/







































































































































































































































































































































































































































































































































































































































































































Author: Адаскевич В.П. Козин В.М.
Tags: кожа наружные покровы тела дерматология кожные болезни венерология медицина инфекционные болезни
ISBN: 5-89677-076-6
Year: 2006




Text
В. П. Адаскевич
В. М. Козин
КОЖНЫЕ И ВЕНЕРИЧЕСКИЕ
БОЛЕЗНИ
Кожные
и венерические
болезни
Учебное руководство
В. П. Адаскевич
В. М. Козин
Москва
Медицинская литература
2006
УДК 616.5 + 616.97
ББК 55.81 + 55.83
А28
Все права защищены. Никакая часть данной книги не может быть воспроизведена в любой форме и любыми
средствами без письменного разрешения владельцев авторских прав.
Авторы, редакторы и издатели приложили все усилия, чтобы обеспечить точность приведенных в данной книге
показаний, побочных реакций и рекомендуемых доз лекарств. Однако эти сведения могут изменяться. Внима-
тельно изучайте сопроводительные инструкции изготовителя по применению лекарственных и технических средств.
Рецензенты.
Семенов В. М. — заведующий кафедрой инфекционных болезней Витебского государственного
медицинского университета, доктор медицинских наук, профессор.
Корсун В. Ф. — доктор медицинских наук, профессор Российского университета Дружбы народов.
Авторы:
Адаскевич Владимир Петрович — доктор медицинских наук, профессор, заведующий кафедрой
дерматовенерологии Витебского государственного медицинского университета. Член Европейской
Академии Дерматовенерологии и Международного Союза по инфекциям, передаваемым половым
путем. Автор свыше 200 научных и учебно-методических работ, в том числе книг «Заболевания,
передаваемые половым путем», «Функциональная морфология и общая патология кожи», «Венери-
ческие болезни и трепонематозы», «Венерические заболевания в судебно-медицинской практике»,
«Аллергические дерматозы», «Неотложные состояния в дерматологии», «Актуальная дерматология»,
«Акне и розацеа», «Актуальные вопросы венерологии», «Алопеция», «Дерматозы эозинофильные и
нейтрофильные», «Гомеопатия в клинической дерматологии», «Воспалительные кожные заболева-
ния», «Диагностические индексы в дерматологии», «Акне вульгарные и розовые», «Системные рети-
ноиды в дерматологии», «Морфофункциональная дерматология».
Козин Владимир Михайлович — доктор медицинских наук, профессор кафедры дерматовенероло-
гии Витебского государственного медицинского университета. Автор свыше 130 научных и учебно-
методических работ, в частности монографий «Псориаз», «Хламидийная инфекция», учебных посо-
бий «Дерматология», «Наружная фармакотерапия дерматозов». Основные научные работы посвящены
патогенезу, терапии псориаза, нейродермита, урогенитального хламидиоза и генитального герпеса.
Автор оригинального и эффективного способа бальнеотерапии ряда хронических дерматозов, вклю-
чающего применение препарата биологически активного «оксидата торфа».
Адаскевич В. П.
А28 Кожные и венерические болезни /В. П. Адаскевич, В. М. Козин. — М.:
Мед. лит., 2006.— 672 с.: ил.
ISBN 5-89677-076-6
В руководстве рассмотрены основные нозологические формы кожных болезней и инфекций, пере-
даваемых половым путем. В доступной форме представлены современные данные о патогенезе, кли-
нике, диагностике, терапии и профилактике кожных и венерических болезней. Руководство содер-
жит 79 цветных фотографий, на которых представлены классические проявления заболеваний
Для студентов высших медицинских учебных заведений, а также для врачей-дерматовенерологов;
может быть полезным для врачей смежных специальностей (аллергологов, косметологов, гинеколо-
гов, инфекционистов, терапевтов, онкологов).
Сайт издательства в Интернете: www.medlit.biz
ISBN 5-89677-076-6
ISBN 985-6333-15-9
УДК 616.5 + 616.97
ББК 55.81 + 55.83
© В. П. Адаскевич,
В. М. Козин, 2006
© изд. Б. И. Чернин,
изд. Ф. И. Плешков, 2006
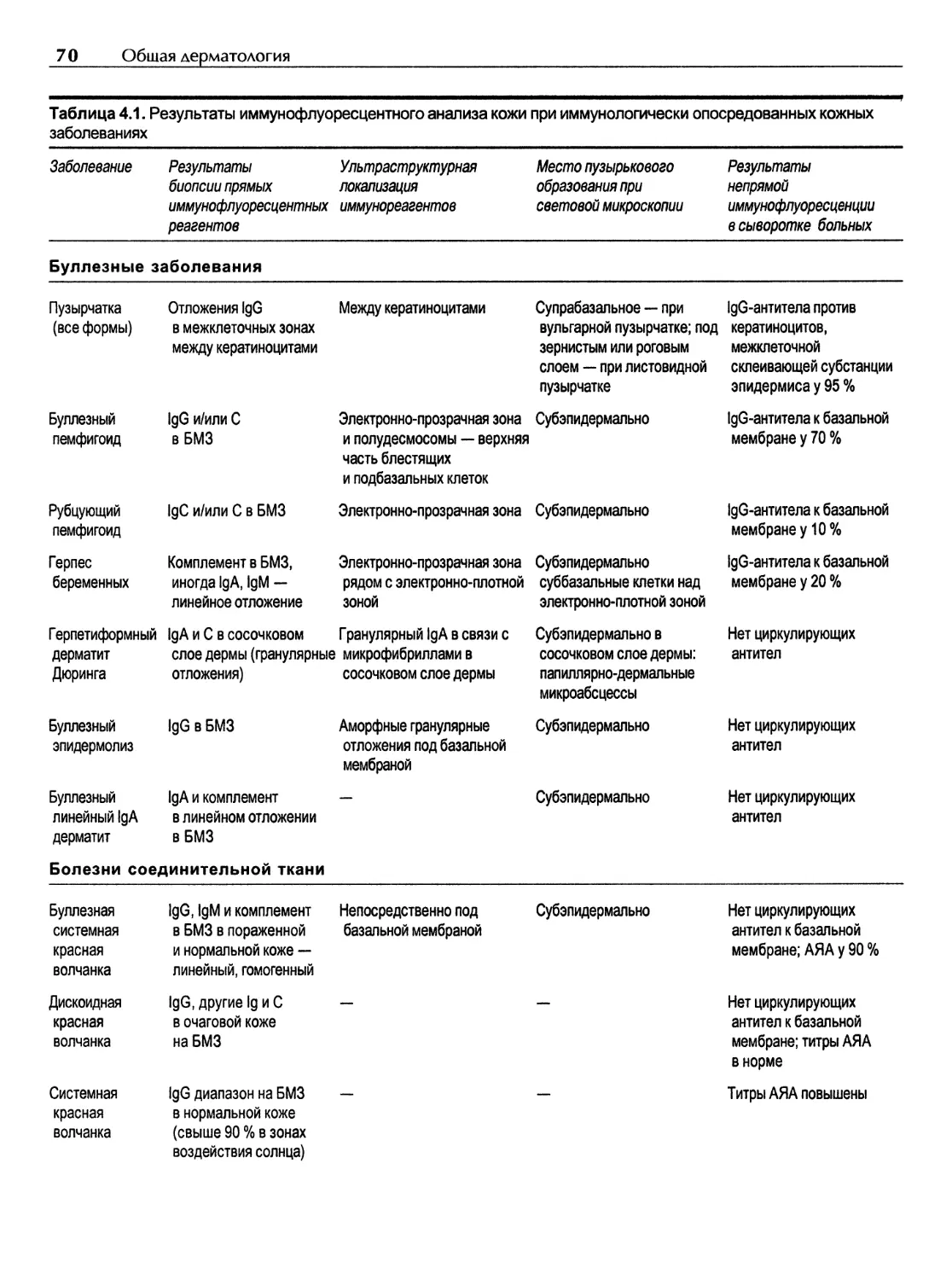
Содержание
ЧАСТЬ 1
ДЕРМАТОЛОГИЯ.......................................................1
Общая дерматология.................................................1
Глава 1
История развития дерматовенерологии.............................2
Глава 2
Анатомия и гистофизиология кожи.................................4
Состав и общий план строения системы кожных покровов...........4
Тонкое строение кожи..........................................10
Строение тонкой и толстой кожи................................47
Иннервация кожи...............................................48
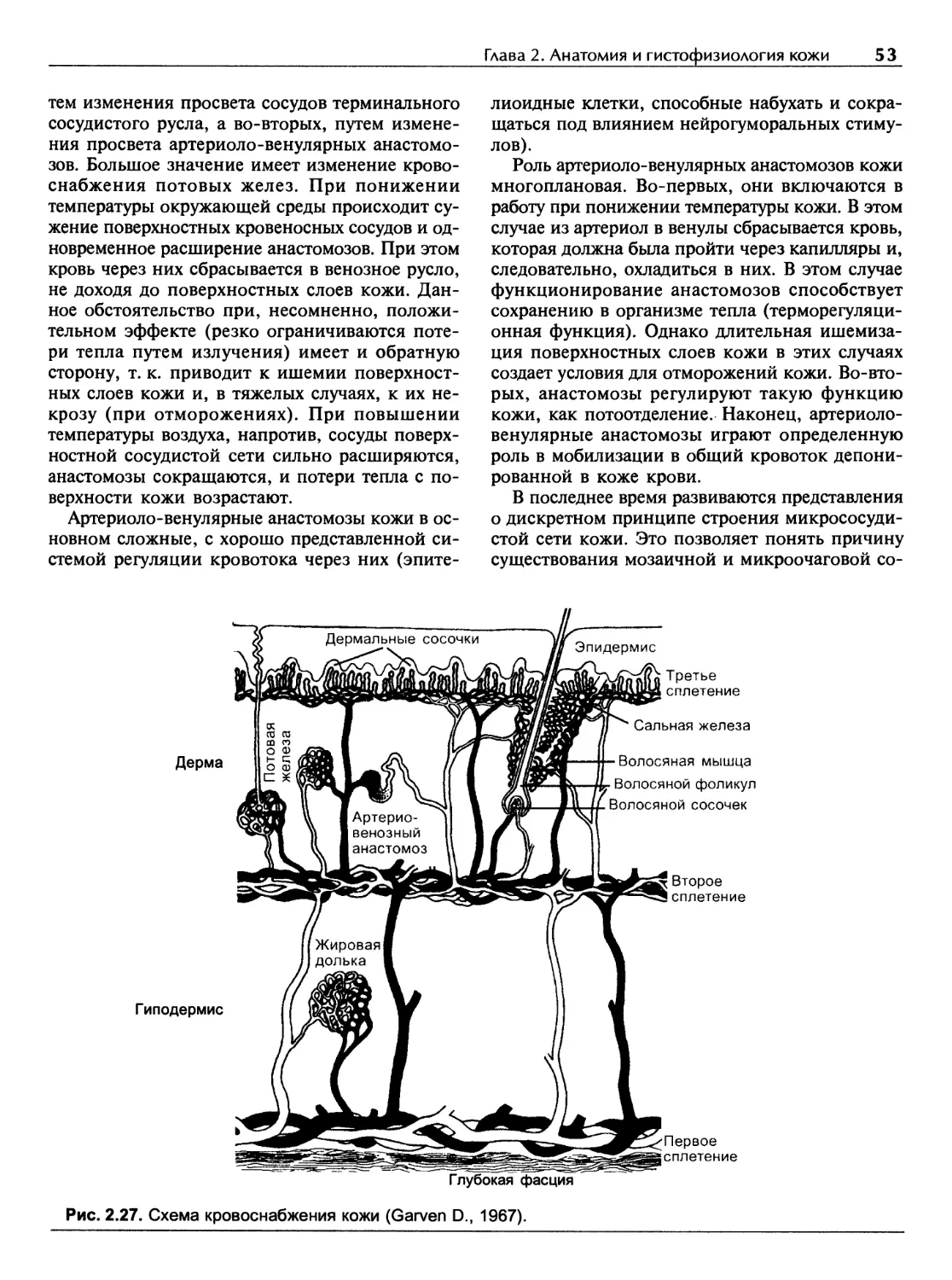
Кровоснабжение кожи...........................................50
Глава 3
Основные функции кожи...........................................55
Глава 4
Общепатологические процессы в коже..............................63
Глава 5
Этиология, патогенез и диагностика кожных болезней.............87
Этиология и патогенез кожных болезней.........................87
Общая симптоматология болезней кожи...........................89
Обследование кожного больного.................................98
Глава 6
Общие принципы лечения кожных болезней........................103
Препараты и методы физиотерапии, применяемые в дерматологии..104
Наружная фармакотерапия дерматозов...........................109
Вопросы деонтологии..........................................115
Гигиена кожи, ногтей, волос..................................116
iv Содержание
Частная дерматология................................................119
Глава 7
Пиодермии (гнойничковые болезни кожи)............................120
Стафилококковые пиодермии......................................121
Стрептококковые пиодермии......................................129
Стрептостафилодермии (смешанные пиодермии).....................132
Хроническая глубокая пиодермия................................ 133
Пиоаллергиды...................................................135
Глава 8
Туберкулез кожи..................................................137
Локализованные формы туберкулеза...............................138
Диссеминированные формы туберкулеза............................143
Глава 9
Лепра............................................................149
Глава 10
Кожный и кожно-слизистый лейшманиозы.............................161
Кожный лейшманиоз Старого Света................................161
Кожный лейшманиоз Нового Света.................................165
Глава 11
Вирусные болезни кожи............................................168
Простой герпес.................................................168
Опоясывающий герпес............................................173
Бородавки......................................................175
Остроконечные кондиломы........................................176
Контагиозный моллюск...........................................176
Узелки доильщиц................................................177
Глава 12
Паразитарные болезни кожи........................................179
Чесотка........................................................179
Педикулез......................................................185
Глава 13
Грибковые болезни кожи (дерматомикозы)...........................189
Разноцветный лишай.............................................190
Микоз крупных складок..........................................191
Микозы стоп....................................................192
Микоз волосистой части головы..................................193
Содержание
v
Кандидоз........................................................196
Онихомикозы.....................................................197
Глубокие микозы.................................................204
Глубокие псевдомикозы...........................................214
Глава 14
Дерматиты........................................................217
Простой ирритантный контактный дерматит.........................218
Аллергический контактный дерматит...............................219
Глава 15
Экзема...........................................................222
Острая экзема...................................................222
Подострая экзема................................................223
Хроническая экзема..............................................224
Экзема ладоней..................................................224
Сухая экзема....................................................225
Монетовидная экзема.............................................227
Дисгидротическая экзема.........................................227
Себорейная экзема...............................................229
Особенности возникновения и течения экземы у детей..............232
Глава 16
Профессиональные болезни кожи....................................235
Глава 17
Кожный зуд........................................................237
Глава 18
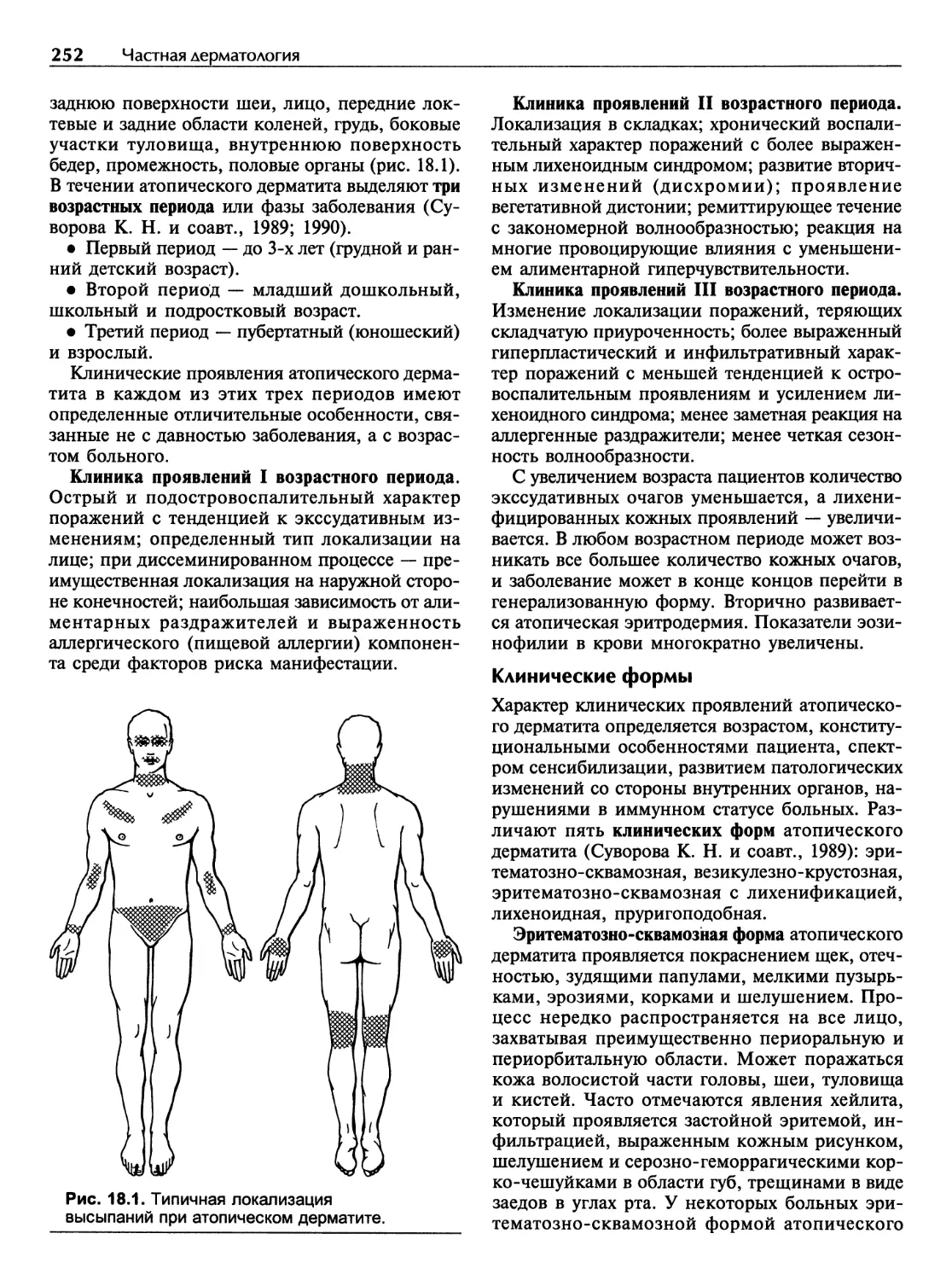
Атопический дерматит.............................................246
Глава 19
Почесуха (пруриго)...............................................264
Пруриго простое острое детское..................................264
Пруриго простое подострое.......................................265
Пруриго узловатое...............................................267
Глава 20
Нейродермит......................................................269
Лихенификация гигантская........................................270
Простой хронический бородавчатый лишай..........................270
Глава 21
Крапивница. Отек Квинке..........................................272
vi Содержание
Глава 22
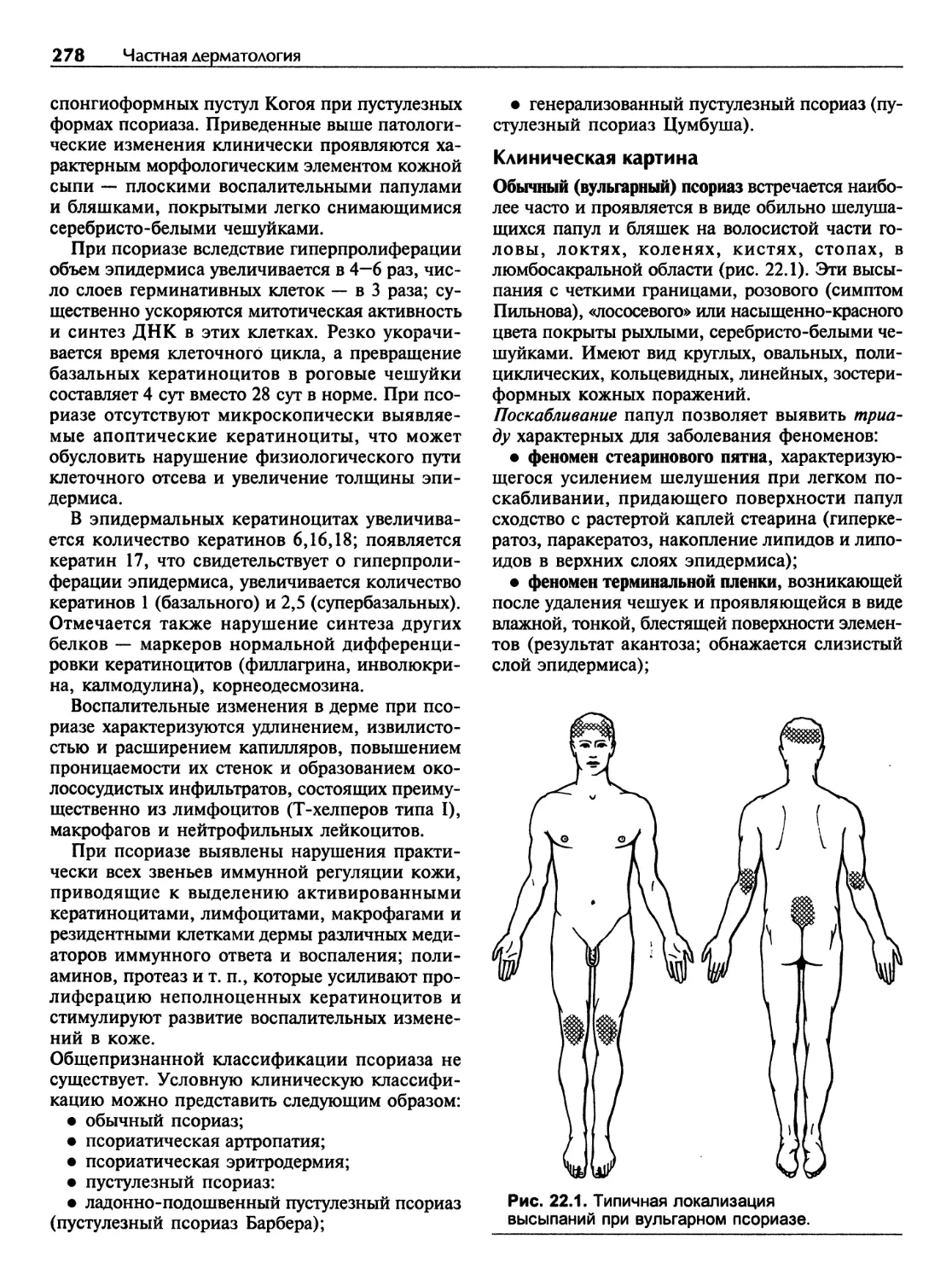
Псориаз..........................................................277
Глава 23
Красный плоский лишай............................................288
Глава 24
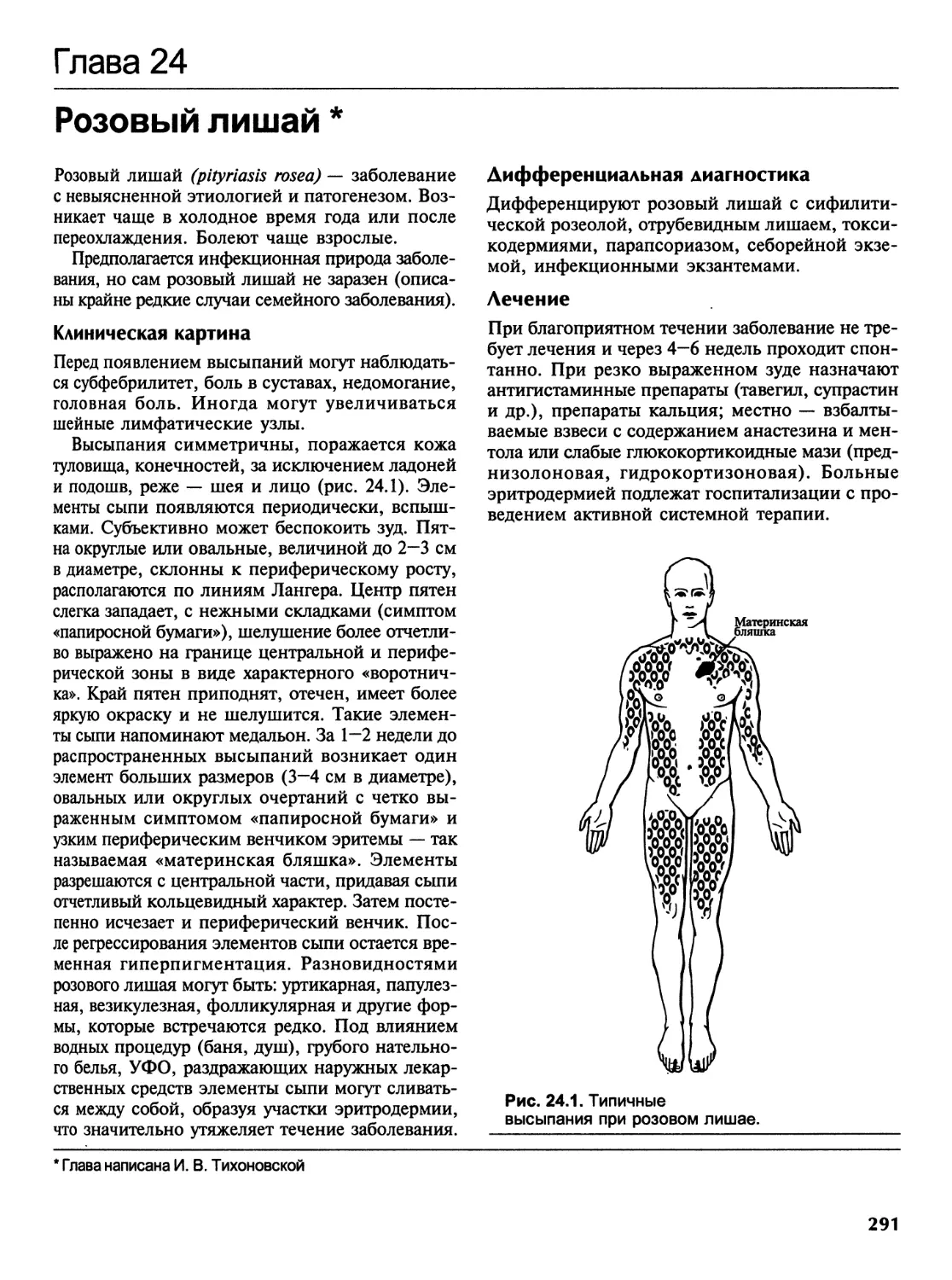
Розовый лишай....................................................291
Глава 25
Болезни с диффузным поражением соединительной ткани..............292
Красная волчанка................................................292
Системная красная волчанка......................................295
Очаговая склеродермия...........................................300
Системная склеродермия..........................................302
Дерматомиозит...................................................309
Глава 26
Многоформная экссудативная эритема...............................314
Глава 27
Клещевой боррелиоз (лайм-боррелиоз)..............................318
Глава 28
Пузырные дерматозы...............................................321
Вульгарная пузырчатка...........................................321
Вегетирующая пузырчатка.........................................325
Листовидная пузырчатка..........................................326
Бразильская пузырчатка..........................................327
Эритематозная пузырчатка........................................328
Паранеопластическая пузырчатка..................................329
Пузырчатка хроническая врожденная семейная......................330
Герпетиформный дерматоз Дюринга.................................331
Буллезный пемфигоид.............................................336
Рубцующийся пемфигоид...........................................338
Глава 29
Болезни придатков кожи...........................................341
Себорея.........................................................341
Акне (вульгарные угри)..........................................342
Розацеа.........................................................345
Гнездная алопеция...............................................351
Содержание vii
Глава 30
Васкулиты кожи..................................................355
Глава 31
Новообразования кожи............................................361
Доброкачественные новообразования кожи.........................362
Предраковые заболевания кожи и слизистых оболочек..............370
Злокачественные новообразования кожи...........................373
Глава 32
Особенности кожных проявлений при СПИДе.........................377
Глава 33
Лимфомы кожи....................................................379
Глава 34
Саркома Капоши..................................................385
Глава 35
Г енодерматозы..................................................390
Ихтиозы........................................................390
Буллезный эпидермолиз..........................................394
Кератодермии...................................................396
Глава 36
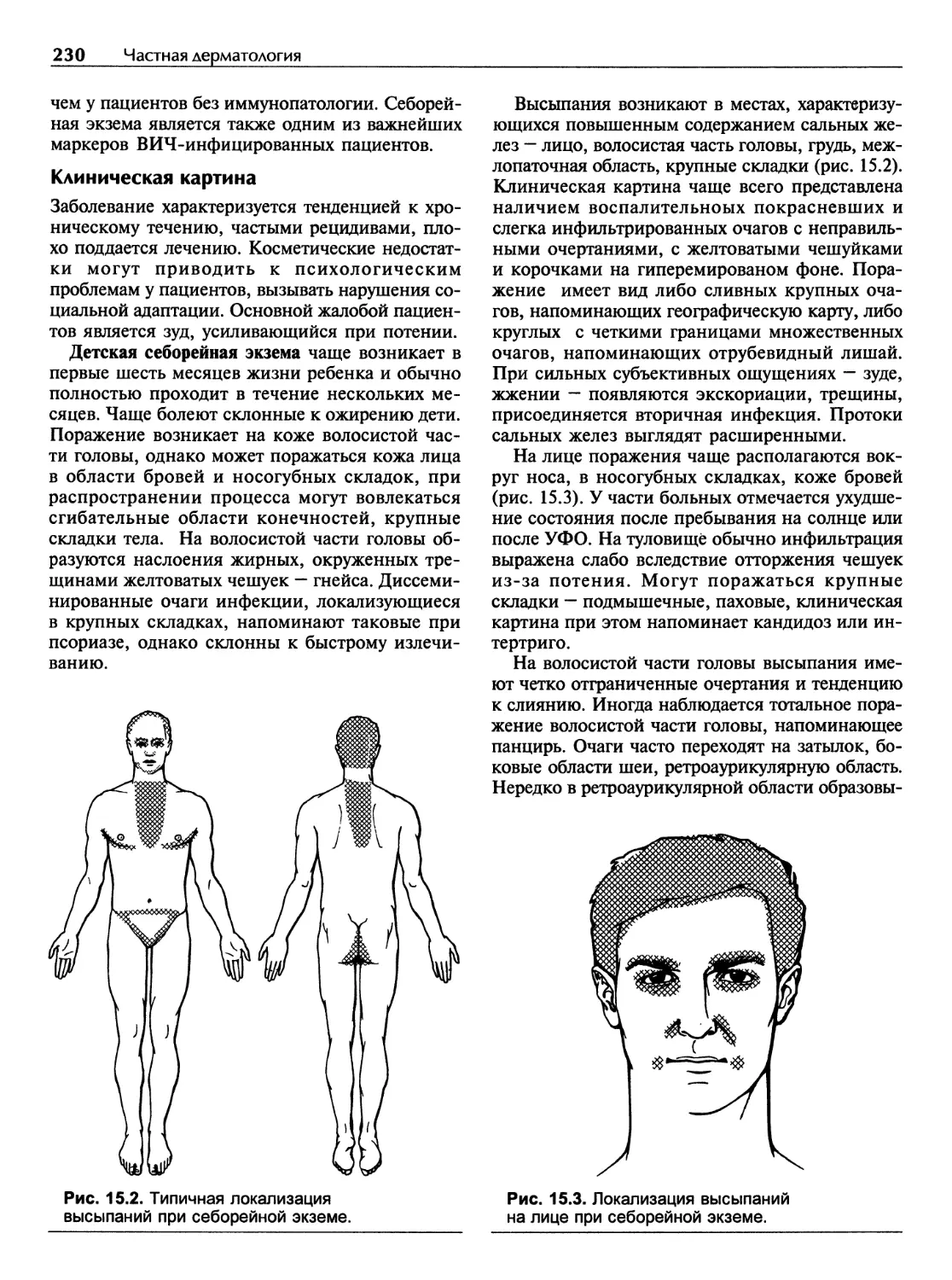
Кожные заболевания в жарких странах.............................398
Тропические пиодермии и смешанные бактериальные инфекции.......398
Тропические микозы.............................................405
Особенности течения специфических дерматозов...................408
Поражения кожи при протозойных болезнях........................409
Кожные поражения, вызываемые гельминтами.......................410
Поражения кожи, вызываемые членистоногими......................415
Поражения кожи, вызываемые ядовитыми змеями,
моллюсками, пиявками, актиниями...............................417
Особенности клиники и течения неинфекционных дерматозов........418
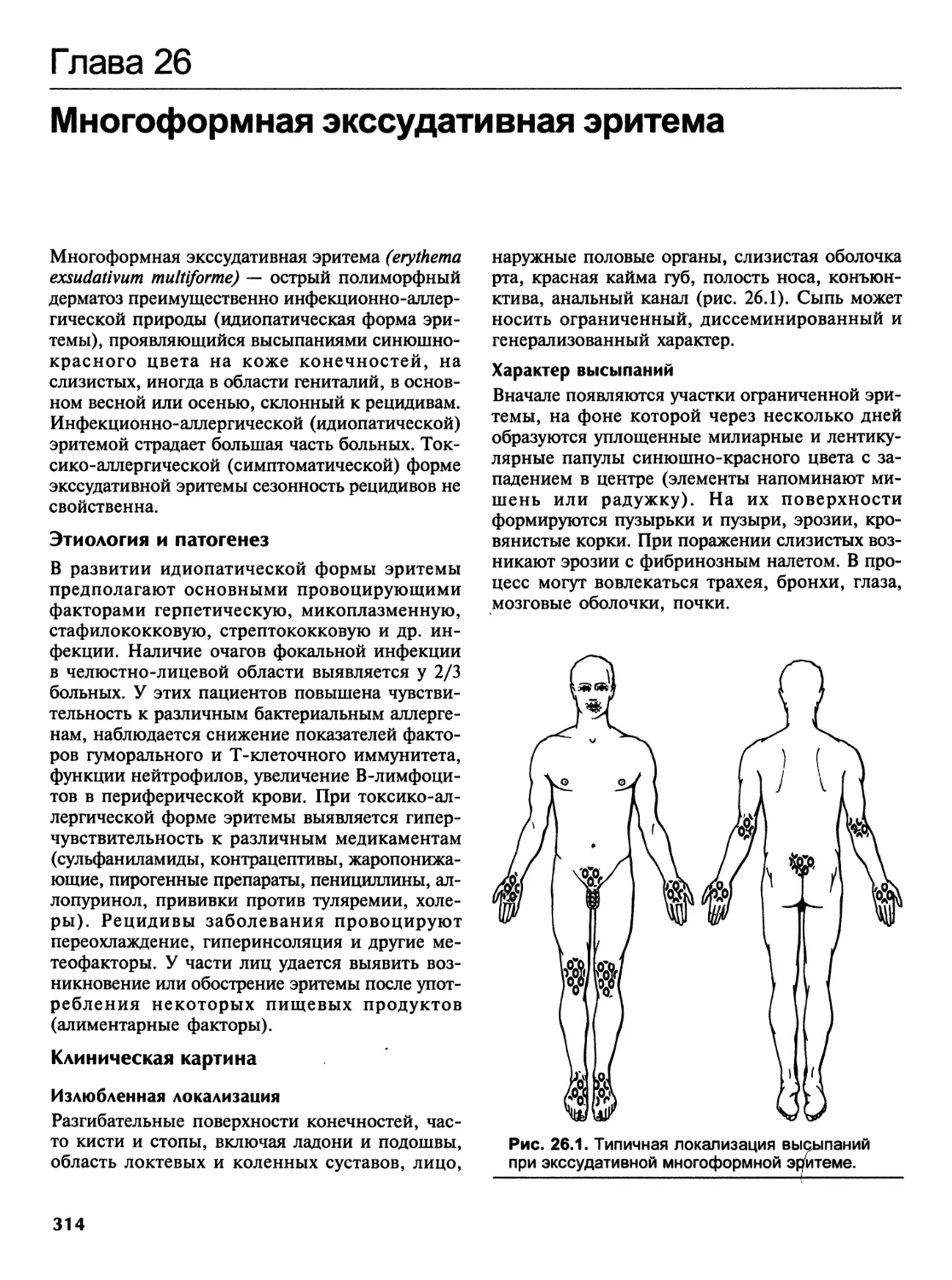
Венерические болезни и эндемические трепонематозы..............419
ЧАСТЬ 2
ВЕНЕРОЛОГИЯ........................................................425
Г лава 37
Основные этапы развития венерологии.............................426
viii Содержание
Глава 38
Сифилис........................................................430
Этиология.....................................................430
Формы сифилиса.............................................. 431
Эпидемиология и общая патология сифилиса......................432
Лабораторная диагностика сифилиса.............................453
Лечение сифилиса..............................................459
Профилактика сифилиса.........................................469
Глава 39
Гонорея........................................................473
Этиология.....................................................473
Классификация.................................................473
Клиника гонореи у мужчин......................................474
Клиника гонореи у женщин......................................477
Клиника гонореи у девочек.....................................479
Диагноз.....................................;.................480
Лечение.......................................................481
Профилактика гонореи у детей..................................496
Глава 40
Шанкроид.......................................................497
Глава 41
Венерический лимфогранулематоз.................................501
Глава 42
Донованоз......................................................506
Глава 43
Трихомониаз....................................................510
Глава 44
Урогенитальный хламидиоз.......................................516
Глава 45
Урогенитальный микоплазмоз.....................................528
Глава 46
Урогенитальный кандидоз........................................533
Глава 47
Генитальный герпес.............................................541
Содержание ix
Глава 48
Вульвовагиниты у девочек различного возраста.............550
Глава 49
ВИЧ/СПИД-инфекция........................................556
ПРИЛОЖЕНИЯ.................................................573
Приложение А.............................................574
Перечень и описание практических навыков по кожным
и венерическим болезням..................................574
Приложение В.............................................585
Ситуационные задачи по дерматовенерологии................585
Приложение С.............................................601
Эталоны ответов к ситуационным задачам
по дерматовенерологии для студентов лечебного факультета.601
Приложение D.............................................609
Тесты по дерматовенерологии..............................609
Приложение Е.............................................647
Ответы к тестам..........................................647
Алфавитный указатель.....................................650
Авторы
Профессор
Адаскевич Владимир Петрович
Профессор
Козин Владимир Михайлович
Введение
Посвящается нашему учителю заслуженному деятелю науки,
лауреату Государственной премии БССР профессору Леониду
Ивановичу Богдановичу
Дерматология и венерология не являются узки-
ми специальностями. Напротив, знание основ
этих дисциплин необходимо врачам любой спе-
циальности, их преподавание обязательно для
подготовки врача общего профиля. В общей
врачебной практике в среднем до 30 % всех па-
циентов страдают заболеваниями кожи. Часто тя-
желые, угрожающие жизни заболевания начина-
ются с внешне безобидных изменений кожи, их
дифференциальный диагноз в это время затруд-
нен, а своевременная и правильно проведенная
терапия имеет решающее значение для судьбы
пациента. Другие распространенные поражения
кожи не представляют угрозы для жизни, хотя
существенно снижают ее качество. И здесь пра-
вильный диагноз поставить сложно, эти кожные
поражения необходимо отличить от банальных
высыпаний при приеме лекарств или от других
видов кожной аллергии. Поэтому следует хоро-
шо знать симптоматику наиболее часто встреча-
ющихся кожных болезней.
Венерические заболевания считаются наибо-
лее распространенными инфекционными забо-
леваниями. В последние годы число инфекций,
передаваемых половым путем, возросло, а все ве-
нерические заболевания успешно лечатся толь-
ко при правильной и своевременной постанов-
ке диагноза.
Все вышеизложенное имеет целью подчерк-
нуть необходимость интенсивной подготовки по
курсу «Дерматовенерология». В рамках цикла
обучения на данную специальность отводится
всего 94 часа. Для достаточной подготовки это-
го времени слишком мало. Тем более поэтому
важно, чтобы будущий врач добросовестно изу-
чил, по меньшей мере, основы дерматологии и
венерологии.
В написании учебного пособия принимали уча-
стие профессор О. Д. Мяделец, доцент, к. м. н.
И. В. Тихоновская, к. м. н. О. С. Зыкова, к. м. н.
В. В. Саларев, В. В. Козловская.
Выражаем благодарность доктору Г. В. Драгу-
ну за любезно предоставленные иллюстративные
материалы.
Выражаем искреннюю благодарность профес-
сору В. Ф. Корсуну и профессору В. М. Семё-
нову за труд по рецензированию данной работы
и высказанные замечания.
Все замечания, предложения по данному по-
собию авторами будут приняты с признательно-
стью и пониманием, а в дальнейшем учтены
при новом издании.
X
Часть 1
Дерматология
Общая дерматология
Глава 1
История развития дерматовенерологии
В истории развития медицины, начиная с ран-
них ее этапов и по настоящее время, кожные
болезни всегда занимали значительное место.
Важные общемедицинские положения и прин-
ципы разрабатывались на примере хорошо изу-
ченных кожных заболеваний. Многочисленные
упоминания о кожных болезнях можно найти в
богословских сочинениях, произведениях лите-
ратуры и искусства с древнейших времен до на-
ших дней.
Первые описания многочисленных кожных
болезней были даны греческими и римскими
врачами. Врачами были греческие философы
Гиппократ, Платон, Аристотель. И сегодня мы
применяем термины древнегреческой медицины,
такие как экзантема, pityriasis, lichen.
Из эпохи Древнего Рима следует, прежде все-
го, упомянуть энциклопедиста Цельсия и Гале-
на, врача гладиаторов и придворного врача им-
ператора. Из применяемых и в настоящее время
терминов римской медицины хорошо известны
erysipel, фурункул, гангрена, карцинома, пусту-
ла, scabies, витилиго.
В средние века (500—1500 гг. н.э.) традиции
греческой и римской медицины развивали пре-
имущественно арабские ученые (Авиценна). Ба-
зой средневековой медицины служили работы
Галена, а также философские теории гумораль-
ных патологий (учение Кразена).
С 1500 года начинается переворот в истории
медицины и кожных болезней. Отсчет медици-
ны нового времени ведется с работ Парацельса
(1493—1541). Первые книги по дерматологии
были написаны на латыни Джироламо Мерку-
риале (1530—1606) и изданы в 1572 году в Ве-
неции, а затем в Тюбингене в 1630 году Самуэ-
лем Хафенреффером (1587-1660).
Развитие естественно-научной медицины от-
крыло новые подходы и к пониманию природы
кожных болезней. Три важных и до сегодняш-
него дня способа анализа и классификации кож-
ных заболеваний предложили:
• Иозеф Якоб Пленк (1735—1807): класси-
фикация кожных симптомов (учение о высы-
паниях) — первая попытка нозологической
классификации заболеваний на основании их
морфологии;
• Фердинанд фон Гебра (1816—1880): класси-
фикация кожных заболеваний на патолого-ана-
томической основе;
• Жан Дарье (1856-1938): классификация
кожных заболеваний по их известной или пред-
полагаемой этиологии с учетом сложных взаи-
мосвязей между морфологией кожных заболева-
ний и их патогенетическими механизмами.
Первый дерматовенерологический журнал под
названием «Сифилидология» издавался в Лейп-
циге (1839-1845). В Париже с 1844 по 1852 г.г.
по заболеваниям кожи выходил журнал, кото-
рый основал А. Казенав.
Учреждение самостоятельных дерматологичес-
ких клиник и кафедр началось в середине XIX
века. Первые кафедры кожных болезней в Рос-
сии были организованы в 1869 году в Москов-
ском университете (в настоящее время Москов-
ская медицинская академия им. И. М. Сеченова),
Санкт-Петербургской медико-хирургической
академии (ныне Российская Военно-медицинская
академия), медицинском факультете Варшавского
университета.
Во второй половине XIX века в разных стра-
нах мира были организованы общества дермато-
венерологов: Американская дерматологическая
ассоциация (1876), Русское дерматологическое
общество (Санкт-Петербург, 9 августа 1885 г.),
Общество итальянских дерматологов и сифили-
дологов (1885), Немецкое дерматологическое
общество (1888), Общество французских дерма-
тологов и сифилидологов (1889). В Париже в
1889 году состоялся первый международный
конгресс дерматовенерологов.
Московская дерматологическая школа сфор-
мировалась на базе медицинского факультета
Московского университета. В 1895 году была по-
строена первая в России клиника кожных и ве-
нерических болезней. Выдающуюся роль в оте-
чественной дерматологии сыграл А. И. Поспелов
(1846—1916), который организовал Московское
общество дерматовенерологов (носит сейчас его
имя) и первый в стране музей муляжей.
2
Глава 1. История развития дерматовенерологии
3
Петербургская дерматологическая школа созда-
валась первым русским профессором дерматоло-
гии А. Г. Полотебновым (1838—1907) и основопо-
ложником русской венерологии В. М. Тарковским
(1837-1906), которые одновременно заведовали
кафедрами в Санкт-Петербургской медико-хи-
рургической академии.
Киевскую школу дерматовенерологов пред-
ставляет М. И. Стуковенков (1843—1897) —
ученик А. Г. Полотебнова. Одним из выдаю-
щихся представителей отечественной дермато-
логической школы является П. В. Никольский
(1858-1940), который вошел в мировую дерма-
тологию с описанным им симптомом при пу-
зырчатке.
Белорусскую школу дерматовенерологов мно-
гие годы возглавлял А. Я. Прокопчук (1896—
1970), который предложил метод лечения боль-
ных красной волчанкой акрихином.
Изменения в условиях жизни людей, а также
изменения общественно-экономических форма-
ций, наряду с развитием диагностических и те-
рапевтических возможностей медицины, преоб-
разили панораму кожных заболеваний.
Дерматология в последние годы переживает
огромный подъем благодаря прогрессу в диагно-
стике (молекулярно-генетические исследования),
разработке новых медикаментов (ретиноиды,
иммуносупрессоры, интерферон), а также в те-
рапии ультрафиолетовыми лучами.
Глава 2
Анатомия и гистофизиология кожи *
Состав и общий план строения системы кожных покровов 4
Тонкое строение кожи 10
Строение тонкой и толстой кожи 47
Иннервация кожи 48
Кровоснабжение кожи 50
Состав и общий план
строения системы
кожных покровов
К системе кожных покровов относятся:
1. Кожа как орган.
2. Производные кожи, имеющие, как прави-
ло, органное строение: железы (потовые, саль-
ные, молочные), волосы и ногти. Кожа как орган
является одним из самых крупных органов че-
ловеческого организма, имея общую площадь
около 2 м2 и массу, составляющую около 20 %
массы тела (без учета массы подкожно-жировой
клетчатки, вместе с которой она может состав-
лять до 50 и более процентов от общей массы
тела).
Кожа как орган представляет собой трехкомпо-
нентную систему, образованную эпидермисом,
дермой и подкожно-жировой клетчаткой, кото-
рые находятся в морфофункциональном един-
стве (рис. 2.1). Таким образом, в коже имеются
эпителиальная и соединительнотканная части.
Эпидермис
Эпидермис является многослойным плоским
ороговевающим эпителием, имеет слоистое
строение и обладает высокими резистентными
и регенераторными свойствами. Это наиболее
сложно устроенный вид эпителия, полидиффе-
ронная ткань, имеющая в качестве основного
клеточного дифферона кератиноциты, произ-
водные кожной эктодермы. Помимо них, в
эпидермисе в качестве дополнительных клеточ-
ных дифферонов с определенной для каждого
вида степенью постоянства находятся клетки
* Глава написана проф. О. Д. Мядельцем
нейроэктодермального (меланоциты, клетки
Меркеля) и мезенхимного (клетки Лангерган-
са, клетки Гринстейна, внутриэпидермальные
лимфоциты) происхождения.
Эпидермис расположен на базальной мембране,
которая служит для него эластической опорой,
прочно связывает эпителий с коллагеновыми во-
локнами дермы и препятствует росту эпителия
вглубь дермы, как это бывает при раковых опу-
холях, когда базальная мембрана разрушается. В
настоящее время считается, что компоненты базаль-
ной мембраны синтезируются как эпителиальны-
ми, так и соединительнотканными клетками.
В эпидермисе более или менее постоянно про-
исходят несколько процессов:
• деление клеток в глубоком слое, выталки-
вание в результате этого части клеток из глубо-
ких слоев по направлению к поверхности;
• превращение клеток, наиболее удаленных от
дермы, в роговое вещество;
• слущивание рогового вещества с поверхно-
сти.
Поскольку при продвижении клеток к поверх-
ности они претерпевают изменения, создается
впечатление, что эпидермис образован несколь-
кими слоями (стратификация эпителия). Эти слои
следующие:
1. Базальный слой 1 Ростковый
2. Слой шиповатых клеток J или мальпигиев слой
3. Зернистый слой
4. Блестящий слой
5. Роговой слой
В последнее время установлено, что эпидермис
млекопитающих организован в структурно-фун-
кциональные единицы, которые называются эпи-
дермальными пролиферативными единицами
4
Глава 2. Анатомия и гистофизиология кожи
5
(ЭПЕ). Они имеют форму вертикальных коло-
нок. В состав колонки входят 6—12 полностью
ороговевших клеток (роговых чешуек), две зер-
нистые, одна шиповатая и 8—12 базальных кле-
ток. Роговые чешуйки имеют гексагональные
очертания. Каждой чешуйке, находящейся на
поверхности, соответствует своя ЭПЕ. ЭПЕ мо-
гут иметь столбчатую и нестолбчатую структуру
в зависимости от функционального состояния
эпидермиса. Организация эпидермиса в эпидер-
мальные пролиферативные единицы позволяет на
оптимальном уровне осуществлять его тканевой
гомеостаз и достаточно успешно адаптироваться
к измененным условиям существования. В ре-
зультате в процессе функционирования и адап-
тации к внешним условиям может наблюдаться
переход ЭПЕ от столбчатой к нестолбчатой фор-
ме организации и наоборот.
Основной функцией эпидермиса является ба-
рьерно-защитная функция, реализуемая прежде
всего достаточно мощным роговым слоем. По-
этому процессы кератинизации, происходящие
в этом эпителии, имеют очень важное значение
и жестко регулируются. Их нарушение лежит в
основе многих заболеваний кожи. Кроме рого-
вого слоя, в защитных реакциях эпидермиса уча-
ствуют присутствующие в нем клетки мезенхим-
ного происхождения.
Дерма
Вторым структурным компонентом кожи явля-
ется дерма. Она придает коже прочность, обес-
печивает питание и дыхание эпидермиса. Явля-
ясь проводником кровеносных сосудов и нервов,
дерма принимает участие в выполнении всех
функций кожи. В дерме различают два слоя:
подэпидермальный, или сосочковый, образован-
ный рыхлой волокнистой соединительной тка-
нью, и глубокий, или сетчатый, состоящий из
плотной неоформленной волокнистой соедини-
Роговой
Мышца,
поднимающая
волос
слой
Мальпигиев
слой
Базальная
Дермальный
сосочек
Тельца
Фатера-Пачини
Рис. 2.1. Строение кожи.
6
Общая дерматология
тельной ткани. Последний без резких границ
переходит в подкожно-жировую клетчатку (ги-
подерму), связывающую кожу с более глубоко
лежащими тканями.
В соединительнотканной части кожи распола-
гаются железы, развивающиеся из эпидермиса,
т. е. имеющие эктодермальное происхождение. В
коже имеются три разновидности желез: саль-
ные, потовые и молочные.
Сальные железы
Это простые альвеолярные железы с разветвлен-
ными концевыми отделами (рис. 2.2). Они по-
чти всегда расположены по соседству с волоса-
ми. Их выводной проток обычно впадает в
волосяную воронку. Один волос имеет одну или
две сальные железы. Размеры сальных желез не
всегда соответствуют величине волос, и иногда
небольшие по размеру волосы сопровождаются
крупными сальными железами. В ряде случаев
протоки крупных сальных желез открываются
непосредственно на поверхности кожи. ЭГо так
называемые свободные сальные железы. Саль-
ные железы функционируют по голокриновому
типу, т. е. при образовании секрета клетки же-
лезы постепенно накапливают липидные капли,
дегенерируют и затем полностью разрушаются,
Рис. 2.2. Сальная железа. * 200.
1 — разветвленный альвеолярный концевой
отдел; 2 — выводной проток.
а их остатки входят в состав секрета (кожного
сала). Кожное сало, выделяясь на поверхность
эпидермиса, смягчает его роговой слой, делает
его более эластичным, а также выполняет тер-
моизолирующую функцию. С секретом кожных
желез на поверхность кожи выделяется лизоцим
(мурамидаза), разрушающий бактериальную
стенку и обладающий в связи с этим бактери-
цидным действием. Компоненты кожного сала
участвуют в формировании так называемой кис-
лотной мантии Маркионини, имеющей pH 5,5 и
губительно действующей на бактериальные клет-
ки.
Потовые железы
Это простые трубчатые железы. Их концевые от-
делы также залегают в дерме (глубже сальных
желез, на границе дермы и гиподермы) в виде
длинных слепых трубок, завитых в клубочек
(рис. 2.3). Основу стенки концевых отделов со-
ставляет однослойный кубический эпителий,
снаружи от которого располагаются миоэпите-
лиальные клетки. Секреторный отдел потовой
железы постепенно переходит в выводной про-
ток, выстланный двух- и трехслойным эпители-
ем. Проток проходит через оба слоя дермы
(трансдермальная часть) и открывается или в во-
лосяное влагалище, выше устья сальных желез,
или, чаще, свободно на поверхность кожи пото-
вой порой. В толстой коже она располагается на
поверхности эпидермального гребешка. При этом
проток спиралевидно проходит через эпидермис,
уплощенные клетки которого формируют трансэ-
пидермальную часть протока. По типу секреции
Рис. 2.3. Строение апокриновых потовых желез,
а. Общий вид апокриновой потовой железы. * 60.
б. Мелкозернистые клетки железы с апокрино-
вым типом секреции. * 120
(Апатенко А. К., 1973).
Глава 2. Анатомия и гистофизиология кожи
7
потовые железы разделяются на мерокриновые
(эккриновые) и апокриновые. Мерокриновые же-
лезы являются преобладающими потовыми же-
лезами у человека и других приматов (в отли-
чие от других млекопитающих, у которых резко
преобладают апокриновые железы). Эти железы
находятся на большей части поверхности кожи
(особенно их много на ладонях и подошвах) и
секретируют без признаков разрушения секре-
торных клеток. Их выводные протоки всегда сво-
бодно открываются на поверхность кожи. Апок-
риновые потовые железы выделяют секрет при
разрушении апикальной части судорифероцитов
(общее название секреторных клеток потовых
желез). Они расположены только в отдельных
частях кожного покрова человека: в области ге-
ниталий, вокруг анального отверстия, на лбу, в
подмышечных впадинах и в некоторых других
участках кожного покрова. Эти железы имеют
размеры, превышающие размеры эккриновых
желез. Секрет апокриновых желез содержит
больше белка, который, разлагаясь на поверхно-
сти кожи, имеет достаточно резкий специфичес-
кий запах. Эти железы особенно интенсивно раз-
виваются в период полового созревания.
Разновидностью апокриновых потовых желез яв-
ляются серные железы кожи наружного слухо-
вого прохода и век.
Волосы
Характеризуются постоянным ростом за счет
деления клеток волосяной луковицы. Все волосы
(рис. 2.4) делятся на длинные (волосы головы,
бороды, усов, кожи лобка, подмышечных впа-
дин); щетинистые (волосы ресниц, бровей, ноз-
дрей, наружного слухового прохода); пушковые
(покрывающие большую часть тела).
Анатомически волос делится на две части.
Часть волоса, выступающая над поверхностью
кожи, называется стержнем, а внутридермальный
отдел — корнем. В области контакта волос с эпи-
дермисом формируется углубление — воронка.
Стержень и корень длинных и щетинистых
волос состоят из трех слоев: коркового, мозгово-
го вещества и кутикулы.
Кутикула — наружная оболочка волоса, обра-
зованная ороговевшими, лишенными пигмента
клетками, дифференцирующимися из камбиаль-
ных клеток волосяной луковицы. По мере диф-
ференцировки и продвижения вверх первона-
чально кубические клетки кутикулы резко
уплощаются и черепицеобразно накладываются
друг на друга. В них образуется твердый кера-
тин, и клетки превращаются в роговые чешуй-
ки. Кутикула обеспечивает эластичность воло-
са, защищает его от вредных воздействий внеш-
ней среды, препятствует разрушению и ломкос-
ти стержня.
Корковое вещество — основное вещество во-
лоса. Оно состоит из нескольких слоев клеток,
дифференцирующихся из камбиальных клеток
луковицы, расположенных медиальнее камбия
для кутикулы. В области луковицы оно образо-
вано кубическими клетками с ядром, тонофиб-
риллами и гранулами меланина, между клетка-
ми лежат меланоциты. При ороговении клетки
в верхних отделах корня превращаются в рого-
вые чешуйки, содержащие твердый кератин, от-
личающийся от мягкого кератина эпидермиса и
других слоев волоса большим содержанием ами-
нокислоты цистина и большей прочностью.
Процесс образования твердого кератина проис-
ходит без промежуточных стадий, т. е. без на-
копления гранул трихогиалина.
Рис. 2.4. Продольный срез через луковицу
и корень волоса. хЮО. (Фалин Л. Е., 1957).
1 — внутреннее, 2 — наружное корневые
влагалища; 3 — внутренняя (стекловидная)
оболочка волосяной сумки; 4 — волосяной
сосочек; 5 — волосяная луковица;
6 — жировая ткань.
8 Общая дерматология
Мозговое вещество занимает центр волоса в его
корне, постепенно исчезая к волосяной ворон-
ке. Образуется из самых центральных клеток во-
лосяной луковицы. В самой нижней части кор-
ня волоса, непосредственно над волосяным
сосочком, мозговое вещество построено из по-
лиморфных малодифференцированных клеток,
в которых имеются тонофибриллы и скопления
гликогена. По мере роста волоса эти клетки про-
двигаются вверх и подвергаются дифференци-
ровке. В них постепенно накапливаются трихо-
гиалиновые гранулы, окрашивающиеся, в
отличие от кератогиалина, не базофильно, а ок-
сифильно. На уровне протоков сальных желез
клетки замещаются мягким кератином (в отличие
от твердого кератина коркового вещества). Клет-
ки полностью теряют воду и превращаются в
клеточные тени — роговые чешуйки. В них и
между ними содержится воздух и небольшое ко-
личество гранул меланина. В пушковых волосах
мозговое вещество отсутствует.
Снаружи корень волоса окружен волосяным
мешком, или фолликулом, состоящим из внутрен-
него и наружного корневых эпителиальных вла-
галищ. Фолликул окружен соединительнотканным
дермальным влагалищем (волосяной сумкой).
Внутреннее корневое влагалище в нижних уча-
стках корня волоса имеет 3 отчетливо различи-
мых слоя:
1. Наружный бледный слой Генле. Образован
одним рядом клеток, не содержащих гранулы
трихогиалина.
2. Средний гранулосодержащий слой Гексли. Об-
разован несколькими рядами клеток с гранула-
ми трихогиалина.
3. Кутикула внутреннего корневого влагалища.
Прилежит к кутикуле волоса. Образована одним
рядом плоских клеток.
Все три слоя внутреннего корневого влагалища
образуются за счет дифференцировки самых пе-
риферических клеток волосяной луковицы. Про-
цесс дифференцировки клеток всех трех слоев
принципиально одинаков, но различается ее ско-
ростью: раньше всего она начинается в слое Ген-
ле, позже всего — в кутикуле. Клетки этих сло-
ев подвергаются ороговению, в процессе
которого в цитоплазме накапливается трихоги-
алин, биохимически сходный с кератогиалином.
Самые мелкие гранулы трихогиалина обнаружи-
ваются в кутикуле, самые крупные — в слое
Гексли. В последующем трихогиалин во всех
слоях превращается в мягкий кератин. В средних
и верхних отделах корня все три слоя сливают-
ся, а на уровне протока сальной железы внут-
реннее корневое влагалище исчезает.
Наружное корневое влагалище состоит из кле-
ток, не подвергающихся ороговению. Оно пред-
ставляет собой как бы дубликатуру эпидермиса
с повернутым наружу ростковым слоем, который
в верхних отделах корня переходит в ростковый
слой эпидермиса. Только здесь клетки приобре-
тают способность к ороговению. Наружное во-
лосяное влагалище построено из 4—5 слоев кле-
ток, в которых содержатся гликоген,
тонофибриллы, кератогиалин и кератиносомы.
Количество слоев минимально (1—2) в самых
нижних отделах корня, где влагалище сливает-
ся с луковицей. По направлению вверх количе-
ство слоев клеток нарастает. На уровне прикреп-
ления мышцы, поднимающей волос, наружное
корневое влагалище утолщается, образуя так на-
зываемую припухлость, или валик. Припухлость
является камбиальной зоной наружного влага-
лища. Размножающиеся здесь клетки мигриру-
ют вверх и вниз, подвергаясь дифференциров-
ке. В эпителии припухлости практически
отсутствуют антигенпредставляющие клетки
Лангерганса и резко снижено содержание анти-
генов главного комплекса гистосовместимости (им-
мунологически привилегированная зона). При
потере этой привилегированности в припухлос-
ти нарастает количество клеток Лангерганса, по-
являются аутоантигены и отмечается воспали-
тельная инфильтрация и аутоиммунная реакция,
что в итоге ведет к выпадению волос (рубцовая
алопеция). Клетки наружного корневого влага-
лища лежат на базальной мембране.
Снаружи от наружного корневого влагалища
находится соединительнотканная капсула — сум-
ка волоса. Она состоит из наружного продоль-
ного и внутреннего циркулярного слоев.
Луковица (матрица) волоса. Волос растет за счет
размножения клеток луковицы — терминальной
части корня волоса. Луковица в виде шапочки
окружает волосяной сосочек. Находящиеся непос-
редственно над волосяным сосочком клетки лу-
ковицы являются камбием для волоса и внутрен-
него волосяного влагалища. Ее расположенные
центрально клетки являются камбием для моз-
гового вещества волоса, кнаружи от них после-
довательно лежат камбиальные клетки для кор-
кового вещества, кутикулы волоса, трех слоев
внутреннего корневого влагалища. Так же как и
припухлость наружного корневого влагалища,
волосяная луковица лишена клеток Лангерганса
и антигенов главного комплекса гистосовмести-
мости. При гнездной алопеции эти качества луко-
вицы теряются, что ведет к аутоиммунной атаке
луковицы и облысению. Наружное корневое вла-
галище и луковица участвуют в посттравматичес-
Глава 2. Анатомия и гистофизиология кожи
9
кой регенерации кожи и в эпителизации раневой
поверхности.
Волосяной сосочек образован рыхлой волок-
нистой неоформленной соединительной тканью
(РВНСТ) с кровеносными сосудами, питающи-
ми волос. У генетически предрасположенных к
облысению мужчин кровеносные капилляры
волосяного сосочка чувствительны к повышению
в крови содержания андрогенов и отвечают на
него стойким спазмом. У таких мужчин при
половом созревании начинается выпадение во-
лос (андрогенетическая алопеция).
К соединительнотканной волосяной сумке под
углом прикрепляется мышца, поднимающая во-
лос. Она образована гладкой мышечной тканью.
Ногти
Ноготь представляет собой роговую пластинку
(ногтевая пластинка), лежащую на эпителии кон-
цевых фаланг пальцев рук и ног (рис. 2.5). Ног-
Дистальная
Рис. 2.5. Схема макро-микроскопического
строения ногтя (Хэм А., Кормак Д., 1983).
тевая пластинка состоит из тела и корня. Тело
ногтя — его видимая часть. Скрытая в ногтевом
желобке проксимальная часть ногтя является его
корнем. Ногтевая пластинка снаружи имеет глад-
кую поверхность, а изнутри покрыта гребешка-
ми и бороздками. Она состоит из роговых плот-
ных чешуек, иногда с остатками ядер,
содержащих твердый кератин. Лежит на ногте-
вом ложе, которое представляет собой толстый
слой эпидермиса без рогового слоя (роговой слой
заменяет ногтевая пластинка). Проксимально
эпидермис ногтевого ложа продолжается в мат-
рицу ногтя, состоящую из эпителиальных кле-
ток онихобластов. Здесь же есть меланоциты,
клетки Меркеля и Лангерганса. У проксималь-
ной границы ногтя роговой слой эпидермиса ок-
ружающей кожи накладывается на ноготь и
прочно к нему прикрепляется. Эта зона ногтя
вместе с роговым слоем называется надногтевой
кожицей, или эпонихием. За счет размножения
онихобластов ноготь растет в длину, а за счет
размножения клеток базального слоя ногтевого
ложа — в толщину.
Подкожная жировая ткань
Подкожная жировая ткань (клетчатка; гиподер-
ма) является третьим, наиболее глубоким слоем
кожи (не все авторы включают ее в состав кожи,
что, на наш взгляд, неверно). Она смягчает дей-
ствие на организм механических факторов, по-
этому особенно хорошо развита в тех участках
кожи, которые подвергаются постоянным меха-
ническим воздействиям (подушечки пальцев,
подошвы и т. д.). Здесь подкожная клетчатка не
исчезает даже при сильном истощении. При
ожирении отмечается не только гипертрофия
жировых клеток, но и увеличение их числа (ги-
перплазия). В результате объем клетчатки может
достигать больших величин. Гиподерма образо-
вана белой жировой тканью, формирующей
жировые дольки, образованные гипертрофиро-
ванными липоцитами, и рыхлой волокнистой
неоформленной соединительной тканью
(РВНСТ). Жировые дольки разделены прослой-
ками РВНСТ с характерным для нее клеточным
составом и обилием кровеносных сосудов
(рис. 2.6).
Там, где кожа малоподвижна, подкожный
слой пронизан туго натянутыми соединительно-
тканными пучками, прикрепляющими ее к фас-
циям и надкостнице. Там же, где кожа более
подвижна, но имеет мало жировых отложений,
подкожная клетчатка образована из соединитель-
нотканных пластинок. В большинстве же учас-
тков подкожно-жировой клетчатки она состоит
10 Общая дерматология
из жировых долек, форма и расположение ко-
торых определяются ходом кровеносных сосудов,
положением соединительнотканных пластинок,
окружающих эти дольки, и механическими фак-
торами.
В жировой ткани гиподермы откладывает-
ся жир, являющийся не только запасом пи-
тательных веществ (трофическая функция), но
одновременно смягчающий толчки и удары
(защитно-механическая функция) и играющий
роль сильного термоизолятора (терморегуля-
торная функция). Кроме того, при расщепле-
нии жира (липолизе) подкожно-жировой
клетчатки образующаяся энергия идет на со-
гревание организма. Распад жира сопровож-
дается также высвобождением большого ко-
личества воды, и поэтому подкожно-жировую
клетчатку рассматривают как своеобразное
депо жидкости.
Рис. 2.6. Гиподерма кожи человека.
Из рисунка видно, что жировая ткань формирует
дольки, между которыми находятся перегородки
из рыхлой или плотной волокнистой
соединительной ткани, содержащие
кровеносные сосуды. Один такой достаточно
крупный сосуд (артерия) виден в правом верхнем
углу, х 100.
Мышцы кожного покрова
К мышцам кожи относятся следующие гладкие
и поперечнополосатые мышцы:
1. Мышца, поднимающая волос. Она представ-
лена небольшими пучками гладких миоцитов,
расположенными косо по оношению к волосу.
Они одним концом прикрепляются к соедини-
тельнотканной сумке волоса ниже впадения в
него протока сальной железы, а другим вплета-
ются в соединительную ткань сосочкового слоя
дермы. У щетинистых волос бровей, ресниц,
носового хода и наружного слухового прохода
мышцы отсутствуют, а у волос усов, бороды и
подмышечных впадин развиты слабо.
2. Диагональные гладкие мышцы. Это гладко-
мышечные пучки, не связанные с волосяными
фолликулами. Они располагаются в коже воло-
систой части головы, щек, лба, тыльной поверх-
ности кистей и стоп, в подмышечной области.
Особенно хорошо развита гладкая мышечная
ткань в коже мошонки, грудного соска и соско-
вого поля, в области заднего прохода, где она
формирует довольно мощные пучки (гениталь-
ные, или дартоидные мышцы).
3. Гладкая мышечная ткань средней оболочки
кровеносных сосудов кожи: артерий, вен, анасто-
мозов.
4. Поперечнополосатые мышцы. Пучки попе-
речнополосатых мышц обнаруживаются в коже
лица (преимущественно в области губ и носогуб-
ных складок), а также шеи и верхней части гру-
ди (m. platisma).
Из мышц кожи могут развиваться доброкаче-
ственные и злокачественные опухоли.
Тонкое строение кожи
После такого краткого рассмотрения общего пла-
на строения кожи далее будут более подробно
рассмотрены детали строения ее составных эле-
ментов.
Эпидермис
Эпидермис имеет слоистое строение и состоит из
5 слоев: базального (stratum germinativum), ши-
поватого (stratum spinosum), зернистого (stratum
granulosum), блестящего (stratum lucidum) и ро-
гового (stratum corneum) (рис. 2.7). Однако на
значительном протяжении кожный покров имеет
4 слоя, т. к. блестящий слой встречается не вез-
де. В составе эпидермиса в настоящее время вы-
деляют следующие виды клеток: эпителиальные,
Глава 2. Анатомия и гистофизиология кожи
11
Рис. 2.7. Схема строения эпидермиса толстой
кожи. Видна нестолбчатая организация рогового
слоя. I — эпидермис; II — дерма. 1 — базальный
слой эпидермиса; 2 — меланоцит;
3 — шиповатый слой; 4 — зернистый слой;
5 — блестящий слой; 6 — роговой слой;
7 — базальная мембрана; 8 — сосочковый слой
дермы (Е. Ф. Котовский).
которые здесь называются кератиноцитами, эти
клетки в различных слоях эпидермиса имеют
различное строение, а в роговом слое превраща-
ются в роговые чешуйки; меланоциты, или пиг-
ментные клетки; клетки Лангерганса, или внут-
риэпидермальные макрофаги; клетки
Гринстейна; клетки Меркеля; в эпидермисе по-
стоянно встречаются внутриэпидермальные лим-
фоциты, а иногда и тучные клетки.
Базальный слой
Наиболее неоднородный по клеточному составу
базальный слой эпидермиса. Именно в нем в
основном встречаются указанные разновиднос-
ти клеток, хотя иногда все эти клетки можно
найти и в слое шиповатых клеток.
Кератиноциты имеют различное строение в
различных слоях эпидермиса. Базальные кера-
тиноциты имеют овальную форму и меньше по
размеру, чем шиповатые клетки. В участках с
утолщенным эпидермисом клетки базального
слоя вытянуты в вертикальном направлении.
Плазмалемма их характеризуется ровными кон-
турами. Ее толщина 7—8 нм.
Между собой кератиноциты соединяются при
помощи десмосом, а к базальной мембране при-
крепляются полудесмосомами.
На электронных микрофотографиях десмосома
выглядит как симметричная структура, образо-
ванная двумя плотными прикрепляющимися
пластинками, расположенными под плазмалем-
мами соседних кератиноцитов (рис. 2.8). С каж-
дой стороны к пластинкам подходят пучки то-
нофиламентов. Плазмалеммы смежных клеток
разделены межклеточным пространством шири-
ной 25-35 нм. Пространство заполнено плотным
материалом, центральная часть которого окраши-
вается более интенсивно, чем периферические
части, прилежащие к мембранам кератиноцитов.
На поперечном срезе центральный слой имеет
зигзагообразную форму, причем от вершин зиг-
загообразных структур отходят тонкие филамен-
ты, пронизывающие плазмалемму и заканчива-
ющиеся на десмосомальных пластинках. В состав
десмосомальных дисков входят белки десмопла-
кины, а пучки тонофиламентов состоят из а-ке-
ратина.
Полудесмосомы являются асимметричными
образованиями, как бы половиной десмосом
(рис. 2.2), однако по химическому составу, им-
мунологически и функционально отличаются от
десмосом.
Ядра базальных кератиноцитов имеют округ-
лую или овальную форму и ровные контуры
12
Общая дерматология
ядерной мембраны с немногочисленными вдав-
лениями. Перинуклеарное пространство опреде-
ляется лишь на отдельных участках. Нуклео-
протеиновые гранулы расположены довольно
равномерно, либо в виде сгущения гетерохрома-
тина вблизи ядерной мембраны, либо рассеяны
по всей нуклеоплазме.
В цитоплазме базальных клеток расположены
фибриллярные элементы диаметром 3—5 нм —
тонофиламенты, а также немногочисленные пуч-
ки тонофибрилл. В переплетениях тонофиламен-
тов расположены структурно связанные с ними
свободные рибосомы. Количество рибосом и
митохондрий значительное. Имеется неплохо
выраженная зернистая эндоплазматическая сеть.
В цитоплазме встречаются также меланосомы —
гранулы, содержащие меланин (последние посту-
пают в эпидермоциты из меланоцитов), и лизо-
сомы, благодаря наличию которых кератиноци-
ты способны осуществлять фагоцитоз. Иногда в
цитоплазме встречаются гранулы гликогена и
пиноцитозные пузырьки, а также липидные кап-
ли. Базальные кератиноциты подразделяются на
две разновидности: с зубчатой и ровной поверх-
ностью (рис. 2.9). Кератиноциты с ровной по-
верхностью являются клетками, способными по-
стоянно делиться. Одна из образующихся клеток
остается в базальном слое и вновь через опре-
деленное время делится, а вторая выталкивает-
ся по направлению к шиповатому слою, где всту-
пает на путь дифференцировки. Кератиноциты
с зубчатой поверхностью являются покоящейся
популяцией клеток, выполняющей резервную
функцию. Они начинают делиться при обшир-
ных повреждениях эпидермиса. Более подробно
об этих популяциях кератиноцитов см. ниже.
В базальных кератиноцитах начинается био-
синтез специфического белка эпидермиса — а-
кератина. Вначале здесь образуется растворимый
предшественник кератина — прекератин. В бо-
лее поверхностных слоях он затем превращает-
ся в фибриллярный нерастворимый кератин. Мо-
лекула кератина состоит из не менее 2-х
субъединиц, скрученных между собой наподобие
волокна
а
б
Рис. 2.8. Схема строения десмосомы.
а. По Arnn I., Staehelin L., 1981.
б. По В. М. Gumbiner. Плазматические мембраны клеток разделены промежутком 20-30 нм, в котором
находятся внемембранные части Са2+-связывающих белков десмоглеина и десмоколлина. К внутрен-
ней (цитоплазматической) поверхности плазматической мембраны прилегает цитоплазматическая
пластинка с вплетенными в нее промежуточными филаментами. В состав этой пластинки входят
десмоплакины, плакоглобин и часть молекулы десмоглеина.
Глава 2. Анатомия и гистофизиология кожи
13
каната. По мере синтеза эти канатоподобные
субъединицы объединяются и образуют керати-
новые филдменты, видимые в электронный мик-
роскоп (рис. 2.3). Они имеют ширину 8—10 нм
и относятся к классу промежуточных филамен-
тов, образующих цитоскелет кератиноцитов и
входящих в состав десмосом и полудесмосом.
Соединяясь, тонофиламенты образуют тонофиб-
риллы, видимые в световом микроскопе. Пре-
кератин, или незрелый кератин, характерен для
всех эпителиальных тканей. Зрелый кератин от-
личается от незрелого прекератина не только
большей молекулярной массой, но и наличием
внутри- и межцепочечных дисульфидных свя-
зей, которые придают молекуле большую нера-
створимость. Зрелый кератин характерен лишь
для кератинизирующихся эпителиев. Он выде-
ляется в наибольшем количестве из рогового
слоя.
Шиповатый слой
В слое шиповатых клеток кератиноциты имеют
полигональную форму и образуют от 2-3 до 10
рядов. Их плазмолемма толщиной 7—8 нм харак-
теризуется очень неровными контурами и обра-
Рис. 2.9. Схема, представляющая гетерогенность
кератиноцитов базального слоя эпидермиса
(Lavker R., Sun Т., 1983). 1 — роговой слой;
2 — зернистый слой; 3 — шиповатый слой;
4 — базальный слой; 5 — кератиноциты
с цитоплазматическими выростами («зубчатые»);
6 — стволовые клетки («гладкие») расположены
на вершине самого глубокого эпидермального
гребешка; 7 — переходные клетки;
8 —- постмитотические клетки.
зует значительное количество шипов, проника-
ющих в соответствующие углубления соседних
клеток. Здесь располагаются многочисленные
десмосомы. Ядра шиповатых клеток округлые и
имеют слегка овальную форму, окружены хоро-
шо очерченной ядерной мембраной. В центре
ядра или эксцентрично располагается хорошо
очерченное ядрышко.
В клетках шиповатого слоя исчезают центри-
оли и пиноцитозные пузырьки, но возрастает
количество свободных рибосом, что свидетель-
ствует об увеличении способности клеток син-
тезировать и накапливать белки, но не секрети-
ровать их. Действительно, по сравнению с
базальным слоем здесь усиливается синтез пре-
кератина. Часть его идет на формирование мно-
гочисленных десмосом, а часть — на образова-
ние тонофиламентов. Здесь также синтезируются
десмоплакины для десмосом.
Количество митохондрий в цитоплазме кле-
ток относительно невелико. Отличительной осо-
бенностью этих клеток является наличие хоро-
шо развитого фибриллярного аппарата,
представленного тонофибриллами и тонофила-
ментами. Они формируют пучки различной про-
тяженности и толщины, образующие концент-
рические сгущения вокруг ядра, от которого
отходят пучки тонофибрилл, направляющихся к
плазмалемме и заканчивающихся в области дес-
мосом. Особой мощности пучки тонофибрилл
достигают в шиповатых клетках эпидермиса сто-
пы и ладоней. Таким образом, тонофибриллы
пронизывают весь объем клетки и формируют
ее внутренний каркас, препятствуя деформации
клетки и особенно ее ядра.
В цитоплазме шиповатых клеток обнаружива-
ются многочисленные округлые везикулы раз-
личного диаметра, канальцы цитоплазматической
сети, а также меланосомы. Десмосомы шипова-
тых клеток имеют форму дисков диаметром от
250 до 700 нм. Впервые их светомикроскопичес-
ки обнаружил в 1870 году Д. Биццоцеро. Он опи-
сал наличие между шиповатыми клетками мос-
тиков или шипов, а в середине каждого из
них — узелка (узелок Биццоцеро), являющего-
ся десмосомой. Десмосомы выполняют две ос-
новные функции: ответственны за адгезию кле-
ток и за поддержание внутренней архитектоники
шиповатых клеток. При усиленном разрушении
десмосом (к ним вырабатываются аутоантитела
при патологических состояниях) соединение
шиповатых клеток нарушается, возникает от-
слойка слоев эпидермиса и формируются пузы-
ри. Это заболевание называется пузырчаткой и
может закончиться летальным исходом. В верх-
14 Общая дерматология
них клетках шиповатого слоя появляются гра-
нулы Одланда (см. ниже).
Базальный и шиповатый слои называются ро-
стковым слоем Мальпиги, т. к. только в них
встречаются митозы (в шиповатом слое — только
при обширных повреждениях эпидермиса) и за
счет этого происходит физиологическая и репа-
ративная регенерация эпидермиса.
Зернистый слой
Клетки зернистого слоя формируют 2—3 ряда.
Они имеют вытянутую форму и ограничены чет-
ко выраженной клеточной оболочкой толщиной
8—12 нм. На боковых поверхностях плазмалем-
ма часто имеет зубчатые контуры. Ядра клеток
зернистого слоя отличаются заметным полимор-
физмом. Одни имеют округлую или овальную
форму, а другие вытянуты параллельно поверх-
ности эпидермиса. У большинства ядер ядрыш-
ко отсутствует. Нуклеоплазма часто выглядит
разреженной, а нуклеопротеиновые гранулы хо-
рошо различимы. В некоторых клетках содержат-
ся лишь остатки ядер. Количество митохондрий
относительно невелико. Одной из характерных
особенностей клеток зернистого слоя является
наличие в них кератогиалиновых масс. Керато-
гиалиновые массы или зерна (отсюда название
слоя) хорошо выявляются обычными красителя-
ми. В электронном микроскопе они выглядят
гомогенными образования и, не ограниченными
мембраной. Число, размеры и форма гранул силь-
но варьируют. Распределены в основном по ци-
топлазме, но иногда встречаются и в ядре. При
своем образовании кератогиалиновые гранулы
имеют размеры 100—200 нм, но, сливаясь, обра-
зуют конгломераты размером до нескольких мик-
рометров. Наиболее часто они покрывают сгуще-
ния коротких тонофибрилл, формируя
тонофибриллярно-кератогиалиновые комплексы,
вокруг которых концентрируются нуклеопроте-
иновые гранулы, митохондрии и рибосомы. В
поверхностных клетках зернистого слоя по мере
накопления кератогиалиновых гранул постепен-
но исчезают органеллы. Однако в нижних рядах
слоя клетки обладают высокой синтетической
активностью. Здесь происходит биосинтез филаг-
рина или белка, агрегирующего филаменты
(раньше его называли гистидинбогатым белком).
Филагрин — основной белок кератогиалиновых
гранул, которые, несмотря на свое название, ни
кератина, ни гиалина не содержат. Филагрин
синтезируется только в зернистом слое при по-
мощи соответствующей матричной (информаци-
онной) РНК. Он объединяется с кератиновыми
филаментами и необходим для ориентации и ста-
билизации их в роговых чешуйках.
В клетках зернистого слоя происходит также
биосинтез кератолинина — белка, накапливаю-
щегося под цитолеммой клетки и вызывающе-
го её утолщение. Специфическая РНК для это-
го синтеза также имеется только в зернистом
слое. В клетках этого слоя продолжается и био-
синтез новых полипептидов прекератина, моле-
кулярная масса которого больше, чем прекера-
тинов, синтезируемых в базальном и шиповатом
слоях. Таким образом, клетки зернистого слоя
достаточно функционально активны, причем ке-
ратогиалиновые гранулы образуются исключи-
тельно в этом слое и не являются продуктом дег-
радации кератиноцитов, как считалось ранее.
Второй характерной особенностью зернистых
клеток является присутствие в их цитоплазме ке-
ратиносом, или гранул Одланда. Эти видоизменен-
ные лизосомы представляют собой электронно-
плотные, округлой формы образования с
ламеллярным содержимым, окруженные однослой-
ной мембраной. В гранулах Одланда обнаружен
целый ряд веществ: гликолипиды, гликопротеины,
свободные стерины, набор гидролитических фер-
ментов. В клетках зернистого слоя кератиносомы
занимают в основном периферическое положение.
Часто можно видеть, как содержимое кератино-
сом выделяется в межклеточное пространство.
Здесь из выделенного материала формируется пла-
стинчатое цементирующее вещество. В процессе
кератинизации происходят выраженные измене-
ния в строении и гистохимии этих межклеточных
пластинок: вначале секретируются гликолипиды,
свободные стерины, которые затем превращаются
в эфиры стеринов. Эти вещества образуют бислой
в промежутках рогового слоя и, являясь гидрофоб-
ными, выполняют главную функцию барьера про-
ницаемости эпидермиса.
Роговой слой
Роговой слой состоит из роговых чешуек (их
иногда называют корнеоцитами), которые соеди-
няются друг с другом с помощью взаимопрони-
кающих выростов оболочки, особенно глубоких
на боковых сторонах, и ороговевших десмосом.
Каждая роговая чешуйка обладает протяженно-
стью до 10 мкм и более, а ее толщина колеблется
от 100 нм до 1 мкм. Снаружи она окружена
электронноплотной оболочкой толщиной 12—
15 нм, а внутри заполнена кератиновыми фиб-
риллами диаметром 7—8 нм, разделенными элек-
тронноплотным аморфным материалом в
соотношении 1:1.
Глава 2. Анатомия и гистофизиология кожи
15
Имеется два типа чешуек: чешуйки с рыхлым
заполнением кератиновых фибрилл (тип Б, они
располагаются ближе к зернистому слою) и че-
шуйки с плотным заполнением (тип А). В отдель-
ных чешуйках, непосредственно примыкающих
к зернистым клеткам, наряду с пучками кера-
тиновых фибрилл, иногда содержится значитель-
ное количество кератогиалиновых гранул и еди-
ничные остатки митохондрий. Эти чешуйки
иногда называют Т-пластинками. По своему
микроскопическому строению эта зона соответ-
ствует блестящему слою, который в настоящее
время многими авторами не выделяется в само-
стоятельный слой эпидермиса. В промежутках
между чешуйками часто содержится электрон-
ноплотная цементирующая субстанция гликоли-
пиновой природы.
Толщина рогового слоя определяется взаимо-
действием двух факторов: скорости размноже-
ния и продвижения в вертикальном направле-
нии кератиноцитов, с одной стороны, и скорости
отторжения роговых чешуек, с другой. При ме-
ханических и некоторых других воздействиях на
кожу активируются митотическая активность
кератиноцитов и их продвижение вверх при нор-
мальной скорости отторжения роговых чешуек.
Так возникает роговая мозоль. В некоторых слу-
чаях кожной патологии нарушаются процессы
отторжения роговых чешуек при нормальных
скоростях пролиферации и дифференцировки
кератиноцитов. Группа заболеваний с подобным
соотношением процессов наработки и потери
роговых чешуек называется гиперкератозами.
В последнее время установлено, что чешуйки
рогового слоя могут иметь столбчатую и нестолб-
чатую организацию. Нестолбчатая организация
характерна для толстой кожи с толстым роговым
слоем. Такая кожа характеризуется высокой ми-
тотической активностью эпидермиса, что препят-
ствует упорядочению роговых чешуек. Столбчатая
организация рогового слоя характерна, напротив,
для тонкой кожи с низкой митотической актив-
ностью эпидермиса. Клетки всех слоев эпидермиса
и роговые чешуйки при столбчатой организации
расположены строго друг над другом. Каждая ро-
говая чешуйка имеет 6-угольную форму и каждой
из 6 сторон контактирует с соседней чешуйкой
(«стеганое одеяло»). Такое расположение как нельзя
лучше обеспечивает связь чешуек и выполнение
роговым слоем барьера проницаемости. При ряде
кожных заболеваний нормальное расположение
роговых чешуек нарушается. Более подробно орга-
низация рогового слоя в колонках будет обсуж-
дена при рассмотрении эпидермальной пролифе-
ративной единицы (ЭПЕ).
Клетки Лангерганса
Клетки Лангерганса, или внутриэпидермальные
макрофаги, впервые были описаны в 1868 году
немецким гистологом Паулем Лангергансом, ко-
торый применил для их выявления метод золо-
чения. Он считал их нервными элементами. В
последующем клетки Лангерганса рассматривали
как стареющие меланоциты. Лишь в последние
годы убедительно показано, что эти клетки имеют
костномозговое (моноцитарное) происхождение и
являются разновидностью макрофагов. Работами
G. F. Murphy и соавт. (1986) была показана транс-
формация дермальных активно фагоцитирующих
макрофагов в эпидермальные клетки Лангерганса
через промежуточные формы, которые совмеща-
ли в себе как свойства моноцитов-макрофагов,
так и типичных клеток Лангерганса. Наряду с фа-
голизосомами в их цитоплазме содержались гра-
нулы Бирбека. Авторы связывают превращение
типичных гистиоцитов дермы в клетки Лангер-
ганса с особенностями эпидермального окруже-
ния, вызывающими эту трансформацию.
Клетки Лангерганса выявляются в эпидермисе
всех млекопитающих, включая человека. Кроме
эпидермиса, они встречаются в многослойных
эпителиях эктодермального происхождения: по-
лости рта, пищевода, роговицы и конъюнктивы
глаза, влагалища и шейки матки. Эти клетки об-
наружены также в лимфоузлах, селезенке, мозго-
вом веществе тимуса. В многорядном реснитчатом
эпителии воздухоносных путей эти клетки в норме
отсутствуют, однако при метаплазии его в много-
слойный эпителий (например, у курильщиков)
появляются там. Подобная же ситуация наблюда-
ется и при метаплазии переходного эпителия мо-
чевого пузыря в многослойный плоский неорого-
вевающий. В последнее время доказано наличие
клеток Лангерганса в многорядном эпителии воз-
духоносных путей в условиях нормы.
Элективными методами выявления клеток
Лангерганса являются методы с солями тяжелых
металлов (хлориды золота и серебра), солями
кобальта, ртути, никеля, хрома. Эти клетки со-
держат такие ферменты, как маннозидаза, АТФ-
аза, гистохимическое исследование которых ши-
роко (особенно АТФазы) также используется для
их визуализации. Наиболее точными являются
методы иммуногистохимической идентификации
клеток Лангерганса, связанные с выявлением
специфических антигенов на поверхности кле-
ток при помощи флуоресцирующих монокло-
нальных антител. Такими антигенами у мышей
являются 1а-антигены, а у человека — HLA-Dr
и Тб. Надежным способом идентификации внут-
16 Общая дерматология
риэпидермальных макрофагов является элект-
ронная микроскопия благодаря характерной их
ультраструктуре.
Количество клеток Лангерганса в эпидермисе,
по данным разных авторов, сильно колеблется от
нескольких десятков (30—40) до 1500 на 1 мм2
площади поверхности кожи. Такие различия мо-
гут быть связаны с различными методами выяв-
ления, возрастом животных или человека, обла-
стью исследования и т. д. Известно, что в толстом
эпидермисе ладоней и подошв человека количе-
ство клеток Лангерганса в 2 раза меньше их числа
в коже туловища и конечностей. По данным
Мядельца О. Д. (1985); Полчаниновой В.В., Мя-
дельца О. Д. (1996), содержание клеток Лангер-
ганса в коже спины половозрелых крыс состав-
ляет 1220 клеток/мм2, а в коже спины и живота
человека — соответственно 330 и 340 клеток/мм2.
В зависимости от возраста число клеток Лангер-
ганса сильно варьирует (Мяделец О.Д., 1985).
Так, у новорожденных животных они не выяв-
лялись, а появлялись в эпидермисе впервые у 3-
суточных крысят (10 кл/мм2). Затем в течение пер-
вого месяца жизни их число постепенно
возрастало и достигало 1000—1100 кл/мм2, что
было характерно и для половозрелых животных.
У старых животных число клеток уменьшалось
почти в 3 раза по сравнению с уровнем половоз-
релых крыс. Нами отмечено также снижение
числа клеток Лангерганса и у пожилых людей.
Имеются половые различия в числе клеток Лан-
герганса у человека и животных, причем в коже
самцов их численность обычно меньше.
Количество клеток Лангерганса может суще-
ственно изменяться под действием различных
факторов внешней среды и факторов, использу-
емых в физиотерапии. Ультрафиолетовое облу-
чение вызывает стойкое снижение численности
этих клеток. Таким же эффектом обладает ла-
зерное облучение. Работами Мядельца О. Д.
(1987, 1993, 1994) показано существенное сни-
жение содержания клеток Лангерганса при дей-
ствии на организм общего глубокого охлажде-
ния. При этом, наряду со снижением содержания
клеток, существенно изменялась форма остав-
шихся в эпидермисе клеток: они теряли харак-
терную отростчатость, уменьшались в размерах,
активность АТФазы в клетках снижалась. Эти
реактивные изменения зависели от возраста
животных и продолжительности холодового воз-
действия. У молодых животных депрессия была
менее выраженной, чем у половозрелых крыс, и
после холодового воздействия достаточно быст-
ро наступала компенсация, а затем и суперком-
пенсация. У половозрелых животных депрессия
количества клеток была более выраженной и
продолжительной в постгипотермическом пери-
оде, наблюдалось лишь восстановление числа
клеток без суперкомпенсации. Еще более резкие
изменения отмечались под воздействием холода
у старых крыс.
Как отмечалось, клетки Лангерганса являют-
ся внутриэпидермальными макрофагами. Однако
они, наряду со сходством (наличие активности
неспецифической эстеразы и кислой фосфата-
зы; наличие на плазмолемме рецепторов к Fc-
фрагменту IgG и СЗ-компоненту комлемента;
способность к активной миграции; способность
к секреции интерлейкина-1; способность к фа-
гоцитозу; экспрессия антигенов гистосовмести-
мости класса II, таких как 1а-антигены у мышей
и HLA-DR-антигены у человека; способность
презентировать антиген и стимулировать лимфо-
циты), имеют и существенные отличия от типич-
ных макрофагов. Эти отличия следующие. Клет-
ки Лангерганса обладают меньшей фагоцитарной
активностью и способностью прилипать к стек-
лянным поверхностям, чем типичные макрофа-
ги. Они не могут поглощать целые клетки или
их фрагменты, а фагоцитируют лишь раствори-
мые и мелкозернистые вещества. У них отсут-
ствует активность типичных для макрофагов
ферментов, таких как p-глюкуронидаза, перок-
сидаза, 5-нуклеотидаза, лизоцим, а-1-антитрип-
син и а-1-антихимотрипсин.
Клетки Лангерганса несут на своей поверхно-
сти отличающийся от макрофагов антигенный
набор. Они в 50 раз сильнее экспрессируют на
своей поверхности HLA-DR-антиген, чем моно-
циты крови. В дополнение к классическим ан-
тигенам макрофагов они несут некоторые мар-
керы Т-лимфоцитов (Тб и Т4). У них выявляется
белок S-100, характерный для нервной ткани и
обычно отсутствующий у макрофагов. Имеют-
ся и ультраструктурные отличия клеток Лангер-
ганса от макрофагов: меньшее содержание лизо-
сом, отсутствие фаголизосом и наличие гранул
Бирбека.
Все эти различия до сих пор вызывают спо-
ры в отношении природы клеток Лангерганса.
Однако все возрастающее количество фактов
подтверждает моноцитарную природу клеток и
заставляет противников этой точки зрения
превращаться в ее адептов. Как отмечалось, вы-
явлены промежуточные клетки, имеющие свой-
ства и типичных макрофагов, и клеток Лангер-
ганса. Было показано, что некоторые макрофаги
также несут белок S-100, который, таким обра-
зом, не является абсолютным маркером нервной
ткани.
Глава 2. Анатомия и гистофизиология кожи
17
В эпидермисе клетки Лангерганса располагают-
ся в базальном слое или супрлбазально. При вы-
явлении АТФазным методом они окрашиваются
в черный или коричневый цвет. Иногда можно
видеть, что ядро клеток не окрашивается или ок-
рашивается слабее, чем перикарионы и отростки.
Поданным Мядельца О.Д. (1985; 1993), Полча-
ниновой В. В., Мядельца О. Д. (1996), клетки
Лангерганса эпидермиса крысы и человека име-
ют похожее строение. В базальном слое или суп-
рабазально находятся лишь перикарионы клеток
Лангерганса. Их ветвистые отростки отходят в
разные стороны. Боковыми отростками клетки
контактируют друг с другом, образуя сложную
сеть. Верхние отростки могут доходить до зерни-
стого слоя, где слепо заканчиваются, иногда пу-
говчатыми утолщениями. Нижние отростки име-
ются не у всех, а лишь у части клеток. Эти
отростки могут уходить в дерму и заканчиваться
там либо слепо, либо контактировать с гемокапил-
лярами. Эта связь отростков клеток Лангерганса
с гемокапиллярным руслом может являться либо
проявлением миграционного процесса, либо отра-
жением какой-то эндокринной функции клеток.
Перикарионы клеток Лангерганса могут иметь ок-
руглую, треугольную или многоугольную форму.
Иногда мы наблюдали, что к клеткам Лангерган-
са подходят из дермы тонкие нервные волокна,
которые заканчиваются в непосредственной бли-
зости от клеток булавовидным утолщением.
Клетки Лангерганса имеют характерную уль-
траструктуру (рис. 2.10). Они характеризуются
лопастным плотным ядром, в котором содержат-
ся среднего размера 1—2 ядрышка. Цитоплазма
светлая и содержит большое количество органелл:
много митохондрий, развитые гранулярную эн-
доплазматическую сеть и комплекс Гольджи.
Обнаруживаются также лизосомы, меланосомы и
микрофиламенты. Характерно наличие особых
гранул Бирбека, имеющих достаточно сложное
строение. Они имеют форму теннисной ракетки
из-за ампулярного расширения конечного отро-
стка. Длина гранул около 200—300 нм, а толщи-
на около 30 нм. Под мембраной гранулы обна-
руживается мелкая зернистость. В «рукоятке»
гранулы две мембраны сближаются. В этом мес-
те между ними обнаруживается линейный элек-
тронноплотный материал с периодической исчер-
ченностью 9 нм. Вопросы происхождения этих
гранул, их функций и содержимого до конца не
разрешены. Ряд исследователей полагают, что они
формируются в комплексе Гольджи и отражают
процесс секреции. По другим предположениям
гранулы формируются в результате пиноцитоза/
фагоцитоза. Авторы этой гипотезы рассматрива-
ют гранулы Бирбека как транспортные органел-
лы, несущие антигены к месту внутриклеточной
переработки или разрушения.
Функции клеток Лангерганса
Роль в иммунологических
реакциях кожи
Наиболее изучена антигенпредставляющая роль
клеток Лангерганса. Они захватывают из внеш-
а
б
Рис. 2.10. Клетка Лангерганса супрабазального слоя эпидермиса, а — общий вид; б — фрагмент
(Михайлов И. Н„ 1979).
а. Ядро лопастной формы с хорошо выраженным гетерохроматином. Цитоплазма содержит многочис-
ленные органеллы: митохондрии (Мх), гранулы Бирбека (ГЛ), лизосомы (Л), цитоплазматическую сеть
(ЦС), пластинчатый комплекс (ПК), гранулы гликогена (Г). х 15500.
б. Характерная структура специфических гранул (в форме «теннисной ракетки») с ампулярным расши-
рением, центральной продольной и поперечными ламеллами в области рукоятки, х 66100.
1 8 Общая дерматология
ней среды антигены, перерабатывают их и экс-
прессируют на своей поверхности. Затем анти-
гены в комплексе с собствеными антигенами (1а-
антигены у мышей и HLA-Dr у человека) и
интерлейкином-1 представляются проникающим
в эпидермис лимфоцитам, в основом Т-хелпе-
рам, которые вырабатывают интерлейкин-2,
который, в свою очередь, индуцирует пролифе-
рацию Т-лимфоцитов. Эти активированные Т-
клетки участвуют в иммунном ответе, например,
при контактной гиперчувствительности кожи.
Клетки Лангерганса могут очень долго удер-
живать на своей поверхности захваченные анти-
гены, участвуя таким образом в иммунологичес-
кой памяти. Они способны презентировать
антигены не только в эпидермисе, но и в дер-
ме. Описана их способность после захвата анти-
гена в эпидермисе транспортировать его по лим-
фососудам из кожи в регионарные лимфоузлы.
Там их обнаруживают в маргинальных синусах
и паракортикальной тимусзависимой зоне. В па-
ракортикальной зоне клетки Лангерганса способ-
ны превращаться в интердигитирующие макро-
фаги, являющиеся обязательными участниками
реакций клеточного иммунитета. Таким образом,
клетки Лангерганса связывают в единую функ-
циональную систему два органа: кожу и лимфо-
узлы. Есть данные, что клетки Лангерганса иг-
рают роль в предотвращении распространения
вирусов внутри эпидермиса и элиминации из
него неопластических клеток.
Секреторные функции
клеток Лангерганса
Эти функции изучены недостаточно, однако
имеющиеся данные не дают оснований сомне-
ваться в реализации их клетками. Во-первых,
сильное развитие в клетках гранулярной эндо-
плазматической сети, комплека Гольджи, нали-
чие гранул Бирбека (возможно, это секреторные
гранулы) позволяет отождествлять эти клетки с
секреторными. Во-вторых, иногда в местах кон-
тактной гиперчувствительности в клетках Лан-
герганса наступает еще большая гиперплазия
указанных органелл и появляются вакуоли.
Из секретируемых клетками Лангерганса ве-
ществ известны интерлейкин-1, у-интерферон.
Сведения о других синтезируемых веществах от-
сутствуют. Можно лишь предполагать, что яв-
ляясь разновидностью макрофагов, клетки Лан-
герганса сохраняют весь диапазон секретируемых
макрофагами веществ (лизоцим, простагланди-
ны, пирогены, ингибиторы плазминогена, фак-
торы регуляции биосинтеза белков в других
клетках, фактор активации тромбоцитов, фак-
тор хемотаксиса нейтрофилов; факторы, инги-
бирующие размножение лимфоцитов, опухоле-
вых клеток и факторы, стимулирующие деление
клеток и др.).
Некоторые авторы показали, что в гранулах
Бирбека клеток Лангерганса содержится эпидер-
мальный кейлон, однако это единичные наблю-
дения и неясно, синтезируют его клетки Лангер-
ганса или лишь депонируют. Наши наблюдения
о связи клеток Лангерганса своими отростками
с сосудами микроциркуляторного русла дермы
дают основание предполагать, что это морфоло-
гические проявления эндокринной функции кле-
ток. По данным I. A. Vega и соавт. (1990), в клет-
ках Лангерганса методом иммуноцитохимии
обнаружено содержание мет-энкефалина — эн-
догенного модулятора боли, одновременно явля-
ющегося сильным стимулятором иммунной си-
стемы.
Роль клеток Лангерганса
в структурно-функциональной
организации эпидермиса
В настоящее время имеются сведения, что клет-
ки Лангерганса участвуют в регуляции числен-
ности популяции кератиноцитов эпидермиса и
его структурно-функциональной организации.
Эти сведения следующие.
Клетки Лангерганса могут концентрировать в
гранулах Бирбека эпидермальный кейлон (а воз-
можно, синтезируют его), подавляющий деление
кератиноцитов. Клетки Лангерганса иницииру-
ют апоптоз кератиноцитов; они осуществляют
лизис кожных папиллом.
Не исключается способность клеток Лангер-
ганса стимулировать дифференцировку керати-
ноцитов. Это подтверждается теми наблюдени-
ями, что при заболеваниях, связанных с
нарушением дифференцировки кератиноцитов
(плоскоклеточный рак и базалиома кожи) чис-
ло этих клеток в эпидермисе резко уменьшает-
ся вместе с нарушением их нормального строе-
ния (Суханов А.Ф., Мяделец О.Д., 1988). Клетки
Лангерганса являются центром эпидермальных
пролиферативных единиц и регулируют их мор-
фологию.
Роль клеток
Лангерганса в патологии
Благодаря своей роли в иммунных реакциях
кожи клетки Лангерганса вовлекаются во мно-
гие иммунопатологические процессы в коже: ал-
лергический контактный дерматит, атопический
дерматит. В последнее время установлено нали-
чие на клетках Лангерганса рецепторов к 1g класса
Глава 2. Анатомия и гистофизиология кожи
19
Е, и через эти и другие рецепторы они включа-
ются в аллергические реакции. При этом они
могут повреждаться, и продукты их распада как
биологически активные вещества могут вызы-
вать повреждение эпидермиса.
Есть сведения о том, что клетки Лангерганса
могут приобретать аутореактивность против соб-
ственных меланоцитов и вызывать их гибель, что
приводит к развитию витилиго.
Популяция этих клеток изменяется при целом
ряде других кожных заболеваний (туберкулез
кожи, лепра, неопластические процессы и др.),
что свидетельствует о их роли в патогенезе дан-
ных заболеваний (Персина И.С., 1985; Тухватул-
лина З.Г, 1994). При гиперпролиферативных
процессах эпидермиса — регенерации, злокаче-
ственном росте — количество клеток Лангерганса
снижается или они исчезают совсем. Число кле-
ток Лангерганса может служить прогностичес-
ким признаком при карциномах. При старении
численность этих клеток также снижается, и с
этим связывают учащение в старости раковых за-
болеваний кожи.
Клетки Лангерганса принимают участие так-
же в патогенезе аутоиммунных заболеваний кожи
(красная волчанка, пузырчатка). Они связыва-
ют собственные антигены эпидермиса и стано-
вятся мишенями для аутореактивных Т-лимфо-
цитов. Число клеток Лангерганса при этом
снижается.
Описано заболевание (гистиоцитоз X), в ос-
нове которого лежит пролиферация и повреж-
дение органов и тканей клетками, имеющими
фенотипические маркеры эпидермальных кле-
ток Лангерганса. В настоящее время гистиоци-
тоз X считают опухолью из клеток Лангерган-
са (Самсонов В.А. и соавт., 1996). Наиболее
распространенная теория заболевания подтвер-
ждает, что клетки Лангерганса на ранних ста-
диях своей дифференцировки повреждаются и
приобретают способность к аккумуляции в фи-
зиологических и атипичных местах, где, про-
дуцируя цитокины (интерлейкин-4 и фактор
некроза опухолей) и простагландины, вызыва-
ют локальные повреждения. Пролиферация
клеток Лангерганса наблюдается в коже, сли-
зистых оболочках, костях, легких, печени и
других органах (Schmid М. Н. и соавт., 1996).
Гистологическая картина зависит от давности
процесса. Вначале обнаруживаются расширение
сосудов, увеличение количества гистиоцитов,
поверхностно расположенный в коже гистиоци-
тарный инфильтрат. Он пронизан лимфоцита-
ми, плазматическими клетками, нейтрофилами
и эозинофилами, что приводит в дальнейшем
к гранулематозному разрастанию. Далее клет-
ки гранулемы накапливают холестерин и час-
тично превращаются в пенистые и гигантские
клетки с центрально расположенным ядром.
Электронно-микроскопическое исследование
позволяет обнаружить гранулы Бирбека.
Принимая во внимание высокую реактивность
клеток Лангерганса, их связь с нервным аппа-
ратом и микроциркуляторным руслом кожи,
можно ожидать изменение их популяции не
только при кожных, но и общесоматических за-
болеваниях организма. Действительно, работами
Мядельца О. Д., Полчаниновой В. В. (1997) по-
казано, что количество клеток Лангерганса и их
морфология отчетливо зависят от тяжести пато-
логического процесса. При таких заболеваниях,
как острый холецистит, грыжа передней брюш-
ной стенки, изменения в популяции клеток Лан-
герганса незначительные или отсутствуют. На-
против, при раковых заболеваниях, длительных
септических процессах количество клеток суще-
ственно снижается пропорционально стадии за-
болевания, а в оставшихся клетках наблюдают-
ся выраженные дегенеративные процессы:
уменьшаются отростчатость, размеры клеток,
снижается активность АТФазы. На наш взгляд,
изучение клеток Лангерганса при ряде общесо-
матических заболеваний дает возможность про-
гнозировать их исход, корректировать лечебные
мероприятия.
Клетки Гринстейна
В 1983 году в эпидермисе мышей обнаружена
популяция дендритических клеток, которая по
ряду показателей, как морфологических, так и
иммунологических, существенно отличалась от
клеток Лангерганса. Эти клетки экспрессиро-
вали Thy-I-антиген, являющийся маркером пе-
риферических лимфоцитов и тимоцитов. С тех
пор этот антиген широко используется в имму-
ногистохимических реакциях для выявления
данного типа клеток. Благодаря этим реакци-
ям было установлено, что клетки (они получи-
ли название Thy-Г-клеток, или клеток Грин-
стейна- Granstein R.D. и соавт., 1983) образуют
в эпидермисе регулярную сеть, соединяясь друг
с другом при помощи отростков. Возникшее
предположение о том, что это клетки Лангер-
ганса, не подтвердилось: при помощи метода
двойной метки было показано, что это две са-
мостоятельные популяции дендритических кле-
ток эпидермиса. На поверхности клеток Грин-
стейна отсутствуют рецепторы к Fc-фрагменту
иммуноглобулинов и С3-компоненту компле-
2 0 Общая дерматология
мента, 1а-антиген, характерные для клеток Лан-
герганса. С другой стороны, они несут мембран-
ные антигены (Lyt-5, упоминавшийся Thy-1-ан-
тиген), отсутствующие на клетках Лангерганса.
Клетки имеют морфологические особенности,
роднящие их с клетками Лангерганса: содержат
в цитоплазме большое количество виментино-
вых филаментов, но совершенно лишены кера-
тиновых тонофиламентов. В клетках отсутству-
ют гранулы Бирбека, но имеется большое
количество специфических гранул с уплотнени-
ем в центре и периферической прозрачной зо-
ной, окруженных мембраной, являющихся уль-
траструктурным маркером клеток. Для клеток
характерно дольчатое ядро с маргинально рас-
положенным хроматином. Количество клеток
Гринстейна составляет от 1 до 3 % всех клеток
эпидермиса, а по некоторым данным, может
колебаться от 5 до 600 кл/мм2. Попытку обна-
ружить подобные клетки в эпидермисе человека
следует, очевидно, считать успешной, т. к. в нем
имеются так называемые неопределенные клет-
ки двух типов, один из которых не содержит
HLA-Dr-антигены, присущие клеткам Лангер-
ганса, и имеет ультраструктурные признаки
мышиных Thy-1+-клеток.
Функция клеток Гринстейна до конца не вы-
яснена. Существуют две точки зрения на их роль
в эпидермисе. Они являются естественными
киллерами, осуществляющими лизис трансфор-
мированных кератиноцитов; Thy-Г дендритичес-
кие клетки являются антигенпредставляющими
клетками для Т-супрессоров, проникающих в
эпидермис (Granstein R. D. и соавт., 1984).
Если это последнее предположение верно (а
фактов, его подтверждающих, появляется все
больше), то в эпидермисе существует своеобраз-
ная регуляторная система иммунных реакций,
действующая по типу «плюс-минус»: один по-
люс этой системы —• клетки Лангерганса и Т-
хелперы, второй — клетки Гринстейна и Т-су-
прессоры. Превалирование одной из систем будет
определять характер иммунной реакции в коже.
Меланоциты
В области базального слоя присутствуют мела-
ноциты (рис. 2.11). Эти клетки также имеют от-
ростчатую форму, но отличаются от клеток Лан-
генгарса наличием пигмента и положительной
ДОФА-реакцией (реакция, выявляющая процес-
сы синтеза меланина). Меланоциты имеют эк-
Рис. 2.11. Меланоциты.
а. Схема расположения меланоцитов в коже. Обратите внимание на то, как их отростки (выделенные
черным) проникают между клетками эпидермиса, доставляя им меланин (Хэм А., Кормак Д., 1983).
б. Схема электрнномикроскопического строения меланоцита (Stuettgen G., 1965): 1 — базальная мембра-
на; 2 — тело меланоцита; 3 отростки меланоцита; 4 — премеланосомы; 5 меланосомы;
6 зерна меланина в межклеточных пространствах; 7 — зерна меланина в цитоплазме кератиноцита.
в. Схема строения эпидермальной меланиновой единицы (Parker F., 1981):
а — кератиноциты; б — меланоцит.
1 базальная мембрана; 2 синтез тирозиназы в гранулярной эндоплазматической сети;
3 — комплекс Гольджи; 4 — премеланосомы; 5 — стадии формирования меланосом (I, II, III, IV);
6 — меланосомные комплексы (у европейца); 7 — отдельные гранулы пигмента (у африканца) по
F. Parker, 1981.
Глава 2. Анатомия и гистофизиология кожи
21
томезенхимное (нейроглиальное) прохождение.
Gramer S. и соавт. (1984) различают 4 стадии
дифференцировки меланоцитов: стадия предше-
ственников меланоцитов в составе нервного греб-
ня; стадия перемещения в дерме от нервных греб-
ней к базальной мембране эпидермиса; стадия
перемещения в эпидермис; дендритическая ста-
дия, когда меланоцит занимает свое место в эпи-
дермисе и формирует отростки.
Окончательная дифференцировка меланоцитов
происходит под влиянием тканевого окружения
и меланотропина гипофиза. Первые меланоциты
в эпидермисе человека появляются на 2-м меся-
це эмбриогенеза, а в дерме — на 2 недели рань-
ше. Их функцией является синтез меланина —
пигмента, придающего цвет коже, избирательно
адсорбирующего ультрафиолетовую часть спек-
тра солнечных лучей. Это является одним из ве-
дущих факторов в защите внутренней среды от
губительного действия на клетки ультрафиолето-
вых лучей. Кроме того, меланин обладает хоро-
шими теплоизоляционными свойствами. Цвет
кожи зависит не от количества меланоцитов, ко-
торое примерно постоянно у людей различных рас,
а от количества пигмента в одной клетке, хотя
доказано, что под действием ультрафиолетовых
лучей пролиферация меланоцитов может усили-
ваться. В отличие от клеток Лангерганса, мела-
ноциты связаны с соседними кератиноцитами по-
средством десмосом, а с базальной мембраной —
полудесмосомами. Отростки соседних меланоци-
тов также связаны десмосомами. Цитоплазма ме-
ланоцитов содержит небольшое количество фи-
ламентов. Цитоплазматическая сеть выражена
хорошо, количество митохондрий значительно.
Пластинчатый комплекс Гольджи развит хорошо.
В цитоплазме содержится множество везикул, ри-
босом, полисом и небольшое количество лизосом.
Ядро имеет неровные контуры мембраны с неглу-
бокими впячиваниями и очень плотной нукле-
оплазмой. Отростки меланоцитов содержат мно-
жество меланосом, рибосом и контактируют с
несколькими кератиноцитами при помощи дес-
мосом. Каждый меланоцит секретирует гранулы
меланина и связанные с ним кератиноциты. Это
партнерство «меланоцит-кератиноцит» называют
меланиновой эпидермальной единицей.
Меланосомы имеют высококонтрастную мел-
козернистую структуру и окружены элементар-
ной мембраной. Механизм образования мелано-
сом до настоящего времени неясен, но считают,
что развитие их протекает в три стадии. С уча-
стием рибосом и пластинчатого комплекса
Гольджи осуществляется синтез белка, фосфо-
липидов и фермента протирозиназы. Эти со-
ставные части будущих меланосом перемещают-
ся в пластинчатый комплекс Гольджи, где об-
разуется промежуточная везикула, отпочковы-
вающаяся от комплекса Гольджи. В эту стадию
из протирозиназы образуется тирозиназа. Из
аминокислоты тирозина при помощи фермен-
тов тирозиназы и ДОФА-оксидазы синтезиру-
ется меланин, и в меланосоме появляются сло-
истые структуры.
Механизм передачи меланина от меланоцитов
к кератиноцитам не совсем ясен. Предполагает-
ся, что это осуществляется путем фагоцитоза
кератиноцитом отдельных гранул или их групп
вместе с частью отростка меланоцита. В этом
процессе принимает участие филаментная система
меланоцита.
Функция меланоцитов, по-видимому, не ог-
раничивается только защитой от ультрафиолета,
т. к. эти клетки в значительном количестве при-
сутствуют в слизистой рта, а также в тех участ-
ках кожи, которые непосредственно ультрафи-
олетовому облучению не подвергаются.
Очевидно, клетки играют определенную, воз-
можно достаточно важную роль в межклеточных
взаимодействиях, но этот вопрос не изучен. По-
казана способность меланоцитов стимулировать
иммунную систему организма.
С патологией связан целый ряд заболеваний
меланоцитов. Меланома — злокачественное пе-
рерождение меланоцитов. Это одно из наиболее
быстротекущих новообразований. Большинство
меланом имеет темно-коричневый или черный
цвет; в некоторых случаях меланомы могут быть
красными, синими. Заболеваемость меланомой
быстро растет и составляет 2,5 % всех вновь вы-
являемых опухолей. Факторами риска являют-
ся: светлый цвет кожи, светлые глаза, белые или
рыжие волосы, солнечные ожоги 1 степени, дан-
ные анамнеза о наличии меланомы у родствен-
ников, врожденный волосяной невус или пиг-
ментная ксеродерма. Альбинизм — заболевание
связано с недостаточностью фермента тирозина-
зы или ее блокированием в меланоцитах, и про-
является недостаточной пигментацией кожи.
Люди с этим заболеванием не переносят солнеч-
ных лучей. Витилиго характеризуется появлением
на коже участков, лишенных пигмента. В таких
участках отсутствуют меланоциты и скаплива-
ются клетки Лангерганса. По современным пред-
ставлениям, заболевание связано с врожденной
неполноценностью меланоцитов, приобретением
ими свойств аутоантигенности. При этом клет-
ки Лангерганса вызывают лизис меланоцитов и
фагоцитируют их. Этой патологией далеко не
исчерпываются нарушения пигментации кожи.
2 2 Общая дерматология
Клетки Меркеля
Эти клетки впервые описаны в 1875 году немец-
ким гистологом Меркелем в эпидермисе морды
крота. В последующем они были обнаружены в
базальном слое эпидермиса у многих млекопи-
тающих как в покрытой волосами, так и в без-
волосой коже. Оволосенные области кожного
покрова содержат очень мало клеток Меркеля,
а в безволосых участках их число, напротив, су-
щественно увеличено. В некоторых, особо чув-
ствительных областях (ладони и пальцы челове-
ка, кожа морды крота и др.) плотность клеток
Меркеля значительно (в несколько раз) превы-
шает их количество в остальных участках тела.
Клетки Меркеля — нейроэндокринные клетки
кожи — по отдельности диссеминированы в ба-
зальном и шиповатом слоях эпидермиса, наруж-
ных слоях волосяного фолликула, а также в не-
которых слизистых. Скопления клеток Меркеля
находятся в так называемых волосяных дисках
Меркеля (Пинкуса), малых механорецепторных
органах волосистой кожи различных млекопи-
тающих, включая человека. Источником разви-
тия клеток Меркеля является нервный гребень.
Морфология клеток Меркеля весьма характер-
на, что отличает их от остальных клеток эпидер-
миса. Осязательные клетки Меркеля, локализо-
ванные в эпителии, округлой или удлиненной
формы. Они крупнее эпителиоцитов, ядро вытя-
нутое и дольчатое, цитоплазма светлее (рис. 2.12).
Клетки Меркеля соединяются с эпителиоцитами
при помощи десмосом и формируют контакт с
нервными терминалями. В цитоплазме равномер-
но распределены митохондрии, лизосомы, муль-
тивезикулярные тельца, вакуоли, микрофиламен-
ты. Эндоплазматическая сеть развита слабо. На
противоположной от ядра стороне расположен
хорошо выраженный комплекс Гольджи.
Типичным признаком клеток Меркеля явля-
ются нейроэндокринные специфические осми-
офильные гранулы размером от 80 до 200 нм и
свободно соединенные в пучки филаменты в ци-
топлазме (рис. 2.12). Биохимически филаменты
состоят исключительно из низкомолекулярных
цитокератинов, которые совершенно отличают-
ся от цитокератин-полипептидов окружающих
кератиноцитов. Поэтому эти цитокератины све-
томикроскопически являются полноценными
маркерами клеток Меркеля, которые впервые
подверглись углубленным исследованиям по их
распределению в коже человека.
В эпидермисе ладоней и подошв имеется от
200 до 400 клеток Меркеля на 1 мм2, в то вре-
мя как на основной поверхности кожи только
около 20 клеток Меркеля на 1 мм2, в феталь-
ном эпидермисе значительно больше (до 1700
на 1 мм2); они появляются уже с 8-й недели.
Интересно, что конгломераты из клеток Мер-
келя находятся в наружном волосяном влага-
лище человека в углублении воронки, в зоне
перешейка и на остальном протяжении воло-
сяного фолликула, а подвергшиеся цикличес-
ким изменениям части фолликулов свободны
от них. Такое необычное распределение и со-
седство клеток Меркеля с базальными клетками
волосяного фолликула, которые, вероятно, ло-
кализуются в утолщении перешейка, говорит
о их функциональном значении при росте во-
лос. В эккринных потовых железах клеток
Меркеля нет.
У взрослого человека клетки Меркеля скап-
ливаются в отдельные группы по 2—3 клетки.
Плотность распределения клеток становится
выше. Среди эллипсовидных появляются от-
Рис. 2.12. Ультраструктура клетки Меркеля.
В — вакуоли; Г — гранулы; БМ — базальная мембрана;
Мх — митохондрии; ПД — полудесмосомы;
Д — десмосомы; ПК — пластинчатый комплекс;
Л — лизосомы (х 13300). (Breathnach A. S., 1971).
Глава 2. Анатомия и гистофизиология кожи
23
дельные отростчатые клетки Меркеля. Изменя-
ется общая ориентация клеток: их продольная
ось становится параллельной поверхности базаль-
ной мембраны. Клетки кожи волосистой части
головы имеют самые крупные размеры. Нейро-
секреторные локализуются в базо-латеральных
отделах цитоплазмы, диффузно или реже фор-
мируют конгломераты, которые встречаются и в
единичных отростках клеток.
Наиболее существенные сдвиги в цитофизио-
логии клеток Меркеля происходят в старческом
возрасте, когда выявляется нехарактерный для
предыдущих возрастных групп тип распределения
клеток Меркеля. Они собираются в островки по
2~3, иногда 4 клетки, причем распределение их
по базальному слою крайне неравномерное, они
в основном располагаются в местах углубления
эпидермиса в дерму. Общее количество клеток
уменьшается, иногда они вообще с трудом выяв-
ляются на фоне общей атрофии эпидермиса.
Продольная ось клеток может быть ориентиро-
вана под углом к базальной мембране. Нейросек-
реторные гранулы перемещаются в апикальные
части клеток, уменьшаются их размеры, количе-
ство этих гранул. Лишь единичные отростки со-
держат гранулы, в основном же многочисленные
отростки от нейросекрета свободны.
В электронном микроскопе в клетках Меркеля
выявляются хорошо развитые комплекс Гольд-
жи и эндоплазматический ретикулум гладкого и
особенно гранулярного типа. Многочисленны
также свободные рибосомы, лизосомы, мульти-
венофиламенты. Специфический признак — на-
личие округлых осмиофильных гранул размером
до 120 нм, которые окружены мембраной. Гра-
нулы нередко сосредоточиваются в месте контак-
та клеток Меркеля с диском Меркеля. В этом
случае формируется подобие синапса. Базальная
часть клетки Меркеля формирует пресинаптичес-
кий полюс и содержит нейросекреторные грану-
лы. Синаптическая щель имеет ширину 30—
60 нм. Внутри щели можно обнаружить
фибриллярные структуры низкой электронной
плотности. Постсинаптический полюс образован
расширением терминали чувствительного не-
рвного волокна (диск Меркеля).
С соседними кератиноцитами клетки Мерке-
ля связаны десмосомами, а с базальной мембра-
ной — полудесмосомами.
Нормальные клетки Меркеля синтезируют
специфические для нервной ткани маркеры
(нейрофиламенты, периферии, нейрональные
клеточные адгезионные молекулы), а также
нейронспецифическую энолазу, протеин-ген-
ный продукт 9,5; хромогранин А и пептиды.
2 Зак. 1357
Оказалось, что они с различной плотностью рас-
пределены по всему человеческому кожному
покрову.
Функции клеток Меркеля следующие.
1. Те из них, которые расположены в волося-
ном диске Пинкуса, вовлечены в медленно адап-
тирующуюся механорецепторную функцию это-
го маленького чувствительного органа кожи
человеческого тела.
2. Эндокринные функции клеток Меркеля ас-
социируются с различными нейропептидами и
факторами роста, которые были локализованы
в клетках Меркеля. Установлено наличие в них
вазоактивного интестинального полипептида
(ВИП), фактора роста нервов, субстанции Р,
бомбезина, мет-энкефалина. В ходе роста пло-
да эти вещества, вероятно, оказывают трофи-
ческое влияние на растущие нервы и придатки
кожи.
3. Клетки Меркеля могут нести позиционную
информацию для определения места окончатель-
ной локализации терминали периферического
отростка чувствительного нейрона в нейрогене-
зе или при регенерации нервов.
4. Есть данные о роли клеток Меркеля в апоп-
тозе кератиноцитов (см. ниже).
На основании присутствия нейропептидов и
специфических гранул в клетках Меркеля их
считают нейроэндокринными клетками кожи и
относят к APUD-системе. Подтверждением этого
предположения является то, что в коже лягуш-
ки типичные элементы APUD-системы обнару-
жены давно, они продуцируют бомбезин. С по-
мощью метода моноклональных антител в
клетках Меркеля человека обнаружен целый ряд
полипептидных гормонов, вырабатываемых
клетками APUD-системы: мет-энкефалин, вазо-
интестинальный полипептид, фактор роста не-
рвов, бомбезин и другие. И. М. Кветной (1981)
указывает на наличие в клетках Меркеля чело-
века эндорфинов.
Роль клеток
Меркеля в патологии
При гипертрофической форме актинического
кератоза с формированием булавовидных зон
пролиферации эпидермиса отмечается увеличение
числа клеток Меркеля на 1 мм длины эпидермиса
(Y. Merot и соавт., 1989). Клетки формировали
скопления и при этом локализовались не только
в базальном слое, как в норме, но и в более вы-
сокорасположенных слоях, вплоть до верхних
рядов шиповатого слоя. Авторы считают, что
клетки Меркеля являются результатом постми-
2 4 Общая дерматология
тотической дифференцировки полипотентных
базальных кератиноцитов.
При хроническом воспалении кожи количество
клеток Меркеля чрезвычайно мало. С другой сто-
роны, в острых стадиях воспаления (острая УФ-
эритема, острая контактная экзема) они, вероят-
но, разрушаются и отторгаются подобно так
называемым «ожоговым» клеткам и в последую-
щем не возобновляются. Значение клеток Меркеля
при воспалительном процессе еще неизвестно. Ус-
тановлено, что иногда они скапливаются в эпи-
дермисе вблизи различных кожных опухолей, та-
ких как базалиома, опухоли придатков кожи и
кожные метастазы, а также в десмопластических
трихоэпителиомах. Трансформированные клетки
Меркеля присутствуют в нейроэндокринных кар-
циномах кожи и трабекулярных карциномах раз-
личной локализации (толстая кишка, легкие). В
подобных опухолях клетки Меркеля сохраняют не-
которые специфичные для них свойства. Так, им-
муногистохимическая реакция на нейронспецифи-
ческую энолазу остается положительной и служит
маркером диагностики опухолей, содержащих
трансформированные клетки Меркеля.
В настоящее время описана карцинома клеток
Меркеля (Toker С., 1972). Карцинома клеток
Меркеля (син. нейроэндокринная карцинома,
трабекулярная карцинома) — это злокачествен-
ная нейроэндокринная опухоль, дифференци-
ровка которой соответствует клеткам Меркеля.
Заболевание встречается редко, однако в после-
дние годы распознается все чаще, преимуще-
ственно у пожилых людей, больше у женщин
(4: 1). Ранее опухоль ошибочно диагностирова-
лась как недифференцированная карцинома,
малоклеточная злокачественная лимфома или
метастатическая бронхиальная карцинома. Ти-
пичная локализация опухоли: лицо, голова, шея,
реже — конечности, редко — туловище. Окра-
шенная в цвет кожи или красноватая, плотная,
экзофитная опухоль безболезненна, быстро ра-
стет и может изъязвляться. Размер ее при поста-
новке диагноза колеблется от 1-^2 до 14 см.
Гистопатологическая картина
Плотные гнезда или широкие трабекулярные ме-
стастазирующие пучки мономорфных малых опу-
холевых клеток с округлыми или овальными яд-
рами пронизывают дерму и достигают гиподермы.
Многочисленные митозы. Эпидермис вначале
остается интактным. Диагностически значимым
является иммунологическое подтверждение ней-
ронспецифической энолазы, нейрофиламентов и
в точечной пробе цитокератина 20. Электрон-
номикроскопическое исследование клеток опухо-
ли выявляет в них сильно развитую гранулярную
эндоплазматическую сеть и мономорфные ней-
росекреторные гранулы «dense core» («плотный
стержень») диаметром 100—200 нм.
Лимфоциты
В эпидермисе всегда находится то или иное ко-
личество лимфоцитов (внутриэпидермальные
лимфоциты, ВЭпЛ). Оно равно у человека 1—4 %
от всех клеток эпидермиса. Располагаются они
либо в базальном слое, либо супрабазально, в
шиповатом, но только в его нижних рядах, ни-
когда не достигая верхних рядов и зернистого
слоя. По данным Ю. К. Скрипкина и Е. М. Лез-
винской (1989), в интактном эпидермисе при
помощи проточной цитометрии обнаружено лишь
0,16 % лимфоцитов по отношению ко всем клет-
кам эпидермиса. При патологии количество лим-
фоцитов в эпидермисе может резко возрастать. По
данным J. Boss и соавт. (1989), самая высокая
инфильтрация эпидермиса лимфоцитами наблю-
дается при псориазе и самая низкая — при ато-
пическом дерматите. В большинстве случаев в
эпидермисе находили супрессоры, цитотоксичес-
кие Т-клетки, клетки памяти. Так называемые
«наивные» клетки были характерны для непов-
режденного эпидермиса. По данным R. Morgolis
и соавт. (1983), внутриэпидермальные лимфоци-
ты являются Т-хелперами и Т-супрессорами.
Морфология внутриэпидермальных лимфоци-
тов не отличается от других телиоцитов (внутри-
эпителиальных лимфоцитов). По описанию К. А.
Зуфарова и соавт. (1979), телиоциты имеют бес-
структурную цитоплазму и плотное ядро. Согласно
данным Мядельца О. Д. (1987), внутриэпидер-
мальные лимфоциты располагаются в базальном
слое или супрабазально. Они имеют округлое или
бобовидное ядро с плотно упакованным хромати-
ном. В ядре часто видны 1—2 ядрышка. Цитоплаз-
ма клеток не воспринимает красители, и ядра лим-
фоцитов окружены светлым ореолом. Иногда
встречаются лимфоциты с неправильными, слож-
ной формы ядрами. Возможно, это лимфоциты на
стадии миграции в эпидермис или из него. Рас-
положенные рядом с лимфоцитами ядра керати-
ноцитов имеют на прилежащей к кератиноциту по-
верхности характерное вдавление.
Функциональное значение внутриэпидермаль-
ных лимфоцитов изучено не до конца. Согласно
одним работам, они входят в состав периферичес-
кой подсистемы иммунологического надзора,
физиологическая роль которой заключается в
элиминации из кожи индуцированных вирусом
или злокачественно перерожденных клеток.
Глава 2. Анатомия и гистофизиология кожи
25
Streilein J. и соавт. (1985), S. Tanako и соавт.
(1985) показали, что у человека плоские бородав-
ки инфильтрируются Т-лимфоцитами и разруша-
ются ими. Н. Pehamberer и соавт. (1983) устано-
вили, что клетки Лангерганса способствуют
превращению Т-лимфоцитов в цитотоксические
клетки, направленные против аллогенных анти-
генов или сенсибилизированных тринитрохлор-
бензолом собственных клеток.
По данным других авторов (Didierjean L. и
соавт., 1980), эпидермис является местом экст-
ратимусной дифференцировки Т-лимфоцитов.
Установлено, что клетки эпидермиса эмбрионов
крыс стимулируют клетки аллогенного тимуса
при их совместном культивировании. Эта сти-
муляция осуществляется при помощи вырабаты-
ваемого кератиноцитами тимопоэтина.
Существует и третья точка зрения на функцию
внутриэпидермальных лимфоцитов. Согласно ей,
лимфоциты эпидермиса выполняют функцию
регуляторов пролиферативной активности кера-
тиноцитов. Эта точка зрения хорошо подтверж-
дается фундаментальными работами А. Г. Баба-
евой (1972, 1985) о важной роли системы
иммуногенеза в управлении процессами нор-
мального и регенераторного роста, а также ря-
дом наблюдений. К. И. Панченко (1978) при
изучении инфильтратов эпидермиса ауто- и ал-
лотрансплантантов полнослойной кожи мышей
показал, что степень инфильтрации эпидерми-
са лимфоцитами при аутотрансплантации прямо
пропорциональна его толщине. Автор высказал
предположение, что лимфоциты стимулируют ре-
генерацию эпидермиса. В. А. Флоренсов (1974),
изучая расположение лимфоцитов в многослой-
ном плоском, однослойном цилиндрическом и
других эпителиях, установил, что проникшие в
толщу тканей лимфоциты в определенном соот-
ношении (от 1 :90 до 1 : 1300) контактируют с
местными клетками ткани. Параметры инфиль-
трации эпителиев лимфоцитами коррелировали
с митотическими индексами, степенью диффе-
ренцировки эпителиальных клеток. На связь
количества внутриэпидермальных лимфоцитов
со степенью дифференцировки эпителиальных
клеток указывают также К. И. Панченко и со-
авт. (1983). D. A. Chambers и соавт. (1984) обна-
ружили на 90 % эпидермоцитов классические
маркеры Т-лимфоцитов. Авторы предположили,
что эти маркеры играют большую роль во вза-
имодействии кератиноцитов и Т-лимфоцитов и
в регуляции пролиферации и дифференциров-
ки кератиноцитов. S. Tanaka и соавт. (1983) по-
казали, что лимфоциты селезенки обладают
сильно выраженной способностью вызывать
спонтанную клеточно-опосредованную цитоток-
сичность кератиноцитов, лимфоциты перифери-
ческой крови проявляют эту способность в ма-
лой степени, а лимфоциты тимуса и лимфоузлов
не обладают такой способностью. Авторы счи-
тают, что спонтанная клеточно-опосредованная
цитотоксичность играет роль в регуляции диф-
ференцировки и пролиферации кератиноцитов.
В. И. Ноздрин (1992) показал, что существует
тесная связь между лечебным действием пальми-
тата ретинола у больных с нарушением орогове-
ния, содержанием в эпидермисе субпопуляций Т-
лимфоцитов, способностью лимфоцитов и
кератиноцитов взаимодействовать между собой и
пролиферативной активностью эпидермиса. А. Н.
Глушков (1989) считает, что есть клоны Т-лим-
фоцитов, специфических к распознаваемым ком-
плексам нелимфоидных клеток. Автор утверждает,
что одной из важных функций Т-лимфоцитов
является способность регулировать состояние
межклеточных контактов, при этом существуют
различающиеся по специфичности клоны лимфо-
цитов для каждого типа контактов.
Тканевые базофилы
В литературе имеются сведения о проникновении
тканевых базофилов в интактный эпидермис. Так,
К. Ш. Баймухамбетов и соавт. (1991) обнаружи-
ли миграцию этих клеток из дермы в эпидермис
у тонкорунных овец. В эпидермисе тканевые
базофилы подвергались дегрануляции, при этом
со временем гранулы, вышедшие из клеток, по-
степенно светлели, вакуолизировались и исчеза-
ли. Есть сведения о наличии тканевых базофи-
лов и в интактном эпидермисе человека. Данные
наблюдения показывают, что сфера функциональ-
ных отправлений тканевых базофилов не ограни-
чивается только соединительной тканью. Ранее
описано проникновение тучных клеток в мерца-
тельный эпителий дыхательных путей и в мно-
гослойный плоский эпителий шейки матки (В. А.
Проценко и соавт., 1987). В работе О. Д. Мядельца
(1993) установлен факт проникновения тканевых
базофилов в многослойный эпителий новообра-
зующихся волосяных фолликулов при органоти-
пической регенерации кожи. При мастоцитозе
тканевые клетки проникают в эпидермис в боль-
шом количестве.
Современные представления
о процессах кератинизации
Морфология процесса кератинизации была опи-
сана достаточно хорошо еще на уровне светово-
2 6 Общая дерматология
го микроскопа. В ходе этого процесса кератино-
циты увеличиваются в размерах (до 30 раз), уп-
лощаются, в них постепенно конденсируется
ядро, в цитоплазме накапливается кератогиалин.
Затем ядро подвергается разрушению, а цито-
плазма гомогенизируется. В клетках накаплива-
ется кератин. В конце концов клетки превраща-
ются в роговые чешуйки. Электронный
микроскоп позволил установить более тонкие
процессы, происходящие при кератинизации.
После миграции базального кератиноцита в ши-
поватый слой увеличивается число десмосом, свя-
зывающих его с другими клетками, а также чис-
ленность тонофиламентов. В верхних рядах
шиповатого слоя в кератиноцитах появляются
пластинчатые гранулы Одланда. Гранулы Одлан-
да занимают периферическое положение и вы-
деляют путем экзоцитоза свое содержимое в
межклеточное пространство, где оно приобрета-
ет пластинчатое строение (межклеточный це-
мент). В цитоплазме зернистых кератиноцитов
выявляются тонофибриллярно-кератогиалиновые
комплексы. Цитолемма клеток заметно утолща-
ется. В ядрах клеток часто видны глубокие ин-
вагинации ядерной оболочки. Хроматин распре-
деляется неравномерно, образуя главным образом
скопления около ядерной мембраны. В нукле-
оплазме нарастают явления разрежения или про-
светления. В конечном итоге ядра клеток разру-
шаются и исчезают. Постепенно редуцируются
органеллы. Матрикс митохондрий разрежается,
в них уменьшается число крист. В последующем
митохондрии разрушаются и исчезают. Исчеза-
ют эндоплазматическая сеть и свободные рибо-
сомы. Кератиноциты превращаются в роговые
чешуйки, склеенные цементирующим материа-
лом и десмосомами. Десмосомы при этом видо-
изменяются: в них теряется межклеточный ком-
пактный слой с последующим слиянием
промежуточных слоев. Роговые чешуйки в ро-
говом слое различаются по распределению кера-
тиновых фибрилл, которое постепенно изменя-
ется по мере созревания роговых чешуек от менее
к более плотному.
Биохимические и иммуноцитохимические ис-
следования лишь в последнее время позволили
понять многие тонкие механизмы кератинизации.
Синтез кератина в форме прекератина начинает-
ся в базальных кератиноцитах. Прекератин имеет
более низкий молекулярный вес, чем зрелый ке-
ратин (40—60 000 дальтон при молекулярной мас-
се зрелого кератиноцита более 60 000 дальтон) и
характеризуется отсутствием внутри- и межцепо-
чечных дисульфидных связей, придающих моле-
кулам кератина прочность и нерастворимость. В
более высоких слоях эпидермиса (шиповатый,
зернистый) биосинтез прекератина продолжается
и даже нарастает, причем молекулярная масса син-
тезируемого прекератина увеличивается. В нижних
рядах рогового слоя прекератин различной моле-
кулярной массы под действием специфических
ферментов превращается в зрелый кератин. При
этом между отдельными полипептидными моле-
кулами и внутри них образуются дисульфидные
связи. Происходит укорочение молекул. Кератин
теряет свойство растворимости в воде и приобре-
тает прочность.
Для упаковки кератина необходим особый
белок филагрин (ранее он назывался «белок, бо-
гатый гистидином». Этот белок вызывает агре-
гацию кератиновых филаментов. Биосинтез фи-
лагрина осуществляется в зернистом слое с
участием специфической и-РНК. Филагрин об-
ладает кислотными свойствами и отличается
сильной степенью фосфорилирования. По мере
синтеза он накапливается в виде кератогиалино-
вых гранул и существует до тех пор, пока плот-
но упакованный кератин не стабилизируется
прочными дисульфидными связями. Как толь-
ко это произойдет, филагрин в корнеоцитах ро-
гового слоя распадается до свободных аминокис-
лот. Интересна их функция в корнеоците:
свободные аминокислоты поддерживают в нем
на высоком уровне осмотическое давление, что
вызывает приток воды, которая сохраняется в
роговых чешуйках даже при понижении влаж-
ности внешней среды. Аккумулированная вода
придает чешуйкам упругость и механичесую
прочность. Кроме того, аминокислота гистидин
превращается в роговых чешуйках в урокановую
кислоту, участвующую в защите кожи от ульт-
рафиолетовых лучей. Все эти превращения фи-
лагрина и входящих в него аминокислот проис-
ходят при участии ферментов, что противоречит
укоренившемуся представлению о роговой че-
шуйке как о мертвой клетке.
В клетках зернистого слоя синтезируется так-
же белок кератолинин или инволюкрин. Этот
белок выстилает внутреннюю поверхность ци-
толеммы зернистых кератиноцитов, утолщая ее.
Утолщенная плазмолемма успешно противостоит
гидролитическим процессам, которые интенсив-
но протекают в верхних рядах клеток зернис-
того слоя и приводят к разрушению кератино-
цитов. В роговом слое молекулы кератолинина
сшиваются глютамил — лизиновой связью при
помощи ферментов трансглютаминаз и дисуль-
фидными мостиками при помощи кальцийза-
висимой сульфгидрилоксидазы. Появление этих
связей в белковых молекулах обусловливает
Глава 2. Анатомия и гистофизиология кожи
27
прочность и нерастворимость их даже в очень
сильных реагентах. В результате корнеоцит при-
обретает прочную толстую так называемую по-
перечно-сшитую оболочку, характерную толь-
ко для ороговевающего эпителия. У ряда
наземных животных не обнаруживаются кера-
тогиалиновые гранулы, а у некоторых водных
млекопитающих, кроме того, не образуется пол-
ноценная поперечно сшитая оболочка. Анало-
гичная ситуация наблюдается при ряде заболе-
ваний кожи.
В процессе кератинизации важная роль при-
надлежит кератиносомам. Эти видоизмененные
лизосомы путем экзоцитоза выделяют свое со-
держимое в межклеточные пространства, где оно
превращается в богатый липидами межклеточный
цемент с пластинчатым строением. Если корне-
оцит содержит лишь 3 % липидов, то межкле-
точный цемент — 80 % (остальные 20 % прихо-
дятся на белки различной молекулярной массы).
Кератиносомы содержат по крайней мере 4 груп-
пы веществ: гликолипиды, гликопротеины, гид-
ролитические ферменты, свободные стерины. По
биохимическим данным в межклеточном матрик-
се обнаруживаются церамиды, холестерин и его
эфиры, свободные жирные кислоты, триглице-
риды, сквалены, белки. Основную массу состав-
ляют церамиды. Все эти вещества обеспечивают
адгезию корнеоцитов, а также, за счет гидрофоб-
ности своих молекул, формируют барьерный
слой эпидермиса.
Таким образом, процессы, происходящие при
кератинизации, заключаются в следующем: об-
разовании нерастворимого белка кератина; ви-
доизменении десмосом с растворением межкле-
точного слоя; лизисе ядер кератиноцитов и их
органелл; экзоцитозе гранул Одланда с образо-
ванием гидрофобного межклеточного цементи-
рующего матрикса; образовании кератолинина,
укреплении его глутамил-лизиновыми и дисуль-
фидными связями с формированием нераство-
римой поперечно-сшитой оболочки; гибели клет-
ки и превращении ее в роговую чешуйку или
корнеоцит.
Структура эпидермальной
пролиферативной единицы
В последнее время установлено, что эпидермис
млекопитающих организован в структурно-
функциональные единицы, названные эпидер-
мальными пролиферативными единицами (ЭПЕ)
(рис. 2.13). ЭПЕ имеют форму вертикальной
колонки. В состав этой колонки входят 6—12
роговых чешуек, две зернистые, одна шипова-
тая и 8—12 базальных клеток. Роговые чешуй-
ки имеют гексагональные очертания и форму че-
тырнадцатигранника (тетрадекаэдра). Именно та-
кая форма наиболее экономна и идеально
укладывается в столбики. Каждой чешуйке, на-
ходящейся на поверхности эпидермиса, соответ-
ствует своя ЭПЕ. ЭПЕ могут иметь столбчатую
и нестолбчатую структуру, в зависимости от
функционального состояния эпидермиса. При
незначительной функциональной нагрузке на
эпидермис он тонкий, а ЭПЕ имеют колонча-
тое строение. При увеличении нагрузки на эпи-
дермис (например, механической) он утолщает-
ся, а ЭПЕ теряют колончатую организацию.
Роговые чешуйки столбчатого эпидермиса
лишены десмосом. Их уплощенные горизонталь-
ные поверхности, обращенные друг к другу, со-
вершенно гладкие. Однако по периметру каж-
дая чешуйка имеет электронноплотную зону,
которой связывается с чешуйками соседних ко-
лонок. Эти зоны называются сквамосомами.
Сквамосомы окружают каждую колонку зам-
кнутым кольцом и связывают соседние колон-
ки рогового слоя в единую систему. В центре
ЭПЕ среди базальных и супербазальных кера-
тиноцитов лежит клетка Лангенгарса, которая,
как полагают, регулирует деление базальных ке-
ратиноцитов. Базальные кератиноциты, лежа-
щие в центре ЭПЕ (3—4 клетки), делятся реже,
чем кератиноциты периферических зон колон-
ки. Из этих 3—4 базальных кератиноцитов одна
является стволовой, способной воссоздавать
структуру ЭПЕ. Эта клетка находится на дне
эпидермальных углублений и содержит много
гранул меланина, т. е. защищена от вредных
факторов местоположением и меланином. Ство-
ловые клетки эпидермиса не имеют на базаль-
ной поверхности выростов цитолеммы и назы-
ваются «гладкими» клетками. Периферические
базальные кератиноциты называются транзит-
ными делящимися клетками. На их базальной
поверхности имеются цитоплазматические вы-
росты («зубчатые» клетки). По мере своего де-
ления эти клетки могут иметь 3 пути развития:
возвращаться в пул стволовых клеток, оставать-
ся транзитными делящимися, приступать к диф-
ференцировке. В последнем случае они вытал-
киваются из базального слоя в вышележащие
слои. Транзитные делящиеся кератиноциты
отличаются от стволовых также тем, что в их
цитоплазме имеются тонофибриллы. В некото-
рых случаях переходные клетки сохраняют спо-
собность совершить 2—4 деления даже после пе-
рехода в супрабазальное положение. В этом
случае картины митоза наблюдаются в нижних
28
Общая дерматология
А
Рис. 2.13. Схема организации эпидермальной пролиферативной единицы.
А. По Е. Ф. Котовскому (1999): 1 — базальный слой; 2 — шиповатый слой; 3 — зернистый слой;
4 — блестящий слой; 5 — роговой слой; 6 — базальная мембрана; 7 — кератиноцит; 7а — митоз;
8 — кератиновые филаменты; 9 — десмосома; 10 — внутриэпидермальный макрофаг (клетка
Лангерганса); 11 — гранулы Бирбека; 12 — кератиносомы; 13 — слой кератолинина; 14 — образова-
ние межклеточного цемента; 15 — межклеточный цемент; 16 — корнеоцит (роговая чешуйка);
17 — отпадающая роговая чешуйка.
Б. По Б. Альбертсу и соавт. (1987): 1 — базальный кератиноцит; 2 — шиповатый кератиноцит;
3 — зернистый кератиноцит; 4 — роговые чешуйки; 5 — отделяющаяся от поверхности эпидермиса
роговая чешуйка.
В. По С Поттену (1974): 1 — базальная мембрана; 2 — «гладкие» кератиноциты; 3 — «зубчатые» керати-
ноциты; 4 — клетка Лангерганса; 5 — шиповатый, 6 — зернистый, 7 — роговой слои эпидермиса;
8 — сквамосомы.
Глава 2. Анатомия и гистофизиология кожи
29
клетках шиповатого слоя. Это, однако, встре-
чается редко и лишь при действии на эпидер-
мис неблагоприятных факторов и патологий. В
норме большинство клеток при переходе в суп-
рабазальное. положение необратимо утрачивают
способность к делению и вступают на путь
дифференцировки.
Эпидермис со столбчатой организацией хоро-
шо предохраняет организм от потерь влаги и
тепла, чему способствует уподобление рогового
слоя стеганому ватному одеялу. При ихтиозе,
когда утрачивается столбчатая организация эпи-
дермиса, проницаемость эпидермиса для воды
существенно изменяется. При всей своей эконо-
мичности столбчатый эпидермис хорошо (лучше,
чем нестолбчатый эпидермис) защищает кожу от
действия ультрафиолетовых лучей.
На подошве у человека эпидермис приспособ-
лен к максимальным механическим нагрузкам и
должен быть резистентным к постоянному тре-
нию. В этом случае (как адаптивное явление)
между кератиноцитами и корнеоцитами сохра-
няются десмосомы.
В столбчатом эпидермисе процессы превра-
щения кератиноцитов в корнеоциты существен-
но замедлены по сравнению с нестолбчатым.
Так, в эпидермисе уха мыши, где ЭПЕ имеют
столбчатую организацию, время превращения
базальных клеток в роговую чешуйку состав-
ляет 24 дня, тогда как в эпидермисе подошвы
мыши, где колонки полностью отсутствуют,
процесс ороговения совершается за 6 суток.
Увеличение митотической активности, вызван-
ное искусственным путем, ведет к увеличению
толщины эпидермиса и потере им столбчатого
строения.
В формировании колончатой структуры эпи-
дермиса участвует фибронектин базальной мем-
браны. Дерма продуцирует вещества неизвестной
природы, которые также способствуют органи-
зации эпидермиса в колонки.
Тканевой гомеостаз в эпидермисе.
Кинетика популяции кератиноцитов
и ее регуляция
Каждая ткань характеризуется определенным
уровнем постоянства своего качественного и
количественного состава. Тканевой гомеостаз
обеспечивается такими фундаментальными про-
цессами, как клеточное обновление, цитодиффе-
ренцировка и клеточная гибель.
В эпидермисе постоянство качественных и
количественных параметров поддерживается эти-
ми же механизмами. Однако в связи с тем, что
эпидермис является пограничной тканью с уни-
кальной особенностью дифференцировки, в его
гомеостазе играют важную роль и такие явления,
как направленная миграция клеток и слущива-
ние корнеоцитов с поверхности.
Таким образом, гомеостаз эпидермиса как
ткани обеспечивают следующие процессы: кле-
точное обновление; целенаправленная миграция
клеток в эпидермальном пласте; цитодифферен-
цировка с превращением кератиноцитов в кор-
неоциты; гибель корнеоцитов, причем в эпидер-
мисе различают две формы клеточной гибели:
гибель в результате процесса кератинизации и
апоптотическая гибель; слущивание корнеоци-
тов с поверхности эпидермиса.
Каждый из этих процессов вносит свою леп-
ту в тканевой гомеостаз и подвержен строгой
регуляции. Ниже мы рассмотрим более подроб-
но каждый из этих процессов и известные к
настоящему времени механизмы их регуляции,
поскольку нарушение их лежит в основе ряда
кожных заболеваний. При этом нарушения ука-
занных процессов могут быть изолированными,
но значительно чаще — комбинированными,
сочетанными.
Клеточное обновление
Клеточному обновлению как наиболее важному
процессу и его регуляции будет уделено особое
внимание.
Эпидермис относится к камбиальным обнов-
ляющимся тканям. Выше отмечалось, что вос-
становление постоянных клеточных потерь как
результата кератинизации и слущивания с
поверхности эпидермиса корнеоцитов происхо-
дит за счет стволовых эпидермальных клеток
(«гладкие клетки»). Они отличаются неспециа-
лизированной цитоплазмой, почти не содержа-
щей признаков дифференцировки; очень низкой
митотической активностью; дают переходную
популяцию (транзитные клетки) пролифериру-
ющих клеток, которые несколько раз делятся,
что приводит к увеличению числа клеток, воз-
никших за счет деления единственной стволовой
клетки; стволовые клетки можно стимулировать
к делению путем искусственного удаления ро-
гового слоя, травм или с помощью специфичес-
ких стимуляторов.
В норме в эпидермисе митотически делящие-
ся кератиноциты локализуются в подавляющем
большинстве случаев в базальном слое и лишь
иногда — в нижних рядах шиповатого слоя. По
данным многих авторов (Лиознер Л. Д., 1966;
30
Общая дерматология
Заварзин А. А., 1967, 1976; Романов Ю. А. и
соавт., 1984 и др.), митотический индекс у мы-
шей может колебаться от 0 до 15 % и зависит
от топографии кожи и времени суток. Он более
низкий в тонком эпидермисе уха и более высо-
кий в коже стопы, а в толстом эпидермисе по-
дошв — максимален. В утренние часы показа-
тель имеет наибольшее значение, а в вечернее
время минимален. У человека наибольшее коли-
чество митозов встречается в интервале 0—4 часа
и снижено (в основном равномерно) в остальное
время суток. Среднесуточные значения митоти-
ческой активности у человека составляют 6 %
(Лиознер Л. Д., 1966).
Скорость обновления кератиноцитов зависит
от возраста животных. Так, у новорожденных
животных она наибольшая, к 5-дневному воз-
расту снижается, а у половозрелых животных ста-
новится еще меньше (Лиознер Л. Д., 1966). По
данным Мядельца О. Д. (1985; 1995), митоти-
ческая активность в эпидермисе спины крыс
имеет максимальные значения у новорожденных
животных и снижается до значений, характер-
ных для половозрелых крыс, в течение первого
месяца жизни, повышаясь в фазу анагена цик-
ла волос.
В связи с чрезвычайной важностью процессов
пролиферации кератиноцитов для нормального
функционирования нс только кожи, но и всего
организма они подвержены строгой многоуров-
невой регуляции. На внутриклеточном уровне
эта регуляция осуществляется разными путями.
Н. И. Епифанова и соавт. (1990) разделили все
эндогенные регуляторы размножения на: 1) ком-
поненты цитоплазматических мембран; 2) цито-
плазматические ингибиторы (триггерные моле-
кулы, в том числе и белки); 3) ядерные
регуляторы. На ядерном уровне митотическая ак-
тивность регулируется генетическими механиз-
мами. Существуют гены, ответственные за вос-
производство клеток. Эти гены кодируют белки,
обеспечивающие деление и дифференцировку
клеток (протеинкиназы, ядерные белки, факто-
ры роста, гуанозинтрифосфатазы). Так, ген
с-егЬВ кодирует рецептор фактора роста эпидер-
миса, а ген егЬА -рецептор стероидных гормонов.
Гены клеточной пролиферации весьма схожи по
строению с вирусными онкогенами и были на-
званы протоонкогенами. При мутации этих ге-
нов они могут спровоцировать опухолевый рост.
Различают так называемые «ранние» и «по-
здние» протоонкогены. Ранние протоонкогены
(myc, fos и др.) создают состояние компетентно-
сти клеток к дальнейшей пролиферации. В при-
сутствии внеклеточных факторов или при дей-
ствии внеклеточных сигналов «компетентные»
клетки начинают экспрессировать «поздние»
гены (ras, myb и др.), которые контролируют про-
хождение клеткой пререпликативного периода.
Таким образом, протоонкогены являются частью
системы генетического контроля клеточного раз-
множения. Наряду с онкогенами существуют ан-
тионкогены, т. е. такие гены, продукты которых
являются супрессорами митотической активно-
сти клеток и, следовательно, развития опухолей.
Из них наиболее известным является ген р53. Он
описан в 1979 году и находится в 17-й хромо-
соме. Ген имеет весьма похожую структуру у всех
позвоночных животных. Активация этого гена
ведет к образованию белка с аналогичным на-
званием (белок р53). Данный белок локализуется
в ядре клетки и в норме очень нестабилен, но
его стабильность резко возрастает при действии
веществ, повреждающих ДНК. В результате это-
го концентрация белка р53 в ядре резко увели-
чивается. Он производит остановку клеточного
цикла, связывается с поврежденным или изме-
ненным участком ДНК и участвует в ее репара-
ции. Если таковая невозможна из-за грубых де-
фектов, белок р53 включает программу апоптоза
измененной клетки, предотвращая репликацию
поврежденного генетического материала и пере-
нос измененной генетической информации от од-
ного поколения клеток к другому. Кроме того,
белок р53 блокирует переход клетки от G]- к S-
периоду, подавляя ферменты, обеспечивающие
этот переход (циклин-зависимые киназы). Эта
функция белка р53 осуществляется путем индук-
ции синтеза продуктов генов р15, р16 и р21. В
связи с описанными функциями белок р53 час-
то называют «сторожем генома», «аварийным тор-
мозом пролиферации».
Нарушение функций гена р53 (его называют
также «диким» типом) в результате мутаций ве-
дет к потере генетического контроля над клеточ-
ным циклом, активной пролиферации клеток-
мутантов и возникновению злокачественных
опухолей (около 50 % опухолей человека связано
с мутацией гена р53). При этом возникает му-
тантный тип белка р53, не способный индуциро-
вать апоптоз измененных клеток. Более того, в
тех случаях, когда одна из аллелей гена сохра-
няется, в опухолевой клетке синтезируются сразу
два типа белка р53, причем мутантный белок
связывается с «диким» и полностью его инак-
тивирует. Это может привести к стимуляции
злокачественного роста. Поэтому существует вы-
ражение, что «... белок р53 как ген-супрессор
опухолей имеет два лица». В отличие от «дико-
го» белка р53 мутантный тип данного белка яв-
Глава 2. Анатомия и гистофизиология кожи
31
ляется стабильным, имеет большой период по-
лужизни, в результате чего его можно легко оп-
ределить иммуногистохимическими методами
при различных опухолевых и предопухолевых
состояниях.
Злокачественные опухоли кожи могут возни-
кать и при мутациях гена р16 (в частности, до
60 % меланом связаны с данной мутацией), а
также гена р15.
Кроме генетического контроля, на внутрикле-
точном уровне митотическая активность регули-
руется также особыми триггерными белками,
которые продуцируются в цитоплазме клеток и
способны включать или выключать в ядре гены,
ответственные за митотическую активность.
К эндогенным ингибиторам размножения от-
носятся также компоненты плазматических мем-
бран, которые, очевидно, участвуют в контакт-
ном торможении размножения.
На межклеточном или тканевом уровне мито-
тическая активность кератиноцитов регулируется
за счет кейлонного механизма. Гипотеза кейло-
нов W. Bullough, Е. Lourence (1962) до сих пор
является наиболее всеобъемлющей теоретической
моделью. Согласно этой гипотезе, дифференци-
рованные клетки эпидермиса вырабатывают бел-
ковые ингибиторы пролиферативной активнос-
ти стволовых клеток эпидермиса или их
ближайших потомков. В настоящее время раз-
личают два типа кейлонов эпидермиса, действу-
ющих в различные фазы митотического цикла:
Q]- кейлон блокирует вступление клетки из Q,-
фазы в синтетическую фазу, а Р2-кейлон инги-
бирует переход от О2-фазы в собственно митоз.
Ингибирование митозов кейлонами усиливает-
ся адреналином (кейлон-адрсналиновый комп-
лекс), что призвано при стрессе предохранять
эпидермис от избыточной потери клеток (Кар-
лсон Б. М., 1986). Митогенные гормоны нейт-
рализуют действие кейлонов.
Из внутриэпидермальных регуляторов мито-
тической активности кератиноцитов следует так-
же назвать у-интерферон, продуцируемый клет-
ками Лангерганса, и систему циклических
нуклеотидов.
у-Интерферон является ингибитором деления
кератиноцитов. Он продуцируется клетками
Лангерганса в достаточно больших количествах.
Обладая противовирусной активностью, интер-
фероны, снижая скорость синтеза ДНК, подав-
ляют также пролиферацию кератиноцитов (Ба-
лаж А., Блажек К., 1982). Кроме у-интерферона,
подавляющего митотическую активность клеток,
аналогичным эффектом может обладать проду-
цируемый клетками Лангерганса ИЛ-1. Во вся-
ком случае, его митозингибирующий эффект
описан для ряда клеточных систем (Sporn М. В.,
Roberts А. В., 1988).
Циклический аденозинмонофосфат (цАМФ)
подавляет митотическую активность кератино-
цитов, а циклический гуанозинмонофосфат
(цГМФ) оказывает противоположный эффект.
Механизм действия циклических нуклеотидов
до конца не изучен. Предполагается, что их
эффективность реализуется путем активации
фосфорилирования белков, регулирующих ак-
тивность генов. Фосфорилирование этих белков
вызывает изменение их конформации и в ре-
зультате — активности. цАМФ активирует пу-
тем фосфорилирования гистоны, которые подав-
ляют активность генов, а цГМФ — кислые
белки — активаторы генов.
Установлено, что кератиноциты могут регули-
ровать собственную митотическую активность
аутокринным путем. Они вырабатывают так на-
зываемый трансформирующий фактор роста а,
стимулирующий митотическую активность других
кератиноцитов (Улумбеков Э. Г. и соавт., 1997).
Митотическая активность кератиноцитов ре-
гулируется на органном уровне. Регуляторные
влияния могут исходить из подлежащей дермы,
с которой эпидермис находится в тесных меж-
тканевых взаимодействиях. В частности, фибро-
бласты дермы вырабатывают фибробластический
фактор роста кератиноцитов, стимулирующий их
митотическую активность (Улумбеков Э. Г. и
соавт., 1997).
На уровне организма митотическую актив-
ность кератиноцитов регулируют эндокринная,
нервная и иммунная системы.
Все гормоны, приводящие к повышению
внутриклеточной концентрации цАМФ, можно
рассматривать как неспецифические ингибито-
ры клеточной пролиферации, в том числе и кс-
ратиноцитов (Балаж А., Блажек И., 1982). Та-
кими гормонами являются АКТГ, глюкагон,
катехоламины (p-адренергическис гормоны),
кортиколиберин, меланоцитстимулирующий гор-
мон, паратирин и некоторые другие гормоны. На-
против, гормоны, снижающие уровень цАМФ в
клетках, активируют митотическую активность
(опиоиды, а2-адренергические гормоны и др.).
Некоторые другие гормоны влияют на мито-
тическую активность эпидермиса нс через сис-
тему циклических нуклеотидов, а через непос-
редственное влияние на генотип клеток.
Подавляют митоз кератиноцитов стероидные, а
стимулируют — тиреоидные гормоны. Обе эти
группы гормонов имеют внутриклеточный ме-
ханизм действия.
32 Общая дерматология:
Стимулирует деление кератиноцитов фактор
роста эпидермиса (ФРЭ). Этот полипептид сек-
ретируется слюнными железами. Механизм дей-
ствия гормона заключается в том, что он ин-
дуцирует активность тирозинкиназы и
фосфорилирование аминокислоты тирозина в
мембранном рецепторе и в некоторых связан-
ных с ним белках. Это фосфорилирование слу-
жит сигналом, опосредующим внутриклеточные
эффекты ФРЭ и приводит к активации деле-
ния кератиноцитов. Интересно, что рецепторы
для ФРЭ имеют поразительное сходство с ре-
цептором для инсулина (который также спосо-
бен стимулировать деление кератиноцитов) и
липопротеинов низкой плотности (Marri R. Е.,
1993).
Так называемый стероидный гормон почек,
или дигидроксихолекальциферол, подавляет син-
тез в кератиноцитах ДНК, их митотическую
активность и стимулирует их терминальную
дифференцировку. На этом основан положитель-
ный лечебный эффект дигидроксихолекальци-
ферола при псориазе, в основе которого лежит
нарушение процессов деления кератиноцитов.
Регулирующее влияние на пролиферативную
активность кератиноцитов оказывает нервная
система. В настоящее время в эпидермисе выяв-
лены два типа симпатических нервных волокон:
у одних медиатором является норадреналин, у
других — ацетилхолин. Норадреналин, стимули-
руя образование в кератиноцитах цАМФ, тормо-
зит их пролиферацию, а ацетилхолин, снижая
уровень этого нуклеотида, стимулирует ее (Кар-
лсон Б. М., 1987; Грабовой А. Н., 1994). Выде-
ляющееся из терминалей чувствительных не-
рвных окончаний вещество Р также стимулирует
митотическую активность кератиноцитов.
В настоящее время доказано регулирующее
влияние иммунной системы на клеточное деле-
ние. Способность Т-лимфоцитов стимулировать
пролиферацию клеток нелимфоидных органов
доказана работами А. Г. Бабаевой (1972; 1985;
1987). С другой стороны, была доказана способ-
ность лимфоцитов подавлять пролиферацию в
этих органах (Донцов В. И., 1990). В настоящее
время показано, что лимфоциты способны регу-
лировать митотическую активность в нелимфо-
идных органах путем секреции медиаторов (Дон-
цов В. И., 1990), однако сведения об этих
медиаторах для нелимфоидных органов весьма
скудны. Вместе с тем хорошо известно, что сте-
пень инфильтрации эпидермиса лимфоцитами
коррелирует с уровнем митотической активности
в нем (Флоренсов В. И., 1986; Панченко К. И.,
1983; Мяделец О. Д., 1985). К иммунной регу-
ляции митотической активности кератиноцитов
следует также отнести подавление ее клетками
Лангерганса, являющимися внутриэпидермаль-
ными макрофагами.
Таким образом, митотическая активность
эпидермиса находится под жестким контролем,
что является одним из условий тканевого го-
меостаза. В дерматологии пролиферация кера-
тиноцитов имеет особо важное значение, так как
подавляющее большинство дерматозов, незави-
симо от их этиологии и патогенеза, на опреде-
ленном этапе своего развития могут характери-
зоваться нарушениями клеточной пролиферации
и дифференцировки, что приводит к таким
морфологическим феноменам, как акантоз, ги-
перкератоз, гипергранулез, дискератоз и др. (Бе-
ренбейн Б. А., 1980). Некоторые авторы
(Voorhees J. J. и соавт., 1975) условно выделя-
ют пролиферативные заболевания кожи, куда
включают псориаз, экзему, ихтиоз, рак и др.
Миграция кератиноцитов
Второй процесс, определяющий тканевой гоме-
остаз эпидермиса — миграция кератиноцитов. О
механизмах перемещения клеток в эпидермаль-
ном пласте известно немного. Многие авторы
рассматривают этот процесс как пассивный, как
результат выталкивания предсуществующих ке-
ратиноцитов в более высокие слои вновь обра-
зующимися клетками. Разумеется, в этом есть
своя логика. Однако нельзя исключить и актив-
ную миграцию клеток за счет деятельности ци-
тоскелета. Способность кератиноцитов к мигра-
ции хорошо доказана при травмах кожи, когда
эти клетки, активно двигаясь, быстро перекры-
вают зону дефекта. Показано, что в ЭПЕ колон-
чатой организации передвижение кератиноцитов
в процессе созревания достаточно сложное, спи-
ралевидное, что трудно объснить с позиций пас-
сивного выталкивания этих клеток. Предпола-
гается, что перемещение кератиноцитов в
неколончатых ЭПЕ также не менее сложное.
Механизмы регуляции перемещения керати-
ноцитов специально не исследовались. Вместе с
тем, очевидно, что скорость перемещения пря-
мо пропорциональна митотической активности
и, следовательно, зависит от всех факторов, вли-
яющих на нее.
Дифференцировка кератиноцитов
События, происходящие в результате дифферен-
цировки кератиноцитов, подробно описаны
выше.
Глава 2. Анатомия и гистофизиология кожи
33
В норме между пролиферацией кератиноци-
тов и их дифференцировкой устанавливается
стабильное равновесие. При этом достаточно ин-
тенсивные процессы превращения кератиноци-
тов в роговые чешуйки уравновешиваются весь-
ма незначительным количеством митозов,
регистрируемых морфологическими методами.
Ситуация, однако, меняется под влиянием гор-
монального, иммунного стимула или механичес-
кого повреждения, а также в результате ряда
кожных болезней (например, при псориазе). При
этом пролиферация кератиноцитов может резко
возрастать, а их дифференцировка — существен-
но нарушаться. В любом из этих случаев ткане-
вой гомеостаз в эпидермисе нарушается.
Регуляция дифференцировки кератиноцитов
тесно связана с регуляцией митотической актив-
ности кератиноцитов. Замедление пролифератив-
ных процессов в эпидермисе вызывает замедле-
ние миграции кератиноцитов к поверхности
эпидермиса, что, в свою очередь, дает возмож-
ность синтезу кератина пройти все стадии, а ли-
зосомные ферменты и содержимое кератиносом
получают возможность оказать свое физиологи-
ческой действие. В результате формируется зре-
лый роговой слой. Внутриклеточными посред-
никами нормального кератопоэза являются
кейлоны и цАМФ, которые замедляют деление
кератиноцитов. Напротив, накопление в клетках
цГМФ вызывает ускорение деления кератиноци-
тов и их дифференцировки. Расположенные на
мембранах кератиноцитов а- и р-адренорецепто-
ры, холинорецепторы, гистаминовые и другие ре-
цепторы играют роль связывающего звена между
гормонами, медиаторами, биологически актив-
ными веществами и внутриклеточным содержа-
нием циклических нуклеотидов. Многие хими-
ческие вещества, применяемые с лечебной целью,
оказывают модулирующее влияние на синтез ке-
ратина и кератинизацию (антибластомные пре-
параты, антикоагулянты, тяжелые металлы, мы-
шьяк и др.). Недостаток некоторых витаминов
(А, С) вызывает гиперкератоз.
Запрограммированная гибель клеток
В эпидермисе наблюдаются две формы гибели
клеток: гибель в процессе кератинизации и апо-
птоз. Обе они являются запрограммированны-
ми. Здесь речь пойдет об апоптозе, т. к. первая
форма гибели описана выше.
В настоящее время под апоптозом понимают
запрограммированное саморазрушение клеток,
строго контролируемое организмом. Апоптоз яв-
ляется функциональной противоположностью
митоза, и эти два процесса направлены на под-
держание тканевого гомеостаза. Так же, как и
способность к делению заложена в генетической
программе каждой клетки, точно также в этой
программе определен исход каждой клетки — ее
смерть путем апоптоза, которая, однако, может
наступить в разные периоды онтогенеза. Реали-
зация этой программы в определенный момент
существования клетки приводит к экспрессии
одного или нескольких генов и образованию про-
дуктов, вызывающих гибель. В некоторых слу-
чаях показано, что смерть клетки путем апоптоза
определяется двумя генами, мутации в которых
предотвращают гибель, и клетка направляется на
путь дифференцировки. Конечный этап клеточ-
ной гибели — деградация хроматина — кодиру-
ется еще одним геном, продуктом которого яв-
ляется эндонуклеаза, разрушающая ДНК. Таким
образом, у каждой вновь образующейся клетки
имеются три возможности развития, заложенные
в ее генетической программе: повторные деления;
дифференцировка; апоптоз. Выбор одного из этих
путей развития определяется конкретными фун-
кциональными запросами ткани; действием в
связи с этим регуляторных механизмов; действи-
ем факторов внешней среды.
В большинстве работ, посвященных апопто-
зу, указывается, что апоптотической гибели под-
вергаются стволовые и быстроделящиеся клет-
ки, тогда как медленно пролиферирующие
клетки проявляют среднюю способность к апо-
птозу. Дифференцированные клетки очень редко
разрушаются путем апоптоза. Поэтому апопто-
тически трансформированные клетки находят
чаще в генеративных зонах тканей. В наших ис-
следованиях (Мяделец О.Д., Адаскевич В.П.,
1994; Мяделец О.Д., 1995) мы также, в основ-
ном, обнаруживали апоптотические клетки в ге-
неративной зоне эпидермиса (базальном, изред-
ка — нижних слоях шиповатого слоя). При
апоптозе кератиноцитов утолщенного в резуль-
тате поражения атопическим дерматитом эпидер-
миса апоптозы могут локализоваться и в верх-
них пластах шиповатого слоя.
Таким образом, две формы гибели кератино-
цитов эпидермиса имеют принципиальное отли-
чие: апоптозу подвергаются недифференциро-
ванные кератиноциты, тогда как гибель клеток
в процессе кератинизации — это последний этап
их специфической дифференцировки.
Установлено, что во многих (если не в каж-
дой) тканях всегда образуется большее количе-
ство клеток, чем их необходимо для нормального
функционирования ткани. Смысл этой избыточ-
ной наработки клеток заключается в том, что-
3 4 Общая дерматология
бы постоянно создавать материальную базу для
функционирования ткани в экстремальных ус-
ловиях, иначе говоря, обеспечивать адаптивные
потенции ткани. Если в данный момент адаптив-
ные перестройки ткани еще не требуются, из-
лишние клетки подвергаются апоптозу. Если
возникает необходимость компенсаторных пере-
строек, клетки направляются на путь дифферен-
цировки. Таким образом, соотношение митоза,
дифференцировки и апоптоза клеток всегда ди-
намичное.
Основным моментом в гибели клеток по ме-
ханизму апоптоза является особое повреждение
хроматина — межнуклеосомная правильная
фрагментация ДНК в результате активации и
действия Ca-Mg-зависимой эндонуклеазы.
Морфология
апоптоза кератиноцитов
Морфология апоптоза описана давно. Еще в 1914
году А. А. Максимов описал маргинацию хрома-
тина в ядрах — характерный признак апоптоза.
В последующем клетки с типичными для апо-
птоза признаками описаны многими авторами и
определялись как «темные» клетки, «тельца Ка-
унсилмена»; использовались также и другие тер-
мины. Следовательно, апоптоз был известен дав-
но, но его сущность и механизм стали известны
лишь в последнее время. Ядра клеток, подвер-
гающихся апоптозу, уплотняются, становятся ги-
пербазофильными, хроматин в них концентри-
руется около ядерной оболочки. Ядрышко не
выявляется. Иногда ядра (особенно в эпидерми-
се) приобретают зазубренный или полулунный
вид. Цитоплазма становится бесструктурной.
Создается впечатление, что ядро окружено «пу-
стым кольцевидным пространством». Иногда в
цитоплазме наблюдаются слабоокрашенные (ок-
сифильные) участки, но никогда не выявляют-
ся гранулы кератогиалина. Такие клетки очень
легко диагностировать, т. к. они резко контрас-
тируют с интактными кератиноцитами. Они лег-
ко поддаются количественной оценке. Локали-
зуются апоптозные клетки в базальном слое либо
супрабазально.
Слущивание клеток с поверхности
эпидермиса (эксфолиация)
С поверхности эпидермиса происходит постоян-
ное слущивание терминально дифференцирован-
ных кератиноцитов (роговых чешуек или кор-
неоцитов). Этот процесс, с одной стороны,
участвует в поддержании тканевого гомеостаза
эпидермиса, а с другой стороны, обеспечивает
постоянное очищение (санацию) кожного покро-
ва. Нарушение слущивания корнеоцитов лежит
в основе некоторых заболеваний (гиперкерато-
за при его снижении и изъязвления эпидермиса
при ускорении).
Десквамация корнеоцитов определяется дву-
мя основными факторами: интенсивностью кле-
точной продукции (митозом) и состоянием меж-
корнеоцитного цемента. Межклеточный цемент,
образующийся за счет деятельности гранул Од-
ланда, создает адгезию корнеоцитов друг к дру-
гу. В основном между митотической активнос-
тью и силой адгезии существует обратная
зависимость: чем выше интенсивность воспро-
изводства клеток, тем ниже сила адгезии корне-
оцитов. Это отмечается, например, при псориа-
зе. Однако это соотношение сохраняется не
всегда. Так, в ранние сроки постнатального он-
тогенеза у крысят при высоких значениях ми-
тотической активности десквамация отсутству-
ет. Не наблюдается она при ихтиозе, когда
митотическая активность кератиноцитов увели-
чивается вместе с повышением силы адгезии
корнеоцитов (Соколов В.Е. и соавт., 1986).
Имеются данные о том, что ключевую роль в
межкорнеоцитной адгезии играет соотношение
холестерин/холестеринсульфат. При некоторых
наследственных заболеваниях, характеризующих-
ся увеличением толщины рогового слоя эпидер-
миса (чешуйчатый эпидермис) имеется недоста-
точность фермента холестеринсульфатазы.
Отсутствие слущивания рогового слоя в ранние
сроки после рождения также может быть связа-
но с недостаточностью или неактивностью ука-
занного фермента.
Таким образом, гомеостаз эпидермиса обеспе-
чивается 5 основными цитологическими процес-
сами: делением клеток, их миграцией, терми-
нальной дифференцировкой кератиноцитов в
корнеоциты, апоптозом и эксфолиацией корне-
оцитов. Все эти показатели тесно взаимосвяза-
ны и подвержены регуляции на клеточном, тка-
невом, органном и организменном уровне.
Дермо-эпидермальное
соединение
Эпидермис прикрепляется к дерме с помощью
специализированного межклеточного матрикса,
названного базальной мембраной (рис. 2.14). При
светомикроскопическом исследовании она вид-
на как тонкая аморфная полоска шириной до
1 нм. Она является ШИК-положительной, т. е.
содержит углеводы. Окраска орсеином на элас-
Глава 2. Анатомия и гистофизиология кожи
35
тические волокна выявляет прямо под эпидер-
мисом тонкие волоконца, идущие к эпидерми-
су от сети эластических волокон дермы. При со-
четанной ШИК-орсеиновой окраске эти
волоконца находятся в области ШИК-позитив-
ной базальной мембраны, поперечно пересекая
ее. Импрегнация серебром ретикулиновых воло-
кон также выявляет в этой зоне волокнистую
сеть. С помощью моноклональных антител им-
мунофлуоресцентным методом в зоне базальной
мембраны обнаружили антиген, антитела к ко-
Шик-реакция
Окраска Импрегнация
эластических ретикулярных
волокон волокон AgNO3
б
Рис. 2.14. Схема строения дермо-
эпидермального соединения по данным:
а. световой микроскопии;
б. электронной микроскопии и биохимических
исследований (Соколов В. Е. и соавт., 1988, с
некоторыми изменениями). I — плазмолемма
базальных кератиноцитов; II — светлая
пластинка; III плотная пластинка; IV — субэпи-
дермальное сплетение. 1 — пиноцитозные
пузырьки; 2 — полудесмосома; 3 — кератино-
вые промежуточные филаменты полудесмо-
сом; 4 — заякоривающие филаменты;
5 — цитолемма кератиноцитов; 6 — Н-линия;
7 — место локализации БПА; 8 — фибронек-
тин; 9 — местолокализации ламинина;
10 — протеогепарансульфат; 11- коллаген IV
типа; 12 — дерма; 13 — микрофиламенты;
14 —- коллагеновые волокна; 15 — заякорива-
ющие филаменты.
торому появляются в коже и в крови у больных
буллезным пемфигоидом. Этот антиген назван
буллезным пемфигоидным антигеном (БПА).
При электронномикроскопическом исследова-
нии области базальной мембраны оказалось, что
та структура, которая в световом микроскопе
представляется базальной мембраной, неоднород-
на и включает 4 достаточно сложных образова-
ния: плазмолемму базальных клеток эпидерми-
са со специализированными клеточными
контактами полудесмосомами; светлую пластин-
ку (электронно-прозрачная зона: lamina lucida);
плотную или темную пластинку (электронно-
плотная зона: lamina densa); субэпидермальное
сплетение волокон.
Чаще всего на электронномикроскопическом
уровне под базальной мембраной понимают со-
вокупность 2 и 3 компонентов, а совокупность
всех четырех компонентов определяют как дер-
мо-эпидермальное соединение.
Плазмолемма базальных кератиноцитов име-
ет сложные цитоплазматические выросты и по-
этому на большом протяжении имеет очень из-
вилистые, неровные контуры. На базальной
поверхности цитолемма кератиноцитов посто-
янно формирует многочисленные пиноцитоз-
ные пузырьки, которые выполняют процессы
эндо- и экзоцитоза низкомолекулярных ве-
ществ, растворимых макромолекул и неболь-
ших коллоидных частиц. Многочисленные дес-
мосомы прикрепляют кератиноциты к
базальной мембране. Они представляют собой
электронноплотные утолщения базальной
плазмолеммы кератиноцитов (пластинки), в
которые внедряются промежуточные кератино-
вые филаменты (тонофиламенты). Эти тонофи-
ламенты, погрузившись в плотные пластинки,
затем вновь возвращаются в цитоплазму. По-
лагают, что полудесмосомы не только механи-
чески прикрепляют кератиноциты к базальной
мембране, но и участвуют в переносе в кера-
тиноцит через дермо-эпидермальное соедине-
ние ряда молекул, а также являются органи-
заторами биосинтеза компонентов базальной
мембраны.
Светлая пластинка. Имеет ширину до 40 нм.
На большем протяжении аморфна, но в местах
расположения десмосом в ней наблюдается плот-
ная Н-линия, идущая параллельно цитолемме ке-
ратиноцита. В этом месте светлая пластинка про-
низана вертикально идущими заякоривающими
филаментами, которые внедряются в плотные
пластинки полудесмосом. В светлой пластинке
на самой границе с цитолеммой кератиноцитов
содержится БПА, в ней обнаружены также ве-
3 6 Общая дерматология
щества, участвующие в адгезии клеток, такие
как ламинин, нидоген/энтактин, фибронектин,
коллаген IV типа.
Ламинин является главным и наиболее посто-
янным адгезивным гликопротеином базальных
мембран. Он не только выполняет роль адгезив-
ного субстрата для различных эпителиальных и
соединительных клеток, но также индуцирует
формирование полярности эпителиальных кле-
ток, регулирует их пролиферацию, дифференци-
ровку и подвижность. Молекула ламинина имеет
крестообразную форму. С ламинином связан в
один комплекс нидоген/энтактин. Чидоген и эн-
тактин ранее считали двумя самостоятельными
молекулами, но по последним данным они иден-
тичны.
Фибронектин, так же как и ламинин, способ-
ствует адгезии и движению эпителиальных и
соединительнотканных клеток, стимулирует про-
лиферацию и цитодифференцировку, участвует
в поддержании цитоскелета. Он синтезируется
как кератиноцитами, так и соединительноткан-
ными клетками (фибробластами, макрофагами,
эндотелиоцитами, нейтрофильными лейкоцита-
ми, а также тромбоцитами). По химической при-
роде фибронектин является гликопротеином.
Особенно в большом количестве фибронектин
вырабатывается при заживлении кожных ран.
Считают, что он также участвует в организации
эпидермальной пролиферативной единицы.
Коллаген IV типа, в отличие от коллагенов
других типов, обнаруживается исключительно в
базальных мембранах, в светлой и темной плас-
тинках. Благодаря своим свойствам молекулы
коллагена IV типа не способны формировать
коллагеновые волокна. Они обеспечивают нера-
створимость и механическую стабильность ба-
зальных мембран.
Б ПА представляет собой гликопротеин. Нахо-
дится в тесном контакте с поверхностью базаль-
ных кератиноцитов. Согласно последним данным,
БПА синтезируется кератиноцитами и может ло-
кализовываться внутриклеточно, в местах соеди-
нения полудесмосом с тонофиламентами.
Плотная пластинка имеет толщину 30—50 нм
и представляет собой непрерывный электронно-
плотный слой разной толщины, состоящий из
микрофибрилл, заключенных в аморфный ма-
териал. В ней обнаружены коллаген IV и V типа,
ламинин, протеогликаны. При этом единствен-
ным изученным протеогликаном базальной мем-
браны является протеогепарансульфат. Протео-
гликаны, благодаря особенностям их молекул,
участвуют в барьерной функции и регуляции
проникновения веществ в кератиноциты.
Субэпидермальное сплетение образовано тремя
типами фибриллярных структур: а) заякорива-
ющими волокнами; б) пучками микрофиламен-
тов, напоминающих микрофибриллы эластичес-
ких волокон; в) одиночными коллагеновыми
волокнами, не формирующими пучки, в отли-
чие от остальных характерных для дермы кол-
лагеновых волокон.
а. Заякоривающие волокна (не путать с заяко-
ривающими филаментами в светлой пластинке)
имеют уникальную структуру, отличающую их от
других субдермальных волокон. В своей цент-
ральной части они имеют нерегулярную попереч-
ную исчерченность, нехарактерную для коллаге-
новых волокон. Концевые части заякоривающих
фибрилл веерообразно расходятся. Верхние кон-
цы их прикрепляются к плотной пластинке, а
нижние уходят глубоко в дерму.
б. Пучки микрофиламент, напоминающие мик-
рофибриллы эластических волокон, прикрепля-
ются к плотной пластинке перпендикулярно или
под наклоном. Они имеют диаметр 8—11 нм,
регулярную периодичность и центральный ка-
нал. Предполагают, что они являются продолже-
нием эластических элементов дермы и способ-
ны окрашиваться орсеином.
в. Коллагеновые волокна легко различимы по
характерной регулярной периодичности. Они
ориентированы случайно и не объединяются в
пучки, что отличает их от коллагеновых воло-
кон более глубоких участков дермы. Они не
прикрепляются к плотной пластинке, но могут
лежать очень близко от нее и попадать в петли
заякоривающих волокон.
Таким образом, дермо-эпидермальное соеди-
нение — гораздо более сложное образование, чем
описанная в световом микроскопе базальная
мембрана. В состав этого соединения входят:
базальная мембрана, состоящая из светлой и тем-
ной пластинок; связь базальной мембраны с эпи-
дермисом осуществляется через полудесмосомы,
заякоривающие филаменты и молекулы адгезии,
а с дермой — через заякоривающие волокна и
пучки дермальных микрофиламентов.
Базальная мембрана выполняет следующие
функции: опорная; скрепление клеток разного
происхождения; регуляция проникновения ве-
ществ в кератиноциты; регулирующая роль в
развитии и морфогенезе.
Различным макромолекулам базальной мем-
браны принадлежит разная роль в выполнении
этих функций. Коллагены IV и V типа стабиль-
ны, нерастворимы и выполняют опорную функ-
цию. Протеогликанам, очевидно, принадлежит
барьерная роль и функция контроля за проник-
Глава 2. Анатомия и гистофизиология кожи
37
новением веществ в кератиноциты. Фибронек-
тин, БПА, ламинин, нидоген/энтактин ответ-
ственны за прикрепление клеток. Осуществле-
ние морфогенетической функции связывают с
ламинином, фибронектином и коллагеном IV
типа. Ламинин и коллаген IV появляются на
самых ранних этапах эмбриогенеза, что указы-
вает на их роль в развитии. Фибронектин игра-
ет важную роль в построении специфической
колончатой структуры эпидермиса и с пере-
стройках эпидермиса при заживлении ран кожи.
Дерма
Дерма подразделяется на сосочковый и сетчатый
слой. Сосочковый слой образован рыхлой, а сет-
чатый — плотной волокнистой неоформленной
соединительной тканью. Оба слоя состоят из трех
компонентов: клеток, волокон и основного ве-
щества.
Сосочковый слой, образованный рыхлой во-
локнистой неоформленной соединительной тка-
нью, в виде сосочков вдается в эпидермис, из-за
чего и получил название. Он не только скрепля-
ет эпидермис и сетчатый слой, но и играет боль-
шую роль в питании эпидермиса, которое осуще-
ствляется за счет капиллярной сети, лежащей под
базальной мембраной. Обилие макрофагов опре-
деляет защитные функции сосочкового слоя, а
находящиеся меланофоры вместе с меланоцита-
ми эпидермиса осуществляют пигментацию кожи.
Клетки сосочкового слоя
Основную массу клеток сосочкового слоя состав-
ляют фибробласты, тучные клетки, макрофаги,
меланофоры и лейкоциты, среди которых наи-
более часто встречаются эозинофилы и лимфо-
циты. В сосочковом слое содержатся также клет-
ки Лангерганса.
Фибробласты
Это наиболее многочисленная популяция в лю-
бой рыхлой волокнистой соединительной ткани,
в том числе и в сосочковом слое дермы. Их функ-
цией является синтез межклеточного вещества:
коллагеновых, ретикулиновых и эластических во-
локон, а также основного вещеста. Популяция
фибробластов неоднородна и включает в себя ма-
лодифференцированные, дифференцированные
фибробласты, фиброциты, фиброкласты и мио-
фибробласты. Кроме этих клеток, в дифферон
(совокупность всех клеток одной линии диффе-
ренцировки от стволовой до конечной) фибро-
бластов включают стволовые клетки и полуство-
ловые клетки-предшественники. Источником
развития всех клетокфибробластического ряда
считают адвентициальные клетки, сопровождаю-
щие микрососуды. В зарубежной литературе ство-
ловой клеткой фибробластов, однако, считают пе-
рициты — клетки, располагающиеся в
расщеплении базальной мембраны гемокапилля-
ров. Эти клетки, делясь, дают малодифференци-
рованные фибробласты — малоотростчатые клетки
с округлым ядром и базофильной цитоплазмой,
в которой развиты свободные рибосомы и слабо
представлены органеллы белкового синтеза: гра-
нулярная эндоплазматическая сеть и комплекс
Гольджи. Данные клетки способны интенсивно
делиться митозом и, дифференцируясь, превра-
щаться в зрелые, дифференцированные фиброб-
ласты, которые могут достигать размеров до 30—
50 мкм. Зрелые фибробласты являются
активными продуцентами межклеточного веще-
ства и имеют характерные черты строения
(рис. 2.15). Они имеют крупные светлые ядра с
1—2 крупными ядрышками. Цитоплазма слегка
базофильна. В ней хорошо развиты гранулярная
эндоплазматическая сеть, комплекс Гольджи,
умеренное количество митохондрий. По перифе-
рии клетки можно видеть микрофиламенты, уча-
ствующие в движении клетки. Постепенно теряя
функциональную активность и старея, фиброб-
ласты превращаются в дефинитивную клеточную
форму — фиброциты, которые имеют веретено-
видную форму, темное вытянутое уменьшенное
ядро. В цитоплазме снижается содержание орга-
нелл.
Функциональными антагонистами зрелых
фибробластов являются фиброкласты. Эти клет-
ки способны фагоцитировать и осуществлять
гидролиз с помощью ферментов лизосом (чис-
ло которых существенно увеличено) компонен-
тов межклеточного вещества. Благодаря функ-
циональной оси «фибробласт-фиброкласт»
межклеточное вещество постоянно обновляется.
Фиброкласты вызывают также инволюцию мат-
ки после родов, обратное развитие и перестрой-
ку соединительнотканных рубцов в коже после
ранений. По современным предствлениям, одна
и та же клетка в зависимости от конкретных ус-
ловий может выступать либо в роли синтезиру-
ющего фибробласта, либо в роли фиброкласта.
Миофиброкласты характеризуются хорошо
развитой гранулярной эндоплазматической сетью
и системой неисчерченных миофибрилл. Эти
клетки способны активно сокращаться и похо-
жи на гладкие миоциты. Они играют большую
роль в контракции краев раны. Их количество
3 8 Общая дерматология
при этом резко увеличивается путем деления
предшественников, а также дифференцировки
из фибробластов. Особое значение принадлежит
этим клеткам в дикой природе, где невозможно
хирургическое сближение краев ран швами или
скобками.
Тканевые базофилы
Тканевые базофилы (тучные клетки, лаброциты,
мастоциты) встречаются во всех слоях дермы и
в подкожно-жировой клетчатке. Существует
мнение, что молодые тканевые базофилы появ-
ляются в сосочковом слое (их предшественни-
ки выселяются здесь из сосудов микроциркуля-
торного русла), а затем, по мере созревания,
смещаются в нижележащие слои дермы и под-
кожную клетчатку, увеличиваясь при этом в раз-
мерах. Источником развития тканевых базофи-
лов является стволовая клетка крови, более
поздний потомок которой единый для них и ба-
зофильных лейкоцитов крови. Содержание туч-
ных клеток в коже у разных видов млекопита-
ющих различно и обратно пропорционально
количеству базофильных лейкоцитов крови.
Чаще всего тканевые базофилы располагают-
ся периваскулярно. В последнее время появились
сведения о том, что эти клетки могут проникать
в интактный эпидермис. Массивное проникно-
вение их в эпидермис описано при мастоцито-
мах кожи.
Тучные клетки отличаются выраженным по-
лиморфизмом. Он проявляется в различной ве-
личине клеток, их форме (от округлой до вытя-
нутой и угловатой), степенью заполненности
цитоплазмы гранулами. Клетки имеют неболь-
шое округлое или овальное, иногда гиперхром-
ное ядро. Отличительной особенностью клеток
является наличие цитоплазматических гранул
размером 0,3-“ 1 мкм. При окраске некоторыми
красителями гранулы проявляют метахромазию
(способность окрашиваться в цвет, отличный от
окраски красителя в растворе). Например, толу-
идиновый синий окрашивает их либо в малино-
вый, либо в сиреневый цвет. Гранулы могут
быть как ШИК-позитивными, так и ШИК-не-
гативными. ШИК-реакция их обусловлена на-
личием углеводов (гепарина) (рис. 2.16).
Ультраструктура гранул различается. Их содер-
жимое может быть зернистым, мелкосетчатым
или слоистым, иногда кристаллоидным. Цито-
и биохимически в гранулах тучных клеток вы-
является белок (60 %), гепарин (30 %), гистамин
(10 %). У некоторых животных, а возможно и у
человека, в гранулах выявляется также серото-
нин. Кроме гепарина, в гранулах тучных клеток
выявлены и другие гликозаминогликаны (хон-
дроитинсульфат и гиалуроновая кислота).
Рис. 2.15. Зрелые фибробласты (коллагенобласты).
а. Коллагенобласт 1 типа с развитыми пластинчатым комплексом (ПК) и цистернами гранулярной
эндоплазматической сети (ГЭР). Около наружной мембраны множество вакуолей (возможно, секре-
торных). В межклеточном пространстве формирование многочисленных микрофибрилл (Мфб) и
отдельных коллагеновых фибрилл (Кф). х17000.
б. Коллагенобласт 2 типа из заживающей раны. Цитоплазма клетки заполнена расширенными цистерна-
ми ГЭР, в просвете которых видно тонковолокнистое содержимое (проколлаген). х45000 (Серов В. В.,
Шехтер А. Б., 1981).
Глава 2. Анатомия и гистофизиология кожи
39
В цитоплазме тканевых базофилов содержат-
ся органеллы: митохондрии, эндоплазматическая
сеть двух видов, рибосомы, микрофиламенты,
комплекс Гольджи, иногда центриоли. Они рас-
полагаются возле ядра, а степень их развития
зависит от зрелости клеток. В молодых клетках
они развиты максимально, а в зрелых благода-
ря содержанию большого количества гранул по-
чти не видны.
Функциональное значение тучных клеток оп-
ределяется содержащимися в гранулах биологи-
чески активными веществами. Они регулиру-
ют проницаемость и тонус микрососудов,
количество в коже воды, поддерживают колло-
идное состояние основного вещества соедини-
тельной ткани. Велика роль тучных клеток в
поддержании оптимального уровня метаболизма
в коже. Тучные клетки участвуют наряду с
фибробластами в биосинтезе межклеточного ве-
щества, в частности гликозаминогликанов. Они
выделяют ряд медиаторов, влияющих на деле-
ние клеток соединительной ткани и эпителия,
а также регулирующих деятельность этих кле-
ток.
Благодаря биологически активным веществам
тканевые базофилы очень часто вовлекаются в
патологические процессы в коже. При этом в них
наблюдается массированная дегрануляция —
выход гранул, содержимое которых может выз-
вать расширение микрососудов, выход из них
клеток крови, в первую очередь зернистых и не-
зернистых лейкоцитов, отек тканей. Медиаторы
осуществляют регуляцию воспалительных и им-
мунных реакций. Заболеванием, связанным с
бластомным поражением тканевых базофилов,
является мастоцитоз, характеризующийся выра-
женными особыми морфологическими измене-
ниями в коже.
Макрофаги
Макрофаги (гистиоциты) имеют неправильную
амебовидную форму, вакуолизированную цито-
плазму и темное, иногда бобовидное ядро. Эти
клетки становятся отчетливо заметными после
инъекции в кожу красителя, что иногда исполь-
зуется для оценки интенсивности фагоцитоза
(рис. 2.17).
В электронном микроскопе гистиоциты име-
ют характерную ультраструктуру. В них разви-
ты органеллы белкового синтеза, но наиболее
многочисленны лизосомы. Часто видны фагосо-
мы — фагоцитированные частицы, окруженные
мембраной.
Функция макрофагов заключается, прежде
всего, в фагоцитозе — захвате и разрушении при
помощи гидролитических ферментов лизосом
чужеродных частиц. Во время фагоцитоза чуже-
родные макромолекулярные вещества или целые
клетки обволакиваются выростами цитолеммы
макрофага, после чего попадают как бы в мешок-
углубление и, наконец, отделяются от цитолем-
мы, оказываясь окруженными частью цитолем-
мы внутри клетки. Такая окруженная мембраной
частица называется фагосомой. К ней устремля-
ются лизосомы, мембраны которых сливаются с
а
Рис. 2.16. Ультраструктура тканевых базофилов по данным световой (а) и электронной (б) микроскопии.
а. Видны просвечивающее ядро, цитоплазма и умеренная дегрануляция клеток. х900 (Фалин Л. И.,
1957).
б. Цитоплазма тучных клеток заполнена специфическими гранулами (Гр) различной структуры.
М — митохондрии, ПК — пластинчатый комплекс (Серов В. В., Шехтер А. Б., 1981).
40 Общая дерматология
мембраной фагосомы. При этом ферменты лизо-
сом изливаются в фагосому и осуществляют гид-
ролиз органических компонентов фагоцитирован-
ной частицы. Слившаяся с лизосомой фагосома
называется фаголизосомой. Дальнейшая судьба
фаголизосомы зависит от содержимого фагосомы.
Если оно полностью расщепляется ферментами,
то фаголизосомы постепенно уменьшаются в раз-
мерах и исчезают. Если содержимое не может
быть разрушено, фаголизосомы превращаются в
длительно присутствующие в клетке остаточные
б
Рис. 2.17. Строение макрофага по данным
электронной (а) и световой (б) микроскопии,
а. 1 — ядро; 2 — фагосома; 3 — лизосома.
Х15000.
б. Микрофотография группы макрофагов,
фагоцитировавших трипановый синий. * 900.
(Хэм А., Кормак Д., 1983).
тельца, содержимое которых может выбрасываться
из клетки путем экзоцитоза.
Фагоцитоз — не единственная функция мак-
рофагов. Они обладают способностью выступать
как антигенпроцессирующие и антигенпредстав-
ляющие клетки. Захватив антигены, которые не
могут быть элиминированы из организма толь-
ко деятельностью фагоцитов, макрофаги пере-
водят их в высокоиммунную форму и в комп-
лексе с собственным антигеном выделяют на
свою поверхность. Здесь с антигеном способны
контактировать лимфоциты, включающиеся в
элиминацию антигенов.
Макрофаги секретируют множество медиато-
ров, активирующих или подавляющих функ-
ции и деление других клеток соединительной
ткани и иммунокомпетентных клеток. Они на-
зываются монокинами. Наиболее известный мо-
нокин — интерлейкин-1, который стимулиру-
ет Т-лимфоциты-хелперы. Второй, не менее
известный монокин — фактор некроза опухо-
лей, подавляющий рост опухолевых клеток и
одновременно являющийся стимулятором деле-
ния фибробластов.
Макрофаги активно участвуют в поддержании
гомеостаза соединительной ткани. В цитоплазме
макрофагов кожи иногда можно обнаружить гра-
нулы меланина и гранулы Бирбека. Такие макро-
фаги, очевидно, являются мигрирующими в эпи-
дермис (и из эпидермиса) клетками Лангерганса.
Плазматические клетки
Плазматические клетки в норме в коже встре-
чаются редко и, как правило, вокруг сосудов.
Они имеют базофильную цитоплазму, эксцен-
трически расположенное ядро, в котором хрома-
тин формирует характерный вид колеса со спи-
цами (рис. 2.18). В цитоплазме развиты
гранулярная эндоплазматическая сеть и комп-
лекс Гольджи. Митохондрии немногочисленны.
Функцией плазмоцитов является секреция анти-
тел. При патологии количество этих клеток в
коже может резко возрастать.
Из других клеток в дерме встречаются лимфо-
циты, моноциты, зернистые лейкоциты. Из лим-
фоцитов преобладают Т-лимфоциты различных
субпопуляций: Т-хелперы, Т-киллеры, Т-супрес-
соры. Обычно лимфоциты имеют одиночное рас-
положение и не формируют, как в других орга-
нах (бронхи, кишечник и т. д.), лимфоидные
фолликулы. Однако Л. В. Чернышенко (1980) с
применением специальных методов показала на-
личие в коже лимфоидных фолликулов. Незави-
симо от того, есть они в коже или их наличие не
Глава 2. Анатомия и гистофизиология кожи 41
будет подтверждено, в настоящее время выделя-
ют так называемую кожноассоциированную лим-
фоидную ткань (КАЛТ), которая составляет важ-
ную часть иммунного аппарата кожи и всего
организма (см. иммунологические функции
кожи).
Межклеточное вещество
Межклеточное вещество дермы состоит из основ-
ного вещества и волокон. При этом в рыхлой
волокнистой неоформленной соединительной
ткани сосочкового слоя преобладает основное
вещество, а в сетчатом слое, который образован
плотной волокнистой неоформленной соедини-
тельной тканью, превалирует волокнистый ком-
понент.
Основное вещество
В его состав входит вода, минеральные вещества,
липиды, углеводы и белки. Основными функ-
циональными компонентами основного (аморф-
ного) вещества являются гликозаминогликаны,
протеогликаны и гликопротеины.
Рис. 2.18. Схема ультраструктуры
плазматической клетки. 1 — гранулярная
эндоплазматическая сеть; 2 — хроматин;
3 — ядрышко; 4 — ядерная мембрана; 5 — пора
в ядерной мембране; 6 — митохондрии;
7 — секреторные гранулы; 8 — центриоли;
9 — комплекс Гольджи; 10 — свободные
рибосомы (Хэм А., Кормак Д., 1983).
Гликозаминогликаны
Гликозаминогликаны (рис. 2.19) представляют
собой длинные неразветвленные цепи из повто-
ряющихся дисахаридных единиц. Один из саха-
ров всегда содержит аминогруппу, что обуслов-
ливает название гликозаминогликанов. Различные
гликозаминогликаны отличаются друг от друга
числом и локализацией углеводных остатков, ти-
пом связи между ними. В коже обнаружены та-
кие гликозаминогликаны: хондроитин-4-сульфат,
хондроитин-6-сульфат, дерматансульфат, гепарин
и гиалуроновая кислота.
Все гликозаминогликаны существуют в соеди-
нительной ткани не в свободном состоянии, а
как правило, связаны с белками. За исключением
гиалуроновой кислоты, которая образует особые
соединения с белком, они входят в состав про-
теогликанов — комплексных молекул, постро-
енных из полипептидного стержня, к которому
по бокам прикреплено большое количество це-
Рис. 2.19. Строение некоторых
гликозаминогликанов, а — хондроитин-6-
сульфат; б — хондроитин-4-сульфат;
в — гиалуроновая кислота.
42
Общая дерматология
почек гликозаминогликанов. Цепочки хондро-
итинсульфатов прикреплены к стержневому бел-
ку группами по 10 цепочек в каждой, а цепоч-
ки дерматансульфатов наиболее многочисленны.
Протеогликановый компонент матрикса со-
единительной ткани в силу своих полианионных
свойств ответственен за гидратацию, прочность
на сжатие, способность деформироваться и пре-
одолевать деформацию и восстанавливать свое
гидратированное состояние — так называемая
«обратимая деформация».
В свою очередь, протеогликаны находятся не
в свободном состоянии, а объединяются в проч-
ные мультиполярные агрегаты с молекулярной
массой до 100 млн дальтон. Эти структуры в
основе имеют молекулу гиалуроновой кислоты,
одна цепь которой связывает большое число
мономерных протеогликанов (до 200). Практи-
чески вся гиалуроновая кислота может быть вов-
лечена в формирование агрегатов.
Гликопротеины
Гликопротеины — это сложные белки, состоя-
щие из полипептидной цепи и короткой развет-
вленной цепи олигосахарида (рис. 2.20). Гликоп-
ротеинами являются стержневые белки
протеогликанов. Кроме них, в основном веще-
стве дермы содержится ряд других гликопроте-
инов, из которых наиболее изучен фибронектин,
о котором уже упоминалось.
Коллагеновые волокна
Коллагеновые волокна обеспечивают механичес-
кую прочность дермы. В сосочковом слое нахо-
дятся мелкие пучки коллагеновых волокон, ле-
жащие под разными углами к поверхности кожи.
В сетчатом слое коллагеновые волокна толстые
и также имеют различную ориентацию.
Коллагеновые волокна окрашиваются кислот-
ными красителями (эозином, кислым фуксином)
в красный цвет.
Каждое коллагеновое волокно имеет достаточ-
но сложную молекулярную организацию и со-
стоит из белка коллагена. Коллаген относится к
семейству фибриллярных белков. Молекула кол-
лагена образована тремя полипептидными цепя-
ми, закрученными по спирали. По аминокислот-
ному составу полипептидные цепи отличаются
друг от друга, в связи с чем различают несколько
(до 12) типов коллагена. Основными разновид-
ностями коллагена дермы являются I и III типы
(в сосочковом слое в основном обнаруживается
коллаген III, а в сетчатом — 1 типа). Последний
формирует толстые фибриллы, является более
прочным и эффективным в выполнении меха-
нических свойств. Коллаген имеет уникальный
аминокислотный состав, т. к. содержит много
редко встречающихся аминокислот: глицин,
пролин, оксипролин, лизин и оксилизин; в виде
следов в нем обнаруживаются тирозин и метио-
нин и полностью отсутствуют цистеин и трип-
тофан (Мазуров В.И., 1974). Остатки лизина и
пролина находятся в коллагене в гидроксилиро-
ванной форме и при помощи своих гидроксиль-
ных групп участвуют в образовании водородных
связей, стабилизирующих тройную спираль кол-
лагена. Это придает коллагену повышенную
прочность. Если искусственно предотвратить
гидроксилирование (гипоксией, дефицитом же-
леза, аскорбиновой кислоты и т. д.), то тройная
спираль не образуется, что ведет к разрушению
коллагена в клетках. В результате кожа и ее со-
суды становятся хрупкими, ломкими.
Полипептидные цепи коллагена синтезируют-
ся на рибосомах гранулярной эндоплазматичес-
кой сети фибробластов. Затем они модифициру-
ются: вначале в цистернах эндоплазматической
сети (гидроксилирование), затем в комплексе
Гольджи (гликозилирование). Только после этих
модификаций происходит сборка а-полипептид-
ных цепей в трехцепочную спираль с помощью
дисульфидных связей. Затем трехцепочные мо-
лекулы тропоколлагена теряют хвостовые участ-
ки, после чего способны к самосборке в фибрил-
Молекула
протеогликана
Неразветвленные
цепи
гликозаминогликанов
Рис. 2.20. Схематическое изображение молекул
протеогликана и гликопротеина (Альберте Б.
и соавт., 1987).
Молекула
гликопротеина
разветвленная
цепь олигосахарида
Глава 2. Анатомия и гистофизиология кожи
43
лы. Это происходит уже вне клетки. Коллагено-
вые молекулы в фибрилле упаковываются в оп-
ределенном порядке, что дает феномен специфи-
ческой поперечной исчерченности (рис. 2.21).
Эластические волокна
Эластические волокна образуют обширную тон-
кую сеть в дерме, несмотря на то, что составля-
ют лишь 2 % от сухой массы кожи человека.
Элективно окрашиваются орсеином, резорцин-
фуксином и железным гематоксилином, причем
не обнаруживают поперечной исчерченности.
Обеспечивают эластические свойства кожи, ее
способность возвращаться к первоначальному
состоянию после растяжения.
Ультраструктурное исследование показало нали-
чие в эластических волокнах двух компонентов:
фибриллярного и аморфного, имеющих разный
химический состав (рис. 2.22). Фибриллярный
компонент образован гликопротеинами фибрилли-
нами. Аморфный компонент состоит из белка эла-
стина. В зависимости от соотношения двух ком-
понентов эластические волокна бывают зрелые и
незрелые (окситалановые и элауниновые). Зрелые
эластические волокна содержатся только в сетча-
том слое дермы, где вплетены между коллагено-
выми волокнами. Содержат до 90 % эластина.
Незрелые эластические волокна содержатся в со-
сочковом слое дермы, где в виде тонких пучков
идут от базальной мембраны вглубь дермы. Здесь
они, возможно, выполняют заякоривающую роль
(см. дермо-эпидермальное соединение). Они име-
ются также вокруг секреторных отделов и прото-
ков потовых желез.
Рис. 2.22. Ультраструктура эластического
волокна.
а. Схема: 1 — центральный гомогенный (аморф-
ный), построенный из эластина компонент;
2 — периферический микрофибриллярный
компонент, состоящий из фибриллина.
б. Электронная микрофотография:
1 — центральный аморфный компонент;
2 — микрофибриллы по периферии волокна;
3 — комплекс Гольджи фибробласта;
4 — эндоплазматическая сеть; 5 — центриоль
фибробласта (Афанасьев Ю. И. и соавт., 1989)
Рис. 2.21. Уровни организации коллагенового
волокна (схемы, А и Б) и электронная
микрофотография коллагеновой фибриллы (В).
А. I — полипептидная цепочка; II — молекулы
тропоколлагена; III — протофибриллы; IV —
фибрилла, у которой становится видна
поперечная исчерченность; V коллагеновое
волокно.
Б. Спиральная структура молекулы коллагена.
Мелкие светлые кружочки — глицин, крупные
светлые — пролин, заштрихованные —
гидроксипролин.
В. Электронная микрофотография коллагеновой
фибриллы, х130000. П — период.
44 Общая дерматология
Эластин отличается высоким содержанием
аминокислот глицина, пролина, валина и алани-
на, десмозина и изодесмозина. Последние две
аминокислоты образуют поперечные сшивки,
обеспечивая нерастворимость эластина.
Синтезируются эластические волокна фибро-
бластами, детали синтеза описаны менее подроб-
но, чем для коллагена, но в общих чертах они
схожи.
Ретикулиновые волокна
Ретикулиновые волокна в виде густой тонкой
сети находятся в верхней части сосочкового слоя
дермы непосредственно под эпидермисом, вок-
руг потовых желез, в соединительнотканных
влагалищах волосяных фолликулов. В сетчатом
слое они сконцентрированы, в основном, толь-
ко вокруг кровеносных сосудов.
Главной отличительной особенностью ретику-
линовых волокон является их способность окра-
шиваться азотнокислым серебром. Аргирофилия
ретикулиновых волокон объясняется наличием
склеивающего аморфного матрикса, состоящего
из гликопротеина с большим содержанием галак-
тозы и маннозы. Фибриллярный компонент во-
локон состоит из коллагена III типа и имеет ха-
рактерную периодическую исчерченность. Это
роднит ретикулиновые волокна с коллагеновы-
ми, особенно с незрелыми. Однако есть между
ними и отличия. Незрелые коллагеновые волок-
на обладают метахромазией и не ветвятся; в ходе
эмбриогенеза коллаген III типа в них замещает-
ся коллагеном I типа. Коллагеновые волокна в
значительно меньшей степени восстанавливают
серебро и окрашиваются в коричневый, а не в
черный, как ретикулиновые, цвет.
Повышение синтеза и распада коллагена на-
блюдается при многих заболеваниях, особенно
при коллагенозах. При склеродермии происхо-
дит повышенный катаболизм коллагена, несмот-
ря на его усиленный биосинтез. В этом случае
гистологически коллагеновые волокна выглядят
утолщенными из-за глубоких нарушений их тон-
кой организации. При склеродермии в фибро-
бластах находят возросшее число рибосом и ми-
тохондрий, что свидетельствует об усилении в
них синтеза белка. В результате увеличения ско-
рости размножения фибробласты не успевают
синтезировать полноценный зрелый коллаген.
Поэтому дальнейшее внеклеточное образование
структуры коллагена на основе самосборки на-
рушается. Заметно изменяется структура колла-
геновых фибрилл, что выражается в уменьше-
нии их колибра, значительной вариабельности
ширины, неполной поперечной исчерченности и
нерегулярности переплетения волокон между со-
бой. В целом, коллагеновые волокна кожи боль-
ных склеродермией напоминают по своему стро-
ению фибриллы коллагена зародыша.
С обменом коллагена связывают старение кожи.
С возрастом увеличивается число межмолекуляр-
ных связей белка, вследствие чего повышается
содержание резистентного к действию энзимов
коллагена. Избыточное количество метаболичес-
ки инертного коллагенового белка приводит к
нарушению обмена в тканях кожи и ее старению.
Группа наследственных заболеваний, характе-
ризующихся нарушением синтеза или метабо-
лизма коллагена, включает различные болезни,
имеющие среди прочих и кожные проявления.
К ним относят синдром Элерса-Данлоса, cutis
laxa (кожа вялая), псевдоксантома эластическая.
Комплексные исследования обмена компонен-
тов соединительной ткани (гликозаминоглика-
нов и коллагеновых волокон) у больных различ-
ными клиническими формами псориаза
показали, что даже при сравнительно благопри-
ятно протекающем ограниченном процессе по-
казатели метаболизма основного вещества уве-
личиваются отчетливо. Нарушение обмена
коллагена является общим метаболическим при-
знаком всех клинических форм псориаза.
Значительная дезорганизация коллагеновых
структур наблюдается при всех формах красной
волчанки, вследствие чего уровень оксипроли-
на в очагах поражения повышается. При гипер-
кератозно-инфильтративной стадии деструкция
коллагеновых волокон выражена еще больше,
чем в эритематозно-отечной стадии, что сопро-
вождается дальнейшим увеличением уровня ок-
сипролина в очагах поражения кожи. В стадии
атрофии, наоборот, наблюдается резкое умень-
шение содержания оксипролина в очагах пора-
жения, что свидетельствует об уменьшенном ме-
таболизме коллагена при регрессировании
процесса и развитии склероза (Мордовцев В.Н.,
Цветкова Г.М., 1993).
Сетчатый слой дермы
Сетчатый слой дермы образован плотной волок-
нистой неоформленной соединительной тканью
и является основной по объему частью дермы.
В нем располагаются самые толстые коллагено-
вые волокна. Они имеют разное направление и
формируют характерную вязь. Между коллаге-
новыми волокнами находится небольшое коли-
чество аморфного вещества и неактивные клет-
ки фиброциты. Коллагеновые волокна сетчатого
Глава 2. Анатомия и гистофизиология кожи
45
слоя состоят из коллагена I типа. Сосочковый
слой проникает в сетчатый вдоль сосудов. Таким
образом, вокругсосудистые зоны сетчатого слоя
образованы рыхлой волокнистой соединительной
тканью с характерными клетками.
Е. В. Виноградова и И. Н. Михайлов (1978) в
зависимости от архитектоники коллагеновых
волокон сетчатого слоя выделили 3 основных типа
переплетения их пучков (вязь).
Пластообразный тип
Пластообразный тип вязи обнаружен в коже
верхних и нижних конечностей и характеризу-
ется расположением пучков и волокон своеоб-
разными пластами, которые лежат параллельно
друг к другу и к поверхности кожи. Между со-
бой пласты соединяются вертикально направлен-
ными пучками фибрилл.
Ромбовидный тип
В этом случае каждая пара коллагеновых пуч-
ков пересекается под разными углами друг к дру-
гу, образуя ромбовидные или треугольные струк-
туры. В местах пересечения пучков имеются
многочисленные анастомозы за счет ветвлений
сети пучков и волокон (кожа груди, колена, по-
дошвы).
Сложнопетлистый тип вязи
Сложнопетлистый тип вязи напоминает ромбо-
видный тип, но при этом замкнутые петли или
ячейки своими острыми или тупыми углами об-
ращены к поверхности кожи. Внутри каждой
петли перпендикулярно к ее плоскости лежат
один или два коллагеновых пучка, анастомози-
рующих друг с другом. Этот тип вязи характе-
рен для сетчатого слоя кожи всей поверхности
спины и надплечья.
В коже шеи, ладони, тыла кистей и стоп, под-
коленной ямки и живота обнаруживается не-
сколько типов вязи, сменяющих друг друга. Так,
в коже живота пластообразный тип вязи в верх-
них отделах сменяется сложнопетлистым в бо-
лее глубоких отделах.
Подкожная жировая
клетчатка — гиподерма
Представляет собой рыхлую соединительную ткань
со скоплением клеток адипоцитов (липоцитов).
Если эти клетки накапливаются в большом коли-
честве и гипертрофируются, то говорят о жировой
ткани, которую относят к соединительным тканям
со специальными свойствами. Гиподерма соединя-
ет дерму с подлежащими тканями.
Подкожная жировая клетчатка неодинаково
развита на различных участках тела. У человека
она предпочтительно развита в нижней части
живота, на бедрах, ягодицах, ладонях, стопах, в
подмышечных впадинах. Жировые клетки в под-
кожной клетчатке скапливаются в дольки, раз-
деленные обычной рыхлой соединительной тка-
нью с ее типичными клетками. Дольки
окружены также кровеносными сосудами.
Различают два вида жировой ткани: белую и
бурую.
Белая жировая ткань
Белая жировая ткань составляет почти всю жи-
ровую ткань приматов и расположена в подкож-
ной клетчатке на всем протяжении кожного по-
крова. Ее структурно-функциональной единицей
является белый адипоцит (рис. 2.23). Цитоплазма
белых адипоцитов занимает узкий ободок по пе-
риферии клетки, содержит свободные рибосомы
и полисомы, митохондрии, слаборазвитые комп-
лекс Гольджи и эндоплазматическую сеть. В
центре клетки в виде крупной капли находится
включение жира, с которым тесно контактиру-
ют митохондрии.
Белая жировая ткань выполняет следующие
функции:
1. Депонирующая функция жировой ткани раз-
нообразна: 1) депо питательных веществ — тро-
фическая функция; 2) депо воды (при распаде
жиров образуется большое количество воды);
3) депо жирорастворимых витаминов А, Д, Е, К,
а также стероидных гормонов, особенно женс-
ких половых (эстрогенов).
2. Энергетическая функция. При распаде жира
образуется большое количество энергии.
3. Терморегуляционная функция: 1. термоизоли-
рующая функция связана с тем, что из-за низ-
кой теплопроводности жира теплоотдача в холод-
ной атмосфере происходит в незначительном
объеме. Животные, обитающие в северных ши-
ротах, имеют хорошо развитую подкожную жи-
ровую клетчатку, а у водных животных таких
широт имеется мощный сплошной слой этой
клетчатки, сохраняющийся на протяжении года.
Подкожно-жировая клетчатка хорошо развита и
у населения северных широт. 2) подкожная жи-
ровая ткань участвует в термопродукции в холод-
ной атмосфере при усилении метаболизма жира.
4. Защитно-механическая и опорная функции со-
стоят в механической защите тех органов, кото-
рые окружает жировая ткань. Располагаясь вдоль
подкожных сосудисто-нервных пучков, подкож-
ная жировая ткань обеспечивает их механичес-
46
Общая дерматология
кую защиту и изоляцию, препятствует смеще-
нию и травматизации.
5. Косметическая функция подкожной жировой
ткани заключается в том, что подкожный жир
участвует в образовании форм человеческого тела.
При этом жировая ткань не только обеспечивает
форму тела, но и ее индивидуальные, половые
особенности и особенности при патологии.
6. В последнее время установлена также эндо-
кринная функция белой жировой ткани.
Во-первых, в ней синтезируются эстрогены.
О возможности такого синтеза свидетельствует
хорошее развитие в липоцитах органелл стерои-
догенеза — гладкой эндоплазматической сети и
митохондрий. Указанная функция жировой тка-
ни очень важна. Показано, в частности, что при
резкой атрофии жировой ткани у женщин мо-
жет наступать аменорея, что связано со значи-
тельным нарушением эстрогенного баланса в
организме.
Во-вторых, липоциты синтезируют гормон, ре-
гулирующий потребление пищи — лептин. Лептин
тормозит секрецию гипоталамусом особого нейро-
пептида (нейропептид Y, NPY), который усилива-
ет потребление пищи, а также стимулирует про-
дукцию инсулина и накопление жира в липоцитах.
При голодании секреция лептина снижается из-за
потери жировой ткани и перехода липоцитов к не-
дифференцированному состоянию. В результате
нарастает секреция NPY, формирующая пищевое
поведение. При насыщении продукция лептина
возрастает, это ведет к снижению секреции NPY
и отказу от пищи. Недостаточная выработка леп-
тина ведет к ожирению (Быков В. Л., 1998).
7. Белая жировая ткань активно задействована
в регенерации кожного покрова, т. к. участвует в
формировании грануляционной ткани (Аничкин
Н. Н. и соавт., 1951; Давыдовский И. В., 1961).
Бурая жировая ткань
Бурая жировая ткань хорошо развита у зимо-
спящих животных и у новорожденных детей. У
взрослого человека она сохраняется в области шеи,
межлопаточной области, вокруг почек, аорты и
в средостении, а на остальных участках заменя-
ется белой жировой тканью. Ее структурно-функ-
циональной единицей является бурый адипоцит,
который меньше белого липоцита (см. рис. 2.23).
Ядро в клетке, в отличие от белого адипоцита, не
Рис. 2.23. Схема строения белой (I) и бурой (II) жировой ткани.
I. а. — липоциты с удаленным жиром в световом микроскопе; б. — ультраструктура липоцитов:
1 — ядро жировой клетки; 2 — крупные капли липидов; 3 — нервные волокна; 4 — гемокапилляры;
5 — митохондрии.
II. а. — липоциты с удаленным жиром в световом микроскопе; б. — ультраструктура липоцитов:
1 — ядро липоцита; 2 — мелко распределенные липиды; 3 — многочисленные митохондрии;
4 — гемокапилляры; 5 — нервное волокно (Афанасьев Ю. И. и соавт., 1989).
Глава 2. Анатомия и гистофизиология кожи
47
смещено на периферию, а находится в центре. В
клетках сильно развиты митохондрии, окружаю-
щие многочисленные диссоциированные жировые
капли. Цитохромы митохондрий обеспечивают ха-
рактерный цвет ткани. Окислительная способ-
ность митохондрий здесь в 20 раз выше, чем в
жировой ткани. При этом энергия не запасается
в виде тепла, а идет на быстрое согревание орга-
низма. Это связано с наличием в митохондриях
особого белка термогенина, разобщающего окис-
лительное фосфорилирование и дыхание. Этот
белок отсутствует в белой жировой ткани.
Функция бурой жировой ткани — терморегу-
ляция и согревание всего организма (при про-
буждении от зимней спячки; у маленьких детей)
или органов, которые она окружает.
Строение тонкой
и толстой кожи
Несмотря на общий план строения кожного по-
крова, отдельные его участки довольно суще-
ственно отличаются друг от друга. Если сравнить
кожу ладоней и подошв, с одной стороны, и ос-
тальную кожу тела, с другой, то даже невоору-
женным глазом можно увидеть разницу между
ними. Еще более отчетливо эта разница видна
под микроскопом.
Строение толстой кожи
К толстой коже относят кожу ладоней и подошв.
Эпидермис толстой кожи имеет все пять харак-
терных слоев, причем роговой слой имеет мак-
симальную толщину, и может по ширине пре-
вышать все остальные слои, вместе взятые. Хо-
рошо развит блестящий слой в виде достаточно
широкой, иногда суживающейся оксифильной
полосы. Зернистый слой представлен несколь-
кими рядами крупных вытянутых клеток с от-
четливо различными гранулами кератогиалина.
Шиповатый слой включает большое количество
клеточных рядов. Клетки базального слоя чаще
округлые, иногда вертикально вытянутые.
Эпидермис кожи имеет отчетливо выраженные
гребешки, видимые невооруженным глазом, меж-
ду которыми находятся бороздки. И гребешки,
и бороздки не изменяются в течение жизни, у
каждого человека имеют свой неповторимый
рисунок. Это позволило в 1880 году шотландцу
Генри Фолдзу предложить дактилоскопирование
как способ идентификации личности.
Гребешки и бороздки развиваются в течение
3—4 месяцев внутриутробной жизни. Их обра-
зование определяется генетическими факторами.
У гомозиготных близнецов характер кожного ри-
сунка очень сходен и похож у членов одной се-
мьи. При хромосомных аномалиях формирова-
ние кожного рисунка существенно нарушается.
У детей с болезнью Дауна, например, эти нару-
шения отчетливы, причем у 70 % таких детей
встречаются комбинации узоров, которые в нор-
ме не обнаруживаются. Поэтому анализ дакти-
лоскопических картин используется для диагно-
стики этих аномалий.
Видимые невооруженным глазом эпидермаль-
ные гребешки (рис. 2.24) образуются благодаря
тому, что эпидермис повторяет дермальные гре-
бешки. Каждый дермальный гребешок расщеп-
ляется на два дермальных сосочка участками
Рис. 2.24. Схемы, иллюстрирующие отношения между эпидермальными и дермальными гребешками
(Хэм А., Кормак Д., 1983).
48 Общая дерматология
эпидермиса, названными межсосочковыми кли-
ньями. Межсосочковые клинья являются местом
впадения протоков мерокриновых желез.
Эпидермальные пролиферативные единицы
эпидермиса толстой кожи характеризуются не-
столбчатым типом организации и более высокой
пролиферативной активностью кератиноцитов и
динамикой клеточной популяции в целом. Ко-
личество клеток Лангерганса в толстом эпидер-
мисе меньше, чем в эпидермисе тонкой кожи.
Толстая кожа всегда гораздо менее пигментиро-
вана, чем тонкая, что связано с более низкой
интенсивностью меланогенеза. Это особенно хо-
рошо заметно у людей негроидной расы.
Дерма толстой кожи может не превышать ее
толщину в тонкой коже (например, дерма кожи
спины имеет очень большую толщину). Однако
характер упаковки и направления коллагеновых
волокон, их толщина и толщина эластических
волокон в толстой коже имеют свои закономер-
ности. В сетчатом слое дермы пучки толстых кол-
лагеновых волокон формируют широкопетлистую
грубую сеть. Клеточный состав в дерме толстой
кожи менее разнообразный, чем в тонкой коже,
и представлен в основном фибробластами, распо-
лагающимися на довольно большом расстоянии
друг от друга. Макрофагов в сосочковом слое мало.
Это, очевидно, связано с резко выраженными за-
щитными свойствами утолщенного эпидермиса.
Уменьшено и количество тучных клеток.
Толстая кожа лишена сальных желез, а содер-
жание в ней мерокриновых потовых желез зна-
чительно выше, чем в тонкой коже.
Подкожно-жировая клетчатка в толстой коже
развита лучше, чем в тонкой и не исчезает даже
при сильном общем истощении организма.
Толстая кожа значительно более богато снаб-
жена чувствительными нервными приборами.
Поэтому кожа рук, например, служит как вы-
сокочувствительный тактильный орган.
Строение тонкой кожи
Эпидермис тонкой кожи, по сравнению с тол-
стой, содержит меньше, слоев, которые тоньше.
Иногда толщина эпидермиса составляет всего не-
сколько клеток. Роговой слой истончен и менее
компактен. Блестящий слой отсутствует. Резко
истончен, иногда до отсутствия или прерывис-
тости, зернистый слой. Шиповатый слой истон-
чается иногда до отсутствия или 1—2 слоев кле-
ток. Клетки базального слоя чаще, чем в толстой
коже, вытянуты в вертикальном направлении.
Характер границы между эпидермисом и дер-
мой в тонкой коже иной. Дерма не формирует
дермальных гребешков, а местами вступает в
эпидермис в виде истинных сосочков, но их на-
личие не приводит к формированию на поверх-
ности кожи видимых гребешков и характерно-
го для толстой кожи узора. Та картина
поверхности эпидермиса тонкой кожи, которая
видна невооруженным глазом, не связана с под-
лежащими дермальными сосочками, а обуслов-
лена в основном линиями, соединяющими вдав-
ленные устья волосяных фолликулов.
В тонкой коже, в отличие от толстой, содер-
жатся многочисленные волосяные фолликулы и
сальные железы. Потовых желез в тонкой коже
меньше, чем в толстой. В тонкой коже некото-
рых участков тела (подмышечные впадины, об-
ласть половых желез) имеются апокриновые
потовые железы.
Иннервация кожи
Кожа связана с ЦНС посредством многочислен-
ных нервных проводников. В зависимости от
выполняемой функции различают чувствительные,
секреторные, сосудодвигательные и моторные не-
рвные волокна кожи. Нервы кожи туловища и
конечностей берут свое начало от спинного мозга
и состоят из пучков эфферентных и афферентных
волокон. Первые являются отростками мотоней-
ронов передних рогов спинного мозга и иннерви-
руют подкожную мышцу (т. platisma), а вторые —
аксонами псевдоуниполярных нейронов спиналь-
ных ганглиев. По мнению большинства исследо-
вателей, вегетативная нервная система кожи при-
надлежит к симпатической нервной системе. При
этом постганглионарные волокна, идущие к саль-
ным и потовым железам, к мышце, поднимающей
волос, являются холинергическими.
Отдельным нервам соответствует определенная
зона иннервации кожи (сегментная иннервация).
Кожа головы иннервируется преимущественно
ветвями тройничного нерва и лишь частично
спинномозговыми нервами. Нервы кожи в составе
крупных стволов по ходу кровеносных и лимфа-
тических сосудов вступают через фасцию в под-
кожную клетчатку и образуют здесь крупное
сплетение. От него в сетчатый слой дермы отхо-
дят более тонкие стволы, где они разветвляются
и формируют глубокое дермальное сплетение. От
этого сплетения еще более тонкие волокна идут
в сосочковый слой и формируют в нем поверх-
ностное сплетение. От каждого из трех сплетений
для иннервации сосудов, придатков отходят тон-
кие нервные волоконца.
Глава 2. Анатомия и гистофизиология кожи
49
Преганглионарные нервные волокна симпа-
тической нервной системы кожи формируются
аксонами нейронов ядер боковых рогов спин-
ного мозга (латеральных промежуточных ядер).
Эти нейроны имеют связь с ретикулярной фор-
мацией промежуточного мозга, корой больших
полушарий и центрами эмоциональной актив-
ности. Аксоны нейронов этих ядер в составе
преганглионарных волокон идут к ганглиям
симпатической цепочки (паравертебральным
ганглиям) и переключаются на нейроны этих
ганглиев. В свою очередь нейроны ганглиев
формируют постганглионарные нервные волок-
на, достигающие смешанных спинномозговых
нервов через серые соединительные ветви. В
составе спинномозговых нервов симпатические
нервные волокна идут в кожу и заканчивают-
ся на сосудах и железах. В коже вегетативные
нервные волокна перемешиваются с чувстви-
тельными нервными волокнами и трудно отли-
чимы без специальных гистоэнзимологических
сплетений от последних. Некоторые авторы
описывают в коже различные типы пептидер-
гических нервных волокон.
Эфферентная
иннервация кожи
Соматическую эфферентную иннервацию кожи
получает только т. platisma. Остальные струк-
туры получают вегетативную эфферентную ин-
нервацию (потовые и сальные железы, сосуды;
мышца, поднимающая волос).
Кровеносные сосуды кожи иннервируются
ветвями нервов, образованных постганглионарны-
ми волокнами нейронов симпатических гангли-
ев. Эти нервные волокна вызывают вазоконст-
рикцию. Особенно богато иннервируются
артериоло-венулярные анастомозы. В последнее
время вазодилатацию сосудов кожи связывают с
пептидергическими нервными волокнами, кото-
рые описаны в коже и в органах с собственной
двигательной активностью и которые относятся,
как известно, к метасимпатическому отделу ве-
гетативной нервной системы. Медиаторами вазо-
дилатации являются гистамин, брадикинин, про-
стагландины, дофамин, ангиотензин, АТФ,
вещество Р. Играет роль в вазодилатации и пе-
риневральная и паравазальная система тучных
клеток. Эта часть тучных клеток лежит между
сосудами и нервами и тесно контактирует с ве-
гетативными нервными окончаниями сосудов.
Она может выполнять роль нейрогормональных
клеток. Вопрос о наличии в коже элементов ме-
тасимпатической системы до настоящего време-
ни не обсуждался и остается открытым, однако
факт присутствия пептидергических нервных
волокон заставляет об этом задуматься.
Нервы, регулирующие работу желез кожи, по
мнению большинства авторов, относятся к сим-
патическому отделу вегетативной нервной сис-
темы. Однако в ряде нервных волокон медиа-
тором является не норадреналин, а ацетилхолин
(характерный медиатор парасимпатической не-
рвной системы). Согласно последним данным,
все виды желез кожи получают двойную иннер-
вацию: холин- и адренергическую. При этом хо-
линергические нервные волокна лежат в виде
густой сети на базальных мембранах секретор-
ных отделов, а тонкие адренергические волокна
заканчиваются непосредственно на секреторных
клетках.
Афферентная
иннервация кожи
Чувствительная иннервация кожи обеспечивается
дендритами псевдоуниполярных нейронов спи-
нальных ганглиев. Они заканчиваются рецеп-
торными нервными окончаниями, которые де-
лятся на свободные и несвободные (рис. 2.25).
Свободные
нервные окончания
Свободные нервные окончания представляют
собой терминальные ветвления дендрита, не
имеющие вспомогательных глиальных клеток.
Этот тип нервных окончаний локализуется в
эпидермисе; в поверхностной нервной сети дер-
мы, расположенной в сосочковом слое; вокруг
волосяных фолликулов в виде корзинчатых
структур.
Внутриэпидермальные нервные волокна у
взрослого человека обнаруживаются чаще всего
в коже век и половых органов. В эмбриональ-
ной коже они многочисленны повсеместно.
Несвободные
нервные окончания
Несвободные нервные окончания подразделяют-
ся на неинкапсулированные и инкапсулированные.
И те, и другие находятся в тесном взаимодей-
ствии с особым видом глиальных клеток (оли-
годендроглия нервных окончаний), которые уча-
ствуют в рецепции. Инкапсулированные нервные
окончания, кроме того, окружены оболочкой из
соединительной ткани.
5 0 Общая дерматология
Неинкапсулированные
нервные окончания
Как пример несвободного неинкапсулированного
нервного окончания некоторые авторы рассмат-
ривают нервные окончания, связанные с клет-
ками Меркеля (см. рис. 2.25). Иногда эти окон-
чания называют дисками Меркеля. Дендрит
псевдоуниполярного нейрона, подходя к клетке
Меркеля, расширяется и формирует своеобраз-
ный диск, который образует синапс с клеткой
Меркеля. Клетка Меркеля содержит нейросек-
реторные гранулы с нейромедиатором. Механи-
ческое воздействие на эпидермис приводит к воз-
буждению клетки Меркеля и к экзоцитозу ее
гранул в синаптическую щель. Затем нейроме-
диатор диффундирует к мембране дисков Мер-
келя и вызывает ее деполяризацию. Диски Мер-
келя находятся в коже высокочувствительных
зон тела (в области ладоней, слизистой рта и губ).
В настоящее время при помощи электронного
микроскопа показано, что гранулы клеток Мер-
келя располагаются вблизи контакта клетки с не-
рвным волокном. Следует отметить, что не все
клетки Меркеля формируют синапсы с нервными
волокнами. Часть из них свободна от этих кон-
тактов. Такие клетки, очевидно, выполняют
только эндокринные функции.
Инкапсулированные
нервные окончания
К инкапсулированным нервным окончаниям кожи
относятся пластинчатые тельца Фатер-Пачини, ося-
зательные тельца Мейснера, концевые колбы Крау-
зе, генитальные тельца Догеля, колбы Руффини и др.
Все они построены по сходному принципу. Расмот-
рим строение инкапсулированных нервных окон-
чаний на примере пластинчатого тельца Фатер-
Пачини, наиболее многочисленного нервного
прибора, имеющего наиболее выраженную соеди-
нительнотканную капсулу (рис. 2.26.). Эти тельца
имеют овальную форму и состоят из трех частей:
слоистой соединительнотканной капсулы, термина-
лей дендрита псевдоуниполярного нейрона и ней-
ролеммоцитов (олигодендроглии). Дендрит при
подходе к капсуле теряет миелиновую оболочку и
внедряется в капсулу, внутри которой покрывает-
ся несколькими слоями нейролеммоцитов. Вместе
с отростком нервной клетки нейролеммоциты фор-
мируют внутреннюю колбу. Наружная колба пред-
ставляет собой соединительнотканную капсулу, ко-
торая состоит из многочисленных концентрических
слоев тонких коллагеновых пластинок с фиброци-
тами между ними. На границе наружной и внут-
ренней колб находится особый вид отростчатых
нейролеммоцитов, контактирующих с осевым ци-
линдром. Давление на поверхность кожи передается
на капсулу тельца Фатер-Пачини, при этом от слоя
к слою многократно усиливается, как в трансфор-
маторе, и вызывает смещение капсулы по отноше-
нию к внутренней колбе. Это вызывает раздраже-
ние отростчатых нейролеммоцитов, которые
передают его на осевой цилиндр. Локализуются
тельца Фатер-Пачини в глубоких слоях дермы и в
гиподерме.
Осязательные тельца Мейснера имеют похожее
строение (см. рис. 2.26), отличаются от телец
Фатер-Пачини своей локализацией (сосочковый
слой кожи волярных поверхностей приматов),
более тонкой наружной капсулой, а также тем,
что осевой цилиндр, войдя в тельце, ветвится и
имеет спиральный ход, а нейролеммоциты лежат
перпендикулярно к осевому цилиндру.
Генитальные тельца Догеля находятся в осо-
бо чувствительных областях кожи, в первую оче-
редь, в коже наружных половых органов. Ок-
ружены очень тонкой соединительнотканной
капсулой. Во внутреннюю колбу проникает од-
новременно несколько осевых цилиндров, явля-
ющихся дендритами нескольких нервных кле-
ток. Это ведет к иррадиации возбуждения при
воздействии на тельце Догеля.
Ранее различным видам рецепторов приписы-
вали свою функцию. Свободные нервные окон-
чания, согласно этим представлениям, отвечают за
ноцицепцию и тактильную чувствительность;
тельца Фатер-Пачини — за чувство глубокого дав-
ления; Мейснера ~ осязание, колбы Краузе — за
терморецепцию и т. д. В настоящее время выска-
зывается мнение, что четыре основных вида чув-
ствительности (тактильную, холодовую, тепловую
и болевую) вряд ли стоит привязывать к конкрет-
ным концевым приборам. Вместе с тем, многие
авторы считают, что поверхностная нервная сеть
дермы отвечает за болевую и температурную чув-
ствительность, а тактильную чувствительность
обеспечивают нервная сеть фолликулов волос,
тельца Мейснера, Фатер-Пачини, свободные не-
рвные окончания эпидермиса и диски Меркеля.
Кровоснабжение
кожи
Артерии, питающие кожу, происходят из трех
групп артерий: кожные ветви артерий (идущие
в межмышечных фасциальных перегородках),
Глава 2. Анатомия и гистофизиология кожи
51
Клетки
Меркеля
Свободное
окончание
афферентного
волокна
Граница между
дермой -
и эпидермисом
Базальная
мембрана
а
Конечный диск
афферентного
волокна
Миелин
Пучки
коллагеновых
волокон
в ядре тельца
Терминальные
веточки
афферентного
волокна
Капсула
Капсула
Базальная
мембрана
Терминальный отросток
афферентного волокна
Субкапсулярное
пространство
Внутренняя
колба
Шванновские
клетки
Наружная
колба
Миелин
Капсула
в
Миелин
Базальная
мембрана
Спиральные
терминали
афферентного
волокна
Уплощенные
шванновские
клетки
Шванновская
клетка
Шванновские
клетки
е
Терминальные
веточки
афферентного
волокна
Рис. 2.25. Схема строения чувствительных нервных окончаний в коже (Хэм А., Кормак Д., 1983).
а. Свободные нервные окончания.
б. Окончание Меркеля.
в. Тельце Фатер-Пачини.
г. Тельце Мейснера.
д. Тельце Руффини.
е. Концевая колба Краузе.
5 2 Общая дерматология
мышечно-кожные артерии, надкостно-кожные
артерии. Артерии, питающие кожу, образуют
широкопетлистую сеть под гиподермой — фас-
циальная сеть (рис. 2.27). Из этой сети в кожу
поднимаются более мелкие ветви, которые на
нижней границе дермы делятся и анастомозируют
друг с другом, образуя вторую глубокую (суб-
дермальную) артериальную сеть, параллельную
первой. Здесь берут начало артерии, питающие
волосяные фолликулы и клубочки потовых же-
лез. Из субдермальной артериальной сети в соб-
ственно кожу направляются микроскопически
малые артерии, которые делятся, образуя длин-
ные артериолы, анастомозирующие друг с дру-
гом. Под эпидермисом из них образуются арте-
риолярные аркады. На границе с сосочковым
слоем располагается поверхностная артериоляр-
ная сеть, от которой отходят терминальные ар-
териолы, идущие к кожным сосочкам. Каждая
терминальная артериола питает группу сосочков,
образуя сосочковые капилляры. Этим объясня-
ется очаговость сосудистых поражений кожи.
Сосочковые капилляры можно исследовать в
клинике при помощи капилляроскопии, при
этом они имеют форму дамских шпилек.
Капилляры кожи, по данным большинства
авторов (Шахламов В.А., 1970; Чернух А.М. и
соавт., 1982 и др.), являются капиллярами сома-
тического типа: имеют непрерывный нефенест-
рированный эндотелий и непрерывную базаль-
ную мембрану, а также развитые перициты.
Из сосочков кровь оттекает в венулы, обра-
зующие поверхностную мелкопетлистую сеть
сразу под сосочками. Несколько глубже распо-
лагается вторая субпапиллярная сеть венул, иду-
щая параллельно первой. В сетчатом слое дер-
мы лежит третья венозная сеть, а в гиподерме
располагается крупноячеистая глубокая веноз-
ная сеть, в которой осуществляется теплообмен.
Эта сеть лежит параллельно расположенной над
ней сети субдермального артериального сплете-
ния, с которым соединяется множеством арте-
риоло-венулярных анастомозов. Это обстоятель-
ство имеет большое значение для регуляции
теплоотдачи. Две сети функционируют по
принципу поворотно-противоточно-множитель-
ного механизма.
В отличие от других органов, микроциркуля-
торное русло в коже выполняет не только транс-
портную и обменную функции, но и очень важ-
ную функцию терморегуляции. Регуляция
температуры тела осуществляется, во-первых, пу-
Рис. 2.26. Схема строения инкапсулированных нервных окончаний кожи Пачини.
а. Тельце Фатер-Пачини (по А. А. Стеллину и соавт.): 1 — слоистая капсула; 2 — внутренняя колба;
3 — дендрит чувствительного нейрона; 4 — спиральные коллагеновые волокна; 5 — фиброциты;
6 — вторичночувствующие клетки с ресничками; 7 — синапсы отростков вторичночувсвующих клеток с
дендритом чувствительного нейрона;
б. Тельце Мейснера (по Р. Кристичу): 1 — капсула; 2 — специальные клетки (леммоциты); 3 — чувстви-
тельные нервные терминали; 4 — миелиновое нервное волокно; 5 — опорные (поддерживающие)
фибриллы; 6 — эпидермис.
Глава 2. Анатомия и гистофизиология кожи
53
тем изменения просвета сосудов терминального
сосудистого русла, а во-вторых, путем измене-
ния просвета артериоло-венулярных анастомо-
зов. Большое значение имеет изменение крово-
снабжения потовых желез. При понижении
температуры окружающей среды происходит су-
жение поверхностных кровеносных сосудов и од-
новременное расширение анастомозов. При этом
кровь через них сбрасывается в венозное русло,
не доходя до поверхностных слоев кожи. Дан-
ное обстоятельство при, несомненно, положи-
тельном эффекте (резко ограничиваются поте-
ри тепла путем излучения) имеет и обратную
сторону, т. к. приводит к ишемии поверхност-
ных слоев кожи и, в тяжелых случаях, к их не-
крозу (при отморожениях). При повышении
температуры воздуха, напротив, сосуды поверх-
ностной сосудистой сети сильно расширяются,
анастомозы сокращаются, и потери тепла с по-
верхности кожи возрастают.
Артериоло-венулярные анастомозы кожи в ос-
новном сложные, с хорошо представленной си-
стемой регуляции кровотока через них (эпите-
лиоидные клетки, способные набухать и сокра-
щаться под влиянием нейрогуморальных стиму-
лов).
Роль артериоло-венулярных анастомозов кожи
многоплановая. Во-первых, они включаются в
работу при понижении температуры кожи. В этом
случае из артериол в венулы сбрасывается кровь,
которая должна была пройти через капилляры и,
следовательно, охладиться в них. В этом случае
функционирование анастомозов способствует
сохранению в организме тепла (терморегуляци-
онная функция). Однако длительная ишемиза-
ция поверхностных слоев кожи в этих случаях
создает условия для отморожений кожи. Во-вто-
рых, анастомозы регулируют такую функцию
кожи, как потоотделение. Наконец, артериоло-
венулярные анастомозы играют определенную
роль в мобилизации в общий кровоток депони-
рованной в коже крови.
В последнее время развиваются представления
о дискретном принципе строения микрососуди-
стой сети кожи. Это позволяет понять причину
существования мозаичной и микроочаговой со-
Рис. 2.27. Схема кровоснабжения кожи (Garven D., 1967).
5 4 Общая дерматология
судистой кожной патологии. Разработаны и обо-
снованы такие понятия, как «кожный ангион»,
«артериоло-капиллярно-венулярные единицы
кожи» (АКВЕК). Выделены субпапиллярные,
парафолликулярные, дермальные и гландуляр-
ные АКВЕК, существенно различающиеся по
структуре и функции.
Из этого краткого освещения можно предста-
вить, насколько сложно и многогранно органи-
зована сосудистая система кожи в структурном
отношении. А ее функциональная организация
несравненно сложнее. Изучение этих вопросов,
несомненно, позволит раскрыть многие механиз-
мы патогенеза кожных заболеваний.
Глава 3
Основные функции кожи
Защитная функция кожи 55
Кожа как орган чувств 56
Участие кожи в терморегуляции 57
Депонирующая функция кожи 58
Дыхательная функция кожи;58
Резорбтивная функция кожи 58
Экскреторная функция кожи 58
Роль кожи в обмене веществ 59
Кожа как эндокринный орган 59
Кожа как орган иммунитета 60
Косметическое значение кожи 61
Экологическое значение кожи 61
Защитная функция кожи
Защитные функции кожи многогранны. Их
можно подразделить на защитно-механические
функции; функции по поддержанию водно-со-
левого гомеостаза; иммунологические функции;
защиту организма от биологических, химичес-
ких, физических факторов.
Определенные структуры в коже, включая
роговой слой, меланин, кожные нервы и дер-
мальную соединительную ткань, выполняют
важные для выживания защитные функции.
Кожа защищает организм от потери важных
жидкостей, от проникновения токсичных аген-
тов и микроорганизмов, вредного воздействия
ультрафиолетового излучения, механических
повреждений и экстремальных температур окру-
жающей среды.
Роговой слой является барьером низкой прово-
димости, который не только замедляет потерю
воды из внутренних эпидермальных гидратиро-
ванных слоев, но и защищает от проникновения
воды из окружающей среды. Барьерные свойства
рогового слоя имеют практическое значение с
нескольких точек зрения:
Во-первых, чрезмерное высыхание или воспали-
тельные реакции в коже ведут к огрублению и
шелушению, поскольку нормально компактные
слои роговых клеток нарушаются. Это ведет к
увеличению трансэпидермальной потери воды и,
если нарушаются обширные зоны рогового слоя
(как при генерализованном эксфолиативном
дерматите, эритродермии или ожогах), тотальная
водопотеря может привести к жидкостному и
электролитному дисбалансу.
Во-вторых, при нарушениях в роговом слое ве-
щества из внешней среды легче проникают в
слой эпидермиса. Так, различные химикаты,
включая медикаменты, при попадании на пора-
женную кожу могут подвергаться системной аб-
сорбции, выступая при этом в качестве гаптенов
или антигенов и увеличивая вероятность аллер-
гического контактного дерматита. Это особенно
часто наблюдается при местном применении
неоспорина, который на хронически воспален-
ной коже провоцирует аллергический контакт-
ный дерматит.
В-третьих, нарушение барьера увеличивает воз-
можность обсеменения кожи патологическими
бактериями, особенно в присутствии экссудатов
тканевой жидкости, которые служат отличной
питательной средой.
55
3 Зак. 1357
5 6 Общая дерматология
В-четвертых, чрезкожная абсорбция различных
медикаментов при местном применении для лече-
ния заболеваний кожи (например, стероидные
мази) может быть усилена путем гидрирования ро-
гового слоя посредством окклюзионных повязок.
Роговой слой служит не только барьером от
проникновения различных бактерий, но и явля-
ется местом обитания ряда аэробных и анаэроб-
ных облигатных механизмов (например,
Staphylococcus epidermitis diphtheroidus,
Propionibacterium acnes и Malassezia). Поврежде-
ние в роговом слое, несоблюдение гигиены и по-
вышенная влажность с мацерацией (особенно в
интертригинозных зонах) способствуют кожным
инфекциям, таким как импетиго, эризепелоид,
фолликулит, фурункулез, эктима.
Вторым структурным компонентом, обеспечи-
вающим защиту, является меланоцит, который
производит пигмент меланин. Меланин — это
крупный полимер, имеющий уникальную спо-
собность поглощать свет в широком диапазоне
с длиной волн от 200 до 2400 нм. Он служит от-
личным экраном против неблагоприятных эф-
фектов солнечного ультрафиолетового излуче-
ния, таких как старение и морщинистость кожи,
а также развитие кожных новообразований. Важ-
ность меланина ярко демонстрируется высокой
степенью вероятности возникновения рака кожи
на подверженных воздействию солнца участках
тела, особенно у тонкокожих, голубоглазых, лег-
ко загорающих индивидуумов и у альбиносов.
Подверженность ультрафиолетовому свету вызы-
вает также старение и морщинистость кожи. Раса
и пол не влияют на количество меланоцитов в
эпидермисе. Кожа негров содержит то же коли-
чество меланоцитов, что и кожа кавказцев, но
пигментация кожи у них более интенсивна в ре-
зультате синтеза большого количества мелани-
на, который распространяется через меланоци-
ты и смежные с ним кераноциты. Черная кожа,
соответственно, менее склонна к новообразова-
ниям и стареет более медленно, чем белая.
Третьим структурным компонентом кожи, иг-
рающим защитную роль, являются дермальные
нервы. Нервные окончания экстенсивно распре-
делены в коже в двух общих морфологических
типах: свободные нервные окончания и специ-
ализированные окончания (тельца Фатер-Пачини
и Мейсснера), которые опосредуют многие ощу-
щения, включая боль, давление и зуд. Боль важ-
на для выживания, поскольку помогает избегать
источника боли и дальнейшей травмы. Потеря
ощущения (например, при диабетической ней-
ропатии) может привести к глубоким трофичес-
ким язвам, образование которых пациент может
не заметить. Повреждения кожных нервов (на-
пример, herpes zoster) может привести к длитель-
ной жгучей боли и постгерпетической невралгии.
Свойства высокоэластичности, упругости,
придаваемые коже волокнистыми протеинами
(коллагеном и эластином), а также основное ве-
щество, составляющее дерму, защищают от ме-
ханического воздействия на кожу. Свойства эла-
стичности и вязкости основного вещества
позволяют сопротивляться сжатию и восприни-
мать определенную форму, что ослабляет точеч-
ное давление на более чувствительные кожные
структуры.
Кожа как орган чувств
Рецепторы различных видов чувствительности
отличаются по своему строению: болевые явля-
ются свободными нервными окончаниями. Дру-
гие виды раздражения воспринимают инкапсу-
лированные нервные окончения (давление —
тельца Мейснера и Фатер-Пачини; температур-
ные — колбы Краузе и тельца Руффини и т. д.).
Кожа является обширным рецепторным полем.
Ни один орган не имеет такой богатой чувстви-
тельной иннервации. Очевидно, это можно
объяснить тем, что эпителиальные образования
кожи и ЦНС имеют единый источник развития
— эктодерму. Рецепторы по кожной поверхно-
сти распределены неравномерно. Подсчитано, что
на 1 см2 кожи приходится 2 тепловых, 12 холо-
довых, 25 осязательных и 150 болевых точек.
Через рецепторы кожи поступает обширная и
сложная информация. В любом участке кожи
имеется сложный комплекс нескольких рецеп-
тивных полей, с каждым из которых связано
множество отдельных рецепторов, объединенных
в определенную анатомо-функциональную сово-
купность.
Наиболее распространенным ощущением и
симптомом в дерматологии является зуд, кото-
рый может возникать в связи с рядом дермато-
логических заболеваний или без клинически
очевидного кожного заболевания. Зуд и боль
проводятся по безмиелиновым С-волокнам,
сформированным дендритами псевдоуниполяр-
ных нейронов. Аксоны этих нейронов входят в
задний рог спинного мозга, переключаются на
вторые нейроны, аксоны которых переходят на
противоположную сторону и в составе спинота-
ламического тракта поднимаются к таламусу.
Затем импульс идет в сенсорную зону задней
центральной извилины коры головного мозга.
Перерезка спиноталамического тракта снимает
боль и зуд (антеролатеральная гемихордотомия).
Глава 3. Основные функции кожи
57
Различные периферические медиаторы стимули-
руют С-волокна и вызывают зуд. К ним отно-
сятся гистамин, трипсин, протеаза, пептиды (бра-
дикинин, вазоактивные интестинальные
пептиды, вещество Р — все возможные высво-
бодители гистамина) и желчные соли.
Простагландины являются скорее модуляторами
зуда, чем первичными медиаторами, понижая
порог зуда, вызванного как гистамином, так и
болью. Центральные модуляторы зуда, такие как
системный морфин, вызывают зуд, облегчая боль
действием на центральные опиатные рецепторы.
Генерализованный зуд в отсутствие первичного
кожного заболевания может быть важным при-
знаком внутреннего заболевания. Такие различ-
ные заболевания, как уремия, желчекаменная
болезнь, лимфомы и миелопролиферативные
заболевания, тиреотоксикоз, диабет, карцинома,
анемии с недостаточностью различного генеза и
психические нарушения могут вызывать силь-
ный зуд. Важная причина зуда — психический
стресс. Некоторые пациенты с психогенным зу-
дом полагают, что зуд вызывается невидимыми
паразитами в коже. Такие пациенты расчесыва-
ются, пока не возникают экскориации и прури-
гинозные папулы в зонах, легко доступных для
пациента (конечности, череп, верхняя часть спи-
ны). Сухая кожа (ксероз) —- распространенная
причина зуда у других индивидуумов. Опреде-
ленные лекарства (аспирин, опиаты) могут выз-
вать зуд без видимой сыпи. Пациенты с истин-
ной полицитемией обладают уникальным типом
зуда, а именно зудом, начинающимся при вне-
запных изменениях температуры, особенно при
выходе из теплой ванны. Зуд по характеру яв-
ляется колючим и длится от нескольких минут
до нескольких часов.
Участие кожи
в терморегуляции
Около 80 % всего количества тепла, вырабаты-
ваемого в организме, выделяется через кожу за
счет испарения, теплопроведения и теплоизлу-
чения. Терморегуляция осуществляется совмес-
тно сосудистой системой и потовыми железами
кожи. Массивная сеть взаимосвязанных кожно-
мышечных артерий и вен, а также капилляров,
артериовенозных шунтов и мелких вен играет оп-
ределяющую роль в поддержании температуры
тела. Основная фракция объема крови кожи со-
держится в больших венозных сплетениях, в
которых кровь движется медленно, вблизи по-
верхности, обеспечивая максимальное распреде-
ление тепла.
Важным для терморегуляции является также
образование эккринного пота, который обеспе-
чивает охлаждение при испарении с поверхнос-
ти кожи. На каждый грамм воды, испаряющей-
ся с поверхности кожи, теряется 580 калорий
тепла. С помощью потоотделения и испарения
через кожу выводится 27 % воды. В обычных
условиях кожа выделяет за сутки 650 г влаги и
около 10 г углекислоты, при усиленном потоот-
делении их количество увеличивается в несколь-
ко раз. Максимально за 1 час может выделить-
ся от 1 до 3,5 л пота, что эквивалентно отдаче
2500—8750 кДж тепла. Огромная работа желези-
стого аппарата кожи во время обильного пото-
отделения поистине благотворна.
Терморегуляция может осуществляться также
с помощью изменения кровотока в коже. При
основных условиях 8,5 % общего кровопотока
или 450 мл в минуту проходит через кожу, кон-
троль потока осуществляется первично симпати-
ческой нервной системой (через адреналин и
норадреналин). При физической нагрузке в теп-
лой окружающей среде кровяной поток может
возрастать до 3 л/мин. Поскольку теплопровод-
ность и удельное тепло крови высоки, большое
количество тепла может распространяться по
коже. Как центральные, так и периферические
терморецепторы стимулируют потоотделение че-
рез периферическую нервную систему, но в слу-
чае потовых желез постганглионарным провод-
ником является ацетилхолин. В терморегуляции
участвуют центральные (расположены в гипота-
ламусе) и периферические (локализованы в
коже) терморецепторы. При этом центральные
терморецепторы примерно в 10 раз эффектив-
нее в отношении вызывания потоотделения.
Ответ на холод начинается тогда, когда охлаж-
денная ниже обычной температуры кровь дости-
гает гипоталамуса, что затем задействует механиз-
мы производства и сохранения тепла в организме.
Симпатическая система сужает кровеносные сосу-
ды и, следовательно, уменьшает перенос тепла к
поверхности тела. Импульсы от гипоталамуса так-
же активируют моторный центр дрожи, который
увеличивает производство тепла на 50 %. Наобо-
рот, если к гипоталамусу притекает кровь теплее
нормальной, центральный механизм теплообразо-
вания отключается, и кожные сосуды расширяют-
ся, позволяя крови собираться у поверхности, а
теплу теряться посредством проводимости и кон-
векции. Вазодилатация, таким образом, происхо-
дит рефлекторно, через прямое нагревание повер-
хности кожи (в теплых средах). Кроме того,
стимуляция гипоталамуса вызывает потение и уве-
личивает испарительную потерю тепла. При пе-
5 8 Общая дерматология
риодическом воздействии тепла или тепла в соче-
тании со стрессом (например, ежедневно 1—2 часа
в течение 10—14 дней) секреторная производи-
тельность эккринных потовых желез увеличива-
ется (акклиматизация). Одним из примеров реша-
ющей роли кожнососудистой системы при
терморегуляции и кардиоваскулярном гомеостазе
являются широко распространенные воспалитель-
ные заболевания кожи, вызывающие эритродер-
мию. При таких заболеваниях, как распространен-
ный дерматит, псориаз, токсикодермии и
лимфомы, воспалительная реакция в коже может
вызвать генерализованную кожную вазодилатацию
с отвлечением от 10 до 20 % общего кровообраще-
ния через кожу. При этом центральный объем кро-
ви может уменьшаться. Для поддержания кровя-
ного давления общее кровообращение должно
возрасти, и у пожилых лиц с дисбалансом карди-
орезерва может произойти сбой в связи с больши-
ми колебаниями температуры и ознобом.
Депонирующая функция кожи
Кожа служит своеобразным кровяным депо, яв-
ляясь как бы второй селезенкой организма. В оп-
ределенных условиях расширенные сосуды кожи
способны вместить более 1 л крови. Это доволь-
но значительная цифра, если учесть, что объем
всей циркулирующей крови равен 5 л. Помимо
воды, в коже депонируется жир, который в свою
очередь является источником воды и энергии.
Дыхательная функция кожи
Через кожу в организм поступает кислород и
выделяется углекислый газ в количествах 2 % от
всего газообмена. Организм человека выделяет
через кожный покров 7—9 г углекислоты и по-
глощает 3—4 г кислорода в сутки. Кожное ды-
хание усиливается при повышении температуры
окружающей среды и тела, во время физичес-
кой работы, при увеличении атмосферного дав-
ления и при остром воспалении кожи. Кожа про-
ницаема и для других газов. Усиленная жировая
смазка на повехности эпидермиса снижает ды-
хание через кожу.
Резорбтивная функция кожи
Кожа не является абсолютно непроницаемой гра-
ницей между внутренней и наружной средой.
Проницаемость может быть различной для водо-
или жирорастворимых веществ, для соединений
с малой или большой молекулярной массой, что
во многом ускоряет или замедляет этот процесс.
На скорость проникновения различных веществ
через кожу влияют разнообразные факторы? ло-
кализация участка кожи, степень его гидратации,
толщина рогового слоя, наличие липидной смаз-
ки. Кроме газов, через кожу могут проникать
токсические, органические и неорганические ве-
щества (глюкокортикоиды, деготь, ртуть, сали-
циловая кислота, димексид). Некоторые вещества
(ртуть, салициловая кислота), проходя через
кожу, повреждают клетки эпидермиса, эластичес-
кие волокна дермы, что приводит к их дезорга-
низации и даже гибели. Вещества, проникающие
через кожу, могут оказать и общетоксическое
действие. В ответ на чужеродное повреждающее
вещество возникают соответствующий иммунный
ответ и аллергические реакции (простой контакт-
ный и аллергический дерматит, токсикодермия).
Изучение и использование проницаемости
лекарственных веществ через кожу представля-
ет собой важное и перспективное направление в
клинической дерматологии. Большинство лекар-
ственных препаратов, вводимых парентерально
или через желудочно-кишечный тракт, не явля-
ются дерматотропными. Для проникновения в
кожу имеются два альтернативных пути: транс-
эпидермальный (через тонкий, относительно не-
проницаемый роговой слой) или через волосяные
фолликулы (включая сальные и/или эккринные
потовые железы). Трансэпидермальный путь
введения лекарственных препаратов может иметь
преимущества перед их введением перорально
или парентерально, так как не зависит от вели-
чины pH содержимого желудка, времени приема
пищи, метаболической инактивации. Трансэпи-
дермальный путь введения лекарственных ве-
ществ обладает важным достоинством — достав-
кой непосредственно к больному органу.
Экскреторная функция кожи
Кожа осуществляет важную функцию выведе-
ния из организма веществ, освобождая его от из-
бытка одних (вода, соли) и от токсического дей-
ствия других (метаболиты, лекарственные
вещества). Это осуществляется потовыми и саль-
ными железами, а также через эпидермис. Ток-
сические вещества, лекарственные препараты,
приносимые кровью в сосочки дермы, вместе с
тканевой жидкостью по межклеточным каналь-
цам проникают в эпидермис, могут оказывать
повреждающее действие на его клетки и уда-
ляться с поверхности. При патологии (например,
при уремии) экскреторная функция кожи рез-
ко возрастает, что носит ярко выраженный адап-
тивный характер.
Глава 3. Основные функции кожи
59
Роль кожи в обмене веществ
Кожа играет важную роль в обмене веществ
организма, что обусловливается ее массой (око-
ло 20 % от массы тела) и положением на грани-
це с внешней средой. Метаболизм кожи объеди-
няется с обменом всего организма с помощью
нервной и эндокринной систем. Давно замече-
на связь обменных нарушений в тканях с дис-
функцией половых желез при таких заболева-
ниях, как себорея, алопеция, акне, дерматозы
беременных. Характерные дистрофические изме-
нения кожи возникают у больных диабетом,
базедовой болезнью, зобом щитовидной железы,
а также при недостаточности надпочечников,
гипофиза, паращитовидных желез.
Выделяют несколько аспектов обмена веществ
в коже и ее участия в общем обмене. Велика роль
кожи в депонировании различных биологичес-
ких жидкостей и твердых веществ: крови, лим-
фы, продуктов тканевого обмена, макро- и мик-
роэлементов. Этому способствует разветвленная
сосудистая сеть кожи. Увеличение остаточного
азота при белковой диете и различных заболе-
ваниях (экзема, атопический дерматит, псориаз,
красная волчанка) показывает, что кожа времен-
но задерживает белковые метаболиты, тем самым
ослабляя их токсическое действие на другие орга-
ны, в частности, на мозг.
Функциональная роль в организме многих
микроэлементов остается недостаточно изучен-
ной, однако для дерматологии наибольший ин-
терес представляют цинк, медь и железо.
У человека при дефиците цинка наблюдаются
различные симптомы и заболевания: энтеропа-
тический акродерматит, алопеция, сонливость,
апатия, эритематозно-сквамозные высыпания,
повышенная чувствительность к различным ин-
фекциям.
Велико значение для кожи меди, так как че-
рез тирозиназу она участвует в синтезе меланина,
через лизилоксидазу — в обмене эластина и
коллагена, через тиолоксидазу — в процессах
кератинизации. Недостаточность меди в орга-
низме приводит к образованию дефектного кол-
лагена, в котором отсутствуют поперечные
сшивки между полипептидными цепями кол-
лагена и эластина, что увеличивает вероятность
их разрыва.
Железо является составной частью гемогло-
бина, миоглобина, цитохромов, каталазы, пе-
роксидазы и этим определяется его важнейшая
роль в организме. У больных псориазом в по-
раженных участках эпидермиса отмечается по-
вышенное содержание железа. При различных
формах фотодерматозов отмечена корреляция
между степенью перекисного окисления липи-
дов и уровнем содержания железа в сыворотке
крови больных.
В коже протекают химические превращения
веществ, находящиеся в связи с обменными про-
цессами других органов, например, процессы ка-
таболизма белков, углеводов и липидов. В ней
содержатся все необходимые для этого фермен-
ты: оксидоредуктазы, трансферазы, гидролазы,
синтетазы, изомеразы и лиазы. В коже осуще-
ствляются специфические для нее превращения:
образование кератина, коллагена, меланина, кож-
ного сала и пота.
Кожа как эндокринный орган
В настоящее время накапливаются сведения,
позволяющие рассматривать кожу как крупный
эндокринный орган. Эти взгляды основывают-
ся на следующих данных.
В коже (очевидно, в эпидермисе) под действи-
ем ультрафиолетовых лучей образуется витамин Д3
(холекальциферол). В последнее время этот ви-
тамин рассматривается как гормон, так как об-
ладает всеми свойствами гормонов: синтезирует-
ся в организме; всасываясь в кровь, достигает
вместе с ней регулируемых органов; действует в
малых количествах; имеет органы-мишени; ак-
тивные формы витамина Д, образуемые в ходе ме-
таболических превращений этого стероида, при-
сутствующего в больших количествах в коже,
обладают многими эффектами стероидных гор-
монов. Таким образом, кожу можно рассматри-
вать как орган, синтезирующий гормон холекаль-
циферол.
Кератиноциты вырабатывают гормон, подоб-
ный тимопоэтину. Он называется также эпидер-
мальным тимоцитактивирующим фактором
(ЭТАФ). Этот фактор имеет молекулярную мас-
су, равную 15000 дальтон. Такую же массу имеет
и интерлейкин-1, вырабатываемый макрофага-
ми. Кроме того, было показано, что интерлей-
кин-1 вырабатывается не только клетками Лан-
герганса, но и кератиноцитами. Интерлейкин-1
кератиноцитарного происхождения и получил
название ЭТАФ. Он так же, как и тимопоэтин,
стимулирует дифференцировку Т-лимфоцитов.
Введенный в организм экзогенный ЭТАФ вы-
зывает лихорадку, нейтрофилию. In vitro он уси-
ливает хемотаксис этих клеток. В отношении
места продукции ЭТАФ нет окончательного
мнения. Показано, что даже в роговом слое
ЭТАФ содержится в больших количествах и с
сохраненными биологическими свойствами.
60 Общая дерматология
В последнее время показана продукция кера-
тиноцитами интерлейкинов-2 и 3. Интерлейкин-
3, как известно, продуцируется стимулирован-
ными лимфоцитами и активирует деление и
дифференцировку тучных клеток.
В коже вырабатываются нейропептиды. Их
продукция связана в основном с клетками Мер-
келя. Нейропептиды, в частности, мет-энкефа-
лин, обнаружены методом иммуногистохимии,
помимо клеток Меркеля, в клетках Лангерган-
са, меланоцитах, нейролеммоцитах чувствитель-
ных нервных окончаний кожи и в пигментных
клетках (меланофорах) дермы.
Клетки Лангерганса продуцируют интерфероны,
которые не только участвуют в иммунных ре-
акциях, но и оказывают системное влияние на
деление клеток различных тканей. Клетки Лан-
герганса продуцируют также интерлейкин-1, ко-
торый наряду с участием в межклеточных взаи-
модействиях клеток иммунной системы обладает
рядом системных влияний (в отношении ЭТАФ).
Адипоциты жировой ткани продуцируют эс-
трогены и лептин.
Таким образом, морфологическим субстратом
эндокринных функций кожи являются керати-
ноциты, клетки Лангерганса, клетки Меркеля и,
возможно, меланоциты. Все эти клетки облада-
ют морфологическими признаками гормонпро-
дуцирующих клеток: имеют развитые органел-
лы биосинтеза, могут контактировать при
помощи отростков с гемомикроциркуляторным
руслом дермы (клетки Лангерганса). Развитие ис-
следований по выяснению эндокринной функ-
ции кожи может оказаться весьма перспектив-
ным не только для теоретической, но и для
практической медицины.
Как важнейшая железа внешней и внутрен-
ней секреции, кожа тесно связана со всеми
внутренними органами, соединительной тка-
нью, гипофизом, надпочечниками и другими
эндокринными железами. Вытяжки из кожи
могут действовать как стимулирующие, сосудо-
суживающие и антиинфекционные средства.
Поэтому нет ничего удивительного в высказы-
ваниях немецкого ученого С. Шмитца, который
считает кожу «самой большой эндокринной
железой».
Многие виды метаболической активности
кожи регулируются гормонами. Действительно,
сальные железы и некоторые волосяные фолли-
кулы не только с готовностью реагируют на ан-
дрогены, но и способны к различным трансфор-
мациям под влиянием стероидных гормонов.
Дигидротестостерон вызывает увеличение саль-
ных желез в период полового созревания, рост
определенного вида волос (волосяного покрова
у лиц мужского пола на груди, верхнем лобко-
вом треугольнике, в носу и ушах), а также рост
и развитие наружных гениталий. Антиандроге-
ны, лекарства, которые блокируют конверсию
тестостерона в дигидротестостерон, осуществляют
это, конкурентно ингибируя либо 5а-редукгазу,
либо цитозолрецепторный протеин. Лекарства,
подобные циметидину и спиронолактону, име-
ют антиандрогенное действие и использовались
для лечения акне и гирсутизма. Вдобавок, тире-
оидные гормоны могут регулировать рост волос,
изменять текстуру кожи (тонкие скудные воло-
сы и гладкая мягкая кожа при гипертиреоидиз-
ме, жесткие грубые волосы и прохладная шер-
шавая толстая кожа при гипотиреоидизме).
Далее, гормоны влияют на образование пигмента
меланина, меланоцит-стимулирующий гормон, и
на эстроген-стимулирующую пигментацию
кожи.
Кожа как орган иммунитета
В настоящее время существует достаточно доказа-
тельств, позволяющих считать кожу органом им-
мунитета. Кожа является высокоорганизованным
периферическим органом иммунной системы и
обладает необходимым составом иммунокомпетен-
тных клеток: эпителиоциты, эпидермальные мак-
рофаги, лимфоциты, гистиоциты, тканевые базо-
филы, Thy-1-положительные клетки. Это дает
возможность коже осуществлять ряд важных для
организма физиологических функций: распозна-
вание антигенного материала и элиминацию его,
внетимусную дифференцировку незрелых клеток
в Т-лимфоциты, иммунологический надзор за опу-
холевыми клетками. Таким образом, кожа явля-
ется одновременно центральным и периферичес-
ким органом иммуногенеза.
Эпидермис и дермоэпидермальная соедини-
тельная зона служат активными участниками
иммунологических реакций. Кожа состоит из
иммунологически важных клеток, включая ке-
ратиноциты, клетки Лангерганса и меланоциты,
а также иммунологические структуры, такие как
lamina lucidum и lamina b as alls, которые вовле-
чены в разнообразные буллезные реакции кожи.
Вероятно, самой важной иммунологической
клеткой в эпидермисе является клетка Лангер-
ганса, составляющая от 2 до 5 % общей эпидер-
мальной клеточной популяции. Клетки Лангер-
ганса играют определенную роль в ряде
иммунологических реакций, включая взаимо-
действие макрофаги-Т-клетки, Т- В-лимфоци-
ты, реакции типа GVH (трансплантат против
Глава 3. Основные функции кожи
61
хозяина), а также отторжение трансплантата.
Клетки Лангерганса синтезируют la-антигены
(антигены класса II, антигены иммунного отве-
та генно-ассоциированные), которые являются
определяющими в переработке и представлении
антигенов Т-лимфоцитам. Лимфокины, произ-
водимые клетками Лангерганса во время имму-
нологических реакций, усиливают и увеличива-
ют их протекание, а также содействуют
сопроводительному воспалительному ответу.
Кератиноциты также играют роль в имму-
нологических ответах, экспрессируя 1а-антиге-
ны на своих поверхностях при таких состоя-
ниях, как GVH-реакция, грибовидный микоз,
аллергический контактный дерматит, красный
плоский лишай, туберкулоидный тип лепры.
При этих состояниях кератиноциты произво-
дят лимфокины, особенно интерлейкин-1,
который обеспечивает второй сигнал, дополня-
ющий макрофаги (клетки Лангерганса) в ми-
тоген- и антиген-индуцированной активации
Т-клеток. Кроме того, эпидермальные клетки
производят другие цитокины, такие, как про-
стагландин Е2 и лейкотриены, которые участву-
ют в воспалительных реакциях кожи. Керати-
ноциты — это иммунологическая мишень в
пемфигусной группе заболеваний, при которых
циркулирующие антитела к межклеточным
антигенам эпидермиса и эпителию мукозной
мембраны вызывают внутриэпидермальный
акантолитический пузырь.
Различные воспалительные заболевания час-
то характеризуются буллезными реакциями, ко-
торые опосредуются иммунореагентами, вклю-
чая IgA и IgМ, и дополнительным скоплением
вдоль дермальной соединительной зоны. Анато-
мическое расположение пузырьковых образова-
ний коррелирует с позицией скопления этих им-
мунореагентов. При некоторых заболеваниях
антигены были изолированы и частично охарак-
теризованы. Использование иммунофлюорес-
центной методики на микроскопическом и осо-
бенно подструктурном уровне очень помогает в
более точном диагносцировании этих буллезных
состояний.
Поскольку кожа является важным звеном еди-
ной иммунной системы, она часто поражается при
первичных и вторичных иммунодефицитных со-
стояниях. В то же время иммунологические
структуры кожи позволяют ей осуществлять не-
которые патологические процессы самостоятель-
но. К ним относятся прежде всего различные
проявления контактной гиперчувствительности,
фотодерматозы, а также лимфопролиферативные
процессы кожи. Выраженная антигенная стиму-
ляция кожи как барьерного органа, другие эндо-
генные и экзогенные факторы могут вызывать
поломки в одном или ряде звеньев иммунной
системы кожи, что приводит к многообразным
иммунопатологическим проявлениям. В коже
могут быть реализованы все типы иммунопато-
логических реакций, что можно наблюдать при
ряде различных дерматозов. Для понимания этих
патологических процессов существенно знать
иммунологические функции здоровой кожи.
Косметическое значение кожи
Косметическое и эстетическое значение кожи
известно с древних времен. Так, например, в
Египте умершего, отправлявшегося в свой пос-
ледний путь, снабжали семью видами проти-
раний и масел, двумя косметическими сред-
ствами типа грима. Ассирийцы и жители
Вавилона красили веки и волосы в черный
цвет соединениями серы и сурьмой. Египетс-
кая царица Клеопатра каждый раз брала с со-
бой в дорогу 500 ослиц, чтобы купаться в ос-
лином молоке. Во времена подъема культуры
Древнего Востока для ухода за кожей исполь-
зовались протирания, масла, благовония, рас-
тительные экстракты.
С возрастом практически все структуры и
функции кожи изменяются. Влияние окружаю-
щей среды, особенно хроническая подвержен-
ность воздействию солнца, ускоряет старение
кожи, проявляющееся в образовании морщин.
Возраст меняет в целом внешний вид кожи: она
грубеет, морщинится, теряет эластичность, имеет
неравномерную пигментацию, разнообразные
доброкачественные и злокачественные пролифе-
ративные очаги.
Экологическое значение кожи
Поскольку человек по своему происхождению
и положению в живой природе является био-
логическим видом, для обеспечения его суще-
ствования требуются такие физические, хими-
ческие и биологические факторы, к которым
он эволюционно приспособлен, и которые ему
жизненно необходимы. В свою очередь, про-
цесс цивилизации оказывает влияние на сре-
ду, и изменяет ее. Воздействия, испытываемые
организмом, и кожей в частности, бесчислен-
ны. Однако биологические и цивилизационные
факторы могут оказывать на человека как бла-
гоприятное, так и неблагоприятное влияние.
Кожа по сравнению с другими органами тела,
существующими в сбалансированной внутрен-
62
Общая дерматология
ней среде организма, находится в очень невы-
годном положении, поскольку она представля-
ет «линию первого контакта» с внешней сре-
дой. Кожа прекрасно приспособлена к
выполнению функции пограничного органа, ее
филогенетическое развитие — лучшее свиде-
тельство такой приспособляемости. Ее вклад в
успешное развитие человека в процессе антро-
погенеза даже выше по сравнению с большин-
ством других органов. Поскольку одним из
главных признаков, отличающих человека от
его ближайших зоологических родственников
и свидетельствующих о совершенстве функций
кожи, является отсутствие на теле волосяного
покрова, можно было бы с таким же основа-
нием называть его «homo nudus», как и «homo
sapiens».
Функции кожи, бесспорно, таковы, что они
позволяют человеческому роду существовать в
самом широком спектре естественного влияния
среды, а в соответствии со своей эстетической
функцией кожа становится и средством регули-
рования межчеловеческих отношений.
Глава 4
Общепатологические процессы в коже *
Патологические процессы в коже могут затрагивать
ее различные структурные компоненты: эпидер-
мис и его производные, дерму, подкожно-жиро-
вую клетчатку. Вместе с тем следует отметить, что
полной изоляции этих процессов не наблюдается,
изменения в эпидермисе неизбежно приводят к
нарушениям в дерме и наоборот.
Патологические процессы в коже по своей
сути аналогичны таковым в других органах.
Однако в связи с гетерогенностью ее структуры
морфологическое выражение некоторых из них
имеет определенное своеобразие. Кроме того, не-
которые из патологических процессов, такие, на-
пример, как нарушение пигментации и некото-
рые другие, характерны только для кожи.
Следует признать весьма удачным изложение
В. Н. Мордовцевым и Г. М. Цветковой (1993) об-
щепатологических процессов в коже, где они
отдельно рассматривают патологические наруше-
ния эпидермиса и дермы. На основании этого мы
сделали попытку создать классификацию обще-
патологических процессов в коже, приведенную
ниже.
Классификация
общепатологических
процессов в коже
А. Общепатологические
процессы в эпидермисе
I. Нарушение клеточной кинетики:
1. Нарушение нормального соотношения про-
лиферации, дифференцировки и апоптоза
кератиноцитов:
• в сторону преобладания пролиферативных
процессов и дифференцировки кератино-
цитов: адаптивные перестройки эпидерми-
са при действии на кожу неблагоприятных
факторов внешней среды (механические
воздействия, ультрафиолетовое облучение
и др.), ведущие к утолщению эпидермиса
* Глава написана проф. О. Д. Мядельцем
и, в особенности, его рогового слоя — про-
лиферативный акантоз;
• в сторону преобладания апоптоза керати-
ноцитов (действие факторов, угнетающих
пролиферацию и активирующих апоптоз
клеток эпидермиса: ионизирующей ради-
ации, цитостатиков и т. д.) с формирова-
нием язв;
2. Нарушение нормального соотношения про-
лиферации кератиноцитов и эксфолиации
корнеоцитов:
• в сторону преобладания пролифератив-
ных процессов и дифференцировки ке-
ратиноцитов — пролиферативный ги-
перкератоз;
• в сторону снижения эксфолиации корнео-
цитов — ретенционный гиперкератоз;
3. Нарушение дифференцировки кератиноцитов:
• замедление ороговения кератиноцитов с
сопутствующим увеличением их пролифе-
ративной активности — паракератоз;
• усиление ороговения кератиноцитов —
дискератоз.
II. Нарушение межклеточных связей кератино-
цитов в эпидермальном пласте — акантолиз,
спонгиоз, гидротическая и баллонизирую-
щая дистрофии.
III. Нарушение дермо-эпидермальных связей,
приводящее к формированию субэпидер-
мального пузыря (при многоформной экс-
судативной эритеме, буллезном пемфигои-
де, красной волчанке и др.).
Б. Обшепатологические
процессы в дерме и гиподерме
I. Патологические изменения соединительно-
тканных волокон.
1. Коллагеновых волокон:
а. гиперпродукция (склероз);
Ь. гипопродукция или усиленный их распад
(коллагенолиз).
2. Эластических волокон: эластолиз, эластоз.
63
64 Общая дерматология
II. Патологические изменения основного веще-
ства соединительной ткани кожи.
1. Соединительнотканные дистрофии кожи —
диспротеинозы:
а. мукоидное набухание;
Ь. фибриноидное набухание;
с. гиалиноз;
d. амилоидоз.
2. Нарушения липидного обмена.
3. Нарушения углеводного обмена.
III. Смешанные дистрофии (нарушения метабо-
лизма, затрагивающие как эпителиальные, так
и соединительнотканные структуры кожи):
1. Нарушения обмена хромопротеидов:
а. гемоглобинные;
Ь. протеиногенные;
с. липидогенные.
2. Нарушения минерального обмена.
3. Патология волосяных фолликулов.
IV. Воспаление кожи.
1. Неиммунное воспаление кожи:
• серозное;
• гнойное;
• продуктивное.
2. Иммунное воспаление кожи:
• аллергическое;
• гранулематозное;
• экзематозное.
VI. Некроз.
VII. Опухоли кожи.
Общепатологические
процессы в эпидермисе
Нарушение клеточной кинетики
Нарушение клеточной кинетики в эпидермисе ле-
жит в основе ряда патологических процессов в
коже. Тканевый гомеостаз в эпидермисе поддер-
живается за счет пяти основных процессов: мито-
тической активности кератиноцитов, их диффе-
ренцировки и миграции в вертикальном
направлении, апоптоза кератиноцитов и эксфоли-
ации корнеоцитов. Нарушение нормального соот-
ношения указанных процессов ведет к нарушению
тканевого гомеостаза эпидермиса и патологии.
Эпидермис как тканевая система весьма четко
и быстро реагирует на нарушения гомеостаза и
также быстро включает указанные выше меха-
низмы своего восстановления. Это хорошо демон-
стрируется, например, при нарушении целостно-
сти эпидермиса в результате травмы. При
действии на кожу продолжительных по времени
неблагоприятных факторов внешней среды про-
исходит стойкое нарушение тканевого гомеоста-
за в эпидермисе. Для его компенсации возника-
ет необходимость перехода тканевой системы на
более высокий уровень функционирования. Про-
исходящие в эпидермисе перестройки носят при
этом адаптивный характер, тем не менее любая
адаптация, по мнению Ф. 3. Меерсона (1981),
имеет свою цену и несет в себе элементы пато-
логии, доля которых может возрастать при нара-
стании интенсивности воздействия. Рассмотрим
эту ситуацию на примере действия на кожу ме-
ханического фактора. Если этот фактор действует
в течение продолжительного времени и не вызы-
вает выраженных повреждений эпидермиса и
кожи в целом, то как ответная реакция эпидер-
миса на механическое раздражение увеличивает-
ся митотическая активность кератиноцитов (оче-
видно, это происходит в результате снижения
продукции Qj-или Р2-эпидермальных хейлонов
и снятия Qj- или Q2- блока митотической актив-
ности), усиливается их миграционная способность
и дифференцировка. Одновременно подавляется
апоптоз клеток. В результате идет наработка кле-
точного материала. Эксфолиация корнеоцитов с
поверхности, очевидно, не претерпевает суще-
ственных изменений. В результате гипер-
продукции клеток и усиления дифференциров-
ки эпидермальный пласт резко утолщается за счет
шиповатого, зернистого и в особенности рогово-
го слоя. Возникает мозоль. Увеличение толщины
эпидермиса в основном за счет шиповатого слоя
называется акантозом. В данном случае речь идет
о так называемом адаптивном пролиферативном
акантозе. Аналогичные процессы происходят и
при действии на кожу других неблагоприятных
факторов внешней среды (ультрафиолетовое об-
лучение, химические вещества, холод и др.).
Гистологически в эпидермисе отмечается уве-
личение толщины указанных выше слоев эпи-
дермиса, иногда и дермы. В сосочковом слое
дермы в редких случаях может формироваться
слабая воспалительная реакция сосудов.
При прекращении действия на кожу вредного
фактора начинает реализовываться принцип
экономизации структурного обеспечения функ-
ции (Саркисов Д.С., 1993): оптимального вы-
полнения функции при минимуме клеточного
материала. В эпидермисе происходят противо-
положные процессы: снижаются митотическая
активность кератиноцитов, их миграция и про-
цессы дифференцировки, могут усиливаться
апоптоз и эксфолиация. Эпидермис как систе-
ма постепенно возвращается в первоначальное
состояние, чтобы повторить цикл при повтор-
Глава 4. Общепатологические процессы в коже
65
ном воздействии. Еще раз следует оговорить-
ся, что все эти процессы укладываются в рам-
ки адаптивных, но могут легко перерасти в па-
тологическое состояние, требующее лечения.
Противоположная ситуация наблюдается при
действии на кожу факторов, резко подавляющих
митотическую активность и дифференцировку
кератиноцитов, их миграцию в вертикальном на-
правлении. Такими факторами являются ионизи-
рующая радиация, цитостатики и цитоцидные пре-
параты. Сюда же можно отнести нарушения
микроциркулярного русла дермы, приводящие к
нарушению трофики эпидермиса. Все эти факто-
ры, наряду с подавлением митотической активно-
сти и дифференцировки кератиноцитов, усилива-
ют их апоптоз. В результате толщина
эпидермального пласта постепенно снижается, он
становится ломким, быстро и легко повреждается
при малейших физических воздействиях, что в
конечном итоге ведет к формированию изъязвле-
ний, эрозий, трещин (например, у рентгенологов).
Нарушение нормального соотношения
пролиферации кератиноцитов
и эксфолиации корнеоцитов
Нарушение клеточной кинетики в эпидермисе
может быть вызвано резким усилением митоти-
ческой активности, миграции и дифференциров-
ки кератиноцитов с избыточным образованием
кератина при одновременном подавлении их
апоптотической гибели. Это состояние ведет к
так называемому пролиферативному гиперкерато-
зу (рис. 4.1). Принципиально он мало чем отли-
чается от пролиферативного акантоза по соотно-
шению гомеостатических механизмов, однако
ряд данных указывают на то, что это уже бо-
лезнь, а не адаптационный процесс. В отличие
от адаптационного пролиферативного акантоза в
данном случае резко утолщаются шиповатый,
зернистый и роговой слои. Формируется папил-
ломатоз — состояние, когда сосочки дермы резко
увеличиваются в длину и ширину и глубоко
проникают в эпидермис, который, в свою оче-
редь, формирует длинные выросты, глубоко
проникающие в дерму (рис. 4.2).
В электронном микроскопе удается наблюдать
увеличение числа десмосом, потерявших связи с
тонофиламентами, изменение структуры тоно-
филл и возрастание количества кератиноцитов в
клетках шиповатого и зернистого слоев. Таким
образом, в основе пролиферативного гиперкера-
тоза лежит нарушение тонофиламентов — дес-
мосомальных связей (Цветкова Г.М., 1993). Про-
лиферативный тип гиперкератоза встречается при
таких заболеваниях, как атопический дерматит,
красный плоский лишай и др.
Вторая разновидность гиперкератоза — ретен-
ционный гиперкератоз. Его развитие связано не
с избыточной продукцией и ороговением кера-
тиноцитов, а с резким снижением процессов
эксфолиации корнеоцитов с поверхности эпи-
дермиса в результате повышения их адгезивно-
сти. Это связано с увеличением синтеза цемен-
тирующей межклеточной субстанции (в
основном гликозаминогликанов). В результате
нарушается разъединение корнеоцитов в роговом
Рис. 4.1. Гиперкератоз
с утолщением зернистого слоя, х 75
(Цветкова Г. М., Мордовцев В. Н., 1986).
Рис. 4.2. Папилломатоз, акантоз и гиперкератоз
с образованием субэпидермальной роговой
кисты (показано стрелкой). х75 (Цветкова Г. М.,
Мордовцев В. Н., 1986).
66 Общая дерматология
слое и снижается их отшелушивание. Имеет
место также нарушение синтеза белков керато-
гиалиновых гранул. Ретенционный гиперкератоз
может быть местным, например, при пигмент-
ной ксеродерме, и общим (при вульгарном их-
тиозе). Характерной гистологической картиной
ретенционного гиперкератоза является увеличе-
ние толщины рогового слоя при атрофии или от-
сутствии зернистого слоя. При ихтиозе могут на-
блюдаться атрофические изменения одних
кератиноцитов с уменьшением их размеров при
одновременном увеличении размеров других ке-
ратиноцитов рогового слоя и их вакуолизация.
При пигментной ксеродерме патологические из-
менения эпидермиса характеризуются также ги-
перкератозом с неравномерной толщиной эпи-
дермиса, истончением росткового слоя, в
котором появляются атрофические изменения.
Изменения нормального процесса дифферен-
цировки кератиноцитов могут происходить как
в сторону их задержки, так и в сторону усиле-
ния. При этом одновременно изменяются и дру-
гие параметры гомеостазирования.
Существенное нарушение, замедление образова-
ния роговых чешуек лежит в основе паракерато-
за (рис. 4.3), который наблюдается при различных
дерматитах, псориазе, экземе, розовом лишае и
других заболеваниях. Замедление дифференциров-
ки кератиноцитов при паракератозе сочетается с
повышением их пролиферации и миграции. Апоп-
тотичсская гибель клеток при этом замедляется.
Очень часто паракератоз сочетается с акантозом.
Рис. 4.3. Паракератоз и акантоз (Цветкова Г. М.,
Мордовцев В. Н., 1986).
В основе паракератоза лежит нарушение способ-
ности клеток эпидермиса вырабатывать такие бел-
ки кератинизации, как филагрин и кератолитин.
Гистологически в очагах паракератоза выявляют
утолщение шиповатого и рогового слоев и одно-
временно резкое истончение (до единичных кле-
ток) или даже полное отсутствие зернистого слоя.
При этом роговой слой представляется незрелым:
в его клетках могут присутствовать ядра, имею-
щие форму палочек, а в цитоплазме сохраняются
органеллы: рибосомы и полирибосомы, а также ли-
пидные включения. Адгезия таких корнеоцитов
нарушена, что связано с дефектом цементирующе-
го межклеточного вещества и является причиной
повышенной десквамации корнеоцитов.
Дискератоз — явление, при котором происхо-
дит преждевременное ороговение кератиноцитов
с усилением их апоптотической гибели. Он мо-
жет встречаться как при доброкачественных (бо-
лезнь Дарье, Хейли-Хейли, Кирле, заразный
моллюск), так и злокачественных заболеваниях
кожи (болезнь Педжета, спино-целлюлярный рак
и др.). При этих заболеваниях отдельные кера-
тиноциты теряют межклеточные связи с соседни-
ми клетками (акантолиз) и приобретают шаровид-
ную форму. Такие кератиноциты могут
располагаться либо группами, либо мозаично. В
дальнейшем клетки претерпевают ряд изменений.
Вначале они имеют интенсивно окрашенные ядра
и базофильную, с зернистостью, цитоплазму,
затем превращаются в клетки с мелкими пикно-
тичными ядрами и резкой ацидофильной цито-
плазмой. Такие клетки располагаются в роговом
слое и называются телами Дарье. При электрон-
номикроскопическом исследовании обнаружива-
ется распад десмосом, потеря их связи с тонофи-
ламентами, последние концентрируются вокруг
ядер клеток. В дальнейшем в клетках происхо-
дит уплотнение и уменьшение количества кера-
тина, в них формируются «пустые» участки и, в
конце концов, клетки превращаются в зерна. При
дискератозе довольно часто формируются надба-
зальный акантолиз и в дальнейшем лакуны, ок-
руженные одним слоем кератиноцитов — интра-
эпидермальный пузырь — как результат потери
десмосомальных связей между кератиноцитами.
Нарушение межклеточных связей
кератиноцитов в эпидермальном
пласте
В эпидермальном пласте кератиноциты объеди-
нены в единую систему при помощи многочис-
ленных десмосом. При ряде патологических со-
стояний эти связи могут разрушаться, что ведет
к формированию внутриэпидермальных полос-
Глава 4. Общепатологические процессы в коже
67
тей, заполняющихся пропотевающей из сосудов
подлежащей дермы жидкостью. Различают пер-
вичную потерю межкератиноцитарных связей,
называемую акантолизом, и вторичную, вслед-
ствие проникновения жидкости в эпидермис в
результате отека — спонгиоз (рис. 4.4).
Акантолиз, в свою очередь, также подраз-
деляется на первичный и вторичный. Первич-
ный акантолиз возникает в результате ауто-
иммунного разрушения межклеточных связей
в эпидермисе. Это происходит в результате
выработки аутоантител класса G и М к ком-
понентам клеточной оболочки кератиноцитов.
В реакции участвуют также компоненты ком-
племента. Инициированная антителами и
комплементом деструкция межклеточных свя-
зей завершается выделяющимися при разру-
шении кератиноцитов гидролитическими фер-
ментами лизосом.
Первичный акантолиз наблюдается при таких
аутоиммунных процессах, как, например, пузыр-
чатка (рис. 4.5). В его основе могут также лежать
генетические нарушения мембран кератиноцитов,
что отмечается при семейной хронической пузыр-
чатке Хейли-Хейли. В данном случае пусковым
механизмом в потере межклеточных контактов
являются экзогенные факторы, например, меха-
нические травмы, бактериальные инфекции и др.
В результате потери межклеточных десмосомаль-
ных связей в шиповатом слое эпидермиса фор-
мируются пузыри, заполняющиеся тканевой
жидкостью — акантолитические пузыри. Ли-
Рис. 4.4. Спонгиотический пузырек при острой
экземе с деструкцией клеток эпидермиса, х 75
(Цветкова Г. М., Мордовцев В. Н., 1986).
шенные связей кератиноциты (акантолитические
клетки) свободно «плавают» в тканевой жидко-
сти. Они приобретают округлую форму из-за
того, что тонофибриллы в них концентрируют-
ся в околоядерном пространстве. В дальнейшем
в клетках существенно снижаются метаболичес-
кие процессы, и они постепенно подвергаются
разрушению.
Вторичный акантолиз связан с повреждением
самих кератиноцитов вирусами (например, ви-
русами герпеса, оспы). В результате этого по-
вреждения в кератиноцитах возникает вначале
внутриклеточный отек, а затем вакуолизация
клеток. Вакуоли могут располагаться перинук-
леарно, а в более выраженных случаях занима-
ют большую часть цитоплазмы, сдвигая ядро на
периферию. Ядро в этом случае приобретает сер-
повидную форму. Такая дистрофия кератиноци-
тов получила название гидропической, или ваку-
ольной дистрофии (рис. 4.6). Обычная ее
локализация — шиповатый и зернистый слои. В
тех случаях, когда гидропической дистрофии
подвергаются клетки базального слоя, происхо-
дит гибель этих клеток, эпидермис отделяется от
базальной мембраны, а в образовавшейся поло-
сти накапливается тканевая жидкость с форми-
рованием субэпидермального пузыря. Этот вид
дистрофии имеет место при красной волчанке,
склероатрофическом лишае и некоторых других
заболеваниях.
Рис. 4.5. Акантолитический
внутриэпидермальный пузырь, в полости
которого находятся акантолитические клетки.
Вульгарная пузырчатка. х75 (Цветкова Г. М.,
Мордовцев В. Н., 1986).
68
Общая дерматология
При резко выраженном отеке эпидермиса раз-
вивается так называемая баллонирующая дистро-
фия эпидермиса с исчезновением межклеточных
контактов на большом протяжении (рис. 4.7).
Акантолитические пузыри крупные, заполнены
серозно-фиброзным экссудатом, в котором на-
ходятся очень крупные, резко отечные керати-
ноциты. Они в последующем превращаются в
гомогенные эозинофильные шаровидные обра-
зования.
Вторичное нарушение межкерацитарных свя-
зей обусловлено первичными изменениями не в
эпидермисе, а в дерме, в сосочковом слое кото-
рой возникает отек. Изменения в эпидермисе
вторичные, связанные с проникновением отеч-
ной жидкости из дермы. Отечная жидкость рас-
ширяет и разрывает межклеточные связи в эпи-
дермисе, и если возникающий вторичный отек
эпидермиса выражен сильно, то часть кератино-
цитов погибает, и на их месте образуются мик-
рополости, заполненные серозной жидкостью.
Эти микрополости могут затем сливаться, фор-
мируя более крупные пузыри. Это явление на-
зывается спонгиозом, а пузыри — спонгиотичес-
кими. При проникновении в них лейкоцитов
образуются спонгиотические пустулы.
Нарушение
дермо-эпидермального соединения
В дермо-эпидермальном соединении находится
большое количество аутоантигенов, способных
выступать в роли факторов аутоиммунной агрес-
сии. Дермо-эпидермальное соединение может
нарушаться при механических воздействиях на
кожу и при воспалительных процессах. Конеч-
ным результатом нарушения этого соединения
является формирование субэпидермального пузы-
ря, но механизм его образования в связи с изло-
женным выше может быть различным. Суб-
эпидермальные пузыри могут быть механической,
воспалительной или иммунной природы.
Субэпидермальные пузыри механической
природы в самом простом варианте могут быть
обусловлены запредельным действием механи-
ческих факторов (длительное трение и др.). В
таких случаях в результате нарушения микроге-
мо- и лимфоциркуляции выходящие из микро-
сосудов кровь или лимфа механически отслаи-
вают эпидермис от дермы. Соответственно
выходящей из сосудов жидкости содержимое пу-
зырей может быть либо серозным, либо гемор-
рагическим. При некоторых заболеваниях (по-
здняя кожная порфирия) целостность
дермо-эпидермального соединения нарушается
при малейших травмах, это же происходит и при
дистрофической форме буллезного эпидермоли-
за. В этих случаях, очевидно, следует говорить
о субэпидермальных пузырях физико-химичес-
кой природы, т. к. их появление связано с фи-
зико-химическими нарушениями дермо-эпидер-
мальной зоны. Так, при дистрофической форме
буллезного эпидермолиза в дермо-эпидермаль-
ной зоне сокращается число (или они исчезают
Рис. 4.6. Вакуольная дистрофия клеток
базального слоя при красной волчанке. х250
(Цветкова Г. М., Мордовцев В. Н., 1986).
Рис. 4.7. Баллонирующая дистрофия клеток
эпидермиса (показана стрелками) с
образованием пузыря. х250 (Цветкова Г. М.,
Мордовцев В. Н., 1986).
Глава 4. Общепатологические процессы в коже
69
полностью) крепящих фибрилл, что, очевидно,
объясняется разрушением их коллагеназой, ак-
тивность которой в этой зоне повышается.
При таких заболеваниях, как красный плос-
кий лишай, многоформная экссудативная эри-
тема, красная волчанка и других дерматозах раз-
рушение базальной мембраны может иметь
воспалительную природу. При этом эозинофиль-
ные и нейтрофильные лейкоциты при помощи
своих циторецепторов связываются с ней и пол-
ностью разрушают ее с помощью своих фермен-
тов, что ведет к отслоению эпидермиса и обра-
зованию субэпидермального пузыря.
При буллезном пемфигоиде механизм образо-
вания субэпидермального пузыря иммунологи-
ческий. В его основе лежит выработка аутоан-
тител к компонентам светлой пластинки
базальной мембраны (Ig G и др.) После специ-
фического связывания аутоантител с базальной
мембраной сюда осаждаются компоненты ком-
племента, которые, в свою очередь, являются
сильными хемотаксическими веществами для
гранулоцитов, в первую очередь эозинофилов,
но также и нейтрофилов. В зоне лизиса базаль-
ной мембраны характерным является также
скопление тучных клеток, которые усиленно
дегранулируют, при этом содержащиеся в гра-
нулах биологически активные вещества активи-
руют гранулоциты и облегчают их движение к
разрушаемой базальной мембране.
Иногда два последних вида субэпидермальных
пузырей определяют как первичные, возника-
ющие в результате первичных нарушений в ба-
зальной мембране. Пузыри чисто механической
природы определяют как вторичные, т. к. в дан-
ном случае вначале возникает внутриэпидермаль-
ный пузырь, и лишь в результате увеличения
размеров последнего он доходит до дермально-
эпидермального соединения, разрушает его и пре-
вращается в субэпидермальный пузырь.
Дермо-эпидермальное соединение
как иммунологическая структура
Различные воспалительные заболевания кожи
характеризуются буллезными реакциями, кото-
рые опосредуются иммунореагентами, включая
IgA и IgM, а также дополнительным скоплени-
ем иммуноглобулинов и комплемента вдоль дер-
мальной соединительной зоны. Анатомическое
расположение пузырьковых образований корре-
лирует с позицией скопления этих иммунореа-
гентов. Антигены при некоторых заболеваниях
были изолированы и частично охарактеризова-
ны. Использование иммунофлуоресцентной ме-
тодики на микроскопическом и особенно ульт-
раструктурном уровне очень помогает в более
точном диагностировании этих буллезных состо-
яний. Они обобщены в таблице (табл. 4.1) вме-
сте с результатами иммунофлуоресцентного ана-
лиза при заболеваниях соединительной ткани.
Общепатологические
процессы в дерме
и гиподерме
Поскольку эти две составные части кожи пред-
ставлены в основном соединительной тканью, то
при общепатологических процессах они реаги-
руют одинаково и логично рассматривать их в
совокупности.
Как известно, соединительнотканная часть
кожи состоит из двух компонентов или ткане-
вых элементов: клеток и межклеточного веще-
ства. Несомненно, что клетки являются главным
тканевым элементом любой ткани, в том числе
и соединительной. Межклеточное вещество —
это лишь продукт жизнедеятельности клеток, оно
создается клетками, с помощью клеток проис-
ходит его перестройка и деградация износивших-
ся компонентов. Однако, несмотря на свое «вто-
ростепенное значение», межклеточное вещество
играет важнейшую роль в жизнедеятельности
соединительной ткани, а в некоторых случаях —
первостепенную, например, в плане трофики
тканей, а также в выполнении опорно-механи-
ческой функции. Если взять сетчатый слой дер-
мы, то межклеточное вещество, в частности,
волокна ответственны за выполнение этим сло-
ем своей функции — механической, а клетки
фиброциты выполняют в большей степени пас-
сивную роль. Поэтому и при патологии соеди-
нительной ткани кожи наибольшие последствия
для организма в конечном итоге будут связаны
с поражением межклеточного вещества.
Поражения
волокнистого каркаса кожи
Изменения со стороны коллагеновых волокон дер-
мы и гиподермы могут быть количественного и
качественного характера, однако полностью разгра-
ничить эти отклонения, очевидно, невозможно.
Относительное увеличение толщины и коли-
чества коллагеновых волокон в коже наблюдается
при различных заболеваниях и определяется как
склероз. Наиболее простым проявлением склеро-
тического изменения кожи является соединитель-
нотканный рубец, возникающий после более или
менее значительных повреждений кожного покро-
ва. В этом случае, как отмечалось ранее, не про-
70
Общая дерматология
Таблица 4.1. Результаты иммунофлуоресцентного анализа кожи при иммунологически опосредованных кожных
заболеваниях
Заболевание Результаты Ультраструктурная Место пузырькового Результаты
биопсии прямых локализация образования при непрямой
иммунофлуоресцентных иммунореагентов реагентов световой микроскопии иммунофлуоресценции в сыворотке больных
Буллезные заболевания
Пузырчатка (все формы) Отложения IgG в межклеточных зонах между кератиноцитами Между кератиноцитами Супрабазальное — при вульгарной пузырчатке; под зернистым или роговым слоем — при листовидной пузырчатке IgG-антитела против кератиноцитов, межклеточной склеивающей субстанции эпидермиса у 95 %
Буллезный пемфигоид IgG и/или С вБМЗ Электронно-прозрачная зона и полудесмосомы — верхняя часть блестящих и подбазальных клеток Субэпидермально IgG-антитела к базальной мембране у 70 %
Рубцующий пемфигоид IgC и/или С в БМЗ Электронно-прозрачная зона Субэпидермально IgG-антитела к базальной мембране у 10%
Герпес беременных Комплемент в БМЗ, иногда IgA, IgM — линейное отложение Электронно-прозрачная зона рядом с электронно-плотной зоной Субэпидермально суббазальные клетки над электронно-плотной зоной IgG-антитела к базальной мембране у 20 %
Герпетиформный дерматит Дюринга IgA и С в сосочковом Гранулярный IgA в связи с слое дермы (гранулярные микрофибриллами в отложения) сосочковом слое дермы Субэпидермально в сосочковом слое дермы: папиллярно-дермальные микроабсцессы Нет циркулирующих антител
Буллезный эпидермолиз IgG в БМЗ Аморфные гранулярные отложения под базальной мембраной Субэпидермально Нет циркулирующих антител
Буллезный линейный IgA дерматит IgA и комплемент в линейном отложении в БМЗ — Субэпидермально Нет циркулирующих антител
Болезни соединительной ткани
Буллезная системная красная волчанка IgG, IgM и комплемент в БМЗ в пораженной и нормальной коже — линейный, гомогенный Непосредственно под базальной мембраной Субэпидермально Нет циркулирующих антител к базальной мембране; АЯА у 90 %
Дискоидная красная волчанка IgG, другие lg и С в очаговой коже на БМЗ — — Нет циркулирующих антител к базальной мембране; титры АЯА в норме
Системная красная волчанка IgG диапазон на БМЗ в нормальной коже (свыше 90 % в зонах — — Титры АЯА повышены
воздействия солнца)
Глава 4. Общепатологические процессы в коже
71
Заболевание Результаты биопсии прямых иммунофлуоресцентнь реагентов Ультраструктурная локализация IX иммунореагентов Место пузырькового образования при световой микроскопии Результаты непрямой иммунофлуоресценции в сыворотке больных
Системная склеродермия Нуклеолярный IgG — Истончение эпидермиса и увеличение коллагена дермы АЯА у 85 %, антицентро- мерные антитела при CREST-синдроме
Смешанные заболевания соединительной ткани (MCTD) IgG/IgM на БМЗ у некоторых больных; нуклеарный IgG в эпидермисе — — Пятнистые АЯА и антитела к ядерным компонентам
Дерматомиозит Негативный — — АЯА в норме
Примечание: БМЗ — базальная мембранная зона, С — комплемент, АЯА — антиядерные антитела.
исходит восстановления нормального первона-
чального строения кожи (органотипической ре-
генерации), а формируется грубый соединитель-
нотканный рубец, покрытый эпидермисом.
Отсутствует и разделение дермы на сосочковый
и сетчатый слои. Такие рубцовые изменения
кожи, если они обширны, не только создают се-
рьезные косметические проблемы, но и суще-
ственно нарушают функции кожного покрова.
Увеличение количества коллагеновой субстан-
ции в дерме наблюдается также при склеродер-
мии. В этом случае происходит не только уплот-
нение и увеличение толщины коллагеновых
волокон, но и их гомогенизация. Дерма утолща-
ется, чему способствует также склероз поверх-
ностных слоев подкожной клетчатки.
Склерозирование дермы кожи наблюдается и
при таких заболеваниях, как склероатрофичес-
кий лишай, пахидермопериостоз, соединитель-
нотканный невус и другие. При соединительно-
тканном невусе количество коллагеновых
волокон значительно увеличивается, и в очагах
поражения формируются выступающие над по-
верхностью кожи узелки.
Единственными продуцентами коллагеновых
волокон являются фибробласты, количество и
функциональная активность которых при скле-
розирующих процессах в коже существенно воз-
растают. Имеет значение тот факт, что при всех
склерозирующих процессах первоначально стра-
дает микроциркуляция и возникает локальная
гипоксия тканей. При этом наряду с нарушением
функции паренхимы, чувствительной к недо-
статку кислорода, происходит активация фибро-
бластов, которые добывают энергию для синте-
тических процессов в бескислородной среде.
Противоположным состоянием, характеризу-
ющимся существенным снижением содержания
в коже коллагеновых волокон, является колла-
генолиз, который может наблюдаться, например,
при epidermolysis bullosa и других кожных забо-
леваниях и характеризуется повышением актив-
ности коллагенолитических ферментов. Актив-
ность этих ферментов может возрастать при
опухолях, при воспалении.
Наряду с усилением коллагенолиза уменьше-
ние размеров, числа и нарушение строения кол-
лагеновых волокон в коже может происходить
при повреждении биосинтеза коллагена в фиб-
робластах. Например, при синдроме Элерса-Дан-
ло (гиперэластическая кожа), имеющем 8 форм
с различным типом наследования, происходят
дефекты различных ферментов, катализирующих
нормальный биосинтез коллагена.
Качественные изменения коллагенового каркаса
кожи отличаются при наследственных заболевани-
ях, связанных с изменениями состава коллагено-
вых молекул и образующихся из них коллагено-
вых фибрилл. Так, например, цепи коллагена
могут обладать неупорядоченной структурой и
быть неспособными к формированию фибрилл
при врожденном дефекте фермента проколлаген-
пептидазы, отщепляющего пептиды с N-концов
цепей коллагена (событие, абсолютно необходимое
для самосборки тропоколлагена).
Изменения эластических волокон дермы и
гиподермы могут проявляться либо разрушени-
ем эластических волокон (эластолиз), либо их
дезорганизацией, склеиванием, огрубением (эла-
стоз). Чаще, однако, эти изменения сочетаются.
Так, при гиперэластической коже или синдро-
ме Элерса-Данло количество эластических воло-
кон в коже относительно увеличивается наряду
с огрубением и одновременно наблюдается фраг-
ментация или комкообразование на месте элас-
тических волокон. При эластической псевдо-
72 Общая дерматология
ксантоме, в основе которой лежит системное по-
ражение эластических волокон кожи и слизис-
тых оболочек, в дерме выявляются скопления на-
бухших и комкообразных эластических волокон,
на которых могут откладываться ионы кальция.
Патологические изменения основного
вещества соединительной ткани
Изменения основного вещества дермы имеют ме-
сто практически при всех воспалительных и опу-
холевых заболеваниях кожи. При системных за-
болеваниях (красная волчанка, дерматомиозит),
склерозирующем лишае и др. происходит накоп-
ление в основном веществе гликозаминоглика-
нов. В некоторых случаях в основном веществе
происходят муцинозные изменения, сопровож-
дающиеся нарушением нормального строения
коллагеновых волокон. При некоторых наслед-
ственных заболеваниях также нарушается состав
основного вещества. Так, при эластической псев-
доксантоме в коже увеличивается содержание
гиалуроновой кислоты, хондроитинсульфатов и
дерматансульфата.
Соединительнотканные дистрофии
Перечисленные ранее поражения соединительной
ткани дермы и гиподермы характеризуются тем,
что патологические изменения происходят непос-
редственно в составляющих компонентах соеди-
нительной ткани. Выделение их в самостоятель-
ную группу общепатологических процессов в
коже является в определенной степени услов-
ным, но целесообразным по двум причинам: во-
первых, с дидактической целью; во-вторых, не-
смотря на то, что подобные изменения
межклеточного вещества встречаются и при дру-
гой патологии, в данном случае все же можно го-
ворить о том, что они являются при рассмотрен-
ных состояниях относительно самостоятельными
и в определенной степени первичными. Во всех
других случаях они вторичные, происходят в ре-
зультате комплекса предшествующих изменений
соединительной ткани. При соединительноткан-
ных дистрофиях в результате нарушения обмен-
ных процессов в соединительной ткани и в со-
судах кожи накапливаются нехарактерные для
них продукты обмена. Они могут приноситься
с кровью, лимфой, но могут быть результатом
нарушения синтеза волокнистых структур или
основного вещества. Мезенхимные дистрофии
могут иметь те же механизмы и возникать при
тех же заболеваниях кожи, что и описанные
выше поражения структурной соединительной
ткани, но могут быть и проявлением специфи-
ческих патологических процессов.
Мукоидное набухание
соединительной ткани
Это патологическое состояние, при котором в
соединительной ткани кожи накапливаются
нехарактерные для нормы гликозаминоглика-
ны. Их накопление и аномальное перераспре-
деление ведет к повышению гидрофильности
основного вещества и изменению сосудистой
проницаемости. В результате из сосудов кожи
выходят белки плазмы, глобулины и гликоп-
ротеины, что ведет к набуханию межклеточного
вещества и коллагеновых волокон. Позднее к
изменениям межклеточного вещества могут
присоединяться клеточные реакции, что ведет
к формированию лимфоцитарных, гистоцитар-
ных и плазмоцитарных инфильтратов. Эти из-
менения носят обратимый характер и могут
исчезать в результате лечения и разрешения
патологических процессов. Встречается муко-
идное набухание при коллагенозах (красная
волчанка, склеродермия), при аллергических
дерматитах и др. Основное вещество при му-
коидном набухании окрашивается базофильно
и может давать метахромазию.
Фибриноидное набухание
соединительной ткани
Фибриноидное набухание соединительной тка-
ни кожи, в отличие от мукоидного набухания,
носит необратимый характер. Оно заключается
в накоплении в ней аномального вещества фиб-
риноида. Его происхождение и состав до насто-
ящего времени являются предметом споров.
Состав фибриноидных масс при различных за-
болеваниях может быть различным. В одних
случаях, например, при аутоиммунных процес-
сах, в фибриноиде преобладают фибрин, комп-
лемент и иммунные комплексы. Этот вид фиб-
риноида называется фибриноидом иммунных
комплексов. В таких случаях иммунные комп-
лексы, привлекая компоненты комплемента, вы-
зывают повреждение микроциркуляторного рус-
ла, паравазальной соединительной ткани, что
затем приводит к выпадению в этом месте фиб-
рина. Это имеет место при аутоиммунных про-
цессах в коже, например, красной волчанке. При
ангионевротических расстройствах, сопровожда-
ющихся плазморрагией, также встречаются от-
ложения фибрина в местах транссудации (фиб-
риноид транссудации). Во всех случаях
отмечается деструкция коллагеновых волокон,
изменение состава гликозаминогликанов основ-
ного вещества, что обеспечивает увеличение про-
ницаемости сосудов кожи и выход из кожи в со-
единительную ткань высокомолекулярных
Глава 4. Общепатологические процессы в коже
73
веществ. Фибриноидное набухание отмечается
при аллергических васкулитах, феномене Артю-
са, коллагенозах.
При гистологическом исследовании очаги с
фибриноидным изменением соединительной тка-
ни дают гомогенную, резко выраженную эози-
нофилию. В начальных стадиях из-за большого
скопления гликозаминогликанов может наблю-
даться их метахромазия при окраске толуидино-
вым синим, несколько позднее отмечается рез-
кое ШИК-позитивное окрашивание.
Исходом фибриноидного изменения соедини-
тельной ткани является резкая деструкция со-
единительной ткани и ее некроз (фибриноидный
некроз) с образованием аморфного детрита.
Гиалиноз
Этот вид мезенхимной дистрофии соединитель-
ной основы кожи представляет собой отложение
в ней гомогенных эозинофильных масс различ-
ного состава — гиалина.
Различают три вида гиалина:
1. простой, образуется в результате выхода из
сосудов неизмененной плазмы крови;
2. липогиалин, в состав которого входят липо-
иды и 0-липопротсины (при сахарном диабете и
других заболеваниях, сопровождающихся нару-
шением липидного состава крови);
3. сложный гиалин.
В его состав входят фибрин, иммунные комп-
лексы и некротизированные участки сосудистой
стенки (например, при коллагенозах). По степе-
ни распространения различают системный гиа-
линоз (коллагенозы и др.) и местный гиалиноз,
который может формироваться в рубцующейся
некротизированной ткани, в местах фибриноид-
ных изменений.
Амилоидоз
Этот вид соединительнотканной дистрофии ха-
рактеризуется образованием в соединительной
ткани нехарактерного для нормы сложного фиб-
риллярного белка — гликопротеина амилоида-
протеина (от латинского amylum — крахмал),
который под действием иода и серной кислоты
дает синий цвет. На этом основании Р. Вирхов
считал амилоид животным крахмалом и дал ему
соответствующее название, т. е. «амилоид».
В настоящее время показано, что амилоид
представляет собой гликопротеин. Его основным
компонентом являются фибриллярные белки (F-
белки или F-компонент). Одни F-белки амило-
ида связаны с легкими цепями иммуноглобули-
нов (так называемыми AL-белками), другие же
не связаны с иммуноглобулинами, но имеют
аналог в сыворотке крови (SAA — белок).
Амилоидоз может быть генерализированным,
или общим, и местным. В зависимости от вы-
зывающей его причины он подразделяется на
первичный (идиопатический), приобретенный,
или вторичный, старческий, наследственный, ло-
кальный (опухолевидный).
Кожные проявления наиболее часты при пер-
вичном, особенно системном, амилоидозе и харак-
теризуются полиморфизмом очагов поражения.
Могут встречаться папулезные или пурпурозные
высыпания, узловатые, ксантомоподобные элемен-
ты. Отложения амилоида могут наблюдаться на
фоне разнообразных хронических дерматозов
(красный плоский лишай, нейродермит и др., вто-
ричный амилоидоз). При первичном амилоидозе
характерно появление узелков (lichen amyloidosis)
и реже пятен (maculas amyloidosis).
Воспаление кожи
Воспалительные заболевания кожи относятся к
наиболее распространенным поражениям данного
органа. Это вытекает из пограничного ее распо-
ложения, что неизбежно ведет к взаимодействию
кожи с самыми разнообразными флогогенными
факторами: механическими (травмы), физичес-
кими (действие высоких и низких температур,
лучевой энергии и др.), химическими (кислоты,
щелочами и т. д.), биологическими (разнообраз-
ные микроорганизмы). Кроме этих экзогенных
факторов, воспалительные процессы в коже мо-
гут быть обусловлены эндогенными причинами
(иммунные комплексы, различные медиаторы,
иммунокомпетентные клетки, продукты азоти-
стого обмена и распада опухолей и др.).
Как известно, воспаление — это сложная
комплексная местная реакция организма на по-
вреждение тканей различными патогенными раз-
дражителями.
Воспалительные процессы в коже отличают-
ся большим разнообразием не только из-за при-
чин, их вызывающих, но и по характеру тече-
ния, площади поражения. Очевидно, трудно
найти другой орган, в котором отмечалось бы
такое разнообразие воспалительных реакций. По-
этому воспалительные поражения кожи доста-
точно трудно классифицировать, и в особенно-
сти это относится к аллергическим
воспалительным процессам. Полной, четкой и
всеобъемлющей классификации воспаления
кожи до настоящего времени не существует.
Многие авторы выделяют иммунное и неиммун-
ное воспаление. Это разделение в определенной
74
Общая дерматология
степени условно, т. к. в каждой из этих двух
форм могут участвовать одинаковые клетки.
Вместе с тем, это разделение имеет большой
смысл, т. к. оно определяет, какие из клеточных
механизмов в данном виде воспаления преобла-
дают. На основании данных, представленных в
фундаментальной работе «Патология кожи»
(1993), учебнике по патологической анатомии
(1995), руководстве «Морфофункциональная
дерматология» (2006), можно представить следу-
ющую классификацию воспалительных заболе-
ваний кожи (табл. 4.2).
Фазы воспалительного процесса
Воспалительный процесс характеризуется отчет-
ливо выраженной фазностью. А. И. Струков
(1990) выделяет следующие фазы воспаления:
1. фаза повреждения или альтерации;
2. фаза экссудации или реакции микроцирку-
ляторного русла с выходом из крови и лимфы в
ткани жидкости;
3. фаза восстановления или пролиферации.
Альтерация
Фаза альтерации характеризуется нарушением
нормального строения клеток и межклеточного
вещества, ведущим к их гибели. При этом ос-
вобождается огромное количество медиаторов
воспаления. Это клеточные медиаторы. Кроме
того, большое количество биологически актив-
ных веществ устремляется к месту повреждения
из плазмы крови (плазменные медиаторы вос-
паления).
Медиаторы воспаления выполняют целый ряд
функций.
• изменение (повышение) проницаемости со-
судистой стенки;
• обеспечение межклеточных взаимодействий
воспалительной реакции; роль хемотоксических
факторов;
• деление и дифференцировка из стволовых
клеток тех клеточных форм, которые играют
основную роль на каждом этапе воспалительного
процесса;
• активация фагоцитоза и иммунных реакций.
К медиаторам воспаления относятся биологи-
чески активные вещества, выделяемые грану-
лоцитами, тканевыми базофилами (дегрануля-
ция), эндотелиоцитами, тромбоцитами и
другими клетками. Сюда относятся гистамин,
серотонин, интерлейкины, лизосомальные гид-
ролазы, простагландины, лейкотриены, систе-
ма кининов плазмы, фактор Хагемана, медлен-
но реагирующие субстанции и целый ряд
других, в том числе и неизученных факторов.
В результате действия этих веществ на сосу-
дистую стенку и основное вещество соедини-
тельной ткани проницаемость микрососудов
резко увеличивается с одновременным резким
расширением их просвета. Сосудистая стенка
подготавливается к следующей фазе воспале-
ния — экссудации.
Экссудация
Экссудация включает в себя такие процессы, как
выход из крови в основное вещество окружающей
соединительной ткани жидкой части плазмы, а
также миграцию через сосудистую стенку в тка-
ни клеток крови, принимающих участие в воспа-
лении. В результате паретического расширения со-
судов и выхода плазмы крови возникает отек тканей.
Отек играет важную роль в системе защитных сил
организма по двум причинам. Во-первых, отечная
жидкость сдавливает кровеносные и лимфатичес-
кие сосуды и локализует флогогенный фактор,
Таблица 4.2. Воспаление кожи
Неиммунное Иммунное
1. Серозное 2. Гнойное 3. Геморрагическое 4. Продуктивное (пролиферативное) 1. Немедленные аллергические реакции (крапивница, некоторые виды лекарственной эритемы) 2. Цитотоксические реакции аутоиммунной природы (пузырчатка, пемфигоид, синдром Лайелла) 3. Иммунокомплексные реакции (многоформная экссудативная эритема, токсикодермия и некоторые другие дерматозы, пурпура Шенляйн-Геноха и др.) 4. Гиперчувствительность замедленного типа (аллергический контактный дерматит, экзема, нейродермит и др.) 5. Гранулематозное воспаление (туберкулезная волчанка, лепра, саркоидоз, грибковые поражения с гранулематозным воспалением) 6. Кожные проявления реакции «трансплантат против хозяина»
Глава 4. Общепатологические процессы в коже
75
препятствует его широкому распространению в
организме. Во-вторых, отечная жидкость умень-
шает концентрацию флогогенного фактора и, сле-
довательно, его патогенное действие (Мусил Я.,
1985). Кроме того, уменьшая плотность межкле-
точного вещества, она способствует миграции кле-
ток к очагу воспаления.
Под действием хемотаксических веществ (ме-
диаторов воспаления) лейкоциты крови, в пер-
вую очередь нейтрофильные лейкоциты, начина-
ют диапедез через сосудистую стенку. Диапедезу
предшествует краевое стояние лейкоцитов, ког-
да они отделяются от общего потока клеток в
сосуде и тяготеют к эндотелиоцитам, в последу-
ющем приклеиваясь к ним. Диапедез нейтро-
фильных лейкоцитов осуществляется интерэн-
дотелиоцитарно, т. е. в местах стыка двух
эндотелиоцитов. Этому способствует образова-
ние так называемых «гистаминовых люков» —
расширенных промежутков между эндотелиоци-
тами из-за сокращения их под действием гиста-
мина, а также деполяризация базальной мемб-
раны под влиянием гиалуронидазы и других
веществ.
Роль нейтрофильных лейкоцитов в воспале-
нии огромна. Под влиянием флогогенных фак-
торов и медиаторов воспаления в них происхо-
дит так называемый респираторный взрыв: в
клетках вырабатывается большое количество ак-
тивных форм кислорода (синглетный кислород,
супероксидный анион, перекись водорода, а
также другие метаболиты), которые оказывают
резко выраженное бактерицидное действие. С
другой стороны, эти вещества губительны для
здоровых тканей и самих нейтрофилов, и при
сильно выраженном воспалительном процессе
происходит их повреждение с развитием гной-
ного воспаления. Таким образом, положитель-
ный эффект нейтрофилов может легко перехо-
дить в отрицательный. Кроме активных форм
кислорода, нейтрофилы продуцируют и секре-
тируют целый ряд других веществ, обладающих
бактерицидным действием. Таковыми являют-
ся содержащиеся в специфических и азуро-
фильных гранулах катионные белки, щелочная
фосфатаза, лактоферрин, нейтральные протеа-
зы, миелопероксидаза.
Лизоцим (мурамидаза) расщепляет бактериаль-
ную стенку, что вызывает осмотическую гибель
бактерий. Лактоферрин конкурирует с бактери-
ями за ионы железа, связывает их, лишая бак-
терии этого важного минерала и действуя бак-
териостатически.
Катионные белки и нейтральные протеазы
вызывают повреждение мембран клеток и раз-
рушение поверхностных белков. Молочная кис-
лота вызывает снижение pH в фагосомах и, кро-
ме того, обладает прямой бактерицидностью.
Комплекс «миелопероксидаза — перекись водо-
рода — галогены» вызывает галогенизацию
(окисление) структурных и функциональных
белков.
Щелочная фосфатаза нарушает структуру
ДНК бактерий, обладая бактериостатическим
действием. Таким образом, нейтрофилы вызы-
вают гибель бактерий еще до фагоцитоза, кото-
рый является вторым этапом их литического
действия. Фагоцитируя бактерии и другие мик-
роорганизмы, продукты некроза тканей, лейко-
циты при помощи лизосом внутриклеточно рас-
щепляют их до низкомолекулярных веществ,
очищая зону воспаления. Это так называемый
завершенный фагоцитоз. Если же имеется функ-
циональная несостоятельность нейтрофильных
лейкоцитов, то микроорганизм может продол-
жать существование внутри лейкоцита и даже
размножаться (незавершенный фагоцитоз). Это
наблюдается, например, при хронической грану-
лематозной болезни детей — генетическом рас-
стройстве, при котором нейтрофильные лейко-
циты и моноциты поглощают, но не могут убить
содержащие фермент каталазу микроорганизмы,
так как теряют способность вырабатывать губи-
тельные для этих микробов активные метаболи-
ты кислорода. Таким образом, как видно из дан-
ного примера, фагоцитарная бактериолитическая
и бактерицидная функции нейтрофилов тесно
сопряжены.
Роль лейкоцитов в воспалительном процессе
настолько велика, что многие авторы (Елисеев
В.Г., 1961; Маянский А.Д., 1991) выделяют са-
мостоятельную лейкоцитарную фазу.
Помимо бактерицидных веществ, нейтрофиль-
ные лейкоциты выделяют целый ряд медиато-
ров иммунных реакций, вовлекая в воспалитель-
ный процесс все новые клетки. По данным
Н. М. Бережной (1989), нейтрофилы выделяют
следующие факторы: ингибирующие дифферен-
цировку В-лимфоцитов, усиливающие пролифе-
рацию тимоцитов, ингибирующие функции Т-
хелперов, Т-супрессоров, фактор хемотаксиса
эозинофилов и др.
При анафилактических реакциях, других ви-
дах аллергии в кожу могут мигрировать эозино-
фильные лейкоциты. Эти клетки участвуют в
элиминации комплексов антиген-антитело. Они
также увеличиваются в количестве при парази-
тарных заболеваниях кожи. Их роль в этом слу-
чае заключается в выделении специфических
белков, которые встраиваются в мембраны кле-
76 Общая дерматология
ток паразитов. Это приводит к формированию
трансмембранных пор и осмотическому повреж-
дению клеток паразита. Одновременно продук-
ты секреции эозинофильных лейкоцитов по-
вреждают ДНК клеток.
Под активирующим влиянием нейтрофиль-
ных лейкоцитов моноциты крови совершают
диапедез через сосудистую стенку и трансфор-
мируются в макрофаги. Эта трансформация
заключается в увеличении размеров клеток,
появлении псевдоподий, накоплении лизосом
в цитоплазме. Это так называемые гематоген-
ные макрофаги. Но под влиянием флогоген-
ных веществ и медиаторов нейтрофилов к оча-
гу воспаления устремляются и макрофаги,
предсуществующие в соединительной ткани
(гистиогенные макрофаги). Лейкоцитарная
фаза воспалительной реакции сменяется мак-
рофагической. По времени она наступает не-
сколько позже лейкоцитарной и достигает
максимума через несколько дней. В активизи-
рованных макрофагах также наступает респи-
раторный взрыв. Механизмы дыхательного
взрыва во всех фагоцитах идентичны: мобили-
зация кислорода начинается после активации
НАДФН-оксидазного ферментного комплекса
в мембранах клеток с образования суперокси-
даниона О 2, который трансформируется в син-
глетный кислород, гидрооксилрадикал ОН".
Медиаторы макрофагов, называемые моноки-
нами, играют очень важную роль в воспалитель-
ной реакции. Они поддерживают в очаге воспа-
ления (при необходимости) содержание
нейтрофильных лейкоцитов путем усиления диф-
ференцировки их из стволовых клеток крови, ак-
тивируют Т- и В-лимфоциты и участвуют в их
взаимодействии, захватывая, перерабатывая и
представляя им антиген в высокоиммунной фор-
ме. Медиаторы макрофагов стимулируют размно-
жение фибробластов, регулируют синтез ими
межклеточного вещества, т. е. инициируют вос-
становительные процессы в очаге воспаления.
Одной из важных функций макрофагов являет-
ся поглощение некротизированных масс и очи-
щение зоны будущей репарации, в том числе и
от погибших лейкоцитов.
Нейтрофильные лейкоциты и макрофаги яв-
ляются основными клетками воспалительных
инфильтратов, особенно на ранних этапах вос-
паления. Кроме них, в состав инфильтратов вхо-
дят Т- и В-лимфоциты, плазмоциты, тканевые
базофилы, а при грануломатозном воспалении
могут обнаруживаться гигантские клетки ино-
родных тел или типа Лангханса-Пирогова.
Т- и В-лимфоциты появляются в инфильтра-
те почти одновременно с макрофагами. Их роль
заключается в регуляции воспалительного про-
цесса и в регенерации. Кроме того, при иммун-
ном воспалении эти клетки могут стать основ-
ными клетками инфильтрата. При этом
В-лимфоциты, превращаясь в плазмоциты, сек-
ретируют специфические антитела, а Т-лимфо-
циты — большое количество медиаторов-лимфо-
кинов, участвующих в разрушении собственных
клеток кожи. Особенно много плазмоцитов на-
капливается в пораженной коже при сифилисе,
паховой гранулеме, риносклероме, глубоких
микозах, хронических фолликулитах.
Роль тканевых базофилов (тучных клеток) в
восстановительном процессе огромна, причем
они являются участниками воспалительной ре-
акции на разных ее этапах. В фазу альтерации
эти клетки усиленно дегранулируют, выделяя ги-
стамин, серотонин, гепарин, являющиеся вазо-
активными веществами, подготавливающими со-
судистую стенку к миграционным процессам
клеток и к выходу из микрососудов жидкой
части крови (отек). В фазу экссудации продол-
жается действие указанных медиаторов, чем под-
держивается необходимое состояние пОйЫШ^М-
ной проницаемости сосудистой стенки. Тканевые
базофилы активно взаимодействуют с макрофа-
гами и другими иммунокомпетентными клетка-
ми, регулируя их активность. Макрофаги, в
свою очередь, фагоцитируют гранулы тканевых
базофилов (Виноградов В.В., Воробьева Н.Ф.,
1972; Юрина Н.А., Радостина А.И., 1991). Туч-
ные клетки играют большую роль и в стимуля-
ции пролиферативных процессов. Так, они сти-
мулируют размножение клеток эпителия,
фибробластов, эпидермиса и его производных
(Проценко А.В. и соавт., 1987).
Пролиферация
После очищения воспалительного ложа от бак-
терий, инородных веществ и разрушенных тка-
ней наступает пролиферация клеток, которая,
как правило, является завершающей фазой вос-
паления (Струков А.И., Серов В.В., 1990). И все
же пролиферативные процессы имеют место и в
самом начале воспалительной реакции, делению
подвергаются эндотелиоциты кровеносных сосу-
дов, адвентициальные клетки, Т- и В-лимфоци-
ты, моноциты. Однако основная интенсивность
пролиферативных процессов приходится на бо-
лее поздние сроки. В это время резко возраста-
ет количество фибробластов как путем диффе-
ренцировки их из адвентициальных клеток, так
и за счет миграции в очаг воспаления фибро-
Глава 4. Общепатологические процессы в коже
77
бластов из окружающих здоровых тканей. В оча-
гах пролиферации фибробластов начинается
фибробластическая фаза воспаления с усиленным
синтезом клетками межклеточного вещества
(коллагеновых и эластических волокон, основ-
ного вещества). Происходит либо ограничение
очага воспаления толстыми коллагеновыми во-
локнами, либо замещение его вновь созданным
межклеточным веществом, часто рубцового типа.
С течением времени в очаге воспаления остаются
лишь фибробласты, постепенно превращающи-
еся в фиброциты.
При воспалительных процессах в коже в них
часто вовлекается эпидермис. Активация в нем
пролиферативных процессов приводит к акан-
тозу и папилломатозу, что встречается при ней-
родермите, хронической экземе и др.
При различных воспалительных процессах в
коже степень выраженности каждого из ком-
понентов может быть различна, что определя-
ется реактивностью организма и флогогенным
агентом. Так, при воздействии на кожу хими-
ческих веществ, токсинов, бактерий, при гипер-
чувствительности немедленного типа преобла-
дают альтеративные процессы, которые могут
сопровождаться фибриноидным изменением со-
единительной ткани (при аллергических васку-
литах, в ревматоидных узелках, при красной
волчанке и др.)
Альтеративные процессы особенно выраже-
ны при образовании тромбов в сосудах кожи,
что наблюдается при васкулитах, в особенно-
сти протекающих с некротическими изменени-
ями (аллергический васкулит кожи Руитера,
узелковый некротический васкулит Вертера-
Дюмлинга).
Преобладание экссудативного компонента в
воспалении кожи имеет место при резко выра-
женной реакции микроциркуляторного русла.
Характер экссудата зависит от количества меди-
аторов, интенсивности их действия, чувствитель-
ности к медиаторам соответствующих рецепто-
ров клеток, а также от степени повреждения
стенок микрососудов.
Серозное воспаление кожи
Характеризуется развитием межклеточного и
внутриклеточного отека в эпидермисе и пораже-
нием мелких сосудов дермы (капилляров, пост-
капилляров, венул).
В эпидермисе развивается спонгиоз с после-
дующим развитием спонгиотических пузырьков
или пузырей. Спонгиоз может сопровождаться
внутриклеточным отеком. В результате форми-
руется ретикулярная дистрофия эпидермиса. Она
всегда сочетается с баллонирующей дистрофией.
При этом из-за резкого внутриклеточного накоп-
ления жидкости во многих кератиноцитах про-
исходит разрыв цитолеммы. Формируются мно-
гокамерные пузыри, в которых камеры
разделяются остатками дегенерирующих керати-
ноцитов. Далее происходит разрушение межкле-
точных контактов кератиноцитов, приводящее к
акантолизу.
Гнойное воспаление кожи
Гнойное воспаление в коже возникает при раз-
личных нарушениях барьерно-защитных сил
организма и, в частности, кожи. Может проте-
кать либо в острой, либо в хронической форме.
Этот достаточно частый вид кожной патологии
может встречаться при гипо- и авитаминозах,
гормональных нарушениях, действии професси-
ональных вредностей и т. д. В. В. Серов (1990)
считает гнойное воспаление результатом сопря-
жения функциональной недостаточности двух
фагоцитарных систем: системы нейтрофильных
лейкоцитов и макрофагов. При этом первичным
является нарушение функции нейтрофильных
лейкоцитов. Происходит массивная их гибель из-
за респираторного разрыва с разрушением здо-
ровых тканей кожи. Фагоцитируя продукты рас-
пада нейтрофильных лейкоцитов и тканей,
макрофаги перегружаются этими продуктами
распада, не успевают их элиминировать и в ре-
зультате оказываются несостоятельными в отно-
шении микроорганизмов.
Таким образом, гнойное воспаление кожи воз-
никает при качественных и количественных из-
менениях динамичного взаимодействия макро-
и микроорганизма. В этих изменениях превали-
руют защитные свойства микрооргнизма, направ-
ленные против бактерицидных свойств макро-
организма.
Вызывается гнойное воспаление гноеродны-
ми бактериями, к числу которых относятся ста-
филококки, гноеродные стрептококки и др.
Эти микроорганизмы имеют ряд факторов,
блокирующих защитные свойства организма и
кожи:
• способность угнетать хемотаксис и опсони-
зацию, противодействовать активности фагоци-
тов и специфических антител (при помощи ак-
тивных веществ и ферментов);
• выработка факторов «агрессии» и «защиты»
(например, гиалуронидаза, выполняя роль фак-
тора распространения, одновременно устраняет
влияние ингибиторов токсинообразования);
78 Общая дерматология
• иммунодепрессивное влияние гноеродных
бактерий на макроорганизм (установлено отно-
сительно недавно): уменьшение количества плаз-
моцитов как продуцентов антител, нарушение
активность Т-лимфоцитов и т. д.;
• способность стрептококков и стафилокок-
ков изменять свои морфологические свойства
(трансформироваться в L-формы, менять серо-
типы), что делает их недоступными для защит-
ных факторов макроорганизма.
Кроме того, в результате изменения антиген-
ных свойств микроорганизмов постоянно напря-
гается иммунная система макроорганизма, за-
трудняется выработка антимикробных и
антитоксических факторов.
Развитию гнойного воспаления кожи способ-
ствует целый ряд изменений со стороны макро-
организма (Глухенький Б.Т. и соавт., 1983):
• снижение реактивных свойств организма;
• заболевания центральной и периферической
нервной системы, голодание, неполноценное пи-
тание;
• развивающееся в результате этого снижение
бактерицидных свойств кожи;
• нарушение целостности кожного покрова в
результате травм и микротравм.
Основным морфологическим элементом при
гнойном воспалении кожи является пустула. Раз-
новидность пустул — пустулы Когоя — много-
камерные пустулы, располагающиеся в верхних
слоях эпидермиса и представляющие собой мел-
кие полости, заполненные гранулоцитами и про-
дуктами разрушения кератиноцитов. Развиты
акантоз и спонгиоз. Встречается при пустулез-
ном псориазе, болезни Рейтера и других кожных
заболеваниях. К хроническому гнойному воспа-
лению относятся пиодермии.
Геморрагическое воспаление кожи
Характерным для этого воспаления является по-
ражение сосудов кожи в виде васкулитов и пос-
ледующий фибриноидный их некроз. В результате
проницаемость сосудов и их прочность настолько
изменяются, что к экссудату любого характера в
большом количестве примешиваются эритроциты
(сибирская язва, геморрагический васкулит Ми-
шера, Шенлейна-Геноха, узелковый некротичес-
кий васкулит Вертера-Дюмлинга). Внешне гемор-
рагическое воспаление характеризуется появлением
геморрагических пятен — петехий и экхимозов.
Иногда при гистологическом исследовании
стенки кровеносных сосудов выглядят полностью
некротизированными, с закрытым просветом,
могут быть инфильтрированы нейтрофилами,
часть ядер которых разрушается (кариорексис). В
дерме — выраженный отек, коллагеновые волок-
на в состоянии фибриноидных изменений, мас-
сивные кровоизлияния. Эти изменения приводят
к некрозу кожи с формированием язв.
Продуктивное воспаление кожи
О продуктивном (пролиферативном) воспалении
говорят в тех случаях, когда в нем преобладает
третья фаза. При этом в очаге возрастает числен-
ность клеток и гистиогенного, и гематогенного
происхождения, а альтерация и экссудация вы-
ражены слабо.
Характеризуется эта форма воспаления кожи
отчетливыми продуктивными изменениями как
в эпидермисе, так и в дерме. В дерме при нерез-
ко выраженных изменениях со стороны сосудов
микроциркуляторного русла формируются кле-
точные инфильтраты различного состава, что
определяется видом возбудителя и состоянием
организма. Инфильтраты могут быть лимфоци-
тарными, макрофагическими, но чаще всего сме-
шанными. Часть клеток инфильтрата затем транс-
формируется в фибробласты, а в последующем
происходит склероз с преобладанием фиброцитов.
В эпидермисе отмечаются акантоз, гиперке-
ратоз, удлинение эпидермальных выростов. Уд-
линенные сосочки дермы при этом могут под-
вергаться фиброзу или инфильтрироваться
клетками воспалительного инфильтрата.
Продуктивное воспаление имеет место при бо-
лезни Дарье, вегетирующей пузырчатке, боро-
давчатой форме туберкулеза кожи. При болез-
ни Дарье и вегетирующей пузырчатке, кроме
отмеченных изменений, могут иметь место акан-
толитические пузыри, содержащие эозинофиль-
ные лейкоциты и акантолитические клетки эпи-
дермиса.
Хроническое воспаление кожи
По механизму возникновения хроническое вос-
паление делят на три вида:
1. хроническое воспаление, возникшее в резуль-
тате внутриклеточной персистенции возбудителей
(глубокие микозы, сифилис, туберкулез и др.);
2. воспаление в результате длительного пребы-
вания в коже не подвергающегося элиминации
пассивного материала (силиций, инородные
инертные тела и др.);
3. аутоиммунное воспаление, при котором ком-
плекс «антиген-антитело» вызывает хронические
изменения в коже (ревматоидный артрит, ауто-
иммунный тиреоидит).
Глава 4. Общепатологические процессы в коже
79
Иммунное воспаление кожи
По мнению А. И. Струкова и соавт. (1990), хотя
при банальном воспалении всегда имеется им-
мунологический аспект, при некоторых заболе-
ваниях в клинико-иммунологической картине с
необычайной яркостью сплетаются в единый
узел иммунный ответ и последовательные вос-
палительные реакции. Авторы считают, что это
не только иммунологический аспект воспаления,
но иммунное воспаление в своей основе.
Основу иммунного воспаления в коже состав-
ляют иммунокомпетентные клетки эпидермиса
и дермы. В эпидермисе это внутриэпидермаль-
ные лимфоциты, клетки Лангерганса и Грин-
стейна. Кератиноциты также не являются пас-
сивными объектами иммунной атаки, а
активными участниками иммунного воспаления,
выделяя разнообразные медиаторы, участвующие
в иммунных воспалительных процессах в коже.
В дерме такими клетками являются лимфоци-
ты, тканевые базофилы, макрофаги, плазмоци-
ты, гранулоциты, в первую очередь эозинофиль-
ные. Кроме этих клеток гистиогенного
происхождения, в иммунном воспалении уча-
ствуют клетки гематогенного происхождения.
В коже реализуются почти все известные виды
иммунопатологических процессов и иммунного
воспаления (Персина И.С., 1993). В этом отно-
шении она является уникальным органом, в связи
с чем используется в многочисленных экспери-
ментальных моделях для изучения иммунопато-
логических феноменов.
Дерматозы, имеющие
в основе немедленные
анафилактические реакции
К этой форме иммунного воспаления в коже от-
носятся крапивница, лекарственные эритемы. Кра-
пивница представляет собой анафилактическую
реакцию кожи на различные биологически актив-
ные вещества, белковые аллергены, лекарственные
препараты, пищевые продукты, а также вещества
эндогенного происхождения, особенно при забо-
леваниях печени, желудочно-кишечного тракта.
Механизм развития крапивницы можно пред-
ставить себе следующим образом (рис. 4.8). В ре-
зультате сенсибилизации под влиянием аллерге-
на в плазмоцитах образуются антитела реагины
или иммуноглобулины класса Е. Реагины явля-
ются необычными антителами, т. к. только им
присуща способность накапливаться в коже и ос-
таваться в ней в течение нескольких недель. При
этом они фиксируются своими Fc-фрагментами
на тучных клетках. При повторном попадании
аллерген, взаимодействуя с Рав — фрагментами
IgE, активирует образование цАМФ в тканевых
Вторичные медиаторы
Аллерген
|дЕ
ТАФ
Тучная
клетка
Базофил
МРС-А (лейкотриены)
ЭХФ
Первичные медиаторы
^Гистамин -----------
Эозинофил
Основной белок
Простагландины
Лейкотриены
Тромбоксан А2
"Арипсульфатоза В
Г истаминаза
Фосфолипаза Д
Анафилактический
шок
Г истамин
Тромбоциты
НХФ
Нейтрофил
Ферменты
гранул
медиаторы
Сокращение
гладкой
мускулатуры
Увеличение
секреции
бронхиальной
слизи
Приступ
бронхиальной
астмы
(бронхоспазм, отек
слизистой,
гиперсекреция
слизи)
Ринит
Конъюнктивит
ПЛФ
Простагландины
Увеличение
сосудистой
проницаемости
Отек Квинке
Крапивница
Кожный зуд
и др.
Стадии:
| ИММУНОЛОГИЧЕСКАЯ
Брадикинин
\ Серотонин
ПАТОХИМИЧЕСКАЯ
♦ ПАТОФИЗИО-
I ЛОГИЧЕСКАЯ
f КЛИНИЧЕСКИЕ
I ПРОЯВЛЕНИЯ
Рис. 4.8. Схема реагинового типа аллергической реакции (Д. К. Новиков и соавт., 1996).
80
Общая дерматология
базофилах. Последний активирует самосборку
микротрубочек в клетках и экзоцитоз их гранул,
содержащих биологически активные вещества:
гистамин, медленно реагирующую субстанцию
(МРС), эозинофильный хемотаксический фактор
(ЭХФ), тромбоцитактивирующий фактор (ТАФ),
нейтрофильный хемотаксический фактор (НХФ),
серотонин и др. Эозинофилы, нейтрофилы и
тромбоциты, в свою очередь, выделяют вторич-
ные медиаторы, которые обеспечивают увеличе-
ние сосудистой проницаемости. В настоящее вре-
мя показано, что у больных атопическим
дерматитом может формироваться высокий уро-
вень IgE как результат нарушения регуляции Т-
лимфоцитами его синтеза.
Клинически крапивница проявляется прехо-
дящими сильно зудящими, слегка возвышающи-
мися, плотными, резко ограниченными от ос-
тальной кожи волдырями от белого до
ярко-красного цвета различной величины и фор-
мы. При распространении воспалительного про-
цесса на гиподерму возникает тестовидный отек
со стертыми границами (ангионевротический
отек, отек Квинке). Гистологическая картина ха-
рактеризуется расширением кровеносных и лим-
фатических сосудов, периваскулярной инфиль-
трацией лимфоцитами и эозинофильными
лейкоцитами. Коллагеновые волокна набухшие,
разрыхлены. В эпидермисе отмечается спонгиоз
и различной степени внутриклеточный отек.
Цитотоксические
реакции аутоиммунной природы
К ним относятся пузырчатка, пемфигоид, син-
дром Лайелла. В патогенезе их ведущую роль иг-
рают аутоиммунные процессы, причем аутоан-
титела образуются против межкератиноцитарной
цементирующей субстанции. Образование комп-
лекса «антиген-антитело» приводит к высвобож-
дению кератиноцитов из связей с соседними
клетками. Морфологическим субстратом этих
реакций являются: вакуольная дистрофия базаль-
ных кератиноцитов, супрабазальный акантолиз
или акантолиз в зернистом слое. При вегетиру-
ющей пузырчатке имеют также место папилло-
матоз, акантоз, в старых очагах — гиперкератоз.
В дерме в сосочковом слое можно обнаружить
незначительный инфильтрат из эозинофилов,
лимфоцитов и нейтрофилов. При рубцующем-
ся пемфигоиде развивается фиброз и далее —
рубцевание.
Иммунокомплексные реакции в коже
К ним относятся многоформная экссудативная
эритема, токсикодермия, пурпура Шенлейн-Ге-
ноха, лейкоцитопластический васкулит, узелко-
вый некротический васкулит Вертера-Дюмлин-
га. По этому механизму развиваются также
системные заболевания соединительной ткани.
Стадии:
ИММУНОЛОГИЧЕСКАЯ
ПАТОХИМИЧЕСКАЯ
Фиксация \ комплемента и активация его компонентов f Сывороточная болезнь
Аллергический альвеолит
Лизосомальные ферменты Васкулит с l альтерацией, _>^ г* экссудацией, пролиферацией Лекарственная и пищевая аллергия Анафилактический шок
Агрегация тромбоцитов Активация калликреинкининовой системы Крапивница Отек Квинке Поствакцинальные аллергические осложнения к
ПАТОФИЗИО- А КЛИНИЧЕСКИЕ
ЛОГИЧЕСКАЯ I ПРОЯВЛЕНИЯ
Рис. 4.9. Схема иммунокомплексных реакций (Д. К. Новиков и соавт., 1996).
Глава 4. Общепатологические процессы в коже
81
При этих заболеваниях образуется большое ко-
личество иммунных комплексов необычных раз-
меров при избытке антигена (рис. 4.9). Это на-
рушает фагоцитоз иммунных комплексов, и они
начинают откладываться в тканях, в первую оче-
редь под базальной мембраной кровеносных со-
судов. Иммунные комплексы содержат иммуно-
глобулины класса G и М и активируют систему
комплемента. Продукты активации комплемента
повышают проницаемость сосудов, вызывают их
расширение, а также привлекают гранулоциты и
макрофаги. Фагоциты, в свою очередь, выделя-
ют вторичные медиаторы, повреждающие ткани.
Основными патогистологическими изменения-
ми являются: артериит, периартериит, флебит и
перифлебит, сопровождающиеся отечностью,
пролиферацией и дегенерацией эндотелия. Ин-
фильтрат пронизывает стенку сосудов и окружа-
ет их. Он состоит из лейкоцитов (нейтрофиль-
ных и эозинофильных), а при васкулите
Вертера—Дюмлинга к ним примешиваются плаз-
моциты. Просвет многих сосудов может быть
закрыт эндотелиальным тромбом или облитери-
рован. В поздней стадии наблюдается гиалиноз
и разрастание соединительной ткани.
Г иперчувствительность
замедленного типа
К заболеваниям, обусловленным данной формой
гиперчувствительности, относятся такие формы
иммунного воспаления кожи, как нейродермит,
красный плоский лишай, аллергический контак-
тный дерматит (АКД), экзема и др. Реакция
отторжения трансплантата кожи также относит-
ся к данному виду гиперчувствительности.
Главную роль в развитии данной формы им-
мунопатологии играют Т-лимфоциты, несущие
специфические рецепторы к антигену
(рис. 4.10). Вначале они сенсибилизируются при
взаимодействии с антигеном, затем претерпева-
ют бласттрансформацию, делятся митозом, уве-
личиваясь в количестве, и дифференцируются в
цитотоксические Т-лимфоциты. Одновременно
лимфоциты выделяют ряд лимфокинов, которые
привлекают в очаг воспаления макрофаги и ней-
трофилы. Формируется очаг воспаления, ин-
фильтрат, клетки которого повреждают чуже-
родные или собственные измененные ткани.
Образующиеся в результате бласттрансформации
Т-киллеры оказывают на антигены прямое по-
вреждающее действие путем выделения перфо-
ринов. Молекулы перфоринов встраиваются в
цитолемму атакуемых клеток. Это ведет к фор-
мированию трансмембранных каналов, мембра-
на теряет свои барьерные функции, и клетки по-
гибают в результате осмотического шока.
Контактная гиперчувствительность как фор-
ма гиперчувствительности замедленного типа
(ГЗТ) вызывается воздействием на кожу низко-
молекулярных веществ, являющихся неполны-
ми антигенами (гаптенами). Соединяясь с бел-
Прямое цитотоксическое
действие
сенсибилизированных
Т-лимфоцитов на клетки
мишени
Т-киллер
Клетка-мишень
Клетка-эффектор \
(сенсибилизированные
Т-лимфоцит)
Аллерген <
Лимфокинсинтезирующий
путь повышенной
чувствительности
замедленного типа (ПЧЗТ)
Стадии:
ИММУНОЛОГИЧЕСКАЯ
Продукты деградации
клеток-мишеней
Медиаторы ПЧЗТ
Г Лимфотоксин
Хемотаксический
фактор
Фактор
ингибиции
миграции
<
Митогенный фактор
Иммунный
интерферон
Кожно-реактивный
фактор
Развитие воспалительной
реакции продуктивного
типа в тканях
Вызывает повреждение или
разрушение клеток
Обусловливает хемотаксис
лейкоцитов в очаг
аллергической реакции
Способствует накоплению
макрофагов и нейтрофилов
в очаге поражения.
Участвует в образовании
гранулем при инфекционно-
аллергических заболеваниях
Стимулирует пролиферацию
и рост лимфоцитов
Обладает противовирусным
действием
Участвует в формировании
кожной реакции в ответ на
внутрикожное введение
аллергена
Контактно-
аллергический дерматит
Инфекционно-
аллергическая
бронхиальная астма
Аутоаллергические и
инфекционно-аллергиче-
ские заболевания
Лекарственная
аллергия
Реакция на
трансплантат
f ПАТОХИМИ-
IЧЕСКАЯ
k а КЛИНИЧЕСКИЕ
ПАТОФИЗИОЛОГИЧЕСКАЯ ПРОЯВЛЕНИЯ
е
Рис. 4.10. Схема реакций гиперчувствительности замедленного типа (Д. К. Новиков и соавт., 1996).
82 Общая дерматология
ками эпидермиса, они образуют полные антиге-
ны. Гаптены являются экзогенным фактором
АКД. К эндогенным факторам относятся наслед-
ственная предрасположенность, обусловленная
изменениями в генах, контролирующих иммун-
ный ответ (Ir-гены) и сцепленных с ситемой
HLA, ответственной за гистосовместимость. Оп-
ределенную роль играет нарушение функций ве-
гетативной нервной системы.
Проникнув в эпидермис, гаптен связывается
не только с кератиноцитами, но и (через неко-
торое время уже в комплексе с эпидермальным
белком) с клетками Лангерганса. И кератиноци-
ты, и клетки Лангерганса способны представлять
антиген Т-лимфоцитам. Однако у кератиноци-
тов такая способность имеется не ко всем аллер-
генам, а только к отдельным (соли кобальта и
никеля) и отсутствует к другим гаптенам, что
может иметь диагностическое значение. Воздей-
ствие указанных гаптенов на кератиноциты вы-
зывает появление у них HLA-Dr-антигенов, ха-
рактерных для клеток Лангерганса.
Количество клеток Лангерганса в месте ап-
пликации гаптена резко увеличивается как в дер-
ме, так и в эпидермисе. Инактивация клеток
Лангерганса ультрафиолетовым облучением пре-
дотвращает развитие сенсибилизации.
Антигены представляются клетками Лангер-
ганса в эпидермисе, дерме либо в регионарных
лимфоузлах, куда они мигрируют и где накап-
ливаются в паракортикальной зоне (Т-зависимой
зоне, основной зоне лимфоузла, где происходит
антигензависимая дифференцировка Т-лимфо-
цитов). Клетки Лангерганса могут оказаться в
очаге АКД клетками-мишенями для воздействия
цитотоксических Т-лимфоцитов и служить ис-
точником вторичных медиаторов.
Патоморфология АКД достаточно полно изуче-
на. Г. М. Цветкова и соавт. (1993) выделяют 3
этапа развития сенсибилизации к гаптенам (ди-
нитрохлорбензол, ДНХБ):
1. первично-контактную реакцию;
2. спонтанную воспалительную реакцию вос-
пламенения;
3. воспалительную реакцию на разрешающее
нанесение ДНХБ.
Гистологическая картина кожи в каждый из этих
этапов различна. Для первой фазы характерен
ограниченный отек, гиперемия кожи, позднее
присоединяется некроз с отслойкой некротизи-
рованного эпидермиса и образованием субэпи-
дермальных пузырей. В дерме отмечается:
• резкое расширение кровеносных и лимфа-
тических капилляров, гемокапилляры полно-
кровны, часто содержат нейтрофилы;
• формирование в верхней трети диффузного
клеточного инфильтрата из нейтрофилов, макро-
фагов; могут присутствовать также лимфоциты и
в значительном количестве тканевые базофилы.
Вторая фаза характеризуется значительными
акантозом и гиперкератозом эпидермиса. В
дерме отмечается расширение и полнокровие
микрососудов, в некоторых наблюдаются ди-
строфические изменения стенок вплоть до пол-
ной деструкции. Вокруг таких сосудов могут
обнаруживаться зоны кровоизлияний, фибри-
ноза и перерождения коллагеновых волокон.
Формируется крупный, на всю верхнюю по-
ловину дермы, клеточный инфильтрат, состо-
ящий из лимфоцитов, макрофагов и базофиль-
ных лейкоцитов, которые иногда могут
преобладать, но редко находятся в состоянии
дегрануляции. Среди лимфоидных клеток ос-
новными являются Т-хелперы и цитотоксичес-
кие Т-супрессоры с преобладанием первых.
Могут встречаться также клетки плазмоцитар-
ного ряда.
Морфологическая картина третьей фазы похожа
на предыдущую, но изменения более выражены
и в эпидермисе, и в дерме. В эпидермисе более
выражены экссудативные изменения, явления
межклеточного отека. Возрастает количество кле-
ток Лангерганса. Изменения в дерме также бо-
лее выражены, особенно в сосудах микроцирку-
ляторного русла, стенки которых утолщаются за
счет гипертрофии и гиперплазии эндотелиоцитов.
В результате просветы суживаются. Базофильные
гранулоциты в инфильтрате уменьшаются в ко-
личестве, большая их часть дегранулирует. В це-
лом состав инфильтрата такой же, как и во 2 фазе.
Атопический дерматит рассматривают как одну
из форм иммунного воспаления в коже IV типа
(Персина И.С., 1993). Его основными симпто-
мами являются кожный зуд, эритема, склонные
к слиянию узелковые элементы, образование
очагов лихенизации.
Патогистологические изменения в коже харак-
теризуются акантозом, гиперкератозом, а при
диффузном поражении кожи в свежих очагах,
кроме того, могут обнаруживаться паракератоз,
спонгиоз и проникновение в эпидермис лейко-
цитов. Характерен также папилломатоз. Базаль-
ные кератиноциты в очаге поражения теряют
меланин, тогда как по периферии, напротив, со-
держание меланина увеличено. В сосочковом слое
дермы и в верхней части сетчатого слоя дермы
обнаруживаются периваскулярные инфильтраты
из лимфоцитов и фибробластов. Отмечается фиб-
роз дермы. В состав инфильтрата могут входить
нейтрофильные и эозинофильные гранулоциты
Глава 4. Обшепатологические процессы в коже
83
(Суворова К.Н. и соавт., 1989), а по нашим дан-
ным (Адаскевич В.П., Мяделец О.Д., 1995) —
также макрофаги и единичные тучные клетки.
Гранулематозное воспаление
Гранулематозное воспаление в настоящее время
относится большинством авторов к разновидно-
стям иммунного воспаления, хотя иммунные
нарушения не всегда могут начинаться с момента
проникновения в организм возбуждающего аген-
та. Этот вид воспаления называют также специ-
фическим (Струков А.И., Серов В.В., 1995;
Струков А.И., Кауман О.Я., 1989). Согласно
данным А. И. Струкова и В. В. Серова (1985),
специфическое воспаление имеет следующие
характерные признаки:
1. каждый вид специфического воспаления вы-
зывается определенным возбудителем;
2. по ходу специфического воспаления проис-
ходит смена тканевых реакций, определяющая-
ся иммунологической перестройкой организма;
3. специфическое воспаление имеет хроничес-
кое волнообразное течение, при котором пери-
оды затихания процесса сменяются периодами
его обострения;
4. при специфическом воспалении преоблада-
ет продуктивная тканевая реакция и происходит
развитие гранулемы — наиболее яркого морфо-
логического признака специфичности;
5. по ходу развития специфического воспале-
ния закономерно появление некроза:
а. первичного, возникающего при преобладании
альтеративной тканевой реакции и без пред-
шествующих клеточных реакций;
б. вторичного, возникающего на фоне предше-
ствующей экссудативной или продуктивной
тканевой реакции.
Все эти признаки специфического воспаления
придают ему особые черты, среди которых наи-
более важные — развитие гранулемы, хрониза-
ция и волнообразность течения, некроз по ходу
воспаления. Это объединяет все виды грануле-
матозного воспаления. Однако строение грану-
лем может несколько различаться при каждом
виде, различными могут быть и темпы воспали-
тельного процесса.
Все виды гранулематозного воспаления делятся
в зависимости от вида вызывающего его возбу-
дителя или агента на:
1. гранулемы инородных тел (вызываются та-
туировкой, солями хрома, кобальта, металлов
бериллия, циркония);
2. гранулемы при туберкулезе;
3. лепрозные гранулемы;
4. гранулематозное воспаление при позднем
вторичном и третичном сифилисе;
5. саркоидозные гранулемы;
6. кандидозные и другие гранулемы.
В зависимости от иммунного статуса организма
различают следующие виды гранулематозного
воспаления (Reyes-Flores О., 1986):
• иммунокомпетентное гранулематозное вос-
паление;
• гранулематозное воспаление с неустойчивым
иммунитетом;
• иммунодефицитное гранулематозное воспа-
ление.
Любое гранулематозное воспаление характери-
зуется образованием гранулем (узелков), возни-
кающих в результате пролиферации и трансфор-
мации способных к фагоцитозу клеток. При
этом гранулема формируется в три стадии:
1. накопление в очаге повреждения ткани юных
моноцитарных фагоцитов;
2. созревание этих клеток в макрофаги и аг-
регация их с образованием зрелой гранулемы;
3. дальнейшее созревание моноцитов и макро-
фагов в эпителиоидные клетки и агрегация их
с образованием эпителиоидной гранулемы.
При этом происходит слияние эпителиоидных
клеток (или макрофагов), что ведет к образова-
нию гигантских клеток инородных тел. Послед-
ние (иногда) превращаются в гигантские клетки
Пирогова-Лангханса, которые могут содержать от
2~3 до 100 и более ядер. В гигантских клетках
инородных тел ядра лежат в цитоплазме равно-
мерно, а в клетках Пирогова-Лангханса — пре-
имущественно по периферии. Фагоцитарная ак-
тивность макрофагов и их производных по мере
их созревания падает.
Отдельные виды
гранулематозного воспаления кожи
Туберкулез кожи — иммунокомпетентное грану-
лематозное воспаление. Проявляется клиничес-
ки сыпью, варьирующей в зависимости от сте-
пени казеозного некроза от пятен до язвенных
поражений. При туберкулезе кожи, как и дру-
гих органов, имеют место различные виды им-
мунного ответа: образование циркулирующих
антител, гиперчувствительность замедленного
типа с формированием гранулемы. Туберкулез-
ная гранулема чаще наблюдается при туберку-
лезной волчанке и состоит из эпителиоидных
клеток, небольшого числа клеток Пирогова-Лан-
гханса, окруженных валом из Т-лимфоцитов.
Строму гранулем образуют аргирофильные во-
локна. В центре гранулемы обычно наблюдает-
ся казеозный некроз.
84 Общая дерматология
Сифилис относится к иммунокомпетентному
гранулематозному воспалению. Гранулемы обра-
зуются в позднем вторичном и третичном пери-
одах. В развитии воспаления при сифилисе на-
блюдается: пролиферативно-инфильтративная
реакция, при которой инфильтрат состоит из
лимфоидных плазматических и эпителиоидных
клеток с наличием продуктивного васкулита
(твердый шанкр); продуктивно-некротическая
реакция с формированием бугорка; экссудатив-
но-некротическая реакция при врожденном си-
филисе. При сифилисе в очаг воспаления в про-
цесс больше, чем при туберкулезе, вовлекаются
кровеносные сосуды (артерииты и флебиты).
При позднем вторичном сифилисе в дерме от-
мечается диффузный, преимущественно лимфо-
цитарный инфильтрат, который имеет тенден-
цию к формированию гранулемы из гигантских
клеток типа Пирогова-Лангханса.
В третичном или гуммозном периоде в коже
развиваются характерные гранулемы, содержа-
щие в центре очаг казеозного некроза, окру-
женный лимфоцитами, плазмоцитами, эпители-
оидными клетками и некоторым количеством
гигантских клеток типа Пирогова-Лангханса.
Лепра относится к иммунокомпетентому гра-
нулематозному воспалению кожи. Однако раз-
личные формы лепры характеризуются различ-
ной устойчивостью организма по отношению к
возбудителю. Туберкулоидная форма характери-
зуется высокой устойчивостью организма к воз-
будителю, лепроматозная форма — низкой, а
промежуточная форма сочетает в себе признаки
первой и второй форм.
Туберкулоидная форма лепры характеризуется
формированием гранулем из эпителиоидных
клеток, окруженных валом из Т-лимфоцитов-
супрессоров. Внутри гранулемы могут находить-
ся Т-хелперы. Гранулематозная реакция может
захватывать весь эпидермис и различные струк-
туры дермы. Микобактерии практически не вы-
являются.
Лепроматозная форма лепры характеризуется
образованием специфических инфильтратов из
макрофагов в состоянии различной по степени
липодистрофии. Развивается при иммунодефи-
цитном состоянии. Лимфоцитов немного. Коли-
чество микобактерий огромное.
При неустойчивом иммунитете развивается
пограничная форма лепры, характеризующаяся
макрофагальной реакцией дермы, иногда с эпи-
телиоидными клетками различной степени диф-
ференцировки. Гигантские клетки отсутствуют.
Окраска по Циль-Нильсону выявляет умеренное
количество микобактерий.
При грибковых заболеваниях кожи также вы-
являют иммунокомпетентные гранулемы, грану-
лемы с неустойчивым иммуни чом и иммуно-
дефицитное гранулематозное воспаление.
К иммунокомпетентному гранулематозному
воспалению при грибковых заболеваниях отно-
сят хромомикоз, вызываемый хромомицетами.
Кожные формы этого заболевания характеризу-
ются изменениями эпидермиса в виде акантоза,
иногда с картиной псевдоэпителиоматозной ги-
перплазии. В дерме формируются гранулемы из
эпителиоидных и гигантских клеток. Примером
гранулемы с неустойчивым иммунитетом явля-
ется кандидоз. Его хроническая форма характе-
ризуется акантозом, папилломатозом, гиперке-
ратозом эпидермиса. В дерме инфильтраты из
лимфоцитов, плазмоцитов, макрофагов, эпите-
лиоидных и гигантских клеток.
Паразитарные гранулемы делятся по такому же
принципу. Гранулематозное воспаление с имму-
нокомпетентной реакцией формируется при
кожном американском мукокутанном лейшмани-
озе. Возбудителем его является Leischmania
brasiliensis. Гистологические изменения в коже
зависят от клинической картины и выражаются
в образовании бляшки, узла, язвы. В дерме на
ранней стадии наблюдается неспецифическая
воспалительная реакция. Инфильтраты при ней
состоят из макрофагов, лимфоцитов, плазмоци-
тов и небольшого количества эозинофильных и
нейтрофильных лейкоцитов. В более старых оча-
гах воспаления макрофаги превращаются в эпи-
телиоидные клетки, формируя окруженные лим-
фоцитами и плазмоцитами гранулемы, которые
могут прорастать в гиподерму. Лейшмании на-
ходятся внутриклеточно, часто вблизи язв. Их
число зависит от интенсивности процесса. В эпи-
дермисе — выраженный акантоз с псевдоэпите-
лиоидной гиперплазией.
Диффузный кожный лейшманиоз является при-
мером иммунодефицитных гранулем, гистологи-
чески напоминающим лепроматозную форму
лепры. При этом в дерме формируется плотный
инфильтрат, состоящий из макрофагов, с высо-
ким содержанием паразитов, видимых даже при
обычных гистологических окрасках. Однако в
отличие от лепры в макрофагах отсутствует жир,
хотя они сильно вакуолизированы. В вакуолях
(фаголизосомы, увеличенные в размерах) содер-
жится множество паразитов. Имеются макрофаги
на разных стадиях превращения в эпителиоид-
ные клетки и большое число лимфоцитов. Эпи-
дермис над гранулемным инфильтратом находит-
ся в состоянии атрофии из-за давления на него
гранулемы.
Глава 4. Общепатологические процессы в коже
85
А. И. Струков и О. Я. Кауфман (1989), наряду
с иммунными гиперчувствительными гранулема-
ми, выделяют неиммунные гранулемы, участие
иммунных механизмов при образовании которых
не доказано. К ним авторы относят неиммунные
инфекционно-токсические гранулемы, возника-
ющие при острых инфекционных заболеваниях,
а также неиммунные токсические гранулемы, к
которым относят как экзо-, так и эндогенные
гранулемы, возникающие вокруг инородных тел.
Согласно определению W. Epstein (1980), грану-
лема инородных тел является неиммунной реак-
цией мононуклеарных фагоцитов на нераство-
римый экзогенный или эндогенный стимул.
Неиммунные гранулемы инородных тел подраз-
деляют на неактивные (доброкачественные) и ак-
тивные или цитотоксические. Первые из них
возникают в ответ на внедрение пластмассовых
гранул, латекса, тентонина и т. д., вторые — на
попадание двуокиси кремния, оболочки стреп-
тококка и т. д. (Epstein W., 1980).
Таким образом, гранулематозное воспаление —
это хроническое воспаление кожи, вызванное
нерастворимыми или медленно разрушающимися
(персистирующими) раздражителями и сопро-
вождающееся образованием гранулем. Последние
представляют собой компактное скопление мак-
рофагов и/или эпителиоидных клеток, а также
их более зрелых потомков (гигантских клеток
инородных тел, гигантских клеток Пирогова-
Лангханса). Оно может сопровождаться рядом
дополнительных признаков (Пауков В.С., Кауф-
ман О.Я., 1995):
• инфильтрацией другими клетками (плазмо-
цитами, лимфоцитами, нейтрофильными или
эозинофильными лейкоцитами);
• наличием фибробластов и развитием скле-
роза;
• развитием деструктивных изменений и не-
кроза.
Условиями развития гранулемы являются два
основных момента (Струков А.И., Серов В.В.,
1995):
1. наличие веществ, способных стимулировать
макрофаги;
2. стойкость раздражителя по отношению к фа-
гоцитам.
Возможны следующие исходы гранулематозно-
го воспаления в коже, как и в других органах
(Пауков В.С., Кауфман О.Я., 1995):
• рассасывание клеточного инфильтрата;
• сухой (казеозный) или влажный некроз с
образованием дефекта кожи;
• нагноение гранулемы с формированием аб-
сцесса;
• фиброзное превращение гранулемы с обра-
зованием фиброзного узелка или рубца;
• рост гранулемы, иногда с образованием псев-
доопухоли.
Реакция «трансплантат
против хозяина»
Реакция «трансплантат против хозяина» (РТПХ)
или «вторичная болезнь» развивается у большин-
ства больных после трансплантации костного моз-
га при тяжелом комбинированном иммунодефи-
ците. Ятрогенная РТПХ возникает как
осложнение при пересадке костного мозга боль-
ным апластической анемией и лейкозами. Она
может быть иногда и следствием переливания
крови иммунодефицитному реципиенту. Более
резкая (врожденная) форма РТПХ является ре-
зультатом взаимодействия лимфоцитов матери с
антигенами гистосовместимости тканей иммуно-
дефицитного плода.
Механизм РТПХ
Реакция развивается при следующих условиях:
1. трансплантат должен обладать иммунологи-
ческой активностью;
2. в тканях реципиента должны содержаться
антигены, отсутствующие в тканях донора (ан-
тигенная чужеродность реципиента для иммуно-
логически активного донора);
3. иммунологическая инертность (пассивность)
реципиента, неспособность отторгать трансплан-
тированные клетки донора.
Клетками-эффекторами при РТПХ являются
цитотоксические Т-киллеры. Кожа является пер-
вичным органом-мишенью при РТПХ. В основе
РТПХ и ряда кожных заболеваний (красная вол-
чанка, склеродермия и др.) лежат общие механиз-
мы. Иммуногистохимически при РТПХ в коже
выявляются супрессорные цитотоксические Т8+-
лимфоциты, локализующиеся среди кератиноци-
тов и клеток Лангерганса (Т-киллеры являются
разновидностью супрессорных Т-лимфоцитов).
Клетками-мишенями в эпидермисе при РТПХ
являются кератиноциты и клетки Лангерганса.
Содержание последних в эпидермисе является
важным критерием степени тяжести РТПХ. РТПХ
может быть острой (10—30 дней после пересад-
ки костного мозга или переливания крови) и
хронической (развивается через несколько месяцев
или даже лет после трансплантации).
Клиника кожных проявлений РТПХ
Острая форма РТПХ проявляется появлением на
коже пятнисто-папулезных высыпаний, преиму-
86 Общая дерматология
щественно на лице, ушных раковинах, верхней
половине туловища, ладонях и подошвах. На
местах давления могут формироваться пузыри.
Такие поражения сходны с токсическим некро-
лизом и часто ведут к смерти больного.
Хроническая РТПХ характеризуется либо ге-
нерализованным, либо локализованным пораже-
нием кожи и делится по типу высыпаний на ли-
хеноидную и склеротическую фазы, следующие
обычно одна за другой. Лихеноидные папулы
имеют фиолетовую окраску, напоминая элемен-
ты красного плоского лишая, часто располага-
ются на ладонях и подошвах, но могут быть рас-
пространёнными и иметь тенденцию к слиянию,
сопровождаются зудом. После себя оставляют
очаги интенсивной гиперпигментации с непра-
вильными очертаниями. При более поздней
склеротической фазе на коже появляются очаги
уплотнения, напоминающие склеродермию.
Придатки кожи при этом атрофируются, часто
формируется алопеция. Кожа в очагах теряет эла-
стичность, с участками атрофии, гипо- и гипер-
пигментации.
Патоморфология РТПХ
Острая РТПХ характеризуется вакуольной дис-
трофией базальных клеток эпидермиса и эпите-
лия волосяных фолликулов, что сочетается с
проникновением лимфоцитов в эпидермис и
небольшими лимфоцитарными инфильтратами в
дерме. Часто отмечается лимфоассоциированный
апоптоз кератиноцитов (такие апоптотически
измененные кератиноциты лежат рядом в тесном
контакте с интраэпидермальными лимфоцита-
ми — сателлитный некроз). При нарастании ва-
куольной дистрофии происходит отслойка эпи-
дермиса с образованием пузырей. По степени
тяжести поражения эпидермиса обычно опреде-
ляют тяжесть течения РТПХ: при РТПХ I сте-
пени возникает вакуольная дистрофия базальных
клеток. II степень характеризуется апоптозом
кератиноцитов с массивным проникновением в
эпидермис лимфоцитов, а при III степени нара-
стает периваскулярная инфильтрация лимфоци-
тов дермы. IV степень РТПХ характеризуется
некрозом эпителиоцитов, возникновением пара-
кератоза, а при эпидермальном некролизе —
отеком сосочков дермы. Тяжесть кожных про-
явлений РТПХ обычно прямо зависит от коли-
чества инфильтрирующих эпидермис и дерму
лимфоцитов.
Хроническая РТПХ. При ранней лихеноидной
хронической РТПХ гистологические изменения
сходны с таковыми при идиопатическом красном
плоском лишае. Отмечаются выраженный гипер-
кератоз, вакуольная дегенерация базальных ке-
ратиноцитов. Под эпидермисом формируется
инфильтрат, в состав которого входят лимфоци-
ты, эозинофильные лейкоциты, некоторое коли-
чество плазмоцитов и клетки, содержащие пиг-
мент (меланофоры). Отличием этого инфильтрата
от инфильтрата при красном плоском лишае яв-
ляется его меньшая плотность и большее разно-
образие входящих в его состав клеток (при крас-
ном плоском лишае в состав инфильтрата входят
в основном лимфоциты).
Поздняя склеротическая фаза хронической
РТПХ характеризуется гистологической карти-
ной, сходной со склеродермией: формируется
атрофия эпидермиса, местами сохраняется ваку-
ольная дегенерация базальных кератиноцитов.
Атрофии подвергаются и придатки кожи. В дер-
ме нарастает количество меланофоров, отмечают-
ся очаговые периваскулярные лимфоцитарные
инфильтраты, которые могут располагаться и
вокруг придатков кожи. Склеротические изме-
нения захватывают в основном сосочковый слой
дермы. В нем изменяется состав коллагеновых
волокон, содержание проколлагена I типа уве-
личивается. Появляются активные фибробласты,
тканевые базофилы, лимфоциты, макрофаги и
меланофоры.
Глава 5
Этиология, патогенез
и диагностика кожных болезней
Этиология и патогенез кожных болезней 87
Общая симптоматология болезней кожи 89
Обследование кожного больного 98
Этиология и патогенез
кожных болезней
Кожа — один из важнейших органов, участву-
ющих в поддержании гомеостаза человека. Яв-
ляясь уникальным полифункциональным мем-
бранным образованием, она через сенсорные и
защитные механизмы осуществляет контакт че-
ловека с внешним миром, отражает качество
функционирования внутренних органов, эндо-
кринной и нервной систем. Кожу сравнивают с
зеркалом, в котором отражается деятельность
организма в целом.
Многие кожные болезни возникают в ре-
зультате взаимодействия внешних (экзоген-
ных) и внутренних (эндогенных) факторов. В
ряде случаев кожные проявления вызывают
сразу несколько факторов, причем один из
них является предрасполагающим, а другой —
непосредственно вызывающим патологические
изменения.
К экзогенным этиологическим факторам отно-
сят химические, физические, инфекционные
агенты, а также воздействие на кожу биологичес-
ких факторов (некоторые растения, животные-па-
разиты, мигрирующие личинки и др.). Наиболее
разнообразны эндогенные этиологические факторы
в возникновении дерматозов. К ним относят за-
болевания желудочно-кишечного тракта, печени,
желчного пузыря, эндокринной, нервной и сер-
дечно-сосудистой систем, нарушения обмена ве-
ществ, гемопоэза, генетические дефекты и др. За-
болевания внутренних органов, эндокринной и
нервной систем приводят к возникновению ин-
токсикации и обменным сдвигам, изменению
висцерокутанных, висцеровегетокортикокутан-
ных связей, что способствует формированию ал-
лергического состояния, играющего важную роль
в патогенезе ряда дерматозов.
Существует множество причин, которые мо-
гут приводить к развитию различных заболева-
ний кожи. В ряде случаев кожные поражения
вызывают так называемые безусловные (обли-
гатные) раздражители, которые в 100 % приво-
дят к определенной реакции со стороны кожных
покровов и являются местной ответной реакцией
организма на повреждающее действие раздражи-
теля. Такое действие оказывают концентрирован-
ные растворы кислот и щелочей, вызывающие
химические ожоги, большие дозы лучистой энер-
гии (рентгеновские лучи, ультрафиолетовое из-
лучение), термические факторы (ожоги, обморо-
жения) и др.
Следует подчеркнуть, что в большинстве слу-
чаев то или иное кожное поражение возникает
при сочетании нескольких причин. Так, напри-
мер, стафилодермии развиваются при наличии ви-
рулентных штаммов стафилококков, нарушении
защитных свойств кожи (изменение водно-ли-
пидной мантии кожи, микротравмы) на фоне
сниженной иммуно-биологической реактивности
организма при гиповитаминозах, диабете, забо-
леваниях внутренних органов и т. д.
Становится понятным, что деление этиологи-
ческих факторов при дерматозах на экзогенные
и эндогенные в ряде случаев следует признать
в достаточной степени условным.
Необходимо также учитывать, что состояние
организма человека часто зависит от социально-
бытовых условий его жизни и труда. Их ухуд-
шение позволяет говорить о болезни как о со-
циальном явлении.
В последние годы в дерматологии возникла
проблема, связанная с неблагоприятным дей-
ствием экологической среды на организм чело-
87
4 Зак. 1357
88 Общая дерматология
века и среду его обитания. Изменение эколо-
гической обстановки провоцирует появление
классических и атипичных дерматозов, как пра-
вило, токсикоаллергического характера, что зат-
рудняет диагностику, лечение и профилактику
этих заболеваний. Врачу дерматологу в своей
практической деятельности необходимо учиты-
вать возможное действие этих антропогенных
факторов (химические производства, радиаци-
онная обстановка и др.).
Рассмотрим подробнее экзогенные этиологичес-
кие факторы кожных болезней.
1. Среди физических экзогенных этиологи-
ческих факторов различают механические, тер-
мические и связанные с действием лучистой
энергии. Длительное давление, трение, ушибы
и другие механические воздействия могут выз-
вать воспалительный процесс различной интен-
сивности, образование пузырей, геморрагии,
омозолелости. В результате воздействия терми-
ческих факторов (высокие и низкие темпера-
туры) могут развиться ознобления, отмороже-
ния, ожоги и т. п. Ультрафиолетовые и
рентгеновские лучи, ионизирующая радиация
могут при воздействии на кожу приводить к
дерматитам различной степени тяжести вплоть
до некротических изменений.
2. Химические экзогенные факторы, способ-
ные вызвать те или иные изменения кожи
(эритема, пузырьки, гнойнички, трещины и
др.), весьма многообразны. Они встречаются на
производствах (профессиональные химические
раздражители), в быту (например, моющие
средства), при назначении наружных дермато-
логических лекарственных средств. В патоге-
незе развития дерматозов, вызванных этими
агентами, важная роль принадлежит степени
сенсибилизации организма, его аллергическо-
му состоянию, что вносит определенные труд-
ности в объяснение механизмов развития дер-
матоза у конкретного пациента.
3. Инфекционные агенты и, в частности, па-
тогенные микроорганизмы служат причиной
развития гнойничковых заболеваний, туберку-
леза кожи, лепры; простейшие — лейшманиоза
и др., вирусы являются возбудителями просто-
го и опоясывающего герпесов, бородавок, ост-
роконечных кондилом, контагиозного моллюс-
ка, узелков доярок.
4. Патогенные грибы (низшие растительные па-
разиты) вызывают трихофитию, микроспорию,
фавус и другие дерматомикозы.
5. Животные паразиты (чесоточный клещ, ли-
чинки овода) проникают в кожу и проходят в ней
цикл развития, или своими укусами (блохи, вши,
клопы, комары, клещи) вызывают зуд, расчесы,
возникновение гнойничковых элементов.
Эндогенные этиологические факторы связыва-
ют с влиянием на кожу патологически функци-
онирующих внутренних и эндокринных органов,
нарушениями со стороны центральной и пери-
ферической нервной системы. Это влияние мо-
жет носить характер рефлекторного процесса,
осуществляться гуморальным или нейрогумо-
ральным путем, в виде метастазирования злока-
чественных опухолей в кожу, распространяться
«по продолжению» (per continuitatem), что отме-
чается при язвенном туберкулезе кожи и тубер-
кулезном поражении лимфатических узлов.
Ряд кожных заболеваний (экзема, нейродер-
мит, красный плоский лишай и др.) возника-
ют под влиянием острых или хронических
стрессовых ситуаций, т. е. в результате дисфун-
кции центральной нервной системы. Функци-
ональные изменения и срывы нервной систе-
мы в ряде случаев являются основным
пусковым фактором развития патологических
процессов в коже. Различные кожные прояв-
ления (перфорированные или трофические
язвы) могут наблюдаться также при органичес-
ких заболеваниях как центральной, так и пе-
риферической нервной системы (сирингомие-
лия, спинная сухотка, алкогольные невриты,
повреждения седалищного нерва и др.).
Различные эндокринопатии являются причиной
разнообразных патологических изменений. Так,
нарушение секреции гипофиза и половых желез
в период полового созревания является причиной
себореи, угревой сыпи, атрофических полос; при
гипофункции щитовидной железы наблюдается
возникновение микседемы (слизистый отек кожи),
при резком снижении функции коры надпочеч-
ников — развитие бронзовой окраски кожи (бо-
лезнь Аддисона) и т. п. Эндокринные нарушения
на фоне невротических расстройств (нейроэндо-
кринные дисфункции) создают условия, при ко-
торых развиваются хронические аллергодермато-
зы (нейродерматозы, экзема).
Возникновение и течение ряда заболеваний
находятся в зависимости от менструального цикла
(рецидивы герпеса), течения беременности (дер-
матозы беременных), развитие кератоза ладоней
и подошв в климактерическом периоде и др.
Нарушения углеводного, жирового, минераль-
ного и водного обменов играют важную роль в
патогенезе некоторых дерматозов. Так, напри-
мер, гипергликемия приводит к развитию фу-
рункулеза, кандидоза, кожного зуда, нарушение
холестеринового обмена способствует возникно-
вению ксантом.
Глава 5. Этиология, патогенез и диагностика кожных болезней
89
Гиповитаминозы могут служить причинами раз-
вития некоторых кожных болезней. При недостатке
в организме витамина С развивается кровоточивость
слизистых и кожные геморрагии (цинга), вследствие
недостатка витамина РР возникает пеллагра (дер-
матит, деменция, диарея). Гиповитаминоз А явля-
ется причиной нарушения ороговения (фринодер-
ма). Себорейный дерматит сопровождается
сниженным содержанием в организме витамина В2.
Установлена связь ряда кожных заболеваний с
фокальной инфекцией (хронический тонзиллит,
гайморит, кариозные зубы), играющей патогене-
тическую роль при таких заболеваниях, как кра-
пивница, многоформная экссудативная эритема,
псориаз, красная волчанка и др. Наличие очагов
хронической инфекции является причиной раз-
вития бактериальной аллергии, клеточного имму-
нодефицита и провоцируется различными фак-
торами (охлаждение, прием лекарственных
препаратов, инсоляции и др.).
Значительную группу кожной патологии состав-
ляют так называемые генодерматозы. Роль наслед-
ственности установлена при таких заболеваниях,
как ихтиозы, буллезный эпидермолиз, кератодер-
мии, атопический дерматит, псориаз и др. От на-
следственных, генетически обусловленных болез-
ней следует отличать врожденные заболевания
кожи, являющиеся следствием нарушения внутри-
утробного развития плода в результате инфекций,
интоксикации у беременных.
Кожа вовлекается в патологический процесс при
таких системных заболеваниях как красная волчан-
ка, склеродермия, Т и В — клеточные лимфомы.
При венозной недостаточности на нижних конеч-
ностях могут развиваться трофические язвы, гипо-
статические экземы, при облитерирующем эндар-
териите — некрозы, язвы, гангрена нижних
конечностей.
В патогенезе многих дерматозов существенная
роль принадлежит аллергической реактивности
организма в результате его сенсибилизации (по-
вышенной чувствительности) к различным веще-
ствам. Повторный контакт с тем или иным аллер-
геном (раздражителем) провоцирует появление
кожных сыпей вплоть до развития отека Квинке,
тяжелых токсидермий. Существует врожденная
повышенная восприимчивость к конкретному ве-
ществу или раздражителю. Такое состояние обо-
значают как идиосинкразия. Из наиболее часто
встречающихся аллергенов следует отметить пище-
вые аллергены, лекарственные средства, химичес-
кие вещества (хром, никель, лаки, краски и др.).
В патогенезе некоторых дерматозов опреде-
ленная роль принадлежит иммунным сдвигам.
Ряд авторов выделяют группу аутоиммунных
дерматозов (красная волчанка, склеродермия, ис-
тинная пузырчатка, дерматоз Дюринга), хотя при
дерматозах неаллергического генеза также выяв-
ляются нарушения клеточного и гуморального
иммунитета. При некоторых гематологических
заболеваниях (лейкоз, лимфогранулематоз, Т и
В-лимфомы, тромбоцитопеническая пурпура,
ДВС-синдром) выявляются характерные и не-
специфические поражения кожных покровов.
У ВИЧ-инфицированных нередко развивает-
ся саркома Капоши (преимущественно у гомо-
сексуалистов), тяжело протекают и плохо под-
даются лечению инфекционные (вирусные,
грибковые, бактериальные, паразитарные) и не-
которые другие дерматозы (себорейный дерма-
тит, псориаз, атопический дерматит и др.).
Разработка новых молекулярных, иммуноло-
гических, генетических, гибридомных методов
позволит продолжить исследования по расшиф-
ровке этиологии, патогенеза и, следовательно,
последующей успешной терапии ряда дермато-
зов установленной и невыясненной природы.
Общая
симптоматология
болезней кожи
Кожная сыпь состоит из отдельных составных
частей, которые называются морфологическими
элементами. Морфологические элементы явля-
ются азбукой дерматологии. Все элементы кож-
ной сыпи рассматриваются обычно в двух боль-
ших группах: первичные и вторичные. Первичные
элементы представлены высыпаниями, которые
появились первично в результате патологичес-
кого процесса в коже. Вторичные элементы яв-
ляются результатом эволюции первичных эле-
ментов сыпи вследствие их обратного развития.
Первичные
морфологические элементы
Различают 8 первичных морфологических эле-
ментов: пятно (macula), узелок (papula), бугорок
(tuberculum), узел (nodus), волдырь (urtica), пу-
зырек (vesicula), пузырь (bulla), гнойничок
(pustula) (рис. 5.1).
Пятно
Пятно (macula) — элемент, характеризующийся
очаговым изменением цвета кожи. Пятно не
90
Общая дерматология
возвышается, не имеет консистенции и по ис-
чезновении не оставляет следов на коже. В за-
висимости от характера пятен различают невос-
палительные и воспалительные пятна. По
происхождению пятна делятся на сосудистые, ге-
моррагические, пигментные и эритематозно-сква-
мозные.
Сосудистые пятна характеризуются расширени-
ем сосудов кожи. Они имеют различные оттенки
Бугорок
Рис. 5.1. Первичные морфологические
элементы кожной сыпи.
красного цвета и исчезают при надавливании.
Обычно сосудистые пятна воспалительного харак-
тера и зависят от временного расширения крове-
носных сосудов. По величине их подразделяют на
розеолы (величиной до ногтя) и эритемы (больше
ногтя). Островоспалительные пятна обычно сопро-
вождаются зудом. К розеолам относятся сифили-
тическая розеола и инфекционная розеола.
Гистологическая картина кожи в зоне розео-
лы характеризуется резким расширением и пере-
полнением кровью сосудов верхней трети дермы,
эндотелий которых чаще всего набухший, иног-
да с явлениями пролиферации. Вокруг сосудов
могут образовываться небольшие инфильтраты,
состоящие преимущественно из лимфоцитов. При
возникновении сифилитической розеолы к лим-
фоцитам примешиваются плазматические клетки.
Эпидермис, как правило, не изменен.
Эритема наблюдается при экземе, дерматите,
красной волчанке, лепре, дерматозе Дюринга,
экссудативной многоформной эритеме. Имеются
невоспалительные сосудистые пятна, зависящие
от стойкого расширения сосудов. К ним отно-
сятся сосудистые родимые пятна и телеангиэк-
тазии.
Гистологическая картина кожи при эритеме
характеризуется более выраженными изменени-
ями воспалительного характера. В зависимости
от этиологического фактора, кроме резкого рас-
ширения сосудов, наблюдается образование пе-
риваскулярного инфильтрата, состав которого
зависит от характера заболевания. Общими при-
знаками эритемы при экземе, атопическом дер-
матите, многоформной экссудативной эритеме
являются резкое расширение сосудов не только
сосочкового слоя дермы, но и более глубоких ее
отделов и выраженная отечность, особенно верх-
ней трети дермы. Инфильтраты, как правило,
небольшие, периваскулярные, состоят в основ-
ном из лимфоцитов, среди которых могут наблю-
даться нейтрофильные лейкоциты и эозинофи-
лы (экзема). Волокнистые субстанции в местах
отека разрыхлены, но их структура и гистохи-
мические свойства не изменены. Эпидермис
чаще утолщен, иногда в состоянии выраженно-
го акантоза (атопический дерматит). При нали-
чии выраженного экссудативного компонента в
эпидермисе может наблюдаться межклеточный
отек, иногда с образованием спонгиотических пу-
зырей (экзема, токсикодермия).
Геморрагические пятна являются невоспали-
тельными и обусловлены выхождением эритро-
цитов из сосудов в ткань. При надавливании не
исчезают, при обратном развитии не шелушат-
ся, но меняют цвет. Вначале они красного цве-
Глава 5. Этиология, патогенез и диагностика кожных болезней
91
та, затем происходит общеизвестное цветение си-
няка. По величине различают петехии (точечные
кровоизлияния), пурпуру (кровоизлияния вели-
чиной до ногтя), экхимозы (кровоизлияния раз-
меров больше ногтя), линейные кровоизлияния и
кровоподтеки.
Гистологическая картина в зоне геморрагичес-
ких пятен характеризуется резким расширени-
ем сосудов, иногда стазами с периваскулярны-
ми кровоизлияниями различной выраженности.
Обычно наблюдается набухание эндотелия сосу-
дистых стенок, местами с очагами деструкции,
в связи с чем в дерме обнаруживают небольшие
инфильтраты, состоящие из полиморфноядерных
лимфоцитов, а также дистрофические изменения
волокнистых субстанций. При экхимозах кро-
воизлияния более обширные, иногда диффузные.
Пигментные пятна, как правило, невоспали-
тельные и возникают в связи с расстройством
пигментации кожи. Пятна, при которых имеет-
ся избыток пигмента, называются гиперпигмен-
тированными. Гиперпигментированные пятна мо-
гут быть врожденными (пигментный невус),
возникать самопроизвольно (веснушки, хлоазма,
пятна при болезни Реклингхаузена), после приема
некоторых лекарственных препаратов (акрихин,
препараты висмута, серебра). Самые мелкие ги-
перпигментированные пятна — веснушки, лока-
лизующиеся на лице.
Гистологическая картина при веснушках ха-
рактеризуется увеличением в пигментном пят-
не количества меланоцитов, меланосомы кото-
рых по структуре длиннее и тоньше обычных.
Отмечается увеличение содержания меланина в
клетках эпидермиса.
Расстройство пигментации, при котором на
гиперпигментированном фоне имеются депиг-
ментированные пятна, называется витилиго.
Депигментированные пятна могут быть врожден-
ными (альбинизм), развиваться на очагах скле-
родермии, псориаза, атопического дерматита.
Депигментированные пятна бывают белыми,
цвета слоновой кости, розовато-белыми. Очаги
поражения, лишенные пигмента, могут быть
диффузными (альбинизм, распространенная фор-
ма витилиго).
Гистологическая картина при витилиго харак-
теризуется следующими изменениями: эпидер-
мис обычной толщины или слегка истончен, от-
ростки его сглажены. Роговой слой чаще всего
утолщен; зернистый слой состоит из одного ряда
клеток; шиповатый слой без особых изменений;
клетки базального слоя не содержат пигмента.
Меланоциты в депигментированной коже встре-
чаются часто, рядом с ними — большое количе-
ство клеток Лангерханса. В дерме могут наблю-
даться набухание и гомогенизация некоторых
коллагеновых волокон, эластическая сеть без
особых изменений. Сосуды расширены, стенки
их набухшие, вокруг них обнаруживаются гнез-
дные скопления фибробластов, гистиоцитов и
лаброцитов. Волосяные фолликулы на участках
депигментации несколько атрофичны, устья их
расширены, заполнены роговыми пробками.
Эритематозно-сквамозные пятна — это пятна
с выраженными явлениями шелушения. При
этом шелушение имеется не только при обрат-
ном развитии, но и в течение всего существова-
ния элемента. Наблюдаются при себорейной эк-
земе, грибковых заболеваниях (трихофития,
микроспория, эпидермофития, руброфития,
эритразма, отрубевидный лишай).
Папула
Папула, узелок (papula) — это бесполостной ком-
пактный элемент, несколько возвышающийся
над уровнем кожи. Возникает вследствие ин-
фильтрации в сосочковом слое дермы (дермаль-
ная папула) или вследствие разрастания эпидер-
миса (эпидермальная папула), или в результате
патологического процесса и в эпидермисе, и в
дерме (смешанная эпидермо-дермальная папула).
По происхождению различают невоспалителъные
и воспалительные папулы. Папулы эпидермо-
дермальные и дермальные — воспалительные, а
эпидермальные папулы — невоспалительные. К
эпидермальным папулам относятся мозоль, бо-
родавки, заразительный моллюск, гиперкератоз
ладоней и подошв, ихтиоз. К эпидермально-дер-
мальным и дермальным папулам относятся эле-
менты при псориазе, красном плоском лишае,
экземе, дерматите, почесухе, атопическом дерма-
тите, ограниченном нейродермите, дерматозе
Дюринга, дискоидной красной волчанке, экссу-
дативной многоформной эритеме, лихеноидном
туберкулезе кожи, лепре, сифилитические папу-
лы и остроконечные кондиломы.
Гистологическая картина. Эпидермальные па-
пулы могут образовываться в результате эпи-
телиальной гиперплазии (бородавки, контаги-
озный моллюск, псориаз, эпителиома), а также
межклеточного и внутриклеточного отека (эк-
зема). В основе эпидермальных папул лежит
пролиферация клеток эпидермиса (псориаз) или
увеличение их объема вследствие отека и дист-
рофических изменений (экзема). При папулах
дермального происхождения (сифилис) наблю-
дается воспалительная инфильтрация дермы.
Инфильтраты в папулезных элементах имеют
периваскулярную локализацию, но иногда ели-
9 2 Общая дерматология
ваются и располагаются более диффузно. Со-
став инфильтрата может быть различным в за-
висимости от этиологического фактора, но в ос-
новном он содержит лимфоидные клетки,
гистиоциты, фибробласты. Эпидермально-дер-
мальная папула (красный плоский лишай) ха-
рактеризуется гиперпластическими процессами
в эпидермисе, гиперкератозом, а также сочета-
ется с выраженной инфильтрацией в дерме. В
этом случае инфильтрат состоит в основном из
лимфоидных элементов.
Папулы различаются по величине: милиарные
(мелкоточечные, величиной с просяное зерно),
лентикулярные (плоские, величиной с горошину
или чечевицу), нумулярные (монетовидные),
бляшки (достигают величины ладони человека).
По форме различают следующие виды папул: ко-
нусовидная, полусферическая, полу шаровидная,
плоская, плоская с вдавлением, плоская с западе-
нием. При обратном развитии папула, как пра-
вило, не изъязвляется, не оставляет после себя
рубцов, а рассасывается, шелушится; на повер-
хности кожи остается временная пигментация
или гиперпигментация.
В ряде случаев на папулах располагаются дру-
гие морфологические элементы: пузырьки (па-
пуловезикулы), пустулы (папулопустулы). При
папулонекротическом туберкулезе кожи папулы
сочетаются с некротическим процессом, а при
некоторых формах васкулитов кожи сопровож-
даются геморрагическими изменениями с изъяз-
влением. В подобных случаях на месте папул
может образоваться рубец. При локализации
папул в складках и местах трения отмечается их
избыточный рост и на поверхности папул раз-
виваются вегетации. Вегетирующие папулы сли-
ваются с образованием кондилом (широкие кон-
диломы при сифилисе).
Бугорок
Бугорок (tuberculum) — бесполостной компакт-
ный неостровоспалительного характера элемент,
отчетливо возвышающийся над уровнем кожи.
Имеет красный цвет с разными оттенками. Ин-
фильтрат при этом скапливается в сосочковом
и ретикулярном слоях дермы, имеет строение ин-
фекционной гранулёмы. Величина колеблется от
просяного зерна до грецкого ореха. В процессе
эволюции бугорок подвергается центральному
некрозу, изъязвлению и завершается образова-
нием рубца. В случаях рассасывания инфильтрата
без изъязвлений на месте бугорка остается руб-
цовая атрофия кожи. Бугорки наблюдаются при
туберкулезной волчанке, бородавчатом туберку-
лезе кожи, язвенном туберкулезе кожи, папуло-
некротическом туберкулезе, бугорковом сифи-
лиде, лейшманиозе, лепре.
Для гистологической картины бугорка харак-
терно наличие гранулем, состоящих из эпители-
оидных клеток с примесью гигантских. Тубер-
кулезной инфекции свойственно образование
вокруг эпителиоидно-клеточной гранулемы лим-
фоцитарного вала и казеозного некроза в цент-
ре бугорка с последующим размягчением и за-
мещением его соединительной тканью. Для
сифилитической гранулемы характерно наличие
большого количества плазматических клеток и
фибробластических элементов, в результате чего
она склонна к рубцеванию. При лепре бугорок
сходен с туберкулезным, однако некроз в нем
никогда не развивается.
Узел
Узел (nodus) — бесполостное воспалительное
образование крупнее бугорка (достигает величи-
ны куриного яйца). Инфильтрат при этом скап-
ливается глубже, главным образом в подкожно-
жировой клетчатке. В процессе эволюции он
обычно изъязвляется (язва будет больше и глуб-
же, чем при разрешении бугорка) и завершает-
ся, как правило, рубцом. В некоторых случаях
узел может рассасываться без изъязвлений и тогда
на месте типичного рубца остается рубцовая ат-
рофия. Узлы наблюдаются при третичном сифи-
лисе, туберкулезе, лепре, кожном лейшманиозе,
индуративной эритеме Базена.
По гистологической структуре узел можно от-
нести к инфекционным гранулемам. Узел при
сифилисе (гумма) представляет собой очаг коагу-
ляционного некроза, окруженный эпителиоидны-
ми и гигантскими клетками (клетки Лангханса-
Пирогова) или гигантскими клетками типа
инородных тел, а также лимфоидными и плазма-
тическими клетками. В грануляционной ткани
имеются сосуды, стенки которых пронизаны клет-
ками воспалительного инфильтрата. Узел при
колликвативном туберкулезе характеризуется об-
разованием неспецифического абсцесса в центре
с последующим расплавлением всех слоев кожи
вплоть до эпидермиса. По периферии очага не-
кроза образуются бугорки с резко выраженными
казеозным некрозом и обширной лимфоцитарной
инфильтрацией. При узловатой эритеме в подкож-
ной жировой клетчатке наблюдается диффузная
инфильтрация лимфоидными клетками с приме-
сью нейтрофильных лейкоцитов, находящихся в
фиброзных перегородках между дольками жиро-
вой ткани и отдельными жировыми клетками. В
инфильтрате обнаруживают небольшое количе-
ство гистиоцитов и эозинофилов, но плазмати-
Глава 5. Этиология, патогенез и диагностика кожных болезней
93
ческих клеток нет. Абсцессы и некробиоз не на-
блюдаются. В сосудах, главным образом капил-
лярах и сосудах среднего калибра, происходят
значительные изменения в виде альтерации их
стенок и пролиферации эндотелия. Среди лим-
фоидных клеток встречаются гигантские клетки
типа инородных тел. Имеются также маленькие
узелки, состоящие из гистиоцитов, располагаю-
щиеся в виде частокола вокруг центральной тре-
щины. Гистологическая структура узла при леп-
ре (лепромы) характеризуется наличием большого
количества гистиоцитов и лепрозных клеток,
среди которых имеются лимфоциты и плазмати-
ческие клетки, а в старых элементах — фиброб-
ласты. В дальнейшем могут произойти распад
инфильтрата и отторжение омертвевших участ-
ков. Для узлов при глубоких микозах характерно
хроническое гнойное воспаление глубоких отде-
лов дермы на границе с подкожной жировой
клетчаткой. Инфильтрат состоит из лимфоидных
и плазматических клеток. В некоторых случаях
среди них могут наблюдаться группы эпителио-
идных клеток, иногда с гигантскими клетками
типа инородных тел или белыми отростчатыми
эпидермоцитами.
Волдырь
Волдырь (urtica) — бесполостной островоспали-
тельный, возвышающийся над уровнем кожи
элемент, возникающий в результате острого ог-
раниченного отека сосочкового слоя дермы.
Имеет цвет от белого до розово-красного. Вели-
чина — от горошины до ладони и больше. Очер-
тания округлые или неправильные. Различают
следующие виды волдырей: нумулярные, ленти-
кулярные, полосовидные, гигантские, гирляндооб-
разные. Волдыри сопровождаются зудом, склон-
ны к периферическому росту, особенно при
расчесах. Исчезают быстро (в течение несколь-
ких часов), не оставляя после себя следов. Вол-
дыри наблюдаются при крапивнице, дерматозе
Дюринга, токсйдермии, детской почесухе, фле-
ботодермии.
Гистологическая картина характеризуется вы-
раженным отеком сосочкового и подсосочково-
го отделов дермы, сопровождающимся резким
расширением сосудов, особенно лимфатических.
Воспалительных инфильтратов обычно не обна-
руживают, но иногда они наблюдаются в неболь-
шом количестве. В эпидермисе может отмечаться
умеренный спонгиоз.
Пузырек
Пузырек (vesicula) — полостной островоспали-
тельный элемент с серозным прозрачным содер-
жимым, развивающийся в результате скопления
экссудата в эпидермисе. Пузырьки могут быть
внутриэпидермальными (располагаются между
слоями эпидермиса), субэпидермалъными (распо-
лагаются под эпидермисом) и юнктиональными
(пограничными, на границе эпидермиса с дер-
мой). Величина от булавочной головки до горо-
шины. Форма полушаровидная. Пузырьки рас-
полагаются изолированно (одиночные) или
группами (герпетиформные). В процессе эволю-
ции пузырьки вскрываются с образованием эро-
зии или ссыхаются в корочку, по отпадении ко-
торой может остаться пигментно-гиперемическое
пятно, которое в дальнейшем исчезает, не остав-
ляя следа. Пузырьки наблюдаются при простом
герпесе, опоясывающем лишае, экземе, герпети-
формном дерматозе Дюринга.
Гистологическая картина характеризуется экс-
судативными изменениями, сопровождающими-
ся спонгиозом и образованием однокамерных
пузырьков (буллезный дерматит, экзема) или
многополостных (герпес). При появлении одно-
камерных пузырьков спонгиоз может быть вы-
ражен в различных отделах эпидермиса. Затем
вокруг пузырька происходит пролиферация эпи-
дермиса, и пузырек, подымаясь кверху, локали-
зуется уже в верхних отделах росткового или под
роговым слоем. Иногда наблюдается акантоз, в
роговом слое — различное количество паракера-
тотических клеток и корки. Воспалительный
инфильтрат в дерме локализуется в основном
вокруг расширенных сосудов и состоит из лим-
фоидных клеток, иногда с примесью нейтро-
фильных лейкоцитов и эозинофилов.
Пузырь
Пузырь (bulla) — воспалительный полостной
элемент с серозным (реже геморрагическим) со-
держимым, более крупных размеров, чем пузы-
рек (до куриного яйца). Пузыри могут быть од-
нокамерные и многокамерные. Различают
эпидермальные, дермальные, пограничные пузыри.
При вульгарной пузырчатке акантолитические
пузыри неостровоспалительного характера. Пу-
зыри наблюдаются при дерматите, эпидермофи-
тии стоп, дерматозе Дюринга, пузырчатке, экс-
судативной многоформной эритеме.
Гистологическое исследование имеет важное
значение в дифференциальной диагностике пу-
зырных дерматозов. Пузырь может находиться
внутри- или субэпидермально. В основе разви-
тия субкорнеальных, спонгиотических пузырей
лежит межклеточный отек (спонгиоз), сопро-
вождающийся инфильтрацией мононуклеарны-
ми элементами, мигрирующими в эпидермис из
9 4 Общая дерматология
верхних отделов дермы, и растяжением десмо-
сом, которые затем исчезают. Акантолитические
пузыри образуются в результате растворения
межклеточной цементирующей субстанции, что
приводит к нарушению связей между отдель-
ными эпидермальными клетками. Акантолити-
ческие пузыри возникают преимущественно в
супрабазальной области (при вульгарной пузыр-
чатке, болезни Дарье) и в субкорнеальной (при
листовидной пузырчатке). Пузыри, образующи-
еся в результате деструктивных изменений ба-
зальных клеток эпидермиса, имеют субэпидер-
мальную локализацию, так же как и пузыри,
развивающиеся вследствие деструктивных из-
менений базальной мембраны. При гистологи-
ческом исследовании таких пузырей отмечается
истончение или отсутствие ШИК-положитель-
ной базальной мембраны, а при электронной
микроскопии — повреждение полудесмосом,
базальной пластинки или крепящих фибрилл.
Изменения в дерме при указанных разновид-
ностях пузырей зависят от заболевания, при ко-
тором они развились. Главную роль в образо-
вании внутриэпидермального пузыря играют
предварительное дегенеративное повреждение
эпителиальных клеток (баллонирующая и ва-
куольная дистрофия), утрата связей между
ними, развитие спонгиоза и акантолиза. Акан-
толиз и основанный на этом явлении феномен
Никольского (отслаивание верхних слоев эпи-
дермиса при механическом воздействии) явля-
ются критериями дифференциальной диагно-
стики истинной пузырчатки и других
пузырных дерматозов. При подэпидермальных
пузырях, образующихся вследствие отека со-
сочкового слоя дермы и нарушения структу-
ры базальной мембраны, скопившийся экссу-
дат отслаивает эпидермис от сосочкового слоя
дермы (экссудативная многоформная эритема,
буллезная токсикодермия, буллезная форма
рожи, пеллагра).
Гнойничок
Гнойничок, пустула (pustula) — полостной вос-
палительный элемент с серозно-гнойным или
гнойным экссудатом. В зависимости от локали-
зации различают поверхностные (эпидермальные)
и глубокие (дермальные) пустулы.
Поверхностные пустулы, при которых процесс
локализуется в эпидермисе, называются импети-
го (при этом инфильтрации нет). Глубокие пу-
стулы, когда гнойный процесс располагается в
дерме, называются эктимой (при этом характе-
рен инфильтрат, при расплавлении которого и
образуется гнойный экссудат. Иногда эктима по-
крывается толстой слоистой коркой, которая на-
поминает раковину улитки и носит название ру-
пия. Импетиго, эктима и рупия относятся к не-
фолликулярным пустулам.
Гистологическая картина при импетиго харак-
теризуется наличием пустулы под роговым сло-
ем, заполненной фибрином и полиморфноядер-
ными лейкоцитами с небольшим содержанием
лимфоцитов. Иногда в ней можно видеть акан-
толитические клетки, образующиеся в результате
воздействия протеолитических ферментов лей-
коцитов. В ростковом слое эпидермиса, под пу-
стулой, выявляют спонгиоз и нейтрофильные
лейкоциты. В верхнем слое дермы расположен
умеренный воспалительный инфильтрат, состо-
ящий из нейтрофилов и лимфоцитов. Гистоло-
гическая картина при эктиме, в отличие от им-
петиго, характеризуется распространением
патологического процесса с эпидермиса на дер-
му с образованием язвенного поражения. Эпи-
дермис по краям язвы, как правило, в состоя-
нии акантоза, соединительная ткань подлежащих
участков дермы может быть некротизирована и
инфильтрирована лейкоцитами. Капилляры в
верхней трети дермы расширены как по пери-
ферии, так и в центре очага. К разновидности
гнойничка по гистологической картине можно
отнести спонгиоформную пустулу Когоя, представ-
ляющую собой многокамерную пустулу, лока-
лизующуюся в верхней части эпидермиса и ха-
рактеризующуюся наличием мелких полостей,
заполненных нейтрофилами, среди которых на-
ходятся деструктивно измененные эпителиаль-
ные клетки в виде остатков оболочек. Эпидер-
мис обычно в состоянии акантоза, местами в нем
выражен межклеточный отек. В дерме отмеча-
ются различные изменения в зависимости от за-
болевания, при котором встречается пустула
Когоя, но не всегда наблюдаются отек, особен-
но сосочкового слоя, расширение сосудов и ин-
фильтрация различной степени выраженности
(пустулезный псориаз, болезнь Рейтера, акродер-
матит Аллопо, герпетиформное импетиго, гоно-
рейный кератоз и кандидоз).
Различают также пустулы фолликулярные,
которые располагаются в области волосяных
фолликулов и называются фолликулитами. Они
могут быть поверхностными, когда нагноение
располагается только в поверхностной части
фолликула, и глубокими, когда процесс захва-
тывает весь или почти весь фолликул. К повер-
хностным фолликулитам относятся обычный
фолликулит и сикоз. К глубоким фолликули-
там относятся фурункул, карбункул, гидраде-
нит. Различают также остиофолликулиты, при
Глава 5. Этиология, патогенез и диагностика кожных болезней
95
которых пустула локализуется в устье волося-
ного фолликула (в центре она пронизана воло-
сом).
При расположении гнойничка вокруг сальных
желез образуется акне (угорь). Угри могут быть
поверхностными и глубокими.
Гнойное содержимое пустул ссыхается в кор-
ки, которые затем отторгаются, остатки инфиль-
трата рассасываются. На месте поверхностных
пустул (импетиго, поверхностные фолликулиты,
поверхностные акне) после отторжения корок
остается быстро исчезающее пигментно-гипере-
мическое пятно, т. е. они разрешаются бесслед-
но. На месте глубоких пустул (эктима, рупия,
глубокие фолликулиты, глубокие акне) остаются
рубцы.
В число первичных элементов А. И. Картамы-
шев (1963) и Л. И. Богданович (1988), кроме
перечисленных, включали опухоль (невоспали-
тельные образования в виде узлов, бугорков и
узелков, величиной от горошины до куриного
яйца, залегающие в дерме и подкожной клетчат-
ке). В качестве первичного элемента К. Н. Су-
ворова и В. Т. Куклин (1996) описывают серо-
папулы (папулы или волдыри с образованием на
их поверхности пузырька).
Вторичные
морфологические элементы
Вторичные морфологические элементы возника-
ют в процессе обратного развития первичных
элементов, т. е. вторично. Различают 10 вторич-
ных морфологических элементов: дисхромия
(dischromia), чешуйка (squama), корка (crusta),
эрозия (erosio), экскориация (excoriatio), трещина
(fissura, rhagas), язва (ulcus), рубец (cicatrix), ли-
хенификация (Uchenificatio), вегетация (vegetatio)
(рис. 5.2).
Дисхромия
Дисхромия (dischromia) — нарушение пигмен-
тации кожи, возникающее вследствие разреше-
ния морфологических элементов сыпи. К вто-
ричным дисхромиям относятся: вторичная
лейкодерма, вторичная гиперпигментация, ге-
мосидериновая пигментация. Вторичные гипер-
пигментированные и депигментированные пят-
на остаются после исчезновения сосудистых
пятен и папул. После бугорков и гумм остает-
ся пигментация рубца либо пигментация вок-
руг рубца, после пузырьков и пустул — слегка
пигментированное пятно.
Гистологическая картина при пигментации
характеризуется увеличением количества пигмен-
та меланина в клетках базального слоя эпидер-
миса, толщина последнего в зависимости от ха-
рактера бывшего элемента может быть различ-
ной. В дерме изменений практически не
Экскориация
Рубец
Рис. 5.2. Вторичные морфологические
элементы кожной сыпи.
Лихенификация
96
Общая дерматология
происходит, за исключением образования руб-
цов при язвенном поражении кожи.
Чешуйка
Чешуйка (squama) — это отторгающиеся клет-
ки рогового слоя эпидермиса. Видимое на глаз
отторжение чешуек называется шелушением.
Цвет чешуек бывает от блестяще-белого до бу-
ровато-желтого и даже серо-черного цвета в за-
висимости от наличия пигмента, кожного сала,
пыли, пота. Различают отрубевидное мелкопла-
стинчатое шелушение (псориаз, отрубевидный
лишай) и крупнопластинчатое (дерматит, токси-
кодермия, скарлатина). Чешуйки образуются на
месте первичных морфологических элементов
сыпи воспалительного характера (пятна, узелки,
бугорки) или являются основным симптомом за-
болевания (ихтиоз).
Гистологическая картина при шелушении
представлена дистрофическими изменениями на
некоторых участках эпидермиса и сопровожда-
ется нарушением кератинизации в виде гипер-
кератоза и паракератоза, а также воспалительны-
ми изменениями в дерме. После снижения
воспаления начинается регенераторный процесс,
в результате которого вновь образованные пла-
сты эпидермиса отодвигают кверху некротизи-
рованные и паракератотические участки, кото-
рые в дальнейшем отторгаются.
Корка
Корка (crusta) образуется в результате ссыхания
содержимого пузырьков и гнойничков, а также
отделяемого язв после бугорков, гумм и злока-
чественных опухолей.
Различают серозные корки (состоят в ос-
новном из фибрина, среди волокон которого
находится небольшое количество лейкоцитов
и деструктивно измененных клеток эпидер-
миса), гнойные (пронизаны большим количе-
ством нейтрофильных лейкоцитов) и кровя-
нистые (находится большое количество
гемолизированных эритроцитов среди масс
фибрина, лейкоцитов и распадающихся эпи-
дермальных клеток).
Корки могут быть тонкими (стрептококковое
импетиго), плоскими, толстыми (сифилитичес-
кая рупия), слоистыми (вульгарное стрептоста-
филококковое импетиго), плотными (вульгарная
эктима, рупиоидный псориаз, гангренозная пи-
одермия), рыхлыми.
Эрозия
Эрозия (erosio) — это дефект кожи в пределах
эпидермиса, который образуется в> результате
обратного развития пузырьков, пузырей, поверх-
ностных пустул.
При гистологической картине дно эрозии
обычно выстлано эпидермисом или частично
сосочковым слоем дермы. Дефект эпидермиса
покрыт фибрином, в петлях которого находят-
ся нейтрофильные лейкоциты и деструктивно из-
мененные клетки эпидермиса. Вокруг эрозии
выражены межклеточный отек и экзоцитоз,
иногда акантоз.
Эрозии повторяют величину и форму пред-
шествовавшего первичного элемента. Эрозии
могут появляться также первично во время
расчесов (экскориаций) при механических раз-
дражениях эпидермиса, а также вследствие
мацерации и трения соприкасающихся повер-
хностей эпидермиса (внутренняя поверхность
бедер, пахово-бедренные складки). Эрозии
возникают на поверхности воспалительных
папул (эрозивные папулы при сифилисе) или
на фоне ограниченного инфильтрата (эрозив-
ный твердый шанкр). Дно эрозии обычно
ярко-красного цвета, иногда покрыто сероз-
ным отделяемым. При заживлении эрозии не
оставляют рубцов.
Экскориация
Экскориация (excoriatio) или ссадина возника-
ет в результате механического повреждения и
нарушения целостности кожного покрова
вследствие расчесов. Различают динейные, по-
лосовидные и округлые экскориации. Экско-
риации могут быть поверхностными (с наруше-
нием целостности эпидермиса и сосочкового
слоя дермы) и глубокими (с проникновением в
глубокие отделы дермы).
Гистологическая картина характеризуется при-
знаками основного заболевания, а также нару-
шением целости эпидермиса с образованием эро-
зий, покрытых толстой коркой, состоящей из
фибрина и эритроцитов. На подлежащем участ-
ке дермы отмечаются расширение сосудов и вос-
палительная инфильтрация различной интенсив-
ности.
Трешина
Трещина (fissura, rhagas) — это линейный де-
фект, возникающий вследствие потери эластич-
ности и инфильтрации участка кожи. Трещи-
ны образуются в местах естественных складок
и в участках растяжения (углы рта, область зад-
него прохода, область пяток). Трещины сопро-
вождаются болезненностью. В зависимости от
глубины трещин различают поверхностные
(эпидермальные), которые после заживления не
Глава 5, Этиология, патогенез и диагностика кожных болезней
97
оставляют следов, и глубокие (дермальные) тре-
щины, после заживления которых остаются руб-
цы. Поверхностные трещины возникают при
повышенной сухости кожи (ксероз), тилотичес-
кой (роговой) экземе кистей и стоп, эпидермо-
фитии стоп, опрелости, заеде. Глубокие трещи-
ны возникают при пеллагре, кератодермиях.
Примером глубоких трещин являются белова-
тые радиальные рубцы Робинсона-Фурнье при
врожденном сифилисе.
Язва
Язва (ulcus) — это глубокий дефект ткани в
пределах собственно кожи, подкожной клетчат-
ки и глубжележащих тканей. Язвы развивают-
ся в результате распада первичных инфильтра-
тивных элементов в глубоких отделах дермы —
бугорков, гумм, узелков, глубоких пустул, зло-
качественных опухолей, некроза при трофичес-
ких и микроциркуляторных нарушениях. Язвы,
как правило, заживают рубцом.
Гистологическая картина дна и краев язвы пред-
ставлена наслоениями некротических масс различ-
ной толщины, отделенными лейкоцитарным валом.
Дно язвы состоит из юной грануляционной тка-
ни, богатой сосудами и содержащей большое ко-
личество клеточных элементов в различных ста-
диях зрелости. При хронических гнойных
воспалительных процессах среди этих элементов
находится большое количество нейтрофильных
лейкоцитов. При различных патологических про-
цессах на дне язвы можно обнаружить опухоле-
вые элементы, инфекционные гранулемы, имею-
щие большую диагностическую ценность.
Язвы имеют различную величину, форму и
глубину. Остро развивающиеся язвы имеют не-
большие размеры. Язвы при хронических па-
тологических процессах (варикозные язвы,
гуммы, хроническая язвенная пиодермия) от-
личаются значительными размерами. Язвы
имеют круглую, резко отграниченную форму
(язвенный твердый шанкр, изъязвившиеся
гуммы, некоторые типы базалиом кожи) или
фестончатые очертания (варикозные язвы, яз-
венные формы туберкулезной волчанки). Края
язвы бывают подрытыми (колликвативный ту-
беркулез кожи, некоторые формы пиодермии,
глубокие микозы), отвесными (сифилитичес-
кая гумма), блюдцеобразными (язвенный твер-
дый шанкр), мягкими (колликвативный тубер-
кулез кожи), плотными (раковые язвы,
варикозные язвы), валикообразными (плоско-
клеточный рак, изъязвившаяся кератоаканто-
ма, распадающаяся опухоль грибовидного ми-
коза). Дно язвы может быть ровным, гладким
(твердый шанкр, мягкий шанкр), неровным,
извилистым (колликвативный туберкулез, кар-
бункул), кратероподобным (сифилитическая
гумма).
Рубец
Рубец (cicatrix) образуется в результате зажив-
ления язв и представляет собой грубоволокнис-
тые соединительнотканные разрастания с разру-
шением дермы и нередко подлежащих тканей.
Рубцы развиваются на месте ожогов, изъязвле-
ний, глубоких пустул, бугорков, узлов. Рубцы
могут формироваться сухим путем, без предше-
ствующего изъязвления, в результате замещения
соединительной тканью (туберкулез кожи, леп-
ра, сифилис). Свежие рубцы имеют розово-крас-
ную окраску, старые рубцы могут быть гипер-
пигментированы или депигментированы.
Различают плоские рубцы (расположены на од-
ном уровне с окружающей кожей), гипертрофи-
ческие или келлоидные (возвышаются над поверх-
ностью окружающей кожи), атрофические
(расположены ниже уровня окружающей кожи).
Гистологическая картина рубца характеризу-
ется атрофией или некоторым утолщением эпи-
дермиса. В дерме обнаруживают разрастание во-
локнистой соединительной ткани, коллагеновые
пучки которой грубые и располагаются беспо-
рядочно. Выявляют участки гомогенизации
коллагеновых волокон, иногда — отложения ги-
алина. Клеточных элементов очень мало, они
представлены в основном фиброцитами. В об-
ласти рубца, как правило, отсутствуют придат-
ки кожи: волосяные фолликулы, сальные и по-
товые железы. При рубцовой атрофии
развивается нежная соединительная ткань в
малом количестве. Поверхность пораженной
кожи при этом лишена характерного для эпи-
дермиса рельефа.
Величина, форма, локализация, цвет и глубина
рубцов имеют важное дифференциально-диагно-
стическое значение. При гуммозном сифилисе
рубцы гладкие, овальной, округлой или звезд-
чатой формы, глубоко втянутые. Бугорковые
сифилиды оставляют после себя пестрые моза-
ичные рубцы с фестончатыми очертаниями.
После колликвативного туберкулеза и конгло-
батных акне рубцы неправильной формы, изъе-
денные, поверхность их перекрыта перемычка-
ми-мостиками. На месте папулонекротического
туберкулеза кожи остаются штампованные по-
верхностные рубцы. Рубцы могут быть симпто-
мом патологического процесса (склеродермия,
эритематоз, атрофическая форма красного плос-
кого лишая, идиопатическая атрофия кожи).
98 Общая дерматология
Лихенификация
Лихенификация (lichenificatio) — утолщение,
уплотнение кожи; усиление кожного рисунка
(кожные борозды и складки резко выражены).
Кожа при этом сухая, шероховатая (шагреневая).
Лихенификация (лихенизация) возникает при
длительно протекающем воспалительном процес-
се, который сопровождается зудом и папулезной
инфильтрацией дермы. Наблюдается лихенифи-
кация при хронической экземе, атопическом
дерматите, ограниченном нейродермите.
Гистологическая картина при лихенификации
характеризуется гиперкератозом и акантозом с
удлинением эпидермальных отростков, иногда
паракератозом и спонгиозом. В дерме обнаружи-
вают хронический воспалительный инфильтрат,
а иногда явления фиброза.
Вегетация
Вегетация (vegetatio) — это разрастание сосочков
кожи и утолщение шиповатого слоя эпидерми-
са, ведущие к возникновению папилломатозных
разрастаний. Вегетации развиваются на поверх-
ности различных патологических процессов: па-
пул, эрозий, воспалительных инфильтратов.
Примером вегетаций являются вегетирующая
пузырчатка, вегетирующие грануляции на дне
язвы, остроконечные кондиломы, папилломы.
По внешнему виду вегетации могут напоминать
цветную капусту (остроконечные кондиломы);
иметь сероватый цвет и быть сухими и плотны-
ми на ощупь (бородавки); могут быть эрозиро-
ваны (вегетирующая пузырчатка).
Гистологическая картина характеризуется
акантозом с удлинением эпидермальных отрос-
тков, которые иногда глубоко погружаются в
дерму. При вегетирующей пузырчатке в аканто-
литическом эпидермисе имеются пузыри с акан-
толитическими клетками и большим количе-
ством эозинофилов. При болезни Дарье в дерме
отмечаются удлинение и утолщение сосочков
дермы, которые покрыты одним или двумя ря-
дами эпителиальных клеток и проникают в пу-
зыри либо лакуны. Лакуны могут быть инфиль-
трированы воспалительными элементами или
фиброзно изменены.
Разделение элементов на первичные и вторич-
ные является в некоторой степени условным.
Так, например, пигментное пятно является пер-
вичным элементом, но может быть и вторичным,
если оно возникает после обратного развития
папулы. При воспалительном процессе элемен-
ты могут переходить один в другой. Например,
сосудистое пятно в результате воспалительного
процесса может инфильтрироваться и превра-
титься в папулу. При остром воспалительном
процессе появившееся вначале сосудистое пят-
но или папула в результате экссудации может
превратиться в пузырек или пузырь. При обрат-
ном развитии бугорка и гуммы образуется язва,
после которой формируется рубец. Таким обра-
зом, рубец является уже третичным элементом
по отношению к вторичному элементу — язве.
Однако в целом это разделение на первичные и
вторичные морфологические элементы являет-
ся принципиально правильным.
Обследование
кожного больного
Анамнез болезни
Вначале выясняют жалобы больного, которые
излагаются в порядке их значимости. Прежде
всего, это могут быть различные субъективные
ощущения, сопровождающие высыпания: зуд,
жжение, боль, онемение, повышенная или по-
ниженная чувствительность, анестезия и др. Ана-
лизируют интенсивность зуда, который бывает
сильным при атопическом дерматите, почесухе,
крапивнице, чесотке, грибовидном микозе и
других дерматозах; менее сильным — при псо-
риазе, пиодермитах, микозах, красном плоским
лишае. Жжение больные отмечают при герпети-
формном дерматозе Дюринга, опоясывающем
лишае, пустулезном псориазе; боль — при глу-
боких пиодермитах, трофических язвах, опоясы-
вающем лишае; анестезию и гипостезию — при
лепре, сирингомиелии; чувство стягивания кожи
отмечается при склеродермии.
Собирают анамнестические сведения об общих
нарушениях, нередко беспокоящих больных
кожными заболеваниями (потеря аппетита, бес-
сонница, снижение трудоспособности, раздражи-
тельность).
Следует отметить, что целиком полагаться на
сведения о субъективных ощущениях, сообщае-
мых больными, нельзя. Оценка и интерпретация
их зависят от психики больного, степени его
восприимчивости, мнительности. Высыпания на
коже всегда ассоциируются у больного с неиз-
бежным зудом, которого на самом деле может и
не быть.
Анамнез заболевания должен включать нача-
ло и давность дерматоза, причинные и провоци-
рующие факторы. Следует указать первоначаль-
Глава 5. Этиология, патогенез и диагностика кожных болезней
99
ную локализацию во время первых признаков
дерматоза. Также необходимо отметить динамику
высыпаний на коже, распространенность, ремис-
сии, рецидивы, сезонность; предшествующее
лечение и его результаты.
Одни заболевания (фиксированная эритема,
простой пузырьковый лишай и др.) склонны к
рецидивам, другие (опоясывающий лишай, ин-
фильтративно-нагноительная трихофития) —
рецидивируют очень редко. Сведения об этих
заболеваниях имеют значение при постановке
диагноза. Выясняют возраст больного к нача-
лу заболевания, что также помогает диагностике.
Например, туберкулезные заболевания кожи на-
чинаются в детстве, красная волчанка — у
взрослых. В ряде случаев имеет значение посто-
янное или временное проживание больного в
той или иной местности (лепра, лейшманиоз,
флеботодермия).
Важно выяснить характер работы, химические,
физические, бактериальные вредности, особен-
но при диагностике профессиональных заболе-
ваний кожи: дерматита, экземы, ознобления, бо-
родавчатого туберкулеза, эризипелоида,
отдельных форм дерматомикозов.
Некоторые кожные болезни поражают людей
определенного пола. Так, например, стафилокок-
ковый сикоз, ринофима, узелковый хондродер-
матит, ромбическая кожа шеи — почти всегда
встречаются у мужчин; розацеа, хроническая
трихофития взрослых, болезнь Фокса-Фордай-
са — чаще у женщин.
Важное значение имеет анамнез жизни больно-
го. Следует расспросить о здоровье родителей, за-
болеваниях в семье (туберкулез, сифилис, онколо-
гические и кожные заболевания). Необходимы
сведения об особенностях перинатального периода
и возрастных этапов развития. Надо учитывать у
больного аллергические реакции на медикаменты,
химические вещества, аллергию или плохую пере-
носимость пищевых продуктов. Имеют значение
при некоторых дерматозах физическая активность
и интеллектуальные нагрузки в различные возрас-
тные периоды, а также психоэмоциональные пере-
живания. Общая характеристика кожи включает
следующие данные:
• цвет (смуглая, покрасневшая, с желтушным
оттенком);
• растяжимость и эластичность кожи; тургор
тканей;
• потоотделение и салоотделение (повышен-
ное, сниженное, умеренное);
• дермографизм (красный, белый, смешан-
ный), быстрота появления, продолжительность,
распространенность;
• мышечно-волосковый рефлекс;
• чувствительность кожи (болевая, темпера-
турная, тактильная);
• подкожно-жировая клетчатка;
• лимфатические узлы (величина, подвиж-
ность, болезненность, спаянность с кожей).
Общие свойства слизистых (конъюнктивы глаз,
век, красной каймы губ, полости рта и носа,
наружных половых органов и ануса) — цвет,
пораженность.
Волосы, ногти, состояние костей и суставов,
мышечная система.
Органы кровообращения: сердце, сосуды,
пульс, артериальное давление.
Органы дыхания: верхние дыхательные пути,
легкие, данные рентгенологического исследования.
Органы пищеварения: полость рта, состояние
зубов, миндалины, желудок, кишечник, печень,
селезенка.
Эндокринная система: щитовидная железа
(увеличение, экзофтальм, тремор рук); надпо-
чечники (пигментация кожи, слабость, утом-
ляемость, АД, глюкоза крови); половые желе-
зы (вторичные половые признаки, развитие,
либидо).
Нервная система: возбудимость, память, сон.
Рефлексы: сухожильные (коленные, ахилловы,
верхних конечностей); зрачковый. Симптом
Ромберга. Черепно-мозговые нервы (при пока-
заниях). Патологические рефлексы.
Органы чувств: зрение, слух, обоняние, вкус,
осязание.
Осмотр больного
1. Осмотр всего кожного покрова является пер-
вым и важным методом обследования больно-
го с заболеванием кожи. Осмотр лучше всего
проводить при рассеянном дневном либо дос-
таточно ярком электрическом свете. Темпера-
тура в комнате должна быть 22—23°С, так как
низкая температура вызывает спазм сосудов
(кожа бледнеет), а более высокая — их расши-
рение (гиперемия), что искажает истинный цвет
кожи. Желательно осматривать всю кожу, об-
ращая внимание и на лицо больного, которое
может выражать различные эмоции и состояния
(раздражительность, страдание, усталость, без-
различие), что свойственно больным с некото-
рыми дерматозами. Иногда при осмотре кожи
возникает необходимость пользоваться лупой,
лучше с увеличением не более чем в 3—5 раз.
Для этого удобна бинокулярная лупа, которую
надевают на лоб. При рассматривании кожи с
помощью лупы лучше различаются характер че-
100 Общая дерматология
шуек, корок, петехиальная и розеолезная сыпь.
Уделяют внимание окраске кожи, которая в
норме может быть бледной, розовой, смуглой.
У здоровых людей она матовая, не имеет жир-
ного блеска, отсутствуют зияющие протоки
сальных желез. При некоторых заболеваниях
кожа меняет свою окраску. Она может быть си-
нюшной при застойных явлениях, сине-бурой —
при акродерматите, коричневой — в начальной
стадии грибовидного микоза, желтой — при
желтухе, серовато-черной — при ихтиозе, мела-
нодермии; бронзовой — при аддисоновой бо-
лезни. При дерматозах, сопровождающихся
нарушением функции вегетативной нервной
системы, кожа бывает влажной; при почесухе
Бенье, атопическом дерматите, ихтиозе — сухой.
2. Пальпация (ощупывание). Пальпаторно оп-
ределяют эластичность и тонус кожи. В норме
эластичность и тонус кожи на разных участках
общего покрова у одного и того же человека не
одинаковы. Учитывают также возрастные изме-
нения кожи: снижение тургора и эластичности
у пожилых и старых людей. Пальпаторно мож-
но определить повышение или понижение тем-
пературы кожи (рожистое воспаление, глубокая
стафилодермия, эритродермия, болезнь Рейно,
склеродермия); расположение элемента (эпидер-
мис, дерма, подкожная основа), его размеры,
форму, консистенцию, спаянность с окружаю-
щими тканями, болезненность. При отеке опре-
деляют его интенсивность, наличие флюктуации
(абсцесс, гидраденит), отсутствие болезненнос-
ти (первичная сифилома).
3. Поскабливание (граттаж). Проводится пред-
метным стеклом или тупым скальпелем. Метод
поскабливания позволяет обнаружить шелуше-
ние кожи, определить его характер (муковидное,
отрубевидное, мелкопластинчатое, крупноплас-
тинчатое), плотность прикрепления чешуек к по-
верхности кожи. Метод применяется при псори-
азе (триада феноменов), разноцветном
(отрубевидном) лишае, парапсориазе. Не реко-
мендуется проведение поскабливания ногтем, так
как при этом возможно заражение (дерматоми-
коз, папулезный сифилид).
4. Диаскопия (витропрессия) — надавливание на
пораженный участок кожи предметным стеклом
либо специальным прибором — диаскопом (про-
зрачная пластмассовая пластинка). С помощью
этого метода определяют характер элемента (со-
судистый, пигментный). Сосудистое пятно при
надавливании побледнеет или исчезнет, а гемор-
рагическое пятно (или пигментное) — не исче-
зает. Этот метод помогает при диагностике тубер-
кулезной волчанки (феномен «яблочного желе»).
Дерматологический статус
Общая характеристика
высыпаний
Распространенность сыпи (ограниченная, рас-
пространенная, диссеминированная, универ-
сальная).
Топографические особенности расположе-
ния элементов сыпи (излюбленная локализа-
ция). Например, при псориазе — разгибатель-
ные поверхности конечностей; при
эпидермофитии стоп — третий и четвертый
межпальцевые промежутки стоп и свод сто-
пы; при красной волчанке — лицо; при ато-
пическом дерматите — подколенные и локте-
вые сгибательные поверхности, задняя и
боковые участки шеи.
Симметричность (истинная экзема, атопичес-
кий дерматит, псориаз) или асимметричность
высыпаний (дерматомикоз, микробная экзема,
кольцевидная гранулема, контактный дерматит);
по ходу нервных и сосудистых стволов (опоясы-
вающий лишай, линейный невус, линейный
красный плоский лишай).
Первичные (пятно, папула, бугорок, узел, вол-
дырь, пузырек, пузырь, гнойничок, опухоль) и
вторичные (пигментные пятна, чешуйки, корки,
эрозии, экскориации, трещины, язвы, рубцы,
лихенизация, вегетации) элементы.
Мономорфная и полиморфная сыпь. Моно-
морфной называется сыпь, состоящая из како-
го-либо одного первичного элемента (папулы при
псориазе или бугорки при туберкулезной волчан-
ке, или пузыри при вульгарной пузырчатке).
Полиморфной называют сыпь, состоящую из
нескольких первичных элементов (при экземе,
экссудативной многоформной эритеме, дермато-
зе Дюринга — пятна, папулы, пузыри, пузырь-
ки, волдыри).
Субъективные явления. Различают зуд,
жжение, болезненность, чувство стягивания
кожи.
Характеристика
первичных элементов
Характер воспалительного процесса: острый,
подострый, хронический. Невоспалительный
процесс. При островоспалительном процессе пре-
обладает экссудация, при неостровоспалитель-
ном — пролиферация.
Цвет с разными оттенками. Характерными
являются фиолетово-лиловый оттенок и воско-
Глава 5. Этиология, патогенез и диагностика кожных болезней
101
видный блеск для красного плоского лишая;
синюшный оттенок для многоформной экссуда-
тивной эритемы; кирпично-красный — для эрит-
размы; желтоватый оттенок — для туберкулез-
ной волчанки.
Величина. Элементы с булавочную головку
или просяное зерно называются милиарными;
с чечевицу или горошину — лентикулярны-
ми, элементы с 2—5-копеечную монету — мо-
нетовидными (нумулярными). При слиянии
элементов образуются бляшки, которые дос-
тигают величины ладони взрослого человека
и больше.
Очертания краев: правильные, неправильные,
овальные, круглые, угловатые, линейные, поли-
гональные, фестончатые.
Границы: резкие, нерезкие, четкие, нечеткие.
Резкие — чаще при хроническом воспалитель-
ном процессе (сифилис, красный плоский лишай,
псориаз), нерезкие границы — при остром вос-
палительном процессе (экзема).
Поверхность: шероховатая, бугристая, гладкая,
блестящая, матовая, зернистая.
Форма: плоская (псориаз), полушаровидная
(лентикулярный сифилид), конусовидная (мили-
арный сифилид), плоская с вдавлением (крас-
ный плоский лишай) или плоская с западением
(экссудативная эритема).
Консистенция: деревянистая (при актиноми-
козе), плотная (при онкодерматозах), плотно-
эластическая (при сифилисе), плотноватая
(при псориазе, красном плоском лишае), тес-
товатая (при липомах), мягкая (при туберку-
лезе кожи).
Глубина поражения: элемент может быть эпи-
дермальный, эпидермально-дермальный, дер-
мальный, суб дермальный.
Отношение к фолликулам: фолликулярные
(обычный фолликулит, сикоз, фурункул), не-
фолликулярные (не связанные с волосяными
фолликулами).
Многие дерматозы имеют специфические при-
знаки (феномены, симптомы), помогающие обо-
сновать диагноз.
Лабораторные и специальные
методы исследования
I. Патоморфология (световая микроскопия:
локализация и характер патологического
процесса, типы клеток, иммунофлюоресцен-
ция, окрашивание специальным методом ок-
раски, электронное микроскопическое и
другие методы исследования).
Гистологическое исследование применяется в
тех случаях, когда с помощью других методов
не удается установить диагноз. При некоторых
диагнозах (опухоли кожи, болезнь Дарье, рети-
кулез, саркоидоз, глубокий микоз) патогисто-
логическое исследование биопсированного ма-
териала является решающим. Гистологическая
картина чаще всего подтверждает клинический
диагноз, то есть на основании патоморфологи-
ческих изменений можно поставить только
предположительный диагноз. В некоторых слу-
чаях биопсию делают для контроля эффектив-
ности лечения.
II. Микробиологическое исследование чешуек
кожи, корок, экссудата, биопсийного мате-
риала, волос, ногтей.
1. Исследование мазков-отпечатков на аканто-
литические клетки.
2. Микроскопия соскоба из пораженного оча-
га.
• Грибы: препарат, обработанный 10 % ра-
створом КОН.
• Бактерии: мазок, окрашенный по Граму.
• Вирусы: проба Тцанка (мазок со дна вези-
кулы).
• Спирохеты: микроскопия в темном поле
зрения.
• Паразиты: чесоточный клещ в соскобе из
чесоточного хода; анализ на клеща Demodex
folliculorum.
3. Посев на среды для выявления бактерий,
грибов, простейших. При наличии грануле-
мы для посева используется измельченный
биопсийный материал.
4. Выделение вирусов в культуре клеток.
III. Исследование крови:
• Посев крови.
• Серологические реакции: отборочные реации
на сифилис; антинуклеарные антитела.
• Общий анализ крови: гематокрит, гемогло-
бин, форменные элементы, лейкоцитарная
формула.
• Биохимический анализ крови: глюкоза
плазмы натощак, азот мочевины крови, кре-
атинин сыворотки, биохимические показа-
тели функции печени.
• Исследование функции щитовидной железы.
IV. Люминесцентная диагностика (осмотр под
лампой Вуда):
• Кожа (in vivo), эритразма (кораллово-крас-
ное свечение), гипопигментрованные пятна
(становятся более темными), синие гипер-
пигментированные пятна при отложении
меланина в дерме (цвет не меняется).
102 Общая дерматология
• Волосы (in vivo): зеленое свечение волос при
дерматофитии волосистой части головы.
• Моча: оранжево-красная при поздней кож-
ной порфирии (добавляют в мочу 1 мл 5 %
соляной кислоты).
V. Проба с уксусной кислотой для выявления
остроконечных кондилом —• появление мел-
ких белых папул после нанесения 5 % ук-
сусной кислоты.
VL Аппликационные, лоскутные пробы (аллер-
гические тесты компрессные).
VII. Лучевая диагностика: рентгенография, уль-
тразвуковое исследование, компьютерная то-
мография, магнито-резонансная томография
(по показаниям).
Глава 6
Общие принципы лечения кожных болезней
Препараты и методы физиотерапии, применяемые в дерматологии 104
Наружная фармакотерапия дерматозов 109
Вопросы деонтологии 115
Гигиена кожи, ногтей, волос 116
Многочисленные кожные заболевания можно ус-
ловно подразделить на следующие основные
группы:
1. Дерматозы с выясненной этиологией (пио-
дермии, туберкулез кожи, лепра, лейшманиоз,
чесотка, вирусные заболевания и др.) и вызван-
ные известными химическими, физическими и
биологическими агентами.
2. Дерматозы с недостаточно ясным патогене-
зом (псориаз, красный плоский лишай, аутоим-
мунные дерматозы, аллергодерматозы, новообра-
зования кожи, Т-клеточные лимфомы и др.).
3. Генетически обусловленные болезни кожи
(генодерматозы).
При большинстве дерматозов проводится общая
и местная терапия. Выбор средств и методов те-
рапии дерматозов зависит от этиологии и пато-
генеза заболевания кожи. При установленной
причине кожной патологии назначается этио-
тропная терапия. Патогенетическая терапия на-
правлена на коррекцию механизмов развития и
течения патологического процесса, выявленных
нарушений со стороны органов и систем, с ко-
торыми может быть связано заболевание кожи.
Важное место при лечении дерматозов уделяет-
ся местной терапии, которая способствует лик-
видации местного процесса, улучшению психо-
эмоционального состояния пациента и, в
известной мере, оказывает положительное вли-
яние на общее состояние организма.
Симптоматическому лечению дерматозов в
настоящее время отводится меньшая роль, чем
этиологической и патогенетической терапии.
Инфекционные дерматозы вызываются биоло-
гическими агентами — бактериями, грибами, ви-
русами, животными-паразитами (чесоточный
клещ, вши, блохи) и некоторыми личинками. Эта
группа дерматозов разнообразна как по клиничес-
ким проявлениям, так и по степени контагиозно-
сти, что объясняется рядом причин: характером
возбудителя и его патогенными свойствами, им-
мунобиологической реактивностью макроорганиз-
ма, санитарно-гигиеническими и социальными
условиями жизни заболевшего человека.
Для лечения инфекционных дерматозов раз-
работаны и внедрены специфические препара-
ты, целенаправленно действующие на возбуди-
теля заболевания. Однако назначение одних
специфических препаратов в виде монотерапии
оказывается эффективным при ограниченном
числе дерматозов — например, при пиококко-
вом импетиго, поверхностных дерматофитиях,
чесотке, лейшманиозе и др. Следует отметить, что
при многих инфекционных дерматозах (лепра,
хронические рецидивирующие формы пиодер-
мии, дерматомикозы, герпесы, глубокие формы
пиодермии и микозов, туберкулез кожи и др.)
только одни этиотропные препараты не дают же-
лаемого терапевтического эффекта. Это связано
с необходимостью подключения препаратов, це-
ленаправленно воздействующих на патогенети-
ческие звенья данных дерматозов. Патогенети-
ческие средства назначаются для коррекции
развившихся тех или иных патологических из-
менений, вызванных непосредственно инфекци-
онным агентом, а также средств, улучшающих
функционирование внутренних органов и сис-
тем (иммунотропные, нейротропные, витамин-
ные, сосудистые средства и др.). Кроме того, в
последние годы в ряде случаев наблюдается фе-
номен лекарственной устойчивости микроорга-
низмов к этиотропным препаратам, что требует
их смены или чередования. Известно, что раз-
личные сопутствующие заболевания, особенно со
стороны желудочно-кишечного тракта, печени,
поджелудочной железы нарушают всасывание и
распределение лекарственных препаратов, замет-
но снижая их терапевтическую эффективность.
103
104 Общая дерматология
Назначая больному лечение, врач обязатель-
но должен учитывать целостность организма, ре-
гулирующую роль центральной нервной систе-
мы, эндокринных органов в поддержании его
гомеостаза. Лечение должно быть индивидуаль-
ным, с учетом пола и возраста пациента, анам-
нестических данных, проводимой ранее терапии,
переносимости лекарственных средств, стадии и
распространенности дерматоза.
Эффективность лечения больных с дермато-
зами во многом зависит от их взаимоотношений
с врачом этического порядка. Авторитет врача
обеспечивается прежде всего его компетентнос-
тью и гуманным отношением к больному. Ин-
дивидуальный психотерапевтический подход к
пациенту в значительной мере способствует бла-
гоприятным результатам фармакотерапии.
Для общей терапии заболеваний кожи в на-
стоящее время используют большое количество
фармакологических средств и методов физиоте-
рапии (табл. 6.1). При лечении больных дерма-
тозами с успехом применяют седативную, гипо-
сенсибилизирующую, гормональную, иммуно —
и химиотерапию, антибиотики, физические, ку-
рортные факторы и др.
В последние годы достигнуты определенные
успехи в терапии различных дерматозов. Разра-
ботка и внедрение новых препаратов (иммуно-
модуляторы, ароматические ретиноиды, цикло-
спорин) позволяют улучшить качество лечения
больных псориазом, атопическим дерматитом,
пиодермией, угревой сыпью, красным плоским
лишаем, многоформной экссудативной эрите-
мой, отдельными генодерматозами.
Арсенал дерматолога пополнился новыми сис-
темными противогрибковыми препаратами (ла-
мизил, орунгал, дифлюкан, низорал), повышаю-
щими эффективность и сокращающими сроки
лечения больных дерматофитиями, онихомико-
зами, трихомикозами, кандидозом.
Для купирования рецидивов герпетической
инфекции применяются препараты ацикловира
(зовиракс, виролекс, цикловир), ганцикловир,
фоскарнет, панавир и др. -
Значительно улучшились перспективы лече-
ния больных псориазом, атопическим дермати-
том, Т-клеточными лимфомами кожи в резуль-
тате разработки метода ПУВА-терапии и ее
комбинирования с ретиноидами, эссенциале.
Активно применяется при лечении дерматозов
облучение крови пациентов ультрафиолетовыми
и лазерными лучами.
При эритродермиях, пустулезном и артропа-
тическом псориазе, пузырчатке, токсидермиях,
тяжелых формах крапивницы и атопического
дерматита внедрены сорбционные методы деток-
сикации (гемосорбция), плазмаферез, а также
пероральный прием энтеросорбентов.
В терапии зудящих дерматозов с успехом при-
меняются методы рефлекторной терапии с по-
мощью ультразвука, лазера, КВЧ-излучения.
Созданы антибиотики новых поколений с
широким спектром активности для терапии яз-
венных и глубоких форм пиодермии (нетроми-
цин, амикацин, тиенам, меронем, зиннат и др.).
Для лечения меланом и лимфом кожи начали
применяться моноклональные антитела, интер-
лейкин-2, лейкинферон, интрон А.
Важное место в лечении и реабилитации боль-
ных хроническими дерматозами отводится баль-
неотерапии (ванны с фитодобавками, солевыми
экстрактами, оксидатом торфа и др.).
Препараты и методы физиотерапии,
применяемые в дерматологии
Таблица 6.1.
Препараты и методы
Показания
Противовоспалительные и сенсибилизирующие препараты
Препараты кальция (хлорид, глюконат, пантотенат кальция и др.)
уменьшают проницаемость сосудов, усиливают выделение
надпочечниками адреналина; при внутривенном введении
(10 % хлорид кальция по 5-10 мл) оказывают некоторое
гемостатическое действие
Аллергодерматозы, псориаз, красный плоский
лишай в острой стадии
Натрия тиосульфат—оказывает противотоксическое,
противовоспалительное и десенсибилизирующее действие.
Вводят внутривенно по 5-10 мл 30 % раствора или по 2-3 г
на прием в виде 10 % водного раствора
Нейродермит, псориаз, экзема, токсидермии,
эритродермии, крапивница
Глава 6. Общие принципы лечения кожных болезней 105
Препараты и методы Показания
Нестероидные противовоспалительные препараты (ибупрофен, напросин и др.) — ингибируют синтез простагландинов Псориатическая артропатия
«Антигистаминные препараты (димедрол, тавегил, фенкарол, перитол, кларитин и др.) — относятся к блокаторам Н1 — рецепторов Аллергодерматозы, псориаз, токсидермии, эритродермии, флеботодермии
Альфа-адреноблокаторы (пирроксан, празозин и др.) — влияют на периферические и центральные адренореактивные системы организма, уменьшают психическое напряжение, зуд кожи Акроцианоз, синдром Рейно, склеродермия, облысение, диффузный нейродермит, хроническая крапивница, псориаз, псориатическая эритродермия, красный плоский лишай
Седативные препараты и транквилизаторы (магния сульфат, бромиды, настойка пустырника и валерианы, диазепам, беллатаминал, беллоид, фенобарбитал, амизил, резерпин, раунатин) — назначают в соответствии с рекомендациями психотерапевта Псориаз, красный плоский лишай, аллергодерматозы, нейродерматозы
Иммунокорригирующие препараты
Иммуностимуляторы — левамизол, тималин, тактивин, нуклеинат натрия, продигиозан, пирогенал, спленин, экстракт плаценты, миелопид, рибомунил Хронические, рецидивирующие и тяжелые дерматозы
Иммунодепрессанты — цитостатики (метотрексат, азатиоприн, меркаптопурин, проспидин и др.), циклоспорин А (сандиммун-неорал), гепарин. Цитостатики лучше переносятся в комбинации с малыми дозами кортикостероидов, витаминами, иммуностимуляторами Красная волчанка (острая, подострая), акантолитическая пузырчатка, тяжелые формы псориаза, эритродермия, предопухолевые и онкодерматозы
Заместительная иммунокорригирующая терапия — интерферон лейкоцитарный, реаферон, интрон А, неовир, циклоферон, антистафилококковая плазма, лейкоцитарная масса, гамма-глобулины и др. Герпесы, экзема Калоши, тяжелые формы псориаза, нейродермита, хроническая язвенная пиодермия
Средства специфической иммунотерапии—стафилококковый антифагин и анатоксин, антистафилококковый гамма-глобулин, антистафилококковая плазма, стафилококковый бактериофаг, вакцины стрептококковая и стафилококковая Хронические фолликулиты, вульгарные угри, фурункулез, рецидивирующий гидраденит, рожистое воспаление, эпидемическая пузырчатка новорожденных, микробная экзема, рецидивирующая эктима
Адаптогены—дибазол, настойка женьшеня, лимонника, оротат калия и другие Хронические дерматозы, герпесы
Гормональные средства
Препараты щитовидной железы (тиреоидин, трийодтиронин, L-тироксин) Применяют при заболеваниях, сопровождающихся микседематозным статусом: псориаз, склеродермия, ихтиоз, нейродермит, экзема, угревая болезнь, эритродермии
«Синтетическиеаналоги глюкокортикостероидов (дексаметазон, триамцинолон, кеналог, дипроспан и другие) — сочетают с препаратами кальция, калия, витаминами С, А, Е, иммуностимуляторами, ограничивают поваренную соль, жиры, углеводы Акантолитическая пузырчатка, дерматоз Дюринга, эритродермии, артропатический псориаз, диффузные формы нейродермита и экземы
Анаболики (ретаболил, нероболил, неробол и другие) Акантолитическая пузырчатка и др. дерматозы с признаками развития кахексии
106 Обшая дерматология
Препараты и методы Показания
° Антибиотикотерапия
Пенициллин и его производные: бензилпенициллин, феноксиметилпенициллин, бензатинпенициллин, бициллины, оксациллин, ампициллин, амоксициллин, карбенициллин и др. Пиодермии, угри, эризипелоид, вирусные дерматозы (тетрациклины), дерматозы с неясной этиологией (склеродермия, красный
Тетрациклины: тетрациклин, метациклин, доксициклин плоский лишай, розовый лишай, многоформная
Макролиды: эритромицин, олететрин, азитромицин (сумамед), рокситромицин (рулид), мидекамицин (макропен) и др. эритема и некоторые другие)
Цефалоспорины: цефазолин, цефалексин (первое поколение), цефуроксим, цефаклор (второе поколение), цефотаксим, цефтазидим (третье поколение), цефепим (четвертое поколение) и др.
Фторхинолоны: ципрофлоксацин (ципробай, квинтор), офлоксацин (таривид), пефлоксацин (абактал), ломефлоксацин (максаквин) и др.
Аминогликозиды: нетилмицин, амикацин, гентамицин, тобрамицин Карбапенемы: тиенам, меронем Монобактамы. азтреонам Линкозамиды: линкомицин, клиндамицин Гпикопептиды: ванкомицин, ристоцетин Комбинированные антибиотики, амоксициллин/клавуланат, котримоксазол
Антибиотики разных групп, фузидин-натрий, хлорамфеникол, рифампицин
Противогрибковые антибиотики: нистатин, леворин, декамин, флуконазол (дифлюкан); гризеофульвин, канестен, кетоконазол (низорал), итраконазол (орунгал), тербинафин (ламизил) Кандидозы, онихомикозы, трихомикозы, распространенные микозы гладкой кожи
Химиотерапевтические средства
Сульфаниламидные препараты (бисептол, сульфадиметоксин, котримоксазол и др.) Пиодермии
Сульфоновые препараты (диуцифон, авлосульфон, сульфатон и др.) Лепра, дерматоз Дюринга, экзема, нейродермит, псориаз, склеродермия, хронические пиодермии
Противомалярийные препараты (резохин, плаквенил, хлорохин, делагил) Красная волчанка, фотодерматозы, порфириновая болезнь
Производные /74Н/С(фтивазид, изониазид, салюзид и др.), рифампицин, дигидрострептомицин Туберкулез кожи
Метронидазол, тинидазол, метрогил Демодекоз, розацеа, периоральный дерматит, себореиды, вульгарные угри
Средства системной терапии и профилактики герпетической инфекции
Аномальные нуклеотиды, ацикловир (виролекс, зовиракс), валацикловир (валтрекс), фамцикловир (пенцикловир), ганцикловир, видарабин, цитарабин, рибавирин, идоксиуридин, трифтортимитидин, фоскарнет Препараты растительного происхождения (панавир) Герпес простой: • первичный с общими явлениями, • осложненный, • диссеминированный, • рецидивирующий Герпес рецидивирующий • опоясывающий лишай • вирус папилломы человека
Глава 6. Общие принципы лечения кожных болезней
107
Препараты и методы Показания
Специфические гамма- и иммуноглобулины, человеческий иммуноглобулин, цитотект, интраглобин, пентаглобин, везикбулин Интерфероны и их индукторы: человеческий лейкоцитарный интерферон, реаферон, ларифан, ридостин, циклоферон, лейкинферон, роферон-А, интрон А, веллферон, камедон, кагоцел, йитамедин Герпетические вакцины живые, инактивированные, рекомбинантные Методы детоксикации Герпес генитальный: • первая атака, • осложненный, • рецидивирующий Герпес опоясывающий Герпес рецидивирующий II и I типов
Пероральные энтеросорбенты (активированный уголь, полифепан, белосорб, энтеросгель и др.) Гемосорбция, плазмаферез. Эффективность методов повышается при сочетании с ПУВА-терапией, кортикостероидами, ретиноидами Аллергодерматозы, псориаз Тяжелые дерматозы, острая красная волчанка, акантолитическая пузырчатка, токсидермии, включая синдромы Стивенса—Джонсона и Лайелла, эритродермии, артропатический псориаз, тяжелые диффузные формы псориаза и нейродермита, крапивница
Кровезаменители (гемодез, полиглюкин и др.^, изотонический раствор натрия хлорида, 5 % раствор глюкозы применяют в виде повторяющихся внутривенных капельных введений Средства метаболической терапии Псориаз, хронические распространенные дерматозы
Витамины группы В, А, Е, С, D, фолиевая и никотиновая кислоты, кокарбоксилаза, пиридоксальфосфат, рутин, витамин F, эссенциале. Применяют в лечебных и профилактических целях как фармакодинамические средства резерва Ароматические ретиноиды (этретинат, изотретиноин, ацитретин, роаккутан) Псориаз, красный плоский лишай, диффузный нейродермит и некоторые другие дерматозы. Витамин D применяют в комплексной терапии туберкулеза кожи Тяжелые формы псориаза, эритродермия, пустулезный псориаз, ихтиозы, кератодермии, кистозные угри
Препараты цинка и меди (цинктерал, цыгапан, витус М, теравит, гравитус, квадевит и др.) Физиотерапевтические методы ^Электрофорез лекарственных веществ (кальция хлорид, Назначают при выпадении волос, гипотрихозах, витилиго Аллергодерматозы, облысение, келоидные
магния сульфат, лидаза, новокаин, бромиды, ихтиол, димексид) рубцы, склеродермия, опоясывающий лишай, очаговый нейродермит, ограниченные формы красного плоского лишая
/Диадинамотерапия (токи Бернара) Экзема, псориаз, нейродермит, красный плоский и опоясывающий лишай
\/Диатермокоагуляция Бородавки, папилломы, сосудистые невусы, телеангиэктазии, келоиды
Рефлексотерапия—воздействие на биологически активные точки (электро-, фоно-, лазеро-, КВЧ-пунктура, иглорефлексотерапия) /Амплипульс-терапия Экзема, нейродермит, псориаз, крапивница, опоясывающий лишай Склеродермия, ограниченный нейродермит, опоясывающий лишай
/Дарсонвализация (искровой разряд) Выпадение волос, угревая болезнь, хроническая экзема, бляшки красного плоского лишая, псориаза, нейродермита, склеродермия, келоиды
108 Общая дерматология
Препараты и методы Показания
Индуктотермия на область надпочечников Экзема, красный плоский лишай, нейродермит, псориаз
Ультравысокочастотные токи (УВЧ) Фурункулы, гидраденит, васкулиты, трофические язвы
Ультразвуковая терапия (местное воздействие) Гидраденит, ограниченный нейродермит, склеродермия, фурункулы
Ультразвуковая терапия (сегментарное воздействие) Хроническая крапивница, универсальный кожный зуд, диффузный нейродермит, склеродермия
Ультрафонофорез (кортикостероиды, витамин А и др.) Ограниченный нейродермит, очаги хронической экземы, псориаза, склеродермии
Фототерапия
Ультрафиолетовое облучение средневолновое общее (275-320 нм) Фурункулез, стрептодермии, псориаз, диффузный нейродермит, себорейная и микробная экземы
Ультрафиолетовое облучение крови Псориаз, диффузный нейродермит, экземы, пиодермии
ПУВА-терапия. Сочетает действие фотосенсибилизатора (пувален) и длинноволнового ультрафиолетового облучения (320-400 нм). Сеансы ПУВА-терапии комбинируют с приемом ретиноидов, эссенциале, кортикостероидов, цитостатиков, иммунотропных средств, гемосорбцией, плазмаферезом, что позволяет повысить эффективность лечения и уменьшить развитие побочных эффектов Тяжелые формы псориаза, экземы атопического дерматита, нейродермита, угревой болезни Грибовидный микоз, ихтиозы
Пограничные лучи Букки (действуют более поверхностно, чем рентгеновские лучи) Сливающиеся и возвышающиеся угри, упорные хронические формы экземы, нейродермита, келоиды, кератодермии
Криотерапия (лечение холодом снежной угольной кислоты и жидкого азота с помощью специальных аппаратов и криоустановок) Папилломы, бородавки, невусы, кератомы, ангиомы, очаги красной волчанки, кольцевидная гранулема, вульгарные угри, очаги псориаза и нейродермита, узловатая почесуха, келоиды
Пелоидотерапия (грязелечение)
Гоязевые аппликации торфа, водорастворимого продукта торфа — оксидата торфа, сапропеля, лиманных грязей Нейродермит, псориаз, облысение, красный плоский лишай, почесуха, келоиды, угревая сыпь, трофические язвы, микробная экзема, подострые дерматиты
Нафталановая нефть (содержит фракции тяжелой нефти — нафтеновые кислоты) Псориаз, экзема, нейродермит вне стадии обострения
Озокерит (воскообразный продукт нефтяного происхождения) Псориаз, нейродермит, экзема, красный плоский лишай (ограниченные формы)
Курортолечение—включает воздушные ванны (аэротерапия), гелиотерапию, морские купания (талассотерапия), минеральные воды для ванн (сульфидные, радоновые, йодобромные, азотные термы, хлоридно-натриевые, рапные и др.) Реабилитация больных хроническими дерматозами
Глава 6. Общие принципы лечения кожных болезней
109
Наружная
фармакотерапия
дерматозов
Многообразие экзогенных и эндогенных причин
и провоцирующих факторов обусловливают по-
лиморфизм кожных сыпей (ответной реакции
организма), которые имеют свои характерные
клинические особенности.
Не вызывает сомнения тот факт, что наряду
с установлением причинно-следственных свя-
зей возникновения дерматозов и применением
современных терапевтических средств и мето-
дов общего воздействия на организм, рацио-
нальное наружное лечение имеет немаловажное
значение для ликвидации кожных проявлений
и улучшения психоэмоционального состояния
пациента.
Кожа весьма доступна и позволяет использо-
вать комбинированное воздействие на различные
участки тела больного. При этом следует учиты-
вать возможность общерезорбтивного действия
и проявления побочных эффектов некоторых
местных препаратов. В связи с этим при назна-
чении нового для больного наружного средства
необходимо проводить пробное лечение.
При наружном лечении дерматозов можно ис-
пользовать разнообразные лекарственные формы
и способы их применения, учитывая при этом
остроту воспалительной реакции, степень, глуби-
ну и распространенность поражения кожи. Лекар-
ственные вещества проникают в организм через
кератиновый слой путем абсорбции, частично
через волосяные фолликулы и сальные железы,
растворяясь в воде и жирах, подвергаясь при этом
сложным физико-химическим изменениям.
Неповрежденный кератиновый слой выступает
в роли депо, из которого лекарственные препа-
раты проникают глубже в кожу, оказывая по-
степенный лечебный эффект. При этом нужно
учитывать возможность кумулятивного токси-
ческого эффекта на организм пациента.
При нарушении кератинового слоя (экзема,
дерматиты и др.) возможно более активное вса-
сывание препаратов и вследствие этого развитие
интоксикаций, аллергических реакций с тяже-
лыми проявлениями (отек Квинке, токсидермии
и др.), требующих интенсивный терапии.
Выбор лекарственной формы (влажные аппли-
кации, кремы, мази и др.) зависит не только от
лекарственного вещества, но и от конкретной кож-
ной патологии, понимания этиологии и патогенеза
заболевания, при котором оно назначается.
Местная терапия кожных заболеваний пред-
ставляет собой, в известной мере, общее воздей-
ствие на организм больного.
Ликвидация кожных проявлений и субъектив-
ных ощущений (зуд, болезненность, нарушение
сна и т. д.) с помощью рациональной фармако-
терапии положительно сказывается на психоэмо-
циональном состоянии пациентов, что в свою
очередь играет важную роль в достижении бла-
гоприятных результатов лечения.
В наружной терапии дерматозов можно выде-
лить следующие основные направления:
1. Устранение причины заболеваний путем при-
менения антибактериальных, противовирусных,
антипаразитарных средств и препаратов, защи-
щающих кожу от внешних вредных факторов
(при профессиональных дерматозах, фотодерма-
тозах, укусах насекомых).
2. Устранение развившегося в коже патологи-
ческого процесса в зависимости от его остроты
(острый, подострый, хронический), распростра-
ненности (единичные очаги, распространенная,
диффузная, генерализованная сыпь), локализа-
ции (лицо, веки, туловище, конечности, наруж-
ные половые органы), сопутствующих субъек-
тивных ощущений.
При проведении наружной терапии дермато-
зов важно соблюдать следующие принципы.
На острый мокнущий воспалительный процесс
назначают водные растворы в виде примочек
или аэрозолей.
Острый воспалительный процесс без мокну-
тия можно приостановить с помощью примочек,
аэрозолей, водных болтушек, присыпок, лосьо-
нов.
При подостром воспалительном процессе при-
меняют жировые болтушки, пасты, присыпки,
кремы, эмульсии, крело.
При хроническом неспецифическом воспали-
тельном процессе целесообразно использовать
мази, согревающие компрессы, пленки, лаки,
пластыри.
При хроническом специфическом процессе
(лепромы, лейшманиомы, люпомы) вводят в
очаги специфические препараты.
При кожных разрастаниях (бородавки, папил-
ломы, вегетации) рекомендуются прижигающие
и цитотоксические средства.
В стадию прогрессирования заболеваний, со-
провождающихся инфильтрацией в очагах (псо-
риаз, красный плоский лишай) показаны кера-
топластические средства в виде мазей, кремов (с
небольшим содержанием действующих веществ):
2 % салициловой кислоты, 2—5 % серы, 5 %
нафталана.
110 Общая дерматология
В стадию стабилизации и регрессирования
инфильтративного процесса назначают керато-
литические средства (с возрастающим содержа-
нием действующих веществ): мази, пленки, пла-
стыри.
При различной интенсивности воспалитель-
ного процесса (острый без мокнутия, подо-
стрый, хронический неспецифический) прием-
лемо назначение местных кортикостероидных
препаратов в виде кремов, мазей, лосьонов,
аэрозолей.
Перед началом фармакотерапии дерматозов не-
обходимо учитывать показания и противопоказа-
ния к использованию того или иного лекарствен-
ного препарата; возможность привыкания к
местным средствам; возраст больного (детская кожа
более активно поглощает лекарственные средства,
например, борную кислоту, серу, салициловую
кислоту, деготь, гормональные препараты); воз-
можность развития аллергических реакций (при
применении антибиотиков, фурацилина, брилли-
антовой зелени и некоторых других средств); воз-
можность побочных явлений, например, при дли-
тельном местном применении кортикостероидов
(атрофия кожи, гипертрихоз, телеангиэктазии,
кушингоид и др.).
Перед назначением местных лекарственных
средств необходимо очистить кожу в очагах от
чешуек, корок, остатков лекарственных препа-
ратов. Обычно с этой целью применяют повяз-
ку с прокипяченным растительным маслом. Ле-
карство наносят на ограниченный участок и
только убедившись в его переносимости паци-
ентом — на остальные участки поражения. Важ-
но соблюдать правильность наложения примо-
чек, лекарственных и мазевых повязок,
смазываний, втираний, аппликаций, временные
интервалы и т. д.
Многие препараты действуют разносторон-
не в зависимости от концентрации, лекарствен-
ной формы и выбранного способа их приме-
нения.
Местные фармакологические средства по ха-
рактеру воздействия на патологический процесс
в коже могут быть разделены на следующие
группы (табл. 6.2).
Дерматологические
лекарственные формы
Пудры
Пудры — порошкообразные вещества, которые
применяют в виде присыпок. Минеральные по-
рошкообразные вещества (оксид цинка, тальк)
и органические (крахмал) смешивают между со-
бой в различных соотношениях. Их обозначают
как индифферентные присыпки.
Действие. Подсушивающее, обезжиривающее,
охлаждающее, сосудосуживающее.
Таблица 6.2. Наружные средства, применяемые в дерматологии
Группа лекарственных средств Лекарственный препарат
Индифферентные Растительные масла, вазелин, ланолин, свиной жир, глицерин, спермацет, силиконовые соединения, порошки (окись цинка, тальк, крахмал, белая глина)
Антисептические (противомикробные) Противовирусные Противовоспалительные Антипаразитарные Анилиновые красители, перманганат калия, пероксид водорода, борная кислота (2-5 %), хлоргексидин, препараты, содержащие антибиотики, сульфаниламиды Препараты, содержащие ацикловир, тромантадин, алпизарин, интерферон и др. Борная кислота (1-2 %), нитрат серебра, танин, ихтиол, кортикостероиды Бензилбензоат, деготь, сера, зеленое мыло, уксусная кислота, ртутная серая мазь, эсдепаллетрин, линдан, кротамитон и др.
Фунгицидные и фунгистатические Деготь, сера, препараты йода, ундециленовой кислоты, клотримазол, кетоконазол, нистатин и др.
Разрешающие (редуцирующие) Сера, нафталан, ихтиол, деготь, дерматол, резорцин, салициловая кислота (2-3 %)
Кератолитические, прижигающие Салициловая, молочная, бензойная кислоты, резорцин, серебра нитрат, подофиллин, «Солкодерм», «Кондилин», «Ферезол»
Противозудные Фотозащитные Ментол, анестезин, димедрол, карболовая, лимонная, уксусная кислоты Салол, хинин, танин, оксид цинка, метилурацил
Глава 6. Общие принципы лечения кожных болезней
111
Показания. Острые и подострые дерматиты
без мокнутия, повышенная потливость и уси-
ленное салоотделение. Для лечения эрозий и
язв в состав присыпок вводят сульфанилами-
ды, ксероформ, дерматол. При наличии при-
знаков пиодермии к ним добавляют антимик-
робные вещества (висмут, дерматол,
сульфаниламиды), для усиления противозуд-
ного эффекта — ментол, анестезин. Для полу-
чения жирных присыпок добавляют нафталан-
скую нефть.
Крахмал следует исключить из присыпок при
повышенной потливости, особенно в кожных
складках (вызывает брожение).
Методика применения. Припудривают участ-
ки поражения 2—3 раза в день.
Примеры присыпок
Rp.: Zinci oxydi Rp.: Urotropini
Talci Acidi borici aa 2,5
Amyli tritici aa 10,0 Zinci oxydi
M. D. S. Присыпка Taici aa 25,0
M. D. S. Присыпка при
потливости стоп
Растворы
Растворы — жидкая лекарственная форма, по-
лучаемая путем растворения в дистиллирован-
ной воде (реже в этиловом спирте) твердого
или жидкого лекарственного вещества. Для
примочек назначают в виде растворов проти-
вовоспалительные, вяжущие, дезинфицирую-
щие вещества.
Действие. Охлаждающее, противовоспалитель-
ное, дезинфицирующее, вяжущее.
Показания. Острое воспаление кожи с мокну-
тием или без явлений экссудации (ограниченные
экземы, острые дерматиты и др.) для уменьше-
ния мокнутия, отечности, жжения, зуда в учас-
тках поражения. Обычно примочки применяют
на ограниченные участки поражения несколько
раз в день (1-3 дня) до уменьшения мокнутия
(островоспалительные явления). При более дли-
тельном воздействии кожа в очагах становится
сухой, могут появляться трещины. При затяж-
ных процессах рекомендуется менять состав при-
мочек. У грудных детей примочки желательно не
применять.
Методика применения. 4—6 слоев марли, смо-
ченной раствором, прикладывают на участок
поражения. Меняют каждые 3—5 мин в течение
часа или более, 2—3 раза в день.
Примеры прописей примочек
Rp.: Sol. acidi borici 2 % Rp.: Sol. Aethacridini lactatis
300,0 0,1 % 500,0
D. S. Для примочек D. S. Для примочек.
Rp.: Sol. Argenti nitratis 0,25 % Rp.: Liquoris Burovi 500,0
200,0
Da in vitro nigro!
D. S. Наружное. Для примочек. D. S. Для примочек.
Лосьоны
Лосьоны — растворимые или суспендируемые
лекарственные компоненты. Комбинируют с
водой (алкоголем), другими физиологическими
растворителями. Жидкая часть, испаряясь, обес-
печивает быстрый охлаждающий и подсушива-
ющий эффекты, лекарственные вещества усили-
вают противовоспалительное действие.
Эмульсии
Эмульсии — жидкости с содержанием масла
30 %. Полужидкие-полутвердые системы вязко-
эластической консистенции. Популярны при
лечении поверхностных воспалительных дерма-
тозов. При нанесении на кожу жидкая часть
эмульсии испаряется, охлаждая кожу, а масло и
лекарственные вещества абсорбируются, усили-
вая противовоспалительный эффект.
Кредо
Крело — липосомальная эмульсия. Липосомы —
ламеллярные жировые кристаллические структу-
ры, чередующиеся с водными «вздутиями». Кре-
ло обеспечивает вместе с вводимыми лекарствен-
ными препаратами (кортикостероидами)
противовоспалительный эффект, гидратацию
кожи и физиологическое испарение воды.
Взбалтываемые взвеси
Взбалтываемые взвеси — мельчайшие порошко-
образные вещества, взвешенные в воде; воде и
глицерине; воде, глицерине и этиловом спирте.
Перед применением взвесь тщательно взбалтыва-
ют («болтушка»). «Болтушка» состоит из 30—40 %
порошковых веществ (цинк, тальк, крахмал) и
60—70 % жидкости (вода, вода и глицерин, гли-
церин и спирт). После испарения воды нанесен-
ные на кожу порошки остаются на ней тонким
слоем. Глицерин их удерживает на коже в тече-
ние многих часов. Спирт способствует ускорению
испарения воды с поверхности очагов поражения.
К этой лекарственной форме можно добавлять
другие препараты: ихтиол, анестезин, серу и др.
В качестве местного противовоспалительного
средства используют масляные болтушки (лини-
112
менты), которые состоят из окиси цинка (30 %)
и растительных масел (70 %).
Действие. Водных болтушек — противовоспа-
лительное, подсушивающее, противозудное.
Масляных взвесей — противовоспалительное,
смягчающее, отшелушивающее.
Показания. Острые и подострые поражения
кожи без мокнутия (дерматиты, аллергические
сыпи и др.). «Болтушки» можно применять без
наложения повязок.
Методика применения. Перед применением
взбалтывают. Наносят тампоном на участок по-
ражения 2—3 раза в день.
Примеры прописей болтушек
Rp.: Zinci oxydi Talci Glycerini Aq. destil. aa 25,0 Rp.: Zinci oxydi Talci aa 30,0 Glycerini Aq. destil. aa 70,0
M. D. S Наружное. Перед применением тщательно взболтать. М. D. S Наружное. Перед применением тщательно взболтать.
Аэрозоли
Аэрозоли — дисперсные системы, состоящие из
газовой среды, в которой взвешены твердые и
жидкие лекарственные препараты. Они обычно
содержат кортикостероиды, антибиотики, анти-
микотики и выпускаются в специальных герме-
тичных баллонах с клапанным устройством.
Действие. По механизму действия близки к
взбалтываемым взвесям.
Показания. Различной степени воспаления
кожи, в том числе сопровождающиеся мокнути-
ем, пиодермией, осложненные воспалением, ми-
козы.
Методика применения. Из баллона орошают
участки поражения 2—3 раза в день.
Мази
Мази — лекарственные формы, основой которых
является жир или жироподобное вещество. При-
менение мази прекращает дыхание кожи, вызы-
вает некоторое повышение ее температуры, раз-
рыхление поверхностных слоев и расширение
сосудов в участках поражения, благодаря чему
происходит всасывание активных веществ, содер-
жащихся в этой лекарственной форме.
В качестве основы применяют желтый вазе-
лин и ланолин (поровну), очищенное свиное
сало, растительные жиры, масла, нафталан, си-
ликоновые соединения.
В мазевые основы вводят серу, деготь, сали-
циловую кислоту, нафталан, ихтиол, резорцин,
при пиодермических наслоениях — антибиоти-
ки, учитывая возможность сенсибилизации.
Мази гидрофобные содержат жировые веще-
ства (вазелин, ланолин, нафталан, свиное сало
и др.), обеспечивают «разогревание» кожи и
глубокое воздействие вводимых лекарственных
веществ на инфильтративный воспалительный
процесс.
Мази гидрофильные содержат синтетические
силиконовые соединения, которые обеспечива-
ют дыхание кожи.
Действие. В небольших концентрациях (до
3 %), например, салициловая кислота, резорцин,
введенные в мазевую основу, вызывают керато-
пластический эффект (отшелушивающий, эпи-
телизирующий, противовоспалительный, легкий
рассасывающий), в больших концентрациях (3—
5 % и более) — кератолитический (отшелуши-
вающий, отслаивающий, выраженный рассасы-
вающий).
Показания. Хронические, подострые воспали-
тельные заболеваниях, наличие в коже воспали-
тельного инфильтрата неинфекционного харак-
тера и выраженное шелушение (псориаз,
нейродермит, красный плоский лишай, хроничес-
кая экзема, ихтиоз, гиперкератозы и др.), лече-
ние грибковых и паразитарных заболеваний
кожи, для ее смягчения и с другими целями.
Методика применения. Рассчитывая на кера-
топластическое действие, используют небольшие
концентрации действующих веществ, смазыва-
ют очаги 2-3 раза в день. Для получения кера-
толитического эффекта используют возрастаю-
щие концентрации действующих веществ.
Применяют смазывание очагов 2—3 раза в день
под повязку, в т. ч. под компрессную 2 раза в
день.
Примеры прописей мазей
Rp.: Acidi salicylic! 2,0 Vaselini ad 100,0 Rp.: Acidi salicylic! 5,0 Sulfuris рр 20,0 Vaselini ad 100,0
М. D. S. Смазывать кожу 2 раза в день (кератопластический эффект). М. D. S. Смазывать очаги 2 раза в день (кератопластический эффект).
Кремы
Кремы отличаются от мазей содержанием воды
в жировой основе. Если частицы воды окруже-
ны маслом («вода в масле»), то образуется жир-
ная мазь (крем), если наоборот («масло в воде»),
то образуется нежирная эмульсионная мазь
(крем).
Глава 6. Общие принципы лечения кожных болезней 113
Чаще применяют крем, состоящий из равных
частей ланолина (животный жир), вазелина (или
растительного масла) и воды.
Нередко в качестве основы для патентованных
кремов применяют синтетические жироподобные
вещества — производные целлюлозы, этиленок-
сидов и др.
Кремы гидрофильные содержат 30 % масла в
воде. Обеспечивают гидратацию, смягчение
кожи, противовоспалительный эффект. Дей-
ствуют непродолжительно (несколько часов).
Кремы, содержащие липобазу — кремы с во-
дой в качестве постоянной фазы, но с более вы-
соким содержанием (70 %) жира, чем в кремах
«масло в воде». Кремы содержат неионизиро-
ванный эмульгатор, который образует ламел-
лярные жировые кристаллические структуры,
чередующиеся с водными пузырьками, форми-
рующими липосомы, и являются наиболее фи-
зиологичными местными противовоспалитель-
ными средствами. На основе липосом создан
кортикостероидный препарат «Локоид-Липо-
крем».
Кремы гидрофобные представляют собой сис-
тему «вода (30 %) в масле». Создают масляную
пленку на коже, предотвращая испарение воды,
регидрируют кожу, способствуют проникнове-
нию лекарственных препаратов, оказывая тем
самым противовоспалительное действие.
В кремы часто вводят противозудные (димед-
рол, ментол), отшелушивающие (салициловая
кислота) вещества, мочевину, витамины А, Е,
кортикостероиды и другие средства.
Действие. При нанесении кремов кожа смяг-
чается и увлажняется, очищается от чешуек, ко-
рочек. При этом сохраняется ее дыхание, про-
являются охлаждающий, сосудосуживающий,
противовоспалительный эффекты.
Показания. Сухая шелушащаяся кожа, трещи-
ны, подострые дерматиты (экземы), зудящие
дерматозы, некоторые инфильтративно-десква-
мативные (псориаз, нейродермиты, ихтиоз) и
другие дерматозы.
Методика применения. Смазывание или легкое
втирание в участки поражения 2—3 раза в день.
Примеры прописей некоторых кремов
Rp.: Lanolini 01. Persicorum Rp.: Triamcinoloni 0,04 Zinci oxydi 3,0
Aquae destil аа 50.0 М. D. S. Смазывать кожу М. D. S. Aq. destil. 7,0 Lanolini 01. Helianthi aa 10,0 Смазывать очаги
2 раза в день. 2 раза в день.
Дерматологические пасты
Пасты дерматологические — это смесь равных
количеств жировых веществ и индифферентных
порошков. Они действуют глубже, чем болтуш-
ки, но более поверхностно, чем мази.
Действие. Противовоспалительное, подсуши-
вающее, охлаждающее, не вызывают прекраще-
ния кожного дыхания.
Показания. Острое и подострое воспаление без
мокнутия (дерматиты, герпесы, подострые эк-
земы и др.). При хроническом инфильтратив-
ном воспалительном процессе (хронические эк-
земы, нейродермиты, псориаз и др.) назначают
жирные пасты, увеличив при этом в лекарствен-
ной форме содержание жировых веществ и ре-
гулируя содержание активных веществ (салици-
ловая, карболовая кислоты, сера, деготь,
нафталан, ихтиол, АСД и др.), можно получить
кератопластический (редуцирующий) или кера-
толитический (рассасывающий) эффекты.
Методика применения. Наносят тонким слоем
на очаги 2—3 раза в день. Не применяют на уча-
стки с обильным волосяным покровом и мок-
нутием.
Накладывают обычно без повязки.
Примеры прописей некоторых
дерматологических паст
Rp.: Pastae Zinci 40,0 Rp.: Acidi salicylic! 10 Zinci oxydi Amyli tritici aa 12,5 Vaselini ad 60,0
D. S. Наносить на очаги М. D. S. Наносить на очаги
2 раза в день 1-2 раза в день (паста Лассара).
Дерматологические лаки
Основа дерматологических лаков — эластичес-
кий коллодий с 10 % содержанием касторового
масла или травмацитин (каучук в хлороформе).
В основу вводят салициловую, карболовую, мо-
лочную кислоты, резорцин, деготь и другие ве-
щества. Лак застывает на коже в виде тонкой
пленки; вещества, содержащиеся в нем, глубо-
ко проникают в кожу. Он удаляется бензином.
Действие. Кератолитическое, отслаивающее,
фунгицидное, антипаразитарное.
Показания. Лечение омозолелостей, бородавок,
ногтевых пластинок при онихомикозах, при
бляшечном псориазе, для отслойки рогового
слоя.
Методика применения. Наносят однократно на
очаг на 3—5 дней, затем применяют мази 1—2
дня, душ; циклы повторяют.
114
Примеры некоторых
дерматологических лаков
Rp.: Ac. salicylici 12.0 Rp.: Ac. Lactici
Ac. lactici 6,0 Collodii elastici ad 100,0 Ac. Salicylici Resorcini aa 10,0 Ol Ricini 3,0 Collodii elastici ad 100,0
M. D. S. Смазывать очаги 1 раз в день. М. D. S. Смазывать 1 раз в день подошвенные бородавки.
Дерматологические пластыри
Основа дерматологических пластырей — воск
или канифоль, обеспечивающие его густую и
липкую консистенцию. Вводят кератолитичес-
кие концентрации салициловой кислоты, моче-
вины, йода, карболовой и уксусной кислот,
свинца оксид и др.
Действие. Активное рассасывающее, отслаи-
вающее, разрыхляющее, дезинфицирующее,
фунгицидное в зависимости от вводимых ве-
ществ.
Показания. Удаление омозолелостей, разрых-
ление ногтей, лечение ногтевого ложа при они-
хомикозах, трихомикозах, веррукозной форме
красного плоского лишая, ограниченном нейро-
дермите, гнойно-воспалительных процессах в
коже.
Методика применения. Перед применением
пластырь нагревают, очаг протирают спиртом
(бензином), меняют 1 раз в 2—3 дня. Пластырь
хорошо удерживается на коже, ногтях.
Примеры некоторых
дерматологических пластырей
Rp.: Ac. Salicylici
Emplastri Plumbi
aa 30,0
M. D. S. Наносить на очаг
1 раз в неделю.
Rp.: Emplastri acidi carbolici
5 % 50,0
D. S. Накладывать
на ногтевые
ложа на 48 часов.
Противозудные средства
Противозудные средства можно разделить на две
группы
Первая группа противозудных средств оказы-
вает только противозудное действие, не влияя на
патологический процесс в коже. К ней относятся
препараты: ментол, анестезин, лимонная и кар-
боловая кислоты, растворы уксуса, антигиста-
минные средства, хлоралгидрат, красители и др.
Эта группа средств применяется чаще в виде
спиртовых растворов для смазываний, обтираний
зудящих участков кожи.
Вторая группа противозудных средств оказы-
вает влияние и на патологический процесс в
коже. Сюда относят препараты серы, дегтя, са-
лициловую кислоту, кортикостероиды и др. Эти
вещества включают преимущественно в состав
болтушек, паст, мазей, кремов.
Показания. Кожный зуд, почесуха взрослых,
крапивница, нейродермит, многоформная экссу-
дативная эритема, укусы насекомых и другие
заболевания, сопровождающиеся зудом.
Методика применения. Смазывание зудящих
участков 2-3 раза в день.
Примеры прописей некоторых
противозудных спиртовых растворов
Rp.: Mentholi 2,0 Sp. aethylici 70 % ad 100 ml Rp.: Ac. Carbolici 2,0 Sp. Aethylici 70 % ad 100 ml
M. D. S. Протирать пораженную кожу. M. D. S. Наружное. Смазывать кожу при зуде.
Местные кортикостероидные средства
Местные кортикостероидные средства эффектив-
но подавляют кожную воспалительную реакцию,
в том числе аллергическую, благодаря сосудосужи-
вающему эффекту и способности тормозить выс-
вобождение цитокинов (интерлейкины, у-интер-
ферон) из лимфоцитов и макрофагов, медиаторов
воспаления из эозинофилов и нейтрофилов, сни-
жать метаболизм арахидоновой кислоты, подавлять
активность гиалуронидазы. Кортикостероиды (осо-
бенно фторированные) снижают скорость деления
клеток, что важно при инфильтративно-десквама-
тивных дерматозах (псориаз, нейродермит, крас-
ный плоский лишай, ихтиоз и др.).
Кортикостероиды для местного применения под-
разделяются на 4 группы:
1. Очень сильнодействующие: беклометазон,
гальцинонид, дифлукортолон, клобетазол.
2. Сильнодействующие: беклометазон, бетаме-
тазон, гидрокортизона бутират, десонид, дифлу-
кортолон, триамсинолон, флуклоролон, флуоци-
нолон, флуоцинонид, флупреднилиден.
3. Обладающие сильной активностью: гидро-
кортизон с мочевиной, дезоксиметазон, флуо-
кортолон, флуоцинолон, флурандренолон.
4. Слабодействующие: гидрокортизон, метил-
преднизолон.
При инфицировании участков поражения их не
применяют без добавления антибактериальных
средств, так как можно усилить воспаление и
диссеминацию процесса.
При нанесении на большие участки пораже-
ния (особенно эрозивно-язвенные) имеется зна-
Глава 6. Общие принципы лечения кожных болезней
115
читальный риск резорбции местных фторирован-
ных кортикостероидов, подавления функции
надпочечников, при этом не исключается их те-
ратогенное действие.
Потому при назначении местных кортикостеро-
идных препаратов следует придерживаться сле-
дующих рекомендаций:
• в начале лечения применяют не очень ак-
тивные препараты группы гидрокортизона, пред-
низолона;
• при их неэффективности назначают высо-
коактивный препарат до получения выраженного
улучшения, после чего переходят на менее ак-
тивный препарат;
• препараты наносят на очаги тонким слоем,
придающим коже небольшой блеск;
• препараты следует выписывать в неболь-
ших количествах, чтобы исключить их пере-
дозировку и контролировать ход лечебного
процесса;
• окклюзионные повязки следует применять
только непродолжительное время;
• высокоактивные препараты назначают при
дискоидной красной волчанке и красном плос-
ком лишае, среднеактивные препараты — при
псориазе, с небольшой активностью (гидрокор-
тизон, преднизолон) — при экземе, дерматитах.
При инфицированных кожных поражениях к
кортикостероидам присоединяют антимикроб-
ные и антимикотические средства (неомицин,
клотримазол, гелиомицин, нистатин и др.), при
этом стероид не предупреждает возможные ре-
акции на эти вещества.
При отсутствии инфекции кортикостероиды
применяют в чистом виде.
Побочные реакции чаще развиваются при ле-
чении сильнодействующими кортикостероидами.
Они выражаются в присоединении инфекции,
атрофии кожи (иногда необратимой), стрий, ме-
стного гирсутизма у женщин молодого возрас-
та, периорального дерматита, местной депигмен-
тации, акне.
Не следует применять сильнодействующие
кортикостероиды на кожу лица (исключение —
дискоидная красная волчанка), век (из-за воз-
можности развития глаукомы).
Противопоказаниями к назначению местных
глюкокортикостероидов (особенно высокоактив-
ных) являются бактериальные, вирусные, гриб-
ковые дерматозы, периоральный дерматит, спе-
цифические инфекции кожи (туберкулез,
сифилис), опухоли кожи, беременность (лакта-
ция), обширные поражения кожи, дерматозы у
детей до года.
Вопросы деонтологии
Медицинская деонтология — наука о долге, мо-
ральной обязанности и профессиональной эти-
ке медицинских работников (врачей, медицин-
ских сестер, младшего медицинского персонала).
Общество предъявляет к врачу значительно боль-
ше требований, чем к представителям каких-либо
других профессий, поскольку в процессе лече-
ния личность врача играет порой большее зна-
чение, чем различные манипуляции и медика-
менты. М.Я. Мудров писал: «...ты изучил
болезнь и знаешь больного, но ведай, что и боль-
ной испытал тебя и знает, каков ты»
Одной из основных задач деонтологической
работы врача является поддержание чувства оп-
тимизма, душевного спокойствия, доверия и
уважения у пациента.
У больного человека наблюдается несколько
типов реакции на свое состояние. Эти реакции
зависят от личностных характеристик и психичес-
кого статуса пациента: адекватная, «уход в болезнь»,
«уход от болезни», недооценка тяжести заболева-
ния, аггравация и симуляция, диссимуляция и
метасимуляция. Молодые пациенты чаще переоце-
нивают свое здоровье и тяжело реагируют на гос-
питализацию, у пожилых пациентов происходит
переоценка тяжести своего состояния.
Врачу любой специальности, в том числе и
дерматовенерологу, необходимо помнить о состо-
яниях, которые могут быть вызваны некоррек-
тным поведением медицинского персонала. К
ним относится ятрогения и соррогения.
Соррогения характеризуется нарушением со-
стояния в результате неправильного поведения
медицинской сестры.
Ятрогения может развиваться в результате
недоучета личностных особенностей пациента и
нарушения правил медицинской этики. Дерма-
тологические больные крайне негативно воспри-
нимают слова «лишай», «короста», «плешивость».
Любые элементы сыпи воспринимаются как
проявление заразного заболевания. Когда врач
сообщает пациенту, что у него хроническое кож-
ное заболевание, например псориаз, у больного
может возникнуть чувство обреченности, особен-
но это часто наблюдается, когда пациенты ви-
дят тяжелые формы заболевания у своих сосе-
дей по палате. Наличие в одной палате пациентов
с различной степенью тяжести дерматозов, раз-
ного интеллектуального и культурного уровня
приводит к тому, что они передают друг другу
искаженную информацию о причине тяжести
своего заболевания и неуспехах в лечении, за-
116 Общая дерматология
частую подрывая авторитет врача, вызывая ре-
акцию эгротогении — ухудшение состояния под
влиянием поведения других пациентов. Профи-
лактика этого состояния требует продуманных
организационных мероприятий по правильному
размещению больных в палатах, постоянного
контроля за психологической атмосферой в от-
делении. В этом деле средний медицинский пер-
сонал неоценимый помощник для врача.
Некоторые кожные болезни являются результа-
том заболеваний внутренних органов. В таких си-
туациях врачу — дерматовенерологу необходимо
назначить дополнительные обследования у врачей
смежных специальностей. Зачастую дерматологи-
ческие больные воспринимают необходимость об-
следования как перебрасывание к другому врачу.
Дерматовенерологу стоит запастись терпением и
тактом, чтобы убедить пациента в необходимости
обследования, не потеряв его доверие.
Дерматологические болезни чаще всего не
приносят сильных физических страданий паци-
ентам, но их психический статус претерпевает
значительные изменения. Кожные заболевания
вызывают у пациентов чувство отверженности
окружающими, венерические — чувство стыда и
вины. Особенно выражены эти реакции у лиц с
высоким социальным статусом и информирован-
ных людей. Пациент в беседе с врачом может
сообщить свои переживания, тонкие подробно-
сти интимной жизни и если эта информация не
является общественно опасной, то ее не следует
никому передавать.
В этих ситуациях врачу очень важно суметь
помочь пациенту справиться с трудной жизнен-
ной ситуацией, вселить надежду в положитель-
ный исход лечения.
Психиатр Балинт писал, что «лекарством, ко-
торым чаще всего пользуются в повседневной
практике, является сам врач... важна та манера,
с которой врач прописывает его больному, и даже
та атмосфера, в которой лекарство назначается
и принимается». При беседе пациента с врачом
важную роль играет материальное окружение:
мебель, освещение, оформление кабинета, вне-
шний вид врача. Экстравагантная с обилием
украшений или неряшливая одежда вызывает
реакцию отчуждения у пациента. Не вызывает
доверия у пациента врач, который во время бе-
седы с ним постоянно пишет или говорит по
телефону. Заваленный бумагами стол врача мо-
жет вызвать подозрение в низкой квалификации
и постоянной черновой работе, пустой стол на-
водит на мысль о безделии. Обращаться к па-
циенту необходимо доброжелательно, демонст-
рировать сочувствие. В процессе общения с па-
циентом врач должен уметь направлять беседу в
нужное русло: в начале беседы позволить паци-
енту свободно излагать свои проблемы, что даст
возможность сориентироваться в его психологи-
ческом статусе, а затем перейти к конкретным
вопросам, касающимся состояния пациента. Ре-
комендации в конце беседы должны быть чет-
кими, излагаться на понятном языке.
Гигиена кожи,
ногтей, волос*
Основной закон личной гигиены — чистота.
Уход за здоровой кожей тела, здоровыми во-
лосами, ногтями и зубами носит название гиги-
енической или профилактической косметики.
Коррекционной косметикой, консервативной и
хирургической, занимаются специалисты — кос-
метологи. Привлекательный внешний вид чело-
века создает хорошее настроение ему самому и
окружающим, способствует предотвращению
некоторых кожных заболеваний. Уход за кожей
должен происходить постоянно, только в этом
случае можно добиться желаемого результата,
выглядеть красивым и оставаться здоровым.
Основными помощниками в этом деле служат
вода и мыло. Уход за телом включает ежеднев-
ный уход, состоящий из утренних и вечерних
процедур и еженедельное общее мытье, предпоч-
тительно в бане. Под влиянием воды из мыла
освобождаются свободные основания, которые
эмульгируют жир рогового слоя, образуют пену,
механически очищая кожу от омертвевших кле-
ток рогового слоя, секрета сальных и потовых
желез, грязи и микробов. От содержания осно-
вания зависит раздражающее действие мыла,
поэтому мыло необходимо выбирать с учетом
индивидуальной чувствительности.
Вода не только очищает кожу, но и обладает
терапевтическим эффектом. Под воздействием
воды возникает рефлекторный ответ со стороны
внутренних органов, изменяется кровоснабжение,
артериальное давление, потоотделение. Быстрая
смена температуры воды (контрастный душ) бла-
готворно влияет на нервные окончания, являясь
своеобразной гимнастикой для кровеносных со-
судов, что в конечном итоге улучшает питание
кожи, приостанавливает ее увядание. Баня иде-
Раздел написан И. В. Тихоновской
Глава 6. Общие принципы лечения кожных болезней
117
ально подходит для таких процедур. Непригод-
ной для ухода за кожей считается жесткая вода.
Вступая во взаимодействие с жиром кожи и
мылом, соли кальция, магния и железа образу-
ют нерастворимые вязкие вещества, которые
способствуют закупорке сальных и потовых же-
лез, вызывают зуд и раздражение кожи. Жест-
кую воду можно смягчить. Самый простой спо-
соб — кипячение. Для здорового человека
полезной считается температура воды 34° по
Цельсию. Для кожи лица рекомендуется комнат-
ная температура воды. Уходу за кожей лица тре-
буется уделять особое внимание, на ней находит-
ся много сальных и потовых желез, лицо
постоянно подвергается действию атмосферных
явлений, солнца, ветра, мороза, пыли.
Гигиена ногтей включает в себя маникюр и
педикюр, которые можно выполнять самостоя-
тельно или в косметических кабинетах.
Маникюр — гигиеническая процедура по ухо-
ду за ногтями рук. Её периодичность 1 раз в 10
дней. Надкожица при маникюре срезается осто-
рожно, не затрагивая кожу ногтевых валиков.
Необходимо помнить, что травматизация ногте-
вых валиков может привести к росту деформи-
рованного ногтя. Свободный край ногтя должен
выступать на 3—4 мм, длинные ногти часто ло-
маются, под ними скапливается грязь и это тре-
бует более тщательного ухода. При ряде профес-
сий длинные ногти недопустимы, например, у
медицинских работников.
Применяемый для маникюра лак может вызвать
воспаление кожи вокруг ногтя. При малейших
симптомах этого явления лак необходимо заменить
или не пользоваться им вообще, поскольку суще-
ствует угроза возникновения экземы.
Педикюр — гигиеническая процедура по уходу
за стопами. Его рекомендуется проводить каж-
дый месяц, но если на коже или ногтях стоп
имеются проявления какого либо заболевания,
то необходимо обратится за помощью к врачу -
дерматологу.
При проведении педикюра удаляют надкожи-
цу ногтевого валика, корректируют длину ног-
тей. Все гиперкератотические наслоения снима-
ются специальными приспособлениями без
травматизации кожи, чтобы не вызвать боль при
ходьбе. Ноготь большого пальца должен быть
длиннее остальных ногтей и слегка выступать
над мягкой частью пальца. В отличие от мани-
кюра, когда свободному краю ногтя кисти мож-
но придавать любую форму, ногти стоп долж-
ны иметь свободный край в виде прямой линии.
При проведении маникюра и педикюра необ-
ходимо соблюдать правила асептики. Незначи-
тельное нарушение целостности кожного покро-
ва может привести к бактериальной и грибко-
вой инфекции. При посещении косметических
кабинетов возникает риск заражения заболева-
ниями, которые могут передаваться через загряз-
ненные кровью предметы (гепатиты В, С, ВИЧ-
инфекция и др.).
При выборе косметических средств для ухода
за волосами необходимо помнить, что все кос-
метические процедуры, проводимые с волосами,
осуществляются в «омертвевшей части» волоса —
стержне. Любые шампуни, исключая лечебные,
применяемые для ухода за волосами, не прони-
кают на глубину залегания волосяного фолли-
кула, следовательно, не могут влиять на рост во-
лоса.
Шампуни состоят из очищающих поверхно-
стно-активных веществ сурфактантов, конди-
ционирующих и функциональных добавок,
консервантов и эстетических добавок. Этими
составляющими и определяется качество шам-
пуня.
Поверхностно-активные вещества состоят из
жирорастворимого компонента, который очищает
волосы от жира и грязи, и водорастворимого
компонента, благодаря которому грязь и жир
растворяются в воде и смываются.
Кондиционеры придают волосам блеск, пре-
дотвращают их спутывание, облегчают расчесы-
вание. Большое значение имеют функциональ-
ные добавки шампуня, регулирующие pH
(важный параметр для поддержания физиологи-
ческой кислотности кожи) и вязкость шампуней.
Для предотвращения развития микроорганиз-
мов в шампуни, среда которых является для них
благоприятной, добавляют консерванты. Эстети-
ческие добавки состоят из отдушек и красителей
и, как правило, безопасны. Следует выбирать
шампуни, на упаковке которых написан состав.
Шампуни могут содержать лекарственные до-
бавки, тогда они применяются соответственно
выявленной патологии волосистой части головы.
После мытья волосы необходимо сушить при
комнатной температуре. Постоянное использо-
вание фена травмирует стержень, в результате
чего волосы становятся сухими и ломкими.
Химические завивки, окрашивание и обес-
цвечивание волос отрицательно влияют на состо-
яние стержня волоса.
В процессе завивки под действием химичес-
ких веществ происходит разрушение дисульфид-
ных связей кутикулы волоса и поверхностных
слоев коркового вещества, благодаря чему воз-
никает возможность изменять форму волоса.
При закреплении химической завивки не все ди-
118 Общая дерматология
сульфидные связи восстанавливаются, следова-
тельно, уменьшается первоначальная прочность
волоса.
Отбеливание волос основано на разрушении
меланина, который располагается в клетках
коркового слоя. Нужно предварительно разру-
шить кутикулу для доступа отбеливателя к ме-
ланосомам. Этот процесс также сопровождается
разрушением дисульфидных связей и сниже-
нием прочности волоса. По данным некоторых
ученых, отбеливатели при длительном исполь-
зовании могут вызывать онкологическую па-
тологию и перекрестные аллергические реак-
ции.
При окраске волос происходит процесс, ана-
логичный отбеливанию, то есть для проникно-
вения красящего вещества в корковый слой раз-
рушается кутикула волоса. Безопаснее проводить
окраску волос полупостоянными красителями,
которые смываются после 6—10 — кратного мы-
тья головы. Они могут добавляться в красящие
шампуни, которые допустимы для применения
при гнездной алопеции.
При здоровых волосах окраску можно прово-
дить не чаще одного раза в четыре месяца. По-
лупостоянные красители меньше разрушают во-
лос, поскольку красящее вещество не проникает
в его глубокие слои. В этом смысле полупосто-
янные красители являются более безопасными.
5 Зак. 1357
Частная дерматология
Глава 7
Пиодермии (гнойничковые болезни кожи)
Стафилококковые пиодермии 121
Стрептококковые пиодермии 129
Стрептостафилодермии (смешанные пиодермии)132
Хроническая глубокая пиодермия 133
Пиоаллергиды 135
Этиология пиодермий
Пиодермии — большая группа дерматозов, вы-
зываемых гноеродными кокками, преимуще-
ственно стафилококками и стрептококками.
Возможна их ассоциация с другими микроор-
ганизмами (вульгарный протей, синегнойная
палочка, микоплазмы и др.). Возникают пио-
дермии как первичные заболевания кожи или
как осложнения других дерматозов (зудящие и
аллергические дерматозы) в результате микро-
травм, расчесов, при попадании на кожу извне
патогенных штаммов или активации микробной
аутофлоры человека.
По данным ВОЗ, больные пиодермиями в раз-
витых странах составляют 1/3 больных инфек-
ционной патологией. Среди дерматозов пиодер-
мии превалируют над другими кожными
заболеваниями.
Пиококки весьма распространены во внешней
среде (пыль, вода, предметы и др.). Высока об-
семененность стафилококком кожи и складок у
здоровых лиц (90—100 %), ногтей со свободно-
го края, носа, зева (выделяются преимущественно
белый и золотистый стафилококки). Стрепто-
кокки обнаруживаются в складках кожи прибли-
зительно у 12 % здоровых людей (50 % состав-
ляет гемолитический стрептококк). Из зева
стрептококки высеваются у 100 % обследован-
ных здоровых лиц (зеленящий, гемолитический,
негемолитический). Следует отметить, что не-
смотря на высокую распространенность стафи-
лококков и стрептококков на коже и слизистых
человека, их патогенное действие проявляется
при нарушении равновесия в системе макроор-
ганизм — микроорганизм при повреждении це-
лостности кожных покровов и снижении имму-
нобиологической сопротивляемости организма.
Предрасполагающие факторы
Кожа обладает защитными свойствами по отно-
шению к различным микробным агентам, одна-
ко, для развития пиодермий необходимы, как
правило, предрасполагающие причины.
К эндогенным предрасполагающим факторам
относят: функциональные нарушения ЦНС,
стресс, сахарный диабет, другие эндокринопатии,
заболевания ЖКТ, гепатобилиарной системы,
хронические инфекции, интоксикации, гипови-
таминозы, местные нарушения кровообращения,
сенсибилизация к пиококкам, первичный и вто-
ричный иммунодефицит, длительный прием кор-
тикостероидов, цитостатиков, иммунодепрессан-
тов, ВИЧ-инфекция.
Экзогенными предрасполагающими факторами
являются: различные травмы и микротравмы, в
т. ч. химического характера, охлаждение, пере-
гревание, гиперинсоляция, загрязнение и маце-
рация кожи, сдвиг pH кожи в щелочную сторо-
ну, зудящие дерматозы.
Пути передачи: воздушно-капельный, меха-
нический перенос пиококков из очагов пораже-
ния, а также через обсемененные пиококками
предметы, белье.
Патогенез пиодермий
Патогенез пиодермий можно представить в виде
схемы (рис. 7.1).
Особенности этиологии и патогенетических
механизмов различных клинических форм пи-
одермий будут рассмотрены ниже.
120
Глава 7. Пиодермии
121
Классификация пиодермий
Группы пиодермий и их клинические разновид-
ности представлены в таблице 7.1.
Стафилококковые
пиодермии
Этиология
Возбудители стафилококковых пиодермий ста-
филококки подразделяются на три вида:
Рис. 7.1. Схема патогенеза пиодермий.
1. Золотистые стафилококки, вызывающие за-
болевания у человека.
2. Эпидермальные стафилококки, принимающие
участие в патологическом процессе у человека.
3. Сапрофитные стафилококки.
Патогенность стафилококков связана с их спо-
собностью образовывать коагулазу. Неконтроли-
руемое применение антибиотиков способствует
трансформации стафилококков в L-формы, ко-
торые утрачивают многие типичные свойства, но
сохраняют способность вызывать атипичные и
хронические формы стафилодермий, бациллоно-
сительство, появление вариантов бактерий с вы-
сокой лекарственной устойчивостью. Например,
белый стафилококк, не коагулирующий плаз-
му, — одна из форм изменчивости патогенных
стафилококков. Эксфолиативный токсин, выра-
батываемый стафилококками второй фагогруппы,
может быть одной из причин пузырчатки ново-
рожденных, буллезного импетиго, скарлатинопо-
добной сыпи и, возможно, синдрома Лайелла. Ми-
коплазменностафилококковая ассоциация может
иметь значение в формировании таких тяжелых
форм, как гангренозная и язвенно-некротическая
пиодермия. Гиалуронидаза стафилококков способ-
ствует инвазии, ДНКаза выполняет функции пи-
тания и защиты от чужеродной ДНК, коагулаза
образует тромбы и фибринозные чехлы вокруг
стафилококков, фибринолизин — лизирует фиб-
рин, препятствуя проникновению фагоцитов в
очаги инфекции.
Стафилококк вызывает гнойное воспаление
кожи, поражая волосяные фолликулы, потовые
апокриновые железы (у детей эккринные) на
различной глубине (табл. 7.2).
Таблица 7.1. Клиническая классификация пиодермий
Гоуппы пиодермий Клинические разновидности
Стафилококковые пиодермии Остиофолликулит, фолликулит, сикоз вульгарный, фурункул, фурункулез, карбункул, гидраденит.
Стафилодермии у детей Везикулопустулез (перипорит), псевдо- фурункулез Фингера (множественные абсцессы у детей), эпидемическая пузырчатка новорожденных (эпидемический пемфигоид), эксфолиативный дерматит Риттера
Стрептококковые пиодермии Импетиго Тильбери-Фокса и его разновидности — кольцевидное, буллезное, поверхностный панариций, щелевидное, слизистых
Атипичные стрептодермии Простой лишай, папулезное сифилоподобное импетиго (пеленочный дерматит), острая диффузная пиодермия
Стрептостафилодермии (смешанные пиодермии) Хроническая глубокая пиодермия Вульгарное импетиго, вульгарная эктима, хроническая диффузная пиодермия Хроническая глубокая язвенно-вегетирующая пиодермия, пиогенная гранулема, шанкриформная пиодермия, гангренозная пиодермия и другие реже встречающиеся формы
122 Частная дерматология
Общая характеристика
стафилококковой пустулы
Фолликулярная пустула болезненная, распрост-
раняется вглубь, с напряженной покрышкой.
Наполнена зелено-желтым экссудатом. Исход —
эрозия, временная пигментация (поверхностные
формы), язва, рубец (глубокие формы).
Поверхностные
стафилодермии
- Остиофолликулит
Острое гнойное воспаление верхних отделов
волосяного фолликула. Наиболее частый возбу-
дитель — золотистый коагулазопозитивный ста-
филококк.
Предрасполагающие факторы: загрязнение,
мацерация кожи, повышенная потливость, по-
верхностные механические травмы, дисэндокри-
нии и др.
Излюбленная локализация: лицо, волосистая
часть головы, конечности.
Клиническая картина
В устье волосяного фолликула появляется красно-
ватый узелок с пустулой, пронизанной волосом,
размером 1—2 мм, с периферическим венчиком
гиперемии до 1 мм. Элементы могут быть единич-
ными или множественными. Пустула в течение 4—
5 дней ссыхается в корочку, которая отпадает,
оставляя временную красноватую пигментацию.
Элементы больших размеров (в диаметре 3—4 мм)
характеризуют как импетиго Бокхарта.
Общие явления отсутствуют.
Субъективные ощущения: небольшая болезнен-
ность, зуд в области элементов.
Течение острое.
Дифференциальный диагноз
Стрептококковое и вульгарное импетиго (флик-
тена неправильных очертаний, корки, склон-
ность к распространению, легкий зуд). Диффе-
ренцируют также с вульгарным сикозом, папу-
ло-пустулезными вульгарными и розовыми уг-
рями, бромистыми и йодистыми угрями, пери-
оральным дерматитом, угревидным сифилидом.
Лечение
Общее лечение обычно не проводится. В случа-
ях частых рецидивЬв назначают аутогемотера-
пию, стимуляторы лейкопоэза (метилурацил,
натрия нуклеинат и др.), биостимуляторы (эк-
стракт алоэ, гумизоль и т. п.), общее ультрафи-
олетовое облучение в субэритемных дозах.
Местная терапия: обтирание здоровой кожи
вокруг очагов 3 % салициловым, 3 % борным,
1 % левомицетиновым спиртом, на очаги — ра-
створы анилиновых красок, пасты, содержащие
антисептики, антибиотики.
' Фолликулит
Острое гнойное воспаление среднего (поверхно-
стный фолликулит) и нижнего (глубокий фол-
ликулит) отделов волосяного фолликула.
Возбудители: различные стафилококки, чаще
золотистый коагулазопозитивный стафилококк.
Предрасполагающие факторы: преимуществен-
но экзогенные; при глубоких фолликулитах —
сахарный диабет, анемии, иммунодефицитные
состояния.
Излюбленная локализация: лицо, волосистая
часть головы, шея, конечности, туловище.
Клиническая картина
В результате внедрения инфекционного агента
в более глубокие отделы волосяного фолликула
развивается острое гнойное воспаление фолли-
кула и частично перифолликулярной ткани. Эле-
мент имеет вид конусовидной папулы красно-
ватого цвета, величиной 5—7 мм, которая часто
превращается в пустулу, пронизанную волосом.
Периферическая зона гиперемии составляет 2-
3 мм. Папула в течение нескольких дней или рег-
рессирует, или абсцедирует. При вскрытии мик-
роабсцесса выделяется сливкообразный гной,
Таблица 7.2. Классификация стафилодермий
Особенности
кожных поражений Клинические формы
Поверхностные формы Остиофолликулит, фолликулит, сикоз
Глубокие формы Фурункул, карбункул, фурункулез, гидраденит
У детей Везикулопустулез (перипорит), псевдофурункулез Фингера (множественные
абсцессы у детей), эпидемическая пузырчатка новорожденных, эксфолиативный
дерматит Риттера
Глава 7. Пиодермии
123
затем образуется небольшая язвочка, заживаю-
щая с образованием рубчика.
Общие явления обычно отсутствуют, однако
при множественных, более крупных пустулах
(более 1 см), захватывающих нижние отделы
фолликулов, появляется выраженная болезнен-
ность, лихорадка, лейкоцитоз, ускоренная СОЭ.
Субъективные ощущения: болезненность обычно
исчезает после естественного вскрытия пустулы.
Течение острое.
Дифференциальный диагноз
Стрептококковое и вульгарное импетиго, вуль-
гарный и паразитарный сикоз, папуло-пустулез-
ные вульгарные и розовые угри, бромистые и
йодистые угри, угревидный сифилид, демодекоз,
кандидозный и герпетический фолликулит,
вросшие волосы, акне-келоид.
Лечение
Общее лечение обычно не проводится. В случа-
ях частых рецидивов назначают аутогемотера-
пию, стимуляторы лейкопоэза (метилурацил,
натрия нуклеинат и др.), биостимуляторы (эк-
стракт алоэ, гумизоль и т. п.), общее ультрафи-
олетовое облучение в субэритемных дозах.
При выраженной общетоксической реакции
назначают антибиотики: диклоксациллин или це-
фалексин по 1—2 г/сут в 4 приема в течение 10 сут;
миноциклин по 100 мг внутрь 2 раза в сутки.
Местная терапия. Обтирание здоровой кожи
вокруг очагов 3 % салициловым, 3 % борным,
1 % левомицетиновым спиртом, на очаги — ра-
створы анилиновых красок, пасты, содержащие
антисептики. Мазь мупироцин 2 раза в сутки на
участки поражения и слизистую носа (носитель-
ство золотистого стафилококка). Антисептичес-
кие мыла, обработка кожи бензоилпероксидом.
Сикоз вульгарный
Хроническое рецидивирующее гнойное воспале-
ние фолликулов в области бороды, усов, реже —
бровей, лобка, подмышечных впадин, волосис-
той части головы.
Возбудители: Золотистый стафилококк или со-
четания различных штаммов стафилококков.
Предрасполагающие факторы: Очаги инфек-
ции в челюстно-лицевой области, хронический
холецистит, простатит, заболевания внутренних
органов, нервно-эндокринная патология, сниже-
ние функции половых желез, злоупотребление
алкоголем, повышенная запыленность производ-
ства, развитие сенсибилизации к стафилококку,
нарушение бактерицидных свойств кожи.
Излюбленная локализация: область бороды,
усов.
Клиническая картина
В области бороды, усов, на других участках ро-
ста коротких толстых волос возникают ости-
офолликулиты и поверхностные фолликулиты,
которые постепенно сливаются. Кожа в участ-
ках поражения приобретает синюшно-бурый
цвет, инфильтрируется и покрывается гнойны-
ми корками. При их снятии выявляется влаж-
ная мокнущая эрозированная поверхность. У
корней волос, удаленных пинцетом из очагов по-
ражения, обнаруживаются стекловидные муфты.
В результате прогрессирования процесса область
роста волос (бороды, усов) принимает характер
единого воспалительного инфильтрата, покры-
того гнойными корками (создается впечатление
человека с неопрятной внешностью).
Общие явления: иногда лихорадка.
Субъективные ощущения: небольшая болезнен-
ность, зуд.
Течение: хроническое, рубцовых изменений в
очагах не остается.
Дифференциальный диагноз
Паразитарный сикоз (положительный анализ на
грибы, острое течение), сикозиформная экзема,
медикаментозный дерматит.
Лечение
Общая терапия. Назначают антибиотики (дик-
локсациллин, аугментин, цефалексин и др.) в
течение нескольких недель (не менее 3-х). Ре-
комендуются: аутогемотерапия, стимулирующие
средства (пирогенал, тактивин, тималин, имму-
нал), по показаниям — половые гормоны.
Местная терапия. Участки поражения обраба-
тывают спиртовыми растворами салициловой и
борной кислот, левомицетина, фукорцина, дру-
гих анилиновых красок, бактробаном. На учас-
тки инфильтрации применяют мази с антибио-
тиками (гентамициновая, линкомициновая,
левомеколь, мупироцин и др.), борно-дегтярную
и серно-дегтярную мази, комбинированные кор-
тикостероиды. Рекомендуется общее ультрафи-
олетовое облучение, на очаги — эритемные дозы
УФО. Запрещается бритье в области пораженных
участков кожи.
Профилактика
Обтирание кожи дезинфицирующими спиртовы-
ми растворами, витаминные препараты, введение
стафилококкового анатоксина; лечение сопут-
ствующих заболеваний; здоровый образ жизни.
124 Частная дерматология
Глубокие стафилодермии
Фурункул
Острое гнойно-некротическое воспаление воло-
сяного фолликула и перифолликулярных тканей.
Возбудитель: коагулазопозитивный золотис-
тый стафилококк, выделяющий большое число
факторов агрессии (гиалуронидаза, коагулаза,
фибринолизин, ДНКаза и др.) и вызывающий
некроз перифолликулярных тканей.
Предрасполагающие факторы: первичный и
вторичный иммунодефициты, снижение бактери-
цидной активности нейтрофилов (хронические
гранулематозы), нарушения хемотаксиса, синдром
гиперпродукции IgE, эндокринная патология (са-
харный диабет, ожирение, нарушение углеводного
обмена), микротравматизм, несоблюдение личной
гигиены, работа на загрязненных производствах,
носительство штаммов патогенных стафилокок-
ков и инфицированная ими одежда; зудящие
дерматозы, чесотка, педикулез и др.
Излюбленная локализация: участки микро-
травм кожи на плечах, ягодицах, бедрах, туло-
вище; чаще — на лице, в области носогубного
треугольника, подбородка, затылка, задней по-
верхности шеи, подмышечных впадин. Обычно
очаги единичные, реже — множественные (фу-
рункулез).
Клиническая картина
Заболевание начинается с остиофолликулита,
который быстро трансформируется в глубокий
фолликулит, а затем в течение 24—48 часов пре-
вращается в резко болезненный, воспалительный
узел размером до лесного — грецкого ореха (ста-
дия инфильтрации). Через несколько дней кожа
над узлом куполообразно выпячивается, истон-
чается, появляются признаки флюктуации. Да-
лее узел вскрывается с выделением гноя, при
этом обнажается омертвевшая ткань зеленоватого
цвета — «некротический стержень» (стадия аб-
сцедирования). В результате процессов демарка-
ции (созревания) некротический стержень вме-
сте с гнойно-кровянистыми массами отторгается
с образованием гранулирующей кратерообразной
язвы, которая достаточно быстро рубцуется (ста-
дия язвы и рубцевания). Эволюция элемента в
среднем составляет около 2-х недель.
Общие явления. При одиночных фурункулах
общее состояние обычно не страдает. При мно-
жественных фурункулах и фурункулезе могут
быть лихорадка, слабость, головные боли; со
стороны периферической крови определяются
лейкоцитоз и ускоренная СОЭ. Посев крови (до
применения антибиотиков) показан при лихо-
радке и других признаках сепсиса.
Возможные осложнения фурункула
У ослабленных больных лимфогенное распрос-
транение инфекции на прилегающие и подлежа-
щие ткани может вызвать появление флегмоны,
остеомиелита, язвенной пиодермии. Проникно-
вение возбудителя в кровь сопровождается сеп-
тическим состоянием, септикопиемией и мета-
стазированием инфекции в различные ткани
(кости, позвоночник, мышцы), внутренние орга-
ны (чаще почки), вызывает бактериальное по-
ражение эндокарда, что в конечном итоге может
привести к летальному исходу. Появление фу-
рункула в области носогубного треугольника,
анатомические особенности венозной сети, а так-
же выдавливание фурункула в начальной стадии,
его срезание, назначение тепловых процедур
могут вызвать тромбофлебит пещеристого синуса
и гнойный менингит (злокачественный фурун-
кул).
Фурункулез — часто рецидивирующие единич-
ные фурункулы или высыпание множественных
Фурункулов. Одиночные фурункулы чаще реци-
дивируют в области задней поверхности шеи, на
ягодицах^ пояснице. Диссеминированные высы-
пания фурункулов наблюдаются по всему кож-
ному покрову. Появление единичных рецидиви-
рующих фурункулов связано, как правило, с
экзогенными причинами, а множественных — с
эндогенными.
Дифференциальный диагноз
Инфильтративно-нагноительная трихофития
(анализ на грибы положительный), рожистое
воспаление, глубокие фолликулиты, карбункул,
гидраденит, скрофулодерма, узловатый васкулит.
Лечение
Единичные фурункулы без нарушения общего
состояния лечат с учетом фазы развития. В ста-
дии инфильтрата применяют УВЧ-терапию,
соллюкс, ультразвук, «лепешки» с чистым их-
тиолом 2 раза в день (на фурункулы области
лица не применяются). При необходимости
волосы вокруг фурункула выстригаются, пери-
фокальные участки кожи обрабатываются сали-
циловым, борным или левомицетиновым спир-
том. Мытье тела запрещается. В стадии
абсцедирования — вскрытие фурункула, дрени-
рование с помощью турунды, смоченной гипер-
тоническим раствором натрия хлорида; стериль-
ные салфетки прикрепляют к коже
бактерицидным пластырем или клеолом (но не
Глава 7. Пиодермии
125
прибинтовывают). Перевязки проводят не ме-
нее 2-х раз в сутки. После отторжения некро-
тического стержня — на язву применяют мази
с антибиотиками, аргосульфан, орошение про-
теолитиками (1 % трипсин, химотрипсин).
При нарушении общего состояния, опасной
локализации фурункула (лицо, шея, волосистая
часть головы) и на фоне иммунодефицита назна-
чаются антибиотики: диклоксациллин до 2,0 г/
сут в 3—4 приема, 10 сут; амоксициллин/клаву-
ланат 20 мг/кг/сут 3 раза в сут, 10 сут; цефалек-
син до 2,0 г/сут внутрь, 10 сут (взрослые), де-
тям — 40—50 мг/кг/сут в 4 приема, 10 сут;
кларитромицин 250—500 мг внутрь 2 раза в сут-
ки, 10 сут; азитромицин 250-500 мг/сут, 5—
7 сут; клиндамицин 150—300 мг внутрь 4 раза в
сутки, 10 сут; миноциклин 100 мг 2 раза в сут-
ки, 10 сут; ципрофлоксацин 500 мг 2 раза в сут-
ки, 7 сут; ванкомицин применяют в тяжелых
случаях и устойчивости 5. aureus к другим ан-
тибиотикам.
Постельный режим при локализации фурун-
кулов на лице, волосистой части головы, шее.
Профилактика фурункулеза
Установить носительство 5. aureus (посевы со
слизистой носа, подмышечных впадин, промеж-
ности, кишечника).
Определить чувствительность возбудителя к
антибиотикам.
Пользоваться антисептическим мылом.
Смазывать слизистую носа и складки тела 2 %
мазью мупироцина.
Антибиотики в среднетерапевтических дозах
применяют до полного исчезновения фурунку-
лов, затем переходят на поддерживающий при-
ем антибиотика 1 раз в сутки в течение длитель-
ного периода.
Для устранения носительства 5. aureus реко-
мендуется прием рифампицина 600 мг/сут в те-
чение 7—10 дней.
Назначается стимулирующая терапия (имму-
нотропные средства, биостимуляторы, витамины
группы В, антиоксиданты, адаптогены), курсы
ультрафиолетовых облучений в субэритемных
дозах.
Обследование ЖКТ, ЛОР-органов, эндокрин-
ной системы, иммунного статуса.
Карбункул
Острое обширное гнойно-некротическое воспа-
ление кожи и подкожной клетчатки в результа-
те слияния нескольких рядом расположенных
фурункулов.
Возбудитель: коагулазопозитивный золотис-
тый стафилококк, нередко в ассоциации со
стрептококками. Патогенное действие инфекци-
онных агентов на фоне резкого снижения им-
мунобиологической реактивности организма
вызывает обширную флегмонозную реакцию
вплоть до фасций и мышц.
Предрасполагающие факторы и излюбленная
локализация: см. «Фурункул».
Клиническая картина
Карбункул образуется в результате слияния меж-
ду собой нескольких фурункулов и проходит те
же стадии развития, однако с образованием ин-
фильтрата более крупных размеров и зоны вы-
раженного перифокального отека. Появление
карбункула сопровождается резкой болезненно-
стью в очаге поражения, повышением темпера-
туры тела, недомоганием, головной болью, изме-
нениями в крови (лейкоцитоз, ускоренная СОЭ).
Характерны множественные, глубокие, многока-
мерные абсцессы, поверхностные пустулы, некро-
тические стержни, после отторжения которых
выявляется симптом «сита» — наличие на повер-
хности очага большого числа отверстий с гной-
ным отделяемым. Далее кожа в очаге расплавля-
ется с образованием глубокой язвы, дно которой
покрыто некротической массой грязно-зеленого
цвета. После ее отторжения язва медленно (в те-
чение 2—4 недель) выполняется грануляциями с
последующим образованием грубого рубца. Тече-
ние карбункула может быть сопряжено с рядом
осложнений (см. «Фурункул»).
Лечение
Лечение проводят в отделении гнойной хирур-
гии. Назначают антибиотики с высоким терапев-
тическим индексом, особенно в случаях опасной
локализации карбункула (амоксициллин/клаву-
ланат, ампициллин/сульбактам, цефтазидим,
цефотаксим, нетилмицина сульфат, меропенем,
ванкомицин), проводят мероприятия по деток-
сикации организма; флюктуирующие зоны кар-
бункула вскрывают и дренируют, вводят проте-
олитики. Для ускорения заживления язвенного
дефекта применяют местные средства, ускоряю-
щие регенеративные процессы (аргосульфан,
мази «СП» и Микулича, ируксол, вулнузан, сол-
косерил, аэрозоли, обладающие антимикробным
и эпителизирующим действием).
'Гидраденит
Острое или рецидивирующее гнойное воспаление
апокриновых потовых желез с преимущественной
126 Частная дерматология
локализацией в подмышечных областях, реже —
в перианальной области, в области половых губ,
лобка, пупка, околососкового ареола.
Возбудитель: коагулазопозитивный золотис-
тый стафилококк, нередко в ассоциации с про-
теем, клебсиеллами, кишечной палочкой.
Предрасполагающие факторы: повышенное по-
тоотделение, недостаточная гигиена указанных
областей, травматизация кожи при подбривании
подмышечных впадин, лобковой области, дис-
функция (гипофункция) половых желез, гипо-
витаминозы, вторичная иммунная недостаточ-
ность.
Излюбленная локализация: аксиллярная об-
ласть.
Клиническая картина
Заболевание начинается с появления в подмы-
шечной области или на других участках кожи,
содержащих апокриновые потовые железы, од-
ного или нескольких болезненных узлов с виш-
невую косточку или лесной орех, которые мо-
гут увеличиваться в размерах до голубиного —
куриного яйца и более. Узлы спаиваются с ок-
ружающей подкожной клетчаткой в болезнен-
ный инфильтрат, имеющий бугристую соско-
видную поверхность, напоминающую «сучье
вымя». В результате абсцедирования кожа над
узлами истончается, размягчается, появляется
флюктуация. Узлы вскрываются, а из образовав-
шихся свищевых отверстий выделяется большое
количество сливкообразного гноя (без некроти-
ческого стержня). Процесс протекает в среднем
2—3 недели и заканчивается рубцеванием. В от-
дельных случаях инфильтраты могут рассасы-
ваться без вскрытия.
Нередко возникают рецидивы гидраденита (на
фоне сниженного иммунитета). При частых ре-
цидивах в гнойный процесс вовлекаются все
новые апокриновые железы. Участки поражения
в таких случаях представлены рубцово-изменен-
ной тканью, свежими воспалительными узлами,
свищевыми отверстиями, из которых при надав-
ливании выделяется жидкая гнойная масса
(иногда с примесью крови).
Общие явления: нередко температура, сла-
бость; выявляются лейкоцитоз, ускоренная
СОЭ.
Течение: острое, без нарушения (с нарушени-
ем) общего состояния; хроническое рецидивиру-
ющее.
Дифференциальный диагноз
Фурункул (инфильтрат, узел с некрозом и стер-
жнем, язва, рубец), лимфаденит при болезни
«кошачьих царапин», скрофулодерма, глубокие
микозы, актиномикоз.
Лечение
Лечение гидраденита практические не отличается
от лечения фурункула. Больным гидраденитом
с рецидивирующим и вялым течением рекомен-
дуют физиотерапевтические процедуры (ультра-
звук, УВЧ, соллюкс, УФО) в сочетании с натив-
ным стафилококковым анатоксином другими
иммунокорректорами (тактивин, тималин и др.).
В некоторых случаях показано иссечение рубцо-
во-измененных тканей.
Стафилодермии у детей
У новорожденных детей до 1 года нередко воз-
никают стафилококковые поражения кожи не-
фолликулярного характера. Наиболее часто про-
являются образованием пустул в области устьев
потовых желез. У ослабленных детей и при не-
своевременном лечении процесс распространяет-
ся вглубь с образованием множественных абсцес-
сов (псевдофурункулез), флегмон, развитием
септического состояния с неблагоприятным ис-
ходом. В некоторых случаях наблюдается обшир-
ная эксфолиативная реакция эпидермиса с тя-
желым токсикосептическим синдромом
(эксфолиативный дерматит Риттера), которую по
тяжести развития и течения сравнивают с синд-
ромом стафилококковой обожженной кожи
(staphylococcal scalid skin syndrome — «SSSS»).
Эксфолиативный токсин у новорожденных об-
разуется в первичных очагах инфекции (гной-
ный конъюнктивит, средний отит, инфекции
носоглотки) и гематогенным путем попадает в
кожу или образуется в самих пузырях (буллез-
ное импетиго). У взрослых SSSS почти не встре-
чается, так как у них эпидермолитический ток-
син быстро разрушается и элиминируется из
организма в связи с более совершенными им-
мунными механизмами.
Везикулопустулез (перипорит)
Гнойное воспаление устьев эккринных пото-
вых желез преимущественно у новорожденных
детей, находящихся на искусственном вскар-
мливании.
Предрасполагающие факторы: развивается у
недоношенных детей, при искусственном вскар-
мливании, нарушении правил пеленания детей
(повышенная потливость).
Излюбленная локализация: волосистая часть
головы, складки кожи.
Глава 7. Пиодермии
127
Клиническая картина
Пустулы с булавочную головку и венчиком ги-
перемии с плотной покрышкой, нередко сгруп-
пированные и быстро распространяющиеся. У
ослабленных детей процесс может прогрессиро-
вать вглубь с образованием абсцессов, флегмон,
лимфаденита, развитием септического состояния,
анемии, отита, бронхопневмонии, пиелонефри-
та (плохой уход, несвоевременно начатое лече-
ние).
Длительность заболевания при благоприятном
исходе 7—14 дней.
Дифференциальный диагноз
Буллезное импетиго.
Лечение
Очаги обрабатывают антисептическими и бакте-
рицидными средствами (0,25—1 % раствор нит-
рата серебра, 2 % раствор бриллиантового зеле-
ного), мази с антибиотиками (эритромициновая,
гелиомициновая, линкомициновая и др.). При
тяжелом течении лечение проводят как при псев-
дофурункулезе.
Псевдофурункулез Фингера
(множественные абсцессы
У детей)
Гнойное расплавление эккринных потовых же-
лез. Развивается у ослабленных, недоношенных,
находящихся на искусственном вскармливании
и недостаточно ухоженных детей с повышенным
потоотделением.
Излюбленная локализация: волосистая часть
головы, задняя поверхность туловища, ягодицы,
бедра.
Клиническая картина
Пораженные потовые железы вначале пальпиру-
ются в виде твердых узлов величиной до виш-
невой косточки синюшно-багрового цвета, ко-
торые затем размягчаются, вскрываются с
выделением сливкообразного гноя, без наличия
некротического стержня, последующим образо-
ванием свищевых ходов и язв, которые при ра-
циональном лечении и уходе рубцуются, инфиль-
траты рассасываются.
При ухудшении состояния ребенка появля-
ются новые воспалительные узлы, абсцессы,
могут развиться флегмоны, менингит, перито-
нит, сепсис, увеличение печени, селезенки, ги-
потрофия. Прогноз в этих случаях неблаго-
приятен.
Дифференциальный диагноз
Везикулопустулез, фурункулез.
Лечение
Обтирают кожу дезинфицирующими спиртовы-
ми растворами. Вскрывают абсцессы и смазыва-
ют их растворами анилиновых красителей, ма-
зями (левомеколь, левосин), пастой с цинка
оксидом и ксероформом (5-10 % ксероформа).
На абсцессы назначается УВЧ, повязки с 10 %
раствором натрия хлорида, общее УФО (10—15
сеансов).
При общетоксических явлениях антибиотико-
терапия, переливание нативной и антистафило-
кокковой плазмы, гемодеза, альбумина, поли-
глюкина, полиглобулина, 20 % раствора
глюкозы; антистафилококковый иммуноглобу-
лин.
Гемотерапия материнской (донорской) кро-
вью, ферроплекс, гемостимулирующие средства,
фитоферролактон, витамины С, В6, А, Е, каль-
ция пантотенат.
Эпидемическая
пузырчатка новорожденных
(эпидемический пемфигоид)
Острая контагиозная поверхностная стафилодер-
мия, возникающая в первые дни жизни ребен-
ка, в основном до 7-10-го дня после рождения.
Возбудитель: золотистый стафилококк, реже —
другие штаммы стафилококков, стрептококки и
иные микроорганизмы. Основное действие на
эпидермис оказывает стафилококковый эксфо-
лиативный токсин, вызывающий поверхност-
ную буллезную реакцию.
Источники инфекции: роженицы, медицинс-
кий персонал. В родильных домах заболевание
может иметь характер эпидемической вспышки.
Предрасполагающие факторы: патогенные ста-
филококки могут попадать на кожу новорожден-
ного из плохо обработанного, инфицированно-
го пупка воздушно-капельным и механическим
путем при уходе за ребенком.
Излюбленная локализация: туловище.
Клиническая картина
Эпидемическая пузырчатка новорожденных ча-
сто начинается с появления пузырей на гипере-
мированном фоне. Покрышка пузыря — бело-
ватого цвета, вялая, дряблая, морщинистая и
возвышается над уровнем кожи. Пузыри увели-
чиваются в размерах из-за выраженного аканто-
лиза в шиповатом слое и легко отслаиваются.
128 Частная дерматология
Содержимое пузырей вначале серозное, затем —
серозно-гнойное.
Пузыри легко разрываются, образуя эрозив-
ные поверхности с неровными, фестончатыми
краями и сочным, розово-красным или бледно-
розовым дном. При неблагоприятном течении
заболевания эрозии сливаются между собой,
быстро захватывая значительную часть кожно-
го покрова.
При благоприятном течении и лечебных ме-
роприятиях эрозии эпителизируются с образо-
ванием временной вторичной пигментации и
шелушения.
Иногда распространенные поражения кожи
могут осложняться импетигинизацией слизис-
тых оболочек полости рта, глаз, половых орга-
нов, тяжелым общим состоянием ребенка. Тя-
жесть заболевания обычно пропорциональна
площади вовлеченных в процесс кожи и сли-
зистых оболочек. Возможны пневмония, оти-
ты, диспепсия, сепсис. В подобных случаях
прогноз для жизни ребенка становится небла-
гоприятным.
Дифференциальный диагноз
Пиококковый пемфигоид новорожденных сле-
дует дифференцировать от сифилитической пу-
зырчатки, при которой пузыри локализуются
преимущественно в области ладоней и подошв.
Диагноз подтверждается обнаружением бледных
трепонем, а также серологическими исследова-
ниями крови ребенка и матери.
Пиококковую пузырчатку новорожденных
необходимо отличать также от врожденной пу-
зырчатки. Возникают пузыри с серозным, сероз-
но-геморрагическим содержимым в области ла-
доней и пальцев кистей, на коже локтевых и
коленных суставов, на подошвах, ягодицах,
реже — на других участках, подвергающихся
трению, давлению.
Лечение
В тяжелых случаях вводятся антибиотики с пред-
варительным определением чувствительности
(цефалоспорины, фузидин-натрий), показано
внутривенное вливание антистафилококковой
плазмы, альбумина, внутримышечное введение
антистафилококкового иммуноглобулина, тима-
лина.
Наружное лечение. Ежедневные ванны с ка-
лия перманганатом 1 : 10 000 и смена белья.
Вскрытие пузырей, обработка здоровой кожи
фукорцином, салициловым спиртом, прокипя-
ченным растительным маслом. На очаги — па-
сты и мази с антибиотиками, цинковое масло,
нафталановый линимент, аэрозоли с антибио-
тиками. Общее УФО. Необходима изоляция
детей в боксы и тщательный уход за ними. Об-
следование матерей и медперсонала йа носитель-
ство патогенных стафилококков. Кварцевание
палат, боксов. Дезинфекция отделений, каран-
тинные мероприятия.
Эксфолиативный
дерматит Риттера
Тяжелая форма эпидемической пузырчатки но-
ворожденных (син.: синдром стафилококковой
обожженной кожи). Характеризуется развитием
диффузной эритемы, образованием крупных
вялых пузырей. Напоминает поверхностную
ожоговую реакцию.
Возбудитель: золотистый стафилококк II фа-
гогруппы, выделяющий эксфолиативный ток-
син, вызывающий массивный акантолиз в эпи-
дермисе и образование крупных поверхностных
пузырей.
Заражение новорожденных происходит в бли-
жайшие дни после рождения от инфицирован-
ных золотистым стафилококком матерей, мед-
персонала. Реже заболевание развивается у детей
старших возрастных групп и взрослых (на фоне
иммунодепрессивной терапии).
Излюбленная локализация: начало в области
пупка и рта с распространением на другие учас-
тки кожи.
Клиническая картина
Различают 3 стадии болезни: эритематозную,
эксфолиативную и регенеративную.
Заболевание обычно начинается в местах из-
любленной локализации с появления быстро
распространяющейся эритемы. На этих участ-
ках возникают пузыри с дряблой покрышкой,
которые быстро вскрываются и образуют эро-
зии с отслаивающимся по периферии эпидер-
мисом (симптом Никольского положитель-
ный). В течение нескольких дней поражается
весь кожный покров с клинической картиной
«обваренной кожи». Отмечается высокая тем-
пература, потеря веса, понос; возможны ослож-
нения (пневмония, отиты, гнойный конъюн-
ктивит, сепсис). В крови лейкоцитоз,
повышение СОЭ, анемия, диспротеинемия.
Прогноз весьма серьезен.
Дифференциальный диагноз
Врожденная пузырчатка, сифилитическая пу-
зырчатка; у взрослых — синдром Лайелла (ле-
карственный).
Глава 7. Пиодермии 129
Лечение
Парентерально вводят антибиотики с предвари-
тельным определением чувствительности (цефа-
лоспорины, фузидин-натрий), показано внутри-
венное вливание антистафилококковой плазмы,
альбумина, внутримышечное введение антиста-
филококкового иммуноглобулина, тималина.
Ежедневные ванны с калия перманганатом 1: 10
ООО и смена белья. Вскрытие пузырей, обработ-
ка здоровой кожи фукорцином, салициловым
спиртом, прокипяченным растительным маслом.
На очаги — пасты и мази с антибиотиками, цин-
ковое масло, нафталановый линимент, аэрозоли
с антибиотиками. Общее УФО. Необходима изо-
ляция детей в боксы и тщательный уход за
ними. Обследование матерей и медперсонала на
носительство патогенных стафилококков. Квар-
цевание палат, боксов. Дезинфекция отделений,
карантинные мероприятия.
Стрептококковые
пиодермии
Этиология
Возбудители стрептококковых пиодермий стреп-
тококки являются преимущественно факультатив-
ными анаэробами. По характеру роста на кровя-
ном агаре подразделяются на гемолитические,
зеленящие и негемолитические; по характеру ге-
молиза — на 4 типа: b, a, al, g; на основании ан-
тигенных свойств выделяют 17 серологических
групп. Наиболее частый возбудитель заболеваний
человека — b-гемолитический стрептококк (51
серологический тип). Для его выделения исполь-
зуют бульоны — сахарный, кровяной, сывороточ-
ный, асцитный. Стрептококки образуют эндо- и
экзотоксины, ферменты. Экзотоксины (гемолизи-
ны, стрептолизины, лейкотоксин, некротоксины,
фибринолизин, токсин летальности, эритроген-
ный) обладают цитотоксическим, иммуносупрес-
сивным, пиогенным действием. Стрептококки вы-
рабатывают дезоксирибонуклеазу, гиалуронидазу,
стрептокиназу, растворяющие сгусток плазмы;
амилазу, рибонуклеазу, протеиназу и другие фер-
менты, обеспечивающие питание, рост и размно-
жение микроорганизмов.
Различные штаммы стрептококков высоко-
чувствительны к пенициллину. Стрептококки,
вызывающие пиодермию, относятся в основном
к серологическим группам А и D и вызывают
типичные поверхностные и глубокие поражения
кожи и слизистых (табл. 7.3).
Обшая характеристика
стрептококковой пустулы
Нефолликулярная пустула (фликтена), распро-
страняется по периферии, покрышка вялая, бе-
рется в складку. Безболезненная, наполнена се-
розно-желтоватым содержимым. Исход —
преимущественно эрозия, красноватая исчезаю-
щая пигментация.
Импетиго стрептококковое
Т ил ьбери—Фокса
Острое контагиозное заболевание кожи, вызы-
ваемое преимущественно патогенными стрепто-
кокками и проявляющееся появлением на теле
и волосяном покрове поверхностных тонкостен-
ных вялых пузырей (фликтен).
Возбудитель: преимущественно р-гемолитичес-
кий стрептококк группы A (Streptococcus
pyogenes). Бактерии вырабатывают экзотоксины,
которые служат причиной эксфолиативной ре-
акции кожи.
Предрасполагающие факторы: детский возраст,
несоблюдение личной гигиены, мелкие травмы,
высокая температура и влажность окружающей
среды, антисанитарные условия, зудящие дерма-
тозы (нейродермит, почесуха, чесотка и др.),
укусы насекомых, глистная инвазия, алкого-
лизм, сахарный диабет.
Излюбленная локализация: лицо, руки, шея.
Таблица 7.3. Классификация стрептодермий
Особенности кожных поражений Клинические формы
Поверхностные формы Импетиго Тильбери-Фокса и его разновидности: кольцевидное, буллезное, околоногтевое, щелевидное (заеда, ангулит), слизистых оболочек
Глубокие формы Атипичные формы Эктима Простой лишай (эритематозно-сквамозная стрептодермия, сухая стрептодермия), папулезное сифилоподобное импетиго (пеленочный дерматит), острая диффузная стрептодермия
130 Частная дерматология
Клиническая картина
На открытых участках тела появляются упло-
щенные поверхностные тонкостенные вялые
пузыри (фликтены) с мутноватым содержимым
(иногда с примесью крови), окруженные венчи-
ком эритемы. Фликтены быстро вскрываются с
образованием эрозий, которые покрываются
чешуйко-корочками или нетолстыми корками.
После их отпадения образуется временная крас-
новатая пигментация.
Течение острое, эволюция обычно длится око-
ло недели; при распространенном поражении —
несколько недель.
Субъективные ощущения: небольшой зуд.
Варианты импетиго
Тильбери-Фокса
Кольцевидное импетиго — пустула, распространя-
ясь по периферии, образует кольцевидные фор-
мы, с периферическим эритематозным венчи-
ком.
Буллезное импетиго — образуются крупные
напряженные или дряблые пузыри с мутноватым
содержимым, которые не имеют воспалительного
венчика. Часто располагаются на пальцах кис-
тей и стоп, вблизи ногтевых пластинок.
Поверхностный панариций (турниоль) — пусту-
лы располагаются в виде подковы вокруг ног-
тевой пластинки по ходу ногтевого валика; при
периферическом росте происходит отторжение
верхних слоев эпидермиса, при этом обнажает-
ся эрозия с серозно-гнойным отделяемым.
Щелевидное импетиго (заеда, ангулит, инфек-
ционный угол) — в углах рта образуется неболь-
шая фликтена, затем эрозия с воротничком ро-
гового слоя, корочка, трещина. Протекает
хронически и сопровождается небольшой болез-
ненностью. Встречается у лиц с привычкой об-
лизывать губы, со слюнотечением. Дифферен-
цируют с кандидозом углов рта (корочка не
образуется), папулезным сифилидом (характер-
на инфильтрация в основании эрозии).
Щелевидное импетиго нередко наблюдается
вокруг носа и за ушными раковинами.
Импетиго слизистых — на слизистых оболоч-
ках рта, носа, конъюнктивы в виде очагов по-
являются дряблые округлые пузыри, быстро
вскрывающиеся; образовавшиеся эрозии быст-
ро покрываются дифтероидным налетом.
Лечение
Спиртовые растворы анилиновых красок, 10 %
борно-дегтярная мазь, мази с антибиотиками
(мупироцин, бактробан, банеоцин и др.), «Дер-
мозолон», «Тридерм», обтирание здоровой кожи
вокруг очагов спиртовыми растворами салици-
ловой (2 %) и борной кислот (1—3 %), левоми-
цетина (1 %).
Эктима
Глубокое язвенное поражение кожи, связанное
с действием стрептококковой инфекции и воз-
никающее на фоне сниженной иммунобиологи-
ческой реактивности организма.
Предрасполагающие факторы: хронические бо-
лезни, алкоголизм, гиповитаминозы, зудящие
дерматозы.
Излюбленная локализация: конечности; может
появиться на любом участке тела.
Клиническая картина
В дерме образуется небольшой узел, на верши-
не которого появляется маленький пузырек, пре-
вращающийся в плоский пузырь с мутно-гемор-
рагическим содержимым, с инфильтрированным
ярко-красным или синюшным основанием. Со-
держимое пузыря вместе с его покрышкой ссы-
хается в плотную коричневую корку, которая
сохраняется около 2-х недель, а затем отпадает,
оставляя рубцово-измененный участок. Обычно
наблюдаются единичные, но иногда множествен-
ные элементы. Эктимы часто осложняются лим-
фангитами, лимфаденитами, реже — флебитами
и абсцессами.
Течение хроническое.
Субъективные ощущения: боль.
Дифференциальный диагноз
Сифилитическая эктима (безболезненность, по-
ложительная RW), скрофулодерма, индуративная
эритема Базена, лейшманиомы.
Лечение
При множественных эктимах — антибиотики,
гамма-глобулин, аутогемотерапия, инъекции ви-
таминов Вр В6 витамины А, С, Р, стимуляторы
лейкопоэза, иммунитета (такгивин, тималин и др.).
Местно — мази с антибиотиками, стимулято-
рами регенерации (мазь Микулича, «СП», ирук-
сол и др.).
Атипичные формы
стрептодермий
В практической деятельности педиатры и дерма-
тологи сталкиваются с неклассическими прояв-
лениями стрептодермий, которые не укладыва-
Глава 7. Пиодермии
131
ются в клинику приведенных выше форм. Не-
редко симптомы сухой формы стрептодермии
(простой лишай), которые встречаются преиму-
щественно у детей на открытых участках тела,
воспринимаются как заболевание со стойкой
потерей кожей пигмента меланина — витилиго.
При неправильном уходе за ребенком в обла-
сти ягодиц и на прилегающих участках могут
развиваться папулоэрозивные элементы с началь-
ными высыпаниями в виде красноватых пятен,
без типичных фликтен (пеленочный дерматит).
Несоблюдение правил гигиенического ухода за
детьми, наличие сопутствующих заболеваний
(сахарный диабет и др.), ослабляющих защитные
функции организма, могут вызвать стрептокок-
ковые поражения кожи в виде обширных диф-
фузных эрозивных очагов с явлениями гипере-
мии и мокнутия, схожими с проявлениями
микробной экземы.
Разновидности атипичных форм стрептодермий:
простой лишай (эритематозно-сквамозная стреп-
тодермия), папулезное сифилоподобное импети-
го (пеленочный дерматит), острая диффузная
стрептодермия.
Простой лишай
Простой лишай — «сухая» форма стрептодермии,
без образования типичных фликтен. Развивает-
ся преимущественно у детей в осенне-зимний
период (функциональный гиповитаминоз, све-
товое голодание, фокальная инфекция и др.).
Излюбленная локализация: лицо, верхние ко-
нечности.
Клиническая картина
Крупные пятна бледно-розового цвета, слегка
шелушащиеся, с подчеркнутым фолликулярным
аппаратом (как бы припудренные). После лече-
ния остаются временные депигментированные
пятна.
Дифференциальный диагноз
Витилиго: депигментированные пятна не шелу-
шатся, имеют четкие границы.
Лечение
Мази с антибиотиками, комбинированные кор-
тикостероиды, содержащие антибиотики (гиок-
сизон, дермозолон и т. п.), 3 % желто-ртутная
мазь с добавлением резорцина и салициловой
кислоты (1 %). При распространенных формах
антибиотики внутрь (эритромицин), поливита-
мины, общее УФО. Рекомендуется ограничить
углеводы, сладости.
Папулезное
сифилоподобное импетиго
(пеленочный дерматит)
Папулезное сифилоподобное импетиго — разно-
видность стрептококкового импетиго у детей до
1 года жизни. Развивается при нарушении пра-
вил гигиены и ухода за ребенком.
Излюбленная локализация: туловище, кожа
ягодиц, реже бедер, голеней.
Клиническая картина
Красноватые пятна до 1 см в диаметре с форми-
рующимися на их поверхности воспалительны-
ми папулами, четко отграниченными от здоро-
вой кожи (розового, красновато-синюшного
оттенков). На поверхности папул возникают
фликтены, которые эрозируются. Эрозии в ди-
аметре меньше, чем папулы, кожа вокруг них
воспалена и инфильтрирована.
Дифференциальный диагноз
Сифилитические папулы: медно-красный цвет,
четкие границы, плотность, безболезненность,
отсутствие фликтен, перифокального воспаления;
RW, РИТ, РИФ положительны; в эрозированных
папулах обнаруживается бледная трепонема.
Лечение
Здоровые участки кожи обрабатывают 1-2 %
спиртовым раствором салициловой или борной
кислоты, пораженные — 0,25 % раствором нит-
рата серебра, 1—2 % раствором резорцина с пос-
ледующими аппликациями мазей и паст, содер-
жащих антибиотики, ксероформ, этакридина
лактат, дерматол.
Острая диффузная
стрептодермия
Острое ограниченное поражение кожи диффуз-
ного характера, вызванное стрептококками, к
которому предрасполагают потливость, мацера-
ция выделениями из гениталий, диабет, плохой
уход и др.
Излюбленная локализация: у взрослых это ко-
нечности вследствие травматизации кожи (раны,
ожоги) — «паратравматическая форма»; у де-
тей —складки (стрептококковая опрелость -
интертриго).
Клиническая картина
Высыпание сливающихся фликтен, образующих
сплошные эрозивные участки с фестончатыми
132 Частная дерматология
очертаниями и венчиком отслаивающегося эпи-
дермиса, явлениями отека, гиперемии, диффуз-
ным мокнутием, серозными корками, возмож-
но образование свежих кольцевидных фликтен
по периферии очага. У ослабленных больных
наблюдаются изъязвления с серозно-гнойным и
геморрагическим отделяемым, ссыхающимся в
корки, явления лимфангита и лимфаденита.
Нередки аллергические сыпи на отдаленных
участках тела.
Дифференциальный диагноз
Микробная экзема (четкие очаги, «серозные ко-
лодца», выраженное мокнутие).
Лечение
Примочки с подсушивающими дезинфицирую-
щими растворами (этакридина лактат 1: 1000,
0,25 % раствор нитрата серебра, 1—2 % раствор
резорцина). После стихания острых воспалитель-
ных явлений переходят на пасты (борно-цинко-
нафталанная, Лассара), аэрозоли (оксикорт, дек-
сакорт и др.), мази с антибиотиками
(гелиомициновая, эритромициновая) и содержа-
щие кортикостероиды (тридерм, лоринден С),
мази и пасты с содержанием дегтя, серы, нафта-
лана, АСД и другие.
Непораженные складки у детей протирают 2 %
салициловым спиртом, припудривают тальком с
содержанием 1 % сульфата меди или 3—5 % бор-
ной кислоты.
При явлениях лимфаденита, лимфангита —
преимущественно антибиотики, при наличии
пиоаллергидов — преимущественно гипосенси-
билизирующая терапия.
Стрептостафилодермии
(смешанные пиодермии)
В ряде случаев течение стрептококковых флик-
тен осложняется присоединением стафилококко-
вой инфекции. Элементы принимают гнойный
характер, склонны к быстрому распространению
по кожному покрову (вульгарное импетиго), по-
крываются гнойно-геморрагическими корками,
изъязвляются (вульгарная экгима), протекают в
виде диффузных эрозированных, инфильтриро-
ванных очагов (хроническая стрептостафилодер-
мия).
Разновидности стрептостафилодермии: вульгар-
ное импетиго (стрептостафилококковое), вульгар-
ная эктима, хроническая диффузная пиодермия.
Вульгарное импетиго
(стрептостафилококковое)
Часто встречающаяся контагиозная форма импе-
тиго, может носить характер эпидемической
вспышки в детском коллективе. У взрослых
встречается как осложнение при чесотке, вши-
вости, зудящих дерматозах.
Излюбленная локализация: кожа лица, верх-
них конечностей, туловища.
Клиническая картина
На фоне гиперемии появляется фликтена с се-
розным содержимым, которое в течение несколь-
ких часов принимает гнойный характер. Осно-
вание пустулы становится инфильтрированным,
окружено венчиком эритемы. Покрышка пусту-
лы вскрывается, образуется эрозия, отделяемое
которой ссыхается в сочные «медовые» корки,
нередко с наличием кровоточащих трещин. В те-
чение недели корки отторгаются, обнажая эри-
тематозные пятна, слегка шелушащиеся и посте-
пенно исчезающие. Элементы импетиго склонны
к периферическому росту, слиянию и могут
принимать характер сплошных очагов.
Дифференциальный диагноз
Многоформная экссудативная эритема, вульгар-
ный сикоз, истинная пузырчатка.
Лечение
Лечение — см. «Стрептококковое импетиго».
При обильной экссудации, гнойных корках про-
водят промывание элементов раствором калия
перманганата 1 : 5000, применяют примочки с
растворами этакридина лактата (1: 1000), резор-
цина, танина (1 %).
После отторжения корок и подсыхания эро-
зий — линименты или масляные взвеси, мази,
содержащие антибактериальные и противовоспа-
лительные средства.
. Вульгарная эктима
Ограниченное язвенное поражение кожи стреп-
тостафилококковой этиологии, возникающее у
лиц со сниженной реактивностью (хронические
болезни, авитаминозы, алкоголизм и др.).
Излюбленная локализация: конечности.
Клиническая картина
На месте фолликулярной пустулы или инфиль-
трированной фликтены возникает пузырь с
гнойно-геморрагическим содержимым, после
вскрытия которого образуется язва округлой
Глава 7. Пиодермии 133
формы с валикообразными краями, кровото-
чащим дном, покрытым гнойно-слизистым
некротическим налетом, ссыхающимся в кор-
ку. На месте отторгшейся корки выявляется
язвенный дефект без признаков регенерации,
который вновь заполняется коркой в резуль-
тате образования некротических масс с при-
месью крови. Процесс может протекать вяло
длительный период. При благоприятном ис-
ходе язва выполняется грануляциями и зажи-
вает рубцом.
Дифференциальный диагноз
Фурункул, сифилитическая эктима, скрофуло-
дерма, чесоточная эктима, язвы при лейшмани-
озе. Дифференцируют также с гангренозной
эктимой, которая в большинстве случаев вызы-
вается синегнойной палочкой (Pseudomonas
aeruginosa). В месте внедрения возбудителя об-
разуется болезненная язва, которая быстро не-
кротизируется в результате поражения сосудов.
В центре язва покрывается черным струпом, а по
ее периферии выявляется красный венчик. Раз-
вивается на фоне резкой нейтропении и длитель-
ной антибиотикотерапии (химиотерапии) у хи-
рургических и онкологических больных;
сопровождается септическим состоянием.
Лечение
При вялом заживлении вульгарных экгим про-
водят их туширование 1 % раствором нитрата се-
ребра, 2 % раствором протаргола; применяют
мази «СП», Микулича, «Солкосерил», «Ируксол»,
«Этоний», «Мадекассол». Проводят общеприня-
тый туалет язвы.
При множественных эктимах назначаются
антибиотики, биостимулирующая терапия, вита-
минотерапия.
Хроническая
диффузная пиодермия
Может развиться из очага затянувшейся острой
диффузной стрептодермий или вялотекущего
вульгарного импетиго (несколько месяцев).
Излюбленная локализация: голени, стопы у
лиц с гипостатическим синдромом (варикоз,
тромбофлебит, перенесенные травмы и др).
Клиническая картина
В очагах развивается выраженная инфильтрация
кожи с синюшным оттенком, эрозированными
участками, мокнутием и слоистыми желтовато-
серыми корками. Контуры очага неправильных
(фестончатых) очертаний с наличием венчика от-
слаивающегося эпидермиса. Очаги могут лихе-
нифицироваться с наличием шелушения и напо-
минать картину бляшечного псориаза.
Дифференциальный диагноз
Микробная экзема («экзематозные колодцы» и
мокнутие в виде капелек росы).
Лечение
Местная терапия не отличается от мероприятий,
применяемых при микробной экземе.
Проводится витаминотерапия (Вр В6, А, Е, С),
назначаются антигистаминные средства, биости-
муляторы, аутогемотерапия, иммуноглобулин,
алоэ, торфот. Местное и общее УФО, лазероте-
рапия, УФОК, лучи Букки в упорных случаях.
Хроническая
глубокая пиодермия
Клиническая картина хронических форм глубо-
кой пиодермии многообразна. Этиологическая
роль пиококков и их ассоциации с другими гно-
еродными возбудителями (протей, синегнойная
палочка и др.), микоплазмами, вирусами в воз-
никновении этих форм пиодермии недостаточ-
но выяснена. Приведенные ниже разновиднос-
ти клинических форм хронической глубокой
пиодермии встречаются редко, однако для них
характерно упорное длительное течение, резис-
тентность к терапии в связи с изменчивостью
биологических свойств пиококков и их устой-
чивостью к антибиотикам, неясностью патоге-
нетических иммуноаллергических нарушений в
организме больных.
Хроническая глубокая
язвенно-вегетирующая
пиодермия
Этиология: из очагов получают рост стафилокок-
ка, стрептококка, реже протея, кишечной, синег-
нойной палочки и др.
Клиническая картина
На местах различных пустул в области волосис-
той части головы, подмышечных и паховых
складок, лобка, голени появляются серпигини-
рующие изъязвленные бляшки синюшно-крас-
ного цвета мягкой консистенции, резко отгра-
ниченные от прилегающей к очагу здоровой
кожи венчиком гиперемии, выступающим над
134 Частная дерматология
их поверхностью, с неправильными округлыми
(овальными) очертаниями, наличием веррукоз-
ных корковых наслоений, язв, вегетаций, узлов,
свищевых ходов, из которых при надавливании
выделяется гнойно-кровянистая жидкость.
Течение: месяцы, иногда годы. Болеют чаще
лица 40—60 лет.
Дифференциальный диагноз
Скрофулодерма, бородавчатый туберкулез кожи,
глубокие микозы.
Пиогенная гранулема
(ботриомикома)
Этиология: в возникновении признается роль ас-
социации стафилококка и вирусной инфекции;
по гистологическим признакам относят к геман-
гиомам.
Клиническая картина
Появление пиогенной гранулемы связывают с
травматизацией кожи области лица, красной
каймы губ, кистей, стоп, слизистых оболочек. На
коже возникает образование полушаровидной
формы темно-красного цвета, умеренно болез-
ненное, мягкой консистенции с множеством
поверхностно расположенных сосудов, дольчатой
(сосочковой) поверхностью, легко кровоточащей,
без тенденции к самостоятельному разрешению
в течение длительного времени.
Дифференциальный диагноз
Вегетирующая пиодермия, йододерма, злокаче-
ственные опухоли, фрамбезиформные сифили-
тические папулы.
Шанкриформная пиодермия
Этиология: стафилострептококковая ассоциация.
Клиническая картина
Встречается у лиц различного возраста и пола
на гениталиях или других участках тела. Эле-
менты чаще единичные. В области вскрывше-
гося пузырька образуется безболезненная эро-
зия или язва в диаметре до лесного ореха,
правильных очертаний, с приподнятыми края-
ми, уплотненным основанием и геморрагичес-
кой коркой. При снятии корки обнажается
ровное дно красноватого цвета, покрытое скуд-
ным гнойным отделяемым. Регионарные лим-
фатические узлы увеличены, плотные, подвиж-
ные, безболезненные.
Дифференциальный диагноз
Твердый шанкр, постскабиозные узелки облас-
ти полового члена и мошонки, укусы насекомых,
плоскоклеточный рак.
Гангренозная пиодермия
(язвенный дерматит)
Этиология: выявляются признаки аллергическо-
го васкулита с сенсибилизирующей ролью различ-
ных микроорганизмов. Сочетается с системными
заболеваниями (язвенный колит, лейкоз и др.).
Клиническая картина
Обширные язвенные поверхности с неровными
подрытыми краями и нависающими обрывками
эпидермиса, приподнятыми валикообразными
краями и периферической зоной гиперемии. Дно
покрыто сочными кровоточащими грануляциями
и гнойно-кровянистым отделяемым с неприятным
гнилостным запахом. Язвы имеют тенденцию к
периферическому росту и рубцеванию. Очаги чаще
располагаются на нижних конечностях, могут быть
на туловище, верхних конечностях, лице. Заболе-
вание рецидивирует, прогноз зависит от эффек-
тивности лечения системного заболевания.
Дифференциальный диагноз
Глубокие микозы, скрофулодерма, гуммозный
сифилид, хроническая язвенно-вегетирующая
пиодермия.
Лечение глубоких пиодермий
Проводится антибиотикотерапия с учетом чув-
ствительности от 2—3 недель до 1—1,5 месяцев (в
зависимости от терапевтического индекса анти-
биотика). Показана специфическая иммунотера-
пия (стафилококковый иммуноглобулин, плазма,
анатоксин). Назначаются иммуномодуляторы,
биостимуляторы, витаминотерапия.
При хронических вегетирующих формах пи-
одермии применяют цитостатики: проспидин в/
м 50 мг (1—2 инъекции), затем по 100 мг, на
курс 1,5—2 г, или циклофосфан (циклофосфа-
мид) — в/м, ежедневно по 100 мг, на курс 1—
1,5 г; кортикостероиды — 20—40 мг в сутки (в
перерасчете на преднизолон) 3—6 недель.
Хирургическая тактика: удаление и кюретаж
участков поражения, их рассечение; электроко-
агуляция свищевых ходов.
Из средств наружной терапии показаны прижи-
гающие средства, дезинфицирующие мази, пласты-
ри. Проводится обработка окружающей здоровой
кожи дезинфицирующими растворами, лосьонами.
Глава 7. Пиодермии 135
Пиоаллергиды
Вторичные высыпания, которые появляются на
кожных покровах в результате сенсибилизации
организма к пиококкам. Наблюдаются чаще при
длительном течении стрептодермий, глубоких
пиодермиях, назначении нерациональной наруж-
ной терапии (примочки с фурациллином, мазе-
вые повязки и др.), некоторых иммуностимули-
рующих препаратов.
Клиническая картина
Вторичные высыпания при обострениях пиодер-
мии проявляются чаще всего в виде распростра-
ненных симметричных пятнистых (розеолезных)
или мелко-папулезных (фолликулярных) высы-
паний, значительно отдаленных от основных оча-
гов поражения. Нередко высыпания могут быть
везикулярного или уртикарного характера. На
ладонях и подошвенных поверхностях стоп пио-
аллергиды проявляются пузырьками и пузыря-
ми с плотной покрышкой и напоминают явления
дисгидроза при экземе и микозах стоп. В неко-
торых случаях обильное высыпание пиоаллерги-
дов сопровождается повышением температуры
тела и другими легкими общетоксическими ре-
акциями. Регресс элементов происходит обычно
в течение 2-х недель путем отпадения мелких
корочек и шелушения, не оставляющих следов.
Дифференциальный диагноз
Инфекционные экзантемы, аллергический дер-
матит, токсидермии, красный плоский лишай,
многоформная экссудативная эритема, микиды,
туберкулиды.
Лечение
Отмена местного средства, применявшегося на очаг
пиодермии, которое могло спровоцировать появ-
ление аллергидов. Проводится гипосенсибилиза-
ция (препараты кальция, антигистаминные сред-
ства). При температурной реакции назначаются
антибиотики. Местно (на участки высыпаний):
индифферентные болтушки, комбинированные
кортикостероиды (тридерм, целестодерм с гарами-
цином, оксикорт и др.) в виде кремов, аэрозолей.
Профилактика пиодермии
При стрептококковом и вульгарном импетиго —
изоляция и лечение больных детей до выздоров-
ления. Не мыть водой пораженные участки
кожи. Кожу вокруг очагов протирать лосьона-
ми, содержащими 1—3 % салициловой и борной
кислоты, резорцина, левомицетина. Избегать
повязок и компрессов.
Предупреждение микротравматизма на произ-
водстве и в быту (санитарно-технические мероп-
риятия).
Санитарно-гигиенические мероприятия (лич-
ная гигиена, чистое белье и спецодежда, уборка
рабочих мест, вентиляция, индивидуальные па-
кеты и др.).
Тщательное обследование беременных, меди-
цинского и обслуживающего персонала на пио-
кокковую инфекцию в родовспомогательных и
детских учреждениях.
Проведение мероприятий, повышающих об-
щую сопротивляемость организма (закаливание,
рациональное питание, уход за кожей, витами-
низация, лечение сопутствующих заболеваний и
очагов инфекции).
Санитарно- просветительная работа.
Наружные средства
для лечения пиодермий
К этой группе относят спиртовые растворы ани-
линовых красителей (пиоктанин, метиленовый
синий, бриллиантовый зеленый) и йода; жидко-
сти, содержащие фуксин, фенол, борную и са-
лициловую кислоты, резорцин, дихлорид ртути,
жидкость Кастеллани, фукорцин, жидкость
Вильчинского; детергенты (зеленое мыло, мыль-
ный спирт); линименты, пасты с содержанием
дегтя, ксероформа, этакридина лактата, нафта-
лана, борной и салициловой кислот; мази, аэро-
золи, в состав которых входят антибиотики (нео-
мицин, гентамицин, эритромицин и др.),
антисептические средства (триклозан, дерматол,
ксероформ и др.).
Для стимуляции заживления «вялых», плохо
регенерирующих язв применяют мазц, содержа-
щие водорода пероксид (мазь «СП»), нитрат се-
ребра и винилин (мазь Микулича), фермент
клостридилпептидазу (мазь «Ируксол»), 5—10 %
растворы азотнокислого серебра, полиметилен-
мета-крезолсульфоновую кислоту («Ваготил»),
раствор «Куриозин»
Действие: антибактериальное, дезинфицирую-
щее, подсушивающее, противовоспалительное,
эпителизирующее, ускоряющее заживление язв.
Показания: первичные инфекции кожи (стреп-
тостафилодермии) .
Необходимо с осторожностью применять при
явлениях импетигинизации и экзематизации в
связи с возможным аллергогенным действием.
Методика применения: приведена в сигнатуре
к рецептам.
136 Частная дерматология
Рецепты наружных средств для лечения пиодермитов
Rp.: D. S. Sol. Pioctanini spirituosae 1 % 50,0 Смазывать очаги 2 раза в день Rp.: Spiritus saponatus 100,0 M. D. S. Смазывать участки поражения 2 раза в день
Rp.: Sol. Viridis nitentis spirit. 1 % 10,0 Rp.: Liquoris Castellani 50,0
D. S. Смазывать элементы сыпи 1-2 раза в день D. S. Смазывать очаги поражения 2 раза в день
Rp.: Saponis viridis 100,0 Rp.: Hydrargyri dichloridi 0,1 (!)
D. S. Для мытья волосистой части головы Ac. salicylic! 1,0
Laevomycetin! 1,5
Rp.: Spiritus saponatus compositus 100,0 Sp. aethylici 96 % 50 ml
D. S. Обрабатывать участки поражения M. D. S. Смазывать очаги 1-2 раза в день
2 раза в день
Rp.: Ol. Ricini 20,0
Rp.: Fucorcini 25,0 Xeroformii 1,2
D. S. Смазывать очаги 1 раз в день Vinylini 1,0
Rp.: Picis Liquidae D. S. Для мазевых повязок
Rp.: Ac. borici
Xeroformii aa 3,0
Ol. Ricini ad 100,0 Naphthalan! aa 0,5
M. D. S. Накладывать на очаг в виде мазевой повязки 1 раз в день Aethacridini lactatis 1,5 Pastae Zinci 15,0 M. D. S. Наносить на очаги 2 раза в день
Rp.: Linim. Streptocidi 5 % 30,0
D. S. Для повязок на раны Rp.: Ung. Xeroformii 5 % 100,0 D. S. Смазывать очаги 2 раза в день
Rp.: Aerosoli «Neogelasol» 60,0
D. S. Для орошения участков поражения Rp.: Ung. Erythromycin! 1 % 10,0 D. S. Смазывать очаги 2 раза в день
Rp.: Ung. Neomycin! sulfatis 2 % 15,0
D. S. Смазывать очаги поражения 2 раза в день Rp.: Ung. Dermatoli 5 % 50,0 D. S. Смазывать очаги 2 раза в день
Rp.: Ung. Heliomycini 4 % 25,0
D. S. Смазывать очаги 2 раза в день Rp.: Streptocidi 1,5
Rp.: D. S. Ung. «Garamycin» 15,0 Смазывать очаги 2 раза в день Sulfadimesini 1,0 Perhydroli 0,2 Lanolini Vaselini aa 23,0
Rp.: Argenti nitratis 0,1
Balsam! Schostakovsky 1,0 M. D. S. Смазать язвы 1 раз в день
Rp.: Sol. Argent! nitratis 10 % 10,0
Vaselini 10,0
M. D. S. Смазать язвы 1 раз в день
Da in vitro nigro!
Rp.: D. S. Ung. Irucsoli 30,0 Смазать язвы 2 раза в день тонким слоем M. D. S. Для прижигания грануляций
Rp.: Vagotili 50 ml
Rp.: Sol. Methyleni coerulei 1 % 10,0 D. S. Прижигать язву в течение 1 -3 мин тампоном,
D. S. Смазывать элементы сыпи 1-2 раза в день насыщенным препаратом, 2-3 раза в неделю
Rp.: lodi puri 5,0 Sp. aethylici 96 % ad 100 ml
M. D. S. Смазывать очаги 1 -2 раза в день ' *
Глава 8
Туберкулез кожи
Локализованные формы туберкулеза 138
Диссеминированные формы туберкулеза 143
Туберкулез кожи — хроническое инфекционное
заболевание, вызываемое Mycobacterium
tuberculosis, характеризующееся разнообразием
клинических форм, и представляющее собой тка-
невую реакцию повышенной чувствительности
микроорганизма на возбудитель.
В основе патологического процесса лежат спе-
цифические гранулемы, образующиеся в коже и
слизистых оболочках. Туберкулез кожи встреча-
ется значительно реже, чем туберкулез внутрен-
них органов, лишь у немногих больных (около
7 %) туберкулезом легких и других органов од-
новременно наблюдаются специфические пора-
жения кожи. Выраженность клеточных иммун-
ных реакций определяет клинические формы и
тяжесть инфекционного процесса. Иммунодефи-
цит способствует развитию тяжело протекающих
форм кожного туберкулеза. Имеются данные о
широкой распространенности туберкулеза кожи
у ВИЧ-инфицированных лиц.
Этиология
Туберкулезные поражения кожи у человека вы-
зываются преимущественно микобактериями че-
ловеческого типа (М. tuberculosis), реже бычьего
(М. bovis) и очень редко микобактериями пти-
чьего типа (Л/, avis). Клиническая картина по-
ражений кожи, за редким исключением, прак-
тически не зависит от типа возбудителя.
Возбудитель М. tuberculosis — это аэробные, не
образующие спор, неподвижные, кислото- и
спиртоустойчивые, грамположительные палочки
длиной 1—4 мк, шириной 0,2—0,6 мк. Характе-
рен полиморфизм микобактерий: различают
кокковидные, фильтрующиеся и L-формы. Наи-
более важной чертой окраски микобактерий яв-
ляется кислотоустойчивость, которую относят на
счет способности задерживать карболовый фук-
син после промывания кислотой или спиртом.
На искусственных питательных средах микобак-
терии туберкулеза растут медленно. Они имеют
сложный антигенный состав, который способен
вызывать Т- и В-клеточные иммунные реакции
макроорганизма. Размножаются микобактерии
поперечным делением и путем сложного цикла
развития. Главную роль при этом играют цитоп-
лазматические зерна, располагающиеся всегда на
полюсе клетки. На определенном этапе разви-
тия они начинают увеличиваться, от них отпоч-
ковываются одно или несколько зерен, при этом
цитоплазма постепенно лизируется, а появивши-
еся из тела микроба зерна могут вести самосто-
ятельное существование. В дальнейшем эти зерна
превращаются в новые палочки. В процессе та-
кого эволюционного развития зерна могут про-
ходить стадию временной потери кислотоустой-
чивое™ (зерна Муха).
Микобактерии туберкулеза сохраняют жизне-
способность и патогенность при очень низких
температурах. При нагревании в течение 5 мин
они могут выдерживать температуру до 80°
С. При кипячении в течение 1—5 мин микобак-
терии погибают. Прямые солнечные лучи уби-
вают их в чистой культуре за 1—1,5 часа.
Патогенез
Специфические поражения кожи развиваюся
лишь у немногих больных, страдающих тубер-
кулезом внутренних органов, что, вероятно, свя-
зано с иммунологическими свойствами барьер-
ного органа. В развитии туберкулеза кожи
важное значение придается путям проникнове-
ния возбудителя в кожу, общему состоянию
организма, состоянию кожи и анатомо — физи-
ологическим особенностям отдельных участков
кожного покрова.
Туберкулезом кожи можно заразиться тремя
путями. Первичное экзогенное внедрение мико-
бактерий туберкулеза в кожу встречается край-
не редко, преимущественно у ослабленных де-
тей. Экзогенная инфекция развивается при
нахождении ее источника вне организма и мо-
137
138 Частная дерматология
жет привести к развитию первичного туберку-
леза кожи в виде туберкулезного шанкра (пер-
вичный инокуляционный туберкулез) или боро-
давчатого туберкулеза кожи.
Второй путь проникновения инфекции —
эндогенное распространение, которое происходит
за счет местного вовлечения кожи (скрофулодер-
ма), распространения по соприкосновению (per
continuitatum) из пораженных туберкулезом лим-
фатических узлов (вторичная скрофулодерма)
или по кровеносным сосудам из очагов в легких
и лимфатических узлах (туберкулезная волчан-
ка, острый милиарный туберкулез кожи).
Третий путь распространения инфекции —
аутоинокуляция возбудителя из очага активно-
го туберкулеза внутренних органов (орифици-
альный туберкулез) встречается при попадании
микобактерий в кожу из мокроты.
Развитию туберкулеза легких способствует
подавление Т-клеточного звена иммунитета, сни-
жение неспецифической защиты организма, ос-
трые бактериальные или вирусные инфекции,
травмы, функциональные нарушения нервной
системы, эндокринные расстройства, недостаточ-
ное и неполноценное питание, гиповитаминозы,
длительная кортикостероидная и цитостатичес-
кая терапия; нарушения жирового, минерально-
го, водного обмена.
Обычно кожа является неблагоприятной сре-
дой для возбудителя туберкулеза из-за особен-
ностей pH, значительной аэрации, поэтому по-
павшие в нее микробактерии, как правило,
погибают. Лишь при нарушении сопротивляемо-
сти кожного покрова М. tuberculosis вызывают
специфическое воспаление, но в этих условиях
из очагов поражения высевают микробактерии
с пониженной вирулентностью, что также гово-
рит о неблагоприятных условиях существования
возбудителя в коже.
Большое значение в возникновении той или
иной клинической формы туберкулеза кожи
имеют анатомо — физиологические свойства не-
которых участков кожного покрова. Бородавча-
тая форма туберкулеза кожи обычно локализуется
на утолщенной и ороговевшей коже стоп и кис-
тей, уплотненная эритема Базена локализуется на
голенях. Бородавчатой формой туберкулеза стра-
дают исключительно мужчины, туберкулезная
волчанка развивается чаще у детей, эритемой
Базена болеют преимущественно женщины.
Патоморфология
На ранней стадии инфицирования макроорга-
низм отвечает неспецифической реакцией врож-
денного иммунитета, которая выражается экссу-
дацией, полиморфно — клеточной инфильтра-
цией. В инфильтрате обнаруживаются микобак-
терии. На последующих стадиях формируется
специфическая клеточная иммунная реакция,
выражающаяся в образовании тканевой туберку-
лезной гранулемы, которая состоит преимуще-
ственно из эпителиоидных и гигантских клеток
Лангханса, фибробластов, лимфоцитов с участ-
ком казеозного некроза в центре бугорка. Воз-
будитель редко встречается в тканевых грануле-
мах. Туберкулиновая проба положительная.
Различные клинические формы туберкулеза
кожи имеют разную степень тканевой реакции,
выражающуюся в изменении клеточного соста-
ва гранулемы, и чувствительность к туберкули-
ну. Так, классическая туберкулезная гранулема
наиболее характерна для туберкулезной волчан-
ки. Другие формы туберкулеза имеют структу-
ру гранулемы, ассоциированной с неспецифичес-
кой воспалительной реакцией.
Клиническая картина
Все клинические формы туберкулеза кожи мож-
но разделить на локализованные и диссеминиро-
ванные. К локализованным формам относятся:
первичный туберкулез кожи, туберкулезная вол-
чанка, колликвативный туберкулез кожи, тубер-
кулез кожи бородавчатый, язвенный туберкулез
кожи, туберкулез слизистых оболочек. Диссеми-
нированный туберкулез кожи объединяет клини-
ческие формы, очаги поражения которых распо-
лагаются рассеянно на различных участках тела:
острый милиарный, папулонекротический, инду-
ративный, лихеноидный. В настоящее время ро-
зацеаподобный туберкулид Левандовского (вол-
чанка лица милиарная) не относят к туберкулезу
кожи.
Локализованные
формы туберкулеза
Первичный туберкулез кожи
Первичный туберкулез кожи развивается при эк-
зогенной инокуляции инфекции и наблюдается
в настоящее время чрезвычайно редко у грудных
детей. Обычно возникает у ранее не инфициро-
ванных лиц при попадании возбудителя через рану
в кожу. Первые случаи первичного туберкулеза
были описаны в прошлом веке при процедуре
Глава 8. Туберкулез кожи
139
обрезания. Если священник, проводивший этот
обряд, страдал туберкулезом слизистой оболочки
полости рта, он становился источником зараже-
ния, поскольку останавливал кровотечение у ре-
бенка, зализывая раневую поверхность.
На месте внедрения возбудителя возникают
воспалительная кожная реакция и лимфаденит.
Необходимым условием является наличие по-
вреждения кожи или слизистой оболочки. Забо-
левание встречается главным образом у детей, но
описаны случаи возникновения этой формы и
у взрослых. Очаги поражения располагаются
обычно на открытых участках кожи (лицо, ко-
нечности), но могут локализоваться на конъюн-
ктиве, в полости рта и на половых органах. Пер-
вичный туберкулез кожи может развиться на
месте татуировки, вакцинации БЦЖ, а также на
месте травм (кошачьих царапин, укусов насеко-
мых, уколов острыми предметами или плохо
стерилизованными иглами, при прокалывании
мочек ушей) у непривитых людей.
Клиника
Через 8—10 дней, а иногда через несколько не-
дель появляется плотноватая папула красно-
коричневого цвета, которая превращается в
бляшку и изъязвляется (туберкулезный шанкр).
Вскоре возникает регионарный лимфаденит,
который имеет тенденцию к изъязвлению. Че-
рез несколько недель наступает заживление, и
лишь увеличенный регионарный лимфоузел
свидетельствует о первичном туберкулезном аф-
фекте.
Патоморфология
В первые две недели отмечается воспалительная
реакция с большим числом полиморфно-ядер-
ных лейкоцитов и микобактерий туберкулеза. В
следующие две недели гистологическая картина
меняется. Лимфоциты и эпителиоидные клетки
замещают полиморфно-ядерные лейкоциты. С
появлением эпителиоидных клеток количество
микобактерий быстро уменьшается. Туберкулез-
ная гранулема формируется не только в месте
внедрения возбудителя, но и в регионарных
лимфатических узлах в течение трех или четы-
рех недель после инфицирования.
Диагностика
Для постановки диагноза первостепенное значе-
ние имеет наличие язвы (туберкулезный шанкр)
и регионарного лимфаденита. При подозрении
на первичный туберкулез следует провести бак-
териоскопическое исследование отделяемого
язвы, в котором обычно легко обнаруживают ми-
кобактерии. Туберкулиновые пробы (реакции
Пирке и Манту) вначале отрицательные, а через
4—6 недель становятся положительными.
Дифференциальный диагноз
Дифференциальный диагноз туберкулезного
шанкра необходимо проводить от шанкроподоб-
ных поражений кожи при грибковых и бакте-
риальных заболеваниях, таких как споротрихоз,
бластомикоз, гистоплазмоз, кокцидиоидоз, но-
кардиоз, сифилис, лейшманиоз, туляремия,
фрамбезия, болезнь «кошачьих царапин». Все эти
заболевания дифференцируются на основании
клинических, бактериоскопических, культураль-
ных и патоморфологических данных.
Туберкулезная волчанка
Туберкулезная (обыкновенная или вульгарная)
волчанка — хроническое заболевание кожи, ко-
торое составляет около 75 % от общего числа
заболеваний туберкулезом кожи. Обычно начи-
нается в детском возрасте. Возбудитель попада-
ет лимфогенным или гематогенным путем из
очагов туберкулеза внутренних органов, а так-
же по соприкосновению.
Клиника
Первичным высыпанием при волчанке являет-
ся бугорок, вначале напоминающий пятно, ко-
торый называют люпомой. Поражаются преиму-
щественно кожа лица, особенно в области носа,
щек, около рта, ушных раковин (мочки), а так-
же шеи, реже конечностей, туловища. Люпома
состоит из множества микроскопических бугор-
ков и представляет собой инфекционную грану-
лему. Люпомы слегка выступают над поверхно-
стью кожи или расположены в толще собственно
кожи. Диаметр люпом составляет 2—5 мм, цвет
желтовато — красный, форма округлая, конси-
стенция тестоватая. При надавливании предмет-
ным стеклом (диаскопия) бугорок приобретает
желто-коричневый цвет — симптом «яблочного
желе». Бугорок часто выглядит прозрачным, при
легком надавливании на его поверхность пугов-
чатый металлический зонд легко проникает и
проваливается в бугорок, имеющий мягкую те-
стоватую консистенцию, обусловленную распа-
дом коллагеновых и эластических волокон. При
воспроизведении этого феномена (симптом про-
валивания зонда Поспелова или проба зондом
Поспелова) возникает кровоточивость бугорка в
месте давления зондом и ощущение болезненно-
сти. Бугорки склонны к смягчению, покрыва-
ются плоскими чешуйками. По периферии оча-
140 Частная дерматология
га можно наблюдать появление новых бугорков.
А в центральной зоне бугорки рассасываются,
оставляя атрофически измененную кожу, напо-
минающую папиросную бумагу, на поверхнос-
ти которой могут появиться новые люпомы. В
ряде случаев при плоской форме волчанки часть
бугорков может некротизироваться с образова-
нием поверхностных изъязвлений с неровными
остроконечными краями и слегка зернистым бо-
лезненным дном.
Выделяют следующие клинические разновид-
ности туберкулезной волчанки.
Язвенная форма (lupus ulcerosa) характеризу-
ется выраженной воспалительной инфильтрацией
с последующим распадом. Обычно развивается
под действием различных неблагоприятных фак-
торов (травм) и характеризуется образованием
поверхностной язвы с ровным, застойно-крас-
ным, покрытым мягкими грануляциями, легко
кровоточащим дном. Края язвы неровные, слегка
подрытые, инфильтрат в основании мягкий. На
месте изъязвления формируются атрофические
рубцы.
Туберкулезная волчанка слизистых оболочек
является наиболее тяжелой формой, при кото-
рой на слизистых оболочках носа, глаз, полости
рта образуются красно-желтые бляшки с зерни-
стой поверхностью, напоминающей рыбью икру.
Переход специфического процесса на хрящи носа
приводит к тяжелому обезображиванию лица —
мутиляции. Мутилирующая волчанка (lupus
mutilans) получила свое название в связи с глу-
бокими, обширными разрушениями подкожной
клетчатки, хрящевой части носа, ушных рако-
вин; отторжениями фаланг пальцев; расплавле-
нием сухожильных влагалищ, надкостницы,
костей.
При плоской форме (lupus planus) очаги не воз-
вышаются над уровнем окружающей кожи, по-
верхность их остается неповрежденной. При на-
личии на поверхности очага большого количества
мелких отрубевидных чешуек говорят о питири-
азиформной волчанке (lupus pityriasiformis).
Эксфолиативная форма (lupus exfoliativus) ту-
беркулезной волчанки характеризуется образо-
ванием крупных чешуек в зоне люпом. Чешуй-
ки беловатого цвета, плотно прилегают,
элементы располагаются на лице в виде «бабоч-
ки», что придает им сходство с дискоидной крас-
ной волчанкой. Иногда очаги волчанки по сво-
ему внешнему виду напоминают высыпания
псориаза (lupus psoriasiformis).
Саркоидоподобная форма туберкулезной вол-
чанки проявляется инфильтрированными эрите-
матозными очагами, возвышающимися над
уровнем окружающей кожи в виде опухолевых
образований.
Люпус-карцинома является самым грозным
осложнением туберкулезной волчанки. Характе-
ризуется малингизацией процесса и высокой
злокачественностью.
В большинстве случаев туберкулезная волчан-
ка возникает в детском или юношеском возрас-
те, чаще у девушек и молодых женщин. Харак-
терная локализация волчанки — кожа лица. Это
обусловлено частыми травмами лица, тесной
связью с лимфатическими сосудами, а также с
поражением шейных узлов туберкулезным про-
цессом. Течение Заболевания торпидное, много-
летнее. Периоды улучшения (особенно в летнее
время) сменяются периодами ухудшения. Наблю-
даются сочетанные поражения кожи и внутрен-
них органов. Туберкулезная волчанка осложня-
ется мутиляцией, рожистым воспалением,
лимфанитами, элефантиазом, импетигинизаци-
ей, люпус-карциномой.
Патоморфология
Для люпомы характерна туберкулезная грануле-
ма с двумя четко выявляемыми зонами: цент-
ральной и периферической. В центральной зоне
расположены казеозный некроз, туберкулезные
бактерии, а также эпителиоидные и гигантские
клетки Лангханса. Периферическая зона пред-
ставлена широким кольцом инфильтрата из лим-
фоидных клеток и фибробластов.
Диагностика
Диагноз туберкулезной волчанки основывается
на выявлении люпом с характерными симпто-
мами («яблочное желе», симптом зонда Поспе-
лова) в виде желтовато-застойных тестоватых
бугорков и бляшек, а также медленного течения
процесса с образованием язв и атрофических
рубцов в местах типичной локализации, резуль-
татов гистологического исследования и специфи-
ческой сенсибилизации к туберкулину.
Дифференциальная диагностика
Дифференциальная диагностика туберкулезной
волчанки проводится с бугорковым сифилидом
третичного периода, мелкоузелковым саркоидо-
зом, дискоидной красной волчанкой, туберку-
лоидной лепрой, туберкулоидной формой лей-
шманиоза, обыкновенными угрями, розацеа.
При сифилисе бугорки темно-красные, при ди-
аскопии не меняют цвет, плотно-эластической
консистенции (нет симптома «зонда»). При рас-
паде сифилитических бугорков язвочки глубо-
кие с плотными краями и дном. Диагноз сифи-
Глава 8. Туберкулез кожи
141
лиса может быть подтвержден серологическими
тестами, обнаружением возбудителя и другими
проявлениями сифилиса.
Дифференциальная диагностика с мелкоузел-
ковым саркоидозом нередко представляет значи-
тельные трудности, так как при диаскопии сар-
коидных узелков также имеется желто-бурый
цвет, но не сплошной, а точечный, пылевидный.
Гистологически саркоидная гранулема похожа на
туберкулезную, но казеозный некроз и микобак-
терии туберкулеза отсутствуют, выражены мел-
кие новообразованные капилляры, в цитоплаз-
ме гигантских клеток определяются тельца
Шаумана, а периферическая зона гранулемы
очень тонкая — «обнаженный бугорок». Реакции
Пирке и Манту при саркоидозе, как правило, от-
рицательны.
Для исключения дискоидной красной волчан-
ки следует ориентироваться на симптом фолли-
кулярного гиперкератоза и результаты патомор-
фологических и иммуноморфологических
исследований биопсии.
Колликвативный туберкулез
кожи (скрофулодерма)
Колликвативный туберкулез кожи (скрофуло-
дерма) представляет собой вторичное поражение,
исходящее из туберкулезного лимфаденита, ко-
стного, суставного или другого туберкулезного
процесса.
Клиника
Процесс начинается с появления в подкожной
клетчатке круглого, подвижного, плотного, без-
болезненного или малоболезненного узла разме-
рами 2—4 см, спаянного с кожей. Вначале кожа
над узлом не изменена, но по мере врастания
узла в кожу и спаивания с ней становится крас-
новатой, а затем синюшно-красной. Узел при
этом несколько выступает над поверхностью
кожи, кожа над ним истончается. Узел расплав-
ляется и вскрывается одним или несколькими
отверстиями, из которых выделяется гной, сме-
шанный с кусочками тканевого распада и кро-
вянистыми массами. Образующиеся при скрофу-
лодерме язвы неглубокие, неправильных
очертаний, их края мягкие, тонкие, подрытые,
вялые с гнойно-некротическим отделяемым.
Язвы нередко имеют между собой фистульные
ходы, открывающиеся свищевыми отверстиями.
При введении в свищевой ход зонда создается
впечатление пустоты («симптом дупла»). Дно
язвы вяло гранулирующее, заживление идет мед-
ленно и неравномерно. Вследствие этого форми-
руются неровные, местами втянутые, местами
возвышающиеся рубцы, между которыми обра-
зуются перемычки и мостики; возможны также
обезображивающие келоидные рубцы. Излюб-
ленная локализация скрофулодермы — зоны
расположения лимфатических узлов в области
шеи: в подчелюстной, надключичной и подклю-
чичной областях, а также зона подмышечных и
кубитальных лимфатических узлов; очень ред-
ко поражаются полость рта, язык.
Патоморфология
Патоморфологически в центре очага обнаружи-
вается неспецифический воспалительный процесс
с преобладанием в инфильтрате нейтрофильных
гранулоцитов. Иногда образуются абсцессы. В
более глубоких отделах дермы, а также по пе-
риферии очага можно обнаружить типичные ту-
беркулезные гранулемы с выраженным некрозом
в центре и значительной инфильтрацией вокруг.
Микобактерии туберкулеза обычно находят в
поверхностных отделах бугорков.
Диагностика
Диагностика скрофулодермы осуществляется на
основании характерных клинических признаков
(типичная локализация, характер рубцов, тубер-
кулезный процесс в лимфатических узлах, сус-
тавах, костях или внутренних органах) и бакте-
риоскопического анализа. Скрофулодерму
следует отличать от третичного гуммозного си-
филиса. Сифилитическая гумма более плотная с
язвой правильной формы, не подрытыми, кру-
тыми краями, с гуммозным стержнем на дне. В
диагностике помогают также серологические ре-
акции на сифилис. Иногда проводят дифферен-
циальную диагностику с гидраденитом и вуль-
гарной эктимой. Скрофулодерму следует
дифференцировать с кожными поражениями при
споротрихозе, бластомикозе, актиномикозе и ту-
ляремии с помощью культуральной, микроско-
пической и гистопатоморфологической техники.
Бородавчатый
туберкулез кожи
Бородавчатый туберкулез кожи возникает в ре-
зультате суперинфицирования и проникновения
возбудителя в кожу ранее инфицированных лю-
дей из экзогенного источника, реже — как след-
ствие аутоинокуляции. Заболевание чаще разви-
вается у ветеринарных работников, мясников,
патологоанатомов, фтизиохирургов, санитаров и
других лиц, работа которых связана с больны-
ми туберкулезом людьми или животными. Обыч-
142 Частная дерматология
но болеют мужчины, но могут инфицироваться
и дети.
Клиника
Бородавчатый туберкулез кожи локализуется пре-
имущественно на тыльной поверхности кистей и
пальцев рук, реже на разгибательной поверхнос-
ти стоп и пальцев ног, а также нижней трети го-
лени. Субъективные ощущения обычно отсутству-
ют. Первые проявления болезни напоминают
бородавку. Первичный бугорок буровато-красный,
плотный, с сероватыми папилломатозными выро-
стами на поверхности. Бугорок постепенно увели-
чивается и превращается в бляшкоподобное обра-
зование с характерными признаками: в центре
очага выражены гиперкератоз, сероватые бородав-
чатые разрастания; вокруг центра располагается
синевато-красное гладкое поле; по периферии
имеется воспалительный ободок лилово-красного
цвета. Расширение очага по периферии происхо-
дит медленно. Одновременно в центральной час-
ти может быть разрешение инфильтрата с разви-
тием атрофии в центре. Длительно существующий
бородавчатый туберкулез кожи может приобретать
злокачественный характер.
Патоморфология
Патоморфологически выявляют акантоз, гипер-
кератоз, папилломатоз. В дерме — сочетание
воспалительной и гранулематозной реакции.
Острый воспалительный инфильтрат состоит из
нейтрофильных гранулоцитов и лимфоцитов.
Микобактерии присутствуют в очаге поражения.
Диагностика
Диагностика обычно не вызывает затрудений.
Диагностике могут помочь, помимо типичной
клинической картины, анамнез, гистологические
данные, положительные туберкулиновые пробы.
Дифференциальная диагностика проводится с
бородавчатой формой красного плоского лишая,
глубоким бластомикозом кожи, йододермой,
бромодермой, вульгарными бородавками. При
бородавчатой форме красного плоского лишая
высыпания чаще локализуются на передней по-
верхности голени, сопровождаются сильным
зудом, цвет бляшек красно-фиолетовый; по пе-
риферии очагов и на других участках кожи мож-
но обнаружить типичные полигональные папу-
лы с пупковидным вдавливанием или сеткой
Уикхема. В случае глубокого бластомикоза кожи
типа Джилкрайста могут помочь бактериоскопи-
ческие исследования. Йододерма и бромодерма
представлены сосочковыми и бородавчатыми
разрастаниями, носят множественный характер,
часто локализуются на лице и волосистой части
головы. Бородавчатый туберкулез кожи отлича-
ется от вульгарных бородавок наличием трех
типичных зон, числом очагов, их величиной.
Язвенный туберкулез
кожи и слизистых оболочек
Язвенный туберкулез кожи и слизистых оболо-
чек развивается при сниженном иммунитете
больных и тяжелом течении туберкулеза горта-
ни, легких, почек, мочевого пузыря, кишечни-
ка и других органов в результате аутоинокуля-
ции микобактерий с мокротой, мочой, калом и
другими выделениями.
Клиника
Очаги поражения располагаются на слизистых
оболочках вокруг естественных отверстий: рта,
ноздрей, половой щели, заднего прохода, отвер-
стия мочеиспускательного канала. Первичным
морфологическим элементом является шаровид-
ный бугорок размером 1—2 мм. Бугорки множе-
ственные, быстро сливаются и изъязвляются.
Язвы резко болезненные, размером 1—1,5 см, с
мягкими подрытыми краями, имеют полицик-
лические мелкофестончатые очертания, их дно
покрыто мелкими узелками — «зернами Треля»
(казеозно перерожденные бугорки). В отделяе-
мом язв содержится огромное число микобакте-
рий туберкулеза. Болезнь протекает на фоне
иммунодепрессии и анемии, поэтому туберкули-
новые пробы отрицательные.
Патоморфология
Патоморфологически вокруг язвы обнаружива-
ется неспецифический воспалительный инфиль-
трат с преобладанием нейтрофильных грануло-
цитов. В глубоких отделах дермы выявляются
специфические туберкулоидные гранулы с казе-
озным некрозом в центре.
Диагностика
Диагностика проводится на основании анамне-
за, клинических проявлений и бактериоскопи-
ческого анализа. При отрицательных туберкули-
новых пробах может возникнуть сомнение.
Однако подтвердить диагноз позволяют обнару-
жение поражений специфическим процессом
гортани, легких, кишечника и большое количе-
ство микобактерий туберкулеза в очагах пораже-
ния. Дифференцируют язвенный туберкулез
кожи и слизистых оболочек от шанкроида, си-
филитической гуммы, венерической лимфогра-
нулемы, язвенной карциномы.
Глава 8. Туберкулез кожи 143
Диссеминированные
формы туберкулеза
Острый милиарный
туберкулез кожи
Острый милиарный туберкулез кожи — редкая
форма туберкулеза. Милиарная диссеминирован-
ная форма туберкулеза кожи развивается у де-
тей грудного и раннего детского возраста.
Клиника
Первичный элемент — милиарный бугорок
плотной консистенции, застойно-красного цве-
та с геморрагическим компонентом. В центре
элементов возникают некротический распад, а
затем — изъязвления. В отделяемом язвы мож-
но обнаружить микобактерии туберкулеза.
Патоморфология
Патоморфологически в поверхностных слоях дер-
мы развивается неспецифическая воспалитель-
ная реакция с явлениями деструкции ткани. Гра-
нулемы отсутствуют или появляются на поздних
стадиях, они единичны и расположены по краю
очагов.
Диагностика
Диагностика затруднена. При этой форме выявля-
ется анергия, и специфические кожные реакции
(Пирке, Манту) могут быть отрицательными. Выс-
казывается мнение, что эта форма не имеет прямой
связи с туберкулезом. Дифференциальную диагно-
стику проводят с саркоидозом, при этом могут по-
мочь гистологические данные и выявление саркои-
доза во внутренних органах и лимфатических узлах.
Папулонекротический
туберкулез кожи
Морфологической основой заболевания являет-
ся аллергический васкулит, вызванный в резуль-
тате сенсибилизации туберкулезными микобак-
териями. Вероятно, патологический процесс в
коже возникает при гематогенном попадании
микобактерий из очагов внутренних органов у
лиц с первичным хроническим туберкулезом.
Заболевание чаще встречается у лиц молодого
возраста (15—25 лет), особенно среди девушек.
Клиника
Высыпания локализуются на заднебоковых по-
верхностях предплечий и плеч, тыльной повер-
хности кистей и стоп, на задней поверхности
бедер, разгибательной поверхности голеней, лок-
тевых и коленных суставов, на ягодицах, на
ушных раковинах. Для этой формы характерно
симметричное расположение плотных, розоватых
или красных с синюшным оттенком папул ве-
личиной с конопляное зерно, чечевицу, чаще
полушаровидной формы. В центре папулы по-
степенно развивается некроз с образованием тем-
но-коричневой плотной корочки, которая через
несколько недель отторгается, обнажая четко
очерченный округлый вдавленный «штампован-
ный» рубчик, вначале гиперемированный, затем
депигментированный, иногда с ободком корич-
невого цвета. Заболевание рецидивирующее,
сопровождается периодическим возникновени-
ем новых высыпаний, чаще весной и осенью.
Клиническая картина часто бывает весьма пес-
трой: наряду с прежними синюшно-красными
папулами (или бугорками), а также темно-си-
нюшными гипер- и депигментированными бе-
лыми штампованными рубчиками на коже име-
ются свежие розовые или ярко-красные бугорки.
Туберкулиновые пробы обычно резко положи-
тельны. Эта форма туберкулеза нередко сочета-
ется с индуративным туберкулезом кожи, реже —
с колликвативным.
Патоморфология
Патоморфологически очаги состоят из эпители-
оидных клеток и лимфоцитов. В узелках отме-
чается некроз центральной части, окруженный
неспецифическим воспалительным, а по пери-
ферии гранулематозным инфильтратом. Отмеча-
ются сосудистые изменения в виде тромбоза мел-
ких вен, эндартериита, эндофлебита.
Диагностика
Диагностика в типичных случаях не представ-
ляет затруднений, если имеются основные кли-
нические признаки. Дифференциальную диаг-
ностику проводят с некротическим акне
(высыпания располагаются на коже лба и воло-
систой части головы), вульгарным акне (более
выраженные воспалительные явления, пустули-
зация и иной характер рубцов), аллергическим
васкулитом (в анамнезе нет туберкулезного про-
цесса внутренних органов), изолированными
бугорками третичного периода сифилиса (кли-
нические и серологические данные), острым
парапсориазом (наличие туберкулеза; отсутствие
полиморфизма высыпаний, характерных для
парапсориаза), узелковым некротическим выс-
кулитом, злокачественным атрофическим пусту-
лезом.
144 Частная дерматология
Лихеноидный туберкулез
кожи или лишай золотушных
Лихеноидный туберкулез кожи может разви-
ваться у лиц любого возраста, однако чаще
встречается у анемичных и гипертрофичных
детей с туберкулезом костей, лимфатических
узлов. Отмечено развитие болезни после инъ-
екций туберкулина у лиц, сенсибилизирован-
ных к туберкулину. Лихеноидный туберкулез
кожи возникает в результате гематогенной дис-
семинации микобактерий туберкулеза (иногда
возможен лимфогенный путь) и всегда соче-
тается с другими туберкулезными поражения-
ми кожи.
Клиника
Высыпания локализуются преимущественно на
боковых поверхностях туловища, спине, пояс-
нице, ягодицах, груди, реже — на коже лица,
бедер, волосистой части головы. Сыпь пред-
ставляет собой папулы величиной с булавоч-
ную головку, желтовато-коричневого цвета,
иногда цвета нормальной кожи. Папулы име-
ют плоскую или остроконечную форму, при-
урочены к волосяным фолликулам, склонны к
группировке, могут сливаться в бляшки раз-
личной формы и размеров, высыпания бесслед-
но регрессируют, иногда остаются маленькие
светло-коричневые пятна, реже — едва замет-
ные поверхностные рубчики. Течение заболе-
вания длительное, хроническое, доброкаче-
ственное. Для лихеноидного туберкулеза не
характерны субъективные ощущения, изредка
бывает незначительный зуд.
Патоморфология
Патоморфологически в перифолликулярных ин-
фильтратах обнаруживаются эпителиоидно —
клеточные гранулемы, как правило, без казеоз-
ного некроза в центре и со слабой лимфоцитар-
ной реакцией вокруг них. Возбудитель не обна-
руживается.
Диагностика
Установить диагноз лихеноидного туберкулеза
позволяют типичная локализация высыпаний,
отсутствие субъективных ощущений. Лихеноид-
ный туберкулез кожи дифференцируют с сифи-
литическими папулами, себорейной экземой,
псориазом, красным остроконечным, плоским и
блестящим лишаями.
Индуративный
туберкулез кожи
Индуративный туберкулез кожи (уплотненная
эритема Базена) развивается при гематогенном
или лимфогенном инфицировании кожи мико-
бактериями туберкулеза. Заболевание часто
встречается у девушек и молодых женщин со
сниженным иммунитетом, старадающих эндок-
ринопатиями, нарушениями периферического
кровообращения, подвергающихся переохлажде-
нию.
Клиника
В подкожной клетчатке преимущественно голе-
ней и вокруг коленных суставов возникают сим-
метричные, глубоко расположенные плотные
узлы, которые постепенно увеличиваются и пре-
вращаются в крупные узловатости уплощенной
формы. Узлы малоболезненные, немногочислен-
ные, изолированные друг от друга. В толще кожи
иногда прощупываются плотные бугристые тяжи,
которые идут от одного инфильтрата к другому.
Узлы медленно разрешаются, кожа над ними
вначале обычного, а затем застойно-синюшного
цвета. При пальпации узлов отмечается умерен-
ная болезненность. Узлы могут существовать
месяцами и полностью регрессировать, оставляя
пигментацию и легкую атрофию. В ряде случа-
ев узлы размягчаются и изъязвляются. Язвы
неглубокие, имеют округлые очертания, желто-
вато-красное дно, покрытое вялыми грануляци-
ями и серозно-кровянистым отделяемым. Пос-
ле заживления язв остается слегка втянутый
гладкий поверхностный рубец с гиперпигмента-
цией по периферии.
Патоморфология
Патоморфологически при индуративном тубер-
кулезе кожи в подкожной клетчатке и дерме
обнаруживаются обширные специфические ту-
беркулезные инфильтраты, состоящие в основ-
ном из эпителиоидных клеток и единичных
гигантских клеток. Отмечаются выраженные
изменения сосудов по типу эндоартерита и эн-
дофлебита; очаги казеозного некроза; неспеци-
фические инфильтраты, состоящие из лимфоци-
тов и плазматических клеток; постепенная
атрофия жировой ткани. Так как гистологичес-
ки при индуративной эритеме Базена в сосудах
очагов поражения находят изменения, близкие
к кожной форме узловатого периартериита, то
ряд авторов полагают, что эта форма не имеет
туберкулезной природы.
Глава 8. Туберкулез кожи 145
Диагностика
При дифференцировании заболевания с узлова-
той эритемой учитывают характерную для пос-
ледней множественность очагов (не только на
голенях), они имеют ярко-красный, синюшный
или коричневатый цвет, симптом «цветения си-
няка». Воспалительные явления выражены, узлы
плотно-эластической консистенции, болезненные.
Заболевание сопровождается общими явлениями
(слабость, лихорадка, недомогание), в крови по-
вышенная СОЭ, лейкоцитоз. От сифилитических
гумм узлы уплотненной эритемы Базена отлича-
ются нерезкой ограниченностью, излюбленной
локализацией на голенях, отсутствием других
признаков сифилиса, отрицательными серологи-
ческими реакциями. Узловатый аллергический
васкулит протекает остро, болезненно; реакции на
туберкулин отрицательные. При тромбофлебите
пораженные вены большей частью прощупыва-
ются в виде резко болезненных тяжей, над ко-
торыми кожа воспалена и отечна.
Лечение туберкулеза кожи
Лечение туберкулеза кожи проводится в соответ-
ствии с общими принципами противотуберку-
лезной терапии и должно быть комплексным,
непрерывным и длительным. Общие принципы
терапии: укрепление общего состояния больно-
го, применение активных противотуберкулезных
препаратов, физиотерапевтические процедуры.
Для получения хорошего терапевтического эф-
фекта от применения противотуберкулезных
препаратов необходимо соблюдать следующие
основные принципы химиотерапии. Лечение
следует начинать сразу же после установления
диагноза туберкулеза и проводить длительно.
Противотуберкулезные препараты необходимо
назначать в оптимальных дозах в комплексе с
другими лекарствами (витаминами, гормонами и
др.). Лечение будет эффективным, если оно
проводится несколькими различными препара-
тами (не менее трех) одновременно, т. е. комби-
нированно и обязательно поэтапно — в стацио-
наре, затем в санатории и, наконец, амбулаторно.
Следует учитывать переносимость больными
антибактериальных препаратов.
На стационарном этапе лечения противотубер-
кулезные лекарства назначаются ежедневно. Наи-
более эффективной лечебной схемой является
сочетание стрептомицина с рифампицином и
изониазидом. После 2—3 месячной терапии этими
препаратами стрептомицин отменяется, и лече-
ние продолжается рифампицином и изониази-
дом. В дальнейшем (на санаторном или амбула-
торном этапе) в зависимости от особенностей
клинического течения процесса, переносимости
лекарств, уже достигнутого терапевтического
эффекта лечение можно проводить интермитти-
рующим методом (через день, 1—2 раза в неде-
лю) двумя противотуберкулезными препаратами.
Противотуберкулезные препараты, применяемые
в настоящее время во фтизиатрической практике,
можно разделить по активности на препараты 1-
го ряда (основные) с высокой противотуберкулез-
ной активностью — гидразид изоникотиновой
кислоты (ГИНК) и его производные (тубазид,
фтивазид, салюзид, метазид); рифампицин; 2-го
ряда (резервные), менее активные. Их использу-
ют при развитии устойчивости микобактерий к
препаратам первого ряда. Это этионамид (проти-
онамид); этамбутол; циклосерин; пиразинамид;
канамицин; флоримицин; вибрамицин. Дозы пре-
паратов подбирают индивидуально, в зависимос-
ти от формы туберкулеза, возраста больного, со-
путствующих заболеваний.
Изониазид (тубазид) — наиболее эффектив-
ный препарат из группы ГИНК, применяется в
суточной дозе для взрослых 10—15 мг/кг, т. е.
0,6—0,9 г; для детей — из расчета 5—8 мг/кг, но
не более 10 мг/кг, или 0,5 г в сутки. Принима-
ют внутрь после еды 2—3 раза в сутки.
Метазид мало токсичен и более эффективен,
чем фтивазид. Назначают внутрь взрослым 1,0 г
в сутки (по 0,5 г 2 раза в день), на курс 100—
200 г. В течение года можно провести 2—4 кур-
са лечения с интервалом 1—2 мес. Следует сле-
дить за переносимостью препаратов этой группы,
так как нередко возникают головная боль, раз-
дражительность, желудочно-кишечные расстрой-
ства, парестезии, пеллагроидный дерматит, ко-
торый можно купировать приемом никотиновой
кислоты, инъекциями витаминов Bt и В6. Про-
тивопоказания для назначения препаратов гид-
разида изоникотиновой кислоты: стенокардия,
сердечная недостаточность, нарушения функций
печени и почек.
Фтивазид применяется в случаях, когда при
приеме тубазида возникают побочные реакции
(головокружение, боли в сердце, нарушения сна,
парестезии, признаки периферических неври-
тов). Суточная доза для взрослых составляет 1—
1,5 г (по 0,5 г 2—3 раза в день), для детей 20—
30 и 40 мг/кг, но не более 1, 5 г.
Салюзид назначают в дозе 0, 5 г 2—3 раза в
сутки, а также вводят парентерально при кол-
ликвативном туберкулезе в область свищей в
виде 5—10 %-ного водного раствора.
Рифампицин — полусинтетический антибиотик
широкого спектра действия. Препарат принима-
146 Частная дерматология
ют внутрь по 0,45—0,6 г/сут в 1 прием за 30 мин
до еды, продолжительность лечения до 1 года.
Суточные дозы для взрослых с массой тела ме-
нее 50 кг — 600 мг, для детей —8—10 мг/кг в
сутки, но не более 450 мг. Рифампицин (бенеми-
цин) является вторым по эффективности проти-
вотуберкулезным препаратом. Побочные эффек-
ты: эозинофилия, токсикодермия, тромбофлебит,
тошнота, диспептические расстройства. Противо-
показания: нарушения функции печени и почек.
В последнее время выпускается растворимый
рифампицин для парентерального введения и
комбинированный препарат (рифампицин + изо-
ниазид).
Стрептомицин используют при острых фор-
мах, при поражении слизистых оболочек и
внутренних органов. Стрептомицин назначается
в комбинации с другими противотуберкулезны-
ми препаратами, чаще всего с изониазидом,
этамбутолом, этионамидом. Суточная доза для
взрослых 1,0 г внутримышечно. Детям и под-
росткам препарат назначают из расчета 15-
20 мг/кг в сутки, но не более 0,5 г детям и
0,75 г подросткам. Побочные эффекты: сниже-
ние слуха, вестибулярные расстройства, альбу-
минурия, гематурия, токсидермия. Больным,
плохо переносящим стрептомицин, старше 60
лет и с массой тела менее 50 кг доза снижается
до 0,5—0,75 г. Стрептомицин применяют так-
же в аэрозолях, его раствор инсталлируют в
плевральную и брюшную полости, в каверны,
бронхи, казеозные лимфатические узлы, свищи.
Во время лечения необходимо обязательно еже-
месячно исследовать слух и вестибулярный ап-
парат, функции почек и сердце больного.
Парааминосалициловая кислота (ПАСК) —
наименее эффективный препарат первого ряда,
применяется внтурь в таблетках 0,5 г: суточная
доза 10—12 г, на курс лечения 400—800 г (60—
90 дней): детям 2—6 г/сут из расчета 0,15—0,
2 г/кг.
Препараты второго ряда (резервные) исполь-
зуются при развитии устойчивости микобакте-
рий к препаратам первого ряда.
Этионаимид обладает бактериостатической
активностью в отношении микобактерий тубер-
кулеза, устойчивых к другим препаратам. Пре-
парат рекомендуется вместо ПАСК в комбина-
ции с тубазидом, циклосерином по 0,25 г 3 раза
в день внутрь после еды. Суточная доза для де-
тей определяется из расчета 10—20 мг/кг, но не
более 0,75 г. Не следует давать этионамид вме-
сте с протионамидом, тиоацетазоном, солютизо-
ном (перекрестная устойчивость микобактерий
туберкулеза). Применение этионамида внутрь
противопоказано при язвенной болезни желуд-
ка и 12-перстной кишки, гастритах и колитах,
тяжелых формах сахарного диабета, при заболе-
ваниях печени, при беременности.
Пиразинамид применяют при устойчивости
микобактерий туберкулеза к препаратам группы
ГИНК. Он назначается вместо них или вместе с
тубазидом по 0,5 г 3—4 раза в день или по 0,75—
1,0 г 2 раза в день. Детям — по 20—30 мг/кг в
сутки, но не более 1,5 г. При интермиттирую-
щем лечении (2—3 раза в неделю) суточную дозу
для взрослых увеличивают до 2,0—2,5 г. Пира-
зинамид противопоказан при нарушении фун-
кций печени, при подагре. С осторожностью
применяют при язвенной болезни, сахарном ди-
абете. Для уменьшения токсического действия на
печень рекомендуется прием метионина.
Циклосерин, или серомицин — антибиотик
широкого спектра действия, заменяет стрепто-
мицин, хотя и менее активен в отношении ми-
кобактерий туберкулеза. Суточная доза для
взрослых 0,75 г (по 0,25 г 3 раза в день), пре-
парат принимают внутрь перед едой. Суточная
доза для детей определяется из расчета 10—20 мг/
кг, но не более 0,75 г. Побочные явления: тош-
нота, рвота. Противопоказания: органические за-
болевания периферической и центральной не-
рвной системы, эпилепсия, психические
расстройства, хронический алкоголизм, нарко-
мания.
Биомицин или флоримицина сульфат, также
обладает широким спектром антимикробного
действия. Препарат вводится внутримышечно по
0,5 г 2 раза в день, суточная доза для взрослых
1,0 г; детям и подросткам назначают из расчета
15-20 мг/кг в сутки (не более 0,5 г детям и
0,75 г подросткам). Препарат применяют в те-
чение 6 дней подряд, на 7-й день перерыв на 2
дня; или в течение 2—3 дней с однодневным
перерывом. Побочные эффекты: нарушение слу-
ха, аллергические реакции, гипокалиемия.
Канамицин — антибиотик широкого спектра
действия, равен стрептомицину по туберкуло-
статической активности, но более токсичен. При
лечении туберкулеза применяют канамицина
сульфат внутримышечно по 1,0 г в сутки од-
нократно или 0, 5 г 2 раза ежедневно с пере-
рывом каждый 7-й день, иногда 2—3 раза в
неделю по 1,0 г в сутки. Суточная доза для
детей и подростков 15—20 мг/кг (не более 0,5 г
для детей и 0,75 г для подростков). Канамицин
нельзя сочетать со стрептомицином и его про-
изводными, флоримицином, мономицином,
неомицином (ото- и нефротоксические антиби-
отики). Лечение канамицином можно начинать
Глава 8. Туберкулез кожи
147
не ранее чем через 10—12 дней после оконча-
ния лечения этими антибиотиками. Канамицин
противопоказан при неврите слухового нерва,
нарушении функции почек и печени, беремен-
ности. Для профилактики невритов назначают
пантеиновую кислоту. Не рекомендуется его
применять совместно с фуросемидом и други-
ми диуретиками.
Этамбутол принимается внутрь однократно
после завтрака: суточная доза для взрослых
25 мг/кг, детям назначают из расчета 20—25 мг/
кг в сутки, но не более 1,0 г. Препарат нельзя
назначать при неврите зрительного нерва и ри-
ните, катаракте, пониженной выделительной
функции почек вследствие опасности кумуля-
ции, а также в первом триместре беременности.
Специфическое лечение туберкулеза следует
начинать с 3-х препаратов основной группы:
стрептомицина и его производных, ПАСК, фти-
вазида или другого препарата ГИНК (салюзид,
метазид, тубазид). Курс лечения продолжается
10—12 месяцев. При разных клинических фор-
мах туберкулеза кожи возможны разные комби-
нации препаратов. Так, при туберкулезной вол-
чанке предпочтительно назначать в комплексе
изониазид или его производные: рифампицин,
стрептомицин, ПАСК, фтивазид, хирургическое
удаление, криотерапию или электрокоагуляцию.
При бородавчатом туберкулезе — изониазид или
его производные, стрептомицин, фтивазид, хи-
рургическое удаление. При миларном язвенном
туберкулезе кожи и слизистых оболочек — изо-
ниазид или его производные, стрептомицин,
фтивазид. При лихеноидном туберкулезе — изо-
ниазид или его производные, стрептомицин,
фтивазид, УФО. При индуративном туберкуле-
зе — изониазид или его производные, стрепто-
мицин, ПАСК, УФО.
Кроме основных специфических средств, в
каждом случае туберкулеза кожи следует назна-
чать витамины D2, Вр В6 и А, никотиновую
кислоту. Витамин D2 применяют при язвенной
форме волчанки и туберкулеза слизистых обо-
лочек. Оптимальная суточная доза витамина D2
при туберкулезной волчанке составляет 100 000
ЕД, при скрофулодерме — от 50 000 до 75 000
ЕД. У детей доза уменьшается до 20 000—50 000
ЕД. Суточную дозу делят на 2—3 приема, вита-
мин D2 принимают внутрь во время еды, рассчи-
тывая дозу в каплях (в 1,0 мл спиртового раство-
ра витамина D2 содержится от 40 до 50 капель).
Курсовая доза при хорошей переносимости со-
ставляет 15—20 млн ЕД (75—100 мл спиртового
раствора) у взрослых и 6—10 млн ЕД (30—50 мл
спиртового раствора) у детей. Курс лечения длит-
ся 6—8 мес. Побочные эффекты: общая слабость,
жажда, ухудшение аппетита, боли в области же-
лудка, головные боли, недомогание, похудание,
повышение температуры.
Наряду со специфическими противотуберку-
лезными препаратами используют адаптогены,
общеукрепляющие, стимулирующие средства —
препараты кальция, железа. В качестве иммуно-
стимулирующих средств рекомендуют фитопре-
параты: экстракты элеутерококка, левзеи, радио-
лы розовой. Антибактериальной активностью
обладают грецкий и манжурский орехи, шалфей,
тысячелистник, зимолюбка, груша, хвощ, вереск,
чабрец. Сборы из овса, гороха, сои, кукурузы,
крапивы, гречихи уменьшают побочные явления
противотуберкулезной терапии, снижают ототок-
сическое действие стрептомицина.
Для лечения туберкулеза кожи используют
физиотерапию, климатические факторы, лечеб-
ное питание, симптоматические средства. Пища
должна быть калорийной, с повышенным содер-
жанием белков, жиров, витаминов; количество
углеводов несколько уменьшают; значительно
ограничивают содержание поваренной соли.
Хорошие результаты дает гелиотерапия или об-
щее и местное облучение искусственными источ-
никами ультрафиолетовых лучей. Светолечение
противопоказано при активных формах тубер-
кулеза легких, туберкулезном поражении нервной
системы.
Физиотерапия проводится с помощью элект-
рофореза 10 % раствора тиосульфата натрия,
общего и местного УФО. При колликвативном
туберкулезе после удаления оперативным путем
очагов поражения применяют криодеструкцию
или диатермокоагуляцию; хорошие результаты
дает фонофорез с 10 %-ной стрептомициновой
мазью, аппликации жидким азотом, противоту-
беркулезных средств (изониазида, стрептомици-
на) в димексиде. При бородавчатом туберкулезе
показаны оперативное лечение, криодеструкция,
диатермокоагуляция. При волчанке туберкулез-
ные бугорки разрушают диатермокоагуляцией,
3 %-ной мышьяковой пастой, 10—50 %-ной пи-
рогалловой мазью, иногда добавляют 10 %-ный
анестезин, 5 %-ную салициловую кислоту. При
индуративном туберкулезе показан фонофорез
10 %-ной стрептомициновой мази в равных ча-
стях с 2,5 %-ной гидрокортизоновой мазью; на
изъязвленные участки накладывают мазь Мику-
лича, солкосерил. При язвенном туберкулезе
слизистых оболочек эффективно применение
эпителизирующих средств (облепиховое масло,
масло шиповника), полуспиртовых и водных
растворов анилиновых красителей.
148 Частная дерматология
Диспансерное наблюдение
Диспансерное наблюдение больных осуществля-
ется после выписки из стационара в противоту-
беркулезном диспансере по соответствующим
группам учета в зависимости от локализации
процесса, давности заболевания и характера кли-
нических проявлений. Для больных внелегоч-
ным туберкулезом предусмотрена V группа ди-
пансерного учета, состоящая из пяти подгрупп:
V «О», V А, V Б, V В, V Г. Подгруппа V А фор-
мируется из больных с впервые выявленными
активными внелегочными поражениями, обо-
стрением или рецидивом заболевания, которым
необходимо длительно проводить лечебные и
социально-профилактические мероприятия, что-
бы добиться клинического улучшения, восста-
новления трудоспособности и оздоровить очаг
туберкулезной инфекции. Срок наблюдения
больных в VA подгруппе определяется продол-
жительностью основного курса химиотерапии.
Если у больного одновременно диагностируется
туберкулез органов дыхания, то сроки наблюде-
ния и частота посещения фтизиатра увеличива-
ются и определяются состоянием легочного про-
цесса. Диспансеризация больных туберкулезом
проводится фтизиатром при постоянном кон-
сультированиии больного врачом-дерматовене-
рологом с учетом его рекомендаций (2—4 раза в
год). После законченного эффективного основ-
ного курса химиотерапии и достижения благо-
приятных клинических результатов, свидетель-
ствующих о затихании специфического процесса,
больной переводится в V Б группу учета. Срок
наблюдения больного в этой группе определя-
ется необходимостью проведения в течение года
2-х курсов противорецидивной химиотерапии
(по 2 месяца каждый), как правило, двумя пре-
паратами. Если в течение года обострений про-
цесса не наблюдалось, он регрессирует и не имеет
клинических признаков активности, больной
может быть переведен в V В подгруппу. В под-
группу V В переводятся больные из подгруппы
V Б, у которых достигнута потеря активности
специфического процесса в коже. Больные этой
группы посещают противотуберкулезный дис-
пансер не реже одного раза в полугодие. Если на-
ступила полная регрессия процесса с малыми
остаточными изменениями или без них, больной
может быть снят с диспансерного учета. При
выраженных остаточных изменениях больной
переводится для многолетнего наблюдения в
подгруппу Г. Подгруппу Г составляют лица, пе-
реведенные из подгруппы В, ранее снятые с уче-
та, впервые выявленные (спонтанно излеченные)
с выраженными остаточными изменениями.
Все больные туберкулезом кожи, а также из-
леченные от него, подлежат клинико-рентгено-
логическому контролю, который участковый
фтизиатр проводит не реже одного раза в 6 ме-
сяцев с целью выявления у них туберкулеза
органов дыхания. При сочетанном туберкулезе
легких и кожи больные учитываются по пора-
жению, проявления которого наиболее выраже-
ны во время учета.
Экспертиза трудоспособности
при туберкулезе кожи
Экспертиза трудоспособности при туберкулезе
кожи основывается на следующих данных. Сред-
няя продолжительность основного курса хими-
отерапии при всех формах туберкулеза кожи
составляет 6—9 месяцев. В это время больные
временно нетрудоспособны (только при стаци-
онарном лечении, а не амбулаторном). В период
ремиссии больным разрешается работать в усло-
виях, исключающих контакт с раздражающими
и загрязняющими кожу веществами, производ-
ственными аллергенами, пылью, а при очагах
волчанки на лице и кистях нежелательно обще-
ние с большим количеством людей. Если пре-
жняя работа противопоказана и трудоустройство
без снижения квалификации невозможно, то
устанавливается группа инвалидности.
Профилактика
туберкулеза кожи
Профилактика туберкулеза кожи аналогична
общим профилактическим мероприятиям при
этой инфекции: улучшение материально-быто-
вых и санитарно-гигиенических условий жизни
населения, выявление начальных форм туберку-
леза внутренних органов и своевременное их
лечение.
Глава 9
Лепра
Синонимы: проказа, болезнь Ганзена, ганзеноз,
скорбная болезнь, черная немочь, болезнь Свя-
того Лазаря, зараат (библейское).
Лепра — хроническое, мало контагиозное
инфекционное заболевание, вызываемое мико-
бактериями Mycobacterium leprae. Возможны рез-
кие обострения. Заболевание считается одним из
бичей человечества и может сопровождаться тя-
желейшими увечьями. При раннем начале лече-
ния возможно выздоровление, но полностью
устранить возбудитель обычно не удается.
Возбудитель лепры
Mycobacterium leprae представляет собой слегка
изогнутую или прямую, не имеющую спор, ма-
лоподвижную кислотоустойчивую палочку ди-
аметром 4—7 или 0,3—0,4 цм. Под микроскопом
возбудитель можно увидеть с помощью окрас-
ки по Ziehl-Neelsen или Fite-Faraco. Часто рас-
положены группами в виде пачки плотно упа-
кованных сигарет. В отличие от родственных
Mycobacterium tuberculosis, Mycobacterium leprae
при окраске нитратом серебра выглядят про-
зрачными, а туберкулезные бактерии — черны-
ми. Культивирование на питательных средах не-
возможно. В качестве подопытного животного
подходит, в частности, броненосец (Armadillo
novemcergtus), у которого развиваются лепромы.
Для культивирования микобактерий лепры ис-
пользуют также других диких животных: мар-
тышкообразных обезьян и шимпанзе. Основ-
ным резервуаром микобактерий лепры является
человек.
Историческая справка
Лепра — это заболевание, распространенное во
всем мире с древнейших времен. Уже в древне-
египетских и древнекитайских рукописях име-
ются тому соответствующие свидетельства. Идет
ли речь о лепре в Библии, в книге Левит пяти-
книжия Моисея, там, где упоминается заболева-
ние под названием «заарат», не совсем ясно,
поскольку в античную эпоху ранее приобретен-
ные знания были временно преданы забвению,
и лишь во времена Александра Македонского их
значение вновь возросло. Благодаря походам
римских легионеров и особенно крестовым по-
ходам эпидемия лепры распространилась в Цен-
тральной Европе. О заразности лепры было из-
вестно уже в средние века. Начиная с 16 века,
эпидемия лепры стала спадать и ограничилась
определенными областями Европы, такими как
южноевропейские страны, Скандинавия и При-
балтика. В настоящее время современные мето-
ды профилактики и леченижтГепры в этих стра-
нах практически привели к исчезновению
заболевания. В Германии регистрируются лишь
принесенные извне случаи болезни (переселен-
цы, гостевые рабочие, приезжие, в том числе и
на лечение).
Проказу нельзя полностью отождествлять с
лепрой. Как прокаженные изолировались ра-
нее из человеческого сообщества не только
больные лепрой, но и, по незнанию, другие
больные с вызывающими отвращение кожны-
ми заболеваниями, такими как пемфигус, гри-
бовидный микоз, псориаз. Термин «лепра»
восходит к греческому слову лепра (от Lepis =
чешуя), а также к индогерманскому слову Lap
(слущивание).
Эпидемиология
Во всем мире количество больных лепрой оце-
нивается сегодня в 10—15 млн человек. Около
5,5 млн из них получают антимикробную тера-
пию. Лепра встречается преимущественно в тро-
пиках и субтропиках, а также и в более прохлад-
ных зонах, таких как Непал и Корея и
предпочитает географический пояс, ограничен-
ный 40° северной и 40° южной широты. Основ-
ные области ее распространения — Индия (около
4 млн больных лепрой), Юго-Восточная Азия,
Южная и Центральная Америка, Центральная
Африка. В США, по данным разных авторов,
количество больных лепрой составляет от 2500
до 6000 человек. Данные о лепре в Китае отсут-
ствуют. Борьба с лепрой принадлежит к первей-
149
150 Частная дерматология
шим задачам медицины, и ВОЗ придает ей осо-
бое значение.
Лепра не наследуется и не передается перина-
тально. Описана, однако, ее ассоциация с HLA-
В8 и (под вопросом) с HLA-A9. Дети восприим-
чивы к лепре. Заражение происходит обычно в
детском или юношеском возрасте (чаще всего в
10—20 лет). В детстве заболевание поражает в
равной мере оба пола, после полового созрева-
ния заболевают преимущественно мужчины. Для
распространения лепры необходимы два факто-
ра: восприимчивость пациента (иммунный ста-
тус) и контакт с открытыми случаями заболева-
ния. К открытым случаям заболевания относятся
богатая возбудителями лепроматозная и димор-
фная форма лепры и редко — реактивная форма
туберкулоидной лепры. Резервуаром бактерий
лепры может быть материнское молоко, еще од-
ним — мышцы. Лепра встречается у людей всех
социальных слоев, однако преимущественно по-
ражает беднейшие слои населения, которые жи-
вут в тесном пространстве при неудовлетвори-
тельных гигиенических условиях. При
достаточной степени гигиены и питания в ци-
вилизованных условиях опасность передачи леп-
ры незначительна. Заболевание истинной лепрой
в животном мире неизвестно. В экспериментах
на животных бактерии могут передаваться внутри
отдельных видов,^например на лапках мышей,
генерализованной лепры при этом не наступа-
ет. У броненосцев наблюдается генерализованная
инфекция лепры.. Броненосцы считаются источ-
никами инфекции; в штате Луизиана вплоть до
нашего времени травмы у охотников, нанесен-
ные броненосцами, или употребление недоста-
точно обработанного мяса броненосцев вызыва-
ли лепру.
Патогенез
Путь передачи от человека к человеку точно
неизвестен. Предполагается, что инфекция пе-
редается в ходе длительного и тесного контакта
с больными открытой формой лепры. Прямая
передача по воздуху невозможна. В жарких стра-
нах косвенную роль могут играть насекомые,
такие как мухи, клопы и блохи, поскольку они
вызывают травмы поверхности кожи, пиодермии
и хронические изъязвления. Первичный очаг,
как, например, при туберкулезе или атипичных
микобактериальных инфекциях, неизвестен.
М. leprae поражают кожу, слизистые, верхние
дыхательные пути и периферические нервы, в
частности, обсеменяют шванновские клетки, т. е.
клетки нейроэктодермального происхождения,
образующие оболочку нервных волокон перифе-
рических нервов (сравнимы с клетками олиго-
дендроглии головного и спинного мозга). Цве-
товое поведение (морфологический индекс)
показывает, жизнеспособны ли бактерии и спо-
собны ли они к размножению или же нет: пол-
ностью окрашенные бактерии (solid staining rods)
способны к размножению; бактерии с отсутстви-
ем окраски в центре или неравномерной диффуз-
ной поляризацией окраски считаются неспособ-
ными к размножению.
Такая дифференциация важна для прогноза и
успеха терапии. Если тест на лепромин негатив-
ный (недостаточная способность к иммунной
защите), то имеется опасность инфекции. Паци-
енты с лепроматозной лепрой в высокой степе-
ни энергичны на антиген Mycobacterium leprae,
слабо реагируют на небактериальные антигены
и лишь редко имеют контактную аллергию.
Тест на лепромин
Это не диагностический тест, а лишь вспомога-
тельный метод для классификации лепры и
оценки прогноза.
Три антигена используются для внутрикожных
инъекций:
• полный лепромин (Mitsuda), смесь из маце-
рированной ткани лепроматозной лепры, кото-
рый содержит 1,6 х Ю8 Mycobacteria leprae/mm\
а также тканевой детрит;
• антиген бациллы (Dharmendra), который со-
держит только М. leprae;
• протеиновый лепромин (Olsmos-Castro), ко-
торый содержит отдельные антигенные фракции
М. leprae.
Результат оценивается через 2 дня и через 3
недели. Через 48 часов может образоваться ин-
фильтрированная эритема размером 10—20 мм
(положительный тест на лепромин). Он указы-
вает на сверхчувствительность, но едва ли име-
ет клиническую значимость. Эта реакция обо-
значается как реакция раннего типа по
Фернандесу (реакция Фернандеса). В дальней-
шем ранняя реакция может перейти в хроничес-
кую воспалительную и преобразоваться в узелок
размером свыше 5 мм. Ткань удаляют и гисто-
логически обрабатывают. При такой поздней ре-
акции Мицуда (реакция на лепромин) имеется
туберкулоидная или саркоидная гранулема.
Положительный тест на лепромин (аллергия)
указывает на туберкулоидную лепру с хорошим
иммунным статусом и благоприятным прогно-
зом, отрицательный тест на лепромин (анер-
гия) — на лепроматозную или диморфную леп-
Глава 9. Лепра
151
ру с плохим иммунным статусом. Лица с отри-
цательным тестом на лепромин в эндемических
областях должны из-за опасности заражения
лечиться профилактически или же эвакуировать-
ся. В свободных от лепры зонах, таких как наша,
тест на лепромин имеет параллель с туберкули-
новым тестом по Манделю-Манту; он положи-
телен у 80 % населения и указывает на иммуно-
логическое столкновение с туберкулезными
микобактериями. В человеческом теле шваннов-
ские клетки считаются клетками-хозяевами ми-
кобактерий лепры; туда бактерии попадают либо
прямо через кожу, либо через кровь.
Клиническая картина
После некоторого точно не определенного инку-
бационного периода (продолжительность от 3 до
20 лет) могут возникнуть совершенно нехарактер-
ные ранние симптомы, которые вариабельны и не
всегда приводят пациента к врачу. В отдаленных
участках планеты люди боятся (даже при обосно-
ванных указующих симптомах, таких как пиг-
ментное посветление кожи, анестезии, атрофии
мышц, хронический насморк или носовые кро-
вотечения) обращаться к врачу, так как диагноз
«лепра» несет с собой их социальное изгнание.
Многолетнее врачебное наблюдение, направление
в лепрозории, расставание с семьей и професси-
ей несет на себе и сегодня печать проказы.
Клинические формы лепры
На основании клинических, бактериологических,
гистологических и иммунологических данных
лепра классифицируется по определенному спек-
тру. Главными ее формами являются два ста-
бильных полярно противоположных типа: энер-
гическая, с большим количеством возбудителей,
лепроматозная лепра и аллергическая, с малым
количеством возбудителей, туберкулоидная леп-
ра. Между этими двумя формами расположены
недифференцированная и пограничная (диморф-
ная) лепра. Особый способ противостояния меж-
ду возбудителем и хозяином приводит к соот-
ветственно выраженной клинической картине.
Кроме приведенной здесь и применяемой ниже
классификации ВОЗ, которая различает лепро-
матозную, туберкулоидную, пограничную и не-
дифференцированную лепру, есть еще и класси-
фикация по Джобу (Job, 1978), который
выделяет лепроматозную, пограничную, погра-
ничную-туберкулоидную, туберкулоидную, не-
дифференцированную и, наконец, чисто не-
вральную лепру.
Лепроматозная лепра
Это злокачественная, анергическая, инфекцион-
ная форма заболевания при плохом иммунитете
и серьезном прогнозе без тенденции к самовыз-
доровлению. Лепроминовая реакция отрицатель-
ная. В проявлениях на коже и слизистых обна-
руживается массовое количество возбудителей.
Такие пациенты могут носить в себе более 1 кг
бактерий. Гистологическое исследование пока-
зывает гранулемы с большим количеством пени-
стых клеток (клеток Вирхова или клеток лепры).
Кожа
Высыпания на коже образуются симметрично и
состоят из пятен, инфильтрированных бляшек
и коричнево-красных, часто цвета кожи ин-
фильтратов, которые быстро становятся узелка-
ми. Эти узелки называются лепромами. Кож-
ные проявления отграничены нечетко. Пятна
круглые или овальные и находятся преимуще-
ственно на закрытых участках тела. Инфильт-
рированные узлы различны по величине, нечет-
ко отграничены, расположенная над ними кожа
сухая, атрофичная или отечная. Сухость, ангид-
роз, алопеция и анестезия — четыре важных
клинических критерия. Папулы и узлы (лепро-
мы) могут выступать из пятен, а также из нор-
мальной кожи. Их размеры от нескольких мил-
лиметров до сантиметра, они бывают разной
консистенции: от эластичной до твердой. Воз-
можно также подкожное расположение. Они
часто располагаются на местах давления (таких
как локти, колени, ягодицы, лицо и уши). Леп-
ромы могут спонтанно изъязвляться (лепрома-
тозные гуммы) — вследствие травмы или в ре-
активных стадиях (лепроматозные реакции). В
этом случае велика опасность вторичной инфек-
ции. Обширные инфильтрации, лепромы и ке-
ратоконъюнктивит составляют феномен Facies
leonina «львиное лицо»: искажение черт лица и
нарушение мимики. Позднее узлы сливаются и
образуют гирляндообразные дуги. Очаги леп-
ры находятся главным образом на прохладных
участках тела, а в теплых зонах, таких как кожа
головы, затылок, подмышечные впадины, па-
ховая область и грудина, они часто отсутству-
ют. На лице лепромы локализуются на носу,
ушах, яремной впадине и лбу. Кожные морщи-
ны остаются нетронутыми: поэтому на лице
бросаются в глаза выступающие морщины. Уве-
личение мочек ушей патогномонично, характер-
ным и ранним симптомом является выпадение
волос на бровях из-за мягкого лепроматозного
инфильтрата (мадароз).
6 Зак. 1357
152 Частная дерматология
При диффузном лепроматозе (Lucio-Latapi),
называемом также лепрой Святого Лазаря, имеет
место генерализованный воскоподобный кожный
инфильтрат, который напоминает микседему.
Руки и ноги отекают, диффузно краснеют с
цианотическим компонентом, слегка гиперпиг-
ментированы и телеангиэктатичны. Сюда же
относятся и алопеция ресниц, бровей и кожи
головы (лепроматозная алопеция). Остальная
кожа сухая и шелушащаяся. Микобактерии леп-
ры массово находятся и в видимо здоровой коже.
В реактивной стадии образуются пузыри и при-
соединяются затем к необычным некрозам кожи
(феномен Lucio, 1852), которые преобразуются
в глубокие изъязвления, а затем, мутилируя,
заживают со шрамами. Поражение слизистой
носа с деформирующим ринитом является одним
из ранних симптомов. Такая форма течения осо-
бенно часто наблюдается в Центральной Америке
(Мексика).
Слизистые оболочки
Слизистая оболочка носа поражается почти все-
гда. Хронический насморк в эндемических об-
ластях вызывает подозрение на заболевание леп-
рой, также как и эпитаксис, нарушение носового
дыхания вследствие лепром и деструкции носо-
вой перегородки с характерным втягиванием
дистальной верхней стенки полости носа (нос в
виде листа клевера по Gay Prieto). В области
гортани лепроматозные инфильтрации могут
привести к гнойному ларингиту и приступам
удушья.
Другие слизистые, такие как губы, ротовая
полость, язык, а также лимфоузлы, яички, пе-
чень, селезенка и эндокринные железы могут
быть также поражены; это приводит к соответ-
ствующим, часто характерным симптомам, таким
как гинекомастия, бесплодие и повреждение
клеток печени.
Глаза
Заболевание часто поражает глаза, причем очень
тяжело. Инфильтрации на веках, склерах, рого-
вице, радужной оболочке и цилиарном теле мо-
гут привести к нарушению зрения или слепоте.
Поражения глаз часто описываются и при леп-
розных реакциях.
Нервы
Изменения нервов выступают на передний план
не так сильно, как при туберкулоидной лепре.
Иногда поражение нервов возникает только при
регрессе кожных проявлений. Вследствие обра-
зования специфических инфильтратов вдоль
периферических нервов наступает паралич, осо-
бенно mm. intreossei, иннервируемой N. ulnaris и
N. radialis, с типичной мышечной атрофией.
Мышечные бугры мизинца и большого пальца
сжимаются, в результате пальцы приобретают
когтеобразное положение. На пальцах рук и ног
вследствие лепроматозных инфильтратов, вторич-
ных инфекций и трофических изъязвлений ча-
сто возникают увечья.
Лепрозные реакции
Острые ухудшения лепроматозной лепры назы-
вают лепрозными реакциями. Erythema nodosum
leprosum возникает не только на ногах, но и на
руках, лице и туловище. Она склонна к реци-
дивам и встречается только при лепроматозной
лепре у половины больных, как правило, в пер-
вые два года после начала лечения. На коже
появляются множественные воспаленные под-
кожные узлы.
Туберкулоидная лепра
Это обычно неконтагиозная, относительно доб-
рокачественная и медленно протекающая лепра
без системного поражения со склонностью к
спонтанной регрессии при хорошем иммунном
статусе организма и в большинстве случаев с
благоприятным прогнозом. С другой стороны,
пациенты могут страдать от тяжелых мутиляций.
Кожные изменения без бактерий или с малым
количеством бактерий. Лепроминовая реакция
резко положительная. Гистологически имеются
туберкулоидные или саркоидные гранулемы.
Туберкулоидная лепра поражает почти исключи-
тельно кожу и периферические нервы, редко
лимфоузлы. Кожные проявления часто незначи-
тельно выражены, асимметричны и нередко ог-
раничены областью распространения одного не-
рва или определенной кожной зоной.
Кожа
Туберкулоидная лепра часто начинается менее
выразительно с одной или нескольких слабо
выдающихся, но всегда резко отграниченных,
красноватых или красно-фиолетовых папул,
которые увеличиваются к периферии, а в цент-
ре заживают, оставляя депигментрованные и
слегка атрофичные очаги. Свежие кожные из-
менения характеризуются гиперстезией. В ходе
заболевания, особенно в центре очага, наруша-
ется чувствительность. Вначале теряется воспри-
ятие температуры, познее тактильных и болевых
ощущений. Ценным диагностическим призна-
ком является сопутствующий ангидроз.
Глава 9. Лепра 153
Нервы
При всех формах туберкулоидной лепры могут
поражаться нервы. Воспалительные гранулома-
тозные изменения клинически выражаются в
напряжении, полосовидном, хорошо пальпиру-
емом утолщении или винтообразном расшире-
нии на периферических нервах и в потере их
функций. Поражение нервов иное, чем при леп-
роматозной лепре, часто тяжелое и асимметрич-
ное. Повреждение нервов приводит к парезам
и вторично к мышечным атрофиям: на лице —
к лицевому парезу, птозу верхнего века и ми-
мической гримасе (Facies antonina — лицо стра-
дающего Святого Антония), к параличу голо-
совых связок с потерей голоса. На руках — к
атрофии mm. interossei, мышечных бугров боль-
шого пальца и мизинца, к контрактурам с ког-
теобразным положением пальцев; на ногах — к
атрофии малых мышц стопы, петушиной поход-
ке (разгибание стопы невозможно, и при ходьбе
больной высоко поднимает ее, чтобы не заде-
вать о землю, синонимы «степаж», перонеапо-
добная походка, бывает при неврите малого
нерва и спинной сухотке), трофическим изъяз-
влениям (mal perforant). Изменения развиваются
относительно медленно. Туберкулоидная лепра
по степени распространенности подразделяется
на две формы.
Минорная форма
При этой форме наблюдаются эритематозные,
слегка выступающие кожные изменения в фор-
ме маленьких папул с неравномерной границей.
Папулы сливаются, образуя очаги, напоминаю-
щие по рисунку географическую карту. Изме-
нения анестезичны, ангидротичны и часто нахо-
дятся вблизи естественных отверстий, особенно
на веках, в области ануса и мошонки. Возмож-
на спонтанная регрессия этой относительно ста-
бильной формы туберкулоидной лепры.
Мажорная форма
Кожные изменения выступают сильнее и выгля-
дят как инфильтрированные, мягкие и эритема-
тозные папулы, узелки или бляшки. Центр их
часто втянут и бледнее по цвету, так что возни-
кают полные и неполные кольца. Туберкулоид-
ные бляшки находятся преимущественно на яго-
дицах, спине, лице, вокруг естественных
отверстий и на дорсальных и боковых участках
конечностей. Бактериологически обнаруживаются
чаще позитивные результаты, в пораженных оча-
гах кожи — примерно у 50 % пациентов, а на
слизистой носа — у 10 %. Бляшки и утолщенные
нервы находятся почти всегда по соседству друг
с другом, опухоли кожи возникают вследствие
анестезии и лагофтальмуса. При этой форме мо-
гут встречаться острые эпизоды в смысле лепро-
идных реакций. Но и здесь возможна спонтан-
ная регрессия, дальнейшее развитие в диморфную
форму, однако чаще развитие происходит в сто-
рону лепроматозной формы. Тогда ранее положи-
тельные реакции становятся отрицательными. При
этой форме речь идет о нестабильной форме те-
чения туберкулоидной лепры.
Неопределенная лепра
Lepra indeterminata, недифференцированная леп-
ра — нестабильная форма лепры, характеризу-
ется клинически ограниченными кожей и нерва-
ми изменениями, бактериологически — малым
количеством или же отсутствием микобактерий
лепры, гистологически — нехарактерными вос-
палительными реакциями, а иммунологичес-
ки — в большинстве случаев слабо положитель-
ным или отрицательным тестом на лепромин.
Кожа
Кожные изменения асимметричны: частично
гипо- или гиперпигментированные с небольши-
ми воспалительными изменениями пятнистые
очаги с анестезиями и ангидрозом. Поражаться
могут ягодицы, шея, туловище и конечности.
Лицо остается, как правило, непораженным.
Нервы
Изменения нервов в смысле лепроматозного
полиневрита с утолщением больных нервов яв-
ляются важной частью этой формы лепры и
приводят к параличам и «когтистой» руке, а также
к местным перфорирующим изъязвлениям на
стопах.
Лепра недифференцированная может длиться
месяцами и годами. Случаются и спонтанные
регрессии. Переход в лепроматозную лепру —
ранее положительный тест на лепромин стано-
вится отрицательным — происходит достаточно
часто, а в туберкулоидную лепру — редко. Леп-
роидных реакций практически не бывает.
Диморфная лепра
Пограничная лепра — редкая форма лепры и
рассматривается как фаза развития мажорной
формы туберкулоидной лепры. Клинически она
характеризуется изменениями в коже и нервах,
бактериологически — большим количеством
микобактерий лепры, гистологически — сосед-
154 Частная дерматология
ствующими одновременно туберкулоидными и
лепроматозными тканевыми реакциями, а им-
мунологически — обычно отрицательным тес-
том на лепромин. Диморфная лепра находится
между двумя полярными типами лепры: лепро-
матозной и туберкулоидной формами. Соответ-
ствующим образом меняются клинические, бак-
териологические, гистологические и
иммунологические показатели. При отсутствии
лечения эта нестабильная форма лепры обыч-
но переходит в лепроматозную форму, при ле-
чении — в туберкулоидную. Небольшие, часто
асимметричные кожные изменения — это сухие
гипопигментированные пятна и отдельные
крупные куполообразные узлы. Типичные вы-
сыпания на коже — анулярные, частично крас-
ные или медного цвета узлы или бляшки с не-
четкой периферической границей и свободными
зонами, выглядящими как бы «вырубленными»
(по типу швейцарского сыра). На ногах встре-
чаются полосовидные или в виде рисунка по
типу географической карты высыпания. Уже в
ранней стадии заметно поражение нервов (асим-
метричный неврит — ранний симптом), а так-
же алопеция бровей.
Поражения органов при лепре
Изменения нервов при лепре
Поражение нервов является важнейшей составной
частью лепры и встречается при всех ее формах.
Поражаются только периферические нервы, а не
ЦНС. Нейрит при лепре определяется как мно-
жественный мононеврит, т. е. одновременное или
последовательное заболевание одного, двух или
нескольких периферических нервов. Речь, следо-
вательно, не идет об истинном полиневрите.
Поражение нервов при лепроматозной лепре чаще
симметрично, при туберкулезной — чаще асим-
метрично. Изменения нервов включают утолще-
ния, нарушения чувствительности, моторные и
трофические поражения.
Утолщения нервов. Пальпация периферических
нервов является неотъемлемой составной частью
любого клинического осмотра при лепре. Пре-
имущественно встречаются следующие локали-
зации: N. ulnaris в эпитрохлеарной впадине,
N. peroneus на головке фибулы, N. auriculas major,
N. occipitalis minor, поверхностные шейные не-
рвы и N. radialis на запястье. Клинически обна-
руживаются цилиндрические шнуроподобные
утолщения, винтообразные расширения и распо-
ложенные в ряд узлообразные утолщения. На
ощупь они фиброзно-твердые, но бывают и
мягкие (псевдоабсцесс). При пальпации часто
ощущается боль.
Нарушения чувствительности. Первыми клини-
ческими признаками могут быть гиперестезия,
парестезия, потом гипостезия и анестезия. Бо-
лее прохладные, расположенные на периферии
участки тела поражаются рано, нарушения чув-
ствительности распространяются в проксималь-
ном направлении. Вначале теряется способность
различать холод и тепло, а самой последней —
ощущать прикосновения и особенно давление.
Поэтому незаметно могут произойти ожоги,
травмы или вторичные инфекции ран. Иногда
потеряна только способность различать один
признак (диссоциированная анестезия). Тест на
гистамин помогает при ранней диагностике.
Лепрозные невриты могут вначале сопровож-
даться непереносимым зудом, парестезиями,
нарушениями потовой секреции и невралгиями.
Моторные нарушения. Следствием заболевания
нервов являются многочисленные мышечные ат-
рофии. Ранним симптомом является мышечная
слабость, например при ходьбе, удерживании
маленьких предметов или при мимике. Дисталь-
ные участки конечностей, ладони и ступни, а
также лицо поражаются в последнюю очередь.
Неврит N. facialis имеет следствием эктропион и
лагофтальмус (незащищенная роговица). При
двустороннем лицевом парезе возникает воско-
вое невыразительное лицо (Facies antonina). Уль-
нарные парезы приводят к атрофии мышц гипо-
тенара, мизинец и указательный палец находятся
в контрактурном положении (когтеобразная
рука). Если поражен также N. medianus, то полу-
чается положение тотальной когтевидной руки.
Парез N. radialis ведет к «висячей руке».
Местные нарушения. В рамках совместного по-
ражения вазомоторных нервов возникают различ-
ные топические нарушения. Кожа вначале сухая,
шелушащаяся, атрофичная. Спонтанно могут раз-
виться пузыри, которые вскрываются и потом
изъязвляются. Характерны выпадение волос, ан-
гидроз, гиперкератозы на подошвах, особенно на
пятке и над 1 и 5-й метадарзальной головкой,
добавляются также пролифирирующие трофичес-
кие изъязвления с опасностью вторичных инфек-
ций. Узуры, фистулы костей (остеомиелит), ре-
зорбтивные процессы на дистальных сегментах
костей ладоней и стоп с типичным безболезнен-
ным искривлением пальцев рук и ног, утолще-
ния ногтей и атрофия ногтей представляют со-
бой тяжелые увечья при лепре. Пальцы могут
полностью мутилировать до коротких культей.
Атрофия альвеолярных отростков верхнечелюс-
тной кости приводит к расслаблению и выпаде-
Глава 9. Лепра
155
нию зубов-резцов (важный антропологический
признак), а также к атрофии костей носа с обра-
зованием седловидного лепроидного носа.
Другие изменения при лепре
Слизистые оболочки. При лепроматозной лепре
очень часто поражается слизистая носа, однако,
не в начальной стадии. Поэтому мазок из носа
на наличие микобактерий лепры не может слу-
жить ранним диагностическим признаком лепры.
Хронические носовые кровотечения (эпистиак-
сис), хроническая простуда (носовая обструкция)
и ринит (напоминающий вазомоторный) являют-
ся часто не распознанными, но очень подозритель-
ными указателями на лепру. Имеют место без-
болезненные перфорации носовой перегородки.
Богатый возбудителями носовой секрет являет-
ся, вероятно, одним из главных источников ин-
фекции при открытой лепре. Инфильтрированы
могут быть небо, язычок и носоглотка.
Глаза. Нелеченная лепроматозная лепра почти
неизбежно приводит к глазным заболеваниям,
вызванным непосредственно микобактериями
лепры или косвенно поражением нервов
(N. trigeminus, N. facialis). Патогномоничным яв-
ляется перикорнеальный безболезненный застой-
ный конъюнктивит, который образует красно-
оранжевое кольцо. Имеет место милиарное
(просовидное) высыпание лепром в склере и
радужной оболочке («жемчужины на радужной
оболочке»). Иридоциклит разрушает цилиарное
тело. Острый ирит может развиться при erythema
nodosum leprosum — угрожающее глазам ослож-
нение с опасностью слепоты. Кератит, язва ро-
говицы и образование паннуса приводят к то-
тальному рубцеванию и слепоте. Птоз век и
лагофтальм косвенно ведут к поражению глаз
(лепрозная панофтальмия).
Печень и селезенка. Печень и селезенка при
лепроматозной лепре поражены у трети пациен-
тов; осложнения — неинфекционный гепатит,
цирроз печени и амилоидоз.
Почки. Примерно у 75 % всех пациентов с
лепрой поражаются почки: альбуминурия, гло-
мерулонефрит, нефротический синдром и ами-
лоидоз. Уремия является наиболее частой при-
чиной смерти при лепре. Вторичный амилоидоз,
кроме печени и селезенки, поражает щитовид-
ную железу и надпочечники.
Половые железы. Болезненный двусторонний
эпидидимит и орхит при лепроматозной лепре
с тестикулярным фиброзом; бесплодие и импо-
тенция не редки. Лепрозная гинекомастия у
мужчин находится во взаимосвязи с этими по-
ражениями. Алопеция бровей дополнительно
придает пациентам евнухоподобный вид. Яич-
ники поражаются очень редко.
Лимфатические узлы. Паховые лимфатические
узлы часто безболезненно увеличены и богаты
бактериями.
Придатки кожи. Выпадение волос по типу
гнездной алопеции на бровях, бороде и коже го-
ловы — ранние и важные признаки лепры. Ногти
останавливаются в росте. Образование пота в
анестезированных лепроматозных очагах почти
полностью прекращается.
Лепроматозные реакции
Так называют эпизодические внезапные острые
изменения хода заболевания, которые могут воз-
никнуть спонтанно после лепроминового теста
или при терапии.
Различают два типа лепроматозных реакций:
Тип I — клеточноопосредуемый — состоит в
воспалительном преобразовании имеющихся
очагов лепры. Он встречается при туберкулоид-
ной, диморфной и лепроматозной лепре.
Тип II — вызывается, вероятно, циркулиру-
ющими иммунными комплексами как реакция
Артюса, но частично рассматривается и как фе-
номен Schwartzman-Sanarelli. Уже существовав-
шие очаги лепры не поражаются, но возникает
erythema nodosum leprosum, или же добавляют-
ся мультиформные очаги эритемы.
При лепроматозной лепре лепроматозные
реакции встречаются намного чаще, чем при
туберкулоидной и течение их очень тяжелое.
Общие симптомы токсического инфекцион-
ного состояния — лихорадка, головная боль,
озноб, адинамия и артралгии. Кроме ухуд-
шения имевшихся и возникновения новых
кожных проявлений, отмечаются невриты,
ухудшение глазной симптоматики с тяжелы-
ми последствиями и висцеральные проявле-
ния в печени, селезенке, почках, яичках или
плевре. У пациентов может наступить ток-
сический шоковый синдром и в этом состо-
янии они могут умереть.
При туберкулоидной лепре системных симпто-
мов не бывает. Лепроматозные реакции ограни-
чиваются ухудшением старых и возникновени-
ем новых кожных проявлений, а также острым
поражением нервов с опуханием нервов и неврал-
гиями.
При недифференцированной лепре лепроматоз-
ные реакции практически не встречаются. Фе-
номен Lucio с некротическими изъязвлениями
кожи встречается только при диффузной лепро-
матозной лепре.
156 Частная дерматология
Диагностика
Диагностические критерии
В подозрительных случаях важен точный анамнез
и вопрос о проживании в зонах лепры. Приведен-
ные здесь мероприятия служат для постановки
диагноза лепры, ее биологической классификации
и распознавания объема поражений нервов. Про-
верка температуры, боли и прикосновения отно-
сится к важнейшим мероприятиям при диагнос-
тике лепры. Диагностика лепры основывается на
оценке дерматологических и неврологических
проявлений заболевания, данных функциональных
и лабораторных исследований, а также учете дру-
гих факторов: пребывание пациента в лепрозных
зонах; длительный контакт с больным лепрой,
особенно лепроматозной формой; длительное те-
чение болезни (годы); подобное заболевание у
других членов семьи.
Лабораторные тесты
Тест на гистамин
Капля гистамина фосфата или гистамина хлор-
гидрата 1:1000 наносится на кожу, и кожа ца-
рапается иголкой; можно также сделать внутри-
кожную инъекцию 0,1 мл гистамина. Нормой
будут трехкратная реакция (Lewis): эритема, из
которой образуется волдырь, за ним следует бо-
лее крупная рефлекторная эритема (аксоновый
рефлекс). При лепре в пораженной зоне волдырь
образуется, но из-за патологических изменений
вазомоторных нервов отсутствует рефлекторная
эритема.
Тест на потоотделение
Отсутствие потовой секреции в лепроматозных
очагах: 0,1 мл пилокарпина хлоргидрата 1: 100
(или мехолил) вводится внутрикожно в лепро-
матозный очаг, обработанный раствором йодного
крахмала. Проводившийся ранее общий тест на
потоотделение теперь из-за возможной нагруз-
ки на систему кровообращения больше не про-
водится.
Доказательства наличия возбудителя
Соскоб из очага поражения. Соскоб материала
ткани (после неглубокого надреза кожи) дела-
ется из мочек ушей и из двух других очагов по-
ражения, затем окрашивается по Циль-Нильсену
или Fite-Faraco. Только при туберкулоидной и
неопределенной лепре бактерии в мазках могут
отсутствовать. Возможна путаница с неспеци-
фическими кислотоустойчивыми сапрофитами.
Мазок или соскоб со слизистой носа в настоя-
щее время применяется редко.
Полимеразная цепная реакция. Молекулярно-
генетическое доказательство в различных секре-
тах из свежих или фиксированных в формали-
не тканей. Идентификация ДНК М. leprae
применяется для диагностики любой формы
лепры и для оценки эффективности проводимой
терапии.
Бактериологическая культура. М. leprae не рас-
тет на питательных средах и в культуре клеток.
М. leprae культивируют, заражая мышей в мякоть
подушечек лапок. Перепрививка на животных
служит лишь научным целям и не может рассмат-
риваться в качестве рутинной диагностики.
Доказательство наличия антител
Антитела на фенольный гликолипид I (PGLI)
положительны в сыворотке пациентов. При
многобациллярных формах лепры (ВС, LL) по-
ложительный результат отмечается у 98 % паци-
ентов, при малобациллярных формах (ТТ, ВТ) —
только у 30—50 % больных. Методика тестиро-
вания — IgN-Elisa, Dot-Elisa и агглютинация
желатиновых частиц.
Биопсия кожи
Важна при лепроматозной лепре, туберкулоидной
лепре (большая форма), а также для отграничения
от диморфной лепры. Лепроматозная лепра: мас-
сивный клеточный инфильтрат, отделенный от
эпидермиса узкой прослойкой коллагена. Придат-
ки кожи разрушены. Макрофаги содержат М. leprae
и имеют «пенистую» вакуолизированную цитоп-
лазму. Такие макрофаги Называют клетками Вир-
хова, или лепрозными клетками. Туберкулоидная
лепра: гранулема из эпителиальных клеток вок-
руг кожных нервов. Кислотоустойчивые палочки
малочисленны или отсутствуют.
Биопсия нерва
Биопсия нерва необходима только тогда, когда
отсутствуют кожные изменения, как при чисто
невральной туберкулоидной лепре или погранич-
ной лепре.
Реакция на лепромин
Проводится главным образом как реакция Ми-
цуды. Лепроминовая реакция отрицательна при
лепроматозной лепре, чаще всего отрицательна
при диморфной лепре, резко положительна —
при туберкулоидной лепре и чаще всего умерен-
но положительна — при недифференцирован-
ной лепре. Положительный тест на лепромин
Глава 9. Лепра 157
(аллергия) указывает на резистентность к ми-
кобактериям лепры и бывает при доброкаче-
ственных формах лепры, у здоровых и контак-
тирующих с больными лиц, а отрицательный
тест на лепромин (анергия) указывает на сни-
женный или отсутствующий иммунитет и встре-
чается при злокачественных формах лепры.
Неврологические исследования
Проверка температурной чувствительности в
очагах поражения (способность различать теп-
ло — холод), ощущения боли (укол иглой) и
прикосновения.
Йодо-калиевый тест
Этот провокационный тест опасен и поэтому
больше не применяется.
Биохимические исследования
Нетрепонемные серореакции на сифилис неспе-
цифически положительны у 30 % пациентов.
Трепонемные реакции — такие как ТРНА-тест
и FTA-ABS-тест — нереактивны. Более частые
отклонения — гиперхолестеринемия, повышен-
ное общее количество липидов, криоглобулине-
мия и гиперглобулинемия.
Дифференциальный
диагноз
При различных формах лепры труден и тре-
бует специального опыта. В эндемичных об-
ластях лучшая рекомендация — постоянно
помнить о лепре. У нас о лепре нужно ду-
мать у людей, проживавших в эндемичных
областях.
Туберкулоидную лепру следует дифференци-
ровать с отрубевидным лишаем, себорейным
дерматитом, витилиго, бляшечной склеродерми-
ей, фрамбезией, алиментарными дисхромиями,
эпидермомикозами.
Лепроматозную лепру дифференцируют с сар-
коидозом, туберкулезом кожи, кожным лейшма-
Таблица 9.1. Бактериологический индекс —
BI (логарифмическая шкала Ридли)
0 Отсутствие бактерий в 100 полях зрения
1+ 1-10 бактерий в 100 полях зрения
2* 1-10 бактерий в 10 полях зрения
3+ 1-10 бактерий в поле зрения
4+ 10-100 бактерий в поле зрения
5+ 100-1000 бактерий в поле зрения
6+ >1000 бактерий в поле зрения
ниозом, липоидным некробиозом, саркомой
Капоши, гранулематозом Вегенера, кольцевид-
ной гранулемой.
Пограничную лепру следует дифференциро-
вать с третичным сифилисом, красной волчан-
кой, грибовидным микозом, кожным лейшма-
ниозом, кольцевидными эритемами.
Лечение
Принципы лечения
Этиотропная антимикобактериальная терапия;
профилактика и лечение реактивных состояний
(лепроматозных реакций); профилактика и ле-
чение неврологических осложнений; обучение
больного: тактика поведения при отсутствии
чувствительности; социальная адаптация.
Общие мероприятия
Состоят в улучшении гигиенических условий,
обеспечении достаточной чистоты и удовлетво-
рительного питания, а также в улучшении жи-
лищных условий.
Медикаменты
Соответственно рекомендациям ВОЗ сегодня ис-
пользуется комбинированная терапия. При этом
схема лечения ориентируется на количество име-
ющихся бактерий. Она имеет также существенное
значение при постановке прогноза и оценке эф-
фективности применяемых медикаментов. В этой
связи используется бактериологический индекс (В!)
и морфологический индекс (MI) (табл. 9.1).
Сульфоны. В настоящее время сульфоны при-
меняются как терапевтическое средство выбора
при лепре. Они действуют бактериостатически,
но не бактерицидно. Средством выбора являет-
ся DADPS (ДДС, дапсон). Обычная доза состав-
ляет 25—50 мг DADPS ежедневно для взрослых,
но прописывают и 100, и даже максимально
200 мг. Лечение Должно проводиться плавно,
особенно при лепроматозной лепре. Начинать
следует с низких доз, чтобы избежать лепроид-
ных реакций. Рекомендуется следующая дози-
ровка (по Canizares и Hartman и Carville-лепро-
зорию в США):
1 месяц: 25 мг DADPS 1 раз в неделю
2 месяц: 25 мг DADPS 2 раза в неделю
3 месяц: 25 мг DADPS 3 раза в неделю
4 месяц: 50 мг DADPS 2 раза в неделю
5 месяц: 50 мг DADPS 3 раза в неделю
6 месяц: 100 мг DADPS 2 раза в неделю
7 месяц: 100 мг DADPS Зраза в неделю
158 Частная дерматология
Рекомендуемая ВОЗ дозировка выше и состав-
ляет 100 мг DADPS 6 раз в неделю. Дапсон
можно вводить внутримышечно по 225 мг; со-
держание его в крови сохраняется достаточно
высоким в течение 2-х месяцев.
При лепроматозной и диморфной лепре в те-
рапии руководствуются бактериологическим
индексом (BI) и морфологическим индексом
(MI). Бактериологический индекс вычисляется
из количества кислотоустойчивых палочек в
мазках из очагов лепры и количественно оцени-
вается в 6+ с большим скоплением бактерий
(> 1000 микобактерий) до 1+ с 1—10 микобак-
терий на 100 полей зрения (при иммерсионной
микроскопии). Морфологический индекс полу-
чают из числа различных гранулярно окрашен-
ных (beaded staining), неспособных к размноже-
нию и неинфекционных микобактерий.
Бактериальные пробы из слизистой носа (бак-
териологический индекс и морфологический
индекс) становятся отрицательными только че-
рез 1—2 года, а из лепром — не ранее, чем через
2 года (5—7 лет). Сульфонотерапия должна по-
этому проводиться много лет в уменьшенных
дозах, вероятно, даже в течение всей жизни.
Рифампицин. Это бактерицидно действующий
туберкулостатик. Дозировка 300—600 мг в тече-
ние нескольких недель. В пределах 1—2 недель
мазок из носа и лепромы будут свободны от
живых возбудителей.
Рифампицин — антибиотик из группы рифами-
цинов; он тормозит бактериальную РНК-полиме-
разу и таким образом действует бактерицидно на
размножающиеся бактерии, особенно на микобак-
терии, но по другому, чем при туберкулезе — про-
блемы резистентности при лепре до сих пор неиз-
вестны. При применении рифампицина у 20 %
пациентов возрастает уровень трансаминаз в сыво-
ротке. При дальнейшем применении показатели
всесторонне нормализуются. Однако при концен-
трации трансаминазы свыше 100 ед/л или повыше-
нии билирубина рифампицин следует отменить, так
как возможна дистрофия печени. В ходе терапии
показаны регулярные проверки показателей пече-
ни. Дополнительно могут быть гастроинтестиналь-
ные нарушения, кожные проявления, такие как зуд
и транзиторная нейтропения (регулярный контроль
крови). Рифампицин может иметь ряд побочных
действий на ЦНС (сонливость, атаксия, нарушение
остроты зрения и мышечная слабость). Редко, в
качестве идиосинкразии, может развиться почечная
недостаточность. Рифампицин способен повлиять
на механизм действия оральных контрацептивов и
антикоагулянтов.
Клофацимин (лампрен) — специфически дей-
ствующий на микобактерии лепры фенациновый
краситель. В лечении лепры применяется либо
дополнительно к дапсону, либо как замена при
резистентности к сульфонам, а также в лечении
лепроматозных реакций (лепрозная узловатая
эритема). При беременности также противопока-
зан, как и рифампицин. Как правило, хорошо пе-
реносится, но часто наступает окрашивание кожи
от красного до коричневого цвета. Характерно
также окрашивание слезной жидкости, пота,
мочи. Иногда бывают ксероз, ихтиоз, фотосен-
сибилизация и акнеподобные высыпания. Часто
отмечаются тошнота, рвота, боли в животе, по-
нос, потеря аппетита и веса, прежде всего при
применении высоких доз в течение длительного
времени (более 3 месяцев). При терапии следует
контролировать функции почек и печени (каж-
дые 4 недели).
В принципе сегодня рифампицин, дапсон и кло-
фацимин рассматриваются как терапевтические
средства первого выбора (табл. 9.2.). При приеме
одного рифампицина бактериологический индекс
уменьшается за несколько недель, а при дапсоне —
Табл. 9.2. Схемы лечения противолепроматозными препаратами
Формы лепры Препарат Дозировка Примечание
Туберкулоидная Рифампицин 600 мг внутрь 1 раз в месяц Продолжительность — 6 мес, после чего все препараты отменяют
Погранично- туберкулоидная Дапсон 100 мг внутрь 1 раз в сутки Наблюдение после лечения — 2 года; обследование каждые 12 мес
Лепроматозная Рифампицин 600 мг внутрь 1 раз в месяц Продолжительность — 2 года, но лучше до тех пор, пока в соскобе из очага
Клофацимин 300 мг внутрь 1 раз в месяц поражения не исчезнут микобактерии
Погранично- лепроматозная Дапсон 100 мг внутрь 1 раз в сутки Наблюдение после лечения — 5 лет; клиническое обследование и бактериологическое исследование
Пограничная Клофацимин 50 мг внутрь 1 раз в сутки каждые 12 мес
Глава 9. Лепра
159
за месяцы и годы. Мазки со слизистой носа бу-
дут негативными только через 1—2 года, а из леп-
ром — только более чем через 2 года (5—7 лет). Мо-
нотерапия сульфонами еще нередко проводится в
наше время с учетом ее относительно недорогой
стоимости, однако она должна проводиться мно-
гие годы, часто пожизненно в уменьшенных до-
зах. Дапсон хорошо действует бактериостатичес-
ки, рифампицин же — бактерицидно. Среди всех
лекарственных веществ, которые применяются в
настоящее время, рифампицин может рассматри-
ваться как самый эффективный. В связи с воз-
можной монотерапией дапсоном следует учесть, что
могут быть резистентные штаммы.
При слабобактериальной лепре (туберкулоидной,
недифференцированной и погранично-туберкуло-
идной) минимум 6 месяцев применяют дапсон по
100 мг ежедневно, а также раз в месяц 0,6 г ри-
фампицина. При сильнобактериальной лепре (леп-
роматозной форме, пограничной форме и погра-
нично-лепроматозном типе лепры) — минимум 2
года до того, как при микроскопии результат ста-
нет отрицательным, прописывают дапсон ежеднев-
но по 0,1 г (дапсон, фатол), а также рифампицин
(рифа) однократно по 0,6 г в месяц и клофаци-
мин (лампрен) ежедневно по 0,05 г и дополнитель-
но однократно в месяц 0,3 г. При лепрозной уз-
ловатой эритеме с тяжелой формой течения
применяется глюкокортикоидная терапия — пред-
низолон по 40—60 мг в сутки с быстрым сниже-
нием дозы, альтернативно — прием клофацими-
на. Он имеет не только антибактериальное, но и
противовоспалительное действие, которое однако
наступает лишь через несколько недель. При бо-
лее легком течении заболевания применяются и
нестероидные противовоспалительные средства,
такие как ацетилсалициловая кислота. Насколько
возможно следует продолжать антимикробную хи-
миотерапию. При иридоциклите местно применя-
ется глюкокортикостероид, а также мидриатик.
Желательные и нежелательные
эффекты действия важнейших
противолепрозных средств
Дапсон тормозит синтез фолиевой кислоты мико-
бактерий лепры. При многолетнем применении в
рамках монотерапии может возникнуть вторичная
резистентность, следует рассчитывать также и на
возможность первичной резистентности возбуди-
теля. В частности, при дефиците глюкоза-6-фос-
фат-дегидрогеназы может наступить угрожающий
гемолиз, поэтому перед терапией нужно провести
соответствующее исследование. К тому же дапсон
в зависимости от дозы приводит к метгемоглоби-
немии, что делает необходимым регулярный кон-
троль гемоглобина. Иногда бывают гастродуоде-
нальные нарушения, периферическая нейропатия
и нефропатия. При терапии лепры дапсоном мо-
жет возникнуть Erythema nodosum leprosum, что
говорит о Id-реакции при распаде возбудителя.
Для купирования могут быть необходимы систем-
ные глюкокортикостероиды. Лечение же дапсоном
можно в большинстве случаев продолжать. Рифам-
пицин снижает уровень дапсона в крови.
Альтернативные препараты
В случае редкой первичной резистентности к
клофацимину или рифампицину применяются
этионамид и протионамид, а также вторая гене-
рация квинолона, такие препараты как ципроф-
локсацин (ципробай).
Реабилитация
Следует использовать все пути семейной, соци-
альной и профессиональной реабилитации паци-
ентов. Физиотерапия, пластическая реконструк-
тивная хирургия (носовая пластика, ушная
пластика, искусственные суставы пальцев, транс-
плантация связок и нервов на ногах и руках для
устранения перфорирующих изъязвлений, кон-
трактуры и малоподвижности суставов).
Лечение лепры сегодня успешно проводится
и может избавить от многих зол, приносимых
ранее этим считавшимся судьбоносным заболе-
ванием.
Профилактика лепры
Относится к важнейшим и самым существенным
задачам по борьбе с лепрой. Она состоит, глав-
ным образом, в учете пациентов с лепрой, селек-
тивной изоляции бактериально-положительных
пациентов, лечении всех известных пациентов с
лепрой, контроля контактирующих лиц в ходе
регулярных обследований, а также в случае не-
обходимости в профилактическом лечении леп-
ромин-отрицательных контактирующих лиц.
Проведенное ВОЗ профилактическое привива-
ние окружающих лиц посредством BCG не дало
положительных результатов. Важную роль играет
разъяснение пациентам сути заболевания, учреж-
дение амбулаторий, а также улучшение гигиени-
ческих условий и питания находящихся под уг-
розой людей. Принудительная изоляция в
последние годы была ослаблена. Пациенты с ту-
беркулоидной лепрой сегодня больше не изоли-
руются. Имеется тенденция изоляции пациентов
с лепроматозной лепрой, требующих ухода, пока
не будут обеспечены достаточно надежный и ре-
160 Частная дерматология
гулярный врачебный контроль и проведение ле-
чебных мероприятий, а также соответствующие
гигиенические условия.
Прогноз
Он определяется в зависимости от типа лепры.
При лепроматозной лепре и феномене Lucio он
хуже, чем при других формах. Некоторые паци-
енты умирают в течение 10—15 лет от постепен-
но развивающихся амилоидоза, интеркуррентных
инфекций, остеомиелита, туберкулеза или сеп-
сиса. При туберкулоидной лепре прогноз более
благоприятен, поскольку существенного умень-
шения срока жизни обычно не происходит.
Здесь прогноз определяется степенью поражения
нервов.
Глава 10
Кожный и кожно-слизистый лейшманиозы
Кожный лейшманиоз Старого Света 161
Кожный лейшманиоз Нового Света 165
Кожный лейшманиоз (leishmaniosis cutanea) —
трансмиссивное протозойное заболевание, энде-
мичное для районов жаркого, тропического и
субтропического климата, развивающееся после
укусов человека москитами, инфицированными
лейшманиями, и характеризующееся поражени-
ями кожи с образованием папул (бугорков), уз-
лов, их изъязвлением и рубцеванием. Различают
кожный лейшманиоз Старого Света (антропоноз-
ный и зоонозный подтипы) и кожный лейшма-
ниоз Нового Света, при котором нередко пора-
жаются и слизистые оболочки (кожно-слизистый
лейшманиоз).
Тяжесть течения лейшманиозов, их исходы во
многом обусловлены степенью снижения клеточ-
ного иммунитета организма.
Этиология
Возбудитель лейшманиоза относится к семейству
Trypanosomatidae, роду Leishmania. Впервые воз-
будителя кожного лейшманиоза выявил Лейш-
ман (1872), подробно его описал русский воен-
ный врач П. Ф. Боровский (1898). Возбудитель
кожного лейшманиоза определен как L. tropica.
Жизненный цикл лейшманий протекает со
сменой хозяев в виде двух стадий — амастигот-
ной (безжгутиковой) — в организме позвоноч-
ного животного и человека и промастиготной
(жгутиковой) — в организме членистоногого.
Амастиготы имеют овальную форму и размеры
3-5 х 1—3 мкм. При окраске по Лейшману или
Романовскому-Гимзе выявляется гомогенная или
вакуолизированная цитоплазма голубого цвета,
центрально расположенное ядро и кинетопласт
рубиново-красного цвета.
Промастиготы имеют веретенообразную форму
длиной до 10—20 мкм и шириной до 4—6 мкм.
При окрашивании наряду с цитоплазмой, ядром
и кинетопластом выявляется жгутик в переднем
отделе простейшего, отходящий от кинетоплас-
та; с помощью жгутика паразит активно двига-
ется. Культивируются на среде NNN (Novi —
Мак-Нила — Николля).
Кожный лейшманиоз
Старого Света
Антропонозный
кожный лейшманиоз
Синонимы: поздно изъязвляющийся, городской
тип, ашхабадка годовик, сухой лейшманиоз
кожи; разновидность — туберкулоидный кож-
ный лейшманиоз.
Возбудитель — Leishmania tropica.
Эпидемиология
Источник инвазии — больной человек; в не-
которых странах (Афганистан, Иран) резерву-
аром возбудителя могут служить также соба-
ки. Переносчики — самки москитов вида
Phlebotomus sergenti. В процессе кровососания
на больном человеке москит инвазируется и
через 6—8 дней становится заразным. Переда-
ча возбудителя человеку происходит при его
укусе москитом.
В месте внедрения в кожу лейшмании размно-
жаются; возникает продуктивное воспаление и
специфическая гранулема (лейшманиома).
Лейшмании могут распространяться лимфо-
генно, иногда образуя бугорки обсеменения,
лимфангиты, лимфадениты. Общая реакция
организма обычно не выражена.
Максимум заражаемости приходится на теп-
лые месяцы, что связано с периодом лёта мос-
китов, однако сезонности в заболеваемости не
отмечается из-за различной продолжительности
инкубационного периода. Антропонозный под-
тип кожного лейшманиоза встречается преиму-
щественно в городах и поселках городского типа,
161
162 Частная дерматология
но иногда выявляется и в деревнях. Восприим-
чивость всеобщая. Эпидемические вспышки не
характерны. После перенесенной болезни фор-
мируется стойкий иммунитет, однако не исклю-
чены и повторные заражения.
Антропонозный кожный лейшманиоз встреча-
ется в странах Ближнего Востока, Западной и Се-
верной Африки, широко распространен на западе
Индии, встречается в городах и поселках сред-
неазиатских стран и Закавказья.
Клиническая картина
Инкубационный период продолжается от 2—8
мес до 1,5 лет и более.
Возможны следующие варианты антропонозно-
го кожного лейшманиоза:
• Первичная лейшманиома, которая проходит
стадии бугорка, изъязвления и рубцевания.
• Последовательная лейшманиома.
• Диффузно-инфильтрирующая лейшманиома.
• Туберкулоидный кожный лейшманиоз.
На месте внедрения возбудителя в кожу воз-
никает первичная гладкая папула (бугорок) ро-
зового цвета, диаметром 2—3 мм. Постепенно
увеличиваясь в течение 3—6 мес, она достигает
1—2 см в диаметре. В центре бугорка появляет-
ся чешуйка, затем корка, которая постепенно
утолщается. После ее отпадения (через 6—10 мес)
образуется неглубокая язва с зернистым дном,
покрытым гнойным налетом. Края язвы неров-
ные, форма ее овальная или круглая. В подкож-
ную клетчатку она не проникает, вокруг язвы
и под ней определяется плотный инфильтрат.
Увеличение размеров язвы происходит вслед-
ствие распада краевого инфильтрата; к 8—12-му
месяцу болезни она может достигать 4—6 см в
диаметре. Отделяемое язвы скудное, серозное
или серозно-гнойное. Через несколько месяцев
происходит рубцевание, при этом грануляции
развиваются как с центра, так и с периферии
язвы. Рубец вначале розовый, затем бледный, ат-
рофичный, имеет характерный «штампованный»
вид. От появления бугорка до формирования
рубца проходит в среднем 1 год («годовик»),
иногда более 2-х лет. В случае присоединения
вторичной инфекции течение язвы осложняет-
ся и удлиняется. Язвы, в большинстве случаев
безболезненные, локализуются обычно на лице
и верхних конечностях, их число от 1—3, реже
до 8—10. Возможно развитие последовательных
лейшманиом: ранних и поздних.
Ранние лейшманиомы развиваются параллель-
но и аналогично первичным лейшманиомам,
поздние — протекают абортивно и не изъязвля-
ются (формируется иммунитет). У пожилых и
ослабленных больных иногда происходит лим-
фогенное распространение лейшманий с разви-
тием диффузно-инфильтративных лейшманиом,
имеющих вид обширных кожных инфильтратов
без наклонности к изъязвлению. Возникающие
язвы неглубокие и оставляют после себя мало
заметные рубцы. Через 5—7 мес инфильтрат на-
чинает рассасываться и постепенно исчезает.
У детей и юношей обычно развивается тубер-
кулоидный кожный лейшманиоз (люпоидный
лейшманиоз, металейшманиоз). Вокруг рубцов,
реже на самих рубцах появляются мелкие (1—
3 мм в диаметре) множественные бугорки, ко-
торые развиваются медленно, к изъязвлению не
склонны, но могут увеличиваться в размерах и
сливаться. Процесс длится до 5—20 лет и разре-
шается рубцеванием.
Диагностика
Клиническая картина заболевания с учетом эпи-
демиологических данных. Диагноз подтверждается
обнаружением лейшманий в материале, получен-
ном из бугорков, со дна язв и краевого инфильт-
рата, где обычно содержится большое количество
возбудителей. Возможно использование биопробы
на белых мышах или хомяках, а также получение
культуры на среде NNN. Серологические реакции
неспецифичны. В мазках-отпечатках с язв и при
аспирационной биопсии при окраске по Райту или
Гимзе обнаруживают амастиготы.
Дифференциальный диагноз
Проводится с эпителиомами, лепрой, сифилисом,
тропическими язвами.
Лечение
Тактика лечения больных, выбор препаратов и
методов зависят от стадии заболевания и тяже-
сти его течения.
В лечении больных кожным лейшманиозом
применяют средства химиотерапии, физиотера-
певтического воздействия, а также хирургичес-
кие методы. Системное химиотерапевтическое
лечение с включением препаратов сурьмы, об-
щеукрепляющая и стимулирующая терапия про-
водится при тяжелом течении заболевания и его
туберкулоидной форме.
Для системной химиотерапии применяют сле-
дующие препараты (табл. 10.1):
В плохо поддающихся лечению случаях при-
меняют амфотерицин В (или стибоглюконат
натрия) в сочетании с у-интерфероном.
Местное лечение. Монотерапию обычно приме-
няют в начальных стадиях кожного лейшманиоза
и при его нетяжелых формах. В очаги инъециру-
Глава 10. Кожный и кожно-слизистый лейшманиозы
163
ют препараты сурьмы в сочетании с кортикосте-
роидами. В лейшманиомы вводят 5 % раствор
мепокрина по 2—3 мл через 3—5 дней № 3-5.
На очаги прикладывают коллагеновую губку
с 30 % мономицина, а также повязки с мазями:
5 % мономициновой, 3 % метациклиновой, 5 %
тетрациклиновой, протарголом (5—10 %), акри-
хином (1—2 %), риванолом (1 %), стрептоцида
(5-10 %), желтой ртутью, борной кислотой,
линиментом по Вишневскому. Применяют при-
мочки с дезрастворами (фурациллин, марганце-
вокислый калий, риванол и др.), анилиновые
красители, присыпки с дерматолом.
Из методов физиотерапии показано воздей-
ствие на очаги поражения углекислотным и ге-
лий-неоновым лазером, жидким азотом (крио-
терапия) и диатермокоагуляция элементов.
По показаниям: хирургическое лечение язв,
рубцовых изменений, в стадии бугорка — их
электрохирургическое разрушение.
Прогноз
Для жизни благоприятный. Летальные исходы
не отмечаются. Остаются косметические дефек-
ты, иногда обезображивающие.
Профилактика
Активное выявление, своевременное лечение
больных. На лейшманиомы накладывают по-
вязки или наклейки для предупреждения за-
ражения москитов. В некоторых странах (Аф-
ганистан, Иран) резервуаром L. tropica могут
служить собаки, поэтому необходимо уничто-
жение инвазированных животных. Проводят-
ся мероприятия по уничтожению москитов ин-
сектицидами и оздоровлению мест их выплода.
Личная профилактика предполагает примене-
ние репеллентов.
В очагах инфекции проводят прививки жи-
вой культурой L. major — возбудителей зооноз-
ного кожного лейшманиоза, создающей перекре-
стный иммунитет к антропонозному кожному
лейшманиозу.
Медикаментозная профилактика лейшманиоза
не разработана.
С целью иммунопрофилактики применяют
рекомбинантную вакцину БЦЖ в смеси с уби-
тыми промастиготами.
Таблица 10.1. Химиотерапия больных кожным и кожно-слизистым лейшманиозом
Препарат Дозировка, схема применения
Солюсурьмин Взрослым: 0,35 мл/кг/сут в 2 введения (утром и вечером), в/в, медленно, курсовая доза 7-8 мл/кг Детям: однократно 0,5 мл/кг/сут, курсовая доза 7,5-9 мл/кг
Стибоглюконат или меглумин-антимониат Мономицин Расчет по сурьме —10-20 мг/кг/сут в течение 10 суток, в/в или в/м; применяется взрослым Взрослым: в/м по 0,25 г в 4-5 мл 0,5 % раствора новокаина каждые 8 часов в течение 10-14 дней, курсовая доза 9—12 г Детям: 4-5 мг/кг/сут в 3 приема
Доксициклин Метациклин По 0,2 г/сут внутрь после еды в течение 15 дней Взрослым: 0,3 г внутрь 2 раза в день Детям: 7,5 мг/кг/сут, курс лечения 10-12 дней, в тяжелых случаях дозу увеличивают в 1,5-2 раза, курс — 15 дней
Кетоконазол (низорал) Метронидазол Взрослым по 400 мг/сут внутрь в течение 28 дней Взрослым по 0,2 г 4 раза в сутки в течение 7 дней, после перерыва (7 дней) по 0,2 г 3 раза в сутки в течение 14 дней. На курс 9,8 г
Аминохинол Взрослым: 0,45-0,6 г/сут Детям: 0,15-0,4 г/сут. Курс лечения 25-30 дней
Фуразолидон Хингамин Взрослым: 0,15-0,2 г 4 раза в день, курс лечения 18-20 дней Взрослым: 0,25 г 3 раза в день в течение, 10 дней или 0,5 г 2 раза в день в течение 7 дней + сульфален 1,0 г в первый день, затем по 0,2 г ежедневно, 10-12 дней
Рифампицин Взрослым: 0,3 г 2-3 раза в сутки за 30-40 мин до еды Детям: по 7,5-10 мг/кг в 2 приема. Курс лечения от 7-10 до 15-20 дней
164 Частная дерматология
Зоонозный
кожный лейшманиоз
Синонимы: остро некротизирующийся, пустын-
но-сельский лейшманиоз, влажный кожный лей-
шманиоз, пендинская язва.
Возбудитель — L. major. Отличается от возбу-
дителя антропонозного подтипа кожного лейш-
маниоза биологическими и серологическими осо-
бенностями.
Эпидемиология
Основным резервуаром L. major являются пес-
чанки, суслики и другие грызуны, ежи и неко-
торые хищные животные. Переносчики — мос-
киты нескольких видов рода Phlebotomus (в
основном Ph. papatasi), они становятся заразными
через 6~8 дней после кровососания на грызунах.
Человек заражается после укусов инвазирован-
ных москитов. Характерна летняя сезонность
заболеваемости, совпадающая с лётом москитов.
Встречается в сельской местности. Восприимчи-
вость всеобщая. В эндемических районах мак-
симальная заболеваемость выявляется у детей и
приезжих, так как большая часть местного на-
селения переболевает в детстве и становится
иммунной. Возможны эпидемические вспышки,
повторные заболевания редки.
При зоонозном лейшманиозе формирование
изъязвлений и рубцевание первичной лейшма-
ниомы происходит заметно быстрее.
Зоонозный кожный лейшманиоз распростра-
нен преимущественно в странах Северной и За-
падной Африки, Азии (Индия, Пакистан, Иран,
Саудовская Аравия, Йемен), Западной Азии,
встречается в Туркмении и Узбекистане.
Клиническая картина
Инкубационный период продолжается в среднем
10—20 дней. Формирование первичной лейшма-
ниомы сходно с развитием гранулемы при ант-
ропонозном варианте. Однако лейшманиома при
зоонозном лейшманиозе крупнее, иногда напо-
минает фурункул с воспалительной реакцией
окружающих тканей, но малоболезненный. Спу-
стя 1—2 недели начинается центральный некроз
лейшманиом, образуются различной формы язвы
диаметром до 10—15 см и более, с подрытыми
краями, обильным серозно-гнойным, часто сук-
ровичным экссудатом, болезненные при пальпа-
ции.
Вокруг первичной лейшманиомы часто возни-
кают обильные мелкие узелки — «бугорки обсе-
менения», которые затем превращаются в язвоч-
ки, сливающиеся в язвенные поля. Лейшманиомы
при сельском лейшманиозе обычно множествен-
ные и локализуются чаще на открытых частях
тела: нижних и верхних конечностях, лице. Че-
рез 2—6 мес начинается регенерация язвы. С мо-
мента появления папулы до формирования руб-
ца проходит не более 6—7 мес. Часто возникают
лимфангиты (определяются в виде узловатых
безболезненных тяжей), лимфадениты, которые
могут изъязвляться и рубцеваться.
Изредка возникает туберкуловдный лейшманиоз
кожи, протекающий годами. Описаны также
лепроматоидная и промежуточная формы лейш-
маниоза. Диффузно-инфильтрирующая лейшма-
ниома практически не встречается. Присоедине-
ние другой микробной флоры осложняет течение
кожного лейшманиоза и задерживает выздоров-
ление.
Диагностика
При зоонозном лейшманиозе в язвах содержит-
ся мало лейшманий, поэтому необходимы повтор-
ные поиски возбудителя в отделяемом язв, со-
скобах и биоптатах кожи.
Прогноз и лечение зоонозного лейшманиоза
аналогичны антропонозному.
Профилактика
Используются различные способы истребления
диких пустынных грызунов. Борьба с моски-
тами проводится по тем же принципам, что и
при антропонозном кожном лейшманиозе. При-
вивки живой культуры L. major проводят в
осенне-зимний сезон (за 3 мес до выезда лиц в
эндемический по зоонозному кожному лейш-
маниозу очаг); развивается прочный, пожизнен-
ный иммунитет.
Суданский кожный
лейшманиоз
Синонимы: египетский кожный лейшманиоз,
нодулярный кожный лейшманиоз.
Возбудитель — L. nilotica — отличается от
возбудителей других типов кожного лейшмани-
оза антигенным строением. Распространен в
Египте, Судане, Кении, Сомали, Чаде, Ливии,
Уганде. Начальные стадии сходны с зоонозным
кожным лейшманиозом. При Суданской форме
лейшманиомы превращаются в келоидоподобные
узлы, существующие неопределенно долго. Ре-
гионарные лимфатические узлы и сосуды не
изменяются.
Диагностика, прогноз, лечение, профилакти-
ка такие же, как при других типах кожного лей-
шманиоза.
Глава 10. Кожный и кожно-слизистый лейшманиозы
165
Кожный лейшманиоз
Нового Света
Возбудителями кожного лейшманиоза Нового
Света являются:
• L. mexicana (5 подвидов): L. т. mexicana,
L. amazonensis, L. т. pifanoi, L. m. garnhami,
L. m. venezuelensis.
• L. braziliensis (3 подвида): L, b. braziliensis,
L. b. guyanensis, L, b. panamensis.
• L. peruviana.
Эпидемиология
Большинство форм этого заболевания являются
природно-очаговыми зоонозами. Источниками
и резервуаром возбудителей могут быть гры-
зуны, сумчатые, многие дикие и домашние жи-
вотные. Основные переносчики — москиты из
родов Lutzomyia и Psychodopygus — нападают на
человека в дневные часы. Болезнь распростра-
нена преимущественно в сельских районах.
Максимальное число случаев заболевания при-
ходится на сезон дождей. Восприимчивость
всеобщая (болеют лица всех возрастов, как ме-
стные, так и приезжие). Регистрируется в стра-
нах Латинской Америки, в южных районах
США.
В Бразилии (бассейн р. Амазонки) выделе-
на L. mexicana amazonensis, поражающая пре-
имущественно диких животных (крысы,
мыши, опоссумы, лисы, паки), обитающих в
лесу на берегах рек и в заболоченных местно-
стях. Человек включается в эпидемический
процесс крайне редко. В случае заражения за-
болевание у человека протекает очень тяжело,
в 30 % случаев не поддается лечению; проте-
кает в форме диффузного кожного лейшмани-
оза и приводит к обезображиванию. В осталь-
ных случаях кожные язвы локализуются
преимущественно на нижних конечностях,
регрессируют самостоятельно.
L. mexicana pifanoi, обнаруженная в лесах Ве-
несуэлы, вызывает у человека неизлечимую обе-
зображивающую форму диффузного кожного
лейшманиоза. Заражение происходит в местах
обитания лесного грызуна рода Heteromys.
Патогенез во многом сходен с кожным лей-
шманиозом Старого Света. Отмечается более
глубокое поражение кожи и нередко распрост-
ранение патологического процесса на слизистые
оболочки (до подслизистой основы) носа, рта,
глотки, гортани, реже половых органов. Имму-
нитет нестойкий и ненапряженный.
Клиническая картина
Инкубационный период продолжается от 2—3
нед до 1—3 мес. Принципиальных различий в
клинической картине кожных поражений при
лейшманиозе Нового и Старого Света нет.
Особенностью кожного лейшманиоза Нового
Света является частое вовлечение в патологичес-
кий процесс слизистых оболочек. Слизистые обо-
лочки поражаются обычно через 1—2 года после
развития язв на коже. Язвенно-некротические
изменения на слизистых оболочках приводят к
глубокой деформации носа, ушей, носовой час-
ти глотки, дыхательных путей, половых органов,
уродующих и инвалидизирующих больных.
Разновидности кожного
лейшманиоза Нового Света
Известно несколько форм кожного лейшмани-
оза Нового Света, которые имеют ряд клинико-
эпидемиологических особенностей и вызываются
определенными видами лейшманий.
Мексиканский кожный лейшманиоз (возбуди-
тель — L. mexicana mexicana) характеризуется
доброкачественным, но затяжным течением и
редким поражением слизистых оболочек. Обра-
зующаяся язва (язва чиклеро) безболезненна, в
большинстве случаев единична, у 60 % больных
локализуется на ушной раковине, при прогрес-
сировании процесса вызывая её деструкцию.
Чаще язва заживает спонтанно в течение не-
скольких месяцев.
Резервуары и источники инвазии — различ-
ные виды лесных крыс и мышей, обитающих
в густых влажных низинных тропических ле-
сах. Заболевание встречается в северной части
Центральной Америки (Мексика, Гватемала,
Гондурас).
Кожный лейшманиоз ута (Перуанский) (возбу-
дитель — L. peruviana), характеризуется добро-
качественным течением. Обычно одиночные без-
болезненные язвы заживают в течение 4—12
месяцев без лечения. Слизистые оболочки пора-
жаются редко, не вызывая тяжелых разрушений
тканей. Встречается в сухих и горных районах
(северные склоны Анд в Перу и Аргентине, в
Боливии), главным образом у детей, которые
являются источником и резервуаром инвазии.
Другим резервуаром возбудителя могут быть
больные собаки.
Еще одна форма кожного лейшманиоза вызы-
вается L. garnhami. Заболевание встречается в
высокогорных районах (800—1800 м над уровнем
моря) западной части Венесуэлы как в сельской
местности, так и в городах. Течение язв добро-
166 Частная дерматология
качественное, выздоровление спонтанное, про-
должается около полугода.
Гвианский кожный лейшманиоз (возбудитель —
L. braziliensis guyanensis)', известен как «лесной
пиан», «лесная Фрамбезия». Протекает иногда с
поражением слизистых оболочек, но без тяжелых
и опасных осложнений, сравнительно доброкаче-
ственно. Заживление безболезненных, как прави-
ло, единичных «сухих» язв происходит спонтан-
но в течение 9 мес. В редких случаях L. b. guyanensis
распространяются лимфогенно, при этом образу-
ются новые язвы с благоприятным исходом.
Резервуаром возбудителя служат щетинистые
крысы, обитающие в лесах северных районов
Южной Америки (Гайана, Суринам, Панама).
Панамский кожный лейшманиоз (возбудитель —
L. braziliensis panamensis) — один из наиболее
тяжелых и самых распространенных форм кож-
ного лейшманиоза Нового Света. Регистрирует-
ся в большинстве стран Центральной Америки.
Заражение происходит в лесных массивах, где
обитают обезьяны, ленивцы, цепкохвостый мед-
ведь, коати, колючий перогнат, хлопковые кры-
сы, являющиеся резервуаром возбудителя.
Спонтанного заживления язв при этом забо-
левании не бывает, нередко наблюдается лимфо-
генная диссеминация лейшманий с образовани-
ем «дочерних» лейшманиом. Поверхность язв
влажная, мокнущая, легко инфицируется, они
нередко болезненны, особенно в случае локали-
зации вокруг рта.
Кожно-слизистый лейшманиоз Нового Света
(возбудитель — L. braziliensis braziliensis). Резер-
вуарами возбудителя служат лесные крысы и
мыши. Заболевание распространено в западных
и северных районах Южной Америки, Бразилии,
встречается в некоторых странах Центральной
Америки.
Кожно-слизистый лейшманиоз протекает тяже-
лее, чем другие формы кожного лейшманиоза. По
внешнему виду кожные язвы похожи на язвы,
вызванные другими представителями рода L. Ь.
braziliensis, но обычно крупнее, а иногда и болез-
ненные. У большинства больных при отсутствии
лечения кожные язвы длительно (более года) не
заживают. Разрушение тканей обусловлено ги-
перчувствительностью к антигенам лейшманий.
Наиболее тяжело заболевание протекает при
одновременном поражении слизистых оболочек
(в 90 % случаев) и образованием кожных язв.
Значительно чаще слизистые поражаются че-
рез несколько лет после заживления кожных
язв (эспундия). Прогрессирующее специфичес-
кое разрушение слизистых оболочек носа («нос
тапира»), мягкого неба, глотки, гортани, тра-
хеи приводит к появлению клинической сим-
птоматики, которая утяжеляется присоедине-
нием разнообразной вторичной инфекции.
Описаны летальные исходы эспундии, чаще
всего от бронхопневмоний, которые не подда-
ются лечению. При относительно благоприят-
ном течении заболевания и проведении специ-
фического лечения возможно выздоровление
больных, однако при этом остаются обезобра-
живающие лицо рубцы, а также тяжелые по-
следствия разрушения носовой перегородки,
голосовых связок.
Диффузный кожный лейшманиоз (L.m. pifanoi)
рассматривается как вариант течения заболевания,
а не отдельная форма. Встречается сравнительно
редко, главным образом у людей с отягощенным
преморбидным фоном, иммунологической недо-
статочностью.
Диффузный кожный лейшманиоз, вызывае-
мый L.aethiopica (эфиопский кожный лейшма-
ниоз), встречается в горных районах Эфиопии
и Кении. Резервуар возбудителей — даманы
двух родов (Procavia и Heterohyrax). Человек
включается в эпидемический процесс редко, при
нападении инвазированных москитов этих ро-
дов на территории обитания даманов.
Течение заболевания длительное (несколько
лет), без тенденции к спонтанному выздоровле-
нию. Инфильтративные высыпания на коже
носят генерализованный характер, практически
никогда не изъязвляются, но и не рассасывают-
ся. Терапия в большинстве случаев неэффектив-
на; некоторым больным помогает пентамидин и
антибиотик амфотерицин В.
Диффузный кожный лейшманиоз, вызывае-
мый Z. mexicana pifanoi, встречается в Венесуэ-
ле. Основной резервуар возбудителей — лесной
грызун рода Heteromys. При случайном зараже-
нии человека кожные изменения сравнительно
Хорошо поддаются обратному развитию под вли-
янием пентостама.
Диффузный кожный лейшманиоз, вызывае-
мый L. mexicana amazonensis, наблюдается в не-
которых районах Бразилии. Резервуары возбуди-
теля — различные виды диких млекопитающих.
Человек заражается случайно при нахождении в
биотопах этих животных (выволоченные леса) в
определенное время года. Считается, что основ-
ные переносчики (Lutzomyia flaviscutellata) на
человека практически не нападают, что делает это
заболевание сравнительно редким. Поддержанию
заболеваемости в человеческой популяции, веро-
ятно, способствует передача лейшманий моски-
тами, инфицированными при нападении на боль-
ного человека.
Глава 10. Кожный и кожно-слизистый лейшманиозы
167
Прогноз при тяжелом течении заболевания
может быть неблагоприятным, особенно в слу-
чае присоединения вторичной инфекции.
Диагностика
Принципы диагностики такие же, как при кож-
ном лейшманиозе Старого Света. Проводят био-
псии тканей с целью поиска амастигот. Исполь-
зуется внутрикожная проба Монтенегро (с
лейшманином), которая, как правило, отрица-
тельная, что объясняется отсутствием иммунно-
го ответа на антигены лейшманий. Из сероло-
гических реакций используются РНИФ, реже
ИФА.
Лечение
Наряду с общей терапией (см. табл. 10.1), при-
меняемой при тяжелых формах кожного лейш-
маниоза Старого Света, обязательно включают
препараты сурьмы — глюкантим (меглумин).
Химиотерапия лейшманиоза с поражением
слизистых оболочек не дает положительного
эффекта. В необходимых случаях применяют
хирургическое лечение (косметические, пласти-
ческие операции).
При диффузном кожном лейшманиозе, вызы-
ваемом L. mexicana amazonensis, химиотерапия
неэффективна, применяются симптоматические
средства, частые горячие ванны на протяжении
ряда месяцев.
Профилактика
Профилактические мероприятия проблематичны,
так как предполагают оздоровление природных
очагов, борьбу с грызунами и другими резерву-
арами лейшманий, москитами. Специфическая
профилактика не разработана. Применяют вве-
дение БЦЖ с убитыми промастиготами, а так-
же вакцинацию рекомбинантной вакциной
БЦЖ, содержащей поверхностный антиген лей-
шманий.
Глава 11
Вирусные болезни кожи
Простой герпес 168
Опоясывающий герпес 173
Бородавки 175
Остроконечные кондиломы 176
Контагиозный моллюск 176
Узелки доильщиц 177
Вирусные болезни кожи составляют значитель-
ную группу дерматозов, которая включает ряд
нозологических форм (табл. 11.1). Среди взрос-
лого населения они регистрируются в 3—4 %, у
детей заболеваемость этой патологией достига-
ет 10 %. Вызываются дерматонейротропными
(герпес простой и опоясывающий) и дермато-
тропными (бородавки, контагиозный моллюск,
узелки доильщиц) вирусами.
Вирусом простого герпеса I типа (ВПГ I) инфи-
цировано около 90 % населения. Персистирует ВПГ
I в сенсорных паравертебральных ганглиях. При
действии неблагоприятных факторов (охлаждение,
стресс и др.) вирус активируется и вызывает в коже
пузырьковую реакцию, чаще в области лица.
Инфицирование вирусом простого герпеса II
типа (ВПГ II) происходит при половом контак-
те с больными клиническими или асимптомными
формами генитального герпеса и дальнейшей
персистенцией вируса в сакральных ганглиях.
Таблица 11.1. Классификация
вирусных дерматозов
Гоуппы кожных поражений Нозологические формы
Герпесы • Простой
• Опоясывающий
Бородавки • Простые
• Плоские
• Подошвенные
• Нитевидные
• Остроконечные кондиломы
• Контагиозный моллюск
Паравакциния • Узелки доильщиц
Реактивация вируса оспы (ОРВИ, снижение
реактивности организма, пожилой возраст и др.)
нередко приводит к возникновению опоясыва-
ющего лишая— более тяжелой кожной формы
герпетической инфекции, сопровождающейся
болями по ходу высыпаний, длительно не исче-
зающими невралгиями, а в некоторых случаях
поражением ЦНС, внутренних органов, органа
зрения.
В группу вирусных заболеваний, передавае-
мых при половых контактах, включены также
генитальные бородавки (остроконечные конди-
ломы) и контагиозный моллюск. Отмечается
тенденция роста этих дерматозов среди лиц мо-
лодого возраста. Заболевание принимает распро-
страненный и упорный характер у инфицирован-
ных ВИЧ, клинической стадией СПИД.
Активация вирусов папилломы человека, вы-
зывающих вульгарные и плоские бородавки,
ворсинчатые папилломы, также связана с разви-
тием иммунодефицитного состояния и снижени-
ем внутриэпидермального иммунологического
контроля (клетки Лангерганса и Гринстейна, Т-
лимфоциты). Наиболее часто папилломатозные
разрастания, кератомы наблюдаются у лиц стар-
ших возрастных групп.
Простой герпес
Часто встречающийся дерматоз, проявляющий-
ся высыпаниями на коже и слизистых оболоч-
ках сгруппированных пузырьков на эритематоз-
ном и слегка отечном основании. Может
168
Глава 11. Вирусные болезни кожи 169
принимать упорный рецидивирующий, диссе-
минированный, иногда системный характер. Ге-
нитальный герпес по частоте значительно пре-
восходит другие инфекции, передающиеся при
половых контактах.
Этиология
Вирус простого герпеса I и II типов (ВПГ I и II)
- дерматонейротропный ДНК-содержащий ви-
рус, размножается в течение 14 часов в ядре и
цитоплазме инфицированных клеток, с последу-
ющей их гибелью и выходом дочерних вирионов.
• Герпес лица и полости рта вызывается ВПГ
I (80—90 %) и ВПГ II (10—20 %).
• Герпес генитальный: ВПГ II (70—90 %) и
ВПГ I (10-30 %).
• Герпетический панариций: ВПГ I выявля-
ется в возрасте до 20 лет, а ВПГ II — старше 20
лет.
• Герпес новорожденных: ВПГ II (70 %), ВПГ
I (30 %).
Пути передачи
Инфицирование ВПГ I происходит при пря-
мом контакте (соприкосновении) с поражен-
ной кожей, слизистой рта, губ больного гер-
песом (обычно в течение первых нескольких
лет жизни). Воздушно-капельный путь зара-
жения возможен, если герпетическая инфек-
ция протекает в форме острого респираторно-
го заболевания или на фоне ОРЗ иной
этиологии. Через предметы обихода заражение
маловероятно, т. к. при комнатной температу-
ре ВПГ быстро погибает.
Антитела к ВПГ обнаруживаются у 80—90 %
взрослого населения.
Передача плоду ВПГ от матери, больной ге-
нитальным герпесом, осуществляется: во время
прохождения через родовые пути (инфицирует-
ся носоглотка, кожа, глаза) — риск заражения со-
ставляет 40 %; через цервикальный канал и да-
лее в околоплодные воды; через плаценту, при
развитии герпетической вирусемии у беремен-
ной.
Пути проникновения
и распространения ВПГ в организме
Внедрение в кожу и слизистые оболочки на ме-
сте контакта; размножение в нижних слоях эпи-
дермиса; гибель зараженных клеток; эритема,
везикулы; дерма; окончания чувствительных и
вегетативных нервов; ганглии (тройничного,
блуждающего нервов, пояснично-крестцовые),
где вирус находится в латентном состоянии —
последующая реактивация.
Факторы риска
Переохлаждение, гиперинсоляция, респиратор-
ные заболевания, стресс, иммунодефициты
(ВИЧ-инфекция, гемобластозы, трансплантация
органов, костного мозга, химиотерапия, лучевая
терапия, системные кортикостероиды, иммуно-
депрессанты), менструация.
Патогенез рецидивов простого герпеса
Снижение противовирусной и неспецифической
резистентности организма, вызванное фактора-
ми риска. Снижение иммунологического конт-
роля в ганглиях. Реактивация вируса в гангли-
ях и его перемещение по периферическим
нервным волокнам в эпителий кожи и слизис-
тые оболочки. Репликация вируса. Своеобраз-
ная пузырьковая реакция.
При иммунодефиците: снижение иммунологи-
ческого контроля; вирусемия; гематогенная дис-
семинапия; поражение ЦНС, печени, легких,
почек, других органов, распространенное пора-
жение кожи и слизистых.
Клиника
Инкубационный период — от нескольких дней до
2—3-х недель.
Продромальные явления: жжение, парестезии,
зуд в месте будущего очага, возможен легкий
озноб, гиперестезия кожи, неинтенсивные мы-
шечные и суставные боли.
Наиболее частая локализация: красная кайма
и кожа губ, крЪлья носа, лоб, щеки (ВПГ I
типа), гениталии (ВПГ II типа).
Клинические симптомы типичной формы просто-
го герпеса. Выявляются единичные очаги ограни-
ченной эритемы на слегка отечном основании, за-
тем возникают сгруппированные пузырьки с
пупковидным вдавлением в центре, диаметром до
2 мм, наполненные вначале прозрачным, затем
мутным и геморрагическим содержимым. Пузырьки
вскрываются, образуя болезненные эрозии поли-
циклических очертаний с мягким дном и влажной
красноватой поверхностью, которые покрываются
буровато-желтоватыми корками. После отпадения
корок образуется красновато-бурая пигментация.
Регресс высыпаний длится 7—14 дней. При инфи-
цировании или травматизации эрозия может пре-
вратиться в неглубокую слегка уплотненную и
отечную язвочку с более длительным периодом за-
живления и наличием регионарного лимфаденита.
Разновидности простого
герпеса по течению
Первичный герпес. У большей части лиц (80—
90 %) инфицирование ВПГ происходит бессим-
170 Частная дерматология
птомно. У части — появляются типичные высы-
пания, возможен регионарный лимфаденит, не-
тяжелые общетоксические явления, которые
нарастают в течение нескольких дней (3—4 сут),
а затем в течение такого же времени постепен-
но исчезают вместе с очагом поражения. У мо-
лодых женщин первичный герпес нередко про-
текает в виде герпетического вульвовагинита.
Первичный герпес у детей чаще проявляет-
ся в виде герпетического стоматита и проте-
кает с выраженными общими явлениями. Раз-
вивается обычно у детей до 1 года,
сопровождается высокой температурой, симп-
томами интоксикации. Наиболее частая лока-
лизация — слизистая щек, десен, внутренняя
поверхность губ, язык. Появившиеся пузырьки
вскрываются, образуя фестончатые резко бо-
лезненные эрозии. Увеличены и болезненны
регионарные лимфатические узлы. Гингивит
сопровождается болезненной отечностью десен
с фиолетовым оттенком, слюнотечением. При
легком течении процесс регрессирует в течение
2 недель. У ослабленных детей возможна дис-
семинация инфекции, развитие тяжелого ме-
нингоэнцефалита, летальный исход.
Рецидивирующий герпес — протекает с менее
выраженными клиническими симптомами, ло-
кализуется на лице, гениталиях, перигениталь-
ной области, возможен кератоконъюнктивит с
частотой рецидивов от 1-3 раз в год до несколь-
ких раз в месяц.
Выраженные физические и психологические
беспокойства у больных вызывает рецидивиру-
ющий генитальный герпес.
Длительно протекающий генитальный герпес
может иметь атипичное течение (без везикуляр-
ных элементов), с наличием эритемы, жжения
и зуда в очаге.
Частые рецидивы генитального герпеса могут
привести к лимфостазу и вызвать слоновость
наружных половых органов.
Атипичные формы простого герпеса
Абортивная — появление слабо выраженных
папул, эритемы, некроза, субъективных ощуще-
ний без типичных высыпаний.
Отечная —- высыпания на веках, губах, сопро-
вождающиеся выраженным отеком.
Зостериформная — болезненные высыпания
наблюдаются по ходу нервов на лице, конечно-
стях, туловище, ягодицах и сопровождаются го-
ловной болью, слабостью.
Диссеминированная — одновременное появле-
ние на различных участках тела очагов сгруппи-
рованных высыпаний.
Мигрирующая — очаги высыпаний появляются
последовательно и занимают новые участки тела.
Геморрагическая и некротическая формы —
определяются кровянистым характером экссуда-
та пузырьков и развитием некроза.
Элефантиазоподобная — длительно сохраня-
ющаяся отечность кожи в местах регрессировав-
шего герпеса.
Хронический кожный простой герпес — неис-
чезающие множественные язвенные очаги диа-
метром до лесного ореха и более у больных с
иммунодефицитом.
Язвенный герпес на гениталиях — развивается
после типичных высыпаний у лиц со снижен-
ной иммунобиологической реактивностью (мяг-
кая язва красноватого цвета, болезненная, воз-
можен лимфаденит).
Рупиоидная форма — развивается на лице в
результате присоединения вторичной инфекции
(слоистые корки, трещины).
Герпетический панариций развивается у медра-
ботников (анестезиологов, стоматологов), кон-
тактирующих со слизистой больных герпесом.
Очаги состоят из единичных, плотных, болез-
ненных пузырьков, иногда с разлитой эритема-
тозной реакцией.
Герпетический фолликулит. Встречается пре-
имущественно у мужчин в области подбородка
и в носогубном треугольнике. В устьях волося-
ных фолликулов образуются везикулы, затем
появляются корки. Процесс сопровождается лег-
ким зудом и жжением.
Герпетиформная экзема Калоши чаще возни-
кает у детей, больных экземой и атопическим
дерматитом (заражаются от родителей, больных
активным герпесом). Сопровождается высокой
температурой, общим тяжелым состоянием, дис-
семинированными высыпаниями на коже, сли-
зистых, множеством элементов с пупковидным
западением, массивными геморрагическими кор-
ками. Возможен летальный исход.
Реже экзема Капоши встречается при болез-
ни Дарье, пузырных дерматозах, ихтиозе, гри-
бовидном микозе.
Генитальный герпес
Заражение происходит путем прямого контакта
от полового партнера с герпетическими высыпа-
ниями на гениталиях или прилегающих участ-
ках, а также в период ремиссии (когда продол-
жается продукция ВПГ) и при асимптомном
вирусоносительстве.
У большей части лиц инфицирование ВПГ II
протекает бессимптомно или без явных клини-
ческих проявлений.
Глава 11. Вирусные болезни кожи 171
В 70 % случаев генитального герпеса выявля-
ется ВПГ II типа, в остальных случаях — ВПГ
_1. Почти половина лиц гомосексуальной ориен-
тации инфицировано ВПГ II, а среди проститу-
ток инфицированность ВПГ II достигает 80 %.
Генитальный герпес является наиболее частой
причиной язвенных поражений гениталий у лиц
молодого возраста.
Первичный герпес гениталий часто сопровож-
дается общей симптоматикой (лихорадка, голов-
ная боль, миалгии, слабость), которая нарастает
в течение 6—8 дней. Высыпания сгруппирован-
ных пузырьков чаще бывают в виде изолирован-
ных небольших бляшек, реже — захватывают
значительную часть полового члена, мошонки,
бедра, ягодицы, а у женщин — половые губы,
клитор, промежность, внутреннюю поверхность
бедер, ягодицы. Характерны зуд, жжение, дизу-
рия, нередко выделения из влагалища, уретры,
явления цистита. Присоединяются явления одно-
стороннего болезненного регионарного лимфаде-
нита, возможны боли внизу живота (тазовый
лимфаденит), явления менингизма, ишиалгия.
Герпетический проктит гомосексуалистов
(возбудитель ВПГ I типа) сопровождается боля-
ми, тенезмами, выделениями из rectum; при ос-
мотре выявляются признаки воспаления прямой
кишки и эрозивно-язвенные участки.
После первичного эпизода генитального гер-
песа у 2 % пациентов рецидивы наблюдаются
ежемесячно, у 13% — каждые 2—11 месяцев,
ежегодно и реже — у 24 %. Прием ацикловира
резко снижает выделение вируса из гениталий и,
следовательно, риск заражения полового партнера.
При рецидивах генитального герпеса симпто-
матика не столь ярко выражена.
Генитальный герпес дифференцируют с ак-
тивными проявлениями первичного и вторично-
го периодов сифилиса (эрозии, язвы, папулез-
ный и пустулезный сифилиды), мягким
шанкром, гонорейными, трихомонадными, хла-
мидийными, неспецифическими бактериальны-
ми эрозиями, язвами; необходимо исключить
ВИЧ-инфекцию.
Герпес беременных, новорожденных
При первичном генитальном герпесе в первом три-
местре беременности риск осложнений у плода до-
стигает 50—80 %. В этих случаях на сроках до 12
недель рекомендуется прерывание беременности.
При возникновении у беременной диссемини-
рованной герпетической инфекции (вовлечение
ЦНС, внутренних органов) в более поздние сро-
ки проводят лечение ацикловиром по стандарт-
ной схеме.
Если за 1 мес до родов у беременной возник
эпизод первичного генитального герпеса или
клинический рецидив инфекции за несколько
дней до родов, показано родоразрешение путем
кесарева сечения.
В случаях подтверждения у беременной носи-
тельства ВПГ (асимптомный генитальный гер-
пес) за 2 недели до родов проводят профилак-
тику ацикловиром по 200 мг 4 раза в сутки —
5 сут.
Риск заражения новорожденного наиболее
высок, если роды совпадают с первичным гер-
песом половых органов.
При рецидивах генитального герпеса у бере-
менной риск заражения новорожденного неболь-
шой и составляет около 3 %.
Дети, родившиеся через инфицированные
родовые пути, подлежат вирусологическому об-
следованию на ВПГ в первые 48 ч жизни (ма-
зок с конъюнктивы, носа, ротоглотки). При
выявлении ВПГ или первых симптомах заболе-
вания проводится лечение.
Возможные осложнения простого герпеса
Герпетический энцефалит, менингоэнцефалит
(при некротической форме летальность достигает
80 %), гепатит, кератоконъюнктивит, рецидиви-
рующее течение инфекции, преканкроз шейки
матки.
При внутриутробной герпетической инфекции
летальность может достигать 80 % в результате
генерализации процесса с поражением ЦНС,
внутренних органов.
Методы диагностики
герпетической инфекции
Проба Цанка. Обнаружение в содержимом све-
жего пузырька (при окраске по Гимзе) гигантс-
ких многоядерных кератиноцитов (у 75 % паци-
ентов — положительна).
Определение антигенов ВПГ содержимом пу-
зырьков с помощью моноклональных антител.
Выделение вируса из патологического матери-
ала (содержимое пузырьков, соскобы с поражен-
ных участков, биопсийный материал) в культу-
ре клеток.
Обнаружение антител к ВПГ в сыворотке
крови является доказательством активного про-
цесса.
С помощью ПЦР выявляется ДНК ВПГ в
патологическом материале.
Гистопатология
Баллонирующая и ретикулярная дегенерация
клеток эпидермиса, акантолиз, наличие внутри-
172 Частная дерматология
эпидермальных пузырьков, гигантских много-
ядерных кератиноцитов с внутриядерными ви-
русными включениями (клеток Цанка). В дер-
ме инфильтрация стенок сосудов нейтрофилами,
периваскулярные инфильтраты (лимфоциты,
нейтрофилы).
Дифференциальный диагноз
Стрептококковое и смешанное импетиго, опоя-
сывающий герпес, первичная сифилома, сифи-
литические папулы, эрозивно-язвенный балано-
постит, чесоточные узелки, язвы, фолликулиты,
фурункул в области гениталий, детская экзема,
осложненная пиодермией (с экземой Капоши),
фиксированная эритема (гениталии), многофор-
мная экссудативная эритема.
Лечение
Первичный герпес (с общими явлениями):
ацикловир 400 мг 3 раза в сутки в течение 5
дней, или 800 мг 2 раза в сутки 5 дней, или
валацикловир 500—1000 мг 2 раза в сутки 5
дней. Витамин С 0,3 г 3 раза в день в течение
3—5 дней.
При отсутствии общих явлений ограничива-
ются применением местных противовирусных
препаратов (ацикловир, ДНК-аза и др.); при
герпетическом стоматите: 0,5 % теброфеновая
мазь, 0,25 % риодоксоловая мазь, мазь цикло-
вир 5 %. Очаги смазывают 5 раз в сутки в те-
чение 5—10 дней.
Рецидивирующий герпес кожи и слизистых, ке-
ратоконъюнктивит: ацикловир по 200 мг 5 раз
в сутки 5—10 дней; или валацикловир 500 мг 2
раза в день в течение 5 дней; или фамцикло-
вир 250 мг 3 раза в день в течение 5 дней. На-
значение этих препаратов в первые 2 дня
уменьшает остроту рецидивов, однако их не
предотвращает.
Панавир 0,004 % раствор в момент рецидива
две внутривенные инъекции по 5,0 мл с интер-
валом 48 часов (табл. 11.2).
Герпес первичный половых органов: ацикловир
200 мг внутрь 5 раз в сутки в течение 10 дней
или 400 мг внутрь 3 раза в сутки 10 дней; или
ганцикловир 5 мг/кг, в/в каждые 12 ч 14—21
день.
Рецидивирующий герпес половых органов:
ацикловир 200 мг внутрь 5 раз в сутки в тече-
ние 5—10 дней или 400 мг внутрь 3 раза в сут-
ки в течение 5—10 дней; или фамцикловир
125 мг 2 раза в день в течение 5—7 дней. Цик-
лоферон по 2 мл (250 мг) в/м через день № 10.
Две внутривенные инъекции 0,004 % раствора
панавир по 5,0 мл с интервалом 48 часов; через
месяц курс рекомендуется повторить.
Простой герпес кожи и слизистых при иммуно-
дефиците: ацикловир 5 мг/кг внутривенно, каж-
дые 8 ч в течение 7 суток, или 200 мг внутрь 5
раз в сутки — 10 дней; или фоскарнет 40 мг/кг
внутривенно каждые 8 часов до исчезновения
высыпаний в течение 10—24 суток (при устой-
чивости к ацикловиру).
Методы физиотерапии: гелий-неоновый ла-
зер на очаг или на область кубитальных вен
(лазерное облучение крови). УФО на область
очага или ультрафиолетовое облучение кро-
ви. Ультрафонофорез на очаг с противови-
русными мазями. Воздействие паравертеб-
рально на область сакральных ганглиев с
помощью ультразвука.
Профилактика первичного герпеса: исключить
тесные контакты с больными, имеющими гер-
петические высыпания на коже и слизистых.
Профилактика рецидивов
Методика непрерывной супрессивной терапии ре-
цидивирующего генитального герпеса: ацикловир
400 мг 2 раза в день в течение одного года (в
случае 6 и более рецидивов в год). Через год
оценивают частоту рецидивов с целью дальней-
шей лечебной тактики.
Инактивированная герпетическая вакцина 0,2—
0,3 мл внутрикожно в сгибательную поверхность
предплечья через 3—4 дня — 5 инъекций; через
2 недели назначают аналогичные инъекции 1 раз
в 5—7 дней № 5; через 3—6 месяцев проводят
ревакцинацию по 5 инъекций с интервалом 7—
Таблица 11.2. Рекомендации к применению противовирусного препарата «Панавир»
Заболевание Методика лечения Ожидаемый эффект
Рецидивирующий генитальный и назолабиальный герпес (ВПГ1,ВПГ2) Две внутривенные инъекции 0,004% раствора Панавир по 5,0 мл с интервалом 48 часов в момент рецидива. Гель защитный Панавир наружно 3-4 раза в сутки. Через месяц курс рекомендуется повторить Сокращение длительности рецидива в 1,5-2 раза, увеличение периода ремиссии в 2 - 2,5 раза. Снятие болевых ощущений, быстрое ранозаживление. Доказанная эффективность более 80%
Глава 11, Вирусные болезни кожи 173
14 дней, через 6—8 месяцев проводят вторую ана-
логичную ревакцинацию. Антиоксидантный ком-
плекс в течение 3—4 недель. Иммуномодулято-
ры назначают после определения состояния
иммунного статуса (тактивин, тималин, циклофе-
рон, амиксин, полиоксидоний, миелопид и др.).
Показаны адаптогены растительного происхож-
дения. Проводят санацию организма. Необходи-
мо соблюдение личной гигиены. Возобновление
половой жизни после регресса высыпаний с при-
менением барьерных контрацептивов.
Опоясывающий
герпес
Острое вирусное заболевание, возникающее в
результате реактивации вируса Varicella zoster
и проявляющееся сгруппированными болезнен-
ными везикулезными высыпаниями по ходу не-
рвов, обычно в пределах одного дерматома. Дети
до 15 лет болеют значительно реже (5 %), чем
взрослые. У лиц старше 60 лет нередким ослож-
нением является постгерпетическая невралгия
(40 % пациентов). Вероятность заболеть опоя-
сывающим лишаем в течение жизни составля-
ет 10-20 %.
Этиология
Дерматонейротропный вирус Varicella zoster (при
первичном инфицировании вызывает ветряную
оспу). Практически все лица старше 20 лет ин-
фицированы этим вирусом.
Факторы риска
Переохлаждение, инфекции и интоксикации,
сахарный диабет, язвенная болезнь желудка и
др. Гиперинсоляция, ионизирующее излучение,
болезни крови, новообразования. Применение
иммунодепрессантов, рентгенотерапии, ВИЧ-
носительство, СПИД; 25 % ВИЧ-инфицирован-
ных страдают опоясывающим лишаем.
Патогенез
Реактивация вируса в черепных и спинно-моз-
говых ганглиях. Первый контакт с вирусом
происходит у переболевших ветряной оспой,
после чего вирус в латентном состоянии пер-
систирует в этих ганглиях. Реактивация вируса
происходит в результате воздействия неблагоп-
риятных экзогенных и эндогенных факторов.
Вирус поражает чувствительные нервы и вы-
зывает явления невритов, затем линейные пу-
зырьковые высыпания, соответственно дерма-
тому. Наблюдается отечность нервных стволов
и их сдавление в межпозвоночных простран-
ствах.
Клиническая картина
Продромальные явления: повышение температу-
ры тела, недомогание, слабость, головная боль,
диспептические расстройства.
Локализация поражения: в зоне межреберных
нервов; по ходу веточек тройничного нерва; на
голове, лице, на конечностях.
Количество очагов: множественные.
Субъективные ощущения: боль по ходу пора-
женных нервов.
Течение: острое, не рецидивирует. Рецидивы
возникают при онкозаболеваниях, болезнях кро-
ви, иммунодефиците (СПИД).
Клинические формы
Обычная форма. Характерно унилатеральное (од-
ностороннее) расположение элементов. Кожные
проявления начинаются с эритемы и папулезных
элементов в виде вытянутых очагов, которые
превращаются в везикулы и пустулы, затем эро-
зии, покрывающиеся корками и последующим
образованием в этих участках буроватой пигмен-
тации. В легких случаях процесс регрессирует в
течение 1—3 недель. Постгерпетическая неврал-
гия (болезненные ощущения и парестезии по
ходу пораженного нерва) может сохраняться до
года и более (результат хронического неврита и
рубцовых изменений нерва).
Везикулярные и эрозивные элементы могут
появляться на слизистой рта, влагалища, моче-
вого пузыря (в зависимости от топографии по-
раженного ганглия и зоны его иннервации).
Возможно развитие регионарного лимфаденита,
расстройств различных видов чувствительности,
пареза мимических мышц.
Абортивная форма: отек, эритема, папулезные
высыпания по ходу нерва, боли в участках по-
ражения.
Язвенно-некротическая форма. В тяжелых слу-
чаях высыпания принимают язвенный и язвен-
но-некротический характер. Регрессирование их
затягивается до 1—1,5 месяцев.
Генерализованная форма наблюдается у час-
ти больных (2—4 %) с болезнями крови, ново-
образованиями, получающих химиотерапию.
Высыпания принимают диссеминированный ха-
рактер в виде множественных, нередко слива-
ющихся папуловезикулезных элементов, бля-
шек, занимающих значительные участки
кожного покрова.
174 Частная дерматология
Гангренозная форма развивается у больных
сахарным диабетом, язвенной болезнью, у пожи-
лых лиц в виде везикул с кровянистым содер-
жимым и образованием на их месте участков
гангренизации с последующим рубцеванием.
Нередко упорные симпаталгии и ганглиониты
(нарушение сердечной деятельности, функции
ЖКТ, мочеиспускания и другие расстройства).
Офтальмогерпес. Поражаются роговица,
конъюнктива, радужная оболочка, цилиарное
тело с развитием светобоязни, блефароспазма,
слезотечения, болями по ходу зрительного не-
рва, наличием высыпаний в области лица, шеи,
волосистой части головы; возможно развитие
менингоэнцефалита.
Осложнения офтальмогерпеса: увеит, керато-
конъюнктивит, ретинит, глаукома, экзофтальм,
деформации века, парез (паралич) глазодвига-
тельных мышц и некоторые другие.
Диагностика
Положительная проба Цанка; определение вирус-
ных антигенов Varicella zoster методом прямой
иммунофлюоресценции в содержимом везикул
или соскобе их дна; выделение Varicella zoster в
культуре клеток; ЭКГ, КТ, МРТ внутренних
органов, легких, спинного и головного мозга
(для исключения их патологии).
Гистопатология
Отек эпидермиса, пузырьки в верхних отделах
мальпигиева слоя, баллонирующая дегенерация
и дистрофия цитоплазмы клеток, гигантские
многоядерные клетки. В дерме небольшая ин-
фильтрация нейтрофилами, тенденция к экзо-
цитозу. Аналогичные изменения выявляются в
структурах головного и спинного мозга и внут-
ренних органах при генерализованной инфек-
ции.
Возможные осложнения. Гангренизация высыпа-
ний, генерализация инфекции с поражением зна-
чительных участков кожного покрова, внутренних
органов, ЦНС, спинного мозга. Постгерпетичес-
кие невралгии, симпаталгии, ганглиониты. Реци-
дивы наблюдаются при иммунодепрессии, ново-
образованиях.
Дифференциальный диагноз
Зостериформный красный плоский лишай, ро-
жистое воспаление, зостериформный простой
герпес, герпетиформный дерматит Дюринга,
контактный дерматит от растений, буллезное им-
петиго.
Лечение
Ацикловир 800 мг внутрь 4 раза в сутки в тече-
ние 7—10 суток. Применяют другие средства: са-
лицилаты 0,5 г 1—3 раза в день, 25 % магния
сульфат по 5—12 мл внутримышечно, мочегон-
ные (фуросемид, лазикс). Пять инъекций 0,004 %
раствора панавир по 5,0 мл. с интервалом 48 ча-
сов, местно — защитный гель панавир 3-4 раза
в сутки (табл. 11.3).
Для купирования болевого синдрома: 1,5 %
раствор ганглерона по 1—4 мл внутримышечно
3—4 раза в день, или пахикарпин 0,05 г 3 раза в
день; новокаиновые блокады, анальгетики
(анальгин, баралгин), индометацин в течение
10—14 дней. Курс инъекций витаминов Вр В12
(500—1000 мкг) № 10—15, аскорбиновая кисло-
та 0,3—0,5 г в сутки. Электрофорез с новокаи-
ном на болевые очаги; паравертебрально диатер-
мия № 6—10 или ультразвук № 10—12.
При офтальмогерпесе и менингоэнцефалите:
ацикловир внутривенно; 40 % раствор уротропи-
на по 10 мл в/в, ежедневно № 5—8.
При иммунодефиците и тяжелых осложне-
ниях-. ацикловир 10—12 мг/кг в/в через каж-
дые 8 ч в течение 7—14 суток; или валацикло-
вир 1,0 г внутрь 3 раза в сутки в течение 7
дней; или фамцикловир 500 мг 3 раза в сутки
7 дней; или фоскарнет в/в; или ганцикловир—
начальная доза 5 мг/кг через 12 ч внутривен-
но в течение 14—21 суток (поддерживающая
доза 6 мг/кг внутривенно 1 раз в сутки 5 раз
в неделю).
Интерферон а-2а в/м по 1 млн ЕД 3 раза в
неделю.
Таблица 11.3. Рекомендации к применению противовирусного препарата «Панавир»
Заболевание Методика лечения Ожидаемый эффект
Опоясывающий лишай Пять инъекций 0,004% раствора Панавир по 5,0 мл с интервалом 48 часов Снятие выраженных неврологических болей, сокращение сроков заживления до 7 - 8 дней. Уменьшение объемов высыпаний
Гель защитный Панавир наружно 3-4 раза в сутки Снятие болевых ощущений, быстрое ранозаживление
Глава 11. Вирусные болезни кожи 175
Местно: ацикловир, дезоксирибонуклеаза,
анилиновые красители, фукорцин, жидкость
Бурова, повязки с физиологическим раствором
на очаги, капсаициновый крем, лидокаиновый
гель, комбинированные кортикостероидные
мази, УФО, ультразвук, лазеротерапия.
Профилактика
Биогенные стимуляторы, адаптогены, грязелече-
ние, сероводородные и радоновые ванны, обсле-
дование на онкозаболевания, ВИЧ.
Бородавки
Появление эпидермальных гиперкератотических
папул с папилломатозной реакцией дермы невос-
палительного характера в результате патогенного
действия вирусов папилломы человека (ВПЧ).
Этиология
Вирусы папилломы человека (содержат ДНК,
состоящую из 2-х цепочек): ПВЧ-1 — подошвен-
ные бородавки; ПВЧ-П — обычные бородавки;
ПВЧ-1П — плоские бородавки.
Пути заражения. Контактным путем от боль-
ных лиц; через инфицированные предметы оби-
хода (полотенца, одежда, обувь и др.).
Предрасполагающие факторы: травматизация
кожи, сухость кожи, снижение общей реактивно-
сти и иммунитета, вегетоневрозы, несоблюдение
личной гигиены, иммунодепрессивная терапия,
дефект клеточного иммунитета, инфицирование
ВИЧ.
Клинические формы
Простые (вульгарные) бородавки — полушаровид-
ные невоспалительные узелки цвета кожи, плотные,
с ороговением, порой с вегетациями, безболезнен-
ные, шероховатые, чаще множественные, локали-
зуются в области кистей, иногда на коленях. В
местах их появления кожный рисунок не визуали-
зируется. При осмотре под лупой выявляются тем-
ные точки (признак тромбоза капилляров сосоч-
ков) — диагностический симптом вульгарных
бородавок. Вокруг удаленной бородавки могут по-
явиться дочерние элементы (изоморфная реакция).
Плоские (юношеские) бородавки — плоские
узелки цвета нормальной кожи или буроватые,
поверхность гладкая, как правило, множествен-
ные; появляются в области лица, на тыле кис-
тей, голенях. Характерна изоморфная реакция.
Подошвенные бородавки — вначале небольшие
блестящие папулы, затем желтоватого цвета об-
разования, покрытые толстыми роговыми на-
слоениями, единичные, болезненные. При сли-
янии мелких бородавок образуется бляшка,
имеющая мозаичный вид. Между пальцами
стоп нередко появляются «целующиеся» боро-
давки, которые необходимо дифференцировать
с меланомой.
Нитевидные (ворсинчатые) бородавки — тонкие
в виде отростков, частично рогового характера раз-
растания с локализацией на веках, подбородке, шее,
подмышечных областях и других участках тела.
Течение хроническое.
Гистопатология
В клетках мальпигиева слоя явления акантоза,
баллонирующей дегенерации шиповатых клеток,
резкое утолщение зернистого и рогового слоев,
вертикальный паракератоз. Характерен койлоци-
тоз — множество эпителиальных клеток с око-
лоядерной зоной просветления. Сосочки дермы
вытянуты, содержат сосуды.
Дифференциальный диагноз
Красный плоский лишай, омозолелость, бородав-
чатый туберкулез кожи, папулезный сифилид, стар-
ческие кератомы, узелки доильщиц (элементы па-
пуло-везикулезного характера с западением в центре
на кистях, запястьях, возможны вегетации).
Лечение
Простые бородавки. Прижигающие средства —
солкодерм, ферезол, 10—20 % салицилово-мо-
лочный коллодий, 10—20 % раствор нитрата се-
ребра, 50 % раствор трихлоруксусной кислоты,
40 % салициловый пластырь. Цитотоксические
препараты — 25 % раствор подофиллина, «Кон-
дилин», колхаминовая мазь (смазывание эле-
ментов 1 раз в неделю). Кератолитические мази,
растворы, лаки. Противовирусные препараты
(5 % теброфеновая мазь, 3 % оксолиновая
мазь).
Криотерапия жидким азотом. Диатермокоагу-
ляция. Углекислотный лазер (1—3 сеанса). Гип-
носуггестивная терапия.
Плоские бородавки. Жженый магния сульфат
per os 0,25—0,5 г 2 раза в день после еды в тече-
ние 3—4 недель; местно на очаги 30 % раствор
трихлоруксусной кислоты, солкодерм.
Подошвенные бородавки. Прижигающие и ке-
ратолитические средства (см. простые бородавки);
криодеструкция; более эффективна электрокоа-
гуляция и углекислотный лазер. Хирургическое
лечение не проводят. Горячие ванны (45 °C) для
стоп в течение 30—45 мин, 2—3 раза в неделю, до
20 процедур.
176 Частная дерматология
Профилактика
Личная гигиена, повышение общей реактивно-
сти организма и иммунитета.
Остроконечные
кондиломы
Очаговые или множественные опухолевидные
сосочковые кожно-слизистые разрастания, чаще
наблюдающиеся в области гениталий, реже — в
области рта и в перианальной области.
Этиология
Вирус папилломы человека VI и XI типов. Про-
онкогенный ДНК-содержащий вирус.
Пути заражения. Половой контакт, инфици-
рованные предметы обихода. У большинства лиц
вирус находится в латентном состоянии. Конта-
гиозность выражена при наличии клинических
проявлений. Новорожденные заражаются от
больной матери.
Факторы риска: несоблюдение личной гигие-
ны, влажность и мацерация кожи, выделения из
гениталий, травматизация, иммунодефицитные
состояния, ВИЧ-инфицирование, заболевания
мочеполовых органов.
Клиническая картина
Инкубационный период: 1—9 месяцев и более.
Преимущественная локализация: у мужчин —
венечная борозда полового члена, внутренний
листок крайней плоти, тело полового члена, пе-
рианальная область (гомосексуалисты); у жен-
щин — половые губы, клитор, вход во влагали-
ще, вокруг уретры, мочевой пузырь, шейка
матки, область прямой кишки, промежность, ро-
тоглотка.
Кожные проявления. Мелкие сосочковые об-
разования остроконечной или конусовидной
формы, мягкой консистенции. При слиянии
образуют дольчатые разрастания в виде цветной
капусты. Имеют ярко-красный цвет, мацериро-
ванную, эрозированную поверхность. При при-
соединении вторичной инфекции появляется
неприятный запах.
Диагностика
На доклинической стадии применяют диагнос-
тическую пробу с 5 % уксусной кислотой. Мар-
левую салфетку с раствором уксусной кислоты
накладывают на исследуемые участки на 5—
10 мин. При осмотре под лупой или кольпоско-
пии кондиломы визуализируются в виде мелких
белых узелков.
Биопсию у женщин проводят при подозрении
на озлокачествление процесса, затем ежегодно —
цитологическое исследование мазков с шейки
матки.
Гистопатология
Выраженный папилломатоз и акантоз, истон-
чение рогового слоя с явлениями паракерато-
за. Вакуолизация эпидермиса. Отек дермы,
расширение капилляров, инфильтрация лим-
фоидными и плазмоцитарными клетками. В
сосочках хронический воспалительный ин-
фильтрат, расширение кровеносных и лимфа-
тических сосудов.
Возможные осложнения
Изъязвление высыпаний, озлокачествление, ге-
нерализация (СПИД).
Дифференциальный диагноз
Широкие кондиломы (медно-красный цвет,
широкое основание, бледная трепонема, RW,
РИТ, РИФ положительны), папулезный сифи-
лид, геморрой, ворсинчатые папилломы, вегети-
рующая пузырчатка, спиноцеллюлярная эпите-
лиома.
Лечение
Ферезол, солкодерм, кондилин. Смазывание
элементов концентрированной трихлоруксусной
кислотой, 20 % раствором ляписа, 25 % раство-
ром подофиллина, 0,5 % колхаминовой мазью.
Кератолитические мази, растворы, присыпки
(резорцин, дерматол поровну), лаки. Противо-
вирусные мази (риодоксоловая, теброфеновая,
бонафтоновая, оксолиновая). Диатермокоагуля-
ция. Криотерапия жидким азотом. Кюретаж. Ла-
зеротерапия.
Профилактика
Соблюдение личной гигиены; использование
презервативов; лечение воспалительных заболе-
ваний мочеполовых органов, опрелостей; диспан-
серное наблюдение.
Контагиозный
моллюск
Вирусное поражение эпидермиса (редко слизис-
тых) в виде полушаровидных узелков розового
Глава 11. Вирусные болезни кожи
177
цвета, величиной до горошины с пупковидным
вдавлением в центре. Болеют преимущественно
дети. Заболевание склонно к самостоятельному
разрешению в течение полугода. У ВИЧ-инфи-
цированных на теле выявляется множество мел-
ких элементов, а на лице — крупных (гигантс-
ких); без лечения прогрессирует. Контагиозным
молюском поражается 5—18 % ВИЧ-инфициро-
ванных.
Распространенное высыпание и рецидивиру-
ющее течение также наблюдается у лиц с врож-
денными нарушениями иммунитета, больных
лимфопролиферативными заболеваниями, ато-
пическим дерматитом, при лечении системны-
ми и местными кортикостероидами.
Этиология
Вирус группы оспы (ДНК-содержащий), се-
мейства поксвирусов. В культуре не выделен.
Различают два типа вируса контагиозного мол-
люска.
Пути заражения. Прямой контакт с больным
человеком. Через инфицированные вирусом
предметы обихода заражение маловероятно. При
половых контактах характерна генитальная ло-
кализация.
Клиническая картина
Инкубационный период: от 2 недель до б меся-
цев.
Локализация: лицо, шея, грудь, подмышечные
впадины, тыл кистей, предплечья, лобковая об-
ласть, наружные половые органы, внутренняя
поверхность бедер.
Кожные проявления. Множественные мелкие
полусферические высыпания, обычно размером
с булавочную головку восковидного телесного
или розового цвета с пупковидным вдавлением
в центре, которое заполнено роговыми массами.
Вдавление лучше выявляется после кратковре-
менной аппликации жидкого азота. Высыпные
элементы располагаются группами или диссеми-
нированно. У лиц с иммунодефицитом может
принять диффузный характер и прогрессировать
даже на фоне лечения.
Атипичные формы
Гигантские моллюски (до 3 см в диаметре); оро-
говевающие, кистозные, изъязвленные, напоми-
нающие милиум, угри, фурункулы, бородавки.
Способ диагностики
При надавливании с боков на элемент пинцетом
выделяется крошковатая масса беловатого цвета
(роговые клетки, жир, моллюсковые тельца).
Гистопатология
Инфекционная акантома. Клетки эпидермиса
увеличены, содержат внутриплазматические
включения (моллюсковые тельца). Отмечаются
признаки легкой воспалительной инфильтрации
дермы.
Дифференциальный диагноз
Диссеминированный контагиозный моллюск —
бородавки (вульгарные, плоские), угревая сыпь,
милиум, фолликулиты, аллергический дерматит;
единичные элементы контагиозного моллюс-
ка — кератоакантома, базалиома, плоскоклеточ-
ный рак. У ВИЧ-инфицированных необходимо
исключить глубокий микоз.
Лечение
Выдавливание элементов пинцетом (поверхность
очага можно предварительно проколоть в не-
скольких местах). Применяется выскабливание
кюреткой и смазывание элемента настойкой
йода, нитратом серебра, ферезолом, ТХУ, проти-
вовирусной мазью. Показаны также: диатермо-
коагуляция, криотерапия, лазерная терапия.
Применяют местно бензоилпероксид, трети-
ноин, кантаридин, подофиллин.
При распространенных высыпаниях назначают
внутрь циметидин (Н2-блокатор с иммуномоду-
лирующими свойствами) — 40 мг/кг/сут в тече-
ние 2-х мес, а также системно изотретиноин.
Профилактика
Соблюдение личной и общественной гигиены.
Дети, больные контагиозным моллюском, до-
пускаются в коллектив после излечения.
Узелки доильщиц
Вызываются вирусом коровьей оспы (паравак-
цины Липшютца). Заболевание возникает при
внедрении инфекции в ссадину кожи с вымени
больных коров. Болеют доярки и другие работ-
ники молочных ферм.
Клиническая картина
На кистях и предплечьях появляются высыпа-
ния в виде узелков величиной до крупной го-
рошины, полушаровидной формы с перифери-
ческим воспалительным венчиком. Вначале
узелки бледно-розового цвета, затем принима-
ют синевато-багровую окраску, в центре появ-
ляется небольшое западение, характерное для
оспы. Узелки могут быть единичные или мно-
178 Частная дерматология
гочисленные. Через 1—2 мес элементы регресси-
руют без признаков рубцевания.
Лечение
На очаги поражения применяют влажные повяз-
ки с растворами калия перманганата, этакриди-
на лактата; аутосанация обычно наступает спон-
танно в течение 1—2 мес.
Профилактика
Ветеринарный надзор за животными, изоляция
больных. Больные лица, не допускаются к здо-
ровым животным. Санитарно-гигиенический
режим.
Глава 12
Паразитарные болезни кожи
Чесотка 179
Педикулез 185
Чесотка
Синонимы: scabies.
Чесотка — инфекционное паразитарное забо-
левание кожи, которое вызывается чесоточным
зуднем (Sarcoptes scabiei) и сопровождается ноч-
ным зудом кожных покровов, расчесами, парны-
ми папуло-везикулами и чесоточными ходами.
Этиология и патогенез
Возбудитель чесотки (Sarcoptes scabiei var.
hominis) относится к типу мелких членистоно-
гих, классу паукообразных, отряду акариформ-
ных, подотряду чесоточных. Клещ имеет неболь-
шие размеры и плохо виден невооруженным
глазом. Строение чесоточного клеща и форма
исследованы с помощью оптического микроско-
па, а затем были уточнены исследованиями с
использованием сканирующего электронного
микроскопа. Тело (или идиома) клеща снабже-
но четырьмя парами конечностей и имеет шаро-
видный вид, напоминающий форму картофели-
ны: на поверхности его спины имеются
многочисленные иголки и поперечная щель. На
брюшке самки также имеется поперечная щель
(токостома), через которую она откладывает яйца.
Яичники после оплодотворения становятся очень
большими и длина самки увеличивается с 200 мк
до 400 мк. Как только взрослая самка прикреп-
ляется к поверхности кожи, она начинается бы-
стро буравить с помощью коготков передних ног
роговой слой эпидермиса, проделывая вначале
едва заметные вертикальные, колодцеобразные
отверстия, затем параллельно поверхности кожи
роет S-образные чесоточные ходы. Клещ имеет
такой телесный объем, который позволяет ему
достичь даже верхних слоёв мальпигиевого слоя,
вызывая при этом альтерацию ороговения, ко-
торое развивается в сторону паракератоза. Сама
самка продвигается в чесоточном ходе с перемен-
ной скоростью, причем этому продвижению
способствуют секреция литических факторов и
интенсивность работы передних ног. По оценке
специалистов, продолжительность жизни пара-
зита может достигать двух месяцев. Спустя не-
сколько часов после начала образования чесоточ-
ного хода самка начинает откладывать яйца (2—3
яйца в сутки: яйца выходят из полости живота
через поперечную щель, находящуюся на брюш-
ке (токостома) и прикрепляются к стенке чесо-
точного хода с помощью аморфного материала.
Через 3—4 дня после кладки яиц из них вылуп-
ляются личинки, которые имеют три пары ног.
Личинки покидают чесоточный ход и возвраща-
ются в эпидермальный слой. Через 2—3 дня у
личинок происходит первая линька, в результате
которой рожцается октоподная нимфа (4 пары
ножек). Нимфа линяет через 2—3 дня после этого
и рожает взрослого самца или самку. Следова-
тельно, весь цикл развития половозрелого кле-
ща, который начинается кладкой яиц и закан-
чивается рождением взрослой особи, происходит
примерно за 10—14 дней. Однако этот цикл раз-
вития характеризуется значительной гибелью
яиц. Всего 10 % всей кладки яиц доживают до
стадии появления взрослых особей. Стенки че-
соточного хода покрыты особыми веществами,
секретируемыми паразитами, и испражнениями.
Эти твердые каловые массы представляют собой
остатки чешуек клеток эпидермиса, принимае-
мых в пищу клещем. Следовательно, вся сово-
купность инородных материалов является для
«хозяина» — носителя паразитов — немаловаж-
ным сенсибилизирующим фактором. Число па-
разитов, которое можно обнаружить на одном
человеке, составляет 11,3 особей. Между числом
паразитов и их «хозяином» устанавливается со-
стояние равновесия, при котором можно иног-
да наблюдать полное и спонтанное исчезновение
паразитов, сопровождающееся клиническим из-
лечением.
При комнатной температуре клещи остаются
живы в течение 5 дней, взрослые особи и незре-
179
180 Частная дерматология
лые формы (личинки-нимфы) живут не более 48
часов при температуре 22°С и степени влажнос-
ти 35 %; при температуре выше 55°С клещ гиб-
нет в течение 10 минут; при кипячении клещи
гибнут моментально.
Эпидемиология
В 95 % случаев заражение происходит при пря-
мом длительном контакте с больным человеком
и требует «благоприятных» условий температу-
ры и влажности. Источником заражения при
чесотке является больной человек. Различают
прямой и непрямой пути заражения чесоткой.
Можно выделить три основных фактора, обес-
печивающих инвазию: прямой телесный кон-
такт, его длительность и обилие возбудителя на
источнике заражения. При прямом пути зара-
жения возбудитель переходит непосредственно
от больного человека здоровому в момент те-
лесного соприкосновения. Основными обстоя-
тельствами, при которых происходит заражение,
являются половые контакты, когда чесотка ста-
новится настоящим источником венерического
заболевания. Непрямая передача заболевания
происходит через предметы личного пользова-
ния (нижнее белье и постельные принадлежно-
сти больного). Типичным примером высокой
вероятности развития эпидемических вспышек
чесотки служат организованные коллективы
(школы, интернаты, детские сады, ясли). При
создании благоприятных условий для инфици-
рованное™ (плохие санитарно-гигиенические
условия, скученность, частая смена половых
партнеров) может произойти резкое увеличение
числа больных чесоткой, особенно среди опре-
деленных групп населения.
За последнее столетие были отмечены три
крупные эпидемии чесотки: 1913—1925 гг.,
1940—1955 гг., начало 1965 года. Эпидемии че-
сотки следуют синусоидальному закону эволю-
ции с общей амплитудой в 30 лет (период 15
лет — эпидемии, период 15 лет — без эпиде-
мий). Однако причины таких колебаний пред-
ставляются комплексными и недостаточно хоро-
шо известными. Причины эпидемий имеют
различное происхождение:
• социологические (бедность, несоблюдение
надлежащих мер санитарии и гигиены, ранняя
половая жизнь, миграция населения, туризм);
• медицинские (ошибки в диагностике заболе-
вания, контагиозность инкубационного перио-
да, атипичная форма чесотки);
• иммунологические (уменьшение иммунных
реакций у людей по отношению к чесоточным
клещам Sarcoptes scabiei hominis).
Факторами роста заболеваемости чесоткой явля-
ются также свобода половых отношений, повы-
шение солнечной активности, недостатки меди-
цинской службы. Существует некоторая
сезонность в течении болезни, а также имеются
данные, что заболеваемость чесоткой в настоя-
щее время находится примерно на уровне забо-
леваемости сифилисом.
Клиническая картина
Инкубационный период при заражении самка-
ми чесоточного клеща отсутствует (процесс вне-
дрения самки в кожу занимает не более одного
часа), а при заражении личинками инкубацион-
ный период составляет от 8 до 12 дней.
Основные клинические симптомы чесотки:
• зуд, усиливающийся в вечернее и ночное
время суток;
• наличие чесоточных ходов;
• полиморфизм высыпаний (папулы, везикулы,
эрозии, геморрагические корочки, экскориации);
• характерная локализация высыпаний (меж-
пальцевые складки кистей, область лучезапяст-
ных суставов, локтевые сгибы, переднебоковая
поверхность живота, поясницы, ягодиц; половые
органы мужчин, молочные железы у женщин).
Зуд является основным субъективным симп-
томом заболевания и обнаруживается практичес-
ки у всех без исключения больных чесоткой.
Преобладает сильный и умеренный зуд, который
имеет два основных характерных признака:
• проявляется в наибольшей степени вечером
и во время сна, но может быть и постоянным;
часто бывает очень сильным и мешает нормаль-
ному сну; зуд может локализоваться на отдель-
ных участках кожи или распространяться по
всему телу за исключением кожи лица и воло-
систой части головы, которые никогда не пора-
жаются;
• является групповым зудом, поражает не-
сколько членов одной семьи или одного коллек-
тива.
Усиление зуда вечером связывают с наличи-
ем суточного ритма активности клещей и уве-
личение его в ночные часы. Механизм возник-
новения зуда при чесотке не выяснен. Считают,
что зуд связан с сенсибилизацией организма к
клещу и продуктам его жизнедеятельности. Дру-
гие исследователи утверждают, что возникнове-
нию зуда способствуют вещества, находящиеся
в экскрементах. Согласно еще одной точке зре-
ния, зуд является результатом раздражения не-
рвных окончаний при движении клеща.
Чесоточный ход является основным диагнос-
тическим признаком чесотки, отличающим ее от
Глава 12. Паразитарные болезни кожи 181
других зудящих дерматозов. Наиболее типичны
ходы, имеющие вид слегка возвышающейся над
кожей линии белого или грязно-серого цвета,
прямые или изогнутые, длиной от 1 мм до не-
скольких сантиметров. Каждый чесоточный ход
роет самка клеща в роговом слое эпидермиса.
Чесоточный ход заканчивается на одном из его
концов (слепом) светопроницаемым вздутием —
акариозным пузырьком, в котором обычно рас-
полагается самка паразита. Различают три кли-
нических варианта чесоточных ходов, в основу
которых положены приуроченность к ходам тех
или иных морфологических элементов и резуль-
таты анализа их содержимого.
Для клинической картины чесотки характер-
ны полиморфизм высыпаний за пределами ходов.
Первичные морфологические элементы представ-
лены фолликулярными папулами, располагающи-
мися на коже туловища и конечностей, а также
изолированными везикулами, локализующими-
ся на кистях. Вторичные морфологические эле-
менты представлены точечными кровянистыми
корочками, эрозиями. Папулы считают вторым
по диагностической значимости признаком чесот-
ки, они значительно преобладают среди других
высыпаний. Характерными признаками везикул
считается их немногочисленность, изолированное
расположение и локализация преимущественно на
кистях. Везикулы в межпальцевых складках счи-
таются патогномоничным симптомом чесотки. В
результате расчесывания (вследствие зуда) появ-
ляются эрозии, кровянистые корочки, линейные
расчесы. Крупные геморрагические корочки, рас-
полагающиеся линейно вдоль конечностей, можно
считать дополнительным диагностическим сим-
птомом чесотки. Типичным для чесотки являет-
ся симптом Горчакова — наличие точечных кро-
вянистых корочек на локтях и в их окружности,
а также признак Арди — импетигинозные эле-
менты на разгибательных поверхностях локтевых
суставов. Высыпания в виде импетигинозных
элементов, пузырьков, корочек, располагающихся
в области межъягодичной складки и переходящих
на крестец, получили название «симптом треу-
гольника».
Излюбленная локализация высыпаний при че-
сотке: межпальцевые складки, лучезапястные су-
ставы, разгибательная поверхность локтевых сус-
тавов; область пупка, переднебоковая поверхность
живота, поясницы, ягодиц; мужской половой
член, молочные железы у женщин (рис. 12.1).
Плотные, красного цвета, пруригинозные папулы,
находящиеся на головке мужского полового чле-
на, являются точными признаками чесотки у муж-
чин. У детей грудного возраста чесоточные ходы
появляются на коже лица и волосистой части го-
ловы, иногда на внутреннем своде стопы.
Атипичные клинические формы
Чесотка без ходов представляет собой начинаю-
щиеся формы заболевания или встречается у
людей, которые соблюдают правила гигиены
тела. Предполагают, что отсутствие чесоточных
ходов возможно на ранней стадии заболевания
у лиц, контактировавших с больными чесоткой
и объясняется заражением личинками. При этом
имеются другие, характерные для чесотки сим-
птомы: зуд, усиливающийся в вечернее время,
папулы и везикулы в местах типичной локали-
зации.
Осложнения
Наиболее распространенными осложнениями
чесотки являются пиодермия и дерматит, реже
наблюдаются экзема и крапивница. Крайне ред-
ко наблюдается поражение ногтевых пластинок
у больных чесоткой, преимущественно у детей
грудного возраста.
Диагностика
Клинический и эпидемиологический диагноз
чесотки должен быть подтвержден лабораторны-
ми методами, цель которых —- обнаружение воз-
Рис. 12.1. Типичная локализация
высыпаний при чесотке.
182 Частная дерматология
будителя. Для лабораторной диагностики чесотки
применяются следующие методы: извлечение
клеща иглой; метод тонких срезов; соскоб без
крови; соскоб до появления крови; соскоб в ми-
неральном масле; щелочное препарирование
кожи; экспресс-диагностика с применением мо-
лочной кислоты.
Лечение
Лечение больных чесоткой направлено на унич-
тожение возбудителя с помощью акарицидных
препаратов. Идеальное противочесоточное сред-
ство должно:
• оказывать одинаково эффективное воздей-
ствие на клещей и их личинки;
• обладать минимальным сенсибилизирую-
щим и раздражающим побочным воздействием
даже в случае регулярного применения;
• быстро выводиться из организма в случае
проникновения внутрь через кожные покровы,
т. е. общая токсичность противочесоточного сред-
ства должна быть ничтожно мала;
• быть простым в применении и способ его
применения должен быть четко указан;
• быть достаточно приятным с косметической
точки зрения: не иметь запаха, не пачкать
одежду.
Для лечения чесотки предлагались различные
препараты: серная мазь, мазь Вилькинсона, мазь
Хельмериха; жидкости Флемингса, Мура, Элерса;
паста Милиана; растворы креолина, лизола; чи-
стый деготь, этиленгликоль, бензойный эфир и
др. Издавна использовали также нелекарствен-
ные средства, например, керосин, бензин, мазут,
автол, сырую нефть, зольный щелок. Начиная
с 1938 года открылась новая эра в лечении че-
сотки благодаря последовательным открытиям
таких препаратов, как бензилбензоат (1936),
Д. Д. Т. (1946), кротамитон (1949), линдан
(1959), спрегаль (1984). Однако в настоящее
время не выработан единый подход к терапев-
тическим методам лечения чесотки. Необходи-
мо также отметить, что в большинстве продава-
емых лекарств дозировка препаратов значительно
превосходит терапевтические потребности. В то
же время необходимо соблюдать некоторые об-
щие правила при лечении больных чесоткой:
• обрабатывать следует всю поверхность тела,
а не только пораженные участки; препарат дол-
жен наноситься тонким однородным слоем; осо-
бое внимание следует уделить обработке рук, ног,
межпальцевых промежутков, подмышек, мо-
шонки и промежности;
• избегать попадания препарата в глаза и на
слизистые оболочки;
• дозировка должна быть не слишком боль-
шой; одновременно с противочесоточными пре-
паратами не следует применять другие местные
средства;
• в случае запущенной чесотки с осложнени-
ями следует вначале обработать кожные пораже-
ния; для лечения вторичной инфекции приме-
няются антисептики и методы общей терапии,
для лечения дерматитов и экзем — смягчающие
препараты местного действия;
• в случае норвежской чесотки необходимо
при помощи кератолитических средств предва-
рительно очистить участки кожи, покрытые
коркой, и немедленно изолировать больного.
Препараты, содержащие серу, уже очень дав-
но применяются для лечения чесотки (мазь
Хельмериха, паста Милиана, серная мазь).
Серная мазь используется наиболее часто (33 %
— для взрослых и 10-15 % — для детей). Пе-
ред началом лечения больной моется теплой во-
дой с мылом. Мазь втирают в весь кожный по-
кров ежедневно в течение 5—7 дней. Через 6—8
дней больной моется с мылом и меняет натель-
ное и постельное белье. Для детей рекоменду-
ется применение 15 % серной мази в 1-й и 4-
й день курса терапии. Недостатки применения
серной мази: длительность лечения, неприят-
ный запах, частое развитие дерматитов, загряз-
нение белья.
Метод Демьяновича основан на акарицидном
действии серы и сернистого ангидрида, выде-
ляющихся при взаимодействии гипосульфита
натрия и соляной кислоты. Лечение заключа-
ется в последовательном втирании в кожу 60 %
раствора гипосульфита натрия (раствор № 1)
и 6 % раствора соляной кислоты (раствор № 2).
Для лечения детей используют более низкие
концентрации — 40 % и 4 % соответственно.
Раствор гипосульфита перед употреблением
слегка подогревают и втирают в кожу в опре-
деленной последовательности: начинают с
кожи обеих кистей, затем втирают в левую и
правую верхние конечности, потом — в кожу
туловища (грудь, живот, спина, ягодичная об-
ласть, половые органы) и, наконец, в кожу
нижних конечностей до пальцев стоп и по-
дошв. Втирание в каждую область длится
2 мин, вся процедура должна занимать не ме-
нее 10 мин. При подсыхании в течение 10 мин
на коже появляется масса кристалликов гипо-
сульфита. После 10-минутного перерыва при-
ступают к втиранию 6 % соляной кислоты, ко-
торое осуществляется в том же порядке по
одной минуте на каждую область 3 раза с пе-
рерывами по 5 мин для обсыхания. По окон-
Глава 12. Паразитарные болезни кожи 183
чании втирания и после обсыхания кожи боль-
ной надевает чистое белье и не моется в тече-
ние 3 дней, но в кисти рук растворы повтор-
но втираются после каждого мытья. Через 3
дня больной моется горячей водой и вновь
меняет бельё. Недостатки метода: трудоёмкость,
часто отмечаются рецидивы, необходимы по-
вторные циклы лечения.
Метод Богдановича основан на применении
полисульфидного линимента (10% концентра-
ция для взрослых и 5 % — для детей). Действу-
ющим началом линимента является полисуль-
фид натрия, для приготовления которого берут
600 мл воды, добавляют 200 г едкого натра
(квалификация «чистый») и сразу же 200 г по-
рошкообразной серы («серный цвет», квалифи-
кация «чистый») и помешивают стеклянной па-
лочкой. Соотношение ингредиентов 3:1:1
(вода: NaOH: сера), содержание полисульфида
в растворе — 27 %. Раствор полисульфида го-
ден к употреблению до 1 года при хранении в
плотно закрытой посуде. Основой линимента
является мыльный гель, для приготовления
которого берут 50 г измельченного мыла (же-
лательно «Детское»), подогревают в 1 л воды до
полного растворения, затем охлаждают в откры-
той посуде при комнатной температуре. Лини-
мент необходимой концентрации готовят сле-
дующим образом: к 100 мл 5 % мыльного геля
добавляют 10 мл (для 10 %) или 5 мл (для 5 %)
раствора полисульфида натрия и 2 мл подсол-
нечного масла. Методика лечения: линимент
втирают в течение 10—15 минут во всю повер-
хность кожи. Повторное втирание проводят на
2-й и 4-й день. Кисти дополнительно обраба-
тываются после каждого мытья рук. Купание
перед первым и третьим втиранием (1-й и 4-й
день) и через 2 дня после последнего третьего
втирания, т. е. на шестой день. Смена белья
после первого втирания и через 2 дня после пос-
леднего втирания (на 6-й день). При распрост-
раненных и осложненных формах заболевания
рекомендуется втирать препарат ежедневно
(один раз в день) в течение 4—5 дней. Недостат-
ки метода: неприятный запах сероводорода,
иногда развиваются дерматиты.
Бензилбензоат применяется в виде 20 % вод-
но-мыльной суспензии, детям до 3 лет — 10 %
суспензии. Суспензией обрабатывают весь кож-
ный покров (кроме головы), а у детей до 3 лет
- и кожу лица. Втирание должно проводиться
в определенной последовательности: начинают
с одновременного втирания в кожу обеих кис-
тей, затем в левую и правую верхние конечно-
сти, потом в кожу туловища и, наконец, в кожу
нижних конечностей. Предложена модификация
методики лечения больных бензилбензоатом:
20 % водно-мыльная эмульсия втирается одно-
кратно только в 1-й и 4-й день курса лечения.
Смена нательного и постельного белья прово-
дится дважды: после первого и второго втира-
ния препарата. Больной не моется в течение
последующих 3 дней, но в кисти рук препарат
втирается повторно после каждого мытья. Че-
рез 3 дня больной моется горячей водой и вновь
меняет белье. У грудных детей вместо втирания
проводят смачивание кожной поверхности ука-
занными растворами, повторяя курс лечение
через 3—4 дня. Недостатки препарата: развитие
дерматита, воздействие на центральную не-
рвную систему, отмечены случаи острой инток-
сикации.
Линдан — препарат применяется в виде 1 %
крема, лосьона, шампуня, порошка, мази. Лин-
дан или гаммабензолгексахлоран — это хлорор-
ганический инсектицид, представляющий собой
изомер гексахлорциклогексана. Препарат нано-
сят на 6—24 часа, затем смывают. Необходимо
строго соблюдать следующие правила: не реко-
мендуется применять линдан для лечения мла-
денцев, детей и беременных женщин; препарат
следует наносить в один прием на холодную
сухую кожу; концентрация препарата должна
быть ниже 1 %. Недостатки препарата: контак-
тная экзема; при попадании внутрь организма
токсичен для нервной системы и крови; прони-
кает через кожные покровы новорожденного;
местная и общая реакция, связанная с присут-
ствием анестетика.
Кротамитон — крем, содержащий 10 % п-
этил-О-кротонилтолудина, эффективен как про-
тивочесоточный и противозудный препарат, не
вызывающий побочных реакций. С успехом
применяется при лечении новорожденных и де-
тей. Кротамитон наносят после мытья дважды с
интервалом 24 часа или четырехкратно через 12
часов в течение 2-х суток.
Тиабендазол. Препарат на основе тиабенда-
зола вначале с успехом применялся для при-
ема внутрь в дозе 25 мг/кг веса в день в тече-
ние 10 дней. Однако в связи с
неблагоприятным воздействием на пищевари-
тельный тракт в настоящее время его приме-
нение ограничено. Последующие исследования
были связаны с наружным примением тиабен-
дазола в виде 5 % крема 2 раза в день в тече-
ние 5 дней, а также в виде 10 % суспензии 2
раза в день в течение 5 дней. Не отмечено
никаких побочных клинических или биологи-
ческих эффектов.
7 Зак. 1357
184 Частная дерматология
Эсдепаллетрин — этот синтетический пирет-
рин используется в качестве действующего на-
чала аэрозольного средства «Спрегаль». Препа-
ратом обрабатывают весь кожный покров, за
исключением лица и волосистой части головы,
распыляя его сверху вниз вдоль туловища, за-
тем покрывая руки и ноги. По истечении 12
часов рекомендуется тщательное мытье с мы-
лом. Обычно достаточно одного цикла обработ-
ки. Зуд и другие симптомы могут наблюдать-
ся в течение 7—8 дней. Если по истечении этого
срока симптомы сохраняются, проводят по-
вторное лечение.
Аэрозольное средство «Спрегаль» может ис-
пользоваться для лечения чесотки у беремен-
ных, кормящих женщин, новорожденных.
Перметрин применяется в виде 5 % мази (или
крема). Методика лечения: мазь тщательно вти-
рается в кожу всего тела от головы к стопам.
Через 8—14 часов принимается душ. Как прави-
ло, эффективно однократное применение препа-
рата.
Ивермектин применяется перорально одно-
кратно в дозе 20 мкг/кг веса пациента. Ивермек-
тин (ivermectin) эффективен и безопасен. Пре-
парат применяется также местно один раз в день,
однако в 50 % случаев требуется повторное ле-
чение спустя 5 дней.
Диэтилкарбамазин используется для лечения
чесотки только перорально. Препарат назнача-
ют по 100 мг 3 раза в день в течение 7 дней. Не-
достаток препарата: невысокая клиническая эф-
фективность (50 %).
Профилактика
Профилактика чесотки основана на особеннос-
тях эпидемиологии этого заболевания и вклю-
чает следующие мероприятия:
• обязательное стационарное или амбулатор-
ное лечение больных;
• установление источника заболевания;
• осмотр всех лиц, с которыми больной имел
бытовой или половой контакт;
• контроль излеченности чесотки осуществля-
ется в течение 2 недель: больных и контактных
лиц осматривают дважды — при первичном об-
ращении и через 2 недели;
• проведение текущей и заключительной де-
зинфекции очага инфекции, одежды и постель-
ных принадлежностей больного.
Обеззараживание постельных принадлежно-
стей, полотенец, нательного белья производят
в 1—2 % растворе соды или любом стиральном
порошке в течение 5-10 мин с момента заки-
пания. Верхнюю одежду (платья, костюмы,
брюки, джемперы, свитера) проглаживают с
обеих сторон горячим утюгом. Часть вещей
(шубы, пальто, плащи, вещи из кожи и замши)
может быть обеззаражена путем проветривания
на открытом воздухе в течение 5 дней. Одежда
и постельные принадлежности, которые нельзя
стирать при температуре выше 55°С, могут быть
продезинфицированы противочесоточным пре-
паратом — аэрозолью A-PAR. В комнате боль-
ного ежедневно проводят влажную уборку по-
мещения 1—2 % мыльно-содовым раствором,
включая мытье пола, протирание предметов об-
становки. Матрацы и одеяла обеззараживают в
сухожаровой камере при температуре +100°С
в течение 1 часа. В зимнее время года при ми-
нусовой температуре эти вещи выдерживают
3—4 часа на открытом воздухе. Заключитель-
ная дезинфекция проводится работником де-
зотделения СЭС после госпитализации боль-
ного, после окончания амбулаторного лечения,
а в детских коллективах дважды: после выяв-
ления больного в группе и по окончании ле-
чения в изоляторе.
Чесотка нераспознанная
Чесотка нераспознанная (инкогнито) развивается
на фоне местного применения кортикостероид-
ных препаратов. Стероиды уменьшают воспали-
тельные явления и подавляют зуд, создавая при
этом благоприятные условия для размножения
клещей, в результате чего образуется большое
количество ходов и повышается контагиозность
заболевания. Чесотка теряет свои специфичес-
кие симптомы, принимает папуло-сквамозный,
папуло-везикулезный, а иногда даже кератозный
характер и становится резистентной, как ни па-
радоксально, по отношению к кортикостерои-
дам.
Узелковая чесотка
Узелковая чесотка (постскабиозная лимфопла-
зия) возникает после полноценного лечения за-
болевания и представлена в виде зудящих узел-
ков. Предполагают, что гранулематозная реакция
кожи может возникать в результате внедрения
чесоточного клеща, вследствие раздражения
кожи при расчесывании или всасывания продук-
тов распада экскрементов. Имеются сообщения
об иммуноаллергическом генезе узелковой чесот-
ки, в пользу чего свидетельствуют гистологичес-
кие данные.
Клинически заболевание выражается появле-
нием округлых, плотных узелков размером до
Глава 12. Паразитарные болезни кожи
185
фасоли, синюшно-розового или буровато-крас-
ного цвета с гладкой поверхностью. Локализа-
ция элементов преимущественно на закрытых
частях тела. Течение доброкачественное, но дли-
тельное (от нескольких месяцев до нескольких
лет). Возможен спонтанный регресс узелковых
элементов и их повторное появление на тех же
самых местах.
Обычная местная и противочесоточная тера-
пия неэффективны. Рекомендуется применение
внутрь антигистаминных препаратов, пресоци-
ла, наружно — стероидные мази под окклюзи-
онную повязку. При длительно персистирующих
узелках применяют жидкий азот, диатермокоа-
гуляцию, лазеротерапию, пиявки.
Псевдочесотка
Псевдочесотка — это зудящий дерматоз, возни-
кающий при заражении чесоточными клещами
животных. Наиболее частым источником зара-
жения человека являются чесоточные клещи со-
бак, реже — других животных: свиней, лошадей,
кроликов, овец, коз, лисиц.
Инкубационный период заболевания очень
короткий и составляет несколько часов. Боль-
ных беспокоит сильный зуд. Клещи не прони-
кают в эпидермис и не образуют ходов. Высы-
пания асимметричные, локализуются на участках
соприкосновения с больным животным. Сыпь
представлена в виде уртикарных и пруригопо-
добных папул, папуловезикул, волдырей с рез-
ко выраженным воспалительным компонентом.
От человека к человеку заболевание не переда-
ётся, поэтому контактных лиц не следует лечить.
Лабораторная диагностика затруднена: выявля-
ются только самки, а неполовозрелые стадии
отсутствуют.
Норвежская чесотка
Норвежская чесотка — заболевание, впервые
описанное при обследовании больных лепрой в
Норвегии. Возбудителем норвежской чесотки яв-
ляется обычный чесоточный клещ. Норвежская
чесотка характеризуется следующими особенно-
стями:
• редкая регистрация заболевания;
• особый контингент больных: болезнь Дау-
на, сенильная деменция, инфантилизм, астения,
иммунодефицитные состояния;
• трудность в постановке диагноза: часто про-
ходит несколько месяцев и даже лет с момента
появления поражений и до установления диаг-
ноза; это объясняется тем, что зуд в течении
болезни часто отсутствует, а поражения затраги-
вают лицо, волосистую часть головы, ногти в
виде корок и гиперкератоза, напоминая другие
заболевани — псориаз, болезнь Дарье, питири-
аз, гистиоцитоз;
• плохо изучен патогенез; основная роль от-
водится иммунодефицитному состоянию; имеет-
ся гипотеза о генетической предрасположенно-
сти организма к развитию гиперкератозных
поражений в связи с уменьшением потребления
витамина А.
Основные клинические симптомы заболева-
ния: массивные корки, чесоточные ходы, поли-
морфные высыпания (папулы, везикулы, пусту-
лы, чешуйки, корочки) и эритродермия.
Излюбленная локализация корок — верхние и
нижние конечности (локти, колени, ладони,
подошвы), ягодицы, лицо, уши, волосистая часть
головы. Поверхность корок шероховатая, покры-
та трещинами или бородавчатыми разрастания-
ми, которые напоминают рупии. Ногти серо-
жёлтого цвета с бугристой поверхностью, легко
крошатся, край изъеден. Выражен ладонно-по-
дошвенный гиперкератоз. Отмечается увеличе-
ние лимфатических узлов. Иногда норвежская
чесотка сопровождается повышением температу-
ры тела, которая держится в течение всего забо-
левания. Заболевание характеризуется высокой
контагиозностью в связи с большим количе-
ством клещей: до 200 на 1 см2 кожи больного.
Педикулез
Возбудители болезни вши относятся к крово-
сосущим насекомым. Вши являются строго спе-
циальными паразитами, которые характерны
для определенных видов животных и человека.
У человека встречается три вида вшей, вызы-
вающих педикулез: головная вошь (pediculus
humanus capitis); платяная вошь (pediculus
vestimentorum); лобковая вошь (pediculus pubis
или phthirus pubis).
Вши относятся к насекомым отряда Anoplura.
Они имеют три пары мощных, снабженных ко-
готками ножек. Оплодотворенные самки посред-
ством нерастворимого в воде вещества, выделяе-
мого добавочной железой яичника, приклеивают
свои 150—300 яиц, овальных гнид длиной при-
мерно 0,8 мм, к волосам на голове или лобке или
же в швы белья. Цикл развития состоит из трех
этапов: яйцо, личинки, половозрелая особь (има-
го). Личинки вшей выводятся примерно через 8
дней, проходят через три превращения и через 2—
186 Частная дерматология
3 недели достигают половой зрелости. Полный
жизненный цикл от яйца до откладывания созрев-
шей самкой яиц длится 15 дней. Каждые два часа
вши сосут кровь, выпивая от 0,7 до 1,2 мл кро-
ви; голодными они могут оставаться лишь не-
сколько дней.
Головной педикулез
Синонимы: pediculosis, вшивость
Головной педикулез — заболевание, вызыва-
емое головной вошью, которая паразитирует на
волосистой части головы и шее.
Этиология и патогенез
Головная вошь (pediculus humanus capitis) имеет
2—3,5 мм в длину и представляет собой особь
серовато-белого цвета. Предпочтительно поража-
ет детей и людей с длинными волосами. Пере-
дача происходит от человека к человеку через
головные уборы или расчески. Недостаточная
гигиена и жизнь в скученных сообществах спо-
собствуют ее распространению. Нередко разви-
ваются небольшие эндемии в школах. Вне хо-
зяина головная вошь может прожить 55 часов.
В отличие от платяной вши, головная вошь не
является переносчиком инфекций.
Клиническая картина
Основное место локализации — голова. Волосы
на бороде и лобке поражаются редко. На воло-
систой части головы чаще всего поражается уча-
сток за ушами. Головные вши впитывают кровь
посредством укуса каждые 2—3 часа. Кожные
проявления становятся заметными только через
несколько часов или дней. При этом развива-
ются ярко-красные уртикарные папулы, которые
из-за проникновения слюны вшей сильно зудят.
На затылке часто возникает типичная экзема
вшивых. Расчесы часто ведут к вторичным бак-
териальным инфекциям и, наконец, к сильно-
му спутыванию волос и образованию колтуна
(трихомы). Вторичной инфекцией обусловлен и
болезненный лимфаденит в затылочной облас-
ти и в области шеи со склонностью к абсцеди-
рованию.
Диагноз
Ставится на основании типичной клиники при
доказательстве наличия вшей. Сильный зуд вши
могут вызвать даже при малом количестве. Если
вшей нет, то ищут гниды — это почковидные,
приклеенные к волосу, овальные, длиной 0,8 мм
яйца вшей, упрятанные в хитиновую оболочку.
Вначале они сидят у основания волоса, а с рос-
том волоса перемещаются к его кончику и к
этому времени уже чаще всего пусты: покрыш-
ка гниды отсутствует. В отличие от перхоти,
гниды не сыплются с волос, а сидят прочно.
Места предпочтительной локализации — участ-
ки за ушами. При осмотре необходимо припод-
нять волосы сбоку над ушами, чтобы обнаружить
там гниды. Вшей найти практически нельзя. Зуд
головы, экзема головы и затылка с импетиги-
низацией должны навести врача на мысль о го-
ловных вшах.
Дифференциальный диагноз
Экзема головы, контагиозное импетиго, псори-
аз волосистой части головы.
Лечение
Следует уничтожать не только вшей, но и их
эмбрионы в гнидах.
Линдан (гамма-гексахлорциклогексан) при-
меняется как 0,3 % гель. Волосы моют шам-
пунем, затем в еще влажные волосы втирают
гель и многократным расчесыванием тщательно
распределяют. Для этого требуется примерно
15 г геля. Гель должен оставаться в волосах три
дня, затем голову моют. При таком лечении
гниды также погибают и удаляются при полос-
кании теплым уксусным раствором (1 часть
6 % пищевого уксуса на 2 части воды) и пос-
ледующем расчесывании частым гребнем. При
беременности и кормлении линдан противопо-
казан, но допустим для грудных младенцев и
дошкольников.
Пара-Плюс (перметрин+малатион). Применя-
ется как спрей и действует также на вшей и гнид.
Препарат распыляют с расстояния 1—2 см, нажа-
тиями продолжительностью в 1—2 сек, распы-
литель снаружи прикрывают рукой. Важно тща-
тельно обработать всё основание волос, включая
виски и затылок. Через 320 мин волосы моют
обычным шампунем, затем полоскают уксусной
водой и расчесывают, как указывалось выше.
Через неделю проводят контрольный осмотр,
при повторном росте вшей или гнид лечение
повторяют. Платки и головные уборы можно
обрызгивать с расстояния 20 см. Препарат про-
тивопоказан во время беременности и кормле-
нии. Применяют его только в больших хорошо
проветриваемых помещениях. Следует обратить
внимание на его особую токсичность для хлад-
нокровных: террариум, аквариум.
Малатион (0,5 % раствор). Этим раствором
(горючий!) полностью покрывают волосы и кожу
головы (10—20 мл). После высыхания на возду-
хе (без фена!) и 12 часов действия волосы тща-
Глава 12. Паразитарные болезни кожи 187
тельно промывают шампунем. Последующий
контроль через 8—10 дней. Малатион поглоща-
ется кератином и поэтому должен иметь опре-
деленный защитный эффект на несколько не-
дель. Он противопоказан грудным детям.
Перметрин (1 % крем-ополаскиватель) вти-
рается во влажные вымытые волосы и через
30 минут смывается. Лечение грудных детей и
дошкольников проводят под наблюдением
врача.
Профилактика
Следует осмотреть всех лиц, контактировавших
с больными (семья, детский сад, школа, дом пре-
старелых) и при необходимости провести лече-
ние. Нередко возникают небольшие эндемии.
Платяной педикулез
Платяной педикулез вызывается платяной во-
шью, которая располагается в швах одежды и
вызывает поражение кожи в виде папул, гипе-
ремированных пятен или волдырей с кровянис-
той корочкой в центре.
Этиология и патогенез
Платяная вошь (pediculus humanus corporis или
pediculus vestimentorum) длиной 3—4,5 мм, т. е.
несколько крупнее головной, кроме того сегмен-
ты задней части туловища не так сильно заост-
рены. Оба вида являются фактически подвида-
ми, которые могут скрещиваться. Платяных вшей
редко обнаруживают при нормальных условиях
жизни. Обычно они распространены среди лиц без
определенного места жительства, во времена нуж-
ды и войн, нередко встречаются вместе с голов-
ными и лобковыми вшами. Платяная вошь рас-
полагается не на теле, а в прилегающей одежде.
Гниды расположены в виде четок в швах одеж-
ды. Платяная вошь очень быстро размножается.
Клиническая картина
Укус вначале остается незамеченным, секрет слю-
ны вшей ведет к покраснению, волдырям и узел-
кам с очень сильным зудом. В местах укусов
возникают гиперемированные пятна, папулы,
мелкие волдыри, в центре которых имеется ге-
моррагическая корочка. Кожа скоро покрывается
штрихообразными эффектами расчесов, которые
часто вторично импетигинизируются. Возника-
ющая таким образом «вагантная кожа» (cutis
vagantium) имеет кроме того многочисленные
светлые шрамики с окружающей их гипер- и
депигментацией. Эта картина в целом достаточ-
но характерна.
Платяные вши переносят риккетсиозы, сып-
ной тиф, возвратный тиф. Волынская (окопная
или пятидневная) лихорадка вызывается риккет-
сиями (R. quintana), которые размножаются в
кишечнике вшей и выделяются с продуктами их
жизнедеятельности. Вши рода Pediculus являются
источником заражения человека эпидемическим
сыпным тифом, причиной возникновения кото-
рого также является риккетсия (Rickettsia
prowaceki). Возвратный тиф вызывается спиро-
хетой (Spirochaeta recurrens), которая попадает с
кровью в желудок насекомого и быстро из него
эвакуируется. Заражение больного происходит
на шестой день после попадания насекомого на
кожу, при его раздавливании и втирании спи-
рохет в поврежденные покровы, ранки, расчесы.
Диагноз
Наличие платяных вшей и гнид в швах нижне-
го белья. Дифференциальный диагноз: герпети-
формный дерматоз Дюринга, экзема, старческий
зуд, диабетический зуд, неспецифические изме-
нения кожи при болезни Ходжкина.
Лечение
Белье кипятится или дезинсектируется. При
необходимости можно применить контактные
инсектициды в форме спрея. Профилактически
опрадано применение пудры «Якутии». Лечение
кожных проявлений проводится по общетерапев-
тическим правилам в зависимости от актуально-
сти и вторичного инфицирования экзематозных
проявлений.
Фтириаз
Синонимы: Лобковый педикулез, площицы,
phthiriasis.
Фтириаз — заболевание, вызываемое лобко-
выми вшами, которые обитают в основном на
лобке, иногда на груди, в подмышечных впади-
нах, на ресницах верхних век.
Этиология
Лобковая вошь (pediculus pubis или phthirus pubis,
площица) на 1,5—2 мм короче головной и платя-
ной, имеет более широкую щитовидную форму.
2-я и 3-я пары ножек заканчиваются выражен-
ными коготками, посредством которых вошь
удерживается у основания волоса. Лобковые вши,
в отличие от других видов, почти не движутся,
поэтому их труднее распознать. Они также мед-
леннее размножаются. Легче найти их гниды.
Передача осуществляется при тесном телесном
контакте, чаще всего при половом акте, а также
188 Частная дерматология
от родителей к детям. Вероятно, возможно рас-
пространение через одежду, постельное белье или
полотенца.
Клиническая картина
Излюбленные места локализации этого вида
вшей — области с апокринными потовыми же-
лезами: лобок, генито-анальная область, подмы-
шечные впадины, а также, при наличии густого
волосяного покрова, грудь и живот. Реже, и чаще
у маленьких детей, их обнаруживают в волоси-
стой части головы, на бровях и ресницах. Зуд при
лобковом педикулезе умеренный, но ночью уси-
ливается. Эффекты расчесов обычно отсутству-
ют. Как следствие укусов лобковых вшей воз-
никают размытые стального синего или серого
оттенка пятна, величиной до размеров ногтя —
maculae coeruleae — в типичных местах локали-
зации. Их происхождение, вероятно, связано с
внутрикожным отложением зеленоватых продук-
тов разложения гемоглобина, происходящим под
действием слюны вшей. При сильном зуде от-
мечаются вторичные изменения в виде экскори-
аций, импетиго, лихенизации. У детей возмож-
но поражение ресниц и бровей в виде серозных
корок, а в некоторых случаях и отека век.
Диагноз
Синюшно-голубые пятна (Maculae coeruleae) с
типичной локализацией, особенно внизу живо-
та или на бедрах являются важными диагности-
ческими признаками. Доказательством служит
наличие вшей или их гнид. Когда пациенты
жалуются на зуд в области гениталий или под-
мышечных впадин, следует учесть возможность
наличия лобкового педикулеза.
Лечение
Осуществляется по той же схеме, что и при го-
ловном педикулезе, причем могут применяться
препараты линдан, аллетрин, пиперонилбутоксид
и экстракт пиретриума. При густом волосяном
покрове на теле эти места также требуют лече-
ния наряду с областью лобка и подмышечных
впадин. Важно осмотреть и провести лечение
контактировавших с пациентом лиц.
Лечение при поражении бровей и ресниц у
детей представляет трудности, поскольку выше-
указанные препараты могут, вследствие токси-
ческого раздражения, вызвать конъюнктивит. По
возможности следует пинцетом удалять вшей с
ресниц. Для местного лечения рекомендуется
простой белый вазелин 5 % водный раствор ма-
латиона, применяется также 1 % желтая ртутная
мазь под повязку на края век 2 раза в сутки в
течение 7—10 дней.
Глава 13
Грибковые болезни кожи (дерматомикозы)
Разноцветный лишай 190
Микоз крупных складок 191
Микозы стоп 192
Микоз волосистой части головы 193
Кандидоз 196
Онихомикозы 197
Глубокие микозы 204
Глубокие псевдомикозы 214
Грибковые заболевания, или микозы (от гречес-
кого «микес» — гриб), относятся к инфекционным
болезням и составляют значительную их часть.
Среди микозов встречаются острые и хронические
(по течению), поверхностные и глубокие (по глу-
бине поражения кожи и слизистых), локализован-
ные и распространенные формы.
Возбудители микозов — растительные парази-
ты, которых насчитывают около 100 000 видов.
Грибы составляют обширную группу низших
растений, лишенных хлорофилла, поэтому для
поддержания существования им требуются гото-
вые органические вещества. Питание осуществ-
ляется двумя способами:
• сапрофитический — питание за счет мертвых
остатков (пример: обычные лесные грибы); от-
части к ним относятся дрожжеподобные грибы
рода кандида;
• паразитический — питание за счет соков жи-
вых растений и животных (примером являются
дерматофиты).
Имеется около 500 видов грибов, патоген-
ных для человека. К этой группе относится
большинство дерматофитов — возбудителей
грибковых заболеваний. Они поражают глав-
ным.образом кожу и ее придатки — волосы,
ногти, в редких случаях другие органы и тка-
ни. Местом их обитания является больной че-
ловек и патологический материал, выделяе-
мый им.
Общим для большинства патогенных грибов
является вегетативное тело — мицелий, или
грибница, состоящая из ветвящихся нитей (ги-
фов). Для определения вида гриба имеют зна-
чение характер и форма концевых ветвей ми-
целия, особенно при выявлении дерматофитов.
Дрожжевые и дрожжеподобные грибы имеют
вид округлых или овальных клеток, эти гри-
бы образуют псевдомицелий, но некоторые
виды — и истинный мицелий. Размножение
патогенных грибов осуществляется половым и
бесполым путем. Бесполое размножение проис-
ходит путем деления, или почкования, которое
бывает почти у всех патогенных грибов, в том
числе у дерматофитов, а также у дрожжеподоб-
ных грибов рода кандида. Патогенные грибы об-
разуют споры как вне мицелия (экзоспоры), так
и внутри его (эндоспоры). Дрожжеподобные
грибы размножаются путем образования экзос-
пор, возникающих в результате почкования ма-
теринской клетки (бластоспоры). Нити, обра-
зующиеся из бластоспор путем почкования,
называют псевдомицелием. Клетки грибов со-
стоят из оболочки, цитоплазмы, ядра с ядрыш-
ками, вакуолей и включений (жир, гликоген,
волютин, реже пигменты и соли органических
кислот).
Грибковые заболевания кожи развиваются в
результате внедрения в нее патогенных грибов.
При обитании вне организма человека или жи-
вотного грибы могут сохранять свою жизнеспо-
собность и заразительность в течение несколь-
ких лет. Поэтому возможно заражение не только
от больного человека, но и путем непрямого
189
190 Частная дерматология
контакта — через предметы обихода, зараженные
чешуйками, кусочками ногтей или обломками
волос, содержащих элементы гриба.
Споры внедряются в организм при наличии
незначительных повреждений кожи. Грибы па-
разитируют на коже, волосах, ногтях, реже — во
внутренних органах человека и животных. В
коже они находят все необходимые для разви-
тия питательные вещества: эпидермис, волосы,
ногти, пот, жир.
Грибковые поражения кожи относятся к са-
мым распространенным заболеваниям челове-
ка. Согласно данным американских ученых, до
90 % американцев болели эпидермофитией
стоп хотя бы раз в жизни. Исследователи от-
дела микологии Центрального кожно-венеро-
логического института (Москва) указывают,
что микозами стоп поражено 30 % населения
России.
По среде обитания дерматофиты подразде-
ляются на антропофильные — поражающие
преимущественно человека; зоофильные — по-
ражающие животных, но иногда и людей; гео-
фильные — находящиеся постоянно в почве и
не имеющие серьёзного эпидемиологического
значения.
Клиническая классификация
грибковых заболеваний
(Н. А. Шеклаков, П. Н. Кашкин)
I. Кератомикозы
• Разноцветный лишай
• Узловатая трихоспория (пьедра)
II. Дерматофитии (дерматомикозы)
• Эпидермофития паховая
• Руброфития (микоз, обусловленный крас-
ным трихофитоном)
• Эпидермофития стоп (микоз, обусловленный
интердигитальным трихофитоном)
• Трихофития (антропонозная, зоонозная)
• Микроспория (антропонозная, зоонозная)
• Фавус
III. Кандидоз
• Поверхностный кандидоз слизистых оболо-
чек, кожи, ногтевых валиков и ногтей
• Хронический генерализованный (гранулема-
тозный) кандидоз
• Висцеральный кандидоз различных органов
IV. Глубокие (висцеральные и системные) ми-
козы
• (гистоплазмоз, кокцидиоидомикоз, бласто-
микозы, криптококкоз, геотрихоз, споротрихоз,
хромомикоз, риноспоридиоз, аспергиллез, пени-
циллинов, мукороз)
V. Псевдомикозы
• (эритразма, подмышечный трихомикоз,
актиномикоз, микромоноспороз, мицетомы, но-
кардиоз)
В настоящее время предложена клиническая
классификация дерматомикозов МКБ-10, которая
основана на локализации грибкового поражения.
Клиническая классификация
дерматомикозов (МКБ-10)
• Микоз волосистой части головы (Tinea
capitis)
• Микоз гладкой кожи (Tinea corporis)
• Микоз крупных складок (Tinea cruris)
• Микоз кистей и стоп (Tinea pedis et manuum)
• Микоз ногтей (онихомикоз)
Разноцветный лишай
Разноцветный лишай (сии.: отрубевидный ли-
шай, pityriasis versicolor) — грибковое заболева-
ние невоспалительного характера с поражением
рогового слоя эпидермиса.
Этиология
Возбудитель — Malassezia spp. — относится к
слабоконтагиозным дрожжеподобным липо-
фильным организмам.
Эпидемиология
Заболевание наиболее распространено в жарких
странах с повышенной влажностью воздуха.
Болеют преимущественно люди молодого возра-
ста. Заболевание имеет хроническое течение и
отличается частыми рецидивами. Контагиозность
его незначительная.
Клиническая картина
Типичная локализация — кожа плеч, груди,
спины и живота; волосистая часть головы (рис.
13.1). Пятна невоспалительные, розоватые, затем
желтоватой и коричневой окраски, шелушащи-
еся. Шелушение напоминает отруби, поэтому
другое название заболевания — отрубевидный
лишай. После загара остаются депигментирован-
ные белые участки кожи.
Диагностика
Характерные высыпания: локализация и вид.
Осмотр под люминесцентной лампой Вуда (жел-
тое, желто-коричневое или бурое свечение).
Микроскопическое исследование (обнаружение
элементов гриба в чешуйке). Бактериологичес-
Глава 13. Грибковые болезни кожи
191
кое исследование (посев на среду Сабуро). Йод-
ная проба Бальцера (при смазывании участков
поражения настойкой йода пятна лишая интен-
сивнее поглощают йод).
Лечение
Основано на примении кератолитических и
противогрибковых средств. Используются шам-
пуни с противогрибковыми препаратами (ни-
зорал); микоспор (крем или раствор 1—2 раза в
день в течение 2—3 недель); ламизил (1 % крем
втирать 2 раза в день в течение 2 недель); бат-
рафен в виде крема или раствора (2 раза в день
10-12 дней). В последующем необходимо в те-
чение 2-3 недель очаги поражения смазывать
2-5 % раствором салицилового спирта 1 раз в
день.
При распространенных формах заболевания с
частыми рецидивами — кетоконазол (низорал)
по 200 мг 1 раз в день в течение 10 дней или
орунгал по 200 мг в сутки в течение 7 дней.
Профилактика
Осмотр всех членов семьи клинически и с по-
мощью люминесцентной лампы. Профилакти-
ческие курсы лечения через 1—2 месяца. Дезин-
фекция одежды, нательного и постельного белья.
Обследование на сифилис.
Рис. 13.1. Типичная локализация высыпаний
при отрубевидном лишае.
Микоз
крупных складок
Грибковое поражение преимущественно круп-
ных складок кожи, в ряде случаев — межпаль-
цевых поверхностей стоп.
Этиология
Возбудитель — Epidermophyton floccosum. В насто-
ящее время термин «эпидермофитон» применим
в отношении микоза, вызываемого единственным
патогенном грибом из рода эпидермофитон. Бо-
леют преимущественно тучные мужчины, страда-
ющие нарушениями углеводного обмена и повы-
шенной потливостью.
Эпидемиология
Заражение происходит при контакте с больным,
но чаще через предметы, инфицированные гри-
бом (одежда, обувь); в спортзалах — коврики,
маты и другой инвентарь; в больнице — белье,
подкладные судна.
Фактора, способствующие заболеванию
Повышенная потливость, опрелость кожи, избы-
ток массы тела, нарушение обмена веществ.
Клиническая картина
Заболевание развивается в области крупных
складок, чаще паховых, реже межъягодичных,
под молочными железами, в подмышечных впа-
динах. Очаги поражения в виде пятен ярко-крас-
ного или красновато-коричневого цвета, с чет-
кими границами, отечным валиком по
периферии, состоящим из пузырьков, пустул и
корочек. Течение острое, сопровождается силь-
ным зудом, жжением и болезненностью. В жар-
кое время года — обострение.
Диагностика
Типичная клиническая картина заболевания.
Микроскопически — обнаружение в чешуйках
кожи из очагов поражения ветвещегося корот-
кого мицелия. Бактериологически —- выделение
культуры гриба эпидермофитон флоккозум с ха-
рактерными микроконидиями в виде гроздьев
бананов.
Лечение
При отсутствии островоспалительных явлений —
серно-салициловая или серно-дегтярная мазь,
2 % настойка йода, мази «Микоспор», батрафен,
микосептин, клотримазол.
192 Частная дерматология
При наличии экзематизации — примочки с
раствором резорцина, борной кислоты, прием
внутрь препаратов кальция, антигистаминных
средств; после разрешения острых воспалитель-
ных явлений — йодно-мазевое лечение.
Противогрибковые кремы и мази с противо-
воспалительным действием: низорал, травокорт,
пимафукорт, ламизил, экзодерил, микозолон,
певарил, тридерм.
Профилактика
Дезинфекция предметов общего пользования.
После разрешения клинических проявлений
микоза профилактически в течение 2—3 недель
следует продолжить лечение.
Микозы стоп
В настоящее время микозы стоп являются самы-
ми распространенными грибковыми заболевани-
ями во всех странах мира. В некоторых странах
ими болеют до 50 % населения. Название «ми-
козы стоп» — собирательное и включает в себя,
в основном, два заболевания:
• микоз, вызываемый грибом красный трихо-
фитон (трихофитон рубрум);
• микоз, вызываемый грибом трихофитон ин-
тердигитале (трихофитон ментагрофитес, вари-
ант дигитале или интердигитале).
Термин «эпидермофития стоп» применим толь-
ко в отношении микоза, обусловленного пахо-
вым эпидермофитоном.
Основным возбудителем в настоящее время
является красный трихофитон, поражающий
также ногти, кожу кистей и других участков
кожных покровов.
Эпидемиология
Заражение микозами стоп и кистей происходит
при непосредственном контакте с больным в
семье, коллективе или через обувь, одежду, пред-
меты обихода (мочалки, маникюрные принад-
лежности, коврики в ванной). Чаще других
грибковому заболеванию стоп подвержены ра-
ботники бань, душевых и прачечных, пловцы,
а также обслуживающий персонал лечебных и
детских учреждений, домов отдыха и санатори-
ев, постоянно пользующийся банями, душами.
Патогенез и факторы,
способствующие заболеванию
Имеет значение узость межпальцевых складок,
плоскостопие, травмы, различные аномалии ко-
нечностей. При наличии сопутствующих заболе-
ваний (сахарный диабет, иммунодефицит, хро-
нические инфекции и интоксикации, поражение
нервной и сосудистой систем) микоз стоп и ки-
стей, обусловленный красным трихофитоном,
приобретает распространенный и генерализован-
ный характер, часто рецидивирует и трудно под-
дается лечению. При этом микозе поражаются
не только эпидермис, дерма, волосяные фолли-
кулы, но и кровеносные и лимфатические сосуды
и регионарные лимфоузлы.
Клиническая картина
Основные возбудители микозов стоп — красный
трихофитон и трихофитон интердигитале — от-
личаются своеобразием вызываемых ими пато-
логических изменений. Красный трихофитон
поражает кожу, ногти стоп и кистей; трихофи-
тон интердигитале — кожу и ногтевые пластин-
ки только стоп.
Клинические проявления, вызываемые этими
грибами, различны.
Рубромикоз (микоз, обусловленный красным
трихофитоном) начинается с кожи межпальце-
вых складок, затем в процесс вовлекается кожа
подошв, боковых и тыльных поверхностей стоп,
ладоней и ногтевые пластинки. Кожа поражен-
ных участков сухая, роговой слой утолщен с
выраженным рисунком кожных борозд и значи-
тельным мукоидным и пластинчатым шелуше-
нием. Для руброфитии характерно множествен-
ное поражение ногтевых пластинок стоп и
кистей по нормотрофическому, гипертрофичес-
кому, атрофическому типу и по типу онихоли-
зиса. Отмечаются и явления лейконихии: появ-
ление пятен или полос белого цвета в толще
ногтя.
Поражение кожи стоп при руброфитии может
быть в виде следующих клинических форм:
интертригинозная, дисгидротическая, сквамоз-
ная, сквамозно-гиперкератотическая, с единич-
ным или множественным поражением ногтей.
При микозе стоп, обусловленном другим воз-
будителем — трихофитоном интердигитале, на-
блюдаются те же клинические формы пораже-
ния, что и при руброфитии, но поскольку гриб
обладает более выраженными аллергизирующи-
ми свойствами, у больных нередко возникают
вторичные высыпания на коже кистей и других
местах кожного покрова, и заболевание проте-
кает с более выраженными воспалительными
явлениями, напоминая экзему. Гриб трихофи-
тон интердигитале поражает кожу 3 и 4 меж-
пальцевых складок стоп, верхнюю треть подо-
швы, боковые поверхности стопы и пальцев,
Глава 13. Грибковые болезни кожи 193
свод. Никогда не поражает кисти. Ногтевые пла-
стинки вовлекаются в процесс у 20—30 % боль-
ных только на 1 и 5 пальцах стоп.
Диагностика
Диагноз устанавливается после лабораторного
обследования больного. Лабораторная диагнос-
тика состоит в микроскопическом исследовании
патологического материала и выделении культу-
ры гриба на питательных средах (Сабуро).
Лечение
Комбинированный метод лечения противогриб-
ковым препаратом общего действия в сочетании
с местным применением противогрибковых и
кератолитических средств, патогенетической те-
рапии дает наиболее эффективные результаты.
Для удаления ногтевых пластинок используют-
ся различные методы:
• по Андриасяну (20 % уреапласт на ногти для
размягчения, затем мазь с салицилово-молочной
кислотой и резорцином);
• по Аравийскому (50 % мазь с йодистым ка-
лием на ланолине на 48 часов с последующей
чисткой лезвием бритвы);
• хирургический метод удаления ногтевых
пластинок (редко применяется), вначале ногте-
вые пластинки размягчают хинозоловым плас-
тырем.
Противогрибковые препараты общего дей-
ствия: гризеофульвин, кетоконазол (низорал)
применяют от 3 до 12 месяцев; тербинафин (ла-
мизил, тербизил) назначают по 250 мг в сутки
в среднем 3 месяца, можно проводить и без уда-
ления ногтей; итраконазол (орунгал) — пульс-
терапия по 7 дней (по 200 мг 2 раза в день пос-
ле еды) с перерывом 3 недели (2 курса при
поражении ногтей пальцев рук и 3 курса при по-
ражении ногтей пальцев стоп).
Комплексное лечение больных с поражением
ногтевых пластинок включает и использование
противогрибковых препаратов для местного при-
менения: нитрофунгин, микосептин, микозолон,
фитекс, батрафен, травоген, травокорт, толмицен,
кетоконазол (низорал, ороназол), клотримазол,
дактарин, певарил, экзодерил, ламизил, лоцерил,
микоспор. Микоспор — крем в наборе для лече-
ния ногтей. В его состав входят крем, 15 полосок
водонепроницаемого лейкопластыря и пилка.
Профилактика
Общая профилактика — гигиеническое содержа-
ние и регулярная дезинфекция бань (полы, по-
ловики, деревянные решетки, скамейки, тазики),
душевых и бассейнов, профосмотры их персона-
ла, своевременное лечение. Личная профилакти-
ка: использование только своей обуви, соблю-
дение правил гигиены кожи стоп, дезинфекция
обуви. Ватным тампоном, смоченным 25 %-ным
раствором формалина, протирают стельку и под-
кладку обуви. Затем помещают в полиэтилено-
вый мешок на 2 часа. Носки, чулки дезинфеци-
руют кипячением в течение 10 мин.
Микоз волосистой
части головы
Микоз волосистой части головы (Tinea capitis)
может быть обусловлен всеми видами грибов
рода Trichophyton и Microsporum, но особую пред-
расположенность к поражению волосистой час-
ти головы имеют возбудители микроспории, три-
хофитии и фавуса (Microsporum audouinii,
Trichophyton violaceum, Trichophyton scholeinii).
Клиническая картина микозов волосистой ча-
сти головы определяется экологией возбудителя
(антропофильные грибы — паразитируют только
на коже человека; зоофильные грибы — основным
хозяином их явялются животные, а человек ин-
фицируется вторично; геофильные грибы — оби-
тают в почве и довольно редко инфицируют че-
ловека), способом расположения элементов гриба
в волосе (эндотриксы, эктотриксы) и степенью
выраженности воспалительной реакции, развива-
ющейся в коже в ответ на внедрение гриба. Зоо-
фильные грибы (эктотриксы) являются возбуди-
телями острых инфильтративно-нагноительных
форм микоза волосистой части головы; антропо-
фильные грибы (эндотриксы) — поверхностных,
слабо воспалительных поражений кожи. Возбу-
дители микроспории, трихофитии и фавуса об-
ладают способностью вовлекать в патологический
процесс не только кутикулу волоса, но и корко-
вое и мозговое вещество волоса. Страдают не
только жесткие волосы волосистой части кожи
головы, ресниц, бровей, верхней губы, подбород-
ка, подмышечных и паховых областей, а также
пушковые волосы гладкой кожи лица, туловища
и конечностей.
Микроспория
Микроспория является высококонтагиозным за-
болеванием с поражением кожи, волос, иногда
ногтей и вызывается грибами рода Microsporum.
Микроспория волосистой части головы отмеча-
ется почти исключительно у детей.
194 Частная дерматология
Этиология
Возбудителями заболевания могут быть как ан-
тропофильные (М. audouinii, М. ferrugineum), так
и зоофильные (М. canis) грибы. Самым распро-
страненным возбудителем микроспории является
зоофильный гриб М. canis (синонимы: собачий,
пушистый, кошачий). Вторым по частоте выде-
ления является антропофильный гриб М.
ferrugineum. Известны также геофильные мик-
роспорумы (М. gypseum), но эпидемиологическое
значение их невелико. Другие виды грибов этого
рода (Microsporum) встречаются редко.
Эпидемиология
Самые главные источники заражения и рапрост-
ранения зоонозной микроспории — животные
(кошки, реже — собаки). Антропонозная микро-
спория, обусловленная ржавым микроспорумом,
передается только от больного человека к здоро-
вому при контакте с ним или через зараженные
предметы. Этот вид микроспории контагиознее
зоофильной, но в настоящее время обнаружива-
ется редко (3—4 % случаев).
Патогенез и факторы,
способствующие заболеванию
Заражение детей чаще всего происходит в возрас-
те от 1 года до 13 лет, а к периоду полового со-
зревания у всех нелеченных больных наступает
самопроизвольное выздоровление, что связано с
гормональной перестройкой организма и активной
работой половых желез. Определенная роль в па-
тогенезе микроспории принадлежит недостаточно-
сти естественной резистентности, снижению фун-
кциональной активности клеток крови,
недостаточности системы лизоцима, комплемента,
пропердина, витаминов, микроэлементов. Распо-
ложение очагов микроспории на определенных уча-
стках кожного покрова определяется микротрав-
мами, недостаточным сало- и потоотделением,
нарушением процессов пролиферации эпидерми-
са, местными нервно-сосудистыми нарушениями.
Клиническая картина
Клинические особенности микроспории зависят
от вида возбудителя, состояния заболевшего че-
ловека, возрастных особенностей, локализации
очагов поражения, глубины проникновения гри-
ба. Клинические проявления заболевания разви-
ваются после инкубационного периода, который
длится 5—7 дней при зоонозной микроспории и
4—6 недель при антропонозной микроспории.
Очаги поражения полициклических очертаний,
округлой формы, с четкими границами, локали-
зуются в краевой зоне волосистой части головй.
В центре этих очагов находятся чешуйки, короч-
ки, а по периферии — папулы и везикулы. Во-
лосы обламываются на высоте 6—8 мм. Зооноз-
ный вариант вызывается Microsporum canis и
характеризуется появлением на гладкой коже
множественных эритематозных, шелушащихся
очагов с четкими границами. На волосистой ча-
сти головы образуются 1—2 крупных, шелуша-
щихся очага также с четкими границами. Воло-
сы обламываются на высоте 5—8 мм.
Диагноз
Диагноз ставят на основании клинической кар-
тины, характерного зеленовато-изумрудного све-
чения волос в лучах лампы Вуда, микроскопи-
ческих и культуральных данных.
Лечение
Лечение предусматривает назначение гризео-
фульвина внутрь с чайной ложкой растительного
масла по 22—25 мг на 1 кг массы тела в сутки в
3 приема ежедневно до первого отрицательного
результата на грибы, затем через день в течение
2 недель. При микроспории волосистой части
головы, единичных очагах на гладкой коже с
поражением пушковых волос сбривают волосы
1 раз в 7—10 дней. Наружно можно использо-
вать микоцидные средства (клотримазол, кето-
коназол, бифоназол, серно-салициловая мазь).
Альтернативным препаратом для лечения мик-
роспории является тербинафин, который прини-
мается в течение 8—12 недель и назначается после
еды по 250 мг 1 раз в сутки (взрослым и детям
с массой тела свыше 40 кг), или 125 мг в сутки
(детям с массой тела от 20 до 40 кг), или 62,5 мг/
сутки (детям с массой тела менее 20 кг).
Клиническое и микологическое излечение
оценивается по разрешению клинических про-
явлений, отсутствию свечения под люминесцен-
тной лампой Вуда и отрицательным результатам
микросопического исследования (3 анализа с 5-
дневными перерывами при поражении волоси-
стой части головы).
Профилактика
Направлена на проведение и соблюдение четких
санитарно-эпидемиологических мероприятий.
Трихофития
Грибковое заболевание кожи подразделяют на
антропонозный и зоонозный типы. При антро-
понозном типе (эндотрикс) споры располагаются
внутри волоса. Возбудителем этой формы явля-
Глава 13. Грибковые болезни кожи
195
ется Trichophyton tonsurans. Возбудителем зооноз-
ной формы трихофитии является Trichophyton
mentagrophytes. Инфицирование человека может
происходить от больных телят (Trichophyton
verrucosum) или от мышевидных грызунов
(Trichophyton gypseum).
Клиническая картина
Выделяют три клинических варианта антропоноз-
ной трихофитии: поверхностную гладкой кожи,
поверхностную волосистой части головы, хрони-
ческую трихофитию взрослых.
Поверхностная трихофития гладкой кожи про-
является округлыми или овальными очагами
бледно-розового цвета с четко очерченным пери-
ферическим ободком и шелушащимся центром.
Поверхностная трихофития волосистой части
головы характеризуется наличием очагов с нечет-
кими расплывчатыми очертаниями и с лабовы-
раженным шелушением. Волосы в очагах пора-
жения обламываются на уровне 1—2 мм.
Хроническая трихофития взрослых развивает-
ся в пубертатном периоде у девочек с эндокри-
нопатиями и характеризуется скудными клини-
ческими симптомами. На волосистой части
головы определяются мелкие очаги в виде про-
плешин с черными точками в теменной или
височных областях.
На коже ягодиц, передних поверхностей го-
леней, реже на туловище отмечается наличие
слегка шелушащихся очагов застойной окраски.
На ладонных поверхностях кистей — муковид-
ное шелушение, ногтевые пластинки тусклые,
грязно-серой окраски, легко крошатся.
При зоонозной трихофитии (эктотрикс) споры
располагаются на поверхности волоса, вызывая
островоспалительные изменения коже. Различа-
ют три формы зоонозной трихофитии: поверхно-
стную, инфильтративную и нагноительную. При
поверхностной трихофитии на коже появляются
крупные очаги с фестончатыми очертаниями и
периферическим валиком из экссудативных па-
пул и везикул. При инфильтративной форме кожа
в области очагов поражения уплотнена, эритема-
тозно-застойной окраски, с отдельными фолли-
кулярными пустулами. Процесс сопровождается
появлением болезненного регионарного лимфа-
денита. Нагноительная форма проявляется обра-
зованием на коже волосистой части лица или
головы болезненных плотных очагов поражения
островоспалительного характера из возвышаю-
щейся застойной эритемы с зияющими фоллику-
лами (керион Цельса). Корни волос расплавля-
ются и выпадают вместе с гнойным субстратом.
Процесс разрешается рубцовой атрофией.
Диагноз
Диагноз ставится на основании характерной кли-
нической картины и микроскопии.
Лечение
Внутрь применяют гризеофульвин из расчета
16—18 мг на 1 кг массы тела в течение 3—4 не-
дель, а наружно 3—5 %-ный раствор йода, 2—
3 %-ную серно-салициловую мазь, 10 %-ный
раствор ихтиола на очаги инфильтративно-нагно-
ительной трихофитии; микоцидные мази, кре-
мы, растворы, лосьоны. Альтернативным препа-
ратом для лечения трихофитии является
тербинафин, который принимается в течение 5—
6 недель и назначается после еды по 250 мг 1 раз
в сутки (взрослым и детям с массой тела свыше
40 кг), или 125 мг в сутки (детям с массой тела
от 20 до 40 кг), или 62,5 мг/сутки (детям с мас-
сой тела менее 20 кг).
Клиническое и микологическое излечение оце-
нивается по разрешению клинических проявлений
и отрицательным результатам микросопического
исследования (3 анализа с 5-дневными перерыва-
ми при поражении волосистой части головы).
Фавус
Фавус — хроническое грибковое заболевание
кожи, волос, ногтей.
Этиология
Возбудитель — Trichophyton schoenleinii, переда-
ющийся от человека к человеку, и Trichophyton
quickeanum Zopf, поражающий кошек, собак,
мышей, овец и редко — человека.
Клиническая картина
Различают типичную (скутулярную) и атипич-
ную (сквамозную и импетигинозную) формы.
При скутулярной разновидности на коже во-
лосистой части головы в зоне волосяных фол-
ликулов появляются эритематозно-отечные,
слегка шелушащиеся пятна. Очаги поражения
имеют тенденцию к периферическому росту,
увеличиваясь в размерах до 1—3 см, инфильтри-
руются в основании. У корня волос формиру-
ются скутулы-корочки, плотно прикрепленные
к коже, за счет прорастания в них грибницы.
Цвет корок становится землисто-серым, волосы
в очагах поражения истончаются, теряют блеск
и эластичность, становятся запыленными, напо-
миная старый парик. Процесс распространяется
на теменно-затылочную область и завершается
рубцовой атрофией.
196 Частная дерматология
При сквамозной форме в очагах поражения у
корня волос формируются чешуйко-корочки,
пронизанные тусклыми волосами. На гладкой
коже отмечается появление рассеянных шелуша-
щихся очагов.
Импетигинозная разновидность встречается
редко и характеризуется наличием инфильтра-
тивно-нагноительных элементов типа кериона.
Ногтевые пластинки поражаются обычно у
взрослых, и в течение длительного периода их
конфигурация не изменяется.
Диагноз
Диагноз ставят на основании клинической кар-
тины, обнаружения фавозных щитков, рубцо-
вой алопеции, септированного мицелия, сохра-
нения нормальных волос по краевой зоне
головы.
Лечение
Лечение проводят гризеофульвином по 18 мг на
1 кг массы тела больного в сочетании с местными
фунгицидными препаратами. Альтернативным
препаратом для лечения фавуса является терби-
нафин, который принимается в течение 5—6
недель и назначается после еды по 250 мг 1 раз
в сутки (взрослым и детям с массой тела свыше
40 кг), или 125 мг в сутки (детям с массой тела
от 20 до 40 кг), или 62,5 мг/сутки (детям с мас-
сой тела менее 20 кг).
Кандидоз
Этиология
Дрожжеподобные грибы рода Candida встреча-
ются повсеместно. В полосе с умеренным кли-
матом обнаруживают Candida albicans, в зонах
с тропическим климатом — С. tropicalis, С.
crusei, С. pseudotropicalis и др. Грибы рода кан-
дида нередко обнаруживаются в ассоциации с
микробами.
Клиническая картина
Кандидозная инфекция поражает слизистые обо-
лочки, кожу и внутренние органы.
Кандидоз слизистых оболочек полости рта яв-
ляется наиболее распространенным поражением
новорожденных детей, подростков и лиц пожи-
лого и старческого возраста, истощенных изну-
рительными болезными.
Кандидозный вагинит и баланопостит — до-
вольно распространенные заболевания, относя-
щиеся в настоящее время к группе заболева-
ний, передаваемых половым путём. В ряде
случаев у 20 % женщин кандида присутствует
в виде каменсел, но в определенных ситуаци-
ях эти каменселы приобретают патогенные
свойства, вызывая боль, отек, воспаление и
обильные выделения.
Кандидозная опрелость локализуется в меж-
пальцевых складках кистей, под молочными
железами, в межъягодичной и паховых склад-
ках. Клинически опрелость характеризуется
наличием очагов поражения в виде мокнущих
поверхностей с обилием творожистого налета
на фоне эритематозно-отечной кожи. На коже
ягодиц, паховых складок могут возникать оча-
ги поражения в виде эритематозно-пемфиго-
идных и везикулезно-пустулезных элементов.
Субъективно больные отмечают боль, зуд,
жжение.
Паронихии и онихии также развиваются под
воздействием кандидамикотической инфекции.
Клинически в околоногтевых валиках формиру-
ются отечность и мокнутие, которые в последу-
ющем приводят к инфицированию и деформа-
ции ногтевых пластинок.
Из внутренних органов в первую очередь по-
ражается желудочно-кишечный тракт. В патоло-
гическиий процесс могут вовлекаться органы
дыхания в виде острых или хронических пнев-
моний, абсцессов легких, бронхитов, ларинги-
тов. У больных, страдающих диабетом, нередко
развиваются циститы, пиелоциститы, пиелонеф-
риты.
Грануломатозный кандидоз — одна из тяжелых
форм поражения кожи и слизистых, развивает-
ся у ВИЧ-инфицированных или страдающих
иммунодефицитом и эндокринопатиями паци-
ентов. Процесс возникает на слизистых оболоч-
ках ротовой полости, красной кайме губ или по-
ловых органах, распространяясь в последующем
на кожу туловища, конечностей и волосистой ча-
сти головы в виде узелковых, бугорковых или
опухолевидных образований с выраженной ин-
фильтрацией и эритемой.
Диагноз
Диагноз ставят на основании клинических и
лабораторных данных.
Лечение
Лечение при орофарингеальном кандидозе про-
водится флуконазолом (дифлюкан, микосист)
по 50—100 мг 1 раз в сутки в течение 7—14 дней.
При вагинальном кандидозе и баланопостите
клиническое излечение достигается однократ-
Глава 13. Грибковые болезни кожи
197
ным назначением флуконазола (дифлюкан, ми-
косист) в дозе 150 мг. При висцеральных и сеп-
тических состояниях доза флуконазола состав-
ляет 400 мг в первые сутки, а далее по 200 мг
в сутки в течение 2—3 недель. Высокоэффек-
тивным является также итраконазол (орунгал).
При системном кандидозе его назначают по
100-200 мг в сутки в течение 3 недель, при по-
ражении только полости рта — по 100 мг в сут-
ки в течение 15 дней, при вульвовагинитах —
по 200 мг ежедневно в течение 3 суток.
Онихомикозы
Онихомикоз — грибковое поражение ногтей
стоп (кистей). Заболевание широко распростра-
нено во всех странах мира. В настоящее время
среди грибковых заболеваний чаще всего встре-
чается микоз стоп, при этом частота поражения
ногтевых пластинок составляет от 18 % до 40 %.
Приблизительно 10 % американцев поражены
онихомикозом, а в Англии насчитывается
1,2 млн больных этой инфекцией. Заболевае-
мость онихомикозом в некоторых профессио-
нальных группах населения России (шахтеры,
металлурги, промышленные рабочие) значитель-
но выше среднепопуляционной. Онихомикоз
часто встречается у пожилых лиц, больных са-
харным диабетом, онкологическими заболевани-
ями, саркомой Капоши, кератодермиями, ихти-
озом.
Онихомикоз является не только косметичес-
кой проблемой. Он представляет собой серьез-
ную угрозу здоровью человека. Исследования
показывают, что грибы, вызывающие онихоми-
коз, в процессе жизнедеятельности могут син-
тезировать токсические продукты. При продол-
жительном грибковом поражении некоторые
токсические метаболиты (ксантомегнин, виомел-
леин) могут привести к поражению внутренних
органов. Токсические продукты жизнедеятель-
ности болезнетворных грибов могут вызывать
гепатопатии. Длительная персистенция грибов в
пораженных ногтях приводит к образованию
даже антибиотикоподобных веществ, которые
могут спровоцировать развитие резистентной
бактериальной флоры. Другая опасность заклю-
чается в том, что при выработке пенициллина
штаммами грибов развивается сенсибилизация
огранизма больного и возникает аллергия к пе-
нициллину. Впоследствии это может привести к
медикаментозной токсидермии и даже синдро-
му Лайелла.
На микозы стоп и кистей приходятся основ-
ные трудопотери в группе больных с инфек-
ционными поражениями стоп, связанными с
наличием у большинства из них онихомико-
зов, которые с трудом поддаются лечению си-
стемными антимикотиками. Число больных
онихомикозом во всем мире растет ежегодно.
Европейская Академия Дерматологии и Вене-
рологии и Европейское общество исследования
ногтей в 1996 году учредили «Ахиллес-проект»,
цель которого состоит в ознакомлении работ-
ников здравоохранения с европейскими эпи-
демиологическими исследованиями в области
микозов и онихомикозов стоп, а также с теми
последствиями, к которым они приводят, в ча-
стности, к снижению работоспособности паци-
ентов, профессиональной непригодности от-
дельных групп населения (спортсмены,
военнослужащие, работники горнорудных и
химических предприятий). В 1997 году началь-
ный скрининг включил 22 000 пациентов в Ев-
ропе, а в 1998 году, кроме 20 стран европейс-
кого континента, проект был осуществлен в
Южной Африке и Израиле, а на следующий
год и в России. Тест на микологическое обсле-
дование прошли около 150 000 человек. В ре-
зультате, более чем у одной трети обследован-
ных (35 %) выявлен микоз стоп, а из них почти
каждый четвертый (23 %) болеет онихомико-
зом.
Этиология и эпидемиология
Возбудителями онихомикоза являются три груп-
пы грибов: дерматофиты (80—94 % случаев),
дрожжевые грибы, плесневые грибы. Наиболее
распространенными дерматофитами считают
Trichophyton rubrum (вызывает поражение ногтей
стоп, кистей, а также гладкой кожи) и
Trichophyton mentagrophytes (поражает ногти на I
и V пальцах стоп и кожу 3—4 межпальцевых
складок). Онихомикоз может быть обусловлен
Epidermophyton floccosum, намного реже — дрож-
жеподобными и крайне редко — плесневыми
грибами.
Дерматофиты являются основными возбуди-
телями онихомикозов — дерматофитный онихо-
микоз. Около 85 % всех случаев онихомикозов
вызвано Trichophyton rubrum. Второй по распро-
страненности возбудитель онихомикозов —
Trichophyton mentagrophytes var. interdigitale. Из
других дерматофитов при онихомикозах чаще
встречается Epidermophyton floccosum (1—2 % слу-
чаев). При эпидермофитийных онихомикозах
поражаются наиболее травмируемые ногти I и V
пальцев обеих стоп.
198 Частная дерматология
После дерматофитов наиболее частыми возбу-
дителями онихомикозов считаются дрожжеподоб-
ные грибы Candida spp., которые преимуществен-
но вызывают поражение ногтевых пластинок
кистей (а в некоторых странах мира кандидоз
поражает чаще ногти стоп). В отличие от дерма-
тофитов, дрожжеподобные грибы вначале пора-
жают кожу, окружающую ноготь, а затем уже
проникают в ткань ногтя.
Плесневые грибы-недерматофиты крайне ред-
ко вызывают онихомикоз. Плесневые грибы не
обладают кератолитической способностью и по-
этому поражают пациентов с кератином, повреж-
денным вследствие травмы или болезни. Около
3 % всех случаев онихомикоза стоп вызывается
плесневыми грибами Scopulariopsis brevicaulis,
Hendersonula toruloidea, Scitalidium hialinum.
Ногтевые пластинки кистей поражают преиму-
щественно грибы рода Fusarium и Altemaria.
Плесневые грибы чаще всего обнаруживают у
пожилых пациентов и у лиц с иммунодефицит-
ным состоянием.
Изолированные онихомикозы встречаются
редко. Их возникновение чаще всего связано с
микозом стоп и кистей, волосистой части голо-
вы, гладкой кожи.
Заражение онихомикозом происходит через
предметы обихода: коврики в ванной, мочалки,
обувь, маникюрные принадлежности; а также
при посещении бассейна, бани, сауны, спортза-
ла, душевой. Вследствие хронического течения
болезни, резистентности к проводимой терапии
онихомикозы являются источником инфекции
для самого больного (распространение на другие
участки тела и генерализация процесса) и для
окружающих людей. Мужчины более чувстви-
тельны к заражению ногтей грибами, однако
наблюдаются онихомикозы чаще у женщин. Это
объясняется особенностями женской обуви, ко-
торая сдавливает стопу и способствует травмати-
зации ногтевых пластинок. Онихомикозы явля-
ются заболеванием взрослых людей, у детей они
встречаются редко. Среди больных онихомико-
зом преобладают люди пожилого и старческого
возраста, что объясняется замедленным ростом
ногтевых пластинок, дистрофическими и гипер-
трофическими изменениями кожи и ногтей;
нарушениями периферического кровообращения,
частой травматизацией кожи стоп и кистей; со-
матической отягощенностью; иммунодефицитом,
обусловленном приемом медикаментозных
средств.
Группами риска являются военнослужащие,
спортсмены, банщики; рабочие, посещающие
душевые; лица, регулярно пользующиеся банями
и саунами; рабочие промышленных предприятий,
шахтеры, рабочие горнорудных производств; ра-
бочие резинотехнической промышленности; стро-
ители туннелей и укладчики.
Источником заражения при дерматофитийном
онихомикозе является кожа стоп самого больно-
го. При руброфитии стоп й ногтей источник
инфекции находится в семье больного. Переда-
ча инфекции происходит через общую обувь,
белье. При эпидермофитии стоп и ногтей зара-
жение чаще происходит в душевых, бассейнах,
банях, саунах, спортзалах. Кандидозный онихо-
микоз чаще встречается у поваров, кондитеров,
прачек, рабочих консервных предприятий. При
кандидозе грибы проникают из кожи самого
больного или из пищевых продуктов, богатых
углеводами. При плесневых недерматофитных
онихомикозах источник заражения находится во
внешней среде, так как многие плесневые гри-
бы обитают в почве и их споры можно встре-
тить на окружающих предметах.
Патогенез
Онихомикозы представляют длительно суще-
ствующий очаг грибковой инфекции, способ-
ный вызвать сенсибилизацию организма. Крас-
ный трихофитон и плесневые грибы могут
выделять токсины. Возникновению онихомико-
зов способствуют следующие предрасполагающие
факторы.
Инфицирование ногтей может произойти
вследствие травмы ногтя, обусловленной слиш-
ком тесной обувью. Длительное давление сбоку
или спереди ведет к отгибанию ногтя, при этом
между ногтевой пластинкой и ложем образуют-
ся идеальные входные ворота для грибков. Пред-
посылками к заболеванию онихомикозом могут
быть анатомические особенности стопы: дефор-
мация пальцев, плоскостопие, узкие межпаль-
цевые промежутки.
Влажная и теплая среда — еще один фактор,
благоприятствующий развитию онихомикоза
стоп и ногтей. Субтропические условия, созда-
ваемые обувью, которая затрудняет воздухооб-
мен, размягчает кожу и ногти, тем самым под-
готавливая почву для заселения грибками.
Более подвержены возникновению и распро-
странению онихомикоза лица, страдающие тяже-
лыми соматическими и эндокринными (сахар-
ный диабет), хроническими инфекционными и
онкологическими заболеваниями, а также имму-
нодефицитами различного генеза; нарушениями
кровообращения; получающие кортикостероид-
ные гормоны, массивную антибиотикотерапию
и иммуносупрессивные препараты.
Глава 13. Грибковые болезни кожи 199
Клинические формы онихомикоза
С патогенетической и клинической точки зре-
ния выделяют различные формы онихомикозов:
дистальный латеральный онихомикоз, белый
поверхностный, проксимальный подногтевой,
тотальный дистрофический.
Дистальный латеральный онихомикоз (дисталь-
ный подногтевой) является самым распростра-
ненным видом заболевания. В 85 % случаев его
возбудителем служит Trichophyton rubrum. Ин-
фекции ногтей, вызванные дрожжеподобными
грибами (в основном Candida albicans), встреча-
ются значительно реже и поражают ногти кис-
тей. Среди плесневых грибов наиболее вероят-
но участие Scopulariopsis brevicaulis. При
дистальном латеральном онихомикозе источни-
ком гриба является инфицированная кожи, от-
куда возбудитель через свободный край ногтя
медленно проникает в ложе ногтя и распростра-
няется по направлению к матриксу. Вначале
ногтевая пластинка остается внешне интактной,
однако вследствие гиперкератоза она постепен-
но отходит от ногтевого ложа и принимает вслед-
ствие этого желтоватую окраску. Может про-
изойти и утолщение ногтевой пластинки.
Бактериальное обсеменение придает ногтевой
пластинке различные оттенки цвета, от зелено-
ватого до грязно-коричневого.
Белый поверхностный онихомикоз встречает-
ся реже всего, примерно в 90 % случаев он вы-
зывается Trichophyton mentagrophytes, реже —-
плесневыми грибками типа Aspergillus spp. В ре-
зультате проникновения грибов в ногтевую
пластинку на поверхности ногтя стопы образу-
ются небольшие белые островки, которые по-
степенно захватывают весь ноготь. Эта форма
онихомикоза обычно наблюдается у пожилых
пациентов, у которых один палец закрывает
ногтевую пластинку соседнего пальца. Ногте-
вая пластинка зачастую становится дистрофич-
ной и крошится, приобретая коричневый или
серый цвет. При этом матрикс и эпителий ног-
тевого ложа не поражаются; воспалительных
реакций со стороны окружающей кожи не на-
блюдается.
Проксимальный подногтевой онихомикоз явля-
ется самой редкой формой онихомикоза. К числу
его возбудителей относятся преимущественно
дерматофиты, прежде всего Trichophyton rubrum,
иногда дрожжи, особенно Candida albicans, но
также и плесневые грибки типа Hendersonula
toruloidea. Проксимальный подногтевой онихо-
микоз возникает при переходе инфекции с кожи
или околоногтевого валика. Оттуда грибы рас-
пространяются по поверхности ногтевой плас-
тинки к матриксу, здесь проникают в ноготь и,
по мере его роста, достигают дистальных отде-
лов ногтевой пластинки. При осмотре можно
увидеть изменения окраски ногтевой пластин-
ки в виде пятен в области лунки и ногтевого
ложа, где довольно быстро может появляться об-
ширная отслойка ногтевой пластинки. Выражен-
ного воспаления ногтевого ложа или матрикса,
как правило, не бывает. При этой форме они-
хомикоза также возможно вторичное бактериаль-
ное инфицирование и последующее потемнение
ногтя.
Тотальный дистрофический онихомикоз встре-
чается как следствие дистального латерального
или реже — проксимального подногтевого; кро-
ме того, он может наблюдаться при хроническом
подкожном кандидозе. В соответствии с механиз-
мом развития заболевания здесь преобладают
эпидермофиты или дрожжевые грибы. При ос-
мотре данная форма онихомикоза характеризуется
поражением всего ногтя, часто его разрушением.
Проксимальные участки ногтевого валика почти
исчезают или патологически утолщаются, поэтому
нормальная ногтевая пластинка больше образо-
вываться не может. Часто видны только ее рас-
крошившиеся остатки.
Диагностика
Поскольку изменения ногтя при онихомикозах
хотя и типичны, но не специфичны, при про-
ведении дифференциальной диагностики забо-
леваний ногтя необходимо прежде всего исклю-
чить следующие заболевания: псориаз, экзему,
гнездную алопецию, красный плоский лишай,
ониходистрофии неясного генеза. Клинический
диагноз грибковой инфекции должен подтвер-
ждаться микробиологически. Для этого прово-
дят микроскопию и обнаружение гриба или
грибковых элементов в пораженных тканях, а
затем выделяют возбудителя из ткани ногтя. Для
идентификации возбудителя необходим посев на
специальные питательные среды (чаще всего на
агар Сабуро).
Диагноз онихомикоза устанавливается на ос-
новании клинических проявлений, обнаружения
гриба при микроскопическом исследовании па-
тологического материала и выделения культуры
гриба на питательных средах.
Для успешного исследования необходим пра-
вильный сбор патологического материала из оча-
гов поражения. Пораженные участки ногтя сле-
дует брать из тех мест, которые не подвергались
местному лечению. Однако при уже проводимом
ранее лечении очаги поражения следует вымыть
с мылом или очистить спирто-эфирной смесью
200 Частная дерматология
и произвести забор материала через 2—3 дня.
Перед взятием материала ногти протираются 70°
этиловым спиртом. Для взятия материала из по-
раженных ногтей применяют маникюрные нож-
ницы, препаровальные иглы, скальпели, пинце-
ты. При часто встречающейся дистальной
подногтевой форме онихомикоза грибы распола-
гаются под ногтевой пластинкой. При поверхно-
стной форме онихомикоза соскобы проводят с
поверхности ногтевой пластинки. Материалом для
исследования должен быть обрезок ногтевой пла-
стинки, соскобы ногтевого ложа и из-под плас-
тинки. Необходимо захватывать и область неиз-
мененного ногтя, так как на границе между
пораженным и непораженным участками ногтя
располагаются самые активные грибы. При па-
ронихиях производят соскоб из проксимального
валика и соприкасающейся поверхности ногтевой
пластинки, а при наличии гноя проводят его за-
бор из-под ногтевых валиков.
Микроскопическое исследование. Микроскопи-
ческое исследование патологического материала
на грибы производят в нативных и окрашенных
препаратах. Материал обрабатывают 10—30 %-
ным раствором КОН (или NaOH), чтобы раство-
рить кератин. На размельченные при помощи
скальпеля ногтевые чешуйки наносят 1—2 кап-
ли раствора щелочи и осторожно подогревают над
пламенем спиртовки до появления нежного бе-
лого ободка из кристаллов щелочи по перифе-
рии капли (не доводить до кипения). После
подогревания каплю накрывают покровным
стеклом и оставляют на 30—40 мин для мацера-
ции и просветления. Приготовленные препара-
ты исследуют под световым микроскопом в те-
чение 2 часов с момента приготовления. Чтобы
лучше были видны части мицелия плесневых
грибов, иногда к раствору КОН добавляют чер-
нила.
При микроскопии обнаруживают нитевидные
гифы грибов или почкующиеся клетки. При
онихомикозе, вызванном красным трихофитоном,
под световым микроскопом отмечают многочис-
ленные полиморфные микроконидии, располо-
женные по обеим сторонам гифы, реже гроздья-
ми; некоторые микроконидии напоминают форму
карандаша. Онихомикоз, обусловленный
Trichophyton mentagrophytes var. interdigitale, харак-
теризуется микроскопически следующей карти-
ной: мицелий образует завитки, может иметь
форму бамбука; микроконидии грушевидной и
овальной формы, макроконидии тупоконечные,
септированные. При онихомикозе, вызванном
Epidermophyton floccosum, микроскопически вид-
ны многочисленные тонко- и гладкостенные,
тупоконечные макроконидии с 2—6 перегородка-
ми, расположенные в виде гроздьев бананов;
микроконидии отсутствуют; ветвящийся мицелий
находят не всегда. В ногтевых пластинках, пора-
женных Candida albicans, при микроскопии вид-
ны круглые или овальные почкующиеся дрож-
жеподобные клетки; наряду с ними постоянно
встречаются удлиненные элементы псевдомице-
лия. При наличии плесневых грибов Scopulariopsis
brevicaulis микроскопически выявляются единич-
ные гифы, многочисленные толстостенные спо-
ры, имеющие форму лимона. Гифы двух грибов
Hendersonula toruloidea и Scytalidium hyalinum
неотличимы друг от друга, но отличаются от гиф
дерматофитов большей вариабельностью шири-
ны и гладкой извилистой поверхностью; они
имеют двойной контур вследствие отражения
цитоплазмы от клеточной стенки гиф.
Однако с помощью микроскопии не всегда
удается идентифицировать различные микроор-
ганизмы, поэтому наиболее точная идентифи-
кация достигается культуральным методом ди-
агностики. Культуральный метод диагностики
позволяет иногда выявить возбудителя при от-
рицательных данных микроскопии. При куль-
тивировании материала можно определить род
и вид возбудителя, что особенно важно для це-
ленаправленной этиотропной терапии и профи-
лактики заболевания. Исследование ногтевых
чешуек проводится стерильной микологической
лопаточкой, увлажненной в конденсате среды,
чешуйки переносят в пробирку на скошенный
сусло-агар в 2—3 точки, прижимая их к повер-
хности среды. Обычно посев делают в 2—3 про-
бирки с последующей инкубацией при темпе-
ратуре 28°-37°С до 5 дней. Некоторые плесневые
грибы, а также дерматофиты, в культуре выра-
стают медленно, за 2—3 недели. При описании
колоний отмечают такие признаки, как время
ее появления, размеры, цвет поверхности и об-
ратной стороны, характер поверхности, форму
и край колонии, ее консистенцию и наличие
врастания в субстрат. При характеристике ко-
лонии указывают ее отличие от типичных штам-
мов, что важно для оценки полиморфизма дан-
ного вида гриба, а также для выявления
атипичных штаммов. Культуры гриба
Trichophyton rubrum развиваются на 8—10-й
день; колонии быстро растут, бархатистые
(иногда мучнистые или складчатые); поверх-
ность белая или розовая, обратная сторона пур-
пурная; пигмент не диффундирует в среду.
Культуры гриба Trichophyton mentagrophytes var.
interdigitale (пушистый) развиваются на 5—7-й
день; колонии плоские, бархатисто-пушистые,
с возрастом — мучнистые, поверхность белая,
обратная сторона желто-коричневая. Культуры
гриба Epidermophyton floccosum развиваются на
4-5-й день; колонии складчатые, куполообраз-
ные, мучнистые, цвет желтый с зеленоватым
оттенком, обратная сторона — от желтой до ко-
ричневой. Выделение плесневых грибов диаг-
ностически значимо только в тех случаях, ког-
да эти грибы получены отдельно, а не вместе с
дерматофитами, если они выделены в 5 из 20
культур и если в патологическом материале об-
наруживается мицелий этих грибов (М. Englich,
1976). Довольно часто могут встречаться сме-
шанные инфекции, вызванные дерматофитами
вместе с другими грибами. При культивирова-
нии материала инфицированных ногтей рост
грибов зачастую отсутствует. До 30 % ногтево-
го материала, заведомо инфицированного дер-
матофитами, могут давать отрицательный ре-
зультат при культуральном исследовании. При
отрицательном микроскопическом и культу-
ральном исследованиях на грибы нужно рас-
смотреть возможность наличия других заболе-
ваний, протекающих под маской онихомикоза.
Лечение больных онихомикозами
При назначении лечения больному онихомико-
зом необходимо учитывать клиническую форму
заболевания, распространенность процесса, ско-
рость роста ногтей, общее состояние, способ при-
менения препарата, финансовые возможности
пациента.
Местная терапия. Местная терапия назначает-
ся больным с дистальным и латеральным подног-
тевым онихомикозом. Методы наружной терапии
назначают также при наличии противопоказаний
к применению антимикотиков системного дей-
ствия. Эффективными препаратами для лечения
ногтей являются кремы микоспор, бифоназол, а
также 5 % лак «Лоцерил» и 0,25 % крем аморол-
фин или 8 % лак циклопирокс (Батрафен) в со-
Глава 13. Грибковые болезни кожи 201
четании с 1 % кремом или 1 % раствором цикло-
пирокс.
Системные противогрибковые препараты. Анти-
микотические препараты системного действия
назначаются пациентам с множественными по-
ражениями ногтей (как правило, свыше 4—5
ногтей) и кожи, с проксимальным подногтевым
онихомикозом, тотальным дистрофическим,
дистальным латеральным с вовлечением в про-
цесс матрикса, а также при отсутствии эффекта
от местной терапии. При гиперкератозе и тоталь-
ном онихомикозе следует удалить пораженные
ногти и роговые наслоения в области ложа с
помощью кератолитических средств.
При выборе системного препарата важным
критерием является спектр его действия. Поэто-
му необходимо знать данные культурального
исследования на грибы. Если этиология возбу-
дителя неизвестна или выделено несколько гри-
бов, то рекомендуются препараты широкого
спектра действия, воздействующие на дермато-
фиты, дрожжеподобные и плесневые грибы.
Одним из таких универсальных препаратов яв-
ляется итраконазол (орунгал). Спектр активно-
сти системных антимикотических препаратов
представлен в таблице 13.1.
При дерматофитном онихомикозе назначают
итраконазол, тербинафин, гризеофульвин. При
кандидозном онихомикозе рекомендуется флу-
коназол, итраконазол. При онихомикозе, выз-
ванном плесневыми грибами недерматофитами,
назначают итраконазол.
В прошлом для системной терапии онихоми-
козов использовали гризеофульвин, а затем ке-
токоназол (низорал). В настоящее время основ-
ными и наиболее эффективными препаратами
являются тербинафин, итраконазол, флуконазол.
Тербинафин (ламизил, тербизил) — фунгицид-
ный (противогрибковый) препарат широкого спек-
тра действия из класса аллиламинов; эффективен
в отношении дерматофитов, дрожжей и плесеней.
Таблица 13.1. Спектр активности системных антимикотических препаратов
Препараты Виды грибов
Дерматофиты Дрожжеподобные Плесневые
Гризеофульвин *** — —
Кетоконазол ** ** —
Флуконазол ★ ** ★
Итраконазол *** ** **
Тербинафин *** ★ *_
Примечание: (***) — очень высокоэффективен; (**) — высокоэффективен; (*) — эффективен; (*-) — эффек-
тивен при повышении дозы; (-) — не эффективен.
202 Частная дерматология
Тербинафин обладает самой высокой среди систем-
ных антимикотиков активностью в отношении
дерматофитов, вызывающих большинство случа-
ев дерматомикозов. Фунгицидные концентрации
препарата сохраняются в коже и ногтевых плас-
тинах в течение 30—36 нед после окончания лече-
ния, т. е. действие препарата в ногте сохраняется
еще до 9 месяцев, что существенно повышает про-
цент полного излечения. Тербинафин назначают
1 раз в сутки по 250 мг (1 таблетка) независимо
от приема пищи. Продолжительность лечения в
среднем составляет: при поражении ногтей на
пальцах кистей — б нед, стоп — 12 нед.
Итраконазол (орунгал) противогрибковый пре-
парат из класса азолов широкого спектра дей-
ствия. Он эффективен у больных онихомикозом,
обусловленным дерматофитами, дрожжевыми и
плесневыми грибами. Итраконазол назначается
методом пульс-терапии: его принимают внутрь
после еды по 2 капсулы по 100 мг 2 раза в день
(400 мг в сутки) в течение 7 дней, затем после
трехнедельного перерыва курс повторяется. При
поражении ногтей на пальцах кистей больные
получают два курса терапии, на пальцах стоп —
3—4 курса в зависимости от формы онихомико-
за, площади поражения, скорости роста ногтей.
Эффективная концентрация препарата в ногтях
на кистях после окончания лечения сохраняется
в течение 3 месяцев, на стопах после 3 курсов —
6—9 месяцев, после 4 курсов — до 1 года.
Флуконазол (дифлюкан, микосист) — препа-
рат из класса триазольных противогрибковых
средств, обладает широким спектром действия.
При онихомикозе, вызванном дерматофитами,
его назначают по 150 мг 1 раз в неделю в тече-
ние 3—12 мес; при поражении ногтей кистей —
3—6 мес, при поражении ногтей стоп — 6—12
мес. После 4-месячного лечения препарат сохра-
няется в ногтях до 6 мес. Флуконазол наиболее
эффективен при онихомикозе, вызванном дрож-
жеподобными грибами. При этом заболевании
его назначают по 200 мг в день после приема
пищи 1 раз в неделю в течение 3—4 мес.
Наиболее распространенные схемы назначения
системных противогрибковых препаратов при-
ведены в таблице 13.2.
Наиболее безопасными в отношении побочных
и токсических эффектов, а также удобными для
пациентов, считаются методики пульс-терапии,
то есть короткие и прерывистые схемы приме-
нения системных антимикотических препаратов.
При всех методах терапии онихомикозов обя-
зательно проводится дезинфекция обуви и пер-
чаток до отрастания здоровых ногтей. В качестве
дезинфектантов рекомендуется 1 % раствор хлор-
гексидина биглюконата и 25 % раствор форма-
лина.
Подробно охарактеризуем три основных систем-
ных антимикотических препарата, которые при-
меняются для лечения онихомикозов: итракона-
зол, тербинафин, флуконазол.
Итраконазол (торговое название — орун-
гал) — триазольный антимикотический препа-
рат; водорастворимый, липофильный; отличается
высокой степенью биодоступности. Препарат
хорошо всасывается в пищеварительном тракте
при нормацидном и гиперацидном состоянии же-
лудочного сока. Орунгал принимают внутрь сра-
зу после еды. Всасывание препарата снижается
при пониженной кислотности желудочного сока:
гипоацидных и анацидных состояниях; приеме
антацидов, Н2-антагонистов, антигистаминных
препаратов; после трансплантации органов. По-
этому препарат целесообразно назначать в ком-
бинации с желудочным соком или соляной кис-
лотой с пепсином. Всасываясь, орунгал быстро
поступает в различные ткани и органы, где на-
капливается в концентрации, превосходящей
плазменную. Орунгал быстро метаболизируется
в печени, распадаясь на большое число метабо-
литов, многие из которых обладают противо-
грибковой активностью. Выводится препарат в
виде неактивных метаболитов в основном с ис-
пражнениями и мочой. Уже через 2—4 часа пос-
ле приема орунгала начинается его накопление
в тканях и постепенное исчезновение из плаз-
Таблица 13.2. Схемы назначения системных противогрибковых препаратов (Ю. В., Сергеев, А. Ю. Сергеев,
1998).
Препарат Схема назначения Суточная доза Продолжительность курса
Ногти рук Ногти ног
Гризеофульвин Стандартная (ежедневно) 1000 мг 4-12 мес 12-18 мес
Ламизил Укороченная (ежедневно) 250 мг 1,5 мес от 3 мес
Низорал Стандартная (ежедневно) 200 мг 4-6 мес 12-18 мес
Орунгал Пульс-терапия (1 день в неделю) 400 мг 2 мес от 3 мес
Дифлюкан Пульс-терапия (1 день в неделю) 150 мг 4-6 мес 6-12 мес
Глава 13. Грибковые болезни кожи
203
мы крови. В ногтях через 7 дней после приема
орунгала в дозе 400 мг/сутки его концентрация
в 10 раз выше, чем после приема в суточной дозе
100 мг. Пик концентрации в ногтях (900 нг/мг)
достигается через 2 месяца после начала терапии.
После прекращения терапии орунгалом его кон-
центрация в ногтях (350—800 нг/мг) сохраняет-
ся в течение 3-6 месяцев, элиминируясь по мере
роста ногтей. Препарат достаточно быстро эли-
минируется из внутренних тканей и органов,
поэтому при лечении больных системными глу-
бокими микозами необходим постоянный при-
ем препарата до клинико-микологического из-
лечения.
Действие орунгала на грибы фунгистатичес-
кое и фунгицидное. Концентрация препарата
0,1 мкг/мл подавляет прорастание спор гриба в
мицелий, а затем вызывает их разрушение, при
этом погибает 94 % штаммов грибов-дерматофи-
тов и до 71 — 100% дрожжеподобных грибов.
Тестированные 6113 видов различных грибов,
играющих важную роль в патологии человека,
оказались чувствительными к орунгалу. С по-
мощью световой и электронной микроскопии
установлено, что концентрация орунгала 10—10—
10-7 подавляет развитие бластопор и превраще-
ние их в гифы, вызывает нарушения в клеточ-
ной стенке, плазматических мембранах,
цитоплазматических везикулах. По механизму
противогрибкового действия орунгал сходен с
другими азольными соединениями. Орунгал из-
бирательно действует на грибковые клетки. На-
капливаясь в фосфолипидном слое грибов, орун-
гал подавляет включение 14С-ацетата в синтез
грибковой клетки. Участие орунгала в 14-а-де-
митилации 24-метилдигидроланостерола способ-
ствует значительному уменьшению и даже ис-
чезновению 14-а-метилтестостерона. Остающиеся
стеролы не обеспечивают мембранный транспорт
и нормальное существование грибковых клеток.
Орунгал действует также на цитохром Р-450 и
С-оксидазу грибковых клеток. Наличие в моле-
куле орунгала длинной боковой липофильной
цепи обусловливает его более высокое сродство
с цитохромом Р-450 в основном грибов, а не
ферментных клеток млекопитающих. Все пере-
численные свойства препарата влияют на гриб-
ковые структуры: они становятся ломкими и
быстро погибают вследствие нарушения осмоти-
ческого давления. При широком спектре проти-
вогрибковой активности, орунгал в 5—100 раз ак-
тивнее низорала в отношении некоторых
дерматофитов, грибов рода кандида, плесневых,
а также возбудителей глубоких микозов, что по-
зволяет достигать терапевтического эффекта при
меньших дозах препарата. Орунгал обладает так-
же иммуномодулирующим и противовоспали-
тельным действием.
Орунгал поступает в кожу и ее придатки тре-
мя путями: экскрецией сальными, потовыми
железами и путем простой диффузии. Орунгал
появляется в дистальных частях ногтевой плас-
тинки в течение недели с начала лечения. Кон-
центрации препарата в ногте повышаются в ходе
лечения. При повышении дозы орунгала со 100
до 200 мг/сут концентрации его в ногте возрас-
тают в 10 раз. Попадая в ногтевую пластинку,
орунгал остается в ней и не возвращается в кро-
воток. Эти свойства препарата позволили разра-
ботать схему пульс-терапии, в которой орунгал
назначают не постоянно, а только одну неделю
в месяц. Препарат в суточной дозе 400 мг/сут
(по 200 мг 2 раза в день) принимают в течение
1 недели с трехнедельным перерывом. Для ле-
чения онихомикозов кистей достаточно 2-х кур-
сов пульс-терапии; для лечения онихомикозов
стоп — 3-х курсов. Орунгал накапливается в под-
ногтевой зоне, основном очаге инфекции, что
считается составляющей частью эффективного
лечения. Препарат накапливается в ногтевом
ложе и гиперкератотических массах даже лучше,
чем в дистальных частях ногтевой пластинки.
При пульс-терапии концентрации орунгала в
плазме крови начинают падать сразу после от-
мены препарата и к началу следующего пульса
становятся практически незаметными. Феномен
сохранения высоких концентраций в ногте при
падении их в крови в настоящее время рассмат-
ривают как предпосылку безопасности лечения
орунгалом.
Для приема внутрь итраконазол выпускается
под торговым названием «Орунгал» в желатино-
вых розово-голубого цвета капсулах, содержа-
щих 100 мг препарата. Блистерная упаковка со-
держит 4 или 15 капсул.
Побочные эффекты приема орунгала чаще
всего связаны с расстройством желудочно-ки-
шечного тракта: дискомфорт в области эпигаст-
рия, чувство переполнения желудка, тошнота,
рвота, диспепсия. У некоторых больных отмеча-
ются преходящие изменения печеночных транса-
миназ. Чрезвычайно редко могут быть высыпа-
ния на коже, кожный зуд. Орунгал не назначают
беременным и кормящим матерям.
Флуконазол (дифлюкан, микосист) — бис-
триазольный антимикотический препарат, во-
дорастворимый, отличается высокой биодос-
тупностью как при приеме внутрь, так и при
внутривенном введении, хорошо всасывается и
накапливается в высоких терапевтических
204 Частная дерматология
концентрациях во всех биологических тканях
организма. Препарат хорошо проникает через
гематоэнцефалический барьер, метаболически
стабилен. Флуконазол угнетает образование эр-
гостерола, основного компонента мембраны
грибов, действуя на фермент 14-а-диметилазу,
входящий в систему цитохрома Р-450. Флуко-
назол высоко специфичен по отношению к за-
висимым от цитохрома Р-450 ферментам гри-
бов.
Флуконазол обладает широким спектром про-
тивогрибкового действия, особенно при кандидоз-
ной инфекции и системных микозах. Активность
флуконазола в отношении разных плесневых гри-
бов меньше, чем в отношении дрожжевых. Флу-
коназол является гидрофильным препаратом; аб-
сорбция его почти полная, биодоступность
составляет 90 %. Препарат не подвергается актив-
ному метаболизму в печени. Флуконазол выводит-
ся с мочой на 70—90 % в неизмененном виде; пе-
риод полувыведения составляет около 20—35 часов.
Флуконазол быстро поступает в ноготь и накап-
ливается там в терапевтической концентрации
1300 нг/г уже в первые сутки лечения. Это свой-
ство препарата позволило назначать его по схеме
пульс-терапии 1 раз в неделю (по 150 мг). При
онихомикозах стоп препарат назначают в течение
12 месяцев, при онихомикозах кистей — до 6 ме-
сяцев. Иногда используется стандартная схема
пульс-терапии онихомикоза стоп с назначением
300 мг препарата в неделю (две капсулы по 150 мг)
в течение 9 месяцев. Из побочных эффектов флу-
коназола отмечают тошноту, чувство дискомфор-
та в эпигастрии.
Тербинафин (ламизил, тербизил) — синтетичес-
кий препарат из класса аллиламинов. По срав-
нению с азольными соединениями тербинафин
действует на ранних стадиях стерольного мета-
болизма, на уровне скваленового эпоксидазно-
го цикла. Подавляя скваленовую эпоксидазу
грибов, тербинафин тормозит формирование
эргостерола — основного компонента клеточной
стенки грибов, препятствуя дальнейшему превра-
щению скваленов.
Действие тербинафина на грибковые клетки
фунгистатическое и фунгицидное. Фунгистати-
ческий эффект связан с недостаточностью эр-
гостерола, вследствие чего цитоплазматические
мембраны становятся пористыми, а сами гриб-
ковые клетки, утрачивая способность расти и
развиваться, только выживают. Фунгицидное
действие тербинафина обусловлено накоплени-
ем скваленов, которые еще больше ухудшают
состояние клеточных мембран, так как подоб-
но своеобразной губке экстрагируют из них ли-
пидные компоненты. При этом усиливается
ущербность стенки гриба и увеличивается объем
липидных гранул. Накапливаясь внутри клет-
ки, постепенно увеличивающиеся липидные
гранулы наконец разрывают ущербную цитоп-
лазматическую мембрану, приводя грибковую
клетку к гибели.
Спектр действия тербинафина включает дер-
матофиты; эффективность препарата по отноше-
нию к недерматофитным плесневым инфекци-
ям неизвестна; дрожжевые грибы (Candida
albicans) устойчивы к тербинафину с МПК око-
ло 128 мг/л. Тербинафин хорошо всасывается в
желудочно-кишечном тракте, в течение 2 часов
абсорбируется более 70 % препарата. Пик кон-
центрации тербинафина в крови достаточно ста-
билен и составляет 0,9 мг/мл при приеме в дозе
250 мг/сут и 1,7—2,0 мкг/мл при дозе 500 мг/сут.
Препарат поступает в ногти медленно: спустя 2—
4 недели его концентрация составляет 0,3—
0,55 мкг/г, через 12 недель — 0,78 мкг/г и да-
лее остается стабильной. Через 55—90 дней после
отмены препарата его концентрация в ногтях ос-
тается 0,1—0,2 мкг/г.
При лечении онихомикозов, вызванных дер-
матофитами, тербинафин применяется в дозе
250 мг в сутки. Курсовая непрерывная терапия
при онихомикозах кистей составляет 6 недель,
при онихомикозах стоп — 12 недель.
Побочные эффекты применения тербинафи-
на — тошнота, рвота, дискомфорт, чувство тя-
жести в желудке. Изредка может развиться
крапивница. Очень редко встречаются наруше-
ния гепатобилиарной системы. Не рекоменду-
ется назначать препарат больным с патологи-
ей печени.
Глубокие микозы
Кокцидиоидоз и гистоплазмоз являются весьма
контагиозными и вызывают тяжелые поражения
кожи, слизистых оболочек и внутренних орга-
нов. Аспергиллез, мукороз, пенициллиоз, спорот-
рихоз не относят к контагиозным глубоким
микозам. Они вызываются условно-патогенны-
ми грибами и развиваются у лиц со сниженной
иммунобиологической реактивностью. Описан-
ные далее глубокие микозы встречаются преиму-
щественно в странах с тропическим и субтропи-
ческим климатом. В зонах с умеренным климатом
некоторые глубокие микозы (споротрихоз, хро-
момикоз) наблюдаются спорадически или не
регистрируются совсем (кокцидиоидоз, гистоп-
Глава 13. Грибковые болезни кожи
205
лазмоз). Споротрихоз, криптококкоз, гистоплаз-
моз, кокцидиоидоз тяжело протекают у больных
в клинической стадии СПИДа.
Кокцидиоидоз
Кокцидиоидоз — глубокий эндемический сис-
темный микоз, протекающий с первичным по-
ражением легких и заканчивающийся в боль-
шинстве случаев аутосанацией. У части лиц
инфекция распространяется гематогенным путем
в кожу, кости, мозговые оболочки, легкие, где
возникают множественные очаги хронического
гранулематозного воспаления. Относится к груп-
пе опасных микозов.
Возбудитель: Coccidioides immitis, диморфный
гриб, обитает в почве в виде мицелия.
Предрасполагающие факторы: цветная раса; бе-
ременность, иммуносупрессивная терапия, ВИЧ-
инфекция.
Пути передачи: воздушно-пылевой: при вды-
хании пыли, содержащей артроспоры грйба, раз-
вивается первичная легочная инфекция. Через
поврежденную кожу возбудитель проникает в
организм редко.
Течение. У 60 % зараженных заболевание про-
текает бессимптомно и наступает аутосанация.
Диссеминированный кокцидиоидоз обычно воз-
никает при нарушениях иммунитета, преимуще-
ственно у ВИЧ-инфицированных. Наиболее
опасно в прогностическом плане поражение моз-
говых оболочек.
Излюбленная локализация. Кожные проявле-
ния: промежность, крестцово-копчиковая об-
ласть, ягодицы, задняя повехность шеи, лицо,
подкрыльцовые ямки.
Клиническая картина
Острый легочный кокцидиоидоз
Острый легочный кокцидиоидоз протекает в
виде гриппоподобного синдрома, бронхита, пнев-
монии, активного туберкулеза. Кожные проявле-
ния: распространенная эритема, кореподобная
сыпь, крапивница, узловатая эритема, полимор-
фная экссудативная эритема.
Диссеминированный кокцидиоидоз
Кожные проявления: папулы, пустулы, бляш-
ки, узлы, абсцессы, флегмона, множественные
свищи, язвы, бородавчатые разрастания, гра-
нулемы, рубцы. Первичное поражение кожи
наблюдается весьма редко и проявляется узлами
и изъязвлениями, лимфангитом и регионарным
лимфаденитом. Поражения костной системы:
остеомиелиты, натечные абсцессы. Нервная си-
стема: менингит.
Лабораторные методы исследования
При микроскопии мокроты или гноя обнаруживают
сферулы возбудителя. Посев проводят на среду Са-
буро; используют гной или биопсийный материал.
Патоморфология кожи. Гранулематозное вос-
паление. В тканях находят сферулы — крупные
круглые спорангии гриба, заполненные спорами.
Диагноз
Обнаружение сферул в мокроте или гное; иден-
тификация колонии Coccidioides immitis при
посеве; результаты биопсии кожи.
Дифференциальный диагноз
Бородавки, пиодермия, узловатая почесуха, ке-
ратоакантома, североамериканский бластомикоз,
криптококкоз, туберкулез кожи, третичный си-
филис. У ВИЧ-инфицированных — фолликулит,
контагиозный моллюск.
Лечение
В тяжелых случаях: амфотерицин В внутривен-
но. При более легком течении: флуконазол, или
итраконазол.
Профилактика
Прием противогрибковых средств в эндемичных
очагах; особенно антимикотики показаны ВИЧ-
инфицированным. При менингите нередко на-
значается пожизненный прием антимикотиков.
Г истоплазмоз
Гистоплазмоз (ретикулоэндотелиальный цитоми-
коз) — глубокий, системный микоз, который
начинается с первичного инфицирования и по-
ражения легких. При гематогенной диссемини-
нации возбудитель попадает в слизистые, кожу,
печень, селезенку и костный мозг. Заболевание
относится к группе опасных глубоких микозов.
Возбудитель: Histoplasma capsulatum, в Афри-
ке — Histoplasma capsulatum var. duboisii. Гриб ди-
морфный; сапрофит красных подзолистых почв,
богатых органическими веществами, содержит-
ся в помете птиц и летучих мышей.
Предрасполагающие факторы: иммунодефицит
(ВИЧ-инфекция, трансплантация органов, хими-
отерапия, гемобластозы), старческий возраст.
Пути передачи: воздушно-пылевой (вдыхание
пыли, содержащей споры гриба).
Течение. При легочном неосложненном гис-
топлазмозе выздоровление обычно наступает са-
206 Частная дерматология
мостоятельно. При хронических формах кавер-
нозного легочного и диссеминированного гистоп-
лазмоза летальность пациентов, не получающих
лечения, достигает 80 %. В ряде случаев при этих
тяжелых формах требуется постоянный прием
системных антимикотиков.
Излюбленная локализация. Кожные проявле-
ния: лицо, конечности, туловище. Слизистые:
узлы, вегетации, болезненные язвы, поражаю-
щие мягкое небо, ротоглотку, надгортанник,
преддверие носа.
Клиническая картина
Острый легочный гистоплазмоз. Гриппоподобный
синдром, который может сопровождаться узло-
ватой эритемой или многоформной экссудатив-
ной эритемой.
Диссеминированный гистоплазмоз. Признаки
интоксикации. У ВИЧ-инфицированных может
развиться сепсис, недостаточность надпочечни-
ков, диарея, боли в животе.
Кожные проявления: красные папулы и узлы,
ороговевающие или некротизирующиеся, эрите-
матозные пятна; фолликулиты, иногда пустулез-
ные или угревидные элементы; бляшки, покры-
тые вегетациями; панникулит; эритродермия.
При поражении надпочечников — диффузная
гиперпигментация в результате надпочечниковой
недостаточности.
Лабораторные методы исследования
При микроскопии возбудитель обнаруживается
в окрашенных по Гимзе мазках-отпечатках с
биопсийного материала, в мазках мокроты и
костного мозга.
Для посева используют кровь, мочу, костный
мозг, биоптаты кожи, слизистой, печени, лим-
фоузлов, легкого.
Определение антигенов Histoplasma capsulatum
в сыворотке больных используют для постановки
диагноза, оценки результатов лечения и предска-
зания рецидивов.
Для выявления антител к Histoplasma
capsulatum в сыворотке больных используют
метод иммунодиффузии и РСК.
При диссеминированном гистоплазмозе в пун-
ктате костного мозга обнаруживают возбудитель.
Диагностика
Клиническая картина, результаты лабораторных
исследований.
Дифференциальный диагноз
Другие глубокие микозы, дерматозы, заболева-
ния легких.
Лечение
В тяжелых случаях и при менингите: амфо-
терицин В в/в. При более легких формах: ит-
раконазол 400 мг/сут внутрь в течение 12 нед,
или флуконазол 800 мг/сут внутрь в течение
12 нед.
Профилактика
Индивидуальные средства защиты (респираторы,
очки, перчатки, защитная одежда) при контак-
те с пометом птиц, летучих мышей и т. п.
ВИЧ-инфицированным назначают постоян-
ный прием итраконазола или флуконазола.
Споротрихоз
Споротрихоз — подостро или хронически про-
текающий глубокий микоз, поражающий кожу,
подкожную клетчатку, лимфатическую систему
и слизистые оболочки. Возможно поражение
печени, почек, легких. Заболевание чаще спора-
дическое, однако возможны эпидемии.
Возбудитель: диморфный гриб Sporothrix
schenckii, обитает в почве, на растениях и т.п. В
тканях существует в виде овальных или сигаро-
образных дрожжевых клеток «челноков» или в
виде их скоплений — астероидных тел.
Предрасполагающие факторы: регионы с тро-
пическим климатом. Сахарный диабет, алкого-
лизм, ВИЧ-инфекция, злокачественные новооб-
разования, иммуносупрессивная терапия.
Угрожаемый контингент: цветоводы, садовни-
ки, сельхозрабочие, лесники и др.
Пути передачи. Заражение происходит при глу-
бокой травматизации кожи (колючки, шипы,
занозы, осколки стекла) или вдыхании, аспира-
ции и пероральном попадании инфицированного
материала (висцеральный споротрихоз). Источ-
ником инфекции могут быть болеющие спорот-
рихозом домашние животные, а также броненос-
цы (после их царапин).
Течение. Инкубационный период в среднем
2—3 нед после травмы кожи. Болевые ощуще-
ния и лихорадка не наблюдаются. Самоизле-
чение не наступает. Возможны частые рециди-
вы после полноценных курсов лечения. Для
ВИЧ-инфицированных характерна торпид-
ность к терапии.
Излюбленная локализация: наиболее частая ло-
кализации споротрихозного шанкра у взрос-
лых — тыл кисти или пальца с наличием лим-
фангита предплечья, у детей — лицо; при
диссеминированной споротрихозной сыпи (узлы
и язвы) ладони и подошвы не поражаются.
Глава 13. Грибковые болезни кожи
207
Клиническая картина
Различают несколько форм споротрихоза:
• поверхностный кожный;
• локализованный подкожный («споротрихоз-
ный шанкр»);
• лимфатический споротрихоидный;
• диссеминированный;
• сочетанный;
• споротрихиды.
Поверхностный кожный споротрихоз: фоллику-
литы, акне, папулы, бляшки, веррукозные, па-
пилломатозные или псориазиформные высыпа-
ния, язвы, корки. Иногда эритематозно-язвенные
и папилломатозные поражения слизистых рта,
носа, зева, гортани.
Локализованный подкожный споротрихоз («спо-
ротрихозный шанкр»)', в месте травмы через не-
сколько недель появляется папула, пустула или
узел, которые спаиваются с окружающими тканя-
ми и изъязвляются. Элемент окружен розовой
эритемой. Формируется споротрихозный шанкр —
болезненная язва на плотном основании, с неров-
ными подрытыми краями. Выявляется увеличение
и воспаление регионарных лимфоузлов.
Лимфатический споротрихоидный споротрихоз
(60 % случаев) возникает вследствие распростране-
ния инфекции из первичного очага (споротрихоз-
ного шанкра) по лимфатическим сосудам. По ходу
дренирующего лимфатического сосуда выявляется
плотный толстый тяж с множественными узлами,
узелками, которые впоследствии изъязвляются.
Диссеминированный споротрихоз развивается
вследствие распространения инфекции гемато-
генным путем. Поражаются кожа (множествен-
ные узлы, язвы, корки), суставы, глаза, мозго-
вые оболочки, внутренние органы.
Споротрихиды: наблюдаются в ряде случаев
споротрихоза в виде аллергических сыпей.
Лабораторная диагностика
При микроскопии мазков материала из очагов
поражения обнаруживаются сигарообразные
клетки.
Патоморфология кожи
Гранулематозное воспаление. Сигарообразные
клетки в материале из очагов поражения можно
выявить только у больных с иммунодефицитами.
Посев. Используют биопсийный материал. В
культуре (t 24° С) вырастает бархатистая белая,
затем чернеющая колония с тонким мицелием и
овальными конидиями; при t 37° С грибы про-
ходят дрожжевую фазу, образуя колонии мягкой
консистенции, гладкие, желтоватые.
Диагностика
Наличие клинической картины, положительные
результаты микроскопии и посева.
Дифференциальный диагноз
Туберкулез кожи; инфекции, вызванные ати-
пичными микобактериями; туляремия; фели-
ноз; первичный сифилис; другие глубокие
микозы; глубокие пиодермии; сибирская язва;
лейшманиоз; мелиоидоз; инородные тела; дер-
матофитии.
Лечение
Препараты йода внутрь (3—4 г/сут); итракона-
зол 200-600 мг/сут; флуконазол 200-400 мг/сут;
кетоконазол 400—800 мг/сут. Амфотерицин В
назначают при диссеминированном споротрихозе
и в случаях поражений легких.
Профилактика
Заключается в предупреждении травматизации
кожного покрова.
Аспергиллез
Аспергиллез — микотическое поражение кожи,
слизистых оболочек, уха, глаза, легких, дру-
гих внутренних органов, вызванное аспергил-
лами.
Возбудитель: грибы Aspergillus fumigatus, А.
flavus, A. niger и некоторые другие. Сапрофиты
почвы и растений, выявляются в воздухе и пыли
помещений на производствах, где аспергиллы ис-
пользуются в качестве продуцентов ферментов,
некоторых органических кислот и др. Носите-
лями аспергилл могут быть домашние животные
и некоторые птицы (например, голуби).
Предрасполагающие факторы: снижение им-
муно-биологической резистентности организма,
тяжелые инфекции (туберкулез), онкозаболева-
ния (рак легкого).
Пути передачи: ингаляторный, эндогенный;
реже — через поврежденную кожу.
Течение: хроническое, без лечения приводит
к тяжелым поражениям легких, печени, почек.
Излюбленная локализация: наружный слухо-
вой проход, кожа ягодиц, ногтевые пластинки,
роговица.
Клиническая картина
Кожные проявления разнообразны: могут про-
текать в виде дерматита, интертриго, везикулез-
ной, папулезной сыпи, эритродермии, хроничес-
ких язвенных и абсцедирующих пиодермий;
208 Частная дерматология
иногда поражаются ногтевые пластинки. Брон-
холегочная форма напоминает бронхит, пневмо-
нию, туберкулез легких, или осложняет течение
туберкулеза, рака легкого. Встречаются аспери-
гиллезные отиты, поражения роговицы (вплоть
до панофтальмита), придаточных полостей носа.
Гистопатология
Некроз тканей, грануляции с гистиоцитами,
фибробластами, гигантскими и эпителиоидны-
ми клетками (гранулема инородного тела).
Диагностика
Обнаружение спор и мицелия гриба в патологи-
ческом материале с дальнейшим выделением
чистой культуры возбудителя.
Дифференциальный диагноз
Туберкулез, саркоидоз, актиномикоз, эумицето-
ма, висцеральные микозы, паразитарные и непа-
разитарные кисты, пылевые фиброзы легких и др.
Лечение
При поверхностных поражениях кожи: местные
антимикотики, фунгицидные жидкости.
При висцеральном аспергиллезе: амфотерицин
В, итраконазол, средства общеукрепляющей те-
рапии.
Аспергилломы легких: лечение хирургическое.
Профилактика
Применение средств защиты у работающих на
вредных производствах (с продуцентами аспер-
гиллами).
Цефалоспориоз
Цефалоспориоз — глубокий микоз кожи, сли-
зистых оболочек, опорно-двигательного аппарата
и внутренних органов.
Возбудитель: грибы рода Cephalosporium, чаще
встречается Cephalosporium acremonium Corda;
природные сапрофиты.
Предрасполагающие факторы: различные трав-
мы кожи. Снижение защитных свойств организма.
Пути передачи: при травме кожи и слизистых
оболочек.
Течение хроническое.
Излюбленная локализация: кожа, слизистые
оболочки.
Клиническая картина
Напоминают дерматиты, пиодермии, везикулез-
но-пустулезные элементы, эрозивно-язвенные
или глубокие очаги типа гумм, язвенно-вегети-
рующих, абсцедирующих элементов, мицетомы
(«мадурской стопы»). Висцеральные поражения
сопровождаются, как правило, септикопиемией.
Гистопатология
Характерной картины не выявляется.
Диагностика
Получение культуры гриба в патологическом ма-
териале, выделение его ретрокультуры, внутрибрю-
шинное заражение лабораторных животных.
Дифференциальный диагноз
Другие глубокие микозы, язвенно вегетирующие
пиодермии, гуммозный сифилид и др.
Лечение
Йодистый калий в нарастающих дозировках
внутрь, пероральные антимикотики (итраконазол,
кетоконазол). Возможна хирургическая тактика.
Местные антимикотики, фунгицидные красители.
Профилактика
Борьба с микротравматизмом в сельском хозяй-
стве и при земляных работах.
Кладоспориоз
Кладоспориоз — глубокий системный микоз с
преимущественным поражением центральной
нервной системы (чаще головного мозга), а так-
же кожных покровов. Заболевание неконтагиоз-
но. Регистрируются спорадические случаи.
Возбудитель: Cladosporium trichoides,
Cladosporium mansonii и др; почвенные и расти-
тельные сапрофиты. Гриб нейротропный.
Предрасполагающие факторы и пути передачи
не известны.
Течение хроническое, прогредиентное.
Прогноз: при поражении мозга прогноз небла-
гоприятный.
Излюбленная локализация: ЦНС (преимуще-
ственно головной мозг), реже — кожа.
Клиническая картина
Поверхностные поражения кожи (эпидермиса)
протекают в виде Tinea nigra, keratomycosis pabnaris.
Головной мозг: головные боли, сонливость,
апатия, лихорадка, гипертензия, связанные с
множественными абсцессами мозга. Встречают-
ся инкапсулированные абсцессы легких.
Гистопатология
В веществе мозга выявляются инкапсулирован-
ные очаги с признаками специфического воспа-
Глава 13. Грибковые болезни кожи 209
ления с сегментированными ветвящимися нитя-
ми гриба.
Диагностика
Обнаружение в тканях возбудителя (пигмен-
тированные коричневые нити, распадающиеся
на споры в виде цепочек), выделение его куль-
туры.
Дифференциальный диагноз
Вульгарные абсцессы мозга; другие глубокие
микозы, поражающие головной мозг (аспергил-
лез, мукороз, криптококкоз).
Лечение
Кожные проявления — см. хромомикоз. Абсцес-
сы мозга — лечение хирургическое.
Профилактика - не определена.
Риноспоридиоз
Риноспоридиоз — хронический глубокий микоз,
встречающийся преимущественно в странах с
жарким климатом.
Возбудитель: Rhinosporidium seeberi; культура
возбудителя не получена.
Предрасполагающие факторы и пути передачи:
заболевание выявляется у лиц, работающих на
рисовых полях и грязных водоемах, которые заг-
рязняются птицами, болеющими риноспориди-
озом.
Течение хроническое.
Излюбленная локализация: слизистая оболоч-
ка носа, влагалища, конъюнктивы, прилегающие
участки кожи.
Клиническая картина
В местах преимущественной локализации разви-
ваются папилломатозные и полипозные разрас-
тания в виде малины или цветной капусты, по-
крытые вязкими слизистыми массами.
Характерна обильная васкуляризация образова-
ний, их легкая травматизация и кровоточивость.
Полипы могут достигать больших размеров. Для
кожных поражений характерны веррукозные ги-
перкератотические разрастания.
Гистопатология
Капиллярная гранулема, явления некроза, выра-
женный инфильтрат из лейкоцитов, эозинофи-
лов, гистиоцитов и ретикулярных клеток.
Диагностика
Выявление в патологическом материале возбу-
дителя.
Дифференциальный диагноз
Полипозные разрастания, ринофима, широкие и
остроконечные кондиломы.
Лечение
Хирургическое удаление или электрокоагуляция
полипозных разрастаний. Внутривенные влива-
ния 5 % раствора солюсурьмина.
Профилактика - не предложена.
Мицетома
Мицетома (эумицетома) (мадурская стопа, маду-
ромикоз, фикомицетома) — хроническое, очаго-
вое, медленно прогрессирующее неконтагиозное
грибковое заболевание, при котором поражает-
ся кожа, подкожная клетчатка, фасции, мышцы,
кости конечностей (стопы, кисти), реже — дру-
гие участки тела. Характерны явления слоново-
сти, деформация конечности с опухолевидными
разрастаниями, образованием свищевых ходов и
гнойным отделяемым.
Возбудитель: настоящие грибы, чаще
Pseudallescheria boydii, Madurella grisea, Madurella
mycetomatis. Почвенные сапрофиты.
Предрасполагающие факторы. Болеют преиму-
щественно жители села (сельскохозяйственные
рабочие) в странах с тропическим и субтропи-
ческим климатом. Развитию заболевания способ-
ствуют несоблюдение личной гигиены, хожде-
ние босиком, травматизация кожи, гипотрофия.
Пути передачи: очаги поражения возникают на
местах травм с последующим инфицированием
почвенными геофилами.
Течение: недели, месяцы, годы.
Излюбленная локализация: наблюдается односто-
роннее поражение голени, стопы, кисти, изредка —
туловища, предплечья, головы, бедра, ягодиц.
Клиническая картина
В месте внедрения возбудителя появляется па-
пула или узел, небольшая болезненность при
пальпации, лихорадка. Конечность становится
отечной и увеличивается в размерах. Узел изъяв-
ляется, из него выделяется гной. Вокруг свище-
вых ходов образуются опухолевидные разраста-
ния. Процесс распространяется на подлежащие
ткани — фасции, мышцы, кости. Возможно уве-
личение регионарных лимфоузлов. Застарелая
мицетома состоит из рубцов и гнойных свищей.
Диагностика
В мазке гноя обнаруживают предполагаемого
возбудителя микоза. Посев позволяет идентифи-
цировать возбудителя.
210 Частная дерматология
Гистопатология
Псевдоэпителиоматозная гиперплазия эпидер-
миса, признаки острого или хронического гной-
ного воспаления с грануляционной тканью и со-
единительнотканной капсулой.
На рентгенограммах костей видны множе-
ственные остеолитические очаги, признаки пе-
риостальной оссификации.
Дифференциальный диагноз
Хромомикоз, североамериканский бластомикоз,
актиномицетома, глубокие пиодермии, инород-
ные тела, дерматофитии, лейшманиоз.
Лечение
Расширенное иссечение небольших очагов. По
возможности избегают ампутации конечностей
или экзартикуляции. Кетоконазол или итрако-
назол per os.
Профилактика
Ношение обуви, защищающей стопу от колючек
и заноз в эндемических зонах. Повышение са-
нитарной культуры населения.
Мукороз
Мукороз — глубокий хронический плесневый
микоз, при котором поражаются кожа, подкож-
ная клетчатка, уши, глаза и внутренние органы
(бронхолегочная система, печень, почки, желу-
дочно-кишечный тракт, центральная нервная си-
стема).
Возбудитель: грибы из родов Мисог, семейства
Мисогасеае. Широко распространены в приро-
де, на продуктах питания, в воздухе.
Предрасполагающие факторы: тяжелые формы
диабета; профессиональные особенности (сбор-
щики апельсинов).
Пути передачи: в основном аэрогенный.
Течение хроническое.
Излюбленная локализация: различные участ-
ки кожного покрова с вовлечением подкожной
клетчатки, ногтевых пластинок и околоногтевых
валиков кистей.
Клиническая картина
Кожные проявления имеют вид хронических
язвенных пиодермий, наружных отитов.
Микотические поражения легких возникают
при пневмониях на фоне лечения антибиотика-
ми. Встречаются мукорозные поражения ЛОР-
органов, желудочно-кишечного тракта, а также
тяжелые менингоэнцефалиты вследствие распро-
странения процесса с придаточных пазух носа.
Указанные проявления характерны для лиц с
тяжелыми формами диабета.
Гистопатология
Гнойно-некротическое воспаление, пролифера-
ция сосудов, приводящая к тромбозу; в дерме
лейкоцитарная и лимфоцитарная инфильтрация,
повышенный процент эозинофилов.
Диагностика
Обнаружение в патологическом материале воз-
будителя и выделение культуры гриба.
Дифференциальный диагноз
Другие глубокие микозы, новообразования, ме-
нингоэннцефалит, пневмония и др.
Лечение
При поверхностных поражениях кожи: местные
антимикотики, фунгицидные жидкости.
При висцеральном мукорозе: амфотерицин В,
итраконазол, средства общеукрепляющей терапии.
Мукороз легких: хирургическая тактика.
Профилактика
Заключается в рациональном применении анти-
биотиков у больных диабетом. Профессиональ-
ный отбор, своевременное лечение онихий и
паронихий у работающих на сборке апельсинов.
Пенициллиоз
Пенициллиоз — поражение ногтей, слизистых
оболочек, внутренних органов, вызванное плес-
невыми грибами рода Penicillium. Заболевание
неконтагиозно.
Возбудитель: сапрофиты кожи и слизистых
оболочек человека. Наиболее патогенными яв-
ляются Penicillium crustaceum Fries, Penicillium
glaucum Link.
Предрасполагающие факторы: снижение за-
щитных сил организма; работа в овощехранили-
щах, на производстве сыров, антибиотиков.
Пути передачи. Заражение — аэрогенное или
при травме кожи и слизистых оболочек.
Течение хроническое на фоне тяжелого общего
заболевания и его нерациональной антибиоти-
котерапии.
Излюбленная локализация: кожа и слизистые
оболочки, редко — бронхолегочный аппарат,
головной мозг.
Клиническая картина
Дерматиты, экземы, хронические язвенные про-
цессы, наружные отиты. Возможны поражения
Глава 13. Грибковые болезни кожи
211
слизистых оболочек, бронхолегочного аппарата,
внутренних органов, развитие абсцесса мозга,
мицетомы.
Гистопатология
Некротическое гранулематозное воспаление, аб-
сцессы, каверны, окруженные зоной некробио-
за с нитями гриба; наличие эпителиоидных,
иногда гигантских клеток.
Диагностика
Обнаружение в гное, мокроте, пунктатах, чешуй-
ках и соскобах кожи, ногтей, слизистых оболо-
чек, наружных слуховых проходов септирован-
ного мицелия с характерным спороношением.
Дифференциальный лиагноз
Другие глубокие микозы, хронические глубокие
пиодермии.
Лечение: см. аспергиллез, мукороз.
Профилактика
Регулярная дезинфекция, эффективная приточ-
но-вытяжная вентиляция овощехранилищ, поме-
щений, связанных с производством сыров (рок-
фор), антибиотиков (группа пенициллинов).
Криптококкоз
Криптококкоз — системный микоз, который
начинается с поражения легких и последующей
гематогенной диссеминации инфекции в мозг и
мозговые оболочки, лимфатическую и костную
систему, у части больных — в кожу и слизистые
оболочки. У ВИЧ-инфицированных криптокок-
коз (криптококковый менингит) — наиболее
частый из глубоких микозов.
Возбудитель: дрожжевой гриб Cryptococcus
neoformans. Является сапрофитом и находится в
почве и помете птиц. Болеют человек и домаш-
ние животные. Для выявления возбудителя ис-
следуют ликвор, гной, мокроту, аутопсийный
материал. Растет на средах, которые применяют
для культуральной диагностики кандидозов и
дерматофитий.
Предрасполагающие факторы: ВИЧ-инфек-
ция, сахарный диабет, лимфосаркома, лимфог-
ранулематоз, саркоидоз, трансплантация органов,
иммунодепрессивная терапия.
Пути передачи: воздушно-пылевой путь; ме-
ханические переносчики инфекции — голуби,
попугайчики, канарейки и др.
Течение. Без лечения процесс принимает хро-
ническое течение и может длиться годами. Поло-
вина больных погибает от менингоэнцефалитов.
Излюбленная локализация. У ВИЧ-инфициро-
ванных поражаются преимущественно лицо и
волосистая часть головы. У небольшой части
больных наблюдаются узловато-язвенные пора-
жения слизистой рта.
Клиническая картина
Элементы кожной сыпи: папулы, узлы, язвы,
корки, под корками папилломатозные разраста-
ния и мелкие пустулы; бляшки с серозно-гной-
ным отделяемым, в центре которых формируется
мягкий рубец. Возможны угревидные элементы,
герпетиформные и везикулезные высыпания,
флегмоны. Кроме мозговых оболочек, легких и
кожи, могут поражаться костный мозг, слизис-
тые мочевых путей и половых органов, предста-
тельная железа. Выявляется гепатоспленомегалия.
Поражения легких клинически и рентгенологи-
чески напоминают туберкулез, абсцедирующую
пневмонию с перфорацией грудной клетки и об-
разованием свищей.
Лабораторные методы исследования
При микроскопии мазков-отпечатков с био-
псийного материала или соскоба с очага пораже-
ния, обработанных калия гидроксидом, обнару-
живается Cryptococcus neoformans.
Патоморфология кожи
Обнаруживают скопления возбудителей, окру-
женные желатиноподобной капсулой или при-
знаки гранулематозного воспаления с неболь-
шим числом возбудителя. В легких казеозный и
гнойный распад очагов, некроз бронхиальных
лимфоузлов.
Для посева обычно используют СМЖ или
биоптат кожи. В культуре возбудитель имеет
дрожжевую или мицелиальную формы с круг-
лыми, овальными или грушевидными кони-
диями. Исследуют также костный мозг, мок-
роту, мочу, секрет предстательной железы,
кровь.
В СМЖ выявляется лимфоцитоз, повышен-
ный белок, сниженный уровень глюкозы. При
микроскопии у значительной части больных
обнаруживают инкапсулированные дрожжевые
клетки.
Проводят определение антигенов Cryptococcus
neoformans в СМЖ, сыворотке крови, моче для
оценки прогноза и эффективности лечения.
Диагностика
Клиническая картина и комплекс лабораторных
исследований, подтверждающих этиологию за-
болевания.
212 Частная дерматология
Дифференциальный диагноз
Пиодермии, контагиозный моллюск (высыпания
на лице у ВИЧ-инфицированных), другие глу-
бокие микозы, флегмоны бактериальной этио-
логии. Туберкулезный менингит, опухоли и аб-
сцессы мозга, энцефалиты. Туберкулез легких,
саркоидоз, бронхоэктазии и др.
Лечение
При менингите: амфотерицин В (2—6 нед) в за-
висимости от тяжести состояния или флукона-
зол; при необходимости — люмбальные пунк-
ции.
При кожных формах: флуконазол 200—400 мг/
сут внутрь или итраконазол 400 мг/сут внутрь.
Профилактика
Раннее выявление и лечение больных в началь-
ной стадии болезни. Текущая и заключительная
дезинфекция. ВИЧ-инфицированным назначают
постоянный прием флуконазола 200—400 мг/сут
внутрь, или итраконазола 200—400 мг/сут внутрь.
Североамериканский
бластомикоз
Североамериканский бластомикоз — хроничес-
кий глубокий микоз, протекающий с образова-
нием в коже, подкожной клетчатке, в легких и
других внутренних органах гнойных, гранулема-
тозных очагов.
Возбудитель: Blastomyces dermatitidis Gilchrist.
Гриб диморфный, в тканях имеет вид дрожже-
вых клеток; размножается почкованием. Природ-
ный сапрофит.
Предрасполагающие факторы: нарушение кле-
точного иммунитета, ВИЧ-инфекция (диссеми-
нированная форма североамериканского бласто-
микоза).
Пути передачи: воздушно-пылевой; контакт-
ный — при повреждении кожи и внедрении в
этот участок возбудителя. Чаще болеют люди,
работающие в сельском хозяйстве, а также со-
баки и лошади.
Течение. Первичная легочная инфекция про-
текает бессимптомно. Высыпания на коже чаще
появляются одновременно с легочной инфекци-
ей; могут поражаться кости, предстательная же-
леза, мозговые оболочки, надпочечники, печень.
Излюбленная локализация: симметричное по-
ражение туловища, реже — лица, кистей, пред-
плечий, часто выявляются множественные очаги.
При контактном заражении очаг локализуется в
месте внедрения возбудителя.
Клиническая картина
Первичная легочная инфекция обычно протека-
ет бессимптомно, в некоторых случаях может со-
провождаться гриппоподобным синдромом или
напоминает пневмонию. Кожные проявления:
узловатая эритема, многоформная экссудативная
эритема.
Диссеминированная инфекция в начальных ста-
диях проявляется в виде папулезно-пустулезных
элементов, далее длительно протекает в виде хро-
нического папилломатозно-язвенного поражения
с последующим образованием рубца, имеющего
вид географической карты. У четверти больных
поражается слизистая рта и носа, иногда горта-
ни.
Поражение легких напоминает туберкулез,
абсцедирующую пневмонию, вплоть до перфо-
рации грудной клетки.
Поражения костно-суставной системы проте-
кают в виде остеомиелитов с образованием сви-
щей, подкожных абсцессов, гнойных артритов.
Мочеполовая система: простатит, эпидидимит.
Лабораторные методы исследования
В мазках гноя и мокроты обнаруживают почку-
ющиеся клетки с толстой двухконтурной стен-
кой, соединенные широкой перемычкой. Для
посева используют мокроту, гной, секрет пред-
стательной железы, биопсийный материал.
Патоморфология кожи
Псевдоэпителиоматозная гиперплазия эпидер-
миса с признаками специфической гранулемы.
В микроабсцессах дермы выявляют почкующи-
еся клетки возбудителя. Определяют антигены
к Blastomyces dermatitidis.
Диагностика
Клиническая картина, подтвержденная резуль-
татами микроскопии мокроты, гноя, мочи и по-
сева биопсийного метериала. Рентгенологическое
исследование легких, костей.
Дифференциальный диагноз
Плоскоклеточный рак кожи, гангренозная пио-
дермия, грибовидный микоз (опухолевая ста-
дия), эктима, бородавчатый туберкулез кожи,
актиномикоз, нокардиоз, мицетома, третичный
сифилис (гумма), донованоз, лепра.
Лечение
При осложнениях менингитом или респиратор-
ным дистресс-синдромом у взрослых — амфоте-
рицин В в/в, итраконазол 200—400 мг/сут (прием
не менее 2 мес), кетоконазол 800 мг/сут.
Глава 13. Грибковые болезни кожи 213
Профилактика
Маловероятна ввиду широкого распространения
возбудителя в природных зонах.
Южноамериканский
бластомикоз
Бластомикоз южноамериканский (бразильский
бластомикоз, паракокцидиоидоз) — глубокий
системный микоз с хроническим прогредиент-
ным течением, вовлечением в процесс слизистых
оболочек щек, носа, кожи, лимфатических уз-
лов, различных внутренних органов (легкие,
печень, желудок, селезенка).
Возбудитель: Paracoccidioides brasiliensis, сап-
рофит растений и почвы.
Предрасполагающие факторы: угрожаемый
контингент: жители сельской местности.
Пути передачи: заражение происходит при раз-
личных повреждениях слизистой оболочки рта
во время чистки зубов веточкой растений. От
человека к человеку не передается.
Течение: без лечения больной погибает в те-
чение 3 лет.
Излюбленная локализация: обычно язвенным
процессом поражаются слизистые оболочки рта
и носа (реже конъюнктивы и прямой кишки),
лимфатические узлы.
Клиническая картина
Язвенный процесс может привести к разрушению
языка, голосовых связок. При переходе на кожу
образуются папулы, пустулы, серпигинирующие
папилломатозные язвы. В процесс вовлекаются
лимфатические узлы, селезенка и легкие. Лим-
фатические узлы могут нагнаиваться, образуя
свищи с грибами в отделяемом. При гематоген-
ной диссеминации поражаются кости, легкие.
Лабораторные методы исследования
обнаружение в патологическом, материале возбу-
дителя и внутрибрюшинное его введение мышам
с последующей микроскопической и гистологи-
ческой идентификацией Paracoccidioides
brasiliensis.
Диагностика
Основывается на клинической картине и обна-
ружении в патологическом материале возбудителя.
Дифференциальный диагноз
Лимфогранулематоз, кокцидиоидоз, североаме-
риканский бластомикоз, актиномикоз, другие
глубокие микозы.
Лечение
Амфотерицин В в/в, итраконазол или кетокона-
зол per os.
Профилактика
Санитарно-просветительная работа среди населе-
ния.
Келоидный бластомикоз
Бластомикоз келоидный — хронический глубо-
кий очаговый микоз кожи, для которого харак-
терно образование в местах поражения келоид-
ных рубцов.
Возбудитель: гриб Glenosporella lobio, близок
к Blastomyces dermatitidis. Во внешней среде не
обнаружен.
Предрасполагающие факторы: встречается у ра-
бочих, добывающих каучук в долине реки Ама-
зонки.
Пути передачи: при травмах, укусах змей, на-
секомых.
Течение длительное (годы), доброкачественное,
без нарушения общего состояния.
Излюбленная локализация: ограниченные вы-
сыпания могут быть на коже лица, ушных ра-
ковин, предплечий, голеней.
Клиническая картина
В участках поражения выявляются бугорки крас-
но-фиолетового цвета, покрытые корочками,
бородавчатые, болезненные и келоидноподобные
образования светло-коричневого цвета, склонные
к слиянию. Лимфатические узлы в процессе не
вовлекаются.
Лабораторные методы исследования
Обнаружение в патологическом материале округ-
лых (овальных) почкующихся дрожжевых кле-
ток с двухконтурной оболочкой. Рост белых бар-
хатистых колоний с септированным мицелием,
эллипсовидными артроспорами (при 25 °C).
Патоморфология
Инфекционная гранулема с элементами гриба;
по строению схожа с акне келоид.
Диагностика
Базируется на характерной клинической карти-
не и обнаружении грибов Glenosporella lobio.
Дифференциальный диагноз
Акне келоид, бородавчатый туберкулез кожи,
бугорковый сифилид, бородавчатый красный
плоский лишай.
214 Частная дерматология
Лечение
Иссечение келоидных рубцов и бородавчатых
разрастаний.
Профилактика
Повышение санитарно-медицинских знаний ра-
ботающих.
Хромомикоз
Хромомикоз (болезнь Пьедрозо, черный бласто-
микоз) — хроническая инфекция кожи и под-
кожной клетчатки, проявляющаяся бородавчаты-
ми разрастаниями на голенях и стопах, нередко
сочетающаяся с поражением внутренних органов
и костей.
Возбудитель: грибы семейства Dematiaceae:
Fonsecaea pedrosoi, Fonsecaea compacta, Phialophora
verrucosa, Cladosporium carrionii, Rhinocladiella
aquaspersa, Botryumyces caespitosus. Продуцируют
меланин; их гифы и конидии имеют темную ок-
раску. Почвенный и растительный сапрофит.
Предрасполагающие факторы: регионы с теп-
лым, влажным климатом в течение года. Наи-
более подвержены возникновению заболевания
сельскохозяйственные рабочие; шахтеры; жи-
тели тропиков и субтропиков, не носящие
обувь.
Пути передачи: возбудитель проникает в орга-
низм через поврежденную кожу и распространя-
ется по лимфатическим сосудам с элементами
аутоинокуляции. Заболевание не передается от
человека к человеку.
Течение: медленный рост очагов отмечается в
течение ряда лет.
Излюбленная локализация: одностороннее по-
ражение голени, стопы, иногда кисти и области
грудной клетки.
Клиническая картина
Различают бугорковую, папилломатозно-язвен-
ную и узловатую формы хромомикоза. В месте
внедрения возбудителя появляется небольшой
шелушащийся узел (бугорок), вокруг которого
(через месяцы или годы) возникают новые по-
добные элементы. Очаг поражения имеет вид ра-
стущих бляшек с бородавчатой поверхностью и
участками здоровой кожи. Бляшки разрешают-
ся с центра и имеют кольцевидную форму. На
их поверхности могут быть опухолевидные раз-
растания в виде цветной капусты, сидящие на
ножке, пустулы, язвочки, «черные точки», кро-
воточащие грануляции. Очаги могут охватывать
всю голень или стопу с признаками слоновости
пораженной конечности.
Гистопатология
Хроническая инфекционная гранулема с субкор-
неальными и интрадермальными микроабсцес-
сами, характерными сферическими тельцами
возбудителя, окруженными лейкоцитами, эпи-
телиоидными и гигантскими клетками.
Лабораторные методы исследования
В мазках гноя (экссудате, корках) видны сфе-
рические тельца (темные крупинки), гифы гри-
бов. На среде Сабуро в течение 4—6 недель вы-
растают полусферические бархатистые колонии,
имеющие цвет от зеленого до черного.
Диагностика
Клиническая картина, подтвержденная резуль-
татами посева, выявление сферических телец в
мазке гноя или в гистологическом препарате.
Дифференциальный диагноз
Бородавчатый туберкулез кожи, другие глубокие
микозы, третичный активный сифилис, фрам-
безия, неинфекционные гранулемы, лепроматоз-
ная лепра, дерматофитии, лейшманиоз, глубокая
пиодермия, плоскоклеточный рак кожи.
Лечение
Мелкие очаги иссекают. Длительный прием ит-
раконазола 200—600 мг/сут (не менее одного
года); кетоконазол 400—800 мг/сут. Термотера-
пия. Противогрибковые мази.
Профилактика
Заключается в предупреждении травматизации
кожного покрова при земляных и сельскохозяй-
ственных работах.
Глубокие
псевдомикозы
Актиномикоз
Актиномикоз — хроническое гранулематозное
гнойное поражение различных тканей и органов,
вызванное бактериями Actinomycetales, которое
сопровождается образованием плотных слегка
болезненных инфильтратов, абсцессов и дли-
тельно не заживающих свищевых ходов.
Возбудитель: Actinomyces Israeli, Actinomyces bovis,
Actinomyces albus Rossi — Doria, Mlcromonospora
parva. Бактерии порядка Actinomycetales (раньше
относили к царству грибов). Сапрофиты кожи,
Глава 13. Грибковые болезни кожи
215
слизистых о.болочек рта, дыхательных путей и
желудочно-кишечного тракта.
Предрасполагающие факторы: снижение за-
щитных сил организма, хронические инфекции,
иммунодефицитные состояния.
Пути передачи: инфицирование преимуще-
ственно эндогенное в результате активации ак-
тиномицетов (нередко после травмы).
Течение: преимущественно хроническое (неде-
ли, месяцы, годы).
Излюбленная локализация: шейно-лицевая и
торакальная области, брюшная стенка и органы
брюшной полости, другие участки кожного по-
крова, слизистые оболочки.
Клиническая картина
Различают первичный и вторичный актиноми-
коз, при котором инфекция распространяется из
основного очага при поражениях различных ор-
ганов и тканей.
Первичный актиномикоз кожи встречается ред-
ко и начинается с появления слегка болезненного
плотного инфильтрата синюшно-красного, затем
багрового цвета, который размягчается, абсцеди-
рует и вскрывается с образованием длительно не
заживающих свищей и плотных втянутых рубцов.
На фоне применения антибиотиков плотный ин-
фильтрат и свищи могут не образовываться.
Возможно образование первичного очага ак-
тиномикоза в виде мицетомы. На месте вне-
дрения возбудителя образуется папула или узел,
отмечается опухание конечности и увеличение
ее в размерах. Далее узел изъязвляется, с вы-
делением из него гноя, в котором содержатся
друзы. Вокруг свища образуются опухолевид-
ные разрастания. Процесс распространяется на
подлежащие ткани и сопровождается рубцева-
нием и наличием гнойных свищей, деформа-
цией конечности. Чаще поражаются стопы, го-
лени, кисти; возможна другая локализация
мицетомы.
При шейно-лицевой форме актиномикоза
инфекция вначале поражает кожу и подкожную
клетчатку, а затем распространяется по фасци-
альным пространствам, разрушая мышечную
ткань, кости, придаточные пазухи, черепную
коробку, проникает в грудную полость, может
вызвать перфорацию грудной клетки и симпто-
матику поражения соответствующего органа.
Актиномикоз брюшной полости чаще возни-
кает в илеоцекальной области и в области пря-
мой кишки с вовлечением брюшной стенки.
Встречается также актиномикоз органов малого
таза, урогенитальной системы, параректальной,
крестцово-копчиковой и ягодичной областей.
Вторичный актиномикоз кожи чаще проявля-
ется в виде гуммозно-узловатой формы, при
которой образуется плотный безболезненный
синюшно-багровый инфильтрат с множествен-
ными свищами.
Лабораторные методы исследования
В гнойно-кровянистом отделяемом элементов ad
oculus выявляются белые или желтоватые зерна,
а при микроскопии видны характерные друзы
актиномицета — сплетения тонкого мицелия с
колбообразными утолщениями и желтоватым
ободком по периферии. В некоторых случаях не-
обходимы повторные исследования.
Гистопатология
Специфическая гранулема, зоны обильной гра-
нуляционной ткани с абсцессами и перифери-
ческой соединительнотканной капсулой. Друзы
обычно выявляются в гное абсцессов.
Диагностика
Клиническая картина, обнаружение друз и иден-
тификация культуры актиномицета. Положи-
тельные кожно-аллергические пробы и реакция
связывания комплемента с актинолизатом.
Дифференциальный диагноз
Новообразования, кокцидиоидоз, другие глубо-
кие микозы, абсцессы, хронические пиодермии,
сифилитические гуммы, туберкулез кожи.
Лечение
Стрептомицин в комбинации с дапсоном или
триметопримом / сульфаметоксазолом. Внутри-
мышечное введение актинолизата по 3—4 мл или
внутрикожно с 0,5 до 2,0 мл 2 раза в неделю (на
курс 20—25 инъекций). Гемотрансфузии, вита-
мины, симптоматические средства. В некоторых
случаях хирургическая тактика.
Профилактика
Предупреждение микротравматизма, санация рото-
вой полости, соблюдение правил личной гигиены.
Нокардиоз
Нокардиоз (атипичный актиномикоз) — острое
или хроническое заболевание внутренних орга-
нов (чаще легких), с нередким вовлечением в
процесс кожи, подкожной клетчатки, слизистых
оболочек, лимфатических узлов. Заболевание не-
контагиозно.
Возбудитель: Nocardia asteroides, Nocardia
madurae, Nocardia brasiliensis Castellani. Аэробы,
8* Зак. 1357
216 Частная дерматология
не образующие спор и капсул, без друз и зерен
в тканях (друзоподобные образования без колб
по периферии). Природные сапрофиты.
Предрасполагающие факторы: заболевание
встречается у лиц, страдающих болезнями кро-
ви, получающих стероидные препараты, прожи-
вающих в субтропиках.
Пути передачи: респираторный, через желудоч-
но-кишечный тракт, травмированную кожу и
слизистые оболочки.
Течение: острое или хроническое.
Излюбленная локализация: внутренние орга-
ны (обычно легкие), реже кожа, подкожная клет-
чатка, слизистые оболочки, лимфатические узлы.
Клиническая картина
Различают легочный (по типу плевропневмонии)
и генерализованный (септический), первичный
и вторичный нокардиоз. Возможны мицетомы
(чаще нижних конечностей), поражающие кожу,
подкожную клетчатку и кости.
Диссеминация инфекции сопровождается сеп-
сисом, поражением внутренних органов, появ-
лением фистулезных ходов в грудной и брюш-
ной полостях, на коже. Возможно развитие
абсцессов головного мозга или менингитов.
Гистопатология
В зависимости от пораженных органов и тканей
напоминает поражение актиномицетами.
Диагностика
Клиническая картина, подтвержденная резуль-
татами посева, обнаружением возбудителя в па-
тологическом материале и экспериментом на
животных.
Дифференциальный лиагноз
Гистоплазмоз, актиномикоз, кокцидиоидоз, ту-
беркулез легких.
Лечение
Стрептомицин, полусинтетические пенициллины
в комбинации с триметопримом/сульфаметокса-
золом или дапсоном, общеукрепляющая терапия.
Профилактика
Предупреждение травм кожи и слизистых обо-
лочек, дезинфекция зараженного материала.
Глава 14
Дерматиты
Простой ирритантный контактный дерматит 218
Аллергический контактный дерматит 219
Острые дерматиты и хронические экземы объе-
диняют иногда под общим названием эпидер-
мальные реакции непереносимости. Эти заболе-
вания встречаются очень часто, ими болеют
15~20 % всех пациентов с кожными заболева-
ниями. Такие заболевания можно отнести к не-
инфекционным, а поэтому незаразным полиэти-
ологическим воспалительным дерматозам, при
которых клиническую картину определяют па-
тологические изменения в эпидермисе и верх-
нем слое дермы.
В случае острого течения этих часто зудящих
кожных проявлений на первый план в клиничес-
кой картине выступают экссудативно-воспали-
тельные процессы с покраснением, отеком, пузы-
рями, мокнутием и корочками, при хроническом
течении — пролиферативно-воспалительные про-
цессы с покраснением, утолщением эпидермиса
(акантозом), шелушением или лихенификацией.
В зависимости от индивидуальной реактивно-
сти организма дерматиты могут вызываться как
известными, так и неизвестными, эндогенно
либо экзогенно действующими провоцирующи-
ми факторами, имеющими токсическую либо
аллергическую природу.
В номенклатуре эпидермальных реакций не-
переносимости до сих пор нет единообразия, по-
скольку отсутствует всесторонне удовлетворяю-
щая их классификация как в этиологическом
плане, так и с точки зрения патологии. Клини-
ческие диагнозы «дерматит» и «экзема» часто
используются в качестве синонимов. С учетом
течения заболевания говорят об остром, под-
остром и хроническом дерматите, но и об ост-
рой, подострой и хронической экземе, имея в
виду одно и то же заболевание. В англо-амери-
канской медицинской литературе для обозначе-
ния такого рода заболеваний чаще употребляет-
ся обозначение «дерматит», В европейской
дерматологии придерживаются распространенно-
го обозначения «экзема». Последним определя-
ют все эпидермальные реакции непереносимос-
ти, которые имеют синхронный полиморфизм
кожных высыпаний и характеризуются выражен-
ным хроническим течением.
В европейской дерматологии в течение уже
многих десятилетий принято называть эпидер-
мальные реакции непереносимости, протекаю-
щие остро и с быстрой способностью к регрес-
су, дерматитом, а протекающие хронически со
слабой тенденцией к спонтанной ремиссии —
экземой.
Однако следует учесть, что в течение заболе-
вания могут происходить самые разные процес-
сы. Например, аллергический контактный дер-
матит при повторном контакте с причинно
значимыми контактными аллергенами может
стать хроническим, т. е. перейти в экзему, а при
хронической аллергической контактной экземе
в случае возобновления воздействия причинных
контактных аллергенов может наступить острое
ухудшение. Различать эти процессы особенно
важно для практического врача. Установление
причины помогает назначению соответствующей
терапии.
С учетом вышеперечисленных определений
принята следующая классификация:
• острый контактный дерматит: простой или
аллергический;
• хронический контактный дерматит: кумуля-
тивно-токсический или аллергический;
• себорейная экзема;
• микробная (нумулярная) экзема;
• атопическая экзема (атопический дерматит).
При однократном экзогенном воздействии
облигатно токсичных, т. е. первично сильно по-
вреждающих кожу веществ, на коже с нормаль-
ной чувствительностью может развиться острый
токсический контактный дерматит. При много-
кратном экзогенном воздействии слабо раздра-
жающих кожу веществ и соответствующей пред-
расположенности кожи может возникнуть
кумулятивно-токсическая экзема. В обоих слу-
чаях речь идет о воспалительных изменениях
217
218 Частная дерматология
кожи с клинически выраженным вовлечением
эпидермиса, причем первичные аллергические
процессы не являются патогенетически ответ-
ственными.
Простой ирритантный
контактный дерматит
Простой ирритантный (раздражительный) кон-
тактный дерматит — это контактный дерматит,
который часто вызывается постоянными контак-
тами с водой, мылом, высокой температурой и
трением. К сильным раздражителям (ирритан-
там) относятся кислоты, щелочи и влажный це-
мент; эти химикаты могут вызывать острые оча-
ги химических ожогов. Хроническое воздействие
слабых раздражителей является наиболее распро-
страненной проблемой, вызывающей экзематоз-
ные изменения. Интенсивность воспаления
обычно связана с концентрацией ирританта и
продолжительностью контакта. Мягкие раздра-
жители jy>i3biBaK)T~сухость, трещиньГгГэ^тему;
сильные раздражающие химикаты могут вызы-
вать немедленную реакцию, которая характери-
зуется жжением, эритемой, отеком и, возмож-
но, изъязвлением кожи. Около 80 % случаев
контактного дерматита представляют собой про-
стой ирритантный контактный дерматит. В от-
личие от аллергического контактного дермати-
та, ирритантный контактный дерматит не
специфичен и не требует сенсибилизации.
Патогенез
Атопический фон (сенная лихорадка, астма или
экзема) предрасполагает к повышенной воспри-
имчивости кожи к раздражению. Восприимчи-
вость к ирританту (раздражителю) индивиду-
альна и может варьировать. Ирритантный
контактный дерматит — наиболее распростра-
ненный тип профессионального кожного забо-
левания, имеющий место в 80 % случаев. Про-
фессиональные и домашние обязанности,
включая уход за ребенком и хобби, являются
критическими составляющими анамнеза. Рабо-
та, при которой руки постоянно контактируют
с влажной средой, например в системе обще-
ственного питания, при уходе за больными и
за маленькими детьми, а также в парикмахер-
ских, предрасполагает к развитию у пациента
простого ирритантного контактного дерматита.
Наиболее распространенные раздражители
включают детергенты, кислоты, щелочные хи-
микаты, масла, органические растворители, ок-
сиданты, химические восстановители и воду.
Грубые волокна, такие как стекловолокно или
дровяные опилки, могут вызвать ирритантный
контактный дерматит. Воздействие раздражите-
ля происходит либо при прямом контакте, либо
по воздушным путям. Примерами ирритантов,
которые могут вызвать контактный дерматит
воздушным путем, являются стекловолокно,
формальдегид, эпоксидные смолы, промышлен-
ные растворители, глютаральдегид и опилки.
Постоянное трение и механическое раздражение
могут вызвать хронический ирритантный кон-
тактный дерматит. Ирритантный экзематозный
дерматит может иметь место и при длительном
контакте со слабыми раздражителями. Когда
порог раздражения достигнут, стойкий дерматит
может развиться и от незначительного контакта
со слабыми раздражителями. Низкая влажность
окружающей среды снижает порог раздражения.
Непрерывное воздействие влаги и цикла «намо-
кание-высыхание» на таких участках тела, как
ладони, ягодичная и пахово-бедренная область
у младенцев, кожа вокруг раны, может иногда
вызывать экзематозное воспаление. Ирритант-
ный дерматит может осложниться аллергией по
мере возрастающего применения наружных ле-
карственных средств, а также в результате более
легкого проникновения аллергенов в поврежден-
ную кожу. Атопики предрасположены к ирри-
тантному контактному дерматиту; дерматит про-
текает у них длительно и лечить его труднее.
Клиническая картина
Кисти поражаются чаще всего. Поражаться мо-
гут как тыльная, так и ладонная поверхности.
Веки являются еще одним участком, подвержен-
ным воздействию раздражителей. Хроническая
привычка облизывать губы может привести к
ирритантному дерматиту вследствие повторяю-
щегося цикла «увлажнение-высыхание». Эрите-
ма, сухость, болезненные трещины и шелушение
типичны. Могут присутствовать и везикулы, но
они не типичны. Симптомы болезненности и
жжения весьма распространены. Жжение часто
преобладает над зудом. Острый ирритантный
дерматит может проявляться мокнущими папу-
лами и/или везикулами на эритематозном пят-
нистом фоне с мокнутием и отеком. Стойкий
хронический ирритантный дерматит характери-
зуется лихенификацией, эритематозными пятна-
ми, трещинами, экскориациями и шелушением.
Гиперкератотическая форма характеризуется
постоянным шелушением, трещинами и слабой
эритемой и может возникать вследствие механи-
Глава 14. Дерматиты 219
ческой травмы, например перекладывания бумаг.
Открытая кожа может «гореть» при контакте с
наружными препаратами, которые в остальных
случаях переносятся нормально.
Лабораторная диагностика
Проводится исследование в растворе КОН для
исключения грибковой инфекции. Проводится
лоскутный тест для оценки роли аллергическо-
го контактного дерматита, если его наличие мож-
но предположить из анамнеза (например, кон-
такт с аллергенами) или если заболевание
торпедное и персистирует, несмотря на лечение
и меры профилактики. Лоскутный тест обычно
проводят, применяя готовые скрининговые се-
рии аллергенов, состоящие из 24 проб. Биопсия
проводится редко и показывает спонгиоз, отек
дермы и воспалительный, преимущественно
лимфоцитарный, инфильтрат.
Дифференциальный диагноз
Аллергический контактный дерматит (везикулы
и зуд часто более типичны для аллергического
контактного дерматита, чем для простого ирри-
тантного контактного дерматита). Атопический
дерматит. Грибковая инфекция (табл. 14.1).
Лечение
Ранняя постановка диагноза, лечение и меры
профилактики могут предотвратить развитие
хронического ирритантного дерматита. Исклю-
чение или уменьшение контактов с раздражите-
лями кожи является решающим для восстанов-
ления эффективности барьерной функции
кожи. Следует уменьшить количество циклов
«намокание-высыхание», имеющих место вслед-
ствие, например, постоянного мытья рук. Хлоп-
чатобумажные перчатки, надетые под виниловые
перчатки, позволяют уменьшить намокание рук
при работе во влажной среде. При работе со спе-
цифическими растворителями или химикатами
следует надевать соответствующие перчатки.
Существенную пользу приносит регулярное ча-
стое применение на пораженную кожу смягча-
ющих эмолентов, таких как вазелин. При про-
стом ирритантном дерматите кистей в течение
нескольких недель дважды в день применяют
топический стероид средней или высокой фар-
макологической эффективности для уменьшения
эритемы, зуда, отечности и болезненности.
Аллергический
контактный дерматит
Аллергический контактный дерматит представ-
ляет собой реакцию замедленного типа, которая
вызывается контактом кожи с аллергеном. Ал-
лергический контактный дерматит является эк-
зематозным дерматитом. Для проявления забо-
левания необходима сенсибилизация, аллергия
является специфичной к определенному хими-
ческому веществу; аллергены обычно имеют
низкий молекулярный вес, являются липидора-
Таблица 14.1. Дифференциальная диагностика простого ирритантного контактного дерматита от аллергического дерматита и истинной экземы
Признаки Простой ирритантный контактный дерматит Аллергический контактный дерматит Экзема истинная
Сенсибилизация Отсутствует Моновалентная Поливалентная
Причина и время развития высыпаний После воздействия раздражителя Развивается через латентный период после контакта с аллергеном Психоэмоциональные стрессы
Локализация высыпаний Непосредственно на месте воздействия раздражителя На месте контакта с аллергеном и за пределами зоны раздражителя Симметричность высыпаний на коже конечностей и лица
Клиническая картина Последовательная смена стадий (эритематозная, буллезная, крустозная, сквамозная) Одновременное появление полиморфных высыпаний Эволюционный полиморфизм (преобладают везикулы, мокнутие)
Прогноз заболевания Быстрый регресс высыпаний после прекращения действия раздражителя При отсутствии контакта с аллергеном высыпания полностью регрессируют После устранения воздействия аллергена регресса высыпаний не наблюдается
220 Частная дерматология
створимыми и обладают высокой реакционной
способностью.
Типичными аллергенами являются металлы,
такие как никель и хром; резиновые добавки в
материал перчаток и обуви (карбаматы, тиура-
мы, меркапто-бензатиазол); консерванты или
другие добавки к продуктам, таким как лосьо-
ны для кожи, солнцезащитные средства и дру-
гие предметы косметики и гигиены; ароматиза-
торы и ароматические добавки; красители;
формальдегид и родственные ему вещества; до-
бавки в лак для ногтей, а также наружные ле-
карственные препараты (например, бактрацин,
неомицин, гидрокортизон).
Патогенез
Первый контакт и первичная сенсибилизация
вызывают клиническое воспаление — как пра-
вило, через 14—21 день после экспозиции. Кон-
такт с химическим веществом необходим для
развития аллергии. Время, необходимое для раз-
вития клинически очевидного воспаления, у
ранее сенсибилизированного человека составляет
12—48 часов, но может колебаться в пределах 8—
120 часов, поскольку высыпания могут несколь-
ко запаздывать, считая от времени контакта.
После однократного контакта высыпания могут
длиться до 3 недель. Аллергия на продукты (кос-
метику, наружные лекарства, аллергены, связан-
ные с профессий пациента) или же на другие
вещества может возникнуть, даже если пациент
ранее длительно и без каких-либо затруднений
контактировал с ними. Высыпания рецидивиру-
ют, если контакты повторяются. Поскольку
многие продукты имеют одинаковые или сход-
ные по химическому составу ингредиенты, за-
мена одного наружного средства другим часто не
приводит к разрешению экзематозного высыпа-
ния. У некоторых пациентов развиваются кон-
тактные аллергии сразу к нескольким аллерге-
нам (множественные).
Тщательный анамнез должен включать следу-
ющее: время начала дерматита, его возможную
связь с работой пациента (а именно: улучшается
ли состояние во время выходных или в течение
длительного отпуска), тип и специфику контак-
та с возможными аллергенами на работе, возмож-
ные контакты при занятиях в свободное время
или работе по дому, а также сведения о том, ка-
кими продуктами ухода за кожей пользуется па-
циент. Некоторые вызывающие аллергию веще-
ства являются фотоаллергенами: для проявления
аллергической реакции необходимо их взаимо-
действие с солнечным светом. У лиц, сенсибили-
зированных к топическим медикаментам или
другим аллергенам, может развиться генерализо-
ванное экзематозное воспаление, если лекарствен-
ные или другие химические вещества поступают
в организм путем употребления внутрь (внутрен-
не-наружная реакция). Например, пациент, сен-
сибилизированный к наружному дифенгидрамину
в форме крема, реагирует генерализованным вы-
сыпанием на прием дифенгидрамина внутрь.
Клиническая картина
Интенсивность экземы зависит от индивидуаль-
ной степени чувствительности, концентрации
аллергена и от величины экспозиции антигена.
Характерными признаками аллергического кон-
тактного дерматита являются везикулы, отек,
краснота и часто интенсивный зуд. Сильные
аллергены, такие как ядовитый плющ, вызыва-
ют появление пузырей. Вначале дерматит распро-
страняется только на область непосредственно-
го контакта. Если этот контакт хронический,
аллергический дерматит может распространить-
ся за пределы области первичного контакта. Зуд
и отек являются ключевыми составляющими
анамнеза и могут указывать на аллергию. Для
простого ирритантного дерматита более типич-
ным является жжение. Дерматит, вызываемый
растениями, часто имеет линейную форму рас-
пространения. Аллергия на наружный препарат,
применявшийся для лица, может проявляться
зудящими пятнами эритемы, расположенными
асимметрично. При поражении лица везикулы
присутствуют редко.
Сильные сенсибилизаторы, такие как ядови-
тый плющ, могут вызывать интенсивное воспа-
ление, несмотря на их низкую концентрацию
или короткий контакт; слабые сенсибилизаторы
могут вызывать только зудящую эритему. Ла-
дони, предплечья и лицо — участки наиболее
типичной локализации аллергического контак-
тного дерматита. Аллергический контактный
дерматит может поражать очень ограниченные
участки кожи, такие как веки, тыльную повер-
хность ладоней, губы, ступни или гениталии.
Аэроаллергены (например, при сжигании ядови-
того плюща) могут вызвать дерматит лица
(включая веки и кожу за ушами), шеи и других
открытых участков тела.
Фотоаллергический контактный дерматит ти-
пично поражает открытые участки тела: лицо,
шею, предплечья и тыльную поверхность ладо-
ней. Участки под подбородком, на верхних ве-
ках и за ушами обычно не поражаются. Профес-
сиональный аллергический контактный дерматит
чаще всего поражает ладони; лицо и веки могут
поражаться, если аллерген воздушный или име-
Глава 14. Дерматиты
221
ет место косвенная передача аллергена от рук к
лицу.
Лабораторная диагностика
Лоскутное тестирование проводится имеющим
опыт специалистом. Оно показано лицам со
стойким или рецидивирующим, несмотря на
адекватную терапию, дерматитом. Лоскутное
тестирование проводится с достаточно широкой
панелью аллергенов из сфер профессиональной
деятельности и досуга, а также связанных с про-
дуктами ухода за кожей. Если набор аллергенов
для лоскутного тестирования ограничен, или же
в него не включены аллергены из сферы про-
фессиональной деятельности пациента, некото-
рые важные аллергены могут быть пропущены.
Правильная методика лоскутного тестирова-
ния требует трех посещений дерматолога. Пер-
вое — для нанесения аллергенного химическо-
го вещества в соответствующей концентрации,
второе — для снятия теста, прочтения и анали-
за результатов, и последнее посещение — для
окончательной оценки отсроченных реакций.
Если последний этап исключить, идентифика-
ция аллергенов может не получиться. Лоскутный
фото-тест следует провести пациентам с распре-
делением поражения по типу фотодерматита.
Дифференциальный диагноз
Ирритантный контактный дерматит (лоскутный
тест на ирритантный контактный дерматит не
проводится; клиническая картина практически
неотличима по клиническим признакам, особен-
но на пораженных ладонях). Атопический дер-
матит (при дифференциации может помочь тип
распределения поражений; у пациентов — ато-
пиков могут быть и часто бывают контактные
аллергии). Целлюлит (отек и эритема могут быть
похожими, но целлюлит обычно болезненный и
без зуда). Заболевание соединительной ткани
(стойкая эритема век; ключ — отсутствие зуда
на пораженной коже и возможная гипертрофия
кутикулы при дерматомиозите). Розацеа (распре-
деление эритемы, приливы крови к коже и на-
личие жжения помогают провести дифференци-
альный диагноз).
Лечение
Для выздоровления необходимо избегать контак-
та с аллергеном. Важно правильно идентифици-
ровать аллерген (ы). Если причина неочевидна,
а это бывает редко, рекомендуется начать лече-
ние, чтобы позже провести обследование и зап-
ланировать лоскутный тест. Как врачи, так и
пациенты обычно плохо угадывают специфичес-
кие для пациента аллергии. Наружное лечение
начинают с упрощения до минимума всего, что
пациент применяет на коже и с назначения то-
пического кортикостероида. Отмените все ув-
лажнители, лосьоны и наружные средства, за
исключением простого вазелина, если он необ-
ходим при сухости кожи. Для лечения дермати-
та назначают кортикостероидную мазь два раза
в день в течение 2—3 недель.
Выбор фармакологической эффективности
кортикостероида зависит от того, какой участок
тела поражен: слабые кортикостероиды приме-
няются для лица, кортикостероиды средней силы
действия — для рук, ног и туловища, а корти-
костероиды с высокой фармакологической эф-
фективностью — для ладоней и подошв. Пред-
почтительнее назначать кортикостероидную
мазь, а не крем, поскольку добавки в крем мо-
гут быть аллергенными. Процедуру ухода за
кожей пациенту необходимо упростить, чтобы
избежать возможного контакта с аллергеном. Во
время лечения нельзя применять никакого дру-
гого наружного средства (за исключением про-
стого вазелина или прописанного кортикостеро-
ида). Гидрокортизон лучше не назначать,
поскольку могут иметь место случаи контактной
аллергии. В случае тяжелого или генерализован-
ного контактного дерматита назначают курс си-
стемных кортикостероидов в течение 3-х недель
с постепенным понижением дозировки. Однако
на повторные курсы такого рода полагаться не
стоит. Некоторые аллергены (такие как в хими-
катах для окраски волос и клеях) могут прони-
кать через резиновые перчатки, так что такая
защита может быть неадекватной. Некоторые
аллергены, такие как никель и хром, ассоции-
руются с хроническим дерматитом, несмотря на
попытки избегать с ними контакта. Следует
уделить Время для подробного объяснения паци-
енту потенциальных источников аллергенных
контактов. Когда аллерген определен в резуль-
тате диагностического лоскутного теста, внима-
тельно изучите список аллергенов и потенциаль-
но связанных с ними веществ, чтобы объяснить
пациенту, чего он должен избегать и какие аль-
тернативы существуют для замены потенциаль-
ных аллергенов. Все это является жизненно важ-
ным для успешного результата лечения.
Глава 15
Экзема
Острая экзема 222
Подострая экзема 223
Хроническая экзема 224
Экзема ладоней 224
Сухая экзема 225
Монетовидная экзема 227
Дисгидротическая экзема 227
Себорейная экзема 229
Особенности возникновения и течения экземы у детей 232
Острая экзема
Острая экзема представляет собой острое экзе-
матозное воспаление, клинически характеризу-
ющееся эритемой, отеком и образованием ве-
зикул, мокнутием очагов, иногда сильным
зудом.
Патогенез
Существует множество причин острой экземы. К
ним относятся контактная сверхчувствительность
к специфическим растительным аллергенам, та-
ким как ядовитый плющ, дуб и к другим аллер-
генам. Никель, наружные лекарственные средства,
такие как бацитрацин, неомицин и бензокаино-
вые ароматизаторы, консерванты в продуктах
личной гигиены и органические вещества в до-
бавках также являются распространенными при-
чинами острого экзематозного воспаления. Дер-
матит раздражения обычно возникает после
повторяющихся контактов с водой, растворами
или растворителями. При так называемой «id»
реакции острая экзема с везикулами возникает в
месте, удаленном от активной грибковой инфек-
ции (например, на ладонях и подошвах). Застой-
ный дерматит, чесотка, реакции раздражения, а
также дисгидротическая и атопическая экземы
могут проявляться в виде острого экзематозного
воспаления.
Клиническая картина
Клинические признаки включают эритему,
отек, образование везикул и мокнутие. Воспа-
ление — от умеренно выраженного до сильно-
го. На поверхности кожи появляются малень-
кие, прозрачные, наполненные жидкостью
везикулы. Могут развиться пузыри. Если про-
воцирующих факторов удается избежать, высы-
пания улучшаются за 7—10 дней, к 3-й неделе
кожа становится полностью чистой. Экскори-
ация предрасполагает к инфекции и вызывает
скопление серозной жидкости, корок и гноя.
Результатом экскориации может стать вторич-
ная стафилококковая инфекция, а также ухуд-
шение дерматита и увеличение его продолжи-
тельности.
Лабораторная диагностика
Следует провести лоскутный тест для оценки
гиперчувствительности замедленного типа, если
локализация экземы предполагает контакт, если
заболевание рецидивирует и не поддается лече-
нию, или же известно о контакте с кожными
аллергенами на работе или при иных привычных
занятиях.
Лечение
Прохладные влажные повязки и наружные сте-
роидные кремы способствуют сужению сосу-
дов кожи, подавлению воспаления и зуда. Чи-
222
Глава 15. Экзема 223
стую ткань смачивают прохладной водой или
раствором Бурова и на 30 минут помещают на
пораженный участок. Затем тщательно втира-
ют соответствующий стероидный крем (груп-
пы II или III). Системные кортикостероиды
применяются лишь в случае тяжелой или ге-
нерализованной экземы. Начальная дозиров-
ка примерно 1 мг/кг/день с постепенным сни-
жением в течение 3 недель. Слишком короткий
курс лечения может вызвать рецидив или ре-
зонансный эффект. Системные антигистамин-
ные препараты первого поколения могут об-
легчить зуд, а их седативный эффект
способствовует улучшению сна. При вторич-
ной инфекции назначается антибиотик против
Staphylococcus aureus (например, цефалексин)
в течение 10—14 дней.
Подострая экзема
Подострое экзематозное воспаление представляет
собой зудящие и шелушащиеся красные пятна,
папулы и бляшки различных размеров и очер-
таний.
Этиология
Контактная аллергия, контактное раздражение,
атопический дерматит, застойный дерматит, мо-
нетовидная экзема, экзема кончиков пальцев и
грибковые инфекции могут проявляться в виде
подострой экземы. Если отсутствует явный ато-
пический анамнез, необходимо искать какой-
либо новый раздражитель кожи или же воздей-
ствие аллергена. Стресс может ухудшить
состояние и способствовать его развитию, но не
является единственной причиной.
Патогенез
Данное состояние может развиться из острой
(везикулярной) экземы. Это наиболее распро-
страненное клиническое проявление атопичес-
кого дерматита. Пациенты жалуются на дерма-
тит, который длится более одной недели.
Интенсивность зуда различна: от слабого до
умеренного и тяжелого. Состояние разрешает-
ся без образования рубцов, когда устраняют про-
воцирующие или способствующие ему факто-
ры. Экскориация и многократное воздействие
ухудшающих состояние условий (воды, чистя-
щих или моющих агентов, ирритантов или дру-
гих распространенных аллергических или раз-
дражающих факторов) переводит заболевание в
хроническую форму.
Клиническая картина
Эритема или шелушение различной формы. Гра-
ницы часто нерезко очерчены. Гиперемия может
быть слабой или интенсивной.
Лечение
Стероидные и нестероидные наружные препара-
ты. Назначаются стероидные кремы группы II —
V дважды в день под полиэтиленовой окклюзи-
ей или без нее. Окклюзия ускоряет разрешение
очагов, усиливая абсорбцию топического стеро-
ида. Продолжительность окклюзии определяет-
ся индивидуально, она должна быть ограничен-
ной и проводиться под контролем. Стероидные
мази применяют два раза в день без окклюзии.
Нестероидный топический иммуномодулятор
пимекролимус (крем «Элидел» 1 %) может при-
меняться дважды в день на пораженные участки
кожи и особенно эффективен при подострой
экземе лица или периорбитальной области. Вна-
чале может возникать ощущение жжения, кото-
рое проходит через несколько дней. Этот вид
терапии эффективен при хроническом течении
подострой экземы у атопиков. Дегтярные мази и
кремы являются альтернативой в случае устой-
чивых к стероидам очагов и оказывают умерен-
но эффективное действие на некоторых пациен-
тов. Влажных компрессов следует избегать,
поскольку они сильно высушивают кожу. При-
менение увлажняющих средств является суще-
ственной частью ежедневной терапии. Увлажни-
тели лучше всего работают, если их применяют
через несколько часов после топических стерои-
дов. Аппликацию следует продолжать в течение
нескольких дней или недель после того, как утих-
нет воспаление. Следует часто применять увлаж-
нители. Увлажнители наиболее эффективны, если
их тщательно втирать в кожу сразу после умы-
вания, предварительно осторожно подсушив её
похлопывающими движениями полотенца.
Кремы простого состава (наприМер,
«Aveeno»), не содержащие ингредиентов, кото-
рые наиболее часто ассоциируются с аллерге-
нами, лучше подходят, чем лосьоны. Простое
вазелиновое желе является отличным увлажни-
телем, преимуществами которого являются
простота состава, отсутствие аллергизирующих
добавок или раздражающих ингредиентов.
Однако приемлемость вазелина для пациентов
ограничена из-за его жирности. При нечастом
мытье кожи с мылом полезнее пользоваться
мягкими кусковыми мылами типа «Dove».
Антибиотики применяются при вторичных
бактериальных инфекциях.
224 Частная дерматология
Хроническая экзема
Пораженная хроническим экзематозным воспа-
лением кожа воспалена, гиперемирована, утол-
щена и подвержена шелушению.
Этиология
Возможные причины — атопический дерматит,
хронический аллергический или ирритантный
дерматит, привычка к расчесам, простой хрони-
ческий лишай, потрескавшиеся подошвы, моне-
товидная экзема, сухая (астеатотическая) экзема,
экзема кончиков пальцев, гиперкератотическая
экзема. Заболевание эволюционирует в резуль-
тате хронического процесса.
Клиническая картина
Интенсивный зуд приводит к экскориациям.
Воспаленная зудящая кожа утолщается, поверх-
ностные кожные линии становятся более выра-
женными. Появляются толстые бляшки с глубо-
кими параллельными кожными линиями
(лихенификация). Наиболее типично поражают-
ся легкодоступные места и зоны складок. Типич-
ная локализация — на задней поверхности шеи,
подколенной впадине, голенях, веках и аноге-
нитальной области. На пораженной коже может
быть гипо- или гиперпигментация.
Лечение
Хроническое экзематозное воспаление часто ус-
тойчиво к лечению; ключ к успеху — прерыва-
ние цикла «зуд-расчесы» в ходе лечения и уст-
ранение причин или источников ухудшения
заболевания. Прохладный влажный компресс на
пораженную кожу в течение 20 минут помогает
успокоить и уменьшить зуд; он может быть
эффективен при ночных позывах к расчесам.
Эффективны стероидные кремы или мази груп-
пы I или II, которые применяются 2 раза в день.
Стероиды группы II —IV применяются с поли-
этиленовой окклюзией в течение 2—8 часов.
Экзема ладоней
Это распространенное, часто хроническое забо-
левание, имеющее множество причинных и спо-
собствующих факторов. Экзему ладоней можно
отнести к таким категориям, как ирритантная
экзема; эксфолиативная экзема; атопическая
экзема; экзема кончиков пальцев; аллергичес-
кая экзема; гиперкератотическая экзема; моне-
товидная экзема; дисгидротическая экзема; про-
стой хронический лишай и «id»- реакция. Каж-
дый из этих типов рассматривается отдельно.
Ирритантная экзема ладоней — самый распро-
страненный тип, за ним следует атопическая
экзема ладоней. Аллергический контактный дер-
матит является причиной экземы ладоней при-
мерно в 10—25 % случаев.
Этиология и патогенез
Женщины болеют чаще, чем мужчины. К фак-
торам профессионального риска относятся кон-
такты с химическими ирритантами, работа во
влажной среде, хроническое трение и работа с
сенсибилизирующими (аллергенными) химика-
тами.
Экзогенные факторы. К ирритантам относят-
ся химические вещества (такие как растворите-
ли, детергенты, щелочи и кислоты), трение, хо-
лодный воздух и низкая влажность. Аллергены
могут иметь связанные с работой и не относя-
щиеся к работе источники аллергенной экспо-
зиции. Немедленная аллергия по типу I может
включать реакции на латекс и пищевые проте-
ины, а более распространенная замедленная ал-
лергия по типу IV может включать реакции на
резиновые добавки, никель, медикаменты (бак-
трацин, неомицин и гидрокортизон) и распрос-
траненные химические ингредиенты в продук-
тах личной гигиены (такие как консерванты,
ароматизаторы, солнцезащитные вещества и дру-
гие добавки). Определенную роль могут играть
и пищевые аллергены. При инфекциях могут
иметь место «id»-реакции, в том числе экзема
ладоней, в качестве реакции на отдаленный очаг
грибковой или бактериальной инфекции.
Эндогенные факторы. Атопический диатез
(сенная лихорадка, астма, атопическая экзема)
часто является предрасполагающим фактором и
может способствовать восприимчивости к забо-
леванию и хронизации процесса, несмотря на
соответствующее лечение и меры предосторож-
ности.
Клиническая картина
Следует тщательно осмотреть все кожные покро-
вы в поисках диагностических ключей и способ-
ствующих заболеванию факторов, а также с це-
лью исключения других дерматозов (например,
псориаза). Данное состояние вариабельно; на-
блюдаются острые, подострые и хронические
экзематозные изменения. Хотя ассоциацию меж-
ду клинической картиной и этиологией устано-
вить с достаточной степенью надежности невоз-
можно, некоторые признаки могут оказаться
Глава 15. Экзема
225
полезными: ксероз, эритема, жжение на тыль-
ной и внутренней поверхности ладоней застав-
ляют подозревать раздражающие факторы. Мо-
нетовидная экзема, тыльные поверхности ладоней
и пальцев говорят о возможности аллергии, раз-
дражения или атопии; иногда виновницей явля-
ется контактная крапивница (аллергия типа I).
Обильные, рецидивирующие, интенсивно зудя-
щие везикулы на латеральных поверхностях
пальцев и ладоней могут указывать на дисгид-
ротическую экзему. При экземе кончиков паль-
цев (сухость, расщепление, болезненность, отсут-
ствие зуда) подумайте о наличии раздражителя,
эндогенного фактора (атопия в зимнее время)
или о фрикционной экземе. При наличии эри-
темы, шелушения, зуда в области основания
пальцев можно предположить атопию.
Если можно определить, с какими ирритан-
тами или аллергенами контактировал пациент и
устранить этот контакт в самом начале заболе-
вания, прогноз полного выздоровления будет
хорошим. Непрерывный или длительный кон-
такт с ирритантами и аллергенами может при-
вести к хроническому процессу. Воздержание от
контакта с провоцирующими факторами и со-
ответствующий уход часто улучшают состояние,
но у некоторых пациентов заболевание полнос-
тью не проходит.
Лечение
Лечение предполагает идентификацию раздража-
ющих факторов, которых следует избегать. Та-
кими факторами могут быть частое мытье рук
и воздействие воды, мыла, детергентов и раство-
рителей. Травма вследствие хронического трения
также является раздражающим фактором, кото-
рый может привести к хроническому рецидиви-
рующему дерматиту. Следует принять защитные
меры (например, виниловые перчатки для работы
с водой или химикатами). Назначают топичес-
кие кортикостероиды средней силы действия
(группа II-IV) два раза в день. Мази предпоч-
тительнее кремов. Можно применить окклюзию
под полиэтиленовой пленкой. Если дерматит не
тяжелый, следует воздержаться от назначения
очень сильных кортикостероидов (группы I).
Топические кортикостероиды для дерматита рук
действуют более эффективно, если их назначать
не постоянно, а с перерывами.
В случае тяжелого дерматита топический кор-
тикостероид с очень высокой фармакологической
эффективностью применяется после влажных
компрессов с раствором Бурова два раза в день в
течение первых 3—5 дней лечения, после этого
назначают кортикостероид средней силы действия
два раза в день в течение нескольких недель.
Можно назначить ванночки для рук с маслом
«Бальнеотар». Два-три колпачка масла растворя-
ют в миске с водой и погружают руки на 15—30
минут. Процедуру проводят 2 раза в день. После
нее применяют топический кортикостероид.
Системные стероиды (преднизон 0,75—1 мг/кг/
день с постепенным снижением дозы в течение
3 недель) могут иногда понадобиться для конт-
роля за сильным и острым воспалением. Состо-
яние большинства пациентов улучшается, если
устраняют раздражители, проводят лечение топи-
ческими кортикостероидами и регулярно и час-
то применяют смягчающие средства. Если есть
подозрение на аллергию (отек ладоней, везику-
лы, зуд и особенно если поражена тыльная сто-
рона ладоней или имеет место экзема кончиков
пальцев) следует провести лоскутное тестирова-
ние, чтобы определить вызывающие или поддер-
живающие заболевание аллергены. В материал для
тестирования нужно включить аллергены, соот-
ветствующие профессиональным занятиям паци-
ента. В случае хронического торпидного заболе-
вания пациент должен находиться под контролем
дерматолога. Другие методики лечения включа-
ют применение псоралена местно в комбинации
с лечением ультрафиолетовым облучением диа-
пазона А и поверхностную короткофокусную
рентгенотерапию. В случаях нарушения трудоспо-
собности еженедельно применяют низкие дозы
метотрексата (5—15 мг еженедельно) или ежеднев-
но низкие дозы циклоспорина внутрь.
Сухая экзема
Сухая (астеатотическая) экзема представляет со-
бой экзематозный дерматит, вызванный избы-
точной сухостью и растрескиванием кожи. Эта
сухая экзема называется также «eczema
craquele». Сухая (астеатотическая) экзема явля-
ется формой подострого экзематозного дерма-
тита, которая имеет тенденцию к медленному
хроническому течению с сезонными вспышка-
ми в зимнее время из-за низкой влажности.
Мужчины и женщины поражаются в равной
степени. Заболевание чаще встречается среди
пациентов с атопическим диатезом, особенно в
поздние годы жизни. У большинства пациен-
тов в анамнезе ранее уже имелись похожие
вспышки заболевания. Заболеваемость достигает
пика к концу зимы и снижается летом, это
особенно касается стран с сухим холодным кли-
матом. Может поражаться любой участок кожи,
226 Частная дерматология
хотя наиболее часто заболевание локализуется
на нижних конечностях. В начале заболевания
пациенты часто замечают, что их кожа выгля-
дит сухой, и ощущают ее сухость. По мере того
как заболевание прогрессирует, зуд и усилива-
ющееся воспаление становятся наиболее выра-
женными симптомами. Пациенты могут ощу-
щать жжение, а в тяжелых случаях образуются
трещины и корки.
Клиническая картина
Клиническая картина типична для подострого
экзематозного дерматита. Ксероз с акцентирован-
ным кожным рисунком является характерным
признаком с самого начала заболевания. Воспа-
ление вначале слабое, но со временем становится
более выраженным. Слабая, с размытыми грани-
цами эритема прогрессирует в ярко красные, ос-
тро-экзематозные папулы, которые сливаются в
широкие бляшки. Везикулы, как правило, не
образуются, а экскориации практически почти
всегда присутствуют. Сухая, тонкая десквамация
прогрессирует с образованием тонких поверхно-
стных трещин до картины, известной как «eczema
craquele», когда кожа становится похожей на трес-
нувший фарфор или пересохшее русло реки.
Кожа очень сухая с мелкими и глубокими тре-
щинами. Она может быть болезненной. Прогрес-
сируя, экзема становится острой, с мокнутием,
образованием корок и интенсивной эритемой.
Следует ожидать сезонных рецидивов в зим-
ние месяцы. Слабые сезонные вспышки с зудом
и ксерозом улучшаются при теплой погоде и при
постоянном применении смягчающих препаратов.
Активное подострое воспаление обычно реагирует
на мази с кортикостероидом средней силы дей-
ствия, а также улучшается с наступлением теп-
лого сезона. Тяжелая локализованная вспышка с
острыми признаками, такими как мокнутие и
корки, также реагирует на индивидуальную на-
ружную терапию, о которой пойдет речь далее.
Тяжелые вспышки следует лечить агрессивно,
поскольку они могут стать генерализованными.
Лабораторная диагностика
Клиническая картина достаточно красноречива,
поэтому биопсия кожи редко нужна для поста-
новки диагноза. Биопсия кожи подтверждает
наличие эпидермального спонгиоза с воспалением
дермы и часто вторичной импетигинизацией.
Дифференциальный диагноз
Дифференциальный диагноз включает другие
подострые экзематозные дерматозы, такие как
застойный дерматит, ирритантный контактный
дерматит, атопический дерматит, аллергический
контактный дерматит и целлюлит. Могут наблю-
даться признаки нескольких дерматозов одновре-
менно. Второй дерматоз может маскировать или
ухудшать первичный экзематозный процесс.
Ирритантный и аллергический контактный дер-
матит могут развиваться как результат собствен-
ных усилий пациента по самолечению. Пациента
следует расспросить о том, что он применяет на
пораженные участки. Застойный дерматит обыч-
но поражает голени у пожилых пациентов. В
анамнезе при этом регистрируется венозная не-
достаточность и отек ног, а также наличие ко-
ричневой пигментации (гемосидероз) кожи.
Лечение
Лечение зависит от стадии сухой экземы (острой,
подострой или хронической) и степени воспале-
ния. Для лечения ксероза предусматриваются ме-
роприятия для чувствительной кожи, а именно:
ограниченное применение только мягкого мыла
и обильное использование смягчающих средств.
В качестве смягчающего средства, не содержащего
консервантов, можно рекомендовать вазелин,
хотя пациенты не всегда согласны его применять.
Увлажняющие средства, содержащие молочную
кислоту, мочевину или гликолиевую кислоту,
также могут быть полезны. Раннее воспаление
лучше всего поддается лечению наружными кор-
тикостероидами средней силы действия преиму-
щественно на мазевой основе. Лечение следует
продолжать до разрешения эритемы и шелуше-
ния. Обильное применение эмолентов следует
продолжить в качестве профилактики от рециди-
вов. Лучше всего применять эмоленты успокаи-
вающего действия, не содержащие ароматических
добавок. Локализованные вспышки с признака-
ми острого экзематозного процесса, такими как
мокнутие и образование корок, следует сначала
лечить как острую экзему. Пациенты требуют
тщательного контроля во время этой стадии, по-
скольку локализованные вспышки могут стать ге-
нерализованными. При рецидивирующих острых
вспышках дерматолог должен провести обследо-
вание на предмет наличия аллергического контак-
тного дерматита. Влажные компрессы с раство-
ром Бурова и наружным кортикостероидом
средней силы действия на основе крема эффек-
тивны для санации раны и уменьшения воспале-
ния. Системные антибиотики могут быть пока-
заны при вторичной импетигинизации, о которой
свидетельствуют липкие корки цвета меда. Ког-
да мокнутие, воспаление и образование корок
регрессируют, влажные компрессы нужно отме-
нить, чтобы избежать избыточного высушивания
Глава 15. Экзема
227
пораженных участков. Мази с кортикостероидом
среднем силы действия (группы II или IV) сле-
дует продолжать применять, пока не пройдут
гиперемия и шелушение, примерно 2—3 недели.
Затем для уменьшения рецидивов применяют
меры ухода за чувствительной кожей, включая
эмоленты. Системные кортикостероиды для ле-
чения сухой экземы применяются редко.
Монетовидная экзема
Монетовидная (нумулярная) экзема - форма
экземы, характеризующаяся наличием часто ге-
нерализованных, сильно зудящих, округлых
(монетовидных) очагов экзематозного воспале-
ния. Взрослые болеют чаще, мужчины чаще, чем
женщины. Начало постепенное, без явного ухуд-
шения и без наличия экземы в анамнезе. Мо-
нетовидная экзема часто начинается с несколь-
ких изолированных очагов на ногах; со временем
появляются множественные очаги без какого-
либо специфического расположения. Очаги ча-
сто разрешаются или улучшаются после приме-
нения топических кортикостероидов, но после
прекращения их применения иногда возвраща-
ются на то же место.
Клиническая картина
Четко ограниченные, шелушащиеся, круглые
экзематозные бляшки появляются на туловище
и конечностях. Мокнутие очагов и везикуляция
характерны для вспышек заболевания. Вторич-
ная инфекция может спровоцировать вспышку
болезния. Корки медово-желтого цвета указы-
вают на вторичную импетигинизацию.
Это одна из наиболее трудных для лечения
форм экземы. Течение заболевания переменчи-
вое и непредсказуемое. Это состояние может
быть хроническим и рецидивирующим в тече-
ние нескольких лет. Сформировавшись, очаги
имеют тенденцию сохранять свой размер и ре-
цидивировать на ранее пораженной коже.
Лабораторная диагностика
Лоскутный тест дает положительный результат
в 1/3 случаев. Посев может выявить наличие
Staphylococcus aureus. Антибактериальное лече-
ние обычно помогает, но часто не приводит к ре-
миссии.
Дифференциальный диагноз
Псориаз (часто более симметричный и «геогра-
фический» характер поражения с серебристыми
чешуйками). Грибковые инфекции (очаги рег-
рессируют в центре и шелушатся по периферии;
исследование с КОН помогает в проведении
дифференциального диагноза). Кожная Т-кле-
точная лимфома на нижних конечностях (её
можно принять за монетовидную экзему; ис-
ключить диагноз поможет биопсия).
Лечение
Следует отменить все необязательные наружные
увлажняющие препараты, принимаемые внутрь
медикаменты, диетические добавки и препараты
из трав на период, как минимум, 3—4 месяца.
Следует провести исследование на наличие мико-
за стоп; грибковую инфекцию, при ее наличии,
лечат противогрибковыми препаратами, так как
иногда генерализованная монетовидная экзема
может быть «1с1»-реакцией на грибковую инфек-
цию. Следует агрессивно применять топический
стероид средней силы действия и смягчающие
эмоленты. Топический стероид наносят на пора-
женную кожу два раза в день в течение 2—3 не-
дель. Лучше всего продолжать лечение неделю или
еще немного дольше, чем необходимо для разре-
шения очагов. Эффективность топического стеро-
ида усиливается при окклюзии под полиэтилено-
вой пленкой или под специальным костюмом для
сауны, а также при приеме увлажняющей кожу
ванны перед нанесением топического стероида или
же при использовании обоих этих методов.
Вторичную инфекцию лечат системными про-
тивостафилококковыми антибиотиками (напри-
мер, цефалексин по 250 мг четыре раза в день).
В случае зуда назначают антигистаминные пре-
параты. Для длительного лечения системные
стероиды не применяются. Фототерапия может
привести к разрешению очагов, если наружная
терапия оказалась неэффективной. Ультрафио-
лет В узкого и широкого спектра лучше всего
подходит для фототерапии; псорален плюс уль-
трафиолет А можно применить, если ультрафи-
олет В оказался неэффективным.
Дисгидротическая
экзема
Для обозначения дисгидротической экземы исполь-
зуются также термины «дисгидроз» и «pompholyx».
В 20—25 % случаев экземы ладоней отмечается
дисгидротическая экзема. Эта форма экземы пред-
ставляет собой характерный хронический рециди-
вирующий экзематозный дерматит неизвестной
228 Частная дерматология
этиологии. Для дисгидротической экземы характер-
ны внезапные высыпания обычно сильно зудящих,
симметричных везикул на ладонях, боковых повер-
хностях пальцев и/или подошвах. В анамнезе у
бальных обычно имеется атопия (в личном или се-
мейном анамнезе — астма, сенная лихорадка или
атопическая экзема). Умеренный или сильный зуд
типично предшествует вспышке заболевания или
его рецидиву. Гипергидроз (избыточное потение)
часто сопровождает данное состояние или ухудша-
ет его. Пик заболеваний у женщин бывает в нача-
ле 2-го десятилетия жизни, а у мужчин в середине
четвертого десятилетия.
Клиническая картина
Везикулы диаметром 1—5 мм представляют собой
мономорфные, глубоко расположенные высыпа-
ния, наполненные прозрачной жидкостью и на-
поминающие крупу. Везикулы появляются вне-
запно и симметрично на ладонях и боковой
поверхности пальцев или на подошвах (рис. 15.1).
Кольца чешуек и шелушение сменяют везикулы
по мере уменьшения зуда. В зависимости от фазы
заболевания клиницист может наблюдать только
коричневые пятнышки. Когда острый процесс
заканчивается, кожа шелушится и открывается
красное потрескавшееся основание с коричневыми
Рис. 15.1. Типичная локализация
высыпаний при дисгидротической экземе.
пятнышками. Коричневые пятнышки — это ме-
ста бывшей везикуляции. Везикулы медленно
разрешаются в течение 1—3 недель. После этого
могут последовать хронические экзематозные
изменения с эритемой, шелушением и лихенифи-
кацией. Неопределенно часто могут иметь место
волнообразные рецидивы с появлением симмет-
рично расположенных везикул. По неизвестным
причинам хронические рецидивирующие высы-
пания иногда со временем проходят.
Дифференциальный диагноз
Пустулезный псориаз ладоней и подошв (основ-
ная жалоба пациентов чаще на боль, чем на зуд).
«Id»- реакция (в результате отдаленного источ-
ника грибковой инфекции). Воспалительная
грибковая инфекция (положительный тест с
КОН на наличие грибов). Острый аллергичес-
кий контактный дерматит. Буллезный пемфиго-
ид (может быть геморрагическим). Кожная Т-
клеточная лимфома (редко).
Лечение
Лечение начинают с применения прохладных
влажных компрессов либо с водопроводной во-
дой, либо с раствором Бурова, затем следует ап-
пликация стероидного крема средней или высо-
кой фармакологической эффективности (группы
I или III). Назначают преднизон 0,5—1 мг/кг/
день с постепенным понижением дозы в течение
1—2 недель. Некоторое облегчение может дать
применение мази с такролимусом («Протопик»
0,1 %), которую чередуют с ежедневным двухра-
зовым применением наружного кортикостерои-
да средней силы действия (группы I-III) в не-
сколько циклов из 3—4 недель. Кортикостероиды
не следует применять многократно или для ле-
чения хронической формы заболевания. Систем-
ные антигистаминные препараты могут облегчить
зуд. Псорален наружно на ладони плюс ультра-
фиолет А — вариант лечения частых торпидных
высыпаний. Назначение дисульфирама
(«Antabuse» 200 мг/день в течение 8 недель) мо-
жет помочь чувствительным к никелю пациен-
там с дисгидротической экземой ладоней. Если
определен отдаленный источник грибковой ин-
фекции и результат исследования с КОН поло-
жителен, следует лечить очаг грибковой инфек-
ции агрессивным наружным противогрибковым
средством (крем «Эконазол» или «Тербинафин»
ежедневно в течение 3 недель) или же коротким
курсом системных противогрибковых препаратов
(«Тербинафин» или «Итраконазол»), подобрав
дозировку и продолжительность лечения, соответ-
ствующие очагу инфекции. Сдерживание или
Глава 15. Экзема
229
устранение стрессов может помочь в лечении,
имеются отдельные сообщения об излечении не-
которых пациентов в таких случаях.
Если устранение контакта с аллергенами, вы-
явленными в ходе лоскутного тестирования, не
дает улучшения, и состояние остается тяжелым,
другие варианты терапии могут включать элек-
трофорез с водопроводной водой, внутрикожное
введение ботулина токсина (100—160 ME), еже-
недельные низкие дозы метотрексата, азатио-
прин (100—150 мг/день для достижения контро-
ля, затем поддерживающие дозы 50—100 мг/
день) и низкие дозы наружной лучевой терапии.
Себорейная экзема *
Себорейная экзема (синонимы: себорейный дер-
матит, диссеборейный дерматит, болезнь Унны)
- хроническое заболевание кожи, в основе ко-
торого лежит нарушение секреторной функции
сальных желез, выявляющееся в богатых саль-
ными железами участках кожи.
Эпидемиология
Средняя встречаемость заболевания среди популя-
ции составляет 3-5%, однако у пациентов с им-
мунодефицитами оно встречается значительно чаще:
от 30 до 80%. Как правило, начинается в период
полового созревания, однако может развиться в
любом возрасте. Большинство пациентов заболе-
вает в возрасте до 30 лет, после 50 лет риск воз-
никновения заболевания вновь возрастает. Муж-
чины болеют чаще. Основная жалоба пациентов —
зуд, усиливающийся при потоотделении. Состоя-
ние чаще ухудшается в зимнее время.
Этиология и патогенез
Этиология и патогенез болезни до настоящего
времени изучены недостаточно. Существенную
роль в ее развитии играют генетические факто-
ры, гиперпродукция сальных желез, патологи-
ческие изменения функции сальных желез, бак-
терии, обнаруживаемые в устьях волосяных
фолликулов и сальных желез, стресс, аллерги-
ческие реакции.
Гиперфункция сальных желез является важным
предрасполагающим фактором. У новорожденных
сальные железы активны вследствие эндогенно-
го образования андрогенов, поэтому себорейная
экзема может развиться у детей до 3 месяцев. В
более позднем возрасте активность сальных же-
* Раздел написан В. В. Валльес-Козловской
лез снижается, поэтому развитие себорейной эк-
земы происходит менее часто. Влиянием андро-
генов объясняется и более частое возникновение
заболевания у мужчин. Качественные изменения
кожного сала доказаны не были.
О роли нервной системы свидетельствуют та-
кие факты, как связь болезни Паркинсона и се-
борейной экземы. При полиомиелите или сирин-
гомиелии кожные изменения часто имеют место
только в области поражения тройничного нерва.
Пациенты также часто подчеркивают, что стресс
усугубляет кожные проявления. Проявления за-
болевания более выражены в зимнее время. При
недостатке цинка или энтеропатическом акродер-
матите повышен риск развития себорейного дер-
матита. Имеет место нарушение обмена эссенци-
альных жирных кислот. Недостаточность витамина
В также может вызвать подобный дерматоз.
В настоящее время широко изучается возмож-
ная роль дрожжей Malassezia (Pityrosporum) в
развитии себорейного дерматита. Связь подтвер-
ждается тем, что при лечении себорейной экзе-
мы противогрибковыми препаратами наблюда-
ется уменьшение проявлений заболевания и
уменьшение колонизации кожи Malassezia. Ко-
личество дрожжевых клеток на поверхности
кожи больного значительно превышает нормаль-
ные показатели (5* 105 см-2 у здоровых и 9,2хЮ5
см'2 у больных себорейной экземой). Мицели-
альная фаза грибов при себорейной экземе встре-
чается у 26% больных (у здоровых — в 6% слу-
чаев). Предполагают также, что себорейная
экзема является своеобразной специфической
реакцией кожи на Malassezia. Изучены различ-
ные нарушения иммунной системы у больных
себорейной экземой как результат деятельнос-
ти дрожжеподобных грибов: отмечена взаимо-
связь титров антител к Malassezia с тяжестью
себорейной экземы волосистой части головы.
Этиологическую роль играют не только дрож-
жи Malassezia. Например, у некоторых младен-
цев, страдающих себорейной экземой, из кала и
с поверхности кожи высеиваются многочислен-
ные колонии Candida albicans, а аппликацион-
ные тесты и реакция трансформации лимфоци-
тов подтверждают наличие сенсибилизации.
Известно также о перекрестных антигенах С.
albicans и Malassezia.
Вероятно, однако, что у разных групп пациен-
тов имеет место свой определенный патогенез
данного заболевания, поскольку, например, у
пациентов с выраженным иммунодефицитом
клетки Malassezia высеиваются значительно реже,
230
чем у пациентов без иммунопатологии. Себорей-
ная экзема является также одним из важнейших
маркеров ВИЧ-инфицированных пациентов.
Клиническая картина
Заболевание характеризуется тенденцией к хро-
ническому течению, частыми рецидивами, пло-
хо поддается лечению. Косметические недостат-
ки могут приводить к психологическим
проблемам у пациентов, вызывать нарушения со-
циальной адаптации. Основной жалобой пациен-
тов является зуд, усиливающийся при потении.
Детская себорейная экзема чаще возникает в
первые шесть месяцев жизни ребенка и обычно
полностью проходит в течение нескольких ме-
сяцев. Чаще болеют склонные к ожирению дети.
Поражение возникает на коже волосистой час-
ти головы, однако может поражаться кожа лица
в области бровей и носогубных складок, при
распространении процесса могут вовлекаться
сгибательные области конечностей, крупные
складки тела. На волосистой части головы об-
разуются наслоения жирных, окруженных тре-
щинами желтоватых чешуек — гнейса. Диссеми-
нированные очаги инфекции, локализующиеся
в крупных складках, напоминают таковые при
псориазе, однако склонны к быстрому излечи-
ванию.
Рис. 15.2. Типичная локализация
высыпаний при себорейной экземе.
Высыпания возникают в местах, характеризу-
ющихся повышенным содержанием сальных же-
лез — лицо, волосистая часть головы, грудь, меж-
лопаточная область, крупные складки (рис. 15.2).
Клиническая картина чаще всего представлена
наличием воспалительноых покрасневших и
слегка инфильтрированных очагов с неправиль-
ными очертаниями, с желтоватыми чешуйками
и корочками на гиперемированом фоне. Пора-
жение имеет вид либо сливных крупных оча-
гов, напоминающих географическую карту, либо
круглых с четкими границами множественных
очагов, напоминающих отрубевидный лишай.
При сильных субъективных ощущениях — зуде,
жжении — появляются экскориации, трещины,
присоединяется вторичная инфекция. Протоки
сальных желез выглядят расширенными.
На лице поражения чаще располагаются вок-
руг носа, в носогубных складках, коже бровей
(рис. 15.3). У части больных отмечается ухудше-
ние состояния после пребывания на солнце или
после УФО. На туловище обычно инфильтрация
выражена слабо вследствие отторжения чешуек
из-за потения. Могут поражаться крупные
складки — подмышечные, паховые, клиническая
картина при этом напоминает кандидоз или ин-
тертриго.
На волосистой части головы высыпания име-
ют четко отграниченные очертания и тенденцию
к слиянию. Иногда наблюдается тотальное пора-
жение волосистой части головы, напоминающее
панцирь. Очаги часто переходят на затылок, бо-
ковые области шеи, ретроаурикулярную область.
Нередко в ретроаурикулярной области образовы-
Рис. 15.3. Локализация высыпаний
на лице при себорейной экземе.
Глава 15. Экзема
231
вается длительно не заживающая трещина, кото-
рая склонна к вторичному инфицированию. В
центре грудины или между лопатками поражение
приобретает вид инфильтрированных гипереми-
рованных очагов.
Себорейная эритродермия является осложне-
нием себорейной экземы и возникает при непе-
реносимости наружного лечения или вследствие
контактной сенсибилизации.
Диагностика себорейной экземы не вызыва-
ет затруднений и основывается на типичной
клинической картине заболевания. Основную
трудность представляет дифференциальная ди-
агностика с вульгарным псориазом, особенно
при поражении волосистой части головы. При
псориазе высыпания располагаются по росту
волос, являются более инфильтрированными,
шелушение более сухое. Себорейная экзема
быстрее поддается терапии, чем псориатичес-
кие поражения. При поражении крупных скла-
док необходимо помнить о кандидозе или ин-
тертриго. При себорейной эритродермии надо
исключить синдром Сезари.
Лечение себорейной экземы может быть как
местным, так и системным, и зависит от тяжес-
ти заболевания. Из-за склонности к рецидивам
лечение является долгосрочным и направлено на
коррекцию себореи. Проводится противовоспа-
лительная и антимикотическая терапия.
При легких формах себорейной экземы с ло-
кализацией высыпаний на гладкой коже ис-
пользуются мазь, крем или раствор противо-
грибкового препарата, которые применяют 1—2
раза в день в течение 2—4 недель. Обезжири-
вание кожи лица производят спиртовыми ра-
створами с добавкой салициловой кислоты (2-
3%) или резорцина (2%). Днем используют
пудру, содержащую серу. Для лица рекоменду-
ются средства с эритромицином (лосьон «Зине-
рит») или кетоконазолом (крем «Низорал»). На
ночь показано подсушивающее лечение: цин-
ковый лосьон с клиохинолом (5%) и/или их-
тиолом (2-5%), а также серой (2-5%). Мокну-
щие очаги хорошо обрабатывать 1% водным
раствором бриллиантовой зелени.
Местные кортикостероиды являются одним из
наиболее эффективных методов лечения себорей-
ной экземы. Кортикостероиды обладают мощным
противовоспалительным действием, однако их
длительное применение ограничено побочными
эффектами — возникновением атрофии кожи,
телеангиэктазий, акне, периорального дерматита.
У детей кортикостероиды следует назначать с
особой осторожностью, учитывая повышенную
всасываемость кожного покрова. На лицо назна-
чают кортикостероидные кремы низкой активно-
сти — преднизолон и гидрокортизон.
Для мытья головы применяют антисеборейные
средства с кератолитическими и антимикробными
добавками: сульфид селена (шампунь Vichy
Dercos с сульфидом селена), салициловую кис-
лоту, деготь («Т-gel», «Friderm-Таг»), цинк
(«Friderm-Zinc»). Показан кетоконазол (шам-
пунь «Низорал»), активный в отношении липо-
фильных дрожжеподобных грибков (2 раза в
неделю). Антисеборейные настойки для волос
содержат серу, салициловую кислоту, резорцин
или нефеминизированные эстрогены. Для крат-
ковременного эффекта показаны спиртовые ра-
створы глюкокортикоидов, иногда с добавкой
дегтя. При сильном воспалительном процессе в
очагах назначают галогенизированные глюко-
кортикоиды. В качестве основ рекомендуются
кремы, лосьоны или гели.
При тяжелом течении заболевания, характе-
ризующемся наличием очагов с выраженным
воспалением и плотным наслоением чешуек, с
целью удаления последних используются кера-
толитики, такие как салициловая кислота или
препараты угольного дегтя. После отшелуши-
вания применяются местные противогрибковые
и кортикостероидные препараты. Дополнитель-
но могут быть рекомендованы антигистаминные
средства, препараты кальция, а в случае присо-
единения бактериальной инфекции назначаются
антибиотики.
При неэффективности наружной терапии по-
казано применение системных антимикотичес-
ких препаратов внутрь в течение одной недели:
кетоконазол (200 мг/сутки), тербинафин (250 мг/
сутки), флуконазол (100 мг/сутки), итраконазол
(200 мг/сутки). Наиболее полно изучено действие
кетоконазола и итраконазола. Флуконазол и тер-
бинафин менее эффективны против Malassezja,
однако также применяются в лечении себорей-
ной экземы.
В особо тяжелых случаях назначаются себосуп-
рессивные препараты, такие как изотретиноин,
уменьшающий активность и размер сальных же-
лез на величину до 90% и обладающий также
противовоспалительным действием. Показано, что
ежедневный прием препарата в суточной дозе от
0,1 до 0,3 мг/кг массы тела приводит к улучше-
нию тяжелой себореи после 4-х недель лечения.
В комплексную терапию включаются антиги-
стаминные препараты, мультивитамины, седа-
тивные средства, препараты для нормализации
функций желудочно-кишечного тракта, а в слу-
чае присоединения вторичной инфекции — ан-
тибактериальные средства и эубиотики.
232 Частная дерматология
Особенности
возникновения
и течения
экземы у детей *
Роль экссудативного
диатеза и наследственности
в развитии детской экземы
В развитии экземы у детей важную роль игра-
ют аномалии конституции — аллергический
(син.: экссудативно-катаральный) и другие ди-
атезы. Как известно, диатез — это особая форма
наследственной предрасположенности организма
к некоторым патологическим состояниям и за-
болеваниям, характеризуется необычной реак-
цией организма на физиологические раздражи-
тели и обычные жизненные условия. Диатез ещё
не является патологическим состоянием или за-
болеванием, а создаёт фон для их развития при
неблагоприятных условиях среды. Диатез дли-
тельный период может протекать скрыто и толь-
ко при неблагоприятных условиях среды (не-
правильный уход за ребенком, питание, режим,
вредно действующие экзогенные факторы), на-
рушениях функционального состояния эндо-
кринной, центральной и вегетативной нервной
систем удаётся выявить аномалии конституции.
Таким образом, сущность экссудативного ди-
атеза заключается в наследственно детерминиро-
ванной полигенной и гетерогенной предраспо-
ложенности с выраженной экспрессивностью
генов, передаваемой как по аутосомно-доминан-
тному, так и по аутосомно-рецессивному типу
наследования, которое в совокупности с общей
неспецифической и иммунологической реактив-
ностью опосредует готовность организма ребенка
к рецидивирующим воспалительно-экссудатив-
ным заболеваниям кожи с затяжным хроничес-
ким течением в ответ даже на обычные эндо- и
экзогенные воздействия.
Установлено, что при наличии аллергических
заболеваний в родословной по отцовской линии
экзема диагностируется у 30 % детей, по мате-
ринской линии — у 50 %, по отцовской и ма-
теринской линии — у 75 % детей. В последнем
случае она развивается в первые недели или
месяцы жизни ребенка и отличается непрерыв-
но-рецидивирующим течением с обширной пло-
щадью поражения кожного покрова. Более того,
достоверно установлено, что у детей, больных
экземой, наблюдаются генетически детермини-
рованные, преимущественно по материнской
линии, и вторичные — приобретенные иммун-
ные нарушения с повышенным содержанием IgG,
IgE и снижением уровня IgM при увеличенном
количестве В-лимфоцитов. Нарушения гумораль-
ной фазы иммунитета сопровождаются сниже-
нием числа функционально активных Т-лимфо-
цитов и снижением содержания Т-клеток.
Установлено, что сенсибилизация и гиперчув-
ствительность немедленно-замедленного типа
ребенка может произойти внутриутробно (транс-
плацентарно) за счет циркулирующих в организ-
ме беременной антигенов.
В постнатальный период у 95 % детей груд-
ного возраста ряд авторов отмечают дисфункции
печени, поджелудочной железы, пищеваритель-
ного тракта, несовершенство или недостаточность
ферментативных систем пищеварительного ап-
парата, повышенную проницаемость слизистых
оболочек желудка, кишечника и нарушение пе-
ченочного барьера. Нельзя не отметить большое
значение, которое имеют в возникновении эк-
земы у детей нарушения диеты матерью, ранний
прикорм, докорм ребенка на фоне обострения
очагов хронической инфекции, ОРВИ, глистной
инвазии, обострения заболеваний внутренних
органов и др.
Клинические особенности
экземы у детей
Экзема у детей раннего возраста имеет ряд
особенностей в клинической картине и тече-
нии. У 72 % детей первые высыпания на коже
появляются в течение 1-го полугодия жизни.
Экзема у детей в возрасте 1—2 лет обычно свя-
зана с экссудативным диатезом и протекает с
мокнутием. Чаще поражаются щеки и лоб (ис-
тинная форма экземы), далее процесс захваты-
вает волосистую часть головы и всё лицо. Кожа
диффузно краснеет, отекает, появляющиеся на
ней мелкие пузырьки быстро вскрываются,
оставляя эрозированные поверхности. Процесс
чаще развивается на 3—6 месяце жизни. Нос и
носогубный треугольник обычно не поражают-
ся. У детей чаще диагностируют обширные,
лишенные рогового слоя мокнущие поверхно-
сти. Процесс имеет тенденцию к распростра-
нению с кожи головы на другие участки по-
крова.
Раздел написан В. В. Саларевым
Глава 15. Экзема
233
Сильный (биопсирующий) зуд постоянного ха-
рактера беспокоит ребенка в течение суток (чаще
при сочетании экземы с заболеваниями органов
пищеварения).
Дети жалуются на нарушение сна, и имеют при
осмотре характерный внешний вид: пастозность
кожи с бледной (бледно-розовой) окраской, пол-
нота с рыхлой, но не упругой жировой клетчат-
кой. Тургор мягких тканей у детей снижен.
У детей чаще диагностируют себорейную (67 %
случаев), импетигинозную (56 % случаев), мик-
робную (49 % случаев) и пруригинозную (23 %
случаев) формы экземы. При присоединении
пиогенной инфекции на участках экземы у де-
тей появляются импетигинозные пустулы или
фолликулиты, корки становятся слоистыми,
желто-зеленого цвета, иногда присоединяется
лимфаденит, нередко повышается температура.
У маленьких детей наблюдается импетигиноз-
ная экзема ягодиц (неопрятное содержание де-
тей, поносы). У детей старшего возраста (от 5 до
14 лет) иногда наблюдается диссеминированное
проявление экземы с локализацией очагов по-
ражения на коже туловища, реже на лице и еще
реже на конечностях. Очаги, как правило, оваль-
ной, неправильной формы в виде пятен или
инфильтрированных бляшек. Зуд сохраняется
постоянно в течение суток.
Гистологические изменения: в острых случаях
экземы характерны явления экссудации, а при
хронической форме — пролиферации. В эпидер-
мисе и дерме отмечается отечность, особенно в
шиповатом слое эпидермиса. Внутри него меж-
клеточный отек раздвигает клетки и формирует
различного размера полости. В мальпигиевом слое
иногда обнаруживаются клетки инфильтрата,
создающие впечатление формирующихся микро-
абсцессов. При хронической экземе в эпидермисе
наблюдается акантоз и нередко паракератоз.
Кровеносные и лимфатические сосуды расши-
рены, некоторые из кровеносных сосудов напол-
нены эритроцитами. В дерме диффузно между
коллагеновыми волокнами, по ходу сосудов и
вокруг придатков кожи располагается инфильтрат.
При острой экземе инфильтрат состоит из поли-
морфноядерных лейкоцитов, а при хронической
экземе в инфильтрате преобладают лимфоциты и
фибробласты, иногда встречаются полиморфноя-
дерные лейкоциты. Эластическая сетка в сосочко-
вом слое находится в состоянии распада; встреча-
ющиеся пучки нервных волокон отечны.
Лечебно-диетические рекомендации
Правильно назначенная диета может оказать
неспецифическое десенсибилизирующее воздей-
ствие и способствовать улучшению состояния
ребенка. В течение первых 3 суток при острой
экземе необходима строгая молочная диета. За-
прещаются острые блюда, сладости, яйца, кофе,
чай и напитки, содержащие алкоголь. Исклю-
чаются из питания (элиминационная диета) про-
дукты, являющиеся аллергенами для данного
ребенка. Питание ребенка, должно строиться с
учётом выявленных заболеваний органов пище-
варения и формы экземы. Так, при истинной
экземе у детей усилен липостаз и выражена бел-
ковая недостаточность, а при себорейной форме
экземы отмечаются липолиз и диспротеинемия.
Поэтому детям I группы рекомендуется диета с
повышенным содержанием животного и расти-
тельного жира и увеличением белка на 10—12 %,
а детям П-ой группы увеличивают потребление
растительного жира. Количество потребляемых
углеводов компенсируется за счет гипоаллерги-
ческих фруктов и овощей. В питание детей не-
обходимо вводить ксилит, за счёт которого сни-
жается потребление сахара и проявляется
выраженное холецистокинетическое действие.
Принципы обшей терапии
1. Этапность и непрерывность лечения (стаци-
онар — поликлиника — санаторий).
2. Комплексность и индивидуальный подход
при построении схем лечения.
3. Обязательная санация очагов хронической
инфекции в ЛОР-органах (тонзиллит, гайморит,
отит), органах пищеварения (хронический пан-
креатит, дисбактериоз кишечника), в бронхоле-
гочной системе, мочевыводящих органах.
4. Антибиотики необходимо назначать с осто-
рожностью и только в условиях стационара по
строгим показаниям.
5. Дезинтоксикационная терапия, элиминация
аллергенов проводится параллельно с санацией
очагов инфекции, преимущественно у детей с
диссеминированным распространением экземы.
6. Дегельминтизация оправдана при наличии
признаков экземы.
7. Показана неспецифическая гипосенсибили-
зация организма, назначение антигистаминных
препаратов II-IV классов и др..
Принципы местной терапии
1. Наружная терапия назначается с учётом ста-
дии экзематозного процесса (обострение, ремис-
сия).
2. Местная терапия проводится с учётом кли-
нической формы экземы.
3. Наружные глюкокортикоиды (преднизоло-
новая или гидрокортизоновая мази, «Элоком»
234 Частная дерматология
или «Апулеин» — крем, мазь) применяют на ог-
раниченные участки поражения кожного покрова
в течение не более 10 дней.
4. Строгое соблюдение методики применения
наружного лечебного средства (примочка, повяз-
ка и др.).
Профилактика детской экземы
Первичная профилактика:
• здоровый образ жизни супругов;
• соблюдение гигиены жилища;
• тренирующий режим жизни ребенка;
• антенатальная профилактика аллергодерма-
тозов: (рекомендации по режиму питания буду-
щей матери; медицинская помощь при токсико-
зах);
• акушерско-педиатрические советы (курсы).
Вторичная профилактика:
• раннее выявление детей, входящих в груп-
пу «риска»;
• полноценная их диспансеризация;
• комплексная консультативная и терапевти-
ческая помощь;
• рациональная наружная терапия.
Глава 16
Профессиональные болезни кожи
Профессиональные дерматозы составляют до
80 % профессиональной патологии и возника-
ют в результате вредного действия различных
производственных факторов.
К профдерматозам относятся кожные заболе-
вания, которые впервые возникли в условиях
производства. Если заболевание появилось до
работы на производстве, а под влиянием произ-
водственных факторов обостряется и рецидиви-
рует, такое заболевание не является профессио-
нальным.
Производственные факторы
1. Химические раздражители (основные) —
кислоты, щелочи, скипидар, синтетические и
эпоксидные смолы, стекловолокно, синтетичес-
кие краски и лаки, соединения никеля, хлора,
ртути, цемент, формалин, ядохимикаты, стреп-
томицин и новокаин.
2. Физические факторы — радиоактивные изо-
топы, лучи рентгена, ультрафиолетовые лучи,
механические и термические факторы.
3. Некоторые инфекции — вирус коровьей
оспы, грибы рода Candida.
Клинические формы
1. Профессиональный простой (контактный)
дерматит ничем не отличается от простого кон-
тактного непрофессионального дерматита, но
возникает в условиях производства.
2. Профессиональный аллергический дерматит
ничем не отличается от непрофессионального
аллергического дерматита, но возникает в усло-
виях производства.
3. Профессиональные фотодерматозы —- фото-
сенсибилизация, возникшая в условиях произ-
водства под действием фотодинамических веществ
(асфальт, гудрон, деготь, креозотовое масло).
4. Профессиональная экзема не отличается от
обычной экземы, но действие аллергена связа-
но с производственным процессом.
5. Профессиональные фолликулиты возникают
при работе с гудроном, дегтем, маслами, керо-
сином. Масла и керосин вызывают раздражение
кожи и появление воспалительньп/фолликули-
тов — красных папул вокруг волрса. Гудрон и
деготь вызывают пролиферацию клеточного эпи-
телия и появление роговых фолликулитов — это
плотные эпидермальные папулы вокруг волоса,
роговые, темно-красного цвета. Локализация —
чаще голень, реже предплечье, иногда туловище.
6. Профессиональные изъязвления отмечаются
при контакте с кислотами, щелочами (сода), ци-
анистыми соединениями. Локализация — тыль-
ная поверхность кистей, ладони в местах повреж-
дения эпидермиса. Язвы маленькие, мелкие,
неглубокие, малоболезненные. Иногда может
быть и большее количество язвочек, покрытых
темными корочками —- симптом «птичий глаз».
7. Профессиональные гиперкератозы и папилло-
матозы возникают при длительном контакте с
продуктами, которые содержат канцерогенные
вещества. Это каменноугольный деготь, синте-
тические смолы, гудрон и др. Локализация —
верхние и нижние конечности. Заболевание воз-
никает через 10—15 лет после контакта с этими
веществами.
Кожные проявления
• Высыпания по типу плоских бородавок —
это эпидермальные папулы, образованные в ре-
зультате пролиферации клеточного эпителия;
• высыпания по типу вульгарных бородавок —-
за счет пролиферации клеточного эпителия и
разрастания сосочков дермы с образование пер-
вичного элемента — эпидермально-дермальных
папул. Клинически выявляются крупные вуль-
гарные бородавки величиной до вишни, оставля-
ющие после себя рубчики;
Дифференциальный диагноз
Верруциформная эпидермодисплазия — поражаются
только открытые участки, наследуется по аутосом-
но-рецессивному типу. Клинически определяются
бородавки-папулы, бляшки желто-коричневого
цвета. Возраст пациентов до 25 лет и менее.
* Глава написана В. В. Саларевым
235
236 Частная дерматология
Папилломы — характерен быстрый рост, болез-
ненны, изъязвляются и нередко перерождаются
в эпителиомы.
Узелки доильщиц — возникают у доярок, зоо-
техников, ветеринарных врачей при контакте с
больными коровами, которые поражены коро-
вьей оспой. Локализация — пальцы, тыл кис-
тей, ладони, лучезапястные суставы, предплечья.
Клинически определяются плотные папулы ве-
личиной с горошину с западением в центре, без-
болезненные, округлые, через 1—2 месяца само-
произвольно исчезают.
Профессиональный кандидоз — развивается
между пальцами кистей у кондитеров, работни-
ков овощехранилищ (перезрелые овощи и фрук-
ты, действие на кожу яблочной, лимонной и
молочной кислот).
Критерии для правильной
постановки диагноза
1. Установление связи с определенным произ-
водственным фактором.
2. Локализация на открытых участках тела.
3. Положительные кожные пробы с предпола-
гаемыми раздражителями.
4. Наличие подобного заболевания у других ра-
бочих, находящихся в тех же условиях.
Лечение
Лечение профдерматозов не отличается от лече-
ния аналогичных кожных заболеваний.
Профилактика
1. Защита кожи от вредных веществ.
2. Механизация, автоматизация, герметизация
производства.
3. Санитарно-технические условия вентиля-
ции, спец, одежда, умывальники, души и др.
4. Санитарно-просветительная работа.
5. Применение защитных средств (мази, пас-
ты, кремы, которые наносятся на участки кожи
до работы):
• при контакте с водными растворами исполь-
зуют силиконовый крем, который защища-
ет руки от вредного действия воды;
• при контакте с органическими растворите-
лями используют биологические перчатки
или казеиновый раствор (казеин, растворен-
ный в спирте);
• при контакте с эпоксидным клеем, смолой
используют для их удаления 96° спирт.
6. Профессиональный отбор рабочих должен
включать методику кожных проб.
Если вредное вещество нельзя заменить, тех-
нический процесс нельзя механизировать, а за-
щитные средства не помогают, проводят профес-
сиональный отбор. Для этого поступающему на
работу человеку ставят кожные пробы с произ-
водственным раздражителем. Если кожная про-
ба отрицательная, рабочий принимается на дан-
ное производство.
Глава 17
Кожный зуд
Зуд (pruritus) — это специфическое для кожи (и
расположенных близко к ней слизистых) чув-
ственное ощущение или ноцицепция, которая
физиологически действует как сигнально-пре-
дупреждающая система для щекочущего раздра-
жения, допороговых болевых ощущений и в
различных качественных и количественных гра-
дациях сопровождает многочисленные кожные
заболевания.
Зуд относится к самым распространенным
жалобам, касающимся кожи. При этом речь идет
о неприятном ощущении, которое сопровожда-
ется непрерывной потребностью в ответном ме-
ханическом раздражении кожи. Хронический
зуд, как и боль, может существенно влиять на
общее состояние пациента и в экстремальных
случаях привести к угрозе самоубийства. Одна-
ко зуд и следующее за ним расчесывание вос-
принимаются негативно в социальном плане и
поэтому, особенно в аногенитальной области, ча-
сто описываются пациентами как жжение или
сухость. Зуд, с одной стороны, это функциональ-
ная ноцицепция, которая служит для удаления
с кожи вредящих ей паразитов, остатков расте-
ний, с другой стороны — это симптом заболе-
вания, которое может привести к повреждению
кожи.
Зуд близкородственен болевому ощущению,
но отличается от него по следующим пунктам:
оба ощущения неприятны, однако боль реали-
зует рефлекс «избегания», а зуд, напротив, по-
чти принудительный рефлекс «обработки», ис-
полнение которого в ходе почесывания, трения
приводят к моментальному (пусть даже кратков-
ременному) удовлетворяющему облегчению. Та-
кая смена ощущений от неприятного к прино-
сящему удовольствие является одной из причин,
почему кожные заболевания воспринимаются
как личностные. К тому же эффекты расчесов
вследствие зуда входят в морфологическую кар-
тину ряда дерматозов и могут со своей стороны
снова через порочный круг вовлекаться в пато-
генез зуда и кожного заболевания.
Общая структурная база боли и зуда иллюст-
рируется многочисленными наблюдениями: от-
сутствие этих ощущений, как врожденное, так
и приобретенное, почти всегда возникает в паре.
Однако зуд не идентичен слабой боли, поскольку
оба ощущения имеют свое собственное качество
и собственный спектр интенсивности, не пере-
ходят друг в друга и диссоциируемы: нагрева-
ние кожи до 4ГС блокирует зуд, но усиливает
болевое ощущение; удаление эпидермиса ведет
к потере ощущения зуда, ощущение боли одна-
ко остается; раздражение токами высокой час-
тоты производит боль с малым зудящим раздра-
жением.
В опосредовании зуда и боли решающую роль
играют свободные окончания волокон типа А
дельта и волокон типа С. Если раньше это под-
тверждалось лишь клиническими наблюдения-
ми, то теперь имеются и нейрофизиологические
исследования, поддерживающие теорию о том,
что волокна зуда и боли представляют собой
единое целое с разным порогом активности.
Спектр восприятия зуда варьирует от легко-
го щекочущего, нагревающе-жгучего, режущего
до тупого, мучительного. Поэтому и рефлексо-
образные «реакции обработки» совершенно раз-
личны: расчесывающие, как при экземах, соот-
ветствуют попытке удалить точечные источники
зуда из кожи (деструктивный акт); осторожно
растирающие, как при красном плоском лишае;
при плохо локализуемом или распространенном
источнике зуда (например, грибовидный микоз
или механическая крапивница) или охлаждаю-
щие (острая крапивница). Расчесы поэтому не
являются само собой разумеющимся следстви-
ем зуда. Вероятно, эта разница в восприятии зуда
уравновешивается многообразием причинных
медиаторов или их сменой.
Помимо физической, химической, биохими-
ческой, антимикробной и иммунологической
барьерных функций кожи в рамках ее роли как
пограничной зоны между телом и окружающей
средой решающую роль в смысле ощущения и
соответствующего реагирования играет иннерва-
ция дермы и эпидермиса. Сегодня уже непри-
емлемо положение о том, что для различных
ощущений существуют отдельные специфичес-
237
238 Частная дерматология
кие рецепторы. В настоящее время исходят из
того, что существуют смешанные рецепторы для
восприятия тепла, холода, боли, зуда и прикос-
новения. Некоторые ноцицепторы реагируют на
химические раздражители, однако их поведение
по отношению к различным химическим веще-
ствам также явно различно. В настоящее время
не существует четко определенных химических
веществ, вызывающих только зуд или только
боль, даже гистамин в зависимости от дозы вы-
зывает либо зуд, либо болевое ощущение.
Как световая, так и электронная микроскопия
не в состоянии полностью и однозначно диффе-
ренцировать эпидермальные сенсорные нейро-
ны. И только методика иммуногистохимии в
комбинации с иммуноэлектронной микроскопи-
ей и применением антител на нейропептиды по-
зволила провести дальнейшую дифференциацию.
В пределах кожных нервных волокон смогли ло-
кализовать такие вещества как субстанция Р,
ген-кальцитониновый пептид, нейротропин и
вазоактивный интестинальный пептид (VIP).
Известно также, что некоторые волокна снабже-
ны комбинацией таких нейропептидов.
С точки зрения физиологии чувств, процесс
развития зуда как симптома зависит от иннер-
вации кожи. Различные кожные рецепторы про-
водят ощущение зуда преимущественно через
полимодальные С и А нервные волокна.
Среди рецепторов кожи различают:
• Тактильные рецепторы: (диски Меркеля, А-
волокна, осязательные тельца Мейсснера, плас-
тинчатые тельца Фатер-Пачини и тельца Голь-
джи-Маццони).
• Температурные рецепторы: (поверхностная
нервная сеть дермы содержит рецепторы для вос-
приятия холода — колбы Краузе, и для воспри-
ятия тепла — тельца Руффини).
• Болевые рецепторы представлены свободны-
ми нервными окончаниями.
Зуд передается преимущественно по безмие-
линовым, медленно проводящим С-волокнам в
центральную нервную систему. Зуд вызывается
механической, термической, электрической или
химической стимуляцией полимодальных С-не-
рвных волокон. Свободные нервные окончания
этих безмиелиновых нервных волокон на границе
эпидермиса и дермы служат как ноцицепторы и
возбуждаются либо непосредственно, либо опос-
редованно путем высвобождения различных ме-
диаторов. К веществам, вызывающим зуд, отно-
сятся амины (гистамин, серотонин), протеазы
(экзогенный папаин, калликреин, трипсин) и
различные пептиды (брадикинин, секретин) и
нейропептиды (вещество Р, вазоактивный инте-
стинальный полипептид), гормон щитовидной
железы — кальцитонин, а также метаболиты
арахидоновой кислоты, интерлейкин-2, факто-
ры роста и различные биологически активные
вещества эозинофилов и тромбоцитов. Проста-
гландины и эндорфины действуют модулирую-
ще на периферическую и центральную нервную
систему. Многие из приведенных веществ явля-
ются потенциальными либераторами гистамина;
другие, как папаин и калликреин, непосред-
ственно вызывают зуд. Гистамин — это важный,
но не единственный медиатор зуда, чем объяс-
няется иногда неудовлетворительная терапевти-
ческая реакция на антигистаминные препараты.
Нервные импульсы, обеспечивающие ощуще-
ние зуда, поступают по афферентным нервным
волокнам в задние рога спинного мозга, где пе-
реключаются на нейроны спиноталамического
тракта, по которому поступают в таламус и да-
лее в сенсорную зону коры головного мозга.
Вследствие перекрестной возбудимости поли-
модальных С-волокон зуд приобретает различные
качества. Например, муканаин, выделенный из
стручков растения Мисипа pruriens, вызывает чи-
стый зуд, а типичное для гистамина ощущение
состоит примерно на 60 % из зуда и на 40 % из
боли. В отличие от этого, горчичное масло вы-
зывает чисто жгучую боль. Стимуляция ноцицеп-
тивных рецепторов брадикинином и, возможно,
кислая тканевая среда при воспалительных дер-
матозах ведут к тому, что электрофоретическое
действие гистамина воспринимается как жжение.
Отдельные медиаторы или комбинация тако-
вых в состоянии активизировать отдельные ре-
цепторы в вышеназванных С-волокнах, вслед-
ствие чего превышается определенный порог
раздражения или же запускается каскад, кото-
рый в ходе нервной стимуляции перерабатыва-
ется в ЦНС интерпретационным сигналом зуда.
Типичный центр зуда в ЦНС до сих пор не
идентифицирован. Посредством функциональ-
ной позитронно-эмиссионной томографии при
обусловленном гистамином зуде кожи смогли
доказать в качестве признака нейрональной ак-
тивации повышение скорости кровотока в обла-
сти поясной извилины. Некоторые исследовате-
ли указывают на то, что этот регион может быть
ответственен за сенсорный аспект обусловленно-
го гистамином зуда, в то время как премотор-
ная зона, вероятно, отвечает за подготовку к
расчесыванию.
Гистамин является наиболее известным веще-
ством, исследовавшимся в связи с зудом. Он яв-
ляется компонентом тучных клеток, а при его
высвобождении в результате их дегрануляции и
Глава 17. Кожный зуд 239
связывании с ^-рецепторами возникают, по Lewis
(1927), три феномена: пятнистая эритема с рас-
ширением капилляров, покраснение без увеличе-
ния массы ткани, развитие волдыря через 60—90
секунд, за которым следует образование неболь-
шого анемичного ареала, вызванного отеком и
связанной с ним компрессией капилляров.
Действие гистамина можно полностью или
частично прекратить блокадой гистаминных ре-
цепторов, применяя Н,-антигистаминные сред-
ства. Поэтому с помощью антигистаминных
препаратов всегда пытались подавить зуд при
различных кожных заболеваниях и самых раз-
ных внутренних болезнях. При этом выяснилось,
что целый ряд форм зуда не реагирует на анти-
гистаминные препараты, поэтому поиск других
медиаторов стал еще более необходимым.
Другой биогенный амин, серотонин, при инъ-
екциях или электрофорезе также способен вы-
зывать зуд и образовывать волдыри. Однако он
является более слабым пруритогеном по сравне-
нию с гистамином. Серотонин не накапливает-
ся в тучных клетках и может производить как
атлетический, так и аналгетический эффекты.
Он, возможно, играет особую роль при уреми-
ческом или гепатическом зуде. Исследования
показали, что капсаицин, хотя и уменьшает ин-
дуцированные серотонином волдыри, не может
повлиять на окружающую их эритему. Протеи-
назы также пруритогенны. Трипсин и химотрип-
син вызывают зуд, его воздействие, однако,
прекращается при применении антигистаминных
препаратов, причем следует принять к сведению,
что модулирование происходит посредством
высвобождения гистамина. Папаин и калликре-
ин, напротив, не вызывают собственных, зави-
сящих от гистамина, эффектов.
Много внимания в последнее время уделяет-
ся взаимосвязи нейропептидов и зуда. Субстан-
ция Р вызывает сильный зуд, частично посред-
ством гистамина. Терапевтическое применение
капсаицина еще больше прояснило этот вопрос.
Местное применив капсаицина на коже приво-
дит к деплеции субстанции Р нейропептидами
вплоть до повреждения немиелинизированных
волокон типа С. Вначале наступает сильное
жжение и боль, а также зуд, затем восприятие
или образование субстанции Р блокируется.
Прояснению природы зуда способствовало
действие опиоидов. Морфин устраняет боль, но
с другой стороны вызывает зуд. Хотя опиоиды,
такие как морфин, вызывают зуд, высвобождая
гистамин из тучных клеток, антигистаминные
препараты не в состоянии прервать его посред-
ством рецепторной блокады.
Простагландины и эйкозаноиды, которые об-
наруживаются в коже в больших количествах в
рамках иммунологических и аллергических ре-
акций, очевидно, также играют какую-то роль
при зуде. После инъекции простагландины мо-
гут вызывать легкий зуд, который, однако, зна-
чительно меньше зуда, вызываемого гистами-
ном, но который, очевидно, гистамином
опосредуется или же гистаминный зуд может
усиливаться простагландином Е2. Лейкотриены,
например типа LTB4, вызывают эритему, одна-
ко после кожной инъекции не дают волдырей.
Ингибиторы синтеза простагландинов, такие как
ацетилсалициловая кислота или индометацин, не
в состоянии отрегулировать этот зуд. С другой
стороны, ацетилсалициловая кислота в лечении
сильного зуда в третьем триместре беременнос-
ти играет решающую роль и более эффективна,
чем Н,-антагонист хлорфенирамин.
Выясняется роль цитокинов и факторов рос-
та в аспекте их возможной сьязи с зудом. В этом
плане особое значение имеет исследование, ко-
торое смогло доказать, что нейротропный фак-
тор нейротропин-4 играет роль в рамках зуда при
атопическом дерматите.
Зуд является наиболее распространенным сим-
птомом в дерматологии, который может возни-
кать в связи с рядом кожных заболеваний или
без клинически видимого кожного заболевания:
ксеродермия (сухость кожи), дерматозоонозы
(чесотка, педикулез, укусы насекомых), дерма-
тит атопический, дерматит контактный, медика-
ментозная таксидермия, красный плоский лишай,
экзема, крапивница, пруриго, герпетиформный
дерматоз Дюринга, солнечный дерматит.
При дерматозах зуд является симптомом и
следствием кожного заболевания. Соответству-
ющий дерматоз диагностируется по типичным
высыпаниям. Многие кожные заболевания со-
провождаются зудом. Интенсивный зуд, приво-
дящий к расчесам и их эффектам, наблюдают
при экземе, атопическом дерматите, некоторых
микозах и паразитарных заболеваниях кожи.
При многих дерматозах (красный плоский ли-
шай, крапивница), несмотря на интенсивность
зуда, нет эффектов расчесов, так как кожу рас-
тирают, а не расчесывают. Такие пациенты име-
ют характерные блестящие ногтевые пластинки.
Для атопического дерматита типичны кризы
зуда. При простом подостром пруриго расчесы
вызывают высыпания, после чего зуд вдруг пре-
кращается, остаются только геморрагические ко-
рочки, но нет следов расчесов. Зуд является так-
же симптомом крапивницы и усиливается
расчесами, но экскориации не встречаются.
240 Частная дерматология
Частые горячие ванны или ежедневный при-
ем горячего душа с применением обезжиреного
мыла и особенно добавок для ванн могут при-
вести к высушиванию кожи, часто с едва види-
мым шелушением, при этом кожа реагирует
сильным зудом. У пожилых людей зудят участ-
ки кожи, бедные сальными железами, особенно
предплечья и голени, преимущественно зимой,
когда вследствие отопления влажность воздуха
в квартире низкая.
Каждый пациент, страдающий от зуда, должен
проверяться на дерматозоонозы (чесотка, укусы
насекомых, педикулез). Чесотка является наибо-
лее частым паразитарным зудящим заболевани-
ем кожи. Зуд при чесотке особенно часто воз-
никает ночью. При зуде волосистой части
головы и ушей следует исключить вшивость;
при зуде в области лобка, промежности, груди,
подмышечных впадин — лобковый педикулез;
при зуде в области поясницы, лопаток, шеи —
педикулез, вызванный платяными вшами.
Зуд — вечный спутник атопического дерма-
тита. Его интенсивность различна; он бывает
диффузным и локализованным, частью ограни-
ченным зоной отдельных высыпаний. Последнее
встречается у пожилых атопиков с пруригиноз-
ными изменениями. Зуд может предшествовать
повторному приступу атопического дерматита.
Обусловленные зудом эффекты расчесов замы-
кают порочный крут, вызывая воспаление, при-
водящее к стафилококковой инфекции, а тем
самым снова к воспалению, что способствует
персистированию заболевания.
Кожный зуд с уртикарными высыпаниями, а
также атопический дерматит вызывает классичес-
кий медиатор гистамин. Многие другие болезни
кожи могут также сопровождаться кожным зудом.
Диагноз «pruritus sine materia» может быть установлен
тогда, когда все диагностические возможности ис-
черпаны, а соматическая причина длительного зуда
не устанавливается. Неосознанные вынужденные
расчесы приводят к линейным полосам на коже.
Иногда дерматологи говорят о «pruritus sine materia»,
когда при осмотре кожа во всем остальном здоро-
ва. Зуд как симптом меньше зависит от гистами-
на, скорее от других медиаторов (серотонин, про-
стагландин и другие вазоактивные вещества).
Хронический зуд без явной причины поражает чаще
пожилых людей, особенно мужчин. При дифферен-
циальном диагнозе следует учитывать pruritus senilis
или общий ксероз (сухость кожи).
Дифференциальный диагноз
При дифференциальной диагностике зуда сле-
дует учесть, что мастоцитоз, пемфигоид или гер-
петиформный дерматоз Дюринга могут вначале
проявляться зудом на клинически здоровой
коже, а себостаз у кожных больных едва ли под-
дается объективизации.
В тех случаях, когда невозможно связать зуд с
каким-либо дерматологическим заболеванием,
следует искать другие причины. Генерализованный
зуд, в отсутствие первичного кожного заболевания,
может быть важным признаком внутреннего за-
болевания: уремический зуд (заболевания почек);
холестатический зуд (механическая желтуха, хо-
лестатический гепатит, первичный билиарный
цирроз); эндокринопатический зуд (сахарный диа-
бет, зуд беременных); паранеопластический зуд
(болезнь Ходжкина, висцеральная карцинома);
неврогенный зуд (неврологические заболевания);
психогенный зуд (психические заболевания); меди-
каментозный зуд (прием некоторых препаратов).
При осмотре тела пациента с жалобами на
зуд следует далее различать зуд с кожными
проявлениями и без них. Однако прежде чем
заключить, что кожных проявлений нет, не-
обходимо провести самый тщательный осмотр
тела пациента, начиная от кожи ступней до во-
лосистой части головы, включая полость носа
и наружный слуховой проход, области ануса,
мошонки и вульвы, а также ногти и межпаль-
цевые пространства. И только когда все эти
участки тела оказываются без изменений, мы
говорим о зуде без кожных проявлений. Да-
лее следует обратить внимание, имеется ли
увеличение лимфоузлов, селезенки, экзофтальм
или признаки сахарного диабета и, конечно,
ксероз.
Верная оценка зуда — наиболее трудная зада-
ча, требующая тщательного обследования паци-
ента (табл. 17.1). Подробный сбор анамнеза о
времени начала, течении и интенсивности зуда
обязателен. Следует всегда проводить опрос о
следующих характеристиках зуда: генерализо-
ванный — локализованный; непрерывный —
пароксизмальный; прогредиентный — затухаю-
щий; зависящий от температуры, ситуации, вре-
мени суток.
Следует выяснить, провоцируют ли зуд или
усиливают его такие факторы, как вода, жара,
сухость или влажность, физическая нагрузка,
охлаждение кожи. Всегда следует осведомиться
о пребывании в тропиках, контакте с животны-
ми, о приеме медикаментов, а также об особен-
ностях питания и любимых лакомствах (краси-
тели, добавки, консерванты). Тщательный
анамнез относительно атопии, а также профес-
сии, социального статуса, вплоть до сексуальной
жизни, также должен присутствовать. Даже без
Глава 17. Кожный зуд 241
Таблица 17.1.
Анамнез заболевания при кожном зуде
• Начало (резкое, постепенное)
• Течение (непрерывное, перемежающееся)
• Характер (колющий, жгучий)
• Продолжительность (дни, месяцы)
• Время (циклический, в течение дня, ночью)
• Степень страдания (влияние на повседневную
жизнь)
• Локализация (генерализованный, ограниченный)
• Провоцирующие факторы (вода, температура,
трение)
• Прием лекарственных средств
• Факторы окружающей среды (профессия, гигие-
на, домашние животные)
• Психотравмирующие ситуации в недавнем про-
шлом
• Аллергии, атопия
• Анамнез поездок (места командировок, отдыха)
• Половой анамнез
• Предшествующая терапия
осмотра кожи пациента, простой вопрос о том,
страдают ли зудом другие члены семьи или парт-
неры, уже может указать на инфекционный ге-
нез. Зуд, который успокаивается с засыпанием,
не приводит к нарушению сна и заметно возра-
стает в интенсивности при вставании, указыва-
ет на напряжение, которое может быть психи-
чески обусловлено. Зуд, который не дает
пациентам уснуть, или же вследствие которого
они просыпаются по ночам, скорее вызван сис-
темным заболеванием. Различные кожные забо-
левания характеризуются непрерывным зудом,
как например атопический дерматит, при кото-
ром только наступивший от истощения сон «при-
крывает» приступы зуда. Ночное потение и суб-
фебрильные температуры, связанные с зудом,
почти патогномоничны для болезни Ходжкина
и часто провоцируются вечерним приемом алко-
голя. Эти немногие примеры заостряют внима-
ние врача на правильной постановке вопросов
при сборе анамнеза у пациента с зудом, прежде
всего без типичных кожных проявлений.
Зуд редко бывает непрерывным. Иногда он
зависит от смены температуры, например, при
входе в теплое помещение после пребывания на
холодном воздухе. Зуд может наступать в фор-
ме кризов днем, но чаще усиливается ночью. При
некоторых дерматозах (например, простое подо-
строе пруриго) ограниченное воспаление кожи
расчесывается до тех пор, пока не пойдет кровь,
только тогда зуд прекращается. Часто при экзе-
мах зуд усиливается при расчесах и отступает,
когда пациент истощается после расчесов. Изве-
стен зуд, отсутствующий днем и оживающий
ночью: типичный анамнез для чесотки.
Определение зуда неясного генеза — это
большая проблема дифференциального диагно-
за. Именно в более позднем возрасте следует ду-
мать об одновременном наличии нескольких за-
болеваний, которые могут дать повод для зуда
(возрастной себостаз в комбинации с гормо-
нальными нарушениями, дефицит питания или
злокачественные опухоли). На практике целе-
сообразно проведение пробного местного лече-
ния индифферентными мазями на жировой
основе. Иногда трудно отграничить вторичные
эффекты зуда на коже от первичных дермато-
зов. Воздействие продолжительного зуда на
психику пациента, вызывающее длительную
бессонницу или неврастению, не должно вести
к ошибочному диагнозу психиатрического за-
болевания. Важно установить, нет ли у больного
лимфаденопатии или гепатоспленомегалии, так
как лимфомы могут сопровождаться зудом. Зуд
без высыпаний на коже иногда служит показа-
телем ВИЧ-инфекции, часто сопровождающей-
ся кандидозом полости рта и лимфаденопати-
ей. Иногда при осмотре кожных покровов
ошибочно расценивают высыпания как причину
зуда, хотя на самом деле они представляют со-
бой его результат.
Следующие вспомогательные средства помогут
поставить диагноз при зуде кожи: увеличитель-
ное стекло (возможно, микроскоп), стеклянная
линейка, пара маленьких щипцов, инструмент с
тупым концом (шпатель), зонд, малый анатоми-
ческий пинцет. Дополнительно к визуальному
осмотру врачу понадобится его чувство осязания
(пальпация, общий осмотр), и в некоторых слу-
чаях — его ноготь. До клинического осмотра
следует как можно более детально собрать анам-
нез у пациента. Для самого осмотра пациента
просят полностью раздеться. В случае зудящих
состояний важно проверить каждый сантиметр
кожи больного, даже если он и отрицает нали-
чие каких-либо видимых кожных поражений
(табл. 17.2).
Помимо сбора анамнеза и физического осмот-
ра следует проводить целенаправленную инди-
видуальную программу обследования (табл.
17.3), так как зуд может предшествовать про-
явлению системного заболевания. Пациентов
следует внимательно контролировать и в даль-
нейшем.
Минимальная лабораторная программа для
определения возможных причин зуда при отсут-
242 Частная дерматология
Таблица 17.2.
План осмотра пациента с кожным зудом
• Общий осмотр (температура, потливость, утомля-
емость, потеря веса)
• Кожа (пигментация, сухость, иктеричность, следы
экскориаций)
• Ногти (изменение окраски, дистрофии, онихоли-
зис)
• Глаза (экзофтальм, изменение окраски склер)
• Эндокринная система (тремор, нарушение тер-
морегуляции, полидипсия, полиурия)
• Система крови (анемия, кровоточивость, лимфа-
денопатия)
• Желудочно-кишечный тракт (тошнота, рвота,
стул, недержание кала и цвет)
• Урогенитальная система (цвет мочи, недержание
мочи, менструация, беременность)
• Нервная система (головные боли, парестезии,
зрительные расстройства)
• Психический статус (настроение, нарушения сна,
галлюцинации, бред)'
Таблица 17.3.
План обследования пациента с кожным зудом
• Общий анализ крови
• Биохимический анализ крови (щелочная фосфа-
таза, билирубин, мочевина, креатинин)
• Т4 (тироксин), ТСГ (тироксинсвязывающий глобу-
лин)
• Анализ крови на глюкозу (натощак)
• Анализ крови на железо, ферритин
• Анализ крови на общий белок и белковые фрак-
ции (а1, а2, ₽, у)
• ВИЧ-серология (ИФА-ВИЧ)
• Анализ кала на скрытую кровь
• Анализ кала на яйца гельминтов
• Анализ мочи (5-гидроксииндолуксусная кислота,
17-кетостероиды)
• Биопсия кожи (гистология, иммунофлуоресцен-
ция, электронная микроскопия)
• Рентгенологическое и ультразвуковое исследо-
вание
• Эндоскопия (фиброэзофагогастродуоденоско-
пия, ректороманоскопия, колоноскопия, лапа-
роскопия)
ствии типичных проявлений какого-либо дерма-
тоза должна включать, кроме определения пара-
метров воспаления (СОЭ и С-реактивный бе-
лок), дифференциальный анализ крови с
подсчетом числа эозинофилов и тромбоцитов,
трансаминаз со щелочной фосфатазой и билиру-
бина, а также трансферрина и железа, мочеви-
ны и креатинина, мочевой кислоты и сахара,
кальция и фосфата. Исследование гормонов,
щитовидной и паращитовидных желез, опреде-
ление общего уровня IgE в связи с доказатель-
ством интестинальных паразитозов завершают
программу.
При подозрении на зуд, ассоциированный с
паранеоплазиями, следует провести соответству-
ющие исследования с использованием опухоле-
вых маркеров, а также минимально инвазивные
исследования, такие как рентген грудной клет-
ки и УЗИ.
Иногда помогает определение уровня гистами-
на, серотонина и триптазы (диффузный масто-
цитоз, нефропатия, гепатопатия). При лихено-
идных поражениях биопсия позволит исключить
гранулематозные дерматозы. Исследования, ас-
социированные с инфекциями, должны всегда
проводиться целенаправленно.
Лечение
Недостаточность знаний патофизиологии зуда
объясняет трудность в подборе эффективного
варианта терапии. На первом месте любых тера-
певтических мероприятий в случае зуда должно
быть лечение основного заболевания. К тому же
следует устранить провокационные факторы, та-
кие как сухость кожи, контакт с раздражающи-
ми веществами, мероприятия по обезжириванию
кожи (спиртовые повязки), употребление опре-
деленных продуктов (алкоголь, специи), а также
температурные перепады окружающей среды.
Следует избегать длительного применения потен-
циальных контактных аллергенов (антигистамин-
ных, местных анестезирующих), а также доксе-
пина (потенциальное антигистаминное), который
успешно применялся местно при атопическом
дерматите, но по своей химической структуре и
вследствие усиления Т-клеточной активности
имеет высокую степень сенсибилизации.
Сопутствующие мероприятия (избегание
стрессов, аутогенная тренировка, помощь психо-
лога, коррекция влияния психосоциального ок-
ружения; соответствующая одежда, прием душа,
влажные обертывания; при необходимости жи-
ровые смазки кожи мочевиной, оказывающей
прямое антипруритическое действие) могут смяг-
чить зуд.
В зависимости от основного заболевания целе-
сообразно включить в рецептуру кортикостеро-
иды, анестезирующие средства (фенол, камфора,
ментол, полидоканол), клиоквенол, резорцин,
деготь в соответствующих основах. В качестве
поддерживающей меры можно применять транс-
Глава 17. Кожный зуд 243
кутанную электрическую нейростимуляцию или
акупунктуру. Новое в терапии зуда — примене-
ние капсаицина. Капсаицин представляет собой
алкалоид, добываемый из растения паприки (пе-
рец).
При терапии зуда следует исходить из трех
аспектов. Вначале — это каузальный (причин-
ный) подход, при котором устраняется специфи-
ческий возбудитель болезни. Если это невозмож-
но или же возможно недостаточно быстро, можно
попытаться смягчить симптомы, влияя, напри-
мер, на высвобождение опосредующих зуд ме-
диаторов. Если и это не удается, можно попы-
таться смодулировать факторы, ухудшающие зуд,
чтобы сделать симптомы переносимыми.
Каузальная терапия может успешно устранить
симптомы острых и средних по длительности
форм зуда, если ясны вызывающие его причи-
ны. Так, в большинстве случаев можно устранить
аллерген, вызывающий острую уртикарию или
другую аллергичесукю экзантему (медикаменты;
аллергены в пищевых продуктах; псевдоаллер-
гены, такие как аспирин и пищевые добавки;
физические раздражители, такие как холод, дав-
ление и УФ-лучи). То же касается и контактных
аллергенов. Паразитов можно также устранить,
применяя соответствующие средства наружно
или внутрь. У некоторых пациентов со злокаче-
ственными опухолями в качестве причины зуда
успешное хирургическое или терапевтическое
лечение приводит к исчезновению симптомов с
их возвратом при рецидивах. При лимфопроли-
феративных заболеваниях, согласно последним
сообщениям, и зуд, и сама боль реагируют на а-
интерферон. Успешное лечение других внутрен-
них болезней (хроническая почечная недостаточ-
ность, болезни печени, сахарный диабет) также
ведет к смягчению или устранению зуда. При
зуде с невротическими или психическими нару-
шениями успешной бывает психотерапия с уст-
ранением стрессовых факторов или лечение тран-
квилизаторами, гипнозом или акупунктурой.
В клинической практике при целом ряде за-
болеваний и при многочисленных патологичес-
ких процессах с зудом неизвестной природы
невозможно устранить основное заболевание или
избежать возбудителя. Здесь начинается врачеб-
ное искусство по смягчению зуда либо путем
влияния на действие медиаторов зуда на целе-
вом органе, свободных нервных окончаниях,
либо путем модуляции передачи зуда по пери-
ферическим и центральным нервным проводя-
щим путям.
Гистамин в экспериментальных и патологи-
ческих условиях является единственным надежно
идентифицированным передатчиком зуда. По-
этому при большинстве заболеваний, обуслов-
ленных тучными клетками, можно лечить зуд и
папулезные высыпания антигистаминными пре-
паратами, однако менее успешно — рефлектор-
ное покраснение. Старые антигистаминные пре-
параты, обладающие центральным седативным
эффектом, по своему действию на зуд и пузырь-
ковые высыпания мало отличаются от новых
неседативных препаратов. Первично обусловлен-
ный гистамином зуд бывает при острой и неко-
торых хронических уртикариях, а также при
некоторых формах физической уртикарии, на-
пример, при механической и пигментной крапив-
нице, у большинства пациентов с холинергичес-
кой уртикарией. Зуд при лечении ран, после
укусов насекомых и контакта с растениями (на-
пример, крапивой) обусловлен тучными клетка-
ми или гистамином.
Неседативные антигистаминные средства у
70 % пациентов с хронической уртикарией пол-
ностью подавляют зуд, а у остальных пациен-
тов наступает улучшение. У больных экземой
в большинстве исследований с хорошим конт-
ролем нет реакции на различные антигистамин-
ные средства. Антигистаминные препараты, ко-
торые к тому же влияют на высвобождение
медиаторов из тучных клеток и миграцию эози-
нофилов, демонстрируют, напротив, определен-
ную эффективность при атопическом дермати-
те (цетиризин, лоратадин). В целом,
антигистаминные средства классифицируются
при лечении зуда в случае экземных заболева-
ний как мало- или неэффективные. Местные
антигистаминные препараты при этих же забо-
леваниях действуют в ограниченном масштабе,
а при лечении детей их следует избегать из-за
потенциального системного побочного действия
(контактной сенсибилизации) при применении
на больших площадях.
Слабое действие антигистаминных средств при
многих воспалительных дерматозах контрасти-
рует с быстрой реакцией зуда на глюкокортико-
иды, причем наряду с симптомами зуда подав-
ляются и другие параметры воспаления. При
отсутствии противопоказаний кортикостероиды
применяются системно при острых заболевани-
ях (острая уртикария, острая контактная экзе-
ма). При хронических заболеваниях они, напро-
тив, противопоказаны, кроме краткосрочного
лечения обострений.
Фотохимиотерапия (ПУВА) может уменьшить
зуд при некоторых заболеваниях, обусловленных
действием тучных клеток и воспалением. В ка-
честве примера служит узловатое пруриго, пара-
244 Частная дерматология
неопластический зуд, пигментная крапивница и
гиперэозинофильный синдром. При световой
крапивнице УФ-терапия применяется скорее в
смысле «закалки» кожи или индукции перено-
симости. Действие УФ-терапии непродолжитель-
но, оно лишь немного превышает длительность
лечения, а кроме того, сама ПУВА-терапия у
некоторых пациентов может стать причиной
зуда.
Циклоспорин А действует уже в малых дозах
(5 мг/кг веса в день) при экземе, крапивнице,
атопическом дерматите, смягчая зуд, но он так-
же небезупречен, поскольку после отмены пре-
парата быстро наступает рецидив. Кроме того,
это потенциально нефротоксичное средство.
При внутригепатическом холестазе при сниже-
нии эндогенных желчных кислот в сыворотке, в
частности, холиевой кислоты, в результате лече-
ния холестирамином или урсодезоксихолевой
кислотой хронический зуд вместе со щелочной
фосфатазой значительно снижается. По после-
дним данным, у взрослых и детей холестатичес-
кий зуд прекрасно реагирует на рифампицин, хотя
следует принимать во внимание его высокую
степень побочного действия, возможное перекре-
стное действие с другими медикаментами и от-
носительно высокую стоимость терапии. Упоми-
нается об относительно хорошем эффекте
холестирамина, действие которого еще можно
повысить одновременным применением УФ-те-
рапии. Умеренно хорошо помогают антагонисты
морфина (налаксон, налмефен) и плазмаферез.
Оперативные мероприятия (отвод желчной жид-
кости — стома, трансплантация печени при соот-
ветствующих показаниях) резко улучшают сим-
птоматику зуда.
При терапии зуда, помимо общих модулиру-
ющих мероприятий, имеет значение снижение
воспаления. При этом не играет решающей роли,
как это будет происходить: путем применения
наружных глюкокортикоидов или, при очень
тяжелом течении, с помощью системной терапии
циклоспорином. Важно уменьшение Т-клеточ-
ной инфильтрации с последующим высвобож-
дением медиаторов воспаления в эпидермис.
Ксероз при атопии является еще одним аспек-
том лечения и требует применения веществ, ко-
торые связывают воду. Поэтому принципиаль-
но показана мочевина, а также деготь, который
смягчает зуд и ослабляет гиперпролиферацию ке-
ратиноцитов и лихенификацию. Зуд при острой
воспаленной коже и при хроническом состоянии
атопического дерматита следует лечить различ-
но. Подострые стадии можно лечить УФ-облу-
чением, однако иногда УФА!-облучение приво-
дит к подавлению воспаления и зуда и в острой
стадии. Если же применяются антигистаминные
средства, то преимущественно вечером и только
седативного типа.
Больным с кожным зудом рекомендуется те-
рапия сопутствующих заболеваний. Назначают
диету с исключением облигатных и индивиду-
альных трофоаллергенов: яичный белок, мяс-
ной бульон, шоколад, пряности, сладости, ал-
коголь; ограничивают применение поваренной
соли, копченых и консервированных продуктов.
Показаны молочно-кислые, растительные про-
дукты.
В качестве симптоматической терапии могут
быть использованы седативные (препараты ва-
лерианы, пустырника, транквилизаторы); анти-
гистаминные (супрастин, фенкарол, диазолин,
эролин, лоратадин); десенсибилизирующие (ге-
модез, препараты кальция, тиосульфат натрия);
анестезирующие средства (0,5 % раствор ново-
каина, 1 % раствор тримекаина); энтеросорбенты
(белосорб, активированный уголь, полисорб, по-
лифепан).
Местная терапия. Местно применяются при-
сыпки, спиртовые и водные растворы, взбалты-
ваемые взвеси, пасты, мази. Противозудный
эффект зависит от лекарственной формы. Про-
тивозудным локальным эффектом обладают сле-
дующие средства различных составов: 0,5—2,0 %
ментол; 1—2 % тимол; 1—2 % анестезин; 1—2 %
фенол (карболовая кислота); спирты (1—2 % ре-
зорциновый, 1—2 % салициловый, камфорный;
30—70 % этиловый); 1—2 % растворы лимонной
кислоты; настои ромашки, травы череды. При
отсутствии эффекта зудящие участки можно не-
продолжительное время смазывать кортико-
стероидными мазями (локоид, элоком, адвантан,
флуцинар, фторокорт).
Противозудное действие оказывают сероводо-
родные ванны; ванны с отваром дубовой коры,
череды (50—100 г), отрубей (300—500 г на ван-
ну); морские купания; ванны с хвойным экст-
рактом, морской солью, крахмалом. Температу-
ра воды 38°С, продолжительность процедуры
15—20 мин, на курс 10—20 ванн.
Показаны гипноз, электросон, акупунктура,
лазеропунктура, магнитотерапия, КВЧ-терапия,
биоритморефлексотерапия, фонофорез гидрокор-
тизона.
Ретиноиды у пациентов с атопической пред-
расположенностью могут провоцировать зуд
вместо его снижения. Однако при красном плос-
ком лишае, склеротическом и атрофическом
лишае зуд исчезает в течение нескольких дней
даже при малых дозах (этретинат или изотрети-
Глава 17. Кожный зуд 245
ноин по 10—20 мг в сутки). Кожные проявле-
ния, напротив, не обязательно реагируют на пре-
парат. То же характерно для местного лечения
2 % эстрогеном или тестостероном на основе
крема.
При лечении широко распространенного пе-
рианального зуда вначале следует устранить при-
чину заболевания и нормализовать гигиену
анальной области. В питании избегать раздража-
ющих веществ: цитрусовых и пряностей. Далее
рекомендуются инъекции 5 % фенола в миндаль-
ном масле в подкожную ткань дистального от-
дела ануса; в 90 % случаев этот метод дает выз-
доровление.
Если причина зуда неизвестна или исчерпа-
ны вышеперечисленные терапевтические возмож-
ности, можно применить мероприятия по сни-
жению зуда. Сюда относится, прежде всего,
регулярный уход за кожей жирными наружны-
ми средствами, особенно в пожилом возрасте. У
пациентов с аквагенным зудом это является
средством выбора.
Приступ зуда может быть существенно сни-
жен самими пациентами путем аутогенного тре-
нинга. Пациентов с зудом следует опросить, как
они привыкли мыться. Слишком частое мытье
горячей водой, чрезмерное употребление мыла
ведет к уменьшению естественной жировой смаз-
ки и сухости кожи, что благоприятствует зуду.
Теплый сухой воздух от нагревательных прибо-
ров и тепло постельного белья являются факто-
рами, ухудшающими состояние больного. Паци-
енты часто сообщают об облегчении ночных
приступов зуда после приема холодного душа.
Следует также дополнительно использовать жи-
ровую смазку на кожу.
Обобщая вышеизложенное, можно сказать, что
зуд контролируется современными фармаколо-
гическими средствами лишь ограниченно. Ис-
ключениями являются хорошо реагирующие на
антигистаминные препараты уртикарные реак-
ции и недавно открытые терапевтические воз-
можности лечения хронического зуда при почеч-
ной недостаточности и заболеваниях печени. Зуд
при хронической экземе реагирует на кортико-
стероиды, но побочные действия неприемлемы
для долговременной терапии. В целом, поиски
причины зуда являются важной базой для успеш-
ной целевой терапии. К тому же зуд у большин-
ства пациентов может быть, по крайней мере,
облегчен умелым применением имеющихся в
настоящее время терапевтических методов и
средств.
Глава 18
Атопический дерматит
Атопический дерматит — наследственное забо-
левание мультифакториальной природы с гене-
тически детерминированным дефицитом функ-
ции Т-лифоцитов супрессоров, одновременной
частичной блокадой p-адренергических рецепто-
ров и В-зависимым IgE-глобулиновым механиз-
мом патологических иммунных реакций. Атопи-
ческий дерматит характеризуется возрастной
вариабельностью, хроническим рецидивирую-
щим течением, зудящими воспалительными по-
ражениями кожи с истинным полиморфизмом
(эритемы, папулы, везикулы), лихенификацией;
симметричной топографией высыпаний, завися-
щей от эволютивной динамики; часто сочетает-
ся с функциональными расстройствами нервной
системы, нарушениями иммунитета, атопически-
ми поражениями респираторных органов.
Терминология
Среди дерматологических заболеваний атопичес-
кий дерматит по причине окончательно невыяс-
ненного этиопатогенеза, хронического течения и
связанных с этим терапевтических проблем за-
нимает особое место в дерматологии. В литера-
туре насчитывается около ста обозначений это-
го заболевания. В отличие от английской и
французской литературы, в которой утвердилось
понятие «атопический дерматит» или «атопичес-
кая экзема», в немецких источниках более час-
то употребляются термины «атопическая экзема»,
«эндогенная экзема», «диффузный нейродер-
мит», «атопический нейродермит». Такой терми-
нологический калейдоскоп затрудняет работу
практических врачей и создает путаницу в иден-
тификации заболевания. Мы рекомендуем при-
держиваться двух равноценных и однозначных
терминов: «атопический дерматит» и «атопичес-
кий нейродермит», хотя в англоязычных руко-
водствах по дерматологии часто употребляют
также наименование «атопическая экзема».
Сложность в применении понятия «атопичес-
кое заболевание» состоит в том, что при аллер-
гическом рините, аллергическом конъюнктиви-
те и аллергической бронхиальной астме речь идет
о IgE-опосредованных аллергических реакциях
немедленного типа (тип I по Coombs и Gell), а
при атопическом дерматите, весьма вероятно,
имеет место комплексное взаимодействие не-
скольких иммунологических и неиммунологичес-
ких факторов, которые частично еще неизвест-
ны. Из этого факта вытекают и существующие
до сегодняшнего дня трудности с терминологи-
ей. Наименование нейродермит, предложенное
Brocq в 1891 году, указывает на предположитель-
ную патогенетическую связь с нервной системой,
поскольку сильный зуд рассматривался как фак-
тор, провоцирующий заболевание. Применяемые
с этим наименованием синонимы конституцио-
нальный или атопический нейродермит указыва-
ют, в частности, на патогенетическое значение
семейных или наследственных факторов, в то
время как названия атопическая экзема, эндоген-
ная экзема или конституциональная экзема ори-
ентируются больше на экзематозные высыпания.
Эпидемиология
Частота атопических заболеваний возрастает.
Они поражают примерно от 5 до 20 % населе-
ния, чаще всего выражаются в виде аллергичес-
кого ринита и атопического дерматита (примерно
50 %) и значительно меньше — в виде бронхи-
альной астмы. Атопический дерматит проявля-
ется в большинстве случаев уже в младенческом
возрасте, часто между 2 и 3 месяцем жизни. За-
болевание может возникнуть и в более позднем
детском возрасте. По оценке ученых, атопичес-
кий дерматит стоит на восьмом месте по часто-
те среди всех дерматозов у лиц младше 25 лет.
Заболевание встречается в грудном возрасте, ран-
нем детском, у подростков и взрослых. Лица
мужского пола чаще болеют в грудном и детс-
ком возрасте, а женщины — в старшем детском
и взрослые. Первичные проявления атопическо-
го дерматита после периода полового созревания
относительно редки.
Тип наследования
Тип наследования во всех деталях пока неясен,
не связан с отдельным геном. Влияние HLA —
системы также, по-видимому, отсутствует. Веро-
246
Глава 18. Атопический дерматит 247
ятность заболевания для ребенка, имеющего од-
ного из родителей с атопией, оценивается в 25—
30 %. Если оба родителя атопики, она существенно
увеличивается и составляет 60 %. Вероятно нали-
чие полигенного типа наследования. Наследует-
ся не определенное атопическое заболевание, а
предрасположенность к атопической реакции
различных систем. Примерно у 60—70 % пациен-
тов имеется положительный семейный анамнез на
атопию. По этой причине тщательный сбор се-
мейного и индивидуального анамнеза с учетом
атопических заболеваний имеет диагностическое
значение для определения атопического дермати-
та. Кроме наследственной предрасположенности,
важную роль играют также экзогенные, индиви-
дуально реализуемые факторы. Среди факторов
окружающей среды, провоцирующих атопические
заболевания дыхательных путей или кишечника,
значение имеют не только ингаляционные (кле-
щи домашней пыли, пыльца растений, шерсть
животных) или пищевые (нередко вместе с аллер-
гической крапивницей) аллергены — такие как
молочный белок, фрукты, яйца, рыба, консерван-
ты, но и такие индивидуальные факторы, как
стресс или сопутствующие психовегетативные и
психосоматические нарушения.
Примерно в 30 % случаев наблюдается вульгар-
ный ихтиоз, с еще большей частотой — сухая
кожа (астеатоз, себостаз) с измененным содержа-
нием липидов и повышенной водопроницаемос-
тью (нарушение барьерной функции). Многие
пациенты имеют типичную ихтиозную ладонь с
сильно выраженным линейным рисунком — ги-
перлинеарность. Витилиго чаще проявляется у
пациентов с атопическим дерматитом, а гнездная
алопеция у таких пациентов имеет неблагопри-
ятный прогноз (атопический тип алопеции). При-
мечательно также образование, хотя и очень ред-
кое, глазных аномалий, таких как атопическая
катаракта, особенно у молодых людей, реже ке-
ратоконус. Имеется связь с дисгидрозом, дисгид-
ротической экземой ладоней и уртикарией. Дис-
кутируется связь с мигренью, однако она не
считается достоверно установленной.
Этиология
Причина атопического дерматита до сегодняш-
него дня не совсем ясна. В частности, не выяс-
нено, почему заболевание начинается уже в мла-
денческом возрасте и так различно протекает у
разных больных. По сути, атопический дерма-
тит в раннем детском возрасте характеризуется
картиной воспалительно-экссудативной экземы
(гиперемия, пузырьки, корочки), а позднее в
клинической картине преобладает воспалитель-
но-пролиферативное и экскориированное
утолщение кожи (лихеноидные папулы, лихе-
нификация и пруригинозные папулы). Этот
макро- и микроморфологический субстрат
нельзя объяснить только аллергической реак-
цией немедленного типа, о которой можно
было бы думать из-за часто повышенного уров-
ня IgE в сыворотке. Определенную роль иг-
рают также замедленные клеточно-опосредо-
ванные реакции экзематозного типа, как это
показали многочисленные исследования.
Точная причина атопического дерматита не-
известна. Предлагается две теории: иммуноло-
гическая гипотеза; гипотеза блокады бета-ад-
ренергических рецепторов.
Иммунологическую теорию подтверждают
несколько фактов: частая ассоциация атопи-
ческого дерматита с другими аллергическими
заболеваниями; существенное повышение сы-
вороточного IgE; положительные реакции на
широкий спектр кожных тестов; склонность к
вирусным и грибковым инфекциям; сочетание
с иммунодефицитными заболеваниями.
Теория блокады бета-адренергических ре-
цепторов предполагает, что дисфункция бета-
рецепторов вызывает гиперреактивность, пус-
ковым механизмом которой могут быть
различные стимулы: физические, иммуноло-
гические, инфекционные, химические, психи-
ческие.
Патогенез
Иммунологическая теория привлекает больше
внимания, но явления, вызывающие реакцию,
еще предстоит идентифицировать. Присутству-
ют аномалии как гуморального, так и клеточ-
но-опосредованного иммунитета. Очевидно, IgE
стимулируется специфическими антигенами.
Он расположен на тучных клетках и вызывает
высвобождение из них воспалительных меди-
аторов. В пользу клеточно-опосредованных
факторов свидетельствует подверженность ви-
русным инфекциям и их рецидивирование,
включая простой герпес, контагиозный мол-
люск и бородавки. Пациенты часто устойчивы
к сенсибилизации динитрохлорбензола. Нали-
чие сниженного количества Т-лимфоцитов
может указывать на нехватку важнейших суб-
популяций Т-клеток, контролирующих произ-
водство иммуноглобулина В-клетками и плаз-
моцитами так, что уровни производства IgE
оказываются высокими. К тому же уменьше-
на фагоцитарная активность и нарушен хемо-
таксис нейтрофилов и моноцитов. Еще одним
фактором, поддерживающим иммунологичес-
9 Зак. 1357
248 Частная дерматология
кую основу, является наличие значительного
количества стафилококков как на больной, так
и на здоровой коже больных атопическим дер-
матитом.
Бета-адренергическая теория поддерживается
рядом аномальных кожных ответов. Они вклю-
чают избыточный констрикторный ответ кож-
ных сосудов, белый дермографизм, замедленное
побледнение на холинергические стимулы и па-
радоксальный ответ на применение никотино-
вой кислоты. Пониженные уровни цАМФ мо-
гут увеличивать выделение медиаторов из
тучных клеток и базофилов.
Нарушения гуморального иммунитета. Лица с
наследственной предрасположенностью к атопии
реагируют на контакт с веществами окружаю-
щей среды (аллергенами) сенсибилизацией не-
медленного типа. Такая сенсибилизация подтвер-
ждается уртикарной реакцией немедленного типа
в ходе интракутанного теста. Иммунологически
речь идет об аллергической реакции немедлен-
ного типа (тип I по Coombs & Gell). Здоровый
человек не реагирует на контакт с такими встре-
чающимися в окружающей среде веществами.
Однако суть атопического дерматита нельзя све-
сти только к одному такому аллергическому
способу реакции атопического организма.
У пациентов уже в период раннего детства
путем кожного теста обнаруживаются позитив-
ные реакции немедленного типа на пищевые и
ингаляционные аллергены. Число положитель-
ных кожных реакций составляет от 50 до 90 %.
Пациенты с аллергической бронхиальной аст-
мой или аллергическим ринитом значительно
чаще имеют положительные интракутанные
реакции на ингаляционные аллергены, в част-
ности домашнюю пыль, клеща домашней пыли
( Dermatophagoides pteronyssinus), пыльцу расте-
ний или животные аллергены (шерсть и перхоть
животных). Перхоть человека и протеины пота
также могут действовать как аллергены. Хотя
причинное значение ингаляционных аллергенов
как провокаторов ухудшения атопического дер-
матита еще не совсем ясно, любому дерматоло-
гу известно, что сезонное обострение аллерги-
ческого ринита сопровождается ухудшением
кожных проявлений, и наоборот. Аллергены
пищевых продуктов (белок в молоке, рыба,
мука, фрукты, овощи) также часто дают поло-
жительные тестовые реакции, хотя они не все-
гда совпадают с клиническими симптомами.
Кроме того, матери часто отмечают тот факт,
что зуд и воспалительные кожные реакции у их
младенцев часто провоцируются определенны-
ми пищевыми продуктами (например, молоком
или цитрусовыми). Перспективные исследова-
ния показывают, что кормление ребенка мате-
ринским, а не коровьим молоком в первые
недели жизни положительно влияет на детей-
атопиков; поэтому в первые месяцы жизни ре-
комендуется именно материнское молоко. Кроме
того, внешний контакт с пыльцой растений
может вызывать воспалительные кожные реак-
ции и провоцировать пыльцевой вульвит у ма-
леньких девочек.
Итак, в целом, несмотря на то, что патогене-
тическое значение реакций немедленного типа
для развития атопического дерматита еще не
полностью оценено, ряд данных говорит в его
пользу. Показаны также соответствующие внут-
рикожные и in vitro — тесты (RAST), причем
тестовые реакции следут рассматривать крити-
чески, во взаимосвязи с общей клинической
картиной, что может послужить поводом для
возможного проведения дальнейших мероприя-
тий, таких как экспозиционные тесты или эли-
минационная диета.
Определение IgE в настоящее время проводит-
ся чаще всего посредством метода PRIST. У
большинства пациентов с тяжелым атопическим
дерматитом уровень IgE в сыворотке повышен.
Повышение уровня IgE регистрируется особен-
но при одновременных проявлениях в дыхатель-
ных путях (аллергическая астма, аллергический
ринит). Однако поскольку у некоторых отдель-
ных пациентов с распространенным поражени-
ем кожи уровень IgE может находиться в пре-
делах нормы, определение его, кроме случаев
подозрения на синдром гипер-IgE, не имеет па-
тогномоничного значения, тем более что и при
других воспалительных дерматозах происходит
повышение уровня IgE в сыворотке. Поэтому
отсутствие IgE в сыворотке не означает, что ато-
пического дерматита нет. Примечательным яв-
ляется также снижение повышенных показате-
лей IgE при ремиссиях заболевания.
В последние годы благодаря современным имму-
нологическим методам достигнуто лучшее понима-
ние регуляции образования IgE. Определенные
цитокины, которые образуются активизированны-
ми Т-лимфоцитами, в частности, интерлейкином-
4 (IL-4) и интерфероном-у (INF-у), вовлечены в
комплексную сеть регуляторных сигналов синтеза
IgE посредством В — лимфоцитов. Дальнейшие ис-
следования в этой области смогли бы показать те-
рапевтические последствия, если бы удалось затор-
мозить перепроизводство IgE.
Методика RAST дает в распоряжение врача
способ доказательства in vitro наличия аллерген-
специфических антител к сыворотке крови па-
Глава 18. Атопический дерматит 249
циента. Этим способом можно доказать наличие
антител класса IgE к ряду ингаляционных и
пищевых аллергенов. При атопическом дермати-
те RAST или САР положительны в большом
проценте случаев; этими методиками можно
доказать наличие циркулирующих антител на
аллергены окружающей среды, которые не были
охвачены интракутанным тестом.
Нарушения клеточного иммунитета. У пациен-
тов с атопическим дерматитом, кроме наруше-
ний гуморального иммунитета, имеет место и
ослабление клеточного иммунитета. Примеча-
тельно, что такие пациенты подвержены вирус-
ным, бактериальным и грибковым кожным ин-
фекциям. Эти инфекции, с одной стороны, у
атопиков встречаются чаще, а с другой стороны,
протекают более тяжело. В качестве осложнений
подобного рода известны eczema verrucatum,
eczema molluscatum, eczema coxsaccium, а также
impetigo contagiosa и tinea corporis. При тяжелом
течении атопического дерматита было доказано
явное уменьшение розеткообразования эритро-
цитов, изменение реакции Т-лимфоцитов на
митогены, снижение стимулируемости in vitro
лимфоцитов с бактериальными и микотически-
ми антигенами и понижение склонности к кон-
тактной сенсибилизации (однако при повышен-
ной распространенности контактной аллергии на
никель), снижение числа или активности есте-
ственных клеток-киллеров. С тяжестью заболе-
вания коррелирует и уменьшение супрессорных
Т-лимфоцитов. Из практики известен тот факт,
что пациенты имеют незначительную тенденцию
к развитию контактного дерматита после мест-
ного применения медикаментов. И, наконец,
доказаны дефекты нейтрофильных гранулоци-
тов (хемотаксис, фагоцитоз) и моноцитов (хемо-
таксис). Эозинофилы крови увеличиваются и
сильнее реагируют на стресс. Очевидно, коли-
чество IgE-несущих лимфоцитов также повыше-
но. Интерпретация этих данных достаточно
сложна. Гипотеза исходит из того, что чрезмер-
ное образование IgE у пациентов с атопическим
дерматитом обусловлено имеющимся, особенно
в первые три месяца жизни, секреторным дефи-
цитом IgA, и оно не может быть компенсирова-
но из-за дефицита супрессорных Т-лимфоцитов.
В этом смысле первопричинный дефект следует
искать в системе Т-лимфоцитов. Можно пред-
ставить, что вследствие нарушения торможения
активности Т-лимфоцитов могут спонтанно раз-
виваться воспалительные изменения на коже,
как это происходит при контактном аллергичес-
ком дерматите. В пользу этой гипотезы говорят
и результаты новейших исследований.
Значительную роль в возникновении кожных
изменений при атопическом дерматите могут
играть также IgE-несущие антиген-презентиру-
ющие клетки в эпидермисе, т. е. клетки Лангер-
ганса. Предполагают, что антигенспецифичные
молекулы IgE, связанные на поверхности эпи-
дермальных клеток Лангерганса через высоко-
аффинный рецептор, аэроаллергены (антигены
клеща домашней пыли с поверхности кожи) и
пищевые аллергены взаимодействуют через кро-
воток. Затем они, подобно другим контактным
аллергенам, презентируются клетками Лангер-
ганса аллергенспецифическим лимфоцитам, ко-
торые вызывают воспалительную аллергическую
реакцию экзематозного типа. Эта новая концеп-
ция патогенеза атопического дерматита образу-
ет мостик между гуморальным (IgE-опосредуе-
мым) и клеточным звеньями иммунного ответа
и находит клиническое подтверждение в том
отношении, что эпикутанные тесты с ингаляци-
онными аллергенами (например, растительной
пыльцой) у пациентов с атопическим дермати-
том, в отличие от здоровых лиц, могут приво-
дить к экзематозной кожной реакции в области
проведения теста.
Нарушения вегетативной нервной системы.
Наиболее известен белый дермографизм, т. е.
сужение сосудов после механической нагрузки
на кожу на ее видимо не измененных участках.
Кроме того, после нанесения эфира никотино-
вой кислоты реактивно возникает не эритема,
а анемия вследствие контракции капилляров
(белая реакция). Инъекция холинергических
фармакологических средств, таких, как ацетил-
холин, также приводит к белой окраске кожи
в месте инъекции. Разумеется, белый дермог-
рафизм нехарактерен для воспаленных участков
кожи. Склонность к контракции сосудов у та-
ких пациентов проявляется и при сравнитель-
но низкой температуре кожи пальцев и силь-
ной контракции сосудов после воздействия
холода. Наверняка неизвестно, идет ли здесь
речь об аномальной чувствительности а-адре-
нергической стимуляции мышечных волокон. В
связи с этим стала известна теория Szentivanzy
о p-адренергической блокаде. Торможение р-
рецепторной активности дает в результате сни-
женный реактивный подъем цАМР клеток с
усиленной тенденцией к образованию медиато-
ров воспаления. Нарушением баланса между а-
и p-адренергическими рецепторами можно, ве-
роятно, объяснить и повышенную чувствитель-
ность гладких мышечных клеток в зоне крове-
носных сосудов и пиломоторов. Отсутствие
цАМР-индуцированного торможения синтеза
250 Частная дерматология
антител может привести к увеличению их об-
разования. Кроме того, в основе фармакологи-
ческих и иммунобиологических нарушений
может лежать общая причина.
Другие функциональные нарушения.
Себостаз (астеатоз). Уменьшение продукции
сальных желез — типичное явление для паци-
ентов с атопическим дерматитом. Кожа сухая и
чувствительная, она склонна при частом мытье
и/или приеме душа к дальнейшему высыханию
и зуду. Отсюда понятна и слабая склонность
таких пациентов к заболеваниям себорейного
круга, таких как вульгарное акне, розацеа или
себорейная экзема. В основе сухости и чувстви-
тельности кожи лежат, вероятно, также нару-
шения образования эпидермальных липидов
(керамидов) или нарушение в обмене веществ
эссенциальных жирных кислот (дефицит 8-6-
дезатуразы), что может иметь иммунологичес-
кие последствия. На аномалии в обмене веществ
эссенциальных жирных кислот опирается реко-
мендуемая диета с содержанием у-линолиевых
кислот.
Нарушения потообразования. Такие нарушения
не доказаны наверняка. Скорее, имеют место
нарушения потоотделения. Многие пациенты
жалуются на сильный зуд при потении. Возмож-
но, потоотделению препятствуют нарушения в
роговом слое (гиперкератоз и паракератоз), так
что пот после выхода из выводных протоков
потовых желез в окружающую кожу инициирует
воспалительные реакции (синдром задержки
потоотделения). Пот также содержит IgE и ме-
диаторы воспаления и может вызвать реакции
рефлекторного покраснения и крапивницы.
Климатические аллергены. В качестве причин
развития атопического дерматита рассматрива-
лись и так называемые климатические аллерге-
ны. В горах на высоте более 1500 м над уров-
нем моря или на побережье Северного моря
пациенты чаще всего чувствуют себя очень хо-
рошо, однако лежащие в основе этого патофи-
зиологические процессы трудно обобщить. По-
мимо аллергологических факторов, значение
могут иметь уровень инсоляции и состояние
психической релаксации.
Нервно-психологические факторы. Они игра-
ют очень важную роль. Действие стресса или
других психологических факторов можно пред-
ставить через систему аденилциклаза-цАМФ. -
Пациенты с атопическим дерматитом часто
представляют собой индивидуумов астеничес-
кого типа, имеют уровень образованности выше
среднего, склонны к эгоизму, неуверенности в
себе, конфликтным ситуациям типа «мать-ре-
бенок», в которых доминирующей является
мать, страдают от фрустрации, агрессии или
подавленных состояний страха. Остается откры-
тым вопрос, что здесь первично, а что вторич-
но. Однако сильно зудящие кожные проявле-
ния могут также участвовать в формировании
личности и чувствительно влиять, особенно у
детей, на их развитие и успехи в школе.
Бактерии. Пациенты с атопическим дермати-
том склонны к стафилококковым поражениям
кожи и могут иметь в сыворотке повышенное ко-
личестве стафилококковых IgE-антител. Патоге-
нетическое значение этого факта не выяснено,
однако его надо учитывать при проведении ле-
чения.
Подводя итог вышеизложенному, следует от-
метить, что современные данные свидетельствуют
об иммунологической основе атопического дерма-
тита. Специфичные для атопии клетки Т-хелпе-
ры могут играть патогенетическую роль, произ-
водя и высвобождая цитокины, релевантные для
аллергического воспаления, такие как ИЛ-4, ИЛ-
5 и другие факторы. Предполагается, что эози-
нофилы играют главную роль в качестве клеток-
эффекторов, опосредующих патогенетически
значимую позднофазную реакцию, которая ас-
социируется со значительными разрушениями
окружающей ткани. Соответственно была обна-
ружена значительная преактивация эозинофилов
периферической крови у пациентов с атопичес-
ким дерматитом, что приводит к повышенной
чуствительности этих клеток к определенным
стимулам, таким, как ИЛ-5. Токсичные проте-
ины, такие как эозинофильный катионный про-
теин, содержащиеся в матрице и ядре вторичных
гранул эозинофилов, могут играть важную роль
в распространении аллергического воспалитель-
ного процесса как опосредованно, так и непос-
редственно, благодаря своим иммуномодулиру-
ющим свойствам.
У больных атопическим дерматитом увеличе-
но содержание «эозинофилов — долгожителей»,
которые in vitro имеют длительный период рас-
пада и менее подвержены апоптозу. Длительный
рост in vitro стимулировался IL-5 и GM-CSF;
содержание обоих медиаторов повышено при ато-
пическом дерматите. Эозинофилы с длительным
жизненным циклом могут быть характерным
признаком атопического дерматита, поскольку
эозинофилы от больных с гиперэозинофильным
синдромом не проявляют подобных свойств in
vitro.
Патогенетическая роль эозинофилов при ато-
пическом дерматите подтверждается обнаруже-
нием протеинов, содержащихся в их гранулах в
Глава 18. Атопический дерматит 251
экзематозной коже пациентов. Более того, совре-
менные данные указывают на значительную
корреляцию между активностью заболевания и
накоплением (отложением) содержимого эозино-
фильной гранулы:
• уровни сывороточного эозинофильного ка-
тионного протеина у пациентов с атопическим
дерматитом были значительно повышены;
• уровни эозинофильного катионного про-
теина коррелировали с активностью заболева-
ния;
• клиническое улучшение ассоциировалось
как с уменьшением клинической оценки актив-
ности заболевания, так и с падением уровня
эозинофильного катионного протеина.
Эти данные четко указывают на то, что ак-
тивизированные эозинофилы участвуют в аллер-
гическом воспалительном процессе при атопи-
ческом дерматите. Следовательно, изменение
активности эозинофилов может быть важным
критерием при выборе фармакологических пре-
паратов для лечения атопического дерматита в
будущем.
Первым и основным аспектом патогенеза ато-
пического дерматита является аллергический дер-
матит. Внутрикожным или накожным введени-
ем различных аллергенов у большинства
пациентов с атопическим дерматитом, имеющих
только кожные поражения, получено 80 % по-
ложительных реакций. Главную роль при ато-
пическом дерматите играют следующие аллерге-
ны: аэроаллергены (клещ домашней пыли,
плесени, шерсть животных, пыльца), живые
агенты (стафилококки, дерматофиты, питирос-
порум орбикуляре), контактные аллергены
(аэроаллергены, никель, хром, инсектициды),
пищевые аллергены. Из всех специфических
аэроаллергенов аллергены клеща домашней пыли
могут вызывать специфическую воспалительную
реакцию у большей части больных атопическим
дерматитом, особенно у лиц старше 21 года.
Пищевые продукты являются важными аллер-
генами при атопическом дерматите в раннем дет-
ском возрасте.
Клиническая картина
Атопический дерматит — мультифакториальное
заболевание, имеющее хроническое рецидивиру-
ющее течение с патологически измененными
иммунными ответами и многовариантной кар-
тиной клинических проявлений. Атопический
дерматит представляет собой довольно разнооб-
разное по морфологическому аспекту и общему
течению воспалительное заболевание кожи с
сильным зудом, которое связано с наследствен-
ными факторами и часто встречается в семье па-
циента или же сопровождается другими атопи-
ческими заболеваниями немедленного типа, та-
кими как аллергический ринит, аллергический
конъюнктивит и аллергическая бронхиальная
астма. Морфологическая выраженность заболе-
вания обычно меняется в зависимости от возра-
ста пациента и остроты кожных проявлений.
Клиника атопического дерматита характери-
зуется зудящими воспалительными поражения-
ми, хроническим рецидивирующим течением,
возрастной динамикой клинической картины, се-
зонностью проявлений, частыми осложнениями
вторичной кожной инфекцией, лимфаденопати-
ей, нередким сочетанием с респираторной ато-
пией и некоторыми моногенными дерматозами,
особенно с вульгарным аутосомно-доминантным
ихтиозом. При этом заболевании имеет место
сложность клинико — морфологического синд-
рома с истинным полиморфизмом сыпи, множе-
ством вторичных элементов, фоновыми патоло-
гическими изменениями кожи и осложнениями,
с наличием ряда клинических форм (эритематоз-
но-сквамозная, эритематозно-сквамозная с ли-
хенификацией, лихеноидная, пруригоподобная).
Характерным признаком атопического дерма-
тита является зуд, который может быть непро-
должительным, легким и мучительным тягост-
ным. Зуд начинается либо в форме кризиса, либо
существует длительно в экзематизированных уча-
стках. На него реагируют сильными расчесами,
поэтому штрихообразные расчесы предсталяют
собой важный симптом. Особенно сильным мо-
жет быть неосознанный зуд по ночам. Отдель-
ные типы высыпаний, такие как пруригинозные
папулы на голове и при пруригоформном ато-
пическом дерматите, расчесываются. Субъектив-
ное чувство облегчения наступает с появлением
кровоточения. При распространенных проявле-
ниях заболевания может возникать дермопати-
ческая лимфаденопатия, а при вторичной бак-
териальной инфекции — также лимфангит и
лимфаденит. Своеобразным признаком клини-
ческой картины атопического дерматита является
постепенно нарастающая лихенификация кожи
и лихеноидные папулезные высыпания. Дли-
тельный срок после начала зуда кожа остается
здоровой на вид и лишь постепенно на ней по-
являются своеобразные изменения, свойствен-
ные атопическому дерматиту. После интенсив-
ного зуда и расчесывания вначале появляются
эритематозные, уртикарные и папулезные высы-
пания, а затем в относительно короткий срок раз-
вивается лихенификация с нечеткими граница-
ми, занимающая симметрично латеральные и
252 Частная дерматология
заднюю поверхности шеи, лицо, передние лок-
тевые и задние области коленей, грудь, боковые
участки туловища, внутреннюю поверхность
бедер, промежность, половые органы (рис. 18.1).
В течении атопического дерматита выделяют три
возрастных периода или фазы заболевания (Су-
ворова К. Н. и соавт., 1989; 1990).
• Первый период — до 3-х лет (грудной и ран-
ний детский возраст).
• Второй период — младший дошкольный,
школьный и подростковый возраст.
• Третий период — пубертатный (юношеский)
и взрослый.
Клинические проявления атопического дерма-
тита в каждом из этих трех периодов имеют
определенные отличительные особенности, свя-
занные не с давностью заболевания, а с возрас-
том больного.
Клиника проявлений I возрастного периода.
Острый и подостровоспалительный характер
поражений с тенденцией к экссудативным из-
менениям; определенный тип локализации на
лице; при диссеминированном процессе — пре-
имущественная локализация на наружной сторо-
не конечностей; наибольшая зависимость от али-
ментарных раздражителей и выраженность
аллергического (пищевой аллергии) компонен-
та среди факторов риска манифестации.
Рис. 18.1. Типичная локализация
высыпаний при атопическом дерматите.
Клиника проявлений II возрастного периода.
Локализация в складках; хронический воспали-
тельный характер поражений с более выражен-
ным лихеноидным синдромом; развитие вторич-
ных изменений (дисхромии); проявление
вегетативной дистонии; ремиттирующее течение
с закономерной волнообразностью; реакция на
многие провоцирующие влияния с уменьшени-
ем алиментарной гиперчувствительности.
Клиника проявлений III возрастного периода.
Изменение локализации поражений, теряющих
складчатую приуроченность; более выраженный
гиперпластический и инфильтративный харак-
тер поражений с меньшей тенденцией к остро-
воспалительным проявлениям и усилением ли-
хеноидного синдрома; менее заметная реакция на
аллергенные раздражители; менее четкая сезон-
ность волнообразности.
С увеличением возраста пациентов количество
экссудативных очагов уменьшается, а лихени-
фицированных кожных проявлений — увеличи-
вается. В любом возрастном периоде может воз-
никать все большее количество кожных очагов,
и заболевание может в конце концов перейти в
генерализованную форму. Вторично развивает-
ся атопическая эритродермия. Показатели эози-
нофилии в крови многократно увеличены.
Клинические формы
Характер клинических проявлений атопическо-
го дерматита определяется возрастом, конститу-
циональными особенностями пациента, спект-
ром сенсибилизации, развитием патологических
изменений со стороны внутренних органов, на-
рушениями в иммунном статусе больных. Раз-
личают пять клинических форм атопического
дерматита (Суворова К. Н. и соавт., 1989): эри-
тематозно-сквамозная, везикулезно-крустозная,
эритематозно-сквамозная с лихенификацией,
лихеноидная, пруригоподобная.
Эритематозно-сквамозная форма атопического
дерматита проявляется покраснением щек, отеч-
ностью, зудящими папулами, мелкими пузырь-
ками, эрозиями, корками и шелушением. Про-
цесс нередко распространяется на все лицо,
захватывая преимущественно периоральную и
периорбитальную области. Может поражаться
кожа волосистой части головы, шеи, туловища
и кистей. Часто отмечаются явления хейлита,
который проявляется застойной эритемой, ин-
фильтрацией, выраженным кожным рисунком,
шелушением и серозно-геморрагическими кор-
ко-чешуйками в области губ, трещинами в виде
заедов в углах рта. У некоторых больных эри-
тематозно-сквамозной формой атопического
Глава 18. Атопический дерматит 253
дерматита присоединяются гнойничковые вы-
сыпания.
Везикулезно-крустозная форма атопического
дерматита формируется на 3—5 месяце жизни и
характеризуется появлением на фоне эритемы
микровезикул с серозным содержимым. Микро-
везикулы вскрываются с образованием серозных
«колодцев» — точечных эрозий, одновременно
отмечается интенсивный зуд пораженных учас-
тков кожи. Процесс наиболее выражен на кож-
ных покровах щек, туловища и конечностей.
Эритематозно-сквамозная с лихенификацией
форма атопического дерматита характеризуется
эритематозно-сквамозными очагами остро- и
подостро-воспалительного характера с наличием
мелких шаровидных фолликулярных и милиар-
ных папул. Кожа сухая, лихенифицированная,
покрытая большим количеством чешуек. Симмет-
ричные поражения кожи Отмечаются в локтевых
сгибах, на тыльных поверхностях кистей, задних
и боковых участках шеи, в подколенных ямках.
При лихеноидной форме атопического дерма-
тита кожа в очагах поражения эритематозная,
отличается сухостью, выраженностью рисунка,
отеком и инфильтрацией. На фоне эритемы рас-
полагаются крупные папулы, сливающиеся в
центре очага и изолированные по периферии.
Папулы покрыты мелкими отрубевидными че-
шуйками. Больных беспокоит мучительный био-
псирующий зуд. Ногтевые пластинки имеют
отполированный вид, блестящую поверхность,
сточенный свободный край. При обострении
заболевания на местах расчесов возникают сса-
дины, трещины и эрозии. Нередко течение дер-
матоза осложняется пиогенной инфекцией, а
затем регионарным лимфаденитом. При этой
форме атопического дерматита часто бывает
эритродермия, захватывающая лицо и шею.
Пруригоподобная форма атопического дерма-
тита характеризуется высыпаниями на фоне ли-
хенифицированной кожи в виде множественных
изолированно расположенных плотных отечных
папул, на вершине которых часто обнаружива-
ются мелкие пузырьки с плотной покрышкой.
Сыпь чаще располагается на разгибательных
поверхностях конечностей, а также на ягодицах
и на туловище. На коже больных обнаружива-
ются глубокие экскориации, атрофические руб-
чики на месте расчесов.
Сопутствующие симптомы. Аллергический
конъюнктивит, аллергический ринит, сенная
лихорадка и/или аллергическая бронхиальная
астма. Эти заболевания встречаются у 30—50 %
пациентов с атопическим дерматитом и разви-
ваются чаще всего после периода детства.
Заболевания глаз. Они могут проявляться в
виде нейродермической катаракты, кератоконуса
или неаллергического кератоконъюнктивита.
Возможен также изолированный атопический
блефарит. Катаракта наблюдается редко (< 5 %)
и чаще у детей и подростков. Иногда описыва-
ют Ablatio retinae.
Увеличение инфраорбитальных складок (линия
Денни-Моргана, инфраорбитальная складка Ден-
ни-Моргана) — важный признак атопического
заболевания (рис. 18.2); при атопическом дерма-
тите складка атопии встречается в 70 % случаев.
Осложнения
Обусловлены главным образом вторичными ин-
фекциями или же неграмотной терапией (жесткой
диетой со вторичными проявлениями дефицита,
побочными действиями глюкокортикоидов). Со-
общается о нарушениях в росте у детей с тяжелым
атопическим дерматитом. При инфекциях опреде-
ленную роль играют нарушения функции лейко-
цитов и лимфоцитов, а также то, что кожные про-
явления у пациентов после многомесячного
лечения наружными глюкокортикоидами стано-
вятся более чувствительны к инфекциям. На коже
таких пациентов часто выявляется Staphylococcus
aureus.
Вторичная бактериальная инфекция. Выража-
ется в импетигинизации очагов, обусловленной
Staphylococcus aureus. Желтые импетигообразные
корочки на кожных проявлениях с неприятным
запахом — типичная картина, которая вместе с
болезненным увеличением лимфатических узлов
позволяет поставить диагноз. Фурункулы, эри-
зипелод и наружный отит достаточно редки.
Рис. 18.2. Типичная локализация
высыпаний на лице при атопическом дерматите.
254 Частная дерматология
Вторичные вирусные инфекции. Нарушенная ба-
рьерная функция кожи у таких пациентов делает
ее более чувствительной и к инфекциям, причиной
которых являются вирусы. Это касается прежде
всего инфекций, вызванных вирусом herpes simplex
(eczema herpeticatum). В настоящее время сообщается
также о переносе вируса кошачьей оспы. Это за-
болевание начинается остро с лихорадки и соответ-
ствующего ухудшения общего состояния. На коже
появляются многочисленные пузырьки в той же
стадии развития. Практически важным является
мазок со дна пузырька для доказательства наличия
эпителиальных гигантских клеток (Tzank-тест).
Иногда наличие возбудителя необходимо доказать
электронной микроскопией, методом негативного
контрастирования, иммунофлуоресценцией, ПЦР
или вирусной культурой. Легко диагностируются
вирусные инфекции, вызванные вирусом Molluscum
contaginosum (eczema molluscatum) или же папилло-
мавирусом человека (HPV) (eczema verrucatum). В
частности, при бородавках в области паронихия и
на подошвах у детей следует думать об атопии.
Вирусная инфекция Коксаки в зоне атопического
дерматита (eczema coxsaccium) встречается очень
редко.
Вторичная грибковая инфекция. Интересно, что
она встречается редко, преимущественно у взрос-
лых, чаще в виде дерматомикоза и наблюдается
тогда, когда более фигуроподобные эритематоз-
но-сквамозные высыпания при соответствующей
терапии глюкокортикоидами не проходят. В на-
стоящее время, в частности, обсуждается пато-
генетическая роль контактной аллергии на
Malassezia spp при атопическом дерматите воло-
систой части головы и затылочной области.
Malassezia spp рассматривается как причина ухуд-
шения состояния при атопическом дерматите в
этой области. В пользу такого значения говорит
успех местного лечения кетоконазолом (низора-
лом).
По распространенности поражения кожи раз-
личают: локализованные поражения (ограничен-
ные поражения в локтевых и подколенных
складках или на кистях и запястьях, периораль-
ная лихенификация); распространенное пораже-
ние; универсальное поражение (эритродермия).
По степени тяжести (тяжелый, средней тя-
жести, относительно легкий) атопический дер-
матит подразделяют на основании распростра-
ненности поражения кожи, длительности
заболевания, частоты рецидивов и продолжи-
тельности ремиссий.
Важнейшими провокационными факторами,
вызывающими при атопическом дерматите обо-
стрение, являются сухая кожа, жара, потение,
холод, физические упражнения, температурные
перепады, инфекции, аллергический контакт-
ный дерматит, волнения, стрессы, пищевая ал-
лергия, аэроаллергены, расчесы, сопутствующие
заболевания (чесотка).
Гистопатология
Гистопатологическая картина заболевания зави-
сит от его вида. При наличии экссудативных
очагов в период младенчества обнаруживают те
же явления, что и при аллергическом контакт-
ном дерматите: спонгиоз и спонгиотические
пузыри, начинающийся акантоз с гипер- и па-
ракератозом и сывороточными включениями, а
также дермальный периваскулярный инфильт-
рат из лимфоцитов и гистоцитов с экзоцитозом.
В лихенифицированных очагах эпидермис акан-
тотически утолщен в 3—5 раз и имеет наруше-
ния ороговения (гиперкератоз); папиллярное тело
гипертрофично и пронизано воспалительными
клетками (лимфоцитами, гистиоцитами). При-
мечательным является также наличие, как и при
псориазе, большого числа тучных клеток, что
объясняется повышенным содержанием гистами-
на в хронических лихенифицированных очагах.
Диагностика
Выделены две группы диагностических крите-
риев (основные или обязательные, и дополни-
тельные или вторичные признаки), которые
помогают в постановке диагноза атопического
дерматита (Hanifin J. М., Raika G., 1980).
Основные (главные или обязательные) крите-
рии:
1. Зуд кожных покровов.
2. Типичная морфология и локализация высыпа-
ний: в детском возрасте — поражение кожи лица,
разгибательных участков конечностей, тулови-
ща; у взрослых — лихенификация на сгибатель-
ных участках конечностей.
3. Атопия в внамнезе или наследственная пред-
расположенность к атопии.
4. Хроническое рецидивирующее течение с обо-
стрениями весной и в осенне-зимнее время года.
Вторичные признаки (дополнительные или
вспомогательные критерии) атопического дерма-
тита — табл. 18.1.
Хотя диагноз атопического дерматита кажет-
ся достаточно прямолинейным, существуют по-
граничные случаи и некоторые другие кожные
состояния у атопических лиц, поэтому важно
придерживаться вышеуказанных диагностичес-
ких критериев. Для постановки диагноза необ-
ходимо, как минимум, наличие трех главных и
трех дополнительных признаков.
Глава 18. Атопический дерматит 255
Таблица 18.1. Дополнительные
признаки атопического дерматита
Клинические признаки
• Ксеродерма или ихтиоз
• Фолликулярный кератоз
• Хейлит
• Потемнение кожи глазниц
• Неспецифический дерматит кистей и стоп
• Кератоконус
• Передняя субкапсулярная катаракта
Иммунологические признаки
• Повышенный уровень общего IgE в сыворотке
• Пищевая непереносимость
• Склонность к кожным инфекциям
Патофизиологические признаки
• Белый дермографизм
• Зуд при потении
• Побледнение или эритема лица
• Непереносимость липидных растворителей и
шерсти
Европейская оперативная группа по атопичес-
кому дерматиту (European Task Force on Atopic
Dermatitis) в 1993 году разработала метод балль-
ной оценки тяжести этого заболевания: индекс
СКОРАД (The SCORAD Index).
При атопическом дерматите диагностика на-
правлена в первую очередь на выявление при-
чинной связи с различными аллергенами, кото-
рые играют ведущую роль в развитии воспаления
кожи. Важным является сбор аллергологического
анамнеза, включающего историю развития по-
ражения кожи, семейный аллергологический
анамнез, наличие атопических респираторных
проявлений, сопутствующие заболевания кожи,
наличие в анамнезе факторов риска (течение
беременности и родов, характер вскармливания,
наличие инфекции в младенческом возрасте,
прием антибактериальных препаратов в раннем
детстве, сопутствующие заболевания и очаги
фокальной инфекции, непереносимость лекар-
ственных препаратов). Аллергологическое обсле-
дование предусматривает постановку кожных
проб (вне обострения и при отсутствии антиги-
стаминной терапии) и провокационных тестов.
При торпидном рецидивирующем течении дер-
матоза и распространенных поражениях кожно-
го покрова проводится определение специфичес-
ких IgE и IgG4 — антител к неинфекционным
аллергенам с помощью МАСТ (множественного
аллергосорбентного теста) или РАСТ (радиоал-
лергосорбентного теста), проводятся также дру-
гие параклинические и специальные инструмен-
тальные исследования (табл. 18.2. (Феденко Е.
С., 2000))
Дифференциальная диагностика
Атопический дерматит необходимо дифференци-
ровать со следующими заболеваниями: ограни-
ченный нейродермит, красный плоский лишай,
пруриго Гебры, грибовидный микоз, хроничес-
кая экзема.
Для ограниченного нейродермита (лишая Ви-
даля) характерно отсутствие атопии в анамнезе,
начало заболевания во взрослом периоде жизни;
Таблица 18.2. Схема обследования
больных атопическим дерматитом
Лабораторные и инструментальные
методы исследования
• Общий анализ крови
• Биохимический анализ крови (общий белок,
билирубин, АлТ, АсТ, мочевина, креатинин, фибри-
ноген, С-реактивный белок, глюкоза)
• Общий анализ мочи
• Иммунологическое обследование (IgE, субпопу-
ляции лимфоцитов)
• Бактериологическое исследование фекалий (на
дисбактериоз)
• Эзофагогастродуоденофиброскопия
• Электрокардиограмма
• Рентгенологическое исследование придаточных
пазух носа
Аллергологическое обследование
• Аллергологический анамнез
• Кожные тесты с атопическими аллергенами
• Определение специфических IgE-антител к
атопическим аллергенам (МАСТ, РАСТ)
• Провокационные тесты (назальные, конъюнкти-
вальные) — при необходимости
Дополнительные исследования
• УЗИ внутренних органов, малого таза — по
показаниям
• Рентгенологическое исследование — по показа-
ниям
• Биопсия кожи — по показаниям
Консультации специалистов
• Аллерголог
• Терапевт (педиатр)
• Гастроэнтеролог
• Отоларинголог
• Психоневролог
• Эндокринолог
256 Частная дерматология
отсутствие зависимости обострений от действия
аллергенов; локализованное поражение; наличие
трех зон в очаге поражения: центральной лихе-
нификации, лихеноидных папулезных высыпа-
ний и дисхромической зоны; сопутствующие
заболевания предшествуют кожным высыпани-
ям; уровень общего IgE в сыворотке крови нор-
мальный; кожные пробы отрицательные.
При красном плоском лишае имеются типич-
ные папулы фиолетового цвета с блестящей по-
верхностью и пупковидным вдавлением в цен-
тре; характерно наличие сетки Уикхема в виде
беловато-сероватых точек и полосок; наблюда-
ется поражение слизистых оболочек.
У больных с пруриго Гебры папулы распо-
лагаются на разгибательных участках конечно-
стей; элементы изолированы друг от друга; уве-
личены лимфатические узлы; нет атопии в
анамнезе.
При грибовидном микозе очаги лихенифика-
ции менее выражены, отсутствуют ремиссии в
летнее время года.
Для хронической экземы характерны поли-
морфизм высыпаний, везикулы, мокнутие, крас-
ный дермографизм.
Лечение
Комплексная терапия атопического дерматита
включает проведение следующих мероприятий:
гипоаллергенная диета (особенно у детей); меди-
каментозное лечение; физиотерапия и курортное
лечение; профилактические меры.
Гипоаллергенная диета включает следующие ос-
новные принципы:
• ограничение или полное исключение из пи-
тания продуктов, обладающих высокой сенсиби-
лизирующей активностью (яйца, рыба, орехи,
икра, мед, шоколад, кофе, какао, алкогольные
напитки, консервы, копчености, горчица, май-
онез, специи, хрен, редис, редька, баклажаны,
грибы; ягоды, фрукты, овощи, имеющие оран-
жевую и красную окраску: земляника, клубни-
ка, малина, персики, абрикосы, цитрусовые,
ананас, морковь, помидоры);
• полная элиминация причиннозависимых пи-
щевых аллергенов;
• обеспечение физиологических потребностей
пациента в основных пищевых веществах и энер-
гии за счет адекватной замены исключенных
продуктов;
• для включения в гипоаллергенную диету ре-
комендуются: ягоды и фрукты светлой окраски,
кисломолочные продукты; крупы (рисовая, греч-
невая, овсяная, перловая); мясо (говядина, не-
жирные сорта свинины и баранины, кролик,
индейка, конина); масла растительные и топле-
ное сливочное; хлеб ржаной, пшеничный второго
сорта; сахара — фруктоза, ксилит. Пища, гото-
вится на пару или отваривается, картофель и
крупы вымачиваются в холодной воде в течение
12—18 часов, мясо дважды вываривается.
Такая диета назначается в острый и подострый
периоды заболевания сроком на 1,5—2 месяца,
затем проводят ее постепенное расширение за
счет введения ранее элиминированных продук-
тов. При отсутствии положительной динамики
от используемой диеты в течение 10 дней сле-
дует диету пересмотреть.
Местная терапия. Для большинства пациентов
с атопический дерматит местная терапия являет-
ся основным способом лечения (табл. 18.3). Ус-
пешный ее результат зависит от многих факто-
ров — мотивации пациента, степени понимания
им метода лечения и его ограничений, прагмати-
ческого подхода врача с точки зрения его уверен-
ности в приемлемости для пациента и терапевти-
ческой эффективности назначаемого им лечения.
Однако для многих пациентов лечение их забо-
левания остается неудовлетворительным, посколь-
ку эффективный контроль за заболеванием тре-
бует многократного применения различных
препаратов на разные участки тела в течение дли-
тельных промежутков времени. Последние раз-
работки топически активных нестероидных им-
муномодуляторов, таких как пимекролимус и
такролимус, потенциально являются настоящим
прогрессом для таких пациентов.
Применение кортикостероидов 50 лет назад
произвело революцию в лечении атопическо-
го дерматита, и для большинства пациентов
они остаются главным средством терапии. Ме-
стные побочные эффекты, такие как атрофия
кожи и риск системной токсичности, исклю-
чают кортикостероиды как оптимальные пре-
параты для лечения тяжелых форм заболева-
ния, особенно при чувствительной коже и у
детей. Однако самый большой барьер на пути
к эффективному лечению представляет собой
боязнь этих побочных действий со стороны
самих пациентов.
Кортикостероиды нового поколения, такие
как негалогенизированные эфиры (например,
предникарбат, метилпреднизолона ацепонат,
мометазона фумарат) обладают высокой проти-
вовоспалительной активностью с более низким
риском системной токсичности. После достиже-
ния ремиссии пациентов следут проинструкти-
ровать о необходимости перехода к более слабому
препарату или же к постепенному уменьшению
частоты применения лекарства.
Глава 18. Атопический дерматит 257
При атопическом дерматите были идентифи-
цированы многочисленные медиаторы воспале-
ния, поэтому вещества, способные блокировать
какой-либо один из медиаторов, вряд ли при-
несут клиническую пользу. Однако некоторые
из антагонистов имеют ценность при атопичес-
ком воспалении (в частности, при астме), что
предполагает доминирующую роль определенных
медиаторных механизмов.
Доксепин, трициклический антидепрессант с
мощной способностью блокировать рецепторы
Нр Н2 и мускариновый рецептор, недавно был
лицензирован в качестве средства местной тера-
пии для контроля над зудом, связанным с ато-
пическим дерматитом.
Макролидные иммуносупрессанты имеют
макролидоподобную структуру и обладают мощ-
ной иммуномодуляторной активностью как in
vivo, так и in vitro. Циклоспорин является, по-
жалуй, наиболее известным из веществ этой
группы, и чрезвычайно активным при систем-
ном применении. Однако некоторые новые пре-
параты, относящиеся к этому классу, демонст-
рируют топическую активность и являются
предметом пристального иследовательского ин-
тереса. Наиболее продвинутых стадий в смысле
разработки для клинического применения дос-
тигли крем «Элидел» (пимекролимус) и мазь
«Протопик» (такролимус).
Пимекролимус (крем «Элидел») разработан
специально для использования в качестве про-
тивовоспалительного наружного препарата для
лечения больных атопическим дерматитом. Пи-
мекролимус относится к группе макролактамных
антибиотиков и является производным аскоми-
цина. Препарат обладает высокой липофильно-
стью, вследствие чего он распределяется преиму-
щественно в коже и практически не проникает
через нее в системный кровоток. Препарат из-
бирательно блокирует синтез и высвобождение
противовоспалительных цитокинов, в результате
чего не происходит активации Т-клеток и мас-
тоцитов, необходимой для «запуска» и поддер-
жания воспаления. В связи с избирательным
действием пимекролимуса на синтез провоспа-
лительных цитокинов Т-лимфоцитами и высво-
бождение медиаторов воспаления тучными клет-
ками, без торможения синтеза эластических и
коллагеновых волокон, его применение исклю-
чает развитие атрофии, телеангиэктазий, гипер-
трихоза кожи. Исходя из данных особенностей
препарата, он может применяться длительно без
риска местных побочных эффектов.
Основным предназначением пимекролимуса
(элидела) является длительное поддержание ре-
миссии без периодического использования на-
ружных кортикостероидов. Препарат применя-
ется в виде 1 % крема и разрешен к
использованию у детей с 3-месячного возраста.
Показанием к назначению элидела является
средняя и легкая степень атопического дермати-
та. Необходимым условием эффективного лече-
ния кремом «Элидел» является его сочетанное
применение с увлажняющими и смягчающими
средствами. Крем «Элидел» можно наносить на
все пораженные участки кожи, включая кожу
лица, шеи, гениталий даже у маленьких детей
при условии неповрежденной поверхности кож-
Таблица 18.3. Местная терапия атопического дерматита
виды Препараты (мази, кремы, гели) местной терапии
Кортикостероиды Антигистаминные Негалогенизированные кремы, мази (элоком, адвантан, локоид, апулеин, афлодерм) Доксепин (крем), фенистил (диметиндена малеат) — 0,1 % гель; совентол (бамипин) — 2 % гель, дифенгидрамин (димедрол) —1-2 % мазь, крем
Иммуносупрессоры Пимекролимус (элидел) — 1 % крем Такролимус (протопик) — мазь 0,1-0,3 %
Нестероидные противовоспалительные Индометацин (5-10 % мазь, 1 % гель), буфексамак (5 % мазь), диклофенак (1 % гель, мазь, крем), ибупрофен (5 % гель, крем), дермарэф (5 % мазь биена), кетопрофен (крем, гель), фенилбутазон (мазь)
Фитопрепараты Препараты дегтя Капсаицин (0,025-0,075 %), ментол (0,1-1,0 %), камфора (0,1-3,0 %) Дёгти древесные: березовый (Oleum Rusci), сосновый (Pix liquida Pini), можжевеловый (Oleum cadini), буковый (Oleum Fagi) Дёготь каменноугольный (Pix Lithanthracis) Дёгти нефтяные: нафталан (паста, мазь, линимент) Дёгти битумные: ихтиол (паста, мазь)
258 Частная дерматология
ных покровов. Эффект от терапии препаратом
отмечается с конца первой недели лечения и
сохраняется в течение одного года. Крем «Эли-
дел» не применяется для лечения больных тяже-
лыми формами атопического дерматита и при
тяжелом обострении заболевания.
Такролимус (мазь «Протопик») является мак-
ролидным соединением 822-Da, первоначально
полученным из ферментационной жидкости
Streptomyces tsukubaensis. Последний экстрагиро-
вали из пробы почвы в Тсукуба (Япония), от-
сюда акроним Т в названии препарата, «акрол»
от термина «макролид» и «имус» от термина «им-
муносупрессант». Такролимус производит целый
ряд действий на различные типы клеток, кото-
рые являются потенциально значимыми для его
терапевтической эффективности при атопичес-
ком дерматите.
Эфирные масла ментола (листья мяты переч-
ной) и камфары (камфорное дерево) проявляют
свое противозудное действие, стимулируя кож-
ные сенсорные рецепторы. Многие пациенты
сообщают о приятном охлаждающем эффекте.
Ментол (0,1—1,0 %) и камфора (0,1—3,0 %) для
местной терапии изготавливаются синтетическим
путем. Для детей лечение этими препаратами не
показано из-за возможного токсического и раз-
дражающего действия.
Капсаицин — вещество, получаемое из струч-
ков перца, применяется для местной терапии
(0,025—0,075 %) болезненных и зудящих дерма-
тозов. Вначале он вызывает жжение в результа-
те высвобождение нейропептидов из перифери-
ческих медленно проводящих С-волокон. При
продолжении применения наступает деплеция
нейропептидов, которой и объясняются проти-
возудный и аналгетический эффекты.
Системная терапия. Конечно, для торпидного
заболевания, особенно распространенного дерма-
тита, системная терапия является наиболее под-
ходящей. Основная проблема терапевтической
дилеммы — недостаточная эффективность безо-
пасных лекарств и большое количество побоч-
ных действий у эффективных препаратов, ис-
пользуемых при системной терапии атопического
дерматита. Остается выбор между пользой и
возможным риском.
Циклоспорин (сандиммун-неорал) является
наиболее изученным из препаратов, применяю-
щихся для системного лечения тяжелых форм
атопического дерматита. Обычная начальная доза
составляет 5/мг/кг/день. Первые терапевтичес-
кие результаты видны уже за период от несколь-
ких дней до недели. Через две недели можно
начинать снижение дозы на 100 мг каждую вто-
рую неделю. Переходить на прием препарата
через день можно в случае, если начальная еже-
дневная доза была 300 мг/кг/день; желаемая
цель — окончание лечения через 3—6 месяцев.
При снижении дозы циклоспорина нужно начи-
нать проводить стабилизирующую терапию,
комбинируя применение ультрафиолетового об-
лучения А и В. Таким образом обеспечивается
возврат к местной терапии, а также профилак-
тика возможного обострения кожного воспале-
ния. Первичные побочные эффекты циклоспо-
рина — нефротоксичность и гипертензия,
поэтому контроль за этими параметрами должен
проводиться до проведения лечения, через 2 не-
дели, через месяц и затем каждый месяц во вре-
мя лечения. Долгосрочные исследования пока-
зали, что при тщательном отборе пациентов и
контроле циклоспорин является безопасной и
эффективной системной терапией в случае тя-
желого торпидного атопического дерматита. По-
скольку стартовую дозу лечения можно выби-
рать, предпочтительно начинают с эффективной
дозировки в надежде сокращения общей продол-
жительности лечения. Некоторые врачи предла-
гают низкую начальную дозировку в 2—3 мг/кг/
день, особенно в педиатрии, где отмечаются слу-
чаи тошноты при более высоких дозах. У взрос-
лых, напротив, для получения ремиссии необхо-
дима более высокая доза в 7 мг/кг/день,
особенно в тяжелых случаях.
Системный препарат такролимус для перо-
рального применения оказался эффективен при
псориазе, но его применение при атопическом
дерматите формально недостаточно исследовано.
В дозах 1—4 мг/день препарат имеет профиль бе-
зопасности и побочных действий, сходный с
таковым у циклоспорина, с которым его можно
взаимозаменять. Это следует особенно учесть для
пациентов, которые неадекватно реагируют на
циклоспорин.
Сейчас разрабатывается новый препарат для
системного применения при атопическом дерма-
тите — пимекролимус. До настоящего времени
изучали местную лекарственную форму этого
препарата, однако недавнее исследование при
псориазе показало, что это лекарство может быть
эффективным при пероральном применении с
более безопасным профилем побочных эффек-
тов, чем у циклоспорина и такролимуса. Ожи-
дается, что эта форма препарата будет эффектив-
на и при атопическом дерматите.
Азатиоприн часто применяют при тяжелых
дерматологических заболеваниях в качестве им-
муносупрессивного средства. Терапевтической
дозой при атопическом дерматите считается 2—
Глава 18. Атопический дерматит 259
2,5 мг/кг/день, причем пациенты должны знать,
что до начала действия препарата может пройти
6 недель. Азатиоприн хорошо переносится, лишь
иногда сообщается о тошноте и рвоте. Регуляр-
ный лабораторный контроль проводится в пер-
вый месяц лечения каждые две недели, а затем
каждый месяц в течение всего срока терапии.
Исследования должны включать полный анализ
крови, анализы на функцию печени и почек, а
также анализ мочи. Длительность терапии, схе-
мы снижения дозировки и необходимость при-
менения стабилизирующей терапии в фазе сни-
жения доз препарата — такие же, как и при
лечении циклоспорином и метотрексатом.
Системные кортикостероиды, включая внутри-
мышечные инъекции триамицинолон ацетонида,
очень эффективны для контроля над симптома-
ми атопического дерматита. Быстрота ответа, хо-
рошая переносимость при кратковременном при-
менении и относительно небольшая стоимость
делают лечение преднизолоном одинаково при-
влекательным как для измученных пациентов, так
и для клиницистов. Однако задокументирован-
ные побочные действия длительной терапии сте-
роидами (например, остеопороз, катаракта) огра-
ничивают их применение при хронических
заболеваниях, к которым относится и атопичес-
кий дерматит. Один-два раза в год, на 6-8 дней
соответственно, преднизолон может применять-
ся для предотвращения тяжелых приступов, при-
чем стероидная зависимость и давление со сто-
роны пациентов повторить преднизолоновую
терапию являются повсеместными. Однако эф-
фект рикошета и снижение эффективности де-
лают повторную терапию кортикостероидами
непривлекательной.
Опыт многих авторов показывает, что пре-
рвать порочный круг от зуда к расчесам при
атопическом дерматите можно с помощью седа-
тивных антигистаминных препаратов. Противо-
воспалительно действующие неседативные анти-
гистаминные препараты нового поколения (при
атопическом дерматите показаны лоратидин,
цетиризин — амертил, парлазин), помимо Hl-
антигистаминного эффекта, уменьшают зуд у од-
ной из подгрупп пациентов с атопическим дер-
матитом.
Пациенты с атопическим дерматитом часто
имеют поверхностную стафилококковую ин-
фекцию, которая, в свою очередь, может выз-
вать обострение дерматита. Системный прием
антибиотиков является базовым в лечении та-
ких пациентов. Стафилококковые культуры не-
изменно устойчивы к пенициллину и обычно
к эритромицину, оставляя нам циклоспорин и
диклоксациллин в качестве препаратов выбора
в дозах 250 мг 4 раза в день для взрослых и
125 мг два раза в день (25—50 мг/кг веса в день,
разделенные на два приема) для младших детей.
Как правило, пустулы разрешаются быстро, и
пациентам редко требуется принимать препарат
более 5 дней. Если у пациентов инфекции ре-
цидивируют, им лучше провести еще один 5-
дневный курс лечения для предупреждения обо-
стрений заболевания. Некоторые пациенты
имеют множественные или непрерывные реци-
дивы, для надежного лечения таковых необхо-
дим курс тетрациклина в течение месяца, что-
бы предотвратить развитие резистентности к
цефалоспоринам (пациенты должны быть стар-
ше 12 лет).
Физиотерапия
Фототерапия. Фототерапия УФ-светом проводит-
ся в целом как мероприятие, дополняющее ле-
чение атопического дерматита, а также для ста-
билизации кожи в конце других терапевтических
мероприятий при выходе заболевания из острой
стадии. Различают терапию селективным УФ-В-
спектром (СУФ), комбинации УФ-В с УФ-А,
ПУВА и новейшую монотерапию «высокодози-
рованным» УФ-А (табл. 18.4).
Таблица 18.4. Фототерапия атопического дерматита
Вид фототерапии Примечание
УФ-А длинноволновые лучи (320-400 нм) 320 -400 нм
УФ-В средневолновые лучи 285-350 нм
УФ-В лучами узкого спектра 311 нм
УФ-А/УФ-В Селективная фототерапия
ПУВА Аутосенсибилизатор+УФА
УФА1 высокие дозы 340-400 нм
Фотодинамическая терапия инфракрасное излучение 600-730 нм + фотосенсибилизатор группы порфиринов
260 Частная дерматология
Недостатком фототерапии является усиленное
высыхание кожи атопика и повышенный риск
раковых заболеваний. Механизм действия фото-
терапии на атопический дерматит еще недостаточно
исследован. Известно, что УФ-В-свет приводит к
торможению клеточно-опосредованных иммунных
ответов, в частности, посредством количественного
уменьшения или ослабления активности клеток
Лангерганса. Новые методы исследования указы-
вают также на то, что УФ-В явно тормозит эксп-
рессию ICAM-1 на кератиноцитах человека и тем
самым может привести к подавлению воспалитель-
ной реакции в коже. Возможно, определенную
роль играет также противомикробный эффект.
Точных данных о специфическом воздействии
ПУВА и отдельно УФ-А-облучения на атопичес-
кий дерматит пока не имеется. Существует мне-
ние, что в качестве действующего механизма имеет
место особое действие УФ-А-излучения на IgE-
несущие клетки Лангерганса. Перед началом ле-
чения следует исключить прием фотосенсибили-
зирующих медикаментов. Рекомендуется
предварительный врачебный осмотр. Дети дош-
кольного возраста меньше подходят для фототе-
рапии, поскольку с учетом их подвижности труд-
но точно определить дозу облучения. Пациенты с
типом кожи I уже при малых УФ-дозировках ре-
агируют тяжелой длительной эритемой, так что
терапевтически эффективные дозы едва ли удается
применить. Противопоказано применение УФ при
одновременно имеющих место провоцируемых
светом дерматозах.
Селективная фототерапия УФ-В. Селективная
УФ-В-фототерапия (СУФ). Начальные дозы
СУФ-облучения (преимущественно 290—320 нм)
должны соответствовать индивидуальной дозе
для минимальной эритемы (МЭД) в диапазоне
УФВ. Во время 2-го сеанса МЭД повышается на
50 %, при третьем — на 40 % и последующих —
на 30 %. Следует стремиться как минимум к 3,
а лучше к 5 сеансам в неделю. При нежелатель-
ном появлении слишком сильной эритемы ле-
чение нужно прервать и применить при необхо-
димости топические кортикостероиды. После
затухания эритемы следует продолжать облуче-
ние в дозе, составляющей 50 % от предыдущего
облучения. При многодневном прерывании те-
рапии лечение продолжают также в дозе, состав-
ляющей половину от назначенной перед преры-
ванием терапии. Побочные действия — это
возможность солнечного дерматита, а также риск
развития эпителиальной или меланоцитарной
неоплазии. При облучении рекомендуется зак-
рывать лицо и область гениталий. В последнее
время при тяжелом атопическом дерматите ре-
комендуются лампы с узким спектром УФ-В
(312 + 2 нм), однако достаточного опыта при-
менения таких ламп еще нет.
Комбинация УФ-В и УФ-А-облучения (УФ-
АВ-терапия). Новейшие исследования позволя-
ют предположить, что комбинация УФ-В (300 +
5 нм) с УФ-А (350 + 30 нм) имеет при атопи-
ческом дерматите лучший эффект, чем только
одно УФ-А или СУФ-облучение. Лечебный эф-
фект при такой комбинации также представля-
ется более продолжительным. Однако этот ва-
риант лечения не применяется как монотерапия,
а только в качестве сопутствующего мероприя-
тия при местном применении кортикостероидов.
Одновременное облучение пациента осуществля-
ется двумя различными источниками света в
одной и той же кабине. Для начала лечения снова
определяется МЭД и при 80 % МЭД начинают
первое облучение. Начальная УФ-А доза долж-
на при этом составлять около 3 Дж/см2, а началь-
ная доза УФ-В дожна составлять 0,02 Дж/см2.
Продолжение облучения осуществляется по ана-
логии с облучением при СУФ. Повышение до-
зировки для обоих видов облучения сответствует
начальной дозе и должно при максимальной
дозировке составлять 6 Дж/см2 для УФ-А и 0,18
Дж/см2 для СУФ. Побочные действия и проти-
вопоказания такие же, как и для СУФ-терапии.
Облучение высокими дозами УФ-Al. Здесь
речь идет о новом варианте, так называемом УФ-
Ар то есть УФ-А-облучении в длинноволновом
диапазоне в 340—440 нм при высоких дозах до
140 Дж/см2 на сеанс. Для этого необходимы осо-
бые источники света. Длительность облучения
составляет 30 мин. Сообщается, что уже через 6-
9 сеансов можно рассчитывать на явный тера-
певтический эффект (улучшение до 50 %), и по-
этому этот вид облучения в некоторых случаях
может успешно применяться и как монотерапия.
Вследствие высоких доз УФ-А, побочное долго-
временное действие которых еще полностью не
изучено, считается обязательно необходимым
проведение подобной процедуры только в ост-
ром периоде тяжелого генерализованного атопи-
ческого дерматита. Их применение в качестве
экспериментальной терапии в настоящее время
ограничено несколькими европейскими универ-
ситетскими центрами. Этот метод применяется
как острое интервенционное мероприятие на
короткое время. Более точное исследование на
более длительный период времени еще предсто-
ит выполнить. Механизм действия неизвестен,
предполагается уменьшение в результате свето-
вого воздействия воспалительных реакций,
включая гамма — интерферон.
Глава 18. Атопический дерматит 261
ПУВА-терапия. Терапия ПУВА показана толь-
ко при обострении атопического дерматита, при
котором имеются противопоказания к примене-
нию кортикостероидов. При этом реакция на
терапию довольно хорошая, однако применение
ПУВА до достижения стабильного результата
требует в целом в два раза больше сеансов, чем,
например, необходимо при псориазе. В одном из
последних исследований указывалась средняя
необходимая кумулятивная доза УФ-А в 118 Дж/
см2, а среднее число необходимых сеансов — 59.
Быстрая отмена часто связана с явлением «ри-
кошета» или реакции подавления после возбуж-
дения. Применив ПУВА у подростков и моло-
дежи должно происходить только по строгим
показаниям и после соответствующих предвари-
тельных обследований. Именно у молодых па-
циентов с атопией к этому виду лечения следу-
ет относиться очень осторожно из-за его еще
неизвестных долговременных эффектов. Для
женщин, желающих иметь детей и беременных,
а также лиц с заболеваниями печени и почек,
ПУВА-терапия противопоказана.
Акупунктура (иглорефлексотерапия). Учитывая
сложность патогенеза и разнообразие клиничес-
ких проявлений атопического дерматита, реко-
мендуется составлять рецептуру точек с учетом
их общего действия и локализации высыпаний
на коже. Лечение начинают с точек общего дей-
ствия, затем включаются местные точки соответ-
ственно локализации процесса и аурикулярные
точки. При наличии сопутствующих заболева-
ний применяют симптоматические точки. В ос-
трой стадии кожного процесса используется I ва-
риант тормозного метода, в подострой и
хронической стадии — II вариант тормозного
метода. Во время проведения процедур исполь-
зуются комбинации и сочетания точек индиви-
дуально для каждого больного с учетом особен-
ностей поражения кожи, выраженности зуда,
наличия сопутствующих заболеваний. Процеду-
ры проводятся ежедневно, 10—12 процедур на
курс. Через неделю назначается повторный курс
лечения, состоящий из 6—8 процедур, проводи-
мых через день. В периоды наиболее вероятных
обострений или рецидивов проводится аурику-
лярная терапия.
Ицдуктотермия на область надпочечников. На-
значается при атопическом дерматите со сниже-
нием функциональности активности коры над-
почечников. Используют высокочастотную
индуктотермию резонансным индуктором
(ЭВТ-1) от аппарата «УВЧ-30». Индуктор рас-
полагают со стороны спины на уровне Т10-Т12,
доза слаботепловая, продолжительность 5—
10 мин, первые 5 процедур ежедневно, затем
через день, на курс 8—10 процедур. Воздействие
на область надпочечников проводится индук-
тотермией СВЧ (диапазона СМВ и ДМВ) от ап-
паратов «Луч-3» и «Ромашка», на курс 10—15
процедур через день.
Магнитотерапия переменным или постоянным
магнитным полем. Переменное магнитное поле от
аппарата «Полюс» рекомендуется в острый и
подострый периоды атопического дерматита с
целью влияния на центральную и вегетативную
нервную систему, на тканевую трофику. Воздей-
ствие осуществляется сегментарно на воротни-
ковую, поясничную области и местно на очаги
поражения кожи. Используются индукторы с
прямым сердечником, режим непрерывный,
форма тока — синусоидальная. Интенсивность
переменного магнитного поля от 8,75 до 25 мТ,
продолжительность 12—20 минут, на курс 10—20
процедур, ежедневно.
Центральная электроаналгезия (ЦЭАН) —
электротерапия и электротранквилизация путем
чрезкожной электростимуляции импульсными
токами. Метод применяется у больных атопичес-
ким дерматитом с неврозоподобными состояни-
ями. Центральной электроаналгезией достигается
изменение поляризационных и электропровод-
ных свойств тканей, что создает благоприятные
условия для нормализующего воздействия на
ЦНС. Импульсное воздействие производится
при лобно-шейном положении электродов аппа-
рата «ЛЭНАР» с частотой от 800 до 1000 гц,
длительностью импульсов от 0,1 до 0,5 мс и
среднем значении тока от 0,6 до 1,5 ма. Продол-
жительность процедуры ограничивается 40 мин,
курс лечения составляет 10—15 ежедневных про-
цедур.
Низкоэнергетическое лазерное излучение. Ле-
чение низкоинтенсивным лазерным облучением
проводится с помощью аппарата «Узор»: импуль-
сный режим 2 Вт, частота импульсов 3000 Гц,
длина волны 0,89 мкм. Курс лечения составля-
ет 12—15 процедур ежедневно.
Лечебное голодание (разгрузочно-диетическая
терапия). Метод показан больным с избыточным
весом, резистентностью заболевания к другим
видам терапии, а также с сопутствующей пато-
логией желудочно-кишечного тракта. Разгрузоч-
но-диетическая терапия (методика Ю. С. Нико-
лаева) продолжается в течение 28—30 дней.
Разгрузочный период длится 14—15 дней, в те-
чение которого при полном воздержании от пищи
больным назначают ежедневные клизмы, прием
минеральной воды до 3 литров в сутки, ежеднев-
ный душ с последующим применением смягча-
262 Частная дерматология
ющих кремов. Восстановительный период продол-
жительностью 14—15 дней начинается с приема
фруктовых соков в первые дни, затем тертых
овощей и фруктов с переходом на специальную
молочно-растительную диету. В дальнейшем для
поддержания достигнутого эффекта больным
рекомендуется строгая гипоаллергенная диета.
Терапевтический эффект разгрузочно-диетичес-
кой терапии обеспечивается очищающим действи-
ем самого процесса голодания путем вымывания
из организма циркулирующих иммунных комп-
лексов, аллергенов, токсинов, санирующим воз-
действием его на функции желудочно-кишечно-
го тракта, а также возможностью поддержания
гипоаллергенного режима питания после процесса
голодания. Метод лечебного голодания противо-
показан больным с сердечно-сосудистой патоло-
гией.
Гипербарическая оксигенация (ГБО). Метод по-
казан больным атопическим дерматитом с явлени-
ями гипотонии, астеникам, а также с сопутству-
ющими заболеваниями, сопровождающимися
симптомами анемии. Сеансы ГБО проводят в од-
номестной барокамере типа ОКА-MT. Давление
кислорода составляет 1,5 атм, продолжительность
сеанса — 40 мин, на курс лечения назначают обыч-
но 10 сеансов. Терапевтическое действие метода
связано с активацией ферментного звена антиок-
сидантных систем, повышением парциального
давления кислорода в пораженных тканях, в час-
тности, в коже, и улучшением микроциркуляции
за счет увеличения скорости кровотока, уменьше-
ния степени агрегации эритроцитов и нормализа-
ции реологических свойств крови.
Плазмаферез. Метод экстракорпоральной де-
токсикации в виде плазмафереза назначают боль-
ным с торпидным течением, эритродермическим
вариантом заболевания, а также при лекарствен-
ной непереносимости. В условиях хирургической
процедурной из локтевой вены кровь эксфузи-
руют в пластикатные контейнеры и центрифу-
гируют при 3000 об/мин в течение 10 мин при
температуре +22°С. Плазму удаляют, а формен-
ные элементы реинфузируют больному в плаз-
мозамещающих растворах. Объем удаляемой
плазмы составляет от 300 до 800 мл, что компен-
сируют таким же или чуть большим объемом
плазмозаместителей. Процедуры проводят обыч-
но 1 раз в 2—3 дня, до 8—12 на курс; при особо
тяжелых формах — ежедневно. При плазмафе-
резе организм освобождается от патологических
метаболитов, циркулирующих иммунных ком-
плексов, очищаются его рецепторы, повышает-
ся чувствительность к различным лечебным, в
том числе медикаментозным воздействиям.
Для лечения больных атопическим дермати-
том применяются также другие методы физиоте-
рапии: пунктурная физиотерапия (фонопункту-
ра, лазеропунктура); миллиметровоЛновая
терапия (КВЧ-терапия); ультразвуковая терапия
(ультразвук паравертебрально и ультразвук на
очаги поражения — ультрафонофорез); эндона-
зальный электрофорез антигистаминных препа-
ратов; диадинамотерапия области шейных сим-
патических узлов.
Таким образом, тяжелый, распространенный
атопический дерматит, не реагирующий на мес-
тную терапию, требует терапии системной. Вос-
паление и зуд в большинстве случаев можно
явно улучшить, применяя описанные вещества,
при этом следует соблюдать равновесие между
его приступообразностью, рецидивирующим и
хроническим течением заболевания, а также ток-
сичностью применяемых веществ. Имеющиеся
в распоряжении системные методы терапии мо-
гут облегчить упорный зуд и должны повсемес-
тно применяться при выраженном и торпидном
течении заболевания. Хорошо продуманное при-
менение дополнительных «стабилизирующих»
видов лечения, — например, УФА/В или агрес-
сивных местных видов терапии — может облег-
чить возврат к применению только лишь топи-
ческой терапии и воспрепятствовать повторному
обострению воспаления.
Санаторно-курортное лечение предусматрива-
ет пребывание в местных санаториях привычного
климата и на курортах с морским климатом (Ев-
патория, Анапа, Сочи, Ялта). Климатотерапия в
теплое время года проводится в виде воздушных,
солнечных ванн и морских купаний. Курорты
позволяют применить сероводородные, родоно-
вые, морские ванны, грязелечение. Лечение
минеральными водами назначается при сопут-
ствующих заболеваниях желудочно-кишечного
тракта и печени.
Прогноз
Вследствие возможных вторичных инфекций у
маленьких детей прогноз нужно делать с осто-
рожностью. В целом интенсивность заболевания
после первого года жизни несколько снижает-
ся. Кожных проявлений становится меньше, и
к 30-му году жизни они почти исчезают. Связь
с другими атопическими поражениями, такими
как бронхиальная астма и аллергический ринит,
индивидуальна и не совсем ясна. Пациенты,
которые дополнительно страдают и этими забо-
леваниями, сообщают, что иногда при спонтан-
ном улучшении кожных проявлений ухудшает-
ся состояние легких или носа и наоборот.
Глава 18. Атопический дерматит 263
Составить прогноз в каждом индивидуальном
случае достаточно сложно.
Профилактика
Генетическая консультациям ограничения в пище
(диетические мероприятия у детей и взрослых для
клинически доказанных случаев на определенный
период времени); избегание аэроаллергенов (исклю-
чить контакт с кошками, собаками, лошадьми,
коровами, свиньями; не заводить домашних жи-
вотных; исключить курение в доме; использовать
вытяжки на кухне; исключить контакт с расте-
ниями, образующими пыльцу); против клещей до-
машней пыли — тщательная чистка ковров и влаж-
ная уборка квартиры; удаление из спальни
ковров, занавесей, собирающих пыль; использо-
вание подушек с полиэстроловым наполнителем,
частая стирка постельного белья; устранение ис-
точников скопления пыли, в том числе телеви-
зора и компьютера); против сухости кожи — сма-
зывание кожи кремами после купания, масла для
купания, увлажнение помещений (поддержание
относительной влажности около 40 %); избегание
перегрева, потения, тяжелых физических упраж-
нений; избегание шерстяной грубой одежды и
синтетических тканей, «непроницаемых» тканей;
диспансерное наблюдение (информация для паци-
ентов с атопическим дерматитом и учет этих боль-
ных); обучение родителей больных атопическим
дерматитом детей.
Глава 19
Почесуха(пруриго) Пруриго простое острое детское 264
Пруриго простое подострое 265
Пруриго узловатое 267
Сопровождающиеся сильным зудом кожные
заболевания называются также пруригинозны-
ми. Под термином «пруриго» (синоним: поче-
суха) объединяют сильно зудящие дерматозы,
первичные высыпания при которых представ-
ляют собой уртикарные серопапулы, уртикар-
ные папулы или персистирующие папулезно-
узловатые элементы, которые чешутся или
расчесываются.
Пруриго простое
острое детское
Синонимы: почесуха детская, строфулюс,
strophulus infantum, prurigo simplex acuta infantum,
urticaria papulosa infantum, lichen urticatus, lichen
simplex acutus.
Речь идет об особо выраженной рецидивиру-
ющей аллергической реакции прежде всего у
детей, которая вначале рассматривается как эпи-
зооноз. Главным образом болеют дети от 2 до 8
лет, преимущественно летом и осенью. Молодые
люди и взрослые редко склонны к такой форме
реакции, отсюда и название strophulus infantum
(почесуха детская).
Этиология и патогенез
Заболевание очень похоже на крапивницу. Не-
которые авторы придерживаются мнения, что им
болеют перекормленные дети вследствие неуме-
ренного потребления шоколада, конфитюров и
сырых, не совсем зрелых фруктов. В качестве
причин предполагаются пищевые аллергии и
реакции на прием медикаментов. Этиологичес-
ки может рассматриваться и прорезывание зубов
у маленьких детей. Упоминаются в качестве
причинных и стрессовые ситуации. Чаще всего,
однако, придерживаются мнения, что это забо-
левание представляет собой не что иное, как
эпизооноз, т. е. заболевание, вызванное укуса-
ми или уколами насекомых-артроподов, таких
как блохи, клещи, комары. В пользу этой вер-
сии говорит тот факт, что частота заболевания
в сельских местностях выше, его сезонность кор-
релирует с периодом укусов насекомых, внутри-
кожный тест с антигенами насекомых значитель-
но более положителен, чем у контрольной
группы и, наконец, при направлении маленьких
пациентов в клинику болезнь обычно проходит
быстро и без какой-либо терапии. При этом
предполагают действие иммунологических меха-
низмов, которые проявляются как реакция на
контакт с антигеном вследствие укуса или уко-
ла в форме комбинированных реакций типа I и
типа IV по Gell и Coombs.
Клиническая картина
На фоне полного здоровья остро возникают
сильно зудящие серопапулы со светло-красным
приподнятым ободком, чаще всего свободно
диссеминированные, но иногда и сгруппирован-
ные. Вначале появляются волдыри размером с
булавочную головку или ноготь, плотные на
ощупь, у которых в центре диаскопически на-
блюдается желтая окраска (выход сыворотки).
Это начало серопапулы, которое далее может
привести к образованию маленького централь-
ного пузырька. При сильно выраженном экссу-
дативном течении пузырьки могут расти и об-
разовывать серопапулы в виде туго наполненных
пузырей (strophulus bullosus). Чаще всего серопа-
пулы через несколько часов теряют свой покрас-
невший ободок и, вследствие увеличивающейся
клеточной инфильтрации, переходят в плотную
папулу. Папула зудит, расчесывается и покры-
вается корочкой. После заживления остается
депигментированное или гиперпигментирован-
264
Глава 19. Почесуха 265
ное пятнышко. Поскольку заболевание может
протекать приступообразно, свежие и более ста-
рые высыпания соседствуют друг с другом и
нередко имеется варицелиформная (ветряная
оспа) картина заболевания. Места излюбленной
локализации — туловище и конечности, часто
участки тела, покрытые одеждой. Иногда в ре-
зультате расчесывания высыпаний наступает
импетигинизация. Слизистая оболочка полости
рта не поражается.
Заболевание может протекать не только при-
ступообразно однократно, но и рецидивировать
или быть хроническим. Вследствие интенсивно-
го зуда дети сильно изнурены. Они плохо спят,
становятся капризными. В летние месяцы неред-
ко развивается импетигинизация вследствие ста-
филококковой инфекции. Смена белья, дезин-
фекция помещения или госпитализация в
кожную клинику чаще всего приводят к выздо-
ровлению.
Гистопатология
Свежая серопапула проявляется как интраэпи-
дермальный, множественно субкорнеальный
пузырек и как отек в папиллярных тельцах на-
ряду с незначительным воспалительным, преиму-
щественно периваскулярным лимфоцитарно-
эозинофильным инфильтратом в верхнем
роговом слое. После расчесывания высыпаний
имеется склонность к реактивному, слегка акан-
толитическому утолщению эпидермиса с короч-
кой в центре и сильной лимфогистиоцитарной
реакцией.
Дифференциальный диагноз
Вследствие пестроты клинической картины,
включающей уртикарные изменения, серопапу-
лы и везикулезные проявления, следует исклю-
чить ветряную оспу. На последний диагноз ука-
зывают изменения на волосистой части головы,
поражение слизистой рта, лихорадка и наруше-
ние общего состояния организма. В сомнитель-
ных случаях помогает электронная микроскопия:
обнаружение вируса в содержимом пузырьков
методом негативного контрастирования или же
обнаружение баллонирующих дегенерировавших
эпителиальных клеток в мазке со дна пузыря
(тест Тцанка). В случае чесотки иная локализа-
ция и зуд, преимущественно в ночное время;
здесь в постановке диагноза решающую роль
играет наличие клещей. В дождливые летние
месяцы следует исключить также тромбидиоз,
при котором поражаются участки кожи в зонах
тесного прилегания одежды (под поясом или
ремнем).
Лечение
Внутрь рекомендуются антигистаминные препа-
раты. Наружно применяют тальк или подсуши-
вающий цинковый лосьон, иногда с 5 %-ным
полидоканолом (тезит), летом — обтирания с
сильно разбавленным винным уксусом или на-
несение спиртового лосьона цинка с 0,5 %-ным
виоформом. Можно применять и антигистами-
новое желе (фенистил, совентол, систрал, таве-
гил). Важен гигиенический уход за ребенком,
смена белья и дезинфекция помещения.
Пруриго
простое подострое
Синонимы: подострое пруриго, почесуха подо-
страя, prurigo simplex subacuta, urticaria papulosa
chronica, prurigo simplex acuta et subacuta
adultorum, strophulus adultorum, lichen Vidal
urticatus.
Острый или хронический воспалительный дер-
матоз с очень типичным зудом в анамнезе и ти-
пичной картиной высыпаний, вероятно полиэти-
ологического генеза. Примерно две трети
пациентов — женщины, которые заболевают в
возрасте от 20 до 30 лет, а затем снова в возрасте
менопаузы. Мужчины болеют чаще после 50 лет.
Этиология
В отличие от пруриго простого острого, эпизо-
онозы не играют причинной роли. Нередко име-
ются психические (невротические) отклонения
со склонностью к артифициальной гиперреак-
тивности на различный зуд. Следует принять во
внимание атопию.
Патогенез
Речь идет, вероятно, о полиэтиологическом ал-
лергическом способе реакции у определенных
людей. В этом отношении имеются принципи-
альные общие моменты с крапивницей. Неко-
торые авторы указывают на фолликулярное рас-
положение первичных высыпаний (спонгиоз
эпителия волосяного фолликула с интра- и пе-
рифолликулярным мононуклеарным инфильтра-
том).
Причинами возникновения и персистирова-
ния пруриго простого подострого и пруриго
простого хронического могут быть различные
факторы: гормональные нарушения (контрацепти-
вы, аллергия на гестагены, дефицит андрогенов
у мужчин, вторичный гиперпаратиреоидизм у
266 Частная дерматология
пациентов, находящихся на хроническом гемо-
диализе); нарушения желудочно-кишечного трак-
та (резко выраженное снижение желудочной
кислотопродукции, гипохлоргидрия, ахлоргид-
рия, хронический гастрит, язва желудка или
двенадцатиперстной кишки, кишечный канди-
доз); очаговые инфекции, нарушения функции пе-
чени (инфекционный гепатит); глистные инвазии
(аскаридоз, оксиуриаз); гинекологические наруше-
ния (предменструальная масталгия, полименорея,
кисты яичников, железисто-кистозная гипер-
плазия слизистой матки, гиперэстрогенемия, на-
рушения менструального цикла).
Патогенетически речь идет об аллергических
проявлениях, а именно о комбинированной ал-
лергической реакции, которая начинается аллер-
гией немедленного типа (реакция типа I по
Coombs и Gell), которая в начале имеет уртикар-
ное выражение, после чего следует аллергия по-
зднего типа (реакция типа IV) с клеточной ин-
фильтрацией уртикарного очага.
Клиническая картина
Преимущественно симметрично поражаются раз-
гибательные поверхности предплечий, пелерино-
подобно — верхняя часть спины, область груди
и наружные поверхности бедер (рис. 19.1). По-
стоянно возникающие новые высыпания распо-
лагаются всегда изолированно, но их сумма, од-
нако, приводит к характерному аспекту общей
экзантемы. Редко имеются такие же высыпания
на лице — это создает картину уртикарного акне.
Первичные высыпания представлены светло-
красными серопапулами, т. е. уртикарные типа
комариных укусов папулы размером 1—5 см, в
центре которых ощущается тугой, похожий на
дробинку, пузырек. Поскольку они очень зудят,
то сразу же после образования расчесываются,
поэтому лечащий врач редко наблюдает их в
первоначальном виде.
Экскориированные первичные высыпания — рез-
ко очерченные с кровавой корочкой папулы или
следы расчесов в виде запавших кровавых коро-
чек размером с булавочную головку или линзу.
Резидуальные (остаточные) высыпания прояв-
ляются как гиперпигментированные или часто
как депигментированные в центре, но по на-
правлению к краю имеются гиперпигментиро-
ванные атрофические рубчики величиной по-
чти с линзу.
У пациентов на местах предпочтительной ло-
кализации обычно наблюдается диморфная кар-
тина, а именно расцарапанные геморрагические
экскориированные высыпания и типичные ат-
рофические с нарушениями пигментации рубчи-
ки. Прочие вторичные высыпания, такие как
лихенификация небольшой или большой повер-
хности кожи, всегда отсутствуют. Отсутствует,
как правило, и импетигинизация. Внутренние
поверхности ладоней и стопы, а также слизис-
тые не поражаются.
Характерен зуд в анамнезе. Пациенты регуляр-
но сообщают, что свежие высыпания мучитель-
но зудят и что зуд сразу же прекращается, как
только высыпание расцарапано и кровоточит.
Расчесываются обычно только высыпания, а не
окружающая их кожа. Этим зуд и последователь-
ность зудящего раздражения при пруриго четко
отличаются от таковых при других дерматозах,
таких как чесотка, атопическая экзема или дру-
гие формы экземы. Поэтому на клинически
здоровой коже эффекты расчесов отсутствуют.
Течение хроническое в течение месяцев и лет.
Только пруриго простое подострое при беремен-
ности быстро заживает после родов, но во вре-
мя второй беременности возникает многократ-
но вновь.
Гистопатология
Гистологическая картина соответствует опреде-
ленной фазе развития заболевания. Первично
образуется серопапула с сильным расширением
Рис. 19.1. Типичные высыпания при почесухе.
Глава 19. Почесуха 267
капилляров в сосочковом слое, наблюдается не-
значительная лимфоцитарная реакция. В даль-
нейшем развиваются суб- или интракорнеальные
пузырьки, которые содержат нейтрофильные, а
иногда и эозинофильные лимфоциты. Экскори-
ированное высыпание характеризуется дефектом
эпидермиса, покрытым корочкой, эпидермаль-
ной регенерацией с боковым акантолитическом
утолщением эпидермиса, сильным расширением
сосудов, отеком и скорее гистоцитарно-фибро-
бластической реакцией.
Диагностика
Типичный зуд в анамнезе, места предпочтитель-
ного расположения и диморфная картина с эк-
скориированными высыпаниями и остаточными
явлениями делают диагноз достаточно простым.
Дифференциальный диагноз
Следует провести разграничение между симпто-
матическими формами пруриго, что может пред-
ставлять некоторые трудности и требует в каж-
дом случае тщательного наружного осмотра и
исследования внутренних органов. Далее нужно
отграничить пруриго от следующих заболеваний.
Пруригинозная форма атопической экземы.
Полиморфная картина, изменения комбиниро-
ванные с типичными проявлениями атопической
экземы. Эта картина заболевания называется
также prurigo mitis или пруриго хроническое
мультиформное.
Пруригинозная форма герпетиформного дер-
матита. Полиморфная герпетиформная картина
с пруригинозными папулами.
Пруриго солнечное. Сильно зудящие и экс-
кориированные пруригинозные папулы локали-
зуются только на открытых солнцу участках
тела. Присутствует сезонность: летнее пруриго.
Лечение
Поскольку при подостром пруриго речь идет о
полиэтиологическом синдроме аллергического
генеза, устранение причин стоит на первом плане.
Только если удается найти причину заболевания
и соответствующим образом его лечить, можно
достигнуть долговременного эффекта. К сожа-
лению, часто причина не выявляется. Внутри-
кожные тесты проводятся как при хронической
крапивнице.
Лечение в любом случае ориентируется на
выявленные нарушения. Применение глюкокор-
тикоидов в средней дозе (40—60 мг эквивален-
тной дозы преднизолона ежедневно) возможно
лишь кратковременно и оказывает в большин-
стве случаев лишь морбостатический эффект.
Вечером показаны антигистаминные препара-
ты с седативным эффектом, транквилизаторы,
так как зуд беспокоит особенно сильно по но-
чам. В отдельных случаях в качестве эффектив-
ного препарата рекомендуется хлорохин (резо-
хин). Если у пациенток начало заболевания
совпадает с началом климакса или менопаузой,
можно применять эстрогены или лютеотропный
гормон; здесь необходима консультация гинеко-
лога. В остальном большинство авторов прида-
ет большое значение санированию желудочно-ки-
шечного тракта. Методика такая же, как при
хронической крапивнице: тетрациклин (2,0 г)
или доксициклин (200 мг) ежедневно в течение
5 дней, затем 5 дней амфотерицин В (4 раза в
день по 100 мг), затем в течение 3 недель нор-
мализация кишечной флоры. При уремическом
зуде морбостатически эффективным является
эритропоэтин. Иногда рекомендуется консуль-
тация психотерапевта.
Наружно рекомендуются мероприятия по
смягчению зуда. Наружные анестезирующие
средства (5 % тезит), ихтиол (5—10 %), Liquor
carbonis detergens (2,0—10,0 %). Обтирания раз-
бавленным винным уксусом, спиртовыми ра-
створами с ментолом (0,25—1,0 %) или ан i и гис-
таминным гелем (фенистил, совентол, сис1рал,
тавегил).
Пруриго узловатое
Синонимы: узловатое пруриго Гайда, узловатая
почесуха, prurigo nodularis Hyde, крапивница
папулезная стойкая, нейродермит узловатый.
Сильно зудящее заболевание, которое харак-
теризуется персистирующими чашеобразными
узлами. Включение этого заболевания в группу
пруриго иногда оспаривается, так как первичное
высыпание при нем не является серопапулой,
иногда его относят к пруриго простому хрони-
ческому.
Распространенность: относительно редкое за-
болевание. Генетические факторы неизвестны.
Болеют преимущественно женщины среднего и
пожилого возраста (40—60 лет). Часто (до 80 %)
отмечается атопический диатез.
Этиология и патогенез
Этиология неизвестна. Неоднократно в качестве
причин указываются стрессовые ситуации. Пре-
имущественно поражаются лица невротического
склада. Имеют место психические (депрессии,
депрессивные испуганные настроения) или психо-
268 Частная дерматология
социальные нарушения. Указывают на связь за-
болевания с нарушением эндокринной системы, а
также наличием гепатита, холецистита. Вследствие
гистологического сходства постоянно предполага-
ется связь с простым хроническим лишаем, и
высыпания интерпретируются как ограниченная
лихенификация, что дало повод французскому
дерматологу Pautrier назвать заболевание
lichenificatlons circonscrites nodulaires chroniques.
Примечательны изменения на кожных нервах с
повышением клеточной пролиферации, особенно
в области шванновских клеток (Schwannom-не-
вринома, шваннома), которые могут дать повод к
шванномаподобным образованиям. Носит ли эта
типичная гипертрофия сосочковых дермальных
нервов причинный характер или же она реактив-
ной природы, т. е. обусловлена сильными расче-
сами, не является принципиальным. В последнее
время патогенетическое значение придавалось
внеклеточным отложениям протеиназосодержащих
эозинофильных гранул из эозинофилов. В пользу
атопии говорят повышение IgE в крови и аллер-
гии немедленного типа на пыльцу растений, до-
машнюю пыль, клещи и т. п. Следует подумать и
о контактной аллергии экзематозного типа (эпи-
кутанный тест).
Клиническая картина
Преимущественно поражаются разгибательные
поверхности конечностей; лицо и туловище в
большинстве случаев остаются свободными от
высыпаний. При билатеральном проявлении
имеют место относительно небольшое количе-
ство изолированно расположенных чашеобраз-
но выпуклых, плотных на ранних стадиях за-
болевания, слегка гиперемированных узлов,
размерами с горошину или фасолину (0,5—
3,0 см в диаметре). Их поверхность при боко-
вом освещении кажется тусклой, цвет ливид-
но-серый или скорее грязно-серый. Так как
высыпания вследствие сильного зуда интенсив-
но расчесываются, они чаще экскориируются на
поверхности или склонны к грязно-серого цвета
кератотическим или верруциформным образо-
ваниям. Окружающая кожа нормальная либо ги-
перпигментирована.
Больных беспокоит очень интенсивный зуд,
который приступообразно может усиливаться до
невыносимого и приводить к суицидальной по-
пытке. Сильные расчесы высыпаний обусловли-
вают эрозии, реже геморрагические корочки, и
способствуют увеличению узлов. Течение забо-
левания упорное хроническое, спонтанная тен-
денция к регрессии отсутствует.
Гистопатология
Гистологическая картина напоминает простой
хронический лишай с утолщением рогового слоя,
нерегулярным гиперакантозом, гиперпапиллома-
тозом и плотным хроническим воспалительным
инфильтратом, состоящим преимущественно из
лимфоцитов и гистиоцитов. Количество тучных
клеток часто увеличено, нередко встречаются
отдельные эозинофилы. Особенно примечатель-
ны изменения нервных окончаний: вначале ги-
пертрофия, затем дегенерация многочисленных
нервных волокон и пролиферация шванновских
клеток, частично с образованием шванномопо-
добных формаций (нейрома Pautrier).
Дифференциальный диагноз
Узловатое пруриго Гайда легко диагностирует-
ся. Дополнительные экзематозные или лихени-
фицированные изменения отсутствуют и таким
образом отличают заболевание от пруригинозной
формы атопической экземы (пруриго хроничес-
кое мультиформное). Часто невозможное отгра-
ничение от пруриго простого хронического осу-
ществляется при наличии первичных высыпаний
(серопапул) и более сильно выраженных воспа-
лительных изменений. Красный лишай — гипер-
трофический или веррукозный — располагается
предпочтительно на голени.
Лечение
Выяснение причины затруднено даже при про-
ведении внутрикожного и эпикутанного тестов.
Во многих случаях рекомендуется сотрудниче-
ство с психотерапевтом или психиатром.
Внутрь назначают транквилизаторы. Антиги-
стаминные препараты эффективны, только если
они имеют седативный компонент. Талидомид
по 100—200 мг ежедневно рекомендуется в самых
тяжелых случаях, но требует тщательного конт-
роля из-за побочных тератогенного и нейроток-
сического действий. Использовались попытки
лечения пимоцидом (орап) или беноксапрофе-
ном, клофацимином (лампреном) или дапсоном
(дапсон-фатол).
Наружно назначают внутриочаговые инъек-
ции триамцинолон ацетонида (волон А кристал-
лическая суспензия 10 мг в растворе 1:5с мес-
тным анестетиком, например, мепивакаином).
При особенно мучительных узлах: экссцизия,
криотерапия посредством жидкого кислорода, ла-
зеро- или электрокоагуляция. Рекомендуется фо-
тотерапия (УФ-В, УФ-Al), бальнеофототерапия
или ПУВА-терапия. Местная слаболучевая (мяг-
колучевая) рентгенотерапия применяется редко.
Глава 20
Нейродермит Лихенификация гигантская 270
Простой хронический бородавчатый лишай 270
Синонимы: ограниченный нейродермит, Lichen
Vidal, лишай Видаля.
Речь идет об ограниченном хроническом вос-
палительном, сильно зудящем кожном заболева-
нии с лихеноидным утолщением кожи, которое
поддерживается постоянным расчесыванием.
Имеется связь с атопическим диатезом.
Заболевание распространено по всему миру,
хотя среди представителей черной расы встреча-
ется реже. Женщины болеют чаще мужчин. Ве-
роятно, существует врожденная предрасположен-
ность к развитию лихенификации в результате
трения или расчесов из-за сильного зуда.
Патогенез
При соответствующей предрасположенности на
ограниченных участках в результате трения или
расчесов возникает воспалительная реакция
кожи, которая характеризуется лихенификаци-
ей, т. е. воспалительным утолщением кожи с
огрублением кожных покровов. С точки зрения
кожных тканей речь идет об утолщении эпидер-
миса с хроническим клеточным воспалением.
Из-за сходства кожных высыпаний с лихени-
фицированными очагами при атопической экзе-
ме постоянно дискутируется связь с этим заболе-
ванием простого хронического лишая. В последнее
время есть предпосылки считать это заболевание
минимальным вариантом атопической экземы.
Некоторые авторы решительно отклоняют такое
мнение и полагают наличие этиопатогенетической
связи с внутренними дисфункциями, например,
нарушениями в работе желудочно-кишечного
тракта (нарушения процесса пищеварения, анацид-
ность, хронический гастрит), болезнями печени,
холецистопатиями. Иногда терапевтический успех
мероприятий, направленных на устранение подоб-
ных нарушений, подтверждает и эту точку зрения.
Важным представляется также исследование
психических факторов, поскольку для возник-
новения простого хронического лишая реша-
ющими являются постоянное трение и расче-
сывание ограниченных участков кожи. Приме-
чательно и наличие признаков нервозности, та-
ких как жевание ногтей, покусывание губ, не-
прерывное курение или наличие конфликтных
ситуаций.
Клиническая картина
Обычно заболевание ограничивается единствен-
ным очагом, реже наблюдают 2—3 очага. У жен-
щин очаги локализуются преимущественно на
затылке. Другими местами предпочтительной
локализации являются разгибательные стороны
предплечий и голеней, верхняя часть спины,
внутренняя поверхность бедра, область крестца,
скротума и вульвы. Часто имеет место себостаз.
Клинико-морфологически речь идет о типич-
ном лихеноидном заболевании. Первичное вы-
сыпание представляет собой солидную папулу,
которая не претерпевает изменений, ее началь-
ные размеры 1—3 мм. Папулы всегда четко ог-
раничены, чаще округлые, реже полигональные,
плоские, с блестящей поверхностью. Цвет серый
или коричнево-красноватый, часто такой же, как
и цвет кожи. Первоначально изолированно рас-
положенные на узком участке папулы собирают-
ся в очаги, которые могут иметь круглую, лен-
тообразную, полосовидную или любую другую
конфигурацию, а по краям вследствие трения
имеет место тенденция к механической гипер-
пигментации. Полностью сформированный очаг
состоит из трех типичных зон:
1. В центре находится первичная плоская ли-
хенификация с воспалительным утолщением
кожи и огрублением кожных покровов.
2. Затем следует зона с лихеноидными папула-
ми, которые расположены плотно рядом друг с
другом и окрашены в цвет кожи или же имеют
серовато-красноватый оттенок.
3. На периферии находится зона гиперпигмен-
тации шириной в несколько сантиметров, кото-
рая нечетко переходит в окружающую кожу и
там теряется.
269
270 Частная дерматология
Иногда заболевание ограничивается одним оча-
гом с изолированно расположенными лихеноидны-
ми папулами без наличия центральной лихенифи-
кации, в других случаях лихенификация может
настолько прогрессировать, что отсутствуют и 2-я,
и 3-я зоны. Примечательно, что иногда может быть
потеря пигмента (ахромия) в пределах очагов про-
стого хронического лишая. Слизистые при этом
заболевании не поражаются.
Типичен сильный, часто пароксизмальный зуд,
который особенно беспокоит по ночам. Все еще
остается неясным вопрос, вызывают ли лихини-
фикацию зуд и расчесы или же сильно зудит
именно лихеноидная папула. Экспериментальные
исследования подтверждают возможность воз-
никновения лихенификации вследствие хрони-
ческой механической нагрузки, какой может
являться расцарапывание.
Заболевание длится несколько месяцев или
лет. Прогноз сегодня с учетом хорошей способ-
ности к излечиванию благоприятный. Тяжелее
бывает устранить психоэмоциональные причин-
ные факторы.
Гистопатология
Эпидермис сильно утолщен с удлинением и утол-
щением клеток, которые часто сетевидно слива-
ются в основании. Умеренный гиперпаракератоз.
Капилляры в сосочковом слое расширены, в вер-
хнем слое обнаруживается преимущественно свя-
занный с сосудами лимфоцитарный инфильтрат.
Гистологическое отличие от красного плоского
лишая и лихенифицированной экземы (более
сильные спонгиотические изменения и экзоци-
тоз) обычно не представляет трудности.
Дифференциальный диагноз
Типичные очаги простого хронического лишая
имеют описанное выше «трехзонное» строение.
Лихенифицированные высыпания при атопичес-
кой экземе чаще всего симметричны, имеют
места предпочтительной локализации, более
сильное воспаление и больший полиморфизм.
Для отличия от отдельных ограниченных очагов,
являющихся поздними проявлениями атопичес-
кой экземы, следует определить уровень
IgE. Другие виды лихенифицированной хрони-
ческой экземы отличаются намного более силь-
но выраженными воспалительными изменения-
ми, «трехзонное» строение всегда отсутствует.
Трудность может быть при отличии от очага
плоского красного лишая, однако высыпания в
этом случае всегда полигональны, часто немно-
го вдавлены в центре и имеют, как правило,
феномен Уикхема.
Лечение
Лечение сопутствующего заболевания. Устранение
зуда с помощью антигистаминных препаратов.
Наилучший успех с учетом устранения зуда
дают глюкокортикоиды в форме кремов или
мазей, применяемые под окклюзионную повяз-
ку. После регрессии лихенификации рекоменду-
ются глюкокортикоидные пластыри или более
слабые глюкокортикоиды (адвантан, дерматоп).
Более мелкие очаги лечат внутриочаговой инъек-
цией ацетонида триамицинолона. После значи-
тельной регрессии переходят на глюкокортико-
иды низкой концентрации в форме кремов или
мазей. Для последующего лечения используют
ихтиол, а также дегти (каменноугольный деготь
2—5 % в мягкой цинковой пасте или чистый ка-
менноугольный деготь) в течение нескольких
дней. В случае устойчивых к терапии очагов по-
казано лечение мягкими рентгеновскими лучами.
Лихенификация
гигантская
Синонимы: Lichenification geante
Встречается главным образом в генито-пахо-
во-анальной области. Здесь очаги не имеют ти-
пичных для простого хронического лишая трех
зон, они представляют собой ограниченные,
поражающие большие половые губы и мошон-
ку высыпания, имеющие более сильную воспа-
лительную лихенификацию с папилломатозны-
ми разрастаниями и мокнутием. Возможно, при
возникновении гигантского лишая болезнетвор-
ную роль играют процессы мацерации в интер-
тригинозных пространствах. Зуд при этой ред-
кой форме заболевания чрезвычайно сильный.
Нередко возникают рецидивирующие рожистые
воспаления.
Дифференциальный диагноз
Гипертрофическая болезнь Хейли-Хейли и ве-
гетирующая пузырчатка.
Простой хронический
бородавчатый лишай
Синонимы: веррукозный нейродермит,
Neurodermitis verrucosa, Lichen simplex chronicus
verrucosus.
Эта форма возникает преимущественно на
голенях пациентов с хронической венозной не-
достаточностью. Поверхность лихенификации в
центре склонна к верруциформным кератозам;
зуд очень сильный. Наверняка сюда же относят-
ся случаи верруциформного кератоза (Вайден-
фельд).
Глава 20. Нейродермит 271
Дифференциальный диагноз
Дифференциальный диагноз проводится в пер-
вую очередь с веррукозным красным плоским
лишаем, амилоидозным лишаем и очень редким
гиалинозом кожи и слизистых.
Лечение
Целесообразно дополнительное лечение комп-
рессионными повязками с кортикостероидами.
Глава 21
Крапивница. Отек Квинке
Крапивница и отек Квинке относятся к острым
аллергическим реакциям или реакциям гипер-
чувствительности немедленного типа. К этим
состояниям также относят синдром Лайелла,
синдром Стивенса — Джонсона, анафилакти-
ческий шок. Перечисленные заболевания пред-
ставляют большую опасность для жизни паци-
ента и некоторые из них могут заканчиваться
летально.
Крапивница является довольно распространен-
ным заболеванием — примерно каждый третий
человек хотя бы раз перенес ее. Заболевание
характеризуется быстрым высыпанием на коже
мономорфной сыпи в виде зудящих волдырей.
Это полиэтиологический дерматоз с мономорф-
ной уртикарной зудящей сыпью. Крапивница
может быть следствием аллергической или псев-
доаллергической реакции при многих заболева-
ниях различной этиологии.
Крапивница и отек Квинке могут возникать
в любом возрасте, но наиболее часто они встре-
чаются в возрасте от 21 года до 60 лет.
Патогенез
Аллергические крапивницы развиваются вследствие
аллергии к экзогенным (инфекционным и не-
инфекционным) и эндогенным (аутологичным)
аллергенам. Патогенез аллергических крапивниц
включает этап сенсибилизации к аллергену и
вторичную гиперергическую реакцию при по-
вторном его попадании в организм. Реакции
часто протекают как типично атопические, в
которых участвуют реагиновые антитела класса
IgE. Наряду с I типом аллергической реакции
крапивница нередко является следствием имму-
нокомплексных реакций III типа, для которых
характерно поражение капиллярной сети. Сре-
ди медиаторов, выделяемых лейкоцитами, важ-
ная роль принадлежит гистамину, который спо-
собен индуцировать волдырь и отек кожи. В
реализации гиперчувствительности немедленно-
замедленного типа у больных разными вариан-
тами крапивницы (особенно хронической) уча-
ствуют и другие физиологически активные
вещества (серотонин, ацетилхолин, брадикинин,
интерлейкины, простагландины), потенцирую-
щие действие гистамина. При хронической кра-
пивнице обнаружены высокие показатели анти-
глобулиновой реакции Кумбса, титры антител
против некоторых бактериальных аллергенов,
снижение уровня Т-лимфоцитов, дисиммуно-
глобулинемия, эозинофилия, тромбоцитопения.
Различают неиммунные (псевдоаллергические)
реакции развития крапивницы и отека Квинке,
которые вызываются нестероидными противовос-
палительными средствами, рентгеноконтрастными
средствами, ингибиторами ангиотензин-превраща-
ющего фермента (АПФ), антагонистами кальция,
а также лекарственными средствами, высвобожда-
ющими гистамин, средствами, действующими на
ЦНС, миорелаксантами, симпатомиметиками, ги-
потензивными средствами, антимикробными пре-
паратами. Нестероидные противовоспалительные
средства частично или полностью подавляют ак-
тивность циклооксигеназы, участвующей в син-
тезе простагландинов. Помимо крапивницы и оте-
ка Квинке, они вызывают риносинусит и
бронхиальную астму. Рентгеноконтрастные сред-
ства вызывают анафилактоидные реакции, изредка
бывают аллергические реакции. Ингибиторы АПФ
могут вызвать (у 0,1—0,2 % больных) быстро на-
растающий отек слизистой носа, глотки, гортани,
голосовых связок, слизистой полости рта и язы-
ка. Отек, вероятно, обусловлен нарушением мета-
болизма кининов. Антагонисты кальция (нифеди-
пин) у 10—30 % больных вызывают отеки голеней
и стоп, которые обусловлены дилатацией артери-
ол и капилляров, а не задержкой жидкости в орга-
низме.
Псевдоаллергические формы крапивниц возни-
кают под влиянием экзогенных или эндогенных
факторов, вызывающих образование и выделе-
ние медиаторов аллергии неиммунологическим
неспецифическим путем. Химические вещества
и лекарственные препараты, биологические аген-
ты, а также физические факторы действуют как
либераторы медиаторов, т. е. индукторы псевдо-
аллергических реакций. Нередко псевдоаллерги-
ческие крапивницы возникают в связи с дисбак-
териозом и ферментопатиями. Различают
272
следующие формы псевдоаллергических крапив-
ниц: холодовая крапивница с двумя типами —
семейная наследственная и приобретенная; ме-
ханическая, тепловая, холинергическая, акваген-
ная, солнечная, дисметаболические крапивницы.
Различные виды крапивницы различаются по
своим взаимодействиям с эозинофилами. При
сывороточной болезни с уртикарными высыпа-
ниями эозинофилии обычно не отмечается.
Эозинофилия характерна для острой крапивни-
цы, которая наблюдается при заражении пара-
зитами, при лекарственной и пищевой аллергии.
Эти ситуации представляют собой ассоциации
аллергии с эозинофилией. У больных холодовой
крапивницей были выделены медиаторы гипер-
чувствительности немедленного типа — гиста-
мин, эозинофильный хемотаксический фактор
анафилаксии и компоненты комплемента. По-
вышение в сыворотке концентрации гистамина
и хемотаксических факторов при холинергичес-
кой крапивнице подобно изменению их у боль-
ных холодовой крапивницей. Исследование у
больных крапивницей хемотаксических факто-
ров позволило подразделить их по молекуляр-
ной массе. Были выделены два хемотаксических
фактора эозинофилов — с молекулярной массой
400-600 и 1500-3000.
Классификация
Различают этиологическую и патогенетическую
классификации крапивницы и отека Квинке.
Однако в связи с неоднородностью патогенети-
ческих механизмов многих видов крапивниц не
всегда представляется возможным отнести их к
той или иной группе.
По этиологическому признаку чаще всего вы-
деляют аллергические, псевдоаллергические и
смешанные крапивницы.
Предлагаем следующую классификацию кра-
пивницы и ее вариантов по их патогенетическим
механизмам.
Патогенетическая классификация крапивницы и
отека Квинке
I. Иммунологически обусловленные крапив-
ницы:
• IgE-зависимая крапивница (классическая
острая или хроническая рецидивирующая
немедленного типа; дермографическая кра-
пивница; холодовая крапивница);
• комплемент-индуцированная крапивница
(наследственный или приобретенный отек
Квинке, анафилактоидная уртикарная реак-
ция, уртикарный тип васкулита).
II. Неиммунологически обусловленные крапив-
ницы:
Глава 21. Крапивница. Отек Квинке 273
• вследствие воздействия физических и хими-
ческих факторов (тепловая, механическая,
световая, аквагенная, холинергическая);
• вследствие различных гистаминолибераторов
(медикаменты);
• вследствие индукторов реакций непереноси-
мости и псевдоаллергических реакций.
III. Пигментная крапивница (увеличение ко-
личества тканевых тучных клеток в коже, с по-
ражением внутренних органов или без него; си-
стемный мастоцитоз).
Клиническая картина
При аллергических реакциях немедленного типа
сенсибилизация обычно развивается в течение
7—14 суток; крапивница может возникнуть как
во время лечения, так и после отмены препара-
та. У сенсибилизированных больных крапивни-
ца появляется через несколько минут или часов.
Иммунокомплексные аллергические реакции
обычно развиваются в течение 7—10 суток. У
сенсибилизированных больных реакция прояв-
ляется через 12—36 часов после начала лечения.
При приеме нестероидных противовоспалитель-
ных препаратов реакция развивается через 20—
30 мин (до 4 час) после введения препарата.
Пациенты предъявляют жалобы на зуд, жжение
ладоней, стоп, ушей; может быть артралгия. При
отеке слизистой дыхательных путей наблюдают-
ся одышка и удушье. При аллергических реак-
циях немедленного типа отмечаются нарушения
общего состояния: приливы, внезапная слабость,
головная боль, головокружение, бронхоспазм,
давящая загрудинная боль, сердцебиение, тош-
нота, рвота, приступообразные боли в животе,
понос.
Клиника острой крапивницы характеризуется
мономорфной сыпью, первичный элемент ко-
торой — волдырь (остро возникающий отек со-
сочкового слоя дермы). Крупные волдыри воз-
никают и разрешаются за несколько часов,
самостоятельно или под действием лекарствен-
ных средств. Они могут появиться вновь на том
же месте. Цвет волдырей — розовый, у круп-
ных элементов — светлый центр и красный
ободок по периферии. Форма волдырей оваль-
ная, дугообразная, кольцевидная, полицикли-
ческая, линейная, мишеневидная. Элементам
свойственен периферический рост и разрешение
в центре. Иногда они сливаются между собой
и образуют причудливые узоры (urticaria
flgurata). При слиянии волдырей в обширные
зоны возможно нарушение общего состояния:
повышение температуры тела, недомогание,
озноб — «крапивная лихорадка», у детей —
274 Частная дерматология
пищеварительные и неврологические расстрой-
ства. Наблюдаются высыпания на слизистых
оболочках губ, языка, мягкого неба, гортани.
Очень редко возникают геморрагические вол-
дыри, оставляющие после себя гиперпигмента-
цию (urticaria cum pigmentosa), а также волды-
ри с пузырями, наполненными серозной или
геморрагической жидкостью (urticaria bullosa).
Разновидностью острой крапивницы являет-
ся дермографическая крапивница (urticaria
factitia, уртикарный дермографизм), характери-
зующаяся возникновением волдырей, чаще ли-
нейной формы, в ответ на механическое раздра-
жение, например, при выявлении дермографизма.
В отличие от обычной крапивницы при уртикар-
ном дермографизме нет зуда.
Холодовая крапивница характеризуется появ-
лением кожного зуда, отека, гиперемии после
воздействия холода. Клинические симптомы
заболевания обычно возникают в холодное вре-
мя года при воздействии пониженной темпера-
туры на открытые участки кожи в сырую влаж-
ную погоду. При глубоком охлаждении кожи
развивается отек Квинке, а быстрое охлаждение
большой поверхности кожи (например, при ку-
пании в холодной воде) может вызвать генера-
лизованную крапивницу и анафилактический
шок.
Тепловая крапивница наблюдается у лиц, ра-
ботающих при повышенной температуре окру-
жающего воздуха. Местные реакции сопровож-
даются головной болью, диареей.
Механическая крапивница встречается в не-
скольких вариантах: от давления на кожу рем-
ней, пряжек и других предметов; от вибрации;
как патологическая форма дермографизма. За-
болевание чаще развивается у лиц с лабильной
нервной системой, у больных вегетативно-сосу-
дистой дистонией,.
Световая (или солнечная) крапивница возни-
кает под действием ультрафиолетового излуче-
ния с длиной волны 290—500 нм. Эта крапив-
ница проявляется в виде зудящих полиморфных
высыпаний или типичной крапивницы и эри-
темы.
Аквагенная крапивница наблюдается при кон-
такте с водой, независимо от ее температуры.
Холинергическая крапивница появляется под
влиянием физической нагрузки, сопровождаю-
щейся повышенным потоотделением, а также
после приема ванны или душа, горячего чая.
Заболевание характеризуется появлением много-
численных мелких (от 1 до 4 мм в диаметре)
зудящих волдырей или папулезных элементов,
окруженных зоной гиперемии. Они могут сли-
ваться и образовывать типичные волдыри, иногда
гигантских размеров. Высыпания локализуются
на лице, шее, верхней части груди, верхних ко-
нечностях.
Отек Квинке (острый ограниченный отек
Квинке, гигантская крапивница) представляет
собой обширный отек кожи и подкожной клет-
чатки, при котором появляется большой, блед-
ный, плотный инфильтрат. При отеке Квинке
поражаются лицо, наружные половые органы и
другие участки тела с рыхлой клетчаткой. Вы-
сыпания могут возникать и на слизистых обо-
лочках губ, век, полости рта (язык, мягкое небо,
миндалины), мошонки. При этом кожа в ука-
занных зонах плотноэластической консистенции,
белого, реже розового цвета. Субъективные ощу-
щения обычно отсутствуют, реже отмечаются
жжение и зуд. Часто наблюдаются нарушения
общего состояния: недомогание, тошнота, голов-
ная боль, повышение температуры тела, боль в
животе, диспепсия.
Особенно опасным является отек Квинке в
области гортани, при котором больные могут
погибнуть от явлений асфиксии. При легкой и
средней тяжести отек гортани продолжается от
часа до суток. При локализации на лице в про-
цесс могут вовлекаться серозные мозговые обо-
лочки с появлением менингеальных симптомов
(ригидность затылочных мышц, резкая головная
боль, рвота, иногда судороги). У некоторых боль-
ных наблюдается склонность к рецидивам, часто
начинающимся без продромальных симптомов, с
затрудненным дыханием и прогрессирующим
удушьем с возможным смертельным исходом.
Наследственный отек Квинке — тяжелое забо-
левание, которое наследуется аутосомно-доминан-
тно. Проявляется отеком лица и конечностей,
гортани и острой болью в животе. Лабораторно
выявляется снижение уровня или активности
ингибитора С1-эстеразы, снижение уровня С4 и
нормальные уровни компонентов комплемента
С1 и СЗ. Отеки развиваются вследствие избыточ-
ной продукции брадикинина.
Периодический отек Квинке с эозинофили-
ей — тяжелое заболевание, которое сопровожда-
ется лихорадкой, увеличением массы тела вслед-
ствие задержки воды, сильным зудом и поражает
лицо, шею, конечности, туловище. Лаборатор-
но выявляются эозинофилия (60—80 % эозино-
филов), высокий лейкоцитоз.
Пигментная крапивница характеризуется оча-
говыми скоплениями тучных клеток в коже и
наличием красновато-коричневых папул и пятен.
Высыпания наблюдаются чаще всего на тулови-
ще или конечностях, реже на лице. Ладони и
Глава 21. Крапивница. Отек Квинке
275
подошвы, волосистая часть головы обычно ос-
таются свободными от высыпаний. Различают
три формы пигментной крапивницы: пятнис-
тую, папулезную, смешанную. При расчесыва-
нии или повреждении появляются эритема и
волдыри вследствие дегрануляции тучных кле-
ток (симптом раздражения или набухания
Унны-Дарье). Этот симптом проявляется при
энергичном потирании шпателем, когда пятно
или папула краснеет, а затем превращается в
плотный, ограниченный, бугристый синюшно-
красного цвета волдыреподобный элемент. В
таком виде элемент сохраняется от нескольких
минут до многих часов и часто сопровождается
ощущением зуда или жжения. Пигментная кра-
пивница наблюдается чаще у детей и может ис-
чезать у подростков. Системный мастоцитоз, на-
оборот, наблюдается чаще у взрослых и
сопровождается инфильтрацией тучными клет-
ками печени, селезенки, лимфатических узлов и
других органов. Под влиянием различных фак-
торов тучные клетки выделяют медиаторы, в
результате возникают приливы крови, свистящее
дыхание, головные боли, отеки и шок.
Патоморфология
Патоморфологически для острой крапивницы
характерны отек сосочкового слоя дермы, рас-
ширение просвета лимфатических кровеносных
сосудов (капилляров), незначительный перивас-
кулярный инфильтрат, состоящий из лимфоци-
тов, иногда выявляют спонгиоз. При хроничес-
кой крапивнице наблюдается периваскулярный
инфильтрат, увеличенное количество лимфоци-
тов. Количество тучных клеток увеличено при-
мерно в десять раз, растет содержание гистами-
на. Почти у половины больных в тканях
обнаруживаются отложения основного щелочно-
го белка эозинофилов.
Диагноз
Диагноз крапивницы не представляет особых
затруднений, обычно достаточно анамнеза и ти-
пичной клинической картины: характерный вид
мономорфной сыпи, быстрое развитие и регресс
элементов. При необходимости проводят био-
псию кожи, при которой выявляют отек дермы
и подкожной клетчатки, дилатацию венул, де-
грануляцию тучных клеток, слабую периваску-
лярную инфильтрацию. Для диагностики холо-
довой крапивницы к коже на 10 мин
прикладывают кусок’ льда или пробирку с ледя-
ной водой. Уртикарный васкулит можно запо-
дозрить в случае сохранения волдырей в течение
24 часов, тогда следует провести биопсию. Сол-
нечную крапивницу диагностируют с помощью
фотопроб с УФ-А, УФ-В и излучением видимо-
го диапазона, когда через 30 мин появляются
волдыри. Уртикарный дермографизм выявляют
при легком штриховом раздражении кожи, что
приводит к немедленному появлению зудящего
волдыря. При тепловой крапивнице положитель-
на проба с теплом: пробирку с теплой водой
(50°С) прикладывают к коже предплечья на
5 мин, гиперемия и отек возникают через 10—
20 мин. Холинергическая крапивница развива-
ется при физической нагрузке до пота (появля-
ются мелкие волдыри) или после внутрикожного
введения 0,1 мл карбохолина (волдырь свыше
15 мм). Адренергическая крапивница возника-
ет при нервных стрессах и при ней в крови об-
наруживают повышенное количество адренали-
на, норадреналина и IgE. Периодический отек
Квинке с эозинофилией проявляется лихорад-
кой, высоким лейкоцитозом (главным образом
за счет эозинофилов) и увеличением веса вслед-
ствие задержки воды). При наследственном оте-
ке Квинке отягощен семейный анамнез, отек
лица и конечностей может быть спровоцирован
незначительной травмой, наблюдаются боли в
животе, снижены уровни компонента компле-
мента С4 и ингибитора С1-эстеразы.
Лечение
При острой крапивнице необходимо выявить
препарат, вызвавший аллергическую реакцию, и
немедленно его отменить. Антигистаминные
препараты назначают после предварительного
очищения желудочно-кишечного тракта или
парентерально в дозе 1—2 мл. Используют диа-
золин по 0,025—0,05 г; тавегил по 0,001 г; дип-
разин по 0,025 г; супрастин по 0,025 г; димед-
рол по 0,025—0,05 г; фенкарол по 0,025 1—2 раза
в день; цетиризин (парлазин) по 10 мг в сутки;
дезлоратадин по 5 мг в сутки.
Применяются десенсибилизирующие средства,
купирующие проявления аллергической реак-
ции, а также дегидратирующие препараты в це-
лях уменьшения отеков. Назначают 25 %-ный
раствор магния сульфата по 1 столовой ложке 3—
5 раз в день (до получения легкого слабитель-
ного эффекта), мочегонные средства (тиазиды,
фуросемид). Внутривенно вводят препараты
кальция (маленьким детям внутрь), натрия тио-
сульфат. Эти средства, обладая выраженным те-
рапевтическим эффектом, обычно снимают ос-
трый приступ крапивницы легкой и средней
степени тяжести.
При отеке Квинке назначают подкожные
инъекции 0,1 % раствора адреналина по 0,3—
276 Частная дерматология
0,5 мл каждые 15—20 мин в зависимости от тя-
жести состояния. При тяжелом состоянии (отек
гортани) назначают внутривенно струйно 60 мг
преднизолона в 20 мл 40 % раствора глюкозы.
Поддерживают проходимость дыхательных пу-
тей и устанавливают венозный катетер. Может
потребоваться искусственная вентиляция лег-
ких и инфузионная терапия. При периодичес-
ком отеке Квинке с эозинофилией показан
преднизон.
При наследственном отеке Квинке показано
длительное лечение андрогенами со слабым мас-
кулинизирующим действием (даназоном и ста-
нозол). При обострениях переливают свежезамо-
роженную плазму или вводят концентрат
ингибитора С1-эстеразы. Эпсилон-аминокапро-
новую кислоту при остром наследственном оте-
ке Квинке вводят внутривенно по 100—200 мл
5 % раствора, затем по 100 мл каждые 4 часа.
Полезно вводить ингибиторы протеиназ и кал-
ликреина (трасилол или контрикал в дозе 30—
40 тыс. ЕД) в 300 мл изотонического раствора
натрия хлорида в течение 3 часов.
В лечении приобретенной холодовой крапив-
ницы особенно эффективен ципрогептадин. При
механической крапивнице (от давления) корти-
костероиды и нестероидные противовоспалитель-
ные препараты предотвращают развитие пораже-
ний.
Лучшим средством лечения адренергической
крапивницы являются b-адреноблокаторы (анап-
рилин, индерал, оксипренолон). При холинер-
гической крапивнице антигистаминные препа-
раты неэффективны, поэтому применяют
холинолитики в комбинации с другими сред-
ствами.
Для лечения больных хронической крапивни-
цей применяют комбинацию Н^Н^антигиста-
минных препаратов: циметидин внутрь по 800 мг
в сутки, димедрол или тавегил, фенкарол, пери-
тол в обычных дозах, с заменой препарата через
10—12 дней.
Профилактика
Больному рекомендуют всегда иметь при себе
список препаратов, вызывающих аллергическую
реакцию. Не рекомендуется использовать рент-
геноконтрастные средства, вызывавшие ранее
побочную реакцию. Если без них нельзя обой-
тись, то за 30—60 мин до исследования назнача-
ют Н,- блокатор и преднизон в дозе 1 мг на кг
веса тела.
Глава 22
Псориаз
Псориаз — хронический дерматоз неустановлен-
ной этиологии, с большой долей генетической
компоненты, характеризующийся воспалитель-
ными шелушащимися папулами и бляшками, с
тенденцией к распространению и утяжелению
кожного процесса в течение жизни больного,
нередко вызывающий патологические изменения
в других тканях и органах (ногти, суставы, по-
звоночник, иногда почки), приводящий к пси-
хической, социальной дезадаптации, иногда к
инвалидизации. История изучения болезни на-
считывает около 200 лет. До этого периода за-
болевших псориазом включали в группу лепроз-
ных («прокаженных») больных.
В развитых странах псориазом страдает 1,2—
2 % населения. Псориазом болеют одинаково
часто мужчины и женщины. Заболевание возни-
кает преимущественно в молодом возрасте (75 %
случаев): у женщин в среднем в 16 лет, у муж-
чин — в среднем в 22 года (I тип), реже
(25 %) — в старших возрастных группах (в сред-
нем в 56 лет) (II тип). Псориаз чаще встречает-
ся у населения с атерогенным профилем пита-
ния; редко — у японцев, эскимосов и жителей
Западной Африки; весьма редко — у севере- и
южноамериканских индейцев.
Факторы риска
Наследственное предрасположение (мультифак-
ториальное) — по аутосомно-доминантному и
рецессивному признаку. У родственников I сте-
пени родства (не страдающих псориазом) выяв-
ляется т. н. латентный псориаз — морфологичес-
ки в коже обнаруживается разрыхление
капилляров, их резкое расширение.
Острый и хронический стресс, функциональ-
ные нарушения ЦНС. Более чем у 40 % больных
можно установить связь возникновения и обо-
стрений псориаза с эмоциональными перегруз-
ками.
Провоцирующая роль обострений очаговых ин-
фекций (стрептококковая, вирусная, стафилокок-
ковая) выявляется при каплевидном псориазе.
Определенную роль в возникновении псори-
аза играют проявления атеросклероза (ИБС,
мозговой инсульт), сахарный диабет, нарушения
обменных процессов (жировой, углеводный),
заболевания печени.
Ряд лекарственных препаратов (кортикостеро-
иды, группы акрихина, соли лития, р-адрено-
блокаторы и др.) могут вызывать псориазифор-
мную реакцию кожи или обострение заболевания.
Травматизация кожи (ссадины, порезы, меха-
ническое растирание) способствует появлению
псориатической сыпи на новых участках.
Псориаз чаще возникает в сезоны минималь-
ной солнечной активности (осенне-зимне-весен-
ний периоды).
Этиология и патогенез
Не ясны. Вирусная (ретровирусная) природа за-
болевания окончательно не доказана.
Псориаз в настоящее время включают в груп-
пу мультифакториальных дерматозов с полигон-
ным наследованием. Доля генетической компо-
ненты составляет 60—70 %, а средовой —
30—40 %. При наличии псориаза у одного из
родителей риск развития заболевания ребенка
составляет 8 %; если псориазом страдают оба
родителя — 41 %. Обнаружена взаимосвязь на-
рушенных звеньев иммунитета с генетическими
маркерами системы HLA. Выявлена связь кож-
ных проявлений с антигенами гистосовместимо-
сти В|3, В|7; кожных проявлений и артропатии с
АГ В17, В27, В33, В4П.
В основе кожных проявлений при псориазе
лежит генетически детерминированное наруше-
ние главной функции кожи — кератинизации,
обусловленное увеличением скорости пролифе-
рации и нормальной дифференцировки эпидер-
мальных кератиноцитов. Этот процесс сопро-
вождается выраженным в разной степени
проникновением нейтрофильных лейкоцитов в
эпидермис и активированных Т-лимфоцитов в
дерму. Гистологически данные процессы харак-
теризуются акантозом, исчезновением зернистого
слоя эпидермиса и паракератозом. Патогномо-
ничным признаком псориаза также является
скопление нейтрофильных лейкоцитов в эпидер-
мисе с образованием микроабсцессов Мунро или
277
278 Частная дерматология
спонгиоформных пустул Когоя при пустулезных
формах псориаза. Приведенные выше патологи-
ческие изменения клинически проявляются ха-
рактерным морфологическим элементом кожной
сыпи — плоскими воспалительными папулами
и бляшками, покрытыми легко снимающимися
серебристо-белыми чешуйками.
При псориазе вследствие гиперпролиферации
объем эпидермиса увеличивается в 4—6 раз, чис-
ло слоев герминативных клеток — в 3 раза; су-
щественно ускоряются митотическая активность
и синтез ДНК в этих клетках. Резко укорачи-
вается время клеточного цикла, а превращение
базальных кератиноцитов в роговые чешуйки
составляет 4 сут вместо 28 сут в норме. При псо-
риазе отсутствуют микроскопически выявляе-
мые апоптические кератиноциты, что может
обусловить нарушение физиологического пути
клеточного отсева и увеличение толщины эпи-
дермиса.
В эпидермальных кератиноцитах увеличива-
ется количество кератинов 6,16,18; появляется
кератин 17, что свидетельствует о гиперпроли-
ферации эпидермиса, увеличивается количество
кератинов 1 (базального) и 2,5 (супербазальных).
Отмечается также нарушение синтеза других
белков — маркеров нормальной дифференци-
ровки кератиноцитов (филлагрина, инволюкри-
на, калмодулина), корнеодесмозина.
Воспалительные изменения в дерме при псо-
риазе характеризуются удлинением, извилисто-
стью и расширением капилляров, повышением
проницаемости их стенок и образованием око-
лососудистых инфильтратов, состоящих преиму-
щественно из лимфоцитов (Т-хелперов типа I),
макрофагов и нейтрофильных лейкоцитов.
При псориазе выявлены нарушения практи-
чески всех звеньев иммунной регуляции кожи,
приводящие к выделению активированными
кератиноцитами, лимфоцитами, макрофагами и
резидентными клетками дермы различных меди-
аторов иммунного ответа и воспаления; поли-
аминов, протеаз и т. п., которые усиливают про-
лиферацию неполноценных кератиноцитов и
стимулируют развитие воспалительных измене-
ний в коже.
Общепризнанной классификации псориаза не
существует. Условную клиническую классифи-
кацию можно представить следующим образом:
• обычный псориаз;
• псориатическая артропатия;
• псориатическая эритродермия;
• пустулезный псориаз:
• ладонно-подошвенный пустулезный псориаз
(пустулезный псориаз Барбера);
• генерализованный пустулезный псориаз (пу-
стулезный псориаз Цумбуша).
Клиническая картина
Обычный (вульгарный) псориаз встречается наибо-
лее часто и проявляется в виде обильно шелуша-
щихся папул и бляшек на волосистой части го-
ловы, локтях, коленях, кистях, стопах, в
люмбосакральной области (рис. 22.1). Эти высы-
пания с четкими границами, розового (симптом
Пильнова), «лососевого» или насыщенно-красного
цвета покрыты рыхлыми, серебристо-белыми че-
шуйками. Имеют вид круглых, овальных, поли-
циклических, кольцевидных, линейных, зостери-
формных кожных поражений.
Поскабливание папул позволяет выявить триа-
ду характерных для заболевания феноменов:
• феномен стеаринового пятна, характеризую-
щегося усилением шелушения при легком по-
скабливании, придающего поверхности папул
сходство с растертой каплей стеарина (гиперке-
ратоз, паракератоз, накопление липидов и липо-
идов в верхних слоях эпидермиса);
• феномен терминальной пленки, возникающей
после удаления чешуек и проявляющейся в виде
влажной, тонкой, блестящей поверхности элемен-
тов (результат акантоза; обнажается слизистый
слой эпидермиса);
Рис. 22.1. Типичная локализация
высыпаний при вульгарном псориазе.
Глава 22. Псориаз
279
• феномен точечного кровотечения, появляю-
щегося после легкого поскабливания терминаль-
ной пленки, в виде точечных, не сливающихся
капелек крови (феномен Auspitz или кровяной
росы А. Г. Полотебнова). Этот феномен обуслов-
лен травматизацией полнокровных сосочков дер-
мы в результате папилломатоза.
Для псориаза характерна изоморфная реакция
(феномен Кебнера) — развитие псориатических
высыпаний на участках кожи, подвергшихся
механическому или химическому действию
(мелкие травмы, расчесы, порезы, капли кислот,
щелочей и др.). Наиболее ярко выражена изомор-
фная реакция в прогрессирующую стадию забо-
левания. Высыпания на местах травматизации
обычно возникают в течение двух недель.
Излюбленные места. Излюбленные места ло-
кализации псориатической сыпи — локти, ко-
лени, волосистая часть головы, реже складки. На
этих участках сыпь может существовать длитель-
но, не распространяясь на другие участки кож-
ного покрова. Такие элементы образно называ-
ют «дежурные» или «сторожевые» бляшки.
Элементы сыпи в ряде случаев располагаются
беспорядочно по кожному покрову, образуя ду-
гообразные, змеевидные, географические фигу-
ры. У некоторых больных элементы могут сли-
ваться в сплошную пораженную поверхность,
напоминающую огромную псориатическую
бляшку, обозначаемую как эритродермия. Эле-
менты сыпи можно обнаружить в области гени-
талий, на ногтевых фалангах, волосистой части
головы, редко в области лица (рис. 22.2).
Обычный псориаз в большей части случаев
проявляется типичными папулезно-бляшечными
высыпаниями. Однако в зависимости от харак-
тера клинического течения и локализации про-
цесса выделяют следующие его разновидности:
себорейный, экссудативный, интертригинозный,
ладоней и подошв, застарелый, рупиоидный,
каплевидный, пятнистый.
Волосистая часть головы является одной из
наиболее частых первоначальных локализаций
псориаза.
Себорейный псориаз как на волосистой части
головы, так и на других «себорейных» участках
имеет своеобразную клиническую картину. При-
чем в таких случаях проявления заболевания
длительно могут существовать самостоятельно в
виде папулезных элементов или бляшек, шелу-
шения на волосистой части головы с типичным
поражением соседних участков гладкой кожи,
особенно лба и заушных областей («псориатичес-
кая корона»). Поражение волосистой части го-
ловы выявляют у 80 % больных псориазом. Для
подтверждения диагноза псориатического пора-
жения волосистой части головы можно приме-
нить прием Картамышева А. И. — при пальпа-
ции с закрытыми глазами появляется ощущение
четких границ по периферии псориатических
бляшек, в отличие от очагов себорейного дерма-
тита.
При экссудативном псориазе выявляются псо-
риатические папулы и бляшки, покрытые кор-
ками и чешуйками, пропитанными желтоватым
экссудатом. Нередко такая форма псориаза раз-
вивается у лиц с избыточной массой тела.
У детей и пожилых пациентов, особенно у
страдающих сахарным диабетом, развивается
псориатическое поражение крупных складок,
т. н. интертригинозный псориаз. При этом шелу-
шение, как правило, не выражено или отсутству-
ет, очаги резко очерчены, их поверхность глад-
кая, насыщенно-красного цвета, иногда слегка
влажная, мацерирована, весьма схожа с прояв-
лениями кандидоза складок, дерматофитиями,
опрелостью.
У ряда больных псориазом, чаще у лиц, заня-
тых физическим трудом, а также при обостре-
нии распространенного псориаза обнаруживаются
поражения ладоней и подошв, которые могут
существовать в виде изолированного дерматоза
(инфильтрация, гиперкератоз, трещины, болез-
ненность).
Рис. 22.2. Инверсный псориаз.
10 Зак. 1357
280 Частная дерматология
Длительное существование бляшек, чаще рас-
положенных на пояснице, ягодицах, бедрах при-
водит к их значительной инфильтрации, иног-
да с папилломатозными и бородавчатыми
разрастаниями (застарелый псориаз), слоистыми
корками, что придает высыпаниям коническую,
рупиоидную форму (рупиоидный псориаз).
При наличии обильных миллиарных и ленти-
кулярных папул по всему кожному покрову
диагностируется каплевидный псориаз, который
развивается остро, вскоре после перенесенных
инфекций (ангина, тонзиллит, синуситы, ОРВИ
и др). В этих случаях из глотки, складок высе-
вается Streptococcus pyogenes.
У 25 % больных псориазом поражаются ног-
тевые пластинки, чаще на кистя, в виде наперст-
ковидной истыканности (симптом «наперстка»).
Реже наблюдаются явления подногтевого гипер-
кератоза, онихогрифоза (симптом «песчаных
волн»), онихолизиса, изменение окраски и на-
личие характерного желтовато-бурого пятна под
ногтем (симптом «масляного пятна» Кейнинга-
Гасенфлюга), онихолизиса, деформация ногте-
вых пластинок («коготь птицы»), подногтевые
петехии (симптом Левенталя), выявляется дефект
эпонихия (признак Попова).
Слизистая оболочка рта (щеки, губы, язык)
поражается у 1—2 % больных вульгарным псо-
риазом. На слизистых выявляются возвышаю-
щиеся плоские папулы серовато-белой окраски,
с четкими границами и розовым периферичес-
ким венчиком. При пустулезном псориазе час-
то наблюдается географический глоссит и более
обширное поражение слизистой.
К тяжелым клиническим формам относят псо-
риатическую эритродермию, артропатический и
пустулезный псориаз.
Эритродермия может возникнуть при посте-
пенном прогрессировании обычного псориати-
ческого процесса, в результате слияния бляшеч-
ных элементов. Кожный покров принимает вид
одной гигантской инфильтрированной бляшки,
с характерным серебристо-белым шелушением,
положительной псориатической триадой. На-
блюдается увеличение лимфатических узлов; во-
лосы не выпадают.
Чаще возникает аллерготоксическая форма псо-
риатической эритродермии. Она развивается дос-
таточно быстро под влиянием провоцирующих
факторов: гиперинсоляции, нерациональной фо-
тотерапии в прогрессирующей стадии заболевания,
в результате быстрой отмены кортикостероидных
препаратов, при лечении сопутствующей патоло-
* Раздел написан О. С. Зыковой
гии с использованием антималярийнЫх препара-
тов, новокаина, пенициллина и некоторых других
препаратов. У больных при этом наблюдаются
признаки общей интоксикации: повышение тем-
пературы тела, головная боль, боль в мышцах, дис-
пепсические расстройства. Кожный покров при-
обретает ярко-красный цвет с бурым оттенком;
отек и инфильтрация на различных участках вы-
ражены неравномерно. Кожа лица натянута, оп-
ределяется эктропион. На волосистой части голо-
вы шелушение отрубевидное, а на других
областях — пластинчатое или крупнопластинчатое.
Типичные для псориаза признаки исчезают либо
выражены незначительно. Увеличиваются лимфа-
тические узлы, выпадают волосы. Больные жалу-
ются на зуд, жжение, стянутость кожи, болезнен-
ность в конечностях при движении.
Псориатическая артропатия. * Артропатичес-
кий псориаз является инвалидизирующей фор-
мой заболевания. Термином «псориатическая ар-
тропатия» обозначается разнообразная патология
опорно-двигательного аппарата у больных псо-
риазом — артралгии, артриты, артрозы и др., а
также их сочетания. В медицинской литературе
для обозначения этого состояния используются
также термины «псориатический артрит», «псо-
риатический спондилоартрит». Псориатический
артрит — воспалительный артрит, который свя-
зан с псориазом и, как правило, негативен по
отношению к ревматоидному фактору (РФ).
Термин «псориатический спондилоартрит» упот-
ребляется при значительном или преимуществен-
ном поражении позвоночника.
Псориатическая артропатия описана у больных
псориазом в возрасте от 9 до 71 года. Средний
возраст больных составляет 30—55 лет. Считается,
что гендерная структура заболевания однородна,
однако некоторые авторы указывают на более
частую встречаемость артропатии у мужчин.
У большинства больных симптомы поражения
кожи предшествуют поражению суставов (око-
ло 70 %), у части больных появляются уже в
первый год после начала псориаза, а у некото-
рых больных предшествуют высыпаниям на
коже. Разные типы псориаза неоднородны в
отношении сроков развития артропатии. Так,
при псориазе I типа поражение опорно-двига-
тельного аппарата развивается по прошествии
нескольких лет после начала кожного пораже-
ния, в то время как у пожилых больных псори-
азом И типа поражения кожи и опорно-двига-
тельного аппарата возникают с большей или
меньшей синхронностью. Обнаружены рентге-
Глава 22. Псориаз
281
нологические изменения в суставах у большин-
ства больных псориазом при отсутствии жалоб
на состояние опорно-двигательного аппарата.
До настоящего времени общепризнанного мне-
ния о механизме развития псориатической арт-
ропатии не существует. Семейственность наблю-
дается не только в отношении псориаза вообще,
но и в отношении псориатической артропатии, и
этот факт наводит на мысль о генетических осо-
бенностях этих больных. Показано, что монози-
готные близнецы при псориазе имеют конкордан-
тность 65—72 %, а дизиготные — 15—30 %. У лиц,
страдающих псориатической артропатией, наря-
ду с «генами псориаза» HLA В17, В13, В16 и др.,
наблюдается повышение частоты HLA В27, В39,
В38. Дополнительную роль в качестве причинных
факторов могут играть инфекции, травмы, стресс.
В патогенезе псориатической артропатии важ-
ную роль играют иммунологические нарушения,
связанные с активацией Т-лимфоцитов. В коже
очагов поражения и синовии поражённых суста-
вов обнаруживаются Т-клеточные инфильтраты
и отложения иммуноглобулинов, повышенные
концентрации провоспалительных цитокинов —
ФНО-альфа, IL-1, IL-15, IL-10. В свою очередь
ФНО-альфа индуцирует синтез ряда других ци-
токинов — IL-6, IL-8, GM-CSF. Большую роль
в развитии периваскулярной инфильтрации иг-
рают морфологические изменения в мелких со-
судах кожи и синовия у этих больных.
Распространённость поражения опорно-двига-
тельного аппарата при артропатическом псори-
азе широко варьирует от моно- и олигоартрита
до поражения нескольких или многочисленных
суставов конечностей или позвоночника, имею-
щих разную степень воспалительной активнос-
ти (рис. 22.3). К характерным клиническим осо-
бенностям псориатической артропатии относят:
• поражение межфаланговых суставов конеч-
ностей;
• энтезоит — воспаление суставов в зоне при-
крепления связок и сухожилий;
• дактилит — воспаление всех тканей пальца;
• асимметричный сакроилеит и спондилит
(под асимметричностью подразумевается не
столько односторонность поражения, сколько
выраженность клинических проявлений сустав-
ной патологии);
• постепенное увеличение количества пора-
жённых суставов с течением времени;
• фазность течения заболевания со сменой пе-
риодов обострения и субремиссии;
• продолжительность и частота ремиссий
обычно уменьшается у больных с полисуставны-
ми поражениями.
Поражение кожи у больных с артропатией бы-
вает распространённым, эритродермическим, экс-
судативным или пустулёзным. Более чем у трети
больных наблюдается поражение ногтевых пласти-
нок, которое, как правило, предшествует артропа-
тии и проявляется в виде напёрстковидной исты-
канности, «масляных пятен», гипертрофии,
гребневидной деформации, онихолизиса и др. При
тяжёлых формах заболевания часто диагностируется
патология внутренних органов — сердечно-сосуди-
стой, мочевыделительной систем, печени, органов
зрения и др., что в свою очередь подтверждает си-
стемный характер псориаза. Выделена злокачествен-
ная форма артропатического псориаза с тяжёлыми
системными проявлениями, быстрым прогрессиро-
ванием и неблагоприятным прогнозом.
При установлении диагноза псориатической
артропатии используют систему дианостических
критериев, которая включает комплекс анамне-
стических, клинических, лабораторных, рентге-
нологических данных.
У 1 % больных наблюдается пустулезный псо-
риаз, который проявляется в виде генерализован-
ной сыпи или ограниченных высыпаний в обла-
сти ладоней и подошв. Пустулы стерильны, хотя
отдельные авторы, в частности Г. Я. Шарапова и
Рис. 22.3. Типичные очаги
поражения при псориатической артропатии.
282 Частная дерматология
соавт., указывают, что у некоторых больных
высевается 5. aureus, другие исследователи обна-
руживали бета-гемолитический стрептококк.
Генерализованный пустулезный псориаз разви-
вается чаще на фоне обычного псориаза, который
впервые описан L. Zumbusch (1910). Большинство
исследователей возникновение пустулезного псо-
риаза связывают с инфекциями, резкими стрес-
совыми ситуациями, применением в прогресси-
рующей стадии антималярийных препаратов,
антибиотиков, гормональными нарушениями, в
том числе вызванными пероральными контрацеп-
тивами, с быстрой отменой кортикостероидов,
нерациональной наружной терапией. Генерализо-
ванный псориаз — это тяжелая, угрожающая
жизни пациента форма заболевания с резким
интоксикационным синдромом. На фоне яркой
эритемы появляются мелкие поверхностные пу-
стулы, сопровождающиеся жжением и болезнен-
ностью, расположенные как в зоне бляшек, так
и на ранее не измененной коже. Вначале разбро-
санные очаги эритемы с пустулизацией быстро
увеличиваются в размерах, сливаются, захваты-
вая обширные участки кожного покрова, так что
псориатические бляшки перестают быть различи-
мыми. Слившиеся пустулы отслаивают поверх-
ностные слои эпидермиса в виде «гнойных озер»,
может развиться эритродермия. У некоторых
больных наблюдаются кольцевидные, серпигини-
рующие и другие фигурные очаги, эритемато-пу-
стулезные, эрозивные, географические высыпа-
ния на слизистой полости рта. Часты
дистрофические изменения ногтей, поражения
суставов, иногда почек.
Пустулезный псориаз ладоней и подошв встре-
чается чаще, чем генерализованная форма. Вы-
сыпания располагаются преимущественно в об-
ласти тенара и гипотенара, свода стоп. Характерна
резкая очерченность эритематозно-сквамозных
бляшек, на фоне которых (реже по периферии)
имеются множественные пустулезные высыпа-
ния. Одни из них ссыхаются с образованием
чешуйко-корочек, другие появляются им на
смену. Пустулезные высыпания могут быть в
зоне бляшек и на других участках кожного по-
крова, чаще всего они развиваются при раздра-
жающем наружном лечении.
Клинические формы и их симптомы
Обычная — эпидермально-дермальные папулы,
серебристо-белые чешуйки, бляшки, иногда зуд.
Локализация — разгибательные поверхности
конечностей, туловище. Клинические изменения
в суставах — у 5~7 % больных, рентгенологи-
ческие — у большинства больных.
Экссудативная — эпидермально-дермальные
папулы, бляшки, чешуйко-корки, часто зуд.
Локализация — различные участки кожного
покрова, нередко поражение складок.
Псориаз волосистой части голбвы — пораже-
ние головы может быть изолированным или со-
четаться с высыпаниями в других областях тела.
Характерен интенсивный зуд; волосы обычно не
выпадают. Четко определяются инфильтриро-
ванные папулы или бляшки с обильным сереб-
ристо-белым шелушением.
Эритродермия — универсальная эритема, ин-
фильтрация, шелушение, зуд, частое увеличение
лимфоузлов, температура, признаки интоксика-
ции; наличие предшествующих типичных папу-
лезных псориатических высыпаний.
Артропатическая (дистальная и аксиальная
форма) — деформация суставов, анкилозы, му-
тиляции, поражение кожи, боль, рентгенологи-
чески — остеопороз, сужение суставных щелей,
остеолизис, остеофиты, кистозные просветления
костной ткани. Ревматологические тесты отри-
цательны.
Поражение ногтей — наперстковидная исты-
канность, разрушение ногтевых пластинок кис-
тей (точечная форма); утолщение, деформация
ногтей с явлениями онихогрифоза (гипертрофи-
ческая форма); истончение и лизис ногтевых
пластинок (атрофическая форма).
Пустулезный генерализованный псориаз Цумбу-
ша — эритематозно-сквамозные бляшки эпидер-
мально-дермального характера с пустулезными
высыпаниями, корочками, гнойными элемента-
ми по типу смешанного импетиго, сливающими-
ся в «гнойные озера»; лихорадка, лейкоцитоз,
ускоренная СОЭ; изменения ногтей, поражения
суставов, иногда почек.
Ладонно-подошвенный пустулезный псориаз
(тип Барбера) — на ладонях и подошвах появ-
ляется множество желтоватых, глубоко располо-
женных в эпидермисе пустул со стерильным
содержимым и последующим образованием на
этих местах красновато-бурых пятен.
Патогномоничные симптомы: «стеариновое пят-
но», «терминальная пленка», «точечное кровоте-
чение». Наиболее четко определяются в прогрес-
сирующую стадию заболевания.
Гистологические признаки обычного псориаза:
гипер- и паракератоз, агранулез, акантоз, папил-
ломатоз. Экзоцитоз лимфоцитов и нейтрофилов
(субкорнеально) — нейтрофильные микроабс-
цессы Мунро. Отек и расширение сосудов в со-
сочках, по ходу скопления лимфоцитов, гисти-
оцитов; в сетчатом слое— расширение сосудов и
инфильтрация менее выражены.
Глава 22. Псориаз 283
Стадии течения
Прогрессирующая — появление новых мелких
высыпаний насыщенно-розового цвета, перифе-
рический рост элементов с наличием эритема-
тозного бордюра, не покрытого чешуйками
(симптом или ободок Пильнова), положительная
изоморфная реакция Кебнера, четко определя-
ется псориатическая триада.
Стационарная — новые элементы не появля-
ются, имеющиеся высыпания сохраняются.
Регрессирующая — разрешение высыпаний и
образование вокруг них псевдоатрофического обод-
ка Воронова (вокруг папул обнаруживается блес-
тящее светлое кольцо слегка морщинистой кожи).
Диагноз
Алгоритм постановки развернутого диагноза
псориаза предлагаем следующий.
Распространенность процесса
Ограниченный — единичные бляшки на локтях,
в области коленных суставов, других участках
тела), волосистой части головы.
Диссеминированный (распространенный) —
множество элементов на различных участках
тела.
Диффузный — слившиеся бляшки, занимаю-
щие значительные участки кожи.
Эритродермия — универсальное поражение
кожного покрова.
Характер папулезных элементов
Вульгарный псориаз — типичная сыпь.
Экссудативный псориаз — с экссудацией, кор-
ками.
Себорейный псориаз — локализация в себорей-
ных участках.
Интертригинозный — в складках.
Пустулезный псориаз — с пустулизацией, эро-
зиями.
Величина элементов
Мелкобляшечный — папулы милиарного, ленти-
кулярного, монетовидного характера. Бляшеч-
ный — бляшки до детской ладони (6—7 см);
Крупнобляшечный — изолированные бляшки с
ладонь взрослого человека (15 см и более);
Диффузные бляшки — сплошные бляшки, за-
нимающие отдельные участки тела (кожу живота,
спины, конечностей);
Универсальное поражение — весь кожный по-
кров представляет сплошную пораженную по-
верхность;
Степень инфильтрации бляшек: умеренная, вы-
раженная (значительная), резкая, с наличием тре-
щин и их кровоточивости.
Тяжесть течения
Обычная (нетяжелая форма) — поражение до 1/3
кожного покрова.
Среднетяжелая форма — поражение составля-
ет 30—50 % кожного покрова.
Тяжелая — поражение 50—70 % кожного по-
крова.
Очень тяжелая — поражение кожи свыше
70%.
Поражение костно-суставной системы: вовле-
чение суставов, позвоночника (с указанием
топографии) и их функциональной недостаточ-
ности.
Поражение придатков кожи: характер пораже-
ния ногтевых пластинок, наличие признаков
алопеции.
Стадии заболевания
Прогрессирующая, стационарная, регрессирую-
щая. Установление стадии играет важную роль
в выборе средств общего и местного воздействия.
Сезонность высыпаний
Зимний тип отличается обострениями в осенне-
зимний период). Летний тип - обострениями в
весенне-летний период). Смешанный тип - вне-
сезонный.
Чувствительность к УФ лучам
Фоточувствительный (УФ лучи оказывают по-
ложительный лечебный эффект, способствуют
регрессу элементов, без раздражения окружаю-
щей их кожи).
Фототоксический с явлениями фотосенсиби-
лизации — выявляется фототоксическое дей-
ствие УФ лучей на пораженную и окружающую
кожу: фотодерматит, болезненность кожи. УФО
таким пациентам проводить не рекомендуется.
Частота рецидивов
Редко рецидивирующий (обострения 1 раз в не-
сколько лет).
Умеренно рецидивирующий (обострения через
1—2 года).
Часто рецидивирующий (ремиссия сохраняет-
ся 1-3 месяца после лечения).
Непрерывно рецидивирующий (торпидный) —
неполный лечебный эффект, обострение заболе-
вания в течение месяца после выписки из ста-
ционара.
Пример описания диагноза
Распространенный вульгарный псориаз, мелко-
бляшечный, обычная форма, прогрессирующая
стадия, зимний тип, фоточувствительный, уме-
ренно рецидивирующее течение.
284 Частная дерматология
Дифференциальный диагноз
Красный плоский лишай — полигональная фор-
ма элементов, восковидный блеск высыпаний,
пупкообразное вдавление в центре папул, сетча-
тый рисунок на поверхности папул.
Папулезный сифилид — (полушаровидная
форма папул, медно-красный цвет, воротничок
Биетта, обнаружение бледной трепонемы, в эле-
ментах RW положительная).
Розовый лишай — наличие материнской
бляшки на туловище, распространение дочерней
пятнистой сыпи с гофрированной поверхностью
по линиям натяжения.
Парапсориаз — феномен «облатки», фено-
мен «пурпуры» при пощипывании элемента;
наличие прикрепленной пластинчатой чешуй-
ки на месте рассосавшегося элемента — сим-
птом «коллодийной пленки»; феномен «скры-
того шелушения».
Себорейный дерматит — отсутствует четко
определяемая инфильтрация кожи.
Дерматофития волосистой части головы —
очаги эритематозно-сквамозных пятен с обло-
манными волосами или пеньками волос (обяза-
тельное исследование на грибы).
Атопический дерматит — неяркая эритема,
шелушение, лихенификация, экскориации в
местах излюбленной локализации (лицо, шея,
верхняя часть груди, локтевые и подколенные
области).
Болезнь Рейтера — уретрит, конъюнктивит,
артрит, псориазиформные высыпания на по-
дошвах, головке полового члена, в полости
рта с тенденцией к экссудации и пустулиза-
ции.
При поражении ладоней и подошв необходи-
мо дифференцировать с дерматофитиями кистей,
стоп, дисгидротической экземой, кератодерми-
ями.
Псориаз кожных складок — дерматофитии,
кандидоз, хроническая экзема, нейродермит.
Псориатическую эритродермию дифференци-
руют с эритродермиями при экземе, таксидер-
мии, розовом лишае, лимфомах кожи.
Артропатический псориаз дифференцируют с
ревматоидным артритом, болезнью Рейтера, ар-
трозами.
Лекарственная таксидермия — псориазифор-
мные высыпания при лечении р-адреноблокато-
рами, метилдофой, препаратами золота.
Бактериемия и сепсис (посев крови) при ге-
нерализованном пустулезном псориазе.
Внезапное начало псориаза может быть свя-
зано с ВИЧ-инфекцией (необходимо обследова-
ние).
Лечение
Проводится с целью получения выраженного
клинического эффекта и его последующего за-
крепления в период ремиссии.
Комплексное лечение больных псориазом
включает: общую и местную терапию, физиоте-
рапию с учетом стадии заболевания, тяжести
клинических проявлений, особенностей локали-
зации кожного процесса, сезонности.
Лечение прогрессирующей стадии начинают с в/
м введения антигистаминных препаратов (супра-
стин, тавегил, пипольфен, димедрол); 30 % тио-
сульфат натрия внутривенно по 3—10 мл ежеднев-
но № 10 или 5 % раствора унитиола (п/к, в/м
3—10 мл через день № 5) или 10 % раствор каль-
ция хлорида 5—10 мл в/в, ежедневно № 4-5,
затем через день № 4—5. Одновременно назнача-
ют седативные средства (бромиды, валериана,
транквилизаторы). Проводят витаминотерапию:
курс инъекций витаминов Вр В6, В12; фолиевая
кислота (0,02 г 3 раза в день), витамин С (до 1,0 г
в сутки). Витамин А 100 000 ME в сутки внутрь
или в/м, «Аевит» 1 капсула 2—3 раза в сутки,
витамин D2 —2000 МЕ/кг в сутки.
При нарушениях жирового обмена — препара-
ты никотиновой кислоты, теоникол (1 таблетка
3 раза в сутки), трентал 0,1 г 3 раза в сутки;
липамид, липоевая кислота, метионин; эссенци-
але по 5—10 мл в/в на аутокрови ежедневно
№ 10, затем по 2 капсулы 2—3 раза в день в
течение 1—2 месяцев; экстракт солянки холмо-
вой по 1 чайной ложке 3 раза в день в течение
1 месяца; полифепан по 1 десертной ложке на-
тощак ежедневно в течение 2-х недель.
В стационарную стадию подключают пироге-
нал 5 мкг в/м, через 1—3 дня дозу повышают в
2 раза, на курс до 100 мкг, или продигиозан —
20—25 мкг (0,5—0,6 мл 0,005 % раствора) в/м,
каждое введение повышают на 20—25 мкг, мак-
симальная разовая доза—100 мкг, инъекции
проводят 2 раза в неделю, на курс № 5—7. На-
значают также экстракт плаценты 1 мл п/к ежед-
невно № 25—30, спленин 1—2 мл ежедневно
№ 10, препараты тимуса (тактивин, тималин и
др.). Проводят курс общего УФО № 10—15, баль-
неотерапию с оксидатом торфа, экстрактом мор-
ской соли, хвойным экстрактом и др.
Приведенные средства являются основными
для лечения обычных форм с поражением до */3
кожного покрова.
При каплевидном псориазе в ряде случаев эф-
фективно назначение антибиотиков (амоксицил-
лин, цефалексин, макролиды) и общего ультра-
фиолетового облучения.
Глава 22. Псориаз
285
Средства и методы
терапии тяжелых форм псориаза
Цитостатики
Метотрексат (антагонист фолиевой кислоты,
подавляет клеточные митозы) по 2,5 мг 2 раза
в день в течение 5 дней с интервалами между
циклами по 3 дня, или 10—25 мг в/в однократ-
но, с недельными интервалами. Курс лечения 4—
5 недель. Возможна комбинация с триамцино-
лоном (16—20 мг в сутки).
Азатиоприн (антиметаболит, структурный ана-
лог пуриновых оснований) 4 мг/кг сутки внутрь
в 3 приема ежедневно; при стабилизации про-
цесса до 50 мг 1 раз в 5 дней до отмены препа-
рата; курс 4—5 недель. Контролируют состояние
периферической крови (количество тромбоцитов,
лейкоцитов), функцию печени.
Ароматические ретиноиды
Ацитретин (неотигазон) — 0,5—1 мг/кг/сут.
После достижения терапевтического эффекта
суточную дозу препарата постепенно снижают и
подбирают поддерживающую дозу. Курс лече-
ния 6-8 недель и более. Наиболее эффективен
при пустулезном псориазе. Эффект лечения уси-
ливается при комбинировании с ПУВА-терапи-
ей. Возможны признаки гипервитаминоза А.
Необходимо наблюдение офтальмолога, невро-
патолога, гинеколога, рентгенография костей.
Циклоспорин А (сандиммун неорал) — подав-
ляет секрецию интерлейкинов и других лимфо-
кинов активированными Т-лимфоцитами, на-
значают 2,5 мг/кг перорально в 2 приема. Курс
лечения около 6 недель, затем подбирают под-
держивающую дозировку препарата. Контроли-
руют артериальное давление, креатинин сыво-
ротки крови, функцию почек 1 раз в неделю.
ПУВА-терапия
(УФО 320—390 нм + метоксален) — цитостати-
ческий эффект. 0,3—0,6 мг/кг метоксалена при-
нимают однократно за 1 ч до облучения. Ритм
облучения 2—4 раза в неделю. Начальная доза
УФО 1,5-2 Дж/см2. Через каждые две процеду-
ры дозу постепенно увеличивают на 1,5—2 Дж,
до максимальной 12—14 Дж/см2. При плотнос-
ти излучения 10—12 мВт/см2 время облучения
составляет 2—18 мин. Поддерживающая терапия
2 раза в неделю в течение 3—4 недель, затем 1
раз в неделю в течение месяца, далее 1 раз в
месяц. Суммарное количество облучений за год
35-50, общая доза облучения 300—600 Дж/см2.
Основной курс лечения 15—20 процедур. ПУВА-
терапию комбинируют с приемом эссенциале
или ароматическими ретиноидами. Возможно
развитие фотодерматита, гипертрихоза, хейлита,
угрей, кандидоза слизистых, фурункулеза, плос-
ких бородавок, кератом, рака кожи, редко—
снижение либидо.
Гепарин
По 10 000 ЕД 2 раза в сутки подкожно в тече-
ние 10 дней, затем 2 раза в сутки по 5 000 ЕД;
после достижения клинического эффекта 1 раз
в сутки по 5000 ЕД в течение 7—10 дней. Осу-
ществляют контроль свертывающей системы
крови.
Методы детоксикации
Гемодез — 200—400 мл в/в капельно с интерва-
лом 2—4 дня, на курс 5—6 вливаний. Контроль
артериального давления.
Гемосорбция проводится с помощью фильтров
«Овосорб», Скн-1к, 2к, 4 м. За одну процедуру
очищают 1,5—2 объема крови пациента. Проце-
дуры повторяют через 5—7 дней, выполняют 3—
5 таких сеансов.
Плазмаферез. В течение курса лечения (в сред-
нем 3 процедуры) у пациента забирается около
4 л плазмы, для компенсации вводится альбу-
мин, кровезаменители. Проводят в отделениях
детоксикации.
Ультрафиолетовое облучение крови. За одну
процедуру воздействию УФО на аппарате типа
«Изольда» подвергается кровь пациента в объе-
ме 2 мл/кг массы тела. На курс 5—7 таких про-
цедур.
Иммуномодуляторы (назначают после имму-
нологического скрининга). Человеческий лейко-
цитарный интерферон. Ингаляции 2—3 раза в
день по 2 500 ЕД в течение 10 дней. Тактивин
по 100 мкг в нижний угол лопатки подкожно в
1, 3, 6, 13 и 20-й день лечения. Тималин — 5 мг/
сут в/м в течение 6—10 дней.
Нестероидные
противовоспалительные средства
Применяют при артропатическом псориазе.
Индометацин по 0,025—0,5 г внутрь 3 раза в
день. Контроль состояния желудка и двенадца-
типерстной кишки (ФГДС).
Напроксен по 0,5—0,75 г внутрь 2 раза в день.
Бруфен по 0,2 г внутрь 3—4 раза в день. Курс
4—6 недель.
Ингибиторы фосфодиэстеразы
Применяют при экссудативном псориазе.
Эуфиллин по 0,015 г 3 раза в день внутрь. Кон-
троль уровня артериального давления, функции
внешнего дыхания и общего самочувствия. Те-
286 Частная дерматология
офиллин по 0,2 г 3 раза в день внутрь. Кавин-
тон по 0,01 г 3 раза в день в течение 2 недель,
затем 2 недели по 0,005 г 3 раза в день. Курс
лечения 3—4 недели.
Для лечения артропатического псориаза исполь-
зуют метотрексат, азатиоприн, циклоспорин А,
которые оказывают иммуносупрессивное дей-
ствие и могут использоваться в комбинациях как
друг с другом, так и с другими противоревмати-
ческими препаратами, а также с сульфосалазином.
Применение этих средств связано с необходимо-
стью контроля функции почек, показателей кро-
ви, возможной активизации микробного факто-
ра и др. Эффективно применение ретиноидов и,
в частности, этретината, во время приёма кото-
рого следует контролировать липидный спектр
крови, состояние печени и учитывать потенци-
альное тератогенное действие препарата.
В настоящее время активно исследуются био-
логические препараты, которые подавляют ак-
тивность провоспалительного цитокина ФНО-
альфа в коже и суставах — этанерсепт и
инфликсимаб, которые, в отличие от нестероид-
ных противовоспалительных средств, приводят
к ослаблению не только суставного, но и кож-
ного патологического процесса. При хорошей пе-
реносимости пациенты могут использовать их в
комбинации с препаратами других групп. Изби-
рательно подавляют активность ФНО-альфа па-
мидронат (бифосфонатный синтетический ана-
лог пирофосфата) и талидомид, которые также
представляют интерес в плане лечения больных
псориатическим артритом.
Для лечения артропатического псориаза при-
меняют противомалярийные препараты (делагил,
плаквенил), дериваты витамина Д, купренил (Д-
пеницилламин), аферез, ПУВА-терапию и её
варианты — с учётом противопоказаний и воз-
можных побочных эффектов. Применение пре-
паратов золота менее эффективно по сравнению
с метотрексатом. Системные глюкокортикоиды
могут применяться лишь в случаях тяжёлого
воспаления суставов и резистентности к терапии.
Из немедикаментозных методов показаны
общие ванны с морской солью, оксидатом тор-
фа, лечебная физкультура и другие мероприя-
тия медицинской реабилитации. При необходи-
мости применяются хирургические методы
лечения — синовэктомия, эндопротезирование
анкилозированных суставов.
Средства и методы наружного лечения
Прогрессирующая стадия: 1—2 % салициловая
мазь, крем Унны, кортикостероидные кремы или
мази (2—3 раза в неделю).
Стационарная стадия: 3 % салициловая мазь,
5—10 % нафталановая мазь, 2—5 % серно-дегтяр-
ная мазь, дитранол (дитрастик), псоркутан, гид-
ротерапия, общее УФО при зимней (смешанной)
форме.
Регрессирующая стадия: 3—5 % салициловая
мазь, 10—20 % нафталановая мазь, 5—10 % сер-
но-дегтярная мазь, мазь Рыбакова (автоловая),
дитранол, псоркутан, псориазин, антипсориати-
кум, общее УФО при зимней, смешанной фор-
мах, бальнеотерапия.
Псориаз волосистой части головы: шампуни,
содержащие деготь или кетоконазол (низорал).
Кортикостероиды в виде лосьонов. При обиль-
ном шелушении и толстых корках: 2—10% са-
лициловая кислота в вазелиновом масле на ночь
под пластиковую шапочку (1—3 процедуры);
0,005 % лосьон с кальципотриолом или шам-
пунь «Скин-кап», элоком-лосьон, дипросалик-
лосьон.
Дежурные псориатические бляшки. Нельзя при-
менять раздражающее лечение (деготь, повышен-
ные концентрации салициловой, молочной, бен-
зойной кислот, резорцина и др.), так как данные
средства могут провоцировать высыпания на
других участках тела. При наличии дежурных
бляшек необходимо исходить из принципа: «Не
буди спящего».
Лечебный эффект оказывают фторированные
кортикостероиды (смазывание или под окклю-
зионную повязку): флуоцинолон, бетаметазона
валерат (дипропионат); наиболее эффективен
клобетазол. В небольшие бляшки можно вводить
внутрикожно триамцинолон. Эффективен так-
же дитранол (крем, мазь, в виде палочек), ко-
торый применяют в соответствии с инструкци-
ей. Постепенный эффект наблюдается при
смазывании бляшек псоркутаном (кальципотри-
ол) 2 раза в день.
Псориаз ногтей: инъекции триамцинолона в
ногтевые валики (2 мг/мл), проводят PUVA-те-
рапию для кистей и стоп, назначают прием ацит-
ретина (0,5 мг/кг), в тяжелых случаях — метот-
рексат, циклоспорин.
Поражение ладоней и подошв: PUVA-терапия
(Re PUVA-терапия), этретинат, метотрексат,
циклоспорин.
Псориаз кожных складок: местные кортикос-
тероиды слабой степени активности, жидкость
Кастеллани, псоркутан.
Профилактика рецидивов
Диета, бедная углеводами и жирами, богатая
овощами и рыбными продуктами. Устранение
или ослабление факторов риска.
Глава 22. Псориаз 287
Проводится коррекция выявленных сопут-
ствующих заболеваний: печени, желчного пузы-
ря, ЖКТ, нарушений липидного обмена; эндок-
ринной сферы — щитовидной и поджелудочной
железы, надпочечников, явлений диэнцефалеза,
гипоталамического синдрома; нарушений гемо-
поэза, иммунной системы; нервно-психической
сферы.
Наружные средства для лечения псориаза
Курсы витаминотерапии, фитотерапии, липо-
тропные средства, УФО, поддерживающая
ПУВА-терапия, прием ретиноидов, неорала.
Бальнеотерапия (оксидат торфа, морская соль,
фитодобавки и др.). Санаторно-курортное лече-
ние.
Rp.: Acidi salicylic! 2,0 Vaselini 100,0 Rp.: D.S. Ung. Sdidoli Ribacovi 40,0 Втирать в очаги поражения 1-2 раза в день.
M. D. S. Смазывать кожу 1 -2 раза в день. Styli Ditrastik 1,5 % №10 № 1. Наносить на очаг на 8 ч.
Rp.: Lanolini Vaselini Rp.: D.S.
М. D. S. Aq. destil. aa 30,0 Смазывать участки поражения 1-2 раза в день Rp.: Acidi salicylici 2,0 Sulfuris рр. 4,0 Zinci oxydi Talciaa22,0 Vaselini ad 100,0 M. f. ung.
Rp.: Linim. Naphthalani 5 % 50,0 D. S. Смазывать очаги поражения 1 раз в день.
D. S. Смазывать бляшки 1-2 раза в день.
Глава 23
Красный плоский лишай
Красный плоский лишай (КПЛ) — хронический
дерматоз, характеризующийся появлением на
коже, реже на слизистой рта или гениталий па-
пул лилового цвета с восковидным блеском.
Типичные папулы красного плоского лишая
можно обнаружить при различных клинических
формах заболевания. Красный плоский лишай
встречается у 0,3—1 % дерматологических боль-
ных; преимущественный возраст 30—60 лет, чаще
болеют женщины.
Концепции этиологии и патогенеза
Иммунно-аллергическая, вирусная (инфекцион-
ная), нейроэндокринная, токсико-аллергическая
(лекарственные препараты и химические соеди-
нения), наследственная. В очагах поражения про-
исходит формирование кожного процесса по
типу гиперчувствительности замедленного типа
Рис. 23.1. Преимущественная локализация
высыпаний при красном плоском лишае.
в результате стимуляции неидентифицирован-
ным антигеном процесса накопления клеток
Лангерганса и их контакта с Т-лимфоцитами и
макрофагами. Характерно разрушение базальных
кератиноцитов (цитотоксический иммунологи-
ческий процесс) и формирование Т-хелперного
(индукторного) инфильтрата.
Предрасполагающие факторы
Стрессовые ситуации, нейроэндокринные рас-
стройства, заболевания ЖКТ, печени, поджелудоч-
ной железы, гипертоническая болезнь, сахарный
диабет, фокальная инфекция в челюстно-лицевой
области, прием медикаментов (тетрациклин, суль-
фаниламиды, ПАСК, стрептомицин, препараты
золота, мышьяк, хлорохин, хлорсодержащие диу-
ретики, ингибиторы АПФ и др.). При поражени-
ях слизистой оболочки рта важную роль играет
патология со стороны зубов, наличие протезов из
пластмассы или их сочетание из различных метал-
лов. Аллергические реакции по типу красного
плоского лишая нередко встречаются у лиц, работа
которых связана с проявителями цветных пленок.
Клиническая картина
Излюбленная локализация
Кожа сгибательных поверхностей предплечий и
голеней, область крестца, наружных половых ор-
ганов, слизистая оболочка рта, губ; нередко по-
ражаются ногтевые пластинки, волосистая часть
головы с развитием рубцовой алопеции (рис.
23.1).
Течение
Острое (появление свежих папул, положитель-
ная изоморфная реакция); хроническое (месяцы,
годы), особенно при бородавчатой форме с по-
ражением голеней. Длительный эрозивно-язвен-
ный процесс на слизистой рта и языка может
привести к трансформации в плоскоклеточный
рак.
У значительной части пациентов высыпания
на коже могут регрессировать самостоятельно в
течение 15 мес.
288
Глава 23. Красный плоский лишай 289
Основные клинические формы
На коже: типичная, бородавчатая, эритематоз-
ная, актиническая, эрозивно-язвенная, атрофи-
ческая, кольцевидная, пузырная, остроконечная,
пигментная, линейная.
На слизистой оболочке ротоглотки и генита-
лий: типичная, экссудативно-гиперемическая,
эрозивно-язвенная (как осложнение первых двух
форм). Поражение слизистой рта без высыпаний
на коже выявляют у 25 % больных.
Поражение ногтей: онихогрифоз, онихолизис,
деструкция ногтевого валика.
По распространенности высыпаний", локализо-
ванная (ограниченная), распространенная.
Типичная форма встречается наиболее часто
(45 % пациентов). На коже возникают папулы
преимущественно милиарного или лентикуляр-
ного характера розовато-фиолетового цвета, плос-
кие, с полигональными очертаниями и воско-
видным блеском, пупкообразным вдавлением в
центре. Восковидный блеск хорошо виден при
боковом осмотре очагов поражения. На поверх-
ности отдельных папул и, в большей мере, бля-
шек выявляется своеобразный сетчатый рисунок
(белесоватая исчерченность, напоминающая кру-
жевной рисунок или листья папоротника), ко-
торый лучше визуализируется при осмотре под
лупой после смазывания пораженных участков
кожи водой, растительным или вазелиновым
маслом (симптом Уикхема). Данный симптом
обусловлен явлениями неравномерного гранулеза
(утолщения) зернистого слоя эпидермиса. В на-
чальной стадии заболевания характерно появле-
ние новых мелких элементов, особенно на уча-
стках травматизации кожи (изоморфная реакция
Кебнера), что указывает на прогрессирование
процесса.
Бородавчатая форма. Фиолетово-красные
бляшки с неравномерными роговыми наслоени-
ями, резкими границами, различной формы и
очертаний, в виде бляшечных элементов, заметно
возвышающихся над поверхностью кожи и со-
провождающихся выраженным зудом. Данная
форма заболевания чаще встречается на нижних
конечностях и доставляет пациентам наибольшие
беспокойства от сильного кожного зуда.
Эрозивно-язвенная форма красного плоского
лишая возникает на местах типичных кожных
высыпаний и характеризуется торпидностью к
терапии (в частности, на подошвенных поверх-
ностях стоп).
У некоторых больных типичным высыпаниям
красного плоского лишая сопутствуют:
• коричневато-буроватая пигментация (пиг-
ментная форма)\
• в участках поражения появляются неболь-
шие пузыри с плотной покрышкой (буллезная
форма)',
• атрофия в области элементов (атрофическая
форма)',
• папулы конической формы в устьях воло-
сяных фолликулов (остроконечная или фоллику-
лярная форма)',
• кольцевидные папулезные элементы (коль-
цевидная форма)',
• преобладает воспалительный сосудистый
компонент (эритематозная форма).
При актинической форме папулезная сыпь рас-
положена на тыле кистей и предплечий и ее
возникновение связано с действием солнечных
лучей.
Для линейной формы красного плоского лишая
характерен унилатеральный полосовидный ха-
рактер сыпи.
Поражения слизистой оболочки ротоглотки и
гениталий (головка полового члена, мошонка,
большие и малые половые губы, слизистая вла-
галища) встречаются у 40—60 % пациентов с
высыпаниями на кожном покрове.
При типичной форме (наиболее частая) в этих
областях обнаруживаются беловато-перламутро-
вые папулы, которые образуют сетчатый рису-
нок.
Эрозивно-язвенные кровоточащие участки на
отечном основании, покрытые фибринозным
налетом, характерны для эрозивно-язвенной фор-
мы красного плоского лишая. В ряде случаев эро-
зивные участки могут захватывать всю слизис-
тую оболочку рта, языка и губ. Эта форма
заболевания протекает наиболее тяжело, склон-
на к длительному течению, быстрым рецидивам
и плохо поддается терапии, несмотря на систем-
ное применение цитостатиков и кортикостеро-
идных препаратов.
Поражения слизистых оболочек могут прояв-
ляться также в виде гипертрофической формы
(плотные белесоватые бляшки с сеткой Уикхе-
ма на их поверхности), атрофической формы
(алые бляшки с сеткой Уикхема на окружающей
их слизистой), буллезной формы (пузыри, эрозии,
белесоватые элементы с сетчатым рисунком).
Поражение ногтей. Выявляется продольная
ребристость ногтя и трещины, уменьшение ног-
тевой пластинки вплоть до ее лизиса, разруше-
ние ногтевого валика (чаще первых пальцев
стоп).
Субъективные ощущения. Для типичной фор-
мы красного плоского лишая, при локализации
высыпаний на коже и гениталиях, характерен
зуд различной интенсивности. На жжение и бо-
290 Частная дерматология
лезненность пациенты предъявляют жалобы в
случаях эрозивно-язвенного процесса на слизи-
стых рта и гениталий.
Патоморфология
В участках пораженной кожи выявляется гипер-
кератоз, гранулез, акантоз, диффузный воспали-
тельный, иногда полосовидный инфильтрат в
дерме, проникающий в эпидермис (экзоцитоз)
и состоящий из лимфоцитов. При бородавчатой
форме преобладают явления гиперкератоза и
неравномерного гранулеза.
Дифференциальный диагноз
Дифференциальный диагноз проводят:
• с сифилитическими папулами (круглая, по-
лушаровидная форма папул медно-красного цве-
та, снимающийся налет, обнаружение бледных
трепонем в элементах сыпи, RW положительная);
• псориазом (триада псориатических симпто-
мов);
• лейкоплакией (сплошной очаг ороговения
серовато-белого цвета, отсутствие характерного
рисунка);
• для исключения вульгарной пузырчатки
слизистой полости рта необходимо исследование
мазков-отпечатков на акантолитические клетки;
• при многоформной экссудативной эритеме
наблюдаются выраженная гиперемия, отек сли-
зистой оболочки рта, пузыри и резко болезнен-
ные эрозии полициклических очертаний с фиб-
ринозным налетом, отсутствие вокруг них
папулезных высыпаний.
Лечение
При первичных распространенных формах — ги-
поаллергенная диета, гипноз и электросон, се-
дативные препараты, включая бромиды. Назна-
чают транквилизаторы, антидепрессанты (азафен
25 мг 2—3 раза в сутки в течение 1—1,5 месяцев).
В некоторых случаях эффективны антибиотики:
пенициллин и его аналоги; или тетрациклин 2 г
в сутки, на курс-12 г; или гризеофульвин по
0,25 г 4 раза в день через день в течение 3 не-
дель (с учетом переносимости данных антибио-
тиков). Показаны также: дезоксирибонуклеаза
по 0,25 г в/м — первые 2 дня, затем по 0,5 г 1
раз в 3 дня — 8 инъекций; никотиновая кисло-
та 0,05 г 2—3 раза в день после еды; теоникол
(компламин) 0,15—0,3 г 3 раза в день после еды;
аскорутин по 1 таблетке 3 раза в день; витами-
ны А, Е, С; курс инъекций витаминов В( и В12;
гистаглобулин по 2 мл подкожно 2 раза в неде-
лю № 10.
При среднетяжелых и тяжелых формах — де-
лагил по 0,25 г 1—2 раза в день в течение 4—6
недель, или ацитреин (этретинат) по 1 мг/ кг/
сут — 2 недели, или преднизолон 25—35 мг в
сутки — 2 недели, с последующим снижением;
при тяжелых формах суточную дозу преднизо-
лона повышают до 40—50, иногда до 70 мг. В
ряде случаев благоприятный эффект оказывают
рефлексотерапия и (или) индуктотермия пояс-
ничной области.
При эрозивно-язвенном поражении слизистой
полости рта назначают циклофосфамид по 50—
100 мг в сутки в течение 5—7 недель. Эффек-
тивны инъекции триамцинолона (3 мг/мл) в
очаги поражений, а также полоскания рта раство-
ром циклоспорина. Рекомендуются ароматичес-
кие ретиноиды (неотигазон по 20-30 мг ежеднев-
но; роаккутан по 0,3 - 0,5 мг/кг массы тела
ежедневно).
При генерализованных формах показаны
ПУВА-терапия и системное применение циклос-
порина в дозе 5 мг/сут до выраженного клини-
ческого эффекта с последующим постепенным
снижением суточной дозы.
Местное лечение. Кортикостероидные мази,
кремы в виде аппликаций и окклюзионных
повязок на очаги поражения. При бородавчатой
форме эффективны инъекции кортикостероидов
в очаг; лазеро-, букки-терапия, криотерапия.
Профилактика
Диспансеризация больных с частыми рецидива-
ми заболевания, санация очагов инфекции, вы-
явление и лечение сопутствующих заболеваний,
запрещение приема медикаментов, способных
провоцировать заболевание, оздоровительные
мероприятия, санаторно-курортное лечение.
Глава 24
Розовый лишай *
Розовый лишай (pityriasis rosea) — заболевание
с невыясненной этиологией и патогенезом. Воз-
никает чаще в холодное время года или после
переохлаждения. Болеют чаще взрослые.
Предполагается инфекционная природа заболе-
вания, но сам розовый лишай не заразен (описа-
ны крайне редкие случаи семейного заболевания).
Клиническая картина
Перед появлением высыпаний могут наблюдать-
ся субфебрилитет, боль в суставах, недомогание,
головная боль. Иногда могут увеличиваться
шейные лимфатические узлы.
Высыпания симметричны, поражается кожа
туловища, конечностей, за исключением ладоней
и подошв, реже — шея и лицо (рис. 24.1). Эле-
менты сыпи появляются периодически, вспыш-
ками. Субъективно может беспокоить зуд. Пят-
на округлые или овальные, величиной до 2—3 см
в диаметре, склонны к периферическому росту,
располагаются по линиям Лангера. Центр пятен
слегка западает, с нежными складками (симптом
«папиросной бумаги»), шелушение более отчетли-
во выражено на границе центральной и перифе-
рической зоны в виде характерного «воротнич-
ка». Край пятен приподнят, отечен, имеет более
яркую окраску и не шелушится. Такие элемен-
ты сыпи напоминают медальон. За 1—2 недели до
распространенных высыпаний возникает один
элемент больших размеров (3—4 см в диаметре),
овальных или округлых очертаний с четко вы-
раженным симптомом «папиросной бумаги» и
узким периферическим венчиком эритемы — так
называемая «материнская бляшка». Элементы
разрешаются с центральной части, придавая сыпи
отчетливый кольцевидный характер. Затем посте-
пенно исчезает и периферический венчик. Пос-
ле регрессирования элементов сыпи остается вре-
менная гиперпигментация. Разновидностями
розового лишая могут быть: уртикарная, папулез-
ная, везикулезная, фолликулярная и другие фор-
мы, которые встречаются редко. Под влиянием
водных процедур (баня, душ), грубого нательно-
го белья, УФО, раздражающих наружных лекар-
ственных средств элементы сыпи могут сливать-
ся между собой, образуя участки эритродермии,
что значительно утяжеляет течение заболевания.
Дифференциальная диагностика
Дифференцируют розовый лишай с сифилити-
ческой розеолой, отрубевидным лишаем, токси-
кодермиями, парапсориазом, себорейной экзе-
мой, инфекционными экзантемами.
Лечение
При благоприятном течении заболевание не тре-
бует лечения и через 4—6 недель проходит спон-
танно. При резко выраженном зуде назначают
антигистаминные препараты (тавегил, супрастин
и др.), препараты кальция; местно —- взбалты-
ваемые взвеси с содержанием анастезина и мен-
тола или слабые глюкокортикоидные мази (пред-
низолоновая, гидрокортизоновая). Больные
эритродермией подлежат госпитализации с про-
ведением активной системной терапии.
Рис. 24.1. Типичные
высыпания при розовом лишае.
* Глава написана И. В. Тихоновской
291
Глава 25
Болезни с диффузным
поражением соединительной ткани
Красная волчанка 292
Системная красная волчанка 295
Очаговая склеродермия 300
Системная склеродермия 302
Дерматомиозит 309
Красная волчанка
Красная волчанка (lupus erythematodes; син.: эри-
тематоз) — аутоиммунное заболевание с пораже-
нием соединительной ткани, при котором обра-
зуются аутоантитела и иммунные комплексы,
откладывающиеся в стенках мелких сосудов и
под базальной мембраной эпидермиса, а также
характеризующееся выраженной фоточувстви-
тельностью и эритематозными высыпаниями на
коже и слизистых.
Этиология и патогенез
Этиология и патогенез заболевания до сих пор
окончательно не выяснены. Существуют следу-
ющие теории: аутоиммунная, вирусная, иммуно-
логическая, генетическая, иммуногенетическая,
нейрогу моральная.
Наиболее распространенной из гипотез проис-
хождения красной волчанки является аутоим-
мунная, согласно которой в организме образу-
ются различные аутоантитела и иммунные
комплексы, откладывающиеся под базальной
мембраной эпидермиса и в стенках мелких со-
судов. Подтверждением аутоиммунного генеза за-
болевания служит часто наблюдаемая ассоциация
красной волчанки с другими аутоиммунными
патологическими состояниями.
Роль генетических факторов в патогенезе
красной волчанки подтверждается семейными
случаями заболевания, развитием болезни у од-
нояйцевых близнецов, ассоциацией патологии с
некоторыми антигенами тканевой гистосовмес-
тимости: HLA—В5, В8, В15, Bw35, Ар А3, А10, А1Р
А18, DR^ DR3.
Иммунологические сдвиги при красной вол-
чанке проявляются угнетением Т-клеточного
иммунитета (Т-супрессоров) и повышенной ак-
тивностью В-клеток. Но указанные изменения
возникают вторично, на базе генетической пред-
расположенности, благодаря чему возникают
соматические мутации стволовых лимфоидных
клеток. На этом фоне различные экзогенные и
эндогенные факторы (ультрафиолетовые лучи,
хроническая очаговая стрептококковая инфек-
ция, лекарственная непереносимость, переохлаж-
дение, вирусная инфекция, аутоаллергия) про-
воцируют включение аутоиммунного механизма
с образованием антител, направленных против
соединительной ткани. .
Предположение о вирусной природе красной
волчанки позволили сделать электронно-микро-
скопические данные, с помощью которых были
выявлены деструктивные изменения в сосудах
кожи, почек, мышц и эндоцитоплазматические
тубулярные вирусоподобные образования, сход-
ные со структурами парамиксовирусов.
Классификация
До настоящего времени общепризнанной клас-
сификации клинических форм красной волчан-
ки не существует. Со времен Капоши в дер-
матологии используется классификация,
основанная на остроте течения процесса. Раз-
личают следующие формы: хроническая крас-
ная волчанка (дискоидная или ограниченная,
диссеминированная или множественная) и ос-
292
Глава 25. Болезни с диффузным поражением соединительной ткани 293
трая (подострая) системная красная волчанка,
лечение которой входит в компетенцию рев-
матологов.
Клиническая картина хронической
красной волчанки
Дискоидная красная волчанка — наиболее часто
встречающаяся форма заболевания. Возникает в
молодом возрасте, у женщин в 2 раза чаще, чем
у мужчин. Процесс развивается в весеннее или
летнее время года после инсоляции, обмороже-
ния, механической травмы. Излюбленная лока-
лизация — кожа открытых участков, особенно
выступающие части лица: спинка носа и приле-
гающие участки щек, лоб, а также волосистая
часть головы, ушные раковины, красная кайма
губ, область декольте (рис. 25.1). Характерна
триада симтомов: эритема, гиперкератоз, атро-
фия.
Течение процесса стадийное:
I. Эритематозная стадия. Появляются мел-
кие, слегка отечные, растущие по периферии
пятна, четко контурированные. Цвет их розовый,
иногда с лиловым оттенком и телеангиэктазия-
ми в центре. Распространяясь и сливаясь, очаги
по очертаниям напоминают бабочку, спинка ко-
торой располагается на носу, а крылья на щеках.
II. Гиперкератозно-инфилътративная стадия.
Основание очагов поражения инфильтрируется,
в центре появляются мелкие, асбестовидные,
плотно сидящие чешуйки. При снятии чешуй-
ки с внутренней стороны видны чешуйки-ши-
пики — выступающие, как гвозди из отрываю-
щегося каблука (симптом «дамского каблука»).
Рис. 25.1. Типичная локализация
высыпаний при дискоидной красной волчанке.
При нажатии на очаг или соскабливании чешу-
ек отмечается болезненность за счет раздраже-
ния шипиками нервных окончаний в фоллику-
ле (симптом Бенье-Мещерского).
III. Атрофическая стадия. В центре очага по-
ражения формируется рубцовая (рубцующийся
эритематоз) атрофия. Кожа истончена, депиг-
ментирована, с множественными телеангиэкта-
зиями. По периферии сохраняются инфильтра-
ция и гиперпигментация. На волосистой части
головы в результате рубцевидной атрофии оста-
ется стойкое облысение.
Патоморфология
Основными гистологическими признаками дис-
коидной красной волчанки являются гиперке-
ратоз, атрофия мальпигиевого слоя, гидропичес-
кая дистрофия клеток базального слоя, отек и
расширение сосудов в верхней части дермы, на-
личие преимущественно лимфоцитарных ин-
фильтратов, располагающихся главным образом
вокруг придатков кожи. В эпидермисе наблю-
даются умеренно выраженный гиперкератоз,
вакуольная дистрофия клеток базального слоя,
резкий отек верхних слоев дермы с периваску-
лярными лимфогистиоцитарными инфильтрата-
ми. В дальнейшем наступает более глубокая де-
зорганизация соединительной ткани —
фибриноидное набухание, в основе которого
лежит деструкция коллагена и межуточного ве-
щества, сопровождающаяся резким нарушением
сосудистой проницаемости.
Диссеминированная красная волчанка характе-
ризуется наличием множественных очагов пора-
жения, клинически не отличающихся от диско-
идной разновидности. При этой форме
заболевания очаги поражения располагаются на
лице, рассеяны по кожным покровам туловища,
волосистой части головы. Чаще, чем при диско-
идной форме, могут наблюдаться признаки по-
ражения внутренних органов с большей вероят-
ностью трансформации в системный процесс.
Выраженность наружных клинических проявле-
ний может быть различной, что и определяет
некоторые разновидности клинических форм.
Различают центробежную, глубокую, гиперке-
ратотическую, гипсовидную, веррукозную, ге-
моррагическую, себорейную, дисхромическую,
пигментную, опухолевую формы.
Центробежная эритема характеризуется отсут-
ствием кардинальных симптомов дискоидной
красной волчанки: фолликулярного гиперкера-
тоза и рубцовой атрофии. Заболевание характе-
ризуется яркой, иногда отечной, резко ограни-
ченной эритемой, располагающейся в области
294 Частная дерматология
лица либо на открытых участках шейно-ворот-
никовой зоны. Воспалительный процесс может
сопровождаться нестойким повышением темпе-
ратуры тела, иногда болями в суставах, костях.
Центробежная эритема чаще других форм транс-
формируется в системный процесс.
Глубокая форма красной волчанки (Ирганга)
относится к редким разновидностям и также
имеет склонность к переходу в системную крас-
ную волчанку. Характеризуется наличием одного
или нескольких глубоко расположенных узло-
ватых образований плотноватой консистенции,
кожа над которыми имеет нормальную или зас-
тойно-синюшную окраску. Очаги располагают-
ся преимущественно в области плеч, лица, яго-
диц. Существуют в течение длительного
времени, иногда кальцифицируются. После за-
вершения процесса остается глубокая атрофия
кожи.
Опухолевая разновидность относится к наибо-
лее редко встречаемым формам красной волчан-
ки. Клинически у больных очаг поражения выг-
лядит как резко очерченный, возвышающийся
над поверхностью плотноватый элемент синюш-
но-красного цвета, покрытый на поверхности
множественными рубчиками. Субъективные
ощущения незначительны.
Эрозивно-язвенная форма красной волчанки с
изолированной локализацией в области красной
каймы губ и слизистой полости рта характери-
зуется выраженным воспалением, на фоне кото-
рого возникают трещины, язвы, покрытые се-
розными, серозно-кровянистыми корками.
Эрозии нередко располагаются на слегка инфиль-
трированном основании с явлениями выражен-
ного гиперкератоза и атрофии по периферии
очага поражения. Больные нередко отмечают
жжение, болезненность, возникающие при раз-
говоре или приеме пищи. Такая форма волчан-
ки может осложниться гландулярным хейлитом
или трансформироваться в рак. Одним из харак-
терных симптомов красной волчанки губ явля-
ется яркое, снежно-голубое свечение в лучах
лампы Вуда.
Диагноз
Диагноз заболевания устанавливают на основа-
нии типичных клинических признаков (эрите-
ма, фолликулярный гиперкератоз, атрофия),
лабораторных данных, гистологического и им-
муноморфологического исследований.
• Клинические признаки: эритема, фоллику-
лярный гиперкератоз, атрофия.
• Вспомогательное значение имеют указания
на повышенную фоточувствительность, прово-
цирующие факторы (инсоляция, УФО, отморо-
жение, травма).
• Лабораторные данные: анемия, лейкопения,
лимфопения, высокая СОЭ, гипергаммаглобули-
немия, гипергаптоглобулинемия, наличие анти-
ядерных антител и LE-клеток).
• В трудных случаях — гистологическое и
особенно иммуноморфологическое исследование.
Дифференциальный диагноз
Дифференциальный диагноз при поражении
кожи лица проводят с псориазом, розацеа, руб-
ромикозом, папуло-некротическим туберкулезом
кожи, лейшманиозом, бугорковым сифилидом.
При локализации очагов поражения на волоси-
стой части головы дифференциальный диагноз
проводят с трихо-микозами, псориазом, очаго-
вой алопецией, псевдопелладой, себорейной эк-
земой.
Лечение
Основными препаратами для лечения кожных
форм красной волчанки являются аминохиноли-
новые — хингамин, резохин, хлорохин, делагил,
плаквенил. Они обладают фотозащитными свой-
ствами, предупреждают полимеризацию ДНК и
РНК, подавляют реакции образования антител
и иммунных комплексов, трансформацию лим-
фоцитов, связывание нуклеопротеинов. Их про-
тивовоспалительное действие основано на подав-
лении активности гидролитических ферментов
(стабилизация мембран лизосом), синтеза воспа-
лительных простагландинов, блокировании хе-
мотаксиса, антигистаминном эффекте. Препара-
ты назначают по 250 мг 2 раза в сутки циклами
по 10 дней с 5-дневными перерывами. По мере
стихания процесса дозу уменьшают до 1 таблет-
ки в сутки. Продолжительность лечения состав-
ляет 2—3 мес, курсовая доза — до 20-25 г.
Применяют различные иммунокорригирую-
щие средства (тактивин, декарис, апилак, мета-
цил, нуклеинат натрия). Тактивин или тималин
вводят подкожно или внутримышечно через день
(на курс 10 инъекций) для повышения иммуни-
тета и достижения противовоспалительного эф-
фекта. Иммуномодулирующий эффект дает апи-
лак по 0,01 г 3 раза в сутки в течение 10 дней.
При нарушениях липидного обмена и патоло-
гии печени — эссенциале внутривенно по 5 мл
на аутокрови ежедневно в течение 15—20 дней.
Наружная терапия: применяют активные кор-
тикостероидные мази — синалар, флуцинар,
фторокорт, целестодерм, клобетазол и др. — на
очаги поражения на ночь. В ряде случаев исполь-
зуют внутрикожное обкалывание очагов гидро-
Глава 25. Болезни с диффузным поражением соединительной ткани 295
кортизоном или 5-10 %-ным раствором резохина
или хингамина; проводят криотерапию жидким
азотом или снегом угольной кислоты. Фотоза-
щитные кремы (мази, пасты) наносят на очаги
при выходе на работу. В яркие солнечные дни
необходимо использовать зонтик или шляпу с
широкими полями, фотозащитные очки.
Профилактика
Больным дискоидной красной волчанкой в ве-
сенне-летнее время года показан профилактичес-
кий прием препаратов хинолонового ряда (дела-
гил по 1 таблетке через день или плаквенил по
1 таблетке ежедневно). Больные должны нахо-
диться на диспансерном наблюдении дерматолога
и ревматолога.
Системная
красная волчанка
Синонимы: lupus erythematosus systemicus, lupus
erythematosus acutus, lupus erythematosus visceralis,
lupus erythematosus integumentalis et visceralis.
Системная красная волчанка — тяжелое ауто-
иммунное полисиндромное системное заболева-
ние соединительной ткани и сосудов, характе-
ризующееся образованием множества антител к
собственным клеткам и их компонентам и раз-
витием иммунокомплексного воспаления с по-
вреждением многих органов и систем (пораже-
ния кожи и слизистых оболочек, люпус-нефрит,
люпус-пневмонит, волчаночный менингоэнце-
фалит, люпус-артрит, полисерозиты — плеврит,
перикардит).
Этиология и патогенез
Этиология и патогенез заболевания до сих пор
окончательно не выяснены. Предложены следу-
ющие теории: вирусная, иммунологическая,
аутоиммунная, генетическая, иммуногенетичес-
кая, нейрогуморальная.
По патогенетическим механизмам системная
красная волчанка относится к аутоиммунным
болезням. В условиях дефицита Т-супрессорной
функции лимфоцитов отмечается продукция
большого количества аутоантител: антинуклеар-
ных, к ДНК, микросомам, лизосомам, митохон-
дриям, форменным элементам крови. Наиболь-
шее патогенетическое значение имеют антитела
к нативной ДНК (нДНК), которые соединяют-
ся с нДНК, образуют иммунные комплексы и
активируют комплемент. Они откладываются на
базальных мембранах различных внутренних
органов и кожи, вызывают их воспаление и
повреждение. Одновременно повышается прони-
цаемость лизосом, выделяются медиаторы вос-
паления, активируется кининовая система.
Клиническая картина
Болеют преимущественно женщины в возрасте
14—40 лет; женщины болеют в 8 раз чаще муж-
чин. Начальные проявления болезни характери-
зуются слабостью, утомляемостью, похуданием,
повышением температуры тела.
Поражения кожи и слизистых оболочек
Эти поражения (встречаются у 85 % больных)
проявляются при системной красной волчанке
многочисленными изменениями:
• эритематозные пятна различной формы и
величины, располагаются на лице по типу «ба-
бочки» на носу и щеках, шее, груди, в области
локтевых, коленных, голеностопных суставов;
• «дискоидные» высыпания — папулы и
бляшки, характерные для дискоидной красной
волчанки (на лице и предплечьях);
• островоспалительные уртикарно-везикулез-
ные, буллезные, геморрагические высыпания,
сетчатое ливедо с изъязвлениями;
• телеангиэктатические или геморрагические
пятна на коже кистей, в области подушечек
пальцев, на подошвах; эритема ладоней;
• трофические нарушения — сухость кожи,
выпадение волос (рубцовая алопеция на месте
«дискоидных» высыпаний, диффузная алопе-
ция), ломкость и хрупкость ногтей;
• люпус-хейлит — выраженное покраснение
губ с сероватыми чешуйками, корочками, эро-
зиями, с последующим развитием атрофии на
красной кайме губ;
• энантема слизистой оболочки полости рта —
участки эритемы с геморрагиями и эрозиями;
• на слизистой оболочке полости рта и носа —
эрозивные, язвенные очаги, белесоватые бляш-
ки, эритематозные пятна, возможна перфорация
перегородки носа.
Поражения костно-суставной
и мышечной систем
Проявляются следующими симптомами:
• выраженная скованность пораженных сус-
тавов по утрам;
• симметричный полиартрит с поражением
проксимальных межфаланговых суставов кистей,
пястно-фаланговых, запястно-пястных, колен-
ных суставов;
• длительные и интенсивные боли в одном
или нескольких суставах;
296
• развитие сгибательных контрактур пальцев
рук вследствие тендинитов, тендовагинитов;
• формирование ревматоидноподобной кис-
ти за счет изменений в периартикулярных тка-
нях;
• возможно развитие асептических некрозов
головки бедренной кости, плечевой и других
костей; миалгии, выраженная мышечная сла-
бость, иногда развивается полимиозит.
Легочные проявления
Со стороны легких выявляются:
• волчаночный пневмонит (легочный васку-
лит), который характеризуется одышкой, сухим
кашлем, иногда кровохарканьем; при рентгено-
логическом исследовании легких выявляются
дисковидные ателектазы в нижних отделах,
иногда видны инфильтративные тени;
• при хроническом течении люпус-пневмонита
наблюдается поражение интерстициальной тка-
ни легких с усилением легочного рисунка;
• сухой или выпотной плеврит (как правило,
двусторонниий) с болями в груди и выраженной
одышкой при накоплении большого количества
жидкости;
• синдром легочной гипертензии;
• возможна тромбоэмболия легочной артерии.
Поражение сердечно-сосудистой системы
Характеризуется развитием панкардита, но чаще
всего наблюдается перикардит, обычно сухой;
иногда развивается тяжелый экссудативный пе-
рикардит. Поражение эндокарда наблюдается
при панкардите и приводит к изменениям кла-
панного аппарата сердца. Чаще формируется
митральная недостаточность, реже — недостаточ-
ность клапана аорты. Поражаются сосуды, пре-
имущественно артерии среднего и мелкого ка-
либра. В патологический процесс могут
вовлекаться подключичная артерия, коронарные
артерии, что иногда приводит к развитию инфар-
кта миокарда. Возникают тромбофлебиты повер-
хностных вен плеча, передней поверхности груд-
ной клетки.
Поражение желудочно-кишечного тракта
Больных беспокоят тошнота, рвота, отсутствие
аппетита. Поражения пищевода сопровождают-
ся его дилятацией, эрозивными изменениями
слизистой оболочки. Нередко обнаруживаются
изъязвления слизистой оболочки желудка и две-
надцатиперстной кишки. Поражение сосудов
брыжейки приводит к сильным болям в живо-
те, преимущественно вокруг пупка (абдоми-
нальный криз), ригидности мышц брюшного
пресса.
Поражение печени (волчаночный гепатит)
Проявляется увеличением печени, желтухой раз-
личной степени выраженности, повышением
содержания в крови аминотрансфераз.
Поражения почек
При поражениях почек (люпус-нефрит) возмож-
ны следующие клинические формы (М. М. Ива-
нова, 1994):
• быстро прогрессирующий волчаночный не-
фрит (тяжелый нефротический синдром, злока-
чественная артериальная гипертензия, быстрое
развитие почечной недостаточности);
• нефротическая форма гломерулонефрита (в
отличие от неволчаночного нефрита, протеину-
рия менее выражена, чаще наблюдаются артери-
альная гипертензия и гематурия, менее выражена
гиперхолестеринемия);
• активный волчаночный нефрит с выражен-
ным мочевым синдромом (протеинурия больше
0,5 г/сут, микрогематурия, лейкоцитурия);
• нефрит с минимальным мочевым синдро-
мом — протеинурия меньше 0,5 г/сут, микроге-
матурия — единичные эритроциты в поле зре-
ния, небольшая лейкоцитурия; артериальное
давление нормальное.
Поражение нервной системы
Наблюдается почти у всех больных и обуслов-
лено васкулитами, тромбозами, инфарктами и ге-
моррагиями в различных отделах головного моз-
га. В изменениях нервной системы важное
значение придается антинейрональным антите-
лам, которые поражают мембраны нейронов.
Поражение нервной системы может иметь сле-
дующие клинические симптомы: головные боли,
психические расстройства, судорожный синдром
(по типу височной эпилепсии), нарушение фун-
кции черепно-мозговых нервов, мононейропа-
тии, полинейропатии, нарушение мозгового кро-
вообращения (вследствие тромбозов,
геморрагий).
Варианты течения
Выделяют три варианта течения системной крас-
ной волчанки: острое, подострое и хроническое.
При остром течении начало болезни внезап-
ное, температура тела высокая, характерны ост-
рый полиартрит с резкой болью в суставах, вы-
раженные кожные проявления, тяжелые
полисерозиты; поражение почек, нервной систе-
мы, трофические нарушения, похудание, резкое
увеличение СОЭ, панцитопения, большое коли-
чество LE-клеток в крови, высокие титры АНФ.
Длительность заболевания 1—2 года.
Глава 25. Болезни с диффузным поражением соединительной ткани
297
Подострое течение характеризуется посте-
пенным развитием, суставным синдромом, нор-
мальной или субфебрильной температурой тела,
кожными изменениями. Активность процесса в
течение значительного времени минимальная,
ремиссии продолжительны (до полугода). Од-
нако постепенно процесс генерализуется, раз-
вивается поражение многих органов и систем.
Хроническое течение проявляется моно- или
малосиндромйостью в течение многих лет. Об-
щее состояние долго остается удовлетворитель-
ным. На ранних этапах наблюдаются кожные
изменения, суставной синдром. Процесс медлен-
но прогрессирует, и в дальнейшем вовлекаются
многие органы и системы.
Лабораторные данные
Для обследования больных системной красной
волчанкой рекомендуются следующие методы:
общие анализы крови и мочи; исследование
крови на LE-клетки; биохимический анализ
крови (общий белок и белковые фракции, фиб-
риноген, фибрин, сиаловые кислоты, гаптогло-
бин, СРП, серомукоид, креатинин, мочевина,
аминотрансферазы, альдолаза); иммунологичес-
кое исследование крови (антитела к нДНК,
АНФ, комплемент, реакция Кумбса, количество
В- и Т-лимфоцитов, субпопуляция Т-лимфоци-
тов, РБТЛ, ЦИК); серологические исследова-
ния крови на сифилис (ИФА, РПГА, РМП),
ЭКГ, ФКГ, эхокардиография, рентгеновское
исследование сердца и легких, суставов; био-
псия кожи, мышц, лимфатических узлов (по
показаниям), почек (по показаниям); спинно-
мозговая пункция, компьютерная томография
головного мозга.
Общий анализ крови. Почти у всех больных
выявляется значительное повышение СОЭ, бо-
лее чем у половины — лейкопения со сдвигом
в формуле крови до промиелоцитов, миелоци-
тов в сочетании с лимфопенией, довольно час-
то — гипохромная анемия, тромбоцитопения.
Патогномонично определение большого количе-
ства LE-клеток (это зрелые нейтрофилы, цито-
плазма которых почти целиком заполнена фаго-
цитированным ядром погибшего лейкоцита,
собственное ядро при этом оттеснено к перифе-
рии). Диагностически значимо обнаружение не
менее 5 LE-клеток на 1000 лейкоцитов. LE-
клетки образуются при наличии антител, реаги-
рующих с комплексом ДНК-гистон и компле-
мент. Единичные волчаночные клетки могут
встречаться и при других заболеваниях.
Общий анализ мочи: при поражении почек —
протеинурия, цилиндрурия, микрогематурия.
Биохимический анализ крови: гиперпротеине-
мия и диспротеинемия. В у-глобулиновой фрак-
ции находится волчаночный фактор, ответствен-
ный за образование LE-клеток, и другие
антинуклеарные факторы. Повышено содержа-
ние сиаловых кислот, фибрина, серомукоида,
гаптоглобина, появляется СРП (С-реактивный
протеин).
В сыворотке крови больных системной крас-
ной волчанкой обнаруживаются антитела к фак-
торам свертывания VIII, IX, XII (повышают риск
кровотечений и вызывают тромбоцитопению), а
также антитела к фосфолипидам (задерживают
выделение простациклина из эндотелия, что
повышает агрегацию тромбоцитов и предраспо-
лагает к тромбозам).
Иммунологический анализ крови: криопреци-
питины, антитела к ДНК, антинуклеарный фак-
тор (АНФ). Антитела к ДНК определяются
РПГА, а также методом радиоиммунного связы-
вания меченой по йоду нДНК и иммунофлюо-
ресценции. Для системной красной волчанки
наиболее характерно периферическое свечение
ядер, обусловленное наличием антител к ДНК,
и высокий титр этой реакции — более 1: 1000.
У 30~40 % больных определяются антитела к
антигену Смита (разновидность антинуклеарных
антител). Рекомендуется исследование компле-
мента СН50 и его компонентов, снижение кото-
рых коррелирует с активностью люпус-нефри-
та. Отмечается снижение функциональной
активности Т-лимфоцитов (в том числе Т-суп-
рессоров), гипер- и дисиммуноглобулинемия
(увеличение содержания в крови IgG, IgM).
Серологические тесты на сифилис: возможны
ложноположительные биологические нетрепо-
немные тесты.
Исследование биопсийного материала почек,
кожи, лимфатических узлов, синовии выявляет
характерные гистологические изменения: фиб-
риноидное набухание или фибриноидный некроз
соединительной ткани и стенок сосудов, воспа-
лительные инфильтраты из плазматических кле-
ток и лимфоцитов, положительные результаты
иммунофлюоресценции.
Патоморфология кожи. Атрофия эпидермиса,
гидропическая дистрофия базального слоя, отек
дермы, воспалительная инфильтрация дермы
лимфоцитами, фибриноидное набухание или
фибриноидный некроз соединительной ткани и
стенок сосудов.
Иммунофлюоресцентная микроскопия. Методом
прямой иммунофлюоресценции выявляют гра-
нулярные отложения IgG, IgM и Clq в виде
непрерывной толстой полосы вдоль границы
298 Частная дерматология
дермы и эпидермиса. Эти отложения присутству-
ют как в пораженной (90 %), так и в здоровой
коже (50—70 %). Отложения иммуноглобулинов
и компонентов комплемента в здоровой коже —
признак люпус-нефрита и дефицита компонен-
тов комплемента.
Исследование спинномозговой жидкости (при
поражении нервной системы): повышение содер-
жания белка (0,5—1,0 г/л), плеоцитоз, повыше-
ние давления, снижение содержания глюкозы,
увеличение уровня IgG (> 6 г/л).
Компьютерная томография головного мозга:
при поражении нервной системы выявляются
небольшие зоны инфаркта и геморрагий в голов-
ном мозге, расширение борозд, церебральная
атрофия, кисты головного мозга, расширение
подпаутинных пространств, желудочков и базаль-
ных цистерн.
Рентгенография суставов: эпифизарный осте-
опороз преимущественно в суставах кистей,
реже — в запястно-пястных и лучезапястных
суставах, истончение субарахноидальных пласти-
нок, мелкие узуры суставных костей с подвы-
вихами.
Биопсия синовиальной оболочки: острый или
подострый синовит с «бедной» клеточной реак-
цией, значительной ядерной патологией и обна-
ружением гематоксилиновых телец.
Исследование синовиальной жидкости: жид-
кость прозрачная, вязкая, с небольшим числом
лейкоцитов и преобладанием мононуклеарных
клеток.
Диагностика
Диагностика системной красной волчанки осно-
вывается на клинической картине (эритема-ба-
бочка с очагами дискоидного типа, синдром Рей-
но, алопеция, изъязвления слизистой оболочки
рта, фотосенсибилизация, артрит без деформации
суставов, полисерозиты, психоз или судороги), а
также на лабораторных данных (LE-клетки, вы-
сокие титры антинуклеарных антител и антител
к нативной ДНК, панцитопения).
Американской Ревматологической Ассоциаци-
ей (1982) разработаны диагностические критерии
системной красной волчанки (табл. 25.1.). При
наличии 4-х признаков из 11 диагноз СКВ счи-
тается достоверным.
Лечение
Для лечения больных системной красной волчан-
кой применяют гормональные иммунодепрессан-
ты, негормональные иммунодепрессанты, имму-
нотерапию, аминохинолоновые препараты,
нестероидные противовоспалительные средства,
антикоагулянты и антиагреганты, эфферентную
терапию.
Гормональные иммунодепрессанты. Глюкокор-
тикоиды считаются препаратами выбора при си-
стемной красной волчанке. Абсолютными пока-
заниями к назначению глюкокортикоидов
являются острая, подострая форма системной
красной волчанки и обострение хронической си-
стемной красной волчанки. Наибольшее значение
из всех глюкокортикоидных препаратов имеет
преднизолон, обладающий высоким терапевтичес-
ким эффектом и вызывающий сравнительно не-
тяжелые побочные реакции при многолетнем
приеме. Лучшей переносимостью и хорошим те-
рапевтическим эффектом обладает метилпредни-
золон (урбазон), который значительно реже дру-
гих глюкокортикоидов вызывает поражение
ЖКТ (стероидные эрозии и язвы). Доза препа-
рата зависит от остроты течения и активности
процесса. При остром и подостром течении пред-
низолон назначают в дозе 40—50 мг в сутки, при
подостром и хроническом течении доза может
быть меньше 30—40 мг, а при первой степени
активности — 15—20 мг в сутки. Критериями
эффективности лечения преднизолоном являют-
ся: уменьшение активности патологического про-
цесса, устранение или значительное уменьшение
выраженности висцеральных проявлений, особен-
но со стороны почек, стабилизация течения за-
болевания. При отсутствии преднизолона можно
применять триамцинолон, дексаметазон, исходя
из расчета: 1 таблетка этих препаратов равна 1
таблетке преднизолона.
При неэффективности пероральной глюко-
кортикоидной терапии и быстром прогрессиро-
вании болезни применяется пульс-терапия ме-
тилпреднизолоном (метипредом), который вводят
внутривенно капельно по 1000 мг в 100 мл изо-
тонического раствора натрия хлорида (в капель-
ницу добавляют 5000 ЕД гепарина) в течение 3
дней подряд. Во время пульс-терапии и после
нее больные продолжают принимать внутрь ту
же дозу преднизолона, что и прежде. После
пульс-терапии рекомендуется применять цикло-
фосфамид по 200 мг в неделю в течение 3—6
месяцев в сочетании с преднизолоном в дозе 20—
25 мг в сутки.
Негормональные иммунодепрессанты (цитоста-
тики). Они обладают противовоспалительным
эффектом, способностью подавлять иммуноком-
плексный противовоспалительный процесс и
аутоантителообразование. Наиболее часто приме-
няют азатиоприн (имуран) и циклофосфамид в
дозе 1—3 мг на 1 кг массы тела, т. е. от 100 до
200 мг в сутки в сочетании с 30 мг преднизо-
Глава 25. Болезни с диффузным поражением соединительной ткани 299
лона. Указанные дозы назначают в течение 2—
2,5 месяцев, а затем переходят на поддержива-
ющие дозы 50—100 мг в сутки в течение мно-
гих месяцев и лет. Цитостатики назначают при
недостаточной эффективности или необходимо-
сти уменьшения поддерживающей дозы предни-
золона, при генерализованном васкулите, быст-
ро прогрессирующем течении и высокой
активности процесса. Цитостатики применяют-
ся также одновременно: азатиоприн + циклофос-
фамид (1 мг/кг в день внутрь) в сочетании с
малыми дозами преднизолона; комбинация аза-
тиоприна внутрь с внутривенным введением
циклофосфамида (по 1000 мг/м2 поверхности
тела каждые 3 месяца). Азатиоприн наиболее
эффективен при диффузном поражении кожи.
Иммунотерапия. Внутривенное введение имму-
ноглобулинов рекомендуется при выраженных
формах системной красной волчанки с тромбо-
цитопенией (по 0,4 г/кг в сутки в течение 5 дней).
Рекомбинантные препараты (а- и у-интерферо-
ны) вводятся внутривенно ежедневно в течение
3 недель, затем дважды в неделю в течение 2
месяцев. При люпус-нефрите применяются анти-
видотипические моноклональные СВ4-антитела.
Аминохинолиновые препараты назначают боль-
ным хронической системной красной волчанкой
прежде всего при поражении кожи, суставов,
люпус-нефрите: делагил по 0,25—0,5 г в сутки,
плаквенил по 0,2—0,4 г в сутки. Лечение про-
должают месяцы, годы.
Нестероидные противовоспалительные средства
(индометацин, вольтарен, ибупрофен, ацетилсали-
циловая кислота) обладают выраженным противо-
воспалительным и умеренным иммунодепрессан-
тным эффектом, стабилизируют лизосомальные
мембраны. Они назначаются при подостром и
хроническом течении системной красной волчан-
ки, преимущественно при наличии артрита. Эти
препараты применяются в обычных терапевтичес-
ких дозах длительно, до исчезновения воспаления
в суставах и нормализации температуры тела.
Таблица 25.1. Диагностические критерии системной красной волчанки
Критерий Характеристика критерия
Высыпания на носу и щеках Фиксированная эритема, плоская или возвышающаяся над кожей, не поражающая носогубные складки
Дискоидные высыпания Эритематозные выступающие пятна с чешуйками и фолликулярными пробками, впоследствии развиваются атрофические рубцы
Фотосенсибилизация Изъязвления в полости рта Кожные высыпания в результате реакции на солнечный свет Изъязвления в полости рта или носоглотке, обычно болезненные, должны наблюдаться врачом
Артрит Неэрозивный артрит двух и более периферических суставов, болезненность, припухание или выпот
Серозит Плеврит — плевритические боли или шум трения плевры (выслушивается врачом или имеются инструментальные доказательства); перикардит—документированный инструментально или выслушиваемый врачом шум трения перикарда
Почечные нарушения Стойкая протеинурия более 0,5 г/сут или цилиндрурия (эритроцитарные, зернистые, смешанные)
Неврологические нарушения Судороги, припадки — при отсутствии приема некоторых препаратов или нарушений метаболизма (уремия, кетоацидоз, электролитный дисбаланс) или психозы
Гематологические нарушения Гемолитическая анемия с ретикулезом или лейкопения (< 4000 в 1 мкл в двух или более анализах) или лимфопения (< 1500 клеток в 1 мкл в двух или более анализах) или тромбоцитопения (< 100 000 в 1 мкл)
Иммунологические нарушения Положительные LE-тест или повышенные титры антител к нативной ДНК или антитела к антигену Смита (он содержит Ц-рибонуклеопротеин-эпитоп и несколько других насыщенных уридином рибонуклеопротеинов) или ложноположительные реакции на сифилис в течение не менее 6 месяцев
Антинуклеарные антитела (АНАТ) Повышенные титры АНАТ, выявляемые с помощью иммунофлюоресценции при отсутствии приема препаратов, способных индуцировать волчаночноподобный синдром
Примечание: дискоидная красная волчанка — кожная форма волчанки,
для которой характерны три признака — эритема, гиперкератоз, атрофия.
300 Частная дерматология
Антикоагулянты и антиагреганты. Из антикоа-
гулянтов рекомендуется гепарин — 10 000—20 000
ЕД в сутки (4 инъекции подкожно в область
живота) в течение нескольких месяцев. В каче-
стве антиагрегантов используют курантил в су-
точной дозе 150—200 мг и трентал — 400—600 мг/
сут в течение нескольких месяцев. Антикоагулян-
ты и антиагреганты применяют в комплексной
терапии при наличии поражения почек, ДВС-
синдрома, при нарушениях микроциркуляции.
Эфферентная терапия — плазмаферез и гемо-
сорбция. Плазмаферез — метод эфферентной,
экстракорпоральной терапии, направленной на
освобождение организма от продуктов метабо-
лизма, циркулирующих иммунных комплексов
путем удаления плазмы крови. Плазмаферез
назначают при тяжелом течении системной
красной волчанки, когда терапия глюкокорти-
коидами, включая пульс-терапию метилпредни-
золоном, а также применение цитостатиков,
оказывается неэффективной. На курс лечения
рекомендуется от 3 до 5 процедур плазмафереза
при однократном удалении 800—1000 мл плазмы.
Гемосорбция — экстракорпоральный метод очи-
щения крови путем пропускания ее через колон-
ку с гранулами активированного угля. Метод
обладает иммунокорригирующим действием, а
также повышает чувствительность клеток кро-
ви и тканей к действию глюкокортикоидов. На
курс лечения рекомендуется от 3 до 5 процедур
гемосорбции, проводимых еженедельно.
Наружная терапия. Кортикостероидные мази
(синалар, флуцинар, фторокорт, целестодерм,
сульфодекортем) применяются на очаги пораже-
ния на ночь. Рекомендуется внутрикожное об-
калывание дискоидных очагов гидрокортизоном
или 5—10 %-ным раствором резохина или хин-
гамина, а также криотерапия жидким азотом или
снегом угольной кислоты.
Прогноз при системной красной волчанке
остается в целом неблагоприятным, хотя адек-
ватное современное лечение значительно увели-
чивает продолжительность жизни больного.
Больные должны находиться под диспансерным
наблюдением ревматолога.
Очаговая
склеродермия
Склеродермия — аутоиммунное заболевание,
которое характеризуется системным поражени-
ем соединительной ткани с преобладанием фиб-
розно-склеротических и сосудистых нарушений
по типу облитерирующего эндартериолита с рас-
пространенными вазоспастическими изменени-
ями, развивающимися преимущественно в коже
и подкожной клетчатке.
Этиология и патогенез
Этиология склеродермии окончательно не уста-
новлена. Предполагается мультифакториальное
наследование. В развитии заболевания имеют
значение переохлаждения, простудные заболева-
ния, травмы, эндокринопатии, заболевания не-
рвной системы.
Патогенез склеродермии связывают с обмен-
ными, сосудистыми и иммунными нарушения-
ми. Нарушение метаболизма соединительной
ткани проявляется гиперпродукцией коллагена
фибробластами, повышенным содержанием ок-
сипролина в плазме крови и моче, нарушением
соотношения растворимой и нерастворимой
фракций коллагена и накоплением в коже меди.
Особое патогенетическое значение при скле-
родермии придают изменениям микроциркуля-
ции. В их основе лежат поражения преимуще-
ственно стенок мелких артерий, артериол и
капилляров, пролиферация и деструкция эндо-
телия, гиперплазия интимы, склероз.
В последние годы склеродермию рассматрива-
ют как аутоиммунное заболевание, при котором
в основе нарушений лежит взаимодействие ауто-
антигенов с лимфоидными клетками.
Классификация
Различают две основные формы склеродермии:
системную (поражаются кожа и внутренние орга-
ны) и очаговую (ограниченную). Наиболее прием-
лемой в практическом плане является классифи-
кация очаговой склеродермии, основанная на
клиническом принципе (С. И. Довжанский, 1979).
• Бляшечная форма и ее варианты: индуратив-
но-атрофическая (Вильсона); поверхностная «си-
реневая» (Гужеро); келоидоподобная; узловатая
глубокая; буллезная (эрозивно-язвенная); гене-
рализованная.
• Линейная форма (полосовидная): саблевид-
ная (по типу «удара саблей»); лентовидная; зос-
териформная.
• Склероатрофический лихен Цумбуша (бо-
лезнь белых пятен).
• Идиопатическая атрофодермия Пазини-Пье-
рини.
Клиническая картина
Склеродермия является хроническим длительно
протекающим заболеванием с постепенной сме-
Глава 25. Болезни с диффузным поражением соединительной ткани 301
ной характерных клинических стадий патологи-
ческого процесса кожи: плотного отека, уплот-
нения вплоть до затвердения или склероза и
атрофии вследствие дегенеративно-склеротичес-
кого поражения соединительной ткани и обли-
терации артериол.
Ограниченная склеродермия проявляется вна-
чале эритематозным отечным пятном бледно-
розового или розовато-лилового цвета {стадия
отека). Размер очага — от просяного зерна до
ладони и больше, границы нечеткие. Затем в
центре пятно приобретает хрящевую или дере-
вянистую плотность цвета слоновой кости {ста-
дия уплотнения). Кожа в очаге блестящая, рису-
нок сглажен, нарушены чувствительность, сало-
и потоотделение, пушковые волосы не растут,
кожа в складку не берется. По периферии очаг
окружен фиолетовым ореолом (сиреневое коль-
цо), что указывает на активность процесса. В
дальнейшем фиолетовый ореол исчезает, умень-
шается уплотнение в очаге, центральная часть
бляшки несколько западает, на месте регресси-
ровавших высыпаний остаются гиперпигмента-
ция и атрофия {стадия атрофии).
Бляшечная склеродермия локализуется на ко-
нечностях, шее, туловище, иногда на лице в виде
очагов неправильной формы (рис. 25.2). Возни-
кает внезапно, незаметно. Обострения сменяются
ремиссиями в течение нескольких месяцев или
лет. В регрессивной стадии на месте поражения
образуются гиперпигментированные пятна. Для
склеродермии характерно наличие телеангиэкта-
зий около очагов.
Линейная (полосовидная) склеродермия харак-
теризуется поражением кожи лба или голени,
напоминает рубец от удара саблей. Располагает-
ся по ходу нервных стволов или рефлексогенных
зон Захарьина-Геда. Чаще болеют дети. Может
сопровождаться гемиатрофией лица Ромберга, при
которой поражается половина лица в виде атро-
фии дермы, гиподермы, жевательных и мимичес-
ких мышц, хрящевой и костной ткани.
Болезнь белых пятен (пятнистая склеродер-
мия, склероатрофический лихен Цумбуша) рас-
полагается на коже шеи, груди, спины, плече-
вого пояса, живота. Встречается чаще у
женщин. Депигментированные атрофические
пятна размером с горошину локализуются вок-
руг волосяных фолликулов, блестящие, мор-
щинистые, рисунок сглажен, могут сливаться
и занимать большие участки кожи. Вокруг
очагов наблюдается эритематозный гиперпиг-
ментированный венчик.
Идиопатическая атрофодермия Пазини-Пьери-
ни (син.: поверхностная ограниченная склеродер-
мия). На туловище, плечах, бедрах образуются
слегка пигментированные цианотичные высыпа-
ния округлой или овальной формы с незначи-
тельной атрофией. Они могут сливаться между
собой и образовывать очаги больших размеров
с фестончатыми краями.
Патоморфология
В стадии отека отмечается утолщение коллаге-
новых волокон, между которыми располагают-
ся умеренно выраженные воспалительные ин-
фильтраты, состоящие преимущественно из
лимфоцитов. При вовлечении в процесс подкож-
ного жирового слоя соединительнотканные пе-
регородки его утолщаются в результате воспале-
ния и новообразования коллагеновых волокон,
которые полностью его заменяют. В стадии уп-
лотнения и склероза воспалительные явления ис-
чезают, и в дальнейшем пучки коллагеновых во-
локон становятся гомогенизированными и
гиалинизированными. В начале процесса они
окрашиваются эозином интенсивно, а затем
бледно. Клеточных элементов и сосудов очень
мало, стенки последних утолщены, а просветы —
сужены. Эпидермис изменен обычно очень мало,
в воспалительной стадии он несколько утолщен,
а в склеротической — атрофичен.
Рис. 25.2. Типичная локализация
высыпаний при ограниченной склеродермии.
302 Частная дерматология
Диагностика
Диагноз склеродермии не вызывает затруднений,
особенно при выраженной клинической симп-
томатике. Диагностические признаки очаговой
склеродермии: ограниченное уплотнение и атро-
фия кожи в виде бляшек, полос или пятен.
Лечение
Современное лечение основывается на патогенезе
заболевания с учетом основных клинических
проявлений активности и варианта течения скле-
родермии.
Базисная терапия — применяют антифиброз-
ные средства:
• D-пеницилламин (купренил, бианодин, ар-
тамин); диуцифон; мадекассол;
• ферментативные препараты — лвдаза, ронцдаза.
D-пеницилламин (купренил) является основ-
ным антифиброзным средством, которое подав-
ляет избыточный синтез коллагена фибробласта-
ми и аутоиммунное воспаление, что проявляется
улучшением состояния кожи (уменьшение плот-
ного отека и пигментации кожи) и облегчением
проявлений синдрома Рейно. Мадекассол (в таб-
летках или в виде мази) ингибирует синтез кол-
лагена и стабилизирует лизосомальные мембра-
ны. Диуцифон — противолепрозный препарат —
оказывает иммуномодулирующее и умеренное ан-
тифиброзное действие. Ферментативные препара-
ты (лидаза, ронидаза) воздействуют на систему ги-
алуроновая кислота — гиалуронидаза и замедляют
фиброзообразование.
Применяют также нестероидные противовос-
палительные средства — вольтарен, бруфен,
пироксикам, индометацин, метиндол-ретард.
Иммунодепрессантные средства — глюкокорти-
коиды, цитостатики (азатиоприн, метотрексат),
аминохинолоны (делагил, плаквенил) — подав-
ляют аутоиммунный воспалительный процесс в
соединительной ткани, тормозят избыточное
фиброзообразование.
Препараты, улучшающие микроциркуляцию,
должны назначаться уже в самом начале заболе-
вания (антагонисты кальция — коринфар, кор-
дафен, верапамил; антиагреганты — курантил,
трентал, аспирин, реополиглюкин; простаглан-
дины — алпростадил; андекалин; солкосерил;
ангиопротекторы — пармидин, продектин, каль-
ция добезилат; никотиновая кислота; мидокалм).
Местная терапия склеродермии применяется
для уменьшения выраженности и прогрессиро-
вания фиброзных изменений кожи и сосудис-
тых нарушений. Назначают следующие виды ме-
стной терапии:
• аппликации ДМСО (диметилсульфоксида);
• электрофорез лидазы (ронидазы);
• фонофорез гидрокортизона, лидазы, теони-
коловой мази;
• гелий-неоновое лазерное излучение;
• теплолечение (аппликации парафина, озоке-
рита; лечебные грязи);
• гипербарическая оксигенация;
• пунктурная физиотерапия (акупунктура,
КВЧ, фонопунктура);
• ванны — сероводородные, радоновые, йо-
добромные;
• массаж, лечебная физкультура.
Эфферентная терапия (гемосорбция, плазма-
ферез) показана при тяжелом течении заболева-
ния и оказывает иммунокорригирующее дей-
ствие, уменьшает выраженность интоксикации
и фиброза.
При рациональном лечении ограниченных
форм болезни обычно наступает клиническая
ремиссия. Прогноз для жизни при очаговой
склеродермии благоприятный, но для полного
излечения сомнительный. Больные склеродерми-
ей должны находиться под постоянным наблю-
дением дерматолога и ревматолога, избегать
физического и нервного перенапряжения, про-
студных заболеваний.
Системная
склеродермия
Синонимы: sclerodermia systemica, диффузная
склеродермия, универсальная склеродермия,
прогрессирующий системный склероз.
Системная склеродермия — диффузное забо-
левание соединительной ткани с преобладанием
фиброза и облитерирующей микроангиопатии,
характеризующееся индуративными изменения-
ми кожи, поражением опорно-двигательного
аппарата, внутренних органов (легких, сердца,
пищеварительного тракта, почек), генерализо-
ванным вазоспастическим синдромом Рейно.
Этиология
Этиология системной склеродермии не выясне-
на. Предполагаются вирусное и наследственное
происхождение заболевания. Системной склеро-
дермии предшествуют инфекционные заболева-
ния или умеренное лихорадочное состояние не-
ясной причины, зябкость, потливость, повторные
ангины, повышенная чувствительность кистей к
холоду, парестезии пальцев кистей (синдром Рей-
Глава 25. Болезни с диффузным поражением соединительной ткани
303
но), боли в мышцах и суставах, диспепсии, го-
ловные боли.
Системная склеродермия диагностируется пре-
имущественно у женщин в возрасте от 35 до 64
лет. Заболевание редко встречается у детей, а
также у взрослых до 30 лет. Заболеваемость си-
стемной склеродермией составляет 6,3—12 слу-
чаев на 1 млн населения.
Патогенез
Патогенез системной склеродермии включает
изменения обмена соединительной ткани (увели-
чение биосинтеза коллагена и неофибриллогене-
за, фиброз тканей), иммунные нарушения (сни-
жение уровня Т-супрессоров при нормальном
содержании В-лимфоцитов в крови, появление
антител к коллагену, иногда антинуклеарных
антител) и поражение микроциркуляторного русла
(цитотоксические лимфоциты повреждают эндо-
телий, что сопровождается адгезией и агрегаци-
ей тромбоцитов, активацией коагуляции, высво-
бождением медиаторов воспаления, увеличением
проницаемости сосудистой стенки с плазматичес-
ким ее пропитыванием и отложением фибрина,
сужением просвета). В развитии фиброза большую
роль играют цитокины и факторы роста, секре-
тируемые лимфоцитами, моноцитами и тромбо-
цитами. Они вызывают гиперпродукцию колла-
гена и макромолекул основного вещества
соединительной ткани с последующим развити-
ем участков фиброза.
Клиническая картина
Системная склеродермия включает кожный и
внекожные синдромы. Кожный синдром: скле-
родактилия; акросклероз; диффузно — генера-
лизованный тип; синдром Тибьержа-Вейссенба-
ха. Внекожные синдромы: мышечно-суставной,
желудочно-кишечный, легочный, сердечно-сосу-
дистый, почечный, неврологический.
Системная или диффузная склеродермия
(универсальная, генерализованная, прогрессиру-
ющая) начинается чаще у женщин в детском или
юношеском возрасте с поражением кожи кистей,
лица, а затем конечностей и туловища. При диф-
фузной склеродермии фиброз кожи локализуется
проксимальнее локтевых или коленных суставов,
включая шею и туловище. У больных может
наблюдаться феномен Рейно в течение первого
года болезни, чаще поражаются внутренние орга-
ны: легкие, почки, сердце. Выделяют также ли-
митированную склеродермию, при которой раз-
вивается фиброз кожи кистей, предплечий и
стоп, но могут поражаться лицо и шея. У боль-
ных данной группы феномен Рейно наблюдает-
ся годами, у них развиваются телеангиэктазии,
кальцификаты в коже, а позднее присоединяется
легочная гипертензия.
Поражения кожи
Поражения кожи являются ведущим признаком
заболевания. Возможно появление акросклеро-
за, иногда образуются многочисленные подкож-
ные известковые узелки с дальнейшим их изъяз-
влением. Уплотнение кожи развивается
вследствие патологической выработки коллаге-
на I типа фибробластами кожи, а также избы-
точных отложений гликозаминогликанов и фиб-
ронектина во внеклеточном матриксе. Внешне у
больного имеются пораженные и непораженные
участки кожи. Однако исследования показали
наличие проколлагена-1 и адгезивных молекул
во всех участках дермы, что свидетельствует о
генерализованном патологическом процессе.
На коже лица, шеи, кистей появляется плот-
ный отек, затем развиваются уплотнение и
склерозирование кожи. Впоследствии наступа-
ет ее атрофия и лицо приобретает маскообраз-
ный вид: складки сглаживаются, мимические
движения отсутствуют, губы истончаются, вок-
руг них образуются радиарные глубокие склад-
ки, похожие на полустянутый кисет, рот не
закрывается, язык не высовывается из-за края
зубов, веки часто не смыкаются; нос истонча-
ется, кожа на его крыльях становится гладкой,
натянутой, блестящей. Голос слабеет, становится
как будто из глубины. Ушные раковины так-
же заметно атрофируются, истончаются. Воло-
сы постепенно выпадают, ногти истончаются,
а затем отделяются от ногтевого ложа. Наблю-
дается выраженная сухость кожи, гиперкератоз
ладоней и подошв. Уменьшается, а затем пре-
кращается секреция потовых и сальных желез.
Процесс нередко захватывает слизистую оболоч-
ку рта, язык, мягкое небо, гортань, пищевод,
легкие и другие внутренние органы, а также
мышцы, суставы, кости. Впоследствии пораже-
ние может распространяться на весь кожный
покров: предплечья, плечи, туловище, бедра,
голени и стопы. Кожа при этом становится
восково-желтой; участки выраженной гиперпиг-
ментации чередуются с депигментированными
очагами; отмечаются множественные телеанги-
эктазии. Подкожно-жировая клетчатка посте-
пенно исчезает, кожа прирастает к подлежащим
фасциям и становится неподвижной над костя-
ми, в складку не берется. В результате насту-
пает одеревенение всего тела. При тотальном
поражении кожи туловища и конечностей раз-
вивается кахексия и мумификация («живые
304 Частная дерматология
мощи»). Резко ограничиваются движения в су-
ставах и даже дыхание, больной испытывает зат-
руднение в приеме пище.
Иногда происходит отложение солей кальция
(кальциноз) в подкожной клетчатке в виде уз-
ловатостей каменистой консистенции, которые
вскрываются с образованием фистул и белыми
крошковидными выделениями. Кальциноз —
это отложения гидроксиапатитов в коже, кото-
рые локализуются премущественно на кистях в
области проксимальных межфаланговых суста-
вов и ногтевых фаланг, вокруг суставов и над
костными выступами (особенно на разгибатель-
ной поверхности локтевого и коленного суста-
вов). Периартикулярное отложение кальция в
виде белых очагов, просвечивающихся сквозь
кожу, носит название синдрома Тибьержа —
Вейссенбаха. Отложения кальция не всегда оп-
ределяются визуально, в этом случае они выяв-
ляются при рентгенографии. Кальциноз сохра-
няется годами и трудно лечится.
Отмечаются множественные телеангиэктазии
(расширенные венулы, капилляры, артериолы),
которые имеют вид овальных или неправильной
формы пятен величиной 2—7 мм в диаметре и
располагаются на кистях, лице, губах и слизис-
той оболочке полости рта. Телеангиэктазии со
временем самопроизвольно исчезают.
Состояние, проявляющееся кальцинозом, син-
дромом Рейно, эзофагопатией, склеродактили-
ей и телеангиэктазиями, получило название
CREST-синдрома (по первым буквам симпто-
мов). Этот синдром считают благоприятно про-
текающим вариантом заболевания и относят к ли-
митированной склеродермии.
Наиболее ранним признаком системной скле-
родермии является синдром Рейно, который ха-
рактеризуется внезапным появлением парестезий
(чувство онемения, ползания мурашек) в облас-
ти II-IV пальцев кистей, стоп, резким их поблед-
нением. По окончании приступа появляются
боли, чувство жара в пальцах, кожа гипереми-
рована. Синдром Рейно при системной склеро-
дермии захватывает не только пальцы рук, ног,
но и губы, кончик языка, части лица. У паци-
ентов с синдромом Рейно возможно подтверж-
дение наличия системной склеродермии на ран-
ней стадии с помощью следующих признаков
наличие антинуклеарных антител, антител к цен-
тромерам или антител к топоизомеразе 1 (анти-
Scl-70-антитела); изменения капиллярного рус-
ла ногтевого ложа — уменьшение числа
капилляров или их дилатация; явления тендова-
гинита (утолщения по ходу сухожилий и шум
трения в области сухожильных влагалищ); плот-
ный отек пальцев рук или ног; сопутствующий
рефлюкс-эзофагит.
Костно-суставной синдром
Костно-суставной синдром может быть одним из
ранних признаков системной склеродермии. Выде-
ляют три основных варианта суставного синдрома:
полиартралгии; склеродермический полиартрит с
преобладанием экссудативно-пролиферативных или
фиброзно-индуративных изменений; периартрит с
развитием контрактур вследствие вовлечения в па-
тологический процесс периартикулярных тканей.
Для поражения костей характерен акросклероз с ос-
теолизом, обычно ногтевых фаланг, укорочение и
деформация пальцев рук и ног. Поражение костей
в основном связано с резорбцией костной ткани.
Часто возникают артралгии и утренняя скован-
ность, наблюдаются анкилозы.
Мышечный синдром характеризуется разными
типами поражения мышц, которые могут быть в
виде интерстициального миозита или полимио-
зита; проявляется болями в мышцах, мышечной
слабостью, ощущением скованности в мышцах.
Первый тип поражения отличается умеренной
слабостью проксимальных мышц вследствие доб-
рокачественной миопатии невоспалительного ге-
неза. При гистологическом исследовании выяв-
ляют атрофию мышечных волокон 2-го типа,
которая связана с обездвиженностью и приемом
кортикостероидов. Концентрация мышечных
ферментов в пределах нормы. Второй тип пора-
жения характеризуется умеренным повышением
концентрации мышечных волокон и наличием
симптома «воскообразной» слабости мышц. При
биопсии обнаруживаются интерстициальный
фиброз и атрофия мышечных волокон. Выявля-
ется умеренная инфильтрация клетками воспали-
тельного ряда. Третий тип поражения проявля-
ется миопатией воспалительного генеза с
повышением концентрации ферментов.
Желудочно-кишечный синдром
Желудочно-кишечный синдром проявляется дуо-
денитом, энтеритом (с развитием синдрома ма-
льабсорбции), колитом (с выраженными запора-
ми, явлениями кишечной непроходимости).
Характерным считается поражение пищевода
(эзофагит), что проявляется дисфагией, диффуз-
ным расширением пищевода, сужением его в
нижней трети, ослаблением перистальтики и
ригидностью стенок, рефлюкс-эзофагитом,
иногда развитием пептических язв, стриктур.
Легочный синдром
Легочный синдром проявляется клиникой фиб-
розирующего альвеолита и диффузного пневмо-
Глава 25. Болезни с диффузным поражением соединительной ткани
305
фиброза с преимущественной локализацией в
базальных отделах легких. У некоторых больных
развивается легочная гипертензия вследствие
поражения сосудов легких. Выраженный пнев-
москлероз иногда сочетается с развитием брон-
хоэктазов, эмфиземы легких, возможно присо-
единение пневмоний.
Сердечно-сосудистый синдром
Сердечно-сосудистый синдром характеризуется
кардиосклерозом с увеличением размеров сердца,
аритмиями, нарушениями сердечного ритма и
проводимости, развитием недостаточности крово-
обращения. Крупноочаговый кардиосклероз мо-
жет имитировать на ЭКГ «инфарктоподобные
изменения». Развитие интерстициального миокар-
дита приводит к появлению клинической симп-
томатики, фактически аналогичной кардио-
склерозу. Поражение эндокарда в области
клапанного аппарата приводит к образованию
склеродермического порока сердца, чаще недоста-
точности митрального клапана. У некоторых
больных формируется пролапс митрального кла-
пана. Возможно развитие перикардита.
Почечный синдром
Почечный синдром проявляется вовлечением
почек в патологический процесс. Различают ос-
трую и хроническую нефропатии. Острая нефро-
патия (истинная склеродермическая почка) про-
является следующими признаками: олигоанурия,
артериальная гипертензия, нарастающие проте-
инурия, микрогематурия, цилиндрурия, ретино-
патия, энцефалопатия. В основе острой нефро-
патии лежит генерализованное поражение
артериол почек и развитие кортикальных некро-
зов, что приводит к острой почечной недоста-
точности. Наиболее частым поражением почек
при системной склеродермии является хроничес-
кая нефропатия. Морфологическим субстратом
этого вида патологии почек считается пораже-
ние сосудов и клубочков почек, а также каналь-
цев и интерстиция* Клиническая симптоматика
соответствует симптоматике хронического гло-
мерулонефрита (протеинурия, цилиндрурия,
микрогематурия, снижение клубочковой филь-
трации, артериальная гипертензия, развитие хро-
нической почечной недостаточности).
Неврологический синдром
Неврологический сивдром характеризуется пора-
жением периферической нервной системы в виде
полинейропатии (боли в руках и ногах, наруше-
ние чувствительности в виде гиперестезии с
последующим развитием гипостезии дистального
типа, снижение сухожильных рефлексов). Поли-
нейропатия приобретает длительное, упорное
течение. Характерен также стойкий, рецидиви-
рующий тригеминит (воспаление тройничного
нерва). Поражение ЦНС наблюдается редко,
описаны энцефалиты, менингоэнцефалиты,
ишемические инсульты, кровоизлияния в мозг.
Эндокринологический синдром
Эндокринологический синдром проявляется наруше-
нием функции щитовидной железы (гипотиреоз,
возможен аутоиммунный тиреоидит, редко — ги-
пертиреоз), надпочечниковой недостаточностью,
снижением функции половых желез. Возможно
сочетание системной склеродермии с сахарным
диабетом. Поражение эндокринных желез обус-
ловлено в первую очередь поражением их сосуди-
стой системы.
При системной склеродермии характерна по-
теря массы тела, вплоть до кахексии, особенно
выраженная при быстропрогрессирующем тече-
нии заболевания и при его обострении.
Течение
Различают три варианта течения системной
склеродермии: острое, подострое и хроническое.
Острое течение характеризуется быстрым (в
течение года) развитием диффузной симптома-
тики, неуклонным прогрессированием пораже-
ния внутренних органов, нарастающим фибро-
зом органов и тканей, развитием склеротической
почки. Острое течение сопровождается выражен-
ными изменениями лабораторных показателей,
которые отражают активность воспалительного
процесса.
Для подострого течения характерна клиника
заболевания с поражением кожи, суставов,
мышц, внутренних органов на фоне нерезко
выраженных вазомоторных трофических нару-
шений, отмечается увеличение СОЭ, фибрино-
гена, а2- и у-глобулинов в крови, наличие рев-
матоидного фактора, антиядерного фактора.
Хроническое течение отличается медленно про-
грессирующими на протяжении нескольких лет
сосудистыми и трофическими нарушениями,
уплотнением кожи, периартикулярной ткани с
образованием контрактур, остеолизом, медлен-
но развивающимися поражениями пищевода,
легких, сердца. Типичные изменения кожи мо-
гут носить очаговый характер и долго оставать-
ся единственным проявлением болезни.
Стадии
Начальная стадия склеродермии проявляется
синдромом Рейно, артралгией, тахикардией, ча-
стыми инфекциями дыхательных путей. Комп-
306 Частная дерматология
лексное лечение в этой стадии приводит к дли-
тельной ремиссии и даже выздоровлению.
Генерализованная стадия протекает со всеми
ранее приведенными симптомами, характеризу-
ется развернутой картиной болезни.
Терминальная стадия наступает при далеко
зашедших изменениях и сопровождается выра-
женной потерей массы тела, недостаточностью
функций одного или нескольких органов. Ле-
чение в этой стадии не дает эффекта.
Активность
По степени активности системной склеродермии
выделяют минимальную (I) степень — вазо-
спастические и трофические нарушения, СОЭ
менее 20 мм/ч; умеренную (II) степень — артрал-
гии, артриты, адгезивный плеврит, кардиоскле-
роз, СОЭ в пределах 20—35 мм/ч; высокую (III)
степень — лихорадка, полиартрит, миокардио-
склероз, нефропатия, СОЭ более 35 мм/ч.
Лабораторные
и инструментальные исследования
Обший анализ крови
признаки гипохромной анемии, лейкопения,
иногда лейкоцитоз, повышение СОЭ.
Анализ мочи
повышение экскреции оксипролина, протеину-
рия.
Биохимический анализ крови
Гиперпротеинемия, увеличение уровня а2- и у-
глобулинов, фибрина, серомукоида, С-реактив-
ного протеина, гаптоглобина, оксипролина (на-
рушение метаболизма коллагена).
Иммунологическое исследование крови
Антитела к эндотелию, антинуклеарные антите-
ла к антигену SCL-70, снижение количества Т-
лимфоцитов-супрессоров, гипер- и дисиммуног-
лобулинемия; обнаружение ревматоидного
фактора (40—45 %), антинуклеарных антител (30—
90%).
Биоптат кожно-мышечного лоскута
Фиброзная трансформация тканей, патология
сосудов.
Патоморфология кожи
На ранних стадиях выявляются небольшие ин-
фильтраты вокруг сосудов дермы и мерокрино-
вых потовых желез, а также инфильтрация под-
кожной клетчатки; на поздних стадиях
отмечается исчезновение межсосочковых клиньев
эпидермиса; утолщенные гомогенные пучки кол-
лагеновых волокон, ярко окрашиваемые эози-
ном; слияние пучков, облитерация и исчезнове-
ние межпучкового пространства; утолщение дер-
мы, замещение жировой ткани (верхних слоев
или всей) коллагеном, гиалиноз; уменьшение
количества сосудов, утолщение стенок и суже-
ние просвета, гиалиноз; атрофия придатков
кожи; потовые железы располагаются в верхних
слоях дермы; отложения солей кальция в скле-
розированной подкожной клетчатке.
Электрокардиограмма
Диффузные изменения миокарда, иногда блока-
да ножек пучка Гиса и атриовентрикулярная.
Рентгенологическое исследование
Участки кальциноза в подкожной клетчатке, пре-
имущественно концевых отделов пальцев рук,
реже — стоп, в области локтевых, коленных и
других суставов. Остеолиз в ногтевых фалангах
пальцев кистей, стоп, дистальных отделах луче-
вой и локтевой костей, задних отделов ребер.
Околосуставный остеопороз, сужение суставных
щелей, иногда единичные эрозии на поверхно-
сти суставного хряща, костные анкилозы. Сни-
жение тонуса и ослабление перистальтики желу-
дочно-кишечного тракта, что приводит к
расширению пищевода, двенадцатиперстной
кишки. Диффузный и кистозный пневмоскле-
роз в базальных отделах и увеличение размеров
сердца.
Диагностика
Американским институтом ревматологии разра-
ботаны диагностические критерии системной
склеродермии (1980):
Главный (большой) критерий — склеродерми-
ческое поражение кожи туловища (проксималь-
нее пястно-фаланговых или плюснефаланговых
суставов — проксимальная склеродермия).
Малые критерии — склеродактилия, рубчики
на дистальных фалангах пальцев, двусторонний
базальный фиброз легких.
Для установления диагноза системной склеро-
дермии требуется наличие главного и двух ма-
лых критериев (табл. 25.2).
Для достоверного диагноза системной склеро-
дермии достаточно наличия любых трех основ-
ных признаков или одного из основных, если им
является склеродермическое поражение кожи,
остеолиз ногтевых фаланг или характерное по-
ражение пищеварительного тракта, в сочетании
с тремя и более вспомогательными признаками.
При меньшем числе симптомов устанавливает-
ся «вероятный» диагноз.
Глава 25. Болезни с диффузным поражением соединительной ткани
307
Лечение
Для лечения больных системной склеродерми-
ей применяют антифиброзные, нестероидные
противовоспалительные, улучшающие микро-
циркуляцию средства, иммунодепрессанты; ме-
стную терапию, массаж.
Антифиброзные средства. Основным базис-
ным препаратом в лечении системной склеро-
дермии является D-пенициллинамин (купренил),
который подавляет избыточный синтез колла-
гена фибробластами; связывает и удаляет медь
из организма, что приводит к активации кол-
лагеназы и вызывает распад коллагена; нару-
шает созревание коллагена и ускоряет его рас-
пад; подавляет аутоиммунное воспаление.
Эффект от применения препарата проявляет-
ся уменьшением плотного отека и пигмента-
ции кожи, уменьшением артралгии и миалгии,
снижением проявлений синдрома Рейно. Вна-
чале препарат назначают по 150—300 мг в день
в течение 2 недель, затем дозу повышают каж-
дые 2 недели на 300 мг до максимальной —
1800 мг. Эту дозу назначают в течение 2 ме-
сяцев, а затем медленно уменьшают до поддер-
живающей — 300—600 мг в сутки. Эффект
действия препарата отмечается не раньше двух
месяцев от начала лечения. Побочные действия
купренила: дерматит, выпадение волос, диспеп-
сия, лейкопения, тромбоцитопения, поражение
почек. При развитии побочных эффектов не-
обходимо уменьшить дозу препарата или от-
менить его. Купренил противопоказан при по-
чечной и печеночной патологии, лейкопении,
тромбоцитопении.
Мадекассол — препарат из экстракта растения
Centella asiatica, содержит азиатиковую и маде-
кассоловую кислоты. Мадекассол ингибирует
синтез коллагена, стабилизирует лизосомальные
мембраны. Назначается по 10 мг внутрь 3 раза
в день в течение 3—6 месяцев.
Таблица 25.2. Диагностические признаки
(основные и дополнительные) системной склеродермии (Н. Г. Гусева, 1993,1997)
Основные признаки Дополнительные (малые) признаки
1. Склеродермическое поражение кожи, проходящее последовательно стадии «плотного» отека, индурации и атрофии с преимущественной локализацией на лице (маскообразность) и в области кистей (склеродактилия), возможно тотальное поражение. Обычно синдром сочетается с пигментацией. 1. Периферические: гиперпигмен- тация кожи, телеангиэктазии, трофические нарушения, синдром Шегрена, полиартрал-
2. Синдром Рейно. гии, полимиалгии, полимиозит.
3. Суставно-мышечный синдром с развитием стойких контрактур, в основе которого — ревматоидноподобный артрит, периартикулярные изменения и фиброзирующий миозит. 2. Висцеральные: полисерозит (чаще адгезивный), хроническая нефропатия, полиневрит,
4. Остеолиз ногтевых, а иногда средних и основных фаланг пальцев рук, тригеминит.
реже — ног, что проявляется укорочением и деформацией пальцев. 3. Общие: потеря массы (более
5. Синдром Тибьержа-Вейссенбаха — отложение солей кальция преимуще- ственно в области пальцев рук и периартикулярно — вокруг локтевых, плечевых и тазобедренных суставов, в подкожной клетчатке, иногда по ходу фасций и сухожилий мышц. 10 кг). Лабораторные: увеличение СОЭ (более 20 мм/ч), гиперпротеине- мия (более 85 г/л), гипергаммаг-
6. Поражение пищеварительного тракта (склеродермический эзофагит с дисфагией, дилатация пищевода, гастрит, дуоденит, нарушение моторики кишечника вплоть до кишечной непроходимости, развитие синдрома мальабсорбции). 7. Поражение сердца по типу первичного крупноочагового кардиосклероза. 8. Поражение легких по типу базального пневмосклероза, кистозного легкого (на рентгенограмме — «медовые соты»). 9. Истинная склеродермическая почка, диагностируется клинически на основании внезапного повышения артериального давления и развития острой почечной недостаточности. 10. Наличие специфических антинуклеарных антител (анти-8с1-70 и антицент- ромерные антитела). 11. Капилляроскопические признаки (по данным широкопольной капилляроско- пии). лобулинемия (более 23 %), антитела к ДНК или антинукле- арному фактору, ревматоидно- му фактору.
308 Частная дерматология
Диуцифон — противолепроматозный препарат,
обладающий также умеренным антифиброзным
и иммуномодулирующим действием. Назначает-
ся по 0,1—0,2 г 3 раза в день внутрь.
Лидаза — ферментный препарат, который
воздействует на систему гиалуроновая кислота —
гиалуронидаза, замедляет фиброзообразование.
препарат применяется при хроническом течении
склеродермии и при очаговой склеродермии.
Противопоказанием к назначению ферментатив-
ных препаратов является высокая активность
процесса и резко повышенная сосудистая про-
ницаемость. Лидаза — препарат, содержащий
гиалуронидазу; вызывает расщепление гиалуро-
новой кислоты до глюкозамина и глюкуроновой
кислоты и уменьшает ее вязкость. Лидаза вво-
дится подкожно или внутримышечно по 64 УЕ
(в 1 мл 0,5 % раствора новокаина), на курс ле-
чения 12—14 инъекций. Препарат противопока-
зан при язвенной болезни, склонности к гемор-
рагиям, почечной недостаточности.
Нестероидные противовоспалительные средства.
Назначают при выраженном суставном синдро-
ме, а также при наличии противопоказаний к
глюкокортикоидной терапии. Нестероидные
противовоспалительные средства применяют при
системной склеродермии в комплексе с амино-
хинолоновыми соединениями или при сниже-
нии дозы глюкокортикоидов. Нестероидные про-
тивовоспалительные средства ингибируют синтез
провоспалительных простагландинов, уменьша-
ют обеспечение энергией участка воспаления и
агрегацию тромбоцитов, оказывают мягкое им-
мунодепрессантное действие. Из препаратов наи-
более часто применяются: вольтарен (диклофе-
нак-натрий, ортофен) по 0,25 г 3—4 раза в день;
вольтарен-ретард по 0,075 г 1—2 раза в день;
бруфен (ибупрофен) по 0,2—0,4 г 3 раза в день.
Иммунодепрессантные средства (глюкокорти-
коиды, цитостатики, аминохинолины) подавляют
аутоиммунный воспалительный процесс в соеди-
нительной ткани, тормозят избыточное фибро-
зообразование.
Глюкокортикоцды назначают при подостром и
остром течении системной склеродермии. Хро-
ническое течение системной склеродермии и I
степень активности не требуют глюкокортико-
идов. Преимущественно назначают преднизолон.
Начальная доза препарата при III степени актив-
ности в среднем 30 мг/сут в течение 1,5—2 ме-
сяцев до достижения клинического эффекта с
последующим снижением поддерживающей
дозы до 20—10 мг/сут. При II степени активно-
сти начальная суточная доза преднизолона со-
ставляет 20 мг/сут в течение 2 месяцев с после-
дующим снижением дозы. Глюкокортикоиды
дают положительный эффект при лихорадке,
полиартрите, кожном синдроме, при висцераль-
ных проявлениях, миозите. Для предупреждения
остеопороза рекомендуется на фоне глюкокор-
тикоидной терапии применять анаболические
препараты (ретаболил, метандростенолон), глю-
конат кальция.
Цитостатики назначают при остром и подо-
стром течении болезни со значительными им-
мунными нарушениями и высокой активностью
процесса, при полимиозите, гломерулонефрите.
Применяются имуран (азатиоприн) или цикло-
фосфамид по 100—150 мг в день, метотрексат —
7,5—10 мг в неделю. Лечение цитостатиками
сочетают с глюкокортикоидами. Лечение цито-
статиками проводят 2—3 месяца.
Аминохинолиновые средства обладают слабым
имммунодепрессантным и противовоспалитель-
ным действием, стабилизируют лизосомные мем-
браны и ингибируют выход из лизосом протео-
литических ферментов. Аминохинолиновые
препараты показаны при любом течении систем-
ной склеродермии в сочетании с базисными пре-
паратами. Назначают делагил по 0,25 г в сутки
или плаквенил по 0,2—0,4 г в сутки непрерыв-
но в течение года, после чего переходят на при-
ем препаратов с перерывом на летний период.
Побочные эффекты аминохинолиновых препа-
ратов: головные боли, головокружение, диспеп-
сия, поражения сетчатки и роговицы.
Средства, улучшающие микроциркуляцию, на-
значают в самом начале заболевания: антагони-
сты кальция, антиагреганты, ацетилсалициловая
кислота, реополиглюкин, каптоприл, простаглан-
дины, андекалин, мидокалм, дилминал, солко-
серил, ангиопротекторы, никотиновая кислота.
Местная терапия. Цель локальной терапии
заключается в уменьшении выраженности и
предупреждении прогрессирования фиброзных
изменений кожи, опорно-двигательного аппа-
рата и сосудистых нарушений. Применяются
аппликации диметилсульфоксида (ДМСО), ко-
торый обладает обезболивающим и противовос-
палительным действием, хорошо проникает
через кожу и проводит через нее различные
препараты, а также способен угнетать проли-
ферацию фибробластов и тормозить развитие
соединительной ткани. На пораженные обла-
сти рекомендуется использовать аппликации
50—70 %-ного раствора ДМСО по 30—40 мин,
курс лечения — от 10 до 30 процедур. Мест-
ное физиотерапевтическое лечение проводит-
ся при отсутствии или минимальной активно-
сти процесса: электрофорез с лидазой или
Глава 25. Болезни с диффузным поражением соединительной ткани 309
ронидазой, фонофорез мадекассола, электрофо-
рез с никотиновой кислотой, димексидом, ла-
зеротерапия, КВЧ-терапия; акупунктура; фо-
нопунктура; аппликации парафина, озокерита.
В комплексное лечение больных системной
склеродермией входит применение лечебной
физкультуры и лечебного массажа, которые на-
значаются в период минимальной активности
процесса под контролем общего состояния и сер-
дечно-сосудистой системы.
Прогноз
Заболевание неуклонно прогрессирует, приводя
к склерозу кожи и внутренних органов. Боль-
ные системной склеродермией с поражением
мышц, костей, суставов, а также внутренних
органов, состоянием которых и определяется
прогноз, нуждаются в длительном лечении, не-
редко становятся инвалидами. В случаях тяже-
лого течения системной склеродермии возможен
летальный исход. Десятилетняя выживаемость
превышает 50 %. Основная причина смерти —
почечная недостаточность; реже — поражение
сердца и легких. Иногда наблюдаются спонтан-
ные ремиссии. Прогноз при синдроме CREST
более благоприятный.
Дерматомиозит
Синонимы: болезнь Вагнера, dermatomyositis.
Дерматомиозит — диффузное прогрессирую-
щее воспалительное заболевание соединительной
ткани с поражением кожи в виде периорбиталь-
ного отека и эритемы век с лиловым оттенком,
эритемы лица, шеи, верхней половины тулови-
ща, сопровождающееся поражением поперечно-
полосатой и гладкой мускулатуры с нарушени-
ем двигательной функции, с частым поражением
внутренних органов. При отсутствии поражения
кожи используется термин «полимиозит».
Дерматомиозит рассматривается как неотлож-
ное состояние, так как заболевание порой раз-
вивается очень быстро и характеризуется высо-
кой смертностью.
Этиология и патогенез
Предполагается роль вирусной инфекции и ге-
нетических факторов. Наибольшее внимание
уделяется пикарновирусам (особенно эховиру-
сам), Коксаки-вирусам. Хроническая вирусная
инфекция персистирует в мышцах и вызывает
вторичный иммунный ответ с развитием поли-
миозита. Имеет значение также антигенная ми-
микрия (сходство антигенной структуры виру-
сов и мышц), обусловливающая появление пе-
рекрестных антител (аутоантител) к мышцам с
последующим образованием иммунных комп-
лексов.
Генетические факторы также играют большую
роль в развитии заболевания. Иммуногенетичес-
кими маркерами дерматомиозита являются оп-
ределенные HLA антигены. При дерматомиози-
те у детей и взрослых наиболее часто встречается
сочетание HLA В8 и DR3, а при полимиозите,
ассоциированном с диффузными болезнями со-
единительной ткани, сочетание HLA В14 и В40,
носительство определенных миозит-специфичес-
ких антител.
Предрасполагающими или триггерными (пус-
ковыми) факторами развития болезни являют-
ся: обострения очаговой инфекции, физические
и психические факторы, переохлаждение, пере-
гревание, гиперинсоляция, вакцинация, лекар-
ственная аллергия.
Основным патогенетическим фактором дерма-
томиозита является аутоиммунный механизм,
появление аутоантител, направленных против
цитоплазматических белков и рибонуклеиновых
кислот, входящих в состав мышечной ткани.
Развитию аутоиммунных механизмов способ-
ствуют дисбаланс в соотношении Т- и В-лимфо-
цитов и снижение Т-супрессорной функции.
Клиническая картина
Различают идиопатический (первичный) и па-
ранеопластический (вторичный) дерматомиозит.
Паранеопластический миозит развивается у лю-
дей старше 55 лет на фоне злокачественных но-
вообразований: рака молочной железы, легких,
яичников, желудка, толстой кишки, предстатель-
ной железы, тела матки, а также при гемоблас-
тозах.
Выделяют три периода заболевания: продро-
мальный (от нескольких дней до месяца), мани-
фестный (с кожным, мышечным, общим синд-
ромами), дистрофический или кахектический
(терминальный).
Начало заболевания обычно постепенное.
Больные жалуются на неуклонно прогрессиру-
ющую слабость в проксимальных мышцах рук
и ног. У некоторых больных мышечная слабость
нарастает в течение 5—10 лет. Менее характерно
острое начало, когда в течение 2—4 недель по-
являются резкие боли и слабость в мышцах,
повышение температуры тела, и состояние боль-
ного быстро ухудшается. Иногда первыми пред-
шествующими симптомами болезни оказываются
кожные сыпи, синдром Рейно, полиартралгия.
310 Частная дерматология
Поражение кожи является характерным призна-
ком дерматомиозита и проявляется следующи-
ми симптомами:
• эритематозно-пятнистая сыпь в области
верхних век (периорбитальный отек и эритема
век с лиловым оттенком — «симптом очков»),
скуловых костей, крыльев носа, носогубной
складки (рис. 25.3);
• эритема и папулезная сыпь в области верх-
них отделов груди, спины, локтевых, коленных,
пястно-фаланговых и проксимальных межфа-
ланговых суставов; возможно образование эро-
зий и язв, после которых остаются причудливые
рубцы;
• симптом Готтрона (плоские лиловые папу-
лы и эритематозные шелушащиеся пятна в об-
ласти проксимальных межфаланговых суставов
пальцев рук; атрофия кожи);
• покраснение и шелушение кожи ладоней;
• поражение ногтевых валиков: околоногтевая
эритема, телеангиэктазии, тромбоз капилляров
сосочков дермы и инфаркты кожи; исчерчен-
ность и ломкость ногтей;
• пойкилодерматомиозит — чередование оча-
гов пигментации и депигментации со множе-
ством телеангиэктазий, истончением кожи, ее су-
хостью и гиперкератозом;
• при длительном течении заболевания кожа
становится атрофичной с участками депигмен-
тации.
Поражение мышц является одним из важных
признаков заболевания. Особенно характерна
резко выраженная слабость проксимальных от-
Рис. 25.3. Очаги поражения
при дерматомиозите.
делов мышц верхних и нижних конечностей, а
также мышц шеи. Больному трудно подняться
с постели, умыться, причесаться, одеться, вой-
ти в транспорт. При вовлечении в патологичес-
кий процесс мышц глотки, пищевода, гортани
нарушается речь, появляются приступы кашля,
затруднение при глотании пищи, поперхивание.
Дистальные отделы мышц рук и ног поражают-
ся реже и в меньшей степени. При исследова-
нии мышц отмечаются болезненность, отечность,
при длительном течении болезни возможна мы-
шечная атрофия. При поражении межреберных
мышц и диафрагмы нарушается вентиляционная
функция легких, что сособствует развитию пнев-
моний Мышцы лица и глазные мышцы поража-
ются чрезвычайно редко.
Поражение слизистых оболочек проявляется
конъюнктивитом, стоматитом, гиперемией, оте-
ком в области неба, задней стенки глотки.
Суставной сицдром малохарактерен для дерма-
томиозита, однако он может наблюдаться. По-
ражаются преимущественно мелкие суставы ки-
стей, лучезапястные, реже локтевые, плечевые,
коленные, голеностопные суставы. Наблюдаются
болезненность, отек, гиперемия кожи, ограни-
чение подвижности указанных суставов.
Кальциноз является характерным признаком
ювенильного дерматомиозита, реже наблюдает-
ся у взрослых. Кальцификаты располагаются
подкожно или внутрикожно и внутрифасциально
вблизи пораженных мышц и в пораженных
мышцах, преимущественно в области плечевого
и тазового пояса. Отложения кальция возмож-
ны также в проекции локтевых и коленных су-
ставов, ягодиц. Массивная кальцификация тка-
ней может привести к инвалидизации больных.
Подкожное отложение кальцинатов может выз-
вать изъязвление кожи и выход соединений
кальция в виде крошковатых масс.
Поражение сердца наблюдается часто у боль-
ных дерматомиозитом и характеризуется следу-
ющими проявлениями: миокардит, миокардио-
фиброз: (тахикардия, умеренное расширение
границ сердца, приглушение тонов, чаще в об-
ласти верхушки, аритмия, артериальная гипотен-
зия, на ЭКГ — атриовентрикулярная блокада
различных степеней, изменения зубца Т, смеще-
ние интервала ST книзу от изолинии); перикар-
дит бывает редко.
Поражение легких обусловлено развитием ин-
терстициальной пневмонии, фиброзирующего
альвеолита, гиповентиляцией (поражение межре-
берных мышц, мышц диафрагмы), аспирацией
при нарушениях глотания и проявляется одыш-
кой, обильной, рассеянной по всем легочным
Глава 25. Болезни с диффузным поражением соединительной ткани
311
полям крепитацией, иногда сухими хрипами,
кашлем. При рентгенологическом исследовании
легких выявляется базальный пневмосклероз.
Поражение желудочно-кишечного тракта про-
является снижением аппетита, дисфагией, неред-
ко болями в животе. В основе этих проявлений
лежат нарушения функции мышц глотки, пище-
вода, васкулиты желудочно-кишечного тракта.
Поражение почек встречается очень редко. У
отдельных больных наблюдается гломерулонеф-
рит, характеризующийся протеинурией, очень
редко — нефротическим синдромом.
Поражение нервной системы малохарактерно. В
отдельных случаях отмечается резко выражен-
ный полиневрит; чаще имеют место выраженные
вегетативные дисфункции.
Поражение эндокринной системы проявляется
снижением функции половых желез, надпочеч-
ников при тяжелом течении заболевания.
Течение болезни
Течение дерматомиозита бывает острым, подо-
стрым, хроническим (А. П. Соловьева, 1980).
Острое течение характеризуется генерализован-
ным поражением мускулатуры вплоть до полной
обездвиженности, дисфагией, эритемой, пораже-
нием сердца и других органов с летальным ис-
ходом через 2—6 мес от начала дерматомиозита.
Подострое течение отличается более медлен-
ным, постепенным нарастанием симптомов, цик-
личностью, через 1—2 года от начала дерматоми-
озита проявляется развернутая клиническая
картина.
Хроническое течение является более благопри-
ятным, циклическим вариантом с умеренной
мышечной слабостью, миалгиями, эритематозной
сыпью, иногда без поражения кожи. Возможно
локальное поражение мышц.
Лабораторные и инструментальные
исследования
Общий анализ крови: признаки умеренной ане-
мии, лейкоцитоз с нейтрофильным сдвигом вле-
во, реже — лейкопения, эозинофилия. СОЭ
увеличивается соответственно активности пато-
логического процесса.
Биохимический анализ крови: в активной фазе
болезни повышение активности креатинфосфо-
киназы (у 65 % больных) — специфический
признак поражения мышц; наблюдается повы-
шение активности фруктозодифосфатальдолазы
(у 40 % больных), трансаминаз, особенно АсАТ,
ЛДГ и альдолазы, что отражает остроту и рас-
пространенность поражения мышц. Возможно
повышение уровня мочевой кислоты.
Анализ мочи: усиление экскреции креатина
(более 200 мг в суточной моче).
Иммунологические исследования: снижение
титра комплемента, в небольшом титре — РФ, в
небольшом количестве и незакономерно — LE-
клетки, антитела к ДНК, снижение количества
Т-лимфоцитов и Т-супрессорной функции, по-
вышение содержания IgM и IgG, снижение IgA;
HLA В8, DR3, DR5, Drw52, высокие титры ми-
озитспецифических антител.
Биопсия кожи: истончение эпидермиса, гидро-
пическая дистрофия базального слоя, отек верх-
них слоев дермы, рассеянные воспалительные
инфильтраты, отложение ШИК-позитивного
фибриноида на границе эпидермиса и дермы и
вокруг поверхностно расположенных капилля-
ров, накопление в дерме кислых гликозамино-
гликанов.
Биопсия мышц: проводят биопсию мышц пле-
чевого или тазового пояса, которые проявляют
слабость или болезненны: (дельтовидная, надос-
тная, ягодичные или четырехглавая мышцы бед-
ра). Гистологическое исследование — восковид-
ный сухой некроз мышечных волокон с утратой
поперечной исчерченности, иногда — регенери-
рующие мышечные волокна; воспалительные
инфильтраты из гистиоцитов, макрофагов, лим-
фоцитов, плазматических клеток.
Электромиография: повышенная возбудимость
при введении электродов, спонтанные фибрил-
ляции, псевдомиотонические разряды (залпы
высокочастотных потенциалов действия после
механического раздражения мышц), положи-
тельные потенциалы (как при денервации). При
нарушении нервной проводимости следует запо-
дозрить злокачественное новообразование.
ЭКГ: признаки миокардита, предсердные и
желудочковые экстрасистолы, атриовентрику-
лярная блокада.
Рентгенография грудной клетки: возможен
фиброзирующий альвеолит.
Спирография: рестриктивная дыхательная не-
достаточность.
Диагноз
См. таблицу 25.3.
Лечение
Пациенты должны быть тщательно обследованы,
а при выявлении опухоли проводится оператив-
ное лечение. Удаление опухоли может привести
к регрессу паранеопластических проявлений дер-
матомиозита, многолетним ремиссиям и даже к
выздоровлению. Препаратами выбора, как при
паранеопластическом (без лечения опухоли эф-
11 Зак. 1357
312 Частная дерматология
фект кратковременный), так и при идиопатичес-
ком дерматомиозите, являются глюкокортико-
иды.
Глюкокортикоиды. Рекомендуется преднизолон
в связи с тем, что он оказывает наименьшее
катаболическое действие на мускулатуру по срав-
нению с другими глюкокортикоидами. При ос-
тром дерматомиозите назначают 80—120 мг в
сутки, при подостром — 60 мг в сутки, при обо-
стрении хронического течения — 30—40 мг в
сутки. Препарат следует принимать в течение 2—
3 месяцев и более до получения клинического
эффекта. Затем дозу очень медленно понижают
до поддерживающей: на 1-м году болезни при
исходном остром или подостром течении она
составляет 30—40 мг в сутки, на 2—3-м — 10—
20 мг в сутки. Преднизолон можно отменить
только при стойкой клинической ремиссии.
Отмена препарата должна производиться посте-
пенно (по */4 таблетки преднизолона в 30 дней).
При очень тяжелом течении острого дермато-
миозита целесообразно на фоне продолжающе-
гося приема преднизолона провести пульс-тера-
пию метилпреднизолоном (вводится внутривенно
капельно 1000 мг метилпреднизолона 1 раз в
день в течение 3 дней) с одновременным исполь-
зованием плазмафереза.
Негормональные иммунодепрессанты. Наибо-
лее часто применяются азатиоприн и метотрек-
сат. Азатиоприн назначается в дозе 2,5 мг/кг в
день, метотрексат — 0,75 мг/кг внутривенно 1
раз в неделю. Цитостатики, как правило, добав-
ляются к преднизолону при отсутствии эффекта
и при необходимости снизить его дозу. Пока-
заниями к назначению негормональных имму-
нодепрессантов являются отсутствие эффекта от
лечения преднизолоном, невозможность исполь-
зования преднизолона вследствие непереноси-
мости его и развития осложнений. Цитостати-
ки применяются в течение 2—6 месяцев, затем
дозу постепенно снижают до поддерживающей,
которую принимают до года. Цитостатики по-
давляют иммунопатологические и аутоиммун-
ные реакции и оказывают противовоспалитель-
ное действие. Перед назначением цитостатиков
следует обследовать пациента для исключения
злокачественной опухоли.
Иммуноглобулин в больших дозах (внутривенно
0,4 г на 1 кг веса тела в сутки в течение 5 дней,
ежемесячно) позволяет снизить дозу кортико-
стероидов, необходимую для достижения и под-
держания ремиссии.
Аминохинолины. Рекомендуется делагил (хин-
гамин) по 0,25 г 1 раз в сутки внутрь в течение
не менее 2 лет. Показания к назначению: хро-
ническое течение дерматомиозита без признаков
активности процесса; при снижении дозы пред-
низолона или цитостатиков для уменьшения
риска возможного обострения.
Нестероидные противовоспалительные средства.
Назначаются при доминирующем болевом и су-
ставном синдромах, а также при хроническом
течении дерматомиозита с малой степенью ак-
тивности при наличии болевого синдрома. Ре-
комендуются индометацин-ретард (по 0,075 г) по
1 таблетке (капсуле) 1 раз в день; вольтарен (ор-
тофен) по 0,025 г 2—3 раза в день; пироксикам
по 0,02 г 2 раза в день.
В комплексной терапии дерматомиозита реко-
мендуются препараты для улучшения метаболиз-
ма в пораженных мышцах: ретаболил (по 1 мл
5 %-ного раствора 1 раз в 2 недели внутримы-
Таблица 25.3. Диагностические критерии дерматомиозита,
предложенные Американской Ревматологической Ассоциацией
Основные диагностические критерии: Дополнительные диагностические критерии: кальциноз, дисфагия.
1. Характерное поражение кожи: периорбитальный отек и эритема (симптом «очков»); телеангиэктазии, эритема на открытых участках тела (лицо, шея, верхняя часть груди, конечности). 2. Поражение мышц (преимущественно проксимальных отделов конечностей), выражающееся в мышечной слабости, миалгиях, отеке и позже — атрофии. 3. Характерная патоморфология мышц при биопсии (дегенерация, некроз, базофилия, воспалительные инфильтраты, фиброз). 4. Увеличение активности сывороточных ферментов — креатин- фосфокиназы, альдолазы, трансаминаз на 50 % и более по сравнению с нормой. 5. Характерные данные электромиографического исследования. 1. Диагноз дерматомиозита достоверен при наличиии трех основных критериев и сыпи или при наличии 2 основных, 2 допол- нительных критериев и сыпи. 2. Диагноз дерматомиозита вероятен при наличии первого основного критерия, при наличии двух остальных из основных критериев; при наличии одного основного и двух дополнительных критериев. 3. Диагноз полимиозита достоверен при наличии четырех критериев без сыпи.
Глава 25. Болезни с диффузным поражением соединительной ткани
313
шечно; на курс 3—4 инъекции); рибоксин (в таб-
летках по 0,2 г; внутрь по 2 таблетки 3 раза в
день в течение 2 месяцев); карнитин (внутрь по
1-2 чайные ложки 20 % раствора 2 раза в день
в течение 2 месяцев); милдронат (внутрь по
0,25 г 2—4 раза в день в течение 10—20 дней или
внутривенно 5 мл 10 % раствора 1 раз в день в
течение 10 дней); АТФ (внутримышечно по 1—
2 мл 1 % раствора 1 раз в день в течение 30
дней); фосфаден (внутримышечно по 2 мл 2 %
раствора 2—3 раза в день в течение 3—4 недель,
а затем внутрь по 0,05 г 2—3 раза в день в тече-
ние 2-3 недель).
Кальциноз в мышцах и подкожной клетчат-
ке лечится путем внутривенного капельного
введения двунатриевой соли этилендиаминотет-
рауксусной кислоты. Разовую дозу препарата
(250 мг) растворяют в 400 мл изотонического
раствора натрия хлорида или 5 % раствора глю-
козы. Указанный раствор вводят ежедневно в
течение 5 дней с 5-дневным перерывом (курс
лечения — 15 вливаний) под контролем содер-
жания кальция в крови, так как возможны бы-
строе его снижение и развитие тетании. Курсы
лечения повторяют 3 раза в год.
Лечебная физкультура играет важную роль в
предотвращении деформаций. В острой фазе за-
болевания следует ежедневно выполнять пассив-
ные движения в суставах в полном объеме, а при
необходимости проводить иммобилизацию для
профилактики деформаций, обусловленных уко-
рочением мышц. Позднее следует перейти к ак-
тивным движениям.
Прогноз
Основными причинами смерти больных дерма-
томиозитом являются: опухоль; вторичная ин-
фекция; дыхательная и сердечная недостаточ-
ность; осложнения кортикостероидной терапии;
общая дистрофия, истощение, перфорации и кро-
вотечения из язв желудочно-кишечного тракта.
Глава 26
Многоформная экссудативная эритема
Многоформная экссудативная эритема (erythema
exsudativum multiforme) — острый полиморфный
дерматоз преимущественно инфекционно-аллер-
гической природы (идиопатическая форма эри-
темы), проявляющийся высыпаниями синюшно-
красного цвета на коже конечностей, на
слизистых, иногда в области гениталий, в основ-
ном весной или осенью, склонный к рецидивам.
Инфекционно-аллергической (идиопатической)
эритемой страдает большая часть больных. Ток-
сико-аллергической (симптоматической) форме
экссудативной эритемы сезонность рецидивов не
свойственна.
Этиология и патогенез
В развитии идиопатической формы эритемы
предполагают основными провоцирующими
факторами герпетическую, микоплазменную,
стафилококковую, стрептококковую и др. ин-
фекции. Наличие очагов фокальной инфекции
в челюстно-лицевой области выявляется у 2/3
больных. У этих пациентов повышена чувстви-
тельность к различным бактериальным аллерге-
нам, наблюдается снижение показателей факто-
ров гуморального и Т-клеточного иммунитета,
функции нейтрофилов, увеличение В-лимфоци-
тов в периферической крови. При токсико-ал-
лергической форме эритемы выявляется гипер-
чувствительность к различным медикаментам
(сульфаниламиды, контрацептивы, жаропонижа-
ющие, пирогенные препараты, пенициллины, ал-
лопуринол, прививки против туляремии, холе-
ры). Рецидивы заболевания провоцируют
переохлаждение, гиперинсоляция и другие ме-
теофакторы. У части лиц удается выявить воз-
никновение или обострение эритемы после упот-
ребления некоторых пищевых продуктов
(алиментарные факторы).
Клиническая картина
Излюбленная локализация
Разгибательные поверхности конечностей, час-
то кисти и стопы, включая ладони и подошвы,
область локтевых и коленных суставов, лицо,
наружные половые органы, слизистая оболочка
рта, красная кайма губ, полость носа, конъюн-
ктива, анальный канал (рис. 26.1). Сыпь может
носить ограниченный, диссеминированный и
генерализованный характер.
Характер высыпаний
Вначале появляются участки ограниченной эри-
темы, на фоне которой через несколько дней
образуются уплощенные милиарные и лентику-
лярные папулы синюшно-красного цвета с за-
падением в центре (элементы напоминают ми-
шень или радужку). На их поверхности
формируются пузырьки и пузыри, эрозии, кро-
вянистые корки. При поражении слизистых воз-
никают эрозии с фибринозным налетом. В про-
цесс могут вовлекаться трахея, бронхи, глаза,
мозговые оболочки, почки.
Рис. 26.1. Типичная локализация высыпаний
при экссудативной многоформной эритеме.
314
Глава 26. Многоформная экссудативная эритема 315
Субъективные ошушения
Зуд, реже жжение и болезненность в местах кож-
ных высыпаний. Поражения слизистой рта все-
гда сопровождаются резкой болезненностью.
Обшие явления
Лихорадка, боли в суставах и мышцах, слабость,
недомогание.
Клинические формы
По тяжести проявлений различают:
1. Простую или легкую форму (папулезная,
обычно протекает без поражения слизистых).
2. Везикуло-буллезную форму (среднетяжелая).
3. Буллезную (тяжелая форма), включая син-
дром Стивенса-Джонсона;
4. Крайне тяжелую форму заболевания (синд-
ром Лайелла). В начальной стадии развертыва-
ния синдрома Лайелла типичные мишеневидные
высыпания на коже, характерные для многофор-
мной эритемы, выявляются у половины боль-
ных. Затем быстро появляются распространен-
ная эритема и явления эпидермального
некролиза.
Простая форма
На коже определяются многочисленные типич-
ные папулы диаметром 1—2 см, с западением в
центре, которые регрессируют в течение 1—2
недель. Возможна пузырная реакция на слизис-
той рта с образованием кровянистых, грязнова-
тых корок, появлением отека, трещин на губах,
болезненности.
Везикуло-буллезная форма
Немногочисленные эритематозные бляшки, с
пузырем в центре и кольцом пузырьков по пе-
риферии. Часто в процесс вовлекается слизис-
тая рта и губы.
Рецидивы вышеуказанных форм многоформ-
ной эритемы связывают с реактивацией вируса
герпеса, так как прием ацикловира у ряда боль-
ных приводит к купированию обострений дер-
матоза.
(/Буллезная форма (синлром Стивенса-Джонсона)
Является тяжелой разновидностью многоформной
экссудативной эритемы или токсико-аллергичес-
кой реакцией на прием лекарственных препаратов.
На слизистой рта возникают обширные пузыри и
кровоточащие эрозии, массивные геморрагические
корки. Симптомы хейлита и стоматита затрудня-
ют прием пищи из-за резкой болезненности. Воз-
можно развитие катарального или гнойного конъ-
юнктивита, изъязвлений роговицы, увеита,
панофтальмита, поражений слизистых гениталий
с вовлечением мочевого пузыря и нарушением
мочеиспускания. На коже обнаруживаются мно-
жественные пятнисто-папулезные высыпания,
пузыри с положительным симптомом Никольско-
го, реже пустулы, иногда возникают паронихии.
Характерна длительная лихорадка, возможно раз-
витие пневмонии, нефрита, диареи, полиартрита,
отита. Без лечения летальность составляет 5-15 %.
Данные проявления необходимо дифференциро-
вать с синдромом Лайелла. Считается, что нали-
чие пузырной реакции более чем на 30 % повер-
хности кожного покрова соответствует клинике
синдрома Лайелла.
Крайне тяжелая форма (синлром Лайелла)
Большая часть случаев возникновения синдро-
ма Лайелла связана с приемом пациентами ле-
карственных препаратов, в меньшей мере упот-
реблением некоторых химических соединений,
а также вирусной и микоплазменной инфекци-
ями, вакцинациями. В некоторых случаях (5 %
пациентов) синдром развивается без предшеству-
ющего назначения медикаментозных средств.
Токсико-аллергическая реакция начинается с
поражения лица и конечностей в виде эрите-
мы, пятнисто-папулезной сыпи или типичных
высыпаний многоформной эритемы. В течение
нескольких часов — суток высыпные элемен-
ты сливаются и занимают весь кожный покров.
Далее происходит образование тонкостенных
вялых пузырей, которые быстро сливаются и
выглядят как беловатые участки, напоминаю-
щие папиросную бумагу. Симптом Никольского
резко положительный. При травматизации уча-
стков поражения некротизированный эпидер-
мис легко отслаивается, обнажая красную эро-
зированную, болезненную поверхность
сосочкового слоя дермы, что соответствует по
клинике ожогу II степени. У большинства па-
циентов эрозивным процессом поражаются сли-
зистая щек, губ, конъюнктивы, кожа и слизи-
стая половых органов, перианальная область.
Эпителизация эрозий начинается с периода
образования пузырей и продолжается в течение
всего периода болезни, который длится около
3—4-х недель. В стадии выздоровления наблю-
дается обильное крупнопластинчатое шелуше-
ние и слущивание не отторгшегося некротизи-
рованного эпидермиса крупными пластами. В
период развертывания основных клинических
проявлений синдрома Лайелла характерна вы-
сокая лихорадка, гриппоподобный синдром.
Нередко развиваются почечная недостаточность,
язвенно-некротические изменения трахеи,
бронхов, желудочно-кишечного тракта, различ-
316 Частная дерматология
ная патология со стороны органа зрения, вплоть
до слепоты. Выявление нейтропении в анализе
периферической крови является плохим прогно-
стическим симптомом. Летальность при синд-
роме Лайелла достигает 30 % и зависит от пло-
щади пораженной кожи и возраста пациентов
(чаще погибают лица пожилого возраста). Воз-
можные причины смерти: сепсис, желудочно-
кишечные кровотечения, выраженные наруше-
ния водно-элекролитного баланса.
Кроме вышеуказанных тяжелых форм токси-
ко-аллергических реакций в практике дермато-
лога достаточно часто наблюдаются кожные ре-
акции на прием медикаментов с более легким
клиническим течением: распространенная и
фиксированная формы многоформной экссуда-
тивной эритемы.
При фиксированной форме поражается слизи-
стая рта, гениталий, перианальная область. Ре-
цидивы обычно возникают на местах бывших
высыпаний. Воспалительная реакция в области
пузырей может отсутствовать. Сезонность реци-
дивов не свойственна.
Для распространенной формы характерны об-
щие явления (обычно температурная реакция) и
возникновение очагов поражения на других уча-
стках кожного покрова. С целью предотвраще-
ния рецидивов заболевания необходимо выявить
аллерген и в последующем выработать рекомен-
дации по устранению контакта пациента с дан-
ным провоцирующим фактором.
Гистопатология
Вакуольная дегенерация в нижних слоях эпидер-
миса, некроз эпидермальных клеток, надбазаль-
ные и субэпидермальные пузыри без акантоли-
за. Лимфогистиоцитарная инфильтрация вокруг
сосудов и в области базальной мембраны, экст-
равазация эритроцитов.
Диагноз
Острое начало, признаки интоксикации, поли-
морфизм и симметричность высыпаний синюш-
но-красного (темно-розового) цвета. Длитель-
ность рецидива обычно составляет 3—4 недели.
Сезонность рецидивов наблюдается при инфек-
ционно-аллергической форме заболевания. При
легком течении дерматоза с наличием везикул и
пузырей симптом Никольского отрицательный.
Дифференциальный диагноз
Пузырчатка вульгарная (отсутствие эритемы,
папул, положительный симптом Никольского,
наличие акантолитических клеток в мазках-от-
печатках с эрозий).
Дерматоз Дюринга (герпетиформность распо-
ложения элементов, чувствительность к йоду,
эффект от применения у пациентов ДДС).
Хроническая мигрирующая эритема Афцелиуса—
Липшютца (кожные проявления Лайм-боррели-
оза). На месте присасывания клеща появляется
синюшно-красное пятно с геморрагической ко-
рочкой в центре, которое, разрастаясь, превра-
щается в кольцо площадью с детскую ладонь и
больше (1 стадия). Через несколько недель в
результате диссеминации спирохет появляется
сердечная и неврологическая симптоматика (2
стадия). Через год и более от момента укуса
клеща (3 стадия) развиваются осложнения: по-
ражение кожи в виде хронического атрофичес-
кого акродерматита, тяжелые изменения со сто-
роны суставов, сердца, ЦНС, периферической
нервной системы.
При установлении диагноза необходимо ис-
ключить также лекарственные токсидермии,
инфекционные экзантемы (скарлатина, корь,
краснуха), дерматомиозит, термические ожоги,
фототоксические реакции, синдром ошпаренной
кожи.
Лечение
Легкая форма
Салицилаты, антигистаминные препараты (таве-
гил, фенкарол, пипольфен, супрастин, эролин,
кларитин, димедрол). Курс в/м инъекций вита-
минов Вр В6, В12, витамины С, Р, препараты
кальция (глюконат, пангамат).
Среднетяжелые формы
Преднизолон 30—60 мг/ сут 5—7 дней с после-
дующим снижением в течение 2—4 недель. Этак-
ридина лактат 0,05 г 3 раза в день в течение 10-
20 дней. Декарис по 150 мг 2 дня с 5-дневными
перерывами в течение 2 месяцев. При необхо-
димости применяют антибиотики с учетом их пе-
реносимости и установления связи возникнове-
ния заболевания с очагами бактериальной
инфекции. Препараты группы ацикловира на-
значают в случаях обострений заболевания, свя-
занных с герпетической инфекцией.
Синдром Стивенса—Джонсона
Начальная доза преднизолона 60—80 мг. Гемодез
(полиглюкин) по 200—400 мл в/в капельно № 3—
5, 30 % раствор тиосульфата натрия 10,0 мл в/в
ежедневно № 6—8, антигистаминные препараты.
Синдром Лайелла
Лечение больных проводят в ожоговом отделе-
нии и (или) отделении реанимации. На началь-
Глава 26. Многоформная экссудативная эритема 317
них стадиях заболевания назначаются большие
дозы кортикостероидов внутривенно капельно;
проводится активная инфузионная терапия с
целью коррекции потерь воды и электролитных
нарушений. Необходимы мероприятия по про-
филактике аспирационной пневмонии, сепсиса
и других инфекционных осложнений. Некроти-
зированные участки кожи удаляют.
Местное лечение
На очаги поражения — фукорцин, 5 % дерма-
толовая мазь, комбинированные кортикостеро-
иды (кремы, аэрозоли). Полоскание рта калия
перманганатом 1: 8000, 2 % раствором борной
кислоты, соды, танина, отваром ромашки, раз-
веденной настойкой календулы, прополиса.
Смазывание эрозий масляным раствором вита-
мина А, дентальной пастой «Солко» с последу-
ющим воздействием гелий-неоновым лазером.
Смазывание пораженных губ препаратами, со-
держащими антибиотики, комбинированные
кортикостероиды.
Профилактика рецидивов
В случаях инфекционно-аллергической приро-
ды заболевания проводится санация очагов ин-
фекции, лечение сопутствующих заболеваний.
Назначаются средства, повышающие неспеци-
фическую резистентность организма: иммуно-
глобулин, витамины, биостимуляторы, аутогемо-
терапия, стафилококковый анатоксин, общее
УФО, закаливание организма. При токсико-ал-
лергической форме многоформной эритемы не-
обходимо выявление аллергена или группы ле-
карственных средств, которые пациент не
переносит (консультация аллерголога). Обяза-
тельное разъяснение больному опасности повтор-
ного применения медикамента. Данные об этих
медикаментах должны быть указаны на опозна-
вательном браслете пациента.
Глава 27
Клещевой боррелиоз (лайм-боррелиоз)
Трансмиссивное инфекционное заболевание,
вызываемое спирохетами рода Borrelia, передаю-
щееся иксодовыми клещами и проявляющееся
хронической мигрирующей эритемой, а в даль-
нейшем поражением нервной системы, опорно-
двигательного аппарата, сердца. Встречается в
зонах ландшафта умеренного климатического
пояса, пик заболеваемости приходится на летние
месяцы. Широко распространен на территории
Европы. Спорадические случаи зарегистрирова-
ны в Азии, Австралии, Северной Америке.
Возбудитель
Спирохеты рода Borrelia. В настоящее время ус-
тановлены несколько видов: Borrelia burgdorferi
(чаще); Borrelia garinii; Borrelia afrelii.
Этиология
Источник инфекции
Дикие позвоночные, крупный рогатый скот.
Пути передачи
Заражение человека происходит при укусе инфи-
цированных боррелиями иксодовых клещей (их
слюна, содержимое кишечника, при удалении кле-
щей с кожи). Не исключается заражение от других
кровососущих, например, слепней, комаров.
Распространение в организме
После попадания в кожу со слюной клеща бор-
релии в течение нескольких дней активно раз-
множаются. В месте внедрения спирохет возни-
кает хроническая мигрирующая эритема
(подобно первичному аффекту при сифилисе).
Далее происходит их гематогенная и лимфоген-
ная диссеминация на другие участки кожи (до-
черние эритемы), внутренние органы (печень, се-
лезенка, почки, сердце, головной мозг, суставы).
Генерализация инфекции сопровождается лихо-
радкой, интоксикацией, органными нарушени-
ями.
Патогенез
На внедрение в организм боррелий развивается
гуморальный и клеточный гипериммунный ответ,
что сопровождается выработкой антител IgM, за-
тем появляются антитела класса IgG, ЦИК, ко-
торые в органах и тканях вызывают воспали-
тельный процесс, лимфоцитарные инфильтраты
(кожа, подкожная клетчатка, лимфоузлы, селе-
зенка, периферические нервные ганглии). Повы-
шаются уровни Тх и Тс; ткани-мишени инфиль-
трированы лимфоцитами и плазматическими
клетками, наблюдаются явления васкулитов,
тромбоваскулитов. Липосахариды боррелий сти-
мулируют выработку мононуклеарами интерлей-
кина-1, активирующего образование простаглан-
динов, коллагеназы синовиальными тканями, что
способствует развитию воспаления суставов. Для
хронического течения болезни характерно раз-
витие аутоиммунных процессов, возможна внут-
риклеточная персистенция возбудителя.
Клиническая картина
Инкубационный период
Инкубационный период — в среднем 1—2 недели
(колебания 24 часа — 180 сут).
Клиническая классификация (стадии):
1. Острая (I период)
2. Промежуточная (II период)
3. Хроническая (III период)
Для боррелиоза характерна стадийность, что
позволяет сравнивать заболевание по течению с
сифилитической инфекцией. В периоде началь-
ных проявлений, который продолжается 2—7
дней, отмечаются головная боль, озноб, тошно-
та, мигрирующие боли в суставах и мышцах,
артралгии, слабость, утомляемость, сонливость,
лихорадка (от субфебрилитета до 40 °C). В мес-
те укуса клеща — зуд, иногда боль, жжение,
эритема, инфильтрация кожи. Пятно или папу-
ла увеличиваются по периферии до 10 см в ди-
аметре и более. Выявляется очаг эритемы округ-
лой или неправильной формы, по периферии
более интенсивной окраски, возвышается над
уровнем здоровой кожи. Центральная часть эри-
темы бледнеет, становится синюшной. Очаг при-
обретает характер кольца (кольцевидная эрите-
ма), которое эксцентрически увеличивается по
318
Глава 27. Клещевой боррелиоз (лайм-боррелиоз)
319
периферии (мигрирующая эритема). На месте
укуса клеща образуется корочка, затем рубец (в
зависимости от локализации очага), выявляется
болезненный регионарный лимфаденит. Возмож-
но образование дочерних эритематозно-кольце-
видных очагов на других участках тела. Субъек-
тивные ощущения в очагах могут отсутствовать.
Обычно эритема исчезает через 2—3 недели. При
этом кольцевидная периферическая зона разра-
стается, фрагментируется, бледнеет и постепен-
но исчезает, т. е. наступает аутосанация. Иног-
да эритема сохраняется в течение нескольких
месяцев (до 18 недель). В очаге может наблю-
даться шелушение, пигментация, возможны па-
рестезии. У некоторых больных период началь-
ных проявлений может протекать без
интоксикации и лихорадки, только с наличием
первичного аффекта на месте укуса клеща, что
соответствует описанию хронической мигриру-
ющей эритемы Афцелиуса — Липшютца, кото-
рая встречается у 80 % больных. Возможно также
развитие доброкачественной лимфоплазии кожи
(саркоид Шинглера-Фендта), которая чаще ло-
кализуется на мочке уха, голове, области мошон-
ки и представляет собой одиночный, безболез-
ненный, красноватый узел (размером до 5 см).
Вторичный период связан с диссеминацией бор-
релий из кожного очага на 4—6 неделе болезни
и поражением нервной системы (менингит, ме-
нингоцефалит, неврит лицевых нервов, радику-
лоневрит), сердца токсико-дистрофического ха-
рактера (изредка возможно развитие
миоперикардита, панкардита). В этом периоде
могут выявляться поражения крупных суставов
по типу реактивного артрита. Без лечебных ме-
роприятий болезнь принимает рецидивирующее
или хроническое течение, что соответствует III
периоду заболевания. Этот период может возни-
кать после длительного латентного периода с раз-
витием поздних осложнений.
Поздние осложнения
Энцефаломиелит, рассеянный склероз, психичес-
кие нарушения, энцефалопатии, нарушения зре-
ния, слуха, глотания, полирадикулоневриты,
полиневропатия, атрофия мышц и другие рас-
стройства; хроническое воспаление 1—2 крупных
суставов с признаками остеопороза, дегенерации
суставных тканей.
Возможно как изолированное поражение
кожи, так и в сочетании с нарушениями орга-
нов и систем. Часто встречается лимфаденоз
кожи Беферштедта: синюшно-красные узелки,
бляшки в области лица, мочек ушей, сосков
молочных желез и регионарная лимфаденопатия.
Эти проявления напоминают очаги красной вол-
чанки, саркоидоза, микоза гладкой кожи, кото-
рые спонтанно исчезают без атрофии. Пораже-
ния кожи могут протекать по типу склеродермии
и хронического атрофического акродерматита
(ХААД). Проявления склеродермии могут быть
в виде бляшечной формы, атрофических полос,
болезни белых пятен, атрофодермии Пазини-
Пьерини, эозинофильного фасциита Шульмана,
глубокой склеродермии. Для ХААД характерна
диффузная цианотично-красного цвета эритема,
отек, инфильтрация, атрофия кожи в виде па-
пиросной бумаги.
Дифференциальный диагноз
Дифференциальный диагноз кожной формы
лайм-боррелиоза проводят с эритематозной фор-
мой рожи (ярко-красного цвета «языки пламе-
ни», болезненность лимфаденит, лимфангит),
аллергическим дерматитом на укусы других на-
секомых (быстро проходящий), центробежной
эритемой Дарье (небольшие множественные оча-
ги, уртикарный характер элементов и их цент-
робежное распространение, появление новых
очагов на месте разрешившихся), а также Т и В-
клеточными лимфомами, ревматоидными узел-
ками, дерматофитиями, узловатой эритемой.
Лабораторные данные
Применяют реакцию непрямой иммунофлюо-
ресценции (нРИФ) с корпускулярным антиге-
ном 1р 21. При наличии в исследуемой сыворотке
специфических антител выявляется яркое жел-
то-зеленое свечение по ходу спирохеты. Устанав-
ливается титр IgM-антител, которые появляют-
ся не ранее 2 недель после первичного
заражения; далее следят за изменением их тит-
ра; возможны серонегативные случаи лайм-бор-
релиоза. Максимальный уровень IgM-антител
выявляется в хронической стадии болезни.
Лучшим способом подтверждения диагноза
является ПЦР, позволяющая идентифицировать
возбудителя в биопсийном материале, перифе-
рической крови, синовиальной жидкости.
Культуру Borrelia burgdorferi можно получить
при посеве биопсийного материала на среду
Келли.
Лечение
Этиотропная терапия антибиотиками проводится
при наличии эритемы в месте присасывания
клеща, лихорадки и симптомов интоксикации
при отсутствии поражения нервной системы,
сердца, суставов: тетрациклин 0,5 г 4 раза/сут
или вибрамицин (доксициклин) 0,2 г 2 раза/сут
320 Частная дерматология
в течение 10 дней. Азитромицин (сумамед) — на
курс 5,0 г. Цефуроксим 500 мг/сут в течение
7 сут. Амоксициллин 500 мг 3 раза в сутки — 10
дней.
Детям до 8 лет: амоксициллин 30—40 мг/сут
в 3 приема или парентерально 50—100 мг/кг/сут
в 4 введения.
При непереносимости тетрациклинов назнача-
ют левомицитин 0,5 г 4 раза/сут — 10 дней. При
выявлении патологии со стороны нервной сис-
темы, сердца, суставов назначают бензилпени-
циллин 500 тыс ЕД в/м 8 раз/сут в течение 14
дней. При клинических признаках менингита
(менингоэнцифалита) суточная доза бензилпени-
циллина увеличивается до 20 млн ЕД, снижает-
ся при нормализации ликвора. Возможно при-
менение пефлоксацина по 0,4 г 2 раза/сут внутрь
или в/в на 5 % растворе глюкозы. Цефтриаксон
назначают по 1,0 г 2 раза/сут в/в в течение 14
дней. При хроническом течении лайм-боррели-
оза курс лечения указанными препаратами про-
должают до 28 дней.
Применяют также экстенциллин 2,4 млн ЕД
в/м один раз в 7 дней, 3 введения. У беремен-
ных: острая стадия — амоксициллин 500 мг 3
раза в сутки, внутрь — 21 сут; при наличии
признаков гематогенной диссеминации — бен-
зилпенициллин до 20 млн ЕД/сут в/в — 14—
28 сут. Одновременно с антибиотикотерапией
при тяжелых формах заболевания проводят де-
токсикационные мероприятия, вводят дегидра-
тационные средства (менингит). Назначают фи-
зиотерапию (невриты, артриты, артралгии), по
показаниям сердечно-сосудистые средства, им-
мунные стимуляторы, при рецидивирующем
артрите — делагил, НПВС.
Прогноз
Без этиотропной терапии болезнь прогрессиру-
ет, хронизируется, приводит к трудопотерям и
инвалидности. Реконвалесценты наблюдаются
специалистами в зависимости от клиники оста-
точных явлений.
Профилактика
Истребление клещей, специальные противокле-
щевые костюмы, одежда, импрегнированная
препаратом «Пермет». Рекомендуется примене-
ние репеллентов. Превентивное назначение ан-
тибиотиков после укусов клещей: тетрациклин
0,5 г 4 раза/сут — 5 дней или экстенциллин 2,4
млн ЕД в/м однократно.
Глава 28
Пузырные дерматозы
Вульгарная пузырчатка 321
Вегетирующая пузырчатка 325
Листовидная пузырчатка 326
Бразильская пузырчатка 327
Эритематозная пузырчатка 328
Паранеопластическая пузырчатка 329
Пузырчатка хроническая врожденная семейная 330
Герпетиформный дерматоз Дюринга 331
Буллезный пемфигоид 336
Рубцующийся пемфигоид 338
Приобретенные и не заживающие спонтанно
заболевания из группы пузырных дерматозов
первично характеризуются акантолитическим,
ведущим к образованию пузырей нарушением
целостности структуры эпидермиса, которое
происходит вследствие расторжения уже имею-
щихся и подавления образования новых десмо-
сомальных межклеточных соединений. Аканто-
литические клетки эпидермиса могут при таких
заболеваниях цитологически обнаруживаться в
основании пузыря (Тцанк-тест) и иметь диагно-
стическую значимость. Ответственными за акан-
толиз являются аутоантитела, которые направ-
лены против адгезионных протеинов на
поверхности клеток эпидермиса и тем самым
препятствуют способности этих клеток к вступ-
лению в контакты на их пути к поверхности
кожи. Такие антиэпителиальные антитела (пем-
фигусные антитела) при пузырных дерматозах
могут обнаруживаться методом прямой иммуно-
флуоресценции в межклеточном пространстве
эпидермиса (пузыри и краевая зона пузырей), а
также многократно подтверждаться методом не-
прямой иммунофлуоресценции в сыворотке па-
циента.
К группе пузырных дерматозов относятся сле-
дующие заболевания: вульгарная пузырчатка,
вегетирующая пузырчатка; листовидная пузыр-
чатка, бразильская пузырчатка, эритематозная
пузырчатка; паранеопластическая пузырчатка;
хроническая пузырчатка доброкачественная се-
мейная, болезнь Гровера.
Вульгарная
пузырчатка
Вульгарная пузырчатка — чаще всего хроничес-
кое заболевание, при котором на имеющей нор-
мальный вид коже и видимых слизистых обо-
лочках высыпают пузыри. Если ее не лечить,
заболевание может иметь смертельный исход.
Пемфигусные антитела, которые направлены
против адгезионных протеинов, обнаруживаются
в сыворотке пациентов и в пораженных участ-
ках кожи.
Заболевание нераспространенное и не связа-
но с полом пациента. Преимущественно пора-
жаются люди в возрасте от 30 до 60 лет, но за-
болевание может возникнуть также и в
детском, и в старческом возрасте. У пациентов
еврейской национальности вульгарная пузыр-
чатка регистрируется чаще, чем у других эт-
нических групп, имеется тесная ассоциация с
серологическим HLA-признаком DR4 или с
DR4/DRw6.
321
322 Частная дерматология
Этиопатогенез
Этиология неизвестна. Заболевание может воз-
никать спонтанно или в связи с УФ и рентге-
новским облучением или же ожогом. Реже воз-
никает в комбинации с другими аутоиммунными
заболеваниями, такими как миастения гравис,
тимома или пернициозная анемия. При предрас-
положенности к аутоиммунной реакции вульгар-
ная пузырчатка может быть вызвана приемом
медикаментов. К ним относятся лекарственные
вещества, содержащие сульфгидрильные группы,
такие как D-пеницилламин, каптоприл и пири-
тинол, а также туберкулостатики, пропранолол,
фенилбутазон, ибупрофен и индометацин.
Аутоиммунная реакция при вульгарной пу-
зырчатке направлена против эпидермального
гликопротеина десмоглеин-3, который принад-
лежит к семейству кадгеринов. Эти связываю-
щие кальций трансмембранные адгезионные
молекулы встречаются в пределах десмосом и на
поверхности кератиноцитов. Антиген вульгарной
пузырчатки десмоглеин-3 имеет молекулярный
вес 130 kD и связан с плакоглобином, еще од-
ним десмосомальным адгезионным протеином с
молекулярным весом 85 kD.
Фиксация пемфигусных антител в сыворотке
на экстрацеллюлярных адгезионных доменах
десмоглеина-3 приводит к нарушению контак-
та между соседними клетками эпидермиса и об-
разованию акантолитической щели. Обсуждается
участие в акантолизе комплементарных факто-
ров, активатора плазминогена или медиаторов
воспаления.
Различные наблюдения показали, что пемфи-
гусные антитела патогенетически активны. Это
подтверждается следующими фактами:
• у большинства пациентов интенсивность за-
болевания, даже с задержкой по времени, кор-
релирует с повышением титра пемфигусных ан-
тител в сыворотке;
• новорожденные от матерей с вульгарной пу-
зырчаткой могут иметь типичные для пузырчат-
ки изменения на коже, которые проходят толь-
ко тогда, когда распадутся переданные через
плаценту материнские IgG-антитела;
• пемфигусные антитела класса IgG в клеточ-
ной культуре эпидермиса могут приводить к на-
рушению внутриклеточной связи с образованием
акантолитической щели без комплементарной си-
стемы или клеток, осуществляющих воспаление;
• в эксперименте на животных после пассив-
ной передачи IgG-пемфигусных антител у ново-
рожденных мышей наблюдались типичные для
вульгарной пузырчатки гистологические изме-
нения.
Клиническая картина
Заболевание начинается незаметно и чаще без
явной причины. В каком-либо участке кожи,
часто в области пупка, возникают прозрачные
дряблые пузыри с серозным содержимым, кото-
рые быстро вскрываются с возникновением по-
краснения и эрозий. Они могут покрываться
корочкой, в то время как край пузыря смеща-
ется далее. В пределах одного очага могут воз-
никнуть новые пузыри. Содержимое пузырей не
имеет геморрагического характера. Примерно в
50 % случаев заболевание начинается в полости
рта. Здесь тонкие пузыри быстро разрываются
вследствие мацерации, поэтому в клинической
картине преобладают болезненные эрозии. Мо-
гут поражаться и глаза (хронический конъюнк-
тивит, хронический блефарит без рубцевания).
В течение заболевания могут поражаться ин-
тертригинозные участки кожи (подмышечные
впадины, субмаммарные участки, паховая об-
ласть), а затем и вся кожа. Участков предпоч-
тительной локализации не существует. Повсеме-
стно на клинически нормальной коже
развиваются различной величины дряблые пу-
зыри с прозрачно-серозным или беловато-мут-
ным содержимым. Пузыри могут быть окруже-
ны воспаленным ободком. Тонкая покрышка
пузырей быстро вскрывается, оставляя эрозиро-
ванные поверхности. В этой фазе заболевание ха-
рактеризуется обширными эрозиями, занимаю-
щими большие участки поверхности кожи. Если
они вторично не инфицируются, эрозии могут
заживать, не оставляя следов, в то время как в
старых областях или на новых участках кожи раз-
виваются новые пузыри. Образование новых
пузырей преобладает, так что все большие и
большие поверхности кожи поражаются эрози-
ями и покрываются корками.
Если на видимо здоровую кожу вблизи пузыря
плотно нажать, смещая ее в сторону, то можно
сместить верхние слои эпидермиса (феномен
Никольского I). Этот признак позволяет уста-
новить, что вследствие акантолиза уменьшилось
сцепление эпидермальных клеток. Если нажи-
мать на пузырь, то можно увидеть, как его со-
держимое смещается дальше в сторону (феномен
Никольского II). К моменту ремиссии оба фе-
номена отсутствуют.
У большинства больных зуд отсутствует. Эро-
зии болезненные, могут трескаться, кровоточить
и покрываться коркой, а также подвергаться
вторичным инфекциям. Особенно болезненны
плохо заживающие эрозии во рту, что резко за-
трудняет прием пищи. Места предпочтительной
Глава 28. Пузырные дерматозы 323
локализации отсутствуют, поскольку речь идет
о заболевании, при котором любой участок кожи
склонен к образованию пузырей. Как показы-
вает феномен Никольского, механические фак-
торы, несомненно, определяют локализацию
пузырей. Именно поэтому кажущимися места-
ми преимущественной локализации являются
паховая область, область ягодиц, спины и учас-
ток под грудными железами.
Предсказать течение заболевания невозможно.
В течение нескольких лет на одних и тех же
участках кожи или слизистых оболочках могут
возникать отдельные пузыри. Приступообразное
высыпание пузырей на коже и слизистых может
происходить внезапно и через несколько недель
снова регрессировать. При отсутствии лечения
через 1—3 года заболевание может привести к
смерти. Так как во многих случаях поражается
слизистая полости рта, процесс приема пищи
затруднен и вызывает сильные физические стра-
дания пациентов. Вовлечение в процесс слизис-
тых оболочек носа, конъюнктивы и анального
отверствия также вызывает болезненные ощуще-
ния. В мазках из старых пузырей и эрозий мно-
гократно увеличено количество бактерий, так что
в любое время возможно возникновение вторич-
ной инфекции.
Если вульгарная пузырчатка длится долго или
протекает тяжело, то, как правило, общее состо-
яние пациентов заметно нарушено из-за плохо-
го самочувствия, потери аппетита и похудения.
Температура тела может быть повышенной, для
конечной фазы характерно резкое повышение
температуры, как при сепсисе. Смерть при ост-
ром течении наступает через несколько месяцев
или же, при перемежающемся течении, через
несколько лет вследствие вторичных осложне-
ний (сепсис, бронхопневмония, кахексия). Час-
то создается впечатление, что причиной смерти
пациента являются не кожные проявления. Кро-
ме того, аутопсия показывает только неспеци-
фические изменения, которые наводят на мысль
о том, что смерть наступила скорее из-за изме-
нений, обусловленных обменом веществ, чем
вследствие органического заболевания.
Прогноз следует делать с осторожностью. Если
пациенты в целях лечения длительное время
принимали высокие дозы глюкокортикоидов и
цитостатиков, следует ожидать проявления их
побочных действий.
Лабораторные данные
Для диагностики пузырчатки эти показатели не
имеют существенного значения. В случаях про-
грессирующего заболевания повышена СОЭ,
имеется вторичная гипохромная анемия, лейко-
цитоз с умеренным смещением влево, иногда
эозинофилия. Вторичны гипо- и диспротеине-
мия (уменьшение содержания альбумина при
увеличении альфа-, бета- и особенно гамма-гло-
булинов), а также изменения электролитов
(уменьшение в сыворотке натрия, хлорида и
кальция).
Цитологическое исследование
мазка из основания пузыря.
Описанная Тцанком в 1947 году акантолитичес-
кая пемфигусная клетка, взятая из основания
свежего пузыря, типична и даже специфична для
вульгарной пузырчатки. После осторожного
вскрытия свежего пузыря из его основания пла-
тиновой петлей или скальпелем берется тканевый
материал, помещается на предметное стекло, как
при анализе крови, и окрашивается (окраска по
Май-Грюнвальду-Гимза). При этом кроме лей-
коцитов или бактерий, свободно расположенных
комплексов эпидермальных клеток, наблюдаются
акантолитические эпидермальные клетки с малым
количеством или же с отсутствием межклеточных
связей (клетки Тцанка). Вокруг базофильного
безструктурного ядра находится зона перинукле-
арного просветления, которая контрастирует с
утолщением базофильной РНК /—/ содержащей
цитоплазмы. Тест Тцанка ценен для первой по-
становки диагноза, но он не может заменить со-
бой гистологического исследования.
Гистопатология
Типичен незначительный межклеточный отек в
нижних слоях эпидермиса с супрабазальным
акантолитическим нарушением целостности и
пузырями. В основе этого лежит ослабление су-
ществующих в межклеточном пространстве в
форме десмосом контактов межу отростками
соседствующих эпидермальных клеток, которое
приводит к внутриэпидермальной трещине и
пузырям. Электронномикроскопические иссле-
дования показали, что в находящейся рядом с
пузырями и на первый взгляд клинически нор-
мальной коже образование десмосомальных кон-
тактов между эпидермальными клетками также
патологически ослаблено. В более старых пузы-
рях часто имеется много нейтрофильных или
эозинофильных лейкоцитов, которые могут
фагоцитировать некробиотические клетки эпи-
дермиса. Изменения в верхнем роговом слое не
характерны и демонстрируют картину незначи-
тельного воспаления, часто с участием эозино-
фильных лейкоцитов. В более поздних прояв-
лениях в клинической картине доминируют
неспецифические воспалительные процессы.
324 Частная дерматология
Иммунологические исследования
Посредством метода прямой иммунофлуоресцен-
ции между эпидермальными клетками в преде-
лах пузыря и его краевой зоны обнаруживаются
сетчатообразные отложения иммуноглобулинов
(IgG, IgA) и продуктов расщепления комплемента
(Clq, С4, СЗ), которые выявляются во всех сло-
ях эпидермиса независимо от уровня образования
трещины. Антитела принадлежат преимуществен-
но к подклассу IgG4, а в области слизистой —
также к классу IgA.
При поражении исключительно слизистых
оболочек прямая иммунофлуоресценция непора-
женной кожи, как и пемфигус-тест, например,
из ягодичной области, уже положительны, что
дает возможность ранней постановки диагноза.
Методом непрямой иммунофлуоресценции в
острой стадии заболевания почти у всех паци-
ентов доказывается наличие в сыворотке ауто-
антител. Подходящим субстратом антигена для
непрямой иммунофлуоресценции являются кри-
остатические срезы пищевода обезьяны; здесь
пемфигусные антитела образуют характерный
сетчатообразный межклеточный образец связы-
вания. Определение пемфигусных антител час-
то коррелирует с активностью заболевания и
может быть использовано для контроля терапии
и течения заболевания. Аутоантитела при лекар-
ственно индуцированной вульгарной пузырчат-
ке при непрямой иммунофлуоресценции демон-
стрируют ту же реакцию, что и спонтанная
форма заболевания.
Примерно в 1 % случаев при непрямой имму-
нофлуоресценции доказательство пемфигусных
антител неспецифично и не ассоциируется с
вульгарной пузырчаткой. Пемфигусподобные
антитела описаны при обширных ожогах, при
синдроме Лайелла, при миастении беременных
и других заболеваниях. Они могут быть также
обусловлены перекрестной активностью антиге-
нов групп крови А и В.
Дифференциальный диагноз
Особенно важно провести дифференциальный
диагноз с буллезным пемфигоидом, герпетифор-
мным дерматозом Дюринга, буллезной токсичес-
кой экзантемой, буллезной формой многофор-
мной экссудативной эритемы и поздней кожной
порфирией. Решающую роль играют анамнез,
гистологические исследования первичного пузы-
ря, иммунологические исследования и наблюде-
ние за течением заболевания. Изменения слизи-
стой рта следует отграничить от афтозных
заболеваний, эрозивной красной волчанки и эро-
зивного красного плоского лишая.
Лечение
Со времени введения глюкокортикоидов и им-
муносупрессивных препаратов летальность от
заболевания значительно снизилась. Поскольку
речь идет о жизненно важном показании, при-
ходится мириться с известными побочными дей-
ствиями этих терапевтических средств. Оправ-
дала себя тактика лечения пузырчатки, при
которой начинают с назначения высоких доз
глюкокортикоидов, чтобы скорее достигнуть ре-
миссии. Начальные дозы при распространенной
пузырчатке составляют от 200 до 250 мг пред-
низолона (декортина) или изодозы других глю-
кокортикоидов, таких как преднизолон, метил-
преднизолон (урбазон) или флюокортолон
(ультралан), при средне-тяжелом течении пузыр-
чатки — 120—200 мг, а при легких формах 80-
120 мг ежедевно до полной ремиссии. Так про-
должают 4—6 недель, после этого высокие
ежедневные дозы относительно быстро уменьша-
ют и подбирают поддерживающие дозы, доста-
точные для поддержания ремиссии. Это часто
удается при 5,0—15,0 мг эквивалента преднизо-
лона в день. При недоборе этих доз снова мо-
жет начаться рецидив. Если поддерживающую
дозу пациент выдерживает неопределенно дол-
гое время без побочных воздействий, то состо-
яние его чаще всего улучшается. К сожалению,
это бывает не всегда. В противных случаях при-
ходится иметь дело с побочными явлениями, та-
кими как сахарный диабет, катаракта, язва же-
лудка, гипертония, тромбоэмболические явления,
остеопороз, активизация органного туберкулеза,
кандидозные инфекции и другие заболевания.
Рекомендуется также комбинация с иммуно-
супрессивными препаратами: азатиоприн (иму-
рек), если он прописывается в начальной фазе
заболевания в ежедневных дозах 100—200 мг в
комбинации с глюкокортикоидами, оказывает
морбостатический эффект. Он может экономить
глюкокортикоиды и редуцировать их побочные
действия и снижает летальность, что показали
долгосрочные исследования. Действие азатиоп-
рина начинается только после латентного пери-
ода в несколько недель. После отмены кортико-
стероидов в случае успешной ремиссии
азатиоприн можно продолжать принимать от-
дельно для профилактики рецидивов. Циклофос-
фамид (эндоксан 50—100—150—200 мг/день),
метотрексат (25—30 г внутривенно или перораль-
но раз в неделю), а также циклоспорин А (сан-
диммун 2,5—5,0 мг/кг веса в день) могут помочь
экономить стероиды, прежде всего в тех случа-
ях, когда заболевание при приеме высоких доз
стероидов перешло в стадию первой ремиссии.
Глава 28. Пузырные дерматозы 325
При самой тяжелой форме течения с успехом
применяется так называемая пульс-терапия:
кратковременное внутривенное введение высо-
ких доз глюкокортикоидов (100—150 мг декса-
метазона внутривенно в течение 3 дней в месяц)
и циклофосфамида (500—1500 мг внутривенно
каждые 2—4 недели, позднее через более длитель-
ные интервалы) общей длительностью 6 месяцев
в комбинации с малыми дозами циклофосфами-
да внутрь с интервалом. При приеме всех имму-
носупрессивных препаратов необходимо следить
за побочными действиями, к которым относят
токсическое действие на костный мозг, печень
и почки, стерильность и тератогенность, подвер-
женность инфекциям; в комбинации с глюко-
кортикоидами и при длительном применении —
опасность развития малигномы. Поэтому реко-
мендуется совместное консультативное ведение
пациента вместе с терапевтом. Комбинационное
лечение глюкокортикоидами и назначаемыми
внутримышечно или внутрь препаратами золота
(ауреотан, аурео-детоксин) примерно у одной
трети пациентов отягощается многочисленны-
ми кожными и системными побочными дей-
ствиями.
В качестве дополнительного мероприятия про-
водится плазмаферез с заменой большого объе-
ма плазмы в целях снижения уровня аутоанти-
тел в сыворотке. Чтобы избежать феномена
отдачи, плазмаферез должен комбинироваться с
приемом высоких доз глюкокортикоидов и им-
муносупрессивных средств. Эта дорогостоящая и
требующая интенсивного ухода за больным со-
путствующая терапия показана пациентам с
быстро прогрессирующей и распространенной
вульгарной пузырчаткой, которые не реагируют
на традиционную терапию.
Вторичные бактериальные и микотические
инфекции требуют лечения антибиотиками или
антимикотическими препаратами, а при усилив-
шейся анемии — повторных переливаний кро-
ви. Следует обратить внимание на достаточное
поступление жидкости, калорийное и богатое
белком питание и прием витаминов (витамины
С, комплекс витаминов В). При долговременной
терапии глюкокортикоидами следует подумать и
о достаточном поступлении в организм калия.
Местное лечение
От наружного лечения кожи и слизистых обло-
чек зависит стабилизация либо ухудшение состо-
яния пациента. Наружные мероприятия носят
лишь поддерживающий характер, они должны
помешать возникновению вторичных инфекций
и способствовать заживлению ожогов. Наружные
глюкокортикоиды не особенно эффективны.
Необходим усиленный контроль бактериального
и микотического обсеменения посредством анти-
биотикограммы. Для лечения эрозий применяются
анилиновые красители, в области слизистой рта
используются многократные полоскания с ис-
пользованием противовоспалительных, противо-
бактериальных и анестезирующих жидкостей.
Полезны также ванночки с добавлением марган-
цовки и детергентов.
Вегетирующая
пузырчатка
Вегетирующая пузырчатка — это особая форма
вульгарной пузырчатки, характеризующаяся ог-
раниченными папилломатозными разрастаниями
у пациентов с относительно хорошей сопротив-
ляемостью заболеванию. Различают два ее типа —
Ноймана (Neumann) и Аллопо (Hallopeau).
Вегетирующая
пузырчатка Ноймана
Это заболевание развивается у больных вульгар-
ной пузырчаткой либо спонтанно, либо при ле-
чении глюкокортикоидами. Начинается обычно
с дряблых беловато-мутных пузырей, которые
быстро надрываются. На эрозированных повер-
хностях заживления, однако, не происходит, а
развиваются папилломатозные разрастания, так
называемые вегетации. Они, в частности, обра-
зуются в интертригинозных участках (таких как
уголки губ, назолабиальная складка, область
вульвы и анального отверстия, подмышечные и
перианальная области), где их развитию способ-
ствуют мацерация и микробное обсеменение.
Часто вокруг бородавчатоподобных мокнущих
или покрытых корочкой вегетаций по краям об-
наруживают остатки пузырей. Высыпания могут
подсыхать и приобретать вид бородавчатоподоб-
ных гиперкератотических изменений с болезнен-
ными трещинами. Заболевание при остром вы-
сыпании пузырей может перейти в вульгарную
пузырчатку. Течение более длительное и терапев-
тически многократно проблематичнее.
Гистопатология
Супрабазальный акантолитический пузырь с
мощным акантозом и папилломатозом. Часто
интраэпидермальные микроабсцессы с эозино-
фильными лейкоцитами.
326 Частная дерматология
Диагноз и дифференциальный диагноз
Диагностические критерии те же, что и при
вульгарной пузырчатке.
Важно избежать неверного диагноза при вто-
ричном сифилисе (широкие кондиломы). Далее
следует подумать о вегетирующей форме буллез-
ного пемфигоида, черном акантозе, а также о
йодо- и бромодерме.
Вегетирующая
пузырчатка типа Аллопо
Речь идет о заболевании, протекающем преиму-
щественно в интертригинозных пространствах, в
частности в подмышечных впадинах, ингвиналь-
ной и периоральной областях, а также на дру-
гих участках кожи, например, на волосистой
части головы. Первичным высыпанием являет-
ся не дряблый пузырь, а желтовато-гнойная
пустула. После разрыва у основания пустулы
образуются верруциформные папилломатозные
пролиферации, которые мокнут и неприятно
пахнут. Типичен рост краев при возникновении
новых пустул. Мокнущая вегетация вызывает
сильные субъективные жалобы (боли) и склон-
на к вторичной бактериальной или микотичес-
кой (Candida albicans) инфекции. Течение хро-
ническое, заболевание может в итоге перейти в
вульгарную пузырчатку и закончиться леталь-
ным исходом.
Дифференциальный диагноз
Дифференциальный диагноз — как при вегети-
рующей пузырчатке типа Ноймана. Следует по-
думать также о других первично пустулезных и
верруциформных заболеваниях.
Лечение
Системное лечение ведется при обоих заболева-
ниях, как и при вульгарной пузырчатке. При
вегетирующей пузырчатке типа Hallopeau из-за
ее выраженной тенденции к выздоровлению
чаще всего достаточно средних доз глюкокорти-
коидов. Для устранения вегетаций используют-
ся хирургическое удаление, терапия слабыми
рентгеновскими лучами или инъекциями крис-
таллической суспензии триамцинолона. Кроме
того, применяются ванны с дезинфицирующи-
ми добавками. Наружно применяют глюкокор-
тикоиды с антимикотическими и антибиотичес-
кими добавками на маложирной основе (лучше
лосьоны или кремы).
Листовидная
пузырчатка
Речь идет в принципе об очень близом к вуль-
гарной пузырчатке заболевании, при котором
нарушение интраэпидермальной целостности
вследствие акантолиза происходит, однако, не
супрабазально, а в верхнем Stratum spinosum или
Stratum granulosum. Поэтому крышка пузыря
очень тонкая и легко надрывается. Клинически
на передний план выступает мокнущее тестооб-
разное шелушение с неприятным запахом. По-
средством прямой и непрямой иммунофлуорес-
ценции пемфигусные антитела обнаруживаются
при этом заболевании в сыворотке, а также в
пораженной и непораженной коже.
Заболевание относительно редкое. Могут болеть
дети, но чаще болеют лица в возрасте между 30
и 60 годами. И в этом случае преимущественно
заболевают лица еврейской национальности.
Этиопатогенез
Примечательно, что провоцирующими фактора-
ми заболевания являются солнечный свет, малиг-
номы (тимомы) или медикаменты (D-пеницил-
ламин). Аутоантитела в сыворотке пациентов с
листовидной пузырчаткой имеют специфич-
ность, отличную от антигенов вульгарной пузыр-
чатки, и реагируют с десмосомальным гликоп-
ротеином десмоглеин-1 (молекулярный вес 160
kD), который, как и антиген вульгарной пузыр-
чатки, относится к семье катедринов. Десмогле-
ин-1 связан с плакоглобином, десмосомальным
протеином с молекулярным весом 85 kD, кото-
рый также относится к антигенному комплексу
вульгарной пузырчатки. Патогенетичность этих
антител листовидной пузырчатки доказана в
экспериментах с новорожденными мышами, у
которых после введения содержащей антитела
сыворотки от пациентов в эпидермисе развива-
лись эрозии и пузыри.
Клиническая картина
Заболевание может начинаться на любом участ-
ке тела, но чаще всего на волосистой части го-
ловы и на лице. Здесь появляются расположен-
ные совершенно плоско дряблые пузыри,
которые вскрываются и развиваются далее в ше-
лушащиеся тестоподобные корочки. Поскольку
самые верхние слои эпидермиса, вследствие суб-
корнеального акантолиза, в некотором роде «пла-
вают» на своем основании, роговой слой припод-
нимается над основанием, что приводит к
Глава 28. Пузырные дерматозы
327
мокнущим клейковато-влажным эрозиям, кото-
рые вследствие выделения бактериального сек-
рета неприятно пахнут. При последовательном
распространении может в конце концов возник-
нуть картина вторичной эритродермии, В этом
случае наблюдается универсальное покраснение
кожи, которая покрыта клейковато-влажными
различного размера тестоподобными шелушащи-
мися отложениями, размягчающимися в интер-
тригинозных участках и дающими картину по-
красневших мокнущих участков. Феномен
Никольского I повсеместно положительный.
Вторичная инфекция может привести к сепси-
су. Изменения на слизистой оболочке полости
рта в форме маленьких поверхностных эрозий
достаточно редкие. Может иметь место конъюн-
ктивит с гнойной секрецией.
Течение заболевания чаще всего хроническое.
У детей заболевание имеет тенденцию к первич-
ному хроническому течению, иногда со спонтан-
ной ремиссией. У взрослых течение обычно хро-
ническое волнообразное. Через месяцы, а иногда
и годы, постепенно прогрессирующая кахексия
у нелечившихся пациентов вместе с общими
симптомами, такими как лихорадка, вторичная
инфекция, бронхопневмония или уремия при-
водит к летальному исходу.
Прогноз у молодых пациентов более благо-
приятен, а у пациентов старше 50 лет — хуже чем
при вульгарной пузырчатке.
Лабораторные данные
Гистопатология
Образование акантолитической щели или пузы-
рей в верхнем эпидермисе, т. е. в верхнем stratum
spinosum или stratum granulosum. Могут добав-
ляться вторичные эпидермальные изменения в
форме легкого акантоза, папилломатоза и гипер-
кератоза, как и дискератотические изменения в
клетках эпидермиса. В роговом слое обнаружи-
вается, как правило, выраженная воспалитель-
ная реакция, иногда с большим количеством
эозинофильных лейкоцитов.
Цитологическое исследование
мазка из дна пузыря
Легко подтверждается наличие пемфигусных
клеток в мазке из основания пузыря на эрозив-
ных участках.
Иммунологические исследования
Посредством прямой и непрямой иммунофлуорес-
ценции листовидную пузырчатку нельзя надежно
отличить от вульгарной. Оба заболевания харак-
теризуются сетчатовидными межклеточными от-
ложениями антител в эпидермисе. При листовид-
ной пузырчатке аутоантитела почти исключитель-
но относятся к подфракции IgG4. Антитела лис-
товидной пузырчатки в сыворотке особенно
хорошо видны на пищеводе морской свинки при
непрямой иммунофлуоресценции. Однако они
имеются не у всех пациентов с листовидной пу-
зырчаткой, поэтому часто необходимо многократ-
ное исследование. Наличие различной антигенной
специфичности у листовидной пузырчатки или же
наличие антител вульгарной пузырчатки в сыво-
ротке удается доказать только с помощью специ-
альной иммунологической методики, такой как
иммуноблотинг или иммунная преципитации с ис-
пользованием экстрактов эпидермальных клеток.
Дифференциальный диагноз
При расположении проявлений заболевания в
центре лица, на коже головы и на груди забо-
левание вначале напоминает себорейную экзему
или подострую кожную красную волчанку. Од-
нако и здесь уже имеется резкий запах в пора-
женных очагах, а поверхностные отложения
вследствие акантолиза легко смещаются.
Лечение
Внутрь рекомендуется вначале монотерапия глю-
кокортикоидами, затем в комбинации глюко-
кортикоиды с иммуносупрессивными препара-
тами, как при вульгарной пузырчатке. Описано
эффективное применение дапсона в ранней фазе
заболевания.
Местно. Ванны с добавлением синтетических
детергентов, средства дезинфекции и противо-
воспалительные (отруби, экстракт овсяной соло-
мы) или вяжущие добавки (отвар дубовой коры).
Можно применять фторированные глюкокорти-
коиды в виде лосьона или крема, при необхо-
димости под окклюзионную повязку. Не реко-
мендуется применять мази. Постоянный
контроль на вторичные бактериальные или ми-
котические инфекции (антибиотикограмма) и со-
ответствующее лечение.
Бразильская
пузырчатка
Синонимы: бразильская листовидная пузырчат-
ка, fogo selvagem.
Речь идет об эндемическом заболевании, ко-
торое очень похоже на листовидную пузырчат-
ку. Это эндемическое заболевание распростра-
328 Частная дерматология
нено в центральной и западной части Бразилии,
а также в северной Аргентине, Парагвае, Боли-
вии, Перу и Венесуэле. Болезнь часто семейная
и поражает преимущественно девушек и моло-
дых женщин до 30 лет. Она встречается преиму-
щественно в сельских тропических лесных рай-
онах. После раскорчевки леса и устройства
лучших жилищ частота заболевания снижается.
Эпидемиологические исследования указывают на
то, что в этих районах заболевание имеет инфек-
ционную этиологию. Вероятно, вызывается и
поддерживается до сих пор пока неизвестными
факторами внешней среды. Возможно, перено-
сится черными мухами (simulium prunosium)\
предполагают также вирусную природу.
Клиническая картина
На лице и волосистой части головы, а также на
груди и спине возникают плоские пузыри, ко-
торые вскоре трескаются и образуют слоистые
чешуйчатые корочки на эрозивной основе. Из-
менения могут сохраняться в течение несколь-
ких лет. Возможен переход во вторичную эрит-
родермию, как при листовидной пузырчатке.
Слизистые не поражаются. При полностью сфор-
мировавшейся картине заболевания пациенты
жалуются на ощущение жжения на коже, как от
огня; отсюда и название fogo selvagem*, дикий
огонь. Феномен Никольского в очагах пораже-
ния всегда положителен.
Летальность при этом заболевании составля-
ет около 5 %. Свыше 55 % пациентов излечи-
ваются при длительном (в течение нескольких
лет) применении глюкокортикоидов, остальные
нуждаются в длительной терапии глюкокорти-
коидами.
Лабораторные данные
Гистопатология
Как при листовидной пузырчатке.
Цитологическое исследование
Цитологическое исследование мазка из основания
пузыря: пемфигусные клетки (клетки Тцанка)
всегда положительны.
Иммунологические исследования
Как при листовидной пузырчатке. Титры пем-
фигусных антител в сыворотке пациентов чаще
всего очень высокие и параллельны тяжести за-
болевания.
Лечение
До появления глюкокортикоидов это заболева-
ние имело очень тяжелый прогноз. В целом от
начала заболевания до наступления ремиссии
требуется 60—90 мг эквивалента преднизолона
ежедневно. Затем дозы постепенно уменьшают-
ся до поддерживающих 5—10 мг ежедневно,
которые принимаются каждые 24 или 48 часов.
Средняя продолжительность терапии составля-
ет 2—4 года. Как и при вульгарной пузырчатке,
применяются и иммуносупрессивные средства.
Вторичные инфекции лечат соответствующим
образом. Небольшие первичные очаги лечат как
при эритематозной пузырчатке.
Эритематозная
пузырчатка
Синонимы: себорейная пузырчатка, синдром
Senear-Usher.
Клинически и гистологически заболевание
представляет собой морфологический вариант
листовидной пузырчатки.
Этиопатогенез
Этиопатогенез соответствует листовидной пузыр-
чатке. Из-за наличия дополнительных, направ-
ленных против базальной мембраны антител и
антинуклеарных антител (ANA) предполагается,
что в основе этого заболевания лежит сосущество-
вание листовидной пузырчатки и хронической
красной волчанки. Провоцируется солнечным
светом, травмой или лекарствами (например, пе-
ницилламином, каптоприлом, пропанололом,
напроксеном, тиазидом).
Клиническая картина
Болезнь очень редкая и ограничивается себо-
рейными участками на лице, голове, груди и
спине. Иногда имеются единичные очаги, но
чаще поражение симметрично. Клиническая
картина на лице напоминает себорейную эк-
зему или поверхностную хроническую эрите-
матозную волчанку. На спине и груди очаги
покрываются корочкой и склонны к поверх-
ностным эрозиям, напоминая в большей сте-
пени о листовидной пузырчатке. На слизистых
изменений нет, а субъективные симптомы не-
значительны.
Заболевание может протекать ограниченно, но
может переходить в листовидую пузырчатку.
Очень редко встречается в сочетании с другими
аутоиммунными заболеваниями, такими как
миастения беременных, тимома или системная
красная волчанка.
Глава 28. Пузырные дерматозы 329
Лабораторные данные
Гистопатология
как при листовидной пузырчатке. В области
базальных кератиноцитов вакуольная дегенера-
ция. Дермальные лейкоциты и периваскулярные
лимфоцитарные инфильтраты, а также утолще-
ние PAS-реактивной базальной мембраны.
Цитологическое исследование
Цитологическое исследование мазка со дна пузы-
ря: пемфигусные клетки, дискератические кера-
тиноциты и много лейкоцитов.
Иммунологические исследования
Посредством прямой иммунофлуоресценции
очагов заболевания доказывают наличие анти-
эпителиальных пемфигусных антител типа IgG
между акантолитическими клетками поражен-
ного эпидермися. IgG и/или составные части
комплемента гомогенно представлены в эпидер-
мальной базально-мембранной зоне, особенно
в подверженных хроническому световому воз-
действию участках кожи, у 50—70 % пациентов.
В сыворотке более 80 % пациентов обнаружи-
ваются антинуклеарные антитела (ANA), а ан-
титела к DNS, Sm или Ro (SSA), как при крас-
ной волчанке, напротив, отсутствуют. Если
исследование на пемфигусные антитела и ANA
в сыворотке пациентов отрицательно, то его
следует повторить через 2—3 недели.
Дифференциальный диагноз
Себорейная экзема, красная волчанка хроничес-
кая поверхностная или подострая кожная крас-
ная волчанка. Диагностически определяющими
являются результаты гистологического, иммуно-
патологического и сероиммунологического ис-
следований.
Лечение
Внутрь глюкокортикоиды назначаются при рас-
пространенной форме заболевания и применя-
ются, как при вульгарной пузырчатке, в сред-
них дозах (60—90 мг эквивалента преднизолона
ежедневно) в комбинации с хлорохином до до-
стижения ремиссии. Иммуносупрессивные пре-
параты применяются также, как при вульгарной
пузырчатке. Рекомендуется дапсон (ДДС по
100-150 мг/день).
Наружная терапия: кремы, содержащие глю-
кокортикоиды, особенно с окклюзионной повяз-
кой; на единичные очаги глюкокортикоидный
пластырь или внутрйочаговая инъекция кристал-
лической суспензии триамицинолона. Защита от
солнца.
Паранеопластическая
пузырчатка
Это заболевание клинически характеризуется
эрозиями на слизистых оболочках с полиморф-
ными изменениями на коже и ассоциируется с
опухолями, чаще всего с лимфомами или тимо-
мами. Гистологически оно начинается с образо-
вания интраэпидермальной супрабазальной
акантолитической щели. Иммунологически до-
казывается наличие пемфигусовидных аутоанти-
тел в эпидермисе и в сыворотке пациентов.
Заболевание встречается редко. Возможно, оно
идентично ранее описывавшимся случаям одно-
временного наличия вульгарной пузырчатки с
миастенией беременных, лимфомой или тимомой.
Этиопатогенез
Дискутируется перекрестная реактивность между
антигеном опухоли и эпителиальным антигеном.
Пассивным переносом аутоантител у новорож-
денных мышей провоцируется образование акан-
толитического пузыря. Протеино-химические
исследования показали, что эти аутоантитела
реагируют с иммунным комплексом, который
помимо десмоплакина I и буллезного пемфиго-
идного антигена с молекулярным весом 230 kD,
содержит еще два эпителиальных антигена с
молекулярными весами 210 и 190 kD.
Сосуществование с опухолями: до настояще-
го времени описаны в качестве лежащих в ос-
нове неоплазий хронические миелотические лей-
кемии, тимомы, доброкачественные гиперплазии
лимфоузлов (опухоли Castleman) и злокачествен-
ные лимфомы.
Клиническая картина
Персистирующие, частично фибринозные, болез-
ненные эрозии в полости рта, на конъюнктивах,
в аногенитальной области, а также на ногтях рук
и ног. Изменения на коже полиморфные и часто
имеют место в верхней части туловища, на руках
и ногах, а также ладонях и подошвах. Здесь при-
сутствуют сливающиеся эритемы, эрозии или па-
пулосквамозные высыпания и подобные аналогич-
ным при многоформной экссудативной эритеме
явления с образованием пузыря в центре.
Течение и прогноз
Изменения на коже и слизистых плохо реаги-
руют на глюкокортикоиды, иммуносупрессив-
ные препараты и плазмаферез. У многих паци-
ентов происходит выздоровление после удаления
330 Частная дерматология
ассоциированной опухоли. Прогноз поэтому за-
висит от опухоли, лежащей в основе заболева-
ния.
Лабораторные данные
Гистопатология
Многообразие гистологических показателей со-
отвествует полиморфное™ гистологической кар-
тины. Супрабазальные акантолитические пузы-
ри, некрозы кератиноцитов и вакуольные
дегенерации базальных кератиноцитов преобла-
дают. В папулосквамозных изменениях может
отсутствовать интраэпидермальный акантолиз.
Поэтому необходимы повторые биопсии.
Иммунологические исследования
При прямой иммунофлуоресценции наблюдаются
отложения IgG между клетками эпидермиса -
как при вульгарной пузырчатке, но более слабые
и неравномерные. Одновременно в базально-мем-
бранной зоне имеют место гранулярные или ли-
нейные преципитаты факторов комплемента. С
помощью непрямой иммунофлуоресценции в
сыворотке пациентов доказывается наличие ауто-
антител, которые по-иному, чем при вульгарной
пузырчатке, связываются с цилиндрическим или
переходным эпителиями в мочевом пузыре, сли-
зистой бронхов, тонком кишечнике и даже с тка-
нями мышц и щитовидной железы
Пузырчатка
хроническая
врожденная семейная
Синоним: болезнь Хейли-Хейли.
Это хроническое семейное заболевание не
имеет никакого отношения к вульгарной пузыр-
чатке, листовидной пузырчатке и их подвидам.
Речь идет скорее о наследуемом по аутосомно-
доминантному признаку кожном заболевании с
различной генной пенетрацией. Встречаются и
единичные случаи. Предполагается ассоциация с
HLA-B8. Заболевание редкое и может провоци-
роваться трением, солнечным излучением, холо-
дом или жарой и микробными инфекциями. Как
показывают исследования генного локуса, нет
связи и с болезнью Дарье.
Этиопатогенез
Указания на связь с вульгарной пузырчаткой,
листовидной пузырчаткой или наследственны-
ми эпидермолизами отсутствуют. Из-за склон-
ности к дискератозу ранее дискутировалось, не
является ли это заболевание везикулезным ва-
риантом болезни Дарье. Акантолиз и дискера-
тоз имеют место при обоих заболеваниях, и оба
они наблюдаются у одних и тех же пациентов.
После обнаружения под электронным микро-
скопом нарушений в синтезе или созревании то-
нофиламентов и десмосомных комплексов, а
также в синтезе межклеточных веществ в пре-
делах эпидермиса, сегодня предполагают нали-
чие генетического дефекта в эпидермальной
дифференциации, который провоцируется тре-
нием, солнечным облучением, холодом, жарой,
контактной аллергией экзематозного типа или
микробными инфекциями (бактериями или
грибками, чаще всего Candida albicans).
Клиническая картина
Преимущественно в местах трения и в интертри-
гинозных или сгибательных участках, таких как
боковые части шеи, подмышечная область, па-
ховая и перианальная области, обычно в позднем
юношеском или раннем взрослом возрасте воз-
никают единичные или сгруппированные и
склонные к слиянию, часто удлиненные дряб-
лые пузырьки, которые, соединяясь, образуют
покрасневшие, покрытые чешуйчатой коркой
экземоподобные очаги, последние, однако, от-
личаются эпидермальными трещинами. Очаги,
имеющие округлые, овальные или кольцевидные
границы, располагаются чаще всего симметрич-
но и зудят. В интертригинозных участках наблю-
даются резко отграниченные, мокнущие поверх-
ности с неприятным запахом и склонностью к
плоским вегетациям и периферическому росту
вследствие вновь вскрывающихся плоских мут-
ных пузырей. Отмечаются временные улучше-
ния, за ними следуют рецидивы. Феномены
Никольского I и II многократно положительны.
Общее состояние хорошее, местно больных бес-
покоит зуд. Иногда на внутренних поверхнос-
тях ладоней заметны кератотические папулы, а
на ногтях пальцев рук — продольные белые или
красновато-коричневые полоски. Течение забо-
левания хроническое рецидивирующее с времен-
ными ремиссиями. Осложнения могут быть
обусловлены вторичными инфекциями.
Лабораторные данные
Гистопатология
Массивный акантолиз в пределах часто слегка
акантолитически расширенного эпидермиса, зах-
ватывающий колбочки сетчатки и продолжаю-
Глава 28. Пузырные дерматозы 331
щийся, в частности, через кончики сосочков меж-
ду базальными клетками. Отсюда нарушение
структуры эпидермального клеточного соедине-
ния («трещина в кирпичной стене»). Кроме акан-
толиза отдельных клеток, в некоторых местах
имеет место дискератоидное преобразование акан-
толитических клеток (эозинофильные гомоген-
ные клетки с пикнотическим клеточным ядром,
в частности, в stratum granulosum). Лейкоцитар-
ный экзоцитоз и эозинофильные лейкоциты в
содержимом пузырей отсутствуют.
Цитология мазка из основания пузыря
Пемфигусные клетки и дискератотические
клетки.
Иммунологические исследования
Пемфигусные антитела отсутствуют и в коже, и
в сыворотке пациента.
Дифференциальный диагноз
Важно отграничение от интертриго, интертриги-
нозной экземы и микоза, в частности, кандидоз-
ного интертриго. Серпигинозный край очага
должен натолкнуть на мысль о микозе гладкой
кожи. Вегетирующую пузырчатку, при которой
могут быть изменения на слизистой рта, следу-
ет исключить путем гистологических и иммуно-
логических исследований. Болезнь Дарье может
представлять диагностическую сложность в ги-
стологическом плане, но не в клиническом, по-
скольку она имеет иные места локализации и
фолликулярную кератотическую папулу как
первичное высыпание.
Лечение
Только при тяжелых формах заболевания оправ-
даны глюкокортикоиды в средних дозах. Анти-
биотики следует выбирать после определения
бактериологической резистентности микроорга-
низмов. Они могут оказать существенную по-
мощь. Можно применять в качестве альтерна-
тивного варианта лечение изотретиноином
(роаккутаном) или ацетретином (неотигазоном).
Комбинированное местное применение фто-
рированных глюкокортикоидов и антибиотиков
(по данным антибиотикограммы) на основе мо-
лочка, крема или пасты является средством вы-
бора. Ванночки из марганцовки и анилиновых
красителей могут также принести пользу. Раствор
кислоты витамина А в концентрации 0,01 % в
виде крема часто бывает эффективным. Важно
избегать провоцирующих факторов, таких как
жара, солнечное облучение, трение или бактери-
альные инфекции. При долговременном местном
применении глюкокортикоидов контроль на
кандидозную инфекцию. Применение СО2-ла-
зера может оказаться эффективным. Дермабра-
зия или полная эксцизия больных участков кожи
с последующим замещением (покрытием) кож-
ным лоскутом у отдельных пациентов определен-
но способствуют выздоровлению.
Г ерпетиформный
дерматоз Дюринга
Герпетиформный дерматоз Дюринга (герпети-
формный дерматит Дюринга) — это доброкаче-
ственное хроническое рецидивирующее поли-
морфное кожное заболевания, которое может
привести к жгучему зуду и образованию гер-
петиформных пузырей и при котором часто
присутствует глютенсенситивная энтеропатия
(целиакия); характерны субэпидермальные гра-
нулярные отложения IgA.
Заболевание редкое (1 на 800 дерматологичес-
ких пациентов) и наступает чаще всего между 30
и 45 годом жизни, но может быть и у детей.
Мужчины болеют чаще женщин (3 : 2). Относи-
тельно часто герпетиформный дерматоз Дюринга
отмечается в Северной Европе, особенно в Скан-
динавии. В пользу наследственной предрасполо-
женности данных нет. Целиакия и герпетифор-
мный дерматит генетически связаны: оба
заболевания более чем у 90 % пациентов имеют
ассоциацию с HLA-класса II- антигенами DR3/
DQ2, более чем у 70 % пациентов имеется ал-
лотип HLA — Al, В8, DR3.
Этиопатогенез
Этиология неизвестна. Иммунологические и
генетические исследования трактуются в пользу
того, что герпетиформный дерматоз представляет
собой особую форму течения целиакии с харак-
терными кожными проявлениями. Глютенсенси-
тивная энтеропатия при герпетиформном дерма-
тозе морфологически, функционально и
клинически идентична энтеропатии при целиа-
кии, хотя и более олиго- или асимптоматична,
поскольку изменения в кишечнике при герпе-
тиформном дерматозе обычно выражены значи-
тельно слабее. При избытке глютена у пациен-
тов с герпетиформным дерматозом наступает
ухудшение симптоматики кожи и тонкого ки-
шечника. При безглютеновой диете, напротив,
у большинства пациентов изменения на коже и
в кишечнике регрессируют, хотя регресс кожных
332 Частная дерматология
проявлений может затянуться от нескольких
месяцев до года. Как при целиакии, так и при
герпетиформном дерматозе, имеются сывороточ-
ные антитела класса IgA, реже IgG или IgM,
которые направлены против глиадина. Антите-
ла тироидных и париетальных клеток могут при
герпетиформном дерматозе указывать на ассоци-
ированные иммунные заболевания.
Специфичными для целиакии герпетиформ-
ного дерматоза являются IgA — антитела к ре-
тикулярной соединительной ткани (ретикулин),
in vitro они реагируют с эндомизием гладкой
мускулатуры, например, в пищеводе (антитела
IgA- эндомизия) или же с субмукозной соеди-
нительной тканью тонкого кишечника (IgA-
Jejenium-антитела). Обнаруживаемые в субму-
козной ткани тонкого кишечника отложения
антител имеют, вероятно, ту же антигенную спе-
цифичность, что и антитела ретикулина и эндо-
мизия в сыворотке. В отличие от них, происхож-
дение и антигенная специфичность связанных в
коже IgA-антител при герпетиформном дерматозе
неизвестны. Возможно, здесь местная иммуно-
комплексная реакция приводит к активизации
системы комплемента (преимущественно альтер-
нативным путем), к хемотаксису и активизации
гранулоцитов и гуморальных медиаторов воспа-
ления, которые вносят свой вклад в образова-
ние субэпидермальных пузырей.
Примечательна особая чувствительность паци-
ентов к галогенам, в частности, к йодиду калия.
В этой связи йодосодержащие медикаменты или
наружные средства (йодоформная марля), при-
менение йодированной поваренной соли или
употребление йодосодержащей морской рыбы
или других продуктов питания приводит к воз-
никновению или ухудшению кожных проявле-
ний.
Клиническая картина
Заболевание развивается либо медленно по на-
растающей линии, либо возникает внезапно и
вызывает субъективно ощущающийся жгучий
или болезненный зуд. Места предпочтительной
локализации обычно симметрично расположен-
ных экзантем — верхний плечевой пояс, область
ягодиц и волосистая часть головы, затем разги-
бательные поверхности предплечий и локти, а
также колени и разгибательные поверхности
голеней. Реже поражаются только локти и ко-
лени. Слизистая полости рта практически никог-
да не поражается.
Для кожных изменений типичен полимор-
физм. Вначале в местах предпочтительной ло-
кализации возникают эритематозные, уртикар-
ные или папулезные изменения кожи. Как
только появляются сгруппированные герпети-
формные пузырьковые высыпания, диагноз
становится ясен. Иногда пузырьки очень мел-
кие и расположены в коже как дробинки, они
лучше обнаруживаются пальпаторно, чем на
глаз. Причудливые пузырьки всегда упругие и
имеют плоскую устойчивую покрышку. У де-
тей и пожилых людей, а также после отмены
лечения могут возникать большие напряжен-
ные пузыри, какие бывают при буллезном
пемфигоиде (крупнопузырный или буллезный
герпетиформный дерматоз). Могут добавляться
и экзематозные, склонные к лихенификации,
изменения. Вторичная гиперпигментация по-
раженных участков кожи присутствует у 50 %
пациентов.
Симптоматически типичен сильный и особо
жгучий зуд в области кожных проявлений. Мас-
сивные расчесы оставляют следы, которые мо-
гут импетигинизироваться. Поэтому болезнь
бывает сложно диагностировать. Рецидивы, эф-
фекты расчесов и импетигинизация создают
картину, заставляющую подумать о cutis
vagantium.
Глютенсенситивная энтеропатия
при герпетиформном дерматозе
Примерно 70 % пациентов имеют в тонком ки-
шечнике изменения, подобные возникающим
при целиакии. Сюда относятся атрофия ворси-
нок различной степени интенсивности, воспали-
тельные инфильтраты из лимфоцитов и наруше-
ние энзиматической активности в эпителии
тонкого кишечника. Однако примерно у 30 %
пациентов слизистая тонкого кишечника гисто-
логически и функционально в норме. При гер-
петиформном дерматозе могут быть получены
следующие данные о желудочно-кишечном трак-
те: глютенсенситивная энтеропатия, атрофия вор-
синок в кишечнике с гиперплазией крипты, руб-
цовое сужение пищевода, дивертикул, стеаторея,
атрофический гастрит, интестинальная липоди-
строфия, ректоколит и лимфома.
Общие симптомы обычно отсутствуют. При-
мечательна частая эозинофилия в сыворотке
крови и содержимом пузыря, а также в спинном
мозге.
Чувствительность к йодидам
Суть гиперсенситивности к йодидам еще не
выяснена. Речь идет о неспецифической прово-
цируемое™ йодом кожных проявлений. Прием
внутрь йодида калия может привести к массив-
ному и иногда небезопасному ухудшению забо-
левания.
Глава 28. Пузырные дерматозы > 333
Герпетиформный дерматоз часто начинается
остро или подостро и протекает приступообраз-
но, причем могут иметь место многомесячные
интервалы без каких-либо проявлений или с
очень незначительными проявлениями заболева-
ния. Поскольку заболевание растягивается на
много лет, больные физически и психически
сильно страдают от кожных проявлений и осо-
бенно жгучего зуда, хотя в принципе речь идет
о доброкачественном заболевании.
Лабораторные данные
Цитология мазка со дна пузыря
Акантолитические пемфигусные клетки отсут-
ствуют, образование пузыря субэпидермальное.
Много эозинофильных лейкоцитов.
Гистопатология
Субэпидермальный пузырь натяжения без акан-
толиза. В содержимом пузыря обнаруживаются
много эозинофильных, а также нейтрофильные
лейкоциты. PAS-реактивная базальная мембра-
на зафиксирована на роговом слое. Отек и вос-
палительный инфильтрат из гистиоцитов, лим-
фоцитов, эозинофильных и нейтрофильных
лейкоцитов в сосочковых тельцах эритематозных
изменений и краевых зон пузырей. Типичны
межсосочковые микроабсцессы в субэпидер-
мальных кончиках сосочков с нейтрофильными
и эозинофильными лейкоцитами, а также остат-
ками клеточного ядра. Субкорнеальные пузырь-
ки встречаются редко.
Электронная микроскопия показывает, что
пузырь начинается в роговом слое, где имеют
место отек, некробиотические изменения колла-
геновых волокон и клеточно-воспалительная
реакция. Изменения, регистрируемые электрон-
ным микроскопом на базальной мембране и
цитоплазматической мембране базальных клеток,
типичны. Так наступает нарушение непрерыв-
ности между клетками эпидермиса и лежащим
под ним роговым слоем.
Иммунологические исследования
Важнейшим диагностическим критерием получа-
емом при герпетиформном дерматозе, посредством
прямой иммунофлуоресценции, являются грану-
лярные отложения IgA, часто вместе с СЗ, в кон-
чиках дермальных сосочков и вдоль базально-
мембранной зоны, реже также и вокруг
дермальных кровеносных сосудов. Почти все па-
циенты с этим гранулярным образцом преципи-
тации имеют изменения в тонком кишечнике в
смысле глютенсенситивной энтеропатии. Отложе-
ния IgA встречаются независимо от интенсивно-
сти заболевания, но в ходе последовательной без-
глютеновой диеты они могут долгосрочно регрес-
сировать. Они относятся преимущественно к
подклассу IgA и существуют в одномерной или
двухмерной формах. Ультраструктурно грануляр-
ные отложения IgA в коже соответствуют так
называемым тельцам герпетиформного дермато-
за, которые можно распознать в сосочковом слое
в окружении коллагеновых волокон или эласти-
ческих микрофибрилл.
При герпетиформном дерматозе в сыворотке
не обнаруживаются аутоантитела к антигенам
эпидермиса или базально-мембранной зоны, хотя
обнаруживаются другие — прежде всего класса
IgA. Сюда относятся: IgA-антитела эндомизия,
IgA-антитела тонкого кишечника и IgA-антите-
ла ретикулина. Они возникают в связи с ассо-
циированной глютенсенситивной энтеропатией
и являются теми самыми серологическими мар-
керами вовлечения тонкого кишечника при гер-
петиформном дерматозе, их наличие доказыва-
ется у 70—80 % пациентов. При соблюдении
безглютеновой диеты они медленно исчезают, а
при повторном отягощении глютеном могут сно-
ва возникнуть. Непрямой иммунофлуоресценци-
ей доказывается наличие этих IgA — антител к
различным антигенным субстратам, что и при-
вело к их единому наименованию; и они — ве-
роятно, с учетом их антигенной специфичнос-
ти, — идентичны. Перекрестной реакции с
глиалиновыми антителами, которые также встре-
чаются при герпетифорном дерматозе, они не
имеют.
Дифференциальный диагноз
Типичным для данного заболевания является
симметричность высыпаний, их полиморфизм,
определенная локализация и интенсивный жгу-
чий зуд (табл. 28.1). Диагностическую помощь
могут оказать и биопсии из свежего пузыря и
края эритемы на гистологическое и иммуноло-
гическое обследование (прямая иммунофлуорес-
ценция). Диагноз можно подтвердить и ех
juvantibus, поскольку герпетиформный дерматоз
хорошо реагирует на сульфоны (дапсон 100—
150 мг ежедневно).
При остром начале заболевания его следует
отграничить от многоформной экссудативной
эритемы и синдрома Свита, в случае клиничес-
кого варианта с крупными пузырями — от бул-
лезного пемфигоида. Субкорнеальный пустулез
с точки дифференциального диагноза рассмат-
ривается только при наличии цирцинарного типа
герпетиформного дерматоза. Если на передний
план в клинической картине выдвигаются пру-
334 Частная дерматология
ригинозные папулы и узелки (пруригинозная
форма герпетиформного дерматоза) следует ис-
ключить пруригинозные формы других дерма-
тозов, таких как атопическая экзема или пру-
риго простое подострое.
Диагноз
Диагноз устанавливается на основании анамне-
за (непереносимость препаратов йода, наличие
глютеновой энтеропатии), клинической карти-
ны (симметрично расположенные на локтях,
коленях, ягодицах, разгибателях предплечий,
голове зудящие высыпания в виде папул, вези-
кул, пузырей), гистологических данных (субэ-
пидермальные пузыри с большим количеством
эозинофилов; на верхушках сосочков дермы
скопления эозинофильных и нейтрофильных
гранулоцитов), реакции прямой иммунофлюо-
ресценции здоровой на вид кожи (отложения IgA
в сосочковом слое дермы).
Интенсивный зуд кожи и жжение, полимор-
физм высыпаний с герпетиформным расположе-
нием везикул, хроническое рецидивирующее
течение позволяют заподозрить заболевание.
Подтвердить диагноз помогают цитологическое,
патоморфологическое, иммунологическое иссле-
дования.
Таблица. 28.1. Дифференциальный диагноз вульгарной
пузырчатки, дерматоза Дюринга, буллезного п пемфигоида
Признаки, симптомы Вульгарная пузырчатка Гэрпетиформный дерматоз Дюринга Буллезный пемфигоид
Возраст больных Старше 40 лет Все возрастные группы Старше 60 лет
Патогенез Аутоиммунный Аутоиммунный Аутоиммунный
Начало со слизистой 60-80% Менее 10 % 30-50%
полости рта
Первичный элемент Мономорфная сыпь (вялые пузыри с серозным содержимым на видимо не измененной коже и слизистых) Полиморфная сыпь (на эритематозном фоне мелкие сгруппированные плотные пузыри, везикулы, уртикарии, папулы, эритемы) Полиморфная сыпь (на фоне отечной эритемы появляются напряженные пузыри, группировка не характерна)
Последующее Рост по периферии, невыраженная Нет склонности к периферическому Нет склонности к
развитие пузырей склонность к эпителизации росту, имеется тенденция к эпителизации периферическому росту, имеется тенденция к эпителизации
Локализация на коже Может быть везде, поражается слизистая оболочка полости рта; группировка не характерна Чаще локти, колени, разгибатели предплечий, ягодицы, волосистая часть головы; герпетиформная группировка высыпаний Склонность к группировке на сгибательных участках конечностей, туловище
Симптом Никольского Положительный Отрицательный Отрицательный, перифокальная отслойка эпидермиса
Поражение слизистой Почти всегда Редко Часто
оболочки рта
Жалобы больных Болезненность в местах эрозий, нарастающая общая слабость Сильный зуд в местах высыпаний Зуд в период свежих высыпаний, иногда повышена температура
Течение Прогрессирующее Приступообразное, толчками Хроническое
Гистология Внутриэпидермальное расположение пузырей, акантолиз супрабазальный Субэпидермальное расположение пузырей, акантолиза нет Субэпидермальное расположение пузырей, акантолиза нет
Цитология Акантолитические клетки (Тцанка) Большое количество эозинофилов, акантолитических клеток нет Лимфоциты, гистиоциты, многочисленные эозинофилы
Прямая РИФ Отложение IgG на десмосомах шиповатых клеток эпидермиса Отложение IgA и СЗ-комплемента на базальной мембране Отложение IgG и СЗ-комплемента в области базальной мембраны
Непрямая РИФ Циркулирующие антитела класса IgG против антигенов эпидермиса Антитела к базальной мембране эпидермиса не определяются Циркулирующие антитела к базальной мембране
Лечение сульфонами Безуспешное Успешное Безуспешное
Прогноз без лечения Неблагоприятный, злокачественное течение с летальным исходом Относительно благоприятный, кроме паранеопластического дерматоза Дюринга Относительно доброкачественный
Глава 28. Пузырные дерматозы 335
Лечение
Рекомендуется комплексное лечение герпети-
формного дерматоза Дюринга, основой которо-
го является безглютеновая диета и назначение
препаратов сульфонового ряда.
Препараты сульфонового ряда
Считаются базисными при терапии дерматоза
Дюринга. Лучше всего оправдали себя такие
сульфоны, как дапсон. Обычно взрослым дос-
таточно доз в 50—150 мг ежедневно для дости-
жения терапевтического контроля над проявле-
ниями заболевания. Однако диапазон
последующих морбостатических доз достаточ-
но большой: для некоторых пациентов доста-
точно 50 мг дапсона через день, другим же нуж-
но более 150 мг ежедневно. При лечении
сульфонами после ремиссии высыпаний необ-
ходимо рассчитать минимальную поддержива-
ющую дозу. Диаминодифенилсульфон (ДДС,
дапсон, авлосульфон) назначают внутрь по 100—
200 мг/сутки. Назначают его производные: ди-
уцифон по 100 мг 2 раза в сутки или димоци-
фон по 200 мг 2 раза в сутки. ДДС (дапсон,
авлосульфон) назначают пятидневными курса-
ми с одно- или двухдневным перерывом. При
достижении клинического эффекта дозу препа-
рата снижают до 25—50 мг в сутки. Рекомен-
дуется проведение 2—4 курсов терапии. Препа-
рат ДДС (реже — его производные) обладает
побочными эффектами в виде общей слабости,
головокружения, головных болей, диспепсии,
тахикардии. Может быть цианоз губ и ногте-
вых фаланг вследствие образования метгемог-
лобина и развития анемии. Возможно развитие
фотосенсибилизации, таксидермии, агранулоци-
тоза, лейкопении, периферической нейропатии.
Для устранения побочных эффектов ДДС-те-
рапии рекомендуется ежедневный прием вита-
мина С в дозе 500—1000 мг.
Важен контроль за уровнем гематологических
побочных действий (образование метгаммаглобу-
лина, цитомегалическая анемия, анемия с нали-
чием телец Гейнца, гемолиз). Механизм действия
сульфонотерапии неизвестен: из-за вероятных
побочных действий необходим постоянный кон-
троль. Поскольку речь идет о морбостатическом
эффекте, лечение проводится годами.
Пациентам без клинических проявлений эн-
теропатии следует также придерживаться стро-
гой безглютеновой диеты, даже если это требу-
ет существенных усилий, а успех терапии может
проявиться только через несколько месяцев,
поскольку благодаря этому можно снизить дозу
лекарств. Еще одна важная рекомендация отно-
сительно диеты: не употреблять йодированной
соли, морской рыбы, йодосодержащих медика-
ментов.
Диета
Исключение из рациона продуктов, богатых
глютеном и/или содержащих соли йода, неред-
ко приводит к исчезновению высыпаний, а так-
же позволяет уменьшить дозы препаратов суль-
фонового ряда. Рекомендуется исключить из
рациона больного продукты, содержащие глю-
тен: все сорта хлеба и другие выпечные изделе-
ния из пшеничной, ржаной, ячменной, овсяной
муки (сушки, бублики, сухари, печенье); изде-
лия из пшеницы, ячменя, ржи, овса, отруби,
добавки солода, изделия из теста (макароны, вер-
мишель, лапша, клецки), изделия из манной
крупы, из перловой и ячневой круп, овсяных
хлопьев; панированные продукты из мяса, рыбы
и птицы; изготовленные с хлебом изделия из
мяса, рыбы, птицы (пельмени, равиоли, чебуре-
ки, вареники); все колбасные изделия с хлебны-
ми наполнителями, включая сосиски, кровяную
колбасу и ливерную колбасу; все супы, которые
содержат запрещенные виды муки и крахмала,
горох, фасоль, чечевицу; капуста, горох, фасоль,
чечевица и злаки; кофейные напитки, содержа-
щие ячмень, рожь, овес; хлебный квас, пиво;
мороженое, пудинги, кремы, шоколад с начин-
кой, марципаны, чурчхела; масло из ростков
пшеницы; готовые супы, соусы, приготовленные
с мукой, сиропы с мукой, продукты с солодом
(Михеев Г. Н. и соавт., 1999). Необходимо
уменьшить употребление поваренной соли (со-
держит соли йода) и полностью исключить про-
дукты, богатые солями йода (морская рыба, каль-
мары, крабы, креветки, водоросли, различные
моллюски) и йодсодержащие медикаменты.
Больным герпетиформным дерматозом Дю-
ринга разрешаются следующие безглютеновые
продукты: хлеб и другие выпечные изделия из
крахмала, кукурузной, соевой и рисовой муки;
изделия из риса, кукурузы, соевой и гречневой
круп (каши, плов, блины, оладьи); все виды
мяса и речной рыбы, домашняя птица, яйца,
цельное молоко (до 0,5 л в день), кисломолоч-
ные продукты (кроме йогурта), творог, нату-
ральный сыр, брынза; отвары и бульоны, супы
овощные и супы-кремы из разрешенных про-
дуктов питания (только с картофельным и ку-
курузным крахмалом); все овощи (кроме капу-
сты, плодов стручковых растений и злаков); все
фрукты; чай, какао, кофе натуральный (не ко-
фейные напитки), минеральные воды без солей
йода и брома; глюкоза, сахароза, мёд, кисель,
336 Частная дерматология
компот, мусс, варенье, повидло, джем, марме-
лад, желе, конфитюр, пудинги и кремы из раз-
решенной муки и крахмала; сливочное и рас-
тительное масло, маргарин, чистый майонез;
зелень, орехи, оливы, пряности, уксус, поварен-
ная соль без солей йода, арахисовое масло (Ми-
хеев Г. Н. и соавт., 1999).
Кортикостероиды
В первые 3—4 недели терапии препаратами
сульфонового ряда (ДДС или его производные)
рекомендуется присоединить кортикостероиды
в средних дозах: преднизолон по 20—40 мг в
сутки.
Сульфапиридин
При непереносимости диаминодифенилсульфо-
на (ДДС, дапсон, авлосульфон) назначают суль-
фапиридин по 1,0—1,5 г в сутки.
Колхицин
Альтернативными препаратами для лечения гер-
петиформного дерматоза Дюринга являются
колхицин по 0,5 мг/день 3 раза в сутки в ком-
бинации с системными кортикостероидами.
Циклоспорин
При торпидных случаях заболевания рекомен-
дуется циклоспорин А (сандиммун) по 5 мг на
кг веса (суточная доза).
Местная терапия
Назначают водные растворы анилиновых краси-
телей, аэрозоли, теплые ванны с перманганатом
калия. Рекомендуется применение 3—5 % поли-
доканола на кремовой основе, а также 1 % гид-
рокортизоновый крем.
Буллезный пемфигоид
Речь идет об обычно протекающем хронически,
относительно доброкачественном, образующем
пузыри заболевании часто ограниченной дли-
тельности, которое характеризуется высыпани-
ем напряженно наполненных пузырей на нор-
мальной или воспаленно покрасневшей коже.
Этнической и половой предрасположенности
нет. Отсутствует ассоциация с HLA-антигенами.
Заболевание встречается чаще всего у пациентов
после 70 лет. Очень редко может возникать в
детском возрасте: ювенильный пемфигоид.
Этиопатогенез
Этиология неизвестна. В основе этого заболева-
ния лежит аутоиммунная реакция с образованием
аутоантител на протеин с молекулярным весом
230 kD, который является нормальной составной
частью базальной мембраны любого многослой-
ного пластинчатого эпителия и синтезируется
базальными кератиноцитами. Буллезный пемфи-
гоидный антиген (ВРА) расположен преимуще-
ственно интрацеллюлярно на цитоплазматичес-
ких пластинках половинных десмосом базальных
клеток и является близким родственником дес-
мосомального адгезионного протеина десмопла-
кин I. Сыворотка у 80—90 % пациентов содер-
жит антитела класса IgG к пемфигиодным
антителам с молекулярным весом 230 kD (ВРА-
1), 30—50 % пациентов имеют антитела, которые
реагируют с еще одним антигеном молекулярно-
го веса 160—180 kD (ВРА2), который, вероятно,
расположен примущественно экстрацеллюлярно
в области lamina lucida базально-мембранной
зоны. Предполагается, что связывание антител
на пемфигоидно-антигенном комплексе посред-
ством активации факторов комплемента, а так-
же действия воспалительных клеток (эозино-
фильных и нейтрофильных гранулоцитов) и
продуктов их секреции вызывает нарушение
целостности связей между эпидермисом и дер-
мой и тем самым образование субэпидермальных
пузырей.
Буллезный пемфигоид встречается и как па-
ранеопластический синдром. Примерно у 16—
30 % преимущественно пожилых пациентов в
связи со вспышкой заболевания или позднее
обнаруживаются злокачественные опухоли (кар-
цинома простаты, грудной железы или бронхи-
альная карцинома). Временная взаимосвязь меж-
ду возникновением опухоли и буллезным
пемфигоидом, однако, очень редко бывает од-
нозначной, так что патогенетическую связь мож-
но лишь предположить.
Возможна также провокация заболевания по-
средством лекарственных средств, например
сульфадимезин, пенициллин, фуросемид или
диазепам. Описано возникновение буллезного
пемфигоида после локального применения 5-
фторурацила, УФ- или рентгеновского облуче-
ния.
Примечательно возможное сосуществование
буллезного пемфигоида с другими заболевани-
ями, при которых имеется или дискутируется
аутоиммунопатогенез: полимиозит, вульгарная
пузырчатка, листовидная пузырчатка, герпети-
формный дерматоз, системная красная волчан-
ка, язвенный колит, нефрит, хронический по-
лиартрит, красный плоский лишай и псориаз. В
пользу инфекционной причины пока свиде-
тельств не обнаружено.
Глава 28, Пузырные дерматозы
337
Клиническая картина
Местами преимущественной локализации симмет-
ричных высыпаний являются боковые участки
шеи, подмышечные впадины, ингвинальные
складки, внутренняя сторона бедер и верхняя
часть живота. На видимо здоровой коже, но чаще
на элевированных эритемах, без причины вдруг
появляются упругие пузыри часто причудливой
формы с прозрачным содержимым. Их диаметр
от нескольких миллиметров до нескольких сан-
тиметров. Вначале высыпания напоминают мно-
гоформную эритему. Типичным является то, что
позднее у части пузырей имеется геморрагичес-
кое содержимое, так как при субэпидермальном
нарушении целостности разрушаются поверхно-
стные капилляры сосочкового слоя. Поэтому воз-
никающие эрозии часто покрываются геморраги-
ческой корочкой. Упругие пузыри гораздо более
устойчивы, чем пузыри при вульгарной пузыр-
чатке, так как их покрышку образует весь эпи-
дермис. Когда они вскрываются, возникают эро-
зии с остатками пузыря по краям, которые
заживают по направлению от края. В интертри-
гинозных участках пузыри вскрываются быстрее
вследствие мацерации, возникают плоские, иног-
да покрытые геморрагическими корочками эро-
зии. Если пузыри причудливой формы возника-
ют сгруппированно, то заболевание может
напоминать герпетиформный дерматит.
Изменения на слизистой полости рта в фор-
ме маленьких пузырей или чаще вторичных, по
форме четко ограниченных эрозий без фибри-
нозных отложений имеют место в 20-30 % слу-
чаев и приводят к болезненным ощущениям. Их
тенденция к выздоровлению незначительна.
Очень редки эрозивные изменения слизистой
полости рта в качестве первичных или исклю-
чительных проявлений буллезного пемфигоида.
Изменения на других слизистых, таких как
гортань, глотка, конъюнктива и генитоанальная
область присутствуют очень редко.
Начинающиеся высыпания могут зудеть, а
эрозии — причинять боль. Иногда зуд являет-
ся единственным симптомом до того, как через
несколько месяцев появятся пузыри. Общее со-
стояние вначале хорошее. В течении заболевания
вследствие рецидивирующих высыпаний, поте-
ри аппетита, снижения веса могут иметь место
общая слабость и температура.
Заболевание протекает хронически с присту-
пами и ремиссиями в течение месяцев и лет. При
отсутствии лечения смертность ниже, чем при
вульгарной пузырчатке. Она оценивается при-
мерно в 40 % и наступает вследствие вторичных
осложнений, таких как бактериальные инфек-
ции — бронхопневмония, сепсис. Прогноз у лиц
пожилого возраста со слабой конституцией сле-
дует делать осторожно.
Лабораторные данные
Типичны вторичные анемия, лейкоцитоз с лег-
кой эозинофилией и повышение СОЭ. Содер-
жание альбумина в крови может быть снижено.
Показатели IgE в сыворотке повышены у 60 %
пациентов. Специфические внутренние измене-
ния не поддаются определению лабораторными
методами.
Гистопатология
По краю пузырей наблюдаются мельчайшие мик-
ропузырьки между эпидермисом и роговым слоем.
В области пузыря его появление вызвано обра-
зованием субэпидермальной трещины. Покрышка
пузыря состоит целиком из эпидермиса, вначале
без дегенеративных изменений. В качестве содер-
жимого пузыря определяются сыворотка с фиб-
риновыми нитями, эозинофильные лейкоциты.
Кожные изменения при буллезном пемфигоиде
невоспалительного типа имеют лишь редкие вос-
палительные инфильтраты из лимфоцитов и не-
скольких эозинофильных гранулоцитов. При вос-
палительном типе буллезного пемфигоида,
напротив, в зависимости от его выраженности в
роговом слое имеются признаки васкулита с оте-
ком эндотелия, утолщением стенок капилляров
и периваскулярным инфильтратом из лимфоци-
тов, нейтрофильных лейкоцитов с незначитель-
ной лейкоцитоклазией и различным количеством
эозинофильных лейкоцитов. Иногда в сосочках
имеют место ограниченные скопления нейтрофи-
лов и эозинофилов, которые позволяют вспом-
нить о межсосочковом микроабсцессе при герпе-
тиформном дерматите. Ведущая к образованию
пузыря трещина при наблюдении под электрон-
ным микроскопом локализуется между цитоплаз-
матической мембраной базальных клеток и
Lamina densa базальной мембраны в пределах
Lamina lucida, места аутоиммунной реакции.
Цитология мазка из основания пузыря
Поскольку пузырь не акантолитический, нет и
пемфигусных клеток. Тест Тцанка отрицательный.
Иммунологические исследования
Прямая иммунофлуоресценция доказывает нали-
чие в области пузыря, в его краевой зоне и в
здоровой коже отложений иммуно-глобулинов и
факторов комплемента в области базально-мем-
бранной зоны (в пределах Lamina lucida), кото-
338 Частная дерматология
рые имеют характерный линейный рисунок.
Чаще всего обнаруживается СЗ вместе с IgG,
главным образом подклассов IgG2 и IgG4, реже
IgA или IgM. И лишь изредка, прежде всего на
ранних стадиях заболевания, подтверждается
наличие отложений исключительно СЗ. Непря-
мая иммунофлуоресценция доказывает наличие
в сыворотке у 70—80 % пациентов циркулиру-
ющих антител в качестве антител базально-мем-
бранной зоны (пемфигоидных антител), которые,
также как и связанные in vivo пемфигоидные
антитела, связываются с базально-мембранной
зоной многослойного эпителия и образуют ли-
нейный узор иммунофлуоресценции. Соответ-
ствующие антисубстраты для непрямой иммуно-
флуоресценции — пищевод обезьяны, или
препарированная в поваренной соли нормальная
человеческая кожа, в которой посредством пред-
варительной инкубации в одномолярным раство-
ре поваренной соли индуцировано образование
трещины (в Lamina luciday, здесь локализуются
IgG — преципитаты чаще на эпидермальной
стороне кожи. Циркулирующие в сыворотке
пемфигоидные антитела относятся преимуще-
ственно к классу IgG, примерно в 25% — к клас-
су IgA. Строгой корреляции между повышени-
ем титра антител и активностью заболевания не
существует.
Дифференциальный диагноз
Проводится в первую очередь с вульгарной пу-
зырчаткой, герпетиформным дерматозом и при-
обретенным буллезным эпидермолизом. Много-
формная экссудативная эритема возникает остро
и очень редко бывает у пожилых людей. Важно
отграничение от мультиформных лекарственных
экзантем. Если напряженные пузыри локализу-
ются ограниченно на ногах, то следует подумать
о буллезном дерматозе при сахарном диабете,
при дисгидрозиформном буллезном пемфигои-
де — о дисгидрозиформной экземе или микозе.
Лечение
Системные глюкокортикоиды можно рекомен-
довать как средство выбора. Поскольку заболе-
вание поражает чаще всего довольно пожилых
людей и его течение относительно доброкаче-
ственное, дозы выбираются меньшими, чем при
вульгарной пузырчатке. Для достижения ремис-
сии обычно бывает достаточно 40—80 мг пред-
низолона ежедневно или же изодозы других глю-
кокортикоидов (метилпреднизолона, урбазона,
фторкортолона, ультралана). Из-за возраста па-
циентов такие побочные действия поддержива-
ющей терапии, как сахарный диабет, остеопороз
или подверженность воздействию инфекций,
могут проявиться раньше и быть более выражен-
ными. При достижении ремиссии доза глюко-
кортикоидов должна быть снижена до необхо-
димой при поддерживающей терапии.
Комбинация глюкокортикоидов с азатиопри-
ном (имурек вначале 100—150, затем 50—100 мг
ежедневно) или хлорамбуцилом (натулан 4—6 мг
ежедневно) для достижения ремиссии при бул-
лезном пемфигоиде требуется редко. Рекомен-
дуется метотрексат (15—50 мг внутривенно раз в
неделю или 5—10 г перорально с 12-часовыми
интервалами 3 раза в неделю) как моносредство
или в комбинации с глюкокортикоидами. Од-
нако у большинства пожилых пациентов следу-
ет учесть возможность медикаментозной имму-
носупрессии и вторичных бактериальных или
микотических инфекций.
Сульфоны, такие как диаминодифенилсуль-
фон (дапсон 50—100 мг ежедневно) эффективны
у большинства пациентов с легким течением бул-
лезного пемфигоида в качестве единственного
препарата или же в комбинации с глюкокорти-
коидами в относительно низкой дозировке (20—
30 мг эквивалента преднизолона ежедневно).
Следует учесть и возможные побочные действия.
В отдельных случаях сообщается о благопри-
ятном эффекте при буллезном пемфигоиде анти-
биотикотерапии тетрациклинами или эритроми-
цином. Плазмаферез при буллезном пемфигоиде
имеет стероидосберегающее действие, но с учетом
пожилого возраста пациентов и отсутствия кор-
реляции между титрами антител и активностью
заболевания показан лишь в небольшом числе
случаев.
Для местного лечения рекомендуется после
вскрытия пузырей использование дезинфициру-
ющих средств с красителем (бриллиантовая зе-
лень 0,1—0,5 % водный раствор), поливидон-йод,
мази с антибиотиками и ванны с антисептичес-
кими добавками; при больших по площади эро-
зиях — накладка марлевых повязок для ран.
Может помочь и современный хирургический
перевязочный материал. При локализованном
заболевании можно использовать глюкокорти-
коиды местно.
Рубцующийся
пемфигоид
Синонимы: доброкачественный пемфигоид сли-
зистых оболочек, буллезный дерматит слизистых.
Глава 28. Пузырные дерматозы 339
Хроническое заболевание с образованием пу-
зырей, которое приводит к рубцеванию и пора-
жает преимущественно конъюнктивы и слизис-
тые оболочки. Заболевание редкое, болеют чаще
женщины (2:1), оно не имеет этнических вза-
имосвязей и проявляется обычно у лиц старше
60 лет.
Этиопатогенез
Имеющиеся в настоящее врямя в распоряжении
морфологические и иммунологические данные
указывают на тесную взаимосвязь с буллезным
пемфигоидом. В пользу этой версии свидетель-
ствует и наличие пациентов, у которых имеют
место генерализованные буллезные высыпания,
очень похожие на буллезный пемфигоид. Одна-
ко неизвестно, направлен ли иммунный ответ при
рубцующемся пемфигоиде против одного анти-
гена или против антигенного комплекса в базаль-
но-мембранной зоне, локализация, распределе-
ние и функция которого отличаются от антигена
буллезного пемфигоида. Почему рубцующаяся
атрофия возникает на слизистых, неизвестно.
Возможно, эти антигены имеют в Lamina lucida
особое функциональное значение для заживле-
ния эпидермальных ран.
Клиническая картина
У 90 % предпочтительно поражаются конъюн-
ктивы и слизистая оболочка полости рта, хотя
заболевание может охватывать и слизистую носа,
глотки, гениталий и анальной области. Кожные
проявления имеют место у 25 % пациентов.
Глаза могут поражаться отдельно. Чаще за-
болевание начинается с одной стороны. Через
1-2 года поражается и другой глаз, вначале при
наличии картины катарального конъюнктиви-
та. При точном наблюдении можно установить
наличие светлых пузырьков, которые быстро
вскрываются. После этого происходит рубце-
видное сморщивание, отсюда и прежнее назва-
ние заболевания: эссенциальное сморщивание
конъюнктивы. Между бульбарной и пальпеб-
ральной конъюнктивами возникают рубцовые
синехии; закрыть веко становится невозмож-
ным. Движения глаз ограничены. Энтропия
ведет к вторичным изменениям роговицы с
возникновением паннуса или же рубцующих-
ся и ведущих к слепоте изъязвлений роговицы.
Вследствие рубцового сморщивания век со сме-
щением выводных слезных протоков и атрофией
бокаловидных клеток конъюнктива высыхает и
образуется ксерофтальмия и панофтальмия,
которая в конце концов приводит к потере зре-
ния.
Слизистая рта: и здесь возникают рецидиви-
рующие пузыри, которые быстро вскрываются
и ведут к болезненным, оставляющим при за-
живлении рубцы, эрозиям. При локализации на
уздечке языка рубцовое сморщивание приводит
к ограничению подвижности языка, при лока-
лизации поражений на мягком небе, дугах мин-
далин и слизистой щек может возникнуть му-
козогенный тризм с ограничении возможности
приема пищи.
На других слизистых, например вульве, мо-
гут также образоваться похожие рубцовые адге-
зии, синехии или стенозы.
В отличие от глаз и слизистой рта, изменения
на коже встречаются только у 25 % пациентов
и протекают легко. На одном или нескольких
местах на эритематозном основании образуются
рецидивирующие и часто геморррагические пу-
зыри, которые имеют прочную покрышку и за-
живают также с образованием атрофических,
чаще всего гиперпигментированных рубцов. При
локализации на волосистой части головы разви-
вается рубцовая алопеция по типу псевдопела-
ды Брока. Очень редко имеют место диссеми-
нированные приступообразные высыпания
пузырей, как при буллезном пемфигоиде.
Течение заболевания часто волнообразное, на
протяжении нескольких лет, без существенного
влияния на общее состояние. Чем быстрее тече-
ние, тем хуже прогноз. Из-за трудностей при
приеме пищи могут развиться дистрофия и ка-
хексия. Слепота наступает у 30—60 % пациентов.
Очень редки случаи развития карциномы на
образующихся рубцовых изменениях слизистой.
Лабораторные данные
Цитология мазка из основания пузыря
Акантолитические пемфигусные клетки отсут-
ствуют.
Гистопатология
Типично образование субэпидермального пузыря
без признаков акантолиза. В верхнем роговом
слое воспалительный инфильтрат из лимфоци-
тов, плазменных клеток, а также эозинофиль-
ных лейкоцитов. Позднее сильная фибробласти-
ческая активность с фиброзом, ангиоплазией и
рубцовым сморщиванием в пределах базально-
мембранной зоны.
Иммунологические исследования
Прямой иммунофлуоресценцией в биопсиях сли-
зистой у 60—80 % пациентов доказывается на-
личие линейных отложений только IgG либо в
комбинации с IgA в базально — мембранной
340 Частная дерматология
зоне. В базальной мембране конъюнктив часто
обнаруживаются отложения СЗ и фибриногена.
Если прямая иммунофлуоресценция дает нега-
тивный результат, следует вновь взять свежую
биопсию из периочаговой эритемы. Посредством
непрямой иммунофлуоресценции в сыворотке
только у 10—20 % пациентов подтверждается
наличие циркулирующих пемфигоидных анти-
тел (антитела антибазально-мембранной зоны)
классов IgG или IgA; их титры, однако, даже при
одновременном вовлечении в процесс кожи чаще
всего очень низкие. Еще не точно установлено,
реагируют ли аутоантитела при рубцующемся
пемфигоиде слизистых с пемфигоидным анти-
геном или с родственным антигенным комплек-
сом в пределах базально-мембранной зоны. При
антигенном картировании методом прямой им-
мунофлуоресценции на дне пузыря связывают-
ся коллагеновые антитела типа IV: юнктиональ-
ный пузырь.
Электронная микроскопия
Электронно-микроскопически при рубцующемся
пемфигоиде имеет место предпочтительное свя-
зывание антител на экстрацеллюлярном участ-
ке пемфигоидного антигена. Некоторые пациен-
ты с клинической картиной рубцующегося
пемфигоида слизистых имеют антитела к эпи-
лигрину; эпилигрин (или ВМ 600) локализует-
ся в lamina lucida и представляет собой адгези-
онный протеин, который опосредует сцепление
базальных кератиноцитов с базально-мембран-
ной зоной.
Дифференциальный диагноз
При начале заболевания очень важно отграни-
чить его от вульгарной пузырчатки, красного
эрозивного лишая, болезни Бехчета и хроничес-
кой красной волчанки. Цитологические, гисто-
логические и иммунологические исследования
играют решающую роль. При возникновении
глазной симптоматики с рубцующимися сенехи-
ями диагноз поставить просто. У пациентов с
вовлечением в процесс кожи трудным является
дифференцировка заболевания с буллезным эпи-
дермолизом, это возможно только при помощи
электронной микроскопии.
Лечение
Комбинированная терапия глюкокортикоидами
и иммуносупрессивными препаратами (азатиоп-
рин или циклофосфамид) приводит к успеху
только при генерализованных буллезных высы-
паниях, а не при хронических изменениях конъ-
юнктив и слизистых. Показано лечение дапсо-
ном по 50—200 мг ежедневно, изотретиноином
(роаккутаном), ацитретином (неотигазоном) или
циклофосфамидом (эндоксаном). Возможно так-
же применение циклоспорина А (сандиммуна)
перорально (2,5—5,0 мг/кг веса ежедневно) или
местно в дополнение к оперативным вмешатель-
ствам, которые часто показаны в поздних ста-
диях рубцевания.
Поражения глаз должен лечить офтальмолог.
Возможно применение внутриочаговых инъек-
ций глюкокортикоидных суспензий, а также
контактных линз. При явлениях на слизистой
полости рта показана симптоматическая терапия
(субкутин, динексан, хервирос, камиллосан, ка-
восан), а также глюкокортикоиды местно (мазь
Волон А, донтизолон, иногда впрыскивание кри-
сталлической суспензии триамицинолона — Во-
лон А 1:4 в растворе с мепивакаином). При раз-
витии тризма — эксцизии зарубцевавшейся
слизистой рта и трансплантация нормальной
кожи, при этом заболевание в соответствующем
участке затухает. Это касается и ограниченных
очагов на коже.
Глава 29
Болезни придатков кожи
Себорея 341
Акне (вульгарные угри)342
Розацеа 345
Гнездная алопеция 351
В этой группе заболеваний придатков кожи (бо-
лезни волос, ногтей, сальных и потовых желез)
будут рассмотрены акне (вульгарные угри), ро-
зацеа (розовые угри) и алопеции (нерубцовые —
гнездная, андрогенетическая, диффузная).
Себорея
Под себореей понимают болезненное состояние
кожи, связанное с гиперпродукцией сальными
железами кожного сала с измененным химичес-
ким составом. Таким образом, патологические
изменения при себорее связаны с количествен-
ными и качественными изменениями кожного
сала.
При повышенном производстве сала на учас-
тках, богатых сальными железами, кожа сильно
блестит. Если провести пальцем по назолабиаль-
ным складкам, лбу, ушным раковинам или лож-
бинке на груди, то ощущается жирная сальная
пленка. Блеск этой сально-потовой пленки ча-
сто косметически нарушает вид лица. При силь-
ной себорее волосы после каждого мытья стано-
вятся жирными, слипаются и теряют свою
форму. Одновременно с повышенной жирнос-
тью кожи головы наблюдается жирное шелуше-
ние. При себорее и недостаточной гигиене ощу-
щается неприятный запах тела. Летом в горячих
климатических условиях себорея особенно вы-
ражена, но заметна лишь из-за сильного поте-
ния. В прохладные зимние месяцы она не так
бросается в глаза.
При себорее существенно изменяется состав
кожного сала. По сравнению со здоровыми ли-
цами, у больных себореей в кожном сале замет-
но повышено количество андрогенов и снижено
- эстрогенов. Однако качественные изменения
кожного сала при себорее касаются прежде всего
концентрации ненасыщенной жирной кислоты —
линолевой. В составе кожного сала изменяется
соотношение между высшими и низшими жир-
ными кислотами в связи с увеличением концен-
трации высших свободных жирных кислот. При
себорее в кожном сале уменьшается концентра-
ция линолевой кислоты, что приводит к увели-
чению pH кожи, изменению проницаемости эпи-
телия, росту микроорганизмов на поверхности
кожи.
Причина себореи неизвестна. Важную роль
играют наследственные факторы, а также дей-
ствие различных гормонов. Определенную роль
в развитии себореи играют эмоциональные фак-
торы, различные расстройства со стороны не-
рвной системы.
Возникновение себореи у женщин связано с
изменениями нормальных соотношений в орга-
низме между андрогенами и прогестероном. У
пациенток чаще всего обнаруживается гиперан-
дрогенемия в сочетании с гипоэстрогенией или
гиперпрогестеронемией. На важную роль гор-
монов в патогенезе себореи у женщин указы-
вают следующие факторы: начало заболевания
в периоде полового созревания, обострение
процесса за неделю перед менструацией (лю-
теиновая фаза менструального цикла), частая
связь заболевания с нарушениями менструаль-
ного цикла, поздним менархе; развитие себо-
реи после беременности, прерывания беремен-
ности, при воспалительных процессах органов
малого таза.
Основной причиной себореи у мужчин явля-
ются изменения соотношений в организме между
андрогенами. Гиперандрогения у мужчин может
быть связана с эндокринным дисбалансом, а
также с андроген-продуцирующими опухолями
(семинома).
341
342 Частная дерматология
Избыточная себорея наблюдается при болез-
ни Паркинсона, болезни Иценко-Кушинга и ле-
таргическом энцефалите. Гиперпродукция кож-
ного сала возможна при длительном приеме
гормонов (глюкокортикоидов, анаболиков, те-
стостерона, прогестерона). Себорея приобрета-
ет значение вследствие тесной связи с рядом
дерматозов, для которых повышенная секреция
сала имеет причинное значение или же играет
хотя бы поддерживающую роль. Причинное
значение себорея имеет при вульгарных угрях,
грамнегативном фолликулите, себорейной эк-
земе и розацеа. Хотя последняя не всегда свя-
зана с себореей. Себорейный статус дает также
благоприятную питательную среду многочис-
ленным патогенным бактериям и грибам, по-
этому себорейная кожа склонна к пиодермиям
и грибковым заболеваниям. Прогностические
оценки при себорее должны быть достаточно
сдержанными, поскольку речь идет о доволь-
но длительном конституциональном состоянии.
Сильная себорея уменьшается только в пожи-
лом возрасте и то незначительно.
Акне
(вульгарные угри)
Самое распространенное заболевание кожи, по-
ражающее до 85 % лиц в возрасте от 12 до 25 лет
и 11 % лиц в возрасте старше 25 лет. Это забо-
левание широко распространено в мире среди
всех рас. Клинически акне достигает пика меж-
ду 16 и 18-м годом жизни и в отдельных случа-
ях может сохраняться до возраста 40 лет и стар-
ше (акне позднего типа). У девушек акне
начинается раньше, чем у юношей. Комедоно-
вое акне чаще всего встречается примерно на 12-
м году жизни; папулезное и пустулезное акне —
на 16-м году жизни; глубокие воспалительные
формы возникают позднее, между 16-м и 20-м
годом жизни. Возрастные различия и меняюща-
яся клиническая картина при акне требуют бо-
лее точных знаний о патогенезе заболевания, а
также клинического опыта при его лечении.
Акне могут привести к обезображиванию и об-
разованию стойких рубцов. Но даже легкие слу-
чаи вызывают значительный эмоциональный
дистресс и имеют серьезные психосоциальные по-
следствия.
Акне представляет собой заболевание волося-
ных фолликулов и сальных желез. В патогенезе
акне имеют значение четыре взаимосвязанных
фактора: патологический фолликулярный гиперке-
ратоз, избыточное образование секрета сальных
желез, размножение Propionibacterium acnes
(Р. acnes) и воспаление. Кроме этого, на харак-
тер и объем секреции сальных желез влияют
андрогены, что также может играть важную роль
в патогенезе акне, хотя данный механизм оста-
ется неясным.
Различают невоспалительные и воспалитель-
ные элементы акне. Невоспалительные элемен-
ты представлены комедонами и милиумами.
Комедоны (acne comedonica) — невоспалитель-
ные элементы угревой сыпи, которые проявля-
ются в двух видах: открытые (черные) и закры-
тые (белые). Открытые комедоны выглядят как
черно-коричневые точки, а закрытые имеют
форму небольшого узелка цвета окружающей
кожи или более светлого цвета. Возникновение
комедонов связано с застоем рогово-жирных
масс в сальных железах в результате гиперпро-
лиферации клеток эпителия выводного прото-
ка. Черно-коричневый цвет открытых комедо-
нов вызван окислением жиров кожного сала под
воздействием кислорода.
Милиумы (микрокомедоны) — единичные, реже
множественные мелкие беловатые, молочно-бе-
лые или беловато-желтоватые полушаровидные
узелки, размером с булавочную головку или
просяное зерно. Они возникают в результате
скопления сального секрета в устьях фоллику-
лов или в протоках сальных желез. Различают
первичные и вторичные милиумы (белые угри).
Первичные милиумы наблюдаются у детей груд-
ного возраста в период гормонального криза на
коже лица (лоб, щеки, нос, носогубные склад-
ки) и в пубертатном периоде. Вторичные мили-
умы образуются на рубцах при дистрофическом,
гиперпластическом и полидиспластическом на-
следственном буллезном эпидермолизе.
Клинические формы
Клинические формы воспалительных акне: папу-
лезные, пустулезные, индуративные, флегмоноз-
ные, конглобатные, узловатокистозные, тропичес-
кие, молниеносные, инверсные, акне-тетрада.
Папулезные акне (acne papulosa) представляют
собой воспалительный инфильтрат у основания
комедонов, в результате чего образуется неболь-
шой узелок ярко-красного цвета, слегка возвы-
шающийся над уровнем кожи.
Пустулезные акне (acne pustulosa) возникают на
месте папулезных. Воспалительный инфильтрат
увеличивается, в центре узелка появляется по-
лость с гнойным содержимым. Пустулы безбо-
лезненны, иногда вызывают умеренный зуд.
Глава 29. Болезни придатков кожи 343
Индуративные акне (acne indurata) формируют-
ся в результате мощного обширного инфильт-
рата с бугристой поверхностью вокруг воспален-
ных сально-волосяных фолликулов. Они
достигают размеров горошины и крупнее. Раз-
решаются очень медленно. Иногда на их месте
образуются небольшие келоиды.
Флегмонозные акне (acne phlegmonosa) — глу-
бокие, медленно развивающиеся фолликулярные
или перифолликулярные дермо-гиподермальные
абсцессы. Флегмонозные акне образуются в ре-
зультате слияния крупных пустул, глубоко за-
легающих и быстро созревающих, склонных к
абсцедированию. Они имеют багрово-красный,
синюшный цвет, достигают размеров сливы.
Конглобатные (шаровидные) акне (аспе
conglobata) — тяжелая форма акне, характеризу-
ющаяся возникновением, помимо комедонов и
атером, крупных полушаровидных узлов, распо-
лагающихся глубоко в дерме и достигающих
верхней части подкожно-жировой клетчатки,
размером от горошины до вишни. Вначале ин-
фильтрат плотный, красно-синюшного цвета,
болезненный. Узлы могут сливаться в конгло-
мераты, формируясь дальше в абсцессы, после
вскрытия которых остаются длительно незажи-
вающие язвы, покрытые грануляциями. После
заживления язв остаются неровные рубцы с пе-
ремычками, свищами. Локализуются конглобат-
ные угри на спине, реже на груди и лице. Тече-
ние этой формы угрей длительное (до 40-летнего
возраста и даже в течение всей жизни) и закан-
чивается формированием обезображивающих
атрофических, гипертрофических, келоидных
рубцов. Некоторые исследователи наиболее ча-
сто отмечают эту тяжелую форму угревой сыпи
у мужчин с дополнительной Y-хромосомой
(XYY).
Узловатокистозные акне (acne nodulocystica)
отличаются выраженными узелками, которые
всегда превышают в размере 1 см и глубоко вда-
ются в дерму. Они, как правило, болезненны.
Часто несколько узелков сливаются, образуя
очаги с абсцедирующими свищевыми ходами,
причем заживающие узлы могут оставлять кис-
тообразные изменения. Так возникает картина
кистозных акне. При этой форме акне может
поражаться только лицо, но иногда также грудь
и спина.
Тропические акне (acne tropicalis) — абсцеди-
рующие конглобатные акне у пожилых людей в
условиях жаркого влажного климата. Заболева-
ние развивается вследствие повышенного содер-
жания влаги в коже, что влечет за собой разрых-
ление стенки фолликула и увеличение
12* Зак. 1357
высвобождения хемоатграктивных веществ с раз-
витием усиленного воспаления кожи. Высыпа-
ния располагаются, в основном, в области спи-
ны и ягодиц, единичные элементы могут быть
на лице, передней части груди. Болеют, как пра-
вило, люди, у которых в молодые годы были
вульгарные угри.
Молниеносные акне (acne fulminans) — мол-
ниеносная угревая сыпь, акне конглобатные и
септицемия — тяжелое заболевание с внезап-
ным началом (от латинского fulmen — молния)
в виде множественных болезненных узелковых
элементов и кистозных элементов с выражен-
ной воспалительной реакцией, быстрым тече-
нием и образованием после вскрытия глубоких,
серпигинизирующих язв. Излюбленная лока-
лизация — верхняя часть груди и спины, бо-
ковые поверхности шеи и плеч. Отсутствуют
высыпания в области лица. Заболевание со-
провождается высокой температурной реакци-
ей, артралгией (боли в коленных и плечевых
суставах), миалгией и лейкоцитозом. Иногда
глубокое расплавление мягких тканей доходит
до костей, что может закончится остеолизисом.
Заболевание встречается преимущественно у
мужчин молодого возраста.
Инверсное акне (acne inversa) — конглобатное
акне с обратной клинической картиной — хро-
ническое воспалительное заболевание сальных
желез и волосяных фолликулов, проявляющее-
ся во всех зонах, кроме лица, груди и спины.
Высыпания представлены в виде узелково-вос-
палительных и фистульных комедонов, свище-
вых ходов в подмышечной, паховой и гениталь-
ной областях, на затылке, а также у женщин под
молочными железами. У пациентов с ожирени-
ем заболевание наблюдается в складках на жи-
воте. Инверсное акне является тяжелой формой
заболевания.
Акне-тетрада (acne-tetrada) — наиболее тяже-
лая форма акне, объединяющая в себе конгло-
батные, инверсные акне, folliculitis nuchae
abscedens et suffodiens (абсцедирующий и подры-
вающий фолликулит Гоффмана), а также пол-
ный или неполный Pilonidalsinus (гнойная по-
лость с пучком волос в области копчика, но
может быть и на других участках тела).
Лечение
Для лечения акне применяется местная и систем-
ная терапия.
Местное лечение
Только местное лечение показано при легких или
средней тяжести невоспалительных акне, легких
344 Частная дерматология
воспалительных акне без образования рубцов, а
также в качестве дополнения к пероральной тера-
пии средней тяжести или тяжелых акне. Местная
антиугревая терапия основана на ее воздействии
на различные патогенетические факторы вульгар-
ных акне. Она включает: местные ретиноиды
(изотретиноин, третиноин, адапален, тазаротен),
азелаиновую кислоту, бензоилпероксид, местные
антибиотики и некоторые другие вещества (сали-
циловую кислоту, цинк и местные эстрогены).
Стандартными препаратами среди местных
ретиноидов являются третиноин (полностью
транс-ретиноевая кислота, кислота витамина А)
и изотретиноин (13-цис-ретиноевая кислота).
Используются кремы, гели и растворы. Ретино-
иды являются универсальными ингибиторами
липогенеза и секреции кожного сала, тормозят
адгезию корнеоцитов в устье сальной железы,
усиливают пролиферацию эпидермоцитов, нор-
мализуют процессы кератинизации, уменьшают
воспалительную реакцию. Наиболее известные
препараты: третиноин (0,01 %; 0,1 %); айрол
(крем, раствор, примочки); изотретиноин (изот-
рекс-гель 0,05 %); атредерм (0,05 %; 0,1 %); ло-
кацид (0,05 %, 0,1 %); ретин-А (0,1 %); клинес-
фар — комбинация 0,025 % третиноина и 4 %
эритромицина. В первые 8—14 дней после при-
менения третиноина, особенно в форме геля или
раствора, происходит обострение акне с усилен-
ным образованием комедонов и пустул. Ретино-
иды применяются местно 1 раз в сутки или че-
рез день в течение 2—3 мес. В России
выпускается 1 % ретиноевая мазь — препарат с
13-цис-ретиноевой кислотой, которая обладает
высокой биодоступностью.
Разработан новый топический ретиноид 4-го
поколения в виде 0,1 %-ного геля — адапален
(дериват нафталиновой кислоты), который об-
ладает противовоспалительным ретиноидоподоб-
ным действием. Адапален воздействует путем
избирательного связывания специфических ре-
цепторов клеточных ядер эпидермиса (а и у-ре-
цепторы ретиноевой кислоты), влияя на норма-
лизацию нарушенного ороговения фолликула.
Активизация у-рецептора ретиноевой кислоты
влияет на дифференциацию кератиноцитов боль-
ше, чем на клеточную пролиферацию, которая
регулируется а-рецептором. В отличие от рети-
ноидов первого поколения адапален не связы-
вается с ретиноидными рецепторами в плазме и
имеет меньшее раздражающее действие на кожу.
Адапален тормозит различные модели воспале-
ния in vitro, а также, благодаря своей структуре,
стабилен против кислорода воздуха и, вероятно,
также против УФ-облучения.
Ретиноиды можно использовать в комбинаци-
ях с местной терапией бензоилпероксидом при
воспалительной папулопустулезной форме акне
перед системным антибактериальным лечением
или приемом внутрь изотретиноина.
Азелаиновая кислота (20 % крем; скинорен) —
это препарат для местного применения с антико-
медогенными свойствами, который предположи-
тельно вмешивается в кератинизацию при обра-
зовании кератинфиламент-аггрегирующих
протеинов. Лечебный эффект препарата связан с
влиянием на основные патогенетические звенья
угревой болезни: гиперкератинизацию стенки
волосяного фолликула, усиленное размножение
микроорганизмов в сальной железе и развитие в
ней воспалительного процесса. Обладая способ-
ностью тормозить биосинтез клеточных белков,
азелаиновая кислота оказывает умеренный цито-
статический эффект, проявляющийся в нормали-
зации процессов кератинизации в стенке волося-
ного фолликула, а также антимикробное действие
(прежде всего, в отношении Propionibacterium acne)
как на поверхности кожи, так и в сальных же-
лезах. Токсикологически азелаиновая кислота бе-
зупречна. Этот препарат можно применять как
молодым людям, так и женщинам во время бе-
ременности. Местно может наблюдаться легкое
покалывание, жжение и зуд, прежде всего в те-
чение первых минут после применения. Контак-
тные аллергии или фотодинамические реакции
едва ли можно ожидать.
Незначительное действие на фолликулярный
гиперкератоз оказывают кератолитики, такие как
салициловая кислота. При акне она применяет-
ся в различных формах, преимущественно в ра-
створах (от 0,5 % до 5 %). Сильнодействующим
антимикробным средством при акне является
бензоилпероксид, который выпускается в гелях,
лосьонах, кремах и растворах для местного при-
менения при папуло-пустулезной форме акне.
Концентрации достигают от 2 до 10 %, а в не-
которых случаях — до 20 %. Азелаиновая кислота,
как и бензоилпероксид, обладает антимикроб-
ным эффектом и особенно действует на Р. acnes
и Staphylococcus aureus. Оба препарата не вызы-
вают резистентности, т. е. изменения фенотипа
возбудителя ни in vivo, ни in vitro. Поэтому они
особенно подходят для длительной антимикроб-
ной терапии.
Системная терапия
Системная терапия акне (часто в сочетании с
местной) показана для лечения больных средней
тяжести или тяжелыми акне, особенно в случа-
ях образования рубцов, дисхромий или значи-
Глава 29. Болезни придатков кожи 345
тельных психосоциальных расстройств. Систем-
ная терапия может быть необходима при непе-
реносимости или неэффективности местного
лечения.
Средством выбора при системной антибиоти-
котерапии при акне средней и тяжелой степени
являются пероральные тетрациклины. Как пра-
вило, несколько месяцев назначают тетрацикли-
на гидрохлорид по 1,0 г в день. Дозу снижают
только после значительного клинического улуч-
шения. Препарат принимают за полчаса до еды,
чтобы обеспечить оптимальную резорбцию. Аль-
тернативно можно назначать доксициклин 50—
200 мг/день. Более благоприятным (из-за мень-
шей степени побочных действий) является
миноциклин по 100 мг/день. Можно применять
эритромицин в дозе 1,0 г/день, причем предпоч-
тительнее лекарственные формы, растворяющи-
еся в тонком кишечнике. Клиндамицин очень
эффективен, но при долгосрочной терапии имеет
Недостаток — может вызвать энтероколит. Си-
стемные антибиотики хорошо сочетаются с ме-
стными препаратами, особенно третиноином,
азелаиновой кислотой и бензоилпероксидом.
Альтернативой антибиотикотерапии акне у
женщин является гормональная терапия с исполь-
зованием препаратов с антиандрогенным действи-
ем. Основной механизм действия антиандроген-
ных препаратов заключается в снижении
циркулирующих в крови активных андрогенов и
в препятствовании образования комплекса анд-
роген-рецептор. К чистым андрогенам относят
препараты, блокирующие андрогеновые рецепто-
ры путем замещения рецепторов андрогенов в
органах-мишенях. Основными представителями
этих нестероидных анитандрогенов являются
спиронолактон (верошпирон) и флутамид.
К блокаторам андрогеновых рецепторов отно-
сится ципротерона ацетат (ЦПА). ЦПА является
сильнодействующим оральным гестагеном. Он
тормозит овуляцию и блокирует рецепторы анд-
рогенов, подавляет секрецию гонадотропных гор-
монов. В виде монопрепарата ЦПА выпускается
под названием «Андрокур-10» и «Андрокур-50».
Чаще ЦПА применяют в форме пероральных
контрацептивов в комбинации с этинилэстради-
олом (ЭЭ) при яичниковой форме гиперандро-
гении. Наиболее известным комбинированным
гормональным контрацептивом является препа-
рат «Диане-35», который содержит 35 мкг ЭЭ и
250 мг ЦПА. Однако этот препарат содержит
относительно высокие дозировки эстрогеновых и
гестагеновых компонентов, что увеличивает риск
развития побочных эффектов. Кроме того, такие
дозировки показаны при выраженных лаборатор-
ных показателях и клинических явлениях андро-
генизации.
Новым оральным контрацептивом с антиан-
дрогенным действием с содержанием в качестве
гестагена диеногеста является препарат «Жанин».
Диеногест сочетает в себе свойства прогестеро-
на и норстероидов и обладает выраженным гес-
тагенным эффектом и антиандрогенным дей-
ствием (40 % от таковых ЦПА).
Среди комбинированных оральных контра-
цептивов использование препаратов с содержа-
нием гестагенов нового поколения и низким
содержанием эстрогеновых компонентов являет-
ся оптимальным. К таким препаратам относят-
ся «Новинет» и «Регулон». В качестве гестагена
они содержат дезогестрел в дозировке 150 мкг,
в качестве эстрогена — этинилэстрадиол в до-
зировке 20 мкг и 30 мкг соответственно.
Наиболее эффективным препаратом для лече-
ния акне среднетяжелой и тяжелой формами
является изотретиноин (роаккутан), который
при системном применении в дозе 0,5 мг/кг веса
тела в сутки вызывает длительные ремиссии или
излечение у большинства больных. Однако пре-
парат обладает рядом побочных эффектов, ко-
торые следует учитывать при его назначении.
Изотретиноин (роаккутан) имеет прямой эф-
фект, воздействуя на пролиферацию и диффе-
ренцировку себоцитов, результатом чего является
выраженная себосупрессия. Себосупрессия зави-
сит от доз, она начинается через 10—12 недель
при дозах 0,1 мг/кг веса тела в сутки, а при дозе
1 мг/кг веса тела в сутки — через 4 недели. При
достаточно высокой общей дозе в течение сред-
него срока лечения (от 6 месяцев до 1 года) ре-
миссии стабильны. Побочные действия вынуж-
дают к осторожному применению системных
ретиноидов для лечения тяжелой степени акне.
В частности, у молодых женщин следует всегда
помнить о тератогенности ретиноидов и приме-
нять пероральные контрацептивные препараты
во время лечения изотретиноином.
Розацеа
Розацеа (розовые угри) — хроническое заболева-
ние кожи лица невыясненной этиологии с про-
гредиентным течением, характеризующееся час-
тым покраснением, персистирующей эритемой,
телеангиэктазиями, эпизодами воспаления, появ-
лением папул, пустул и редко узлов. В более тя-
желых случаях через много лет может развиться
уродующая гипертрофия, особенно в области
346 Частная дерматология
носа — ринофима. Заболевание начинается на
третьем-четвертом десятилетии жизни и достигает
расцвета между 40 и 50 годами. Чаще страдают
женщины, хотя гиперплазия соединительной тка-
ни и сальных желез, приводящие к ринофиме,
наблюдаются исключительно у мужчин.
Патогенез
Причина возникновения розацеа остается неиз-
вестной, хотя относительно происхождения за-
болевания имеется много теорий. В патогенезе
розацеа ведущую роль играют сосудистые нару-
шения, патология желудочно-кишечного тракта,
дисфункции эндокринной системы, психосома-
тические и иммунные нарушения, а также ряд
эндокринных факторов. Генетической предрас-
положенности к розацеа не установлено.
Экзогенные предрасполагающие факторы
Ряд алиментарных факторов (алкоголь, горячие
напитки и пряности) стимулируют слизистую обо-
лочку желудка и за счет рефлекторного действия
вызывают расширение кровеносных сосудов лица.
Полагают, что чрезмерное употребление в пищу
мяса является причиной формирования ринофи-
мы. Однако розацеа развивается и у вегетариан-
цев, что ставит под сомнение это предположение.
Нет достоверной связи между приливами крови к
лицу и приемом пищи с большим содержанием
глютамата натрия, применяемого в качестве вку-
совых добавок в мясные продукты («синдром ки-
тайских ресторанов»). Злоупотребление кофе, чаем
или колой, по мнению некоторых ученых, пато-
генетически связано с розацеа, однако тщательные
исследования показали, что приливы крови к коже
лица вызывает вода с температурой выше 60°С, а
не собственно кофе (J. К. Wilkin, 1979). Тем не
менее авторами в рекомендациях по диете боль-
ным розацеа все же предписывается исключать из
пищи продукты, провоцирующие эритему, такие
как алкоголь, горячие напитки, острые блюда и
цитрусовые.
Одна из наиболее частых причин розацеа —
наличие клещей рода Demodex folliculorum. Об-
наружение при гистологическом исследовании
клещей в отдельных инфильтратах, а также эф-
фективность акарицидных препаратов (гекса-
хлороциклогексан, кротамитон, бензил-бензоат),
метронидазола и серы при розацеа подтвержда-
ют это предположение. Вместе с тем локализа-
ция высыпных элементов розацеа преимуще-
ственно не связана с фолликулярным аппаратом,
a Demodex folliculorum обнаруживается не у всех
больных. Demodex folliculorum — физиологичес-
кий представитель микрофлоры кожи, заселение
им фолликулов значительно увеличивается с
возрастом. Наружное применение серы при ро-
зацеа ведет к клиническому улучшению, но не
к гистопатологической редукции личинок кле-
щей. Большинство авторов связывают возник-
новение экстрафациальной формы розацеа с
инфицированием Demodex folliculorum. У неко-
торых больных обнаруживаются даже специ-
фичные антитела к демодексу.
Генетические факторы
Ни одного приемлемого доказательства генети-
ческой предрасположенности к розацеа пока не
найдено, хотя иногда в семье встречается более
одного случая развития этого заболевания. Кроме
цвета кожи и семейного анамнеза, предрасполо-
женность у людей некоторых национальностей
может выступать как фактор риска в возникно-
вении розацеа.
Заболевания желудочно-кишечного тракта
По мнению большинства ученых, нарушение де-
ятельности желудочно-кишечного тракта не иг-
рает ведущей роли в патогенезе розацеа. У 50-
90 % пациентов с розацеа выявлены клинические
и гистологические признаки гастрита, а у 33 % —
патологические изменения слизистой оболочки
тощей кишки. На основании результатов иссле-
дования желудочного сока патогенетическое зна-
чение придают чаще гипо- или анацидному со-
стоянию, реже — гиперацидному. Сообщается об
ассоциации розацеа с хроническими воспалитель-
ными заболеваниями кишечника. Обращается
внимание на повышенное заселение гастроинте-
стинального тракта больных розацеа спиралефор-
мными бактериями Helicobacter pylori, которые
известны как этиологический фактор хроничес-
кого активного гастрита типа В. На основании
имеющихся исследований взаимосвязь между
розацеа и вызываемым Н. pylori заболеванием
ЖКТ нельзя полностью исключить, хотя данные
при этом значительно противоречивее, чем при
хронической уртикарии.
Патология эндокринной системы
Некоторые исследователи расценивают наличие
сахарного диабета и снижение секреции липазы
на 15 % как доказательство патогенетического
значения нарушения обмена веществ у больных
с розацеа. Овариальная и гипофизарная недоста-
точность, заболевания щитовидной железы и
другие гормональные нарушения не рассматри-
ваются как единые причины розацеа, посколь-
ку этим заболеванием страдают и мужчины, и
женщины разных возрастных групп. Отмечено
влияние надпочечниковой недостаточности на
Глава 29. Болезни придатков кожи 347
формирование розацеа на основе положительного
эффекта от введения гормонов надпочечников.
Психовегетативные нарушения
Психические факторы долгое время считали
первостепенной причиной возникновения роза-
цеа, однако целенаправленные психологические
исследования статистически не подтвердили
предполагаемую связь между розацеа и психи-
ческими факторами. Они скорее указывали на
влияние обезображивающего косметического
дефекта на психику больного.
Сосудистая патология
Клинические и экспериментальные исследова-
ния показывают, что нарушения центральной
регуляции тонуса кровеносных сосудов кожи в
области лица играют в патогенезе розацеа осо-
бую роль. Вследствие этого развиваются замед-
ление перераспределения кровотока и венозный
стаз в области оттока venae facialis sive angularis,
соответствующей наиболее частой топографии
розацеа. В область оттока лицевой вены вклю-
чается также конъюнктива, что объясняет час-
тое вовлечение глаз при этом заболевании.
На основе морфологических и лабораторных
исследований установлено (Е. И. Рыжкова,
1976), что в основе патогенеза заболевания ле-
жит ангионевроз с преимущественным пораже-
нием сосудов лица как одно из проявлений ве-
гетососудистой дистонии.
Роль вазоактивных пептидов
Вазоактивным пептидам желудочно-кишечного
тракта (VIP, пентагастрин) приписывается спо-
собность вызывать приливы. Провоцируемые
алкоголем реакции приливов связаны с его не-
достаточным ферментативным разрушением,
особенно у лиц восточных национальностей. В
качестве причины реакции приливов у больных
розацеа отмечалось также повышение содержа-
ния ряда медиаторных веществ, таких как эн-
дорфины, брадикинин и субстанция Р. Актива-
ция калликреин-кининовой системы и усиление
кининогенеза установлены у многих больных с
розацеа (О. В. Сницаренко, 1988; 1989). Вазоак-
тивная направленность действия кининов, их
способность в чрезвычайно малых концентраци-
ях изменять тонус сосудов и повышать прони-
цаемость капилляров вызывают характерные для
розацеа сосудистые изменения. Преимуществен-
ная локализация на лице, видимо, объясняется
повышенной чувствительностью расположенных
здесь брадикининовых рецепторов. Это являет-
ся подтверждением теории о вазоактивном бра-
дикинине как «эффекторе» розацеа.
Нарушения иммунной системы
У больных розацеа установлено достоверное уве-
личение уровня иммуноглобулинов всех трех
классов: А, М, G. Однако при этом не выявле-
но корреляции между уровнем иммуноглобули-
нов и длительностью и стадией заболевания. С
помощью реакции иммунофлюоресценции на
базальных мембранах иногда обнаруживаются
отложения иммуноглобулинов, реже — компле-
мент СЗ фракция. Ряд авторов при изучении
клеточного иммунитета выявляли достоверное
увеличение абсолютного числа «тотальных» и «ак-
тивных» розеткообразующих клеток и снижение
количества Т-супрессоров.
Таким образом, розацеа представляет собой
заболевание с полиэтиологическими провоциру-
ющими факторами и разнообразной клиничес-
кой картиной. Основное значение в патогенезе
розацеа принадлежит патологической реакции
сосудов, развивающейся под влиянием вазоак-
тивных пептидов, антител, циркулирующих
иммунных комплексов и других эндогенных
раздражителей (ирритантов).
Клиническая картина
В настоящее время в течении розацеа выделяют
три последовательные стадии: эритематозно-теле-
ангиэктатическая, папулопустулезная, пустулез-
но-узловатая. Начало заболевания характеризуется
ливидной приливной эритемой, причиной воз-
никновения которой могут быть многочисленные
неспецифические стимулирующие факторы: ме-
ханическое раздражение кожи, инсоляция, тем-
пературные колебания; употребление горячих
напитков, острой пищи, алкоголя. Вначале про-
должительность эритемы колеблется от несколь-
ких минут до нескольких часов, сопровождается
ощущением жара или тепла и затем бесследно
исчезает, но вскоре под влиянием провоцирую-
щих факторов появляется вновь. Локализация
чаще ограничивается назолабиальной областью
(рис. 29.1). В таком состоянии смены рецидивов
и ремиссий процесс может продолжаться многие
месяцы и годы. В дальнейшем на месте гипере-
мии появляются умеренная инфильтрация, теле-
ангиэктазия. Эритема становится более насыщен-
ной, с синюшным оттенком, нередко переходит
на окружащие нос части щек, на лоб и подборо-
док. При гистологическом исследовании на этой
стадии преобладают лишь расширенные кровенос-
ные и лимфатические сосуды. Позже на фоне
диффузного утолщения пораженной кожи возни-
кают изолированные или сгруппированные вос-
палительные розово-красные папулы, зачастую
покрытые нежными чешуйками. Папулы способ-
348 Частная дерматология
ны к персистенции в течение многих дней или
недель. Наиболее крупные элементы инфильтри-
рованы в основании. Далее большинство узелков
подвергается нагноению, образуя папулопустулы
и пустулы со стерильным содержимым. Пустули-
зация формируется за счет клеточно-опосредован-
ного иммунитета с диффузией большого количе-
ства нейтрофильных гранулоцитов в ответ на
наличие Demodex folliculorum. Поражение распро-
страняется с центрофациальной области на кожу
лба, заушных областей, передней поверхности шеи
и даже предстернального региона. Гистологичес-
ки выявляются периваскулярные лимфогистио-
цитарные инфильтраты в эпидермисе, преимуще-
ственно в области фолликулов сальных желез.
При наличии пустул в гистологической картине
наблюдается спонгиоз фолликулярной воронки,
атрофические изменения фолликулярного аппа-
рата, а также деструкция коллагеновых волокон.
В дальнейшем, вследствие хронического про-
грессирующего течения, патологический процесс
приводит к образованию воспалительных узлов,
инфильтратов и опухолевидных разрастаний за
счет прогрессирующей гиперплазии соединитель-
ной ткани и сальных желез и стойкому расши-
рению сосудов. Эти изменения затрагивают в
первую очередь нос и щеки, реже подбородок, лоб
и ушные раковины, создавая выраженный обе-
зображивающий вид. Некоторые авторы рассмат-
ривают эту стадию заболевания как самостоятель-
ную форму — ринофиму. Ринофима встречается
практически исключительно у мужчин. Форма
носа становится асимметричной, на фоне застой-
но-синюшной эритемы располагаются многочис-
ленные крупные телеангиэктазии. Функ-
Рис. 29.1. Локализация высыпаний при розацеа.
циональная активность сальных желез резко уве-
личена. При надавливании из устьев фолликулов
освобождается беловатый пастообразный секрет.
Аналогичные изменения встречаются и в местах
иной локализации: утолщение кожи на подбород-
ке — гнатофима, подушкообразное утолщение в
области надпереносья и лба — метафима, разра-
стания на мочках ушей — отофима, а также утол-
щение век из-за гиперплазии сальных желез —
блефарофима. Ринофима рассматривается как наи-
более тяжелая форма розацеа, которая развивается
не у всех больных. Это подтверждается подроб-
ными патоморфологическими исследованиями
(Е. И. Рыжкова, 1976).
Некоторыми исследователями (G. Piewig, 1997)
розацеа подразделяется на три степени тяжести,
которые обычно соответствуют стадиям разви-
тия заболевания.
Розацеа I. Персистирующие эритемы и теле-
ангиэктазии. С развитием заболевания эритема
сохраняется от нескольких часов до нескольких
дней (застойная эритема). При этом добавляются
телеангиэктазии, которые расположены главным
образом назолабиально и на щеках и космети-
чески могут мешать больным.
Розацеа II. Папулы, папулопустулы и пустулы.
В центральной части лица встречаются часто изо-
лированные или сгруппированные полусферичес-
кие, воспалительные, гиперемированные папулы
с мелкопластинчатым шелушением, которые на-
блюдаются в течение нескольких дней или не-
дель. Также могут быть папулопустулы, пусту-
лы и легкий отек. Пустулы имеют нормальную
бактериальную флору или являются стерильны-
ми. Комедоны не развиваются. Разрешение вос-
палительных высыпаний не приводит к рубцам.
Впоследствии розацеа может захватывать не толь-
ко центральную часть лица, но и область лба на
границе роста волос, боковые участки шеи, рет-
роаурикулярный регион и предстернальную об-
ласть. Кожа в этих зонах характеризуется усилен-
ным кровообращением и утолщена.
Розацеа III. Воспалительные узлы и бляшки.
При дальнейшем развитии процесса розацеа
может сопровождаться воспалительными узлами,
захватывающими большие участки, а также ин-
фильтратами с тенденцией к воспалению и диф-
фузной тканевой гиперплазии, т. е. образованию
бугров или фим. Это касается особенно щек и
носа (ринофима), реже подбородка (гнатофима),
лба (метафима), или ушей (отофима). Кожа у
пациентов с крупными порами, воспаленно утол-
щена, отечна («апельсиновая кожура»), причем
имеются воспалительные инфильтраты, увели-
чение соединительной ткани, гиперплазия саль-
Глава 29. Болезни придатков кожи
349
ных желез, увеличение кожи в объеме. Могут
возникать утолщения размером с мизинец. На
этой основе может развиться ринофима.
Клинические формы
Различают несколько особых клинических форм
розацеа. Стероидпровоцированная (стероидная)
розацеа развивается у больных, длительное вре-
мя применяющих кортикостероидные мази, осо-
бенно фторированные, по поводу того или ино-
го дерматоза. В результате возникает феномен
«стероидной кожи» с легкой субатрофией и об-
ширной темно-красной эритемой, на поверхно-
сти которой расположены телеангиэктазии и па-
пулопустулезные элементы. После отмены
местных кортикостероидов, как правило, отме-
чается временное обострение.
При люпоидной, или гранулематозной розацеа
на фоне эритемы, локализованной главным об-
разом периорбитально и периорально, густо рас-
положены диссеминированные буровато-красные
папулы или небольшие узлы, при диаскопии
которых порой выявляются желто-бурые пятна.
Прилегая друг к другу, папулы образуют неров-
ную, бугристую поверхность.
Конглобатная розацеа (rosacea conglobata) раз-
вивается на месте уже существующей розацеа и
характеризуется образованием крупных шаро-
видных абсцедирующих узлов и индуцирован-
ных фистул. Подобная аггравация нередко на-
ступает после приема препаратов, в состав
которых входят галогены (йод, бром).
Молниеносная розацеа (rosacea fulminans) на-
блюдается практически только у молодых жен-
щин и представляет собой наиболее тяжелый
вариант конглобатной розацеа. Заболевание на-
чинается остро, высыпания локализуются ис-
ключительно в области лица, отсутствуют при-
знаки себореи. Причины молниеносной формы
розацеа неизвестны. Предполагается влияние
психомоциональных. факторов, гормональных
нарушений, беременности. Молниеносная роза-
цеа начинается внезапно. Общее состояние стра-
дает незначительно. Преимущественная локали-
зация высыпаний — лоб, щеки, подбородок.
Образуются выраженный отек и эритема синюш-
но-красного цвета, папулы и пустулы, узлы по-
лушаровидной и шаровидной форм. Воспали-
тельные узлы быстро сливаются в мощные
конгломераты, появляется флюктуация, образу-
ются синусы и фистулы.
Грамнегативная розацеа характеризуется фор-
мированием многочисленных фолликулитов.
При исследований содержимого пустул обнару-
живаются грамотрицательные бактерии. Грамне-
гативные фолликулиты являются осложнением
длительной, нерациональной терапии розацеа
антибиотиками, преимущественно тетрациклино-
вого ряда.
Розацеа-лимфоэдема (солидный персистирую-
щий отек лица, хроническая персистирующая
эритема и отек лица, болезнь Морбигана) —
редкая форма розацеа. Заболевание проявляется
центрофациально с поражением средней трети
лица. При этой болезни на фоне типичных для
розацеа морфологических элементов наблюдает-
ся крепкий, не сохраняющий ямки при надав-
ливании, отек на лбу, подбородке, веках, носу
и щеках. Речь идет не о собственно отеке, а об
увеличении соединительной ткани и сопутству-
ющем этому фиброзе, возникающих на фоне
хронического воспаления и лимфостаза. Типич-
ным является темно-красного цвета с фиолето-
вым оттенком выраженный отек с переходом в
индурацию. Течение вначале хроническое реци-
дивирующее, а затем персистирующее. Субъек-
тивных симптомов заболевания не отмечено,
однако косметически заметно огрубение конту-
ров лица. Гистопатологически имеется фиброз,
отек с раширением лимфатических сосудов, а
также периваскулярный и перифолликулярный
гистиоцитарный инфильтрат. Характерно увели-
ченное количество тучных клеток. Патогенети-
ческое значение имеют, возможно, хронические
воспалительные реакции с обструкцией лимфа-
тических сосудов, нарушения микроциркуляции
и индуцированный тучными клетками фиброз.
Лечение
Для лечения розацеа применяются различные
препараты, что объясняется многообразием эти-
ологических и патогенетических факторов дер-
матоза, его стадией и клинической формой за-
болевания.
Местная терапия
Местная терапия определяется стадией и фор-
мой заболевания. В период эритематозной ста-
дии применяются примочки 1—2% раствора
борной кислоты, 1—2 % раствора резорцина, ра-
стительные примочки (настойки чая, ромаш-
ки, череды, шалфея, зверобоя, корня алтея).
Холодные примочки оказывают сосудосужива-
ющее действие, устраняют чувство жжения и
жара.
Антидемодикозным действием обладают пре-
параты серы для местного применения, а также
10 % суспензия бензилбензоата, мазь Вилькин-
сона, мазь «Ям», спрегаль, трихополовые мази и
кремы.
350 Частная дерматология
Метронидазол при местном применении в
виде 0,75 % и 1 % геля или крема не только не
уступает пероральному приему, но и конкури-
рует по эффективности с тетрациклинами. От-
мечен положительный эффект использования
примочек раствора клиона, в 100 мл которого
содержится 500 мг метронидазола (Хамаганова
И. В., 1992).
При лечении больных папулопустулезной ро-
зацеа применяют 20 % крем с азелаиновой кис-
лотой. Азелаиновая кислота уменьшает содержа-
ние свободных жирных кислот в липидах кожи
и нормализует процессы кератинизации в про-
токах сальных желез.
Имеются сообщения об успешном примене-
нии при розацеа третиноина в виде 0,025 % кре-
ма и 0,2 % изотретиноинового крема. Длитель-
ное местное использование (32 нед) третиноина
не уступает по эффективности 16-недельному
пероральному приему изотретиноина. Для мес-
тной терапии розацеа не рекомендуется местное
применение кортикостероидов, особенно фтори-
рованных.
Обшая терапия
Общая терапия применяется при осложненных
формах розацеа. Наиболее часто применяемые
препараты — антибиотики, метронидазол, син-
тетические ретиноиды. Антибиотики тетрацик-
линового ряда остаются самыми эффективными
антибактериальными препаратами в терапии
акне. Положительный эффект тетрациклинов
при розацеа объясняется свойством накапливать-
ся в сальных железах, избирательно проникать
в участки воспаленной ткани, угнетать действие
липазы коринебактерий, что снижает уровень
свободных жирных кислот в кожном сале, под-
держивающих перифолликулярное воспаление.
Начальная доза тетрациклина и окситетрацикли-
на в зависимости от массы тела составляет 1000—
1500 мг в сутки и принимается в 3—4 приема.
При клиническом улучшении доза постепенно
снижается до поддерживающей — 250—500 мг в
сутки. Начальная доза доксициклина составля-
ет 200 мг в сутки, поддерживающая — 100 мг,
миноциклина — соответственно — 100 и 50 мг.
Длительность лечения тетрациклинами — 12
недель, что повышает риск развития побочных
действий. Возможны нарушения деятельности
желудочно-кишечного тракта, аллергические
реакции, фотосенсибилизация. Для лечения ро-
зацеа применяются также антибиотики из груп-
пы макролидов — эритромицин (500—1500 мг/
сутки), рокситромицин (150 мг 2 раза в сутки),
кларитромицин (150 мг 2 раза в сутки).
Метронидазол является производным нитро-
имидазола, усиливает защитные и регенератор-
ные функции слизистой оболочки желудка и
кишечника, оказывает выраженное противоотеч-
ное действие. Метронидазол дает бактериостати-
ческий эффект в отношении грамотрицательных
анаэробных палочек, а также антипаразитарный
в отношении Demodex folliculorum. Метронида-
зол в синергизме с пальмитолиеновой кислотой,
содержащейся в коже человека, подавляет функ-
циональную активность нейтрофилов, тем самым
снижая продукцию последними медиаторов вос-
паления (Akamatsu Н. и соавт., 1990). Суточная
доза препарата составляет 1,0—1,5 г; продолжи-
тельность лечения — от 4—6 до 8 недель. Неко-
торые авторы считают, что показанием к перо-
ральному назначению метронидазола является
выраженный демодикоз, а длительность лечения
должна ограничиваться 10 днями (Jansen Т.,
Piewig G., 1996).
Орнидазол — новый имидазольный препарат
(зарегистрирован под названием тиберал) обла-
дает высокой терапевтической эффективностью
и отсутствием устойчивости бактериально-пара-
зитарной флоры к тибералу (М. В. Черкасова,
1997). Препарат тиберал рекомендуется больным
папулезной и папулопустулезной формами ро-
зацеа в суточной дозе 0,5 г (по '/2 таблетки 2 раза
в день) в течение 10 дней.
Ретиноиды применяют для лечения тяжелых
форм розацеа — rosacea conglobata и rosacea
fulminans. Изотретиноин (13-цис-ретиноевая кис-
лота, роаккутан) — синтетический ретиноид —
широко используется также для лечения тяжелых
форм акне. Терапевтическая эффективность пре-
парата связана с влиянием на процессы диффе-
ренцировки и кератинизации клеток эпидерми-
са и сальных желез, что обусловливает
выраженное себостатическое действие. Изотрети-
ноин оказывает противовоспалительное и имму-
номодулирующее действие за счет ингибирования
медиаторов воспаления — лейкотриенов В4. Су-
точная доза изотретиноина составляет 0,2—1,0 мг
на 1 кг массы тела. В особо тяжелых случаях ре-
комендуется назначение до 2 мг/кг веса тела в
сутки. При умеренных проявлениях розацеа при-
менение лишь 2,5—5 мг в день (независимо от
массы тела) приводит к клинической ремиссии.
Длительность лечения изотретиноином составляет
4—6 мес. Улучшение обычно наступает через 1 мес
после начала терапии и продолжается после от-
мены препарата. При необходимости повторный
курс изотретиноина проводится минимум через
8 нед после окончания предыдущего. При лече-
нии женщин возможна комбинация изотретино-
Глава 29. Болезни придатков кожи 351
ина с антиандрогенными препаратами — ципро-
теронацетатом (андрокур) и этинилэстрадиолом.
Глюкокортикоиды применяются при молние-
носной розацеа (pyoderma faciale) в комбинации
с изотретиноином. Разработана эффективная
схема лечения молниеносной формы розацеа:
преднизолон 1 мг на 1 кг массы тела в течение
5-7 дней с последующим снижением дозы и
отменой на 10—14 день; изотретиноин, начиная
с 7-го дня приема преднизолона, по 0,2—0,5—
1,0 мг/кг на протяжении 2—4 мес (Piewig G. и
соавт., 1992). В течение 1-й недели лечения це-
лесообразно применение кортикостероидных
кремов 4-й группы 1—2 раза в день, а в даль-
нейшем наложение теплых компрессов с целью
рассасывания инфильтратов.
Проведено патогенетическое лечение эритема-
тозной и папулезной форм розацеа препаратом
эглонил (по 0,05—0,1 г в течение 1—2 мес, 2—3
цикла в год), который применяется в терапии
астенических и астенодепрессивных состояний,
а также соматических расстройств нейрогенной
природы.
Физиотерапевтические методы. Криотерапия (ап-
пликации жидкого азота) проводится через день
или ежедневно до глубокого отшелушивания.
Метод показан больным независимо от стадии
заболевания. Криотерапия оказывает противовос-
палительное, сосудосуживающее, антидемодикоз-
ное действие. Электрокоагуляция применяется для
разрушения телеангиэктазий, папулезных и папу-
лопустулезных элементов. Количество процедур —
от 20 до 100 — в зависимости от стадии и распро-
страненности процесса. У больных с эритематоз-
ной и ограниченной папулезной стадиями розацеа,
как правило, достигается клиническое излечение.
Лазеротерапия — для деструкции телеангиэктазий
применяются длинноволновые (577 и 585 нм)
лучи. Световой пучок, генерируя тепловой эффект
внутри поверхностного кровеносного сосуда, вы-
зывает разрушение последнего, не затрагивая при-
легающие ткани.
Гнездная алопеция
Алопеция (облысение) — это патологическое
выпадение волос. Различают рубцовую алопецию,
обусловленную разрушением волосяных фолли-
кулов вследствие воспаления, атрофии кожи или
рубцевания, и нерубцовую алопецию, протекаю-
щую без предшествующего поражения кожи. К
нерубцовой алопеции относят гнездную алопецию
(alopecia areata), андрогенетическую алопецию
(alopecia androgenetica), диффузную алопецию
(alopecia diffusa).
Гнездная алопеция (гнездная плешивость,
alopecia areata, круговидное облысение) — хро-
нический дерматоз, характеризующийся патоло-
гическим выпадением волос, обусловленным
различными воздействиями на волосяной фол-
ликул и клинически проявляющийся формиро-
ванием очагов с полным отсутствием волос на
волосистой части головы, в области бороды, бро-
вей, ресниц и туловища.
Этиология и патогенез
Этиология гнездной алопеции точно не выясне-
на. Предполагается наличие аномальной реакции
кератиноцитов волосяного фолликула при гене-
тической детерминированности на эндогенные
или эндогенные влияния. О том, что гнездная
алопеция не является лишь ограниченным забо-
леванием волосяного фолликула, свидетельству-
ют сопровождающие ее нарушения роста ногтей
и обнаруживаемые при гистологическом иссле-
довании изменения на видимо здоровой коже.
В последнее время большинство авторов счи-
тают, что иммунная система играет ведущую
роль в развитии алопеции. У больных гнездной
алопецией наблюдается снижение активности
некоторых лизосомальных ферментов в нейтро-
фильных лейкоцитах, повышение уровня Ig G
и снижение Ig Е. Обнаружены изменения в по-
казателях клеточного иммунитета в виде сниже-
ния субпопуляций Т-л, Т-х, Т-с и повышения
иммунорегуляторного индекса.
Психический стресс может играть большую
роль в развитии и течении гнездной алопеции
и может предшествовать выпадению волос у 87 %
пациентов. Причиной гнездной алопеции может
быть дефицит цинка, нарушения деятельности
нервной системы, изменения со стороны желу-
дочно-кишечного тракта, сверхчувствительность
к бактериальным агентам, нарушения в микро-
циркуляторном русле.
Выделяют 6 типов гнездной алопеции: обыч-
ный, прегипертензивный, атопический, аутоим-
мунный, подростковый.
Обычный тип характеризуется началом в воз-
расте от 20 до 40 лет.
Прегипертензивный тип алопеции возникает у
молодых людей, у которых впоследствии разви-
вается гипертония. Иногда родители пациентов
страдают тяжелыми формами рано возникшей
гипертонии.
Атопический тип — встречается при сочетании
гнездной алопеции с атопическим дерматитом,
бронхиальной астмой и другими атопическими
352 Частная дерматология
состояниями. Такие формы алопеции тяжело
поддаются лечению, особенно если алопеция
возникла до полового созревания, часто разви-
вается тотальная и универсальная алопеция.
Аутоиммунный тип — характеризуется сочета-
нием гнездной алопеции с аутоиммунными за-
болеваниями (тиреоидит Хашимото, витилиго,
аутоиммунный полигландулярный синдром I
типа, гипопаратиреоз, первичная надпочечнико-
вая недостаточность, хронический генерализо-
ванный гранулематозный кандидоз, миастения).
Смешанный тип — чаще возникает у людей
пожилого возраста (старше 40 лет), характери-
зуется торпидностью течения и меньшей часто-
той развития субтотальной и тотальной форм.
Клиническая картина
Клинические проявления гнездной алопеции
весьма характерны: на фоне полного здоровья,
без субъективных признаков (редко может быть
покалывание в месте формирования очага) об-
разуются округлые или овальные очаги, лишен-
ные волос. В безволосых участках кожа вслед-
ствие потери волос и уменьшения луковиц
несколько втянута («утоплена»). Она имеет вид
слоновой кости или же небольшого воспаления.
Признаки атрофии отсутствуют, устья волося-
ных фолликулов сохраняются. Наиболее часто
процесс начинается в затылочной области, что
говорит о заинтересованности в развитии забо-
левания затылочного нерва.
Клинические формы
Выделяют 6 клинических форм проявлений гнез-
дной алопеции: локальная (очаговая), лентовид-
ная (офиазис), субтотальная, тотальная, универ-
сальная, с поражением ногтей.
1. Локальная (очаговая) форма характеризует-
ся изолированными очагами выпавших волос,
округлой формы с практически неизмененной
кожей на месте поражения.
2. Лентовидная форма (офиазис Цельса) про-
является наличием очага поражения, первона-
чально локализующегося в области затылка и
затем постепенно распространяющегося в виде
ленты до ушных раковин или висков. Для этой
формы алопеции характерны переход очага по-
ражения на гладкую кожу и прогностически
неблагоприятное течение. Иногда, сформировав-
шись однажды, очаг не прогрессирует
дальше и не регрессирует, несмотря на проводи-
мое лечение.
3. Субтотальная форма характеризуется обшир-
ными очагами поражения, образовавшимися в
результате слияния более мелких очагов.
4. При тотальной форме волосы отсутствуют на
всей поверхности головы, включая волосы рес-
ниц, бровей, а у мужчин — бороды. Клиничес-
кие формы могут трансформироваться друг в
друга, что особенно выражено при злокачествен-
ном течении заболевания.
5. При универсальной гнездной алопеции воло-
сы отсутствуют на всей поверхности кожи тела
человека.
6. Гнездная алопеция с поражением ногтей. По-
ражение ногтей при гнездной алопеции корре-
лирует с тяжестью процесса и наблюдается в виде
точечных вдавлений, продольной исчерченнос-
ти, онихорексиса, волнообразных ногтей — «пес-
чаные волны», красные луночки.
Течение
В течении гнездной алопеции выделяют три ста-
дии: прогрессирующую, стационарную и регрес-
сирующую. В прогрессирующей стадии при лег-
ком потягивании волос по краю очага происходит
их безболезненная эпиляция на расстоянии 1,5—
2 см от края очага облысения — зона «расшатан-
ных волос». Эпилированные из пограничной
зоны волосы дистрофичны и многократно рас-
щеплены, не имеют корневой щели. Длина кон-
чика зависит от интенсивности дистрофии мат-
рицы. Чем интенсивнее поражена матрица, тем
короче кончик (основание) волоса.
В стационарной стадии зона «расшатанных во-
лос» определяется нечетко или не определяется
вообще.
При регрессирующей стадии заболевания в
очаге облысения начинают расти пушковые во-
лосы (веллюс), которые постепенно утолщаются
и пигментируются. Иногда уже отросшие во-
лосы в очаге облысения отличаются по цвету
от нормальных волос и имеют пеструю окрас-
ку (полиозис). Очень часто наблюдается сле-
дующая картина: отмечается рост пушковых
волос и одновременно определяется зона «рас-
шатанных волос». В таких случаях устанавли-
вается прогрессирующая стадия заболевания,
несмотря на то, что рост волос может опере-
дить выпадение, и очаг поражения в итоге за-
растет волосами. Лечение, особенно местное,
назначается соответственно прогрессирующей
стадии.
Для определения прогноза заболевания у па-
циентов с гнездной алопецией применяется мик-
роскопическое исследование корней волос.
Правила забора волос для микроскопии:
— за две недели до микроскопии с волосами не
должны проводиться косметические процедуры;
Глава 29. Болезни придатков кожи 353
— забор волос проводится на 5-й день после
мытья;
— при очаговом облысении волосы забирают-
ся по краю очага и на участке, противополож-
ном очагу облысения; при диффузном облысе-
нии волосы забирают на 2 см кзади от лобной
линии и на 2 см сбоку от височной линии;
— при длинных волосах перед эпиляцией их
можно остричь на длину до 2 см;
— микроскопию волос производят в день за-
бора волос.
Различают следующие формы корней волос,
которые могут обнаруживаться при микроско-
пическом исследовании: анагеновые волосы,
дистрофические анагеновые волосы, катагеновые
и телогеновые волосы.
При нормальной трихограмме 89 % волос на-
ходятся в анагеновой фазе; до 1 % — в катаге-
новой фазе; до 20 % — в телогеновой фазе. Этим
и объясняется физиологическая потеря волос (до
100) в сутки.
При гнездной алопеции могут обнаружи-
ваться дистрофические анагеновые волосы. По
периферии очага увеличивается количество те-
логеновых волос. Обнаружение телогенового
корня говорит о медленно прогрессирующем
течении; дистрофические волосы характерны
для быстрого прогрессирования процесса. Ана-
логичные изменения обнаруживаются на про-
тивоположном, клинически здоровом участке
кожи.
Для диагностики заболеваний, которые мо-
гут быть причиной или поддерживающим фак-
тором гнездной алопеции и с целью проведе-
ния дифференциальной диагностики пациентам
рекомендуется проводить следующие исследо-
вания: общеклинические исследования (анализ
крови, мочи общий, биохимический анализ
крови), серологическое исследование крови на
сифилис (показано всем больным), исследова-
ние крови ИФА — ВИЧ (в упорных случаях за-
болевания), радиоизотопное исследование крови
для определения уровня гормонов (Т3, Т4, эст-
радиол, кортизол, тестостерон, пролактин,
АКТГ); исследование содержания микроэлемен-
тов (цинк, медь, калий, натрий, железо, маг-
ний); метаболизм кальция, иммунологические
исследования (интерфероны, интерлейкины,
фактор некроза и роста опухолей); при тяже-
лых формах гнездной алопеции необходимо
провести ядерно-магнитный резонанс головного
мозга, УЗИ яичников, надпочечников; реова-
зографию сосудов головного мозга, сосудов вер-
хних конечностей; ультразвуковое исследование
органов брюшной полости.
Дифференциальная диагностика
Дифференциальная диагностика гнездной ало-
пеции проводится с грибковыми инфекциями
(микроспория волосистой части головы, повер-
хностная антропонозная трихофития, хроничес-
кая трихофития волосистой части головы, фа-
вус), с мелкоочаговой алопецией при сифилисе,
трихотилломанией, травматической и андрогене-
тической алопецией.
Лечение
Традиционная терапия
Для лечения гнездной алопеции применяют пре-
параты, улучшающие микроциркуляцию: ксан-
тинола никотинат, мидокалм, пентоксифиллин
(трентал), дипиридамол (курантил); препараты
меди, цинка (цинктерал в таблетках по 0,124 г
и 0,2 г), железа (окись цинка, сульфат меди,
лактат железа) в стандартных дозировках; био-
генные стимуляторы: экстракт алоэ жидкий для
инъекций по 1 мл подкожно ежедневно в тече-
ние 30—50 дней, гумизоль по 1 мл первые 2—3
дня ежедневно, затем по 2 мл 1 раз в день внут-
римышечно в течение 20—30 дней, спленин вво-
дят по 1—3 мл внутримышечно 10—15 дней.
Из ноотропных препаратов — ноотропил в
капсулах по 400 мг три раза в день в течение 1—
1,5 месяцев. Ноотропил положительно влияет на
обменные и энергетические процессы, повыша-
ет устойчивость клеток мозга.
Гормональная терапия
Показанием к назначению глюкокортикоидов
внутрь может быть недавно возникшая алопе-
ция, с поражением волосистой части головы не
более 50 %. При универсальной алопеции и дли-
тельно существующей глюкокортикоиды, как
правило, неэффективны.
Назначают преднизолон по 40 мг в сутки в
течение 6—8 недель с последующим снижением
дозы в зависимости от состояния больного и
динамики заболевания. Некоторыми авторами
предлагается лечение преднизолоном до 6 меся-
цев, но в этом случае дозы препарата составля-
ют 20—30 мг в сутки.
В последнее время широко используется про-
лонгированный глюкокортикоид дипроспан.
Вводят по 1 мл внутримышечно с интервалом 1—
2 недели. На курс лечения 3—5 инъекций.
Местное лечение
Местно применяются различные гормональные
мази в виде смазываний и аппликаций. При
смазывании очагов поражения необходимо зах-
ватывать 1 см видимо здоровой кожи. Гормо-
354 Частная дерматология
нальные мази назначают в прогрессирующей
стадии заболевания. Можно применять глюко-
кортикоиды под окклюзионную повязку (поли-
этиленовую пленку или одноразовую шапочку
для принятия душа) на ночь в течение несколь-
ких недель. Этот метод используется преимуще-
ственно при тотальной алопеции.
Широко используют внутриочаговое введение
глюкокортикоидов. Безыгольным инъектором в
очаг(и) один раз в неделю вводят гидрокорти-
зон в возрастающей дозировке: 0,2—0,4—0,6—
0,8—1 мл и далее по 1 мл. Каждое впрыскива-
ние обеспечивает попадание в кожу 0,1 мл
эмульсии гидрокортизона. Курс лечения состо-
ит из 10 процедур и длится 2,5 месяца.
Дипроспан вводят при помощи инсулинового
шприца в расчете 0,1 мл на 2 см2 очага пораже-
ния с интервалом 1 неделя. На курс лечения не
более 10 инъекций. Рост волос восстанавливается
через 4—6 недель. Иногда в самом очаге пора-
жения отмечается рост волос островками.
Местная иммуносупресивная терапия
Местная иммуносупресивная терапия проводится
в стационарной стадии заболевания с примене-
нием дибутилового эфира скваровой кислоты и
дифенилциклопропенона. Принцип топической
иммунной терапии состоит в интермиттирующем
возбуждении контактной экземы на коже воло-
систой части головы. После проведения сенси-
билизации подбирают такую концентрацию,
которая поддерживает воспалительную реакцию
1—2 суток. После достижения эффекта интервал
между применением увеличивают или совсем
прекращают терапию.
Раздражающая терапия
Раздражающая терапия также проводится в ста-
ционарную стадию. Применяется 0,5—1 % ант-
ралин, 2 % раствор миноксидила, силокаст, ме-
лагинин.
Физиолечение
Гнездной алопеции назначается независимо от
стадии заболевания и включает практически все
виды физиотерапии: гальванизацию, электрофо-
рез, дарсонвализацию, амплипульстерапию, фо-
тохимиотерапию, массаж, криомассаж.
Профилактика гнездной алопеции не разрабо-
тана.
Глава 30
Васкулиты кожи
Васкулитами (ангиитами) кожи называют дер-
матозы, в клинической и патоморфологической
симптоматике которых первоначальным и веду-
щим звеном является неспецифическое воспале-
ние стенок дермальных и гиподермальных кро-
веносных сосудов разного калибра.
Этиология и патогенез
Этиология и патогенез васкулитов во многом еще
остаются неясными. В последние годы все боль-
шее распространение получает теория иммуноком-
плексного генеза васкулитов, связывающая их
возникновение с повреждающим действием на
сосудистую стенку осаждающихся из крови им-
мунных комплексов. Роль антигена в зависимос-
ти от причины васкулита, вероятно, выполняет тот
или иной микробный агент, лекарственное веще-
ство, собственный измененный белок, а антителом
является соответствующий иммуноглобулин. Цир-
кулирующие в крови иммунные комплексы при
благоприятных условиях, исключающих или тор-
мозящих их естественную элиминацию, осажда-
ются субэндотелиально в сосудистых стенках и
вызывают их воспаление. К благоприятствующим
отложению иммунных комплексов моментам от-
носят их усиленное образование, длительную цир-
куляцию в кровяном русле, повышение внутри-
сосудистого давления, замедление тока крови,
нарушения в системе иммунного гомеостаза. Сре-
ди этиологических факторов, приводящих к вос-
палению сосудистой стенки, чаще всего называ-
ют фокальную (реже общую) инфекцию,
обусловленную стафилококком или стрептокок-
ком, микобактериями туберкулеза, энтерококка-
ми, дрожжевыми и другими грибами, вирусом
гриппа. О сенсибилизирующем влиянии инфек-
ции на сосудистые стенки свидетельствуют поло-
жительные результаты внутрикожных проб с со-
ответствующими антигенами. Среди экзогенных
сенсибилизирующих факторов при ангиитах осо-
бое место в настоящее время отводится лекарствен-
ным средствам, в первую очередь антибиотикам и
сульфаниламидам. Существенную роль в патоге-
незе васкулитов могут играть хронические инток-
сикации, эндокринопатии, различные виды обмен-
ных нарушений, а также повторные охлаждения,
психическое и физическое перенапряжение, фо-
тосенсибилизация, артериальная гипертензия, ве-
нозный застой. Васкулиты кожи часто являются
кожным синдромом различных острых и хрони-
ческих инфекций (грипп, туберкулез, сифилис,
лепра, иерсиниоз и др.), диффузных болезней со-
единительной ткани (ревматизм, системная крас-
ная волчанка, ревматоидный артрит и др.), кри-
оглобулинемии, болезней крови, различных
опухолевых процессов.
В настоящее время насчитывают до 50 нозо-
логических форм, относящихся к группе васку-
литов (ангиитов) кожи. Значительная часть этих
нозологий имеет между собой большое клини-
ческое и патоморфологическое сходство, неред-
ко граничащее с идентичностью, что следует
иметь в виду при постановке диагноза. До сих
пор нет единой общепринятой классификации
васкулитов кожи, что объясняется недостаточной
изученностью этиологии, патогенеза, клиничес-
кого течения и ряда других вопросов, связанных
с этой сложной группой дерматозов. Большин-
ство клиницистов пользуются преимуществен-
но морфологическими классификациями кож-
ных васкулитов, в основу которых обычно
берутся клинические изменения кожи, а также
глубина расположения (и соответственно калибр)
пораженных сосудов. Для практических целей
разработана рабочая классификация васкулитов
(ангиитов) кожи (О. Л. Иванов, 1997).
Клиническая картина
Клинические проявления кожных васкулитов
чрезвычайно многообразны. Однако существу-
ет целый ряд общих признаков, объединяющих
клинически эту группу дерматозов:
• воспалительный характер изменения кожи;
• склонность высыпаний к отеку, кровоизли-
янию, некрозу;
• симметричность поражения;
• полиморфизм элементов сыпи (обычно эво-
люционный);
• первичная или преимущественная локализа-
ция на нижних конечностях (чаще на голенях);
355
356 Частная дерматология
Таблица 30.1. Классификация и основные клинические симптомы васкулитов (ангиитов) кожи
Клиническая форма Синонимы, идентичные болезни и синдромы Основные проявления
I. Дермальные ангииты
1. Полиморфный Болезнь Гужеро-Рюитера. Артериолит Рюите-ра. Синдром
дермальный ангиит: Гужеро-Дюперра. Дермальный некротизирующий васкулит. Аллергический кожный васкулит. Гиперсенситивный ангиит
уртикарный тип волдыри Уртикарный васкулит. Неклассифицируемый тип артериолита Рюйтера Воспалительные пятна, стойкие
геморрагический тип Геморрагический васкулит. Анафилактоидная пурпура Петехии, отечная пурпура
Шенлейна-Геноха. Капилляротоксикоз. Геморрагический («геморрагические волдыри»,
тип артериолита Рюйтера. Геморрагический лейкоцитокластический «пальпируемая пурпура»), экхимозы,
микробид Мишера-Шторка. Аллергическая пурпура. Криоглобулинемическая пурпура геморрагические пузыри
папулонодулярный Нодулярные дермальные аллергиды Гужеро Воспалительные узелки и бляшки,
тип мелкие отечные поверхностные узлы
Папулонекротический Некротический нодулярный дерматит Вертера-Дюмлинга. Воспалительные узелки с пустулой или
тип Парапсориаз Мухи-Хабермана. Атрофирующий папулез Дегоса. некрозом в центре, «штампованные» рубчики
пустулезно-язвенный тип Язвенный дерматит. Гангренозная пиодермия Везикуло-пустулы, эрозии, язвы, рубцы
некротически-язвенный Некротическая пурпура. Гангренозная пурпура. Геморрагический некроз,
тип Молниеносная пурпура. геморрагические пузыри, язвы
полиморфный тип Трехсимптомный синдром Гужеро-Дюперра. Чаще сочетание волдырей, пурпуры и
Полиморфно-нодулярный тип артериолита Рюйтера поверхностных мелких узлов. Сочетание элементов любых типов
2. Хроническая Геморрагически-пигментные дерматозы. Пигментно-пурпурный
пигментная пурпура дерматоз. Болезнь Шамберга-Майокки
петехиальный тип Стойкая прогрессирующая пигментная пурпура Шамберга. Болезнь Шамберга. Петехии, пятна гемосидероза
Телеангиэктатический Телеангиэктатическая пурпура Майокки. Дугообразная Петехии, пятна гемосидероза,
тип пурпура Турена. телеангиэктазии
лихеноидный тип Пигментный пурпурозный лихеноидный ангиодермит Гужеро-Блюма Петехии, пятна гемосидероза, телеангиэктазии, лихеноидные папулы
экзематоидный тип Экзематоидная пурпура Дукаса-Капенатакиса Петехии, пятна гемосидероза,
Зудящая пурпура Левенталя лихенификация, чешуе-корки, застойная эритема
II. Дермо-гиподермальные ангииты
1. Нодозный артериит Доброкачественный узелковый периартериит Глубокие узелки по ходу сосудов, узлы
2. Ливедо-ангиит Ливедо-васкулит. Кожная форма узелкового периартериита. Сетчатое ветвистое ливедо, узлы в
Некротизирующий васкулит. Ливедо с узлами. области пятен ливе-до, некрозы, язвы,
Ливедо с изъязвлениями. рубцы
III. Гиподермальные ангииты
1. Узловатая эритема Узловатый васкулит. Узловатый ангиит Узлы (чаще на голенях)
острая Острый узловатый ангиит Отечные ярко-красные узлы артралгии, лихорадка
подострая Мигрирующая узловатая эритема Беверстедта. Плоский узел, растущий по периферии
(мигрирующая) Варибельный гиподермит Вилановы-Пинь-ола. Подострый и разрешающий ся в центре. Узлы
мигрирующий нодулярный панникулит. Болезнь Вилановы подостро-го течения с умеренными общими явлениями
хроническая Узловатый васкулит Рецидивирующие язвы без
выраженных явлений
2. Хронический Нетуберкулезная индуративная эритема. Нодулярный васкулит Плотные узлы с изъязвлением,
узловато-язвенный ангиит Монтгомери-О'Лири-Баркера втянутые рубцы
IV. Смешанные и переходные формы
Глава 30. Васкулиты кожи
357
• наличие сопутствующих сосудистых, аллер-
гических, ревматических, аутоиммунных и дру-
гих системных заболеваний;
• нередкая связь с предшествующей инфек-
цией или лекарственной непереносимостью;
• течение острое или с периодическими обо-
стрениями.
Полиморфный дермальный ангиит
Полиморфный дермальный ангиит, как прави-
ло, имеет хроническое рецидивирующее течение
и отличается чрезвычайно разнообразной морфо-
логической симптоматикой, что и породило его
путанную и обильную номенклатуру. Высыпа-
ния первоначально появляются на голенях, но
могут возникать и на других участках кожного
покрова, реже — на слизистых оболочках (рис.
30.1). Наблюдаются волдыри, геморрагические
пятна различной величины, воспалительные
узелки и бляшки, поверхностные узлы, папуло-
некротические высыпания, пузырьки, пузыри,
пустулы, эрозии, поверхностные некрозы, язвы,
рубцы. Высыпаниям иногда сопутствуют лихо-
радка, общая слабость, артралгии, головная боль.
Появившаяся сыпь обычно сохраняется в тече-
ние длительного времени (от нескольких недель
до нескольких месяцев), имеет тенденцию к
рецидивам. Начало болезни и ее рецидивы за-
частую провоцируются острыми инфекционны-
ми заболеваниями (ангина, грипп, ОРЗ), пере-
охлаждением, физическим или нервным
перенапряжением, реже — приемом каких-либо
медикаментов или пищевой непереносимостью.
В зависимости от наличия тех или иных морфо-
логических элементов сыпи выделяют различные
типы поверхностного дермального ангиита (см.
таблицу 1). Однако нередко разные элементы
комбинируются, создавая картину полиморфного
типа ангиита.
Уртикарный тип, как правило, симулирует
картину хронической рецидивирующей крапив-
ницы, проявляясь волдырями различной вели-
чины, возникающими на разных участках кож-
ного покрова. Однако в отличие от крапивницы,
волдыри при уртикарном ангиите отличаются
особой стойкостью, сохраняясь на протяжении
1-3 суток (иногда и дольше). Вместо выражен-
ного зуда больные обычно испытывают жжение
или чувство раздражения кожи. Высыпаниям
нередко сопутствуют артралгии, иногда боли в
животе, т. е. признаки ситемного поражения.
При обследовании может выявляться гломеру-
лонефрит. У больных отмечают также повыше-
ние СОЭ, гипокомплементемию, повышение ак-
тивности лактатдегидрогеназы, положительные
воспалительные пробы, изменения в соотноше-
нии иммуноглобулинов. Лечение антигистамин-
ными препаратами обычно не дает эффекта. Ур-
тикарным ангиитом болеют чаще женщины
среднего возраста. Диагноз окончательно под-
тверждается патогистологическим исследовани-
ем кожи, при котором обнаруживают картину
лейкоцитокластического ангиита.
Геморрагический тип наиболее характерен для
поверхностного ангиита. Самым типичным сим-
птомом при этом варианте является так называ-
емая «пальпируемая пурпура» — отечные гемор-
рагические пятна различной величины,
локализующиеся обычно на голенях и тыле стоп,
легко определяемые не только визуально, но и
при пальпации, что отличает их от симптомов
других пурпур, в частности от болезни Шамбер-
га-Майокки. Однако первыми высыпаниями при
геморрагическом типе обычно бывают мелкие
отечные воспалительные пятна, напоминающие
волдыри и вскоре трансформирующиеся в ге-
моррагическую сыпь. При дальнейшем нараста-
нии воспалительных явлений на фоне сливной
пурпуры и экхимозов могут формироваться ге-
моррагические пузыри, оставляющие после
вскрытия глубокие эрозии или язвы. Высыпа-
ниям, как правило, сопутствует умеренный отек
нижних конечностей. Геморрагические пятна
Рис. 30.1. Типичная локализация
высыпаний при аллергическом васкулите.
358 Частная дерматология
могут появляться на слизистой рта и зева. Опи-
санные геморрагические высыпания, возникаю-
щие остро после перенесенного простудного за-
болевания (обычно после ангины) и
сопровождающиеся лихорадкой, выраженными
артралгиями, болями в животе и кровянистым
стулом, составляют клиническую картину ана-
филактоидной пурпуры Шенлейна-Геноха, ко-
торая чаще наблюдается у детей.
Папулонодулярный тип встречается довольно
редко. Он характеризуется появлением гладких
уплощенных воспалительных узелков округлых
очертаний величиной с чечевицу или мелкую
монету, иногда больше, а также небольших по-
верхностных нерезко очерченных отечных блед-
но-розовых узлов величиной до лесного ореха,
болезненных при пальпации. Высыпания лока-
лизуются на конечностях, обычно на нижних,
редко на туловище, и не сопровождаются выра-
женными субъективными ощущениями.
Папулонекротический тип проявляется неболь-
шими плоскими или полушаровидными воспа-
лительными нешелушащимися узелками, в цен-
тральной части которых вскоре формируется
сухой некротический струп, обычно в виде чер-
ной корочки. При срывании струпа обнажают-
ся небольшие округлые поверхностные язвочки,
а после рассасывания папул остаются мелкие
«штампованные» рубчики. Высыпания распола-
гаются на разгибательных поверхностях конеч-
ностей и клинически полностью симулируют
папулонекротический туберкулез, с которым
следует проводить самую тщательную дифферен-
циальную диагностику.
Пустулезно-язвенный тип начинается с неболь-
ших везикулопустул, напоминающих акне или
фолликулит, быстро трансформирующихся в
язвенные очаги с тенденцией к неуклонному
эксцентрическому росту за счет распада отечно-
го синюшно-красного периферического валика.
Поражение может локализоваться на любом уча-
стке кожи, чаще на голенях, в нижней полови-
не живота. После заживления язв остаются плос-
кие или гипертрофические рубцы, длительно
сохраняющие воспалительную окраску.
Некротически-язвенный тип является наиболее
тяжелым вариантом дермального ангиита. Он
начинается остро (иногда молниеносно) и прини-
мает затяжное течение (если процесс не заканчи-
вается быстрым летальным исходом). Вследствие
острого тромбоза воспаленных кровеносных со-
судов возникает омертвение (инфаркт) того или
иного участка кожи, проявляющееся некрозом в
виде обширного черного струпа, образованию
которого может предшествовать геморрагическое
пятно или пузырь. Процесс развивается в тече-
ние нескольких часов, сопровождается сильны-
ми местными болями и лихорадкой. Поражение
чаще располагается на нижних конечностях и
ягодицах. Гнойно-некротический струп сохраня-
ется длительное время. Образовавшиеся после его
отторжения язвы имеют различную величину и
очертания, содержат гнойное отделяемое, медлен-
но рубцуются.
Полиморфный тип характеризуется сочетани-
ем различных высыпных элементов, свойствен-
ных другим типам дермального ангиита. Чаще
имеет место сочетание отечных воспалительных
пятен, геморрагических высыпаний пурпурозно-
го характера и поверхностных отечных мелких
узлов, что составляет классическую картину так
называемого трехсимптомного синдрома Гуже-
ро-Дюперра и идентичного ему полиморфно-но-
дулярного артериолита Рюйтера.
Хроническая пигментная пурпура
Хроническая пигментная пурпура (болезнь Шам-
берга-Майокки) представляет собой хронический
дермальный капиллярит, поражающий сосочко-
вые капилляры. В зависимости от клинических
особенностей можно выделить следующие раз-
новидности (типы) геморрагически-пигментных
дерматозов.
Петехиальный тип (стойкая прогрессирующая
пигментная пурпура Шамберга, прогрессирую-
щий пигментный дерматоз Шамберга) — основ-
ное заболевание этой группы, являющееся как
бы родоначальником для других его форм, ха-
рактеризуется множественными мелкими точеч-
ными геморрагическими пятнами без отека (пе-
техиями) с исходом в стойкие буровато-желтые
пятна гемосидероза различной величины и очер-
таний. Высыпания располагаются чаще на ниж-
них конечностях, не сопровождаются субъектив-
ными ощущениями, наблюдаются почти
исключительно у мужчин.
Телеангиэктатический тип (телеангиэктатичес-
кая пурпура Майокки, болезнь Майокки) про-
является чаще своеобразными пятнами — меда-
льонами, центральная зона которых состоит из
мелких телеангиэктазий на слегка атрофичной
коже, а периферическая — из мелких петехий на
фоне гемосидероза (здесь элементы болезни
Шамберга ассоциируются с телеангиэктазиями).
Лихеноидный тип (пигментный пурпурозный
лихеноидный ангиодерматит Гужеро-Блюма)
характеризуется диссеминированными мелкими
лихеноидными блестящими узелками почти те-
лесного цвета, сочетающимися с петехиальными
высыпаниями, пятнами гемосидероза и иногда
Глава 30. Васкулиты кожи
359
мелкими телеангиэктазиями, что наводит на
мысль о болезни Шамберга-Майокки с лихено-
идными высыпаниями.
Экзематоидный тип (экзематоидная пурпура
Дукаса-Капенатакиса) отличается наличием в
очагах, помимо петехий и гемосидероза, явлений
экзематизации (отечности, разлитого покрасне-
ния, папуло-везикул, корочек), сопровождаю-
щихся выраженным зудом.
Ливедо-ангиит
Ливедо-ангиит наблюдается почти исключитель-
но у женщин, обычно в период полового созре-
вания. Первыми его симптомами являются стой-
кие ливедо-синюшные пятна различной
величины и очертаний, образующие причудли-
вую петлистую сеть на нижних конечностях,
реже на предплечьях, кистях, лице и туловище.
Окраска пятен резко усиливается при охлажде-
нии. С течением времени интенсивность ливе-
до становится более выраженной, на его фоне
(преимущественно в области лодыжек и тыла
стоп) возникают мелкие кровоизлияния и некро-
зы, образуются изъязвления. В тяжелых случа-
ях на фоне крупных синюшно-багровых пятен
ливедо формируются болезненные узловатые уп-
лотнения, подвергающиеся обширному некрозу
с последующим образованием глубоких медлен-
но заживающих язв. Больные ощущают зябкость,
тянущие боли в конечностях, сильные пульси-
рующие боли в узлах и язвах. После заживле-
ния язв остаются белесоватые рубцы с зоной
гиперпигментации в окружности.
Узловатый ангиит
Узловатый ангиит (узловатый васкулит) вклю-
чает различные варианты узловатой эритемы,
отличающиеся друг от друга характером узлов и
течением процесса.
Острая узловатая эритема — классический, хотя
и не самый частый вариант заболевания. Она про-
является быстрым высыпанием на голенях (редко
на других участках конечностей) ярко-красных
отечных болезненных узлов величиной до детской
ладони на фоне общей отечности голеней и стоп.
Отмечается повышение температуры тела до 38—
39°С, общая слабость, головная боль, артралгии.
Заболеванию обычно предшествуют простуда,
вспышка ангины. Узлы исчезают бесследно в те-
чение 2—3 недель, последовательно изменяя свою
окраску на синюшную, зеленоватую, желтую
(«цветение синяка»). Изъязвления узлов не быва-
ет. Рецидивов не наблюдается.
Хроническая узловатая эритема — самая час-
тая форма кожного ангиита, отличается упорным
рецидивирующим течением, возникает обычно
у женщин среднего и пожилого возраста, нередко
на фоне сосудистых и аллергических заболева-
ний, очагов фокальной инфекции и воспали-
тельных или опухолевых процессов в органах
малого таза (хронический аднексит, миома мат-
ки). Обострения чаще всего возникают весной
и осенью, характеризуются появлением неболь-
шого числа синюшно-розовых плотных болез-
ненных узлов величиной с лесной или грецкий
орех. В начале своего развития узлы могут не из-
менять окраски кожи, не возвышаться над ней,
а определяться только при пальпации. Почти ис-
ключительная локализация узлов — голени
(обычно их передняя и боковая поверхности).
Отмечается умеренная отечность голеней и стоп.
Общие явления непостоянны и выражены сла-
бо. Рецидивы длятся несколько месяцев, в те-
чение которых одни узлы рассасываются, а на
смену им появляются другие.
Мигрирующая узловатая эритема обычно имеет
подострое, реже хроническое течение и своеоб-
разную динамику основного поражения. Про-
цесс, как правило, носит асимметричный харак-
тер и начинается с появления одиночного
плоского узла на переднебоковой поверхности
голени. Узел имеет розовато-синюшную окрас-
ку, тестоватую консистенцию и довольно быст-
ро увеличивается в размерах за счет перифери-
ческого роста, превращаясь вскоре в крупную
глубокую бляшку с запавшим и более бледным
центром и широкой валообразной более насы-
щенной периферической зоной. Ему могут со-
путствовать единичные мелкие узлы, в том числе
на противоположной голени. Поражение сохра-
няется от нескольких недель до нескольких ме-
сяцев. Возможны общие явления (субфебрили-
тет, недомогание, артралгии).
Узловато-язвенный ангиит
Узловато-язвенный ангиит в широком смыс-
ле можно рассматривать как язвенную форму
хронической узловатой эритемы, но вследствие
весьма характерных клинических особенностей,
сближающих его с индуративной эритемой,
заслуживает обособления. Процесс с самого на-
чала имеет торпидное течение и проявляется
плотными довольно крупными малоболезнен-
ными синюшно-красными узлами, склонными
к распаду и изъязвлению с образованием вяло
рубцующихся язв. Кожа над свежими узлами
может иметь нормальную окраску, но иногда
процесс начинается с синюшного пятна, транс-
формирующегося со временем в узловатое уп-
лотнение и язву. После заживления язв оста-
ются плоские или втянутые рубцы, область
360 Частная дерматология
которых при обострениях может вновь уплот-
няться и изъязвляться. Типичной локализацией
является задняя поверхность голеней (икро-
ножная область), однако возможно расположе-
ние узлов и на других участках. Характерна
хроническая пастозность голеней. Процесс
имеет хроническое рецидивирующее течение,
наблюдается у женщин среднего возраста,
иногда у мужчин. Клиническая картина узло-
вато-язвенного ангиита зачастую полностью
симулирует индуративную эритему Базена, с
которой должна проводиться самая тщательная
дифференциальная диагностика.
Течение и степень активности
При кожных васкулитах можно выделить про-
грессирующую, стационарную и регрессирующую
стадии, особенности которых не требуют допол-
нительной характеристики и легко идентифици-
руются.
Степень активности определяется распростра-
ненностью кожного поражения, наличием общих
явлений и признаков поражения других органов
и систем, а также изменениями лабораторных по-
казателей.
В практических целях выделяют две степени
активности:
I. Низкая;
II. Высокая.
При активности первой степени поражение но-
сит локализованный характер, представлено не-
большим или умеренным числом высыпаний,
отсутствуют общие явления (лихорадка, недомо-
гание, головная боль), нет признаков вовлече-
ния других органов, лабораторные показатели су-
щественно не отклоняются от нормы.
При активности второй степени процесс носит
диссеминированный характер, сыпь обильная,
имеются общие явления (познабливание, лихо-
радка от субфебрильной до гектической, общая
слабость, головная боль), выявляются признаки
системности (артралгии, миалгии, лимфаденопа-
тии, желудочно-кишечные расстройства), наблю-
даются отклонения различных лабораторных
показателей (повышение СОЭ до 20 мм/ч и бо-
лее, лейкоцитоз или лейкопения, эозинофилия,
снижение содержания гемоглобина, тромбоцито-
пения, повышение содержания общего белка, Oj-
или у-глобулинов, изменение содержания отдель-
ных иммуногобулинов, снижение уровня ком-
племента, появление С-реактивного белка, по-
ложительные пробы на ревматоидный фактор,
появление АНФ и антител к ДНК, высокие тит-
ры антистрептолизина-0 и антигиалуронидазы,
снижение уровня фибринолиза).
Лечение основных
форм васкулитов кожи
Лечение кожного васкулита зависит от формы
заболевания, стадии процесса и степени его ак-
тивности. Больные с I степенью активности
подлежат госпитализации или домашнему режи-
му в прогрессирующей стадии заболевания, осо-
бенно при наличии неблагоприятных професси-
ональных условий. При II степени активности
процесса больным необходимо обследование и
лечение в условиях стационара. Первостепенное
значение во всех случаях обострений кожного
ангиита с основными очагами на нижних конеч-
ностях имеет постельный режим, так как у та-
ких больных обычно резко выражен ортостатизм.
В то же время полезна гимнастика в постели для
улучшения кровообращения. Постельный режим
следует соблюдать до перехода в прогрессирую-
щую стадию. Больные, особенно в период обо-
стрения, должны соблюдать диету с исключени-
ем раздражающей пищи (алкогольные напитки;
острые, копченые, соленые и жареные блюда;
консервы, шоколад, крепкий чай и кофе; цит-
русовые). Всем больным следует проводить те-
рапию, направленную на ликвидацию причины
заболевания (санация очага фокальной инфек-
ции, лечение микоза стоп или основного обще-
го заболевания). Необходимо исключить воздей-
ствие провоцирующих обострение заболевания
факторов (переохлаждение, курение, длительные
ходьба и стояние, ушибы, подъем тяжестей). В
острых случаях патогенетически показан плаз-
маферез.
Комплексное лечение больных васкулитами
кожи включает нестероидные противовоспали-
тельные препараты (индометацин по 0,025 г 3
раза в день; напросин по 0,25 г 3 раза в день),
антибиотики (олететрин по 0,25 г 4 раза в день,
метациклин по 0,3 г 2 раза в день, вибрамицин
по 0,1 г в день), хинолины (плаквенил по 0,2 г
2 раза в день; делагил по 0,25 г в день), антиги-
стаминные препараты, витамины (аскорутин).
Местная терапия: троксевазин-гель, бутадионо-
вая мазь, аминокапроновая мазь. У больных II
степени активности показано применение кор-
тикостероидов (преднизолон по 1—2 таблетки 4
раза в сутки), цитостатиков (азатиоприн по 0,05 г
3 раза в день), гепарина по 5000 ЕД подкожно
4 раза в день, гемодеза по 400 мл 2 раза в неде-
лю. При некротически-язвенном типе на I эта-
пе — примочки или мази с протеолитическими
ферментами (химопсин, «ируксол»); на II эта-
пе — дезинфицирующие, анестезирующие мази
и губки, лазерное облучение малой мощности.
Глава 31
Новообразования кожи
Доброкачественные новообразования кожи 362
Предраковые заболевания кожи и слизистых оболочек 370
Злокачественные новообразования кожи 373
Рак кожи из всех видов опухолевых заболева-
ний человека встречается наиболее часто. В США
каждый год регистрируется более 600000 новых
случаев рака кожи с тенденцией роста заболева-
емости и его появлению в молодом возрасте.
Своевременно начатое лечение базальноклеточ-
ных и плоскоклеточных форм рака кожи позво-
ляет получить излечение в 95 % случаев, у лиц
со злокачественной меланомой — в 90 %, на
поздних стадиях меланомы эффективность ле-
чения значительно снижается (у 39 % больных
сохраняются положительные отдаленные резуль-
таты).
Успех лечения рака кожи заключается в ран-
нем выявлении, знании его первых симптомов
врачами различных специальностей, наблюде-
нии за лицами с преканкрозами (пигментная
ксеродерма, актинический кератоз, пигментные
невусы, лейкоплакии и др.) и профилактике
(избегать гиперинсоляции, травм родимых пя-
тен).
Возникновение рака представляет собой мно-
гоэтапный (см. рис. 31.1), а в ряде случаев дос-
таточно длительный процесс, которому предше-
ствуют предраковые состояния (кератозы,
лучевой дерматит, болезнь Боуэна и др.). Эти со-
стояния отличаются от рака отсутствием одного
или нескольких характерных признаков, кото-
рые появляются в результате постепенного про-
грессирования кожного опухолевого процесса.
Результатом предраковых изменений может
быть:
• прогрессирование процесса (радиальный и
вертикальный рост);
• рост без инвазии подлежащих тканей и ме-
тастазов (радиальный рост);
• длительное существование без видимых из-
менений (отсутствие радиального и вертикаль-
ного роста);
• спонтанная регрессия.
Рис 31.1. Схема патогенеза
новообразований кожи.
361
362 Частная дерматология
Первый путь течения процесса является наи-
более опасным и указывает на переход предра-
кового состояния в рак. Прогрессирование пред-
ракового процесса сопровождается, как правило,
длительным или периодическим действием кан-
церогенных факторов. Прекращение действия
этих факторов может приостановить озлокаче-
ствление процесса на любом его этапе.
Морфологические признаки опухолевого про-
цесса в коже по мере его прогрессирования ха-
рактеризуются следующим образом: гиперплазия
- метаплазия - атипическая метаплазия - дис-
плазия - внутриэпителиальный рак (рак in situ)
- инвазивный рак - анапластический рак.
Если при явлениях дисплазии вероятность
возвращения клеток к нормальной пролифера-
ции достаточно велика, то при состоянии ана-
плазии эта вероятность очень низкая.
Классификация
Новообразования кожи подразделяются на пер-
вичные, вторичные и гемодермии.
• Первичные новообразования кожи — очаги
патологической пролиферации её различных
структур врожденного характера (пороки эмбри-
онального развития — невусы) или возникшие
в результате действия активирующих факторов
(приобретенные, эволюционные).
• Вторичные новообразования кожи возника-
ют вследствие метастазирования злокачественных
клеток при раке внутренних органов.
• Гемодермии (лейкемии) развиваются в ре-
зультате пролиферации в коже миелоидных кле-
ток при злокачественных заболеваниях системы
кроветворения.
В прогностическом плане новообразования
кожи подразделяются на доброкачественные,
предраковые (преканкрозы) и злокачественные.
Доброкачественные
новообразования
кожи
• Старческий кератоз
• Кератоакантома (роговой моллюск)
• Дерматофибромы, мягкие фибромы, гипер-
трофические и келоидные рубцы (опухоли фиб-
розной ткани)
• Атеромы (опухоли сальных желез), гипер-
плазия сальных желез
• Сирингомы (опухоли из потовых желез)
• Липомы (опухоли жировой ткани)
• Гемангиомы (сосудистые опухоли)
• Кисты (полости в коже, выстланные эпидер-
мисом или эпителием придатков кожи)
• Отложения в коже (ксантомы, кальциноз)
• Нейрофиброматоз (опухоли нервной ткани)
• Невусы эпителиальные (пороки развития
эпителиальных клеток)
• Невусы пигментные (врожденные, приобре-
тенные)
Старческая кератома
Старческая кератома (старческий кератоз) — наи-
более часто встречающаяся доброкачественная
эпителиальная опухоль наследственного характе-
ра, исходящая из базалиоидных клеток, реже —
из клеток шиповатого слоя эпидермиса. Количе-
ство кератом может достигать у лиц преклонно-
го возраста нескольких сотен (тысяч). Вначале
появляются милиарные папулы, слегка возвыша-
ющиеся над уровнем кожи, обычно гиперпигмен-
тированные. Под лупой на их поверхности вид-
ны множественные мелкие углубления (вид
напёрстка). С течением времени бляшки начина-
ют заметно возвышаться над окружающей кожей,
увеличиваются в размерах (до нескольких санти-
метров в диаметре), их поверхность может при-
нимать бородавчатый характер. При осмотре под
лупой на их поверхности выявляются роговые
кисты. Папулы обычно округлой или овальной
формы, имеют темную или светлую пигментацию.
Локализуются на лице, туловище, руках.
Дифференцируют с вульгарными и плоскими
бородавками, базалиомой, меланомой, пигмен-
тными невусами.
Лечение. Электрокоагуляция, криодеструкция
жидким азотом, солкодерм, 30 % проспидино-
вая и 5 % фторурациловая мази; при множе-
ственных кератомах — внутрь ароматические
ретиноиды.
Кератоакантома
Кератоакантома (роговой моллюск) — доброка-
чественная эпителиальная опухоль в виде полу-
сферического узла с кратерообразным углубле-
нием в центре, заполненным роговыми массами.
Опухоль обычно располагается на лице (редко на
губе, языке), напоминает плоскоклеточный рак
кожи, от которого отличается быстрым ростом
и спонтанным рассасыванием через 3—8 месяцев.
Кератоакантома имеет серовато-красный цвет,
плотную консистенцию, не спаяна с подлежащи-
ми тканями, подвижна, слегка болезненна.
Глава 31. Новообразования кожи
363
В возникновении кератоакантомы играет роль
папилломавирусная инфекция, ультрафиолето-
вое облучение, смолы, нефтепродукты.
Лечение. Единичные элементы иссекают, при
множественных кератоакантомах применяют
ретиноиды и метотрексат. При подозрении на
трансформацию процесса в рак производят ис-
сечение опухоли и её гистологическое исследо-
вание.
Дерматофиброма
Дерматофиброма (узелковый субэпидермальный
фиброз) встречается часто; возникает на конеч-
ностях, туловище и представляет собой внутри-
кожный узел плотной консистенции до 1 см в
диаметре, иногда болезненный. Образование
сливается с окружающими тканями, однако
пальпируется отчетливо. Может куполообразно
возвышаться над кожей или имеет форму вдав-
ления. Кожа над узлом часто не изменена, мо-
жет быть пигментирована или красно-бурого
цвета. Характерен патогномоничный «симптом
ямочки»: при легком сдавлении образования
большим и указательным пальцем узел уходит
вглубь кожи, оставляя западение. Дерматофиб-
ромы обычно единичные, у некоторых лиц их
может насчитываться несколько десятков. Эти
опухоли не малигнизируются. Дифференциру-
ют с меланомой, рубцами, кистами, дерматофиб-
росаркомой.
Лечение. Удаляют при постоянном травмиро-
вании, с косметическими целями и неясном ди-
агнозе. Эффективна также криодеструкция жид-
ким азотом.
Мягкая фиброма
Мягкая фиброма — мягкий полип на ножке,
круглой или овальной формы, цвета нормальной
кожи, или с различными коричневыми оттенка-
ми, размером до вишневой косточки. Выполне-
на рыхлой соединительнотканной стромой и ис-
тонченным эпидермисом. При травматизации
фибромы появляются болезненность, кровоточи-
вость, корки. Мягкие фиброзы возникают в
среднем и пожилом возрасте, чаще у женщин и
тучных людей. Локализуются в подмышечных
впадинах, паховых областях, складках под молоч-
ными железами, а также на шее и веках. С воз-
растом их количество увеличивается.
Дифференциальный диагноз: старческая кера-
тома, пигментные невусы, нейрофиброматоз,
контагиозный моллюск.
Лечение. Электрокоагуляция.
Г ипертрофические
и келоидные рубцы
Гипертрофические и келоидные рубцы появля-
ются в результате избыточного образования со-
единительной ткани в месте повреждений кожи
(раны, ссадины, абсцедирующие угри, прививки,
прокалывание ушей, операционные швы, диатер-
макоагуляция и др.) или спонтанно. Келоидный
рубец, в отличие от гипертрофического, значи-
тельно выходит за пределы травмированного уча-
стка, образуя клешневидные отростки. Форма у
гипертрофических рубцов куполообразная, у ке-
лоидов — линейная; цвет — нормальной кожи
или розово-красный; поверхность — гладкая,
блестящая; консистенция — плотная или твердая.
В келоидных рубцах ткань выполнена коллаге-
новыми волокнами, фибробласты отсутствуют.
Конституциональная склонность к келоидной
реакции кожи наиболее выражена у лиц австра-
лоидно-негроидной расы.
Лечение. Предварительное замораживание руб-
ца жидким азотом и последующее его оттаива-
ние вызывает отек, размягчает рубцовую ткань
и облегчает введение в ткань рубца триамцино-
лона ацетонида (10—40 мг/мл). Такие процеду-
ры проводят 1 раз в месяц. В ряде случаев при-
меняют глубокую криотерапию рубцовой ткани.
Иссечение рубцов не рекомендуется, так как
послеоперационный дефект заполняется ещё
большим количеством рубцовой ткани.
Атерома
Атерома (сальножелезистая киста) обычно лока-
лизуется на волосистой части головы, шее, лбу,
в области копчика, мошонки. Это образование
величиной от горошины до куриного яйца,
круглой формы, телесного или желтоватого цве-
та, подвижное. Образуются атеромы преимуще-
ственно в зрелом и пожилом возрасте. Чаще
возникает как одиночное, возвышающееся над
окружающими тканями плотной или эластичной
консистенции медленно растущее образование,
кожа над которым не изменена. При сдавлении
с боков из образования выделяется сырообраз-
ная масса или густая молочного вида жидкость.
Нередко атерома воспаляется, нагнаивается, ста-
новится болезненной и самопроизвольно вскры-
вается.
Дифференцируют с фурункулом, липомой, гра-
нулемой инородного тела, панникулитами и др.
Лечение. Хирургическое вылущивание с обя-
зательным удалением сумки (капсулы) сальной
железы.
364 Частная дерматология
Гиперплазия сальных желез
Гиперплазия сальных желез часто наблюдается
у лиц пожилого возраста в области лба, висков,
шеи в виде множественных милиарных папул с
пупковидным вдавлением в центре. Элементы
при гиперплазии сальных желез мягкой конси-
стенции, при сдавлении с боков из центра па-
пулы выделяется капелька накопившегося сала.
Лечение. Электрокоагуляция элементов.
Сирингома
Сирингома — доброкачественная опухоль апок-
риновых потовых желез. Высыпания возника-
ют преимущественно в детском возрасте и про-
являются множественными опухолевидными
образованиями величиной с булавочную голов-
ку или просо, слегка возвышающимися над ко-
жей, желтоватой или красноватой окраски и
плотной консистенции. Локализуются в облас-
ти грудной клетки, век, вблизи ключиц, на шее,
животе, на верхних конечностях и наружных
половых органах.
Гистологически выявляются множественные
эпителиальные кисты, состоящие из базальных
клеток.
Лечение. С косметическими целями произво-
дится электрокоагуляция элементов.
Липома
Липома (жировик) — доброкачественное обра-
зование подкожной клетчатки мягкой консис-
тенции, круглой или дольчатой формы, не спа-
янное с окружающими тканями. Липомы могут
достигать больших размеров (до 12 см/ Возни-
кают обычно на шее, туловище, иногда на ко-
нечностях. Кожа над узлами не изменена. Ли-
помы могут быть единичные и множественные.
Состоят из липоцитов, окруженных соедини-
тельнотканной капсулой.
Лечение. Хирургическая тактика.
Сосудистые новообразования
(гемангиомы)
Гемангиомы — это сосудистые опухоли, которые
выявляются при рождении (30 % случаев), или
в течение первого месяца после рождения (боль-
шинство случаев). В основе гемангиом лежит
пролиферация эндотелиальных клеток. Геманги-
омы следует отличать от пороков развития сосу-
дов, таких как пламенеющий невус. При после-
днем заболевании пролиферации эндотелиальных
клеток нет. Все гемангиомы состоят из капилля-
ров и сосудов, просвет которых увеличивается с
возрастом.
Капиллярная гемангиома
Капиллярная гемангиома («земляничный не-
вус») X выступающая над уровнем кожи повер-
хностная сосудистая опухоль, которая появля-
ется вскоре после рождения, а к 5—12 году
жизни самостоятельно регрессирует. Наблюда-
ется у 1—2 % детей. Имеет вид мягкого ярко-
красного или темно-вишневого узла или бляш-
ки до 8 см в диаметре, располагается на голове,
шее, туловище, голенях. Максимальный рост
опухоли обычно наблюдается в течение перво-
го года жизни ребенка. Опухоль обычно рас-
сасывается с центра, возможно её изъязвление,
вторичное инфицирование. На месте регресси-
ровавшей капиллярной гемангиомы остаются
иногда поверхностные рубцовые изменения, ги-
попигментация, атрофия или огрубение кожи,
телеангиэктазии.
Лечение. Обычно наступает спонтанный рег-
ресс опухоли. Иногда прибегают к лазерной те-
рапии, криодеструкции.
Пламенеющий невус
Пламенеющий невус («винное пятно») — порок
развития сосудов дермы в виде красного или
фиолетового пятна неправильной формы. Выяв-
ляется при рождении, самостоятельно не исче-
зает, за исключением невуса Унны. Иногда со-
четается с пороками развития сосудов глаз,
мягкой и паутинной оболочек головного мозга,
варикозом и гипертрофией конечностей, сосудов
спинного мозга, мягких тканей и костей.
Встречается на любом участке тела, но чаще
на лице. Поражение преимущественно односто-
роннее, в пределах одного дерматома (иногда и
прилегающих), на лице — в зоне иннервации
тройничного нерва, нередко с вовлечением конъ-
юнктивы и слизистой рта.
Обычно пламенеющие невусы занимают зна-
чительные участки кожи, имеют четкие грани-
цы. С возрастом на их поверхности возникают
папулы, узлы, усиливающие внешнюю дефект-
ность кожи лица человека.
Невус Унны (врожденная
телеангиэктазия затылка)
Выявляется на задней поверхности шеи пример-
но у трети новорожденных. Иногда невус воз-
никает на веках и в межбровной области.
Лечение. В стадии пятна — маскирующая кос-
метика, в стадии образования папул, узлов — ла-
зерная терапия.
Глава 31. Новообразования кожи
365
Кавернозная (глубокая) гемангиома
Порок развития сосудов кожи (вен, капилляров,
лимфатических сосудов), подкожной клетчатки
или подлежащих тканей. Имеет вид опухолевид-
ного образования полушаровидной формы, мяг-
кой, губчатой консистенции, синюшно-красно-
го или голубоватого оттенков, с дольчатым
строением, нечеткими границами. При надавли-
вании на опухоль пальцем образуется ямка, ко-
торая быстро исчезает. Возникает чаще у детей
различных возрастных групп на конечностях, ту-
ловище.
Кавернозная гемангиома иногда сочетается с
варикозным расширением вен, артериовенозны-
ми свищами и пламенеющим невусом.
Лечение. Давящие повязки, эластичные чул-
ки, методы пневматической компрессии. При
крупных гемангиомах X хирургическая тактика
(иссечение, эмболизация). В случае множествен-
ных гемангиом — большие дозы кортикостеро-
идов.
Старческие гемангиомы
Полушаровидные ярко-красные (фиолетовые)
папулы или многочисленные точечные красные
пятна, похожие на петехии, которые появляют-
ся преимущественно на туловище. Дифференци-
руют с ангиокератомой, венозной гемангиомой,
узловой меланомой и метастазами опухолей
внутренних органов (рак почки).
Лечение. Крупные гемангиомы иссекают, мел-
кие подвергают электрокоагуляции или лазер-
ной терапии (при наличии косметического де-
фекта).
Венозная гемангиома
Венозная гемангиома представляет собой мягкую
папулу темно-синего или фиолетового цвета,
появляющуюся на лице, губах или ушных рако-
винах у лиц старше 50 лет. Обычно венозные
гемангиомы единичные, внешних и субъектив-
ных беспокойств не составляют. Опухоль пред-
ставляет собой полость, ограниченную соедини-
тельнотканной стенкой, с внутренним слоем
эндотелиальных клеток, заполненную эритроци-
тами. Необходимо дифференцировать с узловой
меланомой.
Лечение. При необходимости (косметические
цели) гемангиому удаляют с помощью лазера или
электрокоагуляции.
Паукообразная гемангиома
Паукообразная гемангиома (сосудистые звездоч-
ки, звездчатая гемангиома) — телеангиэктазия,
в центре которой видна мелкая красная папула
(питающая артериола) с радиально расходящи-
мися расширенными извитыми капиллярами-
веточками (напоминает паука). Элементы обыч-
но одиночные, в диаметре до 1,5 см. При диас-
копии радиальные сосуды обескровливаются, а
центральная артериола набухает и пульсирует.
Наиболее частая локализация — лицо (щеки,
нос), предплечья и кисти. Появление паукооб-
разных гемангиом у женщин связывают с избыт-
ком эстрогенов (беременность, прием контрацеп-
тивов), звездочки выявляются при вирусных
гепатитах, алкогольном циррозе печени. Одна-
ко они возникают и у здоровых лиц. Телеанги-
эктазии, появившиеся у женщин после родов, а
также возникшие в раннем детстве, постепенно
исчезают.
Лечение. Удаление с помощью лазера и элек-
трокоагуляции.
Ангиокератома (ороговеваюшая гемангиома)
Чаще встречается ангиокератома Фордайса', на
вульве или головке полового члена, мошонке
обнаруживаются многочисленные темно-красные
гиперкератотические милиарные и лентикуляр-
ные папулы (расширенные венулы).
Редкие формы
Ангиокератома Мибелли
Болеют девочки (аутосомно-доминантный тип
наследования), у которых на локтях, коленях и
тыле кистей выявляются ороговевающие папулы
розового, красного или темно-красного цвета.
Болезнь Фабри (диффузная ангиокератома)
Темно-красные точечные сосудистые опухоли,
которые появляются у мужчин внизу живота, на
половых органах, ягодицах, иногда на губах (ре-
цессивное наследование, связанное с недостаточ-
ностью р-D-галактозидазы в эндотелиальных
клетках, фиброцитах и перицитах). В процесс
вовлекаются сердце, кожа, вегетативная нервная
система.
Телеангиэктатическая гранулема
Телеангиэкгатическая гранулема (пиогенная гра-
нулема) — быстро растущая выбухающая геман-
гиома в виде узла или на ножке. Иногда образу-
ется на месте небольших травм. Обычно это
одиночный конусообразный узел от ярко-крас-
ного до черного цвета, размером до мелкой виш-
ни, который изъязвляется, эрозируется, покры-
вается корками, кровоточит при травмировании
или спонтанно. Возникает в результате разраста-
ния капилляров на лице, пальцах рук, деснах.
Встречается чаще у детей и беременных. Диффе-
ренцируют с узловой меланомой, ангиосаркомой,
избыточными грануляциями.
366 Частная дерматология
Лечение. Иссечение опухоли, электрокоагуля-
ция, импульсная лазеротерапия.
Гломангиома (гломусная опухоль)
Доброкачественная опухоль артериовенозного
клубочкового анастомоза (гломуса). Чаще встре-
чается одиночная гломангиома (после травмы),
которая проявляется лентикулярной папулой
красновато-синего цвета, разной консистенции
и выраженной болезненностью. Локализация —
конечности, преимущественно ногтевое ложе,
редко — шея, лицо, половой член и мошонка.
Одиночная гломангиома представляет собой ин-
капсулированный очаг, состоящий из множества
мелких сосудов, просвет которых выстлан упло-
щенными эндотелиальными и гломусными клет-
ками, или клетками по типу эпителиоидных, а
также множеством немиелизированных нервных
волокон. Множественные гломангиомы чаще
выявляются у детей, наследуются по аутосомно-
доминантному типу.
Дифференциальный диагноз (одиночная гло-
мангиома). Меланома, нейрофиброма.
Лечение. Иссечение, электрокоагуляция, иног-
да эффективен импульсный лазер на красителях.
Кисты
Замкнутые полостные образования кожи и сли-
зистых, высланные эпидермисом или эпителием
придатков кожи и заполненные содержимым раз-
личной консистенции (истинная киста). У лож-
ной кисты эпителиальная выстилка отсутствует.
Эпидермальная киста
Эпидермальная киста — замкнутая полость в
дерме, образованная эпидермисом или эпителием
волосяных фолликулов, которая заполнена ро-
говыми массами и кожным салом. Эпидермаль-
ные кисты чаще встречаются в молодом и сред-
нем возрасте. Представляют собой обычно
одиночный внутрикожный или подкожный узел
с тонкой стенкой в диаметре до 5 см. На поверх-
ности узла видны кожные поры, заполненные
роговыми массами. Преимущественная локали-
зация — лицо, шея, грудь, верхняя часть спи-
ны и мошонка. Постепенное увеличение кисты
приводит к разрыву её стенки, попаданию со-
держимого в дерму и воспалительной реакции,
что сопровождается резкой болезненностью.
Разрыв кисты имитирует гнойный абсцедирую-
щий процесс.
Волосяная киста
Волосяная киста обычно возникает у женщин
среднего возраста; возможны семейные случаи.
Имеет вид гладкого, плотного, куполообразно-
го узла, достигающего крупных размеров (до
5 см). От эпидермальной кисты отличается от-
сутствием центрального отверстия. Волосяные
кисты обычно множественные и локализуются
на волосистой части головы, имеют толстую
стенку, образованную многослойным плоским
фолликулярным эпителием, без зернистого слоя.
Киста содержит плотную массу (кератин с вклю-
чениями холестерина), иногда петрифицируется.
Разрыв кисты сопровождается воспалением и
резкой болезненностью.
Имплантационная киста
Имплантационная киста возникает в результате
травматизации эпидермиса, его внедрения и раз-
растания в дерме с образованием полости, содер-
жащей роговые массы. Стенка кисты образова-
на многослойным плоским эпителием и
полноценным зернистым слоем.
Лечение. Имплантационная киста чаще локали-
зуется на ладонях и подошвах, выглядит как
небольшой внутрикожный узел и подлежит ис-
сечению.
Синовиальная киста
Синовиальная киста — ложная киста, возника-
ющая у пожилых людей на тыле кистей, реже —
стоп, наполненная прозрачной, студнеобразной
жидкостью. Появление кисты у основания ног-
тя приводит к его дистрофии и образованию
продольной борозды шириной 1—2 мм.
Лечение. Хирургическое; введение склерози-
рующих средств, триамцинолона (кеналог, дип-
роспан), наложение давящей повязки на несколь-
ко недель.
Милиум
Милиум (просянки) — мелкие эпидермальные
кисты, содержащие кератин и локализующиеся
на веках, лбу и щеках в виде множественных
милиарных желто-белых папул. Милиум разви-
вается из клеток эпидермиса или эпителия при-
датков кожи. Образуется на неизмененной коже,
нередко на ссадинах, в местах ожогов, при пу-
зырных дерматозах. Возникает в любом возрас-
те, часто у грудных детей.
Лечение. Вскрытие капсулы кисты и выдав-
ливание содержимого.
Отложения в коже
Кальциноз кожи
Кальциноз кожи возникает при нарушениях ней-
роэндокринной регуляции, деятельности паращи-
товидных, зобной, щитовидной и половых желез,
Глава 31. Новообразования кожи
367
а также при некоторых системных (склеродермия,
дерматомиозит, миеломная болезнь, фиброзная
остеодистрофия) и других заболеваниях (опухо-
ли, туберкулез, остеомиелит, гипервитаминоз D
и др). Различают 4 формы кожного кальциноза:
метастатический, дистрофический, метаболичес-
кий и идиопатический. Заболевания связывают
с нарушениями фосфорно-кальциевого обмена.
Клиника. Отложения кальция обнаруживают-
ся преимущественно в коже и подкожной клет-
чатке, реже в мышцах и сухожильных влагали-
щах. В коже возникают как ограниченные
участки кальциевых отложений (преимуще-
ственно в области суставов или конечностей),
так и диффузные отложения кальция, часто со-
провождающие системную склеродермию или
дерматомиозит. Кожа в пораженных участках
слегка цианотична, а в холодное время года на-
блюдаются явления акроцианоза. В начале за-
болевания появляются небольшие ограниченные
уплотнения кожи, которые медленно увеличи-
ваются в размерах. При спонтанном или опе-
ративном вскрытии крупных узлов обнаружи-
вается ограниченная полость в виде кисты,
наполненная густой крошковидной массой бе-
лого цвета с желтоватым оттенком. Как прави-
ло, очаги асимметричны, слегка болезненны при
пальпации и визуализируются рентгенографи-
чески.
Гистологически отложения кальция в дерме и
гиподерме выявляются в виде синих зерен (ок-
раска гематоксилин-эозином).
Лечение. Крупные узлы удаляют оперативным
путем, мелкие — электрокоагуляцией. Нагноив-
шиеся и изъязвившиеся узлы подвергают кюре-
тажу. Назначают УВЧ, соллюкс, диатермию.
Показана диета с ограничением продуктов, со-
держащих кальций.
Ксантомы
Образуются в результате отложения липидов в
коже и других тканях. Это желто-коричневые,
оранжевые или розового цвета образования в
виде пятен (плоские ксантомы), папул (эруптив-
ные ксантомы), бляшек (ксантелазмы), узлов
(бугорчатые ксантомы) или инфильтратов (сухо-
жильные ксантомы). Ксантомы — скопления
пенистых (ксантомных) клеток — макрофагов,
нагруженных липидами. Чаще ксантомы появ-
ляются при нарушениях липидного обмена (пер-
вичных и вторичных гиперлипопротеидемиях),
гистиоцитозах (болезнях, обусловленных проли-
ферацией оседлых макрофагов). Нередко ксан-
телазмы и плоские ксантомы возникают при
нормальном уровне липидов.
Ксантелазма — плоская ксантома с локализа-
цией на коже век. У половины больных ксан-
телазма обусловлена гиперлипопротеидемией (се-
мейная гиперхолестеринемия или семейная
дисбеталипопротеидемия). Болеют лица старше
50 лет. В области век появляются мягкие папу-
лы или бляшки желто-оранжевого цвета, непра-
вильных очертаний. Лечение: после хирургичес-
кого иссечения, электрокоагуляции, лазерной
терапии или прижигания трихлоруксусной кис-
лотой рецидивы наблюдаются редко.
Сухожильные ксантомы — желтые или цвета
нормальной кожи подкожные возвышающиеся
образования (признак семейной гиперхолестери-
немии). При разгибании суставов они движутся
вместе с сухожилием. Лечение: избегают продук-
тов, содержащих насыщенные жирные кислоты
и холестерин. Назначают холестирамин, симва-
статин, правастатин, которые эффективны при
гетерозиготной форме семейной гиперхолестери-
немии.
Бугорчатые ксантомы — желтоватые узлы,
которые располагаются в основном на локтях
и коленях, образуются при слиянии эруптив-
ных ксантом. Появляются при семейной дис-
беталипопротеидемии, семейной гипертригли-
церидемии и семейной недостаточности
липопротеидлипазы. Лечение: лечение основно-
го заболевания.
Эруптивные ксантомы появляются внезапно, в
виде множественных узелковых элементов, со-
провождаются воспалительной реакцией. Обыч-
но локализуются на ягодицах, реже — на лок-
тях, коленях, спине. Наблюдаются при семейной
гипертриглицеридемии, семейной дисбеталипоп-
ротеидемии, декомпенсированном сахарном ди-
абете. Сыпь представлена четкими полусфери-
ческими желтыми папулами с красным
венчиком. Лечение: низкокалорийная диета с
минимальным содержанием жиров.
Плоские ксантомы на ладонях — желто-оран-
жевые плоские или слегка выпуклые полосы в
складках ладоней и пальцев, появляются при
семейной дисбеталипопротеидемии — наслед-
ственном дефекте апопротеина Е, приводящем к
нарушениям связывания ЛПНП с гепатоцитами
и их метаболизма. Лечение: диета с низким со-
держанием жиров и углеводов. Назначают так-
же клофибрат, который подавляет синтез Л ОНП
в печени.
Плоские ксантомы при нормальном уровне ли-
пидов — ограниченные очаги желто-красной
пигментации, которые возникают часто без ви-
димых причин или при болезнях крови — мие-
ломной болезни, лейкозах, лимфомах.
368 Частная дерматология
Нейрофиброматоз
(болезнь Реклингхаузена)
Генетически обусловленный дерматоз с аутосом-
но-доминантным типом наследования, сопровож-
дающийся поражением кожи, нервной системы,
костей и эндокринных желез (врожденные по-
роки, опухоли и гамартомы — дефекты разви-
тия органов). Мутантные гены находятся на 17-
й хромосоме (I тип) и на 22-й хромосоме (II
тип). Нейрофиброматоз, описанный в 1882 г.
Реклингхаузеном, относится к I типу заболева-
ния; встречается с частотой 1:4000. Кожные
проявления в виде пятен цвета кофе с молоком
появляются в первые годы жизни, а нейрофиб-
ромы — к концу периода полового созревания.
Нейрофиброматоз развивается в результате на-
рушения пролиферации и дифференцировки
клеток эмбрионального нервного гребня (мела-
ноциты, шванновские клетки, фибробласты эн-
доневрия).
Основные клинико-диагностические признаки
заболевания:
• Множественные пятна типа веснушек на-
блюдаются по всему кожному покрову, однако
наиболее характерна их локализация в подмы-
шечных областях. Пятна лучше визуализируются
при осмотре под лампой Вуда.
• Выявляются крупные пятна цвета кофе с
молоком — их должно быть более 6 у взрослых
и не менее 5 у детей.
• Характерны нейрофибромы — узлы цвета
нормальной кожи, розовые или коричневые,
мягкой или твердой консистенции, нередко на
ножке. При надавливании на узел возникает
болезненность и углубление, а палец провали-
вается в пустоту, как при вправлении грыжи.
• Выявляются плексиформные нейрофибро-
мы — свисающие, мягкие, рыхлые фибромы,
которыми могут быть усеяны конечности, голова,
часть туловища.
• Необходима консультация окулиста, так как
при I типе нейрофиброматоза выявляются гамар-
томы радужки (узелки Лиша).
• Проводится обследование опорно-двигатель-
ной системы: выявление кифосколиоза, сустав-
ной патологии, искривлений голеней, ложных
суставов, гиперостозов, костных кист; надпочеч-
ников (феохромоцитома); периферической не-
рвной системы и ЦНС (глиомы зрительных не-
рвов).
• В горизонтальных гистологических срезах
обнаруживаются макромеланосомы.
• Определяется наличие у родственников I сте-
пени родства проявлений нейрофиброматоза.
• Больным необходимо проводить медико-ге-
нетическое консультирование и диспансерное
наблюдение, так как с течением времени забо-
левание прогрессирует и возможно злокачествен-
ное перерождение нейрофибром. Этой группе
пациентов требуется социальная поддержка и
адаптация.
Лечение. Лечение заключается в коррекции из-
менений костно-суставной системы, проведении
пластических операций, консультаций невропа-
толога и психиатра; при наличии феохромоцито-
мы — хирургическая тактика; наблюдение за
кожными проявлениями заболевания.
Эпидермальные невусы
Возникают как пороки развития эпителиальных
зачатков (эпителиальные невусы); имеют резкие
очертания, различную форму, оттенки коричне-
вого цвета, поверхность гладкую, мягкую или с
гиперкератотическими плотными разрастаниями
(бородавчатую) и ростом длинных волос.
Односторонний эпидермальный невус
Односторонний эпидермальный невус — порок
развития эпидермиса, который выявляется при
рождении или в грудном возрасте. Наиболее
часто проявляется ограниченным бородавчатым
образованием, линейными веррукозными папу-
лами, полосами полуколец на туловище, эпидер-
молитическим ихтиозом, комедоновым невусом
(фолликулярные папулы, в центре закрытые
роговой пробкой), невусом ILVEN (красные и
псориазиформные высыпания).
Дифференциальный диагноз. Линейное распо-
ложение псориаза, порокератоза, красного плос-
кого лишая, болезни Дарье, acanthosis nigricans,
экземы; недержание пигмента.
Лечение. Кортикостероиды для местного при-
менения, инъекции кортикостероидов в очаг
поражения, криотерапия жидким азотом; мест-
но — дитранол, 5-фторурацил в сочетании с тре-
тиноином, кальципотриол.
Порокератоз Мибелли
Порокератоз Мибелли — заболевание, в основе
которого лежит нарушение ороговения керати-
ноцитов в зоне выводных протоков потовых
желез. Гистологическим признаком порокератоза
является роговидная пластинка — паракератоти-
ческий столбик, состоящий из неправильно рас-
положенных роговых клеток с пикнотичными
ядрами и перинуклеарным отеком. Заболевание
наследуется по аутосомно-доминантному типу.
Поверхностный диссеминированный порокера-
тоз — наиболее частая форма заболевания. По-
Глава 31, Новообразования кожи 369
рокератоз возникает чаще в детском возрасте.
Поражаются конечности, реже — лицо, тулови-
ще и половые органы.
Ладонно-подошвенный порокератоз (точечный)
проявляется мелкими папулами на ладонях и
подошвах, которые затем распространяются на
другие участки тела.
Линеарный порокератоз напоминает линейные
формы эпидермального невуса, псориаза, крас-
ного плоского лишая. В очагах порокератоза
могут возникать плоскоклеточный рак, болезнь
Боуэна и базальноклеточный рак. Риск их воз-
никновения составляет 25 %.
Дифференциальный диагноз
Кольцевидная гранулема, анулярный сифилид в
третичном периоде сифилиса, красная волчанка.
Лечение
Местно: 5-фторурацил, третиноин-гель, криоде-
струкция, иссечение, дермабразия и лазерная
терапия (углекислотный лазер).
Пигментные невусы
Пятнистый невус — доброкачественное пигмен-
тное образование (встречается у 3 % взрослых) в
виде одиночного, обычно крупного пигментно-
го пятна, локализованного на лице, туловище и
проксимальных отделах конечностей. Меланома
на месте пятнистого невуса возникает редко.
Пигментные невусы образуются в результате
скопления в разных отделах кожи так называе-
мых невоидных клеток, которые отличаются от
нормальных меланоцитов отсутствием отростков.
Невусы имеют характер пигментированных,
иногда слегка возвышающихся пятен коричневой
или черной окраски с гладкой или папилломатоз-
ной поверхностью. Количество этих невусов и их
локализация могут быть различными. Пигмент-
ные невусы могут быть гигантскими, обычно они
врожденные, иногда волосатые. В зависимости от
локализации невоидных клеток различают:
• пограничный невоклеточный невус (невус -
ные клетки располагаются преимущественно в
нижнем слое эпидермиса);
• внутридермальный невоклеточный невус,
или «отдыхающий» невус, или родимое пятно
(невусные клетки обнаруживаются в средней и
нижней частях дермы);
• сложный невоклеточный невус (сочетает ги-
стологические признаки пограничного и внутри-
дермального невуса) — промежуточная форма.
Часть пигментных невусов при их травматиза-
ции становятся меланоопасными, т. е. могут транс-
формироваться в меланому. В большей мере это
относится к пигментному пограничному невусу,
который имеет вид плоского узелка (до несколь-
ких сантиметров в диаметре) темно-коричневого,
темно-серого или черного цвета с сухой, чаще не-
ровной поверхностью, на которой отсутствуют при-
знаки роста волос. Эти невусы чаще являются врож-
денными, но могут появиться и у лиц других
возрастных групп. Риск развития меланомы суще-
ствует и при смешанном пигментном невусе.
При гистологическом исследовании первично-
го пятна выявляют признаки акантоза, увеличе-
ние числа меланоцитов в базальном слое, повы-
шенное содержание меланина в меланоцитах и
кератиноцитах, меланофаги в дерме. При гисто-
логическом исследовании мелких пятен, возник-
ших позднее, выявляются признаки погранич-
ных невоклеточных невусов, реже — сложных
или диспластических невоклеточных невусов.
Приобретенные пигментные невусы
Лентиго
К приобретенным доброкачественным пигмен-
тным невусам относится лентиго — одиночные
или множественные, желто-коричневого до ко-
ричнево-черного цвета, плоские или слегка воз-
вышающиеся, размером с чечевичное зерно, пят-
на. Различают юношеское лентиго, которое
появляется в детском возрасте на различных уча-
стках кожи, и старческое лентиго, возникающее
у лиц пожилого возраста на открытых участках
кожи. Эти пятна легко подвергаются криодест-
рукции. Появление на поверхности пятна папулы
или узла указывает на инвазию опухолевых кле-
ток в дерму и переход старческого лентиго в
лентиго-меланому.
Врожденные пигментные невусы
Врожденный невоклеточный невус
Врожденный невоклеточный невус выявляют при
рождении или в грудном возрасте. Это доброка-
чественное пигментное новообразование, разных
размеров (от едва заметного, до гигантского),
состоящее из невусных клеток (производных
меланобластов); может явиться предшественни-
ком меланомы.
В зависимости от размера выделяют гигантс-
кий врожденный невоклеточный невус (диаметр
более 20 см, 1 случай на 20 000 новорожденных);
средний врожденный невоклеточный невус (ди-
аметр 2~20 см, встречается у 0,6 % новорожден-
ных); мелкий врожденный невоклеточный невус
(менее 2 см в диаметре, который выявляется у
1 % новорожденных).
370
Гигантский врожденный невоклеточный невус
головы и шеи часто сочетается с поражением
мягкой и паутинной мозговых оболочек; может
протекать бессимптомно, с клиникой эпилепсии,
неврологической симптоматикой.
На поверхности бляшки, занимающей целую
анатомическую область (туловище, конечности,
голова и шея), нередко встречаются узлы и па-
пулы, темные волосы.
Риск развития меланомы из крупного врожден-
ного невоклеточного невуса в течение жизни
достигает 6,3 % и более. При меланоме, возник-
шей из крупного врожденного невоклеточного
невуса, прогноз неблагоприятный.
У больных с мелкими невоклеточными неву-
сами риск развития меланомы в течение жизни
составляет 1—5 %.
Дифференциальный диагноз
Дифференциальный диагноз при врожденном
невоклеточном невусе проводят с нейрофибро-
матозом (пятна цвета кофе с молоком), монголь-
ским пятном (серовато-синий цвет), невусами
Оты и Ито, эпидермальным невусом, лентиго с
ранним началом и послевоспалительной гипер-
пигментацией.
Риск развития меланомы из гигантского
врожденного невоклеточного невуса в течение
жизни составляет 5—15 %, из мелкого врожден-
ного невоклеточного невуса — до 5 %. Наибо-
лее высокий риск развития меланомы существует
при локализации врожденного невоклеточного
невуса на туловище.
Лечение невоклеточных невусов
При пятнистом приобретенном невусе показано
наблюдение и ограничение инсоляции (возмож-
ность озлокачествления).
Электрокоагуляция сложных и внутридер-
мальных приобретенных невусов с косметичес-
кими целями допускается после биопсии и ус-
тановления доброкачественности невуса.
Мелкие врожденные невоклеточные невусы необ-
ходимо удалить до достижения больными 12 лет.
Гигантский врожденный невоклеточный невус из-
за высокого риска меланомы удаляют в ранние
сроки. При наличии показаний (волосистая часть
головы, подошвы, промежность, слизистые, из-
менение и неравномерность окраски, границ,
очертаний, наличие боли, зуда, кровоточивости)
и гистологическом подтверждении невус иссека-
ют с применением методик трансплантации кожи
(необходимо исключить врожденный невокле-
точный, диспластический и голубой невусы).
Диспластический невус
Диспластический невус — это приобретенное пиг-
ментное образование, возникшее в результате
беспорядочной пролиферации полиморфных ати-
пичных меланоцитов. Возникает на неизмененной
коже или на фоне сложного невоклеточного не-
вуса. Считается предшественником поверхностно
распространяющейся меланомы и фактором рис-
ка меланомы. Вероятность заболеть меланомой в
течение жизни составляет: в популяции — 0,8 %,
при наличии синдрома диспластических невусов
в сочетании с меланомой — 100 %, у лиц с дисп-
ластическими невусами — 18 %. При подтверж-
дении диагноза диспластические невусы подлежат
иссечению в пределах здоровых тканей. Физичес-
кие методы удаления не применяют. Максималь-
но исключается инсоляция. Кожный процесс фо-
тографируют в натуральную величину. Пациентов
осматривают 1—4 раза в год.
Предраковые
заболевания кожи
и слизистых оболочек
• Пигментная ксеродерма
• Актинический кератоз
• Дегтярный и мышьяковый кератозы
• Болезнь Боуэна (рак in situ)
• Лучевой дерматит
• Кожный рог
• Бородавчатый предрак
• Ограниченный предраковый гиперкератоз
• Лейкоплакия
• Абразивный преканкрозный хейлит Манга-
нотти.
Пигментная ксеродерма
Пигментная ксеродерма — гетерогенная группа
генетических заболеваний, обусловленных нару-
шениями репарации ДНК, связанными с дефек-
тами эндонуклеаз. Заболеваемость составляет 1
на 40000—250000 населения; тип наследования —
аутосомно-рецессивный. Проявляется в первые
месяцы жизни повышенной чувствительностью
к солнечному свету, фотофобией и конъюнкти-
витом. Постепенно на пораженных участках
кожи возникают шелушение и множество гипер-
пигментных пятен по типу веснушек, телеанги-
эктазии и гипопигментные пятна с признаками
атрофии кожи, возможно развитие солнечного
кератоза и кератоакантом. В течение первых 6
Глава 31. Новообразования кожи
371
лет жизни у большинства больных пигментной
ксеродермой на пораженных участках кожи раз-
виваются базальноклеточный или плоскоклеточ-
ный рак, меланома, рак передней трети языка,
сочетающиеся с различными поражениями глаз,
неврологическими расстройствами.
Лечение. Максимальное ограничение инсоля-
ции, генетическое консультирование, консуль-
тация офтальмолога и невропатолога. Прием
изотретиноина и этретината в низких дозах.
Регулярное наблюдение.
Солнечный кератоз
(актинический кератоз, сенильная кератома)
Появляется у лиц старше 45 лет на участках, дли-
тельно подвергающихся солнечному облучению,
в виде красноватых пятен и папул, покрытых
грубыми, прочно сидящими чешуйками, резко
болезненными при удалении. Поражается лицо,
волосистая часть головы, ушные раковины, шея,
верхний плечевой пояс. Длительное ультрафио-
летовое облучение способствует трансформации
эпидермальных клеток в анапластические (рак in
situ) и переходу кератоза в плоскоклеточный рак
кожи. Этот переход сопровождается воспалитель-
ной реакцией, усилением явлений гиперкерато-
за и изъязвлением бляшки.
Дифференциальный диагноз проводят с хрони-
ческими кожными формами красной волчанки,
себорейными кератомами, болезнью Боуэна,
базалиомой, дерматомикозами.
Лечение. Иссечение очагов, криотерапия, ла-
зеротерапия, близкофокусная рентгенотерапия.
Мышьяковый и дегтярный кератозы
Мышьяковый и дегтярный кератозы — появле-
ние участков гиперкератоза, ахромии кожи, те-
леангиэктазий у лиц, применяющих с лечебны-
ми целями средства, содержащие эти вещества,
или занимающихся их производством.
Лучевой дерматит
Лучевой дерматит — поражение кожи, развива-
ющееся в результате воздействия ионизирующего
излучения, которое вызывает в коже обратимые
и необратимые изменения. К обратимым изме-
нениям (сохраняются до нескольких лет) отно-
сят эритему, выпадение волос, подавление фун-
кции сальных желез и гиперпигментацию; к
необратимым — острый и хронический лучевой
дерматит и злокачественные опухоли.
Острый лучевой дерматит
Характерно развитие ранней эритемы (держит-
ся около 3 суток) и поздней эритемы (пик че-
рез 2 нед.). По окончании третьей недели появ-
ляется гиперпигментация, а через 5-7 суток
возможно возникновение острого лучевого дер-
матита: эритема (при большой дозе облучения),
пузыри, язвы, после заживления которых оста-
ются стойкие рубцовые изменения.
Хронический лучевой дерматит
При облучении с лечебными целями через 3 нед
после окончания курса лечения возникает глу-
бокая отслойка эпидермиса. После эпителиза-
ции, которая продолжается 3—6 нед., остаются
рубцовые изменения, гиперпигментация, атро-
фируются придатки кожи, эпидермис и дерма.
В последующие несколько лет атрофические из-
менения усугубляются, появляются участки де-
пигментации и гиперпигментации, телеангиэк-
тазии, инъецируются поверхностные венулы.
Характерен гиперкератоз (лучевой кератоз).
При аварийном облучении обычно страдают
кисти, стопы и лицо. Исчезает кожный рисунок
на подушечках пальцев кистей (отпечатки), вы-
падают волосы, развиваются ксеродермия, атро-
фия сальных и потовых желез и гиперкератоз,
иногда образуются болезненные, длительно не
заживающие язвы.
Течение хронического лучевого дерматита
прогрессирующее. На его фоне со временем мо-
жет возникнуть плоскоклеточный или базаль-
ноклеточный рак кожи. Необходимо иссечение
пораженного участка и трансплантация кожи.
Кожный рог
Кожный рог — четко ограниченный очаг гипер-
плазии эпителия с мощным, выступающим кна-
ружи гиперкератозом, схожим с миниатюрным
рогом, нередко возникающий на фоне лейкоп-
лакии, туберкулезной волчанки, в области руб-
цов. У основания кожный рог достигает в диа-
метре 1 см, имеет грязно-серый или
коричневато-серый цвет, плотную консистен-
цию. Течение — хроническое; признаки озлока-
чествления: появление воспаления вокруг обра-
зования, уплотнение его основания, резкое
усиление ороговения. В этих случаях необходи-
мо гистологическое исследование.
Лечение. Иссечение кожного рога в пределах
здоровых тканей.
Бородавчатый предрак
Бородавчатый предрак возникает преимуществен-
но на красной кайме нижней губы (чаще у муж-
чин старше 40 лет) и имеет вид возвышающего-
ся безболезненного узелка полушаровидной
формы в диаметре до 1 см, плотноватой консис-
тенции, застойно-красного цвета или цвета нор-
мальной красной каймы, с плотно сидящими на
372 Частная дерматология
поверхности тонкими чешуйками. Часто напоми-
нает бородавку или ороговевающую папиллому.
Диагноз должен быть подтвержден результа-
тами гистологического исследования биопсийно-
го материала из очага.
Дифференцируют от обыкновенной бородав-
ки (сосочковые разрастания с выраженным ги-
перкератозом), папилломы (наличие ножки,
мягкая консистенция), кератоакантомы (плот-
ный периферический красноватый валик, цен-
тральный кратер, заполненный роговыми масса-
ми), пиогенной гранулемы (наличие ножки,
ярко-красный цвет, мягкая консистенция).
Лечение: своевременное удаление очага пора-
жения в пределах здоровых тканей.
Ограниченный предраковый гиперкератоз
Ограниченный предраковый гиперкератоз красной
каймы губ имеет вид вытянутого, небольшого,
четко ограниченного участка, обычно неправиль-
ных очертаний и с признаками воспаления, плот-
ной консистенции и трудно отделяемыми тонки-
ми чешуйками серовато-белого цвета. Очаг может
быть слегка западающим, или слегка возвышает-
ся над окружающей красной каймой (реже).
Течение. Озлокачествление ограниченного ги-
перкератоза может произойти в течение полугода
после возникновения заболевания. Трансформация
в плоскоклеточный рак занимает длительный пе-
риод времени и сопровождается усилением процес-
сов ороговения, появлением уплотнения в осно-
вании элемента, его эрозированием, изъязвлением.
Определяющую роль в установлении диагноза
играет раннее гистологическое исследование био-
псии из участка поражения.
Лечение. Иссечение очага поражения в преде-
лах здоровых тканей и его гистологическое ис-
следование.
Лейкоплакия
Лейкоплакия рта проявляется четко ограничен-
ными пятнами или бляшками белого цвета на
слизистой, которые при соскабливании не уда-
ляются. Возникает под действием экзогенных
раздражителей (курение, алкоголь, длительное
трение зубных протезов или острых краев зубов
о щеку) и не исчезает неопределенно долго пос-
ле их устранения. Иногда на фоне лейкоплакии
развивается кандидозный стоматит. У 30 % боль-
ных плоскоклеточному раку языка и слизистой
рта предшествует лейкоплакия. При гистологи-
ческом исследовании белых пятен обнаружива-
ется гиперплазия или дисплазия эпителия, либо
плоскоклеточный рак in situ. Гиперпластическая
лейкоплакия нередко спонтанно регрессирует
(злокачественное перерождение — менее 5 %
случаев). Диспластические лейкоплакии (их
доля составляет около 10 %) озлокачествляются
в 10 раз чаще. Клинические признаки диспла-
зии и озлокачествления: появление белых бля-
шек с точечными изъявлениями, темно-красных
пятен с бархатистой поверхностью (эритропла-
кия) или точечных белых папул на фоне атро-
фичного эпителия. В данных случаях проводят
гистологическое исследование.
Клинически различают: плоскую (48 %), вер-
рукозную (39 %), эрозивную (10 %) и лейкоп-
лакию курильщиков Таппейнера (3 % случаев).
Наибольшей потенциальной злокачественностью
обладает эрозивная форма лейкоплакии.
Диагноз. Клинические данные; в сомнитель-
ных случаях — гистологическое исследование.
Дифференцируют лейкоплакию с красным
плоским лишаем, красной волчанкой, сифили-
тическими папулами, кандидозом.
Лечение. Санация полости рта, устранение дей-
ствия раздражающих факторов (прекращение ку-
рения; ликвидация дефектов зубов и их протезов,
часто изготовленных из разнородных металлов).
Показан прием витамина В6, концентрата витами-
на А; местно — препараты, содержащие витами-
ны А и Е. Диспластические лейкоплакии устра-
няют с помощью криодеструкции, назначают
системные ароматические ретиноиды. При выяв-
лении плоскоклеточного рака in situ или инвазив-
ного плоскоклеточного рака необходима оператив-
ная тактика.
Прогноз. Наиболее благоприятный — при
плоской лейкоплакии; при веррукозной и эро-
зивной формах — высокий риск трансформации
лейкоплакии в рак.
Абразивный преканкрозный
хейлит Манганотти
Характеризуется появлением на застойно-гипе-
ремированном фоне красной каймы нижней
губы слегка возвышающихся округлых’ эрозий
с гладкой (полированной) поверхностью, без
признаков инфильтрации, спонтанно эпители-
зирующихся, затем вновь быстро рецидивирую-
щих на тех же или новых участках и резистент-
ных к терапии. Эрозии часто покрываются
корками, легко подвергаются травматизации и
кровоточат. В возникновении хейлита Манга-
нотти определенную роль играют возрастные
трофические изменения тканей (вторичная аден-
тия, разрушение передних нижних зубов, гипер-
инсоляция, острая и хроническая травма, гипо-
витаминоз А, патология желудочно-кишечного
тракта и др.).
Глава 31. Новообразования кожи
373
Диагноз. Клинические данные; цитологичес-
кое, а в упорных случаях гистологическое иссле-
дование.
Дифференциальный диагноз. Эрозивные фор-
мы красной волчанки, красного плоского лишая;
лейкоплакия, пузырчатка, многоформная экссу-
дативная эритема, герпетическая эрозия, эксфо-
лиативный, метеорологический, актинический,
атопический хейлиты.
Лечение. Лечение патологии желудочно-кишеч-
ного тракта, санация полости рта, рациональное
протезирование. Длительное применение витамина
А (до 100 тыс. ЕД в сутки), сосудорасширяющие
препараты (теоникол, компламин, никотиновая
кислота и др.), анаболики, солкосерил (внутримы-
шечно или внутривенно). Местно — аппликации
масляного раствора витамина А, фукорцин, жид-
кость Кастеллани, комбинированные кортикосте-
роидные мази, метилурациловая мазь, солкосерил
(мазь или гель) и др. средства, усиливающие ре-
генеративные процессы.
Злокачественные
новообразования
кожи
• Базалиома (базальноклеточный рак)
• Плоскоклеточный рак кожи
• Меланома
• Болезнь Педжета
Базальноклеточный рак
Наиболее часто встречающаяся злокачественная
опухоль кожи. Базальноклеточный рак обычно
развивается из атипичных (базалиоидных) кле-
ток базального слоя эпидермиса, в том числе из
эпителия волосяных фолликулов, в связи с чем
опухоль редко наблюдается на красной кайме губ
и наружных половых органах. Для опухоли ха-
рактерен медленный местный (радиальный) рост
с разрушением окружающих тканей, метастази-
рует редко (вертикальный рост). Наибольшую
опасность представляет локализация опухоли в
периорбитальной области, носогубных складках,
вокруг наружного слухового прохода и на воло-
систой части головы (прорастание в подлежащие
ткани, разрушение мышц, костей, поражение
твердой мозговой оболочки с последующим кро-
вотечением из разрушенных крупных сосудов
или развитием менингита) (рис. 31.2).
Клинические формы
Узелково-язвенная форма. Чаще базальноклеточный
рак представляет собой полупрозрачный, «перла-
мутровый» узел или папулу, затем язву округлых
очертаний с четкими валикообразными краями,
нередко покрытую коркой (ulcus rodens, разъеда-
ющая язва). Цвет элемента обычно розовый или
красный, поверхность — гладкая, блестящая, ха-
рактерна плотная или твердая консистенция; под
лупой выявляются нитевидные телеангиэктазии.
Склеродермоподобная форма выглядит как
беловатый склерозированный участок с нечетки-
ми границами, иногда с «перламутровыми» па-
пулами на периферии.
При поверхностной форме выявляются множе-
ственные красновато-бурые, слегка шелушащи-
еся бляшки с тонкими валикообразными края-
ми, расположенными на туловище. Необходимо
исключить псориаз, старческую кератому, бо-
лезнь Боуэна и дерматофитию туловища.
Пигментированная форма похожа на узловую
меланому, однако имеет более плотную консис-
тенцию.
Для диагностики требуется хорошее освеще-
ние, осмотр элементов с использованием лупы
и их пальпаторное исследование.
Лечение
Оперативное удаление опухоли. При обширных
послеоперационных дефектах требуется транс-
плантация кожи.
При опасной локализации опухоли (носогубный
треугольник, внутренний и наружный угол глаза,
Рис. 31.2. Типичная локализация
базапиомы на лице.
374 Частная дерматология
ушные раковины и слуховой проход, волосистая
часть головы, склеродермоподобная форма) иссе-
чение проводят с интраоперационной микроско-
пией (для определения объема операции). В неко-
торых случаях применяют лучевую терапию.
При другой локализации базалиомы возмож-
но применение криодеструкции и электрокоагу-
ляции.
Плоскоклеточный рак
Злокачественная инвазивная опухоль кожи и
слизистых, растущая из кератиноцитов, приоб-
ретающих свойства атипии в результате действия
различных канцерогенов (инсоляция, соли трех-
валентного мышьяка, ионизирующая радиация,
лучевая терапия, электронное излучение, лучи
Букки, иммуносупрессивная терапия при транс-
плантации органов, местное применение хлорме-
тина, промышленные канцерогены, фотохими-
отерапия и др.).
Возникновение плоскоклеточного рака рото-
глотки, гортани, вульвы, шейки матки, полового
члена, анального канала, промежности и ногте-
вых валиков связывают с вирусом папилломы
человека.
Плоскоклеточный рак in situ
• Болезнь Боуэна
• Эритроплазия Кейра
• Плоскоклеточный рак in situ, вызванный ви-
русом папилломы человека.
Болезнь Боуэна проявляется медленно расту-
щей, красной, слегка инфильтрированной бляш-
кой с четкими границами. На её поверхности
обнаруживается легкое шелушение и корки. Тон-
кий валик по периферии очага, характерный для
поверхностной формы базалиомы, отсутствует.
Рак in situ головки полового члена или внут-
реннего листка крайней плоти известен как
«эритроплазия Кейра». Метастазирует чаще, чем
болезнь Боуэна. При признаках инвазивного
роста появляются мягкие, сочные, легко крово-
точащие грануляции с корками.
При локализации болезни Боуэна на слизистой
оболочке рта поражаются мягкое нёбо, язычок,
ретромолярная область, язык. Участки поражения
нередко имеют сходство с лейкоплакией и красным
плоским лишаем. Их поверхность может быть эро-
зированной, с сосочковыми разрастаниями, реже
гладкой, иногда слегка атрофичной. При слиянии
очагов формируются бляшки неправильных очер-
таний. Субъективные ощущения не характерны.
Для подтверждения диагноза необходимо ги-
стологическое исследование.
Лечение
Иссечение очага в пределах здоровых тканей или
близкофокусная рентгенотерапия.
Прогноз
Без лечения рак in situ трансформируется в ин-
вазивную форму плоскоклеточного рака кожи.
Плоскоклеточный рак кожи, инвазивный обыч-
но развивается на открытых участках тела из
предраковых заболеваний или рака in situ, скло-
нен к метастазированию (рис. 31.3). Различают
ороговевающий и неороговевающий плоскокле-
точный рак.
Высокодифференцированный плоскоклеточный
рак, ороговевающий (язвенный) тип, вначале
имеет вид плотной папулы, бляшки или узла,
покрытых плотными, с трудом удаляемыми ро-
говыми пластинками. Затем в их центре возни-
кает эрозия, которая превращается в язву с плот-
ными, приподнятыми, ороговевшими краями,
покрытую коркой. Форма новообразования не-
правильная или округлая, цвет — красный, жел-
товатый или обычной кожи, консистенция при
пальпации — твердая. Образования чаще оди-
ночные. При метастазах выявляются увеличен-
ные регионарные лимфатические узлы.
Рис. 31.3. Типичная локализация
высыпаний при плоскоклеточном раке.
Глава 31. Новообразования кожи
375
Плоскоклеточный рак нижней губы развива-
ется на фоне лейкоплакии или актинического
хейлита.
Патоморфологически — признаки анаплазии
кератиноцитов, участки ороговения в глубине
или на поверхности новообразования.
Низкодифференцированный плоскоклеточный
рак, неороговевающий (опухолевидный) тип,
имеет вид папулы или узла, обычно красного
цвета, неправильной формы, с мясистыми гра-
нуляциями (вегетациями), которые легко трав-
мируются, кровоточат, превращаются в эрозии
или язвы с некротизированным дном, мягкими
мясистыми кровоточащими краями и корками
(напоминают цветную капусту).
Опухоль в области наружных половых орга-
нов развивается из эритроплазии Кейра, а на
туловище и лице трансформируется из болезни
Боуэна. Метастазирование в регионарные лим-
фоузлы наблюдается чаще, чем при высокодиф-
ференцированном плоскоклеточном раке.
Патоморфологически — анапластический
плоскоклеточный рак с множественными мито-
зами, низкой степенью дифференцировки кле-
ток, отсутствием ороговения.
Лечение
Хирургическое; объем вмешательства определя-
ется проведением интраоперационной микроско-
пии.
Лучевую терапию применяют у пожилых па-
циентов и при поражении слизистых оболочек.
При раке in situ возможны криодеструкция,
применение 5 % фторурациловой мази.
Меланома
В последние годы заболеваемость меланомой
заметно выросла. Риск возникновения мелано-
мы в течение жизни у лиц белой расы в США
составляет 1: 80 (1,25 %). У больного с мелано-
мой в анамнезе риск развития новой меланомы
составляет 5 %. Играет роль наследственная пред-
расположенность, воздействие внешних канце-
рогенных факторов.
Факторы риска
Основным фактором риска развития меланомы
является наличие диспластического невуса с тре-
мя из пяти следующих признаков:
1. Диаметр более 5 мм, неровные границы,
асимметрия, неравномерная окраска, эритема.
2. Синдром диспластических невусов, особенно
у лиц с отягощенным семейным анамнезом. Риск
меланомы при этом синдроме достигает 100 %.
13 Зак. 1357
3. Гигантский врожденный невоклеточный не-
вус.
4. Избыточная инсоляция в анамнезе.
5. Светлая кожа, рыжие волосы, веснушки,
большое количество пигментных невусов.
Диагноз
Выделяют четыре основных признака мелано-
мы: 1) неправильные границы, 2) появление,
кроме основных (черного и коричневого), дру-
гих оттенков (красного, голубого, белого), 3)
асимметрия, 4) диаметр больше 6 мм. Дополни-
тельную информацию дает эпилюминесцентная
микроскопия. С её помощью в области мела-
номы выявляют «псевдоподии», «лучи» и «чер-
ные точки». Меланома сначала растет в преде-
лах эпидермиса и верхнего слоя дермы (фаза
радиального роста), затем инвазирует глубокий
слой дермы (фаза вертикального роста).
Клинические формы
Выделяют четыре основные формы меланомы.
Наиболее часто встречается поверхностно распро-
страняющаяся меланома (около 70 % всех слу-
чаев). Эта форма меланомы развивается из ме-
ланомы in situ и обладает длительной фазой
радиального роста.
Лентиго-меланома обычно возникает у пожи-
лых людей на открытых участках тела и её
предшественником является злокачественное
лентиго.
Акральная лентигинозная меланома поражает
ладони и подошвы (8 % всех случаев меланомы
у лиц белой расы).
При узловой меланоме фаза радиального рос-
та минимальна, опухоль быстро прорастает в
дерму и метастазирует. Прогноз при этой фор-
ме наиболее неблагоприятен.
Дифференциальный диагноз
Диспластический невус, старческая кератома,
гемангиома и другие новообразования кожи.
Лечение
Показано широкое иссечение опухоли для сни-
жения риска местных рецидивов. Высокие дозы
интерферона-альфа 2р.
Болезнь Педжета
Болезнь Педжета (дискератоз соска) — адено-
карцинома, развивающаяся из одного или не-
скольких ходов молочной железы, поражающая
эпидермис области соска и околососкового
поля с последующей инвазией тканей железы
376 Частная дерматология
и метастазированием в регионарные лимфати-
ческие узлы. Заболевание относится к злокаче-
ственным новообразованиям и развивается из
атипичных клеток протоков молочной железы
(клеток Педжета). Болеют преимущественно
женщины после 40 лет, поражается, как прави-
ло, одна железа.
В начальной стадии клиническая картина на-
поминает экзему соска, которая длительно про-
текает в виде мокнущего, слегка уплотненного
очага. В дальнейшем сосок и прилегающая об-
ласть превращаются в резко очерченную темно-
красную, влажную поверхность, частично по-
крытую корками, резистентную к лечебным
мероприятиям. В течение продолжительного
времени (месяцев, лет) формируется растущая по
периферии эрозия, захватывающая значитель-
ную часть молочной железы. При этом сосок
постепенно втягивается и исчезает с образовани-
ем фиброзного уплотнения. В ткани железы
образуется твердый кожный узел, увеличиваются
регионарные лимфатические узлы, пдртныё й
безболезненные при пальпации. Кожные пора-
жения сопровождаются ощущением зуда и жже-
ния. Возможно образование очагов болезни Пед-
жета в аногенитальной, подмышечной областях,
реже — в области пупка. В этих участках опу-
холевым процессом поражаются апокриновые
потовые железы (экстрамаммарная болезнь Пед-
жета).
Дифференциальный диагноз
Дифференциальный диагноз проводят с экзе-
мой, пиодермией, псориазом, болезнью Боуэна.
Лечение
Объем хирургического вмешательства и других
лечебных мероприятий (химио-рентгенотерапия)
определяет врач-онколог.
Глава 32
Особенности кожных проявлений при СПИДе
Неопластические 377
Инфекционно-паразитарные 377
Дерматозы с неясным патогенезом 378
Общая характеристика
Возникают в различных возрастных группах. Про-
являются атипично, имеют тяжелое течение, пло-
хо поддаются лечению, при наличии лихорадки,
слабости, диареи, потери веса и т. п. являются
плохими прогностическими симптомами и свиде-
тельствуют о развитии клинических проявлений
СПИДа и резком снижении иммунного ответа.
Условная классификация заболеваний:
1. Неопластические (саркома Капоши).
2. Инфекционные (вирусные, грибковые, бак-
териальные, паразитарные).
3. Дерматозы с неясным патогенезом (себорей-
ный дерматит, телеангиэктазии, ксеродерма,
псориаз, атопический дерматит и др.).
Неопластические
Саркома Капоши
Саркома Капоши отмечается преимущественно у
гомосексуалистов. На теле появляются бледно-
розовые пятна и папулы, приобретающие фиоле-
товый или коричневый цвет и увеличивающиеся
по периферии. Кожные проявления напоминают
гемангиому, пиогенную гранулему, дерматофиб-
рому, сосудистые пороки развития, экхимозы.
Высыпания чаще располагаются на лице, животе,
конечностях, на туловище — вдоль ребер. В про-
цесс вовлекаются периферические лимфатические
узлы, из внутренних органов — чаще кишечник
(язвенный колит, кровотечения).
Инфекционно-паразитарные
Кандидоз слизистых оболочек и кожи
Развивается стойкий кандидоз слизистых оболо-
чек рта, нередко с вовлечением пищевода, воз-
никает у молодых людей, не получавших лече-
ния антибиотиками, кортикостероидами, цито-
статиками. Подобные проявления являются ран-
ним симптомом ВИЧ-инфекции. На слизистой
щек, языке образуется сплошная бело-грязная
пленка, плохо удаляемая, в углах рта — канди-
дозная заеда. Реже развивается кандидоз пери-
анальных и паховых складок со склонностью к
слиянию очагов и их изъязвлению.
Простой герпес
Характерно образование пузырьков, очень болез-
ненных эрозий или язв, чаще на губах, половых
органах, голенях, пальцах, в аксиллярной и пе-
рианальной областях (у гомосексуалистов). На-
поминает ветряную оспу или импетиго.
Опоясывающий герпес
Может проявляться в виде легких локализован-
ных форм без постгерпетической невралгии и в
виде тяжелых, диссеминированных, склонных к
рецидивам высыпаний со стойкой лимфадено-
патией. Часто сочетается с саркомой Капоши.
«Волосатая» лейкоплакия
Вероятный возбудитель — вирусы Эпштейна-
Барр или папилломы человека. Частая локали-
зация — краевая зона языка. Возможно распро-
странение на слизистую щек. Очаги поражения
имеют неровную гофрированную поверхность,
покрытую нитевидными разрастаниями эпите-
лия, напоминающими волоски. Спонтанно не
исчезают.
Контагиозный моллюск
Поражение чаще всего локализуется в области
щек, переходит на все лицо и волосистую часть
головы. Имеет тенденцию к быстрому распрост-
ранению на другие участки, слиянию элементов.
Бородавки
У больных наблюдается повышенная склонность
к появлению обычных бородавок, чаще на лице,
377
378 Частная дерматология
во рту. У гомосексуалистов появляется большое
количество обычных бородавок и остроконечных
кондилом в области заднего прохода и на гени-
талиях.
Бактериальные инфекции кожи
Распространенные стафилококковые фоллику-
литы на шее, спине, груди, верхних конечнос-
тях, в подмышечных областях и перианальной
складке. Возможно развитие экгим, фурункулов,
абсцессов, целлюлита, импетиго, элементов, на-
поминающих папуло-некротический туберкулез
кожи, акне.
Дерматомикозы
Могут протекать в виде типичных для дермато-
микозов поражений кожи стоп, кистей, голеней
и других участков. Атипичные проявления встре-
чаются, как правило, на лице и шее и могут
напоминать многоформную экссудативную эри-
тему, себорейный дерматит, розацеа. Руброми-
коз ладоней и подошв имеет характер кератодер-
мии, онихомикозы сочетаются с паронихиями.
Для отрубевидного лишая характерно диссеми-
нированное поражение с тенденцией к развитию
папулезно-бляшечных элементов, инфильтрации
и лихенификации кожи.
Чесотка
Протекает атипично в виде генерализованных,
сильно зудящих папуло-сквамозных очагов по-
ражения. Часто поражаются лицо и волосистая
часть головы. Заболевание весьма контагиозно
в связи с наличием в очагах множества клещей.
Постскабиозный дерматит носит упорный харак-
тер и плохо поддается лечению.
Болезнь Рейтера
Характеризуется тяжелым течением с развити-
ем артрита, уретрита (цервицита), конъюнкти-
вита (увеита). Могут развиваться цирцинарный
баланит, изъязвление слизистой оболочки рта,
кератодермия ладоней и подошв, обнаруживается
энтеропатия.
Криптококкоз, гистоплазмоз, амебная,
микобактериальная инфекция кожи
Относят к редким осложнениям ВИЧ-инфек-
ции. Могут протекать в виде фолликулитов,
папуло-некротических очагов, панникулита, вас-
кулитов, подкожных абсцессов, перианальных
язв, гиперкератозов, напоминать герпес, конта-
гиозный моллюск. Указанные инфекции часто
сочетаются с саркомой Капоши.
Дерматозы
с неясным патогенезом
Себорейный дерматит
Первоначально процесс локализуется преимуще-
ственно на лице (на бровях, около рта, в носо-
щечных складках), волосистой части головы и
разгибательных поверхностях верхних конечно-
стей. На лице иногда приобретает форму бабоч-
ки и напоминает дискоидную красную волчан-
ку. На волосистой части головы характерно
обильное шелушение. На конечностях напоми-
нает нумулярные себореиды. Может принять
характер распространенных сильно зудящих
сливных эритематозных фолликулярных бляшек
с поражением крупных складок.
Телеангиэктазии
Обычно локализуются на груди (чаще над клю-
чицами), реже — на ушных раковинах, ладонях
и подошвах. Развиваются на фоне неяркой диф-
фузной эритемы. Встречаются чаще у наркома-
нов (реакция на амил- и бутилнитриты).
Ксеродерма (сухая кожа)
Развивается у больных СПИДом, страдающих
хронической диареей или синдромом мальабсор-
бции. Напоминает ихтиоз с наиболее частым
поражением лица и голеней.
Аллергодерматозы
Характерна склонность к аллергическим реакци-
ям. Развитие атопического дерматита нередко
происходит в продромальном периоде. Часто
возникает медикаментозная токсидермия.
Псориаз
Тяжелые формы могут быть первым симптомом
ВИЧ-инфекции. Характерно развитие генерали-
зованных пустулезных и эритродермических
форм, поражение ладоней и подошв.
Псевдотромбофлебитический синдром
Сопровождается высокой лихорадкой. Опреде-
ляются отек и болезненность нижних конечно-
стей, напряженные воспаленные болезненные
тяжи, индурация кожи. Тромбофлебит глубоких
вен не характерен. Процесс длится 1—4 недели.
Нарушения питания
Чаще отмечаются у детей. Напоминают пеллагру,
энтеропатический акродерматит, цингу. У некото-
рых больных развивается желтое окрашивание
свободного края ногтевых пластинок и вид «ба-
рабанных палочек» дистальных фаланг пальцев.
Глава 33
Лимфомы кожи
Кожные лимфомы — это клинически и морфо-
логически неоднородная группа неопластических
заболеваний, при которых основой патологичес-
кого процесса является первично возникающая
в коже злокачественная пролиферация лимфо-
цитов. Установлена гетерогенность злокачествен-
ных лимфом в зависимости от вида пролифери-
рующего лимфоцита, его принадлежности к
определенной популяции и субпопуляции. Ис-
пользование иммунофенотипирования оказалось
важным для разделения неходжкинских лимфом
на Т- и В-клеточные группы с учетом степени
злокачественности пролиферации.
Распределение лимфоцитов в коже по фено-
типу у больных с лимфомами такое же, как и у
здоровых: Т-лимфоциты локализуются преиму-
щественно в эпидермисе и верхних слоях дер-
мы, а В-лимфоциты в средних и глубоких сло-
ях дермы. Соответственно Т-клеточные опухоли
в основном занимают верхние слои дермы, а В-
лимфопролиферативные процессы возникают в
глубоких слоях дермы и не носят эпидермотроп-
ный характер. Количество кожных Т-клеточных
лимфом (КТКЛ) существенно превосходит ко-
личество В-лимфом (КВКЛ). Т-лимфомы со-
ставляют 65 % всех вариантов злокачественных
лимфом кожи, В — лимфомы — 25 % и неклас-
сифицируемые лимфомы — 10 %.
Эпидемиология
Относительная редкость КТКЛ и сходство кли-
нических проявлений на ранних стадиях с дру-
гими широко распространенными дерматозами
(экзема, красный плоский лишай, атопический
дерматит, псориаз и др.) затрудняет диагности-
ку и регистрацию заболевания, создает опреде-
ленные трудности в эпидемииологи КТКЛ. В
США в настоящее время КТКЛ поражено 2 %
больных, страдающих всеми формами лимфом,
которые встречаются примерно в 0,3 случаях на
100 000 населения в год. Наиболее часто КТКЛ
регистрируются у пожилых людей, хотя отмеча-
лись единичные случаи заболевания даже у де-
тей. Мужчины болеют в 2 раза чаще, чем жен-
щины.
Этиология
Этиология заболевания не выяснена. В качестве
этиологического фактора развития КТКЛ в на-
стоящее время рассматриваются ретровирусы,
подобные лимфотропному вирусу человека I типа
(HTVL-I), который вызывает Т-клеточную лим-
фому-лейкемию взрослых. Признается роль на-
следственных факторов. Существует связь HLA-
I класса и отдельных нозологических форм
КТКЛ. Например, антигены гистосовместимос-
ти В-5 и В-35 часто встречаются у больных лим-
фомами кожи высокой степени малигнизации,
А-10 — лимфомами, протекающими менеее аг-
рессивно, а В-8 — преимущественно у пациен-
тов эритродермической формой грибовидного
микоза.
Длительно протекающие хронические дермато-
зы, такие как нейродермит, атопический дерма-
тит, псориаз и другие, нередко приводят к нару-
шениям барьерной и иммунологической функции
кожи. Это способствует проникновению различ-
ных инфекционных агентов, в том числе виру-
сов, которые могут усиливать поломки в иммун-
ной системе кожи, что в некоторых случаях
приводит к развитию опухолевого процесса.
Определенное значение в генезе лимфом име-
ют: ионизирующая радиация, УФ-излучения,
различные химические соединения, обладающие
канцерогенными свойствами, и некоторые аллер-
гены. Перечисленные факторы могут привести
к появлению клона «генотравматических» лим-
фоцитов за счет активации протоонкогенов или
инактивации гена опухолевой супрессии и, та-
ким образом, инициировать процесс малигниза-
ции лимфоцитов.
При КТКЛ отмечается преимущественная
пролиферация Т-хелперных лимфоцитов, кото-
рые имеют функциональные и мембранные
свойства, подобные здоровым Т-хелперным лим-
фоцитам. Процесс формирования КТКЛ начи-
нается с активации лимфоцитов под воздействи-
ем различных канцерогенных факторов и
появления доминантного Т-клеточного клона.
При его появлении в коже способность ее осу-
379
380 Частная дерматология
ществдять противоопухолевый контроль в зна-
чительной степени варьирует, определяя дина-
мику процесса у больных КТКЛ — от пятнис-
то-бляшечных очагов до развития крупных
опухолей и летального исхода.
Классификация
Выделяют Т-клеточные и В-клеточные лимфо-
мы, которые отличаются по клинике и прогно-
зу: Т-клеточные лимфомы характеризуются ге-
нерализованным поражением кожного покрова
и более тяжелым течением, В-клеточные лимфо-
мы часто манифестируют одиночными элемен-
тами (табл. 33.1, 33.2).
Таблица 33.1. Кожные лимфомы —
классификация ВОЗ (1996)
1. Кожные Т-клеточные лимфомы
• Грибовидный микоз
• Синдром Сезари
• Педжетоидный ретикулез
• Возрастная Т-клеточная лимфома/лейкемия
2. Кожные В-клеточные лимфомы
3. Кожная плазмоцитома
4. Плеоморфные варианты кожных лимфом
• Иммунобластная Т-клеточная лимфома
• Крупноклеточная анапластическая лимфома
5. Поражение кожи при лейкемиях
Для подразделения лимфом кожи по стадиям
используется классификация TNM (Т — кожа,
N — лимфатические узлы, М — внутренние
органы), которая также имеет определенное про-
гностическое значение (табл. 33.3). Что касает-
ся Т-клеточных лимфом, то прогноз на ранних
стадиях (la — НА), как правило, очень благопри-
ятный и продолжительность жизни составляет
около 10—20 лет. На более поздних стадиях (ста-
дия III, синдром Сезари) прогноз в значитель-
ной степени неблагоприятный. Ожидаемая про-
должительнсть жизни в этом случае составляет
около 3 лет.
Клиническая картина
Для КТКЛ характерен полиморфизм высыпаний
в виде пятен, бляшек, опухолей. Наиболее рас-
пространенной формой КТКЛ является грибо-
видный микоз, в течении которого выделяют три
формы: классическую, эритродермическую и
обезглавленную. Каждая из этих форм имеет
разную степень злокачественности. Например,
классическая характеризуется низкой или сред-
ней степенью злокачественности (КТКЛ I-II), а
обезглавленная — высокой (КТКЛ/-Ш).
Классическая форма
грибовидного микоза
Классическая форма грибовидного микоза по те-
чению подразделяется на три стадии: эритематоз-
но-сквамозную, инфильтративно-бляшечную и
опухолевую. Клинически первая стадия напоми-
нает экзему, себорейный дерматит, бляшечный
парапсориаз, красный плоский лишай. Высыпания
могут существовать годами и спонтанно исчезать.
Диагностическими критериями первой стадии
являются упорный, не проходящий, несмотря на
Таблица 33.2. Классификация рабочей группы EORTC «первичные кожные лимфомы» (1997)
Первичная кожная Т-клеточная лимфома кожи (КТКЛ)
Неагрессивная, медленно протекающая (продолжительность жизни >10 лет) Грибовидный микоз. Грибовидный микоз + фолликулярный муциноз. Педжетоидный ретикулез. Большеклеточная КТКЛ, CD30+ (анапластическая, иммунобластная, плеоморфная). Лимфоматоидный папулез
Агрессивная, быстро прогрессирующая (продолжительность жизни < 5 лет) Синдром Сезари Большеклеточная КТКЛ, CD30- (анапластическая, иммунобластная, плеоморфная)
Переходная Гранулематозная «вялая кожа» (гранулематоз кожи). КТКЛ плеоморфная, мелко-/среднеклеточная. Подкожная панникулитоподобная КТКЛ
Первичная кожная В-клеточная лимфома кожи (КВКЛ)
Неагрессивная, медленно протекающая (продолжительность жизни >10 лет) Лимфома центроцитарная. Иммуноцитома, включая краевые зоны КВКЛ
Агрессивная, быстро прогрессирующая (продолжительность жизни < 5 лет) Большеклеточная В-клеточная лимфома ног
Переходная Интраваскулярная В-клеточная лимфома кожи. Плазмоцитома
Глава 33. Лимфомы кожи 381
лечение, зуд, необычность высыпаний, нехарактер-
ных для доброкачественных дерматозов, возраст
больных — обычно старше 50 лет. Во второй,
инфильтративно-бляшечной стации, очаги пораже-
ния формируются на месте ранее существовавших
пятнистых высыпаний и видимо здоровой коже.
Бляшки резко очерчены, имеют темно-красный
цвет с буроватым оттенком. Их консистенция
напоминает «толстый картон». У более половины
больных отмечается ладонно-подошвенный гипер-
кератоз, лимфаденопатия. В третьей, терминаль-
ной стадии развития болезни появляются безбо-
лезненные опухоли плотновато-эластической
консистенции желтовато-красного цвета. Вид та-
ких опухолей часто напоминает шляпки грибов.
Опухоли могут появляться повсеместно. Возник-
новение опухолей свидетельствует о начавшемся
метастазировании малигнизированных лимфоци-
тов в лимфоузлы и висцеральные органы. Следу-
ет отметить, что экстракутанные поражения явля-
ются менее характерными для грибовидного
микоза, да и для всей группы КТКЛ по сравне-
нию с кожными В-клеточными лимфомами. Они
варьируют от 15 % до 100 %, в среднем составляя
72 %. Наиболее часто поражаются лимфоузлы,
селезенка, печень и легкие. Средняя продолжи-
тельность жизни больных с классической формой
грибовидного микоза от момента постановки ди-
агноза составляет 7,2—8 лет.
Эритродермическая форма
грибовидного микоза
Эритродермическая форма грибовидного мико-
за (синдром Сезари, лейкемический вариант гри-
бовидного микоза, эритродермический вариант
КТКЛ II) характеризуется стремительным раз-
витием эритродермии (1—2 месяца), обычно без
предшествующих высыпаний, неукротимым зу-
дом, отечностью, универсальной гиперемией,
мощным крупнопластинчатым шелушением.
Практически всегда сопровождается ладонно-по-
дошвенным гиперкератозом и диффузной ало-
пецией. Все группы лимфоузлов резко увеличе-
ны. Характерен резко выраженный лейкоцитоз
до 100 000 клеток в 1 мм3, в лимфоцитограм-
мах — не менее 10 % клеток Сезари. Эти клет-
ки представляют собой малигнизированные Т-
хелперы, ядра которых со складчатой
церебриформной поверхностью, с глубокими ин-
вагинациями ядерной мембраны. Летальный
исход наблюдается через 2—4 года.
Обезглавленная
форма грибовидного микоза
Обезглавленная форма грибовидного микоза на-
чинается внезапным появлением опухолей без
предшествующих высыпаний. Эта форма КТКЛ
характеризуется высокой степенью малигнизации
и расценивается как проявление лимфосаркомы.
Летальный исход наступает в течение 1 года.
СОЗО+большеклеточная
Т-клеточная лимфома кожи
Гистологическая картина СОЗО+болыпеклеточной
(плеоморфной или анапластической) Т-клеточной
лимфомы кожи соответствует картине нодальных
CD30+ анапластических лимфом. Модальные
лимфомы с этим фенотипом имеют чрезвычайно
неблагоприятный прогноз. И напротив, прогноз
для CD30+ Т-клеточной лимфомы кожи весьма
положительный. На основании этой ситуации мно-
гие первичные кожные CD30+ лимфомы кожи
лечатся агрессивно, как и нодальные лимфомы,
хотя их биология совершенно отличается. Это
выражается также в отсутствии t (2,5) — трансло-
кации.
Таблица 33.3. TNM-классификация кожных Т-клеточных лимфом (1987)
Т: Кожа ТО Клиническое и/или гистологическое подозрение на грибовидный микоз
Т1 Экзематозные очаги, бляшки: менее 10 % поверхности тела
Т2 Экзематозные очаги, бляшки: более 10 % поверхности тела
ТЗ Опухолевидные образования (более одного)
Т4 Эритродермия
N: Лимфоузлы N0 Лимфоузлы не пальпируются
N1 Лимфоузлы пальпируются; гистологически нет данных за КТКЛ
N2 Лимфоузлы не пальпируются; гистологический инфильтрат КТКЛ
N3 Лимфоузлы пальпируются; гистологический инфильтрат КТКЛ
В: Кровь ВО Атипичные лимфоциты в крови отсутствуют (< 5 %)
В1 Атипичные лимфоциты в крови присутствуют (> 5 %)
М: Внутренние МО Внутренние органы не поражены
органы М1 Гметалогически подтверждается поражение внутренних органо
382 Частная дерматология
В-клеточная лимфома кожи
В отличие от Т-клеточных лимфом, В-клеточ-
ные, которые составляют примерно 25 % всех
кожных лимфом, имеют относительно гомоген-
ную клиническую картину. Клинически чаще
всего имеется быстрорастущая солитарная опу-
холь. Множественные опухоли редки. Экстраку-
танные проявления, как правило, отсутствуют,
поэтому и смертность ниже.
Гистопатология
Основным диагностическим критерием грибо-
видного микоза на ранних стадиях являются:
акантоз с широкими отростками, гиперплазия
и уплотнение базальных кератиноцитов, ваку-
ольная дистрофия части базальных клеток, ати-
пичные митозы в разных слоях эпидермиса,
эпидермотропизм инфильтрата с проникнове-
нием лимфоцитов в эпидермис. В дерме опре-
деляются небольшие инфильтраты вокруг сосу-
дов. Среди клеток инфильтрата наблюдаются
единичные мононуклеарные клетки с гиперх-
ромными ядрами — «микозные» клетки. Во
второй стадии гистологическая картина стано-
вится более отчетливой. Значительно усилива-
ется эпидермотропизм клеток инфильтрата, в
результате чего в эпидермис проникают малиг-
низированные лимфоциты, образуя скопления
в виде микроабсцессов Потрие. В третьей ста-
дии акантоз может быть либо массивным, либо,
наоборот, незначительным из-за атрофии эпи-
дермиса. Усиливается инфильтрация эпидерми-
са опухолевыми лимфоцитами, которые форми-
руют множественные микроабсцессы Потрие.
Пролиферат занимает всю толщу дермы, про-
никая в подкожно-жировую клетчатку, и ста-
новится более мономорфным. Преобладают бла-
стные формы лимфоцитов.
Диагностика
Для диагностики КТКЛ необходимо оценить тип
высыпаний (пятна, бляшки, опухоли), скорость
их появления, данные гистологических, цитоло-
гических, рентгенологических и томографичес-
ких исследований, результаты клинических и
биохимических параметров крови, состояние
лимфатических узлов. Исследования костного
мозга оправданы только в случаях нахождения
опухолевых клеток в лимфоузлах и/или в кро-
ви. Стадийная классификация КТКЛ позволя-
ет с помощью набора классических критериев
(клинических и лабораторных) совершенствовать
оценку тяжести и прогноза заболевания.
Большинство лимфом кожи можно предполо-
жить уже клинически, хотя гистологическими и
иммунологическими исследованиями не стоит
пренебрегать (табл. 33.4). Молекулярно-биоло-
гические методы приобретают в последнее вре-
мя все большее влияние на диагностику и клас-
сификацию злокачественных лимфом кожи, а
также на их отграничение от реактивных лим-
фоцитарных инфильтратов.
Биопсия / гистология. Для диагностики реко-
мендуется пробная эксцизия нелеченного кли-
нически подозрительного участка кожи (иногда
несколько пробных биопсий). В принципе при
подозрении на злокачественную лимфому нуж-
но брать препарат для рутинной парафиновой
гистологии и нефиксированный препарат для
замораживания на микротоме.
Молекулярно-биологические исследования. Моно-
клональность может подтверждаться на DNA-,
mRNA и протеиновом уровне, причем анализи-
руются Т-клеточные рецепторные или иммуно-
глобулиновые гены и генные продукты. У оча-
гов с очень плотным лимфоцитарным
инфильтратом может применяться методика
Southernblot. Однако чувствительность этого
Таблица 33.4. Методы диагностики лимфом кожи
Клиническое обследование Тщательное изучение всех элементов сыпи. Подробное исследование всех лимфатических узлов (пальпация, при необходимости УЗИ лимфоузлов). Пальпация печени и селезенки. Фотодокументирование (вспомогательный метод).
Инструментальная диагностика Лабораторные исследования При В-клеточных лимфомах При Т-клеточных лимфомах УЗИ органов брюшной полости. Рентгенография органов грудной клетки в 2 проекциях. КТ грудной клетки и органов брюшной полости (вспомогательные методы). Полное лабораторное обследование (биохимический анализ крови, общий анализ крови, ферменты печени, почечные пробы, ЛДГ, электролиты, электрофорез белков) Иммуноэлектрофорез сыворотки и мочи. Биопсия КМ грудины. Исследование крови на наличие клеток Сезари. Биопсия пораженного участка кожи. Биопсии увеличенных лимфатических узлов и органов.
Глава 33. Лимфомы кожи
383
метода недостаточна, так что приходится приме-
нять методы с использованием ПЦР.
Лечение
КТКЛ на ранних стадиях, представленных пят-
нистыми и бляшечными высыпаниям, имеют
относительно благоприятный прогноз с выжи-
ванием больных свыше 5 лет в 90 % и 67 % слу-
чаев соответственно. В опухолевой стадии у боль-
ных с синдромом Сезари прогноз значительно
ухудшается, и выжившие пациенты в течение 5
лет составляют примерно 30—35 %, при этом
средняя продолжительность жизни достигает не
более 3-х лет.
Учитывая патогенетические факторы развития
КТКЛ, большинство исследователей предпочи-
тают начинать терапию ранних стадий заболева-
ния относительно мягкими противоопухолевы-
ми методами и препаратами, без выраженных
имуннодепрессивных свойств. В двух крупных
рандомизироиванных исследованиях, сделанных
в национальном институте рака (США) в 1989
году, было показано, что химиотерапия больных
на ранних стадиях КТКЛ в комбинации с элек-
тронно-лучевой терапией приводит к очень вы-
сокому терапевтическому результату, однако не
увеличивает длительность ремиссий по сравне-
нию с менее токсичной наружной химиотерапи-
ей. Основываясь на этих результатах, был сде-
лан вывод, что химиотерапия играет только
паллиативную роль и ее применение целесооб-
разно в поздних стадиях КТКЛ.
Лечение КТКЛ на ранних стадиях (IA и IB),
без лимфаденопатии осуществляется наружными
кортикостероидами, производными азотистого
иприта, фотохимиотерапией, электронно-лучевой
терапией, интерлейкинами, ароматическими ре-
тиноидами. Широкое применение в Европе на-
шли соединения азотистого иприта: эразол и мех-
лорэтамин, позволяющие достичь полной
клинической ремиссии у 70 % больных с пятни-
стыми и бляшечными высыпаниями. Механизм
действия этих препаратов основан на подавлении
пролиферативной активности опухолевых Т-хел-
перных лимфоцитов. Препараты используются в
виде аппликаций на очаги поражения 3—4 раза в
неделю. На курс — 20 аппликаций.
ПУВА-терапия — использование фотосенси-
билизаторов из группы псораленов с УФ-светом
области А — оказалась эффективной на ранних
стадиях заболевания. Полная клиническая ре-
миссия достигается у 76—90 % больных. Эффек-
тивность метода основана на избирательном об-
разовании ковалентных сшивок псораленов с
ДНК в пролиферирующих Т-хелперных клет-
ках, что тормозит их деление. Методика: боль-
ные применяют перорально 8 — метоксипсора-
лен из расчета 0,6 мг/кг массы тела за 2 часа до
облучения. За это время препарат успевает мак-
симально накопиться в коже. Облучение прово-
дится в зависимости от типа кожи, начиная с
0,25—1,0 Дж/см2, с увеличением дозы через каж-
дые 2—3 процедуры на 0,5 Дж/см2. Проводится
30—35 процедур. В опухолевой стадии у больных
с эритродермией эффективность ПУВА-терапии
заметно снижается из-за глубокого расположе-
ния пролиферата. В этих случаях применяются
комбинированные методы: ПУВА + цитостати-
ки (проспидин); ПУВА + интерферон; ПУВА +
ретиноиды (РеПУВА — терапия).
Рекомбинантные интерфероны применяются
в терапи КТКЛ с 1980 года. С успехом исполь-
зуются альфа, бетта и гамма интерфероны. Эти
цитокины играют центральную роль в диффе-
ренцировке клеток. Применяют как высокие (50
MU/м2) дозы, так и низкие (3 MU/м2) по 3 раза
в неделю. Длительность ремиссии при моноте-
рапии интерферонами составляет от 5 до 10 ме-
сяцев. В настоящее время в клинике использу-
ются в основном рекомбинантные альфа
интерфероны: роферон, реаферон (альфа-2а-ин-
терферон) и интрон-А (альфа-2Ь-интерферон),
реже используется гаммаферон (гамма-интерфе-
рон). Комбинирование интерферонов с другими
методами терапии оказывает более выраженный
лечебный эффект (интерфероны + ПУВА, ин-
терфероны + цитостатики, интерфероны + аро-
матические ретиноиды).
Электронно-лучевая терапия (ЭЛТ) проводит-
ся больным с КТКЛ в 2-х вариантах: локально
и тотально. Эта методика применяется в основ-
ном при прогрессировании лимфомы с исполь-
зованием бетатрона или линейного ускорителя
мощностью 6—10 Мэв. Облучение больных про-
водят 1—4 раза в неделю в дозе 1—6 грей. Кур-
совая доза составляет 30—40 грей. Побочные
эффекты лучевой терапии включают дермати-
ты, атрофию кожи, алопеции и сухость кожи.
Фотоферез (экстракорпоральная фотохими-
отерапия) — новый комплексный метод тера-
пии КТКЛ, заключающийся в пероральном
приеме псоралена (8-метоксипсоралена) с эк-
стракорпоральным УФА-облучением перифе-
рической крови больного. Механизм действия
фотофереза объясняют формированием в орга-
низме больного КТКЛ мощного иммунного
ответа на введение поврежденных Т-лимфоци-
тов, то есть проводится своеобразная вакцина-
ция больного против опухолевых лимфоцитов.
Псорален наиболее тропен к малигнизирован-
384 Частная дерматология
ным клеткам. В результате фотохимических
реакций наступает их гибель, тем не менее
маркеры опухолевых клеток сохраняются.
Критериями для проведения фотофереза явля-
ются следующие показатели: уровень CD4 и
CD8 должен составлять не менее 15 % от об-
щего числа лимфоцитов, лейкоцитоз менее 15
ООО, незначительная лимфаденопатия, отсут-
ствие висцеральных поражений, нормальное
количество CD 16. Продолжительность заболе-
вания — менее 2-х лет. Фотоферез проводит-
ся в течение 4—6 месяцев 2 раза в месяц. Оп-
тимальным уровнем 8-метоксипсоралена в
крови считается 100—200 мкг/мл. Полный кли-
нический эффект достигается у 15—25 % боль-
ных. Ремиссии длятся от 6 месяцев до 2-х лет.
Ретиноиды включают витамин А и его про-
изводные (ретинал, ретинол, ретинойную кисло-
ту), а также ароматические аналоги (аротиноид,
13-цис-ретинойную кислоту — изотретиноин,
этретинат). Эти препараты, особенно ароматичес-
кие ретиноиды, способны ингибировать проли-
ферат, замедляя дифференцировку. Изотретино-
ин и этретинат больные КТКЛ принимают в
дозах 0,5—1 мг/кг массы тела в течение 2—3
месяцев. Средняя продолжительность ремиссий
составляет от 3 до 13 месяцев. Наиболее эффек-
тивны ароматические ретиноиды на ранних ста-
диях КТКЛ, особенно у больных с ладонно-по-
дошвенным гиперкератозом. Еще более
выраженной терапевтической эффективностью
обладает комбинация ароматических ретиноидов
с ПУВА-терапией (Ре-ПУВА-терапия).
Монохимиотерапия предполагает применение
различных цитостатиков: алкилирующие препа-
раты (циклофосфан, эмбихин, проспидин, спи-
робромин и др.); винкаалкалоиды (винкристин,
винбластин, этопосид); антиметаболиты (метот-
рексат); противоопухолевые антибиотики (адри-
амицин, брунеомицин и др.); производные пла-
тины (цисплатин); кортикостероидные препара-
ты (преднизолон и др.).
Полихимиотерапия показана в поздних стадиях
КТКЛ как наиболее оптимальный метод лечения
больных лимфомами кожи с высокой степенью
злокачественности. Одновременный прием разных
по механизму действия противоопухолевых пре-
паратов (полихимиотерапия) связан с тем, что
каждый из цитостатиков может ингибировать
конкретный этап синтеза ДНК в опухолевой
клетке на G, М и S-фазе и таким образом при-
вести к более надежному подавлению опухолевой
пролиферации. С этой целью предложены мно-
гочисленные полихимиотерапевтические схемы,
из которых приводим только некоторые: проспи-
дин по 100 мг внутримышечно, на курс 3—4 г;
винкристин по 1 мг внутривенно 1 раз в неделю
в течение 4-х недель и преднизолон в дозе 20—
30 мг в день в течение 3-х недель, с постепен-
ным снижением дозы по 2,5 мг в день. По вто-
рой схеме используется циклофосфан по 600 мг
внутримышечно 1 раз в неделю в сочетании с
винкристином по 1 мл внутривенно 1 раз в не-
делю и преднизолоном по 20—30 мг в неделю в
течение 2—3 недель, с постепенным снижением
дозы. При эффективности первых двух курсов
лечения его повторяют 3—4 раза с перерывами для
первой схемы 4—5 недель между циклами, для
второй — 2 недели между 2 и 3 циклами. При не-
эффективности этих методик можно использо-
вать другие сочетания, например циклофосфан,
адриабластин, винкристин и преднизолон или бле-
омицин, адриамицин и метотрексат, либо цикло-
фосфан, адриамицин, винкристин и этопосид и
другие.
Глава 34
Саркома Калоши
До 1981 года саркома Капоши считалась чрез-
вычайно редкой сосудистой опухолью, которая
типично начинала развиваться у пожилых муж-
чин на коже нижних конечностей. Такие спо-
радически, а в некоторых местностях и эндеми-
чески возникающие опухоли объединяются под
названием классическая саркома Капоши, По-
скольку заболевание начинается в пожилом воз-
расте, а опухоль прогрессирует медленно, она
лишь незначительно влияет на качество и про-
должительность жизни пациента. Кроме того,
саркома Капоши выделяется также при массив-
ной, чаще всего ятрогенной иммуносупрессии
(например, саркома Капоши, ассоциированная с
трансплантациями), ее кожная или лимфатичес-
кая форма наблюдается также эндемично в Цен-
тральной Африке (африканская эндемическая
саркома Капоши).
Тяжелый, быстро прогрессирующий вариант
саркомы Капоши, диссеминированно поражаю-
щий кожу, слизистые, лимфоузлы и внутренние
органы, с начала 80-х годов встречается у моло-
дых ВИЧ-инфицированных мужчин-гомосексу-
алистов. Эта эпидемическая ВИЧ-ассоциирован-
ная саркома Капоши явно зависит от клеточной
иммунной системы и считается злокачественным,
идущим от эндототелия сосудов системным за-
болеванием. Предполагают, что под влиянием
ангиогенетических факторов в любых местах
сосудистой системы могут возникать новые опу-
холи. Метастазирования в собственном смысле
слова не происходит.
Помимо клеточного иммунодефицита, в воз-
никновении опухоли с большой долей вероят-
ности играет роль инфекционный передаваемый
агент. В качестве возбудителей предполагают
открытый в 1994 году герпесный вирус (герпес-
ный вирус человека 8 или герпесный вирус сар-
комы Капоши=К8НУ), а также tat-протеин ви-
руса ВИЧ-1.
Клиническая картина
В настоящее время различают четыре формы
саркомы Капоши (классическая идиопатическая,
эндемическая африканская, ВИЧ-ассоциирован-
ная эпидемическая, ятрогенная иммуносупрес-
сивная), которые отличаются особенностями
клинической картины, течением, эпидемиологи-
ей, прогнозом.
Классическая саркома Капоши
Классическая идиопатическая саркома Капоши
чаще всего начинается на голенях и в типичных
случаях имеет медленно возрастающую прогрес-
сию с поздним вовлечением внутренних органов
(рис734.1)ГТри другие формы саркомы Кало-
ши (африканская эндемическая, ВИЧ-ассоции-
рованная эпидемическая и ассоциированная с ят-
рогенной иммуносупрессией саркома Капоши)
не имеют однозначно предопределенного тече-
ния и могут достаточно рано поражать лимфо-
узлы, слизистые оболочки и внутренние органы
(особенно легкие и желудочно-кишечный тракт).
Рис. 22. Типичная локализация
высыпаний при саркоме Капоши.
385
386 Частная дерматология
При классической идиопатической саркоме Ка-
поши вначале развиваются асимптоматичные си-
нюшно-красные пятна или узлы, располагающи-
еся в направлении кожных линий. Единичные
или немногочисленные опухоли могут годами
оставаться неизменными или же быстро распро-
страняться в течение нескольких недель и воз-
растать в количестве и размерах. Следствием этого
являются сливные бляшки и инфильтративно
растущие узлы, часто сопровождающиеся массив-
ными отеками. Наступает сильный отек всех
конечностей и лица. Типичны контузиоформные
кровоизлияния в области опухоли, имеющие
желто-зеленую окраску. В дальнейшем прогрес-
сирующие отдельные опухоли могут некротизи-
роваться в центре, изъязвляться и кровоточить.
Однако и выраженно гиперкератические формы,
которые полностью скрывают сосудистый харак-
тер опухолей, акцентированно возникают на ниж-
них конечностях. Орально поражается, в частно-
сти, слизистая твердого неба. Здесь развиваются
синюшные эритемы, бляшки и узлы, склонные
к изъязвлениям. Клинический дифференциаль-
ный диагноз типичной ангиоматозной саркомы
Капоши проводится с сосудистыми опухолями,
такими как многоклеточная или тромбозирован-
ная ангиома, телеангиэктатическая гранулома,
ангиокератома и ангиосаркома, а также с бацил-
лярным эпителиоидным ангиоматозом. Опухоли,
имеющие инфильтрат преимущественно из вере-
тенообразных клеток, менее синюшны, поэтому
их можно принять за гистиоцитомы, дермальные
невусы или лимфомы, в редких случаях за мела-
номы или пигментированные базалиомы, а на го-
ленях за акроангиодерматиды.
Африканская эндемическая саркома Капоши
Развивается у жителей Экваториальной Афри-
ки и Гаити. Отмечено, что частота саркомы Ка-
поши резко возрастает у жителей тропической
дождливой зоны, наряду с эндемическими оча-
гами по вирусу Эпштейна-Барра, цитомегалии,
повышенной частоте лепры и туберкулеза. Раз-
личают три основных варианта эндемической
саркомы Капоши. Первый — соответствует клас-
сической форме. Второй вариант — агрессив-
ный — обычно развивается у молодых негров и
может протекать в виде локализованной или
диссеминированной лимфаденопатической фор-
ме. Локализованная агрессивная саркома Капо-
ши характеризуется появлением инфильтриро-
ванных, экзофитных или вегетирующих
опухолей на коже, которые имеют тенденцию к
быстрому распространению в подкожную жиро-
вую клетчатку и окружающие ткани, включая
кости. Третий тяжелый вариант (диссеминиро-
ванная лимфаденопатическая саркома Капоши)
характеризуется высокой заболеваемостью сре-
ди детей и подростков, высокой частотой вис-
церальных поражений, диффузным поражением
лимфатических узлов, часто без кожных прояв-
лений, более частым изъязвлением кожных оча-
гов, сравнительно злокачественным течением с
быстрым летальным исходом, смерть наступает
в течение года практически у всех заболевших.
ВИЧ-ассоииированная
эпидемическая саркома Капоши
ВИЧ-ассоциированная эпидемическая саркома
Капоши встречается в 20—26 % случаев при ВИЧ-
инфекции как основной клинический маркер и
в 7-15 % случаев в сочетании с пневмоцистной
пневмонией. Возбудитель ВИЧ / СПИД-инфек-
ции имеет тропность к клеткам с антигенами
CD4, которые располагаются не только на Т-лим-
фоцитах/хелперах, но и на клетках сосудистого
эпителия, что может иметь существенное значе-
ние в патогенезе заболевания. У ВИЧ-инфици-
рованных больных эта гетерогенная по морфоло-
гической структуре опухоль имеет агрессивное
течение, склонность к генерализации и пораже-
нию внутренних органов. ВИЧ-ассоциированная
саркома Капоши характеризуется появлением
большого количества распространенных высыпа-
ний на коже и слизистой оболочке полости рта.
Очаги могут появляться на любом участке кож-
ного покрова, включая кожу головы (особенно
лица, ушных раковин), шеи, туловища, ладоней,
подошв. Они могут быть единичными и локали-
зованными, в течение более или менее длитель-
ного времени концентрироваться на одном учас-
тке (лицо, живот, конечности), но в последующем
имеют тенденцию к быстрой диссеминации.
Ятрогенная иммуносупрессивная
саркома Капоши
Ятрогенная иммуносупрессивная саркома Капо-
ши встречается на фоне иммуносупресссивной те-
рапии по поводу трансплантации почек, при
врожденных иммунодефицитных состояниях и
патологии тимуса, на фоне глюкокортикоидной
терапии по поводу системной красной волчан-
ки, височного артериита, буллезного пемфиго-
ида, дерматомиозита, полимиозита. Отмена или
уменьшение иммуносупрессивной терапии у
некоторых больных приводит к спонтанной ре-
миссии саркомы Капоши или прекращению
появления новых высыпаний. Ятрогенная сар-
кома Капоши чаще всего локализуется в коже,
но может наблюдаться поражение слизистых
оболочек и внутренних органов.
Глава 34. Саркома Капоши 387
Диагноз
Саркома Капоши обладает рядом клинических
признаков, таких как синюшная окраска, рас-
положение по линиям кожного рисунка, желто-
зеленая околоочаговая окраска (кровоизлияния),
отеки, диссемининация (часто с вовлечением
слизистой), которые уже позволяют поставить
клинический диагноз. Это, в частности, касает-
ся пациентов, у которых уже известно наличие
ВИЧ-инфекции или другой формы иммуноде-
фицита или же присутствуют клинические при-
знаки такового (например Mundsoor, ульцери-
рующий генитально-анальный герпес и т. д.). В
случае единичных опухолей или же в иных со-
мнительных случаях, а также в рамках дотера-
певтического исследования стадийности в зави-
симости от размера опухоли осуществляется
эксцизионная или инцизионная биопсия для ги-
стологического подтверждения диагноза.
Для диагностики распространенности необходим
полный осмотр пациента (включая слизистые),
сонографический статус лимфоузлов, гастродуоде-
но- и ректоскопия, а также рентгенологическое
исследование грудной клетки и абдоминальная
сонография. В случае необходимости проводится
компьютерная томография грудной клетки и
брюшной полости.
Серологический маркер опухоли отсутствует,
однако ПЦР-подтверждение герпесного виру-
са саркомы Капоши (соответствует герпесному
вирусу человека 8) в крови опухолевого обра-
зования у ВИЧ-инфицированных предшеству-
ет Саркоме Капоши или возникает параллель-
но с ней.
Диагноз саркомы Капоши ставится с помощью
рутинной гистологической методики с опреде-
лением пятнистых, бляшкообразных и нодуляр-
ных форм и веретеноклеточного или ангиома-
тозного вариантов. Опухоль вначале развивается
без изменений эпидермиса в среднем или верх-
нем слое эпидермиса. Здесь на нормальных со-
судах и придатках образуются новые щелевид-
ные тонкостеночные сосуды с очаговыми
экстравазатами эритроцитов, отложениями гемо-
сидерина и лимфоцитарным воспалительным ин-
фильтратом.
Следует провести дифференциальный диагноз с
другими сосудистыми опухолями: телеангиэкта-
тической грануломой, воспалением грануляци-
онной ткани, акроангиодерматитом Мали, син-
дромом Stewart-Bluefarb и бациллярным
ангиоматозом.
Прогноз и стадийная классификация
При прогнозе следует также учитывать различия
между классической, африканской, ассоцииро-
ванной с ятрогенной иммуносупрессией и ВИЧ-
ассоциированной саркомой Капоши.
Классическая саркома Капоши считается менее
злокачественной, медленно прогрессирующей
опухолью, носитель которой из-за пожилого
возраста чаще всего умирает от других болезней,
Таблица. 34.1. Стадийная классификация ВИЧ-ассоциированной
эпидемической саркомы Капоши (по ACTG 1)
Ранняя стадия (хороший прогноз) Поздняя стадия (плохой прогноз)
При выполнении всех нижеперечисленных условий При выполнении одного из следующих условий
1. Опухоль
Саркома Капоши ограничена кожей и/или лимфоузлами, во всех случаях оральное вовлечение минимально (нет очагов на твердом небе) Пульмональная или гастроинтестинальная саркома Капоши; распространенное оральное поражение, обусловленные опухолью отек или изъязвления
2. Иммунный статус
Клетки CD4 > 200/р! 3. Симптомы Клетки CD4 < 200/pl
Отсутствие оппортунистических инфекций, отсутствие Mundsoor и В2 -симптоматики ВИЧ-инфекции В анамнезе оппортунистические инфекции, Mundsoor, злокачественная лимфома или ВИЧ-ассоциированные неврологические заболевания, В -симптоматика ВИЧ-инфекции
Примечание:1ACTG — AIDS Clinical Trial Group, 2В-симптоматика — неясная лихорадка, ночные поты или
диарея, длящаяся более 2 недель, потеря массы тела более 10 %.
388 Частная дерматология
прежде чем саркома Капоши достигнет угрожа-
ющего масшатаба.
О саркоме Капоши при ятрогенной иммуносуп-
рессии известно, что при прекращении иммуно-
супрессии может произойти спонтанное ее рег-
рессирование. Агрессивная форма течения у этой
группы пациентов довольно редкое явление.
Возможность спонтанной регрессии опухоли ста-
вит вопрос, идет ли речь в таких случаях дей-
ствительно о злокачественной опухоли или же
о вызванной цитокинами доброкачественной
пролиферации сосудов.
Африканская эндемическая саркома Капоши
имеет как малозлокачественный, подобный клас-
сической саркоме Капоши, характер, так и аг-
рессивное (особенно лимфаденопатическая фор-
ма у детей), быстро приводящее к смерти течение.
ВИЧ-ассоциированная саркома Капоши чрезвы-
чайно вариабельна. Наряду с отдельными узла-
ми и пятнами, которые могут в течение многих
лет оставаться хронически стационарными, име-
ются и быстро прогрессирующие варианты с
диссеминацией и вовлечением лимфоузлов и
внутренних органов. Агрессивный и инфильт-
ративный рост опухоли может в течение не-
скольких недель привести к смерти пациента.
При саркоме Капоши легких доказан злокаче-
ственный клональный рост опухоли. С тех пор
как оппортунистические инфекции при ВИЧ-за-
болеваниях стали лучше поддаваться контролю
(ранняя терапия, длительная профилактика),
диссеминированная эпидемическая саркома Ка-
поши наряду с лимфомами считается наиболее
частой причиной смерти при СПИДе.
Лечение
Общепризнанной стандартной схемы терапии
саркомы Капоши до сих пор нет. ВИЧ-инфици-
рованные пациенты, которые при диагнозе сар-
комы Капоши еще не получали противовирусную
терапию, должны получить ее в любом случае. По
наблюдениям Франкфуртской университетской
клиники, достаточная антивирусная терапия
(уменьшение вирусной нагрузки в сыворотке
ниже диагностического порога, увеличение абсо-
лютного числа СВ4-клеток/ц1) у ряда пациентов
приводит к стабилизации саркомы Капоши и даже
ведет к ее регрессии. Согласно современному
уровню знаний, рекомендуется комбинация од-
ного или двух ингибиторов реверсной транскрип-
тазы с ингибитором протеиназы.
Оперативное лечение
Поскольку саркома Капоши является мультило-
кулярным системным заболеванием, оперативная
терапия ограничивается первичными эксцизион-
ными биопсиями для постановки диагноза и пал-
лиативным устранением мелких опухолей, не-
удобных с косметической точки зрения.
Отсутствие метастазирования и быстрая диссеми-
нация делают любые другие, сверх упомянутых,
оперативные вмешательства нецелесообразными.
Поскольку опухоль часто шире захватывает ок-
ружающие ткани, чем это видно клинически,
могут быть рецидивы in loco. Возможность воз-
никновения новых опухолей после локального
травматизирования (операций, повреждений) в
смысле феномена Кебнера также требует сдержан-
ности относительно оперативных мероприятий.
Лучевая терапия
Саркома Капоши считается очень чувствитель-
ной к лучевой терапии опухолью. Степень рег-
рессии для классической саркомы Капоши и
саркомы Капоши с ятрогенной иммуносупрес-
сией составляет 80—90 %. ВИЧ — ассоциирован-
ная саркома Капоши также успешно лечится
малыми дозами облучения. Поверхностную пят-
нистую и бляшкообразную саркому Капоши
следует лечить Orthvolt-терапией (методика мяг-
кого облучения) или же высокоэнергетически-
ми электронами. Общая доза должна составлять
20—30 Gy с разбитием на фракции (по 3 в не-
делю).
Химиотерапия
Системная химиотерапия в случае классической
саркомы Капоши вследствие малоагрессивного ее
течения, редких и, как правило, поздних орган-
ных манифестаций при пожилом возрасте паци-
ентов применяется лишь в единичных случаях
с висцеральным поражением. На первый план
выдвигаются локальные мероприятия. Для ле-
чения эндемической (ВИЧ — ассоциированной)
саркомы Капоши применяются терапевтические
схемы моно- или комбинационной терапии вин-
каалкалоидами. Саркома Капоши у пациентов с
ятрогенной иммуносупрессией может регресси-
ровать после отмены иммуносупрессивной тера-
пии. Слабо прогрессирующие опухоли без вов-
лечения органов лечатся у таких пациентов как
классическая саркома Капоши.
Лечения эпидемической ВИЧ-ассоциирован-
ной саркомы Капоши с явным удлинением вре-
мени выживания до сих пор нет. У таких паци-
ентов существенным фактором риска является
обусловленная химиотерапией супрессия костного
мозга и, как ее результат, дополнительная иммун-
ная супрессия: угрожающие жизни оппортунис-
тические инфекции при химиотерапии встреча-
Глава 34. Саркома Капоши 389
ются вдвое чаще, чем при нелеченной саркоме
Капоши. Чтобы как можно дольше поддержать
качество жизни пациента, в случае ВИЧ-ассоци-
ированной саркомы Капоши химиотерапию про-
водят только при быстром прогрессировании
опухоли и/или вовлечении внутренних органов.
Иммунотерапия
Для лечения саркомы Капоши применяются раз-
личные интерфероны класса I (IFN-alpha 2а, 2Ь;
IFN-beta) в очень разных дозировках и с разны-
ми интервалами. Классическая спорадическая
саркома Капоши имеет степень ремиссии на ин-
терфероны класса I — 60—70 %. Стандартизиро-
ванной схемы лечения в настоящее время не су-
ществует. Интерферон-гамма приводит к
прогрессии опухоли и противопоказан. Моноте-
рапией интерфероном-альфа и бета достигают
при ВИЧ-ассоциированной саркоме Капоши при
еще функционирующей клеточной иммунной си-
стеме (CD4 > 400/ц1) высоких степеней ремис-
сии. При уже начавшемся иммунодефиците (CD4
200—400/ц1) степень реагирования улучшает до-
полнительная антиретровирусная терапия (AZT,
DDI, DDC). При прогрессирующем иммуноде-
фиците (CD4 < 200/p.l, В-симптомы ВИЧ-ин-
фекции, оппортунистические инфекции) интер-
фероны мало- или неэффективны. Возможно,
причина этому — сильно повышенный в этой
фазе заболевания эндогенный уровень интерфе-
рона-альфа в сыворотке.
Существенными сопровождающими явления-
ми высоких терапевтических доз интерферона
являются выраженные гриппоподобные симпто-
мы, ухудшение общего состояния организма, что
обусловливает необходимость терапии парацета-
молом. К тому же наблюдается токсическое по-
ражение костного мозга с анемией, нейтропенией
и быстро начинающейся тромбоцитопенией.
Следует учитывать и потенциальную гепато- и
кардиотоксичность.
Преимущество местной химио- и иммунотера-
пии, в отличие от системной, заключается в не-
значительных, а то и вовсе отсутствующих побоч-
ных действиях. В опухоли можно достигнуть
высоких концентраций прямо антипролифератив-
но действующих интерферонов и химио-
терапевтических веществ. Местная терапия про-
водится амбулаторно и стоит значительно
дешевле. Варианты местной противоопухолевой
терапии синдрома Капоши приведены в таблице.
Диспансерное наблюдение
Поскольку классическая саркома Капоши пожилых
людей встречается редко, надежных данных о
пользе регулярного диспансерного контроля до сих
пор не имеется. Медленная прогрессия опухоли
позволяет считать целесообразным проведение кли-
нического контроля с интервалом в 6 месяцев.
При ВИЧ-ассоциированной эпидемической
саркоме Капоши интервалы диспансерного кон-
троля определяет течение основного ВИЧ-забо-
левания с присущими ему оппортунистически-
ми инфекциями. Так как в отдельных случаях
даже при пока хорошо сохраняющихся остаточ-
ных функциях клеточной иммунной системы
(CD4 > 400 pl) следует рассчитывать на быструю
прогрессию опухоли и вовлечение других орга-
нов, рекомендуется контроль степени клиничес-
кого распространения через каждые 3 месяца
(кожа, слизистые, лимфатические узлы), а так-
же через 6—12 месяцев контроль легких (рент-
геноскопия грудной клетки) и желудочно —
кишечного тракта (при необходимости соногра-
фия, эндоскопия).
Надежные данные по диспансерному ведению
опухоли, которое привело бы к увеличению сте-
пени ее излечивания благодаря строгому конт-
ролю, для саркомы Капоши при ятрогенном
иммунодефиците, а также для африканской и
ВИЧ-ассоциированной саркомы Капоши до сих
пор отсутствуют.
Таблица 34.2. Местная терапия саркомы Капоши: стандартные методы
в зависимости от величины опухоли
Малые < 1 см2 (пятнистые, нодулярные) Криохирургия. Винкристин внутриочагово. Винбластин внутриочагово Интерферон внутриочагово (эксцизия)
Средние 1-4 см в диаметре (пятнистые, нодулярные) Винкаалкалоиды внутриочагово. Облучение дермопаном (фракционное)
Крупные > 4 см в диаметре (узелковые, инфильтрирующие, оральные) Быстрые электроны, облучение кобальтом (фракционное) на голени: дополнительное лечение компрессами
Все преимущественно пятнистые саркомы Капоши Камуфляж
Глава 35
Генодерматозы
Ихтиозы 390
Буллезный эпидермолиз 394
Кератодермии 396
Ихтиозы
Ихтиозы представляют гетерогенную группу
наследственных заболеваний, характеризующих-
ся чрезмерным шелушением кожи. Термин «их-
тиоз» происходит от греческого «ихти», означа-
ющего «рыба». Наиболее часто встречаются
наследственные формы ихтиозов, но не исклю-
чено возникновение этого состояния вторично
по отношению к другим заболеваниям. Суще-
ствует много форм ихтиоза и целый ряд редких
синдромов, включающих ихтиоз как один из
симптомов. Мы рассмотрим лишь четыре основ-
ные формы, которые встречаются чаще всего:
обыкновенный ихтиоз, X — сцепленный ихти-
оз, ламеллярный ихтиоз, эпидермолитический
ихтиоз (табл. 35.1).
Обыкновенный ихтиоз
Синонимы: вульгарный ихтиоз, обычный ихти-
оз, ichthyosis vulgaris.
Для обыкновенного ихтиоза характерно диф-
фузное поражение кожи, более выраженное на
ногах. Отмечаются шелушение и сухость кожи,
фолликулярный кератоз. Многие больные
склонны к аллергическим заболеваниям.
Патогенез
Для обыкновенного ихтиоза характерен ретен-
ционный гиперкератоз: упрочение связей меж-
ду клетками и задержка отторжения роговых
чешуек приводят к утолщению рогового слоя
эпидермиса. Пролиферация кератиноцитов не
нарушена.
Заболевание начинается в 3—12 мес. Мужчи-
ны и женщины болеют одинаково часто. Тип
наследования — аутосомно-доминантный.
Клиническая картина
Элементы сыпи. Сухость кожи; муковидное,
отрубевидное или мелкопластинчатое шелуше-
ние. Фолликулярный кератоз — ороговение ус-
тьев волосяных фолликулов. На ладонях и по-
дошвах усилен кожный рисунок; изредка —
Таблица 35.1. Клинические и генетические признаки наследственных ихтиозов
Форма ихтиоза Тип наследования и частота Возраст манифестации Тип чешуек Локализация Сопутствующие Гистологическая признаки картина
Вульгарный ихтиоз Аутосомно— доминантный 1:300 В детстве Тонкие Обычно спина и разгибательные участки; ладони, подошвы Атопия, фолликулярный кератоз Может быть диагностической
X— сцепленный ихтиоз X—сцепленный, 1:6000 От рождения или в детстве Большие, темные На шее и туловище Помутнение роговицы Может быть диагностической
Ламеллярный ихтиоз Аутосомно— рецессивный 1:300 000 От рождения Большие, грубье Вся поверхность тела; на ладонях и подошвах кожа утолщена Выворот века Диагностическая
Эпидермоли- тический ихтиоз Аутосомно- доминантный 1:300 000 От рождения Толстые, бородавчатые Большая часть тела; в кожных складках бородавчагость Пузыри Диагностическая
390
Глава 35. Генодерматозы 391
кератодермия. Более половины больных страда-
ют также диффузным нейродермитом. Цвет кожи
не изменен. На коже голеней чешуйки напоми-
нают рыбью чешую. Поражение генерализован-
ное, более выражено на голенях, предплечьях и
спине. Обычно не поражены: подмышечные впа-
дины; локтевые и подколенные ямки; лицо, за
исключением щек и лба; ягодицы; внутренняя
поверхность бедер. У детей фолликулярный ке-
ратоз иногда более выражен на щеках. Редко
кератопатия глаз. Течение и прогноз. С возрас-
том состояние обычно улучшается, кроме того,
улучшение наступает летом и при смене клима-
та на более теплый и влажный. Фолликулярный
кератоз на щеках обычно проходит по мере ро-
ста ребенка.
Диагноз
Обычно достаточно клинической картины. Об-
наружение мелких незрелых гранул кератогиа-
лина при электронной микроскопии окончатель-
но подтверждает диагноз.
Патоморфология кожи
Легкий гиперкератоз, истончение или отсутствие
зернистого слоя эпидермиса, базальный слой без
изменений. Электронная микроскопия: мелкие
незрелые гранулы кератогиалина.
Дифференциальный диагноз
Сухая кожа и шелушение. Сухость кожи, при-
обретенный ихтиоз (может быть паранеопласти-
ческим), лекарственная токсикодермия (при ле-
чениии трипаранолом, который в настоящее
время снят с производства), Х-сцепленный их-
тиоз, ламеллярный ихтиоз, эпидермомикозы.
Лечение
Для увлажнения кожи принимают ванны, после
чего кожу смазывают вазелином. Кератолитичес-
кие средства: эффективны препараты, содержа-
щие пропиленгликоль, глицерин, молочную кис-
лоту. Лечение проводят перед сном. 44—60 %
водный раствор пропиленгликоля наносят на кожу
после принятия ванны и надевают «пижаму» из
полиэтиленовой пленки. После улучшения состо-
яния этот метод лечения применяют один раз в
неделю или еще реже. Салициловая кислота,
мочевина (10-20 %), гликолевая и молочная кис-
лоты применяются в разных лекарственных фор-
мах (мазь «Солкокерасал»). Молочная и глико-
левая кислоты уменьшают шелушение.
Ретиноиды (изотретиноин, ацитретин, этрети-
нат), принимаемые системно, эффективны при
всех четырех формах ихтиоза. В тяжелых случаях
ретиниоды принимают длительно с временны-
ми перерывами.
Х-сцепленный ихтиоз
Синонимы: ихтиоз, сцепленный с полом; ихти-
оз, сцепленный с Х-хромосомой; ихтиоз рецес-
сивный Х-сцепленный.
Этой формой ихтиоза болеют только мужчи-
ны. Вскоре после рождения на шее, конечнос-
тях, туловище и ягодицах появляются крупные
темно — коричневые чешуйки, похожие на
грязь. Заболевание может быть врожденным или
начинается в первые месяцы жизни. Болеют
только мужчины. Тип наследования — рецессив-
ный, сцепленный с Х-хромосомой, локус Хр
22.32. Генетически обусловленный дефект —
недостаточность стеролсульфатазы. Частота: от
1: 2 000 до 1:6 000.
Патогенез
Из-за недостаточности стеролсульфатазы разви-
вается ретенционный гиперкератоз: упрочение
связей между клетками и задержка отторжения
роговых чешуек. Пролиферация кератиноцитов
не нарушена.
Клиническая картина
Элементы сыпи: крупные чешуйки, плотно при-
легающие к коже; муковидное шелушение не-
характерно. Цвет темно-коричневый, напомина-
ющий грязь. Локализация: задняя поверхность
шеи, наружная поверхность предплечий, локте-
вые суставы и подколенные ямки, туловище. В
кожных складках высыпаний меньше. Ладони,
подошвы и лицо не поражены. Отсутствует и
фолликулярный кератоз. Глаза: помутнение ро-
говицы (очаг расположен в строме, по форме
напоминает запятую) выявляют у половины
взрослых больных. Помутнение роговицы иног-
да встречается у женщин, гетерозиготных по де-
фектному гену. Половые органы: крипторхизм —
у 20 % больных. В отличие от обыкновенного
ихтиоза, состояние с возрастом не улучшается.
Заболевание обостряется зимой и при смене кли-
мата на более холодный. Из-за недостаточности
плацентарной стеролсульфатазы рождение боль-
ного ребенка обычно сопровождается слабостью
родовой деятельности.
Диагноз
Клиническая картина и семейный анамнез. Пре-
натальная диагностика: амниоцентез и исследо-
вание ворсин хлориона позволяют выявить не-
достаточность стеролсульфатазы.
392 Частная дерматология
Дифференциальный диагноз
Каммелярный ихтиоз; обыкновенный ихтиоз;
эпидермолитический ихтиоз; наследственные
синдромы, включающие ихтиоз как один из
симптомов.
Патоморфология кожи
Гиперкератоз; зернистый слой эпидермиса не
изменен.
Лечение
Местно назначают смягчающие средства, среди
которых самое простое — вазелин. Кератолити-
ческие средства: пропиленгликоль. Лечение про-
водят перед сном. 44—60 % водный раствор про-
пиленгликоля наносят на кожу после принятия
ванны и надевают «пижаму» из полиэтиленовой
пленки. После улучшения состояния этот метод
лечения применяют 1 раз в неделю или еще реже.
Салициловая кислота, мочевина (10—20 %), гли-
колевая и молочная кислоты в разных лекарствен-
ных формах (мазь «Солкокерасал). Общее лече-
ние: назначают внутрь этретинат или ацитретин.
Суточная доза — 0,5—1,0 мг/кг. После того, как
состояние улучшится, дозу постепенно снижают
до минимальной эффективной. Регулярно про-
водят общий и биохимический анализы крови
(включая липидный профиль), а при длительном
лечении — еще и рентгенографию (обызвествле-
ние мягких тканей, диффузный гиперкератоз).
Ламеллярный ихтиоз
Синонимы: пластинчатый ихтиоз, ichthyosis
lamellaris, ихтиозиформная сухая эритродермия,
пластинчатая эксфолиация новорожденных, кол-
лодийная кожа новорожденных.
Ламеллярный ихтиоз — врожденное заболева-
ние, для которого характерны эритродермия,
пластинчатый гиперкератоз, эктропион, фотофо-
бия. При этом заболевании кожа новорожден-
ного обычно покрыта пленкой, напоминающей
коллодий (это состояние известно как «колло-
дийный плод»). Со временем пленка превраща-
ется в крупные грубые чешуйки, которые покры-
вают все тело, включая кожные складки, ладони
и подошвы. Тело остается замурованным в этот
панцирь в течение всей жизни. Мужчины и
женщины болеют одинаково часто.
Этиология
Патология наследственная, тип наследования —
аутосомно-рецессивный; один из локусов —
14ql 1. Заболевание обычно встречается в близ-
кородственных браках. Популяционная частота
1:300 000.
Патогенез
В патогенезе заболевания преобладает пролифе-
ративный гиперкератоз — гиперплазия базаль-
ного слоя эпидермиса, ускоренное продвижение
кератиноцитов от базального слоя к поверхнос-
ти кожи.
Клиническая картина
Кожа новорожденного обычно покрыта ригид-
ной тонкой пленкой («коллодийный плод»). Со
временем пленка трансформируется в крупные
грубые чешуйки, которые покрывают все тело,
включая кожные складки, ладони и подошвы.
Развивается эритродермия. Через несколько лет
у детей и взрослых все тело покрыто толстыми
чешуйками, напоминающими пергамент. Че-
шуйки трескаются и появляется рисунок, по-
хожий на мозаику или черепицу. На ладонях и
подошвах — кератодермия и трещины. Над су-
ставами возможны бородавчатые разрастания
эпидермиса. Нередки вторичные бактериальные
инфекции. Волосистая часть головы покрыта
обильными чешуйками. Частые инфекции ве-
дут к рубцовой алопеции. Отмечается выворот
век и выворот губ. Ногтевые валики воспале-
ны, наступает дистрофия ногтей. Слизистые
обычно не поражены. Выворот век способствует
развитию инфекций.
Заболевание продолжается всю жизнь. Состо-
яние с возрастом не улучшается. Гиперкератоз
ведет к закупорке протоков мерокриновых же-
лез, из-за чего нарушается потоотделение. Вслед-
ствие нарушенного потоотделения у больных
физические нагрузки и жаркая погода приводят
к гиперрексии (повышению температуры тела до
41°С и выше). Чрезмерные потери жидкости из-
за трещин в роговом слое эпидермиса вызыва-
ют обезвоживание. У детей вследствие ускорен-
ного обновления клеток эпидермиса увеличены
потребности в питательных веществах.
Диагноз
Диагностика основана на типичных клиничес-
ких признаках: эритродермия, пластинчатый
гиперкератоз, эктропион, фотофобия. Диагнос-
тические критерии ихтиозиформной врожденной
сухой небуллезной эритродермии (К. Н. Суворо-
ва и соавт., 1990):
• рождение в «коллодийной» пленке, с трещи-
нами;
• после отшелушивания пленки — выражен-
ная эритродермия и отечность кожи;
Глава 35. Генодерматозы
393
। • многолетнее шелушение на всех поверхностях
тела с наибольшей выраженностью на голенях;
• усиление эритемы при инсоляции, от неко-
торых местных препаратов или общего лечения;
• периодически — интенсивное отшелушива-
ние с временным очищением кожи;
• блестящий коллодийноподобный гиперкера-
тоз гребешковой кожи ладоней, подошв, паль-
цев с трещинами, усилением складчатости; ги-
поплазия эпидермальных гребней на кончиках
пальцев и их лакированный вид;
• слабый дисморфический синдром в виде эк-
тропиона преходящего, наружного блефарофи-
моза, смятости и прирастания завитка ушных
раковин, укорочения и радиальной девиации
мизинцев.
Патоморфология кожи
Гиперкератоз, зернистый слой эпидермиса не
изменен или слегка утолщен, акантоз.
Лабораторные данные
Посев позволяет исключить вторичные бактери-
альные инфекции и сепсис, особенно у новорож-
денных.
Дифференциальный диагноз
Х-сцепленный ихтиоз, эпидермолитический
ихтиоз, синдром Нетертона, трихотиодистрофия.
Лечение
Новорожденных помещают в отделение реани-
мации. В кувезе поддерживают высокую влаж-
ность воздуха. Применяют смягчающие средства.
Наблюдают за водно-электролитным балансом и
признаками инфекций.
Следует предупредить родителей, что физичес-
кие нагрузки, лихорадка и жаркая погода при-
водят к перегреванию и тепловому удару. По-
могают обливания, смачивание кожи,
обертывания во влажную простыню.
Местная терапия. Применяют смягчающие
средства (вазелин), а также кератолитические
средства (пропиленгликоль). Пропиленгликоль
применяется в виде 44—60 %-ного водного ра-
створа, который наносят на кожу после при-
нятия ванны и надевают «пижаму» из полиэти-
леновой пленки. После улучшения состояния
этот метод лечения применяют 1 раз в неде-
лю. Используются также другие средства: са-
лициловая кислота, мочевина (10—20%), мо-
лочная кислота в разных концентрациях (мазь
«Солкокерасал»).
Общая терапия. Эффективны третиноин,
ацитретин и, реже, изотретиноин. Суточная
доза — 0,5—1,0 мг на кг веса. Вследствие тера-
тогенного влияния ретиноидов в период лечения
необходима контрацепция.
Эпидермолитический ихтиоз
Синонимы: врожденная буллезная ихтиозифор-
мная эритродермия Брока, эпидермолитический
гиперкератоз, ихтиоз буллезный, ichthyosis
epidermolytica, буллезный тип ихтиозиформной
эритродермии, эпидермолитический генерализо-
ванный гиперкератоз, буллезный ихтиозиформ-
ный гиперкератоз, врожденный универсальный
акантокератолиз.
Эпидермолитический ихтиоз — очень редкое
врожденное заболевание, для которого характер-
ны эритродермия, буллезные высыпания, гипер-
кератотические линейные образования. Заболе-
вание может быть врожденным или начинается
вскоре после рождения. Характерный признак —
пузыри. Со временем пузыри исчезают, и на
первый план выступает неравномерное орогове-
ние кожи. В кожных складках, локтевых, под-
коленных ямках могут появиться бородавчатые
наслоения.
Этиология
Патология наследственная, тип наследования —
аутосомно-доминантный. Мужчины и женщины
болеют одинаково часто.
Клиническая картина
Заболевание начинается в период новорожден-
ности, плод рождается в «рубашке» или «как
ошпаренный». С рождения у ребенка состояние
влажной эритродермии. Кожа гиперемирована,
влажная, мягкая, податливая. Характерный при-
знак заболевания — пузыри, которые присутству-
ют при рождении или появляются вскоре после
рождения. Отмечается положительный симптом
отслойки эпидермиса (симптом Никольского).
Пузыри вскрываются с образованием эрозий,
которые заживают, не оставляя следов. Со вре-
менем развивается ороговение кожи, вплоть до
бородавчатых наслоений в кожных складках,
локтевых и подколенных сгибах. Чешуйки тем-
ные, плотно прикреплены к коже и образуют
рисунок, похожий на вельвет. Высыпания сопро-
вождаются неприятным зловонным запахом (фе-
тор), который связан с проникновением в кожу
бактерий и развитием инфекций. Повторное
образование пузырей на ороговевшей коже, а
также слущивание роговых наслоений ведет к
тому, что кожа приобретает относительно нор-
мальный вид. Такие островки нормальной кожи
394 Частная дерматология
посреди очагов ороговения — важный диагнос-
тический признак. Волосы не изменены. Воз-
можна деформация ногтей. Слизистые не пора-
жены. Салоотделение и потоотделение не
нарушены.
Эпидермолитический ихтиоз может протекать
в тяжелой форме со смертельным исходом. При
данной форме ихтиоза новорожденный кажется
нормальным: кожные покровы красного цвета,
мягкие, но на отдельных участках заметны уп-
лотнения; ладони и подошвы утолщены, бело-
ватого цвета. Губы мягкие, слизистая оболочка
полости рта нормальная. При трении пальцем
кожи очень легко происходит отслоение эпидер-
миса (симптом Никольского). Вследствие яркой
красноты кожа имеет вид «обваренной кипят-
ком». В данном случае нет пузырей в прямом
смысле, а лишь происходит отторжение эпидер-
миса. Через несколько дней наступает смерть.
Более благоприятное течение при средне-тя-
желой и относительно легкой форме эпидермо-
литического ихтиоза, при которой дети выжи-
вают. Клиническая картина развивается
постепенно и пузыри появляются в более поздние
сроки, на лице поражение незначительно, одна-
ко в носогубных складках отмечается слабо вы-
раженный гиперкератоз. Волосы, брови, ресни-
цы, ногти могут оставаться нормальными. На
коже туловища сохраняется генерализованный
гиперкератоз, более отчетливо выраженный в
складках, с неприятным запахом. Особенно чет-
ко выражен ладонно-подошвенный гиперкера-
тоз; гипергидроз. Эпизодически на различных
участках кожи появляются пузыри. При отно-
сительно легких и среднетяжелых формах эпи-
дермолитического ихтиоза в первые годы жиз-
ни преобладает клиническая картина
эритродермии с развитием пузырей, а в более
поздние сроки происходит нарастание гиперке-
ратоза с заметным уменьшением эритродермии.
Диагностика
Диагностические критерии эпидермолитического
ихтиоза (К. Н. Суворова и соавт., 1990):
• при рождении красная, отечная, мягкая
кожа, места с влажными участками, где содран
роговой слой, нет «коллодийной» пленки;
• появление пузырей с рождения или с пер-
вых дней и недель;
• при заживлении пузырей шелушение, гипе-
ремия, а после 2—4-х лет — сосочковые бородав-
чатые гиперкератозы;
• преимущественное поражение сгибательных
поверхностей и складок;
• фетор (зловоние, резкий неприятный запах).
Патоморфология кожи
В зернистом слое эпидермиса видны гигантские
гранулы кератогиалина и вакуолизация. Лизис
клеток и образование субкорнеальных многока-
мерных пузырей. Папилломатоз, акантоз, гипер-
кератоз.
При электронной микроскопии выявляются
большие зоны перинуклеарного цитоплазмати-
ческого ретикулума, масса рибосом и митохон-
дрий в клетках зернистого и верхней части ши-
поватого слоев, утолщение тонофиламентов,
вертикальная ориентация эпидермоцитов в ро-
говом слое, дефекты кератинизации.
Лечение
Местное лечение — молочная и гликолевая кис-
лоты. Антимикробная терапия. Ретиноиды (эт-
ретинат или ацитретин внутрь) вызывают вре-
менное ухудшение (провоцируют образование
пузырей), однако впоследствии, благодаря нор-
мализации дифференцировки кератиноцитов,
состояние резко улучшается.
Буллезный
эпидермолиз
Буллезный эпидермолиз — группа заболеваний, для
которых характерно спонтанное развитие пузырей
или их появление на месте незначительной трав-
мы, обусловленное генетическими дефектами. Раз-
личают три основные группы этого заболевания:
простой, пограничный, дистрофический (табл. 35.2).
Простой буллезный эпидермолиз с аутосомно-
доминантным типом наследования появляется
при рождении или в раннем детстве. Пузыри
развиваются вследствие малейших травм, зажи-
вают без образования рубца. Отмечается дефект
кератинов 5 и 14.
Пограничный буллезный эпидермолиз — врож-
денное состояние, проявляющееся множеством
пузырей. Полости развиваются в области дермо-
эпидермального соединения и связаны, по-ви-
димому, с дефектом Калинина — молекулы, уча-
ствующей в прикреплении эпидермиса к дерме.
Этот тип эпидермолиза наследуетсядудосомий-
рецессивно.
Дистрофический буллезный эпидермолиз насле-
дуется аутосомно-доминантно или_рецессивно.
Степень выраженности поражений кожи варь-
ирует от легких до тяжелых обезображивающих.
Причина заболевания — дефект дермальных
якорных фибрилл.
Глава 35. Генодерматозы
395
По современным представлениям, группа пу-
зырных наследственных кожных заболеваний
включает в себя более 20 дерматозов.
Этиология
Этиология всех форм буллезного эпидермолиза
определяется многочисленными генными мута-
циями в различных хромосомах. Причем, эта
группа заболеваний генетически неоднородна:
часть из них наследуется аутосомно-доминант-
но, часть — аутосомно-рецессивно.
Патогенез
Патогенез буллезного эпидермолиза изучен не-
достаточно. При всех формах заболевания име-
ются предопределенные структуры нарушения на
уровне базальной мембраны. При ряде форм
буллезного эпидермолиза известны некоторые
ферментные нарушения в эпидермисе.
Клиническая картина
Клинически общими для всех форм буллезного
эпидермолиза являются раннее проявление за-
болевания, чаще прй рождении или с первых
дней жизни, возникновение пузырей и эрозий
на коже и слизистых оболочках в результате не-
значительной механической травмы или гипер-
термии. Цаличие или отсутствие рубцов после за-
живления дает основания для раздёления всех
клинических форм буллезного эпидермолиза на
дистрофические и простые.
Патоморфология кожи
На основании современных данных, полученных
с помощью электронной микроскопии, все фор-
мы буллезного эпидермолиза можно разделить
на три группы: простой, пограничный, дистро-
фический буллезный эпидермолиз.
При простых формах буллезного эпидермолиза
образование пузырей в покровном эпителии
происходит в результате цитолиза базальных
эпителиоцитов, что выявляется на электронно-
граммах в виде резко выраженного отека их
цитоплазмы с разрывом клеточной оболочки.
Покрышкой пузыря является весь отслоившийся
эпидермис, дном — неповрежденная базальная
мембрана.
При пограничных формах буллезного эпидер-
молиза образование пузыря происходит на
уровне светлой пластинки базальной мембра-
ны и обусловлено врожденной неполноценно-
стью полудесмосом базальных эпителиоцитов
и крепящих тонофиламентов. Дно пузыря со-
ставляет плотная пластинка базальной мемб-
раны.
При дистрофических формах буллезного эпи-
дермолиза образование пузыря происходит на
границе базальной мембраны и дермы, связано
с неполноценностью крепящих «якорных» фиб-
рилл — структур, соединяющих базальную мем-
брану с дермой. Таким образом, дном пузыря
является дерма (дермолитический пузырь), что
впоследствии приводит к формированию рубцов.
Таблица 35.2. Классификация
буллезного эпидермолиза
Гоуппа простого буллезного эпидермолиза
(тип наследования — аутосомно-доминантный)
1. Простой генерализованный
2. Поздний локализованный
3. Простой герпетиформный
Гоуппа пограничного буллезного эпидермолиза
(тип наследования — аутосомно-рецессивный)
1. Летальный генерализованный
2. Доброкачественный генерализованный
атрофический
3. Инверсный вариант атрофического пограничного
4. Прогрессирующий вариант атрофического
пограничного
5. Локализованный атрофический пограничный
6. Рубцующий пограничный
Группа дистрофического буллезного эпидермолиза
(тип наследования — аутосомно-доминантный)
1. Атрофический вариант
2. Гиперпластический вариант
3. Альбопапулезный вариант
Гоуппа дистрофического буллезного эпидермолиза
(тип наследования — аутосомно-рецессивный)
1. Дистрофический генерализованный
мутилирующий
2. Дистрофический генерализованный
немутилирующий
3. Дистрофический локализованный инверсный
Лечение
Учитывая наследственный характер заболевания,
существование его в течение всей жизни боль-
ного, активная терапия применяется короткими
курсами во время обострений и возникновения
осложнений, а затем сменяется более длитель-
ным назначением общеукрепляющих и симпто-
матических средств.
Основными задачами лечения являются:
• предупреждение травматизации кожи (раци-
ональный уход, гигиена кожи, правильная про-
фессиональная ориентация) и слизистых оболо-
чек (полноценное питание, богатое растительной
396 Частная дерматология
клетчаткой, хорошо измельченная и мягкая
пища);
• повышение резистентности кожи к травма-
тическим воздействиям;
• ускорение заживления имеющихся высыпа-
ний;
• профилактика и лечение вторичного инфи-
цирования;
• предупреждение и лечение тяжелых ослож-
нений, связанных с рубцеванием.
Патогенетическая терапия. Известно, что в
коже больных дистрофическим буллезным эпи-
дермолизом с аутосомно-рецессивным типом
наследования вырабатывается избыточное коли-
чество структурно измененной коллагеназы. В
связи с этим патогенетически обосновано при-
менение препаратов, ингибирующих выработку
или активность коллагеназы — дифенина (фе-
нитоина), эритромицина, больших доз витами-
на Е и ретиноидов.
Дифенин назначают внутрь 2 раза в день из
расчета 3,5 мг/кг массы тела в сутки у взрослых
и 8 мкг/кг у детей. Положительный эффект
заключается в уменьшении количества пузырей,
ускорении эпителизации эрозий.
Из группы ретиноидов применяются неоти-
газон и ретинола пальмитат (или ретинола аце-
тат). Помимо антиколлагеназного действия, они
обладают способностью ускорять эпителизацию.
Неотигазон назначают в суточной дозе 1 мг/кг
веса тела (равными частями 3 раза в день во
время еды). Неотигазон тератогенен и эмбрио-
токсичен, поэтому противопоказан беременным
и планирующим беременность женщинам. Рети-
нола пальмитат (или ретинола ацетат) применя-
ется в суточной дозе 5 000 МЕ/кг массы тела (в
равных частях после еды утром и вечером). Курс
лечения составляет 1,5—2 мес.
Кератодермии
Кератодермия (син.: кератоз ладонно-подо-
швенный) — группа болезней ороговения на-
следственного и приобретенного генеза. Харак-
теризуется избыточным рогообразованием,
преимущественно в области ладоней и подошв.
По характеру клинической картины кератодер-
мии могут быть диффузными, со сплошным по-
ражением всей поверхности ладоней и подошв
(кератодермии Унны-Тоста, Меледа, Папийона-
Лефевра и др.) и локализованными, при кото-
рых участки избыточного ороговения распола-
гаются очагами (кератодермия Симменса,
кератодермия линейная Фукса, кератодермия
Бушке-Фишера-Брауэра и др.).
Кератодермия Унны-Тоста
Кератодермия Унны-Тоста (кератома врожденная
ладонно — подошвенная) /—/ распространенная
форма наследственной диффузной кератодермии,
для которой характерен кератоз ладоней и подошв
без перехода на другие участки кожи. Тип насле-
дования аутосомно-доминантный. Заболевание
проявляется в первые годы жизни в виде легко-
го утолщения кожи ладоней и подошв. Постепен-
но диффузный кератоз нарастает к 4—5 годам,
редко позднее. К этому возрасту клиническая
картина заболевания формируется полностью.
Роговые наслоения на ладонях и подошвах (иног-
да только на подошвах) гладкие, толстые, жел-
того цвета, с резко очерченным краем, который
окружен эритематозным венчиком шириной 1-
3 мм. Процесс сопровождается локальным гипер-
гидрозом.
Гистологически выявляют ортогиперкератоз,
гранулез, акантоз, в дерме — небольшой пери-
васкулярный воспалительный инфильтрат. Во-
лосы, зубы не изменены. Ногти могут быть
утолщены, но не дистрофичны. Возможны ос-
теопороз и остеолиз фаланг, деформирующий ар-
троз межфаланговых суставов, осложнение про-
цесса грибковой инфекцией.
Кератодермия Меледа
Кератодермия Меледа (кератоз наследственный
трансградиентный) — форма наследственной
диффузной кератодермии, отличающаяся пере-
ходом кератоза с ладонно-подошвенных повер-
хностей на тыл кистей, стоп, области локтевых,
коленных суставов (трансградиентный кератоз).
Описана впервые среди кровных родственников
населения острова Меледа. Тип наследования
обычно аутосомно-рецессивный. Первые прояв-
ления болезни возникают в детском возрасте в
виде стойкой эритемы с шелушением кожи ла-
доней и подошв. В дальнейшем ороговение кожи
усиливается, и к 15—20 годам на ладонях и по-
дошвах видны массивные роговые наслоения
желто-коричневого цвета, лежащие компактны-
ми пластинами, эритема сохраняется лишь в виде
фиолетово-лилового ободка шириной несколь-
ко миллиметров по периферии очага.
Гистологически выявляют ортогиперкератоз,
иногда акантоз, в дерме — небольшой воспали-
тельный лимфоцитарный инфильтрат. Характе-
рен локальный гипергидроз, поверхность очагов
кератоза обычно влажная, с черными точками вы-
водных протоков потовых желез. Роговые наело-
Глава 35. Генодерматозы 397
ения переходят на тыльную поверхность кистей,
стоп, области локтевых и коленных суставов. На
их поверхности образуются болезненные глубо-
кие трещины (особенно в области пяток). Харак-
терны сочетание с атопической экземой, ослож-
нение процесса пиококковой инфекцией,
дистрофия ногтей с их резким утолщением или
койлонихией. Могут отмечаться изменения на
ЭЭГ, умственная отсталость, синдактилия,
складчатый язык, готическое небо.
Кератодермия Папийона-Лефевра
Кератодермия Папийона-Лефевра (синдром Па-
пийона-Лефевра) — наследственная диффузная
кератодермия, сочетающаяся с парадонтозом и
пиогенными инфекциями кожи и десен. Тип на-
следования аутосомно-рецессивный. У больных
отмечается снижение функции щитовидной и под-
желудочной железы, нарушение функциональной
активности лейкоцитов, снижение фагоцитарной
активности нейтрофилов и чувствительности Т- и
В-лимфоцитов к митогенам. Клиническая карти-
на проявляется обычно в возрасте от 1 года до 5
лет (чаще на 2—3 году жизни) в виде эритемы
ладоней и подошв, покрывающихся роговыми
наслоениями, интенсивность которых постепенно
усиливается. Участки кератоза нередко выходят за
пределы ладонно-подошвенных поверхностей на
тыл кистей и стоп, область пяточного (ахиллова)
сухожилия, коленных и локтевых суставов. Харак-
терен локализованный гипергидроз.
Гистологически выявляют гиперкератоз, нере-
гулярный паракератоз, в дерме — небольшой вос-
палительный инфильтрат. В клетках рогового и
зернистого слоев обнаруживают липидоподобные
вакуоли, нарушение структуры тонофибрилл и ке-
ратогиалиновых гранул. Ногти нередко дистро-
фичны (тусклые, ломкие), волосы не изменены.
В возрасте 4—5 лет в результате персистирующе-
го гингивита развивается прогрессирующий па-
родонтоз с образованием и дистрофией альвеоляр-
ных отростков с преждевременным кариесом и
выпадением зубов, аномалией их развития. Воз-
можны кальцификация твердой мозговой оболоч-
ки, арахнодактилия, акроостеолиз.
Лечение
Ретиноиды (неотигазон, этретинат) — по 0,5 мг/
кг в сутки в течение нескольких недель, аевит,
ангиопротекторы (теоникол, трентал). Наружно:
кератолитические мази (20 % салициловая мазь,
мази Ариевича, Уайтфельда), мазь «Солкокера-
сал», солевые ванны, фонофорез с витамином А,
20 % димексид, лазеротерапия. При кератодер-
мии Папийона-Лефевра лечение начинают с
антибиотикотерапии и санации полости рта.
Глава 36
Кожные заболевания в жарких странах
Тропические пиодермии и смешанные бактериальные инфекции 398
Тропические микозы 405
Особенности течения специфических дерматозов 408
Поражения кожи при протозойных болезнях 409
Кожные поражения, вызываемые гельминтами 410
Поражения кожи, вызываемые членистоногими 415
Поражения кожи, вызываемые ядовитыми змеями,
моллюсками, пиявками, актиниями 417
Особенности клиники и течения неинфекционных дерматозов 418
Венерические болезни и эндемические трепонематозы 419
Регионы тропического климата по природным
и социально-экономическим условиям суще-
ственно отличаются от регионов умеренного
пояса: характерна высокая температура возду-
ха, ее незначительное колебание в течение су-
ток, высокая влажность, повышенная солнеч-
ная радиация, наличие двух сезонов года
(сухой — лето и влажный — зима), сезонная
закономерность движения воздушных пото-
ков. В тропиках часто встречаются тепловой
и солнечный удар, солнечный ожог, потница
и др.
Особенности кожных и венерических заболеваний
в странах с тропическим и субтропическим кли-
матом обусловлены:
• предрасполагающими климатическими усло-
виями и природным ландшафтом;
• социально-экономическими факторами (не-
полноценное и недостаточное питание);
• низкой санитарной культурой населения;
• многообразным растительным и животным
миром;
• повышенной функцией меланогенеза в от-
личие от европейцев;
• своеобразным профилем терморегуляции,
водно-солевого обмена, секреторной, защитной
и других функций кожи;
• склонностью кожи к келоидной реакции;
• наличием у трети дерматологических боль-
ных различных гельминтозов и других инфек-
ционных заболеваний.
Тропические
пиодермии
и смешанные
бактериальные
инфекции
В отличие от жителей умеренных поясов, для
населения жарких стран характерны:
• меньшая частота «классических» пиодермий
и слабая выраженность островоспалительного
компонента при возникновении фолликулитов,
сикоза, гидраденита;
• частое возникновение подрывающего фол-
ликулита, акне-келоид, множественных мили-
арных абсцессов у детей, эпидемической пузыр-
чатки новорожденных, рожистого воспаления,
эктим, язв, кожно-костных гангрен;
• частое развитие гнойного процесса после
укусов насекомых, грызунов, пресмыкающих-
ся;
• тяжелое течение у детей и стариков;
398
Глава 36. Кожные заболевания в жарких странах 399
• частые сопутствующие заболевания (гель-
минтозы, хронические инфекции);
• многообразные тропические язвы.
Тропические пиодермии
Пиоз Мансона
Пиоз Мансона характерен для стран с влажным
тропическим климатом (возбудители- стафилокок-
ки: золотистый, гемолитический, белый). Чаще бо-
леют дети старшего возраста, женщины и приез-
жие европейцы. Поражаются крупные складки
(паховые, подмышечные, межъягодичные), в ко-
торых появляются множественные папулы с пе-
риферическим воспалительным венчиком. Затем
возникают небольшие фликтены с серозно-гной-
ным содержимым, которые вскрываются, образу-
ют эрозии и корки, после их отпадения остается
быстро проходящая гиперпигментация. Общее
состояние больных обычно не страдает. У гипот-
рофичных детей процесс может принять универ-
сальный характер, с общетоксическими явления-
ми и сопровождаться подъемом температуры.
Акне-келоид
Акне-келоид (келоидные угри). Частое появление
акне-келоид в области затылка у мужчин афри-
канцев на местах стафилококковых пустул связы-
вают с конституциональной предрасположеннос-
тью лиц австралоидно-негроидной расы к
келоидному диатезу. В результате стафилококко-
вого воспаления волосяных фолликулов и сальных
желез в области задней поверхности шеи, как пра-
вило, на границе гладкой кожи и волосистой ча-
сти головы происходит абсцедирование этих эле-
ментов, их слияние и образование свищевых ходов.
Возникает горизонтальный полосовидный ин-
фильтрат шириной до 5 см, возвышающийся над
уровнем кожи, далее подвергающийся келоидно-
му уплотнению и пигментации. Верхняя граница
очага поражения представлена разряженными пуч-
ками волос, выходящими из свищевых отверстий.
Дифференциальный диагноз проводят с кело-
идным бластомикозом, при котором полиморф-
ные элементы (узелки, бугорки, бородавчатые
разрастания) располагаются преимущественно на
коже лица, предплечий, голеней. Из этих эле-
ментов сыпи выделяется культура гриба
Clenosporella Loboi- Fonseca et Leao.
Тропические угри
Торпидно протекающий дерматоз, по типу вуль-
гарных угрей, развивается у лиц, приехавших
в тропические страны из регионов с умеренным
(холодным) климатом (европейцы, североаме-
риканцы). Возникновению тропических угрей
способствуют повышенная потливость, мацера-
ция кожи, десквамация эпителия, приводящие
к закупорке сальных желез и появлению угре-
подобных элементов. Заболевание развивается
у лиц старше 20 лет, обычно остро, в ближай-
шие несколько месяцев после прибытия в тро-
пики. На спине, груди, животе, пояснице, бед-
рах, реже на лице появляются множественные
изолированные узелковые элементы (напоми-
нают комедо), которые постепенно нагнаивают-
ся, становятся болезненными при пальпации и
в покое. Отдельные пустулезные элементы
могут сливаться, образуя конгломераты по типу
сливных угрей с фистулезными ходами. Кож-
ный процесс носит хронический характер, со-
провождается рубцеванием, келоидизацией,
дисхромией кожи на местах регрессировавших
элементов.
Лечение малоэффективно; выздоровление на-
ступает после возвращения в прежние климати-
ческие условия.
Тропический везикулезный
акродерматит Кастеллани-Челмера
Достаточно редкий тропический пиодермит,
встречающийся в странах с влажным тропичес-
ким климатом (возбудители — вирулентные
штаммы стафилококков, стрептококков и неко-
торых других пиококков). На коже тыла кистей
(реже стоп) появляются болезненный отек и ин-
фильтрация. В течение 2—3 недель кожа в уча-
стках поражения становится синюшной, напря-
женной, блестящей. Общее состояние пациента
ухудшается (небольшая лихорадка, слабость, не-
домогание). Пораженная кожа покрывается мно-
жеством мелких пузырьков с прозрачным или
желтоватым содержимым, которые затем одно-
моментно вскрываются, образуя эрозии округ-
лой формы. Далее эрозии эпителизируются, на
их месте остается пигментация. Одновременно с
инволюцией сыпи постепенно уменьшается отеч-
ность и уплотнение кожи тыла кистей, регрес-
сируют и субъективные ощущения. При отсут-
ствии рациональной терапии возможны
рецидивы пузырьков и формирование слоново-
сти в области кистей (стоп). Иногда болезнь
принимает затяжное течение. В таких случаях
бывает несколько волн появления пузырьков.
После ряда рецидивов и при отсутствии рацио-
нального лечения может сформироваться слоно-
вость кистей (или стоп).
Дифференциальную диагностику проводят с
отеком при лоаозе, эризипелоидом.
400 Частная дерматология
Тропический пиомиозит
(тропический дерматомиозит)
Разновидность глубокой тропической пиодермии,
при которой поражаются кожа, подкожная клет-
чатка и подлежащие ткани (возбудитель — пре-
имущественно стафилококк). Заболевание начина-
ется с внезапного озноба, лихорадки (39° С и
выше), головных болей, болезненности мышц
нижних конечностей. Кожа в области пораженных
мышц становится отечной, гиперемированной,
напряженной, резко болезненной при пальпации.
Обычно через неделю в фокусе затвердения мышц
появляется размягчение и флюктуация (иногда оп-
ределяется несколько таких очагов). Далее абсцессы
спонтанно вскрываются с образованием крупных
язв. У ослабленных больных (сопутствующая ма-
лярия, гельминтозы и др.) болезнь протекает очень
тяжело и может закончиться летальным исходом
без массивной антибактериальной терапии и хи-
рургического вмешательства.
Дифференциальный диагноз проводят с карбун-
кулом, гуммозными язвами, лейшманиомами,
глубокими микозами, дерматомиозитом, глубо-
кими флебитами.
Кожные гангрены
В условиях тропического и субтропического кли-
мата кожные гангрены нередко осложняют тече-
ние кожных миазов, филяритозов, глубоких ми-
козов, фрамбезиом, травматических и других
поражений кожи. Достаточно редко у жителей этих
регионов возникает диабетическая гангрена кожи.
Множественная
гангрена детского возраста
Множественная гангрена детского возраста на-
блюдается у детей, ослабленных серьезной сопут-
ствующей патологией. Начинается заболевание
с появления на коже туловища, конечностей,
реже на лице полиморфных высыпаний (пятна,
волдыри, пурпура, пузыри, пустулы, фурунку-
лоподобные элементы). Через 2—3 дня в центре
элементов появляется сероватое, а затем бурова-
то-черное некротическое пятно. Процесс быст-
ро прогрессирует с формированием глубоких
корок, после снятия которых выявляются язвы
с неровными краями и гнойно-кровянистым
дном. Прилегающие друг к другу язвы могут
сливаться и образовывать обширные очаги не-
кроза с фестончатыми очертаниями, разрушаю-
щими кожу, подлежащие ткани с последующи-
ми мутиляциями носа, ушных раковин, пальцев,
половых органов. Заболевание протекает с вы-
раженным общетоксическим синдромом (лихо-
радка, адинамия, желудочно-кишечные расстрой-
ства, возможны судороги, потеря сознания и др.).
Развитие септицемии может привести к леталь-
ному исходу.
Множественная гангрена взрослых
Множественная гангрена взрослых может быть
первичной и вторичной. Характерна многооча-
говость поражений, тяжелые общетоксические
явления, возможно возникновение сепсиса.
Множественная гангрена взрослых развивается
преимущественно на фоне тропических трепоне-
матозов, донованоза, филяриатозов, американс-
кого и бразильского лейшманиозов, миазов, тро-
пических зудящих дерматозов и др.
Молниеносная гангрена
наружных половых органов
Молниеносная гангрена наружных половых ор-
ганов (некротический фасциит, гангрена Фурнье),
вероятно, связана с аллергизирующим воздей-
ствием золотистого стафилококка, стрептокок-
ков, протея (реже) и развитием ишемии. Забо-
левание начинается внезапно с повышения
температуры до 38—39°С, отека полового члена
и мошонки, лихорадки. Через 1—2 недели раз-
вивается некроз тканей мошонки и полового
члена с их последующей деструкцией в течение
2—3 недель. Яички в процесс не вовлекаются.
Смертность при ган!рене Фурнье достигает 25 %.
Несмотря на своевременно начатое лечение, де-
фекты разрушенных тканей половых органов
полностью не восстанавливаются. У женщин
возможно поражение малых половых губ, обла-
сти клитора, промежности, лобка, бедер.
Дифференциальный диагноз проводят с некро-
тическим и фагеденическим твердым шанкром.
Нома
Нома (водяной рак, гангрена полости рта). Чаще
болеют ослабленные дети 2—4 лет вскоре после
перенесенных инфекционных и паразитарных
заболеваний, на фоне истощения организма.
Возможны эпидемические вспышки номы в дет-
ских больницах тропических стран. В отделяе-
мом язв обнаруживают Вас. perfringens, а также
симбиоз Плаута-Венсана; не исключается вирус-
ная природа заболевания. Болезнь проявляется
быстрым гнойно-некротическим распадом мяг-
ких тканей полости рта (десны, небо, язык),
перфорацией щек, выпадением зубов и нередко
заканчивается тяжелой деструкцией хрящей и
костей лица. Иногда первичный очаг поражения
Глава 36. Кожные заболевания в жарких странах 401
может локализоваться в области шеи, промеж-
ности, половых органов. Процесс сопровождается
икхорозным запахом изо рта, крайне тяжелым
общим состоянием (лихорадка, резкая слабость,
адинамия, судороги, желудочно-кишечные рас-
стройства, пневмония), ранним развитием сеп-
тицемии. При несвоевременном лечении болезнь
часто заканчивается летальным исходом.
Лечение: массивная антибиотикотерапия; об-
калывание очагов поражения противогангреноз-
ной сывороткой (до 20000 АЕ, в первый день),
затем ежедневно по 10000 АЕ в течение 5—7
дней; гамма-глобулин по 2 мл внутримышечно
через день № 4; системные кортикостероидные
препараты; десенсибилизирующие средства, ин-
фузии плазмы крови, поливитамины. Местно —
антисептики, дезинфицирующие, эпителизиру-
ющие средства. Необходимо полноценное высо-
кокалорийное питание.
Мадагаскарская нома
Мадагаскарская нома встречается у детей (1—7
лет) на острове Мадагаскар, прилегающих ост-
ровах, в некоторых арабских странах. Заболева-
ние чаще поражает здоровых детей, эпидемио-
логически не опасно. Для нее характерно более
продолжительное (до 1,5 мес) и относительно
доброкачественное течение по сравнению с
«классической» номой. Наблюдается язвенно-
некротический распад слизистой оболочки десен
с гнойно-кровянистым отделяемым, расшатыва-
ние зубов (иногда их выпадение), поражение
верхней и нижней челюстей, перфорация щек.
Язык, глотка и миндалины в процесс не вовле-
каются. Реакция региональной лимфатической
системы выражена незначительно. Отмечается
лихорадка (38,5-39°С), слабость, желудочно-ки-
шечные расстройства, боли в очагах поражения.
При тяжелом течении возможен сепсис. Исход
болезни — обезображивающие лицо рубцы.
Лечение: см. «Нома».
Тропические язвы
Представляют собой клинически схожие язвен-
ные процессы различной этиологии. В отделяе-
мом язв выявляются микробные ассоциации:
гемолитический стрептококк, стафилококки,
симбиоз Плаута-Венсана и др.
Истинная тропическая язва
(фагеденическая тропическая язва,
джунглевая язва и др.).
Для заболевания характерен «комплекс четырех F»:
Foot (стопа), Filth (грязь), Fusospirillosis (фузоспи-
риллез), Friction (втирание, потертость), его отно-
сят к болезням бедного слоя населения. Излюб-
ленная локализация тропических язв — нижняя
треть голеней, стопы, т. е. места наиболее частой
травматизации, загрязнения, укусов насекомых.
Язвы обычно возникают единичные и поражают
одну конечность. Заболевание начинается с быст-
ро возникающего болезненного пузырька или
фликтены с ободком гиперемии и слегка уплот-
ненным основанием. В течение суток на месте
элемента образуется эрозия, которая затем превра-
щается в язву, растущую по периферии и вглубь,
с формированием на поверхности грязно-серого
струпа с неприятным запахом, под которым вид-
ны розовато-сероватые грануляции. В язвенный
процесс могут вовлекаться фасции, сухожильные
влагалища, иногда мышцы и надкостница. Харак-
терен эксцентрический рост язвы («феномен тя-
жести») в зависимости от преимущественного ста-
тического положения пациента: при вертикальном
положении — к стопе, при горизонтальном — к
икроножной мышце. Общее состояние обычно не
страдает. Без лечебных мероприятий заболевание
может длиться годами и заканчивается формиро-
ванием гладкого и западающего рубца, а у лиц
австралоидно-негроидной расы — образованием
келоида. Редко наблюдается анкилозирование го-
леностопного сустава, гангренизация с ампутаци-
ей конечности; при молниеносной форме — сеп-
тицемия с летальным исходом.
Лечение: обкалывание язвы антибиотиками;
общеукрепляющие средства; местно — дезинфи-
цирующие и эпителизирующие средства, хирур-
гическая обработка язвы.
Тропикадоидная язва (язва Кастеддани,
язва оазисов, песчаная язва и др.)
Встречается в странах Северной и Центральной
Африки, в отдельных регионах субтропическо-
го климата.
Клиническая картина. На коже нижней тре-
ти голени появляются пузырьки или пузыри,
которые затем превращаются в малоболезнен-
ные язвы без икхорозного запаха. В содержи-
мом пузырных элементов обнаруживается
Micrococcus mycetoides Castellani; в язвах пре-
обладают пиококки (стафилококки). Язвы не-
многочисленные, мелкие, окружены неярким
воспалительным венчиком, дно их покрыто
гнойной коркой или псевдопленкой. Течение —
хроническое (более года), без тенденции к руб-
цеванию. При заживлении язвы формируется
поверхностный, нежный, гиперпигментирован-
ный рубец.
Лечение: см. «Тропические язвы».
402 Частная дерматология
Язва пустынь (степная язва,
септическая язва и др.)
Встречается в пустынных и степных районах, на
плоскогорьях стран с сухим тропическим и суб-
тропическим климатом. В язвах выявляются
вирулентные пиококки, нередко — дифтерий-
ные бациллы. Чаще болеют мужчины молодого
и среднего возраста, контактирующие с верблю-
дами и лошадьми.
Клиническая картина: на тыле кистей, пред-
плечьях, локтевых и коленных суставах, голенях
возникают везикулы, везикуло-папулы, которые
превращаются в эрозии и болезненные язвы с
ровным дном, покрытым желтовато-серой плен-
кой или коркой. Через 2—3 недели язвы стано-
вятся малоболезненными. Заживление их длит-
ся до года и более, завершается формированием
мягкого тонкого рубца. При наличии тяжелой
сопутствующей патологии возможны глубокие
некрозы с образованием грубых рубцов, кон-
трактур, инвалидизация.
Лечение: см. «Тропические язвы».
Коралловая язва
Поражает рыбаков и ныряльщиков, контактиру-
ющих с живыми коралловыми рифами (Цейлон,
страны Персидского залива, побережье Красно-
го моря, Япония). На местах контакта кожи с
кораллами появляются болевые ощущения, эри-
тема, инфильтрация, лимфангиты и лимфадени-
ты. Формируется одна или несколько неглубо-
ких язв с подрытыми краями, до 1—3 см в
диаметре, неровным дном, покрытым гнойным
налетом. Вокруг язвы выявляется воспалитель-
ный ободок. Течение язв торпидное (месяцы,
годы); на местах заживших язв остаются гипер-
пигментированные рубцы.
Язва Бурули (язва микобактериальная)
Вызывается Mycobacterium ulcerans. Встречается
в жарких, влажных, болотистых местностях
(преимущественно в Африке, Австралии).
Клиническая картина: на месте травмы кожи,
в среднем через 3 месяца, появляется подкож-
ное уплотнение, которое постепенно увеличи-
вается в размерах и изъязвляется. Язва захва-
тывает подкожную клетчатку, растет по
периферии и может поражать значительные
площади (чаще нижнюю конечность). Язвы
могут существовать месяцы, годы; заживают
рубцеванием; возможны контрактуры, лимфа-
тический отек.
Диагноз подтверждается клинической карти-
ной и результатами посева биопсийного мате-
риала.
Лечение: хирургическая обработка язв, транс-
плантация кожи; антибиотики — рифампицин,
миноциклин, сульфаметоксазол; местно — согре-
вающие компрессы, оксигенотерапия.
Перуанская бородавка
Синонимы: бартонеллез, лихорадка Оройя, бо-
лезнь Карриона — тяжелое инфекционное забо-
левание, которое характеризуется появлением на
коже множественных кровоточащих бородавча-
тых разрастаний и выраженными общими нару-
шениями (интоксикация, лихорадка, ревматоид-
ные боли, гемолитическая анемия).
Возбудитель —- Bartonella baciliformis Barton, гра-
мотрицательные палочки, окрашиваются по ме-
тоду Романовского—Гимзы, выявляются в эрит-
роцитах и ретикуло-эндотелиальных клетках
внутренних органов.
Переносчики инфекции: москиты (Phlebotomus
verrucorrum, Phlebotomus nogushi), возможно кле-
щи Dermacentor andersoni.
Клиническая картина. Инкубационный пери-
од от 2 до 4 недель.
Заболевание может протекать в виде асимп-
томной инфекции; относительно доброкачествен-
ных кожных поражений (перуанская бородавка);
злокачественного тяжелого течения заболевания,
с высокой температурой, анемией и смертель-
ным исходом (лихорадка Оройя).
В стадию клинических проявлений выражен-
ные изменения наблюдаются со стороны внутрен-
них органов: некрозы и разрастание соединитель-
ной ткани в печени, селезенке, кишечнике и
лимфатических узлах. Симптомы общей инток-
сикации приводят к резкому ухудшению обще-
го состояния больного с переходом в кому (бо-
лее чем в 1/3 случаев в течение 2—4 недель
наступает смерть). У 2/3 больных через 3—4 ме-
сяца симптомы болезни стихают, состояние улуч-
шается, т. е. заболевание ограничивается только
острой фазой (лихорадка Оройя). Через 1—2 ме-
сяца болезнь обычно переходит в следующую
фазу — перуанской бородавки. На коже шеи,
лица, разгибательных поверхностей конечностей,
туловища появляются многочисленные мелкие
багрово-красного цвета папулезные высыпания,
сопровождающиеся зудом. Аналогичные элемен-
ты могут выявляться также на конъюнктиве глаз,
слизистой оболочке рта, желудочно-кишечного
тракта, мочеполовых органов. В дальнейшем
высыпания приобретают характер бородавчатых
разрастаний, размером до горошины, с богатой
сетью кровеносных и лимфатических сосудов
(напоминают тутовую ягоду, а по строению —
Глава 36. Кожные заболевания в жарких странах 403
пиогенную гранулему). Эти образования легко
кровоточат, могут изъязвляться и инфицироваться
пиококками. Возникновение сосудистых опухо-
левидных образований в пищеводе и желудке
может явиться причиной дисфагии, кровавой
рвоты, а в урогенитальном тракте — гематурии
и кровотечений из влагалища.
В течение 2—4-х месяцев процесс постепенно
регрессирует. На местах изъязвлений бородавча-
тых элементов остаются рубцы. Эта фаза забо-
левания протекает, как правило, благоприятно,
однако могут наступать рецидивы высыпаний на
коже и поражений внутренних органов.
Лечение. Для профилактики осложнений на-
значается антибиотикотерапия, проводится элек-
трокоогуляция бородавчатых разрастаний. Спе-
цифическое лечение не разработано.
Профилактика. Защита от укусов москитов и
их уничтожение.
Южноамериканская
ложная бородавка
Характеризуется развитием множественных бо-
родавчатых разрастаний, отличающихся от обык-
новенных бородавок пастозно-уплотненным ос-
нованием. Встречается в странах Южной
Америки. Предполагается вирусная природа за-
болевания.
Клиническая картина. На различных участках
тела выявляются множественные (величиной до
горошины) бородавчатые разрастания. У некото-
рых разрастаний определяется основание, глубо-
ко уходящее в дерму и подкожную жировую
клетчатку. Вокруг крупных узлов выявляются
мелкие дочерние бородавки. Появлению боро-
давчатых разрастаний предшествует пастозное
изменение участка кожи, на котором в дальней-
шем появляется ложная бородавка. Заболевание
протекает без субъективных ощущений и изме-
нений состава периферической крови. В течение
неопределенного времени бородавки спонтанно
исчезают.
Лечение. Электрокоагуляция, криотерапия,
прижигающие растворы.
Профилактика. Соблюдение личной гигиены
при контакте с больными.
Аингум
Синонимы: ампутирующая склеродермия.
Спонтанная сухая ампутация мизинцев стоп,
которая встречается у лиц австралоидно-негро-
идной расы. Этиология не ясна.
Процесс чаще захватывает мизинец стопы,
редко — другие пальцы. В основании мизинца
появляется уплотняющийся склеротический тяж,
который постепенно (в течение нескольких лет)
приводит к полному прекращению кровоснаб-
жения мизинца, мутиляции костной ткани и
спонтанной ампутации. Заболевание встречает-
ся у лиц среднего и пожилого возраста.
Лечение. На начальных стадиях — рассечение
склерозированного тяжа; на поздних — хирур-
гическая ампутация мизинца.
Кожные проявления других
бактериальных инфекций
Дифтерия кожи
Достаточно часто регистрируется в странах с
тропическим климатом. Возбудитель —
Corynebacterium diphtheriae (бацилла Клебса-
Леффлера). Дифтерия кожи может быть пер-
вичной и вторичной.
Первичная дифтерия кожи (раневая) развива-
ется в результате инфицирования дифтерийной
палочкой царапин, потертостей, ссадин.
Вторичная дифтерия кожи — результат ино-
куляции инфекции из носоглотки или инфици-
рования сопутствующих кожных сыпей (пиодер-
мия, экзема и др.).
По глубине поражения различают поверхност-
ную и глубокую дифтерию кожи. При поверхнос-
тной дифтерии кожи на очагах экземы или пи-
одермии возникают зеленовато-серые
ограниченные налеты; увеличиваются регионар-
ные лимфоузлы, которые становятся плотными,
болезненными. Течение данной разновидности
дифтерии кожи относительно доброкачественное.
При глубокой форме дифтерии кожи (поражаются
конечности, перианальная область, наружные
гениталии, шейка матки, влагалище, уретра)
возникает болезненная пустула, которая превра-
щается в болезненную язву с ровными краями
и воспалительным венчиком, покрытую влаж-
ной плотно сидящей пленкой различных оттен-
ков (желтый, серый, коричневый). Попытка
снятия пленки сопровождается болью, часто
кровотечением. В таком состоянии язва может
существовать до нескольких месяцев. В некото-
рых случаях развивается флегмонозный или ган-
гренозный дифтерийный кожный процесс, ко-
торый приводит к обширной деструкции кожи
и подлежащих тканей, сопровождается тяжелым
общим состоянием, сепсисом, вплоть до леталь-
ного исхода.
404 Частная дерматология
Диагностика: обнаружение в налете и поражен-
ных тканях дифтерийной палочки; культураль-
ное исследование; патогистологическое исследо-
вание биопсийного материала.
Лечение: противодифтерийная сыворотка, 2—
5 тыс ЕД на 1 кг массы тела в/м; пенициллин
по 1—2 млн ЕД 4 раза в сутки в/м в течение 5—
6 дней, или рифампицин по 0,25 г в/м 2 раза в
сутки в течение 5—6 дней.
Сибирская язва
Тяжелое токсико-инфекционное заболевание,
вызываемое Bacillus anthracis. Заражаются при
контакте с больными животными (овцы, рогатый
скот, лошади, козы), или при обработке их кожи,
шкур, шерсти, весьма редко — от больного че-
ловека. Сибирской язвой болеют преимуществен-
но работники животноводческих ферм, боен,
кожевенной промышленности. Заражение проис-
ходит через травмированную кожу (кожная фор-
ма), реже — при вдыхании спор (легочная фор-
ма) или через желудочно-кишечный тракт при
употреблении инфицированного и недоваренно-
го мяса (кишечная форма). Инкубационный пе-
риод кожной формы сибирской язвы обычно
составляет 2—3 дня. Наиболее частая локализация
кожного процесса — лицо, руки, шея.
Клиническая картина: на месте внедрения воз-
будителя образуется зудящее красное пятно,
которое превращается в папулу с пузырьком на
ее вершине. Пузырек превращается в пустулу,
наполненную кровянистым содержимым; пусту-
ла вскрывается с образованием некротического
струпа черного цвета, растущего по периферии.
Вокруг струпа возникают дочерние пузырьки,
претерпевающие подобную эволюцию. По пери-
ферии очага некроза развивается резкий отек
мягких тканей. Болевые ощущения в очагах
поражения не характерны. Регионарные лимфа-
тические узлы увеличиваются, в некоторых слу-
чаях нагнаиваются.
Течение: сибиреязвенный отек протекает с
выраженной интоксикацией, поражением лег-
ких, кишечника, мозговых оболочек, эндокар-
да, нередко — с септицемией.
Диагностика основывается на клинических
признаках заболевания, обнаружении в мазках
с эрозий, содержимом пузырьков палочки си-
бирской язвы; в периферической крови выяв-
ляется нейтрофильный лейкоцитоз.
Дифференцируют с карбункулом, гангреноз-
ным твердым шанкром, рожистым воспалением,
флегмоной.
Лечение: бензатинбензилпенициллин 2,4 млн
ЕД в/м 1 раз в 7 дней № 3, или ципрофлокса-
цин по 0,5 г внутрь 2 раза в сутки в течение 7—
10 дней, или пенициллин 1 млн ЕД 4 раза в
сутки в/м до исчезновения отека в области
язвы, затем ампициллин 0,5 г внутрь 4 раза в
сутки или оксациллин 0,5 г внутрь 4 раза в сутки
в течение 7—10 дней. Вводят также противоси-
биреязвенную сыворотку; местно — мази с ан-
тибиотиками.
Эризипелоид
Вызывается палочкой свиной рожи (Erysipelothrix
rhusiopathiae). Эризипелоид развивается преиму-
щественно у работников, разделывающих туши
животных, их мясо и мясопродукты и травми-
рующих кожу рук, иногда других участков (по-
резы, уколы острой костью и др.). Различают
кожную, кожно-суставную и генерализованную
формы эризипелоида.
Клиническая картина кожной формы: на кис-
тях, реже — запястье, лице, нижних конечнос-
тях появляется эритематозная бляшка с циано-
тичным оттенком и инфильтрированным
приподнятым периферическим валиком (вид
блюдца). Образуются пузырьки с прозрачным
или геморрагическим содержимым. Признаков
пустулизации, высокой температуры, озноба не
отмечается. Возможны регионарный лимфаде-
нит, лимфангит. Длительность болезни 2—3 не-
дели. Возможна суставная симптоматика (при-
знаки артрита, болезненность, при хроническом
течении — деформация суставов).
Диагноз устанавливается, исходя из данных
анамнеза, наличия клинической картины, полу-
чения культуры возбудителя.
Дифференцируют с рожистым воспалением
(«языки пламени», пузыри, интоксикация), па-
нарицием (признаки нагноения), истинной эк-
земой (мокнутие).
Лечение: пенициллин 1 млн ЕД в/м 4 раза в
сутки в течение 5—7 дней, или эритромицин
0,25 г внутрь 4 раза в сутки в течение 5—7 дней.
Сап
Тяжелое инфекционное заболевание, которым
страдают лица, контактирующие с больными
сапом животными (лошади, ослы, мулы, зебры).
Встречается в Азии, Африке, Южной Америке.
Возбудитель: Pseudomonas mallei — грамотрица-
тельная палочка, напоминающая микобактерию
Коха. Заражение человека происходит при на-
рушении целостности кожных покровов и сли-
зистых оболочек при уходе за больными живот-
ными.
Острая форма сапа. На месте внедрения воз-
будителя (область лица, носа, верхние конечно-
Глава 36. Кожные заболевания в жарких странах 405
сти) после 3—5-дневного инкубационного пери-
ода возникает болезненный островоспалитель-
ный узел, который быстро изъязвляется и сопро-
вождается лимфангитами и регионарным
лимфаденитом. Наблюдается лихорадка, резкая
слабость, мышечные и суставные боли, желудоч-
но-кишечные расстройства. Появляются множе-
ственные высыпания пустул, пузырей, изъязв-
ляющихся узлов, поражающих не только кожу,
но и мышцы. Аналогичные высыпания в носу
затрудняют дыхание и распространяются на сли-
зистую дыхательных путей. При развитии брон-
хопневмонии возможен летальный исход. При
хронической форме узлы образуются на коже и
во внутренних органах; вовлекаются легкие,
суставы, кости. Узлы длительно не заживают,
превращаются в серпигинирующие язвы, возни-
кают фистулезные ходы. Данная форма может
протекать годами с периодическими обострени-
ями. В результате очередной активации процес-
са возможна смерть. Септическая форма: множе-
ство некротических кожных абсцессов, обильная
полиморфная сыпь; однако доминируют пора-
жения нервной системы (менингиты), суставов
(артриты), возникает гепатоспленомегалия, раз-
виваются синуситы, фарингиты, пневмония.
Летальный исход — в 90 % случаев.
Диагностика: клинико-эпидемиологические
данные; обнаружение возбудителя в препаратах
из элементов кожной сыпи и в крови; получе-
ние культуры возбудителя.
Дифференцируют с фурункулами; в стадии
распространенной сыпи — с натуральной оспой,
пустулезным и везикулезным сифилидами.
Лечение: изоляция больных; сульфатиазол 1 г
внутрь 6 раз в сутки в течение 25—30 дней; тет-
рациклины, стрептомицин.
Профилактика: ветнадзор, выявление и унич-
тожение больных животных; изоляция заболев-
ших людей; повышение санитарной культуры,
меры личной и общественной профилактики.
Туляремия
Хроническое инфекционное заболевание. Встре-
чается в жарких странах. Возбудитель: Pasteurella
tularensis — грамотрицательная неспорообразую-
щая коккобацилла, выделяющая эндотоксин.
Носители инфекции — грызуны; передается при
укусах вшей, блох, клещей; возможно инфици-
рование через дыхательные пути и пищевари-
тельный тракт. Отмечаются эпидемические
вспышки.
Клиническая картина: на месте внедрения ин-
фекции, обычно в течение недели, на лице, шее,
кисти возникает некротизирующийся и изъяз-
вляющийся узелок, превращающийся в язву с
мягкими краями. Появление язвы сопровожда-
ется лимфангитом и регионарным лимфадени-
том. Лимфатические узлы нагнаиваются и
вскрываются (или становятся плотными и уве-
личенными) с образованием свищей. По ходу
лимфангита могут появляться нагнаивающиеся
узелки. В начале болезни отмечаются озноб, ли-
хорадка, головная боль. Характерны туляреми-
ды — симметричная полиморфная сыпь, напо-
минающая многоформную экссудативную
эритему. Помимо приведенной язвенно-желези-
стой формы туляремии, встречаются глазо-бу-
бонная, ангиозно-бубонная, железистая, легоч-
ная и генерализованная (без первичной язвы на
месте внедрения возбудителя). При этих разно-
видностях туляремии более выражены общеток-
сические явления и распространенные токсико-
аллергические сыпи.
Течение: хроническое, преимущественно доб-
рокачественное; летальные исходы возможны
при тяжелом течении железистой формы.
Диагностика: клинико-эпидемиологические
данные, идентификация Pasteurella tularensis в
патологическом материале; положительная внут-
рикожная проба с тулярином.
Дифференциальный диагноз: твердый шанкр,
сибирская язва, споротрихоз, колликвативный
туберкулез кожи.
Лечение: живая бактериальная вакцина; стреп-
томицин 7,5—10 мг/кг массы тела в/м каждые 12
часов в течение 7—10 дней; применяют также
тетрациклины, макролиды, левомицетин (сред-
нетерапевтические дозы в течение 2 недель);
местно — комбинированные стероидные мази с
антибиотиками, тепловые физиопроцедуры; при
гнойных лимфаденитах — хирургическое вмеша-
тельство.
Профилактика: своевременное выявление,
изоляция и лечение больных; обеззараживание
воды и пищевых продуктов; вакцинация насе-
ления в эндемичных зонах сухой туляремийной
вакциной. Истребление грызунов в случаях эпи-
зоотий, профилактический ветнадзор.
Тропические микозы
Весьма распространены в жарких странах вслед-
ствие:
• высокой температуры и влажности воздуха,
создающих условия термостата;
• наличия множества растений, обсемененных
грибами сапрофитами;
406 Частная дерматология
• ношения жителями разнообразных головных
уборов;
• традиционных смазываний волос маслами,
молочнокислыми продуктами;
• низкой санитарной культуры населения.
Для грибковых заболеваний в тропиках ха-
рактерны следующие черты: высокая частота по-
ражения ими местного населения и приезжих,
большая контагиозность (заразительность), об-
ширность поражений, большая длительность
заболеваний.
Кератомикозы
Отрубевидный лишай
Встречается значительно чаще, чем у жителей
умеренного климата. В отдельных странах (Ве-
несуэла, Гвиана и др.) поражено до 50 % насе-
ления. Кроме типичных мест локализации, по-
ражаются нижние и верхние конечности, лицо.
Характерны белесоватые, обильно шелушащие-
ся пятна и последующее развитие на этих учас-
тках ахромии.
Черепицеобразный микоз (хронический
фигурный дерматомикоз, тропический
ихтиоз, Токело и др.)
Распространен в тропических странах с высокой
влажностью. Возбудитель: Trichophyton concentricum.
Заражение происходит при контакте с больным и
предметами его обихода. Болеют преимуществен-
но взрослые. Обычно поражаются туловище, раз-
гибательные поверхности конечностей, ягодицы и
другие участки тела, за исключением лица, ладо-
ней, подошв, крупных складок, ногтей, волос.
Клиника: на теле обнаруживаются множе-
ственные светло-коричневые (коричневые) или
белесоватые пятна с эксцентрическим ростом,
шелушащиеся, затем обесцвечивающиеся в цен-
тре, с венчиком нависающих чешуек (колец
шелушения), напоминающих чешую рыбы или
черепичную крышу. На высоте клинических
проявлений кожа выглядит как бы разрисован-
ной узорами, напоминающими песчаные барха-
ны. Характерен зуд различной интенсивности;
возможны импетигинизация, экзематизация,
временные лимфангиты и лимфадениты. Микоз
протекает упорно, годами.
Диагностика: типичная клиническая картина,
обнаружение в чешуйках ветвящегося мицелия
со спорами; при посеве вырастают колонии, на-
поминающие Trichophyton faviforme.
Лечение: системные и местные антимикотики,
кератолитические жидкости и мази. Для предот-
вращения рецидивов необходимы мероприятия
по борьбе с повышенной потливостью, а также
соблюдение личной гигиены.
Ш имбери
По клиническим проявлениям напоминает чере-
пицеобразный микоз, однако с преимуществен-
ной локализацией кожного процесса в области
лица, шеи, на верхних конечностях, реже — на
туловище и нижних конечностях. Предполагае-
мый возбудитель: Endodermophyton Roquettei
Fonseca. Болеют обычно взрослые, не соблюда-
ющие гигиену тела, с повышенной потливостью
и сопутствующими инфекционно-паразитарны-
ми заболеваниями.
Клиническая картина: на указанных участках
появляются мелкие невоспалительные, серовато-
буроватые, слегка шелушащиеся пятна, которые
быстро увеличиваются по периферии и прини-
мают вид овальных или округлых пятен с лег-
кой гипопигментацией в центре, без образова-
ния концентрических кокард, свойственных
черепицеобразному микозу. Возможны пораже-
ния ногтевых пластинок (тусклые, деформиро-
ванные, легко обламываются).
Для постановки диагноза необходимо культу-
ральное доказательство роста колоний, напоми-
нающих возбудителя фавуса.
Лечение: см. «Черепицеобразный микоз».
Черный лишай
Поверхностное грибковое поражение кожи, ха-
рактеризующееся появлением обычно на ладо-
нях и подошвах стоп нешелушащихся пятен
темно-коричневого или черного цвета, не вы-
зывающих субъективных ощущений. Заболе-
вание контагиозно. Возбудитель: плесневый
гриб Exophiala werneckii. Предрасполагающие
факторы: тропический климат. Инкубацион-
ный период длится от нескольких недель до не-
скольких лет. Поражается кожа ладоней и бо-
ковых поверхностей пальцев кистей, а также
подошвы стоп.
Клиническая картина: в излюбленных участ-
ках появляются мелкие буроватые или черные
пятна, постепенно увеличивающиеся по перифе-
рии и сливающиеся в более крупные очаги,
имеющие четкие полициклические очертания.
Шелушение кожи в области пятен обычно об-
наруживается при исследовании с помощью
лупы.
Диагностика: клиническая картина, микроско-
пическое и культуральное исследования.
Дифференциальный диагноз: меланома, пиг-
ментные невусы, лентиго.
Глава 36. Кожные заболевания в жарких странах 407
Лечение: местные антимикотики.
Профилактика: диспансерное наблюдение.
Тропический белый лишай
Тропический белый лишай характеризуется по-
явлением преимущественно на конечностях сим-
метричных полиморфных высыпаний, после
регресса которых остаются участки достаточно
стойкой депигментации неправильных или при-
чудливых очертаний. Предполагаемый возбуди-
тель Trichophyton albicans Nieuwenhuis.
Предрасполагающие факторы: несоблюдение
гигиены тела, повышенная потливость, мацера-
ция кожи, повышенная влажность климата.
Чаще заболевает население Юго-Восточной
Азии.
Клиническая картина: на ладонях, подошвах,
голенях, реже на предплечьях появляются зудя-
щие эритематозно-папулезные высыпания, быс-
тро превращающиеся в пузырьки с серозно-гной-
ным содержимым. Большая часть пустулезных
элементов вскрывается, образуя эрозии с белова-
тым периферическим бордюром отслаивающегося
эпидермиса. Элементы в таком виде могут суще-
ствовать длительно (месяцы, годы). Процесс мед-
ленного регресса высыпаний сопровождается по-
явлением мелких участков депигментации,
которые сливаются и образуют очаги полицик-
лических очертаний, имеющие пестрый вид на
фоне темной кожи. Явления ксероза, гиперкера-
тоза приводят к появлению на подошвах длитель-
но не заживающих трещин. Возможно поражение
ногтевых пластинок.
Лечение: системные антимикотики; антибио-
тикотерапия при выраженной пустулизации;
местно — аппликации красителей.
Прогноз: благоприятный; очаги ахромии но-
сят стойкий характер.
Тропический желтый лишай
Поверхностный микоз, поражающий преимуще-
ственно кожу лица, шеи в виде пятнистых жел-
товато-коричневых зудящих элементов. Возбу-
дитель Hormodendron Fontoynonti. Встречается в
Индокитае, на Цейлоне, в Бразилии, на Кубе,
реже — на Африканском континенте. Болеют
взрослые и дети, проживающие в стесненных
жилищных условиях. Контагиозен для детей.
Клиническая картина: на лице, шее, иногда на
груди появляются мелкие зудящие оранжево-
желтого цвета пятна, которые сливаются в бо-
лее крупные пятнистые элементы полицикличес-
ких очертаний. Вокруг крупных пятен могут
возникать дочерние элементы, которые, слива-
ясь с основными элементами, образуют более
15 Зак. 1357
крупные очаги правильных очертаний в виде
дисков. В результате эволюции пятнистых эле-
ментов остаются участки депигментации с жел-
товато-оранжевым оттенком.
Лечение и профилактика: см. «Черепицеобраз-
ный микоз».
Прогноз: благоприятный.
Пьедра (трихоспория узловатая)
Грибковое заболевание, характеризующееся по-
явлением на поверхности волоса мелких очень
плотных узелков. Возбудитель: плесневый гриб
Piedraia hortai (черная пьедра), Trichosporum
giganteum, Trichosporum cerebriforme и др. (белая
пьедра).
Предрасполагающие факторы: тропический
климат, несоблюдение гигиены волос, их загряз-
нение мучной и сахарной пылью, смазывание
волос растительным маслом, мытье головы с
кисломолочными продуктами, ношение тесных
головных уборов, повязок, сдавливающих воло-
сы, заколок, брошей и т. п.
Пути передачи: заболевание возникает при
указанных условиях.
Течение: без лечения может длиться месяцы
и годы.
Излюбленная локализация: преимущественно
кожа волосистой части головы.
Клиническая картина: на волосах в области
волосистой части головы, реже бороды и усов
образуются множественные (по 30—50 и более)
мелкие (до 0,1—0,2 мм) каменистой твердости
узелки веретенообразной, овальной или непра-
вильной формы, муфтообразно (3—5 мм) окру-
жающие волос и плотно прикрепленные к его
поверхности. Узелки лучше просматриваются
под лупой. Цвет узелков светло-коричневый,
красновато-коричневый или черный. Субъек-
тивные ощущения обычно отсутствуют.
Диагностика: локализация и характерная плот-
ность узелков. При микроскопии обнаружива-
ются элементы гриба в виде нитей мицелия и
мозаично расположенные споры. На среде Сабуро
рост колоний, напоминающих культуру Achorion
(Trichophyton) Schonleinii.
Дифференциальный диагноз: проводится с лож-
ной пьедрой и гнидами головных вшей. Ложная
пьедра вызывается бактериями, для дифферен-
циальной диагностики необходимо культураль-
ное исследование. Яйца головных вшей (гниды)
имеют серовато-белый цвет и плотно приклеи-
ваются к волосам. Заболевание сопровождается
сильным зудом, появлением расчесов, осложня-
ется вульгарным импетиго, фолликулитами,
иногда фурункулами.
408 Частная дерматология
Лечение: бритье волос, местные антимикоти-
ки в виде растворов, кремов, мазей, аэрозолей,
шампуней.
Профилактика: консультации дерматолога,
косметолога по правильному уходу за волосами.
Подкрыльцовый трихомикоз
Относят к псевдомикозам. Бактериальное пора-
жение кутикулы волос подкрыльцовой области
и лобка с образованием на них узелков, без при-
знаков кожных изменений. Возбудитель:
Nocardia (Corynebacterium) tenuis Castellani, час-
то сочетается с пигментообразующими кокками.
Предрасполагающие факторы: повышенная
потливость, нарушение правил гигиены.
Пути передачи: от больного человека, через
загрязненное им белье и предметы обихода.
Течение: без лечебных мероприятий протека-
ет хронически.
Излюбленная локализация: кутикула волоса
подкрыльцовой области и лобка.
Клиническая картина: на поверхности волос
обнаруживаются узелки желтого, иногда красно-
го или черного цвета, мягкой консистенции.
Узелки сливаются и окутывают волос в виде
чехла; на ощупь — слизистые (представлены
кокковыми бациллярными микроорганизмами).
Постепенно волосы становятся хрупкими, обла-
мываются, склеиваются между собой, окраши-
вая пот и белье больного.
Гистопатология: эпидермис и дерма без изме-
нений.
Диагностика: пьедра, трихорексис, монилет-
рикс, лобковые и платяные вши.
Лечение: бритье волос, обработка участков
поражения 0,1% раствором сулемы, 2 % раство-
ром формалина в 70° спирте, 2 % салициловым
спиртом, 1 % настойкой йода.
Профилактика: соблюдение правил личной
гигиены, борьба с повышенной потливостью.
Эпидермомикозы
Руброфития в условиях тропиков и субтропиков
характеризуется распространенным, нередко
универсальным поражением кожного покрова с
вовлечением волос.
Трихофития весьма распространена в жарких
странах. Болеют не только дети, но и взрослые,
у которых, кроме гладкой кожи, поражается
волосистая часть головы. Заболевание в ряде
случаев отличается торпедным течением и пло-
хо поддается лечению.
Глубокий генерализованный трихофитоз — фор-
ма хронической трихофитии, близкая по патогенезу
и клиническим проявлениям к глубоким тропичес-
ким микозам. До периода полового созревания
отличается распространенностью микотического
процесса на коже с переходом в эритродермию,
выраженным гиперкератозом ладоней и подошв,
поражением волосистой части головы, онихомико-
зами. В период полового созревания наблюдаются
гранулематозные поражения глубоких тканей и
внутренних органов, септицемия, описаны энцефа-
литы и менингоэнцефалиты, мицетома стопы, по-
ражения лимфатических узлов, селезенки и пече-
ни. Из пораженных тканей выделяется
преимущественно 7г. Violaceum, Tr. schonleinii.
Фавус часто носит распространенный характер.
Заболеваемость высокая, на уровне трихофитии.
Микроспория по сравнению с перечисленны-
ми эпидермомикозами выявляется реже, отли-
чается доброкачественным течением.
Кандидозы. Наиболее частый возбудитель —
Candida tropicalis, который выявляется на коже,
слизистых, во внутренних органах. Климатичес-
кие условия благоприятствуют большему распро-
странению кандидозов, чем у жителей умерен-
ных поясов.
Плесневые микозы достаточно часто встречают-
ся в жарких странах и вызываются плесневыми
грибами Aspergillus, Penicillium, Acremonium,
Nocardia, Cladosporium и др.
Глубокие микозы — хронические и упорно
протекающие микозы, поражающие кожу, сли-
зистые, подкожную клетчатку и глубжележащие
ткани, внутренние органы, нередко ЦНС. Наи-
более часто встречается южноамериканский бла-
стомикоз, мицетома, споротрихоз, кокцидиои-
доз, гистоплазмоз.
Особенности течения
специфических
дерматозов
Кожный лейшманиоз
В различных регионах с теплым среднегодовым
климатом имеет клинико-эпидемиологические
особенности.
Американский лейшманиоз (бразильский, кож-
но-слизистый, ута, эспундия, лесная фрамбезия,
чиклеро). Возбудитель — L, cutis Americana.
Наряду с поражением кожи, в процесс часто
вовлекаются и при этом разрушаются слизистые
оболочки (до 80 % случаев) губ, носа, рта, глот-
ки, гортани, влагалища с подлежащими тканя-
Глава 36. Кожные заболевания в жарких странах 409
ми и соответствующей симптоматикой. Выявля-
ются хрящевые и костные мутиляции, которые
имеют характер увечий.
Суданский (египетский) кожный лейшманиоз
(возб. L. nilotica). Появляющиеся обычно на
открытых участках тела после укусов москитов
элементы носят характер лентикулярных папул,
которые достаточно быстро (через 1—2 мес)
трансформируются в болезненные узлы с си-
нюшно-багровым воспалительным венчиком
(изъязвляются редко). Далее перифокальное
воспаление вокруг узлов постепенно исчезает,
элементы начинают рассасываться с шелушени-
ем. Характерны рецидивирующие уплотнения
узлов с формированием на этих местах келои-
доподобных опухолевидных образований. Ке-
лоидоподобные узлы могут существовать нео-
пределенно долго, не вызывая субъективных
ощущений. Общее состояние больных обычно
не страдает, лимфатическая система в процесс
не вовлекается; редкое поражение слизистых
оболочек не приводит к распаду тканей и их
деформации.
Другие формы кожного лейшманиоза: эфиоп-
ская (течение 2—3 года); лепроматоидная (Боли-
вия, Венесуэла) напоминают кожные диффузные
инфильтраты при лепре.
Лепра
Основная масса больных регистрируется в стра-
нах Индокитая, Азии, Латинской Америки, на
Африканском континенте. Клиническая карти-
на и течение соответствуют описанным «класси-
ческим» проявлениям заболевания. Важная роль
придается раннему выявлению лепрозных боль-
ных в эндемичных зонах с применением специ-
альных диагностических методов и тестов.
Туберкулез кожи
Туберкулезная волчанка протекает более добро-
качественно; чаще встречается бородавчатая фор-
ма туберкулеза кожи.
Поражения кожи
при протозойных
болезнях
Трипаносомозы
При этом протозойном заболевании поражают-
ся лимфатическая и нервная системы, в началь-
ном периоде выявляются определенные кожные
симптомы и проявления. Различают африканс-
кий и американский трипаносомозы. Возбуди-
тель африканского трипаносомоза (сонная бо-
лезнь) Trypanosoma gambiense и Т. brucei
rhodesiense. Хозяева — позвоночные животные,
человек; переносчики — мухи рода Glossina
(мухи цеце). Возбудитель американского трипа-
носомоза (болезнь Шагаса) — Т. Cruzi- Хозяе-
ва — позвоночные животные, человек; перенос-
чики — триатомовые клопы.
Африканский трипаносомоз
После укуса мухи цеце образуется трипаносом-
ный шанкр — плоский болезненный узел крас-
ного цвета, окруженный белой восковидной зо-
ной, диаметром до 1—3 см, сопровождающийся
лимфангитом и лимфаденитом и разрешающийся
в течение нескольких дней. В дальнейшем воз-
никают эфемерные сыпи на туловище в виде
кольцевидной или полиморфной эритемы, узло-
ватой зудящей сыпи. В последней стадии болез-
ни легко определяются увеличенные лимфати-
ческие узлы в области заднего треугольника шеи
(симптом Winterbottom), что является предвест-
ником сонной болезни (летаргии) и гибели боль-
ного.
Диагноз: обнаружение возбудителя в кро-
ви, пунктате и биоптатах лимфоузлов; про-
водят заражение кровью больного морских
свинок.
Лечение: взрослые — сурамин в/в в дозе 1 г
на 1,3 7, 14, 21 дни (после пробной дозы 0,1—
0,2 г); дети — 20 мг/кг, или пентамидин — 10
в/м инъекций (4 мг/кг на инъекцию) в течение
10—20 дней.
Американский трипаносомоз
На месте внедрения трипаносом в кожу (обыч-
но через неделю) образуется «шагома» — воспа-
лительный инфильтрат темно-красного цвета с
лимфангитом и регионарным лимфаденитом.
При внедрении возбудителя через слизистую
глаза развивается конъюнктивит с отеком век,
лица, лба, височных областей (симптом Рома-
ньи), возможно возникновение паразитарного
зоба. На коже может появляться мелкая пятни-
стая сыпь, сохраняющаяся до 3-х недель. При-
соединяется сердечная недостаточность, гепато-
спленомегалия; развитие менингоэнцефалитов
приводит к гибели больного.
Лечение: в острой фазе — нифуртимокс 8—
10 мг/кг/сут 4 раза в день внутрь в течение 4
мес, или бензнидазол — 5 мг/кг/сут внутрь в 2
приема в течение 2 мес.
410 Частная дерматология
Малярия
Возбудители — 4 вида плазмодиев (Р. vivax, Р.
malariae, Р. falciparum, Р. ovale); источник ин-
фекции — больной человек (паразитоноситель);
переносчики болезни — комары рода Anopheles.
Часто выявляются универсальный зуд и су-
хость кожи (ксероз), при тяжелом течении —
отеки лица и конечностей, иногда всего тела.
Характерны «малярийные пятна» — признаки
геморрагического синдрома в результате воздей-
ствия токсинов возбудителя на сосудистую стен-
ку: геморрагические пятна на коже и слизистых
оболочках, кровотечения из носа, рта, глотки.
Описаны нарушения, напоминающие болезнь
Рейно, акроасфиксию; герпетиформные высы-
пания и по типу узловатой эритемы; дистрофи-
ческие изменения волос и ногтей, дисхромии
кожи и слизистых оболочек, пеллагроидные дер-
матиты на открытых участках («маска маляри-
ка»); возможны пиодермии, абсцессы, гангрены
нижних конечностей, нома.
Диагноз: подтверждается выявлением возбуди-
теля в крови.
Лечение: антималярийные и общеукрепляю-
щие средства.
Токсоплазмоз
Возбудитель — Toxoplasma gondii. Человек ин-
фицируется пищевым путем от животных (ко-
ровы, свиньи, кролики, кошки) и через продук-
ты питания. Со стороны кожных покровов
выявляются: сосудистые и геморрагические пят-
на, узелки, везикулы, волдыри, скарлатинопо-
добное шелушение, сетчатое ливедо, облысение,
генерализованная лимфаденопатия, мышечные
уплотнения (кальцификаты).
Диагноз: подтверждается наличием клиничес-
ких проявлений, результатами гистологического
исследования биоптатов, микроскопией мазков
крови, пунктатов лимфоузлов; серологическими
исследованиями (ПЦР, РСК, реакция с красите-
лем); проводится внутрикожная проба с токсоп-
лазмином и др.
Лечение: наиболее часто используют пириме-
тамин (тиндурин, хлоридин) в сочетании с суль-
фаниламидными препаратами: пириметамин
0,1—0,2 г в сутки внутрь в два приема в тече-
ние 2-х дней, затем 0,025 г внутрь 1 раз в сут-
ки в течение 4—5 недель; сульфадиазин 4—6 г
в сутки внутрь в 4 приема 4—5 недель. В пос-
ледние годы рекомендуется сочетать пиримета-
мин (в первые сутки 0,2 г, затем 0,1 г внутрь
1 раз в сутки) с клиндамицином (0,45 г внутрь
3 раза в сутки или 0,6 г внутривенно 4 раза в
сутки) и фолинатом кальция (0,01 г внутрь 1
раз в сутки) для профилактики угнетения кро-
ветворения. При такой схеме курс лечения со-
ставляет 3—6 недель.
Амебиаз кожи
Возбудитель — Entamoeba histolytica. Обычно
развивается как вторичный процесс в результа-
те аутоинокуляции возбудителя из кишечника,
печеночного абсцесса или прямого контакта с
инфекционным материалом. В перианальной
области, на промежности, ягодицах, реже — на
ногах, туловище, лице возникают серпигиниру-
ющие эрозии, язвы. Язвы глубокие, малоболез-
ненные, с почерневшими краями, некротическим
дном и неприятным запахом. Элементы суще-
ствуют до двух и более лет.
Диагноз: клиническая картина, обнаружение
возбудителя в кале, биоптатах, аспирационной
жидкости.
Лечение: метронидазол (тинидазол), иодохи-
нол, дилоксанида фуроат, паромомицин (моно-
мицин, аминозидин), тетрациклины, дигидро-
эметин.
Кожные поражения,
вызываемые
гельминтами
Шистосомозы (бильгарциоз)
Вызываются несколькими видами червей—со-
сальщиков из сем. Schistomatidae, поражающи-
ми кишечник, кожу и урогенитальный тракт.
Шистосоматоидный (церкариальный) дерматит
развивается в результате проникновения под-
вижных личинок (церкариев) в кожу и слизи-
стые оболочки человека из загрязненных ими
водоемов. Протекает по типу аллергодермато-
за (участки отечной эритемы, волдыри, пу-
зырьки, зуд, лихеноидные папулы), нередко
осложняющегося импетигинизацией, эктима-
ми. Возможно развитие шистосоматидной
эритродермии с выраженной общетоксической
реакцией. При повторных контактах с церка-
риями развиваются явления сенсибилизацион-
ного дерматита.
Мочеполовой шистосомоз весьма распростра-
нен в тропических странах. Обусловлен сенси-
Глава 36. Кожные заболевания в жарких странах
411
билизацией организма человека метаболитами
гельминтов, протеолитическими ферментами
(выделяются яйцами), механическим действием
самих гельминтов, яиц, конкрементов. Созрева-
ние половозрелых форм в воротной и брыжееч-
ной венах и венах мочевого пузыря сопровож-
дается общетоксической реакцией, затем
увеличением печени и селезенки. Появляется бо-
лезненное и учащенное мочеиспускание, терми-
нальная, а затем тотальная гематурия. Дальней-
шая симптоматика связана с воспалением стенок
мочеполовых органов, образованием эрозий, язв
(шистоматозных бугорков — скоплением яиц
возбудителя), папилломатозных разрастаний в
мочевом пузыре, кальцификацией стенок моче-
вого пузыря, мочеточников.
Осложнения: женщины — папилломатозные
разрастания во влагалище, в шейке матки, на на-
ружных половых органах, выраженные нарушения
менструального цикла, аменорея, бесплодие. Муж-
чины — эпидидимит, простатит, везикулит, оли-
гоазоспермия. Оба пола — псевдоэлефантиаз по-
ловых органов, занос яиц в орган зрения, ЦНС,
эмболия легочной артерии, рак мочеполовых ор-
ганов. У детей — задержка умственного и физи-
ческого развития.
Диагностика: клиническая картина, цистоско-
пия, урография, обнаружение яиц шистосом в
моче, эндовезикальная биопсия, РСК, аллерго-
пробы.
Лечение: празиквантел по 20 мг/кг через 4 ч
(2 приема), или оксамниквин — 15 мг/кг, одно-
кратно; общеукрепляющие и симптоматические
средства. Контроль в течение 5—6 мес после ле-
чения; иммунологические реакции становятся
отрицательными не ранее, чем через 3 мес пос-
ле специфической терапии.
Профилактика: массовое обследование и лече-
ние больных в эндемичных зонах; санитарно-
технические мероприятия на рисовых плантациях
и водоемах; строительство колодцев, туалетов и
обезвреживание экскрементов человека; улучше-
ние санитарно-гигиенических условий быта;
ношение специальной одежды и обуви, препят-
ствующих наружному проникновению церкари-
ев в мочеполовые органы и попаданию их на ко-
нечности.
Цистицеркоз кожи
Кроме жарких стран, заболевание регистрируется
в зонах умеренного климата. Развивается при
попадании в организм человека вместе вместе с
плохо обработанным мясом домашнего скота ли-
чинок свиного или бычьего цепня (цистицер-
ков). Возможно заражение от кошек и собак, при
употреблении немытых, зараженных яйцами
гельминтов овощей, фруктов и др. Проникнув
в организм человека (промежуточный хозяин) и
проделывая свой цикл развития, цистицерки
могут оказаться в мышцах, коже и подкожной
клетчатке, головном мозге, органе зрения. В
коже туловища, реже на других участках вокруг
паразита формируются единичные или множе-
ственные опухолевидные образования округлой
или овальной формы, подвижные, плотноватой
консистенции, безболезненные, сохраняющиеся
в течение многих лет. Постепенно узлы могут
рассасываться или развивается их фиброз и
кальцификация.
Диагностика: в биопсии или пунктате узла
обнаруживается сколекс и типичные крючья
цистицерка; проводятся кожные тесты, РСК с
пузырной жидкостью финн животных.
Лечение: тениаринхоз — празиквантел 20 мг/
кг внутрь однократно, или никлозамид, фена-
сал, дихлосал, трихлосал, семена тыквы, экстракт
папоротника мужского; тениоз — празиквантел
5 мг/кг внутрь однократно, или никлозамид.
Вскрытие единичных узлов, аккуратное удале-
ние содержимого узлов (в противном случае
возможна диссеминация на соседние участки
кожи).
Эхинококкоз
Возбудители — личинки Echinococcus granulosus,
поражающие печень и легкие. В коже цисты
образуют невоспалительные узлы различных
размеров, мягкие наощупь, подвижные, безбо-
лезненные. Узлы сохраняются в течение не-
скольких лет, затем подвергаются петрифика-
ции.
Лечение: альбендазол 0,4 мг внутрь с пищей,
28 дней. Методика ПАИР: Пункция, Аспирация
содержимого кисты, Инъекция сколицидного
агента (гипертонического раствора), Реаспирация.
Профилактика празиквантелом.
Спарганоз (дифиллоботриоз)
Гельминтоз, вызываемый личинками (плеоцерко-
идами) широких лентецов, попадающих в кожу,
подкожную клетчатку и мышцы человека. Зара-
жаются при употреблении не прокипяченной
питьевой воды, содержащей процеркоиды (нахо-
дятся в циклопах), или при прикладывании к
коже с лечебными целями инвазированного мяса
птиц, свиней и др. Участки поражения представ-
лены отечными, болезненными узлами, в центре
412 Частная дерматология
которых определяется движущаяся лентоподобная
личинка; возможно развитие множественных аб-
сцессов и септицемии. После гибели личинки со-
храняется выраженное воспаление и деструкция
ткани; характерна эозинофилия. При поражении
лимфатических сосудов конечностей возможно
возникновение ограниченной слоновости.
Лечение: празиквантел 10 мг/кг внутрь, одно-
кратно; никлозамид (вомесан) 2 г однократно,
накануне и в день лечения рекомендуется лег-
коусвояемая пища.
Аскаридоз
Встречается главным образом у детей. Вызыва-
ется паразитированием в организме человечес-
кой аскариды (Ascaris lumbricoides)\ гельминтоз
распространен повсеместно. Клинические прояв-
ления зависят от степени аллергизации организ-
ма. В начальной стадии инвазии кожные прояв-
ления характеризуются множественными
волдырями (иногда везикулами) и выраженным
зудом, в позднем периоде — рецидивирующей
крапивницей, сухостью кожи и легким ее ше-
лушением, элементами по типу центробежной
кольцевидной эритемы.
Лечение: альбендазол 0,4 г внутрь однократ-
но, или мебендазол (вермокс) по 0,1 г внутрь 3
дня, или пирантел памоад И мг/кг однократно
(максимальная доза 1 г); при массивной инва-
зии левамизол (декарис) 0,15 г внутрь однократ-
но (детям 2,5 мг/кг).
Энтеробиоз
Наиболее распространенный гельминтоз челове-
ка в тропических странах (возбудитель —
Enterobius vermicularis). Паразитирует в нижнем
отделе тонкого и толстого кишечника человека.
Обычно через 1—2 нед после инвазии появляет-
ся сильный ночной зуд, чувство жжения, пол-
зания в перианальной области и промежности
(результат миграции самок из кишечника). Эк-
скориации способствуют появлению импетиго,
эктим, фолликулитов, фурункулов, экзематиза-
ции. У лиц женского пола возможно развитие
вульвовагинитов вследствие инокуляции остриц
в наружные половые органы.
Лечение: альбендазол 0,4 г внутрь однократ-
но, прием препарата повторить через 2 недели,
или мебендазол 0,1 г внутрь однократно, повто-
рить через 2 недели. При указанных кожных
осложнениях — мази с антибиотиками, краси-
телями, комбинированные кортикостероиды, со-
держащие антибиотики.
Анкилостомидозы кожи
Возбудители анкилостомидозов Ankylostoma
duodenale и Necator americanus — кишечные па-
разиты. Выделяемые человеком с фекалиями яйца
паразита во внешней среде превращаются в ин-
вазивные личинки. Личинки вызывают кожные
поражения, известные под названием «земляная
чесотка», «водяная чесотка», «сыпь рудокопов» и
др. В местах внедрения личинок, чаще на руках
и ногах, в межпальцевых складках, на суставах
и подошвах появляется зуд или жжение, ограни-
ченная пятнистая, папулезная, папуловезикулез-
ная или везикуло-буллезная сыпь, а в области
лодыжек — отек и уртикарные элементы непра-
вильных очертаний. Высыпания нередко ослож-
няются пиодермией и разрешаются через 2—3 нед.
В поздних стадиях болезни выявляются блед-
ность кожи и слизистых с желтоватым оттенком
(результат анемизации организма, вызванной
сосальщиком), единичные полиморфные элемен-
ты, гипопигментные пятна, признаки общей ин-
токсикации. В кале обнаруживаются яйца гель-
минтов.
Лечение: альбендазол 0,4 г однократно; пиран-
тел памоад И мг/кг/сут 3 дня; левамизол 0,1 г
2 раза в сутки 3 дня; мебендазол 0,1 г/сут 3 дня;
местно: противозудные и противовоспалитель-
ные средства.
Стронгилоидоз
Синонимы: «Почвенный зуд», Larva currens («бе-
гущая личинка»).
Возбудитель — угрица кишечная (Strongeloides
stercoralis Bavayi). На ранней стадии инвазии орга-
низма возникают аллергические реакции; в мес-
тах внедрения личинок — полиморфные зудящие
высыпания, явления дерматита; в крови — эози-
нофилия (необходимо дифференцировать с «по-
чвенным зудом» при анкилостомидозах). В по-
здних стадиях наблюдается ползущая сыпь —
«Larva currens» с изоморфной реакцией. Процесс
начинается с перианальной области и распрост-
раняется на кожу бедер, ягодиц, живота и боко-
вых поверхностей туловища. Голени, стопы, руки,
половые органы не поражаются. Отмечается уме-
ренный зуд, по ходу сосудов линейные, ползу-
щие, валикообразные волдыри, которые могут
распространяться со скоростью до 10 см в час
(активное передвижение личинок в коже).
Лечение: альбендазол 0,4 г/сут 3 дня; тиабен-
дазол 25 мг/кг внутрь 2 дня; ивермектин
200 мкг/кг в день 2 дня.
Глава 36. Кожные заболевания в жарких странах 413
Трихинеллез
Возбудитель — Trichinella spiralis. Заражение
происходит при употреблении сырого или пло-
хо проваренного мяса животных, инвазирован-
ного гельминтом. Заболевание начинается с ли-
хорадки и разлитых мышечных болей. Позднее
отмечается затрудненное дыхание, паралич языка
и челюсти, отек век, лица, окологлоточной клет-
чатки, диффузный отек и напряжение кожи,
напоминающие дерматомиозит. На коже конеч-
ностей возникает преходящая пятнисто-папулез-
ная и геморрагическая сыпь, под ногтями вы-
являются геморрагии. При исследовании
биоптатов кожи часто обнаруживаются парази-
ты. Регресс высыпаний обычно наблюдается
через 2—3 нед.
Лечение: мебендазол 0,3 г внутрь 3 дня, затем
0,5 г внутрь 10 дней и преднизолон 40—60 мг
внутрь в течение 3—5 дней; тиабендазол 25 мг/
кг/сут в 3 приема в течение 7—10 дней.
Синдром Larva migrans
(личинка мигрирующая)
Объединяет болезненные состояния, которые
вызываются мигрирующими в организм челове-
ка личинками некоторых нематод, поражающих
кожу и внутренние органы. Возбудители:
Ancylostoma braziliense; Ancylostoma caninum,
Uncinaria stenocephala (паразиты собак);
Bunostomum phlebotomum (паразит крупного ро-
гатого скота); Ancylostoma duodenale и Necator
americanus (паразит человека). В песчаных по-
чвах, теплых местах яйца гельминтов превраща-
ются в личинки, которые проникают в организм
человека через кожу. Угрожаемые контингенты:
работники сельского хозяйства; дети; отдыхаю-
щие на пляжах.
При Larva currens места внедрения личинок
Strongiloides stercoralis (кишечная угрица) в кожу
и путь их подкожного передвижения сопровож-
дается появлением зудящих папул, папуловези-
кул, волдырей и наличием красноватых извили-
стых («растущих») полос. Извилистость
элементов сыпи и характер их распространения
позволяют заподозрить поражение кожи мигри-
рующими личинками. Заболевание проходит
самостоятельно через 1-2 месяца. Наиболее ча-
сто поражаются: перианальная область, ягодицы,
бедра, живот, спина, плечи. Возможно развитие
дерматитов, экзематизации, вторичной пиодер-
мии. Мигрирующие личинки собачьей и коша-
чьей токсокар и человеческой аскариды поража-
ют внутренние органы. Возможны аллергические
сыпи; характерна эозинофилия; увеличивается
печень; выявляется пневмонит.
Лечение: тиабендазол, внутрь 50 мг/кг/сут
каждые 12 часов — 2—5 сут. Альбендазол внутрь
400 мг/сут, 3 сут; местно кортикостероиды; жид-
кий азот на «продвигающийся» участок кожно-
го хода личинки.
Филяриатозы
Заболевания, вызываемые нематодами семейства
Filaridae. Встречаются, в основном, в тропиках,
поражают значительную часть населения (напри-
мер, на океанском побережье Индии — до 70 %).
Источник инфекции — человек, больной фи-
ляриатозом. Промежуточные хозяева и перенос-
чики личинок паразита (микрофилярий) — ко-
мары, некоторые москиты, слепни, мошки.
После укуса человека микрофилярии разносят-
ся током крови и превращаются в половозрелые
формы в закрытых полостях (грудная, брюш-
ная), подкожной клетчатке, лимфатических со-
судах, сердце, под конъюнктивой, реже — в
других тканях. Кожные проявления наиболее
характерны для следующих форм филяриатозов:
вухерериоза, бругиоза, онхоцеркоза, лоаоза.
Вухерериоз
При этом филяриатозе поражается преимуществен-
но лимфатическая система нижних конечностей и
половых органов. Наиболее частый филяриатоз
жарких стран. Возбудитель (Wuchereria bancrofti)
обычно выходит в периферическую кровь ночью.
После периода инкубации (от нескольких месяцев
до двух лет) появляются лихорадка и уртикарные
элементы, напоминающие сыпь при многоформ-
ной экссудативной эритеме, затем регионарные
отеки кожи и подкожной клетчатки, лимфанги-
ты, лимфадениты, орхит, эпидидимит, фуникулит.
Далее преобладают пролиферативные процессы,
наблюдается расширение лимфатических сосудов
кожи и подкожной клетчатки, лимфадениты. Кожа
утолщается, грубеет, инфицируется пиококками,
изъязвляется, на ней появляются трещины. Вок-
руг паразитов в подкожной клетчатке и мышцах
формируются абсцессы. Для поздних стадий ха-
рактерен элефантиаз (слоновость) нижних конеч-
ностей, половых органов (водянка яичка — лим-
фоскротум), других участков кожи с
экзематизацией, веррукозными разрастаниями,
хронической пиодермией, абсцессами, разрывами
лимфатических сосудов кожи, мочевого пузыря,
кишечника, брюшины, мошонки с явлениями
хилурии, лимфурии, гематохилурии. Течение —
длительное, хроническое.
414 Частная дерматология
Диагностика: оценка клинической картины
болезни с учетом эпидемиологических данных;
обнаружение микрофилярий в периферической
крови, в моче, жидкости полостей. Мазки кро-
ви исследуют методом толстой капли с окраской
по Романовскому-Гимза или гематоксилином.
Кровь для изоляции периодического штамма
берут ночью, субпериодического — днем и но-
чью.
Лечение: диэтилкарбамазин (дитразин, гетра-
зан, нотезин, баноцид): 1-й день 0,05 г внутрь
после еды, 2-й день 0,05 г, 3-й день 0,1 г, 4—21
дни 2 мг/кг в день, курс 21 день.
Бругиоз
Второй по частоте филяриатоз; возбудитель
(Brugia malayi) оседает в крупных лимфатичес-
ких сосудах и внутренних органах. Его микро-
филярии поступают в кровь обычно к вечеру.
Клиническая картина близка к вухерериозу.
Слоновость развивается преимущественно на
нижних конечностях.
Диагностика и лечение: см. «Вухерериоз».
Онхоцеркоз
При онхоцеркозе, обусловленном личинками
гельминта, поражаются кожа, подкожная клет-
чатка, лимфатическая система и орган зрения
(вплоть до слепоты). Ранние симптомы — кож-
ный зуд и полиморфные высыпания без излюб-
ленной локализации (диссеминированные мел-
кие пятнистые и узелковые элементы,
сгруппированные волдыри, сильный зуд («фи-
ляриозная чесотка»), явления хронического дер-
матита, рожистого воспаления с индурацией,
язвочками, корками. В дальнейшем кожа истон-
чается, становится сухой, складчатой, шелушится
(напоминает «старческий дерматит»), появляются
дисхромии, лимфаденопатии. Для клиники он-
хоцеркоза, обусловленного действием половоз-
релого гельминта, характерно формирование в
подкожной клетчатке вокруг паразитов узлов
(онхоцерком) величиной до куриного яйца, плот-
ных, безболезненных, подвижных, локализую-
щихся у африканцев преимущественно на конеч-
ностях, туловище, а у американцев — на голове.
Диагностика: оценка эпидемиологических,
анамнестических и клинических данных. Обна-
ружение микрофилярий в поверхностных био-
птатах (срезах) кожи с очагов поражения и в
содержимом онхоцерком.
Лечение: ивермектин (мектизан) 150 мкг/кг
внутрь, повторить через каждые 6 месяцев, воз-
можно, в течение 10—15 лет. Сурамин (антрипол,
моранил, германии). Вводят внутривенно в виде
10 % раствора на дистиллированной воде или
физиологическом растворе в течение 10 дней.
Доаоз
Возбудитель — небольшая нематода Loa loa.
Окончательный хозяин — человек, у которого
зрелые филярии паразитируют в подкожной
клетчатке, под серозными оболочками и конъ-
юнктивой. Их личинки (микрофилярии) ночью
находятся в сосудах внутренних органов, а
днем — в периферических сосудах кожи. Про-
межуточные хозяева и переносчики — слепни.
При лоаозе поражаются преимущественно кожа,
орган зрения, иногда половые органы. Характер-
ны эфемерные мигрирующие плотные отеки
(калабарская опухоль) кожи и подкожной клет-
чатки на конечностях и лице, сохраняющиеся
несколько часов или дней и спонтанно разреша-
ющиеся. Возможны: выраженный зуд кожи,
уртикарные высыпания, дизурические расстрой-
ства (боли во время мочеиспускания, связанные
с проникновением гельминта в уретру), ощуще-
ния жжения, покалывания, боли, передвижения
(ползания «чего-то» под кожей).
Диагностика: клинические проявления; анам-
нез, характерный для эндемичных зон по лоао-
зу; обнаружение микрофилярий в крови, преиму-
щественно в дневные часы.
Лечение: диэтилкарбамазин (дитразин, гетра-
зан, нотезин, баноцид): 1-й день 0,05 г внутрь
после еды, 2-й день 0,05 г, 3-й день 0,1 г, 4-21
дни 2 мг/кг в день, курс 21 день. Для профи-
лактики — диэтилкарбамазин 0,3 г в неделю.
Взрослых гельминтов удаляют хирургическим
путем.
Дракункулез (Ришта)
Возбудитель — Dracunculus medinensis. Самка
этой круглой нематоды выделяет массу личи-
нок, которые в водоемах заглатываются цикло-
пами (пресноводными рачками). Человек зара-
жается при употреблении не кипяченой воды из
этих водоемов. Через 7-14 мес в местах оседа-
ния личинок в подкожной клетчатке, чаще в об-
ласти голеней и стоп появляется эритема, па-
пулы, волдыри, нарушается общее состояние
(лихорадка, головные боли, слабость, зуд и др.).
Затем в участке локализации головного конца
самки последовательно появляются: выпячива-
ние, воспалительный узел, а в течение несколь-
ких суток — пузырь, в основании которого
прощупывается гельминт в виде плотноэласти-
ческого тяжа. Через несколько дней пузырь
разрывается и формируется колодцеобразная
болезненная язва с головкой гельминта на дне.
Глава 36. Кожные заболевания в жарких странах
415
Для этой стадии характерен симптом «молоч-
ного фонтанчика» — выбрасывание из язвы мо-
лочного цвета жидкости с большим количеством
личинок после ее орошения теплой водой. При
инфицировании подкожного канала, где распо-
ложен гельминт, возможно образование абсцес-
сов, флегмон, развитие гангрены конечности,
септицемии.
Диагностика: клиническая картина, анамнес-
тические и эпидемиологические данные, под-
твержденные гельминтологическими и необхо-
димыми лабораторными исследованиями.
Лечение: дезинфекция кожи и покой для по-
раженной конечности; осторожное механическое
извлечение гельминта (занимает обычно 2—4
дня); на язву — ранозаживляющие средства; мет-
ронидазол 0,4 г/сут 10—20 дней; по показаниям
— антибиотикотерапия.
Профилактика: санитарно-технические мероп-
риятия в населенных пунктах, санитарный над-
зор за гидротехническими объектами, повыше-
ние санитарной грамотности населения.
Поражения кожи,
вызываемые
членистоногими
Ракообразные (морские мелководные рачки
Cymothoidea) вызывают болезненные укусы,
прикрепляясь к рукам или ногам человека. В
местах их прикрепления появляется точечное
кровотечение, в дальнейшем развивается клиника
дерматита, который регрессирует в течение не-
дели.
Скорпионы при укусах человека вызывают
острую боль, припухлость и гиперемию кожи,
вплоть до геморрагий и некроза. От их яда раз-
вивается острая токсическая реакция, которая
(особенно у детей) может привести к коллапсу
и даже к летальному исходу.
Пауки. После укуса каракурта быстро появ-
ляется острая боль и жжение, эритема, отечность,
а затем — папуло-везикулезные элементы и
уплотнение кожи. Через 10—30 мин развивает-
ся общая токсическая реакция, судороги, диссе-
минированная пятнисто-папулезная и везикуло-
пустулезная сыпь. В 3—5 % случаев отмечается
летальный исход.
Укусы тарантула являются не опасными, в
местах их нападения наблюдается только мест-
ная реакция (эритема, отек).
Пузатый клещ вызывает зерновую чесотку.
Обитает в соломе, на злаках, в старых скирдах,
матрацах. После нападения клеща на человека
появляются сильно зудящие, красноватые, мел-
кие папулы или папуловезикулы, склонные к
пустулизации. Аутосанация наступает обычно в
течение 2-х недель. Возможны эпидемические
вспышки.
Личинки клеща-краснотелки (обитают на фа-
соли, плюще, кустах красной смородины, кры-
жовника, злаковых) попадают на человека во
время нахождения в поле, на огороде, в лесу и
внедряются в устья волосяных фолликулов.
Через несколько часов в месте присасывания
появляется небольшая отечная эритема, затем
папула или пузырек, иногда мелкоточечная пе-
техия, возможен лимфаденит. Обычно поража-
ются открытые участки тела. После отпадения
(извлечения) клещей элементы сыпи регресси-
руют в течение непродолжительного времени.
Паразит легко обнаруживается с помощью лупы:
имеет вид красной крупинки, плотно сидящей
в устье волосяного фолликула.
Аргасовые клещи (длиной до 2 см) паразити-
руют на диких и домашних животных, птицах.
После укуса клеща (чаще Ornithodorus) на вто-
рые сутки появляются болезненность и радиаль-
но расходящиеся эритема и отек. В центре эле-
мента часто видна точечная геморрагия или
папула (везикулопустула). В течение последую-
щих 2—3 дней воспаление нарастает, нарушает-
ся общее состояние, появляется распространен-
ная уртикарная и буллезная сыпь. Регресс сыпи
медленный (4— 5 нед).
Гамазовые клещи паразитируют на птицах и
грызунах. В местах укусов (туловище и конеч-
ности) появляются острая боль, жжение, зуд и
геморрагическое пятно, вокруг которого возни-
кают эритематозные пятна, папуло-везикулы и
уртикарии. При наличии повышенной чувстви-
тельности высыпания приобретают распростра-
ненный характер, сопровождаются зудом, пио-
дермическими элементами. Обычно сыпь
регрессирует в течение 3—4 дней (при отсутствии
осложнений).
Иксодовые клещи (обычно «собачий клещ»,
жук-дровосек) являются кровососущими пара-
зитами человека и животных. Через несколько
часов на месте укуса появляется безболезненное
эритематозное пятно, которое постепенно увели-
чивается в размерах, затем приобретает вид эк-
сцентрически растущей эритемы (до 15—20 см в
диаметре) ярко-красного цвета, отечной, зудя-
щей. Через 2—3 нед наступает выздоровление. С
помощью кусательного аппарата клещи внедря-
416 Частная дерматология
ются в складки кожи, область наружных гени-
талий, суставов, живота, ягодиц.
Клещи-железницы паразитируют в волосяных
фолликулах и сальных железах лица, ушных ра-
ковин, слухового прохода, ресниц, грудных же-
лез, гениталий. У человека выявляются две раз-
новидности этих клещей — Demodex folliculorum
и Demodex brevis (длиной 0,2—0,3 мм). Они име-
ют веретеновидное тело, короткие конечности и
колющий ротовой орган. Железницы у животных
вызывают демодикоз, а у человека являются ча-
стой причиной розацеоподобного и периорального
дерматитов (чаще у женщин), блефарита, наруж-
ного отита. У многих людей выявляется бессим-
птомное носительство демодекса.
Постельные клопы — нередко вызывают по-
ражение кожи человека. В месте их укуса (руки,
шея, грудь, наружные гениталии, ягодицы) по-
является красное зудящее пятно, папула или
волдырь с перифокальной эритемой. Характер-
но образование групп сыпи, состоящей из 2—3
элементов, располагающихся линейно. У сенси-
билизированных лиц возможна распространен-
ная кожная реакция по типу папулезной крапив-
ницы и образование пузырьков, пузырей. В
неосложненных случаях элементы разрешаются
в течение нескольких дней.
«Целующие» клопы кусают человека в местах
перехода кожи в слизистые на лице (губы, нос).
На местах нападения могут обнаруживаться па-
пулезные высыпания с точкой в центре, герпе-
тиформно расположенные везикулы, разлитые
уртикарии коричневатого цвета, а также гемор-
рагические и буллезные элементы.
Вши (головные, платяные, лобковые) — кро-
вососущие паразиты человека. Вшивость — сви-
детельство санитарно-гигиенического неблагопо-
лучия человека. В США ежегодно педикулезом
заражаются 6—10 млн детей; широко распрост-
ранен в Африке и других жарких странах.
Гусеницы некоторых бабочек при попаданиц на
кожу человека вызывают явления дерматита.
Через 10—20 мин на месте контакта гусеницы с
кожей появляются жжение, зуд, линейная отеч-
но-эритематозная полоска. Расчесывая зудящий
участок, человек переносит волоски гусениц на
другие места, где возникают новые пятнистые и
уртикарные элементы. Обычно дерматит регрес-
сирует в течение 1—2 нед. При повторных кон-
тактах с гусеницами и наличии сенсибилизации
развивается пятнисто-буллезная сыпь; заболева-
ние приобретает более длительное течение. Ле-
чение симптоматическое.
Поражение кожи некоторыми жуками возни-
кает при их раздавливании и втирании в кожу
содержащейся в теле жука токсической жидко-
сти. В течение суток развивается ответная кон-
тактная кожная реакция в виде эритемы, отека,
различной величины и формы пузырей, имею-
щих тенденцию к периферическому росту. Зуд
незначительный. В течение двух недель высы-
пания разрешаются.
На месте укусов блох (паразитируют на мле-
копитающих, птицах) появляется жжение, затем
пятно, папула, волдырь или пузырек (пузырь)
с геморрагической точкой в центре. При выра-
женной аллергизации организма появляются
папулы, уртикарии, множественные элементы
типа многоформной эритемы с выраженным
зудом. Сыпь чаще выявляется на бедрах, ягоди-
цах, на руках. В Америке, Африке, Азии неред-
ко встречается тунгиаз (саркопсилез) — заболе-
вание, вызываемое песчаной (земляной) блохой,
при котором на половых органах, перианальной
области, бедрах и руках появляются пятна, за-
тем зудящие узелки с пустулой в центре, узлы,
некротические язвы, абсцессы, лимфангиты,
лимфадениты; возможно развитие септицемии,
иногда столбняка, газовой гангрены, самоампу-
тации пальцев.
Среди многоножек для человека имеют значе-
ние травоядные двупарноногие и плотоядные
губоногие. Первые выделяют защитную зловон-
ную жидкость, вызывающую различные пора-
жения глаз, а также кожи (жжение, боль, по-
краснение, отек, волдыри). Губоногие выделяют
яд, вызывающий боль, отек, пурпуры, чаще на
нижних конечностях.
Лечение: извлечение паразита, профилактика
вторичной инфекции; при массивной инва-
зии — тиабендазол 25 мг/кг/сут, или альбенда-
зол 400 мг/сут в течение 3 сут.
Укусы пчел, ос, шершней, муравьев встречаются
часто и обычно сопровождаются жгучей болью,
эритемой и местным отеком кожи. Если не уда-
лить жало пчелы, то на месте укуса может воз-
никнуть длительно существующий гранулематоз-
ный узел (величиной до ореха). При высокой
реактивности к укусам в течение получаса раз-
вивается ангионевротический отек, крапивница,
вплоть до системных нарушений. При повтор-
ных укусах могут быть очень тяжелые реакции
с геморрагической сыпью.
При укусах двукрылых (комаров, москитов,
мошек, слепней, мух-кровососок) появляются
легкая болезненность, эритема, волдыри и в те-
чение суток формируется зудящая папула. У сен-
сибилизированных пациентов могут быть распро-
страненные уртикарные реакции, папулезные и
везикуло-буллезные высыпания.
Глава 36. Кожные заболевания в жарких странах 417
После укусов москитов рода Phlebotomus dermia
появляются флеботодермии и харара (их описа-
ние см. ниже).
Миазы — заболевания, вызываемые черве-
образными личинками некровососущих дву-
крылых (некоторых мух, оводов). Различают
поверхностные и глубокие, облигатные и фа-
культативные миазы. Поверхностные миазы
вызываются мухами, откладывающими яйца в
открытые гноящиеся ссадины и раны. Разви-
вающиеся личинки питаются экссудатом ран.
Глубокие миазы вызываются личинками воль-
фрамовой мухи, поражающими кожу, подкож-
ную клетчатку и слизистые оболочки. Личин-
ки питаются некротизированными и
здоровыми тканями, образуя язвы и глубокие
ходы. При кордилобиазе (африканском миазе),
чаще у детей, образуются узлы, язвы и абсцес-
сы. Дерматобиаз (южноамериканский миаз) ха-
рактеризуется опухолевидным воспалением и
подкожными абсцессами с фистулезными от-
верстиями на поверхности. Личинки оводов
вызывают «линейный мигрирующий миаз»,
проникая в кожу и проделывая в ней извили-
стые ходы.
Лечение: обездвиживание и удаление личинок.
При смазывании фурункулоподобных образова-
ний вазелином личинки покидают свою эколо-
гическую нишу; при укусах двухкрылых — сим-
птоматическое лечение.
Профилактика: меры общественной и личной
гигиены, применение репеллентов.
Поражения кожи,
вызываемые
ядовитыми змеями,
моллюсками,
пиявками, актиниями
Ядовитые змеи
Из ядовитых змей наиболее опасны укусы коб-
ры, очковых змей, гадюк, некоторых морских
змей. Их укусы (обычно — руки, ноги) сопро-
вождаются местной болью, нарастающим оте-
ком пораженной конечности, переходящим
иногда на туловище. В месте укуса выявляются
две красноватые точечные ранки; вскоре вок-
руг них появляются петехии или геморрагии,
которые можно выявить также на слизистых.
Появляется цианоз и резкий отек конечности,
нарастает общетоксическая реакция. В ряде
случаев развивается коллаптоидное состояние.
При укусах тропических змей до 15 % укушен-
ных умирают от сердечной недостаточности,
паралича дыхательного центра на 3—4 день
после укуса.
Лечение: быстрое удаление (отсасывание) яда из
ранок (эффективно в течение получаса); иммо-
билизация конечности; введение противозмеиной
сыворотки в межлопаточную область (20—
100 мл); адреналин, кофеин, кордиамин, эфедрин;
новокаиновая блокада вокруг места укуса; кор-
тикостероиды системно в средних дозах.
Моллюски
Некоторые морские моллюски имеют ядовитый
аппарат и вызывают у купающихся болезненные
ожоги, развитие эрозий, локальных ишемий,
цианоза, онемения, которые быстро могут рас-
пространяться на значительные участки тела.
Контакт с некоторыми видами моллюсков мо-
жет привести к летальному исходу. Описаны
профессиональные аллергические дерматиты от
контактов с кальмарами и мидиями.
Гирудиноз
Вызывается пиявками, которые присасывают-
ся к коже человека в стоячих водоемах, при на-
хождении в джунглях, ходьбе босиком по мок-
рой траве и болотам. Укусы медицинских
пиявок малоболезненны, так как секрет их
слюнных желез содержит анестезирующие ве-
щества. Укусы тропических пиявок вызывают
жжение и резкую боль. После их укусов появ-
ляются кровоточащие ранки, медленно зажива-
ющие, вследствие присоединения вторичной ин-
фекции, вплоть до абсцессов и флегмон (при
сопутствующих истощающих заболеваниях).
При наличии сенсибилизации возникают урти-
карные, буллезные, некротические высыпания;
не исключены анафилактические реакции.
Болезнь ловцов губок (коралловые язвы)
Контактный дерматит, вызванный плавающими
личинками актиний Edwardsiella lineata. На ме-
стах тела, закрытых купальником или плавка-
ми, появляется легкая эритема, мелкие красно-
ватые папулы (папуловезикулы), реже —
пузыри, пустулы. Сыпь держится 1—2 нед. При
повторных контактах с актиниями кожные про-
явления носят более тяжелый характер (до яз-
венно-некротических поражений).
Лечение: кортикостероиды местно или сис-
темно.
418 Частная дерматология
Особенности
клиники и течения
неинфекционных
дерматозов
Аллергодерматозы
Встречаются достаточно часто и обусловлены пре-
имущественно контактом населения с растени-
ями и их продуктами. Они обозначаются как ра-
стительные дерматиты (лесные, джунглевые).
Наиболее часто наблюдаются манговые, анана-
совые, примуловые. полисандровые, буковые, та-
бачные, вызываемые ядовитым плющом и др.
фитодерматиты.
Тропический плоский лишай
Заболевание, которое характеризуется появле-
нием на открытых участках кожи высыпаний,
клинически и морфологически похожих на
классический красный плоский лишай. Возник-
новение заболевания связывают с приемом ан-
тималярийного препарата атебрина, высокой
температурой воздуха, длительным световым
днем, повышенной солнечной радиацией, али-
ментарной дистрофией (дети), авитаминозами,
глистными инвазиями, выпечкой хлеба с помо-
щью тандура. Заболевание обычно возникает в
летние месяцы. Поражаются лицо (лоб, щеки,
губы), открытые участки шеи, тыльные повер-
хности кистей рук и нижних третей предпле-
чий, тыл стоп. Возможно поражение слизистой
оболочки полости рта по типу лейкоплакии, об-
разование пузырьково-эрозивных поражений,
стоматита. Поражения на коже практически не
отличаются от типичных высыпаний красного
плоского лишая (с наличием сетки Уикхема). К
зимнему периоду воспалительные явления, зуд
регрессируют; на местах сыпи остается стойкая
гиперпигментация.
Лечение: исключают прием атебрина или его
заменяют другим противомалярийным препара-
том. Необходима защита кожи от повышенной
температуры воздуха, солнечной радиации. На-
значаются гипосенсибилизирующие и другие
симптоматические средства.
Прогноз: в большинстве случаев благоприят-
ный, однако возможны рецидивы.
Экзема
Характерна слабая экссудативная реакция (мик-
ровезикуляция, мокнутие), больше напоминаю-
щая нейродермит, т. е. «сухое» течение экзема-
тозного процесса.
Нейродермит
Отличается склонностью к диссеминации высы-
паний и слабой их лихенификации.
Псориаз
В условиях тропиков наблюдается реже. Чаще
встречаются инфильтративные формы дермато-
за в виде единичных, «дежурных» сухих бляшек
на местах излюбленной локализации. Изоморф-
ная реакция не характерна. Для подтверждения
диагноза нередко требуется гистологическое ис-
следование.
Эритематоз
Хронические кожные формы заболевания встре-
чаются значительно реже (несмотря на гиперин-
соляцию), чем в странах с умеренным и холод-
ным климатом. Системная красная волчанка
диагностируется несколько чаще, чем в европей-
ских странах.
Тропические зудящие дерматозы
Временная почесуха — появление на разгибатель-
ных поверхностях конечностей зудящих мелких
серозных папул, в последующем покрывающих-
ся корочками серозно-геморрагического харак-
тера. Процесс может протекать до нескольких
месяцев с гриппоподобными явлениями. Отяго-
щают течение заболевания глистные инвазии и
другие сопутствующие заболевания.
Тропическая потница — временный дерматоз,
связанный с закупоркой потовых желез (рого-
вые чешуйки, кожное сало, пыль и др.) на раз-
личном уровне их залегания в коже при работе
в жарких условиях: от прозрачных пузырьков,
наполненных потом, до их пустулизации. Про-
цесс чаще локализуется в локтевых и коленных
сгибах, на груди, животе, других участках, ред-
ко — на лице. Самостоятельно исчезает при пе-
ремене климата.
Тропический помфоликс — носит сезонный
(лето) характер. Явления потницы возникают
только на ладонях и подошвах стоп, боковых
поверхностях пальцев. Внутриэпидермальные
пузырьки вскрываются, эрозируются, затем
быстро эпителизируются, не причиняя субъек-
тивных жалоб. Осложнения — экзематизация,
пустулизация, абсцедирование.
Флеботодермия — распространенный дерматоз,
возникающий после повторных укусов человека
москитами и сенсибилизации организма к секрету
слюнных желез флеботомусов. На открытых учас-
Глава 36. Кожные заболевания в жарких странах
419
тках тела, вскоре после нападения москитов, по-
являются розеолы с точечной геморрагией в цен-
тре, далее превращающиеся в зудящие папулы,
часто покрытые пузырьками. Возможны волдыр-
ная и папуло-буллезная реакции. Появление эле-
ментов сопровождается экскориациями. После
атаки флеботомусов процесс обычно постепенно
стихает. В зависимости от степени сенсибилиза-
ции, наличия сопутствующих заболеваний при
повторяющихся атаках москитов дерматоз может
принять распространенный рецидивирующий
характер с образованием пруригинозных узлова-
тых элементов (до горошины), различных пио-
кокковых высыпаний. Заболевание может затя-
гиваться до нескольких месяцев и дольше. В ряде
случаев необходимо проводить активную тера-
пию, включающую системное применение
кортикостероидов.
Харара (эндемическая многоформная крапив-
ница) — сезонный (лето) зудящий дерматоз,
связанный с повторными укусами человека от-
дельными видами комаров. Чаще встречается в
арабских странах как у приехавших лиц, так и
у местного населения (обоего пола и различно-
го возраста). Наблюдается на открытых участках
тела, особенно в нижней трети голени и тыле
стоп. Характерен полиморфизм сыпи: папуло-
волдыри, серозные папулы, эрозии, серозно-ге-
моррагические корочки, пиококковые элемен-
ты. Возможны общетоксические явления. С
течением времени (1—3 мес) процесс приобретает
«сухой» характер, постепенно стихает, в местах
высыпаний образуется временная гиперпигмен-
тация. В некоторых случаях заболевание может
затягиваться до 1—2 лет (хроническая харара) с
клиникой сухих зудящих узелковых высыпаний
и явлениями лихенификации.
Тропические буллезные дерматозы
Истинная пузырчатка в тропических странах
встречается реже, чем в регионах с умеренным
и холодным климатом.
К тропическим буллезным дерматозам отно-
сят бразильскую эксфолиативную пузырчатку
(БЭП) и онъялаи (onyalai).
БЭП — без продромы на лице, груди, реже на
других участках появляются пузыри, которые
осложняются пиококковым процессом, экзема-
тизацией, иногда веррукозными разрастаниями,
очаговыми дисхромиями. Симптом Никольского
положительный. Слизистые рта не поражаются.
Тяжелое течение наблюдается при молниеносной
(нередко заканчивается летально) и острой фор-
мах БЭП (признаки выраженной интоксикации,
обширные эрозии и участки импетигинизации).
Симптоматика менее выражена при подострой
форме заболевания. Хроническая БЭП имеет
длительное течение и сопровождается клиничес-
ким полиморфизмом с вовлечением костно-су-
ставной системы, зубов, атрофии мышц, груд-
ных желез и другими проявлениями.
Онъялаи — тяжелое заболевание жарких стран
с клиникой буллезной и геморрагической сыпи
на коже и слизистых. Характерен системный
геморрагический синдром, тромбоцитопения,
тяжелое общее состояние и образование крупных
пузырей на слизистых полости рта с гнойными
налетами, напоминающими афты. Прогноз не-
благоприятный при отсутствии интенсивной те-
рапии, включающей кортикостероиды, перели-
вание крови, кровезаменители.
Новообразования кожи
О большей частоте доброкачественных и злока-
чественных новообразований кожи в условиях
жаркого климата единого мнения у исследовате-
лей нет. Имеются данные о редких случаях рака
кожи у населения Китая, Японии, арабских стран,
у негров. Рак полового члена не выявляется у
народностей, у которых традиционно производит-
ся иссечение крайней плоти. У жителей Африки
рак кожи развивается преимущественно на мес-
тах хронического воспалительного процесса, язв,
ранений. Сосудистые невусы у лиц черной расы
часто плохо визуализируется, а волосатые неву-
сы встречаются реже, чем у европейцев. Особен-
ностей со стороны профиля доброкачественных
новообразований (бородавки, остроконечные кан-
диломы, фибромы и др.) не отмечается.
Недостаток витаминов
Гиповитаминоз А встречается достаточно часто,
поражение кожного покрова носит распростра-
ненный или универсальный характер. Хорошо
поддается коррекции витамином А.
Эксфолиативные хейлиты (связанные с арибоф-
лавинозом) отличаются торпидностью к терапии.
Венерические болезни
и эндемические
трепонематозы
Венерические болезни
У лиц, проживающих в условиях жаркого кли-
мата, клинические проявления и диагностика
420 Частная дерматология
сифилитической инфекции имеют некоторые
особенности:
• розеола на темной коже плохо визуализиру-
ется;
• при наличии язвенного шанкра лимфаденит
выражен умеренно;
• на поздних стадиях возникают околосустав-
ные узловатости (локти, колени);
• при установлении диагноза сифилиса необ-
ходимо учитывать возможность ложноположи-
тельных реакций, связанных с другими инфек-
ционными заболеваниями (лепра, малярия,
гельминтозы и др.) и употреблением наркоти-
ков.
Другие классические венерические заболева-
ния (шанкроид, венерический лимфогранулема-
тоз, донованоз) выявляются преимущественно у
жителей жарких стран и некоторых лиц, посе-
тивших эти регионы. Данные заболевания под-
робно описаны в разделе «Венерология».
Эндемические трепонематозы
Для невенерических тропических трепонемато-
зов (фрамбезия, беджель, пинта), являющихся по
сути разновидностями обычного сифилиса, ха-
рактерна высокая заразительность. Передача
инфекции происходит преимущественно через
предметы домашнего обихода (полотенце, по-
стельное белье, посуда и пр.), а также при по-
ловых контактах. Течение этих болезней часто
хроническое. Возбудителями фрамбезии, беджеля
и пинты являются соответственно Г. pertenue,
Т. bejel, Т. carateum.
Их единственный естественный хозяин —
человек.
Фрамбезия
Тропическая инфекционная болезнь, характери-
зующаяся высокой контагиозностью, поражени-
ем кожи, слизистых оболочек, а также костей и
суставов. Типичные кожные папилломатозные
элементы имеют сходство с ягодой малины, (фр.
Framboise).
Этиология
Возбудитель фрамбезии — Treponema pertenue —
по морфологическим и биологическим свойствам
близка к возбудителю венерического сифилиса
(Т. pallidum), однако не вызывает врожденных
поражений, поскольку не проникает через пла-
центу. Т. pertenue быстро гибнет под действием
кислорода, высушивания и нагревания; на пи-
тательных средах не растет, визуализируется в
темном поле микроскопа.
Резервуар инфекции — дети в возрасте от 2 до
15 лет.
Эпидемиология
Фрамбезия передается путем прямого неполового
контакта от человека к человеку (с экссудатом
или сывороткой заразных поражений). Больные
в латентном и третичном периодах практически
Таблица 36.1. Сравнительная эпидемиологическая характеристика трепонематозов
ЭпидемиологическаяСифилис характе ристика Фрамбезия Беджель Пинта
Распространение Спорадическое; преимущественно в городах Эндемическое; сельские районы Эндемическое; сельские районы Эндемическое; сельские районы
Географическое распределение В разных регионах мира Африка, Юго-Восточная Азия, Южная Америка Юго-западная Азия, Сахара Центральная и Южная Америка
Климатические условия Любые Жаркий влажный климат Жаркий сухой климат Жаркий полусухой климат
Возраст больных 18-30 лет 2-10 лет 2-10 лет 15-30 лет
Механизм передачи: -половой -неполовой -врожденный Обычно Редко Иногда Нет Да Нет Нет Да Нет Нет Да Нет
Резервуар инфекции Взрослые Дети 2-15 лет Дети 2-15 лет больные в латентном периоде поражениями кожи Больные 15-30 лет со стойкими
Глава 36. Кожные заболевания в жарких странах 421
не заразны. Распространению фрамбезии способ-
ствуют скученность и низкий санитарно-гигие-
нический уррвень жизни.
Течение заболевания
Инкубационный период — 3—4 недели.
Первичный период — до 0,5 года.
Вторичный период — 1,5—2 года.
Вторичный период характеризуется циклич-
ной сменой активных проявлений и латентных
(скрытых) периодов болезни. На протяжении
всего вторичного периода (1,5—2 года) может
наблюдаться 2—3 клинических рецидива, насту-
пающих после латентных периодов. У многих
больных заболевание на этом завершается.
Третичный период — возникает у 10—30 % не
лечившихся больных через 7—25 лет на фоне
длительного латентного периода, наступившего
после завершения вторичной стадии заболевания.
Клиническая картина
Первичный период — единичная папула, мягкая,
зудящая, бледно-розового цвета, с папилломатоз-
ными разрастаниями (вид малины). На поверх-
ности папулы зачастую формируется неглубокая
язва (фрамбезиома), которая покрывается кор-
кой и заживает рубцом. Фрамбезиомы обычно
располагаются у детей на открытых участках
тела, особенно на лице (губы, ушные раковины,
рот, область носа), кистях; у кормящих матерей
они часто локализуются в области молочных
желез, а у взрослых мужчин — на голенях, сто-
пах, реже на половых органах.
Возможны вторичные фрамбезиомы — шан-
кры-сателлиты.
Вторичный период характеризуется появлени-
ем на туловище, конечностях множественных
зудящих папул — фрамбезидов, имеющих вид
малины, а также сквамозных и эритематозно-
сквамозных пятен, редко — папуло-бугорков,
везикуло-пустул с их изъязвлением. Нередко в
этом периоде выявляется гиперкератоз ладоней
и подошв, который может носить ограниченный
или диффузный характер.
Третичный период — характерно образова-
ние гумм, язв, рубцов, гангозы (мутилирую-
щий ринофарингит), остеопериоститов, сабле-
видных голеней, гиперостоза передней носовой
кости (гунду, гонду), околосуставных узлова-
тостей.
Диагноз
Диагностика фрамбезии основывается на:
• результатах эпидемиологического обследо-
вания очага;
• типичной клинической картине заболевания;
• обнаружении в тёмном поле микроскопа
возбудителя в отделяемом первичной фрамбези-
омы, экссудата из первичного очага поражения,
во вторичных эффлоресценциях;
Таблица 36.2. Дифференциальная диагностика сифилиса и фрамбезии
Сифилис Фрамбезия
Ранний период
Твердый шанкр локализуется в области гениталий Розеолы, папулы, реже пустулы Слизистые оболочки поражаются часто Зуд не характерен Алопеция возможна Поражение глаз —- ирит, хориоидит, ретинит Поражений типа гунду не бывает Поздний период Фрамбезиома локализуется экстрагенитально Папулезно-вегетирующие фрамбезиды Слизистые оболочки поражаются редко Зуд типичен Алопеции не бывает Поражений глаз не бывает Часто бывает гунду
Возможны висцеральные поражения Возможны поражения нервной системы Реакция Вассермана в ликворе положительная при поражении нервной системы Поражение сосудов (облитерирующий эндартериит, тромбоз сосудов мозга) Поражений типа гангоза не бывает Околосуставные узловатости не типичны Наблюдается врожденный сифилис Висцеральных поражений не отмечается Поражений нервной системы не отмечается Реакция Вассермана в ликворе отрицательная Поражений сосудов не наблюдается Нередко бывает гангоза Нередко развитие околосуставных узловатостей Врожденной фрамбезии не бывает
422
Частная дерматология
• гистологическом исследовании элементов
сыпи;
• положительных серологических реакциях
(РВ, РИФ, РИТ).
Лечение
В соответствии с рекомендациями ВОЗ, для
лечения фрамбезии предпочтительно назначе-
ние пролонгированного пенициллина — бен-
затинбензилпенициллина (ББП). Амбулатор-
но проводят одну его инъекцию
внутримышечно в верхний наружный квад-
рант ягодицы в дозе 2 400 000 ЕД, что созда-
ет трепонемоцидный уровень препарата более
чем на 3 недели. Однократное введение ББП
обеспечивает излечение трепонемных болезней
и защиту против реинфекции в течение это-
го периода.
Рекомендуемые профилактические дозы ББП
для лиц, находившихся в контакте с больными
эндемичными трепонематозами:
• в возрасте до 10 лет — 600 000 ЕД бенза-
тинбензилпенициллина;
• для лиц старше 10 лет — 1 200 000 ЕД бен-
затинбензилпенициллина.
В зависимости от распространенности фрам-
безии в эндемичных зонах применяют соответ-
ствующую лечебную тактику (табл. 36.3).
В случаях аллергии к пенициллину больным
фрамбезией, эндемическим сифилисом, пинтой
назначают другие антибиотики:
• тетрациклин или эритромицин по 500 мг
внутрь до еды 4 раза в день, курс лечения не
менее 15 дней;
• детям 8—15 лет назначают половинную дозу
тетрациклина или эритромицина;
• детям до 8 лет назначают только эритроми-
цин в дозах, соответствующих их массе тела.
Препарат следует назначать малыми дозами
каждые 4—6 часов не менее 15 дней.
Не рекомендуется назначать тетрациклин бе-
ременным женщинам, так как он вызывает у них
почечные осложнения и влияет на формирова-
ние костного скелета плода.
Профилактика
• Улучшение экономических, социальных и
бытовых условий жизни населения эндемичес-
ких областей.
• Своевременное выявление больных актив-
ными формами фрамбезии и всех контактиро-
вавших с ними.
• Выявление больных с латентными формами
заболевания, у которых вероятны рецидивы кон-
тагиозных фрамбезидов.
• Проведение этим контингентам соответству-
ющего лечения с целью снижения и ликвида-
ции заболеваемости данным трепонематозом;
• Проведение периодических обследований
жителей сельской местности, где ранее были вы-
явлены больные фрамбезией.
Беджель
Беджель (эндемический сифилис, арабский си-
филис) — хроническая инфекционная болезнь,
встречающаяся главным образом среди детей и
проявляющаяся эритематозно-папулёзными
высыпаниями на коже, поражением слизистых
оболочек, костей, суставов и хрящей.
Этиология
Беджель вызывается Treponema pallidum (bejel)\
некоторые антигенные и патогенные различия
между возбудителями эндемического и венери-
ческого сифилиса отражают лишь штаммовые
особенности этих микроорганизмов.
Резервуар инфекции — дети в возрасте от 2 до
15 лет, а также больные в латентном периоде.
Таблица 36.3. Особенности лечебной тактики при фрамбезии (ВОЗ, 1986)
Распространенность фрамбезии в популяции Рекомендуемое лечение
Высокая — более 10 % населения (гиперэндемическая) Средняя -5-10 % (мезоэндемическая) Назначают бензатинбензилпенициллин всему населению (поголовное массовое лечение) Бензатинбензилпенициллин назначают всем активным больным, всем детям моложе 15 лет и лицам, тесно контактировавшим с заразными больными (массовое лечение подростков)
Низкая — менее 5 % (гипоэндемическая) Бензатинбензилпенициллин назначают всем активным больным, членам семей заразных больных и другим лицам, находившимся также с ними в тесном контакте (избирательное массовое лечение)
Глава 36. Кожные заболевания в жарких странах 423
Эпидемиология
Заболевание передается преимущественно кон-
тактно-бытовым путем, особенно у детей. Это-
му способствуют низкая санитарная культура,
теснота жилищ, питье из одной посуды, риту-
альные омовения. Непрямой путь заражения —
через инфицированную посуду для питья; пря-
мой — через пальцы, загрязненные слюной, со-
держащей трепонемы. Взрослые чаще всего за-
ражаются от своих детей. Переносчиком
заболевания могут быть и мухи.
Течение заболевания
Инкубационный период — 3 недели.
Мимолетные проявления (проходят незамет-
но) — 1 месяц.
Ранний период — от 1 месяца до 1 года.
Иногда латентный период — от 1 до 5 лет.
Поздний период — после 1 года.
Клиническая картина
Мимолетные проявления — на слизистой поло-
сти рта появляются пятна, неглубокие малобо-
лезненные язвы, иногда стоматит и трещины в
углах рта. Через несколько недель эти клини-
ческие проявления исчезают и часто остаются
незамеченными.
Ранний период — возникают диссеминирован-
ные незудящие папулы на туловище и конечно-
стях, в складках — кондиломы (вегетирующие
папулы). Поражаются длинные кости нижних
конечностей, беспокоят ночные костные боли,
т. е. признаки остеопериоститов.
Поздний период — проявляется гуммами кожи,
язвами и рубцами. Характерны гуммы носоглот-
ки, по типу гангоза (мутилирующий ринофарин-
гит), появление костных гумм (остеопериости-
ты), дисхромий (по типу витилиго).
Диагноз
Устанавливается на основании:
• клинической картины заболевания;
• эпидемиологических данных;
• возраста больных (дети);
• обнаружения трепонем в нативных препара-
тах из отделяемого свежих ранних высыпаний (в
тёмном поле микроскопа);
• положительных серологических реакций
(РВ, РИТ, РИФ).
Лечение
Лечение больных эндемическим сифилисом, а
также контактировавших с ними лиц проводится
препаратами пенициллина пролонгированного
действия (бензатинбензилпенициллин, бицил-
лин-1 и др.) в дозах, которые применяются для
лечения фрамбезии. При противопоказаниях к
применению пенициллина лечение проводится
эритромицином или тетрациклином.
Профилактика
• Своевременное выявление больных детей,
изоляция их от здоровых и проведение соответ-
ствующего лечения.
• Клинико-серологическое обследование всех
членов семей и лиц, находившихся в тесном
контакте с больным, для выявления активных
и латентных форм заболевания.
• Превентивное лечение всех контактировав-
ших лиц с больными беджелем.
• Улучшение жилищно-бытовых условий, по-
вышение санитарной культуры населения.
Пинта
Хронический невенерический трепонематоз (эн-
демический обесцвечивающий трепонематоз),
характеризующийся эритематозно-сквамозными
пятнами, склонными к гиперпигментации, затем
их депигментацией и полиаденитом.
Этиология
Возбудитель пинты — Treponema carateum. По
морфологическим и биологическим свойствам
близок к возбудителю фрамбезии и сифилиса.
Резервуар инфекции — больные в возрасте 15—
30 лет со стойкими специфическими поражени-
ями кожи.
Эпидемиология
Непосредственный прямой контакт «поражение-
кожа», а также пользование общей постелью,
полотенцем, посудой.
Течение заболевания
Инкубационный период — 2—3 недели.
Первичный период — до полугода.
Вторичный период — до 3 лет.
Третичный период — наступает после 3-х лет,
может длиться несколько десятилетий.
Клиническая картина
Первичный период — появляются единичные па-
пулы или эритематозно-сквамозные бляшки на
открытых участках тела, беспокоит умеренный
зуд.
Вторичный период — возникают распростра-
ненные зудящие папулы и бляшки (идентичные
начальным), эритематозно-сквамозные пятна
(пинтиды). Они имитируют трихофитию, псо-
424 Частная дерматология
риаз, экзематиды. Со временем их цвет меняет-
ся и становится медно-красным, лиловым, голу-
боватым, фиолетовым, синюшным, свинцово-
серым, серовато-белым.
Третичный период — появляется гиперпигмен-
тация, язвы не образуются. Далее наступает ах-
ромия (прогрессирующая депигментация гипер-
хромных пятен и других элементов); шелушение
пятен, папул, бляшек уменьшается и исчезает.
Иногда возникает гиперкератоз ладоней и
подошв, увеличение лимфоузлов; возможно раз-
витие очагов атрофии кожи («псевдоатрофия»).
Общее состояние пациентов не страдает.
Диагноз
Учитываются:
• эпидемиологический анамнез с учетом пре-
бывания больного в эндемичном очаге;
• наличие на коже дисхромий;
• обнаружение Т. carateum в высыпаниях или
в пунктатах лимфатических узлов;
• положительные серологические реакции
(РВ, РИТ, РИФ).
Лечение
Лечение пинты проводят в соответствии с реко-
мендациями ВОЗ, применяемыми при эндеми-
ческих трепонематозах (см. лечение фрамбезии).
Применяют бензатинбензил пенициллин, эрит-
ромицин и другие антибиотики.
Профилактические мероприятия сводятся к
выявлению и лечению больных и лиц, находя-
щихся с ними в тесном контакте.
Прогноз при этом заболевании благоприятный,
возможны косметические дефекты в виде оча-
гов депигментации, атрофии кожи и белых руб-
цов.
Часть 2
Венерология
Глава 37
Основные этапы развития венерологии
Венерология (лат. Venus — Венера, logos — на-
ука) — раздел медицины, предметом изучения
которого являются инфекционные болезни, пе-
редаваемые от человека к человеку преимуще-
ственно половым путем. Термин «венерические
болезни» был предложен французским врачом
Жаком де Бетенкуром в 1527 году и связан с
именем древней мифологической богини красоты
и любви Венеры.
Каждая из этих болезней — сифилис, мягкий
шанкр, гонорея, хламидийная лимфогранулема
и паховая гранулема (донованоз) вызываются
специфическим возбудителем. Для венеричес-
ких болезней характерны: высокая контагиоз-
ность, быстрое распространение среди некото-
рых групп населения (например, асоциальных),
необходимость в проведении активных проти-
воэпидемических и профилактических мероп-
риятий. Выявление и лечение заболевших на
ранних стадиях, д также их контактов, предот-
вращает развитие тяжелых последствий этих
инфекций.
В последние годы доказано, что при половых
контактах передаются не только указанные выше
«классические» венерические болезни, но такие
инфекции, как генитальный герпес, аногени-
тальные бородавки, кандидоз, контагиозный
моллюск, урогенитальный хламидиоз, лобковый
педикулез, чесотка, эндемические трепонемато-
зы, парентеральные гепатиты и др. Возбудите-
лями этих инфекций являются вирусы, дрожже-
подобные грибы, бактерии, простейшие,
эктопаразиты — всего выявлено более 20 различ-
ных инфекционных агентов.
В соответствии с решением 43-й сессии Все-
мирной ассамблеи здравоохранения и принятым
10-м пересмотром МКБ (1993) все заболевания,
передающиеся половым путем, включая «клас-
сические» венерические болезни, определены как
инфекции, передаваемые преимущественно по-
ловым путем (ИПППП). ИПППП широко рас-
пространены во всех странах мира и приносят ог-
ромный социально-экономический урон.
Наибольшие потери, регистрируемые в высоко-
развитых странах, связаны с сифилисом, гоно-
реей, урогенитальным трихомониазом, хламиди-
озом, генитальным герпесом.
На сохраняющиеся высокие показатели забо-
леваемости ИПППП влияют различные факто-
ры: региональные военные конфликты, низкий
уровень жизни людей, проституция, наркома-
ния, активная миграция населения, сексуальная
революция, неполный учет заболевших и их
контактов, слабая подготовка врачей различных
специальностей по вопросам ИПППП и др.
Краткие сведения
о развитии венерологии
В качестве специального раздела медицины ве-
нерология стала выделяться в конце XV столе-
тия, когда в Европе возникла вспышка сифили-
са. Во многом история венерологии связана с
развитием учения о сифилисе.
До сих пор по вопросу об истории появления
сифилиса в Европе существуют разные мнения
(К. Р. Аствацатуров, 1971; М. В. Милич, 1987).
Одна группа ученых (европеисты) считает, что
сифилис в Европе был известен задолго до на-
шей эры; другие (американисты) утверждают, что
сифилис появился в Европе после открытия Аме-
рики Христофором Колумбом — только в кон-
це XV века. О том, что сифилис существовал в
Европе и Азии задолго до экспедиции Христо-
фора Колумба, свидетельствуют высказывания
Гиппократа, описавшего заболевания гортани и
кондиломатозные высыпания на половых орга-
нах, которые могли соответствовать проявлени-
ям сифилиса. В китайских манускриптах (4535,
2600 гг. до н. э.) имеются описания мягкого и
сифилитического шанкров, бленоррагии, сифи-
лиса у новорожденных, лечения ртутным порош-
ком. В древних индийских, египетских и китай-
ских письменах встречаются описания
заболеваний в области заднего прохода, половых
органов, полости рта и носа, костей, которые
можно трактовать как проявления сифилиса.
Взаимосвязь поражений носа и высыпаний на
половых органах описывается в сочинениях Ди-
оскорида, Галена, Цельса и др., поражения кос-
тей, похожие на сифилитические — у Плутарха
426
Глава 37. Основные этапы развития венерологии
427
и Архигена. В костях, обнаруженных при архе-
ологических раскопках в Забайкалье, выявлены
изменения, схожие с сифилитическими. Кроме
того, при раскопках в Средней Азии в сфероко-
нических сосудах были обнаружены ртутные
смеси, что обосновывает предположение о при-
менении ртути в древние времена для лечения
сифилитических поражений. В трудах таджикс-
кого ученого Авиценны (X—XI век нашей эры)
есть описания среди жителей Средней Азии за-
болеваний, похожих на сифилис. Таким образом,
«сифилис, по-видимому, стар, как само челове-
чество» (Г. В. Вогралик, 1915).
Теорию американского происхождения сифилиса
в Европе подкрепляют следующие доказатель-
ства:
• о болезни в Европе врачам не было извест-
но, пока заболевание не приняло эпидемический
характер и очень быстро распространилось по
всему континенту;
• появление сифилиса в Европе в конце XV
века совпало с возвращением в Испанию экс-
педиции Христофора Колумба (1493), открыв-
шего Америку.
Некоторые ученые считают, что сифилис
возник в результате скотоложства и передачи по-
лового спирохетоза человеку от южно-американ-
ских лам. Вероятно также, что матросы Хрис-
тофора Колумба заразились сифилисом от
местного населения островов Гаити, у которых
сифилис уже существовал задолго до появления
испанцев. После возвращения экспедиции Ко-
лумба в Испанию сифилис стал распространять-
ся по приморским городам, появился в странах,
граничащих с Испанией, а затем — на всем кон-
тиненте Европы.
Большую роль в распространении сифилиса
сыграли частые в то время войны и поход фран-
цузского короля Карла VIII в Италию (1494—
1495), куда сифилис был занесен огромной ар-
мией испанских проституток, которых только в
Риме было около 14 тыс. Карл VIII был вынуж-
ден снять осаду Неаполя из-за «страшной болез-
ни» его наемников из разных стран и распустить
свою армию. После этого инфекция быстро рас-
пространилась по европейскому континенту.
Сифилис стал известен уже не только в Ис-
пании и Италии, но и во Франции, Голландии,
Германии, Дании, Англии, Шотландии, Швей-
царии, Польше, России. В начале XVI века он
появился в Юго-Восточной и Южной Африке,
а затем в Китае и Японии, чему способствовали
быстро развивавшиеся торговые морские связи.
Сифилис стали называть болезнью испанской,
итальянской, французской (галльской) и т. д., по
именам святых, различными терминами (bubas,
lues aetica, pestis inguinalis и др.).
О появлении сифилиса в России в конце XV
века свидетельствует поручение, данное великим
князем Иваном Васильевичем (Иоанн III) в
1499 г. своему литовскому послу Ивану Мамо-
нову: «Пытати ему в Вязьме князя Бориса, в
Вязьму кто не приезживал ли болен из Смолен-
ска тою болестью, что болячки мечются, а сло-
веть франчюжска, а будто из Вильны ее привез-
ли, да и в Смоленску о том пытати, еще ли
болесть есть или нет».
Cockburn (1961) и Hudson (1963) предложи-
ли теорию африканского происхождения сифи-
лиса, согласно которой возбудители тропичес-
ких (эндемических) трепонематозов —
фрамбезии, пинты, беджеля и венерического си-
филиса являются вариантами патогенной тре-
понемы. Трепонематозный паразитизм у чело-
века возник еще в раннем периоде неолита.
Вначале трепонематоз проявлялся как фрамбе-
зия у первобытных людей, живших в Цент-
ральной Африке. При возникновении первых
поселений людей в местности с сухим и более
прохладным климатом трепонематоз начал про-
текать в форме беджеля и пинты. С появлени-
ем городов и ограничением возможности пере-
дачи возбудителя бытовым путем трепонематоз
преобразовался в венерический сифилис. Миг-
рация людей из Африки способствовала распро-
странению трепонематозов в Европу, а через
Аравию в страны Азии. Следовательно, эволю-
цию патогенной спирохеты авторы теории свя-
зывают с изменениями общественно-бытовых
условий жизни людей и, соответственно, харак-
тером клинического течения различных трепо-
нематозов (В. М. Жданов).
Все три упомянутых теории указывают, «что
сифилитическая инфекция по происхождению
уходит в далекое прошлое и дискутируется толь-
ко вопрос о пути проникновения ее в Европу»
(М. В. Милич, 1987).
Наименование «сифилис» принадлежит италь-
янскому врачу, ученому и поэту Gerolamo
Fracastoro, который написал поэму «Syphilis sive
de morbo gallico», изданную в Венеции в 1530
году. Героем поэмы был пастух по имени
Syphilus, которого боги наказали болезнью по-
ловых органов «за дружбу со свиньей» и упре-
ки в сторону богов. От имени пастуха Сифилу-
са (sus — свинья, philos — друг) и возникло
наименование «сифилис», которое не ущемляло
национальное самолюбие разных стран, не по-
рочило святых и благополучно дошло до наших
дней.
428 Венерология
Сифилис как новая, тяжелая и неизлечимая
для того времени болезнь привлекал к себе
внимание многих ученых и врачей (К. Р. Ас-
твацатуров, 1971). Сифилис не сразу выдели-
ли в самостоятельное заболевание. Долгое вре-
мя считали, что сифилис, мягкий шанкр и
гонорея представляют собой различные прояв-
ления одной и той же болезни (сторонники
унитарной теории). Сторонники другой теории
(дуалисты) рассматривали сифилис, гонорею и
мягкий шанкр как самостоятельные заболева-
ния.
Знаменитый парижский венеролог Ricord
(1830), заразив большое число здоровых людей
патологическим материалом от венерических
больных, доказал, что сифилис и гонорея — раз-
личные в этиологическом отношении болезни.
Однако возбудители этих заболеваний еще не
были известны; сифилис не отличали от мягко-
го шанкра, а гонорею — от негонорейных урет-
ритов. В 1879 г. бреславский венеролог A. Neisser
открывает возбудителя триппера — гонококк. В
1887 г. профессор Петербургского университета
О. В. Петерсен сообщил о выявлении возбуди-
теля мягкого шанкра — стрептобациллы, что
позднее было подтверждено работами Ducrey
(1889), Unna (1892) и Krefting (1892).
В 1903 г. И. И. Мечникову и Roux удалось
привить сифилис человекообразным обезьянам.
Вскоре после этих экспериментов в 1905 г.
Schaudin и Hofmann открыли возбудителя вене-
рического сифилиса — бледную спирохету
(Spirochaeta pallida). В 1906 г. немецкий ученый
A. Wasserman совместно с A. Neisser и С. Bruck
предложили серологическую реакцию (РСК) для
диагностики сифилиса, которая и в настоящее
время является важным диагностическим тестом.
Известный немецкий врач и исследователь
Р. Erlich предложил для лечения сифилиса саль-
варсан (1909) и неосальварсан (1912). Сазерак и
Левадити (1921) провели исследования по изу-
чению антисифилитических свойств висмутовых
препаратов. В 1943 г. американцы Mahoney,
Arnold и Harris применили у больных сифили-
сом пенициллин.
Позднее американцы Nelson и Мауег (1949)
разработали более специфическую серологичес-
кую реакцию — иммобилизации бледных тре-
понем (РИБТ), позволяющую исключать лож-
ноположительные сифилитические реакции.
В последние десятилетия разработаны и внедре-
ны в практику ряд новых методов серодиагнос-
тики сифилиса: иммуноферментный анализ —
ИФА (J. Velkamp, A. Viaser, 1975), реакция пас-
сивной гемагглюцинации — РПГА (Т. Rathlev,
1965, 1967; Т. Tomizava, 1966), реакция иммуно-
флюоресценции — РИФ, FTA (Т. Weller,
A. Coons, 1954; W. Deacon, V. Falcone, A. Harris,
1957; В. Н. Беднова и др., 1960), РИФ-абс и её
варианты — IgM-FTA-ABS (1966), 19S-IgM-FTA-
ABS (1977), IgM SPHA (A. Luger и соавт., 1982),
полимеразная цепная реакция (Burstain J. М. и
соавт., 1991) и др.
При лечении больных сифилисом унифици-
рованы методики применения водорастворимо-
го пенициллина, предложены пролонгированные
пенициллины, цефалоспорины, азитромицин,
методы иммунотропной терапии, физической
активации крови (УФО, лазер).
Краткие сведения
о развитии венерологии
в России, Украине и Белоруссии
Основоположником и первым русским профес-
сором кафедры сифилидологии в России (Ме-
дико-хирургическая академия, Петербург), орга-
низованной в 1869 году, является В. М.
Тарновский (1838—1907). Его школой разрабо-
тан ряд важнейших вопросов патологии сифи-
лиса, в том числе и врожденного. В. М. Тарнов-
ский - автор первых русских учебников по
венерическим болезням и работ по изучению
сифилиса и гонореи; инициатор 1 съезда (1897)
по выработке мер борьбы с сифилисом.
Из клиники профессора Т. П. Павлова (ВМА,
Петербург), преемника В. М. Тарковского, выш-
ли ряд оригинальных работ, касающихся дерма-
тологии и венерологии. Его учеником, профес-
сором московской клиники кожных болезней
В. В. Ивановым были проведены обстоятельные
исследования по гистологическому строению
сифилидов. Профессор О. В. Петерсен (Петер-
бург) предложил наружное лечение сифилидов
присыпками висмута.
Важная роль в борьбе с распространением си-
филиса в России в XIX веке принадлежала зем-
ским врачам. Выдающиеся русские клиницисты
Н. И. Пирогов, А. А. Остроумов, И. Е. Дядьков-
ский, С. П. Боткин, М. Я. Мудров, Г. А. Заха-
рьин и др. в своей деятельности большое внима-
ние уделяли вопросам сифилидологии.
Ученик С. П. Боткина, родоначальник рус-
ской дерматологической школы, профессор пер-
вой дерматологической кафедры в Петербурге
А. Г. Полотебнов (1839—1907) применил при ле-
чении сифилитических язв плесневый грибок
(зеленый кистевик, 1872) и изучал действие пре-
паратов ртути в эксперименте на животных.
В. А. Манассеин (1871) и П. В. Лебединский
(1877) также сообщали о возможном лечебном
Глава 37. Основные этапы развития венерологии
429
применении антибиотиков в медицине, в т. ч. у
больных сифилисом.
Под руководством основателя московской
клиники кожных и венерических болезней А. И.
Поспелова (1846—1916) были разработаны воп-
росы лечения сифилиса подкожными впрыски-
ваниями сублимата (1874) и впервые примене-
ны для его лечения препараты мышьяка в виде
подкожных инъекций.
В клинике М. И. Стуковенкова (Киев) углуб-
ленно изучались вопросы лечебного действия
препаратов ртути при сифилисе. Представители
киевской школы профессора П. В. Никольский,
В. К. Боровский, И. Ф. Зеленев также внесли
существенный вклад в развитие дермато-сифи-
лидологии.
Большой вклад в изучение актуальных воп-
росов сифилидологии и организации борьбы с
венерическими болезнями в России внесли
Г. И. Мещерский, П. С. Григорьев, А. А. Студ-
ницин, Н. М. Туранов, Н. М. Овчинников,
В. А. Рахманов, А. И. Картамышев, Н. С. Сме-
лов, Б. М. Пашков, Г. Б. Беленький, Т. В. Ва-
сильев, Л. А. Штейнлухт, Н. А. Торсуев, Л. С.
Резникова и многие другие. Под их руковод-
ством на протяжении ряда лет проводились ис-
следования в области патологии и лечения си-
филиса, экспериментальной сифилидологии,
состояния иммунитета при сифилисе. Многие
важные проблемы венерологии в последние
годы разрабатываются в ЦКВИ М3 РФ (Ю. К.
Скрипкин, В. В. Делекторский, А. А. Кубано-
ва, К. К. Борисенко и др.), а также на профиль-
ных кафедрах и НИИ разных регионов России.
В Белоруссии большое внимание борьбе с ве-
нерическими болезнями уделяли академик
А. Я. Прокопчук, профессора И. И. Богданович,
О. П. Комов, А. Т. Сосновский, Ю. Ф. Коро-
лев, Л. И. Богданович. В настоящее время в об-
ласти ИПППП активно работают профессора
Н. 3. Яговдик, Л. Г. Барабанов, В. П. Адаске-
вич, В. М. Козин, доцент В. Г. Панкратов и др.
Глава 38
Сифилис
Этиология 430
Формы сифилиса 431
Эпидемиология и общая патология сифилиса 432
Лабораторная диагностика сифилиса 453
Лечение сифилиса 459
Профилактика сифилиса 469
Сифилис (syphilis, lues) — общее инфекционное
заболевание с волнообразным течением, со сме-
ной периодов обострения, характеризующихся
поражением кожи, слизистых оболочек, внутрен-
них органов, костей и нервной системы, скры-
тыми периодами, когда клинические признаки
болезни отсутствуют, и только положительные
серологические реакции крови свидетельствуют
о наличии инфекционного процесса.
Этиология
Возбудитель сифилиса — бледная трепонема
(Treponema pallidum). Типичные бледные трепо-
немы представляют собой тонкие спиралевид-
ные микроорганизмы длиной 6—14 мкм, толщи-
ной 0,25—0,30 мкм, с амплитудой спирали
0,5—1,0 мкм. Завитки спирали расположены на
равном расстоянии друг от друга. Трепонемы
очень подвижны и постоянно вращаются вок-
руг продольной оси, иногда изгибаясь и обра-
зуя почти замкнутый круг. Для выявления
бледных трепонем используют темное поле мик-
роскопа или иммунофлюоресцентную окраску.
Бледные трепонемы плохо окрашиваются ани-
линовыми красителями. С помощью электрон-
ной микроскопии установлено, что бледная
трепонема снаружи покрыта чехлом, под кото-
рым находятся трехслойная наружная стенка и
цитоплазматическая мембрана. Она имеет по-
верхностные и глубокие фибриллы, способству-
ющие движению. Трепонемы обычно размно-
жаются поперечным делением. В специальном
растворе и в присутствии редуцирующих ве-
ществ бледные трепонемы сохраняют подвиж-
ность при 25°С в течение 3—6 дней. В цельной
крови или в сыворотке при 4°С микроорганиз-
мы сохраняют жизнеспособность не менее 24
часов, что имеет значение при переливании
крови. Трепонемы быстро погибают при высу-
шивании и повышении температуры до 42°С.
Они мгновенно теряют подвижность и погиба-
ют в присутствии соединений мышьяка, ртути,
висмута. В целом, однако, бледные трепонемы
погибают очень медленно. Объясняется это низ-
кой метаболической активностью и замедлен-
ным размножением этих бактерий (время деле-
ния составляет около 30 часов). При
неблагоприятных условиях существования (воз-
действие антибиотиков, недостаток питания)
трепонемы могут образовывать «формы выжи-
вания» — цисты и L-формы.
В организме человека трепонемы стимулиру-
ют выработку антител, способных окрашивать
бледную трепонему при использовании непря-
мого метода флюоресцирующих антител; вызы-
вать иммобилизацию и гибель живых подвиж-
ных бледных трепонем; связывать комплемент
в присутствии суспензии трепонем.
Трепонемы вызывают также выработку анти-
тел оподобного вещества (реагина), которое дает
положительную реакцию связывания компле-
мента (РСК) и реакцию флоккуляции с водны-
ми суспензиями липидов, экстрагированных из
нормальных тканей млекопитающих. Как реа-
гины, так и противоспирохетные антитела мо-
гут быть использованы для серодиагностики
сифилиса.
430
Глава 38. Сифилис 431
Формы сифилиса
Первичный сифилис
Первичный сифилис (syphilis I primaria) — ста-
дия заболевания, характеризующаяся появлением
твердого шанкра и увеличением регионарных
лимфатических узлов.
Первичный серонегативный сифилис (syphilis I
seronegativa) — сифилис со стойко отрицатель-
ными серологическими реакциями в течение
курса лечения и наличием твердого шанкра.
Первичный серопозитивный (syphilis I
seropositiva) — сифилис с положительными се-
рологическими реакциями и наличием твердого
шанкра.
Первичный скрытый период (syphilis I latens) —
сифилис, характеризующийся отсутствием кли-
нических проявлений у пациентов, начавших
лечение в первичный период болезни и получив-
ших неполноценную терапию.
Вторичный сифилис
Вторичный сифилис (syphilis II secundaria) —
стадия заболевания, обусловленная гематогенным
распространением возбудителей из первичного
очага, проявляющаяся полиморфными высыпа-
ниями (розеолы, папулы, пустулы) на коже и
слизистых оболочках.
Вторичный свежий сифилис (syphilis II
recens) — период сифилиса, характеризующий-
ся многочисленными полиморфными высыпани-
ями на коже и слизистых оболочках, полиаде-
нитом; нередко наблюдаются остаточные
признаки твердого шанкра.
Вторичный рецидивный сифилис (syphilis II
recidiva) — период вторичного сифилиса, кото-
рый проявляется немногочисленными полимор-
фными сгруппированными высыпаниями, а
иногда и поражением нервной системы.
Вторичный скрытый сифилис (syphilis II
latens) — вторичный период заболевания, про-
текающий латентно.
Третичный сифилис
Третичный сифилис (syphilis III tertiaria) — сле-
дующая за вторичным сифилисом стадия, кото-
рая характеризуется деструктивными поражени-
ями внутренних органов и нервной системы с
возникновением в них гумм.
Активный третичный сифилис (syphilis III activa)
проявляется активным процессом образования
бугорков, разрешающихся некротическим распа-
дом, образованием язв, их заживлением, рубце-
ванием и появлением неравномерной пигмента-
ции (мозаичности).
Скрытый третичный сифилис (syphilis III
latens) — период болезни у лиц, перенесших
активные проявления третичного сифилиса.
Скрытый сифилис
Скрытый сифилис (syphilis latens) — сифилис,
при котором серологические реакции положи-
тельные, но отсутствуют признаки поражения
кожи, слизистых оболочек и внутренних орга-
нов.
Ранний скрытый сифилис (syphilis latens
praecox) — скрытый сифилис, с момента зараже-
ния которым прошло менее 2 лет.
Поздний скрытый сифилис (syphilis latens
tarda) — с момента заражения прошло более 2 лет.
Неуточненный скрытый сифилис (syphilis
ignorata) — болезнь с неустановленной давнос-
тью.
Врожденный сифилис
Врожденный сифилис (syphilis congenita) — си-
филис, при котором заражение произошло от
больной матери в период внутриутробного раз-
вития.
Ранний врожденный сифилис (syphilis congenita
praecox) — врожденный сифилис у плода и у де-
тей младше 2 лет, проявляющийся сифилитичес-
кой пузырчаткой, диффузной папулезной инфиль-
трацией кожи, поражением слизистых оболочек,
внутренних органов, костной ткани, глаз.
Поздний врожденный сифилис (syphilis congenita
tarda) ~ врожденный сифилис у детей старше 2
лет, проявляющийся триадой Гетчинсона, а также
поражением кожи, внутренних органов и костей
по типу третичного сифилиса.
Скрытый врожденный сифилис (syphilis congenita
latens) — врожденный сифилис, при котором кли-
нические проявления отсутствуют и лабораторные
показатели спинномозговой жидкости в норме.
Другие формы сифилиса
Сифилис нервной системы (neurosyphilis)'. ранний
(neurosyphilis praecox) — давность болезни до 5
лет, поздний (neurosyphilis tarda) — более 5 лет.
Сифилис висцеральный (syphilis visceralis) —
сифилис, при котором поражены внутренние
органы (сердце, головной мозг, спинной мозг,
легкие, печень, желудок, почки).
432 Венерология
Сифилис бытовой — сифилис, заражение ко-
торым происходит бытовым путем.
Сифилис обезглавленный (syphilis decapitata) —
заражение происходит при попадании возбуди-
теля непосредственно в кровяное русло (через
рану, при исследовании крови); характеризует-
ся отсутствием твердого шанкра.
Сифилис трансфузионный (syphilis
transfusionalis) — заражение в результате перели-
вания крови больного.
Сифилис злокачественный — тяжело протека-
ющий сифилис с массивным поражением внут-
ренних органов и нервной системы, характерным
для третичного сифилиса.
Сифилис экспериментальный — сифилис, воз-
никший у экспериментальных животных (обе-
зьян, кроликов) в результате их искусственно-
го заражения.
Эпидемиология
и общая патология
сифилиса
Пути заражения
и течение болезни
Пути заражения сифилисом подразделяют на
прямой (половой, тесный бытовой контакт,
трансфузионный), косвенный, трансплацентар-
ный.
Наиболее частый путь заражения — половой,
при различных формах половых контактов (per
vaginam, per апит, per os). Заражение происходит
через мелкие генитальные или экстрагенитальные
дефекты кожи либо через эпителий слизистой
оболочки при контакте с эрозивным или язвен-
ным твердым шанкром, эрозивными папулами на
коже и слизистых оболочках половых органов,
полости рта, широкими кондиломами, содержащи-
ми значительное количество возбудителей сифи-
лиса — бледных трепонем. Очень редко инфици-
рование может произойти при тесном бытовом
контакте, в исключительных случаях — через
предметы обихода или при прямом контакте с
экспериментальными животными. Возможно за-
ражение через молоко кормящей женщины, боль-
ной сифилисом. Случаев заражения через мочу и
пот не отмечено. В слюне бледные трепонемы могут
находиться лишь в случае, когда имеются специ-
фические высыпания на слизистой оболочке по-
лости рта. Возможно заражение через сперму боль-
ного при отсутствии у него видимых изменений
на половых органах. При переливании донорской
крови больного сифилисом у реципиента разви-
вается трансфузионный сифилис. Возможно ин-
фицирование медицинского персонала при осмотре
больных сифилисом, проведении лечебных проце-
дур и манипуляций, во время оперативного вме-
шательства, при вскрытии трупов, особенно ново-
рожденных с ранним врожденным сифилисом.
Отмечено внутриутробное инфицирование плода
путем трансплацентарной передачи возбудителя
сифилиса от инфицированной матери. Заражение
может также произойти в момент родов при про-
хождении плода через инфицированные сифили-
сом родовые пути матери.
Для сифилиса характерно волнообразное тече-
ние со сменой периодов заболевания. Традици-
онная схема течения сифилиса включает инкуба-
ционный период, первичный серонегативный и
первичный серопозитивный, вторичный свежий,
вторичный скрытый, вторичный рецидивный и
третичной периоды.
Время от момента инфицирования до появле-
ния первичной сифиломы называется инкубаци-
онным периодом. Продолжительность инкубаци-
онного периода в среднем составляет 3—4 недели.
Однако у отдельных больных он может достигать
4—6 месяцев в связи с бесконтрольным употреб-
лением антибиотиков по поводу различных забо-
леваний, а также под влиянием некоторых дру-
гих факторов. Несмотря на отсутствие
клинических симптомов, в инкубационном пери-
оде диссеминация бледных трепонем из места
внедрения по лимфатическим, периневральным
путям и гематогенно происходит довольно интен-
сивно и завершается формированием первичной
сифиломы — твердого шанкра, сопутствующего
лимфангита и регионарного склераденита. Пер-
вый клинический признак заболевания — твер-
дый шанкр — появляется в среднем через 3-4
недели после заражения на том месте, где блед-
ная трепонема проникла в организм (по образно-
му выражению французов, «при сифилисе пер-
вым наказуется то место, которым согрешили»).
С момента появления твердого шанкра начи-
нается первичный период сифилиса, который про-
должается до возникновения на коже и слизис-
тых оболочках множественных сифилитических
высыпаний. Этот период длится 7—8 недель.
Первичный период сифилиса делится на пер-
вичный серонегативный сифилис, когда стандар-
тные серологические реакции еще отрицательны;
первичный серопозитивный сифилис, когда стан-
дартные серологические реакции становятся по-
ложительными, что происходит в среднем через
Глава 38. Сифилис
433
3-4 недели после возникновения первичной
сифиломы.
При отсутствии лечения естественное течение
первичного периода завершается общими лихо-
радочными явлениями, недомоганием, болями в
костях, суставах и полиаденитом. Эти симптомы
обусловлены генерализацией инфекции и диссе-
минацией бледных трепонем в органы и ткани.
Этот период течения инфекции называется вто-
ричным и наступает через 7—8 недель после по-
явления первичной сифиломы или через 10—12
недель после поражения. Клинически вторичный
период сифилиса характеризуется поражениями на
коже и слизистых оболочках в виде розеолезных,
папулезных и пустулезных высыпаний. В этом
периоде поражаются также внутренние органы,
нервная и костная системы. Высыпания вторич-
ного периода, просуществовав несколько недель,
самостоятельно проходят, и наступает скрытый
(латентный) период заболевания. Через некото-
рое время наблюдается рецидив болезни — на
коже и слизистых оболочках снова появляются
высыпания, характерные для вторичного перио-
да, после чего вновь может наступить скрытый
период заболевания. Вторичный период сифилиса
без лечения может продолжаться 3—4 года.
Таким образом, вторичный период сифилиса
подразделяется на
• вторичный свежий сифилис, когда на коже и
слизистых оболочках впервые появляются
обильные и яркие высыпания при наличии пер-
вичного склероза или его остатков;
• вторичный скрытый сифилис — период си-
филиса без активных проявлений на коже и сли-
зистых оболочках;
• вторичный рецидивный сифилис — период
последующих повторных высыпаний.
Спустя 3—4 года, если больной не лечится или
лечится недостаточно, наступает третичный пе-
риод сифилиса. Этот период характеризуется об-
разованием третичных сифилидов — бугорков и
гумм. Бугорки бывают на коже и слизистых
оболочках, гуммы — в подкожной клетчатке,
костях, внутренних паренхиматозных органах и
нервной системе. Бугорки и гуммы при распаде
могут вызывать деструктивные изменения в
пораженных тканях и органах. Течение сифилиса
в этом периоде также волнообразное, фазы ак-
тивных проявлений могут сменяться фазами
скрытого (латентного) сифилиса.
Третичный период сифилиса подразделяют на
третичный активный сифилис, третичный скры-
тый сифилис. Третичный период сифилиса может
длиться многие годы. У некоторых больных без
лечения или при недостаточном лечении через
10—20 лет и более после заражения могут возник-
нуть поздние формы болезни: сифилис нервной
системы (нейросифилис) — спинная сухотка, про-
грессивный паралич, табопаралич; сифилис внут-
ренних органов (висцеросифилис) — мезаортит,
аневризма аорты, поражение печени, желудка.
Традиционное стадийное течение сифилити-
ческой инфекции наблюдается у значительного
числа больных. Однако в последние годы все
чаще стали выявляться больные с асимптомным
течением болезни, которая диагностируется
только серологически.
Сифилис без клинических проявлений с по-
ложительными серологическими реакциями под-
разделяют на
• ранний скрытый сифилис (до 2 лет после за-
ражения);
• поздний скрытый сифилис (давность болез-
ни более 2 лет).
Реакция организма на внедрение возбудите-
ля сифилиса сложна и многообразна. После кон-
такта с больным сифилисом заражения может не
произойти, а развивается классическое или дли-
тельное бессимптомное течение инфекции.
Рассматривая вопросы общей патологии сифи-
лиса, следует указать, что в ряде случаев после
обычной смены периодов первичного и вторич-
ного сифилиса, несмотря на отсутствие проти-
восифилитического лечения, поздние формы
сифилиса не развиваются и сифилитический
процесс заканчивается самоизлечением, в наступ-
лении которого ведущая роль принадлежит орга-
низму больного, его иммунобиологическим за-
щитным силам.
Первичный период сифилиса
Первичный период сифилиса начинается с мо-
мента образования твердого шанкра и продолжа-
ется 7—8 недель до массивной гематогенной дис-
семинации бледных трепонем, что клинически
проявляется возникновением обильных высыпа-
ний на коже и слизистых оболочках.
Образование твердого шанкра
На месте инокуляции бледных трепонем внача-
ле возникает четко ограниченная эритема округ-
лой формы, которая не беспокоит больного и
быстро превращается в плоскую папулу с незна-
чительным шелушением и небольшим уплотне-
нием ее основания. Через некоторое время на
поверхности папулы появляется эрозия или язва
с уплотненным основанием — первый симптом
первичного периода сифилиса.
434 Венерология
Признаки эрозивного (язвенного) твердого шан-
кра: ярко-красный (медный) цвет «свежего мяса»;
дно эрозии блестящее, лакированное; дно язвы
грязно-желтого цвета; края эрозии четко отграни-
чены; язва блюдцеобразная с покатыми краями;
округлая или овальная форма; отсутствие призна-
ков острого воспаления; безболезненность; нали-
чие инфильтрата у основания эрозии или язвы.
В большинстве случаев твердый шанкр (первич-
ная сифилома) находится в области наружных
половых органов, однако может располагаться на
любом участке кожного покрова или видимых
слизистых оболочек. Для первичного периода си-
филиса возможна экстрагенитальная локализация
очага поражения (слизистая полости рта, область
заднего прохода). Описаны шанкры на пальцах
рук, на конъюнктиве век, на кончике носа, уш-
ных раковинах, волосистой части головы, на коже
затылка, спины, голеней, на подошве.
Несмотря на характерные клинические при-
знаки, необычная локализация и вид первичной
сифиломы затрудняют ее распознавание. К та-
ким атипичным шанкрам относят: шанкр-амиг-
далит; шанкр-панариций; индуративный отек.
Шанкр-амигдалит — специфический амигда-
лит, характеризующийся увеличением и уплотне-
нием небной миндалины без образования эрозии
или язвы, но сопровождающийся болезненностью
при глотании. Одностороннее поражение, отсут-
ствие лихорадки и других нарушений общего
состояния, одностороннее увеличение подчелю-
стных и шейных лимфатических узлов помога-
ют поставить правильный диагноз.
Шанкр-панариций локализуется обычно на
дистальной фаланге указательного пальца, кото-
рый представляется отечным, синюшно-крас-
ным, булавовидным. На тыльной стороне этого
пальца располагается язва с неровными изъеден-
ными краями и дном с некротически-гнойным
распадом. Больного беспокоят резкие, стреляю-
щие боли. Локтевые и подмышечные лимфати-
ческие узлы увеличены и болезненны.
Индуративный отек развивается вследствие
поражения лимфатических капилляров кожи на
половом члене и коже мошонки у мужчин, а
также на малых и больших половых губах у
женщин. Пораженный орган значительно увели-
чивается в размерах, кожа становится плотной,
слегка синюшной. При надавливании ямки не
остается, пальпация безболезненна.
Осложнения твердого шанкра
В результате присоединения вторичной инфекции
и несоблюдения правил личной гигиены у неко-
торых больных развиваются осложнения твердого
шанкра, чаще островоспалительного характера:
• баланит — воспаление эпителия головки
полового члена;
• баланопостит — баланит в сочетании с вос-
палением внутреннего листка крайней плоти;
• фимоз — отек и увеличение в объеме край-
ней плоти, сужение кольца крайней плоти;
• парафимоз — ущемление головки полового
члена и отек крайней плоти, «удавка»;
• гангренизация — язвенно-некротический про-
цесс захватывает всю поверхность твердого шан-
кра с образованием серо-бурого плотного струпа,
по отторжении которого обнажается язвенный
дефект с гнойно-геморрагическим отделяемым;
• фагеденизм — процесс прогрессирует по пе-
риферии и вглубь с обширным, глубоким раз-
рушением окружающих и подлежащих тканей
крайней плоти, головки полового члена, уретры.
Регионарный лимфаденит
Через 5—8 дней после возникновения твердого
шанкра увеличиваются близлежащие лимфати-
ческие узлы (регионарный сифилитический
склераденит), может развиваться специфический
лимфангиит. Регионарный лимфаденит являет-
ся вторым важным симптомом первичного пе-
риода сифилиса. При локализации его в облас-
ти половых органов увеличиваются паховые
лимфатические узлы, на нижней губе или под-
бородке — подчелюстные, на языке — подборо-
дочные, на верхней губе и веках — предушные,
на пальцах рук — локтевые и подмышечные, на
нижних конечностях — подколенные и бедрен-
ные, на шейке матки — тазовые, в области мо-
лочных желез — подмышечные. Регионарный
склераденит проявляется увеличением лимфати-
ческих узлов иногда до размеров лесного ореха.
Признаки регионарного склераденита: увеличен-
ные лимфатические узлы; безболезненность; от-
сутствие изменений цвета кожи; плотноэластичес-
кая консистенция; подвижные узлы; узлы не
спаяны между собой и с подлежащими тканями.
Положительные
серологические реакции
Третьим симптомом первичного сифилиса явля-
ются положительные серологические реакции.
Реакция Вассермана становится положительной
через 3—4 недели после появления твердого шан-
кра, и с этого момента первичный серонегатив-
ный сифилис переходит в стадию первичного
серопозитивного сифилиса.
Глава 38. Сифилис 435
Диагноз
Диагноз первичного периода сифилиса устанав-
ливается на основании следующих данных:
• анамнез заболевания, конфронтация с пред-
полагаемым источником заражения;
• клинические признаки (наличие безболез-
ненной эрозии или язвы со скудным отделяе-
мым и уплотненным основанием, наличие реги-
онарного склераденита);
• в серонегативной стадии — обнаружение
трепонем в отделяемом из очагов поражений;
• в серопозитивной стадии — положительные
серологические реакции.
Диагноз первичного сифилиса, поставленный
только на основании типичной клинической кар-
тины, не подтвержденный обнаружением бледной
трепонемы или положительными серологически-
ми реакциями, не может считаться правомочным.
Однако отсутствие бледной трепонемы в отделя-
емом твердого шанкра при однократном исследо-
вании не исключает сифилис. В таких ситуаци-
ях необходимо трехкратное исследование.
Дифференциальный диагноз
При дифференциальной диагностике твердого
шанкра необходимо отличать его от эрозий и язв
при других заболеваниях, локализующихся
прежде всего в области половых органов.
Эрозия травматического происхождения (при
активных половых актах, травмировании воло-
сами, при введении в кожу инородных тел) име-
ет линейную форму с мягким основанием, со-
провождается островоспалительными явлениями,
болезненна, быстро заживает. Бледные трепоне-
мы не обнаруживаются в отделяемом эрозии.
Мягкий шанкр (шакроид) имеет более корот-
кий, чем сифилис, инкубационный период (1—
5 дней), характеризуется появлением множе-
ственных воспалительных элементов на половых
органах и в перигенитальной области: вначале
пятна, затем — папулы, везикулы, пустулы.
Пустулы изъязвляются. Края язв ярко-красно-
го цвета, отечные, подрытые, с гнойным отде-
ляемым, болезненны. Вокруг язв отмечается ос-
тровоспалительный венчик, инфильтрат в
основании мягкий. В отделяемом язв обнаружи-
вают возбудителя мягкого шанкра: стрептоба-
циллы Дюкрея-Петерсена-Унны. Имеется остро-
воспалительная лимфаденопатия: резкая
болезненность, мягкая консистенция, периаде-
нит, покраснение кожи, очаги флюктуации, об-
разование свищей, гнойное отделяемое.
Трихомонадные и гонококковые язвы характери-
зуются воспалительными явлениями, ярко-крас-
ного цвета с обильным отделяемым, в котором
обнаруживают соответствующие возбудители.
Сопутствующего регионарного склераденита не
имеется.
Простой пузырьковый лишай представлен мел-
кими сгруппированными везикулами с серозным
содержимым на отечном основании и гипереми-
рованной коже. Высыпаниям предшествуют зуд,
жжение, покалывание в участках поражения.
Эрозии простого герпеса множественные, с по-
лициклическими очертаниями без уплотнения у
основания, исчезают бесследно.
Стрептококковая эктима характеризуется ос-
тровоспалительной болезненной язвой с мягким
основанием, дно ее покрыто грязно-желтым
гнойным налетом, отсутствует сопутствующий
склераденит.
Чесоточная эктима и чесоточные папулы
обычно множественные, островоспалительного
характера, сопровождаются зудом и наличием
других типичных симптомов чесотки, подтвер-
ждаются лабораторным обнаружением чесоточ-
ного клеща.
Шанкриформная пиодермия характеризуется
серозно-геморрагической корочкой (покрываю-
щей эрозию или язву) с плотным основанием,
безболезненностью, сопутствующим склерадени-
том. Очаг поражения округлый или овальный
правильных очертаний, края слегка возвышены,
дно ровное, чистое. В отделяемом язвы и пунк-
тате лимфатических узлов бледные трепонемы не
обнаруживаются. Серологические реакции на си-
филис отрицательные.
Туберкулезная язва мягкой консистенции,
неправильной формы, края мягкие, подрытые,
кровоточит. На дне язвы имеются желтоватого
цвета мелкие очаги распада — зерна Треля. Язва
длительно не заживает, не рубцуется. Имеют-
ся другие очаги туберкулезной инфекции.
Острая язва Чаплина-Липшютца на наружных
половых органах встречается у молодых деву-
шек. Заболевание протекает остро, с высокой
температурой тела. На малых и больших поло-
вых губах или в перианальной области возни-
кают резко болезненные язвы округлых или
неправильных очертаний, мягкой консистенции,
на эритематозном фоне. Диагностика основыва-
ется на характерных клинических, бактериоло-
гических и серологических данных.
Эритроплазия Кейра проявляется небольшим
безболезненным очагом на головке полового
члена, четко отграниченным, ярко-красного цве-
та с бархатистой влажной поверхностью без от-
деляемого.
436 Венерология
Базальноклеточная карцинома кожи поражает
людей пожилого возраста, медленно прогресси-
рует, не склонна к метастазам. Край язвы обра-
зован мелкими белесоватыми узелками. Харак-
терны деревянистая плотность у основания,
неровные, несколько размытые, плотные края и
валик* по периферии.
Плоскоклеточный рак кожи встречается чаще у
пожилых людей. Края язвы выворочены, дно изры-
то, покрыто очагами распада, легко кровоточит, дли-
тельно не заживает. Характерен экзо- и эндофит-
ный рост язвы. Лимфатические узлы увеличены,
спаяны друг с другом и окружающими тканями.
Вторичный период сифилиса
Вторичный период сифилиса начинается с появ-
ления первых высыпаний на коже и слизистых
оболочках через 2—3 месяца после заражения или
через 7—8 недель после появления твердого шан-
кра и продолжается без лечения 3-5 лет. При
вторичном сифилисе возникают пятнистые, па-
пулезные, пустулезные, везикулезные высыпания на
коже и слизистых оболочках, а также наблюда-
ются нарушения пигментации, выпадение волос.
Наряду с высыпаниями у 80 % пациентов сохра-
няются твердый шанкр или его остатки, а также
регионарный склераденит. Могут поражаться
внутренние органы, нервная, эндокринная и ко-
стная системы. Течение заболевания во вторич-
ном периоде вариабельно. Через 2—3 месяца вы-
сыпания постепенно исчезают даже без лечения
и сохраняются только положительные серологи-
ческие реакции. Начинается вторичный скрытый
период. Затем наступает рецидив болезни с раз-
нообразным течением.
Общие характерные признаки для всех вторич-
ных сифилидов:
• доброкачественное течение — не разрушают
ткани, не оставляют рубцов, самопроизвольно
проходят через 2—3 месяца;
• удовлетворительное общее состояние, отсут-
ствие субъективных ощущений у больных;
• отсутствие острого воспаления в элементах
вторичного сифилиса; высыпания резко отгра-
ничены от здоровой кожи, не склонны к пери-
ферическому росту и слиянию, располагаются
фокусно;
• полиморфизм высыпаний — одновременное
наличие различных сифилидов (розеолы, папу-
лы, пустулы);
• острозаразность элементов вторичного пери-
ода сифилиса;
• быстрое разрешение вторичных сифилидов
под влиянием противосифилитического лечения.
Для установления диагноза вторичного пери-
ода сифилиса необходимы данные анамнеза,
объективные клинические данные заболевания,
обнаружение возбудителей в очагах поражений,
серологические исследования крови, специаль-
ные функциональные методы исследования.
Пятнистый сифилид
(сифилитическая розеола)
Пятнистый сифилид (сифилитическая розеола)
является самым частым высыпанием во вторич-
ном свежем сифилисе. Сыпь располагается пре-
имущественно на туловище и конечностях. Ро-
зеола появляется постепенно и достигает полного
развития за 7-10 дней. Продержавшись без ле-
чения в среднем 3—4 недели, розеола постепен-
но исчезает. При обострении процесса после
введения пенициллина розеола более выражена
(реакция Герксгеймера-Яриша-Лукашевича) и
может появиться в новых местах.
Признаки пятнистого (розеолезного) сифилида:
пятна розовые и бледно-розовые; обильные, ред-
ко сливаются; расположены беспорядочно, сим-
метрично, фокусно; с округлыми, нерезкими
очертаниями; не возвышаются над кожей; не
шелушатся; не вызывают субъективных ощуще-
ний; при диаскопии исчезают.
Диагностика сифилитической розеолы при вто-
ричном сифилисе затруднений не вызывает.
Дифференциальный диагноз
пятнистого сифилида
При дифференциальной диагностике пятнисто-
го сифилида следует исключить пятнистые вы-
сыпания инфекционных заболеваний (корь,
краснуха, брюшной тиф), розовый лишай, отру-
бевидный лишай, токсикодермию, мраморность
кожи, пятна от укусов лобковых вшей.
При кори пятна обильные, крупные, сливают-
ся. Яркая сыпь сначала появляется на лице, шее,
затем на туловище, конечностях, в том числе на
кистях и стопах. На слизистой оболочке щек,
иногда на губах, деснах возникают точечные
белесоватые пятна Филатова-Кёплика.
У больных краснухой сыпь вначале появляет-
ся на лице, затем на шее и распространяется на
туловище. Высыпания бледно-розовые, размером
до чечевицы, не склонны к слиянию, несколь-
ко выступают над поверхностью кожи, бесследно
исчезают. Иногда беспокоит зуд.
Высыпания при брюшном тифе всегда сопро-
вождаются тяжелыми общими явлениями. Розе-
ола не столь обильна, нередко приобретает ге-
моррагический характер.
Глава 38. Сифилис 437
У больных розовым лишаем на боковой повер-
хности туловища появляется материнская бляшка
в виде крупного овального розово-красного пят-
на, центральная часть которого покрыта пластин-
чатой чешуйкой, подобной смятой папиросной
бумаге. Через 1—2 недели появляются множе-
ственные, мелкие розеолезные пятна на коже
туловища, конечностей, лица, шеи, бедер. Уме-
ренно выражен зуд. Спустя 6—8 недель сыпь
исчезает.
При отрубевидном (разноцветном) лишае пятна
располагаются на коже груди, спины, плеч, шеи.
Пятна невоспалительные, желтовато-бурого цве-
та, разных оттенков, сливаются, шелушатся. При
смазывании раствором йода очаги поражения
окрашиваются более интенсивно по сравнению
с окружающей кожей.
При токсикодермии розеола крупная, красно-
синюшная, сопровождается зудом и жжением,
располагается преимущественно в области круп-
ных складок, отличается острым развитием,
шелушением, склонна к слиянию.
Пятна от укусов лобковых вшей появляются
на животе, лобке, бедрах, боковой поверхности
туловища и отличаются серовато-фиолетовым си-
нюшным цветом с геморрагической точкой в
центре от укуса лобковых вшей. Пятна не исче-
зают при надавливании.
Мраморная кожа возникает в результате про-
свечивания расширенных поверхностных капил-
ляров кожи. В центре их окраска кожи нормаль-
ная, а по периферии — красновато-синюшная.
При дифференциальной диагностике сифили-
тической розеолы с вышеприведенными заболе-
ваниями имеют значение отсутствие других кли-
нических симптомов вторичного сифилиса, а
также результаты серологического обследования
больных.
Папулезные сифилиды
Папулезные сифилиды являются частым прояв-
лением вторичного рецидивного сифилиса. Разли-
чают следующие разновидности папулезных си-
филидов: лентикулярный, милиарный, нумулярный,
мокнущий, ладонно-подошвенный, широкие конди-
ломы.
Лентикулярный папулезный сифилид
Лентикулярный папулезный сифилид представ-
ляет собой разновидность сифилитических па-
пул, которые имеют плотноэластическую кон-
систенцию, округлые, резкие очертания,
полушаровидную форму, без воспалительного
ободка, медно-красного цвета с синющным от-
тенком, размером с чечевицу.
Дифференциальный диагноз
лентикулярного папулезного сифилида
Лентикулярные папулы дифференцируют от вы-
сыпаний при красном плоском лишае, каплевид-
ном парапсориазе, псориазе, папулонекротичес-
ком туберкулезе кожи, контагиозном моллюске.
Красный плоский лишай — плоские, плотные,
полигональные папулы с восковидным блеском,
пупковидным вдавлением в центре, фиолетовые;
излюбленная локализация — внутренняя повер-
хность предплечий.
Каплевидный парапсориаз — мягкие, слегка
возвышаются над кожей узелки, красно-корич-
неватой окраски, покрыты чешуйкой в виде об-
латки; при поскабливании появляются точечные
кровоизлияния на поверхности папулы и на
коже возле нее.
Псориаз — папулы красного и розового цве-
та, покрыты серебристо-белыми чешуйками; при
поскабливании наблюдаются феномены стеари-
нового пятна, терминальной пленки, точечного
кровотечения; элементы склонны к перифери-
ческому росту; расположение симметричное,
преимущественно на разгибательных поверхно-
стях конечностей, в области локтевых и колен-
ных суставов, волосистой части головы.
Папуло-некротический туберкулез кожи ~ па-
пулоподобные красновато-синюшные элементы
с некрозом в центральной части; располагаются
преимущественно на задней поверхности верх-
них и передней поверхности нижних конечнос-
тей, на пальцах, лице; отмечается ложный эво-
люционный полиморфизм, штампованные
рубцы после регресса элементов; часто выявля-
ется туберкулез внутренних органов, костей,
суставов или лимфатических узлов; положитель-
ная реакция Манту; отрицательные серологичес-
кие реакции.
Контагиозный моллюск — небольшие, разме-
ром до горошины папулы полушаровидной фор-
мы, с пупковидным вдавлением в центре, беле-
совато-перламутрового цвета, блестящие, без
воспалительного ободка по периферии; при сдав-
ливании с боков из моллюска выделяется клей-
кая белесоватая густая масса.
Милиарный
(мелкопапулезный) сифилид
Милиарный, или мелкопапулезный, сифилид
встречается редко и располагается преимуществен-
но на коже туловища в виде сгруппированных
438 Венерология
буровато-красных или медно-красных конических
плотных папул величиной с просяное зерно. Вы-
сыпания образуют кольца, дуги, бляшки с зубча-
тыми краями и мелкозернистой поверхностью.
Узелки располагаются вокруг устьев волосяных
фолликул. Иногда милиарные папулы настолько
мелкие, что напоминают гусиную кожу.
Дифференциальный
диагноз милиарного сифилида
Милиарные сифилитические папулы необходи-
мо дифференцировать с лихеноидным туберку-
лезом кожи и микидами:
Лихеноидный туберкулез кожи — мелкие, с бу-
лавочную головку, мягкие, желтовато-розовые,
сгруппированные, фолликулярные папулы, по-
крытые нежной чешуйкой, пронизаны пушком;
расположены на боковой поверхности туловища;
встречаются чаще у детей и подростков, страда-
ющих компенсированным туберкулезом легких,
лимфатических узлов, костей, суставов; сыпь
возникает после перенесенного инфекционного
заболевания — скарлатины, кори, гриппа; поло-
жительная реакция Манту; серологические реак-
ции на сифилис отрицательные.
Микиды — наблюдаются у пациентов с ин-
фильтративно-нагноительной трихофитией; ха-
рактеризуются мелкими, красными, зудящими,
симметрично расположенными папулами.
Нумулярный (монетовидный)
папулезный сифилид
Нумулярный или монетовидный папулезный си-
филид представлен округлыми папулами до
2,5 см в диаметре. Папулы буроватого или крас-
ного цвета, имеют уплощенную полушаровид-
ную поверхность. При рассасывании таких па-
пул долго остается пигментация.
Мокнущий папулезный сифилид
Мокнущий папулезный сифилид обычно пред-
ставлен папулами в местах повышенной потли-
вости и постоянного трения (анальная область,
половые органы, паховобедренные, межъягодич-
ные, подмышечные складки). При этом проис-
ходит мацерация и отторжение рогового слоя с
поверхности папул с образованием мокнущей
эрозии. В серозном отделяемом мокнущих эро-
зивных папул выявляется большое количество
бледных трепонем. Резкие границы каждого
элемента, выступающая над поверхностью эро-
зия, умеренно выраженные ощущения зуда и
жжения позволяют установить диагноз.
Мокнущие папулезные сифилиды имеют сход-
ство с ложносифилитическими папулами, кото-
рые возникают на гребнях больших половых губ
и имеют вид плотных, круглых, возвышающих-
ся над кожей элементов размером с чечевицу,
бледно-розового цвета. Поверхность папул бле-
стящая, сухая, не эрозирована. Бледная трепо-
нема не выявляется.
Ладонно-подошвенный
папулезный сифилид
Ладонно-подошвенный папулезный сифилид
выглядит как резко ограниченные красновато-
фиолетовые или желтоватые пятна с плотной
инфильтрацией у основания. Впоследствии на
поверхности таких элементов образуются плот-
ные, трудно удаляемые чешуйки. Затем роговой
слой в центре папулы трескается, и папула на-
чинает шелушиться, образуя «воротничок Биет-
та». Иногда ороговение на поверхности папул на
ладонях и подошвах приводит к образованию
мозолеподобных утолщений.
Широкие кондиломы
(вегетирующие папулы)
Широкие кондиломы (вегетирующие папулы)
обычно располагаются в области крупных скла-
док, промежности, половых органов, вокруг зад-
него прохода и возникают в результате умерен-
ного длительного раздражения. Они крупные,
возвышаются над уровнем кожи, сливаются,
образуют бляшки. Поверхность широких конди-
лом бугристая, неровная, мацерирована, у неко-
торых больных эрозирована или изъязвлена.
Широкие кондиломы характерны для вторично-
го рецидивного сифилиса, и нередко являются
единственным проявлением болезни.
Дифференциальный
диагноз широких кондилом
Широкие кондиломы следует дифференцировать
от остроконечных кондилом, варикозных гемор-
роидальных вен, вегетирующей пузырчатки.
Остроконечные кондиломы — вегетирующие
опухолевидные разрастания в области крупных
складок, расположенные на узкой ножке, лег-
ко кровоточат, мягкоэластической консистен-
ции; серологические реакции на сифилис отри-
цательные.
Геморроидальные узлы с одной стороны покры-
ты слизистой оболочкой прямой кишки; они
имеют мягкую консистенцию, кровоточат, не
имеют плотноэластического инфильтрата.
Глава 38. Сифилис 439
Вегетирующая пузырчатка — пузыри на неиз-
мененной коже, слизистой оболочке рта, дли-
тельно незаживающие, болезненные эрозии;
положительный симптом Никольского; аканто-
литические клетки в мазках-отпечатках; мягкие
вегетации, возникающие на дне пузырей в об-
ласти крупных складок.
Пустулезные сифилиды
Пустулезные сифилиды обычно возникают при
вторичном сифилисе в результате распада папул
и образования желтовато-коричневых корок,
похожих на пиодермические. В отличие от гной-
ничков, на их периферии имеется валик инфиль-
трата, а не островоспалительный ободок. Пусту-
лезные сифилиды чаще возникают у лиц с
пониженной сопротивляемостью организма,
страдающих сопутствующими заболеваниями
(туберкулезом, малярией, алкоголизмом). Появ-
ление сыпи иногда сопровождается повышени-
ем температуры тела, снижением массы тела.
Разновидности пустулезных сифилидов (угре-
видный, импетигинозный, оспенновидный, экти-
матозный, рупиоидный) зависят от локализации
сыпи, размеров элементов, степени распада.
Угревидный (акнеформный)
пустулезный сифилид
Угревидный или акнеформный пустулезный си-
филид характеризуется мелкими, коническими,
фолликулярными папулами, расположенными на
лице, груди, спине, верхних конечностях. На вер-
шине папул имеется пустула, гнойный экссудат
которой ссыхается в желтовато-коричневую ко-
рочку с пигментированным вдавлением в центре.
Дифференциальный
диагноз угревидного сифилида
Дифференцировать угревидный сифилид следует
с вульгарными угрями, йодистыми и бромисты-
ми угрями, нодулярным аллергическим васкули-
том, папулонекротическим туберкулезом.
Вульгарные угри отличаются островоспалитель-
ными пустулами, болезненностью, наличием
комедонов и ретенционных кист, хроническим
течением с частыми рецидивами.
Йодистые и бромистые угри отличаются ост-
ровоспалительной реакцией, наличием крупных
пустул, отсутствием плотного инфильтрата у ос-
нования, быстрым разрешением высыпаний
после прекращения приема препаратов йода или
брома.
Аллергический узелковый васкулит протекает
длительно, элементы развиваются торпидно и
локализуются на разгибательных поверхностях
конечностей. На месте узелковых высыпаний
остаются «штампованные» рубчики.
Папулонекротический туберкулез кожи харак-
теризуется высыпаниями на задней поверхнос-
ти верхних и передней поверхности нижних
конечностей, эволюционным полиморфизмом,
типичными штампованными рубцами, положи-
тельной реакцией Манту, отрицательными серо-
логическими реакциями на сифилис.
Импетигинозный
пустулезный сифилид
Импетигинозный пустулезный сифилид прояв-
ляется высыпаниями округлых папул с поверх-
ностным распадом в пределах эпидермиса, ко-
торый в дальнейшем ссыхается в грязно-желтую
корочку с красно-синюшным плотным валиком.
Высыпания локализованы на лбу, груди, спине,
передней поверхности верхних конечностей.
Вульгарное импетиго отличается от сифилити-
ческого острым началом, быстрым распростране-
нием, образованием фликтен без уплотнения у
основания, золотистыми или грязно-серыми кор-
ками, слиянием высыпаний в большие очаги.
Оспенновидный
пустулезный сифилид
Оспенновидный пустулезный сифилид пред-
ставляет собой полушаровидные пустулы вели-
чиной с горошину с резко отграниченным ин-
фильтратом медно-красного цвета и
пупкообразным вдавлением в центре. Затем со-
держимое пустулы ссыхается в корку, после от-
торжения которой остаются пигментация и ат-
рофический рубец. Элементы оспенновидного
сифилида расположены обычно на сгибательных
поверхностях конечностей, туловище, лице.
Дифференцировать оспенновидный сифилид
следует с натуральной и ветряной оспой.
Натуральная и ветряная оспа характеризуются
острым началом заболевания с высокой температу-
рой, тяжелым общим состоянием больного, отсут-
ствием у основания пустул плотного инфильтрата,
первоначальным появлением высыпаний на лице,
отрицательными серологическими реакциями.
Эктиматозный пустулезный сифилид
Эктиматозный пустулезный сифилид (сифили-
тическая эктима) возникает в результате глубо-
16 Зак. 1357
440 Венерология
кого распада крупных монетовидных папул.
Распад эктимы прогрессирует вглубь и по пери-
ферии. Возникает вначале темно-красный ин-
фильтрат, в центре которого образуется пусту-
ла, засыхающая в бурую вдавленную корку,
окруженную инфильтратом медно-красного цве-
та. Эктима увеличивается в размерах и после
удаления корки обнажается глубокая язва с от-
весными краями и гладким дном, покрытым
некротическими массами с гнойным отделяемым.
После заживления эктимы остается пигменти-
рованный рубец.
Вульгарная эктима отличается от сифилити-
ческой наличием стрептококковой пустулы с
островоспалительной реакцией окружающей
кожи, отсутствием инфильтрации в основании
пустулы и вокруг нее.
Рупиоидный пустулезный сифилид
Рупиоидный пустулезный сифилид представляет
собой разновидность тяжело протекающих эк-
тим с резко выраженной тенденцией к распрос-
транению и образованием массивных, слоистых
буро-черных корок, похожих на устричную ра-
ковину. При удалении корки обнажается глубо-
кая болезненная язва с инфильтрированными
краями и грязно-кровянистым отделяемым.
После заживления язвы остается глубокий пиг-
ментированный рубец.
Сифилитическая рупия имеет сходство с ру-
пиоидной формой псориаза и рупиоидной пио-
дермией, которые встречаются очень редко.
При рупиоидном псориазе устрицеподобные
наслоения представляют собой не чешуйко-кор-
ки, а только чешуйки без примеси экссудата.
После удаления чешуйки обнажается розово-
красная поверхность псориатической папулы, а
не язва.
Сифилитическая лейкодерма
Сифилитическая лейкодерма (или пигментный
сифилид) возникает во вторичном рецидивном
сифилисе на 6—8 месяце после заражения. По-
ражается преимущественно кожа задней и бо-
ковых поверхностей шеи («ожерелье Венеры»),
иногда — верхняя часть груди, спина, живот.
В очагах поражения на фоне умеренно гипер-
пигментированной кожи выявляются гипопиг-
ментированные пятна округлых очертаний и
различных размеров. Лейкодерма лучше вид-
на при боковом освещении. Пятна отделяют-
ся друг от друга тонкой пигментированной по-
лоской.
Признаки сифилитической лейкодермы: депиг-
мецтированные пятна на гиперпигментирован-
ном или обычном фоне; отсутствуют воспали-
тельные явления; больных не беспокоит зуд; нет
шелушения; пятна стойкие (исчезают через 1—
2 года).
Дифференциальный диагноз
сифилитической лейкодермы
Сифилитическую лейкодерму следует диффе-
ренцировать от витилиго и вторичной лей-
кодермы после отрубевидного лишая, псори-
аза, парапсориаза, экземы, атопического
дерматита.
Витилиго отличается полным отсутствием
пигмента в очагах поражения, более крупными
размерами очагов депигментации, склонностью
к периферическому росту и слиянию.
Вторичная лейкодерма, возникшая на месте
отрубевидного лишая, характеризуется различ-
ной формой и величиной депигментированных
пятен, склонных к слиянию и имеющих фе-
стончатые очертания очагов, а также наличи-
ем шелушащихся элементов молочно-кофей-
ного цвета, выявляемых пробой с йодной
настойкой.
Вторичная лейкодерма после псориаза, парапсо-
риаза, экземы, атопического дерматита имеет
более крупные размеры, различную локализа-
цию. В диагностике помогут анамнестические
данные и отрицательные серологические реакции
на сифилис.
Сифилитическая
алопеция (облысение)
Сифилитическая алопеция (облысение) наблю-
дается чаще у больных вторичным рецидивным
сифилисом. Волосы выпадают в результате нару-
шения питания их корней. Чаще поражаются
височно-теменные и затылочные области. При
тяжелом течении болезни могут выпадать воло-
сы в области бороды, усов, бровей, ресниц, под-
мышечных впадин. На участках поражения кожа
не изменена, нет шелушения и покраснения,
отсутствуют субъективные ощущения. Через 1—
2 месяца рост волос восстанавливается, первона-
чально в старых очагах, а затем в более молодых.
Пораженные ресницы вледствие частичного вы-
падения и последующего отрастания имеют раз-
ную длину (ступенчатообразные ресницы —
признак Пинкуса).
Различают три разновидности сифилитичес-
кой алопеции: мелкоочаговая; диффузная; сме-
шанная.
Глава 38. Сифилис
441
Дифференциальный диагноз
сифилитической алопеции
При дифференциальной диагностике исключа-
ют вторичную рубцовую алопецию после глубоких
язвенных поражений (глубокая пиодермия, трав-
мы, ожоги, инфильтративно-нагноительные
микозы), а также после хронических дерматозов
(красная волчанка, псевдопелада Брока).
При гнездном облысении появляются крупные
округлые очаги, чаще единичные, резко отгра-
ниченные, имеющие блестящую поверхность с
полным отсутствием волос, зону расшатанных
волос по периферии.
При микроспории и трихофитии волосистой
части головы в очагах поражения имеются ше-
лушение и гиперемия, остатки обломанных во-
лос. Лабораторно в пораженных волосах обнару-
живаются споры гриба.
При себорейном облысении волосы жирные или
сухие, тонкие, ломкие, процесс прогрессирует
медленно, сопровождается зудом. Выпадение
волос развивается в лобной и теменных облас-
тях.
Преждевременное облысение имеет наслед-
ственный характер, развивается постепенно, мед-
ленно, волосы выпадают, прежде всего в лобной
и теменной областях. Серологические реакции на
сифилис отрицательные.
Выпадение волос после острых инфекционных
заболеваний (брюшной и сыпной тиф, грипп)
устанавливают на основе анамнестических дан-
ных, диффузного характера облысения, отсут-
ствия признаков сифилиса, отрицательных серо-
логических реакций на сифилис.
Поражения
слизистых оболочек
Поражения слизистых оболочек встречаются ча-
сто у больных вторичным рецидивным сифилисом
и иногда являются единственным симптомом
болезни. Обычно поражается слизистая оболоч-
ка полости рта, губ, языка, глотки и гортани.
Наблюдаются пятнистые и папулезные сифили-
ды слизистых оболочек. Высыпания эрозирова-
ны, мацерированы, заразны и опасны в эпиде-
миологическом плане, регрессируют медленно.
Розеолезные (пятнистые) сифилиды на слизи-
стой оболочке полости рта представляют собой
округлые, красно-синюшные, четко отграничен-
ные розеолы, не возвышаются над уровнем тка-
ней, иногда сливаются, не вызывают субъектив-
ных ощущений, исчезают бесследно. При
сифилитической эритематозной ангине может
быть чувство першения в горле, но боль отсут-
ствует. Сифилитические эритематозные ларин-
гиты могут сопровождаться небольшой охрип-
лостью голоса. При катаральном эритематозном
сифилитическом рините отмечается сухость сли-
зистых оболочек.
Папулезные сифилиды слизистых оболочек яв-
ляются наиболее частыми проявлениями вторич-
ного периода сифилиса.
Признаки язвенных и эрозивных папулезных
сифилидов слизистых оболочек: плотные, плос-
кие, округлые, четко отграниченные; без пери-
ферического воспалительного ободка; ярко-крас-
ного цвета; обычно не беспокоят больных,
иногда болезненны; без острых воспалительных
явлений.
При сифилитической папулезной ангине вы-
сыпания появляются на слизистой оболочке
полости рта, языка, губ. Папулы в области зева
сопровождаются болезненностью, а при их изъяз-
влении — болью при глотании. При поражении
голосовых связок появляются кашель, осиплость
голоса вплоть до афонии.
Папулы на слизистой языка имеют отполи-
рованную, гладкую, блестящую поверхность
(«лоснящиеся» папулы), розовато-синюшный
цвет, неправильные или овальные очертания,
располагаются ниже уровня окружающей сли-
зистой оболочки в виде бляшек «скошенного
луга».
Дифференциальный диагноз
поражений слизистых оболочек
при сифилисе
Сифилитическую папулезную ангину, папулы на
слизистой полости рта и языка дифференциру-
ют с рядом заболеваний.
Язвенно-некротическая ангина Симоновского-
Плаута-Венсана характеризуется островоспали-
тельными явлениями, односторонностью пораже-
ния, сильной болезненностью, язвами с
некротическими массами, гнилостным запахом изо
рта, регионарным лимфаденитом, отрицательны-
ми серологическими реакциями на сифилис.
Обычная ангина сопровождается высокой тем-
пературой тела, нарушением общего состояния
больного, резким отеком и гиперемией зева,
миндалин, дужек, мягкого неба; нечеткими гра-
ницами поражения, сильной болезненностью
при глотании, отсутствием других признаков
сифилиса.
При дифтерии на миндалинах появляется
фибринозный налет грязно-серого цвета, слег-
ка блестящий, плотно прилегающий; отмечают-
ся явления токсикоза.
442 Венерология
Папулы при красном плоском лишае на слизи-
стой оболочке полости рта плотные, белесоватые,
с блестящей поверхностью, полигональные, по-
чти не возвышаются над уровнем тканей. Име-
ются также типичные высыпания на коже внут-
ренней поверхности предплечий. Серореакции
отрицательные.
Афтозный стоматит начинается остро; сопро-
вождается болезненными, желтоватыми эрозия-
ми с ярко-красным ободком; исчезает через не-
делю бесследно, часто рецидивирует.
Плоская лейкоплакия развивается постепенно,
медленно прогрессирует; имеется серовато-белый
налет с шероховатой поверхностью без воспали-
тельных явлений.
Глоссит десквамативный («географический
язык») представлен участками десквамации ярко-
красного цвета, белой каймой по периферии,
фестончатыми очертаниями, отсутствием уплот-
нений в основании, хроническим течением за-
болевания.
Сифилитические поражения гортани, голосовых
связок, слизистой оболочки носа распознаются на
основании клинической картины, лабораторных
данных и характеризуются следующими призна-
ками: безболезненность; длительное течение;
отсутствие острых воспалительных явлений; ус-
тойчивость к традиционной терапии; другие
симптомы сифилиса; положительные серологи-
ческие реакции.
Третичный период сифилиса
Третичный период сифилиса при классическом
течении инфекции развивается спустя 4“5 лет
с момента заражения., В последние десятилетия
третичный сифилис наблюдается редко. Основ-
ными причинами его появления являются тяже-
лые сопутствующие заболевания, физические и
психические травмы, интоксикации, алкоголизм,
туберкулез, ревматизм, неполноценное питание,
иммунодефицитные состояния. Этот период си-
филиса характеризуется поражением кожи, сли-
зистых оболочек, костей, суставов, нервной си-
стемы, внутренних органов (сердце, легкие,
печень, аорта), органов чувств, глаз.
Сифилиды третичного периода сифилиса
имеют следующие особенности: малозаразитель-
ность; мономорфность и асимметричность; мед-
ленное развитие и регресс; отсутствие островос-
палительных явлений; отсутствие субъективных
ощущений; деструктивность очагов поражения;
разрешение рубцевидной атрофией; изъязвление
с формированием рубцов; лимфаденит не типи-
чен.
Различают активную (манифестную) стадию
третичного сифилиса и скрытую (латентную).
Активная стадия сопровождается явными при-
знаками сифилиса, скрытая характеризуется
наличием остаточных признаков активных
проявлений сифилиса (рубцы, изменения ко-
стей).
При третичном активном сифилисе наблюда-
ются дермальные бугорки (поверхностный узло-
ватый сифилид) либо формируются гиподер-
мальные узлы — гуммы (глубокий узловой
сифилид или подкожная гумма).
Кроме основных форм (бугорковый и гуммоз-
ный сифилиды), к третичным поражениям кожи
относится третичная эритема (поздняя третич-
ная розеола). Она локализуется чаще всего сим-
метрично на туловище в виде бледно-розовых
колец диаметром 5—15 см, не шелушится. При
разрешении может оставаться нежная атрофия
кожи. Третичную розеолу необходимо отличать
от кольцевидной мигрирующей, кольцевидной
и экссудативной многоформной эритем, розово-
го лишая, себороида, хронической трихофитии
взрослых.
Бугорковые сифилиды
Бугорковые сифилиды характеризуются высы-
паниями на ограниченных участках кожи в виде
плотных, синюшно-красных, безболезненных
бугорков, залегающих на разной глубине дермы
и не сливающихся между собой.
Различают следующие клинические разновид-
ности бугоркового сифилида: сгруппированный;
серпигинирующий; диффузный; карликовый.
Сгруппированный
бугорковый сифилид
Сгруппированный бугорковый сифилид локали-
зуется на коже разгибательной поверхности ко-
нечностей, носа, лба, в области лопаток, пояс-
ницы. Бугорки располагаются близко, фокусно,
не сливаются, находятся на разных этапах раз-
вития (эволюционный полиморфизм), неболь-
ших размеров, плотные, красно-синюшного цве-
та, могут рассасываться (оставляя рубцовую
атрофию) или изъязвляться. Язва округлая с
плотным валикообразным краем. На дне язвы
располагается некротический стержень, который
через несколько недель отторгается. Язва выпол-
нена грануляциями, рубцуется. Рубец плотный,
глубокий, звездчатый, на нем не бывает реци-
дивных бугорков.
Глава 38. Сифилис 443
Серпигинирующий
(ползучий) бугорковый сифилид
Серпигинирующий (ползучий) бугорковый си-
филид представлен в виде небольшого очага
слившихся бугорков. Постепенно процесс про-
грессирует по периферии, а в центре регресси-
рует. Одновременно с появлением новых бугор-
ков происходит изъязвление и рубцевание старых
элементов. После заживления формируется мо-
заичный рубец с фестончатыми краями и фокус-
ными рубчиками вблизи. Более свежие рубцы ги-
перпигментированы, а более старые
депигментированы. Новые бугорки никогда не
появляются на рубцах.
Диффузный бугорковый сифилид
Диффузный бугорковый сифилид («площад-
кой») представляет собой очаг слившихся бугор-
ков, образующих сплошную уплотненную бляш-
ку размером с ладонь, темно-красного цвета, с
незначительным шелушением, полициклически-
ми очертаниями. Регресс бляшки происходит «су-
хим» разрешением с формированием рубцовой
атрофии либо изъязвлением с последующим
образованием характерного мозаичного рубца.
Карликовый бугорковый сифилид
Карликовый бугорковый сифилид проявляется
бугорками небольших размеров — от булавочной
головки до конопляного зерна. Они никогда не
изъязвляются, регрессируют «сухим» путем, ос-
тавляют после себя рубцевидную атрофию.
Дифференциальный
диагноз бугоркового сифилида
Бугорковые сифилиды дифференцируют с ту-
беркулезной волчанкой, папулонекротическим
туберкулезом кожи, туберкулоидной лепрой, ба-
зальноклеточной карциномой, кольцевидной
гранулемой, розацеа, саркоидозом, эктиматоз-
ной пиодермией, лейшманиозом кожи, липоид-
ным некробиозом, хронической язвенной пи-
одермией.
При туберкулезной волчанке бугорки мягкой
консистенции, красного цвета с желтоватым от-
тенком. При надавливании на бугорок пугов-
чатым зондом длительно сохраняется ямка (сим-
птом Поспелова), язва легко кровоточит
(«симптом зонда»). При диаскопии имеется по-
ложительный феномен «яблочного желе» (изме-
нение цвета очага до оранжевого). Язва дли-
тельно не заживает, отмечается выраженная
болезненность, нет тенденции к рубцеванию.
Края язвы мягкие, неровные, нависающие. Об-
разующиеся рубцы ровные, гладкие, мягкие, на
них часто появляются новые бугорки. Реакции
Пирке и Манту положительные.
При папулонекротическом туберкулезе кожи
высыпания располагаются симметрично на зад-
ней поверхности верхних и передней поверхно-
сти нижних конечностей. Высыпания обильные
с некрозом в центре, впоследствии образуются
штампованные рубцы. Реакция Манту положи-
тельная. Имеются другие очаги туберкулезного
поражения.
При туберкулоидном типе лепры бугорки
буро-красные, кольцевидно расположены, име-
ют запавший атрофический центр и припод-
нятый валикообразный край. В очагах пораже-
ния отсутствуют температурная, болевая и
тактильная чувствительность, а также потоот-
деление.
Базальноклеточная карцинома преимуществен-
но одиночная, локализована на лице, имеет пе-
риферический валик с наличием плотных блес-
тящих узелков наподобие, жемчужин. В центре
базалиомы отмечается слегка кровоточащая эро-
зия, которая длительно существует, медленно
прогрессирует, не рубцуется.
При кольцевидной гранулеме узелки распола-
гаются преимущественно на коже кистей в виде
колец розоватого или нормального цвета, име-
ется легкая атрофия в центре, узелки редко
изъязвляются. Наблюдается у лиц молодого воз-
раста.
Для розацеа характерны островоспалительные
явления в виде венчика гиперемии по перифе-
рии, папулезные и пустулезные элементы, теле-
ангиэктазии, а также обнаружение клеща
Demodex folliculorum.
Мелкоузелковый саркоидоз Бенье-Бека-Шумана
проявляется мелкими множественными, плотны-
ми узелками красно-коричневого цвета. При
диаскопии выявляются мелкие желто-золотис-
тые пылевые точки в виде песчинок. Положи-
тельная внутрикожная проба с антйгеном из
пораженной ткани.
Эктиматозная пиодермия сопровождается ос-
трыми воспалительными явлениями и болезнен-
ностью, развитием лимфаденита и лимфангита,
а также фликтен и фолликулитов.
При лейшманиозе бугорки островоспалительные,
ярко-красного или синюшного цвета, мягкой кон-
систенции. Вокруг бугорков или изъязвлений об-
наруживаются лимфангиты, болезненные при
пальпации. В отделяемом язв находят тельца Бо-
ровского.
444") Венерология
Липоидный некробиоз характерен для больных
сахарным диабетом, сопровождается зудом. Оча-
ги поражения в виде пятнистых или узелковых
элементов розового и красно-синюшного цвета.
Узелки сливаются и образуют бляшки плотной
консистенции с гладкой поверхностью, в цент-
ре которых формируется атрофия и возникают
телеангиэктазии. Бляшки могут изъязвляться и
сопровождаться болезненностью.
Хроническая язвенная пиодермия отличается
значительной болезненностью, мягкими, под-
рытыми и кровоточащими краями язв, дно ко-
торых покрыто гнойным отделяемым и вялы-
ми грануляциями. Серологические реакции на
сифилис и пробное лечение препарами йода или
солей висмута помогут в диагностике заболева-
ния.
Гуммозные сифилиды
Гуммозные сифилиды в настоящее время встре-
чаются редко. Они проявляются отдельными
узлами или разлитой гуммозной инфильтраци-
ей. В развитии гуммозного сифилида выделяют
несколько периодов: образование и рост, размяг-
чение, спаивание с кожей, изъязвление, расплав-
ление и отторжение гуммозного стержня с пос-
ледующим рубцеванием. Возможно также
«сухое» разрешение с рубцевидной атрофией.
Преимущественная локализация гумм — голени,
верхние конечности, реже поражаются грудь,
голова, живот, спина.
Различают следующие клинические формы
гуммозного сифилида: изолированные гуммы; гум-
мозные инфильтраты; фиброзные гуммы («около-
суставные узловатости»).
Изолированные гуммы
Изолированные гуммы возникают в подкожной
основе и представляют собой плотноэластичес-
кие, четко отграниченные, безболезненные, не
спаянные с кожей шаровидные узлы. Постепенно
узлы увеличиваются до размеров лесного ореха,
а иногда куриного яйца, спаиваются с окружа-
ющими тканями и кожей, которая постепенно
краснеет и размягчается. В центре гуммы появ-
ляется флюктуация и гумма вскрывается, выде-
ляется тягучая жидкость грязно-желтого цвета.
Происходит некроз и отторжение гуммозного
стержня с образованием гуммозной язвы. Язва
с плотными, отвесными, валикообразными кра-
ями, четкими границами. После отторжения гум-
мозного стержня дно язвы заполняется грануля-
циями, затем наступает рубцевание. Рубец
гиперпигментированный, глубокий, втянутый в
центре, звездчатый, плотный. Эволюция гуммы
длится от нескольких недель до нескольких
месяцев.
Гуммозные инфильтраты
Гуммозные инфильтрации возникают в резуль-
тате слияния нескольких гумм и представляют
собой очаги с резко отграниченными краями.
Образуется обширная язва с полициклическими
краями, которая заживает рубцом. Характерна
стадийность развития.
Фиброзные гуммы
Фиброзные гуммы («околосуставные узловато-
сти») образуются вследствие перерождения си-
филитических гумм, когда гуммозный инфиль-
трат замещается соединительной тканью и
подвергается фиброзу с последующим отложени-
ем солей кальция. Фиброзные гуммы чаще оди-
ночные и локализуются на передней и задней по-
верхности локтевых и коленных суставов в виде
подвижных, безболезненных образований шаро-
видной формы, хрящевой консистенции. Око-
лосуставные узловатости не изъязвляются.
Дифференциальный диагноз
гуммозного сифилида
Гуммозный сифилид следует дифференциро-
вать с колликвативным туберкулезом кожи,
индуративным туберкулезом кожи, эктимой
обыкновенной, эктимой сифилитической, хро-
нической язвенной пиодермией, плоскоклеточ-
ным раком, базальноклеточным раком, лепрой,
лейшманиозом, узловатой эритемой, узловатым
аллергическим васкулитом, узловатым паннику-
литом, фибролипомой, атеромой.
Колликвативный туберкулез кожи (скрофуло-
дерма) представлен множественными подкожны-
ми узлами по ходу лимфатических сосудов, ко-
торые постепенно сливаются, спаиваются с
кожей и вскрываются с образованием язв. Язвы
неправильной формы с мягкими подрытыми
краями. Содержимое язв гнойное с примесью
крови и творожистых масс. Дно язвы покрыто
вялыми грануляциями, легко кровоточит. Обра-
зуются мягкие рубцы с сосочками по краям и
«мостиками». Проба Манту положительная. Се-
рологические реакции на сифилис отрицатель-
ные. Скрофулодермой болеют преимуществен-
но в детском и юношеском возрасте.
Индуративный туберкулез кожи чаще наблю-
дается у женщин. Плотные и слегка болезнен-
Глава 38. Сифилис 445
ные, спаянные с кожей узлы расположены на
голенях. Над очагами кожа синюшно-красная,
иногда узлы изъязвляются. Дно язвы покрыто
грязно-серыми грануляциями. Течение длитель-
ное, обострения наступают в холодное время
года. Реакция Манту положительная. Серологи-
ческие реакции РИТ, РИФ отрицательные.
Эктима обыкновенная сопровождается остро-
воспалительными явлениями, болезненностью,
гиперемией кожи вокруг очага, отсутствием
плотного валика вокруг язвы и наличием фол-
ликулитов, лимфаденита и лимфангита.
Эктима сифилитическая иногда сопровожда-
ется нарушением общего состояния в виде сла-
бости, утомляемости, головной боли, снижения
аппетита, а также отличается от гуммозного си-
филида мощной коркой серо-бурого цвета.
Хроническая язвенная пиодермия начинается
остро, с импетиго, фолликулита или эктимы.
Впоследствии образуются мягкие, болезненные
язвы, с подрытыми краями и гнойным отделяе-
мым, с вялым хроническим течением.
Плоскоклеточный рак характеризуется язвой
неправильной формы, с плотными «выворочен-
ными» краями, неровным, бугристым дном, от-
сутствием гуммозного стержня, деревянистым
уплотнением у основания.
Базальноклеточный рак имеет валикообразный
край, пронизанный сосудами и содержащий бле-
стящие жемчужные узелки. Дно язвы гладкое,
красного мясного цвета.
Лепромы имеют застойно-красный цвет с бу-
рым оттенком, гладкую блестящую поверхность,
редко распадаются. Отмечается нарушение тем-
пературной, болевой и тактильной чувствитель-
ности. Обнаружение микобактерий лепры в оча-
гах поражения и специфический лепроматозный
инфильтрат при гистологическом исследовании
подтверждают диагноз.
При лейшманиозе кожи имеются характерные
островоспалительные явления с лимфангитами и
лимфаденитами. Пребывание больного в энде-
мичных по лейшманиозу зонах и обнаружение
возбудителя из язв помогают установить диагноз.
Узловатая эритема сопровождается выражен-
ной болезненностью и возникновением в под-
кожной клетчатке узлов с измененным над ними
цветом кожи от ярко-красного до фиолетового;
изъязвлений не наблюдается.
Васкулит узловатый аллергический характери-
зуется развитием болезненных узлов мягкой
консистенции с гиперемированной кожей над
ними. Высыпания сопровождаются общей сла-
бостью, субфебрильной температурой, лейкоци-
тозом, эозинофилией.
При узловатом панникулите гиподермальные
узлы имеют плотную консистенцию с последу-
ющим формированием атрофических участков.
Заболевание отличается хроническим рецидиви-
рующим течением. У больных отмечается нару-
шение общего состояния в виде суставных и
мышечных болей, повышения температуры тела.
Фибролипомы представлены множественными
подкожными мягкими узлами с дольчатым стро-
ением и очень медленным увеличением в разме-
рах; кожа над ними не изменена.
Атеромы представляют собой медленно про-
грессирующую кисту сальной железы плотноэла-
стической консистенции, с четкими границами,
нагнаивающуюся, с творожистым зловонным
содержимым.
Поражения
слизистых оболочек
Поражения слизистых оболочек в третичном пе-
риоде сифилиса встречаются довольно часто. Спе-
цифические изменения наблюдаются на мягком и
твердом небе, слизистой оболочке носа, на задней
стенке глотки и языке. Сифилиды слизистых обо-
лочек отличаются ярким цветом, отечностью
вследствие выраженных экссудативных явлений.
На мягком и твердом небе могут располагаться
гуммозные и бугорковые высыпания. Они
изъязвляются, приводят к разрушению тканей,
иногда к отторжению язычка, а после рубцева-
ния — к деформации мягкого неба.
Гуммы слизистой оболочки носа развиваются
первично или иррадиируют с прилежащих уча-
стков, чаще располагаются в области перегород-
ки, на границе хрящевой и костной частей.
Субъективные ощущения отсутствуют. После
образования язвы слизь из носа становится гной-
ной. При исследовании зондом на дне язвы оп-
ределяется шероховатая поверхность некротизи-
рованной кости. Может произойти разрушение
носовой перегородки и наступает деформация
носа («седловидный нос»).
Третичные сифилиды гортани могут вызывать
перихондриты, поражения голосовых связок
(осиплость, охриплость, афония), кашель с гус-
той грязно-желтой слизью. В результате рубце-
вания язв происходит неполное смыкание голо-
совых связок, и голос навсегда становится
сиплым.
Третичные сифилиды глотки представлены в
виде мелких гуммозных узлов или разлитой гум-
мозной инфильтрации, которые в результате их
изъязвления вызывают боль и функциональные
расстройства.
446 Венерология
Третичные сифилиды языка наблюдаются в
виде ограниченных гумм или диффузных гум-
мозных инфильтраций. В толще языка образу-
ются несколько гумм размером с лесной орех,
которые затем изъязвляются. При диффузном
гуммозном глоссите язык резко увеличен в объе-
ме, плотный, красно-синюшный, легко травми-
руется, малоподвижный, складки сглажены.
После рассасывания инфильтрата язык сморщи-
вается, искривляется, теряет подвижность.
Поражение костей и суставов
Поражение костей и суставов в третичном пери-
оде сифилиса наблюдается часто. Проявляется
это ночными болями разной интенсивности. В
третичном периоде сифилиса чаще страдают ко-
сти голеней, черепа, грудина, ключица, локте-
вая кость, носовые косточки.
Гуммозный периостит проявляется припухло-
стью, болезненностью при надавливании. На
рентгеновских снимках видно сочетание остео-
пороза с остеосклерозом. По краю припухлости
прощупывается плотный костный валик. При
распаде инфильтрата кожа краснеет, образуется
типичная язва с возвышенными, плотными и
отвесно опускающимися на дно краями. После
отхождения гуммозного стержня обнажается
неровная поверхность некротизированной кос-
ти, которая потом секвестрируется. Заживление
происходит глубоким, спаянным с костью, не-
подвижным рубцом.
Гуммозный остит вызывает боль, усиливаю-
щуюся ночью; болезненна даже легкая перкус-
сия. После рассасывания инфильтрата поверх-
ность кости становится неровной. Если он
расплавляется, то образуются большие секвест-
ры с неприятным запахом. При поражении ко-
сти может возникать специфический остеомие-
лит, а при распространении процесса из глубины
кнаружи — остеопериостит с образованием гум-
мозной язвы, дном которой служит омертвевшая
кость. Заживление наступает только после её от-
хождения.
Синовиты наблюдаются преимущественно в
коленных, локтевых, голеностопных и грудино-
ключичных суставах. Припухание, серозный
выпот, ограниченность функции выражены не
очень сильно. При хронических гуммозных си-
новитах имеется сплошное или узловатое утол-
щение синовиальной оболочки. Гуммозный ин-
фильтрат распадается крайне редко, обычно он
замещается рубцовой тканью. При остеоартри-
тах наблюдается сочетание гумм в эпифизах и
хрящах с переходом процесса на синовиальную
оболочку. Поражение развивается медленно, без
лихорадки, иногда с небольшим выпотом в сус-
тавную сумку. Функция суставов обычно не
нарушается.
Скрытый сифилис
Сифилис скрытый (Syphilis latens) —- хроничес-
кое венерическое заболевание, вызываемое блед-
ной трепонемой, характеризующееся положи-
тельными серологическими реакциями на
сифилис и отсутствием признаков поражения
кожи, слизистых оболочек, внутренних органов,
опорно-двигательного аппарата и нервной сис-
темы, а также анамнестических данных о пере-
несенном в прошлом сифилисе.
При скрытом сифилисе заболевание протека-
ет латентно с момента заражения, без клиничес-
ких проявлений, с положительными серологи-
ческими реакциями в крови.
Различают ранний (Syphilis latens ргаесох) и
поздний (Syphilis latens tarda) скрытый сифилис.
Ранний скрытый сифилис — приобретенный
сифилис с давностью инфекции до 2 лет, без
клинических проявлений, с положительными
серологическими реакциями.
Поздний скрытый сифилис — приобретенный
сифилис с давностью инфекции свыше 2 лет, без
клинических проявлений, с положительными
серологическими реакциями в крови и нормаль-
ной спинномозговой жидкостью.
Когда невозможно различить ранний и по-
здний сифилис, ставят диагноз неуточненного си-
филиса (lues latens ignorata), который следует рас-
сматривать как предварительный диагноз,
подлежащий уточнению в процессе лечения и на-
блюдения.
Диагностические признаки
раннего скрытого сифилиса
В анамнезе имеются указания на наличие в те-
чение последних двух лет эрозивно-язвенных
высыпаний на гениталиях, сыпи на коже туло-
вища, выпадения волос, прием антибиотиков по
поводу гонореи и других заболеваний.
Данные конфронтации — обнаружение у по-
лового партнера активного или раннего скрытого
сифилиса.
Данные клинического осмотра — обнаружение
рубца или атрофии кожи, уплотнения на месте
бывшей первичной сифиломы, увеличение па-
ховых лимфатических узлов.
Результаты серологического исследования —
положительные стандартные серологические ре-
акции в высоком титре (1 : 120, 1: 160, 1 : 320)
Глава 38. Сифилис 447
у 2/3 больных, положительная РИФ у всех боль-
ных, положительная РИТ у 1/3 больных.
Реакция обострения (Герксгеймера-Яриша-
Лукашевича) на введение пенициллина может
наблюдаться у 30 % больных.
Относительно быстрая динамика негативации
стандартных серологических реакций в процес-
се лечения.
Ликвор в большинстве случаев нормальный.
При патологическом ликворе преобладает воспа-
лительный компонент (белок, цитоз, глобулино-
вые реакции).
Возраст больных — моложе 40 лет.
Диагностические признаки
позднего скрытого сифилиса
Отсутствие анамнестических данных. Иногда
имеются сообщения о вероятном заражении 2—
3 года назад.
При конфронтации у половых партнеров об-
наруживают поздний сифилис, или у них от-
сутствуют проявления сифилитической инфек-
ции.
При клиническом осмотре на коже и видимых
слизистых оболочках отсутствуют следы разре-
шившихся сифилидов.
Стандартные серологические реакции положи-
тельны в низком титре (1:5, 1: 10, 1: 20, 1: 50)
у 90 % больных. РИФ и РИТ положительны у
всех больных.
Реакция обострения на введение пеницилли-
на не наблюдается.
Динамика негативации стандартных серологи-
ческих реакций в процессе лечения очень мед-
ленная.
Патология ликвора встречается редко. В лик-
воре преобладает дегенеративный компонент
(положительная реакция Вассермана, патологи-
ческая реакция Ланге).
Возраст больных — преимущественно старше
40 лет.
Нейросифилис
В развитии нейросифилиса основную роль иг-
рают отсутствие или недостаточность предше-
ствующего противосифилитического лечения,
черепно-мозговая травма, хронические инфек-
ции, интоксикации, иммунные нарушения. Клас-
сификация поражений нервной системы при
сифилисе несовершенна. С клинической точки
зрения целесообразно выделять сифилис цент-
ральной нервной системы, сифилис перифери-
ческой нервной системы, функциональные не-
рвные и психические явления при сифилисе.
В настоящее время для практических целей
применяют клинико-морфологическую класси-
фикацию. Выделяют ранние и поздние формы
сифилиса нервной системы. Такое подразделение
нейросифилиса условно. Различие между ними
основано на характере патологических изменений
в нервной системе и хронологически не совпадает
с обычной периодизацией сифилиса.
Ранний нейросифилис
Ранний нейросифилис (до 5 лет от момента за-
ражения) называют мезенхимным, так как пора-
жаются оболочки и сосуды мозга, преобладает
мезенхимная реакция.
Различают следующие формы раннего нейро-
сифилиса: скрытый латентный сифилитический
менингит; острый генерализованный сифилити-
ческий менингит; менингоневротическая форма
сифилитического менингита; сифилитическая гид-
роцефалия; ранний менинговаскулярный сифилис;
сифилитический менингомиелит.
Скрытый сифилитический
клинически бессимптомный менингит
Скрытый сифилитический клинически бессимптом-
ный менингит развивается в результате раздраже-
ния оболочек головного мозга либо непосредственно
бледной трепонемой, либо продуктами ее жизнеде-
ятельности. Эта форма раннего нейросифилиса
названа скрытым ранним менингитом потому, что
у большинства больных клинические симптомы
менингита обнаружить не удается. Некоторые боль-
ные могут жаловаться на головную боль («тяжелая
голова»), шум в ушах, снижение слуха и головок-
ружение. Диагноз ставится на основании патоло-
гических изменений ликвора — от значительных
до минимальных. Показателями минимальной па-
тологии считаются белок — 0,4%о, цитоз — 8 кле-
ток в 1 мм3, глобулиновые реакции (Нонне-Апель-
та: ++), реакция Ланге больше двух двоек и
положительная реакция Вассермана.
Острый генерализованный
сифилитический менингит
Острый генерализованный сифилитический ме-
нингит встречается довольно редко. В процесс
вовлекаются все оболочки мозга. Заболевание со-
провождается усиливающейся головной болью,
головокружением, шумом в ушах, рвотой (рво-
та не связана с приемом пищи ~ «мозговая рво-
та»). Объективно отмечаются ригидность затыл-
448 Венерология
ка, положительный симптом Кернита, положи-
тельный нижний симптом Брудзинского. Реги-
стрируются патологические рефлексы Бабинско-
го, Оппенгейма, Россолимо. Ликвор резко
изменен: белок до 1,2%о, цитоз от 200 до 1000
клеток в 1 мм3, реакции Нонне-Апельта и Пан-
ди ++++, реакция Ланге в виде паралитической
или менингитической кривой.
Менингоневротическая форма
сифилитического менингита
Менингоневротическая форма сифилитического
менингита (базальный менингит) характеризует-
ся поражением отдельных участков оболочек го-
ловного мозга, чаще в области основания мозга.
Клиника этого поражения слагается из симпто-
мов менингита и неврита. Кроме головной боли
и головокружения, изредка возникают тошнота
и рвота. Симптомы Кернига, Брудзинского, па-
тологические рефлексы отсутствуют. У больных
отмечается легкий птоз, асимметрия лица, сгла-
женность носогубной складки, отклонение язы-
ка, опущение мягкого неба. В ликворе патология
умеренно выражена: белок до 0,6—0,7%о, цитоз
20—40 клеток в 1 мм3, реакции Нонне-Апельта и
Панди положительные или резко положительные;
реакция Ланге имеет вид менингитической кри-
вой; реакция Вассермана в ликворе положитель-
ная или резко положительная.
Сифилитическая гидроцефалия
Сифилитическая гидроцефалия возникает в слу-
чаях ограниченного воспалительного поражения
мозговых оболочек. Заболевание проявляется
нарастающей головной болью, головокружени-
ем, тошнотой, неукротимой рвотой, спутаннос-
тью сознания. Иногда присоединяются эпилеп-
тиформные припадки, нарушения речи.
Характерным симптомом гидроцефалии является
улучшение общего состояния больного (и голов-
ной боли) после люмбальной пункции. В лик-
воре определяется белково-клеточная диссоци-
ация: много белка (1—2%о) при небольшом
количестве клеток (10—15 в 1 мм3); реакции Нон-
не-Апельта и Панди положительные; реакция
Вассермана в ликворе положительная.
Ранний менинговаскулярный
сифилис
Ранний менинговаскулярный сифилис характе-
ризуется умеренным вовлечением в процесс моз-
говых оболочек. Клиническая картина при этом
разнообразна: головная боль, поражение череп-
ных нервов, нарушение чувствительности, геми-
парезы, альтернирующие параличи, эпилепти-
формные припадки, нарушение памяти, афазия.
Сифилитический менингомиелит
Сифилитический менингомиелит (воспаление мяг-
ких мозговых оболочек и специфический эндарте-
риит спинного мозга) возникает у нелечившихся или
недостаточно лечившихся больных ранним сифили-
сом. Заболевание возникает «внезапно», имеет доволь-
но острое течение и приводит к параплегии нижних
конечностей с глубоким нарушением трофики, сни-
жением или потерей различных видов чувствитель-
ности, нарушением сфинктеров. Диагноз помогают
установить положительные серологические реакции
крови (РВ, РИТ, РИФ), и патология ликвора (РВ
в нем всегда положительная).
Поздний нейросифилис
Поздний нейросифилис (не ранее 6—8 лет после
заражения) называют паренхиматозным в связи
с поражением нейронов, нервных клеток, а также
нейроглии. Изменения носят воспалительно-ди-
строфический характер, мезенхимная реакция не
выражена.
Выделяют следующие формы позднего нейро-
сифилиса: поздний скрытый сифилитический ме-
нингит; поздний диффузный менинговаскулярный
сифилис; сифилис сосудов мозга (васкулярный си-
филис); сухотка спинного мозга (спинная сухотка,
табес); прогрессивный паралич; гумма мозга.
Поздний скрытый (латентный)
сифилитический менингит
Поздний скрытый (латентный) сифилитический
менингит возникает и диагностируется через
много лет после инфицирования сифилисом.
Больные не предъявляют жалоб или иногда от-
мечают нерезкую головную боль, шум в ушах,
снижение слуха и головокружение. Диагности-
ка базируется на патологических изменениях в
ликворе, где воспалительный компонент выра-
жен незначительно. Реакция Вассермана и реак-
ция Ланге, как правило, патологические.
Поздний диффузный
менинговаскулярный сифилис
Поздний диффузный менинговаскулярный сифи-
лис характеризуется умеренным вовлечением в
Глава 38. Сифилис 449
процесс мозговых оболочек. Больные жалуются на
несильную головную боль, реже — на головокру-
жение. Симптомы болезни часто напоминают ги-
пертонический криз. Могут возникнуть наруше-
ния чувствительности, парестезии, гемипарезы,
эпилептиформные припадки, альтернирующие
параличи, расстройства речи и памяти. Серологи-
ческие реакции крови положительны у 50 % боль-
ных. Возможны патологические изменения лик-
вора — невысокое содержание белка (до
0,6~0,7%о), незначительный цитоз (до 20—30 лим-
фоцитов в 1 мм3), слабоположительные глобули-
новые реакции, реакция Вассермана отрицатель-
ная. Нелеченный менинговаскулярный сифилис
может трансформироваться в спинную сухотку.
Сифилис сосудов мозга
(васкулярный сифилис)
Сифилис сосудов мозга (васкулярный сифилис)
характеризуется вовлечением в процесс только
сосудов мозга. Ликвор не изменяется в связи с
отсутствием явлений менингита и поражения
серого вещества мозга. Стандартные серологичес-
кие реакции крови в 60—70 % случаев отрица-
тельны, что затрудняет диагностику. При диаг-
ностике следует учитывать относительную
доброкачественность в начальном периоде забо-
левания, периодичность течения, «разбросан-
ность» симптомов, положительные классические
серологические реакции крови у 30—40 % боль-
ных и, как правило, положительные РИТ и
РИФ. Имеются неврологические расстройства
(моно-, пара-, геми- и диплегии, гемипарезы,
параличи, афазии, нарушения чувствительности,
патологические рефлексы). Могут возникнуть
различные расстройства психической сферы,
эпилептические припадки по типу джексоновс-
кой эпилепсии или общих судорог.
Сухотка спинного мозга
Сухотка спинного мозга (tabes dorsalis), спинная
сухотка, табес. При спинной сухотке пораже-
ние локализуется в задних корешках, задних
столбах и оболочках спинного мозга.
Клинические симптомы сухотки спинного мозга:
• боли стреляющие, кинжальные, сверлящие
и рвущие;
• парестезии — чувство опоясывания, сжатия,
сдавления на определенных уровнях туловища;
чувство онемения, «ползания мурашек», покалы-
вание в ногах;
• расстройство мочеиспускания, дефекации и
появление импотенции;
• парезы черепных нервов и как результат —
птоз, косоглазие, девиация языка, асимметрия
лица;
• зрачковые расстройства — анизокория (из-
менение формы и величины зрачков), миоз (су-
жение зрачков), мидриаз (расширение зрачков),
патогномоничный симптом Аргайлла-Робертсо-
на (отсутствие реакции зрачков на свет или очень
вялая реакция);
• костно-воздушная диссоциация (костная
проводимость снижается, воздушная проводи-
мость не изменяется);
• расстройство координации (атаксия) — по-
явление характерной атаксической походки:
больной сначала становится на пятки, затем на
всю стопу;
• пошатывание в позе Ромберга; нарушения
при пальценосовой и пяточно-коленной пробах;
• расстройства глубокого мышечно-суставно-
го чувства;
• отсутствие коленных и ахилловых рефлек-
сов;
• нарушение трофики, холодовая гиперестезия
в области спины, болевая гипестезия в области
груди.
Стандартные серологические реакции у 25—
50 % больных спинной сухоткой отрицательные.
РИТ положительна более чем в 95 % случаев. У
50 % больных спинной сухоткой спинномозго-
вая жидкость находится в пределах нормы. При
патологических изменениях в ней отмечаются не-
значительное увеличение количества белка (до
0,45—0,55%©), повышение числа лимфоцитов
(20—30 в 1 мм3), положительные глобулиновые
реакции (+++ или ++++) и реакция Вассерма-
на; реакция Ланге имеет характер паралитичес-
кой кривой.
Прогрессивный паралич
Прогрессивный паралич проявляется через 20—
40 лет после заражения у недостаточно или во-
обще нелечившихся по поводу сифилиса. В ос-
нове развития прогрессивного паралича лежит
поражение вещества больших полушарий голов-
ного мозга, чаще в области коры передней доли,
возникающее в связи с воспалительными изме-
нениями капилляров мозга. В острый период за-
болевания отмечаются полный распад личности,
деградация, прогрессирующее слабоумие, различ-
ные формы бреда, галлюцинации, кахексия.
В фазе законченного развития процесса раз-
личают 4 формы прогрессивного паралича: де-
ментная; экспансивная; ажитированная; депрес-
сивная.
450 Венерология
При дементной форме больной теряет инте-
рес к окружающему, быстро наступают апатия,
отупение и прогрессирующее слабоумие. Экс-
пансивная форма характеризуется эйфорией,
бредом величия (мегаломания), психомоторной
активностью с постепенным распадом психики.
У больных ажитированной формой преоблада-
ет возбуждение и склонность к разрушительным
поступкам. Для депрессивной формы характер-
ны подавленность, тревога, ипохондрический
бред.
Клиническая картина прогрессивного парали-
ча слагается из нарушений психической сферы,
различных неврологических симптомов и дан-
ных лабораторного обследования.
К ранним психическим симптомам относятся
следующие:
• изменение личности — вспыльчивость, про-
жорливость, эйфория или вялость, плаксивость,
необоснованное благодушие;
• нарушение памяти — ее снижение на бли-
жайшие события при сохранении на отдельные;
• нарушение счета — связано с нарушением
психики, страдает логика счета;
• нарушение письма — больной пропускает
буквы, слоги, слова;
• нарушение речи — быстрая скороговорка
или, наоборот, замедленная речь; дизартрия —
«спотыкание» на слогах.
К неврологическим симптомам относятся
зрачковые расстройства, симптом Аргайлла-Ро-
бертсона (в 12—20 % случаев), нарушения чув-
ствительности и двигательной сферы, эпилепти-
формные припадки, анизорефлексия (разница в
рефлексах на обеих половинах тела).
Заболевание подтверждается положительными
серологическими реакциями (в 95—97 % случа-
ев), положительными РИТ и РИФ (в 90—94 %
случаев) и изменением спинномозговой жидко-
сти (в 100 % случаев) — реакция Ланге дает
характерную паралитическую кривую.
При наличии у больного симптомов прогрес-
сивного паралича и спинной сухотки ставится
диагноз табопаралича.
Гумма головного
или спинного мозга
Гумма головного или спинного мозга встреча-
ется редко. Обычно она локализуется на выпук-
лой поверхности или в области основания по-
лушарий большого мозга. Неврологическая
картина характеризуется очаговой симптомати-
кой в сочетании с головной болью и повыше-
нием внутричерепного давления. При гумме
спинного мозга могут развиться симптомы пол-
ного его поперечного поражения. В спинномоз-
говой жидкости отмечаются белково-клеточная
диссоциация (повышенное содержание белка
при низком уровне лимфоцитов), положитель-
ные реакции Вассермана, РИТ и РИФ. Кроме
того, РИТ и РИФ положительны в сыворотке
крови.
Профилактикой поздних форм нейросифили-
са является полноценное лечение ранних форм
сифилиса, а также исследование спинномозговой
жидкости при снятии больных с учета. Спинно-
мозговую жидкость необходимо исследовать у
всех больных с клинически выраженными сим-
птомами нейросифилиса с целью диагностики и
для определения качества лечения.
Висцеросифилис
Этим термином обозначают сифилис, при кото-
ром наблюдается поражение внутренних органов.
Наиболее часто сифилитические поражения раз-
виваются во внутренних органах с наибольшей
функциональной нагрузкой (сердце, легкие,
печень, желудок).
Различают ранние и поздние формы висцераль-
ного сифилиса.
Ранние формы
висцерального сифилиса
Ранние формы висцерального сифилиса наблю-
даются в раннем периоде заболевания и при этом
нарушается только функция пораженных орга-
нов. У некоторых больных могут наблюдаться
воспалительные и дегенеративные поражения
внутренних органов.
Для раннего висцерального сифилиса харак-
терны поражения следующих органов и сис-
тем.
Поражения миокарда сопровождаются токси-
ко-инфекционной дистрофией. Отмечаются
функциональные расстройства со стороны сер-
дца (сердцебиение, приглушение тонов).
Изменения показателей функции печени ха-
рактеризуются нарушением протромбино- и про-
теинообразования, реже — пигментной функ-
ции. Могут возникать безжелтушные формы
гепатита с одновременным увеличением уплот-
ненной печени и селезенки.
Поражения желудка регистрируются в виде
сифилитического гастрита и специфической
язвы желудка. Иногда сифилитические пораже-
ния симулируют злокачественную опухоль же-
лудка.
Глава 38. Сифилис 451
Сифилитический гломерулонефрит протекает
доброкачественно, не сопровождается отеками и
повышением артериального давления. Наруше-
ние фильтрирующей способности почек может
проявляться альбуминурией, гематурией.
Диагноз раннего висцерального сифилиса пред-
полагается у больных с острым процессом в том
или ином органе, развитие которого совпадает по
времени с активными проявлениями первично-
го и вторичного сифилиса.
Поздние формы
висцерального сифилиса
Поздние формы висцерального сифилиса наблю-
даются в позднем периоде заболевания. Изме-
нения во внутренних органах сопровождаются
очаговыми деструктивными поражениями, обна-
руживается специфическое гранулематозное вос-
паление.
В настоящее время частота сифилитических по-
ражений при позднем висцеральном сифилисе
выглядит следующим образом:
• 90-94 % случаев — кардиоваскулярный
сифилис;
• 6-4 % случаев — различные поздние пора-
жения печени;
• 1—2% — все остальные поздние висцераль-
ные сифилитические поражения (легкие, почки,
селезенка, желудок).
Поражения
сердечно-сосудистой системы
Поражения сердечно-сосудистой системы (кар-
диоваскулярный сифилис): сифилитический аор-
тит неосложненный; сифилитический аортит, ос-
ложненный стенозом устьев венечных артерий и
недостаточностью клапанов аорты; сифилитичес-
кая аневризма аорты — осложнение сифилити-
ческого аортита; сифилитический миокардит.
Сифилитический неосложненный аортит дли-
тельное время протекает без субъективных ощу-
щений. Одним из ранних симптомов является
постоянная загрудинная боль, давящего или жгу-
чего характера, неинтенсивная, появляется в ноч-
ное время. Характерны расширение восходящего
отдела аорты, мягкий, нерезко выраженный си-
столический шум и акцент II тона у ее устья. В
диагностике аортита имеет значение рентгеноло-
гически определяемое ограниченное расширение
восходящего отдела аорты, а также обнаружение
на рентгенограмме линейного обызвествления по
контуру восходящего отдела аорты.
При сифилитическом аортите в большинстве
случаев поражению подвергаются устья обеих
венечных артерий. Особенность данного процес-
са — медленное развитие, варьирующее от не-
большого сужения до полной облитерации од-
ного или двух устьев. В результате сужения
устьев венечных артерий снижается коронарный
кровоток, что влечет за собой нарушение кро-
воснабжения миокарда. В клинической картине
превалируют симптомы прогрессирующей сер-
дечной недостаточности, что связано с развити-
ем дистрофических и склеротических изменений
в сердечной мышце вследствие прогрессирующе-
го сужения венечных сосудов.
При сифилитической недостаточности аор-
тальных клапанов порок является относительным,
возникающим вследствие расширения поражен-
ной аорты. Недостаточность аортальных клапанов
на ранних этапах протекает бессимтомно. Наибо-
лее характерный признак этого порока — боль в
области сердца типа аорталгии и истинной сте-
нокардии. На аорте одновременно с диастоличес-
ким шумом выслушиваются систолический шум
и звонкий II тон с металлическим оттенком. Важ-
ную роль в диагностике недостаточности аорталь-
ных клапанов играет рентгенологическое иссле-
дование (расширение восходящей части аорты,
увеличение левого желудочка с выраженной его
пульсацией, наличие обызвествления в поражен-
ном отделе аорты).
Клиническая симптоматика сифилитической
аневризмы аорты зависит от локализации и ее
размеров, направления роста, сдавления окружа-
ющих органов, наличия сопутствующего пора-
жения устьев венечных артерий и недостаточно-
сти митральных клапанов.
Основные признаки сифилитической аневриз-
мы аорты:
• пульсирующие участки грудной клетки в
области рукоятки грудины и вправо от нее;
• перкуторное увеличение контуров аорты в
одну или обе стороны от рукоятки грудины;
• разница в пульсе — меньшее наполнение и
запаздывание пульсовой волны на стороне анев-
ризмы;
• своеобразный дующий систолический шум
в зоне сосудистого притупления (выслушивает-
ся иногда самим больным в ночное время);
• симптомы сдавления аневризмой соседних
органов и тканей: верхней полой вены, трахеи,
бронхов, пищевода, нервных стволов;
• симптомы прорыва аневризмы в соседние
органы (в трахею, бронхи, легкие, плевральную
полость, пищевод, средостение, полость пери-
карда);
452 Венерология
• рентгенологически — расширение и отчет-
ливая пульсация сосудистой тени, резко очерчен-
ные и ровные ее края, неотделимость выбухаю-
щей тени от аорты при просвечивании в
различных направлениях.
Сифилис печени
Выделяют 4 формы позднего сифилитического
гепатита: хронический эпителиальный, хроничес-
кий интерстициальный, ограниченный гуммозный
и милиарный гуммозный. Для всех форм харак-
терно длительное течение процесса с постепенным
развитием склерогуммозных изменений, приводя-
щих к циррозу и деформации печени. Диагноз ус-
танавливают на основании анамнеза, специфичес-
ких поражений других внутренних органов,
положительных серологических реакций крови,
результатов противосифилитического лечения.
Сифилис желудка
Сифилис желудка протекает по типу хроничес-
кого гастрита, изолированной гуммы (которая
приводит к язве желудка). Сифилитический
гуммозный инфильтрат в привратнике желудка
вызывает его стеноз. В результате склеротичес-
ких изменений желудок деформируется.
Сифилис легких
Сифилис легких протекает в виде изолирован-
ных гумм или хронической межклеточной сифи-
литической пневмонии. Процесс локализуется в
нижней и средней долях правого легкого. Неред-
ко диагностика проводится только на основании
результатов пробного лечения.
Сифилис почек
Сифилис почек проявляется нефротическим ва-
риантом хронического гломерулонефрита с вы-
раженными отеками, высоким содержанием бел-
ка в моче; появлением гиалиновых, зернистых,
восковидных цилиндров; гипопротеинемией;
гиперхолестеринемией. У некоторых больных
развивается амилоидоз, наблюдаются изолиро-
ванные или диффузные гуммозные поражения.
Основополагающим при диагностике висце-
рального сифилиса является заключение, осно-
ванное на комплексном обследовании внутрен-
них органов и нервной системы. Положительные
серологические реакции в крови и указание в
анамнезе на заболевание сифилисом подтверж-
дают клинический диагноз.
Врожденный сифилис
Врожденный сифилис — это инфекционное за-
болевание, заражение которым происходит внут-
риутробно от больной сифилисом матери. Блед-
ные трепонемы проникают в организм плода
через плаценту, начиная с 10 недели беременно-
сти тремя путями: через пупочную вену; через
лимфатические щели пупочных сосудов; через
поврежденную плаценту с током крови матери.
Согласно принятой классификации, различа-
ют ранний врожденный сифилис (у детей до 2 лет);
ранний врожденный скрытый сифилис (без кли-
нических проявлений с сероположительными
реакциями крови и спинномозговой жидкости);
поздний врожденный сифилис (уточненные либо
проявившиеся признаки через 2 или более лет с
момента рождения); поздний врожденный скры-
тый (без клинических симптомов с положитель-
ными серологическими реакциями и нормаль-
ным составом спинномозговой жидкости).
Ранний врожденный сифилис
Ранний врожденный сифилис диагностируется у
детей до 2 лет и включает: сифилис плода; сифи-
лис детей грудного возраста; сифилис раннего
детского возраста.
Сифилис плода
Сифилис плода заканчивается его гибелью на 6-
7 месяце беременности. Дети, рожденные с ак-
тивными проявлениями сифилиса, оказывают-
ся нежизнеспособными и быстро погибают.
Вероятные клинические признаки сифилиса
плода: плод малого размера, малого веса, с вы-
раженными явлениями кахексии; кожа морщи-
нистая, дряблая, грязно-желтого цвета, мацери-
рованная; новорожденный имеет своеобразный
«старческий вид»; внутренние органы (печень,
селезенка, легкие) увеличены, плотные.
Сифилис детей грудного возраста
Сифилис детей грудного возраста (до 1 года) чаще
всего проявляется в первые 3 месяца жизни и
отличается большим своеобразием, представляет
собой тяжелое заболевание с поражением почти
всех висцеральных органов, центральной нервной
системы, костной системы и специфическими
изменениями кожи и слизистых оболочек.
Вероятные клинические признаки врожденно-
го сифилиса детей грудного возраста: пятнисто-
папулезная сыпь на конечностях, ягодицах, лице;
Глава 38. Сифилис 453
мокнущие папулы вокруг рта и заднего прохо-
да; рубцы Робинсона — Фурнье (беловатые ра-
диальные рубцы вокруг рта); седловидный нос
(вследствие прободения носовой перегородки,
разрушения носовых хрящей и костей); ягоди-
цеобразный череп (развивается в результате ос-
теопериостита лобной и теменных костей и ог-
раниченной гидроцефалии).
Достоверные клинические признаки врожденно-
го сифилиса детей грудного возраста:
• диффузная папулезная инфильтрация (диф-
фузный инфильтрат вокруг рта, на ладонях,
подошвах, ягодицах; вокруг рта образуются глу-
бокие трещины, после которых остаются ради-
альные рубцы);
• сифилитическая пузырчатка (напряженные
пузыри величиной от горошины до вишни с
серозным или серозно-гнойным содержимым на
инфильтрированном фоне в области ладоней и
подошв, нередко на других участках);
• сифилитический ринит (различают три ста-
дии: отек и уплотнение слизистой оболочки носа,
гнойно-кровянистое отделяемое, изъязвление);
• остеохондрит (процесс локализуется в длин-
ных трубчатых костях между эпифизом и диа-
физом, распознается рентгенологически).
Диагноз раннего врожденного сифилиса у
детей грудного возраста должен подтверждаться
серологическими реакциями крови (реакция
Вассермана, РИТ, РИФ), которые при активных
проявлениях заболевания оказываются почти
всегда положительными.
Врожденный сифилис
раннего детского возраста
Врожденный сифилис раннего детского возра-
ста (от 1 года до 2 лет) имеет менее многооб-
разную клиническую симптоматику. На коже
и слизистых оболочках наблюдаются папулез-
ные элементы. Могут отмечаться рубцы Робин-
сона-Фурнье, периоститы, фалангиты, гуммы
костей, орхиты, поражения печени, селезенки.
Поражения нервной системы наблюдаются в
виде умственной отсталости, эпилептиформных
припадков, гидроцефалии, гемиплегии, менин-
гита. В диагностике врожденного сифилиса
раннего детского возраста помогут серологичес-
кие реакции крови, которые у большинства
детей положительные.
Поздний врожденный сифилис
Клиническая симптоматика позднего врожден-
ного сифилиса весьма вариабельна. Выделяют ве-
роятные и патогномоничные (достоверные) при-
знаки позднего врожденного сифилиса.
Вероятные признаки
позднего врожденного сифилиса
Бугорково-язвенные сифилиды и гуммы; ради-
альные рубцы вокруг губ и на подбородке; си-
филитический хориоретинит; ягодицеобразный
череп; «седловидный» нос; «саблевидные» голе-
ни; симметричные синовиты коленных суставов.
Выделяют также дистрофии, которые наблю-
даются не только при позднем врожденном си-
филисе, но и при других заболеваниях. К дист-
рофиям относятся: высокое «готическое» твердое
небо; «инфантильный мизинец»; отсутствие ме-
чевидного отростка грудины; широко расставлен-
ные верхние резцы; «олимпийский» лоб; увели-
чение лобных и теменных бугров; гипертрихоз;
утолщение грудинного конца ключицы (симп-
том Авситидийского).
Обнаружение нескольких дистрофий, сочета-
ние их с одним из патогномоничных признаков
или с несколькими вероятными, с положитель-
ными серологическими реакциями являются
основанием для постановки диагноза позднего
врожденного сифилиса.
Достоверные (патогномоничные)
признаки позднего врожденного
сифилиса (триада Гетчинсона)
• Паренхиматозный кератит (покраснение и
помутнение роговицы, светобоязнь, слезотече-
ние, снижение остроты зрения);
• специфический лабиринтит (шум и звон в
ушах, а затем полная потеря слуха в связи с раз-
витием периостита в костной части лабиринта и
поражения слухового нерва);
• зубы Гетчинсона (дистрофия двух постоян-
ных верхних центральных резцов, которые име-
ют форму долота или отвертки с полулунной
выемкой по режущему краю).
При позднем врожденном сифилисе стандар-
тные серологические реакции положительны у
70—80 % больных. РИТ и РИФ положительны
в 92—100 % случаев.
Лабораторная
диагностика сифилиса
Для установления диагноза необходимы, кро-
ме специальных данных анамнеза и объектив-
ного обследования больного, лабораторные
454 Венерология
методы исследования: бактериоскопическое
исследование; серологические исследования
крови; исследование спинномозговой жидко-
сти.
Сложность клинического течения сифилиса
диктует необходимость сочетанного применения
диагностических тестов. Прямое определение
патогенных Т. pallidum в очагах поражения (пря-
мая визуализация — бактериоскопическое иссле-
дование) позволяет достоверно поставить диаг-
ноз при первичном, вторичном и раннем
врожденном сифилисе.
Бактериоскопическое
исследование
Наиболее убедительным доказательством трепо-
немной природы заболевания является прямая ви-
зуализация трепонем. В настоящее время метод
темнопольной микроскопии для выявления под-
вижных трепонем широко применяется в диаг-
ностике.
При первичном сифилисе на бледную трепо-
нему исследуют отделяемое эрозивно-язвенных
папулезных элементов в области половых орга-
нов, полости рта, в пунктате из регионарных
лимфатических узлов. При вторичном сифили-
се используется материал из различных пораже-
ний кожи и слизистых оболочек. Для диагнос-
тики врожденного сифилиса может быть
использована амниотическая жидкость.
При микроскопическом исследовании бледные
трепонемы имеют вид тонкой спирали с 8—12
равномерными завитками. В темном поле зрения
можно наблюдать поступательное вращение тре-
понемы вокруг оси, ротационное и сгибательное.
Важным признаком является излом в средней
части («приглаженный утюгом»), что отличает
возбудителя сифилиса от других трепонем.
При отрицательных результатах микроскопи-
ческое исследование повторяют либо до обнару-
жения бледной трепонемы, либо до подтверж-
дения диагноза результатами серологического
исследования.
Обнаружение специфических микроорганизмов
с характерной морфологией, подвижностью и ан-
тигенной специфичностью (при исключении та-
ких заболеваний, как пинта и мягкий шанкр)
позволяет поставить диагноз сифилиса при пер-
вичном, вторичном, раннем врожденном, рецидив-
ном сифилисе даже без учета результатов сероло-
гических тестов (Дмитриев Г. А., Брагина Е. Е.,
1996). Следует учитывать, что отсутствие микро-
организмов еще не исключает диагноз сифилиса.
Отсутствие трепонем в типичных очагах пораже-
ния может быть обусловлено длительностью су-
ществования очага, предварительным лечением
пациента или плохой техникой взятия образца или
просмотра материала. При отсутствии возбудите-
лей в очаге поражения следует исключить возмож-
ность других заболеваний (герпес, мягкий шанкр).
Серологические
исследования крови
Серологические исследования имеют важнейшее
значение для диагностики сифилиса, особенно в
латентной и поздней стадиях, а также для оцен-
ки качества проводимого лечения. Однако вслед-
ствие сложности антигенного состава бледной
трепонемы, тело которой содержит два липоид-
ных компонента, протеиновые и полисахаридные
антигены, в сыворотке крови больных отмеча-
ется множественность антител. В различных ста-
диях болезни преобладают то одни, то другие
антитела, относящиеся к разным классам имму-
ноглобулинов. Поэтому в современной венеро-
логии одновременно используется несколько
серологических реакций на сифилис.
Все существующие в настоящее время лабора-
торные серологические тесты можно разделить на
две группы: нетрепонемные тесты, используемые
для отбора; трепонемные подтверждающие тесты.
Нетрепонемные тесты
Нетрепонемные тесты (НТТ) используются в ка-
честве отборочных, они недороги, позволяют
обследовать большое количество больных и не-
обходимы для определения эффективности те-
рапии. Недостатками нетрепонемных тестов яв-
ляются низкая чувствительность при первичном
и третичном сифилисе, снижение чувствитель-
ности при позднем латентном сифилисе, нали-
чие ложноположительных результатов.
Нетрепонемные тесты позволяют обнаружить
в тестируемой сыворотке антитела, реагирующие
с кардиолипин-лецитиновым антигеном. Пер-
вым нетрепонемным тестом был серологический
тест, основанный на реакции фиксации комп-
лемента Борде-Жангу, где в качестве антигена
использовался спиртовой экстракт печени ново-
рожденного, умершего от врожденного сифилиса
(реакция Вассермана).
Все нетрепонемные тесты на сифилис, приме-
няемые в настоящее время, представляют собой
реакции флоккуляции (продукт выпадает в виде
хлопьев), в которых в качестве антигена исполь-
зуются кардиолипин, холестерин и лецитин. Все
Глава 38. Сифилис 455
нетрепонемные тесты направлены на обнаруже-
ние IgG- и IgM-антител к липидам клеточной
стенки трепонем. Вследствие липидной приро-
ды антигена или необычных свойств антител при
этих тестах комплекс антиген-антитело остает-
ся в суспензии, и имеет место флоккуляция, а
не агглютинация или преципитация.
За рубежом (США) используются 4 стандар-
тных нетрепонемных теста в качестве отбороч-
ных реакций на сифилис, среди которых разли-
чают две категории:
• тесты с визуальным определением результатов
реакции — RPR (тест быстрых плазменных реаги-
нов) и TRUST (тест с толуидиновым красным);
• тесты с микроскопическим считыванием
результатов реакции — VDRL и USR (тест оп-
ределения активных реагинов плазмы).
Нетрепонемные тесты с визуальным
определением результатов реакции
RPR (Rapid Plasma Reagins) — тест быстрых
плазменных реагинов и его автоматизированная
модификация — автоматизированный реагино-
вый тест (ART, Automated Reagin Test). Эти те-
сты могут считываться визуально из-за наличия
окрашенных веществ в приготовленном антиге-
не. Тест RPR был разработан для полевых ус-
ловий и сейчас это наиболее распространенный
нетрепонемный тест. Он проводится на пласти-
ковых планшетках с впечатанными кружочками.
При постановке количественной или качествен-
ной модификаций реакции стандартные количе-
ства неразведенной сыворотки или ее серийных
разведений смешиваются с антигеном, содержа-
щим частицы угля. Планшету с антигеном и
сывороткой вращают при 1000 об/мин в течение
8 мин. Антиген не прикрепляется к этим части-
цам, но при наличии в сыворотке антикардио-
липиновых антител происходит флоккуляция
комплексов антиген-антитело, частицы угля как
бы попадают в ловушку в образовавшуюся сеть
и коагулируют. Результаты теста читаются во
влажном месте при ярком свете немедленно пос-
ле проведения реакции. В тесте используется не-
прогретая сыворотка.
TRUST (Toluidin Red Unheated Serum Test) —
тест с толуидиновым красным и непрогретой
сывороткой. В данном тесте стандартные части-
цы азокрасителя толуидинового красного добав-
ляются к основному стабилизированному анти-
гену, причем визуализация выражена лучше, чем
при добавлении частиц угля в RPR. В настоящее
время тест TRUST широко не используется.
Нетрепонемные тесты
с микроскопическим считыванием
результатов реакции
VDRL (по названию лаборатории-разработчика —
Venereal Disease Research Laboratory). В данном
тесте свежеприготовленный антиген и инактиви-
рованная сыворотка смешиваются в стеклянной
лунке механическим вращением. Если в сыворот-
ке присутствуют антитела к кардиолипиновому
антигену, образуются агрегаты в виде коротких
стержневидных структур, выявляемые микроско-
пически. Используется количественная модифи-
кация VDRL до разведений сыворотки 1 : 64.
VDRL является единственным тестом, реко-
мендуемым для исследования спинномозговой
жидкости.
USR (Unheated Serum Reagins) — тест опре-
деления активных реагинов плазмы. Антиген
данного теста немного дороже антигена VDRL,
однако он стабилизирован добавлением холина
хлорида и ЭДТА и не нуждается в ежедневном
приготовлении.
К нетрепонемным тестам относится также ме-
тод непрямого ИФА с кардиолипиновым анти-
геном. В отечественной практике в качестве от-
борочных нетрепонемных тестов применяются
микрореакция с плазмой и инактивированной
сывороткой (МР) и реакция связывания комп-
лемента с кардиолипиновым антигеном (РСК).
Характеристика всех нетрепонемных тестов,
используемых в настоящее время, похожа. Все
они относительно нетрудоемки и просты в ис-
полнении. Чувствительность всех тестов пример-
но такая, как при постановке VDRL (около
70% — при первичном сифилисе и 100% — при
вторичном). Нетрепонемные тесты менее чув-
ствительны при первичном и латентном сифи-
лисе. Антитела, определяемые нетрепонемными
тестами, обычно появляются через 1—4 недели
после формирования шанкра (примерно через 1—
1,5 месяца после инфицирования). У большин-
ства больных нетрепонемные реакции становятся
практически отрицательными в течение перво-
го года после лечения первичного сифилиса.
Трепонемные тесты
Трепонемные тесты (ТТ) обычно не применяют-
ся для отбора, так как они технически сложны и
дороги по сравнению с нетрепонемными. Они
используются для подтверждения результатов
нетрепонемных тестов, особенно при отсутствии
клинически выраженных симптомов заболевания.
Подтверждение диагноза позднего латентного,
456 Венерология
третичного сифилиса при отрицательных резуль-
татах нетрепонемных тестов также проводится с
помощью трепонемных тестов, хотя их чувстви-
тельность уменьшается с течением заболевания.
В будущем, по-видимому, с этой целью будет
использоваться полимеразная цепная реакция.
Развитие методов иммунологии и молекуляр-
ной биологии привело к появлению в течение
последнего десятилетия большого количества
новых тестов, некоторые из них, например, им-
муноферментный анализ (ИФА), прочно заня-
ли место в практике. Такие методики прямого
определения трепонемных антигенов, как по-
лимеразная цепная реакция, метод молекуляр-
ных зондов, основанные на методах молекуляр-
ной биологии, находятся в стадии разработки.
Трепонемные тесты применяются для подтвер-
ждения результатов отборочных реакций, а так-
же в тех случаях, когда скринирующие тесты дают
отрицательные результаты, а клинические данные
позволяют предполагать наличие сифилитической
инфекции. Примерно у 85 % больных после пол-
ноценного лечения сифилиса трепонемные тесты
остаются положительными в течение нескольких
лет, а у некоторых — в течение всей жизни, вслед-
ствие чего эти тесты не рекомендуется исполь-
зовать при контроле излеченности.
В отечественной практике для диагностики
сифилиса применяют комплекс серологических
реакций, включающий стандартные серологичес-
кие реакции: реакцию связывания комплемента;
(реакция Вассермана), реакцию с кардиолипино-
вым антигеном; реакцию с трепонемным антиге-
ном; групповые трепонемные реакции: реакцию
иммунофлюоресценции (РИФ); реакцию иммун-
ного прилипания (РИП); видоспецифические
протеиновые трепонемные реакции: РИТ (реак-
ция иммобилизации бледных трепонем); РИФ-
абс и ее варианты (IgM-FTA-ABS, 19S-IgM-FTA-
ABS); реакцию пассивной гемагглюцинации
бледных трепонем (РПГА).
Реакция Вассермана основана на феномене
связывания комплемента. В постановке реакции
используют как специфические антигены из
бледных трепонем, так и неспецифические ан-
тигены (кардиолипиновый — экстракт из мыш-
цы бычьего сердца). Степень позитивности ре-
акции Вассермана оценивают числом плюсов:
++++ (резко положительная); +++ (положи-
тельная); ++ (слабоположительная); + (сомни-
тельная); — (отрицательная).
Реакция Вассермана (РВ) обязательно приме-
няется в комплексе с двумя осадочными реакци-
ями (Кана и Закс-Витебского). Иммунологичес-
кая сущность этих реакций не отличается от РВ,
но для них готовят более концентрированные ан-
тигены, которые при взаимодействии с реагина-
ми сыворотки дают видимый осадок, оценивае-
мый качественно (++++, +++, ++, +, -).
Комплекс серологических реакций (КСР)
позволяет выявить значительное число больных
сифилисом на разных стадиях заболевания.
Диагноз первичного серонегативного сифилиса
ставят только при отрицательных стандартных
серологических реакциях. Реакция Вассермана
(РВ) становится положительной у 100 % больных
во вторичном серопозитивном периоде сифили-
са. При третичном активном сифилисе КСР по-
ложительны у 65—70 % пациентов. При поздних
формах сифилиса внутренних органов и нервной
системы КСР положительны у 50—80 % больных.
У больных ранним врожденным сифилисом се-
рологические реакции положительны почти в
100 % случаев. При позднем врожденном сифи-
лисе КСР положительны в 75—80 % случаев.
К специфическим серологическим реакциям
относится реакция иммунофлюоресценции (РИФ).
Принцип РИФ основан на выявлении флюоресци-
рующих антител, так как меченые флюорохромом
антитела не теряют способности соединяться с со-
ответствующим антигеном и тем самым обуслов-
ливают свечение препаратов в сине-фиолетовых
лучах ртутно-кварцевой лампы. Отличительной
чертой РИФ, по сравнению со стандартными се-
рологическими реакциями, является более высокая
чувствительность (РИФ бывает положительной уже
в первичном серонегативном периоде сифилиса) при
высокой специфичности. Отмечена ценность РИФ
при выявлении больных со скрытыми и поздни-
ми формами сифилиса при отрицательных резуль-
татах РИТ. Реакция ставится в нескольких моди-
фикациях и наиболее ценной является РИФ-200,
основным назначением которой считается не ран-
няя серодиагностика сифилиса, а распознавание
неспецифических результатов КСР и серодиагно-
стика скрытого сифилиса. Однако РИФ-200 не
обладает абсолютной специфичностью, поэтому
вопрос о наличии или отсутствии заболевания дол-
жен решаться по совокупности клинических и дру-
гих серологических данных. РИФ при вторичном
сифилисе положительна в 100 % случаев. При тре-
тичном активном сифилисе РИФ положительна у
92—100 % пациентов. При поздних формах сифи-
лиса внутренних органов и нейросифилиса РИФ
положительна у 94—100 % больных. У больных
ранним и поздним врожденным сифилисом РИФ
положительна в 100 % случаев. РИФ очень мед-
ленно негативируется в процессе противосифили-
тической терапии. Вследствие этого она непригод-
на для контроля за ходом лечения.
Глава 38. Сифилис
457
Реакция иммунного прилипания (РИП) основа-
на на том, что вирулентные тканевые трепоне-
мы, сенсибилизированные сывороткой больно-
го сифилисом, в присутствии комплемента и
эритроцитов прилипают к поверхности эритро-
цитов и при центрифугировании увлекаются с
ними в осадок, исчезая из надосадочной жидко-
сти. Оценка результатов реакции иммунного
прилипания бледных трепонем следующая: О—
20% — отрицательная; 21—30% — сомнительная;
31-50% — слабоположительная; 51—100% — по-
ложительная.
По специфичности и чувствительности РИП
близка к РИТ и РИФ. РИП применяется в сле-
дующих случаях: когда на основании данных
анамнеза, клиники и результатов КСР и других
исследований не удается подтвердить диагноз
заболевания; при дифференцировании неспеци-
фических результатов КСР; при контрольном
наблюдении после окончания лечения.
Реакция иммобилизации бледных трепонем (РИТ)
является диагностически ценной и признана са-
мым специфичным тестом на сифилис. Однако ее
применение ограничивается из-за дороговизны
исследований и сложности проведения реакции.
Постановка РИТ имеет особое значение при ди-
агностике скрытого сифилиса, распознавании по-
ражений внутренних органов и нервной системы,
при обследованиях беременных с подозрением на
сифилис. Сущность реакции заключается в поте-
ре подвижности бледными трепонемами в присут-
ствии иммобилизинов испытуемой сыворотки и
активного комплемента. Иммобилизины появля-
ются в сыворотке крови больных позднее, чем дру-
гие антитела, и РИТ становится положительной
позже стандартных серореакций и РИФ.
Оценка результатов реакции проводится по
проценту иммобилизации бледных трепонем: до
20% — отрицательная; 21—30% — сомнительная;
31-50% — слабоположительная; 51 — 100% —
положительная.
В первичном периоде сифилиса РИТ у боль-
шинства больных отрицательная или слабопо-
ложительная. При вторичном свежем сифили-
се РИТ дает положительный результат более
чем у половины больных, однако процент им-
мобилизации трепонем невысокий (40—60 %).
Во вторичном рецидивном периоде сифилиса
РИТ бывает положительной у 85—90 % боль-
ных при выраженной степени иммобилизации
(80—100 %). При третичном сифилисе, при спе-
цифических поражениях внутренних органов,
нервной системы, когда РВ часто отрицатель-
ная, РИТ дает положительные результаты в 98—
100 % наблюдений (с высоким процентом им-
мобилизации). В связи с медленной негатива-
цией в процессе противосифилитической тера-
пии РИТ непригодна для контроля за ходом
лечения. При раннем врожденном сифилисе
РИТ положительна у 90—98 % детей, при по-
зднем врожденном сифилисе — почти у 100 %
больных. Диагноз скрытого серопозитивного
сифилиса ставится только по положительным
серореакциям крови с обязательным подтвер-
ждением их РИТ.
Реакция иммунофлюоресценции-абсорбции с
бледной трепонемой (FTA-ABS) становится поло-
жительной примерно в начале 3-й недели после
заражения, а у нелеченных больных она поло-
жительна почти в 100 % случаев. Главные дос-
тоинства реакции — высокая специфичность и
чувствительность. Положительная FTA-ABS
может иметь решающее значение для диагнос-
тики сомнительных случаев, особенно при по-
ложительных реакциях VDRL, выявленных при
скрининге.
После заражения сифилисом уже на 2-й не-
деле вырабатываются трепонемоспецифические
IgM, которых в процессе терапии и после ее
окончания становится все меньше, и в течение
2 лет они перестают определяться. Для сифи-
лидологов представляет интерес динамика этих
антител после лечения. Поэтому для суждения
об эффективности лечения предложены тесты,
направленные на определение специфических
IgM. С помощью методики РИФ с моноспеци-
фическим конъюгатом против IgM стало воз-
можным выявлять трепонемоспецифические
IgM-антитела (IgM-FTA-ABS). Однако при
идентификации антител IgM к бледным трепо-
немам реакцией FTA-ABS с использованием
конъюгата анти-IgM в качестве реагента выяв-
лялись ложноположительные и ложноотрица-
тельные результаты. Реакция 19S IgM-FTA-
ABS в этом отношении является наиболее
точным и специфичным методом. Однако тру-
доемкость и дороговизна теста ограничивают
его применение.
Микрогемагглютинация — Т, pallidum-vQ^r-
глютинационный тест для определения специфи-
ческих антител к Treponema pallidum (микро-
РПГА). В РПГА используются антигены из
трепонем, поэтому он наиболее специфичен.
Чувствительность и специфичность эквивален-
тна РИФ-абс. РПГА позволяет определить на-
личие антител в образцах, не реагирующих в ре-
агиновых тестах. Позитивный образец,
выявленный при скрининге, можно количествен-
но оценить при использовании реагентов ТРНА-
nosticon.
458 Венерология
Современные
методы лабораторной
диагностики сифилиса
Иммуноферментный анализ
Среди множества серологических методов од-
ним из наиболее перспективных для разработ-
ки диагностических тест-систем на сифилис
является иммуноферментный анализ (ИФА). В
настоящее время наибольшее распространение
получил непрямой вариант ИФА, на заключи-
тельном этапе которого используются антиви-
довые АТ к иммуноглобулинам человека,
конъюгированные с пероксидазой хрена или
другими ферментными маркерами.
Метод иммуноблотинга
Одним из современных методов диагностики
сифилиса является иммуноблотинг (Western
blot) для определения IgG либо IgM. При про-
ведении иммуноблотинга Т. pallidum подверга-
ются электрофорезу в градиенте концентрации
додецилсульфата натрия, в результате чего про-
исходит разделение белковых иммунодетерми-
нант на нитроцеллюлозе. Затем производится
обработка разделенных «точек» (blots) исследу-
емой сывороткой и антителами к IgG либо IgM,
меченными ферментами либо радиоактивными
веществами. IgG — иммуноблотинг по чувстви-
тельности и специфичности подобен РИФ-абс.
Применение анти-IgM специфического конъю-
гата (IgM Western blot) при проведении имму-
ноблотинга может служить диагностическим
тестом для врожденного сифилиса, более чув-
ствительным по сравнению с FTA — ABS 19S
IgM (чувствительность и специфичность 83 и
90 % соответственно).
Полимеразная цепная реакция
Полимеразная цепная реакция (ПЦР) применяется
для определения специфических последовательно-
стей ДНК. Метод заключается в многократном
увеличении (амплификации) количества ДНК
диагностируемого микроорганизма. Разработка
вариантов ПЦР для диагностики сифилиса обус-
ловливается недостаточностью стандартных тестов
при раннем серонегативном, врожденном сифилисе
и нейросифилисе. Обнаружение Т. pallidum с по-
мощью ПЦР значительно облегчается выражен-
ным генетическим консерватизмом и практичес-
ким отсутствием рекомбинаций даже между
подвидами. Большим достоинством ПЦР являет-
ся возможность автоматизации реакции путем за-
данного циклического температурного режима.
Применение ПЦР может иметь большое значение
при диагностике врожденного сифилиса (пассив-
ный трансплацентарный перенос антител не вли-
яет на результаты теста), нейросифилиса (чувстви-
тельность существующих тестов около 50 %),
первичного серонегативного (в настоящее время
одним из критериев является прямое микроско-
пическое обнаружение трепонем), а также при ди-
агностике сифилиса у ВИЧ-инфицированных.
Исследование
спинномозговой жидкости
Данные исследования спинномозговой жидкости
(ликвора) используют в следующих случаях: для
определения поражения сифилисом нервной сис-
темы; как критерий качества лечения у лиц с па-
тологическими изменениями в ликворе до лечения;
как один из критериев излеченности больного.
В клинической лаборатории исследуют цитоз,
содержание белка, глобулиновые реакции Пан-
ди и Нонне-Апельта. В серологической лабора-
тории проводят реакцию Вассермана с тремя
разведениями ликвора, реакцию Ланге с колло-
идным золотом, РИТ, РИФ10 и РИФц (реакция
иммунофлюоресценции с цельным ликвором).
Минимальные патологические изменения в
спинномозговой жидкости следующие: содержа-
ние белка, начиная с 0,4%о; цитоз — 8 клеток в
1 мм3; глобулиновые реакции (реакция Нонне-
Апельта + + , реакция Панди + ++); реакция
Ланге — больше двух двоек в цифровом значе-
нии; положительная реакция Вассермана. Пока-
затели выше указанных, даже будучи изолиро-
ванными, свидетельствуют о выраженных
патологических изменениях в спинномозговой
жидкости. Патологической считается также та-
кая спинномозговая жидкость, в которой не-
сколько показателей изменены соответственно
данным минимальной патологии, а реакция Вас-
сермана положительная.
Для обозначения позитивности глобулиновых
реакций и РВ применяется система четырех плю-
сов: резко положительный результат (4+), поло-
жительный (3+), слабоположительный (2+), со-
мнительный (1+).
Использование РИФ также целесообразно
при ликвородиагностике сифилиса. Особенно
высокочувствителен и специфичен тест с цель-
ным ликвором (РИФ-ц). РИТ со спинномозго-
вой жидкостью по постановке, регистрации и
оценке результатов аналогична таковой при ис-
следовании сыворотки крови. Применение РИФ
Глава 38. Сифилис 459
и РИТ расширяет возможности определения
сифилитического поражения нервной системы.
Лечение сифилиса
Специфическое лечение
Специфическое лечение назначается больному
сифилисом после установления диагноза. Диаг-
ноз устанавливается на основании клинической
картины, обнаружения возбудителя заболевания
(при соответствующих клинических проявлени-
ях) и результатов серологического исследования
(КСР, РИФ и в большинстве случаев РИТ).
Могут использоваться и такие специфические
тесты подтверждения диагноза, как иммунофер-
ментный анализ (ИФА), реакция пассивной ге-
магглютинации (РИГА).
Превентивное лечение
Превентивное лечение проводят с целью предуп-
реждения сифилиса лицам, находившимся в
половом или тесном бытовом контакте с боль-
ными заразными формами сифилиса. Превентив-
ное лечение не назначают лицам, находившим-
ся в половом или тесном бытовом контакте с
больными третичным, поздним скрытым, сифи-
лисом внутренних органов, нервной системы. Не
проводится также превентивное лечение лицам,
бывшим в половом контакте с пациентами, ко-
торым назначено превентивное лечение (т. е. при
контактах второго контакта). При выявлении в
детском коллективе больных сифилисом превен-
тивное лечение назначают детям в тех случаях,
когда нельзя исключить тесный бытовой кон-
такт с сотрудниками, имевшими проявления
первичного или вторичного сифилиса на слизи-
стой оболочке рта. '
Профилактическое лечение
Профилактическое лечение проводят беремен-
ным, болеющим или болевшим сифилисом, и
детям, рожденным такими матерями.
Пробное лечение
Пробное лечение можно назначить при подозре-
нии на специфическое поражение внутренних
органов, нервной системы, органов чувств, опор-
но-двигательного аппарата, когда нет возможно-
сти подтвердить диагноз убедительными лабора-
торными данными, а клиническая картина не
позволяет исключить возможность сифилитичес-
кой инфекции.
Больные гонореей с невыявленными источ-
никами заражения подлежат превентивному
противосифилитическому лечению в случае не-
возможности установления за ними диспансер-
ного наблюдения (бомжи, бродяги и др.). Если
у такого больного имеется постоянное место
жительства и работы, то превентивное лечение
против сифилиса ему не проводится, но после
лечения гонореи он должен находиться на кли-
нико-серологическом наблюдении в течение 3
месяцев.
Каждый больной сифилисом проходит в ста-
ционаре тщательное клинико-лабораторное об-
следование. Исследование спинномозговой жид-
кости с диагностической целью проводится
пациентам с клиническими симптомами пораже-
ния нервной системы, а также при скрытых и
поздних формах сифилиса.
До начала лечения необходимо выяснить воп-
рос о переносимости препаратов пенициллина
(или других антибиотиков) в прошлом и зафик-
сировать это в медицинской документации.
Кроме того, за 30 минут до первой инъекции
пенициллина, а также перед каждой инъекцией
дюрантных препаратов пенициллина следует
назначать по 2 таблетке одного из антигистамин-
ных препаратов.
Схемы лечения
больных сифилисом
Превентивное лечение
Превентивное лечение проводится лицам, име-
ющим половой или тесный бытовой контакт с
больными ранними стадиями сифилиса, если с
момента контакта прошло не более 2 месяцев.
Лечение проводят амбулаторно бициллинами
1, 3, 5 в разовых дозах соответственно 1 200 000
ЕД, 1 800 000 ЕД и 1 500 000 ЕД, 2 раза в неде-
лю, на курс 4 инъекции. Бензатинпенициллин
(ретарпен, экстенциллин) вводят однократно по
2 400 000 ЕД внутримышечно двухмоментным
способом. Ретарпен растворяют в 5 мл 0,25 %-
ного раствора новокаина, воды для инъекций,
физиологического раствора. Экстенциллин ра-
створяют в 8 мл растворителя и вводят в каж-
дую ягодицу по 1 200 000 ЕД.
Превентивное лечение реципиентов, получив-
ших кровь больных сифилисом, проводится в
срок не более 2 месяцев после переливания дву-
кратным введением препаратов по 2 400 000 ЕД
с интервалом в 1 нед.
В стационаре проводят лечение натриевой или
калиевой солью пенициллина — по 400 000 ЕД
460 Венерология
на инъекцию 8 раз в сутки (каждые 3 часа) в
течение 14 дней; на курс 44 800 000 ЕД. Возмож-
но применение новокаиной соли пенициллина
по 600 000 ЕД 2 раза в сутки на протяжении 14
дней; на курс — 16 800 000 ЕД.
Лицам, у которых с момента контакта с боль-
ными сифлисом прошло от 2 до 4 месяцев, про-
водится двукратное клинико-серологическое об-
следование (с исследованием КСР, РИТ, РИФ)
с интервалом в 2 месяца. Если с момента контакта
прошло более 4 месяцев, проводится однократное
клинико-серологическое обследование.
Лечение больных первичным
и вторичным свежим сифилисом
Лечение больных первичным и вторичным све-
жим сифилисом проводится по одной из следу-
ющих методик: бициллин-1, 3, 5. Разовая доза —
1 200 000 ЕД, 1 800 000 ЕД, 1 500 000 ЕД соот-
ветственно; число инъекций — 7 (при первичном
серонегативном сифилисе), 8 (при первичном
серопозитивном), 10 (при вторичном свежем си-
филисе). Первая инъекция проводится в непол-
ной дозе — 300 000 ЕД; вторая — в полной ра-
зовой дозе, проводится через сутки; последующие
инъекции проводятся 2 раза в неделю.
Применяется новокаиновая соль бензилпе-
нициллина по 600 000 ЕД 2 раза в сутки на
протяжении 16 дней; курсовая доза составля-
ет 19 200 000 ЕД.
Лечение проводят водорастворимым пени-
циллином, который вводят внутримышечно по
400 000 ЕД через каждые 3 часа в течение 14
дней; курсовая доза — 44 800 000 ЕД (при пер-
вичном серонегативном периоде сифилиса).
При первичном серопозитивном и вторичном
свежем сифилисе лечение длится 16 дней, на 16-
й день через 3 часа после окончания пеницил-
линотерапии однократно вводят бициллин-3 в
дозе 4 800 000 ЕД (по 2 400 000 ЕД вйутримы-
шечно в каждую ягодицу) или бициллин-5 в
дозе 3 000 000 ЕД.
При первичном серонегативном сифилисе пре-
параты бензатинпенициллина вводят однократно
внутримышечно в дозе 2 400 000 ЕД. При первич-
ном серопозитивном и вторичном свежем сифи-
лисе ретарпен или экстенциллин вводят в дозе 2
400 000 ЕД дважды с интервалом в 1 неделю.
Лечение больных вторичным
рецидивным и скрытым ранним
сифилисом бициллином-1, 3, 5.
Для первой инъекции используется доза 300 000
ЕД, для последующих инъекций разовая доза 1
200 000 ЕД, 1 800 000 ЕД, 1 500 000 ЕД соот-
ветственно. Инъекции проводят 2 раза в неде-
лю, число инъекций 14, независимо от того,
какой из бициллинов применяется.
Применяется новокаиновая соль пеницилли-
на по 600 000 ЕД 2 раза в сутки в течение 28
дней.
Лечение проводят водорастворимым пеницил-
лином по 400 000 ЕД 8 раз в сутки в течение 28
дней.
Больным вторичным рецидивным и ранним
скрытым сифилисом производят 3 инъекции
экстенциллина по 2 400 000 ЕД с интервалом в
1 неделю.
При лечении больных вторичным рецидивным
и скрытым ранним сифилисом ретарпеном пер-
вую инъекцию проводят в дозе 4 800 000 ЕД (по
2 400 000 ЕД в каждую ягодицу), вторую и тре-
тью инъекции — по 2 400 000 ЕД с интервалом
в 1 неделю.
Лечение проводят водорастворимым пени-
циллином (натриевой солью) внутримышечно
в разовых дозах 1 000 000 ЕД (пенициллин
разводят в 2 мл физиологического раствора
или дистиллированной воды) 6 раз в сутки в
течение 28 дней; курсовая доза — 168 000 000
ЕД).
При злокачественном течении сифлиса, транс-
фузионном сифилисе и раннем нейросифилисе
предпочтительна последняя методика в комплек-
се с неспецифической и симптоматической те-
рапией.
В тех случаях, когда у больных скрытым ран-
ним сифлисом путем конфронтации, изучения
анамнеза и по данным лабораторных исследова-
ний достоверно установлен срок заболевания,
соответствующий первичному серопозитивному
или вторичному свежему сифилису, лечение этих
больных можно проводить по методикам, реко-
мендуемым для терапии указанных стадий си-
филиса.
Специфическое лечение больных вторичным
рецидивным и ранним скрытым сифилисом це-
лесообразно сочетать с неспецифической терапией.
Принципы лечения больных
сифилисом с сопутствующими
инфекциями мочеполового тракта
Больные сифилисом должны обследоваться на
ВИЧ и другие инфекции, передаваемые половым
путем.
При наличии у больного сифилисом гонореи
лечение проводится антибиотиками, активными
как в отношении гонококка, так и бледной тре-
понемы (препараты пенициллина, доксициклин,
сумамед).
Глава 38. Сифилис
461
При сочетании ранних форм сифилиса и хла-
мидийной инфекции, либо сифилиса, гонореи и
хламидийной инфекции рекомендуется лечение
сумамедом (азитромицином). Лечение проводит-
ся в течение 14 дней, препарат применяется по
0,5 г 1 раз в день (или по 0,25 г 2 раза в день)
через 2 часа после еды или за 1 час до еды. В
первый день лечения доза сумамеда составляет
1,0 г, принимается в один или два приема (ут-
ром и вечером).
При выявлении у больного трихомониаза ле-
чение его проводят одновременно с противоси-
филитической терапией.
При выявлении у больного ВИЧ-антител он
направляется для дальнейшего лечения и посто-
янного наблюдения в регионарный центр по
лечению СПИДа с соответствующими рекомен-
дациями в отношении лечения сифилиса.
При возможности больным сифилисом следует
проводить обследование на другие инфекции
урогенитального тракта, передаваемые половым
путем, с последующим лечением соответствен-
но диагнозу по окончании терапии сифилиса.
Возможно также одновременное лечение сопут-
ствующих инфекций препаратами, высокоэф-
фективными против бледной трепонемы.
Лечение больных
скрытым поздним сифилисом
Лечение начинают с подготовки бийохинолом по
2 мл через день до получения 12—14 мл препара-
та, после чего присоединяют пенициллинотерапию
по 400 000 ЕД через 3 часа в течение 28 дней.
Общую дозу бийохинола доводят до 40—50 мл. В
этой методике бийохинол может быть заменен
бисмоверолом, который применяют по 1 мл через
день или 1,5 мл 2 раза в неделю; на курс 18—20 мл.
Во время пенициллинотерапии целесообразно
применение химотрипсина по 5 мг внутримы-
шечно 2 раза в сутки. Химотрипсин способствует
лучшему проникновению антибиотика в органы
и ткани.
Если имеются противопоказания к назначе-
нию висмутовых препаратов, то лечение прово-
дят двумя курсами пенициллина по 400 000 ЕД
через 3 часа в течение 28 дней. Перед началом
первого курса в течение 10 дней проводят под-
готовку эритромицином, тетрациклином или
олететрином по 0,5 г четыре раза в день. На
втором курсе вместо растворимого пеницилли-
на возможно применение бициллинов. Бицил-
лин- 1 вводят в разовой дозе 1 200 000 ЕД, би-
циллин-3 — в дозе 1 800 000 ЕД, бициллин-5 —
в дозе 1 500 000 ЕД; инъекции 2 раза в неделю;
на курс 7 инъекций.
Специфическое лечение больных поздним
скрытым сифилисом следует сочетать с назначе-
нием неспецифических средств. Лечение боль-
ных скрытым неуточненным сифилисом реко-
мендуется проводить индивидуализированно
(аналогично позднему или раннему скрытому
сифилису).
Лечение больных висцеральным
и третичным сифилисом
Лечение больных висцеральным сифилисом вклю-
чает в себя, помимо специфических, неспецифи-
ческие и симптоматические препараты, и должно
проводиться под наблюдением терапевта.
Специфическое лечение больных висцераль-
ным и третичным сифилисом проводится по схе-
ме позднего скрытого сифилиса с более продол-
жительной подготовкой бийохинолом (до дозы
20 мл). При лечении поздних сифилитических
гепатитов, а также специфических поражений
почек и мочевыводящих путей препараты вис-
мута не назначают. В остальных случаях вопрос
о применении висмута решается индивидуально.
При сифилитическом аортите, осложненном
аневризмой аорты или недостаточностью аор-
тальных клапанов, подготовка бийохинолом
должна начинаться с разовой дозы 1 мл (3 инъ-
екции), с последующим повышением до 1,5 мл
(3 инъекции) и затем до 2 мл. После получения
25—30 мл препарата присоединяют пеницилли-
нотерапию. Последнюю начинают с разовой
дозы 50 000 ЕД через 3 часа. Повышение разо-
вой дозы проводится через день по следующей
схеме: 50 000-100 000-200 000-400 000 ЕД.
Длительность пенициллинотерапии составляет 28
дней. При противопоказаниях к применению
висмута подготовка проводится эритромицином
или олететрином по 0,5 г 4 раза в день в тече-
ние 2 нед. При наличии противопоказаний к
назначению висмутовых препаратов лечение
проводят двумя курсами пенициллина по 400 000
ЕД через 3 часа в течение 28 дней (на втором
курсе с самого начала разовая доза пеницилли-
на составляет 400 000 ЕД).
Если после пенициллин-висмутового курса
остаются клинические симптомы заболевания,
целесообразно назначение дополнительно 2 кур-
сов висмутовой терапии, один из которых про-
водят бийохинолом (40—50 мл на курс), дру-
гой — бисмоверолом (16—20 мл на курс).
Специфическое лечение висцерального сифи-
лиса проводят под контролем функционально-
го состояния пораженного органа (анализы кро-
ви, мочи, биохимические пробы, показатели
свертывающей системы крови, ЭКГ и т. д.).
462 Венерология
Выбор метода лечения при гуммозном пора-
жении зависит от локализации гуммы и общего
состояния больного. При третичном сифилисе с
локализацией гуммы на коже лечение проводится
аналогично позднему скрытому сифилису.
Лечение больных нейросифилисом
При ранних формах нейросифилиса лечение
проводится по следующей методике: водораство-.
римый пенициллин (натриевая соль) внутримы-
шечно в разовых дозах 1 000 000 ЕД 6 раз в
сутки, в течение 28 дней.
Для повышения концентрации пенициллина
в ликворе целесообразно применение препара-
тов, задерживающих выведение антибиотиков из
организма, в частности, пробеницида по 0,5 г 4
раза в день или этамида по 1,05 г (3 таблетки)
4 раза в день в течение 10 дней.
Больные поздним нейросифилисом, за исклю-
чением больных с атрофией зрительных нервов,
подлежат лечению по схемам позднего скрыто-
го сифилиса.
Каждый курс специфической терапии необ-
ходимо дополнять витаминами, общеукрепляю-
щими и стимулирующими средствами. Лечение
следует проводить под наблюдением невропато-
лога и окулиста: первые два курса в стационаре
(а при атрофии зрительных нервов — все 3 курса
в стационаре).
При первичной атрофии зрительных нервов пер-
вый курс лечения начинают с витаминного на-
сыщения:
• прием витамина А по 33 000 ME 2 раза в
сутки в любых перечисленных ниже формах:
драже или таблетки ретинола ацетата; масляный
раствор ретинола ацетата в капсулах; таблетки
или масляный раствор ретинола пальмитата;
• прием комплекса витаминов в виде смеси в
порошке: кислоты аскорбиновой 0,15 г, кисло-
ты никотиновой 0,05 г, кислоты глутаминовой
0,5 г, рибофлавина (витамина В2) 0,025 г;
• внутримышечно назначают инъекции вита-
минов Bj (тиамина хлорид 5% — 2 мл ежеднев-
но № 30, В6 (пиридоксин 5% — 1 мл через день
№ 15) и В12 по 200 мкг ежедневно № 30;
• рекомендуется внутримышечное введение
натриевой соли аденозинтрифосфорной кисло-
ты (АТФ) по 1—2 мл в комбинации с экстрак-
том алоэ (1 мл) или стекловидным телом (2 мл)
№ 30 ежедневно;
• прием препаратов кальция (желательно гли-
церофосфата кальция) по 0,5 г 3 раза в день.
Параллельно начинают пенициллинотерапию
с малых доз (50 000 ЕД), с дальнейшим их по-
вышением на 50 000 ЕД через день (50 000—
100 000—150 000—200 000 ЕД). Пенициллин в
разовой дозе 200 000 ЕД применяют в течение
недели, после чего разовую дозу повышают до
400 000 ЕД. Длительность пенициллинотера-
пии — 28 дней.
В дальнейшем проводят еще 2 курса пеницил-
линотерапии в разовой дозе через 3 часа в тече-
ние 28 дней. Интервал между курсами — 1 мес.
Параллельно со специфической терапией в
каждом курсе следует проводить неспецифичес-
кую, стимулирующую и витаминотерапию, необ-
ходимо систематическое наблюдение окулиста.
Резервные методы
лечения больных сифилисом
При непереносимости препаратов пенициллино-
вого ряда применяются антибиотики широкого
спектра действия: эритромицин, тетрациклин,
олететрин, доксициклин, сумамед.
Для превентивного лечения эритромицин,
тетрациклин и олететрин применяют по 0,5 г 4
раза в сутки; доксициклин в капсулах по 0,1 г
3 раза в день в течение 14 дней.
При свежих формах сифилиса эти антибиоти-
ки в тех же дозах применяются на протяжении
20 дней при первичном периоде сифилиса, 25
дней — при вторичном свежем. При вторичном
рецидивном и раннем скрытом сифилисе реко-
мендуется 2 курса по 30 дней названных анти-
биотиков в указанных дозах, интервал между
курсами 2 недели.
При одновременной непереносимости пени-
циллина, эритромицина и тетрациклинов лече-
ние может быть проведено цефазолином (цефа-
мизином). Препарат применяют внутримышечно
по 1,0 г шесть раз в сутки в течение 14 дней при
первичном серонегативном, 16 дней — при пер-
вичном серопозитивном и вторичном свежем
сифилисе, 28 дней — при вторичном рецидив-
ном и скрытом раннем сифилисе.
При лечении свежих форм сифилиса можно
применять сумамед (азитромицин) перорально
по 0,25 г два раза в день или по 0,5 г один раз
в день в течение 14 дней.
При позднем скрытом сифилисе антибиоти-
ки резерва применяются в объеме 2—3 курсов.
Длительность антибиотикотерапии — 28 дней,
перерыв между курсами — 2 недели.
Неспецифическая
терапия больных сифилисом
Неспецифическая терапия показана при скры-
тых, поздних формах заболевания, сифилисе
нервной системы и внутренних органов, врож-
денном сифилисе, при сопутствующей патоло-
Глава 38. Сифилис 463
гии, включая алкоголизм, при признаках злока-
чественного течения заболевания, при замедле-
нии негативации серологических реакций, серо-
рецидивах и серорезистентности. Целесообразно
назначение неспецифической терапии при вто-
ричном рецидивном и раннем скрытом сифилисе,
а при показаниях — и при свежих его формах.
К методам неспецифической терапии отно-
сятся: пиротерапия, витаминотерапия, биогенные
стимуляторы и средства, влияющие на тканевый
обмен (экстракты алоэ, плаценты, стекловидного
тела, спленина, аспаркам и др.), иммуномодуля-
торы (декарис, метилурацил, натрия нуклеинат,
пирроксан), реинфузии УФ-облученной ауток-
рови. Неспецифическую терапию следует назна-
чать после тщательного обследования больного
с учетом показаний и противопоказаний к тому
или иному препарату.
Пиротерапия приводит к усилению процессов
теплопродукции, улучшению условий крово- и
лимфообращения в пораженных органах и тка-
нях, активации гистиоцитарно-ретикулярной
системы, усилению фагоцитоза, ферментативной
и секреторной деятельности желудка и слюнных
желез. Среди пирогенных препаратов наиболее
апробированы и рекомендуются пирогенал и
продигиозан.
Пирогенал — сложный полисахаридный ком-
плекс — назначается в виде внутримышечных
инъекций в верхненаружный квадрант ягодицы,
в начальной дозе 5—10 мкг, с постепенным по-
вышением на 10—30 мкг на инъекцию, доводя
до 120—150 мкг в зависимости от реакции орга-
низма. Препарат вводится 1 раз в 2—3 дня, все-
го на курс лечения 10—15 инъекций.
Продигиозан — липополисахарид, близкий по
действию на организм к пирогеналу, вводится
внутримышечно 2 раза в неделю в дозах от 25
до 100 мкг; всего на курс 4—6 инъекций.
Биогенные стимуляторы (ФиБС для инъек-
ций, взвесь и экстракт плаценты для инъекций,
спленин, плазмол, стекловидное тело, полибио-
лин) назначают в виде подкожных инъекций по
1 мл ежедневно в течение 10—20 дней. Спленин
вводят внутримышечно ежедневно по 2 мл в те-
чение 10 дней, полибиолин вводят внутримы-
шечно ежедневно по 5 мл раствора (содержимое
флакона — 0,5 г — растворяют в 5 мл 0,25—
0,5 % раствора новокаина) в течение 10 дней.
Витамины С, группы В, аевит применяют од-
новременно со средствами специфической тера-
пии на протяжении всего курса лечения больных
сифилисом. Аскорбиновую кислоту применяют
по 0,2 г 3 раза в день, аевит в капсулах по 1 кап-
суле 3 раза в день. Витамины Вр В6, В12 в виде
ампульных растворов вводят внутримышечно
через день, на курс 10—15 инъекций. Отдельным
больным сифилисом необходимо включать в
комплексную терапию адаптогенные препара-
ты — пантокрин, экстракт элеутерококка, роди-
олы розовой, настойки женьшеня, настойки ли-
монника.
Иммунокорригирующую терапию обычно на-
значают в тех случаях, когда имеются признаки
злокачественного течения заболевания, при на-
личии сопутствующих заболеваний, развиваю-
щихся на фоне иммунодепрессии (кожно-слизи-
стый кандидоз, хроническая пиодермия и др.),
а также больным, страдающим одновременно
хроническим алкоголизмом. Иммунокорригиру-
ющую терапию рекомендуется проводить под
контролем иммунограммы.
Левамизол (декарис) повышает функциональ-
ную активность фагоцитов и Т-лимфоцитов.
Препарат назначается по 150 мг ежедневно в
течение 3 дней с последующим перерывом в те-
чение 4 или 7 дней, всего 2—4 таких цикла (под
контролем количества форменных элементов
крови). Возможны побочные реакции в виде
крапивницы, тошноты, рвоты, токсического дей-
ствия на эритроциты и лейкоциты.
Метилурацил ускоряет процессы клеточной ре-
генерации, стимулирует клеточные и гуморальные
факторы резистентности. Его назначают циклами
по 0,5 г 4 раза в день в течение 10—14 дней с 5—
7-дневными перерывами, всего 2—3 цикла.
Натрия нуклеинат повышает функциональную
активность иммунокомпетентных клеток, стиму-
лирует факторы неспецифической резистентно-
сти организма. Препарат назначают двухнедель-
ными циклами по 0,1 г 3 раза в день с
недельным перерывом.
Пирроксан в комплексной терапии сифилиса
применяется, главным образом, у лиц, страдаю-
щих хроническим алкоголизмом. Он обладает
детоксирующим действием, купирует явления
алкогольной абстиненции, улучшает усвоение
кислорода тканями. Он применяется внутрь в
таблетках по 0,015 г 3 раза в день, 2 цикла по
10 дней, с перерывом в 7—10 дней.
Тактивин и тималин — препараты полипептид-
ной природы, выделенные из тимуса крупного ро-
гатого скота. При иммунодефицитных состояниях
эти препараты нормализуют количественные и
функциональные показатели Т-системы иммуни-
тета, функциональную активность стволовых гемо-
поэтических клеток, усиливают фагоцитоз, норма-
лизуют другие показатели клеточного иммунитета.
Введение иммуномодуляторов целесообразно начи-
нать с 10—12—14-го дня антибиотикотерапии.
464 Венерология
Тактивин вводится подкожно по 1 мл 0,01 %
раствора один раз в день 3 дня подряд, затем 2
раза в неделю; на курс 6—8 инъекций.
Тималин вводится внутримышечно по 10 мг
(разводить в 1—2 мл изотонического раствора
натрия хлорида до получения равномерной взве-
си) через день; на курс 6—8 инъекций.
Тимоген — синтетический пептид — глюта-
милтриптофан. Он нормализует число Т-хелпе-
ров, восстанавливает соотношение иммунорегуля-
торных субпопуляций Т-лимфоцитов. Тимоген
вводится внутримышечно по 100 мкг (растворить
в 1 мл изотонического раствора натрия хлорида)
2 раза в неделю; на курс 5—8 инъекций.
Специфическое,
профилактическое
и превентивное
лечение беременных
При выявлении у беременных ранних стадий
сифилиса лечение проводится по одной из ме-
тодик, изложенных в предыдущем разделе.
При выявлении у беременных первичного или
вторичного свежего сифилиса специфическое ле-
чение проводится пенициллином по 400 000 ЕД
через 3 часа в течение 14—16 дней, при выявле-
нии вторичного рецидивного или раннего скры-
того сифилиса — в тех же разовых или суточ-
ных дозах в течение 28 дней. При установлении
скрытого позднего сифилиса лечение проводится
тремя курсами пенициллина по 67 200 000 ЕД
на курс, в разовых дозах по 400 000 ЕД через 3
часа, с интервалом между курсами 7—10 дней.
Параллельно пенициллинотерапии рекоменду-
ется применение внутрь теоникола по 0,15 г 3
раза в день после еды. Теоникол способствует
повышению концентрации пенициллина в кро-
ви плода.
Лечение беременных ретарпеном или экстен-
циллином производится в соответствии с диагно-
зом в указанных выше дозах, но делают не менее
двух инъекций. Профилактическое лечение бере-
менных проводится 2—3 инъекциями антибиоти-
ка по 2 400 000 ЕД с интервалом в 1 неделю.
Для профилактики врожденного сифилиса
рекомендуется двукратное серологическое обсле-
дование беременных: в первой половине бере-
менности (при явке к акушеру-гинекологу для
взятия на учет по беременности) и во второй ее
половине (на 6—7-м месяце, но не позднее офор-
мления декретного отпуска). При неблагоприят-
ной эпидемиологической обстановке решением
органов здравоохранения может быть введено
трехкратное серологическое обследование бере-
менных на сифилис. Третье обследование про-
водится непосредственно перед родами. При
положительных результатах КСР дифференци-
альный диагноз проводится с помощью РИТ,
РИФ и других специфических серологических
реакций. В случае отрицательных результатов
этих тестов беременная должна находиться на
клинико-серологическом контроле с ежемесяч-
ным исследованием КСР, РИТ, РИФ до родов
и в течение 3 месяцев после них.
В исключительных случаях, при отсутствии
возможности исследования РИТ и РИФ или
других специфических серореакций, у беремен-
ных с резко положительным КСР повторяют
исследование КСР, и при резко положительном
его результате ставят диагноз скрытого сифили-
са. При повторных слабо положительных резуль-
татах КСР беременная подлежит тщательному
клинико-серологическому наблюдению в дина-
мике для дифференцирования биологически
ложноположительных результатов КСР, обуслов-
ленных беременностью.
Женщины, у которых после полноценного
лечения наступила стойкая негативация КСР
(отрицательные результаты, по меньшей мере, в
течение года до наступления беременности), про-
филактическому лечению во время беременно-
сти не подлежат. Исключением могут быть жен-
щины с сохраняющейся резкой позитивностью
РИТ и/или РИФ без тенденции к их сниже-
нию.
Женщинам, у которых в течение года до бе-
ременности отмечались колебания позитивнос-
ти КСР (от отрицательных до положительных
результатов) либо сохранялась позитивность
КСР, профилактическое лечение при беремен-
ности рекомендуется.
Женщины, получившие превентивное проти-
восифилитическое лечение, профилактическому
лечению во время беременности не подлежат.
Профилактическое лечение беременных прово-
дится по одной из следующих методик:
• методика № 1. Водорастворимый пеницил-
лин вводится по 400 000 ЕД 8 раз в сутки в те-
чение 14 дней;
• методика № 2. Новокаиновая соль бензил-
пенициллина вводится по 600 000 ЕД 2 раза в
сутки в течение 14 дней;
• методика № 3. Бициллины-1, 3, 5 вводятся
в дозе 1 200 000 ЕД, 1 800 000 ЕД, 1 500 000
ЕД соответственно, 2 раза в неделю, на курс 7
инъекций.
Если специфическое лечение проведено в пер-
вые месяцы беремености, то профилактическое
Глава 38. Сифилис 465
лечение должно быть начато не позднее 6—7
месяца. Если специфическое лечение проводит-
ся в поздние сроки беременности, то профилак-
тическое следует за ним без перерыва.
Превентивное лечение беременных проводит-
ся в соответствии с рекомендациями, изложен-
ными выше.
При лечении беременных с непереносимос-
тью препаратов пенициллинового ряда нецеле-
сообразно применять эритромицин, так как,
оказывая хорошее клиническое действие у ма-
тери, он недостаточно проникает через плаценту
и не предупреждает возникновения врожденно-
го сифилиса у ребенка. Препараты тетрацикли-
нового ряда предупреждают врожденный сифи-
лис, но откладываются в костной ткани и зубах
плода.
Учитывая это, целесообразно использование у
беременных оксациллина, который вводят внут-
римышечно по 1 000 000 ЕД с интервалом 6
часов, 4 раза в сутки, на протяжении 14 или 28
дней в зависимости от стадии заболевания.
Лечение и профилактика
сифилиса у детей
Превентивное, профилактическое и специфичес-
кое лечение детей проводится препаратами пени-
циллина. Для детей в возрасте до 2 лет исполь-
зуется натриевая и новокаиновая соли
пеницилина, старше 2 лет — также и бицилли-
ны. Суточная доза пенициллина (натриевой и но-
вокаиновой солей) исчисляется из расчета 100 000
ЕД/кг детям в возрасте до 6 мес, 75 000 ЕД/кг в
возрасте от 6 мес до 1 года, 50 000 ЕД/кг — стар-
ше 1 года. Суточная доза делится на 6 равных
разовых доз для водорастворимого пенициллина
и на 2 дозы для новокаиновой его соли.
Бициллины 1, 3 или 5 вводят по 300 000 ЕД 1
раз в сутки. При хорошей переносимости после
нескольких инъекций можно перейти на введе-
ние по 600 000 ЕД 1 раз в 2 суток (по 300 000
ЕД в каждую ягодицу).
Длительность превентивного лечения составляет
2 недели; профилактического — от 2 до 4 недель;
специфического, при раннем врожденном сифи-
лисе — 4 недели; при позднем врожденном — 4
недели антибиотикотерапии в сочетании с висму-
товыми препаратами. Длительность лечения при-
обретенного сифилиса у детей при свежих фор-
мах заболевания составляет 2 недели, при
вторичном рецидивном и скрытом раннем 4 не-
дели.
При непереносимости пенициллина можно
использовать оксациллин и ампициллин.
Оксациллин назначают внутримышечно в сле-
дующих суточных дозах: новорожденным — 20—
40 мг/кг массы тела, детям в возрасте до 3 мес —
200 мг/кг, от 3 мес до 2 лет — 1,0 г в сутки, от
2 лет и старше — 2,0 г в сутки.
Возможно применение оксациллина внутрь, за
1 ч до еды или через 2—3 часа после нее в следую-
щих суточных дозах: новорожденным — 90-
150 мг/кг массы, в возрасте до 3 мес —- 200 мг/кг,
от 3 мес до 2 лет — 1,0 г в сутки, от 2 лет и стар-
ше — 2,0 г в сутки.
Ампициллина натриевую соль применяют внут-
римышечно в следующих дозах: новорожден-
ным — 100 мг/кг, остальным детям 50 мг/кг,
максимально — 2,0 г в сутки. Суточную дозу
делят на 4—6 инъекций.
Ампициллин в таблетках применяют внутрь в
тех же дозах, независимо от приема пищи. Су-
точная доза делится на 4—6 приемов.
В случае непереносимости полусинтетичес-
ких пенициллинов возможно использование
эритромицина детям в возрасте от 1 года до 3
лет — в дозе 0,4 г в сутки, 3—6- лет — 0,5—
0,7 г, 6-8 лет — 0,75 г, 8-12 лет — до 1, 0 г
в сутки. Препарат дают равными дозами 4—6
раз в сутки.
Для предупреждения аллергических реакций
до начала и во время лечения следует назначать
антигистаминные препараты, препараты кальция.
На 2—3 день лечения и в конце курса исследу-
ют кровь на КСР.
Превентивное лечение детей
Вопрос о проведении превентивного лечения
ставится в тех случаях, когда установлена воз-
можность заражения детей при тесном бытовом
или половом контакте с больными заразными
формами сифилиса и ранним скрытым сифили-
сом.
Учитывая особенности ухода за детьми и их
общения между собой, в возрасте до 3 лет про-
ведение лечения, как правило, показано. Для
более старших детей вопрос о лечении решается
индивидуально, с учетом формы сифилиса, ло-
кализации высыпаний и степени контакта с ре-
бенком.
Превентивное лечение проводится, если с
момента последнего контакта с больным прошло
не более 2 мес. При более длительном сроке ре-
бенку необходимо провести полное клиническое
и серологическое (КСР, РИТ, РИФ) обследова-
ние. При отрицательных результатах обследова-
ния лечение не назначается, через 4 месяца про-
водят повторное обследование, после чего
наблюдение прекращают.
466 Венерология
В случаях переливания детям крови от доно-
ров, больных сифилисом, назначается превен-
тивное лечение в сроки до 3 месяцев после пе-
реливания.
Профилактическое лечение детей
Дети, рожденные матерями, болевшими сифили-
сом, не подлежат клинико-серологическому об-
следованию и наблюдению в кожно-венероло-
гическом диспансере в тех случаях, когда у
матери после полноценного специфического ле-
чения произошла стойкая негативация КСР до
беременности (отрицательные результаты КСР в
течение года).
Остальные дети, рожденные матерями, болев-
шими сифилисом или находившимися во время
беременности в тесном контакте с больными
заразными формами сифилиса, должны пройти
клинико-серологическое обследование в первые
месяцы жизни (желательно в возрасте 2,5—3
месяца). Обязательными компонентами обследо-
вания являются: консультация педиатра, дерма-
товенеролога, невропатолога, отоларинголога,
окулиста, исследование крови (КСР, РИФ,
РИТ), рентгенография костей конечностей. При
наличии клинических неврологических измене-
ний показано проведение спинномозговой пун-
кции.
Дети, матери которых подлежали профилак-
тическому лечению во время беременности и
получили его (в том числе и матери с сероре-
зистентностью), при отсутствии у детей клини-
ческих, серологических и рентгенологических
признаков заболевания, не подлежат профилак-
тическому лечению, но остаются под наблюде-
нием кожно-венерологического диспансера в
течение 1 года.
Дети, матери которых подлежали профилакти-
ческому лечению, но не получили его, а также
дети, матери которых получили неполноценное
сифилитическое лечение, подлежат профилакти-
ческому лечению продолжительностью 2 нед.
Дети, родившиеся от нелеченных матерей,
больных сифилисом, подлежат профилактичес-
кому лечению по схеме раннего врожденного
сифилиса длительностью 4 нед, даже при отсут-
ствии у детей клинических, серологических и
рентгенологических признаков заболевания.
При сомнительных результатах обследования
ребенка, рожденного матерью, болевшей сифи-
лисом, вопрос о лечении решается индивидуаль-
но, с учетом данных анамнеза, возраста ребенка
и объема лечения, полученного матерью.
Если ребенок впервые обследуется в возрасте
старше 1 года, то при отрицательных результа-
тах обследования лечение ему не проводится. В
сомнительных случаях рекомендуется пеницил-
линотерапия продолжительностью в 2 нед.
Специфическое лечение детей,
больных врожденным сифилисом
Лечение детей, больных ранним врожденным
сифилисом, проводится натриевой или новока-
иновой солью бензилпенициллина в условиях
стационара (разовые и суточные дозы указаны
выше). Длительность лечения составляет 28 дней.
Лечение детей, больных поздним врожденным
сифилисом, проводится препаратами пеницил-
лина в сочетании с висмутом, по типу позднего
скрытого сифилиса у взрослых. Лечение начи-
нают бийохинолом, вводимым внутримышечно
2 раза в неделю, в возрастной дозировке. При
наличии противопоказаний бийохинол может
быть заменен бисмоверолом.
По достижении */4 курсовой дозы бийохино-
ла его введение прерывают и переходят на инъ-
екции растворимого пенициллина или его ново-
каиновой соли. Суточная доза рассчитывается по
массе тела ребенка. Продолжительность пени-
циллинотерапии 28 дней. По окончании введе-
ния антибиотика продолжают лечение препара-
тами висмута до достижения курсовой дозы.
При непереносимости препаратов пенициллина
назначаются оксациллин, ампициллин или эрит-
ромицин. Одновременно с антибиотикотерапи-
ей рекомендуется прием нистатина и антигиста-
минных препаратов.
Разовые и курсовые дозы препаратов висмута
при лечении детей, больных врожденным сифи-
лисом:
до 3 лет — бийохинол (мл) — разовая доза 0,5—
1,0, курсовая доза 12,0—15,0; бисмоверол (мл) —
разовая доза 0,2—0,4, курсовая доза 4,0—4,8.
от 3 до 5 лет — бийохинол (мл) — разовая доза
1,0—1,5, курсовая 15,5—20,0; бисмоверол — 0,4—
0,6 и 6,0—8,0 соответственно.
от 6 до 10 лет — бийохинол (мл) — 1,0—2,0 и
20,0—25,0; бисмоверол — 0,4—0,8 и 8,0—10,0.
от 11 до 15 лет — бийохинол (мл) — 1,0—2,0
и 25,0—30,0; бисмоверол — 0,6—0,8 и 10,0—12,0.
Лечение приобретенного
сифилиса у детей
Лечение проводится препаратами пенициллина
по принципу лечения сифилиса у взрослых. Рас-
чет суточной дозы антибиотика проводится, как
было указано выше. Длительность терапии при
первичном и вторичном свежем сифилисе — 14
дней, вторичном рецидивном и скрытом ран-
нем — 28 дней. При позднем скрытом приобре-
Глава 38. Сифилис 467
тенном сифилисе лечение проводится так же, как
при позднем врожденном.
Клинико-серологический
контроль после
окончания лечения
Взрослые и дети, получившие превентивное ле-
чение после полового или тесного бытового кон-
такта с больными ранними стадиями сифилиса,
подлежат однократному клинико-серологическо-
му обследованию через 3 месяца после лечения.
Если превентивное лечение проводилось в свя-
зи с переливанием крови от больного сифилисом,
то контроль продолжается в течение 1 года.
Больные первичным серопозитивным и вто-
ричным свежим сифилисом, у которых негати-
вация КСР произошла во время лечения или в
пределах 3 мес после его окончания, должны
находиться на клинико-серологическом контро-
ле в течение 1 года. При замедленной негатива-
ции КСР (в сроки от 3 месяцев до 1 года) сро-
ки контроля продлеваются до 2 лет.
Для больных вторичным рецидивным, третич-
ным, скрытым (ранним, поздним, неуточненным),
висцеральным и нейросифилисом предусмотрен
3-летний срок диспансерного наблюдения. Кли-
нико-серологическое обследование проводят
ежеквартально в течение 2 лет и 1 раз в 6 мес на
третьем году наблюдения.
Если негативация КСР, РИТ и РИФ (под-
твержденная повторно отрицательными резуль-
татами через 6 месяцев) произошла ранее, чем
через 3 года, то пациент может быть снят с уче-
та до окончания этого срока.
Лица с серорезистентностью, при благоприят-
ных данных обследования специалистов, нахо-
дятся на клинико-серологическом контроле 5
лет. По усмотрению врача срок контроля этим
пациентам может быть продлен.
Детям, получившим профилактическое лече-
ние в связи с заболеванием матери, рекоменду-
ется контроль в течение 1 года. Дети, не подле-
жащие профилактическому лечению, также
должны находиться на контроле 1 год.
Дети, получившие специфическое лечение по
поводу как раннего, так и позднего врожденно-
го сифилиса, подлежат клинико-серологическому
наблюдению в течение 3 лет. В течение первых
2 лет КСР исследуют 1 раз в 3 месяца, на тре-
тьем году 1 раз в 6 мес.
Детям, получившим лечение по поводу при-
обретенного сифилиса, клинико-серологическое
наблюдение проводится так же, как взрослым.
При возникновении клинического или серо-
логического рецидива больные подлежат обсле-
дованию терапевта (с рентгенографией органов
грудной клетки), невропатолога, окулиста, ото-
ларинголога; целесообразно произвести спинно-
мозговую пункцию. Лечение проводится в
объеме, предусмотренном для вторичного реци-
дивного сифилиса по любой из методик с на-
значением неспецифических средств.
Серорезистентность после законченного лече-
ния по поводу ранних стадий сифилиса устанав-
ливается в тех случаях, когда в течение года после
окончания терапии РСК с трепонемным и кар-
диолипиновыми антигенами остается стойко
положительной без тенденции к снижению титра
реагинов. В этих случаях назначается дополни-
тельное лечение.
Если через год после полноценного лечения
негативация РСК не наступила, но наблюдает-
ся снижение титра реагинов (по меньшей мере,
в 4 раза) или снижение степени позитивности
РСК от резко положительной до слабо положи-
тельной, эти случаи рассматривают как замед-
ленную негативацию серореакций, и наблюде-
ние продолжают еще 6 мес. Если в течение этого
времени негативация КСР не наступает, прово-
дят дополнительное лечение.
У детей вопрос о серорезистентности целесо-
образно решать через 6 мес после окончания
лечения. При отсутствии к этому сроку сниже-
ния позитивности КСР проводят дополнитель-
ное лечение, при наличии такого снижения ос-
тавляют ребенка без лечения еще 6 мес.
Дополнительное лечение в стационаре прово-
дится одним курсом пенициллина в высоких
дозах в сочетании с неспецифической терапией.
Бензилпенициллин применяют внутримышечно
в дозе 1 000 000 ЕД 6 раз в сутки в течение 28
дней параллельно с назначением неспецифичес-
ких средств. Целесообразно применение 1—2
курсов висмутовых препаратов, если раньше они
не применялись, и при отсутствии противопо-
казаний.
Амбулаторный вариант дополнительного лече-
ния включает 10 инъекций бициллина 1,3 или
5 в дозах 1 200 000 ЕД, 1 800 000 ЕД или 1 500
000 ЕД соответственно, 2 раза в неделю в соче-
тании с неспецифической терапией, например,
инъекциями пирогенала.
Детям дополнительное лечение желательно
проводить в условиях стационара пеницилли-
ном в течение 28 дней (дозы описаны выше),
в сочетании с неспецифической терапией. Же-
лательно также использование висмутовых
препаратов.
468 Венерология
При непереносимости пенициллина дополни-
тельное лечение проводится антибиотиками ре-
зерва. При отсутствии эффекта от дополнитель-
ного лечения повторно дополнительное лечение
не назначается. Срок клинико-серологического
контроля исчисляется от момента окончания
основного лечения.
При сохранении положительного КСР после
полноценного лечения поздних форм сифилиса
вопрос о серорезистентности не ставится и до-
полнительное лечение не проводится.
РИФ и РИТ (РПГА, ИФА) исследуют у боль-
ных всеми формами сифилиса через 6 месяцев
после окончания лечения и затем ежегодно до
снятия с учета.
Снятие с учета
По окончании клинико-серологического на-
блюдения больные всеми формами сифилиса
подлежат рентгенологическому и полному кли-
ническому (осмотр невропатолога, окулиста,
отоларинголога и терапевта) обследованию. Не-
обходимо сопоставить данные этого обследова-
ния с данными, полученными при установле-
нии первоначального диагноза, отраженными в
выписке из истории болезни.
Ликворологическое обследование при снятии
с учета проводится пациентам, лечение которых
было начато по поводу раннего или позднего
нейросифилиса; лицам, у которых в процессе
клинико-серологического контроля появились
какие-то клинические проявления специфичес-
кого поражения нервной системы; лицам с се-
рорезистентностью, сохраняющейся к концу
клинико-серологического наблюдения.
При снятии с учета детей, получивших лече-
ние по поводу врожденного и приобретенного
сифилиса, рекомендуется обследование, включа-
ющее консультации педиатра, невропатолога,
окулиста, отоларинголога, рентгенограммы труб-
чатых костей (при наличии в анамнезе костной
патологии), постановку КСР, РИТ, РИФ
(ИФА).
Критерии излеченности
больных сифилисом
1. Раннее начало противосифилитического ле-
чения.
2. Качество лечения: его соответствие суще-
ствующим схемам, достаточные дозы медика-
ментов, регулярность введения препаратов, про-
должительность интервалов между курсами,
применение методов иммуностимулирующей те-
рапии, соблюдение больными рекомендованно-
го режима.
3. Ближайшие терапевтические результаты:
сроки исчезновения бледных трепонем из сифи-
лидов, время эпителизации или рубцевания
твердого шанкра, сроки исчезновения розеол,
рассасывания папул, негативации серологических
реакций.
4. Отсутствие каких-либо признаков болезни
после окончания лечения и на протяжении все-
го периода диспансерного наблюдения, продол-
жительность которого зависит от стадии болез-
ни к началу лечения.
5. Характер течения заболевания: наличие или
отсутствие рецидивов (клинико-серологических
или только серологических), осложнений во
время лечения, возникновение интеркуррентных
заболеваний.
6. Состояние спинномозговой жидкости через
год после окончания лечения у пациентов, на-
чавших лечение по поводу нейросифилиса, у лиц
со специфическими поражениями нервной сис-
темы, у больных с серологической резистентно-
стью.
7. Состояние внутренних органов и нервной
системы пациентов при переводе на клинико-
серологический контроль и перед снятием с дис-
пансерного наблюдения. Офтальмолог обраща-
ет внимание на состояние зрительного нерва
(сифилитический неврит, первичная табетичес-
кая атрофия); отоларинголог — на воздушную и
костную проводимость; рентгенолог проводит
рентгеноскопию грудной клетки (состояние аор-
ты, неравномерное обызвествление по контуру
восходящего отдела). Терапевт и невропатолог ис-
ключают или подтверждают специфическое по-
ражение внутренних органов или нервной сис-
темы.
Больные сифилисом работники детских уч-
реждений и предприятий общественного пита-
ния допускаются к работе сразу после выпис-
ки из стационара по окончании лечения, а
получавшие амбулаторное лечение — после ис-
чезновения всех клинических проявлений за-
болевания.
Дети, получившие лечение по поводу приоб-
ретенного сифилиса, допускаются в детские уч-
реждения после окончания лечения.
Излеченность конкретного больного являет-
ся сложным вопросом и решается индивидуаль-
но, что требует от врача учета многочисленных
факторов и различных показателей. Заключение
выносится по совокупности критериев.
Глава 38. Сифилис 469
Профилактика
сифилиса
Комплекс профилактических мероприятий при
сифилисе включает в себя лечебно-диагности-
ческие, диспансерные, противоэпидемические,
юридические, в том числе и законодательные,
мероприятия. Учитывая социальную значимость
сифилиса, его профилактика должна быть обще-
ственной и личной.
Общественная
профилактика сифилиса
Общественная профилактика сифилиса предус-
матривает:
• обязательный учет больных сифилисом и
другими инфекциями, передаваемыми половым
путем;
• обязательное привлечение к лечению боль-
ных манифестными и скрытыми формами сифи-
лиса в течение первых суток с момента установ-
ления диагноза; амбулаторное лечение проводится
социально-адаптированным лицам;
• обследование всех лиц, находившихся в по-
ловом и тесном бытовом контакте с заболевши-
ми сифилисом;
• регулярное проведение профилактических
осмотров работников детских учреждений, пи-
щевых объектов, автопредприятий;
• активное выявление больных сифилисом
среди групп риска (проституток, гомосексуали-
стов, наркоманов, лиц без определенного места
жительства);
• обязательное обследование всех доноров с
исследованием крови на серологические реак-
ции;
• серологическое обследование на сифилис всех
взрослых больных, находящихся на обследова-
нии и лечении, в общесоматических стационарах;
• применение единых методов диагностики и
схем терапии больных сифилисом и при пока-
заниях — контактных с больными лиц;
• принудительное освидетельствование и лече-
ние лиц, которым такие меры показаны, но они
уклоняются от добровольного их применения;
• контрольное наблюдение за пациентами
после окончания лечения до снятия их с учета;
• сохранение врачебной тайны;
• проведение мероприятий по повышению
квалификации медицинского персонала лечеб-
но-профилактических учреждений недерматове-
нерологического профиля по вопросам клини-
ки, диагностики и профилактики инфекций,
передаваемых половым путем;
• пропаганда санитарно-гигиенических знаний
по профилактике венерических заболеваний
(внутри- и внедиспансерная, среди населения);
• участие в борьбе с распространением инфек-
ций, передаваемых половым путем, других за-
интересованных ведомств и организаций.
Индивидуальная
профилактика сифилиса
Индивидуальная (личная) профилактика осуще-
ствляется как в пунктах профилактики, так и са-
мостоятельно лицом, имевшим случайную поло-
вую связь и таким образом подвергшимся
опасности заражения сифилисом или другой бо-
лезнью, передаваемой половым путем. Аутопро-
филактика проводится с помощью индивидуаль-
ных (карманных) профилактических средств в
соответствии с прилагаемой к ним инструкци-
ей. К таким средствам относятся хлоргексидин
биглюконат (гибитан), цидипол, мирамистин.
Профилактика в значительной степени обеспе-
чивается использованием барьерных методов
контрацепции — презервативов.
Личная профилактика венерических заболева-
ний у мужчин и женщин проводится в кожно-
венерологических и лечебно-профилактических
учреждениях, которые имеют в своем составе кож-
но-венерологические отделения и кабинеты, сила-
ми среднего медицинского персонала в учрежде-
ниях, имеющих стационар или профилактический
пункт — круглосуточно, а в остальных — в рабо-
чее время. Значимость индивидуальной профилак-
тики зависит от степени ее эффективности и при-
емлемости населением. Профилактика ИППП с
помощью индивидуальных средств, портативных
комплектов, устройств, приспособлений, флаконов,
карманных аптечек наиболее оптимальна, а ее эф-
фективность обеспечивается специфической ак-
тивностью содержащихся в них антисептических
веществ и простотой применения аппаратов.
Методика оказания
профилактической помощи мужчинам
1. Посетитель моет руки, выпускает мочу, за-
тем тщательно обмывает теплой водой с мы-
лом половой член, мошонку, бедра и промеж-
ность.
2. После обсушивания салфеткой те же места
протирают ватным тампоном, пропитанным ра-
створом 0,05 % хлоргексидина биглюконата
(гибитан) или 0,01 % раствором мирамистина
(септиком), либо раствором сулемы 1: 1000.
470 Венерология
3. Из кружки Эсмарха производят промыва-
ние передней части мочеиспускательного кана-
ла 0,05 % раствором хлоргексидина биглюконата
или 0,01 % раствором мирамистина, пропуская
до 0,5 л промывной жидкости.
4. После промывания в уретру вводят 1-2 мл
0,05 % раствора хлоргексидина биглюконата
или 1—2 мл 0,01 % мирамистина, либо с помо-
щью глазной пипетки вводят в уретру 6—8 ка-
пель 2—3 % водного раствора протаргола. Затем
наружное отверстие уретры слегка зажимают на
2—3 минуты и рекомендуют посетителю после
выпускания раствора не мочиться в течение 2—
3 часов.
5. Посетителю выдают стерильную марлевую
салфетку для защиты половых органов от заг-
рязненного белья, которое рекомендуют сменить
при первой возможности.
Методика оказания
профилактической помощи женщинам
1. Посетительница выпускает мочу, тщатель-
но моет руки, а затем обмывает теплой водой с
мылом половые органы, бедра, промежность,
анальную область.
2. После обсушивания салфеткой те же места
протирают ватным тампоном, пропитанным ра-
створом 0,05 % хлоргексидина биглюконата или
0,01 % раствором мирамистина, или раствором
сулемы 1 : 1000.
3. Производят спринцевание влагалища 0,05 %
раствором хлоргексидина биглюконата или
0,01 % раствором мирамистина, при этом во
влагалище впрыскивают 150—200 мл, а в урет-
ру — 1 мл одного из названных растворов. При
отсутствии этих препаратов спринцевание вла-
галища производят раствором перманганата ка-
лия 1:6000 с последующим введением в уретру
с помощью глазной пипетки 8—10 капель 1—2 %
раствора азотнокислого серебра и смазыванием
шейки матки, слизистой влагалища 2 % раство-
ром нитрата серебра.
4. Посетительнице дают стерильную салфетку
для защиты половых органов от загрязненного
белья, которое рекомендуют сменить.
Профилактическая обработка 0,05 % раство-
ром хлоргексидина биглюконата или 0,01 % ра-
створом мирамистина предупреждает развитие
венерических болезней, если она проведена не
позднее, чем через 2 часа после полового сноше-
ния. Растворы хлоргексидина биглюконата и
мирамистина можно использовать не только в
пунктах противовенерической помощи, но и са-
мостоятельно для личной профилактики теми,
кто имел случайные половые связи.
Современный подход
к профилактике сифилиса
Современный подход к профилактике сифили-
са включает в себя три основных направления
(М. Е. St. Louis, 1996):
• Использование элементов или средств-
программ профилактики, таких как скри-
нинг, лечение, уведомление партнеров или
консультирование по вопросам изменения по-
ведения.
• Выбор основных профилактических мероп-
риятий и стратегии их приложения в данной
эпидемиологической ситуации для получения
наибольшего эффекта в отношении распростра-
нения болезни.
• Установление стратегических связей и со-
трудничества с другими пограничными програм-
мами и инициативами органов здравоохранения
для помощи в выполнении реальной работы по
профилактике сифилиса.
Существует много различных путей классифи-
кации потенциальных разделов программы про-
филактики сифилиса (WHO: Geneva, 1985;
W. Cates, S. Meheus, 1990).
Первое направление
профилактики сифилиса
Первое направление — «основные элементы»
профилактики — это первостепенные меропри-
ятия, включающие прямое взаимодействие с
больными ИППП, лицами высокого риска
ИППП или работниками социальных служб для
выявления и лечения инфекций {вторичная про-
филактика) или для влияния на вероятность
передачи или приобретения инфекции {первич-
ная профилактика).
Перечень профилактических мероприятий,
которые могут обеспечить наиболее эффектив-
ное и своевременное лечение (вторичная профи-
лактика) или уменьшить частоту инфицирующих
контактов (первичная профилактика), изложен
ниже.
Основные элементы и мероприятия
программы по профилактике сифилиса
Первичная профилактика
• Санитарное просвещение и информирование
населения об ИППП, их последствиях и профи-
лактике.
• Работа с группами повышенного риска в
сообществах (специфические местности и специ-
фические субпопуляции).
Глава 38. Сифилис
471
• Содействие изменению поведения лиц, от-
носящихся к группам риска (больные нарколо-
гических или ИППП-клиник).
• Уменьшение числа половых партнеров.
• Избегание сексуальных партнеров высоко-
го риска.
• Повышение частоты использования презер-
вативов.
• Увеличение степени доступности презерва-
тивов и их пропаганда.
Вторичная профилактика
• Улучшение качества диагностики и лечения
в специальных учреждениях (ИППП-клиниках).
• Увеличение количества учреждений, работ-
ников социальных служб и/или часов для ока-
зания помощи при ИППП (объем оказанной
помощи, доступность помощи).
• Улучшение доступности услуг ИППП-кли-
ник.
• Улучшение качества, доступности и своев-
ременности лабораторных услуг.
• Эпидемиологическое лечение.
• Массовое лечение.
• Уведомление партнеров.
• Скрининг на инфекцию.
• Обязательная регистрация случаев болезни.
• Содействие лицам с ИППП, нуждающимся
во врачебной помощи.
• Модификация экономических аспектов при
оказании помощи больным ИППП (напр., оп-
лата, материальное поощрение).
Первичная профилактика сифилиса достигает-
ся путем снижения вероятности сексуальных кон-
тактов между инфицированными и неинфициро-
ванными лицами или снижения риска передачи,
если такие контакты происходят, с помощью пре-
зерватива. Задачи первичной профилактики сифи-
лиса идентичны задачам профилактики ВИЧ-ин-
фекции и других ИППП. Мероприятиями,
первичной профилактики, являются просвещение
в области охраны здоровья и акцентирование вни-
мания общества на опасности сифилиса и других
ИППП, их последствиях и их профилактике; ра-
боту с группами повышенного риска в сообществах
(специфические местности и субпопуляции); ин-
дивидуальное или групповое консультирование,
направленное на изменение поведения лиц из
групп риска, обычно в ИППП-клиниках или в
специальных учреждениях (напр., исправитель-
ных). Профилактические мероприятия могут ока-
зать влияние на изменение поведения — от про-
явления желания обратиться за помощью при
возникновении симптомов и признаков заболева-
ния до долговременного изменения сексуального
поведения. Усилия профилактики должны быть
направлены на разъяснение полезности и доступ-
ности презерватива, особенно при сексуальном по-
ведении высокого риска.
Возможно, самым основным элементом кон-
троля и профилактики сифилиса является адек-
ватная терапия инфицированных, которые доб-
ровольно обращаются за лечением. Эта задача
актуальна, в первую очередь, для специализи-
рованных ИППП-клиник. Попытки улучшить
работу клиник могут быть направлены либо на
улучшение качества обслуживания, предлагаемо-
го в ограниченном количестве в местностях с
высоким уровнем ИППП, либо на увеличение
объема помощи больным ИППП (увеличение
количества или типов клинических учреждений,
в которых можно диагностировать и лечить си-
филис, увеличение часов работы, оборудование
мобильных клиник), либо на приемлемость об-
служивания (конфиденциальность, недостаточ-
ное воспитание, осуждение со стороны работни-
ка социальной службы).
Скрининг был и остается необходимым ком-
понентом профилактики сифилиса. Тесты серо-
логического скрининга на сифилис являются
недорогими, чувствительными и довольно спе-
цифичными. Однако крупномасштабный скри-
нинг становится менее эффективным при сни-
жении уровня распространения болезни.
Уведомление и лечение сексуальных партнеров
больных сифилисом является основным сред-
ством контроля сифилиса.
Эпидемиологическое лечение проводится на
основании наличия сексуального контакта с
больным сифилисом или вероятного заражения
сифилисом, особенно в инкубационном перио-
де, который не сопровождается какими-либо
симптомами или положительными лабораторны-
ми тестами. При эпидемиологическом лечении
задача одновременного лечения большого коли-
чества лиц обычно не ставится.
Массовое лечение, т. е. лечение во время ко-
роткого периода времени большого количества
лиц в данной местности с целью снижения рас-
пространения инфекции является необходимой
частью контроля других эндемических трепоне-
матозов. Массовое лечение можно считать край-
ней формой эпидемиологического лечения, и
концепция его применения основана на попыт-
ке сужения резервуара инфекции.
Надзор за подлежащей регистрации болезнью
является одной из основных функций органов
здравоохранения. Без хорошо организованного
надзора контроль и профилактика лишены не-
обходимой информации.
17 Зак. 1357
472 Венерология
Второе направление
профилактики сифилиса
Второе направление профилактики — тактичес-
кое, нацеленное на повышение эффективности
мероприятий путем оптимизации использования
и сочетания основных элементов, а также совер-
шенствования их применения в соответствии с
конкретной ситуацией.
Второе направление профилактических дей-
ствий против сифилиса направлено на опреде-
ление действенности и улучшение стратегии,
описанной выше. Кроме того, мероприятия это-
го направления должны быть наилучшим обра-
зом связаны с действиями общей программы:
• эпидемиологические исследования и анализ
динамики передачи и персистенции сифилиса в
популяции для определения наиболее эффектив-
ных путей вмешательства;
• целевая деятельность медицинских служб,
направленная на отдельные лица, сообщества,
субпопуляции, поддерживающие персистенцию
сифилиса в сообществах;
• участие сообществ в определении и решении
проблем профилактики сифилиса;
• краткосрочные интенсивные местные уси-
лия по контролю, профилактике или ликвида-
ции передачи болезни путем ограниченной по
времени концентрации персонала и средств;
• основные и дополнительные исследования
для разработки более качественных средств, для
диагностики, лечения и профилактики, включая
вмешательство, направленное на изменение по-
ведения.
Третье направление
профилактики сифилиса
Третье направление — «стратегическая связь и
определение позиции» — объединяет действия по
профилактике сифилиса с другими мероприяти-
ями по охране здоровья и социальными иници-
ативами.
Третьим направлением являются стратегичес-
кие связи и действия по профилактике сифили-
са. В программу включают мероприятия по со-
действию в выполнении и выполнению
поставленных задач; по организации связей с
другими программами и инициативами здраво-
охранения (профилактика ВИЧ-инфекции, ох-
рана здоровья матери и ребенка, охрана здоро-
вья меньшинств).
Глава 39
Гонорея
Этиология 473
Классификация 473
Клиника гонореи у мужчин 474
Клиника гонореи у женщин 477
Клиника гонореи у девочек 479
Диагноз 480
Лечение 481
Профилактика гонореи у детей 496
Гонорея — инфекционное заболевание, вызыва-
емое специфическим возбудителем — гонокок-
ком, передающееся преимущественно половым
путем и характеризующееся, в основном, пора-
жением слизистых оболочек мочеполовых орга-
нов. Наблюдаются также гонококковые пораже-
ния слизистой полости рта и прямой кишки, что
выявляется после орогенитальных или гомосек-
суальных контактов.
Источником заражения являются, главным
образом, больные хронической гонореей, преиму-
щественно женщины, так как у них хронический
процесс протекает почти незаметно, более дли-
тельно, труднее диагностируется. Больные острой
и подострой гонореей при наличии островоспа-
лительного процесса обычно избегают половых
связей. Гонорея передается почти исключитель-
но половым путем. В отдельных случаях возмож-
но внеполовое заражение через белье, губки, по-
лотенца, на которых сохранился невысохший
гонорейный гной. Заражение новорожденного
может произойти во время родов при прохожде-
нии плода через родовые пути больной матери.
Этиология
Возбудителем гонореи является Neisseria
gonorrhoeae — грамотрицательный диплококк,
имеющий форму кофейных зерен, обращенных
своей вогнутой поверхностью друг к другу. Го-
нококки имеют хорошо выраженные трехслой-
ную наружную стенку и цитоплазматическую
мембрану, цитоплазму с рибосомами и ядерной
вакуолью. Гонококки обычно располагаются
внутриклеточно в протоплазме лейкоцитов, обыч-
но группами, но иногда можно видеть и внекле-
точных гонококков. Исследования гонококков в
последние годы указывают на изменения их био-
логических свойств (наличие капсул, фагосом, 0-
лактамазы, снижение чувствительности к анти-
биотикам, появление L-форм). Гонококки
поражают слизистые оболочки мочеиспускатель-
ного канала, влагалища, прямой кишки, полос-
тей рта, носа, гортани. Процесс может распрост-
раняться на предстательную железу, семенные
пузырьки, придатки яичек, яички, семявынося-
щие протоки, а у женщин — на матку, яичники,
маточные трубы. Распространяясь по кровяному
руслу, гонококки могут иногда вызывать гоно-
кокковый сепсис и метастазы в различные орга-
ны. При гонококковой бактериемии поражают-
ся суставы, глаза, плевра, эндокард, мышцы,
кости, нервы. У новорожденных поражаются гла-
за, развиваются конъюнктивит, кератит.
Классификация
Классификация гонококковой инфекции, пред-
ставленная в Международной статистической
классификации болезней X пересмотра, отлича-
ется от таковой, принятой в России и странах
СНГ.
473
474 Венерология
Международная статистическая
классификация болезней и проблем,
связанных со здоровьем.
Десятый пересмотр (МКБ-10)
Инфекции, передающиеся преимущественно по-
ловым путем (А50 — А64)
А54 Гонококковая инфекция
AS4.0 Гонококковая инфекция нижних отделов
мочеполового тракта без абсцедирования пери-
уретральных и придаточных желез
Гонококковый: цервицит БДУ*, цистит БДУ,
уретрит БДУ, вульвовагинит БДУ.
Исключена: с: — абсцедированием мочеполовых
желез (А 54.1), периуретральным абсцессом (А 54.1)
AS4.1 Гонококковая инфекция нижних отделов
мочеполового тракта с абсцедированием пери-
уретральных и придаточных желез
Гонококковый абсцесс бартолиновых желез.
А54.2 Гонококковый пельвиоперитонит и дру-
гая гонококковая инфекция мочеполовых органов
Гонококковый (ое): эпидидимит (№ 51.1), вос-
палительное заболевание тазовых органов у жен-
щин (№ 74.3), орхит (№ 51.0), простатит (№ 51.0).
Исключен: гонококковый перитонит (А 54.8).
А54.3 Гонококковая инфекция глаз
Гонококковый конъюнктивит (Н 13.1), ири-
доциклит (Н 22.0).
Гонококковая офтальмия новорожденных.
AS4.4 Гонококковая инфекция костно-мышеч-
ной системы
Гонококковый: артрит (М 01.3), бурсит (М
73.0), остеомиелит (М 90.2), синовит (М 68.0),
теносиновит (М 68.0).
А54.5 Гонококковый фарингит
А54.6 Гонококковая инфекция аноректальной
области
А54.8 Другие гонококковые инфекции
Гонококковый (ая) (ое): абсцесс мозга (G 07),
эндокардит (I 39.8), менингит (G 01), миокардит
(I 41.0), перикардит (I 32.0), перитонит (К 67.1),
пневмония (J 17.0), сепсис, поражение кожи.
Исключен: гонококковый пельвиоперитонит
(А 54.2).
А54.9 Гонококковая инфекция неуточненная
Примечание: * БДУ — без дополнительных уточнений
Клиника
гонореи у мужчин
Гонорейный уретрит — первичная и наиболее рас-
пространенная форма гонококковой инфекции
у мужчин, протекающая в разнообразных фор-
мах. С клинической точки зрения оправдано
выделение переднего уретрита и заднего уретри-
та. По давности заболевания и выраженности
клинической картины выделяют следующие
формы гонореи:
• свежая гонорея (давность заболевания до 2-
х месяцев): острая, подострая, торпидная;
• хроническая гонорея (давность заболевания
свыше 2-х месяцев или неустановленная дав-
ность заболевания).
Свежий острый передний
гонорейный уретрит
Инкубационный период составляет 3—5 дней.
Первичные симптомы — легкий зуд и жжение в
области наружного отверстия мочеиспускательно-
го канала, особенно при мочеиспускании. Затем
появляется отечность, а при надавливании на
мочеиспускательный канал из наружного отвер-
стия появляются скудные слизисто-гнойные вы-
деления. Через 1—2 дня резко усиливаются сим-
птомы болезни: наружное отверстие
гиперемировано, отделяемое становится гнойным.
Крайняя плоть и головка полового члена гипе-
ремированы и отечны. Мочеиспускательный ка-
нал инфильтрирован, при пальпации болезнен-
ный. Пациенты жалуются на режущую, жгучую
боль по всей уретре при мочеиспускании, боле-
вые ощущения при эрекции. При двустаканной
пробе первая порция мочи мутная, вторая про-
зрачная. Объясняется это тем, что первая порция
мочи смывает весь гной в канале и становится
мутной. Вторая порция мочи, проходя по очис-
тившейся от гноя уретре, остается прозрачной.
Свежий острый задний
гонорейный уретрит
Часто гонококковый процесс может перейти на
заднюю уретру. Это происходит при сниженной
сопротивляемости организма, частых половых воз-
буждениях, приеме алкоголя, неправильной мест-
ной терапии (уретроскопия, бужирование, катете-
ризация). С развитием заднего уретрита течение
заболевания осложняется. Кроме клинических сим-
птомов переднего уретрита, у больных появляют-
ся новые признаки и субъективные ощущения:
• частые позывы к мочеиспусканию (каждый
час, а в тяжелых случаях — каждые 15—20 минут);
• болезненность и появление нескольких ка-
пель крови к концу мочеиспускания (терминаль-
ная болезненность и гематурия);
Глава 39. Гонорея
475
• частые эрекции и поллюции с примесью
крови в сперме (гемоспермия), что связано с
наличием воспаления в семенном бугорке;
• моча во всех порциях мутная, в связи с по-
ступлением гноя из предстательной части урет-
ры в мочевой пузырь.
Свежий подострый гонорейный уретрит: призна-
ки такие же, как и при остром гонорейном урет-
рите, но слабее выражены.
Свежий торпидный гонорейный уретрит харак-
теризуется незначительными субъективными
ощущениями. Больные жалуются на чувство
зуда в мочеиспускательном канале, давящую
боль в промежности, иногда у них возникают
болезненные поллюции. Выделения из мочеис-
пусательного канала скудные или совсем отсут-
ствуют.
Хронический
гонорейный уретрит
Наступает после стихания воспалительных про-
цессов при остром и подостром гонорейном урет-
рите. Переходу свежего острого гонорейного
уретрита в хроническую форму способствуют
следующие причины:
• нерациональное лечение, перерывы в лече-
нии, самолечение;
• нарушение режима во время лечения (упот-
ребление алкоголя, погрешности в диете, поло-
вые сношения);
• аномалии мочеиспускательного канала (па-
рауретральные железы, гипоспадия, сужение
наружного отверстия);
• смешанные мочеполовые инфекции (гон-
ококковые и хламидийные, микоплазменныые,
трихомонадные, вирусные);
• различные хронические заболевания (сахар-
ный диабет, туберкулез, анемии, алкоголизм).
Схема обследования
больного хронической гонореей
1. Анамнез — жалобы на зуд, жжение в моче-
испускательном канале, но менее выраженные,
чем при остром заднем уретрите. При пораже-
нии предстательной части мочеиспускательного
канала наблюдаются расстройства мочеиспуска-
ния и половых функций (учащение и усиление
позывов к мочеиспусканию, боль к концу мо-
чеиспускания, эякуляции болезненные, примесь
гноя и крови в сперме).
2. Осмотр выделений — обычно незначитель-
ные и появляются по утрам; могут полностью
отсутствовать; под влиянием различных причин
выделения могут усилиться и симулировать кар-
тину острого уретрита, однако вскоре самопро-
извольно проходят.
3. Осмотр полового члена — обнаруживаются
пораженные парауретральные ходы в виде крас-
ных точек на головке полового члена, из кото-
рых выдавливаются капельки гноя.
4. Двустаканная проба применяется для суж-
дения о состоянии предстательной части моче-
испускательного канала: отделяемое в этой час-
ти незначительно и смывается первой порцией
мочи, поэтому вторая порция может оставаться
прозрачной и не содержать нитей.
5. Пальпация уретры на буже — определяют
наличие или отсутствие в уретре плотных узел-
ков (литтреитов).
6. Пальпация органов мошонки (придаток,
яичко, семенной канатик) с целью выявления
инфильтратов, спаек, болезненности.
7. Пальпаторное исследование простаты и оп-
ределение величины, формы, консистенции,
болезненности, уплотнений.
8. Массаж простаты и семенных пузырьков;
микроскопическое исследование секрета на лей-
коциты, лецитиновые зерна, гонококки, трихо-
монады, флору.
9. Бактериоскопическое исследование выделе-
ний из уретры.
10. Бактериологическое исследование выделе-
ний из уретры.
Уретроскопия производится с целью уста-
новления характера изменений слизистой мо-
чеиспускательного канала при наличии выде-
лений после антибиотикотерапии. Возможные
патологоанатомические изменения в уретре:
мягкий инфильтрат, твердый инфильтрат, пе-
реходный инфильтрат, воспаленные Моргани-
евы пазухи и устья выводных протоков Лит-
треивых желез.
Диагноз
Диагноз гонореи, независимо от типичности
клинических признаков, может быть поставлен
только при лабораторном обнаружении гонокок-
ков.
При наличии клинических и анамнестических
данных, но при отсутствии гонококков таких лиц
учитывают, как подозрительных на гонорею и
проводят лечение, как при гонорее. Также по-
ступают с выявленными источниками заражения
и контактами даже при отсутствии у них кли-
ники и гонококков.
476 Венерология
Осложнения
гонореи у мужчин
Без лечения больные гонореей длительное вре-
мя являются источниками заражения. Кроме
того, у них часто развиваются осложнения, ко-
торые приводят к тяжелым последствиям. Раз-
личают следующие осложнения гонореи у муж-
чин: баланопостит, фимоз, парафимоз,
лимфангиит, кавернит, колликулит, стриктуры
уретры, эпидидимит, деферентит и фуникулит,
орхит, простатит, везикулит.
Гонорея осложняется воспалением головки
полового члена — баланитом и внутреннего
листка крайней плоти — поститом, Обычно эти
воспаления сочетаются, проявляясь баланопос-
титом. При усилении воспалительных явлений
вовлекается вся крайняя плоть, которая отека-
ет и увеличивается так, что не удается ее оття-
нуть назад и обнажить, развивается фимоз.
Иногда воспаленная и отечная крайняя плоть
ущемляет головку полового члена — развива-
ется парафимоз.
В ряде случаев гонококки проникают в лим-
фатические сосуды полового члена и вызыва-
ют их воспаление — лимфангиит. Клиничес-
ки заболевание проявляется плотными
набухшими тяжами в области уретральной
поверхности полового члена. Кожа над ними
иногда гиперемирована и отечна. Лимфанги-
иты могут протекать с повышением темпера-
туры тела, ознобом.
Кавернит — воспаление пещеристых тел поло-
вого члена — развивается при распространении
гонококкового процесса за пределы околожеле-
зистой клетчатки уретральных желез в пещери-
стое тело мочеиспускательного канала. У боль-
ных возникает боль в половом члене при
эрекции, а в пещеристом теле прощупываются
плотные узелки. При острых кавернитах появ-
ляются резко болезненная инфильтрация, боль
и искривление полового члена при эрекции, зат-
рудненное мочеиспускание.
Колликулит ~ поражение семенного бугорка —
характеризуется клиническими симптомами зад-
него уретрита. Это незначительные выделения из
канала, позывы к мочеиспусканию, терминаль-
ная гематурия. Нередко колликулит протекает
асимптомно и диагностируется только при урет-
роскопии. Колликулит может привести к преж-
девременной эякуляции, импотенции, неврасте-
нии.
Стриктуры — сужения мочеиспускательного
канала. Наиболее важным симптомом стриктур
является затруднение мочеиспускания. Струя
мочи тонкая, а в тяжелых случаях моча выделя-
ется по каплям. Диагноз ставят на основании
анамнеза, жалоб больного, инструментального
исследования канала головчатым бужом или
эластическими бужами, а также рентгенографии
мочеиспускательного канала.
Эпидидимит — воспаление придатков яичек —
наблюдается довольно часто и возникает вслед-
ствие проникновения гонококков в придаток из
предстательной части мочеиспускательного ка-
нала через семявыносящий проток. Заболевание
начинается внезапно с появления боли в облас-
ти придатка яичка и в паховой области. У боль-
ных наблюдаются повышение температуры тела
до 39~40°С, озноб, головная боль, слабость. При
пальпации придаток увеличен, плотный и болез-
ненный. Кожа мошонки напряжена, гиперемии
рована. Гонококковые поражения придатков
приводят к образованию рубцов в протоках при-
датков яичек. В результате этого наступают азо-
оспермия и бесплодие.
Деферентит — поражение семявыносящего
протока, и фуникулит ~ поражение семенного
канатика — наблюдаются одновременно с пора-
жением придатков яичек. Семявыносящий про-
ток пальпируется в виде плотного, болезненно-
го тяжа. В ряде случаев включаются фасции и
мышцы семенного канатика, образуя болезнен-
ный тяж толщиной с палец.
Орхит (поражение яичка) встречается редко.
Клинически при этом боль в мошонке значитель-
ная, а ухудшение общего состояния более выра-
жено, чем при эпидидимите. Иногда воспали-
тельный процесс переходит на оболочки яичка,
вызывая в их полости скопление экссудата, ко-
торый пальпируется как флюктуирующее обра-
зование.
Простатит (воспаление предстательной желе-
зы) протекает остро или хронически. По харак-
теру и степени поражения различают простатит:
катаральный, фолликулярный, паренхиматозный.
При катаральном простатите протоки при-
легающих железистых долек предстательной же-
лезы воспалены, инфильтрированы, в их просве-
те имеется значительное количество лейкоцитов,
слизи, эпителия, гонококков. Больные при этом
жалуются на учащенные позывы к мочеиспус-
канию. Отмечается легкое жжение или щекота-
ние в области промежности, незначительное
ощущение давления в области заднего прохода.
Моча прозрачная с примесью единичных нитей
и хлопьев. Пальпаторно в предстательной железе
никаких изменений не выявляется. Простатит
может быть распознан только при микроскопи-
ческом исследовании секрета, в котором повы-
Глава 39. Гонорея
477
шено количество лейкоцитов, а иногда обнару-
живаются гонококки.
Фолликулярный простатит развивается вслед-
ствие дальнейшего распространения воспали-
тельного процесса с закупоркой выводных про-
токов желез и образованием наполненных гноем
изолированных фолликулов. У больных в обла-
сти промежности появляется чувство жара, бо-
лезненность в конце акта мочеиспускания. При
пальпации предстательная железа может быть
увеличена, прощупываются отдельные болезнен-
ные шаровидные уплотнения.
Паренхиматозный простатит возникает при
вовлечении в процесс мышечно-эластической
стромы предстательной железы. Отдельные вос-
палительные инфильтраты, разрушая паренхи-
му железы, сливаются между собой и образуют
вначале ограниченные гнойные полости. Слива-
ясь между собой, гнойники могут образовывать
один большой абсцесс, который захватывает
целую долю и даже всю железу. Субъективные
и объективные симптомы паренхиматозного
простатита соответствуют симптомам заднего
уретрита. Возможно затруднение мочеиспускания
и даже полная задержка мочи. У больных появ-
ляются ощущения давления и полноты в заднем
проходе, боль при дефекации. Боль нередко
иррадиирует в половой член, крестец, таз. При
пальпации обнаруживается увеличение предста-
тельной железы.
Хронический простатит часто приобретает за-
тяжное течение. Появляются различные функ-
циональные расстройства мочеполового аппара-
та: ослабление эрекции, преждевременная
эякуляция, снижение оргазма. Нередко больные
жалуются на быструю утомляемость, снижение
работоспособности, раздражительность. В секрете
предстательной железы отмечается повышенное
содержание лейкоцитов, уменьшение числа ли-
поидных зерен, а иногда их полное отсутствие.
Гонококки в секрете предстательной железы
больных хроническим простатитом содержатся
редко. Простатит може быть вызван и смешан-
ной гонококковой инфекцией: гонококковой и
хламидийной, микоплазменной, трихомонадной
или другой бактериальной.
Везикулит (воспаление семенных пузырьков)
часто сочетается с простатитом и эпидидимитом.
Клинические проявления везикулита обычно
завуалированы симптомами заднего уретрита,
эпидидимита или простатита. Больные жалуются
на выделения, зуд и боль в мочеиспускательном
канале, боль в конце акта мочеиспускания. Воз-
можны терминальная гематурия, повышенная
половая возбудимость, частые поллюции и эрек-
ции, болезненные эякуляции, а также гной и
кровь в сперме. Семенные пузырьки при вези-
кулите пальпируются выше долей предстатель-
ной железы в виде продолговатых болезненных
образований.
Клиника
гонореи у женщин
Особенностями гонорейной инфекции у женщин
являются: вялое, бессимптомное течение (связано
с пониженной функцией яичников; неадекват-
ной дозой сульфаниламидов, которые снижают
вирулентность гонококков; образованием L-
форм гонококка); инфекция часто протекает как
смешанная (гонорейно-трихомонадная, гонорей-
но-хламидийная, гонорейно-микоплазменная,
гонорейно-кандидозная); инфицируется не-
сколько органов (многоочаговое поражение).
Выделяют гонорею нижнего отдела половых
органов (уретрит, парауретрит, бартолинит, ве-
стибулит, цервицит, эндоцервицит) и гонорею
верхнего отдела половых органов, или восходя-
щую гонорею (эндометрит, сальпингит, сальпин-
гоофорит, пельвиоперитонит).
Гонорея нижнего
отдела половых органов
Гонококковый процесс у женщин при пораже-
нии уретры локализуется в эпителии слизистой
оболочки мочеиспускательного канала.
Уретрит сопровождается зудом в уретре, бо-
лью в начале мочеиспускания и учащенными по-
зывами к нему, а также незначительными, сли-
зистыми, почти бесцветными выделениями. При
уретроскопии определяют изменения слизистой
оболочки мочеиспускательного канала в виде
мягкого инфильтрата.
Парауретрит часто сочетается с уретритом го-
нококковой этиологии. Воспалительный процесс
в протоках определяется по гиперемии их усть-
ев и слизистым или слизисто-гнойным выделе-
ниям, которые появляются при выдавливании в
виде капли. В выделениях обнаруживаются го-
нококки.
Вестибулит (поражение преддверия влагали-
ща) обусловлен гонорейным процессом в ямке
и луковице преддверия влагалища, мелких же-
лезках в области клитора, в бороздках между ма-
лыми половыми губами и в промежуточной ча-
сти луковицы преддверия влагалища. Слизистая
478 Венерология
оболочка преддверия влагалища гиперемирова-
на, отечна, покрыта слизисто-гнойными выде-
лениями. В области ямки, луковицы и вывод-
ных протоков малых преддверных желез имеются
воспалительные явления.
Бартолинит (поражение больших желез пред-
дверия влагалища). Гонококки обычно размно-
жаются в цилиндрическом эпителии выводных
протоков больших желез преддверия. Признаком
поражения протока является гиперемированное
пятно величиной с горошину с темно-красной
точкой в центре, соответствующей устью вывод-
ного протока железы. При пальпации воспален-
ной большой железы преддверия из устья вы-
водного протока удается выдавить каплю слегка
мутной слизи. Если выводной проток закрыва-
ется, образуется различной величины болезнен-
ная киста, наполненная патологическим секре-
том и набухшая по направлению к преддверию
влагалища. Иногда в воспалительный процесс
вовлекаются окружающие ткани, что приводит
к образованию болезненного инфильтрата, ко-
торый располагается сбоку у отверстия влагали-
ща. Вокруг воспаленной железы возникают отек,
гиперемия, болезненность. Наиболее тяжелые
формы бартолинита возникают при смешанной
инфекции — гонококковой и хламидийной.
Цервицит и эндоцервицит (воспаление шейки
и шеечного канала матки) — наиболее распрос-
траненное проявление гонококковой инфекции
у женщин. У больных обнаруживается слизис-
то-гнойное отделяемое из канала шейки матки.
При осмотре влагалищной части шейки матки с
помощью зеркал обнаруживают отек, гипере-
мию, эрозии вокруг наружного отверстия шей-
ки матки в виде «красного венчика». Хроничес-
кий эндоцервицит может протекать без
выраженных клинических симптомов и местных
воспалительных процессов.
Гонорея верхнего
отдела половых органов
Эндометрит (воспаление слизистой оболочки
матки) может быть острым или хроническим.
При остром течении гонорейного эндометрита
повышается температура тела, наблюдается лей-
коцитоз. У больных постепенно усиливается боль
внизу живота, нарушаются сроки менструации.
Наблюдаются слизисто-гнойные или кровянис-
тые выделения из канала шейки матки. Матка
увеличена, тестоватой консистенции, болезнен-
на. При хроническом процессе жалобы менее
выражены, боли внизу живота появляются лишь
при физических напряжениях и половых сноше-
ниях, отмечаются нарушения менструального
цикла. При хроническом процессе матка не-
сколько увеличена и плотна.
Сальпингит является наиболее частым проявле-
нием восходящей гонококковой инфекции. Разли-
чают следующие виды гонококкового поражения
труб матки: катаральный сальпингит, глубокий
гнойный сальпингит, нодозный сальпингит, пио-
сальпинкс, гидросалытинкс, аднекстумор.
При катаральном сальпингите поражается сли-
зистая оболочка труб матки (гиперемия, отек
слизистой оболочки и умеренная секреция); тру-
ба отечная, равномерно утолщенная, пальпиру-
ется при исследовании. В ее просвете скаплива-
ется серозно-гнойная жидкость, нередко
образуются внутренние спайки.
Глубокий гнойный сальпингит характеризуется
переходом воспалительного инфильтрата глубже
в подсерозный и мышечный слои трубы, кото-
рая утолщается и стенки ее уплотняются. При
прогрессировании процесса складки труб отека-
ют, препятствуя прохождению секрета, который
при закрытии воронки растягивает стенки тру-
бы, развиваются гидросальпинкс или пиосальпинкс
с гнойным содержимым. Через брюшной конец
трубы гной может вытекать в полость тазовой
брюшины, где под воздействием гонотоксина
выделяется транссудат с большим количеством
фибрина. Между воспаленной трубой, яичником,
маткой, кишечными петлями, сальником обра-
зуются спайки, что приводит к образованию
конгломератной опухоли, в которой невозмож-
но различить отдельные органы (аднекстумор).
Гонококковые сальпингиты протекают подостро,
вяло, с незначительными симптомами. Больные
предъявляют жалобы на ноющие боли внизу
живота, иногда схваткообразные, усиливающи-
еся при физическом напряжении, во время мен-
струации, при дефекации. Заболевание периоди-
чески обостряется вследствие употребления
алкоголя и после половых сношений.
При гонококковом сальпингоофорите больные
жалуются на ноющую боль внизу живота и в
области крестца, усиливающуюся при напряже-
нии брюшной стенки. Иногда повышается тем-
пература тела до 38~39°С, отмечается озноб.
Нарушаются ритм, интенсивность и длительность
менструаций. Гонококки на поверхности яични-
ка вызывают слущивание эпителия, что в пос-
ледующем из-за слипчивого воспаления между
маточной трубой и яичником приводит к обра-
зованию спаек. Вследствие гонококкового саль-
пингоофорита может развиться непроходимость
Глава 39. Гонорея
479
маточных труб, приводящая к бесплодию жен-
щин, внематочной беременности, ограниченно-
му перитониту.
Гонорейный пельвиоперитонит возникает
вследствие инфицирования брюшины гонокок-
ками из брюшного отверстия маточной трубы,
из вскрывшихся пиосальпинкса, пиовариума, а
также проникновения их из подсерозной осно-
вы маточной трубы по лимфатическим сосудам.
Клиническая картина вначале развивается на
фоне симптомов, характерных для сальпингита.
Больных беспокоят резкие, схваткообразные
боли внизу живота, метеоризм, запоры, тошно-
та, рвота, повышение температуры тела до 38—
40°С. Брюшная стенка напряжена, положитель-
ный симптом Щеткина-Блюмберга. При
бимануальном исследовании в прямокишечном
пространстве определяется выпот, выпячиваю-
щий задний свод влагалища и смещающий мат-
ку вперед и вверх. Над симфизом четко опре-
деляется верхняя граница выпота. Гнойный
выпот может самостоятельно вскрыться в сосед-
ние органы (прямую кишку, мочевой пузырь,
влагалище). Диагноз восходящей гонореи и пель-
виоперитонита ставится на основании данных
анамнеза, клиники и после обязательного обна-
ружения гонококков в мазках или в посеве от-
деляемого мочеполовых органов. Одним из по-
следствий гонококкового пельвиоперитонита
является первичное или вторичное бесплодие.
Клиника
гонореи у девочек
У девочек чаще всего встречается нисходящая
гонорея: уретрит, вестибулит, бартолинит (не
ранее 5-летнего возраста), кольпит, эндоцерви-
цит. Гонококк проявляет тропизм не только к
цилиндрическому эпителию (как у взрослых), но
и к многослойному плоскому эпителию половых
путей. С наступлением месячных увеличивает-
ся опасность развития восходящей гонореи (эн-
дометрит, сальпингит, пельвиоперитонит).
Начальные симптомы гонорейного процесса у
девочек выражены в зависимости от общего со-
стояния организма и патогенных свойств возбу-
дителя. Вследствие выделения гонотоксина у
детей на первый план выступают симптомы по-
ражения ЦНС: бессонница, раздражительность,
снижение аппетита, выраженная лихорадка. На-
чало заболевания проявляется чувством боли,
жжения, зуда в аногенитальной области, повы-
шением температуры и дизурическими явлени-
ями. При свежей острой гонорее в процесс вов-
лекаются кожа малых половых губ, слизистая
оболочка входа во влагалище, само влагалище,
уретра и нижний отдел прямой кишки. Разви-
вается яркая диффузная гиперемия и отечность
клитора, больших половых губ и промежности.
Из влагалища на вульву стекает обильное гной-
ное отделяемое, засыхающее в корки, распола-
гающиеся на гребне больших и малых половых
губ. Гименальное кольцо валикообразно утолще-
но, гиперемировано. Местами на слизистой обо-
лочке вульвы видны небольшие эрозии. Пахо-
вые лимфоузлы увеличены и болезненны.
Вагиноскопия при свежей гонорее позволяет
констатировать отечность и гиперемию слизис-
тых оболочек влагалища, особенно выраженные
в области сводов влагалища и гиперемию шей-
ки матки. Одной из характерных черт гонореи
у девочек является многоочаговость поражения,
т. е. помимо половых органов, гонорейный про-
цесс распространяется на уретру, прямую киш-
ку, значительно реже на слизистые оболочки
носа, глаз. У детей почти не бывает гонорейных
артритов и миозитов.
Уретра вовлекается в процесс почти у всех
девочек, страдающих острым гонорейным вуль-
вовагинитом. Поражаются ее передняя и средняя
части: наружное отверстие уретры расширено,
губки гиперемированы и отечны. При надавли-
вании на нижнюю стенку уретры выделяется
гной. Дизурические явления резко выражены,
вплоть до недержания мочи. Обильные гнойные
бели, стекая из влагалища на промежность и
соприкасающиеся поверхности бедер, раздражают
и мацерируют кожу, способствуют инфицирова-
нию прямой кишки.
Гонорейный проктит распространяется на не-
большой (3—4 см проксимальнее сфинктера)
отрезок прямой кишки. При осмотре устанавли-
вается также отечность и гиперемия складок
вокруг заднего прохода, нередко трещины и
гнойное отделяемое.
При вовлечении в процесс цервикального
канала, помимо отечности и гиперемии влагали-
ща и шейки матки, из зева выделяется обиль-
ный гной. Частота эндоцервицитов колеблется в
зависимости от возраста и общего состояния ре-
бенка. Наиболее часто их обнаруживают у дево-
чек от 3 до 7 лет, а также у детей с сопутствую-
щей туберкулезной интоксикацией или очагами
хронической инфекции. В результате лечения
наступает полная инволюция процесса.
Иногда с самого начала гонорейный процесс
принимает торпидное течение. Хроническая гоно-
480 Венерология
рея чаще, чем свежая, осложняется бартолинита-
ми, скеннитами, эндоцервицитами, особенно у
девочек старшего возраста. При хроническом те-
чении большие и малые половые губы могут ос-
таваться неизмененными, но иногда они пастоз-
ны, с очаговой гиперемией на внутренней
поверхности малых половых губ. Симптоматика
поражения бартолиниевых и парауретральных
желез, скенновых ходов незначительна. Уретра
вовлекается почти всегда, течение стертое, дизу-
рические явления выражены слабо или отсутству-
ют. Отмечается гиперемия и отечность слизистой
оболочки задней спайки и складок гимена, где за-
держивается небольшое количество гнойного от-
деляемого. В значительном количестве гной на-
ходят при вагиноскопии в сводах влагалища. На
слизистой влагалища вместо диффузной гипере-
мии, которая характерна для свежей острой го-
нореи, имеются отдельные очажки застойной
гиперемии и пастозности. Во влагалище может
наблюдаться пролиферация сосочков в виде зер-
нистых точек (гранулезный вагинит), слизистая
напоминает «малину». Влагалищная часть шейки
матки бледная, синюшная, иногда по нижнему
краю зева видна эрозия.
Хронический гонорейный эндоцервицит у ослаб-
ленных девочек может распространяться на эн-
дометрий, чему способствует отсутствие внутрен-
него зева и продолжение складок цервикального
канала в полость матки. Однако вовлечение в
процесс вышележащих отделов половых органов
происходит редко. У менструирующих девочек
может развиться восходящая гонорея с пораже-
нием придатков и тазовой брюшины. Иногда
восходящая гонорея сопровождается нарушени-
ем общего состояния, повышением температуры
тела, резкими болями внизу живота, перитоне-
альными явлениями.
Диагноз
Диагностика гонореи основывается на следую-
щих данных: анамнез заболевания, клиническая
картина, обнаружение возбудителя.
Решающее значение имеют лабораторные ме-
тоды исследования. При подозрении на наличие
гонококковой инфекции исследуют отделяемое
мочеиспускательного канала, парауретральных
протоков, большой железы преддверия влагали-
ща, канала шейки матки, стенок влагалища; сек-
рет предстательной железы, семенных пузырь-
ков, желез и лакун мочеиспускательного канала;
промывные воды прямой кишки.
В клинической практике для диагностики го-
нореи чаще применяют бактериоскопический и
бактериологический методы, реже — иммунофлю-
оресцентный, серологический, иммунофермент-
ный. Новыми и перспективными для диагности-
ки гонореи являются молекулярно-биологические
методы: ПЦР (полимеразная цепная реакция) и
ЛЦР (лигазная цепная реакция).
Бактериоскопическое исследование
При типичной клинической картине заболева-
ния, если бактериоскопическое исследование об-
наруживает гонококк, в применении бактерио-
логического метода нет необходимости.
Материалом для бактериоскопического иссле-
дования является отделяемое слизистой уретры
и прямой кишки у мужчин; выделения уретры,
шейки матки, прямой кишки у женщин. Кроме'
того, исследованию подлежат все материалы,
подозрительные на наличие гонококков.
У мужчин отделяемое слизистой оболочки бе-
рут петлей из глубины уретры, предварительно
очистив ее отверстие тампоном, смоченным изо-
тоническим раствором хлорида натрия. При малом
количестве выделений до взятия материала про-
изводят массаж уретры в дистальном направлении.
У женщин патологическое отделяемое из урет-
ры, бартолиновых желез и парауретральных ходов
берут ложкой Фолькмана или петлей, из шейки
матки — влагалищным пинцетом или петлей.
Материал из прямой кишки получают либо
путем промывания ее ампулы теплым изотони-
ческим раствором хлорида натрия с помощью
катетера с двойным током жидкости, либо со-
скабливая его со слизистой оболочки анального
канала ложкой Фолькмана.
При обследовании на фарингеальную гонорею
специально обработанным ватным тампоном
берут материал из лакун миндалин и с задней
стенки глотки выше нижнего края мягкого неба.
Мазки на одном стекле окрашивают 1 %-ным
водным раствором метиленового синего или
0,5 %-ным водным раствором бриллиантового
зеленого в течение 1 минуты, на другом — по
способу Грама. При окраске по Граму гонокок-
ки ярко-розового цвета (грамотрицательные),
остальные кокки фиолетового цвета (грамполо-
жительные). При окраске метиленовым синим на
бледноокрашенном фоне протоплазмы лейкоци-
тов гонококки выглядят темно-синими, ядра лей-
коцитов — синими. В мазках, окрашенных ме-
тиленовым синим, гонококки лучше выявляются,
а в мазках, окрашенных по Граму, они лучше
дифференцируются. Окончательное заключение
дают только на основании окраски по Граму.
Глава 39, Гонорея 481
Бактериологическое исследование
Показаниями к проведению бактериологическо-
го исследования являются неоднократное полу-
чение отрицательного результата бактериоскопи-
ческого анализа при клиническом,
эпидемиологическом и анамнестическом подозре-
нии на гонорею, наличие в мазках из патоло-
гического материала подозрительных на гонококк
микроорганизмов, установление излеченное™
гонореи. Выделения из уретры засевают на сре-
ду асцит-агар или используют безасцитные пи-
тательные среды. Посев проводят не раньше чем
через 5-7 дней после приема антибактериальных
препаратов или местного применения антисепти-
ческих средств. Для выявления гонококков, про-
дуцирующих р-лактамазу, используют чашечный,
йодометрический и ацидометрический тесты.
Серологическая диагностика
Единственной серологической реакцией, исполь-
зуемой при диагностике гонореи, является РСК
(реакция Борде-Жангу), хотя и она применяет-
ся все реже. Реакцию Борде-Жангу рекоменду-
ется проводить при хронической и осложненной
гонорее, особенно в случаях, когда заболевание
не подтверждено бактериологически.
Серологические исследования (реакция Бор-
де-Жангу, реакция определения гонококкового
антигена), а также кожно-аллергическая проба с
гонококковой вакциной имеют лишь вспомога-
тельное значение и непригодны ни для доказа-
тельства гонококковой инфекции, ни для уста-
новления излеченное™.
Иммунофлюореснентный метод
При помощи этого метода вполне удовлетвори-
тельно можно распознать гонококк. Хорошие
результаты получают при прямой иммунофлю-
оресценции, когда фиксированные над пламенем
спиртовки препараты предварительно окрашива-
ют 1 % спиртовым раствором эозина и метиле-
нового синего, а затем обрабатывают флюорес-
цирующей антисывороткой. При таком методе
получается более характерное свечение, позволя-
ющее отличать гонококки от других микроор-
ганизмов.
Для более раннего выявления гонококков в
отделяемом больных гонореей, особенно женщин,
используют замедленный иммунофлюоресцент-
ный метод. В отличие от обычного бактериоло-
гического исследования, этот метод позволяет
распознать гонококки раньше и преимуществен-
но в тех случаях, когда в исследуемом материа-
ле возбудитель находится не в монокультуре, а
в ассоциации с другими микроорганизмами.
Иммуноферментный анализ
Используется метод определения АГ с помощью
ИФА путем исследования мазков из уретры и
проб мочи. Материал больных исследуют с по-
мощью теста Gonozyme, коммерческого набора
для постановки ИФА. Чувствительность и специ-
фичность теста 95,2 % и 100 % соответственно.
Метод ИФА является дополнительным, быстрым
диагностическим методом для обнаружения
Neisseria gonorrhoeae, особенно при неинвазивном
(пробы мочи) обследовании мужчин.
Молекулярно-биологические методы
Полимеразная цепная реакция (ПЦР) применя-
ется в качестве метода коамплификации для оп-
ределения в уретральных и эндоцервикальных
образцах и пробах мочи Neisseria gonorrhoeae и
Chlamydia trachomatis. Обработанные образцы
амплифицируются в одной реакционной смеси,
содержащей праймеры для обоих микроорганиз-
мов. Продукты ПЦР выявляются колориметри-
чески в лунках микропланшета после гибриди-
зации с зондами, специфичными для каждого
патогена. Разрешающая чувствительность обна-
ружения гонококков в уретральных образцах и
моче у мужчин составляет 98,6 % и 94,2 % со-
ответственно. Разрешающая чувствительность
обнаружения гонококков в моче и эндоцерви-
кальных образцах женщин методом ПЦР состав-
ляет 88,9 % и 95,2 % соответственно.
Лигазная цепная реакция (ЛЦР) также может
применяться для одновременного определения в
одном образце Neisseria gonorrhoeae и Chlamydia
trachomatis. Чувствительность ЛЦР при опреде-
лении Neisseria gonorrhoeae в образцах, получен-
ных от женщин и мужчин, составляет 97,3 % и
98,5 % соответственно, а специфичность в обо-
их случаях превышает 99 %.
Эти данные указывают, что двойная ПЦР и
ЛЦР могут использоваться как высоко чувстви-
тельные методы выявления гонококков и хла-
мидий в одном образце мочи или генитальном
мазке, обеспечивая удобный и дешевый подход
к скринингу на наличие нескольких возбуди-
телей.
Лечение
Успех лечения гонорейной инфекции зависит
от рационального использования всех существу-
ющих методов и средств лечения: антибиоти-
ков, сульфаниламидов продленного действия,
специфической (гонококковая вакцина) и не-
482 Венерология
специфической иммунотерапии (пирогенал,
продигиозан, аутогемотерапия, левамизол, ка-
лия оротат, метилурацил, тактивин, тималин,
реинфузия облученной лазером аутокрови),
физиотерапии, местного лечения, соблюдения
режима и диеты.
Назначению лечения должно предшествовать
клиническое и лабораторное обследование боль-
ных с целью выявления сопутствующих заболе-
ваний (сифилис, хламидиоз, трихомоноз и др.)
и одновременного их лечения.
Комплексное лечение включает следующие виды
терапии:
• этиотропная (для уничтожения возбудителя);
• патогенетическая (для повышения сопротив-
ляемости организма к инфекции);
• симптоматическая (для ликвидации местных
патологических изменений в пораженной ткани);
• индивидуальная терапия с учетом пола, воз-
раста больного, клинической формы, тяжести па-
тологического процесса, наличия осложнений;
• соблюдение режима питания, воздержание от
половых контактов и физических нагрузок;
• одновременное лечение половых партнеров
больного.
Основное значение при лечении гонореи име-
ют антибиотики и сульфаниламидные препара-
ты, оказывающие бактерицидное и бактериоста-
тическое действие на гонококки. При острой
свежей гонорее для получения терапевтическо-
го эффекта достаточно этиотропного лечения.
Больным осложненной, торпидной и хроничес-
кой формами гонореи показана комплексная
этиологическая и патогенетическая терапия.
План рационального лечения
разных форм гонореи
1. Свежая гонорея (острая и подострая) — ан-
тибиотикотерапия.
2. Лечение больных хронической, а также све-
жей торпидной гонореей начинают с иммуноте-
рапии и местного лечения, а затем назначают
антибиотики и продолжают местное лечение.
3. Острая восходящая гонорея, острый эпидиди-
мит, простатит, везикулит — постельный режим,
антибиотики, иммунотерапия и местное лечение.
4. Хронический простатит и везикулит —
лечение как при хронической гонорее, с уче-
том особенностей местной терапии; физиоте-
рапия.
При выборе средств антибактериальной тера-
пии необходимо учитывать чувствительность го-
нококков к лекарственным средствам, показания
и противопоказания к их назначению, фармако-
кинетику, фармакодинамику, механизм и спектр
антимикробного действия, а также взаимодействие
с другими лекарственными препаратами.
Воспалительные явления по истечении 10—12
дней после окончания антибиотикотерапии и ис-
ключения гонококков после провокации следует
рассматривать как постгонорейные. Больного
или больную обследуют и, в соответствии с эти-
ологическим и топическим диагнозом, присту-
пают к лечению.
При остропротекающих неосложненных реци-
дивах гонореи следует применять другой анти-
биотик из группы тетрациклинов, макролидов,
аминогликозидов или их комбинации. У боль-
ных с вялым малосимптомным течением реци-
дива, которое наблюдается чаще у женщин, ан-
тибиотикотерапию следует проводить лишь после
местного лечения и иммунотерапии.
Лечение осложненной и восходящей гонореи
в острой стадии заболевания рекомендуют начи-
нать с введения антибиотиков на фоне аутоге-
мотерапии. Лишь после уменьшения островос-
палительных явлений приступают к иммуно- и
физиотерапии. Местное лечение проводят пос-
ле окончания курса антибиотикотерапии.
Мужчинам и женщинам, предполагаемым источ-
никам заражения или половым контактам, у кото-
рых гонококки не обнаружены, необходимо про-
водить лечение по схемам для хронической гонореи.
Больные острой гонореей с невыявленными
источниками заражения в случае невозможнос-
ти установить за ними диспансерное наблюде-
ние (лица без определенного места жительства,
работы и ведущие аморальный образ жизни)
подлежат превентивному противосифилитичес-
кому лечению.
Больным гонореей с невыявленными источ-
никами заражения, но имеющим постоянное
место жительства и работы, превентивное про-
тивосифилитическое лечение не проводится, но
последующий клинико-серологический контроль
(после лечения гонореи) осуществляется в тече-
ние 3 месяцев.
Общая
характеристика препаратов
для лечения гонореи
Для лечения больных гонореей рекомендуют
применять антибактериальные препараты: груп-
пы пенициллина, левомицетин, препараты тетра-
циклинового ряда (тетрациклин, окситетрацик-
лин, метациклин, доксициклин, вибрамицин),
макролиды (эритромицин, олететрин, эрицик-
Глава 39. Гонорея
483
лин), фторхинолоны, аминогликозиды (сизоми-
цин, канамицин), цефалоспорины (цефалексин,
цефтриаксон и др.), рифампицины (рифампицин,
бенемицин), сульфаниламидные препараты про-
лонгированного действия (сульфамонометоксин,
сульфадиметоксин) и комбинированные сульфа-
ниламидные препараты (бисептол, сульфатон).
Антибактериальные препараты
Антибиотики
Антибиотики пеницилинового ряда
Противопоказанием к применению препаратов
пенициллина является наличие в анамнезе непе-
реносимости к этому антибиотику и новокаину
(при растворении пенициллина в новокаине).
Для предупреждения аллергических реакций
рекомендуется до назначения антибиотиков ус-
тановить их индивидуальную переносимость
пациентом путем проведения аллергологических
тестов (внутрикожных, клеточных и др.). Кро-
ме того за 20~30 минут до введения антибиоти-
ка назначают один из антигистаминных препа-
ратов (димедрол, пипольфен, супрастин, тавегил,
фенкарол и др.) по 1 таблетке.
Пенициллины, тетрациклины, аминогликози-
ды, сульфонамиды, макролиды, левомицетин
назначают после определения чувствительности
к ним гонококов.
Бензилпенициллин. У мужчин с острым и
подострым гонорейным уретритом и у женщин
с острой и подострой гонореей нижнего отдела
мочеполовой системы рекомендуется курсовая
доза бензилпенициллина 3,4 млн ЕД.
При свежих торпидных случаях неосложнен-
ной гонореи, острых осложнениях гонореи у
мужчин, восходящей и хронической гонорее
курсовая доза бензилпенициллина должна быть
4,2-6,8 млн. ЕД (в зависимости от тяжести за-
болевания).
При первой инъекции вводится 600 000 ЕД,
а при последующих — по 400 000 ЕД в физио-
логическом растворе с интервалом в три часа без
ночного перерыва.
В исключительных случаях (невозможность
повторного посещения) у мужчин с острым и
подострым уретритом можно воспользоваться
введением одновременно всей курсовой дозы (3
млн. ЕД) бензилпенициллина с 5 мл собствен-
ной крови или дюрантных препаратов пеницил-
лина.
Пенициллин в курсовой дозе 4 000 000 ЕД
может быть также введен внутримышечно одно-
моментно (по 2 000 000 ЕД в каждую ягодицу),
при этом за 30 минут до инъекции и через каж-
дые 6 часов внутрь назначают этамид по 0,7 г,
суммарная доза этамида составляет 2,8 г.
При хронической гонорее у женщин целесооб-
разно использовать регионарное введение бензил-
пенициллина. С этой целью с начала лечения в
толщу мышечного слоя шейки матки вводят од-
нократно 200 000 ЕД препарата. Одновременно
с этим делают внутримышечные инъекции бен-
зилпенициллина в разовых и курсовых дозах,
рекомендуемых при хронической гонорее.
Детям препарат вводится разовыми дозами по
50 000—200 000 ЕД (в зависимости от возраста)
с интервалом в 4 часа круглосуточно.
Бициллин — 1, 3, 5. Мужчинам с острым и
подострым уретритом и женщинам с острой го-
нореей нижнего отдела мочеполовой системы
вводят в виде 6 инъекций по 600 000 ЕД с ин-
тервалом в 24 часа или по 1 200 000 ЕД через
48 часов в курсовой дозе 3,6 млн. ЕД. Больным
с другими формами гонореи назначают 7—10
инъекций по 600 000 ЕД с интервалом в 24 часа,
в курсовой дозе 4,2—6 млн. ЕД.
Инъекции бициллина производят двумомен-
тно — сначала вводят иглу в наружно-верхний
квадрант ягодицы, а затем, при отсутствии кро-
ви из иглы, вводят антибиотик.
Как исключение, только при свежем и по-
достром гонорейном уретрите у мужчин воз-
можны разовые введения бициллина-3 в дозе
2 400 000 ЕД (по 1 200 000 ЕД препарата в
каждую ягодицу). При этом за 30 минут до
инъекции антибиотика больные получают 1,05
этамида (3 табл.). Затем та же доза этамида
назначается через 3, 6 и 9 часов; всего на курс
4,2 г препарата.
Ампициллин. Больным острой и подострой
гонореей назначается 3,0 г, хронической — 8,0 г
препарата (по 0,5 г через каждые 4 часа в день).
Ампиокс. Больным острой и подострой гоно-
реей препарат назначается внутрь по 0,5 г через
4 часа в течение 4—5 дней; при хронической
гонорее — в течение 5—7 дней. Для детей до 12
лет препарат назначается внутрь из расчета 0,1 г
на 1 кг массы тела, старше 12 лет — в тех же
дозах, что и взрослым.
Если препарат назначается внутримышечно,
суточная доза для новорожденных детей и детей
в возрасте до 1 года — 0,1—0,2 г на 1 кг массы
тела; от 1 до 6 лет — до 0,1 г; от 7 до 14 лет —
0,05 г на 1 кг массы тела; взрослым — 2,0 г в
сутки. Суточную дозу разделяют на 3—4 введе-
ния с интервалом 6—8 часов.
Оксациллин. При острой и подострой формах
заболевания назначается внутрь по 0,5 г 5 раз в
484 Венерология
день, на курс 10,0. При других формах гоно-
реи — 14,0 г.
Препараты ампиокс и оксациллин активны и
по отношению к пенициллиназопродуцирующим
штаммам микроорганизмов.
Карфециллин. Назначают по 0,5 г 3 раза в
день, при свежей неосложненной гонорее —
5,0 г на курс лечения, при хронической и ослож-
ненной — 8,0 г.
Аугментин. Назначается при свежей неослож-
ненной гонорее по 375 мг внутрь каждые 8 ча-
сов. На курс — 1,875 г. При осложненной и хро-
нической гонорее: в течение первых трех дней
по 750 мг каждые 8 часов, остальные два дня —
по 375 мг каждые 8 часов.
Сулациллин. Препарат вводится внутримышеч-
но по 1,5 г с интервалом 8 часов, курсовая доза
при свежих формах 6,0 г; при хронических и
осложненных — 9,0 г.
Левомицетин
Мужчинам с острым и подострым уретритом и
женщинам с острой гонореей нижнего отдела
мочеполовой системы левомицетин назначают
внутрь в курсовой дозе 6 г (по 3 г в день), при
других формах гонореи — 10 г (первые два дня
по 3 г, остальные — по 2 г в день). Разовые
дозы в 0,5 г дают в равные промежутки времени
с ночным перерывом в 7—8 часов, за 30 минут
до еды.
Детям левомицетин назначают по 0,2—0,25 г
4 раза в день. Курсовая доза такая же, как и у
взрослых с острой гонореей (6,0 г).
Антибиотики тетрациклинового ряда
У мужчин с острым и подострым уретритом и у
женщин с острой гонореей нижнего отдела мо-
чеполовой системы тетрациклин, хлортетрацик-
лин и окситетрациклин назначают в курсовой
дозе 5 г (5 млн. ЕД).
При острой осложненной гонорее у мужчин,
восходящей — у женщин, а также при хроничес-
кой гонорее курсовую дозу следует увеличить до
10 г. Первые 2 дня назначают по 0,3 г и после-
дующие дни по 0,2 г 5 раз в день через равные
промежутки времени. Ночной перерыв не дол-
жен превышать 7—8 часов. Для предупреждения
развития кандидоза одновременно следует назна-
чать нистатин по 500 000 ЕД 4 раза в день или
леворин по 250 000 ЕД 4 раза в день.
Метациклин (рондомицин). При свежей острой
и подострой неосложненной гонорее назначается
внутрь после еды в дозе 0,6 г (первый прием),
затем по 0,3 г каждые 6 часов; на курс лечения
2,4 г. При остальных формах гонореи на курс 4,8 г.
У мужчин и женщин со свежей неосложнен-
ной гонореей может быть применено одноднев-
ное лечение метациклином (ровдомицином): боль-
ным свежей острой гонореей препарат дается в
2 приема по 1,2 г после еды с 8-часовым пере-
рывом, на курс лечения 2,4 г. Больным свежей
торпидной гонореей назначают в 3 приема по
1,2 г после еды с 8-часовым перерывом, на курс
лечения 3,6 г, а свежей осложненной гонореей —
в 4 приема по 1,2 г после еды с 8-часовым пе-
рерывом; на курс лечения 4,8 г.
Доксициклин (вибрамицин). Больным свежими
неосложненными острыми и подострыми фор-
мами гонореи препарат назначается внутрь по
0,1 г (первый прием 0,2 г) каждые 12 часов, на
курс лечения 1,0 г. При остальных формах за-
болевания препарат назначают по той же мето-
дике, но на курс 1,5 г.
Макролиды
Эритромицин. Мужчинам с острым и подострым
гонорейным уретритом и женщинам с острой
гонореей нижнего отдела мочеполовой системы
эритромицин назначается внутрь круглосуточно
в курсовой дозе 8,8 млн. ЕД (два дня по 400 000
ЕД 6 раз в сутки и последующие дни по 400 000
ЕД 5 раз в сутки). Больным остальными форма-
ми гонореи — 12,8 млн. ЕД по той же методике.
Макропен. Больным с острой и подострой
гонореей назначается по 400 мг 3 раза в сутки
внутрь в курсовой дозе 3,6 г; при остальных
формах гонореи курсовая доза составляет 6,0 г.
Олететрин. Мужчинам с острым и подострым
гонорейным уретритом и женщинам с острой
гонореей мочеполовой системы олететрин назна-
чается внутрь в курсовой дозе 4 млн. ЕД; при
остальных формах гонореи — 7,5 млн. ЕД и бо-
лее. Первый день дают 1 250 000 ЕД (первый
прием 500 000 ЕД и 3 приема по 250 000 ЕД), а
в остальные дни — по 250 000 ЕД 4 раза в сут-
ки через равные промежутки времени.
Эрацин. Назначают внутрь по 400 мг 4 раза в
сутки через 6 часов. Курсовая доза при свежей
гонорее — 8 г (5дней); при хронической —
11,2 г (7 дней).
Эрициклин. Больным с острым и подострым
течением заболевания препарат назначается по
0,25 г пять раз в день после еды, на курс — 3,0 г;
при остальных формах заболевания — 6,0 г (ос-
ложненных формах — 7,0).
Аза л иды
В качестве препарата выбора для лечения гоно-
реи и сочетанных бактериальных инфекций уро-
генитального тракта эксперты ВОЗ рекоменду-
Глава 39. Гонорея
485
ют азитромицин (сумамед) — антибиотик из
группы азалидов.
Азитромицин. При лечении свежей гонореи на-
значается по 1,5—2,0 г однократно либо в 2 при-
ема по 1,0 г. При свежей торпидной и хроничес-
кой гонорее суммарная доза составляет 2,0 г по
следующей схеме: первый прием 1 г, затем каждые
24 часа по 250 мг до достижения курсовой дозы.
Рокситромицин (мидекамицин, джосамицин)
применяют по 1,0 г однократно при лечении
свежей неосложненной гонореи.
Аминогликозиды
Канамицин — антибиотик широкого спектра дей-
ствия. Применяется в виде моно- или сульфата
канамицина, хорошо растворим в воде. При ост-
ром и подостром гонорейном уретрите у мужчин
и острой гонорее нижнего отдела мочеполовой
системы у женщин канамицин назначается внут-
римышечно по 1 млн ЕД через 12 часов в курсо-
вой дозе 3 млн ЕД, при других формах гонореи —
6 млн. ЕД. При длительном применении препа-
рат может оказывать нефро- и ототоксическое
действие. Канамицин нельзя назначать одновре-
менно с другими антибиотиками, обладающими
ото- и нефротоксическим действием.
Цефалоспорины
Цефалексин (кларцеф, цепорекс) — внутрь в пер-
вые 2 дня по 0,5 г 4 раза в день, с третьего дня
по 0,25 г 4 раза в день. Курсовая доза при све-
жей неосложненной гонорее — 5,0 г, при ослож-
ненной и хронической формах — 7,0 г.
Цефтазидим (фортум) вводится внутримышеч-
но в курсовой дозе 2 г при острой гонорее.
Клафоран при остром и подостром гонорей-
ном уретрите у мужчин вводится внутримышечно
по 1,0 г через 12 часов 2 раза в день, на курс
2,0 г. При остальных формах гонореи — курс
лечения 8,0—10,0 г препарата.
Кетоцеф вводится внутримышечно по 1,5 г
однократно при остром, подостром и хроничес-
ком гонорейном уретрите.
Цефтриаксон (роцефин, лонгацеф) вводится
внутримышечно по 1,0 г через 12 часов 2 раза в
день при остром и подостром гонорейном урет-
рите; на курс 2,0 г.
Цефобид вводится внутримышечно по 1,0 г в
день; на курс лечения при свежих формах —
3,0 г; при всех остальных — 5,0 г.
Сульфаниламидные препараты
К сульфаниламидным препаратам пролонгиро-
ванного действия относятся сульфамонометоксин
и сульфадиметоксин. Назначают при непереноси-
мости антибиотиков и после безуспешной пени-
циллинотерапии. Первые 2 дня назначают по
1,5 г 3 раза в сутки через равные промежутки
времени и в последующие дни — по 1,0 г 3 раза
в сутки. Курсовая доза при острой и подострой
неосложненной гонорее — 15,0 г; при других
формах — 18,0 г.
Бисептол, бактрим. Больным свежей острой и
подострой гонореей препарат назначается по 4
таблетки каждые 6 часов; на курс 7,68 г (16 таб-
леток); при других формах — 9,6 г (20 таблеток).
В порядке исключения у мужчин может быть
назначен укороченный (однодневный) метод лече-
ния бисептолом. При острых и подострых фор-
мах гонореи препарат назначается по 5 таблеток
(2,4 г) в два приема после еды с 8-часовым ин-
тервалом; на курс — 4,8 г. При свежей острой
торпидной и осложненной форме — по 5 табле-
ток (2,4 г) в три приема после еды с 8-часовым
интервалом, на курс 7,2 г.
Берлоцид 960, берлоцид 480. При свежей ост-
рой и подострой гонорее назначается по 2 таб-
летки берлоцида 960 (либо по 4 таблетки берло-
цида 480) через 12 часов в течение 2-х дней; на
курс 8 таблеток берлоцида 960 (или 16 таблеток
берлоцида 480). При хронической гонорее кур-
совая доза составляет до 20 таблеток берлоцида
960 либо 40 таблеток берлоцида 480. При необ-
ходимости может быть назначен укороченный
метод лечения берлоцидом: по 2,5 таблетки бер-
лоцида 960 через 8 часов (на курс 5 таблеток),
либо по 5 таблеток берлоцида 480 через 8 часов
(на курс 10 таблеток).
Сульфатон. Больным свежей острой и подо-
строй гонореей, препарат назначают по 4 таблет-
ки (1,4 г) с интервалом 8 часов, на курс — 4,2 г.
При других формах заболевания — 5,6 г (ослож-
ненных формах — 7,0 г).
Фторированные хинолоны. Синтетические
противомикробные препараты широкого дей-
ствия (фторированные производные оксихино-
лонкарбоновой кислоты), обладающие отлич-
ной способностью проникать в ткани и
жидкости.
Применение фторированных хинолонов про-
тивопоказано беременным и детям до 14 лет.
Спектиномицин (тробицин). Для лечения гонореи,
вызываемой p-лактамазообразующими штаммами
гонококков, применяются более эффективные
антибиотики (спектиномицин, розоксацин, цефа-
лоспорины 2-го и 3-го поколений). На этом фоне
положительно выделяется тробицин (спектиноми-
цин) из группы аминоциклитолов. Тробицин
химически отличается от всех остальных антиби-
486 Венерология
отиков, и его антибактериальная активность на-
правлена специфически на Neisseria gonorrhoeae.
На курс лечения неосложненного гонорейного урет-
рита, цервицита или проктита необходима одно-
кратная внутримышечная инъекция препарата
(2,0 г для мужчин и 4,0 г для женщин). Доза 4,0 г
вводится поровну в верхненаружный квадрант
каждой ягодицы. Для лечения детей достаточно
40 мг препарата однократно.
Рифампицин. Больным свежей острой и подо-
строй и неосложненной формой гонореи препа-
рат назначается внутрь по 0,3 г (первый прием —
0,6 г) каждые 6 часов за 30—60 минут до еды,
на курс 1,5 г. При остальных формах гонореи —
по этой же методике на курс 6,0 г. Беременным
рифампицин противопоказан.
Рекомендуемые препараты
и схемы лечения различных
форм гонореи
Свежая острая неосложненная гонорея:
• абактал (пефлоксацин) — однократно 0,6 г;
• максаквин (ломефлоксацин) — однократно
0,6 г;
• нолицин (норфлоксацин) — однократно
0,8 г;
• ципрофлоксацин (цифран, ципробай) — од-
нократно 0,5 г;
• сифлокс — однократно 0,75 г.
Свежая осложненная
и хроническая гонорея:
• абактал — по 0,6 г 1 раз в сутки, на курс
2,4 г;
• максаквин — по 0,6 г 1 раз в сутки, на курс
2,4 г;
• сифлокс — по 0,5 каждые 12 часов, на курс
4 г;
• ципробай (ципрофлоксацин) — первый при-
ем 0,5 г; затем по 0,25 г каждые 12 часов, на курс
1,25 г;
• таривид (офлоксацин) — первый прием
0,4 г, затем по 0,2 г каждые 12 часов, на курс
1,6.
Иммунотерапия
Иммунотерапия является вспомогательным ме-
тодом лечения гонорейных заболеваний и при-
меняется наряду с другими видами терапии с
целью повышения реактивности организма в
борьбе с инфекцией.
Специфическая иммунотерапия
Вакцинотерапия. Применение гонококковой вак-
цины показано больным после безуспешной ан-
тибиотикотерапии, при вялопротекающих ре-
цидивах, при свежих торпидных и хронических
формах заболевания, мужчинам с осложненной
и женщинам с восходящей гонореей (после
уменьшения острых воспалительных явлений).
В амбулаторных условиях вакцинотерапия у
больных гонореей назначается одновременно с
антибиотиками. В условиях стационара антиби-
отики назначаются во время или в конце вак-
цинотерапии. Вакцину вводят внутримышечно
в ягодичную область или внутрикожно. Реак-
ция на введение гоновакцины бывает общей
(недомогание, головные боли, общая слабость),
температурной, очаговой (усиление выделений,
болезненность в пораженных органах, помутне-
ние мочи) и местной (болезненность в области
инъекций).
При осложненной гонорее вакцинацию следует
начинать с 200—250 млн. микробных тел. При
осложнениях, сопровождающихся высокой тем-
пературой, нарушением общего состояния орга-
низма, резкой болезненностью в пораженном
органе, необходимо воздержаться от применения
вакцины.
При торпидной и хронической формах гоно-
реи начальные дозы гоновакцины — 300—400
млн. микробных тел.
Инъекции гоновакцины делают с интервала-
ми в 1—2 дня в зависимости от реакции, а дозы
увеличивают каждый раз на 150—300 млн. мик-
робных тел. Допустимая максимальная разовая
доза не должна превышать 2 млрд, микробных
тел, а количество инъекций — до 6—8.
У женщин с торпидной и хронической фор-
мами гонореи можно применять очаговую
вакцинацию в подслизистую шейки матки,
уретры. В этих случаях вакцинацию начина-
ют с малых доз: от 50 млн. микробных тел,
постепенно повышая до 150—200 млн. Этот
метод вакцинации может сопровождаться
очень бурной общей и температурной реакци-
ей, наступающей через 20—30 минут после
введения вакцины. Очаговую вакцинацию
следует применять только в условиях стаци-
онара и строго учитывать противопоказания
к ней (беременность, менструация, острый
воспалительный процесс).
У детей начальная доза гоновакцины не дол-
жна превышать 50—100 млн. микробных тел.
Высшая однократная доза 500 млн. микробных
тел. Детям до трех лет вакцинация не прово-
дится.
Глава 39. Гонорея
487
Противопоказания: активный туберкулез,
органические поражения сердечно-сосудистой
системы, гипертоническая болезнь, тяжелые за-
болевания почек и печени, истощение, выражен-
ная анемия, аллергические заболевания, менст-
руация.
Все виды иммунотерапии рациональнее про-
водить до назначения антибиотиков. При ам-
булаторном лечении с целью предупреждения
развития восходящего процесса и ввиду эпи-
демиологической настороженности антибиоти-
ки назначаются одновременно с иммунотера-
пией.
Неспецифическая иммунотерапия
Пирогенал. Показания: свежая торпидная и хро-
ническая формы гонореи и негонорейные воспа-
лительные заболевания нижнего отдела мочепо-
ловых органов, а также осложнения (простатит,
эпидидимит, аднексит, периаднексит и др.).
Противопоказания: острые лихорадочные за-
болевания, беременность, гипертоническая бо-
лезнь, активный туберкулез, менструация. Боль-
ным сахарным диабетом и лицам старше 60 лет
препарат назначают с осторожностью, начиная с
низких доз и повышая их в зависимости от ре-
акции.
Препарат назначают, начиная с 2,5—5,0 мкг у
женщин и с 5,0—7,5 мкг у мужчин в виде внут-
римышечных инъекций, которые повторяют
через 1—2 дня. Дозу пирогенала при последую-
щих инъекциях увеличивают на 2,5—5,0—
10,0 мкг (в зависимости от реакции). Макси-
мальная доза (разовая) не должна превышать
100 мкг. Следует добиваться повышения темпе-
ратуры тела в пределах одного градуса. При чрез-
мерно высоком повышении температуры, резком
ознобе, головной боли, рвоте, боли в пояснице
и внизу живота (эти явления продолжаются
обычно 6—8 часов и исчезают без лечения) ре-
комендуется не снижать дозу препарата, а уве-
личить интервал между инъекциями (на 1—2 дня)
или повторить последнюю дозу.
Пирогенал можно применять одновременно с
гоновакциной. При этом начальная доза пиро-
генала 2,5—5,0 мкг, гоновакцины 200—300 млн.
микробных тел. Затем разовые дозы пирогенала
увеличивают на 5,0—15,0 мкг; гоновакцины на
150-300 млн. микробных тел. Максимальная
доза пирогенала 100 мкг, гоновакцины 1,2 млрд,
микробных тел. Препараты вводят в одном
шприце.
Продигиозан. Показания и противопоказания
к применению те же, что и для пирогенала. Дозу
устанавливают индивидуально, в зависимости от
переносимости препарата, начиная с 15 мкг. За-
тем дозу увеличивают на 10—15 мкг в зависи-
мости от реакции организма (общей, местной,
очаговой). На курс лечения 4 инъекции с интер-
валом 4-5 дней. Максимальная разовая доза
75 мкг.
Аутогемотерапия. Показания: гонорейные ос-
ложнения, сопровождающиеся резкой болезнен-
ностью в пораженном органе, высокой темпера-
турой и тяжелым состоянием больного. После
прекращения болей, снижения температуры,
улучшения общего состояния следует перейти к
более активной иммунотерапии вакциной.
Левамизол применяют внутрь курсами. Назна-
чают по 150 мг 1 раз в сутки в течение 3 дней.
После 4-дневного перерыва цикл повторяют. На
курс лечения — 4 цикла. Левамизол назначают
при повторных заболеваниях гонореей, торпид-
ном и хроническом течении гонококковой ин-
фекции, осложненной гонорее.
Калия оротат больным гонореей назначают
внутрь по 0,5 г 4 раза в сутки в течение 20—30
дней.
Метилурацил принимают внутрь после еды по
0,5 г 2 раза в день, курс лечения 10—14 дней.
Миелопид вводится подкожно в среднюю
треть задней поверхности голени в дозе 6 мг,
через день, на курс 18 мг.
Глицирам назначается по 0,05 г 3 раза в тече-
ние 10 дней.
Тактивина 0,01 % раствор вводится по 0,5 мл
подкожно через день, всего 7-8 инъекций (3,5-
4,0 мл) в зависимости от формы заболевания.
Тимактин — иммунокорректор, который при-
меняется у больных с осложненной гонореей.
Назначается сублингвально по 0,1 г один раз в
три дня, на курс 0,5 г.
Реинфузия облученной лазерным светом соб-
ственной крови. Облучение проводится в течение
4—5 минут с помощью установки Л Г-75, длина
волн 633 нм. Мощность 25 мВт. Процедуру по-
вторяют 2—3 раза с интервалом 3—4 дня (на фоне
антибиотикотерапии).
Биогенные
стимуляторы и ферменты
Биогенные стимуляторы и ферменты применяют
для ускорения рассасывания инфильтратов в уретре
и половых железах. Используют экстракт алоэ,
ФИБС, пеллоидодистиллат, стекловидное тело,
трипсин, химотрипсин в обычных терапевтичес-
ких дозировках. Противопоказания: сердечно-со-
судистые заболевания, артериальная гипертензия,
острые желудочные, кишечные расстройства, не-
488 Венерология
фрозонефрит, цирроз печени, беременность свы-
ше 7 месяцев.
Экстракт алоэ назначают в виде ежедневных
подкожных инъекций по 1 мл; на курс лечения
15—30 инъекций.
ФИБС вводят подкожно по 1 мл 1 раз в день;
на курс 15—20 инъекций.
Пеллоидодистиллат вводят подкожно по 1 мл
1 раз в день в течение 15—20 дней.
Стекловидное тело вводят подкожно ежеднев-
но по 2 мл в течение 15—20 дней.
Трипсин или химотрипсин вводят внутримы-
шечно по 0,005-0,01 г (5-10 мг) 1-2 раза в
день; на курс — от 6 до 15 инъекций.
Плазмол вводят подкожно по 1,0—2,0 мл ежед-
невно или через день; на курс 10 инъекций.
Спленин вводят внутримышечно (или подкож-
но) по 2 мл ежедневно в течение 10 дней.
Экстракт плаценты — по 1—2 мл подкожно
ежедневно или через день; на курс 15—20 инъ-
екций.
Особенности лечения
некоторых форм гонореи
Лечение рецидивов гонореи
Антибиотики следует назначать по данным ан-
тибиотикограммы после определения чувстви-
тельности гонококка к антибактериальным пре-
паратам. На фоне применения антибиотика
одновременно внутримышечно вводится трипсин
по 5 мг 2 раза в день или 10 мг 1 раз в день в
течение 5—6 дней.
Доксициклин (вибрамицин) по 0,2 г каждые 12
часов; на курс лечения 1,2 г препарата. Одновре-
менно или за 4 дня до назначения этиотропной
терапии назначают левамизол по 0,15 г ежеднев-
но; на курс 8 капсул или таблеток (1,2 г).
Метациклин (рондомицин) по 0,3 г через 5 ча-
сов с 9-часовым ночным перерывом; на курс
лечения 4,8 г.
Левамизол назначают одновременно или за 3
дня до назначения этиотропной терапии по 1
таблетке ежедневно; на курс 8 таблеток (1,2 г).
Ампициллин по 0,5 г 5 раз в день через рав-
ные промежутки времени с 8-часовым ночным
перерывом; на курс 6,0 г.
Ампиокс по 0,5 гв капсулах через 4 часа 5 раз
в день; на курс 5,0 г.
Цефатоксим (клафоран) по 1,0 г внутримышеч-
но через 12 часов 2 раза в день; на курс 2,0 г.
Кетоцеф внутримышечно однократно 1,5 г.
Цефтриаксон (роцефин, лонгацеф) по 1,0 г 2
раза в день через 8 часов внутримышечно; на
курс 2,0 г.
Лечение гонореи у беременных
Лечение женщин, желающих сохранить беремен-
ность, должно проводиться в условиях стациона-
ра. Бензилпенициллин показан при любом сроке
беременности. Назначение препарата беременным
исключается в случаях индивидуальной неперено-
симости, тогда применяют эритромицин, левоми-
цетин или сульфаниламиды. Препаратами выбо-
ра при беременности являются некоторые
цефалоспорины, макролиды, спектиномицин, бен-
зилпенициллин. Во второй половине беременно-
сти курсовые дозы бензилпенициллина увеличи-
ваются в 1,5—2 раза. Применение препаратов
тетрациклинового ряда противопоказано. Проти-
вопоказаны также фторхинолоны и аминоглико-
зиды.
Из иммунотерапевтических средств при хро-
нической гонорее и при рецидивах назначается
гоновакцина (начиная со второго триместра бе-
ременности) в пониженных дозах — (100—150
млн. микробных тел). Применение имуномоду-
лирующих и биостимулирующих препаратов при
гонококковой инфекции должно быть строго
обоснованным.
Местное лечение уретры, прямой кишки,
больших вестибулярных желез может прово-
диться на всех сроках беременности. Лечение
цервикального канала можно проводить лишь в
виде влагалищных ванночек, без введения лекар-
ственных препаратов непосредственно в церви-
кальный канал (помнить о возможной угрозе
прерывания беременности).
О беременных, перенесших гонорею во вре-
мя беременности, следует сообщать в женскую
консультацию для специального наблюдения за
ними до и после родов и тщательного наблюде-
ния за состоянием новорожденного.
У беременных женщин, больных гонореей,
нарушается процесс биогенеза пептидных и сте-
роидных гормонов, более выраженный при хро-
нической форме заболевания. Для коррекции
выявленных нарушений гормонального звена
фетоплацентарной системы в комплексе лечения
этой группы больных следует назначить но-шпу,
папаверин, димедрол, которые позволяют сни-
зить угрозу беременности, предотвратить после-
родовые осложнения у женщин.
В послеродовом периоде антибиотики назна-
чают по схеме лечения хронической гонореи.
Вакцинотерапия в период лактации не проводит-
ся. Местное лечение уретры и прямой кишки
Глава 39. Гонорея
489
проводится сразу после родов. Местное лечение
шейки матки можно начинать не раньше 5—6
недель после родов (после того, как сформиру-
ется шейка матки). После абортов лечение шей-
ки матки можно начинать не раньше, чем через
10 дней.
К установлению критерия излеченности гоно-
реи приступают через 7—10 дней после заверше-
ния курса лечения. Критериями излеченности
являются: отсутствие субъективных и объектив-
ных симптомов заболевания, отрицательные ре-
зультаты микроскопического и культурального
исследований. При наличии клинико-лаборатор-
ных данных, свидетельствующих о сохранении
воспалительного процесса, рекомендуется по-
вторное обследование с обязательным культу-
ральным исследованием и исключением сопут-
ствующих инфекций. Гонококки, обнаруженные
после проведенного лечения, чаще всего являют-
ся следствием реинфекции. При установлении
рецидива гонореи необходимо осуществлять
культуральное исследование с определением чув-
ствительности гонококка к антибиотикам.
Лечение гонореи у детей
Детям, больным гонореей, лечение следует про-
водить в условиях стационара. Основным анти-
биотиком является бензилпенициллин. В случае
безуспешного лечения бензилпенициллином на-
значают другой антибиотик. У детей старше 3 лет,
больных хронической гонореей, применяют го-
новакцину (начальная доза 50-100 млн. микроб-
ных тел) с последующим назначением антибио-
тиков. Детям до 3 лет иммунотерапия не
проводится. Местное лечение проводят в тех
случаях, когда гонококк исчез, но вялотекущие
воспалительные явления после провокации еще
остаются. В упорно протекающих случаях ваги-
нита рекомендуется вагиноскопия и в дальней-
шем местное лечение в соответствии с обнару-
женными изменениями.
Лечение
экстрагенитальной гонореи
Гонорейный проктит
Встречается у женщин и девочек, больных го-
нореей, и сочетается с поражением мочеполовых
органов. Может быть у мужчин — пассивных
гомосексуалистов. Суъективные симптомы при
гонорейном поражении прямой кишки в боль-
шинстве случаев отсутствуют, однако у некото-
рых больных отмечается зуд, жжение, небольшая
болезненность при дефекации, выделения из
прямой кишки, слизь, гной, кровь в кале. Иног-
да отмечается гиперемия и мацерация анальной
области, утолщение складок сфинктера.
Антибиотики применяют в курсовой дозе, как
при осложненных хронических формах гонореи,
с обязательным последующим проведением ме-
стного лечения: свечи с протарголом (по 0,02 г
проторгола на свечу); введение в прямую киш-
ку из спринцовки или резиновой груши 40-
50 мл 2—5 % раствора протаргола или колларгола
через день, всего на курс 5—6 процедур.
1. Бензилпенициллин в курсовой дозе до 6
млн. ЕД с последующим назначением левоми-
цетина перорально в течение 3 дней, в суммар-
ной дозе 10 г (1 день — 4 г; 2—3-й дни — по 3 г
в сутки).
2. Бициллин-3 в курсовой дозе до 6 млн. ЕД
с последующим назначением 10 г левомицети-
на по вышеуказанной методике.
Орофарингеальная гонорея
У мужчин и женщин поражается преимуще-
ственно слизистая оболочка миндалин и глотки,
наблюдаются гиперемия и отек слизистой, иног-
да на миндалинах отмечается гнойный налет,
регионарный аденит. У некоторых больных
встречаются гингивиты и стоматиты. Назнача-
ют антибиотики в курсовой дозе, как при ослож-
ненной и хронической гонорее.
При всех формах экстрагенитальной локали-
зации гонореи или смешанной инфекции целе-
сообразно использование препаратов из группы
фторированных хинолонов, так как они облада-
ют способностью проникать в ткани и клетки.
Бленорея (гонорея глаз)
Взрослые заражаются при несоблюдении правил
гигиены. Больной гонореей человек может за-
нести инфекцию в глаза загрязненными рука-
ми. У новорожденных гонококки попадают в
глаза во время прохождения через родовые пути
больной матери.
Лечение бленореи должно проводиться
дерматовенерологом совместно с окулистом,
с обязательным внутримышечным введени-
ем антибиотика без учета дозы антибиотика,
примененного местно в глаза. Больных бле-
нореей лечат после консультации окулиста,
согласно его рекомендациям. Снятие с уче-
та больного бленореей проводится после кон-
сультации окулиста.
В целях профилактики гонобленореи всем
детям сразу после рождения глаза протирают
стерильной ватой и закапывают 30 % раствор
490 Венерология
сульфацила натрия; через 2 часа процедуру по-
вторяют. Девочкам одновременно таким же ра-
створом обрабатывают половые органы.
Физиотерапевтические
методы лечения
Физиотерапия гонорейных заболеваний сводит-
ся, в основном, к следующим процедурам: ком-
прессы, местные ванночки, микроклизмы, пара-
фин, озокерит, электрофорез, индуктотермия,
электрическое поле УВЧ, ультразвук, микровол-
новая терапия, фонофорез, диадинамические
синусоидальные токи, электроимпульсная тера-
пия, дарсонвализация.
Ванночки, микроклизмы,
согревающие компрессы
При простатитах и орхоэпидидимитах горячие
клизмочки и ванночки (до 50°С) назначают 2—
3 раза в день, согревающий компресс на вазе-
линовом масле желательно носить постоянно до
стихания острого воспаления в пораженном орга-
не. Сидячие ванночки с калия перманганатом
(1 : 8000) температурой 37—38°С продолжитель-
ностью 10—15 мин. Согревающие компрессы с
вазелиновым маслом на низ живота на 48 часов.
Горячие микроклизмы и ванночки применя-
ют в начальной стадии при острых воспалитель-
ных процессах в предстательной железе, в яич-
ках и их придатках; при восходящей гонорее —
после стихания острых воспалительных явлений,
при парауретритах у женщин. Сидячие ванноч-
ки с калия перманганатом — при бартолинитах,
парауретритах, вульвитах. Согревающие комп-
рессы с вазелиновым маслом — при восходящей
гонорее, после стихания перитонеальных явле-
ний.
Парафино- и озокеритотерапия
Парафинотерапия у мужчин используется по
салфетно-аппликационной методике или мето-
дике наслаивания. Сеансы проводятся ежеднев-
но. Продолжительность процедур от 30 мин до
1 часа. Озокеритотерапия используется по сал-
фетно-аппликационной методике, продолжи-
тельностью процедур 40—60 мин, на курс от 12
до 20 процедур. Озокерит грязи — в виде там-
понов и трусов, 15 процедур.
Парафинотерапия применяется при стихании
острых воспалительных явлений на области су-
ставов и область органов мошонки у мужчин; в
хронической стадии при восходящей гонорее у
женщин. Озокеритотерапия применяется на ос-
троте процесса (экссудативная стадия) при ор-
хиэпидидимитах и простатитах у мужчин, бар-
толинитах у женщин.
Противопоказания: Хронические и подострые
процессы (стадия пролиферации).
Электрическое поле УВЧ
Электрическое поле УВЧ применяется ежедневно
по 10 мин; эффективны первые 3—5 процедур.
Показания: Орхиэпидидимиты и простатиты.
Индуктотермия
Индуктотермия на область мошонки и область
промежности применяется через день с продол-
жительностью 10—20 мин, на курс 10—15 проце-
дур. При артритах продолжительность лечения
увеличивается до 30 мин при том же количестве
процедур. Индуктотермия у женщин применяется
на область промежности или внутривлагалищно
посредством цилиндрического индуктора по 15—
20 мин; количество процедур от 8 до 10.
Показания: острые воспалительные явления,
орхиэпидидимиты, артриты, хронические про-
статиты у мужчин; хронический восходящий
процесс у женщин (хронические эндометриты и
сальпингоофориты).
Противопоказания: острые воспалительные
процессы мочеполового тракта, наличие или
подозрение на опухолевый процесс, склонность
к кровотечениям, повышенная температура тела,
аденома предстательной железы, наличие гемор-
роя, варикоцеле.
Электрофорез
Электрофорез на область полового члена приме-
няется ежедневно или через день от 10—15 мин
до 20-30 мин, на курс 10-12 процедур. Элект-
рофорез на область мошонки проводят ежедневно
или через день от 10 от 20 мин; на курс лече-
ния 10—15 процедур. У женщин используется
вагинально-накожная методика. Применяют
электрофорез препаратов йода (йодистый калий,
йодбромная вода), продигиозана, ронидазы, ли-
дазы, гидрокортизона, совкаина, дикаина, анти-
биотиков, сульфата цинка.
Электрофорез используют после уменьшения
воспалительных явлений при уретритах, орхиэ-
пидидимитах, простатитах, артритах; при хрони-
ческом восходящем процессе у женщин в стадии
обострения, при стриктурах.
Противопоказания: острые воспалительные
процессы, индивидуальная непереносимость
слабого электрического тока, расстройство чув-
ствительности кожи, подозрение на опухолевый
процесс и другие общетерапевтические проти-
вопоказания.
Глава 39. Гонорея
491
Ультразвук
Ультразвук используется по лабильной методи-
ке в непрерывном режиме контактным способом
или посредством ректального излучателя. Про-
должительность процедур 3—10 мин, ежедневно
или через день, на курс 10—12 процедур.
Ультразвук показан при стриктурах уретры,
хронических простатитах и уретритах.
Противопоказания: острые воспалительные
процессы мочеполового тракта, аденома предста-
тельной железы, подозрение на опухолевые про-
цессы и другие физиотерапевтические противо-
показания.
Фонофорез
Фонофорез используется на область промежно-
сти и полового члена по методике ультразвуко-
вого фонофореза при постоянном режиме, ла-
бильно. Продолжительность процедур от 5 до
15 мин через день; на курс 15—20 процедур. В
качестве лекарственных препаратов используют-
ся 1 % гидрокортизоновая мазь, фторокорт, си-
нафлан, целестодерм, белосалик и др.
Фонофорез применяют при стриктурах урет-
ры, хронических простатитах и хронических
уретритах.
Противопоказания: острые воспалительные
процессы мочеполового тракта, аденома проста-
ты, подозрение на опухолевые процессы и дру-
гие физиотерапевтические противопоказания.
Микроволновая терапия (СВЧ)
Микроволновое воздействие осуществляют по-
средством ректального излучателя. Продолжи-
тельность процедуры 8—15 мин, ежедневно или
через день; на курс лечения 10—15 процедур.
Показания: хронические простатиты и везику-
литы.
Противопоказания: подозрение на опухолевый
процесс, аденома предстательной железы, ситем-
ные заболевания крови и другие общие физио-
терапевтические противопоказания.
Диадинамические
токи (синусоидальные)
Для воздействия диадинамическими токами ис-
пользуют ректальный излучатель. Продолжи-
тельность процедуры 10—15 мин, на курс лече-
ния 10—15 процедур и более через день.
Показания: хронические простатиты и везику-
литы.
Противопоказания: подозрение на опухолевый
процесс, аденома предстательной железы, ситем-
ные заболевания крови и другие общие физио-
терапевтические противопоказания.
Электроимпульсная терапия
Электроимпульсная терапия используется на
область промежности. Продолжительность про-
цедур 10—20 мин через день; на курс лечения до
15 процедур.
Показания: хронические простатиты, сопро-
вождающиеся половыми нарушениями; хрони-
ческие везикулиты, наличие геморроя в анамнезе,
варикоцеле.
Противопоказания: острые воспалительные про-
цессы, индивидуальная непереносимость слабого
электрического тока, расстройство чувствительно-
сти кожи, подозрение на опухолевый процесс и
другие общетерапевтические противопоказания.
Дарсонвализация
Дарсонвализации подвергают область промежно-
сти или — с помощью ректального излучателя -
прямую кишку. Воздействие проводят ежеднев-
но или через день, начиная с 5 мин, постепенно
прибавляют по 2 мин, доводя продолжительность
до 15 мин. На курс лечения 12—15 процедур.
Дарсонвализация показана при хронических
простатитах и везикулитах, наличии геморроя в
анамнезе, варикоцеле.
Противопоказания: острые воспалительные
процессы, индивидуальная непереносимость
тока, расстройство чувствительности кожи, по-
дозрение на опухолевый процесс и другие обще-
терапевтические противопоказания.
Местное лечение
гонореи и ее осложнений
Местное лечение применяют при непереносимо-
сти антибиотиков, одновременно с сульфанила-
мидными препаратами, при рецидивах, при све-
жей вялотекущей и хронической гонорее в
период иммунотерапии.
Местное лечение
гонореи у мужчин
Свежий гонорейный передний уретрит
В течение первых 3—5 дней — обильные про-
мывания теплым (37-39 °C) раствором калия
перманганата 1 : 10000—1 : 6000 только пере-
дней уретры. В последующем проводят глубо-
кие промывания.
Хронический гонорейный передний уретрит
При мягком инфильтрате — инстилляции 0,025-
0,5 % раствора нитрата серебра или 2 % раствора
492 Венерология
нитрата серебра или 2 % раствора протаргола
через 1—2 дня в зависимости от местной реак-
ции. На курс 8—10 инстилляций. При отсутствии
эффекта — бужирование через 1—2 дня. Буж
остается в уретре 5 мин, после чего следует про-
мыть уретру 0,05 % водным раствором хлоргек-
сидина биглюконата или нитрата серебра
1: 10000. На курс 8—10 сеансов.
При твердом инфильтрате — бужирование
через 1—2 дня в зависимости от реакции (гной-
ные выделения, помутнение мочи) с последую-
щим промыванием 0,05 % водным раствором
хлоргексидина биглюконата или нитрата сереб-
ра 1 : 10000. На курс 12—15 сеансов.
Если буж не вызывает достаточной реакции,
следует применять тампонаду уретры с 2 % ра-
створом протаргола в глицерине 1 раз в 4—5 дней,
на курс 5—6 сеансов.
При переходном инфильтрате — тампонада
уретры 2 % раствором протаргола в глицерине 2
раза в неделю, на курс 6—8 сеансов.
При уретральном адените — бужирование с
последующим промыванием уретры 0,05 % вод-
ным раствором хлоргексидина биглюконата или
нитрата серебра 1: 10000.
При закрытом адените бужирование должно
сопровождаться массажем уретры на буже. Эф-
фективна в таком случае и тампонада уретры, и
тепловые процедуры (горячие ванночки, индук-
тотермия).
При десквамативном уретрите — инстилляции
1 % раствором сульфата цинка 2 раза в день или
2,5 % взвесью гидрокортизона.
При стриктуре уретры — систематическое и
постепенное расширение стриктуры металличес-
кими бужами. При сужениях ниже калибра бужа
№ 14—16 по Шарьеру, во избежание образова-
ния ложных ходов, применяют эластические
бужи, оставляя их в уретре на 5—10 мин. Бужи-
рование проводят через 48 часов. Если через
сужение с большим трудом проходит нитевид-
ный буж, то его следует оставить на 1—2 суток,
при этом моча проходит мимо бужа. Бужирова-
ние доводят до размера бужа, проходящего че-
рез наружное отверстие уретры. С целью размяг-
чения рубцовой ткани применяют инъекции
стекловидного тела, экстракта алоэ.
При стриктурах, не поддающихся расширению
бужем, показано хирургическое вмешательство.
Свежая гонорея, задний уретрит
При наличии дизурических явлений и болей в
уретре — постельный режим, теплые сидячие
ванночки, тепло на область мочевого пузыря,
ректальные свечи с 0,015 г белладонны 3 раза в
день. При стихании острых явлений — глубокие
промывания уретры по утрам раствором калия
перманганата 1:6000.
Хроническая гонорея, задний уретрит
Инстилляция в заднюю уретру 0,25—0,5 % ра-
створа нитрата серебра или 1—2 % раствора про-
таргола через 1—2 дня. При всех хронических
уретритах обязательно проводится обследование
простаты и семенных пузырьков. При наличии
поражения желез следует проводить одновремен-
ное лечение этих желез и уретры. При грануля-
ционном уретрите — смазывание грануляций
20 % раствором нитрата серебра под контролем
глаза через уретроскопическую трубку 1 раз в 7
дней, всего 5—6 сеансов; при крупных грануля-
циях проводится электрокоагуляция.
Свежая гонорея, парауретрит
Широкие парауретральные ходы промывают (с
помощью шприца с тупой иглой) 30—50 мл ра-
створа нитрата серебра 1: 200—1:1000 с интер-
валом 1—2 дня в зависимости от реакции. При
узком ходе вводят 0,5—1,0 мл 20 % раствора
нитрата серебра или 2—5 % раствора настойки
йода, или проводник от иглы, на который на-
паян чистый нитрат серебра. Эффективна элек-
трокоагуляция ходов. Длинные, извитые, инфи-
цированные ходы, особенно в области крайней
плоти, удаляют хирургическим путем.
Свежая гонорея, эпидидимит
Постельный режим, ношение суспензория, согре-
вающие компрессы, УВЧ. При подостром тече-
нии — горячие ванночки (40—50 °C) 2 раза в
день. При выпоте в оболочках яичка — пунк-
ция с отсасыванием жидкости.
Хроническая гонорея, эпидидимит
Индуктотермия придатка яичка ежедневно по
10—20 мин, 8—9 сеансов. При плохом рассасы-
вании инфильтрата — электрофорез с йодистым
калием, ронидазой.
Свежая гонорея, простатит катаральный
Лечение такое же, как и при заднем уретрите.
После исчезновения дизурических явлений и
просветлении второй порции мочи нужно иссле-
довать секрет простаты.
Хроническая гонорея,
простатит катаральный
Массаж простаты с последующей инстилляцией
в заднююю уретру 0,25—0,5 % раствора нитрата
серебра через день; курс лечения 10—12 сеансов.
Глава 39. Гонорея
493
Лечение проводится под контролем секрета про-
статы.
Свежая гонорея, острый фолликулярный
и паренхиматозный простатит
Горячие микроклизмы 2 раза в день, при болях
в уретре и прямой кишке — свечи с белладон-
ной 2 раза в день. При стихании явлений про-
статита — индуктотермия железы ежедневно, 8—
10 сеансов. При наличии лейкоцитов в секрете
простаты — массаж при наполненном мочевом
пузыре (раствор калия перманганата 1:10000 или
1:6000 или ляписа 1 : 10000, 2—3 раза в неде-
лю), на курс 8—10 сеансов.
Хроническая гонорея, фолликулярный
и паренхиматозный простатит
Регулирование функции кишечника, устранение
застойных явлений в органах малого таза, обус-
ловленных половым воздержанием, сидячим
образом жизни. Индуктотермия простаты с пос-
ледующим массажем 2—3 раза в неделю. После
массажа железы — инстилляция в заднюю уретру
0,25-0,5 % раствора нитрата серебра. На курс
10-12 сеансов. После исчезновения лейкоцитов
из секрета простаты массаж прекращают. При
повторном появлении лейкоцитов снова назна-
чают массаж в сочетании с физиопроцедурами
(ректальная микроволновая терапия или ультра-
звук). Больным показаны также горячие мик-
роклизмы или горячие сидячие ванны, грязеле-
чение.
Свежая гонорея, абсцесс
простаты, парапростатит
При высокой температуре (39 °C), задержке мочи,
болях в прямой кишке и появлении флюктуа-
ции в железе следует вскрыть гнойник через
промежность. Другие мероприятия те же, что и
при остром паренхиматозном простатите.
Острый гонорейный везикулит
Лечение такое же, как и при остром паренхи-
матозном простатите.
Лечение хронических везикулитов такое же,
как и при хронических простатитах.
Местная терапия
при гонорее нижнего отдела
мочеполового тракта у женщин
Свежая острая гонорея, уретрит
Глубокое промывание (после предварительного
мочеиспускания)+инстилляция 1—2 %-ным ра-
створом протаргола или инстилляциями 3 %-ного
раствора колларгола или 0,5 %-ного раствора
нитрата серебра (1 раз в 3 дня).
Свежая подострая гонорея, уретрит
Инстилляция 0,5—1,0 % раствором нитрата сереб-
ра или 3—5 % раствором протаргола.
Хроническая гонорея, уретрит
При полном мочевом пузыре — массаж уретры.
После мочеиспускания смазывание уретры чис-
тым ихтиолом 1 раз в три дня или 1 % раство-
ром Люголя.
Свежая острая гонорея, парауретрит
Горячие сидячие ванночки из раствора перман-
ганата калия 1: 8000 или ромашки (1 столовая
ложка на 2 стакана кипятка). Местная аутогемо-
терапия.
Свежая подострая гонорея, парауретрит
Электрокоагуляция или прижигание ходов зон-
дом с напаянным кристаллическим нитратом
серебра.
Свежая острая гонорея, вульвит
Сидячие ванночки из ромашки (1 столовая ложка
на 2 стакана воды) или из раствора калия пер-
манганата (1: 8000), температура 38 °C, по 10—
15 мин.
Свежая подострая гонорея, вульвит
Смазывание 10 % раствором протаргола в гли-
церине. Коагуляция сопутствующих остроконеч-
ных кондилом или их выскабливание острой ло-
жечкой.
Свежая острая гонорея, бартолинит
Сидячие ванночки из ромашки (1 столовая ложка
на 2 стакана воды) или из раствора калия пер-
манганата (1: 8000), температура 38 °C, по 10—
15 минут, 1—2 раза в день. Местная аутогемоте-
рапия (инъекция 1—3 мл аутокрови над и под
выводным протоком бартолиниевой железы). Ле-
чение проводят 2—4 раза с 2-дневными интер-
валами. При развивающемся абсцессе показано
оперативное лечение.
Свежая подострая
гонорея, бартолинит
Псевдоабсцесс вскрывают разрезом длиной 2 см,
очищают полость, смазывают йодной настойкой,
тампонируют турундой с белым стрептоцидом.
Перед инъекцией полость железы смазывают
йодной настойкой.
494 Венерология
Хроническая гонорея, бартолинит
Местная аутогемотерапия. В условиях стациона-
ра — введение гонококкововй вакцины над и под
выводным протоком железы. При рецидивирую-
щем абсцессе железы показана ее экстирпация.
Лечение восходящей
гонореи у женщин
Эндометрит, метроэндометрит,
сальпингоофорит - острая стадия
Госпитализация, постельный режим. При пель-
виоперитоните — лед на живот на 30—60 мин с
перерывами в 3 часа до исчезновения симптома
Щеткина-Блюмберга. Легкая нераздражающая
пища. Следить за регулярной работой кишечни-
ка. При запорах — микроклизма (1 столовая
ложка глицерина+3 столовых ложки воды или
75 мл прокипяченного подсолнечного масла или
1 чайная ложка слабительной соли натощак).
Ректальные свечи с экстрактом белладонны
(0,02 г) 2 раза в день. При кровотечениях —
внутрь 10 % раствор хлорида кальция по 1 сто-
ловой ложке 3 раза в день. Викасол по 0,01 г 3
раза в день в течение 3 дней. Экстракт калины
по 30—40 капель 3 раза в день. Противопоказа-
ны препараты спорыньи. Аутогемотерапия — от
5 до 10 мл внутримышечно через 1—2 дня 3—4
раза в день. При кровотечениях в межменстру-
альном периоде — прогестерон по 5 мл 1 раз в
день, 5—6 инъекций.
Эндометрит, метроэндометрит,
сальпингоофорит - подострая стадия
На болезненные зоны — согревающий компресс
из вазелина на 48 часов. Через 3—4 дня грелка
на низ живота+позже УВЧ на низ живота и об-
ласть поясницы, ежедневно, 3—4 процедуры.
Затем ионизация с сульфатом цинка (можно
начать при нормальном количестве лейкоцитов,
но даже при СОЭ 40—60 мм/ч по Панченко).
Вакцинотерапия, начиная с 200 млн микробных
тел. В зависимости от реакции последующие
дозы увеличивают, всего 5—6 инъекций с интер-
валом в 2—3 дня, или пирогенал с 2,5 мкг в воз-
растающей дозировке. Максимальная разовая
доза 100 мкг. Доза увеличивается в зависимос-
ти от реакции, на курс 6—8 инъекций, интерва-
лы 1—2 дня. Если реакция слабая, можно вво-
дить гоновакцину с пирогеналом в одном шприце
(начальная доза гоновакцины 150—200 млн м.
т.+пирогенал 5—10 мкг через 2 дня). Дозы уве-
личивают в зависимости от реакции. При запо-
рах ежедневно перед сном 1/2 стакана настоя
трав: коры крушины 1 столовая ложка, корня
валерианы 1 столовая ложка и листа мяты 2 сто-
ловые ложки. Все заваривают в 500 мл кипятка
и настаивают на водяной бане в течение 1 часа.
Эндометрит, метроэндометрит,
сальпингоофорит - хроническая стадия
Грелка на низ живота. Микроклизмы с горячим
изотоническим раствором хлорида натрия по
100 мл 2 раза в день. Введение гонококковой вак-
цины в шейку матки и в подслизистый слой урет-
ры; начальная доза 50—100 млн м. т. или аутоге-
мотерапия от 1 до 3 мл через 2 дня. Ионизация с
калия йодидом или сульфатом цинка на низ жи-
вота и область поясницы; ежедневно, 6—8 проце-
дур. В дальнейшем — индуктотермия по 15 мин,
10 сеансов. Затем назначают озокерит и грязеле-
чение в виде тампонов и трусов, 10—15 процедур.
Проктит - острая стадия
Микроклизмы с 50 мл 1 % раствора протаргола
через день. Свечи с белладонной 2—3 раза в день.
Трещины и экскориации тушируют 3—5 % ра-
створом нитрата серебра.
Проктит - подострая стадия
Микроклизмы с 3 % раствором протаргола. Че-
рез ректальное зеркало осмотр эрозий, исследо-
вание на бледную трепонему, затем туширова-
ние 5—10 % раствором нитрата серебра.
Проктит - хроническая стадия
Тампоны на 24 часа с 2 % раствором пищевой
молочной кислоты.
Эндоиервинит - острая стадия
Влагалищные ванночки с 3 %-ным раствором
протаргола или колларгола. Влагалищные шарики
(протаргола 1,5; белого сахара 4,0; молочного
сахара 3,0; масла-какао 1,0).
Эндоиервииит - подострая стадия
Влагалищные ванночки с перекисью водорода.
Затем влагалище вытереть насухо и вложить там-
пон с 10 % раствором протаргола в глицерине на
24 часа. Чередовать с ванночками из 3 % раство-
ра колларгола. Внутрицервикальное (на глуби-
ну 1,5 см) смазывание 1 % раствором Люголя на
глицерине, 2—3 % раствором нитрата серебра 1
раз в 3—4 дня.
Эндоиервииит - хроническая стадия
При фолликулярной эрозии — прижигание кри-
сталлическим калия перманганатом 1 раз в 2
недели. Ванночки с 2 % раствором нитрата се-
Глава 39. Гонорея
495
ребра через 3—4 дня. Диатермокоагуяяция ретен-
ционных кист шейки матки или вскрытие их
скальпелем.
Местное лечение
гонореи у девочек после
антибиотикотерапии
Свежая гонорея, вульвовагинит
Теплые сидячие ванночки на 10—15 минут 2—3
раза в день из отвара ромашки или раствора ка-
лия перманганата 1: 10000. После ванночек на-
ружные половые органы высушить. Очаги раз-
дражения смазать пастой Лассара. При
экзематозных поражениях кожи — примочки из
3 % буровской жидкости. Смазывание 4 % вод-
ным раствором метиленового синего.
Свежая гонорея, уретрит
Лечение такое же, как при вульвовагините. В
подострой стадии: закапывание глазной пипет-
кой 3-4 капель 0,5—1 % раствора протаргола, 1—
2 % раствора колларгола (чередовать).
Свежая гонорея, проктит или ректит
В прямую кишку вводят 10—20 мл 1—3 % ра-
створа протаргола.
Хроническая гонорея, вульвовагинит
Промывание влагалища через тонкий резиновый
катетер раствором марганцовокислого калия
1: 8000 с последующей инстилляцией через этот
катетер раствора протаргола (1—2 %) или нитрата
серебра (0,25—1 %) в количестве 3—5 мл через
день. В упорно протекающих случаях вагинита —
смазывание влагалища через уретроскопическую
трубку раствором Люголя на глицерине, 10 %
протарголом в глицерине через 2—3 дня.
Хроническая гонорея, уретрит
Закапывание в уретру 3—4 капель раствора про-
таргола (2 %) или нитрата серебра (0,25—0,5 %).
Хроническая гонорея, проктит или ректит
Клизмочки с 2—3 % раствором протаргола по
30—40 мл.
Критерии
излеченности гонореи
Излеченность гонореи устанавливается с помо-
щью клинического, бактериоскопического и
бактериологического исследований.
Критерии излеченности
гонореи у мужчин
Стойкое отсутствие гонококков при микроско-
пическом и бактериологическом исследовании
отделяемого мочеполовых органов; отсутствие
пальпаторных изменений в предстательной же-
лезе, семенных пузырьках, а также нормальное
содержание лейкоцитов (5—10 в поле зрения) в
их секрете при наличии в нем значительного ко-
личества липоидных зерен; отсутствие или не-
резко выраженные воспалительные изменения в
мочеиспускательном канале при уретроскопии.
Излеченность больных, перенесших свежую го-
норею, определяют через 7—10 дней после оконча-
ния лечения. При отсутствии гонококков в иссле-
дуемом материале проводят комбинированную
провокацию: химическую (вводят в мочеиспускатель-
ный канал 4—6 мл 0,5 %-го раствора нитрата сереб-
ра) и одновременно биологическую (внутримышеч-
но 500 млн микробных тел гоновакцины или 200
МПД [20 мкг] пирогенала). Применяют также ме-
ханическую провокацию (в уретру вводят прямой
буж на 10 минут или проводят переднюю уретрос-
копию), алиментарную (дают соленую, острую
пищу, раздражающую слизистые оболочки мочевы-
водящих путей), термическую (прогревание поло-
вых органов индуктотермическим током). Чаще
всего проводят комбинированные виды провока-
ции: химическую, алиментарную и биологическую.
В затяжных, хронических случаях уретрита целе-
сообразно вызвать раздражение слизистой уретры
массажем на прямом буже или тубусе уретроскопа
и внутримышечным введением гоновакцины. Че-
рез 24, 48, 72 часа после провокации для лабора-
торного исследования берут отделяемое из мочеис-
пускательного канала, а при отсутствии такового —
соскоб со слизистой уретры, парауретральных про-
токов, нити из мочи, секрет предстательной желе-
зы и семенных пузырьков. Точность исследования
возрастает, если после провокации отделяемое из
различных очагов инфекции повторно высевают на
искусственные питательные среды.
Если гонококки и другие патогенные микроор-
ганизмы не обнаруживаются, то следующий кон-
троль назначают через 1 месяц. После комбини-
рованной провокации проводят клиническое,
трехкратное лабораторное (для обнаружения гоно-
кокков) и урологическое исследование (уретрос-
копия, микроскопическое исследование секрета
предстательной железы и семенных пузырьков).
Третий, последний контроль проводят анало-
гично через 1 месяц после второго. При отсут-
ствии клинических признаков заболевания и
гонококков при лабораторном исследовании
пациент снимается с учета.
496 Венерология
Лица, перенесшие гонорею и те, у которых не
установлен источник заражения (значит, не ис-
ключено одновременное заражение сифилисом),
подлежат ежемесячному клинико-серологическо-
му контролю в течение 6 месяцев.
Критерии излеченности
гонореи у женщин
Отсутствие гонококков при исследовании выде-
лений из мочеиспускательного канала, канала
шейки матки, прямой кишки; благоприятные
результаты клинического и лабораторного иссле-
дования в течение 2—3 менструальных циклов;
отсутствие боли и явных пальпаторных измене-
ний внутренних половых органов, нормальный
менструальный цикл.
Излеченность определяют через 7—10 дней после
окончания лечения антибиотиками. Выделения из
мочеиспускательного канала, канала шейки мат-
ки, прямой кишки подвергают лабораторному
исследованию на наличие гонококков. При отсут-
ствии их в исследуемом материале проводят ком-
бинированную провокацию: инъекцию гоновак-
цины (500 млн микробных тел) или пирогенала
(200 МПД), смазывание мочеиспускательного ка-
нала и прямой кишки 1 %-ным раствором Люго-
ля, а канала шейки матки — 2—5 %-ным раство-
ром нитрата серебра. Через 24, 48, 72 ч исследуют
отделяемое из уретры, шейки матки и прямой
кишки на наличие гонококков. Во время очеред-
ной менструации снова берут мазки, а после ее
окончания повторяют провокацию с исследовани-
ем отделяемого в течение 3 дней. Такие обследо-
вания проводят в течение 2 менструальных цик-
лов, после чего при благоприятных результатах
клинического и лабораторного исследования боль-
ную снимают с учета. В течение 3 менструальных
циклов следует наблюдать женщин после лечения
восходящей гонореи и женщин, переболевших
гонореей во время беременности.
Больная гонореей находится под наблюдени-
ем врача для установления излеченности после
окончания лечения при свежей гонорее в тече-
ние 2 месяцев, при хронической — 3 месяцев.
Больным с невыявленным источником зараже-
ния срок контрольного наблюдения удлиняют до
6 месяцев. При этом проводят ежемесячные кли-
нические и серологические обследования для
выявления сифилиса.
Критерии излеченности
гонореи у детей
Все девочки дошкольного возраста, посещающие
детские дошкольные учреждения, после оконча-
ния лечения по поводу гонореи остаются в ста-
ционаре в течение 1 месяца для установления из-
леченности. За это время делают 3 провокации
и 3 посева (1 раз в 10 дней). Комбинированная
провокация у детей: инъекция гоновакцины
(150—200 млн микробных тел), смазывание ваги-
ны и вульвы раствором Люголя в глицерине,
закапывание в уретру 2—3 капель 0,5—1 %-ного
раствора нитрата серебра, смазывание нижнего
отрезка прямой кишки раствором Люголя. Через
24, 48 и 72 часа после провокации берут мазки
из уретры, вагины и прямой кишки. Из прямой
кишки лучше исследовать хлопья, выловленные
в промывных водах. Посев проводят через 72 часа.
Критерием излеченности гонореи у детей яв-
ляются нормальная клиническая картина и бла-
гоприятные результаты повторных лабораторных
исследований отделяемого гениталий после 3
провокаций.
Профилактика
гонореи у детей
Для предохранения детей от заболевания гоно-
реей необходимы профилактические мероприя-
тия в семье, детских учреждениях, родильных
домах, своевременное выявление беременных,
больных гонореей, в женских консультациях.
Профилактика в быту: дети должны спать от-
дельно от взрослых, иметь индивидуальный гор-
шок, а также индивидуальные губки и мочалки
для подмывания, индивидуальные полотенца.
Профилактика в детских учреждениях: персонал
должен приниматься на работу после предваритель-
ного обследования врачом-венерологом, а в даль-
нейшем подвергаться таким же обследованиям в
установленные сроки. Медицинским персоналом
должны осматриваться гениталии у детей перед
поступлением в детское учреждение и 1 раз в не-
делю во время нахождения там. В яслях у каждого
ребенка обязательно должен быть индивидуальный
горшок. Подмывание детей производят струей воды
при помощи отдельного ватного тампона на корн-
цанге. Для обтирания половых органов детей пос-
ле подмывания применяют индивидуальные поло-
тенца или салфетки. Персонал медицинских
учреждений должен иметь отдельный туалет.
Заболевшие и подозреваемые на гонорею дети
должны быть изолированы и направлены на
консультацию к венерологу.
С персоналом детских учреждений и матеря-
ми необходимо постоянно проводить санитарно-
профилактическую работу.
Глава 40
Шанкроид
Шанкроид (син.: мягкий шанкр, венерическая
язва, третья венерическая болезнь) — острая
венерическая инфекция, которая вызывается
стрептобактерией Дюкрея-Унны-Петерсена; пе-
редается половым путем, характеризуется болез-
ненными множественными язвами половых ор-
ганов, а также воспалительной аденопатией
лимфатических узлов.
Этиология
Возбудителем шанкроида является стрептобакте-
рия Haemophilus ducreyi, которая представляет
собой короткую тонкую палочку длиной 1,5—2
мкм, толщиной 0,4—0,5 мкм с несколько закруг-
ленными концами и перетяжкой в середине.
Палочки располагаются одиночно или параллель-
но попарно, в виде характерных цепочек (отсюда
их название — «стрептобациллы»). Возбудитель
шанкроида — аэроб, однако растет и при отно-
сительном анаэробизме. Палочки шанкроида
малоустойчивы к высокой температуре (при тем-
пературе выше 40 °C культура погибает), но низ-
кие температуры переносят хорошо. В культуре
1 % раствор фенола убивает возбудителя в тече-
ние 1 минуты, а 0,25 % раствор формалина —
через 40 минут; 0,5 % раствор хинозола губи-
тельно действует на возбудителя в течение не-
скольких минут.
Эпидемиология
Заражение происходит, как правило, половым
путем, в основном от лиц, занимающихся про-
ституцией. В США среди моряков и военнос-
лужащих, имеющих половые контакты пре-
имущественно с проститутками, шанкроид
обнаруживается чаще, чем сифилис. Способ-
ствуют инфицированию повреждения кожного
покрова или слизистых оболочек. Очень ред-
ко, но возможен внеполовой путь заражения.
Имеются сообщения о заражении детей и ме-
дицинских работников.
Шанкроид является эндемичным заболевани-
ем для стран Африки, Юго-Восточной Азии,
Центральной и Южной Америки, а в некоторых
регионах заболевание преобладает над сифили-
сом. Шанкроид регистрируется в Великобрита-
нии, Португалии, Италии, США и других стра-
нах. В США ежегодно сообщается не менее чем
о 1000 случаях заболевания. Во многих крупных
городах Соединенных Штатов шанкроид стал
эндемичным заболеванием; при этом широко
распространена неполная регистрация этой ин-
фекции. В странах бывшего Союза шанкроид
встречается редко, в основном, заражение про-
исходит во время поездок за рубеж.
Шанкроид является кофактором передачи
ВИЧ. Сообщается о высокой распространенно-
сти ВИЧ-инфекции среди пациентов с шанкро-
идом в США. Около 10 % больных с шанкрои-
дом могут быть одновременно инфицированы
Т. pallidum и ВПГ.
Клиническая картина
Инкубационный период шанкроида обычно длит-
ся 3—5 дней, в редких случаях удлиняется до 7—
10 дней. У женщин инкубационный период со-
ставляет 2—8 недель. В первые сутки после
заражения на месте внедрения возбудителя появ-
ляется ярко-красное пятно, которое распростра-
няется по периферии. На следующие сутки в
центре его формируется пустула, после вскрытия
которой, через 3—4 дня после заражения, обра-
зуется очень болезненная язва, быстро увеличи-
вающаяся в диаметре. На 4—5-й день от начала
заболевания язва имеет неправильную округлую
форму, края ее подрыты, окружены отечным
воспалительным венчиком. Через 2—3 недели
развитие процесса прекращается. В неосложнен-
ных случаях заживление наступает через 1—2
месяца после заражения, процесс заканчивается
рубцеванием. Стойкий иммунитет после перене-
сенного мягкого шанкра не развивается.
Обычно язвы шанкроида локализуются на
половых органах: у мужчин — на внутреннем
листке и по краю крайней плоти, в венечной
борозде, на уздечке полового члена; у женщин —
на малых и больших половых губах, клиторе,
реже — на коже лобка, внутренней поверхнос-
ти бедер, вокруг прямой кишки. Описаны слу-
чаи шанкроида полости рта.
497
498 Венерология
Язвы шанкроида имеют неровные подрытые
края и глубокое изрытое дно, покрытое желто-
ватым налетом. Язвы окружены островоспали-
тельным венчиком, но инфильтрация отсутству-
ет, поэтому на ощупь язва представляется мягкой
(отсюда и название «мягкий шанкр»). Величина
язв бывает от размеров булавочной головки до
ладони и больше. Язвы имеют склонность к кро-
вотечению, прикосновение к ним вызывает рез-
кую боль. Отделяемое язвы имеет гнойный или
кровянисто-гнойный характер и содержит боль-
шое количество стрептобацилл. При попадании
обильного отделяемого на окружающие ткани
происходит аутоинокуляция инфекции - один из
важных признаков заболевания. Вследствие это-
го шанкроид становится множественным. По ве-
личине язва обычно не превышает 10—15-ти ко-
пеечную монету. Через 3—4 недели шанкроидная
язва начинает очищаться, уменьшается гнойное
отделяемое, дно язвы покрывается грануляция-
ми и постепенно (через 1—2 месяца) наступает за-
живление с образованием рубца.
Различают следующие формы шанкроида:
• воронкообразный (глубокая язва расположена
в шейке головки полового члена);
• гангренозный (характеризуется гангренозным
расплавлением тканей на головке полового чле-
на в связи с присоединением фузоспириллеза или
анаэробной инфекции);
• дифтероидный (язвы покрыты зеленовато-
серым фибринозным налетом, обусловленным
присоединившимся заражением ложнодифтерий-
ной палочкой);
• импетигинозный (клинически сходен с импе-
тиго);
• серпингинозный (язва заживает рубцом с од-
ного края и увеличивается с другого);
• фолликулярный (возникают множественные
маленькие язвы со слегка приподнятыми края-
ми и глубоким дном, что обусловлено проник-
новением возбудителя в протоки сальных желез
или в волосяные фолликулы);
• смешанный (при одновременном заражении
сифилисом и шанкроидом).
Наиболее частыми осложнениями шанкроида
являются: лимфангиит, лимфаденит, фимоз,
парафимоз. Лимфангиит наблюдается на спин-
ке полового члена и на наружной поверхности
половых губ. Проявляется в виде уплотненного
болезненного тяжа. Кожа над утолщенной повер-
хностью гиперемирована и отечна, пальпация
болезненна. В редких случаях по ходу болезнен-
ного тяжа возникают воспалительные узлы, ко-
торые могут нагнаиваться, а затем превращать-
ся в бубонули (шанкры Нисбета).
Наиболее типичным признаком шанкроида
является регионарный лимфаденит (бубон), воз-
никающий через 2—3 недели после заражения.
Лимфаденит характеризуется остро возникаю-
щим односторонним увеличением одного или
нескольких лимфатических узлов, спаянных
вследствие развития периаденита между собой и
с кожей. Над пораженными лимфатическими
узлами кожа ярко-красного цвета. Постепенно
воспалительные явления стихают и бубон рас-
сасывается, либо в его цецтре появляется флюк-
туация, и он вскрывается. Из полости бубона
выделяется большое количество густого гноя с
примесью крови. Образующаяся язва впослед-
ствие рубцуется. Рубец формируется через 3—4
недели.
Когда развивается отек крайней плоти, может
образоваться фимоз. При этом головку полово-
го члена обнажить не удается, а скапливающийся
под крайней плотью гной выделяется через уз-
кое препуциальное отверстие. В гнойном отде-
ляемом обнаруживают возбудителей шанкроида.
При отеке крайней плоти, находящейся за
обнаженной головкой полового члена, возможен
парафимоз. Его исходом может быть гангрена
головки полового члена вследствие сдавливания
тканей кольцом отечной крайней плоти.
Лабораторная диагностика
Язвенные поражения и гнойное отделяемое
вскрывшихся или невскрывшихся лимфатити-
ческих узлов исследуют на стрептобациллы шан-
кроида.
Используются следующие методы лаборатор-
ной диагностики: микроскопический, культураль-
ный, аутоинокуляционная проба, кожный тест,
иммуноферментный анализ, полимеразная цепная
реакция.
Для микроскопического исследования с целью
обнаружения палочки шанкроида поверхность
язвы очищают от загрязнения изотоническим ра-
створом натрия хлорида. Затем лопаточкой или
острой ложечкой отбирают некротизированную
ткань из края язвы в качестве материала для ис-
следования. Стрептобациллы шанкроида окраши-
вают краской Романовского-Гимзы или метиле-
новым синим, при окраске по Граму они
отрицательны. В препаратах видны ряды цепо-
чек грамотрицательных мелких палочек, распо-
ложенных в виде «стаи рыб» между лейкоцита-
ми. Такое расположение микроба, его
соответствующая окраска, отсутствие другой
флоры характерны для возбудителя шанкроида.
Культуральное исследование используется в
сомнительных и затруднительных случаях.
Глава 40. Шанкроид 499
Возбудитель выделяют, применяя специаль-
ные методы, в частности, посев отделяемого
из очагов поражения на шоколадный агар с
1 %-ным раствором изовиталекса и 5 %-ной
сывороткой барана с добавлением 3 мгк/мл
ванкомицина. Видимый рост колоний обыч-
но появляется через 48 часов инкубации в
атмосфере, содержащей 5 % двуокиси углерода
при 100 % влажности, но иногда для появле-
ния видимого роста может потребоваться 4-
5 суток.
Аутоинокуляционная проба может быть прове-
дена с диагностической целью. Материал, взя-
тый из язвы, вносится в царапину на животе или
бедре самого пациента. Сверху пластырем накле-
ивается часовое стеклышко, дозволяющее наблю-
дать за ходом развития пробы. В течение 1—3
суток развивается типичный шанкроид, в кото-
ром могут быть обнаружены его возбудители. В
настоящее время аутоинокуляционная проба су-
щественного значения не имеет, так как при
подозрении на шанкроид начинают лечение ан-
тибиотиками, которые делают аутоинокуляцию
невозможной.
Внутрикожный тест с использованием инак-
тивированных бактерий (кожный тест Дюкрея)
в повседневной практике считается малонадеж-
ным и применяется редко.
Современная диагностика шанкроида с помо-
щью полимеразной цепной реакции разработана в
последние годы. Предполагается, что ПЦР вско-
ре станет широкодоступным методом диагности-
ки шанкроида.
Дифференциальный диагноз
Шанкроид следует отличать от твердого шанкра,
простого герпеса, эрозивного и гангренозного
баланопостита или вульвита, острой язвы Лип-
шютца, дифтерии половых органов, туберкулез-
ных язв, шанкриформной пиодермии, абсцеди-
рующего пахового лимфаденита, кожно-язвенной
формы туляремии, трихомонадных язв.
Наиболее затруднена диагностика с первич-
ным периодом сифилиса. Необходимы много-
кратные исследования (10—15 раз) серума язв на
бледную трепонему, взятие крови на РВ каждые
5—7 дней, постановка РИФ и привлечение к
обследованию всех половых контактов больно-
го за последние 3—6 месяцев.
Возможно одновременное заражение мягким
шанкром и сифилисом. У больных смешанным
(мягким и твердым) шанкром значительно реже
выявляется палочка шанкроида. В соскобе из-под
края язвы наблюдается картина шанкроида, че-
рез 3—4 недели появляются уплотнение дна язвы
и регионарный склераденит. Шанкроид задер-
живает развитие сифилитической инфекции,
удлиняет инкубационный период и отдаляет
появление вторичных сифилидов.
Первичная инфекция половых путей вирусом
простого герпеса, также как и при шанкроиде,
вызывает болезненную паховую аденопатию.
Вирусную инфекцию можно распознать по осо-
бенностям начала заболевания с везикулярных
очагов на гиперемированном фоне или по нали-
чию в анамнезе данных о возможном заражении
вирусом герпеса и по наличию системных сим-
птомов, таких как лихорадка и миалгия. Шан-
кроид системных симптомов, как правило, не
вызывает.
Вероятный диагноз шанкроида может быть ус-
тановлен, если у пациента обнаруживается одна
(или больше) болезненная генитальная язва,
причем:
а) нет доказательств инфекции, вызванной
Т. pallidum, при исследовании в темном поле яз-
венного экссудата или при проведении сероло-
гических тестов на сифилис, по крайней мере,
через 7 дней после образования язвы;
б) внешний вид и расположение язв, а также
регионарная лимфоаденопатия, если она присут-
ствует, являются типичными для шанкроида, а
результат теста на ВПГ — отрицательный.
Лечение
Лечение шанкроида осложняется трудностью
постановки точного диагноза в связи с трудно-
стью выделения культуры Н. ducreyi. На харак-
тер ответа при лечении может повлиять сопут-
ствующая инфекция, вызываемая Treponema
pallidum или вирусом простого герпеса. Если
бактериологическое подтверждение невозможно,
следует поставить клинический диагноз (пред-
почтительно после исключения при лаборатор-
ном тестировании сифилиса и генитального гер-
песа).
Сульфаниламиды и препараты тетрациклино-
вого ряда считаются активными при лечении
шанкроида, но в ряде случаев лечение оказыва-
ется неэффективным вследствие множественной
лекарственной устойчивости возбудителя. Пер-
воначальную терапию рекомендуют начинать с
сульфаниламидных препаратов, чтобы избежать
маскировки сифилиса:
• котримоксазол (2 таблетки утром и вечером
в течение 12—14 дней);
• мадрибон (по 1,0 г два раза в день в тече-
ние 12—14 дней);
• бисептол (2 таблетки два раза в день в тече-
ние 12—14 дней);
500 Венерология
• триметоприм-сульфаметоксазол (320 мг три-
метоприма и 1600 мг сульфаметоксазола ежед-
невно в течение 12 дней);
• сульфиоксазол (по 3—4 г в день в течение
12—14 дней).
Применяя при шанкроиде антибиотики, сле-
дует учитывать их трепонемоцидное действие.
При этом предпочтение отдают препаратам, ко-
торые не оказывают действия на бледную тре-
понему. С этой точки зрения целесообразно ле-
чение антибиотиками-аминогликозидами
(гентамицин, канамицин, сизомицин, амикацин
и др.):
• канамицин (внутримышечно по 500 000 ЕД
2 раза в сутки, на курс 10—15 млн. ЕД);
• мономицин (внутримышечно по 500 000 ЕД
2 раза в день, на курс — до 10 млн. ЕД);
• гентамицин, хлорамфеникол, тетрациклин
(по 0,5 г каждые 6 часов, на курс до 44 г).
Центр по контролю и предупреждению забо-
леваний (США, 1998) рекомендует следующие
схемы лечения шанкроида:
• азитромицин — 1,0 г однократно;
• цефтриаксон — 250 мг внутримышечно од-
нократно;
• эритромицин по 500 мг внутрь 4 раза в сут-
ки в течение 7 дней;
• ципрофлоксацин по 500 мг внутрь 2 раза в
сутки в течение 3 дней.
Ципрофлоксацин противопоказан беременным
и кормящим женщинам, а также лицам моложе
14 лет. Все четыре схемы эффективны для ле-
чения шанкроида у пациентов с ВИЧ-инфекци-
ей. Азитромицин и цефтриаксон обладают пре-
имуществом, так как их можно применять
однократно. Получены данные из разных реги-
онов мира о выделении нескольких изолятов с
устойчивостью либо к ципрофлоксацину, либо
к эритромицину.
Пациенты должны быть повторно обследова-
ны через 3—7 дней после начала терапии. При
успешном лечении состояние язв улучшается
симптоматически в течение 3 дней и объектив-
но — в течение 7 дней после начала терапии.
Если клинического улучшения не наблюдается,
врач должен рассмотреть следующие возможно-
сти: а) неправильно поставлен диагноз; б) име-
ется смешанная инфекция с другим ЗППП;
в) пациент инфицирован ВИЧ; г) не соблюда-
лась схема лечения; д) штамм Я. ducreyi, явля-
ющийся причиной заболевания, устойчив к на-
значенному препарату. Время, необходимое для
полного излечения, зависит от размера язвы. Для
заживления большой язвы может потребовать-
ся более 2 недель лечения. Кроме того, процесс
заживления проходит медленнее у некоторых
мужчин, не подвергшихся циркумцизии (обре-
занию), у которых язва располагается под край-
ней плотью. Для клинического разрешения
флюктуирующих лимфатических узлов требует-
ся больше времени, чем для заживления язвы и
даже при успешном лечении может потребовать-
ся дренирование. Разрез и дренирование бубо-
нов может оказаться предпочтительнее, чем ас-
пирация содержимого с помощью шприца, так
как после дренирования реже возникает необхо-
димость в применении каких-либо последующих
процедур, хотя аспирацию выполнить проще.
Профилактика
Профилактика шанкроида проводится с учетом
общих принципов предупреждения заражения
болезнями, передаваемыми половым путем. Пос-
ле окончания лечения больных нужно наблюдать
в течение 8—12 месяцев с ежемесячным прове-
дением клинико-серологического контроля.
Лица, имевшие половые контакты с больным
шанкроидом в течение 10 дней до проявления у
этих пациентов клинической картины, должны
быть обследованы и пролечены, даже при отсут-
ствии у них симптомов заболевания.
Глава 41
Венерический лимфогранулематоз
Лимфогранулематоз венерический (лимфогрануле-
матоз паховый, четвертая венерическая болезнь,
болезнь Дюрана-Никола-Фавра, венерическая
лимфопатия) — инфекционное заболевание, вы-
зываемое хламидиями и передающееся, в основ-
ном, половым путем. Заболевание характеризует-
ся преимущественным поражением лимфатических
узлов и мягких тканей урогенитальной области.
Этиология
Возбудитель венерического лимфогранулемато-
за относится к виду Chlamydia trachomatis серо-
логических типов Lj-L3. Он является наиболее
вирулентным представителем хламидий, первич-
но поражающих человека и обитающих в моче-
половых органах, способен размножаться в клет-
ках различных тканей. При естественной
инфекции обладает выраженным лимфотропиз-
мом, определяя лимфопролиферативный харак-
тер заболевания. Может размножаться в ткане-
вых культурах и куриных эмбрионах.
Эпидемиология
Венерический лимфогранулематоз обычно пере-
дается половым путем, но были отмечены слу-
чаи переноса заболевания при контакте через
предметы обихода или в результате случайного
заражения в лабораторных условиях. Чаще забо-
левают лица в возрасте 20—40 лет, ведущие ак-
тивную половую жизнь. Заболевание имеет не-
большие эпидемические очаги в тропических и
субтропических странах Южной Америки, Запад-
ной и Восточной Африки, Юго-Восточной Азии,
в портовых городах Европы. По данным ВОЗ,
венерический лимфогранулематоз составляет в
Индии 6 % от всех заболеваний, передающихся
половым путем, а в Нигерии и Замбии 2 %. В
США ежегодно регистрируется от 500 до 1000
случаев венерического лимфогранулематоза.
Интенсификация коммуникационных связей,
особенно в последние десятилетия, увеличивает
риск возникновения и распространения пахового
лимфогранулематоза в различных регионах мира.
Частота случаев инфицирования Chlamydia
trachomatis значительно меньше этого показателя
для гонококковой инфекции и сифилиса. У
мужчин признаки заболевания выявляют не-
сколько раньше, чем у женщин, которые обра-
щаются к врачу на поздних стадиях болезни. В
США большинство случаев заболевания прихо-
дится на гомосексуальных мужчин, путеше-
ственников, моряков и военнослужащих, возвра-
щающихся из-за границы, а также на лиц,
относящихся к малоимущим слоям населения,
проживающих в регионах с низкой эндемично-
стью на юго-востоке страны. Предполагают, что
главным резервуаром инфекции являются лица
с бессимптомным течением заболевания.
Клиническая картина
Инкубационный период варьирует в пределах 3—
30 дней, хотя имеются данные о длительности
его от 1 до 12 недель. После окончания инку-
бационного периода могут наблюдаться продро-
мальные явления в виде слабости, недомогания,
головных болей, повышения температуры.
При классическом течении заболевания далее
следуют:
• первичный период — на месте внедрения
инфекции появляются первичные симптомы;
• вторичный период — поражение регионарных
лимфатических узлов на фоне общей реакции
организма;
• третичный период — тяжелые проявления бо-
лезни, когда на половых органах, слизистой обо-
лочке прямой кишки и в перианальной области
наблюдаются воспалительно-элефантиатические,
гипертрофические и язвенные процессы с исхо-
дом в стенозирование и рубцевание.
Первичный период
Первичное поражение (лимфогранулематозный
шанкр) у мужчин часто возникает на головке
полового члена, реже — в мочеиспускательном
канале или на коже паховой области, у жен-
щин — во влагалище или на половых губах,
реже — на шейке матки. Первичные поражения
могут иметь вид папул, пустул, поверхностной
язвочки, эрозии или проявляться признаками не-
специфического воспаления мочеиспускательно-
501
502 Венерология
го канала. При орогенитальных, мануальных
половых контактах первичное поражение может
локализоваться на губах, языке, в прямой киш-
ке, на пальцах. Обычно первым признаком за-
болевания является появление единичного пу-
зырька, из которого образуется эрозия или
поверхностная язвочка округлой формы с ров-
ными краями, без инфильтрации в основании,
склонная к самостоятельному заживлению без ле-
чения и образования рубца в течение несколь-
ких дней. Через несколько дней, после появле-
ния первичного поражения, отдельные
регионарные лимфатические узлы начинают уве-
личиваться, возможен даже умеренно выражен-
ный лимфангиит. Возможен и другой вариант
заболевания: шанкр с самого начала выглядит как
отечная папула или безболезненное узелковое пу-
стулезное образование. Иногда он протекает как
эрозивный баланит или баланопостит. В неко-
торых случаях, при локализации эрозии в урет-
ре, могут быть проявления уретрита.
Вторичный период
Спустя некоторое время (от нескольких дней до
2~3 недель, реже до нескольких месяцев) после
появления первичного элемента развивается
наиболее характерный симптом заболевания —
поражение регионарных лимфатических узлов
(вторичный период). Лимфатические узлы уп-
лотняются, становятся болезненными, распрос-
траняется воспалительный инфильтрат. Лимфа-
тические узлы спаиваются между собой. В
паховой области формируется плотный, бугри-
стый инфильтрат, превышающий размеры круп-
ного лимона, появляются боли. Кожа в области
инфильтрата становится болезненной при паль-
пации. Цвет кожи изменяется от розового до си-
нюшно-красного. На инфильтрированной повер-
хности образуются очаги нагноения,
представляющие собой отдельные изолированные
друг от друга многочисленные размягчения. По
мере появления периаденита и гнойного расплав-
ления лимфатических узлов вовлеченная в про-
цесс кожа постепенно истончается и прорывается
с образованием фистульных ходов, из которых
выделяется большое количество желтоватого
клейкого желатиноподобного гноя, иногда с
примесью крови. В паховой области постоянно
возникают новые свищи, при этом более ранние
могут полностью зарубцеваться. Подобная кли-
ническая картина характерна для гетеросексуаль-
ных мужчин, у которых чаще всего наблюдает-
ся паховый синдром, характеризующийся
болезненной паховой лимфаденопатией. У 60 %
больных паховая аденопатия бывает односторон-
ней, и на той стороне, где увеличены паховые
лимфатические узлы, часто пальпируются уве-
личенные подвздошные и бедренные лимфати-
ческие узлы. Чрезмерное увеличение цепочек
паховых лимфатических узлов, расположенных
выше и ниже паховой связки («знак желобка») —
обычное явление при данном заболевании.
У женщин и гомосексуальных мужчин в ре-
зультате инфицирования после анального коитуса
развивается первичный анальный или ректальный
аффект. У женщин ректальное инфицирование
штамами Chlamydia trachomatis может произойти
в результате распространения инфицированных
секретов по промежности или путем распростра-
нения инфекции в прямую кишку через тазовые
лимфатические сосуды. От места первичного
аффекта, расположенного в мочеиспускательном
канале, половых органах, заднепроходном кана-
ле или прямой кишке, микроорганизмы распро-
страняются через регионарные лимфатические
узлы. Инфекция, локализующаяся на половом
члене мужчины, наружных половых органах жен-
щины и в заднепроходном канале, может вызвать
развитие пахового и бедренного лимфаденита.
Инфекция, локализующаяся в прямой кишке,
вызывает развитие подчревного и глубокого под-
вздошного лимфаденита. Инфекция, локализую-
щаяся в верхних отделах влагалища и в шейке
матки, приводит к увеличению обтурирующих и
подвздошных лимфатических узлов.
При вторичном периоде заболевания могут на-
блюдаться общие явления: быстрая утомляемость,
головные боли, тошнота, рвота, возможен озноб и
умеренная интермиттирующая лихорадка. При
лабораторных исследованиях у некоторых больных
обнаруживаются лейкоцитоз, увеличение скорос-
ти оседания лейкоцитов, аномальные результаты
печеночных проб, гиперглобулинемия, смешанная
криоглобулинемия, увеличение активности ревма-
тоидного фактора и повышение уровней содержа-
ния Ig G, Ig A, Ig М.
Третичный период
Без соответствующего лечения болезнь прогрес-
сирует и наступает третичный период, во время
которого развиваются тяжелые деструктивные
изменения не только в лимфатических узлах, но
и в окружающх тканях и органах; увеличиваются
печень и селезенка. Основным признаком тре-
тичного периода является генитоаноректальный
синдром, развивающийся через 1—2 года после
начала заболевания. У женщин генитоанорек-
тальный синдром наблюдается значительно чаще,
чем у мужчин. Это связано с анатомическими
особенностями женских половых органов, не
Глава 41. Венерический лимфогранулематоз 503
только затрудняющими раннюю и своевремен-
ную диагностику заболевания, но и способству-
ющими более быстрому переходу инфекции с
одного органа таза на другой.
Осложнения
К числу ранних осложнений нелеченного генито-
аноректального синдрома относятся: параректаль-
ный абсцесс, свищ заднего прохода, прямокишеч-
но-влагалищный свищ, пузырно-прямокишечный
свищ, седалищно-прямокишечный свищ, язвенное
поражение полового члена, свищевое поражение
полового члена, свищ мочеиспускательного кана-
ла, свищ мошонки.
Поздние осложнения венерического лимфогра-
нулематоза: слоновость половых органов — хро-
нические индурация и отек полового члена или
наружных половых органов женщины вследствие
обструкции лимфатических сосудов, эстиомен —
хронические изъязвления наружных половых
органов женщин, лимфорроиды — гладкие, снаб-
женные ножками перианальные разрастания,
стриктура прямой кишки, стриктура мочеиспус-
кательного канала.
Наиболее ранними признаками развития ге-
нитоаноректального синдрома могут быть уме-
ренно выраженные тянущие боли в области
промежности и скудные сукровично-гнойные
выделения из заднепроходного отверстия. При
ректоскопическом исследовании видны отек
и гиперемия, эрозии, легкая кровоточивость
и болезненность при малейшем растяжении
слизистой оболочки. По мере развития руб-
цово-склеротических изменений в прямой
кишке начинаются процессы фиброзного пе-
рерождения тканей, приводящие к развитию
стриктур, кольцевидно охватывающих пря-
мую кишку. В результате этого просвет киш-
ки постепенно суживается, и она приобрета-
ет форму конуса, обращенного верхушкой
внутрь.
При пальпации в области прямой кишки уда-
ется прощупать концентрически суживающийся
плотный фиброзный тяж, способный привести к
почти полному закрытию просвета кишки, что
довольно часто наблюдается при отсутствии ле-
чения в поздние сроки заболевания. В результа-
те проктита и парапроктита могут развиваться
абсцессы, вскрывающиеся с образованием изъяз-
влений, свищей в области заднего прохода, про-
межности, половых органов. На поверхности
кожи в этих участках в результате мацерации,
трения и других факторов появляются бородав-
чатые разрастания, между которыми скапливают-
ся гнойная жидкость, слущивающийся эпителий,
18 Зак. 1357
грязь. Появляется неприятный запах. Язвенный
распад и последующие рубцевания приводят к
уродующим стриктурам стенок влагалища, осо-
бенно в области входа, значительным деформа-
циям и слоновости малых и больших половых
губ, клитора. Все это может привести к наруше-
нию акта дефекации, недержанию мочи.
При генитоаноректальном синдроме возмож-
на генерализация инфекции, приводящая к се-
рьезным поражениям ЦНС (менингит), сердца
(эндо- и миокардит), легких (пневмония), кожи
(узловатая и многоформная эритема), глаз и
других органов.
Таким образом, лимфогранулематоз венери-
ческий по патогенезу и клиническим проявле-
ниям значительно отличается от других форм
урогенитальных хламидиозов, что во многом
определяется биологией и экологией возбудите-
ля. Клиническими симптомами инфекции мо-
гут быть глубокий инфильтративный язвенный
и склерофиброзный процесс с явлениями эле-
фантиаза области наружных половых органов,
промежности, заднего прохода и прямой киш-
ки. При рубцевании язв возникают тяжелые
осложнения — стриктуры и атрезии уретры и
прямой кишки. Встречаются также папилломы,
вегетации, осложняющие картину пахового лим-
фогранулематоза.
Дифференциальный диагноз
Венерический лимфогранулематоз следует диф-
ференцировать с мягким шанкром, туберкулезом
паховых лимфатических узлов, сифилисом, до-
нованозом, глубоким микозом, туляремией.
Мягкий шанкр начинается, как правило, с раз-
вития язвы. Реакция регионарных лимфатичес-
ких узлов необязательна. Поражение узлов раз-
вивается как осложнение, протекает остро.
Длительно незаживающих фистулезных ходов
обычно не возникает. Мягкий шанкр отличает-
ся аутоинокуляцией инфекции. В сомнительных
случаях имеют значение данные микроскопии.
Сифилис отличается клиническими особенно-
стями твердого шанкра и пораженных лимфати-
ческих узлов. Диагноз сифилиса подтверждает-
ся обнаружением бледной трепонемы, а со второй
половины первичного периода — положитель-
ными серологическими реакциями крови.
При донованозе лимфатические узлы, как
правило, в процесс не вовлекаются. Однако в
поздние периоды заболевания, вследствие рас-
пространения процесса по периферии и пораже-
ния промежности, мошонки возможно развитие
ректовагинальных свищей и слоновости половых
органов. Возбудитель заболевания — простейшие
504 Венерология
(тельца Донована) определяются в соскобах из
пораженных тканей.
При бубонной форме туляремии поражаются
чаще подмышечные, шейные и подчелюстные
лимфатические узлы. Для туляремийного бубо-
на нехарактерно развитие периаденита. Распоз-
наванию помогают положительный результат
туляриновой пробы и другие лабораторные ис-
следования (РСК, заражение животных).
Лимфоретикулез — заболевание, сопровожда-
емое опухолевидными разрастаниями лимфати-
ческих узлов, отличается волнообразной лихорад-
кой, зудом кожи, потливостью и постепенно
нарастающей кахексией. Поражение селезенки,
печени и костного мозга придает этому заболе-
ванию системный характер.
В отличие от лимфогранулематоза с преиму-
щественной локализацией процесса в паховой
области, при четвертой венерической болезни на-
блюдается чаще одностороннее увеличение лим-
фатических узлов с развитием свищей и гной-
ным отделяемым.
Болезнь «кошачьих царапин» вызывается виру-
сом группы Cisticetes. Возбудитель заносится
человеку при укусе или царапаньи кошками или
птицами. Инкубационный период длится от 3 до
28 дней. На месте травмы (лицо, верхние конеч-
ности) возникает пятно или узелок. Общее со-
стояние нарушается (лихорадка, мышечные и
головные боли). В процесс вовлекаются регио-
нарные лимфатические узлы: они увеличиваются,
нередко нагнаиваются, затем развиваются руб-
цово-склеротические изменения. В сыворотке
лиц, перенесших болезнь «кошачьих царапин»,
обнаруживаются комплемент-связывающие ан-
титела, реагирующие с групповыми антигенами
орнитоза и четвертой венерической болезни. У
большинства больных отмечается положительная
внутрикожная проба со специфическим орнитоз-
ным антигеном.
Лабораторная диагностика
Для подтверждения диагноза венерического лим-
фогранулематоза, вызванного С. trachomatis,
используют три типа исследований: прямое мик-
роскопическое исследование соскобов тканей для
выявления типичных внутрицитоплазматических
включений; выделение возбудителя; выявление
антител в сыворотке крови или в местных сек-
ретах.
При микроскопическом исследовании можно
иногда обнаружить крупные хламидии в мазках
гноя или материала, взятого при биопсии. Эле-
ментарные тельца окрашиваются по Романовс-
кому — Гимза в сине-фиолетовый цвет, по
Маккиавелло — в ярко-красный. Однако пря-
мое микроскопическое исследование окрашен-
ных по методу Романовского — Гимзы клеток
характеризуется неприемлемо низкой чувстви-
тельностью, в результате чего может иметь ме-
сто ложноположительная интерпретация резуль-
татов.
Для выделения Chlamydia trachomatis исполь-
зуют бактериологическое исследование посевов
на культурах клеток, заменивших использование
желточного мешка куриных эмбрионов. Чаще
всего используют линии клеток Мак-Коя и
HeLa-229. Положительные результаты посевов
определяют путем визуализации типичных внут-
рицитоплазматических включений, окрашенных
по методу Романовского — Гимза, или йодом,
или же с использованием связанных с флюорес-
цеином моноклональных антител.
При проведении теста с иммунофлюоресциру-
ющими микроскопическими препаратами инфи-
цированные секреты из половых путей наносят
в виде мазка на стекло, фиксируют и окраши-
вают с использованием связанных с флюорес-
цеином моноклональных антител. При исследо-
вании этих мазков с помощью флюоресцентного
микроскопа наличие флюоресцирующих элемен-
тарных телец подтверждает диагноз. Чувстви-
тельность этого теста по сравнению с методом
посевов составляет 85-90 %.
В качестве еще одной альтернативы бактери-
ологическому исследованию посева для выявле-
ния хламидийного антигена разработана методи-
ка твердофазного иммуноферментного анализа.
Согласно сообщениям исследователей, чувстви-
тельность этого теста составляет 67—95 %, а спе-
цифичность в отношении выявления инфици-
рования половых путей — 92—97 %.
Большинство исследователей считают реакцию
связывания комплемента (РСК) наиболее про-
стым серологическим методом выявления анти-
тел к антигенам возбудителя венерического лим-
фогранулематоза. Специфичность при этом
может быть ограничена за счет перекрестной
реакции с антигенами возбудителя пситтакоза.
Положительной реакция становится только спу-
стя 2—4 недели после заражения.
Микроиммунофлюоресцентный тест (микро-
ИФА) с антигенами С. trachomatis более чувстви-
телен, чем РСК. Тест с микроиммунофлюорес-
циирующими антителами выявляет наличие
антител к С. trachomatis почти у всех больных ве-
нерическим лимфогранулематозом с диагнозом,
подтвержденным результатами бактериологичес-
ких исследований посева. Титры микроимму-
нофлюоресцирующих антител в сыворотке кро-
Глава 41. Венерический лимфогранулематоз
505
ви больных составляют обычно > 1 : 512, что
превышает самые высокие значения титров, по-
лученных при хламидийном негонококковом
уретрите.
Лечение
Показаны антибиотики тетрациклинового ряда,
макролиды, а также сульфаниламидные препа-
раты. Рекомендуется прием антибиотиков и
сульфаниламидов в течение 21—23 дней. Этиот-
ропная терапия оказывает хороший эффект, осо-
бенно на ранних стадиях заболевания:
• тетрациклина гидрохлорид — по 250 мг
каждые 6 часов в течение 3 недель или по 500 мг
4 раза в сутки в течение 3 недель;
• доксициклин по 100 мг внутрь 2 раза в сут-
ки в течение 3 недель;
• эритромицин по 500 мг внутрь 4 раза в сут-
ки в течение 3 недель;
• сульфаниламиды (сульфадимезин, норсуль-
фазол) по 1,5 г 3 раза в день в течение 2 недель,
а затем по 1,0 г 3 раза в день в течение 3 недель
(курс лечения 5 недель).
В большинстве случаев после применения ука-
занных препаратов наступает полное излечение.
Резистентности С. trachomatis серотипов Lp L2, L3
к тетрациклинам к настоящему времени не опи-
сано. При наслоении вторичной стафилококковой
инфекции можно применять антибиотики пени-
циллиновой группы, устойчивые к пенициллиназе
(оксациллин, метициллин). Терапия пеницилли-
ном и стрептомицином неэффективна. Активность
азитромицина в отношении С. trachomatis предпо-
лагает, что этот препарат может быть эффективен
при его применении в многократных дозах в те-
чение 2~3 недель, но клинических данных о его
использовании при этом заболевании пока недо-
статочно. Для беременных и кормящих женщин
(больных лимфогранулематозом венерическим)
должна использоваться схема лечения эритроми-
цином.
Для лечения последующих стадий, наряду с
этиотропной терапией, назначают патогенетичес-
кие и симптоматические средства в сочетании с
хирургическим лечением.
При наличии глубоких фиброзных изменений,
в частности стриктур, через 2 недели показано
назначение повторного курса лечения и бужи-
рования в течение длительного времени.
При наличии свищей, уродующих рубцов и
других стойких фиброзных перерождений, по-
мимо сульфаниламидных препаратов и антиби-
отиков, показаны инъекции алоэ, стекловидно-
го тела, лидазы.
По соответствующим показаниям нередко
прибегают к оперативным вмешательствам, на-
значению физиотерапевтических процедур (уль-
тразвук, аппликации парафина).
Через несколько месяцев у большинства боль-
ных, получавших адекватное лечение, происхо-
дит 4-кратное и более снижение титра ФК-ан-
тител или микроиммунофлюоресцирующих
антител. После начала лечения антибиотиками
выделить возбудителя из пораженных участков
не удается. Прогноз венерического лимфограну-
лематоза при полноценном и своевременно на-
чатом лечении можно считать благоприятным.
Заболевание оставляет стойкий иммунитет.
Случаев повторного заражения венерическим
лимфогранулематозом не описано.
Профилактика
Профилактика венерического лимфогранулема-
тоза не отличается от профилактики других ве-
нерических заболеваний. Существенное значение
имеет выявление источников и контактов, а так-
же раннее специфическое лечение и наблюдение
за выявленными больными. Половые партнеры
пациентов с венерическим лимфогранулематозом
должны быть обследованы, тестированы на урет-
ральную или цервикальную хламидийную ин-
фекцию и пролечены, если они имели половые
контакты с больными в течение 30 дней, пред-
шествовавших появлению у последних симпто-
мов венерического лимфогранулематоза.
Глава 42
Донованоз
Донованоз (паховая гранулема, венерическая
гранулема, тропическая язвенная гранулема,
пятая венерическая болезнь, донованоз Бро-
ка) — хроническое, медленно прогрессирующее
инфекционное заболевание, которое передается
преимущественно половым путем, характери-
зуется развитием обширных гранулематозных
изъязвлений кожи, а также поражением лим-
фатических узлов генитальной и перианальной
области.
Этиология
Возбудитель венерической гранулемы
(Calymmatobacterium granulomatis) был открыт
в 1905 году в Мадрасе ирландским врачом
Ч. Донованом, описавшим внутриклеточные
включения в экссудате из очагов поражений
в полости рта больного, у которого наблюда-
лись также поражения в области половых ор-
ганов. Поэтому иногда Calymmatobacterium
granulomatis называют тельцами Донована.
Возбудитель донованоза Calymmatobacterium
granulomatis паразитирует, главным образом,
внутри макрофагов. Это полиморфный грамот-
рицательный микроорганизм овоидной или бо-
бовидной формы, который может иметь форму
кокка или короткой палочки. Палочки обычно
окружены капсулой. В микробных клетках име-
ются одиночные или расположенные биполяр-
но скопления хроматина, что придает клетке ха-
рактерный вид английской булавки. Бактерии
легко выявляются при окраске по методу Рома-
новского — Гимза, при этом тело бактерии ок-
рашивается в голубой, хроматиновые включе-
ния — в темно-голубой или черный цвет, а сам
микроорганизм окружен розовой субстанцией.
При окраске по Райту возбудители венеричес-
кой гранулемы обнаруживаются как бацилляр-
ные тела, окруженные хорошо очерченными
плотными розоватыми капсулами. Другими кра-
сителями микробы окрашиваются плохо. Они
неустойчивы к действию кислот, неподвижны,
грамотрицательны. При электронно-микроско-
пическом исследовании бактерий, выделенных
из пораженных тканей, выявлена фибриллярная
капсула. Размножаются клетки делением попо-
лам.
Calymmatobacterium granulomatis растут на све-
жей среде с яичным желтком, после адаптации —
и на других питательных средах. Тельца Доно-
вана можно выращивать в инфицированном
желточном мешке развивающихся куриных эм-
брионов.
Эпидемиология
Венерической гранулемой болеют только люди.
Заражение происходит, главным образом, поло-
вым путем, значительно реже — бытовым. До-
казательства распространения заболевания поло-
вым путем основаны на следующих факторах:
возрастная специфичность (20—40 лет); частое
сочетание с сифилисом; склонность к пораже-
нию половых органов у гетеросексуалов и к
инфекции в аноректальной области у пассивных
гомосексуалистов. Благоприятствующими фак-
торами распространения инфекции являются
влажный климат и высокая температура окру-
жающей среды. В возникновении и распростра-
нении болезни играют роль несоблюдение пра-
вил личной гигиены и беспорядочные половые
связи. Полагают, что индивидуальная восприим-
чивость к венерической гранулеме играет значи-
тельную роль. Calymmatobacterium granulomatis
характеризуется различной патогенностью и
морфологической вариабельностью как в тканях
пораженных органов, так и при культивирова-
нии, чем и объясняется низкая контагиозность
донованоза по сравнению с другими венеричес-
кими болезнями.
Донованоз — эндемическая тропическая бо-
лезнь, особенно широко распространенная в
Новой Гвинее, среди индусов в Индии, в стра-
нах Карибского бассейна, в Центральной и
Южной Африке, в южных провинциях Китая,
в центральной Австралии. В США венерическая
гранулема встречается редко, большинство случа-
ев заболевания наблюдается в юго-восточных
штатах и преимущественно у гомосексуалистов. В
Бразилии венерическая гранулема занимает 5-е
место среди болезней, передаваемых половым
506
Глава 42. Донованоз
507
путем. В Европе данное заболевание встречает-
ся редко.
Клиническая картина
Инкубационный период донованоза длится от
нескольких дней до 3 месяцев и более. Тельца
Донована проникают через дефекты кожи и сли-
зистых оболочек, вызывая воспалительную реак-
цию в дерме или, реже, в подкожной основе.
Раннее проявление болезни — островоспалитель-
ная дермальная папула с плоской поверхностью,
величиной с горошину. В некоторых случаях
начальное поражение представлено везикуло-па-
пулой с несколько уплощенной поверхностью,
безболезненной, небольших размеров. Папулы
изъязвляются, образуя единичные язвы с барха-
тистой поверхностью, безболезненные, края при-
поднятые. Дно язвы ярко-красного цвета, имеет
несколько зернистую поверхность. Как правило,
поражаются половые органы (головка полового
члена, крайняя плоть, клитор, малые половые
губы), промежность, область заднего прохода,
паховая область, кожа лица, туловища, кисти,
голени, слизистая оболочка носа, полости рта,
гортани, глотки. Типичная донованозная язва
представляет собой увеличивающуюся массу гра-
нуляций, напоминает опухоль, имеет зубчатые
или волнистые края и резко отграничена от ок-
ружающей кожи. Она обычно мягкая на ощупь,
но окружающая кожа может быть отечна и утол-
щена, сочного ярко-красного цвета, дно чистое,
с серозным отделяемым. При вторичной инфек-
ции язва имеет «грязный» вид, покрыта струпь-
ями и некротическими массами. Донованозные
язвы медленно растут по периферии. Выделения
из язв скудные, серозно-гнойные, иногда с при-
месью крови, имеют характерный зловонный
запах. Процесс протекает сравнительно безболез-
ненно, аденопатия незначительна, общее состоя-
ние больного не нарушается. В результате ауто-
инокуляции или переноса заразного материала
руками на участки кожи, могут появиться не-
сколько дополнительных язв. Второе место по
частоте занимает донованоз полости рта. Он от-
личается болезненностью, в процесс вовлекают-
ся зубы. При экстрагенитальном донованозе по-
ражаются губы, десны, щеки, небо, глотка,
гортань, нос, шея, грудь. Системный донованоз,
поражающий кости, суставы, печень и селезен-
ку, возникает редко.
Клинические формы
В зависимости от особенностей клинического
течения и преобладания того или иного симп-
тома заболевания различают несколько клини-
ческих вариантов донованоза: язвенный, веррукоз-
ный, цветущий, некротический, склерозирующий,
слизистый, смешанный.
Язвенная форма
Язвенная форма донованоза встречается наибо-
лее часто. Язвы в области половых органов мо-
гут быть одиночными или множественными и
отличаются склонностью к развитию вегетаций,
серпигинирующему росту, распространению в
паховую область. Язвенные формы заболевания
подразделяются на следующие разновидности:
язвенно-вегетирующая, серпигинозно-язвенная,
язвенно-проникающая, рубцово-келоидная.
Характерной чертой двух первых разновидно-
стей является тенденция к развитию вегетаций
или серпигинирующему росту и распростране-
нию в области паховых складок. Язвенно-про-
никающая форма чаще встречается у женщин,
имеющих обширные поражения, в результате
которых может возникать эстиомен, как при
поздней стадии венерической лимфопатии. При
рубцово-келоидной разновидности отмечена тен-
денция к образованию обширных келоидных
рубцов.
Веррукозная форма
Веррукозная форма характеризуется появлением
на дне безболезненных язв достаточно выражен-
ных, бледно-розового цвета, легко кровоточащих
бородавчатых разрастаний. Выделяющийся в
скудном количестве экссудат засыхает с образо-
ванием серозно-кровянистых, плотно пристающих
к грануляциям корок. Язва малоболезненная,
течение ее торпидное. Разновидностями веррукоз-
ной формы являются 2 типа: гипертрофический,
элефантиазный.
При гипертрофическом типе дно язвы при-
поднято над окружающей кожей и состоит из
крупной, грубой, бледно-розового цвета грану-
ляционной ткани. Язва безболезненна, отделя-
емое из нее скудное, она увеличивается медлен-
но. Элефантиазный тип характеризуется
тенденцией к развитию слоновости наружных
половых органов.
Цветущая форма
Цветущая форма донованоза характеризуется
сочными, ярко-красного цвета грануляциями,
которые имеют бархатистую, влажную повер-
хность. Дно язвы резко болезненное. Кожа
вокруг язвы отечна и инфильтрирована. Вы-
деления из язвы серозно-гнойные, имеют спе-
цифический, неприятный запах. Больные жа-
луются на боль и нестерпимый зуд в области
поражений.
508 Венерология
Некротическая форма
Некротическая форма является наиболее тяжелой
и развивается обычно при хроническом течении
донованоза. Она возникает чаще всего в резуль-
тате осложнения донованозных язв вторичной
инфекцией. На фоне хронического течения вне-
запно возникает острый воспалительный процесс,
при котором розовая грануляционная ткань бы-
стро заменяется серо-коричневой некротической
с обилием густого отделяемого шоколадного цве-
та. Клиническая картина некротической формы
характеризуется появлением увеличивающейся
глубокой язвы с гнойным распадом тканей. По-
ражения быстро распространяются по периферии
вглубь, разрушая ткани вплоть до фасций, мышц
и костей. У женщин наружные половые органы,
промежность и область заднего прохода могут
быть полностью разрушены. У мужчин возможен
некроз полового члена вплоть до его полного
разрушения. Процесс сопровождается лимфанги-
том и лимфаденитом. Появляются симптомы
общего недомогания: слабость, головная боль,
озноб, лихорадка. Может развиться острая вто-
ричная анемия. В результате дальнейшей генера-
лизации процесса возможно метастазирование
инфекции в среднее ухо, печень, селезенку, ко-
сти. При несвоевременно начатом лечении воз-
можно развитие сепсиса со смертельным исходом.
Склерозирующая форма
Склерозирующая форма донованоза обусловли-
вает сужение наружного отверстия мочеиспуска-
тельного канала, ануса и входа во влагалище.
Резко нарушается анатомическое строение жен-
ских половых органов, что приводит к их дефор-
мации.
Смешанная форма
Смешанная форма донованоза характеризуется
наличием симптомов одновременно нескольких
разновидностей заболевания. В зависимости от
превалирования тех или иных симптомов про-
гноз при данной форме может быть различным.
Следствием донованоза могут быть такие ос-
ложнения, как псевдослоновость половых орга-
нов у женщин, стеноз мочеиспускательного ка-
нала, сужение влагалища, фимоз полового члена,
слоновость и некроз полового члена, сужение
заднего прохода, эпидермоидная карцинома.
Диагноз
Диагноз донованоза устанавливается после про-
ведения комплекса лабораторных исследований:
• исследование полученной из язвы серозной
жидкости на бледную трепонему в темном поле
микроскопа;
• исследование мазков отделяемого из язв (ок-
рашенных по Граму препаратов) на стрептоба-
циллы Дюкрея;
• исследование возбудителя донованоза в ок-
рашенных по Лейшману или Романовскому-
Гимза мазках, приготовленных путем раздавли-
вания кусочков ткани краев язв между двумя
предметными стеклами.
Даже при установлении диагноза донованоза
больных необходимо обследовать для исключе-
ния другого сопутствующего венерического за-
болевания.
Дифференциальный диагноз
Донованоз дифференцируют с первичной сифи-
ломой, вторичным и третичным сифилисом, с
мягким шанкром, паховой лимфогранулемой,
раком кожи и слизистых оболочек, филяриозом,
туберкулезными язвами, аногенитальным кож-
ным амебиазом.
Для исключения сифилиса необходимы исследо-
вания на наличие бледной трепонемы и серологи-
ческий контроль. Перианальный донованоз напо-
минает широкую кондидому вторичного сифилиса.
В этом случае, если предполагаемая широкая кон-
дилома не излечивается после применения пеницил-
лина по поводу сифилиса, следует вполне обосно-
ванно предположить наличие донованоза. Это
заключение особенно актуально для стран, где до-
нованоз является эндемической болезнью.
Мягкий шанкр подтверждается коротким ин-
кубационным периодом (3-5 дней), множествен-
ными, болезненными, легко кровоточащими
язвами с подрытыми и неровными краями, изъе-
денным дном, покрытым гнойным налетом, об-
наружением стрептобациллы Дюкрея-Унны-Пе-
терсена в отделяемом из язвы.
При паховом лимфогранулематозе первично
поражаются лимфатические узлы (паховые, по-
ловых органов, прямой кишки). Возбудитель
заболевания — отдельные штаммы Chlamydia
trachomatis, которые выявляют по характерным
внутриклеточным включениям в культуре кле-
ток или в куриных эмбрионах. У больных по-
вышена температура тела, имеется головная боль,
боли в суставах.
Тропический филяриоз вызывает слоновость
половых органов. При этом заболевании язвы не
образуются. В анамнезе больных отмечается на-
личие частых приступов лихорадки. В крови
больных обнаруживаются микрофилярии.
Туберкулез половых органов — туберкулезные
язвы в области половых органов не схожи с яз-
вами при донованозе, при биопсии обнаружи-
вают казеоз, гигантские клетки и микобактерии.
Глава 42. Донованоз
509
Урогенитальный амебиаз — амебиазные пора-
жения наружных половых органов внешне на-
поминают донованозные. Может наблюдаться
одновременное или последовательное заражение
амебиазом и донованозом. Для исключения аме-
биаза исследуют нативные неокрашенные маз-
ки, в которых находят активную, живую
Entamoeba hystolytica.
Лечение
Основными препаратами для лечения доновано-
за являются антибиотики:
• доксициклин по 100 мг 2 раза в сутки, ми-
нимум в течение 3 недель;
• эритромицин по 0,5 г 4 раза в сутки до еды
в течение 3—4 недель;
• тетрациклина гидрохлорид, окситетрациклин
или хлортетрациклин внутрь по 0,5 г во время
или после еды 4 раза в день в течение 14 дней;
• сульфаметаксазол-триметоприм (в одной
таблетке 400 мг сульфаметаксазола и 80 мг три-
метоприма) по 2 таблетки 2 раза в день после еды
в течение 3 недель;
• линкомицин внутрь по 0,5 г за 1—2 часа до
еды или через 2—3 часа после еды 3 раза в день
в течение 14 дней или внутривенно капельно
0,6 г на одно вливание; суточная доза 1,8 г. При
внутримышечном введении разовая доза 0,6 г;
суточная доза 1,8 г;
• ампициллин по 0,5 г 4 раза в день независи-
мо от приема пищи в течение 2—3 недель;
• гентамицин внутримышечно — разовая доза
0,4 мг на кг массы тела; суточная доза 0,8—1,2 мг
на кг массы тела;
• хлорамфеникол (левомицетин) по 0,5 г 4 раза
в день за 30 мин до еды в течение 3 недель;
• хлорамфеникол или тиамфеникол — 500 мг
4 раза в день в течение 3 недель и гентамицин
1 мг/кг внутримышечно 3 раза в день в течение
3 недель;
• азитромицин по 1,0 г 1 раз в неделю в тече-
ние 4 недель или по 500 мг ежедневно в тече-
ние 1 недели;
• ципрофлоксацин 750 мг перорально 2 раза в
день в течение 3 недель.
Стрептомицин применять не рекомендуется,
так как к нему нередко развивается резистент-
ность возбудителя донованоза.
Сульфаниламидные препараты и антибиоти-
ки применяют местно в виде присыпки, примо-
чек на язвы после предварительного их очище-
ния дезинфицирующими растворами. По
показаниям назначают витамины, биостимуля-
торы.
Некоторым больным может потребоваться
хирургическое вмешательство — устранение (ис-
сечение) слоновости, деформирующих, облите-
рирующих рубцов, вульвэктомия, пересадка
кожи. Необходимость в подобных операциях
может возникнуть и у больных с излеченным
донованозом, так как нередко заболевание закан-
чивается увечьем или уродством.
Профилактика
Профилактические меры идентичны таковым
при других венерических заболеваниях. Лица,
имевшие половые контакты с пациентами, боль-
ными паховой гранулемой, должны быть обсле-
дованы и пролечены, если:
а) они имели половые контакты в течение 60
дней до появления у пациентов симптомов;
б) при наличии у них симптомов и призна-
ков данного заболевания.
Прогноз
При своевременно установленном диагнозе и
применении антибиотиков прогноз донованоза
можно считать благоприятным. В случаях по-
зднего установления диагноза и запоздалого на-
чала лечения возможно развитие обширных не-
крозов с последующим разрушением не только
наружных, но иногда и внутренних половых
органов, развитие уродующих рубцовых измене-
ний (сужение, стенозы ануса, уретры, влагали-
ща), слоновости половых органов. Донованозные
язвы подвержены перерождению в плоскокле-
точный рак. Иногда после перенесенного доно-
ваноза возможно развитие вторичной прогрес-
сирующей анемии.
Глава 43
Трихомониаз
Трихомониаз (трихомоноз) — наиболее распро-
страненное, передаваемое половым путем инфек-
ционное заболевание мочеполового тракта, ко-
торое вызывается влагалищной трихомонадой.
Трихомониаз занимает первое место среди
заболеваний, передаваемых половым путем.
Инфекция имеет «космополитический» характер;
ежегодный уровень заболеваемости на земном
шаре обычно составляет около 170 миллионов
человек.
Этиология
Возбудителем мочеполового трихомониаза является
влагалищная трихомонада — Trichomonas vaginalis.
Она относится к простейшим — Protozoa, классу
жгутиковых — Flagellata, роду трихомонад —
Trichomonas. У человека паразитируют 3 вида три-
хомонад: Trichomonas tenax (в полости рта),
Trichomonas hominis (в желудочно-кишечном трак-
те), Trichomonas vaginalis (в мочеполовом тракте).
Урогенитальные трихомонады бывают трех форм:
грушевидной, амебовидной и почкующейся.
Трихомонада представляет собой типичный од-
ноклеточный паразит, признаками которого яв-
ляются (С. В. Батыршина, 1996):
• способность повторять рельеф эпителиаль-
ной клетки (на которой они паразитируют),
проникать между клетками в узкие щели и ин-
вагинировать в клетку хозяина;
• способность урогенитальных трихомонад
фиксировать на своей поверхности большие
количества антитрипсина, что обеспечивает за-
щиту простейших от разрушительного действия
организма;
• вирулентность урогенитальных трихомонад,
которая находится в прямой пропорциональной
зависимости от их гемолитической активности;
• заразительность (развитие заболевания после
полового контакта с больным трихомониазом);
• наличие на поверхности трихомонад проте-
олитических ферментов (гиалуронидаза, амила-
за, каталаза);
• выраженный хемотаксис полиморфноядер-
ных лейкоцитов (на поверхности трихомонад
имеется фибропектин — гликопротеид, способ-
ствующий прикреплению паразита к лейкоци-
там, эпителиальным клеткам, эритроцитам).
Тело трихомонады состоит из оболочки, прото-
плазмы, блефаробласта, ядра, акростиля, четырех
жгутиков, ундулирующей мембраны, фибрилл. В
расширенной передней части тела трихомонады
располагается удлиненное ядро с ядрышком. В
ядре хорошо просматриваются глыбки хроматина.
Обнаружение ядра существенно для идентифика-
ции трихомонад, так как фрагменты цитоплазмы,
кусочки слизи и другие обрывки тканей могут
иметь схожую с трихомонадами окраску. При
электронно-микроскопическом исследовании ус-
тановлено, что все тело трихомонады, включая
жгутики, покрыто трехслойной осмиофильной
мембраной-перипластом; ядро состоит из мелко-
зернистой кариоплазмы; парабазальный аппарат —
из системы нитей, расположенных в один или два
ряда, а в цитоплазме находятся большие пищева-
рительные вакуоли (Н. М. Овчинников, В. В. Де-
лекторский, 1974).
Размножается трихомонада путем продольного
деления. Трихомонады не образуют цист или
других форм устойчивости, обеспечивающих им
сохранение вне человеческого организма.
Влагалищная трихомонада представляет собой
самостоятельный вид, в естественных условиях
обитает только в мочеполовом аппарате челове-
ка и не поражает животных. У женщин местом
обитания Trichomonas vaginalis является влагали-
ще, у мужчин — предстательная железа и семен-
ные пузырьки. Мочеиспускательный канал мо-
жет поражаться у обоих полов.
Трихомонады быстро теряют жизнеспособность
вне человеческого организма. Во внешней среде
при высушивании трихомонады погибают за не-
сколько секунд, губительное действие на них
оказывают температура выше 40 °C, прямЯ&сол-
нечные лучи, медленное замораживание, различ-
ные антисептики. Трихомонады хорошо растут в
анаэробных условиях, оптимум роста их при pH
среды 5,5—7,5 и температуре 35—37 °C. Трихомо-
нады выделяют во внешнюю среду гиалуронидазу,
питаются эндосмотически. Влагалищные трихо-
монады — аэротолерантные анаэробы со своеоб-
510
Глава 43. Трихомониаз 511
разным энергетическим метаболизмом. Гидроге-
носомы (паракостальные или парааксостилярные
гранулы) — органоиды с диаметром от 0,2 до 0,4
мкм, окруженные мембраной с линзовидными
утолщениями и заполненные гомогенным мат-
риксом, являются функциональными аналогами
митохондрий (Е. Е. Брагина и соавт., 1996).
Патогенез
Урогенитальные трихомонады проникают в
организм постепенно per continuitatem, через
межклеточные пространства и затем в субэпи-
телиальную соединительную ткань, а также
лимфогенно через множественную сеть лимфа-
тических щелей. Трихомонады, попадая в урет-
ру, фиксируются на клетках плоского эпителия
слизистой оболочки, проникают в железы мо-
чеиспускательного канала и лакуны. Попадая в
мочеполовые органы, трихомонады либо обус-
ловливают развитие воспаления, либо не вызы-
вают никаких изменений. Умеренно выражен-
ная воспалительная реакция развивается при
наличии большого количества паразитов. Вы-
деляемая трихомонадами гиалуронидаза приво-
дит к значительному разрыхлению тканей и
более свободному проникновению в межклеточ-
ные пространства токсических продуктов обмена
бактерий сопутствующей флоры.
Патогенетические факторы заболевания: ин-
тенсивность инфекционного воздействия; pH
влагалищного и других секретов; физиологичес-
кое состояние эпителия мочеполовой системы;
сопутствующая бактериальная флора.
Трихомонадная инфекция не приводит к раз-
витию выраженного иммунитета. Иммунный
ответ на паразитирование трихомонад изучен
недостаточно. Выявляемые у больных или пере-
болевших трихомониазом сывороточные и сек-
реторные антитела являются лишь «свидетелями»
существующей или ранее перенесенной инфек-
ции, но не способны обеспечить иммунитет.
Эпидемиология
Средняя частота инфицированное™ трихомониа-
зом клинически здоровых женщин детородного
возраста в различных странах и разных социаль-
но-демографических группах колеблется от 2—
10 % (США) до 15—40 % (в тропических странах)
(ВОЗ, 1984). По оценкам специалистов, в США
ежегодно инфицируются 3 млн женщин. При этом
от 30 до, 70 % их партнеров мужчин заражаются
трихомониазом транзиторно. У взрослых девствен-
ниц регистрируется нулевая заболеваемость. Она
достигает 70 % у проституток, у лиц с другими
венерическими болезнями и у половых партнеров
инфицированных больных. Чаще заболевают жен-
щины в возрасте от 16 до 35 лет. Удельный вес уро-
генитального трихомониаза среди первичных не-
гонорейных уретритов достигает 65—80 %. Число
зарегистрированных случаев трихомониаза в мире
за 1995 год составляет 170 млн.
Трихомонада мочеполовая передается при по-
ловом контакте. Хотя возбудитель сохраняет
жизнеспособность в течение 24 часов в моче,
сперме, а также в воде и может выживать в те-
чение нескольких часов во влажном чистом бе-
лье, передача инфекции бытовым путем проис-
ходит редко. Возможность инфицирования
женщин трихомониазом при купаниях в реках,
плавательных бассейнах, а также в банях в на-
стоящее время полностью отвергается.
Клиническая картина
В зависимости от длительности заболевания и
интенсивности реакции организма на внедрение
возбудителя различают следующие формы три-
хомониаза: свежая (острая, подострая, торпид-
ная); хроническая (торпидное течение и давность
заболевания свыше 2-х месяцев); трихомонадо-
носительство (при наличии трихомонад отсут-
ствуют объективные и субъективные симптомы
заболевания). Для хронического трихомониаза
характерны периодические обострения, которые
могут быть обусловлены половыми возбуждени-
ями и сношениями, употреблением алкоголя,
снижением сопротивляемости организма, нару-
шениями функции яичников и изменением pH
содержимого влагалища.
В последнее время предлагается использовать
в практических целях Международную статис-
тическую классификацию X пересмотра: уроге-
нитальный трихомониаз неосложненный, трихо-
мониаз с осложнениями.
Основными очагами поражения при урогени-
тальном трихомониазе являются мочеиспуска-
тельный канал у мужчин, влагалище и уретра у
женщин. В связи с многоочаговостью трихомо-
надной инфекции при постановке диагноза сле-
дует указывать локализацию поражения.
Инкубационный период при трихомониазе
равен в среднем 5—15 дней, но может колебать-
ся от 1—3 дней до 3—4 недель.
Трихомонадный уретрит у мужчин
Трихомонадный уретрит у мужчин по клиничес-
кой картине практически не отличается от урет-
ритов другой этиологии. При острой форме вос-
палительный процесс протекает бурно с
обильными гнойными выделениями из уретры
и дизурическими явлениями. При подостром
512 Венерология
уретрите симптомы незначительные, выделения
из уретры в небольших количествах, серого или
серовато-желтого цвета, моча в первой порции
слегка мутная с умеренным количеством хлопь-
ев. Для торпидной формы трихомонадного урет-
рита у мужчин, которая встречается чаще всего,
характерна скудность объективных и субъектив-
ных симптомов или их полное отсутствие. Иног-
да при хронических и вялотекущих формах три-
хомониаза инфекция распространяется в заднюю
уретру и возникает тотальный уретрит. В таких
случаях появляются учащенные мочеиспускания,
рези и жжение в конце акта мочеиспускания,
тотальная пиурия и терминальная гематурия.
Осложнения при трихомонадных уретритах у
мужчин встречаются часто и представлены в виде
простатита, везикулита и эпидидимита, которые
по клинической картине отличаются от таковых
гонорейной этиологии и протекают более бур-
но. При длительном течении возможно образо-
вание множественных стриктур уретры.
Острый трихомониаз у женшин
Клиническая картина острого трихомониаза у
женщин характеризуется симптомами вагинита,
к которому могут присоединиться вестибулит,
уретрит, эндоцервицит, эндометрит. При острой
форме заболевания женщины жалуются на
обильные, раздражающие кожу выделения и зна-
чительный зуд наружных половых органов. Бес-
покоит также жжение и болезненность при мо-
чеиспускании. При осмотре находят
воспалительные изменения от умеренной гипере-
мии свода влагалища и шейки матки до обшир-
ных эрозий, петехиальных геморрагий и опрело-
сти в области промежности. Весьма характерным,
но не постоянным признаком является наличие
гранулематозных, рыхлых поражений слизистой
оболочки шейки матки красноватого цвета (ма-
линовая шейка матки). В области заднего свода
шейки матки имеется скопление жидких, серо-
вато-желтых, пенистых выделений. Губки урет-
ры нередко отечны и гиперемированы. При вов-
лечении в воспалительный процесс уретры,
можно выдавить небольшое количество гнойных
выделений при массаже. При торпидной форме
трихомониаза субъективные ощущения могут
полностью отсутствовать. Слизистые оболочки
влагалища и наружных половых органов имеют
нормальную окраску или слегка гиперемирова-
ны; выделения из вагины молочного цвета в не-
значительном количестве.
Мочеполовой трихомониаз у девочек
Мочеполовой трихомониаз у девочек чаще все-
го проявляется в виде вульвовагинита или вуль-
вовестибуловагинита. Органы малого таза не по-
ражаются, что связано с недостаточностью эст-
рогенизации у девочек до наступления менархе.
Острый вульвовестибуловагинит проявляется
диффузной гиперемией и припухлостью наруж-
ных половых органов, а также прилегающих
участков кожи бедер и промежности. В области
вульвы, гименального кольца, влагалища и на-
ружного отверстия мочеиспускательного канала
отмечаются эрозии и язвочки. Выделения гной-
ные, жидкие, пенистые с раздражением кожи
наружных половых органов. Нередко в процесс
вовлекается уретра и при надавливании на ее
заднюю стенку можно отметить появление вы-
делений. Больных беспокоят зуд, жжение. От-
мечается неприятный запах отделяемого, расче-
сы в области гениталий. При переходе в
хроническую стадию объективные признаки три-
хомониаза выражены слабее.
Диагноз
Диагноз мочеполового трихомониаза устанавли-
вается на основании клинических признаков
заболевания и обнаружения в исследуемом ма-
териале трихомонад. Для успешной диагности-
ки трихомониаза следует придерживаться следу-
ющих принципов (И. И. Ильин, Е. Н. Туранова,
1991): сочетанное использование различных спо-
собов диагностики; неоднократные исследования
изучаемого материала; забор материала из разных
очагов (уретра, мочевой пузырь, вульва, вагина,
протоки больших вестибулярных желез); пра-
вильная техника забора и транспортировки ма-
териала.
Для лабораторной диагностики трихомониаза
применяются следующие методы: микроскопия
нативного препарата; микроскопия окрашенного
препарата; люминесцентная микроскопия; куль-
туральные; иммунологические.
Микроскопия нативного препарата
В нативных препаратах влагалищная трихомона-
да определяется по грушевидной или овальной
форме тела величиной немного больше лейко-
цита, характерным толчкообразным движениям
и жгутикам, которые особенно хорошо видны
при исследовании в фазовоконтрастном микро-
скопе.
Микроскопия окрашенного препарата
Преимуществом исследования трихомонад в ок-
рашенных препаратах является возможность их
изучения спустя длительное время после взятия
материала. В препаратах, окрашенных метилено-
вым синим или по Граму, трихомонады имеют
овальную, округлую или грушевидную форму с
Глава 43. Трихомониаз 513
хорошо выраженными контурами и нежно-яче-
истым строением цитоплазмы. Для выявления
более тонкого строения трихомонад применяют
более сложные методики окраски препаратов (по
Романовскому-Гимза* Гейденгайну, Лейшману и
др.), которые позволяют рассматривать жгути-
ки, ундулирующую мембрану.
Люминесцентная микроскопия
Метод люминесцентной микроскопии основан
на свечении объекта в ультрафиолетовых лучах
в темном поле. При незначительной собствен-
ной люминесценции материала применяют лю-
минофоры: акридиновый оранжевый.
Культуральный метод
При отрицательных результатах непосредствен-
ного исследования патологического материала
трихомонады могут быть обнаружены с помо-
щью культивирования отделяемого из влагали-
ща, уретры, секрета простаты или спермы. Куль-
туральный метод имеет большую ценность при
распознавании атипичных форм, при диагнос-
тике трихомониаза у мужчин и при контроле за
результатами лечения.
Изучается применимость пробирочного теста
(экспресс-диагностика) количественного опреде-
ления лейкоцитарных пленок (КЛП) для иден-
тификации вагинальной трихомонады. Чувстви-
тельность метода — 100 %, специфичность —
92,3 %.
В настоящее время ведутся разработки мето-
дов молекулярной биологии, которые, возмож-
но, заменят существующие способы диагности-
ки трихомониаза.
Лечение
Современные методы лечения больных трихомо-
ниазом основаны на использовании специфичес-
ких противотрихомонадных средств. Лечению
подлежат все больные, у которых обнаружены
влагалищные трихомонады, независимо от нали-
чия или отсутствия у них воспалительных явле-
ний в половом аппарате, а также больные с вос-
палительными процессами, у которых при
обследовании трихомонады не обнаружены, но
простейшие обнаружены у половых контактов
или источников заражения.
Главным принципом лечения мочеполового
трихомониаза является индивидуальный подход
к больному, основанный на тщательном анам-
незе и всестороннем клиническом обследовании.
Терапия должна проводиться в зависимости от
топического диагноза. Мочеполовой трихомони-
аз является многоочаговым заболеванием и по-
этому лечению следует подвергать все очаги
инфекции — уретру, влагалище, цервикальный
канал, парауретральные ходы. Комплексная те-
рапия больных трихомониазом включает следу-
ющие препараты и методы: этиотропные, имму-
нокорригирующие, биостимуляторы, витамины,
местное лечение.
Этиотропная терапия
Одним из наиболее эффективных препаратов
общего действия при трихомониазе является
метронидазол (трихоцол, флагил). Существует
несколько методик лечения метронидазолом:
• по 0>25 г 2 раза в день в течение 10 дней;
на курс 5,0 г;
• первые 4 дня по 0,25 г 3 раза в день, осталь-
ные 4 дня по 0,25 г 2 раза в день; на курс —
5,0 г;
• в первый день по 0,5 г 2 раза, во второй —
по 0,25 г 3 раза, в последующие 4 дня — по
0,25 г 2 раза в день; на курс — 3,75 г;
• по 0,5 г 4 раза в день в течение 5 дней;
• в первый день по 0,75 г 4 раза, во второй —
0,5 г 4 раза; длительность лечения 2 дня; на курс
лечения — 5,0 г;
• 2,0 г в сутки однократно (американский
метод);
• 2,0 г в сутки в течение 3-х дней (американ-
ский метод);
• по 1,0 г 2 раза в сутки в течение 7 дней (аме-
риканский метод);
• 100 мл раствора метрогила, содержащего
500 мг метронидазола, внутривенно капельно в
течение 20 минут 3 раза в сутки на протяжении
5—7 дней (при хроническом, рецидивирующем
трихомониазе).
Тинидазол (фазижин) назначается по одной их
схем:
• однократно в дозе 2,0 г (четыре таблетки);
• по 0,5 г через каждые 15 минут в течение
часа в курсовой дозе 2,0 г;
• по 0,5 г 2 раза в день ежедневно в течение
недели.
Альтернативные препараты и схемы лечения
Нитазол (аминитразол, трихолавал) в таблетках
по 0,1 г 3 раза в день, курс 15 дней. Одновре-
менно местно 2 раза в день через 12 часов вво-
дят вагинальный суппозиторий с нитазолом
(0,12 г), предварительно очистив влагалище 1 %
раствором натрия гидрокарбоната или 3 % ра-
створом перекиси водорода. Вместо суппозито-
рия можно использовать 2,5 % суспензию ни-
тазола в виде аэрозольной пены 2 раза в день,
вводить во влагалище и уретру в течение 15 дней.
514 Венерология
Курс лечения состоит из 2—3 циклов, назначае-
мых сразу после очередной менструации.
Макмирор — действующее начало препарата
нифуратель. По 1 таблетке 3 раза в день после
еды в течение 7 дней (принимать препарат дол-
жны оба партнера). По 1 свече или 2—3 г мази
вводить во влагалище перед сном в течение 10
дней. Макмирор показан для лечения всех форм
вагинитов, вызванных патогенными микроорга-
низмами: бактериями, трихомонадами, грибка-
ми, инфекциями мочевыделительного тракта.
Клион-D применяют в виде влагалищных таб-
леток 1 раз в день вечером в течение 10 дней.
Препарат содержит 0,5 г метронидазола и 0,15 г
нитрата миконазола, поэтому он особенно эф-
фективен при смешанной кандидозно-трихомо-
надной инфекции.
Эфлоран — это синтетический нитроимидазол,
который действует на грамотрицательные и грам-
положительные анаэробы, а также обладает
сильным противотрихомонадным и лямблицид-
ным действием. Курсовая доза 2,0 г (2 таблет-
ки утром и 3 таблетки вечером после еды). Боль-
ные могут принимать также 5 таблеток (каждая
по 400 мг) однократно.
Атрикан 250 — активным элементом препарата
является тетонитрозол. Принимается по 1 кап-
суле утром и вечером в течение 4-х дней. Пре-
парат не противопоказан беременным.
Наксоджин (ниморазол) — однократный при-
ем четырех таблеток внутрь (2,0 г) при свежем
трихомониазе или по 1 таблетке (500 мг) 2 раза
в день (утром и вечером) в течение 6 дней (при
хронической форме заболевания).
Тиберал (орнидазол) — антимикробное сред-
ство для лечения инфекций, вызванных трихо-
монадами, амебами, лямблиями, анаэробными
бактериями. Препарат содержит действующее
вещество орнидазол. Для лечения трихомониаза
используются следующие схемы.
Лечение однократной дозой: 1-й вариант — 3
таблетки (по 500 мг каждая) вечером; 2-й вари-
ант — 2 таблетки по 500 мг каждая + 1 таблет-
ка по 500 мг вагинально перед сном.
Пятидневный курс лечения: 1-й вариант —
2 таблетки (по 1 таблетке утром и вечером);
2-й вариант — 2 таблетки (по 1 таблетке ут-
ром и вечером) + 1 таблетка вагинально пе-
ред сном.
Для избежания повторного инфицирования
половой партнер во всех случаях должен при-
нимать такие же дозы (вариант 1). Детям на-
значают 25 мг/кг массы тела в виде однократ-
ной дозы. Таблетки всегда следует принимать
после еды.
Тержинан (комбинированный препарат, содер-
жащий производное имидазола — тернидазол)
назначается по 1 вагинальной таблетке в сутки
перед сном в течение 10 дней. Возможно при-
менение препарата в периоды беременности и
лактации по показаниям.
Для лечения трихомонадного вагинита у бе-
ременных применяется клотримазол в вагиналь-
ных таблетках по 200 мг на ночь в течение 2
недель. При недолеченной инфекции во 2—3
триместрах беременности назначают метронида-
зол по 500 мг 2 раза в сутки в течение 7 дней
(эта схема позволяет избежать токсического
уровня препарата в сыворотке).
Для лечения детей от 1 до 5 лет применяют
метронидазол по 1/3 таблетки (которая содержит
250 мг) 2—3 раза в сутки; детям 6—10 лет — «по
0,125 г 2 раза в сутки; 11—15 лет — по 0,25 г 2
раза в сутки в течение 7 дней.
Иммунокорригируюшая терапия
Иммунокорригирующая терапия включает спе-
цифическую (солкотриховак R) и неспецифичес-
кую.
Солкотриховак R — вакцина из специальных
штаммов лактобацилл. Основным показанием к
применению является рецидивирующий трихо-
мониаз и бактериальный вагиноз. Вакцину при-
меняют одновременно с химиотерапией, так как
образование антител длится 2—3 недели. Вакцину
можно также применять как монотерапию прй
торпидном течении трихомониаза и в случаях ре-
зистентности к химиотерапии. Основная вакци-
нация — 3 инъекции по 0,5 мл с интервалом в
2 недели — обеспечивает годичную защиту. Че-
рез год проводится поддерживающая вакцина-
ция — однократно 0,5 мл.
Неспецифическая иммунотерапия проводится
по обычным методикам использования пироге-
нала, аутогемотерапии, лактотерапии; также при-
меняется пчелиный мед (местные аппликации,
влагалищный электрофорез, № 10—15), калия
оротат по 0,5 г 2—3 раза в сутки в течение 20—
30 дней.
Местная терапия
Местное лечение трихомониаза назначается при
наличии противопоказаний к применению мет-
ронидазола (нарушения кроветворения, актив-
ные заболевания центральной нервной системы),
а также у больных с торпидным рецидивирую-
щим течением заболевания.
Местная терапия трихомонадных уретритов у
мужчин сводится к введению различных средств:
раствора серебра нитрата (1: 1000—1: 6000), ртути
Глава 43. Трихомониаз
515
оксицианида (1: 1000—1 : 6000), этакридина лак-
тата (1: 2000) с последующими инстилляциями
в мочеиспускательный канал взвеси осарсола с
борной кислотой и глюкозой. Лечение проводит-
ся ежедневно в течение 8—10 дней.
Женщинам с вульвитом и вагинитом показа-
ны спринцевания, сидячие ванночки с настоем
цветков ромашки, травы шалфея, тампоны с
соком репчатого лука. Эффективна комбинация
препаратов: урогиронин-гексаметилентетрамин
(0,05 г), левомицетин (0,2 г), борная кислота
(0,1 г). Данный состав вводят 1 раз в день во
влагалище или мочеиспускательный канал в те-
чение 10-15 дней.
При непереносимости антитрихомонадных
препаратов рекомендуется мефенаминовая кис-
лота: местные аппликации 1 % водного раство-
ра 1—2 раза в день или 2 % паста; суппозито-
рии бетадина (повидон — йодин).
Критерии излеченности
При установлении критерия излеченности три-
хомониаза необходимо различать этиологичес-
кое и клиническое выздоровление. Под этио-
логическим выздоровлением подразумевается
стойкое исчезновение влагалищных трихомонад
после проведенной терапии. Излеченность мо-
чеполового трихомониаза устанавливается с по-
мощью бактериоскопического и культурального
методов исследования.
После окончания лечения на 7—10 день у
мужчин производят пальпаторное обследование
предстательной железы и семенных пузырьков
и микроскопическое исследование их секрета
(при отсутствии противопоказаний). Спустя 12—
14 дней после окончания терапии назначают
провокацию (алиментарный и механический
способы). Если после провокации в отделяемом
или соскобе со слизистой уретры и в первой
порции свежевыпущенной порции мочи трихо-
монады не обнаружены, а также отсутствуют
симптомы уретрита, больному через месяц про-
водят повторное обследование, уретроскопию и
при необходимости провокацию.
Первые контрольные исследования у женщин
проводят через 7—8 дней после окончания лече-
ния. В дальнейшем исследования производятся
в течение 3-х менструальных циклов. Лабора-
торное обследование у женщин проводят перед
менструацией или через 1—2 дня после ее окон-
чания. Материал для исследования следует брать
из всех возможных очагов поражения.
Больные считаются этиологически излечен-
ными, если после окончания лечения при нео-
днократных повторных обследованиях не удается
обнаружить трихомонады в течение 1—2 месяцев
у мужчин и 2—3 месяцев у женщин. У ряда муж-
чин, несмотря на стойкое этиологическое выз-
доровление, после применения противотрихомо-
надных средств, клинического излечения не
наступает. У больных остаются воспалительные
явления: скудные выделения из гениталий, па-
тологические продукты в моче. Наиболее часто
посттрихомонадные воспалительные процессы
наблюдаются у мужчин с осложненными и хро-
ническими формами трихомониаза.
Профилактика
Профилактические мероприятия при трихомо-
ниазе направлены на своевременное и полное
выявление зараженных трихомонадами лиц:
• выявление и лечение больных трихомониа-
зом;
• выявление, обследование и лечение лиц,
послуживших источником заражения и половых
контактов;
• обследование на трихомониаз всех лиц, стра-
дающих воспалительными заболеваниями моче-
половых органов;
• периодическое обследование на трихомониаз
женщин, работающих в детских коллективах,
больницах, родильных домах;
• санитарно-гигиенические мероприятия
(обеззараживание медицинского инструментария,
обработка резиновых перчаток; влагалищное
исследование только в перчатках, предпочти-
тельно одноразового использования из синтети-
ческих материалов);
• санитарно-просветительная и воспитательная
работа среди населения.
Глава 44
Урогенитальный хламидиоз
Инфекционное заболевание, вызываемое пато-
генными микроорганизмами Chlamydia
trachomatis, передающееся преимущественно при
половых контактах, поражающее мочеполовую
сферу и другие органы, сопровождающееся стер-
той клинической симптоматикой, изначальной
склонностью к хроническому течению, вызыва-
ющее в ряде случаев патологическое течение бе-
ременности, снижение репродуктивной функции
у женщин и мужчин.
Эпидемиология
Урогенитальный хламидиоз (УГХ) — одна из
широко распространенных инфекций, передава-
емых половым путем. В 90-е годы хламидийная
инфекция превратилась из малоизвестной в са-
мое «популярное» заболевание. Очень высокий
уровень распространенности УГХ регистрирует-
ся в развивающихся странах. В отдельных евро-
пейских странах, за счет реализации программ
скрининга, изучения и контроля за ИППП от-
мечено резкое снижение заболеваемости УГХ
(например, в Швеции регистрируется 3 случая
на 100 тыс. населения).
Распространению урогенитального хламидиоза
способствуют: либерализация сексуальных отно-
шений, раннее начало половой жизни, пренеб-
режение средствами барьерной контрацепции,
малосимптомное течение заболевания, а также
использование р-лактамных антибиотиков, гор-
мональных контрацептивов, кортикостероидов,
оперативные вмешательства на генитальной сфе-
ре и др.
Доля хламидийной инфекции среди урогени-
тальных инфекций составляет 3—30 %, а у ги-
некологических больных — 20—40 %.
Негонококковые уретриты хламидийной эти-
ологии регистрируются более чем у 40 % муж-
чин и являются основной причиной постгоно-
рейного уретрита (60—80 %).
У супружеских пар с привычным невынаши-
ванием беременности при хламидийной инфек-
ции 90 % мужчин страдают хроническим проста-
титом, у них снижено количество и подвижность
сперматозоидов (42—50 % от нормы), часто встре-
чаются их дегенеративные формы. У большей
части их жен наблюдаются самопроизвольные
выкидыши (у 82 %) и преждевременные роды (у
10 %).
При выборочном скрининге большой группы
лиц, ведущих активную половую жизнь и не
предъявляющих жалоб на урогенитальную сфе-
ру, хламидии выявлены у 5—10 % обследованных.
Риск заражения после полового контакта с
человеком, инфицированным хламидиями, со-
ставляет в среднем 60 %, а от постоянного парт-
нера — заметно выше.
Урогенитальный хламидиоз протекает с кли-
ническими проявлениями приблизительно у
30 % женщин и 75 % мужчин.
Этиология и патогенез
Согласно классификации, С. trachomatis входит
в семейство Chlamydiaceae, род Chlamydia, 1-й
биовар (серовары А-К, Ba,,Da и 1а) и является
исключительно паразитом человека. Различные
штаммы С. trachomatis способны вызывать тра-
хому, поражения урогенитального тракта, суста-
вов, конъюнктивы и пневмонию у новорожден-
ных. С. trachomatis открыта в 1903 г
Л. Хальберштедтером и С. Провачеком.
Хламидии занимают промежуточное положе-
ние между вирусами и риккетсиями, сочетают
особенности их строения. Имеют ряд признаков,
характерных для грамотрицательных бактерий:
содержат'ДНК и РНК, клеточную оболочку,
размножаются бинарным делением и в этот пе-
риод чувствительны к антибиотикам. У хлами-
дий отсутствует энергетическая система, они не
синтезируют АТФ, размножаются за счет клет-
ки-хозяина и принадлежат к «внутриклеточным
облигатным паразитам». Внеклеточные формы
хламидий (элементарные тельца — ЭТ) не задер-
живаются бактериальными фильтрами (0,2—0,3
мкм). Попадая внутрь эпителиальной клетки, ЭТ
окружаются полимембранной оболочкой, обра-
зуют фагосому, затем через стадию переходных
телец (ПТ) трансформируются в ретикулярные
тельца (РТ — метаболически активные вегета-
тивные формы), которые значительно крупнее
516
Глава 44. Урогенитальный хламидиоз
517
(до 1 мкм) элементарных. Далее ретикулярные
тельца после периода развития, бинарного деле-
ния и ПТ превращаются в элементарные, при
этом апикальная часть эпителиальных клеток
разрушается, а вновь образованные элементар-
ные тельца попадаюгв межклеточные простран-
ства и инфицируют новые клетки. Внутрикле-
точный цикл развития хламидий занимает до 72
часов. Внутрицитоплазматические включения
(«микроколонии») размножающихся хламидий
известны как тельца Провачека-Хальберштедте-
ра. Эпителиальные клетки с внутриклеточными
хламидийными включениями имеют характер-
ный вид — симптом «бычьего глаза».
Элементарные тельца метаболически неак-
тивны и малочувствительны к антибиотикам,
но нейтрализуются антителами, цитокинами,
гамма-интерфероном, фактором некроза опу-
холей, интерлейкином-1 и комплементом.
Однако при хронической хламидийной инфек-
ции выработка этих эндогенных факторов в
организме недостаточна для элиминации мик-
роорганизмов.
Хламидии поражают цилиндрический эпите-
лий уретры, эндоцервикса матки, прямой киш-
ки, конъюнктивы глаз, слизистую оболочку
глотки, а также эпителий влагалища с последу-
ющей воспалительной лимфоцитарно-плазмоци-
тарной реакцией слизистых. Реакция иммунной
системы на внедрение Chlamydia trachomatis не-
стойкая и общая с другими серотипами хлами-
дий, которые вызывают венерическую лимфог-
ранулему, трахому, орнитоз, пневмонию.
Иммунитет после перенесенного хламидиоза не
развивается.
Важным эпидемиологическим фактором хлами-
дийной инфекции является способность возбуди-
теля к персистенции, т. е. к длительному сохране-
нию без существенного роста и размножения в
состоянии равновесия с клетками хозяина. В со-
стоянии персистенции возбудитель не чувствите-
лен к антибиотикам и не передается половым пу-
тем. Однако при различных факторах, приводящих
к снижению иммунной защиты организма, про-
исходит их реверсия в обычные формы и после-
дующее развитие воспалительного процесса в уро-
генитальном тракте. В ряде случаев монотерапия
антибиотиками приводит к внутриклеточному пер-
систированию хламидий (находятся за мембраной
включения) и хронизации заболевания,
У персистирующих форм С. trachomatis изме-
няются морфологические свойства, а также сни-
жается экспрессия хламидийных антигенов (АГ):
основного типоспецифического АГ наружной
мембраны — МОМР, родоспецифического ли-
пополисахаридного АГ (ЛПС), белка клеточной
стенки массой 60 кВа. Усиливается синтез бел-
ка теплового шока (hsp 60 кВа), который запус-
кает вторичный иммунный ответ и вызывает:
гиперпродукцию Ig G и 1g А, развитие реакции
ГЗТ (в результате антигенной перегрузки появ-
ляется инфильтрация подслизистого слоя лим-
фоцитами и моноцитами), возникновение пе-
рекрестных аутоиммунных реакций, торможение
деления ретикулярных телец. Активированные
макрофаги путем выделения цитокинов (ФНО-а,
ИЛ-1) активируют пролиферацию клеток соеди-
нительной ткани, вызывая фибротизацию субэ-
пителиальных тканей.
Сохраняются персистирующие формы хлами-
дий в мембраноограниченных зонах эпителиаль-
ных клеток, нейтрофилах, макрофагах, лимфоци-
тах, эндотелиоцитах, лимфатических капиллярах,
во внеклеточных фагосомах и, вероятно, в лим-
фоидных герминативных центрах.
Пути передачи
Заражение человека генитальной хламидийной
инфекцией происходит преимущественно при
половых контактах. Хламидии заселяют эколо-
гические зоны их обитания у человека (уретра,
шейка матки, иногда прямая кишка). С. tra-
chomatis способны прикрепляться к сперматозо-
идам, которые могут служить переносчиками
инфекции. В связи с этим существует риск ин-
фицирования С. trachomatis не только при поло-
вых контактах, но и при искусственном оплодот-
ворении спермой необследованного донора.
Инфицирование глаз, эпителия дыхательных
путей у новорожденного чаще происходит во
время родов, от больной хламидиозом матери, и
сопровождается развитием конъюнктивита,
реже — пневмонии.
Установлена возможность заражения бытовым
путем от членов семьи, болеющих хламидиозом
(дети, взрослые), с субманифестными проявле-
ниями со стороны урогенитального тракта, не-
ярко выраженными конъюнктивитами, блефаро-
конъюнктивитами («семейный хламидиоз»),
возможно развитие первичного бесплодия у де-
вочек и мальчиков.
Урогенитальный хламидиоз как моноин-
фекция встречается редко. Чаще хламидии ас-
социируются с гонококками, микоплазмами,
трихомонадами, вирусом герпеса I и II типов.
Характерной клиники заболевания не наблю-
дается. Инкубационный период обычно со-
ставляет 2—3 недели, часто установить дав-
ность заражения не удается. Выявляются
признаки малосимптомного, редко подостро-
518 Венерология
го или острого уретрита, цервицита, с обна-
ружением при кольпоскопии на шейке мат-
ки лимфоидных фолликулов (фолликулярный
цервицит). Больных беспокоят различной ин-
тенсивности выделения из уретры или влага-
лища, зуд в области гениталий, боли внизу
живота, в промежности, паховых областях.
Этим субъективным симптомам при сборе
анамнеза следует уделять должное внимание.
При несвоевременной диагностике или нера-
циональном лечении у больных хламидиозом
могут развиться различные осложнения, лече-
ние которых проводят в урогинекологической
клинике.
Клинические формы
урогенитального хламидиоза
Мужчины: уретрит, парауретрит, простатит, ве-
зикулит, эпидидимит, орхиэпидидимит; фарин-
гит, проктит.
Женщины.* цервицит, уретрит, бартолинит,
эндометрит, параметрит, сальпингит, сальпин-
гоофорит, пельвиоперитонит; фарингит, про-
ктит.
Клинические симптомы
урогенитального хламидиоза у женшин:
• Слизисто-гнойный цервицит
• Гнойные вагинальные выделения
• Боли внизу живота
• Посткоитальное или межменструальное кро-
вотечение
• Дизурия
• Признаки воспалительного процесса в ма-
лом тазу с болевым синдромом
Клинические симптомы
урогенитального хламидиоза у мужчин:
• Выделения из уретры
• Дизурия
• Признаки эпидидимита и простатита
Обшие симптомы для женшин и мужчин:
• Аноректальные выделения или дискомфорт
• Конъюнктивит
• Артралгии
Осложнения
Бесплодие, болезнь Рейтера, перигепатит, пери-
аппендицит, перихолецистит, плеврит, спаечная
болезнь кишечника.
У беременных: спонтанный аборт, внематочная
беременность, многоводие, преждевременные
роды, отслойка плаценты, плацентарная недоста-
точность, преждевременный разрыв плодных
оболочек, слабость родовой деятельности, лихо-
радка в родах, изменения последа, послеродовые
эндометриты, замедленная инволюция матки,
субфебрилитет.
У новорожденных: конъюнктивит, пневмония,
офтальмия, асимптомное инфицирование ротог-
лотки, генитального тракта, прямой кишки.
У девочек.* вульвит, вагинит, уретрит.
Клиническая картина
урогенитального хламидиоза у мужчин
Хламидийный уретрит
Для хламидийного уретрита наиболее характер-
но хроническое, реже — острое или подострое
течение болезни.
Частым проявлением уретрита являются выде-
ления йз мочеиспускательного канала слизисто-
го, слизисто-гнойного или гнойного характера,
редко они истекают свободно или появляются при
надавливании на уретру. У большинства пациен-
тов с острым или подострым началом заболева-
ния интенсивность выделений уменьшается в
течение нескольких дней.
Субъективные жалобы больных — неприят-
ные ощущения, зуд, боль в мочеиспускательном
канале, учащенные позывы к мочеиспусканию,
выделения. Заболевание протекает по типу ост-
рого, подострого или хронического воспаления.
При одновременном поражении мочеиспуска-
тельного канала и других половых органов боль-
ные жалуются на боль в области мошонки, про-
межности, заднего прохода, в поясничной и
крестцовых областях, по ходу седалищного не-
рва и в нижних конечностях.
При малосимптомном уретрите с незначитель-
ными жалобами выделения очень скудные, в
виде 1—2 «утренних» капель. Нередко выделе-
ния появляются после длительной задержки
мочи, иногда при дефекации или в конце моче-
испускания.
При незначительном количестве выделений
моча в первой и второй порциях обычно прозрач-
ная и содержит единичные или множественные
слизисто-гнойные нити. При большом количе-
стве выделений моча может быть мутной в пер-
вой или в обеих порциях.
У больных отмечают различную степень выра-
женности воспалительных изменений в области
наружного отверстия мочеиспускательного канала
(гиперемия, пастозность, слипание губок). Неред-
ко удается установить признаки парауретрита
(воспаления парауретральных ходов), диагностика
которого важна для последующей терапии.
При уретроскопии передней уретры у паци-
ентов обычно обнаруживают мягкий инфильт-
Глава 44. Урогенитальный хламидиоз 519
рат или его остаточные явления, переходный
инфильтрат и литтреиты, в предстательной час-
ти канала нередко определяют ограниченные
грануляции и поражение семенного холмика.
Поражение бульбоуретральной железы (хлами-
дийный куперит) может протекать без субъек-
тивных жалоб или с наличием жалоб на перио-
дическую боль в области бедра и промежности.
Воспаленную бульбоуретральную железу обыч-
но пальпируют в толще промежности в виде
плотного болезненного узелка величиной с го-
рошину.
Хламидийный простатит
Хламидийный простатит является частым ослож-
нением хламидийного уретрита, выявляемым у
46-75 % пациентов активного полового возра-
ста.
Хламидийный простатит обычно протекает
хронически, торпидно, периодически обостряясь.
Больные предъявляют жалобы на выделения из
мочеиспускательного канала, иногда во время
акта дефекации или в конце мочеиспускания,
зуд, неприятные ощущения в уретре, прямой
кишке, неинтенсивные боли в промежности,
мошонке, паховой и крестцовой областях, вни-
зу живота, по ходу седалищного нерва. Выявля-
ется слипание губок уретры после длительных
перерывов в мочеиспускании, нередко учащение
мочеиспускания, особенно по ночам. Моча в
первой и второй порциях чаще прозрачная, с
примесью слизистых или слизисто-гнойных
нитей и хлопьев, а в ряде случаев бывает мут-
ной в первой или в обеих порциях (при обостре-
нии процесса).
В зависимости от глубины хламидийного по-
ражения тканей предстательной железы различа-
ют три формы простатита: катаральную (поража-
ются выводные протоки желез), фолликулярную
(поражаются фолликулы желез — секреторно-
инкреторный эпителий ацинусов) и паренхима-
тозную (воспалительный процесс выходит за
пределы железистой ткани с поражением парен-
химы предстательной железы).
При пальпаторном исследовании через пря-
мую кишку предстательная железа при катараль-
ном простатите не увеличена, нормальной кон-
систенции, болезненна. При фолликулярном
простатите предстательная железа не увеличена,
однако при пальпации в ней определяются чув-
ствительные узелки (увеличенные воспаленные
фолликулы) различной консистенции. Увеличе-
ние, болезненность железы или одной из долей,
изменение ее конфигурации и консистенции
определяются пальпаторно при паренхиматозном
простатите. В секрете предстательной железы
выявляются небольшой лейкоцитоз, сниженное
количество (отсутствие) лецитиновых зерен. Для
подтверждения хламидийной этиологии проста-
тита необходима идентификация возбудителя
методом ПЦР или культуральное исследование.
Хламидийный везикулит
Хламидийный везикулит выявляется у 15—16 %
мужчин активного полового возраста при нали-
чии хламидийного уретрита, паренхиматозного
простатита или эпидидимита. Субъективные
жалобы больных аналогичны жалобам, предъяв-
ляемым при уретропростатите. Для хламидийного
везикулита характерно хроническое течение. При
ректальном пальцевом исследовании определя-
ют увеличенные, напряженные и болезненные
семенные пузырьки.
Хламидийный эпидидимит
Хламидийный эпидидимит является наиболее
тяжелым проявлением урогенитального хлами-
диоза у мужчин. Обычно развивается по схеме:
уретрит — простатит — везикулит с последую-
щим проникновением хламидий в придаток яич-
ка каналикулярно, с поражением /эпителия, вы-
стилающего просвет семявыносящих протоков и
развитием у некоторых пациентов признаков
деферентита и фуникулита. Воспаленный семя-
выносящий проток пальпируется в виде слегка
болезненного тяжа. При фуникулите воспален-
ный семенной канатик уплотнен при пальпации
и болезненный.
Эпидидимит, вызываемый хламидиями, может
протекать в острой, подострой и хронической
формах. Хроническая форма может быть само-
стоятельным проявлением хламидийного эпиди-
димита, последствием острой и подострой его
формы.
Для острого эпидидимита характерно повыше-
ние температуры тела до 39°С, сильные боли в
придатке яичка, иррадиирующие по ходу семен-
ного канатика в поясничную и крестцовую об-
ласти. Кожа мошонки гиперемирована и отечна
на стороне пораженного придатка, который уве-
личен в размере, плотный (иногда бугристый) на
ощупь. Отмечается лейкоцитоз и ускорение
соэ.
При подостром эпидидимите отмечаются уме-
ренная боль, субфебрилитет, меньшая выражен-
ность клинических симптомов и признаков ин-
токсикации. Хронический хламидийный
Эпидидимит характеризуется, как правило, от-
сутствием обшей температурной реакции, незна-
чительной болью, умеренным увеличением и
равномерным уплотнением придатка.
520 Венерология
Хламидийный орхиэпидидимит
Воспалительный хламидийный процесс может
распространиться с придатка на оболочки яич-
ка и охватить яичко с развитием клиники ор-
хиэпидидимита. При острой форме заболевания,
помимо клинических симптомов эпидидимита,
наблюдается асимметрия, отечность гиперемиро-
ванной и горячей на ощупь мошонки (на сто-
роне поражения), резкая болезненность при
пальпации и выявляется опухоль с гладкой на-
пряженной поверхностью, дающей ощущение
ложного зыбления. При хроническом (торпид-
ном) течении одностороннего орхиэпидидими-
та, сочетающегося с простатитом, везикулитом и
деферентитом, выявляются нарушения половой
потенции и спермограммы.
Хламидийная инфекция может оказывать суще-
ственное влияние на фертильность мужчин. Воз-
можные механизмы влияния хламидийной инфек-
ции на фертильность мужчин включают
нарушения проходимости каналикулярной систе-
мы половых путей, повреждение эпителиальных
клеток, участвующих в процессе сперматогенеза,
и иммунологические реакции, связанные с обра-
зованием антиспермальных антител. Острый эпи-
дидимит, в особенности осложнившийся орхитом,
может приводить к атрофии яичек и азооспермии.
Клиническая картина
урогенитального хламидиоза у женщин
Хламидийный уретрит
Хламидийный уретрит обычно сопровождается
незначительными субъективными жалобами. При
появлении зуда в мочеиспускательном канале,
болей в начале мочеиспускания и учащенных
позывов обнаруживают легкое раздражение на-
ружного отверстия канала, а также скудные, по-
чти бесцветные выделения, выявляемые чаще
после массажа уретры. Эти явления обычно крат-
ковременны, но у подавляющего числа обследо-
ванных в период ремиссии при уретроскопии
определяются изменения слизистой оболочки
мочеиспускательного канала (мягкий инфильтрат,
остаточные явления мягкого инфильтрата, грубые
складки слизистой оболочки и т. п.).
Нередко хламидийному уретриту сопутствует
парауретрит. Хронический воспалительный про-
цесс в парауретральных протоках определяют по
гиперемии их устьев и слизистому или слизис-
то-гнойному содержимому, выдавливаемому в
виде капли.
Хламидийный бартолинит
Хламидийный бартолинит обычно протекает
вяло, малосимптомно, не сопровождаясь опреде-
ленными жалобами, способными указывать на
локализацию воспалительного процесса.
Хламидии, как правило, размножаются в по-
кровном столбчатом эпителии выводных прото-
ков бартолиниевых желез. Часто признаком по-
ражения протока является гиперемированное
пятно величиной с горошину с центральной тем-
но-красной точкой, соответствующей устью
выводного протока железы.
При пальпации воспаленной бартолиниевой
железы из устья выводного протока удается
выдавить каплю слегка мутноватой слизи. При
закрытии выводного протока он превращается в
большую кисту, наполненную прозрачной жид-
костью. Обычно воспалительные явления в ок-
ружающих тканях не выявляются, но в некото-
рых случаях хламидийного бартолинита
воспалительный процесс распространяется и на
окружающие ткани, что приводит к образованию
болезненного инфильтрата величиной со сливу
и даже с куриное яйцо, расположенного сбоку,
у входа во влагалище. Наиболее тяжелые фор-
мы бартолинита возникают при смешанной ин-
фекции, чаще хламидийно-гонококковой при-
роды.
Хламидийный вагинит
Первичный хламидийный вагинит является ред-
ким проявлением хламидийной инфекции и
практически не встречается у женщин репродук-
тивного возраста с нормальной гормональной
активностью. Хламидии не способны размно-
жаться в нормально функционирующем орого-
вевшем плоском поверхностном эпителии вла-
галища и в свободном состоянии (вне клетки)
проявляют высокую чувствительность к кислой
реакции содержимого влагалища,
В этих условиях, по-видимому, может возни-
кать вторичный хламидийный вагинит — на
фоне эндоцервицита,. в результате мацерирую-
щего действия выделений из канала шейки мат-
ки, а также других факторов (травмы, снижение
эстрогенной активности и др.), влияющих на
эпителий влагалища. Хламидийный вагинит
может развиться у детей (вульвовагинит), пожи-
лых женщин, а также во время беременности —
при структурно-функциональных изменениях
эпителия, связанных с особенностями гормо-
нального состояния. Больные обычно жалуют-
ся на выделения из влагалища, ощущение жже-
ния, зуд.
Хламидийный нервинит
Хламидийный цервицит является наиболее рас-
пространенным первичным проявлением хлами-
дийной урогенитальной инфекции у женщин,
Глава 44. Урогенитальный хламидиоз 521
протекающей в форме эндоцервицита. Заболе-
вание часто не вызывает жалоб. Иногда больные
обращают внимание на появившиеся выделения
из влагалища, редко отмечают неопределенную
тянущую боль внизу живота.
Воспалительный процесс в канале шейки мат-
ки, в столбчатом эпителии которого происходит
размножение хламидий, сопровождается обычно
слизисто-гнойной реакцией.
Острые цервициты характеризуются закупор-
кой шеечного канала, эритемой, появлением
патологического экссудата. Выделения мацери-
руют многослойный сквамозный эпителий сли-
зистой оболочки влагалищной части шейки мат-
ки, вызывают его частичную десквамацию.
Шейка матки становится отечной, гиперемиро-
ванной, вокруг наружного отверстия канала
шейки матки образуются эрозии, часто в виде
«красного венчика», наблюдается эктопия стол-
бчатого эпителия, перемещающегося из канала.
При длительно протекающих хламидийных цер-
вицитах часто возникают гипертрофические эро-
зии. В некоторых случаях в области зева обна-
руживаются везикулы размером с просяное зерно,
наполненные мутным содержимым.
Для хламидийных цервицитов характерно так-
же образование в слизистой оболочке, обычно в
области зева, лимфоидных фолликулов и симп-
том «контактной кровоточивости» — при касании
инструментом стенок цервикального канала по-
является кровоточивость, которая развивается в
результате дисбаланса в простациклин-тромбок-
сановой системе при инфицировании
С. trachomatis. Фолликулярный цервицит не ха-
рактерен для инфекций, вызываемых /V.
gonorrhoeae, Mycoplasma hominis и Ureaplasma
urealyticum.
Хламидийная инфекция шейки матки может
характеризоваться признаками неспецифическо-
го воспаления, не обязательно сопровождаясь
образованием гипертрофических эрозий, лимфо-
идных фолликулов и слизисто-гнойного отделя-
емого. В ряде случаев при персистентной хлами-
дийной инфекции канала шейки матки, кроме
обнаружения возбудителя различными лабора-
торными методами, не удается выявить (в мо-
мент обследования) каких-либо клинических
признаков заболевания.
Хламидийный эндометрит
Хламидии проникают в полость матки из кана-
ла шейки, последовательно инфицируя высти-
лающий матку эпителиальный слой.
Острый хламидийный эндометрит протекает с
жалобами и клиническими симптомами, характер-
ными для эндометритов другой этиологии. У боль-
ных внезапно повышается температура тела до 38—
39°С, определяется лейкоцитоз, ускорение СОЭ,
усиливающаяся боль внизу живота, нарушаются
сроки менструации, отмечаются интенсивные сли-
зисто-гнойные выделения из канала шейки мат-
ки. При гинекологическом исследовании выявля-
ется болезненность матки. Нередко наблюдается
хроническое течение воспалительного процесса и
последовательное поражение маточных труб.
Восхождение инфекции в матку может про-
исходить и без резко выраженных симптомов, а
воспалительный процесс может быть ограничен-
ным и нередко исчезать спонтанно — в период
отторжения функционального слоя эндометрия
при менструации.
Хламидийный сальпингит
Женщины предъявляют жалобы на боль в ниж-
них отделах живота, выделения из влагалища, на-
рушения менструального цикла, дизурические
явления. Боль носит тупой характер, усилива-
ется при физическом напряжении, во время мен-
струации, при затрудненной дефекации, при
гинекологическом обследовании. У больных не-
редко повышается температура тела до 38—39°С,
возникают лейкоцитоз, ускорение СОЭ.
Хламидийная инфекция вначале поражает
слизистую оболочку труб, что выражается гипе-
ремией, отеком и усиленной секрецией. При
катаральном сальпингите труба отечна, равно-
мерно утолщена, иногда пальпируется при гине-
кологическом исследовании. В её просвете скап-
ливается серозно-гнойная жидкость, нередко
образующая внутренние спайки. При прогрес-
сировании процесса складки слизистой оболоч-
ки отекают, препятствуя прохождению секрета,
который при закрытии воронки растягивает
стенки трубы, образуя гидросальпинкс или, при
гнойном содержимом трубы, пиосальпинкс. В
этот период выявляют утолщение трубы до зна-
чительных размеров и ее колбасообразную или
опухолеподобную форму. При хламидийной
инфекции так же, как при гонорее, нередко
наблюдается двустороннее поражение труб мат-
ки. При распространении инфекции через брюш-
ное отверстие маточной трубы обычно возника-
ет поражение яичника.
Хламидийные сальпингиты могут привести к
непроходимости маточных труб и трубной бере-
менности.
Хламидийный сальпингоофорит
При хламидийном поражении верхних отделов
урогенитального тракта у женщин в процесс вов-
522 Венерология
лекаются не только маточные трубы но и яич-
ники. При сальпингоофорите больные наиболее
часто отмечают ноющую боль внизу живота и в
крестцовой области, которая усиливается при на-
пряжении брюшной стенки, нередко появляет-
ся кровотечение в межменструальный период.
Патологический процесс в яичнике при восхо-
дящей хламидийной инфекции вызывает, поми-
мо спаек с окружающими тканями и органами,
воспалительный процесс в яичнике, поврежда-
ющий его внутреннюю структуру.
Хламидийный пельвиоперитонит
В острой стадии болезни на фоне жалоб, харак-
терных для сальпингита, внезапно возникает
резкая боль, вначале локализующаяся внизу
живота. У больных определяют напряжение
брюшной стенки, положительный симптом
Щеткина, повышение температуры тела до 38—
40 °C, учащенный пульс, выраженный лейкоци-
тоз, ускорение СОЭ. Нередко отмечается задер-
жка стула, метеоризм.
С помощью лапароскопии и этиологической
диагностики установлено частое сочетание острого
хламидийного перитонита и острого фибриноз-
ного перигепатита (сийдром Fitz-Hugh-Curtis).
Острые перигепатиты характеризуются развити-
ем болезненных явлений в правой верхней части
живота, обычно появляющихся внезапно и сопро-
вождающихся напряженностью в правом верхнем
квадранте.
Хламидиоз прямой кишки
Хламидиоз прямой кишки у женщин встречается
достаточно часто, нередко в сочетании с хлами-
диозом половых органов. Обнаружение хлами-
дий ассоциируется с наличием видимых измене-
ний в слизистой прямой кишки (фолликулы,
рубцы, застойные явления, слизисто-гнойный
экссудат).
Урогенитальный хламидиоз:
беременность и бесплодие
Прогноз при хламидийных сальпингитах в от-
ношении бесплодия более серьезен, чем при
сальпингитах, вызванных гонококками. По дан-
ным ВОЗ (1990), 25 % женщин, перенесших
острый хламидийный сальпингит, становятся
бесплодными.
Причиной вторичного бесплодия у женщин
могут являться также хронические воспалитель-
ные процессы хламидийной природы в шейке
матки и яичниках.
Урогенитальный хламидиоз у беременных чаще,
чем в популяции, приводит к самопроизвольным
выкидышам (11 %), преждевременным родам
(13 %), несвоевременному отхождению около-
плодных вод (27 %), мертворождению (5,5 %).
Около 40 % рожениц, шейка матки которых была
инфицирована хламидиями, рожали недоношен-
ных детей на 30—37-й неделе беременности с
массой от 1,5 до 2,7 кг. Различные манифестные
формы хламидийной инфекции обнаруживают не
менее чем у 50 % новорожденных, родившихся
от инфицированных хламидиями матерей.
Хламидии у беременных часто локализуются
в цервикальном канале, эндометрии, трубах,
нередко инфицируют децидуальные оболочки,
вызывают хориоамнионит.
Урогенитальный хламидиоз нередко приводит
к послеродовым осложнениям: субинволюции
матки, длительной лихорадке, эндометриту.
К особенностям урогенитального хламидиоза
у женщин относятся: часто первично-хроничес-
кое течение заболевания; рецидивы, сопровож-
дающие его дальнейшее развитие; высокая час-
тота поражений (наряду с уретрой) шейки матки;
выраженное несоответствие между тяжелыми
деструктивными изменениями внутренних поло-
вых органов и умеренно выраженной клиничес-
кой симптоматикой.
Болезнь Рейтера
Синдром, впервые охарактеризованный Н. Reiter
(1916), включает триаду симптомов: поражение
урогенитального тракта (уретрит, цистит, проста-
тит, цервицит, аднексит), конъюнктивит и артрит.
Этиологическим фактором болезни, запускающим
цепь иммунопатологических реакций, признают-
ся хламидии (С. trachomatis), которые обнаружи-
ваются в мочеполовом тракте у большинства боль-
ных синдромом Рейтера. Другие инфекционные
агенты (иерсинии, шигеллы, сальмонеллы, гоно-
кокки, микоплазмы, уреаплазмы), этиологическая
роль которых обсуждается в развитии этого син-
дрома, поражая органы выделения, скорее способ-
ствуют пенетрации в организм истинного возбу-
дителя болезни Рейтера — С. trachomatis. Риск
заболевания высок у лиц с антигеном гистосовме-
стимости HLA В27, с которым связывают форми-
рование слабого иммунного ответа на инфекцион-
ный агент, выработку аутоантител и вовлечение в
патологический процесс суставов.
Клинико-диагностические
признаки и течение болезни Рейтера
Основные признаки
Реактивный асимметричный артрит, ранняя ат-
рофия мышц в области пораженных суставов.
Глава 44. Урогенитальный хламидиоз
523
Мужчины: уретропростатит, простатит с вези-
кулитом, цирцинарный баланит, баланопостит
или ксеротический баланйт.
Женщины: цервицит, аднексит.
Оба пола: офтальмохламидиоз (конъюнкти-
вит, ирит, увеит),
Второстепенные признаки
Кератодермия ладоней и подошв, поражение ног-
тей, диссеминированные псориазиформные вы-
сыпания, узловатая и многоформная эритема,
крапивница, белесоватые эрозии на слизистой рта.
Поражения внутренних органов и систем:
• Сердце (миокардит, недостаточность аорталь-
ных клапанов);
• почки (очаговый гломеролунефрит, пиело-
нефрит, амилоидоз);
• легкие (плевриты);
• нарушения ЖКТ;
• ЦНС (невралгии, невриты, периферические
парезы, менингоэнцефалит).
Течение:
• Острое (до 3-х месяцев);
• подострое (атака до 1 года);
• хроническое (инвалидизирующее).
Диагноз
Диагноз устанавливается при наличии:
• триады симптомов (хламидийный уретрит,
конъюнктивит, артрит)
• или двух основных признаков в сочетании с
двумя второстепенными или двумя лабораторны-
ми признаками (обнаружение хламидий, противо-
хлймидийных антител, высокие титры аутоантител
к ткани предстательной железы, повышенная СОЭ);
• обнаружении у пациента антигена HLA В27.
Диагностика урогенитального хламидиоза
В связи с отсутствием патогномоничной симп-
томатики хламидийной инфекции проведение
лабораторных исследований является определя-
ющим в постановке диагноза и оценке эффек-
тивности терапевтических мероприятий.
Диагностика хламидийной инфекции осуществля-
ется в двух основных направлениях:
1. Обнаружение в исследуемом материале воз-
будителя, его антигенов или специфических
фрагментов ДНК.
2. Выявление специфических изменений имму-
нологического статуса пациента (обследуемого).
С этими целями применяются следующие методы
диагностики:
• Микроскопия (световая, фазово-контраст-
ная, иммуно-флюоресцентная, электронная).
• Культуральный, основанный на выделении
культуры возбудителя (посев материала на клет-
ки Мак-Коя, Не La-229, мышиные фиброблас-
ты L-929, куриные эмбрионы).
• Серологический, позволяющий обнаружить
в сыворотке крови обследуемых специфические
антитела к С. trachomatis и определить динами-
ку их титра.
1. Метод микроскопического обнаружения
хламидий в мазках-соскобах уретрц, цервикаль-
ного канала, прямой кишки, глотки, конъюнк-
тивы путем их окрашивания по Романовскому-
Гимза позволяет выявить ЭТ (окрашиваются в
красный и фиолетово-красный цвет) и РТ (си-
ний и голубой цвет) у 15—20 % мужчин и 40 %
женщин (доступный, недорогой, однако малоин-
формативный).
2. Метод прямой иммунофлюоросценции
(ПИФ) с помощью моноклональных антител
против основного белка наружной мембраны
С. trachomatis, меченных флюоресцеин-изотио-
ционатом, позволяет идентифицировать в маз-
ках-соскобах с помощью люминесцентной мик-
роскопии ЭТ и РТ у 50-80 % обследованных
(доступный и недорогой метод).
3. Культуральное исследование (КИ) позво-
ляет обнаружить жизнеспособные микроорга-
низмы у 50—80 % пациентов. Процедура выра-
щивания хламидий на клетках Мак-Коя
занимает до недели, требует специальных лабо-
раторных условий, дорогостоящих реактивов,
что для широкой практики мало приемлемо.
Данный метод применяют для диагностики
хламидийной инфекции у детей раннего возра-
ста, а также для выявления С. trachomatis в
экстрагенитальных анатомических участках и с
научными целями.
4. Методы амплификации нуклеиновых кислот
(АНК) превосходят по своей чувствительности
(свыше 90 %) другие диагностические методы,
включая КИ, и позволяют обнаруживать мини-
мальное количество элементарных телец
С. trachomatis. Из методов АНК наиболее распро-
странен метод полимеразной цепной реакции
(ПЦР). Исследуются уретральные и цервикаль-
ные соскобы, моча. Для диагностики применя-
ются праймеры, содержащие нуклеотидные пос-
ледовательности, комплементарные генетическому
материалу С. trachomatis. При цикловой темпера-
турной обработке происходит денатурация ДНК
возбудителя и последующая регистрируемая ам-
плификация ее специфического участка компле-
ментарного праймеру.
Поскольку методы АНК позволяют обнару-
живать нежизнеспособные микроорганизмы, для
524 Венерология
анализа могут быть использованы самые различ-
ные биологические материалы, включая мочу и
образцы, самостоятельно взятые женщинами из
преддверия влагалища.
Применение методов АНК для рутинной ди-
агностики ограничивается материальными воз-
можностями лабораторий.
5. Обнаружение специфических антихлами-
дийных иммуноглобулинов класса М и G (ме-
тод иммуноферментного анализа — ИФА) и их
титра позволяет установить первичный (до 1 мес
с момента инфицирования — IgM) и вторичный
(свыше 1 мес — IgG) специфический иммунный
ответ. Эти методы важны для диагностики хла-
мидийной инфекции, в детской практике, при
отсутствии возможности получить материал для
исследования (маточные трубы, яичники, брю-
шина и др.), при подозрении на хламидийное
поражение плода и новорожденного, осложнени-
ях урогенитального хламидиоза, болезни Рейтера,
эпидемиологических и научных исследованиях.
Возможны ложноположительные результаты,
т. к. другие грамотрицательные бактерии также
содержат ЛПС-антиген, вызывающий аналогич-
ный иммунный ответ.
В большинстве случаев доказательством нали-
чия С. trachomatis является исследование методом
ПИФ и ПЦР соскобов (из уретры, шейки мат-
ки, зева, конъюнктивы), мочи, сока простаты,
эякулята и определение антихламидийного IgG в
периферической крови пациентов (обследуемых).
Для диагностики персистирующей хламидийной
инфекции рекомендуются методы:
• Электронная микроскопия (исследуется эпи-
телий половых путей, биопсийный материал из
пораженных тканей — спайки, маточные трубы).
Метод не может быть использован для обследо-
вания широкого круга пациентов.
• Методы АНК (ПЦР, ДНК-зондовой гибри-
дизации) позволяют обнаружить ДНК хламидий
в мазках из уретры и цервикального канала,
фоллопиевых трубах (достаточно редко).
© Реакция непрямой гемагглютинации (РИГА)
с родоспецифическим диагностикумом. У лиц с
персистирующей инфекцией РИГА в 4 раза пре-
вышает диагностический титр. Используется
также реакция непрямой иммунофлюоресценции
(РНИФ) с типоспецифичными антигенами.
• Выявление гиперпродукции Ig А^ и IgG.
Лечение больных
урогенитальным хламидиозом
Лечение больных УГХ, особенно хронических и
персистирующих форм инфекции, является не-
простой задачей. При выборе лекарственных
средств и методов терапии необходимо учиты-
вать следующее:
• к антибиотикам чувствительны только раз-
множающиеся внутриклеточные формы хлами-
дий — РТ;
• ЭТ и персистирующие формы хламидий к
антибиотикам не чувствительны;
• для лечения применяют антибиотики, кото-
рые действуют на внутриклеточные формы мик-
роорганизмов (тетрациклины, макролиды, от-
дельные фторхинолоны, рифампицин);
• монотерапия антибиотиками должна быть
рассчитана на 6~7 циклов развития хламидий (не
менее 2-х недель) и применяется при свежих
формах заболевания и наличии клинических про-
явлений; ?
• препараты пенициллиновой группы, цефа-
лоспорины, левомицетин, сульфаниламиды не
являются этиотропными, при этом способству-
ют трансформации хламидий в персистирующие
и L-формы (аберрантные). Аналогичное дей-
ствие оказывают субтерапевтические дозы про-
тивохламидийных антибиотиков;
• образование аберрантных форм с изменён-
ной морфологической структурой и сниженной
экскрецией ключевых хламидийных антигенов
(МОМР, ЛПС) резко уменьшает транспорт
гидрофильных антибиотиков в хламидийную
клетку;
• иммунотропная терапия проводится при
хронических и персистирующих формах УГХ
(сниженная выработка у-интерферона, факторов
некроза опухоли, ИЛ-2, недостаточность Т и В-
клеточного иммунитета, системы комплемента и
фагоцитоза);
• перед назначением иммунотропных препа-
ратов (в случаях хронической или персистент-
ной инфекции) проводится расширенное имму-
нологическое обследование пациента (включая
профиль CD-антигенов, интерферонов);
• пациенты получают большую лекарствен-
ную нагрузку на организм, не всегда хорошо её
переносят, страдают сопутствующими заболева-
ниями, нередко нарушают лечебный режим;
• однократный прием азитромицина (1,0 г)
эффективен преимущественно при свежих фор-
мах заболевания;
• при хронических и персистирующих формах
УГХ этиотропные препараты назначаются в тече-
ние 3-х недель и более, одновременно проводит-
ся целенаправленная иммунотерапия: пирогенал,
тактивин, тималин, полиоксидоний, индукторы
интерферона (амиксин, циклоферон, неовир, им-
мунофан, ридостин, свечи виферон и др.), ронко-
лейкин;
Глава 44. Урогенитальный хламидиоз
525
• кроме вышеперечисленных препаратов на-
значаются средства для системной энзимотера-
пии (лидаза, трипсин, вобэнзим, флогэнзим),
гепатопротекторы (эссенциале, карсил, легалон,
препараты солянки холмовой), антиоксиданты,
эубиотики (бифидум бактерии, бификол, лакто-
бактерии, энтерол);
• для профилактики кандидоза (кишечного,
генитального, слизистых) назначают нистатин,
леворин, кетоконазол (низорал, канестен), флуко-
назол (дифлюкан), итраконазол (орунгал) в виде
пероральных, мазевых или влагалищных форм;
• показано физиолечение: диатермия, КВЧ,
импульсное тепло, грязевые аппликации, ванноч-
ки с отварами трав и др.
Рекомендуемые антибиотики
и схемы их применения
Доксициклин
Взрослые: при свежем неосложненном хлами-
диозе препарат назначают по 100 мг после еды
2 раза в сутки в течение 7 дней; при остальных
формах и осложнениях — в течение 14—21 дней.
В/в препарат вводят в тех же дозах, медленно,
капельно в течение 1,5—2 ч, с интервалом 12
часов между введениями.
Дети 8—12 лет: внутрь 4 мг/кг/сут — 1-ый день,
затем 2 мг/кг/сут равными дозами через 12 ч.
Азитромицин (сумамед, азикар)
Взрослые: при свежем остром хламидиозе на-
значают однократно 1,0 г за час до еды или че-
рез 2 часа после еды; при остальных формах — в
первый день 500 мг, в последующие — по 250 мг
один раз в сутки в течение 10 дней, общая кур-
совая доза — 3,0 г; или 1,0 г 1 раз в неделю в
течение 14 дней (1, 7, 14 дни), на курс 3,0 г.
Дети старше года: 10 мг/кг 1 раз в день в те-
чение 3-х дней, затем 4 дня по 5 мг/кг 1 раз в
день.
Эритромицин
Взрослые: по 500 мг за 1—1,5 ч до еды 4 раза
в сутки в течение 7 дней или по 250 мг внутрь
4 раза в сутки в течение 14 дней. В/в по 0,5—
1,0 г 4 раза в день в течение 7—14 дней.
Эритромицин
Дети: первых 3-х месяцев жизни по 20—40 мг/
кг массы тела в сутки; от 4-х мес до 18 лет —
30—50 мг/кг/сут равными дозами 4 раза в сут-
ки. Курс лечения 10—14 дней.
Альтернативные антибиотики
и схемы их применения
Тетрациклин
Взрослые: при свежей неосложненной фор-
ме заболевания — по 500 мг после еды 4 раза
в сутки в течение 7 дней; при остальных фор-
мах продолжительность лечения — 14—21
дней.
Дети старше 8 лет: 20—25 мг/кг/сут равными
дозами 3—4 раза в сутки 7—10 дней.
Метациклин (рондомицин)
Взрослые: внутрь после еды в дозе 600 мг
(первый прием), затем по 300 мг через каждые
8 часов в течение 7 дней; на курс лечения при
острых и неосложненных формах — 6,6 г.
Дети старше 8 лет: 7,5—10 мг/кг/сут в 2—4
приема в течение 7—10 дней.
Кларитромицин (клацид, кларикар, фромилид)
Взрослые: при неосложненных формах забо-
левания назначается по 250—500 мг дважды в
день в течение 7 дней, независимо от приема
пищи; при других формах инфекции — по 250—
500 мг 2 раза в сутки в течение 10—14 дней.
Дети: внутрь 7,5 мг/кг/сут в течение 7—10
дней.
Рокситромицин (рулид)
Взрослые: внутрь по 150 мг 2 раза в сутки в
течение 10 дней.
Джосамицин (вильпрафен)
Взрослые: внутрь 0,8—2,0 г в сутки равными
дозами в 3 приема в течение 10—20 дней.
Дети до 14 лет: 30—50 мг/кг/сут в 3 приема
10—14 дней.
Ровамицин (спирамицин)
Взрослые: по 3 млн ME внутрь 3 раза в день
в течение 10 дней.
Офлоксацин (таривид, заноцин)
Взрослые: после еды по 300 мг 2 раза в сутки
в течение 7 дней; или в/в капельно по 200—
400 мг на 0,5 % растворе глюкозы в течение часа
через 12 часов.
Ломефлоксацин (максаквин)
Взрослые: после еды по 400—600 мг один раз
в сутки в течение 7—28 дней.
Рифампицин
Взрослые: первый прием 600 мг, затем 300 мг
2 раза в день в течение 7 дней (свежие формы).
Далацин Ц вагинальный крем
Далацин Ц вагинальный крем (содержит 2 %
клиндамицина фосфата) в тубах по 40,0 г с 7
аппликаторами. Разовая доза 5,0 г крема вводится
во влагалище с помощью аппликатора перед сном.
Не применяют в первые 3 месяца беременности.
Для лечения беременных, больных УГХ,
рекомендуются антибиотики:
Эритромицин. По 500 мг внутрь 4 раза в сут-
ки в течение 7 дней.
Амоксициллин 500 мг перорально 3 раза в сутки
в течение 7—10 дней.
526 Венерология
Джосамицин (вильпрафен) по 750 мг внутрь
2 раза в день в течение 7 дней.
Альтернативные антибиотики:
Эритромицин. По 250 мг внутрь 4 раза в сут-
ки в течение 14 дней.
Эритромицин. По 800 мг внутрь 4 раза в сут-
ки в течение 7 дней.
Эритромицин. По 400 мг внутрь 4 раза в сут-
ки в течение 14 дней.
Азитромицин. По 1,0 внутрь в однократной
дозе.
Кларитромицин. По 250 мг 2 раза в сутки в
течение 10 дней.
Лечение новорожденных
(конъюнктивит, пневмония)
Эритромицин: 50 мг/кг/сут внутрь, в 4 приема,
в течение 10—14 дней
Лечение болезни Рейтера
Продолжительность антибактериального лечения
при болезни Рейтера: от 4-х недель до 4-х меся-
цев в зависимости от тяжести проявлений забо-
левания.
В комплексную терапию при болезни Рейте-
ра, особенно при тяжелых и затяжных формах,
включают:
• нестероидные противовоспалительные сред-
ства — индометацин (метиндол) по 100—150 мг
в сутки, пирабутол или реопирин по 3-5 мл внут-
римышечно в течение 5—7 дней с последующей
заменой другими нестероидными противовоспа-
лительными средствами (метиндол, метиндол-
ретард, пироксикам), вольтарен (ортофен, дикло-
фенак-натрий) по 100—150 мг в день;
• глюкокортикоиды — внутрисуставное вве-
дение гидрокортизона, кеналога; при недостаточ-
ной их эффективности назначают внутрь пред-
низолон 5—20 мг в сутки;
• аминохинолоновые препараты (делагил,
плаквенил);
• при неэффективности глюкокортикоидов и
НПВС назначают иммунодепрессанты (хлорбу-
тин, метотрексат) перорально или парентераль-
но и применяют их внутрисуставное введение;
• препараты золота (криназол, миокризин,
ауранофин);
• физиотерапевтическое лечение, ЛФК, в пос-
ледующем бальнео- и грязелечение.
Контроль излеченности
урогенитального хламидиоза
Всем пациентам после окончания лечения про-
водят клинико-лабораторный контроль в тече-
ние 3—4 месяцев. Первое обследование проводят
через 4—5 недель после окончания лечения, пос-
ледующие 3 контроля — каждый месяц.
Критериями излечения являются: исчезнове-
ние клинических симптомов заболевания, отсут-
ствие морфологических изменений в области
наружных и внутренних половых органов, стой-
кая нормализация лейкоцитарной реакции, эли-
минация возбудителя, т. е. клинико-микробио-
логическое излечение.
Методы провокации урогенитальной хлами-
дийной инфекции не разработаны. У женщин
проведение контрольных исследований желатель-
но приурочить к началу или окончанию менст-
руального цикла.
При оценке клинических показателей, в за-
висимости от локализации перенесенного про-
цесса, используют инструментальные методы
(уретроскопия, уретрография, кольпоскопия,
гистероскопия, малоинвазивная лапароскопия,
УЗИ матки, придатков, предстательной железы).
При оценке цитологической картины соскобов
со слизистой уретры, шейки матки, влагалища и
секрета предстательной железы необходимо при-
менять методы: ПИФ, ПЦР, культуральный; в
сыворотке крови определяют титры противохла-
мидийных антител IgG.
Учитывая половой путь передачи, следует од-
новременно обследовать и лечить половых парт-
неров. Во время лечения и наблюдения половая
жизнь исключается.
Мероприятия по предупреждению
распространения хламидийной
инфекции и её осложнений
Обследование различных контингентов на хла-
мидийную инфекцию с помощью экспресс-ме-
тодов и ПЦР:
• подростков на базе подростковых отделений
поликлиник и при их госпитализации в сома-
тические стационары;
• школьников — с помощью неинвазивных
методов (образцов мочи, смывов из влагалища);
• половых партнеров лиц, инфицированных
С. trachomatis^
• лиц сексуально активного возраста при их
обращении к урологу, а также в соматических
стационарах;
• женщин (особенно моложе 25 лет), состоя-
щих на учете у гинеколога (бесплодие, ВЗПО),
или направленных для прерывания беременно-
сти; *
• женщин перед введением, замещением или
удалением внутриматочных контрацептивов;
• беременных в дородовый период или во
время родов;
Глава 44. Урогенитальный хламидиоз 527
• матерей новорожденных с конъюнктивитом
или пневмонией, вызванных С. trachomatis',
• девочек, больных вульвитами и вульвоваги-
нитами;
• мальчиков с явлениями воспалительного
фимоза;
• взрослых и детей с рецидивирующими конъ-
юнктивитами;
• пациентов с реактивным артритом;
• лиц с длительным лихорадочным состояни-
ем неясной природы;
• пациентов с другими заболеваниями, пере-
даваемыми половым путем;
• лиц, проходящих комиссию медосмотра;
• лиц, планирующих создание семьи.
Глава 45
Урогенитальный микоплазмоз
Урогенитальный микоплазмоз — бактериальная
инфекция, передаваемая половым путем.
Микоплазмозы (уреаплазмозы) широко рас-
пространены среди населения. Они передаются
при половых контактах, и многие исследовате-
ли относят их к заболеваниям, передаваемым
половым путем.
Этиология
Заболевания человека, вызываемые микоплазма-
ми, объединяют в группу микоплазмозов чело-
века. Согласно современной классификации
(N. Romano и соавт., 1984), микоплазмы отно-
сятся к семейству Mycoplasmataceae (порядок
Mycoplasmatales класса Mollicutes). Это семейство
разделяется на два рода: род Mycoplasma, вклю-
чающий около 100 видов, и род Ureaplasma
(urea — мочевина, plasma — принимающий лю-
бую форму), в котором насчитывается пока
только 3 вида. Основным таксономическим при-
знаком уреаплазм является их способность гид-
ролизовать мочевину.
Установлено, что человек является естествен-
ным хозяином, по крайней мере, 14 видов ми-
коплазм. Четыре вида (Mycoplasma pneumoniae,
Mycoplasma hominis, Mycoplasma genitalium,
Ureaplasma urealyticum), в полном соответствии
с постулатами Коха, патогенны для человека.
M.fermentans (шт. incognitas) и М. penetrans, по
данным исследований последних лет, причаст-
ны к развитию СПИДа. М. pneumoniae — воз-
будитель респираторной инфекции, М, hominis,
М. genitalium и U. urealyticum — возбудители
инфекций урогенитального тракта.
Широкое распространение урогенитальных
микоплазм (М. hominis, М, genitalium, U urealyticum)
и их частое выявление у практически здоровых
лиц затрудняет решение вопроса о роли этих мик-
роорганизмов в этиологии и патогенезе заболева-
ний урогенитального тракта. Позиции исследова-
телей по данной проблеме противоречивы.
Часть авторов относят микоплазмы к абсо-
лютным патогенам, ответственным за развитие
конкретных нозологических форм: уретрита,
простатита, послеродового эндометрита, пиело-
нефрита, патологии беременности и плода, арт-
рита, сепсиса.
Другие исследователи считают, что микоплаз-
мы являются комменсалами урогенитального
тракта, способными при определенных услови-
ях вызывать инфекционно-воспалительные про-
цессы мочеполовых органов, чаще в ассоциации
с другими патогенными или условно-патоген-
ными микроорганизмами.
Отличительными особенностями микоплазм и
уреаплазм являются (D. Taylor-Robinson, 1995):
рост в бесклеточной среде; обычно формируют
колонии типа «яичницы-глазуньи»; размер са-
мых маленьких жизнеспособных форм около
200—300 нм; часто сферические, но иногда такие,
как М. pneumoniae или М. genitalium, т. е. имеют
форму бутылки или груши; клеточная стенка
отсутствует, но имеется трехслойная мембрана;
содержат ДНК и РНК; рост подавляется анти-
биотиками широкого спектра действия, однако
устойчивы к пенициллину, что связано с отсут-
ствием клеточной стенки, и рифампицину; мо-
жет развиваться устойчивость к другим антиби-
отикам; рост подавляется антителами, особенно
в присутствии комплемента; тенденция к специ-
фичности в отношении хозяина; некоторые —
иммуностимуляторы, другие — иммуносупрессо-
ры; вызывают заболевания респираторного и
генитального тракта, а также маститы (у живот-
ных), контаминируют (заражают) клеточные
культуры.
Уреаплазмы были описаны как «микроорга-
низмы, при выделении которых следует искать
заболевание». U. urealyticum рассматривается в
качестве возможного этиологического агента
таких заболеваний человека, как НГУ, синдром
Рейтера, простатит, камни мочевого пузыря и
почек, воспалительные заболевания органов ма-
лого таза, артрит и бесплодие. U. urealyticum
может вызывать также такие осложнения бере-
менности, как хориоамнионит, преждевремен-
ные роды, а также перинатальную заболевае-
мость и смертность. В ряде случаев бывает
сложно доказать этиологическую роль
U. urealyticum, поскольку этот микроорганизм ча-
528
Глава 45. Урогенитальный микоплазмоз
529
сто входит в состав нормальной микрофлоры, не
вызывая никаких заболеваний.
Патогенез
Механизм патогенеза урогенитальных микоплаз-
мозов недостаточно ясен. Имеются лишь отдель-
ные сведения о взаимодействии микоплазм с
клетками. Возможным механизмом взаимодей-
ствия уреаплазм и чувствительных клеток может
быть персистенция этих микроорганизмов на
мембране, однако пока не установлено, являются
ли они мембранными паразитами или проника-
ют в цитоплазму. Продукты жизнедеятельнос-
ти этих бактерий могут быть токсичными для
клеток организма и оказывать повреждающее
действие на клеточную стенку. Окисление жи-
ров в результате активности пероксидазы также
способно привести к повреждению стенки клет-
ки. Токсичной для микроорганизма может быть
концентрация аммиака, образующегося в резуль-
тате расщепления мочевины уреаплазмами. Ми-
коплазмы утилизируют компоненты клетки хо-
зяина, такие как холестерин и жирные кислоты.
Механизм прикрепления микоплазм пока недо-
статочно ясен. Если микоплазма прикрепляется
к эпителию уретры, такая связь очень прочная
и не нарушается под действием потока мочи. Од-
нако плотного непосредственного прилипания не
существует между микоплазмами и стенкой клет-
ки хозяина, что установлено при электронно-
микроскопическом исследовании.
Эпидемиология
Урогенитальный микоплазмоз довольно широ-
ко распространен среди разных групп населения.
С наибольшей частотой он обнаруживается у лиц
с повышенной половой активностью, проститу-
ток, гомосексуалистов, при некоторых венери-
ческих заболеваниях, таких как гонорея и три-
хомониаз, и, что особенно важно, во время
беременности.
Заражение урогенитальной микоплазменной
инфекцией происходит преимущественно поло-
вым путем. Возможен непрямой путь заражения
женщин, особенно девочек. Инфицирование
микоплазмами верхних отделов генитального
тракта может осуществляться сперматозоонами —•
носителями микоплазм. Установлено внутриут-
робное инфицирование плода и заражение но-
ворожденных при прохождении через инфици-
рованные микоплазмами родовые пути матери.
Клиническая картина
Клиническая картина микоплазменной инфек-
ции не имеет патогномоничных симптомов.
Описаны клинические симптомы негонококко-
вого уретрита, простатита, орхиэпидидимита,
послеродового эндометрита, спонтанного абор-
та, преждевременных родов, постабортной и
послеродовой лихорадки, артрита у взрослых,
сепсиса и менингита у новорожденных, при ко-
торых выделялись М. hominis, М. genitalium или
U, urealyticum в монокультурах.
Негонококковый уретрит (НГУ) и простатит
Этиологическая роль М. hominis в развитии НГУ
дискутируется. Частота выделения М. hominis при
НГУ варьирует от 13 до 37 %. Показано, что у
10 % лиц с хроническим абактериальным про-
статитом выделяют М. hominis.
Негонококковый или неспецифический урет-
рит — наиболее распространенное следствие уре-
аплазменной инфекции у мужчин. Уреаплазмы
вызывают НГУ, для которого характерно тече-
ние с рецидивами. Инкубационный период
длится 3—5 нед. Заражение происходит половым
путем. При обследовании половых партнеров у
87 % из них заболевание протекало торпидно, а
иногда бессимтомно.
Развитие уреаплазменных простатитов явля-
ется довольно частым проявлением уреаплазмен-
ной инфекции. Присутствие в секрете предста-
тельной железы уреаплазм в количестве 103—104
КОЕ/мл указывает на их этиологическую роль
в развитии простатита.
Часто выявляют колонизацию уреплазмами
прямой кишки мужчин-гомосексуалистов. Выс-
казывается предположение, что U. urealyticum
может быть причиной проктита.
Имеются доказательства того, что М. genitalium
является одним из возбудителей острого НГУ,
есть определенные доказательства ее причастно-
сти к развитию хронического или рецидивиру-
ющего НГУ, однако не получены данные об
участии М. genitalium в развитии хронического
абактериального простатита.
Бесплодие у мужчин
Данные о влиянии микоплазменной инфекции
урогенитального тракта на бесплодие у мужчин
весьма противоречивы. По данным одних иссле-
дователей, такая связь существует, по данным
других, отсутствует. У женщин вторичное беспло-
дие может развиваться в результате воспалитель-
ных процессов, приводящих как к нарушению
овогенеза, так и препятствующих продвижению
яйцеклетки.
Бесплодие у мужчин, вызванное уреаплазма-
ми, может быть обусловлено не только воспали-
тельными процессами (как правило, простати-
том), но и влиянием уреаплазм непосредственно
530 Венерология
на сперматогенез, а также на подвижность спер-
матозоидов.
Не отрицается возможность участия М. genitalium
в развитии бесплодия, особенно у мужчин.
Л/, genitalium обладает выраженной способностью
адсорбироваться на клетках и может прикреплять-
ся к головке или средней части сперматозоида и
изменять его активность.
Диагноз
Общепринятыми для идентификации мико-
плазм являются культуральные методы исследо-
вания, которые постоянно совершенствуются.
Диагностика микоплазменных инфекций при
прямой микроскопии окрашенных препаратов
практически исключена в связи с отсутствием во
всем семействе микоплазм четких морфологичес-
ких характеристик, наличием у них выраженного
полиморфизма, а также мелких размеров.
Для идентификации урогенитальных микоплаз-
мозов используются различные лабораторные
методы диагностики:
• бактериологический (культуральный) метод;
• серологические методы;
• определение антигенов микоплазм методом
РИФ или ИФА;
• метод активированных частиц;
• метод генных зондов;
• метод полимеразной цепной реакции.
Бактериологический метод
Бактериологический метод диагностики (цветная
реакция) основан на расщеплении мочевины,
необходимой для роста уреаплазм, изменении pH
и, как следствие, изменении цвета среды, в ко-
торую добавлен индикатор. Генитальные микоп-
лазмы могут культивироваться как на жидких,
так и на плотных питательных средах. При бак-
териологической диагностике применяют, как
правило, и первый, и второй варианты. Посев
клинического материала на жидкие питательные
среды используется как предварительный тест на
ферменты уреазу (для U. urealyticum) и аргина-
зу (для М. hominis). Под влиянием уреазы про-
исходит расщепление мочевины с образованием
аммиака, что приводит к защелачиванию жид-
кой питательной среды и изменению цвета ин-
дикатора в течение 18—24 часов. Фермент арги-
наза, продуцируемый М. hominis, разлагает
аминокислоту аргинин, что также сопровожда-
ется повышением pH среды.
В последние годы появились специальные
тест-системы, пользуясь которыми, можно од-
ноэтапно определять на жидкой питательной
среде не только наличие того или иного микоп-
лазменного возбудителя, но и его концентрацию
в исследуемом клиническом материале (тест-си-
стема «Mycoplasma DUO» производства фран-
цузской фирмы «Sanofi Pasteur»; тест-система
«Mycoplasma 1ST» производства «bioMeriex»).
Тест-система «Mycoplasma DUO» производства
«Sanofi Pasteur» позволяет культивировать, иден-
тифицировать и дифференцированно титровать
U. urealyticum и М. hominis. Она представляет со-
бой микрометод, который, помимо одновремен-
ного выявления двух микроорганизмов, позво-
ляет определять их содержание. Тест-система
полностью укомплектована и может применять-
ся во всех лабораториях, имеющих ламинар, хо-
лодильник и термостат. В состав набора входят
микроплаты, в лунках которых содержатся и раз-
водятся в момент исследования проб дегидрати-
рованные субстраты, ростовые факторы и селек-
тивные антибиотики, обеспечивающие
реализацию метода (культивирование, иденти-
фикацию, титрование); флаконы с транспортной
средой для сбора, транспортировки и хранения
проб (соскобы, моча, эякулят). Результаты ди-
агностики наблюдают через 24 и 48 ч после за-
ражения сред и инкубации материала в термо-
стате при 37 °C. Аналогичный принцип
культивирования, идентификации и титрования
микоплазм используется и в тест-системе
«Mycoplasma 1ST» производства «bioMeriex». При
использовании данного набора можно, одновре-
менно с выделением и идентификацией
U. urealyticum и М. hominis, определять чувстви-
тельность этих микоплазм к антибиотикам.
При микробиологическом анализе берут про-
бы со слизистой уретры, сводов влагалища, из
канала шейки матки, периуретральной области.
Пробы мочи для выделения микоплазм предпоч-
тительно брать из утренней первой и средней
порции. При подозрении на простатит необхо-
димо получить для посева секрет предстательной
железы. При мужском бесплодии неясной эти-
ологии целесообразно микробиологически иссле-
довать сперму. Микробиологическому обследо-
ванию подлежат также биосубстраты при
лапароскопии, амниоцентезе, а также ткани
абортированных и мертворожденных плодов.
Серологические реакции
Серологические реакции (РСК, РИМ, РПГА,
ИФА) рекомендуется использовать при массовых
обследованиях групп населения, а также для
подтверждения клинического диагноза. Однако
серологическая диагностика микоплазм и уреап-
лазм весьма затруднительна в связи с существо-
ванием большого числа серотипов этих возбуди-
Глава 45. Урогенитальный микоплазмоз
531
телей, а также с недостаточной изученностью
иммунологических изменений при поражении
мочеполовых путей микоплазмами. В настоящее
время одним из общедоступных диагностикумов
является «МикогомоСкрин (G+M)» — выявле-
ние антител классов IgM и IgG методом ИФА
против М. hominis. Четырехкратное нарастание
титра антител в динамике заболевания свиде-
тельствует об инфекционном процессе, обуслов-
ленном М. hominis.
Определение антигенов
микоплазм методом РИФ или ИФА
Разработаны методы выявления антигенов ми-
коплазм и уреаплазм в сыворотке крови и дру-
гих субстратах — РАГА (реакция агрегатгемаг-
глютинации), ИФА, РИФ. Для реализации этих
методов необходимо наличие стандартных набо-
ров антисывороток к разным серотипам возбу-
дителя. Для постановки РИФ используют спе-
цифическую антисыворотку, конъюгированную
с флюорохромом. Среди стандартных диагнос-
тикумов можно отметить тест-систему «УреаС-
лайд» для выявления антигенов U. urealyticum ме-
тодом прямой иммунофлюоресценции и
аналогичную тест-систему «МикоСлайд» для вы-
явления антигенов М. hominis производства НПФ
«ЛАБдиагностика» (Россия). В исследуемых мик-
ропрепаратах микоплазмы выявляются в виде по-
лиморфных структур (зерен, гранул, коккобак-
терий и т. д.), имеющих ярко-зеленое свечение,
на поверхности эпителиальных клеток, в образ-
цах спермы — на сперматозоидах, а также сво-
бодно. Неспецифическая бактериальная микро-
флора окрашивается в оранжевый цвет, а клетки
эпителия, лейкоциты, сперматозоиды — в оран-
жевый и красно-бурый. Качество мазка опреде-
ляется наличием клеток и клеточных элементов
при минимальном количестве слизи, эритроци-
тов. Следует отметить, что выявление антигенов
микоплазм с помощью РИФ практикуется лишь
как качественная реакция на присутствие инфек-
ции и не позволяет определять этиологически
значимое количество возбудителя.
Метод активированных частиц
Метод активированных частиц (диагностикум
«Рохатест») предназначен для выявления
U. urealyticum и М. hominis в мазках. Исследуе-
мый материал из урогенитального тракта боль-
ного наносится на предметное стекло, подсуши-
вается и стеклографом вокруг мазка обводится
поле, в которое наносится нормальная кроличья
сыворотка, предварительно разведенная буфер-
ным раствором. Проводят инкубацию изучаемого
материала (10 мин при комнатной температуре
или 5 мин в термостате при 37 °C). Во время
инкубации аккуратно покачивают предметное
стекло несколько раз. Затем наносится диагно-
стикум Рохатест и растворы смешивают, плав-
но покачивая предметное стекло в течение 2 мин.
По окончании проведенных процедур проводят
учет результатов реакции.
Метод генных зондов
Разработан новый диагностический метод, осно-
ванный на применении генных зондов. Описа-
ны зонды трех классов: зонды из фрагментов
рРНК; зонды из клонированных фрагментов
ДНК, состоящие из последовательностей, специ-
фичных для определенных видов или даже штам-
мов; зонды, сконструированные из плазмид или
вирусов микоплазм.
Метод полимеразной цепной реакции
Для выявления микоплазм разработан метод
ПЦР (полимеразной цепной реакции), который
относится к методам ДНК-диагностики и осно-
вывается на распознавании последовательности
нуклеотидов в генах искомых микроорганизмов
или вирусов. При данном методе исследуются
соскобы из уретры, стенок влагалища и церви-
кального канала.
Для выявления микоплазмозов урогениталь-
ного тракта наиболее подходящими являются
бактериологический метод и ПЦР, а также тест-
система «Mycoplasma DUO». Эффективными
являются также серологические методы диагно-
стики (РПГА и РАГА), особенно при НГУ и
воспалительных процессах нижних отделов ге-
нитального тракта. Рекомендуется использовать
комплекс методов исследования, так как это
повышает достоверность результатов.
Лечение
Подбор лекарственных средств для лечения ми-
коплазмозов определяется особенностями биоло-
гии возбудителей. Они устойчивы ко всем пре-
паратам, действие которых связано с биосинтезом
белков клеточной стенки, и чувствительны к
ингибиторам синтеза мембранных и внутрицитоп-
лазматических белков. Большинство из них чув-
ствительны к антибиотикам тетрациклинового
рада, многим макролидам, линкозамидам, проти-
вогрибковым и противопротозойным препаратам.
Однако некоторые препараты обладают различ-
ной активностью по отношению к разным видам
микоплазм, поэтому требуется информация об ан-
тибиотикоустойчивости штаммов. Особенно это
касается генитальных видов микоплазм. Боль-
шинство микоплазменных инфекций могут быть
излечены соответствующими антибиотиками.
532 Венерология
Трудности могут возникать при лечении больных
с иммунодефицитом.
Критерии назначения этиотропной терапии
при микоплазменной инфекции (В. А. Аковбян
и соавт., 1994)
• Клинические проявления инфекционно-вос-
палительных процессов мочеполовых и других
органов;
• результаты комплексного микробиологичес-
кого обследования, включающего исследование на
наличие патогенных и условно-патогенных мик-
роорганизмов (с количественным определением);
• степень риска предстоящих оперативных
или инвазивных манипуляций (роды, аборт,
введение внутриматочного контрацептива);
• бесплодие, связанное только с микоплазмен-
ной инфекцией;
• у беременных: акушерско-гинекологический
анамнез и течение настоящей беременности.
К препаратам выбора относятся
тетрациклины (доксициклин — 1-й прием 0,2 г;
затем 0,1 г 2 раза в сутки в течение 7—10 дней;
миноциклин — 1-й прием 0,2 г; затем 0,1 г 2 раза
в сутки в течение 7—10 дней);
азалиды (азитромицин — 1-й день 0,5 г; за-
тем 0,25 г 1 раз в день; на курс 1,5 г).
К альтернативным препаратам относятся
макролиды (эритромицин — 0,5 г 4 раза в сут-
ки в течение 7—10 дней; мидекамицин — 0,4 г 3
раза в сутки в течение 5—7 дней; кларитроми-
цин — 0,25 г 2 раза в сутки в течение 5—7 дней;
рокситромицин — 0,15 г 2 раза в сутки в тече-
ние 7—10 дней);
фторхинолоны (пефлоксацин — 0,6 г 1 раз в
сутки в течение 5—7 дней; офлоксацин -0,4 г 1
раз в сутки в течение 5—7 дней; ломефлокса-
цин — 0,6 г 1 раз в сутки в течение 5—7 дней).
Примечание: М. hominis устойчива к макроли-
дам. Как правило, U. urealyticum чувствительна
к макролидным антибиотикам, тетрациклинам и
фторхинолонам. В то же время, применение ами-
ногликозидов и линкозамидов для лечения уре-
аплазменной инфекции обычно оказывается
неэффективным.
При назначении лечения больным микоплаз-
мозом рекомендуется обследовать половых парт-
неров, а при обнаружении возбудителя прово-
дить лечение.
Контрольное обследование больных мико-
плазменной инфекцией осуществляется спустя
2—3 недели после завершения курса антимикроб-
ной терапии.
Профилактика
Меры профилактики, принятые для этой груп-
пы инфекций, должны быть такими же, как и
для других инфекций, передаваемых половым
путем. Планомерное медицинское обследование
на выявление микоплазмозов необходимо начать
в группах «повышенного риска», к которым
большинство специалистов относят проституток,
гомосексуалистов и, главное, всех женщин де-
тородного возраста, страдающих хроническими
воспалительными заболеваниями урогенитально-
го тракта неясной этиологии, а также беремен-
ных с осложненным акушерским анамнезом и
неблагополучным течением настоящей беремен-
ности. Большое профилактическое значение
имеет направленное обследование беременных
женщин на разных стадиях беременности. В слу-
чае выявления у них урогенитального микоплаз-
моза можно предупредить развитие внутриутроб-
ной микоплазменной инфекции путем санации
беременной и ее супруга. Группы повышенного
риска следует выделить среди урологических
больных. В них в первую очередь включают лиц,
нуждающихся в трансплантации почек. Своев-
ременное выявление и лечение урогенитальных
микоплазмозов необходимо для профилактики
восходящей урологической инфекции.
Глава 46
Урогенитальный кандидоз
Урогенитальный кандидоз — грибковое заболе-
вание слизистых оболочек и кожи мочеполовых
органов, вызываемое дрожжеподобными гриба-
ми рода Candida.
Поражение мочеполового тракта является од-
ним из наиболее часто встречающихся проявле-
ний кандидоза. Урогенитальные кандидозы
широко распространены, протекают хронически
и склонны к рецидивам.
Этиология
Возбудителем урогенитальных кандидозов наи-
более часто является Candida albicans. Род кан-
дида насчитывает более 150 видов. Степень па-
тогенности различных штаммов широко
варьирует от безвредных до весьма вирулентных.
Для человека патогенны только С. albicans,
С. tropicalis, С. pseudotropicalis, С. krusei,
С. parakrusei, С. parapsilosis, С. guillermondi. Из
перечисленных дрожжеподобных грибов
С. albicans является наиболее патогенным.
Грибы кандида относятся к условно-патоген-
ным растительным микроорганизмам. Кроме сап-
рофитирования в окружающей среде на субстра-
тах живой и неживой природы, они довольно
часто выделяются с поверхности кожных покро-
вов и слизистых оболочек человека. Дрожжепо-
добные грибы кандида являются одноклеточны-
ми микроорганизмами. Молодые клетки имеют
округлую или яйцевидную форму, зрелые —
удлиненную или округлую. Диаметр клеток ко-
леблется в пределах от 1,5 до 10 мкм. Истинного
мицелия дрожжеподобные грибы не имеют, а
образуют псевдомицелий. Нити его отличаются
от истинного мицелия тем, что не имеют общей
оболочки и перегородок, а состоят из тонких
клеток. В местах сочленения псевдомицелия
дрожжеподобные грибы обладают способностью
отпочковывать бластоспоры, которые располага-
ются группами и образуют так называемые му-
товки, или вертициллы. Установлено наличие у
псевдомицелиальной формы гриба перфоратив-
ного органа, который способствует выраженно-
му повреждающему воздействию его на клетки
хозяина, т. е. формы агрессии (О. А. Голодова,
1982). Этим подтверждено, что в ходе инвазии
бластоспоры дрожжеподобных грибов кандида
трансформируются в псевдомицелий. Клетка
С. albicans имеет мощную шестислойную клеточ-
ную стенку, цитоплазматическую мембрану, ци-
топлазму с розетками гликогена и большим ко-
личеством рибосом, центральную и несколько
мелких вакуолей, ограниченных мембраной, ми-
тохондрии, крупное ядро, ограниченное ядерной
мембраной (Делекторский В. В. и соавт., 1979;
1980).
Дрожжеподобные грибы являются аэробами.
Наиболее благоприятная для роста гриба темпе-
ратура 21—37 °C, оптимальный pH 5,8—6,5. Луч-
шие среды для роста — картофельный и рисо-
вый агар, отвары кукурузы, моркови, а также
пивное сусло. Грибы рода кандида имеют тро-
пизм к тканям, богатым гликогеном. С. albicans
отличается ферментативной активностью, что
проявляется способностью продуцировать про-
теолитические и липолитические ферменты.
Установлено, что С. albicans стимулирует выра-
ботку в организме человека различных анти-
тел — комплементсвязывающих и флюоресци-
рующих агглютининов.
Эпидемиология
Подсчитано, что из 100 млн ежегодных визитов
к врачам по поводу вагинита около 20—25 %
обусловлены вульвовагинальным кандидозом.
Приблизительно 75 % женщин переносят в тече-
ние своей жизни по крайней мере один эпизод
вульвовагинального кандидоза и у 40—50 % из
них развивается по крайней мере один рецидив.
Заболеваемость вульвовагинальным кандидозом
в США насчитывает около 13 млн случаев в год,
что составляет около 10 % женского населения
страны. Генитальный кандидоз наиболее часто
регистрируется в странах с жарким климатом и
низкими санитарно-гигиеническими условиями
(Мексика, Аргентина, Ирак). Урогенитальный
кандидоз чаще встречается у женщин, реже — у
мужчин. Заболевание составляет до 40 % в струк-
туре инфекционной патологии нижнего отдела
гениталий (Прилепская В. Н., Байрамова Г. Р.,
533
534 Венерология
1995). В последние годы появились стертые и
атипичные формы заболевания, а также хрони-
ческие, резистентные к проводимой терапии слу-
чаи урогенитального кандидоза.
Возбудители урогенитального кандидоза распро-
страняются половым путем и при этом опреде-
ленное значение имеют различные факторы:
• физиологические состояния организма (воз-
раст; гормональный статус, например, беремен-
ность);
• наличие эндокринологического заболевания
(диабет, ожирение, синдром Кушинга, полиэн-
докринопатия, гипотиреоз);
• новообразования, инфекционные заболева-
ния, истощение;
• медикаментозные факторы: антибиотики,
кортикостероиды, цитостатики;
• оперативные вмешательства;
• применение гормональных контрацептивов.
Кандидозные грибы нередко попадают в мо-
чеполовые пути при сексуальных контактах
вместе с другими возбудителями половых инфек-
ций, в частности с хламидиями, уреплазмами,
вирусами. Урогенитальный кандидоз может воз-
никнуть и после лечения хламидиоза, микоплаз-
моза, трихомониаза, вирусных инфекций.
Патогенез
Патогенез урогенитального кандидоза довольно
сложен и не вполне изучен. Важная роль в па-
тогенезе этого заболевания отводится состоянию
макроорганизма, его восприимчивости к дрож-
жеподобным грибам и возможности мобилиза-
ции защитно-адаптационных механизмов. Гриб-
ковая инфекция чаще поражает женщин с
ослабленной в результате заболевания или про-
водимого лечения иммунной системой, при сни-
жении естественных защитных сил организма,
нарушениях нормальной бактериальной флоры
и гормонального фона.
Патогенез кандидоза зависит от многочислен-
ных экзогенных и эндогенных факторов (Анто-
ньев А. А. и соавт., 1985).
Экзогенные факторы:
• механическая и химическая травма;
• повышенная влажность и температура;
• лечение антибиотиками, препаратами группы
имидазола, кортикостероидами, цитостатиками;
• побочное действие оральных контрацепти-
вов, антидиабетических препаратов;
• патогенность и вирулентность штамма дрож-
жеподобного гриба.
Эндогенные факторы:
• приобретенная или врожденная иммунная
недостаточность;
• детский и старческий возраст;
• нарушение обмена веществ (углеводного,
белкового, жирового);
• эндокринные заболевания — сахарный диа-
бет, дисменорея, гипотиреоидизм, гипопаратире-
оидизм, гипо- и гиперкортицизм, гипофункция
яичников, полиэндокринная недостаточность;
железодефицитные состояния;
• гиповитаминозы (недостаток рибофлавина,
пиридоксина, никотиновой кислоты, аскорбино-
вой кислоты);
• общие тяжелые заболевания (острые и хро-
нические, инфекционные и неинфекционные),
ослабляющие защитные силы организма;
• заболевания желудочно-кишечного тракта
(дисбактериоз, ахилия, холециститы);
• беременность, заболевания женских поло-
вых органов (аднекситы, кольпиты, эндоцер-
вициты, лейкоплакия, крауроз, нарушения
функции яичников, неопластические заболе-
вания).
Клиническая картина
В настоящее время не существует общепринятой
клинической классификации урогенитального
кандидоза (Плахотная Г. А. и соавт., (1994). Ре-
комендуют различать следующие клинические
формы:
с указанием топического диагноза:
• кандидоносительство;
• острый урогенитальный кандидоз;
• хронический (рецидивирующий) урогени-
тальный кандидоз.
Под кандидоносительством подразумевают
(Антоньев А. А. и соавт., 1985):
• полное отсутствие клинических проявлений
кандидоза;
• постоянное обнаружение псевдомицелия в
биологических средах, в основном почкующих-
ся клеток дрожжеподобного гриба, в незначи-
тельном количестве и только изредка.
Кандидоносители служат причиной инфици-
рования половых партнеров урогенитальным
кандидозом, а также новорожденных в процес-
се родоразрешения.
Увеличение случаев кандидоносительства у жен-
щин обусловлено рядом эпидемиологических
факторов:
• частая передача дрожжеподобных грибов от
матери к ребенку при прохождении по половым
путям;
• возможность инфицирования половым пу-
тем;
• нередкая диссеминация кандидозной ин-
фекции из влагалища в другие органы.
Глава 46. Урогенитальный кандидоз
535
Урогенитальный кандидоз встречается чаще у
женщин, реже — у мужчин. При острой и по-
дострой формах кандидоза длительность заболе-
вания не превышает 2 мес. Воспалительные яв-
ления носят ярко выраженный характер:
гиперемия, отек, высыпания везикул на коже и
слизистых оболочках. При хронических формах
давность процесса составляет свыше 2 мес. На
кожных и слизистых покровах преобладают вто-
ричные элементы в виде инфильтрации, лихе-
низации, атрофичности тканей.
У женщин наиболее часто урогенитальный
кандидозный процесс локализуется в области
наружных половых органов и влагалища.
Кандидозные вульвит и вульвовагинит
Кандидозные вульвит и вульвовагинит без по-
ражения мочевыводящих путей характеризуют-
ся следующими симптомами:
• увеличение количества отделяемого из вла-
галища;
• необычно белый, с «молочными» бляшками
цвет отделяемого;
• зуд, чувство жжения или раздражения в
области наружных половых органов;
• усиление вульвовагинального зуда в тепле
(во время сна или после ванны);
• повышенная чувствительность слизистой к
воде и моче;
• усиление зуда и болезненных ощущений
после полового акта;
• неприятный запах, усиливающийся после
полового сношения.
Кандидозный вульвит
Кандидозный вульвит — поражение дрожжепо-
добными грибами больших половых и малых
половых губ, клитора, преддверия влагалища.
Выделяют следующие основные клинические
формы кандидозных вульвитов (Антоньев А. А.
и соавт., 1985):
• острый и подострый катарально-мембраноз-
ный (типичная форма);
• хронический катаральный (типичная фор-
ма);
• хронический лейкоплакиевидный (атипич-
ная форма);
• хронический краурозоподобный (атипичная
форма);
• хронический пруригинозный асимптомный
(атипичная форма);
• смешанный острый кандидозно-бактериаль-
ный.
Типичные проявления кандидозного вульви-
та характеризуются зудом или чувством жжения
19 Зак. 1357
различной интенсивности. Зуд обостряется в
предменструальный период и исчезает или
уменьшается в менструальную и постменструаль-
ную фазы цикла. Слизистые оболочки наруж-
ных половых органов умеренно или интенсив-
но гиперемированы с багрово-синюшным
оттенком. Возможны высыпания в виде мелких
везикул, которые, вскрываясь, образуют точеч-
ные или сливающиеся между собой эрозии с по-
лицикличными краями и отторгшимся эпидер-
мисом по периферии. Между малыми половыми
губами могут быть наслоения белых крошкова-
тых творожистых пленок. При хроническом те-
чении кандидозного процесса слизистые оболоч-
ки наружных половых органов находятся в
состоянии застойной гиперемии и инфильтра-
ции. Нередко отмечаются лихенификация и су-
хость больших и малых половых губ, имеющих
белесоватую поверхность.
Атипичные формы кандидозного вульвита
диагностируются редко и, как правило, имеют
хроническое течение.
У некоторых больных кандидозный вульвит
распространяется на кожу пахово-бедренных
складок и перианальной области. Этот процесс
наиболее часто наблюдается у больных хроничес-
ким кандидозом, протекающим на фоне диабе-
та, микседемы, гипофункции яичников.
Кандидозный вагинит
Кандидозный вагинит — микотическое пораже-
ние не только слизистых оболочек влагалища, но
и влагалищной части шейки матки. Различают
четыре основные формы кандидозного вагини-
та (Антоньев А. А. и соавт., 1985):
• острый и подострый катарально-экссудатив-
но-мембранозный кандидозный;
• острый и подострый смешанный катараль-
но-экссудативный кандидозно-бактериальный;
• подострый и хронический катаральный (или
мембранозный) кандидозный;
• хронический асимптомный кандидозный
вагинит.
Кандидозный вагинит характеризуется пятни-
стой краснотой слизистой оболочки влагалища с
беловатым густым сметанообразным отделяемым,
а иногда крошковатыми налетами, рыхло сидя-
щими на гиперемированной слизистой оболочке.
Нередко имеются обширные эрозии влагалищной
части шейки матки, эрозированный эктропион и
точечные эрозии на стенках и сводах влагалища.
Бели творожистые, обильные, жидкие или слив-
кообразные, часто пенистые и даже гнойные с
неприятным запахом. Больные ощущают зуд,
жжение и небольшую болезненность.
536 Венерология
Кандидозный эндоиервииит
Кандидозный эндоцервицит как самостоятельное
заболевание никогда не наблюдается, а только
совместно с кандидозным вагинитом. В резуль-
тате влагалищного кандидоза дрожжеподобные
грибы по протяжению проникают в отверстие
шейки матки и в глубь канала. Заболевание на-
блюдается у женщин с неполноценной функци-
ей яичников. При типичной форме эндоцерви-
цита в глубине канала шейки матки имеется
большое количество белых крошковатых комоч-
ков или белое густое слизистое отделяемое. При
малосимптомной форме кандидозного эндоцер-
вицита обнаруживается слизистая пробка, свиса-
ющая на заднюю губу отверстия матки. Возможно
очень скудное молочно-слизистое отделяемое.
Кандидозный уретрит и цистит
Кандидозный уретрит и цистит у женщин ди-
агностируется сравнительно редко, что обуслов-
лено отсутствием патогномоничной симптомати-
ки. При кандидозном уретрите наблюдаются
дизурические расстройства: рези, жжение, болез-
ненность при мочеиспускании, учащенные по-
зывы. Губки уретры отечны и гиперемированы,
у наружного отверстия губок расположены тво-
рожистые крошковатые комочки. При остром
цистите больных беспокоят постоянные тупые
боли в нижней части живота, болезненность при
мочеиспускании, неприятные ложные позывы.
В периоды ремиссий хронического цистита или
уретрита симптомы могут быть весьма незначи-
тельными либо отсутствовать.
У мужчин наиболее частым признаком уроге-
нитального кандидоза является воспаление го-
ловки полового члена (баланит) и внутреннего
листка крайней плоти (постит). Обычно эти
воспаления сочетаются. Баланопостит развива-
ется преимущественно у лиц с узкой и длинной
крайней плотью и особенно часто при несоблю-
дении правил гигиены. Кандидозные баланопо-
ститы полового происхождения у мужчин рез-
ко возросли вследствие роста кандидозов
мочеполовой системы у женщин, что связано с
широким применением антибиотиков, противо-
трихомонадных и гормональных противозачаточ-
ных препаратов. Почти во всех современных ру-
ководствах по венерологии кандидозные
баланопоститы рассматриваются в группе инфек-
ций, передаваемых половым путем.
Кандидозный баланопостит
Кандидозный баланопостит проявляется зудом и
жжением в области головки полового члена.
Клиническая картина заболевания довольно ва-
риабельна. Кандидозный баланопостит может
протекать в эрозивной, мембранозной и эритем-
но-пустулезной формах. Чаще всего на головке
полового члена в области венечной борозды и на
внутреннем листке крайней плоти имеются ги-
перемия кожи, легкая отечность и инфильтра-
ция. На пораженных участках роговой слой ма-
церирован, влажный, покрыт белесовато-серым
налетом, после удаления которого обнажается
эрозивная поверхность красного цвета. На го-
ловке полового члена могут располагаться мел-
кие поверхностные эрозии. Наружное отверстие
уретры становится отечным, краснеет, однако
симптомов уретрита не имеется. На внутреннем
листке крайней плоти очаги поражения покры-
ваются белесоватым псевдомембранозным нале-
том. Крайняя плоть полового члена инфильтри-
рована, обнажение головки затруднено. По краю
препуциального мешка возможно появление
трещин. Из препуциального мешка отделяется
небольшое количество жидкого экссудата, иног-
да гнойного характера - при присоединении
вторичной пиококковой инфекции. При хрони-
ческом течении кандидозного баланопостита
вследствие появления трещин по краю препуци-
ального мешка и последующего рубцевания мо-
жет произойти постепенное рубцовое сужение
крайней плоти полового члена с образованием
стойкого приобретенного фимоза. Возможны и
другие осложнения, такие как гангренизация,
рожистое воспаление полового члена, лимфан-
гиты, лимфадениты, бородавчатые разрастания,
язвы с вегетациями, кандидагранулемы, карци-
номы полового члена.
Кандидозный уретрит
Кандидозный уретрит встречается реже, чем ба-
ланопостит. Заболевание обычно начинается
подостро, течение его торпидное. Инкубацион-
ный период длится в среднем 10—20 дней, иногда
до 2 мес. Выделениям из уретры предшествуют
парестезии в виде легкого зуда или жжения.
Выделения изредка бывают обильными, гнойны-
ми, но чаще скудные, слизистые или водянис-
тые. Для кандидозного уретрита характерно об-
разование белесоватых налетов в виде
творожистых пятен на слизистой облочке моче-
испускательного канала. Уретроскопическая кар-
тина кандидозного уретрита весьма типична:
видны диффузные и ограниченные белесовато-
серые налеты — как при молочнице слизистых
оболочек полости рта. После удаления налетов
обнажается отечная и резко гиперемированная
слизистая оболочка уретры. Кандидозный урет-
Глава 46. Урогенитальный кандидоз 537
рит может осложниться простатитом, эпидиди-
митом, циститом, особенно при смешанной ин-
фекции, обусловленной грибами и другими па-
тогенными микроорганизмами.
При сочетании определенных факторов риска
(недоношенность; искусственное питание; приме-
нение антибиотиков, глюкокортикоидов; упот-
ребление наркотиков; химиотерапия; хирургичес-
кие вмешательства, послеоперационный период;
повреждения кожи и слизистых) в патологичес-
кий процесс вовлекаются различные органы и
наступает генерализация кандидозного процесса.
Генерализованный кандидоз отличается торпид-
ностью течения. Отмечается выраженная лихорад-
ка, реакция со стороны центральной нервной
системы, дисфункция желудочно-кишечного
тракта, изменения со стороны периферической
крови (лейкоцитоз, нейтрофилез со сдвигом вле-
во), изменения гомеостаза.
Диагноз
Принимая во внимание возможность кандидо-
носительства, диагноз урогенитального кандидо-
за устанавливается при наличии клинических
проявлений заболевания и микроскопического
выявления грибов рода Candida с преобладани-
ем вегетирующих форм (почкование, псевдоми-
целий). Наиболее достоверным методом лабора-
торного исследования является культуральный с
количественным учетом колоний грибов. Обна-
ружение грибов при отсутствии симптомов не
является показанием к назначению лечения, так
как они периодически могут выявляться и у
здоровых женщин.
Лабораторная диагностика урогенитального
кандидоза включает следующие виды исследо-
ваний: микроскопия, культуральная диагностика,
серологические реакции, иммунологические мето-
ды, иммуноферментный анализ, полимеразная
цепная реакция.
Микроскопические исследования
Микроскопические исследования являются наи-
более доступными. Материалом для микроско-
пических исследований служат характерные бе-
лесоватые пленки и крошковатый налет с
различных участков слизистой оболочки и кожи
мочеполовых органов (чешуйки кожи, обрывки
эпителия с поверхности губ и слизистых оболо-
чек рта, белые пробки из миндалин глотки, на-
ружных половых органов, стенки влагалища и
шейки матки, соскоб со слизистой уретры или
отделяемое из уретры, осадок мочи). Микроско-
пию проводят сначала при малом увеличении
(хЮО), при котором можно увидеть скопления
дрожжеподобных грибов и псевдомицелия. Об-
наружение псевдомицелия при микроскопии па-
тологического материала является важным под-
тверждением для лабораторного заключения о
дрожжеподобной природе возбудителя. Затем
переходят на большое увеличение (х500), позво-
ляющее видеть отдельные клетки дрожжеподоб-
ных грибов, их форму, расположение. Дрожже-
подобные грибы хорошо окрашиваются
анилиновыми красителями. Применяются такие
методы, как Романовского-Гимза, Грама, Бого-
лепова, Циль-Нильсена. При окраске по Граму
грибы темно-фиолетовые или окрашены нерав-
номерно: периферические слои клетки фиолето-
вые, а в середине — розовые. Окрашенные по
Циль-Нильсену грибы имеют синий цвет с ро-
зово-желтой зернистостью, включениями липи-
дов. В мазках, окрашенных по Романовскому-
Гимза, видны розовато-фиолетовые клетки
дрожжеподобных грибов, красные хроматиновые
образования — темно-фиолетовый волютин.
Наряду с обычными методами микроскопии,
можно использовать люминесцентный метод, по-
зволяющий проводить предварительную иденти-
фикацию и количественный учет грибов. Мазки
изучают при люминесцентном свечении, и степень
свечения микробов оценивают по четырехплюсо-
вой системе. При сопоставлении количества вы-
росших колоний со степенью свечения микробов
установлено, что яркое сияющее свечение перифе-
рии клетки (++++) отмечается у больных с ко-
личеством выделенных колоний от 5000 до 10 000,
умеренное свечение (++) наблюдалось при числе
выросших колоний, равном 1000—3000. Отдельные
светящиеся клетки обнаружены при наличии 100—
250 клеток в поле зрения.
Культуральная диагностика
Культуральная диагностика играет решающее
значение в постановке диагноза урогенитальных
кандидозов. Обнаружение гриба в исследуемом
материале не всегда имеет абсолютное значение.
Необходима оценка по количественным и каче-
ственным показателям (степень обсемененности,
морфологические, биохимические признаки) с
учетом патологического материала и сопоставле-
нием клинической картины заболевания. Для
получения культур Candida применяют твердые
и жидкие питательные среды с углеводами.
Идентификация полученных культур дрожже-
подобных грибов основывается на изучении
морфологических признаков бактериальных
клеток и внешнего вида выросших колоний.
Характерными для каждого вида грибов являют-
ся гигантские колонии.
538 Венерология
Серологические реакции
Для диагностики кандидозной инфекции ис-
пользуются различные серологические реакции:
РА — реакция агглюцинации, РСК — реакция
связывания комплемента, РП — реакция преци-
питации, РИГА — реакция пассивной гемагглю-
цинации. Из комплекса серологических иссле-
дований наиболее существенное значение имеет
РСК с дрожжевыми антигенами. У большинства
больных РСК с дрожжевыми антигенами быва-
ет положительной с небольшим титром антител
(1: 10, 1: 16). Реакция агглюцинации более спе-
цифична при использовании аутоштамма —
культуры гриба, выделенного от больного. Ди-
агностическим титром РА при кандидозах счи-
тается разведение сыворотки более чем 1: 100.
Иммунологические методы
Для исследования состояния клеточного иммуни-
тета при кандидозах применяются реакции с гриб-
ковыми антигенами in vitro: ППН — показатель
повреждаемости нейтрофилов, РБТЛ — реакция
бласттрансформации, реакция Иерне-Клемпарс-
кой — аутоиммунного локального гемолиза или
бляшкообразующих клеток, тест Шелли — моди-
фикация теста дегрануляции базофилов.
Иммуноферментный анализ
Методом ИФА определяют IgE-антитела против
Candida albicans во влагалищных смывах у жен-
щин. Наиболее высоким является титр IgE-ан-
тител к серотипу А, который составляет 83 %
всех штаммов С. albicans.
Полимеразная цепная реакция
С помощью геномной дактилоскопии ДНК в
полимеразной цепной реакции проводится иден-
тификация клинических штаммов Candida
albicans. Метод ПЦР может применяться для
эпидемиологического изучения большого числа
патогенных штаммов.
При постановке диагноза урогенитального
кандидоза микроскопические, культуральные,
серологические, иммунологические, иммунофер-
ментные исследования играют роль только в
комплексе с клинической картиной заболевания.
Лечение
Терапия больных урогенитальным кандидозом
должна быть комплексной, поэтапной, включая
не только этиотропное лечение, но и ликвида-
цию предрасполагающих факторов, а также ле-
чение сопутствующих заболеваний. При комп-
лексном подходе достигается не только
излечение эпизода урогенитального кандидоза,
но и снижение числа рецидивов заболевания. Те-
рапия сопутствующих заболеваний имеет боль-
шое значение в лечении урогенитального канди-
доза. Одним из основных условий на период
лечения является, по возможности, отмена кор-
тикостероидов, цитостатиков, гормональных
эстроген-гестагенных препаратов, антибиотиков;
отказ от вредных привычек.
Лечение острого урогенитального кандидоза
Для лечения острых форм заболевания обычно
применяется местное лечение.
При лечении урогенитального кандидоза исполь-
зуют следующие препараты:
1. Препараты группы имидазола
Миконазол:
Гино-дактанол 0,2 г (вагинальные таблетки);
Гино-дактарин 0,1 г (вагинальные свечи).
Таблетки или свечи вводят в задний свод вла-
галища в положении лежа на спине по 1 шт. на
ночь однократно.
Эконазол:
Гино-певарил 50 (крем 1 %, гранулы 0,05 г;
вагинальные свечи 0,15). Препарат вводится в
задний свод влагалища один раз в сутки на ночь
в течение 2 недель.
Гино-певарил 150 (вагинальные свечи 0,15 г).
Свечи вводят во влагалище один раз в сутки на
ночь в течение 3 дней.
Изоконазол:
Гино-травоген 0,6 г (вагинальные шарики).
Препарат вводится в задний свод влагалища на
ночь.
Травоген 0,01 г (крем). Наносится на пора-
женные участки один раз в сутки до исчезнове-
ния симптомов.
Клотримазол:
Канестен (вагинальные таблетки 0,1 г; мазь
1 %);
Канестен 1 (вагинальные таблетки 0,5 г; мазь
10 %);
Канестен 3 (вагинальные таблетки 0,2 г; мазь
2 %). Курс лечения канестеном 6 дней.
Клотримазол (вагинальные таблетки 0,1 и
0,2 г; крем 1 %, мазь 1 %, раствор 1 %, вагиналь-
ный крем 2 %). Крем, мазь и раствор наносят
тонким слоем на пораженные участки кожи и
слизистых 2—3 раза в сутки и осторожно втира-
ют. Курс лечения в среднем составляет 14 дней.
Вагинальный крем вводится в задний свод вла-
галища по 0,5 г 1 раз в сутки на ночь в течение
3 дней. Вагинальные таблетки вводят 1—2 раза
в сутки в течение 6 дней (целесообразно их при-
менение сочетать с обработкой кремом, мазью и
раствором).
Глава 46. Урогенитальный кандидоз
539
Бутоконазол:
2 % крем «Гинофорт» по 5,0 г интравагиналь-
но однократно.
Тиоконазол:
6,5 % мазь по 5,0 г интравагинально на ночь
однократно.
Терконазол:
Таблетки вагинальные по 80 мг на ночь 1 раз
в сутки в течение 3 дней или 0,4 % крем (5,0 г)
1 раз в сутки на ночь в течение 7 дней.
2. Противогрибковые антибиотики
Натамицин:
Пимафуцин: вагинальные таблетки 0,025 г; ва-
гинальные свечи 0,1 г; раствор во флаконах по
20,0 мл (в 1 мл раствора содержится 0,025 г ната-
мицина), крем в тубах по 30 г (в 1 г крема —
0,02 г натамицина). Крем и раствор наносят на
пораженную поверхность 1—4 раза в сутки. Ваги-
нальные таблетки применяются ежедневно на ночь
или по 1 таблетке 2 раза в сутки в течение 10 дней.
Нистатин:
Мазь по 100 000 ЕД нистатина в 1 г. Наносится
на пораженную поверхность 2 раза в сутки. Курс
лечения от 1 до 2—4 недель. Свечи вагинальные
и ректальные: применяются 2 раза в сутки в
течение 7 дней.
Леворин:
Мазь 500 000 ЕД леворина в 1 г. Применяет-
ся 2 раза в день в течение 1—4 недель.
Амфотерицин:
Мазь, содержащая 30 000 ЕД амфотерицина в
1 г. Применяется 2—3 раза в сутки в течение 1—
2 недель.
Лечение острого вульвовагинального
кандидоза у небеременных женшин
Пероральная терапия острого вульвовагинально-
го кандидоза у небеременных женщин произво-
дится с использованием препаратов;
Итраконазол:
Орунгал внутрь по 200 мг 1 раз в день в те-
чение 3 дней.
Кетоконазол:
Низорал внутрь по 200 мг 2 раза в день в те-
чение 5 дней.
Флуконазол:
Дифлюкан, микосист по 150 мг перорально
однократно.
Лечение хронического
урогенитального кандидоза
При хроническом урогенитальном кандидозе,
наряду с местным лечением, применяется один
из препаратов общего действия.
1. Препараты группы имидазола
Итраконазол:
Орунгал (капсулы по 0,1 г) по 0,2 г перорально
однократно или 0,2 г в сутки в течение 3 дней.
Кетоконазол:
Низорал, ороназол (таблетки 0,2 г). По 1 таб-
летке 2 раза в сутки во время еды в течение 10
дней.
Флуконазол:
Дифлюкан, микосист (капсулы 0,05 г; 0,15 г;
0,1 г; 0,2 г; раствор для внутривенного введе-
ния). Применяется однократно в дозе 150 мг.
2. Противогрибковые антибиотики
Натамицин:
Пимафуцин 0,1 г (таблетки, растворяющиеся
в кишечнике). По 1 таблетке 4 раза в сутки в
течение 7—12 дней. Таблетки следует сочетать с
применением местных лекарственных форм пи-
мафуцина.
Нистатин:
Таблетки 250 000 ЕД, 500 000 ЕД. По 500 000-
1 000 000 ЕД 3-4 раза в сутки.
Леворин:
Таблетки 500 000 ЕД. Применяют по 500 000
ЕД 3—4 раза в сутки в течение 14 дней.
Амфоглюкамин:
Таблетки 100 000 ЕД. Применяют по 200 000 ЕД
2 раза в сутки через 30—40 мин после еды в тече-
ние 10—14 дней.
Одновременно с этиотропной терапией уроге-
нитального кандидоза проводят лечение фоно-
вого заболевания и по показаниям присоединя-
ют препараты, стимулирующие резистентность
организма.
Для лечения беременных предпочтительно ис-
пользовать местную терапию. Рекомендуемые
препараты: клотримазол, миконазол, изоконазол,
натамицин.
Лечение урогенитального кандидоза у детей
Натамицин:
Пимафуцин (раствор для местного примене-
ния во флаконах по 20,0 мл; 1 мл содержит
0,025 мг натамицина. При вагинитах у детей
наносят 0,5—1,0 мл препарата 1 раз в сутки до
исчезновения симптомов. Пероральные формы
применяются по 1/2 таблетки (0,05 г) 2—4 раза
в сутки.
Кетоконазол:
Низорал, ороназол (таблетки 0,2 г). Принима-
ют во время еды 2 раза в сутки из расчета 4-
8 мг/кг массы тела (при массе тела свыше 30 кг
назначают в тех же дозах, что и взрослым).
540 Венерология
Флуконазол:
Дифлюкан, микосист. Назначают детям стар-
ше 1 года из расчета 1—2 мг/кг массы тела в
сутки.
Критериями излеченности урогенитального
кандидоза являются: исчезновение клинических
проявлений заболевания, отрицательные резуль-
таты микробиологического исследования.
Сроки наблюдения устанавливаются индиви-
дуально в зависимости от длительности, харак-
тера клинических проявлений, распространенно-
сти урогенитального кандидоза. Следует
принимать во внимание возможность хроничес-
кого рецидивирующего характера заболевания,
реинфекции, кандидоносительства, а также со-
хранения факторов, способствующих развитию
заболевания. Больных урогенитальным кандидо-
зом следует информировать о том, что их поло-
вым партнерам рекомендуется обследование и,
при необходимости, лечение. Больным рекомен-
дуется воздерживаться от половой жизни до из-
лечения или применять барьерные методы пре-
дохранения.
Профилактика
Профилактика урогенитального кандидоза состо-
ит в соблюдении правил личной гигиены, эф-
фективном лечении больных и уменьшении
контактов с инфицированными пациентами пос-
ле их излечения.
Урогенитальный кандидоз вызывает не толь-
ко местные клинические проявления, но и ге-
нерализацию процесса, а также приводит к ряду
серьезных осложнений.
Для предупреждения развития заболевания
рекомендуются следующие профилактические
мероприятия (Столярова Л. Г., 1995):
• сокращение возможностей инфицирования
(особенно новорожденных) грибами рода кан-
дида;
• настороженность в отношении возможного
кандидоза у больных (особенно в детском и
пожилом возрасте) со сниженной иммунологи-
ческой реактивностью организма;
• отказ от нерациональной и массивной анти-
биотикотерапии, осторожное и только по пока-
заниям применение гормонотерапии;
• применение наиболее эффективных и совре-
менных средств антифунгальной терапии;
• своевременное выявление дисбактериоза и
принятие мер к нормализации микрофлоры в
естественных местах ее обитания в организме
человека;
• разработка и выполнение санитарно-гигие-
нических условий для некоторых медицинских
учреждений, предприятий пищевой промышлен-
ности и производств, связанных с возможнос-
тью заражения грибами кандида и со стимуля-
цией их роста;
• широкая санитарно-просветительная работа
среди врачей разных специальностей, медицин-
ского персонала лечебно-профилактических уч-
реждений, работников пищевых и других пред-
приятий, а также среди населения.
Глава 47
Генитальный герпес
Генитальный герпес — распространенное вирус-
ное заболевание мочеполовых органов, вызыва-
емое вирусом простого герпеса.
Герпес является одной из самых распространен-
ных вирусных инфекций человека и представляет
собой серьезную медико-социальную проблему.
Свыше 90 % людей земного шара инфицирова-
но вирусом простого герпеса (ВПГ) и до 20 % из
них имеют те или иные клинические проявления
инфекции. Обладая нейродермотропизмом, ВПГ
поражает кожу и слизистые оболочки (чаще все-
го на лице и в области половых органов), цент-
ральную нервную систему (менингиты, энцефа-
литы), глаза (конъюнктивиты, кератиты). ВПГ
обусловливает патологию беременности и родов,
нередко приводя к «спонтанным» абортам и ги-
бели плода или вызывая генерализованную ин-
фекцию у новорожденных. Отмечается связь ге-
нитального герпеса с неопплазиями шейки матки
и предстательной железы.
Этиология
Генитальный герпес вызывают два серотипа ви-
руса простого герпеса: ВПГ-1 и ВПГ-2; наиболее
часто ВПГ-2. В начале 60-х годов было установ-
лено, что серологические свойства вирусов различ-
ны. Геном вируса простого герпеса представлен
двуспиральной линейной ДНК, длины которой
достаточно для того, чтобы закодировать еще 60—
70 генных «продуктов» сверх обычной нормы.
Структура генома герпесвирусов отлична от тако-
вой других ДНК-содержащих вирусов: два нук-
леотида, расположенные в специфической после-
довательности, с обеих сторон окружены теми же
нуклеотидами, но расположенными в инвертиро-
ванной последовательности. Эти два компонента
могут меняться местами по отношению друг к
другу, так что ДНК, изолированная от вируса,
состоит из четырех изомеров, различающихся ори-
ентацией двух компонентов. Геномы ВПГ-1 и
ВПГ-2 приблизительно на 50 % гомологичны. Го-
мологичные участки распределены по всей карте
гена. Большинство полипептидов, специфичных
для одного типа вирусов, антигенно связаны с по-
липептидами другого типа. Вирусный геном упа-
кован в капсид правильной икосаэдрической фор-
мы, состоящий из 162 капсомеров. С наружной по-
верхности вирус покрыт липидсодержащей оболоч-
кой, представляющей собой модифицированную
клеточную мембрану, сформировавшуюся при
проникновении ДНК-содержащего капсида
сквозь внутреннюю ядерную мембрану клетки-
хозяина. Между капсидом и двухслойной липид-
ной оболочкой находится еще одна оболочка, со-
стоящая из ряда вирусных белков.
Репликация вируса состоит из ядерной и ци-
топлазматической фаз. Начальные этапы реп-
ликации включают присоединение, слияние
оболочки вируса с мембраной клетки с после-
дующим высвобождением нуклеокапсида в ци-
топлазму клетки, распад нуклеокапсида и вы-
ход вирусной ДНК. После репликации
вирусного генома и синтеза структурных бел-
ков в ядре клетки происходит сборка нуклео-
капсидов. По мере проникновения нуклеокап-
сидов из ядра через внутреннюю ядерную
мембрану в перинуклеарное пространство они
покрываются оболочкой. В некоторых клетках
при репликации вируса в ядре формируются
тельца-включения двух типов: базофильные
тельца типа А, Фельген-положительные, содер-
жат вирусную ДНК; и эозинофильное тельце-
включение, свободное от вирусной нуклеиновой
кислоты или белка, представляющее собой
«шрам» вирусной инфекции. Затем вирионы
транспортируются по эндоплазматической сети
и пластинчатому комплексу к клеточной повер-
хности.
Патогенез
ВПГ, как и некоторые другие герпесвирусы че-
ловека, способен к латентному существованию
с последующей реактивацией, которая может
служить причиной повторяющихся рецидивов
болезни или бессимптомного выделения вируса.
ВПГ внедряется в организм через слизистые
оболочки и кожу и затем быстро захватывается
сенсорными нервными окончаниями при помо-
щи специальных вирусных рецепторов. После
внедрения в аксон нерва, он переносится цент-
541
542 Венерология
ростремительно в нервные клетки дорзальных
корешков ганглиев, где сохраняется в латентном
состоянии. Механизм латентного состояния ви-
руса и его последующей реактивации еще не
вполне ясен. Предполагают, что репликация
вируса блокируется генетическими механизма-
ми, а реактивация, вероятно, достигается устра-
нением этого блокирования.
Латентность и нейровирулентность ВПГ-ин-
фекции — это основные свойства, определяю-
щие патогенез ВПГ-инфекции. Разнообразные
проявления латентности, реактивации и вирулен-
тности у различных пациентов, возможно, оп-
ределяются как свойствами самого вируса, так
и характеристиками хозяина. Латентность — это
свойство вируса персистировать в организме
инфицированного хозяина в отсутствие вирус-
ной репликации и патогенности. Это позволяет
вирусу избегать воздействия иммунного ответа
хозяина. Реактивация может приводить к кли-
нически бессимптомному выделению вируса,
возникновению поражений кожи и слизистых,
или появлению редких неврологических симп-
томов. Среди вирусных факторов, принимающих
участие в реактивации, существенную роль иг-
рает тимидинкиназа, поскольку этот фермент
необходим для репликации вирусной ДНК в
постмитотических нейронах.
Нейроинвазия — это способность вируса при-
обретать доступ из периферических нервных
клеток в ЦНС, а нейровирулентность — способ-
ность вируса реплицироваться в ЦНС и вызы-
вать неврологические заболевания.
Герпес оказывает разрушительное действие на
иммунную систему больного и является причи-
ной вторичного иммунодефицита. Рецидивиру-
ющий генитальный герпес, нарушая нормальную
половую жизнь пациентов, нередко вызывает
нервно-психические расстройства. Тропизм ВПГ
к эпителиальным и нервным клеткам объясня-
ет полиморфизм клинических проявлений гер-
петической инфекции. Поражение кожи и сли-
зистых оболочек — самое распространенное
клиническое проявление болезни, а поражение
ВПГ гениталий относится к наиболее частым за-
болеваниям, передаваемым половым путем.
Эпидемиология
Частота генитального герпеса значительно уве-
личилась в последние годы. По данным амери-
канских исследователей, 30 млн взрослых в США
страдают рецидивирующим генитальным герпе-
сом и каждый год регистрируется еще пример-
но 500 000 новых случаев инфицирования. Та-
ким образом, у каждого пятого жителя США
(20 %) обнаружены серологические признаки
предшествующей инфекции, вызванной вирусом.
Но только у 5 % населения имеются клиничес-
кие проявления, характерные для генитального
герпеса, в остальных случаях он протекает суб-
клинически или бессимптомно. Генитальный
герпес встречается часто в развивающихся стра-
нах и, по-видимому, не всегда регистрируется,
так как многие считают эту инфекцию триви-
альной. Заболеваемость генитальным герпесом
особенно важна для некоторых стран Африки и
Азии, где заболевания половых органов, сопро-
вождающиеся изъязвлениями, вероятно, являют-
ся одним из главных факторов распространения
ВИЧ-инфекции.
Процент инфекций, вызванных двумя типа-
ми вируса, варьирует от 1 до 86, возможно, от-
ражая разницу в популярности орального секса
в различных группах населения. Имеются све-
дения о том, что с ростом осведомленности о
якобы безопасном оральном сексе в определен-
ных популяциях люди применяют его без пре-
зервативов и, как следствие этого, возрастает
процент генитальных инфекций, вызываемых
ВПГ-2 (A. Scoular и соавт., 1990; J. D. С. Ross и
соавт., 1993).
Заболевание передается преимущественно при
сексуальных контактах от больного генитальным
герпесом или носителем ВПГ. Контакт может
быть генито-генитальным, орально-генитальным,
генитально-анальным или орально-анальным.
Заражение может происходить в том случае, ког-
да у партнера, являющегося источником инфек-
ции, рецидив болезни или, что особенно важно,
когда он выделяет вирус, не имея при этом кли-
нических симптомов. Нередко генитальный гер-
пес передается от лиц, не имеющих симптомов
заболевания на момент полового акта или даже
не знающих о том, что они инфицированы
(G. J. Mertz и соавт., 1992; Y. Bryson и соавт.,
1993). Не исключена возможность инфицирова-
ния гениталий при орально-генитальном контак-
те при наличии у партнера герпеса лица, а также
бытовым путем через средства личной гигиены.
Клиническая картина
При определении вида генитального герпеса не-
обходимо учитывать следующие факторы: тип
вируса; инфицирование раньше ВПГ-1 и/или
ВПГ-2; течение инфекции.
Вилы герпетической инфекции
Выделяют следующие виды герпетической ин-
фекции: первичная, непервичная, рецидивиру-
ющая.
Глава 47. Генитальный герпес 543
Первичная инфекция
Первичная инфекция, когда человек впервые
сталкивается с вирусом простого герпеса и не
обладает антителами к нему. Она может сопро-
вождаться симптомами или протекать бессимп-
томно, как это происходит в большинстве слу-
чаев (65 %). Клинические проявления первичной
инфекции длятся 18—22 дня с нарастанием сим-
птоматики в течение первой недели. Заболева-
ние характеризуется длительным выделением
вируса, генитальными и экстрагенитальными
поражениями, сопровождается общими и мест-
ными симптомами, которые проявляются на 2-
14 день. К общим симптомам относятся лихо-
радка, головная боль, недомогание и миалгии. К
местным — зуд, боль, дизурия, выделения из
влагалища, развитие поражений на коже и сли-
зистых, паховая лимфаденопатия. Симптомати-
ка разрешается на 2—3-й неделе. Бессимптомное
слущивание поврежденного вирусом эпителия
наблюдается в течение последующих 2 недель.
Иногда возникают осложнения, требующие гос-
питализации — крестцовая радикулопатия, тя-
желый асептический менингит или диссемини-
рованная инфекция. Таким образом, общая
продолжительность проявлений составляет почти
6 недель.
Непервичная инфекция
Непервичная инфекция наблюдается у пациен-
тов, уже имеющих антитела к одному из типов
ВПГ. Чаще встречается вариант непервичного
генитального герпеса, когда человек, уже име-
ющий антитела к ВПГ-1, впервые инфицирует-
ся ВПГ-2. Непервичная генитальная инфекция
протекает менее выражено: длительность клини-
ческих проявлений приблизительно 15 дней.
Реже наблюдается бессимптомное выделение
вируса, а продолжительность выделения умень-
шается до 8 дней.
Рецидивирующая инфекция
Рецидивирующая инфекция диагностируется у
пациентов, имеющих одновременно симптомы
генитального герпеса и антитела к реактивиро-
ванному типу вируса. Течение ее может быть
бессимптомным, и тогда постановка диагноза
возможна только при использовании очень тон-
ких культуральных или типоспецифических се-
рологических методов исследования. Тяжесть и
продолжительность клинических проявлений
при рецидивах еще менее выражена, чем при
других формах генитальной инфекции, а дли-
тельность слущивания поврежденного эпителия
составляет всего 4 дня. Примерно у 50 % боль-
ных наблюдаются продромальные признаки,
такие как зуд, боль, дизурия и болезненность
лимфатических узлов (L. Corey, 1990). Рециди-
вы при ВПГ-2 проявляются раньше и чаще, чем
при ВПГ-1. Частота и характер обострений у
разных пациентов значительно варьируют и по-
разному влияют на жизнь.
Факторы, способствующие проявлению и/или
рецидивированию генитального герпеса: сниже-
ние иммунологической реактивности; переох-
лаждение и перегрев организма; интеркуррент-
ные заболевания; некоторые психические и
физиологические состояния; медицинские мани-
пуляции (аборты и введение внутриматочной
спирали).
У 10-20 % от общего числа инфицированных
заболевание характеризуется определенными
клиническими проявлениями, способными реци-
дивировать. Первое проявление герпесвирусной
инфекции, как правило, протекает более бурно,
чем последующие рецидивы. У части больных
они непродолжительны. У большинства инфи-
цированных лиц клинические проявления гени-
тального герпеса отсутствуют.
В продромальном периоде больные отмечают
зуд, жжение или боль, затем появляются высы-
пания в виде отдельных или сгруппированных
везикулезных элементов величиной 2—3 мм на
эритематозном основании, имеющих тенденцию
к возвратным проявлениям на том же месте.
Высыпания могут сопровождаться нарушением
общего состояния: недомоганием, головной бо-
лью, субфебрильной температурой, нарушением
сна. В дальнейшем везикулы вскрываются с
образованием эрозивной поверхности, имеющей
полицилиндрические очертания.
Локализация
Характерная локализация генитального герпеса
у женщин: малые и большие половые губы, вуль-
ва, клитор, влагалище, шейка матки; у мужчин:
головка полового члена^ крайняя плоть, уретра.
Клинические формы
Генитальный герпес характеризуется периодичес-
ким появлением на коже и слизистых оболоч-
ках очагов поражения с различной степенью
выраженности и активным выделением ВПГ, что
проявляется в виде|различных клинических
форм: манифестная, атипичная, абортивная, суб-
клиническая. -
Манифестная фбфма /
Для м^ифест^ой фррк^ы рецидивирующего ге-
нитального^герпеса характерно типичное разви-
тие герпетических элементов в очаге поражения.
544 Венерология
Постоянными признаками заболевания являются
пузырьки, эрозии, язвочки, экссудация, реци-
дивирующий характер болезни. Больные гени-
тальным герпесом часто жалуются на недомога-
ние, головную боль, иногда на субфебрильную
температуру, нарушение сна, нервозность. Обыч-
но в начале заболевания отмечается чувство
жжения, зуда и боли в области гениталий. По-
раженное место слегка отекает, краснеет, затем
на гиперемированном основании появляется
группа мелких пузырьков величиной 2—3 мм.
Прозрачное содержимое пузырьков впоследствии
мутнеет, становится гнойным. Через несколько
дней пузырьки вскрываются и начинают засы-
хать в тонкие корочки, которые по мере эпите-
лизации очагов поражения отпадают, оставляя
временную гиперемию или вторичную пигмен-
тацию на месте бывших высыпаний.
Атипичная форма
Среди вариантов атипичной формы рецидиви-
рующего генитального герпеса у женщин встре-
чаются отечная и зудящая формы. Очаг пораже-
ния может быть представлен глубокими
рецидивирующими трещинами слизистой обо-
лочки вульвы и подлежащих тканей малых и
больших половых губ. Трещины самостоятель-
но эпителизируются в течение 4—5 дней.
Абортивная форма
Абортивная форма генитального герпеса обыч-
но встречается у пациентов, получивших ранее
противовирусное лечение и вакцинотерапию.
Очаг поражения при абортивном течении минует
некоторые стадии, характерные для манифест-
ной формы, и может проявляться в виде зудя-
щего пятна или папулы, разрешающихся за 1—3
дня. К абортивно протекающим формам гени-
тального герпеса относят эритематозную, папу-
лезную и пруригоневротическую, при которых
отсутствуют везикулезные элементы. Диагнос-
тика разновидностей генитального герпеса, про-
текающих в стертой абортивной форме, вызы-
вает большие затруднения. Заболевание обычно
распознается в тех случаях, когда типичные ре-
цидивы герпеса чередуются с высыпаниями или
возникают одновременно с ними.
Субклиническая форма
Субклиническая форма генитального герпеса
обычно выявляется при обследовании половых
контактов больных, страдающих ЗППП, или
супружеских пар с нарушением фертильности.
Для этой формы характерна микросимптомати-
ка (кратковременное появление на слизистой
оболочке наружных гениталий поверхностных
трещинок, сопровождающихся незначительным
зудом), или полное ее отсутствие.
Клиническая картина генитального герпеса
находится в прямой зависимости от места пора-
жения, интенсивности воспалительного процесса,
срока заболевания, способности организма к за-
щитным иммунологическим реакциям и виру-
лентности штамма вируса.
Клинические особенности течения у женшин
У женщин герпетические поражения обычно
локализуются на малых и больших половых гу-
бах, в области вульвы, клитора, вагины и шей-
ки матки. Герпетические пузырьки образуют
характерные полициклические фестончатые
фигуры. Впоследствии образуются поверхност-
ные, покрытые сероватым налетом язвочки со-
ответственно числу бывших пузырьков или
сплошная эрозия с гладким дном и неподрыты-
ми краями, окруженными ярко-красным обод-
ком. Язвочки не бывают глубокими и не кро-
воточат. Герпетические изъязвления иногда
очень болезненны. Язвочки и эрозии заживают,
не оставляя рубцов. Герпетические высыпания
на малых губах и вульве у женщин в некоторых
случаях вызывают значительный отек половых
губ. При герпетическом цервиците шейка мат-
ки отечна, часто имеет эрозии. Рецидивы насту-
пают или спонтанно, или после половых сноше-
ний, или после менструации. Часто появление
герпеса гениталий провоцируется другими ин-
фекциями. Герпетическая рецидивирующая ин-
фекция может поражать не только область на-
ружных половых органов, но и слизистую
оболочку влагалища, шейку матки и проникать
восходящим путем в слизистую оболочку мат-
ки, трубы и придатки, уретру и мочевой пузырь,
вызывая в них специфическое поражение.
Клинические особенности течения у мужчин
Герпетические поражения гениталий у мужчин
локализуются в области баланопрепуциальной
бороздки, крайней плоти и на головке полового
члена в виде мелких пузырьков, быстро вскры-
вающихся с образованием поверхностных эрозий.
Клиническая картина генитального герпеса у
мужчин характеризуется постоянством следующих
признаков: наличием герпетических пузырьков,
эрозиями и язвочками, рецидивирующим харак-
тером заболевания. Клинические проявления гер-
петического уретрита у мужчин начинаются с
появления жалоб на боли и чувство жжения, па-
растезии с иррадиацией в область мошонки, по-
лового члена, уретры, в ноги, ягодицы, промеж-
ность. Больные отмечают чувство жжения и боли
Глава 47. Генитальный герпес
545
во время мочеиспускания, появление из уретры
светлых или светло-желтого цвета выделений,
гиперемию и отечность губок наружного отвер-
стия передней уретры. При уретроскопии в об-
ласти передней трети уретры выявляют мелкие,
множественные, сгруппированные, точечные уча-
стки гиперемии. Первичный герпетический ба-
ланопостит сопровождается появлением множе-
ственных пузырьков, после вскрытия которых
образуются обширные болезненные эрозивные
поверхности. После эпителизации эрозий на их
месте остаются эритематозные или пигментиро-
ванные пятна. Хронический рецидивирующий
баланопостит диагностируется примерно у поло-
вины больных генитальным герпесом.
Генитальная инфекция, вызванная ВПГ, ча-
сто вызывает серьезные психологические и
психосексуальные расстройства. Чаще всего
проявляются такие эмоциональные реакции,
как депрессия, душевная боль, гнев, потеря са-
моуважения и враждебность по отношению к
возможному источнику заражения. Для мно-
гих пациентов одной из наиболее сложных
проблем становится извещение о наличии у них
генитального герпеса имеющихся или будущих
половых партнеров. У женщин, как правило,
возникает больше психологических проблем,
чем у мужчин. Исследования пациентов с пер-
вым эпизодом генитального герпеса показы-
вают, что при этом заболевании возникают бо-
лее глубокие психологические проблемы, чем
при других ИППП или хронических дермато-
зах. Существуют значительные разногласия от-
носительно связи стрессовых, жизненных си-
туаций с рецидивами болезни. Некоторые
специалисты полагают, что стресс ускоряет
развитие рецидивов. Однако недавние иссле-
дования позволяют предположить, что у паци-
ентов с частыми рецидивами стресс является
скорее следствием рецидивов, а не их катали-
затором. Частые рецидивы могут накладывать
отпечаток на характер человека. Чрезвычайно
важно сообщать пациентам информацию о
биологии инфекционного агента, его переда-
че, частоте рецидивов заболевания, бессимп-
томном выделении вируса, исходах беременно-
сти и выборе лечения. Для снижения
психологической заболеваемости важное зна-
чение имеют такие аспекты психологической
поддержки, как расширение знаний, а также
поддержка со стороны полового партнера.
Диагноз
В связи с разнообразием клинических проявле-
ний генитального герпеса и частым его сочета-
нием с другими урогенитальными инфекциями,
имеющими сходную симптоматику, важное зна-
чение приобретает идентификация заболевания.
Для диагностики герпетической инфекции ис-
пользуются следующие методы:
• вирусологические методы обнаружения и
идентификации вирусов простого герпеса;
• молекулярно-генетическая диагностика
(ПЦР — полимеразная цепная реакция);
• выявление антигенов ВПГ;
• цитоморфологические методы;
• регистрация иммунного ответа к ВПГ;
• оценка иммунного статуса больных.
В качестве биологического материала для про-
ведения исследования можно использовать со-
держимое везикул, смывы с тканей и органов,
мазки-отпечатки, соскобы, биологические жид-
кости и секреты организма (кровь, слизь, моча,
слезная жидкость, СМЖ).
В настоящее время «золотым стандартом» в
диагностике герпетической инфекции являют-
ся вирусологические методы обнаружения и иден-
тификации ВПГ. Чувствительность этих мето-
дов — 80—100 %, специфичность — 100 %.
Классические вирусологические методы до на-
стоящего времени остаются наиболее важными
и достоверными для окончательной идентифи-
кации ВПГ. Особое значение они имеют при
бессимптомном и хроническом рецидивирующем
течении инфекции. В специализированных ви-
русологических лабораториях используют следу-
ющие методы: заражение клеточных культур,
заражение 12—13-дневных куриных эмбрионов,
заражение экспериментальных животных.
Вирусологические методы
Вирусологические методы складываются из двух
этапов: 1-й этап — размножение и накопление
вирусных частиц и 2-й этап — подтверждение
и идентификация типа ВПГ. С этой целью про-
водят:
1. оценку характерных цитопатических изме-
нений в зараженных клеточных культурах с
помощью световой (образование шарообразных
и многоядерных гигантских клеток с изменением
ядра, хромосомные аберрации) и электронной
микроскопии (выявление внутриклеточных и
внутриядерных вирусных частиц);
2. реакцию нейтрализации на эксперименталь-
ных животных, куриных эмбрионах и в клеточ-
ных культурах с использованием стандартных
противогерпетических антисывороток;
3. идентификацию вирусных антигенов с помо-
щью специфических антител методами иммуно-
флуоресценции или иммуноферментного анализа.
546 Венерология
При учете времени на проведение вирусоло-
гического исследования необходимо учитывать
длительность второго этапа. Продолжительность
вирусологических методов составляет от 2 до 14
дней, диагностическая эффективность (частота
выявления ВПГ) достигает 90—95 %.
К преимуществам вирусологических методов
относят высокую чувствительность, специфич-
ность, а также возможность проведения допол-
нительной дифференцировки выделенных
штаммов ВПГ по антигенным и биологическим
свойствам. Недостатками этих методов являют-
ся высокая стоимость, ограниченная доступность
и относительная длительность.
Молекулярно-генетическая диагностика
Полимеразная цепная реакция (ПЦР) — новый
метод быстрой диагностики генитального герпе-
са. Он высокочувствителен (95 %) и специфи-
чен (90—100 %), достаточно широко применяется
в практике.
Методы выявления антигенов ВПГ
Обнаружение антигенов ВПГ в биологических
жидкостях организма или в клетках является
прямым доказательством активной репликации
вируса и, следовательно, однозначно подтверж-
дает этиологию заболевания. Для выявления
антигенов ВПГ в биологическом материале, по-
лученном от больных, используются серологи-
ческие методы: реакция связывания комплемента
(РСК); реакция нейтрализации (PH); реакция
пассивной гемагглюцинации (РПГА); радиоим-
мунный анализ (РИА); иммуноферментный ана-
лиз (ИФА). Для выявления антигенов ВПГ
широкое применение нашел также метод имму-
нофлуоресцентной микроскопии (ИФ).
Принцип этих методов основан на выявлении
антигенов ВПГ с помощью специфических ан-
тител, либо поликлональных, полученных при
иммунизации животных антигенами ВПГ, либо
моноклональных, полученных с помощью гиб-
ридомной технологии. В основе всех этих мето-
дов лежит реакция специфического связывания
антиген-антитело. Различия методов заключают-
ся в способах регистрации и учета этой реакции.
В настоящеее время классические серологичес-
кие реакции типа РСК, PH, РПГА в силу их
низкой чувствительности, высокой трудоемко-
сти, малой пригодности к автоматизации отсту-
пили на второй план.
Все большее распространение получают методы
ИФА, в которых, как правило, применяются мо-
ноклональные антитела, отличающиеся высокой
специфичностью. При оценке диагностической
ценности следует учитывать, что специфичность
выявления антигенов ВПГ при использовании
коммерческих тест-систем весьма высока, но не
абсолютна. Существуют широкие перекрестные
антигенные связи ВПГ человека 1-го и 2-го ти-
пов с другими герпесвирусами человека (вирусом
Эпштейна-Барр, цитомегаловирусом, вирусом вет-
ряной оспы — герпес зостер), а также герпесви-
русами животных и птиц. Чувствительность ИФА
достаточно высокая, частота подтверждения гер-
песвирусной этиологии заболевания составляет в
остром периоде до 50—75 %, при хроническом ре-
цидивирующем течении — до 35—45 %.
Метод иммунофлуоресцентной микроскопии
(ИФ) широко применяется для диагностики за-
болеваний, вызванных ВПГ. С его помощью
выявляют антигены ВПГ в клетках, полученных
от больных. Принцип метода заключается в ре-
гистрации реакции антиген-антитело с исполь-
зованием коммерческих противогерпетических
антител. Применение метода ИФ возможно в
прямом и непрямом вариантах. При использо-
вании метода ИФ частота подтверждения герпес-
вирусной этиологии достигает, в зависимости от
стадии заболевания, 15—45 %. Длительность
постановки ИФ составляет 1—1,5 часа.
Цитоморфологические методы
Цитоморфологические методы позволяют выя-
вить индуцированные вирусом морфологические
изменения в клетках и тканях пораженных ор-
ганов. Эти методы делятся на две группы: в
одних используется световая, в других — элек-
тронная микроскопия.
При использовании световой микроскопии
в зонах поражения отмечается типичный для
герпеса метаморфоз клеток и их ядер. Размер
клеток увеличен, они имеют крупные, гипер-
хромные ядра, содержащие базофильные
включения, окруженные зоной просветления.
Определяются внутриядерные ДНК-содержа-
щие включения, многоядерные клетки. В ме-
стах поражения тканей отмечается некроз, лим-
фоплазмоцитарная реакция различной степени
выраженности.
Цитоморфологические методы диагностики
являются наиболее доступными и технически про-
стыми. Диагностическая ценность метода может
достигать до 2/3 от эффективности выделения
вируса в культуре тканей. К достоинствам следу-
ет отнести высокую диагностическую значимость
при бессимптомном течении инфекции (цервици-
ты, уретриты). К недостаткам цитоморфологичес-
ких методов с использованием световой микроско-
пии следует отнести их неспецифичность (наличие
Глава 47. Генитальный герпес
547
многоядерных клеток и внутриядерных включе-
ний характерно и для других вирусов, например,
вируса ветряной оспы — герпес зостер), невозмож-
ность дифференцирования первичной инфекции
ВПГ от рецидивирующей, невозможность диффе-
ренцирования типа ВПГ.
Электронно-микроскопическое исследование
позволяет обнаружить в биопробах характерные
по морфологии частицы ВПГ. Дифференциация
ВПГ от других морфологически неотличимых
представителей семейства герпесвирусов может
быть выполнена при комбинации электронной
микроскопии с использованием специфических
антител против соответствующих штаммов ВПГ,
меченных электронно-плотными маркерами —
пероксидазой хрена или ферритином. Все это
позволяет использовать электронную микроско-
пию для быстрой (2—3 часа) дифференциаль-
ной диагностики с другими вирусными заболе-
ваниями. Ограничениями при использовании
электронно-микроскопических методов являет-
ся их неспецифичность, невозможность диффе-
ренцирования первичной и рецидивирующей
инфекции и типа ВПГ, труднодоступность обо-
рудования и отсутствие подготовленного пер-
сонала.
Регистрация иммунного ответа к ВПГ
Для регистрации иммунного ответа к ВПГ ис-
пользуют: выявление циркулирующих в сыво-
ротке или других биологических жидкостях и
секретах организма больного специфических
противогерпетических антител; оценку специфи-
ческого клеточного иммунитета к ВПГ; кожные
пробы.
При инфекции, вызванной ВПГ, организм
отвечает развитием специфического гуморально-
го и клеточного иммунного ответа. При гумо-
ральном иммунном ответе последовательно вы-
рабатываются типоспецифические антитела
классов IgM, IgG, IgA, направленные против ан-
тигенов ВПГ. При первичной инфекции анти-
тела представлены классом IgM. При рецидиви-
рующем герпесе наблюдается нарастание титра
IgM, IgG, IgA. Антитела обладают вируснейтра-
лизующей активностью, которая усиливается в
присутствии комлемента (IgM, IgG).
Выявление у больного высокого титра цирку-
лирующих антител класса IgM может служить
указанием на первичную инфекцию ВПГ у де-
тей и лиц молодого возраста при убедительных
доказательствах отсутствия эпизодов инфекции
ВПГ в анамнезе. Нарастание титра антител в 4
и более раз является характерным признаком
первичной инфекции.
Для выявления антител к антигенам ВПГ ис-
пользуют классические серологические методы
(РСК, РПГА, PH), а также ИФА, чувствитель-
ность которого на 2—4 порядка выше по срав-
нению с классическими серологическими мето-
дами.
К преимуществам методов выявления проти-
вогерпетических антител следует отнести их
высокую специфичность и чувствительность,
техническую доступность, невысокую стоимость,
что имеет значение для диагностики между пер-
вичной инфекцией и активацией персистирую-
щего вируса. К недостаткам следует отнести от-
сутствие диагностической значимости в остром
периоде, выявление антител у большинства кли-
нически здоровых лиц, большая продолжитель-
ность при использовании метода «парных сыво-
роток».
Для оценки клеточного иммунного ответа
предложено использовать различные лаборатор-
ные методы, в том числе реакцию бласттранс-
формации лимфоцитов в ответ на специфичес-
кий антиген ВПГ, продукцию лимфокинов
(MIF/LIF, у-интерферон).
Для оценки специфической иммунной реак-
тивности у больных с предполагаемой герпесви-
русной инфекцией используют внутрикожные
пробы на антигены ВПГ. В качестве антигена
применяют коммерческую противогерпетичес-
кую вакцину. Диагностическая значимость этого
метода отличается широкой вариабельностью и
его применение часто вызывает обострение ин-
фекционного процесса.
Оценка иммунного статуса
Острая или рецидивирующая инфекция, вызван-
ная ВПГ, является следствием имеющегося им-
мунодефицитного состояния у конкретного
больного. Однако развитие острого заболевания,
и тем более переход его в хроническую (реци-
дивирующую) форму, само по себе приводит к
углублению иммунодефицитного состояния.
Поэтому оценка иммунного статуса является
необходимым условием полноценной диагности-
ки при заболеваниях, вызванных ВПГ, так как
позволяет оценить степень тяжести иммуноде-
фицита, назначить адекватную иммунокорриги-
рующую терапию и контролировать ее эффек-
тивность.
При диагностике инфекции, вызванной ВПГ,
рекомендуется проведение повторного исследо-
вания (2—4 раза в течение 5—7 дней) материала
из различных очагов инфекции (у женщин — на
18—20-е дни менструального цикла) для повы-
шения выявляемое™ ВПГ.
548 Венерология
Лечение
В настоящее время химиотерапия и профилак-
тика генитального герпеса заключаются в при-
менении противовирусных препаратов, экзоген-
ного интерферона, индукторов интерферона,
иммуномодуляторов, местных средств. Рекомен-
дуется тщательное клинико-лабораторное обсле-
дование больных и лечение интеркуррентных
заболеваний.
Из противовирусных препаратов базисным
средством является синтетический пуриновый
нуклеозид ацикловир (зовиракс): таблетки
0,2 г; 0,4 г; 0,8 г; суспензия (5 мл — 0,2 и
0,4 г) во флаконах; сухое вещество для инъек-
ций 0,25 г в ампулах. Ацикловир обладает
выраженной активностью против вируса про-
стого герпеса благодаря ингибиции синтеза
вирусной ДНК.
Схемы лечения ацикловиром
Первый клинический эпизод генитального герпеса
— ацикловир 200 мг 5 раз в день в течение 5—
10 дней (или до достижения клинического раз-
решения). Лечение ацикловиром эффективно,
если оно начато в первые часы после появления
первых клинических симптомов заболевания.
Первый клинический эпизод герпетического про-
ктита — ацикловир 400 мг 5 раз в день в тече-
ние 10 дней (или до достижения клинического
разрешения).
Рецидивирующий генитальный герпес — эпизо-
дическое лечение', ацикловир 400 мг 5 раз в день
в течение 5 дней или 400 мг 3 раза в день в те-
чение 5 дней или 800 мг 2 раза в день в течение
5 дней. Лечение целесообразно начинать в про-
дромальном периоде или в первые часы клини-
ческих проявлений.
Супрессивная терапия — для больных, имею-
щих частые рецидивы. Существует перманент-
ный метод лечения рецидивирующего герпеса
ацикловиром (ежедневный прием по 400 мг 2
раза в день или 200 мг 4—5 раз в день в течение
месяцев и даже лет). Ежедневная супрессивная
терапия снижает частоту рецидивов генитального
герпеса у 75 % пациентов. Подтверждена эффек-
тивность и безопасность ацикловира у пациен-
тов, получавших ежедневную терапию в течение
5 лет.
Диссеминированная инфекция. При наличии
диссеминированной инфекции (энцефалит,
пневмония, гепатит и т. п.) следует рекомендо-
вать внутривенное использование ацикловира
каждые 8 часов в течение 5-7 дней из расчета
5—10 мг/кг массы тела.
Другие препараты
Валацикловир (валтрекс) — таблетки по 0,5 г, при-
нимаются внутрь 2 раза в день в течение 5—10
дней. Препарат представляет собой L-валиловый
эфир ацикловира. Валацикловир обеспечивает
высокий уровень всасывания ацикловира при пе-
роральном применении, повышает его биодоступ-
ность в 4—5 раз и сохраняет высокий уровень его
безопасности и переносимости. При генитальном
герпесе, обусловленном ацикловирустойчивыми
штаммами вируса, альтернативными препаратами
являются фоскарнет (фосфономуравьиная кисло-
та) и фамвир (фамцикловир).
Фамцикловир — таблетки по 0,25 г, принима-
ются 3 раза в день в течение 7 дней при острой
герпетической инфекции. Препарат трансформи-
руется в организме в активное противирусное
соедининение — пенцикловир и эффективен в
отношении ВПГ-1, ВПГ-2 и других герпесвиру-
сов. Больным с повышенным риском развития
постгерпетической невралгии рекомендовано
принимать препарат по 0,5 г 3 раза в сутки в
течение 7 дней. В период беременности и лак-
тации препарат можно назначать только по жиз-
ненным показаниям.
Фоскарнет (фоскавир) применяется в виде
аппликаций 3 %-ной мази на места поражений.
При тяжелом течении заболевания возможно
внутривенное, медленное капельное введение
препарата в течение 2 часов по 60 мг/кг массы
тела 3 раза в сутки в течение 10—14 дней. Пре-
парат взаимодействует с ДНК-полимеразой и, в
меньшей степени, с РНК-полимеразой, связы-
вается с пирофосфатом и частично ингибирует
нуклеозидтрифосфат, являясь конкурентным
ингибитором пирофосфата.
Панавир — очищенный экстракт побегов ра-
стения Solanum tuberosum. Препарат способствует
торможению репликации вируса в инфицирован-
ных клетках и приводит к снижению титров
инфекционной активности вируса. Приминяет-
ся в виде двух внутривенных инъекций 0,004 %
раствора по 5,0 мл с интервалом 48 часов в мо-
мент рецидива.
Алпизарин — таблетки 0,1 г; по 1 таблетке 3—
5 раз в день в течение 15—20 дней. Алпизарин
обладает ингибирующим действием на началь-
ных этапах репродукции ВПГ в клетке.
Хелепин — таблетки 0,1 г; по 1 таблетке 3—4
раза в день в течение 15—20 дней.
Гелпин (бривудин) — таблетки 0,125 г по
1 таблетке 4 раза в день (после еды с боль-
шим количеством жидкости) в течение 5—7
дней.
Глава 47. Генитальный герпес 549
Флакозид — флавоноид из листьев бархата
амурского и бархата Лаваля. Применяется перо-
рально после еды по 0,1—0,5 г 3 раза в сутки в
течение 10 дней.
Рибамидил (виразол, рибавирин) — таблетки
0,2 г; по 1 таблетке 3 раза в день в течение 7—
14 дней.
Метисазон — таблетки 0,2 г; по 3 таблетки 2
раза в день в течение 4—6 дней через 1 час пос-
ле еды.
Интерлок 500 000 ME. Применяется внутри-
мышечно 500 000 ME 1 раз в сутки в течение
14 дней.
Полудан — порошок растворяется в стериль-
ной дистиллированной воде (2 мл) в концент-
рации 200 мкг/мл и вводится ежедневно в тече-
ние 10 дней.
Лейкинферон\ ларифан:, 2-интерферон (реафе-
рон), ридостин, комедон (циклоферон).
В качестве индукторов эндогенного интерфе-
рона возможно использование пирогенала, про-
дигиозана, кутизона, зимозана.
В комплексном лечении генитального герпе-
са следует использовать по показаниям иммуно-
модуляторы, биогенные стимуляторы, адаптоге-
ны, симптоматические препараты.
Препараты для наружного использования
Панавир — гель защитный — наружно 3-4 раза
в сутки. Панавир подавляет экспрессию поздних
белков ВПГ 1 типа через 24 часа после зараже-
ния клеток и белков (сверхранних, ранних и по-
здних) ВПГ 2 типа в течение 48 часов после за-
ражения.
Алпизарин — мазь 2 % и 5 %-ная. Апплика-
ции 2 %-ной мази производятся на слизистые
оболочки 4—6 раз в сутки в течение 5—10 дней;
5 %-ная мазь наносится на кожные покровы 4—
6 раз в сутки в течение 5—10 дней.
Вира-МП (видарабин фосфат 10 %) — гель.
Наносится на пораженные поверхности 4 раза в
день в течение 7 дней. Рекомендуется применять
при появлении первых симптомов или первых
герпетических высыпаний.
Пантенол — аэрозольная упаковка, объем —
140 г. Наносится на пораженные участки с рас-
стояния 10 см 4—6 раз в день в течение 2 недель.
Рекомендуется применять в острый период или
в начале возникновения рецидива.
Мегосин — мазь 3 %. Местные аппликации 4—
6 раз в день в течение 5—7 дней.
Хелепин применяется в виде 1 %-ной мази 3
раза в сутки в течение 7—10 дней на поражен-
ные слизистые оболочки; при поражении кожи
применяется 5 %-ная мазь.
Госсипол — линимент 3 %. Местные апплика-
ции 4—6 раз в день в течение 5—7 дней.
Тромантадин в виде 1 %-ной мази наносят на
пораженные участки 3 раза в сутки в течение 7—
10 дней.
Гевизош в виде мази наносится на поражен-
ную поверхность 3—5 раз в день в течение 3—12
дней.
Риодоксоловая мазь 0,25% — 0,5% — 1,0 %.
Применяется при нетяжелом течении герпетичес-
кой инфекции кожи и слизистых оболочек в
виде аппликаций в течение 5—10 дней.
Препараты для наружного лечения следует
применять в продромальном периоде и при раз-
витии рецидива до стадии образования эрозий.
При наличии эрозий или отека необходимо при-
менять примочки с дезоксирибонуклеазой, по-
луданом, 0,5 %-ным раствором сульфата цинка,
а также использовать антисептики, анилиновые
красители.
Профилактика рецидивов заболевания
Вакцины могут использоваться для супрессив-
ной терапии (иммунотерапии) пациентов с гени-
тальным герпесом, предотвращения передачи ви-
руса серонегативным партнерам больных
генитальным герпесом, а также для вакцинации
серонегативных лиц.
Герпетическая вакцина — 0,2 мл внутрикож-
но через 2—3 дня в количестве 5 инъекций не
менее 2 раз в год (при иммунокомпенсирован-
ном состоянии больного).
При иммунодефицитном состоянии у больных
с частыми (1 раз в месяц и чаще) рецидивами
заболевания вакцина применяется по заверше-
нии иммунокоррекции 1 раз в 7—14 дней в ко-
личестве 5 инъекций с ревакцинацией 1 раз в 6—
8 месяцев.
Следует рекомендовать больному воздержи-
ваться от половой жизни до исчезновения кли-
нических проявлений. На этот период использо-
вание презервативов должно быть обязательным
при всех сексуальных контактах. Половых парт-
неров пациентов, имеющих генитальный герпес,
нужно обследовать и при наличии у них герпе-
са — лечить.
Глава 48
Вульвовагиниты у девочек различного возраста *
Вульвовагинит — воспаление преддверия влага-
лища (вульвит) и слизистой оболочки влагалища
(вагинит, кольпит), вызванное различными при-
чинами.
Актуальность изучения вульвовагинитов у
девочек объясняется высокой частотой регистра-
ции этого заболевания, торпидностью течения,
склонностью к рецидивам. В дальнейшем у пе-
ренесших вульвовагинит пациенток возникают
проблемы с репродуктивной функцией: трубное
бесплодие, невынашивание беременности, атопи-
ческая беременность.
Высокая частота заболевания объясняется ана-
томофизиологическими особенностями мочепо-
ловых органов: избыточная складчатость слизи-
стых оболочек, недостаточное смыкание срамных
губ в нижних отделах; низкая эстрогенная на-
сыщенность организма, недостаточная выработ-
ка гликогена, истонченность и замедленная про-
лиферация эндотелия полового тракта,
нейтральная или слабощелочная среда влагали-
ща, преобладание во влагалище кокковой фло-
ры, сниженный местный иммунитет, тонкий
многослойный плоский эпителий преддверия
влагалища, наличие девственной плевы.
Важное значение имеет состояние биоценоза
влагалища, которое определяется функцией яич-
ников, концентрацией лактофлоры. Максималь-
но часто вульвовагиниты возникают в возрасте
3—7 лет, так называемый нейтральный период
полового созревания (85 % пациенток младше 8
лет), поскольку в это время процессы самоочи-
щения влагалища практически отсутствуют.
Одной из причин увеличения числа вульво-
вагинитов у девочек является трудно контроли-
руемый рост урогенитальных инфекций среди
взрослого населения, особенно бесимптомных и
торпидно протекающих.
Этиология и патогенез
Вульвовагиниты делятся на специфические с ус-
тановленным инфекционным агентом (гонокок-
ки, трихомонады хламидии, вирусы, грибы) и
* Глава написана И. В. Тихоновской
неспецифические. Неспецифические вульвоваги-
ниты могут быть первичными и вторичными,
являющимися следствием общего заболевания;
инфекционными и неинфекционными
Неспецифические первичные вульвовагиниты
могут быть вызваны травмой половых органов,
нарушением правил личной гигиены, в том числе
и злоупотреблением гигиеническими процедура-
ми (частое подмывание, спринцевание), перепол-
нением мочевого пузыря, запорами, пребыванием
девочки в запыленном помещении, мастурбацией,
чтением возбуждающей литературы.
Причинами вторичных неспецифических
вульвовагинитов бывают глистные инвазии,
уретровагинальный рефлюкс, атопический дер-
матит, аллергические заболевания (у таких де-
тей обострение вульвовагинита может вызы-
ваться приемом пищевых аллергенов,
обострением аллергического заболевания — в
мазках обнаруживается повышенное количество
эозинофилов); богатая экстративными веще-
ствами и пряностями пища, шоколад; детский
алкоголизм; хронические заболевания ЛОР —
органов, верхних дыхательных путей, органов
мочевыводящей системы; анемия, системные
заболевания крови; сахарный диабет, гломеру-
лонефрит, пиелит, цистит, инфантилиз; имму-
нодепрессивные состояния, возрастная иммуно-
логическая перестройка; скопление крови в
рудиментарных образованиях; длительное ма-
точное кровотечение.
Неспецифические первичные вульвовагиниты
могут вызываться активацией условно-патоген-
ной флоры мочеполовых органов девочки. Это-
му способствует развитие дисбактериоза после
применения антибиотиков, кортикостероидов,
нерациональной витаминотерапии.
В качестве инфекционных причин первичных
неспецифических вульвовагинитов выступают
стафилококки, стрептококки. Вульвовагиниты
могут развиваться в результате заноса во влага-
лище кишечной палочки, энтерококков и дру-
гих представителей флоры кишечника. В этих
550
Глава 48. Вульвовагиниты у девочек различного возраста 551
случаях микробиологическое исследование под-
тверждает диагноз.
В большинстве случаев причиной инфекцион-
ного вульвовагинита являются бактерии и гри-
бы (70 %), в 20 % — вирусы, в 10 % случаев
обнаружить возбудителя не удается.
У половины пациенток наблюдается микст —
инфекция, при которой патогенность каждого
возбудителя увеличивается.
Неспецифические
вульвовагиниты
Клиническая картина
В зависимости от давности заболевания вульво-
вагиниты разделяют на: острые (срок заболева-
ния не более 1 месяца, подострые (от 1 до 3
месяцев) и хронические (свыше 3 месяцев).
Несмотря на различную этиологию, клини-
ческие проявления неспецифических вульвова-
гинитов сходны между собой. Общее состояние
детей, как правило, не нарушается. Отмечаются
жалобы на жжение после мочеиспускания, зуд
и незначительную болезненность наружных по-
ловых органов, иногда боль может иррадииро-
вать в паховые складки. Маленькие дети 4—5 лет
могут жаловаться на боли в бедре или в колене
из-за неумения определить источник боли. Чув-
ство стыдливости детей более старшего возраста
заставляет указывать на боли в животе вместо
гениталий.
Постоянными признаками вульвовагинитов
являются гиперемия и отек наружных половых
органов, бели (водянисто-желтые или серозно-
гнойные). Бели могут наблюдаться не только при
специфической и неспецифической инфекции
мочеполовых органов, но и при общих инфекци-
онных заболеваниях, интоксикациях, заболеваний
сердца и почек, анемии, новообразованиях.
В большинстве случаев неспецифические
вульвовагиниты протекают со скудной симпто-
матикой, не имеют специфических признаков,
обостряются на фоне интеркуррентных заболе-
ваний, переохлаждения; проводимая терапия
(как правило, антибиотики пенициллинового
ряда) приводит к временному улучшению.
Ряд особенностей имеет обследование ребен-
ка с проявлениями вульвовагинита. Дети часто
оказывают физическое сопротивление действи-
ям медицинского персонала, поэтому к осмотру
ребенка нужно подготовиться заранее. Совер-
шенно недопустимо грубое обращение с ребен-
ком, применение насильственных мер. Девочку
нужно отвлечь ласковым обращением, скрыть
приготовленные для обследования инструменты,
позволить держать при себе любимые игрушки,
книги. Применяемые для обследования инстру-
менты должны быть наименее травмирующими,
требуемых размеров. Осматривать девочку мож-
но в положении лежа с ногами, согнутыми в
коленных суставах или с приведенными к жи-
воту. В последнем случае требуется помощь ме-
дицинского персонала. При наличии детского
гинекологического кресла можно воспользовать-
ся им. Вначале осматривается живот, затем на-
ружные гениталии и девственная плева. Забор
мазков производится из преддверия влагалища,
уретры, парауретральных ходов, выводных про-
токов больших вестибулярных желез и прямой
кишки. Для забора материала используются дет-
ские катетеры, пипетки, желобоватый зонд, ло-
жечки различной формы. Если по каким-то
причинам невозможно произвести забор матери-
ала из влагалища, то мазки можно приготовить
из центрифугата утренней мочи. Бимануальное
(прямокишечно — брюшностеночное) исследо-
вание производится у детей до 6 лет мизинцем.
В исключительных случаях допустимо прямоки-
шечно — влагалищное исследование одним паль-
цем. В таких ситуациях необходимо согласие
родителей на обследование, а при непосредствен-
ной процедуре в кабинете должно находится
третье лицо. Иногда для обследования применя-
ется ингаляционный наркоз. Вагиноскопия,
кольпоскопия и цервикоскопия производится по
показаниям.
Диагноз
Диагностика вульвовагинитов включает мазок из
влагалища на наличие микрофлоры, бактерио-
логическое исследование, консультацию нефро-
лога, исследование кала на дисбактериоз, кон-
сультацию гастроэнтеролога, ультразвуковое
исследование внутренних половых органов.
Лечение
Лечение неспецифических вульвовагинитов про-
водится амбулаторно, лишь в торпидных и часто
рецидивирующих случаях показано стационарное
лечение. В легких случаях бывает достаточно со-
блюдать гигиенические требования, наладить
диету. Проводится лечение сопутствующих забо-
леваний, вторично вызывающих вульвовагинит.
Общая противомикробная терапия назначается
редко, только при идентификации возбудителя и
определении чувствительности к нему. При вуль-
вовагинитах аллергического генеза препараты
пенициллина не назначаются, поскольку могут
усугубить процесс из-за выраженного аллергоген-
552 Венерология
ного действия. Местно производится промывание
влагалища растворами фурацилина — 1: 10 000,
риванола — 1 : 5000, 3 % перекисью водорода,
сидячие ванночки с настоем ромашки, шалфея,
листьев эвкалипта. Указанные процедуры назна-
чаются не более чем на 3—5 дней (вымывание
гликогена, снижение факторов местной защиты,
медленное заселение палочками Дедерлейна). В
торпидных и рецидивирующих случаях можно
применить антибиотики местно: назначают инт-
равагинальные свечи с антибиотиками или ороша-
ют влагалище растворами антибиотиков.
Для санации мочевыводящих путей при не-
специфических вульвовагинитах применяются
уросептики. С целью восстановления микрофло-
ры влагалища после проведеной антибактериаль-
ной терапии назначают интравагинально и
внутрь (в возростных дозировках) в течение 10
дней эубиотики: лактобактерии, бифидумбакте-
рин, бактисубтил. В качестве иммуномодулято-
ра можно применять дибазол из расчета 1 мг на
каждый год жизни 1 раз в день в течение меся-
ца. Для лечения вульвовагинитов у детей стар-
ше одного года подключают иммуностимулято-
ры (микробные, химические).
Причиной вульвовагинитов у девочек может
быть энтеробиоз. В таких случаях ребенок пло-
хо спит, просыпается ночью с жалобами на боли
в области наружных половых органов, зуд кожи
промежности. Иногда удается обнаружить остриц
в испражнениях ребенка, на коже в перианаль-
ной области или даже в отделяемом из влагали-
ща. При осмотре обнаруживаются: гиперемия
складок ануса, их утолщение, следы расчесов. В
таких случаях необходимо взять соскоб с периа-
нальных складок на яйца остриц. Для лечения
энтеробиоза применяется пирантель: 10 мг на 1 кг
веса ребенка. Лечение повторяют через месяц.
Членам семьи также необходимо провести анти-
гельминтную терапию и рекомендовать соблю-
дать правила личной гигиены.
Наличие во влагалище инородного тела также
может быть одной из причин вульвовагинита.
Ребенка приводят к врачу с жалобами на кровя-
нисто-гнойные выделения из влагалища; на коже
промежности и бедер могут быть проявления пи-
одермии — результат мацерации. Ведущую роль в
диагностике этого вида вульвовагинитов играет
ректоабдоминальное исследование, вагиноскопия
или осмотр с помощью детских влагалищных зер-
кал. При проведении этих исследований выделе-
ния могут усиливаться за счет травматизации гра-
нуляций, возникших вокруг инородного тела. При
исследовании можно попытаться подтолкнуть
инородное тело к входу во влагалище, где оно более
доступно для захвата зажимом. Мелкие инородные
тела (кусочки ваты, ткань) можно вымыть под дав-
лением жидкости через резиновый катетер, на ко-
торый надевают шприц объемом 20 мм3. После
извлечения инородного тела влагалище промыва-
ют антисептическим раствором, повторяя эту про-
цедуру в течение 2—3 дней.
Специфические вульвовагиниты
Кандидозный вульвовагинит
Переходу условно — патогенных грибов рода
Candida в паразитическую псевдомицелиальную
форму выживания способствует множество фак-
торов: эндокринные заболевания (сахарный ди-
абет, ожирение); недостаточность системы имму-
нитета, применение антибиотиков,
кортикостероидов, дефицит железа в организме,
оперативные вмешательства и т. д.
У девочек кандидозная инфекция урогени-
тального тракта может проявляться кандидоз-
ным вульвовагинитом, катарально-мембранным
вагинитом, кандидозным уретритом. У полови-
ны пациенток поражение половых органов со-
четается с микотическим стоматитом, при ко-
тором выявляется «географический язык». При
кандидозном вульвовагините пациентки жалу-
ются на зуд, жжение, болезненность вульвы.
При осмотре отмечается гиперемия малых и
больших половых губ, клитора, гименального
кольца с творожистыми пленками, эрозиями,
мацерацией. Из-за выраженного зуда могут
наблюдаться экскориации, при длительном те-
чении процесса — лихенизация и мокнутие.
При катарально-мембранном вагините на стен-
ках влагалища обнаруживаются эрозии с легко
травмируемым дном (возникает кровотечение
при соприкосновении с инструментами), по-
крытые творожистыми пленками. В заднем сво-
де влагалища обнаруживается творожистое или
сметанообразное отделяемое. Кандидозный
уретрит характеризуется гиперемией и отеком
губок уретры, сметаноподобным отделяемым из
уретры. С наружных половых органов процесс
может переходить на паховые складки и область
заднего прохода. На гладкой коже очаги пора-
жения имеют полициклические очертания, бле-
стящие, ярко — красного цвета с беловатым
налетом на поверхности; в глубине складок
возникают трещины, мацерация. В результате
сенсибилизации к грибковой инфекции могут
возникать вторичные аллергические высыпа-
ния — микиды, напоминающие высыпания при
острой или подострой экземе.
Глава 48. Вульвовагиниты у девочек различного возраста 553
Диагноз
Диагностика кандидозного вульвовагинита
включает: микроскопическое исследование па-
тологического материала (при окраске мазков по
Граму дрожжеподобные грибы окрашиваются в
темно-фиолетовый цвет, а при окраске по Ро-
мановскому-Гимза — в розовато-фиолетовый),
культуральное исследование, РСК с дрожжевы-
ми антигенами.
Лечение
Лечение кандидозного вульвовагинита включа-
ет: устранение факторов, провоцирующих рост
грибов, местное лечение и системную терапию
противогрибковыми препаратами при торпидно-
сти заболевания к местной терапии.
Местно применяют 5 % раствор бикарбоната
натрия, 3 % раствор перекиси водорода, 1-2 %
водные растворы анилиновых красителей. Пере-
численные растворы вводят во влагалище при по-
мощи резинового катетера. Девочкам старшего
возраста рекомендуются вагинальные таблетки:
пимафуцин по 1 вагинальной таблетке на ночь
в течение 20 дней или по 1 таблетке 2 раза в день
в течение 10 дней; гино-певарил — 50 по 1 таб-
летке на ночь в течение 2 недель или гино —
певарил — 150 по 1 таблетке в течение 3 дней и
кремы «Клотримазол», «Пимафуцин», «Гино-
певарил» по 5 г 1—2 раза в день в течение 6—10
дней; бетадин по 1 суппозиторию 1 раз в сутки
перед сном в течение 14 дней. Для системного
лечения кандидозных вульвовагинитов приме-
няют дифлюкан по 50-150 мг (в зависимости от
возраста и массы тела) однократно или низорал
по 50—200 мг/сут в течении 7—10 дней.
Герпетические вульвовагиниты
Герпетические вульвовагиниты у девочек могут
протекать одновременно с герпетическими гинги-
востоматитами. Инфекция передается от лиц, уха-
живающих за ребенком (родители, обслуживаю-
щий персонал детских учреждений) или путем
инокуляции вируса из других очагов поражения.
Отмечаются жалобы на выраженную болезнен-
ность, зуд и жжение в области вульвы. На вуль-
ве, а иногда на стенках влагалища наблюдаются
мелкие сгруппированные пузырьки на эритематоз-
ном основании. Через 5—7 дней пузырьки вскры-
ваются с образованием эрозий с полициклически-
ми очертаниями или подсыхают с образованием
серозных корочек. За 2—3 дня до появления вы-
сыпаний может наблюдаться изменение общего
состояния — повышение температуры тела, сла-
бость, тошнота, увеличение паховых лимфатичес-
ких узлов, болезненных при пальпации.
Диагноз
При классическом течении заболевания диагно-
стика герпетических вульвовагинитов трудности
не представляет. При атипичных формах реша-
ющее значение в диагностике имеют культураль-
ные вирусологические методы исследования,
ПЦР (полимеразная цепная реакция), ИФА.
Лечение
Лечение включает применение в начальном пе-
риоде болезней противогерпетических мазей: ри-
одоксоловой, Вира-МП и др. При выраженых
симптомах интоксикации назначают ацикловир
перорально в течение 5—7 дней в возрастных до-
зировках.
Гонорейные вульвовагиниты
Многослойный плоский эпителий, как правило,
не подвергается инфицированию гонококками.
Развитие гонорейного кольпита возможно при
беременности, когда разрыхлен эпителий влага-
лища; у детей до полового созревания и у жен-
щин в период менопаузы, когда неполноцен-
ность эпителия влагалища обусловлена
эндокринными особенностями.
Клиническая картина
Клиническая картина острого гонорейного вуль-
вовагинита характеризуется резко выраженной
гиперемией и отеком больших и малых половых
губ, обильным слизисто -гнойным отделяемым,
иногда с примесью крови. При подострой и тор-
пидной форме выделения, гиперемия вульвы и
влагалища незначительны.
При хроническом процессе клинические про-
явления гонорейного вульвовагинита практически
не отличаются от неспецифических вульвоваги-
нитов и характеризуются скудной клинической
симптоматикой.
К. А. Карышева выделила следующие признаки
хронической гонореи у девочек:
• симптом старческой вульвы (дряблость и
сморщенность кожи больших половых губ в
результате хронического воспаления и атрофии
поверхностных слоев эпителия;
• очаговое воспаление слизистой преддверия
влагалища в области нимфогенитальной бороз-
дки, устьев выводных протоков больших желез
преддверия влагалища;
• гиперемия области наружного отверстия
уретры, воспаление параректального венозного
сплетения в области промежности (венозный
узелок Карышевой);
• очаговые воспаления слизистой влагалища с
точечными кровоизлияниями в ней;
554 Венерология
• гиперемия в области наружного зева шейки
матки и гнойное отделяемое из цервикального
канала в виде капли.
У детей с гонорейной инфекцией в крови
могут наблюдаться анемия, эозинофилия, лей-
коцитоз.
Диагноз
Диагностика гонорейного вульвовагинита вклю-
чает бактериоскопическое или бактериологичес-
кое обнаружение возбудителя в отделяемом из
влагалища. При торпидной форме и хроничес-
кой гонорее применяется химическая провока-
ция независимо от возраста путем смазывания
влагалища, прямой кишки 1 % раствором Лю-
голя, а уретры 0,5—1 % раствором нитрата сереб-
ра. Провокация гоновакциной применяется после
трех лет в дозах 150—200 млн. микробных тел с
последующим забором материала из влагалища,
уретры и прямой кишки в течение трех дней.
После наступления половой зрелости физиоло-
гической провокацией служат месячные.
Лечение
Перед назначением антибиотиков необходимо
определить чувствительность к ним гонококков.
Основным средством остаются препараты пени-
циллина. При острой гонорее на курс лечения
требуется 3,4 млн ЕД бензилпенициллина (6 раз
в сутки равными дозами в зависимости от воз-
раста по 50 000—200 000 ЕД), при хронической
гонорее — 4,2—6,8 млн ЕД. При отсутствии эф-
фекта от проводимой терапии применяются
сульфаниламидные препараты, левомицетин.
Вакцинотерапия проводится с трехлетного воз-
раста в виде 4—8 внутримышечных инъекций го-
новакцины с интервалом 2—4 дня (под контро-
лем общего состояния), начиная с 50—100 млн.
микробных тел и повышая последующую дозу
в 1,5 раза от предыдущей. Максимальная доза
для детей — 500 млн. микробных тел. При от-
сутствии острых проявлений местно применяют
сидячие ванночки с раствором ромашки, слабые
растворы перманганата калия (1: 20 000).
Лечение следует проводить в стационарных
условиях. Контроль излеченности школьницам
можно проводить амбулаторно, девочкам дош-
кольного возраста стационарно.
Трихомонадные
вульвовагиниты
Трихомонадные вульвовагиниты у девочек де-
лятся на свежие (острый, подострый и торпид-
ный) — давностью инфекции до 2-х месяцев, и
хронические — свыше 2-х месяцев. Клиничес-
кая картина трихомонадного уретрита характе-
ризуется гиперемией вульвы, иногда бедер и па-
ховых складок. Отделяемое влагалища жидкое,
гнойное, пенистое. Пациентки жалуются на зуд
и жжение. При хроническом течении процесса
развивается эрозивный или фолликулярный
кольпит.
Выделяют трихомонадоносительство, когда
при наличии возбудителя отсутствуют какие-
либо клинические проявления.
Диагноз
Диагностика трихомониаза должна проводиться
многократно, поскольку однократное отсутствие
возбудителя не исключает инфицированности.
Исследуют нативные и окрашенные препараты.
Культуральные исследования показаны в сомни-
тельных случаях и для контроля излеченности.
Лечение
Лечение трихомонадной инфекции проводят
метронидазолом. Девочкам в возрасте 1-5 лет
назначают по 1/3 таблетки (0, 083 г) 2—3 раза в
день в течение 10 дней; 6—10 лет — по 1/2 таб-
летки (0,125 г) 2-3 раза в день; 11-15 лет - по
0,25—2 раза в день в течении 10 дней. Приме-
няют также макмирор, тиберал, нитазол. Мест-
но — вагинальные свечи нитазола в течение 15
дней, для детей младшего возраста — крем Пи-
мафуцин (обладает противогрибковой и проти-
вотрихомонадной активностью).
Хламидийные вульвовагиниты
Хламидийные вульвовагиниты характеризуются
незначительными поражениями вульвы, одна-
ко чаще, чем при неспецифических вульвова-
гинитах, наблюдаются цервициты с петехиаль-
ными кровоизлияниями на шейке матки. При
остром течении отмечатся умеренная гиперемия
наружных половых органов, болезненность
вульвы, слизистые или слизисто — гнойные
выделения из влагалища. При заборе материа-
ла и обследовании при помощи медицинских
инструментов отмечается повышенная рани-
мость оболочек с возникновением кровоточи-
вости. В мазках лейкоцитов обнаруживается
немного, а в крови определяется незначитель-
ное повышение палочкоядерных нейтрофилов
при отсутствии лейкоцитоза. При подостром и
хроническом течении клинические проявления
крайне скудны. Хламидийные вульвагиниты
характеризуются длительным и торпидным те-
чением.
Материал для исследования берут специальны-
ми палочками с дакроновыми тампонами для
Глава 48. Вульвовагиниты у девочек различного возраста
555
забора эпителиальных клеток со стенок влага-
лища, уретры, прямой кишки. Золотой стандарт
обнаружения хламидий — посев на клетки
Мак — Коя. В практике применяется окраши-
вание материала по Романовскому — Гимза и
Папаниколау, прямая реакция иммунофлюорес-
цеяции (ПИФ) и иммуноферментный анализ для
обнаружения антигенов хламидий (ИФА), поли-
меразная цепная реакция (ПЦР).
Лечение
Назначение препаратов тетрациклинового ряда
противопоказано детям до 12 лет, хинолонов —
до 18 лет. Эритромицин назначается в дозе 50 мг/
кг в сутки, в 4 приема в течение 10—14 дней.
Тетрациклин назначается детям старше 12 лет в
дозе 20—25 мг/кг в течение 14 дней, доксицик-
лин применяется у детей старше 12 лет в дозе
взрослых в течение 7—14 дней. По клиническим
показаниям могут применяться в терапевтичес-
ких дозах метилурацил, витамины группы В,
стекловидное тело.
Дифференциальный диагноз
Дифференциальная диагностика вульвовагини-
тов зависит от установленного этиологического
агента(ов). Нередко поражение половых органов
связано с многоформной экссудативной эрите-
мой.
Профилактика
Профилактика вульвовагинитов включает сана-
цию хронических очагов инфекции, лечение
сопутствующих заболеваний, исключение вред-
ных привычек, гигиеническое воспитание девоч-
ки, рациональное питание, контроль заболевае-
мости среди взрослого населения.
Глава 49
ВИЧ/СПИД-инфекция
Синдромом приобретенного иммунного дефици-
та (СПИД) называют заболевание инфекцион-
ной природы, характеризующееся первичным
поражением иммунной системы и развитием
выраженного вторичного иммунодефицита с
различными клиническими признаками. В на-
стоящее время, по рекомендации ВОЗ, принят
термин «ВИЧ-инфекция», а «синдромом приоб-
ретенного иммунодефицита» называют период
выраженных клинических проявлений.
Этиология
Заболевание вызывается вирусом иммунодефи-
цита человека (ВИЧ, англ. HIV — Human
Immunodeficiency Virus), принадлежащим к семей-
ству Retroviridae, подсемейству Lentivirus. Разли-
чают ВИЧ-1 и ВИЧ-2. Вирус иммунодефицита
человека первого типа (ВИЧ-1) был открыт в
мае 1983 года французским исследователем
Л. Монтанье и его американским коллегой
Р. Галло. В 1986 году Л. Монтанье выделил у
жителей Западной Африки вирус иммунодефи-
цита второго типа (ВИЧ-2). Мишени для виру-
сов обоих типов одинаковы: Т-лимфоциты, дру-
гие клетки иммунной системы (макрофаги),
клетки нервной системы (в основном, головно-
го мозга). Эпидемиологические, клинические и
молекулярные характеристики ВИЧ первого и
второго типов показывают, что заражение ВИЧ-
2 происходит реже, инкубационный период у
него больше, он менее вирулентен, чем латент-
ный ВИЧ-1. Наблюдаются случаи двойного
инфицирования (ВИЧ-1 и ВИЧ-2) в среде го-
мосексуалистов, наркоманов, проституток, за-
ключенных. Опасность представляют также рет-
ровирусы, обнаруженные в Западной Африке и
Южной Америке. Они патогенны для человека
и подавляют его иммунитет. Существенным от-
личием ретровирусов от остальных вирусов яв-
ляется наличие у них фермента обратной транс-
криптазы. ВИЧ, проникший в клетки человека
с помощью обратной транскриптазы, трансфор-
мирует затем свой генетический материал в ДНК
за счет клетки-хозяина, а новые клетки, полу-
ченные при репродуцировании, уже содержат
вирусные гены. Развитие СПИДа основывается
на разрушении Т-лимфоцитов-хелперов, а так-
же макрофагов. Как следствие этого возрастает
активность Т-лимфоцитов-супрессоров, и разви-
вается иммунологическая недостаточность. В
настоящее время доказано, что ВИЧ может ин-
фицировать моноциты. Инфекция этих клеток
является продуцирующей. Продуцирование ВИЧ
может активизироваться под действием лимфо-
цитов, при этом активирующим агентом может
быть фактор некроза опухолей. Эти аспекты
проблемы СПИДа объясняют некоторые сторо-
ны его патогенеза.
ВИЧ относится к РНК-содержащим ретрови-
русам, использующим клетки человека для своей
репликации. Патогенные ретровирусы человека
называют «молекулярными пиратами». Среди
них описано 5 различных типов. Первым был
открыт HTLV-1, вызывающий Т-клеточную
лейкемию. Вирус встречается в субтропиках и
является типичным представителем лентивиру-
сов, т. е. «медленных вирусов», равно как ВИЧ-1
и ВИЧ-2.
При репликации ВИЧ внедряется в геном
клеток человека, и РНК с помощью обратной
транскриптазы переходит в ДНК (ретропроцесс,
процесс возврата, аналогично онкогенным виру-
сам — HTLV-1, вирусам, вызывающим Т-кле-
точную лейкемию). Этот вирус также способен
распространяться по организму, имеет те же пути
передачи и вызывает болезнь после длительной
латенции.
Вирус имеет палочковидную или овальную
(реже округлую) форму, диаметр вируса пример-
но 100—140 нм. Он содержит внешнюю липид-
ную оболочку и ядерный протеид (капсид), каж-
дый включает РНК, ассоциируемые с ней белки
и ферменты в центре вирусной частицы. Обо-
лочка ВИЧ на финальной стадии репликации со-
держит белки клеток человека и гликопротеи-
ны, именуемые соответственно их молекулярной
массе gpl20 и gp41. Геном ВИЧ представлен
двумя идентичными однонитчатыми РНК и со-
держит 3 структурных гена: gag, env, pol. Пер-
вый кодирует групповые гликопротеидные ан-
556
Глава 49. ВИЧ/СПИД-инфекиия
557
тигены оболочки (gp41 и gpl20), второй — бел-
ки оболочки (р18 и р24), третий — РНК-зави-
симую ДНК-полимеразу (ревертазу) — фермент,
осуществляющий обратную транскрипцию —
синтез ДНК по матрице РНК вируса. Эта ДНК
встраивается в клеточный геном и называется
протовирусом.
На поверхности ВИЧ образуются многочис-
ленные отростки сложного строения, состоящие
из гликопротеинов. Именно от них зависит спо-
собность вируса проникать в клетку хозяина,
обусловливая ее гибель. Под сферической обо-
лочкой вириона наблюдаются замысловатые
сплетения нитей, образующих симметричную
структуру — «скелет», который удерживает сер-
дцевину вируса в определенном положении. В
конусовидной сердцевине вируса своеобразным
образом «упакованы» тяжи рибонуклеопротеи-
да — геном вируса. Фрагменты генома с помо-
щью обратной транскриптазы встраиваются в
генетическую информацию клеток человека. В
процессе размножения вируса зараженные клет-
ки погибают. В случае поражения клеток (лим-
фоцитов, макрофагов) у человека развивается
поражение иммунной системы. Он становится
беззащитным перед бактериями, другими виру-
сами, грибами. При заражении нервных клеток
(нейроглии) происходит поражение нервной си-
стемы.
ВИЧ-инфекция относится к группе медлен-
ных инфекций. Установлено, что ДНК вируса
СПИДа может находиться в геноме человека как
минимум три года. При этом никакой реплика-
ции вируса не происходит, и антитела в крови
не появляются. ВИЧ длительное время сохраняет
свою жизнедеятельность и в плазме крови, ли-
шенной клеточных элементов. Вероятно, имен-
но с этим связана необычайная «продуктив-
ность» пути передачи инфекции через шприц.
Для этого вируса характерна высокая степень
изменчивости — в один миллион раз больше,
чем у вируса гриппа.
ВИЧ обнаружен во всех жидкостях и тканях
организма. Клетками мишенями являются клет-
ки, имеющие на своей поверхности CD4 анти-
ген, который в наибольшем количестве содер-
жится на Т-лимфоцитах-хелперах. Другие клетки
тоже имеют CD4 антиген, это моноциты и их
тканевые формы — макрофаги, клетки Лангер-
ганса, фолликулярные клетки лимфоузлов, аль-
веолярные макрофаги легких, клетки микро-
глии. Моноциты и макрофаги обеспечивают
диссеминацию ВИЧ, они рефрактерны к цито-
патическому действию вируса, в отличие от CD4-
лимфоцитов.
Устойчивость ВИЧ во внешней среде относи-
тельно невысокая. Установлено, что ВИЧ пол-
ностью теряет инфекционность при нагревании
до температуры +56 °C в течение 30 мин, а так-
же при pH среды меньше 1 или больше 13.
Эффективными дезинфицирующими средства-
ми являются: этиловый и изопропиловый спир-
ты, параформальдегид, перекись водорода, ли-
зол, хлорная известь. Вирус относительно
устойчив к ионизирующему излучению и ульт-
рафиолетовым лучам, достаточно устойчив к вы-
сушиванию.
Патогенез и основные клинические
проявления ВИЧ-инфекции
Репликация ВИЧ начинается с продукции копий
РНК провируса. Это достигается воздействием
нуклеотидных последовательностей LTR (Long
Terminal Repeats), ограничивающих закодирован-
ную вирусную ДНК, каждый конец которой
формирует провирус, с использованием субстра-
тов инфицированных клеток, включая и клеточ-
ную ДНК. Новые вирионы собираются из боль-
шого числа белковых молекул, первоначально
образующих длинные полипептидные цепи бел-
ка-предшественника, которые затем специфичес-
ки раскалываются, представляя вирусные энзи-
мы и структурные протеины новых вирионов.
Энзим, называемый протеазой, содержится в каж-
дой молекуле предшественника, обеспечивая ко-
нечный этап формирования вирусных частиц.
Сначала этот фермент отщепляется от белка пред-
шественника, затем протеаза разрезает полипеп-
тидную цепь предшественника на определенные
сегменты, формируя энзимы. Оставшиеся сегмен-
ты протеинов образуют белковую оболочку, ко-
торая окружает РНК и вирусные энзимы, фор-
мируя внутренний капсид к ядру вирусной
частицы. Структурные протеины и оболочечный
протеин образуются и транспортируются к повер-
хности клеток независимо. Сборка новых вирус-
ных частиц начинается в углах инфицированной
клетки, где соединяются протеины-предшествен-
ники и прикрепляются к клеточной мембране.
Они формируют сферические образования, кото-
рые, разрывая клеточную мембрану, покидают
клетку и содержат внутри себя 2 цепочки РНК.
Последовательность репликации ВИЧ: gp 120
связывается с CD4 рецептором клетки и прони-
кает внутрь клетки.
РНК ВИЧ трансформируется в ДНК с помо-
щью обратной транскриптазы, образуя ДНК
провирус совместно с ДНК клетки.
Провирус транскрибируется обратно в матрич-
ную РНК, которая формирует вирусные части-
558 Венерология
цы для их сборки с помощью фермента протеа-
зы. Новые вирусные частицы покидают клетку.
Механизм биологического взаимодействия
ВИЧ с организмом хозяина представляет особые
трудности, обусловленные типом антител, выра-
ботка которых инициируется этим вирусом.
Поведение вируса в организме инфицированного
человека непредсказуемо. Наблюдения в течение
5 лет не выявили никаких клинических симп-
томов СПИДа более чем у 2/3 инфицированных
лиц. Примерно у 20 % инфицированных после
инкубационного периода наступает прогрессиру-
ющее ухудшение состояния здоровья, либо пе-
риоды вирусной активности сменяются перио-
дами ее затухания.
ВИЧ-1 и ВИЧ-2, вызывая первичную инфек-
цию, не приводят к клинической манифестации
в течение многих лет. Определенно неизвестны
причины длительности бессимптомного перио-
да. Вирусемия держится при этом на низком
уровне, репликация вируса идет медленно. Ухуд-
шение характеризуется интенсивной репликаци-
ей ВИЧ.
ВИЧ-инфекция протекает в виде коинфекции
с другими вирусами. Активная репликация од-
ного или двух из них может иметь значение для
ускорения репликации ВИЧ и прогрессии болез-
ни. Вирусы гепатита В, простого вируса, цито-
мегалии, 6 и 7 типа герпесвирусов, HTLV-1 из-
вестны как стимуляторы репликации ВИЧ в
клеточной культуре. Любой другой антиген,
который вызывает Т-клеточный иммунный от-
вет, стимулируя СО4-лимфоциты, приводит к
активации репликации ВИЧ в этих клетках.
СО4-лимфоциты, стимулированные антигеном,
обычно в ответ пролиферируют, вовлекая транс-
крипцию своего генетического материала в об-
разование новых СВ4-клеток. ВИЧ-провирус,
интегрированный в геном инфицированного
СО4-лимфоцита, вовлекается в процесс проли-
ферации, при этом увеличивается связывающая
способность белка клетки с «Long Terminal
Repeat» ВИЧ. Активация этого протеина, кото-
рый является триггером прогрессии болезни,
способствует репликации ВИЧ.
При ВИЧ-инфекции происходит прогресси-
рующее разрушение иммунной системы организ-
ма. Возбудитель СПИДа имеет выраженный
тропизм к Т-лимфоцитам. Это обусловлено чрез-
вычайно высоким аффинитетом гликопротеина
наружной оболочки вириона к гликопротеино-
вому рецептору Т4, располагающемуся на поверх-
ности Т-хелперов. Вирус размножается сначала
в Т-хелперах, затем распространяется по всему
организму. Поэтому ВИЧ удается выделить не
только из лимфоцитов крови и плазмы, но и из
грудного молока, слюны, мочи, слез, эякулята,
цервикального секрета.
При высокой поглотительной и всасывающей
способности, обильной васкуляризации слизис-
той оболочки мочеполовых путей и прямой киш-
ки легко открывается путь ВИЧ в кровоток.
Содержащиеся в сперме полиамины оказывают
иммуносупрессивное действие, обусловленное ее
трансглютаминазной активностью, и определя-
ют оптимальные условия для проникновения
вируса в слизистую оболочку.
Т-супрессоры являются носителями другого
гликопротеина, не обладающего родством к гли-
копротеину оболочки ВИЧ. Соотношение Т-хел-
перов и Т-супрессоров в норме составляет 2:1,
а при СПИДе —1:5. Известно, что Т-хелперы
стимулируют работу Т-киллеров, способных раз-
рушать мишени-клетки, увеличивать активность
макрофагов и стимулировать продуцирование
антител В-лимфоцитами. Подавление же попу-
ляции Т-хелперов Т-супрессорами ведет к угне-
тению этого процесса.
В-лимфоциты осуществляют свою функцию
только совместно с Т-лимфоцитами при условии
регуляции их взаимодействия белками-регуля-
торами (лимфокинами), секретируемыми Т-хел-
перами. Угнетение функции Т4-лимфоцитов
вызывает ослабление способности организма к
иммунному ответу у больных СПИД. У таких
больных лимфоциты-хелперы в циркулирующей
крови практически отсутствуют.
Установлена тропность ВИЧ и к макрофа-
гам — активным защитникам от инфекций.
Макрофаг способен к фагоцитозу и секреции
Таблица 49.1. Клеточная и вирусная динамика ВИЧ (David Н., 1996)
Жизненный цикл продуктивно инфицированной клетки 22 дня
Полная ВИЧ-1 продукция 10,3x109 вирионов в день
Минимальная продолжительность жизненного цикла ВИЧ-1 in vitro 1,2 дня
Длительность жизни вируса в плазме 0,3 дня
Внутриклеточная фаза 0,9 дня
Среднее время, необходимое ВИЧ-1 для продукции новой генерации 2,6 дня
Глава 49. ВИЧ/СПИД-инфекния
559
антибактериальных белковых веществ, является
помощником Т-киллеров, уничтожающих чуже-
родные субстанции, участвует в дифференциров-
ке тимоцитов и стволовых кроветворных клеток.
Экспериментально доказано, что Т-киллеры не
активны при отсутствии в среде макрофагов.
Макрофаги способны противодействовать росту
опухолевых клеток и осуществлять регуляцию
иммунного ответа.
Когда описанные выше иммунные процессы
снижаются до определенного уровня, человек
становится уязвимым для широкого спектра
оппортунистических инфекций и редко встреча-
ющихся злокачественных новообразований, по-
явление которых служит клиническим показа-
телем лежащего в их основе иммунного
дефицита.
Развитие ВИЧ-инфекции прямо или косвен-
но связано с вирусной инфекцией или реплика-
цией. Распространение ВИЧ-инфекции опреде-
ляется клетками, имеющими CD4 маркер на
своей оболочке. Это CD4-лимфоциты, моноци-
ты и макрофаги, фолликулы лимфоузлов, клетки
Лангерганса, а также другие — астроглия голов-
ного мозга, эндотелиальные клетки сосудов и
другие клетки моноцитарного ряда. Действие
ВИЧ на клетки иммунной системы является
центральным в патогенезе болезни.
На самых ранних стадиях болезни ВИЧ про-
являет тропизм к клеткам моноцитарно-макро-
фагальной линии. Эти клетки, вовлекаясь в
процесс, презентируют антиген. В макрофагах
или моноцитах ВИЧ способен реплицироваться,
продуцируя вирусные частицы и не разрушая эти
клетки. Моноциты/макрофаги способствуют
постепенной диссеминации вируса к другим
клеткам и тканям, однако в процессе вирусной
репликации в моноцитах и макрофагах образу-
ются мутанты вируса, которые быстрее репли-
цируются или формируют синцитиеобразующие
варианты ВИЧ, что имеет важное значение для
повреждения CD4 клеток. Причина этого меха-
низма селекции новых штаммов ВИЧ неизвест-
на, однако в ответ на инфекцию продуцируют-
ся нейтрализующие антитела и вовлекаются в
процесс цитотоксические Т-лимфоциты.
Способность ВИЧ интегрироваться в геном
клеток организма, быстро или медленно репли-
цироваться — важные характеристики, опреде-
ляющие развитие болезни (табл. 49.1). На началь-
ных стадиях болезни наряду с антителами ген nef
может быть выявлен как регулятор медленной
репликации, вирусные гены tat и rev способны
регулировать репликацию ВИЧ много лет. Весь-
ма важное значение для патогенеза имеет спо-
собность ВИЧ проникать в центральную не-
рвную систему через спинномозговой барьер и
спинномозговую жидкость, инфицировать клет-
ки микроглии на ранних стадиях болезни. Мик-
роглиальные клетки, представители моноцитар-
но-макрофагальной линии клеток, продуцируют
цитокины, один из них фактор некроза опухо-
лей (ФНО), играющий важную роль в невроло-
гических нарушениях при ВИЧ-инфекции.
До сих пор окончательно не установлен меха-
низм проявления клинических симптомов забо-
левания и остается неясным развитие СПИДа у
лиц с антителами к ВИЧ. Особую роль при ВИЧ-
инфекции играют заболевания, передающиеся
половым путем (сифилис, герпесвирусная инфек-
ция, хламидиоз, микоплазмоз). В последнее вре-
мя возрос интерес к цитомегаловирусной инфек-
ции в связи с ее большой частотой у больных
СПИДом. Беспорядочные сексуальные связи и
наличие многочисленных половых партнеров слу-
жат причиной того, что у больных ВИЧ-инфек-
цией часто выявляются также разнообразные
болезни, передающиеся половым путем. Особое
практическое значение имеет сочетание СПИДа
и сифилиса.
Стадии развития ВИЧ-инфекции
ВИЧ-инфекция — заболевание, включающее
спектр синдромов и различных ассоциированных
и индикаторных болезней. В раннюю фазу бес-
симптомной хронической инфекции нет ника-
ких признаков заболевания, кроме ПГЛ и лабо-
раторных изменений, присущих ВИЧ-инфекции.
Бессимптомная фаза может длиться много лет.
Продолжительность инкубационного периода ~
до 1 месяца. В это время появляются антитела
к ВИЧ, и только у 5% —- иммунный ответ, анти-
ВИЧ появляются спустя 6 месяцев после зара-
жения.
Острая стадия
Острая стадия развивается у 30—50 % больных,
клинически она напоминает инфекционный
мононуклеоз. Отмечается лихорадка, миалгия,
боли в горле, летаргия, кашель, ночные поты и
гастроинтестинальные симптомы, такие как тош-
нота, рвота, диарея, абдоминальные боли. При
клиническом осмотре выявляются увеличенные
лимфоузлы, гиперемия зева, сыпь. Заболевание
завершается благополучно в течение 1—6 недель.
Полагают, что этот синдром — результат актив-
ной иммунной реакции на ВИЧ. Лабораторные
данные позволяют выявить анти-ВИЧ, но не
постоянно. Не у всех больных анти-ВИЧ мож-
но выявить в начале этого синдрома, чаще — по
завершению его. Надежнее определять р24 ан-
560
Венерология
тиген ВИЧ. В этот период болезни можно об-
наружить транзиторное уменьшение CD4- и по-
вышение CD8-лимфоцитов.
Бессимптомная хроническая инфекция
Бессимптомная хроническая инфекция следует
за острой фазой болезни, но и при отсутствии
симптомов болезни в крови возможно выявле-
ние изолятов ВИЧ как следствие медленной реп-
ликации. Определение иммунологических пока-
зателей, выявление их снижения может иметь
значение для прогноза прогрессии болезни. У
многих больных в бессимптомную стадию пер-
вым клиническим признаком может являться
ПГЛ как результат активации В-лимфоцитов.
Гиперплазия лимфатических фолликулов сопро-
вождается активацией С D8-лимфоцитов, кото-
рые разрушают содержащие ВИЧ клетки. Не-
смотря на то, что отмечается инфекция с
высокой концентрацией ВИЧ в головном моз-
ге, нейрофизиологические симптомы при этом,
как правило, отсутствуют.
Симптоматическая хроническая
фаза ВИЧ-инфекции
Снижение СВ4-лимфоцитов происходит доста-
точно очевидно с появлением первых симптомов
болезни. В симптоматическую фазу в зависимо-
сти от степени снижения СВ4-лимфоцитов и
клинических данных выделяют СПИД-ассоци-
ированный комплекс и СПИД-индикаторные
болезни.
СПИД-ассониированный комплекс
СПИД-ассоциированный комплекс (ARC-AIDS
related complex) характеризуется такими симпто-
мами как профузные ночные поты, прогресси-
рующая слабость, потеря веса, диарея неясной
этиологии. Патогенез этих симтомов связывают
с интерлейкином-1 (ИЛ-1), вырабатываемым
моноцитами-макрофагами в ответ на ВИЧ-ин-
фекцию. Другие исследователи полагают, что это
может быть обусловлено усиленной выработкой
фактора некроза опухолей (ФИО). Вероятно,
имеет значение как усиление экспрессии ФИО,
так и выработка ИЛ-1 макрофагами. Прогнос-
тическое значение имеют также и уровень р24
антигена, IgA, бета-2-микроглобулина, неоптери-
на. Однако решающее прогностическое значение
имеет уровень вирусной нагрузки.
Эпидемиология
ВИЧ передается только при непосредственном
контакте жидкостей или тканей ВИЧ-позитивных
и здоровых людей. Источник инфекции — боль-
ной человек или бессимптомный вирусоноситель.
Особая роль в распространении СПИДа принад-
лежит группам повышенного риска заражения:
гомосексуалисты, бисексуалисты, проститутки и их
клиенты, наркоманы; больные гемофилией, зара-
женные ВИЧ; дети от больных СПИДом матерей,
больные венерическими инфекциями.
Основные пути заражения
• Половой (вагинальное или анальное сноше-
ние с зараженным партнером);
• парентеральный (переливание инфицирован-
ной ВИЧ крови; внутривенное введение лекар-
ственных средств или наркотиков нестерильны-
ми шприцами при повторном их использовании);
• трансплацентарный (внутриутробное зараже-
ние плода от больной матери).
ВИЧ не передается при бытовом контакте, че-
рез слюну и через насекомых. ВИЧ выделен из
многих жидкостей организма, включая слюну и
слезы, но пока неизвестны случаи заражения че-
рез какую-либо среду помимо крови, спермы и
вагинально-цервикального секрета. ВИЧ обнару-
жен в лимфоцитах периферической крови, спер-
ме, спинномозговой жидкости, шеечном и ваги-
нальном секретах, слюне, поте, грудном молоке,
моче инфицированных людей, а также в лимфо-
узлах, тканях головного мозга, клетках костного
мозга и коже. Проникновение ВИЧ в организм
может наблюдаться у медицинских работников
при соприкосновении с инфицированными тка-
нями или кровью, проведении лечебных проце-
дур, при осмотре больных СПИДом, при вскры-
тии трупов.
Во всем мире около 75 % случаев ВИЧ-инфек-
ции передается сексуальным путем. В развитых
странах гомосексуалисты составляют основную
группу больных, в развивающихся странах Аф-
рики и на Гаити преобладают гетеросексуалы.
Незащищенный секс, стадия ВИЧ-инфекции,
уровень вирусной нагрузки (количество РНК
ВИЧ в 1 мл плазмы крови) и состояние поло-
вых путей определяют степень риска сексуаль-
ной передачи. Анальный секс считают самым
рискованным в связи с большой травмирован-
ностью слизистых, так как при этом ВИЧ быс-
трее проникает в кровь. Пассивный партнер при
анальном сексе рискует больше, чем активный,
так как слизистая прямой кишки нежная и лег-
коранимая. При этом инфицируются эпители-
альные клетки слизистой, имеющие маркер CD4.
Травмы слизистой прямой кишки, а также по-
ловые инфекции (генитальный герпес, гонорея,
сифилис) резко повышают риск заражения. Риск
инфицирования при вагинальном сексе колеб-
лется от 7 до 74 %, что объясняется наличием
Глава 49. ВИЧ/СПИД-инфекиия
561
кофакторов: сексуальные болезни, число сексу-
альных партнеров, генетическая восприимчи-
вость, стадия ВИЧ-инфекции, величина вирус-
ной нагрузки. Оральный секс опасен не из-за
слюны (количество ВИЧ в слюне ничтожно
мало), а из-за возможной травматизации слизи-
стой рта и полового члена. Таким образом, риск
сексуальной трансмиссии определяется числом
сексуальных партнеров, видом и характером сек-
са, стадией ВИЧ-инфекции, состоянием генита-
лий, наличием эрозий и язв, сочетанием этих
обстоятельств.
Передача ВИЧ через кровь наблюдается у
инъекционных наркоманов; у пациентов при
использовании нестерильных инфицированных
шприцев и других инвазивных предметов
(ножницы, ножи, щипцы, бронхоскопы, гине-
кологические зеркала); при переливании ин-
фицированной крови и ее продуктов; у работ-
ников здравоохранения при чрезкожной
инокуляции крови и жидкостей инфицирован-
ного пациента.
Только цельная кровь, форменные элементы,
плазма, сыворотка и факторы свертывания опас-
ны, а такие продукты крови, как иммуноглобу-
лин, альбумин, гепатитная В-вакцина не могут
быть причиной инфицирования.
Инъекционные наркоманы считаются второй
группой риска. В некоторых странах (Италия,
Испания, Польша, Россия, Беларусь, Украина)
именно наркоманы определяют эпидемическую
ситуацию по ВИЧ-инфекции. В развитых стра-
нах считают общепринятым заражение через
общие нестерильные шприцы наркомана. При
этом придают значение иммунному статусу нар-
комана, типу наркотиков (наиболее опасный, с
наибольшим риском заражения, крэк-кокаин
с наибольшим риском заражения).
Примерно у 30 % (данные CDC — Center
Diseases Control —- USA) ВИЧ-инфекция связана
с переливанием инфицированной крови или VIII
фактора свертывания крови лицам, страдающим
гемофилией. Посттрансфузионный риск зараже-
ния приближается к 100 %. После внедрения
тестирования крови на ВИЧ риск заражения
снизился до 1 на 150 000 переливаний.
Трансплацентарный путь заражения у 80 %
педиатрического СПИДа ассоциируют с ВИЧ-
инфекцией у матерей-наркоманок. Возможна
перинатальная трансмиссия при грудном вскар-
мливании.
Клинические признаки СПИД
В 1987 году центр по контролю за заболевани-
ями в США (CDC — Center Diseases Control)
объявил вначале о синдроме, а затем о заболе-
вании под наименованием СПИД, наблюдае-
мом у гомосексуалистов и инъекционных нар-
команов. Все остальные сопутствующие или
связанные со СПИД болезни были названы
СПИД-индикаторными. Вначале у гомосексу-
алистов были выявлены необычные для моло-
дых людей пневмоцистная пневмония и сарко-
ма Капоши, наблюдавшиеся ранее только при
тяжелом иммунодефиците и у пожилых. В пос-
ледующем у молодых и «здоровых» гомосексу-
алистов были обнаружены другие возбудители,
вызывающие манифестные и рецидивирующие
заболевания, характерные только для иммуно-
супрессии: кандидоз, простой герпес, цитоме-
галия, токсоплазменный энцефалит, криптос-
поридийная диарея, криптококковый менингит
и большое число других редких болезней.
Спектр этих болезней включал также реакти-
вацию туберкулеза, кахексию, деменцию, лим-
фому головного мозга, атипичный микобакте-
риоз. Все эти патологические состояния
наблюдались при снижении СВ4-лимфоцитов
ниже 200 в мкл крови. Развитию новых синд-
ромов часто предшествовала персистирующая
генерализованная лимфаденопатия (ПГЛ). Это
нередко сочеталось с лихорадкой, диареей,
обильным ночным потоотделением, слабостью,
нарушением работоспособности. Лимфаденопа-
тия, или ПГЛ, рассматривалась как маркер ран-
него заболевания, а в сочетании с другими сим-
птомами и лабораторными данными — позднего
заболевания под названием СПИД-связанный
комплекс. СПИД-связанный комплекс предше-
ствовал развитию СПИД-индикаторных болез-
ней и тяжелой иммуносупрессии, определяемой
как СПИД. Клиническими маркерами СПИД-
связанного комплекса являются: герпес зостер,
лейкоплакия языка, кандидоз рецидивирую-
щий, рецидивирующий простой герпес, себорей-
ная экзема, паротит ЦМВ, гепатоспленомегалия,
ЛюЮрадка неясной причины, нарастающая об-
щая слабость, ночные поты, СПИД-индикатор-
ные болезни. На фоне нарастающей иммуносуп-
рессии развиваются тяжелые прогрессирующие
болезни, которые не встречаются у человека с
нормально функционирующей иммунной сис-
темой. Это болезни, которые ВОЗ определила
как СПИД-маркерные (индикаторные).
ВОЗ подразделяет ВИЧ-индикаторные болез-
ни на 2 группы (табл. 49.2).
Первая группа — это те заболевания, кото-
рые присущи только тяжелому иммунодефи-
циту (уровень СВ4-лимфоцитов ниже 200/мкл)
и поэтому определяют клинический диагноз
562 Венерология
даже при отсутствии анти-ВИЧ или ВИЧ-ан-
тигенов.
Вторая группа — это заболевания, которые
могут развиваться как на фоне тяжелого им-
мунодефицита, так и без него. В этих случаях
необходимо лабораторное подтверждение диаг-
ноза.
Все эти заболевания протекают с поражени-
ем одного или нескольких органов и систем:
головного мозга, легких, печени, желудочно-
кишечного тракта и носят тяжелый прогресси-
рующий характер, представляя угрозу для жиз-
ни. По мере снижения уровня CD4 клеток
патологический процесс принимает необрати-
мое течение: СПИД-индикаторные болезни по-
являются в различных сочетаниях, даже адек-
ватная терапия не приносит ожидаемого
эффекта. '
У детей ВИЧ-инфекция имеет некоторые
особенности. Чаще, чем у взрослых, встреча-
ется интерстициальный лимфоидный пневмо-
нит, гиперплазия пульмональных лимфоузлов,
вторичные бактериальные инфекции. Харак-
терны задержка темпов психомоторного и фи-
зического развития, энцефалопатия. Опухоли,
в том числе саркома Капоши, наблюдаются
редко. Болезнь прогрессирует быстрее, чем у
взрослых.
Значительный полиморфизм клинической
симптоматики ВИЧ-инфекции затрудняет созда-
ние классификации, учитывающей весь симпто-
мокомплекс с вторичными инфекциями, опухо-
лями, состоянием нервной и иммунной систем
и другие параметры.
Согласно классификации центра по контро-
лю за заболеваниями в США, диагноз СПИДа
устанавливается лицам, имеющим уровень CD4-
лимфоцитов ниже 200 в 1 мкл крови, даже при
отсутствии СПИД-индикаторных болезней
(табл. 49.3.).
Таблица 49.2. СПИД-индикаторные болезни (Бюлл. ВОЗ, 1988, 66, 2, 104-110)
Первая группа Вторая группа
• Кандидоз пищевода, трахеи, бронхов.
• Внелегочный криптококкоз.
• Криптоспоридиоз с диареей более 1 мес.
• Цитомегаловирусные поражения различных органов,
помимо печени, селезенки или лимфоузлов у больного
старше 1 мес.
• Инфекция, обусловленная вирусом простого герпеса,
проявляющаяся язвами на коже и слизистых оболочках,
которые персистируют более 1 месяца, а также бронхи-
том, пневмонией или эзофагитом любой продолжитель-
ности, поражающим больного в возрасте старше 1
месяца.
• Генерализованная саркома Капоши у больных моложе
60 лет.
•Лимфома головного мозга (первичная) у больных
моложе 60 лет.
•Лимфоцитарная интерстициальная пневмония и/или
легочная лимфоидная гиперплазия (ЛИП/ЛЛГ комплекс)
у детей в возрасте до 12 лет.
•Диссеминированная инфекция, вызванная атипичными
микобактериями (микобактерии комплекса М. avium
intracellulare) с внелогочной локализацией или локализа-
цией (дополнительно к легким) в коже, шейных лимфа-
тических узлах, лимфатических узлах корней легких.
• Пневмоцистная пневмония.
• Прогрессирующая многоочаговая лейкоэнцефалопа-
тия.
• Токсоплазмоз головного мозга у больного старше 1
месяца.
• Бактериальные инфекции, сочетанные или рецидиви-
рующие у детей до 13 лет (более двух случаев за 2 года
наблюдения): септицемия, пневмония, менингит,
поражения костей или суставов, абсцессы, обусловлен-
ные гемофильными палочками, стрептококками.
• Кокцидиоидомикоз диссеминированный (внелегочная
локализация).
• ВИЧ-энцефалопатия («ВИЧ-деменция», «СПИД-
деменция»).
• Гистоплазмоз диссеминированный с внелегочной
локализацией.
• Изоспороз с диареей, персистирующей более 1 мес.
•Саркома Капоши у людей любого возраста.
• Лимфома головного мозга (первичная) у лиц любого
возраста.
•Другие В-клеточные лимфомы (за исключением
болезни Ходжкина) или лимфомы неизвестного иммуно-
фенотипа:
• мелкоклеточные лимфомы (типа лимфомы Беркит-
та и др.);
•иммунобластные саркомы (лимфомы иммуноблас-
тные, крупноклеточные, диффузные гистиоцитарные,
диффузные недифференцированные).
•Атипичный микобактериоз диссеминированный с
поражением, помимо легких, кожи, шейных или прикор-
невых лимфоузлов.
•Туберкулез внелегочный (с поражением внутренних
органов, помимо легких).
•Сальмонеллезная септицемия рецидивирующая.
• ВИЧ-дистрофия (истощение, резкое похудание).
Глава 49. ВИЧ/СПИД-инфекиия
563
Таблица 49.3. Классификация ВИЧ-инфекции для подростков и взрослых (Центр по контролю за заболеваниями, США, 1993)
Уровень CD4 Клинические категории
А В с
Клеток Бессимптомная острая (первичная) ВИЧ-инфекция или ПГЛ Манифестная, нонеАинеС СПИД-индикаторные состояния
> 500/мкл А1 В1 ci ' '
200-499/мкл А2 В2 С2
< 200/мкл АЗ - - ' ' ' ВЗ • сз - <
— стадия СПИДа
Категория А включает бессимптомных ВИЧ
серопозитивных лиц, лиц с персистирующей
генерализованной лимфаденопатией (ПГЛ), а
также острой первичной ВИЧ-инфекцией.
Категория В включает различные синдромы.
Важнейшие из них: бациллярный ангиоматоз;
орофарингеальный кандидоз; рецидивирующий
кандидозный вульвовагинит, трудно поддающий-
ся терапии; цервикальная дисплазия, цервикаль-
ная карцинома; общеинфекционные симптомы,
такие как лихорадка или диарея более месяца,
волосистая «лейкоплакия» языка; опоясывающий
лишай (один или два эпизода), идиопатическая
тромбоцитопеническая пурпура, листериоз, вос-
палительные заболевания органов малого таза
вплоть до тубоовариального абсцесса, перифери-
ческая нейропатия. Критерии категории В име-
ют приоритет перед критериями категории
А. Например, больная, ранее леченная от канди-
доза полости рта или влагалища (не перешедшая
в категорию С), но без клинических проявлений
в момент осмотра, относится к категории В.
Категория С характеризуется клиническими
состояниями, перечисленными в определении
необходимости эпидемиологического надзора по
поводу СПИДа (табл. 49.4).
При установлении диагноза ВИЧ-инфекции
категории В ее рекомендуется сохранять, даже
если достигнуты успехи в лечении. Вместе с тем,
если есть основание подтвердить диагноз кате-
гории С, то это следует проводить немедленно.
Пересмотренная классификация клинических
и иммунологических проявлений ВИЧ-инфек-
ции создает основу для классификации заболе-
ваний, к развитию которых предрасполагает
ВИЧ и иммуносупрессия, а также помогает оце-
нить влияние этих заболеваний на размер эпи-
демии ВИЧ.
Согласно новой классификации Центра по
контролю за заболеваемостью (CDC, 1993), ди-
Таблица 49.4.
Клиническая классификация ВИЧ-инфекции
1. Стадия инкубации.
2. Стадия первичных проявлений:
А — острая лихорадочная фаза;
Б — бессимптомная фаза;
В — персистирующая генерализованная лимфаде-
нопатия.
3. Стадия вторичных заболеваний:
А — потеря массы тела менее 10 %; поверхностные
грибковые, бактериальные или вирусные поражения
кожи и слизистых оболочек; опоясывающий лишай;
повторные фарингиты, синуситы;
Б — прогрессирующая потеря массы тела более
10 %, необъяснимая диарея или лихорадка более 1
месяца, «волосистая» лейкоплакия, туберкулез
легких, повторные или стойкие бактериальные,
грибковые, вирусные и протозойные поражения
внутренних органов (без диссеминации) или глубокие
поражения кожи и слизистых оболочек, повторный
или диссеминированный опоясывающий лишай,
локализованная саркома Капоши;
В — генерализованные бактериальные, вирусные,
грибковые; протозойные, паразитарные заболевания,
пневмоцистная пневмония, кандидоз пищевода,
атипичный микобактериоз, внелегочный туберкулез,
кахексия, диссеминированная саркома Капоши,
поражения центральной нервной системы различной
этиологии.
4. Терминальная стадия.
агноз СПИДа устанавливается лицам, имеющим
уровень СВ4+Т-лимфоцитов ниже 200 в 1 мкл
крови, даже при отсутствии СПИД-индикатор-
ных болезней.
В России используется преимущественно кли-
ническая классификация ВИЧ-инфекции, пред-
ложенная В. И. Покровским (1989).
564 Венерология
Саркома Капоши
По классификации ВОЗ (1974), саркома Капо-
ши отнесена в группу злокачественных опухо-
лей кровеносных сосудов (ангиоретикулоэндо-
телиоз).
Формы саркомы Капоши
Различают 4 формы саркомы Капоши: споради-
ческая, эпидемическая, иммуносупрессивная,
злокачественная. Наиболее частой формой пора-
жения кожи при СПИДе является злокачествен-
ная форма саркомы Капоши.
Спорадическая или классическая форма сарко-
мы Капоши проявляется багрово-красными,
синюшными, темно-бурыми пятнами, бляшка-
ми, узлами, множественными и часто симметрич-
ными, расположенными в основном на конеч-
ностях, чаще на нижних, реже на коже туловища,
лица, слизистой полости рта. Не связанная со
СПИД саркома Капоши — редкое заболевание.
Болеют чаще мужчины старше 60 лед^Болезнь
протекает медленно (8—10 лет и более), хрони-
чески, с динамикой от пятнисто-узелковых вы-
сыпаний в бляшечные, а затем в опухолевидные.
Эпидемическая форма саркомы Капоши распро-
странена в экваториальной Африке и характери-
зуется высокой частотой висцеральных поражений;
изолированным поражением лимфатических узлов,
которые достигают очень больших размеров, без
сочетания лимфаденопатии с поражениями кожи.
Иммуносупрессивная форма саркомы Капоши
развивается у лиц, длительно получавших имму-
носупрессивную терапию по поводу других за-
болеваний, особенно после трансплантации ор-
ганов и тканей. Отмечается частое поражение
слизистой оболочки полости рта.
Злокачественная, ассоциированная со СПИД,
молниеносная форма саркомы Капоши являет-
ся наиболее характерным проявлением СПИДа,
уступая в частоте только пневмоцистной пнев-
монии. Поражаются лица мужского пола сред-
него и молодого возраста (20—50 лет). Очаги
саркомы Капоши у больных СПИД обычно
множественные, часто симметричные, могут воз-
никать практически на любом участке кожи —
туловище, руках, лице. На лице локализуются на
ушных раковинах, носу, конъюнктиве века, что
вызывает обширный отек на лице и вокруг глаз.
На туловище очаги нередко бывают удлиненны-
ми по ходу кожных борозд и располагаются
вдоль ребер. Возможно раннее поражение сли-
зистых неба, щек, языка, десен, губ и половых
органов. Высыпания иногда имеют вид высту-
пающей над уровнем слизистой оболочки бляшки
с гладкой поверхностью темно-красного цвета;
узла, резко отграниченного, безболезненного,
синюшно-красного или фиолетового цвета, ве-
личиной до лесного ореха. Болезнь характери-
зуется генерализованным поражением кожи,
слизистых оболочек, лимфатических узлов и
внутренних органов, а также резистентным к
терапии течением и быстрым летальным исхо-
дом, главным образом, по причине развития сеп-
сиса или пневмоцистной пневмонии.
Поражения кожи и слизистых оболочек
Различные поражения кожи и слизистых оболо-
чек являются частым проявлением заражения
ВИЧ. Они возникают практически при всех
клинических формах ВИЧ-инфекции, а не толь-
ко в стадии развившегося СПИДа и имеют важ-
ное диагностическое и практическое значение.
Природа поражений кожи и слизистых оболочек
весьма разнообразна, чаще всего встречаются:
себорейный дерматит; вирусные заболевания;
микотические заболевания; пиодермии.
Себорейный дерматит
Себорейный дерматит обнаруживается в период
вирусоносительства у 10 % ВИЧ-инфицирован-
ных при отсутствии других клинических призна-
ков, а в стадии развившегося СПИДа имеет ме-
сто у 46—83 % больных. Возникновение и
развитие себорейного дерматита у ВИЧ-инфи-
цированных связывают с активностью дрожже-
вых грибов, Pityrosporum ovale, увеличением со-
держания в очагах поражения Demodex
folliculorum. У некоторых больных себорейный
дерматит с элементами стрептодермии является
первым клиническим признаком СПИДа, воз-
никающим иногда за 1—2 года до появления
других клинических симптомов заболевания.
Клинические проявления себорейного дермати-
та при ВИЧ-инфекции протекают тяжело и ос-
тро. Высыпания могут выходить за пределы ло-
кализаций классического варианта в связи с
иммунодефицитным состоянием. Кожа приобре-
тает сходство с вульгарным ихтиозом, а высы-
пания из-за сильного шелушения и сухости со-
провождаются выраженным зудом,
преимущественно в области волосистой части го-
ловы. Большое распространение высыпаний и
их переход в эритродермическую форму в виде
сливных эритематозных фолликулярных бля-
шек — неблагоприятный прогностический при-
знак.
Герпес
Из вирусных заболеваний герпес простой и гер-
пес опоясывающий являются наиболее частыми
при СПИДе. Рецидивирующий герпес начина-
Глава 49. ВИЧ/СПИД-инфекиия
565
ется в форме классической инфекции, обуслов-
ленной вирусом простого герпеса, а с присоеди-
нением иммунодефицита приобретает генерали-
зованное или хроническое течение. Высыпания
появляются на губах, в полости рта, на половых
органах и в перианальной области. С присоеди-
нением вторичной инфекции герпес у таких
больных может иметь сходство с ветряной оспой
или импетиго. У инфицированных ВИЧ гомо-
сексуалистов может развиться герпетический
проктит с легким отеком, гиперемией кожи пе-
рианальной области и немногочисленными пу-
зырьками. Герпетические эрозии и язвы на ге-
ниталиях и в перианальной области лишены
тенденции к заживлению, очень болезненны. Ре-
цидивирующий перианальный или генитальный
герпес у больных СПИДом обычно принимает
тяжелую форму со все более длительными и
частыми рецидивами.
По клиническим особенностям у ВИЧ-инфици-
рованных (больных СПИД) можно выделить 4
формы герпетической инфекции:
• латентная — очагов нет, бессимптомное те-
чение;
• локализованная — на коже в виде простого
герпеса, герпетиформной экземы, язвенно-не-
кротического герпеса; на слизистой оболочке
полости рта в виде эрозивного, язвенного или
язвенно-некротического стоматита; на половых
органах — в виде уретрита, вульвовагинита, цеб-
вицита;
• генерализованная форма — в виде менинги-
та, энцефалита, менингоэнцефалита, гепатита;
• смешанная форма — поражаются кожа, сли-
зистые оболочки, внутренние органы, ЦНС.
Клиническим признаком неблагоприятного
прогноза у больных ВИЧ-инфекцией является
Herpes zoster — рецидивирующий опоясывающий
лишай. Полагают, что даже единственный эпи-
зод опоясывающего герпеса следует рассматри-
вать как индикатор ВИЧ-инфекции при возник-
новении его у лиц молодого возраста из группы
риска при отсутствии провоцирующих заболева-
ний и иммуносупрессивной терапии.
Простые бородавки, контагиозный
моллюск, остроконечные кондиломы
Больные, зараженные ВИЧ, имеют повышенную
склонность к появлению простых бородавок,
контагиозного моллюска, остроконечных конди-
лом. Эти высыпания у больных с клинически-
ми проявлениями ВИЧ-инфекции имеют распро-
страненный характер, локализуясь чаще всего на
лице, в полости рта, на половых органах, в пе-
рианальной области. Высыпания резистентны к
лечению и склонны к рецидивам. Папиллома-
вирус чаще обнаруживается у женщин, инфици-
рованных ВИЧ, и приводит к более частому
развитию у них дисплазий в шейке матки, вплоть
до инвазивной карциномы. «Волосатая» лейкоп-
лакия языка описана только у больных, инфи-
цированных ВИЧ. Возбудителем заболевания
считают вирус Эпштейна-Барр или папиллома-
вирусы.
Микотические заболевания
Микотические заболевания при инфекции ВИЧ
чаще всего представлены кандидозом, руброфи-
тией, разноцветным лишаем, паховой эпидермо-
фитией. Общими их особенностями являются
распространенность, упорное течение и стой-
кость к проводимой терапии. Особенно подозри-
тельными на наличие у пациентов ВИЧ являют-
ся стойкий кандидоз у молодых лиц (чаще
мужчин), не получавших антибиотики, цитоста-
тики, кортикостероиды; преимущественное вов-
лечение в процесс слизистых оболочек полости
рта, гениталий и перианальной области; тенден-
ция к образованию обширных очагов, сопровож-
дающихся болезненностью, склонностью к эро-
зированию. Оральный кандй^цоз часто
распространяется на область зева и пищевода.
Этим обстоятельством объясняются затруднения
при глотании, загрудинное жжение и боль во
время приема пищи. Отмечаются клинические
признаки кандидоза в виде поражений красной
каймы губ, углов рта, периоральной области,
паховых складок, туловища и ногтей.
Пиодермия
Вегетирующая, диффузная и, особенно, шанк-
риформная пиодермия считаются частыми при-
знаками ВИЧ-инфекции. Вегетирующая пиодер-
мия поражает преимущественно крупные
складки, клинически симулирует широкие кон-
диломы. Шанкриформная пиодермия, помимо
обычной локализации в области половых орга-
нов, развивается на верхней губе, в области яго-
диц. Шанкриформная пиодермия выглядит как
эрозивно-язвенный дефект кожи диаметром
1,0—1,5 см с округлыми очертаниями и резки-
ми границами. При пальпации выявляется плот-
ноэластический инфильтрат. Для пиодермий на
фоне ВИЧ-инфекции характерны распростра-
ненность процесса и рецидивирующее течение.
Сообщения о поражениях кожи при ВИЧ-
инфекции расширяют представления о клини-
ческих маркерах заболевания. Заслуживают вни-
мания следующие данные: ксеродерма с
выраженным зудом у больных хронической ди-
ареей; телеангиэктазии с локализацией на гру-
566 Венерология
ди, ушных раковинах, ладонях; трофические на-
рушения кожи и ее придатков — пеллагроидный
синдром, преждевременное поседение, гнездная
и очаговая алопеция. У некоторых больных мо-
гут наблюдаться генерализованный зуд и дли-
тельно существующий розовый лишай.
Патология кожи занимает одно из ведущих
мест среди ВИЧ-индикаторных и СПИД-мар-
керных заболеваний. Изменения кожи с упор-
ным течением, особенно у лиц группы риска с
учетом других данных объективного и лабора-
торного обследования всегда настораживают вра-
ча и дают основания для предположения о на-
личии ВИЧ-инфекции.
Диагностика ВИЧ-инфекции и СПИДа
Диагностика СПИДа проводится с учетом дан-
ных эпидемиологического анамнеза, клинических
симптомов, оценки иммунного статуса больных
и обнаружения специфических антител в крови,
а в ряде случаев и получения культуры ВИЧ.
При сборе анамнеза первостепенное внимание
уделяется эпидемиологическим факторам риска.
Учет эпидемиологических особенностей СПИДа
позволяет выявить источник и пути передачи
инфекции. Принадлежность к группам риска или
контакт с ними, переливание крови, другие па-
рентеральные манипуляции во время пребыва-
ния в эндемичных по ВИЧ странах, половые
контакты с гражданами этих стран указывают на
вероятность инфицирования.
Клинические признаки СПИД
Следующие клинические признаки, по мнению
экспертов ВОЗ, могут свидетельствовать в
пользу СПИДа:
• гистологически подтвержденная саркома
Капоши у лиц молодого возраста (30—35 лет и
моложе);
• лимфома головного мозга;
• ангиобластическая лимфаденопатия;
• пневмония (вызванная Pneumocystis carinii
или неясной этиологии), устойчивая к стандар-
тной терапии;
• длительная, необъяснимая лихорадка;
• генерализованная лимфаденопатия неясно-
го генеза;
• продолжительная диарея (более 1-2 месяцев);
• снижение массы тела больного в течение
месяца на 10 % и более;
• длительно не исчезающие поражения кож-
ных покровов и слизистых оболочек;
• бронхиальный и легочный кандидоз;
• эндогенные и экзогенные реинфекции и
суперинфекции.
Иммунологические признаки СПИД
При исследовании иммунологического статуса у
больных СПИДом чаще всего наблюдается:
• снижение общего числа лимфоцитов и Т-
хелперов;
• снижение коэффициента Т-хелперы/Т-суп-
рессоры (< 1,0);
• анемия, или лейкопения, или тромбоцито-
пения;
• уменьшение числа естественных киллеров;
• повышение уровней иммуноглобулинов сы-
воротки крови и циркулирующих иммунных
комплексов;
• снижение бласттрансформации при стиму-
ляции митогенами и уменьшение выработки эн-
догенного интерферона.
Лабораторные данные
Существует несколько методов лабораторной
диагностики СПИДа: выделение вируса, выявле-
ние антител к ВИЧ, обнаружение антигенов ВИЧ.
Выделения ВИЧ
Для выделения ВИЧ могут быть использованы
первичные культуры мононуклеарных лимфоци-
тов и перевиваемые линии Т4- и В-лимфоцитов.
Выделение вируса производится из крови, лим-
фы, пунктата лимфатических узлов, семенной
жидкости, молока кормящей матери, слюны,
мокроты, мочи, цервикального отделяемого,
кала. Чувствительность метода относительно
невысока и он трудновыполним, поэтому метод
изоляции ВИЧ применяется пока в научно-ис-
следовательских целях. Значительные преимуще-
ства имеет метод ДНК-зондов, основанный на
получении клонированных участков вирусного
генома, меченых радиоактивными маркерами
или ферментными зондами. Он позволяет про-
водить молекулярно-биологическое выявление
маркеров ВИЧ без дополнительного культиви-
рования.
Выявление антител к ВИЧ
Для выявления антител к ВИЧ применяется
несколько методов: иммуноферментный анализ,
иммуноблотинг, иммунофлюоресценция, радио-
иммунопреципитация, агглюцинация.
Самым простым, доступным и широко при-
меняемым в практике является иммунофермен-
тный анализ (ИФА) на выявление антител к
вирусу СПИДа в крови больных. По сравнению
с выделением вируса метод ИФА обладает зна-
чительно более высокой чувствительностью,
однако менее специфичен из-за возможности
получения ложноположительных и ложноотри-
цательных результатов.
Глава 49, ВИЧ/СПИД-инфекция
567
Антитела к ВИЧ появляются у 90—95 % ин-
фицированных в течение 3 месяцев после зара-
жения, у 5—9% — через 6 месяцев и у 0,5—1% —
в более поздние сроки. В стадии СПИДа коли-
чество антител может снижаться вплоть до пол-
ного исчезновения.
Забор крови производится из локтевой вены
в чистую сухую пробирку в количестве 3—5 мл.
Материал не следует длительно хранить, так как
наступивший гемолиз повлияет на результаты
анализа. Поэтому после взятия крови желательно
отделить сыворотку, которая может храниться в
холодильнике до 7 дней.
Метод ИФА является скрининговым. В слу-
чае положительного результата анализ в лабора-
тории проводится дважды (с той же сывороткой)
и при получении хотя бы еще одного положи-
тельного результата сыворотка направляется для
постановки подтверждающего теста.
Иммуноблотинг — тестирование на наличие
антител к отдельным вирусным антигенам. Ме-
тод иммуноблотинга используется для потверж-
дения специфичности результата, полученного
методом ИФА. Иммуноблотинг позволяет выя-
вить в сыворотке крови инфицированных обо-
лочечные и регуляторные белки ВИЧ. Однако
и данный метод дает ложноположительные ре-
акции с частотой 1: 4, особенно с белками Р18,
Р24 при исследовании крови больных тропичес-
кой малярией.
По рекомендации Российского центра по про-
филактике и борьбе со СПИД обнаружение ан-
тител к одному из гликопротеинов — gp 41. gp
120, gp 160 следует считать положительным ре-
зультатом. В случае обнаружения антител к дру-
гим белкам вируса результат считается сомни-
тельным и такого человека следует обследовать
еще дважды: через 3 и 6 мес. Отсутствие анти-
тел к каким-либо белкам ВИЧ означает, что
ИФА была ложноположительной. До положи-
тельного результата в иммуноблотинге и при
отрицательном результате человек считается здо-
ровым, противоэпидемические мероприятия не
проводятся.
Метод иммунофлюоресценции относительно
прост, в качестве антигенов в этой реакции ис-
пользуются различные клеточные линии инфи-
цированных ВИЧ.
Радиоиммунопреципитация — один из наибо-
лее специфичных и чувствительных методов
диагностики СПИДа, однако для него требуются
дорогостоящие радиоактивные материалы и обо-
рудование. Кроме того, для постановки реакции
необходимо культивировать инфицированные
вирусом клетки.
20 Зак. 1357
Метод агглюцинации является одним из наи-
более простых, чувствительных и специфичных
для определения антител к ВИЧ.
Обнаружение антигенов ВИЧ
Обнаружение антигенов ВИЧ в исследуемом ма-
териале основано, главным образом, на тех же
принципах, что и выявление антител. Однако су-
ществующие методы не позволяют регулярно об-
наруживать антигены в материалах, полученных
от больных, в связи с низким их содержанием.
В ряде лабораторий некоторых стран изучается
диагностическая ценность новой реакции — эн-
зимной амплификации генов. Эта реакция важна
для уточнения диагностики врожденного СПИ-
Да, так как она позволяет выявлять клетки,
пораженные вирусом, а не антитела.
В последние годы решающее значение для
прогноза и тяжести ВИЧ-инфекции имеет оп-
ределение «вирусной нагрузки» (viral load) —
определение количества копий РНК ВИЧ в плаз-
ме методом полимеразной цепной реакции
(ПЦР). Используется определение и других ви-
русологических маркеров: ВИЧ-антигенемия
(р 24); ВИЧ-виремия в плазме, клетках; ВИЧ
РНК и провирус ДНК качественно и количе-
ственно в плазме методом ПЦР.
Измерение вирусной нагрузки (количества
копий РНК ВИЧ в 1 мл плазмы крови) — наи-
лучший прогностический маркер отдаленного
клинического результата, когда по мере того как
число CD4 клеток падает, а иммунный ответ
слабеет, вирусная нагрузка растет. Однако боль-
ные с низкой вирусной нагрузкой, но с малым
числом CD4 клеток имеют большую склонность
к оппортунистическим изменениям. Если у па-
циентов, не получавших терапии, вирусная на-
грузка находится ниже предела определения (в
настоящее время это менее 5000 копий РНК
ВИЧ в 1 мл плазмы, но разрабатываются мето-
дики, позволяющие считать пределом 20 копий
в 1 мл), это свидетельствует о медленном про-
грессировании или отсутствии прогрессирования
вообще. Существенно, что степень заразности
при этом минимальная.
Вирусную нагрузку определяют методами, ос-
нованными на ПЦР на разветвленной цепи ДНК
и на определении последовательности нуклеино-
вых кислот. К сожалению, измерение вирусной
нагрузки не дает информации о резистентности
вируса, его фенотипе, цитопатичности — фак-
торах, важных для прогноза болезни. Также у
больных с очень высокой вирусной нагрузкой
степень иммунодефицита может быть более вы-
раженной, чем у больных с меньшей вирусной
568 Венерология
нагрузкой. Высокая вирусная нагрузка (напри-
мер, более 100 000 копий в 1 мл у пациентов с
CD4 ниже 300 в 1 мкл) всегда свидетельствует о
прогрессии болезни.
Лечение ВИЧ-инфекции
В настоящее время интенсивно разрабатывается
проблема специфического лечения ВИЧ-инфек-
ции. Весь комплекс используемых лечебных
мероприятий не обеспечивает выздоровления,
удается только облегчить состояние больных,
ослабить выраженность клинических проявле-
ний, продлить жизнь.
Общими принципами терапии ВИЧ-инфек-
ции являются предупреждение прогрессирования
болезни, сохранение состояния хронической вя-
лотекущей инфекции, диагностика и лечение
оппортунистических вторичных болезней. В
комплексную терапию включается полноценная
диета, витамины, психоэмоциональная поддер-
жка больных.
Стратегией и тактикой комплексной терапии
предусмотрена первичная и вторичная профи-
лактика оппортунистических инфекций. Имму-
нодефицит требует первичной профилактики
пневмоцистной пневмонии, криптококковой и
другой грибковой инфекции, цитомегаловирус-
ной инфекции, туберкулеза. Этиотропные пре-
параты с этой целью назначаются при уровне
СГ)4-лимфоцитов ниже 200 в 1 мкл. Вторичная
профилактика проводится этиотропными сред-
ствами для предупреждения рецидивов болезни.
Современные лекарственные средства и ме-
роприятия, применяемые при ВИЧ-инфекции,
можно подразделить на этиотропные, патогене-
тические, симптоматические.
Этиотропные средства
Этиотропные средства воздействуют на вирус
иммунодефицита, подавляя его репродукцию.
Основой назначения анти-ВИЧ противовирус-
ных средств является воздействие на механиз-
мы репликации вируса. ВИЧ интегрируется в
генетический аппарат инфицированной клетки,
ввиду этого идеальное терапевтической средство
должно действовать в нескольких направлени-
ях: предупреждать инфицирование новых кле-
ток организма, разрушать внедрившийся вирус
и/или останавливать его репликацию.
В последние годы разработана комбинирован-
ная противовирусная терапия ВИЧ-инфекции,
включающая ингибиторы обратной транскрип-
тазы и ингибиторы протеазы, воздействующие
на ключевые ферменты, обеспечивающие репли-
кацию ВИЧ.
Нуклеозидные ингибиторы
обратной транскриптазы
Азидотимидин (зидовудин, ретровир) — дидеок-
синуклеотид, 3-азидо, 3-дидеокситимидин —
долгое время был одним из наиболее эффектив-
ных препаратов для лечения болезни. Внутри
клетки азидотимидин (АЗТ) фосфорилируется,
превращаясь в активный трифосфат, аналог ти-
мидинтрифосфата, ингибирует синтез противо-
вирусной ДНК двумя путями: заместительным
путем и прерывая образование цепи. АЗТ — се-
лективный ингибитор репликации ВИЧ-1 и
ВИЧ-2 в СВ4-лимфоцитах, макрофагах, моно-
цитах. Дозы — 200 мг 3 раза или 300 мг 2 раза
в сутки. Принимается с пищей или за 1 час до
или после еды. Побочные эффекты: гематоло-
гические изменения (анемия, лейкопения), ми-
озит, миопатия. Для коррекции анемии назна-
чается гемопоэтин, гранулоцитопении —
нейпоген. АЗТ отчетливо уменьшает реплика-
цию ВИЧ и прогрессию ВИЧ-инфекции, удли-
няет продолжительность жизни и улучшает ее
качество, снижая тяжесть оппортунистических
инфекций. Удлинение жизни и ее качества все-
гда сопровождалось уменьшением тяжести ВИЧ-
оппортунистических инфекций, неврологических
дисфункций, увеличением уровня CD4 клеток
и снижением количества ВИЧ в плазме больных
СПИДом. Побочные эффекты АЗТ в основном
связаны с необходимостью использования боль-
ших доз у тяжелых больных и токсичностью
препарата для костного мозга.
Зальцитабий (2,3 дидеоксицитозин, хивид,
ddC) — пиримидиновый аналог нуклеозида де-
зоксицитидина, в котором гидроксильная группа
заменена в 3-ем положении атомом водорода.
После превращения в активный 5-трифосфат
под действием клеточных киназ он становится
конкурентным ингибитором обратной транс-
криптазы. Дозы — 0,75 мг 3 раза в сутки. На-
значается натощак за 1 час до еды или через 2
часа после еды. Основные побочные эффекты:
головная боль, панкреатит.
Ламивудин (ЗТС, эпивир) — синтетический
аналог нуклеозидов. Внутриклеточно ламивудин
фосфорилируется в активный 5-трифосфат ме-
таболит (L-TP), который находится в клетке до
полураспада от 10,5 до 15,5 часов. LTP ингиби-
рует обратную транскриптазу путем ограничения
синтеза вирусной ДНК. LTP также ингибирует
активность РНК-зависимой ДНК-полимеразы
(обратной транскриптазы). Установлен синер-
гизм действия ламивудина и ретровира. При
комбинированной терапии ламивудином и рет-
ровиром задерживается появление резистентных
Глава 49. В И Ч/СП ИА-инфекция
569
к химиотерапии штаммов ВИЧ. Дозы — 150 мг
2 раза в сутки. Рекомендуется прием с пищей.
Основные побочные эффекты: головная боль,
тошнота; редко — сыпь, нейтропения.
Ставудин (d4T, зерит) — ингибитор обратной
транскриптазы. Дозы — 40 мг 2 раза в сутки
(при весе тела свыше 60 кг). Назначается с пи-
щей, хранить в холодильнике. Основные побоч-
ные эффекты: головная боль, сыпь, нейропатия,
панкреатит.
Видекс (ddl, диданозин, дидеоксиинозин) —
ингибитор обратной транскриптазы. Дозы —
200 мг 2 раза в сутки (при весе тела свыше
60 кг); 125 мг 2 раза в сутки (при весе тела ме-
нее 60 кг). Принимается на голодный желудок
или спустя 2 часа после еды. Основные побоч-
ные эффекты: диарея, боли в животе, тошнота,
рвота, панкреатит.
Ненуклеозидные ингибиторы
обратной транскриптазы
Ненуклеозидные ингибиторы обратной транс-
криптазы — новейший класс антиретровирусных
веществ, останавливают репликацию ВИЧ, при-
крепляясь непосредственно поверх обратной
транскриптазы и предотвращают преобразование
РНК в ДНК. Препараты называются ненуклео-
зидными ингибиторами, потому что они дей-
ствуют на тех же стадиях процесса, что и нук-
леозидные аналоги, но другим образом.
Невирапин (таблетки по 200 мг), принимают-
ся по 200 мг 1 раз в день первые 14 дней, затем
по 200 мг 2 раза в день. Препарат метаболизи-
руется цитохромом, 80 % в неизменном виде вы-
водится с мочой, индуцирует ферменты цитох-
рома.
Делавирдин (таблетки по 100 мг), принимают-
ся по 400 мг 3 раза в день. Препарат выводится с
мочой (51 %) и калом (44 %). Делавирдин подав-
ляет ферменты системы цитохрома, повышает
концентрацию индинавира и инвиразы, при их
совместном назначении необходим перерыв в 1 час.
Ингибиторы протеазы
Ингибиторы протеазы блокируют активность
фермента, который используется ВИЧ для стадии
воспроизведения вирионов. Аспартат протеаза
действует как ножницы, разрезая полоски белков
на зрелые частицы вируса, которые покидают
инфицированную клетку — воспроизводитель
ВИЧ. Ингибиторы протеазы связывают активный
участок фермента, предупреждая образование
новых вирусных частиц. Основные ингибиторы
протеазы проявляют противовирусную активность
в лимфоидных клетках человека, клетках моно-
цитарного ряда, а также в отношении изолятов
ВИЧ, устойчивых к зидовудину. В настоящее
время известны 4 ингибитора протеазы — инви-
раза (саквинавир), индинавир (криксиван), рито-
навир (норвир), нельфинавир (вирасепт).
Инвираза (саквинавир) принимается внутрь по
600 мг 3 раза в сутки (9 капсул по 200 мг в сут-
ки) во время или сразу после еды, содержащей
достаточное количество жира. Побочные эффек-
ты: диарея, тошнота, боли в животе. Препарат
обладает низкой биодоступностью, метаболизи-
руется при первом прохождении через печень под
действием ферментов системы цитохромов. Ин-
дукторы ферментов, рифампицин угнетают его.
Вместе с тем саквинавир проявляет выраженную
противовирусную активность как при моноте-
рапии, так и в сочетании с АЗТ, зальцитабином.
Индинавир (криксиван) принимают внутрь по
800 мг 3 раза в сутки (6 или 12 капсул в сутки
по 200 или 400 мл) натощак, запивая водой в
большом количестве или нежирным молоком (не
менее 1,5 л в сутки). Побочные эффекты: не-
фролитиаз, гиперглобулинемия. При печеночной
недостаточности и нефролитиазе дозу нужно
снизить до 600 мг 3 раза в сутки или отменить
препарат. Индинавир имеет преимущество по
сравнению с ритонавиром из-за низкой связы-
ваемое™ с белками плазмы и, следовательно,
способности достигать более высоких концент-
раций в плазме, тканях и проникать в ЦНС.
Ритонавир (норвир) принимают внутрь по
~600 мг 2 раза в сутки (12 капсул по 100 мг) во
время еды или после еды, запивая молоком.
Побочные эффекты: астения, диарея, тошнота,
рвота. Ритонавир можно использовать для на-
чальной терапии одновременно с ретровиром и
зальцитабином (ddC).
Нельфинавир (вирасепт) принимают внутрь по
750 мг 3 раза в сутки (9 таблеток в сутки) во вре-
мя еды, запивая полноценным необезжиренным
молоком (3,5—4 % жирности). Для детей порошок
разводится молоком или водой. Побочные эффек-
ты: диарея, тошнота, метеоризм. Нельфинавир —
единственный из ингибиторов протеазы, который
рекомендуют для лечения СПИДа у детей.
Ингибиторы протеазы плохо проникают через
гематоэнцефалический барьер, поэтому их целе-
сообразно назначать с ретровиром. При выборе
ингибиторов протеазы следует проявлять осторож-
ность и учитывать последовательность примене-
ния. Длительная монотерапия ингибитором про-
теазы индинавиром может привести к отбору
штаммов ВИЧ с перекрестной резистентностью
к различным классам препаратов. К саквинави-
570 Венерология
РУ формируется особая устойчивость, свойствен-
ная только этому препарату, поэтому он являет-
ся препаратом выбора на старте комбинирован-
ной терапии, и это единственный препарат,
который можно применять после индинавира.
В настоящее время появились данные по ис-
пользованию ненуклеозидных ингибиторов об-
ратной транскриптазы (невира-пин, делавирдин,
лоривид) в сочетании с АЗТ и ингибиторами
протеазы, что замедляет развитие резистентнос-
ти к этим препаратам. Общими выводами для
комбинированной терапии должны быть: лече-
ние ранних стадий ВИЧ-инфекции желательно
начинать в фазу сероконверсии.
Новые данные о быстром развитии резистент-
ности к противовирусной терапии указывают на
необходимость начинать лечение ВИЧ-инфекции
с максимальных доз двух или трех противовирус-
ных препаратов. При этом важна непрерывность
лечения — «привязанность к препаратам», ибо
пропуск, нарушение режима их приема резко
снижает эффективность терапии.
Из этиотропных средств наиболее перспектив-
ным является создание вакцин и лекарственных
препаратов, воздействующих на жизненный
цикл вируса. В настоящее время ведутся интен-
сивные поиски безвредной и эффективной вак-
цины. Для изготовления вакцины используют
убитый вирус, синтетические пептиды, реком-
бинантные вирусы. Однако вариабельность бел-
ков ВИЧ, его быстрая изменчивость затрудня-
ют разработку вакцины. Сложность создания
вакцины определяется также существованием
нескольких типов возбудителя ВИЧ, а также
высокой мутабельностью вируса. Сейчас пред-
принимаются попытки создать вакцины на ос-
нове белка gpl20 и антиидиотипические вакци-
ны на основе антител против CD4. В настоящее
время с помощью генной инженерии в США уже
создана вакцина против ВИЧ и препарат испы-
тывается на больных СПИДом. По мнению спе-
циалистов, даже при успешном результате испы-
таний потребуется 3—5 лет, чтобы вакцина стала
общедоступной.
Патогенетическая терапия
Патогенетическая терапия включает использо-
вание иммуномодулирующих или иммунозаме-
стительных препаратов. К иммуномодулирую-
щей терапии обращаются с целью коррекции
развивающегося при СПИДе иммунодефицита.
Применяют естественные регуляторы иммунных
реакций (гормоны тимуса, интерлейкин, интер-
ферон), синтетические препараты (имутиол,
изопринозин, левамизол, бенамбакс-300). В ка-
честве иммунозаместительной терапии предла-
гаются пересадка костного мозга, зрелые тимо-
циты, фрагменты тимуса. Используют также ги-
пертермию и гипертермическую обработку
лимфоцитов крови больного.
Такие иммуномодуляторы, как интерферон и
амплиген, обладают противовирусным и имму-
номодулирующим действием. Другие — такие
как интерлейкин-6, фактор некроза опухолей,
полипептиды — могут также воздействовать на
ВИЧ репликацию. Некоторые иммуномодулято-
ры способны поддерживать репликацию ВИЧ на
низком уровне, улучшая иммунный статус. Иму-
тиол (дитиокарб) — наиболее изученный имму-
номодулятор. В дозе 5 мг/кг или 200 мг/м2, вве-
денный внутривенно один раз в сутки, он
задерживал прогрессию инфекции, что было
подтверждено клинически и иммунологически,
однако это воздействие носило временный ха-
рактер.
Циклоферон — синтетический низкомолекуляр-
ный индуктор интерферона с иммуномодулирую-
щим эффектом. Препарат выпускается по 250 мг
12,5 % раствора в ампулах по 2,0 мл для внутри-
^мышечных и внутривенных инъекций. Циклофе-
рон как монопрепарат оказывает иммуномодули-
рующее и противовирусное действие при лечении
ВИЧ-инфицированных в тех случаях, когда нет
значительного снижения числа CD4+ клеток. Курс
циклоферона по 4 мл внутримышечно в 1, 2, 4, 6
и 8-й дни лечения. По второй схеме препарат вво-
дится по 4 мл внутривенно в 1 и 2 день, а затем
внутримышечно по 4 мл на 4, 6, 8, 10, 13, 16, 19,
22-й дни лечения. Циклоферон рекомендуется
использовать в сочетании с противовирусными
препаратами у больных с обострениями различных
оппортунистических заболеваний с признаками
активной репликации ВИЧ (высокие показатели
вирусной нагрузки), что позволяет снизить дозы
азидотимидина и уменьшить его токсическое дей-
ствие на организм.
Методы иммунокорригирующей и иммуноза-
местительной терапии могут значительно облег-
чить течение заболевания и продлить жизнь
больного.
Симптоматическая терапия
Симптоматическая терапия направлена на устра-
нение оппортунистических инфекций, неопла-
стических процессов и определяется ведущими
синдромами в клинике заболевания.
Для профилактики пневмоцистной пневмонии
(при CD4 ниже 200 в 1 мкл) и ее лечения ис-
пользуются бисептол, септрин, а при их непе-
реносимости — дапсон.
Глава 49, ВИЧ/СПИД-инфекция
571
При грибковых поражениях используют
кетоконазол (низорал), флуконазол (дифлю-
кан).
Герпесвирусные инфекции являются показа-
нием для назначения лечебных и профилакти-
ческих курсов ацикловира, ганцикловира, фос-
карнета.
Лицам с положительной реакцией Манту про-
водится химиопрофилактика туберкулеза изони-
азидом. На развернутых стадиях туберкулеза
используют сочетания стрептомицина, рифампи-
цина, изониазида, этамбутола. При CD4 менее
100 в 1 мкл, когда повышается вероятность раз-
вития атипичного микобактериоза, назначают
рифампицин.
Бактериальные инфекции требуют комплек-
сной антибиотикотерапии, рассчитанной на ши-
рокий круг условно-патогенных, патогенных
микробов, в том числе анаэробов.
При саркоме Капоши проводится противоопу-
холевая терапия. Раннее выявление, лечение и
профилактика вторичных болезней существен-
но увеличивают продолжительность жизни ВИЧ-
инфицированных пациентов.
В настоящее время известно, что многие про-
тивовирусные агенты могут воздействовать на
репликацию ВИЧ. Среди них глицирризиновая
кислота, тауролитохолевая кислота, кастанб^пер-
мин, рибавирин, амфотерицин В, Д-пеницилла-
мин, папаверин, фузидиевая кислота. В перспек-
тиве целесообразно использовать антисмысловые
олигонуклеотиды.
Однако, несмотря на достаточно большое ко-
личество применяемых препаратов и способов ле-
чения больных СПИДом, результаты терапии в
настоящее время не приводят к полному выздо-
ровлению. Проблема эффективного лечения
СПИДа требует своего дальнейшего изучения и
обоснования.
Профилактика ВИЧ/СПИД
СПИД поставил перед человечеством как науч-
ные проблемы, обусловленные биологическими
свойствами вируса, так и социальные и юриди-
ческие вопросы, связанные с состоянием нравов
и условиями жизни людей.
В настоящее время изучаются и обсуждаются
различные направления в комплексе мер по про-
филактике ВИЧ/СПИД. Главный упор делает-
ся на профилактику заболевания, связанную с
пропагандой средств предупреждения заражения
ВИЧ; изменением образа жизни людей; сведе-
нием к минимуму факторов риска среди лиц,
относящихся к группам повышенной опасности
инфицирования; изучением социальных усло-
вий, способствующих распространению заболе-
вания. Существуют юридические и медицинские
аспекты снижения роста заболеваемости и рас-
пространения ВИЧ-инфекции.
Система профилактики ВИЧ/СПИД включает
следующие основные направления
деятельности медицинских служб:
• раннее, полное и активное выявление боль-
ных СПИДом и ВИЧ-инфицированных лиц;
• информирование и санитарное просвещение
населения;
• организация серологических и вирусологи-
ческих обследований определенных групп насе-
ления и в первую очередь групп риска;
• предупреждение инфицирования ВИЧ при
переливании крови и ее препаратов;
• предупреждение передачи ВИЧ от больной
матери к ребенку;
• профилактические мероприятия среди лиц,
инфицированных ВИЧ;
• предупреждение инфицирования ВИЧ ме-
дицинского персонала;
• разработка средств специфической профи-
лактики СПИДа и методик первичной профи-
лактики.
Следует отметить, что несмотря на длитель-
ный контакт с больными СПИДом, доказанных
случаев инфицирования медицинских работни-
ков при выполнении служебных обязанностей
нет. Это подтверждается многочисленными це-
ленаправленными исследованиями среди меди-
цинского персонала больниц, обслуживающих
больных СПИДом. С целью предупреждения
профессионального инфицирования среди меди-
цинских работников должны проводиться те же
профилактические мероприятия, что и при ви-
русном гепатите.
Клинический и лабораторный персонал,
обслуживающий больных СПИДом, должен:
• работать в специальных халатах, спецодеж-
де, которая снимается при выходе из помещения
или лаборатории;
• после окончания работы, снятия перчаток и
спецодежды необходимо тщательно вымыть руки
и другие открытые участки тела, а случайно за-
раженные кровью поверхности тела предвари-
тельно обработать одним из дезинфицирующих
растворов;
• работа с кровью и биологическими жидко-
стями больных СПИДом без перчаток запреща-
ется;
• предметы и пробирки, загрязненные кровью
больных СПИДом, помещают в специальные
герметические пакеты или мешки с предупреди-
572 Венерология
тельной надписью и обрабатывают так же, как
материал, инфицированный вирусом гепатита В;
• в лабораториях необходимо пользоваться
одноразовыми посудой и наконечниками авто-
матических пипеток;
• избегать случайного ранения острыми инст-
рументами, загрязненными потенциально инфи-
цированным материалом, а также контактов
поврежденной кожи с материалом от больных;
• в случае загрязнения персонала и объектов
окружающей среды (стен, пола) выделениями
больных необходимо провести обработку дезин-
фицирующими растворами;
• лица, имеющие повреждения кожных по-
кровов и слизистых, к работе с больными СПИ-
Дом не допускаются.
Приложения
Приложение А
Перечень и описание практических
навыков по кожным и венерическим болезням
1. Методика телесного осмотра
дерматологического больного
Осмотр лучше проводить при рассеянном днев-
ном либо достаточно ярком электрическом све-
те. Температура в комнате (кабинете) должна
быть 22—23 °C, т. к. низкая температура вызы-
вает спазм сосудов (кожа бледнеет), а более вы-
сокая — их расширение (наступает гиперемия),
что искажает истинный цвет кожи.
Желательно осматривать всю кожу, обращая
внимание и на лицо больного, которое может
выражать различные эмоции и состояния (раз-
драженность, страдание, усталость, безразличие),
что свойственно больным с некоторыми дерма-
тозами. Форма и деформация носа имеют зна-
чение при диагностике сифилиса (седловидный),
туберкулезной волчанки (разрушение мягких
тканей — «бараний нос»), ринофимы.
Уделяют внимание окраске кожи, которая в
норме может быть бледной, розовой, смуглой.
У здоровых людей она матовая, не имеет жир-
ного блеска, отсутствуют зияющие протоки
сальных желез. При некоторых заболеваниях
кожа меняет свою окраску. Она может быть
синюшной при застойных явлениях, сине-бу-
рой — при акродерматите, коричневой — в
начальной стадии грибовидного микоза, жел-
той — при желтухе, серовато-черной — при
ихтиозе, меланодермии, бронзовой — при ад-
дисоновой болезни.
При дерматозах, сопровождающихся наруше-
нием функции вегетативной нервной системы,
кожа бывает влажной; при почесухе Бенье, дис-
семинированном нейродермите, ихтиозе — сухой.
Осматривают волосы, ногти, наружные поло-
вые органы, заднепроходную область.
При осмотре красной каймы губ обращают
внимание на ее окраску, сухость, наличие чешу-
ек, трещин, эрозий, корок. Подлежит осмотру и
слизистая оболочка полости рта, на которой
можно обнаружить высыпания (при кандидозе,
красном плоском лишае, пузырчатке).
2. Специальные методы
обследования кожного
больного
1. Пальпация (ощупывание). Пальпаторно опре-
деляют пластичность и тонус кожи. В норме эла-
стичность и тонус кожи на разных участках об-
щего покрова у одного и того же человека не
одинаковы. Учитывают также возрастные изме-
нения кожи: снижение тургора и эластичности
у пожилых и старых людей. Пальпаторно мож-
но определить повышение или понижение тем-
пературы кожи (рожистое воспаление, глубокая
стафилодермия, эритродермия, болезнь Рейно,
склеродермия); расположение элемента (эпидер-
мис, дерма, подкожная основа), его размеры,
форму, консистенцию, спаянность с окружаю-
щими тканями, болезненность. При отеке опре-
деляют епхинтенсивность, наличие флюктуации
(абсцесс, гидраденит), отсутствие болезненнос-
ти (первичная сифилома).
2. Поскабливание (граттаж). Проводится предмет-
ным стеклом или тупым скальпелем. Метод поскаб-
ливания позволяет обнаружить шелушение кожи,
определить его характер (муковидное, отрубевид-
ное, мелкопластинчатое, крупнопластинчатое),
плотность прикрепления чешуек к поверхности
кожи. Метод применяется при псориазе (триада
феноменов), разноцветном (отрубевидном) лишае,
парапсориазе. Не рекомендуется проведение поскаб-
ливания ногтем, так как при этом возможно зара-
жение (дерматомикоз, папулезный сифилид).
3. Диаскопия (витропрессия) — надавливание на
пораженный участок кожи предметным стеклом
либо специальным прибором — диаскопом (про-
зрачная пластмассовая пластинка). С помощью
этого метода определяют характер элемента (со-
судистый, пигментный). Сосудистое пятно при
надавливании побледнеет или исчезнет, а гемор-
рагическое пятно (или пигментное) — не исче-
зает. Этот метод помогает при диагностике тубер-
кулезной волчанки (феномен «яблочного желе»).
574
Перечень и описание практических навыков по кожным и венерическим болезням
575
3. Методика телесного осмотра
венерического больного
Осмотр половых органов у женщин производят в
положении больной на спине в гинекологичес-
ком кресле. Вначале осматривают большие по-
ловые губы, кожу промежности, заднепроходной
области, обращают внимание на окраску, отсут-
ствие или наличие высыпаний, выделений. Дву-
мя палочками, браншами пинцета или пальца-
ми левой руки в перчатке раздвигают малые
половые губы и осматривают преддверие влага-
лища, обращают внимание на состояние дев-
ственной плевы у девушек или остатков ее у
живших половой жизнью. Затем осматривают
устья выводных протоков больших желез пред-
дверия, обращают внимание на их окраску, вы-
деления. Железы пальпируют большим и указа-
тельным пальцами (левую железу — левой
рукой, правую — правой). При массаже железы
можно получить секрет, который в случае необ-
ходимости берут для микроскопического иссле-
дования. (
Осматривают наружное отверстие и лакуны
мочеиспускательного канала, обращают внимание
на их гиперемию, отечность, выделения. Паль-
пируют мочеиспускательный канал, определяют
состояние его стенок, наличие уплотнений. Паль-
пируя мочеиспускательный канал указательным
пальцем, одновременно производят его массаж
сзади наперед. Полученные выделения исследу-
ют и под микроскопом. Затем больную просят
освободить мочевой пузырь последовательно в два
стакана. Первая порция мочи указывает на состо-
яние мочеиспускательного канала, вторая — на
изменения в вышележащих мочевых путях. У
женщин, живших половой жизнью, осматрива-
ют влагалище с помощью двустворчатого зерка-
ла Куско. При этом определяют величину и фор-
му шейки матки, состояние отверстия матки,
наличие гиперемии, отечности, эрозии, выделе-
ний (их характер и количество). Поворачивая рас-
крытое зеркало, осматривают стенки влагалища,
желобоватым зондом или фолькмановской лож-
кой берут выделения из канала шейки матки для
микроскопического исследования. При наличии
эрозии в подозрительных случаях с ее поверхно-
сти петлей берут отделяемое для исследования на
содержание бледной трепонемы. После удаления
зеркала из влагалища производят ручное иссле-
дование, обращая особое внимание на плотность
шейки матки, что особенно важно при диагнос-
тике первичного сифилиса.
Осмотр половых органов у мужчин. Половые
органы у мужчин осматривают при хорошем
освещении. Больной не должен мочиться на
протяжении 3—4 часов. Вначале осматривают
кожу нижней части живота, внутреннюю повер-
хность бедер, обращая внимание на наличие или
отсутствие высыпаний, состояние паховых и
бедренных лимфатических узлов, а затем — по-
ловой член, наружный и внутренний листки
крайней плоти, головку, край наружного отвер-
стия мочеиспускательного канала. Устанавлива-
ют наличие парауретральных протоков, эрозий,
язв. При обнаружении последних с них снима-
ют налет и пальпируют их основание. Отделяе-
мое берут для исследования на наличие бледной
трепонемы. Осматривая наружное отверстие
мочеиспускательного канала, обращают внима-
ние на его диаметр, наличие воспаления (гипе-
ремия, отечность), выделений, их характер (се-
розные, слизистые, слизисто-гнойные, гнойные,
кровянисто-гнойные). В случае обильных выде-
лений стерильной ватой снимают первую кап-
лю, из последующих капель делают несколько
мазков. После этого пальпируют половой член
по ходу мочеиспускательного канала. При отсут-
ствии выделений легким массажем по ходу мо-
чеиспускательного канала стараются получить
их. В случаях, подозрительных на венерические
заболевания, и при отсутствии выделений пет-
лей или специальной кюреткой делают соскоб
со слизистой оболочки мочеиспускательного
канала. Мазки направляют в лабораторию для
исследования на содержание гонококков или
другой бактериальной флоры. Затем осматрива-
ют мошонку, яички, их придатки, семенные ка-
натики; определяют их размеры, консистенцию,
болезненность. После осмотра ставят двух — или
трехстаканную пробу, при вялых, хронических
процессах в мочеиспускательном канале прово-
дят исследование на буже, уретроскопию.
4. Первичные и вторичные
морфологические элементы
кожной сыпи
Во время осмотра кожи определяют имеющиеся
на ней морфологические элементы — первичные
(пятна, папулы, бугорки, узлы, пузырьки, пу-
зыри, пустулы) и вторичные (пигментация, че-
шуйки, корки, эрозии, язвы, трещины, лихени-
зация, рубцы). При осмотре можно получить
представление о локализации и цвете элемента,
но для более полной его характеристики необ-
ходимо применить пальпацию и поскабливание.
При осмотре определяют мономорфизм (псори-
аз, красный плоский лишай, пузырьковый лишай,
576 Приложение A
вульгарная пузырчатка, крапивница) или полимор-
физм (экзема, герпетиформный дерматоз Дюрин-
га, дерматит) сыпи. Следует дифференцировать
истинный полиморфизм, при котором наблюда-
ется одновременно несколько первичных элемен-
тов (экзема), и ложный, когда имеется один пер-
вичный элемент, а остальные, вторичные,
являются следствием его эволюции (фурункулез).
Обращают внимание на расположение сыпи,
т. к. в ряде случаев тому или иному дерматозу
свойственна излюбленная локализация: красной
волчанке — лицо, псориазу — задние поверхно-
сти локтевых и передние — коленных суставов,
волосистая часть головы, простому герпесу —
губы и половые органы и т. д.
Отмечают особенности расположения сыпи:
фокусное (элементы не сливаются, их окружает
нормальная кожа), диффузное (слияние элемен-
тов в крупные очаги); распространенность —
ограниченная (очаговый нейродермит, очаговая
склеродермия, невус, простой герпес, узловатая
эритема, бородавчатый туберкулез и др.), распро-
страненная (розовый лишай, псориаз), тотальная
(эритродермия); другие особенности — симмет-
ричная (истинная экзема, псориаз), асимметрич-
ная (дерматомикоз, бородавчатый туберкулез
кожи, кольцевидная гранулема, контактный дер-
матит, опоясывающий лишай и др.), систем-
ная — по ходу сосудистых и нервных стволов
(опоясывающий лишай, линейный красный
плоский лишай, линейный невус), фигурная
(псориаз, мигрирующая эритема), серпигиниру-
ющая — ползущая сыпь (туберкулезная волчан-
ка, лепра, бугорковый сифилид и др.).
5. Определение влажности,
сальности, сухости, тургора кожи
Определение тургора и эластичности кожи про-
изводится ощупыванием (пальцами берут кожу
в складку), а также путем поглаживания кожи
и надавливания на нее. О тургоре и эластично-
сти кожи можно судить по времени расправле-
ния складки на тыле кисти. При хорошем тур-
горе она расправляется моментально, при
пониженном и у стариков — держится до не-
скольких минут.
Снижение тургора и эластичности кожи на-
блюдается при различных атрофических процес-
сах, прогерии, повышение эластичности — при
гиперэластической коже.
Исследование потоотделения электрометричес-
ким методом заключается в том, что влажность
кожи обратно пропорциональна сопротивлению
электрического тока.
Специальный наконечник с двумя электрода-
ми плотно прикладывают к поверхности иссле-
дуемого участка кожи. При совершенно сухой
коже слабый постоянный ток, подключенный к
электродам, не может преодолеть сопротивление,
оказываемое сухим роговым слоем.
При влажной коже сопротивление падает и
может быть фиксировано соответствующим при-
бором.
Нарушение потоотделения отмечается при леп-
ре, аллергических зудящих дерматозах, атрофии
кожи, склеродермии.
6. Определение дермографизма
Метод, позволяющий определить состояние ве-
гетативной нервной системы и, в частности, реф-
лекторные реакции сосудистой стенки в ответ на
раздражение кожи.
Тупым концом палочки или ребром шпателя
проводят по коже полосу. Через 10—20 с, стро-
го повторяя движение шпателя, появляется бе-
лая или красная полоса. При белом дермографиз-
ме, характерном, например, для атопического
дерматита, почесухи, полоса исчезает через 2—8
минут. Красный дермографизм (экзема) появля-
ется несколько раньше и держится значительно
дольше, иногда до 1 часа и более. Реже наблю-
даются возвышенный, или уртикарный, дермо-
графизм, особенно типичный для крапивницы,
и рефлекторный, при котором гиперемия появ-
ляется в виде полосы шириной до 3 см.
7. Методика постановки
капельных и компрессных
аллергологических кожных проб
и их интерпретация
Аллергические тесты капельные
Применяются для выявления аллергена при дер-
матозах, чаще профессиональных. Водный или
спиртовой раствор испытуемого вещества слабой
концентрации пипеткой наносят на здоровую
кожу передней поверхности предплечья или над-
чревной области, обводят чернилами или пастой
шариковой ручки. В качестве контроля на сим-
метричные участки кожи наносят капли раство-
рителя и следят за тем, чтобы жидкость полно-
стью испарилась. Реакцию учитывают через 24,
48 и 72 часа. При положительной реакции, в
зависимости от ее интенсивности, в месте нане-
сения растворов возникает эритема, папула, пу-
зырьки, пузырь.
Перечень и описание практических навыков по кожным и венерическим болезням
577
Аллергические тесты компрессные
(лоскутные, аппликационные)
Сложенную вчетверо марлевую салфетку (пло-
щадью 1 х 1 см) смачивают слабым раствором
исследуемого вещества, накладывают на кожу
передней поверхности предплечья или надчрев-
ной области. Салфетку покрывают медицинс-
кой клеенкой площадью 4x4 см, фиксируют
бинтом или лейкопластырем. Наиболее удобно
пользоваться тестопластом, представляющим
собой лист каучукового пластыря с круглым
окном в центре диаметром 0,9 см, где имеется
марля, которую пропитывают с помощью пи-
петки или стеклянной палочки испытуемым
веществом. Центры диагностических окон дол-
жны быть отдалены один от другого на 4,5 см.
Реакцию учитывают через 24, 48 и 72 часа. Для
кожных проб рекомендуется применять хими-
чески чистые вещества. Следует отметить, что
у некоторых лиц иногда сам пластырь может
вызвать аллергическую реакцию или контакт-
ный дерматит.
Положительным считается тест, при котором
в месте воздействия предполагаемого аллергена
возникают: эритема (+); эритема, отек, папула
(++); резко выраженная воспалительная реак-
ция, на фоне которой различаются папулы и
пузырьки (+++); крупные пузыри и некроз
тканей (++++).
8. Лекарственные формы
для наружного применения
при лечении болезней кожи
и показания к их назначению
Краткая характеристика лекарственных
форм, применяемых в дерматологии:
Водные растворы (или с добавлением спирта)
содержат антисептики (борная кислота, резор-
цин, танин, ихтиол, нитрат серебра, риванол и
др.) и применяются в виде примочек, влажно-
высыхающих повязок.
Аэрозоли (пены): взвешенные лекарственные
частицы в среде пропеллента. Удобны в приме-
нении, особенно на эрозированные поверхности.
Пудры: мельчайшие порошкообразные лекар-
ственные вещества (оксид цинка, тальк, крахмал
и др.) в виде присыпок.
Водные «болтушки»: равные части порошков и
жидкостей. Лучше фиксируются к коже при
добавлении глицерина.
Масляные «болтушки» (линименты) содержат
30 % порошкообразных лекарственных веществ
и 70 % масла. Обеспечивают противовоспали-
тельный, противозудный и смягчающий эффект.
Пасты: комбинация порошков (40—50 %) и
жиров. Хорошо удерживаются на коже. Действу-
ют глубже пудр и болтушек.
Пластыри: основа — воск (канифоль) с добав-
лением активных веществ. Хорошо фиксируясь к
коже, обеспечивают продолжительное и глубокое
«кератолитическое» действие активных веществ.
Лаки: основа — эластический коллодий (или
каучук в хлороформе) с добавлением лекарствен-
ных препаратов. Фиксируясь к коже в виде
пленки, обеспечивают глубокое и длительное
«кератолитическое» действие.
Лосьоны: растворимые или суспензируемые
лекарственные компоненты. Комбинируют с
водой (алкоголем), другими физиологическими
растворителями. Жидкая часть, испаряясь, обес-
печивает быстрый охлаждающий и подсушива-
ющий эффекты, лекарственные вещества усили-
вают противовоспалительное действие.
Эмульсии: жидкости с содержанием масла
30 %. Полужидкие-полутвердые системы вязко-
эластической консистенции. Популярны при
лечении поверхностных воспалительных дерма-
тозов. При нанесении на кожу жидкая часть
эмульсии испаряется, охлаждая кожу, а масло и
лекарственные вещества абсорбируются, усили-
вая противовоспалительный эффект.
Крело: липосомальная эмульсия. Липосомы —
ламеллярные жировые кристаллические структу-
ры, чередующиеся с водными «вздутиями». Кре-
ло обеспечивает вместе с вводимыми лекарствен-
ными препаратами (кортикостероидами)
противовоспалительный эффект, гидратацию
кожи и физиологическое испарение воды.
Кремы гидрофильные: содержат масло (30 %) в
воде. Обеспечивают гидратацию, смягчение
кожи, противовоспалительный эффект. Дей-
ствуют непродолжительно (несколько часов).
Кремы, содержащие липобазу: кремы с водой
в качестве постоянной фазы, но с более высо-
ким содержанием (70 %) жира, чем в кремах
«масло в воде». Кремы содержат неонизирован-
ный эмульгатор, который образует ламеллярные
жировые кристаллические структуры, череду-
ющиеся с водными пузырьками, образующими
липосомы, и являются наиболее физиологичны-
ми местными противовоспалительными средства-
ми. На основе липосом создан кортикостероид-
ный препарат «Локоид-Липокрем».
Кремы гидрофобные: вода (30 %) в масле. Со-
здают масляную пленку на коже, предотвращая
испарение воды, регидрируют кожу, способству-
ют проникновению лекарственных препаратов,
578 Приложение A
оказывая тем самым противовоспалительное дей-
ствие.
Мази гидрофобные: содержат жировые веще-
ства (вазелин, ланолин, нафталан, свиное сало и
др.), обеспечивают «разогревание» кожи и глу-
бокое воздействие вводимых лекарственных ве-
ществ на инфильтративный воспалительный
процесс.
Мази гидрофильные: содержат синтетические
силиконовые соединения, которые обеспечива-
ют дыхание кожи.
9. Осмотр больного
под лампой Вуда
Люминесцентную диагностику витилиго прово-
дят в затемненной комнате с помощью лампы
Вуда после адаптации исследователя к темноте.
Метод дает возможность обнаружить участки
кожи в самом начале депигментации. На фоне
темной кожи четко контурируются светлые,
ярко-белые участки различной величины и фор-
мы, невидимые при обычном освещении. Края
пятен резко пигментированы.
Люминесцентная диагностика микроспории:
метод основан на свойстве волос, пораженных
грибами рода Microsporum, давать ярко-зеленое
свечение при облучении его коротковолновой
частью ультрафиолетовых лучей. Источником
последних служит портативная ртутно-кварцевая
лампа специальной конструкции отечественно-
го призводства. Для задержки длинноволновой
части луча используют фильтр Вуда — стекло,
импрегнированное солями никеля. Этим мето-
дом по характерному свечению можно обнару-
жить пораженные грибом волосы головы, а так-
же пушок на гладкой коже.
После смазывания очагов поражения мазями,
5 % спиртовым раствором йода цвет или све-
чение могут искажаться, ослабляться или вооб-
ще исчезать. В этих случаях необходимо тщатель-
но вымыть голову с мылом и повторить
обследование через 3—4 дня. Достоверность из-
ложенного выше метода необходимо обязатель-
но подтвердить микроскопией волос, взятых из
очага поражения.
Более темное свечение, напоминающее мала-
хит, наблюдается при фавусе, а участки бласто-
микоза люминесцируют розово-оранжевым цве-
том.
Люминесцентная диагностика поздней порфи-
рии кожи: у больного собирают суточную мочу
в емкость из темного стекла. Для предупрежде-
ния гнилостных процессов в моче, которые мо-
гут изменить цвет и ее прозрачность, в емкость
добавляют 10—15 мл толуола. Из собранной су-
точной мочи (можно взять разовое количество
мочи после ночного удержания) отливают в про-
бирку 5 мл и помещают ее под люминесцентную
лампу Вуда, лучше в аппарат для люминесцент-
ного анализа витаминов. Реакцию считают по-
ложительной, если исследуемая моча имеет крас-
ную флюоресценцию, у здоровых людей она дает
голубовато-белое свечение.
Люминесцентная диагностика отрубевидного
лишая: метод применяется для обнаружения оча-
гов поражения на волосистой части головы. В
темной комнате освещают лампой Вуда волоси-
стую часть головы. Очаги поражения имеют
золотисто-желтое, желто-коричневое или буро-
ватое свечение.
Выявление участков поражения на волосистой
части головы имеет важное значение для лече-
ния отрубевидного лишая, так как практические
врачи часто забывают об этой локализации, что
приводит в дальнейшем к рецидивам заболева-
ния.
Люминесцентная диагностика эритразмы:
метод применяется для отличия эритразмы от
паховой эпидермофитии, рубромикоза. Очаги
поражения исследуют в лучах лампы Вуда.
При эритразме (очаги поражения предвари-
тельно не должны подвергаться местной тера-
пии) несколько дней наблюдается характерное
кораллово-красное свечение, которое более вы-
ражено в периферической зоне.
10. Проведение пробы Бальзера
со спиртовым раствором йода
Применяется для диагностики отрубевидного
(разноцветного) лишая. Пятна смазывают 5 %
спиртовым раствором йода. Вследствие разрых-
ления рогового слоя в-области высыпаний ра-
створ йода впитывается в эти участки сильнее
и пятно окрашивается интенсивнее, чем окружа-
ющая здоровая кожа. При отсутствии раствора
йода можно пользоваться анилиновыми краси-
телями. Следует отметить, что при остаточной
лейкодерме после отрубевидного лишая, особен-
но у лиц, подвергшихся ультрафиолетовому об-
лучению, проба Бальзера бывает отрицательной.
11. Микроскопическое
исследование
при дерматомикозах
Для исследования можно брать чешуйки, пора-
женные волосы, ногтевые пластинки. От пра-
Перечень и описание практических навыков по кожным и венерическим (юлезням 579
вильного взятия материала во многом зависит
успех микроскопического исследования при дер-
матомикозах. Элементов гриба бывает обычно
больше на свежих, нелеченных, но уже сформи-
ровавшихся участках поражения.
При микозах гладкой кожи (трихофития,
микроспория, микоз стоп, кератомикоз, канди-
доз) для исследования берут чешуйки перифе-
рических участков очага путем соскабливания
скальпелем. У больных дисгидрозом стоп, кис-
тей ножницами или лезвием безопасной бритвы
срезают покрышки пузырьков или бахромки
отслоившегося эйителия.
При дерматомикозах длинные и пушковые
волосы из очагов берут эпиляционным пинце-
том, иногда острием скальпеля, если волос об-
ломан на уровне кожи («черные точки»). При ин-
фильтративно-нагноительных процессах
отбирают с периферии очага волосы, плавающие
в гное; который берут фолькмановской ложеч-
кой и помещают в чашку Петри или на часовое
стекло. Препаровальной иглой вылавливают
пораженный волос и помещают на предметное
стекло. Таким же образом собирают гной из
открытых очагов поражения, свищей, а из зак-
рытых (абсцесс, нагноившиеся лимфатические
узлы и др.) — путем пункции шприцем.
Соскобы из пораженной слизистой оболочки
берут стерильным шпателем, петлей, пинцетом,
фолькмановской ложечкой.
Для взятия материала из пораженных ногтей
используют скальпель, ножницы, маникюрные
щипцы, из более глубоких слоев — бормашину.
При глубоком микозе для исследования берут
мокроту, желчь, мочу, тканевую жидкость и
помещают в стерильные стеклянные банки с
притертой пробкой.
Для микроскопии применяют нативные и
окрашенные препараты. Существует несколько
методов приготовления препаратов в зависимо-
сти от характера исследуемого материала и вида
дерматомикоза. Приводим наиболее доступный
метод лабораторной диагностики грибковых за-
болеваний.
Пораженные волосы, ногтевые пластинки,
плотные роговые массы, чешуйки размельчают
на предметном стекле нагретым скальпелем и
наносят 2-3 капли 10-30 % раствора щелочи
(КОН или NaOH). Для просветления препарат
подогревают на пламени спиртовки, не доводя
до кипения, пока не появится белый ободок по
периферии, а затем, надавливая, накрывают по-
кровным стеклом.
При исследовании чешуек, роговых масс, ног-
тей врач-лаборант может лишь указать, найден
или не найден мицелий («найден мицелий гри-
ба», «найдены группы почкующихся клеток»), а
врач-миколог по клинической картине ставит
диагноз заболевания или дополнительно назна-
чает культуральное исследование.
12. Методика дезинфекции
обуви при грибковых
заболеваниях стоп
Для дезинфекции обуви применяется упрощен-
ная методика В. М. Лещенко: внутреннюю по-
верхность обуви протирают 25 % раствором
формалина или 40 % уксусной кислотой и по-
мещают в полиэтиленовый мешок на 2—4 часа.
Далее обувь сушат и проветривают до исчезно-
вения запаха дезинфектанта.
13. Постановка пробы
Ядассона при герпетиформном
дерматозе Дюринга
Пробы Ядассона с калия йодидом имеют боль-
шую диагностическую ценность для отличия гер-
петиформного дерматоза Дюринга и истинной
пузырчатки. Для герпетиформного дерматоза ха-
рактерна повышенная чувствительность больных
к галогенам, в том числе к йоду. Пробу прово-
дят в двух вариантах.
1. Больной принимает внутрь 1 столовую ложку
5 % раствора калия йодида. Пробу считают поло-
жительной при обострении кожного процесса.
2. На свободный от высыпаний участок кожи
предплечья накладывают 50 % мазь с калия
йодидом, приготовленную на ланолине. Через 24,
реже 48 часов на месте контакта с мазью возни-
кает эритема, иногда пузырьки, папулы, анало-
гичные высыпаниям при герпетиформном дер-
матозе Дюринга, либо наблюдается обострение
основного процесса вне места наложения мази.
14. Симптом П. В. Никольского
при пузырчатке
Имеет диагностическую ценность в основном
при истинной пузырчатке. При потягивании за
обрывок покрышки пузыря пинцетом наблюда-
ется отслойка верхних слоев эпидермиса приле-
гающей видимо здоровой кожи. Обусловлен
акантолизом. Может отмечаться и при хроничес-
кой доброкачественной семейной пузырчатке,
буллезном пемфигоиде, острой лихорадящей пу-
зырчатке, токсическом эпидермальном некроли-
зе Лайелла, вегетирующей пузырчатке и др. Лег-
580 Приложение A
7 кое трение пальцем между двумя пузырями также
вызывает отслойку эпидермиса. В редких слу-
чаях этот симптом может быть положительным
и в участках, отдаленных от пузырей.
При надавливании пальцем на неповрежден-
ный пузырь его площадь увеличивается, так как
давление жидкости приводит к отслойке по-
крышки пузыря по периферии. Это явление
(симптом Асбо-Хансена) наблюдается почти при
всех пузырных дерматозах и по сути является
вариантом симптома Никольского.
Симптом Никольского позволяет отличить
вышеперечисленные дерматозы от неакантоли-
тической пузырчатки, буллезной разновиднос-
ти многоформной экссудативной эритемы, ве-
гетирующей пиодермии Галлопо, болезни
Дарье, болезни Дюринга, субкорнеального пу-
стулезного дерматоза, эритродермии Вильсона-
Брока, доброкачественной неакантолитической
пузырчатки слизистой оболочки только поло-
сти рта.
15. Взятие мазков отпечатков
и их окраска по Романовскому
Гимзе на акантолитические
клетки(Тцанка)
Этот метод незаменим при дифференциальной
диагностике пузырчатки и герпетиформного дер-
матоза Дюринга. С поверхности дна свежего пу-
зыря скальпелем или путем прикладывания и
легкого надавливания кусочком простерилизован-
ной кипячением ученической резинки (метод
отпечатков) берут материал и переносят на сте-
рильные обезжиренные предметные стекла, фик-
сируют в течение 1 минуты метиловым спиртом,
высушивают при комнатной температуре и окра-
шивают по Романовскому-Гимзе; наносят на 20—
25 мин свежеприготовленный раствор азур-эози-
на, затем смывают краситель дистиллированной
водой и высушивают мазки при комнатной тем-
пературе. После приготовления и окраски препа-
раты исследуют под микроскопом при увеличе-
нии 10x40. Акантолитические клетки меньше
нормальных эпителиальных клеток, округлой
формы, с крупным ядром, окрашенным в интен-
сивно-фиолетовый или фиолетово-синий цвет,
занимающий почти всю клетку. В ядре видны два
или несколько крупных более светлой окраски
ядрышек. Цитоплазма как бы оттеснена к пери-
ферии (ободок концентрации), резко базофиль-
на, ближе к ядру — светло-голубая. Количество
клеток различное: от единичных до большого
числа (в виде скоплений).
16. Симптом Бенье-Мещерского
и «дамского каблучка», симптом
«бабочки» при красной волчанке
Симптом сломанного дамского каблучка ис-
пользуется при диагностике дискоидной раз-
новидности красной волчанки. Гиперкератоз,
свойственный этому дерматозу, проникает в
шейку фолликула волоса, образуя конусооб-
разные шипики на обратной стороне чешуйки,
хорошо видные невооруженным глазом. При
энергичном поскабливании участков пораже-
ния больной ощущает некоторую болезнен-
ность в связи с тем, что указанные выше ги-
перкератотические выросты (шипики),
надавливая на нервные окончания, раздража-
ют их (симптом Бенье-Мещерского).
Симптом «бабочки» — наличие эритемы на спин-
ке носа и на щеках (обычно в области скуловых дуг),
по своим очертаниям напоминающей бабочку.
Признак встречается при красной волчанке.
17. Симптом «яблочного желе»
и «пуговчатого зонда» при
туберкулезной волчанке
Симптом проваливания
зонда (пуговчатого зонда)
Бугорки при туберкулезной волчанке, вслед-
ствие разрушения в них эластических и колла-
геновых волокон, приобретают тестоватую,
мягкую консистенцию. Учитывая эту особен-
ность, А. И. Поспелов (1886) предложил симп-
том проваливания зонда: при легком надавли-
вании на бугорок пуговчатым зондом на
поверхности бугорка остается вдавление, кото-
рое очень медленно исчезает. Это явление мож-
но сравнить с картиной, наблюдаемой при на-
давливании пальцем на дрожжевое тесто.
Если на бугорок надавить зондом несколько
сильнее, то он как бы проваливается вглубь
люпомы, при этом появляются легкое кровоте-
чение и незначительная болезненность. Симптом
более выражен при свежей люпоме.
Симптом «яблочного желе»
Диагностический прием при туберкулезной вол-
чанке. При сильном надавливании на люпому
прозрачным шпателем или предметным стеклом
выдавливается кровь из расширенных сосудов
бугорка и появляется буровато-желтая окраска,
напоминающая цвет яблочного желе. Иногда при
этом можно заметить полупрозрачность бугорка.
Перечень и описание практических навыков по кожным и венерическим болезням 581
Данный симптом может быть положительным,
но менее выраженным, при туберкулоидной фор-
ме кожного лейшманиоза.
18. Определение псориатических
феноменов (стеариновое пятно,
псориатическая пленка, кровяная
роса)
Диагностическая триада при псориазе
Почти постоянный симптом при прогрессиру-
ющей и стационарной (реже) стадии псориаза.
При поскабливании псориатических элементов
(папулы, бляшки) шелушение увеличивается и
чешуйки принимают белую окраску, напоми-
ная каплю растертого стеарина (феномен «сте-
аринового пятна»). При дальнейшем поскабли-
вании до зернистого слоя эпидермиса чешуйки
снимаются и обнажается розовая влажная пленка
(феномен «терминальной», или «псориатической»,
пленки). При продолжении поскабливания (до
сосочкового слоя дермы) на поверхности пленки
появляются мельчайшие капельки крови. Число
их зависит от количества поврежденных при по-
скабливании капилляров, входящих в сосоч-
ки, — феномен «точечного кровотечения», или
«кровяной росы».
Поскабливание производят либо предметным
стеклом, либо тупой стороной скальпеля. После
поскабливания ногтем необходимо тщательно
обработать кожу рук во избежание заражения
(псориазиформный папулезный сифилид).
19. Получение сетки Уикхема
при красном плоском лишае
Мелкая сетчатость на поверхности папул; сим-
птом красного плоского лишая, особенно обыч-
ной его разновидности. Папулу или бляшку
смазывают каким-либо жидким маслом (вазели-
новым, подсолнечным и др.). При этом на по-
верхности элемента ясно видны белесовато-опа-
лового цвета точки и пересекающиеся в виде
сетки линии. Этот феномен объясняется нерав-
номерным утолщением зернистого слоя эпидер-
миса (гранулезом), типичным для данного забо-
левания.
При остроконечной, бородавчатой, пигмент-
ной, атрофической и других формах красного
плоского лишая проба менее показательна.
20. Микроскопическое
исследование бледной
трепонемы в темном поле
зрения
Материал для исследования берут с элемента,
подозрительного на первичную или вторичную
сифилому (эрозия, язва, эрозированная папула),
либо путем пункции лимфатического узла.
Элемент очищают стерильным ватным тампо-
ном, после чего прокаленной в пламени горел-
ки и охлажденной петлей слегка поскабливают
его поверхность до получения тканевой жидко-
сти. Последнюю помещают на чистое, обезжи-
ренное предметное стекло с каплей изотоничес-
кого раствора натрия хлорида и накрывают
нетолстым покровным стеклом.
Если получить тканевую жидкость указанным
способом нельзя, то рукой в резиновой перчат-
ке сдавливают сифилому с краев до появления
тканевой жидкости на ее поверхности.
Количество тканевой жидкости и изотоничес-
кого раствора натрия хлорида должно быть не-
большим, так как при большой капле трепоне-
мы будут плавать в жидкости, что затрудняет их
обнаружение.
Для исследования в темном поле зрения при-
меняют специальные приспособления к обыч-
ному микроскопу. Пройдя через эти приспо-
собления (конденсоры), лучи света принимают
косое направление и концентрируются под ос-
трым углом в месте исследуемой капли серу-
ма, не попадая в объектив, чем достигается ис-
следование в темном поле. Для данной
методики можно пользоваться кружком из
плотной черной бумаги (конверт от фотобума-
ги) диаметром 1,5 см, который накладывают
на нижнюю линзу развинченного конденсора
так, чтобы между краями линз и бумаги оста-
вался свободный ободок (просвет) шириной
1,5—2 мм.
Освещение при исследовании в темном поле
зрения должно быть достаточно интенсивным
(электрическая лампа 100—150 Вт).
Рассматривают препарат при объективе
№ 40 и окуляре № 10-15. При этом на тем-
ном фоне можно увидеть большое количество
светящихся движущихся точек (белковые и
коллоидные частицы), а также клеточные эле-
менты, трудно дифференцируемые, что,
впрочем, не входит в задачу исследования.
Бледная трепонема имеет вид светлых што-
порообразных нитей, движущихся плавно
либо маятникообразно. Ее можно спутать с
582 Приложение A
часто встречающейся на половых органах тре-
понемой crassus, однако последняя толще
бледной трепонемы, имеет более широкие за-
витки и движется быстрее.
Данный способ простой и, пожалуй, явля-
ется наиболее надежным. Бледную трепонему
рассматривают in vivo, что позволяет диффе-
ренцировать ее и другие виды спирохет.
21. Методика микроскопического
исследования на трихомонады
и гонококки
Микроскопическое
исследование на трихомонады
Трихомониаз характеризуется многоочаговостью
поражения. Следовательно, материал для иссле-
дования необходимо брать из многих очагов
инвазии.
Выделения, соскоб берут у мужчин из мочеис-
пускательного канала, у женщин — из боковой и
задней частей свода влагалища, мочеиспускатель-
ного канала; у девочек — через отверстие девствен-
ной плевы желобоватым зондом или браншей ва-
гинального пинцета. Перед взятием материала не
рекомендуется туалет наружных половых органов,
больного просят длительное время (не менее 3—
4 ч) воздержаться от мочеиспускания.
Полученный материал помещают на предмет-
ное стекло. Если он густой, предварительно до-
бавляют несколько капель теплого изотоничес-
кого раствора натрия хлорида. Мочеполовые
трихомонады необходимо рассматривать под
микроскопом в свежем и окрашенном состоянии.
Нативные мазки желательно использовать
немедленно. Исследуют при увеличении 10x40
обычным микроскопом или в темном поле зре-
ния фазовоконтрастным. В свежеприготовленных
неокрашенных препаратах мочеполовые трихо-
монады имеют грушевидную форму, величина их
немного больше лейкоцита, движения — толч-
кообразные поступательные. При изучении пре-
парата фазовоконтрастным микроскопом можно
различить в расширенной части трихомонады
жгутики.
При выявлении влагалищных трихомонад в
нативном мазке их необходимо дифференциро-
вать от других видов трихомонад, обитающих в
организме человека (кишечная, ротовая). Поэто-
му, наряду с исследованием материала в натив-
ном состоянии, необходимо всегда производить
микроскопию окрашенных мазков. Мазки под-
сушивают на воздухе и фиксируют. Способ фик-
сации зависит от метода окраски. Для более де-
тального исследования строения трихомонад (об-
наружение ядра, ундулирующей мембраны и
других органоидов) мазок окрашивают по Рома-
новскому-Гимзе. В этих случаях препарат фик-
сируют в смеси спирт-эфир (1:1).
Чаще всего мазки окрашивают 1 % расвором
метиленового синего или по Граму. Препарат
фиксируют обычно над пламенем горелки. Ок-
раска метиленовым синим и по Граму имеет
преимущество перед другими методами, заклю-
чающееся в том, что в мазках можно одновре-
менно исследовать гонококки и трихомонады.
Трихомонады, окрашенные по Граму, красно-
ватого цвета, а метиленовым синим — синего.
Величина их различная. Ядро овальной формы,
расположено эксцентрически и окрашивается в
более темный цвет по сравнению с протоплаз-
мой. Жгутики не определяются. Часто в расши-
ренной части трихомонады можно различить
маленькую щель — цистому, а в цитоплазме —
вакуоли, микроорганизмы, лейкоциты, эритро-
циты, сперматозооны и др. Для диагностики
важным является обнаружение грамотрицатель-
ных диплококков (гонококков). При оценке
препаратов, окрашенных метиленовым синим,
необходимо дифференцировать трихомонады и
эпителиальные клетки, которые больше по раз-
меру, ядро расположено в центре.
Окраска гонококков
метиленовым синим
Перед окраской мазки фиксируют 96 % этило-
вым спиртом в течение 3-х минут или над пла-
менем спиртовки. На высушенный мазок нано-
сят 1 % раствор метиленовой синьки и оставляют
в течение 1 минуты. Остатки красителя смыва-
ют под струей холодной воды и высушивают в
штативе. При исследовании под микроскопом
ядра клеток окрашены в синий цвет, цитоплаз-
ма в голубой цвет различной интенсивности.
Слизь голубого цвета. Бактериальная флора ок-
рашивается в синий цвет различных оттенков.
Гонококки темно-синего цвета, резко очерчены,
бобовидной формы, парные, располагаются
внутри полиморфноядерных лейкоцитов, в слизи
и на эпителиальных клетках.
Окраска гонококков по Граму
Мазок фиксируют над пламенем горелки в те-
чение 1—2 мин и окрашивают каким-либо кра-
сителем трифенилметановой группы (генциан-
виолет, метилвиолет, кристалл виолет),
промывают водой 1—2 мин и заливают раство-
ром Люголя (калия йодид — 2 г, йод кристал-
лический — 1 г, вода дистиллированная —
I
Перечень и описание практических навыков по кожным и венерическим болезням
583
300 мл), а затем фиксируют 96 % этиловым
спиртом в течение 1 мин до появления серо-
фиолетовых струек. После этого препарат про-
мывают водой и дополнительно окрашивают
какой-либо контрастной краской (нейтральрот).
Гонококки грамотрицательны, то есть окраши-
ваются в красный либо розовый цвет. Грампо-
ложительные бактерии фиолетового цвета.
22. Умение собирать
эпиданамнез у больных
венерическими болезнями
Прежде всего, необходимо уточнить жалобы
пациента. Получить сведения о перенесенных
ранее венерических заболеваниях, выяснить
вредные привычки. Далее следует собрать анам-
нестические данные о давности заболевания, дли-
тельности его инкубационного периода, харак-
тере половой жизни (в браке, вне брака, с одним
или разными партнерами). Кроме того, регист-
рируют в медицинской карте сведения о само-
лечении заболевания, о переносимости лекар-
ственных препаратов.
При сборе анамнеза следует обратить внимание
на перенесенные ранее заболевания, развившие-
ся после аборта или родов, развитие воспалитель-
ного процесса вскоре после половой связи, нали-
чие дизурических явлений, появление выделений,
нарушений менструального цикла и бесплодия.
23. Двухстаканная проба при
обследовании больного
уретритом
Двухстаканная проба (проба Томпсона)
Применяется в венерологии и урологии для суж-
дения о месте поражения мочеиспускательного ка-
нала: передней или задней его части. Перед пробой
больной в течение 4—6 ч не должен мочиться, после
чего ему предлагают собрать мочу в два стакана: в
первый — 50—60 мл, во второй — остальную. Если
первая порция будет мутной, а вторая — прозрач-
ной, то процесс локализуется в передней части мо-
чеиспускательного канала. Наличие мути в обоих
стаканах свидетельствует о тотальном поражении мо-
чеиспускательного канала, так как при поражении
задней его части гной может попадать в мочевой
пузырь, в котором будет также мутная моча.
24. Дезинфекция рук
и инструментов при работе
с больными сифилисом,
гонореей и другими
инфекциями, передаваемыми
половым путем
Дезинфекция рук
После осмотра больного с кожными или вене-
рическими заболеваниями врач обеззараживает
руки растворами бактерицидных препаратов. В
качестве средств для дезинфекции рук применя-
ют 70—80 % этиловый спирт, 0,5 %-ный раствор
хлоргексидина в 70 %-ном этиловом спирте,
септоцид Р (антисептик рук).
Дезинфекция инструментов
После осмотра инструменты погружаются в 3 %-
ный раствор хлорамина на 1 час.
25. Методика индивидуальной
профилактики венерических
болезней
Методика оказания
профилактической помощи мужчинам
См. стр. 469
Методика оказания
профилактической помощи женщинам
См. стр. 470
26. Заполнение экстренного
извещения и необходимой
документации на больного
венерическими заболеваниями
и заразными кожными
заболеваниями
При выявлении больного венерическими забо-
леваниями врач, установивший диагноз, незави-
симо от места выявления больного, обязан за-
полнить «Извещение...» (форма № 089/у) и в
трехдневный срок направить его в кожно-вене-
рологический диспансер по месту жительства
больного, а копию в санитарно-эпидемиологи-
ческую станцию.
При выявлении больных чесоткой заполняет-
ся медицинская карта амбулаторного больного
(форма 25/у), куда вносятся сведения об источ-
584 Приложение A
нике заражения и контактных лицах. Сведения
о больном не позднее 12 часов с момента регис-
трации передаются по телефону выявившим уч-
реждением в лечебно-профилактическое учреж-
дение (Л ПУ) по месту жительства или работы и
в территориальный центр гигиены и эпидемио-
логии, а затем в течение 3-х дней дублируются
извещением по форме № 89/у; медработники всех
детских учреждений информируются о выявле-
нии в учреждении больных чесоткой напрямую
из КВД либо через детскую поликлинику.
При выявлении у больного микроспории, трихо-
фитии заполняется извещение по форме № 89/у;
информация по телефону в территориальные
ЛПУ по месту жительства о всех случаях, реги-
страция больных заразными кожными болезня-
ми с указанием места работы, учебы и т. д. При
выявлении зоонозной микроспории и трихофи-
тии копия формы № 89/у дополнительно на-
правляется в территориальные учреждения вете-
ринарии. Проводится выявление и обследование
контактов. В экстренном извещении указывается
фамилия, имя, отчество, возраст, дата обраще-
ния, адрес, подробный диагноз, место работы,
как выявлено заболевание (при обращении, при
профосмотре, при обследовании в стационаре).
Приложение В
Ситуационные задачи по дерматовенерологии
Задача № 1
Больная Я., обратилась к дерматологу с жало-
бами на поражение кожи шеи, груди, спины и
умеренно выраженный зуд в очагах поражения на
протяжении суток. Считает себя больной в тече-
нии четырех месяцев, когда впервые появились
указанные симптомы. Мазь «Синалар», которую
пациентка использовала для лечения, приносила
кратковременный положительный эффект. Пять
месяцев назад больная начала применять новый
импортный дезодорант. Пользуется им ежеднев-
но до настоящего времени.
Дерматологический статус: На коже шеи, гру-
ди, спины имеются розовые пятна различных
размеров, округлых очертаний; папулы, шелуше-
ние, расчесы и кровянистые корочки на повер-
хности очагов.
1. О каком заболевании можно подумать и
почему?
2. Предложите общий план лечения заболева-
ния.
Задача № 2
Утром на станцию «Скорой помощи» посту-
пил вызов от гражданки Н., которая обнаружи-
ла у себя на теле сыпь красного цвета, напоми-
нающую ожоги от крапивы, которая
сопровождалась сильным зудом. По прибытии на
дом врач обнаружил на теле пациентки расчесы
и свежие кровянистые корочки в области внут-
ренней поверхности бедер, спины, груди, лок-
тевых и паховых сгибов. Других элементов сыпи
на коже больной не обнаружено. При опросе вы-
яснилось, что сильный зуд кожи в указанных
областях беспокоил больную во вторую полови-
ну ночи и утром. Перед приездом «Скорой по-
мощи» сыпь и зуд исчезли. Вечером больная
употребляла в пищу апельсины и сухое вино.
1. О каком заболевании может идти речь?
2. Назначьте обследование пациенту.
Задача № 3
Больной П., поступил в кожное отделение с
жалобами на зуд и поражение кожи кистей,
предплечий. Объективно: на коже кистей, пред-
плечий определяется отек, гиперемия, микрове-
зикуляция, мокнутие. Очаги поражения имеют
нечеткие границы.
Установлен диагноз: «Истинная экзема».
1. Определите план общего лечения истинной
экземы.
2. Определите наружное лечение истинной
экземы в соответствии с описанной выше кли-
нической картиной.
Задача № 4
Больная М., поступила в кожное отделение с
жалобами на чувство зуда, жжения кожи груди,
рук, бедер, лица и покраснение кожи в этих
участках. Указанные жалобы появились два дня
назад после втирания в кожу названных облас-
тей спиртовой настойки девясила. В течение двух
дней зуд и жжение усиливались, появилось чув-
ство жара, покраснение кожи, пузырьки и пу-
зыри.
Дерматологический статус: кожа груди, рук,
бедер, лица ярко гиперемирована, отечна, горя-
чая на ощупь. Имеются папулы, пузырьки, пу-
зыри, эрозии.
1. Какой диагноз можно предположить?
2. Назовите признаки, подтверждающие ваше
предположение.
Задача № 5
В кожное отделение поступил больной Д., 16
лет с жалобами на раздражение, болезненность
кожи в паховых областях, зуд кожи спины, рук.
Пять дней назад появился зуд на коже в облас-
ти лобка (связывает его с посещением бани).
Кожу лобка в течение трех дней смазывал серо-
ртутной мазью, после чего на четвертый день
появились высыпания и зуд на коже предпле-
чий, в паховых областях, внутренней поверхно-
сти бедер, половых органов. Еще через сутки
сыпь распространилась на кожу плеч, спины. В
анамнезе — ОРВИ, тонзиллит, ревматический
полиартрит. Периодически принимает индоме-
тацин курсами (два раза в год). Последний раз
принимал индометацин 4 месяца назад. Объек-
тивно: основной патологический процесс лока-
585
586 Приложение В
лизуется в области лобка, пахово-бедренных
складок и половых органов; определяется отек,
гиперемия кожи, островоспалительные папулы.
Очаги эритемы и островоспалительных папул
выявляются также на коже спины и верхних ко-
нечностях. Первичные элементы — воспалитель-
ные пятна и островоспалительные папулы, име-
ющие тенденцию к слиянию.
1. Учитывая анамнез (применение серо-ртутной
мази на кожу лобка, клинику — островоспали-
тельная эритема и островоспалительные папу-
лы), локализацию поражения, изложите свои
предположения о диагнозе.
2. Предложите план лечения больного.
Задача № 6
В клинику поступил больной 19 лет с жало-
бами на зуд кожи и высыпания в области лок-
тевых и подколенных сгибов. Болеет с 6-месяч-
ного возраста (со слов матери). Рецидивы
заболевания наступают после употребления шо-
колада, апельсинов, пряностей. Объективно: на
коже локтевых, подколенных сгибов кожа сухая,
лихенизирована, геморрагические корочки, сле-
ды экскориаций.
1, Установите диагноз, клиническую форму и
стадию заболевания.
2. Предложите план лечения.
Задача № 7
Больной 15 лет жалуется на зуд кожи туло-
вища и высыпания. Объективно: в области лок-
тевых и подколенных сгибов, лица имеются
сквамозные пятна, папулы, шелушение; следы
экскориаций, корочки.
1. Ваша тактика в постановке диагноза?
2. Предложите план лечения.
Задача № 8
К Вам обратился мужчина 23 лет с жалобами
на выпадение волос. Считает себя больным в
течение 4 лет. Из анамнеза стало известно, что
больной злоупотребляет алкоголем, отдает пред-
почтение острой, пряной пище. При осмотре
кожа волосистой части головы повышенной
сальности, покрыта жирными желто-серыми
чешуйками. Волосы диффузно разрежены, мас-
лянистые, блестят. Субъективно — беспокоит
периодический зуд. Патологии внутренних ор-
ганов не выявлено.
1. Предполагаемый диагноз?
2. Ваша тактика по обследованию больного.
3. Ваши предложения по общему и местному
лечению.
Задача № 9
На прием обратился молодой человек 17 лет
с жалобами на высыпания на лице, спине и гру-
ди, появление которых впервые отметил около
двух лет назад. Не лечился, так как у отца в
юношеском возрасте были аналогичные высы-
пания, которые самостоятельно прошли. Из се-
мейного анамнеза известно, что отец в настоя-
щее время страдает сахарным диабетом. При
осмотре: кожа лица, верхней части груди и спи-
ны повышенной сальности, выводные протоки
сальных желез зияют, некоторые закупорены
комедонами. Имеются многочисленные фолли-
кулярные папулы и пустулы, а также ярко-ро-
зовые, болезненные, величиной с крупную го-
рошину, инфильтрированные узлы с пустулой на
вершине, кровянисто-гнойные корочки, округ-
лые рубцы, гиперпигментированные пятна.
1. Ваш предполагаемый диагноз?
2. Тактика лечения.
3. Тактика амбулаторного наблюдения пациен-
та.
Задача № 10
На прием обратилась женщина 53 лет с жало-
бами на сыпь в области лица. Впервые отмети-
ла появление единичных высыпаний около трех
лет назад. Не лечилась, так как высыпания са-
мостоятельно проходили. Однако последние 3-
4 месяца их количество заметно увеличилось,
особенно после ультрафиолетового облучения.
Последний год беспокоят «приливы», менстру-
ации стали нерегулярными. В течение 10 лет
страдает язвенной болезнью желудка. При осмот-
ре: кожа лица (преимущественно щеки, нос и
подбородок) застойно-синюшного цвета, имеется
большое количество телеангиэктазией, мелких
розово-красных папул, единичные гнойнички.
Высыпания сопровождаются умеренным зудом.
1. Ваш предполагаемый диагноз?
2. Укажите факторы, способствующие разви-
тию заболевания, ухудшению течения заболева-
ния в последнее время.
3. Наметьте план лечения и профилактики за-
болевания.
Задача № 11
К дерматологу обратился пациент с жалобами
на утолщение кожи ладоней, болезненные тре-
щины. Временами отмечается незначительный
зуд. Болен на протяжении 2 лет. При осмотре:
выраженный гиперкератоз ладоней, глубокие
трещены.
1. Дифференциальный диагноз.
2. Методы обследования.
Ситуационные задачи по дерматовенерологии
587
Задача № 12
К дерматологу обратилась молодая женщина
с жалобами на поражение кожи туловища. Боль-
на в течение одного месяца. Дерматологический
статус: на коже верхней части туловища, шеи
пятна округлой формы с четкими границами,
кофейного цвета.
1. Дифференциальный диагноз.
2. Методы обследования.
Задача № 13
К пациенту, находящемуся на лечении в ста-
ционаре в связи с дисгидротической экземой,
женщина принесла полукопченую колбасу, огур-
цы, яблоки, шоколадные конфеты.
1. Какие рекомендации по соблюдению дие-
ты необходимо дать больному и посетителям?
Задача № 14
К детскому дерматологу направлен ребенок, 6
лет в связи с выпадение волос в виде единич-
ного очага на волосистой части головы. Впервые
мать заметила очаг около недели назад. Самосто-
ятельно лечила ребенка настойкой чеснока.
Эффекта от терапии не наблюдалось. Дермато-
логический статус: на волосистой части головы
определяется очаг выпадения волос, округлой
формы, с шелушением. В очаге поражения оп-
ределяются обломки волос на уровне 3—5 мм.
Зона расшатанных волос не определяется.
1. Предположительный диагноз.
2. Необходимые исследования для его уточне-
ния.
3. Тактика врача.
Задача № 15
К врачу дерматологу обратилась женщина 33
лет с жалобами на выпадение волос в течение
последнего месяца. Причину указать не может.
Дерматологический статус: на волосистой части
головы очаги выпадения волос размером от 2x2
до 5x5 см. При потягивании волос по краю па-
тологического очага происходит их безболезнен-
ная эпиляция. Кожа в очагах поражения не из-
менена.
1. Ваш диагноз.
2. С какими заболеваниями нужно провести
дифференциальный диагноз?
Задача № 16
На прием к врачу прибыл диспансерный
больной с диагнозом: экзема кистей. При ос-
мотре: на коже кистей единичные очаги вторич-
ной гиперпигментации, застойной гиперемии,
незначительное шелушение. Жалоб пациент не
предъявляет. Около двух месяцев назад полу-
чал лечение в стационарном отделении ОКВД.
Врач назначил лечение: преднизолон 0,005-4
раза в сутки; диазолин 0,1—3 раза в сутки; ти-
осульфат натрия 0,5 г. —3 раза в сутки; мазь
«Гиоксизон».
1. Какие ошибки допустил начинающий врач?
2. Какие препараты и физиопроцедуры необ-
ходимо назначить больному?
Задача № 17
Больная Н., 24 лет, не замужем. Обратилась
к врачу-гинекологу с жалобами на повышение
температуры тела до 38 °C, рези при мочеиспус-
кании, боли внизу живота, гнойные выделения
из половых органов. При осмотре обильное гной-
ное отделяемое из шейки матки, губки уретры
гиперемированы. При бактериоскопическом ис-
следовании гонококк не обнаружен.
1. Какова дальнейшая тактика врача?
Задача № 18
Студент выписал на экзамене рецепты анти-
гистаминных препаратов для лечения острой
экземы. Правильно ли указаны дозы: супрасти-
на — 0,025; кетотифена — 0,001; тавегила —
0,005; димедрола — 0,05.
Задача № 19
Больная С., 24 лет, доярка, обратилась к вра-
чу с жалобами на высыпания в области лица, ко-
торые появились впервые летом 2 года назад пос-
ле длительного пребывания на солнце. При
осмотре: на коже спинки носа и щек имеются
розово-красного цвета пятна, с четкими грани-
цами, напоминающие «бабочку», на поверхнос-
ти их серовато-белого цвета плотно сидящие че-
шуйки. В центре очагов — рубцовая атрофия, по
периферии очагов телеангиэктазии, гиперпиг-
ментация.
1. Поставьте клинический диагноз.
2. Какие лабораторные исследования следует
провести?
3. Назовите заболевания, с которыми необхо-
димо провести дифференциальный диагноз.
4. Наметьте план лечения.
Задача № 20
Больной К., 28 лет, тракторист, обратился к
врачу по поводу высыпаний на коже лица, гру-
ди, умеренного зуда. Заболел впервые 3 месяца
назад. При осмотре на коже щек, волосистой ча-
сти головы, открытой части груди, шеи множе-
ство розово-красного цвета пятен, бляшек вели-
чиной от чечевицы до 5 копеечной монеты,
588 Приложение В
границы очагов четкие. На поверхности боль-
шинства очагов поражения имеются плотно си-
дящие серого цвета чешуйки, которые при по-
скабливании снимаются с трудом, вызывая
болезненность.
1. Какие специальные дерматологические ис-
следования надо провести для подтверждения
диагноза?
2. Поставьте клинический диагноз.
3. Перечислите основные группы медикамен-
тов для лечения этого заболевания.
4. Нуждается ли пациент в рациональном тру-
доустройстве и почему?
Задача № 21
Больная А., 20 лет обратилась к врачу по по-
воду высыпаний на коже лица без субъективных
ощущений. Впервые заболела 2 года назад в
июле после длительного пребывания на пляже.
При осмотре: на коже спинки носа, щек имеет-
ся отечная эритема с синюшным оттенком в виде
«бабочки», границы очагов четкие.
1. Поставьте клинический диагноз.
2. С какими заболеваниями следует провести
дифференциальный диагноз?
3. Назначьте лечение.
Задача № 22
Больная М., 25 лет обратилась к врачу с жа-
лобами на общую слабость, раздражительность,
повышение температуры, боли в суставах и вы-
сыпания на коже. Подобное состояние наступило
через месяц после родов. В анамнезе есть ука-
зание на дискоидную красную волчанку. При
осмотре: температура 37,8 °C, на коже носа, щек,
груди розово-красного цвета бляшки, покрытые
сероватого цвета плотно сидящими чешуйками,
в центре очагов рубцовая атрофия. На ладонной
поверхности ногтевых фаланг имеются пятна
розово-синюшного цвета. Суставы кистей, стоп,
лучезапястные, локтевые отечны, кожа над ними
гиперемирована, функция их нарушена.
1. Поставьте клинический диагноз.
2. Какие лабораторные исследования необхо-
димо провести для подтверждения диагноза?
3. На состояние каких органов следует обра-
тить внимание?
4. С какими заболеваниями следует провести
дифференциальный диагноз?
5. Назначьте лечение.
Задача № 23
Больная Н., 28 лет — каменщица, страдает
дискоидной формой красной волчанки в тече-
ние 3-х лет. Дерматологический статус: на коже
левой щеки бляшка розово-красного цвета, с чет-
кими границами, в центре рубцовая атрофия, по
периферии очаг покрыт плотно сидящими серы-
ми чешуйками.
1. Какие из перечисленных ниже препаратов
назначите больной? Почему? В какой дозе?
а) пенициллин; б) сульфадимезин; в) стафило-
кокковая вакцина; г) делагил; д) преднизолон;
е) димедрол;
2. Какие препараты из выше описанных не
назначаются и почему?
3. Нуждается ли больная в диспансерном на-
блюдении?
4. Перечислите основные звенья диспансерного
наблюдения.
Задача № 24
На прием к врачу обратилась больная 35 лет
с жалобами на высыпания и зуд кожи кистей.
Из анамнеза стало известно, что высыпания
появлялись неоднократно в течение года после
стирки порошком «Лотос». При осмотре: на коже
кистей имеется выраженная эритема, отечность,
множестве узелков и пузырьков, эрозии, мок-
нутие.
1. О каком заболевании можно думать?
2. Какое лечение должно быть проведено
больной?
3. Как можно предупредить возникновение
аналогичного поражения кожи?
Задача № 25
На прием к участковому врачу обратился па-
циент 40 лет с жалобами на недомогание, сыпь
по кожному покрову, сопровождающуюся зудом.
Высыпания на коже появились 2 дня назад пос-
ле приема амидопирина, постепенно увеличива-
ются как в размере, так и в количестве.
1. Можно ли поставить диагноз?
2. Какова ваша дальнейшая тактика?
Задача № 26
Больная К., 27 лет, явилась на прием к вра-
чу по поводу высыпаний на коже лица. Субъек-
тивные ощущения отсутствовали. Больна в те-
чение 1,5 лет. Впервые отметила стойкое
покраснение кожи щек после длительного пре-
бывания на пляже. Последнее обострение насту-
пило в мае этого года. При осмотре на коже
спинки носа, щек имеется «бабочка», границы
очагов четкие.
1. Предположительный диагноз.
2. С какими заболеваниями целесообразно
дифференцировать данный процесс?
3. Наметьте план лечения.
Ситуационные задачи по дерматовенерологии
589
Задача № 27
На прием к врачу обратилась женщина 40 лет
с жалобами на уплотнение кожи живота, кото-
рое появилось месяц назад на месте травмы. Са-
мостоятельно проводила массаж, смазывание пер-
сикбвым кремом — без эффекта. Страдает
гипертиреозом, хроническим тонзиллитом, веге-
то-сосудистой дистонией. При осмотре: на коже
живота справа определяется очаг поражения око-
ло 6 см в диаметре с желтовато-восковидной
блестящей поверхностью, окаймленной фиоле-
товым венчиком, плотный на ощупь (складку в
очаге собрать не удается). Субъективные ощуще-
ния отсутствуют.
1. Диагноз и его обоснование.
2. Ваша тактика лечения и профилактика ре-
цидивов данного заболевания.
Задача № 28
К врачу дерматологу обратился пациент, 36
лет, с жалобами на поражение кожи живота в
течение 6 месяцев, чувство стягивания, онеме-
ния кожи в этом участке. При осмотре: в верх-
ней трети живота справа очаг поражения 7х 8 см,
округлых очертаний, с гладкой, блестящей, жел-
товатого цвета поверхностью, свободной от вы-
сыпных элементов. По периферии очага имеет-
ся сиреневый венчик. При пальпации участка
поражения ощущается его уплотнение, кожу
удается собрать в грубую складку.
1. Ваш диагноз.
2. Какие причины могли вызвать данное за-
болевание?
3. Проведите дифференциальный диагноз.
4. Назначьте лечение.
5. Укажите прогноз заболевания.
Задача № 29
Больная 17 лет обратилась к дерматологу с
жалобами на неприятные ощущения в нижней
трети левого плеча, изменение цвета кожи в этом
участке, которые появились около 3 месяцев
назад после травмы руки. При осмотре: на на-
ружной трети левого плеча имеется участок по-
ражения размером 2x7 см, полосовидной формы,
четких очертаний, кожа в нем плотная, непод-
вижная, холодная на ощупь, цвета слоновой
кости, блестит, безболезненна.
1. О каком заболевании следует подумать в
данном случае?
2. Дайте развернутый диагноз.
3. С какими заболеваниями следует дифферен-
цировать?
4. Назначьте лечение.
Задача № 30
У мальчика, 8 лет, после длительного обмо-
рожения верхних и нижних конечностей появи-
лось уплотнение кожи в этих участках. Через не-
которое время на конечностях развилась атрофия
кожи, на туловище появились эритематозные
уплотненные очаги, окруженные сиреневым вен-
чиком.
1. Какой предположительный диагноз?
2. Проведите дифференциальный диагноз.
3. Составьте план исследования.
4. Укажите необходимые терапевтические ме-
роприятия.
Задача № 31
В дерматологическую клинику направлен
больной 46 лет, с выраженной маскообразностью
лица, ограничением движений. Лицо амимично,
затруднен прием пищи, ротовое отверстие суже-
но. Пальцы рук не сгибаются, тонкие, кожа на
них истончена, цвета слоновой кости.
1. Поставьте диагноз.
2. Какие необходимо провести клинические и
лабораторные исследования больного?
3. Составьте план лечения.
4. Укажите возможный прогноз заболевания.
Задача № 32
У женщины 27 лет развилось уплотнение
кожи в области шеи слева. Высыпных элемен-
тов, зуда, болезненности нет, отмечается неболь-
шое чувство стягивания кожи в этом участке.
1. Ваш предположительный диагноз?
2. Какие причины могут вызвать данное забо-
левание?
3. Какое лечение можно назначить больной?
Задача № 33
К вам обратился больной, 25 лет, с жалобами
на множественные пигментные пятна различной
конфигурации и размеров на верхней половине
туловища, которые появились после загара. На
отдельных участках обнаруживаются единичные
пятна желтовато-бурого цвета с отрубевидным
шелушением на поверхности.
1. О каком заболевании следует думать, и ка-
кие методы исследования нужно применить для
подтверждения диагноза?
2. Назовите методы лечения заболевания.
Задача № 34
К Вам обратилась больная 21 года, с жалоба-
ми на выпадение волос в течение последнего
месяца. При осмотре на волосистой части голо-
вы имеется множество очагов резкого поредения
590 Приложение В
волос величиной до 1 см в диаметре. Выпадение
волос в очагах поражения не полное, фоллику-
лярный аппарат сохранен. Воспалительные яв-
ления и шелушение отсутствуют. На коже шеи
расположены изолированные гипопигментиро-
ванные пятна размером до 1,5 см без воспали-
тельных явлений. Субъективные ощущения от-
сутствуют. Других проявлений на коже и
слизистых нет. Из анамнеза выяснилось, что 3
месяца назад у больной на туловище была сыпь,
которая со временем исчезла без лечения.
1. Укажите, для какого заболевания наиболее
характерна данная клиническая картина.
2. Пути уточнения диагноза.
Задача № 35
К Вам обратился больной 43 лет, у которого
на подошвах имеется муковидное шелушение по
ходу кожных борозд. Все ногтевые пластинки
стоп и кистей серовато-желтого цвета, утолще-
ны, и частично разрушены.
1. Наметьте план обследования, установите
диагноз.
2. Какие препараты следует назначить?
Задача № 36
Больная 70 лет, на протяжении более 1 года
страдает периодически возникающим, иногда
мучительным зудом кожи туловища и частично
конечностей. Объективно: кожа сухая, субатро-
фичная, местами с явлениями лихенификации,
незначительным шелушением и довольно боль-
шим количеством расчесов.
1. Ваш предварительный диагноз и тактика в
отношении больной?
2. Какие обследования необходимо назначить
больной?
Задача № 37
Больная 45 лет обратилась к дерматологу с
жалобами на поражение кожи лица. Больна в
течение трех месяцев. За помощью не обраща-
лась. Злоупотребляет алкоголем. Дерматологичес-
кий статус: на коже щеки справа определяются
бугорки размером до горошины, коричневато-
красного цвета, тестоватой консистенции, с глад-
кой блестящей поверхностью, незначительным
шелушением, располагающиеся изолировано.
1. Ваш предположительный диагноз.
2. Специфическое дерматологическое исследо-
вание и дальнейшая тактика.
Задача № 38
При обследовании рабочих автомобильного
производства выявлено, что антикоррозийные
ациклические аминосоединения нередко являют-
ся причиной дерматозов. Для них характерна
следующая клиническая картина: высыпания
располагаются главным образом на коже меж-
пальцевых складок кистей, сгибательных повер-
хностях кистей, предплечий, реже — на лице.
При этом отмечаются эритематозные, сгруппи-
рованные папуло-везикулезные элементы, еди-
ничные эрозии, серозные корочки. Обращает на
себя внимание выраженная сухость кожи, мел-
копластинчатое шелушение, образование повер-
хностных трещин, значительный зуд.
1. Какой диагноз можно поставить больному
при наличии такой картины?
2. Какое лечение целесообразно при данном
заболевании?
3. Что может подтвердить или отвергнуть про-
фессиональный характер дерматоза?
4. Что является важнейшим мероприятием для
профилактики рецидивов?
Задача № 39
Больной 41 года, 10 лет страдал кожным за-
болеванием. Высыпания постоянные, локализу-
ются на коже локтей, коленей и волосистой ча-
сти головы. В зимнее время высыпания
распространяются на кожу туловища и нижних
конечностей. Последнее обострение — 2 недели
назад. Объективно: на коже волосистой части
головы, туловище и конечностях обильное ко-
личество папул и бляшек ярко-розового цвета,
с четкими границами округлых и фестончатых
очертаний, покрытых серебристо-белыми чешуй-
ками. Субъективно зуд.
1. Ваш предположительный диагноз?
2. Какие дополнительные исследования надо
провести для постановки окончательного диаг-
ноза?
Задача № 40
На приеме у врача больной 18 лет, с жалоба-
ми на наличие обильных ярких высыпаний на
коже туловища и конечностей, сопровождаю-
щихся легким зудом. Болен 3 дня, когда заме-
тил при мытье большое пятно на груди. На сле-
дующий день появилась обильная мелкая сыпь
на туловище и конечностях. При осмотре: дис-
семинированные отечные желто-розовые пятна
овальных очертаний, слегка западающие в цен-
тре и напоминающие форму «медальона», распо-
ложенные по линиям натяжения кожи.
1. О каком заболевании можно думать?
2. Требуется ли в данном случае проводить
дополнительные методы обследования?
3. Возможны ли осложнения?
Ситуационные задачи по дерматовенерологии 591
Задача № 41
Больной 18 лет, обратился к врачу дермато-
венерологу с жалобами на появление язвы в
области головки полового члена. Самолечение
язвы в течение 4 недель различными мазями и
растворами оказалось безуспешным. За месяц до
появления язвы имел случайную половую связь.
1. Какое заболевание можно заподозрить?
2. Какие исследования необходимо провести
для установления диагноза?
Задача № 42
Больная 50 лет, обратилась к врачу по пово-
ду высыпаний на коже лица, конечностей, силь-
ного зуда кожи в очагах. При осмотре: на коже
щек, лба, предплечий, паховых и подмышечных
складках на резко гиперемированном фоне име-
ется множество пузырьков величиной от прося-
ного зерна до горошины, папуло-везикулы рас-
полагаются сгруппировано.
1. Предполагаемый клинический диагноз.
2. Какие специальные дерматологические ис-
следования необходимо провести для подтверж-
дения диагноза?
3. Какие лабораторные исследования следует
провести?
4. С какими заболеваниями необходимо диф-
ференцировать?
5. Назначьте лечение.
Задача № 43
Больной 60 лет, получил курс лечения в ус-
ловиях стационара по поводу крупнопузырной
формы дерматоза Дюринга, трудно поддающейся
терапии. Врач стационара рекомендовал пациен-
ту в амбулаторных условиях продолжить прием
поддерживающей дозы ДДС — 0,05 г. в сутки
и обследование для исключения патологии внут-
ренних органов.
1. Какие лабораторные исследования необхо-
димо регулярно проводить при лечении ДДС?
2. К каким специалистам направите больного
для исключения патологии внутренних органов?
Задача № 44
Больной 55 лет, поступил повторно в стацио-
нар в связи с обострением герпетиформного дер-
матоза Дюринга. При осмотре: на коже разги-
бательной поверхности предплечий, голеней,
живота на отечном, эритематозном фоне сгруп-
пировано располагаются пузыри величиной от
вишни до сливы, эрозии, корки. Субъективно:
выраженный зуд, жжение.
1. Какие из перечисленных ниже медикамен-
тов можно назначить больному? В какой дозе?
а) авлосульфон; б) полькортолон; в) йодистый
калий; г) витамины (групп В, А, С).
2. Какие из перечисленных выше медикамен-
тов назначать нельзя? Почему?
3. Какая диета показана больному?
Задача № 45
К Вам на прием обратилась женщина 21 года
с жалобами на высыпания на коже лица и гру-
ди, которые впервые отметила в 15 лет. При ос-
мотре: температура тела 36,4 °C, кожа лица жир-
ная, выводные протоки сальных желез
расширены, имеется большое количество коме-
донов, депигментированных точечных рубцов. На
щеках и подбородке несколько багрово-красных,
болезненных при пальпации узлов с кровянис-
то-гнойным, вязким экссудатом. Аналогичные
высыпания, не менее выраженные, имеются на
верхней части груди. Со стороны внутренних
органов патологии не выявлено. Сахар крови
5,5 ммоль/л.
1. Ваш предположительный диагноз?
2. Наметьте комплекс лечебных мероприятий?
Задача № 46
На прием к врачу терапевту обратился больной,
65 лет, с жалобами на резкие боли в левой поло-
вине грудной клетки в течении 5 дней; боли по-
степенно усиливаются. При осмотре на коже пра-
вой половины грудной клетки по ходу
межреберных нервов располагается линейный очаг
поражения. Кожа в нем отечна, гиперемирована.
На этом фоне располагаются множественные
сгруппированные везикулы с прозрачным содер-
жимым, отмечается появление свежих высыпаний.
1. О каком заболевании можно думать?
2. Какое необходимо назначить лечение?
Задача № 47
На прием обратился больной, 32 лет, с жалоба-
ми на сыпь и чувство жжения на коже полового
члена. Болен год. За это время аналогичные вы-
сыпания появлялись 6 раз, и возникновению пред-
шествовало переохлаждение. Сыпь, просущество-
вав 7-10 дней, исчезала. При осмотре на коже
полового члена имеется очаг размером с одноко-
пеечную монету. Кожа в нем отечна и гипереми-
рована, имеются мелкие пузырьки и полицикли-
ческие эрозии. Инфильтрата в основании эрозии
нет. Паховые лимфоузлы незначительно увеличе-
ны, слегка болезненны, тестоватой консистенции.
1. О каком заболевании можно думать?
2. С какими заболеваниями следует проводить
дифференциальный диагноз?
3. Какое необходимо провести лечение?
592 Приложение В
Задача № 48
Больная 30 лет, обратилась к врачу по поводу
высыпаний на коже живота. Субъективных ощу-
щений нет. При осмотре: на коже нижней части
живота, лобка имеется множество полушаровид-
ной формы узелков размером до горошины, глад-
ких, блестящих, цвета нормальной кожи, в цен-
тре элементов отмечается пупковидное вдавление.
1. Ваш предварительный диагноз?
2. Какие средства и методы лечения можно
применить после установления диагноза?
Задача № 49
Больной 16 лет, обратился к врачу по поводу
высыпаний в области кистей. Болен 2 года, не
лечился. При осмотре: на коже тыла кистей
множество узелков цвета нормальной кожи,
плоских, округлых очертаний, размером от про-
сяного зерна до чечевицы.
1. Ваш предварительный диагноз?
2. С какими заболеваниями следует провести
дифференциальный диагноз.
3. Какие методы лечения следует применить
при данном заболевании?
Задача № 50
Больная 35 лет, обратилась в поликлинику по
поводу болезненных образований на коже по-
дошв, затрудняющих ходьбу. При осмотре: на
коже подошв (в местах постоянного давления
обувью) имеются желтоватого цвета образова-
ния, покрытые толстыми роговыми наслоения-
ми, болезненные при надавливании.
1. Поставьте клинический диагноз.
2. Наметьте план лечения.
Задача № 51
Врач-хирург госпиталя при лечении ожогово-
го больного 46 лет, бывшего военного летчика,
обратил внимание на отсутствие болевой реакции
у пациента на перевязках. Из анамнеза: служил
в Узбекистане в течение 7 лет, затем был комис-
сован, и переведен на наземную службу в граж-
данскую авиацию, разведен. При исследовании
больного выявлено резкое снижение болевой,
температурной и тактильной чувствительности по
всему кожному покрову утолщение и болезнен-
ность локтевых нервов, легкая атрофия мышц
предплечий и голеней, инфильтраты в надбров-
ных областях и мочках ушей, амимичное лицо.
Последние 4 года страдает импотенцией.
1. Ваш предположительный диагноз?
2. Наметьте план обследования пациента.
3. Ваши лечебные действия после установления
диагноза?
Задача № 52
Для консультации на кафедру дерматологии
обратилась женщина, 68 лет санитарка, с жало-
бами на изменения кожи и костной ткани кис-
тей. В детстве перенесла инфекционную болезнь,
по поводу чего длительно лечилась, в течение
последующей жизни получала курсы профилак-
тической терапии.
Объективно: при осмотре кожного покрова в
области шеи обнаружены мягкие рубцы с сосоч-
ковыми разрастаниями, частичные мутиляции
пальцев кистей, рубцовые изменения кожи ки-
стей, межпальцевые контрактуры.
1. О каком заболевании можно подумать?
2. Какие специфические заболевания могут
вызвать поражения кожи и костной ткани?
3. Какова ваша дальнейшая лечебная тактика?
Задача № 53
В клинику кожных болезней поступил воен-
нослужащий, 22 лет, житель Белоруссии, с диа-
тезом: хроническая пиодермия нижней трети
голени, стоп. Перед поступлением в клинику
лечился в медсанчасти наружными противомик-
робными средствами, УВЧ на область очагов,
получал инъекции витаминов В, и В6, алоэ. Улуч-
шения со стороны кожного патологического
процесса не наблюдалось. Из анамнеза незадол-
го до призыва в армию (1 месяц) летом был у
родственников в Азербайджане. Объективно: на
сгибательных поверхностях нижней трети голе-
ни и тыле стоп инфильтрированные язвочки и
эрозии неправильной формы, покрытые трудно-
отделяемыми корко-чешуйками, окруженные ва-
ликом буровато-красного цвета, слегка болезнен-
ными. Общее состояние удовлетворительное,
имеются явления несколько болезненного пахо-
вого лимфаденита.
1. О каких заболеваниях можно подумать с
учетом эпиданамнеза?
2. Какие исследования необходимо провести
больному для уточнения диагноза?
3. Какие системные и наружные средства не-
обходимо назначить пациенту?
Задача № 54
На туловище у больного множественные ги-
попигментированные пятна, которые появились
после летнего сезона, различной конфигурации
и размеров, расположенные изолированно, на
загорелой коже, местами единичные желтовато-
бурые пятна с отрубевидным шелушением.
1. О каком дерматозе следует думать?
2. Какие наружные средства можно назначить
пациенту?
Ситуационные задачи по дерматовенерологии
593
3. Каковы Ваши рекомендации по профилак-
тике дерматоза?
Задача № 55
К Вам обратился рабочий цементного завода
с жалобами на зуд, покраснение, отечность, гной-
нички, мокнутие кожи вокруг свища от остео-
миелита на правой голени. Границы поражения
резкие, на других участках кожи высыпаний нет.
1. Ваш наиболее вероятный диагноз?
2. Какие наружные средства можно применить
у этого пациента?
Задача № 56
У больной 35 лет после сильного нервного
переживания на тыле кистей появились участ-
ки покраснения и отека с последующим обра-
зованием на этом фоне мелких воспалительных
узелков, пузырьков, эрозий с обильным выде-
лением серозной жидкости. Процесс сопровож-
дается сильным зудом.
1. Ваш наиболее вероятный клинический ди-
агноз?
2. Какие наружные средства необходимо при-
менить при данной стадии заболевания?
Задача № 57
На прием к дерматологу обратился мужчина
с жалобами на зуд кожи, усиливающийся к ве-
черу, появившийся через несколько дней после
возвращения из командировки. Последние два
дня зуд кожи начал беспокоить жену пациента.
Дерматологический статус: на коже живота рас-
пространенная парная папуло-везикулярная
сыпь, следы расчесов. На коже кистей в меж-
пальцевых промежутках, области запястья опре-
деляются точечные узелки.
1. Ваш предположительный диагноз?
2. Назовите методы обследования для его под-
тверждения.
3. Назовите препараты для лечения.
Задача № 58
Больная 17 лет обратилась к дерматологу с
жалобами на поражение кистей. Больна около
недели, не лечилась. Тщательно мыла руки, счи-
тая, что сможет избавиться от «гнойничков».
Дерматологический статус: на коже кистей еди-
ничные пузыри, дряблые, с гнойным содержи-
мым, изолированные друг от друга, некоторые
из них подсыхают в гнойную корочку.
1. Ваш диагноз?
2. Какое лечение назначите и какие рекомен-
дации дадите пациентке.
Задача № 59
В кожно-венерологический диспансер обра-
тился мужчина 22 лет с жалобами на появление
«ссадины» в области полового члена. Согласно
анамнезу 2 месяца тому назад имел однократное
половое сношение с неизвестной женщиной.
Другие половые контакты категорически отри-
цает. Около месяца тому назад обратил внима-
ние на появление язвочки на наружном листке
крайней плоти. При осмотре: в области наруж-
ного листка крайней плоти имеется эрозия, раз-
мером с ногтевую пластинку, безболезненная. В
основании эрозии определяется хрящевидное
уплотнение. Паховые лимфоузлы увеличены.
При исследовании отделяемого эрозии обнару-
жены бледные трепонемы.
1. Ваш диагноз?
2. С какими заболеваниями следует дифферен-
цировать?
3. Какие организационные мероприятия необ-
ходимо провести?
Задача № 60
По поводу ангины больному был назначен
тетрациклин, который он принимал и ранее. На
второй день лечения явления ангины стали раз-
решаться, общее состояние нормализовалось,
температура снизилась, однако на коже появи-
лась обильная, зудящая сыпь в виде розовых
пятен.
1. Ваш клинический диагноз?
2. Какое лечение назначите?
3. Какие рекомендации дадите больному?
Задача № 61
На прием обратился больной с жалобами на
появление высыпаний, сопровождающихся зу-
дом и покалыванием. При осмотре: в области
угла рта, справа на эритематозном отечном фоне
сгруппированные мелкие пузырьки, наполнен-
ные прозрачным серозным содержимым и мел-
кие эрозии.
1. Для какого заболевания характерна указан-
ная выше клиническая картина?
2. Назначьте лечение.
Задача № 62
На приеме у врача больная, 20 лет, с жалоба-
ми на сыпь, сопровождающуюся зудом. Больна
одну неделю. Первое высыпание появилось на
туловище в виде крупного пятна. После мытья
в бане количество сыпи резко увеличилось, она
стала яркой. При осмотре: на коже туловища и
конечностей имеются многочисленные отечные
пятна, розово-желтого цвета, с легким шелуше-
594 Приложение В
нием в центре, овальных очертаний, расположен-
ные по линиям натяжения кожи.
1. Ваш наиболее вероятный диагноз?
2. Какое местное лечение назначите?
Задача № 63
К врачу на прием обратился больной, у кото-
рого два дня назад появились приступообразные
боли в левой половине грудной клетки, общая
слабость, лихорадка и головная боль, высыпания
на коже груди. При осмотре: больного слева по
ходу 4—5 межреберьев на эритематозно-отечном
фоне имеются множественные сгруппированные
пузырьки с напряженной покрышкой, наполнен-
ные прозрачным содержимым, величиной 2—
4 мм в диаметре.
1. Для какого заболевания характерна указан-
ная выше симптоматика?
2. Консультацию каких специалистов необхо-
димо получить?
Задача № 64
На прием обратился больной с жалобами на
болезненные эрозии полового члена. Из анам-
неза выявлено частое появление подобных вы-
сыпаний в течение года. При осмотре, на головке
полового члена сгруппированные пузырьки и
эрозии полициклических очертаний, с четкими
границами, мягкие при пальпации.
1. Для какого заболевания наиболее характер-
на описанная картина?
2. Какие местные препараты можно назначить
пациенту?
3. Укажите средства, применяемые для профи-
лактики заболевания?
Задача № 65
В клинику поступил больной 19 лет с жало-
бами на зуд кожи и высыпания в области лок-
тевых и подколенных сгибов. Болеет с 6-месяч-
ного возраста (со слов матери). Рецидивы
заболевания наступают после употребления шо-
колада, апельсинов, пряностей. Объективно: на
коже локтевых, подколенных сгибов кожа сухая,
лихенизирована, геморрагические корочки, сле-
ды экскориаций.
1. Установите диагноз, клиническую форму и
стадию заболевания.
2. Предложите план обследования и общего
лечения.
Задача № 66
Больная К. 55 лет доставлена в стационар са-
нитарной авиацией. При осмотре: общее состо-
яние больной тяжелое, активные движения из-
за болей затруднены, температура тела 38,3 С.
Из-за болей рот открывается плохо, на красной
кайме губ обилие геморрагических корок. На
коже живота, в складках под грудными железа-
ми, в подмышечных впадинах, пахово-бедрен-
ных множество обширных эрозий, покрытых
грязным налетом; на их поверхности видны пап-
пиломатозные разрастания. По периферии оча-
гов поражения — пузырьки, пузыри на фоне
здоровой кожи. Симптом Никольского положи-
тельный.
1. Поставьте клинический диагноз.
2. Какие лабораторные исследования необхо-
димо провести для подтверждения диагноза?
3. С какими заболеваниями необходимо про-
вести дифференциальный диагноз?
4. Назначьте лечение.
Задача № 67
На прием к дерматологу обратился мужчина
35 лет, животновод по профессии, с жалобами
на поражение кожи лица. Болен в течение не-
скольких дней. Дерматологический статус: на
коже лица в области бороды очаг поражения
округлой формы, резко гиперемирован с боль-
шим количеством гнойных корочек. Из фолли-
кулов выдавливается гной.
1. Предположительный диагноз?
2. Назначьте необходимые методы исследова-
ния для подтверждения диагноза.
3. Тактика врача.
Задача № 68
Больной 48 лет, страдающий сахарным диабе-
том, через несколько недель после посещения
бани заметил у себя в паховой области высыпа-
ния в виде пятен, которые начинают увеличи-
ваться в размерах и вызывают зуд после силь-
ного потоотделения. Объективно: в паховых,
бедренно-мошоночных складках, на внутренней
поверхности бедер имеются крупные очаги, ярко
красного цвета с гиперемированной, мацериро-
ванной поверхностью, приподнятым отечным
краем, наличием корочек, пузырьков. Лабора-
торно при исследовании чешуек из очагов по-
верхности обнаружены нити мицелия гриба.
1. Ваш предположительный диагноз?
2. Какие лабораторные исследования следует
провести дополнительно?
3. План лечения.
Задача № 69
Больная 62 лет неожиданно заметила у себя
эрозии на слизистой оболочке полости рта, ко-
торые вначале эпителизировались и исчезали, а
Ситуационные задачи по дерматовенерологии
595
затем через некоторое время появлялись вновь.
Затем возникли пузыри на внешне неизменен-
ной коже живота, спины. Пузыри крупные с се-
розным содержимым, напряженные, с тенденци-
ей к слиянию между собой. При легкой травме
пузыри быстро вскрываются, образуя эрозии с
сочным красным дном. По периферии очагов
имеются обрывки покрышек пузырей. Эрозии
эпителизируются медленно.
1. О каком заболевании следует подумать?
2. Какие симптомы помогут в диагностике?
3. Какие лабораторные исследования следует
провести?
Задача № 70
Больная 57 лет, жалуется на появление мно-
гочисленных пузырей на коже туловища, спи-
ны, груди. Пузыри крупные с серозным содер-
жимым, дряблые.
После их вскрытия образуются эрозии, которые
медленно эпителизируются. На слизистой полос-
ти рта также имеются эрозии, при приеме пищи
отмечается болезненность. Симптом Никольско-
го положительный. В мазках отпечатках со дна
пузырей обнаружены акантолитические клетки.
1. Ваш диагноз?
2. План лечения.
Задача № 71
Молодой мужчина 22 лет по поводу простуд-
ного заболевания принимал самостоятельно ами-
допирин и сульфамонометоксин. К первоначаль-
ным симптомам заболевания (головная боль,
общая слабость, боли в горле, мышцах, суставах)
через несколько дней после приема лекарств при-
соединились высыпания на коже туловища, сли-
зистой рта, красной кайме губ. Высыпания на
тыле кистей и стоп, разгибательных поверхнос-
тях предплечий и голеней представлены в виде
пятен или плоских папул розово-красного цве-
та, размером 2—3 см. Центральная часть элемен-
тов слегка западает и приобретает синюшный от-
тенок, а периферическая часть сохраняет
розово-красный цвет. Имеются также пузыри с
серозным и единичные — с геморрагическим со-
держимым. Субъективно: жжение, умеренный
зуд. Симптом Никольского отрицательный. В ла-
бораторных показателях существенных измене-
ний нет (незначительное повышение СОЭ).
1. Ваш предположительный диагноз?
2. План лечения.
Задача № 72
Больная 48 лет предъявляет жалобы на мучи-
тельный зуд и поражение кожи в нижней трети
голени. Больна в течение 7 лет, периодически от-
мечаются ремиссии, однако высыпания не рег-
рессируют полностью. Сопутствующее заболева-
ние — холецистит. Дерматологический статус: в
нижней трети передней поверхности левой го-
лени с переходом на тыл стопы имеется бляшка
размером 8x8 см в виде сплошной папулезной
инфильтрации, буровато-красного цвета с оваль-
ными очертаниями, выражена лихенификация.
Кожа в очаге поражения имеет шагреневый вид,
различают три зоны: центральная с лихенизаци-
ей, средняя — с блестящей гладкой поверхнос-
тью и бледно-розовыми папулами, периферичес-
кая зона с гиперпигментацией. Симптом Кебнера
и Ауспитца отрицателен. Сетка Уикхема не оп-
ределяется.
1. Ваш предположительный диагноз?
2. Методы лечения.
Задача № 73
Больной 17 лет жалуется на поражение кожи
волосистой части головы и конечностей в обла-
сти локтевых и коленных суставов, легкий зуд
и шелушение в очагах поражения. Из анамнеза:
симтомы поражения кожи появились около 6 ме-
сяцев назад после спортивных соревнований.
Отец и мать дерматологически здоровы. Двою-
родный брат отмечает периодические высыпания
на коже туловища и конечностей в зимний пе-
риод времени года.
Самостоятельно лечился шампунем «От пер-
хоти», а также смазывал участки поражения на
конечностях 10 % ихтиоловой мазью — эффект
отсутствовал (появлялись новые элементы сыпи).
Дерматологический статус: на волосистой части
головы, преимущественно на границе гладкая
кожа — волосы, за ушными раковинами, в об-
ласти разгибательной поверхности локтевых и
коленных суставов определяются эпидермально-
дермальные папулы, ярко-красного цвета, плот-
новатой консистенции, с резкими границами;
бляшки покрыты серебристо-белыми крупными
чешуйками. Симптом «стеаринового пятна»,
Ауспитца и Пильнова — положительные. Ана-
лиз на грибы с участков поражения — отрица-
тельный (грибы не обнаружены!).
1. Ваш диагноз?
2. На основании каких диагностических при-
знаков может быть выставлен данный диагноз?
Задача № 74
Больная 44 лет поступила в кожное отделение
с диагнозом: Красный плоский лишай. Жалобы
при поступлении: поражение волосистой части
головы, туловища, чувство стягивания и жжения
596 Приложение В
в очагах поражения, шелушение очагов поражения.
Анамнез: страдает заболеванием около 9 лет. Еже-
годно отдыхает на Южном берегу Крыма. Отме-
чает значительное улучшение состояния и стойких
эффект после пребывания на Юге в течение 6—8
месяцев. Последнее обострение наступило после
внезапной смерти отца. Дерматологический статус:
поражение носит распространенный характер с из-
любленной локализацией на волосистой части го-
ловы, разгибательной поверхности коленных и
локтевых суставов, в люмбосакральной области, на
коже спины, груди, боковых поверхностях туло-
вища. Определяются эпидермально-дермапьные
папулы, четко отграниченные от здоровой кожи,
ярко-красного на вид цвета, покрытые рыхлыми
крупнопластинчатыми чешуйками серебристо-бе-
лого цвета, диаметром 2-2 мм, правильной фор-
мы с четким периферическим валиком роста в виде
эритемы. Симптом Ауспитца и Кебнера положи-
тельны.
1. Ваш развернутый диагноз?
2. Перечислите диагностические признаки ста-
дии заболевания.
Задача № 75
Женщина 43 лет обратилась с жалобами на по-
ражение слизистой оболочки рта и кожи верх-
них конечностей, сопровождающееся зудом по-
стоянного характера в течение суток. Эти
симптомы появились около 3 недель после пе-
ренесенной 2-х сторонней пневмонии (лечилась
стационарно). Находится на учете у терапевта по
поводу заболевания ЖКТ и сахарного диабета II
типа. Дерматологический статус: на коже обеих
верхних конечностей (сгибательные поверхнос-
ти) определяется мономорфная сыпь. Первичный
элемент плоская папула с вдавлением в центре,
полигональной формы, размером от 0,5 до 1 см
в диаметре, фиолетово-красного цвета с воско-
видным блеском, плотноватой консистенции.
При смазывании папул (бляшек) вазелиновым
маслом на их поверхности определяются белесо-
вато-опаловые тонкие и пересекающиеся в виде
сетки линии. На передней поверхности голеней
выявляется положительная изоморфная реакция
Кебнера. На слизистой оболочке полости рта
(язык, губы) определяются 2 бляшки до 1 см в
диаметре покрытые серо-белым «напетом».
1. Ваш диагноз?
2. Перечислите диагностические признаки,
подтверждающие ваш диагноз.
Задача № 76
Больная 57 лет поступила в кожное отделение
с диагнозом: Ограниченный псориаз, стационар-
ная стадия. Жалобы на поражение кожи правой
голени, сильный зуд постоянного характера,
шелушение в этих участках. Больна около 13 лет.
Обращалась за помощью в ЦРБ. Лечение вклю-
чало: локальное УФО, преднизолон 0,5 % на
очаг поражения с кратковременным эффектом
и последующим обострением.
Дерматологический статус: поражение кожи
носит ограниченный характер с локализацией на
передней поверхности правой голени в виде
бляшки диаметром 3x5 см, с резкими граница-
ми. В очаге поражения определяется ороговение
белесовато-сиреневого цвета. Симптомы Уикхе-
ма положительный. На коже левого предплечья
(сгибательная поверхность) папулезные элемен-
ты с вдавлением в центре; высыпания фиолето-
вого цвета в виде полосы. Папулы полигональ-
ной формы с пупкообразным вдавлением в
центре, границы четкие, консистенция плотно-
ватая, отсутствует феномен стеаринового пятна,
лаковой пленки и точечного кровотечения. РВ
на сифилис — отрицательная со всеми антиге-
нами.
1. Ваш диагноз?
2. Основные методы лечения.
Задача № 77
Ребенок 10 лет поступил в кожное отделение
по направлению педиатра с диагнозом: стрепто-
кокковое импетиго. Жалобы при поступлении на
наличие сыпи в области нижних конечностей. Са-
мостоятельное лечение кортикостероидными ма-
зями, синтомициновой эмульсией эффекта не
принесло. Единичные элементы сыпи вскрыва-
лись и подсыхали, но одновременно появлялись
новые элементы.
Дерматологический статус: на коже внутрен-
ней поверхности бедер, ягодиц определяются
плотные по консистенции, полушаровидной
формы узелки цвета нормальной кожи величи-
ной от булавочной головки до чечевицы. В цен-
тре узелков — пупкообразное вдавление. При
сдавливании пинцетом из узелков выделяется
белая кашицеобразная творожистая масса.
1. Ваш диагноз?
2. Тактика ведения больного.
Задача № 78
Больной 28 лет поступил с жалобами на по-
ражение головки полового члена и крайней пло-
ти. Из анамнеза выяснено: не женат, ведет бес-
порядочную половую жизнь.
Дерматологический статус: на головке поло-
вого члена и в области внутреннего листка край-
ней плоти маленькие бородавчатые образования
Ситуационные задачи по дерматовенерологии
597
на тонкой ножке, по форме напоминающие цвет-
ную капусту (петушиный гребень). Цвет от блед-
но-розового до интенсивно-красного. Образова-
ния мягкие, не сращены с окружающими
тканями. При эндонуклеарном анализе ДНК,
выделенной из инфицированных тканей — об-
наружен вирус папилломы человека — тип 6, И.
1. Ваш диагноз?
2. Перечислите основные методы лечения.
Задача № 79
Больная 47 лет госпитализирована в кожное
отделение после консультации инфекциониста.
Из анамнеза: жалобы на зуд кожи бессонницу,
снижение трудоспособности, раздражительность.
Больная отмечает возникновение указаных сим-
птомов после употребления блюд из морских
сортов рыб. Состояние резко ухудшилось и вы-
сыпания распространились на кожу всего туло-
вища и конечностей после смазывания кожи в
течение нескольких клинических дней — 5%
настойкой йода.
Дерматологический статус: на коже туловища,
конечностей (бедра, предплечья) определяются
напряженные мелкие пузыри, пузырьки, склон-
ные к группировке, пятна и папулы. В области
поражения на отечном фоне определяются эро-
зии, по периферии которых видны обрывки
покрышек пузырей. Симптом Никольского от-
рицательный.
Лабораторно: в мазках-отпечатках со дна эро-
зии на 100 лейкоцитов определяется 21 % эози-
нофилов. При проведении ИФ-анализа выявле-
ны отложение IgA в дермо-эпидермальной зоне
и частично в сосочковом слое дермы.
1. Ваш диагноз?
2. Тактика ведения больного.
Задача № 80
Больная 19 лет обратилась в кожно-венероло-
гический диспансер. Жалобы на поражение крас-
ной каймы нижней губы, появившееся неделю
назад. При осмотре: в указанной области отме-
чается безболезненная язва, величиной с одно-
копеечную монету, цвета свежеразрезанного
мяса с четкими границами. При пальпации в ос-
новании язвы определяется плотный инфильт-
рат. Подчелюстные лимфоузлы увеличены, плот-
ноэластической консистенции, безболезненные,
подвижные. Кожа свободна от высыпаний. Ре-
акция Вассермана сыворотки крови отрицатель-
ная.
1. Какой должна быть тактика в отношении
больной?
Задача № 81
Пациент 35 лет обратился на прием в связи с
тем, что 16 дней назад имел половой контакт с
женщиной, больной вторичным свежим сифи-
лисом. При тщательном осмотре специфических
высыпаний на коже и слизистых оболочках не
обнаружено. Реакция Вассермана крови и РИБТ
отрицательны.
1. Какие меры следует предпринять?
Задача № 82
Больная 26 лет обратилась на прием к врачу-
гинекологу с жалобами на поражение в области
половых органов. При осмотре: на правой большой
губе располагается язва 0,5 см в диаметре, округ-
лых очертаний, мясо-красного цвета с резкими
границами: при пальпации определяется плотное
безболезненное основание. Язва находится в стадии
рубцевания. На коже туловища, преимущественно
на боковых поверхностях живота обильная розео-
лезно-папулезная сыпь Подчелюстные, подмышеч-
ные, локтевые и паховые лимфоузлы увеличены,
плотноэластической консистенции, подвижные, без-
болезненные при пальпации.
1. Для какого диагноза характерна указанная
клиническая картина?
2. Какой должна быть тактика врача-гинеко-
лога?
3. Какие профилактические мероприятия дол-
жен провести врач-гинеколог?
Задача № 83
Пациентка 40 лет работница столовой прохо-
дила профилактический осмотр у гинеколога.
При осмотре наружных гиниталий обнаружены
папулезные высыпания, резко отграниченные от
здоровой кожи, с неровной поверхностью, часть
элементов эрозированы.
1. О каком диагнозе можно думать?
2. Какие обследования необходимо провести
для подтверждения диагноза?
Задача № 84
Больной 35 лет направлен на консультативный
прием дерматовенеролога с диагнозом: Ранний
скрытый сифилис.
1. Какие изменения могут быть у пациента на
коже, слизистых, со стороны лимфатических уз-
лов?
Задача № 85
Больная 25 лет с диагнозом раннего скрыто-
го сифилиса отказывается от госпитализации и
лечения.
1. Какие меры могут быть к ней приняты?
598 Приложение В
Задача № 86
Больной 24 лет обратился к дерматовенероло-
гу с жалобами на выделения из мочеиспускатель-
ного канала, рези во время и по окончании мо-
чеиспускания. Такие явления возникли через 7
дней после случайной половой связи. При осмот-
ре: в области наружного отверстия уретры — ги-
перемия, отечность, отмечаются свободные уме-
ренные желто-зеленые гнойные выделения.
Пальпация уретры — болезненна. При двухста-
канной пробе Томпсона моча мутная в обеих
порциях. Исследования мазков гноя из уретры,
окрашенных по Грамму: обнаружены гонококки.
1. Установите диагноз.
2. Наметьте план лечения и противоэпидеми-
ческих мероприятий.
Задача № 87
Больной 19 лет обратился к дерматовенерологу
в связи с жалобами на гнойные выделения из
мочеиспускательного канала, а также рези в на-
чале акта мочеиспускания. Заболел через неде-
лю после случайной половой связи. При оцен-
ке двухстаканной пробы Томпсона моча мутная
в первой порции. Бактериоскопическое исследо-
вание выделений выявило расположенные внут-
ри- и внеклеточно гонококки.
1. Установите диагноз и составьте план лечения.
Задача № 88
Больной 27 лет, обратился к дерматовенеро-
логу в связи с жалобами на режущие боли в
конце акта мочеиспускания, а также учащенные
позывы к мочеиспусканию. Заболевание разви-
лось остро, спустя 5 дней после случайной по-
ловой связи, не лечился. При осмотре: гипере-
мия и отек губок наружного отверстия уретры,
гнойно-слизистое желтоватое отделяемое из урет-
ры. При двухстаканной пробе Томпсона моча
мутная в обоих порциях. Повторные исследова-
ния мазков постоянно выявляли лейкоциты,
сплошь покрывающие поле зрения; в каждом
поле зрения большие скопления почкующихся
клеток. Бактерии и трихомонады не определе-
ны. При посеве на среду Сабуро — выраженный
рост С. albicans, а также апатогенного стафило-
кокка.
1. Установите диагноз и предложите лечение.
Задача № 89
Поступил больной с язвочками в области по-
лового члена и мошонки появившихся, 3—4 дня
назад. Возникновение их связывает с внебрач-
ным половым контактом. Объективно: эрозии
округлой формы, величиной до однокопеечной
монеты, в основании — листовидный инфиль-
трат, на поверхности эрозий — «сальный налет».
Регионарные лимфатические узлы не увеличе-
ны, серореакции отрицательные.
1. Укажите план обследования.
2. О каких заболеваниях, следует подумать в
первую очередь?
Задача № 90
Женщина 36 лет обратилась к гинекологу в
связи с безболезненной припухлостью правой
большой половой губы, несколько затрудняю-
щей движение. Больна около трех недель. По
назначению врача-гинеколога принимала стреп-
тоцид внутрь и УФО местно. На фоне лечения
улучшения не наступило. При повторном осмот-
ре замечено увеличение паховых лимфатических
узлов на стороне опухоли и назначено исследо-
вание серологических реакций крови.
1. О каких заболеваниях следует думать?
2. Какие клинические проявления сходны с
бартолинитом?
3. Какие симптомы не характерны для барто-
линита?
4. Какая тактическая ошибка допущена гине-
кологом?
5. Как следовало поступить?
6. Какие результаты серологического исследо-
вания крови ожидаются в подобной ситуации?
Задача № 91
На прием к врачу КВД обратился молодой
человек с явлениями фимоза, развившимся не-
делю назад. Паховые лимфоузлы увеличены до
размера мелкой сливы, плотноэластической кон-
систенции, подвижны, безболезненны, кожа над
ними не изменена. Больной не женат, имел слу-
чайный половой контакт 1 месяц тому назад.
1. Укажите наиболее вероятный диагноз.
2. Какие исследования необходимо назначить
больному?
Задача № 92
Больная 32 лет обратилась к дерматологу по
поводу сыпи на коже туловища. Из анамнеза
стало известно, что три месяца тому назад она
имела половой контакт с неизвестным мужчи-
ной. При осмотре на коже туловища розеолез-
ная сыпь, определяется полиаденит.
1. Какое заболевание можно заподозрить?
2. Какие исследования необходимы для уста-
новления диагноза?
3. Дальнейшая тактика врача?
Ситуационные задачи по дерматовенерологии
599
Задача № 93
Больная 26 лет обратилась с жалобами на уси-
ленное выпадение волос. При осмотре — диф-
фузная алопеция. На коже нижней латеральной
части шеи справа и слева имеются гипопигмен-
тированные пятна овальной формы, напомина-
ющие сетку. При проведении йодной пробы
результат отрицательный.
1. Ваш диагноз?
2. Что необходимо сделать для постановки
диагноза?
Задача № 94
Обратилась беременная женщина 25 лет
(срок беременности 7 недель), у которой при
первичной вассерманизации выявлены резко
положительйые КСР. При осмотре: кожа и
видимые слизистые свободны от специфичес-
ких высыпаний. Однако из анамнеза удалось
выяснить, что у нее на теле существовала ка-
кая-то мелкая и довольно обильная сыпь, не
причинявшая ей никаких субъективных ощу-
щений и которая затем самопроизвольно раз-
решилась.
1. Ваш предположительный диагноз?
2. Тактика в отношении данной больной.
Задача № 95
На прием к дерматовенерологу обратилась
женщина с жалобами на то, что у ее партнера ди-
агностирован сифилис и он по поводу своего за-
болевания находится в венерологическом стаци-
онаре. Половой контакт однократный 10 дней
назад. При обследовании комплекс серологичес-
ких реакций (РВ, РИФ, РИБТ) отрицательный.
При тщательном осмотре больной на слизистых
высыпаний нет. На коже верхней части тулови-
ща имеются пятна размером от лентикулярных
до нуммулярных, темно-коричневого цвета.
Йодная проба положительная.
1. Поставьте диагноз.
2. Показано ли пациентке превентивное лече-
ние?
Задача № 96
На прием к хирургу обратился больной 37 лет
с жалобами на неприятные ощущения в области
заднего прохода. Считает, что заболел 2 недели
назад, когда впервые отметил эти явления. Ме-
стное лечение примочками эффекта не дало.
При осмотре в перианальной области имеются ги-
пертрофические вегетирующие папулы с влаж-
ной, местами мацерированной поверхностью.
Кроме этого, у больного обнаружены папулы на
ладонях и подошвах и диффузное разрежение
волос на голове. Слабо выраженный полиаденит.
1. О каком заболевании можно думать?
2. Каковы пути подтверждения диагноза?
3. Какова должна быть тактика хирурга в дан-
ном случае?
Задача № 97
В клинику госпитализирован мужчина 55 лет
с жалобами на появление язвы на коже правой
голени. Серологические реакции: РВ 3+, РИФ
4+, РИТ—100 %. По данным анамнеза, 4 года
тому назад был установлен диагноз первичного
сифилиса, по поводу которого получил непол-
ный курс лечения. Во время лечения выпивал,
а в конце курса лечение самовольно его прервал
и уехал из города в другую местность, не поста-
вив в известность врачей кожвендиспансера.
Последние три года работал на лесозаготовках.
После незначительной травмы голени образова-
лась язва размером 3x4 см, с плотными отвес-
ными краями и гнойно-некротическим отделя-
емым.
1. Ваш предварительный диагноз?
2. С какими заболеваниями следует провести
дифференциальный диагноз?
3. У каких специалистов следует проконсуль-
тировать больного?
Задача № 98
У больного 52 лет появились высыпания на
коже спины. Объективно: в подлопаточной об-
ласти слева группа бугорков размером с кедро-
вый орешек, медно-красного цвета, плотной
консистенции. Часть элементов изъязвилась. Се-
рологические реакции (РВ, РИФ, РИТ) поло-
жительные.
1. Какой диагноз можно поставить больному?
2. С какими заболеваниями необходимо про-
вести дифференциальный диагноз?
3. У каких специалистов следует проконсуль-
тировать больного?
Задача № 99
Больная 26 лет считает себя больной 7 меся-
цев. Лечилась с диагнозом туберкулезной вол-
чанки слизистой оболочки полости рта. В связи
с отсутствием эффекта была консультирована
отоларингологом. При осмотре: у корня малого
язычка, захватывая небную дужку и прилежа-
щие участки мягкого неба, видна глубокая язва
размером 2x1,5 см с ровными краями, плотной
консистенции, синюшно-красного цвета. Дно
вялое, неровное, покрыто серо-гнойным налетом,
21 Зак. 1357
600 Приложение В
при отторжении которого кровотечение не на-
блюдается. Венерические болезни в анамнезе
отрицает.
1. Возможный клинический диагноз?
2. Необходимые лабораторные исследования?
3. Тактика в отношении бытовых контактов?
Задача № 100
В клинике находится больной, которому ус-
тановлен диагноз: сифилис первичный сероне-
гативный. Из анамнеза установлено, что он не
переносит пенициллин и его производные.
1. Ваша тактика?
Приложение С
Эталоны ответов к ситуационным задачам по
дерматовенерологии для студентов лечебного
факультета
Задача № 1
1. Контактный аллергический дерматит.
2. Десенсибилизирующие препараты (кальция
глюконат, натрия тиосульфат), антигистаминные
препараты (димедрол, тавегил, супрастин); мес-
тно-глюкокортикоидные мази низкой и средней
степени активности.
Задача № 2
1. Крапивница.
2. Консультация аллерголога для установления
причины крапивницы, а следовательно, и про-
филактики рецидивов заболевания.
Задача № 3
1. Антигистаминные препараты (супрастин,
тавегил, димедрол); десенсибилизирующая тера-
пия (кальция глюконат, натрия тиосульфат).
2. Примочки с водными растворами борной
кислоты (1%), риванола (1: 1000).
Задача № 4
1. Контактный дерматит.
2. В анамнезе указывает на контакт с раздра-
жающим веществом; стадийность течения про-
цесса (эритема, пузырь).
Задача № 5
1. Аллергический дерматит.
2. Исключить контакт с серной мазью; внутрь
десенсибилизирующие и антигистаминные пре-
параты; местно — кремы и мази, содержащие
глюкокортикоиды.
Задача № 6
1. Атопический дерматит, эритематозно-скво-
мазная с лихенизацией форма, стадия обостре-
ния.
2. Антигистаминные препараты (пипольфен,
супрастин); десенсибилизирующие препараты
(кальция глюконат, натрия тиосульфат); энтеро-
сорбенты внутрь (белосорб, полифепан). Мест-
но: противовоспалительные мази без глюкокор-
тикоидов (индометациновая, дермарэф).
Задача № 7
1. Собрать подробный анамнез для уточнения
диагноза: Атопический дерматит эритематозно-
сквамозная форма, стадия обострения.
2. Десенсибилизирующие, антигистаминные
препараты, общеукрепляющее лечение, энтеро-
сорбенты внутрь, нормализация функции ки-
шечника. Местно — нестероидные противовос-
палительные мази.
Задача № 8
1. Себорея волосистой части головы. Себорей-
ная алопеция.
2. Обследование эндокринолога, терапевта с
целью исключения патологии внутренних орга-
нов и желез внутренней секреции.
3. Гипоаллергенная диета с пониженным содер-
жимым жиров; нормализация функции кишеч-
ника; витаминотерапия (витамины группы В, А,
С, Е). Местно — шампуни с содержанием дегтя
(«Фридерм»), цинка.
Задача № 9
1. Вульгарные угри папуло-пустулезная, инду-
ративная форма.
2. Посев флоры пустул на чувствительность к
антибиотикам, антибиотики внутрь 1—1,5 мес.,
местно — «Скинорен», мазь «Риодокосол».
3. Обследование у эндокринолога, терапевта
для исключения патологии внутренних органов.
Задача № 10
1. Розовые угри (розацеа).
2. УФО, заболевание желудка, возраст, нару-
шение менструального цикла.
3. Проводить лечение совместно с эндокрино-
логом-гинекологом (устранение нарушений гор-
601
602 Приложение С
монального фона), трихопол — внутрь; мест-
но — метронидазоловый крем.
Задача № 11
1. Роговая экзема, ладонно-подошвенный псо-
риаз, рубромикоз.
2. Бактериологический и бакгериоскопический
анализ на грибы; тщательное исследование дру-
гих участков тела для обнаружения проявлений
дифференцируемых дерматозов.
Задача № 12
1. Отрубевидный лишай, сифилиды, вторичная
лейкодерма.
2. Тщательно собранный анамнез (может указать
на перенесенные дерматологические заболевания,
на факторы риска, способствующие возникнове-
нию (отрубевидного лишая); бактериоскопическое
исследование на грибы; проба Бальцера; анализ
крови на КСР.
Задача № 13
1. Исключить из пищи копчености, шоколад,
кофе, фрукты и овощи красной окраски, со-
ленья и пряности. Допускается принимать в
пищу овощи и фрукты зеленой и желтой ок-
раски.
Задача № 14
1. Микроспория волосистой части головы.
2. Бактериоскопическое исследование волос на
грибы.
3. Госпитализация ребенка в отделение гриб-
ковых инфекций с последующим назначением
гризеофульвина внутрь и применением проти-
вогрибковых мазей местно.
Задача № 15
1. Гнездная алопеция, очаговая форма, обыч-
ный тип, прогрессирующая стадия.
2. Сифилитическая алопеция, грибковые ин-
фекции волосистой части головы.
Задача № 16
1. Необосновано назначено общее лечение,
включая преднизолон внутрь.
2. Необходимо проводить местное лечение с на-
значением мазей, обладающих кератолитическим
действием; применение УФФ с кератолитичес-
кими мазями (ихтиоловая, нафталановая, дегтяр-
ная).
Задача № 17
1. Необходима госпитализация больной в ги-
некологическое (венерологическое отделение),
так как можно предположить развитие эндомет-
рита; углубленное исследование на другие ин-
фекции, передаваемые половым путем; бактери-
ологическое исследование на гонококковую
инфекцию.
Задача № 18
1. Неправильно указана доза тавегила (пра-
вильно — 0,001).
Задача № 19
1. Дискоидная красная волчанка.
2. Исследование на LE — клетки, общий ана-
лиз крови, мочи, иммунограмма.
3. Фотодерматит, рубромикоз, туберкулезная
волчанка.
4. Делагил, витамины В6, В15, аевит, фотоза-
щитные кремы, местные высокоактивные кор-
тикостероиды (клобетазол).
Задача № 20
1. Определить наличие симптомов «дамского
каблучка», Бенье-Мещерского, Хачатурьяна. При
исследовании лампой Вуда — беловато-снежное
свечение очагов атрофии.
2. Диссеминированная форма красной волчан-
ки.
3. Делагил, витамины В6, В15, аевит, фотоза-
щитные кремы, местные высокоактивные кор-
тикостероиды.
4. Работа в закрытых помещениях.
Задача № 21
1. Эритематоз лица, начальная стадия.
2. Фотодерматит, рубромикоз, контактный
дерматит.
3. Местно: высокоактивные кортикостероиды,
фотозащитные кремы, исключить пребывание на
солнце; диспансерное наблюдение.
Задача № 22
1. Диссеминированная красная волчанка.
2. Исследование наЬЕ — клетки, иммунограм-
ма, анализ крови, мочи общий.
3. Необходимо исследовать состояние сердеч-
но-сосудистой системы, легких, почек, печени,
костно-мышечной системы.
4. Пресоцил, витаминотерапия, высокоактив-
ные кортикостероиды местно.
Задача № 23
1. Делагил 0,25 2 раза в день циклами по 5 дней
с пятидневным перерывом (до 20—25 г на курс).
Обладает противовоспалительным и иммуносуп-
рессивным действием.
Эталоны ответов к ситуационным задачам по дерматовенерологии 603
2. Не назначаются вакцины, сульфаниламиды,
так как могут провоцировать волчаночный про-
цесс.
3. Обязательна диспансеризация с клинико-
лабораторным обследованием и профилактичес-
ким лечением 3—4 раза в год.
Задача № 24
1. Контактная экзема.
2. Десенсибилизирующие, антигистаминные,
нестероидные противовоспалительные препара-
ты. Местно — примочки с водными растворами
борной кислоты (1%), риванола (1:1000), танина
(1%).
3. Исключить контакт со стиральным порош-
ком.
Задача № 25
1 Медикаментозная токсикодермия.
2 . Отменить прием медикаментов; обильное
питье; назначить прием внутрь антигистамин-
ных, десенсибилизирующих препаратов, энтеро-
сорбентов. Местно — водная болтушка.
Ежедневный визит к врачу.
Задача № 26
1. Дискоидная красная волчанка.
2. Фотодерматит, рубромикоз, себорейный дер-
матит.
3. После уточнения диагноза: делагил внутрь;
местно — фотозащитные кремы, высокоактив-
ные местные картикостероиды.
Задача № 27
1. Бляшечная склеродермия, стадия уплотне-
ния.
2. Лидаза, пенициллин. Местно: электрофорез
с ронидазой. Профилактические курсы лечения
2—3 раза в год, наблюдение дерматолога, тера-
певта, эндокринолога.
Задача № 28
1. Бляшечная склеродермия, стадия уплотне-
ния.
2. Простудные заболевания, травма, эндокри-
нопатия.
3. Дерматомиозит, глубокий васкулит, дерма-
тофиброма, фиксированная таксидермия.
4. Лидаза, пенициллин. Местно электрофорез
с ронидазой.
5. Профилактические курсы лечения 2—3 раза
в год, наблюдение дерматолога, терапевта, эндок-
ринолога.
Задача № 29
1. Бляшечная склеродермия стадия уплотнения.
2. Дерматомиозит, глубокий васкулит, дерма-
тофиброма.
3. Лидаза, пенициллин. Местно — электрофо-
рез с ронидазой.
4. Профилактические курсы лечения 2—3 раза
в год, наблюдение дерматолога, терапевта, эндок-
ринолога.
Задача № 30
1. Очаговая склеродермия, стадия уплотнения
и атрофии.
2. Болезнь Рейно, диссеминированная красна
волчанка.
3. Обследование ЖКТ, костно-мышечной си-
стемы, легких, почек, глаз; обследование у рев-
матолога.
4. Курс лидазы, аевит. никотиновая кислота;
местно — гидрокортизоновая мазь.
Задача № 31
1. Системная склеродермия, акросклеротичес-
кая форма.
2. Обследование ЖКТ, костно-мышечной си-
стемы, легких, почек, глаз; обследование у рев-
матолога.
3. D-пеницилламин, преднизолон внутрь.
4. Прогноз неблагоприятный.
Задача № 32
1. Бляшечная склеродермия, стадия уплотнения.
2. Простудные заболевания, травма, эндокри-
нопатия.
3. Лидаза, пенициллин. Местно — электрофо-
рез с лидазой.
Задача № 33
1. Отрубевидный лишай; бактериоскопическое
исследование на обнаружение возбудителя; ис-
следование лампой Вуда; йодная проба Бальце-
ра, исследование крови на РВ.
2. Противогрибковые мази, кератолитические
средства.
Задача № 34
1. Сифилис вторичный рецидивный (сифили-
тическая алопеция).
2. Серологическое исследование крови для
подтверждения сифилитической инфекции.
Задача № 35
1. Лабораторное исследование на грибы чешу-
ек кожи и ногтей (микроскопия и посев на сре-
ду Сабуро).
604 Приложение С
2. Ламизил или орунгал внутрь в течение 3—4
месяцев местных противогрибковых средств.
Задача № 36
1 Кожный зуд.
2 . Тщательное терапевтическое обследование,
консультация онколога.
Задача № 37
1. Туберкулезная волчанка.
2. Определение симптома Поспелова и «фено-
мена яблочного желе»; обследование у фтизиат-
ра на туберкулез других органов.
Задача № 38
1. Профессиональная экзема.
2. Устранить контакт; десенсибилизирую-
щие, антигистаминные препараты; местное ле-
чение.
3. Аллергические пробы. Санитарно-гигиени-
ческая характеристика рабочего места.
4. Перевод на другую работу после заключе-
ния профпатолога.
Задача № 39
1. Обычный псориаз, зимний тип, прогресси-
рующая стадия.
2. Граттаж для выявления псориатической три-
ады, симптома Кёбнера.
Задача № 40
1. Розовый лишай.
2. Исследование крови на РВ.
3. При нерациональном местном лечении воз-
можна эритродермия.
Задача № 41
1. Сифилис первичный.
2. Исследование отделяемого в темном поле для
обнаружения бледной трепонемы; серологичес-
кие реакции (РМП, РИФ, РВ).
Задача № 42
1. Дерматоз Дюринга.
2. Проба Ядассона.
3. Исследование эозинофилов крови и пузыр-
ной жидкости.
4. Токсикодермия, пузырчатка, герпес.
5. ДДС, авлосульфон.
Задача № 43
1. Исследование крови и мочи для определе-
ния функции печени, почек, возможной ане-
мии.
2. Терапевт (заболевания ЖКТ), нефролог.
Задача № 44
1. Авлосульфон по 0,05—0,1 г 2 раза в день
циклами по 5—6 дней с перерывом 1 день; все-
го 3—5 циклов.
2. Йодистый калий, потому что вызывает обо-
стрение процесса.
3. Исключить рыбные продукты, продукты,
содержащие глютен (мучные изделия).
Задача № 45
1. Вульгарные угри конглобатные.
2. Посев на чувствительность к антибиотикам;
антибиотики внутрь; ретиноиды внутрь; местные
дезинфицирующие средства.
Задача № 46
1. Опоясывающий лишай.
2. Консультация инфекциониста, невропатоло-
га; ацикловир внутрь; местно — анилиновые
красители, подсушивающие пасты.
Задача № 47
1. Рецидивирующий генитальный герпес.
2. С первичной сифиломой.
3. Учитывая рецидивирующее течение, ацик-
ловир, для купирования обострения. Через 10—
14 дней после исчезновения симптомов — курс
герпетической вакцины.
Задача № 48
1. Контагиозный моллюск
2. Выдавливание элемента пинцетом с после-
дующим прижиганием анилиновыми красителя-
ми; диатермокоагуляция, лазеродеструкция.
Задача № 49
1. Плоские бородавки.
2. Красный плоский лишай, фолликулита кон-
тагиозный молюск.
3. Криодеструкция, лазеротерапия, «Солко-
дерм»; возможно спонтанное исчезновение.
Задача № 50
1. Подошвенные бородавки.
2. Криодеструкция, лазеротерапия, «Солко-
дерм», салициловый пластырь, горячие ванны для
стоп в течении 1,5 мес.
Задача № 51
1. Туберкулез кожи, лепра, неврологическая
патология.
2. Исследование соскобов со слизистой носа,
инфильтратов на М.leprae постановка лепроми-
новой пробы, обследование на туберкулез, кон-
сультация невропатолога.
Эталоны ответов к ситуационным задачам по дерматовенерологии 605
3. При установлении диагноза проводится спе-
цифическое лечение в лепрозории.
Задача № 52
1. Туберкулез кожи колликвативный (перене-
сений в детстве).
2. Сифилис, лепра, лейшманиоз, глубокие
микозы.
3. Назначение профилактического курса про-
тивотуберкулезных препаратов.
Задача № 53
1. Кожный лейшманиоз.
2. Исследование бугорков для обнаружения
возбудителя.
3. Инъекции мономицина; местно — мономи-
циновая мазь 5 %.
Задача № 54
1. Отрубевидный лишай.
2. Противогрибковые мази, отшелушивающие
растворы.
3. Дезинфекция личной одежды, постельного
белья; обследование у терапевта, невропатолога,
эндокринолога, иммунолога.
Задача № 55
1. Паратравматическая экзема, стадия обостре-
ния.
2. Примочки, затем мази с антибиотиками.
Задача № 56
1. Истинная экзема, острая форма.
2. Примочки.
Задача № 57
1. Чесотка.
2. Лабораторное обнаружение чесоточного клеща.
3. Серная мазь, бензилбензоат, метод Богдано-
вича, перметрин.
Задача № 58
1. Стрептококковое импетиго.
2. Анилиновые красители, мази с содержани-
ем антибиотиков (гентамициновая, стрептоцидо-
вая) местно. Исключить контакт с водой.
Задача № 59
1. Сифилис первичный.
2. Эрозии травматического происхождения,
трихомонадные и гонококковые язвы, гениталь-
ный герпес.
3. Лечение пациента; установление личности
женщины и привлечение ее к обследованию и
лечению.
Задача № 60
1. Медикаментозная токсидермия.
2. Отменить прием тетрациклина.
3. В последующем не принимать препараты
группы тетрациклина.
Задача № 61
1. Простой герпес.
2. Анилиновые красители; при начальных сим-
птомах — противовирусные мази.
Задача № 62
1. Розовый лишай.
2. Общее и местное лечение обычно не назна-
чается.
Задача № 63
1. Опоясывающий герпес.
2. Консультация инфекциониста, невропатоло-
га.
Задача № 64
1. Рецидивирующий генитальный герпес.
2. При начальных проявлениях противовирус-
ные мази (оцикловир, риодоксоловая).
3. Герпетическая вакцина 0,2 мл внутрикож-
но через 2 дня 5 инъекций не менее 2 раза в год.
Задача № 65
1. Атопический дерматит, эритематозно-сква-
мозная с лихенификацией форма.
2. Обследование у аллерголога, терапевта; анти-
гистаминные (тавегил, супастин), десенсибилизи-
рующие (натрия тиосульфат), мембраностабили-
заторы (циннаризин, кетотифен); энтеросорбенты
(белосорб), общее УФО.
Задача № 66
1. Истинная пузырчатка, вегетирующая.
2. Мазки-отпечатки для . обнаружения аканто-
литических клеток, прямая иммунофлюоресцен-
ция для выявления фиксированных IgG в меж-
клеточных пространствах.
3. Синдром Стивенса-Джонсона, синдром Лай-
елла, хроническая доброкачественная пузырчатка
Хейли-Хейли, пемфигоид.
4. Преднизолон до 80 мг в сутки; местно —
симптоматическое лечение.
Задача № 67
1. Инфильтративно-нагноительная трихофи-
тия.
2. Лабораторное исследование на грибы (обна-
ружение грибов внутри волоса), при необходи-
мости бактериологическое исследование.
606 Приложение С
3. Госпитализация больного, информация ве-
теринарных служб о случае инфильтративно-
нагноительной трихофитии.
Задача № 68
1. Паховая эпидермофития, стадия обострения.
2. Бактериологическое исследование (посев на
питательные среды); осмотр под лампой Вуда.
3. Антигистаминные препараты внутрь (пи-
польфен, фенкорол или супрастин и др.), мест-
но — растворы анилиновых красителей (для
уменьшения воспалительных явлений); затем —
противогрибковые кремы, растворы, мази (клот-
римазол, канестен, травокорт, экзодерил) в те-
чение 2—3 недель, 1—2 раза в день.
Задача № 69
1. Вульгарная пузырчатка (истинная, аканто-
литическая).
2. Симптомы Никольского, Асбо-Хансена.
3. Мазки-отпечатки со дна эрозий на аканто-
литические клетки; гистологическое исследова-
ние; метод непрямой иммунофлюоресценции.
Задача № 70
1. Вульгарная пузырчатка.
2. Преднизолон 60 мг внутримышечно плюс
60 мг перорально — начальная доза. Для эпите-
лизации эрозий полости рта — масло облепихи
или шиповника; негрубая пища.
Задача № 71
1. Многоформная экссудативная эритема.
2. Преднизолон (дексаметазон) 30—40 мг в сут-
ки; дезинтоксикационная терапия (гемодез, ре-
ополиглюкин) по 400 мл внутривенно капель-
но; обработка слизистой полости рта раствором
KMnO4 1:8000; 2 % раствором борной кисло-
ты, отваром ромашки; на красную кайму губ —
эритромициновая или тетрациклиновая мази с
добавлением кортикостероидов.
Задача № 72
1. Ограниченный хронический нейродермит
Видаля, торпидное, рецидивирующее течение.
2. Гепатопротекторы (карсил, эйконол); эссен-
циале; фонофорез фторированных стероидных
мазей; линимент нафталана.
Задача № 73
1. Ограниченный вульгарный псориаз, про-
грессирующая форма зимний тип, впервые вы-
явленный. Псоиаз волосистой части головы.
2. Диагноз установлен на основании: степени
распространенности процесса, типичных клини-
ческих проявлений, специфических симптомов,
стадии процесса и типа течения заболевания.
Задача № 74
1. Распространенный вульгарный псориаз, про-
грессирующая стадия, зимний тип, непрерывно-
рецидивирующее течение.
2. Ярко-красный цвет папулезных высыпаний,
покрыты серебристо-белым чешуйками, опреде-
ляется периферический валик роста в виде эри-
темы и положительный симптом Кебнера.
Задача № 75
1. Распространенный красный плоский лишай,
прогрессирующая стадия.
2. Папулы полигональной формы с вдавлени-
ем в центре и восковидным блеском; положи-
тельные симптомы Уикхема, Кебнера.
Задача № 76
1. Ограниченный красный плоский лишай,
прогрессирующая стадия.
2. Антигистаминные и антималярийные пре-
параты, гипосенсибилизирующая терапия, ме-
стно-кортикостероиды высокой степени актив-
ности.
Задача № 77
1. Контагиозный моллюск.
2. Изоляция ребенка от детского коллектива,
проведение лечебных процедур (выдавливание
элементов пинцетом с последующей обработкой
5% спиртовым раствором иода); витаминотера-
пия.
Задача № 78
1. Остроконечные кондиломы.
2. Криодеструкция, лазеротерапия, диатермо-
коагуляция, солкодерм.
Задача № 79
1. Герпетиформный дерматоз Дюринга.
2. Госпитализация в кожное отделение, назна-
чение препаратов сульфонового ряда, сульфани-
ламидов, глюкокортикоидов. Обследование у
терапевта, онколога для исключения паренео-
пластического характера заболевания.
Задача № 80
1. Исследование отделяемого язвы в темном
поле для обнаружения бледной трепонемы.
Задача № 81
1. Назначить превентивное лечение как при
первичном серонегативном сифилисе.
Эталоны ответов к ситуационным задачам по дерматовенерологии
607
Задача № 82
1. Для вторичного свежего сифилиса.
2. Исследовать отделяемое язвы в темном поле
на бледную трепонему, назначить исследование
крови на РВ.
3. Направить больную в кожновенерологичес-
кий диспансер.
Задача № 83
1. Вторичный активный сифилис.
2. Исследование отделяемого папул в темном
поле для обнаружения бледной трепонемы; се-
рологические реакции на сифилис.
Задача № 84
1. При это форме заболевания кожа и види-
мые слизистые оболочки свободны от специфи-
ческих высыпаний; иногда выявляется лимфо-
денит; реакция Вассермана резко положительная,
РИБТ 100 %.
Задача № 85
1. Учитывая опасность для половых партнеров,
необходима ее принудительная госпитализация
в венерологическое отделение с привлечением
правоохранительных органов.
Задача № 86
1. Свежий острый тотальный гонорейный урет-
рит.
2. Антибиотики (цефалоспорины, фторхиноло-
ны); клинико-серологический контроль после
излечения гонореи в течение 6 месяцев (так как
источник зарожения не установлен и возможно
инфицирование Т. pallidum).
Задача № 87
1. Свежий острый передний гонорейный урет-
рит; антибиотики (цефалоспорины, фторхиноло-
ны, спектиномицин); клинико-серологический
контроль после излечении гонореи в течение 6
месяцев (не обследован источник заражения, т.е.
не исключено и заражение сифилисом).
Задача № 88
1. Свежий острый тотальный кандидозный
уретрит.
2. Противогрибковые антибиотики внутрь (ин-
траконазол, кетоконазол, флуконазол); в уретру
— инстилляции раствора флуконазола.
Задача № 89
1. Исследование отделяемого язвочек в темном
поле на бледную трепонему, серологические ре-
акции на сифилис (МРП, РИФ, РВ).
2. Активный сифилис, генитальный герпес, го-
нококковые эрозии, травматические эрозии.
Задача № 90
1. Атипичный твердый шанкр при первичном
периоде сифилиса, бартолинит.
2. Припухлость большой половой губы.
3. Отсутствие болезненности и нарушения об-
щего состояния.
4. Назначено лечение без полного обследова-
ния.
5. Взять кровь для серологических реакций на
сифилис.
6. Положительные.
Задача № 91
1. Сифилис первичный (твердый шанкр, ос-
ложненный фимозом).
2. Исследование на бледную трепонему выяв-
ленного твердого шанкра или пунктата увели-
ченных паховых лимфотических узлов; поста-
новка серологических реакций крови на сифилис
(МРП, РИФ, РВ).
Задача № 92
1. Сифилис вторичный свежий. Серологичес-
кие реакции крови на сифилис.
2. Лечение больной после установления диаг-
ноза.
Задача № 93
1. Сифилис вторичный рецидивный (сифили-
тическая алопеция диффузная, пигментный си-
филид).
2. Выяснить анамнез, провести серологическое
исследование крови на сифилис (РВ, РИТ,
РИФ).
Задача № 94
1. Сифилис скрытый ранний.
2. Провести видоспецифические серологичес-
кие исследования на сифилис для подтвержде-
ния диагноза и назначить лечение.
Задача № 95
1. Отрубевидный лишай.
2. Показано.
Задача № 96
1. Сифилис вторичный рецидивный (широкие
кондиломы, ладонно-подошвенные сифилиды,
сифилитическая алопеция).
2. Положительные серологические реакции
крови на сифилис, обнаружение возбудителя в
отделяемом широких кондилом.
608 Приложение С
3. Направить пациента в кожно-венерологичес-
кий диспансер.
Задача № 97
1. Сифилис третичный активный (гуммозный
сифилид голени).
2. Туберкулез, лепра, лейшманиоз, остеомие-
лит.
3. Терапевт, невропатолог, окулист, отоларин-
голог.
Задача № 98
1. Сифилис третичный активный (бугорковый
сифилид).
2. Туберкулез, лепра, лейшманиоз, опоясыва-
ющий герпес.
3. Терапевт, невропатолог, окулист, отоларин-
голог.
Задача № 99
1. Сифилис третичный активный (гуммозный
сифилид малого язычка).
2. Видоспецифические трепонемные серологи-
ческие реакции.
3. В бытовом отношении больной не опасен.
Задача № 100
1. Препараты группы тетрациклина, азитроми-
цин, макролиды (эритромицин, кларитромицин).
Приложение D
Тесты по дерматовенерологии
1. Гистологически в коже выделяют
следующие слои: .
а) эпидермис
б) дерма
в) гиподерма
г) мезодерма
д) периост
2. Эпидермис состоит из следующих слоев
кроме:
а) шиповатого
б)зернистого
в) базальной мембраны
г) блестящего
д) рогового
3. Базальный слой состоит из:
а) одного слоя цилиндрических клеток
б) 3—8 рядов цилиндричеслих клеток
в) одного ряда кубических клеток
г) эластических волокон
д) клеток Лангерганса
4. В состав эпидермиса входят все клетки
кроме:
а) клетки Меркеля
б) меланоциты
в)эритроциты
г) кератиноциты
д) фибробласты
5. Меланоциты входят в состав:
а) базального слоя
б) шиповатого слоя
в) зернистого слоя
г) блестящего слоя
д) рогового слоя
6. В состав дермы не входят:
а) клеточные элементы
б)волокна
в) межуточное вещество
г) базальная мембрана
д) сосуды
7. Дерма состоит из слоев:
а)сосочкового
б)сетчатого
в) базальной мембраны
г) сосудистого
д) рогового
8. В состав гиподермы не входит:
а) соединительная ткань
б) кровеносные сосуды
в) жировые клетки
г) корнеоциты
д) нервные стволы
9. К придаткам кожи относят все кроме:
а) волосы
б) невусы
в) ногти
г) потовые железы
д) сальные железы
10. Сальные железы представляют собой:
а) простые трубчатые образования
б) апокринные железы
в) мерокринные железы
г) эккринные железы
д) голокринные железы
11. Гиперкератоз характеризуется:
а) нарушением процесса меланогенеза
б) потерей связи между клетками
в) утолщением рогового слоя
г) разрастанием сосочков дермы
д) утолщением зернистого слоя
12. Гипергранулез характеризуется:
а) утолщением рогового слоя
б) утолщением зернистого слоя
в) потерей связи между клетками
г) неравномерным утолщением эпидермиса
д) разрастанием сосочков дермы
13. Спонгизэто:
а) образование вакуолей в цитоплазме базаль-
ных эпидермоцитов
б) межклеточный отек шиповатого слоя
в) изменение окраски кожного покрова
г) изменение ядер клеток
д) потеря связи между клетками
609
610 Приложение D
14. Акантолиз это:
а) утолщение рогового слоя
б) утолщение зернистого слоя
в) потеря связи между эпидермальными клет-
ками
г) межклеточный отек
д) увеличение количества рядов клеток шипо-
ватого слоя
15. Акантоз это:
а) потеря связи между эпидермальными клет-
ками
б) утолщение рогового слоя
в) утолщение зернистого слоя
г) неполное ороговение клеток
д) увеличение количества рядов клеток шипо-
ватого слоя
16. К первичным морфологическим
элементам кожной сыпи не относят:
а) пятно
б) бугорок
в) узел
г) волдырь
д) эрозию
17. К полостным первичным
морфологическим элементам кожной
сыпи относятся:
а) пустула
б) папула
в) пятно
г) волдырь
д) везикула
18. Назовите разновидности пятна:
а) розеола
б) бляшка
в) волдырь
г) эритема
д) поверхностное
19. К разновидностям пустулы относят:
а) бляшку
б) микровезикулу
в) фликтену
г) экскориацию
д)корку
20. К полостным первичным
морфологическим элементам кожной
сыпи относят:
а) пустулу
б) папулу
в) пузырек
г) пузырь
д) пятно
21. К бесполостным первичным
морфологическим элементам кожной
сыпи не относится:
а) пятно
б) бугорок
в) волдырь
г) пустула
д) узел
22. К вторичным морфологическим
элементам кожной сыпи не относится:
а) пузырь
б)корка
в)эрозия
г) вегетация
д) экскориация
23. После вскрытия на месте пузырька
остается:
а) язва
б) эрозия
в) корка
г) лихенификация
д) чешуйка
24. Гистологически при волдыре
определяется:
а) ограниченный отек сосочкового слоя
б)спонгиоз
в) гиперкератоз
г) гипергранулез
д)акантолиз
25. Язва это:
а) дефект кожи в пределах эпидермиса
б) глубокий дефект кожи
в) линейный дефект кожи
г) утолщение кожи с усилением ее рисунка
д) изменение цвета кожи
26. Кожа выполняет все функции кроме:
а) иммунной
б)обменной
в) рецепторной
г) секреторной
д) пищеварительной
27. К специальным методам обследования
кожного больного относят:
а) перкуссию
б) граттаж
в) УЗИ внутренних органов
г) диаскопию
д) пальпацию
28. На острый мокнущий воспалительный
процесс необходимо назначить:
а) примочки
Тесты по дерматовенерологии 611
б) мазь
в)аэрозоль
г) лак
29. При подостром воспалительном
процессе показаны:
а) примочки
б) кремы
в) пасты
г) лаки
д) водные болтушки
30. При хроническом неспецифическом
процессе применяют:
а) мази
б) примочки
в) пасты
г) лаки
д) пластыри
31. Пудры применяются при:
а) остром мокнущем процессе
б) остром воспалительном процессе без мокну-
тия
в) подостром дерматите без мокнутия
г) повышенной потливости
д) хроническом воспалительном процессе
32. Примочки применяются при:
а) остром мокнущем процессе
б) остром воспалительном процессе без мокну-
тия
в) подостром дерматите без мокнутия
г) повышенной потливости
д) хроническом воспалительном процессе
33. Примочки оказывают следующее
действие:
а) согревающее
б) противовоспалительное
в) дезинфицирующее
г) кератолитическое
д) иммуномодулирующее
34. Болтушки оказывают следующее
действие:
а) подсушивающее
б) противозудное
в) кератолитическое
г) противовоспалительное
д) кератопластическое
35. Когда применяются мази:
а) в острую стадию при наличии мокнутия
б) при хронических воспалительных дерматозах
в) для смягчения кожи
г) для лечения грибковых заболеваний
д) с целью охлаждения
36. Какое действие оказывают лаки:
а) кератолитическое
б) согревающее
в) отслаивающее
г) противовоспалительное
д) антигистаминное
37. Назовите осложнения
антибиотикотерапии:
а) анафилактический шок
б) дисбактериоз
в)алопеция
г) повышение артериального давления
д) сахарный диабет
38. Назовите препарат, обладающий
наиболее выраженным катаболическим
действием:
а) преднизолон
б) дексаметазон
в) триамцинолон
г) метилпреднизолон
д) бетаметазон
39. Назовите осложнения
глюкокортикоидной терапии:
а) артериальная гипертензия
б) кушингоид
в) анафилактический шок
г) остеолизис
д) деменция
40. ПУВА-терапия это:
а) средневолновое УФО
б) сочетанное действие фотосенсибилизатора и
длинноволнового УФО
в) УФО крови
г) локальное УФО
д) фонофорез фотосенсибилизирующих мазей
41. Антибиотики применяют для лечения:
а) пиодермий
б) бородавок
в) эпидермофитии
г) чесотки
д) сифилиса
42. К противовирусным препаратам
относятся:
а) пенициллин
б)ацикловир
в) ламизил
г) бонафтон
д) фторокорт
43. К противогрибковым препаратам
относятся:
а) ацикловир
612 Приложение D
б) тетрациклин
в) низорал
г) ламизил
д) клотримазол
44. Ароматические ретиноиды это:
а) производные витамина А
б)антибиотики
в)антиоксиданты
г) глюкокортикоиды
д) адаптогены
45. При местном применении
глюкокортикоиды оказывают следующее
действие:
а) противовоспалительное
б) антибактериальное
в) антиэкссудативное
г) противовирусное
д) противоаллергическое
46. Какие формы обладают
кератолитическим действием:
а) 2 % салициловая мазь
б) сернодегтярная мазь 5 %
в) серносалициловая мазь 5 %
г) болтушки
д)примочки
47. К антибиотикам пенициллинового ряда
относятся:
а) ампициллин
б) цефалексин
в)бициллин
г) тетрациклин
д) эритромицин
48. К глюкокортикоидам относятся все
препараты кроме:
а) бетаметазон
б)гидрокортизон
в) метилтестостерон
г) триамцинолон
д) метилпреднизолон
49. Какие из перечисленных лекарственных
форм обладают противовоспалитель-
ным действием:
а) лаки
б) пасты
в) красители
г) мази с глюкокортикоидами
д) прижигающие средства
50. В каких из перечисленных мазей
содержатся глюкокортикоиды:
а) микосептин
б) салициловая
в)оксикорт
г) фторокорт
д)зовиракс
51. Предрасполагающими факторами в
развитии пиодермий не являются:
а) наличие на коже микроорганизмов
б) иммунодефицит
в) повреждение кожи
г) сахарный диабет
д) несбалансированное питание
52. К сгафилодермиям относят:
а) фолликулит
б) простой лишай лица
в) импетиго
г) фурункул
д)сикоз
53. К сгрептодермиям относят:
а) сикоз
б) импетиго
в) простой лишай лица
г) гидраденит
д)карбункул
54. К атипичным пиодермиям относят:
а) пиогенную гранулему
б)сикоз
в)заеду
г) эксфолиативный дерматит Риттера
д) шанкриформную пиодермию
55. Для стафилодермий характерно:
а) фолликулярные пустулы
б) вовлечение в процесс сальных желез
в) потовые железы в процесс не вовлекаются
г) сальные железы в процесс не вовлекаются
д) наличие фликтен
56. Первичным морфологическим элементом
при остиофолликулите является:
а) узел
б) бугорок
в)корочка
г) фолликулярная пустула
д) фликтена
57. Для лечения осгиофолликулита
показано:
а) антибиотики внутрь
б) 2 % салициловый спирт наружно
в) бриллиантовый зеленый
г) мази с глюкокортикоидами
д)ацикловир
58. Вульгарный сикоз локализуется:
а) в области ладоней
Тесты по дерматовенерологии
613
б) в области роста усов, бороды
в) на стопах
г) на голенях
д) на слизистых
59. При лечении карбункула используют:
а) антибиотики внутримышечно
б) сульфаниламиды
в) только местное лечение
г) мази с глюкокортикоидами
д) иммуносупрессоры
60. Наиболее часто гидраденит вызывает:
а) золотистый стафилококк
б) синегнойная палочка
в) стрептококк
г) бледная трепонема
д) кишечная палочка
61. Назовите разновидности
стрептококкового импетиго:
а) заеда
б) стрептококковая опрелость
в) буллезное импетиго
г) остиофолликулит
д) фолликулит
62. Первичным морфологическим
элементом при стрептококковом
импетиго является:
а) фолликулярная пустула
б) фликтена
в)корочка
г) узел
д) волдырь
63. К глубоким формам стафилодермий
относятся:
а) сикоз
б) фурункул
в)гидраденит
г) остиофолликулит
д)карбункул
64. Назовите заболевания, при которых
показан прием антибиотиков внутрь:
а) карбункул
б) остиофолликулит
в) фурункул в области лица
г) заеда
д) простой лишай лица
65. Профилактика пиодермий включает в
себя все, кроме:
а) рациональное питание
б) назначение антибиотиков
в) вакцинация
г) своевременное лечение пиодермий
д) проведение профосмотров
66. Назовите преимущественную
локализацию гидраденита:
а) лицо
б) кисти
в) волосистая часть головы
г) подмышечные ямки
д) голени
67. Назовите 3 стадии эксфолиативного
дерматита Риттера:
а) эритематозная
б) атрофическая
в) эксфоллиативная
г) пойкилодермическая
д) регенеративная
68. Развитию глубоких хронических
рецидивирующих пиодермий
способствует:
а) сахарный диабет
б) длительный прием глюкокортикоидов
в) онихомикоз
г) витаминотерапия
д) прием кетоконазола
69. Для простого лишая характерно:
а) возникает у детей
б) вызывается стрептококком
в) для лечения используют прием антибиоти-
ков внутрь
г) относится к глубоким формам пиодермий
д) острое течение
70. При гидрадените поражаются:
а) апокринные потовые железы
б) голокринные потовые железы
в) сальные железы
г) волосы
д) околоногтевые валики
71. Показанием для назначения
антибиотиков при пиодермиях не
является:
а) длительное хроническое течение заболевания
б) наличие общетоксических явлений
в) наличие регионарного лимфаденита
г) анафилактический шок в анамнезе
д) локализация глубоких пиодермий на лице
72. Назовите осложнения
антибиотикотерапии:
а) кандидоз
б) гипертензия
в) кишечный дисбактериоз
г) остеопороз
д) кушингоид
614 Приложение D
73. Какие из перечисленных препаратов
относятся к иммуностимуляторам:
а) левамизол
б) дексаметазон
в) метотрексат
г) тималин
74. Для эпидемической пузырчатки
новорожденных не характерно:
а) аутосомно-доминантный тип наследования
б) возникает на 7—10 день жизни
в) возбудителем является золотистый стафило-
кокк
г) заболевание начинается с появления пузы-
рей на фоне гиперемии
д)лихорадка
75. Назовите группы препаратов, которые
используют для лечения пиодермий:
а)антибиотики
б) глюкокортикоиды
в) витамины
г) иммуностимуляторы
д) цитостатики
76. Назовите пути проникновения
микобактерий туберкулеза в кожу:
а) экзогенный
б) лимфогенный
в) гематогенный
г) по протяжению
д) все верно
77. Укажите локализованные формы
туберкулеза кожи:
а) туберкулезная волчанка
б) бородавчатый туберкулез
в) скрофулодерма
г) лихеноидный туберкулез
д) папулонекротический туберкулез
78. Укажите диссеминированные формы
туберкулеза кожи:
а) туберкулезная волчанка
б) бородавчатый туберкулез
в) скрофулодерма
г) лихеноидный туберкулез
д) папулонекротический туберкулез
79. Для туберкулезной волчанки
характерны следующие симптомы:
а) триада Ауспитца
б) «яблочного желе»
в)«зонда»
г) «дамского каблучка»
д) зерна Трелля
80. На месте разрешения очагов
туберкулезной волчанки образуется:
а) рубцовая атрофия
б) гиперпигментация
в) вегетации
г) атрофические рубчики
д) депигментированые пятна
81. Каким путем проникают микобактерии
в кожу при бородавчатом туберкулезе:
а) лимфогенно
б) гематогенно
в) по протяжению
г) экзогенным путем
д) путем аутоинокуляции
82. Для скрофулодермы характерно:
а) мягкие бугорки в области лица, склонные
к слиянию и изъязвлению
б) глубокие узлы, склонные к слиянию и раз-
мягчению с последующим абсцедированием, об-
разованием фистул и язв
в) очаг на конечностях, состоящий из трех зон:
атрофической в центре, бородавчатых разраста-
ний и фиолетово-красной каемки по периферии
г) болезненные мелкие язвы вокруг естествен-
ных отверстий с зернами Трелля
д) сплошные глубокие узлы, расположенные
симметрично, преимущественно на голенях
83. Для бородавчатого туберкулеза
характерно:
а) мягкие бугорки в области лица, склонные
к слиянию и изъязвлению
б) глубокие узлы, склонные к слиянию и раз-
мягчению с последующим абсцедированием, об-
разованием фистул и язв
в) очаг на конечностях, состоящий из трех зон:
атрофической в центре, бородавчатых разраста-
ний и фиолетово-красной каемки по периферии
г) болезненные мелкие язвы вокруг естествен-
ных отверстий с зернами Трелля
д) сплошные глубокие узлы, расположенные
симметрично, преимущественно на голенях
84. Для язвенного туберкулеза кожи
Характерно:
а) мягкие бугорки в области лица, склонные
к слиянию и изъязвлению
б) глубокие узлы, склонные к слиянию и раз-
мягчению с последующим абсцедированием, об-
разованием фистул и язв
в) очаг на конечностях, состоящий из трех
зон: атрофической в центре, бородавчатых раз-
растаний и фиолетово-красной каемки по пе-
риферии
Тесты по дерматовенерологии 615
г) болезненные мелкие язвы вокруг естествен-
ных отверстий с зернами Трелля
д) сплошные глубокие узлы, расположенные
симметрично, преимущественно на голенях
85. Для индуративной эритемы Базена
характерно:
а) мягкие бугорки в области лица, склонные
к слиянию и изъязвлению
б) глубокие узлы, склонные к слиянию и раз-
мягчению с последующим абсцедированием, об-
разованием фистул и язв
в) очаг на конечностях, состоящий из трех
зон: атрофической в центре, бородавчатых раз-
растаний и фиолетово-красной каемки по пе-
риферии
г) болезненные мелкие язвы вокруг естествен-
ных отверстий с зернами Трелля
д) сплошные глубокие узлы, расположенные
симметрично, преимущественно на голенях
86. Для папулонекротического туберкулеза
кожи не характерно:
а) экзогенный путь инфицирования
б) сыпь локализуется на разгибательных повер-
хностях конечностей, туловище, лице
в) многочисленные плотные буровато-фиоле-
товые узелки
г) эволюционный полиморфизм
д) положительный симптом Никольского
87. Для лечения туберкулеза кожи
применяют:
а) пенициллины
б) изониазид
в) рифампицин
г) витамин Д2
д) цитостатики
88. Наиболее эффективными препаратами
для лечения туберкулеза являются:
а) изониазид
б) пенициллин
в) стрептомицин
г) эритромицин
д) рифампицин
89. Для лепры характерно:
а) длительный инкубационный период
б) торпидное, затяжное течение
в) обострение при действии УФО
г) поражение кожи, слизистых, нервной сис-
темы, внутренних органов
д) эозинофилия
90. Для микобактерии лепры характерно:
а) окрашиваются по Циль-Нильсону
б) не растут на питательных средах
в) грамотрицательные
г) кислотоустойчивые
д) высокаконтаглозность
91. Длительность инкубационного периода
при лепре:
а) 12 часов
б) до 5 дней
в) 3—10 лет
г) 4—6 недель
д) 3-4 месяца
92. Выделяют следующие типы лепры:
а) недифференцированный
б) лепроматозный
в) смешанный
г) туберкулоидный
д) лейшманиоидный
93. В течении лепры выделяют следующие
стадии:
а) прогрессирующая
б) стационарная
в) регрессивная
г) остаточных явлений
д) все верно
94. Аепроминовая проба положительна при:
а) лепроматозном типе
б) туберкулоидном типе
в) недифференцированном типе
г) всех типах лепры
д) туберкулезе кожи
95. Для лечения лепры используют
следующие препараты:
а) ДДС (дапсон)
б) пенициллин
в) ретиноиды
г) глюкокортикоиды
д) цитостатики
96 Для лепры характерны следующие
симптомы:
а) «львиной морды»
б) «лапы обезьяны»
в) «яблочного желе»
г) «морды тигра»
д) «маски святого Антония»
97. Для лепроматозного типа лепры
характерно:
а) выпадение волос
б) волдыри
в) положительная лепроминовая проба
616 Приложение D
г) потеря болевой, температурной, тактильной
чувствительности
д) атрофия мышц, мутиляция кистей, стоп
98. Для туберкулоидного типа лепры
характерно:
а) отрицательная лепроминовая проба
б) прекращение потоотделения
в) стволовые невриты
г) потеря болевой, тактильной, температурной
чувствительности
д) злокачественное течение
99. Профилактика лепры включает в себя:
а) раннее выявление и лечение больных
б) вакцинация БЦЖ
в) превентивное лечение членам семьи
г) соблюдение правил санитарно-эпидемичес-
кого режима
д) все перечисленное
100. Для подтверждения диагноза лепры
проводят следующие функциональные
пробы, кроме:
а) лепроминовая
б) с никотиновой кислотой
в) Минора
г) гистаминовая
д) Яддасона
101. Источником инфекции при кожном
лейшманиозе является:
а) человек с незажившей лейшманиомой
б) грызуны
в) москиты
г) домашние животные
д) дикие животные
102. Укажите стадии первичной
лейшманиомы:
а) бугорок
б)эрозия
в) язва
г) рубец
д) волдырь
103. Дайте определение туберкулоидному
лейшманиозу:
а) лейшманиоз у больного туберкулезной вол-
чанкой
б) лейшманиоз кожи у больного любой фор-
мой туберкулеза кожи
в) особый вид поздно изъязвляющегося лейш-
маниоза кожи
г) особый вид поздно изъязвляющегося лейш-
маниоза кожи у больного, инфицированного ту-
беркулезом
д) ничего из перечисленного
104. Инкубационный период при
городском поздно-изъязвляюшемся
лейшманиозе:
а) до 1 года
б) до 1 месяца
105. Бугорки при поздно-изъязвляюшемся
лейшманиозе:
а) розово-красные
б) желтовато-бурые
в) красные
г) желтые
д) цвета здоровой кожи
106. Зоонозный (сельский) лейшманиоз
характеризуется, всем перечисленным,
кроме:
а) появления новых бугорков на месте язвы
б) желтовато — бурого цвета бугорков
в) отсутствия феномена «яблочного желе»
г) слабой болезненности бугорков
д) развития стойкого иммунитета
107. Диагноз кожного лейшманиоза
устанавливается с учетом:
а) рентгенологических исследований
б) серологических исследований
в) бактериоскопического и бактериологическо-
го исследований
г) теста Монтенегро
д) периода пребывания в эндемичном очаге
108. Какие из перечисленных элементов
сыпи наиболее характерны для
простого герпеса?
а) гнойнички, язвы
б) узелки, чешуйки
в) папулы, волдыри
г) бугорки, язвы
д) пузырьки, эрозии, корки
109. Укажите наиболее характерный
признак простого герпеса:
а) хроническое длительное течение без перио-
дов ремиссий
б) склонность к рецидивированию
в) возникновение рецидивов после нервного
перенапряжения
г) начало заболевания преимущественно в по-
жилом возрасте
д) провоцирующее воздействие нарушений в
диете
110. Укажите препараты, которые
применяются для лечения простого
герпеса:
а) преднизолоновая мазь
б) целестодерм с гарамицином
Тесты по дерматовенерологии
617
в) салициловая мазь
г) синтомициновая эмульсия
д) крем «Ацикловир»
111. Укажите причины, способствующие
активизации ВПГ:
а) переохлаждение
б) травмы
в) беременность
г) антибиотикотерапия
д) нарушения в диете
112. Укажите способы лечения
хронической рецидивирующей
герпетической инфекции:
а) антибиотикотерапия
б) длительная химиотерапия ацикловером
в) глюкокортикоидная терапия
г) интерферонотерапия
д) специфическая вакцинация
113. Для типичной формы опоясывающего
лишая характерны:
а) эритема
б) отек
в) язвы
г) эрозии
д)некроз
114. К противовирусным препаратам
относятся все перечисленные, кроме:
а) дёгтя
б)ацикловира
в) триоксазина
г) ларифана
д) метисазона
115. Возникновению опоясывающего лишая
чаше всего способствует:
а) переохлаждение
б) перегревание
в) контакт с больным ветряной оспой
г) глюкокортикоидная терапия
д) вторичные инфекции
116. Для опоясывающего лишая
характерно:
а) сгруппированная сыпь
б) диффузная сыпь
в) невралгические боли
г) мышечные боли
д) общее недомогание
117. Укажите препараты, которые
целесообразно включить в обшую
терапию опоясывающего лишая:
а) антибактериальные средства (антибиотики,
сульфаниламиды)
б) анальгезирующие средства
в) кортикостероидные препараты
г) противогерпетические средства
д) витамины Вр В6, В12
118. Укажите разновидности бородавок:
а) лентикулярные
б) обыкновенные
в) подошвенные
г) подногтевые
д) колодцевидные
119. Для лечения бородавок используют:
а) кератолитические средства
б) антисептические растворы
в) противовирусные мази
г) хирургическое лечение
д) физиотерапию
120. Укажите, каким путем осуществляется
заражение остроконечными
кондиломами:
а) трансфузионным
б) трансмиссивным
в) воздушно — капельным
г) прямым половым
д) при тесном бытовом контакте
121. Для остроконечных кондилом
характерны:
а) лентикулярные папулы
б) сосочковые разрастания
в) папулы в виде площадки
г) локализация в области гениталий
д) тенденция к вегетации
122. Укажите необходимые мероприятия
при контагиозном моллюске у
ребенка:
а) изоляция от здоровых детей
б) глюкокортикоидная терапия
в) пенициллинотерапия
г) гемосорбция
д) Пува — терапия
123. Для контагиозного моллюска
характерны, все перечисленные
признаки, кроме:
а) микровезикул
б) узелков
в) бугорков
г) выделения творожистой массы при сдавли-
вании элементов пинцетом
д) изъязвления
124. При эритразме происходит
повреждение:
а) гладкой кожи
618 Приложение D
б) кожи в крупных складках
в) придатков кожи
г) слизистой оболочки полости рта
д) кожи волосистой части головы
125. Какие лекарственные препараты
назначают для лечения эритразмы?
а) дезинфицирующие мази и кремы
б) противогрибковые мази
в) антибиотики (эритромицин)
г) глюкокортикоиды
д) дезинтоксикационные препараты
126. Укажите, какими путями
осуществляется заражение чесоткой:
а) трансфузионным
б) трансмиссивным
в) при тесном бытовом контакте
г) воздушно — капельным
д) при половых контактах
127. Укажите, к какой группе этиологических
факторов относится возбудитель
чесотки:
а) бактерии
б) вирусы
в) простейшие
г) клещи
д) гельминты
128. Укажите, в каком слое кожи самка
клеша прокладывает чесоточный ход:
а) роговой
б) блестящий
в) зернистый
г) шиповатый
д) базальный
129. Для чесотки характерны:
а) точечные зудящие пузырьки и узелки
б) геморрагии
в) лентикулярные папулы
г) волдыри
д) полигональные папулы
130. Распространенность сыпи при чесотке
зависит от всех перечисленных
факторов, кроме:
а) состояния иммунитета
б) количества клещей попавших на кожу в
момент инфицирования
в) половых и возрастных особенностей кожи
г) индивидуального чувства зуда
д) гигиенических навыков
131. Укажите наиболее характерный
признак неосложненной чесотки:
а) экзематизация в очагах поражения
б) инфильтрация в очагах поражения
в) полиаденит
г) вечерний и ночной зуд
д) импетигинизация в очагах поражения
132. Укажите клинические проявления на
коже, патогномичные для
неосложненной чесотки:
а) эктима
б) чесоточный ход
в) папула
г) розеола
д) волдырь
133. Какие участки тела у взрослых не
поражаются чесоткой?
а) межпальцевые складки кистей
б) молочные железы
в) генитальная область
г) лицо
д) кожа бедер
134. Укажите наиболее частое осложнение
чесотки:
а) вторичная пиодермия
б) рубцовая атрофия кожи
в) сепсис
г) аллергический дерматит
д) сухость кожи
135. Лечение больного чесоткой
предусматривает:
а) исключение острой пищи
б) применение бензилбензоата
в) назначение УФО
г) применение салициловой мази
д) применение 33 % серной мази
136. Для осложненной чесотки характерны:
а) экзематизация
б) лимфанглит, лимфаденит
в) симптом Арди-Горчакова
г) симптом Послерова
д) ночной зуд
137. Укажите места типичной локализации
чесотки у взрослых:
а) межпальцевые складки кистей
б) волосистая часть головы
в) лицо и шея
г) живот
д) ладони и подошвы
138. Какие разновидности вшей поражают
человека?
а) головные
б) платяные
в) площицы
Тесты по дерматовенерологии 619
г) бытовые
д)половые
139. Какие методы применяются при
диагностике педикулеза?
а) посев на питательные среды
б) микроскопия
в) осмотр ad oculus
г) проба с йодной настойкой
д) граттаж
140. Выберите наружные средства для
лечения лобкового педикулеза:
а) флуцинар
б) лосьон «Дипросалик»
в) метилурациловая мазь
г) цинковая паста
д) лосьон «Ниттифор»
141. Грибковые заболевания кожи
вызываются всеми перечисленными
возбудителями, кроме:
а) эпидермофитонов
б) трихофитонов
в) палочек Ганзена
г) микроспорумов
д) дрожжеподобных грибов
142. Дайте определение локализации
патологического процесса при
дерматофитии:
а) локализуется в собственно коже и подлежа-
щих тканях, внутренних органах с явлениями
воспаления
б) локализуется на коже, слизистых, придатках
кожи — ногтях, а также внутренних органах
в) локализуется в поверхностных слоях эпидер-
миса, роговом слое или кутикуле волоса, воспа-
лительные явления отсутствуют
г) локализуется в более глубоких слоях кожи
с развитием воспалительной реакции, могут по-
ражаться также придатки кожи
д) локализуется в глубоких слоях кожи с раз-
витием воспалительной реакции, придатки кожи
не поражаются
143. Профилактика грибковых заболеваний
кожи включает:
а) выявление источников заражения, их изо-
ляцию и дезинфекцию помещения
б) ветеринарный надзор за животными
в) обследование контактов
г) периодический медицинский осмотр персо-
нала, подверженного грибковым заболеваниям
(детские и бытовые учреждения)
д) все перечисленное
144. Какие препараты применяют для
наружного лечения микозов?
а) кетоконазол (низорал)
б) интраконазол (орунгал)
в) тербинафин (ламизил)
г) азитромицин (сумамед)
д) микосептин
145. Какие изменения в очагах поражения
характерны для разноцветного лишая?
а) отрубевидное шелушение
б) лихенификация
в) расчесы, поражение волос
г) отсутствие воспалительных явлений
д) пузырьки, эрозии
146. Назовите методы диагностики
отрубевидного лишая:
а) осмотр под лампой Вуда
б) граттаж очагов поражения
в) йодная проба Бальцера
г) микроскопическое исследование чешуек
д) все перечисленное верно
147. Отрубевидный лишай необходимо
дифференцировать от:
а) сифилитической (истинной) лейкодермы
б) лишая Жибера
в) фавуса
г) сифилитической розеолы
д) эритразмы
148. Какие заболевания обусловлены
грибами рода трихофитон:
а) фавус
б) эритразма
в) вульгарный сикоз
г) паразитарный сикоз
д) опоясывающий лишай
149. Какими грибами наиболее вероятно
заразиться от крупного рогатого скота?
a) Trichophyton rubrum
б) Trichophyton shoenleini
в) Trichophyton verrucosum
г) Trichophyton canis
д) Trichophyton mentagrophytes
150. Поверхностная трихофития
вызывается антропофильными
трихофитонами:
а) фавиформным
б) фиолетовым
в) гипсовым
г) кратериформным
д) ментаграфитным
620 Приложение D
151. Клиническими особенностями
инфильтративно — нагноительной
трихофитии являются:
а) эритема и отек
б) фолликулярные абсцессы
в) инфильтративные бляшки
г) экзематозные колодцы
д) медовые соты Цельса
152. Для хронической трихофитии
волосистой части головы характерны:
а) крупные очаги, сплошное поражение волос
б) мелкие очаги с симптомом «черных точек»
в) белая муфта у основания волос
г) четкие очертания очага
д) выраженные воспалительные явления в очагах
153. Для микроспории гладкой кожи
характерно перечисленное, кроме:
а) пятен
б) нечетких границ и быстрого самопроизволь-
ного разрешения
в) пузырьков по краю очагов
г) отрубевидного шелушения
д) пустул
154. Для поражения волос при
микроспории характерно
перечисленное, кроме:
а) обламывания волос в очаге поражения на
уровне 1—2 мм от поверхности кожи
б) обламывания волос в очаге поражения на
уровне 5—8 мм от поверхности кожи
в) муфтообразных «чехликов» на волосах
г) синеватого свечения волос при облучении
лампой Вуда
д) рубцовой атрофии
155. Волосы обламываются на уровне 5-
8 мм от поверхности кожи:
а) при микроспории
б) при фавусе
в) при поверхностной трихофитии
г) при трихофитии инфильтративно-нагнои-
тельной
д) при алопеции
156. К клиническим формам фавуса
волосистой части головы относятся
все перечисленные, кроме:
а) скутулярной
б) импетигинозной
в) сквамозной
г) эритематозной
д) везикулезной
157. Для скутулярной формы фавуса
характерны все перечисленные
признаки, кроме:
а) рубцовый атрофии
б) очаговой алопеции
в) амбарного запаха
г) тусклых, безжизненных волос
д) обломанных волос
158. Для паховой эпидермофитии
характерно все перечисленное, кроме:
а) лихенификации
б) чешуек и зуда
в) вегетации
г) кораллово — красного свечения при облу-
чении лампой Вуда
д) излюбленной локализации
159. Развитию паховой эпидермофитии
благоприятствуют:
а) плоскостопие
б) повышенная потливость
в) травмы
г) избыток массы тела
д) нарушение обмена веществ
160. Укажите клинические разновидности
онихомикоза:
а) сквамозно — гиперкератотический
б) атрофический
в) интертригинозный
г) гипертрофический
д) нормотрофический
161. В патогенезе онихомикозов имеют
значение:
а) применение кортикостероидов
б) недостаточная гигиена стоп и их травмати-
зация обувью
в) юношеский возраст
г) эндокринопатии
д) недостаточное и неполноценное питание
162. Укажите клинические симптомы
онихомикоза:
а) онихолизис
б) онихошизис
в) изменение цвета ногтевых пластинок
г) лейконихия
д)паронихия
163. Для лечения онихомикоза, вызванного
Т. rubrum, применяются следующие
препараты, кроме:
а) гризеофульвина
б) нистатина
в) кетоконазола (низорала)
Тесты по дерматовенерологии
621
г) итраконазола (орунгала)
д) тербинафина (ламизила)
164. К глубоким микозам относятся
следующие заболевания, кроме:
а) бластомикоза
б) актиномикоза
в) гистоплазмоза
г) нокардиоза
д) хромомикоза
165. Укажите клинические формы
бластомикоза:
а) болезнь Лутца — Сплендоре — Алмейда
б) болезнь Педрозо
в) болезнь Гилкриста
г) болезнь Дарлинга
д) болезнь Буссе — Бушке
166. К факторам, способствующим
развитию кандидоза кожи, относятся
все перечисленные, кроме:
а) гиповитаминоза
б) гиперинсоляции
в) дисбактериоза
г) иммунодефицитного состояния
д) эндокринных и обменных нарушений
167. Укажите клинические формы
кандидоза:
а) кандидоз складок кожи
б) кандидоз слизистых оболочек
в) висцеральный кандидоз
г) хронический генерализованный кандидоз
д) все перечисленное
168. Для местного лечения кандидоза
применяют:
а) раствор буры (10 %-20 %) в глицерине
б) мази: нистатиновую, амфотерициновую, де-
каминовую
в) мази: бонафтоновую, оксолиновую, тебро-
феновую
г) клотримазол
д) гиоксизон
169. При хроническом генерализованном
кандидозе наиболее целесообразно
применять следующие препараты:
а) гризеофульвин
б) кетоконазол (низорал)
в) нистатин и леворин
г) амфотерицин В
д) пимафуцин
170. Укажите клинические формы
дерматита:
а) солнечный дерматит
б) горшочный дерматит
в) простой контактный дерматит
г) аллергический контактный дерматит
171. Артифиииальный дерматит
характеризуется всем перечисленным,
кроме:
а) четких границ
б) локализации на месте действия раздражителя
в)бугорков
г) розеол
д) жжения
172. Аллергический контактный дерматит
характеризуется всем перечисленным,
кроме:
а) развития сенсибилизации
б) мономорфной сыпи
в) наличия аллергидов
г) полиморфной сыпи
д) вегетаций
173. Аллергический дерматит развивается в
результате воздействия следующих
этиологических факторов:
а) синтетических тканей
б) концентрированных кислот и щелочей
в) высоких и низких температур
г) моющих средств
д) солей никеля
174. Для аллергического контактного
дерматита характерны:
а) эритема
б) волдыри
в)эрозии
г) пузырьки
д) пустулы
175. При развитии аллергического
дерматита от стирального порошка
уместны следующие рекомендации:
а) прекратить использовать стиральный поро-
шок
б) 30 % раствор тиосульфата натрия pes os или
в/в
в) местно: примочки
г) антигистаминные препараты: димедрол per os
д) все перечисленное верно
176. Какой из перечисленных ниже
препаратов является эффективным
при аллергическом контактном
дерматите в подострой стадии:
а) 2 % борно-нафталановая мазь
б) 1 % салициловая мазь
в) 10 % серно-салициловая мазь
622 Приложение D
г) мазь «Флуцинар»
д) водно — цинковая взвесь
177. В обшую терапию аллергического
контактного дерматита включают
следующие препараты, кроме:
а) антибиотиков
б) энтеросорбентов
в) препаратов кальция
г) сульфаниламидов
д) тиосульфата натрия
178. Укажите клинические особенности
простых дерматитов:
а) нарушение общего состояния
б) быстрое разрешение процесса после устране-
ния действия раздражителя
в) возникают под действием только внешних
факторов
г) эозинофилия
д) резкие границы
179. Укажите факторы, способствующие
развитию токсикодермии:
а) контрацептивы, содержащие металлы (медь,
серебро)
б) наследственное предрасположение
в) аллергические заболевания
г) применение антибиотиков, кортикостероидов
д) все перечисленное верно
180. Укажите разновидности
токсикодермий по этиологии:
а) профессиональные
б) непрофессиональные
в) механические
г) лекарственные
д) пищевые
181. Токсикодермия характеризуется:
а) бурной воспалительной реакцией кожи
б) большой распространенностью, вплоть до
эритродерм ии
в) нарушением общего состояния
г) быстрым регрессом после устранения кон-
такта с аллергеном
д) отрицательными кожными тестами с аллер-
геном
182. Для распространенной токсикодермии
характерно все перечисленное, кроме:
а) поражения слизистой рта, губ
б) хронического рецидивирующего течения
в) некролиза
г) бугорков
д) гумм
183. Для фиксированной эритемы
характерно все перечисленное, кроме
а) жжения
б) угрей
в) гумм
г) эритемы
д) пузырей
184. Синдром Лайелла характеризуется
следующими клиническими
симптомами:
а) эозинофилией
б) лихорадкой
в) благоприятным исходом
г) положительным симптомом Никольского
д) симптомами обезвоживания
185. Какие из перечисленных ниже
мероприятий производятся при
медикаментозной токсикодермии,
развившейся в результате перорального
приема антибиотиков?
а) отмена лекарственных препаратов
б) рациональная диета, прием жидкости
в) десенсибилизирующая общая терапия
г) противовоспалительная местная терапия
д) все перечисленное верно
186. Укажите общие признаки экземы:
а) симметричность высыпаний
б) дермографизм белый
в) дермографизм красный
г) атопический синдром
д) эволюционный полиморфизм
187. Какое патоморфологическое
изменение является характерным для
экземы:
а) папилломатоз
б) паракератоз
в) агранулез
г) спонгиоз
д) акантоз
188. Укажите стадии экземы и их
симптомы:
а) эритематозная (яркая эритема)
б) мокнутия (эритема, пузырьки, экссудациия)
в) рубцевания
г) везикулезная (эритема, пузырьки)
д) эритематозно-сквамозная (эритема, шелуше-
ние, папулы, пузырьки, эрозии)
189. Какой первичный морфологический
элемент наблюдается при экземе?
а) пятно
б) узелок
в) пузырек
Тесты по дерматовенерологии 623
г) волдырь
д)бугорок
190. Какие признаки характерны для
экземы?
а) нервно-аллергический характер процесса
б) рецидивирующие течение
в) полиморфизм сыпи
г) обострение нередко без видимой причины
д) все перечисленное верно
191. Укажите первичный морфологический
элемент не характерный для экземы:
а) микровезикула
б)бугорок
в) гнойничок
г) папула
д)пузырек
192. Какая из перечисленных стадий
экзематозного процесса является
наиболее ценной для подтверждения
диагноза экземы?
а) корковая
б) сквамозная
в) мокнутия
г) эритематозная
д) везикулезная
193. Патогномичными для экземы являются
следующие морфологические
элементы сыпи:
а) корки, чешуйки
б) эритема, пустулы
в) эрозии и язвы
г) микровезикулы и мокнущие эрозии
д) папулы и волдыри
194. Какие признаки характерны для
истинной экземы?
а) нечеткие границы
б) обострение без видимой причины
в) определяются «экзематозные колодцы»
г) симметричность расположения очагов
д) четкие границы
195. Какие пищевые продукты можно
употреблять больному экземой в
стадии обострения?
а) творог, молоко
б) копченую колбасу
в) цитрусовые
г) шоколад и кофе
д) овощи
196. В стадии мокнутия для наружной
терапии экземы применяются только:
а) присыпки
б) кремы
в) пасты
г) холодные примочки
д) взбалтываемые взвеси
197. Общую кортикостероидную терапию
при экземе назначают в случаях:
а) диссеминации процесса
б) поражении кожи туловища
в) пустулизации в очагах
г) резистентности к общей терапии
д) торпидных распространенных формах эк-
земы
198. Какие лекарственные препараты
назначают при острой мокнущей
экземе?
а) кортикостероиды
б) антигистаминные препараты
в)антибиотики
г) энтеросорбенты
д) сульфаниламиды
199. Наружная терапия при хронической
экземе вне обострения включает:
а) 1 % раствор резорцина
б) 0,025 % мазь синафлана
в) 10 % серно — дегтярную мазь
г) аэрозоль “’’Оксикорт”
д) флуцинар с добавлением мази Вилькинсона
200. Лечение больного острой экземой
предусматривает:
а) исключение молочно — растительной
пищи
б) общие теплые ванны с отваром ромашки
в) антигистаминные препараты
г) назначение УФО
д) гипосенсибилизирующие препараты
201. Для профессиональной экземы
характерно:
а) элементы сыпи не исчезают при устранении
контакта с аллергеном
б) резкие, четкие границы очагов поражения
в) поражение кожи выходит за границы кон-
такта с аллергеном
г) поражаются преимущественно волосистая
часть головы, лицо, грудь
д) заболевание начинается с появления пиодер-
мического элемента
202. Для профессиональной токсидермии
характерно:
а) развивается у лиц с индивидуальной непе-
реносимостью химических веществ, которые
проявляют повреждающее действие преимуще-
624 Приложение D
ственно ингаляционным или пероральным пу-
тем
б) проявляется мономорфными уртикарными
элементами
в) возникает при непосредственном контакте с
кожей кислот, щелочей
г) локализация поражения соответствует мес-
ту действия раздражителя
д) при повторном контакте с аллергеном про-
цесс рецидивирует и протекает более тяжело
203. К профессиональным стигмам
относятся:
а) узелки доильщиц
б) омозолелости
в) поздняя рентгеновская лучевая язва
г) эризипелоид
д) сибирская язва
204. Для подтверждения диагноза
профессионального заболевания кожи
используют пробы:
а) Ядассона
б) компрессная
в) Бальцера
г) капельная
д) скарификационная
205. Профессиональные дерматозы могут
вызываться факторами:
а) механическими
б) инфекционными
в) физическими
г) психоэмоциональными
д) наследственными
206. Диагноз профдерматоза
подтверждается:
а) положительными результатами кожных
проб с веществом, применяющимся на произ-
водстве
б) данными санитарно-гигиенической экспер-
тизы условий труда
в) обширным поражением кожи, тяжелым те-
чением заболевания
г) наличием аналогичных заболеваний у дру-
гих работников предприятия
д) положительным симптомом Никольского
207. Зуд кожи не характерен при
следующих заболеваниях:
а) сахарный диабет
б) лимфогранулематоз
в) хроническая почечная недостаточность
г) цирроз печени
д) микроспория
208. При кожном зуде для уточнения
диагноза не назначают следующие
лабораторные исследования:
а) глюкозы крови
б) биохимический анализ крови
в) на LE-клетки
г) кала на яйца глист
д) спинномозговой жидкости
209. При кожном зуде не характерно
наличие:
а) полированных ногтей
б) волдырей
в) экскориаций
г) лихенификаций
д) мокнутия
210. При лечении атопического дерматита
не назначают:
а) гипоаллергенную диету
б) цитостатики
в) энтеросорбенты
г) антигистаминные
д) препараты кальция
211. Наиболее вероятный возраст
возникновения атопического
дерматита:
а) до 2 лет
б) 15 лет
в) 25 лет
г) 50 лет
д) внутриутробно
212. К клиническим формам атопического
дерматита не относят:
а) эритематозно-сквамозную
б) пруригоподобную
в) инфильтративную
г) пигментную
д) лихеноидную
213. Больному атопическим дерматитом не
назначают:
а) санаторно-курортное лечение
б) кисломолочные продукты
в) седативные препараты
г) прием препаратов кальция
д) диету, богатую фруктами, ягодами, овоща-
ми
214. В классификации атопического
дерматита выделяют:
а) 5 клинических форм
б) 3 степени тяжести
в) 3 возрастных периода
г) 2 степени активности
д) 5 форм по этиологическому фактору
Тесты по дерматовенерологии
625
215. В патогенезе атопического дерматита
не играет роль:
а) блокада В-адренергических рецепторов
б) аллергия к микробным антигенам
в) аллегия к пищевым аллергенам
г) врожденный дефицит ингибитора С-3 фак-
тора комплемента
д) снижение количества Т-лимфоцитов
216. Диагноз атопического дерматита
подтверждается:
а) гистологическим исследованием пораженно-
го участка кожи
б) определением уровня содержания IgE в крови
в) определением количества Т-лимфоцитов
г) скарификационными пробами на пищевые
аллергены
д) достаточно клинических данных
217. Основные диагностические критерии
атопического дерматита:
а) зуд
б) начало в детском возрасте
в) белый дермографизм
г) склонность к кожным инфекциям
д) семейная или индивидуальная атопия в
анамнезе (бронхиальная астма, крапивница)
218. Аллергены, играющие роль в развитии
атопического дерматита:
а) контактные
б) аэроаллергены
в) живые агенты
г) пищевые аллергены
д) все перечисленные
219. Типичная локализация высыпаний при
атопическом дерматите:
а) в детском возрасте — поражение разгиба-
тельных участков конечностей, туловища, лица
б) у взрослых — поражение разгибательных
участков конечностей, туловища, лица
в) в детском возрасте — лихенификация на
сгибательных участках конечностей
г) у взрослых — лихенификация на сгибатель-
ных участках конечностей
д) распространенная сыпь без четкой локали-
зации
220. Белый дермографизм наиболее
характерен при следующих
заболеваниях:
а) крапивнице
б) аллергическом дерматите
в) атопическом дерматите
г) истинной экземе
д) кожном зуде
221. Признаки атопического лица:
а) симптом псевдо-Хертога
б) линии Дени, складки Моргана
в)хейлит
г) врожденные дисплазии
д) признак Пинкуса
222. Первичный элемент при ограниченном
нейродермите:
а) пятно
б) папула
в) бугорок
г) волдырь
д) пузырь
223. Для очага поражения при
нейродермите характерно наличие
зон:
а) центральной лихенификации
б) зоны лихеноидных папулезных высыпаний
в) зоны застойной гиперемии
г) периферического валика
д) дисхромической зоны
224. Для местного лечения ограниченного
нейродермита применяются мази,
содержащие:
а) глюкокортикоиды
б) деготь, нафталан, АСД
в)антибиотики
г) противогрибковые препараты
д) цитостатики
225. При детской почесухе Вы посоветуете:
а) употребление для кормления больного ре-
бенка разбавленного грудного молока
б) раннее включение в рацион кисломолочных
продуктов
в) диету, богатую ягодами и фруктами
г) диету, не содержащую злаковых (рис, пше-
ница), иод (морская рыба)
д) прием глюкокортикоидов внутрь
226. Для детской почесухи не характерно:
а) сильный зуд с невротическими расстрой-
ствами
б) серозные корки, экскориации
в) поражение ладоней и подошв
г) поражение разгибательных поверхностей ко-
нечностей
д) увеличение периферических лимфатических
узлов
227. Для узловатой почесухи характерно:
а) поражение лица
б) увеличенные плотные лимфатические узлы
в) пузыри с серозным содержимым
626 Приложение D
г) узелки и узлы с гладкой или бородавчатой
поверхностью
д) поражение нижних конечностей
228. Первичный элемент при острой
крапивнице:
а) пятно
б) волдырь
в) пузырь
г) узелок
д)бугорок
229. Препарат, оказывающий наиболее
выраженный эффект при отеке
Квинке:
а) пипольфен
б)адреналин
в) тиосульфат натрия
г) кетотифен
д) преднизолон парентерально
230. Для острой крапивницы характерно:
а) сильный зуд
б) белый дермографизм
в) лихенификация в области локтевых сгибов
г) волдыри
д) геморрагические пятна темно-красного цвета
231. Для отека Квинке характерно:
а) постепенное развитие в течение 5—7 дней
б) одностороннее поражение лица
в) относится к угрожающим жизни состояниям
г) могут поражаться половые органы
д) после регресса высыпаний остается стойкая
гиперпигментация
232. Для отека Квинке не характерно:
а) гиперемия с четкими границами
б) повышение температуры, озноб
в) острое начало
г) зуд, жжение
д) сочетание с парезом лицевого нерва
233. Для псориаза характерно:
а) мономорфность высыпаний
б) биопсирующий зуд
в) кирпично-красный цвет элементов
г) наличие пузырей по краю элементов
д) образование серебристо-белых чешуек на
элементах
234. Предпочтительная локализация
высыпаний при вульгарном псориазе:
а) разгибательные поверхности конечностей
б) ладони и подошвы
в) лицо
г) межлопаточная область
д) слизистые
235. Первичные элементы при псориазе:
а) узлы
б) волдыри
в) папулы
г) бугорки
д) пятна
236. Для псориаза патогномоничны
симптомы:
а) облатки
б) терминальной пленки
в) стеаринового пятна
г) яблочного желе
д) Никольского
237. Для псориаза патогномоничны
симптомы:
а) Кебнера
б) повышенной кровоточивости
в) точечного кровотечения
г) Ауспитца
д) Ядассона
238. Для прогрессирующей стадии
псориаза характерно все, кроме:
а) появления свежих ярко-красных папул
б) изоморфной реакции Кебнера
в) положительной псориатической триады
г) чешуек покрывающих всю поверхность па-
пулы
д) наличие псевдоатрофического воротничка
Воронова вокруг очагов
239. В стационарную стадию псориаза не
выявляется только:
а) шелушение всей поверхности папул
б) псевдоатрофический ободок Воронова вок-
руг очагов
в) прекращение периферического роста очагов
г) положительная изоморфная реакция Кебнера
д) свежие высыпания отсутствуют
240. Для регрессирующей стадии псориаза
характерно только:
а) разрешение элементов из центра
б) положительная изоморфная реакция Кебне-
ра
в) биопсирующий зуд
г) признак Пелькова
д) положительная псориатическая триада
241. Для псориаза волосистой части головы
не характерно:
а) зуд
б) выпадение волос
в) бляшки покрыты толстыми, трудно отделя-
емыми чешуйками
Тесты по дерматовенерологии 627
г) мокнутие за ушными раковинами
д) шелушение серебристо-белыми чешуйками
242. Для наружной терапии
прогрессирующей стадии вульгарного
псориаза назначите:
а) 1—2 % салициловую мазь
б) 5 % серно-дегтярную мазь
в) 5 % салициловую мазь
г) 10 % ихтиоловую мазь
д) наружное лечение не показано
243. Для лечения прогрессирующей стадии
псориаза применяется все
перечисленное, кроме:
а) кремов с глюкокортикостероидами
б) седативных препаратов
в) витаминов группы В
г) общее УФО
д) гепатопротекторов
244. Триада Ауспитна включает в себя
следующие феномены:
а) стеаринового пятна
б) терминальной пленки
в) точечного кровотечения
г) облатки
д) масляного пятна
245. При псориазе поражаются:
а) потовые железы
б) ногти
в) волосы
г) суставы
д) мозг
246. Феномен Кебнера выявляется в
следующих стадиях псориаза:
а) прогрессирующей
б) стационарной
в) регрессирующей
г) ремиссии
д) во всех стадиях
247. К атипичным формам псориаза
относятся:
а) бляшечный
б) себорейный
в) лентикулярный
г) ладоней и подошв
д) нуммулярный
248. Виды пустулезного псориаза:
а) экссудативный
б) Цумбуша
в) рупиоидный
г) Барбера
д) артропатический
249. При лечении псориаза не используют:
а) седативные препараты
б) ретиноиды
в) ПУВА-терапию
г) иглорефлексотерапию
д) антибиотики
250. Пути передачи возбудителя болезни
Лайма:
а) воздушно-капельный
б) трансмиссивный
в) половой
г) контактно-бытовой
д) алиментарный
251. Для лечения болезни Лайма
обязательно назначите:
а) системные глюкокортикоиды
б)антибиотики широкого спектра дей-
ствия
в)делагил
г) ретиноиды
д) цитостатики
252. При болезни Лайма выявляют:
а) узлы красно-бурого цвета
б) мигрирующую эритему
в) геморрагические пятна
г) сгруппированные бугорки
д) папулезные элементы лилового цвета с пуп-
ковидным вдавлением
253. При красном плоском лишае типична
локализация сыпи:
а) на коже предплечий и голеней
б) на ладонях и подошвах
в) на лице
г) на половых органах
д) на слизистых оболочках
254. При красном плоском лишае
поражаются:
а) суставы
б) слизистые оболочки
в)волосы
г) ногти
д) сальные железы
255. Для первичного элемента при красном
плоском лишае характерно:
а) восковидный блеск
б) пупкообразное вдавление в центре элемен-
та
в) полигональность
г) симптом облатки
д) феномен скрытого шелушения
628 Приложение D
256. Первичный элемент при красном
плоском лишае:
а) пятно
б) пузырь
в) папула
г) волдырь
д)бугорок
257. Для розового лишая характерно
появление:
а) пузырей
б) пустул
в) волдырей
г) материнской бляшки по типу медальона
д) пятен лилового цвета
258. Для местного лечения розового лишая
примените:
а) ванны с морской солью
б) мази с анестетиками
в) мази с кортикостероидами
г) мазь с дегтем
д) мазь с серой
259. Для лечения красной волчанки
применяются:
а) антибиотики широкого спектра действия
б) системные глюкокортикоиды
в) антистафиллококковый иммуноглобулин
г) противомалярийные препараты
д) противовирусные препараты
260. Излюбленная локализация очагов
поражения при красной волчанке:
а) ладони и подошвы
б) голени и бедра
в) щеки и нос
г) предплечья
д) слизистые
261. Дискоидная красная волчанка
проявляется всем перечисленным,
кроме:
а) рубцовой атрофии кожи
б) эритемой
в) фолликулярным гиперкератозом
г) инфильтрации
д) пигментация бугорков
262. Больному красной волчанкой не
назначают:
а) поддерживающую терапию преднизоло-
ном
б) санаторно-курортное лечение, с солнечны-
ми ваннами
в) диспансерное наблюдение
г) диету с исключением злаков
д) ношение широкополой шляпы
263. Для прогрессирующей стадии
склеродермии характерно:
а) положительный симптом Никольского
б) наличие отечного розово-лилового пятна
в) бордюр из отслоившегося эпидермиса по пе-
риферии очагов
г) положительный симптом Бенье
д) положительный симптом яблочного желе
264. Какая из перечисленных форм
склеродермии не относится к
очаговым формам:
а) линейная
б) белый лихен Цумбуша
в) акросклеротическая
г) атрофодермия Пазини-Пьерини
д) бляшечная
265. Укажите клинические формы
ограниченной склеродермии:
а) бляшечная
б)линейная
в) болезнь белых пятен
г) акросклеротическая
д) CREST-синдром
266. При начальной стадии ограниченной
склеродермии рационально назначить
комбинацию препаратов:
а) пенициллин+тигазон+перитол
б) ангиопротекторы+купренил+перитол
в) пенициллин+лидаза+ангиопротекторы
г) купренил+тигазон
д) тигазон+лидаза
267. Для дерматомиозита не характерно:
а) поражение кожи в виде отека, эритемы, «оч-
ков»
б) миозит, миалгии, креатинурия
в) поражение желудочно-кишечного тракта
г) фолликулярный гиперкератоз
д) резкое снижение массы тела
268. Излюбленная локализация высыпаний
при многоформной экссудативной
эритеме:
а) разгибательные поверхности конечностей
б) туловище
в) лицо
д) волосистая часть головы
д) кожа ладоней и подошв
269. Для многоформной эксссудативной
эритемы не характерно:
а) острое начало
б) зуд и жжение в местах поражения
в) уплощенные папулы синюшно-красного
цвета с западением в центре
Тесты по дерматовенерологии 629
г) нарушение общего состояния (лихорадка,
боли в суставах)
д) шелушение элементов рыхло сидящими че-
шуйками
270. Для лечения многоформной
экссудативной эритемы не применяют:
а) салицилаты
б) антигистаминные
в) препараты кальция
г) преднизолон
д) метотрексат
271. Типичная локализация высыпаний при
вульгарной пузырчатке:
а) туловище, слизистая полости рта
б) волосистая часть головы и туловище
в) ладони и подошвы
г) гениталии и лицо
д) гениталии и слизистая полости рта
272. Выделяют следующие формы
пузырчатки:
а) бразильская
б) норвежская
в) листовидная
г) лентикулярная
д) себорейная
273. Для пузырчатки характерны
следующие симптомы:
а) Никольского
б) Асбо-Хансена
в) Арди-Горчакова
г) Поспелова
д) «яблочного желе»
274. Первичный элемент при пузырчатке:
а) пятно
б) микровезикула
в) папула
г) пустула
д) пузырь
275. Для подтверждения диагноза
пузырчатки наиболее важно:
а) обнаружение акантолитических клеток в
мазках-отпечатках
б) эозинофилия в эрозий пузырной жидкости
в) выраженный лейкоцитоз в периферической
крови
г) обнаружение LE-клеток
д) выраженная тромбоцитопения
276. Для лечения пузырчатки наиболее
эффективны препараты:
а) антибиотики
б) сульфаниламиды
в) глюкокортикоиды
г) делагил
д) фтивазид
277. К осложнениям
глюкокортикостероидной терапии не
относят:
а) остеопороз
б) гипертензию
в) пептическую язву
г) тромбоцитопению
д) сахарный диабет
278. Преимущественная локализация
себорейной пузырчатки:
а) голова, грудь, спина
б) ладони и подошвы
в) половые органы
г) разгибательные поверхности конечностей
д) складки кожи вокруг естественных отверстий
279. При дерматозе Дюринга
противопоказаны препараты:
а) Лоринден С
б) препараты, содержащие иод
в)дапсон
г) преднизолон
д)унитиол
280. Для дерматоза Дюринга характерны:
а) полиморфизм высыпаний
б) эозинофилия в экссудате пузырей
в) поражение слизистой рта
г) обнаружение акантолитических клеток в
мазках-отпечатках эрозий
д) положительный симптом Никольского
281. Для подтверждения диагноза
дерматоза Дюринга проводится проба
на переносимость:
а) сульфаниламидов
б)антибиотиков
в) глюкокортикоидов
г) иодидов
д)новокаина
282. Для лечения дерматоза Дюринга
наиболее эффективны:
а) препараты сульфоновоо ряда
б)антибиотики
в) ацикловир
г) кетоконазол
д) препараты иода
283. Вульгарные угри возникают:
а) в детском возрасте
б) в период полового созревания
в) в период лактации
630 Приложение D
г) в старческом возрасте
д) в период новорожденное™
284. Укажите препараты, вызывающие
медикаментозные акне:
а) иодиды
б) глюкокортикоиды
в) анаболические стероиды
г) антибиотики
д) сульфаниламиды
285. Для лечения вульгарных угрей
применяются следующие препараты:
а) мази с глюкокортикоидами
б) Бензакне-гель 5—10 %
в) местные ретиноиды
г) клиндамицин в виде геля
д) примочки с фурацилином
286. Излюбленная локализация высыпаний
при вульгарных угрях:
а) лицо, грудь, спина
б) голени, ягодицы
в) ладони, подошвы
г) половые органы
д) волосистая часть головы
287. В этиологии розацеа имеет значение:
а) наличие хеликобактерной инфекции
б) вегетоневроз
в) заболевания легких
г)гипотиреоз
д) атеросклероз
288. Излюбленная локализация высыпаний
при розацеа:
а) лицо
б) спина, грудь, лицо
в) грудь, спина
г) разгибательные поверхности конечностей
д) кожа вокруг естественных отверстий
289. Для лечения розацеа применяют
препараты:
а) «Метрогил» гель
б) глюкокортикоиды внутрь
в) цитостатики
г) кремы с глюкокортикоидами
д) метронидазол внутрь
290. Для себорейного дерматита
характерно:
а) бляшки, покрытые жирными чешуйками
б) поражение носощечных, носогубных складок
в) пятна лилового цвета на уплотненном осно-
вании
г) поражение ладоней и подошв
д) положительный симптом Ауспитца
291. Больному себорейным дерматитом
рекомендуется:
а) диета, богатая жирами и углеводами
б) шампунь «Низорал»
в) общее УФО ежедневно, 15—20 процедур
г) глюкокортикостероиды внутрь по схеме
д) Тигазон внутрь по схеме
292. К поверхностным васкулитам кожи
относят:
а) болезнь Шамберга
б) мигрирующую узловатую эритему
в) острую узловатую эритему
г) геморрагический васкулит Шенлейн-
Геноха
д) узелково-некротический васкулит
293. К глубоким васкулитам кожи относят:
а) болезнь Шамберга
б) мигрирующую узловатую эритему
в) острую узловатую эритему
г) геморрагический васкулит Шенлейн-
Геноха
д) узелково-некротический васкулит
294. Назовите общие признаки,
характерные для кожных васкулитов:
а) воспалительный характер изменений кожи
б) преимущественная локализация высыпаний
на нижних конечностях
в) преимущественная локализация высыпаний
на лице
г) связь начала заболевания с перенесенной ин-
фекцией
д) проявляются в виде мономорфной папулез-
ной сыпы
295. Различают следующие формы
геморрагического васкулита, кроме:
а) кожно-суставной
б) абдоминальной
в) почечной
г) злокачественной
д) дементной
296. Для острой узловатой эритемы
характерно все перечисленное, кроме:
а) повышение температуры тела до 38—39 гра-
дусов
б) локализация высыпаний на голенях
в) узлы исчезают бесследно в течение 2—3 не-
дель
г) поражаются только поверхностные сосуды
кожи
д) проявляется образованием ярко-красных бо-
лезненных узлов
Тесты по дерматовенерологии 631
297. Для хронической узловатой эритемы
не характерно:
а) упорное рецидивирующее течение
б) локализация высыпаний преимущественно
на ладонях
в) обострение заболевания в осенне-зимний
период
г) образование синюшно-розовых плотных
безболезненных узлов
д) рецидив длится несколько месяцев
298. Для пемфигоида характерно:
а) наличие крупных напряженных пузырей на
неизмененном фоне или на фоне эритемы
б) наличие дряблых пузырей на уплотненном
основании
в) содержимое пузыря серозное или серозно-
геморрагическое
г) группировка высыпаний (герпетиформ-
ность)
д) симптом Асбо-Хансена положительный
299. Морфологические признаки
пемфигоида:
а) субэпидермальное расположение пузырей
б) интраэпидермальное расположение пузырей
в) выраженный гиперкератоз
г) инфильтрат в дерме, состоящий из гистио-
цитов и эозинофилов
д)акантолиз
300. Для лечения пемфигоида применяют:
а) цитостатики
б) глюкокортикоиды
в) сульфоновые препараты
г)антибиотики
д) противовирусные препараты
301. К факультативным предраковым
заболеваниям кожи и слизистых не
относят:
а) кожный рог
б) сенильную кератому
в) папилломатоз языка
г) ксантому кожи
д) лейкоплакию вульвы
302. Факультативным предраковым
заболеванием не являются:
а) хронический дерматит от воздействия хими-
ческих факторов
б) поздняя рентгеновская язва кожи
в) хронический фотодематит
г) хроническая язвенная пиодермия
д) хроническая профессиональная экзема
303. Облигатным предраковым
заболеванием не является:
а) эритроплазия Кейра
б)болезнь Боуэна
в)крауроз вульвы
г) дерматофиброма
д) лейкокератоз языка
304. Признаком малигнизаиии
предракового заболевания не
является:
а) эрозирование или изъязвление
б) появление вегетаций
в) уплотнение основания
г) отсутствие болезненности
д) подострое воспаление кожи
305. Для болезни Кейра не характерно:
а) резко ограниченная эритематозная бляшка
б) выраженное шелушение
в) локализация в области половых органов
г) бархатистая поверхность
д) имеет вид полициклической эрозии или по-
верхностной язвы
306. Болезнью Кейра называется
поражение, локализующееся:
а) в ротовой полости
б) на открытых участках кожи
в) на наружных половых органах
г) на красной кайме губ
д) ареолах сосков
307. Укажите неверный ответ. Для болезни
Боуэна характерно:
а) происходит из плоскоклеточного эпителия
б) представляет собой ограниченную эритема-
тозную, с непостоянным шелушением бляшку
в) может располагаться в любом месте кожно-
го покрова
г) представляет собой неглубокую, слегка кро-
воточащую при травматизации язву
д) имеет несколько клинических форм
308. Какова тактика врача при
установлении диагноза облигатного
предракового заболевания:
а) диспансеризация
б) назначение наружного лечения
в) назначение системной терапии цитостатичес-
кими препаратами
г) направление к онкологу
д) назначение витаминов и биогенных стиму-
ляторов и наблюдение
309. К доброкачественным опухолям кожи
не относят:
а) фиброму
22 Зак. 1357
632 Приложение D
б) липому
в) базалиому
г) гемангиому
д) лимфому
310. Укажите неверный ответ. Для
меланомы кожи характерно:
а) происходит из меланоцитов и невусных кле-
ток
б) развивается только на открытых участках
кожи
в) может иметь различные оттенки цвета
г) иногда развивается под ногтевой пластинкой
д) наиболее злокачественная опухоль
311. Укажите признак, не характерный для
базалиомы кожи:
а) наиболее часто встречающаяся злокачествен-
ная опухоль кожи
б) характерен медленный рост
в) редко метастазирует
г) происходит из пигментных невусов
д) чаще всего поражает кожу лица, височных
областей, ушей
312. Злокачественной опухолью не
является:
а) базалиома
б) пигментный невус
в) плоскоклеточный рак
г) меланома
д) саркома
313. Укажите неверные ответы. Среди
невусов различают:
а) сосудистые
б) пигментные
в) соединительнотканные
г) лимфоидные
д) келоидные
314. Признаками активизации невуса
являются:
а) быстрый рост
б) усиление пигментации
в) ослабление пигментации
г) появление субъективных ощущений
д) верно все
315. Саркома кожи — злокачественная
опухоль, которая происходит из:
а) клеток плоского эпителия
б) клеток железистого эпителия
в) фибробластов
г) моноцитов
д) меланоцитов
316. Для фибросаркомы не характерно:
а) стадийность течения
б) образование метастазов
в) залегание в толще кожи
г) появление «отсевов»
д) первичный очаг может располагатся вне
кожи
317. Саркома Капоши чаше встречается
среди:
а) мужчин
б) женщин
в) детей
г) частота встречаемости не зависит от пола
д) частота встречаемости не зависит от воз-
раста
318. Для лечения саркомы Капоши не
применяют:
а) цитостатические препараты
б) антибиотики
в) иммуномодуляторы — препараты интерфе-
рона
г) хирургические методы лечения
319. Укажите неверный ответ. Лимфомы
кожи — это опухолевые заболевания
кожи, которые:
а) характеризуются злокачественным течением
б) происходят из клона лимфоцитов
в) требуют немедленного оперативного лечения
г) иногда являются исходом хронической эк-
земы
д) могут поражаться внутренние органы
320. Среди лимфом кожи не выделяют:
а) низкой степени злокачественности
б) высокой степени злокачественности
в) В — типа
г) Т — типа
д) эозинофильную
321. Диагностика лимфом не включает:
а) выделение клинической формы
б) морфологическое подтверждение
в) клинико — лабораторное обследование
г) генетическое консультирование
д) инструментальное обследование внутренних
органов
322. Для грибовидного микоза
нехарактерны стадии:
а) эритематозно — сквамозная
б) инфильтративно — бляшечная
в) опухолевая
г) атрофическая
д) стационарная
Тесты по дерматовенерологии
633
323. Для прогноза тяжести течения
грибовидного микоза не имеет
значения:
а) поражение лимфоузлов
б) поражение внутренних органов
в) поражение кожи грибковой инфекцией
г) состояние иммунной системы
д) распространенность поражения
324. Для лечения грибовидного микоза
применяют всё, кроме:
а) цитостатических препаратов
б) глюкокортикостероидов
в) системных антимикотиков
г) ПУВА-терапии
д) общеукрепляющих средств
325. Укажите правильное определение
лимфомы кожи:
а) поражение, локализующееся только в коже
б) поражение лимфатической системы с пер-
вичным образованием в коже клона опухолевых
клеток
в) доброкачественная опухоль кожи
г) лимфоцитарная инфильтрация кожи при
хронических воспалительных заболеваниях
д) поражение кожи при хроническом лимфо-
лейкозе
326. При поддерживающей
глюкокортикостероидной терапии
необходимо:
а) постоянный прием витаминов группы «В»
б) прием препаратов кальция
в) прием препаратов калия
г) прием мочегонных средств
д) прием биогенных стимуляторов
327. Гнездная алопеция является:
а) рубцовой
б) нерубцовой
в) травматической
г) дистрофической
д) инфекционно-зависимой
328. Прогноз при гнездной алопеции
неблагоприятный, если у больного
имеется:
а) множественные телеангиэктазии
б) изменения ногтевых пластинок
в) язвенная болезнь желудка
г) проявления атопии
д) ихтиоз
329. Обязательным лабораторным
исследованием при алопеции является:
а) исследование крови на глюкозу
б) исследование крови на сифилис
в) исследование крови на тиреоидные гормоны
г) исследование крови на эозинофилию
д) исследование крови на циркулирующие им-
мунные комплексы
330. Аля лечения гнездной алопеции не
применяют:
а) глюкокортикостероиды
б) местно-раздражающие вещества
в) ретиноиды
г) биогенные стимуляторы
д) седативные препараты
331. Врожденной кератодермией
называется:
а) поражение кожи при авитаминозе А
б) наследственное нарушение ороговения
в) гиперкератоз вследствие микотической ин-
фекции
г) гиперкератоз вследствие постоянной травма-
тизации
д) выраженная форма ихтиоза
332. Кератодермией Папийона — Лефевра
называется:
а) сочетание диффузной ладонно-подошвенной
кератодермии с поражением коленных суставов
б) сочетание диффузной ладонно-подошвенной
кератодермии с поражением ногтей
в) сочетание диффузной ладонно-подошвенной
кератодермии с пародонтозом и гнойничковы-
ми заболеваниями кожи
г) сочетание ихтиоза со слабоумием, остеопо-
розом и карликовостью
д) сочетание ихтиоза с атопией
333. Кератодермией Меледа называется:
а) сочетание диффузной ладонно-подошвенной
кератодермии с поражением коленных суставов
б) сочетание диффузной ладонно-подошвенной
кератодермии с поражением ногтей
в) сочетание диффузной ладонно-подошвенной
кератодермии с пародонтозом и гнойничковы-
ми заболеваниями кожи
г) сочетание ихтиоза со слабоумием, остеопо-
розом и карликовостью
д) сочетание ихтиоза с атопией
334. Кератодермией Унны — Тоста
называется:
а) сочетание диффузной ладонно-подошвенной
кератодермии с поражением коленных суставов
б) сочетание диффузной ладонно-подошвенной
кератодермии с поражением ногтей
в) сочетание диффузной ладонно-подошвенной
кератодермии с породонтозом и гнойничковы-
ми заболеваниями кожи
634 Приложение D
г) сочетание ихтиоза со слабоумием, остеопо-
розом и карликовостью
д) сочетание ихтиоза с атопией
335. К наследственным формам ихтиоза не
относится:
а) ламеллярный ихтиоз
б) X —- сцепленный
в) простой
г) сенильный
д) врожденный эпидермолитический
336. Укажите неверный ответ. Для
обыкновенного ихтиоза характерно:
а) состояние ухудшается с возрастом
б) ретенционный гиперкератоз
в) усилен кожный рисунок ладоней и подошв
г) коллоидный плод
д) поражение ногтевых пластинок
337. Укажите неверный ответ. Для X —
сцепленного ихтиоза характерно:
а) болеют только мужчины
б) обусловлен рецессивным геном
в) обусловлен доминантным геном
г) грубые, темного цвета чешуйки
д) проявление заболевания с детского возраста
338. Укажите неверный ответ. Для
эпидермолитического ихтиоза
характерно:
а) образование пузырей в период новорожден-
ное™
б) гиперкератоз в местах образования пузырей
в) образование пузырей на месте ороговевшей
кожи
г) аутосомно-рециссивный тип наследования
д) состояние улучшается с возрастом
339. Укажите неверный ответ. Для
врожденного буллезного эпидермолиза
характерно:
а) образование пузырей при механических
травмах
б) гиперкератоз на месте образования пузырей
в) для лечения применяется ретиноиды
г) образование пузырей в период новорожден-
ное™
д) аутосомно-рециссивный тип наследования
340. Для ламеллярного ихтиоза характерно:
а) ретенционный гиперкератоз
б) пролиферационный гиперкератоз
в) акантоз
г) папилломатоз
д) амилоидоз
341. К ЗППП не относят:
а) гонорею
б) хламидиоз
в)трихинеллез
г) чесотку
д) герпес II типа
342. К осложнениям различных
венерических заболеваний относят:
а) аднексит
б) простатит
в) артрит
г) мезаортит
д) все верно
343. Для какого заболевания характерны
высыпания на коже ладоней и подошв:
а) сифилис
б) герпес
в) микоплазмоз
г) гонорея
д) шанкроид
344. Для сифилиса характерны следующие
пути передачи:
а) контактный
б) вертикальный
в) трансфузионный
г) все перечисленное верно
д) не верно б
345. Укажите правильные ответы. Для
профилактики заражения ЗППП могут
применяться следующие методы:
а) барьерной контрацепции
б) применение антисептических средств
в) прерывание полового акта
г) прием атибиотиков
д) все верно
346. Какой из видов трепонем является
возбудителем сифилиса:
а) 7г. refringens
б) 7г. pallida
в) 7г. balanitidis
г) 7г. buccalis
д) Tr. dentinum
347. Возбудитель сифилиса наиболее
чувствителен к следующим агентам:
а) высоким температурам
б) низким температурам
в) химическим веществам
г) УФ-излучению
д) все верно
Тесты по дерматовенерологии
635
348. Аля лабораторного определения вида
спирохеты наименее важно:
а) подвижность
б) способность окрашиваться
в) количество завитков
г) форма завитков
д) характер движений
349. Под влиянием неблагоприятных
условий существования бледная
спирохета может переходить в
следующие формы:
а) цисты
б) элементарные тельца
в) L — формы
г) Грам-положительные формы
д) все верно
350. Укажите неверные ответы. Антигенная
структура Tr. pallida состоит из:
а) гликопротеидного АГ
б) липопротеидного АГ
в) кардиолипинового АГ
г) липополисахаридного АГ
351. Твердый шанкр при первичном
сифилисе чаше всего возникает через:
а) 3—4 недели
б) 3—4 дня
в) 1—2 недели
г) 1—2 месяца
д) 3—4 месяца
352. Укажите правильные ответы.
Первичный период сифилиса
проявляется:
а) появлением сыпи на туловище и конечнос-
тях
б) повышением температуры тела, слабостью,
головными болями
в) образованием твердого шанкра
г) увеличением всех групп лимфоузлов
д) увеличением регионарных лимфоузлов
353. Аля твердого шанкра не характерно:
а) отсутствие болезненности
б) инфильтрация основания
в) медно-красный
г) подрытые края язвы
д) регионарный лимфаденит
354. К атипичным шанкрам не относят:
а) шанкр-амигдалит
б) шанкр-панариций
в) муцинозный отек
г) индуративный отек
д) мягкий шанкр
355. Укажите правильные ответы. Обычно
твердому шанкру сопутствует:
а) регионарный склераденит
б) регионарный отек
в) регионарный лимфангит
г) повышение температуры тела
д) реакция Гексгеймера
356. Для первичного сифилиса не
характерно:
а) самопроизвольное рубцевание язвы
б) удовлетворительное общее состояние боль-
ного
в) образование сифилитической гуммы
г) поражение регионарных лимфоузлов
д) поражение лимфатических сосудов
357. Укажите правильные ответы. К
характерным признакам
специфического сифилитического
аденита не относится:
а) безболезненность
б) образование свищей
в) подвижность лимфоузлов
г) образование «пакетов» узлов
д) плотно — эластическая консистенция лим-
фоузлов
358. Какое обязательное условие для
постановки диагноза первичного
серопозитивного сифилиса:
а) эрозия (язва) в области половых органов
б) эрозия (язва) в области половых органов в
сочетании с лимфаденитом
в) обнаружение в отделяемом эрозией (язвы)
бледных спирохет
г) положительные классические серологические
реакции
д) положительные РИФ, РИБТ
359. Укажите правильные ответы. При
первичном сифилисе выделяют:
а) серонегативный период
б) серопозитивный период
в) скрытый период
г) инкубационный период
д) все верно
360. Осложнениями генетального твердого
шанкра у мужчин может быть все,
кроме:
а) гангренизации
б) фимоза
в) фагеденизации
г) регионарного лимфаденита
д) парафимоза
636 Приложение D
361. Укажите неправильное утверждение:
а) твердый шанкр может быть покрыт грязно-
желтым налетом
б) твердый шанкр локализуется только в об-
ласти естественных отверстий
в) твердый шанкр может локализоваться на
любом участке кожи и слизистых
г) имеет атипичные формы
д) твердые шанкры могут быть множественны-
ми
362. Укажите правильные ответы. Твердый
шанкр может быть в форме:
а) эрозии
б) папулы
в) язвы
г) вегетации
д) индуративного отека
363. Вторичный период сифилиса
подразделяют на:
а) свежий
б) скрытый
в) умеренно активный
г) рецидивный
д) суб-манифестный
364. Во вторичной рецидивной стадии
сифилиса редко поражаются:
а) кожа и слизистые
б) центральная нервная система
в) костно-мышечная система
г) волосистая часть головы
д) лимфатическая система
365. Укажите неверные ответы. Во
вторичном периоде сифилиса
поражение волосистой части головы
может быть в виде:
а) диффузной алопеции
б) гнездной алопеции
в) мелкоочаговой алопеции
г) смешанной алопеции
д) субтотальной алопеции
366. Укажите неверный ответ. Во
вторичном периоде сифилиса
выделяют следующие разновидности
сифилидов:
а) папулезный
б) пигментный
в) пятнистый (розеолезный)
г) бляшечный
д) пустулезный
367. При вторичном рецидивном сифилисе
не развивается:
а) сифилитическая ангина
б) сифилитический ларингит
в) сифилитический ринит
г) полиаденит
д) спинная сухотка
368. Для сифилитической ангины наиболее
характерно:
а) односторонность поражения
б) безболезненность (малая болезненность)
в) налеты
г) затруднение глотания
д) яркая гиперемия
369. Для вторичного рецидивного
сифилиса характерно:
а) обильные яркие множественные высыпа-
ния
б) немногочисленные, бледные, склонные к
слиянию высыпания
в) сифилиды разрешаются с образованием руб-
ца
г) сифилиды полностью не разрешаются без
лечения
д) поражение лимфоузлов
370. При вторичном свежем сифилисе
поражение кожи наиболее часто
бывает в виде:
а) пустул
б) розеол
в)алопеции
г) широких кондилом
д) твердого шанкра
371. Широкие кондиломы возникают
обычно вследствие:
а) ослабленного иммунитета
б) присоединения вторичной инфекции
в) длительного раздражения кожи в складках
г) неадекватного местного лечения
д) повторного инфицирования
372. Пустулезный сифилид возникает
обычно вследствие:
а) пониженной сопротивляемости организма
б) присоединения вторичной инфекции
в) механического раздражения кожи
г) неадекватного местного лечения
д) повторного инфицирования (реинфекции)
373. Укажите неверный ответ.
Разновидности папулезного сифилида
при вторичном сифилисе:
а) лентикулярный
б) мокнущий
в) широкие кондиломы
г) остроконечные кондиломы
д) верно все
Тесты по дерматовенерологии 637
374. Поражение слизистых оболочек
при вторичном сифилисе чаше
бывает в виде высыпаний:
а) розеолёзных
б) папулезных
в) язвенных
г) пустулёзных
д) лейкодермы
375. Наличие у больного сифилисом
эрозивного поражения на половом
члене и пятнисто-папулезной сыпи на
коже туловища свидетельствует о
следующей стадии заболевания:
а) вторичном свежем
б) вторичном рецидивном
в) первичном серопозитивном
г) третичном активном
д) манифестном
376. Признак Пинкуса наблюдается при
сифилитическом поражении:
а) слизистых
б) кожи
в)волос
г) ногтей
д) красной каймы губ
377. Укажите правильные ответы. Наиболее
тяжелыми разновидностями
пустулезного сифилида являются:
а) сифилитическое импетиго
б) широкие кондиломы
в) сифилитическая эктима
г) сифилитическая рупия
д) сифилитическая гумма
378. Выберите стадию (форму) сифилиса,
при которой часто поражаются
слизистые:
а) первичный серопозитивный
б) третичный активный
в) вторичный рецидивный
г) висцеральный
д) вторичный скрытый
379. Для сифилической розеолы не
характерно:
а) розовый, бледно-розовый цвет
б) исчезновение при диаскопии
в) зуд
г) обычно не шелушится
д) разрешается бесследно
380. Во вторичном периоде сифилиса
встречаются редко:
а) пятнистые и папулезные сифилиды
б) пигментные и пустулезные сифилиды
в) папулезные сифилиды и алопеция
г) широкие кондиломы и алопеция
д) розеола и остатки твердого шанкра
381. Для третичного активного периода
сифилиса характерным:
а) папулы ладоней и подошв
б) сгруппированные бугорки
в) диффузная алопеция
г) полиаденит
д) пустулезный сифилид
382. При висцеральном сифилисе чаше
всего поражается:
а) почки
б) аорта
в) брыжейка тонкой кишки
г) легкие
д) пожелудочная железа
383. Для третичного периода сифилиса не
характерны:
а) гуммозные поражения костей
б) бугорковые поражения кожи
в) бугорковые поражения легких
г) мезаортит
д) гуммозные поражения кожи
384. Укажите правильные ответы. В течении
третичного периода сифилиса
выделяют следующие стадии:
а) активную
б) латентную
в) прогрессирующую
г) подострую
д) стационарную
385. Развитию третичного сифилиса
способствуют:
а) алкоголизм
б) иммунодефицитные состояния
в) тяжелая сопутствующая патология
г) все верно
д) все не верно
386. Для бугорков при третичном сифилисе
не характерны:
а) плотная консистенция
б) зуд
в) разрешение с образованием рубца
г) медно-красный, с синюшным оттенком цвет
д) склонности к группировке
387. Для гуммозного сифилиса не
характерно:
а) плотный узел
б) разрешается путем некроза
в) при разрешении образуется рубец
638 Приложение D
г) островоспалительные явления
д) множественность поражения
388. Сифилитическая гумма поражает:
а) надкостницу и хрящи
б) костно-мышечную систему
в) внутренние органы
г) подкожную клетчатку
д) все верно
389. Для сифилидов третичного периода не
характерно:
а) отсутствие островоспалительных явлений
б) образование рубцов
в) высокая контагиозность
г) отсутствие субъективных ощущений
д) медленная эволюция
390. Укажите неверные ответы. Для
диагностики активного третичного
сифилиса используют следующие
критерии:
а) выявление бугорков или гумм
б) обязательное обнаружение бледной трепоне-
мы в отделяемом сифилидов
в) положительные КСР, РИФ, РИБТ
г) анамнез
д) общий анализ крови
391. Укажите не достоверный клинический
признак врожденного сифилиса
грудного:
а) диффузная папулезная инфильтрация
б) седловидный нос
в) сифилитическая пузырчатка
г) сифилитический ринит
д) остеохондрит
392. Достоверный признак позднего
врожденного сифилиса:
а) сифилитический ринит
б) высокое «готическое» нёбо
в) зубы Гетчинсона
г) сифилитическая пузырчатка
д) все верно
393. Ранний врожденный сифилис
включает следующие периоды:
а) сифилис раннего детского возраста
б) сифилис плода
в) сифилис детей грудного возраста
г) вторичный сифилис
д) все верно
394. Вероятными клиническими
признаками врожденного сифилиса
детей грудного возраста не являются:
а) седловидный нос
б) рубцы Робинсона-Фурнье
в) ягодицеобразный череп
г) диффузная алопеция
д) все перечисленное верно
395. К достоверным клиническим
признакам врожденного сифилиса
детей грудного возраста относят:
а) высокое «готическое» нёбо
б) сифилитическую пузырчатку
в) остеохондриты
г) сифилитический ринит
д) диффузную папулезную инфильтрацию
396. К достоверным клиническим
признакам позднего врожденного
сифилиса относят:
а) паренхиматозный кератит
б) специфический лабиринтит
в) зубы Гетчинсона
г) сифилитический ринит
д) недоразвитие мечевидного отростка грудины
397. При врожденном сифилисе раннего
детского возраста выявляют:
а) папулезные сифилиды
б) широкие кондиломы
в) поражения трубчатых костей
г) зубы Гетчинсона
д) паренхиматозный кератит
398. Поздний врожденный сифилис
развивается:
а) на поздних сроках беременности
б) в возрасте до 1-го года
в) в возрасте до 2-х лет
г) после 2-х лет жизни
д) в период новорожденное™
399. Исключите признак, не характерный
для сифилитической пузырчатки:
а) напряженные пузыри
б) инфильтрация в основании пузыря
в) отсутствие субъективных ощущений
г) положительный симптом Никольского
д) локализация на ладонях и подошвах
400. Укажите признаки сифилиса плода:
а) паренхиматозный кератит
б) дистрофия подкожной жировой клетчатки
в) увеличение печени
г) триада Гетчинсона
д) увеличение плаценты
401. Какой срок от момента заражения
характеризует ранний скрытый
приобретенный сифилис:
а) менее 2 лет
Тесты по дерматовенерологии
639
б) более 3 лет
в) 3 года
г) 5 лет
д) 4 года
402. Какой срок от момента заражения
характерен для позднего скрытого
сифилиса:
а) более 2 лет
б) до 2 лет
в) 5 месяцев
г) 6 месяцев
д) 1 год
403. При скрытом раннем сифилисе
реакция Вассермана:
а) отрицательная с трепонемным антигеном
б) резко положительная с титрами 1:320—1:160
в) резко положительная с титрами 1: 20—1:10
г) слабо положительная
д) отрицательная со всеми антигенами
404. Наиболее специфической реакцией на
сифилис является:
а) РМП
б) RW
в) РИТ
г) РИФ
д) VDRL
405. Какое количество «плюсов» при
постановке реакции Вассермана
указывает на резко положительный
результат:
а)1
б) 10
в) 4
г) 5
Д)3
406. По истечении какого срока после
появления твердого шанкра становятся
положительными классические
серологические реакции:
а) 1 месяц
б) 5—7 дней
в)2 недели
г) 3 месяца
д) 2-3 дня
407. Через какой срок после заражения
сифилисом становится положительной
реакция Вассермана:
а) 7—8 недель
б)1 неделя
в)3 месяца
г) 6 месяцев
д) 2 недели
408. К современным методам лабораторной
диагностики сифилиса относятся:
а) ИФА
б) бактериологический метод
в) метод иммуноблоттинга
г) ПЦР
д) микроскопический метод
409. На какие группы в настоящее время
можно разделить серологические
тесты на сифилис:
а) трепонемные
б) бактериологические
в) нетрепонемные
г) вирусологические
д) микроскопические
410. При каком проценте иммобилизации
бледных трепонем РИТ считается
положительной:
а) до 20 %
6)51-100 %
в) 21-50 %
г) 5-10 %
Д) 1-5 %
411. К стандартным серологическим
реакциям относят:
а) РСК
б) РИФ
в) РИП
г) РИТ
д) реакцию с трепонемным антигеном
412. К групповым трепонемным реакциям
относят:
а) РИП
б) РИФ
b)RW
г) РМП
Д) РИТ
413. К нетрепонемным тестам с визуальным
определением результата относят:
а) КСР
б) RPR
в) TRUST
г) РИТ
д) РИФ
414. Какой основной препарат
применяется для лечения сифилиса:
а) доксициклин
б)пенициллин
в) бисептол
г) эритромицин
д) цефазолин
640 Приложение D
415. Какие препараты пролонгированного
действия применяются для лечения
сифилиса:
а) бициллины
б) экстенциллин
в) тетрациклин
г) азитромицин
д) сульфадиметоксин
416. Какие препараты применяются для
неспецифической терапии сифилиса:
а) пирогенал
б) ретиноиды
в) витамины
г) биогенные стимуляторы
д) глюкокортикоиды
417. Какие препараты кроме антибиотиков
применяются при поздних формах
сифилиса:
а) препараты висмута
б) фторхинолоны
в) препараты иода
г) сульфаниламиды
д) глюкокортикоиды
418. При непереносимости пенициллинов
применяют:
а) экстенциллин
б) доксициклин
в) эритромицин
г) метронидозол
д) азитромицин
419. Какое наиболее серьезное осложнение
может возникнуть при лечении
препаратами пенициллина:
а) анафилактический шок
б) расстройства ЖКТ
в) кожный зуд
г) повышение температуры тела
д)полиневриты
420. Первичная профилактика сифилиса
включает:
а) санитарное просвещение
б) использование презервативов
в) избегать случайных половых контактов
г) работу с группами риска
д) все верно
421. Что является основой профилактики
врожденного сифилиса:
а) своевременное выявление и лечение боль-
ных сифилисом женщин
б) трехкратное исследование крови на RW во
время беременности
в) профилактические курсы лечения беремен-
ных у женщин, перенесших сифилис
г) прерывание беременности
д) витаминотерапия
422. Общественная профилактика
сифилиса предусматривает:
а) обязательный учет больных сифилисом и
другими ИППП
б) обязательное привлечение к лечению боль-
ных скрытыми формами сифилиса
в) обязательную флюорографию 1 раз в 6 ме-
сяцев
г) обследование всех лиц, находившихся в по-
ловом и тесном бытовом контакте с больным си-
филисом
д) использование презерватива
423. Для личной профилактики после
полового контакта применяют
следующие препараты:
а) 0,05 % раствор хлоргексидина
б) 0,01 % раствор мирамистина
в) 1 % бриллиантовый зеленый
г) 0,5 % раствор хлоргексидина
д) 20 % спиртовой раствор
424. Возбудитель шанкроида:
a) Tr. Pallidum
б) Trich. vaginalis
в) стрептобактерия Haemophillus Ducrei
г) Chlamidia trachomatis
д) Gardnerella vaginalis
425. Назовите наиболее частые
осложнения шанкроида:
а) лимфангиит
б) лимфаденит
в) фимоз
г) рожистое воспаление
д) перитонит
426. Какой препарат рекомендуется
применять для начальной терапии
шанкроида:
а) бисептол
б) экстенциллин
в) азитромицин
г) доксициклин
д) офлоксацин
427. Возбудителем четвертой венерической
болезни являются:
а) простейшие
б) хламидии
в) дрожжеподобные грибы
г) Грамм-отрицательные диплококки Нейссера
д) пневмококки
Тесты по дерматовенерологии 641
428. Какие осложнения венерического
лимфогранулематоза относят к
поздним:
а) слоновость половых органов
б) полиневриты
в) лимфорроиды
г) стриктуры прямой кишки и мочеиспуска-
тельного канала
д) периоститы
429. Какие препараты показаны для
лечения венерического
лимфогранулематоза:
а) тетрациклины
б) пенициллины
в) макролиды
г) сульфаниламиды
д) фторхинолоны
430. Возбудители гонореи:
а) пневмококки
б) менингококки
в) кампилобактерии
г) хламидии
д) диплококки Нейссера
431. Каким путем не заражаются гонореей:
а) при генитально-генитальные контактах
б) при генитально-анальные контактах
в) при генитально-оральные контактах
г) воздушно-капельным путем
д) при пользовании предметов общего обихо-
да
432. Гонококки поражают:
а) слизистую уретры
б) слизистую глотки и миндалин
в) слизистую 12-перстной кишки
г) слизистую протоков предстательной железы
д) слизистую ободочной кишки
433. Какой материал берут для диагностики
гонореи:
а) выделения из уретры
б)соскоб коньюнктивы
в) кровь из вены
г) пунктат лимфатического узла
д) соскоб слизистой прямой кишки
434. Какай метод окраски мазков
используют для диагностики гонореи:
а) бриллиантовым зеленым
б) по Граму
в) метиленовым синим
г) по Романовскому-Гимзе
д)по Шику
435. Укажите возможные варианты течения
гонорейного уретрита у мужчин:
а) свежий острый гонорейный уретрит
б) свежий подострый гонорейный уретрит
в) хронический гонорейный уретрит
г) асимптомный гонорейный уретрит
д) все верно
436. Какие симптомы характерны для
свежего острого переднего
гонорейного уретрита:
а) мутная моча в двух порциях
б) прозрачная моча в двух порциях
в) обильные гнойные выделения в течение дня
г) учащенное болезненное мочеиспускание
д) мутная моча в первой порции
437. Какие осложнения могут возникать у
мужчин при гонорее:
а) простатит
б) орхоэпидидимит
в) водянка яичка
г) баланопостит
д) семинома
438. Какие экстрагенитальные осложнения
возможны у мужчин при гонорее:
а) артрит
б) эндокардит
в) полиневрит
г) коньюнктивит
д) генерализованный кожный зуд
439. Какие симптомы характерны для
диссеминированной гонорейной
инфекции:
а) артралгии
б) диффузная алопеция
в) септицемия
г) кожные поражения
д) мезаортит
440. Назовите симптомы, характерные для
хронического гонорейного уретрита:
а) наличие выделений из уретры по утрам
б) наличие экссудата при взятии материала из
уретры ложечкой Фолькмана
в) обильные гнойные выделения в течение дня
г) наличие мутной мочи в первой порции
д) прозрачная моча в двух порциях с наличи-
ем нитей
441. Какие среды используют для
культурального исследования на
гонорею:
а) Сабуро
б) агар-агар
в) асцит-агар
642 Приложение D
г) мясо-пептонный бульон
д) рисовый агар
442. Какой препарат является препаратом
выбора для лечения гонореи:
а) цефтриаксон
б)пенициллин
в) эритромицин
г) азитромицин
д) бисептол
443. При хронической гонорее
применяются следующие методы
лечения:
а) антибиотики
б) цитостатики
в) глюкокортикоиды
г) иммуномодуляторы
д) местное лечение
444. Какие виды провокаций используют
для диагностики и определения
критериев излеченности гонорейного
уретрита:
а) механическая
б) биологическая
в) химическая
г) алиментарная
д) все верно
445. Какой основной критерий
излеченности гонорейного уретрита:
а) отсутствие клинических проявлений
б) отсутствие жалоб больного
в) отсутствие роста возбудителя на питательной
среде
г) верно а, б, в
д) лечение полового контакта
446. Назовите возбудителя трихомониаза:
a) Tr. Pallidum
б) Trich. vaginalis
в) стрептобактерия Haemophilias Ducrei
г) Chlamidia trachomatis
д) Gardnerella vaginalis
447. Укажите формы течения
урогенитального трихомониаза:
а) острая
б) подострая
в)хроническая
г) малосимптомная
д) все верно
448. Укажите наиболее достоверный метод
микроскопической диагностики
урогенитального трихомониаза:
а) окраска по Граму
б) окраска метиленовым синим
в) окраска бриллантовым зеленым
г) окраска по Романовскому-Гимзе
д) обнаружение трихомонады в нативной капле
449. Назовите препарат выбора для
лечения урогенитального
трихомониаза:
а) доксициклин
б) азитромицин
в) ципробай
г) метронидазол
д) кетоконазол
450. Какие методы (препараты) провокации
применяют для выявления
урогенитального трихомониаза:
а) плазмол 1,0 подкожно
б) массаж уретры на буже
в) массаж простаты
г) алиментарный метод
д) все верно
451. В какие сроки проводится
лабораторный контроль излеченности
трихомониаза:
а) через месяц после первого контроля
б) через 7 дней
в) через 2 недели
г) через 5 дней
д) через 3 месяца
452. Укажите возбудителя микоплазменной
инфекции:
а) атипичные бактерии
б) вирусы
в) простейшие
г) грибы
д) эктопаразиты
453. Какие лабораторные методы
применяются для диагностики
микоплазмоза:
а) бактериологический метод
б) ПЦР
в)серологический метод
г) прямая микроскопия
д) исследование спинномозговой жидкости
454. Какие осложнения может вызвать
микоплазменная инфекция у мужчин:
а) простатит
б) орхиэпидидимит
в) сепсис
г) параметрит
д) болезнь Рейтера
Тесты по дерматовенерологии
643
455. Укажите основные группы препаратов
для лечения урогенитального
микоплазмоза:
а) пенициллины
б) фторхинолоны
в) сульфаниламиды
г) макролиды
д) тетрациклины
456. Препарат выбора для лечения
микоплазменной инфекции,
вызванной U.urealiticum
а) доксициклин
б) азитромицин
в) бисептол
г) ретарпен
д) офлоксацин
457. Через какой срок проводится
контроль излеченности урогентального
микоплазмоза:
а) 10 дней
б) 2 месяца
в) 2—3 недели
г) 6 месяцев
д) месяц
458. Возбудителем урогенитального
кандидоза являются:
а) трихомонады
б) хламидии
в) микоплазмы
г) гонококки
д) дрожжеподобные грибы
459. Какие факторы способствуют
возникновению кандидозного уретрита
у мужчин:
а) остеомиелит
б) длительный прием антибиотиков, кортико-
стероидов
в) сахарный диабет
г) иммунодефицит
д) плоскостопие
460. Какие методы диагностики применяются
при урогенитальном кандидозе:
а) микроскопическое исследование
б) бактериологическое исследование
в) ПЦР
г) ИФА
д) реакцию Видаля
461. Укажите возможные проявления
урогенитального кандидоза у мужчин:
а) уретрит (острый, подострый, хронический)
б) бессимптомное носительство дрожжеподоб-
ных грибов
в) баланопостит
г) полиаденит
д) высокая температура
462. Какие системные препараты
применяют для лечения кандидозного
уретрита мужчин:
а) флуконазол (дифлюкан)
б) кетоконазол (низорал, оранозол)
в) итраконазол (орунгал)
г)доксициклин
д)дипроспан
463. Какие сроки проведения контроля
излеченности урогенитального
кандидоза у мужчин после окончания
лечения:
а) через 2 недели
б) через месяц
в) через неделю
г) через 6 месяцев
д) 3 раза в течение 3 месяцев
464. Острый гонорейный вульвовагинит у
девочек развивается после заражения
через:
а) 5—7 дней
б) 3 дня
в) месяц
г) 2 недели
д) полгода
465. В виде каких воспалительных
процессов проявляется хламидийная
инфекция у девочек:
а) менингит
б) вульвовагинит
в)уретрит
г) цервицит
д)хейлит
466. Какой препарат рекомендуется для
лечения хламидийного вульвовагинита
у девочек до 8 лет:
а) доксициклин
б) эритромицин
в)пенициллин
г) бисептол
д) аспирин
467. Какие лабораторные методы
применяют для подтверждения
диагноза гонореи у девочек:
а) микроскопический
б)серологический
в)бактериологический
г) РИФ
д) ИФА
644 Приложение D
468. К клиническим формам генитального
герпеса не относят:
а) манифестную
б) атипичную
в) терминальную
г) абортивную
д) субклиническую
469. Какие факторы способствуют
ренидивированию генитального
герпеса:
а) снижение иммунологической реактивности
б) переохлаждение
в) ВИЧ-инфекция
г) инъекции витамина В]
д) прием элеутерококка
470. Укажите возможные клинические
варианты урогенитальной
герпетической инфекции:
а) поражение наружных половых органов
б) герпетические кольпиты, цервициты и урет-
риты
в) герпетический эндометрит, сальпингит или
цистит
г) генерализованная сыпь
д) менингоэнцефалит
471. Какие методы используют для
диагностики герпетической инфекции:
а) ПЦР
б) РИТ
в) КСР
г) цитоморфологический метод
д) выявление антигенов ВПГ
472. Какие серологические методы
используют для выявления антигенов
ВПГ:
а) реакция связывания комплемента
б) ИФА
в) РМП
г) КСР
д) РПГА
473. Какой препарат является базисным для
лечения генитального герпеса:
а) пенициллин
б)бисептол
в)ацикловир
г) флуконазол
д) рифампицин
474. Выделите общие черты ВИЧ-инфекции
и возбудителей венерических
болезней:
а) быстрая гибель во внешней среде
б) половой путь передачи инфекции
в) распространение заболеваний в группах риска
г) способность вызывать различные степени тя-
жести клинические проявления и осложнения
д) все верно
475. Основные пути заражения ВИЧ:
а) половой
б) бытовой
в) парентеральный
г) трансплацентарный
д) воздушно-капельный
476. Какие клинические признаки могут
указывать на ВИЧ-инфекцию:
а) длительная лихорадка и диарея
б) генерализованная лимфаденопатия
в) саркома Капоши у молодых лиц
г) кандидозное поражение слизистых и кожи
д) все верно
477. Выделяют формы саркомы Капоши,
кроме:
а) доброкачественной
б) злокачественной
в) спорадической
г) эпидемической
д) иммуносупрессивной
478. Какие поражения кожи и слизистых
наиболее характерны для ВИЧ-
инфекции:
а) себорейный дерматит
б) вирусные заболевания
в) микотические заболевания
г) паразитарные заболевания
д) трофические заболевания кожи
479. К стадии первичных проявлений ВИЧ-
инфекции относятся:
а) острая лихорадочная фаза
б) прогрессирующая потеря массы тела
в) бессимптомная фаза
г) персистирующая генерализованная лимфа-
денопатия
д) генерализованные грибковые поражения
480. Какие инфекционные заболевания
кожи характерны для ВИЧ-инфекции:
а) опоясывающий лишай
б) контагиозный моллюск
в) простой пузырьковый лишай
г) красный плоский лишай
д) широкие кондиломы
481. Какой препарат является наиболее
эффективным для лечения грибковых
инфекций у ВИЧ-инфицированных
а) дифлюкан
Тесты по дерматовенерологии 645
б)ацикловир
в) макропен
г) офлоксоацин
д) гризеофульвин
482. Какие методы применяются для
выявления антител к ВИЧ
а) ИФА
б) ПЦР
в) РИТ
г) иммунофлюоресценция
д) иммуноблоттинг
483. Какие лекарственные препараты нельзя
применять ВИЧ-инфицированному
человеку
а) системные противогрибковые антибиотики
б) сульфаниламиды
в) иммуностимуляторы
г) витамины
д) противовирусные препараты
484. Возбудителем урогенитального
хламидиоза является:
a) 7r. pallidum
б) Trich. vaginalis
в) стрептобактерия Haemophilias Ducrei
г) Chlamydia trachomatis
д) Gardnerella vaginalis
485. Каким путем происходит заражение
урогенитальным хламидиозом:
а) воздушно-капельным
б) при переливании крови
в) при иньекциях
г) новорожденным в период родов от инфици-
рованной матери
д) половым
486. Chlamydia trachomatis это:
а) простейшее
б) риккетсия
в) вирус
г) атипичная бактерия
д) грамположительная бактерия
487. Какие формы хламидий являются
инфекционными:
а) ретикулярные тельца
б) хламидийные включения
в) элементарные тельца
г) промежуточные (переходные) тельца
д) внутриклеточные микроколонии этих орга-
низмов
488. Какой материал берут для
исследования методом прямой РИФ на
хламидии:
а) выделения из уретры, шейки матки
б) соскоб из уретры, шейки матки
в) эйякулят
г) секрет предстательной железы
д) выделения из влагалища
489. Укажите варианты течения
урогенитального хламидиоза:
а) острое
б) подострое
в) малосимптомное
г) хроническое
д) все верно
490. Какие симптомы характерны для
острого переднего хламидийного
уретрита:
а) обильные серо-белые или гнойные выделе-
ния из уретры
б) рези при мочеиспускании
в) гематурия
г) пенистые светлые выделения
д) скудные слизистые выделения
491. Какие осложнения могут возникнуть
при урогенитальном хламидиозе:
а) простатит
б) эпидидимит
в) реактивный артрит
г) гнездная алопеция
д) острый панкреатит
492. Для хламидийного уретрита у мужчин
характерно:
а) скудные выделения из уретры
б) боли в промежности, мошонке, прямой киш-
ке
в) предстательная железа пальпаторно уплотне-
на, увеличена, болезненна
г) предстательная железа не изменена
д) обильные гнойные выделения из уретры
493. Какие основные системы поражаются
при болезни Рейтера:
а) орган слуха
б) ЖКТ
в) мочеполовая система
г) опорно-двигательная система
д) орган зрения
494. Методы диагностики урогенитального
хламидиоза:
а) ПЦР
б)МРП
646 Приложение D
в) РИФ
г) реакция Борде-Жангу
д) ИФА
495. Какие группы антибактериальных
препаратов применяют для лечения
урогенитального хламидиоза:
а) макролиды
б) препараты нитрафуранового ряда
в)пенициллины
г) препараты тетрациклинового ряда
д) фторхинолоны
496. Наиболее эффективный препарат для
лечения урогенитального хламидиоза
из группы макролидов:
а) эритромицин
б) рокситромицин
в) кларитромицин
г) эрацин
д) олеандомицин
497. Какова минимальная
продолжительность цикла
антибиотикотерапии при хроническом
хламидиозе:
а) 7 дней
б) 14—21 день
в) 2 месяца
г) 21—28 дней
д) 7—10 дней
498. В течение скольких циклов развития
хламидий рекомендовано проводить
антибиотикотерапию:
а) 2—3 цикла
б)16 циклов
в)7 циклов
г) 4—5 циклов
д) без учета циклов
499. Какой вид провокации применяют
больному урогенитальным
хламидиозом:
а) массаж предстательной железы
б) массаж уретры на буже
в) инстилляцию 0,05 % раствором хлоргекси-
дина
г) плазмол подкожно 1,0
д) инстилляцию 0,5 % раствором нитрата се-
ребра
500. Через какой срок после окончания
лечения проводится первый
лабораторный контроль излеченности
хламидиоза:
а) 7 дней
б) 2 месяца
в)2 недели
г) 1 месяц
д) 6 месяцев
Приложение Е
Ответы к тестам
1. а, б, в 46. б, в 91. В 136. а, в, д
2. в 47. а, в 92. а, б, г 137. а, г
3. в 48. в 93. д 138. а, б, в
4. в, д 49. б, г 94. б, в 139. в
5. а 50. в, г 95. а 140. д
6. г 51. а 96. а, б, д 141. в
7. а, б 52. а, г, д 97. а, г, д 142. г
8. г 53. б, в 98. б, в, г 143. Д
9. б 54. а, д 99. д 144. а, в, д
10. д 55. а, б 100. д 145. а, г
11. в 56. г 101. а, б 146. д
12. б 57. б, в 102. а, в, г 147. а, б, г
13. б 58. б 103. в 148. а, г
14. в 59. а, б 104. а 149. в, д
15. Д 60. а 105. б 150. б, г
16. д 61. а, б, в 106. в 151. б, в, д
17. а, д 62. б 107. В, Г, д 152. б
18. а, г 63. б, в, д 108. д 153. б, д
19. в 64. а, в 109. б 154. а, г, д
20. а, в, г 65. б, в НО. д 155. а
21. г 66. г 111. а, б, в 156. г, Д
22. а 67. а, в, д 112. б, г, д 157. б, д
23. б 68. а, б 113. а, б, г 158. а, в
24. а 69. а, б 114. а, в 159. б, г, д
25. б 70. а 115. а, в, г 160. б, г, д
26. д 71. г 116. а, в, д 161. а, б, г
27. б, г, д 72. а, в 117. б, г, д 162. а, в, г
28. а, г 73. а, г 118. б, в 163. б
29. б, в 74. а 119. а, в, г 164. б, г
30. а, г, д 75. а, в, г 120. г 165. а, в, д
31. б, в, г 76. д 121. б, г, д 166. б
32. а 77. а, б, в 122. а 167. д
33. б, в 78. г, Д 123. а, в 168. а, б, г
34. а, б, г 79. б, в 124. б 169. г
35. б, в, г 80. а, г 125. а, б, в 170. а, б, в, г
36. а, в 81. г 126. в, д 171. в, г
37. а, б 82. б 127. г 172. б, д
38. в 83. в 128. а 173. а, г, д
39. а, б, г 84. г 129. а 174. а, в, г
40. б 85. д 130. г 175. д
41. а, д 86. а, д 131. г 176. г
42. б, г 87. б, в, г 132. б 177. а, г
43. в, г, д 88. а, д 133. г 178. б, в, д
44. а 89. а, б, г 134. а 179. Д
45. а, в, д 90. а, б, г 135. б, д 180. а, г, д
647
648 Приложение Е
181. а, б, в 236. б, В 291. б, в 346. б
182. б, г, д 237. а, в, г 292. а, г, д 347. а, в
183. б, в 238. г, д 293. б, в 348. б
184. б, г, д 239. г 294. а, б, г 349. а. в
185. Д 240. а 295. д 350. в
186. а, в, д 241. б 296. г 351. а
187. б, г, д 242. а 297. б 352. в, д
188. а, б, г 243. г 298. а, в, д 353. г
189. а, б, в 244. а, б, в 299. а, д 354. в, д
190. Д 245. б, г 300. а, б, в 355. а, в
191. б, в 246. а 301. г 356. в
192. в 247. б, г 302. г, Д 357. б, г
193. г 248. б, г 303. г 358. в, г
194. а, б, в, г 249. д 304. д 359. а, б
195. а, д 250. б 305. б 360. г
196. г 251. б 306. в, а 361. б
197. а, г, д 252. б 307. г 362. а, в, д
198. б, г 253. а, г, Д 308. г 363. а, б, г
199. в, д 254. б, г 309. в, д 364. в
200. в, д 255. а, б, в 310. б 365. б, д
201. а, в 256. в 311. г 366. г
202. а, д 257. г 312. б 367. д
203. б 258. в 313. г, д 368. б
204. б, г, д 259. б, г 314. д 369. б, д
205. а, б, в 260. в 315. в 370. б
206. а, б, г 261. д 316. г 371. в
207. Д 262. б, г 317. а 372. а
208. д 263. б 318. б 373. г
209. б, д 264. в 319. в 374. а
210. б 265. а, б, в 320. д 375. а
211. а 266. в 321. г 376. в
212. в, г 267. г 322. г, д 377. в, г
213. д 268. а 323. в 378. в
214. а, в 269. д 324. в 379. в
215. г 270. д 325. б 380. б
216. д 271. а 326. в 381. б
217. а, б, д 272. а, В, д 327. б 382. б
218. г 273. а, б 328. б, г 383. в
219. а, г 274. д 329. б, в 384. а, б
220. в 275. а 330. в 385. г
221. а, б, в 276. в 331. б 386. б
222. б 277. г 332. в 387. г
223. а, б, д 278. а 333. а 388. д
224. а, б 279. а, б 334. б 389. в
225. а, б 280. а, б 335. г 390. б, д
226. в 281. г 336. г 391. б
227. б, г, д 282. а 337. в 392. в
228. б 283. б 338. Г, Д 393. а, б, в
229. д 284. а, б, в 339. б 394. д
230. а, г 285. б, в, г 340. б, в 395. а
231. в, г 286. а 341. в 396. а, б, в
232. а, б, д 287. а, б 342. д 397. а, б, в
233. а, д 288. а 343. а 398. г
234. а 289. а, д 344. г 399. г
235. в 290. а, б 345. а, б 400. б, в, д
Ответы к тестам 649
401. а 426. а 451. а, б 476. д
402. а 427. б 452. а 477. а
403. б 428. а, в, г 453. а, б 478. а, б, в
404. в 429. а, в, д 454. а, б, д 479. а, в, г
405. в 430. д 455. б, г, д 480. а, б, в
406. в 431. г 456. д 481. а
407. а 432. а, б, г 457. в 482. а, г, д
408. в, г 433. а, б, д 458. д 483. в
409. а, в 434. б, в 459. б, в, г 484. г
410. б 435. д 460. д 485. г, Д
411. а, в, д 436. В, г, д 461. а, б, в 486. г
412. а, б 437. а, б, г 462. а, б, в 487. в
413. б, в 438. а, б, г 463. б, в 488. б
414. б 439. а, в 464. а 489. Д
415. а, б 440. а, б, д 465. б, в, г 490. а, б
416. а, в, г 441. в 466. б 491. а, б, в
417. а, в 442. а 467. а, в 492. а, г
418. б, в, д 443. а, г, д 468. в 493. в, г, д
419. а 444. д 469. а, б, в 494. а, в, д
420. д 445. г 470. а, б, в 495. а, г, д
421. а, б, в 446. б 471. а, г, д 496. в
422. а, б, г 447. д 472. а, б, д 497. б
423. а, б 448. д 473. в 498. в
424. в 449. г 474. Д 499. а, б, г
425. а, б, в 450. д 475. а, в, г 500. б
Алфавитный указатель
А
Абразивный преканкрозный хейлит Манганотти 372
Акантолиз 67
Акне (вульгарные угри) 342
акне-тетрада 343
инверсное акне 343
индуративные акне 343
комедоны 342
конглобатные (шаровидные) акне 343
милиумы 342
молниеносные акне 343
папулезные акне 342
пустулезные акне 342
тропические акне 343
узловатокистозные акне 343
флегмонозные акне 343
Альбинизм 21
Альтерация 74
Ангиокератома Мибелли 365
Антибиотикотерапия 106
Апоптоз 33
Атерома 363
Атопический дерматит 246
везикулезно-крустозная форма 253
лихеноидная форма 253
пруриго Г ебры 256
пруригоподобная форма 253
сетка Уикхема 256
эритематозно-сквамозная
с лихенификацией форма 253
эритематозно-сквамозная форма 252
Африканская эндемическая саркома Капоши 386
Афферентная иннервация кожи 49
Аэрозоли 112
Б
Базальноклеточный рак 373
Базальный слой 11
Беджель 422
арабский сифилис 422
эндемический сифилис 422
Белая жировая ткань 45
Биццоцеро узелок 13
Болезнь
белых пятен 301
Боуэна 374
Дарье 78
Дюрана-Никола-Фавра 501
Фабри (диффузная ангиокератома) 365
Бородавки 175
клинические формы 175
Бородавчатый предрак 371
Бразильская пузырчатка 327
бразильская листовидная пузырчатка 327
Бугорок 92
Буллезный
пемфигоид 336
эпидермолиз 394
Бурая жировая ткань 46
в
В-клеточная лимфома кожи 382
Васкулиты кожи 355
Вегетация 98
Вегетирующая пузырчатка 325
Ноймана 325
типа Аллопо 326
Венерическая
гранулема 506
лимфопатия 501
Венерический лимфогранулематоз 501
лимфогранулематозный шанкр 501
Венозная гемангиома 365
Взбалтываемые взвеси 111
Вирус простого герпеса 168
Витилиго 21
ВИЧ-ассоциированная
эпидемическая саркома Капоши 386
ВИЧ/СПИД-инфекция 556
простые бородавки, контагиозный моллюск,
остроконечные кондиломы 565
рецидивирующий опоясывающий лишай 565
саркома Капоши 564
себорейный дерматит 564
СПИД-ассоциированный комплекс 560
СПИД-индикаторные болезни 562
Волдырь 93
Волосы 7
Волосяной сосочек 8
Воспаление кожи 73
Врожденные пигментные невусы 369
Врожденный невоклеточный невус 369
Вторичные морфологические элементы 95
Вульвовагиниты у девочек 550
герпетические 553
гонорейные 553
кандидозный 552
неспецифические 551
специфические 552
трихомонадные 554
хламидийные 554
Вульгарная пузырчатка 321
Высокодифференцированный плоскоклеточный рак 374
650
Алфавитный указатель 651
Г
Гексли средний гранулосодержащий слой 8
Геморрагическое воспаление кожи 78
Генитальный герпес 541
абортивная форма 544
атипичная форма 544
манифестная форма 543
непервичная инфекция 543
первичная инфекция 543
рецидивирующая инфекция 543
субклиническая форма 544
Генле наружный бледный слой 8
Герпетиформный дерматоз Дюринга 331
Гигантские клетки
инородных тел 83
Пирогова-Лангханса 83
Г игантский врожденный невоклеточный невус 370
Гигиена кожи, ногтей, волос 116
Гиперплазия сальных желез 364
Г ипертрофические и келоидные рубцы 363
Гиперчувствительность замедленного типа 81
Гликозаминогликаны 41
Гликопротеины 42
Г нездная алопеция 351
гнездная алопеция с поражением ногтей 352
лентовидная форма (офиазис Цельса) 352
локальная (очаговая) форма 352
нерубцовая 351
рубцовая 351
субтотальная форма 352
тотальная форма 352
универсальная 352
Гнойничок 94
Гнойное воспаление кожи 77
Гонорея 473
диагноз 480
иммуноферментный анализ 481
иммунофлюоресцентный метод 481
кожно-аллергическая проба с гонококковой
вакциной 481
лигазная цепная реакция (ЛЦР) 481
молекулярно-биологические методы 481
полимеразная цепная реакция (ПЦР) 481
реакция Борде-Жангу 481
реакция определения гонококкового антигена 481
классификация 473
клиника гонореи у девочек 479
восходящая гонорея 479
нисходящая гонорея 479
клиника гонореи у женщин 477
бартолинит 478
вестибулит 477
гидросальпинкс 478
гонококковый сальпингоофорит 478
гонорея нижнего отдела половых органов 477
парауретрит 477
пельвиоперитонит 479
пиосальпинкс 478
сальпингит 478
уретрит 477
цервицит и эндоцервицит 478
эндометрит 478
клиника гонореи у мужчин 474
баланит 476
баланопостит 476
везикулит 477
деферентит 476
кавернит 476
колликулит 476
лимфангиит 476
орхит 476
осложнения гонореи у мужчин 476
парафимоз 476
постит 476
простатит 476
свежий острый задний гонорейный уретрит 474
свежий острый передний гонорейный уретрит 474
свежий подострый гонорейный уретрит 475
свежий торпидный гонорейный уретрит 475
стриктуры 476
фимоз 476
фуникулит 476
хронический гонорейный уретрит 475
эпидидимит 476
лечение 481
критерии излеченности гонореи 495
лечение гонореи у беременных 488
лечение гонореи у детей 489
лечение рецидивов гонореи 488
лечение экстрагенитальной гонореи 489
местное лечение гонореи и ее осложнений 491
профилактика гонореи у детей 496
этиология 473
Гормональные средства 105
Г ранулематозное воспаление 83
Г ринстейна клетки 19
д
Дерма 5, 37
Дерматиты 217
аллергический контактный 219
простой ирритантный контактный 218
Дерматозы с неясным патогенезом 378
Дерматологические
лаки 113
пасты 113
пластыри 114
Дерматологический статус 100
Дерматомикозы 189
актиномикоз 214
аспергиллез 207
гистоплазмоз 205
глубокие микозы 204
глубокие псевдомикозы 214
кандидоз 196
келоидный бластомикоз 213
кладоспориоз 208
клинические формы онихомикоза 199
кокцидиоидоз 205
криптококкоз 211
микоз волосистой части головы 193
микоз крупных складок 191
микозы стоп 192
мицетома 209
652 Кожные и венерические болезни
мукороз 210
нокардиоз 215
онихомикозы 197
пенициллиоз 210
поверхностная трихофития
волосистой части головы 195
гладкой кожи 195
разноцветный лишай 190
риноспоридиоз 209
североамериканский бластомикоз 212
споротрихоз 206
трихофития 194
фавус 195
хромомикоз 214
хроническая трихофития взрослых 195
цефалоспориоз 208
южноамериканский бластомикоз 213
Дерматомиозит 309
идиопатический 310
паранеопластический 310
Дерматофиброма 363
Дермо-эпидермальное соединение 34
Диморфная лепра 153
Дискератоз 66
Диспластический невус 370
Дистрофический буллезный эпидермолиз 394
Дисхромия 95
Доброкачественные новообразования кожи 362
Догеля тельца 50
Донованоз 506
веррукозная форма 507
гипертрофический тип 507
некротическая форма 508
склерозирующая форма 508
смешанная форма 508
цветущая форма 507
элефантиазный тип 507
язвенная форма 507
донованоз Брока 506
3
Защитная функция кожи 55
Зернистый слой 14
Злокачественные новообразования кожи 373
и
Иммунное воспаление кожи 79
Иммунокорригирующие препараты 105
Инкапсулированные нервные окончания 50
Иннервация кожи 48
Ихтиозы 390
к
Кавернозная (глубокая) гемангиома 365
Капиллярная гемангиома 364
Кератин 7
мягкий 8
твердый 7
Кератоакантома 362
Кератодермия
меледа 396
Папийона-Лефевра 397
Унны-Тоста 396
Кисты 366
волосяная 366
имплантационная 366
милиум 366
синовиальная 366
эпидермальная 366
Классическая саркома Капоши 385
Классическая форма грибовидного микоза 380
Клетки
Вирхова 151
Лангерганса 15
лепры 151
Меркеля 22, 50
сосочкового слоя 37
Тцанка 323
Клещевой боррелиоз (лайм-боррелиоз) 318
Кожные заболевания в жарких странах 398
Кожный зуд 237
Кожный лейшманиоз Нового Света 165
гвианский кожный лейшманиоз 166
диффузный кожный лейшманиоз 166
кожно-слизистый лейшманиоз Нового Света 166
кожный лейшманиоз ута 165
мексиканский кожный лейшманиоз 165
панамский кожный лейшманиоз 166
Кожный лейшманиоз Старого Света 161
антропонозный кожный лейшманиоз 161
зоонозный кожный лейшманиоз 164
Суданский кожный лейшманиоз 164
Кожный рог 371
Коллагеновые волокна 42
Коллодийный плод 392
Контагиозный моллюск 176
Корка 96
Крапивница 272
аквагенная 274
дермографическая 274
механическая 274
острая 273
световая 274
тепловая 274
холинергическая 274
холодовая 274
Красная волчанка 292
LE-клетки 294
глубокая форма 294
дискоидная 293
диссеминированная 293
опухолевая разновидность 294
симптом Бенье-Мещерского 293
системная 295
центробежная эритема 293
эрозивно-язвенная форма 294
Красный плоский лишай 288
Краузе колбы 50
Крело111
Кремы 112
Алфавитный указатель 653
Кровоснабжение кожи 50
Кутикула 7
л
Ладонно-подошвенный порокератоз 369
Ламеллярный ихтиоз 392
ихтиозиформная сухая эритродермия 392
пластинчатая эксфолиация новорожденных 392
Лейкоплакия 372
Лейкоцитопластический васкулит 80
Лентиго 369
Лепра 149
лепроматозная 151
лепроматозные реакции 155
Ливедо-ангиит 359
Лимфомы кожи 379
Лимфоциты 24
Линеарный порокератоз 369
Липома 364
Листовидная пузырчатка 326
Лихенификация 98
Лишай Видаля 269
Лосьоны 111
Луковица волоса 8
Лучевой дерматит 371
м
Мази 112
Макрофаги 39
Мальпиги ростковый слой 14
Маникюр 117
Маркионини кислотная мантия 6
Медицинская деонтология 115
Межклеточное вещество 41
Мейснера тельца 50
Меланиновая эпидермальная единица 21
Меланома 21, 375
Меланоциты 20
Мелкие врожденные невоклеточные невусы 370
Местные кортикостероидные средства 114
Метод
Богдановича 183
Демьяновича 182
Методы детоксикации 107
Микроспория 193
Многоформная экссудативная эритема 80, 314
инфекционно-аллергическая (идиопатическая) 314
Мышьяковый и дегтярный кератозы 371
Мягкая фиброма 363
н
Невус Унны (врожденная телеангиэктазия затылка) 364
Неинкапсулированные нервные окончания 50
Нейродермит 269
лихенификация гигантская 270
простой хронический бородавчатый лишай 270
Нейрофиброматоз (болезнь Реклингхаузена) 368
Неопределенная лепра 153
Несвободные нервные окончания 49
Низкодифференцированный плоскоклеточный рак 375
Новообразования кожи 361
Ногтевая пластинка 9
Ногтевое ложе 9
Ногтевой желобок 9
Ногти 9
о
Обезглавленная форма грибовидного микоза 381
Обыкновенный ихтиоз 390
Ограниченный нейродермит 269
Ограниченный предраковый гиперкератоз 372
Односторонний эпидермальный невус 368
ОнихобластЭ
Опоясывающий герпес 173
Основное вещество 41
Остроконечные кондиломы 176
Острый лучевой дерматит 371
Отек Квинке 80, 272, 274
гигантская крапивница 274
наследственный 274
острый ограниченный 274
периодический с эозинофилией 274
пигментная крапивница 274
симптом Унны-Дарье 275
Отложения в коже 366
бугорчатые ксантомы 367
кальциноз кожи 366
ксантелазма 367
ксантомы 367
плоские ксантомы на ладонях 367
плоские ксантомы при нормальном
уровне липидов 367
сухожильные ксантомы 367
эруптивные ксантомы 367
Очаговая склеродермия 300
бляшечная склеродермия 301
идиопатическая атрофодермия Пазини-Пьерини 301
линейная (полосовидная) склеродермия 301
ограниченная склеродермия 301
склероатрофический лихен Цумбуша 301
п
Папула 91
Паракератоз 66
Паранеопластическая пузырчатка 329
Паукообразная гемангиома 365
Педикулез 185
головной 186
платяной 187
фтириаз 187
Педикюр 117
Пелоидотерапия (грязелечение) 108
Пемфигоид 80
Первичные морфологические элементы 89
Пигментная ксеродерма 370
Пигментные невусы 369
Пинта 423
хронический невенерический трепонематоз 423
Пиодермии 120
гангренозная (язвенный дерматит) 134
654 Кожные и венерические болезни
пиоаллергиды 135
стафилококковые 121
везикулопустулез (перипорит) 126
гидраденит 125
глубокие стафилодермии 124
карбункул 125
остиофолликулит 122
поверхностные стафилодермии 122
псевдофурункулез Фингера
(множественные абсцессы у 127
сикоз вульгарный 123
стафилодермии у детей 126
фолликулит 122
фурункул 124
эксфолиативный дерматит Риттера 128
эпидемическая пузырчатка новорожденных
(эпидемический пемфигоид) 127
стрептококковые 129
атипичные формы стрептодермий 130
импетиго стрептококковое Тильбери—Фокса 129
острая диффузная стрептодермия 131
папулезное сифилоподобное импетиго 131
простой лишай 131
эктима 130
стрептостафилодермии (смешанные пиодермии) 132
вульгарная эктима 132
вульгарное импетиго (стрептостафилококковое) 132
хроническая диффузная 133
хроническая глубокая 133
пиогенная гранулема (ботриомикома) 134
шанкриформная 134
Плазматические клетки 40
Пламенеющий невус 364
Плоскоклеточный рак 374
Поверхностный диссеминированный порокератоз 368
Пограничный буллезный эпидермолиз 394
Подкожная жировая клетчатка — гиподерма 9,45
Полиморфный дермальный ангиит 357
Порокератоз Мибелли 368
Потовые железы 6
Почесуха(пруриго) 264
нейродермит узловатый 267
пруриго Г айда 267
пруриго простое острое детское 264
пруриго простое подострое 265
пруриго узловатое 267
узловатая почесуха 267
Предраковые заболевания
кожи и слизистых оболочек 370
Приобретенные пигментные невусы 369
Проба
Бальцера 191
Тцанка 101
Продуктивное воспаление кожи 78
Пролиферативный гиперкератоз 65
Пролиферация 76
Простой буллезный эпидермолиз 394
Простой герпес 168
абортивная форма 170
атипичные формы простого герпеса 170
беременных, новорожденных 171
геморрагический и некротический 170
генитальный 170
герпетиформная экзема Капоши 170
герпетический
панариций 170
фолликулит 170
диссеминированная форма 170
зостериформная форма 170
мигрирующая форма 170
осложнения 171
отечная форма 170
первичный герпес 169
рецидивирующий герпес 170
рупиоидная форма 170
хронический кожный 170
элефантиазоподобная форма 170
язвенный герпес на гениталиях 170
Простые формы буллезного эпидермолиза 395
Противовоспалительные
и сенсибилизирующие препараты 104
Противозудные средства 114
Профессиональная экзема 235
Профессиональные
гиперкератозы и папилломатозы 235
дерматозы 235
изъязвления 235
фолликулиты 235
фотодерматозы 235
аллергические дерматиты 235
простые (контактные) дерматиты 235
Псориаз 277
генерализованный пустулезный 278
дежурные бляшки 279
застарелый 280
изоморфная реакция Кебнера 283
интертригинозный 279
ладонно-подошвенный пустулезный 278
нейтрофильные микроабсцессы Мунро 282
обычный (вульгарный) 278
псориатическая артропатия 278
псориатическая эритродермия 278
пустулезный генерализованный Цумбуша 282
пустулезный 278
рупиоидный 280
себорейный 279
феномен
auspitz или кровяной росы 279
Кебнера 279
стеаринового пятна 278
терминальной пленки 278
точечного кровотечения 279
экссудативный 279
эритродермия 279
Пузырек 93
Пузырчатка 80
хроническая врожденная семейная 330
болезнь Хейли-Хейли 330
Пузырь 93
Пурпура Шенлейн-Г еноха 80
Пятно 89
655
Р
Растворы 111
Реакции Мицуда 150
Реакция
«трансплантат против хозяина» 85
Фернандеса 150
Ретенционный гиперкератоз 65
Ретикулиновые волокна 44
Роговой слой 14
Розацеа 345
грамнегативная 349
гранулематозная 349
конглобатная 349
люпоидная 349
молниеносная 349
ринофима 348
лимфоэдема 349
стероидпровоцированная 349
Розовый лишай 291
Рубец 97
Рубцующийся пемфигоид 338
буллезный дерматит слизистых 340
доброкачественный пемфигоид
слизистых оболочек 340
Руффини колбы 50
с
Сальные железы 6
Саркома Капоши 377, 385
Свободные нервные окончания 49
Себорея 341
Серозное воспаление кожи 77
Сетчатый слой дермы 44
Симптом
ГоттронаЗЮ
Пильнова 283
Уикхема 289
Синдром
larva migrans 413
senear-Usher 328
Лайелла 80, 315
Рейно 302, 304
Сезари 381
Стивенса-Джонсона 315
Элерса-Данло 71
Сирингома 364
Системная склеродермия 302
диффузная 303
критерии 306
главный 306
малые 306
лимитированная 303
Сифилис 430
атипичный шанкр 434
баланит434
баланопостит 434
бугорковые сифилиды 442
диффузный 443
карликовый 443
сгруппированный 442
серпигинирующий (ползучий) 443
висцеросифилис 450
кардиоваскулярный сифилис 451
поздние формы 451
ранние формы 450
сифилис желудка 452
сифилис легких 452
сифилис печени 452
сифилис почек 452
врожденный 431,452
поздний 453
ранний 452
вторичный 431, 436
гангренизация 434
гуммозные сифилиды 444
гуммозные инфильтраты 444
изолированные гуммы 444
фиброзные гуммы 444
индуративный отек 434
лабораторная диагностика 453
RPR тест 455
TRUST тест 455
USR тест 455
VDRL тест 455
бактериоскопическое исследование 454
иммуноферментный анализ 458
метод иммуноблотинга 458
метод непрямого ИФА с кардиолипиновым
антигеном 455
микро-РПГА 457
микрореакция с плазмой и инактивированной
сывороткой 455
нетрепонемные тесты 454
осадочные реакции Кана и Закс-Витебского 456
полимеразная цепная реакция 458
реакция Вассермана 456
реакция иммобилизации
бледных трепонем (РИТ) 457
реакция иммунного прилипания (РИП) 457
реакция иммунофлюоресценции (РИФ) 456
реакция иммунофлюоресценции-абсорбции
с бледной трепонемой 457
реакция связывания комплемента с
кардиолипиновым антигеном 455
серологические исследования крови 454
трепонемные тесты 455
лечение 459
клинико-серологический контроль
после окончания лечения 467
критерии излеченности больных сифилисом 468
лечение и профилактика сифилиса у детей 468
превентивное лечение беременных 467
профилактическое лечение беременных 467
снятие с учета 468
специфическое лечение беременных 464
лимфаденит регионарный 434
нейросифилис 447
гумма головного или спинного мозга 450
менингоневротическая форма сифилитического
менинги 448
острый генерализованный сифилитический
менингит 447
поздний диффузный менинговаскулярный
сифилис 448
656 Кожные и венерические болезни
поздний нейросифилис 448
поздний скрытый сифилитический менингит 448
прогрессивный паралич 449
ранний менинговаскулярный сифилис 448
ранний нейросифилис 447
сифилис сосудов мозга 449
сифилитическая гидроцефалия 448
сифилитический менингомиелит 448
скрытый сифилитический клинически
бессимптомный менингит 447
сухотка спинного мозга 449
неуточненный сифилис 446
общая патология 432
папулезные сифилиды 437
ладонно-подошвенный 438
лентикулярный 437
милиарный (мелкопапулезный) 437
мокнущий 438
нумулярный (монетовидный) 438
широкие кондиломы 438
парафимоз 434
первичный 431, 433
поражение костей и суставов 446
поражения слизистых оболочек 441, 445
профилактика 469
индивидуальная 469
общественная 469
современный подход 470
пустулезные сифилиды 439
импетигинозный 439
оспенновидный 439
рупиоидный 440
угревидный 439
эктиматозный 439
пятнистый (сифилитическая розеола) 436
сифилитическая алопеция 440
сифилитическая лейкодерма 440
склераденит регионарный 434
скрытый 431, 446
твердый шанкр 433
третичный 431, 442
фагеденизм 434
фимоз 434
формы 431
шанкр-амигдалит 434
шанкр-панариций 434
эпидемиология 432
этиология 430
Склеродермоподобная форма 373
Смешанные бактериальные инфекции 398
Солнечный кератоз (актинический кератоз,
сенильная кератома 371
Сосудистые новообразования (гемангиомы) 364
Спонгиоз 67
Среда NNN (Novi — Мак-Нила — Николля) 161
Средства метаболической терапии 107
Средства системной терапии и профилактики
герпетической инфекции 106
Старческая кератома 362
Старческие гемангиомы 365
Стратификация эпителия 4
Субэпидермальное сплетение 36
т
Т-клеточные и В-клеточные лимфомы 380
Тела Дарье 66
Телеангиэктатическая гранулема 365
Тест на лепромин 150
Тканевые базофилы 25, 38
Токсикодермия 80
Трещина 96
Трихогиалин7
Трихомониаз 510
мочеполовой у девочек 512
острый у женщин 512
трихомонадный уретрит у мужчин 511
Тропическая язвенная гранулема 506
Тропические пиодермии 398, 399
аингум 403
акне-келоид 399
амебиаз кожи 410
анкилостомидозы кожи 412
аскаридоз 412
бартонеллез 402
бацилла Клебса-Леффлера 403
болезнь Карриона 402
дифтерия кожи 403
истинная тропическая язва (фагеденическая
тропическая язва, джунглевая язва и др.) 401
кератомикозы 406
кожные гангрены 400
кожные проявления других бактериальных
инфекций 403
кожный лейшманиоз 408
американский лейшманиоз 408
суданский (египетский) кожный лейшманиоз 409
коралловая язва 402
лепра 409
лихорадка Оройя 402
мадагаскарская нома 401
малярия 410
множественная гангрена
взрослых 400
детского возраста 400
молниеносная гангрена наружных половых
органов 400
недостаток витаминов 419
гиповитаминоз А 419
эксфолиативные хейлиты 419
новообразования кожи 419
нома 400
отрубевидный лишай 406
перуанская бородавка 402
пиоз Мансона 399
подкрыльцовый трихомикоз 408
Пьедра (трихоспория узловатая) 407
сап 404
сибирская язва 404
спарганоз (дифиллоботриоз) 411
стронгилоидоз 412
токсоплазмоз 410
трипаносомоз 409
американский 409
Алфавитный указатель 657
африканский 409
трихинеллез 413
тропикалоидная язва (язва Кастеллани, язва
оазисов) 401
тропические буллезные дерматозы 419
бразильская эксфолиативная пузырчатка 419
истинная пузырчатка 419
Онъялаи 419
тропические зудящие дерматозы
временная почесуха 418
тропическая потница 418
тропический помфоликс 418
флеботодермия 418
харара 419
тропические микозы 405
тропические угри 399
тропические язвы 401
тропический белый лишай 407
тропический везикулезный акродерматит
Кастеллани-Челмера 399
тропический желтый лишай 407
тропический пиомиозит (тропический
дерматомиозит) 400
туберкулез кожи 409
туляремия 405
цистицеркоз кожи 411
черепицеобразный микоз 406
черный лишай 406
шимбери 406
шистосомозы 410
энтеробиоз 412
эпидермомикозы 408
эризипелоид404
эхинококкоз 411
южноамериканская ложная бородавка 403
язва Бурули (язва микобактериальная) 402
язва пустынь (степная язва, септическая
язва и др. 402
Туберкулез кожи 137
диссеминированные формы туберкулеза 143
диспансерное наблюдение 148
индуративный 144
лечение 145
лихеноидный или лишай золотушных 144
острый милиарный 143
папулонекротический 143
профилактика 148
экспертиза трудоспособности 148
локализованные формы туберкулеза 138
бородавчатый 141
колликвативный (скрофулодерма) 141
люпус-карцинома 140
первичный 138
плоская форма 140
саркоидоподобная форма 140
симптом Поспелова 139
симптом «яблочного желе» 139
туберкулезная волчанка 139
туберкулезная волчанка слизистых оболочек 140
эксфолиативная форма 140
язвенная форма 140
язвенный туберкулез кожи и слизистых
оболочек 142
Туберкулиновый тест по Манделю-Манту 151
Туберкулоидная лепра 152
У
Узел 92
Узелки доильщиц 177
паравакцина Липшютца 177
Узелковый некротический васкулит
Вертера-Дюмлинга 80
Узловато-язвенный ангиит 359
Урогенитальный кандидоз 533
кандидозный
баланопостит 536
вагинит 535
вульвит 535
уретрит 536
уретрит и цистит 536
эндоцервицит 536
Урогенитальный микоплазмоз 528
Урогенитальный хламидиоз 516
болезнь Рейтера 522
хламидийный
бартолинит 520
вагинит 520
везикулит 519
орхиэпидидимит 520
пельвиоперитонит 522
простатит 519
сальпингит 521
сальпингоофорит 521
уретрит 518, 520
цервицит 520
эндометрит 521
эпидидимит 519
хламидиоз прямой кишки 522
ф
Фатер-Пачини тельца 50
Феномен Lucio 152
Феномен Никольского 1322
Феномен Никольского II322
Фибробласты 37
Филяриатозы413
аллергодерматозы 418
аргасовые клещи 415
болезнь ловцов губок (коралловые язвы) 417
бругиоз414
вухерериоз 413
вши 416
гамазовые клещи 415
гирудиноз417
гусеницы некоторых бабочек 416
дерматобиаз (южноамериканский миаз) 417
дракункулез (Ришта) 414
жуки 416
иксодовые клещи 415
клещи-краснотелки 415
клещи-железницы 416
кордилобиаз (африканский миаз) 417
лоаоз414
миазы 417
658 Кожные и венерические болезни
многоножка 416
моллюски 417
нейродермит 418
онхоцеркоз 414
пауки 415
песчаная (земляная) блоха 416
постельные клопы 416
псориаз 418
пузатый клещ 415
ракообразные 415
скорпионы 415
тропические зудящие дерматозы 418
тропический плоский лишай 418
тунгиаз (саркопсилез) 416
укусы блох 416
укусы двукрылых 416
укусы пчел, ос, шершней, муравьев 416
«Целующие» клопы 416
экзема 418
ядовитые змеи 417
Формы меланомы 375
акральная лентигинозная меланома 375
лентигО’Меланома 375
поверхностно распространяющаяся 375
узловая меланома 375
Фототерапия 108
Фрамбезия 420
Функции кожи 55
X
Х-сцепленный ихтиоз 391
Химиотерапевтические средства 106
Хроническая пигментная пурпура 358
Хронический лучевой дерматит 371
Хроническое воспаление кожи 78
ч
Чесотка 179
норвежская 185
псевдочесотка 185
узелковая 184
нераспознанная 184
чесоточный ход 180
Чешуйка 96
ш
Шампуни 117
Шанкроид 497
мягкий шанкр 497
воронкообразный 498
гангренозный 498
дифтероидный 498
импетигинозный 498
серпингинозный 498
смешанный 498
фолликулярный 498
Шиповатый слой 13
э
Экзема 222
дисгидротическая 227
ладоней 224
монетовидная 227
острая 222
подострая 223
себорейная 229
сухая 225
хроническая 224
Экскориация 96
Экскреторная функция кожи 58
Экссудация 74
Эластические волокна 43
Эмульсии 111
Эндемические трепонематозы 420
Эпидермальная пролиферативная единица 27
Эпидермальные невусы 368
Эпидермальный тимоцитактивирующий фактор 59
Эпидермис 4, 10
Эпидермолитический ихтиоз 393
врожденная буллезная ихтиозиформная
эритродермия Брока 393
ихтиоз буллезный 393
Эпонихий 9
Эритематозная пузырчатка 328
эритродермическая форма грибовидного микоза 381
Эритроплазия Кейра 374
Эрозия 96
Эфферентная иннервация кожи 49
я
Язва 97
Язва чиклеро 165
Ятрогенная иммуносупрессивная саркома Капоши 386
Иностранные термины
A. flavus 207
A. niger207
Actinomyces albus Rossi — Doria 214
Actinomyces bovis 214
Actinomyces israeli 214
Ancylostoma braziliense 413
Ancylostoma caninum 413
Ankylostoma duodenale 412
Ascaris lumbricoides 412
Aspergillus f umigatus 207
Bacillus anthracis 404
Bartonella baciliformis Barton 402
Blastomyces dermatitidis Gilchrist 212
Borrelia afrelii 318
Borrelia burgdorferi 318
Borrelia garinii 318
Botryumyces caespitosus 214
Brugia malayi414
C. crusei 196
C. pseudotropicalis 196
C. tropicalis 196
Алфавитный указатель 659
Calymmatobacterium granulomatis 506
Candida albicans 196, 533
Cephalosporium acremonium Corda 208
Chlamydia trachomatis 501,516
Cladosporium carrionii 214
Cladosporium mansonii 208
Cladosporium trichoides 208
Coccidioides immitis 205
Corynebacterium diphtheriae 403
CREST-синдром 304
Cryptococcus neoformans 211
Demodex brevis 416
Demodex folliculorum 416
Dracunculus medinensis 414
Edwardsiella lineata 417
Enterobius vermicularis 412
Epidermophyton floccosum 191,197
Erysipelothrix rhusiopathiae 404
Exophiala werneckii 406
Filaridae413
Fonsecaea compacta 214
Fonsecaea pedrosoi 214
Glenosporella lobio 213
Haemophilus ducreyi 497
Hendersonula toruloidea 198
Herpes zoster 565
Histoplasma capsulatum 206
Human Immunodeficiency Virus 556
L. braziliensis 165
L. cutis Americana 408
L. mexicana 165
L. nilotica 409
L. peruviana 165
Leishmania tropica 161
Loaloa414
Madurella grisea 209
Madurella mycetomatis 209
Malassezia spp. 190
Micromonospora parva 214
Microsporum audouinii 193
Mucoraceae210
Mycobacterium leprae 149
Mycobacterium tuberculosis 137
Mycoplasma genitalium 528
Mycoplasma hominis 528
Necator americanus 412
Neisseria gonorrhoeae 473
Nocardia (Corynebacterium) tenuis Castellani 408
Nocardia asteroides 215
Nocardia brasiliensis Castellani 215
Nocardia madurae 215
Ornithodorus415
P. falciparum 410
P. malariae410
P. ovale 410
P. vivax410
Paracoccidioides brasiliensis 213
Pasteurella tularensis 405
Pediculus humanus capitis 185
Pediculus pubis или phthirus pubis 185
Pediculus vestimentorum 185
Penicillium crustaceum Fries 210
Penicillium glaucum Link 210
Phialophora verrucosa 214
Piedraia hortai (черная пьедра) 407
Pseudallescheria boydii 209
Pseudomonas mallei 404
Rhinocladiella aquaspersa 214
Rhinosporidium seeberi 209
Richomonas vaginalis 510
Sarcoptes scabiei 179
Schistomatidae410
Scitalidium hialinum 198
Scopulariopsis brevicaulis 198
Sporothrix schenckii 206
Strongeloides stercoralis Bavayi 412
T. bejel 420
T. carateum 420
T. pertenue 420
Treponema carateum 423
Treponema pallidum 430
Treponema pallidum (bejel) 422
T rich inella spiralis 413
Trichophyton albicans Nieuwenhuis 407
Trichophyton concentricum 406
Trichophyton mentagrophytes 197
Trichophyton quickeanum Zopf 195
Trichophyton rubrum 197
Trichophyton schoenleinii 195
Trichophyton scholeinii 193
Trichophyton tonsurans 195
Trichophyton violaceum 193
Trichosporum cerebriforme (белая пьедра) 407
Trichosporum giganteum 407
Uncinaria stenocephala 413
Ureaplasma urealyticum 528
Varicella zoster 173
Wuchereria bancrofti 413
Endodermophyton Roquettei Fonseca 406
Entamoeba histolytica 410
Mycobacterium ulcerans 402
Hormodendron Fontoynonti 407
Phlebotomus dermia 417
Toxoplasma gondii 410
Язвенная ниодоркия
Сгорлтсakcihciuou икпетиго
Кандидоз кожи области ко. юч>-их желез
Микроспория гладкой кожи
Контактные Сул, >езный дерматит
Ко-1гад1ньй буллезный дезмятит
Солнечная экзема
Микробная экзема
Тилотическая экзема
Хроническая экзема кистей
Дисгидротическая экзема
Контактный буллезный дерма шт
Обыкновенные бородавки
Аногенитальные бородавки Ошроконечные кондиломы
Генитальный герпес
Яадонмая илерлимеармость
(больной атопическим дерматитом)
Лихенизадея кожи (атопический дерматит)
Белый дермографизм
у больного атопическим дерматитом
Атопический жейлит
Пруриголодобная форма
атопического дерматита
Токсццермия, буллезная форма
Ограниченный нейродермит
половых органов
Фотодерматит
Хроническая рецидивирующая крапивница
Токсидсрмия
Фоп>дерма|И1 лица и груди
Пеорияшчаская бляшка
Пустулезный псориаз ладоней
Пеория1ич«ская эритродермии Псориатическая эритродермии
Каплевидный псориаз
Псориатическая эритродермии
Красный плоский лишай
Красный плоский лишай
Красный плоский лишай, линейная форма
Линейная склеродермия по типу «удара саблей»
Дискоидная красная волчанка
Эритематоз волосисюй части головы
Дискоидная красная волчанка
Ограниченная склеродермия, бляшечная форма
Многоформная экссудативная эритема
Многоформная экссудативная эритема
Хроническая мигрирующая Гнрпетиформный дерматит Дюринга
зриIнма {Лайм-боррелиоз) --------------------
Герпетиформный дерматит Дюринга I нрлетиформный дерматит Дюринга
Г эрпетиформный дермагиг Дюринга
Пузырчатка вульгарная
Пузырчатка вульгарная
Жирная себорея лица
Лкнс ю юшсскис, узловато-кистозная форма
Периорапьный дерматит
Акне юношеские, узловато-кистозная форма,
вторичные дисхромии. атрофические рубцы
Диффузная алопеция
Андрогенетическая алопеция
Гнездная алопеция
Розацеа
Периоральпый дерматит
Базальноклеточный рак Невус Беккера
Плоскоклеточный рак
Саркоидоз кожи щек, базапиома
области носа (после лечении)
Базалиома
Базалио м а
Липоидный некробиоз
Голубой невус
Пигментный невус
Подошвенная кератодермия
Врожденная ихтиозиформная эритродермия
Врожденный обыкновенный ихтиоз
Болезнь Дарье
Келоидные рубцы
Кольцевидная гранулема
Учебное издание
Адаскевич Владимир Петрович
Козин Владимир Михайлович
Кожные и венерические болезни
Редакторы: Б. И. Чернин, Ф. И. Плешков, Н. П. Базеко
Оригинал-макет: П. С. Скакун
Обложка: И. В. Курильчик
Подписано в печать с готовых диапозитивов 02.06.2006. Формат 84х 108*/|б.
Печать офсетная. Бумага офсетная. Гарнитура «Тип Таймс».
Усл. печ. л. 70,56. Уч.-изд. л. 52,97. Тираж 2000 экз. Заказ 1357.
ООО «Медицинская литература». Лицензия ЛР № 065380.
117071, г. Москва, ул. Орджоникидзе, д. 13/2.
При участии издателя Б. И. Чернина.
Лицензия №02330/0133039 от 30.04.04.
210001, г. Витебск, ул. Кирова, 1-3.
Республиканское унитарное предприятие
«Издательство «Белорусский Дом печати».
220013, Минск, пр. Независимости, 79.