дугу или между ней и верхней задней дугой впереди сегмента проходит отверстие для выхода переднего, или брюшного, корешка спинномозгового нерва, а между верхней передней и верхней задней или через заднюю дугу — отверстие заднего, или спинного, корешка спинномозгового нерва.
В переднем отделе позвоночника верхние и нижние дуги в каждом сегменте смыкаются и утрачивают границы; слияние выражено между сегментами тем полнее, чем ближе они к черепу. Весь комплекс сросшихся позвонков постепенно переходит в затылочный отдел черепа, составляя с ним единое целое — затылочпо-позвоноч-ный отдел черепа (regio occipito-vertebralis). Количество позвонков, вошедших в состав этой области черепа, можно установить по выходу спинномозговых нервов.
Ребра (costa, ct.) стерляди, как и у акуловых рыб, прикрепляются к поперечным отросткам нижних дуг и лежат в горизонтальных септах, поддерживая верхнебоковые отделы стенки брюшной полости. Они хорошо развиты в переднем отделе туловища и постепенно уменьшаются к его заднему отделу. В средней своей части ребра окостеневают, а концы их остаются хрящевыми.
В дополнение к рассмотренному на целом скелете разбирают строение осевого скелета па его поперечном разрезе, выясняют относительное положение хорды, элементов позвоночника, продольной позвоночной связки и спинного мозга.
Скелет непарных плавников (рис. 76)
Внутренний скелет с п и н н о г о плавника (pnn.d.) состоит из ряда хрящевых лучей (radialia, rad.), каждый из которых расчленен и состоит, как правило, из двух члеников. Основания лучей
соединительной тканью связаны с остистыми отростками верхних дуг позвонков. На этих лучах начинаются мускульные пучки, прикрепляющиеся к коже лопасти плавника и к основаниям его лепи-дотрихий. Наружный скелет плавника представлен кожными костными, главным образом расчлененными, лучами — лепидотри-хиями (1р.).
В анальном плавнике (рпп. а.) внутренний скелет составляют также хрящевые расчлененные лучи, проксимальные концы которых соединительной тканью укреплены с гемальными отростками соответствующего отдела осевого скелета. Наружная лопасть плавника поддерживается расчлененными лепидотрихиями, как и в спинном плавнике.
Рис. 76. Скелет хвостового отдела позвоночника и непарных плавников стерляди Acipenser ruthenus (из Гудрича):
bd.—базидорзалиа; bv. — базивеитралиа; ch. — хорда; f. — фулькра; id. — интердор-залиа; iv. — иитервентралиа; 1р. — лепидотрихии; рпп. а.—анальный плавник; рпп. с. — хвостовой плавник; рпп. d. — спинной плавник; рг. sp. d. — дорзальный остистый отросток; рг. sp. v. — вентральный остистый отросток; rad. — хрящевые лучи
Далее следует рассмотреть внутренний скелет гетероцеркального хвостового плавника (рпп. с.). Хорда (ch.) здесь постепенно утончается и поворачивает в его верхнюю лопасть. Сопровождающие ее верхние хрящевые дуги позвонков (bd.) с хрящевыми дорзальными остистыми отростками (рг. sp. d.) постепенно уменьшаются в размерах. Нижние хрящевые дуги с вентральными остистыми отростками (рг. sp. v.) в передней части плавника крупные с сильным разрастанием гемальных дуг, служащих в значительной мере основанием для вентральной лопасти хвостового плавника.
В ряду верхних и нижних дуг позвонков хвостового плавника промежуточные элементы (интердорзалиа и иитервентралиа) прослеживаются трудно.
Наружный скелет верхней дорзальной лопасти представлен вильчатыми, раздвоенными при основании косточками — фуль-крами (fulorum, f.) и с боков — ромбическими чешуйками. В начале плавника фулькры укороченные и широкие, в средней его части они длинные, узкие, к концу лопасти плавника — значительна
но уменьшаются и утончаются. На всем протяжении дорзальная поверхность фулькр выпуклая. Они лежат черепицеобразно, сохраняя незначительную подвижность относительно друг друга.
Вентральная лопасть хвостового плавника значительно больших размеров и в передней части имеет вырезку. Ее внутренний скелет составляют слабо выраженные хрящевые лучи — радиалии (rad.), в значительной мере прирастающие к геМальным остистым отросткам. Наружный скелет особенно хорошо выражен в переднем отделе нижней лопасти хвостового плавника. Его составляют здесь, как и в других плавниках, расчлененные лепидотрихии.
Скелет парных плавников (рис. 77)
Грудные плавники и их пояса (pinna pectoralis). Скелет грудных плавников осетровых рыб состоит из: 1) хрящевого первичного пояса грудных плавников, одетого с наружной стороны покровными костями кожного происхождения, при помощи которых он прикреплен к черепу, и 2) скелета свободных плавников, представленных хрящевыми радиальными лучами и наружными костными лучами кожного происхождения — лепидотрихиями.
Хрящевой пояс грудных плавников представлен единым хрящом сложной конфигурации без четкого разделения на отделы. При рассмотрении его с внутренней стороны можно выделить три части. Вентральная, вытянутая вперед часть — коракоидный отдел (pars, coracoidea, р. со.). Дорзальная, вертикально стоящая и несколько расширенная часть — лоп ат о ч н ы й от д е л (pars scapu-laris, р. sc.). Их соединяет также идущий вертикально и имеющий направленный вперед вырост м езо кор а ко ид (mesocoracoideum, ms со.). Последний имеет на задней поверхности сочленовую фасетку для причленения плавника. Над и под мезокоракоидом располагаются два обширных (верхнее и нижнее) отверстия (foramen superior et inferior) для мускулатуры, нервов и сосудов плавника. Над лопаточным отделом лежит небольшой вытянутый надлопаточный хрящ (cartilago suprascapularis, ssc.).
Вторичный костный пояс грудных плавников представлен системой покровных окостенений кожного происхождения, являющихся, как было указано при описании внешних признаков, производными передних чешуй — костных жучек наружного скелета туловища. В своей форме они сохраняют структуру кожных чешуй с гребнем посередине и радиальным расположение,м наружных бугорков. Рассматривать их необходимо в последовательном порядке, начиная с вентральной стороны.
Ключица (clavicula, cl.)—парная мощная покровная кость, производная передних жучек брюшного ряда. Ключицы сохраняют строение кожных чешуй и прикрывают коракоидную часть хрящевого пояса. На вентральной стороне они срастаются друг с другом (см. рис. 74).
Клейтрум (cleithrum, clt.) покрывает снаружи и спереди среднюю часть хрящевого пояса и соответствует переднему элемен
ту бокового ряда жучек туловища, также сохраняя структуру кожных чешуй (см. рис. 74).
Верхняя клейтрум (supraclcithrum, sclt.) и задняя клейтрум (postcleithrum, pelt.) лежат над клейтралыюй костью и прикрывают снаружи верхнюю часть лопаточного отдела хрящевого пояса и его надлопаточный хрящ. Верхняя клейтрум своим верхним краем причленястся к задневисочной (posttemporale), или верхней, или надвисочной (supratemporale, stp.) покровной кости дермокраниума (см. рис. 74).
Рис. 77. Скелет парных плавников стерляди Acipenser ruthenus. Л — грудной плавник, правый с медиальной стороны; Б—брюшной плавник, правый с вентральной стороны:
bspt. — базиптеригиум; cl.— ключица; сП. — клейтрум; 1р.—лепидотрихии; msco. — мезокоракоид; mtpt. — метаптеригиум; pelt.—постклейтрум; р. со. — коракоидная часть; р. sc. — лопаточная часть; rad. — хрящевые лучи; ssc. — надлопаточный хрящ; sclt. — надклейтрум;
I — 1-й луч — шип
Скелет свободного грудного плавника в основании состоит из восьми хрящевых радиальных лучей (radialia, rad.) образующих внутренний скелет. Хрящевые лучи членистые и неодинаковой длины. Проксимальными концами 1, 2, 3, 4 и 8-й лучи подвижно сочленяются с мезокоракоидпой частью первичного пояса, а 5, 6 и 7-й включены между 4-м и 8-м и не доходят до пояса. Последний, 8-й луч увеличен, на дистальном конце он ветвится. Этот луч в соответствии с хрящевым скелетом грудного плавника акуловых рыб называется метаптеригием (metapterygium, mtpt.).
К дистальным концам хрящевых лучей причленяются нижний и верхний ряды костных, как правило, расчлененных кожных лучей — лепидотрихий (1р.), составляющих наружный скелет плавника,
поддерживающий наружную лопасть последнего. По переднему краю плавника проходит массивный перасчленспнып костный луч — шип (/), образовавшийся путем слияния передних лепидо-трихий.
Брюшные плавники (pinna abdominalis, рис. 77, Б). Скелет брюшных плавников стерляди, как и других осетровых рыб, сохраняет много примитивных черт организации. Пояс брюшных плавников с осевым скелетом туловища нс связан и представлен двумя хрящевыми лопатовидными пластинками, так называемыми ба-зиптеригиями (basipterygium, bspt.). Их дистальные концы расширены и направлены латералыго. Они расчленены и служат местом прикрепления хрящевых лучей внутреннего скелета свободного брюшного плавника.
Внутренний скелет свободного брюшного плавника, как и грудного, состоит из хрящевых, у старых форм частично окостеневающих радиальных лучей (radialia rad.) в количестве не более 10, каждый из них расчленен па 2—3 элемента.
Лучи от переднего к заднему увеличиваются в длину. В последних лучах члепистость выражена полнее. Сравнительное сопоставление базиптеригиума и лучей убеждает в формировании базиптеригиума за счет срастапия члеников проксимальных отделов передних плавниковых лучей.
Наружный скелет брюшных плавников построен лепидотрнхия-ми, как и в других плавниках.
Осевой хрящевой череп, или нейрокраниум (рис. 78)
Мозговая часть черепа, нейрокраниум (neurocranium), или осевой череп, представляет сплошную массивную хрящевую коробку, окружающую головной мозг и высшие органы чувств — обоняния, зрения, слуха и равновесия.
В черепе стерляди можно выделить следующие друг за другом отделы: ростральный, рыло, или рострум (rostrum), обонятельный, или этмоидный (regio ethmoidalis), глазничный, или орбитальный (г. orbitalis), слуховой (г. otica) и затылочно-позвоночный (г. occi-pito-vertebralis).
Основание черепа па всем протяжении подослано непарными покровными окостенениями — парасфеноидом и базальными ростральными окостенениями — базиростралиями.
Парасфеноид (parasphcnoidcum, psph.) представляет массивную покровную кость, простирающуюся под основанием черепа от обонятельной области до переднего отдела позвоночника и таким образом подстилающую глазничный, слуховой и весь затылочно-позвоночный отделы хрящевого черепа. По всей длине он пологосводчатый. Боковые края его слегка загибаются дорзалыю. На уровне заглазничных отростков хрящевого черепа он дает боковые отростки (processus lateralis),’ или крылья, восходящие дорзолатерально и подстилающие основание и заднебоковые поверхности заглазничных отростков.
Задний конец парасфеноида имеет медиальную вьфезку, латерально от последней он переходит в задние боковые отростки— крылья (processus lateralis posterior, pr. lat. post.).
Базиростральные окостенения (basirostrale) располагаются впереди парасфеноида, как бы продолжая его вперед и подстилая основание рострума до переднего его конца. На вентральной стороне базиростралии несут несколько медиальных выростов, уменьшающихся к переднему концу.
пг. в t. orb-
Рис. 78. Хрящевой череп (осевой и висцеральный) стерляди Acipenser ruthenus.-: b. br. — базибранхиале; b. rst. tn. — среднеростральное окостенение; c. br. — цератобранхиа-ле; с. h.—цсратохиалс; d.—зубная кость; е. br. — эпибранхиале; f. с. ini. — отверстие для-внутренней сонной артерии; h. br. — гипобранхиале; hh. — гипохиале; hind. — гномандибу-ляре; ih. — интерхиале; тх.+ртх.— максила-прсмаксила; ph. br.—фарингобранхиале: pl. — небная кость; pr. lat. post. — задний боковой отросток парасфеноида; рг. ос. I. боковой затылочный отросток; рг. ос. т. — средний затылочный отросток; pr. plb. I. — боковой, небно-основной отросток; pr. plb. tn. — средний небно-основной отросток; pr. porb.—пред-глазничпый отросток; pr. pt. orb.—заглазнвчный отросток; psph.—парасфеноид; pt. —- крыловидная кость; sy. — симплектикум; /, II, III, IV— VII, IX, X — отверстия для выхода головных нервов
Ростральный отдел, или рострум (rostrum, рис. 78, rst.), представляет собой передний, непарный, конической формы вырост хрящевого черепа, кзади постепенно переходящий в расширенный обонятельный отдел. Передний конец рострума заострен. На его вентральной поверхности медиально на всем его протяжении наподобие киля проходит продольный хрящевой валик. Последний является как бы непосредственным продолжением среднего небно-оспов-пого отростка черепа (processus palato-basialis medialis, pr. plb. m.). Вентрально-хрящевой валик подстилается покровным среднеростральным окостенением (os basirostrale mediale, b. rst. m.). По бокам от медиального рострального гребня проходят желобообразные продольные впадины. Они тянутся сзади от нижней части предглазничного отростка (processus praeorbitalis, pr porb.) вперед к концу рострума.
Желобообразные впадины служат для прохождения в них ветвей тройничного и лицевого нервов и кровеносных сосудов, иннер-240
вирующих и питающих органы чувств и другие системы вентральной стороны рострума.
Дорзальная поверхность рострума выпуклая, спадающая к его краям. Топография (рельеф) дорзальной стороны рострума неровная. Здесь проходят продольные бороздки и желобки с тонкой и сложной сетью перемычек и отверстий, ведущих в толщу хряща и служащих для прохождения нервов и сосудов. Кроме этого, дорзальная сторона рострума несет отпечаток прилегающих к нему покровных костей.
Обонятельная, или этмоидная, область (regio ethmoidalis). Эта расширенная часть черепа позади основания рострума является скелетной капсулой органа обоняния. Задняя граница этой области пройдет по переднему краю глазниц и по заднему краю обонятельных капсул. Характерным для этой области в отличие от таковой хрящевых рыб является латеродорзальное положение обонятельных капсул с обширными наружными отверстиями. Обонятельная капсула вмещает обонятельный мешок, дно ее, направленное назад, пронизано системой отверстий для выхода обонятельных нервов (n. olfactorius).
Глазница — орбита (orbita). Представляет обширную глубокую впадину в хрящевом черепе позади обонятельной области. Спереди она ограничена предглазничным отростком (рг. ргае-orbitalis.pr.porb.), сзади — за глазничным (pr. postorbitalis, pr. pt. orb.), сверху — латеральным краем крыши хрящевого черепа, часто в виде навеса глазницы.
Вентрально глазница подходит к основанию черепа, подостланному парасфеноидом, а вперед продолжается до бокового небноосновного отростка черепа (pr. palato-basalis lateralis, pr. plb. I.). Глазница служит вместилищем органа зрения — глаза, его мускулатуры, сосудов. В пей проходят нервы, снабжающие ряд органов передней части головы, а также прикрепляются и располагаются мышцы челюстного аппарата.
В глазнице находят отверстия для выхода нервов и кровеносных сосудов. Главнейшие из них: 1) отверстие для выхода зрительного нерва (for. n. optici, II)—большое отверстие, располагается в передне-нижнем отделе глазницы и переходит в расширяющийся желобок по направлению передп'е-ннжпего угла орбиты; 2) отверстие для выхода глазодвигательного нер-в a (foramen n. oculomotorii, III) — небольшое округлое отверстие, несколько позади первого, на том же уровне; 3) отверстие для выхода блокового нерва (foramen n. trochlearis, IV)—маленькое отверстие в верхней части глазницы, выше двух первых и посередине расстояния между ними; 4) в заднем нижнем углу глазницы на линии отверстий зрительного и глазодвигательного нервов лежит с неправильными контурами обширное отверстие для выхода тройничного, отводящего и лицевого нервов и кровеносных сосудов (for, nn. trigemini, abducentis, facialis, V, VI, VII)-, 5) в середине основания глазницы, несколько выше латерального края парасфеноида, почти под отверстием глазодвигательного
нерва видно округлое отверстие для выхода внутр синей сонной артерии (for. a. carotis interna, f. с. int.).
Кроме этого, топография глазницы усложнена системой других отверстий и желобков для прохождения нервов и сосудов и перихондральными окостенениями, особенно у старых экземпляров. Из последних Встречаются окостенения, гомологичные костям костных рыб: боковая клиновидная кость (laterosphenoideum) около выхода III, V, VII нервов, глазнично-клиновидное (orbito-sphenoideum) окостенение около выхода IV нерва и в передней стенке глазницы впереди выхода зрительного нерва — этмоидальное окостенение (ethmoideum).
Слуховой, или височный, отдел (regio otica, s. temporalis). Представляет расширенную часть черепа позади заглазничных отростков и кроме большей части головного мозга заключает внутри органа слуха и равновесия. При рассмотрении сверху у молодых форм видны просвечивающие через прозрачную хрящевую стенку полукружные каналы внутреннего слухового лабиринта. Внизу эта область подостлана парасфеноидом и его передними боковыми крыльями, назад она суживается вплоть до обширного отверстия выхода блуждающего нерва (п. vagus, X). Позади заглазничных отростков и передних боковых крыльев парасфеноида располагается сочленовная впадина подвеска (fossa glenoidalis). Под пей между хрящевой стенкой слуховой области и основанием крыла парасфеноида проходит канал для челюстно-подъязычной ветви лицевого нерва (ramus hyomandibularis, VII), кзади переходящий в желобок, в котором ниже сочленовной впадины и несколько отступя от заднего края основания крыла парасфеноида хорошо видно отверстие для выхода языкоглоточного нерва (for. n. glossopharyngei, IX).
Нижнебоковая стенка слуховой области между сочленовной впадиной подвеска и отверстием блуждающего нерва (X) в своей средней части служит местом укрепления передних жаберных дуг.
Затылочно-позвоночная область (regio occipito-vertebralis) черепа стерляди расположена кзади от выхода блуждающего нерва. Она постепенно расширяется в стороны и переходит в парные боковые затылочные отростки (рг. occipitalis lateralis, рг. ос. lat.), а медиально образует средний затылочный отросток (рг. occipitalis medialis, рг. ос. т.), направленный назад, и, наконец, переходит в передний отдел позвоночного столба из сросшихся хрящевых позвонков.
Задняя граница затылочно-позвоночного отдела черепа стерляди лежит на уровне заднего конца парасфеноида. Между медиальным и латеральным затылочными отростками этой области черепа располагаются глубокие задние затылочные впадины, открытые сверху и сзади. Эта впадины служат местом начала туловищной мускулатуры. Здесь по отверстиям выхода корешков спинномозговых нервов хорошо прослеживается количество сегментов позвоночного столба, вошедших в состав затылочно-позвоночного отдела черепа.
к боковым затылочным отросткам прикрепляются верхние отделы плечевого пояса, а к среднему — продольная позвоночная связка и дорзальная часть туловищной мускулатуры.
Висцеральный скелет, или спланхнокраниум (см. рис. 78 и 81)
Для того чтобы рассмотреть висцеральный скелет стерляди (splanchnocranium), следует при очистке скелета не отчленять висцеральный скелет от осевого черепа, а составить его при черепе па связках. Внутрь висцерального черепа рекомендуется через ротовое отверстие вставить тампон ваты и расправить на нем висцеральные дуги.
Висцеральный скелет стерляди состоит из семи висцеральных дуг, охватывающих с обеих сторон ротоглоточную полость: из челюстной дуги, подъязычной дуги и пяти жаберных дуг.
Челюстная п подъязычная дуги образуют характерный для осетровых рыб челюстно-подъязычный аппарат в виде выбрасываемой вперед выдвижной воронки, что является важным биологическим приспособлением для донных рыб, собирающих пищу со дна. Остальные висцеральные дуги выполняют преимущественно дыхательную функцию, а также участвуют в сложных глотательных движениях. Все они подвижно сочленены своими дорзальными элементами со слуховой и затылочно-позвоночной частями черепа, а на вентральной стороне подвижно связаны между собой межжаберными элементами, а через последние прикреплены связками к вентральному элементу подъязычной дуги. Таким образом, при совершении хватательных, глотательных, дыхательных и других движений весь висцеральный скелет морфофункционально является единым целостным .аппаратом.
Челюстная дуга (arcus mandibularis). Челюстная дуга состоит из верхней и нижней челюстей, преобразованных в вытяжной ротовой аппарат. Первичная верхняя челюсть представлена сложным небно-крыловидно-квадратным хрящом. Первичная нижняя челюсть представлена меккелевым хрящом.
Верхняя челюсть — неб по-кры л овидно -квадратный хрящ (cartilago palato-pterygo-quadrata, рис. 81, pl. pt. qu.) и его окостенения у осетровых рыб имеют сложное строение. На каждой стороне верхняя челюсть имеет форму веерообразного хряща. Медиальный край ее расширен и несколько выпуклый сверху, каудальный — короткий, утолщен и несет сочленовную площадку для меккелева хряща и симплектикума. Медиально верхние челюсти противоположных сторон соединены узким симфизисом. На вентральной стороне каждая половина челюсти подостлана веерообразной покровной крыловидной костью (pterygoi-deum, рис. 78, рt.). Между передними и задними крыльями ее медиальной части у старых стерлядей встречается небольшое покровное кожно-небное окостенение (dermopalatinum). Передний край хрящевой верхней челюсти до сустава с первичной -нижней челюстью одет покровной комплексной вторичной верхней
челюстью—челюстио-предчелюстной костью (maxillo-praemaxillare, тх.-\-ртх.), соответствующей двум костям вторичной верхней челюсти большинства костных рыб и наземных позвоночных— челюстной (maxillare) и предчелюстной (praemaxillare).
Дополнительный хрящ верхней челюсти — не бно - кр ы л о-видная пластинка (cartilago palato-pterygoidea accessoria, рис. 81, pl. pt. асе.) —непарная хрящевая пластинка ромбовидной формы, лежит непосредственно позади небно-крыло-квадратпого хряща и образует задний выступ крыши выдвижного челюстного аппарата.
Нижняя челюсть (mandibula) состоит из парных первичных хрящевых челюстей— меккелевых хрящей (cartilago Meckeli, рис. 81, с. М.) и вторичных костных челюстей—покровный зубных костей (dentale, d.), одевающих снаружи большую часть меккелева хряща, который в его суставном отделе сохраняется хрящевым. У старых осетров на внутренней стороне меккелева хряща встречаются дополнительные покровные кости — пластинчатая (spleniale) и угловая (angulare). Медиально нижние челюсти дают симфиз.
Подъязычная, или гиоидная дуга (arcus hyoideus). Это вторая висцеральная дуга, выполняет функции подвеска челюстного аппарата к осевому черепу и поддерживает жаберную крышку. Верхний отдел подъязычной дуги, соответствующий гиомандибу-лярному хрящу акуловых рыб, состоит из двух подвижно сочленяющихся элементов: собственно подвеска — гиомандибулярпого хряща и симплектикума. Вентральный отдел — гиоид расчленен на три элемента и на вентральной стороне причленяется к жаберному скелету.
Гиомандибулярный хрящ (hyomandibulare, рис. 78, hmd.) подвешивает выдвижной челюстной аппарат к черепу в слуховой Области. От лопатовидной формы значительная часть его окостеневает. На дорзальном конце хряща помещается сочленовная головка. Нижний расширенный конец образует подвижное сочленение с симплектикумом. Спереди и сзади к нему прикрепляются мощные мышцы, связывающие хрящ с осевым черепом, а прочная соединительнотканная связка идет к верхней челюсти.
Сим'плектикум (symplecticum, рис. 78, sy.), второй элемент подвеска, направлен по отношению к гиомандибулярному элементу вперед и вниз. Передний расширенный конец его сочленяется с задним концом верхней челюсти.
Гиоид (hyoideum, рис. 78), вентральный отдел подъязычной дуги, соединяет челюстной аппарат с жаберным скелетом. Он состоит из трех хрящей, связанных между собой плотной соединительной тканью: 1) и н те р х и а л ь н ы й хрящ (interhyale, ih.)— верхний элемент, соединяющийся передним концом с симплектикумом, а задним — с гиоидом; 2) цератохиальный хрящ (ceratohyale, с. h.) — основной элемент гиоида, одетый в средней части костной муфтой. На его дистальном конце имеется крыловидный вырост, от которого простирается мандибулярная связка к 244
нижней челюсти; 3) г и п о х и а л ь н ы й хрящ (hypohyale, hh.) — небольшой вентральный хрящевой элемент гиоида, сочленяющийся с непарной копулой жаберного аппарата.
Скелет жаберного аппарата. Состоит из пяти жаберных дуг (arcus branchialis), связанных друг с другом соединительнотканными связками в единую систему. Жаберный скелет прикрепляется дорзальными элементами к основанию осевого черепа, а вентральными концами — к непарным хрящам, копулам, соединяющим жаберные дуги на брюшной стороне. Первые три жаберные дуги расчленены на четыре скелетных членика. На основных (средних) дорзальных и вентральных члениках жаберных дуг с внутренней стороны, спереди и сзади сидят два ряда жаберных тычинок— треугольных хрящиков, образующих цедиальный аппарат. С наружной стороны основных члеников жаберных дуг прикрепляются два ряда жаберных лепестков, отделенных друг от друга жа-, верными перегородками (см. жаберный аппарат, стр. 206). При препарировании скелета они удаляются от скелета жаберных дуг. Скелет жаберных дуг хрящевой, но основные дорзальные и вентральные членики частично окостеневают и вокруг хрящевых стержней перихондрально развиваются костные манжетки.
На примере первых трех жаберных дуг разбирают строение отдельной дуги, рассматривая жаберный скелет с брюшной стороны (рис. 78).
Копулы, или б а з и б р а х и а л ь ны е хрящи (basibran-chiale, b. br.),— непарный ряд хрящей, плотно сросшихся друг с другом, — лежит на брюшной стороне жаберного скелета: к ним справа и слева причленяются жаберные дуги. Первые три жаберные дуги причленяются к слившимся первой и второй базибранхиале; четвертая и пятая дуги причленяются к слившимся двум задним базибранхиальным элементам.
Поджаберные, ил и гипобранхиальные, членики (hypobranchiale, п. br.) — короткие, почти кубические хрящики, которыми жаберные дуги сочленяются с копулами.
Рожковидно-жаберные, цератобранхиальные, членики (ceratobranchiale, с. br.) —основные вентральные части жаберных дуг, самые крупные элементы жаберного скелета. Они дугообразной формы, выпуклая сторона их обращена кнаружи, верхними концами наклонены назад.
Надглоточно-жаберные, или эпибранхиальные, членики (epibranchiale, рис. 78, е. br.) — основные дорзальные элементы жаберной дуги. От места сочленения с цератобранхиале они направлены вверх и вперед, но они более короткие, чем вентральные элементы.
Глоточно-жаберные, или ф а р и н г о б р а н х и а л ьп ы е, хрящики (pharyngobranchiale, рис. 78, ph. br.) —двойные дорзальные элементы, которыми жаберные дуги причленяются к слуховому и затылочно-позвоночному отделам осевого черепа. Передний хрящик связывает заднюю дугу с передней, а задний — с осевым черепом.
Четвертая и пятая жаберные дуги неполные: у четвертой нет глоточного элемента, и она причленяется эпибранхиальным элементом к глоточному элементу третьей дуги; пятая дуга имеет только вентральный — церато-бранхиальный элемент.
Мускулатура
Мускулатура осетровых рыб сохраняет типичные черты мускульной системы рыб, заключающиеся в сохранении сегментального строения из последовательного ряда мышечных сегментов-миомеров лишь с началом распределения на отдельные мышцы.
Рис. 79. Мускулатура головы, 'переднего отдела туловища (сбоку) и грудных плавников (сверху) у стерляди Acipenser ruthenus:
hmd. — подвесок; тт. — миомеры; т. ad. р. — мускул, поднимающий грудной плавник; т. consir. sv. — поверхностный вентральный сжиматель; т. obi. ех. — наружная косая мышца; т. ор. — крышечная мышца; т. pr. ft. — протрактор подвеска; т. pr. р. d. — дорзальный протрактор грудного плавника; т. рг. а— вентральный протрактор грудного плавника; т. г. ab. — прямая мышца живота; т. rt. ft. — ретрактор подвеска; т. rt. р. d.— дорзальный ретрактор грудного плавника; ms. — миосепта; s. ft.—горизонтальная септа
Только в голове имеется сложная система отдельных мышц челюстного и жаберного аппаратов, а также мускулатуры глаз.
Туловищная мускулатура (рис. 79) с наружной стороны имеет вид зигзагообразно изломанных поперечных лент — миом еров (тт.), отделенных друг от друга соединительнотканными прослойками— миосептами (ms.). На обеих сторонах тела туловищная мускулатура делится продольной перегородкой — горизонтальной септой (s.h.) — в виде желобка на две группы. Над горизонтальной септой обособляется надосная — эпаксиальная группа спинной мускулатуры. Под горизонтальной септой выделяет-246
ся группа миотомов боковых стенок брюшной полости, называемая подосной — гипаксиальной группой. Поверхностный слой этой группы обособляется в наружные косые мышцы живота (m. obliquus abdominis externus, т. obi. ex.). На вентральной стороне также обособляется продольная полоса прямой мышцы живота (m. rectus abdominis, т. г. ab.).
Мускулатура непарных плавников. Состоит из отдельных сегментальных мускульных пучков, отчленившихся от дорзальных концов миотомов, проксимальным концом прикрепляющихся к радиальным лучам, а дистальным — к лепидотрихиям.
Мускулатура грудных плавников. Подразделяется па дорзальную и вентральную. Дорзальная группа мускулов поднимает плавник, эти мышцы называются аддукторами (m. adductor рес-toralis). Вентральная группа мускулов опускает плавник, они называются абдукторами (m. abductor pectoralis). Кроме того, имеются мышцы, растягивающие плавник и оттягивающие его,— это п р о т р а к т о р ы и р е т р а к т о р ы. Соответственно функции они носят названия: m. protractor pectoralis dorsalis et. m. retractor pectoralis dorsalis; m. protractor pectoralis vontralis et m. retractor pectoralis ventralis.
Мускулатура головы. Мускулатура головы разделяется на соматическую и висцеральную. Соматическая мускулатура в голове представлена затылочными мышцами, соединяющими череп с туловищем, и поджаберной—гипобранхиальной — мускулатурой челюстного и жаберного аппаратов, идущей от плечевого пояса к подъязычному аппарату, а также глазными мышцами. Висцеральная мускулатура челюстного и жаберного аппаратов достигает сложного функционального расчленения на систему мышц, совершающих самостоятельные акты: захватывание пищи, дыхательные и глотательные.
Затылочные мышцы являются продолжением эпакси-альной группы мышц миомеров туловища, заполняющих затылочные впадины затылочного и затылочно-позвоночного отделов черепа.
Поджаберная — гипобрапхиальная — мускулатура представлена тремя мышцами, идущими от плечевого пояса вперед к вентральным отделам челюстного и жаберного аппаратов. Соответственно местам прикрепления от коракоидного отдела пояса грудных плавников к жаберным и подъязычной дугам они называются: m. cartilago arcuata anterior, т. cartilago arcuata posterior, m. branchio-mandibularis.
Мускулатура челюстной и подъязычной дуг •представляет группу мышц, тесно связанную в единую систему выдвижения челюстного аппарата для захватывания пищи со дна. Мышцы, соединяющие верхнюю челюсть с осевым черепом, отсутствуют.
Дорзальный отдел мускулатуры челюстной дуги образует мощную мышцу — протрактор гиомандибулярного хряща (m. protractor hyomandibularis, рис. 79, т. pr. h.), тянущий че
люстной аппарат вперед. Эта мышца оканчивается на переднем крае гиомандибулярного хряща, а не на верхней челюсти, как у акул.
Вентральный отдел челюстных мышц образует челюстной аддуктор, (m. adductor mandibularis, рис. 81, т. add. md.), прикрепляющийся одним концом к верхней, а другим — к нижней челюсти.
Рпс. 81, Мускулатура челюстного ап-парата стерляди Acipenser ruthenus: с. т. — меккелев хрящ; d.—зубная кость; т. add. md. — мускул, поднимающий нижнюю челюсть; т. add. md. а. — сочленовная порция мускула, поднимающего инжнюю челюсть; т. add. md. s.—симфнзиальная порция мускула, поднимающего нижнюю челюсть: тх,+ртх. — верхнечелюстно-
предчелюстная кость; pl. pt. асе.—дополнительная пебно-крыловндная пластинка; pl. pt. qu. — небно-крыловидно-квадратный хрящ
Рнс. 80. Мускулатура головы и грудного плавника стерляди Acipenser ги-thenus (снизу):
т. ab. р. — мускул, опускающий грудной плавник; т. br. md. — жаберио-чслюстной мускул; т. const, sv. — поверхностный вентральный сжиматель; т. сог. аг. — мускул коракондно-дужный; т. рг. h. - протрактор гломандибулярного хряща; т. рг. V. — вентральный протрактор грудного плавника; т. rt. и. — вентральный ретрак-_тор грудного плавника; 1 1-й луч —шип
Дорзальный отдел мускулатуры подъязычной дуги обособляется в две сильные мышцы, оттягивающие челюстной аппарат назад: передняя мышца носит название ретрактора (m. retractor hyomandibularis, рис. 79, т. rt. h.), а задняя — оперкулярной мышцы (m. opercularis, рис. 79, т. ор.).
Вентральный отдел мускулатуры подъязычной дуги носит общее название вентрального сжимателя — констриктора (m. constrictor ventralis), который распадается на четыре мышечных пучка, имеющих самостоятельные названия (рис. 79 и 80, т. constr. vs.).
Дорзальные мышцы жаберных дуг образуют ряд мышц — под-нимателей дуг (m. levator arcus branchialis).
Вентральные мышцы жаберных дуг образуют три системы мы-
щечных пучков: поддужные прямые (m. subarcuata recti), вентральные косые (m. obliquus ventralis) и вентральные поперечные (m. transversus ventralis).
Литература по осетровым рыбам (хрящевые ганоиды)
Берг Л. С. Система рыб. «Труды Зоологического ин-та», т. 5, вып. 2. М,— Л., 1940.
Гиммельрейх Г. А. Висцеральный аппарат осетрообразных как орган приема пищи. «Труды ин-та Морфологии животных им. А. Н. Северцова». АН СССР, вып. 38, 1963.
3 о г р а ф Н. Ю. Материалы к познанию организации стерляди. «Известия об-ва любителей естествознания», вып. 52. М„ 1887.
Северцов А. Н. Положение хрящевых ганоидов в системе рыб. «Русский зоологический журнал», т. 3, 1922.
Суворов Е. К. Основы ихтиологии. Л., 1948.
Goronowitsch N. Das Gehirn und die Cranialnerven von Acipenser ruthe-nus. Morph. Jahrb. Bd. I, H. 4, 1888.
Jonston J. B. The Brain of Acipenser. A contribution to the morphology of the xrertebfal brain. Zool. Jahrb. Abt. Anat., 15, 1901.
Luther A. Beitrage zur Kenntniss des Kopfskelett der Knorpelganoiden. Acta Soc. Scient Fenn., 41, n. 8, 1913.
Maschkowzeff A. A. Der Genitalapparat der Acipenseridae. Zool. Jahrb., Bd. 58, 1934.
Meinel W. Zur Morphologie und Functi on der Splanchnocranium von Acipenser. Zeitschr. f. Morph. Okol. Tiere, Bd. 51, 1962.
Parker W. B. On the structure and development of scull in Sturgeon Acipenser ruthenus and Acip. sturio. Phil. Trans. R. Soc. London, 22, 1881—1883.
Rako w W. Zur Frage der Verknocherungen der Knorpelschadels der Acipen-seriden. Zool. Jahrb. Abt. Anat. n. Ontog., H. 3, 1933.
КЛАСС КОСТНЫЕ РЫБЫ — OSTEICHTHYES
ПОДКЛАССЛУЧЕПЕРЫЕ-ACTINOPTERYGII
Надотряд костистые рыбы — Teleostei
Общая характеристика надотряда
К надотряду костистых рыб относится большинство (около 95%) современных рыб. Они населяют всю толщу океана до самых больших глубин, все реки и другие пресноводные водоемы.
Разнообразные формы, объединяемые в этот падотряд, обладают следующими общими признаками. Чешуя костная — циклоидная или ктсноидная. Позвонки амфицельные (двояковогнутые) и полностью окостеневшие. В затылочной области черепа имеется верхпезатылочпая кость. Нижняя челюсть состоит из трех костей: dentale, angulare, articulare. Срединные гулярныс пластинки (gularia) утрачиваются, но па их месте развиваются связанные с гиоидом лучи, поддерживающие жаберную перепонку. Грудные плавники располагаются по бокам тела, за головой, и очень подвижны. Положение брюшных плавников сильно варьирует. У разных форм они могут прикрепляться к брюшной поверхности тела в разных местах и часто лежат впереди грудных плавников. Хвостовой плавник гомоцеркальный (вторично равнолопастной) или ги-фоцеркальный (образовавшийся в результате слияния спинного и анального, например луна-рыба). Артериальный конус редуцирован, на его месте имеется большая луковица аорты. Спиральный клапан отсутствует. Плавательный пузырь у примитивных форм в течение всей жизни соединен особым протоком, или шнуром, с дорзальной стороной пищевода. '
Представители надотряда костистых рыб
К этому падотряду относятся около сорока отрядов. По степени примитивности они объединяются в три группы. Из первой группы здесь рассматривается щука — отряд щукообразные (Esociformes), из второй — сазан (карп)—отряд карпообразные (Cypriniformes), из наиболее прогрессивной третьей группы — окунь и судак — отряд окунеобразные (Perciformes).
Изучение скелета костистых рыб удобнее вести на судаке, череп которого отличается достаточно крупными размерами и довольно полным набором окостенений. В этом разделе даются ссылки и на особенности скелета других рассматриваемых в руководстве представителей.
За основной вид, по которому будет вестись изучение всех систем органов костистых рыб (кроме скелета), взята щука (Esox 250
lucius L.)—вид, широко распространенный по всему Советскому Союзу. Щука — наиболее удобный объект для работы по своим размерам. Для двух других также широко распространенных представителей костистых — окуня и карпа — описываются только отличительные особенности, ссылки на которые даны непосредственно в тексте, после описания соответствующей системы щуки.
Щука Esox lucius L. (рис. 82,Л) принадлежит к семейству щуковых (Esocidae), одному из четырех семейств отряда щукообразных (Esociformes). Этот вид населяет почти все водоемы нашей страны: реки, озера, водохранилища. Щука — типичный хищник, питающийся в основном рыбой. Крупные щуки могут хватать утят, гусят, водяных крыс, лягушек и т. д. Стоя неподвижно среди зарослей подводных растений, около коряг, камней, под обрывами берега, опа подстерегает добычу и при ее появлении сильным рывком бросается вперед и хватает почти без промаха. Схватив добычу, опа проплывает с ней некоторое расстояние и только потом начинает заглатывать. Заглатывается рыба всегда головой вперед. Половозрелыми щуки становятся обычно на четвертом году жизни. Самки откладывают, главным образом па растительность, от 100 000 до 1 000 000 икринок.
Окунь обыкновенный — Perea fluviatilis L. (рис. 82, В)—представитель семейства окуневых (Percidae). Это семейство вместе почти с 40 другими входит в отряд окунеобразных (Perciformes).
Окунь —широко распространенная рыба, живет в реках, озерах, водохранилищах. Здесь он скрывается в зарослях травы, в корягах, под нависшими деревьями, в ямах и т. и. Такие места служат для пего защитой и засадой, откуда он бросается на свою добычу. Так же, как и щука, окунь — хищник, по в соответствии с меньшими размерами (рыба весом в 1 кг считается уже крупной) питается и более мелкими объектами. В отличие от щуки окунь упорно преследует свою добычу, пока не овладеет ею. Половоззрелыми окупи становятся па третьем году жизни. Одна самка откладывает от 12 000 до 30 000 икринок.
Судак обыкновенный — Lucioperca lucioperca L. (рис. 82, Б) так же, как окунь, — представитель семейства окуневых, но относится к роду судаков (Lucioperca). Это довольно распространенная у нас рыба, населяющая реки бассейнов Балтийского, Черного, Каспийского и Азовского морей. В южных морях живет и в солоноватой воде. Кроме рек, судак населяет большие и малые озера, водохранилища. Судак — типичный хищник, ведет ночной образ жизни. Питается в основном мелкими рыбами других пород, причем так же, как и окунь, активно преследует добычу. Половозрелыми рыбы становятся в возрасте более трех лет. Одна самка откладывает от 200000 до 1 000 000 мелких (1—1,3 мм в диаметре) икринок. Самец активно охраняет икру.
Сазан — Cyprinus carpio (рис. 82, Г)—представитель отряда карпообразных (Cypriniformes), или костнопузырных (Ostariophy-si). Этот довольно богатый представителями отряд объединяет 4 подотряда. Сазан относится к подотряду карповидных (Cyprinoidei),
впп. d.
CCCpI^A-Z^Ew^uc^ (ИЗ кииг” <п.Ромысловые рыбы
J - Г — ^S^Pl^rCa;
а'^п^п^
семейству карповых (Cyprinidae). Он распространен в реках бассейнов Черного, Азовского, Каспийского и Аральского морей, а также в реках бассейна Тихого океана. Живет обычно в водоемах с замедленным течением. В отличие от видов, описанных выше, — это «мирная» рыба, питающаяся донными организмами. Половозрелым становится на 4-м — 5-м году жизни при общей длине около 40 см. Самка откладывает от 96 000 до 1 810 000 икринок. На занятиях может быть использована и культурная форма сазана —карп.
Карп является основным объектом прудового разведения костистых рыб. Для этой цели в результате селекционной работы выведен ряд ценных домашних пород, отличающихся высокой скороспелостью при применении интенсивных форм ведения хозяйства (удобрение прудов, подкормка и т. д.).
Для изучения костистых рыб желательно иметь па каждого студента по два экземпляра рыб: один — для отработки задания ПО' вскрытию, другой — по мускулатуре и нервной системе.
Изучение скелета проводится по заранее приготовленным скелетам судака. Приготовление скелетного материала описано в разделе методики и в соответствующем разделе по скелету. В данном случае возможен следующий порядок отработки заданий: вскрытие, скелет, мускулатура и периферические головные нервы, головной мозг. В таком порядке изложен материал и в настоящем практикуме. Если готового материала по скелету не имеется, то на одном экземпляре производят вскрытие и смотрят головной мозг, другой экземпляр после изучения мускулатуры идет на изготовление скелета. При наличии готового материала можно ограничиться и одним свежим или фиксированным экземпляром на каждого студента. Порядок работы тогда будет следующий: скелет, мускулатура и периферическая нервная система, вскрытие, головной мозг.
Вскрытие
Внешние признаки
Тело щуки (рис. 82, А) сильно вытянуто в длину и сжато с боков, почти одинаковой высоты на всем протяжении. Спереди оно-заканчивается довольно крупной головой, а сзади — суженным округлым хвостовым стеблем. Ясных границ между отдельными частями тела нет. Можно считать, что голова кончается на уровне заднего края жаберной крышки, за ней до уровня анального отверстия следует туловище, далее назад — хвост.
Вытянутая форма тела с заостренным передним концом — приспособление к стремительному движению в воде во время броска на добычу. Ровная прямая форма спины позволяет щуке плавать или стоять под самой поверхностью воды и на мелких местах, оставаясь незамеченной. Этому же способствует и окраска, которая хорошо видна на свежей рыбе. Как у огромного большинства рыб, верхняя часть тела щуки окрашена в темные тона, нижняя — светлая. В воде рыба обычно освещена светом, падающим сверху.
В этом случае па брюхе возникает демаскирующая тень, подчеркивающая объемность тела рыбы. Светлая окраска брюха смягчает эту тень, и вместе с темной окраской спины делает рыбу однотонно окрашенной. Тело рыбы как бы теряет свой объем и становится менее заметным. Покрывающие бока более светлые и темные пятна, объединяющиеся у некоторых особей в продольные или поперечные полосы, хорошо маскируют рыбу в зарослях водной растительности.
В отличие от щуки тело окуня (рис. 82, В) более высокое и компактное, довольно сильно сжатое с боков, с большим или меньшим горбом на спине. Спереди оно 1приостренное, но с относительно тупым концом, а сзади плавно переходит в хвостовой стебель. Спина темпоокрашенная, брюхо светлое. По бокам тела ряд темных поперечных полос, играющих маскирующую роль.
Тело сазана (карпа) (рис. 82, Г)—малоподвижной рыбы — плотное, вальковатое, с относительно слабым изгибом спины и почти прямой линией брюха. Верхняя часть темно, нижняя светло окрашены.
Покровы. Туловищный и хвостовой отделы костистых рыб покрыты настоящей костной чешуей, отсутствие ее — явление вторичного характера, что наблюдается в различной степени у разных форм карпа. Чешуя местами заходит и на голову: в затылочной области, на жаберной крышке и па «горле». Отдельные чешуйки располагаются па теле косыми рядами, причем передние чешуйки налегают на задние, так что с поверхности видны только небольшие задние края их. Наиболее крупные чешуи лежат по средней линии с боков тела. На спине, брюхе, хвосте и верхней части головы они значительно мельче.
Для того чтобы ознакомиться со строением чешуи рыбы, выдергивают несколько чешуек пинцетом, очищают их задние края от покрывающего эпидермиса, промывают и рассматривают под лупой. Рассматривать следует в воде: подсохшие чешуйки становятся менее прозрачными. Можно изготовить временный препарат, поместив чешуйку в каплю глицерина и зажав ее между двумя предметными стеклами. У костистых рыб имеется два типа костных чешуй: циклоидная (округлая) и ктеноидпая (гребенчатая).
Чешуя щуки — циклоидная, однако форма ее не строго округлая. Задний, выступающий наружу край округлый; передний, сидящий в коже, разделен на лопасти, частично налегающие друг на друга (рис. 83, 1). Вся внешняя поверхность чешуи покрыта кольцевыми, склеритовыми ребрышками. Расстояние между склеритами различно. Считается, что весной и летом в период интенсивного роста склеритовыс кольца закладываются па большом расстоянии друг от друга. Осенью и зимой склериты закладываются близко друг к другу пли совсем не закладываются. Граница между тесно расположенными склеритами осеннего роста и широко раздвинутыми склеритами весенне-летнего роста называется годовым, или годичным кольцом (рис. 83, г. к.). По числу годовых колец определяется возраст рыбы. Рост чешуи идет за счет образования повой
тонкой костной пластинки па ее внутренней поверхности, причем эта новая пластинка по своим размерам несколько больше имеющейся старой. При дальнейшем росте снизу закладывается еще одна большая пластинка и т. д. В результате такого нарастания центральная часть чешуи становится более толстой, чем ее края. Кроме круговой исчсрченности, в чешуе видны также радиально идущие полосы — это каналы, служащие для питания чешуи. С внутренней прилегающей к телу стороны на чешуе имеется тонкий блестящий серебристый слой, содержащий 'кристаллики гуанина и извести и придающий чешуе блеск.
У сазана (карпа) чешуя,
как и у щуки, циклоидная, но все края ее ровные и общая форма чешуи неправильно квадратная. На этой крупной чешуе хорошо заметно, что внутренняя и наружная поверхности выступающего края покрыты эпидермисом. На выступающей части склеритовые ребрышки не образуют правильных линий, но образуют выросты и выступы неправильной формы. Слой гуанина имеется только на выступающей части чешуи.
У окуня чешуя ктеноид-ная (рис. 83, 3) и имеет ско-
Рнс. 83. Чешуя щуки Esox lucius (/ и 2) и окуня Perea fluviatilis (3 и 4) (2 и 4 — чешуйки из боковой линии): г. к. — годовое кольцо
рсс квадратную, чем округ-
лую, форму. Верхняя поверхность заднего, выступающего наружу края чешуи покрыта большим количеством расположенных радиальными рядами выступов и зубчиков и напоминает гребешок. Передний, сидящий в коже край, разделен па 6—8 округлых лопастей. Границы между лопастями могут доходить почти до заднего покрытого зубчиками участка. Однако в отличие от щуки отдельные лопасти нс налегают друг па друга. Что касается расположения склеритов и годовых колец, то к ктеноидпой чешуе можно отнести все ранее сказанное о циклоидной с той лишь разницей, что кольца склеритов прерываются в гребешковой части чешуи.
Микроскопическое строение кожи. Кроме рассмотрения внешне-
го вида чешуи, желательно ознакомиться и с тонким строением покровов костистых рыб по готовым микроскопическим препаратам поперечных разрезов кожи. Знакомство с микроскопическим строением кожи можно перенести па другое время, так как оно займет довольно много времени, по свежий материал, подготовленный для вскрытия, следует обработать в первую очередь.
При достаточном увеличении микроскопа на препарате хорошо видны оба слоя, составляющие наружные покровы рыб. Верхний —
относительно тонкий, окрашивающийся по Маллори обычно в розоватый цвет и содержащий крупные клетки, — эпидермис. Ниже лежит отделенный от него пограничной пластинкой обычно темносиний с хорошо различимыми волокнами кориум. Последний делится на две части: нижнюю, волокна в которой тянутся в продольном направлении и плотно прилегают друг к другу, и верхнюю, в которую вдаются сидящие наклонно костные пластинки чешуй. Здесь кориум имеет вид небольших прослоек между чешуями. Ниже, под кожей, на препарате видны волокна туловищной мускулатуры. Чтобы разобраться в тонком строении кожи, необходимо рассмотреть ее отдельные слои под большим увеличением микроскопа (Х140).
Эпидермис состоит из многих слоев эпителиальных клеток. Нижний, прилегающий к пограничной пластинке слой (мальпигиев) состоит из одного ряда вытянутых в высоту клеток, он является ростковым слоем и дает начало всем остальным клеткам эпидермиса. Верхний, наружный, слой состоит из нескольких рядов уплощенных клеток. Между верхним и нижним слоями лежит несколько слоев многочисленных клеток, форма которых постепенно, по мере приближения к поверхности, меняется от вытянутой к плоской. Между мелкими клетками эпидермиса можно видеть крупные железистые клетки, которые на некоторых участках кожи так многочисленны, что почти полностью вытесняют собственно эпидермальные клетки. Среди них можно различить железистые клетки трех типов: 1. Слизистые клетки, которые развиваются из клеток базального слоя, постепенно оттесняются кверху, превращаются в кубковидные и опорожняют свой секрет на поверхность тела, 2. Серозные железистые клетки, содержащие или зернистый, или гомогенный секрет. Они могут открываться на поверхность при помощи узких канальцев, 3. Колбовидные клетки. Одни из них могут быть связаны с базальным слоем эпителия тонким стебельком, другие эту связь потеряли. Они также постепенно вытесняются наружу и открываются на поверхность. Особенно сильного развития эти клетки достигают у рыб со слабо выраженным или вовсе отсутствующим чешуйчатым покровом.
На границе эпидермиса с кориумом видны отдельные группы пигментных клеток, а также перерезанные кровеносные сосуды, заполненные эритроцитами.
Кориум, или собственно кожа, — довольно толстый слой, лежащий под эпидермисом. Однако большая его часть занята сидящими в нем чешуями. В той области, где сидят чешуи, кориум представлен в виде небольших прослоек между ними. Его волокна тянутся здесь параллельно чешуям. Нижний слой собственно кожи как бы подстилает основания чешуй, здесь волокна тянутся в продольном направлении. При внимательном изучении можно заметить, что слой кориума, лежащий непосредственно под эпидермисом, между верхними концами чешуй, более рыхлый, чем глубже лежащие слои. Таким образом, можно говорить о двух слоях кориума — наружном рыхлом и внутреннем плотном.
Под кожей иногда бывает видна рыхлая подкожная, или субку-танеальная, ткань. В обоих слоях кориума видны кровесные сосуды, заполненные окрашенными в оранжевый цвет эритроцитами.
Следует более детально рассмотреть поперечные срезы чешуй. Хорошо видно, что чешуйки сидят в коже наклонно, причем передняя чешуйка почти полностью прикрывает заднюю. Большая часть чешуи погружена в кориум и только верхний конец вдается в эпидермис, который также покрывает все чешуйки сверху. При большем увеличении можно рассмотреть строение отдельной чешуйки. Каждая чешуйка состоит из двух слоев: нижнего базального и верхнего покрышечного, причем покрышечный слой образует характерные выступы — кольцевые, или -склеритовые, ребрышки.
Плавники. Костистые рыбы имеют хорошо развитые парные и непарные плавники. К непарным плавникам относятся спинные (их может быть несколько), анальный и хвостовой. Парные плавники представлены грудными и брюшными (причем брюшные плавники у некоторых видов рыб могут лежать впереди грудных). Свободная лопасть плавника представляет кожную складку, поддерживаемую плавниковыми лучами. В отличие от акуловых рыб лучи плавников не хрящевые, а костные. Счистив эпидермис с одной стороны плавника (лучше спинного) или рассматривая его па просвет, можно убедиться, что перо плавника поддерживается лучами трех типов.
Ближе к каудальному концу плавника лежат мягкие ветвистые лучи. Верхняя часть таких лучей разделена на отдельные лучики и имеет вид кисточки.
Ближе к переднему краю лежат неветвистые лучи, которые могут быть разделены на две группы. Одни неветвистые лучи разделены по длине на отдельные членики, они мягкие и способны гнуться. Такие лучи называются членистыми. Другие лучи не имеют члеников, обычно они твердые, с острой вершиной. Такие лучи называются колючими. Особенно хорошо различные типы лучей видны в спинном плавнике окуня. Количество ветвистых и неветвистых лучей в плавниках служит систематическим признаком.
Спинной пл а в н и к (pinna dorsalis рис. 82, А, рпп. d.) у щуки один, сильно сдвинут назад и сидит на верхней части начала хвостового отдела. Как и все плавники у свежих экземпляров, он окрашен в желто-оранжевый цвет с темными пятнами. Плавник расправляют и рассматривают составляющие его костные лучи и тонкую перепонку, соединяющую их. Первые 6—10 лучей плавника неветвистые, следующие 13—16 лучей в верхней части разделены па 2—3 ветви.
У окуня два спинных плавника (рис. 82, В, рпп. d.), они занимают почти всю среднюю линию спины. Все лучи переднего плавника колючие (перасчлененные); в заднем плавнике первые 1—3 луча членистые, а остальные 13—15 ветвистые. В отличие от щуки лопасти плавников менее мясистые, так как лучи далеко отстоят друг от друга и соединяющая их перепонка очень тонкая.
У сазана (карпа) спинной плавник (рис. 82, Г, рпп. d.) длинный и расположен на уровне середины тела. Первые 3—4 луча у него 9—3467 257
твердые. Первый очень короткий, а последний превращен в мощный, зазубренный по заднему краю шип. Остальные лучи ветвистые и число их может варьировать от 16 до 22.
По дхвостовой, или анальный, плавник (pinna analis, рис. 82, А, рпп. а.) расположен у щуки на вентральной стороне начала хвостового отдела. Строение, форма и функция era очень сходны с таковыми спинного, но количество лучей меньше: неветвистых 6—7, ветвистых 10—13. Следует обратить внимание на то, что анальный и спинной плавники располагаются друг против друга на том участке тела, где туловище резким сужением переходит в хвост. Они также сближены с хвостовым плавником. Такая концентрация плавников на хвостовом отделе дает щуке возможность резко набирать скорость при броске на добычу.
У окуня анальный плавник (рис. 82, В, рпп. а) лежит на уровне второго спинного. Передние 2 луча колючие, остальные 8—9 — ветвистые.
У сазана (карпа) подхвостовой плавник (рис. 82, Г рпп. а.) лежит на уровне заднего края спинного. Спереди он несет мощный шип, а перед ним еще два более коротких колючих луча. Остальные лучи (5—6) ветвистые.
Хвостовой плавник (рис. 82, А, рпп. с.). Лучи хвостового' плавника сидят вдоль заднего края, покрытого чешуей до самого конца хвостового стебля, а также отчасти переходят на верхний и нижний края последнего. В хвостовом плавнике различают две лопасти, примерно одинаково развитые, — верхнюю и нижнюю. Такой тип плавника называется наружноравнолопастным (гомоцеркальным), так как внутренний скелет распределен в лопастях не одинаково; конец позвоночника, как и у акулы, заходит в верхнюю лопасть. Верхняя и нижняя лопасти плавника поддерживаются членистыми лучами. Причем лучи, лежащие по верхнему и нижнему краю, мощные и неветвистые. Они обычно и наиболее длинные. К середине плавника лучи становятся короче и сильно ветвистыми. Деление на ветви начинается почти от основания луча. Перед крупными краевыми лучами сидит несколько мелких лучей, начинающихся от верхнего и нижнего края хвостового стебля. Тонкая кожная складка между лучами может быть покрыта у основания плавника чешуей.
Грудные плавники (pinna thoracalis, рис. 82, А, рпп. th.) у щуки лежат на вентральной стороне тела, на уровне заднего края жаберной крышки. Оттянув наружный край плавника вперед, можно убедиться, что плавник расположен горизонтально. Такое положение указывает на роль плавников как рулей глубины.
Брюшные плавники (pinna abdominalis, рис. 82, Д, рпп. ab.) у щуки лежат также на брюшной поверхности, примерно на уровне середины туловища, и расположены горизонтально.
У окуня грудные плавники располагаются на боковой части туловища, непосредственно за жаберной щелью. Плоскость плавников вертикальная. Такое положение объясняется их функцией — удержанием высокого сплющенного с боков тела рыбы в нормальном
-вертикальном положении. Брюшные плавники расположены горизонтально. Они сильно сдвинуты вперед и лежат на вентральной стороне тела, почти на уровне грудных.
У сазана (карпа) грудные плавники лежат на вентро-латеральной поверхности на уровне заднего края жаберной крышки. Первый луч у них очень мощный и в нижней части нерасчлененный. Брюшные плавники расположены на середине туловищного отдела и лежат горизонтально.
Теперь следует рассмотреть органы, располагающиеся на голове.
Ротовое отверстие (rima oris, рис. 84, г. о.) щуки очень велико. Начинается оно на конце вытянутого рыла (конечный рот) и доходит почти до уровня переднего края глаза. Сверху и снизу оно ограничено челюстями. Нижняя челюсть (рис. 84, тп..) немного длиннее верхней. На боковых частях нижней челюсти находятся высокие конусовидной формы зубы, вершины которых направлены вверх и немного назад. Зубы, сидящие на переднем конце, мелкие и напоминают терку. Такие же мелкие зубы сидят и по краю верхней челюсти, а также на крыше ротовой полости. В задней части верхней челюсти следует рассмотреть выступающий край верхнечелюстной кости (рис. 84, тх.). Он соединен с черепом только при помощи кожной складки, что дает ему возможность сильно опускаться и образовывать боковые стенки ротового отверстия при открывании рта. В то же время он не мешает сильному отведению вниз нижней челюсти. На свежей рыбе нижнюю челюсть следует отвести вниз, для этого оттягивают назад мускулатуру, расположенную на нижней части головы и соединяющую нижнюю челюсть с плечевым поясом. Обращают внимание на то, что в таком положении ротовая полость щуки напоминает большой сачок с широким отверстием, очень удобный для захвата крупной добычи.
Ротовая щель окуня более короткая, зубы в связи с питанием мелкой добычей значительно меньше. Задний край верхнечелюстной кости может не только опускаться вниз, но и выдвигаться вперед, отчего передняя часть ротовой полости при открывании рта несколько удлиняется. На нижней челюсти имеются мясистые губы, которые обеспечивают плотное замыкание ротовой щели.
У сазана (карпа) ротовая щель короткая, но широкая, окруженная толстыми мясистыми «губами», так что челюстные кости снаружи незаметны. Если у свежей рыбы приоткрыть рот, оттянув нижнюю челюсть, то видно, что при этом одновременно выдвигается вперед верхняя челюсть и образуется подобие трубки, направленной вниз (приспособление к питанию бентосом). На уровне углов рта отходят крупные усики (органы осязания), а над ними еще по одному мелкому.
Наружные жаберные отверстия парные, расположенные по одному с каждой стороны. Они имеют вид длинной узкой щели, ограниченной спереди и снаружи задним краем жаберной крышки, а сзади и изнутри — стенкой тела. Снизу щель тянется далеко вперед и только немного не доходит до переднего конца нижней челю
сти. Передние концы жаберных щелей правой и левой сторон соединяются друг с другом.
У сазана (карпа) жаберная щель почти вертикальная, и нижний ее конец не заходит далеко вперед, как бы ограничивая конец головы снизу.
Жаберная крышка (operculum) прикрывает сбоку жаберную область, образуя заднюю боковую поверхность головы. Она состоит из четырех крышечных костей (см. раздел «Скелет»), покрытых кожей, и лежит впереди от жаберной щели. Ее задний край оторочен тонкой, но довольно широкой складкой кожи, которая
Рис. 84. Голова щуки Esox lucius (А) и голова окуня Perea jluviati-lis (Б):
ж, п. — жаберная перепонка; к. с.— кожная складка; л. ж. п,—лучн жаберной перепонки; can. ino. — подглазничный канал; can. I. — боковая линия иа теле; сап. ор. т. — гномаиднбулярный канал; сап. ро.—заглазиичный канал; can. spo.— надглазничный канал; can. st. надвисочный канал; тп. — нижняя челюсть; тх. —верхнечелюстная кость; паг. — ноздря; ос.—глаз; г. о.—ротовое отверстие
переходит на нижний край и далее тянется вперед вдоль ветви нижней челюсти. Эта складка носит название жаберной перепонки.
Жаберная перепонка (рис. 84, ж. п.) переходит с заднего края жаберной крышки на ее нижний край, а затем продолжается вперед вдоль ветви нижней челюсти. В нижней части она поддерживается особыми костными образованиями — лучами жаберной перепонки (рис. 84, л. ж. п.), внутренние концы которых сидят на гиоиде (подробнее см. раздел «Скелет», стр. 323). В наличии лучей легко убедиться, если растянуть жаберную перепонку. Количество лучей у щуки в связи с удлиненной мордой велико (13—16), их меньше у окуня (7) и сазана (3).
Отвернув жаберную крышку, необходимо обратить внимание на гладкий участок стенки тела, расположенный под ее задним краем и образованный костями плечевого пояса. К нему при приведении жаберной крышки плотно прилегает кожная складка, что играет немаловажную роль при дыхании. Следует рассмотреть также лежащие под жаберной крышкой четыре жаберные дуги с сидящими на них в два ряда темно-красными жаберными лепестками.
Глаза (oculus, рис. 84, ос.) у щуки сравнительно крупные, расположены по бокам и несколько выдаются кверху над общим уровнем головы. Зрачок относительно велик и округлой формы. Как и у всех рыб, глаза лишены век, но по краям имеется кожная складка, срастающаяся с передней стенкой глазного яблока, не мешающая его движению.
У окуня и сазана положение глаз боковое.
Подробно со строением глаза необходимо познакомиться при изучении органов чувств, поэтому следует сохранить один или оба глаза, осторожно вычленив их у рыбы и поместив в 10%-ный формалин.
Ноздри (naris, рис. 84, паг.) лежат несколько впереди глаз и направлены вверх и в стороны. Характерно, что каждая ноздря открывается двумя отверстиями. У щуки переднее из них округлое, а заднее — полулунной формы с приподнятым передним краем. У окуня оба отверстия округлые, а задний край переднего приподнят и образует подобие раковины.
У сазана (карпа) переднее отверстие ноздри округлое и имеет вид трубочки. Заднее, несколько вытянутой формы, расположено чуть позади и латеральнее переднего, в нем видны складки слизистой оболочки дна обонятельного мешка. Мостик между отверстиями ноздрей образует вертикальную пластинку, которая обеспечивает попадание воды в переднюю ноздрю и выход ее через заднюю даже при медленном движении рыбы.
Непосредственно под обоими отверстиями ноздрей лежит полость обонятельной капсулы. При движении рыбы вода входит в переднее отверстие, попадает в обонятельную капсулу и выходит через заднее отверстие; тем самым достигается непрерывная смена воды в органе обоняния.
Повернув рыбу брюшной стороной вверх, следует рассмотреть открывающиеся несколько впереди от анального плавника анальное, половое и мочевое отверстия.
Анальное отверстие (anus, рис. 85, ап.)—самое переднее из трех отверстий. Оно округлой формы и заметно очень хорошо.
Половое отверстие (foramen genitale, рис. 85, f. g.) располагается непосредственно за анальным на небольшом кожном возвышении. У рыб в период размножения кожная складка несколько припухает и заметна очень хорошо.
Мочевое отверстие (foramen uretricum, рис. 85, f. иг.) открывается на дне полулунной складки, которая зажата между возвышением полового отверстия и основанием анального плавника. Само отверстие имеет вид узкой поперечной щели и плотно прилегает к основанию плавника, так что рассмотреть его не всегда удается. Иногда половое и мочевое отверстия открываются в одну общую складку: первое —в ее переднюю, а второе — в заднюю часть. У сазана (карпа) перед анальным плавником только два отверстия: переднее — анальное и заднее, в которое открываются мочевые и половые протоки.
рп. ves.pn. gon. г.
Рис. 85. Общая топография внутренних органов: А — щуки Esox lucius; Б — окуня Perea fluviatilis-, В — карпа Cyprinus carpio:
ов.— место расположения овала; a. cl. т. — чревно-брызжеечиая артерия; ап. — анальное отверстие; арр. р. — пилорические придатки; atr. — предсердие; br. — жабра; bul. ао. — луковица аорты; с. г. — место расположения газовой железы; d. chol. — желчный проток; d. cyst.—проток желчного пузыря; d. h. — печеночный проток; d. рп. — соединение плавательного пузыря с пищеводом; f. g. — половое отверстие; /. иг. — мочевое отверстие; gast. — желудок; gon. половая железа; h. — печеиь; ints. — кишечник; I. — селезенка; oes. — пищевод; рп — головная почка; рпег. — поджелудочная железа; psbr. — ложная жабра; г. — почка; ret. — прямая кишка; г. о. — ротовое отверстие; ventr. — желудочек; ves. f. — желчный пузырь; ves. рп. — плавательный пузырь; ves. иг. — мочевой пузырь; р. h. — воротная вена печени
Сеисмосензорная система (органы чувств боковой линии) (рис. 84)
Изучение системы органов чувств боковой линии правильней отложить до рассмотрения других органов чувств, после изучения головного мозга. Однако располагающиеся поверхностно, под кожей, каналы боковой линии обычно удаляются при снятии кожи или частично разрушаются при вскрытии черепной коробки. Поэтому лучше всего их рассмотреть на свежем экземпляре.
Сейсмосензорная система представлена у большинства костистых рыб системой каналов, идущих внутри стенок тела и посылающих ответвления к поверхности. Концы таких ответвлений могут открываться на поверхность или, наоборот, быть затянутыми тонкой кожной перепонкой. На дне каналов отдельными группами располагаются чувствующие клетки, воспринимающие колебания водной среды. Сейсмосензорная система способна воспринимать колебания, возникающие вследствие движения тела самой рыбы, и колебания давления и движения струй воды, возникающие от движения других животных. Точно так же она воспринимает те колебания, которые производятся самими животными, а затем отражаются от соседних твердых тел, что позволяет рыбам обходить встречающиеся на пути препятствия и вообще ориентироваться в водной среде, не пользуясь зрением; ослепленная щука безошибочно схватывает проплывающую мимо нее рыбу.
Большая часть каналов концентрируется на голове, где они у большинства костистых рыб занимают совершенно определенное положение. Кроме того, обычно один длинный канал тянется вдоль всего тела рыбы по его боковой поверхности, откуда и произошло название всей системы — органы чувств боковой линии.
Пользуясь лупой, постараемся более подробно познакомиться с расположением каналов сейсмосензорной системы. Сначала целесообразно рассмотреть каналы, концентрирующиеся на голове, а затем — каналы туловищной области.
У всех костистых рыб расположение каналов органов боковой линии на голове очень сходно. Однако не у всех видов они прослеживаются четко, так как могут проходить в толще покровных костей и судить о их местоположении тогда можно только по отверстиям, которыми они открываются наружу.
У щуки, как уже указывалось выше, каналы открываются наружу рядом отверстий, поэтому ход каналов можно вполне хорошо проследить по этим отверстиям. Они довольно крупные и сразу бросаются в глаза. Сами каналы проходят недалеко от наружных отверстий, но идут под кожей или даже в костях.
Надглазничный канал (canalis supraorbitalis, рис. 84, сап.). Ход его отмечен пятью отверстиями, лежащими на верхней стороне головы. Они начинаются за глазом, огибают его; одно из них располагается чуть медиальнее ноздри и еще одно — впереди на рыле. Канал продолжается и дальше — вперед от переднего отверстия и хорошо заметен, так как имеет вид неглубокого желобка
на верхней стороне рыла. Каналы правой и левой сторон соединяются у своих передних концов перемычками. Чувствующие клетки этого канала иннервируются веточками поверхностной глазничной ветви VII нерва.
Подглазничный канал (canalis infraorbitalis, рис. 84, сап. ino.) прослеживается по ряду (7) отверстий, огибающих глаз снизу и продолжающихся вперед по боковой поверхности рыла. Переднее отверстие капала лежит под задним отверстием ноздри. Иннервируется канал веточками, отходящими от щечной ветви VII нерва.
Задние концы двух вышеописанных каналов сливаются в заглазничной области.
Заглазличный канал (canalis postorbitalis, рис. 84, сап. ро.) тянется назад от места слияния двух предыдущих каналов вдоль верхнего края жаберной крышки. Ход этого канала у щуки проследить трудно, так как отчетливо видны только три отверстия у верхнего копца жаберной щели. Канал иннервируется латеральной ветвью VII нерва и X нервом.
Надвисочный капал (canalis supratemporalis, рис. 84, can. st.). На уровне затылочных костей, на верхней стороне головы видны короткие затылочные каналы, тянущиеся в поперечном направлении. Они имеют вид двух небольших немного изогнутых желобков.
Гиомандибулярный, или крышечно-челюстной, канал (canalis hyomandibularis, s. operculo-mandibularis, рис. 84, can. op. m.) тянется вдоль заднего края предкрышки и по нижней поверхности нижней челюсти. Шесть его отверстий видны на пред-крышке и пять — на нижней челюсти. Верхним концом канал сливается с заглазничной ветвью. Иннервируется канал ветвями VII нерва.
Надглазничная комиссура (commissura supraorbitalis), соединяющая па уровне глаз'надглазничные каналы обеих сторон, у щуки снаружи видна плохо.
Рассмотреть ход каналов у щуки можно также, выламывая их наружную костную стенку. Обычно они располагаются недалеко от отверстий.
У окуня каналы сейсмосензорных органов, расположенные на голове, заметны довольно плохо, так как сами каналы лежат внутри костей или в особых костных трубочках, а их ответвления к поверхности обычно не открыты, как у щуки, а затянуты кожной перепонкой, правда, очень тонкой. Проследить ход каналов полностью удается только после их наполнения какой-либо окрашенной жидкостью, например черной тушью. Инъекцию лучше всего проводить через ответвление надглазничного канала, которое открывается отверстием в кости на верхней стороне головы на уровне переднего края глаза. Для того чтобы его обнаружить, достаточно счистить кожу в этой области.
Вторым отверстием для инъекции может служить отверстие, которым открывается нижнечелюстная ветвь крышечно-челюстного 264
канала на вентральной стороне нижней челюсти, недалеко от ее переднего конца. Частично расположение каналов можно проследить и не прибегая к помощи инъекции, в этом случае полезно вводить в канал щетинку или темноокрашенную леску.
Сняв кожу с заднего края предкрышечной кости, можно обнаружить верхнюю часть крышечно-челюстного канала (рис. 84, сап. ор.-т.), который лежит в костной трубке, внутри предкрышечной кости, и дает ряд ответвлений и отверстий к ее заднему краю. Дальнейший ход канала вперед можно проследить, если счистить скальпелем кожу с вентральной поверхности одной из ветвей нижней челюсти.
Сняв кожу с верхнего угла головы за глазом, необходимо проследить идущий в толщине кости заглазничный канал (рис. 84, сап. ро.). Он тянется назад, делая резкий изгиб на уровне заднего края предкрышки, и сливается с каналом, идущим по боковой поверхности тела. В месте перегиба от него отходит надвисочный канал (рис. 84, can. st.), который тянется в поперечном направлении по верхней поверхности головы и сливается с одноименным каналом другой стороны.
Продолжением заглазничного канала вперед является надглазничный канал (рис. 84, can. spo.), который делает изгиб на уровне заднего края глаза, переходит на верхнюю сторону головы и оканчивается несколько впереди от переднего отверстия ноздри. В районе изгиба от него отходит надглазничная комиссура, которая является вторым соединением между каналами двух сторон.
Подглазничный канал (рис. 84, can. ino.) становится видимым, если снять кожу с боковой поверхности головы позади глаза. Эта ветвь отходит от места слияния двух предыдущих и тянется вдоль заднего края глазницы и под ней. Она оканчивается несколькими расширениями, лежащими над дорзальным краем верхнечелюстной кости.
Боковая линия (canalis lateralis, рис. 84, can. I.). Сейсмо-сензорные органы в туловищной области представлены одним каналом, который проходит по боковой поверхности тела. Наружу канал открывается отдельными канальцами, проходящими через чешуи. Чешуйки, прободенные канальцами, довольно хорошо отличаются от общего чешуйчатого покрова, их ряд и образует характерную боковую линию. Боковая линия начинается непосредственно позади верхней части жаберной крышки, тянется вдоль стенки тела, делая более или менее крупный изгиб вниз, и в районе хвостового стебля идет строго по средней линии его боковой поверхности. Выдернув одну чешуйку из боковой линии, рассматривают ее строение так же, как рассматривали строение обычной чешуи. У щуки каждая чешуйка несет по наружному краю довольно глубокую вырезку, через которую проходит канал к поверхности тела (рис. 83, 2). У окуня канал проходит через особую трубочку, лежащую вдоль средней линии чешуи (рис. 83, 4).
Общая топография внутренних органов
Кладут рыбу на спину и, придерживая левой рукой, делают ножницами небольшой поперечный разрез брюшной стенки тела несколько впереди от анального отверстия. Затем вставляют в разрез тупой конец ножниц и проводят продольный разрез вдоль средней линии брюшной стороны тела вперед до уровня грудных плавников. При проведении разреза следует как можно сильнее оттягивать брюшную стенку на себя, чтобы не повредить лежащие под ней внутренние органы. Перерезка костного пояса брюшных плавников требует некоторого усилия.
Продолжают разрез вентральной стенки также назад до переднего края анального плавника. При этом необходимо обойти лежащие перед ним отверстия по левой боковой стенке тела. Затем можно полностью удалить левую стенку полости тела. Для этого делают дугообразный разрез, начиная от переднего края анального плавника, вверх к спине и затем вдоль боковой линии, несколько ниже ее, до заднего края жаберной крышки, точнее до заднего края пояса передних конечностей, который здесь располагается. Положение пояса передних конечностей легко прощупать, так как большая его часть лежит прямо под кожей. Верхний разрез должен пройти чуть ниже верхней границы полости тела, примерно рядом с линией прикрепления к стенке тела плавательного пузыря. Для этого все время контролируют направление внутренней части разреза, оттягивая в сторону отрезаемую стенку тела.
У сазана внутренние органы, а особенно большие жировые скопления в брюшной полости связаны спайками с боковой стенкой тела, поэтому прежде чем провести разрез по боковой стенке тела, следует оттянуть ее и, перервав связки, отделить от нее внутренние органы. После этого разрез можно провести на уровне верхней трети тела. Соединяют передний край верхнего разреза с передним концом разреза, проведенного вдоль средней линии брюха. Для этого проводят разрез вдоль заднего края пояса передних конечностей. У щуки и окуня можно целиком удалить левую часть плечевого пояса, но на свежей рыбе это необходимо делать очень осторожно, чтобы не повредить лежащие под ней крупные венозные сосуды и задние отделы сердца. У сазана (карпа) под верхней частью плечевого пояса, кроме того, располагается крупная головная почка, богато снабженная кровеносными сосудами, поэтому у данного вида на свежих экземплярах удалять плечевой пояс не следует.
Теперь очень осторожно подводят под середину вентральной части пояса передних конечностей тупой конец ножниц и перерезают пояс, а затем продолжают разрез вперед до внутреннего края переднего конца нижней челюсти. Особенно внимательно следует проводить разрез непосредственно под жаберной крышкой. В этой области под толстой мускульной стенкой лежат сердце и крупные тонкостенные сосуды. Поэтому, перерезав пояс грудных плавников, оттягивают вентральную стенку жаберной области и 266
через имеющееся отверстие рассматривают сердце. Дальше продолжают вскрытие небольшими разрезами, все время контролируя положение внутреннего конца ножниц относительно сердца. Только в этом случае можно гарантировать сохранность сердца и сосудов. Раздвинув края разреза, рассматривают общее расположение внутренних органов.
Все внутренние органы туловища лежат в полости тела — целоме (coelom), которая тонкой поперечной перегородкой, лежащей в области пояса грудных плавников, разделяется на две неравные части: небольшую переднюю — околосердечную сумку, где помещается сердце, и заднюю — брюшную, в которой лежат все внутренние органы. Поперечная перегородка состоит из двух листков, которые разделяются у стенок тела. Передний листок выстилает стенки перикардиальной, задний — брюшной полости.
Непосредственно по латеральным частям перегородки проходят венозные сосуды, несущие кровь в сердце.
Листок, выстилающий стенки брюшной полости, носит название париетального листка и имеет вид тонкой блестящей пленки. Его легко обнаружить, рассматривая внутреннюю сторону вырезанной стенки тела. На дорзальной стенке брюшной полости тела листок переходит на внутренние органы, которые он одевает, и с его помощью органы подвешаны к стенке тела. Здесь он уже называется висцеральным листком.
Вся верхняя часть брюшной полости занята плавательным пузырем (рис. 85, ves. рп.). По бокам и несколько ниже плавательного пузыря тянутся половые железы — гонады (рис. 85, gon.). В зависимости от времени года они могут быть или небольшими лентовидными образованиями, или (в период размножения) огромными мешками, занимающими, особенно у самок, почти все пространство брюшной полости.
Под половой железой лежит длинный относительно узкий ж е-лудок (рис. 85, gast.), который тянется от передней верхней части брюшной полости к ее нижнему заднему концу. Желудок рыб может очень сильно растягиваться, поэтому форма его зависит от степени наполнения пищей. Ниже желудка в брюшной полости лежат петли кишечника (рир. 85, ints.). Далее кишечник тянется назад вдоль вентральной стенки брюшной полости и заканчивается анальным отверстием.
В петле между желудком и началом кишечника лежит жироподобное тело— поджелудочная железа (рис. 85, рпог.).
К заднему концу желудка прилегает буровато-красная селезенка (Uen, рис. 85, /.). Она имеет треугольную форму и уплощена в дорзовентральном направлении. С пищеварительной системой этот орган связан только топографически, так как является органом кроветворения.
Передняя и нижняя части брюшной полости заняты печенью (рис. 85, h.). Оттянув вверх задний конец половой железы, рассматривают лежащий между каудальным концом плавательного
пузыря и задней частью кишечника мочевой пузырь (рис. 85, ves. иг.).
У окуня в отличие от щуки желудок (рис. 85, gast.) более короткий и имеет вид слепого выроста, так как места впадения пищевода (рис. 85, oes.) и выхода кишечника (рис. 85, ints.) лежат рядом. Непосредственно около желудка от кишки отходят три слепых выроста — пилорические придатки (рис. 85, <W- р.).
Сам кишечник имеет большее количество изгибов и более длинный, чем у щуки.
Печень (рис. 85, h.) располагается не снизу, а слева от желудка. Буроватая селезенка (рис. 85, I.) лежит в петле кишечника под желудком. Поджелудочная железа рассеяна по брыжейке в виде микроскопических долек. Половая железа (рис. 85, gon.) у самцов парная, а у самок — непарная. Мочевой пузырь (рис. 85, ves. иг.) лежит в левой половине задней части брюшной полости, сбоку от половой железы.
У сазана (карпа) внутренние органы покрыты большим количеством жира, который приходится удалять, чтобы их рассмотреть. Это следует делать очень осторожно, так как вместе с жиром можно удалить и доли печени, на свежей рыбе слабо отличающиеся по цвету. Для некоторого уплотнения жира рекомендуется детальную препаровку пищеварительной системы вести на объектах, пролежавших несколько часов в слабом растворе формалина, а пока ознакомиться только с общей топографией внутренних органов.
В верхней части полости тела виден плавательный пузырь (рис. 85, ves. рп.), вентральная часть которого прикрыта большим жировым телом. Из-под последнего выступает нижний край гонады (рис. 85, gon.). Половая железа может быть по-разному развита в зависимости от возраста рыбы и сезона. Вентральная часть полости занята петлями сравнительно длинного у карповых рыб кишечника (рис. 85, ints.), между ними лежат беловатые скопления жира. От беловатого жира отличаются буроватокрасным цветом доли печени (рис. 85, h.). Печень включает в себя также ткань поджелудочной железы и поэтому носит название гепатопанкреас. Она имеет значительные размеры и на препарате видны несколько ее лопастей. Две лопасти печени, имеющие вертикальное направление, видны на поверхности кишечника в передней части полости и одна — крупная, идущая горизонтально,— в средней части кишечника под гонадой. Проследить весь ход кишечника из-за большого количества покрывающего его жира трудно, поэтому знакомство с ним лучше отложить до изучения пищеварительной системы.
Особое внимание при вскрытии сазана (карпа) следует обратить на головную почку. У взрослой рыбы она является лимфоидным органом. Это тело темно-красного цвета, лежащее вне полости тела под париетальным листком брюшины. Ее задний конец виден, если оттянуть назад переднюю стенку плавательного пузыря. Передняя часть уходит под плечевой пояс и спускается почти до уровня верх
него края грудного плавника, располагаясь дорзальнее перикардиальной полости (рис. 85, ph.), поэтому она хорошо видна при рассмотрении сердца. Через нее проходит крупный кровеносный сосуд — задняя кардинальная вена. Необходимо запомнить форму и положение головной почки.
Теперь раздвигают края разреза вентральной стенки жаберной полости и рассматривают находящуюся впереди от диафрагмы околосердечную полость и лежащее в ней сердце. Околосердечная, или перикардиальная, полость — обособленный передний отдел вторичной полости тела, имеет форму треугольника, вершина которого направлена вперед. Так же, как и брюшная полость, стенки ее выстланы париетальным листком, который здесь носит название перикардия (pericardium). Переходя на сердце, этот листок одевает его отделы снаружи и называется эпикардием (epicardium). Ближе всего к препарирующему в вентральной части перикардиальной полости лежит небольшой мускулистый желудочек (рис. 85, ventr.). С двух сторон из-под него выступают темно-красные от заполняющей их крови края предсердия (рис. 85, atr.).
У сазана (карпа) основная часть предсердия располагается с левой стороны перикардиальной полости, закрывая почти целиком желудочек, который выступает из-под него только небольшим вентральным участком.
К задней части предсердия примыкает тонкостенный венозный синус, в который впадают кювьеровы протоки. Вперед от сердца тянется брюшная аорта — крупный расширенный у основания влуковицу аорты (рис. 85, bul. ао.) артериальный сосуд.
Щитовидная железа (glandula thyroidea, рис. 90, gl. thr.), как правило, разбросана у костистых рыб в виде отдельных долек вдоль брюшной аорты, которые анатомически плохо различимы. У некоторых рыб дольки могут образовывать заметные скопления. Из описываемых рыб такое скопление можно наблюдать у щуки. Оно расположено перед передним концом брюшной аорты.
Органы дыхания
Органы дыхания костистых рыб так же, как и большинства водных позвоночных, приспособлены к обмену газами между кровью и водой и представлены жабрами. В отличие от рассмотренных выше хрящевых рыб костистые имеют только четыре полные жаберные дуги (пятая редуцируется), а следовательно, и четыре полные жабры.
Так же, как и у акул, каждая жабра состоит из двух полужабр. Однако разделяющая две полужабры межжаберная перегородка редуцируется так, что концы лепестков остаются свободными в большой или меньшей степени.
Каждая отдельная полужабра — довольно сложно устроенный орган, основной функцией которого является обмен кислородом и углекислотой между кровью и внешней средой. В связи с этим жабры состоят из многочисленных тонкостенных жаберных лепестков,
обладающих большой поверхностью. К каждому лепестку подходят кровеносные сосуды, разветвляющиеся здесь на сеть капилляров. Через тонкие стенки капилляров и жаберных лепестков в окружающую среду выделяется углекислый газ, а в кровь поступает кислород. Окисленная, богатая кислородом кровь оттекает из жабр по выносящим жаберным сосудам, а затем разносится по всему телу.
Дыхательная система костистых рыб характеризуется общим для костных рыб (Osteichthyes) строением жаберного аппарата, приспособленного к оперкулярному типу дыхания, при котором акт дыхания совершается за счет движения жаберной крышки (operculum). Этими движениями вода, входящая в ротовую полость через рот, всасывается через жаберные щели и жабры в околожаберную полость, а затем выталкивается из нее через общее оперкулярное отверстие наружу (подробное описание акта дыхания см. ниже). Весь висцеральный аппарат, включая челюстной аппарат, подъязычный и жаберный, представляет сложную двигательную систему, объединенную в одно целое связками и мышцами.
Жаберную крышку оттягивают или отрезают ее задний край совсем и рассматривают лежащие под ней жабры.
Жабры (branchia, рис. 85, Ьг.) в количестве четырех пар лежат в переднем отделе пищеварительной системы, стенки которой они, собственно, и составляют.
Между отдельными жабрами, а также между первой жаброй и жаберной крышкой находятся отверстия — жаберные щели, через которые полость глотки сообщается с полостью под жаберной крышкой. Последняя жаберная щель между жаброй и стенкой тела неполная, так как почти по всей длине затянута соединительнотканной перепонкой.
Каждая жабра состоит из двух частей: верхней, более короткой, тянущейся назад почти горизонтально, и нижней, длинной, идущей косо вперед и вниз и прикреп'ляющейся своим передним концом к непарному элементу, который соединяет жаберные дуги двух сторон. Познакомьтесь более детально со строением отдельной жабры, для этого вырежьте небольшой ее участок и рассмотрите его под лупой.
Жаберная дуга (arcus branchialis, рис. 86, arc. br.) лежит в основании жабры. Как и у акуловых рыб, она состоит из четырех элементов, однако здесь они уже костные, а не хрящевые. На поперечном разрезе жаберная дуга имеет V-образную форму, заостренный конец ее направлен вперед и внутрь. Наружные поверхности угла покрыты относительно крупными костными пластинками — жаберными тычинками (рис. 86, ж. т.), разделенными друг от друга соединительнотканными прослойками. При помощи соединительной ткани эти пластинки причленяются и к жаберной дуге. Внешняя поверхность пластинок покрыта многочисленными зубчиками, острые концы которых направлены в сторону глотки. Сверху пластинка покрыта эпителием, так что наружу торчат только острия зубчиков.
У окуня тычинки имеют форму небольших выростов, так что здесь хорошо видно их расположение в два ряда. Под лупой четко
видны острые зубчики, выступающие на их поверхности.
У карпа жаберные тычинки мощные, имеют вид вертикально
поставленных лопастей. Их край, направленный в Тычинки являются своеобразным фильтром, который препятствует вымыванию пищи из глотки через жаберные щели вместе с водой. Поэтому их форма и число значительно варьируют у разных видов в зависимости от рода пищи. Щука, питающаяся крупной добы- чей, имеет тычинки, которые скорее играют роль зубов, способствующих удержанию пищи.
Во внутренней части V-образпой жаберной дуги лежат два крупных кровеносных сосуда. Приносящая жаберная артерия (рис. 86, a. br. ad.) лежит более каудально, т. е. ближе к основанию жаберных лепестков. По этому сосуду венозная кровь притекает к жаберным лепесткам. Выносящая жаберная артерия (рис. 86, a. br. г.) лежит ближе к передней части, т. е. непосредственно прилегает к костной жаберной дуге. Мелкие капилляры несут в этот сосуд артериальную «кровь из жаберных лепестков.
Ветвление сосудов в жаберных лепестках видно только на инъецированном материале.
Жаберные лепестки (filum branchialis, рис. 86, fit. br.) —тонкие выросты экто-дермического происхождения, сидящие вдоль заднего края жаберной дуги. Как уже говорилось выше, они богато снабжены кровеносными сосудами, отчего цвет их у свежей рыбы темно-красный. Именно в жаберных лепестках происходит обмен кислородом и углекислым газом между кровью и водой.
При детальном рассмотрении среза жабры под лупой можно видеть, что каждый лепесток имеет примерно треугольную форму. Основанием треугольника является задний край жаберной дуги, а вершина направлена назад,
глотку, рассечен.
Рис. 86. Схема поперечного разреза жаберной дуги щуки Esox lucius:
ж. т. — жаберные тычинки; х. т, —скелетная основа жаберного лепестка; a. br. ad. — приносящая жаберная артерия; a. br. г. — выносящая жаберная артерия; arc. br. — жаберная дуга; fit. br. — жаберные лепестки
причем вершины
лепестков поочередно отогнуты больше в правую или левую сторону, так что фактически на жаберной дуге лепестки сидят в два
ряда.
Очистив отдельный лепесток от покрывающих его мягких тканей, можно рассмотреть тонкий хрящевой тяж (рис. 86, х. т.), составляющий скелетную опору лепестка.
Ложная жабра (pseudobranchia, рис. 85, psbr.). У окуня на внутренней части жаберной крышки, против верхнего конца пер-
вой жаберной дуги, сидит небольшая дополнительная жабра с хо-рошо заметными отдельными жаберными лепестками. Для того чтобы ее рассмотреть, необходимо отвернуть жаберную крышку в сторону.
У щуки положение ложной жабры почти такое же — у переднего верхнего конца первой жаберной дуги, но снаружи, особенно на фиксированном материале, она почти не видна. На свежих экземплярах ее можно видеть как просвечивающее красноватое пятно.
У сазана (карпа) ложная жабра имеет такой же вид и расположена на внутренней поверхности переднего верхнего угла крышечной кости.
Функция ложной жабры еще не совсем ясна. Как орган дыхания, она функционирует только у мальков рыб. У взрослых — ложная жабра снабжается артериальной кровью и функционирует как железа внутренней секреции и орган, способствующий дополнительному выделению СО2 из крови и аккумулирующий кислород, который используется через кровеносную систему тканями головы при недостатке О2 в воде.
Акт дыхания у костистых рыб совершается следующим образом. При отведении жаберной крышки в стороны тонкая складка, лежащая по заднему ее краю, плотно прилегает к стенке тела и не дает воде проходить в жаберную область через наружную щель. В результате создавшегося в полости под жаберной крышкой отрицательного давления вода из ротоглоточной полости попадает в полость под жаберной крышкой, омывая жаберные лепестки. При сжимании жаберных крышек жаберные лепестки каждой жабры расходятся в стороны до соприкосновения с лепестками соседней жабры и препятствуют выходу воды в полость глотки. Под давлением жаберной крышки вода поднимает кожную складку и выходит наружу через щель за жаберной крышкой.
В связи с оперкулярным типом дыхания развиваются особые мышцы, приводящие и отводящие жаберную крышку. Они подробно разбираются при изучении мускулатуры.
Пищеварительная система
Строение пищеварительной системы костистых рыб отличается большим разнообразием. Построенная у всех рыб по одному плану, она варьирует в деталях. Эти отличия пищеварительной системы в первую очередь связаны с характером питания. Причем изменения можно отметить во всех отделах. Например, положение ротового отверстия — нижнее, верхнее или конечное — указывает на добывание рыбами корма у дна, под поверхностью или в толще воды. У рыб, питающихся моллюсками, а также растительностью (все карповые), лежащие в задней части ротоглоточной полости глоточные зубы имеют своеобразную форму жерновов, приспособленных для перетирания пищи. Разнообразное строение имеет и пищеварительный тракт, соответственно разному характеру пищи. У хищных рыб (щука, судак) желудок громадный, приспособлен-272
ный к вмещению крупной добычи. У карповых желудок совсем отсутствует. Кишечник у мирных рыб более длинный и имеет много изгибов, соответственно всасывающая поверхность его велика. Увеличение всасывающей поверхности идет также за счет образования у многих видов слепых выростов — пилорических придатков. У хищников кишечник более короткий.
Пищеварительная система начинается ротовым отверстием (rima oris, рис. 85, г. о.), описание которого было дано выше при внешнем осмотре. Ротовое отверстие ведет в ротовую полость, на дне которой помещается язык. Чтобы рассмотреть последний, необходимо сильно оттянуть вниз нижнюю челюсть.
Язык (lingua, s. glossa) рыб — образование, не гомологичное языку вышестоящих позвоночных, так как он лишен мускулатуры и представляет собой покрытый слизистой оболочкой передний конец нижнего отдела скелета жаберного аппарата. В частности, он поддерживается небольшой косточкой glossohyale (см. раздел «Скелет»).
Ротовая полость (cavum orale) без резких границ переходит в глотку (pharynx), так что у рыб можно говорить от общей ротоглоточной полости. Чтобы рассмотреть эту полость, необходимо сделать разрез одной из ее стенок, лучше левой. Для этого вставляют один конец ножниц в ротовое отверстие, проводят его через ротоглоточную полость и выводят в нижней части первой жаберной щели. Разрезают боковую стенку полости и остаток жаберной крышки. Если работа ведется на фиксированном материале или на свежем, но долго лежавшем, так что кровь уже успела свернуться, можно также перерезать и жаберные дуги. Разрез жаберных дуг лучше всего вести по местам их перегиба. Края разреза раздвигают в стороны и рассматривают ротовую полость. Вся полость выстлана слизистым эпителием и довольно гладкая.
Зубы (dens). У большинства рыб в ротоглоточной полости имеются многочисленные зубы. Форма, количество и размеры их различны у разных видов и зависят от рода пищи.
У щуки наиболее крупные зубы сидят на нижней челюсти. Форма их вытянутая, конусовидная, как и у большинства других зубов. Вершинами они направлены назад. Такие зубы могут выполнять только функцию удержания добычи, а не ее обработки. Несколько рядов более мелких зубов сидят и на верхней челюсти, а именно, на межчелюстных костях. Зубами покрыта также передняя часть нёба, здесь они связаны с сошником, предчелюстными и нёбными костями. На этих костях зубы сидят в несколько рядов и имеют вид терки. Точно такие же мелкие зубы сидят двумя рядами на языке.
У окуня зубы располагаются на тех же местах, но они значительно мельче в связи с питанием более мелкой пищей, и отсутствуют на языке.
У сазана (карпа) —мирной рыбы — зубы в ротовой полости отсутствуют.
Перерезанные жаберные дуги оттягивают вниз и рассматривают задний отдел ротоглоточной полости. Видны внутренние отвер-
«стия жаберных щелей, жаберные тычинки, сидящие на жаберных дугах, а также верхние и нижние глоточные зубы.
Верхние глоточные зубы у щуки располагаются в задней дорзальной части ротоглоточной полости и сидят на глоточных (pharyngo-branchiale) элементах третьей и четвертой жаберных дуг. Они имеют вид небольших площадок, покрытых многочисленными мелкими зубчиками.
Нижние глоточные зубы сидят на рожковых (cerato-branchiale) элементах редуцированной пятой дуги и располагаются в вентральной части заднего конца ротоглоточной полости. Отдельные зубные пластинки располагаются также и на дне глотки. Задняя часть глотки, резко суживаясь, переходит в пищевод.
У окуня верхние глоточные зубы связаны со 2, 3 и 4-й жаберными дугами, нижние — также с рудиментами 5-й жаберной дуги. Жаберные тычинки окуня кроме наружных тычинок 1-й жаберной дуги, которые удлинены, имеют вид отдельных шероховатых бугорков, сидящих по наружной и внутренней поверхности жаберных дуг. Причем при сведенных жаберных дугах бугорки передней дуги заходят в межбугорковые пространства задней дуги, так что жаберная щель становится очень узкой.
У сазана (карпа) верхние глоточные зубы отсутствуют. Нижние глоточные зубы развиты очень сильно. Они видны на дне самого заднего отдела ротоглоточной полости в виде двух рядов крупных белых бугорков с продольной исчерченностью. Количество глоточных зубов у карпа больше, чем это видно на препарате, но более подробно с их строением лучше ознакомиться при изучении скелета. Следует обратить внимание на то, что на крыше ротоглоточной полости, против глоточных зубов располагается твердая роговая подушка — жерновок, о который и происходит перетирание пищи.
Жаберные тычинки соседних дуг, как и у окуня, заходят друг за друга, образуя цедильный аппарат.
Для дальнейшего ознакомления с пищеварительной системой необходимо отпрепарировать все ее отделы. Для этого достаточно перерезать препаровальной иглой довольно рыхлые брыжейки, которые соединяют отдельные части пищеварительного тракта друг с другом и с другими органами. При препаровке брыжеек следует не повредить крупных кровеносных сосудов. Наиболее крупные сосуды лежат на дорзальной стороне печени. Поэтому начинают с того, что аккуратно перерезают брыжейку, соединяющую печень с желудком и пищеводом. Отворачивают левый край печени вниз и рассматривают проходящую по средней части дорзальной поверхности печени чревно-брыжеечную артерию (рис. 85, a. cl. т.). Она тянется по брыжейке справа от пищевода, затем проходит вдоль правой стороны желчного пузыря и далее идет в направлении селезенки сначала в жировом веществе и затем в веществе печени. Хорошо видны только ее начальный отдел до желчного пузыря и сосуды, отходящие от нее к печени и кишечнику. Параллельно сосудам, отходящим от вышеназванной артерии, а также
вдоль нее самой тянутся венозные сосуды, сливающиеся в воротную вену печени (рис. 85, v. р. h.), которая входит в печень у ее заднего края между двумя намечающимися долями.
Большие гонады, особенно сильно развитые у экземпляров, пойманных в период нереста, необходимо аккуратно отвернуть вверх. У самок щуки можно даже удалить одну из них, предварительно рассмотрев сосуды, идущие по брыжейке к половой железе.
У окуня для того, чтобы рассмотреть пищеварительную систему, необходимо отвернуть печень вперед, так как она почти полностью прикрывает желудок и передний отдел кишечника. Кроме того, чревная артерия идет по дорзальной стороне желудка и затем переходит на кишечник, давая ответвления к печени.
У сазана (карпа) изучение пищеварительной системы, как уже указывалось, затруднено из-за большого количества жира, покрывающего все внутренние органы, многочисленных связок, соединяющих отдельные петли пищеварительной трубки друг с другом и с внутренними органами, а также из-за многодольчатой печени, между долями которой проходят петли кишечника. Все органы, включая и жировые тела, богато снабжаются кровеносными сосудами, которые приходится перерывать при препаровке, поэтому, как указывалось выше, перед изучением пищеварительной системы необходимо несколько часов или всю ночь подержать вскрытую рыбу в слабом (1:19) растворе формалина.
Сложность и рыхлость внутренних органов карпа приводит к тому, что при препаровке пищеварительной системы приходится перерывать большое количество мелких кровеносных сосудов, однако необходимо сохранить наиболее крупные.
Длинный кишечник карповых рыб (желудок у них отсутствует) делится па три отдела: начальный, идущий от пищевода до заднего конца полости тела; средний, представленный участком кишки, возвращающимся от каудальной части полости тела к головному концу, и конечный, идущий от головного конца к анальному отверстию. У сазана средний и конечный отделы тянутся параллельно друг другу и имеют дополнительные изгибы, как это видно на рис. 87.
Прежде чем приступить к выделению пищеварительной трубки, необходимо срезать до половины высоты правую стенку тела, что облегчит доступ ко всем петлям кишечника. Кроме того, у крупных половозрелых особей удаляют большую часть левой гонады, оставив только ее самый задний конец, а также большое жировое тело, расположенное с левой стороны на уровне вентральной части плавательного пузыря.
Теперь следует рассмотреть пищевартительную систему. Вент-ральней средней и задней части плавательного пузыря тянется конечный отдел кишечника, а параллельно ему, чуть ниже, средний. Перед ними лежит левая доля печени, от которой отходит тонкий, идущий вниз отросток, доходящий до вентральной части полости тела. Этот отдел печени очищают от скоплений жира. После этого, начав от заднего конца, отпрепарируют все изгибы конечного и
среднего отделов кишечника, ориентируясь по приведенному рис. 87. При этом стараются не отделять параллельно идущие отделы друг от друга. Отвернув вправо, к хвостовой части, эти отделы стараются сохранить их естественную форму. После этого открывается более толстый начальный отдел кишечника. Отпрепарируют его, отделив от окружающего вещества печени. Теперь можно
Рис. 87. Часть пищеварительного тракта рыб. А — окуня Perea fluviatilis, Б — карпа Cypri-nus carpio:
арр. р. — пилорические придатки; d. chai. — желчный проток; gast. — желудок; h. — печень; ints, — кишечник; I. — селезенка; ves. f. — желчный пузырь
приступить к более детальному ознакомлению с пищеварительной системой.
Расправив пищеварительный тракт, рассматривают его строение.
У щуки органы пищеварения состоят из следующих отделов.
Пищевод (oesophagus, см. рис. 85, oes.).. Глотка, резко суживаясь, переходит в области плечевого пояса в толстостенную трубку — пищевод, внутренняя поверхность которого у щуки покрыта продольными складками. Следует вскрыть стенку переднего отдела пищеварительной системы и найти место окончания пищевода, там где его продольные складки переходят в извилистые складки желудка.
Внешне граница между лищеводом и желудком не выражена.
У окуня задней границей пищевода можно счи
тать место, которое лежит несколько впереди от места выхода из желудка кишечника.
У сазана (карпа) пищевод очень короткий и лежит на уровне заднего края верхнего отдела перикардиальной полости. Чтобы его вскрыть, необходимо продолжить разрез ротоглоточной полости дальше назад так, чтобы он захватил и передний отдел кишечника. При этом приходится перерезать плечевой пояс. Кроме того, разрез проходит через головную почку и кювьеров проток, что нежелательно проделывать до изучения мочевой и кровеносной систем. Поэтому лучше ограничиться разрезом левой стенки переднего отдела кишечника. Промыв просвет кишки под струей воды, можно
видеть задний отдел пищевода, отличающийся продольной складчатостью слизистой выстилки. Мускульная оболочка пищевода образует род сфинктера. Если он находится в сокращенном состоянии, то просвет пищевода выглядит как небольшое отверстие.
Желудок (gaster, см. рис. 85, gast.) у щуки внешне не отличается от пищевода и является его непосредственным продолжением. Стенки желудка толстые мускулистые, но довольно эластичные. Наполненный пищей желудок сильно растягивается и может занимать большую часть брюшной полости. В сокращенном состоянии у щуки он доходит примерно до середины брюшной полости. Вскрыв стенку желудка, можно убедиться в наличии извилистых складок на его внутренней поверхности.
У окуня желудок (рис. 85, 87, gast.) значительно меньше и имеет форму слепого выроста. Складки его внутренней стенки более крупные и тянутся в косом направлении.
У сазана (карпа) желудок отсутствует.
Кишечник (intestinum, см. рис. 85, ints.) у щуки, как и у всех хищников, короткий и имеет немного изгибов. Он начинается от заднего конца желудка и отграничен от последнего сфинктером. Начальный его отдел тянется вперед параллельно желудку и пищеводу. Недалеко от переднего конца брюшной полости кишечник круто поворачивает назад и идет вдоль вентральной стенки полости до анального отверстия. Кишку трудно дифференцировать па отделы, так как она не имеет сколько-нибудь заметных утолщений, ширина просвета ее постепенно убывает спереди назад. Однако по аналогии с высшими позвоночными небольшой участок, лежащий сразу за желудком, в который впадают протоки желчного пузыря и поджелудочной железы, называют двенадцатиперстной кишкой. Весь кишечник, как уже указывалось, подвешен на брыжейках, которыми соединены и его отдельные части. Наиболее плотные брыжейки находятся в самом заднем отделе, по ним подходят и крупные кровеносные сосуды. Внутренняя поверхность всей кишки выстлана многочисленными мелкими ворсинками — выростами слизистой оболочки, в чем можно убедиться, вскрыв ее продольным разрезом.
У окуня кишечник (рис. 85, 87, ints.) более длинный и имеет значительное количество петель. По выходе из желудка он сначала спускается вниз, затем тянется назад примерно до середины брюшной полости, потом снова вперед до желудка, делает поворот и продолжается назад вдоль нижней стенки брюшной полости, открываясь паружу самостоятельным анальным отверстием.
У сазана (карпа) (рис. 87) кишечник представлен средней кишкой, которую трудно разделить даже на условные отделы, поскольку она представляет гистологически однородную трубку. Как уже указывалось, условно его можно разделить на начальный, средний и конечный отделы. Лучше употреблять именно эти термины, чтобы не возникали ассоциации с передней, средней и задней кишкой других позвоночных. Следует проследить местоположение каждого из отделов. Начальный отдел несколько расширен, поскольку явля-
ется аналогом желудка. Он начинается от пищевода и идет по правой стороне полости тела вниз и назад, доходя почти до каудального конца полости тела. Восстанавливают первоначальное положение остальных отделов кишечника, уложив их опять в полость тела. Средний отдел па своем пути вперед делает несколько изгибов. От конца начального отдела кишечная трубка делает крутой изгиб, переходит на левую сторону полости тела, доходит до уровня ее передней трети и здесь круто поворачивает назад. Дойдя до уровня задней трети полости тела, кишечник снова делает поворот и тянется вперед по правой стороне до передней границы полости тела. Здесь он несколько поднимается вверх, переходя на левую сторону, и после изгиба продолжается в конечный отдел, который повторяет все изгибы среднего отдела в обратной последовательности. Его конечный отдел лежит вентральнее плавательного пузыря и тянется в каудальном направлении, заканчиваясь анальным отверстием.
Продольными разрезами вскрывают разные отделы кишечника и убеждаются в том, что все они имеют одинаковую выстилку, образующую характерный сетчатый рисунок.
Пилорические придатки (appendix pylorica, рис. 85, 87, арр. р) — слепые выросты, сильно варьирующие у разных видов рыб по числу и форме. Из описываемых видов слепые выросты имеются только у окуня в количестве трех. Здесь они располагаются в начале кишечника, непосредственно под желудком (рис. 87). Пилорические выросты по всей вероятности служат для увеличения всасывающей поверхности кишечника и, может быть, для выделения фермента.
Прямая кишка (rectum, см. рис. 85, rd.)—задний отдел кишечника — носит название прямой кишки. Прямая кишка заканчивается анальным отверстием.
Анальное отверстие (anus, см. рис. 85, ап.) лежит на вентральной стенке тела перед 'анальным плавником, впереди от мочевого и полового отверстия.
Печень (hepar, см. рис. 85, h.) у щуки однолопастная и занимает левую и отчасти вентральную часть переднего отдела брюшной полости. Кроме отмеченных проходящих по дорзальной поверхности печени сосудов, обращают также внимание на печеночную вену — синеватый сосуд, просвечивающий через вещество печени с ее вентральной стороной. Печеночная вена впадает в венозный синус, в чем легко убедиться, оттянув назад передний край печени, тогда натянется и выходящий из нее сосуд.
Желчный пузырь (vesica fellea, см. рис. 85, ves. f.) у щуки— довольно крупный тонкостенный мешок вытянутой формы, лежащий на дорзальной стороне печени в передней ее части справа от средней линии. С печенью желчный пузырь связан при помощи брыжейки. Для того чтобы его рассмотреть, отворачивают левую часть печени вниз и в сторону. У свежих экземпляров он интенсивно зеленого цвета. Зеленоватый цвет сохраняется и при фиксации. Задний конец желчного пузыря, постепенно сужаясь, переходит в 278
проток желчного пузыря (ductus cysticus, см. рис. 85, d. cyst.). На некотором протяжении в него впадают выходящие прямо из вещества печени печеночные протоки (ductus hepa-ticus, см. рис. 85, d. h.)\ приняв их, проток продолжается назад уже под названием желчного протока (ductus choledochus, см. рис. 85, d. chol.). Желчный проток на свежем материале заметен довольно хорошо благодаря заполняющей его темно-зеленой желчи. Если проток плохо заметен, осторожно надавливают па желчный пузырь, желчь заполнит проток, и его ход легко проследить. Каудальная часть протока, начиная от конца печени и до места его впадания в начальный отдел кишечника, лежит в веществе поджелудочной железы и плохо видна. Следует отпрепарировать место впадения желчного протока в кишечник. Для этого осторожно подрезают с нижней (вентральной) стороны брыжейку, соединяющую поджелудочную железу с кишечником. Оттянув поджелудочную железу, можно видеть конец желчного протока и место его впадения в кишечник. Более точно место впадения можно установить, счистив сосочковый слой с внутренней стороны кишки через уже проведенный разрез (особенно удобно это делать на фиксированных экземплярах). Наличие зеленоватого пятна желчи на внутренней стенке указывает место впадения протока. После очистки эпителия впадение желчного протока точно обозначается небольшими сосочком.
По джелудочная железа (pancreas, см. рис. 85, рпег.) располагается в брыжейке, соединяющей кишечник с печенью, т. е. фактически лежит в петле между желудком и кишечником. По виду она напоминает жир, которым бывает покрыта. Однако если осторожно счистить жир, то обнаруживается и само вещество железы. Оно более плотное, чем жир, и в отличие от последнего имеет серовато-коричневый оттенок.
Протоки поджелудочной железы (ductus рапегеа-ticus) открываются в передний отдел кишечника и проходят по брыжейкам, соединяющим железу с кишечником, рядом с кровеносными сосудами. Обычно они плохо заметны.
У окуня печень (рис. 85, 87 h.) также однолопастная и занимает левую переднюю часть брюшной полости. Повернув рыбу брюшной стороной вверх, отворачивают печень вперед, а переднюю часть кишечника немного назад и рассматривают лежащий на каудальной стороне печени желчный пузырь (рис. 87, ves. f.). Он грушевидной формы и у свежей рыбы красноватого или буровато-янтарного цвета, на фиксированном материале это тонкостенный мешок с прозрачной жидкостью. Довольно короткий желчный проток (рис. 87, d. chol.) впадает в кишечник у основания переднего пилорического придатка. Так же, как и у щуки, он проходит по брыжейке среди густого переплетения кровеносных сосудов. Чтобы точнее определить место его впадения, вскрывают продольным разрезом пилорический придаток, очищают его от эпительной выстилки. Место впадения обозначено небольшим беловатым сосочком.
Поджелудочная железа у окуня диффузно рассеяна по стенкам кишечника.
У карповых рыб печень имеет сложное строение (рис. 87). Как правило, она состоит из трех долей: левой, правой и средней, которые несут дополнительные выросты и отростки. Кроме того, в состав печени входит также ткань поджелудочной железы и образуется единый орган — «гепатопанкреас». У некоторых карповых в состав печени входит и селезенка (lien), но у карпа она является самостоятельным органом, лежащим по средней линии тела под вентральной стенкой плавательного пузыря, прикрытая справа и слева долями печени. Хотя селезенка рыб является кроветворным органом и относится к кровеносной системе, ее лучше рассмотреть при препаровке долей печени. Селезенка — вытянутый в длину орган, начинающийся в передней части полости тела и доходящий до уровня перетяжки плавательного пузыря. От печени на свежих препаратах она хорошо отличается темно-красным цветом.
У сазана (карпа) средняя доля печени отсутствует. Левая доля располагается слева от начального отдела кишечника, занимает верхнюю переднюю часть полости тела и кроме вышеописанного отростка дает еще небольшой вырост, лежащий в петле кишечника в области перехода среднего отдела в конечный. Правая доля более крупная, располагается справа от начального отдела кишечника. Чтобы ее рассмотреть, необходимо повернуть рыбу брюхом вверх и оттянуть кишечник на левую сторону рыбы. При этом становится видно, что доля занимает весь правый передний отдел полости тела. Ее длинный отросток направляется назад, проходя вдоль вентральной стороны плавательного пузыря почти до заднего конца полости тела. Затем он через вентральную область переходит на левую сторону и входит в виде лопасти в направленную вперед петлю конечной и средней кишки.
Желчный пузырь у сазана (карпа) (рис. 87) довольно крупный и лежит на средней линии тела между двумя лопастями печени. Чтобы его рассмотреть, следует отпрепарировать и оттянуть вниз и в сторону начальный отдел кишечника. Повернув теперь рыбу брюшной стороной кверху, можно видеть расположенный на уровне передней трети брюшной полости крупный, у свежей рыбы зеленоватого цвета желчный пузырь. Каудальный конец пузыря оканчивается слепо, а краниальный продолжается в довольно широкий проток желчного пузыря. На фиксированной рыбе можно аккуратно счистить ткань печени по бокам протока и тогда становятся видны впадающие в него от обеих долей печеночные протоки. Отпрепарируют продолжение протока, которое после впадения в проток печеночных протоков будет называться желчным протоком. Место впадения желчного протока в кишечник можно точно установить, найдя его отверстие па внутренней стенке кишечника. Впадение протока сразу за пищеводом является еще одним подтверждением отсутствия желудка у карповых рыб.
Плавательный пузырь (vesica natatoria, s. vesica pne-umatica, см. рис. 85, ves, pn.) генетически связан с пищеваритель
ной системой, так как представляет не что иное, как выпячивание верхней стенки переднего отдела пищеварительной трубки, однако функционально не имеет никакого отношения к пищеварению. У щуки плавательный пузырь имеет вид длинного, полупрозрачного, наполненного газом мешка, занимающего всю дорзальную часть брюшной полости. На поверхности пузыря хорошо видны тонкие кровеносные сосуды, подходящие сюда от спинной аорты. Параллельно им идут венозные сосуды, несущие кровь от стенок плавательного пузыря к задним кардинальным венам. У щуки, как и у остальных открытопузырных рыб, плавательный пузырь в течение всей жизни сохраняет связь с пищеводом через небольшой воздушный канал.
Воздушный канал (ductus pneumaticus, см. рис. 85, d. рп.). Чтобы рассмотреть этот канал, необходимо отпрепарировать передний конец плавательного пузыря. Для этого отворачивают вверх, насколько это возможно, левую половину половой железы, оттягивают вниз желудок и подрезают брыжейки, соединяющие его и пищевод с плавательным пузырем, начиная сзади и постепенно продвигаясь вперед. Вдоль верхней стороны желудка и пищевода проходит узкий жировой тяж. Примерно па уровне заднего края жаберных лепестков он поворачивает вверх и прикрепляется к дорзальной стенке полости тела. Примерно под местом его изгиба и лежит проток плавательного пузыря. Рядом с протоком в брыжейке проходят сосуды, несущие кровь от чревно-брыжеечной артерии к газовой железе плавательного пузыря, а также вены, так что хорошо отпрепарировать проток, не повредив сосудов, невозможно. Поэтому лучше всего рассмотреть его, оттянув пищевод вниз, а переднюю часть плавательного пузыря — кверху. В таком положении проток растягивается и хорошо виден. Через этот проток у щуки, как и у остальных открытопузырных рыб, из плавательного пузыря удаляется избыток газов. Газы для наполнения пузыря выделяются из крови специальной железой, лежащей внутри пузыря. Делают продольный разрез передней части плавательного пузыря, немного не доведя его до переднего конца. Разрез проводят ниже брыжейки, соединяющей пузырь с половой железой. Стенки пузыря относительно толстые и состоят из нескольких слоев. Наружный слой наиболее плотный, а внутренний тонкий эластичный. Раздвигают края разреза и рассматривают газовую железу.
Газовая железа, или красное тело (corpus ruber, см. рис. 85, с. г.), занимает у щуки весь передний конец плавательного пузыря. Она красноватого цвета (на фиксированном материале — бурая) от большого количества капиллярных сосудов и представляет как бы утолщение мягкой внутренней оболочки плавательного пузыря. Железа вырабатывает секрет, который, очевидно, и способствует выделению газа из крови. Деятельность железы регулируется ветвями блуждающего нерва.
У сазана (карпа) плавательный пузырь (рис. 88) состоит из двух отделов, разделенных перетяжкой, по сообщающихся друг с другом. Перетяжка проходит, примерно, в средней части пузыря, на
уровне покрывающей его лопасти туловищной почки. Отпрепарируют эту лопасть, стараясь при этом не повредить отходящих от нее
кровеносных сосудов, приподнимают ее ,и рассматривают место сужения плавательного пузыря. Сазан, как и щука, относится к открытопузырным рыбам, но воздушный капал начинается от
задней половины плавательного
Рис. 88. Схема веберова аппарата карповой рыбы (из Гирсберга и Ритчела):
int. — вставочная косточка; h'g.—связки; limph. — лимфатическая полость; scaph. — чашевидная косточка; sin. е. с.—общий эндолимфатический синус; sus. — os suspensorium; in. ex. — наружная оболочка плавательного пузыря; tr. — трехногая косточка; /, 11, 111, /V—тела позвонков
пузыря. Чтобы его рассмотреть, следует отпрепарировать вентральную стенку плавательного пузыря в области его сужения. Практически на уровне сужения, почти на медиальной линии вентральной стенки задней камеры, выходит тонкостенная трубка воздушного канала. У небольших рыб стенки канала тонкие, прозрачные и видно, что канал заполнен воздухом, это хорошо отличает его от кровеносных сосудов. У крупных рыб стенки капала более плотные, канал тянется вперед вдоль вентральной стенки плавательного пузыря. Препарирование его задних отделов не представляет особых трудностей. При выделении передней части приходится отпрепарировать и оттянуть впиз левую долю печени, а также раздвинуть и оттянуть крупные кровеносные сосуды и головную почку. Желательно при препарировании не повредить кровеносных сосудов, особенно на свежей рыбе. После препаровки видно, что воздушный проток открывается в средний отдел пищевода, со стороны его дорзальной стенки. Перед впадением в пище
вод канал несколько расширяется и стенки его утолщаются.
После рассмотрения воздушного хода следует отпрепарировать передний отдел плавательного пузыря. Оттянув переднюю стенку назад, убеждаются, что дорзальная ее часть в правом и левом углу соединены при помощи связок с задним концом одной из косточек веберова аппарата, прощупав эту кость иголкой. Подробное ознакомление с веберовым аппаратом правильнее было бы отложить до знакомства с другими органами чувств, но он тесно связан с плавательным пузырем, поэтому его описание приведено после опи
сания плавательного пузыря.
Рассмотрев форму плавательного пузыря, вскрывают одну из его стенок и убеждаются, что она состоит из плотной двухслойной
наружной оболочки и тонкой внутренней. Газовая железа у карпа располагается в переднем отделе вентральной стенки задней камеры пузыря. Макроскопически она слабо заметна, но ее положение легко установить по подходящему сюда крупному артериальному сосуду, тянущемуся вдоль воздушного канала.
У окуня, относящегося к закрытопузырным рыбам, взрослые формы не имеют протока, соединяющего пузырь с пищеводом. Сам пузырь более тонкостенный. Газовые железы (см. рис. 85, с. г.) лежат на вентральной стенке 'передней части пузыря и имеют вид небольших лопастей, центральная часть которых занята сплетением кровеносных сосудов, а края образованы веществом железы. К отдельным долькам подходят веточки кровеносных сосудов, идущие от крупного общего сосуда.
Для выделения газов из пузыря служит специальное образование— овал.
Овал (см. рис. 85, ов.) — особое образование у закрытопузыр-ных рыб, лежащее на дорзальной стороне задней части плавательного пузыря. Это отверстие во внутренней оболочке пузыря имеет по краям мускулатуру и поэтому способно изменять размеры. Через овал газы имеют доступ к среднему богатому венозными сосудами слою,, где и происходит их поглощение кровью. Венозная кровь затем поступает в задние кардинальные вены. Так 'как внутренняя оболочка плавательного пузыря, вероятно, непроницаема для газов, то изменение величины отверстия овала регулирует количество газов, уходящих из плавательного пузыря.
Веберов аппарат, характерный для карповых рыб, — сложно устроенный орган, передающий изменения давления внешней среды в полость внутреннего ухода. В его образовании участвуют плавательный пузырь, скелетные элементы первых позвонков и вырост перилимфатической полости внутреннего уха. Таким образом он связан с несколькими системами, при изучении которых можно было бы познакомиться с его строением. Удобнее рассмотреть его при изучении скелета. Однако не все его части удается сохранить при очистке скелета. Как правило, они бывают утрачены и па готовых препаратах, поэтому лучше всего этот аппарат рассматривать при вскрытии, пользуясь приведенной схемой его строения (рис. 88) и обязательно контролируя положение костных его частей на готовом хорошо очищенном скелете сазана (карпа).
Сазана кладут на правый бок и удаляют у него большую часть эпиаксиальной мускулатуры позади головы, срезав ее скальпелем от затылочной части до начала спинного плавника. Оставшуюся часть мускулатуры выщипывают пинцетом так, чтобы были хорошо видны остистые отростки, тела позвонков и верхние концы ребер. При этом бросается в глаза очень мощное ребро третьего позвонка, идущее параллельно заднему краю жаберной крышки. Из-под ребра на латеральную поверхность выходит одна из косточек веберова аппарата — tripus (трехногая). Некоторые авторы называют ее также malleus или malleoides. Передний ее отдел налегает с дор-
зальной стороны на поперечный отросток второго позвонка. Аккуратно очистив эту довольно крупную кость, но не вычленяя ее, можно видеть, что она имеет вытянутую форму с отростком в средней части, который направлен внутрь и упирается в тело третьего позвонка. Косточка может вращаться вокруг точки опоры, что легко проверить, пошевелив ее кончиком иголки. По своему происхождению эта косточка образовалась из поперечного отростка третьего позвонка. Задний суженный отдел при помощи связки соединен с соединительнотканным наружным слоем (tunica externa) плавательного пузыря, что уже отмечалось при вскрытии. От ее переднего конца в дорзокраниальном направлении также тянется относительно крупная связка. Примерно в середине ее длины она как бы прерывается довольно тонкой второй косточной веберова аппарата— вставочной или соединительной (intercalate). Для нее употребляются и другие названия: incus, intercalarium, incudoideum. Косточка имеет вытянутую форму. Каудальный ее конец разделен на две ножки, каждая из них упирается в тело второго позвонка, над поперечным отростком. Вставочная кость развивается из верхней дуги второго позвонка.
Впереди связка оканчивается на третьей косточке, а именно на scaphium (чашевидной). Применять два других названия (stapes и stapedoideum) нежелательно, так как ими обычно обозначают также связанное с внутренним ухом стремечко наземных позвоночных, которому описываемая кость не гомологична. Она образуется из верхней дуги первого позвонка, сохраняет ее положение, прикрывая собой сбоку спинномозговой канал в районе первого позвонка. На очищенном скелете хорошо видно, что верхние дуги и остистый отросток у первого позвонка отсутствуют. На препарате это подчеркивается тем, что спинной мозг в этой области остается открытым сверху.
Оттянув чашевидную кость на себя, можно видеть задний конец перилимфатического синуса, который представлен отверстием (стенка удаляется вместе с костью), лежащим нейтральнее спинного мозга.
В состав веберова аппарата у карповых рыб входят еще две косточки. Первая лежит рядом с чашевидной, носит название claustrum (запор) и является производной остистого отростка первого позвонка; вторая — в области заднего отростка tripus и носит название os suspensorium. Эти косточки отпрепарировать довольно трудно.
Функция веберова аппарата. Функция этого аппарата заключается в передаче воспринимаемых плавательным пузырем звуковых колебаний и изменений наружного давления. Передний отдел плавательного пузыря, или «веберовская воздушная камера», специализирован для этих функций и отделен от задней гидростатической части узким протоком, который может замыкаться. При увеличении давления происходит сжатие стенок плавательного пузыря (рис. 88), что приводит к изменению положения заднего отростка tripus и передается через серию косточек на непарный перилимфа-284
тический синус и затем на эндолимфу, которая и вызывает раздражение чувствующих окончаний в са'ккулюсе (см. «Органы чувств»).
Функции плавательного пузыря. Функции плавательного пузыря весьма разнообразны. Издавна считалось, что плавательный пузырь — гидростатический орган. Изменение его объема меняет удельный вес рыбы, что позволяет ей всплывать, погружаться или оставаться на определенной глубине без значительной работы локомоторных мышц. Связь плавательного пузыря с веберовым аппаратом указывает на его функцию как органа, воспринимающего-звуковые колебания и участвующего в определении глубины погружения рыбы.
Некоторые рыбы с помощью плавательного пузыря могут издавать звуки. Кроме того, плавательный пузырь может являться дополнительным органом дыхания.
Мочеполовая система
Мочеполовая система костистых рыб отличается от таковой низших рыб обособлением от пищеварительной системы (нет общей клоаки) и полным разделением выделительной системы (почки, мочеточники) от половой; последняя имеет у самцов и самок самостоятельные выводные каналы, открывающиеся наружу особыми отверстиями (см. «Внешние признаки»).
Половая система
Половые железы (gonas, рис. 85, gon.) у самцов и самок щуки парные, они располагаются по бокам и несколько книзу от плавательного пузыря, к которому прикреплены брыжейкой.
Выводные протоки половых желез у костистых рыб — новообразования, они не связаны в своем происхождении ни с мюллеро-вым, ни с вольфовым каналами.
Семенники (testis, рис. 89, test.) имеют вид нешироких лент молочно-белого или чуть желтоватого цвета. Передний конец их лежит на уровне плечевого пояса в верхней части брюшной полости. Отсюда семенники несколько опускаются вниз и тянутся вдоль середины брюшной полости. Задний отдел доходит почти до заднего конца плавательного пузыря. В этой части семенники опускаются в нижнюю часть брюшной полости. На всем их протяжении по брыжейке к ни.м подходят кровеносные сосуды.
Семяпроводы (ductus spermaticus, рис. 89, d. sp.) идут вдоль верхнего края семенника, за задним концом которого они круто поворачивают вниз и имеют здесь вид довольно толстых тяжей, состоящих из рыхлой соединительной ткани, в которой трудно различить основной просвет семяпровода. Отверстие семяпровода наружу очень невелико (рис. 89, d. sp.') и открывается на задней стороне выступа степки тела позади анального отверстия. Срежьте левую стенку конца семяпровода, найдите общую полость, которая
-образуется в результате слияния правого и левого семяпровода и, введя в нее щетинку или леску, постарайтесь пропустить последнюю через наружное половое отверстие.
Яичники (ovarium) щуки парные. Они занимают почти то же самое положение, что и семенники. У половозрелых экземпляров, особенно перед нерестом, они сильно увеличены и занимают почти всю брюшную полость; через тонкие стенки яичника просвечивают отдельные темно-желтые икринки, обусловливая цвет всей железы. Задние концы яичников, несколько суживаясь, переходят в яйцеводы.
Яйцеводы (oviductus) занимают то же положение, что и се-
мяпроводы у самцов, и открываются самостоятельным половым отверстием. Так же, как и семяпроводы, яйцеводы — самостоятельные образования, не связанные по происхождению с мюллеровыми каналами.
Половая система карпа по своему строению принципиально не отличается от таковой щуки, если не считать, что в связи с корот
ким телом половые железы меньше вытянуты в длину.
У окуня половая си-
Рис. 89. Выводные каналы мочеполовой системы самца щуки Esox lucius:
ап. — анальное отверстие; г. anst. — анастамоз между хвостовой и кардинальной венами; a. caud. — хвостовая артерия; а. т. inf. — нижняя брыжеечная артерия: ао. d.—спинная аорта; d. sp. — семяпровод; a. sp.' — наружное отверстие семяпровода; ints. — кишечник; г.— почка; test. — семенник; иг. — мочеточник; иг/ — наружное отверстие мочеточника; v. caud. — хвостовая вена; v. crd. р.— задняя кардинальная вена (правая); ves. иг. — мочевой пузырь;
у. р. г. — воротная вена почек
стема отличается от таковой карпа и щуки тем, что у самок половая же-леза непарная и образует общий икряной мешок с полостью внутри, куда и выделяются икринки при нересте, а затем уже по яйцеводу — наружу.
Выделительная система
Почки (геп, рис. 85, г.) костистых рыб являются настоящими туловищными (мезонефрическими) почками. Головная почка, как правило, функционирует только у личинок, а потом в ней развивается лимфоидная ткань. У щуки почки лежат на дорзальной стенке полости тела, над плавательным пузырем, и отделены от нее брюшиной,
так что па самом деле они находятся вне брюшной полости. Для
того чтобы их рассмотреть, следует осторожно отделить плавательный пузырь от стенки тела почти по всей его длине, кроме самого
переднего отдела. Для этого необходимо перерезать брыжейку,, соединяющую пузырь со стенкой тела, хотя бы с одной стороны. Повернув рыбу на спину, оттягивают на себя пузырь вместе с внутренностями и рассматривают темно-красные почки, которые тянутся в виде двух лент по бокам от позвоночника. Почки начинаются непосредственно за жабрами и идут вдоль всей брюшной полости, заходя задним .концом даже за брюшную полость, в переднюю часть хвостового отдела. В задней части правая и левая почки сливаются, образуя хвостовую почку, в средней они лежат раздельно, так что между ними виден позвоночник и проходящая по его средней линии опинная аорта. Передние, также слитые и несколько расширенные концы образуют головную почку, особенно хорошо этот отдел выражен у окуня. Анатомическое строение почек, кровеносных сосудов и выводящих путей у сазана (карпа) отличается от такового щуки, поэтому их описание будет дано отдельно.
Мочеточники (ureter, рис. 89, иг.) у самцов и у самок — эта вольфовы каналы. Лучше всего они видны в задней части почек.
Одновременно с препарированием мочеточников можно проследить и начало правой задней кардинальной вены (рис. 89, v. crd. р.) в каудальном конце почек.
На фиксированных экземплярах ход мочеточников легко проследить; для этого, положив рыбу на правую сторону, осторожно, пинцетом выщипывают вещество почек, начиная от края и постепенно углубляясь к средней линии. При этом довольно легко обнажаются не очень толстые, ветвящиеся, сероватого цвета мочеточники. Они тянутся вдоль вентральной поверхности почки, ближе к средней линии, несколько прикрытые веществом почки.
Медиальнее от задней части левого мочеточника, который отпрепарирован, в веществе почки лежит начальный отдел правой задней кардинальной вены. При тщательном препарировании можно видеть, что этот сосуд начинается в результате слияния большого количества мелких сосудов, отходящих от вещества почки в ее заднем конце. Кроме того, применяя лупу, удается найти один-два анастомоза, соединяющих правую заднюю кардинальную вену с хвостовой веной.
На свежем материале выщипывать вещество почки рискованно, так как рядом с мочеточниками проходит правая задняя кардинальная вена и ее легко повредить.
Примерно на уровне заднего конца плавательного пузыря мочеточники выходят из вещества почек, сливаются вместе и далее тянутся вниз, вдоль задней стенки брюшной полости в виде непарного протока. В месте их слияния, между ними из вещества почки па ее вентральную поверхность выходит правая задняя кардинальная вена. Она тянется некоторое время почти по средней линии, постепенно расширяясь, а затем переходит на правую почку. В левой почке соответствующий сосуд появляется примерно в средней части.
Вскрывают левую стенку непарного мочевого канала недалеко от места слияния мочеточников, раздвигают края разреза и рас-
сматривают лежащее на передней стенке отверстие мочевого пузыря.
Мочевой пузырь (vesica urinaria, рис. 89, ves. иг.) представляет вырост передней стенки непарной части мочеточника и имеет вид вытянутого в краниальном направлении тонкостенного мешка. Он лежит непосредственно под задним концом плавательного пузыря и между половыми протоками. Передняя часть его несколько смещена влево.
Вскрывают непарный мочевой проток по его левой стенке вплоть до наружного мочевого отверстия, для чего вводят скальпель или ножницы в уже имеющийся разрез и продолжают его дальше вниз.
Наружное мочевое отверстие (рис. 89, иг.) имеет вид узкой поперечной щели, прилегающей к стенке тела непосредственно перед основанием анального плавника. Мочевое и половое отверстия открываются на дне небольшого углубления непосредственно одно за другим и разделены только узкой перегородкой. Они достаточно малы, поэтому иногда при внешнем осмотре рыбы бывает видно только два отверстия; анальное и общее углубление, па дне которого открываются два вышеназванных отверстия.
У сазана (карпа) туловищная почка имеет наиболее развитую среднюю часть. В этой области, как уже указывалось, она спускается почти до вентральной стенки плавательного пузыря. У карпа, как и у всех позвоночных, почки лежат вне полости тела, прикрытые париетальным листком брюшины, но при вскрытии полости тела он может удаляться вместе со стенкой тела, а не оставаться на органах. Тогда создается впечатление, что почки лежат в полости тела. Чтобы рассмотреть форму туловищной почки целиком, отворачивают средний ее участок к спинной стороне рыбы. При этом следует обратить внимание на то, что от переднего нижнего его угла отходит крупный кровеносный сосуд. На свежей рыбе его нужно перерезать, предварительно наложив две лигатуры. Затем оттянуть вниз плавательный пузырь и, повернув рыбу брюшной стороной вверх, рассмотреть форму почек. От расширенной части почки тянутся назад в виде узкой непарной ленты, идущей сначала под позвоночником, а затем по задней стенке полости тела. По левой части ленты проходит обычно хорошо заметный широкий сосуд— приносящая вена почек, который выходит на вентральную поверхность почек на уровне задней границы полости тела. Впереди он распадается па два сосуда, входящих в правую и левую лопасти почки.
Вперед от средних долей тянутся сильно редуцированные передние отделы почек. Правая и левая почки здесь лежат отдельно и между ними хорошо видна спинная аорта. Эти отделы доходят до переднего конца полости тела. В правой и левой почках проходят тонкостенные венозные сосуды — задние кардинальные вены, причем в связи с редукцией воротной системы в левой почке сосуд начинается почти от переднего края боковой лопасти.
На переднем и заднем участках почек хорошо видно, что они расположены вне полости тела и прикрыты париетальным листком 288
брюшины. Оттянув почки вниз, обращают внимание также па то, что в почки с их дорзальной стороны входят многочисленные сегментарные кровеносные сосуды.
Головная почка, являющаяся преобразованным в лимфоидный орган пронефросом, у карпа выражена хорошо. Ее положение над верхней частью перикардиальной полости было подробно описано при вскрытии.
Мочеточники. Поскольку у сазана (карпа) функционирующими являются средние части почек, то мочеточники начинаются от них и имеют вид двух беловатых прозрачных трубочек. В их начальных отделах хорошо видны составляющие их отдельные канальцы. Далее мочеточники тянутся по обеим сторонам заднего отдела почек и сливаются друг с другом в месте его окончания. Образующийся после слияния общий проток расширяется в небольшой мочевой пузырь, который открывается наружу позади полового отверстия, на дне общего углубления.
Кровеносная система
Изучение кровеносной системы лучше всего вести на отдельном, предварительно инъецированном экземпляре. В первую очередь можно рекомендовать инъекцию желатиновой массой. Вполне достаточна инъекция только артериальной системы, которую лучше всего проводить через выносящую жаберную артерию. Для этого примерно на середине любой из жабр накладывается лигатура. Жабра перерезается выше лигатуры и в выносящую жаберную артерию вводится инъекционная игла, которая также фиксируется лигатурой. После инъекции верхняя лигатура также затягивается.
При работе со свежим или фиксированным материалом наиболее удобным объектом следует считать щуку, так как благодаря значительным размерам на ней легко можно проследить большинство деталей строения кровеносной системы. Окунь менее пригоден для препарирования в силу своих небольших размеров. Не очень удобен для изучения кровеносной системы и сазан, особенно если вскрытие и изучение кровеносной системы ведется на одном и том же экземпляре и многие сосуды повреждаются при разборе пищеварительной системы. Однако карп может быть использован в качестве объекта, так как его легко добыть. Поэтому здесь приводится подробное описание кровеносной системы щуки со ссылками па отклонения в строении карпа. Необходимо учесть, что описание ведется но инъецированным объектам, поэтому на объектах с незаполненной кровеносной системой не все описанные сосуды будут видны достаточно хорошо. Во всех случаях для рассмотрения мелких сосудов рекомендуется пользоваться бииокулярньъм микроскопом.
Более подробное знакомство с кровеносной системой рыб лучше всего начать с главного органа кровообращения—сердца.
Сердце
Сердце (cor, рис. 90, cor.) костистых рыб, как и акуловых, называется двухкамерным, но состоит из трех отделов: венозного синуса, предсердия и желудочка. Имеющийся у акуловых и осетровых рыб четвертый отдел — артериальный конус — у костистых рыб
Рис. 90. Кровеносные сосуды жаберной области щуки Esox lucius:
a. br. ad. — приносящие жаберные артерии; а. сог. — венечная артерия; a. h. т. — средняя поджаберная артерия; а. о. о. — брюшная аорта; atr. — предсердие; bul. ао,—луковица аорты; d. С. — кювьеров проток; gl. thr. — щитовидная железа; sin. о. — венозный синус; ti. crd. а.—передняя кардинальная веиа; о. crd. р. ~ задняя кардинальная вена; ventr. — желудочек; ti. h. — печеночная вена; v. j. inf. — нижняя яремная вена
отсутствует. Как и у огромного большинства водных позвоночных, имеющих только один круг кровообращения, через сердце проходит чисто венозная кровь. Последняя по венам собирается в венозный синус, затем из него попадает в предсердие и оттуда — в желудочек. Сокращением мощных мышечных стенок последнего кровь проталкивается в брюшную аорту. Положение сердца в вентральной части жаберной области и окружающая его перикардиальная
полость были описаны в общей топографии внутренних органов. Для того чтобы 'подробно ознакомиться с отделами сердца, нужно снять покрывающую их оболочку (эпикардий). Поэтому ее захватывают пинцетом лучше всего в том месте, где под ней лежит желудочек, и делают ножницами небольшой надрез. Расширив его вперед и в стороны, освобождают отделы сердца.
Желудочек (ventriculus, рис. 85, 90, 92, ventr.) располагается наиболее вентрально относительно всех других отделов сердца. Стенки его мускульные и довольно толстые, на фиксированном материале почти белого, а на свежем — розового цвета.
Предсердие (atrium, рис. 85, 90, 92, atr.) лежит дорзальнее от желудочка (т. е. под ним — для препарирующего) и частично охватывает последний с боков. Стенки его сравнительно тонкие. Темно-красный цвет предсердия зависит от цвета просвечивающей через них крови. Оттянув на себя задний конец желудочка, следует рассмотреть место соединения его с предсердием. Дорзальнее и несколько сзади от него лежит место соединения предсердия с венозным синусом.
Венозный синус (sinus venosus, рис. 90, 92, sin. v.) лежит каудальнее от предсердия и примерно на его уровне. Он очень тонкостенный и имеет вид треугольной полости, лежащей поперек геля.
У сазана (карпа) предсердие несколько смещено в левую сторону. Место его соединения лежит на вентральной стенке желудочка. Венозный синус короче, чем у щуки. Сверху в него впадают кювьеровы протоки, имеющие вид тонкостенных широких вертикально стоящих трубок. В дорзальную часть протока впадает крупная задняя кардинальная вена, выходящая из вещества головной почки. Если оттянуть вверх плечевой пояс, то хорошо видна и несущая кровь от головы передняя кардинальная вена, также впадающая в верхнюю часть кювьерова протока. Повернув рыбу брюшной стороной вверх и освободив заднюю стенку венозного синуса, можно видеть впадающую в него печеночную вену. Оттянув сердце влево, следует найти идущую справа от желудочка, по дорзальной стенке перикардиальной полости, правую нижнюю яремную вену, проследив ее вплоть до места впадения в венозный синус. Вышеописанные сосуды у карпа желательно рассмотреть именно при изучении сердца, так как они могут быть легко повреждены при ознакомлении с деталями строения артериальной системы.
На фиксированном материале можно вскрыть сердце, разрезав его вдоль, и рассмотреть отверстия, соединяющие его отделы, и лежащие в них клапаны. Если материал свежий, то эту операцию необходимо провести только после ознакомления со всей кровеносной системой.
Чтобы познакомиться с внутренним строением сердца, разрезают его на две половины. Разрез следует провести строго по медиальной линии. Лучше всего его сделать лезвием безопасной бритвы или остро отточенным скальпелем. При этом необходимо разрезать 10* 291
и венозный синус, и начальную часть брюшной аорты, так чтобы задний конец разреза прошел непосредственно перед передним концом печени, а передний доходил до переднего конца луковицы аорты. Разрез можно провести или на вычлененном сердце, или не вычленяя последнего. Чтобы вычленить сердце, перерезают брюшную аорту, примерно чуть отступая назад от места отхождения последних приносящих жаберных артерий; кювьеровы протоки перерезают примерно на уровне латеральных сторон предсердия, а также отделяют последние и венозный синус от печени; приподняв передний край сердца, перерезают соединительнотканные связки, прикрепляющие венозный синус и предсердие к дорзальной стенке перикардиальной полости, и вынимают сердце.
Если имеется свежий препарат, то необходимо промыть обе половинки под струей воды. На фиксированном препарате приходится очищать полости венозного синуса, предсердия и желудочка, выбирая из них сгустки крови пинцетом. После этого можно рассмотреть внутреннее строение отделов сердца.
Венозный синус на разрезе имеет вид небольшой округлой полости, прилегающей к задней части предсердия. На правой и левой половинах видны открывающиеся в него отверстия кювьеровых протоков. С полостью предсердия венозный синус сообщается широким отверстием. Обычно при разрезе, прошедшем строго по медиальной линии, оно целиком остается при левой половине сердца. На дорзальной и вентральной сторонах синусо-артериального отверстия сидят относительно плотные полулунной формы клапаны. Действие этих клапанов препятствует обратному току крови из предсердия в венозный синус при сокращении предсердия. На продольном разрезе хорошо видно, что стенки предсердия относительно тонкие и в то же время рыхлые. Внутренняя поверхность имеет вид сложно переплетенных балок с отверстиями между ними, ведущими в полости большей или меньшей величины. В месте соединения предсердия и желудочка лежит атриовентрикулярное отверстие, соединяющее эти два отдела 'сердца. В просвете атриовентрикулярного отверстия находится пара атриовентрикулярных клапанов. Они имеют вид карманов, открытая сторона которых направлена в сторону желудочка. Клапаны прирастают к передней и задней стенкам. При поступлении крови из предсердия в желудочек клапаны прижимаются к стенкам желудочка и не препятствуют току крови. При сокращении желудочка клапаны под давлением крови расправляются, их медиальные стенки плотно соединяются друг с другом и закрывают крови путь в предсердие. При строго медиальном 'разрезе каждый из клапанов режется примерно пополам, поэтому, чтобы полностью представить себе их форму, необходимо рассмотреть их части на правой и левой половине сердца.
Хорошо видна на продольном разрезе и толщина стенок желудочка. Мышечная ткань составляет основную его массу. Полость желудочка очень невелика и по ширине немногим превосходит просвет брюшной аорты. Краниальным концом полость желудочка соединяется с полостью начального отдела брюшной аорты — луко-292
вицей аорты. На границе между желудочком и брюшной аортой располагаются два полулунных клапана. Они имеют вид довольно плотных образований полулунной формы, которые прикреплены наружными краями к правой и левой стенкам полости желудочка и частично к стенке луковицы аорты. Свободные края, соприкасаясь друг с другом, свешиваются в просвет полости желудочка. При прохождении крови из желудочка клапаны прижимаются к стенкам, образуя просвет. При обратном токе крови клапаны расправляются и препятствуют попаданию крови из луковицы аорты в желудочек. Место, где располагаются клапаны, считается рудиментом артериального конуса, который хорошо развит у других рыб.
Артериальная система
Артериальная система костистых рыб состоит из целого ряда кровеносных сосудов, которые несут кровь от сердца и доставляют ее ко всем частям тела. Эти сосуды носят название артерий, хотя не во всех из них течет артериальная кровь. Отходящий непосредственно от сердца крупный сосуд — брюшная аорта — несет венозную кровь, которая, пройдя по приносящим жаберным артериям к жабрам и там окислившись, превращается в артериальную. Артериальная кровь по выносящим жаберным артериям собирается в два параллельных сосуда — корни спинной аорты, которые сливаются в идущую вдоль всего тела спинную аорту. От последней берет начало большинство сосудов, несущих кровь к различным органам.
Препарирование начальных отделов артериальной системы требует большой осторожности, так как тонкостенные сосуды легко рвутся, и, кроме того, мелкие сосуды, идущие в жабры или выходящие из них, довольно плохо заметны. Поэтому в процессе работы необходимо все время пользоваться лупой. Для того чтобы легче было вести препаровку, необходимо расправить и несколько растянуть ротоглоточную полость, для чего вводят в нее большой ватный тампон. Препарируя артериальную систему в области головы, следует не повредить проходящие обычно рядом с артериями венозные стволы.
Вперед от желудочка тянется довольно крупный сосуд — брюшная аорта. Ее часть, прилегающая непосредственно к желудочку, расширена в луковицу аорты.
Луковица аорты (bulbus aortae, см. рис. 85, 90, 92, bul. ао.) в отличие от артериального конуса хрящевых рыб не имеет внутри клапанов и не является отделом сердца. Это только утолщение стенки брюшной аорты.
Брюшная аорта (aorta ventralis, см. рис. 90, 92, ао. о.). Осторожно сняв поджаберную мускулатуру, лежащую впереди от плечевого пояса, прослеживают ход брюшной аорты. Она тянется вперед как продолжение луковицы аорты по средней части вентральной стороны жаберного отдела между нижними частями
жаберных дуг. Примерно на уровне нижнего конца четвертой жаберной дуги брюшная аорта делает резкий изгиб в дорзальном направлении, и передний ее отдел идет значительно глубже, под мускулатурой. Брюшная аорта заканчивается спереди, распадаясь на. два приносящих жаберных сосуда первой жабры.
У сазана (карпа) брюшная аорта лежит более поверхностно и не делает резкого изгиба, по в средней части ее охватывают костные отростки третьей жаберной дуги, удаление которых требует некоторых усилий.
Нижняя яремная вен a (v. juqularis inferior, см. рис. 90„ 92, V. j. inf.) —тонкостенный непарный венозный сосуд, идущий параллельно брюшной аорте и прилегающий непосредственно к ее дорзальной стороне, но несколько смещенный от нее влево (вправо от препарирующего). Этот сосуд относится к венозной системе и упомянут здесь потому, что при дальнейшем препарировании его легко повредить. Примерно на уровне луковицы брюшной аорты нижняя яремная вена распадается на два сосуда, которые проходят справа и слева от сердца по латеральной части дорзальной стенки перикардиальной полости и впадают в кювьеровы протоки на уровне заднего края предсердия.
У сазана (карпа) передний отдел нижней яремной вены обычно-прослеживается с трудом.
Приносящие жаберные а ртер и и (a. branchialis adve-hens, рис. 90, 92, a. br. ad.). На своем протяжении брюшная аорта* отдает вправо и влево три пары приносящих жаберных артерий. Первая и вторая пары сосудов, считая от переднего конца головы, снабжают кровью первую и вторую жабры. Третий сосуд, отходящий от аорты примерно в месте изгиба, сразу распадается на два, несущих кровь к третьей и четвертой жаберным дугам. Чтобы полностью рассмотреть приносящие жаберные сосуды, необходимо отпрепарировать их. Для этого осторожно при помощи препаровальной иголки с крючком на конце и маленького пинцета очищают сосуд от окружающей его соединительной ткани, и прикрывающих сверху мышц. Начинать препарирование следует от места отхождения сосуда от брюшной аорты, а затем продвигаться вдоль него вплоть до вхождения его в жабру. На фиксированных экземплярах толстый слой мышц частично срезают ножницами.
Для того чтобы проследить ход приносящих жаберных сосудов в жаберной дуге, нужно снять внутренний ряд жаберных лепестков. Тогда обнажатся два сосуда: идущий ближе к основанию жабры выносящий жаберный сосуд и лежащий более каудально приносящий сосуд, на последнем обычно видны небольшие отверстия сосудиков, идущих в жаберные лепестки.
Войдя в жабру, сосуд делится на две части: одна, меньшая, идет вперед и снабжает кровью передний вентральный конец дуги, другая — назад и несет кровь к остальной, большей части жабры. Только сосуд четвертой жаберной дуги не делится, так как входит в передний конец вентральной части четвертой жабры.
У сазана (карпа) вентральные концы жабр небольшие, и приносящие сосуды входят в них так, как в четвертую жабру щуки.
Кроме упомянутых сосудов в нижней части жаберной области иногда бывают также видны сосуды, питающие вентральные отделы жаберного аппарата, они образуются в результате слияния нижних концов выносящих жаберных артерий и обычно проходят вентральнее брюшной аорты и яремной вены.
Наиболее крупный из них — средняя поджаберная артерия.
Средняя поджаберная артерия (a. hypobranchialis medialis, рис. 90, a. h. т.) бывает хорошо видна на крупных или инъецированных экземплярах. Она тянется вдоль вентральной поверхности брюшной аорты и является результатом слияния вентральных концов выносящих сосудов третьей жаберной дуги. Примерно на уровне переднего конца луковицы аорты артерия распадается на три сосуда. Два парных идут по наружному краю околосердечной полости вдоль пояса передних конечностей и сливаются на боках тела с подключичной артерией своей стороны. Один, непарный, тянется по луковице аорты к стенкам сердца, которое и снабжает кровью. Он носит название коронарной или венечной артерии (a. coronalis, рис. 90, а. сог.).
Чтобы проследить дальнейший ход сосудов артериальной системы, необходимо отвернуть дно ротовой полости и нижнюю часть глотки. Для этого разрезают правую стенку ротовой полости и перерезают жаберные дуги так же, как это было сделано с левой стороны, а затем отворачивают назад нижнюю часть головы.
Осторожно снимают выстилку крыши полости глотки между жаберными дугами и находят лежащие непосредственно на парасфеноиде парные сосуды — корни спинной аорты. Справа и слева в них впадают выносящие жаберные артерии первой и второй дуг.
У сазана (карпа) в связи с наличием жерновка и толстой выстилкой ротоглоточной полости ход препаровки выносящих жаберных сосудов несколько иной. Удаляют выстилку ротоглоточной полости непосредственно перед жерновком. При этом обнажается непарный сосуд — спинная аорта — и впадающие в него справа и слева мощные сосуды, образованные в результате слияния выносящих жаберных артерий двух последних жабр. Следуя вперед по ходу спинной аорты, можно обнаружить и ее корни. При этом приходится удалять верхние глоточные элементы костных жаберных дуг.
Выносящие жаберные артерии (a. branchialis ге-vehens, рис. 91, a. br. г.). Эти сосуды, как показывает само название, выносят артериальную кровь из жабр. Они начинаются в вентральной части жаберной дуги обычно двумя или большим числом тонких сосудов. От одного из них в каждой жабре отходит уже упомянутый выше сосуд к вентральной части жаберного аппарата. Примерно на границе нижней трети жабры парные сосуды сливаются в один, который идет вверх по жаберной дуге, постепенно утолщаясь. Однако на неинъецированном материале начальные отделы сосудов видны плохо и обычно удается рассмотреть только
Рис. 91. Схема артериальной системы головы щуки Esox lucius: a. br. rt — a. br. r4 — выносящие артерии жаберных дуг; а. с. с. — общая сонная артерия; а. с. е. — наружная сонная артерия; а. с. е.' — собственно наружная сонная артерия; а. с. I, — внутренняя сонная артерия; a. cl. tn. — чревнобрыжеечиая артерия; a. hy. — гиоидная артерия; ао. d. — спинная аорта; а. ог.-п. — глазнично-носовая артерия; a. psbr. — псевдобранхиальная артерия; a. scl. — подключичная артерия; rd. ао. — корни спинной аорты
часть сосуда, которая идет по верхней половине жабры. Мелкие кровеносные сосуды, идущие от жаберных лепестков и впадающие в выносящий жаберный сосуд, тоже видны только при заполнении кровеносной системы специальной массой. Хорошо видны выносящие жаберные артерии на поперечном срезе жабры (см. рис. 86).
Выйдя из жабры, выносящие артерии переходят на дно черепной коробки и впадают в корни спинной аорты (артерии первой и второй жаберных дуг) или в спинную аорту (артерии третьей и четвертой жаберных дуг).
Корпи спинной аорты (radix aortae, рис. 91, rd. ао.) — два крупных парных сосуда, проходящие но дну черепа вентральное от парасфеноида и по бокам от пего. Продолжаясь вперед,, корни аорты входят в отверстия в. кости парасфеноида и там сливаются. Сзади на уровне четвертой, жаберной дуги корпи аорты также сливаются вместе, образуя спинную аорту. Таким образом, образуется так называемый г о-л о в н о й круг.
У сазана (карпа) топография сосудов несколько иная, поэтому их расположение будет описано после описания сосудов головы, щуки.
Общие сонные артерии (a. carotis communis, рис. 91, а. с. с.) являются непосредственным продолжением корней спинной аорты вперед от места впадения в них выносящих артерий первой жаберной дуги. Общая сонная артерия — очень короткий сосуд, который почти сразу делится на два.
Наружная сонная а р т ер и я (a. carotis externa, рис. 91,. а. с. е.) отходит от дорзальной стороны общей сонной артерии, чуть-чуть впереди места впадения выносящей жаберной артерии первой дуги и идет вглубь от препарирующего. Вскоре она разветвляется на два сосуда. Собственно наружная сонная артерия (рис. 91, а. с. е.) идет вперед, подходит под ложную жабру, выходит в область глазницы и тянется вдоль ее заднего края по внутренней стороне. На своем пути она отдает много мел
ких сосудов и заканчивается на нижнем крае глазницы примерно на уровне середины глазного яблока. Гиодпая артерия (a. hyoidea, рис. 91, a. hy.) отходит в сторону от наружной сонной, идет по внутренней стороне гиомандибулярпой кости, а затем переходит на внутреннюю часть середины жаберной крышки.
Внутренняя сонная артерия (a. carotis interna, рис. 91, а. с. i.) является непосредственным продолжением общей сонной артерии. Продолжаясь вперед, внутренние сонные артерии правой и левой сторон входят в хрящевое основание черепа, несколько дорзальней от парасфеноида, и там сливаются друг с другом.
Перед самым вхождением в хрящ внутренние сонные артерии отсылают в стороны небольшие а р т е р и и к ложной жабре (a. pseudobranchiae, рис. 91, a, psbr.). Эти артерии почти тотчас же распадаются на целую серию мелких сосудов, большинство из которых входят в ложную жабру. Однако два из них (по одному с каждой стороны) продолжаются вперед вдоль боковых сторон парасфеноида, проходят через хрящевые предглазничпые отростки и заканчиваются в носовой области. Это — г л а з и и ч н о - но со в а я артерия (a. orbito-nasalis, рис. 91, а. ог.-п.). Разворачивают в стороны правую и левую половины плечевого пояса (если последняя не была удалена). Перерезав пищевод и немного оттянув внутренности назад, отпрепарируют место перехода корней аорты в непарный сосуд — спинную аорту — как раз над началом пищевода. Непосредственно за этим местом в спинную аорту впадают парные сосуды, образовавшиеся в результате слияния выносящих жаберных сосудов двух последних жаберных дуг (рис. 91).
У сазана (карпа), как и у щуки, сосуды образуют головной круг, но наружная и внутренняя сонные артерии отходят у пего отдельно.
Внутренняя сонная артерия отходит от вентральной стороны первой выносящей жаберной артерии, недалеко от места ее выхода из жаберной дуги. Она идет в медиальном направлении и видна только на коротком участке, так как сразу же входит в полость черепа. Чтобы проследить ее дальнейший ход, нужно выломать дно черепа при помощи большого пинцета. В полости черепа артерии противоположных сторон сливаются при помощи тонких сосудов, образуя головной круг. Основные стволы идут вперед, покидают полость черепа рядом с крупными обонятельными трактами. Они соответствуют глазнично-носовым артериям щуки.
Наружная сонная артерия отходит также от первой выносящей, но чуть медиальпей внутренней и от оральной стенки сосуда. Опа направляется к области глазницы, где проходит по ее нижнему краю, и отдает многочисленные мелкие сосуды. Почти у самого основания наружной сонной артерии от ее дорзальной стенки, вглубь от препарирующего, отходит толстый сосуд, который сразу же распадается на два. Сосуд, направляющийся краниалыю, соответствует гиодной артерии. Проследив его ход, можно видеть, что он переходит на внутреннюю поверхность жаберной крышки и дальше
продолжается внутри предкрышечной кости. Второй сосуд тянется в каудальном направлении по верхнему краю внутренней поверхности жаберной крышки.
Подключичные артерии (a. subclavia, рис. 91, a. scl.) —парные сосуды, отходящие или в самом начале спинной аорты, перед впадением последних приносящих жаберных сосудов,, или от конечных участков корней аорты. Каждая артерия проходит вдоль переднего, а затем наружного края головной почки, переходит на внутреннюю сторону плечевого пояса, вдоль которого и тянется. На своем пути артерия отдает сосуды к грудным плавникам и к стенкам тела. Вентральный ее конец, как уже указывалось, сливается с сосудами поджаберной области.
У сазана (карпа) перед впадением в спинную аорту общего ствола двух последних выносящих жаберных артерий, топографически на том месте, где у щуки отходит подключичная артерия (рис. 91), отходит крупный сосуд к пятой жаберной дуге, которая несет глоточные зубы и имеет мощную мускулатуру. Подключичная артерия отходит от спинной аорты каудальнее. Прежде чем ее рассмотреть, приходится проделать дополнительные операции. От места впадения последних выносящих жаберных артерий спинная аорта тянется назад, проходя у карпа через канал в отростке основной затылочной кости. Необходимо уточнить положение этого канала на готовом препарате скелета черепа сазана. Для того чтобы рассмотреть спинную аорту, необходимо вскрыть канал, удалив нижнюю часть отростка вместе с жерновком, аккуратно срезав ее ножницами. После этого открываются отходящие здесь от спинной аорты парные подключичные артерии и непарная чревно-брыжеечная артерия.
Чревно-брыжеечная артерия (a. coeliaca-mesenterica рис. 91, a. cl.-m.) отходит от вентральной стороны спинной аорты, непосредственно за местом впадения выносящих сосудов последних жаберных дуг. В своем начале она круто поворачивает вправо (влево от препарирующего) и тянется вдоль брыжейки, соединяющей печень и передний отдел кишечника, отдавая многочисленные мелкие сосуды к печени, стенкам желудка и кишечника.
В самом начале от артерии отходит очень короткий сосуд, который в свою очередь распадается на множество мелких ветвей. Часть этих ветвей подходит к дорзальной поверхности передней части желудка, часть — к переднему концу плавательного пузыря. Из сосудов, подходящих к плавательному пузырю, некоторые входят в него и питают газовую железу.
Чтобы проследить дальнейший ход спинной аорты, нужно аккуратно отделить дорзальную стенку плавательного пузыря от верхней стенки брюшной полости, для чего необходимо подрезать брыжейку и с правой стороны (слева пузырь был отпрепарирован, когда рассматривались почки). Следует еще раз обратить внимание на то, что к стенкам плавательного пузыря, а по ним и к половым железам идут тонкие кровеносные сосуды. Отвернув плавательный?
пузырь, рассматривают почки и проходящий между ними широкий сосуд — спинную аорту.
Спинная аорта (aorta dorsalis, рис. 91, ао. d.) идет вдоль позвоночника, непосредственно прилегая к нему. На уровне межпозвоночных пространств она узкая, на уровне тел позвонков расширяется не только в боковые стороны, но и входя в специальные углубления на вентральной стороне позвонков, так что вся она -имеет четкообразный характер. От дорзально-боковых частей аорты отходят сегментарные сосуды. Одни из них идут к мускулатуре и почкам. Другие, отдав веточки к почкам и мускулатуре, переходят на плавательный пузырь и входят в вещество половой железы. “Спинную аорту можно проследить примерно до половины брюшной полости. Дальше она прикрыта от препарирующего почками, так что отходящая от нее на уровне заднего конца плавательного пузыря нижняя брыжеечная артерия как бы выходит из вещества почек.
Нижняя брыжеечная артерия (a. mesenterica inferior, рис. 89, а. т. inf.) снабжает кровью заднюю часть плавательного пузыря и конец кишечника.
Хвостовая артерия (a. caudalis, рис. 89, a. caud.). Спинная аорта в хвостовом отделе идет внутри гемального канала и носит название хвостовой артерии. Увидеть ее можно, отрезав заднюю часть хвостового отдела и рассмотрев поперечный срез. Хвостовая артерия лежит под телом позвонка над тонкостенным сосудом — хвостовой веной.
Венозная система
Венозная кровь собирается от всех частей тела в венозный синус по системе венозных сосудов тремя путями.
1. От головы кровь течет по передним кардинальным и нижней яремной венам.
2. Кровь, идущая от хвостового отдела, проходит через воротную систему почек и затем собирается в задние кардинальные вены, которые, сливаясь с передними, образуют кювьеровы протоки. В отличие от хрящевых рыб воротная система почек у костистых рыб неполная. Одна из кардинальных вен, обычно правая, соединяется с воротной веной почек большим или меньшим количеством сосудов, так что часть крови, проходя по ним, минует воротную систему почек.
3. Основная масса крови, идущая от пищеварительной системы, проходит через воротную систему печени, а затем по -печеночной вене попадает в венозный синус.
Дальнейшее описание воротной системы почек и указания по ее препарированию относятся только к щуке. У сазана (карпа) функционируют в основном только средние расширенные отделы почек. Сосуды, приносящие к ним кровь и выносящие ее, хорошо видны и без специальной препаровки и детально были описаны при разборе мочеполовой системы. Ход отдельных венозных стволов у
карпа проследить также трудно, поэтому можно видеть только впадение наиболее крупных из них в венозный синус. Описание мест их впадения было дано при ознакомлении со строением сердца.
Сначала следует рассмотреть со-
sin. и
a.br. at
V-crtp.
ansi.
v.p.r.
ao. v.
v. crd. a.
baLa, v.j. inf. ventr. atr.
d.C.
v.h.
v. caud..
Рис. 92. Схема венозной системы щуки Esox lucius:
a. br. ad. — выносящие жаберные артерии; anst. — анастомоз между воротной веной почек и правой задней кардинальной веной; ао. и. — брюшиая аорта; atr. — предсердие; but. а.—луковица аорты; d. С.— кювьеров проток; sin. v. — венозный синус; v. caud. — хвостовая веиа; v. crd. а. — передняя кардинальная вена; v. crd. р. — задняя кардинальная вена; ventr.—желудочек; v. h. — печеночная вена; v. j. inf. — нижняя яремная вена; v. р. h. — воротная вена печени; v. р. г. — воротная вена почек
суды, несущие венозную кровь от задних частей тела.
Хвостовая вена (v. cauda-lis, рис. 89, 92, v. caud.) проходит в хвостовом отделе по нижней части гемалыюго канала, под хвостовой артерией. Ее положение на поперечном разрезе мы уже видели.
Снимают мускулатуру с левой половины хвостового стебля, вплоть до вскрытой части брюшной полости. Для этого проводят разрез скальпелем вдоль внутренних частей костных лучей анального плавника, вплоть до тел позвонков. Отворачивают образовавшийся лоскут и удаляют его совсем, отделив от позвоночника. Очищают от остатков мускулатуры, соскоблив их скальпелем. Осторожно, начиная сзади, вскрывают гемальный капал и находят в нем хвостовую вену.
Выйдя из гемального канала,, хвостовая вена идет по дорзальной стороне почек, принимая от них небольшие сосуды. Для того чтобы увидеть вену, необходимо отделить-верхнюю часть почек от спинной стенки тела и оттянуть их немного вниз. Примерно на уровне переднего края спинного плавника, или, точнее, на уровне слияния мочеточников, хвостовая вена делится на две
воротные вены почек.
Воротные вены почек (v. portae renalis, рис. 89, 92, и. р. г.) так же, как и хвостовая вена, проходят по дорзальной стороне почек, постепенно истончаясь^ так как разветвляются в последних на многочисленные мелкие сосуды. Оканчиваются они примерно па уровне заднего конца желудка или селезенки. Чтобы их рассмотреть, надо оттянуть вниз вещество почек.
Задние кардинальные вены (v. cardinalis posterior, рис. 89, 92, v. crd. р.) проходят но вентральной стороне почек.
Правая задняя кардинальная вена начинается путем слияния мелких венозных сосудов в самой задней части почек, которая лежит уже в хвостовом отделе. Выщипав пинцетом вещество почек в их заднем конце, рассматривают анастомозы, соединяющие кардинальную вену с воротной веной почек (рис. 92, anst.). На поверхность почек вена выходит между мочеточниками, как раз перед местом их слияния. Примерно на уровне разделения почек па две она идет почти строго по средней линии, а затем переходит па правую почку и тянется вдоль ее внутреннего вентрального края. На своем пути кардинальная вена принимает многочисленные вены от почек и постепенно утолщается. Таким образом, кровь, поступающая в правую заднюю кардинальную вену, частично проходит через воротную систему почек, однако некоторое количество крови минует эту систему, так как поступает прямо из хвостовой и воротной вен почек через имеющиеся анастомозы (рис. 89, 92, anst.).
Левая задняя кардинальная вена проходит по вентральной стороне левой почки. Кровь, поступающая в нее из левой воротной вены почек, целиком проходит через воротную систему. Начинается эта вена небольшими сосудиками на уровне задней трети желудка.
Каждая из кардинальных вен проходит через вещество головной части ночек, выходит из его передней боковой части и сразу же по выходе сливается с передней кардинальной веной, образуя кювье-ровы протоки.
Передние кардинальные, или верхние яремные, вены (v. cardinalis anterior, s. v. jugularis superior, рис. 92, v. crd. a.)—парные сосуды, несущие венозную кровь от верхних частей головы. Начало сосуда каждой стороны следует искать в области глазницы. Сосуд идет вначале параллельно наружной сонной артерии по заднему внутреннему краю глазницы вдоль наружной прямой мышцы глаза. Затем сосуд поворачивает назад, тянется вдоль основания черепа, прободает отросток переднеушной кости. Около места прободения в сосуд вливается г иодная вена (v. hyioidea), идущая параллельно гиодной артерии от внутренней поверхности жаберной крышки. Далее передняя кардинальная вена идет параллельно корню спинной аорты, затем поворачивает латеральнее и, пройдя вдоль переднего края почки, сливается с задней кардинальной веной.
Кювьеровы протоки (ductus Cuvieri, рис. 91, 92, d. С.) —парные сосуды, лежащие на уровне заднего конца сердца и образовавшиеся в результате слияния кардинальных вен соответствующих сторон. Протоки вливаются в вышеописанный (см. раздел «Сердце») венозный синус.
Нижняя яремная вена (v. jugularis inferior, рис. 91, 92, v. j. inf.) несет кровь от нижних частей головы и, как уже указывалось, тянется параллельно брюшной аорте, проходя дорзальнее
нее. На уровне переднего конца околосердечной сумки она делится на два сосуда, каждый из которых, пройдя вдоль наружной стенки околосердечной сумки, впадает в соответствующий кювьеров проток.
Воротная вена печени (v. portae hepatis, рис. 92, v. р. h.), собирающая кровь от внутренних органов, и печеночная вена (v. hepatica, рис. 91, 92, v. Ji.), выносящая кровь из печени и впадающая в венозный синус, были описаны выше.
Скелет
Для изучения скелета костистых рыб в первую очередь можно рекомендовать судака Lucioperca lucioperca, так как у него скелет окостеневает полностью, а границы между костями (особенно в черепе) видны хорошо. Желательно дополнительно познакомиться со строением черепа щуки — представителя отряда щукообразных Eso-cifotmes. Щука принадлежит к низшим костистым рыбам со значительным количеством хряща в черепе, поэтому различия в расположении хрящевых и покровных костей у нее достаточно наглядны. Интерес представляют также и особенности строения черепной коробки в связи с удлинением ее переднего отдела.
При необходимости судака можно заменить треской достаточно крупных размеров. Ссылки на особенности строения скелета трески даны в тексте. Иногда целесообразно изучение скелета тех видов, на которых проводилось вскрытие, поэтому в тексте даются ссылки на особенности скелета окуня и карпа, но только в тех случаях, если различия с судаком будут очень велики. Кроме того, даже при изучении скелета на препаратах судака желательно познакомиться по готовым препаратам с особенностями его строения и у других описываемых видов.
Скелет костистой рыбы лучше приготовить из свежего, нефиксированного материала. Для этого удаляют все внутренние органы, а также мускулатуру и другие мягкие части. Перед препарированием необходимо подержать рыбу в горячей воде минут 5—10 или положить ее в холодную воду и нагревать до кипения (ни в том, ни в другом случае не кипятить!). Затем следует отчленить пояса конечностей с плавниками. Следует обратить внимание на место причленения грудного пояса к черепной коробке и на положение брюшных плавников. Счищают скальпелем мускулатуру с непарных плавников и рассматривают их положение по отношению к осевому скелету. При очистке позвоночника в туловищном отделе следует быть очень внимательным: ребра и сидящие на них мышечные косточки легко отделяются. Рекомендуется детальную очистку передних позвонков производить при их изучении. Отчленять череп от позвоночника следует скальпелем, так как соединение очень прочное.
В черепе прежде всего удаляют «щечную» мускулатуру, расположенную в пространстве между глазом и жаберной крышкой, за
тем удаляют глаз, окруженный кольцом тонких о к о л о г л а з н и ч-ных косточек (orbitale). Чтобы их не растерять, выделяют эти косточки с самого начала, точно заметив положение, и наклеивают их на бумагу. Передняя из них, наибольшая, называется слезной (lacrimale, см. рис. 99, lacr.). В дальнейшем, руководствуясь рисунком, отчленяют от черепной коробки весь висцеральный аппарат (челюсти, жаберную крышку, подъязычную и жаберные дуги); отделяют жаберные дуги, предварительно отметив их положение по отношению к остальной части висцерального скелета.
Для лучшей очистки черепной коробки ее следует дополнительно продержать в горячей воде минут пять. Рекомендуется найти носовые кости (nasale, см. рис. 99, nas.) и снять их, так как они слабо соединены с черепом и легко теряются. Следует обратить также внимание па межглазщичную перегородку.
Череп не рекомендуется отбеливать в пергидроле или обезжиривать в бензине, так как после такой обработки границы между костями видны хуже.
Общий обзор скелета
Прежде чем изучать строение отдельных частей скелета, необходимо провести общий осмотр целого скелета, пользуясь музейным препаратом или недоочищенным скелетом судака.
Скелет костистых рыб, как и всех рыб, подразделяется на три отдела:
1. Осевой скелет, или позвоночник, состоит из костных позвонков, соединенных друг с другом телами позвонков в позвоночный столб. В туловищном отделе позвонки несут ребра;
2. Скелет непарных и парных плавников, состоящий из костных лучей. Из непарных плавников у судака имеются два спинных, хвостовой и анальный; из парных — грудные плавники с плечевым поясом и брюшные с тазовым поясом; то же количество плавников и у окуня. У щуки только один спинной плавник, а у трески три спинных и два анальных;
3. Скелет головы — череп — состоит из двух отделов: нейрокра-ниума, или черепной коробки, и спланхнокраниума, или висцерального скелета (челюстной аппарат и жаберный аппарат, покрытый жаберной крышкой).
Кости в скелете различаются по происхождению. Одни из них эмбрионально развиваются из хрящевой ткани на месте хряща, вытесняя его. Такие кости называются хрящевыми (хондральными), основными или замещающими (преформированными). Другие кости развиваются из соединительной ткани кожных покровов, минуя хрящевую стадию. Поэтому они занимают поверхностное положение относительно хрящевых костей, часто легко от них отделяются и обычно имеют плоскую форму. Такие кости называются кожными или накладными, или покровными. В ходе онтогенетического развития некоторых типичных покровных костей процесс окостенения перекидывается на подстилающий их хрящ нейрокраниума. Таким
образом, в них появляется замещающий компонент. Эти кости смешанного происхождения потенциально имеют два названия, из которых употребляется одно или другое в зависимости от того, какой компонент превалирует.
Хрящевые и покровные кости вполне сходны по гистологическому строению.
Осевой скелет
Осевой скелет судака представлен костным позвоночным столбом, или позвоночником, состоящим из отдельных двояковогнутых (амфицельных) позвонков. Внутри позвоночника на всем его протяжении проходит хорда. В средней части тела позвонка она тонкая, а в пространстве между амфицельными телами позвонков расширяется, так что в целом имеет четкообразный характер. Убедиться в этом лучше всего на заранее приготовленном препарате продольного разреза небольшого участка позвоночного столба.
В осевом скелете различают два характерных для всех рыб отдела: туловищный и хвостовой. Позвонки туловищного отдела несут ребра, защищающие вместе со стенками тела внутренние органы животного. Позвонки туловища более массивны и менее подвижно сочленены между собой, чем хвостовые, так как принимают на себя всю силу поступательного движения, создаваемую хвостовым плавником. Позвонки хвостового отдела не имеют ребер, их замыкающиеся нижние дуги образуют гемальный канал, в котором проходят кровеносные сосуды — хвостовая артерия и хвостовая вена. Начало гемалыгого канала служит границей между туловищным и хвостовым отделами. Задний конец позвоночника образует первичный скелет хвостового плавника (рис. 94).
Вычленяют отдельно туловищный и хвостовой позвонки и рассматривают строение каждого из них.
Туловищный позвонок. Состоит из тела позвонка, верхних и нижних дуг.
Тело позвонка (corpus vertebrae s. centrum, рис. 93, c. vrt.) имеет цилиндрическую форму с сильно вогнутыми передней и задней поверхностями (амфицельный тип позвонка), между позвонками в виду студенистой массы находятся остатки хорды. На нижней поверхности каждого тела позвонка имеются продольные бороздки, образующие в целом позвоночнике желоб, в котором помещается спинная аорта. От тела позвонка отходят верхние и нижние дуги.
Верхние дуги (arcus neuralis, s. neurapophysis, рис. 93, arc. n.) отходят в виде широких пластинок от верхней поверхности тела позвонка вверх: постепенно сужаясь, они срастаются друг с другом в непарный верхний остистый отросток (processus spinosus superior, рис. 93, рг. s. s.), который служит для прикрепления мышц. У низших костистых рыб (Malacopterygii— мягкоперых и Esoces — щукообразных) верхние концы дуг в туловищной области не срастаются и образуют парные остистые отростки,
pr s.s.
аге. 77.
.^p.art. а.
с. vrt.
А
print.
....pr. art. р.
f.n.
Рис. 93. Туловищный позвонок судака Lucioperca lucioperca. А — вид спереди; Б — вид сбоку:
аге. п, — верхняя дуга позвонка; с. vrt. — тело позвонка; f. п. — отверстия для корешков спинномозговых нервов; pr. ast. а. — передний сочленовным отросток; pr. art. р.—задний сочленовный отросток; pr. lat. — нижние дуги; pr. s. s. — верхний остистый отросток
но и у них в заднем отделе тела, позади спинного плавника, верхние дуги образуют непарные остистые отростки. Верхние дуги вместе с остистыми отростками образуют на целом позвоночнике канал, в котором помещается спинной мозг. Брюшной и спинной корешки спинномозговых нервов проходят через два отверстия, расположенные одно над другим в середине расширенной части верхней дуги (рис. 93, f. п.). Основания верхних дуг вытянуты вперед в небольшие сочленовные отростки (processus articularis anterior, s. praezygapophysis, рис. 93, pr. art. a.).
Передние сочленовпыс отростки сочленяются с задними сочленовными отростками (processus articularis posterior, s. po-stzygapophysis, рис. 93, pr. art. p.), которые у судака сидят на заднем крае верхней дуги, а у щуки, трески, сазана, окуня — на теле позвонка. Эти отростки служат для придания позвоночному столбу в целом большей прочности и меньшей подвижности. Поэтому они особенно сильно развиты
на передних туловищных позвонках, 'которые вместе с головой принимают на себя всю силу сопротивления поды при движении рыбы вперед. В подвижном хвостовом отделе эти отростки слабо выражены.
Нижние дуги (parapophysis, s. processus lateralis, рис. 93, pr. lat.) отходят от нижней поверхности тела позвонка и направлены косо вниз и назад. В туловищной области к ним прикрепляются ребра, и они называются боковыми отростками позвонка. У судака, окуня, карпа они одинарные. У щуки можно заметить, что отростки двойные; особенно хорошо эта двойственность видна на боковых отростках трески. На первых восьми позвонках судака эти отростки отсутствуют.
Ребра (costa)—топкие, изогнутые кости, ограничивающие полость тела, прикрытые изнутри только париетальным листком брюшины (нижние ребра). У судака на первых двух туловищных позвонках ребер нет, па последующих шести, где отсутствуют боковые отростки, ребра причленяются к заднему краю позвонка.
Мышечные косточки по форме сходны с ребрами, но значительно тоньше и короче их. Они имеются на передних десяти позвонках, начиная с первого, и прикрепляются к заднему краю соответствующего ребра, ближе к его проксимальной части. На первых двух позвонках, где ребра отсутствуют, мышечные косточки
прикрепляются непосредственно к телу позвонка у основания верхних дуг. Мышечные косточки соответствуют верхним ребрам древних костных рыб.
Всего туловищных позвонков у судака двадцать три.
На переднем позвонке спереди следует рассмотреть фасетки, которые имеются с каждой стороны под сочленовным отростком. Они сочленяются с соответствующей фасеткой, образованной боковой затылочной костью черепа (см. описание черепа).
Позвонки хвостовой области. На целом позвоночнике видно, что боковые отростки туловищных позвонков ближе к хвостовому отделу все больше сближаются и, наконец, срастаются друг с другом, образуя гемальный канал, где проходят хвостовые вена и артерия. Внизу канал замыкается нижними остистыми отростками (processus spinosus inferior), образованными за счет ребер. Пространства между остистыми отростками затянуты тонкой перепонкой.
В конце хвостового отдела осевой скелет сильно изменен. Верхние и нижние дуги позвонков здесь превращены в расширенные пластинки, поддерживающие наружные лучи хвостового плавника. Измененные нижние дуги носят название гипуралий (hypura-lia, рис. 94, hyp.), а верхние уроневралий (uroneuralia, рис. 94, игоп.).
У всех рассматриваемых видов, кроме трески, тела последних позвонков сливаются в вытянутую палочковидную косточку — уростиль (urostyl, рис. 94, urst.), направленную в верхнюю лопасть хвостового плавника. Таким образом, большая часть плавника поддерживается гипуралиями, произошедшими от нижних дуг позвонков. Тем самым внешне симметричный хвостовой плавник поддерживается несимметрично расположенными элементами осевого скелета. Такой тип хвостового плавника называется гомоцеркальным. У трески скелетные элементы в хвостовом плавнике расположены симметрично. Верхняя лопасть поддерживается уроневралия-ми, а нижняя — гипуралиями. Столь полная (внешняя и внутренняя) симметрия плавника вторична, поскольку у мальков он гомоцеркален, и поэтому плавник такого строения называется вторично дифицеркальным или изоцеркальным.
Скелет плавников
У судака, как и у акулы, имеются непарные и парные плавники. Непарные плавники располагаются всегда по средней линии тела на спине и задней части брюха, парные грудные и брюшные плавники занимают латеральное положение и причленяются всегда к поясам конечностей — плечевому и тазовому.
Непарные плавники. У судака имеются два непарных дорзальных плавника, один вентральный (подхвостовой, или анальный) и хвостовой плавник. У трески — три спинных и два анальных. Каждый из этих плавников состоит из наружного и внутреннего скелета. Наружный скелет состоит из костных плавниковых лучей кож-306
ного происхождения— лепидотрихий (lepidotrichia), которые поддерживают тонкую кожную складку.
Внутренний скелет непарных плавников представлен внутренними радиальными лучами (radialia) —тонкими косточками, заостренными внизу и расширяющимися наверху. Верхний конец внутреннего луча имеет шаровидную форму. К нему прочно, но подвижно прикрепляются лепидотрихии. Таким образом, луч внутреннего скелета становится носителем наружного кожного луча — птери-гиофором (pterygiophora). Чтобы рассмотреть эту структуру,
выделяют часть плавника, кипятят его, очищают и расчленяют на отдельные лучи. Птеригиофоры нижними заостренными концами вклиниваются между соответствующими остистыми отростками позвонков и Связаны с ними общей соединительнотканной перепонкой.
Как общее количество, так и количество лучей разных групп (см. раздел «Внешний вид») у разных видов очень
Рис. 94. Скелет хвостового плавника судака Lucioperca lucioperca: hypur. — гипуралин; urst. — уростиль;
uron. — уровневралия
различно и во многих случаях служит систематическим признаком.
Хвостовой плавник (рис. 94). Его строение описано при изучении заднего отдела позвоночника. Различие между его строением и строением остальных непарных плавников сводится к тому, что наружные плавниковые лучи (лепидотрихии) поддерживаются не птеригиофорами, которые у костистых рыб в хвосте отсутствуют, а при помощи своеобразно измененного заднего отдела позвоночника.
Парные плавники, или конечности (рис. 95 и 96), представлены грудными и брюшными плавниками, которые поддерживаются поясами конечностей — плечевым и тазовым. У судака, окуня, трески тазовый пояс с брюшными плавниками расположен не на середине брюшной поверхности тела, а отодвинут вперед и лежит непосредственно под грудными плавниками. У щуки и карпа брюшные плавники, как и у акулы, помещаются непосредственно перед анальным отверстием.
Если при первичной обработке скелета поясов сохранилась связь грудного пояса с тазом, то следует отчленить их друг от друга и очистить от оставшихся мышц. Для удобства так же расчленяют обе половины грудного пояса.
Пояс передних конечностей образован как хрящевыми костями, возникшими внутри хрящевого пояса (гомология с хрящевым плечевым поясом акулы), так и покровными, расположенными одна над другой, из которых верхняя причленяется к заднему отделу черепа. Таким образом, весь плечевой пояс с плавниками опирается на осевой череп.
Замещающих костей, гомологичных хрящевому поясу акул, всего две: лопатка и коракоид. Они примыкают сзади к нижней части крупной покровной кости —клейтрума.
Лопатка (scapula, scap.)—небольшая кость, занимающая дорзальное положение. Она пронизана большим отверстием, через которое проходит нерв. У щуки в связи с горизонтальным расположением плавников опа расположена не выше, а латеральнее коракоида.
Рис. 95. Плечевой пояс и грудной плавник судака:
сап. Z. — каиал боковой линии; clthr.— клейтрум; сгс.— коракоид; 1р.—лепн-дотрихии; pr. 1 — верхний отросток задневисочной кости; рг. 2 — нижиий отросток задневисочной кости; pst.—задневисочная кость; rad. — радиальные элементы; scap. — лопатка; sclethr. — подХлейтрум; spclthr. — иадклейтрум
Коракоид (coracoideum, сгс.) лежит под предыдущей костью на вентральной стороне. Сверху он имеет вырезку, прикрытую покровной костью.
К задней поверхности лопатки и коракоида причленяется свободная конечность — грудной плавник. Основную же часть плечевого пояса составляют четыре покровные кости: клейтрум, надклейтрум, подлейтрум и задневисочная кость.
Клейтрум (cleithrum, clthr.) —большая изогнутая кость. Она располагается тотчас за жаберной крышкой, ограничивая сзади жаберную полость. Обе кости правой и левой сторон сходятся внизу.
Надклейтрум (supracleithrum, spclthr.) лежит над предыдущей костью и покрывает ее верхний конец. Она значительно меньше клейтрума.
Задневисочная (posttemporale, pst.)—у судака небольшая кость с двумя отростками, лежит поверхностно, так что покрыта сверху кожей. Задний край зазубрен, чем напоминает чешуйку. 308
Имеет два отростка: верхний, большой, прикрепляется к верхне-ушной кости черепа (pr. 1), нижний, меньший (рг. 2), — к небольшому выступу заднеушной (см. описание черепа). На ее наружной поверхности выступает валик с каналом (can. I.), внутри него проходит канал органа боковой линии, который направляется вперед и лежит на верхней стороне черепа. Форма этой косточки у рас-
сматриваемых видов сильно варьирует. Она очень короткая у карпа, тогда как у трески два ее отростка сильно вытянуты.
У щуки на месте этой кости лежат две, верхняя из которых носит название задневисочной, а нижняя—надвисочной кости (supratemporale).
Подклейтрум (subcleithrum, sclethr.) расположена на внутренней стороне плечевого пояса, имеет вид тонкой пластинки или палочки и расположена вертикально. У трески она лежит ме-диальнее лопаточно-ко.ракоидного отдела, слабо связана с клейтрумом и может быть легко утеряна при препаровке. К клейтруму кость прикрепляется верхним концом с внутренней стороны.
Грудные плавники у судака в отличие от акул занимают вертикальное положение (см. описание внешней формы). Основные элементы (basalia), имевшиеся у акул, отсутствуют.
Радиальные элементы плавника (radialia, rad.) представлены только четырьмя косточками. Они прикрепляются к
Рис. 96. Тазовый пояс и брюшные плавники судака Lucioperca lucioperca:
Ip. — лепидотрихии; pel. — таз
заднему краю лопатки и кора-
коида.
Лепидотрихии (lepidotrichium, 1р.)—наружные костные лучи, так же как и в непарных плавниках, расположены кнаружи от радиальных элементов.
Брюшные плавники, или задние конечности, причленяются к тазовому поясу.
Таз (pelvis, рис. 96, pel.) у судака парный. Он состоит из двух удлиненных треугольных пластинок, к задним расширенным основаниям которых причленяются брюшные плавники.
У других видов пластинки могут быть более узкими (сазан) или иметь неровный внутренний край '(треска).
Брюшные плавники представлены наружными костными лучами — лепидотрихиями. Радиальные и базальные элементы в них отсутствуют.
Скелет головы (рис. 97)
Череп судака полностью окостеневает. Он образован хрящевыми, покровными костями и костями смешанного происхождения. Как и у акулы, череп судака состоит из двух отделов: осевого черепа (neurocranium) и висцерального скелета (splanchnocra-nium).
Нейрокраниум представляет собственно черепную коробку, окружающую головной мозг, с приросшими к ней скелетными капсулами органов чувств—обоняния и слуха.
Висцеральный скелет состоит из челюстного аппарата, подвижно прикрепленного к черепной коробке, жаберного скелета из пяти жаберных дуг с сидящими на них жабрами, и жаберной крышки, покрывающей жаберный скелет снаружи.
Черепная коробка
Черепная коробка (neurocranium) судака имеет узкое основание, так как стенки глазниц сближаются в результате развития крупных глаз, и между ними остается только тонкая перегородка. По той же причине головной мозг располагается позади глазниц, а не между глазами, как у акуловых рыб. Такой тип черепа с узким •основанием называется тропибазальным.
Из рассматриваемых видов только у карпа череп имеет широкое основание, а глазницы отделены друг от друга. Такой тип черепа, как уже указывалось, носит название платибазального. Черепная коробка гомологична хрящевой черепной коробке хрящевых рыб, но образована отдельными замещающими костями. С наружной стороны она покрыта покровными костями, развивающимися поверх хрящевой ткани; они обычно имеют плоскую форму и легко отделяются при кипячении черепа.
Изучение черепной коробки судака следует начать с покровных костей; кости окологлазничного кольца уже были рассмотрены и удалены раньше, при очистке черепа.
Крыша черепной коробки. Крышу черепной коробки образуют три пары костей: 1) носовые (nasale, см. рис. 99, nas.)—две парные маленькие косточки, лежащие впереди над обонятельным отделом; 2) лобные (frontale, fr.), расположенные за носовыми. Они образуют почти всю крышу черепа и имеют характерную скульптуру, напоминающую чешую осетровой рыбы. Носовые и лобные кости на внутренней поверхности имеют трубку, в которой проходит канал органа боковой линии (его необходимо проследить на черепе, руководствуясь рисунком). Эта трубка начинается на переднем заостренном конце носовой кости; дойдя приблизительно до ее середины, выходит на поверхность, переходит на передний 310
Рис. 97. Осевой череп судака Lucioperca lucioperca. А — вид сбоку; Б — вид сверху; В — вид сзади:
Ъосс. ~ основная затылочная кость; bsph. — основная клиновидная кость; can. I.— канал боковой линии; ecteth. — боковая обонятельная кость; epot. — верхнеушная кость; ехосс. — боковая затылочная кость; f. о. т. — большое затылочное отверстие; fr. >— лобная кость; Isph. — боковая клиновидная кость; meth. — срединная обонятельная кость; opot. — задиеушиая кость; par.—теменная кость; parsph. — парасфеноид; pr. epot. — отросток верхнеушиой кости; prot. — переднеушиая кость; ptot. — крыловидиоушиая кость; г. hmd. VII — отверстие для выхода подъязычночелюстной ветви лицевого нерва; sphot. — клииовидноушиая кость; space. — верхнезатылочная кость; vom. — сошник; V, VII, IX, X — отверстия для выхода головных нервов
конец лобной кости, доходит до уровня конца межглазничной перегородки и открывается наружу отверстием. Через это отверстие канал органа боковой линии выходит из лобной кости и продолжается назад в открытом желобе, образованном крыловидноушной костью (см. «иже описание слухового отдела, стр. 313). Затем он переходит в канал, расположенный на задневисочной кости плечевого пояса; 3) теменные кости (parietale, par.) лежат позади лобных и значительно меньше их по размерам. Они имеют на задней стороне выемку, ограничивающую впадину между верхнеушной и крыловидноушной костями.
По сравнению с судаком форма носовых костей наиболее сильно отличается у щуки. У нее носовые кости имеют вытянутую форму и на большом протяжении плотно соединяются с так же сильно вытянутыми отростками лобных костей. Кроме того, у щуки при осевом черепе обычно остаются две надглазничные косточки из покровных костей окологлазничного кольца. Одна лежит на уровне середины глазницы и имеет округлую форму. Вторая лежит перед ней, имеет сильно вытянутую форму, так что передним концом достигает носовой кости и медиальным краем прилегает к отростку лобной. Ее можно считать предлобной костью (praefron-tale). У карпа носовые кости короткие, округлые и лежат по бокам от сошника и мезэтмоида. У карпа имеется также одна надглазничная (supraorbitale) косточка, лежащая на середине 'верхнего края глазницы.
Правая и левая лобные кости трески срастаются друг с другом по средней линии, так что границу между ними провести трудно.
Теменные кости карпа отличаются почти квадратной формой.
Дно черепной коробки образовано двумя непарными покровными костями: 1) парасфеноид (parasphenoideum, parsph.)—характерная для рыб покровная кость, подстилающая дно черепа. Простирается от заднего конца черепа до переднего края глазницы; 2) сошник (vomer, voni.) лежит впереди от парасфеноида и значительно меньше его по размерам. Спереди сошник расширен и несет заостренные зубы, задний его край входит в выемку парасфеноида.
У карпа сошник короткий и зубы па нем отсутствуют.
Далее следует рассмотреть замещающие кости черепной коробки, при этом следует обратить внимание на прочное соединение этих костей друг с другом. Как и у акулы, первичная черепная коробка судака состоит из четырех отделов: затылочного, слухового (окстенений слуховых капсул), глазничного и обонятельного (окостенений обонятельных капсул).
Затылочный отдел. Этот отдел черепной коробки образован четырьмя затылочными костями, которые окружают большое затылочное отверстие (foramen occipitale magnum, f. о. m., рис. 97).
Основная затылочная кость (basioccipitale, Ьосс.) — большая непарная кость, лежащая под большим затылочным отверстием: сзади она имеет вогнутую поверхность, которой соеди-312
няется с телом первого позвонка. Снизу частично прикрыта покровной костью — парасфеноидом. У карпа основная затылочная кость, несет мощный отросток, идущий в каудальном направлении. На нем с вентральной стороны располагается широкая вогнутая площадка жерновка. Основание отростка имеет отверстие, отграничивающее его от тела основной затылочной кости. Через это отверстие проходит спинная аорта.
Боковые затылочные кости (exoccipitale, s. occipitale laterale, exocc.)—парные кости, лежат по бокам затылочного отверстия, образуя его боковые стенки, смыкаются друг с другом в верхнем углу большого затылочного отверстия. Внизу над основной затылочной костью они образуют по фасетке, которые сочленяются с такими же поверхностями, находящимися под сочленовными отростками первого позвонка. Над каждой фасеткой имеется отверстие, через которое выходит блуждающий нерв (X). При рассмотрении черепа сбоку видно, что боковая затылочная кость лежит над основной затылочной, а вверху граничит с верхней затылочной. Боковые затылочные кости пронизаны отверстием для выхода языкоглоточного нерва (IX). Его можно найти около шва заднеушной кости.
У щуки, трески и карпа боковые затылочные кости не образуют фасеток для сочленения с позвоночником. У трески к ним при помощи шва причленяются верхние дуги первого позвонка. У карпа боковые затылочные кости имеют большие латеральные крылья, к медиальной стороне которых через клейтрум причленяется пояс передних конечностей. Кроме того, у карпа эти кости пронизаны двумя крупными отверстиями.
Верхнезатылочная кость (supraoccipitale, spocc.) непарная, замыкает сверху костное кольцо вокруг большого затылочного отверстия. Сбоку она имеет вид тонкой пластинки, ее передняя часть образует расширение, вклинивающееся между теменными костями.
Верхнезатылочная кость образует по-разному развитый у рассматриваемых видов гребень, служащий для прикрепления мышц. У щуки верхнезатылочная кость относительно крупная (рис. 98, socc.) и округлой формы, однако края ее прикрыты теменными костями, так что кость имеет вид узкого вклинивающегося между ними отростка. У карпа она, наоборот, широкая и доходит только до заднего края теменных костей.
Окостенения слуховой области. Отдел слуховых капсул располагается впереди от затылочного отдела, справа и слева от мозговой полости. Каждая капсула окостеневает пятью ушными костями (ossa periotica, о. per.).
Переднеушная кость (prooticum, prot.) большая, трапециевидной формы, занимает переднюю часть ушного отдела. Верхний край ее рострального выступа утолщен и на середине переднеушной кости образует перемычку, нависающую над двумя отверстиями; через переднее, более крупное, выходит лицевой нерв (п. facialis, VII), через заднее — его гиомапдибулярная ветвь
(г. hyomandibularis VII, г. hmd. VII). На границе с лежащей впереди костью находится отверстие, через которое проходит тройничный нерв (п. trigeminus, V).
У щуки переднеушная кость занимает большую часть боковой поверхности слуховой области. В связи с неполным окостенением черепной коробки она выделяется четко, так как отграничена от соседних костей прослойками хряща (рис. 98, prot.).
preth.
socc.
epot.
sphot. ptot.
ecteth.
Isph.
prot.
exocc.
toco.
Рис. 98. Осевой череп щуки Esox lucius. А — вид сверху; Б — вид снизу:
Ьосс. ~ основная затылочная кость; ecteth. ~ боковая обонятельная кость; epot. — верхнеушная кость; ехосс. — боковая затылочная кость; Isph. — боковая клиновидная кость; preth. ~ предобонятельная кость; prot. ~ переднеушная кость; ptot. — крыловндноушная кость; socc. — верхнезатылочная кость; sphot. ~ клиновидноушная кость
У карпа эта кость также крупная, почти в центре нее лежит крупное отверстие для выхода гиомандибулярной ветви VII нерва. Сзади она граничит с боковой затылочной костью, от которой отделена имеющимся здесь отверстием, и с основной затылочной. Верхняя ее граница проходит в своей передней части по средней линии суставной впадины для гиомандибулярной кости. Передний край ее отделен четко.
У трески переднеушная кость лежит также в передней части слуховой области. Вместо отверстия имеется глубокая вырезка на ее переднем краю. Чтобы установить ее верхнюю границу, следует
найти два отверстия, расположенных опереди и сзади от клиновид-ноушной кости, граница проходит по их нижним краям. Между отверстиями граница с клиновидноушной костью зигзагообразна.
Заднеушная кость (opisthoticum, opot.) лежит позади переднеушной и значительно меньше ее по величине. Нижним краем она граничит с боковой затылочной костью, суженным передним налегает на переднеушную.
У судака к заднеушной кости прикрепляется нижний отросток задневисочной кости.
У щуки заднеушная кость отсутствует.
У трески заднеушная кость развита сильно, имеет округло-вытянутую форму и отверстие, примерно в центре. Снизу она на большом протяжении граничит с основной затылочной, а впереди с парасфеноидом, сзади — с боковой затылочной, и граница проходит по отверстию для IX нерва. Ее верхняя граница с крыловидноушной легко прослеживается по стыку различно направленных выступов скульптуры костей. Заднеушная кость составляет нижнюю часть направленного назад отростка, и здесь к ней прикрепляется ножка заднетеменной кости.
У сазана заднеушная кость очень мала, лежит между боковой затылочной и крыловидноушной и составляет переднюю часть вертикального пребня, образованного боковой затылочной костью.
Крыловидноушная кость (pteroticum, ptot.) является окостенением заднего выступа слуховой капсулы. Сверху она граничит с покровной теменной и лобной костями; на ее поверхности имеется желоб, где проходит надглазничный канал боковой линии (can. I.). Крыловидноушная кость образует боковой слуховой отросток (processus pteroticus), особенно хорошо заметный, если смотреть на череп сзади.
Границы крыловидноушной кости у щуки видны довольно четко.
У карпа крыловидноушная кость составляет верхний край большой впадины в слуховой области. Снизу и спереди она граничит с переднеушной и клиновидноушной костями. Наиболее четко граница видна в суставной впадине для причленения гиомандибулярной кости. Сверху небольшим участком крыловидноушная кость соприкасается с лобной и далее назад с теменной и верхнеушной.
У трески границы крыловидноушной кости менее четки. Снизу она на большом протяжении граничит с заднеушной костью. Передняя граница с клиновидноушной проходит по отверстию между этими костями. Верхняя граница с верхнеушной костью идет по щели, далее вперед линия границы с теменной костью изогнута почти под прямым углом. Впереди крыловидноушная кость граничит с узким выростом клиновидноушной.
Клиновидноушная кость (sphenoticum, sphot.) лежит впереди от предыдущей, над переднеушной костью. Она имеет форму ромба, вытянутого вперед. Сверху на нее налегает суженная часть крыловидноушной кости. В задней части клиновидноушная
кость образует выступ в виде округлоизогнутого гребня, являющегося продолжением костной перемычки переднеушной кости. На задней поверхности выступа клиповидноушпой кости имеется сочленовная впадина, которая продолжается назад на крыловидпоушную кость. В этом месте челюстной аппарат причленяется к черепу при помощи верхнего элемента подъязычной дуги — подвеска.
У щуки крыловидноушпая кость невелика, расположена в передневерхнем углу слухового отдела и сверху почти полностью прикрыта лобной и крыловидпоушной костями.
У карпа нижний отдел клиновидноушпой кости образует часть суставной поверхности для гиомандибуляре. Вперед от этого отдела отходит мощный отросток кости, сочленяющийся с лобной костью. Верхняя граница с лобной и крыловидноушной проходит в вершине впадины, образованной тремя этими костями.
У трески клиновидноушная кость имеет форму горизонтальной пластинки, выходящей на дорзальную поверхность черепа, и идущей от нес вниз под углом ножки. Ножка имеет зигзагообразную границу с переднеушной костью, а верхняя пластинка настолько прикрыта лобной, крыловидноушпой и теменной костями, что имеет вид узкого отростка.
Верхнеушная кость (epioticum, epot.) хорошо видна при рассмотрении черепа сзади, сбоку же виден только ее отросток. Она занимает верхнее положение среди всех ушных костей и образует надслуховой отросток (processus epioticus) по бокам верхнезатылочной кости. Отметим, что к отросткам верхнеушной и заднеуш-пой костей причленяется покровная кость плечевого пояса — задневисочная (posttemporale, см. рис. 95, pst.).
У щуки и трески положение верхнеушной кости такое же, как у судака. У щуки она хорошо видна при рассмотрении черепа сзади. Ее границы с боковой и верхпезатылочной костями очень четки. У трески верхнеушная кость на большом протяжении граничит с боковой затылочной. Эта граница проходит чуть ниже и медиальное гребня, образованного верхнеушной костью. Переднебоковая граница с крыловидноушной костью проходит по щели, а передняя граница с темной — примерно в средней части гребня.
Глазничный отдел. Глазничный отдел нейрокраниума образован громадными глазничными впадинами, настолько сближенными друг с другом, что между ними проходит только тонкая соединительнотканная перегородка. Кости, развивающиеся в этом отделе, называются клиновидными или сфеноидами. У большинства костистых рыб их бывает три, у судака же две, так как в межглазничной перегородке не развиваются глазоклиновидные кости (orbitosphenoideum); таким образом, здесь имеются только боковые клиновидные и основная клиновидная кости.
Из рассматриваемых видов глазоклиновидная кость имеется только у карпа.
Боковая клиновидная кость (laterosphenoideum) парная, лежит над выступом переднеушной кости, гранича с ней снизу, а сверху — с клиновидноушной и с отростком лобной кости.
У щуки и карпа боковая клиновидная кость лежит впереди от переднего отверстия в переднеушной кости. Границы ее видны довольно четко. У карпа ее задний верхний угол входит в состав сочленовной впадины для гиомапдибуляре.
У трески маленькая боковая клиновидная кость имеет вид узкой пластинки, которая отделена щелью от лежащей каудальнее клиновидноушной кости.
Основная клиновидная кость (basisphenoideum) — непарная V-образная кость, расположенная у судака внутри черепной коробки. Если посмотреть на черепную коробку сбоку, видна ее передняя часть (bsph.). Целиком же кость можно хорошо увидеть при рассмотрении мозговой коробки спереди или же сделав продольный распил черепной коробки. У других видов она отсутствует.
Глазоклиновидная кость (orbitosphenoideum)—крупная кость, лежащая у карпа в середине глазничного отдела. Кость имеет трапециевидную форму. Поверхность ее покрыта большим количеством углублений. Широкое основание кости граничит с парасфеноидом. Сверху она граничит с лобной костью. Задняя граница с боковой клиновидной идет почти вертикально, как и передняя с боковой этмоидной.
Между основной клиновидной костью и покровным парасфеноидом проходит назад канал — м ио до м (myodom), ® котором помещаются прямые мышцы глаза.
Обонятельный отдел. Самый передний отдел в черепной коробке— обонятельный отдел. Обонятельные капсулы окостеневают тремя обонятельными костями — этмоидами.
Средняя обонятельная кость (mesethmoideum, meth.) крупная, непарная, занимает среднюю часть этого отдела. Снизу она граничит с сошником, по бокам от нее выступают парные боковые обонятельные кости (ectoethmoideum, ecteth.). Они образуют значительные выступы, ограничивающие переднюю часть глазницы.
Границы между обонятельными костями заметны хорошо, так как в этой области сохраняется много хряща. Следует проследить их сбоку, сверху и опереди.
У карпа средняя обонятельная кость имеет широкие, расходящиеся в стороны крылья, которые лежат впереди лобных костей на дорзальной поверхности черепа.
У щуки мезетмоид отсутствует, но по бокам расширенной передней части рыла выступают из-под покровных носовых костей небольшие предобонятельные кости (praeethmoideum, рис. 98, preth.). Боковые обонятельные кости располагаются у щуки по бокам основания рострума (рис. 98, ecteth.).
У трески боковые обонятельные кости сильно расширены и имеют вид крыльев, лежащих впереди от лобных .костей. Ниже средней обонятельной кости остается большой участок хряща.
Закончив изучение строения черепной коробки, следует отметить, что у низших рыб некоторые из перечисленных костей могут
заменяться покровными, и тогда они носят другие названия. Так, в обонятельной области средняя обонятельная кость (meseth-moideum) может развиваться как покровная. Тогда она называется верхнеобонятельной (supraethmoideum). Если боковые обонятельные кости (ectoethmoideum) являются покровными, то они называются предлобными (praefrontale). Из окостенений слуховой капсулы постоянно хрящевыми костями сохраняются переднеушная (prooticum), верхнеушная (epioticum) и заднеушная (opisthoticum). Последняя обычно бывает мала или может не развиваться совсем (у щуки). Что касается клиновидноушной (sphe-noticum) и крыловидноушной (pteroticum) костей, то у некоторых рыб они могут быть покровными. В таком случае клиновидноушная называется заднелобной (postfrontale), а крыловидноушная — чешуйчатой (squamosum).
Продольный разрез нейрокраниума
После изучения черепной коробки судака с внешней стороны делают продольный распил (при помощи лобзика) и рассматривают ее строение с внутренней стороны. Границы между костями видны хорошо, следует их проследить.
Основная клиновидная кость видна полностью. Снаружи она покрыта выростом парасфеноида, сверху граничит с боковой клиновидной, сзади—с переднеушной костью. Между ней и парасфеноидом виден миодомный канал.
Боковая клиновидная кость видна полностью, в то время как снаружи ее нижняя граница частично прикрыта выростом переднеушной кости.
Интересно отметить, что ушные и затылочные кости образуют углубления (камеры) во внутренней полости черепной коробки, заметные на сагиттальном распиле. Границы между камерами вдаются внутрь мозговой коробки широкими перегородками, которые служат опорой для центральной части перепончатого лабиринта, в камерах же размещаются полукружные каналы. Камер пять. Верхняя образована только верхней ушной костью, в ней размещается задний полукружный канал. Под ней расположены две камеры, из которых передняя образована только крыловидноушной костью, а задняя — заднеушной и отчасти боковой затылочной. В стенке, разделяющей обе камеры, имеется отверстие, через которое из одной камеры в другую проходит латеральный полукружный канал. Четвертая камера ограничена гребнями, отходящими от внутренних стенок переднеушной и клиновидноушной костей. Здесь помещается передний полукружный канал. Пятая, нижняя, камера имеет овальную форму; она образована спереди переднеушной костью, а сзади — основной и боковой затылочными. Здесь помещается круглый мешочек с крупным отолитом.
На сухом черепе перепончатый лабиринт не сохраняется. Его можно рассмотреть на сагиттальном распиле головы, фиксированной формалином.
Осевой череп щуки (рис. 98)
У щуки, которая является представителем примитивного отряда, в осевом черепе сохраняется довольно большое количество хряща. Наряду с ним появляются хондральные (замещающие) кости, которые здесь особенно хорошо видны благодаря наличию между ними хрящевых прослоек. Поэтому желательно, если изучение скелета ведется на судаке, дополнительно рассмотреть осевой череп щуки.
Чтобы отпрепарировать осевой череп, голову щуки варят дольше, чем это необходимо для изготовления общего скелета головы. На таком разваренном черепе покровные кости легко снимаются, обнажая осевой череп. Очищенный череп следует покрасить ализарином, тогда основные кости окрашиваются в красный цвет и отчетливо выступают на фоне почти белого хряща. Приготовленный таким образом осевой череп щуки следует рассмотреть и разобрать положение замещающих костей, пользуясь рис. 98 и указаниями на особенности их строения у щуки, приведенными выше.
Висцеральный скелет
Висцеральный аппарат (splanchnocranium) костистых рыб выполняет две важные функции. Челюстной аппарат выполняет функцию захвата пищи и имеет разнообразное строение в соответствии с различными способами питания. При этом большую роль играет вторичная верхняя челюсть, образованная челюстной и предчелюстной костями. Жаберный отдел вместе с висцеральным аппаратом в целом несет дыхательную функцию.
У хищников, какими являются щука, окунь, судак, ротоглоточная полость усажена многочисленными зубами, покрывающими не только челюсти, но и другие кости ротовой полости. У мирных рыб, к которым относятся карповые, зубов на челюстях нет, но зато имеются сложные глоточные зубы на цератобранхиальных элементах пятой жаберной дуги. Они служат для перетирания захваченной пищи.
Висцеральный аппарат в целом обеспечивает так называемый оперкулярный тип дыхания за счет действия нагнетательного и разрежающего насосов, образованных жаберной крышкой, подкрышечными бранхиостегальными лучами, жаберным и челюстным аппаратом.
Соответственно этим функциям висцеральный аппарат имеет сложно дифференцированную мускулатуру, образованную как висцеральной мускулатурой челюстного и жаберного аппаратов, так и вентральной поджаберной, или подъязычной, мускулатурой туловищного происхождения.
Висцеральный скелет судака, как и акулы, состоит из отдельных дуг, развивающихся в переднем отделе пищеварительной трубки. Они в виде полудуг охватывают ротоглоточную полость и соединяются на брюшной стороне. Всего висцеральных дуг у судака
семь: челюстная, подъязычная и пять жаберных. Кроме того, у судака развивается костная жаберная крышка, прикрывающая жаберные дуги. Жаберные дуги в связи с развитием жаберной крышки смещены вперед и располагаются под осевым черепом (в отличие от акул).
Все элементы висцеральных дуг развиваются из хряща и полностью окостеневают. В челюстной дуге помимо окостенений небноквадратного хряща и меккелева хряща акулы появляются покровные кости.
Челюстная дуга (arcus mandibularis). Состоит из верхней и нижней челюстей. Первичная верхняя челюсть образована костями, замещающими небно-квадратный хрящ, а также непосредственно связанными с ним покровными костями. Это небная кость впереди, квадратная сзади и несколько крыловидных костей между ними.
Квадратная кость (quadratum, рис. 99, q.) располагается в заднем отделе первичной верхней челюсти и, вопреки названию, имеет треугольную форму. Направленная вперед и вниз вершина ее несет сочленовную поверхность для нижней челюсти.
Небная кость (palatinum, рис. 99, pl.) —смешанное окостенение в переднем отделе небно-квадратного хряща. По ее наружному краю сидят зубы, а на передневерхнем конце имеется вырост в виде крючка, прилегающий к сошнику. Рассмотреть эту кость удобнее с внутренней стороны челюсти, так как снаружи она частично прикрыта двумя покровными челюстными костями. Отметим, что в отличие от акул, передние концы небно-квадратных элементов (в данном случае небные кости) не соединены друг с другом по средней линии.
Форма небных костей у других рассматриваемых видов отличается довольно сильно.
У щуки эта кость сильно вытянута в длину и уплощена. На ее нижней поверхности располагается целая щетка зубов. Кость тянется параллельно роструму и своим задним концом соединяется с боковой обонятельной, а передним — с челюстной и межчелюстной костями.
У карпа и трески эти кости короткие. У первого она как бы состоит из трех отростков. Передний связан со слезной, задний с боковой этмоидной, медиальный — со срединной этмоидиой костями.
Задняя крыловидная кость (metapterygoideum, рис. 99, metpt.) является хондральным окостенением небно-квадратного хряща. Она имеет округлую форму, лежит сверху и несколько сзади от квадратной кости. У трески кость имеет вытянутую форму.
Наружная крыловидная кость (ectopterygoideum, рис. 99, ectpt.) — покровная кость, узкая, изогнутая, ограничивающая наружный край верхней челюсти между небной и квадратной костями. От нее назад отходят два отростка; один упирается в квадратную кость, другой — в заднекрыловидную.
У других описываемых видов она может быть более короткой, но легко различается по положению вдоль переднего края квадратной кости.
Внутренняя крыловидная кость (entopterygoideum, рис. 99, ent pt.) — очень тонкая покровная пластинчатая кость, лежит ковнутри от предыдущей, образуя дно глазницы.
По, положению кости медиальнее наружной крыловидной ее легко найти и у других видов. Правда, у щуки ее форма изменена,
Рис. 99. Череп судака Lucioperca lucioperca сбоку:
ang. — угловая кость; art. — сочленовная кость; dent. — зубная кость; ectpt. — наружная крыловидная кость; entpt. — внутренняя крыловидная кость; hma. — гио-мандибулярная кость; 1пор. — межкрышечная кость; lacr. — слезная кость; metpt. — задняя крыловидная кость; тх. — верхнечелюстная кость; nas. — носовая кость; ор. — крышечная кость; pl. — небная кость; ргтх. — предчелюстная кость; prop.—предкрышечиая кость; symp.—симплектикум; q.— квадратная кость;
sop. — подкрышечная кость
и снаружи виден только небольшой участок, вклинивающийся сверху между задней крыловидной и отростком квадратной кости.
Из окостенений небно-квадратного хряща истинно хрящевыми являются только квадратная и заднекрыловидная кости. Небная кость имеет смешанное происхождение, а наружная и внутренняя крыловидные развиваются как покровные кости на небно-квадратном хряще.
Вторичная верхняя челюсть образована с каждой стороны двумя покровными костями, ограничивающими сверху ротовое отверстие — предчелюстной и верхнечелюстной.
Предчелюстная, или межчелюстная (praemaxillare, s. intermaxillare, рис. 99, ргтх.), — длинная узкая кость, занимает переднее положение, на нижней ее поверхности сидят зубы. Ее передний конец сильно расширен и образует два отростка, направленные вверх. При помощи них кость прочно прикреплена своим передним концом к осевому черепу.
У трески форма и положение предчелюстной кости примерно соответствует таковым судака.
У карпа предчелюстные кости срастаются верхними концами и не несут зубов.
У щуки предчелюстные кости короткие, лежат по бокам от передней части рострума и снизу усажены мелкими зубами. К заднему краю каждой кости причленяется передний конец верхнечелюстной кости.
Верхнечелюстная кость (maxillare, рис. 99, тх.) лежит у судака над предыдущей, задний конец ее расширен в виде тонкой пластинки, на переднем образуется сочленовная впадина, сочленяющаяся с бугром второго отростка предчелюстной кости.
У карпа эта кость широкая и имеет сложную форму.
У щуки верхнечелюстная кость длинная и является основной костью, образующей вторичную верхнюю челюсть, поскольку, как мы видели раньше, предчелюстная кость очень короткая и причленяется к переднему концу верхнечелюстной.
Нижняя челюсть. В заднем отделе нижней челюсти лежит сочленовная кость (articulare, рис. 99, art.). Она имеет треугольную форму, острием направлена назад. На заднем конце можно увидеть суставную поверхность, при помощи которой нижняя челюсть подвижно сочленяется с квадратной костью верхней челюсти. От сочленовной кости вперед отходит сильно редуцированный меккелев хрящ. Он проходит по внутренней стороне впереди лежащей кости. На сухой челюсти виден слабо, поэтому размачивают ее в теплой воде.
Покровных костей в нижней челюсти две: зубная и угловая.
Зубная кость (dentale, рис. 99, dent.) образует всю переднюю часть нижней челюсти. Верхняя ее сторона усажена зубами. У карпа зубы на этой кости отсутствуют. Она простирается назад дальше половины длины всей челюсти и соединяется с сочленовной костью. На ее внутренней стороне проходит остаток меккелева хряща.
Угловая кость (angulare, рис. 99, ang.) невелика, располагается на задневнутреннем углу нижней челюсти.
Сравнивая строение челюстного аппарата акуловых рыб с таковым костистых, можно увидеть большие различия. Хватательным аппаратом у акул служат первичные челюсти, представленные небно-квадратным и меккелевым хрящами. У костистых рыб, кроме первичных челюстей, гомологичных этим хрящам, появляются вторичные челюсти, ограничивающие ротовое отверстие и образованные покровными костями — предчелюстной, верхнечелюстной и зубной. Они и образуют в основном хватательный аппарат. Отметим, однако, что элементы небно-квадратного и меккелева хрящей не потеряли полностью функции челюстей, так как эти кости находятся еще в составе челюстного аппарата, и небная кость несет зубы.
Подъязычная дуга (arcus hyoideus). Подъязычная дуга — вторая висцеральная дуга. Она, как и у акул, состоит из двух парных
отделов: подъязычно-челюстного, или гиомандибулярного, и подъязычного, или гиоидного.
П о дъ я з ы ч н о - че л ю ст н а я кость (hyomandibulare, рис. 99, hmd.) выполняет функцию подвеска, связывая челюстной аппарат с осевым черепом. Верхний ее край связан с клиновидно- и крыловидноушной костями. Сзади к ней прикрепляются крышечная и предкрышечная кости. Спереди она граничит с metapterygoideum. Наиболее сложной формы кость с многочисленными отростками имеется у трески.
Собственно подвесок (symplecticum, рис. 99, symp.) — небольшая косточка, отходящая от нижнего края гиомандибуляре и по происхождению являющаяся ее отчленившейся частью. Эта вытянутая косточка направлена вперед и соединяет гиомандибуляре с квадратной костью. Таким образом, как и у акулы, имеется гиостилия, т. е. первичная верхняя челюсть подвешена к черепной коробке при помощи подвеска, состоящего у судака из двух элементов— hyomandibulare et symplecticum.
Нижний отдел подъязычной дуги и жаберные дуги можно рассматривать только с внутренней стороны. Для этого снимают все кости челюстной дуги с одной стороны и рассматривают противоположную подъязычную дугу с внутренней стороны.
Межгиоид (interhyale, s. stylohyale, рис. 100, sth.)—небольшая кость, отходящая от нижней части гиомандибуляре, занимающая почти вертикальное положение и связывающая гиомандибуляре с гиоидом. По своему происхождению она является окостеневшей связкой.
Гиоид (hyoideum, рис. 100)—вентральный отдел подъязычной дуги — состоит из четырех парных частей: верхнегиоидной (epihyale, eph.), рожковидно-гиодной (ceratohyale, cerh.) и двух маленьких нижнегиодных (hypohyale, hyph.). Следует отметить, что приводимые обозначения условны, так как эти элементы не гомологичны соответствующим элементам жаберных .дуг. Нижние элементы гиоидной дуги на вентральной стороне соединены непарной удлиненной костью, называемой основной подъязычной, или подъязычной (basihyale, s. glossohyale, рис. 100, bhy) \ она направлена вперед и поддерживает слизистую оболочку дна ротовой полости. От верхней и рожковидной частей гиоида отходят тонкие изогнутые косточки, налегающие друг на друга своими нижними краями, — это лучи жаберной перепонки (radius branchiostegi). Как указывалось выше, число их у разных видов различно. Они проходят в жаберной перепонке, которая является продолжением кожи, покрывающей жаберную крышку. Таким образом, перепонка с лучами образует как бы продолжение жаберной крышки на вентральной стороне и соответственно участвует вместе с ней в акте дыхания.
От вентральной стороны гипогиальных элементов отходит непарная заднеподъязычная кость (urohyale, s. basibran-chiostegale). Она направлена назад и при помощи связок соединяется с плечевым поясом. Окостенения подъязычной дуги являют-il* 323
ся замещающими, за исключением лучей жаберной перепонки и заднеподъязычной кости.
Жаберные дуги (arcus branchialis). В количестве пяти пар жаберные дуги расположены позади подъязычной дуги. В отличие от акул они тесно сдвинуты под задним отделом черепа, так как жаберные щели стали очень узкими и близко прилежат одна к другой. Каждая жаберная дуга, как и у акулы, расчленена на четыре парных элемента, а внизу дуги соединены непарными копулами (basi-
Рис. 100. Жаберный аппарат судака Lucioperca lucioperca:
bhy. — основная подъязычная кость; cbr. — рожковидиожаберный элемент; cerh. — рожковидно-гиоидный элемент; сор. — копула; ebr. — верхнежаберный элемент; eph. — верхнегиоидный элемент; hbr. — поджаберный элемент; hyph. — вижнегнондный элемент; phbr^ 4 — глоточно-жаберные элементы; sth. — межгиоидная кость
branchiale). Полностью представлены только три первые жаберные дужки. Все окостенения жаберных дуг только хрящевые. Чтобы рассмотреть границы между частями, надо очистить жаберные дуги от жаберных лепестков, тычинок и прочих мягких тканей.
Дорзальные элементы жаберных дуг называются глоточножаберными (pharyngobranchiale). На 1-й жаберной дуге этот элемент представлен небольшой косточкой (рис. 100, phbr\.). Фарингоэлементы 2, 3 и 4-й жаберных дуг расширены, тесно сближе
ны между собой и на нижней поверхности (исключая карпа) усажены массой мелких зубов, которые называются верхними глоточными зубами (рис. 100, phbrx, 2, з, <)• На 5-й жаберной дуге этот элемент отсутствует.
Второй дорзальный элемент называется верхнежаберным (epibranchiale, рис. 100, ebr.). На верхнем конце он раздвоен, на 5-й жаберной дуге отсутствует.
Третий элемент, расположенный на вентральной стороне глотки, называется р о ж ков и д н о -ж а б е р н ы м (ceratobranchiale, рис. 100, cbr.). В 5-й жаберной дуге это единственный сохранившийся элемент. На нем находятся мелкие зубы, называемые нижними глоточными зубами.
У карпа этот элемент развит наиболее сильно. Сидящие на нем зубы расположены в три ряда, и по своей форме и структуре соответствуют зубам других позвоночных.
Самые нижние парные элементы имеются только на первых трех жаберных дугах. Они называются поджаберными (hypobranchiale, рис. 100, hbr.).
Непарные основные вентральные элементы, или копулы (basib-ranchiale, s. copula, рис. 100, cop.), имеются только у первых трех жаберных дуг. 4-я и 5-я жаберные дуги при помощи соединительнотканного тяжа прикрепляются к заднему концу последней копулы.
Жаберная крышка (operculum). Жаберная крышка покрывает снаружи жаберные дуги; она образована четырьмя покровными костями. Передняя из них предкрышечная (praeperculum, рис. 99, prop.) имеет изогнутую форму. Передний ее край соединяется с задним краем гиомандибулярной кости. Нижняя часть переднего края соединяется с квадратной костью. Задний край выпуклый. В верхней части выпуклости отверстие, через которое входит в кость крышечно-челюстной канал органа боковой линии, а ниже него — отверстие, через которое канал сообщается с внешней средой.
Крышечная кость (operculum, рис. 99, ор.) прилегает к верхнему заднему краю предкрышечной кости и верхним передним углом связана с гиомандибулярной костью.
Подкрышечная кость (suboperculum, рис. 99, sop.) — небольшая плоская кость, лежащая под вышеназванной.
Межкрышечная кость (interoperculum, рис. 99, inop.) лежит впереди от подкрышечной и граничит со всеми тремя костями жаберной крышки. Передним своим краем она связана при помощи сухожилий с угловой костью нижней челюсти.
Мускулатура
Для изучения мускулатуры лучше всего взять целый экземпляр рыбы. Поскольку на том же препарате будет изучаться и нервная система, то полезно после промывки материала или без нее подвергнуть его частичной декальцинации, для чего его следует поме
стить в 5—7%-ный раствор азотной кислоты на 1—2 дня, затеи хорошо промыть в течение суток в проточной воде. Одновременно* с изучением мускулатуры можно знакомиться и с отдельными элементами периферической нервной системы, поэтому при препаровке необходимо тщательно сохранять лежащие часто поверхностно* ветви головных нервов.
Снимают кожу с передней половины левой стороны тела рыбы. Для этого делают два небольших разреза. Один разрез .проводят вдоль средней линии брюха, второй перпендикулярно первому по боковой стенке тела. Затем, захватив угол образовавшегося лоскута, постепенно снимают кожу в направлении снизу сзади — вперед вверх. При таком направлении она лучше всего отделяется от мышц. Однако для того чтобы часть мышц не была снята вместе с кожей, необходимо все время отделять их от последней скальпелем. По мере отделения кожи продолжают начатые разрезы вперед и вверх, а затем проводят и продольный разрез вдоль спины. Снимают так кожу с латеральной и медиальной сторон плавников и с головы. Снимать кожу с боковой поверхности головы, а также жаберной крышки следует осторожно, так как расположенные здесь, небольшие мышцы плотно связаны с кожей и легко могут быть удалены. Начинать снимать кожу с головы лучше всего, счистив ее скальпелем с костных частей, а затем, сняв частями при помощи пинцета и скальпеля, уже и с мускулатуры. Для ознакомления с мышцами и нервами, расположенными в глазнице, необходимо вынуть глазное яблоко. Для этого отделяют его от кожи, оттянув наружу переднюю часть и, перерезав глазные мышцы у места их прикрепления к глазу и глазной иерв, удаляют глаз совсем.
Глаз следует сохранить, положив его в сосуд с формалином. Строение глаза будет рассмотрено при изучении органов чувств.
В общих чертах мускулатура костистых рыб сходна с мускулатурой акуловых, однако здесь наблюдается дальнейшая дифференцировка и появление особых мышц, связанных с движением жаберной крышки. Описание мускулатуры будет дано в основном по* щуке со ссылками на отличия, которые имеются в мускулатуре окуня.
Мускулатура туловищной области костистых рыб отличается еще слабой дифференцировкой и имеет сходное строение у разных видов. Несколько отличается ее строение в передней части туловища у видов, брюшные плавники которых передвинуты далеко вперед. В области головы значительной дифференцировки достигают мышцы, участвующие в движении жаберной крышки. Однако набор этих мышц у различных видов сходен, а их форма во многом зависит от формы головы и жаберной крышки. В связи с этим нет необходимости подробно описывать мускулатуру у всех видов. Ее описание дается только для щуки со ссылками на особенности строения у окуня как рыбы с более коротким туловищем и головой. Пользуясь этим описанием, легко разобраться в строении мускулатуры и у других видов.
Туловищная мускулатура
Туловищная мускулатура костистых рыб так же, как и у хрящевых, делится продольной горизонтальной септой (septum horisontale, рис. 101, 103, spt. h.) на спинную (эпаксиальную) и брюшную (гипаксиальную). Каждая из этих частей состоит из отдельных миомеров (рис. 101, 103, тт.), имеющих изогнутую форму и разделенных тонкими соединительнотканными прослойками — миосептами (рис. 101, 103, ms.). Такое простое строение миомеров, видимое с поверхности, не соответствует их строению в толще мус-
Рис. 101. Мускулатура и нервы щуки Esox lucius (вид сбоку):
а. а. р. — приводящий мускул небной дуги; т. ad. т. — приводящий мускул нижней челюсти; tn. d. о. — мускул, отодвигающий жаберную крышку; т. long. d. — длиннейшая мышца спины; т. I. а. р. — подниматель небной дуги; tn. I, о. — подниматель жаберной крышки; пип.— миомер; т. о. а. е. — наружная косая мышца живота; т. о. а. «.’-’внутренняя косая мышца живота; пг. s. — миосепта; tn. tr.—трапециевидная мышца; г. 6. V// —щечная ветвь VII нерва г. hmd. V//— гиомандибуляриая ветвь VII нерва; г, hy. VII — гиоидная ветвь VII нерва; г. md. е. — наружная нижнечелюстная ветвь; г. md. i. — внутренняя нижнечелюстная ветвь; г. md. V—нижнечелюстная ветвь V нерва; г. тх. V — верхнечелюстная ветвь V нерва; г. opht. pr. V—глубокая глазничная ветвь V нерва; г. opht. s. VII ~ поверхностная глазничная ветвь VII нерва; г. pl. V//— небная ветвь VII нерва; spt. h.— горизонтальная септа; // — зрительный нерв; /// — глазодвигательный нерв; IV — блоковый нерв
кулатуры, где миосепты проходят наискось, вследствие чего миомеры имеют вид конусов, вложенных друг в друга, а на поперечном разрезе — концентрически расположенных колец. Причем, конусы спинной порции направлены вершинами назад, а брюшной — вперед. Сложное строение конусов лучше всего рассмотреть на поперечном разрезе. Для этого вполне достаточно отрезать хвостовой отдел или воспользоваться препаратом, на котором рассматривались внутренние органы, и хвостовой отдел уже отрезан.
После рассмотрения поперечного разреза следует подробней разобрать расчленение туловищной мускулатуры на отдельные
слои — мышцы туловища. Так же, как и у акуловых рыб, в туловищной мускулатуре щуки намечается дифференцировка на от-
дельные самостоятельные мышцы.
Длиннейшая мышца спины (m. longissimus dorsi, рис. 101, т. long, d.) очень хорошо видна у щуки благодаря резкому сужению и изгибу дорзальных частей спинных миомеров. Здесь
миомеры сначала направляются
Рис. 102. Мускулатура щуки Esox lucius (вид снизу):
т. g. h. у.— подбородочно-подъязычная мышца; т. г. а. — прямая мышца живота; т. sthy. — грудинно-подъязычная мышца
назад, а затем снова вперед, причем резкость перегиба постепенно уменьшается по направлению к хвостовому отделу. В передней части от мускула отходят длинные сухожилия, прикрепляющиеся к верхней части плечевого пояса и затылку. У окуня эта порция мышц не обособлена.
Между длиннейшими мышцами обеих сторон у многих рыб проходит неширокий мускульный тяж. Это остатки мускулатуры редуцированных передних отделов спинного плавника.
В вентральной части также можно различить несколько слоев или порций мышц.
Наружная косая мышца живота (m. obliquus abdominis externus, рис. 101, т. о. а. е.) занимает у щуки поверхностную дорзальную часть гипаксиальной мускулатуры. Она хорошо разли-
чима по направлению волокон. Отдельные мышечные волокна в миомерах тянутся косо, начинаются сверху и направляются вниз и назад.
Внутренняя косая мышца живота (m. obliquus abdominis internus, рис. 101, т. о. a. i.) видна на боковой поверхности брюшной стенки, несколько ниже вышеописанного мускула. Волокна ее так же, как и в пре
дыдущей мышце, направлены косо, но в противоположном направ-
лении, т. е. начинаются сверху сзади и тянутся вперед и вниз. Мышца занимает всю боковую стенку брюшной полости, но верхняя ее часть прикрыта наружной косой мышцей. В этом легко убедиться, сняв волокна наружной косой мышцы между двумя миосептами и рассмотрев под ними идущие в другом направлении волокна внутренней косой мышцы.
Прямая мышца живота (m. rectus abdominis, рис. 102, т. г. а.) занимает у щуки самую среднюю часть вентральной поверхности туловища. От косых мышц она отличается раоположе-
нием волокон, которые идут не косо, а вдоль продольной оси тела. Особенно хорошо эта мышца видна в передней своей части между грудными плавниками.
У окуня все слои гипаксиальной мускулатуры выражены не отчетливо.
Соматическая мускулатура в голове представлена мышечными пучками, тянущимися от плечевого пояса вперед к гиоиду. Повернув рыбу на спину и раздвинув в стороны жаберные крышки, рассматривают отдельные мышцы.
Грудинноподъязычиая мышца (m. sternohyoideus, s. coracohyoideus, рис. 102, т. sthy.) у окуня начинается в виде парного образования от передней части плечевого пояса, а у щуки — от латеральных сторон клейтрума и тянется вперед в виде непарной мышцы, но с двумя сухожилиями на переднем конце, которыми прикрепляется к urohyale.
Подбородочно-подъязычная мышца (m. geniohyoi-deus, рис. 102, т. g. hy.) —парная мышца, идущая от латеральной стороны передней части гиоида к внутренней поверхности переднего конца нижней челюсти, к которым они прикрепляются несколькими пучками.
Кроме того, имеется еще несколько мышц соматического происхождения, но связанных с подъязычным аппаратом, однако они относительно мелки, и мы на них останавливаться не будем.
Мускулатура плавников
Мускулатура парных и непарных плавников в связи с их большей подвижностью, чем у акуловых рыб, усложняется. Она расчленилась на отделньые пучки, или слои, выполняющие различные функции.
Мускулатура непарных плавников состоит из отдельных пучков, идущих к плавниковым лучам, которые поднимают или опускают последние. Чтобы рассмотреть отдельные мускульные пучки, необходимо сделать продольный разрез кожи на одной стороне у основания спинного плавника, а затем снять лоскут кожи с одной стороны тела, осторожно отделив ее от мускулатуры при помощи пинцета и скальпеля.
Рассматривают мускульные пучки, отходящие от оснований лучей спинного плавника.
Опускатели (m. depressor)—небольшие мышцы, прикрепляющиеся к каудальной стороне луча и служащие для опускания лучей плавника.
Подниматели (m. erector). Волокна этого (у щуки двойного) мускула прикрепляются к передней части плавникового луча и при сокращении поднимают лучи.
Кроме двух вышеназванных групп мышц можно видеть также небольшие мышечные пучки (особенно развитые у щуки) m. incli-nater, идущие от луча к коже и к соединительнотканному футляру туловищной мускулатуры.
Впереди от спинного плавника плавниковые мышцы претерпевают некоторую редукцию. Здесь они тянутся в виде особого длинного мышечного тяжа, передняя часть которого прикрепляется к затылочному отделу и, очевидно, участвует в движении головы.
Этот мышечный тяж хорошо выражен у щуки, у окуня он очень короткий, так как спицной плавник почти вплотную подходит к голове.
Парные плавники. Парные плавники в связи с подвижностью, в частности с самостоятельной подвижностью переднего и заднего краев, обладают дорзальной и вентральной группами мышц, расчлененными на отдельные пучки, оттягивающие и прижимающие передние и задние лучи плавника. Очищают основания грудных плавников от кожи как с наружной, так и с внутренней стороны, а также снимают кожу в области брюшных плавников и рассматривают отдельные мышечные пучки, участвующие в движении плавников. Сначала рассматривают мускулатуру грудных плавников.
Отводящий поверхностный мускул (abductor superficialis, рис. 103, tn. ab. s.) отводит грудной плавник. Он начинается на внутренней поверхности клейтрума и оканчивается на основаниях лучей плавника, занимая почти всю латеральную поверхность основания грудного плавника.
М. arrector ventralis виден дорзальнее от вышеописанного; основная часть этой мышцы лежит под отводящим поверхностным мускулом, так что на препарате видна только ее небольшая задняя часть. Она начинается на клейтруме и коракоиде и оканчивается на основании переднего, несколько утолщенного луча. При сокращении он так же, как и m. arrector dorsalis (который будет описан ниже), расправляет первый луч плавника.
Отводящий глубокий мускул (m. abductor profudus, рис. 103, т. ab. р.) лежит под поверхностным мускулом, и его нижняя часть выступает из-под вентрального края поверхностного мускула. Оттянув поверхностный мускул вверх, убеждаются, что глубокий отводящий мускул начинается на коракоиде и оканчивается на латеральной поверхности оснований лучей плавника.
Оттянув грудной плавник вперед и отпрепарировав соединительную ткань, соединяющую медиальную сторону плавника со стенкой тела, рассмотрите мышцы, приводящие грудной плавник. Общий план расположения их такой же, как и отводящих, но все они лежат с медиальной стороны первичного плечевого пояса.
Приводящий поверхностный мускул (m. adductor superficialis) отличается почти вертикальным расположением волокон, которые идут от верхней части клейтрума к основаниям лучей плавника. У щуки он развит слабо.
Приводящий глубокий мускул (ш. adductor profundus) лежит вентральнее вышеназванной мышцы, его волокна тянутся более или менее горизонтально. Он начинается на медиальной поверхности коракоида и оканчивается на основаниях лучей.
М. arrector dorsalis у щуки слабо отделен от m. adductor profundus и лежит вдоль дорзального края медиальной стороны 330
коракоида. Под вышеназванными мускулами можно иногда рассмотреть небольшие мышцы m. coracoradialis, идущие от коракоида к лучам плавника.
Брюшные плавники имеют те же мышцы, что и грудные.
Висцеральная мускулатура
Висцеральная мускулатура костистых рыб, как уже указывалось, имеет более сложное строение, чем у хрящевых. В связи с появлением жаберной крышки исчезает система жаберных констрикторов, зато усложняются производные констрикторов челюстной и подъязычной дуг, в частности, появляются мышцы, приводящие в движение жаберную крышку.
'Рис. 103. Мускулатура и периферическая нервная система окуня Регса Jluviaiilis (вид сбоку): т. а. а. р. — приводящий мускул вебной дуги; т. ad. т. — приводящий мускул нижней челюсти; т. ab. рг.~ отводящий глубокий мускул; т. ab. s. — отводящий поверхностный мускул; т. d. э. — мускул, отодвигающий жаберную крышку; ml. а. ф1. — подниматель небной дуги; т. I. о. — подниматель жаберной крышки; тт. — миомер; ms. — мносепта; т. d. г. — трапециевидная мышца; г. hmd. VII— гиоманди-•булярная ветвь VII нерва; г. hy. V// —гиоидная ветвь VII нерва; г. тх. V — верхнечелюстная ветвь V нерва; г. md. V — нижнечелюстная ветвь V нерва; г. md. I. VII — внутренняя нижнечелюстная ветвь VII нерва; г. pl. V// —небная ветвь VII нерва; spt. h. — горизонтальная септа; / — обонятельный нерв; // — зрительный нерв: /// — глазодвигательный нерв; IV — блоковый нерв
Сначала следует рассмотреть лежащие поверхностно мышцы, участвующие в движении жаберной крышки.
Подниматель жаберной к р ы ш к и (m. levator operculi, рис. 101, 103, т. I. о.) имеет обычно 3 порции, которые неясно отграничены друг от друга. Мускул начинается на латеральной стороне pteroticuni и затем его волокна, веерообразно расходясь, прикрепляются вдоль верхнего края крышечной кости.
У окуня деление мускула на порции более отчетливо, чем у щуки. Этот мускул иннервируется ветвью VII нерва, является про-изводны.м сжимателя гиоидной дуги (m. constrictor hyomandibula-ris dorsalis) акуловых рыб и, как указывает название, поднимает жаберную крышку.
Мускул, отодвигающий жаберную крышку (ш. dilatator operculi, рис. 101, 103, т. d. о.), лежит впереди вышеописанного и имеет вид треугольника, направленного вершиной вниз. Верхней широкой частью мышца прикрепляется к наружному краю pteroticum, а заканчивается на (верхнем переднем углу крышечной кости. По происхождению она является отделившейся частью ш. levater arcus palatini и, как и последний, иннервируется, ветвями V нерва и является производным дорзального констриктора челюстной дуги. При сокращении этой мышцы задняя часть жаберной крышки отводится в сторону.
Отвернув вперед жаберную крышку, рассматривают лежащий на ее внутренней поверхности мускул, приводящий ее к телу. Обычно, чтобы увидеть место его отхождения и прикрепления, приходится настолько сильно оттягивать крышку, что волокна мускула обрываются.
Мускул, пр и во д я щи й жаберную к р ы ш к у (m. adductor operculi), начинается на боковой поверхности затылочной области черепа, так что место его отхождения обычно скрыто под туловищной мускулатурой. Волокна его тянутся в поперечном направлении и прикрепляются ко внутренней поверхности крышечной кости. Мускул иннервируется VII парой нервов и является производным m. constrictor dorsalis акуловых рыб.
Впереди от жаберной крышки лежит мускулатура, связанная с движением костных элементов верхней и нижней челюстей. Она образует так называемые «щеки» рыб.
Приводящий мускул нижней челюсти (m. adductor mandibulae, рис. 101, 103, т. а. т.) лежит впереди предкры-шечной кости. Волокна его расходятся веерообразно по направлению снизу вверх. При детальном изучении в этой мышце можно различить три порции. Мышца начинается на боковой поверхности мозгового черепа, в частности на sphenoticum, и на передней поверхности предкрышки, оканчивается на задней части нижней челюсти и заднем верхнем крае верхнечелюстной кости. Она иннервируется нижнечелюстной ветвью V нерва, является гомологом одноименной мышцы акуловых рыб и при сокращении закрывает рот, приводя нижнюю челюсть, а также оттягивает назад задний конец верхнечелюстной кости. Осторожно отпрепарировав мышцу и стараясь не повредить проходящие здесь нервы, разрезают ее порции в средней части, поперек хода волокон, отворачивают два мышечных края в стороны. Рассматривают мышцы, лежащие у щуки под приводящими мускулами.
Подниматель небной дуги (m. levator arcus palatini, рис. 101, 103, tn. I. a. p.). У щуки большая часть волокон этой мыш-чы лежит под m. adductor mandibulae и видна только после удаления этой мышцы. У окуня в связи с меньшим развитием приводящего мускула верхняя часть поднимателя небной дуги обнажена и хорошо видна с поверхности. У обоих представителей передний край этого мощного мускула участвует в образовании заднего края глазницы, так что небольшую его часть можно видеть со стороны 332
глазницы. Мускул начинается на передней части боковой поверхности слуховой области мозгового черепа и оканчивается на латеральной поверхности гиомандибуляре. Соответственно с таким расположением его еще называют гл. levator hyomandibulare. Иннервируется мышца ветвями V нерва и является производной под-нимателя небно-квадратного хряща акуловых.
Прив одящий мускул небной дуг и (m. adductor arcus palatini, рис. 101, 103, т. а. а. р.) лежит еще глубже, под вышеназванным мускулом. Чтобы его увидеть целиком, необходимо снять поднимающую мышцу, но при этом легко повредить нервы, идущие по этому мускулу. Поэтому детальное изучение мест прикрепления этого мускула следует отложить до окончания рассмотрения периферических нервов. Однако небольшая часть мускула видна и с поверхности, в заднем нижнем углу глазницы. Мускул начинается на боковой части мозгового черепа, а оканчивается на гиоманди-булярной и крыловидных костях.
За неимением времени не будем рассматривать большого количества мелких мышц, приводящих в движение висцеральный аппарат, в частности отдельные жаберные дуги. Из висцеральной мускулатуры отметим еще только трапециевидную мышцу.
Трапециевидная мышца (m. trapezius, рис. 101, 103, т. tr.)—небольшая мышца, лежащая над верхним краем жаберной крышки. Она как бы окружена со всех сторон костными образованиями. По бокам спереди находятся отростки pter- и epioticum; сзади она входит в вилочку, образованную костями плечевого пояса — post- и supratemporale. Таким образом, так же, как и у акулы, эта мышца связывает плечевой пояс с затылочной областью черепа и так же, как у последней, иннервируется самостоятельной ветвью X нерва, гомологичной нерву XI пары у млекопитающих.
У карпа в связи с плотным срастанием вышеназванных костей эта мышца отсутствует.
Глазные мышцы
Необходимо рассмотреть также шесть глазных мышц (четыре прямые и две косые), располагающихся в области глазницы. Их расположение, наименование и иннервация у костистых рыб такие же, как и у всех вышестоящих позвоночных, и полностью соответствует таковым акуловых рыб, поэтому подробно на них можно не останавливаться. Отличие заключается только в том, что четыре прямые мышцы медиальными частями плотно охватывают зрительный нерв и вместе с ним входят в особую трубку (myodom) в костях черепа, на дне которой и прекрепляются.
Нервная система
Нервная система всех костистых рыб имеет типичное строение для первично водных позвоночных, какими являются все подклассы костных рыб. Однако каждый подкласс отличается ясно выраженными особенностями биологии.
Костистые рыбы отличаются слабым развитием переднего отдела головного мозга, имеющего перепончатую крышу, лишенную нервных клеток. В дне промежуточного мозга обособляется особый отдел— сосудистый мешочек, свойственный только рыбам. Средний мозг сложно дифференцирован, в связи с ведущей ролью органов зрения в добывании пищи он расчленен на зрительный и слуховой отделы и имеет сложно дифференцированную стволовую часть сетевидную формацию как важный центр координации нервной деятельности. Продолговатый мозг также сложно дифференцирован в связи с большой ролью в биологии органов стато-акустики и кожных органов чувств. Мозжечок, ведающий передвижением рыб при плавании и несущий сензорные функции, достигает крупных размеров, закрывая собой лежащие под ним отделы головного мозга. Головные нервы подчинены функциям моторики жаберного аппарата (захватывание пищи, дыхание).
Знакомство с нервной системой лучше всего начать с периферических головных нервов; они относительно хорошо видны на препарате, по которому изучалась мускулатура. Отхождение головных нервов от мозга у костистых рыб мало отличается от такового акуловых. Однако в связи с меньшим размером мозга разобрать места отхождения и проследить начальный путь нерва значительно труднее. Поэтому рассматривают только периферические части нервов, а места отхождения будут разобраны при изучении головного мозга. Эту операцию следует проводить на одной стороне объекта, оставив ненарушенной картину расположения мускулов на другой стороне.
Головные нервы
Обонятельный тракт, или нерв (tractus (nervus) olfactorius, рис. 103, 104, /). В отличие от акуловых рыб, у которых обонятельная луковица непосредственно прилегает к обонятельной капсуле и связана с мозгом его длинным выростом — обонятельным трактом, у костистых рыб могут наблюдаться два различных варианта. У одних видов обонятельная луковица также непосредственно прилегает к обонятельной капсуле, и нервный тяж, соединяющий ее с мозгом, также является обонятельным трактом (карп). У других обонятельная луковица непосредственно прилегает к переднему мозгу, и тогда нервные волокна, идущие от обонятельной капуслы, являются отростками рецепторных клеток и образуют обонятельный нерв (щука, окунь).
Обонятельный нерв щуки и обонятельный тракт карпа проходят внутри черепной коробки и видны только после ее вскрытия. Обонятельный нерв окуня виден со стороны глазницы и выглядит как толстый белый тяж, проходящий по ее верхней части.
Зрительный нерв (n. opticus, рис. 101, 103, 104, 11) отходит от зрительного перекрестка на дне промежуточного мозга и имеет вид толстого белого тяжа, окруженного соединительнотканной оболочкой; его небольшой кусочек торчит между четырьмя 334
прямыми глазными мышцами. Дистальный 'конец, входящий в глазное яблоко, соответственно удаляется при удалении последнего.
Глазодвигательный нерв (n. oculomotorius, рпс. 101, 103, 104, 111), как и у всех позвоночных, иннервирует три прямых глазных мышцы и одну (нижнюю) косую; он берет начало от дна
Рис. 104. Схема головных нервов и их ветвей у костистой рыбы:
т. b. VII— щечная ветвь VII нерва; г. hmd. VII — гиомандибулярная ветвь VII нерва; г. hy. VII — гиоидная ветвь VII нерва; г. hypo.—подъязычная ветвь; г. ini. X — внутренностная ветвь X нерва; г. lat. X — боковая ветвь X нерва; г. mand. V—нижнечелюстная ветвь V нерва; г. max. V — верхнечелюстная ветвь V нерва; г. md. е. VII — наружная нижнечелюстная ветвь VII нерва; г. md. i. W/— внутренняя нижнечелюстная ветвь VII нерва; г. opht. s. VII — поверхностная глазничная ветвь VII нерва; г. pl. VII — небная ветвь VII; г. pl. IX — небная ветвь IX нерва; г. prtr. — преджаберные ветви; г. pstr. — зажаберные ветви; I, II, III, IV, V, VIII, X, XII — головные нервы
среднего мозга. На препарате обычно бывает хорошо видна одна его ветвь, идущая к нижней косой мышце со средней части медиальной стенки глазницы, остальные ветви частично удаляются, а частично скрыты мышцами глаза.
Блоковый нерв (n. trochlearis, рис. 101, 103, 104, IV) иннервирует верхнюю косую мышцу глаза и у щуки тянется по углу глазницы между дорзальной и медиальной стенками, вместе с кровеносным сосудом и рядом с глазничной ветвью V нерва, от которой отличается большей толщиной и с которой может давать анастомозы. Начинается блоковый нерв от крыши среднего мозга на
границе с мозжечком, у окуня и карпа он тянется совершенно самостоятельно вдоль верхней стенки глазницы и заканчивается, как и у щуки, на верхней косой мышце глаза.
Тройничный нерв (и. trigeminus, V) отходит мощным корешком от боковой поверхности продолговатого мозга и имеет большое количество ветвей. Он является производным двух слившихся вместе нервов, образующих при выходе из головного мозга 2 ганглия: глазничный и гассеров (ganglion trigemini I, s. g. profundi, ganglion trigemini II, s. g. Gasseri). Оба эти ганглия обычно сливаются вместе.
От первого ганглия начинаются две ветви: поверхностная и глубокая глазничные; от второго — челюстная ветвь. Поверхностная глазничная ветвь обычно слабо развита и идет вместе с волокнами ветви VII нерва, имеющей то же название. Глубокая глазничная хорошо выражена у современных костных рыб, а у Polypteri является самостоятельным нервом.
У рассматриваемых представителей по выходе из ганглия, который у щуки сохраняет двойственность, V нерв распадается на две ветви: глубокую глазничную и собственно тройничный нерв.
Глубокая глазничная ветвь (n. trigeminus I, s. г. ophthalmicus profundus V, см. рис. 101, г. opht. pr. V.) хорошо видна в области глазницы. У щуки она тянется параллельно IV нерву и кровеносному сосуду, входит в переднюю стенку глазницы, прободает ее, выходит на поверхность и иннервирует кожу верхней части головы.
У карпа это тонкая ветвь, тянущаяся параллельно вентральному краю поверхностной глазничной ветви VII нерва.
Собственно тройничный нерв (n. trigeminus II) по выходе из черепа отдает от себя тянущуюся назад ветвь к m. dilatator operculi. Эта небольшая веточка на препарате не видна. Далее нерв разделяется на два крупных ствола.
Нижнечелюстная ветвь (г. mandibularis, рис. 101, 103, 104, г. md. V) у щуки тянется вниз, вдоль заднего края глазницы под двумя верхними порциями m. adductor mandibulae, поэтому ее начало можно видеть только после удаления этих порций. Затем она выходит на поверхность, проходит по переднему краю вышеназванного мускула и подходит к заднему верхнему краю нижней челюсти. Здесь нерв распадается на целую серию мелких веточек, которые теряются в мягких тканях и костях нижней челюсти. Эти ветви иннервируют кожу, выстилку ротовой полости и отчасти мышцы, связанные с нижней челюстью. Кроме того, в начальной своей части, еще до разветвления, нижнечелюстная ветвь отдает многочисленные, разной величины нервы к челюстной мускулатуре (m. adductor mandibulae). Одна такая ветвь, отходящая на уровне средней части заднего края глазницы и иннервирующая глубокий слой мышцы, хорошо видна. У окуня основная часть нерва идет под m. adductor mandibulae или непосредственно в нем. Поэтому, чтобы ее рассмотреть, следует аккуратно выщипать переднюю часть этой мышцы. Правда, небольшой участок нерва, проходящий по ЗЗВ
заднему краю глазницы, виден и с поверхности, но здесь его трудно отличить от ветвей V и VII нервов, рядом с которыми он проходит.
У карпа в связи с небольшими размерами ротового отверстия нижнечелюстная ветвь проходит по нижней части глазницы вместе с верхнечелюстной V и щечной ветвью VII нервов. Она достигает значительной толщины и лежит не непосредственно у края глазницы, а несколько в глубине. На уровне середины глазницы она делает крутой изгиб вниз, направляясь к нижней челюсти.
Верхнечелюстная ветвь (г. maxillaris, рис. 101, 103, 104, г. тх. V), так же как и вышеописанная ветвь, проходит сначала по заднему наружному краю глазницы, затем переходит на нижнюю ее сторону, где тянется вместе с щечной веткой VII нерва. Эти ветви довольно легко разделить, если аккуратно снять одевающую их общую соединительнотканную оболочку. Челюстная ветка V нерва более мощная, и ее часть, идущая по заднему краю глазницы, лежит более медиально, чем ветвь VII нерва; последняя в этом месте лежит как раз между двумя ветвями V нерва. Пройдя глазницу, верхнечелюстная ветвь выходит на боковую часть верхней поверхности головы и располагается здесь прямо под кожей, так что ее легко рассмотреть, если снять последнюю. Этот нерв своими ветвями иннервирует кожу верхней поверхности головы и верхней челюсти, а также внутреннюю поверхность верхнечелюстной кости.
У окуня эта ветвь занимает то же положение, но в связи с небольшими размерами объекта ее трудно отделить от ветви VII нерва.
У карпа эта ветвь также идет вдоль нижней стороны глазницы вместе с указанными выше ветвями V и VII нервов. Дойдя до переднего края глазницы, она идет вперед под крупной слезной костью и далее распадается на ряд ветвей, иннервирующих верхнюю челюсть.
Отводящий нерв (n. abducens, VI) иннервирует наружную (заднюю) косую мышцу глаза и на препарате не виден.
Лицевой нерв (n. facialis, VII) имеет много ветвей, отходящих от продолговатого мозга позади тройничного нерва целым пучком; большинство из них связано с органами боковой линии и поэтому у костистых рыб, как и у всех водных форм, хорошо развиты.
Поверхностная глазничная ветвь (г. ophthalmicus superficial is, VII, рис. 101, 104, г. opht. s. VII), проходит вдоль медиальной части верхнего края орбиты, параллельно ветвям IV, V нервов и кровеносному сосуду. На своем пути она отдает ряд ветвей к надглазничному каналу органа боковой линии, который и иннервирует. Прибегнув к помощи бинокулярного микроскопа, эту ветвь можно рассмотреть у окуня и карпа.
Небная ветвь (г. palatinus, VII, рис. 101, 103, 104, г. pl., VII) проходит у щуки вдоль латерального края основания черепа, примерно по средней линии глазницы. Чтобы ее рассмотреть, следует снять выстилающую глазницу соединительнотканную перепонку, и тогда у основания черепа обнажится нерв. Пройдя по глазни
це, он выходит через ее переднюю стенку и распадается на большое количество ветвей, которые иннервируют выстилку ротовой полости в небной части.
У окуня и карпа она занимает то же положение, но у последнего достигает значительной толщины.
Щечная ветвь (г. buccalis, VII, рис. 101, 103, 104, г. b., VII) у щуки тянется тараллельно верхнечелюстной ветви V нерва вдоль заднего, а затем нижнего края глазной орбиты. По выходе из нее в области переднего нижнего угла он отклоняется кверху и переходит на наружную поверхность головы, где идет вдоль латеральной стороны рострума. Здесь ее легко обнаружить, если спять кожу с верхней части рыла. Эта ветвь у щуки иннервирует переднюю и заднюю части подглазничного канала органа боковой линии.
У окуня и карпа нерв занимает примерно то же положение, и у последнего легко отличим, так как имеет значительные размеры и оканчивается на поверхности переднего края глазницы в области слезной кости несколькими крупными ветвями.
Дополнительная щечная ветвь (г. buccalis accessorius) иннервирует среднюю часть подглазничного канала и у щуки видна в виде тоненького нерва, проходящего вдоль вентрального края m. levator arcus palatini, и прикрыта снаружи m. adductor mandibulae, так что ее рассмотреть можно только в том случае, если последний удалей. Эта ветвь подходит к медиальной стороне ряда подглазничных косточек н иннервирует подглазничный канал.
Гиомандибулярная ветвь VII головного нерва (г. hyomandibularis VII, рис. 101, 103, 104, г. hmd. VII) — крупный нервный ствол, являющийся частью лицевого нерва и идущий вдоль переднего края жаберной крышки. От него отходит вышеназванная добавочная щечная ветвь.
Нерв проходит по латеральной поверхности гиомандибулярной кости, подстилающей предкрышечную кость, поэтому, чтобы его рассмотреть, необходимо аккуратно снять предкрышечную кость, тогда обнажается основной ствол нерва.
Гиомандибулярная ветвь затем распадается на несколько ветвей. Три из них, наиболее крупные, можно легко рассмотреть. Распадение на них гиомандибулярной ветви происходит у разных рыб на разном уровне. У щуки это деление происходит на уровне верхнего края жаберной крышки. У окуня и карпа это деление происходит на уровне средней части крышечной кости, но две передние ветви вначале одеты общей соединительнотканной оболочкой и расходятся только на уровне нижнего края крышечной кости.
Гиоидная ветвь (г. hyoideus VII, рис. 101, 103, 104, г. hy. VII) раополагается наиболее каудалыю то сравнению с другими ветвями. Она идет вниз почти строго по вертикали, достигает заднего края гиоидной кости, затем круто поворачивает вперед и проходит вдоль оснований лучей жаберной перепонки. Иннервирует внутреннюю поверхность предкрышки и некоторые мышцы нижней челюсти.
Внутренняя нижнечелюстная ветвь (г. mandibu-
laris internus, s. chorda tympani, рис. 101, 103, 104, r. md. i.) — наиболее краниально отходящая ветвь. Она обнажается только после удаления всех слоев m. adductor mandibulae. Первоначально проходит по наружной поверхности metapterygoideum, затем идет по внутренней поверхности квадратной кости и входит в нижнюю челюсть. Эта ветвь иннервирует выстилку нижией части ротовой полости.
Наружная нижнечелюстная ветвь (г. mandibularis externus, рис. 101, 104, г. md. е.) отходит между двумя предыдущими, идет сначала параллельно chorda tympani по передней поверхности подвеска, затем поворачивает вниз, проходит вдоль квадратной кости и входит в нижнюю челюсть с внутренней стороны. Этот нерв иннервирует нижнечелюстной и гиомандибуляр-ный каналы боковой линии.
Слуховой нерв (n. acusticus, рис. 104, VIII) оканчивается в слуховой капсуле и иннервирует внутреннее ухо; в связи с этим он короток и на препарате не виден.
Девятый и десятый нервы отходят вместе от продолговатого мозга, позади слухового нерва. Чтобы рассмотреть их ветви, необходимо сильно отвернуть вперед жаберную крышку так, чтобы обнажился верхний конец первой жаберной дуги. При этом приходится обрывать отводящую мышцу жаберной крышки (m. adductor operculi).
У окуня и карпа в связи с укорененным по сравнению со щукой жаберным аппаратом недостаточно только отвернуть жаберную крышку. Обычно приходится также отламывать пинцетом заднюю часть гиомандибуляре. При этом почти всегда удаляется и верхний отдел гиомандибулярной ветви VII нерва. Чтобы его сохранить, необходимо сначала проследить его ход вдоль внутренней стороны заднего края гиомандибуляре, а затем удалить последнюю, стараясь не повредить нерва.
Языкоглоточный нерв (n. glossopharyngeus, рис. 104. IX). Над верхним концом ряда жаберных лепестков первой жаберной дуги просвечивает под соединительнотканной пленкой темный кровеносный сосуд и лежащий от него каудальнее светлый нерв. Сняв пленку, обнажают языкоглоточный нерв и затем отпрепарируют его. Он идет вдоль основания первой жаберной дуги и фактически является зажаберной веткой (г. posttremticus) IX нерва.
У окуня и особенно карпа этот нерв лучше обнаружить, если на фиксированной рыбе удалить покрывающие ткани с латеральной поверхности первой жаберной дуги, тогда обнажается выносящая жаберная артерия и идущая рядом с ней ветвь IX нерва.
У карпа этот нерв, так же как и ветви следующего X нерва, развит очень сильно. Здесь особенно хорошо видна в отличие от щуки и окуня идущая вперед, к крыше ротовой полости, его небная ветвь (г. palatinus, IX, рис. 104, г. pal. IX).
Блуждающий нерв (n. vagus, X, рис. 104, X) дает многочисленные ветви, которые подходят к жабрам, внутренним ор
ганам и органам боковой линии. Снимают соединительнотканную выстилку дальше назад от IX пары нервов. Примерно под местом прикрепления к черепу мышцы, приводящей жаберную крышку, обнажается целый пучок ветвей X пары головных нервов. Первые три ветви подходят к жаберным дугам, которые и иннервируют. Каждая из этих трех жаберных ветвей (г. branchialis), приуроченная, как и у акулы, к определенной жаберной щели, делится на преджаберную и зажаберную ветви, точнее пред-щелевую и защелевую (г. ргае-, posttrematicus). Первая ветвь, обычно более тонкая, иннервирует жабру впереди, а вторая позади жаберной щели.
Разделение на два ствола наблюдается у первых двух жаберных ветвей примерно на половине их пути к жабрам. Преджабер-ная ветка (г, praetrematicus) третьего жаберного нерва отделяется очень рано и в виде самостоятельной ветви подходит к верхнему концу четвертой жаберной дуги, где резко, почти под прямым углом, поворачивает вперед и идет к третьей жабре. Зажа-берная ветвь (г. posttrematicus) третьего жаберного нерва тянется сначала вместе с другими ветвями X нерва в каудальном направлении, затем поворачивает налево и подходит к медиальной поверхности четвертой жабры рядом с выносящим жаберным сосудом.
Примерно параллельно зажаберной ветке (г. posttrematicus) третьей жаберной ветви тянутся еще две крупные ветви X нерва. Нижняя из этих двух — глоточная (г. pharyngeus.)—поворачивает за четвертой жаброй перед плечевым поясом вниз, а затем на уровне верхнего конца жабры — внутрь в глотку. Перед поворотом внутрь она отдает небольшую предщелевую веточку (г. praetrematicus) к четвертой жаберной дуге. Основная ветвь иннервирует область глотки, а отдельные ее веточки — также и сердце.
Верхняя кишечная ве-твь X нерва (г. intestinalis, рис. 104, г. int. X) идет в каудальном направлении примерно параллельно оси тела и скрывается под дорзальным концом плечевого пояса. Эта ветвь иннервирует внутренние органы.
Наконец, наиболее дорзально из всех ветвей проходит боковая ветвь X нерва (г. lateralis, рис. 104, г. lat. X). Она тянется сначала почти горизонтально, потом поворачивает немного вверх, подходит под задний край заднетеменной кости (posttem-porale) плечевого пояса и продолжается дальше к хвостовому отделу вдоль боковой поверхности тела, иннервируя боковую линию.
У окуня и карпа в связи с более короткой головой нервы не делают значительных изгибов, а каждый идет прямо к соответствующей жаберной дуге. У окуня нервы довольно тонки, имеют отчетливые ветви, и в них легко разобраться, следуя описанию, данному для щуки. Отличительной особенностью является то, что слабо выраженная зажаберная часть четвертой ветви практически не видна. Внутренностная ветвь отходит как бы от четвертой жа
берной ветви. Чуть выше нее прямо назад отходит боковая ветвь.
У карпа в связи с обработкой пищи в ротоглоточной полости каждая из жаберных ветвей X нерва очень мощная и над верхними концами жабр образует утолщение, от которого отходит целый пучок нервов. У этого вида хорошо выражена ветвь, идущая к пятой жаберной дуге, которая, как говорилось выше, несет глоточные зубы. Позади нее несколькими ветвями отходит внутренностная ветвь, а чуть сзади нее — боковая ветвь.
Головной мозг (рис. 105 и 106)
Изучение головного мозга и отхождения от него периферических нервов желательно проводить на экземплярах, предварительно выдержанных в течение 2—3 суток в 5%-ном водном растворе азотной кислоты для размягчения костей. Такая обработка не затрудняет изучения и мускулатуры. Однако в черепе щуки имеется довольно большое количество хряща, который легко удаляется и без предварительной декальцинации.
Знакомство с головным мозгом костистых рыб можно разделить на три этапа: изучение верхней поверхности головного мозга, изучение нижней поверхности головного мозга и отхождения головных нервов и изучение разрезов головного мозга.
Для того чтобы вскрыть черепную коробку, необходимо срезать ее крышу. Поэтому большим скальпелем делают несколько» тонких срезов верхней части головы, начиная от уровня заднего края глаза и затем назад до конца черепа. Позади черепа разрез можно провести глубже—так, чтобы срезать мускулатуру туловища вплоть до оснований верхних дуг позвонков. Рассмотрение головного мозга можно вести на тех же экземплярах, на которых, изучалась мускулатура.
У щуки крыша черепа между глазными яблоками хрящевая, поэтому после нескольких срезов можно вскрыть ее в этом месте пинцетом. Расширяют образовавшееся отверстие, постепенно выламывая пинцетом кусочки хряща и продвигаясь назад и в стороны. Примерно на уровне заднего края предкрышечпой кости на крыше черепа видна крестообразная фигура. Темным пигментом обозначен путь полукружных каналов в хрящах черепа. При вскрытии черепной коробки в этой области следует не повредить полукружных каналов и осторожно вычленить их верхние отделы из хряща. Сняв мускулатуру в передней части туловища по бокам позвоночника, срезают ножницами верхние части дуг позвонков и затем при помощи пинцета снимают их боковые части и обнажают спинной мозг. Продвигаясь вперед, вскрывают переднюю часть спинномозгового канала и далее заднюю стенку черепной, коробки. Во время последней операции необходимо оставить неповрежденными полукружные каналы.
У окуня и карпа вскрытие черепа на недекальцинированном материале несколько затруднительно из-за плотности костей кры-
fes.rh.
A
bul. о.
erst.
lob.o.
lab.l
- em.yr. er crb.
I
ch.o.
Ш. infd. -
о
Б — вид снизу, и F _neDeKpecT зрительных нер-bul о — обонятельные луковицы, полосатые тела; ет. gr. — бо
ши черепа. После срезания покровных костей видно пенистое вещество, которое закрывает мозг и является ничем иным, как мозговой оболочкой. Полукружные каналы лежат, как и у щуки, примерно на уровне предкрышечной кости, однако заметны хуже. Поэтому при препаровке лучше просто удалить их с одной стороны. Подрезав с одной стороны стенку черепной коробки на половину высоты, удаляют оболочки мозга. У щуки оболочка мозга довольно сильно пигментирована и богата кровеносными сосудами. У окуня и карпа все пространство между мозгом и внутренней частью черепной коробки заполнено пенистой массой, которую необходимо убрать, а мозг промыть под слабой струей воды.
Головной мозг сверху
Головной мозг костистых рыб состоит из типичных для большинства позвоночных пяти отделов, однако сверху видны только четыре. Рассматривают эти отделы в порядке их расположения, двигаясь спереди назад.
Передний мозг (telencephalon) у костистых рыб по сравнению с другими отделами головного мозга очень мал. Крыша переднего мозга щуки не содержит нервных клеток, а состоит из тонкой эпителиальной перепонки, которая обычно при препаровке снимается вместе с оболочкой мозга. Поэтому на препарате видно дно первого желудочка, разделенное глубокой продольной бороздой на два полосатых тела. Точно так же легко крыша переднего мозга снимается и у карпа, но у окуня тонкая крыша может сохраняться.
Полосатые тела (corpus striatum, cr. st.) состоят из двух отделов. Дорзальный отдел (epistriatum) более широкий и прикрывает вентральный отдел, который является прямым продолжением обонятельных луковиц. Деление на два отдела можно видеть при рассмотрении мозга сбоку. У карпа вентральные отделы увеличены и деление переднего мозга видно сверху (рис. 106).
Обонятельные луковицы (bulbus olfactorius, bul. о.) непосредственно прилегают к переднему краю переднего мозга. Они невелики и имеют продолговато-овальную форму. Вперед от них отходят обонятельные нервы. У карпа в отличие от щуки и окуня обонятельные луковицы (рис. 106, bul. о.) прилегают непосредственно к обонятельным капсулам. Идущие вперед обонятельные тракты (tractus olfactorius, рис. 106, tr. olf.) являются отделами мозга и несут в себе полости. Поскольку верхняя стенка их оснований перепончатая и ее легко удалить вместе с крышей переднего мозга, в этом месте они имеют вид желобков. Промежуточный мозг сверху не виден, так что непосредственно за передним мозгом лежит средний, нависающий сверху над ним.
Средний мозг (mesencephalon). Сверху видна только его дорзальная часть, имеющая вид двух крупных овальных, вытянутых по длине зрительных долей.
зрительные opticus s. tectum
Рис. 106. Головной мозг карпа Cyprinus carpio. А—вид сверху; but. о. — обонятельные луковицы; crb. — тело мозжечка; cr. st. — полосатые тела; ет. gr.— боковые выступы мозжечка; ер. — эпифиз; fos. rh. — ромбовидная ямка; I. ас. — слуховые доли; I. im.—медиальный бугор; lob. о. — зрительные доли; I. V. — вагальные доли; tor. sem. — полулунные тела; tr. o/f.—обонятельный тракт; v. сег.— мозжечковая заслонка
доли, или зрительная крыша (lobus opticum), — парные образования, отделенные друг от друга глубокой продольной бороздой. Зрительные доли являются зрительными центрами; в них оканчиваются волокна зрительного нерва. Дно полости среднего мозга носит название покрышки (tegmentum). В нем имеется сложная система ядер и проводящих путей, носящая название сетевидной формации (formatio reticularis), которая является важным центром координации нервной деятельности рыб. Ниже располагаются мощные продольные пучки волокон, которые связывают продолговатый мозг с промежуточным и называются н о ж к а м и мозга (pedunculus cerebri).
У карпа зрительные доли достигают значительного развития, а крыша среднего мозга (tectum opticum) хотя и состоит из мозгового вещества, но достаточно тонка и легко может быть повреждена при удалении мозговых оболочек. В этом случае хорошо видны вдающиеся в полость среднего мозга утолщения его нижнего отдела — полулунные тела.
Если вскрыть зрительные доли среднего мозга, то видно, что в полости зрительных долей от мозжечка отделяется складка мозжечка, носящая название мозжечковой заслонки (valvula cerebelli). Последняя особенно сильно развита у карпа (рис. 106, V. сег.). По бокам от нее в дне полости среднего мозга выделяются два бобовидных возвышения, называемые полулунными телами (torus semicircularis), которые служат важными центрами органов чувств боковой линии. Нижняя же поверхность крыши полости сред
него мозга образует возвышения, называемые продолговатыми утолщениями (torus longitudinalis). Они четко видны на тонкой крыше среднего мозга карпа.
Мозжечок (cerebellum) лежит непосредственно за зрительными долями и у щуки имеет округлую, а у окуня и карпа почти шарообразную форму. Задним краем он прикрывает продолговатый мозг. Выступающая вверх часть является телом мозжечка (corpus cerebelli, cr. crb.). Оттянув его в сторону, можно видеть два боковых выступа (eminentia granularis) на его вентральной части. Они отделены от тела неглубокой бороздой. Мозжечок служит центром точной регуляции всех моторных иннерваций, связанных с плаванием и схватыванием пищи.
Продолговатый мозг (myelencephalon) передним отде
лом заходит под мозжечок, а сзади без видимых границ переходит в спинной мозг. У щуки по бокам задней части продолговатого мозга, непосредственно к нему прилегая, лежат два полупрозрачных пузырька — части стато-акустического органа, содержащие отолиты. При дальнейшей (препаровке необходимо их сохранить.
Чтобы рассмотреть передний отдел продолговатого мозга у щуки, необходимо отвернуть вперед тело мозжечка. У окуня и карпа мозжечок невелик, и передний отдел продолговатого мозга хорошо виден. Крыша этого отдела мозга представлена эпендимой. Поэтому ее легко удалить, и на препарате открывается большая ромбовидная ямка (fossa rhomboidea), которая служит полостью четвертого желудочка. У щуки ромбовидная ям'ка расширена на переднем конце, а сзади переходит в узкую медиальную щель. У окуня она продолговатая.
В утолщенных краях ямки — вагальных долях (lobus va-galis, 1. v.) залегают ядра X пары черепномозговых нервов. У карпа вагальные доли развиты очень сильно (рис. 106), а между ними со дна четвертого желудочка поднимается медиальный бугор (lobus impar, рис. 106, I. im.), отчего передние концы вагальных долей сильно раздвинуты в стороны, а ромбовидная ямка имеет почти округлую форму. Впереди от вагальных долей по бокам от медиального бугра видны утолщения — слуховые доли (lobus acusticus, рис. 106, I. ас.), включающие в себя ядра акустического нерва. Вагальные доли у карпа продолжаются и назад на спинной мозг, называясь здесь задними долями (lobus posterior, рис. 106, I. р.).
Продолговатый мозг служит местом отхождения большинства головных нервов, а также проводящим путем, связывающим различные центры передних отделов головного мозга со спинным. На дне продолговатого мозга лежит дыхательный центр.
Рассмотрев головной мозг сверху, вычленяют его из полости черепной коробки. Для этого необходимо срезать или обломать пинцетом одну из боковых сторон черепной коробки вплоть до уровня вентральной поверхности головного мозга. При этом одновременно стараются отпрепарировать внутреннее ухо с системой полукружных каналов и отолитом. (Сохраняют его в сосуде со слабым раствором формалина.) Лучше всего эту операцию проводить под бинокулярным микроскопом или штативной лупой с достаточно большим увеличением. Проведя скальпелем под вентральной поверхностью мозга, перерезают отходящие от него нервы. Ту же операцию проделывают и со стороны оставшейся стенки черепной коробки. Спереди перерезают обонятельные нервы, а сзади — спинной мозг. Вынимают головной мозг из черепа и помещают его в сосуд с водой. Очищают вентральную поверхность мозга от покрывающей ее богатой кровеносными сосудами оболочки. Особенно осторожно следует снимать оболочку с боковых поверхностей продолговатого мозга, стараясь сохранить корешки отходящих здесь головных нервов. Рассматривают строение головного мозга с нижней стороны.
Головной мозг снизу
Первое, что бросается в глаза па нижней поверхности головного мозга,— это толстые зрительные нервы, которые образуют перекрест зрительных нервов (chiasma nervorum optico-rum, ch. о.), располагающийся па границе между дном переднего и промежуточного отделов головного мозга. Впереди переднего мозга видны прилегающие к нему обонятельные доли и идущие вперед от последних обонятельные нервы. За зрительным перекрестом видны отделы промежуточного мозга.
Непосредственно примыкая к заднему краю перекреста, лежит нижняя мозговая железа — гипофиз (hypophysis, hyp.). У щуки эта железа плотно входит в специальное углубление в дпе черепа и обычно при препаровке обрывается; тогда хорошо видна воронка (infundibulum, infd.). У окуня сохранить гипофиз при мозге значительно легче. По бокам и сзади от гипофиза располагаются относительно крупные овальной формы нижние доли (lobus inferior, lob. i.) промежуточного мозга. Между ними у окуня виден полупрозрачный сосудистый мешок (saccus vasculosus). У щуки он отсутствует.
Что касается остальных отделов, то их легко разобрать, сопоставляя с темп частями, которые видны сверху.
Рассматривая мозг снизу, можно различить и места отхождения некоторых нервов. От латеральной стороны передней части продолговатого мозга отходят три круглых корешка. Первый, лежащий наиболее краниальпо, принадлежит V и VII нервам, второй, средний корешок — только VII нерву и, наконец, третий корешок, лежащий каудалыю, является VIII нервом. За ними, также от боковой поверхности продолговатого мозга, несколькими корешками отходят вместе IX и X нервы. Остальные нервы тонки и обычно обрываются при препарировании.
Органы чувств
Сейсмосензорные органы. Эти органы были описаны при внешнем осмотре.
Орган обоняния. Представлен парными мешками, лежащими на верхней поверхности головы впереди глаз. Наружу каждый из мешков открывается двумя отверстиями ноздрей.
Чтобы познакомиться с внутренним строением органа обоняния, необходимо срезать верхнюю стенку, на которой располагаются ноздри, тогда откроется полость обонятельного мешка. Дно носовой полости выстлано складками, на которых у окупи, как и у большинства рыб, помещаются обонятельные клетки. У щуки обонятельные клетки, наоборот, лежат небольшими группами на дне обонятельной полости, между складками. Складки у щуки и у окуня располагаются по радиусам от центра мешка. Отделив нижиюю часть мешка от стенок и отвернув ее в сторону, можно видеть отходящий от органа обоняния обонятельный нерв.
Орган слуха, или стато-акустический орган. У рыб этот орган построен относительно сложно. Он соответствует внутреннему уху высших животных. Вычлененный при препаровке мозга орган слуха помещают в воду (если при препаровке он был сильно поврежден, то вычленяют этот орган с другой стороны головы). Располагают его в ванночке так, чтобы наружная сторона была сверху, и рассматривают отделы. Орган слуха состоит из двух частей: овального мешочка (utriculus), несущего полукружные каналы, и расположенного под ним круглого мешочка (sacculus). От овального мешочка отходят три полукружных канала (canalis semicircularis), которые лежат во взаимно перпендикулярных плоскостях: два из них вертикальные, один горизонтальный. Каждый из каналов несет на одном конце расширенную ампулу; причем ампулы переднего вертикального и горизонтального каналов лежат на их передних концах, задний вертикальный канал имеет расширение на заднем конце. Безампульные концы вертикальных каналов сливаются вместе. К этому общему каналу с латеральной стороны подходит и горизонтальный канал. При 6—10-кратном увеличении лупы в каждой ампуле можно различить похожие по форме на катушку от ниток места окончания ветвей слухового нерва, носящие название слуховых гребней (crista acustica). Под ампулами горизонтальных и вертикальных каналов находится расширение, в котором лежит один из трех отолитов (lapillus). На дне расширения видны скопления чувствующих клеток, называемые слуховыми пятнами (macula acustica). Под овальным мешочком (utriculus) лежит вторая часть перепончатого лабиринта — круглый мешочек (sacculus), обычно снабженный слепым мешкообразным выростом — у л и т к о й (laqena).
Почти весь круглый мешочек (sacculus) занят самым крупным из трех отолитов (saqetta). Третий отолит (asteriscus) располагается у щуки также .в каудальной части круглого мешочка; у окуня и других костистых он лежит в улитке. С медиальной стороны к круглому мешочку подходят веточки слухового нерва, которые оканчиваются на его оральном и каудальном краях.
Все части перепончатого лабиринта заполнены эндолимфой. Снаружи между стенкой лабиринта и стенкой полости, в которой он лежит, находится перелимфа.
Органы вкуса. Органы вкуса имеют строение чувствительных почек микроскопической величины, рассеянных не только в ротовой полости, но и по всему телу костистых рыб. Особенно они развиты у донных форм, располагаясь на внешней поверхности головы, брюхе и усиках. Чувствительные почки расположены в чувствительных ямках, выложенных длинными опорными клетками, между которыми лежат чувствительные клетки.
Органы зрения. Органы зрения представлены парными глаза-м и, расположенными на боковых поверхностях головы так, что рыба смотрит каждым глазом отдельно. Зрение играет большую роль при добывании пищи, особенно у хищных рыб (щука, окунь). У донных рыб, например сомов, глаза маленькие.
Глаз имеет шарообразную форму. Чтобы рассмотреть строение глаза, его падо вынуть из глазницы и острым скальпелем разрезать пополам. Глаз состоит из нескольких слоев. Наружный слой — склера (sclera) играет роль склетной капсулы. Снаружи она соединительнотканная, внутри хрящевая. Спереди склера становится прозрачной и называется роговицей (cornea). С внутренней стороны склера выстлана серебристой оболочкой (arqen-tea) —тонким слоем клеток, содержащих игловидные кристаллы гуанина, придающего им блеск. Следующий слой — сосудистый (chorioidea). В основании глаза, там где в глаз входит глазной стебелек — зрительный нерв, расположена характерная для глаз рыб сосудистая железа (qlandula chorioidea). На наружной стороне глаза сосудистая оболочка переходит в радужную оболочку (iris), которая окружает крупный шаровидный хрусталик (lens). С внутренней стороны глазного яблока к хрусталику прикрепляется серповидный отросток (processus falciformis), при сокращении которого хрусталик передвигается через зрачковое отверстие в глубь глаза, чем достигается наводка •на фокус (аккомодация). Зрение у рыб близорукое (около 1 м). Внутренний слой глазной стенки выстлан сетчаткой (retina), представляющей собой три ряда нервных клеток, разделенных двумя волокнистыми слоями. Самый наружный слой сетчатки представлен чувствующими клетками, воспринимающими световые раздражения. Чувствительные клетки двух родов, как и у всех других позвоночных, — палочки и колбочки. Своими чувствующими отростками палочки и колбочки направлены к наружной стороне сетчатки, одетой с наружной стороны тонким пигментным слоем мозгового происхождения, играющим роль экрана, отражающего свет.
Движение глазного яблока совершается типичным для позвоночных шестью глазными мышцами. Четыре из них прямые мыш-ц ы (m. rectus) — верхняя, нижняя, наружная и внутренняя —прикрепляются в основании черепа в длинном канале, называемом мио до мом. Две косые (m. obliquus) —верхняя и нижняя — прикреплены к боковой поверхности глазницы.
Литература по классу костных рыб (костистые рыбы)
Беркос П. А., Ингеницкий И. Окунь и щука. СПб. 1899.
Пучков И. В. Физиология рыб. Пищепромиздат, 1954.
Суворов Е. К. Основы ихтиологии. «Советская наука», 1948.
В г о n n Н. G. Dr. Н. G. Bronn’s Klassen und Ordnungen der Tierreichs. Bd. 6, Abt. I. Pisces (Fische), Lif. 16—20, 1905; Lif. 21—22, 1906.
• Grasse P. Traite de zoologie anatomle system, biologie. T. 13, Fasc. 1, 2, 3, .1958.
G r-o d z i n s k i Z. Anatomia i anbriologia Ryb. Panst Wydawn Polnicze i. Leske, Warszawa, 1961.
Panschin B. A. Die peripheren Nerven des Hechtes. Anat. Anz., Bd. 35, 1910.
Rauther M. Echte Fische, II Кар., Skelett. In «Bronn’s Klassen und Ordnungen der Tierreichs. Echte Fische», Teil I, 1929.
ОГЛАВЛЕНИЕ
Предисловие..........................................................
Введение ............................................................
Общие методические указания к ведению большого практикума............
Методика и техника приготовления анатомических препаратов............
Оборудование.........................................................
Техника вскрытия ....................................................
Подготовка материала.................................................
Приготовление скелетного материала....................................
Инъекция кровеносной системы.........................................
Литература по технике приготовления препаратов ......................
Тип хордовые — Chordata
Общая характеристика хордовых....................................
Система типа хордовых........................................, . .
Сравнительный метод исследования и типы сопоставления органов при сравнении .......................................................
Родословное древо хордовых ......................................
Литература по зоологии и сравнительной анатомии позвоночных . . . .
Подтип личиночно-хордовые, или оболочники, — Urochorda, s. Tunicata
Общая характеристика подтипа.....................................
Класс асцидии — Ascidiae.............................................
Общая характеристика класса......................................
Представители класса асцидий. Род Styela (Flemming, 1822), s. Go-niocarpa (Hunstman)..................................................
Внешний осмотр.......................................................
Вскрытие асцидий ....................................................
Строение личинки асцидии ............................................
Стр.
3
7
7
9
9
12
13
16
19
25
26
29
32
35
36
38
38
38
39
40
42
50
Подтип бесчерепные — Acrania
Класс головохордовые — Cephalochorda.................................51
Общая характеристика класса......................................51
Представитель класса — ланцетник (Branchiostoma lanceolatum Pallas, s. Amphioxus lanceolatus).........................................52
Внешний осмотр...................................................... 52
Вскрытие............................................................ 53
Изучение просветленных тотальных препаратов......................... 55
Изучение поперечных разрезов ланцетника............................ 59
Поперечный разрез ланцетника в жаберной области (59). Поперечный разрез ланцетника позади атриопора (64)
Литература по низшим хордовым..........................................65
Подтип позвоночные — Vertebrata
Раздел б ес ч е л ю с т и ы е — A g п a t h а Класс круглоротые — Cyclostomata....................................
Общая характеристика класса....................................
Представитель класса круглоротых — невская минога — Lampetra fluviati-lis L..............................................................
Вскрытие ...........................................................
Внешние признаки...............................................
Общая топография внутренних органов............................
Дыхательная система............................................
Органы пищеварения.............................................
Мочеполовая система............................................
Кровеносная система............................................
66 66
68
69
69
73
74
77
78
79
Сердце (81). Артериальная система (82). Венозная система (84) Поперечные разрезы......................................................85
Разрез туловищной области..........................................85
Разрез хвостовой области..........................................87
Разрез жаберной области..........................................87
Продольный разрез...............................................89
Мускулатура.........................................................91
Нервная система................................................... 93
Спинной мозг (94). Головной мозг сверху (95). Головные нервы (97). Головной мозг снизу (99)
Органы чувств......................................................100
Скелет..............................................................ЮЗ
Осевой скелет..................................................104
Скелет плавников...............................................105
Скелет ротовой воронки..........................................ЮЗ
Скелет языка....................................................Ю7
Осевой череп....................................................ЮЗ
Висцеральный скелет.......................................... НО
Литература по классу круглоротых и бесчелюстных....................111
Раздел челюст поротые — Cnathostomata
Надкласс рыбы — Pisces.............................................
Класс хрящевые рыбы — Chondrichthyes...............................
Общая характеристика класса....................................
Представители класса — колючая акула иокотиица, или катран (Squalus acanthias L.), и шиповатый скат (Raja clavata L.)..................
Внешний осмотр акулы...........................................
Внешний осмотр ската...........................................
Вскрытие.......................................................
Общая топография внутренних органов ...........................
Пищеварительная система . . . '................................
Жаберная область...............................................
Органы дыхания.................................................
Ротоглоточная полость .........................................
Кровеносная система............................................
112
112
112
113 ИЗ По 118 118 120 124 125 127 127
Артериальная система (131). Веиозиая система (136) Мочеполовая система самцов хрящевых рыб..........................140
Мочеполовая система самок хрящевых рыб...........................143
Скелет хрящевых рыб..................................................145
Общий обзор скелета акулы катраиа................................146
Осевой скелет — позвоночник акулы................................147
Скелет плавников (150). Скелет головы (154). Осевой череп — иейрокраниум (154) Висцеральный скелет..............................................158
Мускулатура хрящевых рыб..............................................ЮЗ
Мускулатура туловища..............................................ЮЗ
Мускулатура головы................................................Ю6
Нервная система хрящевых рыб..........................................Ю9
Кожные органы чувств боковой линии...............................170
Головной мозг хрящевых рыб.......................................172
Головной мозг ската сверху (172). Головной мозг ската снизу (175). Головной мозг акулы (177)
Головные нервы хрящевых рыб......................................178
Отхождение головных нервов (178). Разветвление головных нервов (179) Спиппой мозг и нервы..............................................184
Органы чувств хрящевых рыб............................................185
Органы зрения.................................................... 185
Стато-акустический орган (187). Орган обоняния (188) Литература по классу хрящевых рыб.....................................189
Надкласс рыбы — Pisces
Класс костные рыбы — Osteichthyes
Общая характеристика класса............................................190
Подкласс лучеперые — Actinopterygii........................................191
Надотряд хрящевые ганонды — Chondrostei....................................191
Характеристика представителя осетровых рыб — стерляди (Acipenser ruthenus L.).........................................................
Вскрытие.............................................................
Внешний осмотр.................................................
Общая топография внутренних органов..............................}9о
Пищеварительная система стерляди.................................198
Мочеполовая система..............................................
Ротоглоточная полость ...........................................
Кровеносная система..............................................
Артериальная система (209). Венозная система (212)
Нервная система...................... ...............................^1°
Общая топография нервной системы черепной полости (217). СИхождение от мозга головных нервов (218)
Головной мозг....................................................21®
Сравнительная характеристика головного мозга разных видов осетровых рыб (222)
Органы чувств....................................................223
Скелет...............................................................
Общий обзор скелета..............................................
Наружный, или костный, скелет....................................
Наружный скелет туловища (228). Наружный скелет головы — дермокраниум (230) Внутренний скелет................................................233
Осевой скелет туловища (234). Скелет непарных плавников (235). Скелет парных плавников (237)
Осевой хрящевой череп, или нейрокрапиум...........................239
Висцеральный скелет, или сплаихнокраниум..........................243
Мускулатура............................................................246
Литература по осетровым рыбам (хрящевые ганонды).......................249
Класс костные рыбы — Osteichthyes
Подкласс лучеперые — Actinopterygii....................................250
Надотряд костистые рыбы — Telcostei....................................250
Общая характеристика надотряда....................................250
Представители надотряда костистых рыб..................................250
Вскрытие...............................................................253
Внешние признаки..................................................253
Ссйсмосензориая система (органы чувств боковой линии).............263
Общая топография внутренних органов...............................266
Органы дыхания....................................................269
Пищеварительная система...........................................272
Мочеполовая система...............................................285
Половая система (285). Выделительная система (286)
Кровеносная система...............................................289
Сердце (290). Артериальная система (293). Венозная система (299)
Скелет.................................................................302
Общий обзор скелета...............................................303
Осевой скелет.....................................................304
Скелет плавников ................................................ 306
Скелет головы.....................................................310
Черепная коробка (310). Продольный разрез ненрокраниума (318). Осевой череп щуки (319). Висцеральный скелет (319)
Мускулатура............................................................325
Туловищная .мускулатура...........................................327
.Мускулатура плавников............................................329
Висцеральная мускулатура ........................................ 331
Глазные мышцы.....................................................333
Нервная система ................................................. 333
Головные нервы .................................................. 334
Головной мозг.....................................................341
Головной мозг сверху (343). Головной мозг снизу (346) Органы чувств............................................................346
Литература по классу костных рыб (костистые рыбы)........................348
И. Н. Гуртовой, Б. С. Матвеев, Ф. Я- Дзержинский
ПРАКТИЧЕСКАЯ
ЗООТОМИЯ
ПОЗВОНОЧНЫХ
Редактор В. С. Капышева. Художник А. Е. Коленков Художественный редактор Т. А. Коленкова. Технический редактор Э. М. Чижевский. Корректор С. К. Марченко
Сдано в набор 20/VI—75 г. Подп. к печати 9/1—76 г.
Формат 60Х90*/и Бум. тип. № 2 Объем 22 печ. л. Уел. п. л. 22 Уч.-изд. л. 25,49 Изд. № Е—263 Тираж 18.000 экз. Цеиа 1 р. 12 к. План выпуска литературы издательства «Высшая школа» (вузы и техникумы) иа 1976 г. Позиция № 82. Москва, К-51, Неглинная ул., д. Й/14, издательство «Высшая школа».
Московская типография № 8 Союзполиграфпрома при Государственном комитете Совета Министров СССР по делам издательств, полиграфии и книжной торговли. Хохловский пер., 7. Зак. 3467.
Н.Н,ГУРТОВОЙ, Е/С МАТВЕЕВ, Ф.Я, ДЗЕРЖИНСК ИЙ
ПРАКТИЧЕСКАЯ ЗООТОМИЯ ПОЗВОНОЧНЫХ